/





























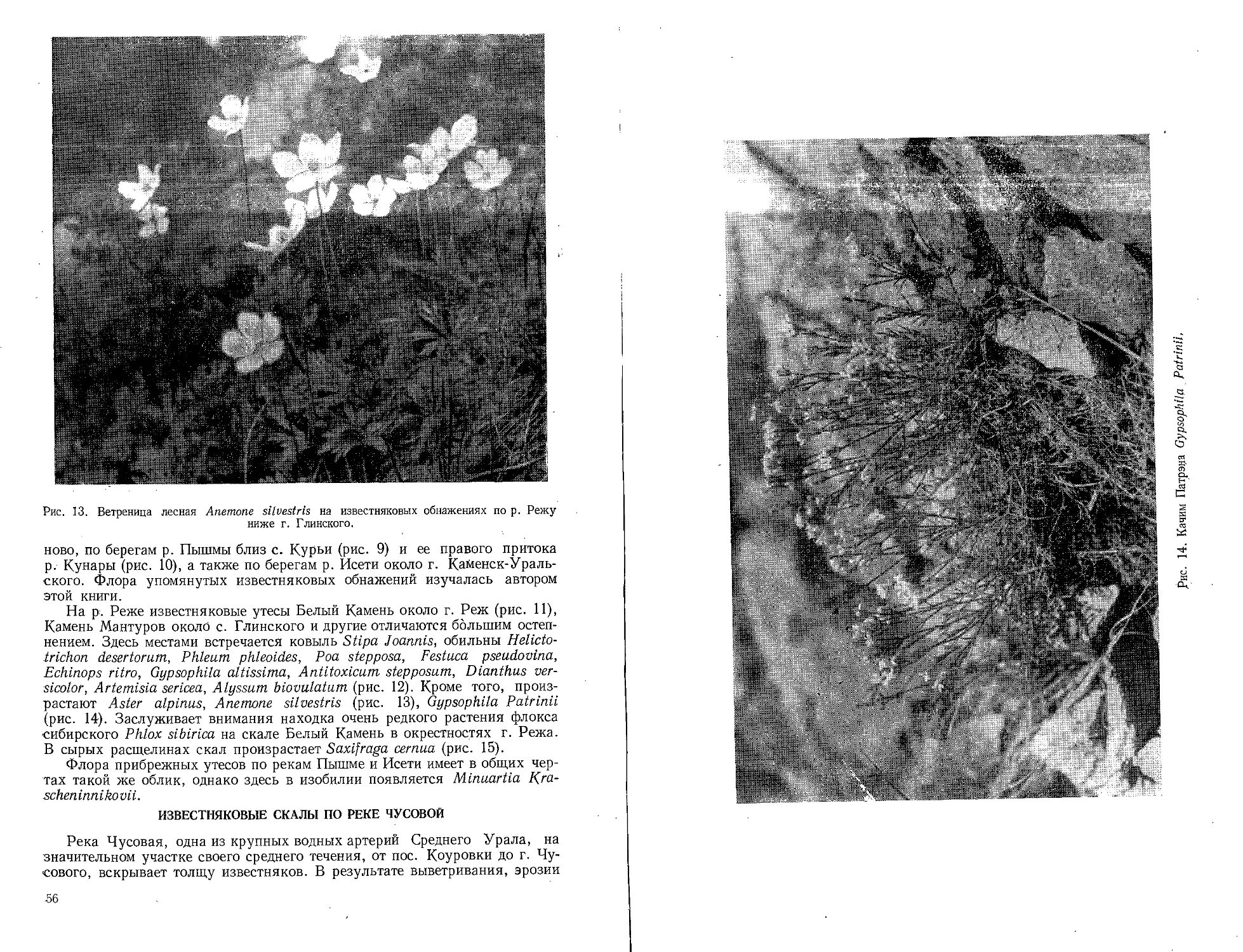


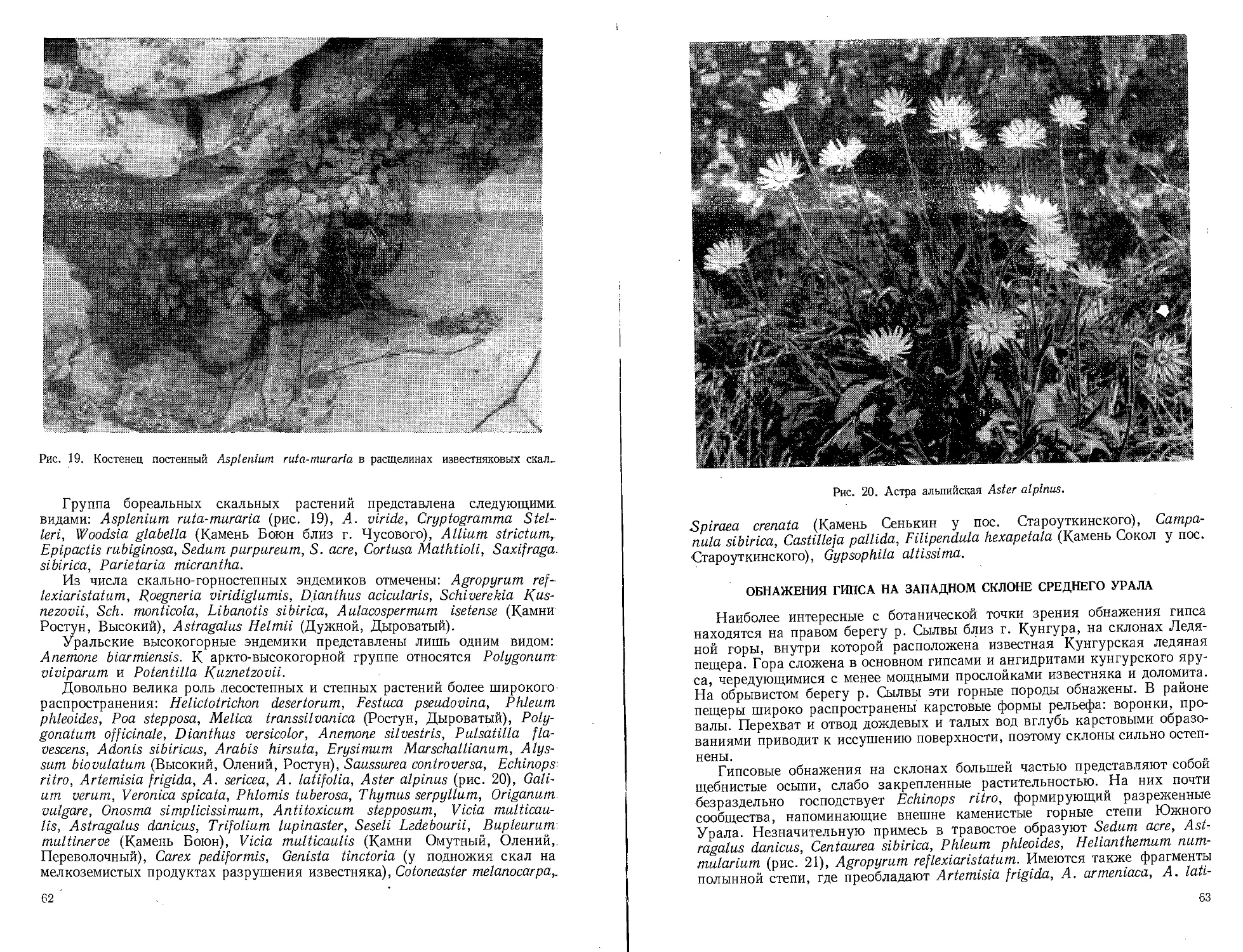




































































































Text
П. Л. ГОРЧАКОВСКИЙ
ОСНОВНЫЕ
ПРОБЛЕМЫ
ИСТОРИЧЕСКОЙ
ФИТОГЕОГРАФИИ
УРАЛА
СВЕРДЛОВСК
1969
АКАДЕМИЯ НАУК СССР
УРАЛЬСКИЙ ФИЛИАЛ
П. Л. ГОРЧАКОВСКИЙ
ОСНОВНЫЕ
ПРОБЛЕМЫ
ИСТОРИЧЕСКОЙ
ФИТОГЕОГРАФИИ
УРАЛА
СВЕРДЛОВСК
1969
Труды Института экологии растений и животных
УФАН СССР (выпуск 66)
Печатается по постановлению
Редакционно-издательского совета
Уральского филиала АН СССР
Ответственный редактор С. А. Мамаев
ВВЕДЕНИЕ
Для ботанико-географа Урал интересен во многих отношениях. Это свое-
образная горная страна, протянувшаяся почти в меридиональном направлении
более чем на две тысячи километров и пересекающая ряд растительных зон,
от тундровой на севере до степной на юге, с ярко выраженной поясностью
растительности.
В течение своей длительной геологической истории, особенно в кайнозое,
Урал был ареной сложных преобразований растительного покрова, связанных
с процессами горообразования, трансгрессиями морей, четвертичным оледе-
нением, изменениями климата, а затем и воздействием человека. Пограничное
положение Урала и значительная протяженность с севера на юг определили
еще одну его особенность: это область столкновения различных флорогене-
тических элементов — сибирских, европейских, арктических, среднеазиатских.
На поверхности Уральских гор большую площадь занимают выходы
разнообразных горных пород, находящихся в разных стадиях выветрива-
ния и закрепления их растительностью. Вследствие эпейрогенических под-
нятий и эрозии поверхность горных пород непрерывно обнажается, образуя
свободный субстрат, более или менее пригодный для заселения растениями.
На скалистых обнажениях деревья произрастать не могут или встречаются
лишь одиночно. Отсутствие конкуренции со стороны древесной раститель-
ности, разнообразие условий увлажнения и освещения благоприятствовали
сохранению на скалистом субстрате нескольких наслоений реликтовых ком-
плексов растений.
В неогене и антропогене Урал явился ареной формообразования, центром
формирования флористического эндемизма и расселения растений. Появлению
эндемиков благоприятствовало разнообразие субстрата и ряд исторических
причин.
Все эти особенности определили пестроту и сложную картину распреде-
ления растительного покрова. Выявление закономерностей распределения
растений на Урале, изучение эндемиков и реликтов проливает свет на исто-
рию формирования растительного мира этой горной страны. А это необходи-
мо для лучшего понимания современного растительного мира, его рациональ-
ного использования и преобразования.
История развития растительного мира Урала, как и других территорий,
изучалась методами палеоботаники. Однако палеоботанические данные сами
по себе, вследствие их неизбежной неполноты, недостаточны для решения
всех вопросов истории флоры и растительности. К тому же палеоботаниче-
ские исследования проводились обычно в отрыве от историко-фитогеографи-
ческих.
Для познания генезиса флоры и растительности большую ценность
представляет выявление закономерностей современного распространения рас-
тений (Вульф, 1932, 1944; Лавренко, 1938; Попов, 1923, 1927). При этом,
конечно, ареалогические данные должны быть, увязаны с палеоботаниче-
скими.
Впервые некоторые специфические черты флоры Урала были отмечены
классиком отечественной фитогеографии С. И. Коржинским (1894), указав-
3
шим на встречаемость здесь ряда эндемичных и реликтовых растений.
В дореволюционное время вопросы генезиса флоры Урала освещались также
в работах А. Я. Гордягина (1895).
Яркий след в познании происхождения флоры и растительности Урала
оставили работы выдающегося ботаника и географа И. М. Крашенинникова
(1937, 1939), уделившего много времени и внимания изучению этой горной
страны и сопредельных территорий. Труды И. М. Крашенинникова послужили
толчком для дальнейших исследований в этом направлении. Исторический
подход к современной флоре и растительности Урала и примыкающих рав-
нин нашел отражение в работах М. М. Ильина (1922), И. В. Сюзева
(1921, 19236), Б. Н. Городкова (1926, 1929, 1939, 1947, 1952), В. С. Го-
ворухина (1929), Ю. Д. Клеопова (1941), В. Б. Сочавы (1929, 1945),
К. Н. Игошиной (1943, 1964, 1966), А. Н. Пономарева (1938, 1945, 1948,
1949), Ю. П. Юдина (1963), П. Л. Горчаковского (1951, 1954а, б, 1955,
1956, 1957а, б, 1958а, б, 1960а, 1962а, б, 1963, 1964, 1966, 1967) и др.
Несмотря на то, что многочисленными исследователями уже накоплен
большой фактический материал по фитогеографии Урала, он еще недостаточ-
но обобщен и интерпретирован. Многие вопросы разработаны слабо, а факти-
ческая аргументация недостаточна.
К числу основных проблем исторической фитогеографии Урала и сопре-
дельных территорий на современном этапе относятся: а) выявление специ-
фики и происхождения скальной флоры; б) эндемизм уральской флоры, его
уровень, время и пути возникновения эндемиков; в) реликты во флоре Урала,
их возраст и происхождение; г) этапы формирования флоры и раститель-
ности (по материалам изучения современного распространения растений с
учетом палеоботанических данных).
Обсуждение этих вопросов и составляет содержание предлагаемой книги.
В основу ее положены итоги исследований, проводимых автором в течение
ряда лет (1946—1967 гг.) на территории Урала и прилегающих к нему
территорий Восточно-Европейской и Западно-Сибирской равнин и Казахстана.
Предварительные результаты освещались в ряде ранее опубликованных работ.
Здесь эти материалы обобщены, изложены более полно, с учетом накопив-
шихся за последнее время дополнений и поправок, а также новых литера-
турных данных.
Автор посвящает свой скромный труд памяти выдающихся ботаников —
С. И. Коржинского и И. М. Крашенинникова, заложивших основы истори-
ческого подхода к познанию флоры и растительности Урала.
Если эта книга пробудит у начинающих ботаников интерес к вопросам
флорогенетики и послужит стимулом для дальнейших изысканий в этой
области, автор будет считать свою задачу выполненной.
ГЛАВА I
ОСНОВНЫЕ ЭТАПЫ ФОРМИРОВАНИЯ ФЛОРЫ
И РАСТИТЕЛЬНОСТИ УРАЛА В КАЙНОЗОЕ
(ПО ПАЛЕОБОТАНИЧЕСКИМ ДАННЫМ)
В палеоботаническом отношении Урал оказался изученным сравнительно
хорошо, так как здесь производились большие работы по поискам месторож-
дений полезных ископаемых и изучению геологических структур, а 'поэтому
для стратиграфии потребовались сборы и определение большого количества
растительных остатков. На протяжении многих лет эта горная страна прив-
лекала внимание крупнейшего палеоботаника А. Н. Криштофовича (1933,
1935, 1938а, б, 1950, 1957 и др.). Позднее ряд публикаций по третичной
и четвертичной флоре Урала дали П. И. Дорофеев (1950, 1952) и
Т. Д. Колесникова ( 1957, 1963, 1966). Г. А. Благовещенский (1940, 1943)
и В. Н. Сукачев (Сукачев и Поплавская, 1946) изучали пыльцу растений
в болотных и озерных отложениях, что способствовало расшифровке исто-
рии растительности Зауралья в плейстоцене и голоцене. Для определения
возраста и условий образования различных более древних, в том числе
третичных отложений Урала и Предуралья, пыльцевой анализ применяли
И. М. Покровская (1950), А. А. Чигуряева (1952) и их сотрудники. Накоп-
ленная палеоботаническая документация (отпечатки и полуразложившиеся
остатки листьев, окаменевшая древесина, остатки плодов, семян, спор,
пыльцы и т. п. в последовательно сменяющихся стратиграфических горизон-
тах на большой территории) имеет огромную познавательную ценность. Она
дает возможность объективно судить об изменениях флоры и растительности,
происходивших в кайнозое на территории Урала и прилегающих равнин.
ТРЕТИЧНЫЙ ПЕРИОД (ПАЛЕОГЕН И НЕОГЕН)
В течение третичного периода в основном установились современные
очертания материков (Съюорд, 1936; Криштофович, 1941; Страхов, 1948;
Марков, 1951). Североатлантический материк, соединявшийся вначале с Ев-
ропой, с середины эоцена отделился от нее Атлантическим океаном. Позд-
нее, в конце нижнего олигоцена, Северная Америка и Европа, вероятно,
вновь кратковременно соединились и снова создалась возможность для
обмена флорами. В Европе в палеогене было два погружения, сопровождав-
шихся трансгрессией моря. Первое из них совпало с палеоценом, второе — с
концом эоцена и началом олигоцена. Во время второго погружения море
залило области Альп, Карпат, Южных Пиренеев, Русской платформы и ши-
роко распространилось в Западно-Сибирской низменности. С середины оли-
гоцена море стало быстро отступать. В продолжение почти всего третичного
периода Азия в районе современного Берингова пролива и южнее его смы-
калась с Северной Америкой. Великое средиземное море Тетис в палеогене
занимало территорию Южной Европы, юг европейской части СССР и значи-
тельную часть Центральной Азии. В миоцене площадь Тетиса значительно
5
сократилась. D вистичнии tilu HaviH iu.u __ —r.
море, занимавшее территорию Черного и Каспийского морей и Северного
Кавказа. Позднее, в плиоцене, обособились Черноморский и Каспийский
бассейны. Современные Средиземное, Черное и Каспийское моря—остатки
некогда огромного Тетиса.
С первой половиной третичного периода — палеогеном — связано образование
молодых складчатых сооружений: Альп, Пиренеев, Карпат, Крыма, Кавказа,
Копет-Дага и других. Позднее, в неогене, возникли многочисленные разло-
мы земной коры в Азии. Подъем некоторых глыб, отграниченных от сосед-
них расколами, привел к «омоложению» рельефа и формировнию гор
Тянь-Шаня, Тарбагатая, Алтая, Саян, Яблонового и Станового хребтов.
Большого и Малого Хингана, Сихотэ-Алиня и ряда хребтов Центральной
Азии. Формирование рельефа этих молодых азиатских глыбовых поднятий
продолжалось и в течение следующего, четвертичного периода.
Урал в палеогене омывался с востока водами Тургайского моря, занимав-
шего тогда всю территорию Западно-Сибирской низменности. Лишь в конце
олигоцена море, в связи с резким поднятием материка, отступило на юг,
к Аральскому бассейну. Поднявшаяся с морского дна суша представляла
собой слегка всхолмленную равнину с многочисленными замкнутыми водое-
мами (озерами, болотами.)
К западному склону Урала и в палеогене примыкала суша. В жаркую
эоценовую эпоху в Предуралье от Мугоджар до р. Вишеры были распрост-
ранены подвижные пески (Наливкин, 1943). Режим суши сохранился здесь
и в неогене. Лишь в конце плиоцена вдоль рек Камы и Белой в Предуралье
вторгся залив так называемого Акчагыльского моря, представляющего собой
разросшийся и значительно продвинувшийся к северу Каспийский бассейн.
Вековые процессы разрушения горных пород привели к тому, что Урал
ко второй половине третичного периода превратился в равнину с разбросан-
ными по ней небольшими всхолмлениями. Горы, возникшие в конце палео-
зоя, были почти сглажены. Поверхность пересекали медленно текущие реки,
развившиеся позднее в ныне существующую речную сеть. Многочисленны
были озера, заросшие по краям густой растительностью.
В конце третичного периода происходило постепенное поднятие древних
палеозойских массивов. Поверхность усиленно размывалась речными руслами,
рельеф стал более расчлененным. Еще большей интенсивности молодые глы-
бовые поднятия достигли в начале антропогена.
Третичный период ознаменовался особенно широким распространением
покрытосемянных растений. К этому времени сформировались уже все основ-
ные систематические группы покрытосемянных, но географическое распрост-
ранение растительности еще имело очень мало общего с наблюдаемым в
настоящее время. В первой половине третичного периода (палеоген) на тер-
ритории Европы и Северной Азии существовали, цо А. Н. Криштофовичу
(1936, 1957), два основных флористических комплекса:
1) «Полтавская» флора. Субтропическая, а южнее даже тропическая,
в основном вечнозеленая. Для нее характерно присутствие мангровой пальмы
Nipa, пальмы Sabal, секвойи Sequoia Couttsiae, циннамомума Cinnamomum.
2) «Тургай с кая» флора. Умеренная листопадная лесная с большей
или меньшей примесью хвойных. В ее состав входили граб (Carpinus), бук
(Fagus), ольха (Alnus), орех (Juglans), комптония (Сотpt onia), ликвидам-
бар (Liquidambar), орешник (Corylus), листопадные виды тополей (Populus)
и др., а также некоторые специфические виды хвойных Taxodium distichum,
Sequoia Langsdorfii).
В палеогене эти комплексы занимали всю территорию тогдашней суши
в пределах СССР и соседних стран. Граница между ними проходила пример-
но по линии, соединяющей место, где теперь находится устье р. Вислы
6
£ устьем р. Янцзы в Китае; к югу от этой линии простиралась область пол-
тавской флоры, а к северу — тургайской. Таким образом, в палеоцене —
эоцене территория Западной Европы и юга европейской части СССР была
занята полтавской флорой, а во всей северной Азии от впадины Тургайского
пролива до побережья Тихого океана была распространена очень однообраз-
ная тургайская флора.
Полтавская флора не была однородной. В. С. Корнилова (1955) делит
ее на следующие провинции: казахстанскую—-субтропическую с умеренны-
ми элементами, туркменскую — аридную с Dryandra, Palibinia и собственно
полтавскую — субтропическую с тропическими элементами (например Nipa).
В восточном направлении полтавская флора обеднялась субтропическими
элементами.
В неогене по мере похолодания на большей части Европы и Западной
Азии тургайская флора постепенно продвигалась на запад, оттесняя пол-
тавскую. На востоке Азии изменение границ между северной листопадной и
южной вечнозеленой флорами было менее существенным.
Процесс постепенного оттеснения на Урале полтавского флористического
комплекса тургайским хорошо иллюстрируется серией ископаемых флор.
Ознакомимся с основными палеоботаническими документами, характери-
.зующими изменение и развитие растительности Урала в третичном периоде.
ПАЛЕОЦЕН
Позднепалеоценовая флора Мугоджар. Изучалась Г. И. Водорезовым,
А. А. Петренко и А. Н. Криштофовичем, однако полных материалов
обработки еще не опубликовано. Из предварительных сообщений (Петренко,
1939, Криштофович, 1932) известно о находке в верхнепалеоценовых отло-
жениях Мугоджар дуба Quercus diplodon, девальквеи Dewalquea grandifolia,
калины Viburnum giganteum, кизила Cornus mugodsharica, циннамомума
Cinnamomum sp., кипарисовника Chamaecyparis belgica, папоротника, близ-
кого к Osmunda, и каштанодуба Dryophyllum sp. К каштанодубам относятся
вымершие древесные породы, по характеру листвы занимающие промежуточ-
ное положение между современными дубами и каштанами. Листья у них
вечнозеленые, зубчатые по краю, с резко выдающейся сетью жилок; плюска,
окружающая плод, сильно развитая, полуоткрытая.
Наряду с другими растениями здесь найдены отпечатки листьев деваль-
квеи (Dewalquea grandifolia и, возможно, других видов этого рода) — вечно-
зеленых кустарников с плотными кожистыми листьями, состоящими из 3—5
долек. Девальквея была обычным растением палеоценовых лесов из кашта-
лодуба; не исключена вероятность, что ее отдаленным потомком является
зимовник кавказский Helleborus caucasicus— современное травянистое расте-
ние из сем. лютиковых, встречающееся на Черноморском побережье Кав-
каза в широколиственных лесах (с господством каштана и дуба). Отмечено
•большое сходство мугоджарской флоры с ископаемой флорой саратовского
яруса Поволжья, хорошо изученной в районе г. Камышина (Баранов, 1959).
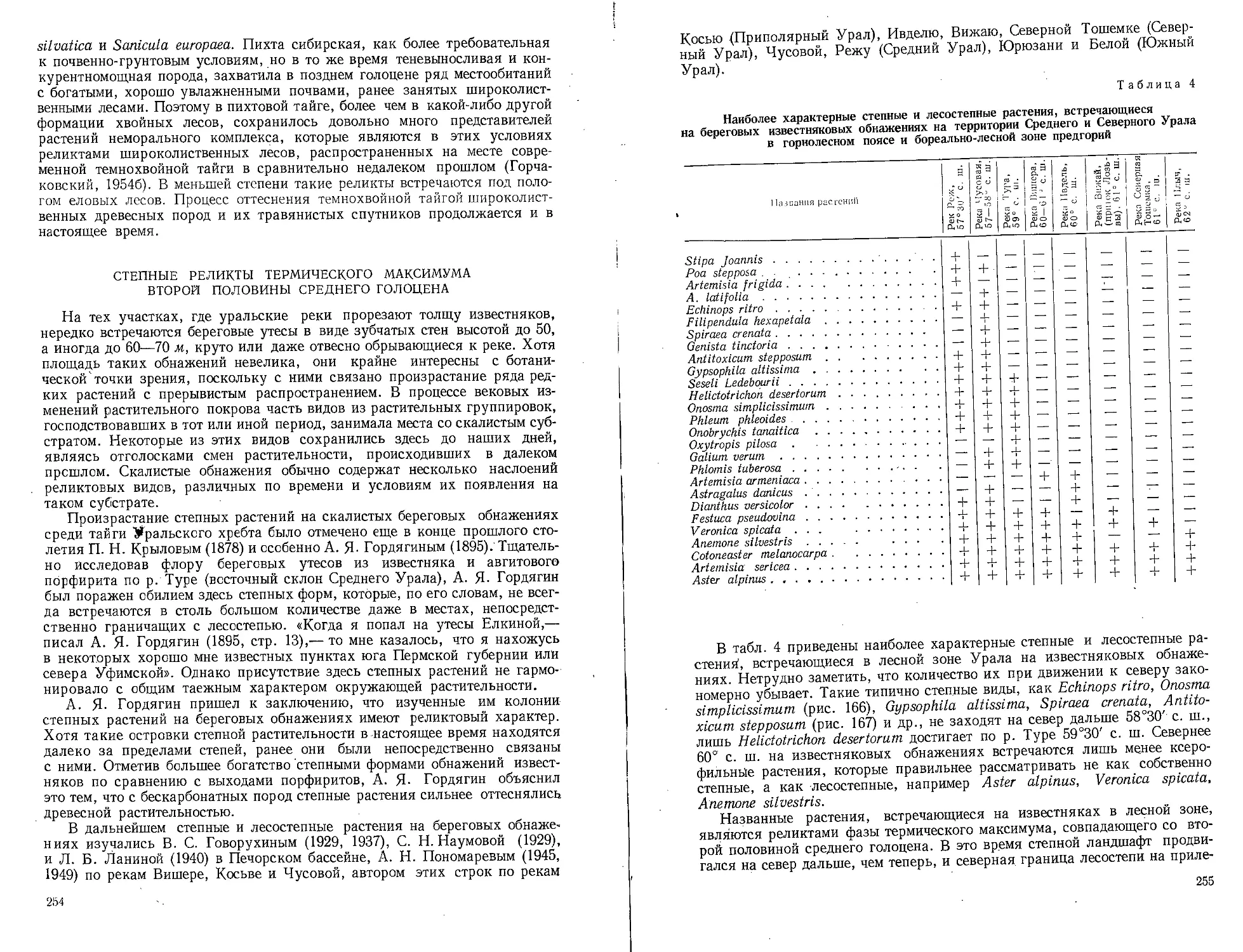
Вероятно, в палеоцене Мугоджары являлись восточным пределом распрост-
ранения флоры камышинского типа.
Палеоценовая (или эоценовая) флора из района г. Орска. На Южном
Урале, в бассейне р. Ори, в морских палеогеновых отложениях (глины с
прослоями глауконитового песка) А. Л. Яшнин (1948) собрал коллекцию
отпечатков растений, из которой А. Н. Криштофович определил Chamaecy-
paris belgica, Dewalquea grandifolia, Quercus diplodon, Viburnum giganteum,
Dryophyllum Dewalquei, Dryandra Schrankii, Magnolia sp., Ficus sp. Мор-
ские отложения перекрыты континентальными образованиями — плотными
кварцитовидными песчаниками, содержащими близ пос. Халилово отпечатки
7
вечнозеленых растений: Sabal sp. fcf. Haeringiana), Andromeda protogaea^.
Dryophyllum furcinerve, Quercus cf. bifurcata, Dryandra Schrankii, Cinna-
momiun sp., Sequoia Couttsiae, Pimela sp., Podocarpus sp. Эта ископаемая
флора имеет в целом палеоценовый облик. Однако, по мнению А. Л. Яшни-
на, возраст изученных им морских отложений ранне- или среднеэоценовый,,
а континентальных — позднеэоценовый.
На левобережье р. Ори в урочище Романкуль (район пос. Батамшинского)
А. А. Петренко (1939) в толще темно-красных железистых кварцитовидных
песчаников собрал отпечатки Lauras Omalii, Quercus furcinervis (Syn. Dryo-
phyllum furcinerve), Celastrophyllum Benekenii, Aralia transversinervia. Andro-
meda protogaea, Sequoia Couttsiae, Glyptostrobus parisiensis, Phyllites-
sp.
На склонах к ручью Романкуль в глауконитовых песчаниках А. А. Пет-
ренко (1939) собрал отпечатки, определенные А. Н. Криштофовичем как
Dryandra Schrankii, Dewalquea grandifolia, Magnolia sp. Здесь же найде-
на окаменевшая древесина Chamaecyparis belgica. Позже это обнажение
привлекло внимание Н. Д. Василевской, а затем В. И. Баранова, собравших
здесь коллекции отпечатков. Предварительная обработка (Баранов, 1949)
показала, что в состав этого комплекса входят Dewalquea gelindenensis,
Aralia cf. venulosa, Cornus cf. sezanensis, Grewiopsis uralensis, Viburnum
giganteum var. uralense, Daphnogene Vasilevskajae, D. gigas, D. crassifolia,
Cornus Janschinii, C. Baranovii, C. angusta, Ficus uralica и др.
Эта флора содержит еще много видов с широкими листьями, что отли-
чает ее от типичной эоценовой флоры, в составе которой преобладают узко-
листные формы. Поэтому В. И. Баранов (1959) определяет ее как палеоце-
новую. Точная датировка возраста этого ископаемого флористического комп-
лекса— дело будущих исследований.
Палеоценовые растения из района г. Челябинска. А. Н. Криштофович;
(1934) в палеоценовых песчаниках в районе оз. Смолино, вблизи Челябинска,,
обнаружил окаменелую древесину беловатого цвета и своеобразные плоды
типа Oxycarpia bifaria. Плоды имеют округлую, ромбовидную или почти’
почковидную форму, в верхней части они утолщены, снабжены острием.
При увеличении снаружи заметна кремневая пленка, возникшая, вероятно, в
результате окаменения скорлупы плода или кожуры семени. Такие же плоды
ранее были найдены в отложениях саратовского яруса палеоцена. Г. Траут-
шольд, впервые описавший эти окаменелости, считал их плодами каштано-
дуба. А. Н. Криштофович, не решая окончательно вопроса об их видовой,
принадлежности, указывает, что это скорее семена из какого-либо плода
или отдельные плодики из соплодия. В палеоценовых песчаниках из окрест-
ностей г. Еманжелинска отмечены, кроме того, остатки листвы Phyllites sp.,.
относящейся, возможно, к какому-нибудь сережкоцветному дереву или кус-
тарнику. Точному определению она не поддается.
Соплодие Steinhauera subglobosa в палеоценовых отложениях Орского'
Урала. Т. Н. Байковская (1951) указала на находку остатка этого соплодия^
на Южном Урале, в палеоценовых отложениях по р. Романкуль, левому
притоку р. Ори. Отпечаток сочетается с минеральным выполнением полостей:
и минеральным же замещением частей соплодия. Соплодие имеет почти ша-
ровидную форму, плодики овальные, длиной около 1 см, снабженные тол-
стой оболочкой, двугнездные, в каждом гнезде содержится по одному семени.
Систематическое положение растения, которому принадлежали такие соплодия,,
точно не установлено. Многие авторы относили эти образования к хвойным,,
сближая их с шишками Abies, Cedrus, Araucaria или Sequoia. Другие иссле-
дователи предполагали, что это соплодие покрытосемянных. Уральский;
образец соплодия Steinhauera subglobosa, по заключению Т. Н. Байковской;
(1951), наиболее близок по своему строению к соплодиям современных.
«.
.представителей рода Syncarpia из сем. миртовых, сохранившегося до на-
стоящего времени во флоре Австралии.
Пыльцевые спектры палеоценовых отложений Урала. Для них, по
И. М. Покровской (1950, 1954), характерно большое количество пыльцы
Taxodiaceae (Taxodium и др.) и Myrtaceae. Кроме того, встречается пыль-
ца широколиственных древесных пород из сем. Aceraceae, Juglandaceae,
Fagaceae, Betulaceae (роды Alnus, Betula), Salicaceae (род Salix). Здесь
.присутствует еще пыльца некоторых форм, более свойственных меловому
периоду (род Sequoia).
ЭОЦЕН
Эоценовая флора Южного Урала. Судя по отпечаткам растений, собранным
.А. Л. Яншиным (1948) на Южном Урале в районе г. Орска и в других
местах и определенным А. Н. Криштофовичем, в эоценовую эпоху здесь
произрастала теплолюбивая тропическая растительность ксероморфного обли-
ка— Andromeda protogaea, Dryophyllum furcinerve, D. sp., Eucalyptus sp.,
пальма из рода Sabal, Sequoia Couttsiae и др. Особенно примечательно при-
сутствие в этом комплексе отпечатков листьев веерной пальмы Sabal, най-
денных в песчаниках Таналыкского района, в окрестностях. г. Орска (Криш-
тофович, 1935). Эта пальма растет в настоящее время в Центральной Аме-
рике, в тропическом климате. В третичное время представители этого рода
были распространены в Японии, в Индокитае, Индии и в европейской час-
ти СССР. На территории Северной Азии от Урала до побережья Тихого
океана ископаемых остатков пальмы Sabal не обнаружено. По-видимому,
Южное Предуралье (под 51° с. ш.) являлось в эоцене восточным пределом
•распространения этой пальмы в составе вечнозеленого полтавского комплекса.
Более подробные данные об эоценовой флоре Южного Урала были опуб-
.ликованы М. Д. Узнадзе-Дгебуадзе (1948). Обработанная им большая кол-
лекция ископаемых остатков была собрана А. Л. Яншиным и другими гео-
логами близ хутора Баки на Южном Урале в континентальных эоценовых
•отложениях (глыбы плотного кварцитовидного песчаника).
Растительные остатки представлены отпечатками листьев и пустотами,
•оставленными разложившимися веточками. К числу изученных отпечатков
растений относятся: Myrica angustata, М. uralica, Quercus аросуnophyHum,
Q. bifurcata, Q. elaena, Q. sp. (два вида), Dryophyllum furcinerve (Syn.
Quercus furcinervis), Ficus multinervis, F. sp., Cinnamomum sp., Laurus sp.,
Terminalia sp., Apocynophyllum helveticum, Myrtophyllum. Warderii, Myr-
.sine doryphora, Andromeda protogaea, Dalbergia cf. bella. Здесь же найде-
ны отпечатки пальмы Sabal sp. и хвойных Podocarpus eocenica, Sequoia
Sternbergii, S. Couttsiae.
В этой коллекции преобладают узкие, большей частью мелкие листья
покрытосемянных древесных растений. Они чаще удлиненноланцетовидной
формы, кожистые, с завернутыми вниз краями. Ксероморфное строение
листьев, их вечнозеленость свидетельствуют о произрастании лиственных по-
род эоценового комплекса Южного Урала в условиях теплого субтропи-
ческого климата, но с засушливым периодом в летнее время.
По мнению Д. В. Наливкина (1943), в эоцене в районе Южного Урала
расстилалась жаркая тропическая пустыня, покрытая чистыми сыпучими бар-
ханными песками. В понижениях рельефа временами возникали водные бас-
сейны— оазисы, окаймленные зарослями вечнозеленых засухоустойчивых
растений, преимущественно с узкими кожистыми листьями.
Трудно сказать, насколько прав Д. В. Наливкин в своих реконструкциях
эоценового ландшафта. Во всяком случае, в результате анализа рассмотрен-
ной ископаемой флоры можно прийти к заключению, что на Южном Урале
зв эоцене произрастали низкорослые и, вероятно, разреженные леса из вечно-
9
зеленых древесных пород (узколистные дубы, лавры, циннамомумы и др.)
с примесью некоторых хвойных (подокарпус, секвойя) и пальм (Sabal). Под-
лесок в них составляли вечнозеленые ксероморфные кустарники. Южнее,
по-видимому, были распространены пустынные ландшафты.
Отпечатки листьев и ископаемая пыльца из тасаранской свиты Му-
годжар. В коллекции образцов, собранных Г. С. Трошиным на западном-
склоне Мугоджар в кварцево-глауконитовых песчаниках, В. И. Баранов (1953)
обнаружил отпечатки Fagus deucalionis, Quercus kamyschinensis, Q. odonti-
phylla, Q. diplodon, Q. cf. platania, Q. sp., Ficus Gordjaginii, Dryandra
karaculensis, D. sp.. Litsaea Troschinii, Hackea sp., Rhus turcomanica, Myr-
tophyllum Warderii, Cornus sp., Andromeda protogaea, Viburnum giganteum
и др. Из некоторых образцов, относящихся к тому же горизонту, выделена
пыльца Araucariaceae, Cupressaceae, Taxodiaceae, Podocarpus, Cedrus, Picea,
Pinus, Myricaceae, Quercus, Juglans, Carya, Acer, Ilex, Nyssa, Rhus. По
А. Л. Яншину (1952), тасаранская свита относится к среднему эоцену и к
нижней половине верхнего эоцена. В состав ископаемой флоры из этой сви-
ты входят дубы и каштанодуб, характерные для палеоцена Поволжья, но
в то же время здесь есть несколько растений, близких к жестколистному
эоценовому комплексу Южного Урала. Присутствие Rhus turcomanica сбли-
жает флору тасаранской свиты Мугоджар с эоценовой флорой из района •
озера Ер-Ойлан в Бадхызе (Туркмения).
Морская трава посидония в эоценовых отложениях восточного склона-
Урала. Ныне живущая Posidonia oceanica представляет собой травянистое
растение из сем. болотниковых (Helobiae), распространенное в Индийском-
океане и Средиземном море. На восточном склоне Урала, в окрестностях
с. Колчеданского, в серых каменистых глинах (опоках) А. Н. Криштофович
(19386) нашел остатки более древнего представителя посидоний — Posidonia
parisiensis. В ископаемом состоянии этот вид известен из палеоценовых и
эоценовых отложений Западной Европы. Находка А. Н. Криштофовича ука-
зывает на чисто морские условия образования опок. Факт распространения
зарослей посидонии в теплом море, омывавшем в конце палеоцена и начале
эоцена восточный склон Урала, свидетельствует о сравнительной однородно-
сти климатических условий указанного отрезка третичного периода. Эта одно-
родность прослеживалась на значительной территории от Западной Европы
до Зауралья.
Ископаемая пыльца из эоценовых отложений Южного Урала. В суль-
фидоносных эоценовых глинах Айдырлинского никелевого месторождения
на восточном склоне Южного Урала обнаружена пыльца Pinus subgen. Нар-
loxylon, Taxodiaceae, Betula, Salix, Juglans, Carya, Castanea, Quercus, Acer,
Rhus, Myrtaceae (Eucalyptus, Eugenia) и Ericaceae (Покровская, 1950). В це-
лом для эоценовых спорово-пыльцевых комплексов Урала, как и палеоцено-
вых, характерно значительное участие пыльцы Myrtaceae и Taxodiaceae (По-
кровская, 1954).
ОЛИГОЦЕН
На территории Урала ископаемых флор, достоверно относящихся к оли-
гоцену (за исключением шишки вымершей ели и отпечатков листьев из рай-
она Мугоджар), пока не найдено. Поэтому судить о растительном покрове
Урала в олигоцене приходится в основном по данным пыльцевого анализа.
Вымершая ель, близкая к тяньшанской, из юго-западного Приуралья..
И. В. Палибин (1931) описал по ископаемым остаткам шишки ныне вымер-
ший вид ели. Шишка ели типа Piceostrobus Neustruevi была найдена
С. С. Неуструевым в районе Общего Сырта (юго-западное Приуралье) на го-
ре Точильной в кварцевых песчаниках, по-видимому, олигоценового возраста.
Шишка длиной 8 см, шириной 1,1 см отнесена к формальному роду Pi~
10
ceostrobus, соответствующему современному роду Picea. Эта ископаемая ель
наиболее близка по строению шишек к тяньшанской ели Picea Schrenklana,
но отличается от последней меньшим размером их и более отчетливо вы-
дающимися средними жилками на семенных чешуях.
Отпечатки листьев из верхнеолигоценовых отложений Мугоджар. В
железистых песчаниках западного склона Мугоджар Г. С. Трошин собрал
коллекцию отпечатков растений. Согласно определению В. И. Баранова (1953),
,в ее состав входят Sequoia sp., Taxodium sp., Cyperites sp., Quercus Alexe-
evii, Ulmus appendiculata, U. Braunii, U. plurinervia, Corylus turgaica,
Oreodapime mugodsharica. Этот комплекс, включающий ряд широколиствен-
ных деревьев, близок к ископаемой флоре самой верхней песчаниковой сви-
ты континентальных отложений тургайской серии Приаралья и Тургайской
низменности.
Ископаемая пыльца из олигоценовых глин Среднего и Южного Урала.
По данным И. М. Покровской (1950), в палинологическом спектре тем-
но-серых глин с оз. Синара отмечены Pinus subgen. Haploxylon, Cedrus,
Sequoia, Alnus, Betula, Salix, Carya, Pterocarya, Quercus, Tilia, Nyssa,
Ericaceae. Из серых глин Александровского лога Висимской депрессии отмы-
та пыльца Pinus subgen. Haploxylon, Pinus subgen. Diploxylon, Picea, Abies,
Taxodium, Sequoia, Tsuga, Alnus, Betula, Corylus, Salix, Carya, Pterocarya,
Quercus, Tilia. В огнеупорных глинах у дер. Пеньки (Красноуфимский район
Свердловской области) содержится, наряду с пыльцой сосен, елей и пихт,
пыльца Tsuga, Taxodiaceae, Alnus, Betula, Carpinus, Corylus, Salix, Carya,
сем. Juglandaceae, Tilia, Acer, Fagus, Rhus, Liquidambar, сем. Saxifraga-
ceae, сем. Compositae. Обращает на себя внимание богатое содержение пыль-
цы различных широколиственных древесных пород. Бурые олигоценовые гли-
ны Башкирского Приуралья содержат также довольно разнообразную пыль-
цу хвойных и лиственных деревьев; в числе последних отмечен падуб (Ilex).
Характерная черта олигоценовых спорово-пыльцевых комплексов Урала
и Западной Сибири — богатый набор широколиственных влаголюбивых пород,
преобладающих в спектре, некоторое участие Taxodiaceae и Pinaceae; еди-
ничная встречаемость вечнозеленых субтропических пород (Покровская, 1954).
На основании всех этих материалов можно заключить, что на Урале
в олигоцене были распространены леса, состоящие из хвойных и особенно
широколиственных древесных пород. Хвойные были представлены родами
Sequoia, Taxodium, Pinus, Picea, Abies, Tsuga. Наиболее характерными ком-
понентами тепло- и влаголюбивого широколиственнолесного комплекса были
Carpinus, Acer, Tilia, Fagus, Juglans, Quercus, Ulmus, Corylus, Carya, Pte-
rocarya.
МИОЦЕН
Уже с конца олигоцена в связи с похолоданием климата вечнозеленая
субтропическая (с тропическими элементами) полтавская флора стала оттес-
няться умеренной листопадной тургайской. Этот процесс в основном завер-
шился на территории Урала в миоцене, о чем свидетельствует ископаемая
флора из Башкирской АССР.
Растения из ранне миоценовых буроугольных отложений Башкирии.
В пластах бурого угля основной угленосной свиты южноуральского буро-
уголы-юго бассейна (Молокановское, Хабаровское, Тюльганское и Бабаевское
месторождения) найдены остатки растений в виде трухи, содержащей плоды,
семена, мегаспоры, хвоинки, обломки древесины. Эти остатки хорошей со-
хранности, без признаков переотложения. Из них Т. Д. Колесникова (1960)
определила следующие виды: Sequoia Langsdorfii, Glyptostrobus europaeus,
Chamaecyparis sp., Myrica Suppanii, Alnus cf. cordata, Aralia sp., Ampe-
dopsis cf. rotundata, Caldesia proventitia, Epipremnum ornatum, E. crista-
11
turn, Brasenia tuberculata, B. Schreberi, Decodon gibbosus, Diclidocarya
Menzelii, Proserpinacea reticulata.
В другой, позднее опубликованной работе Т. Д. Колесникова (1961) сооб-
щила о находке в образце серой элевритовой глины из угленосной свиты;
Ворошиловского буроугольного месторождения Башкирии остатков других
растений. В их числе остатки семян Liriodendron geminata, известного по
палеокарпологическим данным из нижнего плиоцена Западной Европы и оли-
гоцена Западной Сибири, эндокарп растения из рода Ilex, створки коробочки,,
принадлежащей представителю сем. Ericaceae. Особый интерес представ-
ляет находка фрагмента шишки ископаемого вида сосны Pinus Thomasiana.
Современные аналоги этого вымершего вида — средиземноморские сосны нз
секции Eupitys Schpach: Pinus nigra, P. taricio, P. Pallasiana. Другая за-
служивающая внимания находка — остатки семян по-видимому вечнозеленого
растения Distylium uralensis и сем. Hamamelidaceae. Ныне живущие пред-
ставители рода Distylium — вечнозеленые деревья и крупные кустарники с
кожистыми листьями, они распространены в субтропических районах Восточ-
ной и Юго-Восточной Азии.
Анализ всего этого ископаемого комплекса показывает, что во время
образования угленосной толщи на территории Башкирии существовала расти-
тельность, характерная для областей с умеренно теплым климатом. Леса
состояли в основном из хвойных и лиственных (с опадающей листвой) пород,
в их состав входили кустарники и лианы. Участие вечнозеленого субтропи-
ческого элемента было незначительным.
Рассмотренная ископаемая флора имеет как бы переходный характер-
между олигоценовой и миоценовой, хотя более близка к последней. В. Л. Яхи- '
мович (1958) относит основную угленосную свиту Башкирии к нижнему мио-
цену. С этой датировкой в какой-то мере согласуются результаты споро-
во-пыльцевого анализа (Чигуряева, 1951).
Кинельская флора бассейна р. Камы. В кинельских слоях, относимых
геологами к среднему плиоцену, но, возможно, соответствующих нижнему
плиоцену, на левом берегу р. Камы близ пос. Набережные Челны и Камские-
Поляны найдены остатки (семена, плоды, мегаспоры, хвоя, веточки) Abies sp.,
Larix sp., Pinus sp., Betula sp., Morus cf. nigra, Morus sp., Acer sp., Cornus cf.
alba, Sambucus sp., Weigela Kryshtofovichiana и др. По мнению П. И. До-
рофеева (1957), обработавшего эту коллекцию, здесь произрастала темно-
хвойная тайга с миоценовыми реликтами, несколько напоминавшая ситхинские-
или нижнеамурские леса. В состав тайги входили многие широколиственные
деревья, подлесок был разнообразным, имелись лианы. Но таксодиевых
(Tsuga) и некоторых широколиственных деревьев — ведущих компонентов,
олигоценово-миоценовых флор — в составе лесов уже не было. П. И. До-
рофеев полагает, что темнохвойная тайга к кинельскому веку стала господ-
ствующим типом растительности на Русской равнине.' В это время уже офор-
мились основные ландшафтные зоны равнин: арктическая, таежная, зона
открытых пространств (к югу от широты Воронежа и Куйбышева); однако
они заметно отличались от современных по составу и строению.
Кинельская флора Башкирии. В кинельских (в основном пресноводных)
отложениях, относимых к среднему плиоцену, на территории Уфимского и.
Нуримановского районов Башкирской АССР найдены макроскопические ос-
татки (хвоя, веточки, плоды, семена и другие части) Picea sp., Abies sp.,.
Larix sp., Pinus cf. silvestris, Tsuga sp., Juniperus sabina, Betula bashklrica,
B. sp., Alnus bashkirica, Cornus cf. alba, Weigela sp. и др. На основании;
анализа этого ископаемого комплекса П. И. Дорофеев (1962) пришел к вы-
воду, что в кинельский век на территории Башкирии были распространены.!
леса таежного типа, содержащие в своем составе впоследствии исчезнувшую*
тсугу.
12
В 1943 г. П. А. Никитин изучил остатки растений из пяти образцов
кинельских отложений, вскрытых двумя скважинами на левом берегу р. Уфы
близ с. Базилевки. Здесь найдены Azolla pseudopinnata, Salvinia tuberculata,
Najas lanceotata, N. fbveolata, Potamogeton planus, Dulichium vespiforme,
Epipremnutn crassum, Decodon globosus, Nautnburgia subthyrsiflora, Hyperi-
cum septestum, Sparganium noduliferum и другие (Яхимович, 1958; Дорофе-
ев, 1960). В этом списке преобладают водные и прибрежноводные растения.
Миоценовая флора юго-западного Приуралья. Г. В. Вахрушев в районе
Стерлитамака (Башкирская АССР) около дер. Кинзябаевой по р. Суракай
и близ дер. Ромодановки (Осиновой) по р. Карташлы собрал растительные
остатки, которые были изучены А. Н. Криштофовичем (1932). В этой кол-
лекции оказались отпечатки листвы платана Platanus aceroides, каштана
Castanea Kubinyii, граба Carpinus grandis, бука Fagus Antipofii, гинкго
Ginkgo adiantoides, глиптостробуса Glyptostrobus europaeus. Бук, кроме того,
представлен отпечатками плюски плода. >
В окрестностях дер. Сидтикмуллиной (теперь дер. Берекмяк), по балке
Шкатлы (Ушкатлы), притоку р. Аургазы (бассейн р. Белой), было открыто
местонахождение остатков третичных растений. Анализ этой флоры показы-
вает ее близкую связь с ископаемой флорой Приаралья, с одной стороны,
а также украинского сармата и западноевропейского верхнего миоцена — с
другой. Из собранной здесь коллекции А. Н. Криштофович (1938а) опреде-
лил Taxodium distlchum miocaenum, Glyptostrobus europaeus var. Ungeri,
Phragmites oeningenensis, Alnus Kefersteinii, Betula macrophylla, B. prisca,
Comptonia oeningenensis, Fagus Antipofii, Hicoria fSyn. Carya) riphaeica,
Quercus cf. Nimrodi, Q. neriifolia, Q. cf. Olafsenii, Liquidambar europaeum,
Crataegus Fominii, Diospyros brachysepala, Myrsine doryphora, Rhamnus
Graeffii, Jug Ians acuminata, Corylus Macquarrii, Ulmus cf. longifolia. Кроме
того, в отложениях по р. Ташлаиру найден отпечаток Populus balsamoides.
Относящаяся к сем. восковниковых Comptonia oeningenensis, по А. Н. Криш-
тофовичу, — тип, чрезвычайно характерный по своей листве и свойственный
всем тургайским флорам от Аляски, Японии и Сахалина до Приуралья. На-
ходка комптонии в третичных отложениях Башкирии очень интересна.
В 1960 г. обнажение в овраге Ушкатлы у дер. Берекмяк (в 10 км к
юго-востоку от районного центра Толбазы) обследовала Т. Д. Колесникова
(1963) и собрала там коллекцию отпечатков растений. Всего отсюда было
определено 23 вида растений, в том числе некоторые из них отсутствовали
в списке А. Н. Криштофовича: Pseudolarix fossilis, Myrica vindobonensis,
M. cf. Mairei, Pterocarya castaneifolia, Ulmus carpinoides, Sassafras Fer-
retianum, Vitis zaisanica, Tilia sp., а три вида были описаны как новые
для науки: Alnus hirsutiformis, Zizyphus uralensis и Acer baschkirica. В то
же время некоторые виды, указываемые А. Н. Криштофовичем, не были
найдены.
Ископаемые растения из окрестностей дер. Кинзябаевой, Ромодановки
и Берекмяка (Сидтикмуллиной) несомненно относятся к одному комплексу.
Сопоставление этой флоры, которая в целом по месту нахождения может
быть названа Стерлитамакской, с третичной (тургайской) флорой Приаралья
и с сарматским комплексом Украины (флора Крынки) вскрывает их значи-
тельное сходство.
Тургайская флора Приаралья почти лишена вечнозеленых форм. Стерли-
тамакская флора, существовавшая в условиях умеренного климата, еще со-
держит некоторые вечнозеленые узколистные формы, к которым относятся
Myrsine doryphora, Quercus neriifolia и Myrica vindobonensis.
По-видимому, Стерлитамакская флора генетически связана с аральской.
А. Н. Криштофович (1938а, стр. 81) по этому поводу пишет: «Весьма ве-
роятно, что, распространяясь через Урал с востока и северо-востока, тургай-
13
екая флора, вторгшись в Стерлитамакский район, застала там еще остатки
вечнозеленой растительности, которые и сохранились в ней до времени отло-
жения слоев Шкатлы в виде довольно заметного элемента. Отмечая поэтому
одновременность Стерлитамакской и аральской флоры или несколько более
поздний возраст первой, мы .все же должны считать ее, т. е. Стерлитамак-
скую флору, значительно более древней, чем сармат». Проследив ход рас-
селения листопадной тургайской флоры, А. Н. Криштофович пришел к вы-
воду, что Стерлитамакские слои можно считать нижним миоценом или первым
средиземноморским (бурдигальским) ярусом, оставляя некоторое уточнение
в этом отношении до следующего этапа исследований.
Т. Д. Колесникова (1963), проанализировавшая этот комплекс с учетом
данных других исследователей и результатов иалеокарпологических исследо-
ваний, считает, что он относится к верхнему олигоцену или нижнему мио-
цену. В это время на территории Башкирии водораздельные пространства
и склоны гор были покрыты широколиственными лесами из каштана Cas-
tanea atavia и бука Fagus Antlpofii с примесью других деревьев — Pseudo-
larix fossilis, Sequoia Langsdorfii, Betula macrophylla, Ulrnus carpinoides,
Zelkova Ungeri, * Liriodendron geminata1, Sassafras Ferretianum, Acer ba-
schkirica, Tilia sp. Подлесок образовывали Comptonia dryandroides, * Myri-
ca Suppanii, M. vindobonensis, * Distylium uralensis, Ilex sp. и Aralia sp.
Из лиан встречались Vitis zaisanica, * Ampelopsis cf. rotundata и ароидные
Epipremnum ornatum и * E. cristatum. По берегам водоемов и в долинах
сообщества образовывали Alnus hirsutiformis, Quercus neriifolia и Liquidambar
europaeum, а на заболоченных местах — Glyptostrobus europaeus, Taxodium
dubium, Typha sp. и * Decodon gibbosus. В неглубоких стоячих водоемах
росли * Caldesia proventitia, * Brasenia tuberculata и * Proserpinacea reticu-
lata.
Ископаемые древесины хвойных из района оз. Смолино. Прогрессирующее
похолодание климата в неогене сопровождалось постепенным отмиранием
теплолюбивых видов древесных растений, отступанием их к югу. Верхнемио-
ценовые флоры Зауралья имеют уже явно обедненный характер. На Урале
в связи с возрастанием суровости климата создались предпосылки для более
широкого распространения хвойных древесных пород, а местами, вероятно,
возникли хвойные леса. Отчасти это подтверждается находкой ископаемых
лигнитизированных древесин хвойных. Образцы древесины были собраны
в 1914 г. И. М. Крашенинниковым в каолиновых глинах в районе оз. Смо-
лино, около Челябинска. Обработка, произведенная А. В. Ярмоленко (1934),
показала, что среди этих образцов имеется древесина можжевельника Juni-
peroxylon uralense, трех видов таксодия Т axodioxylon distichum, Т. sequoia-
num, Т. Krasheninnikova, глиптостробуса Glyptostroboxylon tenerum и
древесной породы из группы сосновых (Pityoxylon Paxii).
Этот комплекс представлен породами, теперь на. территории Урала вымер-
шими, но сохранившимися в более или менее измененном виде в других
пунктах земного шара. Род таксодий (Taxodium) содержит два ныне живу-
щих вида в Северной Америке, а близкий к нему род глиптостробус (Glypto-
strobus) — один вид в юго-восточном Китае. Древовидные можжевельники
в современной флоре представлены приблизительно 40 видами, широко распро-
страненными в северном полушарии от Субарктики до горных массивов тро-
пической зоны.
Позднее образцы из этой же коллекции изучил Р. Худайбердыев (1958).
Он установил, что древесина, описанная А. В. Ярмоленко как Taxodioxylon
distichum, фактически относится к Pseudolaricixylon firmoides. Кроме того,
1 Звездочкой * в этом списке отмечены виды, присутствие которых установлено по па-
леокарпологическим данным. ;
!4
из этой же коллекции определена древесина Podocarpoxylon uralense. Род
Pseudolaricixylon — формальный, соответствующий по строению древесины
современному роду Pseudolarix. В настоящее время этот род монотипный,
единственный его представитель Pseudolarix amabilis произрастает в восточ-
ном Китае. Podocarpoxylon uralense близок по анатомическим признакам
древесины к современному роду Dacrydium. К нему относится около 20 видов,
произрастающих в Австралии, Новой Зеландии, Тасмании, Новой Каледонии,
Новой Гвинее, на Филиппинах, Целебесе, Борнео, Суматре и доходящих до
Малайского п-ова, Аннама и Хайнаня.
Ископаемый комплекс древесин хвойных из района оз. Смолино А. В. Яр-
моленко (1939), а вслед за ним и В. И. Баранов (1959) датируют миоценом.
Р. Худайбердыев (1958) считает, что он относится к верхнему олигоцену или
нижнему миоцену.
Ископаемая флора из верховьев р. Тавды. Представление о характере
растительности, распространенной в начале неогена на восточном склоне
Урала и в прилегающих частях Западно-Сибирской низменности можно полу-
чить, рассмотрев данные изучения ископаемого комплекса из верховьев
р. Тавды, вблизи поселков Белоярки и Антропово. В этом районе Л. В. Введен-
ским (1933) найдены отпечатки листьев магнолии Magnolia spectabilis, мирта
Myrica Torreyi, дубов Quercus groenlandica, ильма Ulmus appendiculata,
нескольких видов ольхи Alnus nostratum, A. Kefersteinii, A. sp., березы Betula
prisca, B. macrophylla, лещины Corylus Macquarrii, клена Acer cf. laetum,
тополя Populus balsamoides, таксодия Taxodium distichum miocenum и секвойи
Sequoia Langsdorfii.
Тавдинский ископаемый комплекс, как и сходная ископаемая флора из
окрестностей г. Томска, имеет уже в основном тургайский облик. Возраст
ее следует датировать, по-видимому, миоценом.
Ископаемая пыльца из миоценовых отложений Урала. По данным
И. М. Покровской (1950), пыльцевые спектры уральского миоцена характе-
ризуются следующим составом: Picea, Pinus subgen. Dipioxylon, Pinus
subgen. Haploxylon, Abies, Juglans, Carya, Pterocarya, Tilia, Carpinus, Fagus,
Quercus, Acer, Rhus. В миоценовых комплексах усиливается (по сравнению с
олигоценом) значение Picea и Abies, уменьшается участие сем. Juglandaceae,
почти исчезают Taxodium, Sequoia.
Отличительная особенность миоценовых спорово-пыльцевых комплексов
Урала и Западной Сибири — преобладание пыльцы различных сосен (Pinus),
существенное участие пыльцы многих широколиственных пород, особенно из
сем. Juglandaceae и Fagaceae (Покровская, 1954).
В миоцене, как об этом свидетельствуют ископаемые листовые флоры,
окаменелые древесины и спорово-пыльцевые комплексы, на Урале были
распространены хвойные и широколиственные леса (а также, вероятно, сме-
шанные хвойно-широколиственные) с небольшой, примесью вечнозеленых узко-
листных деревьев и кустарников.
ПЛИОЦЕН
Непосредственно на территории Урала ископаемые листовые флоры плио-
ценового возраста пока изучены слабо. Поэтому о растительности Урала
в плиоцене приходится судить, главным образом, по палинологическим данным,
а также на основании палеоботанической документации, относящейся к смеж-
ным областям.
Плиоценовая флора низовьев р. Камы. В низовьях реки Камы, между
поселками Рыбной Слободой и Горицами, в 1913 г. В. А. Чердынцевым
была отмечена встречаемость в третичных отложениях ископаемых остатков
растений. Это обнажение детально изучал в 1947 г. В. И. Баранов, собрав-
ший вместе со своими сотрудниками богатую коллекцию отпечатков растений,
15
заключенных в плотные глинистые конкреции сидерита. Сходное местонахож-
дение растительных остатков выявил В. А. Полянин около с. Татарского
Бурнашево на р. Свияге; собранные здесь отпечатки также поступили в рас-
поряжение В. И. Баранова. Результаты обработки коллекции, дополненные
спорово-пыльцевым анализом, были опубликованы В. И. Барановым в виде
специальной статьи (1949) и вошли в его сводную работу по третичным
флорам СССР (1959).
Из наиболее интересных ископаемых остатков, найденных в плиоценовых
отложениях низовьев р. Камы, следует отметить: Tsuga еигораеа (шишка),
Quercus roburoides, Q. Kryshtofovichiana, Carpinus betulus, Corylus avellana,
Betula Noinskii, В. Tscherdincevii, Alnus glutinosa, Pterocarya sp., Juglans
cf. acuminata, Populus balsamoides, P. cf. lalior, P. cf. mutabilis, P. tremula,
P. Komaroviana, Salix acutissima, S. angusta, S. Integra, S. varians, Acer
Myrtovae, A. Palibinii, A. Pojarkovae, Tilia cordata, Fraxinus cf. excelsior,
Ulmus carpinoides, U. longifolia, U. minuta, Zelkova Richardii, Rhus querci-
folia, Myrica carpinifolia. В этих отложениях обнаружена пыльца ряда
хвойных (сосна, пихта, ель) и лиственных древесных пород.
При анализе этой ископаемой флоры обращает на себя внимание присут-
ствие в ней, наряду с вымершими растениями, ряда ныне живущих древес-
ных пород (ольха черная, лещина обыкновенная, липа мелколистная и др.).
В. И. Баранов (1959) приходит к заключению, что в среднем плиоцене в
низовьях р. Камы существовали сравнительно богатые по своему флористи-
ческому составу широколиственные леса из дуба, граба, вяза, липы, клена
с участием хвойных древесных пород (тсуга, ель, пихта, сосна). Некоторые
теплолюбивые породы (падуб, сумах) сохранялись в этих лесах, по-видимому,
на положении реликтов.
Растения из плиоценовых отложений Среднего Урала. В урочище Новый
Лог Висимского района Свердловской области еще в 1937 г. был взят обра-
зец серо-коричневой плиоценовой глины, содержащей растительную труху.
Из детрита, удалось выделить обломки хвои, шишек, веточки, семена, плоды
или их обломки. П. И. Дорофеев (1952) определил в этой коллекции остат-
ки следующих растений: Tsuga sp., Picea sp., Pinus sp. (пятихвойная, видимо,
из subgen. Haploxylon), Metasequoia sp. (?), Chamaecyparis cf. nootkatensis,
Liriodendron tulipifera, Alnus sp., Corylus cf. avellana, Dulichium spathaceum
(современный североамериканский вид), Epipremnum cristatum (вымерший вид;
другие виды этого рода сохранились лишь в Юго-Восточной Азии и в Австра-
лии), Carex sp. Возраст ископаемого комплекса—-средний или поздний плио-
цен. По составу он близок к среднеплиоценовой флоре Западных Карпат
(Кросьценко на Дунайце). В это время на территории Среднего Урала суще-
ствовали леса из хвойных и листопадных древесных пород. По составу они
значительно отличались от современных, так как включали, наряду с предста-
вителями родов, вошедших в современную флору • (Pinus, Picea, Alnus,
Corylus), ряд растений, чуждых современной флоре Европы, но сохранивших-
ся в Северной Америке и Юго-Восточной Азии (Tsuga, Metasequoia, Chamae-
cyparis, Liriodendron).
Плиоценовая флора Южного Урала и Предуралья. П. И. Дорофеев (I960)
изучил остатки растений в образцах’ миоценовых отложений из скважин,
пробуренных в Башкирии и Оренбургской области. Близ пос. Иглино найдены
остатки Picea sp., Tsuga sp., Larix sp., Salix sp., Betula sp., Cornus cf.
alba, Selaginella pliocenica, Chenopodium rubrum. Polygonum lapathifolium,
Hypericum coriaceum, Taraxacum tanaiticum и др.
В долине р. Нугуша в озерных отложениях обнаружены остатки Salvinia
cf. glabra, S. cf. tuberculata, Najas marina, Potamogeton vaginatus, P. colo-
ratus, Typha pliocenica, Alisma plantago-pliocenica, A. plantago-minima.
Ranunculus sceleratoides, Decodon globosus, Vaccinium pltocenicum.
16
Скважина на левом берегу р. Мендыма у с. Ново-Усмановки вскрыла
остатки Weigela Rryshtofovichiana, Zannichellia pliocenica, Typha pliocenica,
Hypericum coriaceum, Najas marina, N. lanceolata, Pinus sp., Picea sp.
В толще озерных зеленовато-серых глин найдены остатки Aldrovanda
Eleonorae, Potamogeton bashkiricus, P. palaeocompressus, P. trichophyllus,
P. pectinatus, P. filiformis, Decodon globosus и др.
Из числа растений, остатки которых найдены в прослойках супесей в толще
серых слоистых глин у с. Кумурлы в Архангельском районе Башкирской АССР,
заслуживают упоминания Pinus sp., Picea sp., Tsuga sp., Betula sp., Morus
sp., Cornus sp., Satnbucus sp., Weigela Kryshtofovichiana, Sagittaria sagit-
iifolia, Butomus umbellatus, Cyperus glomeratus, C. fuscus, Heleocharis pa-
lustris, Rumex maritimus, Hypericum coriaceum, Zannichellia pliocenica.
По заключению П. И. Дорофеева, эти ископаемые комплексы воспроиз-
водят лесную, точнее таежную флору, представленную в основе Pinus, Picea,
Tsuga, Larix, Salix, Betula, Morus, Cornus cf. alba, Vactinium pliocenicum,
Weigela, Sambucus.
Плиоценовая тайга, по сравнению с современной евро-сибирской, была
намного богаче видами. По составу компонентов она несколько напоминала
современную тайгу Дальнего Востока или юга Канады.
Наряду с растениями, которых нет в современной европейской флоре
(Tsuga, Weigela), этот комплекс включал ряд видов, близких современным
(например, Typha pliocenica соответствует современной Т. latifolia) или
совершенно им тождественных.
Пыльца из плиоценовых отложений восточного склона Среднего Урала.
Пыльцевой анализ образца черной пластичной глины с р. Мраморной на
восточном склоне Среднего Урала дал, по И. М. Покровской (1950), такие
результаты: здесь обнаружено много пыльцы Alnus (73%), в меньшем коли-
честве пыльца Salix (10%), Betula (8%), Pinus (5%), единично пыльца
Abies, Picea, Larix, Carpinus, Acer, Tilia, Quercus, Pterocarya. Кроме того,
единично отмечена пыльца травянистых двудольных растений и. в ничтожном
количестве — споры папоротников.
Плиоценовый спорово-пыльцевой комплекс Среднего Предуралья. В глинах
и торфе долины Пра-Камы (Палео-Камы) найдена, по И. М. Покровской
(1950), пыльца Pinus subgen. Diploxylon, Pinus subgen. Haploxylon, Picea,
Abies, Tsuga, Taxodium, Betula, Alnus, Corylus, Salix, Quercus, Ulmus, Tilia,
Carpinus, Rhus, Pterocarya, сем. Ericaceae, пыльца травянистых однодольных
(сем. Сурегасеае), пыльца травянистых двудольных. В небольшом количестве
примешаны споры папоротников.
На территории Урала в плиоцене наиболее распространенными сообще-
ствами были хвойные и широколиственные леса. По составу они уже в ос-
новном приближались к современным, однако содержали представителей
некоторых впоследствии исчезнувших на Урале родов Taxodium, Tsuga,
Carpinus, Fraxinus (Покровская, 1950).
Характеризуя спорово-пыльцевые спектры кинельского времени (ранний и
средний плиоцен) Башкирского Предуралья, В. К. Немкова (1964) считает
пыльцу наиболее теплолюбивых древесных пород — Juglans, Fagus, Liqui-
dambar, Pterocarya, Nyssa, Sequoia, Taxodium переотложенной, вторично
захороненной в кинельских отложениях. Судя по пыльцевым спектрам из
второго чебеньковского горизонта, на территории Кармаскалинского района
Башкирской АССР в это время существовали сосново-еловые и листопадные
леса, в состав которых входили пихты, тсуги, ели (в том числе из секции
Omorica), липы, вязы; реже встречались дубы и березы. В бассейне Палео-
Сакмары, по данным анализа пыльцы из третьего чебеньковского горизонта,
преобладала лесостепь и степь, листопадных лесов из дуба, вяза с примесью
липы, шелковицы, ореховых было немного. Хвойные леса, по-видимому, росли
2 Заказ X? 251 17
восточнее, на склонах Уральских гор. Севернее, в долине р. Сима в это
время произрастали вязовые леса с дубами и липами (Tilia cordata, Т. cf.
atnurensis, Т. cf. mandshuricd).
Постепенное поднятие в плиоцене древних палеозойских массивов сопро-
вождалось усилением эрозии и расчленением рельефа. Хвойные леса сосре-
доточились преимущественно в горных районах, а широколиственные —
в предгорьях и на склонах в южной части Урала. Площадь широколиственных
лесов на Урале в конце плиоцена, по-видимому, значительно превышала,
современную.
П. И. Дорофеев (1962) предполагает, что растительность плиоцена была
уже во многом сходна с современной. К концу плиоцена сформировались
все основные ландшафтные зоны, которые и по внешнему облику и по за-
нятой ими территории были близки к современным зонам на территории СССР.
Развитие плиоценовой флоры шло постепенно и без существенных отклонений
в одном направлении — по линии общего ее обеднения третичными элемен-
тами, модернизации за счет появления новых видов, сформировавшихся в это
время, и усиления контрастности зон.
Плиоценовые спорово-пыльцевые спектры восточной окраины Русской
равнины. По данным палинологического анализа (Ананова, 1962), уже с са-
мого раннего плиоцена на Русской равнине была выражена зональность
растительности: на территории от Приазовья до Нижнего Дона и Прикамья
с юга на север сменяли одна другую степная зона, лесостепь, зона темно-
хвойных лесов с участием широколиственных деревьев, а в более северных
пределах Русской равнины располагалась таежная зона. Прямых доказательств
существования в плиоцене тундр не обнаружено, хотя можно предполагать,
что на северной окраине Русской равнины началось становление безлесных
тундровых ландшафтов.
О растительности раннего плиоцена к северу от нижнего течения р. Камы
дает представления пыльцевой спектр нижнекинельских отложений. Здесь
преобладает древесная пыльца, главным образом ели из секции Eupicea.
Много пыльцы сосен из подрода Haploxylon. Небольшую примесь составляет
пыльца Abies, Tsuga, Pseudotsuga, Betula, Alnus. Широколиственные породы
представлены единичными пыльцевыми зернами Quercus, Ulmus, Juglans,
Nyssa, Liquidambar. Южнее, в районе Самарской Луки, в палинологических
спектрах преобладает пыльца сосен из подрода Diploxylon с небольшой
примесью сосновой пыльцы, относящейся к подроду Haploxylon. Постоянно,
но в небольшом количестве встречается пыльца Picea, Abies, Tsuga, а также
некоторых таксодиевых, не определенная до рода. Имеется и пыльца листвен-
ных: Carya, Juglans, Pterocarya, Carpinus, Liquidambar, Fagus и др.
На основании этих пыльцевых спектров (Ананова, 1962) можно заклю-
чить, что в раннем плиоцене на востоке Русской равнины в пределах совре-
менной лесной зоны преобладали еловые леса с примесью пихты, тсуги и
псевдотсуги, а на территории современной лесостепи — светлохвойные сосно-
вые леса с примесью ели, пихты, тсуги и других хвойных. Широколиствен-
ные породы образовывали примесь в хвойных лесах или формировали местами
самостоятельные сообщества.
В среднем плиоцене в районе нижнего течения р. Камы и в Среднем
Поволжье пыльцевые спектры свидетельствуют о преобладании пыльцы ели
из секции Eupicea, большой доле пыльцы сосен из подродов Haploxylon и
Diploxylon, присутствии пыльцы Tsuga и Abies. Миоценовые реликты — Li-
quidambar, Nyssa, Juglans, Carya в среднем плиоцене постепенно исчезают.
Значительное участие в спектре принимает пыльца Ulmus и Tilia (12—20%),
в небольшом количестве встречается пыльца Carpinus, Fagus, Pterocarya,.
Weigela (Ананова, 1962). По-видимому, здесь преобладали еловые леса с
примесью сосен, тсуги, пихты. Липа, ильмовые и другие широколиственные-
18
образовывали заметную примесь в хвойных лесах или даже преобладали в
благоприятных для них местах.
В Прикаспии в это время произошла трансгрессия Каспийского моря, по-
крывшего всю равнинную часть вплоть до отрогов Общего Сырта. О расти-
тельности повышенных участков Общего Сырта в эту эпоху можно судить
по пыльцевым спектрам акчагыльских отложений. Здесь преобладает пыльца
древесных растений, главным образом хвойных (от 40—60 до 80%). Пода-
вляющая часть ее принадлежит соснам из обоих подродов; постоянную при-
месь образуют ели (два вида), тсуга, пихта. Из лиственных отмечены ольха
(два вида), береза (два вида), липа (два вида), вяз, дуб.
В палинологических спектрах позднего плиоцена северной части Русской
равнины преобладает пыльца Picea sect. Eupicea, Pinus cf. sibirica, P. cf.
silvestris, значительна примесь пыльцы Abies sibirica, Tsuga canadensis.
Пыльца широколиственных {Tilia, Ulmus, Acer, Quercus, Corylus) более обиль-
на, чем в акчагыльских спектрах. Из водных растений отмечены теплолюби-
вые Brasenia, Trapa, Salvinia. В средней полосе в спектрах преобладает'
пыльца сосны, близкой к Pinus silvestris. Некоторое потепление климата
в это время повлекло за собой, по мнению Е. Н. Анановой (1962), оттесне-
ние темнохвойных лесов к северу и продвижение за ними степей, примерно
до их современных границ. Возросла роль широколиственных пород, особенно
ильмовых и липы; несколько увеличилось значение тсуги и пихты в составе
темнохвойных лесов.
ОБЩИЙ ОБЗОР ТРЕТИЧНЫХ ИСКОПАЕМЫХ ФЛОР УРАЛА
После ознакомления с основными находками ископаемых растений на тер-
ритории Урала, следует выделить самые важные, решающие моменты в исто-
рии развития третичной флоры. Палеоценовые ископаемые комплексы евро-
пейской части СССР с магнолиями, падубом, девальквеей имеют субтропический
или почти тропический облик. Восточным вариантом этого флористического
комплекса является охарактеризованная ранее палеоценовая флора Южного
Урала и Мугоджар; в состав ее входили: каштанодуб Dryophyllum, цинна-
момум Cinnamomum, девальквея Dewalquea, ныне вымершие виды дуба Quercus,
кипарисовника Chamaecyparis, калины Viburnum, кизила Cornus, магнолии
Magnolia, лавра Laurus, фикуса Ficus. В эоцене климат стал еще более
теплым, но в ряде областей (в частности, на Южном Урале) с засушливым
периодом в летнее время. В Европе, включая и европейскую часть СССР,
была распространена пышная теплолюбивая вечнозеленая флора с мангровой
пальмой Nipa, пальмой Sabal, вечнозелеными представителями родов Laurus,
Cinnamomum, Oreodaphne и др. На Южном Урале, где летний засушливый
период был выражен сильнее, флора в эоцене была представлена вечнозеле-
ными деревьями и кустарниками с узкими листьями ксероморфного строения
(из родов Myrica, Ficus, Laurus, Dryophyllum, Quercus, Cinnamomum, Andro-
meda, Myrsine и др.); здесь же отмечены пальма Sabal, хвойные были
представлены родом Sequoia. Судя по данным пыльцевого анализа, на восточ-
ном склоне Южного Урала в это время произрастали представители родов
Pinus subgen. Haploxylon, Betula, Salix, Juglans, Carya, Castanea, Quercus,
Acer, Rhus, а также сем. Myrtaceae и Ericaceae и др. На севере и северо-
востоке Русской .платформы развивалась, по-видимому, более умеренная флора.
Урал в эоцене омывался с востока Тургайским морем и был отделен от
Ангариды. Поэтому его растительность формировалась тогда под влиянием
западных элементов. На северо-востоке Азии, в пределах Ангарского материка,
в эоцене развивалась умеренная флора с тополем, ольхой, листопадными
видами дуба и другими, но в то же время в ее составе сохранились неко-
торые более древние теплолюбивые формы (Криштофович, 1941; Баранов,
2*
19
1959). В конце олигоцена в результате медленного поднятия Западно-Сибир-
ской низменности Тургайский пролив исчез. Перемычка суши между Уралом?
и Ангаридой, возможно, возникла и несколько раньше. В связи с похолода-
нием климата полтавская флора стала отступать к западу и юго-западу. По-
пятам за ней из Сибири через Урал шла сформировавшаяся на территории
Ангариды умеренная листопадная флора тургайского типа. Олигоценовая флора
Урала представлена очень бедной палеоботанической документацией. Здесь
найдены отпечатки Sequoia, Taxodium, Quercus, Ulmus, Corylus, Oreodaphne.
Данные пыльцевого анализа свидетельствуют, что в олигоцене на территории
Урала произрастали леса из хвойных (Pinus subgen. Diploxylon, Р. subgen.
Haploxylon, Taxodium, Sequoia, Tsuga, Picea, Abies, Cedrus) и листопадных
лиственных древесных пород (Alnus, Betula, Salix, Quercus, Tilia, Cary a,
Pterocarya, Carpinus, Acer, Fagus, Corylus и др.), наряду с ними сохранились,
еще некоторые вечнозеленые породы (например, Ilex'). Постепенное оттеснение
полтавского комплекса тургайским, происходившее в конце палеогена и на-
чале неогена, хорошо иллюстрируется серией ископаемых флор из Башкирии,
Подолии и окрестностей Харькова (Криштофович, 1938а, 1946). Миоценовая
ископаемая флора из Башкирии имеет уже в основном тургайский облик.
Здесь указываются различные виды Betula, Alnus, Quercus, Fagus, Carpinus,.
Juglans, Corylus, Ulmus, Castanea, Hicoria, Acer, Platanus, Diospyros, Rham-
nus, Liquidambar, Comptonia, Sassafras и др. Из числа голосемянных отме-
чены Ginkgo, Glyptostrobus, Taxodium. Лиственные деревья и кустарники
представлены, главным образом, формами с опадающей листвой, но наряду
с ними в этом комплексе сохранились некоторые вечнозеленые, преимуще-
ственно узколистные растения (Myrsine doryphora, Quercus neriifolia, Myrica.
vindobonensis).
Ксерофилизированные потомки представителей полтавского флористиче-
ского комплекса, по-видимому, еще долго сохранялись в Средней Азии
(ископаемая флора из района озера Ер-Ойлан-Дуз в Туркмении, описанная
Е. П. Коровиным); в Средиземноморье они в измененном и обедненном виде
дожили до настоящего времени, смешавшись с некоторыми компонентами:
тургайской флоры, приспособившимися к местным климатическим условиям.
Изменение климата в сторону похолодания продолжалось и в плиоцене. На.
Урале в плиоцене были распространены леса из хвойных (Pinus subgen.
Diploxylon, Р. subgen. Haploxylon, Picea, Abies, Larix, Tsuga, Taxodium,.
Metasequoia (?) и летнезеленых лиственных древесных пород (Quercus, Tilia,
Acer, Ulmus, Carpinus, Fraxinus, Salix, Corylus, Pterocarya, Liriodendron,
Betula, Alnus. Наряду с представителями родов, сохранившихся в совре-
менной флоре Урала, в этот комплекс входили растения, впоследствии вы-
мершие на территории Европы, но сохранившиеся на юго-востоке Азии и в
Северной Америке (виды из родов Metasequoia, Chamaecyparis, Tsuga,
Liriodendron).
ЧЕТВЕРТИЧНЫЙ ПЕРИОД (АНТРОПОГЕН)
Четвертичный период, при своей очень небольшой продолжительности,,
очень богат событиями. В это время произошли значительные изменения
физико-географических условий и растительного покрова. В течение этого
периода в результате продолжавшегося поднятия и расчленения поверхности
эрозией окончательно сформировался горный рельеф Урала. ‘
ПЛЕЙСТОЦЕН
Плейстоцен на .территории Евразии ознаменовался оледенением. В евро-
пейской части СССР обнаружены следы не менее трех ледниковых эпох
(Лаврова, 1952, Москвитин, 1952). В последнее время (Марков, Лазуков,
20
Николаев, 1965) предлагается следующая их номенклатура: окская, днепровская:
(включая московскую стадию, выраженную не везде) и валдайская. На севере-
Западной Сибири геологи установили существование двух ледниковых эпох
(Нагинский, 1946). В северной части Средней и Восточной Сибири, вслед-
ствие континентальности климата, сплошной ледниковый покров отсутствовал.
Здесь существовали только местные разобщенные горные ледники, местами,
спускавшиеся на равнины (Сакс, 1947, 1952).
На Урале в плейстоцене происходили эпейрогенические поднятия. Оледе-
нению подвергались полярная, приполярная и северная части хребта, а не-
большие леднички существовали и в южной части (Варсапофьева, 1934;
Сирин, 1947; Боч и Краснов, 1946; Колоколов и Львов, 1945; Каменский,
1957). Центром наиболее интенсивного оледенения был Приполярный Урал —
самая расширенная часть хребта, где сосредоточены крупнейшие горные-
вершины. На Урале, по мнению большинства исследователей, было два;
оледенения, рассматриваемых ими как покровные: первое, интенсивное, по-ви-
димому, совпало с днепровским, второе, более слабое — с валдайским. Следы
этих оледенений прослеживаются достаточно четко (Боч и Краснов, 1946;
Дибнер, 1953; Троицкий, 1966). По данным С. Г. Боча и И. И. Краснова,,
южная граница наибольшего оледенения на Урале проходила несколько север-
нее г. Перми, смещалась к югу и пересекла р. Чусовую на широте г. Ниж-
него Тагила, затем', перевалив через хребет, шла на север по восточному
склону через г. Серов и на широте Денежкина Камня резко поворачивала,
на восток. Некоторые специалисты допускали возможность, кроме того,,
одного-двух горных оледенений. Так, М. С. Калецкая и А. Д. Миклухо-
Маклай (1958) приводят данные, свидетельствующие о том, что на Полярном
Урале было три оледенения, из них два покровных и одно горное, не рас-
пространившееся далеко за пределы гор. Л. С. Троицкий (1966) полагает,
что решающую роль в четвертичном оледенении горных районов Урала играла:
трансгрессия Ледовитого океана. Он отмечает, что выделение стадий ураль-
ского оледенения и их датировка в настоящее время не могут быть проведенье
в полной мере
Раннеплейстоценовая ископаемая флора Северного Урала. О раститель-
ности Урала в период, непосредственно предшествовавший наступлению оле-
денения, дают представление опубликованные П. И. Дорофеевым (1950)
результаты обработки ископаемых остатков из торфа, образец Которого был
взят из карстовой воронки в южной части Северного Урала. В торфе найдена;,
хвоя ели из секции Omorica, близкой к Picea orientalis2, обломки хвои дру-
гого вида ели из секции Eupicea (быть может, Р. obovata или Р. excelsa),
орешки березы, напоминающие Betula папа, и орешки двух других видов
берез, систематическое положение которых неясно. Кроме того, здесь же
обнаружены остатки Selaginella selaginoides, Ranunculus sceleraius, Androme-
da polifolia, Menyanthes trifoliata, Polygonum ex sect. Avicularia, Potentilla-
cf. Kuznetzowii, Scirpus sp., Carex sp., Bryales.
На основании этого списка можно заключить, что в момент образования
изученного ископаемого торфа на юге Северного Урала произрастали хвойные-
леса, составленные двумя видами елей (одна из них, относящаяся к секции
Omorica, была очень близка к современной Picea orientalis') и двумя видами
берез. Для нижних ярусов были характерны березка, близкая к карликовой,,
андромеда, селагинелла, осоки, камыш, несколько представителей разнотравья
и зеленые мхи.
Этот комплекс растений, в котором еще сохранилась вскоре исчезнувшая-
ель из секции Omorica, непосредственно предшествовал на Северном Урале-
2 П. И. Дорофеев сближает эту ель с кавказской Р. orientalis, хотя и отмечает сход-
ство изученных образцов с хвоей Р. omorica. Видовое определение подлежит уточнению,.
2Ь
наступлению ледника (начало плейстоцена или конец плиоцена). Возможно,
что он относится ко времени, соответствующему первому оледенению в Во-
сточной Европе. Флористический состав этого комплекса, его общая обеднен-
ность, отсутствие в нем широколиственнолесных видов свидетельствуют
о наступлении суровых климатических условий.
Ископаемая плейстоценовая флора Башкирии. В юго-западной и южной
части Башкирии, в бассейнах рек Белой и Сакмары собрана и обработана
• коллекция плодов и семян (Колесникова, 1957). Более богатый и древний
карпологический материал получен из озерно-старичных отложений, вскры-
вающихся у основания вторых террас. Здесь найдены остатки древесных
растений: Alnus incana, A. glutinosa, Betula alba, Quercus robur, Tilia cordala,
Salix aurita, S. cinerea, S. viminalis, Populus nigra, P. sp., Ulmus sp.,
Picea sp., Juniperus sabina, Padus racemosa, Frangula alnus. Остатки ольхи
и березы встречались постоянно, в большом количестве; вяза, ивы ушастой
и тополя Populus sp. — часто; остальных пород — редко. Из травянистых
обнаружены главным образом озерно-болотные виды: Potamogeton natans,
Hippuris vulgaris, Zannichellia palustris, Nymphaea alba, Nyphar luteum, Najas
marina, Menyanthes trifoliata, Scirpus lacustris и др.; в меньшем количестве
луговые — Rumex crispus, Polygonum lapathifolium, Filipendula ulmaria,
Thalictrum angustifolium, Lycopus europaeus и др.
Несколько иной комплекс характерен для буровато-коричневых аллювиаль-
ных лёссовидных суглинков, слагающих основную часть вторых террас. Со-
став древесных пород здесь обеднен за счет выпадения дуба, вяза и других
(более обычны Alnus glutinosa и Betula папа); из травянистых растений ис-
чезают Menyanthes trifoliata и Filipendula ulmaria, но появляются в большом
количестве сорняки или растения открытых местообитаний — Urtica dioica,
Thlaspi arvense, Neslia paniculata.
T. Д. Колесникова, работая совместно с геологами из Башкирии, предва-
рительно отнесла этот комплекс к нижнему плейстоцену. Здесь уже нет
многих теплолюбивых видов деревьев, характерных для плиоцена. Выпадение
их можно связать с приближением ледниковой эпохи, с общим возрастанием
суровости и континентальности климата. По-видимому, он был несколько
более холодным, а может быть, и более влажным, чем современный.
В. Л. Яхимович (1958), производившая геологические исследования в этом
районе, датирует рассматриваемый ископаемый комплекс эпохой, непосред-
ственно предшествовавшей оледенению.
Однако позднее, после обработки всех материалов и, по-видимому, под
влиянием взглядов П. И. Дорофеева, Т. Д. Колесникова (1957) отнесла
изученную ею ископаемую флору к среднему плейстоцену. Отсутствие в ней
таких реликтов плиоценовой эпохи, как Brasenia Schreberi, Aldrovanda vesi-
culosa и др., можно связать с более выраженным внутриконтинентальным
положением территории. Т. Д. Колесникова высказала предположение, что
растительность Башкирии от среднего плейстоцена к голоцену развивалась
спокойно, без резких перестроек, по линии постепенного выпадения более
теплолюбивых видов в связи с усилением континентальности климата. Дати-
ровка этого комплекса еще нуждается в уточнении; не исключено, что
первоначальное определение, совпадающее с заключением геологов, было
более правильным.
Раннеплейстоценовые пыльцевые комплексы Башкирского Предуралья.
В южной части Башкирского Предуралья в конце плиоцена и начале плейстоце-
на, судя по материалам спорово-пыльцевого анализа, выполненного И. Г. Воро-
ниной (1965), была распространена преимущественно степная и лесостепная
растительность. Преобладали травянистые сообщества, лесов было, мало, они
отличались однообразным видовым составом, слагались, главным образом,
сосной обыкновенной и березой бородавчатой с примесью лиственницы, липы,
вяза и дуба. Ель образовывала примесь к сосне или формировала местами
небольшие лески.
Основывалась на результатах изучения ископаемой пыльцы, И. Г. Воро-
нина (1960) пришла к выводу, что в плейстоценовое время на территории
Башкирии липа была представлена большим числом видов, чем теперь.
Наряду с Tilia cordata здесь произрастала Т. cordifolia.
Растительность перигляциальной зоны. Характер так называемой пери-
гляциальной растительности, располагавшейся близ окраины ледникового
покрова, еще достоверно не установлен. Особенно много споров вызывает
вопрос о существовании в перигляциональной зоне степной раститель-
ности.
Возможность существования в плейстоцене степей в непосредственной
близости от окраины ледника признают Е. М. Лавренко (1938, 1946),.
И. М. Крашенинников (1939) и ряд специалистов по спорово-пыльцевому
анализу (Гричук, 1945, 1950а, и др).
Е. М. Лавренко (1938) считает, что на окраине ледников располагались
гольцово-тундровые группировки. «На некотором отдалении от последних
(возможно на расстоянии нескольких десятков или даже сот километров),
на более повышенных местах, не страдавших от влияния ледниковых (флю-
виогляциальных) разливов, располагались группировки перигляциальных степей
(или лесостепи), где степные ксерофиты были перемешаны с альпийскими
и аркто-альпийскими формами» (стр. 296). По мнению этого автора, среди
перигляциальных степей по склонам речных долин, вероятно, встречались,
рощицы сосны, а в Сибири—-лиственницы, ели и других древесных пород.
Таким образом, в ледниковые эпохи к югу от материковых льдов характер-
зональности растительного покрова резко отличается от современного.
В позднее опубликованной работе Е. М. Лавренко (1946) отмечает, что
в пределах Евразии и сейчас существуют аналоги перигляциальных ландшаф-
тов. «Это, с одной стороны, высокогорные степи Тянь-Шаня, где степные
группировки подходят вплотную к альпийским лугам и где в этих высоко-
горных степях перемешаны степные и альпийские формы. В Монголии тоже
неоднократно описывался контакт между группировками высокогорными и
степными, причем в последних опять-таки имеется большая примесь альпий-
ских форм. Для Забайкалья и бассейна Верхнего Енисея описаны группировки,
где дриас сочетается со степными растениями (Г. И. Поплавская и В. В. Ре-
вер датто). В знаменитой Чуйской степи Алтая чрезвычайно пестро перемешаны
группировки степные, пустынные, бореально-болотные и даже лесные, т. е.
имеется тот комплекс, который возможно предполагать для перигляциаль-
ной полосы. Подобные явления очень характерны для северных частей внут-
ренней Азии, где климат континентальный, с очень холодной зимой и весной,
но жарким летом» (стр. 380).
' И. М. Крашенинников (1939) полагал, что по периферии ледника был
развит особый ландшафт плейстоценовой сосново-лиственнично-березовой
лесостепи, в состав которого входили наряду с лесами из сосны, лиственни-
цы и березы, участки холодной плейстоценовой степи.
В. П. Гричук (1946) приводит результаты пыльцевого анализа отложений,
эпохи максимального оледенения (образцы были взяты в пунктах, находив-
шихся на расстоянии 200—300 км от периферии ледникового покрова).
Общий состав: древесная пыльца 5—14%, травяная 64—89%, споры 1—22%.
Состав травяной пыльцы: злаки 15—-71 %, осоки 0—5%, лебедовые 0—42%,.
полыни 0—10%, Statice 1—4%. Древесная пыльца представлена сосной,
березой, а в некоторых пунктах в небольшом количестве — елью. Из этого
В. П. Гричук делает вывод, что на некотором расстоянии от края ледника,
на территории Русской равнины существовали степи с островными сосно-
выми и березовыми лесами.
23
В другой, позднее опубликованной работе, В. П. Гричук (1950а,
стр. 114—115) высказывает предположение, что в эпоху максимального
оледенения Предуралье (южная часть) и Южный Урал «представляли собой
сосново-березово-лиственничную лесостепь с переменной ролью древесных
насаждений, площадь которых временами сильно сокращалась. Здесь темно-
хвойные, в частности еловые, леса несомненно играли известную роль, осо-
бенно в западных предгорьях Южного Урала и по среднему течению р. Урала.
В последнем районе существовали также, но, вероятно, в весьма угнетенном
состоянии, элементы широколиственной флоры в виде смешанных, хвойно-
широколиственных лесов. Основной фон ландшафтов создавала травянистая
ксерофильная растительность степного облика, в составе которой значитель-
ную роль играли как злаковые, так и полынные группировки». В отличие от
И. М. Крашенинникова (1939), В. П. Гричук считает, что в плейстоценовом
лесостепном ландшафте Южного Урала и Предуралья степные участки были
не вкраплены на фоне лесов, а преобладали в растительном покрове; роль
же лесов в этом ландшафте была второстепенной.
Взгляды И. М. Покровской (1950) относительно приледниковых ландшаф-
тов Урала в плейстоцене несколько расходятся с высказываниями В. П. Гри-
чука. И. М. Покровская утверждает, что в эпоху максимального оледенения
на Урале зона лесной растительности, особенно лиственных лесов, сильно
•сузилась, и степная растительность продвинулась довольно далеко на север
и северо-запад. Степная растительность существовала в плейстоцене в усло-
виях резко континентального климата, характеризующегося очень низкими
•зимними и очень высокими летними температурами. В качестве одного из
.доказательств правильности своих предположений И. М. Покровская приводит
данные о том, что в аллювиальных отложениях третьей надпойменной тер-
расы р. Чусовой, образование которой геоморфологи относят к эпохе днепров-
'ского оледенения, обнаружен бедный пыльцевой комплекс с преобладанием
пыльцы травянистых растений.. Из этого же горизонта В. И. Громовым опре-
делены кости антилопы — сайги.
В противоположность взглядам упомянутых исследователей, Б. Н. Город-
жов в ряде своих работ (1935, 1939, 1948, 1952) развивает иную, точку
зрения. Он доказал, что представления о так называемых аркто-степных
дизъюнкциях ареалов ряда растений основаны на ошибках, незнании экологии
•отдельных видов и путанице в этикетках. Теперь на равнинах нет физико-
географических условий, позволяющих существовать вперемежку степным и
тундровым ландшафтам. Точно так же нет оснований предполагать, что такие 1
условия могли существовать в прошлом, в плейстоцене. Пыльцевой анализ не
.дает материала для решения вопроса, существовали ли степи по окраине
оледенения. Нельзя забывать, что пыльца ветроопыляемых растений может
переноситься на огромные расстояния (до 1000—1500 км). Принято истол-
ковывать присутствие в приледниковых отложениях пыльцы злаков, Artemisia
как доказательство существования здесь степей; это неверно, так как злаки
произрастают и на лугах, и в лесах, а среди полыней есть немало лесных,
тундровых и сорных видов. Распознавание даже рёцентной пыльцы полыней
и определение ее до вида очень затруднительно; поэтому не всегда можно
доверять приводимым в литературе указаниям на встречаемость ископаемой
пыльцы степных видов полыни в приледниковых отложениях. На основании
этих соображений и анализа палеогеографических данных, а также изучения
закономерностей распределения современной растительности в континенталь-
ных областях СССР, ' Б. Н. Городков пришел к выводу, что по периферии
ледника была распространена не степная, а тундровая растительность, со-
державшая в своем составе много высокогорных видов (и этим напоминавшая
современные тундры северной части Уральского хребта). Южнее располагалась
лесная зона, быть может, более узкая, чем теперь; она отделяла приледни-
24
ковую тундру и лесотундру от' степной зоны. «Никаких степей в перигля-
циальной области, — пишет Б. Н. Городков (1939, стр. 46),—со смешанной,
аркто-таежной фауной не могло быть в условиях ледникового климата, но в,
более южных территориях лесной зоны мы допускаем слабое местное остеп-
нение долинных луговых террас и южных склонов гор, особенно под воз-
действием усиленного выпаса стад копытных».
Отрицает возможность существования перигляциальных степей и Б. А. Ти-
хомиров (1946, стр. 308). Он пишет; «... мы полностью согласны:
с Б. Н. Городковым и не допускаем возможности развития по краю ледника
степных формаций и ландшафтов, во всяком случае в западных участках
евразиатской Арктики».
В последнее время ископаемая пыльца и макроскопические остатки расте-
ний из перигляциальных отложений были найдены в окрестностях г. Москвы.
На основе обработки собранных материалов В. Н. Сукачев (Сукачев и др.,
1959) дает следующую реконструкцию ландшафтов перигляциальной зоны.
Сплошного густого травяно-кустарничкового покрова, а тем более сколько-
нибудь значительного торфянистого слоя на почве тогда не было, здесь,
произрастали Betula папа, ивы, Selaginella selaginoid.es, сфагновый покров,
отсутствовал. На относительно возвышенных местах было столь сухо, что.
там мерзлотные явления не наблюдались, а развитая почва отсутствовала;,
здесь росли Ephedra, представители сем. Chenopodiaceae и рода Artemisia.,
Леса из ели, сосны и березы занимали небольшую площадь.
Ознакомимся с основными палеоботаническими данными, характеризующими;
растительность перигляциальной зоны на территории Урала и прилегающих
равнин.
Ископаемая арктическая флора Зауралья. В ледниковых отложениях
Западно-Сибирской низменности, в районе с. Демьянского на р. Иртыше и
дер. Шкариной на р. Васюгане В. Н. Сукачев (1938) нашел остатки расте-.
ний арктического комплекса, представленного полярной и травянистой ивами?
Salix polaris, S. herbacea, карликовой березой Betula папа, дриадой Dryas.
octopetala, пахиплеурумом альпийским Pachypleurum alpinum, осокой Carex
sp., голубикой Vaccinium uliginosum и др. Пыльцевой анализ показал, что в<
этих отложениях имеется пыльца сосны, лиственницы, ели, пихты, березы,
ольхи и ивы; найдены также древесина и кора сосны, ели, березы, хвоя-
пихты, ствол ели с корнями, листья пушистой березы.
Встречаемость в одних и тех же слоях ископаемых остатков арктических;
и лесных растений привела В. Н. Сукачева (1938) к выводу, что по перифе-
рии плейстоценового ледника, по крайней мере местами, к растительности^
арктического облика подходила более южная, в том числе и лесная расти-
тельность. Смещение зон растительности в плейстоцене определялось наступ-
лением или отступанием ледников. В ледниковые эпохи с их суровым Клима-,
тическим режимом арктическая растительность, свойственная окраине ледника,,
более или менее значительно продвинулась на юг. На территории, покрытой;
ледником, и на некотором расстоянии от его края лесов не было.
Ископаемая арктическая флора Северных Увалов в Предуралье. В При-,
лузском районе Коми АССР в районе Северных Увалов (вытянутая в широт-
ном направлении возвышенность, достигающая высоты 250 м над ур. м.}
геологическая партия двумя скважинами вскрыла местонахождение арктиче-.
ской флоры. Из этих образцов Т. Д. Колесникова (1966) определила следую-,
щие растительные остатки: Dryas octopetala (листья), Salix herbacea (листья
и коробочки), 8. polaris (листья), Arctous alpina (лист), Armeria arctica (ча-.
шечка с остатками плода), Betula папа (листья, плодики и чешуи), Arctosta-.
phylos uva-ursi (семена), Viola palustris (семена), Coryospermum Marschallii,
Menydnth.es trifoliata (половина семени), Potamogeton natans, P. pusillus,
P. filiformis (эндокарпии), Heleocharis palustris (орешек), Hippuris vulgaris;
25
•(эндокарпии), Selaginella selaginoides (мегаспоры), Picea obovata (семена и
хвоя).
Этот ископаемый комплекс найден под 59°30' с. ш. более чем в 500 км
•к западу от Уральских гор. Современная растительность района представлена
хвойной тайгой. Наиболее интересна находка в этом комплексе арктических
и аркто-высокогорных растений — дриады, травянистой и полярной ивы,
•арктоуса альпийского, армерии арктической. В современной флоре Прилузского
района эти растения отсутствуют. Ближайшие их местонахождения располо-
жены в горах Урала, а некоторых видов (Dryas octopetala) на известняковых
скалах по р. Ижме в Ухтинском районе Коми АССР и на р. Каме близ
пос. Полазны (севернее г. Перми); и те и другие отделены от места находки
-ископаемой флоры расстоянием не менее 450 км.
Ископаемый комплекс Северных Увалов говорит о распространении в этом
районе в один из периодов плейстоцена перигляциальной флоры и раститель-
ности, существовавшей в условиях более холодного климата и включавшей
наряду с тундровыми (арктическими и аркто-высокогорными) элементами ряд
лесных, болотных и водных растений. Т. Д. Колесникова предполагает, что
в плейстоцене тундровые сообщества существовали на таких возвышенностях,
как Северные Увалы, которые в то время, возможно, были более высокими,
чем теперь. Остатки тундровых растений могли сноситься водными потоками
в речные и озерные долины, имевшие более мягкий микроклимат (поэтому
они оказались в одном комплексе с остатками лесных, болотных и озерных
видов). Не исключено, что в то время к этому району ближе подступало
морское побережье (хотя бы его мелководные заливы)3.
Ископаемый, спорово-пыльцевой комплекс типа перигляциального из
низовий р. Камы. В нижнем течении р. Камы близ пос. Набережные Челны
в толще нижнечетвертичных осадков Е. Н. Ананова (1959) обнаружила пыль-
цу 65 наименований (родов или видов растений). Содержание древесной
пыльцы в среднем составляет 10%, недревесной — 84%, остальное приходит-
ся на долю спор папоротников и мхов. Из древесных растений отмечены
Picea excelsa, Pinus Silvestris, Betula cf. pubescens, В. папа, Alnus incana,
A. fruticosa, Tilia cor data. Недревесные растения представлены в основном
пыльцой маревых и полыней. Здесь можно выделить несколько комплексов:
а) древесные растения лесной зоны; б) кустарники и травы тундры и лесо-
тундры (Alnus fruticosa, Betula папа, Selaginella selaginoides, Cassiope)-
в) растения степей и пустынь (Eurotia, Kochia, Salsola, Artemisia, Echinops,
Ephedra и др.); г) сорные растения; д) водные и прибрежно-водные растения.
Е. Н. Ананова рассматривает этот спектр как перигляциальный. В это
время, по ее мнению, в низовьях р. Камы господствовал открытый, безлес-
ный ландшафт, в сложении которого принимали участие растения степной,
лесной и тундровой зон. Хотя в его состав входили степные элементы,
Е. Н. Ананова не склонна называть этот ландшафт степным. Возможно, что
наибольшее место в растительности занимали группировки ксерофильных полу-
кустарников. Лесные сообщества играли подчиненную роль и располагались
в пониженных местах.
Спорово-пыльцевой спектр аллювиальных отложений р. Сакмары близ
устья р. Ика. Из отложений основания третьей надпойменной террасы
В. П. Гричук (1950) выделил два комплекса пыльцы и спор: 1) лесной
3 Последнее заключение Т. Д. Колесникова (1966, стр. 735) основывает на находке
Potamogeton filiformis и Armeria arctica: Первый вид в настоящее время в Коми АССР
не встречается, он обитает главным образом в водоемах приморского и морского происхож-
дения. Что касается Armeria arctica, то ее Т. Д. Колесникова характеризует как вид, свя-
занный в своем распространении «исключительно с морскими побережьями севера европейской
части СССР». Это заключение неверно, так как Armeria arctica в горах Урала заходит на
юг до Косьвинского Камня (Горчаковский, 1966).
26
(пыльца Picea, Pinus, Betula, Tilia, Abies'), связанный с сообществами еловых
и елово-липовых лесов, и 2) степной (пыльца Artemisia, Ephedra, Chenopo-
diaceae), связанный co степями и сообществами остепненных солончаковых
пойм. На спорово-пыльцевой диаграмме можно различить две зоны: зону А
с переходными типами спектров, соответствующую фазе некоторой облесен-
ности ближайшего района, и зону Б со спектрами степного и переходного,
типов, соответствующую фазе развития ксерофильной растительности полу-
пустынно-степного облика и исчезновению местных лесов. В. П. Гричук счи-
тает, что зону Б нужно сопоставить со стадией наибольшего приближения
льдов днепровского оледенения, а зону А — с предшествующей стадией
более далекого расположения материковых льдов.
* *
*
Рассмотренные результаты палеоботанических исследований показывают,
что в северных районах Предуралья и Зауралья в непосредственной близости:
от окраины ледникового покрова существовала тундроподобная растительность,
представленная в основном арктическими и бореальными элементами.
Южнее, например, в низовьях р. Камы и в бассейне р. Сакмары, судя
по данным спорово-пыльцевого анализа, господствовали открытые безлесные
сообщества, растительность которых включала ряд степных и полупустынных,
элементов. Леса занимали подчиненные положения. Однако нет никаких
оснований рассматривать эту растительность как приледниковую, так как на.
Южном Урале оледенение было лишь незначительным и не выходило за
пределы горных районов его центральной части. Видимо, разногласия между
ботаниками по вопросу о перигляциальной растительности во многом объяс-
няются разным пониманием интенсивности плейстоценового оледенения, неоди-
наковой трактовкой ширины приледниковой полосы и даже разным смыслом,
вкладываемым в само понятие «перигляциальная зона».
Если понимать под перигляциальной зоной территорию, непосредственно
примыкающую к окраине оледенения, то не приходится сомневаться в том,
что ее растительность имела тундроподобный характер, хотя и включала ряд.
болотных, лесных и луговых элементов. Здесь были, по-видимому, распро-
странены тундры, болотца, заросли кустарников, приснежные и приручьевые-
лужайки. Общий характер зональности растительного покрова на равнинах
приближался к современному, хотя лесная зона была, вероятно, уже, чем:
теперь.
Эпоха максимального (днепровского) оледенения. По мнению И. М. Кра-
шенинникова (1939), похолодание климата накануне максимального оледенения
привело к тому, что из состава лесов на Урале более быстро (по сравнению,
с окружающими равнинами) стали выпадать теплолюбивые растения. Этот
процесс еще более усилился в днепровскую ледниковую эпоху, когда северная
часть. Урала подвергалась оледенению, а на Среднем Урале и в некоторых,
районах Южного Урала местами сложилась физико-географическая обстановка,
характерная для перигляциальной зоны.
По склонам гор Среднего и Южного Урала и в предгорьях были распро-
странены лиственничные, сосновые и березовые леса; местами на Южном:
Урале, главным образом в речных долинах, произрастали темнохвойные-
древесные породы — пихта и ель. По периферии лесной зоны, которая имела,
по-видимому, значительно меньшую ширину, чем теперь, были распространены
лиственничные, сосновые и березовые лески, перемежавшиеся с участками тра-
вянистой растительности из холодо- и засухоустойчивых видов. Флорогенети-
ческий анализ этого комплекса растений, распространенного на Урале в плейсто-
цене (Крашенинников, 1937, 1939), показал, что составляющие его виды истори-
27
чески связаны с флорой лиственничных, сосновых и мелколиственных' лесов
Восточной Сибири. Такие леса в восточной Азии очень широко распространя-
лись с конца третичного периода, и в них вырабатывались холодоустойчивые
•формы травянистых растений, оказавшихся наиболее приспособленными к су-
ровым условиям плейстоцена. Широколиственные леса в эпоху максимального
оледенения сохранились в некоторых районах Южного Урала и Предуралья.
Здесь смогли пережить суровое время максимального оледенения дуб, клен,
липа, вяз и некоторые сопутствующие им кустарники и травянистые растения.
Небольшой островок крайне обедненной неморальной флоры в это время,
по-видимому, существовал и в Зауралье (вяз, липа).
В палинологических спектрах второй половины среднего плейстоцена
Башкирского Предуралья (Яхимович, 1958) преобладает пыльца древесных
пород—-67%. В том числе на долю хвойных приходится 43,8% (главным
образом пыльца сосны, меньше — пыльца ели и пихты). Из лиственных обиль-
но представлена ольха, в меньшем количестве — береза, липа, еще меньше —
:вяз, дуб, лещина. В более южных районах Предуралья (бассейн р. Сакмары,
левобережье р. Белой) в пыльцевых спектрах возрастает роль лиственных
(содержание пыльцы липы достигает 12%, а в некоторых разрезах 70%).
Юбильно встречается пыльца вяза, лещины, березы. Судя по этим данным,
во второй половине среднего плейстоцена в горах Южного Урала и на
Уфимском плато преобладали хвойные леса, южнее же, как в Предуралье,
так и на южных отрогах Урала, были распространены лиственные (главным
образом широколиственные леса). На юге Предуралья леса чередовались
с открытыми степными пространствами, общий характер ландшафта был лесо-
степным.
Днепровско-валдайская межледниковая эпоха. Отступание днепровского
ледника сопровождалось значительным изменением растительного по-
крова.
И. М. Покровская (1950) сообщает интересные данные, характеризующие
•спорово-пыльцевые комплексы межледниковых отложений Урала, которые
предположительно относятся к днепровско-валдайскому межледниковью.
В бассейне р. Камы (в Колво-Вишерском и других районах) в межледниковой
толще определена пыльца Pinus, Picea, Betula, Alnus, Salix и других дре-
весных пород таежной зоны. Широколиственные породы в этом комплексе
представлены пыльцой Quercus, Ulmus, Carpinus, Fagus, Tilia. На p. Вычегде
у дер. Шубино в песчаных отложениях отмечена пыльца Pinus subgen.
Diploxylon, Р. subgen. Haploxylon, Picea, Abies, Larix, Fraxinus. Как видно,
:В эту межледниковую эпоху в состав лесов Предуралья наряду с обычными
элементами хвойной тайги входили некоторые широколиственные древесные
породы; не исключено, что пыльца бука и ясеня переотложенная.
Р. Е. Гитерман (1953) приводит результаты анализа пыльцы в погребен-
ном древнем торфянике, расположенном на третьей надпойменной террасе
в низовьях р. Чусовой. В подстилающих торфяник голубовато-зеленых глинах
господствует пыльца широколиственных пород: липы (свыше 50%), дуба (до
12%), вяза (до 5%), лещины (до 20%), встречаются также единичные пыль-
цевые зерна граба. Пыльца хвойных содержится в сравнительно небольшом
количестве. Пыльцевой спектр указывает на преобладание в растительном
покрове широколиственных лесов. Анализ пыльцы в различных горизонтах
торфа, перекрывающих отложения озерных глин, показывает, что в дальней-
шем широколиственные леса сменились еловыми, с незначительной примесью
широколиственных пород, а еще позднее — хвойными лесами, в составе кото-
рых господствовала ель, но также принимали участие сосна и пихта. Хотя
Р.. Е. Гитерман относит отложение подстилающих торфяник озерных глин
к миндель-рисскому межледниковью, по-видимому, более правильно датиро-
вать ее термическим оптимумом днепровско-валдайского межледниковья.
.28
В Днепрэвско-валдайскую межледниковую эпоху, в связи с потеплением
климата, верхняя граница леса повысилась, а водораздельная полоса Среднего
Урала облесилась. Сильно сократилась площадь горных тундр Южного и
Среднего Урала и, вероятно, прервалась их связь с зональными тундрами
севера. На склонах Уральских гор была распространена, в основном, темно-.
хвойная тайга из ели и пихты, лишь на восточном склоне Среднего и Южного
Урала, где климат был более континентальным, преобладали сосновые и
лиственничные леса. Широколиственные древесные породы вышли из своих
убежищ и распространились в предгорьях и на западном склоне Южного, а
отчасти и Среднего Урала. В Зауралье увеличилась площадь вязовых и ли-
повых лесов.
Эпоха валдайского оледенения. Валдайская ледниковая эпоха не сопро-
вождалась таким интенсивным оледенением, как предыдущая, днепровская.
Тем не менее в это время существовал ледниковый покров в северной части
Уральского хребта. Режим холодного континентального климата, установив-
шийся во время этого оледенения, вновь вызвал сокращение пло-
щади широколиственных лесов на территории Урала и прилегающих к
нему равнин. В хребтовой полосе расширилась площадь горных тундр.
На склонах гор стали господствовать лиственничные, сосновые и березовые
-леса.
Пыльцевые спектры конца позднего плейстоцена на территории Башкир-
ского Приуралья были изучены Н. Я- Хвалиной (1963, 1965). В бассейне
р. Сакмары на широтном участке ее течения установлено девять спорово-
пыльцевых комплексов, в том числе один лесной, шесть лесостепных и два
степных. Лесной комплекс содержит 84,8% пыльцы древесных пород, в том
числе 49,7% пыльцы Pinus, 24,4% —Alnus, 11,8%—Betula. Доля участия
других, в основном лиственных пород, не превышает 4%. В лесостепных
комплексах из древесной пыльцы преобладает пыльца Pinus silvestris, Р. sp.,
Betula и Alnus. Довольно обильна пыльца Tilia, в небольшом количестве
.встречается пыльца Ulmus, Carpinus, Corylus, Acer, Quercus.
Примерно такой же характер имеют спорово-пыльцевые комплексы конца
позднего плейстоцена с левобережья р. Белой. В противоположность этому,
комплексы правобережья (горная часть Башкирской АССР) характеризуются
преобладанием пыльцы древесных над пыльцой травянистых растений. По-ви-
.димому, в горах Урала в это время преобладали леса.
В отложениях конца позднего плейстоцена Н. Я. Хвалина (1965) обна-
ружила пыльцу граба (Carpinus betulus). Эта пыльца найдена в ряде мест
башкирской и оренбургской частей Предуралья, большей частью на востоке
этой территории. Крэме того была обнаружена пыльца Tilia platyphyllos,
встречающейся теперь лишь на крайнем юго-западе СССР. Пыльца Carpinus
betulus и Tilia platyphyllos залегает в средней части аллювия вторых речных
террас и свидетельствует, по мнению Н. Я- Хвалиной, о сравнительно мяг-
ком климате того времени, когда отлагались эти слои.
Находка пыльцы граба в отложениях конца плейстоцена хорошо согласу-
ется с литературными данными о произрастании этой древесной породы на
Общем Сырте вплоть до середины прошлого столетия.
В конце плейстоцена по мере отступания ледников и потепления климата,
леса продвигались вверх по склонам, покрывая до вершин средневысотные
горы. В верхних поясах особенно крупных гор еще долго продолжали су-
ществовать изолированные местные леднички; позднее и они растаяли, а
освободившиеся ото льда пространства заселились горнотундровой раститель-
ностью. Снизу горные тундры, отступавшие в наиболее повышенные участки
Уральского хребта, теснились лесами. В результате, некогда единая гольцовая
область Урала распалась в южной части хребта на ряд изолированных остро-
вов, связанных с наиболее крупными горными массивами.
29
ГОЛОЦЕН
Наиболее богатый материал, характеризующий изменение раститель-
ности в послеледниковое время, получен методом пыльцевого анализа. Изу-
чение ископаемой пыльцы из торфяных и сапропелевых отложений Урала
было начато Д. А. Герасимовым (1926), опубликовавшим лишь некоторые
предварительные данные о результатах своих работ. Исследования в этом
же направлении были продолжены Г. А. Благовещенским (1940, 1943).
В первой работе (1940) он изложил результаты пыльцевого анализа иско-
паемых и современных торфяников водораздельной территории между
рр. Пышмой и Исетью (Среднее Зауралье). Материалом для следующей
статьи (1943) послужили результаты исследования ископаемой пыльцы из
озерных и торфяных отложений восточного склона Уральского хребта
(в южной части Среднего Урала и северной части Южного Урала).
Г. А. Благовещенский исследовал горные озера (Тургояк, Увильды, Змеи-
ное, Сугомак), озера предгорий восточного склона хребта (Малое Миассо-
во, Таткуль) и группу равнинно-степных озер, находящихся в 14 юн к се-
веро-востоку от г. Челябинска.
В 1942 и 1943 гг. микропалеонтологическим исследованием озерно-
иловых (сапропелевых) отложений Среднего Урала занимались В. Н. Су-
качев с сотрудниками (Сукачев и Поплавская, 1946). Район работ охваты-
вал среднюю и южную части Свердловской области и северную часть Че-
лябинской и Курганской областей между 55°28' и 58°2' с. ш. и 60°00' и
66°35' в. д. - ,
В ходе этих работ были исследованы как горные озера в пределах хреб-
товой полосы Урала, так и озера, расположенные на абразионно-эрозион-
ной равнинё восточного склона Среднего Урала и на территории Западно-
Сибирской низменности. Исследования В. Н. Сукачева и Г. И. Поплав-
ской дали возможность реконструировать историю растительности вос-
точного склона Среднего Урала в течение всего голоцена, начиная с его
самых ранних стадий.
Представляет интерес сопоставление стадий развития растительности,
установленных В. Н. Сукачевым для Среднего Урала, с аналогичными
периодами, установленными Д. А. Герасимовым для Татарской АССР1
и М. И. Нейштадтом для средней полосы европейской части СССР. Соотно-
шение этих этапов послеледникового изменения растительности иллюстри-
руется табл. 1.
На основе изучения Горбуновского торфяника В. Н. Сукачев попытался
определить абсолютную продолжительность выделенных им стадий. Для
суждения о времени, потребовавшемся для накопления сапропелевой тол-
щи, была использована слоистость сапропелей, обнаруживающаяся после
их промораживания. Установлено, что эта слоистость есть результат се-
зонной периодичности отложения сапропелей. В. Н. Сукачев подсчитал,
что средняя мощность годичного слоя сапропеля равна 100 р; следователь-
но, прослойка сапропеля толщиной в 1 см образовалась за 100 лет. Отсюда
можно заключить, что вся толща сапропеля в ГорбуновскоМ торфянике
образовалась за 11 000 лет.
Время, потребовавшееся на накопление слоя торфа, перекрывающего
сапропель, исчислено В. Н. Сукачевым более ориентировочно, исходя из
археологических данных. В Горбуновском торфянике найдены следы оби-
тания первобытного человека так называемой «шигирской» культуры (ос-
татки свай и деревянной стлани, поплавки из сосновой коры, грузила, на-
конечники стрел, деревянный идол и другие предметы). Археолог Эдинг
определил, что эта стоянка существовала приблизительно 4 тыс. лет тому
назад. Археологические находки приурочены к пограничному горизонту,.
30
Таблица 1
Соотношение основных этапов истории растительности в голоцене
(по Сукачеву и Поплавской, 1946)
Периоды по Блитту и Сернандеру История ландшафтов
Средняя полоса европей- ской части СССР (по Нейштадту) Татарская АССР (по Герасимову) Средний Урал (по исследованиям Сукачева)
С убатлантический Суббореальный Атлантический Максимум ели, мно- го сосны, меньше бе- резы, мало широко- листненпых Березово-елово-сосно- вые леса. Уменьшение * (по сравнению с преды- дущим) широколист- венных пород Максимум широко- лиственных лесов, не- много березовых и со- сновых, мало ели. Сосна Сосновые и широколиствен- ные леса Широколист- венные леса. Со- сна и береза Стадия со- сновых лесов
Бореальный Безусловное господ- ство сосново-березовых лесов. Других пород ничтожное количество Преобладание сосны, много березы Стадия бере- зовых лесов
Субарктический Арктический Нижний максимум ели и ивы. Сосны и березы немного Господство ивы и березы. Мало ели и сосны Нижний мак- симум ели. Бе- реза, ива Стадия ело- во-лиственич- ных лесов •Стадия лесо- тундры
* В работе В. Н. Сукачева и Г. И. Поплавской (1946) ошибочно напеча-
тано «увеличение».
который залегает почти в середине очень уплотненного слоя торфа. Следо-
вательно, для накопления всей толщи торфа потребовалось не менее
8000 лет, а вернее — 9000 лет.
Пыльцевая диаграмма, составленная для Горбуновского торфяника,
показывает, что сосновая стадия, включая и березово-сосновую подстадию,
которая южнее соответствует березовой стадии, охватывает слой торфа,
а также слои коричневого и серого сапропеля. Это означает, что она дли-
лась 17 000 лет, из которых 8000 лет приходится на березово-сосновую под-
стадию. Елово-лиственничной стадии соответствует толща розового и зе-
леного сапропеля, отложившаяся за 3000 лет.
В результате этих расчетов В. Н. Сукачев предлагает следующую хро-
нологию:
Сосновая стадия............. 9000 лет
Березово-сосновая подстадия,
соответствующая южнее
березовой стадии......... 8000 лет
Елово-лиственничная стадия . 3000 лет
Всего. . . 20 000 лет
Конечно, полученные данные приблизительны, они дают лишь общее
представление о соотношении (во времени) отдельных стадий.
Исследованиями В. Н. Сукачева и Г. И. Поплавской, к сожалению,
не был охвачен западный склон и более северные районы Уральского хреб-
та. Поэтому предложенное указанными авторами название последней ста-
• 31
дии («стадия сосновых лесов») отражает особенности изменения раститель-
ности восточного склона Урала (исключая северную его часть), но не всего
Уральского хребта в целом. На западном склоне Урала этот последний от-
резок голоцена ознаменовался широким распространением темнохвойной
тайги.
По-видимому, на восточном более континентальном склоне Среднего и
Южного Урала в последнюю стадию голоцена были менее значительно вы-
ражены климатически обусловленные изменения в составе лесов, чем на
территории Русской равнины и на западном склоне Уральского хребта.
В частности, в этом районе очень слабо выражена фаза максимума широко-
лиственных древесных пород. Трудно установить, имела ли здесь место
так называемая ксеротермическая фаза; можно допустить, что она действи-
тельно существовала, мало отразившись на пыльцевых спектрах, харак-
теризующихся явным преобладанием сосны. С. Н. Тюремнов (1956) считает,
что пограничный горизонт отлагался в условиях климата с меньшим коли-
чеством осадков и несколько повышенной средней температурой воздуха
по сравнению с этими же показателями предшествующей фазы развития
торфяника. Наиболее резко этот горизонт выражен на тех торфяниках,
в водном режиме которых основную долю составляют атмосферные осадки
или грунтовые воды, лежащие близко к поверхности. Торфяники, питаемые
потоками грунтовых вод повышенного дебита и более глубокого залегания,
воздействия ксеротермического периода не испытали.
Описывая ход заторфовывания Горбуновского водоема близ г. Нижнего
Тагила, В. Н. Сукачев указывает: «Среди толщ торфа замечается более
разложившийся горизонт с крупными пнями сосны. Нет основания не ви-
деть в нем аналога пограничному горизонту, установленному для торфя-
ников более западных областей» (Сукачев и Поплавская, 1946, стр. 16). Пе-
риод более сухого и теплого климата, по его мнению, приходится прибли-
зительно на середину стадии сосновых лесов. В нижних частях залежей
сапропелей Г. А. Благовещенский (1940, 1943) и А. П. Жузе (1939) отме-
чают слои с остатками солоноводных диатомовых водорослей. В одном из
озер на глубине 6,5 м обнаружена прослойка угля (Жузе, 1939). Г. А. Бла-
говещенский связывает образование этих слоев с периодом усыхания озер,
когда климат был более сухим и жарким.
Учитывая, что номенклатура стадий изменения растительности, пред-
ложенная В. Н. Сукачевым, относится лишь к восточному склону Среднего
Урала и не имеет универсального значения, мы вслед за М. И. Нейштадтом
(1952, 1955, 1957) принимаем в дальнейшем изложении подразделение голо- •
цена на древний, ранний, средний и поздний.
Древний голоцен. Вначале, сразу же после отступления ледника, климат
оставался некоторое время суровым, но .не сухим. На территории Север-
ного, а отчасти и Среднего Урала еще была распространена растительность
лесотундрового характера, представленная островными лесами и безлес-
ными участками тундры с карликовой березкой, арктическими ивами и.
другими растениями. В островных лесах на севере преобладала ель, на
юге — лиственница. Затем появилась пихта, а лиственница стала шире
распространяться в северной части Урала. Отсутствие сплошного лесного
покрова содействовало развитию в это время эрозионных процессов.
В горах Южного Урала уже с начала древнего голоцена господствовал
лесной ландшафт, причем леса были не островными, а сплошными или почти
сплошными. И здесь во многих районах преобладающей древесной поро-
дой была лиственница. Однако местами наряду с лиственничными лесами,
на западном склоне хребта существовали еловые и обедненные по составу
широколиственные леса, а на восточном склоне — сосновые и березовые.
На территории Башкирского Предуралья, как об этой свидетельствуют-
пыльцевые диаграммы (Рябова, 1965), в период древнего голоцена продол-
жается формирование хвойных лесов, начавшееся еще в позднем плейсто-
цене. Преобладают сосновые леса с примесью березы, ольхи, дуба, липы.
В северной части района в составе лесов довольно большую роль играет
лиственница, редко встречается пихта (в предгорьях, не проникая далеко
на запад).
Во второй половине древнего голоцена в средней, а затем и в северной
части Урала ландшафт лесотундры сменился лесным ландшафтом с господ-
ством лиственничных и еловых лесов (ель принимала большее участие в
растительном покрове северных районов). Продолжала расселяться пихта,
а в составе лесов более южных районов несколько возросла роль сосны.
Эрозия стала менее интенсивной.
Ранний голоцен. В раннем голоцене климат стал несколько теплее.
В южной части Урала началось оттеснение лиственницы темнохвойными
древесными породами и сосной; широколиственные породы стали посте-
пенно расселяться и проникли по речным долинам на Средний Урал, а
липа — и в прилегающую часть Западно-Сибирской низменности. Зоны ра-
стительности на прилегающей к Уралу части Западно-Сибирской низмен-
ности несколько сместились к северу. На восточном склоне Среднего Урала
климат стал более континентальным, произошло некоторое потепление.
Лиственница и ель стали вытесняться березой бородавчатой и отчасти сос-
ной. Ландшафт, присущий этой стадии, по В. Н. Сукачеву, представлял
собою березовую, а местами березово-сосновую сухую лесостепь, вероят-
но, с более теплым климатом. Из сопоставления пыльцевых спектров от-
ложений разных озер можно заключить, что сосна в начале этой стадии от-
сутствовала, а затем стала распространяться, начиная с юга. Уменьшилась
роль пихты, а в ряде пунктов эта порода совсем исчезла. В это же время леса
из ели, березы и других пород продвинулись далеко на север. В Западной
Сибири появился кедр.
На севере Башкирского Предуралья в начале раннего голоцена в со-
ставе лесов лиственница сохраняет большую роль, но постепенно ее вы-
тесняют лиственные породы. На большей части остальной территории со-
сновые леса сменяются сосново-еловыми, сосново-березовыми, иногда бе-
резово-сосновыми. Из широколиственных шире распространился дуб (Ря-
бова, 1965). Южнее господствовали злаково-разнотравные степи с участ-
ками березовых и сосновых лесов; местами встречалась галофитная расти-
тельность (Чигуряева и Хвалина, 1961).
Средний голоцен. Первая половина среднего голоцена характеризуется
более мягким, относительно теплым и влажным климатом. В это время
широколиственные леса занимали значительную площадь на западном
склоне Южного и Среднего Урала. Так, в погребенном торфянике на вто-
рой надпойменной террасе р. Чусовой, образование которой относится к
послеледниковому времени, в одном из горизонтов, по данным Р. Е. Ги-
терман (1953), на долю пыльцы широколиственных пород (липы, в меньшей
степени — вяза, дуба и лещины) приходится 48%. Форпосты широколист-
венных лесов проникли по западному склону далеко на север, вплоть до
Приполярного Урала. Р. В. Федорова (19516) изучала пыльцу, содержа-
щуюся в образцах погребенного торфа на террасе р. Маньи, притока Ляпина
(Приполярный Урал). В одном из горизонтов отмечено до 3% пыльцы ши-
роколиственных пород (Ulmus, Tilia, Quercus, Corylus). Аналогичные
данные приводят Г. А. Чернов (1940) для верховьев р. Печоры и В. С. Го-
ворухин (1929) для бассейна Илыча (Северный Урал). По речным долинам
отдельные виды широколиственных пород (липа сердцелистная, вяз) уси-
ленно расселялись и в прилегающей к Уралу части Западно-Сибирской низ-
менности.
3 Заказ № 251
33
В Башкирском Предуралье в это время в составе лесов преобладали
лиственные породы — дуб, вяз, клен, липа, береза, ольха, ива, лещина.
Леса были главным образом смешанные — сосново-березово-широколист-
венные или березово-широколиственные. За счет оттеснения широколист-
венными породами уменьшилась роль сосны; в некоторых спектрах возра-
стает значение ели (Рябова, 1965).
Реликтами этой фазы активизации неморального комплекса в первой
половине среднего голоцена являются широколиственные древесные по-
роды — липа мелколистная и вяз в некоторых пунктах восточного склона
Южного Урала и Зауралья. В горных районах северной части Уральского
хребта продолжалось оттеснение лиственницы темнохвойными древесными
породами. На восточном склоне Северного, Среднего и Южного Урала
сосна оттесняла лиственницу и березу. С этого времени на восточном склоне
Среднего Урала начинают преобладать сосновые леса, удерживающие здесь
свое господство до настоящего времени. В это же время, по-видимому, не-
сколько повысился предел леса в горах и сильнее распространились
высокотравные луга в подгольцовом поясе.
Во второй половине среднего голоцена климат, оставаясь теплым, стал
более сухим. Слои торфа, время отложения которых совпадает с этой фазой
(«пограничный горизонт»), обладают темной окраской и нередко содержат
в себе пни и стволы деревьев; следовательно, торф тогда быстро разлагался,
торфяники подсыхали и зарастали лесом (Сукачев и Поплавская, 1946).
В ранней стадии развития некоторых озер восточного склона Среднего
и Южного Урала установлена солоноводная фаза, характеризующаяся
специфическим комплексом диатомовых водорослей. Так, например, в
озере Малый Шарташ Свердловской области в глинах, подстилающих са-
пропели с пресноводной флорой, найден солоноводный комплекс диатомей
(Кордэ, 1949). Ранняя стадия осолонения озер совпала с засушливой эпо-
хой среднего голоцена. В этот период многие озерные котловины стали
бессточными и поэтому подверглись осолонению. В средней и южной части
Зауралья в то время уровень озер понизился, усыхание их сопровождалось
осолонением, о чем свидетельствуют остатки солоноводных диатомовых
водорослей в озерных отложениях (Благовещенский, 1940, 1943). В степ-
ной полосе восточного склона Урала и Зауралья многие озера, вероятно,
совсем осушились (Жузе, 1952).
Изменение климата повлекло за собой смещение растительных зон в се-
верном направлении. Площадь лесов на юге сократилась, а к северу про-
двинулась степная растительность. В это время существовал крупный моно-
литный остров лесостепи в районе городов Кунгура, Красноуфимска и
Бирска; участки горной лесостепи на Южном Урале (например, в районе
гор Сугомак и Егозинская и хребта Южный Крака) сливались с лесостеп-
ными пространствами равнин.
Фрагменты степных растительных сообществ на известняковых холмах
в пределах западных предгорий Среднего Урала (например, Александров-
ские сопки близ г. Красноуфимска), а также на некоторых известняковых
горах Южного Урала (горы Егозинская и Сугомак в окрестностях г. Кыш-
тыма), где безлесье поддерживается интенсивными процессами эрозии,
следует рассматривать как реликтовые растительные сообщества сухой
и теплой фазы второй половины среднего голоцена.
В это время на территории Башкирского Предуралья участки
лугово-степной растительности чередовались лесами. В составе их наме-
тилась тенденция замены лиственных пород хвойными, главным образом
сосной (Чигуряева и Хвалина, 1961).
Поздний голоцен. В растительном покрове северной и средней части
Урала с поздним голоценом совпадает увеличение роли таежного, элемента
34
за счет оттеснения елью и пихтой широколиственных древесных пород. Как
наследие этой экспансии до наших дней на значительном пространстве
Среднего и Южного Урала среди горной темнохвойной тайги сохранились
небольшие островки липовых, реже дубовых лесов, а в древостое или под-
леске ельников и пихтачей встречаются более или менее угнетенные экзем-
пляры липы мелколистной, клена остролистного, ильма, лещины обыкно-
венной и бересклета бородавчатого. В состав травяного покрова темно-
хвойных лесов входят многочисленные неморальные реликты: Asarum еиго-
paeum, Asperula odorata, Sanicula europaea, Dryopteris filix-mas, Actaea
spicata, Stachys silvatica и др. На восточном склоне Среднего и Южного
Урала продолжалось оттеснение лиственницы сосной.
На Среднем Урале, в северной части Южного и в прилегающих равнин-
ных районах в результате расселения березовых лесов по окраине луговых
степей горная и предгорная лесостепь распалась на отдельные изолирован-
ные острова. Процесс наступления леса на степь приостановился в ряде
мест лишь в самое последнее время под влиянием хозяйственной деятель-
ности человека. Там, где влияние человека не столь интенсивно, лес про-
должает наступать на степь. Так, например, в Троицком районе Челябин-
ской области (Троицкое учебно-опытное лесное хозяйство Пермского уни-
верситета) и в настоящее время площадь березовых колков расширяется
за счет участков луговых степей. Явные признаки наступления листвен-
ничного леса на степную растительность обнаруживаются на склонах гор
Сугомак и Егозинская.
Иная картина наблюдается в более южных районах, на предгорьях
западного склона Южного Урала и на низких уровнях гор. Здесь в позднем
голоцене леса постепенно уступали место лесостепи, а южнее — степи.
В составе лесов преобладали мелколиственные породы (береза, ольха и др.),
сосна и ель. Широколиственные породы в пыльцевых спектрах представ-
лены главным образом липой. Во второй половине периода в ряде районов
еще более сокращается площадь лесов (в результате истребления их чело-
веком и иссушения климата), возрастает роль безлесных открытых про-
странств.
Остатки вымерших или редких водных растений в озерных отложениях.
Д. А. Герасимов (1926) указывал, что на Среднем Урале в торфяниках озер-
ного происхождения в верхнем слое сапропеля в изобилии найдены семена
Najas flexilis (семена определены В. С. Доктуровским). В. Н. Сукачев и
Г. И. Поплавская (1946), более детально изучившие уральские сапропели,
отмечают в них семена не только N. flexilis, но и N. marina.
Najas marina теперь на территории Урала не встречается, a N. flexi-
lis крайне редка. По «Флоре СССР», современный ареал N. marina охва-
тывает Северную и Южную Америку, Северную Африку, Западную Евро-
пу, южную и северо-западную части европейской территории СССР, Кав-
каз, Среднюю Азию, западную и Восточную Сибирь, Дальний Восток СССР,
Центральную, Южную и Юго-Восточную Азию, Австралию. В пределах
этого вида в Евразии можно выделить по крайней мере три эколого-геогра-
фические расы (Колесникова, 1965). Одна из них, к которой относится тип
вида, распространена вдоль северного морского побережья Европы от
Финского залива до Великобритании и Франции, а также вдоль атланти-
ческого побережья Северной Америки и встречается только в солоноватых
водоемах лагунного типа или в морских мелководных заливах. Вторая раса
встречается в солоноватых озерах Средней Азии и Казахстана и по побе-
режью Каспийского и Азовского морей; третья (У. marina var. brachycarpa)
известна только из оз. Алаколь в Казахстане. Вид N. marina — строго
приморский, поэтому по находкам его семян в ископаемом состоянии можно
судить о существовании здесь в прошлом или окраин морских бассейнов,
3*
35
или солоноводных водоемов морского происхождения. Ареал AC flexilis
более ограничен. Это растение произрастает в Северной Америке, Средней
Европе, Скандинавии, западной и северо-западной частях европейской
территории СССР. Изолированные местонахождения известны на Южном
Урале (в Оренбургской области и в Башкирской АССР), в Павлодарской
области, Целинном крае, в бассейне Оби в Западной Сибири (Нарым) и
на Байкале. И. М. и В. М. Крашенинниковы (1908) упоминают о находке
Najas sp. (вероятно, N. flexilis) в небольшом пресном озерке с илистым дном
у дер. Могильной на границе между б. Челябинским и Курганским уезда-
ми. Имеются также сведения о встречаемости A. flexilis в Башкирии, в
старице р. Уфы. Т. Д. Колесникова (1965) отмечает, что оторванные место-
нахождения N. flexilis связаны с водоемами ледникового происхождения.
В ископаемом состоянии семена двух указанных видов Najas, как от-
мечают В. И. Сукачев и Г. И. Поплавская (1946), начинают встречаться
даже в отложениях глины и розового сапропеля; в последнем особенно
часто находились семена N. marina. Семена того и другого вида встреча-
ются и в выше расположенном горизонте — кремнистом оливковом сапро-
пеле; здесь уже N. marina попадается реже, а вскоре и совсем исчезает.
Семена N. flexilis находились еще в ряде последующих горизонтов
сапропеля, а иногда встречались всего лишь на глубине 35—40 см от верх-
ней границы сапропеля. По мнению В. Н. Сукачева и Г. И. Поплавской,
оба вида Najas были широко распространены в уральских озерах в течение
стадии елово-лиственничных леров, березовых и большей части стадии со-
сновых лесов, а затем вымерли на Урале; причем вначале исчезла N. marina,
а затем N. flexilis. Цитируемые авторы указывают, что причины вымира-
ния на Урале обоих видов Najas очень загадочны. Остатки N. marina най-
дены Т. Д. Колесниковой (1957) в четвертичных отложениях Башкирии.
Справедливо предположение (Колесникова, 1965), что вымирание этих
видов связано с существенными изменениями физико-географической об-
становки, происшедшими в течение четвертичного периода, в частности —
с увеличением континентальности климата и уменьшением обводненности
территории.
ГЛАВА II
СКАЛЬНАЯ ФЛОРА УРАЛА
В различных частях Урала и Приуралья на дневную поверхность вы-
ходят обнажения разнообразных горных пород. Повсеместно, от северной
оконечности хребта до его южных окраин, встречаются обнажения извест-
няков в виде причудливых прибрежных утесов. Более ограниченное рас-
пространение имеют обнажения некоторых метаморфических пород —
гнейсов, сланцев и др. В хребтовой полосе Урала обнажаются магматиче-
ские горные породы—ультраосновные (перидотит, дунит), основные
(габбро, диабаз), средние (диорит, сиенит) и кислые (гранит).
Хотя площадь таких обнажений относительно невелика, они служат
местом обитания многих редких и интересных растений, в том числе энде-
миков и реликтов. Своеобразие скальной флоры отметил еще X. Лессинг
(Lessing, 1834), выделявший на Южном Урале особую скальную область —
Regio lapidea. Впоследствии скальная флора Урала привлекала внимание
многих исследователей (Крылов, 1878; Гордягин, 1895; Поле, 1907, 1912;
Говорухин, 1929, 1937; Ланина, 1940; Игошина, 1966; Пономарев, 1938,
1949, 1951; Грюнер, 1960; Юдин, 1963; Горчаковский, 1951, 1964 и др.).
Автор этой книги занимался в течение ряда лет изучением скальной
флоры Урала и Предуралья. Большой материал был собран во время ло-
дочных маршрутов по рекам Северной Тошемке, Вижаю, Вишере, Режу,
Сылве, Чусовой, Юрюзани, Белой, экспедиций и экскурсий по Приполяр-
ному, Северному, Среднему и Южному Уралу с прилегающей равнинной
территорией. На основе этого материала, с учетом опубликованных резуль-
татов работ других исследователей, можно сделать общий обзор скальной
флоры Урала и Предуралья, выявить некоторые закономерности ее состава
и распределения. Флора скальных обнажений высокогорий в излагаемом
далее обзоре не рассматривается, так как ей посвящен особый раздел в ра-
нее опубликованной монографии (Горчаковский, 1966).
ИЗВЕСТНЯКОВЫЕ СКАЛЫ НА ЗАПАДНЫХ ПРЕДГОРЬЯХ
СЕВЕРНОГО И ПРИПОЛЯРНОГО УРАЛА
Правые притоки р. Печоры — Илыч, Подчерем, Щугор и Уса с относя-
щимися к ее бассейну реками Б. Сыня и Кожим (рис. 1), выходя с гор на
предгорную всхолмленную равнину, пересекают сложенные известняками
продольные увалы, называемые местным населением «пармами» (Высокая
Парма, Ыджид Парма, кряж Чернышева). Здесь часто встречаются обна-
жения известняков в виде утесов или сплошных каменных стен. Раститель-
ное население этих известковых обнажений в общем однотипно, хотя и
варьирует в зависимости от их размеров и площади, географической широгы
и положения по отношению к горной цепи. Наиболее богатый и лучше изу-
ченный скальный комплекс связан с утесами по р. Илычу (Говорухин,
1929, 1937, Ланина, 1940, Юдин, 1963), поэтому он может быть принят за
эталон скальной флоры этого района. Скальные комлексы по другим упо-
37
Рис. 1. Известняковые утесы на берегу р. Кожима.
мянутым рекам несколько обеднены по сравнению с илычским, хотя они
иногда содержат некоторые виды, отсутствующие на р. Илыче (в этом слу-
чае местонахождения видов указаны в скобках).
Мы не будем упоминать довольно многочисленные виды обычных лес-
ных, луговых и болотных растений, произрастающих на скалах, а остано-
вимся на специфических растениях, характерных именно для скал и от-
сутствующих в соседних зональных и интразональных растительных со-
обществах. Эти растения могут быть подразделены на следующие группы:
а) бореальные скальные виды — Dryopteris Robertiana, Cystopteris
fragilis, Woodsia glabella, W. alpina (реки Щугор, Подчерем), W. il vensis
(реки Щугор., Подчерем), Cryptogramma crispa, C. Stelleri, Asplenium vi-
ride, A. ruta-muraria, Polystichum lonchitis, Polypodium vulgare, Allium
strictum, Epipactis rubiginosa, Dendranthema Zawadskii, Cortusa Matthiolv,
38
б) эндемичные высокогорные виды — Scorzonera Ruprechtiana, Gypso-
phila uralensis (p. Щугор);
в) аркто-высокогорные виды с более широким ареалом—Salix reti-
culata, Minuartia verna, Saxifraga oppositifolia, S. nivalis, S. caespitosa,
Potentilla nivea, P. Ruznetzowii, Dryas octopetala, D. punctata, Gypsophila
uralensis (Щугор), Oxytropis sordida (Щугор), Viola biflora, Arctous alpina
(Щугор), Draba hirta (Щугор, Б. Сыня); Rhodiola rosea (Щугор);
г) лесостепные виды — Aster alpinus, Artemisia sericea.
Нетрудно заметить, что в этом комплексе широко представлена группа
бореальных скальных видов, велика роль аркто-высокогорных видов,
распространенных в горах Урала выше, границы леса, но встречающихся
в реликтовом состоянии на известняковых обнажениях в предгорьях.
Характерно присутствие, хотя и в небольшом количестве, высокогорных
эндемиков, также имеющих здесь реликтовую природу. Доля участия лесо-
степных видов незначительна, хотя сам факт произрастания здесь этих
растений в столь значительном удалении от основной области их распро-
странения очень интересен.
ОБНАЖЕНИЯ ИЗВЕСТНЯКОВ НА ЗАПАДНЫХ ПРЕДГОРЬЯХ
СЕВЕРНОГО УРАЛА. В БАССЕЙНЕ РЕКИ ВИШЕРЫ
В месте пересечения р. Вишерой и ее правым притоком р. Колвой По-
людова кряжа и других предгорных возвышенностей на некоторых участ-
ках русла обнажаются известняками в виде крупных прибрежных утесов.
Наиболее интересны с ботанической точки зрения Камень Дивий на
р. Колве (севернее пос. Ныроб), Камни Ветлан и Говорливый на р. Вишере
(первый — выше г. -Красновишерска, второй — ус. Говорливого).
Первым ботаником, посетившим эти обнажения, был П. Н. Крылов
(1878). Впоследствии здесь проводили исследования А. Н. Пономарев (1949)
и автор этой книги.
На обнажениях известняков в этом районе произрастают бореальные
скальные виды Aspleniutn ruta-muraria, Sedum purpureum, Allium strictum,
высокогорный эндемик Gypsophila uralensis (Ветлан),. аркто-высокогорные
виды Saxifraga caespitosa и Minuartia verna (оба на Ветлане), скально-гор-
ностепные эндемики Libanotis sibirica, Dianthus acicularis, Scorzonera
Ruprechtiana, Astragalus Helmii (Ветлан, Говорливый), Silene baschkirorum
(Ветлан). Здесь же встречается довольно много лесостепных и степных ра-
стений, причем большая часть их сконцентрирована на утесах по р. Вишере,
особенно на Говорливом Камне: Artemisia sericea, A. armeniaca, A. borea-
lis, Festuca pseudovina, Aster alpinus, Pulsatilla flavescens, Bupleurum mul-
tinerve, Astragalus danicus, Alyssum biovulatum, Anemone silvestris, Achy-
rophorus maculatus, Vicia multicaulis.
Для этих обнажений характерно сочетание степняков с арктовысоко- .
горными видами. Однако эти несходные по экологии растения занимают
разные местообитания: в то время как степняки сосредоточены на сухих
освещенных склонах южной экспозиции, аркто-высокогорные виды про-
израстают в сырых тенистых расщелинах, откуда сочится вода.
ПРИБРЕЖНЫЕ ИЗВЕСТНЯКОВЫЕ УТЕСЫ НА ВОСТОЧНОМ СКЛОНЕ СЕВЕРНОГО
УРАЛА В БАССЕЙНЕ РЕК ЛОЗЬВЫ И СОСЬВЫ
Река Лозьва и ее притоки Ивдель, Вижай и Северная Тошемка (рис. 2)
прорезают на восточном склоне Северного Урала толщу известняков. По
берегам этих рек встречаются многочисленные известняковые утесы, даю-
щие приют многим интересным растениям.
39
Рис, 2, Река Северная Тошемка и известняковая скала на ее берегу — самый северный пункт встречаемости P/iloy. sibirica,
Обнажения девонских известняков по р. Ивделю (60°42' с. ш.) были
обследованы автором на 5-километровом участке выше порогов Пильное
(окрестности г. Ивделя). Долина реки здесь глубоко врезана. Береговые
обнажения представляют собой крупные и нередко почти отвесные зубча-
тые стены высотой до 60—70 м, сверху поросшие редкими низкорослыми
•соснами. Стены береговых обнажений покрыты несомкнутой травянистой
растительностью, ютящейся в расщелинах скал и по каменистым уступам.
На голых скалах и в расщелинах растут мхи Anomodon viticulosus, Timmia
austriaca, Thuidium abietinum, Rhytidium rugosum, Lophozia quinqueden-
tata, Hylocomium splendens, Pleurozium Schreberi. Очень обилен на скалах
папоротник Asplenium ruta-muraria-, кроме него, в глубоких расщелинах
встречаются CystopterisDickieana, С. fragilis, на задернованных участках
растет Athyrium crenathum. Из цветковых растений, произрастающих на
открытых, сильно освещенных и хорошо прогреваемых местах, характер-
ны Eritrichium uralense, Dianthus acicularis, Alyssum biovulatum, Schi-
verekia Kuzrtezovii, Thymus serpyllum, Scorzonera Ruprechtiana, Aster
alpinus, Artemisia armeniaca, Artemisia sericea, Veronica spicata, Astragalus
danicus, Agropyrum reflexiaristatum, Campanula rotundifolia var. linifolia,
Dendranthema Zawadskii (Leucanthemum sibiricum).
В затененных местах и расщелинах отмечены Sedum purpureum, Cor-
tusa Matthioli и Equisetum scirpoides. В местах накопления мелкозема
(на плоских уступах и замоховелых площадках вблизи низкорослых сосен)
произрастают Libanotis sibirica, Cypripedium guttatum, Vicia cracca, Arc-
tostaphylos uva-ursi, а также кустарники Cotoneaster melanocarpa и Juni-
perus communis
По p. Вижаю (61°10z с. ш.) известняковые прибрежные скалы (рис.З)’.
распространены от низовьев до устья р. Кула. Местами известняковые
утесы, смыкаясь друг с другом, образуют высокую, иногда отвесную камен-
ную стену.
Флористически обнажения известняков по р. Вижаю близки к ивдель-
«ским. Однако здесь местами на крупных, обращенных к югу утесах доми-
нируют полыни Artemisia sericea и A. armeniaca. Нередко в расщелинах
встречаются дернинки Minuartia Helmii. На некоторых утесах растет
Dianthus repens.
Вдоль русла р. Северной Тошемки (61°20' с. ш.) обнажения девонских
известняков осмотрены начиная с пятого км от устья вверх на протяжении
10 км. Скалистые уступы здесь не столь высоки, не составляют сплошной
-стены и сильнее заросли сосновым и лиственничным лесом. Эти обнажения
•флористически более обеднены по сравнению с ивдельскими известняками.
Здесь в больших количествах встречаются Aster alpinus и Scorzonera.Rup-
rechtiana, но наряду с ними появляются новые виды — Oxytropis uralensis
и Phlox sibirica.
Флора известняковых обнажений по рр. Ивделю, Северной Тошемке
и Вижаю по своему характеру очень неоднородна. Если исключить из рас-
смотрения лесные растения и ряд видов с широкой экологической ампли-
тудой и менее определенными фитоценотическими связями, ее можно под-
разделить на следующие эколого-фитоценотические группы:
1. Уральские скальные горно-степные эндемики. Сюда относятся Mi-
nuartia Helmii, Dianthus acicularis, Agropyrum reflexiaristatum, Schi-
verekia Kuznezovii, Eritrichium uralense.
2. Бореальные скальные виды. В эту группу входят широко распро-
страненные в пределах бореальной зоны скальные растения Sedum purpu-
reum, Asplenium ruta-muraria и Cystopteris Dickieana.
3. Уральские высокогорные эндемики. Единственный представитель —
Scorzonera Ruprechtiana.
41
i
i
Рис. 3, Река Вижай на Северном Урцле,
4. Аркто-высокогорные виды. Растения, свойственные арктическим
тундрам и высокогорьям, но встречающиеся в реликтовом состоянии на
прибрежных известняковых скалах низких уровней гор. Эту группу пред-
ставляет Dianthus repens.
5. Лесостепные и степные виды. К этой группе принадлежат растения,
широко распространенные в горной лесостепи и степи Южного Урала, а
нередко и в лесостепной и степной зонах равнин. По обнажениям известня-
ков они проникают довольно далеко на север, в глубь бореально-лесной
зоны. Представителями этой группы являются Veronica spicata, Artemisia
sericea, A. armeniaca, Phlox sibirica, Alyssum biovulatum, Aster alpinus,
Dendranthema Zawadskii.
Для многих из упомянутых лесостепных, степных видов и уральских
эндемичных скально-горностепных видов — Phlox sibirica, Eritrichum
uralense, Dianthus acicularis, Veronica spicata — местонахождения на из-
вестняковых скалах по рр. Ивделю, Вижаю и Северной Тошемке являются
самыми северными на Урале или, по крайней мере, на его восточном склоне.
Сходный, но значительно более обедненный комплекс скальных расте-
ний обнаружен на правом берегу р. Сосьвы близ с. Тренькино под 60°25'
с. ш. (в районе бывшего заповедника, ныне промыслового хозяйства «Де-
нежкин Камень»). Известняковые скалы здесь невысокие (до 10 м). На них
в тенистых расщелинах растут папоротники Cystopteris fragilis, Crypto-
gramma Stelleri, Asplenium ruta-muraria, Dryopteris Robertiana, а на более
освещенных местах Dendranthema Zawadskii, Carex alba, Epipactis rubi-
ginosa, Pulsatilla flavescens, Adonis sibiricus. В тенистых местах у под-
ножия скал, где сочится вода, растет Cortusa Matthioli.
ГРАНИТНЫЕ ОСТАНЦЫ НА ВОСТОЧНОМ СКЛОНЕ СРЕДНЕГО УРАЛА
На восточном склоне Среднего Урала, например, в радиусе 50—70 км
ют г. Свердловска, среди леса, преимущественно на возвышенных местах,
встречаются многочисленные выходы кислых магматических горных
пород—гранитов (содержание кремнезема около 70%) в виде останцев и
глыб. Гранитные останцы достигают значительной высоты (до 15—18, а
иногда до 40 м), и очертания их порой очень причудливы. Многие выходы
скал не имеют собственных названий и объединены под общим наимено-
ванием «каменных палаток» или «шиханов».
К северо-западу от г. Свердловска, в пяти километрах от пригородной
станции Исеть, на вершине горы возвышается над сосновым лесом нагро-
мождение скал Чертово городище (рис. 4). Оно находится на высоте 349 м
над ур. м., представляет гряду гранитных башен-останцев, тесно смыкаю-
щихся друг с другом и образующих огромную каменную стену. Каждая
башня сложена из плоских гранитных глыб, как бы наложенных стопкой
друг на друга. Некоторые башни вверху грибообразно расширены, другие,
напротив, сужаются на вершине. Наибольшую высоту (18 м над поверх-
ностью) имеют каменные башни, составляющие центральную часть стены;
к краям же их высота постепенно снижается. Вся зубчатая гряда, выгну-
тая на вершине дугой, ориентирована своей длинной стороной в западно-
северо-западном направлении.
В районе станции Исеть менее значительные гранитные скалы выходят
на поверхность во многих других местах. Это скалы Петра Гронского, ска-
листые обнажения на горах Мотаиха и Толстиха.
К западу от г. Свердловска, в 200—250 м от ж.-д. станции Северка,
находятся очень живописные скалы, возвышающиеся как огромный утес,
над межгорной котловиной, где протекают реки Северка и Решетка.
В 2—3 км от этих скал на запад-северо-запад, по левому берегу небольшой
43
Рис. 4. Гранитные скалы Чертово городище в окрестностях г. Свердловска.
горной речки Северки (приток р. Решетки), находятся гранитные скалы,
имеющие общее название Соколиный Камень.
Около ж.-д. станции Палкино (в 1 км к юго-западу) и села того же
названия расположены Палкинские каменные палатки. Скалистый гребень
на вершине увала протянулся с юго-запада на северо-восток на 300—500 м.
Самые высокие скалы находятся в юго-западной части гребня. Они имеют
вид устремленных кверху гранитных зубьев, ячеисто изъеденных вывет-
44
риванием. Высота скал с южной стороны достигает 12—15 м, а с северной
5—6 м. Скалы сильно разрушены, крупные глыбы гранитов округлились.
Гранитные скалы Семь братьев венчают купол горы Семибратской
(442 м), расположенной к востоку от северного конца Верхнейвинского пруда.
Это самые высокие гранитные скалы в окрестностях г. Свердловска. Длина
каменной гряды, вытянутой по гребню горы Семибратской с северо-востока
на юго-запад, более 500 м. Относительная высота отдельных столбов до 30 ж,
а самый большой центральный столб имеет высоту 42 м. Преобладающая
форма залегания гранитов — столбы и матрацевидные плиты, расчленен-
ные серией вертикальных и горизонтальных трещин. Основные башни-
столбы отчленены друг от друга только в верхней своей части, а у основа-
ния скалы имеют общий цоколь. Скалы совершенно отвесны на северном
склоне и ступенчаты на южном. Зубчатый гребень в плане слегка дугообраз-
но изогнут; западная его часть ориентирована в северо-западном направ-
лении, центральная — в широтном, а восточная — в северо-восточном.
У основания южного склона нагромождены многочисленные каменные
обломки. Кроме семи самых крупных столбов, имеются менее значитель-
ные выходы гранитных глыб.
Шарташские каменные палатки расположены к югу от озера Шартащ,
в 0,8 км от берега. Скалы, венчающие вершину горы (301 м над ур. м.)
образуют каменную стену, вытянутую с юго-запада на северо-восток. Дли-
на этой каменной гряды около 200 м, ширина до 6 м. Гряда расчленена на
три более крупных выхода скал. Высота скал с южной стороны около 10 м,
с северной стороны 8 м. Граниты представлены здесь матрацевидными пли-
тами, наложенными друг на друга. Местами скалы образуют ниши, карни-
зы, сквозные отверстия. Скалы расположены среди березово-соснового
леса, сильно измененного человеком, а скальная растительность во многих
местах полностью уничтожена.
Обнажения гранита на восточном склоне Среднего Урала обычно резко
выступают над окружающей местностью в виде столбов, монолитных глыб
или нагромождения плоских плит с характерной матрацевидной отдель-
ностью. Гранитные останцы почти всегда находятся среди леса и лишь
в редких случаях превышают по высоте уровень окружающих деревьев.
Восточные, южные и западные склоны их в большей или меньшей степени
затенены древесными кронами; крутые и отвесные северные склоны сов-
сем не получают прямых солнечных лучей. Вследствие плохой прогревае-
мости, затененности и защищенности от ветров, гранитные скалы пред-
ставляют собой в общем довольно холодный и увлажненный субстрат, что
благоприятствует поселению на нем лишайников, мхов, а в местах, где
скапливается мелкозем, и некоторых сосудистых растений. Из накипных
лишайников, поселяющихся непосредственно на гранитных глыбах, наи-
более характерны Lecanora Garovaglii, L. muralis, Crocynia membranacea
и Lepraria chlorina. Очень обильны на гранитных скалах листоватые лишай-
ники Umbilicaria pennsylvanica и Gyrophora Milhlenbergii, поселяющиеся
преимущественно на отвесных северных стенах останцев и местами оде-
вающие их почти сплошь, а также Parmelia molliuscula, Р. omphalod.es и
Р. conspersa. Кустистые лишайники, например Cladonia destricta и С. chlo-
rophaea, встречаются лишь в местах скопления мелкозема в расщелинах
и на горизонтальной поверхности гранитных глыб. Мхи на гранитных глы-
бах представлены такими видами, как Ulota curvifolia и Leucodon sciuroides;
в расщелинах на мелкоземе растут Hedwigia ciliata и Thuidium abietinum,
а в нишах у основания скал на дресве — Pohlia nutans. Кроме того, на гра-
нитные скалы заходят и многие виды типично лесных мхов.
Для расщелин гранитных глыб и оснований останцев очень характерны
папоротники Polypodium vulgare (рис. 5), Cystopteris fragilis, Dryopteris
45
Рис. 5. Многоножка обыкновенная Polypodium vulgare в расщелинах гранитных скал.
Robertiana (рис. 6), D. Linnaeana, D. filix-mas. В местах, где накапли-
вается больше мелкозема — на плоских поверхностях уступов и на вер-
шинах гранитных плит •— растут деревья Pinas silvesrtis, Р. sibirica (редко,
только на Чертовом городище), Abies sibirica, Picea obovata, Betula verru-
cosa и кустарники Juniperus communis, Rubus idaeus. В нижней части гра-
нитных останцев и по расщелинам произрастают Actaea erythrocarpa, Lin-
паеа borealis, иногда Ledum palustre. У основания гранитных останцев на
скоплениях мелкоземистых продуктов разрушения гранитов в хорошо
освещенных местах встречаются Viola rupestris, Androsace septentrionalis,
Euphorbia Seguieriana и Astragalus Clerceanus. Затененные места в глубоких
Нишах и у основания скал с гранитной дресвой дают приют Parietaria mi-
crantha.
Комплекс сосудистых растений, обитающих на обнажениях гранита,
заключает в себе следующие основные группы: 1) бореальные скальные
виды, куда относятся Sedum purpureum, Parietaria micrantha и влаголюби-
вые папоротники Woodsia ilvensis, Cystopteris fragilis, Dryopteris Robertia-
na, Polypodium vulgare, обитающие в расщелинах; 2) уральские скально-
горностепные эндемики, единственный представитель — Astragalus Cler-
ceanus, иногда встречающийся с южной стороны у подножия гранитных
останцев на мелкоземистых продуктах их разрушения (Чертово горо-
дище, Палкинские каменные палатки); 3) лесные виды, характерные для
46
окружающих скалы лесов (Pinus silvestris, Picea obovata, Juniperus com-
munis, Dryopteris Linnaeana, Rubus idaeus, Linnaea borealis, Vaccinium
vitis-idaea и др.); 4) болотные виды (Ledum palustre), заходящие на скалы,
расположенные в низких сырых местах.
. i Ряс. 6, Щитовник Роберта Dryopteris Robertiana на гранитных скалах.
Нетрудно заметить, что холодные сырые скалы кислых горных пород —
гранитов населены в основном бореальными (лесными, болотными, скаль-
ными} растениями и флористически мало специфичны.
ОБНАЖЕНИЯ ДУНИТА НА УКТУССКИХ ГОРАХ В ОКРЕСТНОСТЯХ
СВЕРДЛОВСКА
Очень интересное с флористической точки зрения обнажение ультра-
основных магматических пород — дунитов (содержание SiO2 около 35%,
MgO около 43%) находится на Уктусских горах (328 ж над ур. м.) близ
южной окраины г. Свердловска. Уктусский габбро-пироксенитовый массив
залегает среди вулканогенно-осадочных пород предположительно силурий-
ского возраста. С севера и запада Уктусские горы ограничены небольшой
речкой Патрушихой (притоком р. Исети), а с востока — Нижне-Исетским
прудом. С южной стороны к Уктусским горам примыкает более высокий
Елизаветинский массив.
Уктусские горы представляют собой останцевый массив, возвышаю-
щийся над размытой поверхностью Зауральского пенеплена. Горы сло-
жены интрузивными основными и ультраосновными породами: в запад-
ной части по склонам их к долине р. Патрушихи обнажаются дуниты, на
северо-востоке местами на поверхность выходят габбро, а центральная
часть массива состоит преимущественно из пироксенитов и перидотитов.
Уктусские горы покрыты сосновыми лесами, среди которых кое-где на
поверхность выходят обнажения различных горных пород. Из них наи-
47
Рис. 7. Обнажение дунита на Уктусских горах.
Рис. 8. Костенец зеленый Asp!enitim viride на дунитсвых скалах.
больший интерес представляют обнажения дунита, хорошо выраженные
в северо-западной части массива на крутых склонах к речке Патрушихе.
Сосновый лес, окружавший в недалеком прошлом эти обнажения, сильно
расстроен рубками, а местами полностью уничтожен.
Обнажения дунита (рис. 7) имеют вид скалистых гребней и уступов на
вершинах и на крутых, иногда почти отвесных, склонах гор. Такие обна-
жения обычно не затенены кронами деревьев, хорошо освещены, сильнее
прогреваются и более подвержены действию ветров. По химическому со-
ставу дуниты резко отличаются от гранитов, большим содержанием окиси
магния за счет уменьшения количества кремнезема. В целом как скалы
дунита, так и скопления мелкоземистых продуктов их разрушения пред-
ставляют собой, по сравнению с гранитом, значительно более теплый и
сухой субстрат для поселения растений. Видовой состав лишайников,
встречающихся на этих обнажениях, значительно беднее; для дунитовых
скал характерны виды, обычно не встречающиеся на граните: Caloplaca
elegans, Physcia caesia, Candelariella vitellina. Моховой покров на таких
скалах практически отсутствует.
Из сосудистых растений непосредственно на обнажениях дунита растут
скальные бореальные виды — папоротники Asplenium viride (рис. 8) и
A. ruta-muraria, а также такие лесостепные и степные виды, как Festuca
4 Заказ № 251
49
sulcata, Echinops ritro, Centaurea sibirica, Euphorbia Seguieriana и Allium
rubens.
В этом скальном комплексе, в отличие от флоры гранитных останцев,
отсутствуют лесные и болотные растения. На восточном склоне Среднего
Урала на соответствующей широте обнажения дуниты представляют собой
наиболее сухой, хорошо прогреваемый субстрат. Поэтому с ним связаны
некоторые из наиболее северных местонахождений степных растений (Cen-
taurea sibirica, Echinops ritro'). Кроме того, на Уктусских горах (на крутых
щебнистых склонах логов, спускающихся к долине речки Патрушихи, на
взлобках у пос. Елизавета и на Арамильском холме) встречаются неболь-
шие участки степных растительных сообществ. В их состав входят, по на-
блюдениям автора и В. И. Прокаева, Filipendula hexapetala, Potentilla
argentea, P. humifusa, Onosma simplicissimum, Aster alpinus, Echinops ritro,
Centaurea sibirica, Veronica spicata, Sitene baschkirorum, Gentiana barbata,
Eoeteria gracilis, Helictotrichon desertorum, Stipa Joannis, Artemisia sericea,
Inula hirta, Antitoxicum stepposum, Trifolium montanum, Polygala comosa,
Phtomis tuberosa, Spiraea crenata, Genista tinctoria.
ОБНАЖЕНИЯ СИЕНИТА, ДИОРИТА И ГАББРО НА ВОСТОЧНОМ
СКЛОНЕ И В ВОДОРАЗДЕЛЬНОЙ ПОЛОСЕ СРЕДНЕГО УРАЛА
На Среднем Урале, кроме гранита и дунита, нередки обнажения других
магматических горных пород — интрузивных (сиенит, диорит, габбро) и
эффузивных (диабаз). Сиениты и диориты относятся к средним породам
(содержание SiO2 от 55 до 65%), а габбро — к основным (содержание SiO2
около 45%). Диабазы близки по составу к габбро, так как представляют
собой эффузивное производное габбровой магмы.
Сиениты. Долина р. Тагила на некоторых ее участках врезана в толщу
сиенитов. Самое крупное обнажение сиенита — Медведь-Камень, распо-
ложенный на р. Тагиле близ устья р. Баранчи (в 12 км от г. Нижнего Та-
гила). На этих скалах произрастают папоротники Cystopteris fragilis, Wo-
odsia ilvensis и Polypodium vulgare, а из цветковых—Allium strictum,
Sedum purpureum, Arabis hirsuta, Phtomis tuberosa, Libanotis sibirica,
Polygonatum officinale, Trifolium lupinaster, Sanguisorba officinalis, Atra-
gene sibirica, Pulsatilla flavescens, Dianthus versicolor, D. acicularis, Mi-
nuartia Helmii, Helictotrichon desertorum, Agropyrum reflexiaristatum.
Габбро-диориты. На водоразделе между pp. Чусовой и Тагилом, в ок-
рестностях пос. Висима (на территории бывшего Висимского заповедника)
находится гора Старик-Камень (753 м над ур. м.). Скалистый гребень горы,
сложенный габбро-диоритами, безлесен. На вершине горы растут Woodsia
ilvensis, Polypodium vulgare (в расщелинах), Polygonatum officinale, Sedum
purpureum, Gypsophila uralensis, Dianthus acicularis, Veronica spicata, Gali-
um boreale, Solidago virga-aurea, Trifolium, lupinaster, Chamaenerium
angustifolium, Vaccinium vitis-idaea, V. myrtillus.
Габбро. В относительно пониженной части Среднего Урала на широте
г. Свердловска в водораздельной полосе возвышается гора Волчиха (525 м
над ур. м.). Она находится на правобережье р. Чусовой. Вершина, горы,
сложенная габбро, куполообразная, увенчанная скалистым гребнем.
По глубоким расщелинам скал до самой вершины взбираются сосны, реже
лиственницы. Отвесные скалы и обращенные на юг каменистые склоны
безлесны, здесь растут Polypodium vulgare, Rubus idaeus, Vaccinium vitis-
idaea, Calamagrostis arundinacea, Dianthus acicularis, Aster alpinus, Echi-
nops ritro, Veronica spicata, Trifolium lupinaster и другие растения.
Скальный комплекс гор осевой полосы Среднего Урала, сложенных
габбро и габбро-диоритами, сильно обеднен, здесь растут обычные лесные
растения, к которым примешиваются скальные бореальные виды (Sedum
50
purpureum, Woodsia ilvensis и др.), степные и лесостепные (Veronica spicata,
Echinops ritro и др.), уральские скально-горностепные эндемики (Dianthus
acicularis'). Характерная черта — появление на более высоких горах (Ста-
рик-Камень) уральских высокогорных эндемиков (Gypsophila uralensis').
Диабазы. В левобережной части бассейна верхнего течения р. Чусовой,
на восточном склоне меридионального Уфалейского хребта неподалеку от
г. Полевского находится гора Азов (588 м над ур. м.). Склоны ее покрыты
сосновым лесом, а вершина представляет собой зубчатый гребень диаба-
зовых скал, расчлененных глубокими трещинами. Здесь растут Atragene
sibirica (цепляется за молодые деревца), Rubus idaeus, Dianthus acicularis и
другие виды.
ОБНАЖЕНИЯ [НЕКОТОРЫХ МЕТАМОРФИЧЕСКИХ ГОРНЫХ ПОРОД
НА СРЕДНЕМ УРАЛЕ
В хребтовой полосе Среднего Урала местами на дневную поверхность
выходят обнажения некоторых метаморфических пород (амфиболитов,
гнейсов, змеевиков, сланцев). Они образуют невысокие береговые о .рывы,
обычно с рыхлой корой выветривания.
Амфиболиты и гнейсы. На восточном берегу озера Исетского близ
г. Средне-Уральска имеется скалистый обрыв высотой до 10 м, где вскры-
вается толща амфиболита и гнейса. Она слагает восточное крыло Верх-
Исетского антиклинория, контактируя с гранитным массивом. Амфиболиты
и гнейсы возникли здесь за счет метаморфизма пироксеновых и плагиоклаз-
пироксеновых порфиритов и их туфов. На этих обнажениях произрастают
Allium strictum, Dianthus versicolor, Euphorbia Seguieriana, Libanotis sibirica.
Змеевики. Обнажения змеевиков сосредоточены по берегам р. Тагила
ниже г. Нижнего Тагила. На этих прибрежных утесах произрастают обыч-
ные скальные папоротники Cystopteris fragilis, Dryopteris Robertiana,
Asplenium viride, A. ruta-muraria, Polypodium vulgare и многие виды цвет-
ковых растений, в том числе Festuca sulcata, Agropyrum reflexiaristatum,
Phleum phleoides, Helictotrichon desertorum, Artemisia sericea, A. armeniaca,
Dendranthema Zawadskii, Echinops ritro, Veronica spicata, Galium verum,
Minuartia Helmii, Dianthus versicolor, Sedum purpureum, Pulsatilla flaves-
cens, Polygonatum officinale, Libanotis sibirica, Antitoxicum stepposum.
Здесь же растут кустарники Cytisus ruthenicus, Genista tinctoria, Cotone-
aster melanocarpa.
Сланцы. По берегам p. Серебрянки, притока p. Чусовой, разбросаны
обнажения сланцев. Они служат местом обитания следующих видов:
Phleum phleoides, Festuca pseudovina, Agropyrum reflexiaristatum, Poten-
tilla.humifusa, Sedum acre, S. purpureum, Dendranthema Zawadskii, Veronica
spicata, Galium verum, Pulsatilla flavescens, Arabis hirsuta, Libanotis
sibirica, Astragalus danicus, Origanum vulgare, Dracocephalum Ruyschia-
na, Antitoxicum stepposum, Allium strictum.
Сопоставление этих списков показывает, что для амфиболитов и гней-
сов характерен наиболее обедненный скальный комплекс, что, возможно,
обусловлено меньшей площадью обнажений. Обращает на себя внимание
отсутствие обычных скальных папоротников на обнажениях сланцев. Наи-
более остепнены обнажения змеевиков.
ИЗВЕСТНЯКОВЫЕ ПРИБРЕЖНЫЕ УТЕСЫ НА ВОСТОЧНЫХ
ПРЕДГОРЬЯХ СРЕДНЕГО УРАЛА (РЕКИ ТУРА, РЕЖ, ПЫШМА, ИСЕТЬ)
Некоторые реки бассейна р. Тобола, берущие начало в горах Урала,
врезаются на восточных предгорьях хребта в толщу известняков. Здесь
по берегам в окружении лесной растительности встречаются известняко-
вые утесы.
>4*
51
Рис. 9. Обнажения известняков на р. Пышме близ с. Курьи,
Рис. 10, Известняковые скалы на р. Кунаре, притоке р. Пышмы,
Рис, 11. Вид на р. Реж с Белого Камня.
Рис. 12. Бурачок двусемянный Alyssum biovulatum на Мантуровой Камне (р. Реж).
Прибрежные скалы по р. Туре (выше г. Верхотурья, близ дер. Коре-
линой и Елкиной) в свое время привлекли внимание А. Я. Гордягина
(1895), опубликовавшего описание их растительности. Против дер. Елки-
ной правый берег р. Туры представляет собой довольно высокую извест-
няковую гряду, круто обрывающуюся к реке. Системой оврагов эта гряда
расчленена на отдельные утесы, обращенные крутой стороной на запад
и юго-запад. Другая гряда длиной около 1,5 км (Дыроватый Камень) рас-
положена на левом берегу р. Туры в 3 юн выше дер. Корелиной; она имеет
форму дуги, разбитой оврагами на ряд утесов, обращенных своей открытой
стороной на одном конце дуги на юго-восток, а на другом — на северо-за-
пад. Высокие берега реки покрыты сосновым бором, редеющим к обрыви-
стой стороне утесов; отдельные сосны растут по карнизам и на обрывах.
Если исключить обычные лесные растения, иногда поселяющиеся на
утесах, флору обнажений известняка по р. Туре можно подразделить на
следующие группы: а) скальные бореальные растения — Cystopteris fra-
gilts, Asplenium. ruta-muraria, Parietaria micrantha, Sedum purpureum:,
б) скально-горностепные эндемичные для Урала—Agropyrum reflexiari-
statum, Dianthus acicularis, Schiverekia sp., Libanotis sibirica-, в) скально-
горностепные с более широким распространением — Helianthemum пит-
mularium, Carex alba, Allium strictum, Epipactis rubiginosa, Aster alpinus,
Dendranthema Zawadskii, Seseli Ledebourii; г) лесостепные и степные—-
Phleum phleoides, Helictotrichon desertorum, Oxytropis pilosa, Astragalus
danicus, Vicia multicaulis, Onobrychis tanaitica, Veronica spicata, Galium
verum, Onosma simplicissimum, Artemisia sericea, Phlomis tuberosa, Thesium
rejractum.
Несколько южнее известняковые утесы встречаются по берегам р. Режа
(приток Ницы, впадающей в Пышму) на участке между г. Режем и с. Миро-.
55
Рис. 13. Ветреница лесная Anemone silvestris на известняковых обнажениях по р. Режу
ниже г. Глинского.
ново, по берегам р. Пышмы близ с. Курьи (рис. 9) и ее правого притока
р. Кунары (рис. 10), а также по берегам р. Исети около г. Кайенск-Ураль-
ского. Флора упомянутых известняковых обнажений изучалась автором
этой книги.
На р. Реже известняковые утесы Белый Камень около г. Реж (рис. 11),
Камень Мантуров около с. Глинского и другие отличаются большим остеп-
нением. Здесь местами встречается ковыль Stipa Joannis, обильны Helicto-
trichon desertorum, Phleum phleoides, Poa stepposa, Festuca pseudovina,
Echirtops ritro, Gypsophila altissima, Antitoxicum stepposum, Dianthus ver-
sicolor, Artemisia sericea, Alyssum biovulatum (рис. 12). Кроме того, произ-
растают Aster alpinus, Anemone silvestris (рис. 13), Gypsophila Patrinii
(рис. 14). Заслуживает внимания находка очень редкого растения флокса
сибирского Phlox sibirica на скале Белый Камень в окрестностях г. Режа.
В сырых расщелинах скал произрастает Saxifraga сегпиа (рис. 15).
Флора прибрежных утесов по рекам Пышме и Исети имеет в общих чер-
тах такой же облик, однако здесь в изобилии появляется Minuartia К.га-
scheninnikovii.
ИЗВЕСТНЯКОВЫЕ СКАЛЫ ПО РЕКЕ ЧУСОВОЙ
Река Чусовая, одна из крупных водных артерий Среднего Урала, на
значительном участке своего среднего течения, от пос. Коуровки до г. Чу-
сового, вскрывает толщу известняков. В результате выветривания, эрозии
56
Рис. 14. Качим Патрэна Gypsophila Patrinii,
Рис. 15. Камнеломка поникшая Saxifraga cerniia на скалах по р. Реж.
и процессов карстообразования здесь сформировались многочисленные
прибрежные утесы, нередко причудливой формы, с крутыми склонами или
отвесными обрывами, обращенными к реке, глубокими трещинами, нишами
и пещерами (рис. 16—18). На некоторых участках утесы как бы сливаются
друг с другом, образуют сплошную каменную стену протяжением до
1—1,5 км. Самые крупные прибрежные скалы («камни», «бойцы»), напри-
мер Камни Смутной, Олений, достигают высоты 70—75 м над уровнем реки.
В нижнем течении р. Чусовой также встречаются утесы, но они меньшей
высоты и отстоят далеко один от другого.
Скальный флористический комплекс бассейна р. Чусовой в целом един,
хотя и состоит из разнородных элементов. Наблюдается некоторое варьи-
рование видового состава в зависимости от размера утесов, их положения
и экспозиции склонов. Более крупные, смыкающиеся друг с другом утесы
с отвесными стенами (Камни Олений, Омутный, Высокий, Дужной, Пере-
волочный и др.) дают приют большему числу видов растений, чем мелкие
изолированные скалы. Склоны южных и западных экспозиций значитель-
но сильнее остепнены по сравнению с северными и восточными склонами,
на карнизах и вершинах утесов степные злаки Hetictotrichon desertorum
и Festuca pseudovina образуют фрагменты сообществ, физиономически сход-
ных со степями.
58
Рис. 16. Камень Толстяк на р. Чусовой близ пос. Коуровки,
Рис. 17. Одна из башен Камня Столбы на р. Чусовой ниже устья р. Чизмы.
Ботаническое изучение известняковых скал по р. Чусовой производи-
лось К- Н. Игошиной (1966), А. Н. Пономаревым (1949), Н. М. Грюнер
(1960) и автором этих строк. На основании этих данных можно привести
список наиболее характерных и интересных растений, слагающих скаль-
ный флористический комплекс прибрежных известняков (для более редких
растений в перечне указываются места встречаемости).
60
Рис. 18. Камень Шайтан на р. Чусовой против с. Нижнего.
Рис. 19. Костенец постенный Asplenium ruta-muraria в расщелинах известняковых скал..
Группа бореальных скальных растений представлена следующими,
видами: Asplenium ruta-muraria (рис. 19), A. viride, Cryptogramma Stel-
ler i, Woodsia glabella (Камень Боюн близ г. Чусового), Allium strictum,
Epipactis rubiginosa, Sedum purpureum, S. acre, Cortusa Mathtioli, Saxifraga
sibirica, Parietaria micrantha.
Из числа скально-горностепных эндемиков отмечены: Agropyrum ref-
lexiaristatum, Roegneria viridiglumis, Dianthus acicularis, Schiverekia Kus-
nezovii, Sch. monticola, Libanotis sibirica, Aulacospermum isetense (Камни
Ростун, Высокий), Astragalus Helmii (Дужной, Дыроватый).
Уральские высокогорные эндемики представлены лишь одним видом:
Anemone biarmiensis. К аркто-высокогорной группе относятся Polygonum-
viviparum и Potentilla Kuznetzovii.
Довольно велика роль лесостепных и степных растений более широкого
распространения: Helictotrichon desertorum, Festuca pseudovina, Phleum
phleoides, Poa stepposa, Melica transsilvanica (Ростун, Дыроватый), Poly-
gonatum officinale, Dianthus versicolor, Anemone silvestris, Pulsatilla fla-
vescens, Adonis sibiricus, Arabis hirsuta, Erysimum Marschallianum, Alys-
sum biovulatum (Высокий, Олений, Ростун), Saussurea controversa, Echinops
ritro, Artemisia frigida, A. sericea, A. latifolia, Aster alpinus (рис. 20), Gali-
um verum, Veronica spicata, Phlomis tuberosa, Thymus serpyllum, Origanum
vulgare, Onosma simplicissimum, Antitoxicum stepposum, Vicia multicau-
lis, Astragalus danicus, Trifolium lupinaster, Seseli Ledebourii, Bupleurum
multinerve (Камень Боюн), Vicia multicaulis (Камни Омутный, Олений,.
Перевод очный), Carex pediformis, Genista tinctoria (у подножия скал на
мелкоземистых продуктах разрушения известняка), Cotoneaster melanocarpa,.
62
Рис. 20. Астра альпийская Aster alpinus.
Spiraea crenata (Камень Сенькин у пос. Староуткинского), Campa-
nula sibirica, Castilleja pallida, Filipendula hexapetala (Камень Сокол у пос.
Староуткинского), Gypsophila altissima.
ОБНАЖЕНИЯ ГИПСА НА ЗАПАДНОМ СКЛОНЕ СРЕДНЕГО УРАЛА
Наиболее интересные с ботанической точки зрения обнажения гипса
находятся на правом берегу р. Сылвы близ г. Кунгура, на склонах Ледя-
ной горы, внутри которой расположена известная Кунгурская ледяная
пещера. Гора сложена в основном гипсами и ангидритами кунгурского яру-
са, чередующимися с менее мощными прослойками известняка и доломита.
На обрывистом берегу р. Сылвы эти горные породы обнажены. В районе
пещеры широко распространены карстовые формы рельефа: воронки, про-
валы. Перехват и отвод дождевых и талых вод вглубь карстовыми образо-
ваниями приводит к иссушению поверхности, поэтому склоны сильно остеп-
нены.
Гипсовые обнажения на склонах большей частью представляют собой
щебнистые осыпи, слабо закрепленные растительностью. На них почти
безраздельно господствует Echinops ritro, формирующий разреженные
сообщества, напоминающие внешне каменистые горные степи Южного
Урала. Незначительную примесь в травостое образуют Sedum acre, Ast-
ragalus danicus, Centaurea sibirica, Phleum phleoides, Helianthemum num-
mularium (рис. 21), Agropyrum reflexiaristatum. Имеются также фрагменты
полынной степи, где преобладают Artemisia frigida, A. armeniaca, A. lati-
63
Рис. 21. Солнцецвет монетный Helianthemum. nummularium на обнажениях гипса
Ледяной горы близ г. Кунгура.
folia, A. macrantha. В местах, где накопилось больше мелкоземистых про-
дуктов разрушения гипса, формируются более сомкнутые сообщества, в
состав которых входят Helictotrichon desertorum, Phleum phleoides, Festuca
pseudo vina, Stipa Joannis, Veronica spicata, Anthemis tinctoria, Onosma
simplicissimurn, Centaurea sibirica, Gypsophila altissima, Salvia stepposa,
Artemisia sericea, Oxytropis pilosa, Astragalus danicus, A. sulcatus, Campanu-
la sibirica, Potentilla humifusa, Genista tinctoria и др.
64
На берегах р. Сылвы в окрестностях г. Кунгура (близ дер. Токареве
у подножия Подкаменной горы и на Спасской горе) на мелкоземистых
продуктах разрушения гипса и ангидрита встречается очень редкий энде-
мичный вид астрагала — Astragalus kungurensis.
Гипсовые прибрежные утесы встречаются также в низовьях р. Чусовой.
Некоторые из них, например Сиролова гора, примечательны тем, что на
склонах северной экспозиции, в местах, где из расщелин сочится вода,
произрастает Dryas punctata.
ОБНАЖЕНИЯ ИЗВЕСТНЯКОВ НА ЗАПАДНОМ СКЛОНЕ ЮЖНОГО УРАЛА
В БАССЕЙНЕ РЕКИ БЕЛОЙ
Русло р. Белой на значительном протяжении, от г. Белорецка до г. Ме-
леуза, врезано в толщу известняков, по берегам возвышаются известня-
ковые утесы, по размерам и красоте не уступающие чусовским (рис. 22).
Известняковые скалы нагромождены и по другим рекам бассейна Белой —
правым ее притокам Зигану, Зилиму, Симу с впадающим в него Инзером
(рис. 23), Уфе (с впадающими в нее Юрюзанью и Аем), а также по берегам
более мелких речек этого бассейна. В районе распространени известня-
ков нередки карстовые формы рельефа.— воронки, провалы, пещеры
(в том числе крупная Капова пещера на правом берегу р. Белой). Большая
часть обнажений известняка находится в горнолесном поясе, и лишь не-
многие из них, в низовьях упомянутых притоков р. Белой, располагаются
в лесостепных районах. Обычно пологие склоны утесов и их вершины об-
лесены, тогда как обрывистые склоны, обращенные к реке, безлесны и дают
приют многим видам растений. Многие растения скального комплекса бас-
сейна р. Белой распространены в пределах этого района почти повсемест-
но, но некоторые виды встречаются редко, и их местонахождения будут
оговорены.
Бореальные скальные виды и здесь составляют примечательный эле-
мент скального комплекса, однако на сухих склонах они отступают на
задний план или совсем выпадают. К этой группе относятся: Cystopteris
fragilis, Asplenium ruta-muraria, A. trichomanes (p. Сим, гора Жукова
Шишка — рис. 24), Dryopteris Robert tana, Cortusa Matthioli (в тенистых
расщелинах, у основания скал, под сводами пещер, в нишах), Parietaria
micrantha (в тени), Allium strictum, Sedum hybridum (рис. 25), Saxifraga
sibirica (p. Белая выше пос. Узян), Helianthemum rupifragum (низовья p.
Ая), Epipactis rubiginosa. Богато представлена группа скально-горностеп-
ных эндемиков: Dianthus acicularis, Agropyrum reflexiaristatum, Scutel-
laria oxyphylla, Minuartia Krascheninnikovii, 7И. Helmii (pp. Ай, Юрю-
зань), Astragalus Kfarelinianus (p. Белая у с. Биккузино), Aulacospermum
isetense, Silene baschkirorum, Schiverekia berteroides, Sch. monticola
(pp. Ай, Юрюзань), Dianthus uralensis (p. Белая).
Очень многочисленны лесостепные и степные растения широкого рас-
пространения: Роа stepposa, Helictotrichon desertorum, Festuca sulcata,
Koeleria sclerophylla, Melica transsilvanica, Stipa Joannis, Artemisia sericea,
A. campestris, A. austriaca, A. frigida, A. santolinifolia, Dianthus versicolor,
Pulsatilla flavescens, Trifolium lupinaster, Aster alpinus, Centaurea sibirica,
Echinops ritro (рис. 26), Antitoxicum stepposum, Gypsophila altissima,
Veronica spicata, Thymus serpyllum, Onosma simplicissimum, Allium rubens,
Phlomis tuberosa, Origanum vulgare, Euphorbia Seguieriana, Salvia step-
posa, Clausia aprica, Thalictrum foetidum, Polygonatum officinale, Thymus
Marschallianus, Dendranthema Zawadskii (низовья p. Ая), Galium verum,
Pimpinelld titanophila (Нугуш), Asperula petraea, Alyssum tortuosum, Bup-
leurum multinerve, Dianthus versicolor (рис. 27). Нередко на скалах обра-
5 Заказ №251 (?5
Рис. 22. Известняковый утес Шершияр на берегу р. Белой близ д. Агапкиной,
*
Рис. 23. Скалистые берега р. Сим.
Рис. 24. Костенец волосовидный Asplenium trichomanes.
Рис. 25. Очиток гибридный Sedum hybridum.
Рис. 26. Мордовник обыкновенный Echinops ritro.
зуют заросли или встречаются одиночно степные кустарники: Caragana
frutex, Cytisus ruthenicus, Genista tinctoria, Cerasus fruticosa, Spiraea crenata,
Amygdalus папа, Cotoneaster melanocarpa.
Характерно появление в этом скальном комплексе некоторых туранских
пустынно-степных видов: Ferula caspica (скалы по р. Зилиму), Atraphaxis
frutescens (скалы по рекам Зилиму, Белой близ с. Иргизла), Ephedra
distachya. На скалах по рр. Белой и Зилиму встречается Juniperus sabina.
ГОРЫ СУГОМАК И ЕГОЗИНСКАЯ НА ВОСТОЧНОМ
СКЛОНЕ ЮЖНОГО УРАЛА
На северной окраине Южного Урала, близ г. Кыштыма, расположен
Сугомакский горный узел, включающий относительно высокие змеевико-
вые массивы — горы Сугомак и Егозинскую (609 м над ур. м.). Основу
этих массивов слагают змеевики, а местами на поверхность выходят пла-
сты мрамора. Склоны гор покрыты сосновым и сосново-лиственничным ле-
сом, но на вершинах и южных скатах (не ниже высоты 500 м над ур. м.)
располагаются участки горной степи.
Степная растительность этих гор (рис. 28) изучалась П. Н. Крыловым
(1878), В. Б. Сочавой (1945) и автором этих строк. Здесь распространены
сообщества типчаково-разнотравной степи (доминанты — Festuca sulcata,
Koeleria gracilis, Thalictrum foetidum, Clausia aprica, Carex pediformis,
Artemisia frigida), кустарниковой степи (с преобладанием Spiraea crenata,
7«
Рис. 27. Гвоздика разноцветная Dianthus versicolcr.
Cerasus fruticosa, Cotoneaster tnelanocarpa, Cytisus ruthenicus. Genista tin-
ctorial и каменистой степи. Сообщества каменистой степи развиты на зме-
евиковых гребнях гор. В их состав входят эндемики: Dianthus acicularis,
Minuartia Krascheninnikovii, Eritrichium uralense, Agropyrum reflexiari-
statum, Schiverekia monticola, а также степные и лесостепные виды, имею-
щие обширный ареал: Aster alpinus, Helictotrichon desertorum, Orostachys
spinosa, Goniolimon speciosum (рис. 29), Asperula petraea, Thlaspi cochleari-
,forme, Veronica spicata, Potentilla humifusa, Centaurea sibirica, Gypsophila
altissima, Allium rubens, Alyssum biovulatum, Echinops ritro.
Сходные фрагменты горных степей встречаются несколько севернее
Сугомакского горного узла, на склонах Вишневых гор, сложенных миаски-
тами и сиенитами (Пономарев, 1959). Однако список скально-горностеп-
ных эндемиков здесь пополняется еще тремя видами: Astragalus Clerceanus,
A. Karelinianus и Oxytropis spicata.
71
№
Ptjc. 28. Фрагменты горной степи на горе Егозинской.
72
Рис. 29. Гониолимон красивый Gcniolitnon speciosum на вершине горы Егозинской.
КАМЕНИСТЫЕ СТЕПИ НА ХРЕБТЕ ИРЕНДЫК
(ВОСТОЧНЫЙ СКЛОН ЮЖНОГО УРАЛА)
К числу районов наибольшего развития каменистых горных степей от-
носится хр. Ирендык, расположенный на восточном склоне южной оконеч-
ности уральских гор на водоразделе между рр. Сакмарой и Уралом. Этот
хребет вытянут с севера на юг, общая протяженность его около 120 км,
наибольшая высота 987 м над ур. м., сложен разнообразными изливши-
мися, осадочными и метаморфическими горными породами, главным обра-
зом кварцитами и известняками.
Склоны хр. Ирендык, особенно в его северной половине, покрыты в ниж-
ней части лесами, взбирающимися языками по долинам рек и логам. Более
высокие уровни заняты разнообразными степными группировками. На ска-
листых гребнях формируются сообщества каменистой степи, в состав кото-
рых входят эндемики: Dianthus acicularis, Scutellaria oxyphylla, Minuartia
Helmii, Schiverekia berteroides, а кроме того, многие более широко распро-
страненные степные и лесостепные виды: Helictotrichon desertorum, Н.
Schellianum, Роа stepposa, Festuca sulcata, Koeleria gracilis, Onosma simp-
licissimum, Achillea nobilis, Salvia stepposa, Galatella angustissima,
Asperula petraea, Artemisia frigida, A. campestris, Echinops ritro, Centaurea
sibirica, Aster alpinus, Orostachys spinosa, Thymus mugodzaricus, Th. ser-
pyllum, Th. Marschallianus, Campanula sibirica, Carex pediformis, Alyssum
tortuosum.
73
СТЕРЛИТАМАКСКИЕ ШИХАНЫ
На фоне окружающей равнинной местности в окрестностях г. Стерли-
тамака резко выделяются четыре одиночных горы, обычно называемых
шиханами. Эти горы, сложенные известняками, находятся на правобережье
р. Белой. Они располагаются цепочкой с севера на юг, причем самая се-
верная называется Юрак-Тау, а следующие Куш-Tay, Шак-Тау (Селеук>
и Тра-Тау.
Существует несколько гипотез о происхождении этих оригинальных
форм рельефа: тектоническая, эрозионная и рифовая. В последнее время
большинство геологов склоняется к мысли, что это остатки рифовых со-
оружений, возникших в позднепермское время на грани между глубоко-
водной и мелководной зонами. Впоследствии большинство рифовых мас-
сивов, располагавшихся в виде прерывистой цепочки вдоль западного борта
предуральского прогиба, было погребено под мощной толщей гипсов,
ангидритов и каменной соли, и только некоторые участки рифовой зоны,
в том числе близ г. Стерлитамака, были приподняты и подверглись эрозии,,
в результате чего рифовые массивы оказались . обнаженными (Шамов и
Микрюков, 1960).
Самая северная гора — Юрак-Тау (рис. 30), почти коническая, длина.
1 км, ширина 850 м, высота 336 м над ур. м. (220 м над поймой р. Белой)..
Западный ее склон, обращенный к реке, крутой, остальные более пологие.
Гора Куш-Тау имеет вид небольшого трехвершинного меридиональ-
но вытянутого хребта длиной 3,5 км, шириной 1 км, высотой 371 м над.
ур. м. Западный склон более крутой, скалистый. На южном склоне, рас-
члененном промоинами на несколько отрогов, имеются скалистые обнаже-
ния.
Форму короткого хребтика длиной 1,3 км, шириной 1 км, высотой 336 At-
над ур. м. имеет гора Шак-Тау. Западный склон, омываемый речкой Се-
леук, крутой, скалистый, резко расчлененный эрозией. Гора промышлен-
но разрабатывается, поэтому естественный ландшафт ее сильно изменен..
Самая южная гора Тра-Тау — куполообразная, наибольшая высота
ее 402 м над ур. м., длина 1,2 км, ширина 0,8 км. Склоны ее довольно по-
логие, с переломом в средней части. Склоны западной экспозиции круче,
здесь имеются красивые скалистые гребни и осыпи.
Северные и отчасти восточные склоны шиханов сплошь или только в.
нижней части покрыты низкорослым порослевым широколиственным (Tilia-
cordata, Acer platanoides, Quercus robur) или осиновым (Populus tremulcfy
лесом и зарослями кустарников. Западные и южные склоны почти совер-
шенно безлесны, лишь местами по оврагам и промоинам взбираются вверх
языки низкорослого порослевого широколиственного или осинового леса.
В ложбинках и на уступах развиты заросли кустарников — Caragana fru-
tex, Cerasus fruticosa, Amygdalus папа. По окраинам лесков произрастают
Lavatera thuringiaca, Melandrium album, Galium boreale, Aconitum anthora^
Centaurea ruthenica и другие виды. Значительную площадь на склонах и
вершинах гор занимают щебнистые осыпи, обнажения известняковых
скал, участки каменистой степи. Избыток солей кальция в почве, сухость
субстрата, сильный нагрев поверхности в жаркие летние дни определяют
преобладание здесь кальцефитов и ксерофитов.
Для каменистых степей характерны следующие виды: Veronica spicata,.
Dianthus acicularis, Onosma simplicissimum, Thalictrum foetidum, Achillea;
nobilis, Echinops ritro, Campanula sibirica, Filipendula hexapetala, Libano-
tis sibirica, Agropyrum imbricatum, A. reflexiaristatum, A. desertorum,
Helictotrichon desertorum, Poa stepposa, Melica transsiluanica, Oxytropis,
pilosa, O. spicata, Astragalus Helmii, A. Karelinianus, Tanacetum uralen-
74
Рис. 30. Шихан Юрак-Тау на берегу р. Белой близ г. Стерлитамака.
se, Antitoxicum cretaceum, Gypsophila paniculata, G. altissima, Silene baschki-
rorum, Dracocephalum thymiflorum, Salvia stepposa, Berteroa incana, Aula-
cospermum isetense, Trinia muricata, Silene nutans, Anthemis tinctoria. Ha
осыпях растут Alyssum tortuosum, Euphorbia Seguieriana, Asperula petra-
ea, Chelidonium majus, Thymus Marschallianus, Th. mugodzaricus, Hedy-
sarum grandiflorum, Carduus nutans, Centaurea sibirica, C. Marschalliana,
Leonurus glaucescens и Limonium caspicum. На скалистых обнажениях
встречаются Aster alpinus, Artemisia sericea, Alyssum tortuosum, Minu-
artia Erascheninnikovii, Zygophyllum macropterum, Trinia muricata.
Скальная и горностепная растительность шиханов характеризуется
необычной концентрацией эндемиков; эти редкие растения, обыкновенно
рассеянные по удаленным друг от друга местонахождениям, здесь встре-
чаются вместе и в большом количестве: Dianthus acicularis, Silene basch-
kirorum, Oxytropis spicata, Astragalus Helmii, A. /(arelinianus, Aulacosper-
mum isetense, Tanacetum uralense, Agropyrum reflexiaristatum, Libanotis:
sibirica. Следует обратить также внимание на присутствие некоторых пу-
стынно-степных (туранских) видов, например, Zygophyllum macropterum.
Это редкое растение найдено нами на шихане Шак-Тау (Селеук). Для тер-
ритории Урала оно указывалось также из окрестностей г. Орска. Кроме
того, нами обнаружено местонахождение Limonium caspicum на шихане
Юрак-Тау — второе и самое северное из известных в Башкирии (ранее
этот вид находили в Хайбуллинском районе у сел Ново-Зиргана и Макан-
Николаевки).
Сопоставление скальных флористических комплексов показывает, что
наиболее богаты реликтовыми и эндемичными растениями обнажения из-
вестняков и гипсов. Для северных районов характерно произрастание на
скалах высокогорных эндемиков и ар кто-высокогорных видов. По мере
движения на юг эти элементы постепенно выпадают из состава скального
комплекса, причем отдельные их представители достигают Среднего Урала.
Одновременно возрастает роль скально-горностепного эндемичного элемен-
та, а также лесостепных и степных видов, имеющих более широкое распро-
странение. Колонии степных растений и фрагменты степных сообществ на
известняковых утесах по берегам рек Северного и Среднего Урала напоми-
нают по флористическому составу и внешнему облику каменистые горные
степи Южного Урала. На южной оконечности уральских гор в состав скаль-
ного комплекса входит туранский пустынно-степной элемент.
Наиболее резкий контраст между растительностью известняковых скал
и соседствующих с ними зональных типов растительного покрова наблю-
дается на Северном Урале. При продвижении к югу этот контраст в какой-
то степени стирается, и физиономически растительность скал приближается
к зональным степным сообществам окружающей горной территории. Од-
нако растительность обнажений скал и каменистых степей в значительно
большей степени, чем растительность окружающих сообществ, насыщена
эндемичными скально-горностепными видами. Это служит основанием для
предположений о ее более древнем возрасте и реликтовой природе основного-
ядра флоры скальных и каменистостепных растительных сообществ.
ГЛАВА III
ЭНДЕМИКИ УРАЛЬСКОЙ ФЛОРЫ
В течение длительного времени считалось, что флора Уральского хребта
мало отличается от флоры равнин, прилегающих к нему с запада и востока.
Высказывалось мнение о том, что уральская флора совсем не содержит
древних таксонов, а образовалась в результате смешения элементов евро-
пейского и азиатского происхождения, столкнувшихся здесь на путях свое-
го расселения.
С. И. Коржинский (1894), проводивший ботанические исследования на
территории Среднего и Южного Урала с прилегающей частью Предуралья
и хорошо знакомый с флорой и растительностью этих мест, впервые взял
под сомнение эту концепцию. Имевшиеся в его распоряжении геологиче-
ские данные свидетельствовали о том, что южная часть Уральского хребта
не была затронута оледенением; в эпоху наибольшего распространения
ледников, а отчасти в послеледниковый период Каспийское море занимало
значительно большую площадь, простираясь в виде широкого залива до
рек Камы и Белой; Западно-Сибирская равнина, прилегающая к Уралу
с востока, представляла собой обширный водный бассейн4. Исходя из этих
данных, С. И. Коржинский считал, что Южный Урал явился своеобраз-
ным островом, где смогли сохраниться остатки древней, доледниковой
флоры. Остатками этой древней флоры являются некоторые эндемичные
виды, свойственные Уралу и Приуралью, например, Mulgedium hispidum
(Syn. Cicerbita uralensis), Knautia tatarica, а также ряд европейских расте-
ний, имеющих на Южном Урале изолированные (реликтовые) участки аре-
алов, например, Sanicula europaea, Cephalanthera longifolia, Circaea lute-
tiana и др. В статье С. И. Коржинский приводит разбор двух эндемиков и
семи реликтовых видов.
Относительно европейских растений широколиственнолесного комплек-
са, встречающихся на Южном Урале в значительном удалении от основной
области их распространения, С. И. Коржинский (1894, стр. 27) писал: «Оби-
тание таких растений на Урале нельзя считать, так сказать, за аванпосты
их современного распространения. Расстояние между Уралом и восточной
границей их европейского распространения слишком велико, чтобы можно
было делать предположения о каком-либо случайном их занесении. При-
том совершенно необъяснимо было бы их отсутствие в широкой промежу-
точной полосе, которая ни по климатическим особенностям, ни по харак-
теру местности не может препятствовать обитанию этих форм. Современ-
ный характер распространения названных растений мы можем объяснить
себе лишь таким образом, что они существуют на Урале с отдаленных вре-
мен, когда растительность в большей части Европейской России была унич-
4 Согласно взглядам современных палеогеографов, единого обширного водного бассейна
на территории Западно-Сибирской низменности в то время не существовало, но поверхность
ее была испещрена огромным количеством озер.
77
тожена ледниками. С другой стороны, эти же формы сохранились также
в разных пунктах Южной Европы и, по минованию ледникового периода,
стали распространяться оттуда к северу на восток... Мы застаем их в на-
стоящее время в разных стадиях переселения; в то время как одни виды
находятся еще лишь у западной окраины России, другие уже глубоко про-
никли в ее равнину; третьи и, быть может, самые многочисленные, уже
дошли до Урала и соединили свои прежде разделенные ареалы».
Важным этапом в познании генезиса уральской флоры явились содер-
жательные статьи И. М. Крашенинникова (1937, 1939), занимавшегося в
течение многих лет флористическими исследованиями на Южном Урале
и в прилегающих к нему частях Западно-Сибирской низменности и Север-
ного Казахстана. Будучи хорошо осведомленным в палеогеографии и гео-
морфологии, И. М. Крашенинников умело использовал данные этих наук
для своих флорогенетических построений. Он кратко проанализировал
22 известных в то время на Урале эндемичных вида и 24 вида, являющихся,
по его мнению, реликтами в составе уральской флоры. Большинство рас-
смотренных этим исследователем реликтов имеет основной ареал в горах
Сибири и Монголии, отсутствует в Западно-Сибирской низменности и
представлено на Урале изолированными фрагментами ареалов или
' серией изолированных местонахождений5. На основе проведенного анализа
И. М. Крашенинников пришел к выводу о существовании в плейстоцене
непосредственной связи флоры Урала с флорой горных массивов Алтая,
Саян, Забайкалья и Северной Монголии. По его мнению, в то время непо-
далеку от окраины ледникового покрова сформировался особый тип расти-
тельности — «сосново-лиственнично-березовая плейстоценовая лесостепь».
Этому типу растительности был свойствен специфический набор видов,
довольно разнообразных по экологии, или «плейстоценовый флористиче-
ский комплекс», основу которого составляли холодостойкие травянистые
растения — дериваты более древних (доледниковых) форм, свойственных
сибирским светлым лиственничным, сосновым, березовым лесам, лугам и
растительным группировкам скалистых обнажений, а также выходцы из
Арктики и высокогорных областей Азии. Современная лесостепь Южного
Урала, как отмечал И. М. Крашенинников, сохранила много черт, прису-
щих «холодной плейстоценовой лесостепи», от которой она берет свое на-
чало, а большинство уральских реликтовых растений относится к реликтам
«плейстоценового флористического комплекса».
Высоко оценивая классические исследования И. М. Крашенинникова,
связанные с восстановлением основных этапов развития флоры и расти-
тельности Урала, не следует забывать, что этот исследователь исходил из
научных данных своего времени и в уральском цикле работ базировался
почти исключительно на ботанических материалах, относящихся к хорошо
ему известной южной части хребта. В последующие годы, преимуществен-
но в процессе работы над созданием многотомной «Флоры СССР», совет-
скими ботаниками выявлено и описано немало новых эндемичных для Урала
и Приуралья видов растений. Уже после опубликования упомянутых ра-
бот И. М. Крашенинникова на Урале проведен ряд углубленных ботанико-
географических исследований, накопились новые материалы (особенно по
флоре северных и высокогорных районов хребта), выявлены вновь или
более детально изучены некоторые реликтовые растения, внесены коррек-
тивы в прежние представления о распространении отдельных эндемичных
и реликтовых видов.
6 Почти одновременно с И. М. Крашенинниковым и независимо от него такого рода
дизъюнкции в ареалах некоторых растений были отмечены шведским ботаником Э. Хульте-
ном (Hulten, 1937).
78
В свете новых данных по экологии, распространению и происхождению
эндемичных и реликтовых элементов уральской флоры возникает необхо-
димость критического пересмотра или уточнения некоторых положений,
высказанных И. М. Крашенинниковым. Увлеченный идеей о тесных исто-
рических связях флоры Урала с флорой горных районов Сибири и север-
ной Монголии, И. М. Крашенинников недооценивал древность эндемичного
ядра уральской флоры, относя его становление к плейстоцену и включая
его в состав «плейстоценового флористического комплекса» сибирского
происхождения. Подчеркивая связи эндемиков Урала с их сибирскими
родичами, он в отдельных случаях не уделял должного внимания фактам,
свидетельствующим об их западни.., европейских, связях. Заслуживают
обсуждения и пересмотра безоговорочно принимавшиеся многими пред-
ставления И. М. Крашенинникова о характере перигляциальной расти-
тельности и о местах сохранения широколиственных лесов и свойственной
им флоры в плейстоцене.
В этой работе дается обзор эндемиков уральской флоры, по возможности
устанавливается их происхождение и возраст, предлагается их классифи-
кация. Приведенные характеристики распространения и условий произ-
растания эндемичных (а в следующем разделе и реликтовых) растений
Урала основаны на литературных данных (Флора СССР, 1934—1960; Фло-
ра юго-востока европейской части СССР, 1927—1936; Флора Казахстана,
1956—1966; Коржинский (Korshinsky), 1898; Крылов, 1927—1964; Hegi,
1906—1931, и др.), ознакомлении с гербарными материалами (гербарии
Ботанического института АН СССР, Института экологии растений и жи-
вотных Уральского филиала АН СССР и др.), а также на результатах фло-
ристических исследований, проводившихся автором в течение ряда лет на
территории Урала и прилегающих равнин.
Изучение эндемичных растений дает материал для суждения о специ-
фических чертах флоры той или иной территории, о происхождении совре-
менного растительного мира. Однако анализ эндемизма затрудняется тем,
что в ботанической литературе под названием «вид» нередко кроются кате-
гории различного таксономического ранга — начиная от крупных видов
в понимании К- Линнея и кончая подвидами, расами и т. п. Некоторые
виды описаны по скудному гербарному материалу из одного пункта, а мор-
фологические особенности, экология и . географическое распространение
их еще недостаточно выяснены (например, Cerastium Porphyrii, С. Gorod-
kovianum, С. Krylovii, Impatiens uralensis, Bromus vogulicus, B. Julii и др.).
По такому интересному полиморфному роду, как Hieracium, находя-
щемуся в расцвете формообразования и представленному на Урале рядом
эндемичных форм (Юксип, 1959 а, б), накопленные материалы настолько
бедны, что в большинстве случаев еще невозможно достоверно судить о
подлинном таксономическом значении и сродстве отдельных форм, описан-
ных в ранге видов. Еще мало изучены уральские манжетки, несмотря на
то, что они в течение ряда лет привлекали внимание знатока рода Alche-
mz7/a С. В. Юзепчука (1954, 1955 б). Иногда при выделении новых видов по
гербарным образцам, без полевых наблюдений, некоторыми авторами ис-
пользовались формальные критерии, не учитывалась встречаемость в при-
роде целой гаммы растений с переходными признаками, в результате чего
в ранг видов были возведены экологические формы (например, Rhodiola
iremelica).
Поэтому в предлагаемом обзоре исключен из рассмотрения ряд недо-
статочно изученных или сомнительных эндемичных форм, хотя некоторые
из них очень интересны. Наибольшее внимание уделено эндемичным так-
сонам, достаточно морфологически и географически обособленным и доста-
точно изученным.
79
Общее представление о количестве и систематическом составе описан-
ных для территории Урала эндемичных форм растений дает табл. 2. Как
видно из таблицы, из 116 описанных в ранге видов форм автором проана-
лизировано 94 таксона. Исключены из рассмотрения главным образом
малоизученные эндемичные формы из родов Alchemilla и Hieracium, один
вид Impatiens (/. uralensis) и один вид Rhodiola (Rh. iremelica).
Таблица 2
Систематический состав эндемиков уральской флоры
Семейство
Всего
описано
видов
Из них
рассмат-
ривается
в этой
работе
Семейство
Всего
описано
видов
Из них
рассмат-
ривается
в этой
работе
Gramineae
Liliaceae
Caryophyllaceae
Ranunculaceae
Cruciferae
Crassulaceae
Rosaceae
Leguminosae
Linaceae
8
1
11
2
3
1
36
(в tom
числе из
рода Al-
chemilla
34)
11
1
8
1
11
2
3
23
Balsaminaceae
Onagraceae
Umbelliferae
Polemoniaceae
Boraginaceae
Labiatae
Scrophulariaceae
Dipsacaceae
Compositae
1
1
2
1
1
7
2
1
26
(в tom
числе из
рода
Hieraci-
um 21)
1
2
1
1
7
2
1
19
Всего . . 116 94
К уральским эндемикам мы относим виды, распространенные исключи-
тельно или преимущественно в пределах Уральского хребта и его пред-
горий и, по всей вероятности, сформировавшиеся на этой территории. Ра-
стения, в равной степени распространенные и на Урале, и на прилегающих
равнинах, заходящие далеко на восток и запад (например, Pedicularis ura-
lensis), нами не рассматриваются.
Эндемики уральской флоры могут быть подразделены по их экологи-
ческим особенностям и ценотическим связям на три основные группы:
а) высокогорные — обитают выше границы леса (в подгольцовом, горно-
тундровом поясах и поясе холодных гольцовых пустынь) в сообществах
горных тундр, мелколесий, высокотравных подгольцовых и низкотравных
околоснежных лугов; б) скально-горностепные — обитают на скалистом,
чаще щелочном субстрате в средней и нижней частях склонов хребта
(в каменистых горных степях Южного Урала, а севернее — на береговых
известняковых или гипсовых обнажениях в пределах горнолесного пояса);
в) широколиственнолесные — свойственны дубовым, липовым, кленовым,
ильмовым и смешанным широколиственным лесам западного склона южной
части Уральского хребта и прилегающей равнины.
Высокогорные и скально-горностепные эндемики более многочисленны,
из них первые теснее связаны в своем распространении с Уральским хреб-
том. Группа эндемиков широколиственных лесов включает лишь несколько
видов.
ВЫСОКОГОРНЬЕ ЭНДЕМИКИ
Среди эндемиков высокогорной флоры Урала есть виды, сравнительно
широко распространенные по хребту, и виды узколокальные, встречаю-
щиеся только в его отдельных частях. Различаются они по своему проис-
80
Рис. 31. Лаготис уральский Lagotis uralensis на берегу снегового ручейка на горе
€ Заказ № 251
хождению и характеру родст-
венных связей с другими ви-
дами. Рассмотрим группу
высокогорных эндемиков,,
имеющих ряд близкородст-
венных викариантов в дру-
гих горных областях.
Типичный представитель
этой группы Lagotls uralensis-
(рис. 31, 32) — высокогорный
эндемик, произрастающий в
горнотундровом и подголь-
цовом поясах Северного и
центральной части Юж-
ного Урала — Ялпинг-Ньер
(рис. 33), Ойка-Чахль, Чи-
стоп, Кумба, Хоза-Тумп,
Муравьиный, Денежкин, Кон-
жаковский, Семичеловечный,
Вогульский Камни, Иремель.
Наиболее характерные места
его обитания — лужайки по
берегам ручейков, берущих
свое начало в снежниках;
встречается он также в гор-
ных тундрах. [Наиболее бли-
зок к лаготису уральскому
L. intergifolia, произрастаю-
щий в горах Азии (Алтай,
Саяны, горы Забайкалья,
Монголии и Средней Азии).
Другие родственные виды рас-
пространены в Арктике (L..
minor), на Камчатке и бли-
жайших островах (A. glaucct),.
в горах Средней [Азии (А.
decumbens, L. Ikonnikovii).
Другой весьма характер-
ный для высокогорий Урала
вид — Gypsophila uralensis-
(рис. 34) — обычно произ-
растает на высоких горных
вершинах в горнотундровомг
и подгольцовом поясах, ти-
пичные места его обитания —
скалистые останцы и некото-
рые типы горных тундр..
Очень обилен на горах Народ-
Рис. 32. Ареал лаготиса уральского Lagotis НОЙ, Манарага, хр. Сабля'
uralensis. (рис. 35), горах Ишерим, Ял-
пинг-Ньер, Чистоп, Денеж-
кин, Конжаковский Камни, Таганай, Иремель и Яман-Тау. Кроме-
того, это растение встречается в горнолесном поясе на известняковых-
береговых обнажениях по рекам Щугору, Манье, Хулге, Вишере и Лобве.
Распространен качим уральский (рис. 36) почти по всему Уралу от-
82
Рис. 33. Вид на гору Ялпинг-Ньер.
*6*
1
Рис. 34. Качим уральский Gypsophila uralensis.
Полярного (верховья рек Малой Харуты, Лопты, Нельки, Хобе-Ю и др.)
до Южного (Губерлинские горы).. Изолированное местонахождение извест-
но в районе Тиманского кряжа, где на известняковых обнажениях это
растение собирал еще Р. Р. Поле (см.. Юдин, 1946).. Близкие виды, относя-
щиеся к тому же ряду Imbricaria Fenzl, произрастают на Кавказе (в высо-
когорьях — Gypsophila tenuifolia, на скалах в ниже расположенных поя-
сах— G. Steupii) и в арктической части Сибири на обнажениях (G. Sam-
bukii). Кроме того, на красных пермских песчаниках по р. Пинеге в
Архангельской области произрастает очень близкий к G. uralensis вид—•
G. pinegensis (Перфильев, 1941). Этот вид произошел от общего с G. uralensis
предка, проникшего сюда с Урала, вероятно; в плейстоцене. Широкое
распространение лесов на территории севера европейской части СССР,
последовавшее после отступления оледенения, вызвало отмирание исход-
ной формы на равнине, за исключением некоторых мест обнажений горных
пород по рекам бассейна Северной Двины. Географическая изоляция и осо-
бые условия среды на обнажениях по Пинеге способствовали формиро-
ванию здесь местного эндемичного вида Gypsophila, который, по-видимому,
правильнее рассматривать как особую разновидность G. uralensis.
К этой же группе эндемиков относится Polemonium nudipedum — мел-
кий вид из европейской субарктической группы ряда Humilia V. Vassi.
(Клоков, 1955). Произрастает он в горах на щебнистых осыпях горнотунд-
рового пояса и в предгорьях на речном аллювии, на лугах и среди болот
на кочках. Ареал (рис. 37) расположен на Приполярном Урале (хр. Сабля,
рис. 38, и верховья р. Хобе-Ю), на Полярном Урале и в его предгорьях
84
(бассейн притоков Усы-Ад-
зьвы и Ельца, бассейн
рек Кары и Хадаты),
по северной оконечности
Уральской складчатой
системы доходит до о.
Вайгач. Этот вид близок к
Р. pulchellum, распростра-
ненному на скалистых и
щебнистых склонах в вы-
сокогорных поясах Алтая
и Саян, а также к двум
мелким эндемичным видам,
встречающимся на евро-
пейском севере — Р. 1ар-
ponicum (Кольский полу-
остров, о. Кильдин) и Р.
onegen.se (берега Онеж-
ского озера).
Происхождение Р. nudi-
pedum представляется нам
в таком виде. Исходная
предковая форма, давшая
начало широко распрост-
раненному виду Р. pulchel-
lum и близким к нему
узколокальным видам из
северных районов евро-
пейской части СССР, воз-
никла в горной р части Си-
бири. В конце плиоцена и
в начале плейстоцена она
стала распространяться на
северо-запад, достигнув
Карелии и Кольского
п-ова. Расселению пред-
ковой формы по равни-
нам благоприятствовало не
только похолодание кли-
мата, вызвавшее сокраще-
ние лесистости, но и оби-
лие обнаженного субстрата
(речные и флювиогляциаль-
'Рис. 36. Ареал качима уральского Gypsophila ига- ные наносы) В начале
lensis- голоцена в связи с интен-
сивным облесением тер-
ритории севера европейской части СССР и Западной Сибири предковая
форма вымерла почти везде на равнинах, но сохранилась в горных
и предгорных районах северной части Уральского хребта, а также на
берегах оз. Онежского и на Кольском п-ове. В результате географической
изоляции и своеобразия местных условий среды здесь обособились энде-
мичные формы.
Для анализа эндемичного элемента высокогорной флоры Урала пред-
ставляет интерес Linum boreale (рис. 39), растущий в высокогорных поясах
(горнотундровом и подгольцовом) на щебнистых склонах, на скалах и по
86
берегам рек на галечниках. В горах он чаще обитает на’основных и ультра-
основных горных породах. Быть может, преимущественной связью этого-
вида со щелочными горными породами объясняется расчлененность его аре-
ала (рис. 40) на две части: полярноуральскую (верховья рек Войкара,.
Соби, Крнгора, Лопты и Лире-Югана) и североуральскую (Чистоп, Денеж-
кин, Конжаковский, Косьвинский, Серебрянский Камни). На Приполяр-
ном Урале пока не найден. Вне высокогорий встречен на обнажениях из-
вестняков по рекам Усе и Большой Сыне. Родственные виды из ряда Регеп-
Рис. 37. Ареал синюхи голоногой Polemonium nudipedum.
nia Ju?., подряда Extraaxillaria Juz. растут в высокогорьях Алтая,.
Монгольского Алтая с прилегающей частью гор Средней Азии (L. altaicum), на^
высокогорных лугах Средней Азии (A. atricalyx) и высокогорных поясах
на Балканах и в Карпатах (L. extrdaxillare)-, лишь один вид из этого под-
ряда произрастает на более низких уровнях — на лугах в Восточной Си-
бири (L. Komarova).
К названным выше видам по условиям обитания, распространению и ха-
рактеру родственных связей примыкает Anemone biarmiensis (рис. 41), об-
ладающая более широкой экологической амплитудой. Ветреница пермская-
произрастает в горнотундровом и подгольцовом поясах на скалах, в гор-
ных тундрах, на подгольцовых лугах, околоснежных лужайках и в разре-
женных редколесьях и криволесьях. Кроме того, она встречается, хотя
и реже, в горнолесном поясе в светлых лиственничных, сосновых и березо-
вых лесах, а также на скалистых обнажениях. Распространена ветреница,
пермская (рис. 42) почти на всем протяжении Урала — от Полярного до-
Южного. В северной части хребта теснее связана с высокогорными поясами,.
87:
Рис. 38. Скалистый гребень хр. Сабля.
Становится редкой в его средней, пониженной части, где встречается на
известняковых обнажениях, например по р. Чусовой6. На Южном Урале
заходит в горные светлохвойные и березовые леса (по полянам), на при-
брежные галечники и даже в каменистые горные степи (хребты Крыкты
и Ирендык).
Этот вид относится к ряду Narcissiflorae Juz., многочисленные предста-
вители которого произрастают на Дальнем Востоке с прилегающей к нему
6 Предположение И. М. Крашенинникова (1937) о полном отсутствии ветреницы перм-
ской в средней части Урала не подтвердилось.
.88
Рис. 39. Лен северный Linum boreale.
частью Северной Америки, в Восточной Сибири, на Алтае, Саянах, в горах:
Средней Азии, на Кавказе и в горных районах Европы.
Из числа близких к перечисленным эндемикам форм в болыпинстве-
случаев едва ли можно назвать такие, которые явились их непосредствен-
ными предками. Однако близкородственные им виды имеются во многих,
других горных областях; в совокупности с названными эндемиками они
образуют серии географически замещающих видов. Уральские эндемики
из родов Lagotis, Gypsophila, Polemonium, Linum, Anemone, как и викари-
рующие виды в других горных странах, возникли в результате экологиче-
ской, морфологической и географической дифференциации ранее широко,
распространенных исходных видов, свойственных, по всей вероятности,
растительным группировкам лесной зоны на равнинной или слабо всхолм-
ленной территории. Горообразовательные процессы и связанные с ними
изменения условий среды способствовали возникновению эндемичных ви-
дов в различных горных областях, в том числе и на Урале; это обособление-
совпало с интенсивным эпейрогенезом конца третичного и начала четвертич-
ного периодов. Изменение климата и растительного покрова в четвертич-
ное время во многих случаях вызвало оттеснение исходных форм на равни-
нах, раздробление первичных обширных ареалов исходных видов на не-
сколько частей, связанных с различными горными системами. Возникшая’
89
Рис. 40. Ареал льна северного Linum boreale.
Рис. 41. Ветреница 'пермская Anemone biarmimsis.
«К*
Рис. 42. Ареал ветреницы пермской Anemone
biarmiensis.
географическая изоляция еще
более способствовала обособ-
лению эндемичных высокогор-
ных видов на Урале.
Среди уральских высокогор-
ных растений выделяется группа
эндемиков, связанных тесными
узами родства с другими, более
широко распространенными
высокогорными видами, кото-
рые могут с полным основа-
нием рассматриваться в качестве
их непосредственных предков.
К этой категории относится
Epilobium uralense, произра-
стающий в горах, преимущест-
венно выше границы леса (в
горнотундровом и подгольцовом
поясах) по берегам ручьев, но
особенно характерный для при-
ручьевых лужаек около тающих
снегов. Иногда по долинам
ручьев это растение спускается
и в прилегающую часть горно-
лесного пояса. Ареал этого
вида (рис. 43) расположен в
основном в высокогорьях Поляр-
ного (гора Енганепе), Приполяр-
ного (хр. Сабля, верховья рек
Хугли и Хобе-Ю) и Северного
Урала (Яны-Емпты, Мань-Хачет-
Чахль, Ялпинг-Ньер, Кваркуш,
Чувал, Тулым, Конжаковский
и Косьвинский Камни). Южнее
не был обнаружен, кроме не-
скольких экземпляров, собран-
ных Ю. Шеллем в горной Баш-
кирии, в Стерлитамакском райо-
не у дер. Сеитовой (образец
хранится в Ботаническом ин-
ституте АН СССР, определен
Р. Р. Поле). Наиболее близкий
к кипрею уральскому вид Е.
alpinum распространен в горах
Европы, в арктической части
Скандинавии, на европейской
территории СССР и в Сибири, а
также в горах Кавказа и Алтая.
На Урале типичный Е. alpinum
отсутствует7, замещаясь Е. ura-
lense.
’ Для Урала Е. alpinum приводился всего лишь из одного пункта — гора Ялпинг-
Ньер на Северном Урале («Флора Западной Сибири», вып. 8; это указание перешло и в не-
которые другие руководства, в том числе во «Флору СССР»). Как указывает Е. И. Штейн-
берг (Флора СССР, т. XV), у Е. uralense семена густо покрыты кругловатыми сосочками,
92
Рис. 43. Ареал кипрея уральского Epilobium uralense. За пре-
делами карты — оторванное местонахождение в Стерлитамакском
районе Башкирской АССР.
(верхние листья заостренные, нечасто мелкозубчатые, на нижней стороне и по срединной
жилке неясно и густо опушенные. В отличие от этого у Е. alpinum семена голые (без со-
сочков), все листья тупые, цельнокрайние, голые, стебли обычно несколько короче. Наши
сборы с горы Ялпинг-Ньер показали, что здесь в подгольцовом поясе и в нижней части
горнотундрового произрастает типичный Е. uralense, выше же по ручьям, близ тающего
снега, встречаются низкорослые особи, габятуально напоминающие Е. alpinum с тупыми,
почти цельнокрайними листьями. Однако и у них семена с сосочками. П. Н. Крылов
{1927—1954) для отграничения Е. uralense от Е. alpinum не приводил наличие сосочков на
семенах; распознавание этих видов он основывал только на форме листьев. Надо полагать,
что собранные на Ялпинг-Ньере низкорослые экземпляры Е. uralense с почти цельнокрайни-
ми листьями он отнес к Е. alpinum. Из сказанного следует, что типичный Е. alpinum на
урале,' по-видимому, не встречается, а здесь его замещает эндемичный вид Е. uralense.
93
Есть основания считать Е. alpinum непосредственным предком Е. ига-
lense. Первичным ареалом исходного вида, по-видимому, были горы Средней
Европы, Балкан, Малой Азии и Кавказа. В плейстоцене это растение ши-
роко расселилось в евразиатской Арктике. Проникнув из Арктики на Урал,.
оно преобразовалось здесь под.
влиянием местных условий сре-
ды в Е. uralense.
Другой пример — Gagea sa-
mojedorum — высокогорный эн-
демик северной части Ураль-
ского хребта (Сабля, Тельпос-
Из и верховья р. Сортыньи,
притока Ляпина), характерный
для приручьевых околоснежных
лужаек (растет обычно выше
границы леса). В последнее вре-
мя оторванное местонахожде-
ние этого растения обнаружено-
Л. И. Красовским и А. К - Сквор-
цовым (1959) в подгольцовом-
поясе Еловского Урала, в райо-
не Денежкина Камня (рис. 44).
Близкородственный] вид G. fis-
tulosa распространен в горах
Средней и Южной Европы. Off
мог проникнуть на Урал через:
полосу тундр на севере Восточ-
ной Европы, которая в эпоху
наибольшего плейстоценового-
оледенения непосредственно смы-
калась с горными тундрами Се-
верной Европы и Урала. Впо-
следствии это растение вымерло-
в промежуточных пунктах на
севере Русской равнины, но
сохранилось на Урале, где-
преобразовалось в G. samojedo-
rum.
Рис. 44. Ареал гусиного лука ненецкого Gagea
samojedorum.
Указанные выше эндемики возникли в результате преобразования в-
местных условиях исходных высокогорных или аркто-высокогорных видов,,
мигрировавших на Урал из других горных областей через полосу равнин-
ных тундр. В обоих случаях исходные формы таких эндемиков могут быть
названы совершенно определенно; время их проникновения на Урал дати-
руется плейстоценом (вероятно, эпохой наибольшего оледенения).
Отметим ряд высокогорных эндемиков, обособившихся от широко рас-
пространенных видов, свойственных равнинам и невысоким горам в пре-
делах тундровой, лесной или степной зон. Сюда, в частности, относится
Cerastium K.rylovii, высокогорный эндемик Южного Урала, описанный по
экземплярам, собранным автором этой книги на Яман-Тау (Шишкин, 1951).
Ясколка Крылова ранее собиралась на Яман-Тау другими исследователями'
(Ю. К. Шелль, Д. И. Литвинов, С. Ю. Липшиц), но неправильно отождест-
влялась с С. alpinum, с которой она имеет некоторые черты габитуального
сходства. По данным наших исследований, С. Krylovii распространена
(рис. 45) на крупных вершинах Южного Урала (Яман-Тау, Иремель, Зи-
гальга, Нары), заходит также на Северный Урал (Мани-Хачет-Чахль, Де-
94
нежкин, Конжаковский (рис. 46),
Косьвинский, Вогульский Камни).
Обитает выше границы леса в гор-
ных тундрах и на травянистых
лужайках. Этот вид относится к
секции Orthodon Ser., подсекции
Perennia Fenzl., ряду Leiostemona
Fenzl. и близок к широко рас-
пространенной С. caespitosum, про-
израстающей, в частности, на Юж-
ном Урале на более низких уров-
нях гор (леса, лесные опушки,
.луга).
Еще один эндемичный вид яс-
колки из этого же ряда — Ceras-
tium Gorodkovianum (Шишкин,
1951). Обитает на гольцах и по
берегам рек близ верхнего предела
леса. Ареал его (рис. 47) — При-
полярный и Северный Урал (р.
Манья, верховья р. Нанксорыньи,
подножие горы Сумах-Ньер в бас-
сейне р. Щугора).
Эндемик горных тундр Поляр-
ного Урала Cerastium Porphyrii
известен по сборам из бассейна
р. Сыни (перидотитовая гора в
верховьях р. Лопты), из верховий
р. Конгор и с горы Рай-Из (рис. 48).
Относится к подроду Eucerastium
Boiss., секции Orthodon Ser., под-
секции Perennia Fenzl., ряду
Arvensia Schischk. Близок к С. ar-
vense, от которой отчленился, при-
способившись к произрастанию на
оливиновых горных породах в
каменистых тундрах. Подлинный
таксономический ранг этой формы
еще не выяснен.
Недавно Е. Г. Победимова
(1968) описала новый эндемичный
.для Урала вид ясколки Cerastium.
Igoschinae, принадлежащий к тому
же ряду Arvensia Schischk. Он
Рис. 45. Ареал ясколки Крылова Cerastium
Krylovii.
встречается на Конжаковском Камне в верховьях р. Южного Иова и на
«плече» Косьвинского Камня (в обоих случаях на обнажениях дунита),
а также на Полярном Урале близ ж.-д. станции 106 км. Как и преды-
дущий эндемик, С. Igoschinae, вероятно, представляет собой обособив-
шуюся эдафическую расу более широко распространенного вида С. arvense.
Senecio Igoschinae растет главным образом в высокогорных поясах —
в горных тундрах и на каменных россыпях, но изредка заходит и в горно-
лесной пояс, где произрастает на скалистых обнажениях (рис. 49). Встре-
чается на Северном, Среднем и Южном Урале (Ишернм, Кумба, Денежкин,
Вогульский, Конжаковский и Косьвинский Камни, .Юрма, Таганай, Ире-
мель, Яман-Тау, Машак, Зигальга, Шатак, известняковые обнажения па
95
Рис. 46. Конжаковский Камень.
Рис. 47. Ареал ясколки Городкова Cerastium Gorodkovianum.
Рис. 48. Ареал ясколки Порфирия Cerastium Porphyrit.
7 3»кы № 251
Рис. 49. Косьвинский Камень (вид с Конжаковского Камня)
р. Сиказе — рис. 50). Этот
вид наиболее близок к S. inte-
grifolius, имеющему широкий
евразиатский ареал и рас-
пространенному на прилегаю-
щих к Уралу равнинах.
Преимущественно выше
границы леса (в подголь-
цовом и горнотундровом
поясах) на каменных россы-
пях, в горных тундрах и
редколесьях, а также иногда
на скалистых береговых об-
нажениях в горнотаежном
поясе произрастает Scorzo-
nera Ruprechtiana (Syn.—S.
glabra, S. austriaca ssp. gla-
bra). Этот вид (рис. 51) рас-
пространен . на Северном,
Среднем и Южном Урале.
Встречается на гольцах Ял-
пинг-Ньера, Чистопа, Кумбы,
Денежкина, Куроксарского,
Чувальского, Конжаковского,
Косьвинского Камней, на
вершинах гор Егозинской,
Сугомака, Ильменских, Ну-
рали, Крака, Крыкты, а так-
же на известняковых утесах
по рекам Илычу, Северной
Сосьве, Лозьве, Ивделю, Ви-
шере, Каме и Чусовой
(рис. 52). За пределами Ура-
ла он известен как реликт
на известняковых и гипсовых
обнажениях по рекам Север-
ной Двине, Пинеге, Мезени,
Сухони и Сотке. Ближайшим
родичем S. Ruprechtiana яв-
ляется широко распростра-
ненный степной вид S. aus-
triaca s. str. (южная часть
Европы и вся степная зона
Азии), от которого уральская
форма, по-видимому, и обо-
собилась, приспособившись
к произрастанию на скали- „
стом субстрате, преимущест- Рис‘ 50' Ареал К™ Игошинои Senecta
венно в высокогорных поясах.
Самый низкорослый из уральских тимьянов — Thymus paucifolius —
распространен (рис. 53) в высокогорной части Северного Урала, где оби-
тает в горных тундрах и на каменных россыпях (по нашим сборам, опре-
деленным М. В. Клоковым, он известен с хр. Чистоп, гор Денежкин и Кон-;
жаковский Камни). Относится к секции Euserpyllum Klok., к ряду Euser-
pylla Klok. et Schost., близок к Thymus sibiricus, распространенному на.
7*
99
Рис. 51. Основной ареал козлеца Рупрехта
Scorzonera Ruprechtiana.
скалах в Западной Сибири и на
предгорьях Алтая, а в несколь-
ко меньшей степени Th. serpyl-
lum s. str.
По мнению M. В, Клокова
(личное сообщение), родиной
тимьянов Еявляется древнее
Средиземноморье, где они сфор-
мировались в сообществах фри-
ганоидного типа. Азиатский ряд
Asiatici Klok. при расселении
его представителей на запад дал
начало ряду Euserpylla Klok. et
Schost. В процессе расселения
исходного азиатского типа на
запад от него обособился Thy-
mus paucifolius', этому обособ-
лению благоприятствовали эпей-
рогенические поднятия и сопря-
женные с ними изменения усло-
вий среды. Th. paucifolius в ряде
Erusepylla Klok.et Schost. зани-
мает крайнее положение не
только в связи с тем, что он
наиболее специализирован по
линии приспособления к высоко-
горным условиям, но и потому,
что он в большей степени, чем
другие представители этого ряда,
обнаруживает генетические свя-
зи е исходным рядом Asiatici
Klok.
Обитателем горных тундр и
лиственничных ^редколесий По-
лярного и Приполярного Урала
(верховья рек Щучьей и Вой-
кара, гора Восточная Енганепэ,
бассейн р. Грубе-Ю, притока
Хулги, бассейн р. Кожима) явля-
ется Bromus vogulicus (рис. 54),
встречающийся также в восточ-
ной части Болыпеземельской
тундры. По всей вероятности,
непосредственным предком его
является широко распространен-
ный вид В. sibiricus.
Другой, близкий к костру
мансийскому эндемичный вид —
В. Julii, видимо, имеет сходное
с ним происхождение. Это расте-
ние известно с Приполярного
Урала (верховья р. Грубе-Ю,
притока Хулги), произрастает оно выше границы леса в расщелинах скал.
Еще один эндемичный представитель этого рода — В. uralensis — про-
израстает на Приполярном и Северном Урале (верховья рек Лемвы и Уньи),
ЮО
Рис. 52. Козелец Рупрехта Scorzonera Ruprechtiana.
на высокотравных подгольцовых лугах. Он также обнаруживает черты
сродства с В. sibiricus. Таксономический ранг всех этих эндемичных форм
костра неясен, они подлежат критическому изучению.
Упомянутые эндемики (Cerastium Rrylovii, Senecio Igoschinae, Scorzonera
Ruprechtiana, Thymus paucifolius, три вида из рода Bromus) возникли в
результате отбора и приспособления популяций широко распространенных
низинных или низкогорных видов к высокогорным условиям. В настоящее
время^эти эндемичные формы, обычно сильно варьирующие по морфологи-
ческим признакам, обособлены от широко распространенных исходных
видов не столько географически, сколько экологически по условиям
обитания. Их ареалы вкраплены в виде островов на фоне более обширного
ареала исходного вида или прилегают к нему, но располагаются на более
высоких уровнях гор.
В высокогорной флоре Урала имеются и эндемики гибридного происхож-
дения. К ним относится Saussurea uralensis, произрастающая в подгольцо-
вом и горнотундровом поясах Уральского хребта, чаще в горных тундрах,
реже в расщелинах среди каменных глыб или, в редколесьях (рис. 55). На
Северном Урале отмечена на горах Ойка-Чахль (рис. 56), Муравьиный Ка-
мень, Чистоп, Кумба, Денежкин, Конжаковский, Косьвинский, Семиче-
ловечный Камни, на Среднем — на горе Качканар и на Южном — на го-
101
Рис. 53. Ареал тимьяна малолистного Thymus paucifo-
lius.
pax Иремель и Яман-Тау. Этот вид, по мнению С. Ю. Липшица (1954),
возник в результате естественной гибридизации S. controversa и S. alpina.
Однако он довольно широко распространен в высокогорной части Урала
и представляет не случайное гибридогенное явление, а строго локализован
Рис. 54. Ареалы уральских эндемичных видов рода Bromus.
/—костер мансийский В. vogulicus, 2 —костер Юлия В. Julii,
3 — костер уральский В. uralensis.
географически. Возникновению этого эндемичного вида благоприятствовал
не только непосредственный контакт ареалов двух исходных форм, произ-
растающих иногда совместно, но и появление в высокогорной части Урала
^(вскоре после освобождения от ледникового покрова) «жизненного про-
103
Рис. 55. Ареал сосюреи уральской Saussurea
uralensis.
странства», еще мало заселен-
ного растительностью, представ-
ляющего простор для отбора*
подходящих для этих условий
форм.
В высокогорных поясах Ура-
ла произрастает довольно боль-
шое количество апомиктических
эндемичных видов Alchetnilla.
Наиболее характерные места их
обитания — околоснежные лу-
жайки, луга близ ручьев, вы-
текающих из снежников, высоко-
травные подгольцовые луга.Ряд.
видов (Alchetnilla semispoliata^
A. Gortschakowskii, A. auricu-
lata, A. cunctatrix. A. malimon-
tana) описан С. В. Юзепчуком
(1954, 19556) по нашим сборам.
Некоторые высокогорные виды,
описаны по сборам Е. А. Сели-
вановой-Городковой (1963) и
А. М. Овеснова. Закономерности,
географического распределения
манжеток на Урале на основе*
гербарных материалов разных
лиц изучала К- Н. Игошина
(1966).
С. В. Юзепчук (19556) насчи-
тывал на Урале 35 эндемичных
видов манжеток, в том числе
один пока не описанный. Из них
21 вид характерен для высоко-
горий, остальные обитают в ниж-
них поясах гор на лугах и в,
разреженных лесах. Эндемич-
ные для Урала высокогорные*
манжетки принадлежат к под-
роду Pes Leones Juz., к секциям
Pubecentes Buser, Calicinae Buser
и Vulgares Buser.
Из секции Pubescentes Buser
известны три эндемичных ман-
жетки Alchetnilla exul, A. Hele-
nae и A. consorbina, произра-
стающих на высокотравных под-
гольцовых (иногда несколько-
остепненных) лугах на Южном
Урале на высоте 1200 м над
ур. м. (хр. Шатак). Первый ив.
упомянутых видов относится к циклу Sericatae Juz., два других — к
циклу Pseudosericatae Juz. Остальные виды из этих циклов обитают на Кав-
казе.
Секция Calcinae Buser представлена на Урале двумя эндемичными вы-
сокогорными видами, обитающими на подгольцовых лугах и в редколесьях:
104
Рис. 56. Гора Ойка-Чахяь.
A. Haraldi (Иремель, Яман-Тау, Зигальга, Шатак) и A. pycnoloba (горы
Басеги, Ослянка, Верх-Яйвинские поляны, Острая гора, Растесский
Камень).
Значительно больше высокогорных эндемиков (17 видов), обитающих,
на подгольцовых лугах и в редколесьях, содержится в секции Vulgares
Buser. В том числе группа Hursutae Lindb. fil представлена следующими
видами: Alchemilla crassicaulis (Кваркуш, Еловский Урал, Лопьинский
Камень, Чувал, Цепельские поляны, Басеги, Ослянка), A. Gortschakowskii
(Сабля), A. cinerascens (Чувал, Харюзный Камень, хр. Ильменский),
A. malimontana (Яман-Тау), A. hyperborea (верховья рек Щугора, Сыпи,
хр. Сабля, горы Чувал, Еловский Урал, Харюзный Камень, Ослянка, Ба-
сеги), A. macroclada (Шатак), A. riphaea (Конжаковский Камень, Юрма,.
Уреньга, Басеги, Иремель, Яман-Тау), A. kvarkushensis (Кваркуш),
A. iremelica (Ослянка, Иремель), A. semispoliata (Сабля), A. amphipsila
(Еловский Урал), A. auriculata (Манарага, верховья р. Нанксортыньи,.
Ялпинг-Ньер) и A. parcipila (верховья р. Большой Харуты, Танью, Кок-
пела, Ослянка, Басеги, Растесский Камень).
Другая группа — Subglabrae Lindb. — той же секции заключает энде-
мичные для Урала высокогорные виды Alchemilla obtusiformis (Манарага,.
верховья рек Сыни, Усы, Хулги, Маньи, Польи, Щугора), A. cunctatrix-
(горы Пай-Пудына, Сабля, Ялпинг-Ньер, Семем-Ю-Из, Чистоп, Конжаков-
ский Камень), A. paeneglabra (Ослянка, Басеги) и A. glabriformis (истоки
рек Щокурьи, Кось-Ю, Сыни, Лозьвы, Хус-Ойка, Чувал, Басеги).
Всем, упомянутым видам присущ облигатный апомиксис. По мнению*
С. В. Юзепчука (1954, 1955), возникновение этих форм относится в основ-
ном к плиоцену, а формирование их ареалов продолжалось в плейстоцене'
и голоцене. «Агамные комплексы» видов, по его мнению, возникли в ре-
зультате усиленной межвидовой гибридизации, сопровождаемой возник-
новением полиплоидии и закономерным, хотя и постепенным переходом
к тотальному апомиксису. Число узкоареальных «апомиктов» наиболее-
велико в центре возникновения такого комплекса и закономерно убывает
к периферии. Урал наряду с Альпами, Кавказом, горами Средней Азии и
Алтаем явился одним из центров видообразования в роде Alchemilla..
Манжетки северной части Уральского хребта генетически связаны с фло-
рой Северной, а через нее и Западной Европы. Южноуральские манжетки.
(A. Helenae, A. Haraldi и др.), напротив, непосредственно связаны с кав-
казскими. Основным центром видообразования, как полагал С. В. Юзеп-
чук, был Кавказ, откуда манжетки проникли в Западную Европу (через;
Малую Азию и Балканы), а также на север. С Урала они расселились в;
пределы Сибири, а оттуда в Среднюю Азию.
В высокогорьях Урала, по исследованиям А. Я. Юксипа (19596), встре-
чается не менее 14 эндемичных форм ястребинОк (род Hieracium). Распро-
странение их и экология выявлены еще недостаточно, а таксономический’
ранг большинства из них остается неясным, хотя все они по традиции,,
установившейся у исследователей этого рода, описаны в качестве видов.
Эндемичные представители ястребинок входят в подрод Euhieracium
Torr, et Gray. Представителем секции Prenanthoidea Koch, является Hie-
racium suberectum—хорошо отличимый вид, свойственный лугам и ред-
колесьям подгольцового пояса. Распространен от хр. Сабля и верховьев;
р. Хулги на Приполярном Урале до гор Иремель, Яман-Тау, Зигальга
и Шатак на Южном Урале (рис. 57).
Секция Alpina Fr. содержит следующие эндемичные высокогорные виды:
Hieracium iretnelense (горные тундры Северного, Среднего и Южного Урала).,
Н. Ijapinense (подгольцовые луга и прибрежные галечники в верховьях
рек Хулги и Щугора), Н. pyrsjuen.se (верховья р. Пырсь-Ю), Н. uralen.se
106
(осыпи и прибрежный галечник в
районе хр. Сабля, лиственничные
редколесья пор. Балбан-Ю), Н. kuro-
ksarense (гора Куроксар), Н. Socza-
vae (верховья р. Хадаты), Н. steno-
piformae (горы Пай-Ер и Сале-Урр-
Ойка), Н. Gorodkowianum (реки Боль-
шой Ханмей, Уса, Адзьва, Манья),
Н. manifestum (р. Малая Хобе-Ю, в
лиственничном редколесье).
Из секции Vulgata Fr. известны
эндемичные высокогорные формы:
Hieracium wologdense (хр. Сабля, бас-
сейн рек Усы и Щугора), Н. cuspi-
dellum (Сабля), Н. kosvinskien.se (Кось-
винский Камень) и Н. konshakov-
skianum (Конжаковский Камень).
Таким образом, можно придти к
заключению, что уральские высоко-
горные эндемики подразделяются
в зависимости от их происхождения
на ряд групп: 1) эндемики, возник-
шие в результате эколого-морфологи-
ческой дифференциации и раздробле-
ния ареала первичного низинного
предка с образованием серии вика-
рирующих видов в разных горных
областях ((время обособления ураль-
ских ареалов совпадает с эпейрогенезом
конца третичного и начала четвер-
тичного периодов); 2) эндемики, воз-
никшие в результате преобразования
в местных условиях высокогорных
видов, проникших на Урал в плейсто-
цене из других горных областей;
3) эндемики, возникшие в результате
отбора и приспособления к высоко-
горным условиям популяций низин-
ных видов, распространенных и ныне
на предгорьях и низких уровнях
гор; 4) эндемики, возникшие в связи
с явлениями межвидовой гибри-
дизации с последующим переходом
к апомиктическому воспроизведению
ее продуктов; 5) эндемики, сформиро-
вавшиеся на базе апомиксиса; 6) энде-
мики гибридного происхождения.
Более древними (плиоцен — плей-
стоцен) являются первая и четвертая
группа эндемиков; несколько позже
(в плейстоцене) сформировались вто-
рая и третья группы. Пятая и шестая
группы представлены молодыми,
Рис. 57. Ареал ястребинки почти-прямой
Hieracium suberectum.
прогрессивными эндемиками, возникшими уже после освобождения высо-
когорной части Уральского хребта от -ледникового покрова.
107
ГОРНОСТЕПНЫЕ И СКАЛЬНЫЕ ЭНДЕМИКИ
Группа горностепных и скальных эндемиков на Урале очень своеобраз-
на; анализ ее представляет интерес для выяснения происхождения ураль-
ской флоры.
В составе уральской горностепной и скальной флоры содержится не-
сколько эндемичных астрагалов. Пока лишь из двух пунктов (Спасская
гора близ г. Кунгура Пермской области и Подкаменная гора в окрестностях,
дер. Токарево, ниже г. Кунгура) известен Astragalus kungurensis (рис. 58).
Это растение, описанное А. Г. Борисовой (1950) по сборам А. Н. Понома-
рева, произрастает на крутых склонах правого коренного берега р. Сылвы.
Характерные места его обитания —
участки обнаженного мелкоземи-
стого субстрата (продукты разру-
шения гипса и очень близкого к.
нему по химическому составу ангид-
рита). Такие участки, вкрапленные
среди соснового леса, но не облесен-
ные, почти лишенные травянистой
растительности, располагаются
обычно в нижних частях склонов,,
куда сносятся мелкоземистые про-
дукты разрушения гипса.
Этот астрагал очень близок к
A. longiflorus, распространенному
на песках Предкавказья, нижнего^
течения Дона, Нижнего Поволжья,.
Западного Казахстана и Прибал-
хашья. По-видимому, A. longiflo-
rus и A. kungurensis имели об-
щего, довольно ксерофильного*
Рис. 58. Ареал^страгала^кунгурского Astra- предка. Впоследствии популяции,.
g us и guren s. связанные с песками в аридных,
районах юга европейской части:
СССР и Средней Азии, обособились от популяций, свойственных мелкозе-
мистым продуктам разрушения горных пород на Южном и Среднем Урале..
Это привело к появлению двух видов — A. longiflorus и A. kungurensis..
В последние стадии голоцена, в связи с оттеснением степей лесной расти-
тельностью и зарастанием лесом многих обнажений горных пород и про-
дуктов их разрушения, ареал астрагала кунгурского сильно сократился.
Сохранению этого вида именно в районе Кунгура благоприятствовало-
широкое распространение здесь выходов гипса и других богатых известью,
горных пород в сочетании с карстовыми образованиями. В таких местах
создаются особые эдафические условия — мелкоземистый, довольно под-
вижный субстрат, сильно обезвоженный как благодаря высокой водопро-
ницаемости мелкоземистых продуктов выветривания гипса, так и вслед-
ствие отвода грунтовых вод, перехватываемых карстовыми образованиями,.,
в более глубокие горизонты.
Другой эндемичный астрагал — Astragalus Clerceanus — встречается
довольно редко в горной каменистой степи и на скалистых обнажениях
Южного и отчасти Среднего Урала (рис. 59). Он произрастает на извест-
няковых утесах по р. Белой близ дер. Иргизла, восточнее г. Стерлитамака
на р. Малом Шешеняке и горе Уаткан-Таш, на горах Вишневых близ;
г. Кыштыма и на гипсовой скале Шалашный Камень в низовьях р. Чусовой.
Кроме того, он встречается на выходах гранита в окрестностях г. Сверд-
108
Рис. 59. Ареал астрагала Клера Astragalus
Cl ere eanus.
ловска — на Палкинских каменных .палатках около станции Палкино и
на Чертовом городище около станции Исеть. Здесь он растет на южных хо-
рошо освещенных склонах у основания гранитных останцев, где скапли-
ваются мелкоземистые продукты
разрушения гранитов. Родствен-
ный ему вид из ряда Elati Gon-
tsch. — A. sulcatus — распрост-
ранен преимущественно в степ-
ной зоне, на степных лугах и
солонцах; ареал его:—Средняя -
Европа, Балканы, европейская
часть СССР, Западная и Восточ-
ная Сибирь, Средняя Азия.
Другой . родственный вид — А.
consanguineus, ^относящийся к
тому же ряду,— распространен
в Средней Азии, где растет на
солончаковых почвах.
Для горных (преимуществен-
но каменистых) степей и обна-
жений скал характерен Astra-
galus Karelinianus. Большая
часть местонахождений этого
вида сосредоточена в бассейне
р. Большого Ика, притока
р. Сакмары, на хр. Наказ, на
р. Белой у с. Биккузино и на
шиханах близ г. Стерлитамака.
Значительно севернее располо-
жены оторванные его место-
нахождения на Вишневых горах
в районе г. Кыштыма и на бере-
говых. утесах по р. Туре в
окрестностях г. Верхотурья,
откуда он описан (рис. 60). По-
всюду этот астрагал тесно связан
и известняковым субстратом.
Принадлежит он к секции Xiphi-
dium Bge., к ряду Macrotropis
Vass. A. Karelinianus— единст-
венный представитель ряда Macrotropis Vass, на Урале; другие представители
этого ряда встречаются в Средней Азии, на Алтае и его предгорьях, на
Кавказе, реже в европейской части СССР, в Западной и Восточной Сибири.
Подобно предыдущему виду, Astragalus Helmii свойствен горным каме-
нистым склонам и скалистым обнажениям. Ареал этого астрагала (рис. 61)
дизъюнктивный. Область более или менее сплошного его распространения
охватывает Южный Урал, северную часть Мугоджар, Бугульминско-Бе-
лебеевскую возвышенность, Общий Сырт и достигает на западе Жигулей.
Наиболее обильно и часто встречается на Южном Урале — на хр. Ирен-
дык, в Губерлинских горах, в бассейне р. Сакмары и ее правого притока
Большого Ика, на известняковых шиханах близ г. Стерлитамака и в бас-
сейне р. Демы, левого притока р. Белой. Самое южное местонахождение
этого вида — близ аула Кызыл-флот на южном щебнистом склоне к
р. Кара-Бутак в Актюбинской области. В удалении от основной части ареала
астрагал Гельма встречается на известняковых скалах в среднем течении
109
Рис. 60. Ареал астрагала Карелина Astragalus Kare-
linianus.
В систематическом отношении к названному
Чусовой (Камни Дужной
и Дыроватый) и в низовьях
Вишеры (Камень Говорли-
вый и гора Сыпучая)8. Еше
более оторваны от основ-
ного ареала местонахожде-
ния этого растения в об-
ласти Казахского мелко-
сопочника (окрестности
г. Кокчетава и оз. Боро-
вого). Кроме астрагала
Гельма в секцию Helmia
Bge входят очень близкий
к нему алтайско-саянский
вид A. depauperatus и два
вида из гор Средней Азии
(A. Polakovii и A. heptapo-
tamicus).
Описанный Д. И. Лит-
виновым (Litvinov, 1892)
Astragalus uralensis обитает
на каменистых (преимуще-
ственно известняковых и
гипсовых) обнажениях.
Этот вид распространен
(рис. 62) на Южном Урале
(гора Косотур в окрест-
ностях г. Златоуста), на
Среднем Урале (известня-
ковые скалы по р. Туре у
дер. Елкиной, обнажения
гипса на Сироловой и
Сокольей горах по р. Чусо-
вой и на Подкаменной горе
по р. Сылве), на Северном
Урале (известняковые уте-
сы по рекам Северной
Сосьве и Вижаю, притоку
Лозьвы) и достигает По-
лярного Урала (прибреж-
ный галечник в верхнем
течении р. Соби). По-види-
мому, к этому же типу от-
носятся растения, найден-
ные на севере Русской рав-
нины, на береговых обна-
жениях в устье р. Пинеги.
астрагалу очень близок А.
australis, обитающий в высокогорных поясах Центральной и Южной
Европы (Альпы, Пиренеи, Апеннины, Карпаты). Другая близкая форма,
еще недостаточно изученная, распространена на Алтае, в Саянах, в горах
8 С. И. Коржинский (Korshinsky, 1898) растения из наиболее северных пунктов (они
Й1ЛИЛИЗ)еСКНЫЬП0 Сб°Рам из бассейна Вишеры) относил к особой форме var. permiensis
ПО
I
Рис. 61.. Ареал астрагала Гельма Astragalus Helmii. За пределами карты —
оторванные местонахождения в Казахском мелкосопочнике.
Забайкалья и Северной Монголии. Весь этот цикл форм (европейских,
уральских и сибирских) объединялся некоторыми авторами (Hegi,
1906—1931; Гончаров и Борисова в обработке астрагалов для «Флоры
СССР») в сборный вид A. australis s. I. Однако уральская форма не тожде-
111
Рис. 62. Ареал астрагала уральского Ast-
ragalus uralensis.
Рис. 63. Ареал остролодочника близкого
Oxytropis approximata.
ственна европейской и обособлена географически, поэтому есть основания
согласиться с трактовкой ее Д. И. Литвиновым как самостоятельного вида.
Горностепная и скальная флора Урала содержит также несколько энде-
мичных остролодочников. Из них три вида относятся к ряду Sordidae Vass.
Oxytropis approximata в пределах Урала имеет две изолированные об-
ласти распространения (рис. 63). Одна из них приходится на Южный Урал
(Ильменские горы, Шатак, хр. Малый Ирендык, бассейн р. Большого Ика),
Рис. 64. Ареал остролодочника Гмелина Oxytropis Gmelini.
где это растение произрастает на каменистых, преимущественно остепнен-
ных склонах. Другой участок ареала находится в районе Конжаковского
и Денежкина Камней (здесь этот вид встречается на низких уровнях гор
по каменистым руслам рек и на прибрежных галечниках), а также на из-
вестняковых скалах по р. Сосьве. На северном пределе своего распростра-
нения О. approximata, по-видимому, связан гаммой переходных форм с
О. sordida, встречающимся в горных тундрах, и трудно отличим от него.
О. Gmelini произрастает на каменистых местах в горных и предгорных
степях Южного Урала в Башкирской АССР и Челябинской области
(рис. 64) — в верховьях рек Урала и Уя, на хребтах Крыкты и Ирендык,
в верховьях р. Сакмары, в бассейне р. Большого Ика.
О. Hippolytii (рис. 65) характерен для каменистых мест в степной зоне
западной, предгорной части Башкирии (бассейн Ика, притока Камы, и
бассейны рек Чермасана и Демы, притоков р. Белой).
Все три названных желткоцветковых эндемичных для Урала вида свя-
заны тесными узами родства друг с другом и с аркто-высокогорным О. sor-
dida, проникающим довольно далеко на юг по горным вершинам Урала.
8 Заказ № 251 113
Другие родственные виды произрастают в Сибири, на Дальнем Востоке,
в горах Средней Азии, Кавказа и Европы.
А. Г. Борисова (1936) считает, что упомянутые эндемичные для Урала
остролодочники, как и родственные им виды из Северо-Восточной Азии и
Европы, имели общих предков азиатского (сибирского) происхождения.
В процессе расселения по Азии и Европе они дифференцировались на ряд
близких друг к другу видов с мелкими изолированными ареалами.
В целом эта полиморфная группа родственных форм нуждается в даль-
нейшем углубленном изучении.
Эндемик Oxytropis spicata из ряда Songoricae Vass, произрастает
в каменистых и луговых степях, на обнажениях известняка и мергеля.
Рис. 65. Ареал остролодочника Ипполита Oxytropis Hippolytii.
Распространен (рис. 66) в основном на Южном Урале (хр. Ирендык, Губер-
линские горы, бассейн рек Сакмары, Большого Ика, Салмыша, шиханы
близ г. Стерлитамака, верховья р. Урала, Ильменские горы). В Пред-
уралье встречается на Бугульминско-Белебеевской возвышенности, на се-
вер доходит до Красноуфимской лесостепи (Александровские сопки, рис. 67),
на юг — до северных острогов Подуральского 'мелового плато. Оторван-
ные самые западные местонахождения известны в Приволжье (близ горо-
дов Серноводска, Бугуруслана, между дер. Шиловкой и Липовкой, между
селами Нижним и Верхним Шунгутом, между селами Сидоровкой и Коз-
ловкой). Этот вид близок к азиатскому степному О. songorica, распро-
страненному в горах южной Сибири (Алтай, Саяны) и прилегающей части
Средней Азии (хр. Тарбагатай), от которого он, по-видимому, и произошел.
Тесно связан с обнажениями пород, содержащих углекислый кальций
(известняков, мергелей, мергелистых глин и мела), эндемик южного
Предуралья — Hedysarum Razoumovianum. Он произрастает на каменистых
вершинах холмов и склонах, преимущественно обращенных на юг. Ареал
этого растения (рис. 68) расположен на Южном Урале, Бугульминско-
Белебеевской возвышенности, Общем Сырте, Подуральском меловом плато
и частично вклинивается в Приволжскую возвышенность. На север заходит
114
до с. Давлеканово на р. Деме (склон горы Ярыш-Тау). Самое южное место-
нахождение этого копеечника отмечено на меловых горах Акшатау (Уиль-
ский район Актюбинской области Казахской ССР). Относится он к секции
Multicaulia Boiss., куда во флоре Советского Союза входят не менее 35
Рис. 66. Ареал остролодочника колосистого Oxytropis spicata.
видов, обитающих на обнажениях скал, каменистых склонах и т. п., пре-
имущественно в Средней Азии и на Кавказе. К числу ближайших его род-
ственников принадлежат широко распространенный в степях Сибири, Сред-
ней Азии и Монголии Н. Gmelini, а также Н. ucrainicum (меловые горы
в бассейне Северного Донца в районе г. Старобельска Луганской области
УССР), Н. cretaceum (меловые холмы Средне-Русской возвышенности
и Заволжья).
И. И. Спрыгин (1934) рассматривал Н. Razoumovianum как обособив-
шуюся эдафическую форму Н. Gmelini. В этом случае остается необъяснен-
ным генезис близких видов — Н. ucrainicum и Н. cretaceum.
Однако с не меньшим основанием можно полагать, что Н. Razoumovi-
anum, как и названные виды с меловых обнажений европейской части СССР,
произошли от общего пришедшего из Азии предка, близкого к Н. Gmelini.
Уменьшение площади различных обнажений горных пород на Восточно-
8* 115
Рис. 67. Красноуфимская лесостепь, вид с Александровских сопок,
Европейской равнине в ходе их заселения лесной растительностью привело,
к раздроблению западного крыла ареала первичного вида и обособлению,
южноуральской, среднерусской и северодонецкой форм, из которых пер-
вая связана в основном с известняками, мергелями и мергелистыми гли-
нами, а остальные — с выходами мела. Здесь мы видим пример эдафиче-
ского эндемизма, возникшего в условиях географической изоляции на ре-
ликтовой основе.
С сухой и теплой фазой голоцена совпала активизация распространения,
Н. Gmelini, расселившегося довольно далеко на запад, по Южному Уралу
Рис. 68. Ареал копеечника Разумова Hedysarum Razoumovianum.
и прилегающим к нему областям Восточно-Европейской равнины. Поэтому
Н. Razoumovianum оказался включенным в западное крыло ареала1
Н. Gmelini, но остался обособленным от последнего эдафически.
Характерным растением горных и предгорных степей, сухих камени-
стых склонов и особенно прибрежных известняковых обнажений является^
Scutellaria oxyphylla. Ареал ее (рис. 69) компактный, ограниченный Южным,
Уралом и примыкающей частью Предуралья в пределах Башкирской АССР,.
Оренбургской и Челябинской областей (бассейн рек Белой, Инзера, Сима,
Уфы, Юрюзани, Урала, Сакмары, Большого Ика). На севере доходит до
Месягутовской лесостепи, на юге — до Губерлинских гор. Самое западное-
местонахождение — окрестности г. Оренбурга, самое восточное — окраина
бора Джабык-Карагай близ пос. Анненского (рис. 69). Принадлежит этот
вид к ряду Supinae Juz., причем наиболее близок к алтайско-саянско-мон-
гольскому S. supina, с которым раньше отождествлялся, и к предволжскому
117.’
~S. cisvolgensls. Кроме названных растений, в этот ряд входит несколько
видов, встречающихся на меловых и известняковых обнажениях европей-
ской части СССР, а также в горах Средней Азии, на Алтае и в Саянах.
Недавно описанный Л. П. Сергиевской вид Eritrichium uralense (Кры-
лов, 1927—1964, том 12) был в течение длительного времени известен на
Урале только из двух пунктов — из каменистой степи на вершине горы
Нурали (Южный Урал) и с обнажений габбро на склоне Семичеловечного
Рис. 69. Ареал шлемника остролистного Scutellaria oxyphylla.
Камня близ верхней границы леса (Северный Урал). Нами обнаружены
(Горчаковский, 1951) еще два местонахождения незабудочника уральского:
на известняковых прибрежных скалах по р. Вижаю и по р. Ивделю (при-
токи р. Лозьвы, восточный склон Северного Урала). Позднее А. К- Сквор-
цов (1959) нашел его на отроге Денежкина Камня — Вересовом Увале
(рис. 70). Близкородственный вид, с которым это растение раньше отождест-
влялось, — Е. pectinatum — произрастает на скалах, щебнистых и каме-
нистых склонах гор и холмов, преимущественно в степном, реже в лесном
поясе горной части Сибири. Ареал этого вида, понимаемого здесь в узком
смысле, охватывает, согласно обработке М. Г. Попова во «Флоре СССР»,
предгорья Алтая, Салаирский кряж, Саяны. По-видимому, уральский эндемик
Е. uralense возник на реликтовой основе, в результате изоляции и
118
приспособления к местным усло-
виям популяций исходного вида
Е. pectinatum, проникших на Урал
в плейстоцене.
Aulacospermum isetense растет
в каменистых горных степях, на
открытых склонах гор и скали-
стых обнажениях главным образом
на Южном Урале с его пред-
горьями и в прилегающей части
Среднего Урала: Белебеевская,
Месягутовская и Красноуфимская
лесостепь (рис. 71), хр. Крыкты,
степные склоны на хр. Крака, бас-
сейн р. Большого Ика и другие
места. В Предуралье проникает
на север в глубь [лесной зоны по
известняковым утесам р. Чусовой
(рис. 72). Кроме того, указывается
для Жигулей. Относится к ряду
AnomalaVjSchischk., к которому
также принадлежат A. anomalum
(горы Алтая, Саян и Средней
Азии) и A. darvasicum (альпийский
пояс гор Средней Азии).
Горные ковыльные степи и ска-
листые обнажения остепненных
склонов — характерные местооби-
тания Potentilla Eversmanniana.
Эта лапчатка пока найдена всего
лишь в нескольких пунктах Юж-
ного Урала: в Оренбургской об-
ласти близ дер. Андреевки (низовья
Большого Ика, притока Сакмары)
и в пределах Башкирской АССР,
как в центральной горнохребтовой
части Южного Урала на хр. Ава-
лях, так и на восточном склоне
в окрестностях с. Баймака гна [хр.
Ирендык (рис. 73). Относится
Р. Eversmanniana к секции Multif-
idae Rydb., куда входит около 20
видов. И. М. Крашенинников
(1937), следуя монографу рода
Potentilla Т. Вольфу (Th. Wolf),
сближал Р. Eversmanniana с Р, ти-
Рис. 70. Ареал незабудочника уральского-
Eritrichium uralense.
Itifida. Однако, по мнению С. Ю. Юзепчука (личное сообщение), наиболее
близким к Р. Eversmanniana видом является Р. sericea — горно-
степное и скальное растение, произрастающее в южной части Сибири, гор-
ных районах Средней Азии и Монголии, К Р. Eversmanniana также доволь-
но близка описанная С. В. Юзепчуком Р. volgarica — реликтовый вид,,
встречающийся на известняковых степных склонах в окрестностях г. Хва-
лынска (Саратовская область); этот вид описан и издан в «Гербарии флоры
СССР» за № 3410, русский диагноз опубликован в восьмом издании «Флоры»
П. Ф. Маевского (1954).
119
Еще очень слабо изучен описанный С. В. Юзепчуком (1955а) эндемич-
ный вид Potentilla Gordiaginii, известный по гербарным экземплярам ра-
стений, собранных А. Я- Гордягиным на предгорьях восточного склона
Среднего Урала в Шадринском районе Курганской области на степных,
склонах (с ковылем) к рр. Караболке и Синаре. Эта лапчатка относится
Рис. 72. Ареал бороздоплодника исетского Aulacospermum
isetense (кроме местонахождения в Жигулях).
к группе Argentea Th. Wolf и близка к евросибирскому виду Р. argentea..
Габитуально она напоминает ещё более широко распространенный евра-
зиатский вид Р. canescens. По всей вероятности, Р. Gordiaginii возникла,
в результате гибридизации Р. argentea с Р. canescens.
Эндемичный уральский вид мокричника — Minuartia Helmii — оби-
тает на скалистых береговых обнажениях в горнолесном поясе, на камени-
стых вершинах невысоких гор, а на юге — на тенистых скалах в горных,
й предгорных степях. Распространен на Южном, Среднем и отчасти Север-
ном Урале (рис. 74). В пределах Южного Урала встречается в горных сте-
пях на хр. Нурали, Ирендык, на обнажениях известняков по рекам Белой,.
Сиказе, Юрюзани, Аю, в Ильменских горах и на скалистых берегах оз.
Увильды. На Среднем Урале очень характерен для известняковых утесов-.
121
по рекам Чусовой, Усьве, Уфе, Режу, Кунаре, Тагилу, Исети, Туре. Захо-
дит на Северный Урал, где встречается на известняковых утесах в бассейне
Камы (по р. Вишере) и в бассейне Лозьвы (по Вижаю и Северной Тошемке),
а также на обнажениях дунита, габбро и пироксенита некоторых невысоких
Гор — Вересовой увале (отрог Денежкина Камня), горе Кумба, Сухогор-
ском и Семичеловечном Камнях. Родственный вид М. Kjyloviana произра-
Рис. 73. Ареал лапчатки Эверсманна Potentilla Euersman-
niana.
стает на Алтае и в горах Средней Азии на скалах и каменистых склонах
в нижней части гор; другой близкий вид этого же ряда — М. Biebersteinii—
обитает на Кавказе в альпийском и субальпийском поясах на скалистом
субстрате.
В каменистых степях, в горной части лесостепи и на прибрежных обна-
жениях известняка растет Minuartia Krascheninnikovii (рис. 75). Большая
часть ее ареала находится на Южном Урале — на хр. Ирендык, в верховьях
р. Большого Ика, на шиханах близ г. Стерлитамака, в верховьях р. Урала,
в бассейне р. Уй, в Месягутовской лесостепи, в Ильменских горах, на го-
рах Сугомак, Егозинской и Вишневых. Кроме того, этот вид заходит на
Средний Урал, где встречается на прибрежных известняковых скалах по
рекам Исети, Пышме и Режу и по р. Уфе близ г. Красноуфимска (рис. 76).
Этот эндемичный вид мокричника встречается значительно чаще на более
122
Рис. 74. Ареал мокричника Гельма Minuartia Helmii.
Рис. 75. Мокричник Крашенинникова Minuartia Krascheninnikovii.
I
континентальном (по климатическим условиям) восточном склоне Южного
и Среднего Урала. Наиболее близкий к мокричнику Крашенинникова
вид — М. setacea, обитающий на скалах, каменистых склонах, галечниках
и опушках сосновых лесов, распространен в Приатлантической и Средней
Рис. 76. Ареал мокричника Крашенинникова Minuartia
Krascheninnikovii.
Европе, на Балканах, в средней и южной полосе европейской части СССР
(не заходит восточнее Волги).
Более широким ареалом, по сравнению с предыдущим эндемиком, об-
ладает Dianthus acicularis (рис. 77), растущая в каменистых степях, на
каменистых склонах и скалах, а иногда в южных сухих сосновых борах на
песчаной почве. Гвоздика иглолистная не является уральским эндемиком
в узком смысле этого слова, так как встречается и на прилегающих к нему
равнинах. Однако центр обилия этого вида, несомненно, находится на
Урале (рис. 78). Названное растение распространено на Южном Урале
преимущественно в горных степях (горы Сугомак, Егозинская, Вишневые,
хребты Крака, Ирендык, Губерлинские горы, степи в районе г. Орска, Бе-
лебеевская лесостепь), на шиханах близ г. Стерлитамака и на обнажениях
известняков по берегам рек Белой, Ая, Сима. На Среднем Урале становит-
ся_более редкой, встречается на горе Азов, на известняковых скалах по ре-
125
Рио. 77. Гвоздика ’иглолистная Dianthus acicularis.
Рис. 78. Ареал гвоздики иглолистной Dianthus acicularis. Не показаны изо-
лированные местонахождения в Западной Сибири и Казахстане.
кам Исети, Пышме, Режу, Туре, Чусовой, Яйве, Ваграну, Уфе. По извест-
няковым скалам заходит в южную часть Северного Урала до р. Вишеры
и до р. Ивделя (последнее местонахождение — самое северное; Горчаков-
ский, 1951). Кроме того, по обнажениям дунитов и реже габбро встречается
на южных склонах гольцов у верхней границы леса (Косьвинский, Семи-
человечный, Золотой Камни, Вересовый увал — отрог Денежкина Камня).
Рис. 79. Ареал гвоздики уральской Dianthus uralensis.
Оторванные от основного ареала местонахождения имеются в области Ка-
захского мелкосопочника (до Омска, Петропавловска и Кокчетава)9. При-
надлежит гвоздика иглолистная к ряду Arenarii Schischk., куда, кроме
того, входят близкий вид D. arenarius (растет на лугах с песчаной почвой,
в песчаных сосновых борах в Скандинавии, Средней Европе и европейской
части СССР), а также D. squarrosus (на песках в степной зоне европейской
территории СССР).
Другой эндемичный вид — D. uralensis — произрастает в скалах и в
каменистых степях преимущественно в степных и отчасти в лесостепных
районах Южного Урала (Башкирия, Оренбургская, частично Челябин-
ская области). Местонахождения этого вида сосредоточены в районах
9 По данным Б. К. Шишкина (Флора СССР, т. VI, 1936), D. acicularis в значитель-
ном удалении от основного ареала встречается на берегах Волги, в Жигулях. Однако
С. В. Юзепчук (1950, а также заметки на этикетках в Гербарии Ботанического института
АН СССР) установил, что растения из Жигулей и окрестностей г. Сызрани относятся
к особому, близкому к D. acicularis виду, названному им D. volgicus. Следовало бы так-
же тщательнее сравнить растения из Казахского мелкосопочника, относимые к D. acicula-
ris, с аутентичными уральскими экземплярами. М. В. Клоков (личное сообщение) намечал
к выделению из D. acicularis s. 1. особой расы D. toboliensis, произрастающий в Зауралье
и Казахстане.
428
хр. Ирендык, верхнего течения р. Урала, бассейна р. Сакмары и его правого
притока р. Большого Ика, а также на известняковых скалах по р. Белой
(рис. 79). Относится он, согласно обработке Б. К. Шишкина (Флора СССР,
т. VI) к ряду Pallidiflorae Schischk., куда входит несколько видов, обитаю-
щих на каменистых склонах, меловых обнажениях, в степях, на речном га-
лечнике и песках: D. rigidus (степная зона европейской части СССР, Запад-
ной Сибири и Казахстана), D. pallidioflorus (степи европейской части
СССР), D. ramosissimus (от степей Западной Сибири до полупустынь Сред-
ней Азии), D. kirghizicus (Тянь-Шань). Эта эндемичная уральская форма
гвоздики еще мало изучена, поэтому нет достаточных данных о ее происхож-
дении. И. М. Крашенинников (1937) считал, что D. uralensis является
примером более молодого, прогрессивного эндемизма, относящегося к го-
лоценовым явлениям формообразования. Доказательств в пользу этого
суждения он не приводил, но, вероятно, опирался на мнение С. И. Коржин-
ского (Korshinsky, 1898), допускавшего возможность гибридного проис-
хождения названного вида. Однако более вероятно, что D. uralensis
отчленилась от широко распространенного степного предка, близкого
к D. rigidus, чему способствовали особые эдафические условия Южного
Урала и его предгорий. Таким образом, возраст этого вида, как и боль-
шинства других горностепных эндемиков, более древний.
Более обширный ареал имеет Silene baschkirorum, произрастающая в ка-
менистых степях и на обнажениях известняка и мергеля. Центр обилия смо-
левки башкирской расположен на Южном Урале (рис. 80). Здесь это расте-
ние встречается в горных степях Губерлинских гор, Урал-Сакмарского водо-
раздела, бассейна р. Большого Ика, верхнего течения р. Урала, на шиханах
близ г. Стерлитамака, известняковых утесах по рекам Белой, Уфе, Юрюзани,
Аю, в Ильменских горах. Заходит на Средний Урал, где отмечено на извест-
няковых обрывах по р. Синаре, на дунитовых обнажениях Уктусских гор
близ г. Свердловска, на Александровских сопках в Красноуфимской лесосте-
пи и на обнажениях гипса в низовьях р. Чусовой у с. Куликово. В Башкир-
ском Предуралье этот вид встречается на Бугульминско-Белебеевской возвы-
шенности. К западу от основного ареала имеется серия оторванных местона-
хождений на Приволжской возвышенности (Белые горы близ с. Криуши,
окрестности городов Вольска, Хвалынска) и в Кинельских горах (близ г. Ки-
нель).
Этот вид принадлежит к подроду Otites (Adans.) Schischk., ряду Wolgen-
ses Schischk., куда, кроме него, входят несколько видов, обитающих в сте-
пях, на луговых склонах, на известняковых скалах, галечниках и песках:
S. wolgensis (Евразия), S. densiflora (Балканский п-ов, Крым, Причерно-
морье, Кавказ), S. pseudotites (Средняя и Восточная Европа), S. polaris
(евросибирская Арктика, север европейской части СССР и Западной Сибири).
Serratula Gmelini связана преимущественно с обнажениями известняка и
мела в лесостепной и степной зонах, но встречается также в луговых степях,
на лугах, лесных опушках и в зарослях степных кустарников. Распростране-
на серпуха Гмелина (рис. 81) главным образом на Южном Урале (в верховь-
ях р. Урала, в бассейне рек Сакмары, Белой с ее притоками Уфой и Юрю-
занью), но заходит также в прилегающую часть Среднего Урала. Самые
южные местонахождения — окрестности г. Оренбурга и пос. Кувандыка, са-
мое северное — Александровские сопки в Красноуфимской лесостепи. На вос-
ток заходит до гор Егозинской и Сугомак. Западная граница основного ареа-
ла проходит в Предуралье, однако, кроме того, имеется ряд изолированных
местонахождений на Бугульминско-Белебеевской возвышенности, в районе
Бузулукского бора, в Приволжье, в Жигулях, в окрестностях городов Сара-
това, Хвалынска, Сызрани, Вольска. Этот вид относится к секции Klasea
(Cass.) DC., ряду Centauroides Boriss., он близок к S. centauroides (Восточ-
9 Заказ № 251
129
ная Сибирь, и Монголия), к S. radiata (Средняя Европа, юг европейской
части СССР и Кавказ) и S. tanaitica (меловые горы в Волгоградской облас-
ти). Указания на встречаемость серпухи Гмелина на меловых горах Средне-
Рис. 80. Ареал смолевки башкирской Silene baschkirorum. За
пределами карты — оторванные местонахождения в Заволжье.
русской возвышенности могут относиться к одному из двух последних упомя-
нутых видов.
На обнажениях скал, щебнистых обрывах и в каменистых степях обитает
эндемик Cerastium uralen.se (Грубов, 1968). Пока этот вид найден лишь в
130
двух пунктах Башкирской АССР: в Караидельском районе на берегу р. Уфы
близ пос. Красный Ключ и в Нуримановском районе близ дер. Ельдяк.
Ясколка уральская относится к подроду Eucerastium Boiss., секции Orthodon
Ser., подсекции Perennia Fenzl., ряду Arenaria Schischk. Близка к широко
распространенному в северном полушарии виду С. arvense и, по-видимому,
представляет собой его эдафическую расу, возникшую в результате отбора в
Рис. 81. Ареал серпухи Гмелина Serratula Gmelini.
популяциях, приспособившихся к произрастанию на каменистом субстрате в
степной зоне Южного Урала.
Libanotis sibirica (L. uralensis) — эндемик Урала, обитающий главным
образом в лесостепи в светлых лиственничных или сосново-лиственничных ле-
сах и на лугах. Связан со щебнистыми маломощными почвами, формирующи-
мися на известняках. В северной части своего ареала встречается на извест-
няковых обнажениях по берегам рек, а также на южных склонах гор близ
верхней границы леса. Распространен на Южном Урале в бассейнах рек Бе-
лой, Демы, Уфы, Ая, Юрюзани, Сима, на шиханах близ г. Стерлитамака,
на хр. Крыкты, горах Шатак, Ильменских, Вишневых. На Среднем Урале
встречается на Уктусских горах близ г. Свердловска, на известняковых уте-
сах по р. Уфе около г. Красноуфимска, близ рудника Новоасбест, на горах
Басеги, Качканар, на известняках по р. Косьве около станции Губаха,
р. Чусовой около пос, Кына и на гипсах по р. Сылве около г. Кунгура.
Заходит на Северный Урал, где встречается на склонах горы Кумба, Кон-
жаковского, Косьвинского и Денежкина Камней, а также на известняковых
утесах по рекам Вижаю и Тошемке (рис. 82).
9* 131
Рис. 82. Ареал порезника сибирского Libanotis sibirica.
Б. К. Шишкин (Флора СССР,
т. XVI) относит Л. sibirica10 к ряду
Seseloides Schischk., куда входят,
кроме него, L. seseloides (Восточная
Сибирь, Дальний Восток, Манчжу-
рия) и L. Schrenkiana (горы Сред-
ней Азии). В. Н. Тихомиров (лич-
ное сообщение) сближает L. sibirica
с L. condensata (Сибирь, Дальний
Восток), отнесенному Б. К. Шиш-
киным к ряду Condensatae Schischk.
К этой же группе эндемиков
относится Agropyrum reflexiaris-
tatum, обитающий в горных каме-
нистых степях на Южном Урале (хр.
Ирендык, горы Нурали, Сугомак и
др.), а по скалистым обнажениям
заходящий на Средний Урал и юж-
ную часть Северного. В северной
части своего ареала (рис. 83) этот
вид встречается на обнажениях из-
вестняка по рекам Чусовой, Косьве,
Язьве, Ивделю, Вижаю, Северной
Сосьве, Илычу и другим и на обна-
жениях гипса по р. Сылве близ
г. Кунгура. По обнажениям дунита
заходит в высокогорья до верхней
границы леса (Желтая сопка массива
Денежкин Камень, дунитовое «плечо»
Косьвинского Камня, водосборная
воронка р. Южного Иова на Кон-
жаковском Камне). Наиболее близок
к нему A. strigosum, произрастаю-
щий в горах Крыма; другие родст-
венные виды распространены в гор-
ных районах Средней Азии, Сибири
и Дальнего Востока.
На скалах и щебнистых скло-
нах Южного Урала произрастает
эндемик Agropyrum pruiniferum.
Этот вид распространен (рис. 84)
в бассейне р. Урала, его притока
р. Сакмары с впадающей в нее
р. Большим Иком, а также в Иль-
менских горах. Изолированное место-
нахождение отмечено в Жигулях.
К ряду Stipifoliae Nevski, в кото-
рый входит пырей инееватый, принад- Рис. 83. Ареал пырея отогнутоостиого Agro-
лежит еще более десятка видов, pyrum reflexiaristatum.
произрастающих на юго-западе евро-
пейской территории СССР, в Крыму, на Кавказе, в Иране, в горах Средней
Азии, на Алтае и в Саянах на скалах, осыпях, каменистых склонах.
10 Название L. sibirica не соответствует его распространению; в Сибири он не растет.
В первой полозине XIX столетия, когда этот вид был описан К- Мейером, Урал было
принято считать частью Сибири. В. Н. Тихомиров предлагает новое название L. Krylov И.
133
Эндемик Южного Урала Roegneria uralensis распространен (рис. 85) пре-
имущественно на восточных и западных предгорьях хребта, в поясе горной
и предгорной лесостепи (хр. Ирендык, верховья р. Сакмары, верховья
р. Большой Сурени в бассейне р. Большого Ика, верховья р. Урала, бас-
сейн р. Большого Кизила,
Ильменские горы, бассейн
р. Демы). Характерные
места его обитания —
луговые степи и разно-
травные луга (Невский,
1930). R. uralensis при-
надлежит к ряду Subse-
cundae Nevski, который
включает еще два близ-
ких вида — R. tianscha-
nica (растет на лугах и
склонах в горах Средней
Азии) и R. K.omarovii
(на лугах, в долинах рек
и по склонам на Алтае
с прилегающей частью
среднеазиатских гор, в
Саянах и Северной Мон-
голии).
Другой эндемичный
вид Roegneria viridiglu-
mis (рис. 86) произра-
• стает на береговых из-
вестняковых обнажениях
по р. Чусовой (Камень
Гилев) и в лесостепных
березовых колках восточ-
ного склона Южного
Урала (окрестности дер.
Аджитаровой между горо-
Рис. 84. Ареал пырея инееватого Agropyrum pruiniferum. дами Златоустом и Челя-
бинском). Этот малоизу-
ченный вид принадлежит к ряду Viridiglumes Nevski, куда входит еще
один вид — R. taigae, распространенный в Мариинско-Чулымской тайге Запад-
ной Сибири.
Koeleria sclerophylla—-эндемик Южного Урала с Предуральем (водораз-
дел рек Малый Чуран и Ток, район озера Асли-Куль, окрестности пос. Кунд-
рава) и Приволжья (Жигули и смежные места). Произрастает на обнажениях
мела и других горных пород, на каменистых степных склонах. Относится к
секции Caespitosae Dornin, ряду Cristatae Dornin, включающему несколько
азиатских, сибирских, дальневосточных и европейских видов. Этот еще мало-
изученный мелкий эндемичный вид крайне близок к келерии Талиева /С. Та-
lievii, распространенной на меловых обнажениях Среднерусской возвышенно-
сти (Курская, Воронежская, Белгородская, Пензенская области и Мордов-
ская АССР).
К числу скально-горностепных эндемиков относятся три мелких вида из
рода Schiverekia (рис. 87). Одна из шиверекий — Sch. berteroides — обитает
на скалистых обнажениях и в каменистых степях на вершинах невысоких
гор Южного Урала (от окрестностей г. Златоуста и горы Уреньга до окрест-
ностей г. Оренбурга и Губерлинских гор). Этот вид наиболее близок к Sch.
134
podolica s. str. (Алексеенко, 1946, 1950), ареал которой расположен на По-
дольско-Волынском плато и в Восточной Галиции.
На Среднем Урале с примыкающими к нему частями Северного и Южно-
го Урала на скалах (чаще известняковых) и в каменистых горных степях
произрастает Sch. monticola. Близкие формы, встречающиеся по р. Донцу и
на Приволжской возвышенности, М. И. Алексеенко раньше рассматривала
как особый подвид этой шиверекии (Sch. monticola ssp. mutabilis),' однако
позднее она сочла целесообразным выделить их в самостоятельный вид Sch.
Рис. 85. Ареал регнерии уральской Roegneria uralensis.
mutabilis. На Южном Урале Sch. monticola обитает в каменистых горных
степях на горах Сугомак, Егозинской, Вишневых, на хребтах Крака, Крыкты
и Ирендык. Встречается на обнажениях известняков по берегам рек как на
Южном Урале (реки Белая, Сиказа, Уфа), так и на Среднем (реки Чусовая,
Косьва, Яйва, Пышма, Исеть и Тура). Заходит в подгольцовый пояс на скло-
нах гор Юрма, Басеги, Армия и Куроксар.
На скалах как в горнолесном, так в горнотундровом и подгольцовом поя-
се Северного и отчасти Среднего Урала произрастает Sch. Kusnezovii, отли-
чающаяся большей изменчивостью по сравнению с другими уральскими вида-
ми шиверекий. Шиверекия Кузнецова нередка на известняковых утесах по
рекам Илычу, Вишере, Колве, Лозьве, Ивделю, Сосьве. Встречается близ
верхней границы леса на некоторых горах Северного Урала по обнажениям
дунита (водосборная воронка в верховьях Южного Иова на Конжаковском
Камне, «плечо» Косьвинского Камня).
Уральский ряд шиверекий, по мнению М. И. Алексеенко (1950), обосо-
бился от подольского еще в третичное время. В неогене на обнажениях мас-
сивнокристаллических горных пород в южной части Урала произрастала ис-
ходная форма уральского ряда шиверекий, проникшая сюда из горных райо-
нов Европы по средней и южной полосе европейской части СССР. К этой
исходной форме наиболее близка современная Sch. berteroides. Эпейрогениче-
135
ские поднятия в конце третичного и
начале четвертичного времени способст-
вовали дифференциации этой исход-
ной формы, обособлению Sch. monti-
cola, а затем и Sch. Kusnezovii. Из
всех уральских видов этого рода Sch.
Kusnezovii — самый молодой, образо-
вавшийся уже в плейстоцене и рас-
селившийся на горных вершинах и ска-
листых склонах Северного Урала вскоре
после освобождения их от ледникового
покрова.
Со скалистыми (преимущественно
известняковыми) обнажениями Ураль-
ского хребта связаны некоторые энде-
мичные j представители рода Thymus.
В их число входит, в частности, Th.
Talievii (рис. 88), встречающийся на
Среднем Урале на обнажениях извест-
няков по рекам Чусовой и Серге, на
Южном Урале — по рекам Юрюзани
и Белой, на горе Егозинской, а также
на прилегающей к Уралу равнинной тер-
ритории Башкирии (ареал указывается
по Клокову и Шостенко, 1936, с
дополнениями по нашим данным). Этот
тимьян относится к секции Euserpyl-
lum Klok., ряду Asiatici Klok., однако
имеет признаки, связывающие его с
рядом Euserpylla Klok. et Sehost.
Другой представитель ряда Asiatici
Klok. ; et Shost?—Th. hirticaulis —
является эндемиком Полярного Урала,
известен из его западных предгорий
в районе кряжа Чернышева (Адак),
где обитает на обнажениях извест-
няка. Этот вид наиболее близок к
Th. asiaticus, хотя прослеживаются
некоторые черты его сходства с. Th.
stbiricus, относящимся уже к ряду
Euserpylla Klok. et Sehost. К ряду
Asiatici Klok. принадлежат два новых
вида тимьяна, намеченных к описанию
М. В. Клоковым по нашим сборам.
Один из них встречается на известня-
ковых обнажениях Приполярного Урала
(р. Косью), а другой довольно широко
распространен по различным каменис-
тым обнажениям на Среднем Урале.
Рис. 86. Ареал регнерии зеленочешуй- Оба они родственны Th. asiaticus Serg.
чатой Roegneria viridiglumis. особенно второй, являющийся его средне-
уральским викариантом. s,V“
Секция Kotschyani Klok. также представлена на Урале несколькими энде-
мичными видами. Сюда принадлежат Th. baschkiriensis (рис. 89) из ряда
Suffruticosi Klok. et Sehost. — эндемик Южного Урала (верховья р. Сакма-
136
Рис. 87. Ареал эндемичных уральских видов рода Schitie-
rekia.
1~ шиверекия Кузнецова Sch. Kasnezotwf 2 — шиверекия горн ая
Sch. mC'titicola’, 3—шиверекия икотниковидная Sch. berteroides.
ры, бассейн р. Большого Кизила) и прилегающей части Предуралья (Бугуль-
минско-Белебеевская возвышенность), где он растет в каменистых степях и
на обнажениях известняка и мергеля.
Более ограниченное распространение имеет другой эндемичный вид из той
же секции — Thymus guberlinensls, пока достоверно известный только из Гу-
берлинских гор на Южном Урале, где он растет в каменистых степях и на
обнажениях скал.
Thymus mugodzharicus — вид, близкий к двум предыдущим, распростра-
нен в каменистых степях, на обнажениях известняка и мела на Южном Ура-
Рис. 88. Ареал тимьяна Талиева Thymus Talievii.
ле, в Предуралье и Мугоджарах. На север доходит до окрестностей г. Стер-
литамака, р. Ай и хр. Крыкты. Центр обилия этого вида расположен на
Южном Урале. Изолированные местонахождения известны в области Казах-
ского мелкосопочника.
Особняком в секции Kotschyani Klok. стоит Th. binervulatus, обнаружен-
ный пока только на известняковой скале Жукова Шишка, находящейся на
берегу пруда близ г. Сима в Челябинской области (Клоков и Шостенко-Де-
сятова, 1936). Это растение было найдено впервые С. Ю. Липшицем;
в 1959 г. здесь же оно было собрано автором этих строк (определил
М. В. Клоков). Th. binervulatus пока является единственным представителем
особого ряда Binervulati Klok. et Shost.
138
Как уже упоминалось, род Thymus сформировался, вероятно, в фригано-
идных сообществах древнего Средиземноморья. Позднее его представители
широко расселились в Азии. Секция Kotschyani Klok., в том числе ряд Suf-
fruticosi Klok. et Schost., генетически связана с Ираном. Ряд Binervulati Klok.
et Schost., вероятно, является производным от ряда Suffruticosi Klok. et
Рис. 89. Ареалы некоторых уральских эндемичных видов
рода Thymus.
1 — тимьян мугоджарский Th. mugodzharleas; 2 — тимьян губерлинский
Th. guberlinensis', 3 —тимьян башкирский Th. baschkiriensts; 4 — тимь-
ян двужилковый Th. binervulatus.
Schost. Ряд Asiatici Klok. из секции Euserpyllum Klok. связан узами родст-
ва с китайскими и гималайскими тимьянами из секции Verticillati Klok., а
также с рядом Suffruticosi Klok. et Schost. из секции Kotschyani Klok., име-
ющим иранское происхождение. Уральские эндемичные представители ряда
Asiatici Klok. являются его крайними звеньями, возникшими при расселении
азиатских тимьянов на запад. Все упомянутые эндемичные виды рода Thy-
139
mus на Урале, в понимании М. В. Клокова, являются обособившимися гео-
графическими расами.
Недавно описан новый эндемичный вид Onosma guberlinense (Доброчаева
и Виноградова, 1966). Растения были собраны в Губерлинских горах, на степ-
ном склоне правого коренного берега р. Урала в 5 км выше устья р. Губер-
ли летом 1964 года. В. М. Виноградова, участница экспедиции, работавшей на
Южном Урале под моим руководством, обратила внимание на то, что расту-
щая здесь оносма не похожа на обычный вид 0. slmplicissimum, резко от-
личается от него желтой окраской венчика. Исследование, проведенное.
В. М. Виноградовой совместно с Д. М. Доброчасвой, показало, что это но-
вый вид из секции Euonosma, подсекции Haplotricha Boiss. (группа Stepposa
М. Pop.), ряда Arenaria. Onosma guberlinense наиболее близка к 0. trans-
rhymnense, произрастающей на каменистых склонах и в степях от Зауралья
до оз. Байкал. Интересно, что в этом же районе (между дер. Подгорной и
с. Губерля) такие же растения были собраны в 1874 г. С. М. Смирновым
и намечены к описанию на этикетках гербария Ботанического института
АН СССР под наименованием О. Trautvetteri. О. guberlinense — локальный
эндемик, отчленившийся от широко распространенного исходного вида, близ-
кого к О. transrhymnense и давшего начало, кроме того, нескольким видам,
встречающимся на Украине — О. borystenicum и О. graniticola.
В группе горностепных и скальных эндемиков к числу немногих примеров
молодого, прогрессивного эндемизма относится Tanacetum uralense (Pyreth-
rum uralense'), встречающийся на предгорьях Южного Урала в каменистых,
реже разнотравных степях, на обнажениях известняка и песчаника как в степ-
ной зоне, так и в лесостепи. Самые северные местонахождения пижмы ураль-
ской— окрестности дер. Кандры в Белебеевском районе и гора Балкан-Тау в-
Учалинском районе Башкирской АССР, самое южное — Губерлинские горы
(рис. 90). Этот вид очень близок к Т. millefoliatum, распространенному в
степях Восточной Европы и Казахстана, и, в сущности, является особой эда-
фической формой последнего.
Как видно, рассмотренные нами эндемичные виды в большинстве случаев-
распространены на Южном Урале в каменистых степях и на скалах, некото-
рые из них заходят на Средний и отчасти Северный Урал, где встречаются
на каменистых, чаще известняковых обнажениях. Распространение этих ви-
дов прерывисто, особенно в северных частях ареала, у некоторых из них
имеются оторванные изолированные местонахождения за пределами Урала
(Жигули, Подуральское меловое плато, Мугоджары, Казахский мелкосопоч-
ник).
Генетические связи рассмотренных здесь скальных и горностепных энде-
миков Урала различны. У некоторых из них (например, Minuartia Krasche-
ninnikovii и местные виды из рода Schiverekia) родственные виды распрост-
ранены в Европе. У других (например, Aulacospermum isetense, Roegneria
uralensis, большинство уральских эндемиков из рода Thymus') они произрас-
тают в горах Азии. Наконец, ряд эндемиков связан с серией родственных
видов, обитающих на территории Русской равнины, в Крьгму, на Кавказе, на
горных хребтах Средней Азии, на Алтае, в Средней и Восточной Сибири.
Эти родственные виды произрастают преимущественно в горах на скалистом
субстрате (чаще на низких уровнях, значительно реже в высокогорных поя-
сах), а в равнинных условиях-—на песчаных почвах, наносах галечника и
песка и различных обнажениях (мел, известняк, мергель и т. п.). Большей
частью ареалы видов, родственных уральским скальным и горностепным энде-
микам, находятся на значительном расстоянии от Уральских гор.
В третичное время Уральские горы были уже сильно снижены и разруше-
ны. Урал представлял тогда слабо всхолмленную местность, где, несомненно^
были широко распространены обнажения различных горных пород и продук-
140
тов их разрушения (скалы, щебень, песок и т. д.). С западной стороны к
Уралу в течение всего третичного периода примыкала суша; с восточной сто-
роны в палеогене он омывался морем, и лишь в неогене здесь установился
режим суши.
Проникновение исходных форм скальных и горностепных растений на
Урал из других горных районов (Крым, Кавказ, Средняя Азия и др.) мож-
но объяснить, допуская, что в плиоцене, а может быть, даже еще в миоцене,
на юге европейской части СССР, в южной части Западной Сибири и в Се-
Рис. 90. Ареал пижмы уральской Tanacetum uralense.
верном Казахстане существовал открытый безлесный или слаболесистый
ландшафт.
По всей вероятности, в плиоцене в южной части Урала на скалистых
и щебнистых склонах холмов, особенно обращенных на юг, существовали
своеобразные слабо сомкнутые сообщества травянистых растений — ксе-
рофитов. Флора этих своеобразных сообществ формировалась главным об-
разом за счет прастепных растений, а также растений периодически обна-
жающегося субстрата — эрозиофилов. Путями проникновения на Урал
эрозиофилов по равнинам европейской территории СССР и Западной Си-
бири были сосновые боры на песках в южной части лесной зоны, прибреж-
ные обнажения различных пород и продуктов их разрушения, а также бе-
чевник по берегам рек.
Эпейрогенические поднятия в конце плиоцена и начале плейстоцена
повлекли за собой усиление эрозионной деятельности на Урале, возник-
новение на значительно большей площади обнажений различных горных
пород, первоначально не заселенных или слабо заселенных раститель-
ностью. Некоторые пришлые растения, способные расти на каменистом и
щебнистом субстрате при недостаточном водоснабжении, нашли здесь для
себя подходящую обстановку и широко расселились.
141
Надо заметить, что оголенность субстрата или маломощность почвы на
нем в горных районах поддерживается, вследствие интенсивной эрозии,
более регулярно, чем на равнинах, а площадь как отдельных обнажений,
так и всех их в совокупности несравненно больше. Поэтому в горах и на
предгорьях создаются более благоприятные условия для растений, способ-
ных произрастать на незакрепленном или слабо закрепленном субстрате,
но обычно не выдерживающих конкуренции с компонентами более сфор-
мировавшихся сообществ. На равнинах же такие растения по мере раз-
вития почвенного слоя на обнажениях и формирования сомкнутого расти-
тельного покрова легко вытесняются.
В ходе смен растительного покрова, происшедших в конце плиоцена
и в плейстоцене, ареалы ряда достигших Урала исходных ксероморфных
видов оказались разорванными, так как эти растения вымерли на равни-
нах, за исключением немногих мест (например, меловые обнажения на
Русской равнине), но сохранились в горах и на предгорьях. Своеобразие
эдафических условий в отдельных районах и географическая изоляция
способствовали образованию новых форм. Так, в частности, экологически
и морфологически обособились эндемичные для Урала растения горностеп-
ных и скальных местообитаний.
В конце плейстоцена и в голоцене, по мере накопления мелкозема на
горных склонах, формирования более развитого слоя почвы и закрепления
склонов древесной растительностью, площадь скалистых обнажений в юж-
ной части Урала значительно уменьшилась. Следовательно, сократилась
и территория, пригодная для произрастания горностепных и скальных
видов. Все это вместе с многократными изменениями климата и смещением
растительных зон и поясов привело к сокращению ареала таких растений.
Горностепные и скальные эндемики уральской флоры представляют со-
бой фитоценотически единую группу, тесно связанную с былым ландшафтом
древних горных каменистых степей или степеподобных группировок травя-
нистых растений-ксерофитов. Об этом свидетельствует то, что многие пред-
ставители этой группы распространены в основном в горных и предгорных
степях Южного Урала, а севернее изредка встречаются в реликтовом со-
стоянии на береговых известняковых обнажениях. Закономерности их
распределения говорят о том, что в прошлом был период, когда сообщества
первичных каменистых горных степей заходили по Уралу дальше на се-
вер, чем теперь проникают современные степи.
Таким образом, уральские горностепные и скальные эндемики явля-
ются своеобразными реликтами флористического комплекса первичных
каменистых степей — древних ксерофильных растительных сообществ,
широко распространенных в южной части Урала в конце третичного пе-
риода на каменистых склонах и различных продуктах разрушения горных
пород.
ЭНДЕМИКИ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
К этой группе относятся эндемичные виды, связанные с областью рас-
пространения широколиственных лесов в южной части Уральского хребта
(рис. 91) и на прилегающей территории Русской равнины.
Примером может служить Lathyrus Litvinovii — растение лиственных
(в особенности широколиственных) лесов и кустарниковых зарослей. Энде-
мик Южного Урала и Предуралья. Наибольшее число известных его место-
нахождений сосредоточено в бассейне р. Белой и ее притоков Демы и Уфы,
а также в бассейне р. Сакмары и ее притока Большого Ика на территории
Башкирской ССР и в прилегающих частях Оренбургской и Челябинской
областей (рис. 92). Самые северные местонахождения отмечены в районе
г. Бирска и в среднем течении рек Юрюзани и Ая. Наиболее восточный
142
Рис. 91. Дубовый лес на Зилаирском плато (Южный Урал).
пункт встречаемости — верховья р. Белой, самый южный
пос. Кувандыка на р. Сакмаре. На западе доходит до г. Бугуруслана Это
мелкий вид, трудно отличимый от родственных ему L. miniatus, °^та^
щего на Кавказе, и L. rotundifolius, распространенного в Крыму, Малой
Азии (горы по южному побережью Черного моря) и Иране. Однако он рез
ко обособлен от них географически. Вследствие недостаточной изученное?
143
пока еще трудно судить о правомерности выделения уральских растений
в самостоятельный вид; не исключена возможность, что они окажутся под-
видом кавказской L. miniatus. Ареал L. Litvinovii в основном совпадает
с областью распространения широколиственных лесов на Южном Урале
и в Предуралье, тогда как ареал L. miniatus — с областью широколиствен-
ных лесов на Кавказе. Разрыв ареала первичного предка в плейстоцене
повлек за собой обособление кавказской, крымской и уральской форм.
Более широкий ареал имеет K,nautia tatarica — характерное растение
широколиственных или смешанных с широколиственными породами ле-
Рис. 92. Ареал чины Литвинова Lathyrus Lituinovii.
сов, опушек, лужаек и зарослей кустарников. Этот вид распространен на
западном склоне Среднего и Южного Урала и в прилегающей части Пред-
уралья (рис. 93). На севере граница его ареала проходит через с. Осу, по
р. Чусовой через пос. Чизму и с. Чусовое; на востоке — через поселки
Билимбай, Бердяуш, г. Белорецк. Самые южные местонахождения зареги-
стрированы в районе с. Ташлы в бассейне р. Большого Ика и с. Юлуково
в бассейне р. Сакмары. Близ западной границы Башкирии этот вид уже
встречается редко. Отдельные оторванные местонахождения отмечены
С. И. Коржинским (Korshinsky, 1898); из этих мест сохранились лишь не-
многие гербарные экземпляры, собранные в смежных с Башкирией и Перм-
ской областью районах Татарской и Удмуртской АССР. Согласно литера-
турным данным, пока не подтвержденным новейшими сборами, К. tatarica
встречается также в Жигулях близ оз. Елгуши (коллектор В. И. Смирнов,
сбор 1904 года).
Крайне близкий вид К- montana обитает на лесных опушках и лугах
в лесном поясе Кавказа и Малой Азии (Восточная Анатолия). Кавказские
растения отличаются более короткими семянками, лишенными, оторочки;
в остальных признаках они сходны с К., tatarica, поэтому некоторыми ис-
следователями отождествляются с последней.
Такого же рода эндемиком является Cicerbita uralensis (С. Gmelini),
свойственная лиственным лесам (преимущественно широколиственным
144
или с участием широколиственных пород), лесным опушкам и лесным лу-
гам. Область распространения ее (рис. 94) охватывает западный склон
Южного и Среднего Урала (Башкирская АССР и смежные районы Челя-
бинской области, Пермская область, юго-запад Свердловской области),
Рис. 93. Основная часть ареала короставника татарского Knautia
tatarica на Урале и в Предуралье.
а также восточную часть средней полосы европейской территории СССР от
Предуралья до Московской, Ивановской и Тульской областей. Трудно от-
личима от близко родственного вида С. grandis, произрастающего в лесах
Кавказа, но хорошо отчленена от него географически.
Вероятно, С. uralensis и С. grandis имели общего предка, распростра-
ненного в доледниковое время на Кавказе и во всей полосе широколиствен-
ных лесов Русской равнины и Урала. В эпоху наибольшего оледенения
первичный ареал исходного вида оказался разорванным; сохранились
лишь кавказская и южноуральская его части. На Южном Урале это рас-
тение смогло сохраниться в убежищах, где пережили оледенение широко-
Ю Заказ № 251
145
Рис. 94. Основной ареал цицербиты уральской Cicerbita uralensis на Урале
и в Предуралье.
лиственные леса. Географическая изоляция способствовала обособлению
уральских популяций в самостоятельный вид. В послеледниковое время
уральская форма расселилась довольно далеко на запад по полосе широко-
лиственных лесов.
К рассмотренным здесь эндемикам примыкает Anemone uralensis, про-
израстающая на сырой, богатой перегноем почве в горных долинах, по бе-
регам речек, заросших кустарниками, на заливных и заболоченных лугах
и в лиственных лесах. Эта ветреница распространена (рис. 95). на Сред-
Рис. 95. Ареал ветреницы уральской Anemone uralensis.
нем Урале (с. Ильинское, окрестности г. Красноуфимска, пос. Билимбая,
городов Нижнего Тагила и Свердловска) и в северной части Южного Урала
(окрестности г. Златоуста, р. Ик, приток Камы). Она относится к серии
близкородственных форм, объединившихся ранее в сборный вид А. гапип-
culoides s. ampl., куда С. И. Коржинский (1892) включал ее в ранге под-
вида, названного им ssp. uralensis. Теперь A. uralensis рассматривается
обычно как самостоятельный вид. К нему очень близки A. ranunculoides
(Европа, включая европейскую часть СССР, Кавказ, Средний и Южный
Урал), A. coerulea (Алтай, прилегающая к нему часть Западно-Сибирской
низменности и западная часть Саянской горной системы, на востоке не до-
ходя до Енисея), A. jenisseensis (Саяны и их предгорья от южной части
Красноярского края до окрестностей г. Иркутска). Самая характерная,
отличительная особенность A. uralensis состоит в окраске ее цветков, ле-
пестки обычно бывают розово-красными, белыми, но иногда и голубыми
с целой гаммой промежуточных оттенков между ними. Хотя уральские
растения с голубыми цветками крайне близки к А. coerulea, их не следует
относить к последней, так как они, по мнению С. И. Коржинского (1892),
представляют собой уклонение от уральской расы. Ареал A. uralensis
смыкается с ареалом желтоцветковой A. ranunculoides, а на их контакте
наблюдается масса гибридных форм с переходными признаками.
10*
147
Все эти близкородственные виды анемон, по-видимому, происходят от
одного общего предка, который был широко распространен в доледниковое
время (плиоцен) от Европы до Восточной Сибири в лиственных (преиму-
щественно широколиственных) лесах с опадающей на зиму листвой. При-
способившись к цветению ранней весной, когда деревья еще не оделись
листвой и под полог проникает много света, этот исходный вид выработал
своеобразную ритмику развития, выражающуюся в раннем появлении
листьев, быстром зацветании и созревании семян. Как и у других предста-
вителей группы весенних раноцветущих растений флоры плиоценовых
широколиственных лесов, у этого вида выработалась способность цвести-
при относительно низких температурах. Похолодание климата, проис-
шедшее в плейстоцене, повлекло за собой деградацию широколиственных
лесов и раздробление ареала исходной формы названных анемон, вымира-
ние ее во многих местах, где она прежде была распространена. Поскольку
в Европе широколиственные леса и в плейстоцене были распространены
на больших площадях, сформировавшаяся здесь A. ranunculoides сохра-
нила свою преимущественную связь с ними. Уральская, алтайская и сред-
несибирская (енисейская) формы образовались в горных и предгорных
районах в результате приспособления исходного вида к произрастанию
в светлых лиственных (березовых, осиновых, ольховых) лесах в условиях
более сурового климата. Хотя в предгорьях Южного Урала, Алтая и Саян
были местные центры сохранения теплолюбивой неморальной флоры,
вновь возникшие виды анемон обособились уже вне фитоценотической связи
с ними и вне их территориальных границ. В голоцене, когда площадь ши-
роколиственных лесов расширилась, ареалы A. uralensis и A. ranunculoides
на Урале сомкнулись и началась интенсивная гибридизация между ними.
Таким образом, A. uralensis генетически была связана с широколиствен-
нолесными формациями, но в ходе изменений состава лесов Урала, проис-
шедших в плейстоцене, приспособилась к произрастанию в мелколиствен-
ных лесах.
Рассмотренные уральские неморальные эндемики в большинстве слу-
чаев морфологически не резко обособлены от родственных им видов, оби-
тающих в лесах Кавказа, Крыма и Малой Азии, хотя изолированы от них
географически. Поэтому можно спорить о том, следует ли возводить эти
формы в ранг видов или лучше рассматривать их как подвиды. Однако сам
факт, что в составе уральской неморальной флоры имеются эндемичные расы
растений, представляет большой интерес. Он служит поводом для предпо-
ложений, что широколиственные леса южной части Уральского хребта не
всегда были связаны с аналогичными лесами Русской равнины, а для
истории их развития характерны свои самобытные черты. И действительно,
изучение эндемичных, а также реликтовых (см. следующую главу) расте-
ний неморального комплекса позволяет детализировать некоторые момен-
ты истории широколиственных лесов Восточной Европы.
В конце третичного периода, в плиоцене, широколиственные леса за-
нимали видное место в сложении растительного покрова европейской
части СССР, Кавказа, Урала и Сибири. В плейстоцене же, особенно в лед-
никовые эпохи, площадь таких лесов значительно сократилась и они смогли
сохраниться только в немногих убежищах.
Представляет интерес вопрос о том, существовали ли на Урале такие
убежища, в которых хотя бы обедненные по составу широколиственные
леса могли пережить эпоху наибольшего оледенения. Решение этого во-
проса затрудняется тем, что широколиственные леса западного склона
Урала в настоящее время смыкаются с полосой аналогичных лесов, протя-
нувшейся через Русскую равнину к горным районам Средней Европы, а
некоторые древесные породы широколиственного комплекса (Тilia cordata,
П8
UImus laevis), расселяясь на восток после эпохи максимального оледене-
ния, перевалили через хребет и проникли в прилегающую к Уралу часть
Западно-Сибирской низменности.
С. И. Коржинский. (1894) склонялся к тому, что не только отдельные
представители лесной флоры, но и вообще широколиственные леса Южного
Урала не представляют результата современной миграции, а ведут свое
начало из отдаленной эпохи. Близкие взгляды высказывали Е. М. Лав-
ренко (1938) и Ю. Д. Клеопов (1941). По мнению И. И. Спрыгина (1936),
липа пережила ледниковый период в таких убежищах, как Урал и При-
волжская возвышенность.
Однако некоторые ботаники исключают возможность существования
в плейстоцене убежищ широколиственнолесной растительности на Урале.
Г. Э. Гроссет (1935, 1962) утверждает, что реликтовые широколиственные
леса пережили ледниковые эпохи в таких удаленных от ледника странах,
как Южная Европа, Крым, Кавказ и Дальний Восток. По его мнению,
реликты широколиственных лесов, рассеянные по всей средней европей-
ской части СССР, на Урале, а возможно, и на Алтае, являются реликтами
уже послеледниковой, теплой и влажной эпохи, когда эти тепло- и влаго-
любивые виды имели более широкое распространение по сравнению с со-
временным. И. М. Крашенинников (1939) также допускал возможность
полного исчезновения на Южном Урале в эпоху наибольшего оледенения
широколиственных лесов вместе с большинством сопутствующих им тра-
вянистых растений. Он считал, что территории, лежащие к западу от гор
Урала, между Уральским хребтом и Волгой, в особенности повышенные
пространства Общего Сырта с прилегающими с севера не затронутыми оле-
денением частями Заволжской возвышенности, были более надежными
убежищами для широколиственнолесной растительности.
Однако нужно иметь в виду, что предположение о полном исчезновении
в плейстоцене широколиственных лесов на Южном Урале и его западных
предгорьях не согласуется с многими ботанико-географическими и палео-
ботаническими фактами. В связи с этим уместно вспомнить высказывание
Н. Я. Каца (1952) о том, что сторонники гипотезы дальних миграций не
учитывают относительной кратковременности периода, прошедшего после
эпохи наибольшего оледенения и не увязывают свои построения с реаль-
ными темпами естественного расселения древесных пород.
Ботанико-географические данные (Горчаковский, 1968) свидетель-
ствуют в пользу предположения о сохранении широколиственных лесов
в некоторых пунктах предгорий и нижней части склонов Южного Урала,
где они в настоящее время являются преобладающим элементом раститель-
ного покрова. В горах и на предгорьях, где рельеф более расчленен, созда-
ется значительно большее разнообразие почвенно-грунтовых условий и
термического режима, чем на равнинах, а следовательно, более велика
вероятность местных благоприятных сочетаний условий среды для сохра-
нения теплолюбивой растительности. Именно с предгорьями связаны ос-
новные местонахождения липы сибирской (Tilia sibirica) в Сибири (пред-
горья Алтая, Кузнецкий Алатау и Салаир, а также предгорья Восточного
Саяна близ устья р. Маны на Енисее южнее г. Красноярска). Третичная
реликтовая природа указанных местонахождений липы сибирской в на-
стоящее время не вызывает сомнений. Надо полагать, что если липа сибир-
ская пережила ледниковые эпохи в предгорных районах в условиях суро-
вого, более континентального климата Сибири, то и крайне сходная с ней
по морфологическим признакам и экологическим свойствам липа мелко-
листная и другие широколиственные породы смогли пережить ледниковые
эпохи на западном склоне и в предгорьях Южного Урала. Нужно иметь
в виду, что западный склон Южного Урала по сравнению с прилегающими
149
равнинными районами отличается повышенной влажностью воздуха и более
обильными атмосферными осадками, что влечет за собой уменьшение
континентальности климата.
Еще недостаточно изучены морфологические и экологические особен-
ности широколиственных древесных пород, произрастающих на Урале.
Весьма вероятно, что некоторые из них представлены здесь особыми,
менее требовательными к теплу формами, сформировавшимися в условиях
относительно сурового климата, когда уральские фрагменты их ареалов
были изолированы в эпоху наибольшего оледенения.
В. П. Гричук (1949), опираясь на микропалеонтологические данные,
высказывает интересные мысли о времени и соотношении тех миграцион-
ных потоков теплолюбивой флоры, из которых в периоды климатических
оптимумов плейстоцена и голоцена складывались широколиственные леса
Восточно-Европейской равнины. Пыльцевые диаграммы показывают, что
в лихвинско-днепровскую межледниковую эпоху неморальная флора по-
лучила чрезвычайно широкое распространение, причем очень велика была
роль южноуральского центра ее консервации. В следующую, днепрово-
валдайскую межледниковую эпоху прежде всего сказалась роль балкан-
ского центра консервации; радианты из этого центра первыми широко рас-
пространились по Восточно-Европейской равнине. Миграционные потоки
из других центров сохранения — центральноевропейского и южноураль-
ского — распространились по равнине лишь позднее, в середине днепров-
ско-валдайского межледниковья, вытесняя и поглощая существующую
уже здесь кверцетальную флору. В голоцене, в период климатического
оптимума, радианты из всех трех центров консервации (центральноевро-
пейского, балканского и южноуральского) появляются почти одновременно,
хотя балканский миграционный поток слегка превалирует.
Результаты палинологических исследований (Хвалина, 1965; Рябова,
1965) свидетельствуют о том, что последнее плейстоценовое оледенение не
распространилось на территорию Башкирского Предуралья. В конце плей-
стоцена и в самом начале голоцена здесь существовали вполне сформиро-
вавшиеся леса. Однако похолодание климата в период валдайского оледе-
нения вызвало увеличение роли хвойных пород в составе лесов (листвен-
ница, сосна) и сокращение, а местами отступление широколиственных.
В древнем голоцене Башкирского Предуралья, как это установила
Т. П. Рябова (1965), в пыльцевых спектрах отмечается присутствие пыльцы
широколиственных пород (в небольшом количестве), тогда как в соседних
районах Русской равнины пыльца широколиственных появляется лишь
в раннем голоцене. Это говорит в пользу существования рефугиумов широ-
колиственных лесов на западных предгорьях Южного Урала в раннем го-
лоцене.
О вероятном сохранении на Урале и в Предуралье в ледниковые эпохи
плейстоцена массивов широколиственных лесов свидетельствует встречае-
мость в южной части хребта и смежных районах Предуралья эндемичных
рас некоторых травянистых растений, относящихся к неморальному ком-
плексу. Образование этих уральских эндемиков способствовало раздроб-
ление ареалов исходных неморальных видов в результате вымирания их на
территории Русской равнины.
В эпоху наибольшего оледенения уральская часть ареала исходных
видов оказалась обособленной от основного ареала названных растений,
охватывающего горные и предгорные районы Европы и Кавказа. Особые,
более суровые, условия среды, характерные для Южного Урала, и геогра-
фическая изоляция способствовали образованию на Урале местных энде-
мичных форм растений. Эти эндемики являются географическими викари-
антами неморальных видов, произрастающих в Европе, на Кавказе и в
150
Малой Азии. Эндемики, сформировавшиеся в связи с обособлением ураль-
ской части ареала от более широкой области распространения неморальных
предков, в большинстве случаев (например, Lathyrus Litvinovii, Knautia
tatarica, Cicerbita uralensis') пережили ледниковые эпохи в очагах сохране-
ния широколиственных лесов на Южном Урале, а некоторые из них
(Anemone uralensis) уже вне этих очагов. В послеледниковое время созда-
лись предпосылки для расселения ряда широколиственнолесных ураль-
ских эндемиков (Cicerbita uralensis, Knautia tatarica) на запад.
НЕКОТОРЫЕ ЧЕРТЫ ЭНДЕМИЗМА УРАЛЬСКОЙ ФЛОРЫ
Хотя Урал имеет свою давнюю геологическую историю, современная
флора его мало специфична и формировалась главным образом под влия-
нием флор других, более или менее близких к нему областей с издавна
сложившимся самобытным ходом развития. Объясняется это тем, что в ре-
шающий период «века антофитов», когда вырисовывались контуры совре-
менных флористических областей Евразии (в третичное время), древние
палеозойские поднятия на территории Урала были сильно пенепленизи-
рованы и как по строению поверхности, так и по другим физико-географи-
ческим условиям мало отличались от прилегающих равнин. Возрождение
и окончательное формирование Уральских гор произошло в конце третич-
ного и начале четвертичного периода. Лишь с этого времени начался этап
относительно самостоятельного развития уральской флоры. Эпейрогени-
ческие поднятия происходили сравнительно быстро, поэтому только немно-
гие представители местной равнинной флоры смогли столь же быстро при-
способиться к изменившимся условиям среды, преобразовавшись в энде-
мичные уральские виды. Возникшие «экологические ниши» на поднимав-
шихся горных цепях заполнялись преимущественно за счет пришлых ра-
стений из смежных областей, которые по экологическим свойствам более
или менее соответствовали новым условиям среды.
Поэтому Урал не явился местом развития резко обособленной, специ-
фической флоры, влияние которой простиралось бы далеко за пределы этой
горной страны. Хотя во флоре Урала прослеживаются некоторые черты
самобытности, развитие ее шло главным образом за счет неуральских по
своему происхождению элементов. Местные новообразования сыграли в
в этом развитии лишь второстепенную роль.
В составе уральской флоры содержится сравнительно небольшое число
эндемичных видов и разновидностей. Эндемичные формы более высокого
таксономического ранга, чем вид, во флоре Урала отсутствуют.
Характерно, что ни один из уральских эндемиков не играет ведущей,
определяющей роли в сложении растительных сообществ, не является эди-
фикатором. Обращает на себя внимание полное отсутствие на Урале энде-
мичных видов древесных пород; древесный ярус лесов, занимающих столь
значительное место в растительном покрове Уральских гор, слагается ис-
ключительно пришлыми породами, сформировавшимися за пределами этой
горной страны. Эдификаторами в растительных группировках на терри-
тории Урала являются виды сибирского или европейского происхождения,
широко распространенные восточнее или западнее Уральского хребта.
Возникновение уральского эндемизма флоры было тесно связано;
а) с обилием на территории Урала выходов разнообразных горных пород и
продуктов их разрушения, остающихся вследствие эрозии непригодными
для произрастания древесной растительности; б) с эпейрогеническими
поднятиями в конце третичного и начале четвертичного периодов и связан-
ными с ними изменениями условий среды; в) с преобразованиями раститель-
ного покрова в связи с плейстоценовыми оледенениями и наличием в
151
южной части Уральского хребта и в некоторых пунктах на прилегающих
к нему равнинах рефугиумов широколиственных лесов, где могли пережить
эпохи оледенения отдельные представители плиоценового неморального
комплекса.
Эндемичные виды придают лишь своеобразный оттенок уральской фло-
ре; не являясь активными фитоценозообразователями, они не определяют
характера растительного покрова Уральского хребта. В большинстве слу-
чаев уральские эндемики встречаются редко, распространены прерывисто
и часто связаны со скалистым субстратом, где ослаблена конкуренция со
стороны других видов растений, особенно деревьев и кустарников.
Значительная часть, если не большинство уральских эндемиков, — это
виды, которые в прошлом были распространены гораздо шире, но теперь
находятся в состоянии угасания и сохранились в немногих местах с осо-
быми эдафическими условиями. Таким образом, на Урале ярко выражен
реликтовый эндемизм.
Лишь небольшая группа уральских эндемиков относится к молодым
видам, которые сформировались сравнительно недавно, отличаются зна-
чительным варьированием и обнаруживают признаки более или менее ин-
тенсивного расселения в современной обстановке11. Интересно, что даже
среди молодых уральских эндемиков есть виды, сформировавшиеся на ре-
ликтовой основе.
Высокогорные эндемики встречаются главным образом в северной
части Уральского хребта, где лучше выражен высокогорный ландшафт;
они обычно (за редкими исключениями) не выходят за пределы Уральской
горной системы. Горностепные и скальные эндемики, напротив, распро-
странены преимущественно в южной части хребта и встречаются на приле-
гающих к нему равнинах. К ним тяготеют некоторые виды, в основном
связанные с Уралом, но имеющие изолированные местонахождения далеко
за пределами — в Жигулях и в области Казахского мелкосопочника.
Эндемики широколиственных лесов встречаютсяв южной части Урала —
в хребтовой полосе и на западном склоне, а некоторые из них довольно
далеко заходят в пределы Русской равнины. Закономерности территориаль-
ного распределения эндемиков свидетельствуют о тесной генетической связи
отдельных их групп с определенными элементами растительного покрова;
эти закономерности определяются как былым, так и современным распре-
делением растительности на Урале и примыкающих к нему равнинах.
НЕОБХОДИМЫЕ МЕРЫ ПО ОХРАНЕ ЭНДЕМИЧНЫХ РАСТЕНИЙ
Как было показано выше, эндемичные растения Урала представляют
большой интерес для науки, так как их изучение проливает свет на исто-
рию формирования флоры и растительности этой территории. Некоторые
эндемики (например, Anemone biarmiensis, Minuartia Helmii, Gypsophila
uralensis) могут быть использованы в цветоводстве в качестве декоративных
растений. Несомненно, что в процессе дальнейшего изучения эндемиков
выявятся и другие полезные свойства отдельных видов.
В связи с тем, что многие эндемики уральской флоры встречаются ред-
ко, а некоторые из них уже сейчас находятся под угрозой уничтожения,
встает вопрос об охране этих растений от истребления.
Охрана уральских эндемичных растений неразрывно связана с сохра-
нением в неприкосновенности растительных сообществ, в составе которых
11 Вторичная активизация расселения наблюдается и у некоторых реликтов и эндемиков
(свойственных обнаженному каменистому, щебнистому или песчаному субстрату) в связи
с разрушением естественного растительного покрова под влиянием деятельности человека.
152
они обитают, а также соответствующих этим сообществам элементов гео-
графического ландшафта. Приведенные выше данные свидетельствуют о
том, что основными местами локализации эндемичных растений являются:
а) высокогорные пояса (горнотундровый и подгольцовый), б) утесы и об-
нажения скал по берегам рек, в) каменистые горные степи и г) широколи-
ственные леса Южного Урала. С этими же стациями связаны и многие
реликты во флоре Урала.
Единственным местом на Урале, где в какой-то степени охраняются
ландшафты и растительные сообщества высокогорий, является промысло-
вое хозяйство «Денежкин Камень». Назрела необходимость создания за-
поведников на Приполярном и Южном Урале в районе наиболее крупных
горных вершин, где сконцентрированы местонахождения эндемичных вы-
сокогорных растений.
Самые интересные береговые утесы и обнажения скал, представляю-
щие большую ценность для ботаники и геологии, не говоря уже об их эсте-
тическом значении, должны быть объявлены памятниками природы.
Ландшафты и растительность горных степей на Южном Урале пока еще
должным образом не охраняются. Следовало бы заповедать или объявить
заказниками наиболее интересные и лучше сохранившиеся участки горных
степей на хр. Ирендык в Башкирской АССР и горах Сугомак и Егозинской
в Челябинской области.
Дубравы и вообще широколиственные леса Южного Урала, имеющие
большое и разностороннее народнохозяйственное значение (благодаря их
водоохранным, почвозащитным свойствам и т. п.), также должны бережно
охраняться. В наиболее типичных их участках следует установить режим
заповедников или заказников. Этим будет также обеспечена и охрана свой-
ственных широколиственным лесам эндемичных и реликтовых растений.
Кроме того, не следует забывать еще об одном способе сохранения для
науки специфических и редких растений уральской флоры — разведении
их в культуре, особенно в ботанических садах. К сожалению, Ботаниче-
ский сад Института экологии растений и животных Уральского филиала
АН СССР из всего разнообразия эндемичных растений Урала пока предла-
гает для обмена семена только двух эндемиков —• Anemone biarmiensis и
Gypsophila uralensis. Необходимо расширить ассортимент семян эндемич-
ных уральских растений, предлагаемых для обмена, и усилить работы по
введению эндемиков в культуру (особенно видов, ценных в декоративном
отношении).
ГЛАВА IV
РЕЛИКТЫ ВО ФЛОРЕ УРАЛА
ВОПРОСЫ МЕТОДИКИ ИЗУЧЕНИЯ РЕЛИКТОВ НА ПРИМЕРЕ
НЕСКОЛЬКИХ КЛЮЧЕВЫХ РАСТЕНИЙ
В ботанике термин «реликт», как известно, понимается двояко: в систе-
матическом и фитогеографическом смысле. С точки зрения систематики,
реликт — это представитель какой-либо в основном вымершей системати-
ческой группы, оказавшийся обособленным и занимающий изолированное
положение в системе. С точки зрения фитогеографа, реликт — это вид,
в прошлом более широко представленный во флоре данной территории,
но впоследствии, в связи с изменениями условий среды, вымерший в зна-
чительной части своего прежнего ареала и сохранившийся лишь в немно-
гих, обычно изолированных местообитаниях, где условия для него были
более благоприятными. В дальнейшем речь пойдет о реликтах в фитогеогра-
фическом смысле этого слова.
Выявление реликтовой природы тех или иных видов в составе флоры
определенной территории, выяснение времени и условий их вхождения в
местную флору, установление причин, вызвавших образование дизъюнк-
ций, сопряжено со значительными трудностями. Решение этих вопросов
должно базироваться на тщательном изучении закономерностей современ-
ного распространения видов, их экологии, условий произрастания в раз-
ных частях ареала, анализе разнообразных факторов — палеогеографи-
ческих, экологических и антропогенных, — оказавших влияние на форми-
рование ареалов. Эти положения будут проиллюстрированы на примере
трех ключевых растений, представляющих разные генетические наслое-
ния в современной флоре Урала и Приуралья — вереска обыкновенного Cal-
luna vulgaris, дазиформы кустарниковой Dasiphora fruticosa и можже-
вельника казацкого Juniperus sabina.
ВЕРЕСК ОБЫКНОВЕННЫЙ CALLUNA VULGARIS
Единственный представитель монотипного рода Calluna — вереск
обыкновенный С. vulgaris — относится к числу наиболее обычных и широко
распространенных растений в Европе. Этому виду и образуемым им расти-
тельным сообществам посвящено немало ботанических работ, в том числе
и монография Бейеринка (Beijerinck, 1940). К сожалению, как в упомяну-
той монографии, так и в других зарубежных источниках недостаточно
охарактеризована восточная граница ареала вереска, проходящая по тер-
ритории СССР, и не освещены особенности поведения этого растения на
восточном пределе его распространения. Наши же отечественные ботаники
не пытались проследить в деталях восточную границу вереска и выяснить
причины, ее определяющие. Хотя в ряде источников вскользь упоми-
нается о произрастании вереска в Зауралье и в Сибири, за пределами его
154
основного ареала, этот интересный факт не привлекал внимания фито-
географов и не получил должной оценки. Однако изучение вереска — од-
ного из немногих типично европейских растений, проникающих за Урал
в Сибирь, — именно на восточном пределе его распространения может ос-
ветить некоторые моменты генезиса современной флоры нашей страны.
Важнейшие черты экологии вереска в оптимальных для него
климатических условиях
Вереск растет лишь в местах, где нет резких колебаний влажности воз-
духа и почвы. Наибольшего расцвета образуемые им растительные сооб-
щества, по Бейсринку (Bcijerinck, 1940), достигают в условиях влажного,
мягкого, прохладного океанического и субокеанического климата (Бри-
танские о-ва, Нидерланды, северная часть ФРГ и ГДР, Дания, юго-запад-
ная часть Норвегии и Южная Швеция). Это типичный олиготроф, произ-
растающий на бедных почвах, содержащих небольшое количество раство-
римых минеральных солей. Для населяемых им почв характерна кислая
или близкая к нейтральной реакция (pH 3,5—6,7). На щелочных почвах
он не встречается. Избегает известнякового грунта, хотя в области своего
климатического оптимума иногда встречается на почвах, подстилаемых
известняком, но лишь при наличии поверхностного кислого слоя перегноя,
в котором укореняется.
В районах с оптимальным для него климатом вереск обладает довольно
широкой эколого-фитоценотической амплитудой, входит в состав различ-
ных, иногда широко распространенных растительных сообществ, а в неко-
торых из них доминирует (Beijerinck, 1940; Bocher, 1943). Сообщества с
господством вереска — низинные и горные верещатники или вересковые
пустоши (Heide немецких авторов, heath, moor англичан) — занимают в
Северо-Западной Европе большие площади и местами имеют ландшафтное
значение, придавая всей местности, особенно в период массового цветения
этого вида, характерный колорит. Кроме того, вереск встречается на
закрепленных растительностью песчаных дюнах, более устойчивых каме-
нистых осыпях, на торфяных сфагновых болотах, а также в негустых со-
сновых, березовых, иногда дубовых лесах. В наиболее северных и высоко-
горных районах вереск произрастает лишь там, где зимой имеется доста-
точно глубокий снежный покров, под прикрытием которого он перези-
мовывает. Он требует достаточного освещения, поэтому под пологом сильно
сомкнутых тенистых лесов и в других затененных местах не произрастает.
В северо-западных районах европейской части СССР вереск в блап>
приятных для него почвенно-грунтовых условиях в изобилии разрастается
на вырубках и гарях, образовавшихся на месте сведенных сосновых лесов,
По мере восстановления древостоя участие вереска в живом напочвенном
покрове уменьшается, но он сохраняется в качестве характерного и до-
вольно обильного растения в некоторых типах леса (например, в сосняке
лишайниковом с вереском). Беглые низовые пожары и выборочные рубки
усиливают позицию этого растения в лишайниковых борах.
Основной ареал вереска в Европе и тяготеющие к нему островные и
изолированные месторождения
Основной ареал вереска (рис. 96) охватывает большую часть Европы
за исключением ряда ее восточных и юго-восточных районов. Западная гра-
ница ареала проходит по западному побережью Пиренейского п-ова, се-
верная — от северной окраины Пиренейского п-ова идет по северо-запад-
ному побережью Франции, побережью Бельгии, Нидерландов, ФРГ и
155
ГДР, Дании и по северной окраине Скандинавии. Как видно, на западе
и северо-западе Европейского материка не существует климатических пре-
град, ограничивающих распространение вереска, но есть физическая пре-
града — море. В процессе своего расселения это растение повсюду достигло
морского побережья, поэтому западная и большая часть северной границы
ареала вереска совпадают с очертаниями береговой линии континента.
Однако в самой северной части Норвегии граница распространения вереска,
судя по карте Хультена (Hui ten, 1958), местами начинает отступать от
береговой линии. На о. Магерейя близ побережья Северной Норвегии
Рис. 96. Основной ареал Cdlluna vulgaris. По Хультену (Hulten,
1958) с исправлениями автора для территории СССР.
1—сплошное распространение; 2 — изолированные местонахождения.
(71°5'.’с/ш.) зарегистрировано самое северное из всех известных местона-
хождений вереска. Далее граница постепенно начинает смещаться к югу.
Она пересекает побережье залива Варангер-Фьорд (Beijerinck, 1940), п-ов
Рыбачий, затем проходит по северному побережью Кольского п-ова (Чер-
нов, 1959) и направляется к п-ову Канин. В Северной Норвегии, на Коль-
ском п-ове и на п-ове Канин вереск заходит в тундровую зону. В Хибинах
места обитания вереска (Мишкин, 1953) еще довольно разнообразны: он
встречается в сосновых лесах, на моховых болотах; образует значительную
примесь в долинных ерниковых тундрах в пределах лесного пояса и захо-
дит в нижнюю полосу горных тундр.
Южная граница ареала вереска проходит вдоль морского побережья
южной и восточной части Испании (на востоке несколько отходит от бере-
говой линии), затем по южному побережью Франции, Северной Италии,
Балканскому п-ову и, огибая с юга Восточные Карпаты (Hulten, 1958),
вступает в пределы СССР. На Украине (Барбарич, 1957) граница ареала
пересекает с. Залещики на Днестре, проходит немного севернее г. Винницы
(близ с. Калиновки), севернее г. Черкасс (у станции Золотоноши), близ
с. Великой Рублевки северо-восточнее г. Полтавы, севернее г. Харькова
(около станции Золочев). В горной части УССР, в Карпатах, вереск под-
нимается до верхней границы леса, заходя на полонины; в равнинных рай-
онах характерными местами его обитания являются сосновые, реже дубово-
2
156
сосновые леса, а иногда он встречается и по краям торфяных болот. Далее
граница пересекает с. Дмитриевку в южной части Курской области, идет
близ городов Воронежа, Усмани и подходит к Балашову (Маевский, 1954).
Здесь вереск произрастает только в сосновых лесах.
К югу от основной области распространения вереска известен неболь-
шой изолированный участок его ареала на Средиземноморском побережье
Африки в Марокко, близ городов Танжера и Тетуана, под 35°40' с. ш. (это
самые южные пункты встречаемости рассматриваемого вида). Отдельные
местонахождения отмечены на о-вах Корсика и Сардиния в Средиземном
Рис. 97. Восточное крыло ареала Calluna vulgaris в СССР.
1 — восточная граница сплошного распространения; 2 — островное распространение за пределами
сплошного ареала; 3 — изолированные местонахождения.
море12. Кроме того, вереск встречается в ряде пунктов южного и юго-за-
падного побережья Черного моря (от г. Бургаса на западе до устья р. Офа
на востоке). В юго-западной части черноморского побережья, на стыке
Европы и Малой Азии, вереск произрастает на обращенных к морю скло-
нах гор в зарослях кустарников, близких к маквису (Endriss, 1921; Matt-
feld, 1929; Stojanoff, 1930). На причерноморских склонах Понтийских гор
в Турецком Лазистане (например, близ г. Офа) вереск встречается в сосно-
вых лесах (Шишкин, 1930). Как видно, на юге распространение вереска
ограничивает сухость климата. Изолированные места его встречаемости,
находящиеся южнее основного ареала и более или менее удаленные от
него, располагаются на островах и участках морских побережий, где ре-
жим увлажнения более благоприятен для произрастания этого растения
(устойчивая повышенная влажность воздуха, более обильное увлажнение
почвы).
Рассмотрим теперь восточную границу ареала (по данным Коржинского
Korshinsky, 1898; Федченко, 1936; Спрыгина, 1941, и материалам гербария
Ботанического института Академии наук СССР). От п-ова Канин (рис. 97)
она резко смещается к юго-востоку, все более и более отходя от побережья
Северного Ледовитого океана, идет к Тиманскому кряжу, вдоль него к
р. Елве, притоку Выми, и затем к междуречью Бадья — Визяха в бассейне
12Хультен (Hulten, 1958) указывает, что Calluna vulgaris приводилась для Сицилии
(коллектор Pojero), но произрастание вереска на этом острове не упоминается другими
итальянскими авторами.
157
р. Черной, притока Весляны. Далее граница направляется к р. Тимшеру,
притоку Южной Кельтмы, и выходит на левобережье Вишеры между де:
ревнями Губдором и Татарской. Здесь расположен самый восточный вы-
ступ ареала вереска. От этого пункта граница идет уже в юго-западном
направлении. Она пересекает р. Долог, приток Косы, неподалеку' от
с. Юксеево, потом идет к южной окраине Вятского Увала (дер. Морки,
р. Петьялка к северо-западу от Казани), Раифской лесной даче, далее к
г. Зеленодольску на Волге, пос. Алатырю на р. Суре и оттуда к восточной
окраине Приволжской возвышенности (верховья р. Алая к северо-западу
от г. Вольска) и, наконец, к Балашову. Везде на восточном пределе своего
распространения вереск произрастает только в сосновых лесах. Встреча-
ется он здесь редко, и отдельные его местонахождения нередко значительно
удалены друг от друга.
В северо-восточной части европейской территории СССР главными фак-
торами, ограничивающими распространение вереска, являются континен-
тальность климата и заболоченность (возрастающие в восточном направ-
влении), а также появление вечной мерзлоты в ряде районов. Поэтому гра-
ница ареала этого вида при движении на восток резко смещается к югу.
Распространен вереск на северо-восточном пределе ареала крайне преры-
висто. Отдельные пункты его нахождения связаны с сосновыми лесами,
растущими на относительно повышенных местах (Тименский кряж, более
мелкие продольные увалы, носящие местные названия «парм» и «чугр») или
на древних песчаных наносах близ речных долин. На таких участках, рас-
полагающихся обычно среди обширных заболоченных пространств, усло-
вия дренажа более благоприятны, почва лучше прогревается, а вечная
мерзлота отсутствует или уровень ее понижен. На юго-востоке и юге евро-
пейской части СССР распространение вереска ограничивают уже другие
факторы — сухость воздуха и почвы. Самые южные местонахождения его
связаны с сосновыми борами (чаще лишайниковыми), располагающимися
на песчаных террасах рек на южной окраине подзоны широколиственных
лесов и в лесостепи. Вереск здесь обитает только под защитой древесного
полога при условии достаточного увлажнения почвы. В степную зону оц
не заходит.
При движении от области климатического оптимума на восток в связи
с возрастанием континентальности климата наблюдается постепенное су-
жение экологической амплитуды вереска; в восточных районах европей-
ской части СССР он уже не выступает в роли доминанта растительных со-
обществ, не встречается на открытых местах (сфагновые болота, вырубки,
гари13), а сохраняется лишь в сосновых лесах под прикрытием их древес-
ного яруса, где выше влажность воздуха и не столь резки температурные
колебания. Все более неблагоприятными для него становятся суровые ус-
ловия севера и засушливый климат юга. Ареал вереска, столь широкий в
центральной части (от 35°40' до 71°5' с. ш.), постепенно сужается и в виде
острого клина подходит к предгорьям Урала на южной окраине лесной
зоны.
Кроме Европейского материка, вереск распространен на Британских
о-вах, где встречается почти повсюду, за исключением немногих мест, ин-
тенсивно освоенных человеком, а также участков со щелочными почвами.
Произрастает вереск также на Шетландских и Фарерских о-вах. В Ислан-
дии он встречается повсеместно, за исключением северо-западной и цент-
ральной части острова (Ostenfeld a. Grontved, 1934; Hulten, 1958). Кром&
13 Видимо, самым восточным районом, где вереск еще обильно встречается на гарях,
и вырубках, является бассейн левых притоков Волги — Ветлуги и Б. Кокшаги в Марийской:
АССР (см. Васильков, 1933).
158
того, в значительном удалении от основного ареала, вереск произрастает
на Азорских о-вах, особенно на Пику и Терсейра (Guppy, 1917).
Что касается высотных пределов вереска, то в Европе он распространен
почти от уровня моря до субальпийского и альпийского поясов в горах.
В Альпах вереск поднимается в горы до 2680 м над ур. м. (Hegi, 1906—1931),
подходя довольно близко к линии вечных снегов, но нигде ее не достигая.
Однако, как указывает Фаварже (Favarger, 1956), в Альпах вереск осо-
бенно характерен для контактной полосы между субальпийским и альпий-
ским поясами, где он, ассоциируясь с Nardus stricta, образует монотонные
сообщества, простирающиеся иногда на протяжении нескольких километ-
ров. В центральной Скандинавии вереск достигает высоты 1200 ж, в южной
Испании — 350 м над ур. м. (Beijerinck, 1940). На Кольском п-ове он за-
ходит в нижнюю полосу пояса горных тундр (Мишкин, 1953), т. е. до высо-
ты 600 м над ур. м. На Британских о-вах доходит до уровня 1095 м, а как
доминант растительных сообществ — до 853 м (Gimingham, 1960). В группе
Азорских о-вов, на о-ве Пико, вершина которого достигает высоты 2351 м
над ур. м., в вертикальном направлении сменяются пояса мирта, лавра,
можжевельника, вереска азорского Erica azorica, вереска обыкновенного
Calluna vulgaris, еще выше располагается пояс скал. Вереск обыкновен-
ный доминирует на высоте 1600—2100 м над ур. м., где отсутствует конку-
ренция со стороны лавра благородного Lauras nobilis и можжевельника
Juniperus oxycedrus ssp. brevifolia, этот же вид встречается и на более низких
уровнях, но только во вторичных растительных сообществах (Danse-
reau, 1966).
Вереск в Северной Америке
Для Северной Америки впервые14 Calluna vulgaris была указана Гуке-
ром (Hooker, 1840). Ссылаясь на сведения, полученные от Де ля Пиле, он
отметил, что этот вид произрастает на о. Ньюфаундленд. Однако это сооб-
щение в свое время не привлекло внимания ботаников. Зато целую сенсацию
произвел экспонировавшийся в 1861 г. на выставке, организованной Мас-
сачусетским обществом садоводства, экземпляр С. vulgaris, снабженный
такой этикеткой: «Дикий вереск естественно произрастает в 20 милях от
Бостона» (Gray, 1861; Clarkson, 1958). Никто не сомневался, что это именно
С. vulgaris, но утверждение о естественном произрастании этого вида было
принято с недоверием. Через несколько дней члены специално назначен-
ной комиссии встретились с участником выставки садоводом Дж. Даусоном,
который провел их на заброшенное поле близ Тьюкбери (штат Масса-
чусетс), где вереск рос на площади, несколько превышающей 0,2 га, на до-
вольно болотистой почве вместе с Chamaedaphne calyculata, Azalea viscosa
и другими растениями. Путем расспросов было установлено, что вереск
произрастал на этом участке еще до распашки, 50 лет тому назад. Аза Грей
(Gray, 1862), посетивший это местонахождение осенью того же года, также
пришел к заключению, что вереск здесь, как и на о. Ньюфаундленд, явля-
ется, по-видимому, аборигенным растением. Вскоре в распоряжение бота-
ников стали поступать гербарные образцы С. vulgaris с о-вов Ньюфаунд-
ленд и Кейп Бретон и с п-ова Новая Шотландия, а затем и из ряда пунктов
Новой Англии. Лаусон (Lawson, 1854), сообщивший о встречаемости
С. vulgaris в заливе Святой Анны на о. Кейп Бретон, писал: «Я не нашел
14 Более 130 лет тому назад в «Эдинбургской энциклопедии» (Giesecke, 1830) вереск
был упомянут в числе растений, произрастающих в Гренландии. По-видимому, это указание
основано на ошибке, так как никто из ботаников впоследствии названного растения там не
собирал и в новейшей сводке по гренландской флоре (Bocher, Holmen a. Jakobsen, 1957)
оно не приводится. Тем не менее, в некоторых источниках (см., например, Барбарич, 1957;
Чернов, 1959) вереск до последнего времени ошибочно указывается для Гренландии.
159
ни одного факта, хотя бы в малейшей степени указывающего на то, что
Calluna была завезена в залив Святой Анны искусственно. Напротив, я
убежден, что это настоящий туземный вид». По мере того, как устанавли-
вались новые местонахождения вереска, в печати стали появляться проти-
воречивые суждения о природе этого растения в Северной Америке. В не-
которых пунктах вереск был найден неподалеку от человеческого жилья
в таких условиях, когда нельзя было исключить возможности преднаме-
ренного посева его семян или случайного заноса их из Европы. Многие бо-
таники пытались доказать, что это растение было первоначально интроду-
цировано в Северную Америку, а затем здесь натурализовалось. Привер-
женцев гипотезы аборигенной природы этого вида становилось меньше.
Мнения некоторых ботаников по этому поводу менялись в разные годы
в зависимости от поступающих в их распоряжение фактов.
Можно смело сказать, что ни одно из растений, распространенных по
обе стороны Атлантического океана, но отсутствующих в тихоокеанской
части Земного шара (а таких «амфиатлантических» видов в атласе Хультена
(Hulten, 1958) насчитывается 278), не привлекало к себе столь большого вни-
мания в Северной Америке и не вызывало таких оживленных и длительных
дискуссий. Эго в значительной степени объясняется, вероятно, тем, что
вереск был хорошо знаком многим жителям Канады и США еще по Европе,
где он широко распространен, и находка его в Америке в виде немногих
экземпляров или небольших клочков зарослей воспринималась как явле-
ние необычное и ненормальное. Высказывались самые различные догадки
о возможных средствах заноса семян вереска в Америку. Одни считали
«виновниками» заноса скандинавских мореплавателей — викингов, ве-
роятно, достигавших берегов Америки в VIII—XI вв., другие — шотланд-
ских солдат, имевших обыкновение подметать вересковыми метлами свои
лагеря; иные полагали, что семена вереска могли быть завезены из Европы
вместе с семенами древесных пород, с подстилкой для скота или в почтовых
посылках. Все эти догадки имели одну цель: дать простое объяснение ка-
завшемуся крайне необычным факту встречаемости европейского расте-
ния в Америке. Конечно, никаких прямых доказательств не приводилось
и, видимо, не могло быть приведено. Местонахождение вереска в приатлан-
тической части Северной Америки при этом рассматривалось и объяснялось
вне связи с тем, что имеются другие, очень отдаленные от основного ареала
местонахождения этого растения на островах Атлантического океана,
между Европой и Северной Америкой — в Исландии и на Азорских о-вах.
Между тем в свете аналогичных фактов амфиатлантического распростра-
нения растений встречаемость вереска в Северной Америке не выглядит
уже столь необычной.
Всего в настоящее время по Кларксону (Clarkson, 1958) достоверно из-
вестно 25 пунктов нахождения вереска (рис. 98) на о-вах Ньюфаундленд
и Кейп Бретон, на п-ове Новая Шотландия в Канаде и в штатах Мэн, Нью-
Гемпшир, Массачусетс, Род-Айленд, Коннектикут, Нью-Джерси, Нью-
Йорк и Западная Виргиния в США15.
Несмотря на то, что во многих «флорах» и определителях растений ве-
реск традиционно приводится для Северной Америки как интродуцирован-
ное, впоследствии натурализовавшееся растение, до сих пор в печати по-
являются высказывания исследователей, наблюдавших этот вид в полевых
условиях, в пользу его аборигенной природы (Grew, 1936; Le Gallo, 1945).
Так, Дру сообщает, что, по его наблюдениям, вереск на о. Ньюфаундленд
16 Приводимый Бейеринком (Beijerinck, 1940) пункт нахождения вереска близ Маркетта
на южном побережье оз. Верхнего (штат Мичиган) в обзоре Кларксона (Clarkson, 1958) не
указывается; по-видимому, эти сведения не подтверждены последующими исследованиями.
160
занимает естественные, не нарушенные хозяйственным воздействием ме-
стообитания, где вместе с ним или неподалеку произрастают другие расте-
ния европейского типа, как например: Ranunculus hederaceus, Potamogeton
polygonifolius, Glyceria fluitans, Sieglingia decumbens, Nardus stricta, Pedi-
cularis silvatica.
При оценке изложенных выше данных следует, как нам кажется, иметь
в виду, что многие из первых находок вереска в приатлантической части
Северной Америки относятся к местам, тогда еще мало затронутым хозяй-
Рис. 98. Пункты встречаемости Calluna vulgaris в Северной
Америке. По Р. Кларксону (Clarkson, 1958).
ственной деятельностью человека. У ботаников, обнаруживших и обсле-
довавших в шестидесятых и семидесятых годах прошлого столетия эти
места обитания вереска, не возникало сомнений в том, что они имели дело
с туземным растением. Однако позднее, по мере возрастания воздействия
человека на первобытную природу, выявились местонахождения вереска
в местах, где естественный растительный покров был более нарушен. В на-
стоящее время не наблюдается тенденции к расселению вереска из мест,
где он был первоначально найден. Напротив, в ряде пунктов его распро-
странение сократилось, а кое-где он был истреблен в связи с хозяйственным
освоением территории. Нельзя также забывать, что вереск встречается
в промежуточных пунктах между Европой и Америкой — в Исландии и на
11 Заказ № 251 161
Азорских о-вах, находящихся на линии крупных, смыкающихся друг
с другом подводных хребтов — Рейкьянеса и Североатлантического.
Былое соединение Азорских островов перемычкой суши с Исландией, а через
нее и с материками северного полушария более удовлетворительно объяс-
няет присутствие на этих островах вереска, чем другие гипотезы, тем более,
что едва ли кто-нибудь станет серьезно доказывать возможность заноса
С. vulgaris в эти португальские владения норманнами или шотландскими
стрелками. Как видно, нет оснований отбрасывать гипотезу о том, что ве-
реск в приатлантической части Северной Америки является не заносным,
а аборигенным растением реликтового характера, хотя п не имеется до-
статочно данных для ее безоговорочного принятия.
Распространение и условия произрастания вереска на Урале,
в Зауралье и Сибири
Как уже упоминалось, граница основного ареала Calluna vulgaris на
востоке Русской равнины немного не достигает предгорий Урала. Собствен-
но на Урале (понимая этот термин в физико-географическом смысле как
наименование горной системы) он встречается лишь в одном пункте —
в окрестностях оз. Таватуй (в 40 км к северо-западу от г. Свердловска).
Но после перерыва, приходящегося на территорию почти всего горного
Урала, вереск вновь появляется в Зауралье и в Сибири (рис. 99). Некоторые
сибирские дизъюнктивные местонахождения вереска упоминаются в ряде
ботанических работ (Словцов, 1891; Сиязов, 1892; Крашенинниковы, 1908;
Буш, 1919; Кудряшов, 1924; Крылов, 1927—1964, и др.). Однако это
разрозненные и, в сущности, случайные данные, они не раскрывают
полной картины распространения и особенностей экологии вереска в Си-
бири. Наше изучение вереска в Зауралье и в Западной Сибири проводилось
в течение ряда лет, а в. 1961 г. районы его наибольшего распространения
были пересечены сетью специально подобранных маршрутов. В процессе
этих исследований обнаружено много новых, ранее неизвестных место-
нахождений вереска и установлено, что в некоторых боровых массивах он
широко распространен. Кроме того, использованы все доступные гербарные
материалы и литературные источники, а также получена ценная информа-
ция от отдельных лиц.
Подавляющее большинство сибирских местонахождений вереска скон-
центрировано в бассейне Иртыша. Но вдоль самого Иртыша это растение
не встречается, а распространено по его левым притокам — Конде, Тоболу
с впадающими в него реками и Вагаю. На Конде вереск произрастает в со-
сновых борах от пос. Шаима до устья р. Мулымьи и по самой Мулымье
в 15—20 км от устья (сообщение Е. П. Смолоногова). В бассейне Тобола
и его притоков Тавды, Туры с Пышмой и Исети удалось выявить и нанести
на карту много островных и точечных местонахождений этого вида. На Тав-
де вереск встречается близ дер. Тагильцы. По Туре, кроме давно извест-
ного местонахождения близ дер. Бабихино (Н. С. Попов, 1804), он обнару-
жен около с. Туринской Слободы. Ниже по Туре и на междуречье Туры и
её правого притока Пышмы вереск встречается уже очень часто. Крупные
островные участки ареала имеются на междуречье Тура — Пышма близ
Бахметского болота, затем к югу и юго-востоку от г. Тюмени, а также на
правом берегу Туры у ее устья. Собственно по Пышме вереск единично
встречается близ с. Елани, пос. Ертарского и дер. Первомайской, в то вре-
мя как на левом берегу близ г. Тугулыма и на правом берегу на участке
от дер. Луговой до устья имеются крупные островные местонахождения
этого вида. В бассейне Исети на левобережье отмечен изолированный пункт
встречаемости вереска по речке Ичкиной близ дер. Воденникове, а кроме
162
того, островной участок ареала близ устья Исети. На правобережье Исети,
точнее, на междуречье между нею и Тоболом, имеются крупные островные
местонахождения вереска в верховьях речки Нияп, близ с. Боровлянки
и у с. Исетского. На левобережье Тобола вереск особенно широко распро-
странен к западу, северо-западу и северу от г. Кургана. Отдельные пункты
его встречаемости отмечены севернее пос. Юргамыша в районе озер Уби-
енного и Окуневского. Островные местонахождения располагаются между
Рис. 99. Распространение Calluna vulgaris в Зауралье
и Западной Сибири.
1 -- сплошное распространение; 2 — изолированные местонахожде-
ния (за пределами карты — местонахождения в Кокчетавских горах*
близ пос. Бердска на Оби, в Хакассии и около г. Канска).
оз. Чаша и станцией Кособродск, к северо-западу от с. Введенского, к се-
веру от пос. Старого Просвета, близ с. Б. Чаусово и западнее с. Упорово.
Близ р. Тобола вереск произрастает изолированно юго-восточнее с. Бело
зерского, островные же местонахождения выявлены по речке Емуртле,
а также около пос. Заводоуспенского и г. Ялуторовска. На р. Вагай вереск
найден в сосновых лесах у Истятских юрт и у пос. Бегитино (сборы
Б. Н. Городкова).
Кроме того, известны изолированные местонахождения вереска из более
удаленных районов. В Ботаническом институте АН СССР хранится гер-
барный образец вереска, собранный в сосновых лесах Кокчетавских гор
близ. пос. Борового в восьмидесятых годах прошлого века фармацевтом
11*
163
любителем ботаники А. М. Абжолтовским, 1901 г. (гербарий БИНа). В кол-
лекции, собранной лесоустроительной экспедицией в борах Джебашской
лесной дачи (Хакасская автономная область), Л. П. Сергиевская обнару-
жила и определила экземпляр С. vulgaris-, эти сведения вошли во «Флору
Западной Сибири», а затем во «Флору СССР». В последнее время вереск
найден еще в одном пункте Красноярского края — в окрестностях с. Браж-
ного Канского района (любезное сообщение Л. П. Сергиевской, образец
собран учителем Барсуком; экземпляр хранится в гербарии Томского уни-
верситета) .
Наши наблюдения показали, что экологический ареал вереска в Запад-
ной Сибири до предела сужен, и это растение, занимает здесь строго опре-
деленные стации (рис. 100). Вереск произрастает в более крупных сосно-
А
Рис. 100. Экологические профили, показывающие наиболее характерные места обитания
Calluna vulgaris в Зауралье. Д—в районе Бахметского болота на древних песчаных
наносах вдоль р. Пышмы; Б — на берегу оз. Тулубаево близ Тюмени.
1 — сфагновое болото с низкорослой сосной; 2 — сосняк-брусиичник с вереском; 3 — сосняк-бруснич-
ник с напочвенным покровом, восстанавливающимся после низового пожара; 4 — сосняк лишайнико-
вый с ^покровом, восстанавливающимся после пожара; 5 — заросли камыша; 6 —заросли кустарни-
ковых ив; 7^—березовая согра.
вых массивах на юге лесной зоны и в лесостепи. Эти боры располагаются
на древних песчаных речных наносах. Наиболее характерные места его
обитания — сосновые леса по берегам озер, а также крупных болот с от-
крытой водной поверхностью, представляющих собой разные стадии за-
растания озер. Встречается он иногда и в сосняках на песчаных «островах»
(гривах) среди болот мезотрофного типа. Почвы здесь песчаные и супесча-
ные; нередко на глубине от 40 до 100 см имеется прослойка ортзанда или
суглинка. Так, например, на древней песчаной террасе р. Пышмы в районе
огромного Бахметского болота вереск произрастает в сосняке-бруснич-
нике на контакте с торфяным болотом, поросшим низкорослой сосной
(сосновый «рям»). В травяно-кустарничковом покрове сосняка-бруснич-
ника преобладают Vactinium vitis-idaea, Trifolium lupinaster, Solidago
virga-aurea, Lycopodium clavatum и Rubus saxatilis. Моховой покров обра-
зует Pleurozium Schreberi. В этой ассоциации Calluna vulgaris встречается
с обилием sp.— сор., в полосе шириной 20—25 м от кромки ряма. Выше
располагается восстанавливающийся после низового пожара сосняк-брус-
ничник, где вереск отсутствует за исключением немногих поврежденных
огнем экземпляров, а на вершине всхолмления — фрагмент лишайнико-
вого сосняка также со следами пожара. В сосновом ряме наряду с сосной
обыкновенной произрастает Betula pubescens. В ярусе кустарников пре-
обладает Ledum palustre и Chamaedaphne calyculata, а в травяно-кустарнич-
ковом покрове — Carex globularis и Vaccinium vitis-idaea. Моховойпокров
164
аиз Polytrichum commune и Aulacomnium palustre с примесью сфагновых
.мхов.
На берегу оз. Тулубаево, в 19 км к югу от г. Тюмени, вереск довольно
обилен в узкой, шириной 10—15 л, полосе сосняка-брусничника, примы-
кающей непосредственно к высокой террасе этого озера. В этом сосняке
сохранился напочвенный покров из зеленых мхов и нет следов пожара.
Выше располагается тот же тип леса, но напочвенный покров нарушен
многократно повторяющимися низовыми пожарами. Вереска здесь нет.
Ниже, на озерной террасе, распространены березовая согра и заросли
кустарниковых ив, а в мелководной прибрежной полосе озера — заросли
камыша озерного Scirpus lacustris.
Низинное положение таких местообитаний, а также присутствие в почве
труднопроницаемого для влаги горизонта определяет неглубокое залегание
грунтовых вод или верховодки. Крупные озера и болота оказывают увлаж-
.няющее и смягчающее влияние на климат непосредственно прилегающей
к ним местности. Здесь в течение всего вегетационного периода поддержи-
вается повышенная влажность воздуха, часты туманы, роса. Почва в со-
сняках по кромкам озер и болот отличается устойчивым повышенным увлаж-
нением. Пожары такие местообитания почти на затрагивают. Все это соз-
.дает наиболее благоприятные условия для произрастания и сохранения
вереска в лесах по окраинам озер и болот, где он встречается наиболее
обильно.
В Сибири с ее континентальным климатом вереск не способен заселять
•обширные открытые пространства среди леса (вырубки, гари), так как не
выдерживает присущих им резких колебаний температуры и влажности
•воздуха и почвы. Поэтому он растет здесь лишь под прикрытием древес-
ного яруса, оказывающего умеряющее влияние на режим тепла и влаги
приземного слоя воздуха. Беглые низовые лесные пожары, повреждающие
живой напочвенный покров и подрост, но не затрагивающие древостоя,
не всегда вызывают полную гибель вереска. От сохранившейся в толще
лесной подстилки нижней части стебля у него могут возникать вегетатив-
ные побеги. Имеются данные, что семена вереска, находящиеся в почве,
могут сохранять способность к прорастанию и после пожара (Kujala, 1926).
Кроме того, на участки, где прошел низовой пожар, заносятся семена ве-
реска из тех мест его обитания, где он не пострадал от огня. Поэтому после
низовых пожаров в сосновых лесах бассейна Тобола вереск нередко стано-
вится обильным, а иногда и приобретает роль доминанта в травяно-кустар-
ничковом ярусе растительных сообществ. Эго послужило поводом для вы-
деления <П. И. Чудниковым (1930) в Припышминском сосновом массиве
«верескового бора» (по аналогии с типом леса, распространенным в запад-
ных районах Европейской части СССР). Однако площадь таких вересковых
боров в Западной Сибири ничтожна, а доминирование вереска в них яв-
ляется непосредственным результатом низовых пожаров. Эти кратко-
временнопроизводные ассоциации впоследствии быстро возвращаются к ис-
ходным типам бруснично-зеленомошных или лишайниковых сосняков.
Вблизи крупных населенных пунктов под влиянием интенсивного вы-
паса скота и часто повторяющихся беглых низовых пожаров вереск в сос-
новых лесах вымирает. Так, в некоторых пунктах в непосредственной бли-
зости от г. Тюмени, где ботаники находили его в конце прошлого века, он
исчез совершенно.
Причины возникновения евро-сибирской дизъюнкции ареала вереска
В систематическом отношении род Calluna наиболее близок к роду
Erica, пр'едставленному в Южной Африке (Капская флористическая об-
ласть) более чем 50 видами и всего лишь несколькими видами в Европе.
165
Не приходится сомневаться в третичной древности монотипного рода Cal-
luna. К сожалению, прямые палеонтологические доказательства этого,
почти отсутствуют: известно лишь, что пыльца этого типа найдена в плио-
ценовых отложениях Богемии (Rudolph, 1936). По-видимому, родиной
Calluna vulgaris были приатлантические или присредиземноморские районы.
Европы, откуда этот вид в плиоцене проник в Северную Африку, Малую
Азию, а также в Западную Сибирь (через Урал, тогда пенепленизирован-
ный). Вероятно, в плиоцене условия обитания вереска в Сибири были более
разнообразными, и он встречался не только в сосновых лесах, но и в дру-
гих лесных и нелесных типах растительности. На восточном крыле своего'
ареала вереск пережил ледниковые эпохи плейстоцена где-то на юге Запад-
ной Сибири или в примыкающей к ней части Северного Казахстана, но вы-
мер на Урале в связи с эпейрогеническими поднятиями в этой области и свя-
занным с ними возрастанием суровости климата.
Как было показано, вереск в Сибири теперь наиболее распространен
в левобережной части Тобола в низовьях его притоков — Туры (с впадаю-
щей в нее Пышмой) и Исети. Всюду здесь он встречается в сосновых лесах,,
произрастающих на песчаных почвах. Материнской породой для этих, почв
являются песчаные наносы, широко развитые вдоль рек и на междуречьях.
Песчаные наносы особенно мощны и широки в низовьях Туры и Исети.
Местами они полностью перекрывают междуречья, достигая ширины
50—60 км. Толща песчаных наносов подстилается глинистыми отложениями.
Эти рыхлые песчано-глинистые образования, для которых характерна го-
ризонтальная и волнистая слоистость, отлагались на дне крупных мелко-
водных озер и рек (Введенский, 1933). Озера служили сточным бассейном
многоводных рек, берущих начало на восточном склоне Урала, или же
были проточными; в них накапливалось огромное количество песчаных
продуктов разрушения горных пород. Сами реки в разных частях русла
также отлагали песчаные наносы. Вероятное время отложения этих осад-
ков — нижний миоцен или олигоцен. В плейстоцене речные русла углу-
бились на несколько десятков метров, и базис эрозии понизился. Песча-
ные наносы подверглись размыву и переотложению, поверхность их рас-
членилась. Водная эрозия была, по-видимому, особенно интенсивной
в плювиальные эпохи плейстоцена. В сухие же периоды, особенно в южных
районах, происходило перевевание песков ветром (свидетельство этого —
косая слоистость песков близ г. Ялуторовска и пос. Заводоуковского). Так
сформировались обширные, возвышающиеся над современными речными
долинами пространства песчаных наносов, имеющих всхолмленную по-
верхность и испещренных многочисленными озерами.
На многих участках песчаные наносы представляли собой на первых
порах свободный, незаселенный растительностью субстрат, пригодный для
поселения растений-олиготрофов. Вероятно, климат южной части Западной
Сибири в конце плейстоцена, когда в основном сформировался современ-
ный рельеф песчаных наносов, оставался некоторое время влажным и про-
хладным. Вереск, несомненно, был в числе первых растений, поселившихся
на приречных песках. Вероятно, именно в это время он широко распро-
странился в Западной Сибири и примыкающей к ней части Северного Ка-
захстана (Кокчетавские горы) и продвинулся на восток, достигнув бассей-
на Енисея. Дальнейшее заселение песчаных террас основными лесами выз-
вало в голоцене оттеснение вереска из ряда типов местообитания, особенно
в травяных борах, где при задернованности поверхности почвы он не вы-
держивает конкуренции с травянистыми растениями. В северных районах
возможность дальнейшего расселения вереска уменьшалась, а ареал его
сокращался под влиянием прогрессирующего заболачивания лесных пло-
щадей (это впоследствии повлекло за собой прерывистость распростране-
166
мия вереска на севере Западной Сибири). Озера на песчаных террасах
-внутри сосновых массивов постепенно заторфовывались. В сухие периоды
голоцена распространение вереска в Сибири на песчаных почвах сокра-
щалось, а местами он совершенно исчез. Наиболее подходящими стациями
для сохранения вереска в засушливые периоды были участки сосновых
лесов по окраинам крупных озер и болот; не исключена вероятность, что
вереск тогда заселял кое-где и подсыхающие торфяники. Окраины озер
и болот были не только местами сохранения этого растения в засушливые
периоды, но и центрами, откуда оно впоследствии расселялось, когда кли-
мат становился более влажным.
В период активного воздействия человека на природу главным факто-
ром, определяющим сокращение распространения вереска в Сибири, стали
.лесные пожары. Однако пожары обычно не затрагивают или мало затра-
гивают периферическую часть сосновых лесов, непосредственно примы-
кающую к озерам или крупным, но полностью заторфованным болотам.
Здесь вереск сохраняется и впоследствии отсюда вновь расселяется в при-
мыкающие участки сосновых лесов, где почвенно-грунтовые условия для
.него благоприятны.
Участки соснового леса на более увлажненных местах по окраинам бо-
лот и озер, следовательно, являлись своеобразными резерватами, где ве-
реск сохранялся в сухие периоды и во время лесных пожаров и откуда он
затем вновь распространялся. В пределах лесостепи Притоболья вереск
’отсутствует в небольших островных сосновых борах. Вероятно, в некото-
рых из них он раньше произрастал, но впоследствии исчез под влиянием
.повторяющихся лесных пожаров.
ДАЗИФОРА КУСТАРНИКОВАЯ DASIPHORA FRUTICOSA
бб * * 9* Род Dasiphora, по данным обработки его для «Флоры СССР», произве-
денной С. В. Юзепчуком (1941), представлен в нашей стране шестью вида-
ми, из которых наиболее распространена дазифора кустарниковая —
.Dasiphora fruticosa (Syn. Potentilla fruticosa). Как вид, дазифора кустарни-
ковая (другие русские названия: курильский чай кустарниковый, лап-
чатка кустарниковая) ранее понималась довольно широко. Однако в по-
следнее время из этого вида sensu laio исключены морфологически и геогра-
фически обособленные формы из Средней Азии и Сибири, возведенные в
ранг самостоятельных видов: D. phyllocalyx (Заилийский Ала-Тау),
D. dryadanthoides (Памир) и D. parvifolia (Памир, Тянь-Шань, Северное При-
балхашье, Джунгарский Ала-Тау, Алтай, Забайкалье, Монголия). Но и
в такой более суженной трактовке D. fruticosa остается сильно изменчивым
видом. Как отмечает С. В. Юзепчук, в ходе дальнейшего изучения в пре-
делах этого типа могут быть выделены другие, более дробные единицы.
Дазифора кустарниковая характеризуется широким географическим
распространением, но пространственное распределение ее на суше север-
ного полушария неравномерно и прерывисто. Ареал Dasiphora fruticosa
состоит из нескольких неравноценных по своей величине частей, значи-
тельно разобщенных друг от друга.
Наиболее крупным и почти монолитным является азиатский участок
ареала дазифоры кустарниковой, охватывающий Алтай, Саяны, Средне-
Сибирское плоскогорье, Забайкалье, горную область северо-востока Азии
от Верхоянского хребта до Чукотки, Охотское побережье, Амурскую об-
ласть, Приморский край, Камчатку, Сахалин, Курильские о-ва, Японию,
Северную Корею, Китай, Монголию и отчасти горы советской Средней
Азии. От этой основной области распространения резко обособлены фраг-
: 167
менты ареала дазифоры кустарниковой на Урале, Кавказе, в Европе, а
также ареал ее в Северной Америке.
Дазифора была знакома мне по наблюдениям в природной обстановке--
в Сибири (Восточные Саяны, Хакассия). Когда впоследствии я столкнулся
с этим кустарником на Урале, мне бросилось в глаза, что он распространен
здесь крайне прерывисто, экологическая амплитуда его резко сужена, и
он встречается почти исключительно на крупных горных вершинах или в
непосредственно прилегающих к ним местах. В последующие годы мне-
удалось наблюдать и коллекционировать/}, fruticosa в ряде пунктов Урала,
изучить особенности ее экологии, ознакомиться с гербарными экземпля-
рами, а также получить от других исследователей ценные сведения о рас-
пространении и условиях произрастания этого кустарника. Здесь излага-
ются основные выводы, вытекающие из анализа всех этих данных и лите-
ратурных источников.
Некоторые эколого-биологические особенности дазифоры кустарниковой
Dasiphora fruticosa — светлолюбивый кустарник, нередко (во всяком?
случае, в периферических частях его ареала) произрастающий в местах,,
где ослаблена конкуренция со стороны других видов кустарников и де-
ревьев. Условия, ограничивающие произрастание других древесных ра-
стений, могут быть эдафическими (каменистость субстрата) или климати-
ческими (резкая сокращенность вегетационного периода, свойственная вы-
сокогорьям).
Дазифора кустарниковая часто растет на слабо задернованных каме-
нистых местах — на прибрежном галечнике, каменистых склонах и камен-
ных россыпях. Поселившись на таком субстрате, она быстро заселяет его,,
разрастаясь с помощью подземных побегов, которые пронизывают гальку
и щебень, причем впоследствии на поверхности субстрата образуется гу-
стая щетка надземных побегов. Заросли дазифоры, задерживая мелкозем,
способствуют закреплению подвижного каменистого субстрата и, следо-
вательно, подготавливают почву для поселения других растений. Таким?
образом, дазифора кустарниковая относится к числу растений-пионеров,
заселяющих и закрепляющих каменистый субстрат. Однако в дальнейшем,,
по мере развития мелкоземистого слоя почвы, дазифора кустарниковая
может быть полностью оттеснена более теневыносливыми и сильными в кон-
курентном отношении деревьями и кустарниками.
Нетрудно заметить тесную связь дазифоры с высокогорьями, где она
способна подниматься значительно выше границы леса. Основным факто-
ром, исключающим возможность обитания здесь деревьев и ограничиваю-
щим произрастание кустарников, является сильная сокращенность веге-
тационного периода. Однако, по сравнению со многими другими кустар-
никами, D. fruticosa s. 1. обладает существенным преимуществом, облег-
чающим ее существование в высокогорных условиях,— способностью за-
кладывать цветы в зимующих почках еще в конце предыдущего вегетацион-
ного периода. Так, по исследованиям Т. Н. Кишковского и 3. Т. Артюшен-
ко (1951), проведенным в высокогорьях Памира, в зимующих почках:
D. fruticosa s. I.16 содержатся уже в основном сформировавшиеся цветы;
в них хорошо развиты оба круга околоцветника, имеются многочисленные
тычинки и начинают дифференцироваться наружные круги плодолистиков,
однако завязь находится еще в зачаточном состоянии. Наличие зачатков
16 Т. Н. Кишковский и 3. Т. Артюшенко приводят для изучавшегося ими растения
название D. fruticosa, однако их наблюдения, вероятно, относятся к D. dryadanthoid.es.
Желательно провести такие же наблюдения над D. fruticosa s. str. из высокогорных мест
обитания.
168
^цветов в зимующих почках сокращает подготовительный период, необхо-
димый для окончательного формирования и распускания цветов, следо-
вательно, значительно увеличивает шансы успешного созревания плодов
в условиях резко сокращенного периода вегетации, характерного для
высокогорий.
Знание указанных эколого-биологических особенностей D. fruticosa
помогает лучше понять историю расселения этого растения, закономер-
ности его современного распространения и позицию его в тех растительных
сообществах, в состав которых оно входит.
Распространение и условия произрастания дазифоры кустарниковой
в пределах основного ее ареала в Азии
На Алтае, по В. И. Баранову (1954), Dasiphora fruticosa особенно обиль-
на в восточной части — в истоках р. Чулымшана (оз. Джулу-Куль или по
.другой транскрипции Джавлукуль) и в бассейне его правых притоков
(реки Шавла, Чульча). Дазифора кустарниковая образует заросли вместе
с кустарниковыми видами берез (Betula humilis, В. rotundifolid) или же со
•стелющейся ивой и спиреей (Salix depressa, Spiraea salicifolia). «Вообще
же,— указывает В. И. Баранов (1954, стр. 265),— она входит в состав
большой группы фитоценозов, представляющих последовательный эколо-
гический ряд от прибрежно-ручьевых с развитым моховым покровом до
остепненных17, сопровождаемых подушками кладоний». Тот же исследо-
ватель (Баранов, 1931) отмечает D. fruticosa в составе кобрезиевых луговин
в пограничной с Монголией юго-восточной части Алтая (хр. Сайлюгем и
смежная с ним южная часть хр. Чихачева) на высотах порядка 2500—2600 м
над ур. м. В гербарии Центрального Сибирского ботанического сада
•СО АН. СССР есть просмотренные мною экземпляры D. fruticosa из следую-
щих пунктов Алтая: 1) хр. Нолго, окрестности Каракольского озера,
южный щебнистый склон на высоте 2000 м над ур. м., 2) Чуйская степь,
в 18 км от Кош-Агача, пойменный луг по р. Чуе и 3) Кош-Агачский аймак,
верховья р. Джазатора, горная тундра. Согласно личному сообщению
Е. И. Лапшиной, в юго-восточной части Кош-Агачского аймака Горно-
алтайской области по долине р. Кок-Су, притока Аргута, заросли дази-
форы кустарниковой тянутся из лесного пояса в подгольцовый и отчасти
внедряются в гольцовый пояс. Для Чуйской степи имеются и литератур-
ные данные А. В. Калининой (1948), указывающей, что D. fruticosa произ-
растает там всюду на заболоченных лугах, в ивняках и в лиственничных
.лесах.
Заросли дазифоры кустарниковой встречаются и в Восточном Казах-
стане— на отрогах Алтая, хребтах Саур и Тарбагатай (Быков и Степа-
нова, 1953). На хребте Джунгарский Алатау18 D. fruticosa произрастает
на склонах в альпийском и субальпийском поясах (Рубцов, 1941а). О про-
израстании дазифоры кустарниковой в горах Тянь-Шаня и других райо-
нах советской Средней Азии нет достоверных сведений из-за недостаточ-
ности гербарного материала, а также в связи с тем, что предыдущие иссле-
дователи не отграничивали ее от других близких среднеазиатских видов из
рода Dasiphora.
17 Не исключена вероятность, что экземпляры из степных мест обитания относятся не
к собственно D. fruticosa, а к £>. parvifolia (Прим. наше. — П. Г.).
11 По данным гербария Института ботаники Академии наук Казахской ССР, на хреб-
'тах Саур и Тарбагатай растет типичная D. fruticosa (характерные места ее обитания —
‘сухие русла рек и ущелья), а растения из Джунгарского Алатау имеют переходные приз-
наки между ней и D. parvifolia, но более близки к последней.
169
На восточном склоне Кузнецкого Алатау, в долине реки Улень, притоке-
Белого Июса (Хакасская автономная область), в полосе парковых лист-
венничных лесов, смыкающихся снизу с остепненными лугами и степями,,
встречаются кобрезиево-кустарниковые луга, мало гармонирующие с со-
временным растительным покровом. В кустарниковом ярусе их преобла-
дают Dasiphora fruticosa, Betula humilis, Salix, depressa v. cinerascens и
S. Kochiana. Доминантой травостоя является Cobresia Bellardi. Обращая
внимание на присутствие здесь растений, характерных обычно для высо-
когорий (например, Cobresia Bellardi), а также некоторых степняков на
буграх, К- А. Соболевская (1941) считает эту ассоциацию реликтовой и
рассматривает ее как наследие ледниковой эпохи. В долине Белого Июса.
по нашим наблюдениям, заросли дазифоры кустарниковой широко распро-
странены; этот кустарник встречается также в подлеске лиственничных
лесов. Неподалеку от этих мест D. fruticosa отмечена в притеррасных ча-
стях поймы р. Уйбата, притока Абакана, и в Июсо-Ширинских степях, по
окраинам озер, на каменистых склонах, с близким залеганием грунтовых,
вод (Тарчевская, 1941; Тарчевский, 1941 и личное сообщение указанных,
авторов).
В Западном Саяне (Ревердатто, 1946) D. fruticosa нередко встречается
в подлеске высокогорных лиственничных лесов; отдельные ее кустики от-
мечены и среди подгольцовых лугов. В верхней части горнолесного пояса,
в несколько заболоченных расширенных речных долинах распространеньг
заросли низкорослых кустарников — ерники. В них преобладают D. fru-
ticosa, Betula rotundifolia, В. humilis. Такие заросли, в частности, отме-
чены в долине р. Большой Арой близ слияния с Малым Ароем на высоте
около 1400 м над ур. м. и в широкой плоской долине р. Каратош на высоте-
около 1580 м над ур. м.
В Тувинской автономной республике, согласно К- А. Соболевской (1953),.
Dasiphora fruticosa является одним из наиболее широко распространенных
растений и «имеет ландшафтное значение». Она образует сплошные заросли,
в подгольцовом поясе, но спускается и значительно ниже по долинам гор-
ных рек. Произрастает дазифора кустарниковая обычно с Betula humilis,.
В. rotundifolia, а в юго-восточной Туве образует вместе с Caragana 'jubata:
характерные ерниковые формации среди лиственничного редколесья.
Dasiphora fruticosa отмечалась нами в пределах Восточного Саяна в до-
линах рек Маны и Большой Дербиной. Ал. А. и Ан. А. Федоровы (1951)
находили это растение в ряде пунктов высокогорной части Восточного-
Саяна. В частности, характеризуя растительность троговой долины р. Ма-
лого Агула, указанные авторы писали: «Плоские берега реки заросли
сплошными, крайне однообразными на большом протяжении и довольно-
унылыми зарослями курильского чая, круглолистной березки и мелкой
ивы по покрову лишайников и мхов» (Ал. А. и Ан. А. Федоровы, 1951,
стр. 144). Для некоторых троговых долин Восточного Саяна D. fruticosa-
отмечена также И. М. Забелиным (1952). В среднем течении р. Сигача (при-
ток Большого Агула, бассейн р. Казыра) в Восточном Саяне на узких ниж-
них террасах среди разреженных лиственничных лесов разбросаны поляны-
ерников, в которых преобладают D. fruticosa, Betula rotundifolia (Петров,
1948). В верховьях р. Оки, в расширенном днище ее долины, D. fruticosa:
также образует заросли совместно с кустарниковыми ивами и березовым^
ерником (Назаров, 1935).
На Енисейском кряже D. fruticosa вместе с Betula humilis и В. fruticosa-
входит в состав ерников, развитых в днищах каньонов (Игошина, 1951).
В Прибайкалье дазифора кустарниковая встречается как в гольцовом
поясе Хамар-Дабана, так и на более низких уровнях гор (М. Г. Попов,.
1957). В Забайкалье D. fruticosa произрастает повсеместно по берегам рек.
170
и в поймах среди кустарников, по каменистым склонам и обнажениям (Гру-
бов, 1949).
На Буреинском хребте (Шмидт, 1874) выше полосы зарослей кедрового
•стланика (Pinus pumila) D. fruticosa растет на каменной россыпи среди
глыб, покрытых лишайниками. Кроме того, дазифора кустарниковая от-
мечена на песчаных берегах Бурей и на скалах южного отрога Сихотэ-Алиня
(Комаров, 1950). В средней части Сихотэ-Алиня D. fruticosa встре-
чена на седловине между двумя вершинами горы Шайтан, на высоте около
1650 м над ур. м., в группировках кустарниково-моховых горных тундр
на более увлажненных местах (личное сообщение Б. П. Колесникова).
В Монгольской Народной Республике D. fruticosa встречается как в
лесном поясе, где она обитает в лиственничных лесах, березовых колках,
на лесных опушках, по лощинам, днищам и склонам падей, по берегам и
долинам рек, так и в высокогорном поясе, где она связана с галечниками
и россыпями, кобрезниками и болотистыми лужайками (Грубов, 1955).
’Заросли низкорослых кустарников в высокогорных районах Монголии рас-
полагаются обычно близ верхней границы леса и внедряются в него в виде
отдельного яруса. Характерным компонентом таких зарослей, наряду
с низкорослыми березками (Betula rotundifolia, В. exilis) является D. fru-
ticosa (Юнатов, 1950).
На плато Пей-Шань и Корейском хребте D. fruticosa растет одиночными
кустами и небольшими зарослями по тальвегам горных ручьев (Комаров,
1950). В северо-западном Китае, в горной системе Кунь-Луня (Куэнь-Луня),
произрастает в высокогорных поясах на высоте 3450—4500 м над ур. м.
(Ильинский, 1937). А. А. Юнатов, изучавший растительность северо-за-
падного Китая, сообщил мне, что в восточном Наньшане и западной части
Ланьчжоу на высотах порядка 3000 м над ур. м дазифора кустарникова'я
образует заросли на северных склонах, обычно в мелких долинках. В юго-
западном Китае (Юньанань, Гуйчжоу, Сычуань) она растет также в высо-
когорных поясах, поднимаясь до высоты 4400—5000 м над ур. м. (Иль-
инский, 1937).
В Якутии дазифора кустарниковая встречается в лесной и лесотундро-
вой зонах и в высокогорных поясах. Обычно она встречается в лиственнич-
ных и березовых лесах и редколесьях по берегам и долинам рек, по лощи-
нам, в ерниках и на кочковатых болотах (Караваев, 1958). В частности,
в долине р. Далдына (Олекминско-Вилюйское плато) В. Б. Сочава (1957)
сделал описание разнотравно-кустарниковых лиственничников с подлес-
ком из D. fruticosa, Betula fruticosa и Junuperus sibirica.
В так называемом Анадырском крае (на Анадырском, или Чукотском,
хребте) D. fruticosa растет в горах в тундрах и зарослях кедрового стланика
Pinus pumila, а в равнинной части края — в типичных тундрах по водо-
разделам и долинам рек, в долинных кустарниковых зарослях и на галеч-
никах (Васильев, 1956, и личное сообщение).
На Камчатке, по В. Л. Комарову (1929), D. fruticosa произрастает оди-
ночно или небольшими группами на сухих террасах со скудной почвой, на
моховых болотах и на сухих каменистых склонах. Кроме того, она встре-
чается в гольцовом поясе в сухих лишайниковых тундрах и на осыпных
склонах (отмечена, в частности, на Ключевской сопке на высоте 1080 м над
ур. м). В группе Курильских о-вов D. fruticosa указывается только для
Шумшу, Парамушира и Шикотана. На последнем из упомянутых островов
дазифора кустарниковая, по Д. П. Воробьеву (1956), является широко
распространенным растением, местами образующим небольшие заросли.
На Сахалине, по А. И. Толмачеву (1950, 1956), D. fruticosa произрастает
в различных, обособленных друг от друга местах на юге и в центральной
•части острова, отчасти высоко в горах. Она отмечена, например, в районе
171
грязевого вулкана Магунтан и на Восточно-Сахалинском хребте, где на?
горе Лопатина растет в гольцовом поясе на высоте более 1300 м над ур. м.
Из сказанного можно заключить, что D. fruticosa является преимуще-
ственно горным растением, хотя на северо-востоке Азии она спускается
и в равнинные тундры. Чем южнее, тем отчетливее выступает в Азии связь,
дазифоры кустарниковой с высокими горными вершинами. Наиболее
обильна в горах Алтайско-Саянской горной системы, где образует заросли
как в верхней части горнолесного пояса, так и в высокогорных поясах, нс,
в виде единичных кустиков спускается вниз до предгорий.
Распространение и условия произрастания дазифоры кустарниковой
в Северной Америке
Естественным продолжением основного азиатского ареала D. fruticosa
является область распространения ее в Северной Америке, отделенная
Беринговым проливом.
На островах Канадского Арктического архипелага (Porsild, 1957) и.
в Гренландии (Bocher, Holmen, Jabobsen, 1957) это растение не было най-
дено; прежние указания о встречаемости его в Гренландии (Крылов, 1927—
1964) ошибочны.
На Аляске дазифора кустарниковая распространена почти повсеместно’
и заходит на север до 70° с. ш., почти до побережья Северного Ледовитого
океана. Так, например, на арктическом склоне Аляски, по Л. Спетцману-
(Spetzman, 1959), дазифора растет на приморской равнине, у подножия,
гор и в горах до 1200 м над ур. м. Типичные ее местообитания — берега’
и поймы рек, где она характерна для начальных стадий зарастания обна-
женного субстрата.
В западной части Канады дазифора кустарниковая распространена на>
север до побережья Северного Ледовитого океана (устье р. Макензи). Се-
верная граница ее распространения (по данным гербариев Национального»
музея и Департамента земледелия в Оттаве) от устья р. Макензи идет-
к заливу Коронейшен, затем смещается на юг к оз. Большому Невольничье-
му и Саскачеван, после чего поворачивает на северо-восток, подходит к;
Гудзонову заливу, огибая его южную оконечность на участке от Черчилла:
до Грейт-Уэйл-Ривер. Далее граница резко уходит на юг, к району оз..
Онтарио, откуда направляется к низовьям р. Св. Лаврентия, затем к Бернт-
Крику и выходит к побережью Атлантического океана в районе оз. Мел-
вилл.
В Канаде дазифора произрастает как в горах, так и в равнинах в раз-
личных местообитаниях: по песчаным, галечниковым и каменистым бере-
гам рек и озер, по берегам ледниковых ручейков, в зарослях кустарников,,
на сырых замоховелых лугах близ ручьев, по окраинам торфяников, на,-
травяных и лесных болотах, в расщелинах глыб на скалистых обнажениях..
В Соединенных Штатах Америки дазифора проникает дальше всего на
юг по горным цепям Сьерра Невада (штат Калифорния) и Скалистых гор
(штат Нью-Мексико). Южная граница от штата Калифорния идет через,
штаты Аризона, Нью-Мексико, Южная Дакота, Миннесота (северная
часть), Айова, Иллинойс, Индиана, Огайо, Пенсильвания и Нью-Джерси
(Fernaid, 1950). Кроме того, есть указания (Small, 1933), что этот вид.
встречается в штате Теннеси.
На севере этот кустарник еще растет на равнинах или на слабо всхолм-
ленной местности. Так, в районе Великих Озер (округ Окленд в штате-
Мичиган) дазифора кустарниковая произрастает на высотах порядка
400—550 м над ур. м. по окраинам торфяных и травяных болот, на сырых;
172
лугах и в смешанных сырых лесах (Bingham, 1945). Кроме того, в северной
части Соединенных Штатов она встречается в речных долинах, на камени-
стых склонах холмов и на обнажениях известняка и гранита.
Однако близ южного предела своего распространения дазифора стано-
вится горным и даже высокогорным растением. В Скалистых горах,
в штате Колорадо, она распространена с высоты 2000 м над ур. м.. на об-
ширных открытых субальпийских площадях с довольно щебнистым суб-
стратом по берегам рек и ручьев вместе с кустарником Betula glandulosa,
а также на лугах (личное сообщение В. Вебера). В горах Сьерра Невада
дазифора произрастает близ верхней границы леса на высоте от 2450 до
3660 я над ур. м., встречается от округа Мадепа до округа Лассен (Jenson,
1925).
На территории Северной Америки известны следующие разновидности
и формы Dasiphora fruticosa: var. tenuifolia, var. parvifolia, var. grandiflora,
f. villosissima, f. humilis. Rydberg, 1898, считает, что крупноцветная форма
не заслуживает возведения в ранг разновидности.
Предпринятое недавно Р. Боуденом (Bowden, 1957) сравнительное ци-
тотаксономическое изучение дазифоры кустарниковой показало, что дико-
растущие особи этого вида в Северной Америке диплоидны (2п = 14), в то
время как растения из Англии (долина р. Тис) и Швеции (о. Эланд) являются
тетраплоидами. Исследованные образцы дазифоры кустарниковой из Се-
верной Америки сильно варьируют морфологически, наблюдается варьиро-
вание и в европейском материале, которым располагал автор. Однако если
сравнить друг с другом много образцов, трудно найти устойчивые морфо-
логические различия между растениями, происходящими из Европы
(Англия, Швеция) и из Северной Америки.
Заслуживает внимания тот факт, что дазифора более обычна в западной,
более гористой, части Северной Америки (Аляска, западная часть Канады,
Сьерра Невада, Скалистые горы в США). В Канадских провинциях Бри-
танская Колумбия и Альберта она встречается почти исключительно в го-
рах (Береговой хребет, Скалистые горы) и крайне редка на равнине. В юж-
ной части Саскачевана большинство местонахождений сосредоточено в го-
рах Сайпрес Хиле. Если на Аляске дазифора кустарниковая доходит почти
до побережья Северного Ледовитого океана, то по мере движения на восток
ее северная граница смещается к югу, а количество местонахождений
уменьшается. Крайние пункты встречаемости этого растения по побережью
Гудзонова залива отделены дизъюнкцией от.мест ее более широкого рас-
пространения в районе Великих Озер и в гористой части бассейна р. Св.
Лаврентия и п-ова Лабрадор.
Эта закономерность в какой-то степени связана с историей расселения
дазифоры. По мнению Д. Гопкинса (Hopkins, 1959), Берингийско-Чукот-
ская платформа была сушей в течение большей части третичного периода,
за исключением относительно коротких промежутков времени в середине
эоцена, середине и в конце плиоцена, когда здесь существовал морской про-
лив. В плейстоцене Азиатский и Американский континенты были разде-
лены морским проливом в течение каждого межледниковья и соединялись
с сушей в течение каждой ледниковой эпохи. В последний раз перемычка
суши исчезла 10—11 тыс. лет тому назад. Дазифора кустарниковая, ве-
роятно, проникла в Северную Америку из Азии через перемычку суши в
районе Берингова пролива. Первоначально она расселилась в северо-за-
падной части Американского материка и лишь позднее продвинулась на
восток и по горным хребтам на юг. В ходе послеледниковых смен расти-
тельности дазифора была оттеснена из некоторых мест обитания на равни-
нах, что вызвало раздробленность ее ареала в северо-восточной части
Северной Америки.
173
Дазифора кустарниковая в Европе и на Кавказе
Распространение Dasiphora fruticosa в Европе крайне прерывисто.
Согласно литературным данным (Hegi, 1919; Hulten, 1937; Clapham, Tutin,
Warburg, 1952), здесь насчитывается четыре основных фрагмента ареала
этого растения:
1. Острова и побережье Балтийского моря (острова Эланд, Готланд,
побережье Балтики в Эстонской и Латвийской ССР);
2. Британские о-ва (Пеннины, район Камберлендских гор, графства
Мейо, Голуей и Клэр в западной части Ирландской республики);
3. Пиренеи;
4. Приморские Альпы.
Фрагменты ареала дазифоры кустарниковой в Европе, так же как на
Кавказе и на Урале, отделены от основного ареала и друг от друга значи-
тельными дизъюнкциями. Возникает вопрос о причинах столь прерывистого
распространения ее вне основного ареала. Является ли эта прерывистость
следствием неравномерного распространения зачатков или же следствием
последующего оттеснения растения из некоторых мест на периферии ареа-
ла? Еще А. Гризебах (1874) считал более вероятным второе предположение.
Он писал: «Некоторые случаи спорадического распространения (растений—
П. Г.) едва ли могут быть разъяснены иначе как предположением о вытес-
нении их с прежних мест пребывания. Сюда, например, относится появле-
ние одного сибирского кустарника из розоцветных (Potentilla fruticosa)
в альпийском поясе Пиреней, на Британских островах, на Эланде и в Рос-
сии, при полном отсутствии его в германской флоре, пока, впрочем, как бы
последние следы его не были отысканы в Баварском Ризе» (стр. 186—187).
Последняя находка, как указывает А. Гризебах в примечании (1874,
стр. 495), относится к пункту между подошвой Баварской Юры у Вемдинга
(на восток от Нердлингена) и Верница, где «на луговом болоте» Dasiphora
fruticosa росла в сопровождении ряда растений с далекого северо-востока
(Pedicularis sceptrum-carolinum, Veronica tongifotia, Polenwnium coeruleum,
Iris sibirica) в условиях, где трудно допустить возможность ее заноса под
влиянием человеческой культуры.
На островах и на побережье Балтики местонахождения дазифоры рас-
полагаются почти у самого уровня моря, тогда как на Британских о-вах,
в Пиренеях и Приморских Альпах этот кустарник заходит довольно высоко
в горы.
В Центральной Европе (Австрия, ФРГ и ГДР и прилегающие к ним части
Польши и Франции) дазифора издавна разводится в садах как декоратив-
ный кустарник и местами одичала.
На Эланде, по данным Р. Штернера (Sterner,, 1938), дазифора кустарни-
ковая обыкновенна в южной части острова, изредка встречается в средней
и совсем отсутствует в северной. В своем распространении она связана с так
называемыми альварами. В Швеции альварами называют пастбищные
угодья низкой продуктивности, где выпас скота препятствует поселению
древесной растительности. Почвы альваров маломощны, обычно подсти-
лаются известняком, растительный покров представлен кустарниками,
травами, мхами и лишайниками. Особенно обильна дазифора по окраинам
так называемого Большого Альвара, где образует густые заросли в пони-
женных, слегка всхолмленных местах, на достаточно увлажненных извест-
няковых моренных почвах. Встречается и в расщелинах выветривающихся
скал. За пределами Большого Альвара она произрастает преимущественно
в недостаточно дренированных местоположениях с моренными почвами.
На Готланде дазифора известна только в приходе Хейнум (в двух пунк-
тах) и близ Калльтутбурга. В Эстонской ССР она встречается в приморской
174
части материка (северо-запад республики). Наибольшее количество место-
нахождений сгруппировано западнее Таллина и севернее районного центра
Кейла, в нижнем течении рек Кейла и Ваэна. Кроме того, южнее имеются
оторванные местонахождения близ сел Энге и Охукотсу. Произрастает
дазифора на влажных альварах и на почти обнаженных известняках, ме-
стами образует густые заросли (Вильбасте, 1953; Флора Эстонской ССР,
1956). Другое изолированное местонахождение D. fruticosa известно в Лат-
вийской ССР — в долине р. Абава близ г. Кандавы.
В восточных прибалтийских районах дазифора кустарниковая была
известна в дикорастущем состоянии давно. По мнению ряда исследовате-
лей, ранее она была распространена здесь шире, но в ходе последующих
смен растительности была оттеснена из других местообитаний, сохранив-
шись на альварах. К- Купфер (Kupffer, 1925) считал дазифору реликтом
растительности бореального периода, Г. И. Вильбасте (1953) отмечает, что
в Эстонии наблюдается тенденция к активному расселению дазифоры на
пастбищах и залежах, причем границы распространения этого кустарника
в Эстонии за последние 150 лет несколько расширились. Это побудило
Г. И. Вильбасте высказать предположение о возможном заносе или искус-
ственном переселении дазифоры в Эстонию с о-вов Эланда и Готланда, хотя
он не располагает какими-либо фактическими данными, подкрепляющими
это предположение. На указанных островах дазифора является несомнен-
ным реликтом и произрастает в таких же местообитаниях, как и в Эсто-
нии,— на альварах. Приведенные Г. И. Вильбасте данные об активизации
распространения дазифоры ни в какой мере не противоречат представлению
о реликтовой природе ее первичных местообитаний в Прибалтике, в том
числе и на альварах Эстонии. Являясь на европейском крыле своего ареала
реликтом, дазифора активно расселяется там, где этому благоприятству-
ет появление обнаженного каменистого или щебнистого субстрата вслед-
ствие хозяйственной деятельности человека (вырубка лесов, выпас скота
и т. п.).
На Британских о-вах D. fruticosa встречается в Англии и в Ирландской
республике.
В Англии она известна в Тисдейле (графства Дургам и Северный Йорк-
шир), а также в Уосдейле и Эннердейле (графство Камберленд).
В Тисдейле, на восточном склоне северных Пеннин19, дазифора произ-
растает по берегам р. Тис на отрезке ее верхнего течения протяженностью
в 16 км в высотных пределах от 230 до 380 At над ур. м. Здесь она всегда свя-
зана со скалами основных пород, известняками каменноугольного воз-
раста или базальтовыми скалами. Экземпляры дазифоры произрастают по
берегам реки в расщелинах скал, на речном наносе и продуктах разруше-
ния горных пород, где это растение образует пятна до 5—6 м в попереч-
нике, а иногда и на небольших скалистых островках.
В графстве Камберленд (Камберлендские горы, Озерный округ) имеется
два изолированных месторождения дазифоры. Одно из них — в Уосдейле,
где это растение встречается в глубокой лощине, врезанной в обнажение
известковых скал на высоте 430 м над ур. м. Другое местонахождение от-
мечено в Эннердейле на крутом северо-западном склоне на высоте 610 м.
В Ирландии дазифора кустарниковая произрастает лишь в западной
части острова, близ побережья. Ее распространение ограничено двумя
районами. Один из них — в графстве Клэр и прилегающей юго-восточной
части графства Голуэй; другой — на границе между северо-восточной
частью графства Голуэй и графством Мейо. В этих местах дазифора встре-
19 Интересно, что в Пеннинах дазифора кустарниковая представлена популяцией с четко
выраженной двудомностью.
175
чается довольно обильно. Растет она в сырых местах, периодически зали-
ваемых водой, исключительно лишь на известковых почвах, подстилаемых
известняками каменноугольного возраста. В северных пунктах типичными
местами ее обитания являются берега больших озер (например, Лох Кор-
риб), в южных — края карстовых депрессий (furloughs), которые зимой
в дождливые периоды времени заполняются водой. Все эти ирландские ме-
стонахождения дазифоры расположены невысоко над уровнем моря.
К- Пиготт (Pigott, 1956) рассматривает флору Верхнего Тисдейла,
включающую Dasiphora fruticosa, Dryas octopetala и ряд других видов с
прерывистым распространением, как реликтовую, являющуюся отголос-
ком растительности позднеледникового времени. Он отмечает ее сходство
с богатой реликтовой флорой о. Эланда. В Ирландии дазифора также трак-
туется как реликт флоры бореального типа, покрывавшей остров в поздне-
ледниковое время. Эта флора на больших пространствах вымерла в голо-
цене в связи с распространением лесов и болот, но отдельные ее предста-
вители смогли сохраниться в известняковой области Западной Ирландии,
где, по-видимому, лесной покров никогда не был сплошным.
В ископаемом состоянии дазифора кустарниковая была найдена на Бри-
танских о-вах дважды, в обоих случаях на территории Англии. Ископае-
мые остатки ее были определены из позднеледниковых отложений близ
Нейзинга, Эссекс (семянки этого растения вместе с фрагментами Dryas
octopetala), и из ледниковых отложений близ Барнуелла, Кэмбриджшир.
В Пиренеях этот кустарник изредка встречается в восточной части на
каменистых местах в пределах альпийского и субальпийского поясов.
Дазифора кустарниковая произрастает в нескольких пунктах Примор-
ских Альп на территории южной Франции и примыкающей части Италии
(Burnat, 1896). Кроме того, она упоминается А. Фиори (Fiori, 1924) для
других мест Италии: Пьемонт, окрестности Турина (Monte delle Cafarse)
и Апеннины, Абруццо, район Гран Сассо, близ приюта di Campopericoli
(в последнем случае, по мнению этого исследователя, возможно, происхо-
дит от культуры). На о. Сицилия дазифора кустарниковая, вопреки упоми-
нанию П. Н. Крылова (1927—1964), не встречается.
На Кавказе дазифора известна из Предкавказья, Главного Кавказ-
ского хребта и Закавказья (северная часть Армянского нагорья, Талыш-
ские горы). Она произрастает на скалистых и каменистых склонах от пред-
горий до альпийского пояса включительно (Гроссгейм, 1952; Флора Арме-
нии, 1958). Однако наибольшее количество известных местонахождений
сосредоточено в высокогорной части Большого Кавказа на участке между
Эльбрусом и Казбеком и в Дагестане. Кроме того, дазифора кустарниковая
заходит и в смежные с Кавказом районы Турции и Ирана.
Распространение и условия произрастания дазифоры кустарниковой
на Урале
На основании анализа гербарного материала, хранящегося в Ботани-
ческом институте Академии наук СССР и в Институте экологии растений
и животных Уральского филиала АН СССР (включая сборы автора), а так-
же литературных источников можно составить значительно более полное
и точное представление о распространении дазифоры кустарниковой на
Урале (см. табл. 3) по сравнению с тем, что сообщалось до сих пор в фло-
ристических сводках. В свете этих данных распространение D. fruticosa на
Урале вырисовывается в таком виде (рис. 101).
На Приполярном Урале дазифора кустарниковая встречается на галеч-
никах по берегам рек, преимущественно выше границы леса. Она отмечена
как на западном склоне в бассейне р. Кожима, притока Кось-Ю, так и на
восточном склоне, в бассейне р. Ляпина (по речке Манье).
176
Таблица 3
Местонахождения дазифоры кустарниковой на Урале
Место сбора
I
Коллектор или литературные I Местонахождение гербар-
источники ных экземпляров
Бассейн р. Кожима, притока р. Ко-
сью, галечник на берегу ручья, выше
границы леса
Тобольская губерния, Березовский
уезд, бассейн р. Северной Сосьвы,
р. Манья (приток Ляпина), между
устьем р. Хальмезы-Я и р. Сорпинг-Я,
берег реки
По р. Щокурье в 12 км выше устья
р. Польи
По р. Щугору
Бассейн р. Северной Сосьвы, горы
в верховьях р. Уольи (Вольи), в комп-
лексе с высокогорными растениями
Бассейн р. Северной Сосьвы, по
р. Яны-Манье, под 63°15' с. ш.
(у Крылова и Говорухина ошибочно
указано 64°40' с. ш.)
Бассейн р. Северной Сосьвы, на бе-
регу р. Няысь, травянистая лужайка
Хребет Чистоп, гольцовый пояс, на
Лоуньинской сопке, у основания ска-
листого останца
Река Вижай, приток Лозьвы, на бе-
реговых галечниках в ряде пунктов от
низовий до устья Кула
Ивдельский район, бассейн р. Юж-
ной Тошемки (приток р. Ивделя), вбли-
зи пос. Тошемки, в 3 км вверх по
р. Фадеевке, на сфагновом болоте
В 50 км к северо-западу от г. Ив-
деля, у пос. Плотинка, заросли по бе-
регу ручья, притока р. Ивделя
Денежкин Камень
Денежкин Камень, долина Сухого
Шарпа, прибрежная заросль кустарни-
ков в полосе подгольцового листвен-
ничного леса
Денежкин Камень, горная тундра
на перевале близ истоков Сухого
Шарпа
Денежкин Камень, горная тундра в
истоках р. Быстрой
Гора Кумба, подгольцовый пояс,
скалы на вершине горы
Конжаковский Камень, гольцовый
пояс, южные скалистые склоны сопки
Сторож
Конжаковский Камень, гольцовый
лояс, южные склоны перевала, среди
скал
Конжаковский Камень, выше грани-
цы леса, в днище каровидного углуб-
ления на каменной россыпи
Тылайский Камень, юго-западный
склон, мшистые поляны среди скал
Косьвинский Камень, выше границы
леса на восточном склоне, в районе
прииска Кытлым
Косьвинский Камень, южный склон,
у границы леса
В. Б. Сочава,
17/VII 1927
Б. Н. Городков,
10/VII 1915
И. Д. Кильдюшевский
Р. Р. Поле
Штаб-лекарь Селиванов,
по Ф. И. Рупрехту (1856)
По П. Н. Крылову
(1927—1964)
В. Б. Сочава,
21/VIII 1928
П. Л. Горчаковский,
5/VIII 1950
П. Л. Горчаковский,
12- 14/VII 1951
А. С. Валитова,
5/IX 1945
К. Н. Игошина
16/VIII 1943
П. Н. Крылов, 1878
П. Л. Горчаковский,
15/VIII 1948
П. Л. Горчаковский,
15/VIII 1948
П. Л. Горчаковский,
16/VIII 1948
К. Н. Игошина,
17/IX 1941
К- Н. Игошина,
28/VII 1925
К. И. Игошина,
29/V 1925
П. Л. Горчаковский,
13/VII 1953
А. А. Черданцев,
19/VII 1906
М. М. Сторожева,
27/VIII 1947
С. Г. Шиятов,
15/IX 1956
Ботанический инсти-
тут АН СССР
Там же
»
»
Неизвестно
Неизвестно; возмож-
но, в Томском универ-
ситете
Ботанический инсти-
тут АН СССР
Институт экологии
растений и животных
УФАН СССР
Там же
Ботанический инсти-
тут АН СССР
Институт экологии
растений и животных
УФАН СССР
Там же
»
»
Ботанический инсти-
тут АН СССР
Там же
Институт экологии
растений и животных
УФАН СССР
Там же
»
»
»
»
12 Заказ № 251
177
Таблица 3 (окончание)'
Место сбора Коллектор или литературные источники Местонахождение гербар- ных экземпляров
Сухогорский Камень И. И. Пастухов, 2-я по- ловина июля 1868 Институт экология растений и животных
Сухогорский Камень П. Н. Крылов, УФАН СССР Ботанический инета-
Семичеловечный Камень, скалы, ка- 6/VII 1876 К. Н. Игошина, тут АН СССР Там же
менистая горная тундра Окрестности прииска Кытлым, гора 5/VIII 1925 С. Г. Шиятов, Институт 'огпп
Третий Камень, горная тундра 5/VII 1956 растений и животных
Южный Урал, Миньярский завод Южный Урал, Башкирская АССР, М. В. Редикорцева УФАН СССР Там же
А. Л. Лыпа и Д. К. Зе- Ботанический сад
Белорецкий район, окрестности Яман- ров, 21/VIII 1942 Башкирского филиала
Тау, на берегу безымянной речки, близ АН СССР
бывших Куянтавских углевыжигатель- ных печей
Южный Урал, в верховьях р. Тир- К. Н. Игошина, (1966) Ботанический инсти-
ляна, на берегу безымянной речки, тут АН СССР
в зарослях Salix pyrolifolia
Губерлинские горы И. Г. Борщов (1865) Неизвестно
Большая часть местонахождений D. fruticosa сосредоточена на Северном
Урале (к югу от широтного отрезка р. Щугора). Она найдена в бассейне
Северной Сосьвы в верховьях Вольи (Уольи) и ее притока Яны-Маньи,
а также по р. Няысь (Няйс). По наблюдениям М. М. Сторожевой (личное;
сообщение) по р. Няысь, ниже устья речки Няысь-Маньи, дазифора кустар-
никовая образует заросли на прибрежных галечниках. Затем после пере-
рыва она появляется, согласно нашим исследованиям, в горнотундровом’
поясе хр. Чистоп и по р. Лозьве и ее притокам — Северной Тошемке, Ви-
жаю, Талице, Ивделю и некоторым другим. По-видимому, верхнее течение-
Лозьвы с ее притоками является центром наиболее обильной встречаемости
дазифоры кустарниковой в пределах уральского фрагмента ее ареала.
Она произрастает не только в высокогорьях, но по долинам рек спускается:
и в горнотаежный пояс, где образует прибрежные заросли на галечниках.
Обычно такие заросли встречаются в глубоких долинах, куда стекают со
склонов массы охлажденного воздуха. Благоприятствует произрастанию
этого кустарника наличие галечникового субстрата, периодически обна-
жающегося вследствие размывающей деятельности речного русла. Напри-
мер, по Вижаю (рис. 102) заросли дазифоры кустарниковой тянутся узкой
полосой вдоль реки на протяжении многих километров (почти от самых ни-
зовий до устья Кула). В прибрежных зарослях дазифоре кустарниковой
сопутствуют Betula humilis, некоторые виды ив, а из травянистых расте-
ний — Allium schoenoprasum, Pinguicula vulgaris, Ligularia sibirica, Po-
lygonum bistorta, Sanguisorba officinalis, Parnassia palustris и др. На Де-
нежкином Камне (Горчаковский, 1950) D. fruticosa встречается лишь в
высокогорной части — в горных тундрах и по прибрежным галечникам
выше границы леса (до высоты 1100—1200 м над ур. м). Встречена она так-
же и на горе Кумба. На Тылайско-Конжаковско-Серебрянском массиве
местонахождения ее также связаны с высокогорьями (горные тундры, ка-
менные россыпи, галечники выше границы леса). Отмечена она в этом райо-
не и на Сухогорском Камне.
На территории Среднего Урала, характеризующейся значительной по-
ниженностью рельефа, дазифора кустарниковая не встречается. Но в самой
центральной части Южного Урала, в районе его крупнейшей вершины —
178
горы Яман-Тау, находится изолиро-
ванное местонахождение D. fruti-
cosa. По сведениям, полученным от
Е. Л. Лыпы, в этом районе она произ-
растает на берегу безымянной реч-
ки — близ бывших Куянтавских угле-
выжигательных печей. Приблизи-
тельно здесь же ее собирал Р. А. Еле-
невский. Кроме того, в этом же райо-
не дазифора была найдена в зарослях
Salix pyrolifolia на берегу безымян-
ной речки, впадающей в р. Тирлян
(Игошина, 1966). Указанные место-
нахождения отделены от южных пунк-
тов встречаемости D. fruticosa в райо-
не Конжаковского Камня расстоянием
почти 600 км. Не исключена вероят-
ность нахождения этого растения и
в некоторых других пунктах цент-
ральной части Южного Урала. В
гербарии Института экологии расте-
ний и животных Уральского филиала
Академии наук СССР есть экземпляр
D. fruticosa, собранный М. В. Редикор-
цевой в районе Миньярского завода
(ныне г. Миньяр). По этикетке, запол-
ненной слишком лаконично, нельзя
судить о точном местонахождении и
условиях произрастания собранного
экземпляра. В 300 км к югу от Яман-
Тау И. Г. Борщов (1865) нашел дази-
фору кустарниковую на Губерлинских
горах. К сожалению, последующие
исследователи не повторили сборов
И. Г. Борщова из этого пункта,
находящегося у самой южной оконеч-
ности Южного Урала.
Как видно, на Урале дазифора
кустарниковая встречается редко, рас-
пространена прерывисто, причем боль-
шая часть ее местонахождений рас-
положена в центральной горно-хреб-
товой полосе на крупных горных вер-
шинах или в непосредственной бли-
зости от них. Ясно заметна преиму-
щественная связь дазифоры с высоко-
горными поясами — подгольцовым и
горнотундровым, хотя по долинам
Рис. 101. Распространение дазифоры кустар-
никовой (Dasiphora fruticosa) на Урале.
Местонахождения: 1 —по гербарным экземпля-
рам, 2 — по литературным данным.
рек она может спускаться и в горнолесной пояс.
На Урале D. fruticosa произрастает на прибрежных галечниках, в гор-
ных тундрах, на каменных россыпях и в редких случаях по окраинам сфаг-
новых болот. Она очень тесно связана с каменистым (нередко периодически
обнажающимся) субстратом, произрастая в слабо задернованных местах,
где ослаблена конкуренция со стороны других кустарников и деревьев.
Таким образом, экологическая амплитуда D. fruticosa на Урале значитель-
12*
179
Рис. 102. Дазифора кустарниковая Dasiphora fruticosa на прибрежном галечнике по р. Вижаю;
но уже, чем в центральных районах ее основного азиатского ареала. Местом
наиболее обильной встречаемости D. fruticosa в пределах Уральского хпеб-
та является восточный склон Северного Урала - горы и речные долины
верхнем течении р. Лозьвы, а также район Денежкина Камня и Тылай-
ско-Конжаковско-Серебрянского горного массива. В других местах она
встречается лишь спорадически. Вероятно, в прошлом D. fruticosa была
распространена на Урале гораздо шире, но впоследствии подверглась от-
теснению лесными и кустарниковыми фитоценозами, сохранившись лишь,
на каменистых местах, преимущественно в высокогорных поясах, где эко-
лого-биологические особенности данного вида дают ему некоторые преиму-
щества в борьбе за существование с другими кустарниками и деревьями..
Промежуточные местонахождения дазифоры кустарниковой
между Уралом и Алтаем
(Кокчетавский горнолесной массив)
Глубоко в степной зоне Северного Казахстана в окрестностях оз. Боро-
вого (область Казахского мелкосопочника) находится исключительно ин-
ересныи Кокчетавский горнолесной массив, где сосновые леса произра-
стают на склонах невысоких гранитных гор и сопок.
Производя геоботанические исследования в б. Кокчетавском уезде в
' г., а. н. 1 ордягин описал сфагновый торфяник, обнаруженный им..
180
в юго-восточной части Кокчетавского массива в окрестностях оз. Чортан-
Куль (близ Щучинска) среди соснового леса. Торфяник расположен в лож-
бине, идущей, по словам местных жителей, к Чортан-Кулю; по ней струит-
ся ручеек, теряющийся в торфянике. В пониженной части торфяника на
фоне покрова из Sphagnum teres и некоторых гипновых мхов возвышаются
небольшие бугорки из Sphagnum fuscum, на которых растут низкорослая
сосна, береза бородавчатая и «много красивой Potentilla fruticosa» (Гордя-
гин, 1901, стр. 301). Как на бугорках, так и в понижениях между ними
отмечены Oxyccccus quadri pel alas, Menyanth.es trifoliata, Eriophorum angu-
stifolium и другие бореальные виды растений.
Необычный факт встречаемости в степной зоне ряда не свойственных,
ей растений побудил А. Я- Гордягина (1916) вновь посетить район Кок-
четавских боров в 1901 г. Экскурсируя здесь, А. Я- Гордягин обнаружил
еще несколько торфяников, причем в торфянике у оз. Карасьего Dasiphora
fruticosa росла наряду с такими бореальными растениями, как Drosera го-
tundifolia и Rhynchospora alba. Кроме того, А. Я. Гордягин познакомился
с небольшой коллекцией растений, собранных Н. П. Лоскутовым на вер--
шине горы Синюхи (887 м над ур. м.), высшей точке Кокчетавских гор.
В числе растений, собранных на скалистой вершине этой горы был изуро-
дованный низкорослый экземпляр Dasiphora fruticosa. Впоследствии в ни-
зине на южной стороне оз. Светлого D. fruticosa была встречена также на
открытом осоково-гипновом болоте на более плотных участках дернины-
(Жаркова, 1930).
По наблюдениям Л. Н. Соболева (1937), дазифора кустарниковая най-
дена кроме того на северо-западном склоне Кокчетавского хребта (Кокче-
Тау), обращенном к Чебачьим озерам. Здесь она произрастает по долинам;
в березняках и кустарниковых зарослях.
Из этих данных явствует, что в районе Кокчетавских горных боров
D. fruticosa20 встречается, главным образом, в пониженных местах, на тор-
фяниках, но была отмечена также и на скалистой вершине горы Синюхи.
А. Я- Гордягин (1916), не ограничиваясь констатацией приведенных им-
интересных фактов, высказал предположение, что D. fruticosa, как и другие
бореальные растения, проникла в район Кокчетавских боров, по-видимому,
не с севера, а с востока, из района Алтая и прилегающих к нему мест. Это-
проникновение было возможным при иных, более суровых, чем теперь,
климатических условиях.
Этот исследователь не высказывался определенно о возможном времени
внедрения во флору Кокчетавского массива отмеченных им бореальных ра-
стений. По экологии и условиям проникновения он не отграничивал D.
fruticosa от глубоколесных и болотных видов. Это нельзя признать пра-
вильным. Необходимой предпосылкой для внедрения в район Кокчетав-
ских гор глубоколесных и болотных растений является сплошная или более
или менее сплошная облесенность территорий между этими горами и
Алтаем. Однако D. fruticosa в отличие от собственно лесных и болотных,
растений могла проникнуть сюда и тогда, когда сплошной облесенности не-
было. Наиболее вероятным временем проникновения дазифоры кустарни-
ковой в район Кокчетавских гор является плейстоцен.
20 К сожалению, мы не располагаем гербарными экземплярами дазифоры из района-
оз. Борового. По-видимому, отсутствуют они и в гербарии Ботанического института Ака-
демии наук СССР. Однако, судя по условиям произрастания дазифоры в этом районе (пре-
имущественно сфагновые болота) в комплексе с другими бореальными растениями, находя-
щимися здесь на южном пределе своего островного распространения, можно предположить,,
что здесь встречается D. fruticosa s. str. Старые сборы этого кустарника из б. Петропав-
ловского округа и Кокчетавского уезда, хранящиеся в гербарии Института ботаники Ака-
демии наук Казахской ССР, относятся к D. fruticosa.
181!
Об истории расселения дазифоры кустарниковой на европейском крыле
ареала и реликтовой природе ее уральских местонахождений
Дазифора кустарниковая в настоящее время наиболее распространена
в горных районах Азии (горы южной части Сибири, Монголия, Китай).
В основной азиатской части своего ареала это растение более изменчиво.
Здесь же оно отличается наибольшей высотной амплитудой (от подножия
гор до высокогорий) и наибольшим размахом приспособленности к раз-
личным условиям среды. По-видимому, именно в горных районах Азии
О. fruticosa сформировалась как вид и затем стала расселяться в различных
направлениях. Существование в прошлом связи между материками Восточ-
ной Азии и Северной Америки обусловило возможность проникновения
-ее на североамериканский континент. Это произошло, вероятно, в конце
третичного периода.
Похолодание климата, начавшееся в Европе в конце плиоцена и уси-
лившееся в начале плейстоцена, вызвало деградацию теплолюбивой дре-
весной растительности и замену широколиственных лесов светлохвойными
(лиственничными и отчасти сосновыми) и мелколиственными (березовыми,
осиновыми). В Западной Сибири, на Урале и на севере европейской части
СССР особенно широко распространились лиственничные леса (Крашенин-
ников, 1939). В лиственничных лесах с их разреженным светлым пологом
дазифора кустарниковая, отличающаяся светолюбием, находит благо-
приятные условия, тогда как в тенистых широколиственных и темнохвой-
ных лесах она расти не может. Распространение лиственничных лесов и
вообще светлых разреженных лесов благоприятствовало проникновению
Дазифоры кустарниковой, наряду с другими сибирскими растениями, че-
рез Западно-Сибирскую низменность и область Казахского мелкосопочника
на Урал, а затем через Русскую равнину в Европу. По северной части
Европы дазифора расселилась вплоть до Британских о-вов, куда смогла
проникнуть по перемычкам суши, неоднократно соединявшим их с мате-
риком в плиоцене и в плейстоцене (Вульф, 1944).
Есть основания предполагать, что в период наибольшего (рисского,
днепровского) оледенения D. fruticosa произрастала в Европе и на Урале
в кустарниковых зарослях по берегам речек и ручьев в перигляциальной
полосе. В последовавшую межледниковую эпоху, по длительности пре-
восходившую весь голоцен, дазифора кустарниковая стала расселяться
особенно активно. Этому благоприятствовало обилие обнаженного каме-
нистого, галечникового и песчаного субстрата (лишенные растительности
флювиогляциальные и речные наносы, освободившиеся от оледенения ка-
менистые склоны гор и россыпи). Соглашаясь с предположением
Э. Хультена (Hulten, 1937), что в течение великого межледниковья D.
fruticosa распространилась на большей части Европы, можно добавить,
что это в равной степени относится к Уралу и к смежной с ним территории.
Второе, менее интенсивное оледенение едва ли могло существенно повлиять
на распространение этого кустарника.
Изменение климата, а вследствие этого и растительности в голоцене
повлекло за собой оттеснение D. fruticosa из многих районов Европы и пол-
ное вымирание ее на территории Русской равнины и Западно-Сибирской
низменности. Самую существенную роль в этом процессе сыграло сокра-
щение площади обнаженного субстрата (вследствие заселения раститель-
ностью) и широкое распространение темнохвойных и вообще тенистых
лесов. D. fruticosa смогла удержать свои позиции в ряде районов Европы
(преимущественно в горах или на альварах побережья и островов Балтий-
ского моря) и в горах Урала. На Урале она сохранилась на каменистых
или щебнистых местах, главным образом в высокогорьях или в некоторых
182
речных долинах горнотаежного пояса, где скапливаются холодные массы
воздуха, а субстрат периодически обнажается вследствие размывающей
деятельности рек.
Из сказанного можно заключить, что D. fruticosa на Урале является/
плейстоценовым реликтом, сохранившимся лишь в местах, где ослаблена
конкуренция со стороны других кустарников и деревьев. Своеобразные
заросли этого растения в долинах рек бассейна Лозьвы на Северном Урале,.
где оно растет вместе с Betula humilis, очень близки к аналогичным расти-
тельным сообществам, характерным для гор Южной Сибири (например,.
Алтай, Саяны). Такие кустарниковые заросли, вероятно, были широко'
распространены на Урале в плейстоцене, когда здесь установился клима-
тический режим, сходный с современным климатом верхних горных поясов
Алтая и Саян. Таким образом, заросли дазифоры кустарниковой на Урале-
являются отголосками перигляциального ландшафта; это, в сущности, ре-
ликтовые растительные сообщества плейстоцена.
Предположение о том, что в плейстоцене, когда ботанико-географиче-
ские зоны были смещены к югу, D. fruticosa произрастала на территории-
Западно-Сибирской низменности в прилегающей к ней части Казахского
мелкосопочника, подтверждается как палеоботаническими документами,,
так и ареалогическими данными.
П. А. Никитин (1935) приводит описание растительных остатков, най-
денных в толще глин на обрывах по р. Оби и ее притокам (Западно-Сибир-
ская низменность). В образце глины на левом берегу р. Чулыма, близ дер.
Елань, обнаружены семянки D. fruticosa. Описанную им ископаемую флору
П. А. Никитин датирует коссожской фазой, соответствующей отступлению-
днепровского ледника. В то время, по мнению указанного исследователя,.,
здесь была распространена перигляциальная растительность, в составе ко-
торой преобладали кустарниковые заросли из D. fruticosa, Alnus frutico-
sa и кустарниковой березки, близкой к североамериканской Betula pumila.
В послеледниковое время D. fruticosa вымерла на территории Западно-Си-
бирской низменности, но сохранилась лишь в Кокчетавском горнолесном-
массиве в области Казахского мелкосопочника, где, как и на Урале, она-
является плейстоценовым реликтом. Материалы по изучению реликтов —
этих «живых ископаемых» — удачно подкрепляются палеонтологической
документацией. Это служит лишним подтверждением плодотворности со-
временных методов исторической географии растений, позволяющих вос-
станавливать историю расселения растений на основе изучения их совре-
менного распространения.
МОЖЖЕВЕЛЬНИК КАЗАЦКИЙ JUNIPERUS SABINA
К числу малоизученных, но интересных в ботанико-географическом от-
ношении представителей уральской дендрофлоры относится казацкий мож-
жевельник Juniperus sabina. Ареал этого вида — евразиатский дизъюн-
ктивный. Он разобщен на несколько частей, связанных с отдельными гор-
ными массивами; кроме того, имеются небольшие фрагменты ареала и изо-
лированные местонахождения на внутриконтинентальных возвышенных?
равнинах. Этот стелющийся кустарник распространен в горах Южной и
отчасти Средней Европы (Альпы, Пиренеи, Апеннины, Карпаты, горы
Балканского п-ова), в высокогорной части Кавказа, в горах на юге Крыма,
в некоторых пунктах юго-востока Восточно-Европейской равнины, на;
Южном Урале, в горах Средней Азии (Тянь-Шань, Джунгарский Алатау,.
Тарбогатай, Саур) с прилегающей к ним равнинной или слабо всхолмлен-
ной территорией Казахстана, в горах Кузнецкого Алатау, Алтая, Запад-
ного Саяна (бассейн р. Усы) и в смежной с Алтаем горной части севера МНР'
(Крылов, 1927—1964; Комаров, 1934; Hegi, 1906—1935; Малеев, 1949).
18&
В Европе казацкий можжевельник произрастает на солнечных склонах
гор, на скалах и в подлеске разреженных сосновых лесов, поднимаясь в
Швейцарских Альпах до высоты 3000 м над ур. м. (Hegi, 1906—1935). На
Кавказе его распространение ограничено верхним лесным, субальпийским
и альпийским поясами (в пределах 1500—3000 м над ур. м.); обитает он на
щебнистых и скалистых склонах и осыпях, где нередко образует чистые
заросли или растет совместно с другим сланцевым можжевельником J. dep-
ressa. В восточных районах Большого и Малого Кавказа J. sabina входит
в состав формаций нагорных ксерофитов (Васильев и Сахокиа, 1959).
Местонахождения этого вида в горной части Средней Азии связаны преиму-
щественно со скалами на низких и средних уровнях гор (Рубцов, 1941а,б;
Поляков, 1951; Степанова, 1962), но изредка он заходит и в высокогорья,
в субальпийский пояс (Головкова, 1959). В горах Южной Сибири казацкий
можжевельник произрастает по открытым южным склонам холмов и невы-
соких гор в степной и смежных частях лесной зоны (Крылов, 1927—1964).
Таким образом, при движении на восток наблюдается снижение вертикаль-
ных пределов его распространения.
Большой интерес представляют изолированные местонахождения
J. sabina на юге и востоке Восточно-Европейской равнины (Кеппен, 1885;
•Литвинов, 1891). Этот можжевельник указан в среднем течении р. Дона на
участке между станицами Казанской и Сиротинской на меловых холмах
‘(где он местами очень обилен), на рыхлых меловых песчаниках по берего-
вым обрывам, в лесистых оврагах и на песчаных дюнах правобережья.
Кроме того, он известен на меловых холмах по левому притоку Дона —
р. Иловле на участке ниже с. Ольховки. Отдельные его местонахождения
ютмечены на мелах в низовьях притока Дона — р. Медведицы, в окрест-
ностях г. Камышина у с. Чухонастовки (Литвинов, 1891) и в Жигулях
(Сукачев, 1927). Ф. П. Кеппен (1885) приводит интересное указание о на-
ходке М. Твардовским казацкого можжевельника в Белоруссии, в б. Пин-
ском уезде близ дер. Велесницы; никем из последующих исследователей
эти данные не были подтверждены. Вероятно, в некоторых изолированных
•местонахождениях на юге СССР казацкий можжевельник в последнее
время бесследно исчез в результате безжалостного уничтожения на топливо.
В Западном Казахстане, в бассейне р. Урала, J. sabina местами встре-
чается на бугристых песках, причем по В. В. Иванову (1958), является
теперь весьма редким видом из-за интенсивного истребления. Восточнее он
известен в области Казахского мелкосопочника (от северного побережья
•оз. Балхаш на юге до Кокчетавских гор и горы Акнет в окрестностях Баян-
Аула на севере), где растет на гранитных сопках, иногда поросших сосной,
и реже на глинистых обрывах по берегам рек21. Л. Н. Грибанов (1960) от-
мечает этот вид в подлеске степных сосновых боров (в том числе из группы
скальных) Казахского мелкосопочника, Калбинского хребта, зоны Тур-
тайского пролива и Обь-Иртышского междуречья.
Литературные данные о распространении казацкого можжевельника
.'на Южном Урале очень скудны. Они, в сущности, ограничиваются мате-
риалами известной монографии С. И. Коржинского (Korshinsky, 1898),
где дан перечень немногих известных в то время местонахождений назван-
ного вида без указаний на условия произрастания.
Автором обнаружен и описан ряд новых местонахождений J. sabina на
Южном Урале (в Башкирской АССР, в Оренбургской и Челябинской
областях в период с 1957 по 1963 гг.), а Б. П. Колесниковым — в Джабык-
Карагайском бору Челябинской области в 1962 г. Эго дало возможность
21 Сведения о распространении казацкого можжевельника в Казахстане приводятся
с учетом данных гербария Института ботаники Академии наук Казахской ССР.
-184
составить с учетом имеющихся гербарных материалов и литературных,
данных карту распространения казацкого можжевельника (Горчаковский
и Колесников, 1964), существенно дополняющую прежние представления
об уральском фрагменте его ареала (рис. 103); большая часть приведенных,
в ней местонахождений ранее оставалась неизвестной.
Рис. 103. Распространение можжевельника казацкого Juniper us sa-
bina на Южном Урале.
По характеру условий среды южноуральские местонахождения казац-
кого можжевельника могут быть подразделены на пять следующих групп..
1. Известняковые утесы и обнажения по берегам рек в увалисто-хол-
мистой полосе западного склона Южного Урала. Местонахождения этой
группы располагаются на высоте 300—400 м над ур. м. и связаны с ланд-
шафтом широколиственных или широколиственно-хвойных лесов. Казац-
кий можжевельник найден по р. Белой в 20 км выше пос. Авзяна, в 3 км-
ниже дер. Старомунасыпово, между деревнями Ишдавлетово и Миндегу-
лово и в 18 км выше с. Биккузино; по притокам Белой: р. Зилиму против,
устья речки Кат в 3 км от дер. Сарала, у дер. Куйли-Тамак и близ с. Имен-
дяшево, по р. Зигану близ устья р. Сиказы, р. Нугушу (рис. 104, 105) у
устья речки Урюк и по р. Кане в 10 км к северу от с. Кананикольского.
Особенно обильно встречается казацкий можжевельник на прибрежных,
известняковых утесах по р. Белой (рис. 106), на участке между пос. Авзя-
ном и с. Биккузино, где его распростертые кусты образуют довольно гу-
стые стланиковые заросли. Склоны утесов, обращенные к реке, круты,
иногда отвесны. Лишь плоские вершины и более пологие склоны покрыты/
сосновым лесом. Кроме того, отдельные низкорослые экземпляры сосны:
185.
Рис. 104. Река Ну гут близ устья речки. Урюка.
Pinus silvestris и березы Betula verrucosa ютятся в лощинках, в ущельях,
и на выступах крупных каменных глыб; у основания утесов нередко встре-
чается вяз UImus laevis. Заросли казацкого можжевельника занимают
обычно обращенные на юг взлобки утесов и прилегающие к ним участки,
осыпей. Стволики можжевельника распростертые, прижатые к субстрату,.
Рис. 105. Можжевельник казацкий на известняковых скалах по р. Нугушу.
диаметром до 8—12 см, длиной до 10—15 м. Высота отходящих от них зе-
леных ветвей не превышает 50—70 см. Самый старый экземпляр казацкого.^
можжевельника, найденный в бассейне р. Ну’гуша, имел возраст 260 лет.
Травяной покров в можжевеловых зарослях не сомкнутый, состоит в основ-
ном из скальных видов Agropyrum reflexiaristatum, Sedum hybridum, Schi-
verekia berteroides, Asperula petraea и степняков Echinops ritro, Centaurea
sibirica, Onosma simplicissimum, Thymus Marschallianus, Antitoxicum-
stepposum и др. Встречаются также и мезофильные растения — папорот-
ник Cystopteris fragilis и Asplenium ruta-muraria (в глубоких тенистых,
расщелинах, Polygonatum officinale, Thalictrum foetidum и др. Из кустар-
ников отмечены Cerasus fruticosa, Caragana frutex, Spiraea crenata, Rham-
nus cathartica, Genista tinctoria, Cotoneaster melanocarpa. За выступы скал.;
и ветки кустарников цепляются длинные гибкие стебли Rubus caesius и
Humulus lupulus.
Местами, например на склоне к р. Нугушу близ устья речки Урюк, ка-
зацкий можжевельник встречается не только на прибрежных обнажениях..
187
i
106. Распластанные экземпляры Juniperus sabina на прибрежных известняковых утесах по р. Белой.
известняка, но изредка и в периферической части редкостойных камени-
стых дубняков.
По прибрежным обнажениям известняков на западном склоне Урала
J. sabina заходит дальше всего на север, вплоть до бассейна р. Зилима под
54° с. ш. Известняковые скалы по Зилиму интересны и в том отношении,
что с ними связаны самые северные местонахождения крайне редких на
Южном Урале пустынно-степных растений — Atraphaxis frutescens и Fe-
rula caspica.
2. Обнажения и осыпи известняка на склонах сыртов в пределах Зила-
ирского плато. Отмечены местонахождения около пос. Зилаира, южнее
его близ устья р. Большого Шора, в бассейне р. Большого Ика (у деревень
Кузьминовки, Нукаево, Ургенкп и Андреевки), в бассейне его правого
притока — р. Большой Сурени в 3,5 км выше дер. Тазларово и близ дере-
вень Кугарчи и Байдавлетово, а также в бассейне р. Сакмары у дер. Бляу-
Тамак.
Казацкий можжевельник встречается одиночными экземплярами и за-
рослями на крутых южных склонах плосковершинных холмов на высоте
от 300 до 550 м над ур. м. В западной части Зилаирского плато его место-
обитания располагаются среди горных каменистых степей, а в восточной —
в окружении остепненных луговых полян, дубовых лесов с примесью сосны,
а также сосновых лесов. Заходит этот кустарник иногда (например, близ
пос. Зилаира) и в окраинную часть сосняков.
На сыртах близ дер. Тазларово казацкий можжевельник образует на
площади около 3 га густые заросли распластанных кустов, нередко смы-
кающихся друг с другом. Кусты можжевельника хорошо видны даже из-
дали, так как резко контрастируют своей темно-зеленой окраской с более
светлым фоном окружающих обнажений и степных участков. Травяной
покров в зарослях казацкого можжевельника развит слабо, состоит он из
обычных степных и скально-осыпных растений (Asperula petraea, Chelido-
.nium majus, Schiverekia berteroides и др.) co значительной примесью сорня-
ков (вследствие интенсивного выпаса скота).
Значительный интерес представляет находка нескольких экземпляров
-J. sabina на второй (надпойменной) террасе р. Большой Сурени в 500 м
выше дер. Тазларово. Распластанные довольно старые кусты казацкого
можжевельника растут здесь на песчаной почве в редкостойном осокорнике
(из Populus nigra). Нет никаких данных, говорящих в пользу искусствен-
ного разведения здесь этого вида. Вероятно, такие местообитания являются
вторичными; казацкий можжевельник перешел на песчаные наносы реч-
ной террасы со своих основных стаций — известняковых склонов сыртов.
3. Скалистые гребни лесистых хребтов восточного склона Южного Ура-
ла. Стланиковые экземпляры казацкого можжевельника растут на
хр. Крыкты, в верхней части скалистого склона ущелья, пропиленного не-
большой речкой, вытекающей восточнее вершины Шершил-Тау, среди
лиственнично-сосновых и производных от них березовых лесов. Это место-
нахождение в отличие от предыдущих расположено значительно выше в
горах (на высоте около 800 м над ур. м.) и не связано с карбонатными гор-
ными породами.
4. Обнажения различных горных пород среди горных и предгорных
степей восточного склона Южного Урала. Сюда относятся местонахожде-
ния казацкого можжевельника на хр. Ирендык (к западу от дер. Файзули-
но, по дороге на пос. Баймак и по р. Таналыку), в Губерлинских горах (по
р. Губерле у дер. Аджалямовой, на склонах к р. Уралу в 5 км ниже устья
р. Шошки и в 18 км выше устья р. Губерли), а также в области зауральского
пенеплена в бассейне верхнего течения р. Урала (по р. Сосновской, в ни-
зовьях р. Ташлы и по р. Уралу выше устья р. Ташлы).
189
На хр. Ирендык J. sabina встречается преимущественно на известня-
ках, на склонах южной экспозиции среди каменистых степей на высоте до
400—750 м над ур. м. Растет он одиночными экземплярами, жмущимися
к скалам, или небольшими группами. Травяной покров на таких участках
бедный, несомкнутый; для него особенно характерны Helictotrichon deser-
torum, Festuca sulcata и Orostachys spinosa. В Губерлинских горах казацкий
можжевельник сохраняет связь с южными экспозициями, но встречается
уже на бескарбонатных горных породах (кристаллические сланцы и др.)
на высоте 300—400 м над ур. м. Среди распластанных кустов можжевель-
ника здесь растут Sedum hybridum, Centaurea sibirica, Veronica incana,
Galium verum и другие виды. По данным С. А. Никитина (1956), в Губер-
линском мелкосопочнике казацкий можжевельник иногда заходит в "бере-
зовые колки среди типчаковых степей с кустарниками. В области заураль-
ского пенеплена встречается на скалах по склонам ущелий, образован-
ных небольшими, пересыхающими летом речками — притоками р. Урала.
5. Островные сосновые боры на гранитах Урал-Тобольского между-
речья (Джабык-Карагайский бор). На возвышенной Зауральской равнине
пока описано одно местонахождение казацкого можжевельника и по сосед-
ству с ним, по данным местных жителей, указывается еще два. Они распо-
лагаются в центральной части Урал-Тобольского междуречья (бассейн
верховий р. Караталыаята), достигающего здесь абсолютной высоты около
400 м и характеризующегося многочисленными выходами на поверхность
гранитных интрузий палеозойского и мезозойского возраста. Для выходов
гранитов типична плоская матрацевидная отдельность плит и глыб. Опи-
санное местонахождение казацкого можжевельника расположено в
149 квартале Анненского лесничества, вблизи дороги из с. Анненского
в дер. Ольховку, в центре крупного лесного массива Джабык-Карагай-
ского соснового бора22.
Единственный хорошо развитый куст казацкого можжевельника (вы-
сота его около 1,5 м, поперечники площади, занятой распростертыми вет-
вями, около 3 и 4 м) растет здесь на окраине обширной поляны с несо-
мкнутым покровом из степных злаков (Helictotrichon desertorum, Festuca
sulcata, Stipa sp., Phleum phleoides) полыней (Artemisia sericea, A. dracun-
culus, A. laciniata) и разнотравья (Pulsatilla flavescens, Gypsophyla pani-
culata, Dianthus versicolor, Filipendula hexapetala, Medicago falcata, Vicia
tenuifolia, Onosma simplicissimum, Galium verum, Salvia stepposa). Лес-
ные мезофильные растения отсутствуют совершенно, а лугово-лесные ксе-
ромезофилы представлены видами: Calamagrostis epigeios, Fragaria viridis,
Galium boreale и др. Характерны пятна лишайников (Cladonia mitis, С. cor-
nuta, C. pyxidata и др.), связанные с окраинами гранитных плит. Повсюду
разбросаны группы степных кустарников Cerasus fruticosa, Spiraea crenata,
а также отдельные экземпляры Cotoneaster melanocarpa, Genista tinctoria,
Cytisus ruthenicus. Среди поляны попадаются одиночные старые сосны,
сосновый подрост и группы молодой березы Betula verrucosa. Через узкую
полосу вейниково-осокового луга с группами Salix sibirica эта поляна гра-
ничит с молодым березняком, в составе которого встречаются старые сосны.
Такие степные поляны с одиночными соснами очень характерны для
Джабык-Карагайского и других степных островных каменистых боров
Зауралья (Брединские боры, отчасти Санарский); они, несомненно, обра-
22 Этот бор входит в систему островных каменистых боров Зауралья, протянувшихся
вдоль восточного склона Южного Урала через всю лесостепную зону Челябинской области
на юг до засушливых степей Оренбуржья и Восточного Казахстана (Соловьев, 1960; Ко-
лесников, 1961). По многим своим особенностям островные каменистые боры Зауралья
напоминают аналогичные боры Казахстанского мелкосопочника, описанные В. Н. Сукаче-
вым (1948) и Л. Н. Грибановым (1960).
190
зовались в результате разрушения лесными пожарами теперь редко встре-
чающихся сухих сосновых редколесий с покровом из лишайников, степ-
ных злаков, разнотравья и кустарников. Их бегло описал Ф. А. Соловьев
(1960) под названием мелкозлаковых сосновых редколесий.
Кроме этого местонахождения, где казацкий можжевельник, видимо,
случайно уцелел от лесного пожара, в Джабык-Карагайском бору, по дан-
ным сотрудников Аненского лесничества, имеется еще два экземпляра этого
кустарника — один в западной части бора, а другой — в южной, па за-
брошенном мусульманском кладбище в районе казахского пос. Акмолда.
Ветви этого можжевельника используются местным населением на венки
гщп похоронах, поэтому не исключена возможность, что в последнем место-
нахождении можжевельник культурного происхождения -:i.
Приведенные материалы позволяют высказать несколько соображений
об особенностях ареала казацкого можжевельника и его ботанико-геогра-
фических позициях на Южном Урале и в смежных районах.
Казацкий можжевельник следует относить к группе мезоксерофитов
горных местообитаний. Коренные его стации — участки открытого ланд-
шафта с выходами массивно-кристаллических пород. Значительно реже этот
вид встречается на песках (такие местообитания, по-видимому, являются
вторичными). Экология и история расселения казацкого можжевельника
•определили сильную раздробленность его ареала и четкую связь его от-
дельных участков с крупными горными странами или с древними остан-
цевыми возвышенностями на внутриконтинентальных равнинах. Южно-
уральский участок ареала казацкого можжевельника явно изолирован Ьт
•остальных смежных участков (приволжско-донского, казахстанского).
В западной части уральского фрагмента ареала местообитания этого кус-
старника связаны с выходами карбонатных пород, к востоку он переходит
на выходы кристаллических сланцев, гранитов и других горных пород,
не содержащих известь. Высоко в горы на Урале J. sabina не заходит, ха-
рактерен для ландшафтов широколиственных лесов западного склона Юж-
ного Урала, сосновых лесов восточного склона, а также для лесостепи и
степи нижнего горного пояса и предгорий. Казацкий можжевельник отно-
сится к числу древних доледниковых элементов флоры Южного Урала.
Обособление уральской части ареала от смежных частей и разобщение
местонахождений внутри уральского фрагмента ареала произошло, по-ви*
димому, в неогене и плейстоцене и усилилось в историческое время под
влиянием хозяйственной деятельности человека (лесные и степные пожары,
рубки). Это дает основание рассматривать местонахождения казацкого
можжевельника на Южном Урале как реликтовые. Они заслуживают
охраны в качестве памятников природы.
ОБЩИЙ ОБЗОР РЕЛИКТОВ
Реликтовые элементы во флоре Урала подразделяются нами в зависи-
мости от их возраста на три категории: а) доледниковые (плиоценовые),
б) плейстоценовые и в) голоценовые.
ДОЛЕДНИКОВЫЕ (ПЛИОЦЕНОВЫЕ) РЕЛИКТЫ
К этой категории относятся преимущественно реликты флоры долед-
никовых широколиственных лесов. С современными широколиственными
лесами Урала и Предуралья связаны фрагменты ареалов некоторых евро-
пейских неморальных видов.
23 Несколько хорошо развитых культурных экземпляров J. sabina растут в дендрарии
Боровского лесного техникума в пос. Бармашино Кокчетавской области.
191
Как показано в одной из предыдущих работ автора (Горчаковский,
1968), травянистые растения, встречающиеся на восточном фланге европей-
ских широколиственных лесов (на территории Урала и Предуралья) и тесно
связанные с этой группой формаций, по типу ареалов подразделяются на
следующие группы.
1. Эндемики Урала и Предуралья. Большая часть ареала находится
на Урале, отчасти вклинивается в смежные районы Восточно-Европей-
ской равнины (Lathyrus Litvinovii, Knautia tatarica, Cicerbita uralensis).'
2. Европейские. Ареал в основном в Центральной и Восточной Европе,
отчасти в Малой Азии и на Кавказе:
а) со сплошным распространением в Восточной Европе вплоть до Урала
(Festuca gigantea, Carex pilosa, Convallaria majalis, Aristolochia clematitis,
Mercurialis perennis, Pulmonaria officinalis ssp. obscura, Ajuga reptans,
Campanula tracheliumy,
б) дизъюнктивные, отсутствующие в ряде районов Русской равнины,
но вновь появляющиеся на Урале и в Предуралье {Cephalanthera longifolia,
Laser trilobum, Scutellaria altissima, Scrophularia Scopolii, Veronica maxima).
3. Евро-южносибирские дизъюнктивные. Ареал в основном в Европе,
иногда отчасти в Малой Азии, на Кавказе и реже в горах Средней Азии.
Изолированная часть ареала на Алтае, реже на юге Красноярского края:
а) со сплошным распространением в Восточной Европе вплоть до Урала
и дизъюнкцией на Западно-Сибирской равнине {Dryopteris filix-mas, Carex
silvatica, Actaea spicata, Asarum europaeum, Viola mirabilis, Circaea lute-
tiana, Stachys silvatica Bromus Benekeni);
б) с основным ареалом в Центральной Европе, дизъюнкциями на Вос-
точно-Европейской равнине и в Западной Сибири {Festuca silvatica, Gera-
nium Robertianum, Sanicula europaea, Digitalis grandiflora).
4. Евро-дальневосточные. Основной ареал разобщен на две части.
Одна, большая, находится в Центральной и Восточной Европе, другая :— в
Приморском крае, на Сахалине, в Китае (Маньчжурия) и Японии. Изоли-
рованные фрагменты ареала имеются в промежуточной области между
этими двумя частями ареала — на Алтае, иногда в некоторых горных райо-
нах Средней Азии и Восточной Сибири {Asperula odorata).
Реликтами в современной флоре Урала и Предуралья являются пред-
ставители 26 и За групп, имеющие здесь изолированные фрагменты ареалов
(карты распространения см. Горчаковский, 1968).
Рассмотрим вначале кратко несколько характерных представителей
группы растений 26 с основным ареалом в Центральной и Восточной Евро-
пе, имеющих более или менее изолированные фрагменты на Урале и в при-
легающей к нему части Русской равнины.
Интересным примером этой группы реликтов является Cephalanthera
longifolia, характерная для лиственных (главным образом широколиствен-
ных) лесов. Это растение распространено в горах Средней и Южной Евро-
пы; по Восточно-Европейской равнине заходит на восток Калининской,
Московской, Орловской и Тульской областей. Встречается, кроме того, на
Кавказе и в Иране; достоверных данных о нахождении этого вида в Сред-
ней Азии нет. Серия оторванных местонахождений отмечена на Урале
(окрестности Красноуфимска, близ Златоуста, между Златоустом и Уфой
и около пос. Катав-Ивановского).
Со светлыми широколиственными (главным образом дубовыми) лесами
связан шлемник высочайший Scutllaria altissima. Основной ареал его на-
ходится в южной и юго-восточной частях Европы, где этот вид местами
культивируется и встречается в одичалом состоянии. В СССР этот шлемик
распространен в Крыму, на Кавказе и в средней полосе европейской части
СССР, по которой, постепенно редея, доходит на восток до Волги. После
192
дизъюнкции он вновь появляется на Южном Урале, в Башкирской АССР,
где это растение найдено у с. Привольного в низовьях р. Нугуша, в бывшем
Стерлитамакском уезде в окрестностях дер. Кубячев, между деревнями
Пестровкой и Подлесной и в Мелеузовском районе на водораздельном
хребте, к западу от с. Воскресенского на правобережье р. Белой, а также
на правобережье р. Белой у с. Сибашево. Типичные места обитания шлем-
ника высочайшего в Башкирии — смешанные липово-ильмово-кленовые
леса.
В пределах своего основного ареала Scrophularia Scopolii растет в лесах
(преимущественно горных широколиственных), заходит также в субаль-
пийский пояс, где встречается на лугах с каменистой почвой. Распростра-
нена в Приатлантической и Средней Европе, в Карпатах, заходит в рав-
нинную часть Украины, встречается в Крыму, на Кавказе, в Иране и, по-
видимому, в западной части Гималаев. Редка в юго-восточной части Восточ-
но-Европейской равнины (Молдавия, Закарпатье, Крым). После дизъюнк-
ции вновь появляется в Приволжье (окрестности г. Саратова) и на Урале,
в юго-западной части Свердловской области в окрестностях городов Ниж-
ние Серги и Красноуфимска, а также в Башкирии (близ пос. Чандар). Здесь
норичник Скополи обитает в широколиственных лесах или в хвойных
лесах с примесью широколиственных древесных пород.
Veronica maxima произрастает преимущественно в горных широколист-
венных лесах на сухой каменистой почве на лугах, в ущельях, а также на
скалах и осыпях. Распространена в горах Европы от Пиренеев до Польши
и Балканского п-ова. В Альпах встречается на высоте 1650—1800 м над
ур. м., преимущественно на известняковой почве. В пределы СССР основ-
ной ареал вклинивается в закарпатской части Украины. Небольшой изо-
лированный фрагмент ареала находится на Урале. Здесь это растение про-
израстает на опушках и открытых известняковых склонах среди елово-
пихтовой тайги с липой в бассейнах рек Вишеры (район Полюдова Камня,
Камень Ветлан, устье р. Улса) и Яйвы. В горнохребтовой полосе встре-
чается на склонах гор Косьвинский, Конжаковский и Сухогорский Камни,
на горе Ослянка, близ поселков Кизела и Губахи, на облесенных известня-
ковых склонах по р. Чусовой у пос. Кына и близ устья этой реки, а также
по р. Вижаю около пос. Пашия. Ю. Шелль (1881—1885) приводил это ра-
стение для подножия горы Иремель на Южном Урале, однако гербарный
экземпляр, по-видимому, не сохранился, и это указание, не подтвержден-
ное последующими сборами, остается сомнительным.
А. Г. Борисова относила V. maxima с Урала к особой разновидности
var. uralensis, отличающейся несколько укороченными тычинками, почти
равными венчику, и яйцевидными, несколько островатыми на верхушке
семенами 1 мм длиной, 0,75 мм шириной. Насколько устойчивы эти при-
знаки — пока не выяснено.
Laser trilobum (Syn. Siler trilobuni) свойствен преимущественно широко-
лиственным лесам. Этот вид распространен в Приатлантической, Средней и
Южной Европе, на Балканах, в Малой Азии, Иране, Прикарпатье, Крыму,
на Кавказе; по средней полосе европейской части СССР он серией более
или менее разобщенных местонахождений доходит до Волги (Спрыгин,
1936), самые восточные местонахождения — на левобережье Волги в рай-
оне Самарской Луки. Относительно обособлена уральская часть ареала
лазурника трехлопастного в бассейне р. Камы и ее притоков Сылвы и Белой
(с притоком Уфой). Более обычен лазурник на западном склоне и пред-
горьях Южного Урала и на Бугульминско-Белебеевской возвышенности.
Заходит отчасти и на западный склон Среднего Урала с прилегающей к нему
равниной (в районе городов Красноуфимска, Кунгура, Оханска и с. Осы).
В Осинском районе Пермской области это растение местами образует до-
13 Заказ № 251
193
вольно густые заросли в ложках, где имеется подрост вяза, клена и ильма.
Заросли лазурника трехлопастного наблюдались нами в нижней части
гипсовых склонов по берегам р. Сылвы близ г. Кунгура. Нередко встре-
чается это растение по окраинам карстовых воронок.
Другая не менее интересная группа реликтов, связанных с широко-
лиственными лесами — растения с основным ареалом в Центральной
Европе и дизъюнкциями на Восточно-Европейской равнине и в Западной
Сибири.
Сюда относится, в частности, Festuca silvatica— обитатель тенистых
лиственных, преимущественно широколиственных лесов европейского
типа. Распространена в Европе за исключением северной части Скандина-
вии и Пиренейского п-ова. В европейской части СССР довольно обычна на
юге бореально-лесной зоны и в широколиственно-лесной зоне, заходит на
юге до Предкавказья. К востоку от Волги заметно редеет, но вновь стано-
вится обильной в Предуралье и на Урале. Самые северные местонахожде-
ния в Предуралье отмечены в лесах с липой близ с. Усть-Косы по р. Каме
и у подножия Полюдова Камня по р. Вишере в островных липняках с иль-
мом. В горах Урала заходит на север до гор Басеги и Колчимский Камень,
где встречается в елово-пихтовых лесах, нередко вместе с ильмом и липой.
На востоке распространена до горы Таганай и смежных гор, а самый южный
пункт встречаемости на Урале — бассейн р. Большой Сурени, притока
р. Большого Ика.
На территории Западно-Сибирской низменности овсяница лесная не
встречается. После дизъюнкции вновь появляется в горах Кузнецкого
Алатау и на Алтае, в районе обитания реликтовой липы Tilia sibirica.
Другой вид—Geranium Robertianum — произрастает в широколиствен-
ных и хвойных лесах с примесью широколиственных древесных пород,
а также на скалах в тенистых влажных местах. Широко распространен в
Приатлантической, Средней и Южной Европе, встречается в Средиземно-
морье и в Иране. В СССР растет как в горах, так и на равнинах. Заходит
также в горные районы Кавказа и Средней Азии (до Зеравшанского хребта
и северного Тянь-Шаня). По широколиственнолесной зоне и южной части
бореально-лесной зоны Восточно-Европейской равнины герань Роберта
доходит на востоке до Приволжья (Жигули, Самарская Лука, окрестности
г. Саратова). Отмечена в б. Бугурусланском уезде у дер. Сосновки. После
слабо выраженной дизъюнкции вновь появляется в Предуралье и на Урале.
В более северных районах (по рекам Вишере, Яйве и Чусовой) встречается
лишь на обнажениях известняка, поросших хвойным лесом с примесью
липы, иногда ильма, нередко вместе с другими травянистыми растениями
широколиственнолесного комплекса. Южнее, в Башкирской АССР, встре-
чается значительно чаще в каменистых дубняках, смешанных ильмово-
кленово-липовых лесах и на обнажениях известняка среди широколиствен-
ных лесов.
На территории Западно-Сибирской равнины герань Роберта отсутствует,
но встречается в значительном удалении от основного ареала в горах Куз-
нецкого Алатау и Алтая в черневых лесах с липой сибирской и некоторыми
другими реликтовыми растениями.
Очень интересным примером растений этой группы может служить
Sanicula еигораеа. Обитает в тенистых широколиственных лесах, а также
в. хвойных лесах с примесью широколиственных древесных пород. В Запад-
ной Европе подлесник европейский распространен почти повсюду за
исключением северной части Скандинавии и южной части Пиренейского
п-ова.
В пределах СССР этот вид произрастает на Кавказе и в полосе широко-
лиственных лесов западной части европейской территории, где, постепенно
194
редея к востоку, доходит до района г. Горького, северной части Тамбовской
области и окрестностей г. Малмыжа на юге Кировской области. После
дизъюнкции вновь появляется на Урале, где отмечен в окрестностях
пос. Сима в Челябинской области и с. Тастубы в Башкирской АССР, около
дер. Субботино близ г. Перми, около с. Поташки в Артинском районе Сверд-
ловской области, а нами найден (Горчаковский, 1956) в Саранинской лесной
даче Красноуфимского лесхоза. Разобщенные местонахождения в Башкир-
ской АССР, Пермской и Свердловской областях в совокупности образуют
уральский фрагмент ареала подлесника европейского, резко обособленный
от основного (европейского) ареала этого вида. Еще более удалены от ос-
новного ареала изолированные местонахождения в горах Кузнецкого Ала-
тау и на Алтае, где подлесник европейский встречается в черневых елово-
пихтовых лесах вместе с другими реликтами широколиственных лесов,
в том числе нередко с липой сибирской {Tilia sibirica).
Ю. Д. Клеопов пытался возвести уральские популяции S. еигораеа в
ранг особого вида S. uralensis, наметив его к описанию на этикетках гер-
бария Ботанического института АН СССР. Однако фактически уральские
особи подлесника не отличимы от растений из Кузнецкого Алатау или из
Западной Европы.
Еще один несколько обособленный вид — Digitalis grandiflora. Свя-
зан, в основном, с широколиственными и смешанными лесами, но встре-
чается также в зарослях кустарников, на лесных опушках, вырубках, за-
дернованных и каменистых склонах, реже на лугах. Распространен в Бель-
гии, Франции, на Пиренейском п-ове, в Италии, ФРГ и ГДР, Швейцарии,
Австрии, Румынии, в северной части Балканского п-ова. В европейской
части СССР идет на восток до западной части Харьковской области, Кур-
ской, Орловской и Тульской областей. Далее после разрыва ареала этот
вид вновь появляется в Приволжье, а также в Предуралье, на Среднем и
Южном Урале и в прилегающей части Западно-Сибирской равнины. На
Урале и в прилегающей равнинной территории наперстянка крупноцвет-
ковая проявляет себя как светолюбивое растение, произрастает в светлых
сосновых, березовых и смешанных лесах, иногда с примесью липы в древо-
стое и подлеске, а также на лесных опушках и лужайках.
Отсутствует на большей части Западно-Сибирской равнины к востоку
от р. Тобола. Далеко от основного ареала встречается на западных отро-
гах Алтая, например, близ курорта Белокуриха (Крылов, 1927—1964).
Другой заслуживающий внимания штрих в истории широколиствен-
ных лесов южной части Уральского хребта — их былая связь в плиоцене
с широколиственными формациями Дальнего Востока — раскрывается
в результате анализа своеобразного ареала Galium paradoxum. Это ра-
стение— типичный представитель мезофильной широколиственнолесной
флоры Юго-Восточной Азии. Морфолого-биологические черты его (тонкие,
нежные, широкие листья темно-зеленой окраски, длинные, тонкие корне-
вища, располагающиеся в лесной подстилке) свидетельствуют о приспособ-
ленности к существованию в условиях повышенной влажности воздуха
под пологом тенистых лесов. Произрастает G. paradoxum преимущественно
в смешанных хвойно-широколиственных лесах и в хвойных лесах с при-
месью неморальных травянистых растений.
Основной ареал расположен в Китае (Ганьсу, Сычуань, Хубей, Юнь-
нань, Сикан и северо-восточные провинции), в.Непале и Сиккиме (Гималаи),
на п-ове Корея, в Японии и в южной части советского Дальнего Востока
(Приморский край, некоторые районы Хабаровского края). Разобщенные
реликтовые местонахождения этого подмаренника известны в Сибири
(побережье оз. Байкал, район Телецкого озера на Алтае) и на Южном
Урале.
13*
195
Реликтовая природа местонахождения G. paradoxum в Сибири была от-
мечена впервые М. А. Мартыненко (1941), обнаружившим его на Алтае,
близ Телецкого озера (в 8—9 км от юго-восточной оконечности), на высоте
около 1000 м над ур. м., в разреженном пихтовом лесу с незначительной
примесью кедра. Необходимо отметить, что с пихтовой тайгой этого района
Алтая связаны другие третичные широколиственные реликты как европей-
ского (Asperula odorata, Asarum europaeum, Sanicula europaea), так и
дальневосточного происхождения (Qstnorhiza aristata).
На Урале было зарегистрировано только одно местонахождение Galium
paradoxum (Lipschitz, 1929). Это растение С. Ю. Липшиц нашел в 1927 г.
на горе Жукова Шишка (Челябинская область), где оно произрастало в
елово-пихтовом лесу по склону горы на скалах. Не будучи знаком с G. pa-
radoxum, он принял найденное им растение за новый вид и описал его под
названием G. Syreitschikovii. Однако при обработке рода Galium для
«Флоры СССР» (т. XXIII) Е. Г. Победимова установила, что растение,
собранное С. Ю. Липшицем на Южном Урале, является типичным G. pa-
radoxum и ничем не отличается от экземпляров из Сибири и Дальнего
Востока.
В 1959 г. автор этой книги обследовал Жукову Шишку, но не обнаружил
G. paradoxum, так как елово-пихтовый лес на ее склоне уже вырублен.
По-видимому, это растение здесь навсегда исчезло, остается только наде-
яться, что оно еще будет найдено где-нибудь в смежных местах Южного
Урала.
G. paradoxum как вид сформировался, очевидно, в широколиственных
и смешанных хвойно-широколиственных лесах Юго-Восточной Азии.
В плиоцене вместе с листопадными широколиственными лесами он рассе-
лился по Южной Сибири, достигнув Урала. Редукция широколиственных
лесов, происшедшая в плейстоцене, особенно в эпоху максимального оле-
денения, вызвала вымирание этого теплолюбивого растения на большей
части территории западного (урало-сибирского) крыла его ареала. Но под-
маренник парадоксальный смог сохраниться как реликт в некоторых
пунктах Урала и Сибири, где условия для него были более благоприятными.
Заслуживает внимания, что оба сибирских реликтовых местонахождения
G. paradoxum находятся в непосредственной близости от крупных озер
(Байкал, Телецкое озеро), оказывающих влияние на климат прилегающей
местности, в целом отличающийся континентальностью.
Все три оторванные местонахождения подмаренника парадоксального
(Южный Урал, Алтай, Прибайкалье) совпадают с центрами сохранения
третичных широколиственнолесных реликтов.
Находка G. paradoxum в реликтовом состоянии на Урале свидетель-
ствует, что в состав флоры широколиственных лесов, существовавших в
плиоцене на Южном Урале, входили элементы' не только европейского,
но и дальневосточного происхождения (из Юго-Восточной Азии).
Как видно, число реликтов, которые с достоверностью могут рассматри-
ваться как реликты доледниковых широколиственных лесов Урала и Пред-
уралья, сравнительно невелико. По всей вероятности, на Урале было
фактически значительно больше реликтовых неморальных видов, пережив-
ших эпоху максимального оледенения в местных широколиственнолесных
рефугиумах, с которыми тогда совпадали обособленные части ареалов. Од-
нако впоследствии, когда площадь широколиственных лесов стала увели-
чиваться, вместе с ними распространилось и большинство сопутствующих
им видов травянистых растений. Поскольку на равнине европейской части
СССР образовалась сплошная полоса широколиственных лесов (от Карпат
до Южного Урала), уральские участки'ареалов многих неморальных видов
полностью или почти полностью сомкнулись с основными их ареалами на
196
западе. Этим, видимо, и объясняется тот факт, что местонахождения неко-
торых неморальных видов (например, Bromus Benekeni, Campanula tra-
chelium, Circaea lutetiana и др.) на территории Восточно-Европейской рав-
нины заметно редеют к востоку; эти растения становятся крайне редкими
на участке между р. Волгой и Башкирией, а затем обильно появляются
в широколиственных лесах на западном склоне Южного и отчасти Среднего
Урала, а также на их предгорьях. По мере приближения к горам возра-
стают численность и концентрация местонахождений этих растений, а так-
же увеличивается их роль в сложении лесных растительных сообществ.
Есть основания предполагать, что такие виды, как Asarum europaeum,
Stachys siloaiica, Sanicula europaea, Geranium Roberlia/ium и др., сохранив-
шиеся в качестве доледниковых реликтов на предгорьях Алтая и Кузнец-
кого Алатау, смогли пережить ледниковую эпоху и на западном склоне
Южного Урала, где климатические условия в плейстоцене, вероятно, не
были суровее, чем в южной части Сибири.
Обособленность уральской части ареалов сохранилась как наследие
прошлого лишь у некоторых травянистых растений широколиственных
лесов, которые в силу тех или иных причин расселялись медленнее осталь-
ных. Уральские фрагменты ареалов этих видов так и не успели слиться с
основными их ареалами на западе.
Горные и предгорные районы Южного Урала, а также возвышенности
и сырты востока Русской равнины характеризуются более мощным и устой-
чивым снежным покровом по сравнению с равнинной территорией, лежа-
щей к западу, где снежный покров маломощен и неустойчив. Поэтому
условия для перезимования травянистых растений дубравного комплекса
здесь более благоприятны теперь и, вероятно, были более благоприятны
в прошлом, в ледниковые эпохи. Видимо, этим в какой-то степени объяс-
няется возникновение дизъюнкций в распространении некоторых широко-
лиственнолесных видов между Карпатами, Кавказом и Уралом.
Анализ восточного фланга ареалов древесных и травянистых растений
широколиственнолесного комплекса (Горчаковский, 1968) позволил уста-
новить области экологического оптимума, где условия для этих видов
более благоприятны и где они встречаются чаще и обильнее.
Области экологического оптимума европейской широколиственнолес-
ной флоры на восточном фланге ее ареала — основные: 1) предгорья и
увалисто-холмистая полоса западного склона Южного Урала, 2) Бугуль-
минско-Белебеевская возвышенность, 3) Общий Сырт; второстепенные:
4) бассейн рек Тавды и Туры и 5) район оз. Медвежьего в лесостепи То-
бол-Ишимского междуречья и 6) Мугоджары. В основных областях широ-
колиственные леса еще образуют сплоченный комплекс видов, тогда как
во второстепенных встречаются лишь отдельные компоненты, ведущие са-
мостоятельное существование.
Эти области экологического оптимума неморальной растительности и
флоры вместе с тем являются, по-видимому, центрами ее сохранения в те
эпохи плейстоцена и голоцена, когда климатические условия для широко-
лиственных лесов были неблагоприятными.
Вследствие иссушения климата и интенсивного хозяйственного воздей-
ствия человека Мугоджарский центр сохранения неморальной флоры в по-
следнее время оказался сильно обедненным, и растения этого комплекса
находятся на грани исчезновения. Однако, несомненно, что такие виды, как
Convallaria majalis, могли появиться на Мугоджарах лишь в условиях со-
ответствующей ценотической среды, то есть тогда, когда здесь существо-
вали широколиственные леса.
Обособленное положение среди доледниковых реликтов занимает мож-
жевельник казацкий Juniperus sabina, связанный не с широколиственными
197
лесами, а с открытыми каменистыми склонами. Распространение этого вида
уже рассмотрено.
ПЛЕЙСТОЦЕНОВЫЕ РЕЛИКТЫ
В течение сравнительно короткого исторического отрезка времени —
в четвертичном периоде — растительный покров Урала подвергся боль-
шим изменениям. Особенно значительные изменения произошли в плейсто-
цене. В начале плейстоцена в основном завершились мощные эпейрогенй-
ческие поднятия, начавшиеся на территории Урала еще в конце третичного
периода. В плейстоцене ряд горных районов Уральского хребта, особенно
в его северной части, а также равнины, прилегающие к хребту с севера,
неоднократно подвергались оледенению. Эти обстоятельства и далеко иду-
щие последствия их вызвали не только коренную перестройку раститель-
ности на территории Урала и прилегающих к нему равнин, но способство-
вали большей дифференциации растительности горных и равнинных обла-
стей. Поэтому плейстоценовые реликты в уральской флоре наиболее мно-
гочисленны. Они могут быть подразделены на ряд эколого-генетических
групп, а именно: 1) перигляциальные реликты арктической флоры;
2) перигляциальные реликты, проникшие из высокогорных районов Азии;
3) перигляциальные реликты горно-европейского происхождения;
4) скальные и горностепные реликты горноазиатского происхождения,
проникшие на Урал в конце плейстоцена и в начале голоцена; 5) ре-
ликты азиатского происхождения, свойственные светлым лесам (листвен-
ничным, сосновым, березовым) и лесным лужайкам; 6) реликты европей-
ского происхождения, связанные с приморскими верещатниками, сосно-
выми лесами и окраинами болот (вереск обыкновенный Calluna vulgaris,
уже рассмотренный в начале этой главы).
ПЕРИГЛЯЦИАЛЬНЫЕ РЕЛИКТЫ АРКТИЧЕСКОЙ ФЛОРЫ
К этой группе реликтов относятся растения, широко распространенные
в настоящее время в арктическом секторе Евразии или даже циркумполяр-
но, но встречающиеся изредка в реликтовом состоянии далеко за пределами
их основного ареала. Они входили в состав комплекса растений, произра-
ставших вблизи окраины ледникового покрова, а в период наибольшего
оледенения продвинулись довольно далеко на юг по Уральскому хребту
и его предгорьям. Такие растения в зависимости от положения современных
реликтовых мест их обитания в зонально-поясной системе растительного
покрова могут быть подразделены на две подгруппы: а) встречающиеся
на береговых обнажениях в предгорьях Урала и на прилегающих к нему
равнинах (в пределах таежной подзоны или горнотаежного пояса);
б) встречающиеся на гольцовых вершинах некоторых южноуральских гор.
Перигляциальные реликты арктической флоры на скалистых береговых
обнажениях
На береговых обнажениях известняка, гипса, а реже других пород в
значительном удалении от гор, поднимающихся своими вершинами выше
границы леса, местами произрастают растения, характерные для высоко-
горных и арктических областей.
К ним относится, в частности, Dryas punctata, свойственная в пределах
основного своего ареала арктическим и горным тундрам, а также скали-
стым обнажениям. Этот вид распространен в Арктике европейской части
СССР, Сибири и Дальнего Востока вплоть до Чукотки, а также на гольцах
Урала, Восточной Сибири и Северной Монголии. В европейской части СССР
это растение найдено также на известняках по р. Пинеге, в 3—8 км от ее
198
устья (Толмачев, 1938). На Урале D. punctata растет в высокогорной части
(Хус-Ойка, Муравьиный Камень, Кумба, Иремель и другие горы), как
реликт встречается в нижнем течении р. Чусовой на обнажениях известня-
ка|(Камень Боюн, или Плакун) или гипса (Сиролова гора), а также по
р. Каме на обнажениях гипса около пристани Полазны близ Перми.
На Чусовой и Каме это растение встречается на скалах, экспонированных
на север, в местах, где из расщелин сочится вода24.
Другой вид этого же рода — D. octopetala — близок к предыдущему
по экологии, распространен в арктической части Евразии (кроме крайнего
дальневосточного сектора), в Исландии и Гренландии, в высокогорьях
Западной Европы и Урала (изредка от северной оконечности хребта до
горы Иремель). Этот вид отмечен на гипсах по р. Пинеге, неподалеку от
бывшего Красногорского монастыря (Архангельская область). Реликтовую
природу имеют местонахождения дриады восьмилепестной на обнажениях
известняка в лесной зоне Предуралья по рекам Белой Кедве (Кирпични-
ков, 1947), Щугору, Илычу, Унье.
Дриады (£). octopetala, D. punctata) в плейстоцене следовали за ледни-
ками, когда те отступали с территории Европы, севера Русской равнины
и северной части Урала. Однако впоследствии, когда климат стал более
благоприятным для древесной растительности и широко распространились
леса, D. octopetala и D. punctata на значительной части их прежнего ареала
вымерли вследствие смены тундровых сообществ лесными. Лишь на скали-
стом субстрате и высоко в горах эти растения смогли сохраниться, причем
их местонахождения на прибрежных скалах и горных вершинах, удален-
ных от области сплошного распространения горных или равнинных тундр,
являются реликтовыми.
К этой же группе реликтов относится Saxifraga caespitosa, свойствен-
ная арктическим и горным тундрам. Эта камнеломка распространена цир-
кумполярно в Арктике, заходит также в высокогорную часть Урала, Вос-
точной Сибири и Северной Америки. По высокогорьям Урала ареал ее
вклинивается на юг до Конжаковского и Косьвинского Камней. Реликтовые
местонахождения камнеломки дернистой обнаружены вдали от высоких
гор, на отвесных известняковых утесах по рекам Большой Сыне, Щугору,
Илычу, Сосьве, Ивделю, Вишере (Камень Ветлан), Усьве, Косьве (у стан-
ции Губаха) и по Чусовой (Камень Поныш).
Характерным аркто-высокогорным растением является произрастаю-
щая в тундрах на сырых скалах и близ снеговых ручейков Pinguicula alpina.
Она распространена на горах Европы, в Европейской и Сибирской Арк-
тике, встречается на гольцовых вершинах Полярного, Приполярного и
Северного Урала (на юг от Чистопа), в Саянах, в горах Якутии и Гима-
лаях. Как реликт эта жирянка отмечена в лесной зоне Приуралья на из-
вестняках по р. Белой Кедве, в бассейне р. Ижмы, притока Печоры (Кир-
пичников, 1947), и на известняковых скалах по р. Большой Сыне, притоку
Усы.
К числу аркто-высокогорных растений принадлежит также Dianthus
repens, распространенная в Арктике, Евразии и западной части Северной
Америки25 и заходящая в высокогорную область Приполярного и Север-
24 Особенно обильно вода сочится из Камня Плакун, который именно по этому приз-
наку получил свое название. На р. Чусовой отвесные тенистые скалы, обращенные на
север, плохо прогреваются даже в жаркие летние дни. В глубоких расщелинах долго со-
храняется лед, поэтому местные рыбаки используют их как природный холодильник для
хранения рыбы.
25 Вопреки указанию Б. К. Шишкина (Флора СССР, т. VI), это растение в Гренлан-
дии не встречается (Bocher, Holmen, Jakobsen ,1957); нет его и на Канадском арктическом
архипелаге (Porsild, 1957).
199
него Урала (на юг до Конжаковского Камня), Забайкалья (бассейн р.Тун-
гира) и Дальнего Востока (горы Охотского побережья, Камчатка и другие
места). За пределами высокогорий на Урале эта гвоздика встречается на
обнажениях известняка по р. Вижаю, притоку Лозьвы.
Представляет интерес еще один реликт из этой группы — Potentilla
Kuznetzowii, распространенная в Скандинавии, на Новой Земле, в аркти-
ческой полосе европейской территории СССР и Сибири (западная часть),
в горных тундрах северной части Уральского хребта (от Полярного до Се-
верного). Как реликт встречается на известняковых скалах по рекам Ви-
жаю, притоку Лозьвы, по Косьве и Чусовой (на Камнях Балабан, Гардым
и ниже дер. Еквы на Камне Синем).
В эпоху максимального оледенения перечисленные здесь растения
вместе с другими компонентами в основном тундроподобного перигляци-
ального комплекса широко расселились по Уралу, но затем отступили
к горным вершинам и на север. Однако некоторые представители этого
комплекса в реликтовом состоянии сохранились на береговых известня-
ковых и гипсовых обнажениях, главным образом на склонах северной
экспозиции.
Интересно, что Dryas punctata, например, не встречается на скалах
в верховьях горных рек, в непосредственной близости от высоких горных
вершин. Отсутствует она и на скалистом субстрате в пределах Южного
Урала. Местонахождения дриады точечной на береговых обнажениях из-
вестняка и гипса располагаются в удаленных от хребта предгорьях (ни-
зовья Чусовой, р. Кама около Полазны). Приблизительно здесь же прохо-
дит южная граница распространения ряда других ледниковых реликтов.
Эти местонахождения примерно совпадают с границей наибольшего оле-
денения. Конечно, такое совпадение не случайно. Названные растения
с полным основанием можно считать реликтами тундроподобной расти-
тельности, характерной для окраины ледникового покрова в период его
наибольшего развития.
Наряду с перигляциальными реликтами в пределах тех же крупных
скалистых обнажений иногда встречаются и степные растения. Однако по
условиям обитания перигляциальные реликты резко обособлены от послед-
них. Они связаны с тенистыми, круто обрывающимися к северу склонами,
где из расщелин сочится вода; уступы таких склонов нередко порастают
елью. Степные растения, напротив, занимают сильно инсолируемые, обра-
щенные к югу склоны, которые совершенно безлесны или дают приют оди-
ночным низкорослым соснам.
Перигляциальные реликты арктической флоры на гольцах Южного Урала
В центральной, наиболее повышенной, час'ти Южного Урала в настоя-
щее время основу растительного покрова составляют леса. Лишь вершины
самых крупных гор (Таганай, Яман-Тау, Машак, Иремель, Зигальга,
Нары), поднимающиеся выше предела леса, покрыты растительностью
высокогорного типа. Здесь распространены горные луга, тундроподобные
• группировки лесных и болотных кустарничков (yaccinium myrtillus„
V. vitis-idaea, V. uliginosutn) и небольшие фрагменты горных тундр. Эти ми-
ниатюрные гольцы, затерявшиеся среди моря лесов, отделены от ближайших
гор, где хорошо выражен горнотундровый ландшафт (район Конжаков-
ского Камня на Северном Урале), расстоянием 550—600 км. Между голь-
цовыми вершинами Южного и Северного Урала располагается понижен-
ная, лесистая часть хребта — Средний Урал.
Поэтому большой интерес представляют изолированные местонахожде-
ния на гольцах Южного Урала некоторых арктических и арктовысокогор-
200
Рис. 107. Дриада восьмилепестная Dryas octopetala в тундре на горе Ире-
мель.
них растений, широко распространенных в северной части хребта. Эти ме-
стонахождения отделены от области более или менее сплошного" распро-
странения указанных видов на Уральском Севере ясной дизъюнкцией,
приходящейся на наиболее пониженную часть Уральского хребта — Сред-
ний Урал.
К числу таких видов относятся: Dryas octopetala (рис. 107), D. punctata
(Иремель, рис. 108), Lloydia serotina (Иремель), Arctous alpina (горы Машак,
рис. 109, и Таганай, рис. НО), Androsace Bungeana (Яман-Тау, Иремель),
Pedicularis Oederi (Иремель), Р. verticillata (Юрма, Таганай, Иремель,
Зигальга, Шатак), Salix reticulata (Иремель), Polygonum viviparum (Тага-
най, Уреньга, Иремель, Машак), Calamagrostis lapponica (Юрма), Diant-
hus repens (Зигальга) и некоторые другие.
Высокие горы Южного Урала (Таганай, Яман-Тау, Иремель и др.) на-
ходятся значительно южнее предположительной границы максимального
оледенения на равнинах. Поэтому арктические и аркто-высокогорные ра-
стения могли проникнуть сюда лишь с севера, по горному хребту. Вероят-
ное время этого проникновения — эпоха наибольшего плейстоценового
оледенения, когда на Южном Урале существовали местные леднички,
а верхний предел леса был значительно снижен. Водораздельная полоса
201
Рис. 108. Гора Иремель,
Рис. 109. Гора Машак.
Рис. 1 IQ.-Гора Тагадай.
вши
Среднего Урала в то время была, по-видимому, покрыта горнотундровой
растительностью26 и могла служить своеобразной трассой для расселения
к югу характерных высокогорных растений. Надо полагать, что период,
когда возможность такой миграции существовала, был кратковременным,
поэтому в гольцовую флору Южного Урала смогли внедриться лишь немно-
гие растения этой экологической группы.
ПЕРИГЛЯЦИАЛЬНЫЕ РЕЛИКТЫ, ПРОНИКШИЕ НА УРАЛ ИЗ ВЫСОКОГОРНЫХ
РАЙОНОВ АЗИИ
Во флоре высокогорной части Урала содержится своеобразная группа
реликтовых растений, основной ареал которых расположен в горах Азии
(Алтай, Саяны, горы Северной Монголии и Средней Азии). В высокогорьях
Рис. 111. Распространение первоцвета Палласа Primula Pallasii на Урале.
Урала эти растения встречаются изредка, причем уральские местонахож-
дения отделены от основного ареала значительной дизъюнкцией, прихо-
дящейся на территорию Западно-Сибирской низменности и Северного
Казахстана.
Примером может служить Primula Pallasii — высокогорное растение,
обитающее в горах на лужайках, близ верхней границы леса и выше ее
26 Не исключена возможность, что водораздел Среднего Урала или отдельные вершины
на нем достигали в то время несколько большей высоты, чем теперь; позднее их уровень
понизился вследствие эрозии.
205
у пятен тающего снега. Основной ареал этого вида располагается в Саянах,
на Алтае и в прилегающей к нему части среднеазиатских гор (Тарбагатай).
Изолированный фрагмент ареала расположен на Кавказе, в Иране и Ту-
рецкой Армении. Кроме того, изолир ованные местонахождения известны
на западном склоне Северного и Средн его Урала (горы Ишерим, Яны-Ёнки,
Лопьинский Камень, Куроксар, Чувал, Ослянка, Кваркуш), где это ра-
стение произрастает на лугах под-
гольцового пояса (рис. 111)’.
К этой же категории реликтов
принадлежит Phlojodicarpus villosus,
произрастающий в горах выше гра-
ницы леса (в горных тундрах и на
каменистых местах). Основной ареал
этого растения — Алтай, горы Сред-
ней и Восточной Сибири, Дальнего
Востока; заходит оно и в арктичес-
кую часть Восточной Сибири. Бли-
жайшее к Уралу местонахождение —
район г. Норильска, на северо-запад-
ной окраине Средне-Сибирского пло-
скогорья. Отсутствуя на территории
Западно-Сибирской низменности, взду-
топлодник мохнатый вновь появ-
ляется в высокогорьях северной части
Уральского хребта (рис. 112) на горе
Пас-Ньер в верховьях р. Няысь, в
верховьях р. Ляпина, на горе Ялпинг-
Ньер в верховьях р. Малой Сосьвы.
Характерные места обитания Оху-
graphis glacialis — горные тундры,
расщелины скал, околоснежные лу-
жайки и берега снеговых ручейков
(нередко вблизи края тающего снега).
Это высокогорное азиатское растение
распространено в горах Сибири, Сред-
ней Азии, Северной Монголии и, по-
Рис. 112. Распространение вздутоплодника видимому, в Гималаях, заходит также
мохнатого Phlojodicarpus villosus на в арктическую часть Сибири (Тай-
уРале' мыр, Северная Якутия, о. Врангеля).
На территории Западно-Сибирской
низменности оно отсутствует, но после перерыва вновь появляется на По-
лярном (перевал между реками Хуутой и Байдаратой, верховья рек
Щучьей, Нёмур-Югана, Лонгот-Югана, Латта-Ю и Харбея), на Приполяр-
ном (верховья р. Хулги, гора Селем-Ю-Из), а также на Северном Урале
(Денежкин Камень). В горах Урала (рис. 113) чаще всего встречается на
высоких седловинах и перевалах в пятнистых тундрах (непосредственно
на глинисто-щебнистых пятнах).
Rhodiola quadrifida обитает в высокогорных поясах, горных тундрах,
на скалах и осыпях около ледников. Основной ареал этого вида находится
на Алтае, в Саянах, в горах Забайкалья, южной Якутии и северной Мон-
голии. Не встречается в пределах Западно-Сибирской низменности. Изоли-
рованная часть ареала (рис. 114) располагается в высокогорьях Полярного
(гора Пай-Ер, верховья рек Холонг-Югана, Соби, Хууты, Войкара), При-
полярного (горы Народная, Манарага, Колокольня и др.) и Северного
Урала (Чистоп, Пахна, Денежкин, Конжаковский, Косьвинский и Семи-
206
Рис. 113. Распространение оксиграфиса ледникового Оху gra-
phic glacialis на Урале.
человечный Камни). На Урале
этот вид обычно произрастает
в сухих тундрах и на камен-
ных россыпях.
Основной ареал Swertia
obtusa (рис. 115) занимает
высокогорные пояса Алтая,
Кузнецкого Алатау, Саян,
Джунгарского Алатау, гор-
ных хребтов Северной Мон-
голии, Забайкалья, южной
Якутии и Дальнего Востока.
Здесь это растение выступает
преимущественно как высоко-
горное (растет на лугах, в
том числе и болотистых, по
берегам рек и ручьев), однако
внедряется и в нижележащий
пояс горных лесов. Кроме
того, этот вид встречается в‘
полярно-арктической области
Сибири (Бреховские о-ва в
низовьях Енисея). Уральские
местонахождения S. obtusa
(рис. 116) значительно отор-
ваны от основного ареала.
Они сосредоточены главным
образом в высокогорной части
Южного Урала (Яман-Тау,
Машак, Иремель, Зигальга,
Бакты (рис. 117), Аваляк,
Шатак, но по речным доли-
нам это растение спускается
в горнотаежный пояс и даже
в горную лесостепь восточ-
ного склона, где встречается
на болотистых лугах и боло-
тах. Интересно, что И. И. Ле-
пехин (1804) приводил свер-
цию тупую для района Кон-
жаковского Камня, но ни-
Рис. 114. Распространение родиолы четырехчленной кому ИЗ последующих ИС-
Rhodiola quadrifida на Урале. следователей найти здесь ЭТО
растение не удалось.
Thlaspi cochleariforme (рис. 118) — преимущественно высокогорный
вид; обитает обычно на лугах и на скалах выше границы леса, но встре-
чается, на каменистых склонах в нижележащих поясах. Ареал ярутки ло-
жечной охватывает Алтай, Саяны, Забайкалье, горы Якутии, горную
часть Таймыра, горы Средней Азии и северной части МНР. Изолированные
местонахождения (рис. 119) известны в северной части Уральского хребта
(преимущественно перидотитовые горы в бассейнах рек Соби, Войкара,
Сыни, Большой Харуты и Лопты), где это растение произрастает на камен-
ных россыпях выше границы леса, по сухим склонам древних ледниковых
морен и в лиственничных редколесьях, а также на Южном Урале (горы
Егозинская и Сугомак в окрестностях г. Кыштыма).
208
Рис. 115. Сверши тупая Sweriia obtusa.
Ясно выраженная дизъюнкция прослеживается и в распространении
Pedicularis compacta. Его сплошной ареал охватывает Алтай с прилегаю-
щими горами Средней Азии (Саур, Тарбагатай), Восточную Сибирь и Се-
верную Монголию. Произрастает этот мытник в высокогорных поясах
(в горных тундрах, на лужайках, скалах и каменных россыпях), а иногда
в прилегающей части горнолесного пояса. Изолированный участок ареала
расположен на Урале (от северной оконечности хребта до центральной
14 Заказ № 251
209
части Южного Урала —• рис. 120) и в прилегающей к нему северо-восточ-
ной части Русской равнины (Большеземельская и Малоземельская тунд-
ра). В уральской части ареала Р. compacta проявляет себя преимуществен-
но как высокогорный вид, произрастая на лугах и в тундрах выше границы
леса, но иногда заходит и в горнотаежный пояс.
Carex Ledebouriana свойственна высокогорным поясам, где обитает на
сырых лугах и в горных тундрах. Основной ареал ее расположен в горах
Рис. 116. Распространение сверции тупой Swertia obtusa
на Урале.
Алтая, Восточной Сибири, Дальнего Востока (до хребтов Анадырского
и Джугджур) и Северной Монголии. Значительной дизъюнкцией отделены
местонахождения осоки Ледебура в высокогорьях северной части Ураль-
ского хребта (рис. 121). Здесь она встречается на Полярном (Пай-Ер,
верховья рек Соби, Сыни, Войкара, Харбея), Приполярном (верховья рек
Хулги, Хобе-Ю, Маньи, гора Сале-Ур-Ойка) и Северном Урале (Денежкин
Камень, сборы автора, определенные Б. Н. Городковым).
Такая же закономерность наблюдается в распространении Carex saby-
nensis. Это преимущественно высокогорный вид, свойственный горным
тундрам, лугам, моховым болотам и берегам ручьев; в Восточной Сибири
и на Дальнем Востоке встречается и в лесном поясе. Ареал этой осоки —
Алтай, горы Восточной Сибири, Дальнего Востока, Северной Монголии
и Северо-Восточного Китая. Изолированный фрагмент ареала (рис. 122)
расположен на гольцовых вершинах Полярного, Приполярного и Север-
ного Урала (на юг до Косьвинского Камня). Значительно южнее собрана
Л. А. Соколовой на хр. Бакты. Кроме того, С. sabynensis найдена в приле-
гающей к Уралу части Западно-Сибирской низменности, близ г. Ханты-
Мансийска на р. Оби.
Сходные черты прослеживаются в ареале еще одной осоки — С. саиса-
sica. Это растение высокогорных лугов, распространенное в горах Средней
210
Рис. 117. Хребет Бакты.
14*
Рис. 118. Ярутка ложечная Thlaspi cochleariforme на вершине горы
Егозинской.
Азии, Кавказа, Ирана и Малой Азии. Изолированная часть ее ареала на-
ходится на Северном, Среднем и Южном Урале (от Чувальского Камня и
Куроксара на севере до Яман-Тау на юге), где этот вид обитает на высоко-
травных лугах и в разреженных лесах подгольцового пояса (рис. 123).
Местами на Южном Урале это растение встречается на более низких уров-
нях гор на лесостепных лугах (например, на хр. Крыкты).
К числу типично высокогорных растений относится Festuca Kryloviana,
свойственная Алтаю, Саянам, горам Средней Азии и Северной Монголии,
где она обычно произрастает в горных тундрах, на лугах и каменистых
212
Рис. 119. Распространение ярутки ложеч-
ной Thlaspi cochleariforme на Урале.
Рис. 120. Распространение мытника
компактного Pedicularis compacta на
Урале.
склонах выше границы леса. Обнаруженные нами (Горчаковский, 1954а)
реликтовые местонахождения этой овсяницы (определение подтверждено
В. В. Ревердатто) в горных тундрах и на каменных россыпях гольцов Юж-
ного Урала (Яман-Тау, Машак, Иремель, Зигальга — рис. 124, Шатак)
также отделены огромной дизъюнкцией от основного ареала (рис. 125).
Характерное растение высокогорных лугов Alopecurus glaucus распро-
странено на Алтае, хр. Тарбагай и в Саянах (Тункинские белки). Изоли-
рованная часть ареала расположена на Урале (рис. 126). Здесь это растение
Рис. 121. Распространение осоки Ледебура Carex Lede-
bouriana на Урале.
произрастает главным образом в подгольцовом поясе на влажных высоко-
травных лугах (горы Ялпинг-Ньер, Муравьиный Камень, Кваркуш, Чу-
вал, Яны-Енки, Ослянка, Харюзный, Растесский, Кырьинский Камни,
Таганай, Ильменский хребет, Иремель, Яман-Тау, Зигальга), но по скло-
нам Южного Урала местами спускается и в расположенные ниже пояса.
Кроме того, к этой же группе реликтов относится рассмотренная ранее
дазифора кустарниковая Dasiphora fruticosa.
Названные растения горноазиатского происхождения, вероятно, про-
никли на Урал в плейстоцене, особенно в эпоху наибольшего оледенения,
по перигляциальной полосе, имевшей тундроподобный облик. Позднее они
были полностью или частично оттеснены видами, входившими в другие
растительные группировки из промежуточных местонахождений на тер-
ритории Западно-Сибирской низменности, но остались в качестве реликтов
в высокогорных поясах Урала, где до сих пор сохранились условия среды
и фитоценозы, соответствующие их экологическим особенностям.
Перечисленные выше растения в основной части своего ареала, нахо-
дящегося далеко за пределами Урала, характеризуются, как правило, до-
вольно большим размахом приспособляемости к условиям среды; некото-
рые из них встречаются не только в высокогорных, но и в нижерасполо-
женных поясах растительности, в разнообразных растительных сообщест-
214
Рис. 122. Распространение осоки шабинской Carex saby- Рис. 123. Распространение осоки
nensis на Урале. кавказской Carex caucasica на
Урале.
Рис- 124. Хребет Зигальга.
вах. На территории Урала имеются лишь небольшие фрагменты их ареалов.
Здесь экологическая амплитуда этих растений значительно уже, они тес-
нее связаны с растительными сообществами высокогорий и обычно не выхо-
дят за пределы горнотундрового и подгольцового поясов.
Такая закономерность объясняется, по-видимому, тем, что в плейсто-
цене из горных районов Азии на Урал смогли проникнуть лишь те немно-
гие популяции названных видов, которые были наиболее приспособлены
Рис. 125. Распространение овсяницы Крылова Festuca
Kryloviana на Южном Урале.
к суровым условиям эпохи максимального оледенения, в то время как нд
родине сохранился более разнообразный набор их экологических форм,
ПЕРИГЛЯЦИАЛЬНЫЕ РЕЛИКТЫ ГОРНОЕВРОПЕИСКОГО ПРОИСХОЖДЕНИЯ
Реликтовые элементы западного, горноевропейского происхождения;
на Урале немногочисленны. К ним относится Alchemilla glabra — европей-
ский вид, По происхождению альпийский. Встречается названная манжет-,
ка в Средней и Приатлантической Европе (ГДР, ФРГ, Франция, Швей-,
цария, Англия), а также на Аландских о-вах, в Финляндии и на Карель-
ском перешейке. После значительного разрыва в ареале этот вид появля-
ется на берегах Печоры (в 25 км выше дер. «У камешка») и в горах Припо-
лярного и Северного Урала (хр. Сабля, гора Койп, Конжаковский Камень),
Надо полагать, что A. glabra вместе с некоторыми другими альпийцами,
происходящими из гор Европы (например, Bartsia alpina) расселилась,
в плейстоцене на восток по перигляциальной полосе Восточно-Европей-
ской равнины вплоть до северной части Уральского хребта. В послелед-.
никовое время, главным образом в связи с расселением лесов на террито-
рии, ранее занятой перигляциальной растительностью, это растение вымер-
ло на Восточно-Европейской равнине, но сохранилось на положении ре-
ликта на Урале (рис. 127), преимущественно в высокогорных поясах.
217
Рис. 126. Распространение лисохвоста Рис. 127. Распространение манжетки голой Alche-
сизого Alopecuras^ glaucus на Урале. milla glabra на Урале.
Рис. 128. Степь на южных отрогах Уральских гор близ пос. Кувандыка.
Рис. 129. Горноколосник колючий Orostachys spinosa на хр. Ирендык.
Хотя участие европейско-альпийского элемента в высокогорной флоре
Урала невелико, сам факт флористических связей Урала с Европейскими
Альпами представляет интерес и заслуживает внимания.
скальные и горностепные реликты горноазиатского
ПРОИСХОЖДЕНИЯ, ПРОНИКШИЕ НА УРАЛ В КОНЦЕ ПЛЕЙСТОЦЕНА
И НАЧАЛЕ ГОЛОЦЕНА
К этой группе реликтов относятся растения, распространенные в горах
Азии (Алтай, горы Средней и Восточной Сибири, Монголия, горные районы
Средней Азии) на каменистом субстрате, чаще в поясе горных и предгор-
ных степей, а также на каменистых обнажениях в выше расположенных
поясах. Отсутствуя на территории Западно-Сибирской низменности, они
вновь появляются в южной части Уральского хребта (рис. 128). В свое
время на эту группу реликтов обратил внимание И. М. Крашенинников
(1937). Мы значительно дополняем список реликтов, относящихся к этой
группе, уточняем экологию и географическое распространение отдельных
видов.
Хорошим примером таких реликтов может служить Orostachys spinosa
(рис. 129). Этот суккулент характерен преимущественно для горных каме-
нистых степей, каменистых и щебнистых склонов, встречается в равнинных
степях на солонцеватых и песчанцсшх почвах, а также по окраинам раз-
реженных степных боров. Ареал этогомзида охватывает Восточную Сибирь,
22Q
южную часть советского Дальнего Востока, Северо-Восточный Китай, се-
верную часть п-ова - Корея, Японию, Монголию, западный Тибет, горы
Средней Азии, Алтай и вклинивается на территорию Западно-Сибирской
низменности и Казахского мелкосопочника (самые западные местонахожде-
ния в районе г. Кокчетава). В отрыве от основного ареала горноколосник
колючий встречается на Южном Урале и в прилегающей части Зауралья
(рис. 130) в пределах Башкирской АССР, Челябинской и Оренбургской
Рис. 130. Распространение горноколосника колючего Orostachys spinosa
на Урале.
областей (в каменистых степях на хребтах Нурали, Ирендык, в Губерлин-
ских горах, в районе пос. Халилово в бассейне верхнего течения р. Сак-
мары, в бассейне р. Большого Ика, на горе Балкан-Тау на восточной окраи-
не Белебеевской возвышенности, на окраине Джабык-Карагайского бора,
в окрестностях г. Троицка,' на скалистых обнажениях по р. Миассу близ
г. Челябинска).
Сходный разрыв ареала прослеживается у Linaria altaica, обитающей
на степных каменистых склонах и скалах, реже в равнинных степях. Ос-
новной ареал этого растения — Алтай и его предгорья. В области Казах-
ского мелкосопочника' известно местонахождение льнянки алтайской в
районе Улутау. После перерыва она появляется вновь на Южном Урале
(в Орском районе Оренбургской области^надр. Ирендык, в Губерлинских
горах, в бассейне р. Сакмары и ее прщ^’1а'"'Большого Ика), на южных от-
221
рогах Общего Сырта близ с. Кииделя и в Мугоджарских горах27
(рис. 131).
Такая же закономерность наблюдается в распространении Linaria debi-
lis. Это растение, свойственное каменистым степям Алтая (Чуйская степь),
отсутствует на территории Западно-Сибирской низменности и Северного
Казахстана, но вновь появляется на Южном Урале (рис. 132) на горе Его-
зинской, хр. Нурали и Крыкты, Губерлинских горах и в других местах.
Подобная же дизъюнкция характерна для ареала Phlox sibirica (рис. 133).
Характерные его местообитания — каменистые степи и сухие лужайки,
главным образом в поясах горной лесостепи и степи, иногда и в горнолес-
Рис. 131. Распространение льнянки алтайской unaria altaica на
Урале.
ном поясе. Основной ареал его — Алтай (Чуйская степь), южная
часть Средней и Восточной Сибири; отдельные местонахождения известны
в более северных районах Восточной Сибири (центральная Якутия, бас-
сейн Вилюя и дальше на север, почти до границы тундры) и на Аляске.
Обособленная часть ареала (рис. 134) расположена на Урале. Этот вид
встречается главным образом в лесостепи Южного Урала и Предуралья
(на хр. Ирендык, в верховьях р. Урала, в Месягутовской лесостепи,
рис. 135, на Бугульминско-Белебеевской возвышенности). Обычно он вхо-
дит в состав ассоциаций каменистой степи. На Ирендыке и в Предуралье,
в районе оз. Асли-Куль, наблюдается сопряжённость местонахождений
флокса сибирского с местами единичной встречаемости лиственницы. Най-
ден также на Борзовских горах близ г. Кыштыма. Нами это растение было
обнаружено значительно севернее основного района концентрации его ме-
стонахождений: на Среднем Урале, на известняковой скале Белый Камень
по р. Режу близ г. Режа, а также в южной части Северного Урала на из-
вестняковых обнажениях по р. Северной Тошемке, притоку Лозьвы, где
он растет в верхней части скалы на открытом южном склоне в окружении
лиственниц. Последняя находка неожиданна и очень интересна.
27 М. И. Котов (1946) предлагал уральские популяции этого растения возвести в ранг
особого вида — L. uralensis. По мнению Л. А. Куприяновой (личное сообщение), этого
делать не следует, так как они практически не отличимы от казахстанских и алтайских.
Описывая этот вид, М. И. Котов, по-видимому, не имел возможности сравнить уральские-
растения с алтайскими.
222
Sedum hybridum — скально-горностепной азиатский вид. Распрост-
ранен на юге Сибири (Алтай, горы Средней и Восточной Сибири и Забай-
калья), в горах Средней Азии (Тянь-Шань), в Туве и северной части Мон-
голии. Западная граница основного ареала достигает г. Томска, с. Локоть
Рис. 132. Распространение льнянки слабой Linaria
debilis на Урале.
на р. Алее. Изолированные местонахождения известны в Омской области
на холмах Бестюбе под 60° с. ш., в’Целинном крае (окрестности г. Цели-
нограда), в некоторых других районах Казахского мелкосопочника (Се-
веро-Казахстанская и Кокчетавская области). Обособленная часть ареала
находится на Южном Урале и в Приуралье (рис. 136). Здесь очиток гиб-
ридный растет на обнажениях известняка, кварцита, песчаника, на щеб-
нистых склонах гор и в каменистых степях. Основные его местонахождения
сосредоточены в бассейнах рек Урала, Сакмары, Большого Ика, Белой,
Нугуша, Юрюзани, Миасса, в степях пенеплена восточного склона Южного
Урала — в бассейнах рек Тобола, Уя, Тогузака. На север этот вид идет
223
Рис. 133. Флокс сибирский Phlox sibirica в каменистой степи на вершине хр. Ирендык.
Рис. 134. Распространение флокса сибирского Phlox sibirica на Урале.
Рис. 135. Каменистая степь на склонах к р. Юрюзани.
до г. Челябинска, на юг до Губерлинских гор. Встречается также в Мугод-
жарах.
Поразительная дизъюнкция прослеживается в распространении Allium
himenorrhizum. Лук плевокорневищный произрастает в горах — на лужай-
ках, на травянистых болотах и на сырых солонцеватых лугах. Основной
ареал его — Алтай, Тарбагатай, Джунгарский Алатау, Заилийский и
Рис. 136. Распространение очитка гибридного Sedum hybridum на Урале.
Кунгей Алатау, Тянь-Шань, Кашгария, Иран. В значительном удалении
от основного ареала этот лук найден на Южном Урале (рис. 137) — на
хр. Ирендык в долине р. Таналыка и в верховьях р. Большого Сапсала,
на лугу.
Artemisia santolinifolia произрастает в каменистых горных степях и
на щебнистых склонах. Основная область ее распространения охватывает
горную и предгорную степь и лесостепь Алтая, Саян, Памиро-Алая, Тань-
Шаня, Тарбагатая, Джунгарии, Забайкалья и Монголии. Этот вид от-
сутствует на территории Сибири и Казахстана: После дизъюнкции, он по-
15*
227
является на Южном Урале (гора Тра-Тау в окрестностях г. Стерлитамака,
бассейн р. Урюка, известняковые скалы по р. Сиказе у хутора Кук-Караук,
каменистые степи в бассейне р. Сакмары и ее притока р. Большой Ик, из-
вестняковый утес Соколья гора на р. Уфе близ г. Нязепетровска, рис. 138).
Другой вид полыни Artemisia bargusinensis характерен для горных
степей и каменистых, чаще остепненных склонов. Основной ареал распо-'
ложен на Алтае (Сайлюгемская степь, долина рек Чулышмана, Сайгомы-
ша, Чарыша, Карлыка), на предгорьях Саян, в Забайкалье и Якутии. На
территории Западно-Сибирской низменности этот вид отсутствует. Изоли-
рованная часть ареала находится на Среднем и Южном Урале (Александ-
ровские сопки в Красноуфимской лесостепи, окрестности оз. Чебаркуль
в Челябинской области и озера Асли-Куль в Башкирской АССР, рис. 139).
Интересный представитель этой группы Patrinia sibirica (рис. 140),
произрастающая на скалистых обнажениях, 'щебнистых и каменистых
склонах гор в различных поясах, начиная от горностепного и кончая гор-
нотундровым. Распространена патриния сибирская на Алтае, в Средней
и Восточной Сибири, в Забайкалье, Приморье, Северной Монголии, Северо-
Восточном Китае и Японии. Обособленный участок ее ареала расположен
на Южном Урале (рис. 141). Здесь это растение встречается на каменистом
субстрате в лесостепных предгорьях восточого склона (окрестности
с. Балбука, с. Ахуново в Карагайском лесничестве), на низких уровнях
гор (хребты Ильменский, Нурали, Малый Ирендык), а также выше гра-
ницы леса на скалистых останцах и в каменистой горной тундре (горы Ире-
мель, Зигальга, Машак, Уреньга, Шатак, Ирендык, Крыкты и др.).
Внимание ряда исследователей уже привлекло распространение Вир-
leurum multinerve. Этот вид произрастает на каменистых, чаще степных
склонах, на сухих степных лугах, реже по опушкам лесов; иногда в горах
встречается выше границы леса на скалистых обнажениях. Основной ареал
володушки многожильчатой — Алтай с примыкающей к нему частью
Западно-Сибирской низменности, хр. Саур, южная часть Средней и Восточ-
228
ной Сибири, Забайкалье, Монголия. Изолированный фрагмент ареала
находится на Южном, Среднем и отчасти Северном Урале (рис. 142). В пре-
делах Урала В. multinerve произрастает в каменистых горных и предгор-
ных степях (на территории Месягутовской и Красноуфимской лесостепи.
Рис. 138. Распространение полыни сантолинолистной
Artemisia santolinifolia на Урале.
на горах Егозинской и Сугомак, на хребтах Уреньга, Нурали, Крыкты,
Ирендык), а севернее — на известняковых обнажениях вдоль рек Чусо-
вой, Усьвы и Вишеры. Кроме того, встречается близ верхней границы леса
на обнажениях дунита, которые обычно дают приют более ксерофильным
растениям (Желтая сопка на Денежкином Камне, водосборная воронка
в верховьях р. Южного Иова на Конжаковском Камне — рис. 143, «плечо»
Косьвинского Камня). К западу от Урала это же растение встречается как
реликт на Средне-Русской возвышенности, в Воронежской области.
229
к числу горностепных реликтов относится и Vieta multicaulis, расту-
щая обычно на открытых каменистых склонах, реже на степных лугах.
Распространена на Алтае, в Средней и Восточной Сибири, на Дальнем
Востоке, в Северной Монголии и Северо-Восточном Китае, заходит в Арк-
тику по р. Анадырю. На территории Западно-Сибирской низменности,
кроме прилегающей к Алтаю и Салаиру части, отсутствует. Далеко ото-
рванная часть ареала (рис. 144) находится на Урале (скалы по рекам Вишере,
Рис. 139. Распространение полыни баргузинской Artemisia bargusinensis на
Урале.
Туре, Колве, Чусовой, Сылве, Красноуфимская лесостепь, горная лесо-
степь на хр. Крыкты).
Chamaerhodos erecta произрастает на каменистых горных склонах и
обнажениях, щебнисто-глинистых осыпях, прибрежных галечниках, ред-
ко на ровных степных местах и по окраинам сосновых боров на песчаной
почве, преимущественно в пределах степной зоны и лесостепи, а реже в при-
легающей части лесной зоны. Основной ареал этого растения расположен
в Монголии, Китае, северной части п-ова Корея, в южной части советского
Дальнего Востока, в Восточной Сибири, южной части Средней Сибири,
на Алтае и в прилегающих к нему районах Западно-Сибирской низмен-
ности. Западная граница более или менее сплошного его распространения
от окрестностей Томска идет к Салаирскому кряжу, к с. Локоть на Оби
и оттуда к западным предгорьям Алтая. Несколько оторванных местона-
хождений этого растения известно в области Казахского мелкосопочника
й в равнинных степях на территории Западно-Сибирской низменности
(горы Джаван-Тюбе близ устья р. Селеты, Кокчетавские горы в окрест-
ностях оз. Борового, близ дер. Ярки на р. Ишиме). После перерыва
Ch. erecta вновь появляется на Урале (рис. 145). В пределах Среднего Урала
он отмечен в Красноуфимском районе Свердловской области на остепненном
230
Рис. 140. Патриния сибирская Patrinia sibirica.
Рис. 141. Распространение патринии сибирской Patrinia sibirica
на Урале.
Рис. 142. Распространение володушки многожиль-
чатой Bupleurum multinerve на Урале.
Рис. 143. Обнажение дунита в верхней части водосборной воронки в истоках р. Южного Иова на Конжаковском Камне.
Рис. 144. Распространение горошка
многостебельного Vida multicaulis на
Урале.
склоне увала у с. Кадочникове и на
восточном склоне хребта по скалистым
берегам рек Режа (Глинский Камень у
с. Глинского), Исети (у сел. Смолино
и Щербаковского) и ее притока Синары
(устье р. Караболки). На Южном Урале
этот вид найден в Троицком районе
Челябинской области по р. Ую (близ
дер. Осиповки), около пос. Кизильского
в устье р. Большого Кизила, притока
р. Урала, и в Оренбургской области
в степи у станицы Березовской.
Potentilla sericea — обитатель каме-
нистых горных степей, каменистых
склонов и обнажений скал. Основной
ареал охватывает Алтай, Саяны, гор-
ные районы Забайкалья, Монголии
и Средней Азии (Джунгарский Ала-
тау, Тарбагатай, Заилийский и Кунгей
Алатау). Ясно выраженной дизъюнк-
цией отделен фрагмент ареала этого
вида на Южном Урале (рис. 146) — на
Борзовских горах близ г. Кыштыма,
в Ильменских горах, в верховьях рек
Уя, Урала и Белой. Между Уралом
и Алтаем известно лишь несколько изо-
лированных местонахождений лапчатки
шелковистой: близ пос. Тюкалинска на
р. Иртыше, г. Тобольска и в Северо-
Казахстанской области неподалеку от
пос. Бишкульского.
Silene altaica распространена, в ос-
новном, на Алтае и его предгорьях,
в Саянах, в Джунгарском Алатау и на
хр. Тарбагатай, заходит в Китай, из-
редка встречается в некоторых райо-
нах Казахского мелкосопочника и
Прибалхашья (северо-восточная часть
Семипалатинской области, Семиречен-
ская область, южная часть Целин-
ного края). Это растение произрастает
на каменистых и щебнистых склонах
сопок, обнажениях скал, на степных
и солонцеватых лугах в степной и
пустынной зонах и аналогичных им
высотных поясах.
На Южном Урале смолевка алтай-
ская встречается на обнажениях из-
вестняков, песчаников, конгломератов
и в каменистых степях в районе
г. Орска, в Губерлинских горах и на хр. Ирендык, в бассейне рек Сакмары
и Большого Ика. Кроме того, встречается в Мугоджарах. От основной об-
ласти распространения южноуральско-мугоджарский фрагмент ареала
(рис. 147) отделен дизъюнкцией, приходящейся на Тургайскую впадину
и прилегающую часть Западно-Сибирской низменности.
234
Рис. 145. Распространение хамеродоса прямого Chamaerho- Рис. 146. Распространение лапчатки шелковистой Potentil-
dos erecta на Урале. . sericea на Урале.
Подобно предыдущим видам, Alyssum biovulatum обитает в горных сте-
пях, по обнажениям скал и каменистым склонам, изредка встречается
в равнинных степях на песчаных почвах и по окраинам южных сосновых
боров. Основная область его распространения — Алтай с прилегающими
к нему районами Западно-Сибирской низменности и Средней Азии, горы
Рис. 147. Распространение смолевки алтайской Sile-
ne altaica на Урале.
Восточной Сибири (вплоть до Крайнего Севера) и Дальнего Востока, Север-
ная Монголия. К западу от основного ареала отмечено несколько изо-
лированных местонахождений этого вида в области Казахского мелко-
сопочника (Кокчетавские горы) и на песчаной почве в окрестностях горо-
дов Омска, Тюмени и Ишима. На Урале (рис. 148) A. biovulatum снова
становится обильным, здесь он встречается в каменистых степях на горах
Егозинской, Сугомак, Вишневых, а также на обнажениях известняка по
рекам Вишере, Чусовой, Ивделю и Режу. В печорском Приуралье он до-
стигает полярного круга (известняковые обнажения по рекам Большой
Сыне, Адзьве, Усе). Найден также на Полярном Урале (верховья рек Соби,
Сыни, Войкара, Малой Харуты, гора Рай-Из), где обитает на обнажениях
оливиновых горных пород и на моренах древних ледников.
236
Oxytropis uralensis s. I.28
в Сибири является обычным
лесостепным растением, про-
израстающим на участках
северных луговых степей, на
сухих лугах, каменистых
склонах и остепненных сос-
новых борах. Распространен
этот вид от юго-востока Запад-
но-Сибирской низменности
(от района Томска) и пред-
горий Алтая на восток по
островным степям Средней и
Восточной Сибири вплоть до
Забайкалья, где постепенно
редеет. Глубоко в таежную
зону он не проникает, но в
Якутии по степным островкам
и по примыкающим к ним
участкам светлых листвен-
ничных лесов заходит на
север до Якутска и устья
Вилюя. На большей части
Западно-Сибирской низмен-
ности это растение не встре-
чается, но после перерыва
вновь появляется на Урале
(рис. 149), где произрастает
в южных районах в степях
и по окраинам остепненных
лесов (Ильменские, Вишневые
горы, шиханы в районе
г. Стерлитамака, степи Баш-
кирии и Оренбургской об-
ласти), а севернее — на обна-
жениях гипса по р, Сылве и
известняковых береговых об-
нажениях (например, по ре-
кам Ваграну, Лозьве, Ив-
делю, Тошемке и др.). Кроме
того, изолированное место-
нахождение этого остролодоч-
ника отмечено на территории
Русской равнины по р. Сухо-
не (на обнажениях извест-
няка).
Еще одно интересное ра-
стение — Gentiana decumbens.
Это обитатель каменистых и
щебнистых склонов, степных,
иногда солонцеватых лугов.
Рис. 148. Распространение бурачка двусемянного
Alyssum biovulatum на Урале.
28 Мы понимаем здесь таксономию и ареал О. uralensis так, как это вытекает из ре-
зультатов исследований Б. А. Юрцева (1959). В обработке рода Oxytropis для «Флоры СССР»
А. Г. Борисова относила к О. uralensis только растения с Урала.
237
Рис. 149. Распространение остролодочника ураль-
ского Oxytropis uralensis на Урале.
Названная горечавка распро-
странена в степном и горно-
лесном поясе, но иногда по
каменистым местам прони-
кает и выше границы леса.
Основной ее ареал охватывает
Алтай и прилегающую к нему
часть среднеазиатских гор,
Саяны, южную часть Средне-
Сибирского плоскогорья, За-
байкалье и некоторые районы
Северной Монголии. На тер-
ритории Западно-Сибирской
низменности это растение не
встречается. После перерыва
вновь появляется на Южном
Урале (на восточном склоне
хр. Ирендык в долине р. Джу-
валы-Узяк, между хр. Ирен-
дык и р. Сакмарой и в верхо-
вьях р. Урала), где произра-
стает на солончаковых лугах
и на солончаках (рис. 150).
Как видно, изолирован-
ные части ареалов названных
растений расположены в боль-
шинстве случаев на Южном
Урале, отдельные же местона-
хождения их известны и
севернее (Средний Урал, юж-
ная окраина Северного Ура-
ла). Если на Южном Урале
такие растения произрастают
главным образом в горных и
предгорных каменистых сте-
пях, то севернее они встре-
чаются лишь на обнажениях
известняка и дунита. Инте-
ресно,- что некоторые из пред-
ставителей группы скально-
горностепных реликтов встре-
чаются и выше границы леса
на каменистом субстрате. Все
реликты этой группы рас-
пространены на Урале пре-
рывисто и встречаются редко.
Наиболее вероятное время проникновения скальных и горностепных
растений азиатского' происхождения на Урал — плейстоцен (после от-
ступления днепровского оледенения) и начало голоцена. Активное рас-
селение их по равнине возможно лишь при наличии незадернованного
субстрата. Благоприятная обстановка для расселения таких растений на
запад, с предгорий Алтая на Урал, создалась сразу же после отступления
ледников, когда на равнинах на значительной площади еще были распро-
странены флювиогляциальные и речные наносы, не заселенные или слабо
заселенные растениями. По песчаным, галечниковым и другим наносам,
238
а также по обнажениям коренных горных пород эти растения интенсивно
расселялись на запад. Расселение их в этом направлении продолжалось
и в начале голоцена, пока сплошное облесение отсутствовало.
Основной путь расселения горноазиатских растений проходил через
юг Западно-Сибирской низменности29 и Северный Казахстан, о чем свиде-
тельствуют изолированные местонахождения некоторых из них в промежу-
точных пунктах (Казахский мелкосопочник). Достигнув Урала, эти имми-
гранты нашли для себя в горах благоприятную обстановку, так как после
Рис. 150. Распространение горечавки стелющейся
Gentiana decumbens на Урале.
отступления ледникового покрова здесь на огромной площади были рас-
пространены обнажения горных пород и продуктов их разрушения, а так-
же различные ледниковые и речные наносы, еще лишенные сомкнутого
растительного покрова. Расселяясь по Уралу, некоторые виды проникали
в высокогорные пояса, так как сплошной облесенности горных склонов
еще не было. Позднее, в связи с развитием лесов на горных склонах, рас-
пространение этих растений на Урале сильно сократилось, и они сохрани-
лись почти, исключительно на каменистых местах. Облесение песчаных на-
носов сосной и другими древесными породами вызвало оттеснение скально-
горностепных растений из большинства промежуточных пунктов на тер-
ритории Западно-Сибирской низменности, привело к обособлению ураль-
ской части их ареалов от основного ареала в горных районах Азии.
29 Своеобразной трассой для такого расселения мог служить, в частности, бечевник
вдоль русел Оби и ее многочисленных притоков. В отдельных случаях расселению мог
благоприятствовать перенос зачатков растений речными водами.
239
Рис. 151. Распространение горечавки борода-
той Gentiana barbata на Урале.
Возможно, что вторичная акти-
визация этих скальных и горно-
степных азиатских растений про-
изошла в одну из последующих
фаз голоцена, характеризующуюся
более сухим и теплым климатом.
РЕЛИКТЫ АЗИАТСКОГО
ПРОИСХОЖДЕНИЯ,
СВОЙСТВЕННЫЕ СВЕТЛЫМ
ЛЕСАМ (ЛИСТВЕННИЧНЫМ,
СОСНОВЫМ, БЕРЕЗОВЫМ) И
ЛЕСНЫМ ЛУЖАЙКАМ
Основной ареал растений, вхо-
дящих в эту группу реликтов,
расположен в северной части Ази-
атского материка (Сибирь, за
исключением ее западной, низмен-
ной части, Дальний Восток, Север-
ная Монголия, иногда горы Сред-
ней Азии). Эти растения входят в
состав светлохвойных (главным
образом лиственничных), а иногда
и мелколиственных (березовых)
лесов, растут на лесных опушках
и луговых полянах среди леса.
Изолированные части ареалов этих
видов находятся на Урале.
В качестве примера назовем
Gentiana barbata, растущую по
лесным опушкам, на лесных и
высокогорных лугах. Распростра-
нена эта горечавка на Алтае,
в Средней и Восточной Сибири, на
Дальнем Востоке, в горах Сред-
ней Азии, Монголии и северной
части Китая. В Сибири западная
граница ее сплошного ареала про-
ходит на долготе г. Томска. Обособ-
ленная же часть ареала горечавки
бородатой (рис. 151) расположена
на Южном, Среднем и отчасти
Северном Урале. В пределах Юж-
ного Урала этот вид встречается
в Губерлинских горах, на хребтах
Ирендык, Уреньга, Бакты (Бехты),
Ильменских горах близ оз. Белого
в Учалинском районе, в окрест-
ностях пос. Тирлян и в других
местах. На Среднем Урале он найден на Уктусских горах в окрестностях
г. Свердловска, близ пос. Билимбая, на горе Качканар и на р. Косьве
около станции Губаха. Группа североуральских местонахождений сосре-
доточена в районе Денежкина, Конжаковского и Сухогорского Камней
(здесь этот вид произрастает близ верхней границы леса, чаще на обнаже-
ниях дунита, реже габбро) и вблизи этих гор на восточном склоне хребта.
240
К уральской части ареала примыкают изолированные местонахождения
в районе городов Тобольска, Тюмени, Тары и пос. Усть-Ламенского.
Рис. 152. Распространение чины.Гмелина
Lathyrus Gmelini на Урале и в Приуралье.
Другой пример — Lathyrus Gme-
lini, обитающая в светлых лесах,
в негустых хвойных (особенно
лиственничных) и смешанных ле-
сах, а также на лугах в пределах
горнолесного и подгольцового поя-
сов. Основной ареал этого расте-
ния— Алтай с прилегающей к нему
частью Западно-Сибирской низ-
менности (самое западное место-
нахождение — р. Чая около устья
Иксы), южная часть Средней и
Восточной Сибири, Забайкалье,
горы советской Средней Азии и
Кульджинские горы на северо-за-
паде Китая. Обособленный участок
Рис. 153. Распространение зигаде-
нуса сибирского Zygadenus sibiricus
на Урале.
ареала находится на Урале (рис. 152), где этот вид обитает главным
образом в светлых (с хорошо развитым травяным покровом) листвен-
ничных, сосновых, березовых и смешанных лесах, на полянах и лес-
16 Заказ № 251
241
Рис. 154. Распространение сосюреи спорной Saussu-
rea coniroversa на Урале.
ных лугах. Повсеместно
встречается в горах Юж-
ного и Среднего Урала
от района горы Яман-Тау
на юге до г. Нижнего Таги-
ла на севере. В более север-
ных районах чина Гмелина
становится редкой, здесь
она связана преимущест-
венно с лесными полянами
на южных известняковых
склонах по берегам рек
Чусовой, Косьвы, Яйвы,
Южной Сосьвы. На юг
заходит до хребтов Алатау,
Южный Крака, Крыкты и
до северной части хр. Ирен-
дык. К уральскому фраг-
менту ареала примыкают
изолированные местона-
хождения этого растения
в Зауралье (между Тю-
менью и дер. Кулаковой,
а в бывшем Туринском
уезде между дер. Носовой
и пос. Большой Кырты-
ньей).
Для светлых лесов (сос-
новых, а особенно листвен-
ничных) характерен Zyga-
denus sibiricus, реже он
встречается по сухим лу-
гам и на скалистом субст-
рате. Основной ареал его—
Алтай, Средняя и Восточ-
ная Сибирь, Дальний Вос-
ток, Северная Монголия
и Северо-Восточный Китай.
Отсутствуя на территории
Западно-Сибирской низ-
менности, это растение
вновь появляется на Урале
(рис. 153) близ городов
Кыштыма и Нижние Серги,
по берегам р. Уфы в окрест-
ностях г. Красноуфимска
и с. Караидель, на Уфим-
ском плато, в районе
г. Бирска, в нескольких
пунктах по рекам Уфе, Аю
и Юрюзани, по р. Белой
близ устья р. Иргизлы,
около деревень Биккузи-
ной и Кутаюновой, а также
на восточном склоне хр.
242
Ирендык близ пос. Баймака. На Урале это растение чаще всего встречается
на щебнистых известняковых склонах, травянистых лужайках среди сос-
новых (нередко с примесью лиственницы) лесов.
Такого же рода закономерность наблюдается в распространении Sau-
surea controversa. Она растет в лиственничных, березовых и сосновых ле-
сах, на каменистых склонах и на лугах, главным образом в пределах гор-
нолесного пояса, но местами поднимается почти до верхней границы леса,
а также проникает в горную и равнинную лесостепь. Основной ареал со-
Рис. 155. Распространение первоцвета кортузовидиого
Primula cortusoides на Урале.
сюреи спорной — Алтай с прилегающей к нему частью Западно-Сибир-
ской низменности, горы Средней Азии (Джунгарский Алатау, Памиро-
Алай), южная часть Средней Сибири, Восточная Сибирь, северная часть
Монголии и Китая. Изолированный участок ареала находится на Урале
(рис. 154). На Южном Урале этот вид распространен почти повсеместно
к северу от хр. Ирендык (в верхнем течении р. Урала, а бассейне рек Бе-
лой, Уфы, Ая, Юрюзани, на хр. Крыкты, горах Шатак, .Яман-Тау, Машаке,
хр. Уреньга, горе Юрма). На Среднем Урале встречается реже, главным
образом по берегам рек Чусовой, Камы и на горе Качканаре. Местонахож-
дения в пределах Северного Урала связаны со светлыми лесами по склонам
гор Конжаковский, Косьвинский, Семичеловечный, Денежкин, Вогуль-
ский Камни, Басеги, Кумба, Куроксар, Ялпинг-Ньер и по берегам рек
Лозьвы, Ивделя, Вишеры.
Другой вид сосюреи — Saussurea parviflora — обитает в редкостойных
лесах, на лесных опушках, лесных лугах, окраинах болот и на камени-
стых склонах, иногда заходит в высокогорья, где растет в тундрах и на
альпийских лугах. Основной ареал охватывает горы южной части Сибири
(Алтай, Саяны, Забайкалье) и Монголии, Среднесибирское плоскогорье.
В пределы Западно-Сибирской низменности сосюрея малоцветковая вкли-
16*
.243
нивается лишь незначительно, достигая на западе пос. Бердского на
р. Оби. Изолированный участок ареала расположен на Урале и в При-
уралье, где этот вид на севере достигает бассейна р. Няысь, а на юге — бас-
сейна р. Белой; самые западные местонахождения его известны близ г. Сык-
тывкара и в приуральской части Башкирской АССР, а самые восточные —
в окрестностях г. Тюмени. На некоторых горах Урала (Ойка-Чахль, Яны-
Енки, Сухогорский Камень)
достигает верхней границы
леса.
В пределах своего основ-
ного ареала, расположенного
на Алтае, в Тарбагатае, на
Саянах и в Монголии, Pri-
mula cortusoides произрастает
в редкостойных светлых ле-
сах, на каменистых склонах,
на лугах, иногда заходит в
березовые колки предгорий.
Западная граница основного
ареала достигает городов
Томска, Мариинска, пос. Су-
зуна на р. Оби. На большей
части Западно-Сибирской низ-
менности первоцвет кортузо-
видный не встречается. Изо-
лированный фрагмент его
ареала находится на Южном
Урале (рис. 155), где это
растение встречается в лист-
венничных и смешанных ле-
сах на известняковых скло-
нах. Основные пункты встре-
чаемости этого вида — берега рек Ая, Юрюзани, Сима, Уфы, Нугуша, Белой,
окрестности г. Златоуста, пос. Усть-Катава и с. Макарово. И. Георги
(Georgi, 1775) упоминает о встречаемости Р. cortusoides на берегах р. Чусо-
вой, но после него никто этого растения здесь не находил.
К предыдущим видам близка по своей экологии Trigonella platycarpos.
Она растет в разреженных березовых, осиновых, лиственничных, а также
в сосновых и пихтово-еловых лесах, содержащих примесь березы, на опуш-
ках и лесных лугах. Основной ареал этого вида расположен в восточной
части Западной Сибири, на Алтае, в горах Средней Азии (хр. Терскей Ала-
тау и некоторые горные районы Восточного Казахстана), в Саянах, в юж-
ной части Якутии, в Приморском крае, в горных районах Монголии и в
Китае (Большой Хинган30 и на юг до Пекина — горы Похуашань). Запад-
ная граница ареала на территории Западно-Сибирской низменности про-
ходит по линии, соединяющей с. Парабель на р. Оби, с. Кыштовку на
р. Таре и пос. Колывань на Оби.
Западнее основного ареала есть несколько изолированных местонахож-
дений. Одно из них находится в низовьях р. Иртыша (с. Уват, пос. Ханты-
Мансийск и другие смежные пункты). Другой пункт встречаемости отме-
чен В. Ф. Семеновым в Кокчетавской области. Кроме того, Т. platycarpos
обнаружена на Южном Урале, в Башкирии (рис. 156). Впервые это расте-
30 И. Т. Васильченко (1953) считает, что правильность отнесения имеющихся сборов
пажитника из района Большого Хингана к Т. platycarpos сомнительна.
244
ние было найдено здесь А. Н. Богдановым и С. Ф. Курнаевым в 1935 г.
в Бурзянском районе Башкирской АССР, в долине горной речки Кужа близ
с. Старосубханкулово. Гербарные экземпляры, определенные А. Н. Богда-
новым, были просмотрены И. М. Крашенинниковым, подтвердившим опре-
деление. На этом основании И. М. Крашенинников (1937) упомянул
Рис. 157. Распространение аконита противоядного
Aconftum anthora на Урале.
Т. platycarpos в числе растений, имеющих на Урале изолированные части
ареалов. Позднее Е. В. Кучеров (1954) обнаружил это растение в том же
Бурзянском районе около с. Исламбаево, на опушке осиново-березового
леса с примесью сосны, неподалеку от лесной сенокосной поляны Тукмак
(приблизительно на высоте 600 м над ур. м.).
Довольно широкой экологической амплитудой обладает Aconitum an-
thora. Произрастает на степных, реже заливных лугах, в разреженных гор-
ных (чаще лиственничных) лесах, по травянистым и каменистым склонам
гор. Заходит в высокогорья, где встречается на лугах и в горных тундрах,
а также на моренах близ ледников. Ареал этого вида раздроблен на ряд
изолированных частей. Основная его часть — азиатская, охватывающая
245
Алтай с его предгорьями и прилегающей территорией Западно-Сибирской
низменности и среднеазиатских гор (Джунгарский Алатау, Тарбагатай)
и Саяны. Изолированная часть ареала находится на территории Среднего
и Южного Урала (рис. 157); граница ее проходит приблизительно через го-
рода Красноуфимск и Свердловск на севере, Кыштым и Миасс — на восто-
ке, Оренбург — на юге, Бирск и Белебей — на западе. Уральские место-
Рис. 158. Распространение герани ложносибирской Geranium pseudosibiri-
сит на Урале.
нахождения связаны главным образом с лесостепью гор и предгорий, а
также с прилегающими степными и лесными районами, где этот аконит
растет на скалистых вершинах невысоких хребтов, на остепненных лугах,
в луговых степях, у подножия известняковых склонов, в карстовых ворон-
ках, на сухих опушках березовых, сосновых и широколиственных лесов.
Небольшие фрагменты ареала имеются в субальпийском поясе Кавказа
и на Крымской Яйле. Затем следует довольно большой участок ареала,
приходящийся на территорию Средней и Южной Европы (лесной и субаль-
246
пийский пояса), равнинную
лесостепь Украины и приле-
гающую часть Средне-Рус-
ской возвышенности31.
Geranium pseudosibiricum
произрастает в светлых ле-
сах, по их опушкам и лужай-
кам. Основной ареал охваты-
вает Тарбагатай, Джунгар-
ский Алатау, Алтай, Восточ-
ную Сибирь (от Забайкалья
на юге до низовьев р. Ени-
сея и центральной части Яку-
тии на севере), Монголию.
Совершенно обособлен ураль-
ский фрагмент ареала герани
ложносибирской (рис.- 158).
Самые северные ее место-
нахождения здесь — хр. Бас-
сеги и р. Чусовая в районе
пос. Чусового, самые юж-
ные — бассейны рек Боль-
шого Ика и Таналыка. На
запад она доходит до Беле-
беевского района в Баш-
кирии, а на восток — до
окрестностей г. Свердловска
и бассейна р. Уя в Челябин-
ской области. Область дизъ-
юнкции населена другим ви-
дом — G. bifolium-, по мне-
нию Е. Г. Боброва, автора
обработки рода Geranium для
«Флоры СССР», это образова-
ние молодое и явно произ-
водное.
Thalictrumfoetidum харак-
терен для каменистых скло-
нов, обрывов, зарослей кус-
тарников и окраин разрежен-
ных лесов на щебнистой „
v, Рис. 159. Распространение василистника вонюче -
почве. Распространен В основ- г0 phalictrum foetidum на Урале,
ном на Алтае с прилегаю-
щими районами Западно-Сибирской низменности, в Восточной Сибири, на
юге Дальнего Востока, в Монголии, в Китае (Тибет), в Торах Средней
Азии, на Кавказе, в Иране. В пределах Западно-Сибирской низменности
западная граница основного ареала проходит от устья р. Иксы, притока
р. Чаи, к с. Кожевникове на р. Оби, южнее г. Колывани (Крылов, 1927—
1964). Западнее этой линии известно лишь несколько изолированных
местонахождений (в Барабе близ с. Кыштовского и близ г. Омска).
Уральский фрагмент ареала (рис. 159) ясно обособлен, здесь этот вид
31 В последнее время некоторые систематики сохраняют название A. anthora лишь за
растениями, произрастающими в Средней Европе, в то время как растения из Сибири, гор
Средней Азии, Кавказа и европейской части СССР относят к A. nemorosutn.
247
распространен от г. Чусо-
вого на севере до бассейна
р. Большого Ика на юге,
на запад он идет до г. Стер-
литамака, на восток до
окрестностей г. Кыштыма.
Обособленные фрагменты
ареала имеются в Цент-
ральной Европе и на Бал-
канах.
Carex alba растет в лес-
ной зоне на скалах, песча-
ных обрывах и по окраи-
не разреженных лесов.
Основной ареал — Алтай,
Саяны, Средне-Сибирское
плоскогорье, Забайкалье.
Изолированные фрагменты
ареала имеются на Урале,
на Кавказе (Дагестан) и
в Центральной Европе.
На Урале (рис. 160) осока
белая растет преимущест-
венно на обнажениях из-
вестняков по рекам. Више-
ре, Каме, Чусовой, Сылве,
Туре. На значительном
расстоянии от Уральских
гор этот вид найден на Рус-
ской равнине (на Тиманс-
ком кряже и по р. Пинеге).
Allium obliquum произ-
растает в лесном и лесо-
степном поясах, а иногда
и в поясе высокогорных
редколесий, главным обра-
зом на горных лугах,
в светлых лесах и по бере-
гам рек. Основной ареал
Рис. 160. Распространение осоки белой Carex alba на его горноазиатский, ОХ-
Урале. ватывающий горные мас-
сивы Средней Азии от
Тянь-Шаня (Ферганская
долина) до Джунгарского Алатау и Саура, Алтай, юго-восточную часть
области Казахского мелкосопочника, Кузнецкий Алатау и Саяны. Ясно
выраженной дизъюнкцией, приходящейся на территорию Западно-Сибир-
ской низменности и Казахстана, отделены местонахождения A. obliquum
на Южном Урале (рис. 161) — в Башкирской АССР близ с. Воскресенского
на р. Зюрганке и на хр. Ирендык, в Оренбургской области в Губерлинских
горах (рис. 162), в окрестностях Оренбурга и у с. Спасского на р. Боль-
шом Ике. Отмечено это растение в смежных с Башкирией районах Пред-
уралья (окрестности пос. Сергиевска и в Бузулукском бору). Кроме того,
указывается еще более удаленное местонахождение A. obliquum на извест-
няковых скалах в центральной Трансильвании.
По сравнению с ранее рассмотренными видами несколько более тене-
248
вынослива Cerastium pauciflorum. Растет она в лиственных, смешанных:
и хвойных (лиственничных, сосновых, иногда разреженных еловых) лесах,
на лесных и высокотравных подгольцовых лугах. Основной ареал ясколки
малоцветковой — Алтай и прилегающая к нему часть Западно-Сибирской
низменности, Тарбагатай, Средняя и Восточная Сибирь, южная часть.
Дальнего Всстока, Северная Монголия, северо-восточный Китай, север-
Рис. 161. Распространение лука косого Allium obliquum
на Урале.
ная часть п-ова Корея. Резко обособлен уральский фрагмент ее ареала
(рис. 163). В пределах Южного Урала этот вид встречается на горах Яман'
Тау, Машак, Иремель, хр. Зигальга, горе Юрма, Ильменских горах, в бас-
сейнах рек Ай, Юрюзань, Уфа и близ г. Златоуста. На Среднем Урале он
известен из окрестностей городов Свердловска, Ревды, Билимбая,
Невьянска и Красноуфимска, пос. Павды. Заходит на Северный Урал в.
район Конжаковского и Денежкина Камней и в бассейн р. Ваграна.
К этой же группе реликтов может быть отнесена Anemone reflexa, свой-
ственная светлым березовым, лиственничным, иногда разреженным сосно-
вым лесам, а также негустым темнохвойным лесам с примесью березы. Ареал
ее охватывает юго-восточную часть Западно-Сибирской низменности (Том-
ская область, особенно березовые леса в низовьях Чулыма), южную часть
Средней и Восточной Сибири, южную часть Дальнего Востока, северную
часть п-ова Корея и Северную Монголию. В отрыве от основного ареала
ветреница отогнутая встречается (рис. 164) в Предуралье (Липовая гора
в окрестностях г. Перми), в горнохребтовой полосе Среднего Урала (по,
р. Казачьему Шишиму, на Бардымском хребте, в окрестностях пос. Бисера
и станции Арасланово) и на восточном склоне Среднего Урала (Верхне-
Туринское лесничество).
П. В. Сюзев (1923а, б) считал Anemone reflexa на Урале доледниковым
реликтом. Мотивом для этого вывода послужило то обстоятельство, что
в обнаруженном им впервые на Урале местонахождении на Липовой горе
близ Перми ветреница отогнутая произрастала в лесах с участием липы и
на опушках. Однако, по наблюдениям А. А. Хребтова (1925, 1941), на Ли-
повой горе она чаще растет на разреженных участках елово-пихтового
леса, по окраинам массивов, не заходя в глубь леса более чем на 6—8 м.
249
Рис. 162., Река Урал в районе Губер,пинских гор.
I
Впоследствии это же растение было
найдено в хребтовой полосе Сред-
него Урала и на восточном склоне,
где оно не связано ни с липовыми
лесами, ни с отдельными место-
нахождениями липы. Основной
сибирский ареал A. reflexa рас-
положен вне области распростра-
нения широколиственных лесов.
В заключение упомянем еще
один реликт, принадлежащий к
этой же группе,— Dentaria tenui-
folia. Она растет на лугах, в за-
рослях кустарников и в негустых
лесах. Основной ареал ее распо-
ложен на Алтае с прилегающими
районами Западно-Сибирской низ-
менности, в горах Восточной Си-
бири и Дальнего Востока. Резко
обособлены местонахождения з'у-
бянки узколистной на Среднем
Урале (рис. 165) близ дер. В. По-
пово и пос. Билимбая на р. Чусо-
вой и около станции Кузино. Кро-
ме того, ряд изолированных место-
нахождений этого вида отмечен
в Тульской области (близ г. Ве-
нева и по р. Осетру, у сел. Мохов-
ского и Лидино).
Установившийся в плейстоцене
режим сурового континентального
климата благоприятствовал широ-
кому распространению на терри-
тории, не затронутой оледенением,
светлых, главным образом листвен-
ничных, а в меньшей степени сос-
новых и березовых лесов, пере-
межавшихся луговыми полянами.
Лиственничные леса в плейстоцене
и голоцене занимали видное место
в сложении растительного покрова
не только в Средней и Восточной
Сибири, где они сохранили свое
господство до наших дней, но и на
территории Западно-Сибирской
низменности и Урала (Крашенин-
Рис. 163. Распространение ясколки малоцвет-
ковой Cerastium pauciflorum на Урале.
ников, 1939; Горчаковский, 1953). Это создало предпосылки для миграции на
запад, из Сибири на Урал, многих травянистых светолюбивых и холодо-
стойких растений, свойственных светлым лесам, лесным опушкам и лугам.
Лиственничные леса были широко распространены на Урале и в Западной
Сибири и в начале голоцена. Однако в последующие стадии голоцена лист-
венничные леса на территории Западно-Сибирской низменности, а также в
ряде районов Урала стали усиленно оттесняться темнохвойными лесами.
Это привело к полному исчезновению некоторых светолюбивых лугово-лес-
ных растений на территории Западно-Сибирской низменности, где площадь
251
лиственничных и вообще светлых лесов сильно сократилась, и они были
полностью оттеснены темнохвойной тайгой, за исключением северной
Рис. 164. Распространение ветреницы отогнутой Ane-
mone reflexa на Урале.
Рис. 165. Распространение зубянки узколистной Denta-
ria tenuifolia на Урале.
27
окраины лесной зоны. Однако на Урале в силу большего разнообразия ус-
ловий среды эти растения в ряде мест смогли сохраниться (на каменистых
склонах, в разреженных светлых лесах, а также на лужайках в подгольцо-
252
вом, горнолесном и лесостепном поясах). Так возникли урало-сибирские
дизъюнкции ареалов упомянутых здесь растений.
Следовательно, эти растения являются реликтами флористического
комплекса лиственничных и вообще светлых лесов и лесных лужаек, ко-
торый имеет сибирское происхождение и особенно широко распространился
на территории Урала и Западно-Сибирской низменности в плейстоцене и
в начале голоцена.
Характерно, что обособленные уральские фрагменты ареалов рассмот-
ренных выше растений расположены главным образом в южной и средней
частях Урала, где в сложении растительного покрова видное место зани-
мают лиственничные, сосновые и мелколиственные леса. В более северных
районах, где в горнолесном поясе господствует темнохвойная тайга, эти
виды не смогли сохраниться.
Ознакомление с плейстоценовыми реликтами во флоре Урала пока-
зывают, что они неодинаковы по экологии, фитоценотическим связям, ха-
рактеру их ареалов и возрасту.
ГОЛОЦЕНОВЫЕ РЕЛИКТЫ
Голоцен был во много раз короче плейстоцена, а происходившие в те-
чение его изменения физико-географических условий не были столь сущест-
венными. Однако в течение послеледниковья растительный покров Урала
подвергся довольно значительной перестройке, постепенно приближался
к его современному облику. Индикаторами происшедших в это время смен
растительности являются реликтовые растения и даже реликтовые расти-
тельные сообщества. Наибольший интерес среди голоценовых реликтов
представляют неморальные реликты фазы максимального распростране-
ния широколиственных лесов и степные реликты, соответствующие фазе
наибольшего продвижения степной растительности на север.
ШИРОКОЛИСТВЕННЫЕ РЕЛИКТЫ ПЕРВОЙ ПОЛОВИНЫ
СРЕДНЕГО ГОЛОЦЕНА
В четвертичное время первое значительное расширение пределов широ-
колиственных лесов совпало с климатическим оптимумом днепровско-
валдайской межледниковой эпохи, после чего площадь лесов сократилась.
В первой половине среднего голоцена широколиственные леса вновь рас-
пространились на Урале на довольно значительной территории. В позднем
голоцене, когда климат стал более прохладным (по сравнению со сред-
ним голоценом), темнохвойная тайга стала частично оттеснять широколист-
венные леса с занятых ими площадей. Лишь в некоторых местах
с наиболее благоприятными условиями широколиственные породы и свой-
ственные им травянистые растения могли удержаться на положении ре-
ликтов.
Южная граница зеленомошной темнохвойной тайги бореального типа
проходит на Урале, согласно К- Н. Игошиной (1943), по линии, соединяю-
щей Чермоз с Кизелом, затем смещается на юг вдоль хребта, отсюда идет
к востоку через станции Теплая Гора, Хребет Уральский и Кушву, а за-
тем направляется к Верхотурью. Далее к югу зеленомошная темнохвойная
тайга уступает место южной темнохвойной тайге с примесью некоторых
широколиственных древесных пород и их травянистых спутников. Наи-
более велика примесь растений неморального комплекса в пихтовой тайге;
из древесных пород здесь встречаются Tilia cordata, Acer platanoides,
Ulmus scabra (последние две породы с явными признаками угнетения),
а из травянистых растений — Asarum europaeum, Asperula odorata, Stachys
253
silvatica и Sanicula europaea. Пихта сибирская, как более требовательная
к почвенно-грунтовым условиям, но в то же время теневыносливая и кон-
курентномощная порода, захватила в позднем голоцене ряд местообитаний
с богатыми, хорошо увлажненными почвами, ранее занятых широколист-
венными лесами. Поэтому в пихтовой тайге, более чем в какой-либо другой
формации хвойных лесов, сохранилось довольно много представителей
растений неморального комплекса, которые являются в этих условиях
реликтами широколиственных лесов, распространенных на месте совре-
менной темнохвойной тайги в сравнительно недалеком прошлом (Горча-
ковский, 19546). В меньшей степени такие реликты встречаются под поло-
гом еловых лесов. Процесс оттеснения темнохвойной тайгой широколист-
венных древесных пород и их травянистых спутников продолжается и в
настоящее время.
СТЕПНЫЕ РЕЛИКТЫ ТЕРМИЧЕСКОГО МАКСИМУМА
ВТОРОЙ ПОЛОВИНЫ СРЕДНЕГО ГОЛОЦЕНА
На тех участках, где уральские реки прорезают толщу известняков,
нередко встречаются береговые утесы в виде зубчатых стен высотой до 50,
а иногда до 60—70 м, круто или даже отвесно обрывающиеся к реке. Хотя
площадь таких обнажений невелика, они крайне интересны с ботани-
ческой точки зрения, поскольку с ними связано произрастание ряда ред-
ких растений с прерывистым распространением. В процессе вековых из-
менений растительного покрова часть видов из растительных группировок,
господствовавших в тот или иной период, занимала места со скалистым суб-
стратом. Некоторые из этих видов сохранились здесь до наших дней,
являясь отголосками смен растительности, происходивших в далеком
прошлом. Скалистые обнажения обычно содержат несколько наслоений
реликтовых видов, различных по времени и условиям их появления на
таком субстрате.
Произрастание степных растений на скалистых береговых обнажениях
среди тайги Уральского хребта было отмечено еще в конце прошлого сто-
летия П. Н. Крыловым (1878) и особенно А. Я- Гордягиным (1895). Тщатель-
но Исследовав флору береговых утесов из известняка и авгитового
порфирита по р. Туре (восточный склон Среднего Урала), А. Я- Гордягин
был поражен обилием здесь степных форм, которые, по его словам, не всег-
да встречаются в столь большом количестве даже в местах, непосредст-
ственно граничащих с лесостепью. «Когда я попал на утесы Елкиной,—
писал А. Я- Гордягин (1895, стр. 13),— то мне казалось, что я нахожусь
в некоторых хорошо мне известных пунктах юга Пермской губернии или
севера Уфимской». Однако присутствие здесь степных растений не гармо-
нировало с общим таежным характером окружающей растительности.
А. Я- Гордягин пришел к заключению, что изученные им колонии
степных растений на береговых обнажениях имеют реликтовый характер.
Хотя такие островки степной растительности в настоящее время находятся
далеко за пределами степей, ранее они были непосредственно связаны
с ними. Отметив большее богатство степными формами обнажений извест-
няков по сравнению с выходами порфиритов, А. Я- Гордягин объяснил
это тем, что с бескарбонатных пород степные растения сильнее оттеснялись
древесной растительностью.
В дальнейшем степные и лесостепные растения на береговых обнаже-
ниях изучались В. С. Говорухиным (1929, 1937), С. Н. Наумовой (1929),
и Л. Б. Ланиной (1940) в Печорском бассейне, А. Н. Пономаревым (1945,
1949) по рекам Вишере, Косьве и Чусовой, автором этих строк по рекам
254
Косью (Приполярный Урал), Ивделю, Вижаю, Северной Тошемке (Север-
ный Урал), Чусовой, Режу (Средний Урал), Юрюзани и Белой (Южный
Урал).
Таблица 4
Наиболее характерные степные и лесостепные растения, встречающиеся
на береговых известняковых обнажениях на территории Среднего и Северного Урала
в горнолесном поясе и бореально-лесной зоне предгорий
Stipa Joannis.........................
Роа stepposa . ........................
Artemisia frigida............................
A. latifolia.................................
Echinops ritro...............................
Filipendula hexapetala.......................
Spiraea crenata..............................
Genista tinctoria............................
Antitoxicum stepposum........................
Gypsophila altissima ........................
Seseli Ledebourii............................
Helictotrichon desertorum ...................
Onosma simplicissimuin .........
Phleum phleoides.............................
Onobrychis tanaitica ........................
Oxytropis pilosa....................... . .
Galium verum ................................
Phlomis tuberosa........................
Artemisia armeniaca..........................
Astragalus danicus ..........................
Dianthus versicolor..........................
Festuca pseudovina...........................
Veronica spicaia ... ...............
Anemone silvestris................... ...
Cotoneaster melanocarpa......................
Artemisia sericea............................
Aster alpinus. ..............................
В табл. 4 приведены наиболее характерные степные и лесостепные ра-
стения, встречающиеся в лесной зоне Урала на известняковых обнаже-
ниях. Нетрудно заметить, что количество их при движении к северу зако-
номерно убывает. Такие типично степные виды, как Echinops ritro, Onosma
simplicissimum (рис. 166), Gypsophila altissima, Spiraea crenata, Antito-
xicum stepposum (рис. 167) и др., не заходят на север дальше 58°30' с. ш.,
лишь Helictotrichon desertorum достигает по р. Туре 59°30' с. ш. Севернее
60° с. ш. на известняковых обнажениях встречаются лишь менее ксеро-
фильнЫе растения, которые правильнее рассматривать не как собственно
степные, а как лесостепные, например Aster alpinus, Veronica spicaia,
Anemone silvestris.
Названные растения, встречающиеся на известняках в лесной зоне,
являются реликтами фазы термического максимума, совпадающего со вто-
рой половиной среднего голоцена. В это время степной ландшафт продви-
гался на север дальше, чем теперь, и северная граница лесостепи на приле-
255
Рис. 166. Оносма простейшая Onosma simplicissimum па известняковом
утесе Высоком на р. Чусовой.
тающих к Уралу равнинах достигала, вероятно, 59—59°30' с. ш. Неко-
торые степные и лесостепные растения смогли тогда в предгорных райо-
нах по южным склонам известняковых холмов, прибрежным обнажениям,
а также по песчаным и галечниковым наносам проникнуть в южную часть
лесной зоны. В результате последовавшего смещения зон к югу и связан-
ного с ним интенсивного расселения таежной растительности в этих райо-
нах степные и лесостепные растения были оттеснены из многих занимаемых
ими мест обитания. Лишь на некоторых экспонированных на юг известня-
ковых обнажениях они смогли сохраниться в реликтовом состоянии.
256
Рис. 167. Ластовень степной Antitoxicum siepposutn на известняковой
скале по р. Режу близ Мантурова Камня.
ЗАКЛЮЧЕНИЕ
Во флоре Уральской горной страны и примыкающих частей сопредель-
ных равнин содержится около 5% эндемичных растений. Флористический
эндемизм проявляется здесь лишь на видовом и внутривидовом уровне;
все эндемики представлены травянистыми растениями. По своим эколого-
центоническим связям эндемичные растения Урала и Приуралья подраз-
деляются на три группы: а) высокогорные (горнотундровые и горнолуго-
вые), б) скальные и в) горностепные и широколиственнолесные.
17 Заказ № 251
257
Среди эндемиков имеются формы, тесно связанные в своем распростра-
нении с орографической областью Урала (эндемики в узком значении этого
слова), и формы, основная область распространения которых расположена
на Урале, где они встречаются чаще и обильнее, но периферические части
ареалов или серии изолированных местонахождений вклиниваются в со-
предельные равнинные территории (эндемики в широком смысле). К пер-
вым относится большинство высокогорных эндемиков, а ко вторым — зна-
чительная часть скально-горностепных и широколиственнолесных. Встре-
чаемость ряда эндемичных форм уральского происхождения не только
на Урале, но и на повышенных'местах прилегающих к нему равнин (При-
волжская возвышенность, Общий Сырт, холмы Казахского мелкосопоч-
ника и др.) является следствием как экологического сходства некоторых
местообитаний и типов растительных сообществ, так и былого флористи-
ческого обмена между этими территориями. Поскольку некоторые физико-
географические, биохорологические и видообразовательные явления и
процессы, приведшие к формированию флористического эндемизма, нередко
развертывались на более обширной, чем Урал, территории (включая и
прилегающие части равнин), при анализе уральского эндемизма необхо-
димо понимать его достаточно широко, не ограничиваясь лишь узко-
локальными эндемиками.
Формирование флористического эндемизма было тесно связано со спе-
цифичностью горных пород древней уральской складчатой области, эпей-
рогеническими поднятиями в неогене и плейстоцене и сопровождавшими
их процессами эрозии, а также плейстоценовым оледенением и сопутство-
вавшим ему изменением климата. Эти явления повлекли за собой на Урале
и в Приуралье деградацию неогенового широколиственнолесного комплек-
са (очаги которого сохранялись на Южном Урале и Общем Сырте даже
в эпоху максимального оледенения), образование обширных скально-щеб-
нистых пространств как на высоких уровнях гор, так и на южной оконеч-
ности хребта.
В разные эпохи неогена и плейстоцена в разных частях Урала, на раз-
ных высотных уровнях возникали (вследствие эрозии, вызванной эпейро-
геническими поднятиями, в связи с отступлением ледникового покрова
и т. п.) скально-щебнистые пространства, лишенные или почти лишенные
растительности. Эти пространства постепенно заселялись растениями, про-
никавшими сюда из других областей (из Сибири, Средней Азии, Европы).
На таких местообитаниях, крайне гетерогенных по химизму горных пород,
физическим свойствам коры выветривания, режиму увлажнения и осве-
щения, термическому режиму, шел отбор форм, приспособленных к спе-
цифическим условиям среды, создаваемым Уральской горной ^страной;
здесь происходила интогрессивная гибридизация, возникали новые поли-
плоидные формы. Разнообразные обнажения крупных каменных глыб и
щебня были главной ареной видообразования на Урале. Эндемизм ураль-
ской флоры.— в основном эндемизм эдафический.
Основными путями образования уральских эндемиков были: а) раз-
дробление ареала широко распространенного низинного предка (в связи
с появлением горных поднятий и другими изменениями условий среды),
отчленение уральской части ареала и последующая эколого-морфологи-
ческая: специализация; б) постепенное изменение низинного предка по мере
изменения условий среды, вызванных эпейрогенезом; в) преобразование
исходных- форм, .-проникших из других районов, путем приспособления
к, новым условиям среды; г) гибридизация, иногда с последующим пере-
ходом; к апомиктическому воспроизведению ее продуктов.
-от Дзучение экологических особенностей эндемиков, их родственных,-свя-
зей, выявление путей расселения исходных, форм дает возможность уста-
258 ...
Таблица 5-
Классификация эндемичных растений Урала
Флороценогенетические группы и их подразделения Время образования
Плиоцен Плейстоцен - Голоцен
Возникшие в результате эко- лого-морфологической диффе- ренциации и раздробления аре- ала первичного низинного пред- ка в ходе эпейрогенеза конца третичного и начала четвертич- ного периода с образованием серии викарных видов в разных горных областях Lagotis uralensis, Gypsophi- la uralensis, Polemonium nudi- pedum, Linum boreale, Anemo- ne biarmiensis
Возникшие в результате пре- образования в местных услови- ях высокогорных видов, проник- ших на Урал в плейстоцене из других горных областей Epilobium uralense, Gagea samojedorum
Высокогорные эндемики Возникшие в результате отбо- ра и приспособления к высоко- горным условиям популяций низинных видов, распространен- ных и ныне на предгорьях и низких уровнях гор - Cerastium Krylovii, C. Gorodkovianutn, C. Porphyrii, C. Igo- schinae, Scorzonera Ruprechtiana, Thy- mus paucifolius, Bro- musvogulicus, B. Ju- lii, B. uralensis, Se- necio Igoschinae
Возникшие в связи с явления- ми межвидовой гибридизации с последующим переходом к апо- миктическому воспроизведению ее продуктов Alchemilla crassicaulis, A. cinerascens, A. semispoliata, A. Gortschakovskii, A. auriculata, A. hyperborea, A. kvarkush- ensis, A. iremelica, A. parci- pila, A. tnalimontana, A. mac- roclada, A. obtusiformis, A. cunctatrix, A. glabriformis, A. amphipsila, A. Helenas, A. consorbina, A. exul, A. Haral- di, A. picnoloba, A. paene- glabra
Возникшие на базе апомик- сиса Hieracium sub- erectum, H. ire- melense, H. Ija- pinense, H. ura- lense, H. kurok- sarense, H. So- czawae, H. ste- nopiforme, H. Gorodkovianutn, H. manifestum, H. wologdense, H. cuspidellutn, H. kosvinskiense, H. konshakov- skianum, H. pyrsjuense
Гибридного происхождения Saussurea ura- lensis
17*
259
Таблица 5 (продолжение)
Флороценогенетические группы и их подразделения Время образования
Плиоцен Плейстоцен Голоцен
Скальные и горностепные эндемики j Возникшие в процессе становления флористического комплекса древних (доледниковых) горных каменистых степей или степе- подобных растительных сообществ Генетически связан- ные с серией родствен- ных видов, обитаю- щих на Русской рав- нине, в Крыму, на Кавказе, в горах Сред- ней Азии, на Алтае, в Средней и Восточ- ной Сибири Minuartia Helmii, Dianthus uralensis, A gr ору rum reflexia- ristatum, A. pruiniferum, Koe- leria sclerophylla, Scutellaria oxyphylla, Astragalus kungu- rensis, A. Clerceanus, A. Ka- relinianus, A. uralensis, Hedy- sarum Ra-oumnvianum., Oxytro- pis approximata, 0. Gmelini, 0. Hippolytii, Thymus baschkiri- ensis, Th. guberlinensis, Th. mugodzharicus, Th. binervula- tus
Генетически связан- ные с родственными видами, обитающими в Европе Schiverekia berteroides, Sch. monticola, Sch. Kusnezovii, Mi- nuartia Krascheninnikovii, Di- anthus acicularis, Silene basch- kirorum
Генетически связан- ные с родственными видами, обитающими в Азии Roegneria uralensis, R. viri- diglumis, Thymus Talievii, Th. hirticaulis, Aulacospermum ise- tense, Potentilla Eversmannia- na, Astragalus Helmii, Oxytro- pis spicata, Eritrichium uralen- se, Libanotis sibirica, Onosma guberlinense
Возникшие в ходе новейшего формообразования Tanajcetum uralense
Гибридного происхождения Potentilla Gor- diaginii
Эндемики широколиственных лесов Обособившиеся от третичного неморального предка в связи с раздроблением его ареала (по мере деградации широколист- венных лесов в плейстоцене) и сохранившие доныне связь с ши- роколиственными лесами Lathyrus Litvino- . vii, Knautia tatari- ca, Cicerbita uralen- sis
Обособившиеся от третичного нёморального предка в связи с раздроблением его ареала (по мере деградации широколист- венных лесов в плейстоцене) и переходом к обитанию в свет- лых лиственных лесах Anemone uralensis
новить время их возникновения (табл. 5). Относительно более древний,
неогеново-плейстоценовый возраст имеет преобладающая часть скально-
горностепных эндемиков. Многие высокогорные эндемики сформировались
в конце неогена и в плейстоцене в связи с эпейрогеническими поднятиями
и оледенением. Плейстоценовый возраст имеют местные широколи-
ственнолесные эндемики. Однако среди уральских эндемичных растений
встречаются и молодые, недавно возникшие формы.
260
Уральские эндемики, независимо от того, насколько велики их морфо-
логические отличия от других родственных форм, строго локализованы
географически. Поэтому ботаники, в том числе и авторы «Флоры СССР»,
следуя в основном концепции вида В. Л. Комарова (1940), хотя и понимая
ее по-разному, традиционно описывали их в ранге видов. Фактически под
этими видами скрывается серия различных по своему таксономическому
значению форм, заслуживающих тщательного изучения.
Существующие диагнозы эндемичных уральских растений неполны,
а экологическая характеристика их далеко не достаточна. Это объясняется
тем, что новые виды (большинство уральских эндемиков выявлено в период
подготовки «Флоры СССР») описывались чаще всего по гербарным экзем-
плярам, собранным случайными коллекторами, без тщательного изучения
растений в природе. Нередко авторы располагали лишь немногими гербар-
ными экземплярами, хромосомные числа большинства эндемиков еще
неизвестны. Опубликованные ранее диагнозы не отражают реаль-
ную природу эндемичных растений, не показывают амплитуду варьиро-
вания основных признаков. Необходимо дальнейшее изучение уральских
эндемичных растений с применением методов биосистематики и цитотак-
сономии.
Уральская флора содержит в своем составе несколько наслоений ре-
ликтовых видов. Некогда эти растения были широко распространены на
территории Урала или отдельных его частей, но впоследствии, в связи с
изменением условий среды, их распространение в пределах горной страны
значительно сократилось, а на прилегающих равнинах они вымерли пол-
ностью. В результате этого возникли дизъюнкции, причем уральские фраг-
менты ареалов таких видов оказались обособленными от основных ареалов,
располагающихся в значительном удалении от Уральских гор.
Реликтовые растения в зависимости от времени их былого процветания,
когда они играли существенную роль в сложении растительного покрова
Урала, можно подразделить на три категории: доледниковые (плиоценовые),
плейстоценовые и голоценовые.
К числу доледниковых реликтов относятся главным образом растения
европейских широколиственных лесов с разорванными ареалами: Scutella-
ria altissima, Scrophularia Scopolii, Veronica maxima, Cephalanthera longi-
folia, Laser trilobium, Festuca silvatica, Geranium Robertianum, Sanicula
europaea, Digitalis grandiflora, а также некоторые растения открытых ка-
менистых местообитаний (Juniperus sabina).
Очень многочисленны и разнообразны плейстоценовые реликты. К ним
принадлежат, в частности, реликты арктической и горной флоры, связан-
ные в свое время с перигляциальным ландшафтом. Перигляциальные ре-
ликты — выходцы из Арктики, сконцентрированы или на береговых из-
вестняковых и гипсовых обнажениях (Dryas punctata, D. octopetala, Saxi-
fraga caespiiosa, Pinguicula alpina, Dianthus repens, Potentilla Ruznetzowii)
или на гольцах Южного Урала (Lloydia serotina, Arctous alpina, Andro-
sace Bungeana, Pedicularis Oederi, P. verticillata, Salix reticulata, Polygonum
viviparum, Dryas octopetala, D. punctata, Calamagrostis lapponica, Dianth-
us repens') в значительном удалении от области основного распространения
этих видов на севере Урала и Приуралья. Перигляциальные реликты гор-
ного происхождения представлены главным образом выходцами из высо-
' когорных районов Азии (Primula Pallasii, Phlojodicarpus villosus, Oxy-
graphis glacialis, Rhodiola quadrifida, Swertia obtusa, Thlaspi cochleariforme,
Pedicularis compacta, Carex Ledebouriana, C. sabynensis, C. caucasica, Fes-
tuca Rryloviana, Alopecurus glaucus, Dasiphora fruticosa) и в значительно
меньшей степени — выходцами из высокогорий Европы (Alchemilla glabra).
Все эти растения проникли на Урал в плейстоцене, главным образом в
261
эпоху максимального оледенения, и сохранились доныне в некоторых наи-
более благоприятных для них местообитаниях.
В течение плейстоцена на территории Сибири и Урала широкое рас-
пространение получили светлые лиственничные, березовые и сосновые ле-
са, мезофильные луга, а в более южных районах — степи.
Сразу же после отступления ледников в горах Урала освободилась
значительная площадь оголенных, лишенных растительности скалистых
обнажений, а на прилегающих к нему равнинах — флювиогляциальных
и речных песчаных наносов. Климат тогда был еще относительно холод-
ным, континентальным, но сухим. Оголенный, незадернованный субстрат
стал заселяться достаточно холодостойкими и малотребовательными к поч-
венно-грунтовым условиям растениями — выходцами из различных со-
обществ (горные степи, группировки скалистых и песчаных обнажений).
Это были преимущественно травянистые растения, расселение которых
происходило быстрыми темпами. Травянистые растения значительно опе-
режали появление на оголенном субстрате древесных пород, которые посе-
лились здесь позже, когда климат стал менее суровым, а почва была под-
готовлена в результате жизнедеятельности травянистых растений.
Поскольку наиболее холодостойкие ксерофильные формы растений
были сосредоточены в горах Азии (особенно в южной части Сибири), на
флювиогляциальные и песчаные речные наносы Западно-Сибирской низ-
менности хлынули преимущественно горноазиатские виды. Расселяясь
на территории Западно-Сибирской низменности, они достигли гор Урала,
где нашли для себя подходящие условия обитания на скалистых обнаже-
ниях. Таким образом, песчаные и другие наносы (особенно вдоль р. Оби
и ее притоков) явились своеобразной трассой для проникновения из горных
районов Сибири на Урал многих горностепных и скальных ксерофильных,
относительно холодостойких видов. В послеледниковое время такие сибир-
ские иммигранты полностью (или почти полностью, за исключением отдель-
ных местонахождений) вымерли на равнинной территории Западно-Сибир-
ской низменности и Северного Казахстана, так как здесь распространились
растительные сообщества, в составе которых они произрастать не могут.
Но эти растения смогли сохраниться в качестве реликтов в горах Урала.
К числу реликтов горноазиатского происхождения, связанных со
скальными и горностепными местообитаниями, относятся Orostachys spi-
nosa, Linaria altaica, L. debilis, Phlox sibirica, Sedum hybridum, Artemisia
santolinifolia, A. bargusinensis, Patrinia sibirica, Bupleurum multinerve,
Vicia multicaulis, Chamaerhodos erecta, Potentilla sericea, Silene altaica,
Alyssum biovulatum, Oxytropis uralensis и Gentiana decumbens.
Плейстоценовые реликты азиатского происхождения, связанные со
светлыми лиственничными, сосновыми, березовыми лесами и лесными лу-
жайками, представлены такими видами, как Gentiana barbata, Lathyrus
Gmelini, Zygadenus sibiricus, Saussurea controversa, S. parviflora, Primula
cortusoides, Trigonella platycarpos, Aconitum anthora, Geranium pseudosi-
biricum, Thalictrum foetidum, Carex alba, Allium hymenorrhizum, A. obli-
quum, Cerastium pauciflorum, Anemone reflexa и Dentaria tenuifolia. Особое
положение среди пледстоценовых реликтов во флоре восточного склона
Урала и Зауралья занимает Calluna vulgaris—вид европейского про-
исхождения, связанный в области своего основного ареала с приморскими
верещатниками, сосновыми лесами и окраинами болот.
Для послеледниковой истории растительности особенно характерны
два события, наложивших наиболее яркий отпечаток на современный ра-
стительный покров Урала и Приуралья: значительное усиление роли ши-
роколиственных лесов в первой половине среднего голоцена и инвазия
степных элементов в эпоху термического максимума второй половины сред-
262
него голоцена. Реликтами былого продвижения широколиственных лесов
на север и в горы являются растения неморального флористического ком-
плекса (Asarum europaeum, Asperula odorata, Siachys silvatica, Sanicula
europaea и др.), сохранившиеся местами в темнохвойной тайге, впослед-
ствии пришедшей на смену этим лесам. О происшедшем некогда продвиже-
нии степной растительности на север свидетельствуют колонии степных и
лесостепных растений (Veronica spicata, Anemone Silvestris, Echinops ritro,
Onosma simpilicissimum, Gypsophila altissima, Artemisia armeniaca,
A. sericea, Phlomis tuberosa, Galium verum, Helictotrichon desertorum,
Phleum phleoides и др.) глубоко в лесной зоне на прибрежных известня-
ковых утесах.
Изучение эндемичных и реликтовых растений, их распространения,
истории формирования ареалов и экологии дает ценный материал для ре-
шения ряда важных вопросов исторической фитогеографии Урала и смеж-
ных территорий. Накопленные данные, в частности, служат фактической
основой для следующих историко-флористических и палеогеографических
выводов.
1. Появление растительности степного типа относится не к плейсто-
цену, как полагал И. М. Крашенинников (1939), а восходит к более древ-
нему периоду геологической истории — к неогену. В конце неогена, в пли-
оцене, на сухих каменистых склонах гор уже существовали своеобразные
общества травянистых растений — ксерофитов. С этой растительностью
преемственно связана большая группа горностепных и скальных эндеми-
ков Урала. Их исходными формами явились ксерофиты из других горных
областей, проникшие на Южный Урал по прилегающим к нему равнинам
(где в южной части, вероятно, уже существовал ландшафт лесостепного и
степного типа) и впоследствии преобразовавшиеся под влиянием местных
эдафических условий.
2. В плейстоцене комплекс широколиственнолесной растительности
не был полностью разрушен, хотя распространение его сильно сократи-
лось. Очаги неморальной растительности смогли сохраниться и пережить
эпоху максимального оледенения в ряде убежищ на территории западного
склона Южного Урала и Приуралья.
3. Местонахождения реликтов приледниковой арктической флоры и
реликтов скально-горностепной флоры азиатского происхождения отчет-
ливо отграничены территориально и редко смыкаются друг с другом.
Южнее местонахождения реликтов арктической флоры примерно совпа-
дают с предполагаемой границей максимального оледенения. На западных
предгорьях Урала ледниковые реликты встречаются южнее, чем на восточ-
ных, что также находится в соответствии с положением границы леднико-
вого покрова. Во всех случаях растения этих флорогенетических групп
занимают совершенно разные местообитания. Судя по данным современ-
ного распространения растений, перигляциальный комплекс на Урале и
в Приуралье был представлен, в основном, тундроподобными сообщест-
вами; степной элемент в нем отсутствовал. Отдельные степные виды могли
встречаться местами в южной части перигляциальной зоны, но они не при-
ближались к окраине ледникового покрова.
*
Эндемичные и реликтовые растения представляют большой научный
интерес для восстановления основных этапов формирования раститель-
ного покрова Урала. В связи с непрерывно возрастающим освоением при-
263
родных ресурсов Урала характерные стации ряда эндемиков и реликтов
подвергаются усиленному хозяйственному воздействию, а следовательно;
возникает опасность уничтожения таких растений. Поэтому в комплексе
неотложных задач по охране природы Урала важное место занимает воп-
рос об охране эндемичных и реликтовых растений в характерных местах
их обитания.
S и M М A R Y
THE PRINCIPAL PROBLEMS OF HISTORICAL PHYTOGEOGRAPHY
OF THE URALS
P. L. GORCHAKOVSKY
1. The flora of the Urals is but slightly specific. The proportion of the
endemics does not overpass 5% of the total number of vascular species. The
endemism of the Ural flora manifests itself only at the specific and
interspecific level. Not a single of the endemics plays a leading determinant
role in the formation of plant communities (none of them may be regarded
as a constructive or building species). There are no endemic arboreous species
in the Urals. Although the endemic plants do impart a peculiar speciality to
the flora of the Urals, nevertheless, not being constructive species, they do
not determine the main features of the vegetational cover of the Ural moun-
tains. In most cases the endemic plants of the Urals occur rarely, their dist-
ribution is discontinuous and they are frequently confined to the rocky sub-
stratum where the competition with the other plant species is relatively sli-
ght. Probably they were much more widespread in the past, but now they
are in a state of gradual extinction, persisting in but a few localities with
special edaphic conditions. Thus the endemism in the Urals is of a pronoun-
ced relict character and only a small proportion of endemic plants of the
Ural flora may be regarded as young endemics that have appeared relati-
vely recently, are highly variable and possess some characters indicating their
more or less intense dispersal under present conditions.
2. The endemic plants of the Ural flora may be subdivided into the fol-
lowing three groups: a) the high-mountain endemics; b) the mountain-steppe
and rock-inhabiting endemics and c) the endemics of broad-leaved forests.
3. The group of high-mountain endemics includes: a) the endemics that
have originated as a result of the ecological and morphological differentiation
of the formerly widespread plain-inhabiting ancestral species, and the conse-
quent development of a series of vicarious species in separate mountain regi-
ons (the disjunctions of areas were concurrent with the epeirogenesis of the
end of the Tertiary and the beginnining of the Quatenary period); b) the
endemics which have originated as a result of the transformation under the
local conditions of the high-montain species that had immigrated to the Urals
from other mountain regions in the Pleistocene; c) the endemics that have
developed from the populations of plain-inhabiting species (up to the present
time dwelling at lower altitudes and in the foothills of the Urals) due to
natural selection and adaptation to the high-mountain conditions; d) the en-
demics of hybrid origin (sometimes with the consequent transfer to the apo-
mictic reproduction); e) the endemics developed on the basis of apomixis.
4. Some of the mountain steppe ang rock-inhabiting endemics of the
Urals are of European kinship, a few other species have descended from the
ancestors immigrated from the mountain regions of Asia, whereas a certain
number of them have a series of allied species inhabiting the Russian Plain,
the Crimea, the Caucasus and some mountain areas of Asia.
265
As early as the end of the Tertiary period there existed peculiar steppe-
like communities of herbaceous xerophytes inhabiting rocky hill slopes, espe-
cially those facing to the south. Those communities were formed mainly by
wide-spread species. The epeirogenic upheavals at the end oi the Tertiary and
at the beginning of the Ouatenary period resulted in the further intensification
of denudative erosion in the Urals, in the appearance of new areas of bare
rocks. These conditions were favorable for the ecological and morphological
isolation of rock-inhabiting plants, for the appearance of the Ural mountain-
steppe and rock-inhabiting endemics. Later on the areas of these endemic
species became reduced.
5. The endemics of broad-leaved forests are represented by a few species
of European origin which have separated due to isolation of the Ural popula-
tions of their ancestral forms during the glacial epochs of the Pleistocene.
6. The main groups of relict elements of the Ural flora are as follows:
a) the relicts of the pre-glacial broad-leaved forests; b) the Pleistocene relicts
(periglacial, mountain-steppe and rock-inhabiting relicts, as well as relicts pe-
culiar to larch and birch forests; all of them mostly of Asiatic origin) and
c) the Holocene relicts.
7. On the basis of the analysis of the endemics and relicts in the light of
all available paleobotanical evidences the main features of the development of
the flora and vegetation of the Urals (from the Tertiary period up to the pre-
sent time) are revealed.
ЛИТЕРАТУРА
Алексе? вко М. И. Новые виды рода Schiverkia Andrz.—Бот. мат-лы гербария Бот.
ин-та АН СССР, т. 9. АТ- - Л,, Изд-во АН СССР, 194G.
Алексеенко М. И. О некоторых эндемиках известняковой флоры Европейской части
СССР.— Труды науч.-исслед. ин-та биологии Харьковского ун-та, 1950, т. 13.
А н а н о в а Е. И. Флора типа «перигляциальной» из древнечетвертичных отложений
Камы.— Пробл. ботаники, вып. 4. М.—Л., Изд-во АН СССР, 1959.
А н а н о в а Е. Н. О флоре и растительности Русской равнины в плиоцене (по палино-
логическим данным).— К. первой международной палинологической конференции
(Таксон, США). Доклады советских палинологов. М., Изд-во АН СССР, 1962.
Байковская Т. Н. К палеоценовой флоре Южного Урала (Steinhauera subglobosa
Presl.) — Бот. ж., 1951, т. 36.
Баранов В. И. Высокогорная тундра в юго-восточном Алтае.— Двадцать пять лет
педагогической и общественной деятельности академика Б. А. Келлера. Воро-
неж, 1931.
Баранов В. И. К истории лесов Волжско-Камского края.— Изв. Казанского фил.
АН СССР, серия биол. и с.-х. наук, 1949, № 1.
Баранов В. И. Новая ископаемая флора из третичных и верхнемеловых отложений
западного склона Мугоджар. Докл. АН СССР, 1953, т. 92, № 5.
Баранов В. И. Этапы развития флоры и растительности СССР в третичном периоде.
Часть третья. Итоги изучения ископаемых третичных флор и проблема реликтов
в современной растительности СССР.—Уч. зап. Казанского гос. ун-та, т. 114, кн. 1
(ботаника), 1954.
Баранов В. И. Этапы развития флоры и растительности в третичном периоде на тер-
ритории СССР. М., «Высшая школа», 1959.
БарбаричА. И. Calluna.— Флора УССР, т. 8. Киев, Изд-во АН УССР, 1957.
Благовещенский Г. А. Об ископаемых в современных торфяниках среднего
Зауралья.— Сов. ботаника, 1940, № 3.
Благовещенский Г. А. История лесов восточного склона Среднего Урала (на осно-
вании изучения пыльцы в озерных отложениях и торфяниках).— Сов. ботаника,
1943, № 6.
Борисова А, Г. О видах рода Oxytropis в пределах Урала.— Сов. ботаника, 1936,
№ 4.
Борисова А. Г. Новые бобовые (Chesneya, Astragalus) флоры СССР.— Бот. мат-лы
гербария Бот. ин-та АН СССР, т. 13. М.—Л., Изд-во АН СССР, 1950.
Б о р щ о в И. Г. Материалы для ботанической географии Арало-Каспийского края. При-
ложение к VII тому Записок имп. Академии наук, СПб., 1865.
Боч С. Г. и Краснов И. И. К вопросу о границе максимального четвертичного оле-
денения в пределах Уральского хребта в связи с наблюдениями над нагорными
террасами.— Бюлл. ком. по изуч. четв. периода, 1945, № 8.
Б у ш Е. A. Ericaceae. — Флора Сибири и Дальнего Востока, т. 3, 1919.
Буш Е. А. Вереск — Calluna.— Флора СССР, т. 18, 1952.
Быков Б. А. и Степанова Е. Ф. Кустарниковые степи как тип растительности.—
Изв. Всесоюз. геогр. о-ва, 1953, № 1.
Варсанофьева В. А. О следах оледенения на Северном Урале.— Тр. Ком. по изуч.
четв. периода, 1934, т. 3, вып. 2.
Васильев В. Н. Растительность Анадырского края. М.—Л., Изд-во АН СССР, 1956.
Васильев А. В. и Сахокиа М. Ф. Juniperus.— Дендрофлора Кавказа. Тбилиси,
Изд-во АН Груз. ССР, 1959.
Васильков Б. П. Вересковые пустоши в Марийской автономной области.—Уч. зап.
Казанского ун-та, серия ботаника, т. 6, 1933.
Васильченко И. Т. Обзор видов рода Trigonella.— Труды Бот. ин-та АН СССР,
серия 1 (Флора и систематика высших растений), вып. 10, 1953.
Введенский Л. В. Геологический очерк западной части Западно-Сибирской низмен-
ности.— Тр. Всесоюз. геол.-развед. объединения, 1933.
Вильбасте Г. И. Распространение курильского чая в Эстонской ССР.— Юбилейный
сборник Общества естествоиспытателей при Академии наук Эстонской ССР
(1853—1953). Таллин, 1953.
267
Воробьев Д. П. Материалы к флоре Курильских островов.— Труды Дальневосточ-
ного филиала Академии наук СССР, сериябот., 1956, т. 3 (5).
Воронина И. Г. Липа (Tilia L.) из четвертичных отложений Башкирии.— Сборник
студенческих работ. Изд. Саратовского гос. ун-та, 1960.
Воронина И. Г. Флора и растительность Башкирского Предуралья в верхнеплиоце-
новое— нижнеплейстоценовое время (по палинологическим данным).— Автореф.
дисс. Саратов, 1965.
Вульф Е. В. Введение в историческую географию растений. Л., Изд-во Ин-та расте-
ниеводства НКЗ СССР, 1932.
Вульф Е. В. История флор Земного шара. М., Изд-во АН СССР, 1944.
Герасимов Д. А. Геоботанические исследования торфяных болот Урала.— Торф,
дело, 1926, № 2—3.
Г и т е р м а н Р. Е. Некоторые данные по истории растительности низовьев р. Чусовой
в четвертичное время.— Бюлл. Ком. по изуч четв. периода, 1953, № 17.
Говорухин В. С. Растительность бассейна р. Илыча (Сев. Урал)—Тр. О-ва изу-
чения Сибири, Урала и Дальнего Востока, 1929, вып. 1.
Говорухин В. С. Флора Урала. Свердловск, Свердлгиз, 1937.
Головкова А. Г. Растительность центрального Тянь-Шаня. Фрунзе, Изд-во Кир-
гиз. ун-та, 1959.
Гордягин А. Я. Растительность известковых скал на реке Туре в Пермской губер-
нии.— Труды О-ва естествоисп. при Казанском ун-те, 1895, т. 28, вып. 2.
Гордягин А. Я- Матералы для познания почв и растительности Западной Сибири.—
Труды О-ва естествоисп. при Казанском ун-те, 1901, т. 35, вып. 2.
Гордягин А. Я. К флоре Акмолинской области.— Ежегодник Тобольского губерн-
ского музея, 1916, вып. 27.
Городков Б. Н. Полярный Урал в верхнем течении рек Соби и Войкара.— Изв.
АН СССР, 1926, № 9.
Городков Б. Н. Полярный Урал в верховьях рек Войкара, Сыни и Ляпина.— Мат-лы
комис. экспед. иссл. АН СССР, серия Урал, вып. 7, 1929.
Городков Б. Н. Растительность тундровой зоны СССР. М.—Л., Изд-во.АН СССР,
1935.
Городков Б. Н. Есть ли родство между растительностью степей и тундр? — Сов. бо-
таника, 1939, 6—7.
Городков Б. Н. Ботанико-географический очерк Крайнего Севера и Арктики
СССР.— Уч. зап. Ленингр. гос. пед. ин-та им. Герцена, т. 49, 1947.
Городков Б. Н. Приледниковые ландшафты плейстоцена на севере Азии. — Докл.
АН СССР, нов. серия, 1948, т. 9, № 3.
Городков Б. Н. Происхождение арктических пустынь и тундр.— Тр. Бот. ин-та
АН СССР, серия 3 (геоботаника), 1952, вып. 8.
Горчаковский П. Л. Высокогорная растительность заповедника «Денежкин
Камень». Свердлгиз, 1950.
Горчаковский П. Л. О реликтовой флоре известняковых береговых обнажений по
рекам Ивдель и Тошемка на Северном Урале.'—Докл. АН СССР, 1951, т. 81,
№ 4.
ГорчаковскийП. Л. История развития растительности Урала. Изд. 2, Свердл. кн.
изд-во, 1953.
ГорчаковскийП. Л. Высокогорная растительность Яман-Тау — крупнейшей вер-
шины Южного Урала. — Бот. ж., 1954а, т. 39, № 6.
Горчаковский П. Л. Пихтовая тайга Среднего Урала.— Зап. Урал, отдела геогр.
о-ва СССР, вып. 1. Свердл. кн. изд-во, 19546.
Горчаковский П. Л. Растительность горных тундр Урала.— Зап. Урал, отдела
геогр. о-ва СССР, вып. 2. Свердл. кн. изд-во, 1955.
Горчаковский П. Л. Важнейшие типы горных еловых и сосновых лесов южной
части Среднего Урала. — Сб. тр. по лесн. х-ву, вып. 3. Свердл. кн. изд-во,
1956.
Горчаковский П. Л. История растительности Урала в плейстоцене и голоцене в
свете изучения реликтовых элементов флоры.— Делегатский съезд Всесоюз. бот,
о-ва. Тезисы докл., вып. 4 (секции флоры и растительности), ч. 1. Л., 1957а.
ГорчаковскийП. Л. Высокогорная растительность хребта Чистоп на Северном Ура-
ле;— Землеведение, 19576, т. 4.
Горчаковский П. Л. Растительность хребта Сабли на Приполярном Урале.— Расти-
тельность Крайнего Севера СССР и ее освоение, вып. 3. М.—Л., Изд-во АН СССР.
1958а.
Горчаковский П. Л. Растительность Свердловской области.— Природа Свердлов-
ской области. Свердл. кн. изд-во, 19586.
Горчаковский П. Л. О распространении и условиях произрастания дазифоры
кустарниковой (Dasiphora fruticosa (L.) Rydb.) в связи с реликтовой природой
ее уральских местонахождений,—Зап. Свердл. отд. Всесоюз. бот. о-ва, вып. 1,
1960а.
268
Горчаковский П. Л. Об охране реликтовых растений и уникальных растительных
сообществ на Урале.— Охрана природы на Урале, вып. 1. Свердловск, 19606
(УФАН СССР).
Горчаковский П. Л. Границы распространения вяза гладкого (Ulmus laevis Pall.)
на Урале и реликтовый фрагмент его ареала в Западной Сибири.— Пробл. бота-
ники, 1962а, т. 6.
Горчаковский П. Л. К географии, экологии и истории формирования ареала
вереска.— Бот. ж., 19626, т. 47, № 9.
Горчаковский Ц. Л. Эндемичные и реликтовые элементы во флоре Урала и их
происхождение.— Материалы по истории флоры и растительности СССР, вып. 4.
М—Л., Изд-во АН СССР, 1963.
ГорчаковскийП. Л. Реликтовое местонахождение липы мелколистной в лесостепи
Тобол-Ишимского междуречья и генезис сибирского крыла ее ареала.— Бот. ж.,
1964, т. 49, № 1.
Г о р ч а к о в с к и й П. Л. Флора и растительность высокогорий Урала. Труды Ин-та био-
логии УФАН СССР, 1966, вып. 48.
Горчаковский П. Л. Красноуфимская лесостепь — ботанический феномен Пред-
уралья.— Бот. ж., 1967, т. 52, № II.
Горчаковский П. Л. Растения европейских широколиственных лесов на восточном
пределе их ареала. Труды Ин-та экологии растений и животных УФАН СССР,
1968, вып. 59.
Горчаковский П. Л. и Архипова Н. П. Растительный мир обнажений гра-
нита, дунита и других горных пород на восточном склоне Среднего Урала.—
Зап. Свердл. отд. Всесоюз. бот. о-ва, 1964, вып. 3 (УФАН СССР, Свердл. отд.
Всесоюз. бот. о-ва).
ГорчаковскийП. Л. и Колесников Б. П. Распространение казацкого мож-
жевельника Juniperus sabina L. на Южном Урале.— Бот. ж., 1964, т. 49, № 10.
Грибанов Л. Н. Степные боры Алтайского края и Казахстана, М., Сельхозгиз, 1960.
Гризбах А. Растительность Земного шара согласно климатическому ее распределе-
нию. Перев. с нем. с прим. А. Бекетова. СПб, 1874.
Г р и ч у к В. П. К истории растительности европейской части СССР в четвертичном пе-
риоде.— Труды Ин-та географии АН СССР, 1946, т. 37.
Гричук В. П. Основные моменты истории широколиственных лесов Восточно-Евро-
пейской равнины в четвертичном периоде.— Бот. ж., 1949, т. 34, № 1.
Гричук В. П. Растительность Русской равнины в нижне- и среднечетвертичное вре-
мя.—Труды' ин-та географии АН СССР, 1950а, т. 46.
Г р и ч у к. М. П. Основные этапы истории растительного покрова Восточно-Европейской
равнины, в четвертичном периоде.— Труды Конф, по споровопыльцевому анализу
1948 года. М., Изд-во МГУ, 19506.
Гричук М. П. О растительном покрове межледниковой и ледниковой эпох в средней
части бассейна р. Оби.— Вести. МГУ, 1960, № 5
Гроссгейм А. А. Флора Кавказа, т. 5. Тбилиси, Изд-во АН Грузинской ССР, 1952.
Гроссет Г. Э. О возрасте реликтовой флоры равнинной европейской части СССР
(критический анализ реликтового вопроса).— Землеведение, 1935, т. 37, вып. 3.
Гроссет Г. Э. Возраст термофильной реликтовой флоры широколиственных лесов
Русской равнины, Южного Урала и Сибири в связи, с палеогеографией плейсто-
цена и голоцена.— Бюлл. МОИП, отд. биол., 1962, т. 6, № 3.
Грубов В. И. Rosaceae— розоцветные.— Флора Забайкалья, вып. 5. Л., Изд-во
АН СССР, 1949.
Грубов В. И. Конспект флоры Монгольской Народной Республики.— М.—Л., Изд-во
АН СССР, 1955.
Грубов В. И. Новый вид ясколки из Башкирии. Новости систематики высших расте-
ний, вып. 3. М.—Л., «Наука», 1968.
Грюнер Н. М. Скальная флора притагильской части Среднего Урала в связи с петро-
графическими и топографическими условиями.— Труды Свердловского областного
краеведческого музея, вып. 1. Свердл. кн. изд-во, 1960.
Дибнер В. Д. О следах двухкратного горного четвертичного оледенения на Конжа-
ковском Камне.— Изв. Всесоюз. геогр. о-ва, 1953, т. 85, вып. 5.
Добр оч а ев а Д. М. и Виноградова В. М. Новый вид оносмы (Onosma
guberlinense Dobrocz. et Vinogr.) из Южного Приуралья. Украинский бот. ж.,
1966, т. 23, № 3.
Дорофеев П. И. Ископаемая ель Picea orientalis (L.) Link, на юге Северного Ура-
ла.— Бот. ж., 1950, т. 35, № 3.
Дорофеев П. И. Плиоценовые растения Урала.— Бот. .ж., 1952, т. 37, № 6.
Дорофеев П. И. Новые данные о плиоценовой флоре Камы.— Докл. АН СССР, 1957, .
т. 117, № 3.
Дорофеев П. И. О плиоценовой флоре Башкирского Приуралья.— Вопр. геологии
восточной окраины Русской платформы и Южного Урала, вып. 5. Уфа, 1960.
(Башкирский фил. АН СССР).
269
Дорофеев П, И. О плиоценовой флоре Башкирии. Бот. ж., 1962, т. 47, № 6.
Жаркова А. М. К изучению торфяников Боровской лесной дачи б. Кокчетавского у.
Акмолинской области (предварительное сообщение).— Изв. Зап.-Сиб. отдела
Русск. геогр. о-ва. Оме, 1930, т. 7.
Жузе А. П. Палеогеография водоемов на основе диатомового анализа. — Труды
Верхне-Волжской экспедиц., № 4, 1939 (Географо-экон, ин-т Ленингр. гос. ун-та).
Жузе А. П. Диатомовые водоросли в отложениях четвертичного возраста.— Мат-лы
по четвертичному периоду СССР, вып. 3. М., Изд. АН СССР, 1952.
Забелин И. М. Особенности распределения растительности в ледниковых и эрозион-
ных долинах в Восточном Саяне.— Вестник Моск. гос. ун-та, 1952, № 2.
Иванов В. В. Степи западного Казахстана в связи с динамикой их покрова.— Зап.
Геогр. о-ва СССР, т. 17, 1958.
Игошина К. Н. Остатки широколиственных ценозов среди пихтово-еловой тайги
Среднего Урала.— Бот. ж СССР, т. 28, № 4. 1943.
Игошина К. И. К изучению растительности Енисейского кряжа.— Тр. Бот. ин-та
АН СССР, серия 3 (геоботаника), 1951, вып. 7.
Игошина К. Н. Растительность Урала.— Труды Бот, ин-та АН СССР, серия 3 (гео-
ботаника), 1964, вып. 16.
Игошина К. Н. Флора горных и равнинных тундр и редколесий Урала. Растения
севера Сибири и Дальнего Востока. М.—Л., «Наука», 1966.
Ильин М. М. К реликтовой флоре Южного Урала.— Изв. Гл. бот. сада, 1922, т. 21,
вып. 4.
Ильин М. М. Растительность Среднего Урала.— Природа Урала, Свердлгиз, 1936.
Ильинский А. П. Растительность Земного шара. М.—Л., Изд-во АН СССР’ 1937.
К а л е ц к а я М. С. и А. Д. Миклухо-Маклай. Некоторые черты четвертичной
. истории восточного склона Печорского бассейна и западного склона Полярного
Урала. — Мат-лы по геоморфологии и палеогеографии СССР, вып. 20 М., Изд.
АН СССР, 1958.
Калинина А. В. Растительность Чуйской степи на Алтае.— Тр. Бот. ин-та АН СССР,
серия 3 (геоботаника), 1948, вып. 5.
Каменский А. И. К геоморфологии Южного Урала. О следах древнего оледенения
на Южном Урале.— Уч. зап. Моск, городского пед. ин-та, т. 46. Тр. геогр. фак.,
вып. 5, 1957.
Караваев М. Н. Конспект флоры Якутии. М.—Л., Изд-во АН СССР, 1958.
К а ц Н. Я. О ледниковых убежищах и расселении широколиственных пород на Восточ-
но-Европейской равнине в послевалдайское время.— Бюлл. Моск, о-ва испыт. при-
роды, отд. биол., 1952, т. 57, вып. 6.
Кеппен Ф. П. Географическое распространение хвойных дерев в европейской Рос-
сии и на Кавказе.— Зап. имп. Акад, наук, т. 50 (прилож.), 1885.
Кирпичников М.'Заметка о флоре известняков по реке Белой Кедве.— Бот. ж.,
т. 32, № 5, 1947.
Кишковский Т. Н. и Артюшенко 3. Т. К биологии высокогорных растений
Памира. — Бот. ж., 1951, т. 36, № 5.
Клео по в Ю. Д. Основные черты развития флоры широколиственных лесов европей-
. ской части СССР. — Мат-лы по истории флоры и растительности СССР, вып. 1.
М.—Л., Изд-во АН СССР, 1941.
К л о к о в М. В. Новые виды рода Thymus L. в СССР.— Бот. мат-лы гербария Бот. ин-та
АН СССР, т. 16. М.—Л., Изд-во АН СССР, 1954.
Клоков М. В. Евразиатские виды рода Polemoniutn L.— Бот. мат-лы гербария Бот.
ин-та АН СССР, т. 17. М.—Л., Изд-во АН СССР, 1955.
Клоков М. В. и Шостенко-ДесятоваН. А. Новые виды рода Thymus L.
из европейской части Союза. — Ж. Ин-та ботаники АН УССР, 1936, № 9 (17).
Колесникова Т. Д. Четвертичные флоры Башкирии.— Бот. ж., 1957, т. 42, № 6.
Колесникова Т. Д. Третичные растения из буроугольных отложений Башкирии.—
Бот. ж., 1960, т. 45, № 1. ' .
Колесникова Т. Д. О двух новых ископаемых растениях для третичной флоры
Южного Урала.— Бот. ж., 1961, т. 46, № 12.
Колесникова Т. Д. Новые данные о третичной флоре Башкирии.— Бот. ж., 1963,
т. 48, № 10.
Колесникова Т. Д. Современное и прошлое распространение видов рода Najas L.
в СССР и их значение для палеогеографии четвертичного периода.— Бот. ж.,
1965, т. 50, № 2.
Колесникова Т. Д. О новых находках ископаемой арктической флоры на севере
европейской части СССР.— Бот. ж., 1966, т .60, № 5.
Колоколов А. А. и Львов К. А. О следах оледенения на Южном Урале (гео-
морфологический очерк хребта Зигальга). — Изв. Всесоюз. геогр. о-ва, 1945,
№ 1—2.
Комаров В. Л. Флора полуострова Камчатки, т. 2. Л., Изд-во АН СССР, 1929.
Комаров В. Л. Juniperus.— Флора СССР, т. 1. М.—Л., Изд-во АН СССР, 1934.
270
Комаров В. Л. Учение о виде у растений. М.—Л., Изд-во АН СССР, 1940.
Комаров В. Л. Флора Маньчжурии, ч. 2. Избранные сочинения, т. 4. М.—Л., Изд-во
АН СССР, 1950.
Кордэ Н. В. История альгофлоры некоторых озер Среднего Урала.— Тр. Лабор. сапро-
пелевых отложений Ин-та леса АН СССР, вып. 3. М., Изд-во АН СССР, 1949.
Коржинский С. И. Северная граница черноземностепной области восточной полосы
Европейской России в ботанико-географическом и почвенном отношениях.—
Тр. о-ва естествоисп. при Казанском ун-те, 1891, т. 12, вып. 6.
К о р ж и п с к и й С. И. Флора востока Европейской России в ее систематических и гео-
графических отношениях, т. 1. Томск, 1892.
Коржинский С. И. Следы древней растительности на Урале.— Изв. ими. Акад, наук,
№ 1, 1894.
Коржинский С. И. Об Урале как центре распространения растений.— Труды СПб.
о-на естествоисп., отд. бот., т. 25, протоколы заседаний, 1895.
Корнилова В. С. О полтавской флоре Казахстана. Докл. АН СССР, 1955, т. 104,
вып. 1.
К о т о в М. И. Linaria uralensis Kotov sp. п. Бот. ж. (АН УССР), № 3—4, 1946.
Красовский Л. И. и СкворцовА. К. Флора сосудистых растений района запо-
ведника «Денежкин Камень».— Труды заповедника «Денежкин Камень», вып. 1.
Свердловское кн. изд-во, 1959.
Крашенинников И. М. Анализ реликтовой флоры Южного Урала в связи с исто-
рией растительности и палеогеографией плейстоцена.— Сов. ботаника, 1937,
№ 4.
Крашенинников И. М. Основные пути развития растительности Южного Урала
в связи с палеогеографией Северной Евразии в плейстоцене и голоцене. — Сов.
ботаника, 1939, № 6—7.
Крашенинниковы И. М. и В. М. О новинках для флоры Оренбургской губернии.—
Русск. бот. ж., № 5—6, 1908.
Криштофович А. Н. Третичные растения с западного склона Урала.— Мат-лы по
геологии Башкирии. Труды Совета по изучению производительных сил, серия баш-
кирская, вып. 1, 1932.
Криштофович А. Н. Развитие ботанических провинций Европы с конца мелового
периода и связь их'с современным распределением растительности. Мат-лы Все-
союз. геогр. съезда, вып. 1. Л., 1933.
Криштофович А. Н. Остатки Oxycarpia в палеоценовых песчаниках Челябинска.—
Изв. АН СССР, 1934, серия 7, № 7.
Криштофович А. Н. Пальмы в третичных отложениях Южного Урала,—Природа,
1935, № 2.
Криштофович А. Н. Развитие ботанико-географических провинций северного полу-
шария с конца мелового периода.— Сов. ботаника, 1936, № 3.
Криштофович А. Н. Миоценовая флора Украины и ее связь через Урал с третич-
ной флорой Азии.— Сборник памяти акад. А. В. Фомина, 1938а.
Криштофович А. Н. Находка эоценовой Posidonia parisiensis Desm. на Восточном
склоне Урала. Изд. АН СССР, М., серия геол., № 3, 19386.
Криштофович А. Н. Каталог ископаемой флоры СССР. М.—Л., Изд-во АН СССР,
1941.
Криштофович А. Н. Материалы к третичной и меловой флоре Средней Азии.—
Бот. ж., 1941, т. 26, № 2—3.
Криштофович А. Н. Эволюция растительного покрова в геологическом прошлом и
ее основные факторы.— Мат-лы по истории флоры и растительности СССР, т. 2.
М.—Л., Изд-во АН СССР, 1946.
Криштофович А. Н. Эволюция растений по данным палеоботаники.— Пробл. бота-
ники. 1950, вып. 1.
Криштофович А. Н. Палеоботаника. 4-е изд. Л., Гостоптехиздат, 1957.
К р ы л о в П. Н. Материалы к флоре Пермской губ.— Труды о-ва естествоисп. при
Казанском ун-те, 1878, т. 6, вып. 1.
Крылов П. Н. Флора Западной Сибири. Тома 1—12. Изд. Томского ун-та, Томск,
1927—1964.
Кудряшов Б. В. Результат обследования торфяных болот близ села Ожогина Тюмен-
ского уезда в 1921 г.— Зап. Тюменского науч, о-ва изуч. места, края, 1924.
Кучеров Е. В. Пажитник плоскоплодный на Южном Урале.— Мат-лы Первого Все-
союз. совещ. ботаников и селекционеров (24—27 марта 1950 г.). М.—Л., Изд-во
АН СССР, 1954.
Лавренко Е. М. История флоры и растительности СССР по данным современного
распространения растений.— Растительность СССР, т. I. М.—Л., Изд. АН СССР,
1938.
Лавренко Е. М. Некоторые итоги обсуждения на конференции вопросов истории
флоры й растительности.— Труды ин-та географии, вып. 37. Проблемы палеогео-
графии четвертичного периода, М.—Л., 1946.
271
Лаврова М. А. Основные этапы истории четвертичного периода севера европейской
части СССР.— Мат-лы по четвертичному периоду СССР, вып. 3. М., Изд-во
.АН СССР, 1952.
Ланина Л. Б. Флора цветковых и сосудистых споровых растений Печорско-Илычско-
го заповедника.— Труды Печорско-Илычского заповедника, вып. 3, 1940.
Лепехин И. И. Дневные записки путешествия академика медицины доктора Ивана
Лепехина по разным провинциям Российского государства, ч. III. СПб. 1804.
Липшиц С. Ю. Новые виды рода Saussurea.— Бюлл. Моск, о-ва испыт. природы, от-
дел биол. 1954, т. 59, вып. 6.
Литвинов Д. И. Геоботанические заметки о флоре европейской России. — Bull. Soc.
Imper. natur. Moscou. Аппёе 1890, 1891.
Маевский П. Ф. Флора средней полосы европейской части СССР. Изд. 8. М., Сель-
хозгиз, 1954.
Малеев В. П. Juniperus.— Деревья и кустарники СССР. М.—Л. Изд-во АН СССР,
1949.
Марков К- К. Палеогеография. М., Географгиз, 1951.
Марков К. К-, Лазуков Г. И. и Николаев В. А. Четвертичный период (ледни-
ковый период — антропогеновый период). Том 1, территория СССР. Изд. МГУ,
1965.
Мартыненко М. А. Новый представитель реликтовой флоры Алтая.— Докл.
АН СССР, нов. серия, 1941, т. 31, вып. 9.
Мишкин Б. А. Флора Хибинских гор, ее анализ и история. М.—Л., Изд-во АН СССР,
1953.
Москвитин А. И. Схема палеогеографии плейстоцена европейской части СССР на осно-
ве новых представлений о стратиграфии четвертичных отложений.— Материалы
по четвертичному периоду, вып. 3. Изд. АН СССР. М., 1952.
Нагинский Н. А. К истории максимального оледенения Западно-Сибирской низмен-
ности.— Проблемы физической географии, вып. 12. М.—Л., Изд-во АН СССР,
1946.
Назаров М. И. Очерк растительности Окинского края в Восточном Саяне.— Изв. Гос.
геогр. о-ва. 1935, т. 67, вып. 1.
Нали'Вкин Д. В. Геологическая история Урала. Свердлгиз, 1943.
Наумова С. Н. Ботанико-географические исследования 1926 года в предгорьях Север-
ного Урала (р. Б. Сыня и Б. Оранец).—Тр. О-ва изучения Урала, Сибири и Даль-
него Востока. 1929, т. 1, вып. 1.
Невский С. А. О новом виде Agropyrum Gaerth.— Изв. Глав. бот. сада СССР, 1930,
т. 29, вып. 1—2.
Нейштадт М; И. О подразделении позднечетвертичной (послевалдайской или голо-
ценовой) эпохи в СССР и Европе.— Мат-лы п'о четв. периоду СССР, вып. 3, 1952.
Нейштадт М. И. К палеогеографии территории СССР в голоцене.— Изв. АН СССР,
серия геогр., 1955, вып. 5.
Нейштадт М. И. История лесов и палеогеографии СССР в голоцене. М.—Л., Изд-во
АН СССР, 1957.
Немкова В. К- Растительность южного (Башкирского) Предуралья в кинельское
время. Автореф; дисс. Саратов, 1964.
Никитин П. А. Коссожские флоры Западной Сибири.— Труды Биол. науч.-исслед.
ин-та при Томском ун-те, т. 1, 1935.
Никитин С. А. Лесорастительные условия низовий Урала.— Тр. Ин-та леса, т. 24,
1956.. .
Палибин И. В. Новые хвойные из неогена Урала и Кавказа.— Изв. Гл. бот. сада. Л.,
т. 30, 1931.
Перфильев И. A. Gypsophila pinegensis Perf. sp. п. Бот. ж., 1941, т. 26, № 1.
Петренко А. А. Геологический очерк Орско-Домбаровско'го промышленного района.—
Зап. Всесоюз. минерал, о-ва, 1939, т. 68, вып. 3.
Петров Б. Ф. Ландшафты центральной части Восточных Саян.— Землеведение,
т. 2 (42). Изд. Московского общества испытателей природы, 1948.
Победи мова Е. Г. Новый вид ясколки с Урала. Новости систематики высших рас-
тений, вып. 3. М,—Л., «Наука», 1968.
П о к р очв с к а я И. М. Основные этапы развития растительности Урала в течение мезо-
зоя и кайнозоя по данным пыльцевого анализа.— Труды конференции по спорово-
пыльцевому анализу. Изд. МГУ, 1950.
Покровская И. М. Основные этапы в развитии растительности на территории СССР
в третичное время (по данным палинологического анализа).— Бот. ж., 1954,
т. 39, № 2.
Покровская И. М. и др. Пыльцевой анализ. М., Госгеолиздат, 1950.
Поле Р. Р. Материалы для флоры Северной России.—'Изв. СПб. бот. сада, 1907, т. 7,
вып. 1; 1912, т. 12, вып. 5—6.
Поляков П. П. Растительность Джунгарского Алатау в районе рек Саркана и Бак-
сана.— Мат-лы исслед. растительности Казахстана. Изд-во АН Казах. ССР', 1951.
272 ' .
Пономарев А. Н. О местонахождениях Dryas punctata Juz. в западном При-
уралье.— Бот. ж., 1938, т. 23, № 2.
Пономарев А. Н. Об относительной стенотопности калькофитов флоры Среднего
Урала в связи с их реликтовым характером. — Докл. АН СССР, 1945, т. 48, № 3.
Пономарев А. Н. Лесостепной комплекс северной окраины Кунгурской лесостепи.—-
Изв. Естеств.-науч. ин-та при Пермском ун-те, 1948, т. 12, вып. 6.
Пономарев А. Н. О лесостепном флористическом комплексе Северного и северной
части Среднего Урала.— Бот. ж., 1949, т. 35, № 4.
Пономарев А. Н. К флоре Среднего Урала.— Изв. Естеств.-науч. ин-та при Перм-
ском ун-те, 1951, т. 13, вып. 2—3.
Пономарев А. Н. Степная растительность Вишневых гор Среднего Урала.— Докла-
ды пятого всеуральского совещания по географии и охране природы Урала.
Пермь. 1959.
Попов Н. С. Хозяйственное описание Пермской губернии. 1804.
Попов М. Г. Флора пестроцветных толщ (краснопесчаниковых нагорий) Бухары.
Фрагмент к истории флоры Туркестана.— Труды Туркестанского науч, о-ва, т. 1.
Ташкент, 1923.
П о п о в М. Г. Основные черты истории развития флоры Средней Азии.— Билл. Средне-
азиатского гос. ун-та, вып. 15. Ташкент, 1927.
Попов М. Г. Флора Средней Сибири, т. 1. Изд. АН СССР, 1957.
Ревердатто В. В. Очерк растительности Западного Саяна.— Изв. Зап.-Сиб. филиала
АН СССР, серия биол., 1946, № 1.
Рубцов Н. И. Растительность Джунгарского Алатау в районе верхнего течения рек
Аксу и Биень.— Мат-лы исследований растительности Казахстана. М.—Л., Изд-во
АН СССР, 1941а.
Р у б ц о в Н. И. Геоботанические исследования в бассейне р. Малой Алмаатинки.—
Мат-лы иеследований растительности Казахстана. Алма-Ата, Изд-во АН Казах.
ССР, 19416.
Рупрехт Ф. И. Флора Северного Урала. О распространении растений на Северном
Урале, по результатам географической экспедиции 1847 и 1848 годов.
В кн.: Э. Гофман «Северный Урал и береговой хребет Пай-Хой». т. 2. СПб.,
1856 (прибавление).
Рябова Т. П. Развитие растительности Башкирского Предуралья в голоцене.— Науч,
докл. высшей школы (биол. науки), № 1, 1965,-
С а к с В. Н. Четвертичное оледенение севера Сибири.— Природа. 1947, № 4.
Сакс В. Н. Опыт восстановления истории развития Сибири в четвертичный период.—
Мат-лы по четвертичному периоду СССР, вып. 3. Изд-во АН СССР, М., 1952.
Селиванова-Городкова Е. А. Особенности распространения видов рода
Alchemilla на Южном Урале.— Труды Башкирского гос. заповедника, 1963, вып. 2.
С и р и н Н. А. О следах двух оледенений на Приполярном Урале.— Бюлл; Ком. по изуч.
четв. периода, № 10, 1947.
СиязовМ. К. К ботанической характеристике Ялуторовска и его ближайших окрест-
ностей.— Зап. Зап.-Сиб. отд. Русск. географ, о-ва, 1892, т. 13, вып. 1.
Скворцов А. К. Новые флористические находки в районе Денежкина Камня.— Бот.
мат-лы гербария Бот. ин-та АН СССР, т. 19, 1959.
Словцов И. Я. Материалы к фитогеографии Тобольской губернии, Зап. Зап.-Сиб.
отд. Русск. геогр. о-ва, т. 12, 1891.
Соболев Л. Н. К характеристике растительного покрова заповедника «Боровое» на
фоне природных ландшафтов.— Землеведение, 1937, т. 39, вып. 4—5.
Соболевская К- А. Реликтовые ассоциации ледниковой эпохи в Хакассии.— Изв.
Всесоюз. геогр. о-ва, 1941, т. 73, вып. 3.
Соболевская К. А. Конспект флоры Тувы. Изд. Западно-Сибирского филиала
АН СССР, Новосибирск, 1953.
Соловьев Ф. А. Материалы к типологии островных сосновых боров Притоболья и юж-
ной части Челябинской области.— Природные условия и леса лесостепного
Зауралья. Труды Ин-та биологии УФАН СССР, 1960, вып. 19.
Сочава В. Б. О происхождении ареала некоторых растений уральской флоры.—
Ж- Русск. бот. о-ва, 1929, т. 14, № 5.
Сочава В. Б. Фрагменты горной степени на Среднем Урале. — Сов. ботаника, 1945,
№ 3.
Сочава В. Б. Тайга на северо-востоке Средне-Сибирского плоскогорья. Бот. ж., 1957,
т. 42, № 9.
Спрыгин И. И. Выходы пород, татарского яруса пермской системы в Заволжье как
один из центров видообразования в группе калькофильных растений (предвари-
тельное сообщение).— Сов. ботаника, 1934, № 4.
Спрыгин И. И. О некоторых лесных реликтах Приволжской возвышенности.— Уч. зап.
Казанского ун-та, т. 96, кн. 6 (ботаника), 1936.
Спрыгин И. И. Реликтовые растения Поволжья.— Мат-лы по истории флоры и
растительности СССР, I, 1941.
18 Заказ № 251 273
Степанова Е. Ф. Растительность и флора хребта Тарбагатай. Алма-Ата, Изд-во
АН Каз. ССР, 1962.
Страхов Н. ЛА. Основы исторической геологии, ч. I—II. М.—Л., Госгеолиздат, 1948.
Сукачев В. Н. Juniperus.— Флора юго-востока европейской части СССР, т. I, 1927.
Сукачев В. Н. История растительности СССР во время плейстоцена.— Раститель-
ность СССР, т. 1, 1938.
Сукачев В. Н. Очерк лесной растительности заповедника Боровое.— Труды заповед-
ника Боровое, 1948, т. 1.
Сукачев В. Н. и Поплавская Г. И. Очерк истории озер и растительности Сред-
него Урала в течение голоцена по данным изучения сапропелевых отложений.—
Бюлл. комиссии по изуч. четвертич. периода, № 8, 1946.
Сукачев В. Н., Горлова Р. Н., Недосеева А. К. и ЛА е т с л ь ц г з а Е. П.
О растительности перигляциальных зон центральной части Русской равнины.—
Докл. АН СССР, 1959, т. 125, № 2.
Съюор Д А. Ч. Века и растения. ЛА.—Л , ОНТИ, 1936.
Сю зев П. В. Конспект флоры Урала в пределах Пермской губернии. М., 1912.
Сюзев П. В. О флоре Среднего Урала. Дневник 1-го Всеросс. съезда русских бота-
ников в Петрограде в 1921 году. Пг., 1921.
Сюзев П. В. Краткий обзор рода Anemone L. на Урале. — Изв. Биол. науч.-исслед.
ин-та и биол. станции при Пермском ун-те, т. 1, вып. 3—4, 1923а.
Сюзев П. В. О находке Anemone reflexa Steph, в окрестностях Перми.— Изв. Глав,
бот. сада, 19236, т. 27, вып. 2.
Тарчевская 3. И. Растительность верхнего течения системы реки Уйбата.— Учёные
записки (Вологодский педагогический институт), вып. 2. Вологда, 1941.
Тарчевский В. В. Июсо-Шаринская степь.— Ученые записки (Вологодский педаго-
гический институт), вып. 2. Вологда, 1941.
Тихомиров Б. А. Пути формирования растительного покрова арктической Евразии
в четвертичном периоде.— Сов. ботаника, 1946, т. 14, № 5.
Толмачев А. И. О характере аркто-альпийских и некоторых локально связанных с
ними элементов флоры в районе рек Пинеги и Сотки.— Мат-лы по истории разви-
тия флор Европейского севера СССР. Изд. Северной базы АН СССР. Архангельск,
1938.
Толмачев А. И: О высокогорной флоре горы Лопатина (о. Сахалин).— Бот. ж., 1950,
т. 35, № 4.
Толмачев А. И. Деревья, кустарники и деревянистые лианы Сахалина.— ЛА.—Л.,
Изд-во АН СССР, 1956.
Троицкий Л. С. История оледенения Урала. — Оледенение Урала. М., «Наука»,
1966.
Тюремнбв.С. Н. К вопросу о пограничном горизонте.— Академику В. Н. Сукачеву
к 75-летию со дня рождения. Сборник работ по геоботанике, лесоведению,' палео-
географии и флористике. ЛА.—Л., Изд-во АН СССР, 1956.
У з н а д з е - ДТ е б у а д з е ЛА. Д. Эоценовая флора Южного Урала.— Тр. Геол, ин-та
АН Грузинской ССР, серия геол., т. 4 (9), 1948.
Федорова' Р. В. К вопросу о распределении граба в послеледниковое время на тер-
ритории европейской части СССР.— Пробл. физ. географии, вып. 16, 1951а.
Федорова Р. В. Материалы палеоботанического исследования погребенных торфяни-
ков Приполярного Урала.— Изв. Всесоюз. геогр. о-ва, 19516, т. 83, вып. 6.
Федоров Ал. А. и Федоров Ан. А. Два года в Саянах. М., Географгиз, 1951.
Федченко Б. A. Ericaceae.—Флора юго-востока европейской части СССР, т. 6, 1936.
Флора Армении, т. 3. Ереван, 1958.
Флора Казахстана, тома 1—9. Изд. АН Казахской ССР. Алма-Ата, 1956—1966.
Флора СССР. Тома I—XXV, XXX. М.—Л., Изд-во АН СССР, 1934—1960.
Флора Эстонской ССР, т. 2. Таллин, 1956.
Флора юго-востока европейской части СССР, тома 1—6. Л., 1927—1936.
Хвалина Н. Я- О растительности бассейна р. Сакмары во второй половине верхнего
плейстоцена.— Науч. докл. высшей школы (биол. науки), № 4, 1963.
Хвалина Н. Я. Растительность конца верхнего плейстоцена Башкирского Пред-
уралья (по данным спорово-пыльцевого анализа). Автореф. канд. дис. Саратов,
1965.
X р е б т о в А. А.. Об устройстве заповедника на «Липовой горе» (вблизи г, Перми).—
Экономика, 1925, № 7 (26) (Пермь).
Хребтов А. А. Полезные и вредные растения Урала. Свердл. обл. изд., 1941.
Худайбердыев Р. Ископаемые древесины из окрестностей оз. Смолино. — Бот. ж.,
1958, т. 43, № 5.
Чернов Г. А. Аллювиальные отложения верхней Печоры и Илыча.— Тр. Печорско-
Илычского заповедника, т. 1, 1940.
Ч е р н о в Е. Г. Вереск Calluna.— Флора Мурманской обл., т. 4, 1959.
Чигуряева А. А. Этапы развития растительности южного Предуралря в третичном
периоде на основании изучения спор и пыльцы. Автореф. дисс. Л., 1951.
274
Чигуряева А. А. Материалы к эоценовой растительности правобережья реки Дона.—
Уч. зап. Саратовского гос. ун-та, 1952, т. 35.
Чигуряева А. А. и Хвалина Н. Я. Материалы к флоре и растительности Башкир-
ского Предуралья в голоцене.— Науч. докл. высшей школы (биол. науки), № 1,
1961.
ЧудниковП. И. Талицкое учебно-опытное лесничество (исторический и естеств. исто-
рич. очерк).— Труды по лесн. опыта, делу Талицк. лесн. техникума, 1930.
Ш а м о в Д. Ф. и Микрюков М. Ф. Стерлитамакские шиханы — ценные памятники
неживой природы.— Состояние и задачи охраны природы Башкирии. Изд. Баш-
кирского филиала АН СССР, Уфа, 1960.
Шелль Ю. Материалы для ботанической географии Уфимской и Оренбургской губер-
ний.— Труды о-ва естествоисп. при Казанском ун-те, 1881, т. 9, вып. 5; 1883, т. 12,
добавление к вып. 4.
Ill и ш к и н Б. К. Ботанико-географический очерк Приморского склона Понтийского
хребта.— Закавказский краеведческий сборник, серия А, № 1. 1930.
Шишкин Б. К. Новые виды рода ясколки с Урала.— Труды Томского ун-та, т. 116,
сер. биол. Томск, 1951.
Шмидт Ф. Б. Амгунско-Буренская флора.— Труды Сибирской экспедиции Русского
географического общества, физ. отдел, 2, ботаническая часть. СПб., 1874.
Юдин Ю. П. Распространение и характер произрастания липы в бассейне р. Конды.—
Геоботаника, 1940, вып. 4.
Юдин Ю. П. О нахождении Gypsophila uralensis Less, на Тимане.— Бот. ж., 1946, №6.
Юдин Ю. П. Реликтовая флора известняков северо-востока европейской части СССР.—
Мат-лы по истории флоры и растительности СССР, вып. 4. М.—Л., Изд-во
АН СССР, 1963.
ЮзепчукС. В. Род Dasiphora Raf.— Флора СССР, т. 10, 1941.
Юзепчук С. В. Песчаные гвоздики Среднего Поволжья.— Бот. мат-лы гербария Бот.
ин-та АН СССР, т. 13. М.—Л., Изд-во АН СССР, 1950.
Юзепчук С. В. Новые манжетки европейской части СССР, Сибири и Средней Азии.—
Бот. мат-лы гербария Бот. ин-та АН СССР, т. 16. М.—Л., Изд-во АН СССР, 1954.
ЮзепчукС. В. Десять новых лапчаток.— Бот. мат-лы гербария Бот. ин-та АН СССР,
т. 17. М,—Л, Изд-во АН СССР, 1955а.
ЮзепчукС. В. Новые виды и список манжеток уральской флоры.— Бот. мат-лы гер-
бария Бот. ин-та АН СССР, т. 17. М.—Л., Изд-во АН СССР, 19556.
Ю к с и п А. Я. Новые виды ястребинок Советского Союза.— Бот. мат-лы гербария Бот.
ин-та АН СССР, т. 19. М.—Л., Изд-во АН СССР, 1959а.
ЮксипА. Я. Таблица для определения видов Hieracium L., обнаруженных на Урале.—-
Изв. Томского отд. Всесоюз. бот. о-ва, т. 4. Томск, 19596.
Ю н а т о в А. А. Основные черты растительного покрова Монгольской Народной Респуб-
лики.— Труды Монгольской комиссии, 1950, вып. 39.
Ю р ц е в Б. А. Материалы к систематике арктических остролодочников. — Бот. мат-лы
гербария Бот. ин-та АН СССР, т. 19, М.—Л., Изд-во АН СССР, 1959.
Яншин А. Л. Геологическая история Орского Урала в третичном периоде.— Изв.
АН СССР, серия геология, 1948, № 4.
ЯншинА. Л. Геология Северного Приаралья. Автореф. дисс. М., 1952.
Ярмоленко А. В. Значение ископаемых древесин для стратиграфии осадочных
пород.— Сов. ботаника, 1934, № 2.
Я хи м о в ич В. Л. Кайнозой Башкирского Предуралья, т. 1, ч. 2 и т. 2, ч. 1. 1958 (Баш-
кир. фил. АН СССР).
Beijerinck W. Calluna: a monograph on the Scotch heather. Verhandelingen der
Koninklijke Nederlandsche Akademie van Wetenschappen, Afdeeling Natuurkunde.
Tweede Sectie, v. 38, № 4, Amsterdam, 1940.
Bingham M. F. Flora of Oakland county, Michigan. Cranbrook Institute of Science,
Bulletin № 22, 1945.
В 6 ch er T. W. Studies on the plant geography of the North-Atlantic heath formation, 2.
Danish dwarf shrub communities in relation to those of Northern Europe. Det Kgl.
Danske Videnskab. Sebskab. Biol., v. 2, № 7, 1943.
В б c h e r T. W., K. Holmen, K. Jakobsen. Groenlands Flora. Kobenhavn, 1957.
Bowden Wray M. Cytotaxonomy of Potentilla fruticosa, allied species and cultivars. J.
of the Arnold Arboretum, 1957, v. 38, № 4.
В u r n a t E. Flore des Alpes Maritimes, v. 2, 1896.
Clapham A. R., Tutin T. G. a Warburg E.F. Flora of the Britisch Isles. Cam-
bridge, 1952.
Clarkson R. B. Schotch heather in -North America.—Castanea. J. South. Appalach.
Bot. Club., 23, 1958.
Dansereau P. Etudes macaronesiennes. III. La zonation altitudinale.— Naturaliste
Canadien, 1966, v. 93, № 6.
Drew W. B. The North American representatives of Ranunculus, Batrachium. Rhodora,
v. 38, 1936.
18*
275
E n d r i s s Z. Das Pflazenleben der Bithynischen Halbinsel.— Beihefte zum Bot. Cen-
tralbl., Bd 27, № 2, 1921.
Fava r ger CL Flore et vegetation des Alpes, V. 1, 1956.
Fernaid M. L. Gray’s manual of botany.— 8-th. edition, N. Y., 1950.
Fiori A. Nuov. Fl. anal. ItaL, v. 1, 1924.
Georgi J. G. Bemerkungen einer Reise in Russischen Reich in Jahren 1772—1774.
St.-Petersb., 1775;
Giesecke С. C. L. The Edinburg Encyclopedia, v. 10, pt 2, 1830.
Gimingham С. H. Biological flora of the Britisch Isles. Calluna vulgaris (L.) Hull.—
J. Ecology, v. 48, № 2, 1960.
Godwin H. The history of the British flora. A factual basis for phytogeography. Cam-
bridge. 1956.
Gray Asa. Calluna vulgaris and Aira caryophylles in the United States.— Amer. J. Sci.,
ser. 2, v. 32, 1861.
Gray Asa. Editor’s note.— Amer. J. Sci., ser. 2, v. 33, 1862.
Guppy H. B. Plants, seeds and currents in the West Indies and Azores, 1917.
H e g i G. Illustrierte Flora von Mittel-Europa, В. 1—7, Miinchen, 1906—1935.
Hooker W. Flora Boreali-Americana, v. 2, 1840.
Hopkins D. M. Cenozoic history of the Bering Land Bridge.— Science., v. 129, jNs 3362,
5 June, 1959.
H u 11 ё n E. E. Outline of the history of arctic and boreal biota during the Quaternary
period. Stockholm, 1937.
H u 11 e n E. The ampli-atlantic plants and their phytogeographical connections, 1958
Jepson W. L. A manual of the flowering plants of California, 1925.
Korshinsky S. Tentamen Florae Rossiae orientalis. Mem. de I’Academie Imp. des
Sciences de St.-Petersbourg, 1898, v. 7, № 1.
Kujala V. Untersuchungen fiber den Einfluss von Waldbranden auf die Waldvegetation
in Nord-Finland.— Commun. ex Instit. quast. forest. Finland., v. 10, 1926.
Kupffer K. R. Grundziige der Pflanzengeographie des Ostenbaltschen Gebietes, Riga,
1925.
Lawson G. Notice of the occurence of heather {Calluna vulgaris) at St. Ann’s Bay,
Cape Breton Island. Proc, and Trans. Nova Scotia Inst. Sci., v. I, 1864.
L. e Gallo S. La Calluna vulgaris L. dans 1’Amerique du Nord.— Naturaliste Canadien,
1945, v. 72, № 11—12.
Lessing Chr. Fr. Beitrag zur Flora des Stidlichen Ural und der Steppen — Linnaea,
Bd 9, 1834.
Lipschitz S. Galium Syreitschikovii — eine neue Art aus Stid-Ural Gebirge. Fedde,
Repert. sp. novarum, Berlin, 1929.
Litvinov D. Astragalus uralensis species nova.— Bull, de Societe Imper. des Natura-
listes de Moscou, v. 4, 1892.
Mattfeld J. Pflanzengeographische Gliederung Ost-Thraciens.— Verhandl. des Bot.
Vereins der Provinz Branderburg, Bd 7, 1929.
Ostenfeld С. H. a. Grontved J. The flora of Iceland and the Faroes, 1934.
Pigott C. D. The vegetation of Upper Teesdale in the North Pennines.— J. of Ecology,
1956, v. 44, № 2.
P or si Id A. E. Illustrated flora of the Canadian Artic Archipelago. Museum of Canada,
Bull. № 146, Ottawa, 1957.
Rudolph K. Microflor. Untersuchung tert. Ablagerungen im nordl. Bohmen. Beih. zum
Bot. Centralblatt, Bd 54, Abt. B., 1936.
Rydberg A. A monograph of the North American Potentillae. Memoirs from the De-
partment of Botany of Columbia University, v. 2, N. Y„ 1898.
Rydberg A. Dasiphora Raf. North American flora, v. 22, pt 4, N. Y., 1908.
Stojanoff N. Die Verbreitung der mediterranen Vegetation in Siidbulgarien.— Bot.
Jahrbiich. fur Systematik, Pflanzengeschichte und Pflanzengeographie, Bd 60, 1930.
S m a 11 J. K. Manual of the Southeastern flora, 1933.
Spetzman L. A. Vegetation of the arctic slope of. Alaska. Washington, 1959.
Sterner R. Flora der Inseln Oland. Acta phytogeographica suecia, v. 9, Uppsala, 1938.
Stojanoff N. Die Verbreitung der mediterranen Vegetation in Siidbulgarien. Bot.
Jahrbiich., v. 60, 1930.
УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ, УПОМЯНУТЫХ В ТЕКСТЕ
Abies Mill 8, И, 12, 15, 17, 18, 20, 27, 28
Abies sibirica Ledeb. 19, 20, 28, 46
Aceraceae Lindl. 9
Acer L. 10, 11, 12, 15, 17, 19, 20, 29
A. baschkirica Kolesn. 13, 14
A. lactim C. A. Mey 15
A. Myrtovae Baran. 16
A. Palibinii Baran. 16
A. platanoides L. 74, 253
A. Pojarkovas Baran. 16
Achillea nobilis L. 73, 74
Achyrophorus maculatus (L.) Scop. 39
Aconitum anthora L. 74, 245, 247, 262
A. nemorosum M. B. 248
Actaea grythrocarpa Fisch. 46, 48
A. spicata L. 35, 192
Adonis sibiricus Patr. 43, 62
Agropyrum desertorum (Fisch.) Schult. 74
A. imbricatum (M. B.) Roem. et Schult.
74
A. pruiniferum Nevski 133, 134, 260
A. reflexiaristatum Nevski 41, 50, 51, 55, 62,
63, 65, 71, 74, 76, 133, 187, 260
A. strigosum (M. B.) Bbiss. 133
Ajuga reptans L. 192
Alchetnilla L. 79, 80, 104, 106
A. amphipsila Juz. 106, 259
A. auriculata Juz. 104, 106, 259
A. cinerascens Juz. 106, 259
A. consorbina Juz. 104, 259
A. crassicaulis Juz. 106, 259
A. cunctatrix Juz. 104, 106, 259
A. exul Juz. 104, 259
A. glabra Neygenf 217, 218, 261
A. glabriformis Juz. 106, 259
A. Gortschakowskii Juz. 104, 106, 259
A. Haraldi Juz. 106, 259
A. Helenae Juz. 104, 106, 259
A. hyperborea Juz. 106, 259
A. iremelica Juz. 106, 259
A. kvarkushensis Juz. 106, 259
A. macroclada Juz. 106, 259
A. malimontana Juz. 104, 106, 259
A. obtusiformis Alech. 106, 259
A. paeneglabra Juz. 106, 259
A. parcipila Juz. 106, 259
A. pycnoloba Juz. 106, 259
A. riphaea Juz. 106
A. semispoliata Juz. 104, 106, 259
Aldrovanda Eleonorae Nikit. 17
A. vesiculosd L. 22
Alisma -plantago-minima (Nikit.) Dorof.
16
A. plantago-pliocenica (Nikit.) Dorof. 16
Allium hymenorrhizum Ledeb, 227, 228,
262
.4. obliqutim L. 248, 249, 262
A. rubens Schrad. 50, 65, 71
A. schoenoprasum L. 178
A. strictum Schrad. 38, 39, 50,51, 55, 62,65
Alnus Gaertn. 6, 9, 11, 15, 16, 17, 18, 20,
28, 29
A. bashkirica Dorof. 12
A. cordata (Loisl.) Desf. 11
A. fruticosa Rupr. 26, 183
A. glutinosa (L.) Gaertn. 16, 22
A. hirsutiformis Kolesn. 13, 14
A. incana (L.) Moench. 22, 26
A. Kefersteinii (Goeppert) Unger, 13, 15
A. nostratum Ung. 15
Alopecurus glaucus Less. 214, 218, 261
Alyssum biovulatum N. Busch. 39, 41,
43, 55, 56, 62, 71, 236, 237, 262
A. tortuosum Waldst. et Kit. 65, 73, 76
Ampelopsis rotundata Chandl. 11, 14
Amygdalus папа L. 70, 74
Andromeda L. 19
A. polifolia L. 21
A. protogaea Ung. 8, 9, 10
Androsace Bungeana Schischk. et Bobr.
201, 261
A. septentrionalis L. 46
Anemone L. 89
A. biarmiensis Juz. 62, 87, 91, 92, 152,
153, 259
A. coerulea DC. 147
A. jenisseensis (Korsh.) Kryl. 147
A. ranunculoides L. 147, 148
A. reflexa Steph. 249, 251, 252, 262
A. silvestris L. 39, 56, 62, 255, 263
A. uralensis Fisch. 147, 148, 151, 260
Anomodon viticulosus (Hedw.) Hook, et
Tayl. 41
Anthemis tinctoria L. 64, 76
Antitoxicum cretaceum Pobed. 76
A. stepposum Pobed. 50, 51, 56, 62, 65,
187, 255, 257
Apocynophyllum helveticum Heer. 9
Arabis hirsuta Scop. 50, 51, 62,
Aralia L. 11, 14
A. transversinervia Sap. et Mar. 8
A. venulosa Sap. et Mar. 8
Araucaria Juss. 8
Arctostaphylos uva-ursi (L.) Spreng. 25,
41
Arctous alpina (L.) Niedenzu 25, 39, 201,
261
Aristolochia clematitis L. 192
Armeria arctica (Cham.) Wallr. 25, 26
Artemisia L. 24, 25, 26, 27
A. armeniaca Lam. 39, 41, 43, 51, 63, 255,
263
277
A. austriaca Jacq. 65
A. bargusinensis Spreng. 228, 230, 262
A. borealis Pall. 39
A. campestris L. 65, 73
A. dracunculus L. 190
A. frigida Willd. 62, 63, 65, 70, 73, 255
A. laciniata Willd. 190
A. latifolia Ledeb. 62, 63, 255
A. macrantha Ledeb. 64
A. santolinifolia Turcz. 65, 227, 229,
262
A. sericea Web. 39, 41, 43, 50, 51, 55, 56,
62, 64. b5, 76, 190, 255, 263
Asarum europaeum L. 35, 192, 196, 197,
253, 263
Asperula odorata L. 35, 192, 196, 253, 263
A. petraea V. Krecz. 65, 71, 73, 76, 187,
189
Asplenium ruta-muraria L. 38, 39, 41, 43,
49, 51, 55, 62, 65, 187
A. trichomanes L. 65, 68
A. viride Huds. 38, 49, 51, 62
Aster alpinus L. 39, 41, 43, 50, 51, 55, 56,
62, 63, 65, 71, 73, 76, 255
Astragalus australis (L.) Lam. 110, 111
A. Clerceanus Iljin et Krasch. 46, 71,
108, 109, 260
A. consangiuneus Bong, et Mey 109
Л. danicus Retz. 39, 41, 51, 55, 62, 63, 64,
255
A. depauperatus Ledeb. 110
A. Helmii Fisch. 39, 62, 74, 76, 109, 111,
260
'A. heptapotamicus Sumn. 110
A. Karelinianus M. Pop. 65, 71, 74, 76,
100, 109, 110, 260
A. kungurensis Boriss. 65, 108, 260
A. longiflorus Pall. 108
A. Polakovii M. Pop. 110
A. sulcatus L. 64, 109
A. uralensis Litv. 110, 112, 260
Athyrium crenatum Rupr. 41
Atragene sibirica L. 50, 51
Atraphaxis frutescens (L.) Koch. 70, 189
Aulacomnlum palustre (Hedw.) Schwaegr.
165
Aulacospermum anomalum Ledeb. 119
A. darvasicum (Lipsky) Schischk. 119
A. isetense (Spreng.) Schischk. 62, 65,
76, 119, 121, 140, 260
Azalea viscosa L. 159
Azolla pseudopinnata Nikit. 13
Bartsia alpina L. 217
Berteroa incana (L.) DC. 76
Betulaceae S. Gray 9
Betula L. 9, 10, 11, 12, 16, 17, 18, 19, 20,
25, 27, 28, 29
B. alba L. 22
B. bashkirica Dorof. 12
B. exilis Sukacz. 171
B. fruticosa Pall 170, 171
B. glandulosa Michx. 173
B. humilis Schrank. 169, 170, 178, 183
B. macrophylla (Goeppert) Heer. 13, 14,
15
В. папа L. 21, 22, 25, 26
B. Noinskii Baran. 16
B. prisca Ettingsh. 13, 15
B. pubescens Ehrh. 26, 164
B. pumila L. 183
B. rotundifolia Spach. 169, 170, 171
В. T scher dincevii Baran. 16
B. verrucosa Ehrh. 46, 187, 190
Brasenia Schreb. 19
B. Schreberi J. F. Gmel. 12, 22
B. tuberculata C. et E. M. Reid. 12, 14
Bromus L. 101
B. Benekeni (Lange) Trimen. 192, 197
B. Julii Gowor 79, 79, 100, 103, 203, 259
B. sibiricus Drob. 100, 101
B. uralensis Gowor. 100, 103, 259
B. vogtilicus Socz. 79, 100, 103, 259
Bupleuruni inultinerve DC. 39 62, 65, 228,
229, 232, 262
Butomus umbellatus L. 17
Clamagrostis arundinacea (L.) Roth. 50
C. epigeios (L.) Roth. 190
C. lapponica (Wahl.) Hartm. 201, 261
Caldesia proveniitia Nikit. 11, 14
Calluna Salisb. 154, 160, 165, 166
C. vulgaris (L.) Hull. 154, 156, 157, 159,
161, 162, 163, 164, 166, 198, 262
Caloplaca elegans (Link.) Th. Fr. 49
Campanula rotundifolia L. var. linifolia
Wahl. 41
C. sibirica L. 63, 64, 73, 74
C. trachelium L. 192, 197
Candelariella vitellina (Hoffm.) A. Z. 49
Caragana frutex (L.) C. Koch. 70, 74, 187
C. jubata (Pall.) Poir. 170
Carduus nutans L. 76
Carex L. 16, 21, 25
C. alba Scop. 43, 49, 55, 248, 262
C. caucasica Stev. 210, 215, 216, 261
C. globularis L. 164
C. Ledebouriana C. A. Mey. 210, 214, 261
C. pediformis C. A. Mey. 62, 70, 73
C. pilosa Scop. 192
C. sabynensis Less. 210, 215, 261
C. silvatica Huds. 192
Carpinus L. 6, 11, 15, 17, 18, 20, 28, 29
C. betulus L. .16, 29
C. grandis Unger 13
Carya Nutt. 10, 11, 13, 15, 18, 19, 20
Cassiope D. Don. 26
Castanea Mill. 10, 19, 20
C. atavia Ung. 14
C. Kubinyii Kovats. 13
Castilleja pallida (L.) Kunth. 63
Cedrus Link. 8, 10, 11, 20
Celastrophyllum Benekenii Sap. et Mar. 8
Centaurea Mars chailiana Spreng. 76
C. ruthenica Lam. 74
C. sibirica L. 50, 63, 64, 65, 71, 73, 76,
187, 190
Cephalanthera longifolia (Huds.) Fritsch.
77, 192, 192, 261
Cerastium alpinum L. 94
C. arvense L. 95, 131
C. caespitosum Gilib. 95
C. Gorodkovianum Schischk. 79, 95, 97
259
C. Igoschinae Pobed. 95, 259
C. Krylov ii Schischk. et Gortschak. 79,
91, 94, 95, 101, 259
C. pauciflorum Stev. 249, 251, 262
C. Porphyrii Schischk. 79, 95, 97, 259
C. uralense Grubov 130
278
Cerasus fruticosa (Pall.) Woronov 70,71,
74, 187, 190
Chamaecyparis Spach. 11, 16, 19, 20
Ch. belgica Sap. et Mar. 7, 8
Ch. noontkatensis (Lamb.) Spach. 16
Chamaedaphne calyculata (L.) Moench.
159, 164
Chamaenerium angustifolium (L.) Scop.
50
Chamaerhodos erecta (L.) Bge 230, 235,
262
Chelidonium majus L. 76, 189
Chenopodiaceae Vent. 25, 27
Chenopodium rubrum L. 16
Cicerbita grandis (C. Koch.) A. Schchi-
an. 145
C. Gmelini Beauverd. 144
C. uralensis (Rouy) Beauverd. 77, 144,
145, 146, 151, 192, 260
Cinnamomum Blume 6, 7, 8, 9, 19
Circaea lutetiana L. 77, 192, 197
Cladonia chlorophaea (Flk.) Spr. 45
C. cornuta Nyl. 190
C. destricta Nyl. 45
C. mitis Sanst. 190
C. pyxidata (L.) Fr. 190
Clausia aprica (Steph.) Korn. 65, 70
Cobresia Bellardi Degl. 170
Compositae Geseke 11
Comptonia Banks 6, 20
C. dryandroides Ung. 14
C. oeningenensis A. Br. 13
Convallaria majalis L. 192, 197
Cornus L. 10, 17, 19
C. alba L. 12, 16, 17
C. angusta Baran. 8
C. Baranovii Vasilevsk. 8
C. Janschinii Vasilevsk. 8
C. mugodsharica Krysht. 7
C. sezanensis Long. 8
Cortusa Matthioli L. 38, 41, 43, 62, 65
Corylus L. 6, 11, 16, 17, 19, 20, 29, 33
C. avellana L. 16
C. Macquarrii (Forbes.) Heer. 13, 15
C. turgaica A. Pojark. 11
Coryospermum Marschallii Stev. 25
Cotoneaster melanocarpa Lodd. 41, 51,
62, 70, 71, 187, 190, 255
Crataegus Fominii Krysht. 13
Crocynia membranacea (Dicks.) A. Z. 45
Cryptogramma crispa (L.) R. Br. 38
C. Stelleri Prantl. 38, 43, 62
Cupressaceae F. W. Neger 10
Cyperites sp. 11
Cyperus fuscus L. 17
C. glomeratus L. 17
Cypripedium guttatum Sw. 41
Cystopteris Dickieana Sims. 41
C. fragilis (L.) Bernh. 38, 41, 43, 45, 46,
50, 51, 55, 65, 187
Cytisus ruthenicus Fisch. 51, 70, 71, 19Q
Dacrydium Soland. 15
Dalbergia bella Heer. 9
Daphnogene crassifolia Baran. 8
D. gigas Baran. 8
D. Vasilev ska'] ae Baran. 8
Dasiphora Raf. 167, 169
D. dryadanthoides Juz. 167, 168
D. fruticosa (L.) Rydb. 154, 167, 168,
169, 170, 171, 172, 173, 174, 175, 176,
178, 179, 180, 181, 182, 183, 214, 261,
262
D. parvifolia (Fisch.) Juz. 167, 169
D. phyllocalix Juz. 167
Decodon gibbosus E. M. Reid. 12, 14
D. globosus (E. M. Reid.) Nikit. 13, 16, 17
Dendranthema Zawadskii (Herbich.)
Tzvel. 38, 41, 43, 51, 55, 65
Dentaria tenuifolia Ledeb. 251, 252, 262
Dewalquea gelindenensis Sap. et Mar. 8
D. grandifolia Krassnow 7, 8
Dianthus acicularis Fisch. 39, 41. 43, 50,
51, 55, 62, 65. 71, 73, 74, 76, 125, 126.
127, 128, 260
D. arenarius L. 128
D. kirghizicus Schischk. 129
D. pallidiflorus Ser. 129
D. ramosissimus Pall. 129
D. repens Willd. 41, 43, 199, 201, 261
D. rigidus M. B. 129
D. squarrosus M. B. 128
D. toboliensis Klok. 128
D. uralensis Korsh. 65, 128, 129, 260
D. versicolor Fisch. 50, 51, 56, 62, 65, 71,
255
D. volgicus Juz. 128
Diclidocarya Menzelii E. M. Reid 12
Digitalis grandiflora Mill. 192, 195, 261
Diospyros L. 20
D. brachysepala A. Br. 13
Distylium uralensis Kolesn. 12, 14
Draba hirta L. 39
Dracocephalum Ruyschiana L. 51
D. thymiflorum L. 76
Drosera rotundifolia L. 181
Dryandra R. Br 7
D. karaculensis Baran. 10
D. Schrankii (Sterb.) Heer. 7, 8
Dryas octopetala L. 25, 26, 39, 176, 199,
201, 261
D. punctata Juz. 39, 65, 198, 199, 200,
201, 261
Dryophyllum Dewalquei Sap. et Mar. 7,
19
D. furcinerve (Rossm.) Heer 8, 9
Dryopteris filix-mas (L.) Schott. 35, 46,
192
D. Linnaeana C. Chr. 46, 47
D. Robertiana (Hoffm.) C. Chr. 38, 43,
45, 46, 47, 51, 65
Dulichium spathaceum Rich. 16
D. vespiforme C. et E. M. Reid 13
Echinops L. 26
E. ritro L. 50, 51, 56, 62, 63, 65, 70, 71,
73, 74, 187, 255, 263
Ephedra L. 25, 26, 27
E. distachya L. 70
Epilobium alpinum L. 92, 93, 94 .
E. uralense Rupr. 92, 93, 94, 259
Epipactis rubiginosa Crantz. 38, 43, 55,
62, 65
Epipremnum crassum C. et E. M. Reid 13
E. cristatum Nikit. 11, 14, 16
E. ornatum Reid et Chandler 11, 14
Equisetum scirpoides Michx. 41
Ericaceae Juss. 10, 11, 12, 17, 19
Erica L. 165
E. azorica 159
279
Eriophorum angustifolium Roth. 181
Eritrichium pectinatum (Pall.) DC. 118,
119
E. uralense Serg. 41, 43, 71, 118, 119, 260
Erysimum Marschallianum Andrz. 62
Eucalyptus L’Her. 9, 10
Eugenia Michx. 10
Euphorbia Serguieriana Neck. 46, 50, 51,
65, 76
Eurotia Adans. 26
Fagaceae Dum. 9, 15
Fagus L. 6, 11, 15, 17, 18, 20, 28
F. Antipofii (Abich) Heer. 13, 14
F. deucalionis Ung. 10
Ferula caspica M. B. 70, 189
Festuca gigantea (L.) Vill. 192
F. Kryloviana Reverd. 212, 217, 261
F. pseudovina Hack. 39, 51, 56, 58, 62,
64, 255
F. silvatica (Poll.) Vill. 70, 192, 194, 260
F. sulcata Hack. 49, 51, 65, 70, 73, 190,
192, 194
Ficus L. 7, 9, 19
F. Gordjaginii Baran. 10
F. multinervis Heer 9
F. uralica Krysth. 8
Filipendula hexapetala Gilib. 50, 63, 74,
190, 255
F. ulmaria (L.) Maxtni. 22
Fragaria viridis Duchartre 190
Frangula alnus Mill. 22
Fraxinus L. 17, 20, 28
F. excelsior L. 16
Gagea fistulosa (Ram.) Ker-Gawl. 94
G. samojedorum Grossh. 94, 259
Galatella angustissima (Tausch.) Novo-
pokr. 73
Galium L. 196
G. boreale L. 50, 74, 190
G. paradoxum Maxim. 195, 196
G. Syreitschikovii Lipsch. 196
G. verum L. 51, 55, 62, 65, 190, 255, 263
187, 190, 255
Genista tinctoria L. 50, 51, 62, 64, 70, 71,
187, 190, 255
Gentiana barbata Froel. 50, 240, 262
G. dectimbens L. f. 237, 239, 262
Geranium bifolium Patz. 247
G. pseudosibiricum J. Mayer. 246, 247,
262
G. Robertianum L. 192, 194, 197, 261
Ginkgo L. 20
G. adiantoides (Unger) Heer. 13
Glyceria fluitans (L.) R. Br. 16, 161
Glyptostroboxylon tenerum. Krausel 14
Glyptostrobus Endl. 14, 20
G. europaeus (Br.) Heer 11, 13, 14
G. parisiensis Brongn. 8
Goniolimon speciosum (L.) Borss 71, 73
Grewiopsis uralensis Vasilevsk. 8
Gypsophila L. 84, 89
G. altissima L. 56, 62, 63, 64, 65, 71, 76,
255, 263
G. paniculata L. 76, 178, 190
G. Patrinii Ser. 56, 57
G. pinegensis Perf. 84
G. Sambukii Schischk, 84
G. Steupii Schischk. 84
G. tenuifolia M. B. 84
280
G. uralensis Less. 39, 50, 51, 82, 84, 86,
152, 153, 259
Gyrophora Milhlenbergii Ach. 45
Hackea sp. 10
H amamelidaceae R. Br. 12
Hedwigia ciliata (Ehrh.) Hedw. 45
Hedysarum cretaceum Fisch. 114, 115
H. Gmelini Ledeb. 114, 115, 117
H. grandiflorum Pall. 76
H. Razoumovianum Fisch. 114, 115, 117,
260
FI. ucrainicum Kaschm. 114, 115
Heleocharis palustris A. Br. 17, 25
Helianthemum nummularium (L.) Dunal.
55, 63, 64
H. rupifragum A. Kerner. 65
Helictotrichon desertorum (Less.) Pilger
50, 51, 55, 56, 58, 62, 64, 65, 71, 73, 74,
79, 106, 190, 255, 263
H. Schellianum (Hack.) Kitagawa 73
Helleborus caucasicus A. Br. 7
Hicoria Raf. 13, 20
H. riphaeica Krysht. 13
Hieracium cuspidellum Pohle et Zahn.
107, 259
H. Gorodkowianum Juxip 107, 259
H. iremelense Juxip 106, 259
H. Ijapinense Juxip 106, 259
H. konshakovskianum Juxip 107, 259
H. kosvinskiense Juxip 107, 259
H. kuroksarense Juxip 107, 259
H. manifestum Juxip 107, 259
H. pyrsjuense Juxip 106, 259
H. Soczavae Juxip 107, 259
H. stenopiforme Pohle et Zahn. 107, 259
H. suberectum Schischk. et Steinb, 106,
107, 259
H. uralense Elfstr. 106, 259
H. wologdense Pohle et Zahn. 107, 259
Hippuris vulgaris L. 22, 25
Humulus lupulus L. 187
Hylocomium splendens (Hedw.) Br. et
Sch. 41
Hypericum coriaceum Nikit. 16, 17
H. septestum Nikit. 13
Ilex L. 10, 11, 12, 14, 20
Impatiens uralensis Skvortz. 79, 80
Inula hirta L. 50
Iris sibirica L. 174
Juglandaceae A. Rich 9, 11, 15
Juglans L. 6, 10, 15, 17, 18, 19, 20
J. acuminata A. Braun. 13, 16
Juniperoxylon uralense Jarm. 14
Juniperis communis L. 41, 46, 47
J. depressa Stev. 184
J. oxycedrus L. 159
J. sabina L. 12, 22, 70, 154, 183, 184,
185, 188, 189, 190, 191, 197, 261, 262
J. sibirica Burgsd. 171
Knautia montana (M. B.) DC. 144
K. tatarica (L.) Litv. 77, 144, 145, 151,
192, 260
Kochia Roth. 26
Koeleria gracilis Pers. 50, 70, 73
K. sclerophylla P. Smirn. 65, 134, 260
K. Talievii Lavr. 134
Lagotis Gaertn. 89
L. decumbens Rupr. 82
L. glauca Gaertn. 82
L. Ikonnikovii Schischk. 82
L. integrifolia (Willd.) Schischk. 82
L. minor (Willd.) Standi. 82
L. uralensis Schischk. 81, 82, 259
Larix Mill. 12, 16, 17, 20, 28
Laser trilobum (L.) -Borkn. 192, 193, 261
Lathyrus Gmelini (Fisch.) Fritsch. 241,
262
L. Litvinova Iljin. 142, 144, 151, 192, 260
L. miniaius M. B. 143, 144
L. rotundifolius Willd 143
Laurus L. 9, 19
L. nobilis L. 159
L. Omalii Sap. et Mar. 8
Lavateru tliuringiaca L. 74
Leconora Garovaglii (Kerb.) A. Z. 45
L. muralis (Schreb.) Rabh. 45
Ledum palustre L. 46, 47, 164
Leonurus glaucescens Bge 76
Lepraria cklorina Ach. 45
Leucodon sciuroides (Mill.) Garcke 45
Libanotis condensata (L.) Crantz. 133
L. Schrenkiana C. A. Mey. 133
L. seseloides (Fisch, et Mey.) Turcz. 133
L. -sibirica (L.) C. A. Mey. 39, 41, 50, 51,
55, 62, 74, 76, 131, 132, 133, 260
L. uralensis Nevski. 39, 50, 131
Ligularia sibirica (L.) Cass. 178
Limonium caspiutn (Willd.) Gams. 76
Linaria altaica Fisch. 221, 222, 262
L. debilis Kupr. 222, 223, 262
L. uralensis Kotov 222
Linnaea borealis L. 46, 47
Linum L. 89
L. altaicum Ledeb. 87
L. atricalyx Juz. 87
L. boreale Juz. 86, 89, 90, 259
L. extraxillare Kit. 87
L. Komarovii Juz. 87
Liquidambar L. 6, 11, 17, 18, 20
L. europaeum A. Br. 13, 14
Liriodendron L. 16, 20
L. geminata Kirchn. 12, 14
L. tulipifera L. 16
Litsaea Troschinii Baran. 10
Lloydia serotina (L.) Reichb. 201, 261
Lophozia quinquedentata Cogn. 41
Lycopodium clavatum L. 164
Lycopus europaeus L. 22
Magnolia L. 7, 8, 19
M. spectabilis Kn. 15
Medicago falcata L. 190
Melandrium album (Mill.) Garcke 74
Melica transsilvanica Schur. 62, 65, 74
Menyanthes trifoliata L. 21, 22, 25, 181
Mercurialis perennis L. 192
Metasequoia Miki 16, 20
Minuartia Biebersteinii (Rupr.) Schischk.
122
M. Helmii (Fisch.) Schischk. 41, 50, 51,
65, 73, 121, 123, 152, 260
M. Kraschenlnnikovii Schischk. 56, 65, 71,
76, 122, 124, 125, 140, 260
M. Kryloviana Schischk. 122
M. setacea (Thuill.) Hayek. 125 ,
M. verna (L.) Hiern. 39
Morus L. 12, 17
M. nigra L. 12
Mulgedium hispidum DC. 77
Myricaceae Blume 10
Myrtaceae Juss. 9, 10, 19
Myrica L. 19
M. angustata Schimp. 9
M. carpinifolia Heer. 16
M. Mairel Zevelle 13
M. Suppanii Kirch. 11, 14
M. Torreyi Lesq. 15
M. uralica Uzn.-Dgeb. 9
M. vindobonensis (Ett.) Heer. 13, 14, 20
Myrsine doryphora Ung. 9, 13, 20
Myrtophyllum Warderii Sap. 9, 10
Najas L. 35, 36
N. flexilis (Willd.) Rostk. et Schmidt 35,
36
N. foveolata A. Br. 13
N. lanceolata C. et E. M. Reid 13, 17
N. marina L. 16, 17, 22, 35, 36
N. marina L. var. brachycarpa Trautv. 35
Nardus stricta L. 159, 161
Naumburgia subthyrsiflora Nikit. 13
Neslia paniculata (L.) Desv. 22
Nipa Thunb. 6, 19
Nuphar luteum (L.) Smith 22
Nymphaea alba L. 22
Nyssa L. 10, 11, 17, 18
Onobrychis tanaitica Spreng. 255
Onosma borystenicum Klok. 140
O. graniticola Klok. 140
O. guberlinense Dobrocz. et Vinogr. 140,
260
O. simplicissimum L: 50, 55, 62, 64, 65,
73, 74, 140, 187, 190, 255, 256, 263
O. transrhymnense Klok. 140
Oreodaphne mugodsharica Baran. 11
Origanum vulgare L. 51, 62, 65
Orostachys spinosa (L.) C. A. Mey. 71,
73, 190, 220, 221, 262
Osmorhiza aristata (Thunb.) Makino et
Vabe 196
Osmunda L. 7
Oxycarpia bifaria Trautschold 8
Oxycoccus quadripetalus Gilib. 181
Oxygraphis gracialis (Fisch.) Bge 206,
207, 261
Oxytropis DC. 237
O. approximata Less. 112, 113, 260
O. Gmelini Fisch. 113, 260
O. Hippolyti Boriss. 113, 114, 260
O. pilosa (L.) DC. 55, 64, 74, 255
O. songorica (Pall) DC. 114
O. sordida (Willd.) Pers. 39, 113
O. spicata (Pali.) O. et B. Fedtsch. 71,
74, 76, 114, 115, 260
O. uralensis (L.) DC. 41, 237, 238, 262
Pachypleurum alpinum Ledeb. 25
Padus racemosa (Lam.) Gilib. 22
Palibinia 7
Parietaria micrantha Ledeb. 46, 55, 62, 65
Parmelia conspersa Ach. 45
P. molliuscula Ach. 45
P. omphalodes (L.) Ach. 45
Parnassia palustris L. 178
Patrinia sibirica (L.) Juss. 228, 231, 262
Pedicularis compacta Steph. 209, 210, 213,
261
P. Oederi Vahl. 201, 261
P. sceptrum-carolinum L. 174
P. silvatica L. 161
281
uralensis Vved. 80
P. verticillata L. 201, 261
Phleum phleoides (L.) Simonk. 51, 55, 56,
62, 63, 64, 190, 255, 263
Phlojodicarpus villosus Turcz. 206, 261
Phlomis tuberosa L. 50, 62, 65, 255, 263
Phlox sibirica L. 40, 41, 43, 56, 222, 224,
225, 262
Phragmites oeningenensis (A. Br.) Heer
13
Phyllites sp. 8
Physcia caesia (Hoffm.) Hampe 49
Picea Dietz. 10, 11, 12, 15, 16, 17, 18, 19,
20, 22, 27, 28
P. excelsa Link. 21, 26
P. obovata Ledeb. 21, 26, 45, 46, 47
P. omorica Purk. 21
P. orientalis (L.) Link. 21
P. Schrenkiana Fisch, et Mey. 11
Piceostrobus Neustruevi Palib. 10
Pimela sp. 8
Pimpinella titanophila Woron 65
Pinguicula alpina L. 199, 261
P. vulgaris L. 178
Pinaceae Lindl. 11
Pinus L. 10, 11, 12, 15, 16, 17, 19, 27, 28,
29
P. laricio Poir. 12
P. nigra Arn. 12
P. Pallasiana Lamb. 12
P. pumila (Pall.) Rgl. 171
P. sibirica (Rupr.) Mayr. 19, 46
P. Silvestris L. 12, 19, 26, 29, 46, 47, 187
P. Thomasiana (Goepp.) E. Reich. 12
Pityoxylon Paxii Krausel 14
Platanus L. 20
P. aceroides (Goeppert). Heer 13
Pleurozium Schreberi Mitt. 41, 164
Poa stepposa (Kryl.) Roshev. 56, 62, 65,
73, 74, 255
Podocarpoxylon uralense Chudajb. 15
Podocarpus L’Her. 8, 10
P. eocenica Ung. 9
Pohlia nutans (Hedw.) Lindb. 45
Polemonium coeruleum L.’ 174
P. lapponicum Klok. 86
P. nudipedum Klok. 84, 86, 87, 259
P. onegense Klok. 86
P. pulchellum Bge. 86
Polygala comosa Schkuhr. 50
Polygonatum officinale All. 50, 51, 62,
65, 187
Polygonum L. 21
P. bistorta L. 178
P. lapathifolium L. 16, 22
P. viviparum L. 62, 201, 261
Polypodium vulgare L. 38, 45, 46, 50, 51
Polystichum lonchitis (L.) Roth. 38
Polytrichum commune Hedw. 165
Populus L. 6, 22
P. balsamoides Goepp. 13, 15, 16
P. Komaroviana Baran. 16
P. latior A. Br. 16
P. mutabilis Heer. 16
P. nigra L. 22, 189
P. tremula L. 16, 74
Posidonia oceanica L. 10
P. parisiensis Fritel. 10
Potamogeton bashkiricus Dorof. 17
282
P. coloratus Vehl. 16
P. filiformis Pers. 17, 25, 26
P. natans L. 22, 25
P. palaeocompressus Dorof. 17
P. pectinatus L. 17
P. planus Nikit 13
P. polygonifoius Pourr. 161
P. pusillus L. 25
P. trichophyllus Dorof. 17
P. vaginatus Turcz. 16
Potentilla argentea L. 50, 121
P. canescens Bess. 121
P. Eversniiintiiana Fisch. 119, 122, 260
P. frulicosn L. 167, 174, 181
P. Gordiaginii Juz. 121, 260
P. humifusa Willd. 50, 51, 64, 71
P. Kusnetzowii (Gowor.) Juz. 21, 39, 62,
200, 261
P. multifida L. 119
P. nivea L. 39
P. sericea L. 119, 234, 235, 236, 262
P. volgarica Juz. 119
Primula Pallasii Lehm. 205, 261
P. cortusoides L. 243, 244, 262
Proserpinacea reticulata C. et E. M. Reid
12, 14
Pseudolaricixylon firmoldes Chudajb. 14,
15
Pseudolarix Gord. 15
P. amabilis (Nels.) Rehd. 15
P. fossilis Jarm. 13, 14
Pseudotsuga Cirr. 18
Pterocarya Kunth. 11, 15, 16, 17, 18, 20
P. castaneifolia (Goepp.) Schlecht 13
Pulmonaria officinalis L. ssp. obscura
Murb. 192
Pulsatilla flavescens (Zucc.) Juz. 39, 43,
50, 51, 62, 65, 190
Pyrethrum uralense H. Krasch. 140, 261
Quercus L. 9, 10, 11, 15, 17, 18, 19, 20,
28, 29, 33
Q. Alexeevii A. Jojark. 11
Q. apocynophyllum Ett. 9
Q. bifur cat a Wat. 8, 9
Q. diplodon Sap. et Mar. 7, 10
Q. elaena Ung. 9
Q. furcinervis (Rossm.) Heer. 8, 9
Q. groelandica Heer. 15
Q. kamyschinensis Goepp. 10
Q. Kryshtofbvichiana Baran. 16
Q. neriifolia Ung. 13, 14, 20
Q. Nimrodi Ung. 13
Q. odontiphylla Sap. 10
Q. Olafsenii Heer. 13
Q. platania Heer. 10
Q. robur L. 22, 74
Q. roburoides Bereng. 16
Ranunculus hederaceus L. 161
R. sceleratus L. 21
R. sceratoides Nikit. 16
Rhamnus L. 20
Rh. cdthartica L. 187
Rh. Graeffii Heer. 13
Rhodiola L. 80
Rh. iremelica Boriss. 79, 80
Rh. quardrifida (Pall.) Fisch, et Mey.
206, 208, 261
Rh. rosea L. 39
Rhus L. 10, 11, 15, 17, 19
Rh. quercifolia Goepp. 16
Rh. turcomanica (Kryscht.) Korovin 10
Rhytidium rugosum Lindb. 41
Rhynchospora alba (L.) Vahl. 181
Roegneria Komarovii Nevski 134
R. taigae Nevski 134
R. tianschanica (Drob.) Nevski 134
R. uralensis Nevski 134, 135, 140, 260
R. viridiglumis Nevski 62, 134, 136, 260
Rubus caesius L. 187
R. idaeus L. 46, 47, 50, 51
R. saxatilis L. 164
Rumex crispus L. 22
R. maritimus L. 17
Sabal Adans 6, 8, 9, 10, 19
Sagittaria sagittifolia L. 17
Salicaceae Mirbel. 9
Salix L. 9, 10, 11, 16, 17, 19, 20, 28
S. acutissima Goepp. 16
S. angusta A. Br. 16
S. aurita L. 22
S. cinerea L. 22
S. depressa L. 169, 170
S. herbacea L. 25
S. Integra Goepp. 16
S. Kochiana Trautv. 170
S. polaris Wahl. 25
S. pyrolifolia Ledeb. 178, 179
S. reticulata L. 39, 201, 261
S. sibirica Pall. 190
S. varians Goepp. 16
S. viminalis L. 22
Salsola L. 26
Salvia stepposa Schost. 64, 65, 73, 76, 190
Salvinia Micheli 19
S. glabra Nikit. 18
S. tuberculata Nikit. 13, 16
Sambucus L. 12, 17
Sanguisorba officinalis L. 50, 178
Sanicula europaea L. 35, 77, 192, 194,
195, 196, 197, 254, 261
S. uralensis Kleop. 195
Sassafras Ness. 20
S. Ferretianum Massal. 13, 14
Saussurea alpina (L.) DC. 103
S. controversa DC. 62, 103, 243, 244,263
S. parviflora (Poir.) DC. 244, 263
S. uralensis Lipsch. 101, 104, 260
Saxifragaceae Juss. 11
.Saxifraga caespitosa L. 39, 199, 262
S. cernua L. 56, 58
S. nivalis L. 39
S. oppositifolia L. 39
S. sibirica L. 62, 65
Schiverekia Andrz. 137, 140
Sch. berteroides Fisch. 65, 73, 134, 137,
187, 189, 261
Sch. Kusnezovii Alex. 41, 62, 134, 136,
137, 261
Sch. monticola Alex. 62, 65, 71, 134, 13Q,
261 \
Sch. monticola Alex. ssp. mutabilis Alex.
134, 137
Sch. mutabilis Alex. 134
Sch. podolica Andrz. 134
Scirpus L. 21
S. lacustris L. 22, 165
Scorzonera austriaca Willd. 99
S. glabra Rupr. 99
S. Ruprechtiana Lipsch. et Krasch. 39, 41,
99, 100, 101, 160
Scrophularia Scopolii Hoppe 192, 262
Scutellaria altissima L. 192, 262
S. cisvolgensis Juz. 118
S. oxyphylla Juz. 65, 73, 117, 118, 261
S. supina L. 117
Sedum acre L. 51, 62, 63
S. hybridum L. 65, 69, 187, 190, 224,
228, 263
S. purpureum (L.) Schult. 39, 41, 46, 50,
51, 62 ,
Selaginella pliocenica Dorof. 16
S. selaginoides (L.) Link. 21, 25, 26
Senecio Igoschiuue Schischk. 95, 99, 101,
260
S. integrifolius (L.) Clairv. 99
Sequoia Endl. 8, 9, 11, 15, 17, 19, 20
S. Couttsiae Heer. 6, 8, 9
S. Langsdorfii (Br.) Heer. 6, 11, 14, 15
S. Sternbergii (Goeppert) Heer. 9
Serratula centauroides L. 129
5. Gmelini Tausch. 129, 131
S. radiata (Waldst. et Kit) M. B. 130
S. tanaitica P. Smirn. 130
Seseli Ledebourii G. Don. 62, 256
Sieglingia decumbens (L.) Bernh. 161
Silene altaica Pers. 235, 237, 263
S. baschkirorum Janisch. 39, 50, 65, 76,
129, 130, 161
S. densiflora D’Urv. 129
S. nutans L. 76
S. polaris Kleop. 129
S. pseudotites Bess. 129
S. wolgensis (Willd.) Bess. 129
Siler trilobum Scop. 193
Solidago virga-aurea L. 50, 164
Sparganium noduliferum C. et E. M. Reid.
13
Sphagnum fuscum (Schimp.) Klinggr. 181
Sph. teres (Schimp.) Angstr. 181
Spirata crenata L. 50, 63, 70, 187, 190, 255
S. salicifolia L. 169
Stachys silvatica L. 35, 192, 197, 253, 263
Statice L. 23
Steinhauera subglobosa Presl. 8
Stipa L. 190
S. Joannis Celak. 50, 56, 64, 65, 120, 255
Swertia obtusa Ledeb. 208, 209, 210, 261
Syncarpia Ten. 9
Tanacetum uralense (Krasch.) Tzvel. 74,
76, 140, 141, 260
T. millefoliatum (L.) Tzvel. 140
Taraxacum tanaiticum Dorof. 16
Taxodiaceae E. W. Neger. 9, 10, 11
Taxodioxylon distichum (Merckl.) Jarm.
14
T. Krascheninnikovii Jarm. 14
T. sequoianum Krausel 14
Taxodium Rich. 9, 11, 14, 15, 17, 20
T. distichum (L.) Rich. 6
T. distichum miocaenum Heer. 15
T. dubium (Sternb.) Heer. 14
Terminalia L. 9
Thalictrum angustifolium L. 22
T. foetidum L. 65, 70, 74, 187, 247, 262
Thesium refractum C. A. Mey. 55
Thlaspi arvense L. 22
283
Th. cochleariforme DC. 71, 208, 212, 213,
261
Thuidium abietinum Br. et Sch. 41, 45
Thymus L. 136, 139, 140
Th. asiaticus Serg. 136
Th. baschkiriensis Klok. et Schost. 136,
139, 260
Th. binervulatus Klok. et Schost. 138,
139, 260
Th. guberlinensis Iljin 138, 139, 260
Th. hirticaulis Klok. 136, 260
Th. Marschallianus Willd. 65, 73, 76, 187
Th. mtigodzaricus Klok. et Schost. 73, 76,
138, 139, 2G0
Th. paucifolius Klok. 99, 100, 101, 102,
259
Th. serpyllum L. 41, 62, 65, 73, 100
Th. sibiricus Klok. et Schost. 99, 136
Th. Talievii Klok. et Schost. 136, 138, 260
Tilia L. 11, 13, 14, 15, 17, 18, 19, 20, 27,
28, 29, 33
T. amurensis Rupr. 18
T. cordata Mill. 16, 18, 22, 23, 26, 74,
148, 253
T. cordifolia Bess. 23
T. mandshurica Rupr. 18
T. platyphyllos Scop. 29
T. sibirica Fisch. 149, 194, 195
Timmia austriaca Hedw. 41
Trapa L. 19
Trifolium lupinaster L. 50, 51, 62, 65, 164
T. montanum L. 50
Trigonella platycarpos L. 244, 246, 262
Trinia muricata Godet. 76
Tsuga Carr. 11, 12, 16, 17, 18, 20
T. canadensis (L.) Carr. 19
T. europaea Mensol. 16
Typha L. 14
T. latifolia L. 17 ,
7". pliocenica Dorof. 16, 17
Ulmus L. 11, 17, 18, 19, 20, 22, 28, 29,
33
U. appendiculata Heer. 11, 15
U. Braunii Heer. 11
U. carpinoides Goepp. 13, 14, 16
U. laevis Pall. 149, 187, 250
U. longifolia Ung. 13, 16
U. minuta Goepp. 13, 16, 17
U. plurinervia Unger. 11
U. scabra Mill. 253
Ulota curvifolia (Wahlenb.) Brid. 45
Umbilicaria pennsylvanica Hoffm. 45
Urtica dioica L. 22
Vaccinium myrtillus L. 50, 200
V. pliocenicum Dorof. 16, 17
V. uliginosum L. 25, 200
V. vitis-ldaea L. 47, 50, 51, 164, 200
Veronica incana L. 190
V. longifolia L. 174
V. maxima Mill. 192, 193, 261
V. spicata L. 19, 41, 43, 50, 51, 55, 62, 64,
65, 71, 74, 255, 263
Viburnum L. 19, 41, 43, 50, 51, 62,64,65,
71, 74, 256, 264
V. giganteum Sap. et Mar. 7, 10
,V. giganteum Sap. var. uralense Baran.
7, 8
Vida cracca L. 41
V. multicaulis Ledeb. 39, 55, 62, 230, 234,
235, 262
V. tenuifolia Roth. 190
Viola biflora L. 39
V. mirabilis L. 192
V. palustris L. 25
V. rupestris L. 46
Vitis zaisanica Baik. 13, 14
Weigela Thunb. 12, 17, 18
W. Kryshtofovichiana Dorof. 12, 17
Woodsia alpina (Bolt.) S. F. Gray. 38
W. glabella R. Br. 38, 62
W. ilvensis R. Br. 38, 46, 50
Zannichellia palustris L. 22
Z. pliocenica Dorof. 17
Zelkova Richardli Mich. 16
Z. Ungeri Kovats. 14
Zizyphus uralensis Kolesn. 13
Zugadenus sibiricus (L.) A. Gray 241,
242, 262
Zygophyllum macropterum C. A. Mey.
76
ОГЛАВЛЕНИЕ
, Стр.
Введение ...................................................... 3
Глава I. Основные этапы формирования флоры и растительности Урала
в кайнозое (по палеоботаническим данным)......................... 5
Третичный период (палеоген и неоген).........................—
Палеоцен . .............................................. 7
Эоцен................................................... 9
Олигоцен................................................ 10
Миоцен....................................................И
Плиоцен ............................................... 15
Общий обзор третичных ископаемых флор Урала..............19
Четвертичный период (антропоген)............................20
Плейстоцен .............................................. —
Голоцен..................................................30
Глава II. Скальная флора Урала.......................................37
Известняковые скалы на западных предгорьях Северного и Припо-
Обнажения известняков на западных предгорьях Северного Урала
в бассейне реки Вишеры .........................................39
Прибрежные известняковые утесы на восточном склоне Северного
Урала в бассейне рек Лозьвы и Сосьвы ..........................—
Гранитные останцы на восточном склоне Среднего Урала . . . . 43
Обнажения дунита на Уктусских горах в окрестностях Сверд-
ловска . . ........................................... 47
Обнажения сиенита, диорита и габбро на восточном склоне и в водо-
раздельной полосе Среднего Урала................................50
Обнажения некоторых метаморфических горных пород на Среднем
Урале.........................................................51
Известняковые прибрежные утесы на восточных предгорьях Сред-
него Урала (реки Тура, Реж, Пышма, Исеть).....................—
Известняковые скалы по реке Чусовой ............................56
Обнажения гипса на западном склоне Среднего Урала...............63
Обнажения известняков на западном склоне Южного Урала в бас-
сейне реки Белой ............................................65
. Горы Сугомак и Егозинская на восточном склоне Южного Урала. 70
Каменистые степи на хребте Ирендык (восточный склон Южного
Урала)........................................................73
Стерлитамакские шиханы. ........................................74
Глава III. Эндемики уральской флоры....................,..............77
Высокогорные эндемики...........................................80
Горностепные и скальные эндемики...............................108
Эндемики широколиственных лесов................................142
Некоторые черты эндемизма уральской флоры ...................151
Необходимые меры по охране эндемичных растений.................152
Глава IV. Реликты во флоре Урала.....................................154
Вопросы методики изучения реликтов на примере нескольких ключе-
вых растений................................................ —
Вереск обыкновенный Calluna vulgaris..................... —
Дазифора кустарниковая Dasiphora, fruticosa..............167
Можжевельник казацкий Juniperus sabina...................183
Общий обзор реликтов...........................................191
Доледниковые (плиоценовые) реликты ... —
Плейстоценовые реликты..................................198
Перигляциальные реликты арктической флоры ........ —
Перигляциальные реликты, проникшие на Урал из высокогорных
районов Азии........................................ 205
Перигляциальные реликты горноевропейского происхождения .217
Скальные и горностепные реликты горноазиатского происхожде-
ния, проникшие на Урал в конце плейстоцена и Начале
голоцена .............................................. 220
285
Реликты азиатского происхождения, свойственные светлым лесам
(лиственничным, сосновым, березовым) и лесным лужайкам 240
Голоценовые реликты . ..................................253
Широколиственные реликты первой половины среднего голоцена —
Степные реликты термического максимума второй половины сред-
него голоцена ......................................... 254
Заключение.................................................. 257
Summary...................................................... 265
Литература....................................................267
Указатель латинских названий растений, упомянутых в тексте . . . 277
ПАВЕЛ ЛЕОНИДОВИЧ ГОРЧАКОВСКИЙ
ОСНОВНЫЕ ПРОБЛЕМЫ
ИСТОРИЧЕСКОЙ ФИТОГЕОГРАФИИ УРАЛА
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стра- ница Строка Напечатано Следует чит
154 15-я снизу дазиформы дазифорь
165 1-я снизу 50 500
263 14-я снизу Южнее Южные
269 29-я сверху Гризбах Гризеба?
274 18-я снизу Тома I—XXV, Тома I—X.
XXX
Заказ № 251
РИСО УФАН СССР
Свердловск, К-49
Первомайская, 91
Редактор изд-ва Л. В. Богданович.
Техн, редактор Н. В. Семенова Корректоры/7. В. Винокурова, М. И. Зубринская
РИСО УФАН СССР № 6/3—12/68/ НС 23501. Подписано в печать 9/1 1969 г.
Печ. л. 18,0. Уч.-изд. л. 22,09. Бумага типографская №1. Формат 70х 1081/1в.
Заказ 251. Цена 1 р. 70 коп. Тираж 1400.
Типография изд-ва «Уральский рабочий», г. Свердловск, проспект Ленина, 49.