/





















































































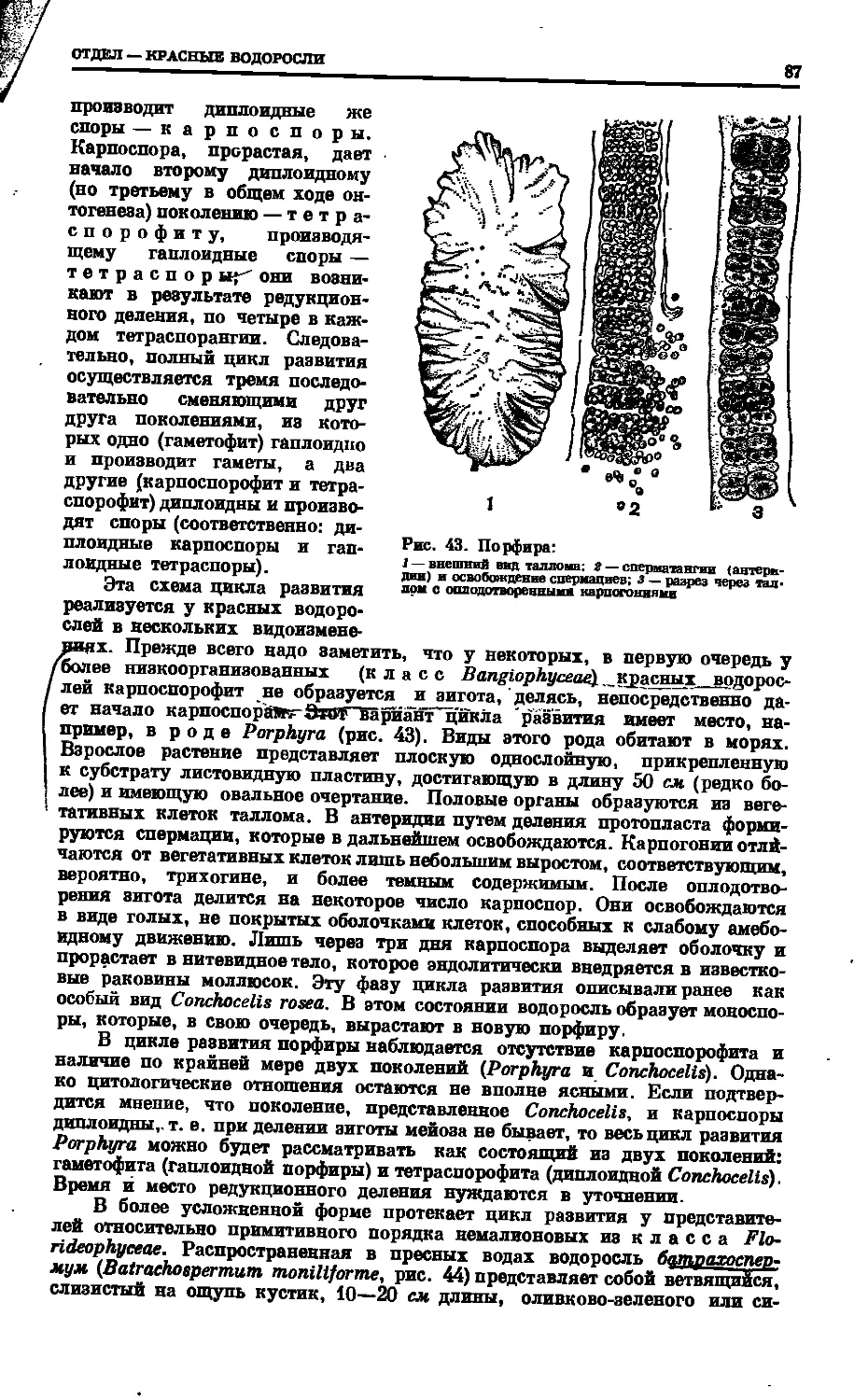






























































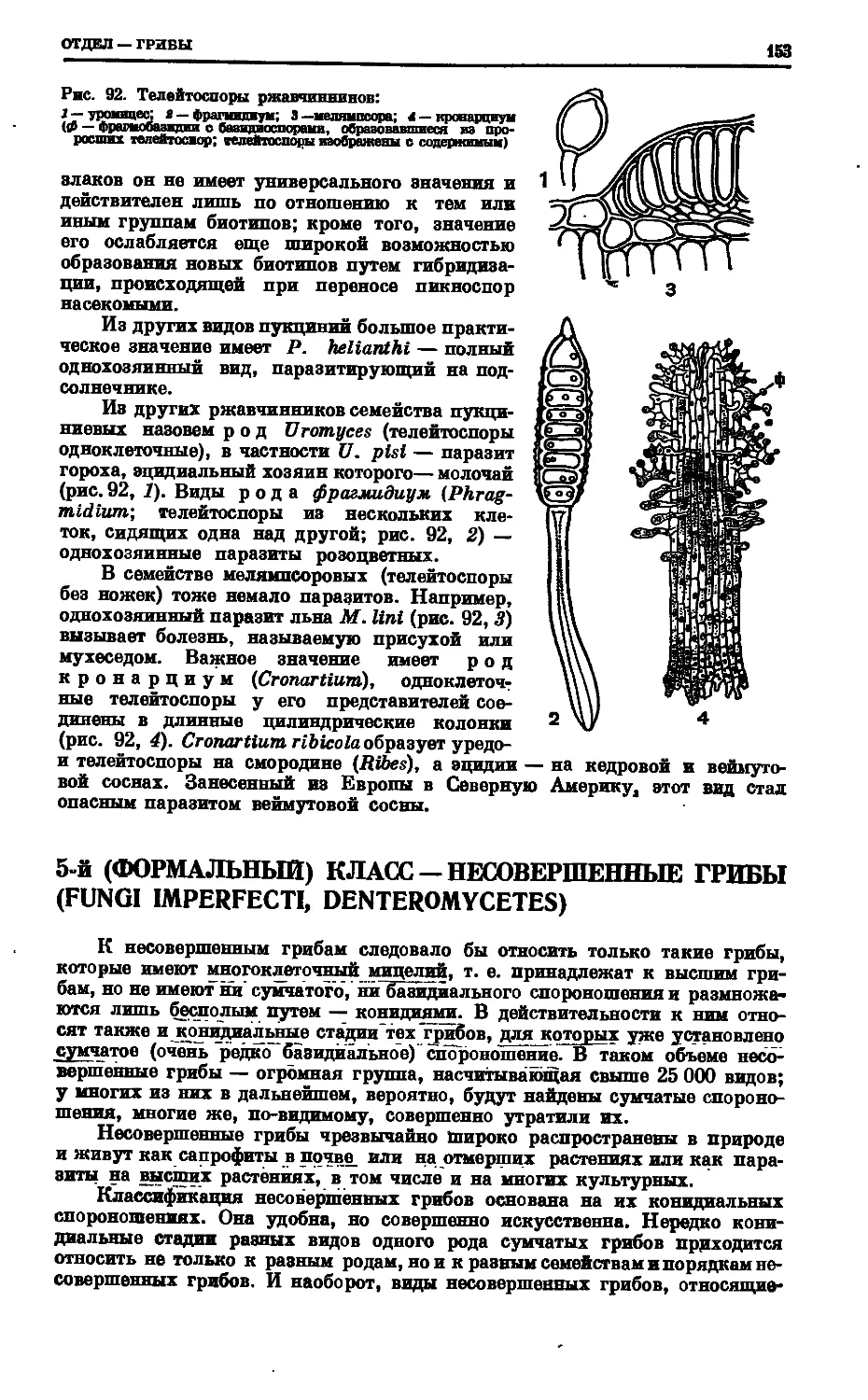
















































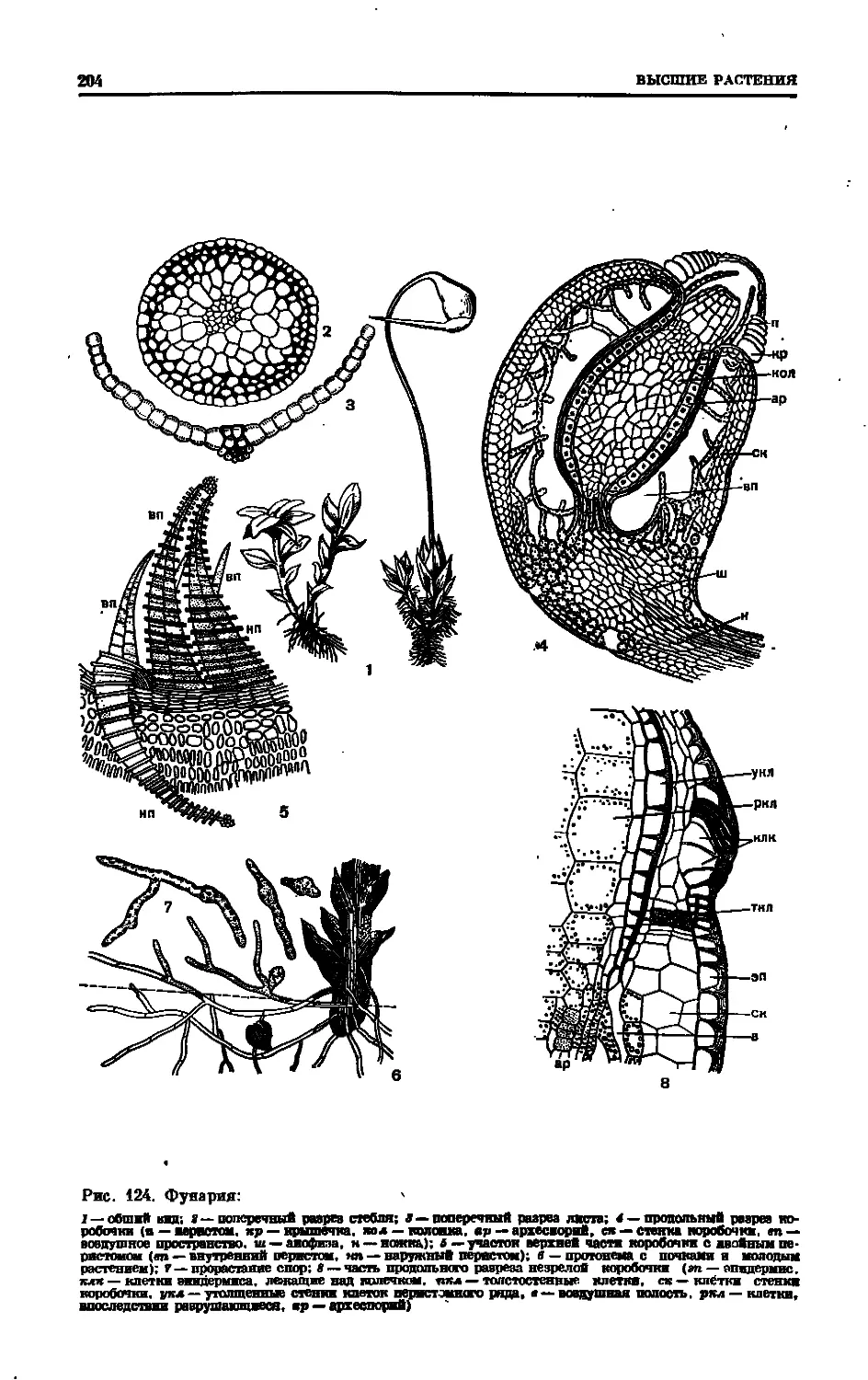









































































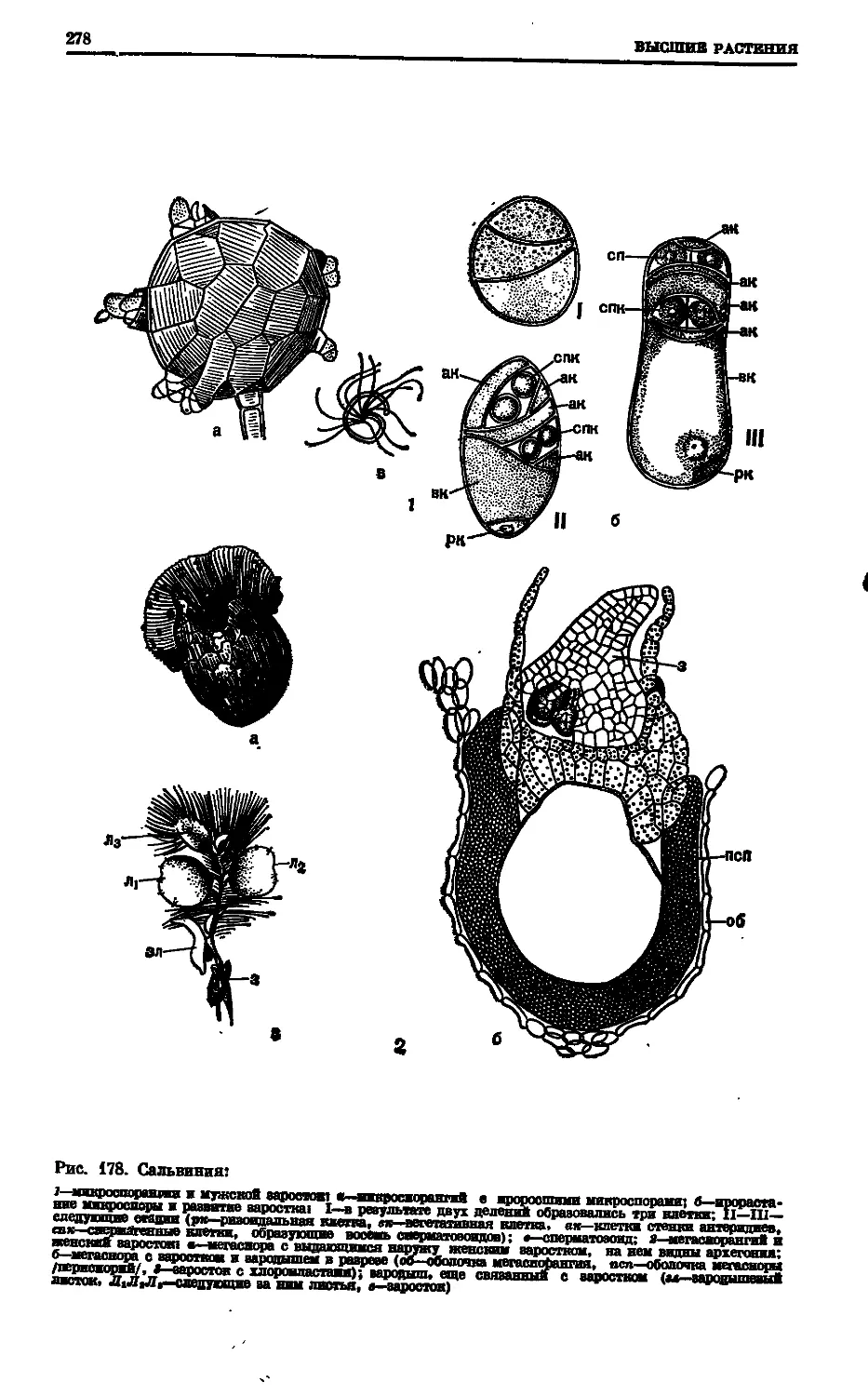






























































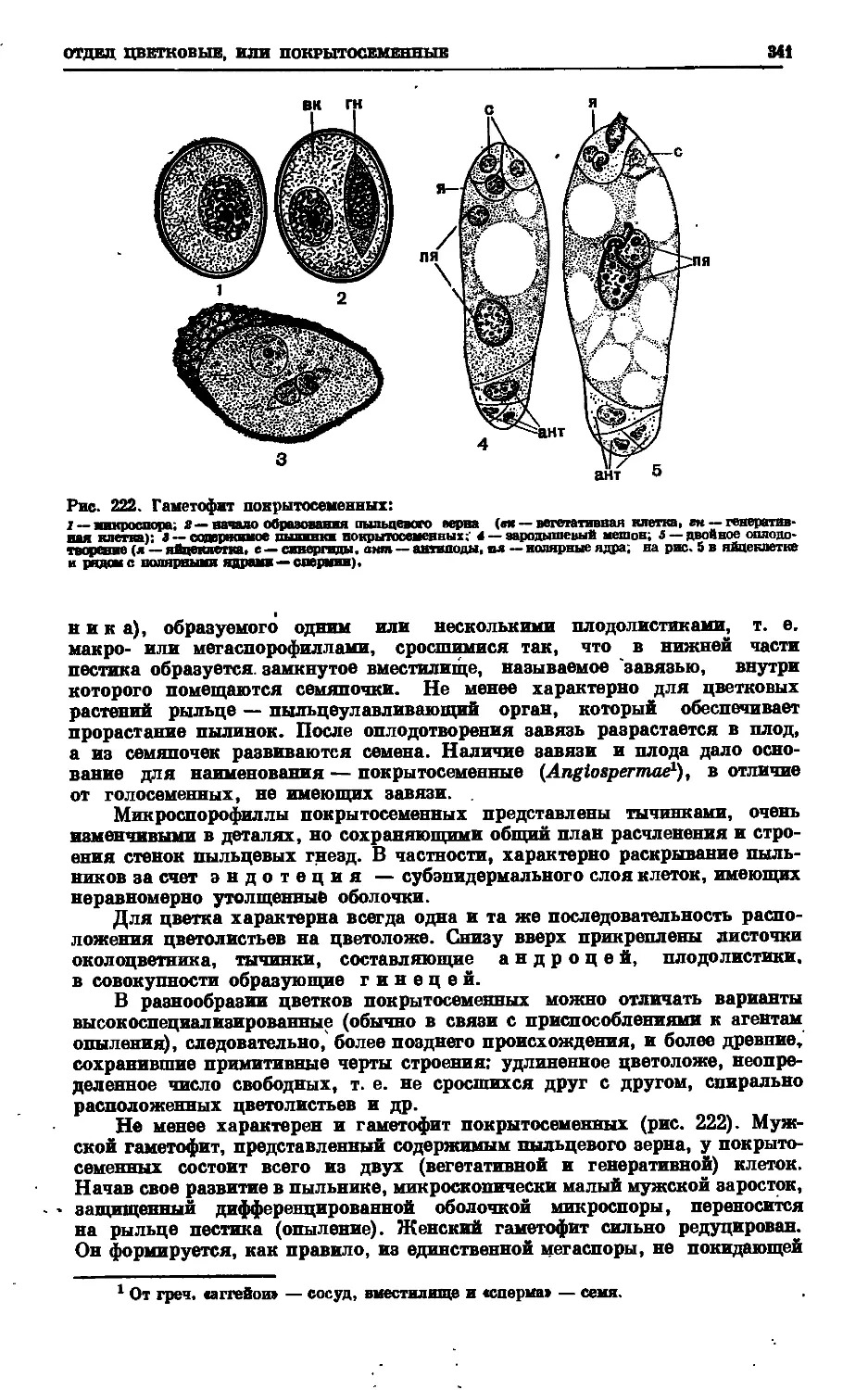












































































































































































































































































Text
Н. А.КОМАРНИЦКИЙ
Л. В. КУДРЯШОВ
А.А.УРАНОВ
М О С ICB А
БОТА Н И КА
СИСТЕМАТИКА РАСТЕНИЙ
ИЗДАНИЕ 7-е, ПЕРЕРАБОТАННОЕ
Допущено
Министерством просвещения СССР
в качестве учебнике? для студентов
биологических факультетов
педагогических институтов
ПРОСВЕЩЕНИЕ* 1975
58
К63
Комарницкий Н. А. др.
К63 Ботаника (систематика растений), изд. 7-е, пере-
раб. М., «Просвещение», 1975.
608 с. « ил.
Перед загл. an.: И. А. Комарницкий, Л. В. Кудряшов,
А. А. Уранов.
Настоящее яадаяве учебника еначнпиыю переработано ио сраанеик»
е преяывушжм ваданнем. Многве разделы напоены заново (введвнне,
характерястнкв отдельных грунп ннзшнх и высвжх рдеишвй). Учебвжк
соответствует ныне пействупщеа прогршпе но бонмпм в пииопнео*
ах ннснвгутх.
Шестое нзданне вышло в 1962 в.
0069В—616
----------- 80—71
108(09)—7&
✓
ПРЕДИСЛОВИЕ
В настоящее издание внесено много нового в соответствии
с современным состоянием науки.
В группе водорослей наиболее капитальной переработ-
ке подверглись пиррофитовые, бурые и сине-зеленые
водоросли; красные написаны заново. Существенно из-
менены многие страницы, посвященные грибам. Описание
низших растений кончается кратким филогенетическим
очерком.
В систематике лиственных мхов большее внимание обра-
щено на разнообразие зеленых мхов и развитие перистома..
В отделе «Папоротникообразные» изъяты некоторые порядки
из класса «Клинолистовидные», внесены изменения в текст
по строению стебля хвощей, упрощен материал по разно-
споровым папоротникам. В отделе «Голосеменные» обновлен
материал по хвойным. Очень большой перестройки потре-
бовал отдел «Цветковые растения», так как опыт препода-
вания показал, что педагогически удобно деление двух
классов этого отдела ха подклассы. Описание многих по-
рядков и семейств сформулировано более лаконично, чем
в предыдущем издании. Некоторые таксоны исключены по
той причине, что составляют дополнительный материал к
программе.
Введение, «Низшие растения» и «Покрытосеменные рас-
тения» переработаны и дополнены А. А. Урановым, «Архе-
гониальные» — Л. В. Кудряшовым. «Низшие растения»
в предыдущем издании были каписаны Н. А. Комарн
Ведущей тенденцией настоящего издания было сохранение
по возможности авторского теиста.
При работе учтены пожелания рецензентов и многочис-
ленные замечания преподавателей пединститутов, универ-
ситетов и студентов. Авторы выражают признательность
всем, кто своими критическими -замечаниями способствовал
улучшению этой книги.
ВВЕДЕНИЕ
Огромная роль, которую играют растения в жизни человека, доставляя
ему основную массу органических веществ, необходимых для питания и удов-
летворения других насущных потребностей, общеизвестна. Но не все растения
в этом отношении равнозначны, и с этим фактом, несомненно, при-
ходилось считаться уже первобытному человеку, тем более что наряду с
растениями полезными существует немало и вредных. Отсюда, вероятно, и
возникла у человека потребность отличать растения друг от друга, а в связи
с. этим — и необходимость давать им названия. Возрастание числа расте-
нии, которые становились в том или ином отношении известными, должно
было привести к первым попыткам классификации. Каковы были принципы
этих первичных классификации — неизвестно; они возникали, конечно, не-
зависимо у разных народов и, по всей вероятности, уже в глубочайшей
древности.
До нашего времени дошли сравнительно поздние образцы классифика-
растений, относящиеся ко времени расцвета древнегреческой культуры
•in
и периоду римского владычества.
Греческая античная наука достигла большого развития в трудах Арис-
тотеля (384—322 гг. до н. э.). Он создал классификацию животных, за-
ложив этим основы зоологической систематики, и авторитет его в этой области
оставался незыблемым в течение длительного времени. Ботанические труды
Аристотеля погибли, но по сохранившимся его работам можно заключить,
что он интересовался и вопросами ботаники.
Ближайший друг и ученик Аристотеля — Теофраст (370—285 гг.
до и, э.), отличавшийся, как и его учитель, разносторонностью научных инте-
ресов, специально занимался растениями. Его ботанические труды1 имели
столь большое значение и оказывали столь большое влияние на воззрения бо-
таников вплоть до XVII в., что Теофраста и до наших дней справедливо
называют «отцом ботаники». В центре его внимания были общие вопросы бо-
таники, в первую очередь проблемы специфичности строения и жизненных
отправлений растений. Он, несомненно, осознавал необходимость классифи-
кации растений. По Теофрасту, «первыми и самыми главными видами,
охватывающими почти все растения или большинство из них, будут деревья,
кустарники, полукустарники и травы»1 2. В составе наземной флоры он выде-
лял растения вечнозеленые и листопадные, а в флоре водной — пресноводные
и морские растения. Теофраст тесно увязывал знания о растениях с вопроса-
ми их практического использования.
1 Сохранились следующие ботанические сочинения Теофраста: «Причины растении»
в 6-ти книгах и «Исследования о растениях» в 9-ти книгах. См.: Теофраст. Исследо-
вания о растениях. Пер. с древнегреч. и прим. М. Е. Сергеенко. Изд-во АН СССР, 1951,
Серия «Классики науки». /
2 Т еоф р а с т. Исследования о растениях, кн, I. Издт&о АН СССР, 1951, стр. 18,
ВВЖ)
В дальнейшем утилитарное направление в изучении растении на долгое
время стало господствующим. Оно получило отражение, например, в трудах
римского ученого Плиния Старшего (23—79.) 1.
Много путешествовавший и знавший растения по личным наблюдениям
Диоскорид (жил в I в. н. э.), грек по происхождению и популярный в
Риме врач-практик, был автором сочинения «О лекарственных средствах», со-
державшего описание более 500 видов растений и сведения об их местонахожде-
нии и распространении. Диоскорид пользовался авторитетом не только у
современников, но и у ботаников средних веков и эпохи Возрождения. Од-
нако труд Диоскорида не имел существенного значения для разработки прин-
ципиальных основ классификации растений.
В течение первых веков нашей эры и почти всего периода средневековья,
в том числе и в эпоху господства арабской культуры, когда ботаника обога-
тилась многими сведениями, главным образом о лекарственных растениях,
вопросы классификации растений, насколько известно,ни разу не были постав-
лены на базе широкого обобщения знаний о природе растений, как это было
сделано Теофрастом.
Возрождение ботаники началось с конца XV в. Гуманисты Италии на-
чали отыскивать в окружающей их природе растения, упоминавшиеся бота-
никами древности. Это движение получило свое дальнейшее развитие в XVI в.
и по северную сторону Альп. Изобретение книгопечатания в Европе (XV в.)
и развитие искусства резьбы по дереву (гравюра) способствовали обмену све-
дениями о растениях между ботаниками. Стали появляться целые сборники
изображений растений, так называемые травники. Все это привело к тому,
что число растительных форм, известных науке, быстро возрастало. Кроме
того, великие путешествия конца XV — начала XVI в. приносили сведе-
ния о растениях, ранее неизвестных, совершенно необычных для Европы.
Иноземные растения, особенно полезные в том или ином отношении, стали г
разводить в особых садах, получивших название ботанических. Наиболее
древний ботанический сад основан в Салерно (Италия) в 1309 г., вто-
рой (по времени основания) — в Венеции в 1333 г. Они еще мало отличались
от монастырских садов, где преимущественно была сосредоточена культура
лекарственных и душистых растений. Позднее перед ботаническими сада-
ми встала задача изучения флоры путем культуры местных и чужеземных
растений, описания и классификации их. Такого типа ботанические сады ста-
ли появляться лишь в XVI в. Из них известны ботанические сады в Ита-
I, основанные в Падуе (1525) и Пизе (1544).
В России первые аптекарские огороды для разведения лекарственных рас-
тений появились в первой половине XVII в. В начале XVIII в. число их
сильно возросло. Аптекарский огород, заложенный в Москве в 1706 г., был в
начале XIX в. (1805) преобразован в Ботанический сад Московского уни-
верситета. В 1714 г, основан Аптекарский огород в Петербурге; в 18^3 г.
он переименован в Ботанический сад, входящий в настоящее время в состав
крупнейшего ботанического учреждения — Ботанического института
им. В. Л. Комарова Академии наук СССР.
В середине XVI в.-было положено начало составлению коллекций расте-
ний*— гербариев, которые способствовали развитию систематики растений.
Инициатором этого начинания считают Луку Гини, первого директо-
ра Пизанского ботанического сада, и его учеников — Альдро ванди и
11SIS
SIU
SIU
SIU
siTT
I fl
Ботанические сады, гербарии и травники имели большое значение и в по-
знании разнообразия растении. При наличии образцов уже известных растении
или хотя бы их рисунков было сравнительно нетрудно путем сравнения уста-
1 П л и н в й Старший, Гай Секунд — автор капитального труда «Ес-
тественная история» в 37 книгах, представлявшего энциклопедию естествознания того
времени. Для составления этого труда автор использовал до 2000 книг, большинство ко-
торых не сохранилось. Книги 12—27 «Естественней истории» Плиния относятся к ботанике.
ILi
г. г,
ir
ГГТТг
Г, 11
во вить, известно ли науке данное растение или оно является новым, еще ни**
кем не описанным. Но для использования накопленного материала стало со-
вершенно необходимо систематизировать его, расположить в известной по-
следовательности. Попытки использовать для этой цели алфавитный порядок
названии, как это делали в травниках, приводили к неудовлетворительно-
му результату. Не подходили и «утилитарные системы», которыми пользова-
лись некоторые естествоиспытатели древности.
В связи с накоплением огромного фактического материала, начиная с
XVI и кончая приблизительно первой половиной XVIII в., задача построения
системы растений становится одной из центральных в ботанике. Так, один
из наиболее авторитетных ученых того времени, лейденский медик и естест-
воиспытатель Б о э р г а в (1688—1738), еще на пороге XVIII в. определял
ботанику как часть естествознания, «посредством которой удачно и с
меньшим трудом распознаются и удерживаются в памяти названия растений».
Одной из наиболее выдающихся ранних (в 1583 г.) попыток создания
системы растений была система итальянского ботаника Андреа Чезальпино
(1519—1603).
Чезальпино заслужил почетную репутацию «Аристотеля XVI в.», что
соответствует широте и глубине взглядов этого ученого и его пытливости в
познании природы. В сочинении «О растениях» (1583) он развивает взгляд
на растение как целостный живой организм (книга 1) и описывает около
1500 видов растений, половину которых он сам же собрал, открыв при этом
много новых видов.
Чезальпино принимал традиционное деление растений на древеснеющие
(деревья и кустарники) и травянистые (полукустарники и травы). Он понимал,
что для классификации нужна целая система соподчиненных единиц и сфор-
мулировал понятие сходного, выражая его в следующих словах: «По за-
кону природы сходное всегда рождает сходное и такое, что относится к тому
же самому виду». К этой мысли естествоиспытатели впоследствии возвраща-
лись не раз в связи с развитием учения о виде. Принципиально важной за-
растаний было то, что он считал
необходимым опираться на признаки, принадлежащие растению — объ-
екту исследования — и важные для него самого, а не на такие, которые пред-
ставляют лишь тот или иной интерес для человека. Из свойств, присущих ор-
я, Чезальпино стремился выделить наиболее существенные
и подходил к этому вопросу с точки зрения своих воззрений на растение. По
его мнению растение должно поддерживать свое индивидуальное и родовое
существование. Первое осуществляется через питание, для чего служит ко-
рень; второе — через развитие плодов и семян, образуемых стеблем. Образо-
вание семян является как бы конечной целью индивидуального существова-
ния растения и началом новой жизни. Поэтому Чезальпино в основу своей
классификации положил строение плодовместилищ семян и сами семена.
Он выделял 14 классов на основании строения плода, числа гнезд и семян в
нем; более мелкие группы выделялись с учетом строения цветка. Особое по-
ложение в системе Чезальпино занимает 15-й класс, куда были отнесены мхи,
папоротники, хвощи, водоросли, грибы и ... кораллы. Растения 15-го класса
Чезальпино считал наименее совершенными, занимающими промежуточное
место между растительным миром и неживой природой.
. Некоторые группы системы Чезальпино оказались вполне естественными.
Например, в 11-м классе объединены бурачниковые и губоцветные, сближае-
мые иногда и современными систематиками, в 10-м классе сосредоточены все
сложноцветные. Но все же в целом эта система, основанная на изменчивости
немногих признаков (нлод, отчасти цветок), имела и с к у с с т в ен ны й
характер. В большинстве классов оказались растения^ резко различные.
Тем не менее система Чезальпино, несовершенная с современной точки
зрения, была важнввс этапом в развитии систематики. Она создана, подобно
. классификации Теофрабта, на базе широкого анализа природы растения и по-
ложила начало длительному периоду искания методов построения системы,
ИГ
5
и»
I ;
^шедшими
«г»
1:1
1:1
r.i
без чего, ввиду огромности все возраставшего материала, становилось поло-
жительно невозможно ориентироваться в разнообразии растений.
Работа ряда ботаников XVI—XVIII вв., создавших новые системы
растений, получила известное завершение в трудах К. Линнея (1707—
1778). Отказавшись от деления растений на деревья, кустарники и травы,
предложенного Теофрастом, Линней избрал в качестве основного система-
тического признака орган размножения, но не плод, как было сделано Чезаль-
ю, а цветок, точнее, строение андроцея. Растительный мир он разделил
на 24 класса, а классы — на 116 отрядов.
Система Линнея оказалась практически чрезвычайно удобной. Для каж-
дого нового растения в ней было легко найти вполне определенное место, а
если требовалось узнать наименование уже известного растения, то неслож-
ный анализ цветка сразу показывал, в каком классе его следует искать. Таким
образом, проблема классификации, имеющей чисто служебное значение,
облегчающей ориентировку в многообразии растений, получила удовлетво-
рительное решение.
Успеху системы Линнея, помимо простоты, способствовало еще и то
что при построении ее он последовательно придерживался определенного
метода описания и наименования растений. До Линнея растения часто опи-
сывали по сравнению с другими, ранее известными. Например, олеандр опи-
сывался как растение «с листьями, как у лавра, и с цветками, как у розы»,
и т. п. По таким описаниям часто было трудно представить себе облик расте-
ния. Описания, которыми пользовался Л:
но иной основе — здесь каждый орган описывается в терминах, имеющих
вполне определенное значение. Л
значил точными терминами в первую очередь листья растений,
благодаря чему все описания растений получили новый вид и освещение1.
Весьма важным нововведением в систематику растений была бинар-
ная номенклатура растений, последовательно применявшаяся
в биологии Линнеем. Сущность бинарной номенклатуры состоит в том, что
каждому растению присваивается название, состоящее из двух слов, первое
из которых представляет название рода, к которому относится данный вид, а
второе есть так называемый видовой эпитет, который вместе с
родовым названием служит для обозначения вида.
Очевидно, использование такого способа наименования растений могло
возникнуть не ранее, чем в ботанике выработалось понятие о р о д е и
виде растений.
Уже в середине XVI в. естествоиспытатели в большой степени прибли-
зились к понятию о виде как систематической категории. В начале XVII в.
швейцарский ботаник Каспар Баугин (Бозн, 1560—1624) в ре-
зультате сорокалетнего труда осуществил критическое описание около 6000 ра-
стений. Обладая исключительной эрудицией и знанием древних и новых
языков, К. Баугин устранил, насколько это было возможно, беспорядок в
наименованиях растений, проистекший из того, что нередко одному и тому
же растению приписывались разные наименования. Важной заслугой Баугина
было то, что он, распределив растения по родам, подчинил последним более
мелкие систематические единицы. Родовые названия растений у Баугина со-
стоят из одного или большего числа слов, названия же подчиненных роду ка-
тегорий — обычно из нескольких (иногда до 20) слов; но довольно часто,
особенно в небольших родах, обе части названий были однословными, а все
название растения становилось таким образом двусловным (биноминальным).
Однако как Баугин, так и последующие ученые предпочитали названия поли- -
номинальные, многословные, так как из них были непосредственно видны
иг
ГI
же время и кратким описанием его. Историки естествознания отмечают, что
К. Баугин достиг в краткости описания растений большого искусства.
I
Е. Г. Боб р о в. Ливией, его жизнь а труден. М., Изд-во АН СССР, 1957, стр. 157.
10
ВВЕДЕНИЕ
Однако понятие о роде и виде Баугином определено не было. Определе-
ние этих понятий относится к более позднему времени. Так, Джон Рэй в
«Истории растений» (1686) очертил понятий вида на основе происхождения
относящихся к нему особей. По словам Д. Рэя, «у растений нет надобности
в каких-либо других доказательствах видовой одинаковости, кроме происхож-
дения из семян растений, специфически или индивидуально идентичных. Фор-
мы, которые по отношению к виду различны, сохраняют зту свою видовую
природу различной, ни одна из них не происходит из семян другой или на-
оборот»1.
К. Линней считал, что отличия родов можно достаточно выявить, опи-
раясь на признаки генеративных органов. Что касается видов, то он понимал
объем этой единицы в смысле, близком к определению Д. Рэя, но счел необ-
ходимым отделить виды от разновидностей, чего не делали его предшест-
венники.
К. Линней первоначально пользовался полиноминальными названиями
растений, стремясь, однако, предельно сократить их, выбирая из многих
отличий вида наиболее существенные. Однако в сочинении «Виды растений»,
первое издание которого вышло в 1753 г., Линней систематически приписы-
вал всем видам и бинарные (двусловные) названия. Прежние многословные
названия сохранили при этом значение кратких описаний (диагнозов) соот-
ветствующих видов; двусловные же названия, или, по Линнею, «простые наи-
менования», приобрели характер названия-клички. Произведенная Линнеем
реформа номенклатуры растений оказалась практически удобной, и с тече-
нием времени двусловные названия растений вошли во всеобщее употребле-
ние, ими пользуются и до настоящего времени.
При всех достоинствах система Линнея имела и существенный недостаток,
так как отдельные растения, явно сходные между собой строением андроцея,
утопали в различные классы.
Линней сделал опыт построения фрагмента естественной системы, выде-
лив 67 естественных порядков, которые, однако, не исчерпывали всего разно-
образия растений.
При построении искусственных систем ботаники в качестве основы из-
бирали какой-нибудь один признак или узкий комплекс признаков (у Лин-
нея — некоторые особенности строения цветка, у Чезальпино — строение
плода и число семян и т. п.) и по градациям или вариациям таковых разделя-
ли растения на группы. В разработке естественной системы принятие только
одного-двух априорных признаков, которые служили бы базой классифика-
ции, исключалось. Растения должны быть объединяемы по их «общему сходст-
ву», т. е. по сходству в возможно большем числе признаков. Но трудности,
возникавшие на этом пути, заставили Линнея сомневаться, возможно ли во-
обще построение завершенной естественной системы. Тем не менее «естест-
венному методу» он посвятил немало лет своей деятельности.
Важным этапом в разработке естественной системы были труды Бер-
нара Жюссье (1697—1777) и его племянника Антуана Жюс-
сье (1748—1838). Б. Жюссье расположил растения в ботаническом саду в
Трианоне (Версаль) в особом порядке. Но единственным печатным отраже-
нием этой большой работы оказался садовый каталог. Спустя 30 лет, в 1789 г.,
А. Жюссье опубликовал «Роды растений». Значение этого сочинения состоя-
ло в том, что в нем сформулированы диагнозы (краткие описа-
ния) сравнительно некрупных групп растений — порядков (порядок
Жюссье соответствует по объему примерно семейству современной системати»
ки). Это было существенным шагом вперед сравнительно с Линнеем, который
описания выделенных им 67 порядков не дал; кроме того, число порядков
в классификации А. Жюссье возрастало до 100, т. е. в 1,5 раза. Сформулиро-
вать диагнозы порядков можно было только на основе глубокого изучения
входящих в каждый из них более мелких единиц. А. Жюссье сделал и следую-;
1 В. Л. Комаров. Избр. cost., т, L М,—Л., Изд-во АН СССР, 1945, стр. 138,
ввк
пи
ЕГИЕ
If
щй шаг: он объединил порядки в
классы (числом 15), а эти последние — в
н г.
еще более крупные единицы. Таким образом, все разнообразие раститель-
ного мира, начиная от водорослей и грибов и кончая цветковыми растениями,
в системе А. Жюссье представлено в виде категорий разного объема, иерар-
хически подчиненных друг другу. Создание естественной системы А. Жюссье
было переломным моментом в развитии научной систематики. В течение более
чем 2000 лет, считая с Теофраста, задача классификации растений сводилась
к тому, чтобы как можно резче отграничить одни группы растений от других.
В системе А. Жюссье впервые классификация была построена так, что систе-
матические единицы располагались во взаимной связи, как бы сливаясь друг
с другом, несмотря на наличие достаточно четких отличий между ними.
Прогрессивная по идее система А. Жюссье, однако, даже в эпоху ее по-
явления в ряде положений была спорной. В последующее время другие бота-
ники выступали со своими предложениями естественной классификации;
среди них наиболее известны системы О. Декандолля (1819),
Ст. Эндлихера (1836—1843), А. Броньяра (1843), А. Б р а у -
н а (1864).
Разработка системы растенийАв конце XV111 — первой половине XIX в.
ознаменовалась рядом важных исследований, сыгравших в развитии ботаники
вообще и систематики в частности выдающуюся роль. Прежде всего следует
отметить морфологические труды О. Декандолля. Им были заложены основы
сравнительной морфологии, опиравшейся на учение о симметрии и о плане
строения органов. <Цлан строения гвыясняется, по Декандоллю, ио мере того
как устанавливаются абортирование (выпадение), дегенерация, срастание час-
тей — вообще вторичные преобразования. Используя сравнительно-морфоло-
гический метод, Декандолль в сущности вплотную подошел к эволюционной
концепции, но, как ни странно, продолжал оставаться на метафизических
позициях неизменяемости видов. Особое значение имели работы английского
ботаника Р. Броуна (1773—1858) по истолкованию морфологии цветка в
ряде трудных случаев (злаки, орхидные, ласточниковые, раффлезиевые и др.).
Он также внес существенный вклад в изучение строения семяпочки, образова-
ния эндосперма и перисперма, исследовал «цветки» саговников и хво:
установил систематическую особенность голосеменных (которые до этого со-
единяли с двудольными). Им же были открыты архегонии («корпускулы»)
в семяпочке голосеменных. /Л I -<
Привлечение результатов морфологических (в широком смысле слова)
исследований к решению вопросов близости или, наоборот, разобщенности
систематических (классификационных) групп растений со времен О. Декан-
долля и Р. Броуна становится все более необходимым. В этом отношении
совершенно исключительную роль сыграли знаменитые труды В. Г о ф-
мейстера (1824—1877). Он придал своим «сравнительным исследовани-
ям» онтогенетическое направление, оказавшееся исключительно плодотворным
и прочно вошедшим в дальнейшем в практику построения и совершенст-
вования системы растительного мира. Гофмейстером было показано, что у мо-
хообразных, у равноспоровых и разноспоровых папоротникообразных и у
голосеменных растений наблюдается один и тот же ход индивидуального
развития, характеризующийся ритмичным чередованием гаметофита и споро-
фита. Этим решалась одна из труднейших задач систематики.
Еще в произведениях Линнея фигурировал термин «тайнобрачные» рас-
тения — это 24-й класс системы, объединивший водоросли, грибы, лишайники,
мхи, папоротникообразные — словом, все так называемые споровые растения.
При построении естественной системы одной из трудных проблем было на-
хождение связи между тайнобрачными и явнобрачными. Исследованиями
Гофмейстера эта проблема была решена, если иметь в виду мохообразные и
папоротникообразные как тайнобрачные, а семенные, в лице голосеменных,
в качестве явнобрачных. Те же исследования определили место голосеменных,
которые долгое время помещали среди двудольных. Голосеменные заняли по-
ложение между папоротникообразными и покрытосеменными. Следовательно,
; 1 l4 i М;
закрывалась брешь, нарушавшая единство той непрерывности растительных
существ, искание которой было, по К. А. Тимирязеву, основной идеей систе-
мы А. Жюссье. Этот факт непрерывности требовал объяснения, и оно выра-
батывалось в общебиологических теориях, шаг за шагом приближавшихся
к эволюционному взгляду на живую природу.
Весьма существенная роль в том отношении принадлежит Ж.-В. Ла-
марку (1744—1829) — автору цервой научно обоснованной теории эво-
Отказавшись от принципа неизменяемости видов и рассматривая пос-
ледние как результат естественного эволюционного развития, Ламарк писал
про естественную систему, что она «есть не что иное, как исполненный чело
веком набросок пути, по которому следовала природа в своих произведениях».
Появление теории Чарлза Дарвина (1809—1882) было началом
новой эпохи в развитии всех биологических наук, в том числе и науки о клас-
сификации организмов — систематики.
вися в XVI столетии и закончившийся пост-
роением системы К. Линнея в середине XV111 в., был периодом искусствен-
ной систематики. Система Жюссье положила основание естественной система-
тике. Переворот же, совершенный Ч. Дарвином, открыл новый, третий период
истории систематики — период эволюционн odi, или фило-
;Ти
Си
ii
I Г, I *,
генетической, систематики. Признание факта эволюции
привело к необходимости при построении классификации' объединять расте-
ния, единые по происхождению, а не просто сходные по боль-
шинству признаков, как это делалось в период естественных систем. Эволю-
ционная основа требовала такого взаимного расположения классифика:
онных единиц в системе, чтобы оно соответствовало путям эволюционного
развития.
Построение системы растительного мира на эволюционной основе потребо-
вало углубленного изучения самих растений, их организации и соотношений
с окружающей средой. Сопоставление растений на основе внешних черт стро-
ения, знание признаков и свойств только взрослых форм, использование толь-
ко организмов, живущих в современную нам геологическую эпоху, не могли
удовлетворить ботаника-эволюциониста. В течение второй половины XIX в.
и в XX в. круг фактов, используемых систематикой растений, постепенно
возрастал, чему особенно способствовало совершенствование методов, ис-
пользуемых частными ботаническими дисциплинами. Авторы современных
систем растительного мира используют огромный фактический материал, на-
копленный бурно развивавшейся в последние десятилетия палеонтологией
растений. Много ценного для понимания действительного хода эволюции да-
ли сравнительная морфология, сравнительная анатомия, эмбриология и он-
тогенетика растений. Многие вопросы филогенетической истории видов, ро-
дов и семейств цветковых растений разрешаются на базе изучения их географи-
ческого распространения. Наконец, за последние 50 лет в систематику все
больше проникают данные физиологии и биохимии растений, причем в этих
экспериментальных отраслях ботаники в настоящее время развивается особое
эволюционное направление.
Необходимость учитывать все сведения о растениях, с одной стороны,
облегчает задачу построения системы, так как всесторонний анализ растений
страхует от ошибочных выводов, но, с другой стороны, этим создаются и чрез-
стоящее время лишь в отношении ограниченного числа растительных форм,
. Авторы эволюционных систем вынуждены восполнять недостаток материала
более иля менее вероятными, но по необходимости субъективными гипотезами.
Этим и объясняется, что, хотя со времени выхода в свет основного произве-
дения Ч. Дарвина — «Происхождение видов путем естественного отбора»
(1859) — прошло более ста лет, все же единой общепринятой филогене-
тической системы растений до настоящего времени пока еще нет. Но фраг-
менты системы разработаны, и нередко достаточно детально; это касается
; 11
J___
BBBJ
ни:
is
как мелких групп (роди, семейства), так в некоторых случаях и более круп-
ных подразделений (порядки и классы).
Наиболее распространенным в настоящее время вариантом филогенети-
ческой системы является система А. Энглера (1844—1930). Несмотря на
то что система во многих отношениях несовершенна, ею продолжают широко
пользоваться, так как это пока единственная современная система, охватыва-
ющая весь растительный мир и разработанная с детализацией до семейств,
родов и подродов, а в некоторых случаях — до видов. Поэтому ею удобно
пользоваться как достаточно детальной основой для расположения групп
растении в определенной последовательности. Например, в большом коллек-
тивном труде «Флора СССР», изданном Ботаническим институтом имени
В. Л. Комарова Академии наук СССР, группы растений расположены по
этой системе.
За истекший период текущего столетня накопилось много фактических
данных, в свете которых предпринимались попытки пересмотра более ран-
них систем и создания новых. В большинстве случаев это касалось отдель-
ных крупных групп растений, как покрытосеменные, голосеменные, водо-
росли и т. п. В частности, в качестве примера можно указать ряд систем,
принадлежащих советским ученым: Н. И. К у 8 н е ц о в у, Н. А. Бушу,
Б. М. Козо-Полянскому, А. А. Гроссгейму, А. Л. Тах-
таджяну; из зарубежных систем, принципиально отличающихся от сис-
тем А. Энглера иР. Веттштейна, широко известны системы Галли-
ра, Бэсси, Гетчинсона.
Основной единицей филогенетической классификации является вид
(а р е с i е s). Понятие о виде, несомненно, возникло под влиянием, с одной
стороны, факта сходства между многочисленными особями растений, в особен-
ности между теми* которые находятся между- собой в непосредственном ге-
нетическом родстве (родители — дети — внуки и т. д_), а с другой — факта
несходства, явного различия во всей организации и поведении многих орга-
; I: I;
низмов, в первую очередь тех, которые имеют происхождение от разных, явно
несходных родителей.
• Понятно о виде выражает, по- меткому определению В. Л. Комарова,
явление «преемственной повторяемости живых существ».
Ив многочисленных определений, которые давали виду биологи после-
да рвинского периода, отметим сформулированное В. Л. Комаровым на осно-
ве богатого опыта непосредственного изучения видов в природе и анализа
истории развития учения о виде. По В. Л. Комарову, «вид есть совокуп-
ность поколений, происходящих от общего предка и под влиянием среды и
борьбы за существование обособленных отбором от остального мира живых
существ; вместе с тем вид есть определенный этап в- процессе эволют
(1945)1.
Условия жизни растений подчинены в своем распределении по земной
поверхности известной закономерности, они географичны. Отсюда вытекает,
чтовид может обитать только на той ограниченной части земной поверхности,
где есть необходимые для его существования условия. Территорию, занимае-
мую видом, называют ареалом. Определяя принадлежность данного
растения к тому или иному виду, ботаник опирается 1) на сходство во всех
. существенных признаках, 2) на сходство экологических условий и 3) на
общность ареала.
Признаки, сходство в которых учитывается при отнесении данного рас-
тения к тому или иному виду, неодинаковы в различных группах растений.
Например, в систематике цветковых растений предпочтение оказывается внеш-
морфологическим чертам строения, которые в той или иной степени бы-
вают коррелятивно связаны и с анатомическими и с биологическими видовы-
ми особенностями. Но в других группах, например у бактерий, внешние мор-
фологические признаки решающей роли не играют, так как разнообразие
II: 1
> :Г । ’
1 В. Л. Комаров. Иабр. сон., т. 1. М.—Л., Иад-во АН СССР, 1945, стр. 873.
14
ВВЕДЕНИЕ
внешней формы в этом случае очень велико и при одной и той же внешности
бактерии могут проявлять себя как существенно различные организмы.
Здесь ведущее значение имеют не признаки строения, а их биологические и
биохимические особенности. То же в известной степени относится и к не-
которым группам грибов.
Нередко виды подразделяются на более мелкие категории. Главнейшие
из них следующие:
Подвиды (subspecies). Они менее резко отграничены друг от друга,
чем виды, и нередко между подвидами имеются переходные формы, но каж-
дый подвид имеет свою обособленную от других подвидов того же вида или
лишь частично с ними совпадающую область распространения, свой ареал.
Разновидности или вариации (varietas), Еще менее рез-
ко отличаются друг от друга. Кроме того, не имеют и своего обособленного
ареала.
Как подвиды, так и разновидности имеют отличительные особенности,
вполне наследственно закрепленные. Но нередко наблюдаются собрания
особей вида, которые хотя и отличаются морфологически от других, но приз-
наки их отличия еще не стали постоянными и потому легко изменяются при
изменении жизненных условий. Такие группы особей называют формы
или морфы (forma., morfa).
В качестве мелких систематических подразделений вида выделяются еще
специальные формы и биотипы.
Понятие о специальных формах применяется преимущественно в систе-
матике грибов. Здесь, главным образом среди паразитных грибов, известны
случаи, когда в пределах вида выделяют несколько более мелких наследствен-
но стойких единиц, которые практически сходны морфологически, но резко
отличаются приуроченностью к различным хозяевам. Эти подразделения ви-
да и называют специальными формами.
В растениеводстве широко применяется попятив сорт, которому в жи-
вотноводстве соответствует термин порода. Сортом называют группу
особей в пределах какого-либо ботанического вида, подвида или разновид-
ности культурного растения, которая отличается по некоторым мелки1£, но
наследственно более или менее постоянным признакам от других особей то-
го же вида, подвида или разновидности. Наряду с морфологическими от-
личиями или признаками сорта главную роль играют те его свойства, кото-
рые определяют его хозяйственную ценность.
Морфологические признаки сорта, особенно важные для хлебных зла-
ков,—окраска зерновок, остистость или безостость, степень опушенности или
отсутствие ее и т. д. Величина, форма и окраска плодов характеризуют сор-
та у плодовых деревьев и ягодных кустарников; величина, форма, окраска
продуктивных органов положены в основу различия сортов клубне- и корне-
плодов. Окраска и размер цветков, махровость, высоко- или низкорос-
лость и т, д, используются как сортовые признаки декоративных растений.
Физиологическими, биохимическими и определяемыми ими хозяйственны-
ми признаками сорта считают различную степень урожайности, холодостой-
кости, засухоустойчивости, сахароносности, крахмалоносности, устойчи-
вость к болезням, скороспелость или позднеспелость, вкусовые качества,
лежкость, пригодность к транспортировке и т, п,
В сельском хозяйстве сорта имеют исключительно важное значение.
Для культуры в разных районах рекомендуются разные, так называемые
районированные сорта.
Близкородственные виды, имеющие общего предка, объединяют в более
крупные систематические категории — роды (genus). Последние по тому
же принципу общности происхождения соединяют в семейства (jarhi-
lia), семейства — в порядки (ordo), порядки — в классы (class is).
Наконец, классы на той же основе общности происхождения объединяют в
отделы (divisio). Каждую из этих систематических, или таксономиче-
ских, единиц можно подразделить, в целях облегчения обзора, на более мел-
ВВЕДЕНИЕ
15
кие, обозначаемые теми ясе словами с приставкой «под» («sub») — подотдел
(subdivision подкласс (subclassis), подпорядок (subordo) и т. д.
Кроме этого, в семействах и подсемействах выделяются еще трибы (tribus),
а в родах и подродах — секции (sectio).
Систематические категории любого ранга обозначают термином таксон
(taxon — ед. число, taxa — мн. число).
Выделяемые в современных системах отделы растительного мира явля-
ются отраясением очень ваясного итога почти векового развития филогенети-
ческой систематики, суть которого в том, что историческое развитие расти-
тельного мира осуществлялось в форме не единственного прогрессивного
усложнявшегося эволюционного ряда, а в виде нескольких более или
менее параллельных эволюционных рядов.
Единство всего растительного мира с точки зрения современной филоге-
нетической системы выражается в общности происхождения всех растений от
каких-то древнейших и простейших существ. Последние дали начало не од-
ному, а нескольким стволам эволюционного развития, в каждом из которых
независимо от других происходило усложнение организации (или, наоборот,
в отдельных ветвях — морфологическое упрощение, например в эволюции
паразитов).
Самостоятельность отдельных эволюционных стволов, т. е. их генетиче-
ская независимость друг от друга, и дает одно из существенных оснований для *
выделения наиболее крупных классификационных единиц — отделов.
Однако не всякий ствол эволюции рассматривается как один отдел. В фи-
логенезе некоторых стволов могли иметь место периоды резких переломов, оп-
ределяемые глубокими изменениями всей организации растений в связи с
приспособлением к новой среде существования, несвойственной более древ-
ним представителям данного ствола. Так, например, считают, что один из
отделов водорослей, эволюционируя, дал некогда начало приспособившим-
ся к жизни на ^уше листостебельным растениям. Конкретные формы расте-
ний, знаменовавшие своим появлением наступление этого переломного момен-
та в эволюции, вместе с происшедшими от них потомками, все лучше осваи-
вавшими сухопутную среду, также заслуживают выделения в особый отдел.
Подобные глубокие перемены в организации, вообще говоря, каждого эволю-
ционного ствола могут повторяться. Как правило, конкретная группа орга-
низмов, давшая начало тому или иному отделу, остается неизвестной, и таким
образом эта крупнейшая единица классификации оказывается в системе изо-
лированной от других.
Под именем отдела в современной систематике, следовательно, вы-
деляют либо целиком изолированные от других стволы эволюции со всеми их
- разветвлениями, либо части (отрезки) этих стволов, соответствующие длитель-
ным этапам исторического развития и отграниченные от предшествующих
и последующих этапов коренным изменением организации в связи с приспо-
соблением к жизни в новых специфических условиях существования.
В основу расчленения всего растительного мира в этом руководстве поло-
жена классификация А. Энглера в ее последней (1954) редакции со следую-
щими отступлениями. Обширный отдел Chrysophyta расчленен на три;СЛгу-
sophyta в узком смысле, Xanthophyta (или Heterocontae) и Bacillariophyta
(или Diatomeae). Отдел Glaucophyta^ самостоятельность которого продолжает
оставаться гипотетичной, исключен. В таком виде система включает 18
отделов, для облегчения обзора которых может послужить следующая
таблица (стр. 16).
Неоднократно обращалось внимание на то, что пиррофитовые, золотис-
тые, желто-зеленые, диатомовые и бурые водоросли выделяются среди про-
чих незеленой окраской, связанной с преобладанием у них ксантофиллов над
хлорофиллом а, хлорофилл Ъ у них отсутствует. Это дает основание некото-
рым ботаникам соединять названные отделы (в ранге классов) в один отдел
Chromophyta^ причем для каждой из объединяемых групп сохранять значе-
ние независимой от других линий эволюционного развития. Объединение их
1 16
ввж
КИ И .1.
отдел, следовательно, лишь отражает предположение о возможном
всех (или почти всех) названных водорослей от общего нптл.
готического предка. Но, с другой стороны, своеобразие даже бурит, диато-
новых и золотистых водорослей столь велико, что с классификационной точки
зрения их целесообразнее рассматривать как особые отделы. Еще в большей
степени это относится к пиррофитовым и желто-зеленым водорослям.
Развитие систематики растении' как науки происходило начиная с*
XVIII в. при непосредственном и активном участии наших отечественных!
ученых. Вклад нашей отечественной науки в построение системы рястителк»-
кого мира достаточно велик.
Построение филогенетической системы требует проработки ее во всех
звеньях, начиная с основной единицы — вида — и кончая наиболее круп-
ными подразделениями. На огромной территории Советского Союза про-
израстает более 17,5 тыс. видов, относящихся к 160 семействам одних только
г II J
сосудистых растений, т. е. папоротников, хвощей, плауновидных, голо-.
и покрытосеменных. Многие из них общие с растениями пограничных и бо- :
лее отдаленных стран, но многие ограничены в своем распространен
it
ТОЛЬ-1
ко нашей территорией. Выявление видового разнообразия нашей флоры —
. дело отечественных ученых. t Уже в XVIII в. наряду с заграничными учены-
ми, которых царское правительство приглашало для работы в России, вы-
Вметав» «ли лвстостебельные»
растявя» свособные к фото-
синтезу1
« о Cyanophyta1 * 3 * * * * 8 (сине-зеленые Bacteriophyta (бактерии)
g водоросли)
Euglenophyta (эвгленовые
водоросли)
Pyrrhophyta (пиррофито-
вые водоросли)
Chrysophyta (золотистые
водорослн)
Xanthophyta (Heterocontae,
желто-зеленые, или раз-
ножгутиковые, водоросли)
Bacillariophyta (Diato-
шеае, диатомовые водо-
росли)
Chlorophyta (зеленые во-
доросли)
Chaiophyta (харовые водо-
рослн, лучивд)
Phaeophyta (бурые водо-
рослн)
Rhodophyta (кра
доросли)
Myxophyta (Миксомице-
ты, слизевики)
Mycophyta (Fungi, грибы)
Lichenophyta (лишайники)
Bryophyta (мохообразные)
Pteridophyta (папоротни-
кообразные)
Gymnoepermatophyta (го-
лосеменные)
Angiospennatophyta (по-
крытосеменные, или цвет-
ковые)
1 Кроне немногих, вторично утративших хлорофилл.
9 Кроме некоторых бактерий.
3 Названия отделов водорослей CftryMpfcyta, CMoropXyta, Pltaeopfcyta нт. п.*в дослов-
ном переводе на русский язык будут соответствовать «золотистые растения», «зеленые
растения», «бурые растения» и т. п. Согласно постановлениям последних международных
ботанических конгрессов, названия отделов желательно составлять так, чтобы они отра-
жали характерные их особенности. Поэтому иногда рекомендуют оканчивать названия
отделов водорослей на „PXycopXyie** («водорослевые растения»), и в таком случае типы
водорослей должны называться С&гумрйрсорйугв, CXbropkycopJbyta, Phaeophycophyta и т. щ
17
, 11 i
делились талантливые исследователи из русских людей, начавшие накопле-
ние флористического материала. Одним из первых русских ботаников был
С. П. Крашенинников (1713—1755). Начиная с первой половины
XVIII в. число ботаников-флористов неуклонно возрастало, как возрастала
их научная продукция. В середине XIX в. на основании накопленного к
тому времени материала оказалось возможным издание сводной флористи-
ческой работы, выполненной профессором Юрьевского университета
К. Ф. Ледебуром,— tFlora Rossie а*. Этот четырехтомный труд, объ-
единяющий описание более 6500 видов растений1, сделал эпоку в изучении
нашей флоры, как сводка, заключающая не только перечень видов и их ха-
рактеристику, но и свод всех литературных данных о флоре России. tFlora,
Rossica* чрезвычайно облегчила дальнейшее изучение растительного мира
нашей страны, и она до сего времени является одним из важных справочных
источников для ботаников-систематиков.
Накопление флористического материала осуществлялось в ряде круп-
ных научных центров: во всех университетах, в Академии наук, в Петербург-
ском ботаническом саду и других ботанических учреждениях. Уже к концу
XIX в. ^Floras Ледебура нуждалась в крупных пополнениях вследствие на-
копления массы новых данных и в связи с присоединением к России средне-
азиатских территорий, растения которых не вошли во tFlora Rossica*. Коли-
чество растений, известных как произрастающие в России, намного возрос-
ло, и требовалось создание новой сводки. Однако выполнение этой работы
было не под силу одному лицу. Коллективную обработку отечественной флоры
удалось организовать лишь после Великой Октябрьской социалистической
революции, когда было начато составление обширного сводного труда по
цветковым, голосеменным и папоротникообразным растениям большим кол-
лективом ботаников, который возглавил академик В. Л. Комаров. Пер-
вый том «Флоры СССР» был издан в 1934 г. Завершено издание в 1964 г.
«Флора СССР» нужна прежде всего для практических целей. Самые раз-
нообразные учреждения и предприятия, имеющие дело с растительным сырь-
ем, нуждаются в сводном флористическом сочинении.
В связи с выявлением видового состава флоры отечественные исследова-
тели участвовали в разработке понятия о виде и методов видовой системати-
ки. Важное общенаучное значение получили теоретические обобщения
С. И. Коржинского (1861—1900), который обосновал понятие о расе.
Коржинский относил к одной расе все формы, «которые при обладании извест*
иыми морфологическими отличиями представляют собой ареал распростране-
ния». Понятие о расе как географическом явлении легло в основу морфоло-
го-географического метода систематики, которым Коржинский стал пользо-
ваться первым и который в настоящее время применяется во всех флористи-
ческих монографиях и солидных обзорах высших растений. Коржинский
развивал и обосновал этот метод ранее Веттштейна, которому в зарубежной
литературе обычно приписывают приоритет в этом отношении. Те же вопросы
разрабатывал в своих трудах академик В. Л. Комаров, обогативший науку
монографией «Учение о виде у растений», в которой критически проанализи- -
ровал понятие о виде и сформулировал его определение, исходя из теории
Ч. Дарвина и основных положений диалектического материализма.
Б. А. Келлер (1874—1945), развивая идею морфолого-географи-
ческого метода, выступил с обоснованием эколого-морфологического метода,
который он применял в систематических изысканиях, целеустремленно на-
правленных на изучение эволюции растений.
Работа по изучению отечественной флоры как высших, так отчасти и низ-
ших растений влекла за собой необходимость критического пересмотра опи-
саний видов, объема и положения их в системе. В результате такие крити-
г. I:
иг.
1 Интересно сравнить эту цифру с числом видов, которые указывались Линнеем в
ere „Specie plantarum" для России,— 350. Общее число видов флоры Земли Линней оце-
нивал 8—10 тыс,
18
ВВЦ
ческие пересмотры выливались нередко в монографические обработки целых
родов. В монографиях на основании широких обобщений устанавливались
родственные связи между частями рода (секциями) и отдельными группами
видов. Таким образом выяснялись отдельные* звенья филогенетической сис-
темы. Плоды монографических обработок, выполненных русскими учены-
ми (например, Н. И. Кузнецовым, В. Л. Комаровым,
А. А. Бунге, А. А. Еленкиными многими другими), вошли в об-
щий фонд мировой систематики растений.
Построение системы растений в целом и разработка ее принципов всегда
вызывали живе:
й интерес и активный отклик со стороны русских ученых.
1111 \
Уже в начале XIX в. были изданы сочинения, посвященные критике господст-
вовавшей тогда системы Линнея (Т. А, Смеловский, 1808). Позднее
М. А. Максимович выступил с диссертацией «О системах растительного
царства», в которой он, характеризуя содержание естественной системы и ее
отличия от искусственных, устанавливает различие между понятиями срод-
ства и сходства мещду растениями, оценивает систему А. Жюссье скорее как
искусственную, чем естественную, возражает против линейного расположе-
ния растений в системе и против представления о мире растений как о сово-
купности звеньев, расположенных в порядке возрастающего совершенства
строения. Некоторые русские ученые выступали как авторы своих систем
цветковых и других крупных групп растений, а также и всего растительного
мира: Н. И. Кузнецов (1864—1932), Н. А. Б у ш (1869—1941),
А. А. Гроссгейм (1888—1948), А. Л. Т а х т а д ж я н, X. Я. Гоби
(1847—1919), Б. М. Козо-Полянский (1890—1957).
Чрезвычайно важное значение имели многочисленные исследования
русских ученых в области эволюционной морфологии. Многие из достиже-
ний в этом отношении оказали глубокое влияние на развитие филогенети-
ческой систематики растений. К числу таких работ относятся исследования
И. Н. Горожанкина (1848—1904) по онтогенезу некоторых зеленых
водорослей (вольвоксовых) и по оплодотворению голосеменных растений,
И. Н. Го рожа нк ин предложил продуманное и глубоко обоснованное деле-
ние всего растительного мира на три больших отдела: оогониальных, архего-
ниальных и пестичных (цветковых) растений. Два последних принимаются и
сейчас некоторыми систематиками.
Работы В. И. Беляева (1855—1911) по изучению развития мужских
заростков разноспоровых папоротникообразных и голосеменных растений
внесли много нового в теорию филогенетической связи между тайнобрачны-
ми и явнобрачными растениями. Они дали толчок к исследованию микроспор
низших голосеменных и привели к открытию сперматозоидов у саговников
и гинкго, чем окончательно установились связи между голосеменными и
папоротникообразными растениями.
Крупным ученым, сыгравшим своими эмбриологическими исследования-
ми большую роль в установлении своеобразия важнейшей группы растений—
цветковых, — был С. Г. Навашин (1857—1930). Ему принадлежит от-
крытие двойного оплодотворения (1898) — оригинальной особенности по-
крытосеменных, — открытие, вместе с последующими работами доставившее
С. Г. Навашину репутацию ученого с мировым именем.
Построение филогенетической системы растений в нашей стране осущест-
влялось на базе критического восприятия идей эволюционного учения Ч. Дар-
вина. Наряду с такими крупнейшими ботаниками, как А. Н. Бекетов,
П. Ф. Горянинов, К. А. Тимирязев, В. Л. Комаров,
Б. М. Козо-Полянский и многие другие, в этом отношении боль-
шую роль сыграли и русские зоологи. Труды В. О. Ковалевского,
А. О. Ковалевского, М. А. Мензбира, Н. А. Северцева,
А. Н. Северцева, И. И. Мечникова и многих других не толь-
ко способствовали распространению у нас идей эволюционного учения, но
и внесли много принципиально нового в теорию Ч. Дарвина, т. е. развива-
ли дарвинизм как общебиологическую науку.
НИЗШИЕ РАСТЕНИЯ
НИЗШИЕ РАСТЕНИЯ
ОБЩАЯ ХАРАКТЕРИСТИКА
Понятие «низшие растения» включает 14_отделов системы. Общей для
них чертой является отсутствие расчленения вегетативного тела на характер-
ные для большинства высших растении органы вегетативной жизни: побег,
лист и корень. Тело низших растений, таким образом, представляет так назы-
ваемое слоевцо, слоевище или таллом. Поэтому для обозначения
: Г,i
мени применяетсярабочее название Thallophyta.
ИнотДЁГу низших растений наблюдается конвергентное сходство с выс-
шими благодаря образованию листо-^или стеблеподобных органов. Но ^спо-
соб их формирования в онтогенезе осительная простота строения резко
отличают_хакие_ участки тела слое низщцх^растений от внешне сходных '
органов высщих. Низшие растени^>тличаются от высших также и строени-
ем органов размножения. Их гвдцнзцн^в (в случае оогамного полового
процесса — оогонии и антеридии!. и спорангии почти всегда, бывают одно-
клетрчвдми. v----'
Низшие растения подразделяются по способу обеспечения органически-
ми веществами на две обширные экологические группы» Для одной, так на- .
зываемых водорослей, типично наличие хлорофилла. Водоросли, следова-
тельно, организмы автотрофные. Их дополнительные пигменты иной, чем
хлорофилл, химической природы, имеют не зеленую, а иную окраску; преоб-
ладание в пигментном комплексе тех или иных красящих веществ обуслов-
ливает окраску водоросли.
Вторую группу низших растений составляют бесхлорофилльные, потому
в основном они нуждаются для поддержания своей жизни в готовом органи-
ческом веществе. Растения этой группы — грибы, миксомицеты, бактерии —
гетеротрофны. Исключение — две небольшие группы бактерий:, в одну вхо-
дят немногочисленные бактерии, способные к фотосинтезу^ в другую — свое-
образные бактериальные организмы, способные к синтезу органических ве-
ществ на основе использования энергии химических процессов (не света!),
С гетеротрофными сближают лишайники — комплексные растения, образо-
ванные грибом и водорослью, с преобладанием обычно первого.
По широкому распространению в природе и по численности индивидуу-
мов низшие растения превосходят высшие. Роль их в природных процессах
и в жизни человека очень велика, и с течением времени оценка значения низ-
: растительных организмов все повышается. Название «низшие» указы-
вает лишь, на простоту дх морфплппгай^кпй организации и на болыпуХгисто- -
, рическую древность. ------ - - -
иг.»'
НИЗШИЕ РАСТЕНИЯ
ОТДЕЛ —БАКТЕРИИ (BACTERIOPHYTA)
Бактерии — преимущественно одноклеточные, иногда нитчатые, обыч-
но неокрашенные1 низшие растения, не имеющие типичного клеточного ядра и
размножающиеся вегетативно поперечным делением клетки, реже — спо-
рами. Отдел бактерий содержит около 3000 видов. Его можно разделить
на 4 класса.
1-й КЛАСС — ТИПИЧНЫЕ БАКТЕРИИ (EUB ACTERIA)
Строение и размножение. К типичным бактериям относятся мельчайшие
растительные организмы, в подавляющем большинстве одноклеточные, не
имеющие хлорофилла, размножающиеся поперечным делением. В зависи-
мости от формы клетки, постоянной для отдельных видов, бактерии носят
разные названия: шаровидные называются кокками (или микрококка-
ми); палочковидные прямые — бациллами; имеющие форму запятой —
вибрионами; спирально закрученные с одним или немногими завит-
ками — спириллами (рис. 1). Кокки, располагающиеся попарно,
называют диплококками, располагающиеся цепочкой — стрепто-
кокками, собранные в гроздь — стафилококками. Клетки
у шаровидных бактерий большей частью достигают 0,5—1 мкм в диаметре,
у палочковидных 2—5 мкм длины и 0,4—0,8 мкм ширины. Некоторые ните-
видные бактерии видны даже невооруженным глазом.
Клетки бактерий имеют плотную оболочку, которая становится замет-
ной при плазмолизе; целлюлозы и хитина в ней нет. В составе оболочки есть
аминосахара и аминокислоты. Многие бактерии вокруг клетки образуют сли-
зистые капсулы (азотобактер, пневмококк и др.), нередко превосходящие
размеры самих клеток. В протоплазме у старых бактерий находятся неболь-
шие вакуоли. Запасные продукты — жир, гликоген, волютин (полифосфаты
с рибонуклеиновой кислотой), крахмал а. нет* * В протопласте содержится 40%
и более нуклеопротеидов, т. е. больше, чем в клетках любого другого орга-
низма. Нуклеопротеиды образуют скопленияшаровидной, эллипсоидальной,
почковидной или нитевидной формы, которые нередко принимают за ядра;
их называют нуклеоидами *. Нуклеоиды отличаются от ядер отсутствием
Рис. 1. Форма бактерий:
I — кокки, 2 — диплококки; 3 — стафи-
лококки; 4 — стрептококки; б — бацил-
лы; « — кибриокы; 7 — спирилла; в —
спирохета; 9 — нитчатая . бактерия;
Ю — бактерии со слизистыми капсулами
1 Содержащийся в клетках окрашенных бактерий бактериохлорофилл а отличен от
хлорофилла а водорослей и высших растений. Своеобразие фотосинтеза окрашенных бак-
терий в том, что в результате его не выделяется свободный кислород.
* От лат. «нуклеус» — ядро и грея, «эйдос» — вид.
ОТДЕЛ — БАКТЕРИИ
23
Рис. 2. Расположение жгутиков у
подвижных видов бактерий
ядерной оболочки и ядрышек; ядерное
веретено при делении клеток не фор-
мируется.
В нуклеоидах сосредоточена ДНК
и в ряде случаев обнаружена редупли-
кация ее нитей, предшествующая деле-
нию бактериальной клетки. Делятся
нуклеоиды поперечно или продольно.
Электронно-микроскопически в ци-
топлазме установлено наличие телец,
которые по составу и размерам подобны
рибосомам. У немногих окрашенных
бактерий найдены и мельчайшие но-
сители пигментов, величиной около
0,05 лскл€. Они содержат зеленый бак-
териохлорофилл и каротиноиды. H(f
сители пигментов имеют ламеллярное
строение, но не имеют своей мембраны,
чем и отличаются от пластид. В цитоплазме же, вблизи наружной ее мембра-
ны, прилегающей к клеточной оболочке изнутри, помещаются базальные
тельца жгутиков.
Благодаря жгутикам многие бактерии подвижны. Жгутики бактерии
очень тонки. Располагаются они или по одному на конце клетки (монотри-
хальные бактерии), или пучком — тоже на конце клетки (лофотрихальные),
или, наконец, по всей поверхности ее (перитрихальные бактерии) (рис. 2).
У многих бактерий число жгутиков не постоя5щбГизменяется иногда в зави-
симости от условий питания. Клетка может сбрасывать и вновь восстанавли-
вать жгутики. Строение-жгутаков у бактерий отлично от таковых у других
организмов, где они составлены из 9 + 2 двойных фибрилл. У бактерий жгу-
тик имеет как ^ы-влштообразное (с поверхности) строение.
Размножаются бактерии простым делением клетки, которое происходит
поперечно, у форм нешаровидных — перпендикулярно к продольной оси клет-
ки. При благоприятных условиях деление может происходить через каждые
полчаса, т. е. чаще, чем у любых других организмов. Этим объясняется чрез-
вычайная4 иногда быстрота размножения некоторых бактерии. На основании
чисто арифметических подсчетов установлено, что потомство одной бактери-
альной клетки через 5—6 дней могло бы заполнить все моря и океаны на Зем-
ле, а через 10 дней дать потомство, равное по объему всему земному шару.
Конечно, в действительности этого никогда не может быть, так как бактерии
очень быстро используют необходимые для роста и размножения питатель-
ные вещества, отравляют среду продуктами своей жизнедеятельности и по-
гибают от неблагоприятного действия внешних факторов. Но подсчеты
подобного рода характеризуют потенциальную энергию размножения.
У многих палочковидных и немногих шаровидных бактерии образуются
эндогенные споры, по одной в .клетке. Они формируются в центре или на кон-
це клетки путем сжатия и уплотнения цитоплазматического содержимого
клетки; вокруг образующейся споры выделяется новая плотная оболочка
(рис. 3). Споры обеспечивают возможность переносить неблагоприятные усло-
вия. Они выдерживают длительное высыхание, кипячение и действие других
неблагоприятных факторов. Попав в благоприятные условия, споры набуха-
ют и прорастают, образуя новую вегетативную клетку бактерии.
Вопрос о цикле развития типичных бактерий нельзя'считать вполне изу-
ченным. В простейшем случае клетка делится, и дочерние клетки, вырастая
до размера материнской, повторяют деление. Но такое простое следование
поколений бывает не' у всех бактерий. Так, например, у сенной палочки
(Bacillus subtilis, рис. 4) подвижная жгутиковая клетка, размножаясь, дает
начало нити, состоящей из подвижных клеток, которые далее теряют жгутики;
в этих клетках в дальнейшем развиваются споры, последние освобождаются
I
НИЗШИМ РАСТЕНИЯ
Рис. 3. Разные типы распо-
ложения спор у бактерий
и при соответствующих условиях прорастают, давая начало новым подвиж-
ным клеткам. В этом случае в цикле развития организм проходит несколько
закономерно сменяющих друг друга этапов. Описывались и более сложные
циклы развития бактерий.
До недавнего времени оставалось совершенно неясным, существуют ди
у бактерии половые явления. Применением главным образом генетического
анализа сейчас установлено, что бактерии способны к объединению генети-
ческой информации, подобно тому как это бывает у организмов, имеющих
половой процесс. Однако у бактерий явления этого рода совершаются в свое-
образной форме, даже если передача информации происходит путем непо-
средственного контакта между конъюгирующими клетками. Выяснено также,
что передача наследственной информации у бактерий может осуществляться
и без прямого контакта клеток — частицами бактериофага. Наконец, иногда
изменение наследственных качеств клетки совершается за счет поступления
в нее ДНК, находящейся в среде в свободном (например, вследствие разру-
шения отмерших клеток) состоянии.
Распространение бактерий. Бактерии чрезвычайно широко и в огромных
количествах распространены в природе. В 1 а черноземных почв находится
около 5—6 млрд, бактерий, в 1 а подзолистых почв — около 2 млрд., в 1 а пес-
ка — около полумиллиарда. В 1 см8 воды вдали от населенных мест содер-
жится несколько сотен и даже тысяч (2—4) бактерий; в загрязненной органи-
ческими веществами воде рек, вытекающих из больших городов, — десятки
тысяч бактерии. В 1 м* воздуха в больших городах содержится до 8 тыс. бак-
терий, а в местах большого скопления людей — десятки и даже сотни тысяч
бактерий. В1 см8 молока отличного качества содежится 10—15 тыс., а в моло-
ке хорошего качества —. до 500 тыс. бактерий.
Физиология и образ жизни бактерий. Основным методом изучения био-.
логических свойств отдельных видов (и более мелких таксонов) бактерий,.
как и многих других низших растении, являются так называемые чистые
иг
культуры, т. е. такие, в которых находится только один вид (или более мел-
Рнс. 4. Цикл развития сейной палочки
Цр ОТДЕЛ—БАКТЕРИИ 25
gss?-'
кая таксономическая единица — раса и т. п.). Наилучшая гарантия чистоты
культуры — получение ее из одной клетки, что достигается специальными
£&•-• микробиологическими методами.
I;;" По характеру питания бактерии бывают автотрофными и ге-
теротрофными. Автотрофных бактерий немного, они могут сами
синтезировать органическое вещество из неорганического путем фотосин-
теза или хемосинтеза. К фотосинтезу способны зеленые и пурпурные бак-
1Г
терпи.
Хемосинтез у некоторых бактерий был впервые открыт в конце XIX в.
русским микробиологом С. Н. Виноградским. Этот процесс осу-
ществляется без участия света, за счет энергии, которая получается при
окислении различных неорганических соединений. Нитрифицирующие бак-
терии окисляют: одни — аммиак до азотистой кислоты (Nitrosomonas, Nit-
rosococcus), другие (Nitrobacter} — азотистую кислоту до азотной. Железобакте-
рии (Leptothrix ochracea, Crenothrix polyspora и др.) окисляют закисные соеди-
нения железа в окисные. Существуют бактерии, которые окисляют водород,
метан, окись углерода и т. д. Усвоения углерода путем хемосинтеза у других
одгянмямпв не встречается. ” ---
Тетеротрофные бактерии питаются как сапрофиты или как паразиты
человека, животных и растений. Многие гетеротрофшде бактерии вызыва-
ют различные виды брожений. Некоторое продукты брожении действуют
угнетающе на развитие других бактерий в их антагонистических межвидо-
вых взаимоотношениях. С брожениями, которые приводят к накоплению мо-
лочной, масляной и уксусной кислот, приходится считаться в повседневной
жизни. Это молочнокислое, маслянокислое, уксуснокислое и другие брожения.
При пектиновом брожении разлагаются пектиновые вещества, находящиеся
в межклеточных пластинках у растений.
Разложение бактериями азотистых веществ, в основном белковых соеди-
не!^, наэиявшбТ7^«шё^Кбнечные^ародукты4ниения" разнообразны; обыч-
но в числе их, имеются дурно пахнущие вещества: скатол, индо л,се ров одо-
род и др.; некоторые вещества, выделяющиеся при гниении, могут вызвать
отравление. Гниение вызывается многими бактериями, причем процесс мо-
жет проходить ряд последовательных этапов. Продукты, получающиеся
вследствие разрушения исходного вещества одними бактериями, подверга-
ются дальнейшему разрушению другими, третьими.
Как в теоретическом, так и в практическом отношении представляют
большой интерес азотофиксирующие бактерии, способные усваивать сво-
бодный азот атмосферы. Среди них широко известны так называемые клу-
беньковые бактерии (Rhizobium leguminosarum. Bacterium radicicola}, живу-
щие в почве. Внедряясь в корни бобовых растений, они вызывают на них не-
большие местные разрастания в форме клубеньков. От бобового растения эти
бактерии получают преимущественно углеводы, сами же они усваивают азот
атмосферы, проникающий в межклетники корня, и срадащт за счет его белко-
вые вещества своего тела. При частичном отмирании этих бактерий в клетках
корЯя их белковые вещества усваиваются бобовым растением. Пожнивные
.остатки бобовых и корни их при разложении обогащают почву азотистыми
соединениями, почему бобовые растения обычно и включают в севооборот.
На 1 га клубеньковые бактерии связывают 100—400 кг азота. Существует
несколько различных рас этих бактерий^ Приспособившихся к сожительству
с теми или иными видами бобовых. Взаимоотношения бобового растения и
бактерий можно рассматривать как (взаимовыгодный) симбиоз, а вернее,
как обоюдный паразитизм.
Другие азотофиксирующие бактерии живут свободно в почве. Важней-
м среди них является азотобактер (Azotobacter chroococcum), имеющий
форму коротких палочек, окруженных толстыми слизистыми капсулами. Уг-
лерод он получает из органических веществ почвы. По отмирании азотобак-
тера его тела разрушаются другими бактериями с образованием неоргани-
ческих соединений азота* усвояемых зелеными растениями. На 1 га почвы азо-
nr,
26
НИЗШИЕ РАСТЕНИЯ
тобактерии связывают 10—30 ха свободного азота. Для лучшего развития
азотобактера необходима хорошая аэрация почвы и наличие в ней достаточ-
ного количества безазотистых органических соединений.
Энергию, необходимую для жизненных процессов, бактерии, как и дру-
гие живые существа, покрывают за счет экзотермических процессов, составля-
ющих сущность акта дыхания. У бактерий дыхание проявляется в разнооб-
разных формах. У большинства оно, как и у человека, животных и подавля-
ющего большинства растений, состоит в окислении свободным кислородом
воздуха различных органических веществ внутри их тела. Это так называе-
мые аэробные1 бактерии. У огромного большинства их при этом выде-
ляется углекислота; у некоторых реакция окисления не доходит до конеч-
ного предела и углекислота не выделяется, как, например, у уксуснокислых
бактерий, окисляющих этиловый спирт лишь в уксусную кислоту. К аэроб-
ным бактериям относятся также хемосинтезирующие (нитрифицирующие и
др.) бактерии.
Другой своеобразной по физиологии группой бактерий являются ана-
эробные бактерии. Они могут жить при отсутствии свободного кисло-
рода; энергию для жизненных процессов большинство их черпает из процес-
сов расщепления сложных органических соединений до более простых, без
участия свободного кислорода, при разных типах брожений. Брожения, по-
добно дыханию, являются экзотермическими процессами, и выделяющаяся
при этом энергия используется бактериями для их жизненных процессов,
подобно энергии, получающейся при кислородном дыхании. Среди анаэроб-
ных бактерий различают облигатные. т. е. нежизнеспособные в присутствии
свободного кислорода или переносящие лишь небольшое его количестве в ок-
ружающей среде (маслянокислые бактерии, бактерии столбняка и др.),
и Факультатив. могущие жить и при наличии свободного кислорода, и
без него (молочнокислые бактерии, возбудители сибирской язвы., брюшно-
го тифа, различных нагноений и т. д.).
Классификация бактерий. Вследствие того что амплитуда морфоло-
гических различий у бактерий в общем ограничена, в классификации их ши-
роко используются различные физиологические свойства — подвижность,
образование капсулы, спорообразование и др. Важные признаки — форма и
окраска колоний, образующихся в результате размножения одной клетки,
а также влияние бактерий на окружающую среду, а именно характер раз-
жижения застывшей желатины, воздействие на различные продукты выделе-
ния специфических веществ и др. При диагностике применяют специальную
окраску, предложенную датским врачом Граммом; в зависимости от
результатов ее отличают грамположительные и грамотрицательные бак-
терии.
Класс Eubacteria делят на 4—5 порядков. Наиболее крупный по-
рядок Eubacteriales включает одноклеточные неветвящиеся виды, распре-
деляемые по форме клеток и другим признакам в 6 или более семейств: ша-
ровидные — семейство Соссасеае и др.; палочковидные
без спор — семейство Bacteriaceae; палочковидные, образу-
ющие споры,—с емейство Baccilaceae; изогнутые или спираль-
но извитые с одним или многими завитками — семейство
Spirtllaceas.
К порядку микобактерий (Mycobacteriales, Corynebacteriales) от-
носятся одноклеточные палочковидные, иногда слабоветвящиеся неподвижные
бактерии, например Mycobacterium tuberculosis —возбудитель туберкулеза,
Corynebacterium diphtherias—возбудитель дифтерии, Mycobacterium lepras —
возбудитель проказы, ряд сапрофитных видов. Многие микробиологи отно-
сят микобактерии к актиномицетам.
В порядок хламидобактерий, или трихобактерий (Chlamy-
dobacteriales, Trichobacteriales)* объединяют многоклеточные нитчатые бакте-
1 От лат. <аэр» — воздух,
ОТДЕЛ — БАКТЕРИИ
27
Рис. 5. Серобактерия беджиатоа:
1 — с серой; 2 — без серы (видны перегородки между клетками)
рии—сферотилюс, или кладотрикс (Sphaerotilus natans,
Cladothrixnatans)^ образующий в загрязненных водоемах
беловатые космы, состоящие из множества ложноразветв-
ленных нитей, железобактерии—лептотрикс (Leptothrix) и
кренотрикс (Crenothrix), серобактерию беджиатоа (Beg-
giatoa) и др. (рис. 5).
В вопросах классификации зубактерий между ми-
кробиологами существуют разногласия, поэтому в насто-
ящее время существует несколько классификаций бакте-
рий.
2-й КЛАСС — МИКСОБАКТЕРИИ (MYXOBACTERIA)
Миксобактерии — подвижные палочковидные клетки, не имеющие жгу-
тиков и передвигающиеся путем отталкивания слизью, выделяющейся из
конца клетки. Клеточная оболочка, если она* есть (существование ее у мик-
со бактерий подвергается сомнению), эластична; при движении клеток их
форма изменяется. У большинства миксобактерий в клетке имеется ядро-
подобная хромативная структура, принимаемая нередко за ядро. Большей
частью у миксобактерий образуются ярко окрашенные, так называемые пло-
довые тела, состоящие из слизистой ножки и головки; последняя сложена
из цист, содержащих по нескольку бактериальных клеток. Миксобактерии
встречаются в почве, навозе, воде. Как и типичные бактерии, миксобактерии
играют большую роль в круговороте веществ в природе. К миксобактериям
относится один порядок, который делится на 5 семейств.
3-й КЛАСС—СПИРОХЕТЫ (SPIROCHAETAE)
Спирохеты представляют собой длинные (от 4 мк до 0,5 мк) гибкие изви-
тые клетки с многочисленными завитками и заостренными концами. В цент-
ре клетки находится волнистый центральный тяж, окруженный снаружи
цитоплазмой. Наружная мембрана обвита пучком тонких и гибких фибрилл.
У многих спирохет вдоль тела идет перепонка (сходная с ундулирующей
мембраной трипаносом). Представители спирохет — бледная спирохета
(Treponema pallidum) — возбудитель сифилиса, Borrelia recurentis — воз-
будитель возвратного тифа, безвредная спирохета (Borrelia buccalis)t часто
встречающаяся в налете на зубах. Один порядок и одно семейство.
4-й КЛАСС — АКТИНОМИЦЕТЫ, ИЛИ ЛУЧИСТЫЕ ГРИБЫ
(ACTINOMYCETES)
Актиномицеты — своеобразная группа микроорганизмов, имеющих при-
знаки отчасти бактерий, отчасти грибов. Типичные актиномицеты предста-
вляют собой очень тонкие (0,3—1,5 мкм) нити, моноподиально ветвящиеся и
часто лучеобразно распространяющиеся во все стороны, как у грибов1. Попе-
речные перегородки между клетками нитей обычно не обнаруживаются. Сос-
тав оболочек, по-видимому, близок к бактериальному. Типичного ядра в
клетках не обнаружено. Вегетативные нити (гифы) находятся в субстрате,
1 Название «актиномицеты» происходит от греч. «актис» (род. падеж «актинос») —
луч и «микос» — гриб.
на поверхности субстрата и приподнимаются над ним; воздушные гифы у мно-
гих актиноиицетов закручены в спираль. Ветвление нитей настоящее. Сово-
купность гиф называют, как и у грибов, мицелием.
Размножаются актиномицеты легко обламывающимися кусочками ми-
целия, а также спорами; половой процесс неизвестен. Споры образуются на
ветвях воздушного мицелия простых прямых, реже — мутовчато-разветвлен-
ных, или у многих — на концах закрученных спирально (рис. 6). Споры
образуются двояко: 1) путем распадения цитоплазмы гифы на шаровидные^
реже — продолговатые, участки, оформляющиеся далее как споры. Пустые
участки гифы, остающиеся между спорами, позднее ослизняются; 2) путем
распадения специализированной гифы — спороносца — на короткие цилинд-
рические членики (споры). Споры актиномицетов хорошо выдерживают высу-
шивание, но в противоположность спорам бактерий погибают при кипячении
воды. Прорастают споры, как и у грибов, одним или несколькими выростами,
вытягивающимися в г и ф ы.
Все актиномицеты грамположительны. Они стойки к щелочам, но, как и
настоящие бактерии, очень неустойчивы к кислотам. Большинство их аэроб-
ны. Многие выделяют пигменты и летучие пахучие вещества.
Живут актиномицеты в почве, в воде, постоянно встречаются в воздухе;
их выделяли из молока, сыра, овощей, фруктов, живых и мертвых растений
и т. п. В 1 а почвы встречаются>десятки, сотни тысяч и миллионы актиномице-
тов; они обусловливают запах земли (Actinomyces odorifer и др.). Актиномице-
ты участвуют в разложении органических веществ в почве и в процессах кру-
говорота веществ. Некоторые актиномицеты вызывают опасные заболевания
человека и домашних животных — актиномикозы. Парша клубней картофеля
(непорошистая) вызывается активомицетами (Actinomyces scabies и др.). По не-
которым датаым, актиномицеты, способные усваивать атмосферный азот,
. образуют клубеньки на корнях ольхи и лоха. *
<; ; Особенно большое внимание привлекли к себе актиномицеты в три пос-
ледних десятилетия, когда выяснилось, что многие из них вырабатывают
антибиотики. В лабораториях многих стран из почв ежегодно выделяют сот-
ни тысяч культур актиномицетов, тщательно изучаемых и исследуемых на
антибиотические свойства. Из актиноиицетов получают стрептомицин, био-
ИН
гг
ИН
Классификация актиноиицетов находится в настоящее время в состоя-
нии разработки. Одни исследователи придают главное значение морфологи-
ческим признакам (строение спороносцев и др.), другие — культуральным
признакам (образование пигментов и т. п.), третьи учитывают совокупность
признаков.
ЗНАЧЕНИЕ БАКТЕРИЙ И АКТИНОМИЦЕТОВ
В ПРИРОДЕ И В ЖИЗНИ ЧЕЛОВЕКА
Роль Bacteriophyta в природе очень велика. Благодаря их деятельности
происходит разложение и минерализация (превращение в неорганические
соединения) трупов и выделений (мочи, кала) животных и различных расти*
тельных остатков (в разложении последних значительную роль играют и гри-
бы). Минерализация органических веществ представляет важное звено круго-
ворота веществ на Земле, без которого была бы невозможна жизнь на ней.
Ря(утопия, содержащие хлорофилл, синтезируют различные органические со-
единения из неорганических, потребляя при этом в общем ограниченный запас
элементов минерального питания, находящийся в поверхностном слое земной
коры. Животные, поедающие растения или других животных, также синте-
зируют органические вещества. Вся эта масса органического материала под .
влиянием бактерий (отчасти и грибов) вновь превращается в неорганиче-
ские соединения, поступающие в землю и становящиеся вновь доступными
растениям. Белковые вещества различными гнилостными бактериями посте-
ГчЛ
ГчЛ
Рис. 6. Актиномицеты:
J—wnnoma актшомацота; 2 — 4 — рамочные форма аворооосцав; S — абрааовашю спор; а — Фпагммип-
цяоивое, б — ссгмеатацкоавос; 6 — црорвсммшо спор
30
НИЗШИЕ РАСТЕНИЯ
пенно разлагаются с образованием аммиака и других продуктов распада.
Аммиак нитрифицирующими бактериями переводится в нитриты, а затем —
в нитраты, последние используются зелеными растениями, и этим замыка-
ется круговорот азота. Так называемые динитрифицирующие бактерии, жи-
вущие в почве и воде, могут восстанавливать нитраты до молекулярного азота,
который азотофиксирующими бактериями снова переводится в связанные
формы.
При гниении белков образуется также сероводород, который ассимили-
руется серобактериями и переводится в соли серной кислоты, доступные для
питания других растений. Таково участие бактерий в круговороте серы.
Фосфороорганические соединения при гниении белков расщепляются
с образованием фосфорной кислоты, которая, соединяясь с основаниями поч-
вы (кальции, магний, железо, алюминий и др.), переходит в труднораство-
римые ди-, три- и тетрафосфаты; последние под действием различных бак-
терий-кислотообразователей переходят в монофосфаты, усвояемые зелеными
растениями.
Углерод, связанный растениями при фотосинтезе в сложные органиче-
ские соединения, при некоторых брожениях, а также при гниении освобожда-
ется в виде углекислого газа, снова используемого растениями, содержа-
щими хлорофилл. Приведенные, а также многие другие примеры указывают
на огромную роль бактерий в круговороте веществ в природе, в создании и
поддержании плодородия почвы. Бактерии вместе с лишайниками и водо-
рослями разрушают горные породы, участвуя этим в начальных стадиях поч-
вообразовательных процессов.
Для жизни человека бактерии и актиномицеты имеют и отрицательное
и положительное значение. Многие опасные заболевания человека и домаш-
них животных вызываются бактериями, таковы туберкулез, дифтерия, холера
(возбудитель Vibrio choleras), брюшной тиф (возбудитель Salmonella typhosa),
паратифы (другие виды Salmonella), чума (возбудитель Pasteurella pestis), го-
норрея (возбудитель Neusserria gonorrhoeas), сепсис (возбудители—некоторые
виды Streptococcus, Staphylococcus), сибирская язва (возбудитель Bacillus
anthracis), сап (возбудитель Coryneeacterium mallei) и др. Гнилостные бак-
терии, находящиеся в толстых кишках, образуют там вредные для орга-
низма вещества (токсины), поступающие через стенки кишечника в кровь и
лимфу. Актиномицеты вызывают у человека и домашних животных особые
заболевания — актиномикозы.
При заболеваниях, вызываемых бактериями, после попадания бактерий
в организм (инфекция) проходит некоторый так называемый инкубационный
период, в течение которого происходит размножение их в организме, но види-
мых признаков заболевания еще нет. Наступающие далее различные местные
и общие расстройства организма происходят главным образом от отравле-
ния токсинами, выделяемыми бактериями в организм или освобождающимися
из клеток бактерий по отмирании и разрушении их.
Ряд заболеваний культурных растений, так . называемые бактериозы, вы-
зываются многими фитопатогенными бактериями.
Порча различных пищевых продуктов: мяса, рыбы, яиц, скисание*молока,
прогорклость масла, образование колбасного яда в мясе и рыбе — вызывает-
ся бактериями. Для предотвращения этой порчи применяют различные спо-
собы: 1) замораживание или действие низкой температуры (ниже 0°) в хо-
лодильниках и ледниках; 2) маринование продуктов в уксусной кислоте,
которая подавляет развитие бактерий гниения; 3) солку, так как повышенная
концентрация поваренной соли подавляет развитие бактерии. При солке
огурцов, помидоров, грибов, квашении капусты консервирование произво-
дится главным образом не солью, добавляемой лишь в небольшом количест-
ве, а молочной кислотой, которая образуется в результате деятельности мо-
лочнокислых бактерии, сбраживающих сахара, находящиеся в этих продук-
тах, и препятствует развитию других бактерии, не выносящих кислой среды.
Подобное образование молочной кислоты оказывает консервирующее деист-
ОТДЕЛ —БАКТЕРИИ
31
вне и при силосовании кормов; 4) высокие концентрации сахара, например,
при изготовлении варенья, джема, повидла, что вызывает плазмолиз в клет-
ках бактерии и прекращает их развитие; 5) сушку и копчение, в комбина-
ции с предварительной солкой; 6) действие высокой температуры.
Для уничтожения бактерий, не образующих спор (молочнокислые, бакте-
рии тифа, паратифа, туберкулеза), достаточно простого кипячения или да-
же нагревания до 65°С в течение получаса (пастеризация). Для уничтоже-
ния спорообразующих бактерии применяют кипячение при температуре
около 120°С и повышенном давлении в автоклавах с последующей гермети-
ческой упаковкой продуктов, исключающей попадание в них бактерий.
Положительная роль бактерий в хозяйственной деятельности человека
также очень велика. Молочнокислые бактерии участвуют в приготовлении
различных молочнокислых продуктов: простокваши, ацидофильного молока,
кумыса, кефира, масла, сыра. Они же, как уже указывалось, содействуют
консервированию продуктов при солке, мочении, силосовании.
Уксуснокислые бактерии применяют иногда при изготовлении столового
уксуса из вина. Бактерии, вызывающие брожение пектиновых веществ, ис-
пользуются в процессах мочки льна, конопли, где они разрушают межклеточ-
ное вещество и способствуют выделению прядильных волокон. Эти бактерии, а
также бактерии, сбраживающие целлюлозу, постоянно находятся в пище-
варительном тракте травоядных животных и значительно способствуют их
пищеварению: они мацерируют пищу, переводят целлюлозу в другие усвоя-
емые соединения и, кроме того, разрушая клеточные оболочки, делают дос-
тупным для ферментов животного содержимое клеток.
На заводах, используя бактерии, получают молочную, масляную и про-
пионовую кислоты, ацетон, бутиловый спирт, некоторые ферментативные
препараты и другие продукты.
Некоторые бактерии применяют в виде бактериальных удобрений при
выращивании сельскохозяйственных растений.-Культуры клубеньковых бак-
терий в смеси со стерильной почвой, мелом и песком применяют под названием
нитрагина для предпосевной обработки семян бобовых растений, особенно на
вновь освояемых землях или там, где долго не возделывались бобовые расте-
ния. Азотобактерин, или азотоген, — препарат азотобактера — прибавля-
ют к посевному или посадочному материалу некоторых небобовых растении.
Фосфоробактерин — препарат бактерий, способствующих минерализации ор-
ганических соединений фосфора в почве, — также вносят в почву вместе с по-
севным или посадочным материалом. Препарат АМБ представляет собой про-
известкованный торф, в котором размножен комплекс различных бактерии,
способствующих образованию веществ, необходимых для корневого питания
растений.
Актиномицеты, в меньшей степени настоящие бактерии, продуцируют
различные антибиотики, применяемые в медицине, и роль их в этом отношении,
несомненно, будет возрастать.
Приведенные примеры далеко не исчерпывают разнообразной роли бак-
терий и актиномицетов в природе и в жизни человека.
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ БАКТЕРИЙ
Четыре класса, входящие в состав отдела Bacteriophyta, естественно,
распределяются в две группы. Одна объединяет спирохеты и миксобактерии,
сходные в том отношении, что клетки их при движении изгибаются, изменяя
свою форму (флексибильные клетки). Два других класса — собственно бак-
терии и актиномицеты, наоборот, имеют клетки устойчивой формы благодаря
наличию жесткой оболочки (ригидные клетки). В этом случае, если бактерия
подвижна, она обладает одним или несколькими жгутиками. Нет оснований
думать, что флексибильные и ригидные бактерии связаны друг с другом как
потомки с предками. Онил вероятно, представляют две независимые линии
г, Л
ИЯ
u:i:
развития, связанного происхождением от общего предка. Возможно, его
была примитивная клетка, еще не имевшая твердой оболочки, безъядерная
и способная к изменению формы.
Принимая во внимание резкие различия в строении клеток спирохет и
миксобактерий, следует допустить, что эти два класса тоже развивались не-
зависимо друг от друга, как две параллельные линии эволюции. Надо заме-
тить, что спирохеты имеют некоторые общие черты с трипаносомами (Protozoa)
и вполне вероятно, что они вообще будут исключены из состава бактерио-
фигов. Что касается миксобактерий, то среди них имеются формы, не отлича-
ющиеся по организации ядерного материала от типичных бактерий, а также
и такие, у которых крупные хроматиновые структуры становятся, по-видимо-
му, постоянными. Принимая во внимание наличие переходов между этими
крайностями, можно считать вероятным, что в этом классе происходила вы-
работка органоидов, приближавшихся к типичному ядру.
В пределах второй группы классов, эволюционное начало которой было
положено выработкой у общего предка всех бактериофитов жесткой оболоч-
ки, дальнейшая эволюция, вероятно, осуществлялась несколькими парал-
лельными рядами. В их числе наиболее ясны два. В одном на известном этапе
развития возникло ветвление клеток (некоторые Mycobacteriales), которое,
усиливаясь, привело к выработке мицелиальных актиномицетов1. В дру-
гом ряду ведущим признаком было появление фотосинтеза. Если принять
во внимание формы жгутования, способность к образованию спор и биохи-
мические особенности, то намечаются еще по крайней мере три эволюцион-
ных ряда, параллельные двум указанным выше.
В этих независимых направлениях эволюционного процесса, вероятно,
- повторялись одни и те же формы внешней морфологии клеток: от шаровид-
ной к палочковидной и к извитой, от безжгутиковых клеток к монотрихаль-
ным или перитрихальным. Нитевидные бактерии следовали, конечно, за од-
jii
л
Покрытый оболочкой одноклеточный общий предок эубактерий и акти-
номицетов мог быть также родоначальником сине-зеленых водорослей.
Ji
ВИРУСЫ
иг
В открытии вирусов и в истории развития учения о них определяющую
роль сыграли два фундаментальных факта, установленных в конце XIX —
начале XX в. Первым было наблюдение «самопроизвольного» лизиса (раст-
' ворения) бактерий в культуральной жидкости, которое наблюдал Н. Ф. Га-
малея (1898); вторым — установление Д. О. Ивановским (1892)
факта заражения здоровых растений табака соком пораженных мозаичной
болезнью. Клеточный возбудитель инфекции в виде бактерии не был обнару-
жен — патогенное начало не удавалось задержать мелкопористыми бактери-
альными фильтрами. Для возбудителя (фактора), вызывающего лизис бакте-
рий, впоследствии было предложено название бактериофага (фага*), для
вовбудителя мозаичной болезни — фильтрующегося вируса (вируса1 * 3). Впо-
следствии оказалось, что и фаги, и вирусы имеют принципиально одну и ту
же природу; если вирусы — возбудители болезней и вообще нарушений
жизненных процессов макроорганизмов, то бактериофаги играют ту же роль
по отношению к бактериям.
Вирусы представляют собой частицы ничтожного размера: от десятых
до 2—5 сотенных долей миллимикрона4, причем частицы каждого вируса со-
вершенно однородны по их величине. Они имеют разную форму: шарообрн.
г-
v
1 Наряду с этим существует и другой взгляд на актиномицеты — их представляют
как вторичные формы, происшедшие от низших грибов путем деградации их мицелия*
1 От греч. «фагус» — пожиратель, обжора.
3 От лат. «Игн«> — яд.
4 Миллимикрон — тысячная доля микрона, ши 10"7
IU 'i
III
11: i:
зную, овальную, палочковидную, в виде головки с хвостиком и т. д, В составе
вирусной частицы обязательно содержится нуклеиновая кислота и облекаю-
щий ее покров белковой природы — так называемый капсид. В свободном
состоянии, т. е. находясь вне клетки растения или животного, вирусные
частицы, насколько известно, не обладают собственным обменом веществ.
Они способны вводить свою нуклеиновую кислоту внутрь живых клеток,
где она репродуцируется с использованием синтезирующего аппарата
зараженной клетки, что и приводит в итоге к созданию новых частиц виру-
са, способных после освобождения заражать новые объекты. Если считать
вирусы организмами, то они облигатные паразиты. Одни из них поражают
ограниченный, другие — более широкий круг организмов. Многие из них мо-
ей длительное время в почве, воде и других средах; многие
высушивание, нагревание выше 100°С, действие спирта, кар-
боловой кислоты и других веществ, убивающих живые клетки.
Значение вирусов велико. Они являются возбудителями таких болезней,
как грипп, полиомиелит (детский паралич), оспа, корь, бешенство, желту-
ха шелкопряда; от них часто зависит пятнистая окраска, курчавость и дру-
гие деформации листьев, карликовость и скручивание стеблей, уродливость
и позеленение цветков и многие другие патологические явления.
Бактериофаги могут быть использованы для борьбы с бактериальной ин-
Вирусы нельзя причислить ни к растениям, ни к животным. Разнообра-
зие взглядов на их природу сводится к двум основным концепциям. Сто-
ронники одной из них видят в вирусах результат регрессивной эволюции
паразитных организмов, утративших практически все характерные элемен-
ты клеточной структуры и существующих в виде вуклеинового аппарата,
использующего ферментативную и другие системы клеток хозяина для само-
поддержания. Противоположное мнение рисует вирусы как пережиток от-
даленнейших времен, когда жизнь только формировалась. Возникавшие при
особых уровнях среды того времени нуклеиновые соединения частью вошли
в состав клеток и сделались их постоянными компонентами, другие же со-
хранились, входя во временный контакт с клеточными телами и обеспечивая
себе таким способом неограниченно длительное существование.
n ir
>11:1
ВОДОВОСЛИ (ALGAE)
Водоросл^^сак уже указано, представляют группу слоевцовых расте-
ний, объединяемую способностью к фотосинтезу. Большинство водорослей —
растения, живущие в воде, но есть немало и приспособившихся к жизни вне
воды, при более или менее обильном, хотя бы и эпизодическом, увлажнении.
В систематическом отношении они делятся на 10 самостоятельных отделов,
различающихся по происхождении^ окраске^ а у некоторых — и по органи-
зации клетки? .
Структура водорослей. Большое морфологическое разнообразие ве-
; гетативного тела водорослей можно свести в несколько категорий, образую-
щихся по возрастанию сложности. Такие ряды повторяются в различных по
происхождению отделах водорослей и по общепринятому мнению отражают
главнейшие ступени морфологической эволюции. Ввиду этого считают веро-
ятным, что эволюция водорослей протекала в виде независимых, но более
или менее параллельных рядов.
Простейшая, структура — монадная1 — характеризуется наличием
у клеток жгутиков, обусловливающих активное движение. Форма клеток
большей частью продолговато-овальная. Водоросли монадной структуры
могут быть одноклеточные или колониальные. Жгутики обычно (но не
у всех) отходят от переднего конца клетки. Электронно-микроскопическими
лиг,
s*
1 От грея, <монао (род. падеж «монадосЦ
2—602
J t • i:
34
НИЗШИЕ РАСТЕНИЯ
наблюдениями установлено, что жгутик образован 11-ю двойными волокон-
цами, из которых одно находится в центре, другие — по периферии; с поверх-
ности жгутик покрыт сократимой плазмой1. В цитоплазме клетки в основании
жгутика находится так называемое базальное тельце в виде зернышка, кото-
рое у многих форм особой нитью связано с клеточным ядром. Жгутики боль-
шей частью направлены вперед, по движению клетки. Для многих монадных
форм характерно наличие красного глазка, или стигмы1 2. Глазок, или
стигма, представляет пигментное пятно красного цвета на переднем,
несущем жгутики конце клетки, реже он занимает иное положение. Строе-
ние глазка разнообразно. В наиболее сложном случае в глазке различают
две части: переднюю, неокрашенную, двояковыпуклую (аналогична хруста-
лику глаза), и заднюю, более или менее чашевидную, содержа]^» пигмент,
близкий к каротину. Глазок служит акцептором света, ориентирующим дви-
жение клетки (фототаксис).
Немногие простейшие водоросли имеют амебоидную (ризоподиаль-
ную) структуру. В этом случае клетки лишены твердой оболочки, не имеют
жгутиков и передвигаются, как амебы, выпуская псевдоподии разной фор-
мы — толстые, короткие, тонкие, длинные. Такая структура есть у немногих
jii
водорослей (незеленых).
Пальмеллоидная структура — соединение нескольких кле-
ток, погруженных в общую слизь, но независимых друг от друга. Эта
структура может быть иди постоянной, или временной.
Коккоидная3 * структура — неподвижные клетки, одиночные или
разной формы (но ненитчатые); распространена
объединенные в колов
1ST.
широко.
Нитчатая (трихальная) структура характеризуется соединением
безжгутиковых клеток в простые или разветвленные нити. Клетки обычно
одинаковые, но иногда выделяются базальные, служащие для прикрепления,
и 11:15
• s j
и верхушечные клетки.
Пластинчатая структура — тело водоросли представляет плас-
тинку из клеток, расположенных в один и более слоев. При делении клеток
да трех направлениях пространства могут возникать паренхимати-
ческие структуры разнообразных внешних форм.
Сифональная*, или сифоновая, структура — водоросли формаль-
но одноклеточные, но клетки очень крупные (иногда до нескольких десятков
сантиметров). Единый протопласт содержит огромное число ядерс
Размножение. Вегетативное размножение у одноклеточных водорослей
происходит делением клеток, у колониальных — распадения колоний или
образованием новых колоний внутри материнской особи (рольвоксовые, мно-
гие протококковые); у нитчатых водорослей — случайным разрывом нитей или
распадением их на небольшие участки; у немногих водорослей образуются
специальные органы вегетативного размножения (клубеньки у _ харовых
я др.). ' ------
Бесполое размножение у водорослей осуществляется образова-
нием специальных клеток — спор, как правило, покидающих материн-
скую клетку. У большинства они подвижны, имеют монадную структуру, не
одеты оболочкой и называются зооспорами5; образуются они по
нескольку, по многу, реже по одной в зооспорангиях, большей частью не от-
личающихся, реже (например, у бурых водорослей) отличающихся от вегета-
тивных клеток. Зооспоры после непродолжительного движения теряют жгу-
тики, выделяют клеточную оболочку и прорастают в новую водоросль. У ря-
1 Такое же строение жгутиков наблюдается и у других организмов: у монадных форм,
зооспор и сперматозоидов низших грибов; у сперматозоидов мхов, папоротникообразных,
моллюсков, лягушек, человека и др. Отличаются в этом отношении только бактерии,
2 От греч. «стигма» — знак, метка,
8 От греч. «коккос» — зерно.
« От греч. «сифон» — трубка.
8 От греч, «зоон» — животное,
«
л
ВОДОРОСЛИ
35
да водорослей споры бесполого размножения неподвижны (не имеют жгути-
ков); они носят общее название — апланоспор1.
У водорослей различают следующие Формы полового процесса: х о л о-
г а м*и ю8 -— слияние двух одноклеточных особей; и з о г а м и юд, при
которой сливаются специально вырабатываемые водорослью голые жгутико-
вые половые клетки — гаметы, одинаковые по величине и строению: J Q Хо-
рога м и ю 4 с участием в половом акте, как и в предыдущем случае, под-
вижных гамет, но разной величины (более крупную считают женской);
о о г амию а — в этом случае относительно крупная "неподвижная клетка
(яйцо, яйцеклетка) оплодотворяется подвижной жгутиковой мужской
гаметой — сперматозоидом. Есть еще особая, форма полового процес-
са — ко н ъ ю га ция ь, когда две вегетативные клетки соединяются
няпрявленньйии друг Другу выростами и протопласты этих клеток сливают-
ся через йойуляцирнный канал, КЛёТки водоросли, b кёторых^ырабатывает-
ся гамета, обозначают общим названием г а мет а н г и е в. Гаметангии
ijiT
яйцеклетки)"и амхеридиями—(ЯРОй&водяЩие Сперматозоиды).
Если копулируют Друг с другом гамугы, рйЗЬИЙШИёСЗГна одной особи, то
такие виды водорослей называют гомоталличными7; если копули-
ровать могут только гаметы, развившиеся на разных особях, то такие виды
называют хетероталличными. Внешних морфологических отли-
чий у гетероталличных особей разных полов не бывает, и если их гаметы тоже
неразличимы (изогаметы), то такие особи, различающиеся лишь физиологи-
чески, по полу, обозначают как + (плюс) и — (минус) особи (соответственно:
+ (плюс) и — (минус) гаметы). Продукт полового процесса во всех случаях —
зигота. Она покрывается оболочкой и прорастает: или немедленно у
морских водорослей, или (главным образом у пресноводных) после периода
покоя. Прорастание зиготы состоит либо в развитии прямо из нее нового рас-
тения, либо в образовании зооспор, из которых развивается новое поколение
особей.
Слияние клеточных ядер в половом процессе влечет за собой образо-
вание диплоидного ядра. Переход от диплоидного числа хромосом к
гаплоидному совершается, как известно, во время редукционного
деления водорослей. Редукционное деление в разных систематических груп-
пах происходит в различные моменты их жизненного цикла. У одних водо-
рослей правильное чередование бесполого поколения (спорофита) и полово-
го (гаметофита), а также редукционное деление происходит, как у высших
растений перед образованием на спорофите спор. У очень многих водорослей
первое деление ядра в зиготе редукционное, и поэтому весь жизненный цикл
водорослей происходит в гаплоидной фазе; диплоидна лишь зигота, в этом
случае говорят не о чередовании поколений, а лишь о смене ядерных фаз.
Одна лишь смена ядерных фаз, без чередования поколений, имеет место и в
случае, противоположном предыдущему, когда жизнь всей водоросли про-
ходит в диплоидной фазе и гаплоидны лишь гаметы, перед образованием ко-
торых происходит редукционное деление.
Подавляющее большинство водорослей обитает в воде, где они находят-
ся во взвешенном состоянии или свободно плавают в верхних слоях воды,
входя в состав планктона, а другие, входя в состав бентоса, при-
крепляются ко дну или к разным подводным предметам, реже свободно лежат
на дне. Немногие водоросли живут на суше — на стволах деревьев, на почве;
ряд водорослей типичных обитателей водной среды могут встречаться в воде и
во влажной почве.
1П
11 и
UH
1 От
* От
8 От
1 От
6 От
• От
’ От
греч, «аплаяес» — неблуждающий, неподвижный,
греч. «холос» — цельный и «гамос» — брак,
греч. «изоо — равный.
греч. «гетерос» — другой, иной*
греч. еоон» — яйцо.
латин. <югум> — ярмо. u
греч. «гомойос> — одинаковый, равный.
НИЗПШВ P ACT
a;t
ОТДЕЛ—СИНЕ-ЗЕЛЕНЫЕ, ИЛИ ЦИАНОВЫЕ,
ВОДОРОСЛИ (CYANOPHYTA)
Сине-зеленые водоросли получили свое название ^а окраску действи-
тельно обычно сине-зеленую, но она бывает также почти черной, желтовато-
иди оливковато-зеленой. Часть водорослей этого отдела одноклеточные, боль-
шинство же колониальные и нитчатые.
Клетки сине-зеленых часто мелкие, цилиндрические, округлые, бочко-
видные и других форм (рис. 7). Вокруг отдельных клеток или их скоплег
и так называемых трихомов1 имеется обычно слизист!<й.че^ол, кофорьн1 мо-
жет превышать диаметр самих клеток. Такие чехлы то бывают^горошо очер-
чены, то имеют расплывчатый характер. Масса чехла состоит главным
образом из пектиновых в
ной обо л о ЧАД" Или выдал;
r.i:
ществи образуется в результате ослизнения клеточ-
ется протопластом через узкие поры в ней. У мно-
ывздртхжружевы осо-
згемицеллюлоэыи целлюлозы/Стрдё-
а состоит из^ пектиновых
цитоплазматическими м
бым,
оболочки
о
довольно ело
ЧI '> 11 i
III.
ранами, что дает основание говор
На, имея волнистый характер, примыкает к самой оболочке в определенных
точках, где посредством пор осуществляется ее контакт с внутренней мемб-
раной.
В протопласте клеток сине-зеленых водорослей оптически различимы
две области: наружная, окрашенная — хроматоплазма — и внут-
енная окраски — центроплазм а (центральное тело).
Хроматоплазма имеет сложную с^бмикро<жацическую структуру, ос-
новными элементами которой являются волнообразно изогнутые пластины —
носители пигментов — и разделяющие их слои цитоплазмы. Эта ламел-
лярная структура сходна со строением хроматофора у более высокоорга-
низованных растений, отличаясь, однако, отсутствием индивидуализации
носителей пигментов. У сине-зеленых не только нет особой мембраны, отгра-
ничивающей окрашенные образования, но, более того, хроматоплазма посте-
пенно нереходихивинеокрашевдую центроплазму. ------"
^ствешЁуво"роль играе^хлорофилл д,
хлурофняя Ъ отоутечвует. Зеленый пигмент сопровождается карбТШиидШи,
среди которых есть специфические ксантины (миксоксантин, миксородин).
Очень характерно присутствие фикобилинов, ШЙ'ШИНМ вёлковой Природа
(хромопротеиды). Синий фикобилин (фикоциан) присутствует всегда, второй
же — красный (фикоэритрин) — констатирован не у всех сине-зеленых и,
по-видимому, является необязательным. Ламеллярная структура не распрост-
раняется на центроплазму за исключением некоторых зернистых образоваг
В составе центроплазмы было установлено наличие нуклеиновых кислот
(ДНК и РНК). Субстанция, содержащая таковые, представлена в виде зер-
нышек или палочек, образующих сеть, или состоит из расположенных более
или менее параллельно нитей. Вся совокупность этих телец составляет так
it.
рым данным, нитевидные "Структуры удваиваются, подобно хромосомам.
В^Ю^ЕШтроплазму можно рассматривать как Физиологический эквивалент
нет отдели-
ядра. Отличия от последнего сост
тпнртй ее пт остал^ной плаамы мем!
нет здесь и ядрышка, а при деле!
гг
я тпгпгпа ПД уттпппттпг.т. кпттртяту..
реликтовое, подобное тому, что, возможно, было у древнейших организмов,
nirr
1 Термин «трихом» в морфологии сянмелеиых водорослей равнозначен нити у водо-
рослей других отделов. Нитью же у сине-зеяевых обычно называют трихом или несколько,
трихомов вместе с окружающим их влагалищем, и
ОТДЕЛ — СИНЕ-ЗЕЛЕНЫЕ ВОДОРОСЛИ 37
— 7 ....... . - -
2
Рис* 7. Строение клетки у сине-зеленых водорослей:
1 — хрооккок; f — аяабеяа; >—осцвллягоряя. В центральной части водны окрашенные (черные) хро-
ыапновые одсютты
не имевших специализированных клеточных органоидов. Это — протопласт
предъядериого (прокариотического) уровня.
В клетках сине-зеленых водорослей отсутствует цшхральпая^дакуоля.
Вйтеулдияяди1дг ттпяпттм а лгяриут отмирающихклетках. Сама же
плазма имеет^повутп^шую вязкость, и даже под влиянием сильных нлазмо-
лизирующих средств^ддмо^бйластплохЬ~~отделяетсяот оболочки. С другой
стороны, пляямя г.ттаияйлдтгтлх обладает "хорошо выраженной способностью.
гг,
отдавая воду при засухе, вновь энергично поглощать ее при выпадении дож-
дя. При отсутствии обычной вакуолизации для клеток преимущественно
планктонных синй-яялдтот тярятстарнп пйряяпнянив меЛКИХ газовых BflKVO-
лей, наполненных, по- некоторым экспериментальным данным, азотом, а по
друппГДВняым угЛбКИСЛЫМ гааом/ТЬяовыевакуоли концентрируются близ
границы хромато- и паптпопляямы. Если их много то благотатж сильному
лучепреломлению, ощи дрндятт матка томную окраску. Значение газовых
_ *> • о _ -***“—^'—1 . О
есть
хроматоплазму в качестве продуктов запаса откладываются хшощ-
ген,пбй£ейхарид, окрятпп^яюпттгйг.я пт йппя в бурый цвет, а также шдюхин=-
Оогатоефосфоромвещество, соЭержашее РНК и красящееся метиленовой сцнью
в сший цветУ наконед, в хроматоплазме содержатся так называемые ЦЙаЙсь)
красящиеся от нейтральной красной в
красный цвет; ноне дающие цветной реакции с метиленовой синью. Запасные
7
глюкопротеиды и углеводы' (анабенин).
Клетки сине-зеленых" водорослей редко существуют одиночно, чаще
они образуют колонии или нити. В колониях (хроококкальныи'"тйп стро-
ения), по-видимойу, каждая клетка физиологически вполне самостоятельна.
То же приписывают и там нитчатым, у которых клетки одеты толстыми обо-
лочками (плеврокапсальное строение). У других нитчатых в поперечных
ругойг имеется по одной поре,
1 (гормогональный
первичная пёрйаоШа которой процжзана!
'1 Некоторые специалисты отрицают существование плазмодесм.
38
Ci
тип строения). В этом случае, следовательно, протопласты клеток находят-
ся во взаимной связи и вся нить является физиологически единой.ч^м
нитчатых форм в разных местах на протяжении нити или в основа!_______
образуются особые клетки ~ г е т е р о ц и с т ы, — более^светлые^сводя;
нистым содержиъщм^._утолщённъиш25бШГОЧКЗй4И~Й закупоренными порами.
Биологичёско^значение их не выяснено. Гормогональные сине-зеленые иног-
да ветвятся; ветвление редко бывает настоящее, обычно оно ложное. В по-
следнем случае участки трихома между гетероцистами, плотно прилегаю-
щими к влагалищу, не могут расти в длину и пробивают одним концом боко-
вую стенку влагалища, образуя ложную ветвь (рис. 8, 6).
Половой процес с^у_ишн&ашшиых--чохсутспует. Размножение
преимущественно вегетативное. У одноклеточных форм оно состоит в деле-
ний материнской клетки на две дочерние. При этом.оболочка-матерюской
клетки может набухать и дочерние остаются, окруженные пбряяпяя шпейс я
слизью^При повторных делениях происходит то же с клетками следующих
поколе! __________________
вляется гормогониями (рис. S, 7). Это короткие, состоящие из нес-
кольких (многих)I клеток участки материнского трихома, часто отдАля^уцяя-
ся пр гетероцистам; в дальнейшем они растут и^ становятся взрослыми осо-
бями. "У бентосных представителей образованию гормогонией предшествует
возникновение в клетках большого количества газовых вакуолей, благо-
даря чему гормогонии способствуют не только размножению, но и распро-
странению водоросли.
Рост, естественно, связан с делением клеток. При этом на.внутренней
поверхности клеточной оболочки возникает кольцевидный вырост, увеличи-
вающийся перпендикулярно к длине нити; отверстие такого кольца посте-
пенно суживается и, наконец, замыкается. Делению клетки предшествует
далдтшй ттантрптнгяямм И СОДержяртитгп п-плй'ур^'ятпнпппплбптдк СТруКТУТЬ
Как средство переживания неблагоприятных условий, главным обра-
зом у нитчатых форм сине-зеленых водорослей, возникают п о к о я ш и е-
Г, IS
s I: ।
• sis
1:1
ее' утолщается и в плазме накапливаются запасные питательные вещества.
Прорастает покоящаяся спора гормогонием. Период неблагоприятных усло-
вий некоторые сине-зеленые переживают иногда в виде гормоцист.
Так называют целые веточки, покрывающиеся общей плотной оболочкой.
Несмотря на отсутствие полового процесса у сине-зеленых водорослей
можно наблюдать закономерную последовательность развития. Так,
у некоторых нитчатых вид?в ФКйУадос*- апгшижиыдг выделитьщ^тыре сменя-
ющие друг друга- спг.тлсттптст Перезимовавшие споры прорастают весной Ц),
после чего наступает вр^ея^активного "вегетативного размножения гормо-
гонияшЦ2)^ сменяющегося периодом нарастания мит<8ЙД^Идшу^(Э)г сезон-
ное развитие заканчивается отмиранием вегетативных частей и массовым
образованием спор (4).
К сине-зеленым водорослям относится около 1400 видов, классифици-
руемых по-разному. В системе, предложенной А. А. Еленкиным, выделено
3 класса.
1-й класс — хроококковые (Chroococceae) — содержит одиночные и
колониальны^ виды. Хроококк (Chroococcus) — обычно одиночные округлые
клетки. Глеокапса (Gloeocapsa) — округлые клетки с толстой слоистой сли-
зистой оболочкой, нередко соединенные в колонии (рис. 8, 2).
2-й класс — хамесифоновые (Chamaesiphoneae) — нитчатые или
ложнопаренхимные, размножающиеся эндоспорами. Большинство живет
в морях на скалах, раковинах или как эпифиты.
3-й класс — гормогониевые (Hormogoneae) — нитчатые, эндоспор
нета у многих образуются гормогонии. Представители этого класса гораздо
более многочисленные, чем двух предыдущих.
Осцилляторил (Oscillatoria) — однорядные, неветвящиеся трихомы из
плоских клеток, без влагалищад без спор и гетероцист (рис. 8Д 5). Нити обла-
i1 s
ir
Sill
-ЗЕЛЕНЫЕ ВОДОРОСЛИ
ОТДЕЛ —
т
Рис. & йдие-зеленые водоросли:
2—дйовддж;g — микроцистис; 4 —анабешц 4—ностокз отдельная ввть» в—колония нихсВ; 4—-
(И! ЦВМ жиооиц 4 шцмощрико; f — мингОвв о Рормогониями
i
38
дают характерным поступательным скользящим движением. Лин&бия (Lyng-
bya) близка к осциллятории, но имеет влагалище около нити и гормогонии.
Анабена (Anabaena) — нити из округлых клеток, прямые или спирально
извитые, со спорами, гетероцистами (рис. 8., 3). Носток (Nostoc, рис. 8, 4, а,
б) — нити, похожие на анабену, но погруженные в общую слизь, имеют форму
пленок, подушечек или округлую в виде мелких или крупных шариков;
Афанизомвнон (Aphanizomenoft) — короткие инти, соединены в скопления
в виде чешуек.
Разные виды сине-зеленых водорослей широко распространены .в планк-
тоне и бентосе пресных водоемов и морей- Многие живут в мд^уяу^ загряя-
_____________ г . _________________2 _ 9 т. е.
синтезируя, а также используя и готовое органическое вещество. Некоторые4
виды ностоков (и другие) часто встречаются—да почве полупустынь нашего
Юго-Во&тдка».-ера.ТГпочве находили сине-
зеленые водоросли^ла глубинб до 1 ле.Л1екоторые почвеннцсийДН'-могут
— ~ ’ 'нтересна способность некоторых сине-зеле-
ных жить в горячих источниках, гейзерах при температуре 65—93°С.
летом. Красное море получило свое название от желтовато-красноватой
окраски воды, нередко наблюдаемой в разных местах его и обусловленной
массовым развитием планктонной сине-зеленой водоросли триходесмиум
(Triehodesmium erythraeum), имеющей желтовато-красноватую окраску (о*5
содержания фикоэритрина).
Носток и еще-некоторме другие сине-зелбйые встречаются в качестве
симбионтов во многих лишайниках. Находят сине-зеленые также в некото-
рых простейших животных (Protozoa), в корнях саговников и т. п. Следы
может быть, даже архейской эры, образовавшихся от 1,5 до 2,5 млрд, лет
назадлЭ™ организмы, сд
III
По строениюНЛбтки сине-зеленые вбдореели~нееколько схбдны с бакте7
риши, но прямые филогенетические связи их маловероятны: у многих
бактерий есть жгутики, споры у них образуются иначе. Связей с другими
группами растений сине-зеленые водоросли не имеют и сохранились в своей
примитивной форме с древнейших времен, не дав начала каким-либо выше-
стоящим группам растений.
ОТДЕЛ — ЭВГ ЛЕНОВЫЕ ВОДОРОСЛИ
(EUGLENOPHYTA)
Эвглеяовые водоросли (рис. 9) отличаются от других отделов водорос-
лей чисто-зеленой окраской хроматофоров, содержащих хлорофиллы <х и б.
каропш и ксантофилл. Форма- клеток у этих водорослей большей частью
удлиненная — эллипсовидная, веретеновидная и т. и. Размеры микроско-
пические. Целлюлозной оболочки нет; ее роль играет наружный упл< тлен-
ный слой цитоплазмы, так называемый перипласт или пелликула; если он
мягкий, гибкий, то клетки способны метаболически изменять форму тела.
t
тайный солями железа, не прилегающий плотно к. протопласту. Число и
форма хронатофоровраздцдны. Крахмала нет; запасной продукт — угле-
\ - - вод памща» —"близок к крахмалу, но от йода не изменяет окраску" или
слегка желтеет; он откладывается в виде блестящих зерен в хроматофорах
или вне их. На переднем конце тела находится воронковидио^'угяублввие,
называемое часто (но ошибочно) «глоткой»; оно является выводным концом
для системы сократительных вакуолей, находящихся возле него. От дна
«глотки» отходит у болы
иг»
1 От трвч. «миксис» — смешение ж строфе» — питание,
1И
отдал—ЭВГЛЕНОВЫЕ ВОДОРОСЛИ
ся
далеко наружу. У некоторых видов водоро-
этого отдела жгутиков два. У боль-
ва имеется красный глазок. Движут-
вгленовые водоросли за счет метаболи-
х изменений формы тела и плавают при
помощи жгута, одновременно вращаясь во-
круг продольной оси. Еммнажахшш__про-
чаше- -ди-шшодвижном состояния. Подгпвпй
процесс у ник с достоверностью не уста-
новлен.
Живут эвгленовые водоросли преиму-
щественно в небольших пресных водоемах,
богатых органическими веществами, где мо-
гут питаться миксотрофно, т. е. нутем фо-
тосинтеза и усвоения растворенных в воде
Органических веществ. Некоторые живут в
солоноватых водоемах.
Эвгленовых водорослей насчитывают
около 400 видов (25 родов).
Очень широко распространены предста-
вители большого рода эвглена (Euglena).
Иногда, размножаясь в массовых количест-
II
водоемах в зеленый или кроваво-красный
цвет («цветение» воды); последнее обуслов-
зается эвгленами, содержащими в прото-
плазме пигмент гематохром- Нередко встре-
чаются виды факуса (Phacus).
Отроение протопласта,' наличие одного
жгутика, отсутствие крахмала, замененного
парамилоном, резко отличают эвгленовых
от зеленых водорослей; это слепая ветвь
эволюции.
Рис. 9. Эвгленовые:
1 — эвглена; Я — трахешшонас; 4—фа-
ОТДЕЛ —ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA)
В большой отдел зеленых водорослей объединяют все водоросли expo-
III:
лом в качестве запасного продукта, с зооспорами, где они имеются, или с
гаметами, снабженными двумя, реже четырьмя равными жгутиками. Внеш-
ний вид, строение и размножение зеленых водорослей весьма разнообразны.
________________________ . _ _____иства водорос-
лей этого типа птгипятгерные. Хроматофоры весьма различной формы, чаще
довольно крупные, по одному или по нескольку в клетке, но у некоторых —
мелкие, зернистые, многотасленные^1я^яд1^^ хлоро^ллы а и 5. каро-
тин, ксантофиллы (лютеин и др.)£В£^атнвное размножение происходит
путем далёкий клеток у одноклеточных, распадом колоний или нитей и обра-
зованием новых колоний внутри клеток старых колощДЕ.^есполрК размно-
жение происходит посредством зооспор или (при утрате жГ^тййов) аплано-
спорами. Головое размпожёнйе. в отличие от предыдущих типов водорослей,
очень широко распространено и происходит в форме хологамии, изога-
мии. гетерогамии, оогамии и конъюгации. Зигота у бояыпинства~одевается
толстой оболочкой и прорастает после длительного периода покоя; у мень-
иг.
I fl. 1
шинства, преимущественно у морских видов зеленых водорослей, зигота.
прорастает немедленно.
РАСТЕНИЯ
Особи зеленых водорослей чаще гаплоидны, и диплоидное состояние
представлено в этом случае только зиготой, при прорастании которой со-
вершается редукционное деление, ^еже вегетативное тело зеленой водоросли
дщплоидно. и тогда редукция происходит при образовании гамет. Наконец,
у ряда зеленых водорослей полный цикл развития осуществляется двумя
последовательно сменяющими друг друга поколениями: половым (гаплоид-
ным поколением — гаметофитом) и бесполым (диплоидным поколением —
спорофитом), правильно сменяющими друг друга (антитетическая смена
поколении). В этом случае редукционное деление происходит при образо-
вании (на спорофите) зооспор. Эти зооспоры, отличные по способу образо-
вания от зооспор обычного бесполого размножения (м и т о с п о р1), было
предложено назвать м ей а спор а м и*.
Большинство зеленых водорослей живет в пресных водоемах, некого-
рые группы — в морях; немногие живут ласырой земле, скалах^коре де-
ревьев, обнаж^яоЙ древесиде (яябпрях и т. п.)
Отдел зеленые водоросли делят на 2 класса.
1-й КЛАСС—РАВНОЖГУТИКОВЫЕ ВОДОРОСЛИ
(CHLOROPHYCEAE, ИЛИ ISOCONTAE)
Самый большой класс зеленых водорослей, в котором представлены
очень разнообразные их формы, начиная от одноклеточных, подвижных в
течение всей вегетативной жизни, до многоклеточных пластинчатых или
мешковвдных. Конъюгац
1ГГ
не бывает. Этоткласс делят на несколько по-
рядков.
1-Я ПОРЯДОК — ВОЛЬВОКСОВЫЕ (VOLVOCALES)
111 л \
11 г. г
Одноклеточные и колониальные водоросли в основном подвижные.
Передвигаются с помощью жгутиков в вегетативном состоянии. Из одно-
клеточных широко распространен большой р о д. хламидомонас (Chlamydo-
monas), содержащий около 320 видов, встречающихся преимущественно
в мелких водоемах, лужах, особенно загрязненных органическими веще-
ствами. "
Хламидомонады имеют овальную, или грушевидную, или шаровидную
.форму с небольшим бесцветным носиком на переднем конце, от которого
отходят два равных по длинежгутика." Ободочка состоит из^п^^щщщх
веп^еств и гемит^еллюлозы и часто несколько отстает от протопласта, особен-
но сзади Хрис- 10, 7—5). В задней части протопласта находится зеленый
хроматоДорд имеющйй форму^чаши. в более толстом дне которой помещается
крупный пиреноид. В цитоплазме, заполняющей углубление чашевидного
хрома то фора, йа?&дится ядро^ В передней, бесцветной части протопласта
видны две пульсирующие вакуоли и красный глазок. Хламидомонады быст-
ро (в сравнении с размерами их тела) движутся в воде, вращаясь при этом
вокруг своей продольной оси. Благодаря положительному фототаксису
(при не очень ярком освещении) и аэротаксису они скапливаются у поверх-
ности воды в более освещенной ее части.
При 6jb с п о л о м_ р а з м и о жен и и. происходящем большей
частью ночью, хламидомонада останавливается, теряет жгутики и содержи-
мое ее после предварительных кариокинетических делений ядра Распада-
ется на 2—4, реже на Sjmocnop, вырабатывающих жгутики И оболочку еще
внутри материнской“оболочки и выходящих после ослизнения ее в виде
новых хламидомонад, только меньших размеров. При недостатке кислорода
или пересыхании водоема хламидомонады теряют жгутики, выделяют
иг
г< I'
1 От греч. «митос» — нить.
8 От греч. «мейозио — уменьшение.
ОТДЕЛ — ЗЕЛЕНЫЕ ВОДОРОСЛИ
43
Рис. 10. Одноклеточные вольвоксовые:
1— хламидомонада: а — вегетативная особь» б — формирование зооспор» в — пальмеллевидная стадия;
2 — сферелла
слизь, но при этом сохраняют способность размножаться делением кле-
ток, благодаря чему образуется пальмеллевидная стадия
(рис. 10,1 в). При наступлении благоприятных условии клетки их выраба-
тывают жгутики и возвращаются в стадию подвижных хламидомонад.
При половом раз мио ж е н и и (рис. 11) у большинства видов
хламидомонад в клетках образуются одинаковые по величине гаметы (изога-^
меты), похожие на зооспоры, но меньших размеров и в Колышем "числе (8—
16, иногда 32 — 64). Для очень немногих видов характерна г е т е р о г а-
м ия или (для других) рога ми я, отмеченные впервые у хламидомонад
русским ботаником И. IL F орожанкиным. При гетерогамии обе
гаметы подвижны и содержиздре_меньшей переливается в другую, большую:
при^ оогамии‘маленька я мужская гамета сливается с крупной неподвижной
женской, вкоторую превращается вся вегетативная клетка. Во всех случа-
ях зигота одеваетсятолсгай^ цёллюлознойо1болочкой. напол-
няется запаснымивеществами и приобретает красную окраску от гемато-
хрома — пигмента каротиноидной природы, растворенного в масде .И харак-
терного вообще для покоящихся . состояний „щщорродей. После периода
покоя.при наступлении благоприятных условий зигота прорастает, образуя
в 'результате редукционного деления ее диплоиднбгоПядра обычно четыре
HI
Более примитивные одноклеточные вольвоксовые не имеют углеводной
оболочки, а лишь уплотненный наружный слой цитоплазмы. Они размно-
жаются продольным делением в подвижном состоянии. Половой процесс у
, них происходит в форме х о д о гам, и и — слиянием цельных вегетатив-
t ных особей без образования специальных гамет. Примером подобных водо-
Рис. 11. Половой процесс
у хламидомонад*.
J — образование ивогамет;
9 —изогаметы к их копуляция;
а —гетерогамия; 4—оогамия;
4—зигота
я
рослей может быть дунализлла
(Dundliella salina), живущаяГв
соленых озерах. Она имеет
красноватую окраску от гема*
нередко красноватую окраску
самосадочной соли и рапы1»
Из подвижных ^солуриаль-
вых—-нодьвоксовых гониим
Рис. 12. Колониальные вольвоксовые:
2— пандорина; а — евдорина: а — вегетативная коло-
ния» б — оплодотворение, в — формирование дочерних
колоний
состоящих из р1 ji
го и 4 клеток
У другого вида, расположенных
в один слой. Клетки имеют в
общем строение, подобное хла-
мидомонадам, и направлены
жгутиками в одру сторожу.
Пандорина (Panaorina) и
эвддрина (Hudorina) — микро-
скопические колойиальные во-
' доросли. У первой в колонии
обычно 16 обратнояйцевидных
клеток,"^ТБсно прилегающих
друг к другу; у второй, обра-
зующей полый шар, — боль-
шей частью 32 шаровидные или
овальные клетки, располагаю-
щиеся по шаровидной поверх-
ности на некотором расстоянии
друг от друга (рис. 12). Отдель-
ные клетки их колоний имеют
строение, сходное с хламидо-
монадами.
форму (0,5 — 2 мм в диаметре)
г I
Вольвоксы (Volvox) имеют шаровидную форму (0,5 — 2 мм в диаметре)
и состоят Из Многих, от сотен до нескольких десятков, клеток. ряг.ттплпж^т-
ных по периферии шара, внутренность которого заполнена слизью. Отдель-
ные клетки с поверхности имеют форму плотно соприкасающихся шести-
угольных ячеек; протопласты сосеутот ^четок^хоединены п ротонд аз ма ти-
не сними тяжами. проходящими через разбухшие слои оболочки (рис.13).
/ размножение колониальных водорослей порядка воль-
/воксовых осуществляется колониями, образующимися в клетках материн-
СК°Д особи путем последовательных продольных^ делений тгроттглястлп
этих клеток. При этом у всех вначале образуется плоская табличка (так
называемая готическая стадия), которая затем к вдавливается внутрь;
позднее чаша такая выворячтгапдлт>д пашшяшгу (рис. 44), после чего края ее
смыкаются. Таким образом, концы клеток, вырабатывающие по два жгутика,
оказываются обращенными наружу. Дочерние колонии у Пандорины и звдо-
рины освобождаются из расплывающейся слизи материнской особи.
У вольвокса дочерние колонии выпадают внутрь материнского шара и осво-
бождаются лишь после разрушения его. В дальнейшем клетки новых коло-
ний только увеличиваются в размере, но вегетативных делений клеток в них
не бывает. У гониума, пандорины и звдорины все клетки колонии образуют
'дочерние особи; у вольвокса к такому размножению способны лишь 8 —
5 клеток, находящихся в задней части шара, отличающихся своими более
II
г. 1 .
: I i 11 \ r
Вода, насыщенная солю в солевых озерах*
ОТДЕЛ — ЗЕЛЕНЫЕ ВОДОРОСЛИ
Рис. 13. Вольвокс:
1 — колония с дочерними шарами; я — часть колонии (я — яйцеклетки, е — пучки сперматозоидов);
а —вил участка колонии с поверхности (a); d - ан же в разрезе (6).
Рис. 14. Вольваке. Последовательные стадии развития дочерней колонии
46
НИЗШИЕ РАСТЕНИЯ
; t: ।
ж ________________w____________ ____х____а изогамное
или нерезко гетерогамное. Всё клетки колонии путем последовательных де-
лении образуют 16 — 32~голые двужгутиковые гаметы, сливающиеся по-
парно, причем водоросли гетероталличтпд в копулируют лишь гаметы из
разных особей, ^др^жпа. .раздельнопола: в мужских колониях каждая
клетка (кроме четырёх передних) дает по 64 удлиненных желтоватых спер-
матозоида; они оплодотворяют яйцеклетки женских колоний, в которые
превращаются там все клетки (кроме четырех передних), увеличивающиеся
в размерах, но не теряющие жгутиков, так что здесь имеется промежуточ-
ный тип между гетерогамией и оогамией. У вольвокса —~ типичная оргдуия,
причем одни виды обоеполы, другие — раДдел^норгрды. От 5 до 100 клеток
колонии превращаются в антеридии, содержимое которых делится на 64
11:1;
желтоватых сильно удлиненных двужгутиковых сперматозоида. От 5 до
15 клеток (редко до 30) превращаются в шаровидные крупные оогонии, в
каждом из которых лежит по одной темно-зеленой неподвижной яйцеклет-
ке, оплодотворяемой подплывающим сперматозоидом. Зиготы красноватые.
После периода покоя они прорастают, при этом происходит редукционное
деление; у гониума образуются 4 зооспоры, в каждой из которых затем фор-
мируется по одной колонии; у прочих 3 зооспоры отмирают и лишь в од-
ной, выживающей образуется новая колония или у вольвокса новая коло-
ния может образоваться прямо в зиготе (ооспоре). Следовательно, вольвок-
совые гаплоидны в вегетативной жизни и диплоидна у них лишь зигота.
К порядку вольвоксовых можно отнести еще ряд родов, для~5сбторых
характерна подавленность жгутиковой стадии. Крупные мешковидные сли-
зистые колонии, взвешенные в воде, с расположением клеток часто по 4,
образует тетраспора (Tetraspora); подобные же, но прикрепленные груше-
видные колонии образует апиоцистис (Apiocystis). Некоторые альгологи
выделяют их в самостоятельный порядок Tetrasporales или даже в класс.
Филогенетически они интересны как показатели одной из возможностей
эволюции подвижных вольвоксовых в неподвижные водоросли.
К вольвоксовым относится несколько десятков родов, представители
которых почти все живут в пресных водоемах, большей частью мелких? не-
редко временных (ямах, лужах), чистых или загрязненных, многие встре-
чаются в планктоне, а некоторые нередко — во влажной почве.
2-Й ПОРЯДОК — ХЛОРОКОККОВЫЕ, ИЛИ ПРОТОКОККОВЫЕ (CHLOROCOCCALES,
ИЛИ PROTOCOCCALES)
Водоросли порядка хлорококковых отличаются от вольвоксовых от-
сутствием жгутиков и в связи с этим неподвижностью в вегетативном
состоянии. Хлорококковые водоросли npeAC¥5SSKfifFl!36dH^H6o одиночные
клетки, либо колонии разнообразного внешнего вида и сложения, но~ не-
нитчатые. Общее с вольвоксовыми то, чт& при бесполом ^размножении но- *
вые клетки возникают внутри материнских клеток й в дальнейшем растут
I'm
ъ за счет увеличения размеров составляющих их клеток, а не путем их
деления. Одни хлорококковые способны к образованию зооспор, у других
эта форма спор отсутствует. Будучи главным образом Организмами гапло-
1,1,1
идными, хлорококковые могут размножаться и половым путем. У них почти
всегда имеет место изогамия и только в виде редкого исключения половой
процесс <зд$вдый (открыт 71. А. Коршиковым у Golenkinia1 и еще
у некоторых представителей порядка).
Наиболее примитивные одноклеточные хлорококковые очень похожи
на хламидомонады, потерявшие подвижность: шаровидные клетки с пекти-
ново-целлюлоаной-оболочкой, чашевидным хроматофором с одним пирено-
идом и с ядром в выемке хроматофора; у немногих имеются’ дне дульсярую-
щир вякуодпия-инлгдя деже красный гляяокг характерные вообще для под-
1 Названа в честь московского ботаника М. И. Г о л е и к и и а.
ОТДЕЛ — ЗЕЛЕНЫЕ ВОДОРОСЛИ
47
Рис. 15. Протококковые:
1 — хлорококк: а — общий вид (одна клетка с зооспорами). б — зооспора; а — хлорелла] а — общий вид»
о—формирование автоспор; Л — сценедесмус: б — анкистродесмус; 5 — голенкиния; в — яедиаструм
(ва — выход зооспор)
вижных форм. Такую форму и строение, но без пульсирующих вакуолей
и глазка имеет ждорококк (Chlorococcum)\ часто встречающийся в пресных
водоемах и на сырой земле (рис. 15,1). По внешнему виду на хлорокок>Г
похож цистококк, или требуксия (Cystococcus, или Trebou&ta), однако
резко отличающийся массивным хроматофором, занимающим центральную
часть клетки. Цистококк живет на сырой земле, в нижней части стволов
деревьев, на пнях и, кроме того, входит как водорослевый компонент в со-
став большинства лишайников. Беспол о е р аз.мн о ж е я и я, у этих
родов происходит путем образования в клетках последовательными деления-
ми 8 — 32 голых двужгутиковых зооспор. По ло_н о й п р о ц е —
копуляция двужгутиковых изогамет, похожих по форме на зооспоры.
Примером цолоздщдьяых^. многоядерных хлдрококковых может слу-
жить водяная сеточка* ш гидрддикйион (Hudrodictuon}. нередко встреча-
ющийся в пресных водоемах со стоячей или медленна текущей водой, бога-
той азотистыми солями- Он имеет вид. замкнутого сетчатого мешкд, дости-
гающего нескольких десятков сантиметров в длину и 10 — 15 см в ширину.
Стенки мешка образованы, цилиндрическими клетками до 1 см длиной с ту-
поконусовидными концами, которыми они соединяютея- на З— 4^17образуя
пяти-шестйугольные ячейки (рис. 16, 1 а,б). В постенном слое, цитоплазмы
находится __ бледно-зеленый.^ сложно расчлененный.. хроматофор со многими
1Г.1
пиреноидами, а под ним — многочисленжме-ядра • (до нескольких тысяч),
видные только после окраски. Срединная часть клетки представляет^огром-
иую вакуоль с вдетоддымс, шшом. Принёс п ол ом размножении
протопласт раскалывается на огромное количество (7000 — 20 000),двужгуг_.
тиковых зооспор, содержащих каждая одно ядро и участок хроматофора.
Они не покидают оболочки материнской клетки, недолго движутся, как бы
1 Раньше хлорококк называли протококком (Protococcus), во так как это название да-
вали и многим другим, более или менее сходным с ним водорослям, относимым в настоящее
время к другим родам, то название протококк теперь не употребляется. В соответствии
с этим и весь порядок следует называть не протококковые, а хлорококковые, •
48
НИЗШИЕ MOW
Рис, 16. Гидродикциов и протосифон:
7 —гццродвкцааи! а—молодая сетка внугря
• — образование в нем сетки; 2 — нротосифов
материаской клетки.
б — ячейка .сетки» в — шшиэдвр.
«танцуют» на одном месте, затем втягивают жгутики и складываются в новую
маленькую сетку, покрываясь при этом оболочками. Возникшая таким обра-
зом юная сеточка освобождается после расплывания оболочки материнской
клетки и постепенно увеличивается в размерах только за счет роста (но не
увеличения числа) составляющих ее клеток, становящихся при этом много-
ядерными. При п о л овом размножении в клетках образуется
множество (до ЗО 0иО) мелких двужгутиковых гамет. Они выходят из мате-
ринской клетки и кппУАйруют тпяпил (ияогамияк Шаровидная -вигота
прорастает цосде периода покоя, образуя 4 зооспоры, чему, вероятно, пред**
шествует редукционное деление ее диплоидного ядра. После некоторого
периода движения зооспоры останавливаются и каждая прапрятяятс.я в
мипгоугдл^рую. ляяая^ятуот" "Апятут оболочкой Клетку. которую
11I;
описывали раньше как самостоятельный род под название^ полиэдеп (Ро1и~
edera рис. 16,1 в, а). Полиэдер некоторое впамя увеличивается в размерах.
затем содержимое его распадается на зооспоры, пп«_жа штрпянчст.
тельной» движения внутри полиэдра складывается в сеточку, освобождаю-
щуюся после разрыва оболочки полиэдра.
astrum), часто встречающиеся в планктоне.
Многоядёрность клеток, кроме водяной сеточки, - можно наблюдать и
у одноклеточных хлорококковых, например у впотосшЦон (Pnlotiphon,
рис. 16, 2). Р. botryoldes живет вне воды. Его клетка имеет в наземной части
до 1,5 мм длины и 0,5 мм ширины; от нее отходит, внедряясь в почву, бес-
цветный неветвистый ризоид. Многочисленные ядра и сетчатый хроматофор
находятся в постенной части. Сифональное строение дало повод к предаю-
к
отдвл—звлкныв ВОДОРОСЛИ
вожению о родственной связи порядка сифоновых (си. ниже) с хлорокок-
новыми.
У большинства хлорококковых стадия зооспор утрачена. Бес п о лор
р аз м н о ж е и и е_ в этом случае осуществляется последовательньйиде^
лением протопласта внутри материнской оболочки на несколько неподвиж-
ных клеток — Так называются клетки, которые, возникнув
из протопласта материнской клетки путем его деления, еще внутри мате-
ринской клетки покрываются оболочкой и вырабатывают все особенности,
характерные для взрослых особей, а у колониальных форм здесь же скла-
дываются в новую колонию, которая освобождается после разрыва или
ослизнения материнской оболочки. Такой способ размножения, по-види-
мому, связан с планктонным образом жизни этих водорослей, при котором
нет надобности в зооспорах для расселения. Вероятно, в связи с этим у очень
многих хлорококковых нет полового процесса и размножение их происхо-
дит только автог.ттрями Ия Tgyffv YjrnpnwfVKwnтаиу следует отметить хлореллу
(ChUxrella)~ имеющую вид зеленого шарика (рис. 15, 2 а — б), анкистроЗ&6~
мус (Ankistrodesmus) — одиночные или соединенные в рыхлые пучки иголь-
чатые или веретеновидные клетки (рис. 15,4), гиенедесмис (Scenedesmus) —
продолговатые клетки, соединенные в 1—2 ряда в пластинки по 4—8, реже
до 16 клеток (рис. 15,3).
К порядку хлорококковых водорослей относятся свыше 100 родов
и свыше 700 видов. Они широко распространены в пресных водоемах (иног-
да вызывают «цветение» воды), а также в почве. Нередко они вызывают
позеленение воды при длительном
стоят
ее в сосудах, куда
попадают
с пылью из воздуха. Некоторые из хлорококковых живут симбионтами в
лишайниках, в зеленой гидре (Zoochlorella
Хлорелла (Chlorelld^ легко культивируемая в лабораторных услови-
ях, служит нередко о&ьектом для изучения фотосинтеза. Выяснено, что
хлорелла использует от 10 до 12% световой энергии (против 1—2% у назем-
ных растений). В клетках хлореллы содержится большое количество пита-
тельных веществ,• а также бактериостатический антибиотик хлореллин.
Хлоредд^, а также сценедесмус начинают разводить (Н. С. Гаевская}
в массовыхколичаствят. ня корм ддд рачкпв да служащих, в свою оче-
редь, кормом для мальков рыб на рыбных заводах? Ведутся опыты по раз-
ведению хлореллы — легко получаемого и дешевого углеводного корма в
промышленных масштабах.
8-Й ПОРЯДОК — УЛОТРИКСОВЫЕ (ULOTH RICHALES)
К улотрякеовым относятся водоросли, имеющие нитчатое строениег
а также некоторые пластинчатые и мешковидные, имеющие в начале онто-
генеза нитчатую форму. В отличиёот предыдущих порядков талломы во
время вегетативной жизни нарастают путем деления клеток. Почти у всех
улотриксовых клетки одноядерные. П о л о в о й пр о п есс — изогамия.
н иещюисходит посредством овальных четырех-, реже др
<шо
гетерогамия и оогамия.
*"л5е’7Гйничнйм представителем может служит^ цлотрикс (Ulothrtx)^ имею-
щий форму простых неряяпяттигдппых цдудД, пддйдеплшрщцхся бесцветной
к^^^о_й кдбткои к подводным предметам в пресных водоемах (рис. 17).
Прочие клетки нити все одинаковые, коротко цилиндрические, с постенным
тгр дояска е
размноже- '
тутиковых зоо-
азаЯ^Йбиэ При
разующихся по нескольку в любой клетке,
причем копулировать могут лишь гаметы из разных нитей (гетероталлизм).
. Пе
тонласт ев делится на~~четыре иди <
споры), прорастающие в дальне
прорастанием зиготы в ней происходит редуцдиппное дмдчрд клро-
JII
ем в нити.
50
низ:
игл;
РАСТЕНИЯ
Рис. 17. Улотрикс:
2— участок вегетативной инти; 2 — образование зооспор; 8— зооспора; 4— прорастание зооспора
4 — образование гамет; « — гамета; 7. 8 — копуляция гамет
У большинства улотриксовых перед прорастанием зиготы происходит
редукционное деление ядра,, и вся жизнь водоросли, как у вольвоксовых и
хлорококковых, происходит в гаплоидной фазе. Исключением из этого
являются водоросли семейства улъвовых (Ulvaceae), живущие
в основном в морях, и немногие другие. Улъеа(Шуа) имеет форму крупной
широкой двуслойной пластинки с гофрйрованными краями (рис. 18, 4).
Пластинчатая форма ее получается в результате продольных делений кле-
ток в первоначальйбЙ^йИти. В цикле развития ее осуществляется изоморф- '
ная смена двух поколений, одинаковых по внешности, но размножающихся
одно^Хспорофит)“зооспорами, а другое (гаметофит) половым путем — копу-
ляцией изогамет. Зигота, образующаяся в. результате копуляции, прорастает
без периотта покоя и без редукционного деления. Ия нее развивается дипло-
идный спорофит, у которого потенциально любая клетка может функциони-
ровать как зооспорангий. Перед образованием зооспор происходцт^редук-
ционное деление ядра. Из гаплоидных зооспор развиваются гаплоидные
гаметофиты, по внешности не отличающиеся' от спорофитов. Гаметофиты
Особое место среди улотриксовых занимают нижеследующие и неко-
торые другие^ здесь не упоминаемые роды, которые выделяют либо как под-
порядок Chaetophorinae, либо как особый порядок Chattophorales. JJ этих
растений тело ветвистое, часто гетеротрихальное, состоящее из нитей, как
прилегающих к субстрату^ так и поднимающихся вверх. Некоторые имеют
длинные щетинки.
У видов рода драпарналъдия (Draparnaldia), обитающих в пресных
водах, интересна дифференцировка на стволик и мутовки боковых ветвей,
сложенные из разных по строению клеток (рис. 18,1). У индийской драпар-
налъдиопсис (Drapamaldtopslsj клетки стволика дифференцированы на круп-
ные клетки «междоузлий» и мелкие клетки «узлов», от последних отходят
боковые ветви, часть их обвивается вокруг стволика, образуя на нем кору.
В лице этого рода иногда видят предшественника рассматриваемых харо-
вых водорослей.
«С
ОТДЕЛ — ЗЕЛЕНЫЕ ВОДОРОСЛИ
' Рис. 18. Улотриксовые:
j—драпнрняльдаи; 2“трентеполия; ж— плеврококк; 4 —ульва: « — разрез таллома, б —общий вид
НИЗШИЕ PACT
nt
Виды рода колеохете (Coleochaetae) икивуг в пресных водоемах эпифит-
но на высших растениях. Они имеют вид подушечек, состоящих из радиаль-
но расходящихся разветвленных нитей, или вид плоских пластинок, обра-
- зованных нитями, сросшимися боками. Бесполое размножение
у них осуществляется двужгутиковыми зооспорами, а половое — оога-
Некоторые улотриксовые живут на суше, и у них выработался ряд
приспособлений к наземному образу жизни.
Род фричиелм (Pritschiella) представлен одним в F. tuberosa
(Индия, Африка), приспособленным к наземному с
тело этой водоросли представляет сист
ванню тем, что
от которых отходят надземные ответвл ; последние при выходе на поверх-
ность обильно ветвятся, образуя н лыпой кустик, и служат органами фо-
тосинтеза. Зооспоры и гаметы возникают только из клеток подземных нитей.
Фричиелла имеет изоморфную смену поколений. Водоросли такого строения
и образа жизни, по мнению некоторых ботаников, могли быть предками
наземных высших растений.
Трешпепмия (Trentepohlia^ рис. 18, 2) образует кирпично-красные '
налеты на коре деревьев. Под микроскопом она имеет вид коротких, слабо-
разветвленных нитей, состоящих из толстостенных клеток. Зеленая окраска
11: i
растворенным в масле, пропитывающем цитоплазму. Размноже-
ние происходит большей "частью Путем разламывания нити на отдельные
части. Кроме того^ сохранилось ^бесполое размножепио дву- или четырех-
жгутиковыми йубспораад и половое — двужгутиковыми.изогамёТ&щ. Зоо-
спорангии развиваются на приподнимающихся веточках, они могут отры-
ваться от нити и переноситься ветром, что рассматривается как особое при-
способление для расселения в условиях наземной жизни.
Еще дальше зашло приспособление к наземной жизни у плевроко
(Pleurococcum), образующего зеленые налеты в нижней части стволов деревь-
ев, на заборах и т. п. Под микроскопом имеет вид одиночных клеток или
чаще пакетиков, состоящих в основном из четырех клеток, иногда из корот-
ких нитей (рис. 18,3). Размножается только делением клетод.
Порядок улотриксовых,' вероятно, возним—от тсЯИЖ^то примитивных
одноклеточных хлорококковых.
H11;
ti;i
4-Й ПОРЯДОК — ЭДОГОНИЕВЫЕ (OEDOGONIALES)
К водорослям порядка эдогониевых относится всего одно семейство, но-
сящее то же название и содержащее 3 рода и около 400 видов. Наиболее
крупный р од (свыше 300 видов) — эдогониим (OedogoniwtC). Это прикреп-
ленные базальной клеткой простые неразветвленные бледно-зеленые нити,
сложенные из о^^адерЕРых ^еток с постёнЫм 1цииндрическим."11оЖДм,
сетчатым хгюмайм^кфом^ " ' ~
Характерной особенностью эдогониевых является наличие на апикаль- •
t А ном конце некоторых клеток одного или чаще нескольких узких колечек,
г ц так называемых колпачков^ образующихся (каждый) в результате деления
\ 7 клетки (рис. 19). Переддёлепием внутренний слой оболочки образует блмя
апикального конца кольцевой валик, влаю^и^ся в полость клетку. После
деления ядраГ наружшт сдой оболочки над валиком разрывается, а валик
поД напором содержимого клетки растягивается, покрывая разрастающий-
ся в длину протопласт. У нижнего края разрыва материнской оболочки
образуется поперечная перегородка, и из старой клетки образуются две.
Нижняя одета оболочкой материнской клетки. Верхняя одета новой
оболочкой, образовавшейся из растянувшегося валика, и лишь на вер-
е ее находится небольшое колечко материнской оболочки; оно
несколько шире диаметра новой клетки и выступает на ней в виде кол-
пачка. По числу колпачков можно судить о числе делений клетки. Б е с п о-
i: ir.
in
111: i
мдал—звлвныв водоросли
Рис. 19. Эдоганжлс
54
urn:
РАСТЕНИЯ
л о е размножение происходит своеобразными зооспорами, образую-
щимися по одной в клетке; они имеют овальную или шаровидную форму и на
переднем конце под широким бесцветным носиком — венец жгутиков.
П о л о в о й п р.о. д е с с оогамный. В коротеньких антеридияд^лаС"
полагающихся по нескоЛБК^ в ряд,оёразуетёя"в каждом подважелтоватых
сперматозоида, похожих по форме на зооспоры, но более мелких. Клетки,
. превращающиеся н оогонии, принимают бочонктшдную форму и содержат
Л по одной- шаровидной или овальной яйцйкдоткд. Сперматозоид проникает
, в оогоний через отверстие, образуюшеесяГ^ГёТО^йболочке; после оплодотво-
рения яйцеклетка разрастается и превращается и_ддстдру прорастающую
после периода покоя, длящегося в течение года и более. Перед прораста-
нием ооспоры происходит редукционное делена^и „образуются- i зооспоры.
У - (2;ВоёЬ’б)разный способ деления клеток и особенная форма зооспор и спер-
матозоидов дают основание не включать эту группу в состав порядка улот-
риксовых, как это иногда делается, а выделять эдогониевые водоросли в
особый порядок, родственные связи которого с другими порядками остаются
неясными.
5-Й ПОРЯДОК — КЛАД ОФОРОВЫЕ (CLADOPHORALES) 1 ’
ни
К кладофоровым относятся нитчатые, большей частью ветвистые Формы.
Хро-
матофор в виде сетчато-продырявленного цилиндра со многими пиреноида-
'raT^pacno^oSceHHoro в постенном слое цйтбВлазмы, или~в виде отдели2
пых дисков. Бе с п о л ое размножение происходит четырех-, реже
таужгу тиковыми зооспорами." иногда^ ядраноспо^ами; половое — копу-
ляцией двужгутиковых jggmmgT, ^>еже У многих кладофоро-
вых цикл развития проходит, как у ульвп'шдх, с и?р^ррфнпй смяпой поко-
лений — бесполого (спорофита) н полового (гаметофита).
В пресных водах и в морях распространены виды (около 200) рода
кл(Cladophora), образующие мацррскщщч^дкие. темно-зеленце^
_ кустики^ дерновинки или довольно длинные пряди, состоящие из разветвлен-
ных нитей с крупными толстостенными клетками. В конечных клетках
нитей развивается много зооспор или гамет. В озерах Западной Сибири н
Казахской ССР кладофоры развиваются в огромных количествах, занимая
иногда у берегов площади в несколько квадратных километров. Такие скоп-
ления кладофор могут быть использованы для ИЯглтоплгаитЛт бумага м кяр-
Некоторые виды кладофоры образуют шары, достигающие иногда 10 —
20 см в диаметре, свободно плавающие в воде. Размножаются они только
разламыванием шаров.
Близок к кладофоре риаоклониум (Rhizoclonium)t часто встречающийся
в пресных водах в виде длинных косм или скоплений другой формы.
К кладофоровым относят около 10 родов, обитающих в пресных водах
и в морях. Филогенетически кладофоровые водоросли развивались, вероят-
но, из улотриксовых»
6-Й ПОРЯДОК —
н;
ФОНОВЫЕ, (SIPHONALES, ИЛИ (BRYOPSIDALES)
В порядок сифоновых объединяют: а) водоросли, имеющие сифональ-
ную или ^еклеточную структуру (стр. 34) — ценоцит с множеством ядер
и зернистым хроматофором; б) многоклеточные впдорпслц, состоящий из
не. дешшшимдтерйн-
j i: г
ской клетки на две равноведтше пплптптн. а особым образом: у одних во
взроадщ_кл£тках.отадевяются перегородкам» пабольтие^сепуент^!, "jguree
разрастающиеся (валоияя и др.); у других отделяются перегородкашГуже
развитые/но ранее не~ отделенные ветви (дазикладус, ацетабулярия и др.).
Отдел— зеленые водоросли
55
—кауддрпа (общий вид): a — Caulerpa macrodiaca; б —
Рис. 20,
низшие РАСТЕНИЯ
Некоторые альгологи выделяют многоклеточные сифоновые в отдельный
порядок или подпорядок.)
В целом тело сифоновых водорослей бывает достаточно крупным, мо-
жет достигать нескольких дециметров.
Сифоновые содержат около 60 ныне живущих родов и несколько десят-
ков ископаемых. Все сифоновые обитают в морях и океанах, преимущест-
венно теплых. Они очень разнообразны по ^Ьорме: неоольшиё" шарики, раз-
ив из множества переплетающихся нитей
I п г
(кобиулс стелющиеся по дну нити с приподнимающимися от них
перистыми веточками (бриопсис — Bryopsls), группы крупных пузыревид-
ных клеток (валония — Valonia), стволики с мутовками боковых ветвей
(давикладус — Dasycladus, ацетабулярия — Acetabularia) и др. (рис. 20).
Каулерпа (Cauterpa, около 60 видов) расчленена на стеблевидную или сте-
лющуюся корневищевидную часть до 1 м длины, листовидные части, у не-
которых очень крупные (до 30 см длины) и корневидные ризоиды. Внутри
водоросли имеются 1шогочисленные целлюлозные балки, отходящие от обо-
лочек, но не делящие еена отдельные। вдетая. Они повышают механическую
прочность, служат средством передвижения веществ и увеличивают осмо-
тически действующую поверхность протопласта (рис. 20, 4а,б,а).
Оболочку многих сифоновых водорослей пропитываются известью, в
таком ‘Состоянии они хорошо сохраняются с древнейших геологических
времен.
Некоторые ископаемые сифоновые известны из нижнесилурийских
отложений, имеющих возраст около 400 млн. лет. Из остатков ископаемых
сифоновых в значительной мере сложены триасовые известняки Альп, Гима-
лаев. Некоторые ныне живущие сифоновые участвуют в образовании корал-
ловых рифов в тропиках.
I: г. г.
.111
II!
Г, I
.ш
Зооспоры известны лишь у очень немногих. Некоторые могут размножаться
вегетативно отрывающимися или распадающимися при разрастании участ-
ками теда, легко регенерирующими: оболочка в местах отрыва очень быстро
I восстанавливается.
i Половое размножение изогамное или гетерогамное. Дву-
жгутиковые гаметы образуются в гаметангиях, отделяющихся йёрегород-
ками, или, например у каулерпы, в отдельных участках тела, не отделен-
ных перегородкой. Многие сифоновые раздельнополы. Зиготы прорастают
без редукционного деления, так что водоросли в вегетативном состоянии
ддплондны; редукционное деление происходит* при боразова
^гапйоадная * представлена лишь гаметами.
У некоторых имеется гетероморфная смена поколений (Derteaia).
Неклеточные сифоновые водоросли филогенетически можно выводить
из хлорококковых, подобных протосифону (стр. 54Х Многоклеточные
сифоновые могли развиться из неклеточных. По-видимому, в отдаленные
геологические периоды сифоновые водоросли были представлены более
обильно.
liMi
2-й КЛАСС—КОНЪЮГАТЫ, ИЛИ СПЕПЛЯНКИ
(CONJUGATOPHYCEAE)
t Г J >
К конъюгатам относятся одноклеточные и многоклеточные нитчатые
водоросли. Характерные особенности их: полнот щсутедвяе жтуттеямж
£Х жгутиками гамет) и* особый тип полового
(стр, 35/ 57)._Клетки конъюгат олиояпеттыя?
хроматофоры крупные, пластинчатые или лептляпттпые.Ввегетативном
состоянии все вдвддрдиШг- диплоидны лирр, зиготы, перед прорастанием
которых происхоД^редукционное деление.
i:
ОТДЕЛ — ЗЕЛЕНЫЕ ВОДОРОСЛИ
Б7
К конъюгатам относится около 4500 видов, живущих не прикреплен*
ними к субстрату в пресных водоемах или, значительно режеа в сырых мес-
тах в земле. Их делят обычно на 3 порядка.
1-И ПОРЯДОК — ЗИГНЕМОВЫЕ (ZYGNEMATALES)
иг.
так называемой лестничной к о н ъ*
Рве. 21. Спирогира:
Многоклеточные в форме нитей, почти всегда неветвящихся и покрытых
слизью. Они имеют вид ватообразных скоплений, лежащих на дне или пла-
вающих на поверхности воды, в стоячей иди медленно текущей воде, обра-
зуя так называемую тину; Роды (около 10) различаются главным образом
по форме хроматофоров.
Наиболее широко распространен род спирогира (Spirogyra), содержа-
щий около 275 видов. Ее легко узнать по хроматофорам, которые в виде
1—2 или нескольких довольно широких спиральных лент лежат в тонком
постенном слое цитоплазмы в каждой клетке. В хроматофорах в виде свет-
лых бляшек видны пиреноиды с окружающим их крахмалом. Большую часть
объема клетки занимает вакуоля с клеточным соком, пронизанная тяжами
цитоплазмы, отходящими от ее постенного слоя и пересекающимися в цент-
ре клетки, образуя плазменный мешочек, в котором помещается ядро с
крупным ядрышком (рис. 21). Все клетки спирогиры способны к делению,
происходящему обычно ночью. Вегетативное размножение
осуществляется путем разрыва нитей на отдельные участки, иногда даже
на отдельные клетки. При половом размножении обычно две
нити располагаются параллельно друг другу. Супротивные клетки образу-
ют выросты, направленные друг к другу и срастающиеся концами. Оболочки
их в месте соприкосновения растворяются и получается сквозной канал,
через который сжавшееся содержимое одной клетки в течение нескольких
минут~йеремещается в другую и сливается с ее протопластом, тоже сжав-
мся к этому времени. Кроме
ю г а ц и и, изредка наблюда-
ется (иногда у тех же видов)
боковая
через носредство бокового ка-
нала над поперечной перего-
родкой содержимое клетки пе-
реходит в соседнюю клетку той
же нити. Зиготы шаредз^щой
или овальной формы одеты тол-
стой трехслойной обблочкой; в
содержимом их много - мййла
гематохрома. После периоца по-
коя, во время которого разру-
шаются оболочки клеток нити,
зиготы прорастают; этому пред-
шествует редукционное деление
диплоидного клеточного ядрдз
из 4 получающихся гаплоид-
ных ядер 3 отмирают, а одно
останется ядром единственного
проростка, выходящего через
разрыв наружных слоев оболоч-
ки зиготы.
. Спирогира, довольно легко
культивируемая, часто служи-
ла объектом различных физио-
логических и общебиологиче-
ских экспериментов. В част-
1
58
НИЗШИЕ РАСТЕНИЯ
Рис. 22. Зигнемовые:
2 — зигнема; 2 — мужоция: а — клетка с хроматофо-
ром в плане, б — клетка с хроматофором в профиль,
« — конъюгация
ности, на спирогире русский
ботаник И. И. Герасимов
проводил свои первые в исто-
рии биологии эксперименты по
выявлению значения ядра в
жизни клетки1, получая путем
воздействия на делящиеся клет-
ки холодом или анестезирую-
щими веществами безъядерные
и двуядерные клетки.
Часто встречаются еще ви-
ды двух других родов зигне-
мовых: зигнема (Zygnema) и
мужоция (Mougeotia). У зигне-
мы в ее короткоцилиндрических
клетках симметрично располо-
жены два звездчатых хромато-
фора с пиреноидом в центре
каждого; между хроматофораии
лежит ядро с крупным ядрыш-
ком, окруженное цитоплазмой
(рис. 22, I). У мужоции —
один пластинчатый хроматофор
с несколькими пиреноидами,
расположенный в с рединной
плоскости клетки по длинному
диаметру ее (рис. 22, 2). Он
может менять свою ориента-
цию. Зиготы у мужоции часто
образуются в копуляционном
канале.
2-Я ПОРЯДОК — ДЕСМИДИЕВЫЕ (DESMIDIALES)
Внешний облик представителей десмидиевых водорослей характеризу-
ется резко выраженной двухсторонней симметрией клеток. У подавляющего
большинства клетка разделена, перетяжкой на две симметричные половин-
ки; у немногих родов, не имеющих перетяжки (пениум, клостериум), двух-
I! 1I
сторонняя симметрия в расположении и строении хроматофоров. фболочка
клетки состоит нз двух половинок, такллотно соединенных, что это обнару-
живается лишь йб время деления клеток. В оболочке находятся поры. Че-
рез них выделяется слизь, окружающая клетки рчень“тонким. слоем. Выде-
ление слизи -через особые широкие поры на концах клеток дает возмож-
ность им медленно передвигаться, отталкиваясь от.субстрата. Это'хорошо
заметно у клостерцу^ов. К десмидиевым водорослям относится около 20 ро-
дов и 3900 видов. Йз числа десмидиевых, не имеющих перетяжки в середине
клетки, часто встречаются представители родов пениум (Penium) и клос-
териум (Closterium). Клетки клостериума полулунные. В каждой половине
клетки находится по одному хроматофору, который состоит из нескольких
радиально расположенных пластинок, соединенных в общем центральном
продольном стержне. Пиреноиды видны в виде светлых телец в хромато-
форе. Ядро находится в цитоплазме между хроматофораии (рис. 23, I).
Виды рода космариум (Gosmariurr^ имеют форму двух полукруглых
или слегка удлиненных половинок, соединенных перетяжкой (рис. 23, 2).
У микрастериас (Micrasterias) каждая половинка округлой клетки
сильно сплюснута и изрезана на круглые лопасти и мелкие дольки; плас-
1 Результаты работ опубликованы в 1890—1905 гг.
ОТДЕЛ —ь ЗЕЛЕНЫЕ ВОДОРОСЛИ
59
б
Рис. 23. Десмидиевые:
клетки у космарин; е
— общий виз; б — конъюгация к
— микрасте-
образование
60
НИЗШИВ PACT
и: г
Ill
тинчатый осевой хроматофор образует выросты, заходящие во все лопасти
клетки. Микрастериас — одна из красивейших пресноводных водорослей
(рис. 23, 3).
Десмидиум (Desmidium) имеет вид нитей, состоящих из коротких тре-
угольных клеток. Третий угол треугольника вместе с заходящим в него
хроматофором проецируется в виде темно-зеленой полоски на клетке: так
как вся нить скручена вокруг продольной оси, то вдоль по нити видна тем-
но-зеленая зигзагообразная полоса, идущая от одной стороны к другой
(рис. 23, 7).
При размножении клетки десмидиевых делятся в плоскости симметрии,
в результате получаются 2 клетки, соответствующие каждая не цельной
материнской, а только половине ее. Каждая из «неполных» дочерних клеток
«достраивает» себе вторую симметричную половинку, и лишь после этого две
получившиеся дочерние клетки (у одноклеточных видов) разъединяются
(рис. 23, 2).
При половом размножении две сблизившиеся клетки окру-
жаются слизью. У некоторых представителей десмидиевых выросты клеток
образуют копуляционный канал, у большинства половинки оболочек клеток
просто раздвигаются и выходящие из них протопласты сливаются (рис. 23, 66).
У нитчатых десмидиевых клетки большей частью разъединяются перед
конъюгацией. Зигота прорастает после периода покоя,
образующихся в ней после редукционного деления, два
два ядра становятся ядрами двух проростков.
Десмидиевые водоросли широко распространены в
(в бентосе и планктоне), а также в почве, многие живут в торфяных водоемах,
бедных известью: избыток извести вызывает выпадение железа, а десмиди-
евые — жедезодюбы. Некоторые роды десмидиевых водорослей содержат
по нескольку сотен видов (клостериум — около 200 ввдовх космариум —
свыше 800).
Из четырех ядер,
ядра отмирают, а
пресных водоемах
।; । \
3-Я ПОРЯДОК— МЕЗОТЕНИЕВЫЕ (MESOTAEN1ALES)
Почти все мезотениевые (6—7 родов, 50 — 75 видов) — одноклеточные
водоросли цилиндрической формы с округленными концами и сплошной
оболочкой. Хроматофоры у разных родов похожи на хроматофоры зигне-
мовых — спиральный у спиротекии (Sptrotaenia)^ осевая пластинка у
мезошекиужа (Mesotaenium)t в виде двух звездочек у цилиндроцистиса
(Cylindrocystis) ит. д. Вегетативное размножение осуществля-
ется делением клеток, и о л о в о е — конъюгацией. При прорастании пос-
ле редукционного деления все четыре ядра остаются жизнеспособными и из
зиготы вырастают четыре проростка.
Мезотениевые водоросли являются исходными для всех других поряд-
ков конъюгат. Одна линия их эволюции дала начало нитчатым зигнемовым
с уменьшением числа проростков, образующихся из зиготы, до одного.
Уменьшение численности потомства компенсируется тем, что один пророс-
ток получает всю цитоплазму и запасные вещества зиготы и выходит в жизнь
более мощным. Вторая линия эволюции от мезотениевых дала начало десми-
диевым, у которых эволюция шла по пути усложнения конфигурации клет-
ки, выработки своеобразного способа дел
ростков из зиготы до двух.
L Г Г
[il.T.
ФИЛОГЕНЕТИЧЕСКИЕ СВЯЗИ ЗЕЛЕНЫХ
ВОДОРОСЛЕЙ
Наличие стадии зооспор в начале онтогенеза у очень многих зеленых
водорослей указывает на происхождение их от организмов монадной струк-
туры. Предковой группой зеленых водорослей могли быть примитивные
<пдвл—ЗЕЛКНЫВ ВОДОРОСЛИ
61
гл;
ill
в
1TIJ
ципг.ц
вожьвоксовые, не имевшие плотной оболочки иного состава, чем цитоплав-
иа, размножавшиеся продольным делением в подвижном состоянии и имев-
шие половой процесс в форме хологамии. Эволюция самих фол^воксовых
шла по пути формирования оболочки, образования колониальных форм —
таблитчатых и шаровидных — с некоторой специализацией клеток, по пути
усложнения полового процесса (оогамии); другим направлением развития
была утрата подвижности (тетраспоровые).
Из сравнительно простых одноклеточных вольвоксовых типа хламидо-
монад при утрате ими подвижности могли получить свое начало хлорокок-
ковые. Дальнейшая эволюция их выразилась, как и вольвоксовых, в по-
явлении колониальных форм, в утрате у многих подвижных стадий и поло-
вого процесса, что, вероятно, связано с их планктонной жизнью.
Из неспециализированных хлорококковых развились нитчатые улот-
рнксовые. В пределах улотриксовых эволюция выразилась в появлег
ветвления, дифференцировке на распростертую и приподнимающуюся (вер-
тикальную) часть таллома, деление клеток не только в поперечном, но и
в продольном направлении, что привело к появлению пластинчатых форм,
разнообразии форм полового процесса.
Образование многоядерных клеток у улотриксовых могло привести
развитию кладофоровых.
Появление многоядерности у простейших хлорококковых (вроде про-
тосифона), вероятно, повело к развитию своеобразной группы неклеточных
сифоновых, по разнообразию своих форм превосходящих все прочие зеленые
водоросли.
Из простейших хлорококковых, возможно, развились и простейшие
конъюгаты — мезотениевые. Эволюция этой группы выразилась в разнооб-
разии очертаний клеток, форм хроматофоров, в развитии нитчатых форм и в
уменьшении числа новых особей, возникающих из зиготы.
Родственные связи эдогониевых неясны.
Дали начало зеленые водоросли вышестоящим зеленым растениям или
нет, сказать трудно. Однако несомненно, что многие группы зеленых водо-
рослей представляют линии эволюции, закапывающиеся слепо (сифоновые,
эдогониевые, кладофоровые, конъюгаты). Некоторые ученые предполагают,
что высшие наземные зеленые растения могли возникнуть из улотриксовых
водорослей, в частности от хетофоровых, расчлененных на горизонтально
распростертую и вертикально стоящую части таллома.
Что касается происхождения конъюгат, то можно предположить,
что они представляют боковую ветвь эволюции равножгутиковых, отделив-
шуюся когда-то от простейших одноклеточных хлорококковых. В конъ-
югирующих протопластах (гаметах) спирогиры найдены пульсирующие
вакуоли — атавистический остаток их когда-то жгутиковой стадии. С дру-
гой стороны, у некоторых хламидомонад при копуляции гамет, одетых обо-
лочками, между ними образуется что-то вроде популяционного канала.
Г; hi
I: i' । '
ОТДЕЛ—ХАРОВЫЕ ВОДОРОСЛИ,
ИЛИ ЛУЧИЦЫ (CHAROPHYTA)
Харовые — крупные пресноводные донные- водоросли, достигающие
20 — 30, иногда 50 см и более высоты и несколько похожие по внешнему
виду на хвощи (рис. 24). Они имеют мутовчато разветвленное тело, главная
ось которого явственно расчленена на длинные междоузлия и узлы. Узел
состоит из коротких клеток, а каждое междоузлие образовано одной очень
длинной (до 5 см) многоядерной клеткой. С поверхности она большей частью
бывает покрыта корой" йзодного слоя клеток. Клетки коры берут начало
от базальных клеток боковых ветвей, образующих мутовки, несколько на-
поминающих хвою и называемых «листьями». От «листьев» двух соседних
узлов образуются выросты, сначала одноклеточные, затем делящиеся. Они
НИЗШИЕ PACT
IE III ‘
Рис. 24. Хара:
I — общий вид; 2 — часть «листа» с осгоннек
(вверху) и антеридием (внизу); 8 — щиток ан-
_ ,,,л со сперматогенными нитями; 4^.
сперматозоиды
направляются навстречу друг другу и сталкиваются посередине междоуз-
лия. Мутовчато расположенные боковые ветви — «листья» — построены,
как и главная ось, отличаясь от нее меньшей длиной и ограниченным на-
растанием. В пазухе одного из «листьев» мутовки может возникнуть боковая
ветвь того же строения, как и главная ось растения, и также способная
к длительному нарастанию. В грунт водоема отходят бесцветные нитевид-
ные ризоиды. На вершине растущей оси находится верхушечная клетка.
Располагающиеся рядом с ней молодые междоузлия еще коротки, благода-
ря чему узлы очень сближены, а листья, загнутые вверх, образуют почку;
в ней и скрыта верхушечная клетка стебля. Она отчленяет от себя пооче-
редно клетки, становящиеся клетками междоузлий и узлов.
Бесполого размножения спорами у харовых нет. Вегета-
тивно они размножаются: 1) клубеньками на ризоидах и на нижних стеб-
левых узлах; 2) укореняющимися ветками, отходящими от нижних узлов.
Половой процесс оогамный. На узлах «листьев», у большинст-
ва на одном и том же растении, развиваются сидящие рядом многочислен-
ные оогонии и антеридии. В оогонии удлиненная яйцеклетка окружена
пятью спирально обвивающими ее нитевидными клетками (нитями), отчле-
няющими на вершине 5 (у хары) или 10 (у нителды) клеток, образующих
так называемую коронку. Антеридии — оранжевые шарики, видные нево-
оруженным глазом. Стёнка их образована 8 плоскими клетками, так назы-
ваемыми щитками. От них внутрь антеридия отходят особые подставки, на
которых развиваются длинные свернутые в клубок сперматогенные нити.
Каждая нить состоит из 100—300 плоских клеток, в которых развивается
fill
г. г
ни
иг
ОТДЕЛ — ХАРОВЫЕ ВОДОРОСЛИ
63
г, г.
по одному сперматозоиду. Всего в антеридии образуется 20—50 тыс. спер-
матозоидов. Щитки антеридия при созревании его разъединяются, и сперма-
тозоиды освобождаются. После оплодотворения яйцеклетка разрастается,
покрывается плотной бурой оболочкой и превращается в зооспору («споро-
почку»).
Ооспора прорастает после периода покоя. Прорастанию предшествует
редукционное деление диплоидного ядра ее, так что все растение харовых
гаплоидно.
К хяровым относится 6—7 родов и свыше 200 видов, растущих в чис-
тых спокойных пресных водах всего света. В ископаемом состоянии харо-
вые известны с девонского периода (около 300 млн. лет назад).
Происхождение и филогенетические связи харовых неясны,
ву пигментов лучицы могут быть сближены с зелеными
. их и относят иногда в ранге класса.
есть некоторое сходство с родом драпарнальдиопсис из
вых (стр. 49—50); у последнего также имеются короткие клетки узлов и
длинные клетки междоузлий: от узлов отходят мутовки боковых ветвей^ а
часть ветвей оплетает в виде коры стебель.
По соста-
водорослями, куда
По вегетативным органам у харовых
порядка улотриксо-
<:
I г I".
,|н
ОТДЕЛ — ПИРРОФИТОВЫЕ ВОДОРОСЛИ
(PYRRHOPHYTA)
Пиррофитовые водоросли обычно имеют монадную, реже коккоидную (
организацию,-л только отдельные виды имеют строение, ^ сходное с нитча-
тыми. клетки у них не радиально-,, а двухсторонне-симметричные или асим-
метричные, дорзовентральные, покрытые оболочкой или без нее. Харак-
терно наличие двух жгутиков неравной длины, обеспечивающих движение^
реже движение амебоидное, или клетки неподвижны. Клетка покрыта часто
сложно устроенной оболочкой или (у примитивных форм) лишена ее. Ядра
довольно крупные, с различными в нем четковидными хроматиновыми
нитями. Хроматофоров — несколько желто-бурых, содержащих хлорофил-
лы а и (немного) с^дкаротин и ксантофилл; у бесхлорфилльных форм есть
леййопласты/Продукты запаса — крахмал и масло. Размножаются деле-
нием, причем в случае наличия оболочки каждая из дочерних клеток полу-
чает одну половину материнской, а вторую вырабатывает вновь. В других
случаях внутри материнской клетки делением протопласта образуются две
двужгутиковые зооспоры или апланоспоры. Иногда, при неблагоприятных
условиях, формируются цисты. Половой процесс известен только у единич-
ных представителей.
Богатый видами отдел (около 1100) пиррофитовых водорослей делят на
4 класса, из которых наиболее распространен класс перидиней (Peridi-
пеае, или Dinophyceae), содержащий до 1000 видов. Для распространенных
у нас и вообще типичных перидиней характерна целлюлозная оболочка,
состоящая, как панцирь, из многих щитков. Щитки соединены друг с другом
швами. Очень характерно наличие двух бороздок, или ложбинок:. попе-
речная ложбинка опоясывает клетку посередине или смещена к одному из
концов и делит оболочку на две части — переднюю и заднюю; продольная—
находится только на вогнутой брюшной сторона jk летки и большей частью
лишь в задней половине клетки. "Ложбинки тоже выстланы щитками. На
брюшной стороне, в месте пересечения кольцевой и продольной ложбинок,
кольцевой ложбинке, опоясывая клетку и волнообразно изгибаясь; дру-
гой жгутик, нитевидный, лежит в продольной ложбинке и выступает за пре-
делы клетки с ее заднего конца.
Из широко распространенных в пресных и морских водах перидиней
отметим род церациум (Ceratium). Для этого рода (рис. 25, 2) очень харак-
терно наличие одного длинного переднего рога и двух-трех рогов на заднем
EPACT
Рис. 25. Перидинеи;
1— верцадниум; Я — церацнум; л — нокти шока.' а — общий вид. б — эоосжора
конце тела. При размножении церациума протопласт делится на две особи,
а панцирь раскалывается косой трещиной на две части; каждая новая особь
получает половину старого панциря, а недостающую половину вырабаты-
вает заново. У очень немногих первдиней описан половой процесс в форме
копуляции двух гаметуподи£ш(бвых по форме и величине.
Более примитивные перидинеи голые. Известны среди них и непод-
вижные представители. К неподвижным перидинеям относится ночесветка,
или поктилюка (Noctiluca, рис. 25, 3)9 широко распространенная в морском
планктоне, где она является одним из главнейших возбудителей ночного
, свечеция моря1.
Пиррофитовые водоросли представляют самостоятельную линию эволю-
, двойного развития, не имеющую прямых связей с другими отделами расти-
тельного мира. Практически они важны как массовые продуценты органи-
ческого вещества. Теоретически эти растения многосторонне интересны.
Здесь можно найти переходы от клеток, не имеющих оболочки, к имею-
щим таковую. Формы, лишенные оболочки, способны к животному типу
литания, который не чужд и формам, имеющим хлорофилл. Пластинки, кото-
рые* слагают покрывающий панцирь, пронизаны многочисленными отверс-
тиями, и сквозь них цитоплазма выступает наружу в' виде тончайших нитей:
они улавливают бактерии и другие микроорганизмы.
Большой интерес представляет ядро. Его оболочка пронизана порами,
а хромосомы, в виде четкообразных нитей, различимы и в покоящемся
ядре (в интерфазе). При делении ядерное веретено не образуется.
Эти морфологические, а также некоторые химические отличия дали повод
рассматривать организацию ядра в данном случае как промежуточную меж-
ду ядерной структурой эукариотических и прокариотических растений.*
11 ,п ।
ШИ
1 Свечение моря вызывают также некоторые перидиниумы, бактерии и другие орга-
низмы. Свет бывает серебристо-белый или слегка голубоватый. Он выделяется в результате
окислительных процессов внутри клеток. Механическое раздражение и усиленная аэ-
рация усиливают свечение.
i
ОТДЕЛ — ХРИ30М0НАД0ВЫВ ВОДОРОСЛИ
ев
ОТДЕЛ—ХРИЗОМОНАДОВЫЕ, ИЛИ ЗОЛОТИСТЫЕ^
ВОДОРОСЛИ (CHRYSOPHYTA)
Характерные особенности представителей этого отдела — монаддая струк-
тура^ золотисто-желтая, желто-зеленая или бурая окраска, радиальная.
симметрия клеток. Подавляющее большинство — одноклеточные подвиж-,
ные формы, значительно меньше колониальных, тоже подвижных; немно-
гие — неподвижные^ коккоидной или нитчатой структуры». Окраска зави-
сит от хроматофоров, распбложенныхТУбЛЬпгей^чГОЖ^ двух в наруж-
ном сдре цитоплазмы и имеющих корытовидную или дисковидную форму;
В хроматофорах, кроме хлорофиллов а и с, находятся еще каротиноиды —
желтый лютеин и буроватый фуккоксантии. Крахмада-яет- Запасные про-
дукты — масло и углевод хризоламинарии (хризоза, лейкозин) — хризо-
монадовыб водороСЛй'откЛадЦваюг^в виде одной или нескольких капель*
сильно преломляющих свет. Клетки у одних голые, способные к метаболи-
ческим изменениям формы тела, у других одеты оболочкой из целлюлозы
и пектиновых веществ. У многих в оболочке или в наружном слое голой
клетки находятся известковые или окремнелые пластинки, у некоторых с
иглами. Движение осуществляется при помощи одного или двух одинако-
вой или разной длины жгутиков.
Размножаются хризомонадовые водоросли продольным делением кле-
ток, большей частью не прекращая движения. Размножение происходит
также путем'образования зооспор с одним-двумя жгутиками. Колониаль-
ные формы размножаются распадением крупных колоний на несколько
меныпих. У многих хризомонадных водорослей при наступлении неблаго-
приятных условий образуются эндогенные цисты: протопласт выделяет под
материнской оболочкой новую, толстую, окрашенную, нередко скульптур-
ную оболочку, снабженную одной порой, закрытой пробочкой. После пери-
ода покоя циста прорастает, образуя зооспоры.
Большинство хризомонадовых — планктонные водоросли чистых, не
загрязненных органическими веществами пресных водоемов, где они встре-
чаются преимущественно ранней весной и поздней осенью. Немногие живут
нг.
в морях.
Отдел золотистых водорослей содержит менее 500 видов (60—80 родов).
В стоячихпресных.^одоемах нередко встречаются хиомулины (Chro-
mulina) — одшоЙЛетоЦйыеТ с-жгутаком, не имеющие обособленной оболочки
(рис. 26, 2). У малломонад fМаПотошде)7 обитающих в пресных чисйХх во-
доемах, в оболочке находятся окрашенные чешуйки, снабженные длинными
кремневыми иглами (рис. 26, 2). Из планктонных подвижных колониаль-
ных родов хризомонадовых водорослей следует отметить силуру и диноб-
рион. Стура (Synura) — красивые золотисто-желтые шарики, крутящиеся
1Г1
пл
как волчок и состоящие из обратно-яйцевидных клеток с двумя неравными
жгутиками и окремнелыми чешуйками в оболочке (рис. 26, 5). Динобрион
(Dinobryori) — изящные, активно движущиеся кустики, состоящие из про-
зрачных бокальчиков (панцирей); в каждом из них сидит прикрепленный
стебельком ко дну цилиндрический или веретеновидный протопласт с дву-
мя неравными жгутиками. Колонии разрастаются путем последовательных
делений протопластов: из двух получающихся протопластов один остается
в старом домике, а другой выходит наружу, прикрепляется на краю старого
бокальчика, после чего вырабатывается новый бокальчик (рис. 26, 4).
В холодных, быстро текущих речках и ручьях живет гидру рус (Hydrurus)
в виде ветвистых слизистых буроватых кустиков, прикрепленных осно-
ванием к камням, бревнам и т. п. В слизь погружены округло-эллиптичес-
кие клетки без жгутиков, размножающиеся делением, иногда дающие нача-
ло новым колониям (рис. 26, 5)- Из нитчатых, утративших подвижность
хризомонадовых водорослей можно указать феотамнион (Phaeothamniori)t
гшъупуй эпифитно на нитчатых пресноводных водорослях и имеющий вид
3—602
66
НИЗШИЕ РАСТЕНИЯ
5
Рис. 26. Хризомона довые:
1 — хромулина: а — общий вид* б — циста; а — малломонас; з — синура; 4 — динобрион; 5 — гидруруо;
а — общий вид. б — кончик ветви» в — зооспора; в — феотамнион
однорядных разветвленных нитей, прикрепленных базальной клеткой к
субстрату (рис. 26, б); размножается он зооспорами.
Хризомон адовые водоросли — древняя группа: некоторые из них из-
вестны из кембрийских отложений. Можно предположить некоторые род-
ственные связи хризомонадовых (происхождение из общих предков)
с диатомовыми и разножгутиковыми водорослями.
Менее вероятны родственные связи хризомонадовых с бурыми водо-
рослями.
ОТДЕЛ — РАЗНОЖГУТИКОВЫЕ, ИЛИ
ЖЕЛТО-ЗЕЛЕНЫЕ, ВОДОРОСЛИ — ГЕТЕРОКОНТЫ
(HETEROCONTAE, XANTHOPHYTA)
Разножгутиковые отличаются тем, что два жгутика у их зооспор не-
одинаковы. Они прикреплены несколько сбоку на переднем конце зооспо-
ры. Один жгутик, более длинный, направлен вперед и несет на оси перисто-
расположенные отросточки; другой гладкий, короткий, направлен назад
I
ОТДЕЛ — РАЗНОЖГУТИКОВЫЕ ВОДОРОСЛИ 67
(рис. 27). Оболочка клетки разножгутиковых спддргедт много^пектиновых П V -
веществ и большей частью не дает реакции на целлюлозу. У многих ошгсо-
стоит из двух равных или неравных половинок и при разрыве клеток распада-
ется на Нтобразные участки. Хроматофоры обычно зернистые: они содержат
хлорофиллы дми с, а также одного каротиноидов и потому имеют у многих
желтовато-зеленоватую окраску. Хроматофоры при обработке крепкой
соляной кислотой становятся голубовато-зелеными, тогда как у зеленых
водорослей они при такой обработке не изменяются или желтеют. Крахмал
не образуется, и запасным продуктом являетсяjgacno. Большинство разно-
жгутиковых—неподвижные, одноклеточные, колониальные и нитчатые рас-
тения.
. Вегетативное размножение происходит делением кле-
ток, распадом колоний или нитей. Бесполое размножение —
зооспорами, автоспорами. Половой процесс, известный у очень
немногих родов, обычно изогамный, реже — оогамный (Vaucheria). Рас-
пространены разножгутиковые в планктоне jh бентосе пресных вод и морей, у г
в почве, на сырых стенах строений, на камнях. ' '
Классификационные подразделения разножгутиковых аналогичны и
параллельны в основном порядкам равножгутиковых зеленых водорослей.
Есть порядки подвижных в вегетативном состоянии водорослей (Hetero-
chlorldales), неподвижных коккоидной структуры — одноклеточных и коло-
ниальных (Heterococcales), нитчатых (Heterothrichales) и неклеточных (Hete-
rosiphonales).
Из отдельных представителей разножгутиковых можно отметить хло-
рамебы (Chloramoeba из порядка Heterochloridales) — голые, сильно мета-
боличные организмы, с псевдоподиями и жгутиками, живут (разные виды)
в пресной воде и в морях; трибонемы (Tribonema из порядка Heterothri-
chales), раньше известные как конфервы (Conferva), — неразветвленные
нити из слегка боковидных клеток, оканчивающиеся в местах разрыва дву-
зубой вилочкой — половиной оболочки оторвавшейся клетки; ботридиум
(Botrydium из порядка Heterosiphonales) — наземную водоросль неклеточ-
2
Рис. 27. Разножгутиковые:
1 — хлорамеба; 2 — трибонема: а — участок нити, б — распадение нити и выход зооспор, • — зооспора!
S— ботридиум: а — общий вид. б — зооспора (после специальной окраски жгутиков)
3*
1
,r±d»j>'
РАСТЕНИЯ
кого строения, растущую на ей-
рой земле, обычц£иц^ больших ко*
личествах. гВ1Ргпипичм. имеет вид
зеленых грушевидных пузырей до
1 жж, от которых в почву отхо-
дят ветвистые бесцветные ризо-
иды; после дождя он образует
огромное количество зооспор, а в
сухую погоду содержимое пузыря
переходит в ризоиды и распадает-
ся на цисты, хорошо выдержива-
ющие высыхание. В последнее вре-
мя к порядку Heterosiphonales от-
дно-Зелешм... слаборазветвленные
толстые нити (рис. 28), достйгаю-
рор ддины. Хро1датофоры зерни-
стые, без пиреноидов, крахмал.
не ^обрайуетсСГзапаснои .про- х i
Рис. 28. Вошеряя:
1 — нить с оогониями
с находящей зооспорой: Л — антеридий (слева) и оого- р д
““ ДИТ
____ зооспор, образующихся'по одной
происхо-
лые, имеют на поверхности многочисленные пары жгутиков; под каждой
парой жгутиков в цитоплазме находится ядро. Внутри зооспоры имеется
большая вакуоля с клеточным соком. Существует предположение, что зоо-
спора вошерии образована многими неразъединившимися двужгутико-
выми зооспорами.
-~М-д_д n n n ft п р п п п г Г, у «лтпррии оогамим
коротких ветвях ее образуются как боковые вырос
|' и J <
1
рцдий развивается в виде изогнутого рога и в месте перегиба отделяется пе-
регородкой от базальной части. Перед оплодотворением оболочка оогония в
верхней части разрывается и небольшая часть цитоплазмы выходит на-
ружу. Она, по-видимому, действует хематоксически на сперматозоиды,
* которые выходят из разорвавшегося кончика антеридия; сперматозоиды
почти бесцветные, овальные или грушевидные, с двумя жгутиками, из кото-
рых один перистый. После оплодотворения в оогонии развивается ооспора
с толстой пбпзпапой. срдоржмпаи много масла и гематохром. После периода
покоя в ней происходит редукционное деление, и она прорастает в новую
1'1'.
Виды вошерии (около 40) широко распространены в различных пресных
водоемах, немногие — в морях; некоторые виды развиваются на сырой зем-
ле, образуя зеленый войлок.
Ранее^в течение долгого времени, Потеряю относили к порядку сифо-
новых отдела зеленых водорослей. Но этому противоречат гапдоидность,
половой процесс, отсутствие крахмала .и наличие масла как запасного про-
дукта. Наличие у вошерии пигмента гетероксантина, отсутствие хлорофилла с
Ь, неодинаковое строение и характерное положение жгутиковых у спер-
матозоидов сближает этот род с сифональными разножгутиковыми. Неко-
торые альтологи сближают разножгутиковые с хризомонадами, предпола-
гая, что они могли иметь общих предков.
нг<
t.
I
ОТДЕЛ — ДИАТОМОВЫЕ ВОДОРОСЛИ
ев
ОТДЕЛ—ДИАТОМОВЫЕ ВОДОРОСЛИ, ДИАТОМ
И
(DIATOMEAE, ИЛИ BAC1LLARIOPHYTA)
Диатомовые водоросли — микроскопически мелкие одноклеточные,
колониальные или нитчатые водоросли буровато-желтого цвета. Оболочка
их содержит большее (иногда до 50% веса клетки) или меньшее (порою все-
го около 4%) количество кремнезема, кремневый панцирь представляет
основу оболочки и с гтпрптт ттпкрыт слоем опганетеского вещества.
В целом оболочка состоит из двух половинок, налегающих одна на другую;
одна из них, несколько большая (элитш^)» ^ялегает на меньшуюП^юдотеку),
как крышка налегает^на коробку. В свою очередь, гипотека и эпитека со-
стоят из двух частей: плоской с немного загнутыми краями, называемой
створкой, и плотно соединенного со створкой кольца, называемого пояском.
Поясок эНитеки плотно надвинут на поясок гипотеки. У некоторых диато-
мовых таких поясков с каждой стороны бывает по нескольку. Если диато-
мовая водоросль обращена под микроскопом к наблюдателю пояском, то
в оптическом разрезе она имеет очертание приблизительно прямоуголь-
v ное и, во всяком случае, отличное от вида со стороны створки (рис. 29).
В кремневом панцире створок находятся сквозные поры, в виде то-
чек, а также камеры, или пустоты, открывающиеся внутрь или наружу.
Все они придают створкам особую
и имеющую очень большое значе-
ние в систематике диатомовых.
В зависимости от конфигура-
ции клетки одни диахрмовые-ше^.
ют радиальную СИф*Дтрит СТВО-
РОК. Другие — януугтпрлтиптп.
Первые объединяют в класс цр*-
трических (Centrical, вторые — в
класс перистых, или иенндтных
г - --------Г
У многих диатомовых из
класса перистых в створках (в
обеих или в одной) находится так
называемый ^шойч— продольная
щель в оболочйегта концах шва
и посредине его находятся утол-
щения оболочки, называемые соот-
ветствецжГ'полярными (концевы-
ми) ^^центральным узелками^ >
Шов и особенно узелки имеют
довольно сложное строение. Цито-
плазма движется по шву и узел-
кам. Трение движущейся цитоплаз-
мы (или выделяемой ею слизи) о
воду или о субстрат создает дви-
жущую отталкивающую силу, в
результате чего клетка движется
в направлении, противоположном
движению цитоплазмы. Диатомо-
вые водоросли, не имеющие шва,
не могут двигаться.
Цитоплазма расположена в
клетках тонким постенным сдоем,
а у перистых еще и в середине
клетки, в виде мостика, в котором
помещается ядро. Остальная., ббль-
структуру, видимую под микроскопом
Рис. 29. Пиннулярвя:
вид со створки; 2—вид с пояска; а—поперечны*
раврев и — 8 изображена только оболочка); 4—ан*
со створки с содержимый; 4 — поперечный раврев обо-
лочки с протопластом; оболочка бесцветная, прото-
илазыа светло-серая» хроматофор темно-серийг -р—
увещав. ш’—шов. о—створки. а —пояски* > — ве-
йка. • — ншотека
70
НИЗШИЕ РАСТЕНИЯ
Рис. 30. Навикуля:
1 — «5 — схема деления клетки (вид с пояска); 4 — дочерние клетки с новыми гипотеками
шая часть клетки заполнена клеточным соком. Хроматофоры, находящи-
еся в цитоплазме, бывают в разном числе (1, 2, 4 — много) и различной
формы — зернистой, пластинчатой; у некоторых есть пиреноиды. Пигмен-
ты в хроматофорах — хлорофиллы а и с, каротин, бурый фукоксантин и
еще несколько ксантинов, два из которых вместе раньше называли диато-
мином. После отмирания водорослей бурые пигменты извлекаются во-
дой, а хлорофилл остается и водоросль посмертно зеленеет. Крахмал не
образуется. Запасные продукты — масло, валютин и хризоза — отлагаются
как в плазме, так и в вакуолях, но без видимой связи с пиреноидами-_
При вегетативном делении клеток протопласт после митотического
деления ядра раскалывается на две половины в плоскости, параллельной
створкам. Створки несколько раздвигаются, унаследованная от материн-
ской клетки створка, становится эпитекой, гипотеку же каждый дочерний
3
Рис. 31. Схема полового процесса центрической диатомеи мелозиры
1 — вегетативная нить (с пояска) а — развитие и выход сперматозоидов, мейоз в женской клетке, оп
лодотворение и начало формирования ауксоспоры; 3 — нить с ауксоспорами
ОТДЕЛ — ДИАТОМОВЫЕ ВОДОРОСЛИ
71
протопласт вырабатывает заново и у оболочки; после этого две получив-
шиеся дочерние клетки разъединяются (рис. 30). Так-какобо-
лочки клеток не способнырастягиваться, то при каждом делении одна из
дочернйх~~клеток оказывается несколько^моиьше материнской. Уже после
третьего деления лишь одна из восьми получившихся клеток имеет размеры
первоначальной, Материнской. В результата большого количества делений
размеры многих^клэтОК уменьшаются в 2 — 3 раза. Такое измельчение кле-
ток в действительности не всегда достаточно выражено. Экспериментально
показано, что у некоторых диатомовых его практически нет. Это объясняют
более частым делением клеток, сохраняющих размер исходных, некоторой
растяжимостью пояскового кольца, т. е. возможностью хотя бы ограничен-
ного роста дочерних клеток, и другими причинами. Заметному сниже-
нию среднего размера особи противопоставляется увеличение их размеров
благодаря половому процессу. Но оплодотворение имеет место и у неизмель-
ченных клеток.
Оплодотворению у диатомовых. предшествует^-редукционное деление
ядра. При сравнительно недавно открытой в этом отделе^оогамии в одних
клетках образуются четыре сперматозоида с одним,”'но7 возможно, и с двумя
жгутиками; в других же три ядра после мейоза дегенирируют и протопласт
такой клетки остается с одним гаплоидным ядром; он и оплодотворяется
сперматозоидом. Зигота докрывается тонкой пектиновой оболочкой, быстро
растет (за что такую" зиготу йЗЗЫваЮТ а у7к с о <Гп о р о й1) и в дальнейшем
вырабатывает' створки панциря. Так протекает половой процесс, например*
у Melo str a varians (рис. 31). Ранее
у других видов Melosira, а также
у Chaetocerus было описано обра-
зование ауксоспор, происходящее
в результате автогамии, В этом
случае два из четырех ядер ре-
дукционного деления дегенериру-
ют, а два оставшихся сливаются
друг с другом. У __ центрических
диатомовых наблюдалось образо-
вание так называемых микроспор,
которые, рассматривали та как
изогаметы, то как зооспоры. После
открытия сперматозоидов стало
вероятным, что эти, снабженные
жгутиками клетки суть сперма-
тозоиды.
Оогамный половой процесс
известен и у перистых диатомовых
(Rhabdonema adriaticum) в фор-
ме, принципиально сходной с
вышеописанной, но отличной в
деталях (рис. 32). Отметим толь-
ко, что микрогаметы в данном
случае не имеют жгутиков и дви-
гаются амебообразно. Однако у
перистых диатомовых давно извест-
на и, по-видимому, более' типична
Рис. 32. Рабдонема. Оплодотворение (я —*
яйцеклетка <с многочисленными пластидами;
яя — ядро яйцеклетки; яяд — дегенерирующее
ядро яйцеклетки; она — антеридий с двумя
микрогаметами; ан — антеридий без гамет;
е — микрогаметы. пяа — переход ядра гаметы
в оогоний; мяд — дегенерирующее мужское
ядро)
иная форма оплодотворения.
Еще в конце прошлого века
было установлено, что у Rhopalo-
dia gibba образованию ауксоспор
предшествует образование безжгу-
От греч. «ауксо»— увеличиваю,
расту.
НИЗШИЕ PACT
2
Рмо. 33. Схема полового процесса перистых диатомовых:
У—цкмбелипз •» б, •—редукционное далекие ядер и дегенерация двух гаплоидных ядер в каждое кв
вмпзведукмцих клеток, •—образование двух гаме* разно* величавы, б — слияние пикет, в — две пово-
дов знгвш; ж—ауксоспоры; л—гомфонемви •—гаметы и редукция их ядер, б —в — образование двух
виток
отдал—ДИАТОМОВЫЕ ВОДОРОСЛИ
ГТ.
тиковых гамет. Две копулирующие клетки, часто неравные, лежащие ря-
дом, склеиваются друг с другом выделяемой ими слизью. В каждом про-
топласте происходит редукционное деление ядра, но из четырех мейотиче-
ских ядер два дегенерируют, что проявляется в уменьшении их размера
сравнительно с двумя другими. Протопласты копулирующих клеток делятся
на две безжгутиковые гаметы, в каждой из которых оказываются по два
ядра: одно нормальное и одно дегенерирующее. Гаметы, принадлежащие
разным клеткам, двигаясь амебообразно, выходят из клеток, сливаются
попарно, причем происходит слияние и их нормальных ядер, дегенери-
рующие разрушаются,
Аналогично происходит обравдванне.-ауксосоцр у Cymbella lanceolate
(рис. 33.2). При таком течении полового процесса у других перистых диато-
мей (Gomphonema parvalum) констатировано, что одна из гамет может быть
активно подвижной, оплодотворяющей, т- е. мужской, другая, остающая-
ся неподвижной, играет роль оплодотворяемой, т. е. женской. Образовав-
шая зиготы покрываются пектиновой оболочкой, растут и, позднее,
вырабатывают типичную двухстворчатую окремневающую оболочку. По срав-
нению с типичной оогамией эту форму полового процесса можно рассматри-
вать как редуцированную (утрата гаметами жгутиков). У некоторых пе-
ристых диатомовых из четырех ядер редукционного деления дегенерируют
три; в этом случае копулируют цельные протопласты, участвующие в по-
ловом процессе особей, ж формируется только одна ауксоспора (зигота). У
перистых диатомовых известны также случаи самооплодотворения, когда
сливаются две гаметы, возникшие из протопласта одной клетки, а также
образование ауксоспор бесполым путем.
Сказанное о половых явлениях у диатомовых водорослей позволяет
иг.
г. г.
У центрических диатомовых водорослей известно образование покоя-
щихся клеток — цист. Цисты формируются эндогенно, путем сокращения
протопласта с последующим выделением толстой окремневающей оболочки.
Значение их — в переживании неблагоприятного состояния среды.
К диатомовым относится около 170 —190 родов и 10 000 (по’
другим данным, 12 000 — 15 000) видов. Их делят большей частью. на
2 класса — перистые и центрические.
1-й КЛАСС — ПЕРИСТЫЕ, ИЛИ ПЕННАТНЫЕ,
ДИАТОМОВЫЕ (PENNATAE)
- Через створки можно провести 1 — 2 плоскости симметрии, или они
асимметричны. У многих есть шов. Половой процесс — своеобраз-
ная изогамия, несколько сходная с конъюгацией у конъюгат.
Представители этого класса: пинмулярия (Pinnularia) — одиночные
клетки, имеющие со створки удлиненно-эллипсоидальную форму, с перис-
той структурой и швом, с пояска — форму удлиненного прямоугольника
(см. рис. 29); навикуля (Navicula) близка к пиннулярии, -но со створки име-
ет лодочковидную форму с сильно суженными концами (рис. 34,2); табел-
лярия (Tabellaria, рис. 34, 4) — таблитчатые с пояска клетки.
2-й КЛАСС—ЦЕНТРИЧЕСКИЕ ДИАТОМОВЫЕ
(CENTRICAE)
Через створки можно провести не менее трех, у большинства — много
плоскостей симметрии; шва нет; половой процесс, по последним
данным, оогамный.
Представители этого класса: циклотелля (CycloteUa) — обычно одиноч-
ные, почти дисковидные клетки, со створки круглые, с пояска в виде удои-
74
НИЗШИЕ РАСТЕНИЯ
Рис. 34. Певватиые диатомовые:
1 — вавикуля; 2 — цимбелла; Л — синедра; 4 — табеллярия; S — астарнттляя
1
Рис. 35. Центрические диатомеи:
1 — циклотедля; а - вид со створки;
б — вид с поиска; а — хетицерос
отдал — БУРЫЙ ВОДОРОСЛИ 75
пенного прямоугольника; мелозира (Melosira) — нитчатые колонии из
клеток, более или менее бочонковидных с пояска и круглых со створки;
хетоцерос (Chaetocervs) — клетки короткоцилиндрические с пояска, эллип-
тические со створки, с длинными щетинками, соединенными в нитевидные
колонии (рис. 35).
Диатомовые водоросли широко распространены в бентосе и планктоне
морей и пресных водоемов (разные виды там и здесь). Нередко бентосными
видами обрастают подводные камни, ветки, образуя на них буроватые бах»
ромки. Некоторые виды живут эпифитами на других, более крупных водных
растениях. Нередко встречаются они в почве и на влажных скалах. Во влаж-
ных тропических странах есть диатомовые, селящиеся (вместе с сине-зеле-
ными) на листьях деревьев.
Бентоносных видов больше, но по количеству особей планктонные не-
измеримо превосходит их. В морях в океанах они являются одними из
главных продуцентов органических веществ.
Ископаемые диатомовые известны с юрского периода (около 110 —
150 млн. лет назад). Оболочки планктонных диатомовых, отлагавшиеся
после отмирания их на дне мелководных морских и больших пресноводных
бассейнов в третичном и четвертичном периодах, образовывали там нередко
мощные отложения, известные под названием диатомита и Трепела. Их
используют для тепловой и звуковой изоляции, как материал для шлифовки
металлов, как фильтрующее вещество.
Филогенетически диатомовые водоросли сближают обычно с хриэомо-
надовыми водорослями. Об этом говорят наличие жгутиковой стадии (спер-
матозоидов) и пульсирующих вакуолей у некоторых центрических диато-
мовых, сходство пигментов в обоих отделах, наличие у диатомовых масла .
и хризоэы как запасных продуктов, нахождение кремнезема у ряда хризо-
монад, образование цист. Центрические диатомовые более древние, из них
через вымершие промежуточные формы развились перистые.
ОТДЕЛ —БУРЫЕ ВОДОРОСЛИ (PHAEOPHYTA)
Бурые водоросли почти все живут в морях как донные, эпифитные и,
реже, вторично планктонные организмы. Как указывает само название,
прежде всего они характеризуются окраской в разные оттенки бурого цве-
та, от оливково-бурого.
Все бурые водоросли^щцгошеточные; ердди них нет ян оппптслаточиых,
ни колониальных. Размеры, их--варьируют от нескольких миулпметЬов до
несколькйх "метров, а у единичных видов.до.десятков.(60)метров. По внеш-
нему виду это ветвистые кустикиг корочкиг пластинки- шнурыт ленты' слотк-
нцрасчлененныё-' на стеблевидны§__ии_листовидные органы. Анатомическое
чдроопис У-мпотйх75шжнеё^.чем у зеленых.водо<юслей^41изп1иа.представи-
телц. отдела —ветвистые_однорядные, или. мшщорядные^нитц^ вышеорга-
низованные имеют тканевое строение вследствие деления клеток в разных
плоскостях; у высших форц^естьрасчленение ^а^специадизированные тка-
ни — ас£ймилЯЦИбйЙые^ запасающие, механические, проводящие. Обо-
лочка клеток снаружи ослизнённая, состоит из пектиновых веществ и внут-
реннего целлюлозного слоя. Слизь защищает от ударов волн и т. п., а у
водорослей, обнажаюшихся во время отлива, удерживает воду и защищает
от высыхания. В оболочках много камедеподобных веществ и альгиновой
кислоты (CgHgO#)^ образующей соли с металлами (альгинаты) и белково-
альгиновые комплексы. Цитоплазма содержит крупную вакуоль с клеточ-
ным соком щелочной (а не кислой, как бывает в большинстве случаев) реак-
ции. Кроме того, в плазме имеются мелкие вакуоли — ф и з о д ы, разме-
ром порядка 4 мкм, содержащие дубильные вещества; сокфизодов называ-
ется фукоэаном. Физоды наиболее характерны для меристематических*
ассимилирующих и репродуктивных клеток.
л
Ядро в клетке одно. Зерна хроматофора почти всегда в постенном, слое
цитоплазмы, большей частью многочисленные, их называют ф е о п л а с-
т а м и. На них обнаружены занимающие периферическое положение пире-
^поиды. Окраска феопластов зависит от ряда пигментов: хлорофиллов а и с,
каротинов, ксантофиллов и еще ряда бурых каротиноидов (ксантинов), в
частности фукоксантина, долгое время считавшегося специфичным лишь
для бурых водорослей. Эти пигменты, по современным представлениям,
играют существенную роль в фотосинтезе, способствуя утилизации главным
образом коротковолновой радиации видимой Части спектра.
Запасные питательные вещества не скапливаются вокруг пиреноидов,
переходя в раствор. Крахмала нет. В клетке накапливаются маннит (шести-
атомный спирт) и растворимые углеводы, в Частности полисахарид лами-
нарии, обычно присутствует масло. Из оболочек бурых водорослей экстра-
гируется углевод фукоидин, который входит в состав слизистой массы, за-
щищающей клетку от высыхания во время отлива.
По продолжительности жизни одни виды однолетние, другие многолет-
ние. Рост происходит у одних при помощи одной, реже нескольких верху-
шечных кдеток; у других — интеркалярно.
Вегетативное размножение может происходить случайно
оторванными участками таллома, лишь редко (например, у сфацеллярие-
вых) образуются специальные ветви вегетативного размножения. Беспо-
лое размножение имеется во всех группах, кроме фукусовых и других
порядков группы циклоспоровых^ У большинства оно осуществляется зоо-
спорами, развивающимися*“помногу в одноклеточных (иногда в много-
клеточных) зооспорангиях; зооспоры грушевидной формы; они имеют в
задней, более широкой, части один или несколько хроматофоров, а в перед-
ней — красно-бурый глазок и два отходящих сбоку неравных жгутика:
передний длинный, с перистыми ответвлениями, задний короткий. В поряд-
ке диктиотовых споры бесполого размножения, неподвижные (аплано-
споры), называемые тетраспорами, потому что они образуются по
четыре в каждом тетраспорангии.
Половое размножение изогамное, гетерогамное и оогамное.
Изо- и гетерогаметы образуются в многоклеточных гаметангиях, разделен-
ных поперечными и продольными перегородками; антеридии и оогонии
одноклеточные.
— Все бурые водоросли, за исключением циклоспоровых, имеют чередо-
I ванне поколений; на бесполом поколении (спорофите) в зооспорангиях (или
/ тетраспорангиях) после редукционного деления образуются зооспоры (или
’ тетраспоры); иэ них вырастают гаплоидные половые растения (гаметофиты),
\ раздельнополые или обоеполые. После оплодотворения зиготы без периода
' покоя развивается в новое бесполое диплоидное растение (спорофит). У цик-
лоспоровых вся жизнь водоросли проходит в диплоидной фазе и гапло-
идны лишь сперматозоиды и яйцеклетки, перед образованием которых про-
исходит редукционное деление.
К бурым водорослям относится около 1500 видов, входящих в состав
40 семейств, объединяемых в 11 порядков.
В зависимости от характера чередования поколений и смены ядерных
фаз бурые водоросли делятся на 3 группы порядков.
ГРУППА ПОРЯДКОВ — ИЗОГЕНЕРАТНЬГК fTSOGENERATAEJ ^
У большинства водорослей этой группы спорофит и гаметофит одинаковы
по форме и величине; у некоторых они различаются по форме. Господствует
гаметофит.
Широко распространены в морях представители большого рода экто-
корпус (Ectocarpus); они имеют вид маленьких кустиков или дерновинок, со-
стоящих из обильно ветвящихся однорядных нитей, оканчивающихся не-
редко бесцветными многоклеточными волосками. Одни нити стелются по
ОТДЕЛ —ВУРЫЕ ВОДОРОСЛИ
77
Рис 36. Эктокарпус:
J —общий вид; 9 — зооспорангий; J — зооспора; < —ветвь с гаметангиями
субстрату, другие поднимаются вертикально (рис. 36). Рост нитей-иятер-
кялярный. Сбоку на ветвях бесполых особей образуются одноклеточные
зооспорангии. В них происходит редукционное деление и еще ряд делений кле-
точного ядра и образуется много зооспор; они выходят в воду и прорастают
в новое, такое же по внешности, но гаплоидное растение — гаметофит. На нем,
на коротких боковых веточках, образуются многочисленные многоклеточные
гаметангии (рис. 36, 4). Гаметы одинаковы по величине и форме, но отлича-
ются поТёдением. Одни, которые можно считать женскими, скоро останавли-
1Г.1
ваются; .другие, которые можно считать мужскими, окружают женскую га-
мету; после некоторого времени, когда они еще продолжают движение, одна
мужская гамета сливается с женской. Зигота без периода покоя прорастает
в новую диплоидную нить с зооспорангиЯШУ ОшишШьхй цикл развития мо-
жет усложняться.' Так йНОТДана спорофите, кроме обычных одноклеточных
зооспорангиев, образуются еще многоклеточные (многокамерные) и в них
зооспоры без редукционного деления. Такие диплоидные зооспоры дают
начало новому поколению спорофитов. Иногда и гаметы ведут , себя как зо-
I
ОТДЕЛ — БУРЫЕ ВОДОРОСЛИ
79
оспоры — из гаметы, минуя половой процесс, может развиваться новая га-
плоидная особь. Наконец, известно, что у однопг-из видов эктокарпуса
(Ectocarpus confervoides) с гетероморфной сменой поколений, нормальные
(гаплоидные) зооспоры, не развиваясь в половые особи, копулируют, т. е.
ведут себя как гаметы.
Весьма интересна кутлерия (Cutleria), широко распространенная у евро-
пейских берегов Атлантического океана и в Средиземном море. Это расте-
ние имеет вид дихотомически разветвленных многослойных лент от 20 см
длины, оканчивающихся волосками. У основания волосков на одних экзем-
плярах развиваются крупноклеточные макрогаметангии, дающие двужгу-
тиковые макрогаметы; на других экземплярах — мелкоклеточные микро-
гаметангии, содержащие микрогаметы. После оплодотворения зигота без
периода покоя развивается в новое диплоидное растение (спорофит), имеющее
форму многослойной пластинки или корочки, плотно прижатой к субстра-
ту. Она настолько не похожа на предыдущее, что была описана под особым
родовым названием аглаозонии (Aglaozonia). На верхней стороне ее развива-
ются одноклеточные зооспорангии, в которых после редукционного деления
формируются гаплоидные зооспоры, из последних снова развивается гап-
лоидная нитевидная кутлерия (гаметофит).
Диктиота (Dictyota) и близкий к ней дилофус (Di tophus), растущие и у
нас в Черном море на небольших глубинах, имеют изоморфное чередова-
ние спорофита и гаметофита, одинаковых по внешности лентовцдных, ди-
хотомически разветвленных, имеющих вид плоских кустиков длиной 10—
20 см у диктиоты (рис. 37, 7). На спорофите из поверхностных клеток раз-
виваются крупные шаровидные тетраспорангии, в которых после редук-
ционного деления образуются 4 гаплоидные неподвижные (безжгутиковые)
голые тетраспоры. Из них развиваются такие же по форме и величине, но
уже гаплоидные мужские и женские растения (гаметофиты). На первых из
поверхностных клеток образуются группами (сорусами) многоклеточные анте-
ридии: из каждого соруса формируется около 150—300 тыс. сперматозои-
дов. На женских растениях образуются сорусы одноклеточных оогониев,
содержащие в каждом оогонии по одной яйцеклетке, при созревании вы-
падающей из него. После оплодотворения зигота развивается в новый спо-
рофит. Такой же цикл развития имеет падина (Padina pavonia), развиваю-
щаяся у нас в Черном море во второй половине лета в виде изящных вееро-
образных желтовато-серых пластинок, суженных к основанию; гаметофит
ее однодомен (рис. 37, 2). 1 ‘
ГРУППА ПОРЯДКОВ — ГЕТЕРО ГЕНЕРАТНЫЕ (HETEROGENERATAE)
В цикле развития водорослей этого класса чередуются макроскопи-
ческий (у многих мощно развитый) спорофит и микроскопический мелкдй
нитевидный гаметофит, нередко называемый по аналогии с папоротниками
заро&тком. Пол о «о й п р о ц е * с—ияогямтттли или оогамный.
Высшие--представители этой группы относягся^тг^чиц^ря д к у лами-
нариевых (Laminariales). Почти у всех родов (кроме одного)таЗгяем^разделен
на базальную_часть (пластинку или когтевидные выросты, служащие для
прикрепления), цилиндрическую стеблевидную часть (или «черешок») и
одну или несколько крупных листоподобных'пластинок (рис. 38). На по-
перечных разрезах стеблевидной (черешковидной), и листовидной, частей
МОЖНО видеть три зоны: периферическую "-ясстгуиляштлппут-дя кЛеТОК,
богатых хроматофорами, под ней крупноклеточную! В центре так
называемую сердцевину, образованную рыхлым сплетением нитей, идущих
продольно. Для~многих ламинариевых характерны ситовидные трубци;
они имеют густое содержимое, а поперечные перегородки их продырявлены
многочисленными сквозными отверстиями. Предполагавшаяся проводящая
функция их подтверждена методом меченых атомов. На границе между стебле-
видной и листоподобной частями таллома находится меристема, 8а счет де-
Рис. 38. Ламинариевые:
2 — Laminaria eaccbarina; а — L. digitate (смена листа); Л — Macrocystls; 4 — Lessonia
Рис. 39. Цикл развития ламинарии:
Л— спорофит: 2—часть соруса^зооспоранги» (ап—зооспорангии, п — парафизы, сл — слизь); в— зоо-
окры (общий вид) и прорастание; 4—мужской гаметофит: S — женский гаметофит с оогонием и на-
чиважицими разбиваться спорофитами; <—молодой спорофит
отдал—БУРЫЕ ВОДОРОСЛИ
st
пи
ления клеток которой происходит нарастание обеих^Чрстей. У многолетних
вадов листовидная часть ежегодно сбрасывается (в северных широтах —
в январе — феврале) и за счет меристемы нарастает новая пластинка.
Из периферических клеток — «листьев»,— перед сбрасыванием их, обра-
зуются группы (сорусы) одноклеточных зооспорангиев. В каждом зооспо-
рангии ядро редукционно делится, и дальнейшими делениями число ядер
увеличивается до 16—64; соответственно плазма разделяется на такое же
число одноядерных участков, которые, приобретая жгутики, становятся
зооспорами. На одном экземпляре водоросли образуются миллиарды зооспор.
После нвдолгого периода движения зооспоры прорастают в микроскопически
. мелкие нитевидные мужские (более ветвистые) и женские гаметофиты (за-
ростки). Мужские заростки (рис. 39) образуют на некоторых клетках вы-
росты, развивающиеся в антеридии, в каждом из которых формируется по
одному сперматозоиду. В оогоний может превратиться любая клетка женского
заростка. Ее содержимое через отверстие в верхней части оболочки выхо-
дит наружу и сидит на оогонии в виде голой яйцеклетки. После оплодотво-
рения из зиготы развивается постепенно новый мощный спорофит.
В наших северных и дальневосточных морях на глубинах 2—40 м и бо-
лее растут виды ламинарий (Laminaria) с цельными или пальчато-рассе-
ченными листовидными пластинками, достигающими 1—5 м и более длины
и несколько десятков сантиметров ширины, на черешках 5—30 см длины.
Оригинальную форму имеет ряд тихоокеанских родов. Пальмовидная постель-
сия (Postelsia palmaefornus) на вершине стебля около 60 см рдяны несет кро-
ну ланцетовидно-изогнутых «листьев». Лессония (Lessonia) имеет ствол до
4 ж высоты с многочисленными разветвлениями на вершине, оканчивающими-
ся ланцетовидными «листьями»А^ак^рщшви^с (Macrocystis), растущая на глу-
бине 10—20 ле, имеет тонкие, гибкие длинные стебли, на которых с одной
стороны сидят листья, снабженные каждый у основания пузырем, наполнен-
ным воздухом. Эти стебли и листья, плавающие у поверхности воды, при
скоплении макроцистов давали основание прежним мореплавателям при-
писывать этим водорослям 200 и даже 400 м длины1.
г
г. г
. 9i гл ’
ГРУППА ПОРЯДКОВ — ЦИКЛОСПОРОВЫЕ (CYCLOSPOREAE)
Сюда относятся бурые водоросли,, не имеющие чередования поколений,
а лишь смену ядерных фаз: вся водоросль диплоидна, гаплоидны лишь га-
меты. Бесполого размножения нет. Эта систематическая единица содержит
всего один порядок — фукусовые (Fucales).
Кожистые, оливково- или желтовато-бурые, дихотомически разветвлен-
ные ремневидные талломы фукусов (рис. 40) достигают 0,5—1,0 м длины,
1—5 см ширины; у некоторых имеются вздутия, наполненные воздухом.
Растут при помощь одной верхушечной клетки. Ко времени полового размно-
жения на концах ветвей образуются бобовидные желтоватые вздутия, в стен-
ках которых находятся округлые полости, скафидии, сообщающиеся с на-
ружной средой только узким отверстием (рис. 41, 1—2). В скафидиях среди
многоклеточных волосков развиваются оогонии и анте'ридии — или в одном
скафидии, или в разных, или на разных экземплярах (двудомность). После
редукционного деления в оогониях образуется 8 яйцеклеток, в антеридиях —
64 двужгутиковых Сперматозоида (рис. 41, У). Развивающаяся слизь вытал-
кивает оогонии и антеридии из скафидия наружу (рис. 41, 4); здесь иэ них
освобождаются половые клетки. Сперматозоиды окружают яйцеклетку, каса-
ются ее своими жгутиками и приводят во вращательное движение (масса яй-
цеклетки в 30 000 pas'больше массы сперматозоида); затем один спермато-
зоид внедряется в нее (рис. 41, 5). Оплодотворенная яйцеклетка сейчас же
одевается оболочкой и без периода покоя прорастает в новое растение.
кг,
ш 200—270 ж, другие, ссылаясь на специальные измерения, указывают всего 60 м*
НИЗШИЕ РАСТЕНИЯ
В Черном море в прибрежной
полосе широко распространена
самая крупная черноморская
водоросль — цистозира (Cyst-
oseira barbata), имеющая вид
сильноразветвленных кустиков
до 1 ле высоты. В оогониях ее
по одной яйцеклетке, так как
7 из 8 ядер, образующихся
после мейоэа, отмирают. По
одной яйцеклетке в оогониях
имеют и виды саргасымя (Sar-
gassum)—большого рода (250 ви-
дов), распространенного глав-
ным образом в теплых морях
и океанах. Расчленение саргас-
сумов напоминают высшие ра-
стения : на ци л инд рических
«стеблях» сидят «листья» и на
особых веточках — плаватель-
ные пузырьки, напоминающие
по внешности ягоды. В Атлан-
тическом океане между 25—35°
с. ш. и 38—60° в. д.,в области
затишья между Гольфстримом
и Северным Экваториальным
течением, находится Саргассово
море, в котором на площади
около 4,4 млн. клеа живет глав-
ным образом два вида саргассу-
мов, общая масса которых со-
ставляет около 20 млн. т.
Первичная родина этих во-
дорослей находится не здесь, а,
вероятно, где-то у берегов ближайших материков.
Почти все бурые водоросли, за единичными исключениями, распростра-
нены только в морях и океанах1, преимущественно холодных. Крупные ла-
минариевые образуют на больших площадях подводные леса:
Многие крупные виды водорослей широко используются на корм скоту
и на удобрение полей (богаты калием). Некоторые виды ламинарий (так
называемая морская капуста), алария (Alaria esculenta) и другие использу-
ются в пищу. Кроме пищевого, они имеют и диетическое значение, так как
богаты йодом. Количество этого элемента в морской воде составляет деся-
тые доли процента, в теле же ламинарий концентрация йода достигает 0,3%,
т. е. повышается примерно в 1000 раз. Это делает возможным добывание
йода из морских водорослей в промышленных масштабах. В значительных ко-
личествах ламинариевые используются для получения альгина, который об-
ладает высокой клеющей способностью и применяется в промышленности.
Достоверные остатки ископаемых бурых водорослей известны не ра-
нее триасового периода (около 185 млн. лет назад). Вероятнее всего родство
бурых водорослей с хризомонадовыми, у которых также имеется пигмент
фукоксантин и встречаются ветвистые формы, как, например, феотамнидн
(Phaeothamnion).
Три группы порядков Phaeophyta соответствуют трем ступеням эволю-
ционного развития онтогенеза этих растений: от изоморфной смены поко-
1 Известны только три рода: Pleurocladia, Zithoderma и Bodenella. Все они из груп-
пы Isogeneratae, ящца которых (и то не исключительно) обитают в пресных водах.
ОТДЕЛ — БУРЫЕ ВОДОРОСЛИ
83
Рис. 41. Фукус:
™ ч?рва скь*ИДЙЙ с оогониями; 2 — разрез через скафидий о антеридиями; з — неточна с анте-
ридиями. « — выход сперматозоидов из антеридия; з — яйцеклетка вскоре после оплодотворения (я —
ядро яйцеклетки, ял — ядро сперматозоида, я — хроматофоры) "чрс *
м
ленвй у изогенератных ж преобладанию в онтогенезе диплоидного спорофкта
у гетерогенератвых, у которых гаметофит сведен к микроскопически мало-
му здростку, и, наконец, к полной поте1№ гаметофита у циклоспоровых.
Эволюция жизненного цикла у бурых водорослей осуществлялась, вероятно,
более чем в одной линии эволюции. В частности, можно, как это и делали
некоторые ботаники, полагать, что от общего предка бурых водорослей до-
статочно рано возникли две эволюционные линии. В одной нарастание ве-
гетативного тела осуществлялось апикально, в другой — интеркалярно,
ж в каждой совершенствование онтогенеза совершалось путем перехода от
изоморфной смены поколении к гетероморфной с преобладанием спорофита
и, наконец, к полной морфологической редукции гаметофита.
Существует и получила широкое распространение гипотеза происхожде-
х растений от бурых водорослей. Предполагается, что по выходе
бурых водорослей на сушу периферические клетки их многоклеточных га-
метангиев стерилизовались, чем была обеспечена защита клеток, произво-
дящих гаметы, от иссушения, и гаметангии могли превратиться таким обра-
111'
зом в многоклеточные антеридии и архегонии высших наземных споровых
растений. Одним из затруднений для принятия этой гипотезы является рае»
личие в пигментах и в запасных веществах у бурых водорослей и высших
растений. В настоящее время чаще придерживаются мнения о происхожде-
l высших растений от земных водорослей.
it.
ОТДЕЛ—КРАСНЫЕ ВОДОРОСЛН, ИЛИ БАГРЯНКИ
(RHODOPHYTA)
Почти все красные водоросли многоклеточны. Это главным образом
простые или ветвящиеся нити, разветвленные кустики, пластинки, иногда они
бывают расчленены на стебель и листовидные органы. Красные водоросли
не бывают столь крупными, как бурые, лишь единичные виды достигают 2 м.
Клетки красных водорослей покрыты оболочкой, в которой различимы
внутренний, целлюлозный и наружный, пектиновый ослизняющийся слой.
Получаемый из последнего студень (агар-агар) содержит, кроме пектина,
сахара и белки. Оболочка может быть инкрустирована СаСО8 и (в меньшем
количестве) MgCO8 во всю толщу, исключая только самую внутр©
часть, непосредственно прилегающую к плазме. Инкрустация иногда быва-
ет настолько полной, что водоросль становится как бы окаменевшей. Цито-
плазма отличается повышенной вязкостью и трудно отделима от оболочки;
она располагается постенным слоем, окружая центральную вакуоль. Ядро,
как правило, одно, но в старых клетках бывает и несколько ядер. Хромато-
• i-rc
тшстый бед пиреноидов. Зерна хроматофора, за их обычно красный цвет, на-
зывают родопластами*. Как и у других растений, окраска пластид и всего
тела красной водоросли обусловлена сочетанием нескольких пигментов:
хлорофиллов а и d (причем последний известен только в этом отделе расти-
тельного мира) каротина, ксантофилла, в том числе лютеина, содержащего-
ся также в клетках зеленых и некоторых других водорослей и у высших
растений. Особую группу пигментов представляют фикобилины, названные
так за химическое сходство с пигментами желчи* 8. Фикобилины — хромо-
протеиды, т. е. белковые соединения, имеющие в своем составе пигментную
группу. Из фикобилинов для красных водорослей особенно характерен крас-
ный фикоэритрин, а у некоторых в небольшом количестве есть и синий —
фико циан. Фикобилины передают поглощенную ими световую энергию хло-
рофиллу а. Доля энергии, поглощенной фикобилинами и используемой в-
процессе синтеза органического вещества, может превышать, как показали
1 От греч. «родос» — красный
8 От лат. «bilis» — желчь. 4
отдел—КРАСНЫЕ ВОДОРОСЛИ
85
__ *
Рис. 42. Споронотения краевых водорослей!
Д—б—тетрагаторянгии (разные типы); 4-* цистодаранн (в разрезе)! «—* наллитамтгон. б — шшненфо-
нви
некоторые исследования, энергию, поглощенную непосредственно хлоро-
филлом.
Относительные количества перечисленных элементов пигментного ком-
плекса определяют реальную окраску красной водоросли — от светло-
розовой до красной и темно-фиолетово-красной, иногда — желтоватую,
оливково- иди синевато-зеленую (стальную).
Запасные питательные вещества —-маслои особый полисахарид, более
близкий к гликогену, чем к крахмалу, называемый багрянкодым
Он откладывается вне связи с хроматофором, в цитоплазме; окрашивается
йодом в винно-красный цвет.
У подавляющего большинства красных водорослей в перегородках,
отделяющих генетически связанные клетки, находится по одной крупной
поре. Такая пора представляет неутолщенное место клеточкой оболочки.
Ее первичная перепонка пронизана множеством мельчайших, канальцев, за-
полненных протоплазмой (плазмодесмы). По порам устанавливается гене-
тическая связь между клетками. Оказалось, что самые разнообразные ва-
рианты анатомического строения красных^ водорослей почти всегда могут
быть сведены к системе ветвящихся нитей. Однородная нитчатая структура
в неизменном виде также встречается у некоторых красных водорослей (на-
пример, у Calliihamnton^ см. рис. 45, 2). Более сложные структуры характе-
ризуются наличием центральной оси, состоящей из одной нити или из пучка,
образованного несколькими параллельными нитями с верхушечным нараста-
нием, и отходящих от них ветвей, располагающихся более или менее
мутовчато. Боковые ветви ограниченно нарастают, но обильно ветвят-
ся. Конечные разветвления боковых ветвей одной и той же и соседних
мутовок могут тесно смыкаться, переплетаться и даже склеиваться друг с
другом цементирующим веществом, выделяемым самой водорослью. Таким
i ' 7 7 » / • . . л-f ♦ / / -. *
1
86
НИЗШИЕ РАСТЕНИЯ
образом формируется кора, у одних достаточно отдаленная от центральной
нити или пучка центральных нитей, у других — приближенная к ней.
Наружные клетки коры более мелкие, богаты родопластами и составляют ас-
симилирующую ткань. Таким образом паренхимное строение тела красной
водоросли в самом деле оказывается ложнопаренхимным. Только у наиболее
низкоорганизованных красных водорослей при паренхимном строении нет
ни плазмодесм, ни пор, а нарастание таллома у них почти исключительно
диффузное, не верхушечное.
Вегетативное размножение для красных водорослей
нетипично. Оторванные участки тела большей частью погибают, но иногда
наблюдается образование выводковых почек, которые представляют видо-
измененные вегетативные ветви, отламывающиеся от материнского тела и
. вырастающие в новые особи (например, у Melobesia из порядка Cryptone-
\miales).
В размножении красных водорослей спорами и в половом процессе,
достаточно разнообразном, имеются общие черты. Из них в первую очередь
надо отметить отсутствие жгутиковых клеток. Ди споры, ни гаметы квасных
водорослей никогда не образуют жгутиков. Перемещение их осуществляет-
ся главным ибрааом пассивно, водой. Б некоторых случаях споры способны
к амебоидному движению.
Споры красных водорослей возникают внутри споропроизводяшей клет-
ки — спорангия — по одной (моноспоры), по четыре (тетраспоры; или по
нескольку (полиспоры). В тетраспорангиях споры могут располагаться по
углам тетраэдра в один ряд или по две клетки в два ряда. Спорангии разме-
щаются на растении одиночно или группами, иногда на особых ветвях
\ (рис. 42).
пн
Половое воспроизведение красных водорослей всегда
оогамное Мужские половые органы -^.антеридии — развиваются из
конечных клеток нитей или из поверхностных коровых клеток. Антеридий
одноклеточен, его содержимое, покрытое тончайшей оболочкой, разрывая
клеточную стенку, выходит наружу. Это—мужская гамета, называемая
ен жгутиков, содержит крупное ядро и немного
цитоплазмы." Жёйёкий половой орган красных водорослей называют и а р-
п о г о ц о м. Он развивается из конечной клетки короткой, 3—4-клеточной,
«каршцонной» нити, нередко погруженной в окружающую ее ткань. У более
примитивных красных водорослей карпогоном может стать обычная—клетка
таллома. Типично развитый карпогон имеет нижнюю расширенную (брюш-
ную) часть и верхнюю нитевидновытянутую — трихогину. Содержи-
мое брюшной части состоит из пл дамы, хроматофора и ядра и соответствует
яйцеклетке. Трихогина молодого карпогона имеет ядро (за исключением
некоторых относительно примитивных форм), ко,времени рплодотворения
дегенерирующее. У примитивных форм трихогины может и не быть.
Спермаций, пассивно переносимый водой, встречаясь с трихогинои. при-
кипает к ней, и его содержимое переливается внутрь карпогона через.отвер-
/ стиву-возникающее в оболочке трихогины. Спермаций достигает брюшной
I части карпогонаГ^гдГ и осуществляется Оплодотворение. Последующие фазы
1 цикла развития развертываются без периода покоя ян гот.ы; в некоторых слу-
I чаях развитие может быть непосредственно из зиготы; обычно же более слож-
; ным и длительным путем смены поколений возникают, в конечном счете, *
1 гаплоидные споры, из которых вырастает новая половоспособная особь,
чем и завершается цикл развития водоросли.
Циклы развития красных водорослей разнообразны и изучены _йце не-
достаточно. На основании имеющихся данных можно считать основной,
т. е. наиболее распространенной, следующую схему: гацлоищ^ш_гамето-
фит ряявинрет половые органы и гаметы; после оплодотворения из дипло-
идной зиготы (точнее — из карпогона) формируется второе поколение —
карпоспорофит, отличный по строению от гаметофита, живущий
на последнем, питающийся за его счет. Будучи диплоидным, карпоспорофит
ОТДЕЛ — КРАСНЫЕ ВОДОРОСЛИ
87
производит диплоидные же
споры — карпоспоры.
Карпоспора, прорастая, дает
начало второму диплоидному
(но третьему в общем ходе он-
тогенеза) поколению - т е т р а'
спорофиту, производя-
щему гаплоидные споры —
тетраспор ы^ они возни-
кают в результате редукцион-
ного деления, по четыре в каж-
дом тетраспорангии. Следова-
тельно, полный цикл развития
осуществляется тремя последо-
вательно сменяющими друг
друга поколениями, из кото-
рых одно (гаметофит) гаплоидно
и производит гаметы, а два
другие (карпоспорофит и тетра-
спорофит) диплоидны и произво-
дят споры (соответственно: ди-
плоидные карпоспоры и гап-
лоидные тетраспоры).
Эта схема цикла развития
1 ©2
Рис. 43. Порфира:
J —внешний вид таллома: Я — сперматангии (антери-
дии) и освобождение спермациев; з — разрез через тал-
лом с оплодотворенными карпогониями
реализуется у красных водоро-
слей в нескольких видоизмене-
ниях. Прежде всего надо заметить, что у некоторых, в первую очередь у
более низкоорганизованных (класс Bangiophyceae) красных, водорос-
лей карпоспорофит не образуется и зигота, * делясь, непосредственно да-
ет начало карпоспорсГ^^йтоТ^вариант^цикла развития имеет место, на-
пример, вроде Porphyra (рис. 43). Виды этого рода обитают в морях.
Взрослое растение представляет плоскую однослойную, прикрепленную
к субстрату листовидную пластину, достигающую в длину 50 см (редко бо-
лее) и имеющую овальное очертание. Половые органы образуются из веге-
тативных клеток таллома. В антеридии путем деления протопласта форми-
руются спермации, которые в дальнейшем освобождаются. Карпогонии отли-
чаются от вегетативных клеток лишь небольшим выростом, соответствующим,
вероятно, трихогине, и более темным содержимым. После оплодотво-
рения зигота делится на некоторое число карпоспор. Они освобождаются
в виде голых, не покрытых оболочками клеток, способных к слабому амебо-
идному движению. Лишь через три дня карпоспора выделяет оболочку и
прорастает в нитевидное тело, которое эндолитически внедряется в известко-
вые раковины моллюсок. Эту фазу цикла развития описывали ранее как
особый вид Conchocelis rosea, В этом состоянии водоросль образует моноспо-
ры, которые, в свою очередь, вырастают в новую порфиру,
В цикле развития порфиры наблюдается отсутствие карпоспорофита и
наличие по крайней мере двух поколении (Porphyra и Conchocelis). Одна-
ко цитологические отношения остаются не вполне ясными. Если подтвер-
дится мнение, что поколение, представленное Conchocelis, и карпоспоры
диплоидны,, т. е. при делении зиготы мейоза не бывает, то весь цикл развития
Porphyra можно будет рассматривать как состоящий из двух поколений:
гаметофита (гаплоидной порфиры) и тетраспорофита (диплоидной Conchocelis).
Время и место редукционного деления нуждаются в уточнении.
В более усложненной форме протекает цикл развития у представите-
лей относительно примитивного порядка немалионовых из класса Flo-
rideophyceae. Распространенная в пресных водах водоросль батлахоспер-
мум (Batrachospermum moniliforme, рис. 44) представляет собой ветвящийся,
слизистый на ощупь кустик, 10—20 см длины, оливково-зеленого или си-
11J11
невато-зеленого цвета, каждая ветвь которого имеет в основе однорядную
клеточную нить с отходящими от нее более или менее мутовчатыми боковыми
ответвлениями — ассимиляторами. Группы последних образуют как бы
бусы, нанизанные на нитку. Вдоль старых осевых клеток от мутовок отхо-
дят продольные нити, образующие как бы зачаточную кору. На ассимиля-
торах образуются карпогоны и антеридии. После оплодотворения ядро зи-
готы делится митотически1. Получившиеся таким образом диплоидные ядра
входят вместе с плазмой в выросты брюшка карпогона, которые, несколько
* вырастая в длину, становятся ветвящимися однорядными клеточными ни-
тями. Совокупность этих нитей представляет маленькое растеньице —
новое, уже дицлоддрое поколение цикла развития, живущее на гаметофите,
т. е. кдлно^Ь?о$ихЗ Его периферические клетки становятся. спорангиями,
производящими каждый по одной диплоидной споре — Щфпбспоре. По-
следняя к дает начало * третьему4 поколению батрахоспермУЙГ^ особи,
имеющей вид неправильно ветвящихся нитей ,’ЧасЮТйо'-стелющихся по
субстрату, частично поднимающихся над ним. Эту фазу развития первона-
чально считали особой, относя ее к роду Chantrangia, затем сталипри-
давать ей значение предростка, подобного протонеме мха. В настоящее вре-
мя шантрансию, как образование диплоидное, считшт1г:хехра^щ£^д0ви
В отдельных клетках его совершается редукционной’деленйе, но др обра?
зования тетраспор процесс не доходит, вследствие чего мейоз долгое время
не был замечаем. Из гаплоидных клетой, возникающих в итоге Фор-
мируется новый гамеЮфит, остающийся' в связи Ъ^шантрансией —’тетрас-
порофитом. ч ’
,11
11 if.
1 Долгое время существовало мнение о редукционном деления ядра зиготы, что но-
вейшими исследованиями не подтвердилось.
отдал—КРАСНЫЕ ВОДОРОСЛИ
89
Аналогично протекает цикл развития у видов /рода Lemanea. Эти
растения — жители чистых пресных вод, относит о холодных и быстро
зиготы
рпоспорофит, образуемые
небольшие тетраспоро-
е деление в атомвдучае про-
г. г.
текущих. У них, так же как у Batrachospermumfwpb прорастая
редукционное деление не происходит и поэтому
им карпоспоры и, наконец, развивающиеся из
фиты типа шантрансии диплоидны. Редукцио
исходит в верхушечной клетке тетраспорофита. fVCd&GCUZ
У водорослей порядка немалионовых иногда тетраспорофит может быть
очень небольшим, как у Liagora farinosa, где он* был открыт только в куль-
туре. ,С этим интересно сопоставить, что у другого вида того же рода —Lt
tetrasporifera — тетраспорофит не развивается вовсе и редукционное деление
совершается в клетках карпоспорофита, становящихся тетраспорангиями.
Деление ядра зиготы без редукции хромосом свойственно вообще крас-
ным водорослям из класса Feorideophyceae^ но ход их онтогенеза бы-
вает обычно более сложным, чем у описанных выше представителей нема-
л ио но вых.
Одним из усложняющих обстоятельств надо считать то, что в противог
положность описанным выше случаям, нити карпоспорофита, называемые
ообластемными или с п о р о г е н н ы м и, растут по направлению
к так называемым ауксилярным1 клеткам, сливаются с ними
и на местах слияния формируются кучки спор — цистокарпии. Ауксйляр-
ные клетки могут помещаться как вблизи карпогона, так и в отдалении от
него. Такого рода отношения можно пояснить конкретным примером во-
дорослей *дюртш и пмтомы (Dudremaya coccinea и Platoma из порядка
криптонемиевых, рис. 45,
Дюренея образует небольшие кустики, ветви которых имеют осевую нить,
несущую так называемые ассимиляторы — боковые ветви, из которых обра-
зуется кора. Иэ оплодотворенного карпогона дюренеи возникает несколько'
ообластемных нитей. Каждая из них сливается с одной (или более) клеткой
карпогонной ветви, это — нетипичные ауксилярные клетки, имеющие зна-
чение только для питания карпоспорофита. Слияние с ауксилярной клеткой
карпогонной ветви не прекращает роста ообластемной нити; она направля-
ется к ауксилярным клеткам других ветвей таллома, выделяющимися гу-
стой плазмой и редуцированным хроматофором (типичные ауксилярные клет-
ки), и, слившись с одной такой клеткой, направляется далее к новым аук-
силярным клеткам. Этот процесс может повториться несколько раз. Слияние
клеток ообластемной нити с ауксилярными клетками состоит в соединен
k их плазм, ядра же сохраняют свою самостоятельность. Надо подчеркнуть,
что ядро ауксилярной клетки, как принадлежащей половому растению, га-
плоидно; ядра же клеток ообластемных нитей, возникших из оплодотво-
ренного карпогона, диплоидны.
Против места слияния с ауксилярной клеткой клетка ообластемной ни-
ти образует выпячивание, куда и переходит ее ядро. Эта выпуклость от-
деляется перегородкой и становится основой для образования ц и с т о-
иг.
Следовательно, у дюренеи на основе оплодотворенного карпогона фор-
' мируется диплоидшй карпоспорофит в виде совокупности диплоидных
ообластемных нитей и цистокарпиев. Отличия же от хода развития немалио-
новых состоят в том, что во-первых, карпоспорофит у дюренеи имеет воз-
можность использовать дополнительное питание от ауксилярных клеток и,
во-вторых, вследствие однократного оплодотворения формируется не один,
а много (по числу ообластемных нитей и слияний их с ауксилярными клет-
ками) цистокарпиев.
У растений, онтогенез которых протекает по типу дюренеи, из дипло-
идных карпоспор развиваются диплоидные же рутения, представляю
третье поколение цикла развития — тетраспорофи|. На нем формируются
х0т лат, «ауксилиарисг-^вспомогательвыйг---
lit:
90
НИЗШИЕ РАСТЕНИЯ
Рис. 45. Цикл развития; половой процесс и развитие карпоспор у красных водорослей:
1 — немалион: а — антеридия и карпогон (ан — антеридии, бк — брюшная часть карпогона* т-трихо-
гина. сп — спермапий), б — в — слияние мужского и женского ядер в брюшной части карпогона, я — О —
развитие карпоспор (кс — карпоспора); 8 — развитие ообластемных нитей у дюренеи (т — трихогина.
ся — спермации. о — ообластемные нити, at — а« — аукснлярные клетки); 8 — развитие ообластеиных
нитей и последовательность образования цистокарпиев у платимы (к — карпогон, о — ообластемная нить»
а— аунсилярные клетки, ц — цистокарпии); 4— цикл развития красных водорослей
ОТДЕЛ — КРАСНЫЕ ВОДОРОСЛИ
91
тетраспорангии и в них — тетраспоры, при этом осуществляется мейоз.
Будучи гаплоидными, тетраспоры развиваются в новые гаплоидные (и вместе
с тем половые) растения.
В подобных случаях, следовательно, полный цикл развития слагается
из трех поколений: гаплоидного гаметофита, диплоидного карпоспорофита
и диплоидного тетраспорофита.
Разнообразие циклов развития красных водорослей этим не исчерпы-
вается. Например, в порядке Gigartinales в слоевище образуется одна
ауксилярная клетка, которая связывается с карпогоном очень короткой
ообластемной нитью. В порядке церамиевых (Ceramiales) ауксилярные клет-
ки образуются лишь после оплодотворения, а клетки, из которых они фор-
мируются (так называемые материнские клетки ауксилярных клеток),
вместе с четырехклеточной карпогонной' ветвью и карпогоном составляют
характерный комплекс, называемый прокарпием.
Из совокупности сведений о ходе цикла развития красных водорослей
можно сделать некоторые обобщающие выводы. Карпоспорофит развивается
не у всех представителей этой группы. Он не известен в классе Вап-
giophyceae. Это как бы промежуточное между гаметофитом и спорофитом
поколение может быть достаточно крупным, как у Dudresnaya, или скромно
развитым, как у Batrachospermum* или, наконец, сильно редуцированным,
как у церамиевых. Иногда, при полной редукции тетраспорофита (Liagora
tetrasportfera), карпоспорофит может производить тетраспоры.
Что касается гаметофита и тетраспорофита, то в большинстве случаев
они бывают сходно построены и не отличаются друг от друга по величине.
В этом случае при достаточно глубокой редукции карпоспорофита цикл раз-
вития в целом принимает форму, сходную с изоморфной сменой поколений.
Однако, как уже было упомянуто выше, иногда тетраспорофит бывает сильно
редуцирован и даже вовсе выпадает из цикла развития. Более редко редук-
ции подвергается гаметофит, сохраняющийся в виде крохотных, 2—3-кле-
точных, растеньиц (Rhodochorton из Nemalionales).
Красные водоросли почти исключительно жители морей. Приблизи-
тельно из 4000 относящихся сюда видов только около 50 живут в пресных.
водах и на земле.
В отделе различают два класса.
1-й КЛАСС — БАНГИЕВЫЕ (ВANGIOPHYCEAE)
В этот класс входит небольшая группа низкоорганизованных красных
водорослей. Встречаются одноклеточные, нитчатые и пластинчатые формы.
Хроматофор у большинства звездчатый. Поры между клетками встречаются
лишь как редкое исключение. Карпогон л
сосочками вме-
сто трихогины.
Типичный представитель — порфира (Porphyra* стр. 87).
2-й КЛАСС—ФЛОРИ ДЕИ (FLORIDEOPHYCEAE)
Этот класс включает всегда многоклеточные различного облика — от вет-
вящихся нитей до сложноустроенных, иногда внешне напоминающих выс-
шие растения. Рост верхушечный, между клетками имеются поры. Карпо-
гон с трихогиной. Цикл развития из трех, реже (вследствие редукции) двух
поколений. Гаметофит и тетраспорофит у большинства изоморфны.
В пресных водах с чистой, хорошо аэрированной водой нередко встре-
чается батрахоспермум (Batrachospermum, стр. 87—88).
В морях на скалах в области прибоя растет немалион (Nemalion) —
слизистые бледно-розовые шнуры 10—20 см длины и 2—5 мм толщины
(рис. 46, 5).
г< г.
92
низшие растения
Рис. 46. Красные водоросли:
1—каллитамнион (мд под микроскопом); е—гигартина; 4—нвиадшош 4—дитотамнисн; 5—дазжя;
< — допессерия
ОТДЕЛ — КРАСНЫЕ ВОДОРОСЛИ
93
Каллитамнион (Callttamnton corymbosum, рис. 46, 7), обычный в Чер-
ном море, образует изящные кустики ярко-розового цвета до 10 см высоты,
состоящие из однорядных, сильно разветвленных нитей.
У представителей большого рода полисифония (Polysiphonia) тал-
лом нитевидный, 3—20 см высоты, с многоклеточной ветвящейся осью. Цис-
токарпии окружен незамкнутым покровом, формирующимся еще до опло-
дотворения и имеющим отверстие наверху. *
У делессерий (Delesseria) на короткой стеблевидной части растения си-
дят листовидные розовые пластинки, у некоторых с перистым жилкованием
(рис. 46, фиг. 6). Эти своеобразные органы, представляющие замечательный
пример конвергенции, по данным онтогенеза могут быть сведены к системе
ветвистых сросшихся нитей.
В семействе кораллиновых (Corallinaceae) в оболочках клеток обиль-
но отлагаются углекислый кальций и углекислый магний. Кораллина (Со-
rallina) имеет вид ветвистых кустиков: в сочленениях веточек нет отложенид
солей, благодаря чему водоросль обладает некоторой гибкостью и поэтому мо^
жет расти в местах сильного прибоя. Некоторые виды родов литотам-
ниона (Lithothamnium рис. 46, 4) и литофиллюма (Lithophyllum) имеют фор-
му кораллоидных кустиков или как бы окаменелых клубней; они вместе с ко-
раллами участвуют в образовании коралловых рифов в тропических морях.
Все морские красные водоросли входят в состав бентоса. Большинство
видов встречается в теплых морях. Красные водоросли могут расти на до-
вольно значительной глубине, так как красный пигмент их участвует в фо-
тосинтезе, используя зеленые, голубые и синие лучи солнечного спектра,
проникающие в воду гораздо глубже красных.
Многие красные водоросли находят практическое применение. В стра-
нах Восточной Азии, на Гавайских и других островах, ряд видов употре-
бляют в пищу. Порфиру (Porphyra) даже разводят в Японии. Из видов
гелидиума (Gelidium), грацилярии (Gracilaria), а в СССР из анфельции(АНп/е1-
tia) получают агар-агар (см. стр. 84), широко применяемый для изготовле-
ния твердых питательных сред в микробиологической практике, а также в
пищевой, в особенности в кондитерской промышленности. Из филлофоры
(Phyllophfora), которая в Черном море занимает на глубине 30—60 м око-
ло 10 000 км*, добывали йод.
Происхождение красных водорослей неясно. Достоверные ископаемые
остатки их встречаются в отложениях мелового периода, менее достоверно
их существование в палеозойской эре (силур, девон). Полное отсутствие жгу-
тиковых стадий, высокоспециализированный половой процесс ставят крас-
ные водоросли в изолированное положение среди других водорослей. Сход-
ство пигментов с пигментами сине-зеленых водорослей, может быть, указы-
вает на каких-то очень отдаленных общих предков их, но совершенно иное
строение клетки и способов размножения у сине-зеленых не дают возмож-
ности установить между ними более близкие родственные связи.
i:r,
гл.
ins
III
ЭКОЛОГИЯ И РАСПРОСТРАНЕНИЕ ВОДОРОСЛЕЙ
* Распространение водорослей в глубину зависит от света и, следовательно,
прозрачности воды. Морская вода прозрачнее пресной, и поэтому массо-
вое распространение водорослей в морях наблюдается глубже — до глуби-
ны 30 ж; более теневыносливые заходят до глубины 100—200 ж, а отдельные
виды — даже до 500 м, Глубина, которой. достигают водоросли в пресных
водах, меньше.' Все же в Болотовском озере на глубине 5—6 м находили
вошерию. В Байкале, имеющем очень прозрачную воду, драпарнальдия и
эгагропила (кладофора) живут на глубине до 10—16 ж, а диатомовые —
да 35 ж. В морях наиболее глубоко задодят бурые и особенно красные во-
доросли. Объяснение этому следует искать в том, что уже на глубине 10 м
остается лишь 1—2% красных лучей солнечного спектра и около 4% си-
•л I
.in
94
НИЗШИЕ РАСТЕНИЯ
них и близких к нему по длине волн. Следовательно, на глубине фотосин-
тетическая деятельность хлорофилла, использующего главным образом
красные лучи, затруднена. Фикоэритрин — красный пигмент красных во-
дорослей — поглощает дополнительные к его цвету синие и голубые лу-
чи спектра, дальше проникающие в глубину, и передает поглощенную им
световую энергию хлорофиллу. Это до некоторой степени объясняет встре-
чаемость красных водорослей на больших глубинах, а также более интен-
сивную красную окраску у глубинных экземпляров одного и того же вида
по сравнению с поверхностными.
Температурные границы, в которых возможна жизнь водорослей,
очень
ин
роки. Многие водоросли, вмерзшие в лед, по оттаивании оживают.
В полярных и высокогорных странах нередко на отдельных участках встре-
чается красный снег: окраска его зависит от живущей в нем одноклеточной
вольвоксовой водоросли — снежной хламидомонады (Chlamydomonas ntvar
lis), окрашенной гематохромом; при таянии снега эта водоросль начинает
двигаться. Встречаются также участки снега, окрашенные другими, преи-
мущественно одноклеточными водорослями в зеленый, реже желтый или
буроватый цвет. Описано свыше 70 видов водорослей, живущих в снегу.
В высоких широтах на севере у бурых ламинариевых водорослей в январе
начинается прорастание из листовидных талломов при —0,65°С. Отмечали
вегетацию водорослей в морях и при —3,3°С.
В то же время живые сине-зеленые водоросли встречаются в горячих
источниках при +85°С и даже при 4-93°С. Интересно отметить, что боль-
шинство этих водорослей — те же виды, какие встречаются и при обычных,
относительно низких температурах.
Питание. Общая соленость в пресных водоемах очень невелика —
0,001—0,05%, в морях (кроме опресненных) и океанах в среднем 3,4—3,5%.
С уменьшением солености в морях происходит обеднение (по числу
видов) их бентоносной водорослевой флоры.
В континентальных водоемах соленость может доходить до 20% и выше.
Число видов водорослей в таких водоемах невелико — это главным обра-
зом одноклеточные сине-зеленые и вольвоксовые, но развиваются они в
огромных количествах, окрашивая воду и рапу.
Углерод водоросли берут из свободного углекислого газа, растворенного
в воде. При падении его содержания некоторое количество растворенных в
воде бикарбонатов расщепляется на карбонат, выпадающий в осадок, угле-
кислый газ и воду: Са(НСОз)2 — СаСОз + СО2 + Н2О; освобождаю-
щийся углекислый газ используется водными растениями. Наоборот, при
увеличении содержания СО2 в воде свыше пределов, необходимых для под-
держания равновесия с бикарбонатами, часть СО2 становится активной и
расщепляет карбонаты, переводя их в растворимые бикарбонаты.
Кроме автотрофного питания, некоторые водоросли, особенно
сине-зеленые, прококковые и другие, могут усваивать органические вещества,
питаясь теми и другими смешанно, или миксотрофно. Некоторые
водоросли даже утратили хлорофилл, живут в местах, богатых органичес-
кими соединениями, и целиком питаются гетеротрофно, как са-
профиты. Таковы, например, астазия (Astasia) из эвгленовых, одноклеточ-
ная политома (Polytoma) из вольвоксовых, прототека (Prototheca) из про-
тококковых, некоторые диатомовые и др.
Водоросли, питающиеся миксотрофно, могут служить показателями
степени загрязнения водоема органическими, особенно азотистыми, вещества-
ми. В пресных водоемах различают группы видов (не роды!) водорослей:
полисапробные, а-меэосапробные, р-м еэосапроб-
ные, олигосапробные. Две первые группы водорослей исполь-
зуют непосредственно органические соединения; третья группа характерна
для водоемов с малым содержанием органических соединений и использует
неорганические продукты их распада; водоросли четвертой группы живут
в водоемах с практически чистой водой.
ОТДЕЛ — КРАСНЫЕ ВОДОРОСЛИ
95
Sill
Некоторые зеленые и сине-зеленые водоросли живут в симбиозе
с другими растениями или низшими животными. Широко известный пример
такого симбиоза — лишайники. Хлорококковая зоохлорелла живет симбио-
тически с некоторыми инфузориями, гидрой, губками, червями. Некоторые
сине-зеленые водоросли живут внутриклеточно в обесцветившихся зеленых
водорослях, выполняя у них функции исчезнувших хроматофоров.
ЭКОЛОГИЧЕСКИЕ ГРУППИРОВКИ ВОДОРОСЛЕЙ
Распределение водорослей в природе в значительной степени зависит от
местных экологических условий, в связи с чем создаются характерные при-
родные экологические группировки, такие, как планктонные, бентоносные
и почвенные.
Фитопланктон. Планктоном1 называют совокупность микроско-
пических или очень мелких макроскопических организмов, взвешенных в
воде, не обладающих собственным движением или хотя и подвижных, но не
могущих сопротивляться движению воды. Планктонный образ жизни ве-
дут все водоросли, обладающие жгутиками, очень многие диатомовые и си-
не-зеленые, многие протококковые, некоторые десмидиевые. Размеры таких
растений обычно измеряются микронами, и только редко они достигают
1—2 жж, а масса их ничтожно мала, составляя в целом миллионные доли
миллиграмма. Кроме ничтожной абсолютной массы возможность жить во
взвешенном состоянии облегчается снижением относительной массы (замена
крахмала маслом, уменьшение окремнения оболочки у диатомовых, газовые
вакуоли у сине-зеленых), а также выработкой приспособлений, увеличи-
вающих трение о воду (длинные иглообразные отростки у церациума и
голенкинии, лентовидная, как у фрагиллярии, или парашютовидная, как
у педиаструма, форма тела) иногда — слизистой оберткой, выделяемой во-
дорослью и т. д.
В морях с прозрачной водой обильное развитие планктона наблюдается
не глубже 100 ж, а в пресных водоемах — не глубже 10 ж. У самой поверх-
ности, где весь азот и фосфор часто бывают израсходованы, фитопланктон
нередко развит слабее, чем на некоторой глубине, куда происходит подток
этих соединений со дна. В течение суток имеет место вертикальная мигра-
ция фитопланктона: днем водоросли вместе с прилипшими к ним пузырь-
ками выделяемого кислорода поднимаются кверху, а ночью, когда фотосин*
теза и обильного выделения газа нет, медленно опускаются вниз. Состав
фитопланктона изменяется по временам года. В озерах средней полосы Со-
ветского Союза весной в планктоне преобладают диатомовые, затем появ-
ляются перидиниевые и хризомонадовые, позднее — зеленые и еще позднее,
в наиболее жаркое время,— сине-зеленые водоросли. К осени снова преобла-
дают диатомовые, частично сохраняющиеся в планктоне и зимой.
В холодных морях и океанах количество индивидов в равном объеме во-
ды бывает в 5—10 раз больше, чем в теплых. Это зависит от происходящей в
холодных морях вертикальной циркуляции воды: охлаждающиеся поверх-
ностные слои воды опускаются вниз, снизу поднимается вода, более богатая
соединениями азота и фосфора, поступающими в нее из разлагающихся рас-
тительных и животных остатков.
Нередко массовое развитие одного или 2—3 видов планктонных водо-
рослей вызывает «цветение» воды — окрашивание ее в зеленый, оливко-
вый, сине-зеленый, реже буроватый или красноватый цвет. Цветение на-
блюдается как в пресных водоемах, так в морях и в океанах, иногда на зна-
чительной площади. При сильном цветении воды в 1 сж3 ее находятся сотни
тысяч и миллионы особей планктонных водорослей (до 300 000 нитей Апа-
baena flos aquae, до 10 000 000 особей Ankis trodesmus faleatusv. Scene desmus
1 От греч. «планктосэ — блуждающий.
г
НИЗШИВ РАСТ
мл
obliquus, до 40 000 000 одноклеточной Chlorella vulgaris). Цветение воды вле-
чет нежелательные последствия. Водоросли забивают сети рыбаков, за-
купоривают фильтры водопроводных станций. При таком интенсивном раз-
витии водоросли скоро потребляют в небольшом водоеме необходимые для
них питательные вещества и начинают отмирать. При гниении водорослей
в воде сильно уменьшается количество кислорода, потребляемого бактери-
ями, вызывающими их разложение; появляются продукты гниения их (серо-
водород, аммиак и др.). Все это может вызвать гибель рыбы (замор) в во-
доёме. В настоящее время, главным образом в нашей стране, разработаны
способы прогнозирования приближающегося цветения воды и меры по его
предупреждению.
Фитобентос1. К бентосным водорослям относятся виды, растущие на
дне; отчасти сюда можно отнести водоросли, растущие эпифитами на других
водорослях и прочих водных растениях, а также перифитон, т. е.
водоросли, которыми обрастают различные подводные сооружения, подвод-
ные части судов и т. п. Бентос морей и океанов состоит главным образом из
бурых и красных, в меньшей степени из зеленых — сифоновых, кладофоро-
вых, ульвовых— и некоторых сине-зеленых водорослей. Они растут, прикреп-
ляясь к каменистому грунту; на побережьях с песчаным, илистым, мелко-
галечным или ракушечным грунтом их не бывает. В песке, иле закрепляются
каулерпы; так же растут пеннатные, бентосные, диатомовые, распро-
страненные также на каменистом грунте и среди крупных растущих там во-
дорослей. В пресных водоемах в бентосе на различном грунте встречаются
разнообразные зеленые водоросли (улотриксовые, эдогониевые, кладофоро-
вые, конъюгаты, харовые), пеннатные, диатомовые, сине-зеленые. Они или
прикреплены к субстрату, или лежат свободно на дне, или (конъюгаты и др.)
образуют ватообразные скопления, поднимающиеся к. поверхности вместе
с пузырьками кислорода, выделяемыми при фотосинтезе.
Основными факторами распределения морского фитобентоса по верти-
кали является глубина и зависящее от нее освещение. Различают 3 пояса
растительности по вертикали, которые дополнительно можно подразделять
на горизонты. Литоральный пояс, или литораль* *, — пояс от высшего *
уровня прилива до нижнего уровня отлива. В наших северных морях в ли-
торали распространены из крупных водорослей главным образом фукусы;
на них (эпифитно) и среди них встречаются более мелкие бурые, красные и
зеленые водоросли. Условия существования в литорали весьма своеобразны.
Водоросли должны выносить удары волн, обсыхание и яркое освещение во
время отлива, временами пресную воду дождей, низкие температуры
зримой.
1
Сублитораль — пояс от низшего уровня отлива до глубины 4О'лс.
Здесь состав водорослей наиболее разнообразен. В морях холодной и умерен-
ной зоны в сублиторали очень широко распространены крупные ламинариевые
водоросли, образующие подводные луга или леса; в более нижних горизон-
тах наиболее распространены различные красные.
Элитораль — пояс от 40 до 100 ж (редко более) глубины. Бенто-
сная растительность здесь небогата, и в ней в связи с цветностью освещения
(см. стр. 84) могут преобладать и часто действительно преобладают красные
водоросли. В связи с температурой в развитии бентосной растительности
наблюдается известная сезонная цикличность, более резко выраженная в
холодных и умеренно холодных морях. Продолжительность жизни у много-
летних видов большей частью не превышает 2—4 лет, у немногих достигает
10—12 лет.
Фитобентос холодных и умеренно холодных морей по видовому соста-
ву водорослей беднее фитобентоса теплых морей, но превышает его по раз-
мерам своих представителей, количеству их и плотности зарослей. В теп-
1 От греч. «бентос» — глубина.
* От лат, «литоралио — береговой,
ОТ ДЕЛ —КРАСНЫЕ ВОДОРОСЛИ
II Г, ► 1
лых морях значительно увеличивается количество красных и зеленых (си-
фоновых) водорослей.
Биомассы бентосных водорослей могут быть значительные. Так, для
наших северных морей они колеблются от 4,4 до 30 кг сырых водорослей на
1 лса. Принимая в среднем около 10 кг на 1 ж3, получим сухой массы около
1 кг на 1 ле3 и около 10 m на 1 га. Так как эта масса у многолетних водорослей
получается за 3—4 года, то годовой прирост составляет около 2,5 т
на 1 га.
Обрастание водорослями подводных частей судов отрицательно ска-
зывается на судоходстве, так как, развиваясь в значительных количествах,
они уменьшают скорость их хода. Для предотвращения этого подводные части
судов покрывают специальными ядовитыми красками. Вредно отражается
обрастание и на внутренней поверхности водопроводных труб, на. защитных
сетках, где с ними борются периодическим пропусканием слабых растворов
хлора и медиого купороса или обрабатывают горячей водой.
Наземные и почвенные водоросли. Есть целый ряд водорослей, живущих
вне воды. Некоторые из них поселяются на коре деревьев (плеврококк, трен-
теполия, реже хлорококк, хлорелла и др.). Источником влаги для них яв-
ляются дожди и роса. Значительную часть жизни они проводят в состоянии,
близком к анабиозу, из-за отсутствия воды или из-за низких температур
зимой. На поверхности влажных скал нередко поселяются различные сине-
зеленые водоросли, образующие слизистые скопления, в сухую погоду почти
превращающиеся в черные крошащиеся корочки. На почве в сухих степях
и полупустынях часто встречается один вид ностока (Nostoc commune); при
увлажнении эта водоросль покрывает землю своими крупными (3—4 см) сли-
зистыми лопастными талломами. На Подсыхающем дие водоемов и по бере-
гам их на обнаженной почве встречаются ботридиум, иногда некоторые виды
вошерии. Защитным приспособлением от высыхания у этих водорослей яв-
ляются толстые слизистые обвертки (у сине-зеленых), толстые оболочки к ле-
ток (у трентеполии), переход содержимого в подземные ризоиды и образо-
вание цист (у ботридиума); важнейшую роль играет, конечно, физиологи-
ческая способность цитоплазмы выносить длительное высыхание, низкие
температуры, солнечный нагрев й т. п.
Водоросли могут проникать и в глубь почвы на несколько сантиметров,
иногда даже до 50 см и более. Здесь можно встретить зеленые (хламидо-
монады, хлорококки, мелкие улотриксовые и др.), сине-зеленые и диатомо-
вые водоросли; всего отмечено до 700 видов водорослей, встречающихся в
почве. Они живут не только в окультуренных или луговых и других более
или менее увлажненных почвах, но и в сухих местах Сахары, в сыпучих
песках Средней Азии. В засуху они образуют цисты, выделяют много
слизи; перистые диатомовые уползают вглубь.
Образуя в почве органическое вещество, водоросли могут способство-
вать развитию там азотобактера — бактерии, усваивающей атмосферный
азот, что положительно сказывается на урожае культурных растений.
ИЗО
ЗНАЧЕНИЕ ВОДОРОСЛЕЙ В ПРИРОДЕ И В ЖИЗНИ
. ЧЕЛОВЕКА
ь *
' В водоемах, особенно морских, водоросли являются главным проду-
центом органического вещества, необходимого для всего животного насе-
ления их. Первенствующая роль в этом принадлежит океаническому план-
ктону, продукция которого превышает бентосную в сотни и даже тысячи
раз. Это' объясняется значительно большей площадью открытых водных
пространств, сравнительно с узкими полосами прибрежий, большой вод-
ной толщей, насыщаемой планктоном, и, наконец, огромной раздроблен-
ностью зеленой массы планктона, а следовательно, исключительно большой
поверхностью восприятия света.
4—602
98
НИЗШИЕ РАСТЕНИЯ
Отми^)яютдие планктонные и отчасти бентосные водоросли образуют
на дне нёгдубо^^ отложения мягкрго^студне-
образного сапропеля, из -которого послвь соответствующей обработки полу-
чают ряд^цениых,веществ; смолу, бензин, керосин, техническое масло, лаки
и др. Сапропеля прошлых геологических времен дали начало некоторым
сортам каменных углей^^богхедов), горючих сланцев, быть может, нефти.
Лечебные грязи, представляющие сложный комплекс веществ, получа-
микробами органичес-
ются в значительной степени благодаря разложен
ких веществ, образованных водорослями.
Многие бентосные морские водоросли находят различное применение
в хозяйственной деятельности человека. Из-нескольких видов красных во-
дорослей получают плотный студень агар-агар, используемый в конди-
терской промышленности, при „ изготовлении лекоторых^сортон^конфет, в
пищевой — для добавления в хлеб, который при этрхмедл еннее^черствеет,
в бумажной й" тейстильнойГ—"для придания бумаге и тканям^ прртаости, и
глянцаТ Агар-агар находит, широчайшее применение в Лабораторных _био-
логических работах, как твердая среда, на которой с дпбямгдимАм
ных питательных веществ культивируют грибы, водоросли, бактерии. Из
ряда крупных бурых водорослей добывают ддьхин (натриевую соль, аль-
гиновой кислоты), который обладает большой клеящей способностью, при-
меняется для отделки .тканей, в производстве мороженого и кремов, может
использоваться для получения пластмасс, искусственного волокна и т. д.
Сбор водорослей производится вручную, а также драгами. В США, где растут
особенно крупные бурые водоросли, уборка их производится специальными
жатвенными машинами.
Из золы крупных морских водорослей можно добывать соли калия, лат-
рия, а также^йрд (в нашей стране — из черноморской" Красной водоросли
филлофоры)?
В приморских странах выбросы морских водорослей,.богатых калием и
отчасти азотом, употребляют как удобрение под культивируемые растения.
В этих же странах морские водоросли используют на корм скоту.
В Китае употребляют в пищу некоторые виды востока. Ъ ряде стран
Восточной Азии, на океанических островах, применяют в~пищу ряд водо-
рослей: бурые ламинарии («морская капуста»), порфиру, родименйю, ульву
• (Ulva lactuca — «морской салат»), энтероморфу и др. В Японии вблизи
устьеврек и в заливах в значительных количествах разводят порфиру.
Водоросли эти содержат много углеводов, небольшое количество белков,
мало жиров и некоторые витамины, богатые йодом. В диетической"отноше-
нии употребление водорослей полезно тем, что они улучшают перистальти-
ку кишечника, а большое содержание в них йода предохраняет человека от
заболевания зобом и развития атеросклероза.
РАЗМНОЖЕНИЕ ВОДОРОСЛЕЙ, ЧЕРЕДОВАНИЕ
ПОКОЛЕНИЙ И СМЕНА ЛДЕРНЫХ ФАЗ
Следование поколений и сопутствующее ему размножение осуществляются
у водорослей различно. У одних новые поколения возникают только ве-
гетативным путем; таковы многие сине-зеленые водоросли, хризомонады,
эвгленовые, большинство пиррофитовых. У других наряду с вегетативным
имеется и специализированное бесполое размножение, осуществляющееся
посредством особых клеток размножения — спор, преимущественно зооспор.
Возникновение новых поколений бесполым путем может повторяться иногда
в течение неопределенно длительного времени. Так, в опыте Гармана в
течение нескольких лет из культуры эвдорины было получено 1300 поколений,
возникших бесполым путем. Образование зооспор в некоторой мере зависит
от внешних условий. В частности, по экспериментальным данным Г. К л е-
б с а, спорообразование можно вызвать перенесением водоросли из пита-
^111
ОТДЕЛ — КРАСНЫЕ ВОДОРОСЛИ
99
тельного раствора вчистую воду, помещением культуры в темноту, что вы-
зывает повышение содержания СО2, переносом водорослей, живущих в про-
точной воде, в стоячую воду, а наземных — в водную среду. Размножение
одноклеточных водорослей спорами по сравнению с вегетативным делением
клетки повышает энергию размножения. Появление специальных клеток
бесполого размножения расценивается как прогрессивный шаг в эволюции
цикла развития водорослей.
Но наряду с вегетативным и собственно бесполым размножением во-
доросли имеют и половой процесс, иллюстрации различных форм которого
приведены выше. Существенно, что при морфологическом тождестве гамет
(изогамия) они могут отличаться своим половым знаком. В сближении га-
мет водорослей известную роль играют химические агенты (половые ве-
щества), в единичных случаях даже определена их химическая природа как
производных каротина. Установлено, что они могут оказывать свое дей-
ствие в ничтожных концентрациях — порядка 10“и.
Половой процесс наступает обычно при неблагоприятных условиях пи-
тания, температуры и т. п. Образующиеся в результате полового процесса
зиготы ведут себя различно.
У одних водорослей (например, у большинства зеленых) они покрыва-
ются толстой оболочкой и переходят в состояние покоя, по окончании кото-
рого прорастают, чаще всего образуя споры. При этом ядра зиготы делятся
редукционно. Ив таких, образующихся при прорастании зиготы, мейоспор1
вырастают особи нового поколения, способные, как и материнские, к раз-
множен
сначала, обычно, бесполым путем, а позднее и половым. В этом
случае, следовательно, из всех состояний, проходимых водорослью (взрослая
особь, митоспоры, гаметы, зигота, мейоспора), только одно — зигота — ха-
рактеризуется диплоидностью, все остальные гаплоидны.
У других водорослей зигота не имеет периода покоя и становится пер-
вой клеткой нового поколения. Так как при этом редукционного деления не
происходит, особь, выросшая из зиготы, оказывается диплоидной. С дипло-
идностью часто коррелятивно связана неспособность таких особей произво-
дить гаметы. Но любая клетка диплоидной особи потенциально способна
производить споры, при этом совершается редукционное деление, и зооспо-
ры оказываются гаплоидными. Вырастающие из них новые особи водо-
росли, естественно, тоже гаплоидны, и это, в свою очередь, коррелятивно
связано со способностью производить гаметы. Таким образом, в этом слу-
чае наблюдается правильное чередование поколений, функционально от-
личающихся друг от друга: половое поколение сменяется бесполым, бес-
полое — половым. Такое следование поколений может осложняться, на-
пример, тем, что диплоидные особи наряду с мейоспорами производят
диплоидные митоспоры, возникающие без редукционного деления; в других
случаях гаметы (всегда гаплоидные!) могут вести себя как зооспоры, прора-
стая без полового процесса в гаплоидные растения. Такие отношения харак-
терны, например, для эктокарповых из бурых водорослей и ряда других.
. Этого рода явления имеют немалое биологическое значение. Образование
диплоидных спор диплоидным же спорофитом делает последний способным
к размножению помимо полового процесса, что ускоряет появление новых
спорообразующих особей и, следовательно, должно привести к увеличе-
ин
количества мейоспор.
Прорастание -гамет в гаплоидные половые особи
способствует увеличению численности последних, а также количества гамет,.
чем повышается вероятность половых слияний.
В процессе эволюционного развития форм следования поколений воз-
никали, с одной стороны, дальнейшие усложнения, примером чего может
служить наличие третьего, вставочного поколения у красных водорослей»
так называемого карпоспорофита (см. стр. 86). С другой стороны, возника-
ли и упрощения. Так, вследствие редукции гаплоидного поколения произ-
1 От грея, «мейоз» — редукционное деление.
100
РАСТЕНИЯ
водство гамет стало функцией диплоидных растений у фукусовых. Весьма
вероятно, что в этом случае гаметами стали мейоспоры, которые, не превра-
щаясь во взрослые растения, сами осуществляют половую функцию.
По отношению к этому ряду форм циклов развития водорослей занимают
как бы боковое положение некоторые, довольно примитивно организованные
водоросли, проводящие всю свою жизнь в диплоидном состоянии, так что
у них гаплоидны только гаметы. Это диатомовые и некоторые зеленые во-
доросли. Такая форма цикла развития могла возникнуть как видоизменение
При этом диплоидная зигота из покоящейся
онтогенеза гаплоидных растег
клетки превратилась в основное (по длительности) состояние растения.
Только перед оплодотворением на основе мейоза у него вырабатываются гап-
лоидные клетки, выполняющие роль гамет. Таким образом здесь происхо-
дит лишь смена ядерных фаз и нет чередования поколений.
С появлением чередования спорофита и гаметофита произошла существен-
ная перестройка онтогенеза в первую очередь в том отношении, что число мей-
О8ов, происходящих в результате единичного слияния гамет, резко Возрос-
ло, так как в этом случае редукционное деление может совершаться почти
во всех (или, по крайней мере, во многих) клетках спорофита. Если учесть,
что при редукционном делении могут возникать новые комбинации призна-
ков, то следует заключить, что возрастание числа мейозов должно иметь
следствием увеличение генетического разнообразия потомства, и, следова-
тельно, повышение способности вида сохраняться в условиях среды, из-
менчивых как в пространстве, так и во времени.
ОТДЕЛ — МИКСОМИЦЕТЫ,
(MYXOPHUTA)*
и
ИЛ
СЛИЗЕВИКИ
Миксомицеты — своеобразный отдел низших растений, имеющих не-
которые черты животной организации.
Для слизевиков особенно характерно, что их вегетативное тело пред-
ставляет голую, т. е. не покрытую оболочкой, плазменную массу,
содержащую множество ядер.
В результате более или менее продолжительного вегетативного существо-
вания плазмодий переходит к спороношению. Споры слезевиков покрыты
оболочками и прорастают или амебоподобными, или голыми жгутиковыми
клетками. И те и другие представляют тот материал, из которого в конечном
счете формируется новый плазмодий.
Слизевики в основном сапрофиты. Они питаются, воспринимая необхо-
димые вещества осмотически, но могут заглатывать и твердые частицы, со-
держащие питательные материалы. Редко встречаются паразиты.
Около 500 видов (60 родов), составляющие отдел Myxophyta^ распре-
деляются в два класса. Ниже рассматривается один их низ — Myxomycetes.
К классу Myxomycetes относится подавляющее большинство слизе-
виков и у них особенно ясно выражены все характерные признаки этого от-
дела (pncJ 47). Вегетативное тело их — плазмодий — развивается на от-
мерших растениях, опавшей листве, гнилой древесине, коре и т. п. Размеры
плазмодия 1 мм и больше (иногда до 1 ле) в диаметре. Окраска их неяркая —
серая, белая, желтая, красноватая и т. д.,— довольно постоянная для каж-
дого вида. Наружные участки плазмодия и его тяжей имеют более или ме-
нее однородную консистенцию; внутренние участки состоят из более жидкой,
мелкозернистой протоплазмы, находящейся в постоянном движении. Кон-
туры тяжей протоплазмы постоянно меняются, и весь плазмодий обладает
поступательным движением, перемещаясь по какому-нибудь направлению и
отступая (втягиваясь) с противоположного конца. Во время вегетативной
1 Этот отдел в некоторых руководствах по систематике обозначается как Myxomycefa9
MyxothaUophniat МусеЬшъ
ОТДЕЛ — МИСКОМИЦЕТЫ
101
Рис. 47. Миксомицеты:
1 — часть плазмодия; 2 — спорангии трихии; S — спорангии стемонитеса; 4 — споры
трихин; s — прорастание споры; в — зооспора
и нити капиллиции
. жизни плазмодий обнаруживает отрицательный фототаксис и положитель-
ный гидротаксис, т. е. передодрает в менее освещенные и более влажные
•' места. Поэтому в таком состоянии он обычно незаметен, так как находится
пбд опавшей листвой, между корой и древесиной в гнилых пнях и тГ п .
-Эапасннмй продуктами в нлаямидии бываюг-ясиры, углевод, глико-
^ген. очень много белков (до 30% от общего количества органических веществ “
и выше); крахмала не бывает. Пигменты плазмодия мало изучены; они рас-
творены в каплях жира или в самой протоплазме. В небдшюприязиых-ушо-
виях плазмодий может одеваться оболочкой, превращаясь в склероций- из
которого вновьмогут образоваться плазмодии.
После' некоторого периода чисто вегетативного развития наступает спо-
рообрааодание. К этому времени таксисы плазмодия изменяются иоИ вы-
ползает на более освещенное и cvxftfl ЦОРТП, ня торцовую поверх-
ность пней. Плазмодии почти лестГраспадается на большое количество спо-
рангиев, одетых плотной оболочвой7~располагающихся обычно тесными
группами, имеющих большей частью округлую или овальную форму, к осно-
ванию обычно сужецшни-в^щц^су; размеры их у большинства измеряются
немногими^миллиметрами. У довольно многих миксомидетов сМОрашни "со-
единяются вмёстев более крупные <улпр<1нотпстмят в форме горошин или зна-
чительно более крупных подушковидных образований; их назыйЯбт эТТа-
л и я м"иГ1ЭтАЛШ1 бывают одетыЪбщей оболочкой на наружной повер^йбсти;
от боковых же оболочек составляющих их спорангиев остаются лишь от-
дельные пластинки, тяжи, или они вовсе исчезают.
Содержимое спорангиев и эталиев распадается на одноклеточные ми-
кроскопически мелкие споры/одетые каждая своей плптрпй ^ii Л ин и uii У (цспт
многих миксомидетов в спорангиях и эталиях образуется, кроме спор, еще
102
I It', IУ
НИЗШИЕ РАСТЕНИЯ
так
вых
ние
ы: l
актера, а чисто нсгогдтицпон. так кцщышяния ядер
Плазмодии питается как растворенными веществами,
телами, например бактериями и т. п., обволакивая и постё-
Диплоидные ядра в плазмодиях ряямттлясдштг.я
сомиНетов, как видно из вы-
вязываемый ^к a-м и и й1 в виде отдельных волоконец, сплот-
или полых внутри, или в виде связанной сети нитей.
Перед образованием спор в плазмодии происходит одновременное деле-
ядер с редукцией числа хромосом.
По созревании спорангие^и эталиев оболочка их разрывается и споры
постепенно вкидываются наружу, чему способствуюд^гигроскопические нити
капиллиция.
' В^сухом виде споры могут долго сохраняться При наличии воды они
прорастают; при этом из споры выходят!, чаще 2. иногда 4 амебрвддннд-Чзес-
---—jy прлтпттласта, превращающихся скоро в^ооогпррч с пульсирующей
Вакуолью и двумя житакамд^на^переднем конце (одним длинным, другим
очень1 короткии)7^бчень часто из спор непосредственно выходят сформиро-
вавшиеся двужгутиковые зооспоры. Зооспоры могут размножаться делени-
ем, прекращая на это время движение. Вскоре зооспоры теряет жгутики и
превращаются в ам ные о ____________ называем^^е/в отличие от настоя-
щих амеб, ; они тоже размножаются делением. Затем мИкеаме-
бы (а иногда зооспоры) попарно копулируют. причем сейчацдщсливаются и
их^ядраГУ некоторых мдксомицетов при этом обнаружена гетероталличность.
Непрокоцулировавщие гаплоидные миксамебы недолговечны; их могут пое-
дать диплоидные миксамебы. При наличии в одном месте нескольких или мно-
гих диплоидных^миксамеб они шпэд^сливаться вместе, образуя-болеетгруп-
gjjft пляй^одийлторпхт границы отдельных ммксамеб неразличимы. Это
слияние не подового ха
при этом^нелцоисходит.
так и твердыми^
пенно переваривая их>
митотическим делением. В ддкде развитие __ _________
шеизложенного;—преобладает ципдоипттяя фаза; гаплоидны лишь споры,
^и^гтосамебы до их копуляпии. ' ‘ '
К K^acS^~Mlyxomycetes относится более 400 видов. На гниющих пнях во
второй половине лета и осенью часто встречаются фулиго (Fuligo) в виде
крупных подушковидных эталиев с рыжеватой или сероватой оболочкой;
ликогаля (Lycogala) — группы эталиев величиной с горошину вначале крас-
новато-розовые, затем серые; ретикулярия (Reticularia) — подушковидные
эталии с блестящей серебристой оболочкой; стемонитис (St emonit is) — тесно
скрученные длинноцилиндрические спорангии на ножках с центральной
колонкой и отходящей от нее сеткой капиллиция — и многие другие виды.
К отделу Myxophyta иногда относят еще класс Phycomyxi-
пеае — паразитную группу плазмодиофоровых, положение которых оспа-
ривается — многие систематики помещают ее среди грибов. Однако двужгу-
тиковость подвижных стадий, наличие плазмодия и диплоидность в веге-
тативном состоянии сближают эту группу со слизевиками. Вегетативное
протоплазматическое тело Phycomyxineae^ не одетое оболочкой, заполняет
клетки растений-хозяев; там же оно распадается на многочисленные споры,
освобождающиеся после разрушения тканей хозяина.
Наибольшее практическое значение среди класса Phycomyxineae име-
ет плазмодиофора (Plasmodiophora brassicae, рис. 48), вызывающая килу
корней капусты и других крестоцветных. Возбудитель болезни впервые
был описан в 70-х годах XIX в. русским микологом М. С. В о р о н и н ы м.
На корнях пораженных растений "образуются вначале небольшие вздутия,
постепенно разрастающиеся в очень крупные опухоли с неровной поверх-
ностью. На разрезах этих опухолей видны под микроскопом группы гипер-
трофированных паренхимных клеток хозяина, заполненных густой прото-
плазмой паразита. На более поздних стадиях в этих клетках можно обнару-
жить огромное количество мелких бесцветных шаровидных спор паразита,
освобождающихся после сгнивания опухоли. Цемотря на большое количество
1 От лат. «капиллюс» — волос.
ОТДЕЛ — мискомицвты
.103
иг
Рис. 48. Нлазмодиофора в корне капусты:
1 — вегетативная стадия в корневом волоске; г — в клетках корня; Л — споры
исследований этого паразита, производившихся в разных странах; цикл раз-
вития его недостаточно выяснен и описывается исследователями различно.
По-видимому, из спор в почве выходят двужгутиковые зооспоры1, превра-
щающиеся в мик с амебы; они копулируют попарно и после этого проникают
в корневые волоски или в клетки эпиблемы корня, а оттуда — в более глу-
боко лежащие клетки растения-хозяина. Так протопласт паразита раз-
растается, становится многоядерным, отчленяет новые плазмодии, проникаю-
щие в соседние клетки. К пораженным участкам обильно притекаю^ пласти-
ческие вещества хозяина, клетки его усиленно делятся и гипертрофируются,
в результате чего получаются крупные опухоли. Перед образованием спор
описано редукционное деление ядер паразита, и, по-видимому, большая часть
его жизни проходит в диплоидной фазе.
Пораженные корни плохо разрастаются; заболевшие растения капусты
или гибнут, или, во всяком случае, не дают кочанов. В борьбе с этим пара-
зитом применяют плодосмен и известкование почвы, так как паразит зара-
жает растения при кислой реакции почвы.
Спонгоспора (Spongospora solani) вызывает порошистую паршу клубней
картофеля: на их поверхности образуются небольшие язвочки, позднее от-
крывающиеся; в них находятся споры паразита, собранные в губчатые шары.
Предполагают, что предками миксомицетов были какие-то бесцветные
жгутиковые организмы. У некоторых жгутиковых наблюдается образование
небольших плазмодиев. У миксомицетов состояние плазмодия стало постоян-
ным.
1В течение долгого времени зооспоры как сапрофитных, так и паразитных миксоми-
цетов описывались как одножгутиковые. В последнее время обнаружено, что они имеют
один длинный жгутик, другой короткий.
104
ОТДЕЛ—ГРИБЫ (FUNGI, ИЛИ MYCETES,
ИЛИ MYCOPHYTA)
иг
111 л
ч II \
4^—1_____.___
.111
III'.
Грибы в противоположность водорослям составляют единый отдел низ-
[х растений, насчитывающий около 100 000 видов* От водорослей они резко
отличаются отсутствием хлорофилла, от бактерий —• наличием типичны*
ядер.
То, что в общежитии называют грибами, представляет только небольшую
часть этой обширной группы, и притом гриб в бытовом понимании представ-
ляет лишь так называемое плодовое тело; вегетативная часть растения обычно
бывает скрыта в субстрате.
Вегетативное тело грибов состоит изобильно разветвленных, у боль:
ства бесцветных, нитей, так называемых г и ф1, образующих грибницу,
или мицелий1 2. У подавляющего большинства грибов мицелий находит-
ся в субстрате (почва, растительные остатки, живые растения, древесина
и т. д.), а на поверхность его выступают лишь плодовые тела или другие споро-
ношения грибов в виде шляпок на ножках, копытообразных тел, корочек,
пушка, порошковатой массы (плесени и др.), шариков, точек и т. д.
Обильно разветвленный мицелий имеет огромную поверхность, через
которую осмотически поглощаются вода и растворенные в ней питательные
вещества.
У самых низших грибов мицелия нет, и вегетативное тело их представ-
ляет'Тббоимикроскопически малый комочек протоплазмы, голый или одетый
оболочкой. У довольно большого числа этих безмицелиальных грибов имеют-
ся как бы зачатки мицелия в виде коротких, разветвленных, чрезвычайно
тонких безъядерны* нитей д так называемого р и з о и иц е л и я, отходя-
шетки.
_ мицелий у низших грибов (фикомицетов) без поперечных пе-
регородок (н е с ё п тированный), содержит огромноекдлйчёство кле-
точных. ядщи т. е. имеет так называемое неклеточное строение, внешне не-
сколько сходное со строением сифоновых водорослей. У высших грибов, к
которым относится огромное их большинство, мицелий разделён поперечными
перегородками на клетки, содержащие по 1—2 и несколько ядер (с е и т и -
рованный м й ц е л и й ).
Лишь у немногих грибов (оомицетов) в оболочке клеток содержитец цел-
людщщ. У всех прочих оболочка состоит из других полисахаридами, кроме
того, содержит некоторое количество хитина, тождественного с хитином на-
секомых. У всех прочих растений хитин не встречается. В поперечных пере-
городках между клетками находятся отверстия (сквозные поры) -
Протоплазма во взрослых клетках располагается вдоль клеточных сте-
нок; тяйй2 ’ВеЛйУрЬсекают клетку по разным направлениям и иногда имити-
руют поперечные перегородки. Хроматофоров и вообще никаких пластид в
клетках грибов нет; крахмал не образуется; запасными продуктами являют-
ся жиры и гликоген, большей частью "пропитывающий протоплазму. Как
запасное вещество часто встречается еще волютин (соединение нуклеиновых
кислот с полифосфатами и другими органическими веществами).
Окраска грибов обусловливается пигментами различной химической при-
роды, находящимися в оболочках, цитоплазме, вакуолях или в виде экскре-
тов, выделяемых на поверхность клеток.
Образование новых клеток и нарастаИие мицелия происходит обычно пос-
редством верхушечных клеток. Боковые ветви мицелия по строению не отли-
чаются от тех, которые их производят. Но у большинства паразитных грибов
некоторые боковые ветви, проникающие в клетки растения-хозяина, дальше
не растут, а превращаются в шаровидные, грушевидные или разветвленные
1 От греч. «гифе» — ткань.
2 От греч, «микео — гриб,
ОТДЕЛ — ГРИВЫ
105
Рис. 49. Гифы мицелия разных видов грибов с гаусториями в корнях растения-хозяи-
на (протоплазма в клетках растения-хозяина не изображена)
г а у с тлели и1, поглощающие питательные вещества из клетки хозяина
(рис. 49)7
Настоящие ткани, образующиеся в результате деления клеток по всем
направлениям пространства, встречаются у грибов очень редко. Компакт-
ные плодовые тела шляпочных и других грибов состоят из плотного переп-
летения тех же грибных гиф.
Из грибных гиф, идущих параллельно и тесно сближенных, состоят и
мицелиальные тяжи, отходящие, например, в почву от плодового тела шля-
почных грибов (в общежитии называемые корешками), а также р изо-
мор ф ы — плотные шнуры в несколько миллиметров толщиной и иногда
в несколько метров длиной, снаружи большей частью темные, встречаю-
щиеся, например, у опенка, у домового гриба и др.
Из сильно переплетающихся гиф с толстыми оболочками образованы и
с клероли и — твердые, округлые, овальные или рожковидные образо-
вания, служащие у ряда грибов для переживания неблагоприятны^ условий
(например, «рожки» спорыньи). Хотя они на разрезах нередко производят
впечатление паренхимной ткани, но, так как и здесь гифы делятся только по?
перечне, а не во всех направлениях, эту ткань называют п с е в й опаре н -
х и м о й или плектенхимой2. Плектенхимное строение имеет и
кожица на шляпочных грибах.
У ряда грибов в специальных гифах, млечных трубках, находится бе-
лый или оранжевый млечный сок (рыжики, белянки и т. п.).
Размножение у грибов, как и у водорослей, бывает вегетативное, бес-
, полое и половое, причем ни в одной группе растений, кроме грибов7нет такого
разнообразия "органов,джзмншкшш* на них главным образом и основыва-
ют классификационные подразделения грибов.
Вег е т а т и в и о е. ,p,aje Рледв е^ происходит случайно от-
/ рывающимися участками мицелия, почкованием клеток (например, у дрож-
*. жей). артроспорами (оилиями) и хла^ЛР1ШГПШ1И^1А.ртроспо рами или оидиями
' называют клетки с тонкой оболочкой, на которые может распадаться мице-
лий у ряда грибов. Хламидоспорами называют клетки с толстой, обычно бу-
роватой оболочкой, на которые распадается мицелий.
1 От лат. «гаустор» — яерпатель.
« От грея, «плекгос» — сплетенный, онхима» — налитое, в ботанике — ткань*
106
НИЗШИЕ РАСТЕНИЯ
Рис. 50. Органы спорообразования у грибов:
1 — зооспорангий сапролегнии: а — молодой, б —
вскрывшийся: а — спорангий мукора (в оптическом
разрезе): л — конидиеносец пеницилла: 4 — сумка с
аскоспорами: 5 — базидия с базидиоспорами
гиями, если в них образу-
Бесполое размно-
жение в узком смысле осу-
ществляется посредством спор,
образующихся на особых вет-
вях мицелия. Споры могут со-
стоять из одной, двух или
нескольких клеток и образу-
ются эндогенно или экзогенно.
Э идогенные споры
бесполого размножения, харак-
терные для большинства низ-
ших грибов, образуются внутри
специализированных клеток,
отделяющихся обычно перего-
родкой от несущей их гифы и
называемых з о о с п о р а н-
ются подвижные заооспоры, или
просто спорангиями,
если в них образуются непод-
вижные спорангиоспо-
р ы (рис. 50).
Экзогенные споры бесполого размножения в^бод^щцнсхве случаев носят
название конидии1. Образуются они по одной или цепочками (иногда ску-
чены в головки) на вершине или также по бокам большей частью специальных
гиф мицелия, простых или разветвленных конидиеносцев, поднимающихся
обычно вертикально.
Половое р*а змножение у самых низших грибов бывает
в форме хологамии, изогамии, гетерогамии; у вышеорганизованных, нр тоже
низших грибов с неклеточным мицелием "оно бывает в форме оогамии или
зигогамии, напоминающей конъюгацию у водорослей конъюгат. Зигота у
большинства низших грибов некоторое время находится в состоянии покоя.
Перед прорастанием (или, реже, задолго до него) в ней происходит редук-
ционное деление^после чего образуются зооспоры или формируется короткая
гифа с зооспорангием, спорангием или Конидией. Всяжизнь многих низших
грйб<5в~проходит в гаплоидной фазе, диплоидна у них лишь зигота.
У многих высших грибов, имеющих многоклеточный мицелий', половое
размножение осуществляется в виде слияния не дифференцированного на
отдельные гаметы содержимого двух различных по внешности половых орга-
нов (гаметангиогамия, т. е. слияние гаметангиев, а не гамет).
У ряда высших грибов происходит оплодотворение женского полового органа
мелкими неподвижными клетками — спермациями, переносимыми воздуш-
ными течениями или насекомыми. У очень многих высших грибов произош-
ло угасание типичного полового процесса и оплодотворение осуществляется
переползанием протопласта из одной вегетативной клетки в другую (псев-
домиксис, или, лучше, соматогамия) или попарным соединением
ядер в одной клетке (автогамия). Почти во всех случаях у высших
грибов при половом размножении происходит вначале слияние цитоплазм
(плазмогамия). Слияние ядер отделено большим или меньшим про-
межутком времени, в течение которого ядра противоположных полов размно-
жаются делением и очень часто бывают соединены в пары, так называемые
дикарионы. После полового слияния ядер (кариогамия) диплоид-
ное ядро, не проходя длительного периода покоя, делится редукционно, и по-
лучающиеся гаплоидные ядра становятся ядрами спор полового размноже-
ния. Таким образом, в цикле развития подавляющего большинства высших
грибов чередуются закономерно три фазы: гаплоидная, дикарионтическая
1 От греч. «копие» — пыль.
ОТДЕЛ — ГРИБЫ
107
и диплоидная, причем диплоидная обычно кратковременна, а гаплоидная и
дикарионтическая в разных группах грибов различны по продолжительности.
Споры, образующиеся в результате полового процесса, возникают или
эндогенно, или* экзогенно. Спорообразующии орган в обоих случаях фор-
мируется из двухъядерной клетки, в ней совершаются кариогамия и редукци-
онное деление ядра. При эндогенном спорообразовании вслед за мейоти-
ческим делением обычно происходит еще одно деление и каждое из восьми
ядер становится ядром одной из спор, сама же произведшая их клетка —
спорообразующим органом, называемым с у MtK о jL или а с к о м. При
экзогенном образовании спор после кариогамии редукционного деления
формируются четыре гаплоидных ядра, а спорообразующии орган, называе-
мый здесь баз и дней, образует выросты по числу ядер, куда эти послед-
ние и переходят. В дальнейшем выросты, содержащие ядра, отшнуровывают-
ся и становятся спорами. Эндогенные_1щрры называются аскоспорами, а
экзогенные — базидиоспорами (см. рис 50).
" Сумки и базидии, играющие существенную роль (в процессе их форми-
рования совершаются кариогамия и мейоз) в цикле развития, у подавляющей
части грибных организмов могут возникать прямо на мицелии, но гораздо
чаще они образуются в связи с так называемыми плодовыми телами (внутри
их или на них). Плодовые тела формируются из переплетений гиф. Они имеют
определенную форму и строение, типичные для систематических единиц то-
го или иного ранга.
Обычно бесполое и половое спороношения закономерно сменяются в
цикле развития грибов, причем половое спороношение завершает жизненный
цикл. X.довольно многих .видов грибов имеется несколько различных беспо-
лых спороношений. Так как о внешнем виде грибов обычно судят по их спо-
роношениям, то эти различные спороношения описывали под разными родо-
выми названиями, и даже позднее, когда была установлена их связь, опи
нередко продолжают фигурировать в систематике грибов каждая, под своим
первоначальным именем. Это дает основание говорить о плеоморфиз-
ме1, т. е. о многоформенности (многообличье), грибов.
Большинство грибов — сапрофиты, питающиеся отмершими органи-
ческими остатками, главным образом отмершими растениями, разложение
и минерализация которых производятся преимущественно грибами; значи-
тельное меньшинство сапрофитов живет на трупах животных. Паразитные
грибы в подавляющем большинстве (свыше 10 000 видов) живут на живых
растениях, меньшинство (около 1000 видов) — на животных или человеке.
^Многие грибы большую часть своего развития проводят как паразиты па
живых растениях и образуют в это время копидиальные спороношения, но
после отмирания растепия-хозяина они продолжают жить на нем как сапро-
фиты и образуют через некоторое время половое, обычно сумчатое, спороно-
шение. Довольно многие сапрофитные грибы могут поселяться на ослабленных
больных растениях, на их органах, находящихся в состоянии ослаблен-
ной жизнедеятельности (например, на плодах, корне- и клубнеплодах), и пи-
таются ими уже как паразиты; такие грибы называют факультативны-
юй-п аразитами. В лабораторных условиях удается культивировать па
различных питательных средах очень многие паразитные грибы как сапро-
фиты, но некоторые группы паразитных грибов (мучнеросные, ржавчинники)
не удается культивировать как сапрофиты, и их называют облигатны-
м и, т. е. обязательными паразитами.
Предпочтение грибами растительных субстратов объясняется наличием
у ник ферментов, расщепляющих клеточные оболочки и углеводы растении,
а также способностью грибов выносить кислую реакцию растений, подавля-
ющую развитие большинства бактерий. На субстратах животного проис-
хождения, богатых белками, грибы не выдерживают конкуренции с бакте-
риями, которые подщелачивают среду, что неблагоприятно для грибов.
1 От греч. «плеон» — много. '
108
НИ8ШИВ РАСТЕНИЯ
Большинство грибов требует высокой влажности субстрата и воздуха,
так как у них почти нет приспособлений для проведения воды и защиты от
испарения* Однако в жидкой среде (вода и др.) большинство грибов не разви-
вается, так как она недостаточно аэрируется. При искусственном разведении
грибов в жидкой среде (например, на пенициллиновых заводах) в нее необхо-
димо продувать воздух.
Продолжительность жизни грибов у разных видов весьма различна —
от немногих дней у одних до нескольких десятков лет у других*
Грибы можно разделить на 5 классов. Два класса, у представителей ко-
торых мицелий совсем не развивается (Archimycetes) или он неклеточный, не-
септированный (Phycomycetes), объединяют в группу низших грибов.
Два класса высших грибов имеют септированный мицелий, при-
чем класс Ascomycetes характеризуется половым спороношением — сумкой, а
класс Basidiomycetes — базидией. К высшим грибам относится и формальный
класс несовершенных грибов (Fungi imperfecti или Dcnteromycetes), у кото-
рых неизвестно половое спороношение.
1-й КЛАСС - АРХИМИЦЕТЫ (ARCHIMYCETES)
В класс архимицеты объединены микроскопически мелкие грибы,
_ - .имеющие мицелия. живущие как внутриклеточные паразиты в виде голой плаз-
менной массы. Бесполое размножение этих грибов осуществляется зооспора*
ленным сзади и имеющим характерное строение. Его проксшальная часть
относительно толстая, покрытая тонкой оболочкой, видимой посредством
электронного микроскопа, заканчивается очень тонким хвостовидным придат-
ком; составляющим дистальную часть. Пплрвпй прдцесс ияо^^ц^й или ге-
терогамяый,
К архимицетам относится несколько сотен видов. Большинство живут в
воде, паразитируя на водорослях, на водных грибах и высших растениях.
Ряд видов паразитирует на высших наземных растениях, заражение которых
зооспорами происходит большей частью в то время, когда их проростки
проходят через почву. Некоторые сапрофиты живут в воде на растительных
остатках или в почве.
На капустной рассаде в парниках часто встречается ыипидиим (Olpidium
brassicae), вызывающий заболевание, известное под наямииеъсяерцдД ножки:
рассада у корневой шейки утончается, чернеет и погибает. В клетках парен-
химы первичной коры иди в эпидермисе находится вегетативное трло паразита
в виде голого одноядерного комочка протоплазмы (рис. 51). Позднее он ста-
новится мкогояддрщ^<т эятем одевается оболочкой и целиком превра-
щается взооспорангий. Развившиеся в нем зооспоры с одним задним
жгутиком выходят наружу через длинную выводящую шейку, развивае-
мую зооспорангием и прободающую оболочки клеток хозяина. В тканях пора-
женного растения наблюдаются еще и толстенные цисты, одетые звездчатой
оболочкой и содержащие в молодом возрасте по два ядра. Как и у некоторых
других, еще ранее изученных видов ольпидиума, эти цисты являются зиго-
тами: зооспоры в случае голодания цх (или по другим причинам) ведугд&бя
как гаметы — сливаются попарно и дают двужгутиковую подвижную зиго-
^гу^которая заражает хозяина, одевается там толстой оболочкой и прорастает
ъ после сгнивания тканей.
Основные меры борьбы с черной ножкой уничтожение сырости в пар-
никах, их проветривание и обеззараживание почвы. При появлении болез- *
ни полезно насыпать наГпочвУ СДой Шёбка, что обеспечит развитие придаточ-
ных корней из стеблей рассады выше пораженного места.
К базмицелиальным архимицетам относится также большой род
V сцяяутрмух (Sync AyiНшп), содержащий около 150 видов, паразитирующих на
высших наземных растениях; зооспоры у него тоже имеют один задний жгу-
.ш
1111
иг
ОТДЕЛ —ГРИБЫ
109
Рис. 51. ОлБпидиум в клетках корневой шейки
капустной рассады:
1 — зооспорангии; 2 —голые протопласты: а —паразита,
б —зиготы; в — зооспоры
Рис- 52. «Рак» на клуб-
не картофеля
тик, но, в отличие от олипидиума, синхитриумы
развивают не oj
и: ।
спо-
рангий, а труппу (сорус) зооспорангиев. У растеЙВЙ=ХРЭЯ№" синхитриумы
вызывают сильное увеличение размеров клетки, в которой поселился паразит,
а у многих еще и ашогократиое делёйтс>1клёток, окружающих пораженную.
В результате образуются галлы в виде бородавочек или более крупных на-
ростов. Большое практическое ’значение ^He&r^Synchytrium endcbioticum9
вызывающий большие бугристые наросты на клубнях (реже на других ор-
ганах) картофеля — заболевание, известное под названием рака картофеля
(рис. 52), приводящее к гибели урожая. Кроме бесполого размножения зосн
спорами для синхитриума характерно и половое — копуляпяя
Рак картофеля, широко распространенный в Северной Америке и Западной
Европе, до недавнего времени не наблюдался в СССР; заносу его препятство-
вали мероприятия, осуществляемые Государственной карантинной службой.
Занесенный в западные и некоторые другие районы СССР в период времен-
ной оккупации их во вторую мировую войну, он уничтожается обеззаражива-
нием почвы химикатами^ также посевом ракоустойчивых сортов картофеля.
О происхождении архимицетов и их филогенетических связях единого
мнения нет. Можно предположить, что они произошли от бесцветных жгути-
ковых организмов (флагеллят)
II’•
2-й КЛАСС— ФИКОМИЦЕТЫ (PHYCOMYCETES)
Большинство фикомицетов характеризуется хорошо развитым некле- -
точным мицелием, содержащим огромное количество ядер;' поперечные п£~
йёпиюдки в л»птк при пдяйУМИУи'опганрв раамнояго^
$ия. У примитивных форм мицелий развит слабо, представляя тонкие гифы,~~\
не*содержащие ядер и отходящие от одноядерной или многоядернойтсяетки. I
Наличие клеточной оболочки, равно как образование органов размножения \
не из всего тела гриба, а из специализироваюшх'екГчаСТ^. ЗЫЯТЯГОЦйо \
отграничивает этот класс от а пхимипетов.~ Бесполое размножение осущесяь*
ляется спорами (сццры, покрытые оболочками, безжгутиковые — спорая-
гиплппрн) нплпйлй процесс разнообразный: от изогамии до оогдуни вгтяпдг
тангиогамци. В вегетативном состоянии фикомицеш'обычно гаплоидны, в та-
ком слуУаадицяьид яа толькоаиготя, в которой перед прорастанием происходит
редукционное деление. Но у некоторых фикомицетов известен я более слож-
ный цикл развития в форме изоморфной смены поколений. 4
no
НИЗШИЕ РАСТЕНИЯ
1-Я ПОРЯДОК — ХИТРИДИЕВЫЕ (CHYTRIDIALES)
.в
Вегетативное тело хитридиевых обычно небольшое. Бесполое размно-
жение зооспорами, с одним, задним по положению, жгутиком. Шловои?прУ
цесс в виде копуляции подвижных гамет иди, гаметангйевП(гаметангиогамия).
Нередко встречается pusofiuduySr^Rhizophidium pollinis) на пыльце
сосны, попавшей в воду. - Его зооспора, имеющая один жгутик сзади, садится
на пылинку, покрывается оболочкой и пускает внутрь пылинки очень тон-
кий слабоветвящийся безъядерный ризомицелий (рис. 53, 7). После некото-
рого периода роста ядро делится и содержимое пузыревидной клетки распа-
дается на зооспоры, которые выходят через отверстие в ее оболочке. Наблю-
дались и толстостенные покоящиеся клетки, у некоторых видов они являют-
ся зиготами, образующимися в результате копуляции двух сидящих рядов
одетых оболочками клеток.
На неподвижных эвгленах нередко паразитирует полифаг (Polyphagus
euglenae), Его пузыревидное, покрытое оболочкой вегетативное тело дает
много отростков ветвящегося ризомицелия, которые могут захватить несколь-
ко десятков эвглен (рис. 53, 2). Этот гриб размножается зооспорами, причем
спорангий формируется из центральной клетки. Одножгутиковая спора ос-
танавливается среди эвглен и, покрываясь оболочкой, дает начало новому
ризомицелию. Половой процесс у полифага в виде копуляции ветвей ризоми-
целия. Образующаяся в результате оплодотворения покоящаяся зигота про-
растает в зооспорангий.
2-Й ПОРЯДОК — БЛАСТОКЛАДИЕВЫЕ (BLASTOCLADIALES)
Бластокладиевые представлены водными и почвенными грибами, питаю-
щимися как паразиты или сапрофиты. У них имеется хорошо развитый ми-
целии. Зооспоры с одним (задним) жгутиком. В анизогамном половом про-
цессе участвуют жгутиковые гаметы. Часто наблюдается изоморфная смена
поколении.
3-Й ПОРЯДОК — МОПОБЛЕФАРИДОВВДЕ (МО NO В БЕРНА R IDA LES)
— * г ь
Зооспоры и сперматозоиды, у представителей моноблефаридовых с од-
ним задним жгутиком. Это единственная группа грибов^в которой, опло-
дотворение осуществляет сперматозоид.
' Мицелий моноблефаридовых развивается по преимуществу весной и
осенью" в чистых водоемах ввиде пушка на сучьях и других раститель-
ных остатках.
2
Рис. 53. Хитридиевые грибы:
j— ризофидиум на ныльое сосны (м«— зачаточный мицелий, а — зооспоры, выходящие из зооспорангия):
> — полифаг. паразитирующий на евгленах
ОТДЕЛ — ГРИВЫ
Ш
Наиболее крупный род — моноблефарис
(Monoblepharis).lfa концах гиф слаборазвитого
мицелия грибовэтого рода отчленяетсд_дере-
горрдкоц по одному удлиненному зооспоран-
гию, где формируются .овальдые или, шаро-
видные зооспоры. Они выходят через отверстие
на в ер хупткег спорангия. Прорастает зоосццрдх.
двух концов, образуя & одной
q другой — гифу.
На мицелии* позднее развиваются оогонии
и антеридии, отделяющиеся друг от другаПиТЯГ
'вШЦелия поперечными перегородками (рис. .54).
В каждом антеридии Формируется несколько
сперматозоидов, похожих на зооспоры, но бо-
лее мелких. В оогонии образуется только одна
яйцеклетка. Прй^вскрыванииЪогЬциямз.него
выделяется слизь, на которую реагируют спер-
матозоиды (хемотаксис). Сперматозоиды двйга-
^ся^амебообрдддр, отчасти с помощью жгу-
тика. После оплодотворения яйцеклетка оде-
вается оболочкой (ооспоЬ^ и прорастает только
после периода -ишокшь Моноблефаридовые,
Рис. 54. Моноблефарис:
1 — зооспорангий; s — оплодотворе-
ние сперматозоидом яйцеклетки в
оогонии (ан— антеридий, о — оого-
ний)
вероятно, возникли
ковых
с другими грибами не имеют.
от каких-то одножгути-
филогенетической связи
4-Й ПОРЯДОК — САПГО ЛЕТНИЕ ВЫЕ (SAPROLEGNIALES)
К сапролегниевым относятся врдные^грмбь^, живущие главным обра-
зом сапрофитно на трупах насекомыхи других мелких животных. Для них
характерны двужгутиковые зооспоры, отсутствие сперматозоидов, образо-
вание в^оогонии цо нискольку яйцеклеток (лишь у некоторых по одной), на
формирование которых идет все содержимое оогонии. У большинства в
субстрате находятся лишь сильноветвящиеся ризоиды, а мицелий находит-
ся снаружи.
Один из наиболее широко распространенных родов — сапролезния
(Saprolegnia). Ее легко получить в культуре, набросав в чашки или аквариу-
мы с водой, взятой из природного водоема, свежие или сушеные «муравь:
ин
яйца». Через несколько дней на некоторых из них появляется белый пуптпр
мицелия. Вскоре на концах гиф образуются длинные пилиттдртптг.и.тн> .wu
спорангии, с густой цитоплазмой, отделяющиеся перегородкой от гифы. В
них формируется много грушевидных зооспор с двумя одинаковыми жгути-
ками, из которых один перистый, а другой простой.
После приблизительно получасового движения зооспор^^останавливает-
ся, одевается оболочкой, и примерно черед час_из каждой зооспоры выходит
новая’голая зооспора, но уже почковидной формы и с двумя жгутиками^
Прикрепленными в выемке сбоку: передний йз них перистый, а задний прос-
той (рис. 55). Это явление называют дji п л а н е т_ид д о м; оно известно
и у очень немногих архимицетов. Вторичные зооспоры могут плавать доль-
ше; привлекаемые хемотаксцческц^КЗтруппам я^ивотныхТоёй подплывают' к
ним, одеваются оболочкой и прорастают в^мицелий. Позднее на том же ми-
целии, который образовал зооспорангии^ формируются половые—органы:
оогонии и антеридии. Они возникают на одной и той же ветви или на разных
и" отделяются от них перегородками. Оогоний имеет шаровидную форму;
внутри него образуется несколько одноядерных яйцеклеток, на формирова-
ние которых идет все содержимое оогония. Антеридии образуется на конце
гифы, подрастающей и прикладывдющейся/к оогонию; он имеет короткоци-
линдрйческую форму, содержит густую цитоплазму и много ядер. Из анте-
112
п ш J
PACT
и; I;
Рис. 55. Сапролегния:
1 — выход воосподы та воосоо-
рангод; я — вервиям вооспора;
*2" вторичная аоосрора; 4 —оою-
ввй с антсридиями
поры) внутрь его врастают
ггйеЬья; пбСЛе разрыва обо-
лочки на вершине этих выростов часть цитоплазмы антеридиев с одним ядром
сливается с яйцеклеткой и оплодотворяет ее. Оплодотворенные' яйцеклетки
одеваются оболочками и превращаются7 вросподу. прорастающие после пе-
риода покоя в короткую гифу, дающую вскоре зооспорангий. Перед прорас-
танием ооспоры в неи происходит редукционное деление ее диплоидного ядра.
ридия черев более тонкие места оболочки оогония
трубчатые выросты; пни направляются к яйцеД
У ряда родов сапролегниевых наблюдается подавление первичных гру-
шевидных зооспор. У дажш (Achlya) они, не образуя жгутиков, выползают
амебообразно из зооспорангвйт и здесь Же одеваются оболочками, а затем
образуют вторичные зооспоры., У Ашствмрусд (Dtctyuchus) первичные зоо-
споры одеваются оболочками уже внутри ^вооспорангия, и из них выходят вто-
ричные зооспоры, каждая через свое отверстие, а опустевший зооспорангий
остается многокамерным,
Сапролегния, ахлия и некоторые другие могут при подходящих для
этого условиях жить и как паразиты на рыбьей икре, мальках рыб, поранен-
ных рыбах и т. п., нанося этим ощутимый вред на рыбных заводах и в ак-
вариумах.
Виды рода афаномицес (Aphanomyces) паразитируют на водорослях
и на корнях некоторых высших растений. Лептомшпус (Leptomitus lac-
teas) — характерный обитатель сильно загрязненных клоачных вод.
5-Й ПОРЯДОК — ПЕРОНОСПОРОВЫЕ (PERO NO SPO RALES)
— । iw .iw.iiiiw 1 ........
ин
ъ очень немногие пероноспоровые живут в воде; подавляющее
CZL’J
^люразом_"паразитами на высших растениях. “
г В разнообразии пероноспоровых можно найти переходы от примитив-
ных к эволюционно вышестоящим формам. Например, прослеживается пе-
реход от сапрофитов к факультативным и далее к облигатным паразитам;
переход от мипедшя’. находящегося рнрсу^рм^ впутрйклетпйндмуЦр-
целию без ^гаусториев, быстро убивающему клетки растения-хозяина, и
далее ^межклеточному мицелию с гаусториями, характерному для более
типичных паразитов, медленно вызывающих отмирание растений-хозяев.
При бесполом размножении лишь у немногих пероноспоровых образуются
цилиндрические зооспорангии, не отделяющиеся от произведшей их гифы;
\ у большинства же зооспорангии резко отличаются от несущих их гиф, от-
\ падают от них и переносятся воздушными течениями, как конидии. Поэто-
му и адаД™ на которых они образуются, называют кон hjjmj^h о о
....... ' ------------------- ..............
Г
г г
ОТДЕЛ —ГРИБЫ
<13
ц а м и. У одних пероноспоровых зооспорангии-конидии вырабатывают,
какТгравило, зооспоры; у других в зависимости от внешних факторов могут
развивать зооспоры или прорастать прямо в гифу; наконец, у наиболее
приспособившихся к наземному образу жизни спорангии проржууяшт всдг-
да в.гифу, не образуя зооспор и, следовательно ~не нуждаясь в капельно-
жидкой воде. Зооспоры пдрпттпртопоных ПОЧКОВИДНОЙ ЙОРМЫ с ДВУМЯ ЖГУ-
г г
~~"11ри половом размножении, кая и у сапролегниевых, к шаровидному
оогонию прикладывается антеридий, образующийся почти у всех на" том же
г. г
________________________________________________________ пероноспо-
ровых дифференцируется на яйцевую (внутреннюю) и периферическую (п е»
р и п л а 8 м у)1. Таким образом на образование гамет идет не вся цито-
плазшг
К пероноспоровым относится несколько сотен видов.
У большого рода питиум (Pjjthium) шаровидные иди грушевидные
зо&аюрашяж, образуются они на концах спорангиеносцев, мало отличаю-
щихся от ветвистых гиф. Зооспорангии, отрываясь, могут переноситься
пр врядуту, как конидии» ипорястяк» ДНИ, ряяпивая ЗООСПОРЫ. Ряд ВИДОВ —
сапрофиты. Многие виды растут на корнях сеянцев высших растений, в
том -числе и культурных (свекла, капуста, редис, салат, табак, горчица,
кукуруза и др.), вызывая у них заболевание, называемое корнеедом.
В роде (йитофтора (Phytophthora) ахо^мятяевпе^а больше отли-
чаются от гай мнцряия. чем у питиума, ветвятся симпрдиально.
Наиболее известный вид этого р о д a—PhytopKifiora infestans (рис. 56)—
вызывает опасное заболевание ботвы и клубней картофеля. Заметное
развитие болезни проявляется осенью, когда картофельная ботва чернеет и
И: 17 И
*-Рко. 56. Фитофтора (Phytophthora infestans):
1 — разрез листа картофеля с выходящими из устьица но-
нидненосцами; 2 — конидия (зооспорангий); 3 — зооспора;
4 — прорастание конидии зооспорами: & — прорастание
жоиидви гифой
Рис. 57. Конидиеносцы:
1 — олазмопара; г — пероноснора
1 От греч, «пера» — вокруг, около,
114
НИЗШИЕ РАСТЕНИЯ
Рис. 58. Альбуго:
1 — конидиеносцы в листе растения-хозяина
(часть разреза листа); 2 — прорастание кони*
дай; 3 — зооспоры; 4 — прорастание зооспоры;
5 — оогоний (с яйцеклеткой и периплазмой) и
антеридий; в — ооспора в оогонии
левания) картофеля. Борьба с ней
ских мероприятий, опрыскиванием
I быстро отмирает. Зооспорангии, по-
1 падая на почву и прорастая, зара-
жают клубни картофеля, особенно
при копке его. Ооспоры гриба в
растении не образуются, но в почве
могут развиваться ооспоры и геммы,
а кроме того, мицелий может зимо-
вать на растительных остатках. Пер-
вичное заражение картофеля прои-
сходит через почву; заболевание
может передаваться также через
клубни картофеля, у которых зара-
жена часть глазков; после развития
здоровой ботвы из здоровых глазков
развиваются больные побеги из за-
раженных глазков; они являются
источником новых заражений, и
этим отчасти объясняется позднее мас-
совое развитие картофельной болез-
ни. Phytophthora infestans^ зане-
сенная в Европу из Северной Аме-
рики в ЗО-х годах XIX в., нередко
вызывала эпифитотии (массовые забо-
ведется как при помощи агротехниче-
мстении бордоской жидкостью, так и
выведением и распространением в тутРурс устпйэдртых сортов- _—
Plasmopara viticola (рис. 57) — опасный паразит виноградной, .лозы,
вызывающий ее заболевание, называемое милдью.. Гриб, завезенный из Сё-
верной Америки в Европу в 70-х годах прошлого века, поражает листья,
усики и ягоды; зимой он сохраняется мипелием в ветвя^ р почках, а также
в виде ооспощ которые в больших количествах (до 200 на 1 мм3^образуются
в листьях. Борьба ведется путем предохранительных опрыскиваний расте-
ний бордоской жидкостью.
Род пероноспора также содержит паразитные виды. Одним из наи-
более обычных видов является Peronospora parasitica — сборный вид, по-
ражающий многие к^естоодезище—настоящее время этот вид разбивают на
несколько (около бО)оолее мелких видов, каждый с достаточно узкой специа-
лизацией относительно хозяев. На луке паразитирует Р. schleideni, зиму-
ющая в луковицах; на свекле — Р. schachtii, на клевере — несколько ви-
дов пероноспор.
Род цистопус, или алъбуго (Cystopus, или Albugo), резко отличается
от прочих пероноспоровых своим конидиальншг .сдомнощением (рис. 58).
Его конидиенрсцы, представляющие собой удданенцо-вздутые окончания
гиф, располагаются довольно тесным сдоемпод эпидермисом растения-хо-
зяина и здесь базипетально отчленяют цепочки конидий. На "растениях-хо-
зяевах в этих местах видны белые волдырики. Под напором конидий эпидер-
мис, разрывается, и_разъединяющиеся конидии разносятся воздушными
течениями. Прорастают они почти всегда .жюсцоравд.
Albugo Candida часто встречается на различных крестоцветных.
Порядки Monoblepharidales^ Saprolegniales и Peronosporales на осно-
вании сходства в оогамности иногда объединяют в подкласс 0^тисМИмг
который противополагают подклассу Zuzomucetidae (зигомицетам) с харак-
терным половым процессом — з и г о га м и е и я. Зооспор у зигомцдетов
1 Phytophthora Infestans — гриб . гетероталличнып. В Мексике мицелии обоих
полов встречаются одинаково часто, образование ооспор не представляет редкости»
В США. Канаде, Западной Европе и в СССР образованию ооспор, по-видимому, препят-
ствует отсутствие мицелия одного из полов.
2 От греч. «зигон» — ярмо, перекладина
ОТДЕЛ — ГРИБЫ
115
Sill
никогда не бывает. При бесполом размножении образуются спорангиоспо-
ры, реже конидии? В оболочках клеток зигомицетов есть хитин и нет целлю-
лозы. Почти" всё зигомицеты — наземные видь^.
К зигомицетам относится около 500 видов, распределяемых между
4 порядками. Ниже рассматриваются два: мукоровые (Mucorales) и энтомо-
фторовые (Entomophthorales), “
- МУКОРОВЫ* _ШЗ) ™
Мицелий у мукоровых хорошо развит, обычно неклеточный. При бес-
полом . размножении образуются неподвижные споры, у подавляющего
большинства в спорангиях (спорангиоспоры).
Большинство мукоровых живётТв" почйе, на растительных остатках, в
навозе; иногда они образуют так называемые головчатые плесени на пищевых
продуктах, некоторые паразитируют на других мукоровых, на человеке и
животных. В 1 г почвы содержатся тысячи зачатков мукоровых, и грибы
эти наряду с другими почвенными грибами и бактериями участвуют в почво-
образовательных процессах.
Род мукор (Мисог) — наиболее крупный по числу видов. Пред-
ставители его повсеместно распространены в почве и в навозе. Обильно раз-
ветвленный мицелии мукора находится в субстрате, а наверх — в воздух —
он направляет отрицательно геотропичные длинные спорангиеносцы, рас-
ширяющиеся в зачаток спорангия (рис. 59). Плазма вместе с -многочислен-
ными ядрами в ^юлодомспорангвм перемещаете^ фбсцовном .к.его перифе-
рии и "образует многочисленные периклйнные вакуоли. Слиянием послед-
них образуется щель, ограничивающая внутреннюю, почти безъядерную
часть цитоплазмы. Эта внутренняя плазма выделяет свою оболочку. В ре-
зультате внутри спорангия образуется так называемая колонка. Содержимое
спорангия раскалываетсялна огромное количество мелких частей, содержа-
щих по нескольку ядер, одевающихся' тэболочкамйГИ"СТй^№яЩих^ спорами
различной формы: шаровидными,'' эдШшсовд^ьными и ^ругидш у разных
видов. Оболочка зрелых спорангиев расплывается, споры освобождаются.
* ----------------_ . ____
t
Рис. 59. Мицелий мукора со спорангиенос-
цами
Рис. 60. Мукор:
1 — подвески, отходящие от гиф мицелия; 8 — подвес-
ки и гаметангии; 3 — зигота о подвесками; 4 — про-
растение зиготы
116
НИЗШИЕ РАСТЕНИЯ
Рис. 61. Пилобояюс:
1 — спорангиеносец со спо-
рангием; 2 — отбрасывание
спорангия
том же мицелии.
^Рдамноакавие мукоровых происходит главным
образом бесполым путем* Половой процесс наблюда-
ется у них редко. <)то зависит от того, что" большин-
ство видов их гетероталличны, а половой процесс
возможен лишь в случае, если близко окажутся два
мицелия противоположных половых знаков. В таком
случае на двух соседних гифах мицелиев противо-
положного пола образуются короткие выросты, ;
растущие навстречу друг другу (рис. 60). На рас-
ширенных соприкясяттцихся концах ^ЛЩДРНЯетСя
^^егородадмя пп-ддипй мппгодддрцпй
йазываемые г а м е т а н гни : гаметы в них не
дифференцируются. Оболочки гаметангиев в месте
Соприкосновения" растворяют^яГ'^ддержтде^дбоих
гаметангиев сливается; под старой оболочкой их
образуется но вря пну х-трехгл пиная бурая оболочка
получившейся зиготы: в ней остаются те короткие
участки мицелиев, на которых образовались гаме-
тангии, их называют мп две сдам и или суспензорами. У го-
моталличных видов, копулирующие гаметангии образуются на одном и
4 Ядра ..гаметангиев BJBHTOTe сливаются* После длительного периода по-
коя она прорастает, чему предшествует редукционное деление диплоидных
ядер. Из зиготы вырастят* w еттпраотитг на вершина- Этот
первый после полового процесса спорангий называется аа Он
имеет обычное строение, но существенно отличается от сппрянгиев беспплп-
го размножения генетической разнородностью спор. Половой заряд (+ или—)
у гетероталличных мукоровых определяется генотипически, вследствие че-
го в зародышевом спорангии оказываются споры + и — полового заряда, в
то время как в спорангиях бесполого размножения они все имеют одинако-
вый половой знак.
Пилоболюс (Pilobolus), растущий на навозе, имеет короткий, в верхней
части сильно грушевидно-вздутый спорангиеносец. с маленьким .черным
спорангием (рис. 61). Вследствие сйльновоэрастающего тургорного давле-
ния спорангиеносец разрывается на вершине и со струей жидкости отбрасы-
вает весь спорангий до 1,8 м в высоту.
Среди мукоровых наряду с развивающими многоспоровые спорангии
есть и такие, у которых спорангии содержат мало и даже одну спору; пос-
ле,
е равнозначны
конидиям.
Вероятно, эволюция бесполого размноже-
ния в порядке Mucorales шла по этому пути.
У некоторых родов мукоровых имеется гетерогамная зигогамия: один
из сливающихся гаметангиев и его подвесок крупнее другого.
У мортьереллы (Mortierella) вокруг зиготы развивается защитная кора
из вегетативных гиф и получается нечто вроде маленького плодового тела.
У некоторых зигомицетов, кроме того., зигоспорой становится не про-
дукт слияния гаметангиев, а его вырост.
Ряд видов мукоровых выделяет амилолитические ферменты, гидроли- '
зующие крахмал; они применяются вместо солода в спиртокурении. Неко-
торые мукоры (Mucor javanicus, М. rouxianus hl др.) могут еще далее сбра-
живать эти сахара с образованием спирта; они применяются вместо дрож-
жей в странах Юго-Восточной Азии для получения алкогольных напитков.
Некоторые виды мукора разлагают пектиновые вещества и являются
активными участниками так называемой росяной мочки льна и конопли.
nigricans) часто образует черную плесень (спорангии
у него темные) на овощах, фруктах, коробочкахи^врдокДДХЗлддчзднИВяд
прорастаюншх сшденах. '
“Некоторые мукоры и еще чаще виды абсидии (Absldia) вызывают иног-
да заболевания бронхов и легких у человека.
ОТДЕЛ —ГРИБЫ
117
Ряс* 62. Эмпуза:
1 — пораженная йуха; 2—кожв-
ца мухи о конидиеносцамк — жн;
Л — конидненосец. отчленяющий
конидию; 4 —развитие мицелия
на конидия; л —мицелий» выде-
ленный из жировых масс мухи
7-Й ПОРЯДОК — ЭНТОМОФТОРОВЫЕ (ENTOMOPHTHORALES)
зрелом
К
' состоянии имадт^пддрретаиа перетурудки^или распадается на отдельные мно-
; гоядерные или „одноядерные, почкующиеся или делящиеся^ клетки. Беспо-
лое размножение происходит только кощдияш. Наиболее крупные поГчисй
лу видов р оды амш/ад (Emnusa} и &нтомо^)тора (Entomophthora), парази-
тирующие на насекомых. На комнатных мухах часто встречается, особенно
осенью, Ттрпза пшзсае (рис. 62). Мицелий гриба распадается в жир^ром
тедемухйна_отдельные ппчкуютци^ся! илиделящиесяклетки. Изних вы-
растают слегка булавовидное конидиеносны, высовывающийся из дыхатель-
ных отверстий и через тонкие места хитинового покрова мухи; благодаря
Нередко осенью на стеклах окон можно видеть мертвых мух, окруженных
белым порошком конидий. Половой процесс, описанный у некоторых дру-
гих видов, состоит в копуляции при помощи коротких выростов двух кле-
ток и в образовании покоящейся толстостенной зиготы.
Некоторые энтомофторовые паразитируют на гусеницах шелкопрядов,
на саранче и других насекомых. Неоднократно делались попытки культи-
вировать их и применять для борьбы с этими вредными насекомыми, но пока
практически ценных результатов получить не удалось.
3-й КЛАСС — АСКОМИЦЕТЫ, ИЛИ СУМЧАТЫЕ ГРИБЫ
(ASCOMYCETES)
Большой класс аскомицетов, содержащий около 25 000 видов, характе-1
ризуется унпгокдеточным мицелием и наличием сумо к или асков
(стр. 107). Наиболее распространенная форма цикла развития сумчатых
грибов слагается из следующих этапов. Спора, возникшая в сумке, прорас-
118
НИЗШИЕ РАСТЕНИЯ
4 б в
Рис. 63. Эре маску с (Eremascus fertilis);
I — 6 — последовательные стадии полового процесса и
развитие сумки
тает и дает начало гаплоидному
МИЦелИЮ, СОСТОЯЩему ИЗ мипгпя-
дерныхили одноядерных клеток.
Это — вегетативное тело гриба.
У подавляющего большинства оно
находится в субстрате. Мицелий
гриба способен к размножению
бесполым путем. У сумчатых гри-
бов щирдао распростр£цены кошь
дйальные .спороношения.
"Половой процесс £
разных „групп сумчатых грибов
протекает различно." У простейших
сумчатых он сходен с половым процессом зигомицетрд. и состоит в слиянии
двух клеток мицелия, сопровождающемся слиянием их ядер (рис. 63). Зигота
в этом случае не переходит в состояние пойоя, как у зигомйцетов, а^коцуля-
ционное ядро ее делится редукционное в результате трех последовательных
делений образуются 8 гаплоидных ядер и далее столько же спор. Зигота,
таким образом, становится сумкой (аском),
У большинства аскомицетов в половом процессе происходит слияние
недифференцированного на гаметы содержимого женского и мужского по-
ловых органов. При этом сливаются только плазмы, слияние же ядер не про-
исходит. Вместо этого каждое из женских ядер сближается с мужскимг бля-
годаря чему образуются пары 'ядер — д и к а р и о н ы 1. Из женского
полового органа, где возникли дикарионы, вырастаюгособые нити, называе-
мые а с к о г е нны aljl куда и входят дикарионы. Между
дикарионами в аскогенной гифе появляются поперечные перегородки, бла-
годаря чему она оказывается состоящей из двухъядерных клеток. На концах
аскогенных гиф из двухъядерных клеток довольно сложным путем образу-
ются сумки. В клетке, представляющей зачаток сумки, осуществляется вто-
рой этап полового процесса — кариогамия. Получившееся диплоидное яд-
ро делится трижды, причем происходит редукция числа хромосом. Вокруг
восьми (уже гаплоидных) ядер обособляется цитоплазма, она покрывается
оболочкой и этим завершается образование сумчатых спор (аскоспор).
Таким образом, в результате единичного полового акта образуется много
сумок и сумчатых спор.
Итак, если в цикле развития низших аскомицетов, как и у фикомицетов,
доминирует гаплоидная фаза и диплоидна лишь зигота, то у подавляющего
большинства аскрмипетов между длтельдой гаплоиднои фазой и диплоид-
ной есть еще кратковременная дккАрплнтштег.кйя фяяя^ когда в клетках ас-
_ кргв!нных нитей находятся пары несливщихся ядер.— дикарионы Днередко
ее приравнивают к диплоидной фазе, что не совсем правильно).
У большинства аскомицетов образуются плодовые тела, в одних слу-
чаях — непосредственно на мицелии, в других — на поверхности или внут-
ри особых специальных плотных переплетений гиф, называемых с т р о -
м а м и или л ожа м й. Стромы" Рбё&печивШбт лучшее питание и защиту,
развивающихся сумокГ
У многих высших сумчатых грибов половой процесс более или менее ре-
дуцирован. Но и в таких случаях все-таки происходит формирование дика-
рионов, развитие аскогенных гиф, и в формирующейся сумке совершается
слияние ядер, редукционное деление и формирование аскоспор.
Характер цикла развития, в частности наличие или отсутствие дикари-
онтической фазы, отсутствие или наличие плодовых тел и расположение в
них сумок дают материал для суждения об относительной высоте организа-
ции той или иной группы сумчатых грибов и отчасти об их родственных свя-
зях. В тех же целях используется и строение сумок. У одних аскомицетов
1 От греч. <дис» — дважды, вдвое, «карион» — орех, ядро.
ОТДЕЛ — ГРИБЫ
119
II
сумка является органом, лишь производящим споры. Оболочка такой сумки
разрушается или ослизняется, благодаря чему и происходит освобождение
спор. Такие сумки, следовательно, не участвуют в рассеивании спор. Сум-
чатые грибы этого рода некоторые микологи характеризуют как протуни-
катные1 и противополагают их эутуникатным. У последних сумки—аппа-
рат не только продуцирующий споры, но активно их разбрасывающий.
Большинство аскомицетов живет сапрофитдо _на отмерших травянистых
и древесных растениях, в почве, на пищевых продуктах (плесени), в саха-
ристыхйстечениях растении (дрожжи) {многие являются паразитами на
растениях, реже—на животных и человеке. Многие виды аскомицетов в тече-
ние вегетационного периода живут как паразиты на живых растениях и раз-
множаются в это время конидиями; после отмирания растений-хозяев разви-
тие гриба не прекращается, но он питается уже сапрофитно и к весне обра-
зует сумчатые спороношения.
Аскомицеты делят на 2 подкласса.
1-Й ПОДКЛАСС - ГОЛОСУМЧАТЫЕ (PROTOASCOMYCET1DAE,
ИЛИ CYMNOASCOMYCET1DAE)
Подкласс делится на 2 порядка.
(>Й ПОРЯДОК>Л1ЕРВИЧНОСУМЧАТЬЩ ИЛИ ЭНДОМИЦЕТОВЫЕ (PROTASCALES,
ИЛИ Е NDOM YCETALES)^
1
Первичносумчатые живут преимущественно ^сахаристых выделениях
растений. Не у всех первичносумчатых развивается многоклеточный ми-
целии, ряд относящихся сюда грибов — одноклеточные растения. У мице-
лиальных форм бесполое размножение осуществляется почти всегда оиди-
ями. При половом процессе соседние клетки образуют копуляционные
{ветви, и в итоге оплодотворения образуется специализированное спороноше-
ние — сумка. В деталях половое воспроизведение в разных семействах
протекает неодинаково,
В теоретическом отношении интересны половые явления всемейст-
ве диподаско&ых (Dipo^zscaceae), в частности у pipodasc^s albus (рис. 64)—
грибка, обитающего в слизистых истечениях деревьев. В этом случае
соседниеклетки мицелия образуют копуляционные ветви, отчленяющиеся
перегородками, л становящиеся таким образом 10—12-ядерными гаметан-
гиями. Они, соприкасаются друг с другом, и в месте контакта возникает
отверстие, через-которое ядра одного гаметангия, принимаемого ааимужскоц,
переходят внутрь- другого» „женского. Однако сливаются. только по одному t .
ядру каждого из гаметангиев. Женский гаметангий образует длинный вырост, / -
который становится .сумкой. Копуляционное ядро.многократно.делится, при
этом, вероятно, происходит редукцияхромосом. Многочисленные потомки
копуляционного ядра становятся центрами образования^ сумчатых хлор.
Прочие ядра впоследствии погибают. Число аскоспор велико- и неопреде-
ленно. Они освобождаются "через отверстие на верхушке спорангия. У дру-
гого вида диподаскуса (D. uninucleatus) K3&XWL. мицелия., и^. гаметангии
одноя^рные.ПЭднбядерность гаметангиев является постоянным признаком
в семействе ямдомицетовые (Endomycetaceae); сумка, образующаяся
в этом случае непосредственно шГзиготы,. содержит фиксированное (8 или 4)
число аскоспор.
Большое практическое значение и наибольшее распространение среди
грибов этого порядка имеют дрожже^ыегрибы. или сяжяролиц^ты^ выделя-
емые в семейство Saccharomycetaceae. У дрожжевых грибов нет ми-
1 От лат. «рго» — пред, перед, «tunica» — род одежды.
1»
штатив pact!
; I;
Я
Рис. 64. Диподаскуег
1 — формирование налогах органов; 2 —а —цено-
вой нроцесс; 4 — 7 — формирование сумки (0—де-
генерирующие гаплоидные ядра)
не л ия, а лишь одиночные боль-
шие овальны^ цветки, размио-
жающиеся по^ованиемиали у не-
которых (главным образом род
Schizosaccharomuces, распростра-
ненный в тёплых странах) делани-
ем. Если Клетки быстро почку-
ются и не успевают разъед
ся, то получаются короткие не-
прочные депочки клёми (вттб5у.
При недостатке питания _ и
большом доступе кислорода обра-
зуются аскоспоры по 4, реже по
в клаткдгРяттктпа предполагали,
что они образуются без полового
процесса, однако в настоящее вре-г
мя установлено, что у них прои-
сходит>_ копуляция двух клеток, У
одних видов это происходит толь-
ко перед спорообразованием; у
других копулируют аскоспоры, и
поэтому вся жизнь гриба прохо-
дит в дипольной фазе; у третьих,
ДЕржжей, ramgmrn^t^TK?, во-
зникшие из аскоспор, размножа-
образуя ряд
поколений", дбёле чего попарно
копулируют (со ПйпкгашйГядер);'
получившиеся диплоидные клетки
снова почкуются в течение очень
длительного времени, так что
здесь имеет место нечто вроде
чередования гаплоидного и дипло-
идного поколений с сильным прео-
бладаниемсВторого.
Дрожжевые грибы в сахаристых средах вызывают спиртовое брожение.
Наибольшее значение среди дрожжей имеет род сахаромицес (Sасе ha romyces),
в особенности два вида этого рода, содержащие йбкЯый пбГ многу -рае, — это
пивные, или хдебогаеипркые, дрожжи (5. cerevislae), применяемые в пивова-
рении, хлебопечении, спиртокурении, и винные дрожжи (S. pint, или
5. ellipsoidal*), применяемые в виноделии.
Первый вид существует только в культуре,' второй — живет на поверх-
ности и в соке различных сочных плодов1. При хлебопечении важно выде-
I s I:
I s I:
ление углекислого газа, который разрыхляет тесто и делает его лучше
пропекаемым и усваиваемым, спирт же улетучивается. При спиртовом'
брожении спирт накапливается в бродящей жидкости в количестве до 8—43%
(редко до 19%); дальнейшее выделение его подавляет развитие дрожжей.
В быту нередко разводят в подслащенном отваре чая так называемый
11'
lire
поверхности в виде плоских слизистых лепешек и образующий кисловатый
напиток, имеющий диетическое значение. Чайный гриб представляет со-
четание аспорогенных (не образующих спор) дрожжевых клеток и уксусно-
кислых бактерий- Последние, имеющие мощйб развитая капсулы, и при-
дают всему комплексу слизисто-хрящевидную консистенцию.
1 Советский специалист по дрожжевым грибам В. И. Кудрявцев считает, что
и винные дрожжи встречаются в соках ягод только в условиях виноделия.
отдел—грибы
121
Рже. 65. Дрожжи!
л—Saccharomyces оегетШае: а — от-
дольная клетка, б—псякованке» в —
вдгшш клеток, не разъединившихся
после почкования, а — аскоспоры в
сушее; в— Schlzosaccbaromyceg octoepo-
rus (копуляция и образование аскоспор)
Цервичиосумчатые являются» по-видимому, примитивными сумчатъь
ми грибАМИ, Д|ЮЯУошедиЙ1ДГ бТЧШКИХчи вмгемнцеяев, ^у*°КОТОрых зигота
^црорЯЗШШ непосредственно вслед за половым процессом. Такая зигота
ем преобразоваться в сумку. Дрожжевые грибы, по
и могла в дальне:
1111
всей вероятности, вторично утеряли мицелий» перейдя к жизни в форме
обособленных клеток. Эта морфологическая деградация сопровождалась,
ное использование ресурсов среды обитания.
2-Й ПОРЯДОК — ТАФРИНОВЫЕ ИЛИ ЭКЗОАСКОВЫЕ (TAPHRINALES,
ИЛИ EXOASCALtiS)
Небольшой порядок тафриновых характеризуется расположи™»*
дптюльнп плотным cjjpfrMj но без образования плодовых тел. Почти все таф-
риновые — паразиты на древесннЗГушУгениях, прштуШебтвенно из семейств
йОЗовыхДподсемейотва сливовых), березовых, буковых, Ивовых и на папорот-
никах* Мицелий их помещается у одних видов под кутикулойпоуяжвятпат
органов», у других проникает в глубже, лезкащие ткян^Ппряжептгё органы
растения-хозяина характерным образом видоизменяются, и на поверхности
их под кутикулой, образуются .сумки (рис. 66).
ТГо л о вой процесс и ядерный цикл у тафринопых отлуп1я«г>тс.я от боль-
Uli
ii:
Г1м,ия, а лдра образуют дикарион, который при прорастаншГдвухъядер-
ной клетки входит в гифу й размножается благодаря одновременному де-
лению обоих ядер; у других дикарионы образуются делением ядра, не соп-
ровождающегося делением клетки, или в аскоспоре, или в клетках гиф.
К a D и о г^ мия наступает только при образовании сумки. Таким образом
почти весь жизненныйцикл или большая часть его проходйт в дикарионти-
ческой фазе; диплоидны лишь развивающиеся сумки непосредственно эа
кариогамией; гаплоидны только аскоспоры и у некоторых — молодой ми-
целий.
Почти все виды тафриновых относятся к одному роду ткбрина
(Taphrina). Tanbrina deformans вызывает «курчавость» листьев у персика,
а и 11
122
НИЗШИЕ РАСТЕНИЯ
Рис. 66. Тафрина (Taphrina pruni
var. pad! (на черемухе):
1 — ветка с настоящими в деформиро-
ванными плодами; е — сумки на поверх-
ности дутого плода (в сумках почк уютен
аскоспоры)
пустыми внутри (дутые сливы, кармашки); разновидность ее (Var.paai) пора-
жает черемуху; Т. cerasl вызывает у вищнедцх деревьев. образование ведь-
миных метел. Мерами борьбы с возбудителями этих болезней является
удаление пораженных органов и опрыскивание фунгицидами.
Положение тафриновых в системе аскомицетов неясно.
2-Й ПОДКЛАСС — ПЛОДОСУМЧАТЫЕ (EUASCOMYCET1DAE,
ИЛИ CARPOASCOMYCET1DAE)
пи
Для плодосумчатых, как указывает их название, характерно образо-
вание плодовых тел, причем сумки образуются обычно в большом количест-
ве на" концах аскогенных гиф.
К плодосумчатым относится подавляющее большинство аскомицетов;
они отличаются разнообразием полового процесса, морфологией спороноше-
ний и образом жизни. Как характерный для многих тип полового процесса
принимают описанный во всех деталях лишь в 1912 г. немецким ботаником
Клауссеном половой процесс у пиронемы (Pyronema omphalodes),
раньше называвшийся Р. confluens* * — сапрофитного гриба из группы дис-
комицетов, нередко встречающегося на почве — на местах бывших костров1.
Половые органы у пиронемы образуются на мицелии группами. Каждая груп-
па состоит из нескольких пар мужских и женских половых органов^рис. 67).
Женский половой орган а дэ х иц ajut состоит иэ двух многоядерных кле-
ток:нШКПЯй,^бЬлее крупная, пузыревидно вздутая, называемая а с к 6 г о н о _м®,
и верхняя,, цилицдрическая. называемая тр и х о г и ной. Таметы цё даф-
ференцируются. Агсгеридий прикладывается к верШИЙУ ‘Грихогины, и его_со-
де£жимое переливается в аскс^н^через трихогину; в перегородке между
трихогииой и аскогоном образуется отверстие, содержимое трихогины
к этому времени дегенерирует. В аскогоне происходит слияние.венской
(принадлежащей аскогону) не мужской ^пришедшей из антеридия) прото-
плазмы, что рассматривают как первыи~этап полового процесса — njiaji-
могямию. Ядра при этом не сливаются, а располагаются попарно —
мужские и женские, образуя -дииариетан- Непосредственно 8а этим из
аскогона развиваются ветвистые выросты — аскогенные гифы.
1 Нахождение его на таких местах объясняют тем, что там уничтожены конкуренты,
подавляющие его развитие в других местах.
* От греч. «аскос» — мешок, сумка и «гонос» — рождение.
ОТДЕЛ — ГРИБЫ
123
В них переходят цитоплазма и дикарионы из аскогона, и дикарионы делятся,
причем оба ядра каждой пары делятся одновременно, и оси деления ядер
одной пары располагаются при этом параллельно. В результате получаются
новые пары, состоящие каэедая опять из мужского (по происхождению)
и женского ядра. Этого рода деления ядер повторяются; в аскогенных ги-
фах„образуются_ дюперечные._де^егородки, делящие их на двухъядёрные и
МНОГОЯДерныа-участки. Наконец, на концах аскогенных гиф довольно слож-
ным способом образуются сумки. Концевая клетка аскогенной гифьГ'из-
габается" крючком, и два её ядра располагаются в месте перегиба. Эти ядра,
составляющие дикарион, делятся одновременно, причем оси деления распо-
лагаются под некоторым углом друг к другу. В результате получаются че-
тыре ядра: два несестринских в месте перегиба крючка, одно в кончике
крючка и одно в основании делящейся клетки, ниже перегиба. Образую-
щиеся далее две поперечные перегородки отделяют базальную одноядерную
клетку в основании крючка, конечную, тоже одноядерную клетку кончика
крючка и среднюю двухъядерную клетку в месте перегиба. Последняя ста-
новится в дальнейшем сумкой: она вытягивается в длину, увеличивается в
размерах и оба ее ядра сливаются, что представляет заключительный этап
полового процесса — к a jj Qj^a-мию ^Затем получившееся диплоидное
9
Рис. 67. Оплодотворение и образование сумок и аскоспор у пировемы (Pyronema от-
phalodes):
I — группа поповых органов (а — аскогон, ™ — трихогина. ан — антеридий); 2 — содержимое антеридия
перешло через трахогину в аскогон; 3 — развитие аскогенных гиф (аса — аскогенные гифы); 4 — схема
образования крючков и сумок (лс — мужское, ядро; ж — женское); 5 — крючок, двукъядерная молодая
сумка и сумка о одним копуйяционным ядром (к — модуляционное ядро); * в — 3 — деление ядер в сумке
(первое, второе, третье); а — формирование аскоспор
124
; r Ki 111 s
В РАСТЕНИЯ
ядро делится трижды с редукцией числа хромосом, я вокруг получающихся
восьми гаплоидных ядер формируются особым образом восемь спор: лучи,
отходящие от центросомы на одном из полюсов ядра, заТЯвЙВв^ЧН&реэают»
из окружающей цитоплазмы вокруг ядер участки, приобретающие затем
оболочку»
Одноядерный кончик крючка и базальная его клетка сливаются, но их
ядра сохраняют свою самостоятельность.- Оти_дш^стринскиа-ядра_образуют
новый дикарион. Из получившейся таким образом двухъядерной клетки
йпрнстЗёт^выйкрючокили более длинная гифа, образующая на'концё-евое^с
крючок; этот процесс может "повторяться. Благодаря образованию аскоген-
ных гиф, делению дикарионов, образованию крючков сильно увеличивается
число сумок, возникающих вследствие единичного слияния содержимого
антеридия и аскогона.
В сумке не вся цитоплазма идет на обпааованиеспор. В неиспользованной
часттГ^'ёёГ^ и И1тл аз м е ко времени созревания сумки происходит фер-
ментация гликогена и образование сахара. Она поглощаетвбду,~~Ж^в^ней
возрастает тургорндё~ давление, В результате оболочка сумки на вершине
разрывается Дилй у некоторых вскрывается крыпВДШбйХ айсоспоры "выб-
расываются. на расстояиие^до 45 см.
Таким образом, в цикле развития плодосумчатых закономерно чере-
дуются три ядерные фазы: продолжительная — гаплоидная, в течение ко-
торой многие сумчатые грйбы размножаются бесполым путем; непродолжи-
некая; очень кратковременная — пиплоиддая (моло-
дая сумка с диплоидным ядром).
г~От указанного типа полового процесса наблюдается много отклоню
Сопоставление конкретных форм, в которых осуществляется половой процесс у
плодосумчатых, приводит к выводу, что в этой группе наблюдается тенденция
Г. I.
пируют или вовсе не развивается трихогина и антеридии; у некоторых
половые органы совсем отсутствуют,b таких случаях образование дикари-
онов происходит в пределах одного аскогона путем ассоциации его ядер.
Если половые органы отсутствуют, вегетативные клетки мицелия соединяют-
ся анастомозами и путем ассоциации ядер образуются дикарионы. Так или
иначе почти у всех плодосумчатых возникают аскогенные гифы, образуются
дикарионы и в формирующейся сумке происходит кариогамия.
Для некоторых аскомицетов характерен иной тип полового процесса —
I Г Г;
ми клетками типа конидий, содержимое которых через трихогину переходит
;|ш
i 1 Л ?
в аскогон.
Плвдовыа_~хела плодосумчатых аскомицетов имеют сложную морфоло-
гическую и цитологическую природу. Они образуются иаболее или менее
плотного переплетения гиф мицйд^яг. принадлежащих гаплоидной фазе
гриба; находящиесявйлЬдбвыхтелаХ аскогенные гифы и молодые сумки
до слияния в них ядер принадлежат к дикарионтической фазе, сумчатые
споры — снова к гаплоидной фазе.
Основные типы плодовых тел: к л ейстокар п и и* 1 (или клейсто-
теции) — вполне замкнутые, округлые; if ер и те g и грушевидные,
овальные, округлые, с узким отверстием на вершине; а п о те ди и 3—блюд-
цевидные, чашевидные, у некоторых лопатовидные или в виде шляпки на
ножке; сумки расположены тесным слоем — гимением на верхней открытой
поверхности их (рис. 68).
В развитии плодового тела сумчатых грибов выявлено два существенно
различных пути, коррелятивно связаных с некоторыми особенностями сумок
и их расположением.
1 От греч. «клейстоо — запертый и «карпою — плод.
8 От греч. «пери» — вокруг, около и «гене» — ящик, сумка*
1 От греч, «апо» — вдали от и «тек©» —> ящик, сумка.
t
ОТДЕЛ —ГРИБЫ
125
Рис. 68 Плодовые тела аскомицетов:
1 -- клейстокарпий; т — перитеций; 3 — апотеций (а — общий виц. б — разрез)
1. В большинстве случаев плодовое тело формируется за счет гиф,'
развивающихся из базальных клеток^женскогд полового органа <апхикарпа)
соучастием вегетативных гиф. Вся эта совокупность сплетенных гиф становится
ложем половых органов, аскогенных гиф и сумок и окружает их, не только
обеспечивая питание дикарионтической фазы и сумок, но и защищая их.
При таком развитии плодового тела сумки занимают некоторое общее про-
странство, ограниченное стенками плодового тела. У протуникатных пло-
составляя пучок или протяженный гимениальный слой. Кроме сумок
в полость плодового тела — между сумок — врастают окончания тонш
вететаташныхгиф, называемых п а р а ф и з д„мл!Г*06олочка сумки в этом
случае не дифференцируется на слой на всем протяжении, кроме верхушки,
где в связи с формированием сложного аппарата раскрывания наблюдается
расчленение оболочки на два слоя. Сумки в этом случае однопокровные
ГК
г, г,
Аскомицеты, у которых плодовое тело развивается описанным способом,
называются а с к о г КМЛ н_и (см. рис. 73).
2. По-иному формируется плодовое тело а с к о л о к улярны ха
(см. рис. 72) сумчатых грибов с двупокровными (битуникатнымн) сумками.
Оболочка сумки в этом случае на всем протяжении состоит из двух хорошо
НаружныислоУ ттнтаВ^*ж^ткии, нерастяжи-
мыд^ только ко времени созревания спор на верхушке растр^йШйЮЩйЙбя.
Йнутрркрдй "СДОЙ толстый и сильно растджирт^й. Сквозь отверстие на вер-
хушке сумки он высовывается в виде цилиндрического мешка и через апи-
кальное верхушечное отверстие выбрасывает одну за другой созревшие аско-
споры. Эта особенность сумок обычно связана с процессом формирования
плодового тела. Еще до того, как возникнут половые оргаш^_мкцедяй
образует плотноеисддедсрие,. ^внутри которого заклададзаются половые органы,
цосла.яего_ это строматическое1оздаз^аниепртобр^а1»^АряктйртоРттриярй>
™ jffPfloporo тсрта данного грибаТТГак развивающиеся плодовые тела было
предложено называть а с к о с т рома м и (или, сокращенно, аскомами).
Аскостромы, как видно из сказанного, существенно отличаются от плодовых
тел аскогимениальных аскомицетов. Если у аскогимениальных Сумчатых
грибов вйутри плодового тела в типичном случае имеется одна общая по-
лость, где помещаются сумки, то у асколокулярных каждая сумка или
иг,
1 От грея, «пара» — возле и ефиаис» — происхождение, рост.
8 От греч. «аскос» — мешок, сумка и лат. «докулюо — местечко, камера.
126
НИЗШИЕ РАСТЕНИЯ
которой является результатом сдавливания и растворения аскогенными
гифами и сумками окружающей их ткани. Несмотря на существенно иной
способ образования, строение аскостром может быть очень сходно со строе-
нием настоящих плодовых тел унитуникатпых сумчатых грибов.
Подкласс плодосумчатых аскомицетов подразделяют на порядки на
основании строения и развития плодовых тел и отчасти по строению и рас-
положению в них сумок. Число порядков в различных классификациях
неодинаково (до 20). Несколько обособленное положение занимает порядок
плектасковые, характеризующийся протуникатными сумками. В остальных
порядках они эутуникатные. Среди эутуникатных преобладают однопокров-
ные (унитуникатные), В нижеследующем обзоре некоторые порядки, фило-
генетически близкие или хотя бы конвергентно сходные, объединены в груп-
пы и отдельно не рассматриваются.
1-Я ПОРЯДОК — ПЛЕКТАСКОВЫЕ, ИЛИ ДСПЕРГИЛЛОВЫЕ (PLECTA SCALES,
ИЛИ ASPERGILLALES)
или
Цлодовн^^еда.— клейстокарпии с беспорядочно расположенными в них
среди бесплодных клеток округлыми сумками (рис. 69,26). Сумкипротуникат-
-^-^щде. Оболочки сумокраноразрушаются, аскоспоры л ежат в полости плодовых
. туг в освобождаются послесгшвания перидия (стенки клейстокарпия).
У,более примитивных плодовые тела зачаточные, с рыхлым перидием.
, Наибольшее практическое значение имеет семейство аспергилло-
вых (Asp#rgillaceae), к которому относятся обычнейшие плесневые грибы,
живущие в большинстве сапрофитно в почве, а также на разных пищедыхдро-
дуктах и других предметах, в обстандВКё, созданной человеком. Особенно
распространены многочисленные виды двух родов — аспепзилл (Aspergillus)
и (Penicillium), обычно встречающиеся в конидиальной стадии.
Сходные по внешнему макроскопическому облику, эти роды легко различа-
ются ™ ^пттъгтщеносцам, образующим вместе с конидиями характерные по-
/рошковатые налеты плесеней сизого, зеленоватого, желтого, черного
/ и других цветов. У _§сцергилла конидиеносец одноклеточный, шаровидно
I или обратногрушёвидно вздутый—на , вершине. ПоГповершости всего этрго
\ вздугшгфасположейы короткие цилиндрические или остроконечные клетки,
у-с т ер и г м ы, отшнуровывающие базипетальные цепочки одноклеточных
конидий (рис. 69, 2). У пеницилла конидиеносец многоклеточный, на верх-
нем конце дважды или трижды разветвленный в виде кисточки; конечные
клетки веточек, стеригмы, тоже базипетально отшнуровывают*"цепочки од-
ноклеточных конидий~(рис. 69,2).
Рис. 69. Плесневые плектасковые грибы:
I — асиергилл; а — конипденосец» б — клейстокарпий; s —•пешпшлл (конкдвеиосец)
отдал —грибы
127
Вредное значение плесневых грибо^ вызывающих порчу разнообразных
продуктов, и материалов. пбщеиянастшк Некоторые виды аспёргиллов
(A. bronihiales, A. malignis и др.) вызывают у человека заболевания дыха-
тельных^уэганов и ушных проходов — аеттёргиллр^тд Ряд видовимеетполез-
ное практическое применение; Aspergillus niger, Л. wentii выращиваются
в промышленных масштабах для получения лимонной кислоты; A. oryzae,
A. flavus и другие аспергиллы дают много амилазы, осахаривающей крах-
мал; Penicillium roqueforti и Р. camemberti используются в производстве
сыровдюкфора и камамбера. Громадное значение имеет Penicillium notatum
иобв&енно рГ chrysogenum как продуценты антибиотика пенициллина.
На лечебное значение зеленой плесени указывали еще *в конце 60—70-х
годов прошлого века русские врачи М а на с се и н и П о л от ебно в.
Однако на это не было обращено внимание. В 192& г. англичанин И. Ф л е-
м и н г обратил внимание на свойство Penicillium notatum подавлять рост
и размножение оолотастого, стафилококка — возбудителя нагпоенчч Лишь
во время второй мировой войны совместными работами Флеминга, Флёри,
Чайна и других было организовано в производственных масштабах полу-
чение пенициллина — одного из величайших достижений пауки за последнее
время. С тех пор во всех крупных странах ведется большая исследователь-
ская работа по селекции наиболее продуктивных штаммов пенициллина,
усовершенствованию способов получения и применения его, по изысканию
новых антибиотиков. Развилась новая отрасль промышленности — произ-
водство антибиотиков. Выход пенициллина на единицу продуцента увели-
чился по сравнению с начальным почти в 1000 раз. Лечебное действие
пенивдлдица, выделяемого притоп w«ir п среду, заключается
в подавлении способности бактерий - возбудителей сепсиса* .пневмонии, си-
бирской язвы, гонорреи, сифилиса и др» —ПГ^размшмиенанэ (бактериоста-
тическое действие).
Филогенетически аспергилловые (плектасковые) связывают с первично-
сумчатыми. По сравнению с последними, аспергилловые отличаются нали-
чием дикарионтической фазы в виде совокупности аскогенных гиф и нали-
чием плодовых тел.
2-Й ПОРЯДОК — ПЕРИСИОРИЕВЫЕ, ИЛИ ЭРИЗИФОВЫЕ (PERISPORIALES,
ИЛИ ERYS1PHALES)
'4и
У периспориевых, в отличие от плектасковых, сумки расположены в
клейстокарпия* не беспорядочно, а пудком; суженные основания сумок
обращены к нижней части клейстокарпия, а вершины — к верхней стороне
его. Это дает возможность активного разбрасывания спор. По созревании
их под напором возрастающего тургорного давления сумок ободочка .клей-
стокарпия в верхней части нлодового-теларазрывается, сумки высовывают-
ся и из них выбрасывяются споры- Мицелий у подавляющего большинства
периспориевых эктотрофный, живущий на поверхности листьев и других
органов растений.
Наибольшее практическое значение имеет семейство паразитных
мучнеросных грибов (Erysiphaceae). В пределах семейства есть виды, мице-
лий которых находится внутри тела хозяина, и только органы размножения
развиваются снаружи. У других (как у Phyllactinia), мицелий живет как
эктопаразит, но часть его гиф через устьица врастает в паренхиму поражен-
ного органа. У большинства же представителей этого семейства мицелий
находится только на поверхности наружных органов растений — листьев,
стеблей, реже плодов. Внутрь эпидермических клеток он пускает только
гаустории, при помощи которых гриб прикрепляется и питается. На мицелии,
на коротких приподнимающихся неразветвленпых конидиеносцах, разви-
ваются базипетально цепочки овальных конидий (рис. 70). Они переносятся
ветром и вызывают новое заражение; часть их падает на месте образования
и вместе с беловатым мицелием создает., мучнистый налет на поражен-
12В
НИЗШИЕ РАСТЕНИЯ
Рис. 70. Мицелий с гаусто-
риями
клетках хозяина),
эпидермически
ковндие-
носцами и конидиями мучне-
росного гриба микросферы
к названию этих грибов мучнеросными. Позд-
ных органах, давший основание
нее, в результате полового процесса, образуются клейстокарпии в виде малень-
ких, еле видных невооруженным глазом шариков, вначале рыжеватого, за-
тем черно-бурого цвета. От перидия клейстокарпиев отходят или обыкновен-
ные гифы, или более характерные нитевидные придатки, так называемые
подвески, загнутые на концах крючком, разветвленные вроде оленьих рогов
или иной формы (рис. 71). Форма этих придатков используется в систематике
мучнеросных. Аскоспоры выбрасываются из клейстокарпиев вышеуказанным
способом осенью или после перезимовки следующей весной. К мучнеросным
относится около 150 видов. В сапрофитных условиях они не живут и явля-
ются облигатными паразитами.
На листьях культурных и дикорастущих злаков паразитирует вид
(Erysiphe gramims). Подвески клейстокарпиев у него похожи на
'ТЙфы;в*клейстокарпиях — по нескольку сумок.
Р о д c^epemfna^(Sphaerotheca) тоже имеет подвески, похожие на гифы.
В каждом клестокарпии развивается лишь одна сумка. Spjiaerotheca mors
uvae, завезенная из Северной Америки в Западную Еврбпу и Россию-в
первом десятилетии XX в., очень быстро распространилась и вызвала мас-
совую гибель крыя^овтшка. Гриб поражает завязи, ягоды, молодые ветви,
образуя на них вначале белый, затем вскоре буреющий налет (буреют гифы
мицелия). Борьба с этим заболеванием заключается в обрезке и уничтожении
пораженных частей кустарника, опрыскивании его поздней осенью и ранней
весной (до распускания почек) 3%-ным раствором железного купороса или
л:»
'.Hi
Рже. 71. Клейстокарпии мучнеросных грибов:
1 — уницииула; 2 — микросфера; я — сферотека (носов разрыва перидия выеоапается едннспеивая у
•того рода сумка с аскоспорами)
ОТДЕЛ — ГРИБЫ
129
Г If
весной до цветения 0,1—0,2%-ным раствором мышьяковокислого кальция
или 0,3—0,5%-ным раствором кальцинированной (бельевой) соды. В нас-
тоящее время есть сорта крыжовника, сравнительно устойчивые к поражег
сферотекой.
Sphaerotheca pannosa имеет две разновидности: одна поражает розы,
другая — персики.
В роде унцинула (Uncinula) подвески клейстокарпием на кпппа спи-
рально загнуты. Uiicinuld necator завезена из Северной Америки в Европу
в середине ХТХ в.; это опасный паразит виноградной лозы, вызывающий
ее заболевание, известное под названием оидиум1 или пене лица. Борьба—опы-
ливание препаратами серы.
Урода лшкуосфеъа (Microsphaera) подвески клейстокарпиев на концах
дихотомически разветвленные, вроде оленьих рогов. Microsphaera
alphitoides, тоже завезенная в Европу из Северной Америки в начале нашего
века, поражает дубы, покрывая листья и молодые ветви белым налетом.
Особенно опасна она для сеянцев и молодых дубков. Борьба, как и с боль-
шинством мучнеросных грибов, — опыливание препаратами серы.
В филогенетическом отношении периспориевые можно рассматривать
как результат дальнейшей эволюции плектасковых. По сравнению с пос-
ледними у них выработалась способность к активному выбрасыванию аско-
спор благодаря 'Изменению формы сумок (более удлиненных) и их располо-
жению параллельным пучком.' Согласно другому мнению, клейстокарпий
рассматривается как результат превращения открытого плодового тела (апо-
теция) в закрытое. В этом случае замкнутость плодового тела расценивается
как вторичное явление, а параллельное расположение сумок как унаследо-
ванное от предков. Что касается эволюции внутри порядка, то она состоит
в приспособлении к зктопаразитическому существованию мицелия и к вы-
работке подвесков, непохожих на гифы, вероятно играющих некоторую роль
в распространении плодовых тел, а следовательно, и сумчатых спор.
Ilf
иг.
ГРУППА ПОРЯДКОВ — ПИРЕНОМИЦЕТЫ (PYRENOMYCETIIDAE)
К пиреномицетам относят аскомицеты с плодовыми телами перитециями,
имеющими грушевидную, кувшиновидную, овальную форму и открывающими-
ся на вершине узким отверстием. На дае их пучком расположены сумки, ко-
торые поочередно подрастают и в отверстие активно выбрасывают споры.
Оболочка сумки, выбросившей споры, спадает, шГее'месте подрастает дру-
гая, и т. д. Плодовые тела находятся непосредственно на мицедии. У дру-
гих оци помещаются на поверхности или, чаще, внутри особого плотного де-
реплетениягиф, -так-называемой стромы. (или ложа), имеющей различную
величину, форму и окраску и обеспечивающей сумкам лучшее питание и за-
щиту. Стромы не следует путать с плодовыми телами. Кроме сумчатогоьспо-
ррношения, у очень «"р^^миц^т^г раятшуяются предшествующие
ему конидйальные спороношения.
пиреномицетам относится около 14 000—15 000 видов. Кроме сапро-
фитов, среди пиреномицетов очень много паразитов. Уитхлх_лидав~в~д$че-
ние вегетационного периода гриб живет как паразит и образует в это время
Лги
которых частей его, например листьев) гриб продолжает на нем и дальше раз-
виваться, уже как сапрофит, и к весне образует сумчатое спороношение.
Пиреномицеты включают несколько порядков, одни из которых харак-
теризуются асколокуляркостью, а другие — аскогимениальностыо (стр. 125).
^Аскожжуляриые. Вентурия (Venturia порядок Pseudospo^
riales), возбудитель партпи^ябдокДУ. inaequalis) и груш (V. pyrtna) — од-
ного из самых распространенных заболеваний этих культур. Летом и осенью
гриб встречается в конидиальной стадии, носящей (как несовершенный гриб)
:м
1 Oidium — название конидиалъного спороношения мучнеросных грибов
5—602
I
।
t
130
НИЗШИЕ РАСТЕНИЯ
на яблонях название Fusicladium dendriticum, на грушах — Т. pyrlnum
(рис. 72). На верхней стороне листьев и наПйлодах образуются буровато-
зеленоватые пятна с бархатистым налетом конидиеносцев. На плодах под
ними образуется защитный сероватый или буроватый слой пробки, часто
обнажающийся вследствие сбрасывания наружного пораженного слоя. Ткань
плода в этих местах становится деревянистой, при сильном поражении пло-
ды недоразвиваются, растрескиваются и товарные качества их резко сни-
жаются. На опавших листьях с осени развиваются аскостпомы (плодовые те-
л§) и половые органы в них; в это же время происходит переход содержи-
мого В ЦДДдгон.ТУйразование аскогенных гиф осутцествляется^шдц>
следующей^весной; они врастают в молод^ГасйдСТроиу, где Образуются сум-
йии аскоспоры. В сформированном видеасЦПГСтр^^ округ-
лая, сосочковидным отеерстием она открывается наружу Г В' нёи1нахо дятся
суМкиг расположентге^параллельно друг другу. bl псевропдряфтаы. Все
плодовое тёЛ и нигружшо в уже "мертвую ткань листа. В целом оно напоми-
нает перитеции аскогимекиаЗГЁЯ^'ёумчмых грибов. На одном листе может
Phd. 72. Парша яблонь, вызываемая вентурией (Venturia inaequalis):
1 — ва даете; я — на плода; 3 — поперечный разрез пятна с конвдиальным спороношением гриба н от-
цельно конивданосцы с воевднямн; 4—псевдопернтедай в тканях лнста
ОТДЕЛ —ГРИБЫ
131
2
Рис. 73. Перитеции:
1 — плевраге (перитедиа в разрезе); в—зпплов. часть стромы (на влагалище ялава) с погруженными
нее перитециями
г.г,
образоваться до 2—3 млн, двухклеточных аскоспор; выбрасывание аскоспор
и первичное заражение ™и мпгртдлдтьг.я вте^ятоМ^«щя тжЯпдгйа. Борьба—
уборка и уничтожение пораженных листьев; опрыскивание растении весной
фунгицидами. ?
^МшсоаЬереллл (Mpeosphaerella) — большой р о д из того же порядка
(но из другого семейства), содержащей свыше 1000 видов. Конидиальны?
стадии вызывают пятнистость на листьях высших растений. /
Аскогимениальные. Сордари& (Sordaria) и илеадага (Рleurage, или
Podospord) — широко распространенные копрофильные грибы из п о р я д-
к a SyhaerialeS' живущие на экскрементах травоядных животных. Их тем-
но-бурые перитеции, расположенные прямо на мицелии, без стромы, има-
ют грушевидную форму с оттянутым и немного согнутым носиком (рис. 73, I).
Нектария (Nectria cynabarina) из порядка Sphaeriales — широко рас-
пространили сапрофит, который обита'й’гнн шмврших ВБТвях лисхвецных
древесных пород. Может быть и паразитом. Другой, реже встречающийся вид
нектрии —ЦУ. galllgena — вызывает на стволах плодовых деревьев незара-
стающие раны с утолщенными краями, называемые раком.
Эпиялоэ (Epichloe typhina) из "пГЬ pFff’f —паразит
луговых злаков, вызывающий так называемую чехловидную болезнь. В на-
чале лета на влагалищах верхних листьев образуете^7цилиндртеская до
5 см длиной беловатая строма, одевающая их как бы чехлом; на ней разви-
. ваются одноклеточные эллиптические конидии; позже строма становится
золотисто-желтой и в ней развиваются целиком погруженные в нее много-
численные перитеции (рис. 73, 2). Соцветия на пораженных стеблях не раз-
виваются. Гриб приносит вред при культуре луговых злаков на семена.
Спорынья (Clavicevs purpurea) — паразит ржи и многих дикорастущих
злаков/ лорошо заметной и общеизвестной формой этого гриба являются
иг.
изогнутые, фиолетово-коричневые, внутри белые. После воздействия низ-
ких зимних температур склероции в соответствующих условиях прорастают;
из них развиваются 10—30 пурпуровых стром в виде шадовддяых.головок,
сидящих на довольно длинных (1,5—4 см) ножках. На периферии головки в
132
НИЗ
пг>
В РАСТЕНИЯ
нее погружены эллиптические, суженные кверху перитеции; в них находят-
ся узкоцилиндрические сумки с восемью нитевидными аскоспорами в каж-
- дой (рис. 74). По созревании сумки высовываются из отверстия перитеция и
выбрасывают аскоспоры, которые разносятся ветром и заражают рыльца
злаков. Мицелий проникает в завязь, разрушает ее и образует конидиальное
спороношеиие, описывавшееся раньше как самостоятельный несовершенный
гриб сфацелия (Sphacelta segetum). При этом выделяется сладкая жидкость
(медвяная роса), на которую прилетают насекомые. Перелетая на другие
растения, они переносят туда и мелкие бесцветные конидии, вызывающие но-
вые заражения. Мицелий гриба, начиная с нижней части завязи, постепен-
но плотно сплетается и образует к концу лета склероций. Вред от спорыньи
состоит не столько в снижении урожая (большей частью не превышающем
1%), сколько в ядовитости склероциев. Хлеб из муки, содержащей свыше
0,06% размолотых склероциев, вызывает заболевание, называемое эрготиз-
мом, которое проявляется в двух формах: конвульсивной в виде судорог
(«злые корчи»), болей головы, желудка; при более редкой, гангренозной
форме, кроме головных болей и других симптомов, происходит еще прогрес-
сивно развивающееся омертвение выдающихся частей тела — грудей, фа-
ланг пальцев, носа («антонов огонь»), которое может кончиться отпадением
их. Обе формы заболеваний могут привести к смерти. Борьба со спорыньей
ведется очисткой посевного материала и рядом агротехнических мероприя-
тий. В настоящее время благодаря успешной борьбе со спорыньей отравления
ею наблюдаются редко. Содержащиеся в спорынье ядовитые вещества —
эргометрин, эрготамин и др., — вызывающие сужение кровеносных сосу-
Рис. 74. Спорынья (Claviceps purpurea):
1 — колос ржи со склероциями; 2 — склероций со строками; Л — верхушка стромы с перитециями (про-
дольный разрез); 4 — перитеций с сумками (в строме); S — сумка с нитевидными спорами; в — раскрыв-
шаяся сумка с торчащими из нее спорами; 7 — конидиальное спороиошение
отдел—грибы
133
Рве. 75. Формы апотециев:
Л — 4 — общий вид; S — продольный разрез одного из них
дов, сокращение некоторых мышц, обусловливают применение склероциев
спорыньи в медицине (особенно в акушерстве) как кровоостанавливающего
средства.
Другой вид того ясе рода — Claviceps paspali,— паразитирующий в
Закавказье на дикорастущем злаке — пальчатой траве, или паспалюм
(Paspalum),— вызывает у скота расстройства координации движения («пья-
ная болезнь» или «бандала»).
Филогенетически пиреномицеты, вероятно, могут быть выведены из
плектасковых грибов как две самостоятельные эволюционные ветви —
асколокулярная и аскогимениальная. Асколокулярные не достигли такой
высоты организации, как аскогимениальные. У высших представителей по-
следних выработались стромы, дифференцированные на стерильную и пло-
дущую части, у некоторых весьма своеобразные, напоминающие плодовые
тела базидиомицетов.
ГРУППА ПОРЯДКОВ — ДИСКОМИЦЕТЫ (DISCOMVCETIIDAE)
Для дискомицетов характерно, что в их плодовых телах {апотециях)
ко времени созревания спор гимений лежит весь открыто на поверхности бес-
плодной части плодового телят. ТТ молодости у одних дискомицетов гимений
бывает прикрыт или полуприкрыт бесплодной тканью, которая разрушается
или разрывается по мере разрастания апотеция; у других он открыт с са-
мого начала. По способу развития плодовых тел дискомицеты являются
а^когимениальтттгя у подавляющего большинства дискомицетов апотеции
имеют форму диска, блюдца^ или чаши, на верхней, открытой поверхности
которой находитсянгимёний — сумкйУпарафдаы (рис. 75776)ГУ некоторых
апотеции булавовидные, лопатрвидные илив^виде шляпки на ножках. Раз-
виваются они у ~ большинства дискомицетов непосредственно на ^мицелии,
у некоторых — на склероциях, у немногих — на стромах. Крнидиальные
спороношения встречаются у сравнительно небольшого числа видов.
К дискомицетам относится около 6000 видов, не считая дискомицетных
компонентов лишайников. Большинство их сапрофиты в почве или на расти-
тельных остатках. Паразитов меньше, чем среди пиреномицетов*
Дискомицеты делятся на две большие группы: о пе р к у л ятные1,
у которых сумка вскрывается крышечкой, и инопер кулятные,
1 От лат, «оперкулуш — крышка.
«34
низшие РАСТЕНИЯ
f
I*
встреча
ziza) x
или гелоциевые, у которых сумка вскрывается простым отверстием на
вершине; этому соответствует различное строение оболочки сумки на ее
вершине. /
К оперкулятнкм дискомицетам, объединяемым в порядок пеци-
цешх (Pezizale^{ относится 5 семейств и около 60 родов. Уже упоминавшаяся
пиронема Ругъпета omphalodes или Р, confluens с ярко-розовыми блюдце-
ециями, 1—3 лсл в диаметре, часто сливающимися, нередко
на земле, на выжженных местах. Большой р о д пецица (Ре-
теризуется блюдцевидными или чашевидными мясисадми^апоте-
разно^ощета^растущюйЯГбольшей частью на земле; у некоторых ви-
д)ыГапотеции на ножках.
* К семейству гельвелловых относятся „съедобные сморчки истрмлш,
появляющиеся йГлесах рано весной, некоторые —^осенью. Их крупные
мясистые плодовые тела состоят из ножки, "у" некоторых гельвелло-
вых очень укороченной, и шляпки у большинства видов сильно морщи-
нистой, покрытой складками и ячейками (роды Morchella, Gyromitra,
Verpa, Helvetia и др.). Сморчки содержат ядовитую гельвелловую кислоту,
111 J
растворяющуюся в кипятке и разрушающуюся при сушке; поэтому перед
употреблением в пищу в свежем виде их предварительно необходимо обва-
рить кипятком и через 5—7 мин слить его.
К иноперкулятным дискомицетам, объединяемым в порядке ге-
мциетх (Helotiales), относится около 10 семейств и около 300 родов. Боль-
шое практическое значение имеют некоторые паразитные ввды рода скле-
ротиния (Sclerotinta) с апотециями в виде чашечек на ножках.. В пики раз-
cierolinia) склероций образованы
род Stromatinia) склероции
ар
г
Рис. 76. Схематичный разрез небольшого
апотеция (с—сумки на разных стадиях
развития, п—парафизы, пер—перидий,
ар—остатки архикарйа)
цев. У одних видов (п ok Р о д Eus-
о грибными гифами, у других ( п о д-
зуются из м^тфицдр^пйдтот 1Т^Г°*
дов, пронвиавдыхгрибными гифа-
ми. . Sclerotinia fructigena (из
под рода Stromatinia) в своей
конидиальной стадии вызывает гниль
яблок и груш, мякоть которых
становится дряблой и окрашивается
в светло-коричневый цвет. Для
заражения необходимо повреж-
дение эпидермиса; большую _ „роль
в заражении играет жук-ка&арка,
ip)
да и переносящий при этомйтайдии
гриба. На ЪораженныХ“йлодах обра-
зуются нередко кощентрическими
кругами сероватые подушечки
(рис. 77) — конидиальное споррноше-
циегриба, которое раныпедтносили к
особому роду несовершенных грибов
монил^(Моп|/ш7ггссЬ"£епа). Гниение
распространяется и в хранилищах.
Осенью часть плодов (яблок), остав-
шихсяпвисеть на деревьях?" мумифи-
цируетсяjutpHPf&eraeT" Зрюверхно-
сти синевато-черную блестятцую^о'к-
раску. Весной на них" обрсюуются
конидии или апотеции^д^вытдцуты-
ми -иияшими;—вПрбчем^^мчатое
спороношенйе у них" почти никогда
не развивается. Аналогичное^забо-
левание на сливах, абрикосах, виш-
нях вызывает вид£. cinered, Поражаю-
ОТДЕЛ —ГРИБЫ
135
Рве. 77. Фруктовая гниль:
> 1 — яблоко с конидидльными подушечками; 2 — часть мовпдпальвой шщушечкн (яродольный раарев)
nt
[ий плоды, цветки, ветви. Мицелий зимует в ветвях. Меры борьбы — си*
стоматическое уничтожение пораженных плодов, веток и опрыскивание
плодовых деревьев бордоской жидкостью.
Бемю гниль овощей (в лежке), а также подсоднечника. махорки и ряда
других равФемий-(в-веявБЫХ^С31бвиЙХ^ВД8ывдет^?7715йгЬа?га (из н о д р о-
д a Eusclerotlnia). Поверхность пораженных частей растений покрывается
пышно развитым белым мипелием. на котором образуются /черные склера-
ции, состоящие только из грибных гиф. После перезимовки на них вырастают
апотеции. К онид£и~не" развиваются; размножается S. libertiana аскоспора-
ми и мицелием, который не теряет жизнеспособности и тогда, когда высы-
хает и превращается в пыль. Серую гниль овощей, пвет™^ ттлпддм ппень >гч<>-
гцх ^вштенйй-Хвинограда, малины, земляники, клубники, тыквенных и др.)
вызывает S. fuckeliana. Серая окраска поражецныхластей^зависит от обиль-
ного развития древовидно разветвленных конидшнеецен,—несущих головки
сероватых одноклеточных"щшидий;" эта стадия как несовершенный гриб
носит название Bbtrytis cinerea. Из мелких черных склероциев, особенно
часто развивающихся на капусте, после периода покоя образуются апо-
теции.
Филогенетически дискомицеты, возможно, происходят на^првмигив-
ных аскомицетов. Вероятно, самостоятельно развивались два эволюционных
ряда — оперкулятные и иноперкулятные. Эволюция шла в направлен
укрупнения и усложнения плодовых тел, разделения их на бесплодную ножку
и спороносную верхнюю часть.
*: I;
3-Й ПОРЯДОК — ТРЮФЕЛЕВЫЕ (TUBERALES>
Небольшой порялоктрщфелевтд^ содержащий uirinio 200 видов, харак-
^клубневидными плоловыш телами, замкнутыми
в зрелости. В начальных стадиях развития они бывают открытыми вроде
блю)Щбвидных апотециев. Затем, по-айримому. в рвщш с пппятгатом образом
ЙШЩИИ1 край' ИХ “завертываютсяисмыкаются. а бывшая верхддд етррйня
несущая гнмени§7образует кнутри плошшпгр теля дилядки и щелевидные
подости с сумками. Этот онтогенез трюфелевых был объяснен главным обра-
зом работами микологов — швейцарского Эд. Фишера и русского
Трюфелевые грибы растут в лесах. Некоторые виды в^око пенятся как
съедобные. Лучший —- перигорский трюфель (Tuber melanosporunl) вели-
чиной с кулак и более/с черно-фиолетовой ароматной мякотью, растет
136
НИЗШИЕ РАСТЕНИЯ
" 11 ' -» —мС"" " " 1
и дубовых леде, у которых сумка вскрыругие виды — зимний трюфель
{Tuber brumale)\ соответствует различное с*)» растущий в СССР — на Укра-
ине, Черноморскч в других местах. Белый, или
полъскийЛтрюфель щ дискомицетам —отпали Ch. meandrijormis) имеет,свет-
ло-желтые плодовые ^сится 5 до***6 по форме, величине и окраске на клуб-
ни картофеля. Этот вид трюфеля встречается и в центральных областях
СССР (например, в Московской); ценных вкусовых качеств не имеет.
Аскоспоры трюфелевых грибов разносятся животными, поедающими
их плодовые тела, а также после сгнивания плодовых тел — подземными
токами вод.
Филогенетически трюфелевые можно выводить из дискомицетов.
4-й КЛАСС—БАЗИДИОМИЦЕТЫ, ИЛИ БАЗИДИАЛЬНЫЕ
ГРИБЫ (BASIDIOMYCETES)
^Базидиомицеты — второй класс высших грибов, содержащий около
15 ООО видощ^в числе- которых ^находятся почти все шляпочные грибы. У ба-
^зидиомицетов, как и у аскомицетов, мицелий многоклеточп1ыги) но споры
полового размножения экзогенные, образующиеся обычно по четыре ^на
баЗИДИЯХ. БаЗИДИЯ ГОМОЛОГИЧНа Сууке: И та И Друга я вначале двухъ стдерные;
й в той и в другой происходит заключительный.этап полового процесса —
кариогамия (у некоторых' базидиомицетов она происходит немного раньше),
за которой* следует редукционное деление липлоидногхЕдюа: у базидиоми-
цетов оно делится дважды и получившиеся четыре гарлоидвых ядра входят
по одному в образ ующиеся базид иоспоры. Но половых органов у базидиомп-
- ДвУов нет1 половой процесс и смена ядерных фаз протекают несколько иначе,
чйГ’уТйж омицетов.
Рис. 78. Строение аскомицета и базждиомицета:
1 — сморчок; 2 —баваавоимцет. Оба гриба гетеротадяичны: ядра разных волов воображены кружками—
светлима с червой точкой. У аскомицета динариовтиаацая происходит неоднократно в оводовых те-
лах. гае образуются аскогенные гифы, у бявидипмцт однажды, в начале онтогенеаа
ОТДЕЛ —ГРИБЫ
137
Рис. 79.
Образование «пряжки» на
мицелии гименомицета
nt
I! 1
Из гаплоидной базидиоспоры у подавляющега болыпинства базидиоми-
цетов развивается мицелии1 состоятцигй дз клцтлк. лдттпму
гаплоидному ядру. Но этот первичный мицелий недолговечен и заменяется
вторичным, сложенным ив двухъядерных клеток. В типичном случае это
происходит путем анастомозов клеток и перехода содержимого из одной в .
другую, причем ядра и последней не сливаются, а образуют дикарион/рис.76Y.
У грмо^алличн^щ. видов анастомозируют клетки одного и того же мицелия;
у гетероталличных, которых большинство, анастомозы происходят при встре-
че мицелиев противоположных полов^+и —), что легко осуществимо, так
как базидиоспоры образуются в огромных" йдЛйчествах. Образование дика-_
рионов происходит и-иными способами: 1) при переносе гаплоидных. корт- J
дий с мццелия одного знака нампцелии противоположного знака, 2) при ко-
пуляции базидиоспор или даже 3) путем деления ядра в базидиоспоре и'
развития из нее сразу мицелия с двухъядерными клетками.
При делении клеток, в которых возникли дикарионы, оба ядра дика-
риона делятся одновременно: таким путем образуется мицелий, сложенный
из клеток, содержащий дикарионы. Этот дшщрионтический мицелий и есть
основной здцелии базидибмицетовУ живущий нередко очень долго. Следо-
вательно, у базйдйо^цетотгв цикле развития доминирует дикарионтическая
фдоа (см. рис. 78). У аскомицетов дикдрионтическая фаза недолговечна,
представлена только аскогенными гифами. В цикле развития доминирует *
гаплоидная фаза.
При размножении клеток дикарионтического мицелия баяидиомицетов
обд.ядра дпуарисшт, как уже сказано, делятся однпвреметтп (синхронно),
причем оси.чделения располагаются пярялглшгкно и между двумя-тшушт-
шимися парайГЭЗДёр" образуются деретюродкйГ У других базидиомицетов
деление клетки происходит сложнее. Перед делением дшсдршша яацротив
него клетка образует маленький блкпнрй вмрпг.т t загибающийся к ее ба-
зальной части--(рис. 79). Оси делящихся ядер дикариона располагаются
под некоторым углом, так что веретено одного делящегося ядра заходит в
этот вырост. Когда ядра дикариона разделятся, то два несестринских ядра
отойдут в верхний (дистальный) конец делящейся клетки; одно ядро из
другой пары несестринских ядер будет в нижней (базальной) части,
а второе ядро этой пары будет в боковом выросте. Затем образуется попе-
речная перегородка, отделяющая одноядерную базальную клетку, и вто-
рая перегородка, отделяющая боковой вырост от дистальной части клетки.
После этого боковой вырос^иаагибае«ся_и^одноядерной базальной клетке;
между ними появляется отверстие, через которое ядро из выроста переходит
в базальную клетку, и в ней таким путем оказываются два несестрияских
ядра. Эти боковые выросты, слившиеся с гифой, называют пряжками. Та-
138
нияшии PACT
и; i:
Рис» 80» Развитие сумок и базидий:
1 _ ю — сумки; и —19 — базидии. «Крючки» у аскомицетов соогаетстпуют «пряжкам» в основании ба-
аидий у базидномицетов
и™ образом ив одной двухъядерной клетки возникают две, я следом со-
вершившегося деления остается пряжка. Значение этого сложного процесса
остается невыясненным. Замечено, что деление с образованием пряжек бы-
вает чаще в узкопросветных гифах. Вообще же, образование пряжек из-
вестно примерно у половины базидиомицетов. Морфологически образование
пряжек соответствует образованию крючков в аскогенных гифах при обра-
зовании сумок у аскомицетов. Такие же пряжки образуются и при развитии
базидии (рис. 80), что подтверждает гомологию между сумкой и базидией.
Интересно, что у некоторых аскомицетов (Helvetia crlspa9 Sclerotinia trl~
foliorum) на аскогенных гифах также образуются пряжки.
На дикарионтическом мицелии у немногих базидиомицетов (из порядка
I экзобазидиальных) прямо-иоаникают базц^и^.. базидиоспорами; у других
(головневых, ржавчинных) на нем образуются покоящиеся споры, из ко-
торых позднее развиваются ба^ддим; у болыпинстШГже на дикарионтическом
мицелии образуются плодовыолед@г и уже на них или в них развиваются
(базидии с базидиоспорами. В отличие от аскомицетов эти плодовые телф сложе-
ны целиком из дикарионтических гиф. Форма и размеры плодовых тел весьма
разнообразны и широко используются как систематические признаки.
Базидиомицеты по строению и развитию базидий делятся на 2 под-
! класса.
i
1-Й ПОДКЛАСС - ХОЛОБАЗИДИОМИЦЕТЫ
(HOLOBASIDIOMYCETIDAE)
Базидии одноклеточные (рис. 81, lt2), называются ^холрбазцдцдми1.
Холобазидиомидетьь^г более^, обшцршы[й_^подкласс базидиомицетов.
К нему относятся около % всего видового состава класса, чт<Г Составляет
по крайней мере 8000 видов.
1 От греч, «холо» — цельный
отдал—грибы
1-И ПОРЯДОК — ЭКЭОБАЗИДЦАЛЬНЫЕ (EXOBASIDIA.LES)
Экзобаэидиальные —=- небольшой порядок, содержащий окодо 30 видов
грибов, паразитирующих на выстпх ряг.теипих- (1снопной отличительный
признак состоит в отсутствии плодовых тел, Каяидии ряспрлягяютги рых-
лым слоем меж ~
часто встречается ркзобааидиуль (Exobasidium vaccinii) на бруснике. реже
на других видах УасЫЯНМ, где он вызывает гипертрофию листьев и молодых
стеблей, пршппгдтптуих беловатую или рпаовАтуи^^кряеку. ТртБ'ЧйПНйш в
1861 г. русским микологом М. С. В орониным, впервые открывшим эту
своеобразную группу базидиомицетов.
ГРУППА ПОРЯДКОВ — ГИМЕНОМИЦЕТЫ (HYMEHOMYCETIIDAE)
Гименомицеты — большая группа, насчитывающая около 7000 видов.
Это наиболее типичные и широко распространенные базидиомипеты.с имею-
щие плодовые,тела различной консистенпци и формы с гимением, лежащим
открыто на поверхности плодового тела или обнажающимся еще до созре-
вания ббльшинитъатдор. ? ’ ' . . .. . .. - 1
Tlo консистенции плодовые тела у более примитивных родов паутинис-
тые или плотновойлочные, у вышеорганизованных — кожистые, мясистые.
дередянисхые. По форме плодовые тела эволюционировали от плоских, рас-
простертых по субстрату (ресупинатных), до пластинчатых, горизонтально
отстоящих от субстрата, копытообразных или шляпочных (в виде шляпки на
ножке), У распростертых по субстрату гимений находится на верхней сто-*
роне плодового тела; у_ллочих он в процесса. эволюд^и переместился на
Нйжнюю, что способстау££лучп1ему распространению спор.базидйомице-
тов тургорное дарениепчепь^узкий канал в
стеригме? Поэтому базидиоспоры отбрасываются с баз^гдта (ппочепепно)
на ничтожное расстоянй©7ТОГ0,1—(у фрагмобазидиомицетов до 0,6—
0,8 лме). В этом случае биологически выгодно расположение гимения на ниж-
ней стороне плодового тела, так как при этом споры, цядцщщие очень мед-
ленно, могут быть подхвачены конвекционными токами воздуха, возникаю-
щими у поверхностр "умеющими ничтожные скорости. Строго вер-
тикальное падение спор обеспечивается гертропическими изгибами плодовых
тел. У растуших на зёмле^шляпочааых мясистых гименомицетов, когда споры
все же очень^скрр_р достигают земли; они нёредко распространяются живот-
ами насекомых, улитками, млекопйтающими)7вПпб0дай)щими
плодовые тела и разносящими с экскрементами непереваренные ими споры.
it; П1
Рис, 81. Базидии:
1 f- ходобшда; Л — а—фрагмобазидия
140
НИЗШИЕ РАСТЕНИЯ
Рис. 82. Строение пластинчатого гименофора агарикового гриба (т— бесплодная тра-
ма; а — гимений, б — базидии, л — псевдопарафизы, ц — цистиды)
Hi'.
II*:
Поверхность плодовых тел, несущая гимений, называется гимено-
фором1, ------------------—------------------ ------------------------
ИИСб’Г'форму зубпов, пласт^ттркг трубочек (вмстлятшит внутри гимением),
что значитеЬвяи увеличивает его повеохность и, тем самым количество про-
дуцируемых спор. Форма гименофора используется в систематике гимено-
мицетов.
В гимениид_кроме базидий (обычно цилиндрической формы), находятся
еще похожие на них, но не несущие спор п с е в д о п а р а ф и з ы, отли-
чающиеся от парафиз а<йсомщетовгнб~т5лько формой, но и двухъядерностью
клеток. У многих видовв’гимении находятся еще д и с т и д ы® — клетки,
различной, но наследственно постоянной формы, более крупные, чем ба-
зидии (рис. 82). Большинство гименомицетов живет сапрофитно в почве и
на отмершей древесине. Ряд видов паразитирует на живых деревьях. Мно-
гие почвенные сапрофиты живут в симбиозе с корнями древесных пород,
образуя микоризу3.
о: *:
2-Й ПОРЯДОК — ПОРИЕВЫЕ, ИЛИ АФИЛЛОФОРОВЫЕ (PORIALES,
ИЛИ APHYLLOPHORALES)
является щф
способностью
Плодовые тела афиллофоровых разнообразны^как по общей форме (в
виде корочки консолевидные, редко в виде шляпкина ножке),так "И по кон-
систенции (кожистые, пробкрватые, деревяни&гыет но нёмягкомясистые}^
Гименофор тожё^разноЗбр^^Ь га^щшйгЛ^цде. складок, валиков, зубцов
ущест5ениой особенностью гимениального слоя
ь составляющих его элементов. Это связано со
расти впшрь неопределенно длительное время.
телефоротлх (Telephoraceae) с их распростертыми по субсчету(ресупиПЭт*-
* От греч. «форео» — несу. “ ч
® От греч. «кюстио — пузырь.
,8 Микоризу могут образовывать и другие грибы, в том числе некоторые фикомидеод.
ОТДЕЛ —ГРИВЫ
141
нГц
ными1) или приподнятыми над субстратом плодовыми телами и гладким
гименофором. Сюда относится кониофора (Coniophora cerebella) — разреши-
ал и крепежного лесав_щахтах. В том же семействе находят место
(Cartfharellus) с врронковидными плодовыми телами и небольшими
складками на нижней поверхности* Большинство телефоровых — сапро-
фиты на растительных остатках;
Оригинальны ветвистые с гладаим гименофором плодовъимтела
mujSQjsax (семейство С1аиаг1меае)"^почвеяш1е лесные сапрбфйтыГ
ИIII t
ватый — на
верхней стороне ресупинатных и на нижнеи у шляпочных ило-
г»
довых тел.
Важное практическое значение имеют (семейство
Meruliaceae) и оддапдеикащв (семейство Polyporaceae). У мерулиевых
Цйодовые тела чаще ресупинатные. Гименофор в виде беспорядочно анасто-
мозирующих, большей частью невысоких складок. Наиболее известер
мтй гриб (Serpula, или Merulius^ eacrymans) — сапрофит, опасный раз-
рушитель деревяниых^застей^достроек. Мицелии гриба проникает в древе-
сину, извлекая из оболочщСклеток в первую очеоедь^нерлюлозу/ вызывая
разрушение древесины. Мццелцй. развивается тшрке..п.JUL З^йерхности^ре-
весины в виде белых ^ватообразных скоплении, приора спядяюпщтся в пепель-
но^ещ^в-^пленкй. Узкие пепельно-серые тяжи мицелия могут «переходить»
йзэтажа в этаж по каменным стенам и в подходящих условиях снова разви-
ваться в древесине. Плодовые тела образуются нечасто и имеют форму плос-
ких jmanggeK желто-бурого, на периферии белого цвета: сверху они несут
гименофор .виде невысоких перепл^аюпщхся складок. Распространяется
гриб как спорами, так и участками мицелия/ легко" переносимыми людьми
на ногахг вдструментах и т. п. Так как грв^рёйййО^дГлиптъ j сырой дре-
весине (19—20% воды от сухого вещества), то профилактические меры борьбы с
ним состоят в использовании для построек сухой /щевесины й в‘ соблюдении
надлежащих санитарно-строительных правил,'"обеспечивающих хорошую
вентиляцию. При появлении гриба необходимо удаление и сжигание пора-
женных и ближайших к ним на вид здориБ^х^аете^^бряботка здоровых
частей антисептическими пастамш^(Г"фторисТНм^натрием,сланцевыми смо-
лами (препарат ЗАГ-2) и другими химикатами; -
В семействе трутовиковых плодовые тела плотные» твердые, к старости
не загнивающие, ресупинатные, копытообразные или другой формы, ~ -от-
стоящие от субстрата, с гименофором на нижней стороне (рис. 83). Сюда
Рис. 83. Трутовик:
1 — общий вид плодового тела; г — поперечный разрез трубочек и гименофора
1 От лат. «ресупинатус» — положенный навзничь, на спину.
I
143
НИЗШИЕ РАСТЕНИЯ
относятся два вида^
vaillantii), объедини:
домового гриба (Coriolus vaporarius и Fibuloporia
г рййёб ТИД общий названием Porta vaporaria.
На многолетних плодовых телах видов, паразитирующих на стволах де- .
ревьев, ежегодно нарастает новый слой трубочек гименофора; таковы»
например, настоящий трутовик (Fomes fomentarlus) с копытообразными
серыми плодовыми телами, р^и^ще употреблявшимися для^ изготовления
igniarius) с темно-серыми растрескиваю-
щимися деревявдстыми плодовыми тел^дщ^./^рн^^^^1^ (Fomitopsis аппо-
sa), вызывающая гниль корней хвойных деревьев. Под именем «чага» известны
черные растрескивающиеся, не образующие спор наросты на березах, обра-
зуемые грибом Inonotus obliquus. Они применялись в народной, а в послед-
нее время и в научной медицине как средство, улучшающее в некоторых слу-
чаях самочувствие больных с опухолями различной локализации.
Многие виды трутовиковых имеют мясистые плодовые тела, например
базовый трутовик (Plptoporus betultnus).
3-Й ПОРЯДОК — АГАРИКОВЫЕ (AGARICALES)
Плодовые тела агариковых однотипные, почти всегда состоящие из
пенька (ножки! ишляпки. причем ножка прикреплена-в середине шляпки.
НзЙдовое тело у этих грибов в определенный момент времени завершает свой
рост, вследствие чего элементы гимениального слоя практически одновоз-
растны. рименофор в типе состоит ив плястинок. наход
' стороне шляпки и расходящихся радиально. У некотои
меноФоо трубчатый. д
слойкой рыхлой тааи (чего не бывает у трутовиковых из по рядка ftonwtes,
нмеюЩзбГ тоже трубчатый пшвнофор)./В целом плодовые тела мягкомясис-
1МДДГ у ~ (Ш&ИШШЬШ, бССННИИ ОПННОК,
некоторые масленки Идр.). При еднорп/дрсуу плпдппух тед о^и и пяаяи-
ваются сходно. Плодовые тела заклдутиттея а мпа еплвтендя^гаФ округ-
лой или од^^н^^^ТПЯйГТ2)7Вэтомзеддймш^щффщ)£нцируетсяяаруж-
пый слойобразующий так называемое общее по к р ы в а л о. В не-
которых случаях^ оно представляет
исчезающее, в других^ “
; i I i I•
ir.i
таточно прочное. Если находящиеся
Рис. 84. Развитие плодовМх тел:
1—шашшньок с частным покрывалом; а—« — бледная поганка с общин ж частным покрывалом
ОТДЕЛ —ГРИБЫ
143
валп оййошщется.ПКо^ времени. хозр$ваяия
плодового^ тела прочное^ общее покрывало
разрывается, и остатки" его сохраняются в
ОГО
►ан-
шляпки (мухомор) в в виде
влагалища у
ней фазе разв
вознйЖ58твше
поверхности которой и шюисхопит Формиро-
вание гименофора. Позднее на уровне этой
полости образуется кольцевая трещина, ко-
торая отделяем шляпку от ножкйГТаким
образом гименофор вначалесвоегосуше-
ствовання лкяянв^ется прорытым так на-
i 11 s 1
Часть его у многих грибов остается на нож-
ке в виде кольца.
Подавляющее больппшство агариковых—
почвенныесапрофйты в лесах, реже на лу-
гах, болотах, встепях; вшогиё —jmhkoph3O-
образователи.
Из относящихся к этому порядку се-
мейств остановивши на двух:
или^дацшкоше в узком смысле (yigariea-
сеое),с пластинчатым гименофором и болту-
созыв (Boletaceae) с трубчатым гименофором.
Семейство пластинниковые (Agari
сасеае). Гименофор -л—виде- -пластинок на
нижней стороне шлянки плодового тела* _ ос _
Пласттаки расположены отвесно и у боль- L?c‘ °5" Е*®** веРтякальН0Г0 Р^"
" W реза ПДДСТЩД0К ШЛЯПОЧНОГО гриба
шинства имею? ^в jpaspeep из семейства о ла ститгтги пиит
Фдому». суживающуюсякшвзУ, 4'Ак что (стрелками показаны траектории
споры с верхней части пластинки падают, ведения базидиоспор)
не ЗКДёшя нижних (рис. 85). Серединная,
большая часть пластинок состоит из бесплодных грф, образующих бесплод-
* “ покрывает пласэдку-. .гдордп и
низ^шдщшсхяюмещухках между пластинками. Между трамой и гимтщрм
находится мелкоклеточный псевдопаренхимттг^й суб г и м"е"НЖУлУн ы й
слой, в шию^м закладываются ^азипии. ; - -
пластинйбвым относится не менее 10000 видов, живущих в большин-
стве сапрофитно в лесах на почве; многие образуют микоризу с корнями
древесных пород; паразитов немного.
Виды лактариуса (Lactorius) содержат млечный сок, белый, реже оран-
жевый илй краиШЛШтый; из них съедобны рыжик (L. deliciosus), груздь (L.
resimus), волнушка (L. torminosus), чернушка (L. necator) и др. К лакта-
риусу сийтемаТйчески близок большой ро д сыроежек (RusauZu), содер-
жащий много съедобных, но малоценных грибов, jtfacmoaupiu, или осенний,
оленек (Atmilariella mellea) — неплохой съедобайпГТфйбТ’Таёредко парази-
тирующий на деревьях, чаще же сапрофит на отмершей древесине; гифы его
мицелия, находящиеся в гнилой древесине, могут в теплую влажную пого-
ду светиться. Съедобен также ^епиш& пмшж (Pholiota mutabilis) н так на-
зываемый лреоеой оунок (Maramtus oreades). растущий на лугах (не около
пней), где он часто, как, впрочем, в многие другие шляпочные грибы, обра-
зует «ведьмины кольца» (плодовые тела расположены в виде круга вследствие
равномерного роста по всем радиусам многолетнего мицелия). К роду
I ’> i;
1 От лат. «трама» — утдк в тканье, ткань.
144
РАСТЕНИЯ
шампиньонов (Agaricus или Рsailiota) относится ряд съедобных видов; один
из них, двуспоровый шампиньон (4. bisporus), культивируют в специальных
теплицах, где он образует плодовые тела в течение зимы и ранней веской,
когда нет других свежих грибов. Гриб предпочитают выращивать из 'спе-
циально разводимых чистых культур, дающих плодовые тела лучшего! ка-
чества. У рода ^опринус (Coprinus) плодовые тела многих видов по сорре-
вании распЛываются ‘% быстро чернеющую жидкость (автолиз), стекающую
вниз и содержащую споры. К роду аманита (Amanita) относится ряд
ядовитых видов: красный мухомор (4. muscaria)', смертельно ядовитые
бледные поганки (4. phalloides, 4. тарра и др.), которые можно узнать по
белым пластинкам^ наличию кольца на ножке и влагалища, свободного
идти приросшего к основанию ножки. Шляпка у них бледно-зеленоватая,
желтовато-зеленоватая или белая, иногда (но не всегда!) с хлопьями на
верхней сверена. К* этому же роду относятся и очень вкусные съедобные
грйбйг-- 4? caesarea, растущий в Закавказье и Южной Европе, ,4. caesa-
reoides, растущий на Дальнем Востоке.
Плодовые тела у грибов семейства болетусовые (Boletaceae) мя~
Чщстые. Гименофор трубчатый, легко отделяющийся от мякоти и шляпки.
Рай^ыце эти грибы включали в сейейство трутовиковых, но уже ука-
занные>эсобенности и рйд переходных форм между болетусовыми и пла-
лее близкими друг к другу,
являются почвенными-сапрофи-
бразуют эктоэндЬтрофную микоризу с корнями дре-
крупный род — (юлетус (Boletus), разби-
более мелких родов (названия их указаны
видов: белый гриб— Boletus edulis,
^jcabrJj, подосиновик — В. versi-
(TutitpOrm) luridus, маслен-’
nutatus и др., моховик — В. (Xerocomus)
стинниковьпйг^дадут основание считать
чём к трутовик а м.^Пб5тиь*все^о лету со
тами в лесах; мно
весиых растений. Наи
ваемый в по
в скобках), сод
1____ ,_____1>ко
ки — В. (ZsoS
subtometosus и др
»:r.
аТу,1шогОч^ъед
ее время на
го
s) luteus, В
:in
ГРУППА ПОРЯДКОВ — ГАСТЕРОМИЦЕТЫ (GASTEROMYCETIIDAE)
У гастеромицетов, в отличие от гименомицетов, плодовые тела^долго
остаются совершенно замкнутымиПй'~вск‘рываются лишь после окончатель-
ного созревания базидиоспор? Плодовые тела различной, формы и размеров
состоят из о^)ол очкщназываемой пефл-дд ем, и внутренней части, называ-
емой^ гл е б ой. В глебе находятся щцщсти, округлые и вытянутые, щеле-
видные, в которые вдаются, базидии, расположенные^.правильный гимени-
альным слоем .или беспорядочно. Между полрсхями.-лаходнхсл бесплод-
ная тканьтрамд. В зрелисти сохраняется вся трама или большая ее часть
разрушается и от нее остаются только некоторые волокнистые гифы — к а -
Базидии у многих видов в зрелости тоже разрушаются. Базидиоспо-
ры у большинства темные, и поэтому глеба, вначале белая, по мере со-
зревания базидиоспор;йзмбНяе¥Л)краску.
Смена ядерных фаз, изученная у немногих гастеромицетов, сходна со
сменой их у гдмжошщетов. У некоторых гаплоидная^^фаад_чрездычайно
кратковременна, так как. дикарион образуется уже в, базидиоспоре путем
деления ее ядра ня два. Диплоидная фаза, как и у гименомицетов, очень
краТйовременяа и представлена молодой лишь базидией с диплоидным
ядром.
К гастеромицетам относится около 1000 видов. Почти все гастеромице-
ты — почвенные сапрофиты с надземными или подземными плодовыми тела-
ми} многие, вероятно, микоризообразователи; некоторые живут на отмершей
древесине; практического значения не имеют.
На лугах, выгонах, полянах, лесных опушках встречаются дождевики
(рис. 86, 2, 2), относящиеся к родам бовиста (Воvista)t ликопердон (Lyco-
ОТДЕЛ — ГРИВЫ
145
perdon), кальвация (Calvatia). Их
шаровидные или булавовидно-
грушевидные плодовые тела, в
молодости белые и упругие,
могут употребляться в . пищу
(до изменения окраски мяко-
ти). При созревании они стано-
вятся снаружи серовато-буроваты-
ми, перидий разрывается на вер-
шине или распадается (у кальва-
ции) и шоколадно-бурая масса
спор постепенно рассеивается.
У гигантского дождевика (Calvatia
gigantea) плодовые тела достигают
в диаметре 30 см и более и раз-
вивают триллионы спор.
В семействе гнездов-
ковых (Nidulariaceae) плодовые
тела, растущие на древесине,
имеют вид миниатюрных, в
зрелости открытых вазочек, в ко-
торых лежат лепешечки — участ-
ки трамы (перидиолы), со-
держащие внутри полости с бази-
дия
У растущего в лесах фаллюса>
или веселки (Phallus impudicus),
плодовое тело в молодости белое
Рис. 86. Гастеромицеты:
шт
овальное, ИЛИ яйцевидное. При 1 — ликопердон: а — общий вид, б — базидии*, г—
, пп^тчт^п/1,., бовиста; s — гнездовка; 4 —фаллюс
созревании его перидии разрывает-
ся и наружу выходит быстро удли-
няющаяся (до 10—20 см) белая губчатая ножка, несущая на вершине ячеистую
тупоконическую «шляпку» (рис. 86,4); последняя покрыта зеленоватой слизью,
содержащей базидиоспоры. Гриб издает неприятный запах падали, привле-
кающий мух, откладывающих яйца в гниющее мясо. Ползая по зеленоватой
слизи, мухи попутно захватывают базидиоспоры и разносят их. Этот род и
близкие к нему другие, преимущественно тропические, роды с ярко окра-
тленными плодовыми телами и энтомохорным распространением спор стоят*
на наиболее высоких ступенях эволюции гастеромицетов и грибов вообщеу^
2-Й ПОДКЛАСС — ФРАГМОБАЗИДИОМИЦЕТЫ
(PHRAGMOBAS1D1OMYCET1DAE)
У большинства фрагмобазидиомицетов базидия разделена поперечными
перегородками, у немногих — продольными, на четыре клетки и называется
фрагмобазидией. Их делят на четыре порядка: два с плодовыми те-
лами и два без них.
i-Й ПОРЯДОК — ДРОЖА Л НОВЫЕ (TREMELLALES)
Плодовые тела дрожалковых слизистой или студенистой консистенции,
что особенно заметно во влажную цогоду. Базидии разделены продольными
взаимно перпендикулярными перегородками на четыре бок о бок располо-
женные клетки; у'некоторых перегородки идут вкось или почти поперечно,
что является переходом к следующему порядку. Преимущественно сапро-
фиты на древесных сучьях, пнях. Включают 3 семейства, свыше 100 видов».
РАСТЕНИЯ
Рис. 87. Прорастание го-
ловневых спор:
1 — устнляго (б — базидия,
бсп — базидиоспоры); г —тмл-
люцвя (изображены копули-
рующие базидии и одна не-
копулирующая)
2-Й ПОРЯДОК — АВРИКУЛЯРИЕВЫЕ (AURICULARIALES)
Плодовые тела аврикуляриевых тоже слизистые или студенистые, но
базидии разделены поперечными перегородками на 4 клетки. Большинство
видов тоже сапрофиты на древесине. Включают 250 видов. Плодовые тела так
называемого иудина уха (Auricularia auricula judae) в разбухшем состоя-
[ напоминают ушную раковину человека. У некоторых родов (Septobasi-
dium, Uredinella) фрагмобазидии развиваются не прямо из гиф плодового те-
ла, а из особых толстостенных клеток, что, как правило, имеет место у двух
следующих порядков.
1515
З^Й ПОРЯДОК — ГОЛОВНЕВЫЕ (USTILAGINALES)
Все головневые — паразиты на высших растениях, вызывающие забо-
левания их, известные под названием головни. Особенно вредоносны и заметны
они на хлебных злаках, где темные (коричневые) споры гриба развиваются
у большинства в более или менее разрушенных соцветиях, приобретающих
как бы обожженный вид. Заражение у многих видов происходит в поч-
ве во время прорастания растения-хозяина. У культурных злаков головне-
вые споры, распыляющиеся и прилипающие к зерну во время обмолота, по-
падают вместе с ним в почву при посеве. Здесь в головневой споре сливаются
бывшие в ней два ядра (дикарион). Обряяовяицтяеея липлоипиод ядро
делится редукционно в споре или в развивающейся из нее бааитди. Н& послеп-
ней образуются базидиоспоры, причем в каж-
дую входит по одному 1йВ1йиДйбМУ‘”вдру
(рис. 87). Базидиоспоры, еще находясь на
базидии или отпав с нее, могут почковаться
и давать одноядерные гаплоидные клетки,
тоже, в свою очередь, почкующиеся (п о ч-
2
Рис. 88.
8
Копуляция у головневых
грибов:
одноа базидиоспоре, в которую пере-
шло содержимое другой баяйдиоспоры,
резвилась двухъядерная конидия)
жет развиться слабый гаплоидный мице-
лии, но он Не способен заразить растение.
Базидиоспоры или их потомство, получаю-
щееся почкованием, могут копулировать,
образуя выросты, направленные друг к другу
(рис. 88,7). Содержимое одной клетки пере-
ходит в другую, но ядра не сливаются, и по-
лучается дикарион. При копуляции обнару-
живается гетероталличность гриба. Из кле-
ток с дикарионами развивается дикарионти-
ческий мицелий, и только он способен вы-
звать заражение, очень молодого проростка
высшего растения, имеющего тонкие оболоч-
ОТДЕЛ —ГРИБЫ
147
Рио. 89. Голозпя:
J — шцеипды (п-^х^ная); а — пшеницы (твердая); Л — овса; 4 — проса; 5 — кукурузы
ки клеток. Мицелий гриба быстро проникает в конус нарастания растения-
хозяина и растет вместе с ним, оставаясь вср4ремя в дикарионтическом состо-
янии благодаря синхронным делениям дв^ ядер дикариона. Больные расте-
ния по внешнему виду почти неотличимы 0т здоровых. Незадолго до выкола-
шивания (или выметывания метелки) мицелий гриба на*
гл:
ает особенно сильно
разрастаться в области соцветия, чему способствуют обильно поступающие
сюда питательные вещества. Ткани несформировавшегося соцветия разруша-
ются; в клетках .мицелия под старой оболочкой образуется новая, более тол-
стая коричневая, старые же оболочки ослизняются, имвделвд распадается на
множество округлых двухъядерных клеток типа хламидоспор, называемых
головне hjm ми спорами. В таком состоянииразрушенное соцветие
и выходит из влагалища верхнего листа (рис. 89). По такому типу происхо-
дит развитие головни проса (Sphacelotheca panici miliacei1), овса (JJstilago ave-
nae и U. 1еи1в)^лчмнл (Ustilago hordei), твердой, или вонючей* головни пиле-
кащ» LTiZZf Ito дЮйМУ^ричем в этом последнем случае околоплодник зернов-
ки пшеницы не разрушается, но внутри находится не эндосперм и зародыш, а
темная масса спор; у стеблевой годооти pacu (Urocustis occulta} головневые
споры образуются не вГ соцветии, а в нижних частях верхнего междоузлия,
но зерна в колосе обычно не вызревают. Меры борьбы с головней этого типа
заключаются в обеззараживании посевного материала, так как источник ин-
фекции (головневые споры) находится на поверхности высеваемых зерновок.
Обеззараживание производится путем обработки посевного материала ела-
1 Раньше этот гриб относился^ роду Ustilago', в настоящее время некоторые мико-
логи относят его ж р о д у Sorosportum*^
/
148
НИЗШИЕ РАСТЕНИЯ
быми растворами формалина в воде (1 часть формалина на 300 частей воды)
или опыливанием его мышьяковистыми, ртутными или сероорганическими
препаратами (НИУИФ-2 — гранозан, протарс, ТМТД и др.). Эти очень эф-
фективные меры борьбы свели потери урожая от головни, которые раньше
достигали 10—12%, а в некоторых местах 30—50% и более, до 1% и
менее.
У немногих видов головневых грибов имеет место другой способ зара-
жения, характерный, например, для пыльной золоти пшеницы (Ustilago tri-
tici) и пыльной головни ячменя (U. пи da). Здесь заражение происходит во-
время цветения злака. Головневые споры, разносимые воздушными течения-
ми, попадают на рыльца и прорастают, давая фрагмобазидии. Базидиоспо-
ры на них не образуются, а гаплоидные клетки фрагмобазидии копулируют
попарно (рис. 88, 2)\ клетки, в которых получаются дикарионы, про-
растают в мицелий, проникающий в завязь и развивающийся внутри форми-
рующейся зерновки — в эндосперме и зародыше, но не разрушающий их.
Пораженные зерна почти не отличаются от здоровых. Когда они прорастают,
находящийся в них мицелий тоже трогается в рост, проникает в конус нарас-
тания и перед выколашиванием распадается на распыляющиеся головневые
споры. Борьба путем поверхностного протравливания посевного материала
фунгицидами (протравителями) здесь невозможна. При небольших посев-
ных площадях можно использовать обработку посевного зерна, при которой
семена выдерживают в течение четырех часов в воде при температуре 28—
32°C; мицелий за это время трогается в рост, а зародыш не успевает на-
чать расти. Затем зерно погружают в воду с температурой 52—53°С на 8 —
7 мин, Эта температура убивает тронувшийся в рост мицелий, но не вредит
зародышу зерна. При больших размерах посевов такие процедуры затруд-
нится ьны, в этом случае посевной материал надо брать с полей, где не было
пыльной головни. В последнее время ведутся поиски протравителей для борь-
бы с возбудителями этой головни.
Третий способ заражения описан для головни кукурузы (Ustilago zeae).
В этом случае возможно местное заражение в любом месте растения, где
находится молодая ткань с тонкими оболочками клеток (цветки, основания
междоузлий, листовые пластинки). Заражение вызывается ппокопулировав-
шими базидиоспорами или кпнидпямттт переносимыми с почвы ветром, В ре-
зультате получаются местные опухоли, достигающие иногда огромных раз-
меров (с головуребенка} и наполненные головневыми спОрамн^МёрЕГ борьбы
возможны главным образом* профилактические: удалений больных расте-
ний до распыления спор, плодосмены и т. п,; рекомендуется и протравлива-
ние посевного материала препаратом гранозан (НИУИФ-2). В последнее вре-
мя появились предположения, требующие проверки, как будто у кукурузы
заражаются молодые проростки еще в почве и диффузно распространяющий-
ся внутри всего растения мицелий дает споров ошения в местах с особенно
обильным притоком питательных веществ.
Из описанного видно, что паразитизм головневых грибов достиг высокого
совершенства, так как зараженное растение не погибает, а долгое время
вегетирует; лишь в конце жизненного цикла растения обнаруживается его
' поражение головней и развивается спороношеиие гриба. Кроме того, углуб-
ленное изучение практически важных головневых грибов показало, что.
ботанический вид гриба состоит из некоторого количества морфологически 1
неотличимых рас (или биотипов), каждая из которых может пора-
жать только определенный сорт культурного растения. Такие биотипы, ин-
тересные и в практическом, и в теоретическом отношении, имеются и у дру-
гих паразитных грибов (ржавчинников и др.).
В отношении смены ядерных фаз в цикле развития головневых гри-
бов чередуются: 1) кратковременная гаплоидная фаза — базидиоспоры и
почкующиеся от них конидии; 2) дикарионтическая фаза, занимающая поч-
ти весь цикл развития — от копуляции базидиоспор или. конидий до слия-
ния ядер дикариона в головневой споре; 3) очень кратковременная дипло-
ОТДЕЛ — ГРИВЫ
149
идная фаза — головневая спора от слияния ядер дикариона до редукцион-
ного деления его.
К головневым грибам относится около 700 видов, распределенных меж-
ду двумя семействами: устиллгиновые (Ustilaginaceae), имеющие ти-
пичную фрагмобазидию с поперечными перегородками (рис. 87, 1) и бази-
диоспорами, образующимися по бокам ее, и тиллециевые (Tilletiaceae), имею-
щие базидию, не разделенную перегородками и образующую базидиоспоры
на своей вершине (рис. 87, 2). При формальной, искусственной классифи-
кации их следовало бы отнести даже в разные подклассы, но все черты строе-
ния, развития и образа жизни представителей этих двух семейств настолько
сходны, что не может быть сомнения в их филогенетической близости.
4-И ПОРЯДОК — РЖАВЧИННЫЕ, РЖАВЧИННИКИ (UREDINALES)
Все ржавчинные грибы, как и головневые, являются паразитами высших
растений из различных семейств, вызывают заболевания их, называемые ржав-
чиной за ржавую окраску некоторых спороношений. В отличие от головне-
вых, которые можно культивировать как сапрофиты и которые небольшую
часть жизненного цикла в природе живут сапрофитно (базидиоспоры и поч-
кующиеся от них клетки), ржавчинники — облигатные паразиты1 и в га-
плоидной и в карионтической фазах своего развития. В отличие от головневых
подавляющее большинство ржавчинников вызывает не диффузное^ местное
поражен^a, и ттд ттрпрпсткпв. а взрослых растений. В цикле развития подав-
ляющего большинства' ржавчинникдв зйкинимирнъ сменяют друг друга раз-
личные спороношения, обычно на вегетативных органах растений (листьях,
стеблях), и у многих видов ржавчинников не на одном, а на двух растениях-
хозяевах, совершенно различных в систематическом отношении (смена расте->
ний-хозяев).
Как пример разнохозяйственного ржавчинника с полным циклом раз-
вития обычно описывают один вид пукцинии (Puccinia graminis), вызываю-
щий линейную ржавчину злаков. Весной развитие гриба начинается на листьях.
барбариса, ГДиЧяЯЗ^фЯ^айтся небольшие гипертрофированные оранжевые'
участки ткани, пронизанные межклетным мицелием с гаусториями. Мице-
лий этот состоит из одноядерных клеток с гаплоидными ядрами; оранжевый
ri:i
ются два погруженных в ткань спороношения гриба: на верхней стороне
листа — пикнид ы, или сперМагонии, на нижней стороне —
э ц и д и и. ПикнидИ имеют кувшинообразную форму (рис. 90, 2); в полость
их от СТ₽Нбк врастают короткие конидиеносцы, отчленяющие мелкие одно-
ядерные конидии,называемые пикноспорами или спермация-
ми. Наружу из пикв-иды высовывается пучок гиф и выделяется пахучая
сахаристая жидкость, в которую погружены пикноспоры; вызвать новую ин-
фекцию пикноспо{$ы~не могут^Эцидий закладывается в'глубине "Листа в виде
клубка-гиф из одноядерных клеток (рис. 90,2). Затем (не вполне ясно, каким
путем) в базальной части его образуется компактная площадочка из двухъ-
ядерных цилиндрических, так называемых базальных клеток, которые от-
членяют базипетальные цепочки двухъядерных, материнских клеток cnops
каждая из последних делится на верхнюю — эцидиоспору с оранжевым-ео-
лержимьш—и нижнюю. маленькую, позднее разрушающуюся промежуточную
клетку (рис. 90,3). Периферические базальные* клетки образуют подобным же
образом .оболочку эцидия — перидий,—окружающую его с боков; наружные
эцидиоспоры превращаются в клеткиГобразующие’крышку перидия,-смыкаю-
щуюся с его боковыми стенками. ЭцЙ/^ии отчасти выдаются йад нижней поверх-
ностью. листа; перидий эцидид на морфологической вершине его, или крыше,
разрывается, и лопасти его в виде зубчиков отворачиваются наружу, так что
1 Недавно удалось заставить один ржавчинник постепенно перейти в лабораторных
условиях на сапрофитное питание.
ISO
н гл >
РАСТЕНИЯ
Рис* 90. Пикниды и эцидии пукцинии (Рпс-
cinia gtaminis):
1—пикниды на листе барбариса; 2—мрщнй; «^мо-
лодая цепочка эцидиоспор с базальной и промежу-
точными клетками
эцидии получает вид вазочких на-
полненной эцидиоспорами. По-
следние разъединяются вследствие
разрушения промежуточных кле-
ток, выпадают вниз из эцидия и
разносятся воздушными течения-
ми. Они не могут заражать вновь
барбарис, а заражают. листовые
влягя^типа и стебли' злаков. Там
они раанивяют межклеточный мц-
целий, состоящий из клеток с
дикарионами. Через 5—7 дней на
нем под "Эпидермисом растения-
хозяина образуется новое спора-
ношение гриба—собрание оваль-
ных одноклеточных двухъядерныу
уд^Д^о сЪ о /р^сидящих каждая
*йа ножке \^и6. pl, I). Эпидермис
растения-хоз^инй раскрывается, и
обнажаются • ( f редоспоры, со-
бранные в заметные невооружен-
ным глазом/ржцвые полоски; не-
сколько похожие на ржавчиду на
железе. Окмсжа зависит от оран-
жевыхНадаль масла в клетках
уредоспор/^Разносимые воздушны-
ми течениями, уредодйд^Ьг про-
растают и вызывают (рис, И1,
новое заражеййё злаков; такие
заражения повторяются 5—6 раз
в лето, так что от первичного за-
ражения злака одной эцидиоспорой
потенциально к концу лета может образоваться 1015—10м уредоспор. К кон-
цу лета на том же мицелии, который давал уредоспоры, образуется новое
спороношение — т_а д е й т о с п о р ы: они также сидят на ножках и состоят
каждая иц ппут лиухъядернЫХ тслЕгок с толстой темно-бурой оболочкой,
(рис. 91, 3). На злаках скопления телейтоспор заметны в виде черных про-
долговатых полосок на влагалищах листьев и на стеблях. Тмейтосдоры про-
растав только-следующей весной, после перезимовки (рис. Sl,4)e, Перед
1Г, Г’
этим ядра дикарионов сливаются; получившиеся пйплоидтае ядра делятся
редукционной И па тгяжттпи тдпдйтлр.ттлрм «трдетддт квтмрйтклетрчг-
ная Апагмобямтпжя; на каждой клетке фрагмобазидии образуется по одной
гаплоидной одноядерной базидиоспоре. Ботатгыпртт/Т^т> рпппоспмыо воздуш-
ными течениями, заражают листья барбариса, образуя в них гаплоидный
мицелий, и цикл развития повторяется.
Весь цикл развития Puccinia graminis и смена ею растений-хозяев были
впервые выяснены работами немецкого миколога А. Де-Бари в 1864—
1865 гг. В течение долгого времени была неясна роль пикноспор, и только в
1927 г. канадский миколог и фитппятллпг К р э г и значение»
Его опыты обнаружили, что ржавчиннцки гетероталличны и что на гаплоид-
ном мицелии (+ или —), развивающемся из заражения одной базидиоспо-
рой, не могут образоваться эцидиоспоры, содержащие но 2 ядра (дикарионы)*
Для образования их необходимо наличие е£це, ржавчинника противополож*
114
ного полового знака. В природе это осуществляется или при срастаг
лиев противоположных знаков, получившихся на одном листе барбариса
вследствие заражения несколькими (хотя бы двумя) разнозначными бази-
1-1'
диоспорами, или при переносе насекомыми пикноспор одного знака в
пикниды мицелия противоположного знйка. Лучшему улавливанию этих
отдал—гр
л м
Рис. 91. У ре до- и телейтоспороношение пукцинии (Puccinia graminis):
1 — тредоспоровошеяме; s — проросшая урепоспора; а — телейтоспороношение; 4 — проросшая телейяо-
спора
пикноспор способствуют особые длинные извилистые - гифы, высовы-
вающиеся из пикнид {рис» 91, 7): к ним прилипают пикноспоры, пе-
реносимые насекомыми. Содержимое пикноспоры переходит в эти гифы и
далее по ним в мицелии. Ядра пикноспор, очевидно, размножаются делением
и через некоторое время образуют с ядрами мицелия, куда они попали, дика-
рионы, после чего становится возможно образование двухъядерных эцидио-
спор. Цитологические подробности
выяснены1.
этой дикарионтизации еще недостаточно
Таким образом, в полном цикле развития ржавчинного гриба имеются:
1) гаплоидная фаза, начинающаяся с базидиоспор и представленная далее
гаплоидным мицелийм с пикнидами и зачатками эцидиев (только зачатками,
но без эцидиоспор!); 2) дикарионтическая фаза, начинающаяся незадолго
до образования эцидиоспор, длительно продолжающаяся затем через боль-
шую часть цикла развития с повторным образованием дикарионтического
мицелия с уредоспорами и заканчивающаяся в телейтоспорах перед слия-
нием их ядер; 3) диплойдная фаза в виде телейтоспор с диплоидными яд-
рами до редукционндГб ДёЛййия их". " ~~~
У других родов и видов ржавчинников много отличий от описанных
морфологических особенностей и цикла развития Puccinia graminis. Пик-
ниды, кроме кувшинообразных, бывают еще плоские. Эцидиальные споро-
1У гименомицетов при анастомозах клеток мицелия противоположных знаков ди-
карионы тоже образуются не сразу: ядро, перешедшее в клетку другого мицелия, может
перейти в последующие нлетки, размножаясь при этом делением, пока, нанонец, на неко-
тором расстоянии от места первичного анастомоза не ассоциируется с ядрами другого по-
лового знака в дикарионы. Канадский миколог Беллер путем экспериментов и наблю-
дений определил, что скорость перемещения ядра равна приблизительно 4,5 мм в час.
452
НИЗШИЕ РАСТЕНИЯ
ношения, кроме типичных эцидиев в форме вазочки, бывают в виде пузырей
неправильной формы, разрывающихся лоскутами (так называемый пери-
д е р м и й), или в виде довольно длинных рожков, вскрывающихся часто
продольными трещинами (рестелия), или в форме плоского спороно-
шения, не имеющего оболочки (перидия) и окруженного лишь венцом булаво-
видных парафиз (ц е о м а)1. Телейтоспороношения представляют наиболь-
шее количество различии и кладутся в основу подразделений на роды. У до-
вольно многих видов телейтоспоры могут прорастать без перезимовки.
Многие роды являются однохозяинными, т. е. проходят весь цикл
развития на одном растении-хозяине. Предполагают, что это позднейший
признак и что все примитивные ржавчинники были разнохозяинны.
Далеко не все ржавчинники имеют такой полный цикл развития, как
Puccinia graminis. Существует очень много так называемых неполных видов
(у них нет тех или иных спороношений), которые считают вторично упро-
щенными.
Описано около 4500 видов ржавчинников. Обычно их делят на 2 се-
мейства: пукциниевые (Pucciniaceae) с телейтоспорами на ножках и
мелямпсоровые (Melampsoraceae) с телейтоспорами без ножек.
Самый крупный род семейства пукциниевых (Pucciniaceae) — пук-
циния (Puccinia), содержащий свыше 1800 видов (в СССР—459). Кроме опи-
санной уже линейной ржавчины (Puccinia graminis), широко распространены
бурая ржавчина ржи (Р. disperse,), образующая эцидии на некоторых дико-
растущих бурачниковых; бурая ржавчина пшеницы (Р. triticina), образую-
щая эцидии в Европе на видах василистника (Thalictrum), а в Сибири на
Leptopyrui^fumartoides-, желтая ржавчина (Р, glumarum); корончатая ржав-
чина овса (Р. coronifera)9 образующая эцидии на крушине слабительной
(Rhamnus cathartica).
Убытки от ржавчинников вряд ли меньше, чем от головни, вследствие
, их массового распространения. В результате разрушения паразитом хлоро-
. филлоносной паренхимы и более интенсивного дыхания больных растении
снижается вес стеблей и глаййЫМ'"ЦС разим зеина. Уменьшается содержание
белков в зерне. Убытки во всем мире от ржавчины хлебных злаков составля-
ют ежегодно около 1,5 млрд, долларов. Столь огромные потери вызвали необ-
ходимость тщательного изучения ржав*
яснилось, что ботанический вид у этих ржавчиг
я хлебных злаков. При этом вы-
кг.
ков является довольно
сложным комплексом., более мелких наследственно постоянных, но морфоло-
гически неотличимых форм. Так, например, линейная ржавчина (Р, graminis)
имеет ряд специализированных форм (formae spectates), или биологических
видов, паразитирующих только на определенных злаках, — пшеничная,
ржаная, овсяная и другие специальные формы* В свою очередь, эти специаль-
ные формы состоят из большого количества еще более мелких, но тоже наслед-
ственно постоянных единиц — биологических (или физиологических) рас,
или биотипов, способных поражать только сорта или группы сортов. Так,
например, у пшеничной специальной формы линейной ржавчины (Р. grami-
nis /. sp. tritici) установлено около 200 биотипов, у бурой ржавчины пшеницы
(Р, triticina) — свыше 100 биотипов, у корончатой ржавчины овса (Р, со-
ronifera) — около 40 биотипов.
|> и
пи
Бороться с ржавчиной хлебных злаков труднее, чем с головней. Унич-
тожение промежуточных хозяев, например барбариса, установленное в не-
которых странах как обязательное в законодательном порядке, дает поло-
жительный эффект. Рекомендуется опыливание полей с самолетов молотой
серой. Большое значение имеют выведение и распространение в культуре им-
мунных или ела бопоражаемых сортов (метод, широко применяемый и в борь-
бе с другими паразитическими грибами). Однако для ржавчины хлебных
1 Различные наименования разных спороношений ржавчинников, нока не было выяс-
нено, что они являются стадиями развития одного вида, употреблялись как самостоятель'
ные родовые названия,
ОТДЕЛ — ГРИБЫ
153
Рис. 92. Телейтоспоры ржавчинников:
1 — уромицес; в — фраппщиум; 3 —мелямпсора; 4 — кронарциум
((3 — фрагмобазидии с базидиоспорами, образовавшиеся из про-
росших телейтосиор; телейтоспоры изображены с содержимым)
злаков он не имеет универсального значения и
действителен лишь по отношению к тем или
иным группам биотипов; кроме того, значение
его ослабляется еще широкой возможностью
образования новых биотипов путем гибридиза-
ции, происходящей при переносе пикноспор
насекомыми.
Из других видов пукциний большое практи-
ческое значение имеет Р. helianthi — полный
однохозяинный вид, паразитирующий на под-
солнечнике.
Из других ржавчинников семейства пукци-
ниевых назовем род Uromyces (телейтоспоры
одноклеточные), в частности £7. pisi — паразит
гороха, эцидиальный хозяин которого— молочай
(рис. 92, 1). Виды рода фрагмидиум (Phrag-
т id turn; телейтоспоры из нескольких кле-
ток, сидящих одна над другой; рис. 92, 2) —
однохозяинные паразиты розоцветных.
В семействе мелямпсоровых (телейтоспоры
без ножек) тоже немало паразитов. Например,
однохозяинный паразит льна М. lint (рис. 92, 3)
вызывает болезнь, называемую присухой или
мухеседом. Важное значение имеет род
кронарциум (Cronartium), одноклеточ-
ные телейтоспоры у его представителей сое-
динены в длинные цилиндрические колонки
(рис. 92, 4). Cronartium ribicola образует уредо-
и телейтоспоры на смородине (Ribes), а эцидии — на кедровой и веймуто-
вой соснах. Занесенный из Европы в Северную Америку* этот вид стал
опасным паразитом веймутовой сосны.
5-й (ФОРМАЛЬНЫЙ) КЛАСС — НЕСОВЕРШЕННЫЕ ГРИБЫ
(FUNGI IMPERFECTI, DENTEROMYCETES)
К несовершенным грибам следовало бы относить только такие грибы,
которые имеют многоклеточный мицелий, т. е. принадлежат к высшим гри-
бам, но не имеют"ни сумчатого, ни базидиального спороношения и размножа-
ются лишь бесполым путем — конидиями. В действительности к ним отно-
сят также и конидиальные стадиитех 'грибов, для которых уже установлено
сумчатое (очень редко базидиальное) спороношение.Втаком объеме несо-
вершенные грибы — огромная группа, насчитывающая свыше 25 000 видов;
у многих из них в дальнейшем, вероятно, будут найдены сумчатые спороно-
шения, многие же, по-видимому, совершенно утратили их.
Несовершенные грибы чрезвычайно Широко распространены в природе
и живут как сапрофиты в почве или на отмерших растениях или как пара-
зиты на высших растениях, в том числе и на многих культурных.
Классификация несовершенных грибов основана на их конидиальных
спороношениях. Она удобна, но совершенно искусственна. Нередко кони-
диальные стадии разных видов одного рода сумчатых грибов прдходится
относить не только к разным родам, нои к разным семействам и порядкам не-
совершенных грибов. И наоборот, виды несовершенных грибов, относящие*
НИЗШИЕ PACT
х: । i
Рис. 93. Спороношеиия несовершенных грибов:
1 — гифошщегы: а — отдельные конидвеносцы; б "-коремий, »— ыелянконнешАе (лаке); Л — сфера-
вскдные (пикнида)
ся в искусственной системе к одному роду, могут быть конидиальными ста-
диями грибов, относящихся к разным порядкам (например* из пиреномице-
тов и из дискомицетов).
В распространенной искусственной системе несовершенных грибов, раз-
работанной итальянским микологом П. Саккардо* выделяют три по-
рядка.
1-Й ПОРЯДОК — ГИФОМ ИЦЕ ТЫ (HYPHOMYCETALES, ИЛИ HYPHALES,
ИЛИ MONILIALES)
Конилиенпстпл у нит расположены поодиночке Па. мицелии или собраны
^ небольшие пучки — коремии (рис. 93, 7);
Из гифомицетов можно отметить вертациллиум (Verticillium). Некото-
рые виды его, поселяясь в сосудах вышних растении (хлопчатник, картофель
и др.), вызывают увядание их, известное под названием вилт или вертиллез.
Алыпернария (Alternaria) — сапрофиты и паразиты на картофеле, табаке и
других растениях. Фузариум (Fusarium) —Тюлыпо? род с сапрофитными и
паразитными виДами;~последние вызывают так называемые фузариозы рас-
тений, иногда увядание (вилт); некоторые ядовиты и при употреблении пора-
женных ими растений в пищу («пьяный» хлеб, «пьяное» льняное масло) вызы-
вают симптомы, сходнее с опьянением; F. nivale вызывает иногда в сырых
местах гибель озимых от «снежной плесени». Кандида (Candida albicaus) —
возбудитель молочницы у грудных детей и других^заболеваний (во всех воз-
растах) слизистых оболочек и кожи* носящих общее название кандида-
микозов.
ОТДЕЛ — ГРИВЫ
2-Й ПОРЯДОК — MEЛЯНКОНИЕВЫЕ (MELANCONIALES)
Конириеноспы собраны плотным слоем. тдпа гимения, на поверхности
плоского переплетения гиф-т-ложа иди стромы (рис. 93, 2).
Из мелянкониевых можно отметить глеоспориум (Gleosporium) —
большой род преимущественно паразитных грибов, вызывающих пятни-
стости, так называемое антракнозы, на листьях, стеблях и плодах ра-
стений (у винограда, фасоли, тыквенных и др.).
3-Й ПОРЯДОК — СФЕРОПСИДНЫЕ (SPHAEROPSIDALES)
Конидиеносцы внутри шаровидных шш яйцевидных споровмеетилшдт
имеющих отверстия на вершине, -- пикнид. Споры, называемые также пикно-
спорами или стилоспорами, выходят наружу через отверстие, часто в тяже
разбухающей слизи Трис. 93, 3).
' Из сферопсидных можно отметить фома (Phoma) — большой род
(свыше 2000 видов). Многие — паразиты на винограде,, капусте, свекле и
других растениях. Септария (Septoria) — свыше 1000 паразитных видов.
Паразиты на груше, томатах, конопле, кендыре и других высших растениях.
ФИЛОГЕНЕЗ ГРИБОВ
Единого мнения о происхождении и филогенезе грибов нет. Некоторые счи-
тают их группой полифилетической и выводят отчасти из бесцветных жгу-
тиковых, отчасти, из разных групп водорослей — разножгутиковых, конъю-
гат (зигомицеты), сифоновых (оомицеты), красных (аскомицеты). Однако
связи с водорослями маловероятны. Полное отсутствие не только пластид-
ных пигментов, но и самих пластид, иное положение жгутиков и ха-
рактер зооспор, иные запасные продукты (у грибов—гликоген, отсутствую-
щий у водорослей) и ряд других особенностей противоречат сближению гри-
бов с какими-либо водорослями.
Вероятнее монофилетическое происхождение грибов от каких-то бес-
цветных жгутиковых. С этой точки зрения наиболее примитивными должны
считаться архимицеты, не имеющие даже зачаточного мицелия. Из них воз-
никли фикомицеты, причем уже очень рано образовались два эволюционных
ИГ
ряда — с одножгутиковыми и двужгутиковыми спорами; мицелий, вначале
зачаточный (ризрмицелий), стал затем типичным неклеточным. Одножгутико-
вые не получили широкого распространения и большого развития; наибо-
лее высоко организованы среди них моноблефаридовые. Двужгутиковые
дали большое количество представителей, и притом более высокоорганизо-
ванных; вершиной их эволюции являются пероноспоровые, уже вполне при-
способившиеся к наземному образу жизни. Однако они, как моноблефаридо-
вые, представляют слепые ветви в эволюции грибов.
Из каких-то примитивных мицелиальных форм фикомицетов, имевших
зигогамный половой процесс (вроде полифага), могли развиться зигомицеты.
Из зигомицетов нетрудно вывести некоторые первичные аскомицеты
(например, диподаскус, эремаскус), имеющие сходный половой процесс и
гг
отличающиеся от зигомицетов лишь многоклеточностью и немедленным про-
растанием зиготы, становящейся сумкой.
Эволюция аскомицетов выразилась в увеличении числа сумок (что было
достигнуто развитием аскогенных гиф) и в образовании плодовых тел. В фор-
мировании последних видны два направления. Одно состояло в появлении
первично замкнутых плодовых тел — клейстокарпиев — с беспорядочно рас-
положенными сумками. По мнению некоторых микологов, клейстокарпии
дали начало перитециям, открывающимся узким отверстием, через которое
аскоспоры активно выбрасываются из сумок, расположенных более или ме-
нее параллельно и обращенных вершиной в сторону отверстия плодового
тела. Другое направление выразилось в развитии апотециев, содержащих
156
НИЗШИЙ РАСТЕНИЯ
гораздо большее число сумок и еще лучше приспособленных к активному
разбрасыванию аскоспор. Это направление также могло привести к образо-
ванию полузамкнутых перитециев. Не исключена возможность, что клейсто-
карпии в некоторых линиях развития (Erysiphales) также могли возникнуть
вторично, как замкнувшиеся перитеции. Изучение онтогенеза трюфелевых
дает основание предполагать, что они в качестве особой эволюционной линии
развились из дискомицетов; вследствие подземного образа жизни плодовые
тела их стали замкнутыми и грибы утратили способность активно выбрасы-
вать аскоспоры.
Половой процесс у аскомицетов развивался вначале в направлении ус-
ложнения половых органов и дифференцировки их на мужские и женские
(архикарпы и антеридии). Затем многократно в разных эволюционных ли-
ниях происходила утрата антеридиев. Оплодотворение стало осуществлять-
ся или ^пермациями (микроконидиями, вторично приобретшими эту функ-
||'М
), или клетками вегетативных гиф, или же в пределах самого аскогона
путем попарной ассоциации его ядер. Дальнейшее угасание типичного поло-
вого процесса у аскомицетов привело к полной утрате половых органов и к
соматогамии (псевдомиксису) — половому слиянию вегетативных кле-
ток мицелия.
Происхождение базидиомицетов довольно неясно. Одни микологи вы-
водят их из зигомицетов, другие — из аскомицетов, в частности из диско-
мицетов. При этом приходится допускать, что базидиоспоры имеют, в сущ-
ности, эндогенное происхождение: на гомологичной сумке базидии после
редукционного деления диплоидного ядра образуются четыре наружных выпя-
чивания; в каждое выпячивание входит ядро и цитоплазма, вокруг которых
образуется новая оболочка (как вокруг аскоспор), отделяющая их от канала
стеригмы; иногда эта оболочка бывает заметна, так как неплотно прилегает
к наружной оболочке (выпячиванию) базидии. Аналогия такому образованию
спор имеется у некоторых зигомицетов, где сходно развиваются односпоро-
вые спорагиоли (хетокладиум и др.).
Половой процесс у базидиомицетов морфологически крайне простой и
выражается у большинства в форме анастомозов двух вегетативных клеток
мицелия. Специальных половых органов ни у кого из представителей базидио-
мицетов нет; зато дикарионтическая фаза существует гораздо более дли-
тельно, чем у аскомицетов (кроме тафриновых) и продолжается почти в те-
чение всего цикла развития.
Наиболее примитивны среди базидиомицетов простейшие холобазидио-
мицеты с зачаточным плодовым телом в виде плоского рыхлого сплетения
гиф, имеющим гладкий гименофор. Дальнейшая эволюция их выразилась в
образовании более массивных, различно устроенных плодовых тел, с сильно
увеличенной поверхностью гименофора, развивающих огромное количество
базидиоспор.
Связь гастеромицетов с гименомицетами неясна; возможно, что они не-
сколькими эволюционными рядами развивались из низших гимеиомицетов; с
другой стороны, есть немногочисленные сторонники теории происхождения
агариковых из гастеромицетов.
их холобазидиоми-
Фрагмобазидиомицеты рано отделились от просто:
. ш
цетов. Среди них порядки с плодовыми телами, вероятно, первичны.
Различное положение перегородок в фрагмобазидиях у дрожалковых
(Tremellales) и аврикуляриевых (Auriculariales) рисует эти два порядка как
две самостоятельные линии эволюции. Но переходы в формах фрагментации
базидий приводят к выводу, что эти порядки могли возникнуть от общего
предка. Таковым, вернее всего, был какой-то таксон из холобазидиальных
грибов, близкий к примитивным Aphyllophorales. Что касается головневых
(U stilaginales) и ржавчинных (Uredinales), то, учитывая поперечное положе-
ние перегородок в их базидиях, можно говорить о родственной связи с Auri-
culariales. Это подтверждается и другими, более специальными особенностя-
ми организации. Головневые и ржавчинные* возникнув от общего предка*
ОТДЕЛ — ГРИБЫ
*
157
эволюционировали независимо друг от друга. В связи с паразитным су-
ществованием они утратили плодовые тела, которые были у их предков, но
зато выработали много специальных приспособлении как в образе жизни
(у ржавчинников), так и в морфологии. Конидиальные формы у многих выс-
ших грибов, особенно у аскомицетов, в связи с регрессом или утратой поло-
вого процесса приобрели физиологическую и морфологическую независи-
мость; в систематике грибов это дало основание к выделению их в отдельную
таксономическую группу несовершенных грибов.
ЭКОЛОГИЯ ГРИБОВ
Ранее уже указывалось, что грибы питаются готовыми органическими
соединениями как сапрофиты, паразиты или как симбионты, причем между
всеми этими группами имеется ряд переходов. Первоначальным является
сапрофитное питание, свойственное и в настоящее время подавляющей части
грибов. Большинство сапрофитов поселяется на разнообразных органи-
ческих субстратах и не отличается большой требовательностью к определен-
ному их составу. Но есть и специализированные экологические группы
сапрофитов, например грибы, живущие в воде на трупах животных (сапролег-
ниевые), копрофильные грибы, поселяющиеся преимущественно на экскре-
ментах животных, особенно травоядных (мукоровые, некоторые аскомицеты
и базидиомицеты), кератинофильные грибы, живущие на рогах, копытах, во-
лосах, перьях, ногтях животных и содержащие много особых белков — ке-
ратинов. Большую отрицательную роль в природе и в хозяйстве человека иг-
рают дереворазруш'ающие грибы, питающиеся древесиной и вызывающие
разрушение (гниение) ее (трутовиковые и др.)- Большинство дерево разру-
шающих грибов — чистые сапрофиты; у многих грань между сапрофитиз-
мом и паразитизмом неясная и нерезкая: они поселяются на живых деревь-
ях,но питаются одревесневшими оболочками клеток и продолжают развиваться
на этих деревьях и после их отмирания. Гнили, вызываемые ими, различа-
ются по окраске (белая, красноватая, бурая и др.), по расположению —
в центральных частях дерева (сердцевинная — у большинства трутовиков)
или на периферии его. В зависимости от выделяемых ферментов одни дерево-
разрушающие грибы разрушают прежде всего лигнин клеточных оболочек
(коррозионная гниль), другие (домовый гриб и др.) — целлюлозу (деструк-
тивная гниль), многие действуют одновременно на оба этих компонента кле-
точных оболочек.
Очень велика группа почвенных сапрофитов, среди которых имеются как
виды без плодовых тел или с микроскопически мелкими плодовыми телами
(почвенные микрофиты), так и с крупными плодовыми телами (почвенные
макрофиты). К почвенным микрофитам относятся многие сотни видов; наи-
более часты среди них аспергилловые, мукоровые, многие гифомицеты, ак-
тиномицеты, некоторые пиреномицеты, В лесах их больше, чем в степях или на
лугах. В 1 г почвы содержатся в том или ином виде десятки и сотни тысяч
зачатков грибов. К почвенным макрофитам относятся главным образом гиме-
номицеты, в меньшем количестве гастеромицеты и некоторые крупные аско-
мицеты (ряд дискомицетов). Особенно богаты ими леса, что объясняется оби-
лием в их почве органических веществ, высокой влажностью почвы и возду-
ха, а также и тем, что очень многие почвенные макрофиты живут в симбиозе
с корнями древесных пород, образуя микоризу, и в других условиях не раз-
вивают плодовых тел. Бедны почвенными макрофитами обработанные почвы, а
также почвы песков, пустынь, хотя и на них имеется некоторое количество
специально им свойственных видов. Интересно образование плодовыми те-
лами почвенных макрофитов, так называемых «ведьминых колец»: плодо-
вые тела гриба располагаются на небольшом расстоянии друг от дру^а,
образуя явственный круг. Объясняется это равномерным центробежным
ростом мицелия из какого-либо пункта по всем радиусам; на периферии тако-
158
низ:
и»:
СВ РАСТЕНИЯ
н»
го многолетнего мицелия, постепенно отмирающего в старых частях, ежегод-
но образуются плодовые тела. Диаметр «ведьминого кольца» ежегодно рас-
ширяется на 8—50 см и может достигнуть нескольких десятков метров. Осо-
бенно часто наблюдаются «ведьмины кольца» у лугового опенка (Maramiut
oreades), шампиньонов, масленков и других грибов.
Узкий слой почвы, непосредственно прилегающий к корням высших
растений, называемый ризосферой, бывает особенно обильно насе-
лен микроорганизмами — бактериями и грибами, использующими корне-
вые выделения и продукты частичного отмирания корней. Некоторые компо-
ненты ризосферы, возможно, играют роль в питании высших растений, пе-
реводя сложные органические соединения почвы в более простые, усвояемые
корнями, а также связывая атмосферный азот и обогащая им почву (азото-
бактер и др.).
Кроме сапрофитных грибов, в почве находится еще много паразитных,
но не в вегетирующем состоянии, а в виде спор и других покоящихся образо-
ваний. При подходящих условиях они могут вызвать инфекции растений, и
микрофлора почвы может быть источником инфекции.
Переход от сапрофитизма к паразитизму осуществлялся, вероятно, пос-
тепенно в разных линиях эволюции, и, как уже ранее указывалось, между ти-
пичными сапрофитами и типичными паразитами существует ряд промежу-
точных форм взаимоотношений.
Биологическая выгода паразитизма заключается в том, что паразит ухо-
дит от конкуренции с сапрофитами, не могущими питаться за счет живого
организма. Эволюция паразитизма шла, вероятно, от полифагии к монофа-
гии, т. е. от способности паразитировать на растениях различных видов,
родов и даже семейств к паразитизму только на видах одного рода, или на
одном виде, или даже на определенных сортах его, кйк это наблюдается у био-
типов многих ржавчинников, головневых и др. Эволюция паразитизма выра-
жалась также в разном воздействии на ткани растения-хозяина: на низших
ступенях паразитизма паразит быстро убивает находящиеся возле него клет-
ки растения-хозяина; на высших ступенях паразит вначале не только не
убивает клетки хозяина, но у некоторых даже вызывает разрастание их, об-
разование опухолей, галлов, «ведьминых метел» или, как у многих головне-
вых, при диффузном поражении мицелий паразита пронизывает все растение
или отдельные побеги его и проявляет свое разрушительное действие лишь в
конце вегетации. От этой высокой взаимоприспособленности паразита и хо-
зяина возможен был переход к мутуалистическому симбио-
зу1, т. е. такому сожительству двух организмов, при котором они извле-
кают из этого какую-то пользу. Так как оба симбионта находятся все-таки в
антагонистических отношениях друг к другу, то мутуалистический симбиоз
нередко называют сбалансированным паразитизмом
или аллелопаразитизмом1 * 3, здесь имеет место не «дружба», а
равновесие в борьбе. Примерами таких отношений являются лишайники и
микориза3.
Микоризой называют сожительство гиф гриба с корнями высшего рас-
тения. Различают два типа микориз — эктотрофную и эндотроф-
ную . У растений с эктотрофной4 микоризой гифы гриба,оплетают снаружи
короткие молодые боковые корешки, образуя вокруг них плотный чехол.
Корешки эти перестают расти в длину и начинают ветвиться, иногда виль-
чато; корневой чехлик и корневые волоски у них не развиваются. Функции
1 Симбиозом в широком смысле этого Слова цожно называть всякое сожительство
двух организмов при наличии непосредственного обмена веществами между ними, в том
числе и паразитизм. В более узком смысле термин «симбиоз» применяется обычно для
обозначения мутуалистического симбиоза (от лат. «мутус» — взаимный, обоюдосторонний 1
и от греЧ. «симбиоз» — живущий вместе).
*От греч. «аллелон» — друг друга, взаимно.
3 От греч. «микео — гриб и «ридза» — корень.
4 От греч. «эктос» — снаружи и «трофе» — питание,
ОТДЕЛ —ГРИБЫ
159
корневых волосков выполняют гифы гриба, отходящие от грибного чехла и
пронизывающие почву. С другой стороны, часть грибных гиф отходит от *
грибного чехла внутрь корня, в первичную кору, где они характерным об-
разом располагаются, образуя сеть между клетками («сеть Гартига»). От-
ветвления этих гиф нередко проникают внутрь паренхимных клеток первич-
ной коры и в дальнейшем частично растворяются и «перевариваются» ими.
Следовательно, эктотрофнаямикориза не целиком наружная, и ее часто на-
зывают эктоэндотрофной1.
. Эктоэндотрофные микоризы образуются у большинства наших древесных
хвойных и лиственных растений. Грибы, образую
иг
е их, принадлежат глав-
ным образом к различным гименомицетам, съедобным и несъедобным, в мень-
шей степени — к гастеромицетам и высшим дискомицетам. Узкой приурочен-
ности отдельных видов грибов к определенному виду дерева у большинства
грибов, по-видимому, нет, хотя некоторое предпочтение все же имеется,
что отражается в названиях грибов, например подберезовик, подосиновик.
Вне леса грибы, образующие
микоризу, не растут, возможно, потому, что
сожительство с корнями деревьев для них необходимо. Гриб получает из
корней растворимые углеводы; при помощи меченых атомов углерода в по-
следнее время доказана передача их из высшего растения в микоризный гриб.
В свою очередь, высшие растения при посредстве .гиф гриба, пронизываю-
щих почву, получают из нее воду и различные элементы минерального пи-
тания. Гриб разлагает в почве сложные органические соединения, недоступ-
ные высшим растениям; при помо
ИГ.
меченых атомов экспериментально до-
казано поступление в корни через микоризу азота, фосфора. Возможен так-
же взаимный обмен между высшим растением и грибом ростовыми веществами
и витаминами. Одни виды древесных пород, как сосна, ель, лиственница,'
дуб, особенно нуждаются в микоризе; другие, как береза, ива, липа,
слабомикотрофны; бобовые и некоторые другие растения вообще не имеют
микоризы.
При искусственном лесоразведении углубленное изучение вопросов ми-
коризообразовапия имеет большое значение, так как без микоризы многие
древесные породы гибнут или плохо растут.
Эндотрофная микориза обнаружена у очень большого количества тра-
вянистых растений из самых различных семейств и у некоторых древесных
(клен и др.), причем новые исследования все увеличивают число растений с
такой микоризой. Существует несколько типов эндотрофных микориз. У од-
них, например у ладанниковых, гриб пронизывает все растение и передает-
ся с семенами; у большинства он находится только в клетках коры подзем-
ных органов. Мицелий гриба проникает внутрь клеток и образует там у од-
них клубки гиф, у других древовидные разветвления (арбускулы),
пузыревидные вздутия (везикулы). Клетки корня «переваривают» про-
никший в них гриб, что иногда сравнивают с фагоцитозом у животных.
У некоторых растений с эндотрофной микоризой (например, у грушанковых)
гриб образует наружный чехол на корнях, у подавляющего большинства
чехла нет; у некоторых (например, орхидных) гифы гриба выходят наружу в
почву, у других наружные гифы наблюдаются редко. Орхидные в естествен-
ных условиях и при культуре в теплицах не могут развиваться без гриба, и
отношения высшего растения с грибом должны характеризоваться как а л -
лелопаразитизм (обоюдный паразитизм) или у бесхлорофилльных
орхидных (гнездовка и др.) как умеренный паразитизм высшего растения
на трибе. Во многих других случаях, по-видимому, имеет место паразитизм
гриба на высшем растении, причем высшее растение частично переваривает
своего паразита (толерантный — переносимый — паразитизм,
т. е. прятаняпптций малый ущерб). Грибы эндотрофных микориз относятся
к разным систематическим группам. Они могут быть и зигомицетами, и
1 От греч. «эндон» — внутри.
160
НИЗШИЕ РАСТЕНИЯ
ri:i
высшими грибами с многоклеточным мицелием (некоторые гименомицеты и
др.), в большинстве своем не дающими спороношений и поэтому точно не
определяемыми.
Впервые микориза была описана русским ботаником Ф, М. Камен-
ским в 80-х годах XIX в. Несколько позднее вышел ряд работ немецкого
ботаника А.-Б. Франка, предложившего и самый термин «микориза»;
в настоящее время имеется уже свыше 1000 работ по изучению микориз.
Широкое и повсеместное распространение грибов в значительной степе-
ни объясняется огромным количеством производимых ими спор и легкостью
их распространения. В одном спорангии у многих мукоровых образуются
тысячи и десятки тысяч спор (у фикомицесс — 70 000 — 80 000). Колония
пеницилла диаметром 2,5 см дает около 400 000 конидий. В листе ябло-
ни, пораженной вентурией, могут образоваться 2—3 млн. аскоспор. На ме-
телке овса, пораженной головней, находится несколько миллионов головне-
вых спор. На одном кусте барбариса могут быть миллионы эцидиоспор ли-
нейной ржавчины злаков. Шампиньон за 6 дней спороношения образует около
16 млрд, базидиоспор; настоящий трутовик (Fomes fomentarius) за 20 дней
образует более 7,5 трлн, базидиоспор.
Распространение спор подавляющего большинства грибов происходит
воздушными течениями (анемохория), что вполне понятно, так как
вес одной споры обычно составляет миллионные или десятимиллионные
доли миллиграмма. Споры могут распространяться на километры, десятки
и даже сотни километров. Особенно велико количество спор грибов в возду-
хе в летнее и раннеосеннее время, когда они даже бывают причиной ал-
лергических заболеваний у человека (заболевания от введения чужеродного
белка).
Споры разносятся также животными (зоохория): млекопитающими,
поедающими плодовые тела шляпочных грибов; несекомыми, питающимися
плодовыми телами, сахаристыми выделениями (конидии у спорыньи и др.),
спорами (у ржавчинников) или откладывающими яйца в плоды (казарка на
яблоках и др.), в гниющее мясо (мухи на фаллюсах и др.) и т. д. Некоторую
роль в распространении спор у грибов играет вода (головневые и др.).
НРАВТИЧЕСВОЕ ЗНАЧЕНИЕ ГРИБОВ
Грибы играют большую роль в круговороте веществ в природе и, в част-
ности, в почвообразовательных процессах: они разрушают и минерализуют
органические вещества (главным образом остатки отмерших растении), т. е.
возвращают в почву элементы минерального питания, делая их вновь доступ-
ными для высших растении.
В хозяйственной жизни человека грибы играют как положительную, так
и отрицательную роль. Среди первых, пожалуй, наибольшее значение имеют
дрожжевые грибы, производящие спиртовое брожение; они необходимы для
призводства спирта, вина, пива, кваса, в хлебопечении (разрыхление теста).
Дрожжи, содержащие много белков и витаминов, применяются, кроме того,
и как пищевой продукт, и с лечебными целями, и для добавления в корм сель-
скохозяйственным животным. Дрожжевидные грибы (Torulopsis) участвуют
в производстве кефира, кумыса. Для получения спиртных напитков приме-
няются еще, особенно в Восточной Азии, некоторые виды мукоровых грибов,
вызывающие спиртовое брожение.
Съедобные шляпочные грибы в ряде стран в значительных количествах
употребляются в пищу, причем круг использования, кроме состава грибной
флоры, зависит еще от укоренившихся привычек населения. В СССР растет
свыше 150 видов грибов, пригодных для употребления в пищу, но широкое
использование имеют лишь несколько десятков. Однако пищевое значение
грибов несколько преувеличивают, называя их «растительным мясом». На де-
ле плодовые тела съедобных шляпочных грибов содержат в свежем состоя»
ОТДЕЛ — ГРИБЫ
161
нии около 80—90% воды, белков ясе — около 2—4%; кроме того, около 15—
45% белков не усваивается организмом человека. Жиров в свежих съедоб-
ных грибах менее 1%; углеводов (кроме оболочек) часто еще меньше, чем
белков. Таким образом, по питательности грибы стоят не выше овощей, ио с
добавлением их в пищу содерясащиеся в них экстрактивные вкусовые вещест-
ва вызывают большее выделение пищеварительных ферментов и улучшают
ркус блюд.
Особые виды пенициллов применяют при изготовлении сыров (рокфор,
камамбер). В медицине издавна применяют спорынью, а в последнее время
огромное значение приобрел Penicillium chrysogenum как продукт антибио-
тика пенициллина.
Некоторые мукоровые и несовершенные грибы, разлагающие пектино-
вые вещества, способствуют отделению прядильных волокон льна при так
называемой росяной мочке (расстил) его.
Многие грибы причиняют большой вред человеку и его хозяйству. Раз-
; нообразные болезни культурных растений, вызываемые паразитными гри-
бами, значительно снижают уроясаи (в среднем на 10—30%), а иногда вызы-
вают и полную их гибель. Неисчислимые потери народному хозяйству нано-
сит вызываемое грибами гниение древесины в лесах на корню, в постройках,
' гниение шпал на железных дорогах, телеграфных столбов и т. д. Большой
ущерб наносит гниение овощей и фруктов в лежке, плесневение и порча раз-
личных пищевых продуктов, а во влажных тропических странах разруше-
ние грибами и других предметов домашнего обихода.
Ряд заболеваний человека и домашних животных, главным образом бо-
лезней кожи и волосяных покровов (дерматомикозы), вызывается некоторыми
несовершенными грибами. Отравления вызываются не только при употреб-
лении в пищу ядовитых шляпочных грибов (бледные поганки, некоторые
сморчки, красный мухомор и др.), но и хлебом, испеченным из муки с боль-
шой примесью рожков спорыньи или из зерна,. пораженного Fusarium grami-
пеагит. В последние десятилетия наблюдались серьезные заболевания чело-
века, лошадей, вызывающиеся употреблением в пищу зерна или кормов,
пораженных* некоторыми сапрофитными несовершенными грибами.
АНТАГОНИЗМ. АНТИБИОТИКИ
При совместном произрастании двух или нескольких микроорганизмов
они часто оказывают заметное влияние друг на друга, причем один или как
бы благоприятствует другому, или, наоборот, подавляет его развитие. Пер-
вые явления называют ассоциативными, вторые — антагонистическими. Ас-
социативные взаимоотношения мало изучены, хотя, по-видимому, они широ-
ко распространены в природе. В лабораторных условиях они выражаются,
например, более обильным ростом мицелия одного гриба в присутствии
другого или образованием органов спороношения у одного при наличии ми-
целия другого, при этом один гриб вырабатывает какие-либо вещества, типа
ростовых или витаминов, не синтезируемые другим.
Более изучены антагонистические отношения. При совместном произ-
растании микроорганизмы в ряде случаев вырабатывают вещества, называе-
мые антибиотиками, и некоторые из них применяются в медицине как лечеб-
ное средство, так как они подавляют развитие различных патогенных микроор-
ганизмов. При антагонистических отношениях механизм подавления одним
видом другого (или других) бывает в разных случаях различен: быстрое раз-
множение и перехватывание питательных веществ, изменение реакции среды
в сторону, неблагоприятную для другого вида, выделение специфических
веществ — антибиотиков1, делающих невозможным развитие некоторых дру-
гих микроорганизмов, и др.
1 От греч. «анти» — против, «биос» — жизнь.
6—602
162
и г. I;;
РАСТЕНИЯ
III
На антагонистические отношения микроорганизмов обращали внимание
еще во второй половине XIX в., в том числе и русские ученые В. А. М а-
нассеин, А.Г. Полотебнов, И. И. Мечников и другие* * В теку-
щем столетии число таких наблюдений значительно увеличилось, но особен»
но большой размах работы по антибиотикам получили после того, как во
время второй мировой войны был получен пенициллин, спасший жизнь мил»
лионам людей. Пенициллин является кислотой состава R — C9H11O<N2S,
где R означает радикал — углеводород, различный у разных вариантов
пенициллина. Применяется пенициллин в виде натриевой соли. Он мало ток-
сичен для человека и весьма эффективен для лечения ряда опасных заболе-
ваний, вызываемых некоторыми бактериями. Он действует на последние
бактериостатически, т. е. подавляет у них способность поглощать из окру-
жающей среды необходимые для синтеза белков аминокислоты, вследствие
чете бактерии йе могут давать новые жизнеспособные поколения. Однако пе-
[иллин безвреден для грамотрицательных бактерий (например, кишечно-
тифозных и др.), так как они сами могут синтезировать внутри клетки необ-
ходимые им аминокислоты.
Открытие пенициллина было мощным стимулом для поисков новых
антибиотиков в разных группах низших растений. Были исследованы тысячи
видов, и довольно многие среди них оказались производящими антибиоти-
ческие вещества, но лишь очень немногие нашли практическое применение,
так как подавляющее большинство антибиотиков оказывается токсичным для
человека.
II!
и к'
.вой получен из* бактерии Bacillus brevis var. G—В. антибиотик совет-
ский грамицидин (грамицидин С), получивший широкое применение для про-
филактики и лечения нагноений. В 1944 г. американским микробиологом
3. А. Ваксманом был получен антибиотик стрептомицин из акти-
. номицета Actinomyces (Streptamyces) globisporus streptomycini1. Актиномице-
ты оказались вообще наиболее перспективными как продуценты антибио-
тиков, пригодных для применения в медицине. Из них получены еще биоми-
цин, тетрациклин, террамицин, левомицетин, нистатин и другие антибиотики.
Кроме применения в медицине, антибиотики используются также в жи-
вотноводстве, ветеринарии и в борьбе с некоторыми болезнями растений.
ОТДЕЛ ЛИШАЙНИКИ (LICHENOPHYTA)
IH
Лишайники — своеобразная группа низших растений, образованных
грибом и водорослью, соединенными вместе в новые комплексные йли симбио-
тические организмы с иными морфологическими, физиологическими и эколо-
гическими свойствами. Их иногда смешивают с мхами, что отражается в
названии некоторых лишайников (олениймох, исландский мох). Однако они
легко отличимы от мхов отсутствием расчленения на стебель и листья и иной,
не зеленой, окраской. Они бывают сероватые, серовато-ведецоватые, корич-
невые, реже желтые, оранжевые, иногда* почти чёрЕсые^, По форме различают
3 основных типа лишайников (рис?94): 1) до р ков ые , или накип-
ные, имеющие вид налетов или корочек, плотно сросшихся с субстратом и
неотделимых от него без разрушения таллома; 2) лист овитые, имеющие
форму дорзовентральных пластинок, сильно распростертых по субстрату;
они срастаются с ним главным образом пучками грибных гиф, называемых
ризинами, и сравнительно легко могут быть отделены от субстрата.,
3) кус т и с т ы е, имеющие форму столбиков или лент, обычно разветвлен*
, f: ।: ।
иг,
।
1 3. А. Ваксман считал, что продуцентом стрептомицина является Actinomyces
griseus, отнесенный им к роду Streptomyces^^ это название часто встречается в литера- >
* туре. Но советский ученый Н. А. Красильников показал, что продуцентом стреп-
томицина является вид Actinomyces globisporus streptomycint*
ОТДЕЛ — ЛИШАЯНИКИ
163
3
Рис. 94. Талломы лишайников:
1 — горновые, соирикасакиаиеся друг с другом; а — кортовый с перитециями (черные точки); з — листо-
ватый с апотециями; 4 — кустистый
ных и срастающихся с субстратом только основанием, а в остальной части
отходящих от него. Эти три морфологических типа не имеют большого сис-
тематического значения и в некоторых родах, например, встречаются и листо-
вые и кустистые виды. Имеются и промежуточные формы, например в центре
накипные, а на периферии почти "листоватые.
Uli
Грибные компоненты лишайников относятся главным образом к сумча-
тым грибам — пиоеномицетам и дискомицетам, но наряду с этим уже дав-
но известны лишайники с участием базидиомицетов из рода телефора, рас-
пространенные в тропической зоне и насчитывающие 1х/2 — 2 десятка видов.
В настоящее время выяснено, что базидиальные лишайники растут и в уме-
ренной зоне, причем в качестве грибного компонента в них участвуют клава-
риевые и агариковые гименомицеты. Если у ранее известных базидиальных
лишайников водорослевыми компонентами считались только _сине-зеленые
водоросли, то в настоящее время круг их расширен за счетзеленых.ТбЧЕГая
164
НИЗШИЕ РАСТЕНИЯ
Рис. 96. Разрез гомеомерного таллома
коллемы
Рис. 97. Разрез гетеромерного таллома:
к — верхняя и нижняя' кора; в — водоросли (гониди-
альный слой); с — сердцевина; р — ризина
: 111
: 111
: 111
: 111
идентификация лишайниковых грибов с отдельными видами или родами
самостоятельно живущих грибов обычно невозможна.
Водоросли (сине-зеленые и зеленые), входящие в состав лишайников,
мало изменились по сравнению со свободно живущими водорослями. Зеле-
цы&у подавляющего большинства относятся к роду цистококк (Cystococcus,
или Trebouxia, рис- 95) из порядка хлорококк о в ы~х. Это одно-
клето’ЩШ! микроскопически ^мелкие шарики с массивным . центральным,
слегка лопастным хроматофоромГ (рисГ 9бу. У?некоторых лишаиииков водоро-
слевый компонент^™ трёнтеполия (Trentepohlia) из цорядка улотриксовых;
у немногих — еще некоторые другие роды.
Jis сине-зеленых водорослей в лишайниках встречается иосток (No st ос),
реже хроркокк (Chroococcus) и глеокапса (Gloeocapsa). У некоторых находят
водоросли из других родов сине-зеленых.
Каждому виду лишайников свойственна водоросль только одного опре-
деленного рода. Имеются ли видовые различия между водорослями в раз-
личных видах и родах лишайников, сказать трудно.
По анатомическому строению лишайники бывают гомеомерные1
и е-р н_ы_е1 2. У гомеомерных лишайников во всей толще равно-
мерно расположены грибные гифы и водоросли. Обычно те и другие погру-
жены в слизью выделяемую водорослью (рис. 96). Этот тип строения более
примитивный. При гетеромерном строении на поперечном разрезе лишайника
сверху находится так называемаярерхняя кора. Она имеет вид плектенхимы
(псевдопаренхимы), образованной переплетающимися и тесно смыкяттц^мрг-
ся грибными гифами (рис. 97). Полней грибные гифы лежат 6плрр рыхло, и
Между ними находятся водоросли. Этот слой называют гонидиаль-
нТьГм^ я все “водорослей’ в лишайниках— г о и и д и я м"и"2 — название,
данное им в то время, когда лишайник считали не комплексным, а единым,
цельным организмом, а зеленые или сине-зеленые клетки его (водоросли) счита-
ли органами его размножения. Под гонвдиальным слоем грибные гифы распо-
лагаются еще более рыхло, с бсшьшимипромежутками между ними, заполнен-
ными воздухом; эту;часть называют сердцевиной^Под ней находится нижняя
кора, по строению сходная д верхней/Через нее из сердцевины проходят пуч-
ки гиф (ризины), прикрепляющие лишайник к субстрату. У корковых лишай-
: 111
ил!
: 111
: 111
: 111
1 От греч. «гомойосж — одинаковый, равный, «мерос» — часть, доля,
2 От греч. «гетерос» — другой, иной, отличающийся.
• От греч. «гоноо — рождение.
ОТДЕЛ — Л
гл 11
АЙНИКИ
165
Рис. 98. Разрез таллома лишайника с пери-
тециями
ли
’.III
Рнс. 99. Разрез лецидневого я леканорового
апотеция:
1 — лецидиевый; 2 — леканоровый
ли
ли
ли
ников нижней коры нет и грибные
гифьГСердцевиньгпрямо срастают-
ся с~субстратдм'.’“ У~"Кустистых,
радиально построенных гетеро-
мерных лишайников на периферии
поперечного раз рез а нах о дится
кора, под ней — гонидиальный
слой, а внутри — сердцевина.
Характерной особенностью
айников является образование
। особых органических соединений,
; называемых лишайниковыми ки-
i слотами и, как долгое время счи-
/ та л ось, не встречающихся нигде,
$ кроме лишайников. В настоящее
время установлено, что некоторые
лишайниковые кислоты встреча-
ются у грибов. Большинстволи-
" тайниковых кислот имеют кислую
реакцию, многие горьки иа вкус.
Они отлагаются нагифахв виде
зернышек" палочек, кристаллов и
т. п., преимущественно на поверх-
ности коры, отчасти и глубже.
Многие из них окрашены и обу-
словливают окраску лишайников.
С рядо|с химических реактивов
многие лишайниковые кислоты да-
ют цветные реакции, что исполь-
зуется при определении лишайни-
ков. Биологическая роль лишай-
никовых кислот не вполне ясна.
Возможно, что они отчасти служат
для защиты от сильной инсоля-
ции. Предполагают также, что, отлагаясь на оболочках особых гиф, они де-
лающих не смачивающимися водой и этим достигается наличие внутренней
атмосферыв~лйшайнйке,нужной для развития гриба. Установлено, что
лишайниковые кислоты обладают антибиотическим действием. В медицин-
ской практике используется натриевая соль усниновой кислоты, оказываю-
щая влияние на грамположительные бактерии. Используется при лечении
ран, ожогов, трещин.
Водоросли в лишайнике размножаются простым делением клеток -иди
путем образования дБтоспд^ (йепрдвижные споры внутри старой клетки),
айников и культивируемых отдельно цистококков вос-
станавливается способность образовывать зооспоры и даже гаметы, утрачен-
ные ими в лишайнике.
Грибы лишайников образуют характерные для них спороношения. Пе-
ритеции погружены в таллом лишайника и заметны йа~нем липйьв виде тем^
ных точек—устьев их (рис. 98). Апотеции даходятся обычно на поверхнос-
ти лишайника ввидемаленьких дисков, блюдечек йлй подуЩечёк, окрашен-
ных в тот же цвет, что и таллом, или чаще инауе.
Апотеции бывают двух наследственно постоянных типов: лецидиевые и
леканоровые (названия даны по латинским названиям двух родов — лецидея
и леканора). Лецидиевые образованы только грибом и имеют весь диск одно-
цветный (рис. 99, 7). В леканоровых апотециях под субгименйальным слоем,
а также на периферии диска находятся и водоросли, обусловливая несколь-
ко иную окраску диска (рис. 99, 2). Образованию сумчатых спороношенийу
ряда лишайников предшествует половой процесс. Он происходит в виде оп-
: 111
: 111
ли
ли
и;г,
: 111
ли
ли
л: । л
166
НИЗЩИВ РАСТЕНИЯ
2
таллома лишайника из
гриба, захватывающих
Рис. 100. Развитие
прорастающих спор
водоросли:
1 —начальная стадия (с —спора, а —гифа
4—водоросль); в—более поздняя стадия
Вегетативное размножение лишай*
Рпс. 101.
виков:
1 — разрез таллома е сералей; г— соредии
лодотворения аскогона., сперма-
циями через трихогину, отходя-
щую от аскогона и немного вы-
совывающуюся из л
Сперма ц и и — одноклеточ-
ныеспоры, называемые иначе
51П
развиваются внутри особых
спороношений — с п е р м о -
г о н и е в, или п и к и ид,
имеющих то ясе строение, что и
у сумчатых (или несовершен-
ных сферопсидных) грибов.
У многих л:
сумчатых грибов, типичный по-
ловой -процесс редуцирован, но
всегда все ясе происходит ассо-
(II
5111
511!
Аскоспоры, выбрасывае-
мые, как и у сумчатых грибов?
из сумок, прорастая'в*п6дходя-
щих условиях, образуют миле-
лий; ftrjrwr поблизости находятся
соответствующие водоросли, то
гифы гриба оплетают их и посте-
пенно образуется таллом , ли-
шайника (рис. 100).
айников очень широ-
ко распространено вегетативное
размножение отламывающимися
участками таллома, способными
к регенерации, а также особы-
ми с о р е д и я м и1 и и зи-
д и я м й. Соредии, широко
распространенные у многих лис-
товых и кустистых Л]
представляют собой одну или
несколько клеток водоросли,
оплетенных немногими грибны-
ми гифами (рис. 101). Соредии
образуются в гон1^иальйом слое
и через разрыв _ верхней коры
выходят на поверхность, обра-
зуя порошкодатые скопления.
называемые соралями. Соредии
разносятся ветром или потоками
дождя и, попав в подходящие ус-
ловия^ развиваются в новый тал-
лом лишайника. Йзидии, встре-
чающиеся у гораздо меньшего
числа лишайников, чем соредии,
представляют собой небольшие
выросты на талломе, состоящие
из водорослей а грибных гиф
5111
1 От греч. «сорейя» — скопле-
ние, куча.
ОТДЕЛ — Л
।; * । i
АЙНИКИ
чет
(рис. 102), одетые корковым слоем; эти вырос-
ты отламываются и, также прорастая, дают
новый таллом.
Интересно, что лишайники, у которых
развиваются соредии или изидии, почти утра-
тили способность образовывать сумчатые" спо-
роношения^ менее обеспечивающие размноже-
ние f лишайников. Способность образовывать
соредии или изидии — свойство, Приобретен-
ное лишайниками в процессе эволюции.
Двойная морфологическая природа лишай-
ников была открыта в 60-х годах XIX в. немец-
ким ботаником С. Щв енденером, но
получила общее признание лишь после продол-
: 111
: 111
|||
: 111
Рис. 102. Разрез таллома, с изи-
диями
:п!
жительной борьбы мнений. Доказательствами ее ^служат: а) тщательные
морфолого-анатомические исследования, доказывающие тождество гони-
айниках с водорослями, бесцветных элементов лишайников с
гифами грибов и тождество спороношений лишайников со спороношени-
ЛИ
ли
i I 'Л 11
1ЛГ
грибов, а также отсутствие всякой генетической связи между г«ни-
дия
ПИ
и гифами; б) получение отделы
культур водоросли и гриба;
в) получение синтетически таллома лишайников из спор, взятых из сумчатых
ли
плодовых тел и проращенных в мицелий, и чистых культур водорослей.
Взаимоотношения гриба и водоросли в л:
ли
айниках часто характеризу-
ют как мутуалистический симбиоз, при котором гриб берет от водоросли вы-
pa6prari№6 Сю углеводы, а ей доставляет главным"образом воду~~м|инепяэт^-
ные соли. а также защиту от сильной инсоляции. высыхания и т7 п. Но такая
гармония вряд ли вообще существует в природе. Имеется наблюдение, что
гриб образует гаустории, у некоторых лишайников прпникающия в клетку
водоросли и явно вредящие ей, у большинства же лишайников гаустории
лишь прободают оболочку и только соприкасаются с протопластом водорос-
ли
ли
ли
ли. Поэтому правильнее считать, что у лишайников наблюдается или толе-
рантный (переносимый) паразитизм гриба на водоросли, или даже обоюдный,
тоже телерантный паразитизм их друг на друге (aj л ё л о п а р аз ития м).
Воду лишайниктЪглощают всей поверхностью тела из атмосферных
осадков, и отчасти из водятдх паров атмосфёрыУбсобенпбпрйбЬльшом на-
^ышении^зе-имиГ^УдёржИвается'и^передвйгаетсявода внутрйтелал^
КоВ ГЛаЪНым образом по чисто физическим законам карцллярно дуеж
ли
Л II
миг а также--в"фязпухающих оболочках некоторых гиф. Углекислый газ
поглощают из атмосферы. Питательные вещества (неорганические и органиче-
тру-р iiHj^rrrar^.r через ловерхноеть-тела-лз водных растворов. Ис
никами-их являются субстрат и атмосферная пыль, оседающая на лиша
ки. Роль атмосферной пыли в питании лшпайциков возрастает JoT норковых
лишайников к листоватым и особенно к кустистым. Для последних субстрат
служит большей частью лиШь-местом-прикрепления (особенно у лишайников,
растущих на скалах и деревьях), и все питание они получают из воздуха,
атмосферных осадков и пыли.
Большинство лишайников легко переносит полноевысыхание. Фотосин-
тез и все питание у них в это время прекращаются. Накопление органиче-
ского вещества у них происходит очень медленно, чем и объясняются^нёзна-
чительный прирост лишайникбВГ норковые лишайники нарастают в среднем
на 1—8 год; листовые и кустистые — на. I м— 35 жж.~~ ~......
По отношению к субстрату среди лишайников имеются различные эколо-
гические группы: ряг.тутиА па питав, дв даревктгт ня скалах и т. п. Внутри
* их различаются еще более мелкие труппы: растущие на известковых или на
кремнистых горных породах, на коре деревьев, обнаженной древесине, на
листьях (у вечнозеленых растений) и т. п. На обрабатывамых землях ли-
‘ шайники не встречаются вследствие их очень медленного роста. Они очень
требовательным чистоте воздуха и, за единичными исключениями, не выпо-
иг.
ли
ли
ли:
168
НИЗШИЕ РАСТЕНИЯ
по-видимому, сернистых газов фабрично-за-
особенно,
: 111
сят дыма, копоти
водских районов.
Известно около 18 000—20 000 видов лишайников. Классификация круп-
ных подразделений их основывается на плодовых телах грибов. Более мел-
кие подразделения основываются на особенностях грибных спор, типе водо-
рослей, внешнем облике таллома и т. п. Лишайники делят на2 кл accaj
1) сумчатые (Ascolichenes), к которым относится основная масса лишайни-
ков, й 2^^азидиальные (Basidiolichenes). '
Сумчагаёлишаиники делят на 2 подкласса: 1) циреномицетные
: 111
: 111
: 111
: 111
.in
Discolichenes).
Из дискомицетных лишайников-модсно отметить следующие. Большие
роды лецйдея (Lecidea) и леканора (Lecanbra), содержащие первый около 1500,
второйсвышё 1100 корковых видов" Клеканорам относят и виды так называ-
емой лишайниковой манны (Lecanord esculentd и др.) — угловатые или округ-
лые бугорчатые комсЯши, растущие нёйрикрепленно к почве в степях и по-
лупустынях юго-востока СССР, Передней Азии, Северо-Западной Африки.
Сильными ветрами они могут переноситься на значительные расстояния.
Пельтигера (Peltigerd) — крупнолистовые серые или буроватые почвенные
айники. Пармелия (Parmelia, около 800 видов) и цетрарця. (Cetraria,
свыше 70 видов) в большинстве листоватые; С. islandica — кустистая, расту-
щая на почве в тундрах, в борах, на песках и называемая исландским мхом;
при недостатке хлеба можно использовать в пищу; раньше применялась и в,
медицине. Ксанп&ория (Xanthoria parietina) — широко распространенный в
виде желтовато-оранжевых листовых розеток на стволах деревьев (особенно на
осинах и тополях), на заборах, скалах. Некоторые виды алектории (Alectoria)
тллснеи (Usnea) растут в виде длинных сероватых или' чёрно-бурых косм
(бородатые лишайники) на ветвях и стволах деревьев и характерны для ланд-
шафта некоторых лесов. Эверния (Еиегта prunastri), растущая в виде неболь-
ОТДЕЛ —I
rin
169
t
иг,
ших зеленовато-сероватых кустиков на лиственных деревьях (рис. 103, 7),
применяется в парфюмерии при изготовлении духов (как, например,
«Шипр»). Морфологически интересен и практически важен род кладония
(Cladonia, свыше 300 видов). Он имеет двойной таллом: первичный накип-
ной .иди У большинства видонмелкоч^^готятгЖивт^ичньгй/вырястятптпуй
ga первом в виде^более крупных вертикальных выростов — простых шиловид-
ных, бокальчатых-или более или мёнёёсильно~^разветвленных. кустистых.
Некоторые кустистые виды кладоний (С. rangiferina, С. sylvatlca, С, alpestris
и др.), известные под названием оленьего мха или ягеля (рис. 103, 2),
покрывают огромные пространства почвы в тундрах и в~сосновых борах и ха-
рактерны для их ландшафта; на севере эти виды имеют большое практиче-
ское значение, так как зимой являются единственной пищей северных оле-
ней. Из некоторых видов кустистой (АдесеПа), растущей на скалах
по берегам Средиземного моря, Атлантического и других океанов, добывают
лакмус, раньше из них получали и краски для тканей.
Происхождение лишайников полифилетическое. В разное время из гри-
бов разных систематических групп, соприкасавшихся с различными водо-
рослями, постепенно в результате длительного процесса взаимоприспособ-
ления образовались новые, своеобразные комплексные организмы с новыми
морфологическими и физиологическими свойствами и новыми закономернос-
тями филогенетического развития. Эволюция лишайников выразилась глав-
ным образом в изменении таллома — в переходе от корковых форм к листо-
ватым и кустистым, что влекло за собой развитие большей поверхности,
поглощающей углекислый газ, уменьшение зависимости от субстрата и пере-
ход к существованию в воздухе. Размножение соредиями, изидиями, образо-
вание леканоровых апотециев также выработались в процессе эволюции.
ЭВОЛЮЦИОННЫЕ ОТНОШЕНИЯ НИЗШИХ
РАСТЕНИЙ
Эволюционные отношения низших растений можно представить в виде
схемы, изображенной на рисунке 104.
Рис. 104. Происхождение и пути эволюции низших растений
170
НИЗШИЙ РАСТЕНИЯ
И1
В схеме отображены следующие общие положения:
1. Наличие среди низших растений двух существенно различных уров-
ней организации клетки — прокариотического и эукариотического.
2. Параллельное существование в продолжение всей известной истории
развития растений двух типов питания — автотрофного и гетеротрофного,
дополнявших друг друга и обеспечивающих постоянство малого, или биоло-
гического, круговорота элементов минерального питания. Этим определяется
целостность растительного мира и его относительная автономность, хотя в
разрушении органического вещества всегда немалую роль играли и
животные.
3. В разнообразии фототрофных низших растений отражена группиров-
ка их по составу пигментов, имеющих отношение к фотосинтезу: группа
зеленых, содержащих хлорофиллы а и Ь; группа содержащих повышенное
количество каротиноидов и в то же время не имеющих хлорофилла Ь9 он заме-
няется обычно хлорофиллом с; наконец, особняком стоящая группа, с обяза-
тельным участием в пигментном комплексе фикобилинов. Генеалогическое
единство каждой из трех «цветных» группировок считать доказанным еще
нельзя, так как не исключена возможность конвергентного возникновения
пигментов.
4. Среди гетеротрофных эукариотических низших растений предпола-
гается существование двух эволюционных стволов, вероятно развивавшихся
независимо, миксомицетов и грибов. Эволюция первых базировалась на со-
хранении не покрытого оболочкой плазменного тела (плазмодий); более же
мощная грибная ветвь эволюции развилась на основе клетки, покрытой
оболочкой, с ясной тенденцией к максимальному соприкосновению тела с
окружающей средой (мицелий), т. е. в том же направлении, что и у фототроф-
ных растений.*
Высшие, грибы вместе с континентальными бактериями и миксомицета-
ми составили иа суше фототрофно-гетеротрофный ансамбль с высшими расте-
• НИЯМИ* .
Г.1
ОБЩАЯ ХАРАКТЕРИСТИКА
Высшие растения произошли от низших. Они, в отличие от низших,
характеризуются рядом признаков более высокой организации. Прежде
всего это растения наземные, обитатели воздушной среды, утратившие не-
посредственную связь с первичной водной обстановкой.
Выйдя на сушу, растения оказались в иной, совершенно любой обста-
новке. Развиваясь при периодическом, а не постоянном увлажнении, неред-
ко при наличии продолжительных сухих периодов, эти растения вырабо-
тали ряд специфических признаков, отличающих их от низших растений.
Тело высших растений расчленено на стебли и листья. В связи с этим
их называют/лл ртостебельными р гГст’ен й я м и (Cormophyta),
Громадное большинство высших растений имеет корень. Такое сложное
расчленение отсутствует у низших растений, тело которых не расчленено
на стебель и листья, а представлено талломом или слоевищем.
Появление сложного расчленения, образование органов — стебля,
листа, корня — вызвано было переходом растений в новую, наземную
среду.
Для ассимиляции в новых условиях требовалась перестройка фото-
синтезирующего аппарата. Растение должно было, образно говдря, припод-
няться над землей и развить большую поверхность, что и было достигнуто с
появлением и развитием листьев и стебля.
Органы высших растений имеют сложное строение. Вышедшие на сушу
растения не могли воспринимать воду всей поверхностью тела. Первоначаль-
но с функцией всасывания воды и минеральных солей и прикрепления к
субстрату справлялись ризоиды — выросты поверхностных клеток расте-
ния. С появлением листостебельного строения и увеличения размеров у на-
земных растений возникли корни (они, таким образом, возникли позднее),
лучше справляющиеся с задачей. Корни обеспечили минеральное питание,
а листья — воздушное питание высших растений.
Для нормального функционирования двух разделенных пространст-
вом органов — корня и листа — необходимо быстрое передвижение воды
к листьям и органических веществ из листа в другие части растения. Это
вызвало к жизни развитие проводящей системы внутри тела рас-
тения в виде специальных клеток — трахеид, а затем трахей или сосудов,
а также ситовидных трубок — образований, зачатки которых мы встреча-
ем у низших растений (например, ситовидные трубки у бурой водоросли
Macrocystis).
В дальнейшем проводящие элементы группируются в закономерные
сочетания — пучки. Возникает центральный цилиндр —'с“т"ё~л ь," "сначала—
в виде простейшей протостели. Затем в связи с увеличением стебля и листо-
вого аппарата вырабатываются более сложные типы стели.
174
ВЫ
(uni;
® РАСТЕНИЯ
В наземной среде у растений развилась сложная система п о к р о в-
н ы х тканей, защищающих их от чрезмерной потери воды, отТГОНйжен-
ных температур*? т. д. (эпидермис, перидерма, корка), а также развился
сложный устьичный аппара т, обеспечиб&жИЦий газообмен и регу-*
лирующий испарение воды при транспирации. Затем получили мощное раз-
витие мех аническ и е т^к_а н и, надобность в которых отсутствовала,
при жизйи растений в водной среде, более плотной, чем воздух.
Все эти органы и ткани появились и развились, конечно, не сразу. Рас-
сматривая современные нам высшие растения, мы находим среди них и вы-
сокоорганизованные формы, хорошо приспособленные к жизни на суше,
и целый ряд примитивных представителей, не расчлененных на стебель и
листья, не имеющих корней, лишенных проводящей системы, и т. д.
Изучение вымерших ископаемых растений открывает нам целый ряд
промежуточных форм, ^доказывающих возникновение и этапы развития тех
или иных органов, тканей^ а также отдельных групп высших растений.
Сопоставляя данные, полученные при детальном изучении ныне живу-
щих растений, с данными па леЬцто логин, мы довольно уверенно восстанав-
ливаем историю развития высших растений. Для уяснения ее первенствую-
щее значение имеют органы размножения, их возникновение и эволюция. ’
Половые органы высших растений: мужские — антеридии и жен-**
Sill
hi;
ские — архегонии — всегда многоклеточны, в отличие от однокле-
точных половых органов большинства низших растений (лишь у харовых
: 111
имеются многоклеточные оогонии и у некоторых бурых водорослей — мно-
гоклеточные гаметангии).
Антеридии — небольшие овальные или шаровидные тельца, оде-
тые снаружи бесплодными клетками (стенка антеридия), расположенными
в один или несколько рядов.
В антеридии развиваются спермагенные клетки, из которых затем воз-
никают мужские гаметы1 — подвижные сперматозоиды, снабженные жгу-
тиками. При созревании антери-
дии вскрываются — стенка их ло-
пается, и тогда сперматозоиды
выходят наружу. Активно двига-
ясь в капельно-жидкой воде, спер-
матозоиды подплывают к архе-
гониям.
Архегонии — небольшие
бутылкообразные или колбообраз*
ные тельца. Архегонц?- состоит
из нижнейт дДЕйГиренной части—
брюшка й верхней, суженной —
шейки (рис. 105J. Снаружи архе-
гоний окружен бесплодными клет-
ками (стенка архегония), защища-
ющими от высыхания его содер-
жимое. В брюшке помещается
неподвижная женская гамета —
яйцеклетка. Над яйцеклеткой рас-
положена брюшная канальцевая
клетка.
. .Внутри шейки расположены
1 2 8
Рис. 105. Архегоний:
1—молодой невскрывшийся архегоний (в брюшке:
о — яйцеклетка, о — брюшная канальцевая клетка;
в шейке: ш — шейковые канальцевые клетки* их окру-
жают стерильные /бесплодные/ клетки); 2—зрелый
вскрывшийся архегоний (канальцевые клетки расплы-
лись); Л — оплодотворенный архегоний (шейка аавя-
Ввет, оплодотворенная яйцеклеша разделилась)
шейковые канальцевые клетки.
К моменту созревания яйцеклетки
канальцевые клетки (как шейко-
вые, так и брюшная), а также
верхние клетки стенки ослизняют-
ся и архегоний на верхушке
вскрывается. От его вершины к
ОБЩАЯ ХАРАКТЕРИСТИКА
«75
Рис. 106. Происхождение антврвдиев и ар»
хегониев и промежуточные образования:
1,3,5 J — происхождение архегоннн; 2.4ЛЛ — про-
исхождение антеридия; а — проденсутечние обра-
аовапв (антеридве-архегоняй) у печеночного мха
нцрсмшде (д — яйцеклетка» с— соермагеннан ткань)
образовавшейся из
расплыв-
иг.
яйцеклетке ведет канал, заполненный слизью,
шихся клеток. По этой слизи сперматозоид проникает в брюшко архего-
ния, где и сливается с яйцеклеткой, производя оплодотворение.
Такие антеридии и архегонии являются исходными и имеются у более
простых высших растёнйи.ПВ процессе эволюции высших' растений они прё-
терревают упрощение, редукцию.
1 Архегонии отсутствуют у наиболее высокоорганияованиых покрыто-
семенншграстений,' такжетупдекбТОрых" голосеменных (у Gnetum, Wei-
witschia).
Антеридии и архегонии произошли, вероятно, из многоклеточных га-
метангиев водорослей, подобных тем, что имеются у бурых' вбдоррслей (на-
Пример" "у EctocafpiLsyrРйЭвНйаибьГв^водйои^среде, тяметанг лгиодирисл ей
не имеют защитного слоя. Все их клетки образуют гаметы.
При выходе растений на сушу появилась необходимость в защите га-
метангиев от высыхания. Наружные слой клётбк такйх'ТЛМбТйНГИев стано*
вились бесплодными’(стерилизовались), теряли способность к образованию
гамет и образовали стенку гаметангия — защитный слой. Первоначально
такие гаметангии, вероятно, содержали одинаковые или слабо различав-
еся подвижные гаметы (подобно мужским и женским гаметангиям у бу-
рой водоросли Cutleria). Со временем половая дифференцировка станови-
лась все более резкой. Из таких гаметангиёв и выработались антеридии и
архегонии. В антеридии, который и внешне очень сходен с многоклеточны-
ми гаметангиями бурых водорослей, развивалось большое количество мел-
ких подвижных гамет — сперматозоидов.
В.архегониях число гамет уменьшил:рсь,, нолши становились более круп-
ными и теряли подвйжность. Вначале еще все они были способны к оплодо-
творению, затем стала оплодотворяться лишь одна, наиболее крупная Гаме-
та — яйцеклетка. Другие стали стерильными, но приобрели новую функ-
ПС
176
выс
1(15
[Е РАСТЕНИЯ
цию — проводят сперматозоиды к яйцеклетке. Так сформировались
брюшная и шейковые канальцевые клетки.
Цроисхожденйе антеридиев и архегониев из многоклеточных гаме-
тангиев показано на^рисунке 106.
О гомологичности антеридиев и архегониев, юб их происхождении из
одного цсточника свидетельствует наличие промежуточных образовании —
антерйдиев-архегониев; Они найдены у многих печеночников, лиственных
м£ов; у папоротников и даже у голосеменных.
Так, у печеночника корсинии (Corsinia) в нижней части такого проме-
жуточного образования, имеющего общую форму архегония, развивалась
яйцеклетка, а в верхней части вместо шейковых канальцевых клеток была
развита спермагенная ткань (рис. 106, 9).
Характерной чертой высших растений является наличие правильной
смены поколений в цикле их развития. Растение здесь представлено двумя
фазами развития, закономерно ' сменяющими друг друга, — гаметофитом
и спорофитом. Гаметофит — половое поколение, на котбром образу-
ются половые органы — антеридии и архегонии (у папоротникообразных
мужской и женский гаметофиты являются самостоятельными растения-
ми), — сменяется бесполым поколением — спорофитом, на котором
формируются органы бесполого размножения — спорангии-
В спорангиях образуются споры. Спорангий одет оболочкой из одного
или нескольких слоев клеток. Внутри спорангия возникает 1шогоклеточная
спорогенная ткань — археспорий.
При делении археспориальных клеток образуются материнские клетки
спор. Последние приступают к редукционному делению, в результате кото-
рого каждая из них образует(четыре гаплоидные клетки — тетраду
спор. Спорофит всегда диплоиден,—каж©я его клетка содержит двойной
набор хромосом (2х).
У всех высших растений редукционное деление происходит при образо-
вании спор в спорангии. Спора гаплоидна, и от нее начинается развитие
гаметофита.
Гаметофит всегда гаплоиден, все его клетки имеют одинарный (z) набор
хромосом. Переход от гаплоидного состояния к диплоидному наблюдается
при оплодотворении. Зигота диплоидна. В дальнейшем из нее развивается
спорофит.
Высшие растения произошли от водорослей. Это более молодая группа
растений. Выход растений на сушу совершился лишь после того, когда
уже в первичной, водной среде водоросли выработали довольно сложное
строение. Это были морские водоросли.
Выход растений на сушу осуществлялся в раннем палеозое, вероятно,
в кембрийском периоде палеозойской зры.
Какие именно группы водорослей дали начало высшим растениям, мы
достоверно hF знаем.' 1Г<Тсбйр0мёНнЫм представлениям, это были, вероятнее
всего, бурые водоросли. Именно среди них мы встречаем наиболее сложно-
расчленённые формы, у бурых водорослей имеются и сложные многоклеточ-
ные гаметангии^
НекоторыТГботаники выводят высшие растения от зеленых водорослей.
Их роднит наличие зеленого пигмента хлорофилла, но среди зеленых водо-
рослей нет сложнорасчлененных форм.
Высшие растения произошли не из современных, а из каких-то прими-
тивных, давно вымерших бурых водорослей, имевших более простое
^строение.
Водоросли, вышедшие на сушу, обладали ясно выраженной сменой
поколений. Уже в водной среде у водорослей, особенно у бурых, сложились
различные типы смены поколений. Так, у ламинарии в цикле развития пре-
обладает спорофит, а гаметофит представлен микроскопическими растеньи-
цами; у фукуса гаметофит совсем выпал из циила развития, и растение
представлено лишь спорофитом; у кутлерии, наоборот, преобладает гаме-
с 111
II
ОБЩАЯ ХАРАКТЕРИСТИКА
<77
тофит; наконец, у диктиоты оба поколения — гаметофит и спорофит — раз-
виты одинаково, внешне совершенно сходны, отличаясь, однако, цитологи-
чески по набору хромосом и по наличию различных органов размножения*
Первые наземные растения, вышедшие из воды на сушу, обладали сме-
ной поколений, вероятно, типа диктиоты с ^одинаковым развитием гамето-
фита и спорофита.
Простейшие из известных нам наземных растений — псилофиты,
остатки которых впервые встречаются в силурийских отложениях, были
уже высокоорганизованными растениями, хорошо приспособленными к
жизни на суше. Они имели проводящую систему в виде пучков (стел ь),
покровные ткани с устьицами, спорангии, одетые толстой оболочкой, и
т. д. (стр. 219).
Несомненно, псилофитам предшествовал длинный ряд растений более
простого строения, приближавшихся к водорослям.
Нам известны лишь спорофиты псилофитов. Каковы были их гамето-
фиты, мы не знаем* Однако детальное исследование над близкими к ним
Psilotales показали в последнее время, что гаметофиты, например, Ттез*
ipteris имеют некоторое сходство с молодыми их спорофитами. В тканях их
гаметофита находят остатки стели в виде рядов и групп трахеид (стр. 223).
Это подтверждает соображения о вероятном сходстве гематофита и споро-
фита у водорослевых предков высших растений.
С момента выхода на сушу высшие растения_£азвивались в^цвухоснов-
ных направлениях^они" обрайОвалн две болыпие зволюпионные ветви —
гаплоидную и диплоидную > Первая из них представлена мохообразными
(Bryophyta). Эта группа характеризуется прогрессивным развитием гаме-
тофита. Гаметофит сообразно основной его функции — обеспечения по-
лового процесса, требующего капельно-жидкой воды, — жался к земле и
первоначально был талломным растением. Эти формы постепенно совершен-
ствовались, усложнялись, увеличивая ассимиляционную поверхность, при-
обретая расчленение в связи с необходимостью обеспечить питание и обра-
зование спор развивающемуся на нем спорофиту.
Спорофит (спорогон), наоборот, претерпевал редукцию. Будучи связан
с гаметофитом, получая от него питательные вещества, спорофит мохообраз-
ных при небольшом размере и слабом морфологическом расчленении вполне
обеспечивал достаточную продукцию спор. В процессе эволюции у споро-
фита мохообразных выработались различные приспособления для лучшего'
рассеивания спор.
Развитие мохообразных шло от талломных к листостебельным растениям.
Вторая эволюционная ветвь представлена всеми остальными высшими
растениями. Развитее этой группы связано с совершенствованием споро-
фита. Спорофит в наземных условиях оказался значительно более жизне-
способным. Именно эта группа растений достигла наибольших успехов
в завоевании суши. Спорофит у них нередко имеет крупные размеры, обла-
дает сложным внутренний и внешним строением.
Гаметофит у этой группы, наоборот, претерпевал упрощение, редукцию*
У более простых форм (у споровых растений) гаметофит еще имеет самосто-
ятельное существование и представлен зеленым заростком (Pteropsida,
Lycopsida, Sphenopsida), но уже у разноспоровых представителей этих клас-
сов гаметофит сильно упрощен, редуцирован. У более организованных —
семенных растений — гаметофит утерял способность к самостоятельной
жизни и развивается в тканях спорофита, а у наиболее совершенных — по-
крытосеменных (цветковых) — растений сведен почти на нет.
Постепенно совершенствуясь в новой обстановке,, наземные растения
вскоре дали начало ряду самостоятельных групп (отделов) растений, при-
спосабливавшихся к разнообразным условиям жизни на суше.
Высшие растения, насчитывающие
настоящее время свыше 300 000
видов, господствуют на Земле, населяя ее от арктических областей до эква-
тора, от влажных тропических лесов до сухих пустынь. Они образуют
178
ИГЛИ
ВЫСШИВ РАСТЕНИЯ
леса, луга, болота, заполняют водоемы* Многие из них достигают гигант-
ских размеров (например, эвкалипты — из цветковых — достигают 90 —
100 м высоты, секвойи — гигантские хвойные — 110 ж и более), другие
представлены маленькими растениями в несколько миллиметров (напри-
мер, ряски, многие печеночники, мхи и т. д.).
Однако при всем огромном разнообразии внешнего вида И внутреннего
строения все высшие растения сохраняют известное единство в строении.
Среди высших выделяются четыре отдела. Они сравнительно легко увязы-
ваются друг с другом, что свидетельствует о единстве происхождения выс-
ших растений.
ВЫСШИЕ РАСТЕНИЯ
С ПРЕОБЛАДАНИЕМ В ЦИКЛЕ
РАЗВИТИЯ ГАМЕТОФИТА
ОТДЕЛ —МОХООБРАЗНЫЕ (BRYOPHYTA)
К этому отделу относятся высшие растения, обычно связанные в своей*
жизни с достаточным, часто избыточным увлажнением. Очень немногие
мохообразные произрастают в засушливых областях, например виды рода
зеленого мха — Tor tula desertorum и др., многие виды Riccia. Эти мхи в сухой
период находятся в состоянии покоя и возобновляют жизнедеятельность
при выпадении осадков.
'Менее организованные представители мохообразных не -имеют расчле-
нения на стебель и лист, и тело их представлено талломом, или слоевищем..
Более организованные являются настоящими листостебельными растения-
ми. Корни у всех мохообразных отсутствуют.
Всасывание воды и прикрепление к субстрату осуществляют ризои-
ды — выросты поверхностных клеток.
Проводящая.сдстема в виде пучков отсутствует, и только у некоторых
представителей (в семействе Polytrichaceae^ среди лиственных мхов) появля-
ются проводящие элементы, сходные с трахеидами и ситовидными трубка-
ми. Они собраны в образования, напоминающие стелярное строение других
высших растений.
Наиболее характерный признак мохообразных, резко отличающий
их от остальных высших растений, — преобладание.в^цикле развития гаме-
тофита (полового поколения) и более слабое-развитие спброфйтггДбесполого
поколения). При этом спорофит всегда связан с гаметофитом, х.
>ёпорофит, называемый у мохообразных спорогоНом, состоится коро-
ббчки со спорами и ножки, нижняя часть которой утолщена в виде присос-
ки — гаустории. При ее посредстве спорогон внедряется в тело гаметофита
И.получает от него необходимые питательные вещества.
Из споры у мохообразных развивается протонема (первичная нить,
предросток) — нитчатое или пластинчатое образование, напоминающее
водоросль. По внешнему виду протонема резко отличается от взрослого
растения и иногда развита очень сильно. Протонема или непосредственно
дает начало растению мха, или образует почки, развивающиеся далее во
взрослое растение.
На взрослом растении образуются антеридии и архегонии (чаще на раз-
ных особях)^ В антеридии развиваются двужгутиковые сперматозоиды.
После оплодотворения из зиготы развивается спорогон.
Bryophyta — древняя группа растений. Ископаемые остатки мо-
хообразных известны из каменноугольного периода, но это были уже высо-
коорганизованные растения, несомненно проделавшие длинный путь эво-
люции, так что появление мохообразных должно быть отодвинуто на зна-
чительно более раннее время — девон или силур.
ОТДЕЛ — МОХООБРАЗНЫЕ
179
У некоторых псилофитов (у Homeophyton) известны признаки, сходные
с мохообразными (спорангий напоминает строение коробочки сфагнума).
, Это указывает на отдаленное родство и общность происхождения мохообраз-
ных и известных нам псилофитов, являющихся родоначальной группой для
остальных высших растений.
Мохообразные и пснлофиты возникли от простейших наземных расте-
ний, имевших- перо ятапт одинаковое развитие" м
В дальнейшем у BryopKyta совершенствовался преимущественно гаме-
трфит, у остальных же высших растений прогрессивно развивался грлрл-
фит, а гаметофит* претерпевал редукцию. Таким образом, oryogT^ita являют-
ся самостоятельной линией эволюции. Отдел. ВгцдркуЫ уапделяиц^на
^класса..
1-й КЛАСС—ПЕЧЕНОЧНЫЕ МХИ, ПЕЧЕНОЧНИКИ
(HEP ATICOPSIDA, -НЕРАТГСАЕГ
Представители этого класса характеризуются дорзовентральным стро-
ением тела. Наряду с талломными здесь имеются и листостебельные расте-
ния, которые, однако, и при расчленении тела на стебель и лист сохраняют
дорзовентральность. В некоторых клетках печеночников (как талломных,
тат^ и листостебельных]гимшотся масляные тельца — одно, два или несколь-
ко. Они пока не обнаружены у сферокарпуса и лнтоцррртбвых.^'
Масляные тельца представляют собой дмдед^аия. клеток (секреты). По
химическому' составу они являются смесью эфирных масел и, вероятно,
служат растодцю_-ддя лащиты от поедания животными.
Имеется очень небольшая группа листостебельных* печеночников с ра-
пулкним строением тела (Haptomitriales, см. стр. 190). /
В коробочке спорофита, кроме спор, содержатся пружинки (элатеры)»
имеющие важное значение для рассеивания спор.
Протонема' у печеночников развита слабо и представлена большей
частью короткой нитью или пластинкой.
В классе печеночников выделяют 4 порядка.
МАРШАНЦИЕВЫЕ (MARCHANT
Обширный порядок маршанциевых насчитывает 33 рода и 400 видов,
Все они являются талломными растениями, но имеют сложное анатомиче-
ское строение. В качестве представителя этого порядка мы рассмотрим мар-
шанцию обыкновенную (Мar chant ia polymorpha) — один из наиболее рас-
пространениях в нашей флоре печеночников. Маршанция встречается
Рис. 107. Маршанция:
таллом смужскими аодэтавкамя; 3 — таллом с женскими подставками
180
ВЫСШИЕ РАСТЕНИЯ
2
ас
Рве. 108. Маршанция:
1 — поперечный разрез таллома: а — при малом увели*
чении, о — при большом (с анфигэстрии» р — ризо-
иды» де ассимиляторы» у — устьице); 2 — ризоиды:
в — простой, б,в — язычковые; з — поверхностный срез
(сверху) таллома (к —> воздушные камеры» у — устьица)
очень широко — йо болотам, в
лесах, на местах пожарищ. Ленто-
видные fift дихотомически
ветвятся, cTflJT,a,w*q
^стигают 5—20 в длину и 1—2 см
в ширину (рис. 107 J7~
Верхушка талломов выемча-
тая, и здесь, в выемке, находится
точка роста, елртпрщяд дд
меристематических^ клеток. Делё-
нном их. осуществляется рост _тал-
Верхняя сторона таллома тем-
но-зеленого цвета. На неб хорошо
заметна «средняя жилка», идущая
вдоль таллома и являющаяся
углублением; ложбинкой.
Верхняя поверхность таллома
имеет как бы сетчатую структуру
и разбита на многоугольные уча-
стки (воздушные ,камеры) с тем-
ными точками в каждом (рис. 108).
Темно-зеленые участки отделяются
друг от друга светлыми линиями.
Нижняя сторона таллома бурова-
то-красная. Вдоль средней жилки
располагаются многочисленные4
одноклеточные ризоиды, которые
отходят также" и в стороны от сред-
ней жилки таллома, но там онй
плотно к нему прижаты.
Кроме ризоидов, на верхней
( поверхности хорошо заметны (в
лупу) особые выросты таллома —
однослойные брюшные ^ешуйки,
или амфигастрии. Они тянутся в два ряда вдоль средней жилки, а
также вдоль краев таллома (рис. 108, 2).
Ризоиды у маршанции двух родов — простые и язычковые (рис. 108,2).
Простые ризоиды имеют более широкий просвет. Внутренние их стенки
гладкие. Расположены они главным образом вдоль средней жилки. Простые
ризоиды направляются вертикально вниз, врастают в землю и всасывают
воду с растворенными в ней минеральными веществами, а также прикрепля-
ют таллом маршанции к почве.
Язычковые ризоиды в просвете ^же простых и имеют на внутренней
поверхности выросты в виде сосочкой, язычков. Язычковые ризоиды спле-.
таются в тяжи, протягивающиеся вдоль краев таллома". Они прижимаются
к таллому амфигастриями. По язычковым ризоидам и между ними, как по
фитилю, происходит движение воды вдоль таллома. Такие тяжи язычковых
ризоидов направляются также в различные выросты таллома (подставки,
см. ниже). 4
Анатомическое строение таллома маршанции довольно сложно. На
поперечном разрезе таллома хорошо заметны верхний и нижний эпидерми-
сы и ткани, заключенные между ними.От нижнего эпидермиса отходят ризо-
идКг^и-^ьмфиысхщшА, В верхнем эпидермисе местами уУТЯЩтг-
Они имеют своеобразную, бочонкообразную форму и состЬйт из четырехря^
дов клеток, расположенных друг над другом. Нижние клеши ус1ьица не-^
редко глубоко, врастают в полость устьица; ~1 ТйКйЯйХ «амыкающих*^слеток
ОТДЕЛ — МОХООБРАЗНЫЕ
181
здесь нет, и устьица служат лшпьдля ^вентиляции таллома, а не для регу-
лировки испарения (рис. 108, 26).
Под эпидермисом находятся воздушное. камера т полости,^отграни-
ченные друг от друга бесцветными крупными клетками^расположенными
в 1 — 2 этажа^ Со дна воздушных камер поднимаются ветвистые нити —
а с с и м и л я т о рju, составленные округлыми клетками, содержащими
хлорофилловые зёрна/ Они являются главным ассимиляционным аппаратом
маршанции и обусловливают темно-зеленую окраску верхней стороны ее
таллома. Светлые полосы соответствуют клеткам, разделяющим камеры.
Ткань таллома под воздушными камерами состоит из широких бесцвет-
ных паренхимных клеток. Они нередко заполнены крахмалом. Местами
встречаются клетки, в которых находятся крупные темные включения —
масляные тельца, содержащие масло. Изредка попадаются слизевые
ходы — полости, образовавшиеся от разрушения соседних клеток и запол-
ненные слизью. Наличие масляных телец и слизевых ходов является ха-
рактернрй .особенностью всех печеновдиков.
Нижний эпидермис нерезко отграничен от паренхимной ткани. От него
отходят в виде выростов язычковые и простые ризоиды. Здесь же прикреп-
ляются и амфигастрии, которые на поперечном разрезе представляются
однорядными пластиночками*
Внутри таллома маршанции, таким образом, нет проводящей ткани.
Прохождение воды вдоль к растущим частям растения обеспечивается здесь
как бы наружной проводящей системой в виде язычковых ризоидов.
Вегетативное размножение маршанции осуществляется образованием
mi.
выводковых почек-, развивающихся в выводковых корзиночках, которые
возникают на верхней стороне таллома (рис. 109).
Сформировавшаяся выводковая почка имеет вид овальной пластиночки
с выемками по бокам, где помещается точка роста. От ночки отходят ризо- ‘
иды. На разрезе через корзиночку можно всегда видеть большое количест-
во почек, находящихся на разных стадиях развития.
Во время сильного дождя "выводковые почки (расположенные на одно-
клеточной ножке, от которой легко отламываются) вымываются из корзи-
ночки водой, попав на землю, нарастают, при помощи точек роста и посте-
пенно формируются в новые талломы маршанции.
При половом размножении маршанции образуются антеридии и архе-
гонии. Маршанция — растение двудомное. На одних ее талломах возникают
Рис. 109. Таллом маршанция и выводковая почка:
1 — таллом маршанция с паводковыми корзинками; в— развитие выводковой ночки; з — выводковая ши-
ка в плане; 4 — ома же в разрезе (н — ножка, о — место опадения почки, р — зачатки ризоидов)
182
ВЫСШИЕ РАСТЕНИЯ
* Рис. 110. Маршанция:
j — вертикальный разрез черев
мужскую полтвку (в некоторых
антервдвдАьных полостях нахо*
датой антеридии); £ —антери-
дий в аитервднальясй полости
(н — ножка антеридия); я — дау*
ст жгутиковый сперйатовоид; 4 —
вертикальный разрез черев жен-
скую подставку (п — веряхепий*
р — пучок ризоидов, м — моло-
дой архегоний, ст —старый,
оплодотворенный. с -увядающей
шейкой. Вокруг каждого архего-
ния возвышается перианцай)
ill
своеобразные выросты в виде многолопастного диска, расположенного на
ножке. Это мужские, антеридиальные_додетадки. На вертикальном разре-
зе через диск в верхней его части заметны антерцдиальцые полости, нц дне
которых расположено по одному антеридию Дрис. 110, 7). Антеридий —
овальное тельце с однослойной стенкой, прикрепляется ко дну полости не-
большой ножкой (рис. 110, 2). Содержимся антеридия состоит из многочис-
ленных спермагенных клеток; каждая из них образуетдва двужгутиковых
сперматозоида (рис. 110, 3). По созревании антеридий вскрывается наверху
щедыо; сперматозоиды во время дождя через канал антеридиальной полости
выходит наружу и, активно двигаясь в воде, подплывают к архегониям..
На других" талломах образуются выросты, имеющие ножку, увенчан-
ную многолучевой звездой. Это женские, архегониальные подставки. На ;
вертикальном разрезе через нее мы заметим архегонии, группами сидящие :
между лучами; они шейками обращены вниз (рис. 110, 4). Каждая группа :
архегониев покрыта общим покровом (перихецием), а каждый архегоний
одет частным покровом (перианцием), возникающим в виде кольцевидного
валика у его "основания. Перианций затем сильно разрастается. Перихеций
и перианций защищают архегонии, а затем и спорогоны от высыхания.
Ко времени созревания архегония шейка на вершине вскрывается,
шейковые и брюшная канальцевые клетки ослизняются, и сперматозоиды
проникают внутрь архегония. Здесь один .из них сливается с яйцеклеткой,
производя оплодотворение. г
Лучи молодой женской подставки плотно прижаты к ножке, а затем
по мере развития они принимают сначала горизонтальное положение, а
ко времени созревания спорогонов загибаются кверху,(рис. 112).
Ткань подставок имеет строение таллома, в ней есть воздушные камеры
с ассимиляторами, устьицами. Ножка подставок состоит из паренхимных
клеток. Вдоль ножки (внутри ее) протягиваются два тяжа язычковых ризо-
идов — от нижней стороны таллома до диска или ‘до лучей подставки
(рис. 110, 4). Молодые подставки расположены на коротких ножках. В даль-
нейшем ножка начинает усиленно расти и выносит антеридии и архегонии
высоко над талломом.
Из оплодотворенной яйцеклетки развтаается бесполое поколение мар-
шанции — спорофит, или спорогон. И
Сформировавшийся спорогон состоит из шаровидной коробочки, корот-
кой ножки, расширенная нижняя часть которой называется гаустори-
е й. При помощи гаустории спорогон внедряется в ткань подставки (гаме-
тофита) и воспринимает из ее клеток питательные вещества, необходи-
мые для его развития (рис. 111). Коробочка содержит споры и пружинки
(элатеры). Пружинки представляют собой удлиненные клетки, стенки
ci:ir.
гл ЛI
’.lit
1111 г.
ОТ^ЕЛ — МОХООБРАЗНЫЕ
183
которых имеют спиральные утолщения в виде одной или двух спиралей,
идущих навстречу друг другу.
Первоначально спорогон заключен р гбрттикп^рхегонпя.стенка кото-
рого разрастаетсявслед за растущим спорогономи обраау^СЗ^вг лгазызЙПЕР-
мый к о дцимц к*(рНС. 111,112). Сйа®5пнйй**ёа6рогон окружен такжесиль-,
но разросшимся перианцием (рис. Ill, 1). По созревании коробочкде^пПЗкка
ее на^верхушке и выносится ножкой за Остаток
брюшка архегония расположен у основания ножки в виде оторочки (колпа-
чок, калиптра). Коробочка закрывается на верхушке створками, тюсле чего
споры высыпаются наружу. Пружвдки очень Т1^оскопичны. Скручиваясь
-1 j
в сухую погоду и раскручиваясь в сырую, они разрыхляют споровую массу,
способствуя равномерному рассеиванию спор из коробочки (рис. Ill, 1, 2).
Оплодотворенная яйцеклетка тотчас же делится поперечной перегород-
кой на две клетки. Из нижней клетки благодаря последующим делениям
формируется ножка спорогона. Из верхней клетки развивается коробочка.
Верхняя клетка делится двумя взаимно перпендикулярными перегородка-
ми на четыре клетки, а последние благодаря делению в тангентальном на-
правлении отчленяют наружные клетки, создающие стенку коробочки,' и
внутренние, образующие археспорий. '
Архёспориальиые клетки делятся миогократнЬ и образуют материнские
клетки спор. Часть материнских клеток спор приступает к делению. Первое
их деление — редукционное. Образовавшиеся две клетки делятся далее
обычным кариокинетическим способом еще раз, и в результате образуется
тетрада (четверка) клеток спор. Таким образом, споры гаплоидными образсь
I' I;
; II
вались в результате редукционного деления, которое у всех высших расте-
ний всегда происходит при образовании спор (рис. Ill; 36). j
Другие материнские клетки спор вытягиваются и благодаря неравно-
мерному утолщению оболочки спирально закручиваются. Каждая такая
Рис. 111. Маршанция:
2 — молодой споротой (м — коробочка, н — короткая ножка. * — гаустории, бр — брюшко архегогшя,
ft— 1разросшийся перманций); в—часть более старого спорогона (видны споры и пружинки); J — мате-
рашжяэкпеткн спор: а—два рнддматерннских клеток спор и часть пружинки (удлиненная клетка),
б—шлеркнскмв юга стаде делятся с образованней тетрад, л — споры и пружинка
184
РАСТЕНИЯ
Рис. 112. Маршанция:
1 — зрелая женская подставка со
споротонами (справа более зре-
лая» лучи подставки поднялись);
2 — ножка спорогсна удлинилась»
коробочка вскрылась, высышдот-
сн споры и пружинки (с — стен-
ка коробочки* п — нерианций»
к — колпачок — калиптра); з —
споры; 4 — пружинка; з — про-
растание спор и молодые талломы
Спора одета двумя. оболочками — наружной, более толстом (экзо-
спорном) и внутренней, тонкой (э и д о с п о р и е м). Эндоспорий
облегает содержимое споры, состоящее из плазмы и ядра. Спора, попав
на почву, прорастает. Наружная оболочка ее лопается, а содержимое, окру-
женное эндоспорием, вытягивается в короткую нить. На ее конце отчле-
няется клетка, которая делится, вновь отчленяя
клетки, продолжающие
делиться в разных направлениях. В результате формируется небольшая плас-
тинчатая протонема, далее развивающаяся в таллом маршанции (рис. 112).
У маршанции, таким образом, имеется смена поколений (генераций): поло-
вое поколение (гаметофит) сменяется бесполым (спорофитом), спорофит —
гаметофитом. В цикле развития преобладает гаметофит —^растение маршан-
ции с развивающимися на. нем мужскими и женскими половши* органами —
антеридиями и архегониями. Все клетки его гаплоидны. Гаметофит берет
начало от гаплоидной споры. Спорофит (спорогон) имеет небольшие разме-
ры, он питается за счет гаметофита, к которому прикрепляется при помощи
х^^стории. Все клетки спорогона диплоидны, так как развиваются из ди-
плоидной зиготы. Переход от спорофита к гаметофиту совершается в резуль-
тате редукционного деления.
Порядок маршанциевых разделяют на 12 семейств. Наиболее обширным
из них является семейство j^apuu^^4£SMS~^Marchantiaceae)9 куда от-
носится рассмотренная нами маршанция.
Из близкого к нему семейств а п нашей флоре ши-
роко распространен коноцефалцм (Confcephsrtum conicum). Встречается это
растение вдоль уреза воды по лесным ручьям. Внешне оно похоже на маршан-
цию, но имеет более четкий рисунок камер на верхней стороне таллома. Вывод»
ОТДЕЛ — МОХООБРАЗНЫЕ
185
Рис. 113. Печеночники:
1 — коноцефалум (Conocephalum conlcum); 3 — рмччин (Riccia glauca): а — общий вид таллома, б — по-
перечный разрез (внутри архегония — молодой спорогон), в — разрез спорангия с тетрадами спор;
3 — риччиокарпус Ricciocarpus natans); 4 —риччия (Ricda fluitans)
ковых корзиночек у него не бывает. Антеридиальные подставки не имеют но-
жек и помещаются прямо на талломе. Женские подставки конической формы
(рис. 113, 1).
Упомянем еще о семействе Это наиболее
совершенное семейство в порядке маршанциевых. Содержит 4 рода и около
200 видов. Наиболее распространены роды Riccia и Ricciocarpus.
Талломы 'их не имеют^ хорошо выраженных воздушных камер. Подставок
на талломах не развивается, а антеридии и архегонии погружены в ткань
таллома. Спорогон также скрыт в талломе. Он состоит из коробочки, ножка
не развивается. В коробочке развиты только споры, пружинок не образует-
ся. По созревании спор стенка коробочки разрушается. Риччия (Riccia)
распространена во всех частях света и представлена большим числом видов.
Маленькие талломы ее дихотомически ветвятся и расходятся радиально от
центра во все стороны. В нашей стране особенно часто R. fluitans; она пла-
вает на поверхности воды, при высыхании водоемов растет на сырой почве
(рис. 113, 4). Другие виды (R. glama, R.sorocarpa, R. ciliata) растут на сырых,
илистых и глинистых почвах, на пашнях и по обочинам канав. Большинст-
во же видов^Ддеаа — растения засушливых, (арцдцых) областей, произрас-
тающие на обнаженных почвах. Род риччиокарпус (Ricciocarpus) пред-
ставлен у нас широко распространенным видом R. natans. Это маленькое
растение, плавающее на поверхности водоемов с непроточной водой или
Ill',
186
вы
Hili'
Л РАСТЕНИЯ
Рис, 114. Пеллия:
1—общий вид; 2—поперечный разрез таллома (а—антеридий); л — архегонии; 4 — молодой споротой
(н —полоса» а — гаустория. * — коробочка «у птуймы. а — апатофор» бр — брюшко архегонии)
иг.
поселяющееся на илистой почве. Кожистые талломы его достигают 5 —
10 жм длины и 3 — 8 мм ширины. На их верхней поверхности хорошо
заметен срединный дихотомически разветвленный желобок (рис. 113, 3).
2-Й
— ЮНГЕРМАННИЕВЫЕ ( JUNGE RMANNMcffiE)
Юнгерманниевые отличаются от маршанциевых простотой анатомиче-
ского строения. Среди них есть как талломные, так-йлйотеетебеДьные рас-
тения; имеются также переходные между ними формы.
Это наиболее обширный порядок печеночников, насчитывающий 195
родов и около 7800 видов. Большинство их произрастает в тропических об-
ластях, но они широко развиты также в умеренных и холодных зонах.
Порядок разделяют на 3 подпорядка. .
1-й подпорядок — Анакрогинные юнгерманниевые (Anacrogynales)
В этот порядок входят главным образом талломные печеночники,
имеются и облиственные.
но
У цсОх анацрогинных юнгерманний точка роста не идет на образ ова-
вне архегониев и таллом продолжает нарастать" после югобразовапия. Под-
ставок никогда не образуется. К подпорядку принадлежит 10 семейств,
20 родов и 560 видов, распространенных по всей Земле. ***’....
ОТДЕЛ — МОХООБРАЗНЫЕ
1©
Рис. 115, Юнгерманиии анакрогинные и калобрневые:
Л — фоссомброняя; я—блявия: а — общий ищ растения со спорогоноы. б — таллом снизу (аж — амфи-
гастрин, епл — спинные листая, ди — лвстовыв ушки); У — кцдобриум: а — общий вид (верхушка муж- .
ского гаметофита). б — продольный разрез верхушки мужской веточки (видны антеридии) t в - продоль-
ный paepep верхушки женской веточки (видны архегонин); 4—гапломитриум (женские растеши со
спороншами) -
В качестве представителя талломных юнгерманний мы рассмотрим
лию (Pellia epiphyIla), широко распространенную у нас по сырым Лесным
оврагам, около ручьев и канав (рис. 114). Тонкие лентовидные узкие талло-
мы пеллии дихотомически ветвятся. Они светло-зеленые. в центре много-
слойные, по краям однослойные. Таллом и^йэт ппаик прплтпп анатомиче-
ское строение. Он одетГверхним и нижним эпидермисами и состоит из немно-
гих слоев паренхимных клеток. От клеток нижнего эпидермиса, нерезко
отграниченного от основной ткани таллома, пч1юдр,р прпстде рдзоиды. JBo3-
душных камедмиарс^мы^д^румд а хлорофилловые зерна имеются во'
всёх кдетках тал^ма^рис. 114, 2).
Подставок на образуется антеридии и архегонни расположены прямо
на талломе. Они возникают, несколько отступя от точки роста, которая
продолжает свою деятельность, наращивая таллом.
ной формы, сидят на короткой ножке и погружены в ткань таллома.
Над антеридием эпидермис прерывается и несколько возвышается,
образуя отверстие, идущее в антеридиальную полость (рис. 114, 2).
В антеридии развиваются двужгутиковые сперматозоиды. Архегонни
возникают группами в небольшом углублении, вблизи точки роста
(рис.114, 3). Они имеют обычное строение.
188
ВЫСШИЕ РАСТЕНИЯ
г г.
Спорогон состоит из коробочки* ножкит гаустории которая внедряется
в ткань талломаПТ^рЁдначально спорогон заключен в брюшке архегония.
Затем, ко времени созревания спор, ножка спорогона сильно удлиняется
(обычно это б^ваёт^весной следующего года) выносит коробочку высоко
над талломом (рис, 114, 4). У основания ножки со^рййя'ется колпачок (оста-
ток "брюшка архегония). Коробрчк^^вскртпается^на,_верхушке__четырьмя
створками, и из нее высыпаются споры и пружинки. Пружинки пеллии
собраны в пучок, прикрепляющийся в центре коробочки (так называемый
Спора, прорастая, образует короткую протонему, вскоре развивающую-
ся в новый таллом пеллии.
Из других родов упомянем метцгерию (Metzgeria). Около 120 видов
Metzgeria растут на гниющей древесине, преимущественно в тропиках, а
также у нас на Кавказе, в Карпатах. Антеридии и архегонии образуются
на коротких боковых ветвях.
Листостебельными являются всего лишь 2 рода: фоссомброния (Fossom-
bronia, рис. 115, 2) и трейбия (Treubia). Имеются также переходные между
талломными и листоСТёбИХЬШЗйй^формы из родов Riccardia (Апеига),
Hymenophyton.
Очень интересна бл^зия маленькая (Blasia pusilia}, растущая у нас по
сырым глинистым дорогам; в лесах — по канавам и т. д. (рис. 115, 2). Ее
распластанные на земле курчавые ярко-зеленые талломы (1 — 3 см дл
и около 5 мм ширины) имеют краевые лопасти, являющиеся редуцирован-
ными листьями. На нижней стороне таллома, по обе стороны от жилки,
развиваются шарообразные, полые внутри ушки, в которые вдается слизис-
тый волосок. Выводковые почки двух родов — звездчатые на талломе и
шарообразные в особых вместилищах (колбочках). Антеридии и архегонии
развиваются на верхней стороне таллома. Блязия произошла, вероятно, от
листостебельных предков в результате редукции. Впрочем, многие ботани-
ки считают краевые лопасти таллома бЛязйи зачаточными листьями.
tn
,1:1
ill
2-й подпорядок — Акрогинные юнгерманнии (Acrogynales)
К этому подпорядку относятся исключительно листостебельные пече-
_цсрюки, талломных растений среди них нет.
Архегонии возникают на верхушках стеблей и веточек. Точка роста их
идет на образование .архегониев, в связи с чем рост веточек^прёк^^цЖРСя?
Отсюда и название подпорядка: акрогинные — верхушечно-плодные.
К акрогинным юнгерманниям принадлежит 18 семейств, 180 родов и
около 7500 -видов печеночников. Они распространены всюду, но наиболь-
шего развития и разнообразия достигают в тропических влажных лесах,
где играют важную роль в растительном покрове. Акрогинные юнгерманнии
одевают сплошным покровом почву, а также стволы, ветви деревьев (э п и-
ф и т ы), часто развиваются на листьях (эпифилл ы), и столь обильно^
что одевают их пластинку сплошь и затрудняют фотосинтез. В тропиче-
ских лесах печеночники достигают наибольших размеров. “
Много печеночников распространено и в умеренных областях. Здесь
они произрастают главным образом в лесах, на нижних частях стволов де-
ревьев, на отмерших ветвях или на почве — среди травы, а также на лугах,
болотах среди других мхов. В холодных областях они также представлены
большим числом видов, и иногда на почве покрывают значительные площади
(например, Ptilidium в тундре).
'Большинство акрогинных юнгерманний имеют лежачие дорзовентраль-
ные побели, Стебель несет пбытоп ури ряда два рядгПй
один ряд брюшных листьев — амфигастриев. Обычно амфигястрци
развиты слабее, а у некоторых видов и совсем атрофируются (у Radula}.
Спинные ^Листья цельные, реже на верхушке. двузубчатые, иногда со-
стоят из лопастей различной величины. В этом случае обычно маленькая
11.1:1:1
ОТДЕЛ — МОХООБРАЗНЫЕ
189
Рис. 116. Радула и хилосодфус:
радула: а —общий вид (на верхушке одной веточки виден колокольчатый псрихедий). б — антери-
дии Ндархегонии. в — молодой спорогон (н — ножка, г — гаустория, л — коробочка со спорамв и пружин*
ками), а — масляные тельца* я — хилосцифус: а — общий вид. о — амфигастрин
ЛОЛ
вод
тос:
жсть загибается на нижнюю сторону стебля и служит для .удержания
ы, в то время как крупная верхняя (спинная) лопасть является фо-
аитезирующей.
У некоторых печеночников, маленькие лопасти приобретают вид кув-
пиков, в которых также собирается вода.
игл
Иногда листья сильно рассечены на многократно ветвистые мелкие доли
(Ptiltdium). Анатомическое строение стебля и листьев акрогинных юпгер-
манний очень простое. Стебель состоит из однородных удлиненных тонко-
стенных клеток. Листья однослойные, без средней жилки.
От стебля и ^ТисгСев- отходят ризоиды, исключительно простые. Они
служат для прикрепления растения к субстрату и всасывания водыГФорма
листьев, взаимное их расположение на стебле имеют большое систематиче-
ское значение.
Для ознакомления с этой группой печеночников мы рассмотрим два
рода — радулу и хилосцифус.
Радум (Radula) насчитывает,J27O видов — тропических и субтропи*
.in
ческих эпифитов на коре деревьев и эпифиллов на листьях. У нас широко
распространена на стволах липы, дуба радула распластанная (Radula
complanata). Ее дорзовентральные побеги 2 — 4 см длиной плотно прижаты
190
ВВДШИВ РАСТЕНИЯ
к коре деревьев. Вычлененное из дернинки растение имеет ветвистый сте-
бель, Главный стебель и боковые веточки имеют два ряда листьев. Каждый
лист состоит из двух лопастей.
Спинная лопасть крупная, брюшная мельче, загнута вниз и прижата
к стеблю. Однослойные листья своими краями налегают друг на друга.
В результате растение имеет ясно дорзовентральное строение (рис. 116).
Брюшные листья (амфигаствии> у радулы отсутствуют.
От нижних лопастей листа отходят ризоиды. Ветвление радулы, как и
всех юнгерманний, происходит очень своеобразно: ветви выходят не из па-
зух листьев, а несколько ниже своего листа, с нижней стороны стебля. Ра-
дула — однодомный печеночник: антеридии и архегонни развиваются на оД-
ном растении.
Архегонни группами располагаются на верхушке веточек, причем точ-
ка роста идет на их образование и рост данной веточки на этом прекраща-
ется. Архегонни обычного строения (рис. 116, 26).
Листья, ближайшие к архегониям, отличаются от других листьев по
форме и размерам. Они срастаются между собой, образуя вокруг архегониев
колпакообразное покрывало — перйхеций (рис. 116, 1а).
Ацтеридии находятся .несколько ниже архегониев и расположены каж-
дый ЩГдлйшойножке (рис. Нб, 2BJ. Кнэтраз^^^^ Двужгутиковые
сперматозоиды.
После оплодотворения из зиготы развивается спорогон. Зрелый споро-
гон состоит из коробочки, ножки и гаустории (рис. 116, 7а). При помощи
гаустории спорогоний внедряется в ткань верхушки стебля, получая от
гаметофита питательные вещества. Коробочка имеет двухслойную стенку.
В коробочке развиваются споры и пружинки. Ко времени созревания спор
ножка спорогона сильно удлиняется и выносит коробочку вверх, за преде-
лы перихеция. Коробочка вскрывается четырьмя створками на верхушке.
Споры при этом высыпаются и падают на землю.
Из спор вырастает короткая пластинчатая протонема, развивающая
далее облиственное растение.
Хилосцифус (Chylosciphus) широко распространен у нас в лесах — у
основания стволов, на опавших гниющих ветках (Ch. pallescens) или на
глинистой почве среди трав (Ch. polyanthus^ рис. 116, 2). Он образует рых-
лые тонкие дерновинки, прижатые к субстрату или приподнимающиеся,
и достигают 2 — 6 см длины при 2 — 3 мм в поперечнике. Листья трехряд-
ные. Спинные листья (два ряда) широкие цельные или же двуразднльные
наверху (у Ch. pallescens). Брюшные листья (один ряд) — амфигастрии,
глубоко двураздельные. Строение антеридиев, архегониев и спорогона такое
же, как и у радулы.
3-Й подпорядок — Гапломитриевые (Haplomitriales)
Это небольшая группа олиственных печеночных мхов, имеющих прямо-
стоячие стебли, внизу переходящие в «корневище». Листья однослойные,
без средней жилки, расположенные трехрядио. Листья или все одинаковые,
или иногда брюшные листья (амфНгастрин) более мелкие. Растения лишены
ризоидов. Сюда принадлежат 2 рода — калобриум (Calobryim) и еапмн
митриум (ffaplomitrium).
Калобриум представлен пятью видами, растущими на гнилой древеси-
не, произрастают они в Восточной Азии, Новой Зеландии, Центральной и
Южной Америке. Это акрогинный печеночник. У него антеридии и архего-
нии (на разных особях) развиваются группами на верхушке стеблей, и точка
роста идет на их образование, в связи с чем прекращается дальнейший рост
стебля (рис. 115, 3).
Гапломитриум представлен одним видом Н. Hookert, растущим на сы-
рой почве в верещЗгаиках Западной Европы. У нас отмечен, только для
- ОТДЕЛ — МОХООБРАЗНЫЕ
191
ли
Карпат. Это анакрогиниый печеночник. Растение двудомное. Антеридии
развиваются по 2 — Зв пазухах листьев. Архегонии в пазухах листьев по
3 — 4 расположены близ верхушки стебля, которая не идет на их образова-
ние и продолжает свой рост (ряс. 115, 4). В коробочках растений обоих ро-
дов развиваются споры и пружинки. Гапломитриевые стоят изолированно
среди печеночников.
Интересно отметить выработку прямостоячих форм среди акрогинных
и анакрогинных юнгерманний. Многие крупные ботаники (Лейт г е б,
Гебель) считали Haplomitriales за переходную группу — между ана-
крогинными и акрогинными юнгерманниями. Вместе с тем они являются
специализированной группой. Отсутствие ризоидов связано, вероятно, с
сапрофитным образом жизни на гнилой древесине (калобриум имеет сильно
разветвленное корневище).
1.1
3-Й ПОРЯДОК — СФЕРОКАРПОВЫЕ (SPHAEROCARPALES)
К порядку принадлежат 2 семейства — сферокарповые и риеллиевые.
Семейство сферокарпотх (Sphaerocarpaceae) представлено двумя
родами: родом Sphaerocarpus, насчитывающим 12 видов в северном полу-
шарии, из коих в Западной Европе распространен Sph. micheli^ в СССР
он отсутствует, иродом Geothallus с одним видом G. California^ произ-
растающим в Калифорнии (США).
Сферокарпус образует маленькие пластинчатые талломы, состоящие из
однородных паренхиматических клеток, на большем протяжении одно-
слойные.
В средней части талломы состоят из нескольких слоев клеток. К земле
талломы прикрепляются простыми, ризоидами.
Архегонии и антеридии развиваются на разных растениях. Они скрыты
внутри бокальчатых выростов таллома, имеющих наверху отверстия
(рис. 117).
Архегонии и антеридии (последние сидят на коротких ножках на дне
антеридиальных вместилищ) по строению и развитию очень сходны с поло-
выми органами маршанциевых, как и споро гоны. Спорогон состоит из луко-
вицеобразно утолщенной ножки, внедряющейся в ткань таллома, и коро-
бочки. Коробочка имеет однослойную стенку. В коробочке образуются споры,
соединенные в тетрады (четверки), и питательные клетки, лежащие сре-
ди спор. Последние гомологичны пружинкам других печеночников. Споры
освобождаются лишь после сгнивания стенки коробочки. Из четырех спор
тетрады обычно две развивают мужские, а две другие — женские тал-
ломы.
Сферокарповые объединяют признаки, свойственные маршанциевым
и юнгерманниевым. Их предки, еще более просто устроенные, произошли,
вероятно, из водорослей или первичных наземных растений, сходных с во-
дорослями.
I: i i 1
4 И ИОРДДОКи АНТОЦЕРОТОВЫЕ (ANTHOСЕ ROTATES)
I
Представители этого небольшого порядка резко * отличаются от других
печеночников своеобразным и наиболее примитивным гаметофитом и очень
сложно устроенным спорофитом, способным к длительномуj росту при по-
мощи особой меристемы. 4 * * * В
В строении гаметофита имеется ряд признаков, сближающих антоце-
ротовые с водорослями, в строении же спорогона — признаки, свойствен-
ные лиственным мхам.
К порядку антоцеротовых относятся 2 семейства: Anthocerotaceae и
Notothylaceae. К семейству Anthocerotaceae относятся 3 рода, из кото-
рых наиболее распространенным является ашпоцерос (Anthoceros), насчи-
192
выс
11Г1>
РАСТЕНИЯ
Ряс. 117. Сферокарпус:
2 — общий вид женской особи; £ — поперечный разрез мужского таллома (антеридии расположены
внутри полостей); 3 — спорогон со спорами и нитателышми клетками; 4 — три крупные тетрады спор
и две питательные клетки
тывающий около 200 видов, встречающихся в умеренных областях обоих
полушарии, а также в тропиках.
Другие 2 рода встречаются в тропиках и в южном полушарии. К се-
мейству Notothylaceae относится только род нототилас (Notothylas)
с 10 видами, растущими в тропических областях, и лишь один вид N. orbicu-
laris произрастает в Западной Европе иСеверной Америке.
В СССР встречается только р о д/j^jijfeocgrbij, представленный двумя
видами — A, punctatus и A. laevis.
Они растут на сырой глинистой почве ~ на полях, по сырым канавам.
Талломы антоцероса рпяетковидтпуе; Х),5~ — 3 см в поперечнике, по краям
надрезайШг-^лотгасТные, темно-зеленые. В середине они плотно прижаты к
субстрату и состоят из 6 — 10 слоев клеток, по краям же однослойные и.
часто загнуты вверх (A. punctatus). К земле талломы прикрепляются, при
помощи простых ризоидов. Брюшных чешуек (амфигастрий) нет. Нарастают
талломы при помощи верхушечных клеток, расположённых меЖДу лопастя-
ми слоевища (рис. 118).
Таллом имеет очень простое анатомическое строение. Он состоит из
и
однородных паренхиматических клеток. Эпидермис не выражен. В клетках
таллома, как и у водорослей, имеются пластинчатые'хроматофоры (по одно-
му в клетке), внутри которых находится пиреноид.
В талломе имеются слизевые полости, открывающиеся наружу. В эти
полости врастают нити' сине-зеленой водоросли носток (Nostoc). Колое
иостока, нарастая, увеличиваются в размерах, а вместе с ними разрастаются
। г
и полости, внутренние стенки которых образуют выросты в виде волосков.
На внутренних стенках полостей находятся устьица. Полости заполнены
ОТДЕЛ — МОХООБРАЗНЫЕ
193
Рис. 118. Антоцеротовые:
1 — антоцерос: а — общий вид растения со сппрагонамв, б — антеридии, в — сперматовояд, е — архегонииа
д —спорогон (ст — створки, код — колонка» чш — чашевидно разросшаяся часть таллома); е —* имжцян
часть спорогона (на границе гаустории и ножки заметна меристематическая ткань, клетки ее уменьшаются
кверху и книзу. Выше — створка» колонка и два ряда аухеспорнальных клеток); ж — продольный раз*
рев средней части спорогона (с — споры» пр — пружинки); г — нонинлас: а — общий вид растения со
спорогонамн, б ** спорогон, е — продольный разрез спорогона (пол — колонка» ар — археспорий» а — гау-
стория)
слизью. Слизь выступает наружу и защищает точки роста таллома от
высыхания.
Очень своеобразны выводковые падки, служащие для вегетативно го
размножения антоцероса. Они образуются внутри клеток таллома, как апла-
носпоры у водорослей. Содержимое некоторых клеток отстает при этом
от стенок и окружается оболочкой. Благодаря ряду последовательных
. делений образуется небольшое тельце, освобождающееся по ослизнении сте-
нок произведшей их клетки. Освободившись, эти тельца разрастаются и
дают новые талломы.
шается, антеридии вскрываются и двужгутиковые сперматозоиды выплы-
вают наружу. Аутегпттмд тяу]ие внутри ткани таллома. Они
не имеют собственнои"стенки и состоят из четырехклетной шейки, брюшной
канальцевой клетки и яйцеклетки. Сверху архегонни одеты поверхностными
клеткамиталлома, которые ослизняются ко времени созревания архегония
(рис» 118, Канальцевые клетки (шейковые и брюшная) расплываются
7—602
194
в слизь, по которой сперматозоид проникает к яйцеклетке. Из оплодотво-
ренной яйцеклетки развивается весьма своеобразный споротой (спорофит).
Зрелый споротой имеет вид довольно-толстой щетинки, возвышающейся над
поверхй№ТЙ&‘1,мл^а на 2 — 3 см, в нижней Частной расширяется в гауо-
торию, внедряющуюся в ткань таллома (ряс. 118, 1 5,а). Ц верхней части
споротой вскрывается двумя продольными створками сверху вниз, обнажая
при этом серединную колонку, протягивающуюся вдоль щетинки и состоя-
j [|; ।
щую из бесплодной ткани.Между ^пппнкпй и.створкмд Pacnon<ngffaHM ГЛА-
ры ..и. пружжшсообразные рВЗй^вленные клетки, гомологичные элатерам
других печеночников (рис. 118, 1ж). Ниже спорогон сомкнут,'и здесь можно
наблюдать, следуя сверху вниз, постепенное формирование спор и пружинок.
В месте перехода щетинки в гаусторию в спорогоне развивается мери-
стематическая ткань, в результате деятельности которой и происходит на-
растание щетинки вверх и ее формирование (рис. 118, 1е).
Клетки створок имеют по 2 хроматофора. Для эпидермиса створок
характерно наличие нормально функционирующих устьиц с замыкающими
клетками. В створках развита ассимиляционная ткань, и спорогон антоце-
роса в значительной мере обеспечивает себя питательными веществами.
Вокруг образующегося спорогона ткань таллома чашевидно разраста-
ется в виде узкой длинной обвертки (рис. 118, 1д). При прорастании споры
ГГ,
образуется тонкая пластинчатая протонема, на ней обозначается много то-
чек роста. Нарастая, они формируют лопастной курчавый таллом анто-
цероса.
У некоторых видов антоцероса (например, у-американского Л. fusel-
formis) спорогоны сильно разрастаются, достигая 6 — 16 см ддптм, внедря-
ются в почву, добывая оттуда воду и минеральные вещества, и становятся
на. некоторое время самостоятельными растениями.
Своеобразию.строение имеет спорогон Notothylas (из семейства
Notothylaceae). Коробочка у него обычно не возвышается над обверткой.
Она вскрывается или двумя лопастями, илж продольной щелью. Внутри
заметна короткая колонка. Спорогон напоминает коробочку Sphagnum
(стр. 195). Стенки коробочки без устьиц, элатеры у него неразветвленные
(рис. 118, 2) ,
2-й КЛАСС—ЛИСТВЕННЫЕ МХИ (BRYOPSIDA, MUSCI)
Лиственные мхи всегда расчленены на стебель и листья. Стебли имеют
радиальное строение, на них обычно тесной спиралью расцодорсины листья.
В коробочке развиваются ТОЛЬКО mnn" irnvwcmre» ("ЛЗТМНг) "°
''реже пластинчатая. В коробочке большинства' мхов развивается п ej> и с-
то м — особый аппарат, способствующий, рассеиванию’’снор_. {перистома
нет у Sphagnum и Andreaza). *
Лиственные мхи разделяют на 3
W-Я ПОРЯДОК — СФАГНОВЫЕ, ИЛИ ТОРФЯНЫЕ (БЕЛЫЕ).
МХИ (SPHAGNA£BS)(<jtau
К этому порядку принадлежит одно семейство Sphagnaeeae с един-
ственным родом сфагнум (Sphagnum}, насчитывающим свыше 350 видов1.
Наиболее широко сфагновые мхи распространены на равнинах в умеренных
и холодных областях северного полушария, образуя сплошные покровы на
болотах, и яаболочйпиых лесах и т. п., занимая огромные пространства в
Евразии и Срлпрной' Ампрнпп В горных областях сфагны расТут на влаж-
1 Некоторые авторы понимают вид в пределах рода очень широко и уменьшают
;ов Sphagnum до 100.
ОТДЕЛ — МОХООБРАЗНЫЕ
195
Рис. 119. Сфагнум:
1-явемшЛ мд: г— верхушка ветви со своротами; л—часть (увеличено) атчиосю листа (хля — хяо>
рофйнлснвепйе клетки, е* — водоносные клетки, п — норы); 4 — поперечный разрез листа; 9 — протоне-
ма; * — часть стебля (айв — антериямльные ведшим. аре — еркегоннаяьные веточки, л — ембааше
листья)
why амязгядг Довольно широко сфагновые мхи представлены в умеренных
областях южного полушария — на болотах Новой Зеландии, Тасмании,
Огненной Земли и Патагонии.
В тропических областях сфягионыа мхи встречаются только в горах
на известной высоте, нередко в большом числе видов, но не образуют боль-
ших зарослей. В СССР наибольшего развития сфагны достигают на севере
ЧВДИПГ В тундровойjjone роль сфагновых мхов резко „снижается, ме-
F;»; ।:
г,*
1; ।:
ном из дярнняы растении хорошо вамегао листост^ел^|рр^дш1еиие. Сте-
бель одет' стеблевыми листьями. В ^^Л^Яжмяюрагг та стт ряттатря^икг
ястиндсобранные пучками, по 2 — 7 ветвей в каждом. На веточках располо-
жены веточные листья. Ветви имеют различный вид в разных участках стеб-
ля. Ц& вершине стебля находятся короткие веточки, плотно скученные в
головку. Листья на них сидят тесно, рассто
очень ^короткие — ^ур^дддчные ветви, Ниже по стеблю пучки располо-
жены на заметном расстоянии друг от друга. В каждом пучке здесь 2 —
4 ветви, отстоящие горизонтально; с более редким расположением листьев, ~
торчащие ветви; остальные ветви пучка — очень длинные, с редко располо-
ЙенПЫмиТтЙсЙВгми — свешиваются вниз вдоль стебля, плотно к нему при-
жимаясь, — свисающий ветви, (рис. 119).
В конце лета одна ив верхушечных ветвей начинает расти вверх, про-
должая рост сфагнума. Нижний конец стебля постепенно отмирает.
Анатомическое строение стебля очень простое. Центральная часть стеб-
ля занята сепдшвиной, обпаяпняинпй шитюмиыи тгяпАятиынымЙ клетками.
ТОК С
I»
ЫДЛИ-ШШЙВДггцвет»
ШЩ^МгЛридает проч-
196
ВЫСШИЕ РАСТЕНИЯ
iiiua]
ность стеблю. Внешня%кора состоит из нескольких слоев широких мерт-
вых прозрачных клеток, стенки-которых имеют спиральные утолщенйя и
крупные отверстия---joBlFIphc. 12(Jt Л-Снаружи стебель^бдёт^шщермИ-
/•ОМ, клтт-ноторого чмоюттакое ж» дтр^ение. [Внешнюю кору й5шидермйс
часто называют мтогпследя5мэпидеомисом(гй а одер ми с ом). Че-
рез поры соседние^иацшш "гиадцДёрММшьсорбгпяш Дпй— Лии лег-
ко всасывают воду, которая быстро пододвигается вдолхНяёбля.
Веточные листья сфагнума лапцетйыеили яйцевидно-ланцетные. У них
очень своеобразное_строенпет Листья состоят из одного слояклеток, не име-
ют средней жилки и поэтому очень сходны "слистьями лцстостебедьных
юнгермяннйи Г...
Листья имеют клетки двух родов — хлорофиллоносные и водоносные, или
гцаддновые. Хлорофиляпносныё~Пклетки узкие, червеобразные, содержат
хлорофилловые зерна. Они зажаты между широкими бесцветными гиалино-
выми клетками. Последние заполнены водой. '
Стенки водоносных клеток имеют спирадьиыа^ухрдщенмяиифрдыряв-
лены отверстиями — порами. Поры ^шличцо^^едизины^и^формы^-дхЕ
мелких, едва заметйыхГтрСрчень широких. Поры могут быть простыми и
окаймленными. Число их, как и расположение в клетке, различно (вдоль
боковых стенок, в углах и т. д.). Эти признаки строго постоянны для опре-
деленных видов сфагнума.
На долю водоносных клеток приходится до ВА поверхшм»™ пиптй Такое
анатомическое строение обеспечивает сфагнуму быстрое всасывание и про-
ведение воды. Установлено, что отдельные виды могут поглощать воды в
37,5 раза более своего веса.
Высокой гигроскопичностью объясняется быстрое заселение
территории и развитие болота в тех местах,
сфагнами
где они появляются.
Рис. 120. Сфагнум:
1 — поперечный разрез стебля: а — многослойный эпидермис, б — древесинный цилиндр. < — сердцевина;
я — веточка с антеридиями; 3 — вскрывшийся антеридий; 4 — двужгутиковый сперматозоид; 4 — группа
архегониев; 4 — спорогон (ш — остатои шейки архегония, хр — крышечка, сп — снорангвй. км — колон-
ка, н — ножка спорогона, лх — ложная ножка); 7 — дифференцировка клеток листа
ОТДЕЛ — МОХООБРАЗНЫЕ
. 197
III.
ТГТ
Описанным выше строением объясняется и белесоватый u цветсфагнума,
так как на долю зеленых кдетпк приходится лишь */а ппворхнпдтй'^тйт^п
Листья разных ветвей отличаются фо рмой, величиной, количеством1
и расположением пор. .........................
Ассимилирующими являются верхушечные и торчащие ветвв^свисаю-.
же пбряаушт ипируг стебля" как бк чёхолТфитиль, способствуя быстрому ~
прпдяижепит- ппды тадплтк стебля.
На стебле сфагнума расположены стеблевые листья. По внешнему_вцду
и строению они обычно несколько отличаются пт веточных листьев. Однако
они такзкб-^дпопяойттыо» бея средней жилки и состоят из хлорофиллоносных
и гиалиновых клеток. Гтаблепня строго постоянны в своих призна-
ках у разных видов и имеют большое значение для систематики сфагно-
вых мхов. ~ "
Uli
Дернины сфагнума ежегодно отмирают в нижней части, и одна из верху-
шечных ветней^продолжает рост стебля вверх. Отмершие части сфагнума
вследствие пбвпд^еиия^и нехватки кислорода не разлагаются до конпа. а
образуют слои полуразложившихся остатков — в вине торфа. При этом
образуются органические кислоты, которые еше^бодее угнетают бактериаль*.
ную флору и задерживают раяложение торфа
В верхней части стеблей образуются антеридии и архегонии. Антеридии
возникают на оси верхушечных веточек, гпе они чередуются с листочками.
Такие янтдрпдяялгктдА пдтатж пдмупид тдяапм Лии шмЧМПНЛ удли-
ненные имеют буроватую _ окраску. Антеридии строением и способом
вскрывания ияплуштяшт ятгтертгдии плиствепных юнгерманнии^
Антеридии — овальные или шаровидные тельца, расположенные на.
длинных ножках Вскрываются они в верхней части путем разрыва его стен-
ки на ряд лопастей, которые заворачиваются при этом наружу.
(рис 120, 2. 3).
В антеридии образуется большое количество двужгутиковых спермато-
* зондов.
Архегонии образуются на верхушке коротких веточек (верхушечных
и некоторых торчащих) в числе 3—5 (рис. 120, 5). Они имеют обычное
строение.
В результате развития оплодотворенной яйцеклетки образуется споро-
гон. Спорогон имеет очень своеобразное строение. Он состоит из шаровид-
ной коробочки, которая ниже, после небольшой перетяжки, переходит в
ножку (рис. 120, 6), Ножка внедряется в верхушку стебля. Ко времени со-
зревания спорогона верхушка стебля удлиняется и выносит коробочку над
покровными листьями.
Эта удлиненная часть стебля называется ложной ножкой (так
как она принадлежит гаметофиту). Граница между настоящей и ложной
ножкой хорошо заметна (рис. 120, б).
Вверху коробочка ^покрыта округлой крышечкой, которая отграничена
от остальных частей коробочки несколькими слоями тонкостенных мелких
клеток. Внутри коробочки имеется поднимающаяся со дна колонка. Клетки
колонки паренхиматические, широкие. Сверху колонки расположен купо-
лообразный спорангий. В нем образуются споры. Стенка коробочки много-
слойная. В эпидермисе находятся редуцированные устьица,/^оторые не име-
ют щели и состоят из двух замыкающих клеток.
Все клетки эпидермиса, кроме устьичных, в молодом состоянии содер-
жат хлорофилловые зерна, и, таким образом, спорогон сфагнов питается
отчасти самостоятельно.
Молодой спорогон скрыт в брюшке архегония; по мере роста брюшко
архегония сначала растягивается, а затем разрывается в середине. Верхняя
часть брюшка вместе с остатками шейки сбрасывается (колпачок); нижняя
‘ его часть остается в виде небольшой оторочки у основания коробочки.
ill:
198
(ыиггН
PACT
j и: г
Рис. 121. Андреевы! box (Andreaea ru pestrig):
1—растенав co свдюгапш (слева — наработка, вскрывающаяся четырьмя щелями); х—верхушка
стебля с ацтервдинмя; 3 — арквгамй; 4-* споротой (мм — неловка, сп —спорангий со спорам*, ар —
остаток архегевмя)
К этому времени разрушаются колонка и стенка спорангия. Споры оказыва-
ются в полости коробочки. Затем сбрасывается крышечка, отрываясь от
коробочки с большой силой.
Споры при этом выбрасываются наружу. Периетома у Syhagnales нет.
Попав на почву, споры прорастают, образуя при этом пдлвтдпмв<|днп тгрёйго-
яему. На протонеме образуются ризоиды (отсутствующие у взрослого рас-
тения) и почки, развивающиеся далее в олиственные побеги. Первые листья
молодого растения состоят из однородных клеток. Дифференцировка кле-
ток листа на хлорофиллоносные и водоносные начинается позднее*
г *
I:
1 ГЛ:
f 2-Й ПОРЯДОК — АНДРЕЕВЫЕ МХИ (ANDREAEALES)
ственное в порядке семейство Andreaeaceae представлено тре-
мя родами и насчитывает 92 вида. Все они являются горными и тундровыми
растениями, образующими небольшие дернинки, прижатые к камням.
Наиболее распространен род андреа (Andreaea), представленный
90 видами в обоих полушариях Земли. Это преимущественно горные виды,
заходящие в Арктику и Антарктику. Остальные два рода встречаются в
Антарктике и Южной Америке в Андах, на Огненной Земле.
Стебди Andreaea обильями многократно ветвятся (рис. 121) и к суб-
страту прикрепляются нитчатыми или пластинчатыми ризоидами. Стебли и
ветви густо одеты листьями. Листья однослойные, со средней жилкой»
Антеридии и архегонии образуются яа "верхушках разных ветвей: ТГо стро-
и развитию они напоминают половые органы следующего порядка —
Bryales.
Сцорогон bq, многом напоминает спорогон сфагнума. Он состоит из ко-
робочки и ножки. В центре коробочки находится колонка и куполообраз-
ный спорангий. Колонка не доходит до верхушки коробочки. Как и у сфаг-
Гг
ГII]
ОТДЕЛ — МОХООБРАЗНЫЕ
ИЦ
пума, верхушка стебля ко времени созревания спорогона удлиняется, обра-
зуя ложную ножку.
Вскрывается коробочка очень своеобразно — четырьмя створками.
При этом споры высеиваются наружу. Створки остаются соединенными навер-
ху. Крышечки и перистома здесь нет (рис. 121, 1). Колпачок маленький,
развивается на верхушке коробочки и рено опадает.
При прорастании споры образуется пластинчатая стелющаяся протоне-
ма, хорошо приспособленная к произрастанию на камнях. Она обильно
ветвится. На протонеме возникают почки, развивающиеся далее в облист-
венные побеги.
Andreaeales занимают промежуточное положение между Sphagnales и
Bryales. Со Sphagnales их сближают строение спорогон! и наличие ложной
ножки, а с Bryales — тешняИ облик и развитие антеридиев и архегониев.
3-Й ПОРЯДОК — ЗЕЛЕНЫЕ, ИЛИ БУРЫЕ МХИ (BRYALES)
К этому наиболее обширному порядку относится свыше 135 000 видов,
всей^ Земле—от Арй-
ИГ.
тики до Антарктики, поднимаются в горы до крайних пределов раститель-
ности и наиболее обильны и разнообразный тропиках.
В сложении растительного покрова^Вемли зелёные мхи принимают
большое участие. Они образуют сплошные покровы на болотах, где многие
из них являю^явщце^шшш торфообразователя»^ (главным образом на
дат зеленых мхов, в отличие от сфагновых и андреевых, характерна
хорошо развитая^
цдаддаи В коробочке
аратл обеспечивающий
Ill
и»;
:! I
По строению вегетативных органов^ как и по устройству перистома^ зе-
леные мхи очень разнообразны.
Рассмотрим в качестве примеров, иллюстрирующих важнейшие при-
знаки растений этого порядка, два рода зеленых мхов — Polutrichum и
Funaria.
' Го д кукушкин лен (Polytrichum) тгяггептыияотЭЗ видяд произрастаю-
_ :х обоих полушарий и в тропиках.
В СССР встречается13. кукуптппна лтдяГНяибплее распространен-
ными в забелачивающихся^хвоЙных лесах и по окраинам болот явл
Р. gnmmang-H Р. На болотах по кочкам растет Р. strletum*
В сухих, сосновых лесах на песке растет маленький Р. pilifesrwn, Все
виды Polytrichwn обрелут плотные, густые дернинки.
Обычно простой, неветвистый, прямостоячий стебель обыкновенного
кукушкина льна (Р. commiw) достигает 30 — 40 см в длину. Стебель густо
покрыт.линейшьшнд<шидШ|Шиишсхьамм.*Нвер^ей и средней частях стебг
ля листья рииггпгты^ веллпыт; в шинней части стебля лис^я более
мелкие, бурые (рис. 122). Ниже стебель Дурум* (<корневшпей.
Здесь от стебля отходят многочисленные токаво-бурые многоклеточные
v Стебель Polytrichwn имеет ело
"стебля^распомжан ко
В середин©
точная
; продольные
вых клеток с тонкими сотенными по
стенки их Д ьно толстые (рис. 122, 2).
ахевдам более оргаЕпгапвяшшх высших рас-
тений. Однако морфологически~ не являются трахеидами, так_нак не
.имеют пор, обеспечивающих сквозное сообщение клеток дпугйм
За ксилемой в 1—2 слоя расположено кр^мд^ов^нм__^дугадише^
Клетки его- заполнены крахмальными зернами- Ва крахмальным вдагалий
200 .
ВЫСШИЕ РАСТЕНИЯ
Рис. 122, Кукушкин лен:
1 растение со спорогон&ми; г — аиввреявиЛ раярез стебля (яс — ксилема, ки — кратна лкww* влагали-
ще. 0 — флсама, лс — листовые следы, кор — кора, а — эпщйрмис); з — поперечный разрез листа (а —
ннидармис. ас*- ассимиляторы, лие — механические клетки); 4 — продольный разрез соорашя аитвдндиев
(ок — авдцшдой, пр — парафизы, ал — верхушечные краевые листья); 5 — продольный разрез собрания
щем расположена кольцом в нескор"" И1гп,"П" флошп Она состоит
ИЗ ЖИВЫХ удлиненных клеток С ГУСТН™ т»7<»п.му„„^тм лпддряйдпМ»,. Kjwtkm
видные трубки, которым и сОлтпетг.твутпт функционально. Они проводят
ПдастИёбййе вещества- Однако клетки эти не имеют ситонмдчых пластинок
илиндр. М JJJjytta сквозных отверстий ни на поперечных, ни на продольных
стенках.
Пуяпк окружает кора. В ней лнтпты листовые следы — ответвления от
централыюи стеЯИ, идущие в листья. Снаружи стебель покрыт эпидер-
и сост
Н ПО дрцип-»*«».
еэ весь лист.У,
На верхшУГ
щие из одного т» _ _______ ._______________
ДЁЁЯМдддвфы. ga поперечном разрезе они имеют вид столбиков из ' 3 —
Дщ££2Б, отходящих почти от каждой клетк^верхней стороны листа. 'Кдетки
ассимиляторов врдеяые, с^ддржат хлорофилловые верна.
I
ОтДют — МОХООБРАЗНЫЕ
201
Совокупность ассимиляторов резко увеличивает ассимиляционную по-
верхность л иста.
В жилке имеются ксилема, флоэма, механические клетки (рис. 122, 5).
Кукушкин лен — раст^цие двудомное. На верхушке стеблей^ обра-
зуются антеридии ^ли архегпнщь, Те~^ДРДГйр собрапы группами и окруже-
ны верхушечными ЛИСТКЯМД^
” Антеридии имеют^мешковидную^форму. Они расположены на короткой
многоклеточной ножкр.^и одеты .однослойной-СФенкой. В антеридии~~ббразу-
ется большое количество спермагенных клеток. Каждая из них дает начало
одному двужгутщшвдму сперматозоиду Г
Антеридий вскрывается в дождливую.
этом сперматозоиды выуо^яумупунцг Среди янтериппоп рягтто^^отгкт
~фйзъг — выросты стебляУ^гмеюшиевид-однорядных нитей или расшипен-
"^^Верхушечные листья мужских растений, окружающие в виде розетки
собрания антеридиев, величиной и формой отличаются от остальных листь-
ев. Они обычноишрашены в коричневый или красный цвет, 'f ’ 1
Точка роста стебля не идет'нар^аз6ваниё!антёрйдиев. и стебель в даль-
пейшем прорастает, образует новый побег (рис. 122, 2). На стебле кукушки-
на льна можно видеть нередко 5 — 6 таких розеток. Архегонии имеют
обычное строение, состоят из брюшка и длинной шейки. Листья, окружаю-
щие группы архегопиев, ничем не отличаются от обычных вегетативных
листьев.
После оплодотворения из зиготы развивается спорогон. Зрелый споро-
гон Polytrichum состоит из коробочки и длинной ножки. При помощи
гаустории спорогон внедряется в ткань верхушки стебля (гаметофита), от
которого и получает необходимые для его развития питательные вещества.
Первоначально спорогон скрыт внутри брюшка архегония, по затем
ножка сильно удлиняется и_коробочка разрывает брюшко архегония.
Верхняя часть брюшка выносится вверх в виде волосйстоШЗПлдачка
(kjML-И пт р ы), одевающего коробочку и защищающего ее .от высыхания.
Ко времеии созревания "сйор~~в""йОрбббЧКе кол5ачок сбрасывается!
Зрелая коробочка имеет сложное строение. Она состоит из средней,
расширенной части ~ урн о ч к п. расположенной у основания коробоч-
ки апофиз ы Ошейки) и крышечки (рис. 123, 2). Внутри от основания
коробочки вверх поднимается колонка — .тяж бесплодных паренхимных
клеток. Колонка в верхней части расширяется, образуя на границе крышеч-
ки и урночки тонкую пластинку — а п и ф р а г_м у (рис. 123, 2).
Вокруг колонки расположен спорангий, подвешенный на тонких ни-
тях. Эти нити прикрепляют спорангий к колонке и стенкам урночки.
В спорангии развиваются многочисленные мелкие споры. Клетки стенки ко-
робочки содержат хлорофилловые зерна. Снаружи коробочка одета эпидер-
мисом; На.границе урночки и шейки в эпидермисе находятся многочислен-
ные устьица. ОВД ймеют две замыкающие клетки и щель между ними. Та-
ким образом, сподогон у Ро/р/псТштГкаК и ^ЙЦОГИХ Др^шУ^ВЯЙньДЯ Тйхов,
имеет ассимиляционную, ткань _и устьица й питается отчасти с^мпстгш-
тельно (отличие jolr печеночников). ~ - х
Вверхней части урночйй, на гряддце у п, гртдптечкой, развивается
тъп[ образованное несколькими* рядами мелких клеток, отличаюйях^
ся _по ф»рме и размерам от соседних! Клетки колъпа" имеют неравномерно
утолщенные стенки. При подсыхании коробочки по кольцу происходит от-
деление крышечки от урночки, она сбрасывается. По опадании крышечки
П23, Л). Па хрднице
совокупность зубцов,
расположенных вокруг всей урночки* У Polytrlchum перистом однорядный.
Зубцы его на верхушке короткие, тупые. Они o6jm»rtwfl™ нисколькими диздн-
тттлигСТ- крипте ИМеЮШИМИ £ЯЛЬНО;уЗЮЛП1еННЫе ~СТёнЯИГ“
Кенпд^ти^аятщ у основания (рис. 12Э, молодой коробочке соседние"
зп зп зп
Рис. 123. Кукушкин лен:
л — продольный разрез коробочки (гр — удочка, ш — шейка, хр — крышечка, кол — колонка, сп —спо-
рангий со спорами, &лф — шифрагма, а» — зачаточный перистом); я — поперечный разрез коробочки; з—
коробочка с акмфрапюй. закрывающей вход в урночку; 4 — перистом; л — часть продольного разреза не-
зрелой коробочки (хлч — колечко, хлп — клетки перистома* апф — эпифрагма); в — часть поперечного
разреза незрелой коробочки (за — зубцы перистома)
зубцы перистома отделяются несколькими рядаьш_паренхимшг1х клеток,
котерые^дрятгпее разрушаются (рис. 123, 5, о). 'Между зубцами перистом*
име^тсдГотверстия. Ко времени созрр^я^дяд-^п^олонкДГ~сте|4ка_С1шиан^
дз№~раарушаются. Врезультате^стъдры -д^амгааштея рястплдзцд»пыми и „
ПОЛОСТИ КЦДОООФШ. ВысмпйНШгГспор ЦАРУКУ цлмлгяйт ПерИСТО|£.
ЗубцЫшУристима очень^гйгроскопичны. Во влажную^погоду они ия бу-
хают и заворачиваются,внутрь коробочки, прижимая на^ухшу>оэпифрагму ,
к стенкам урночта^р результате чего вход вХоробочку закрывается.
Вода в коробочку не попадает, что предохраняет споры от преждевре-
менного прорастания внутри коробочки.
В сухую погоду зубцы перистома теряют влагу, выпрямляются и от-
гибаются наружу. Эпифрагма ссыхается. Тогда через отверстие между эпи-
фрагмой и зубцами перистома при покачивании коробочки ветром споры пор-
циями высеиваются наружу (рис. 123, 3). Спора, попавшая на влажную
почву, прорастает, давая начало нитчатой протонеме. Протонема обильно
на поверхности земли^ ледет^ють^фугт"* нити пртгникают-в верхний слой
Так заканчивается тткл У него, как и у всех
Bryophyta, ^колениом^вллоъся^гаметофит — протонёмаг '
само листостебельное растение с развивающимися на нем антёриДИкмМ и ар^
хегониями. Спорофит в виде и ^аметоФитом.
Он развит и расчленен значительно слабее/Прп образованиихлор вроисхо^.
дит редукцИотое^Деление, и спора, таким образом,гахшоидна.
ОТДЕЛ — МОХООБРАЗНЫЕ
ИГ.
Большинство Bryales имеют гораздо более простое строение- Типичным
примером таких мхов может служить фунария.
- , Фунария (Funaria hygrometrtca} широко распространена у нас на боло-
тах, в лесах, чаще на местах костров. Растение (гаметофит) достигает 1 —
3 см высоты (рис. 124, 1).
Фунария образует небольшие светло-зеленые дернинки. Стебель ее по-
крыт спирально расположенными листьями. От стебля вниз отходят в почву
многочисленные ризоиды. В центре стебля фунарии находится прпдтгартрагй
имеют клетки с утолщенными стенками (рис. 124, 2).
Листья фунарии "однослойные на большом протяжении и только в об*
ласти средней жилкП состоят из"нёскольких слоев клеток. 'Вк летках лйста
находятся хлорофилловые зёрна. ЛйстовыМследы в стебле никогда не_дохо-
дят до центрального пучка (рис. 124, 3).
Антеридии и архегонни собраны группами на верхушках побегов и
имеют обычное строение. ' у ------------
Спорогон состоит из коробочки и ножки. Ножка на
, разно изогнута, и коробочка"фунарйи свешивается вниз,
изогнутая, несимметрична я. Аппфияя (шейка) нерезко отп _
шг. Вверху коробочка снабжена крышечкой. В середине коробочки рас-
положена массивная колонка, вверху -тюрятпдтцрст паплпррдствеиро в
ке ДУГ°°б-
ючкаг* косо
летающий к ней спорангии. Таким образом, в отличие от Polytrichum^ у
фунарии между колпятспй л-дпдрянгием нет - -
-.Стенкутгбррйо^^ с пл таг ее образует вырос-
1П
»т.
(е урночки и шейки в эпидермисе развчааютг.я усткяттЯ| спорогон фуна-
рии также способе^ частично к самостоятельному питанию.
Йй границе урночки и крышечки развиваются -клетки кольца, по кото-
рым происходит отрыв И сбряеирдтаа тертлтдики КлвТКИ КПУТПШй.
После сбрасывания крышечки на верхушке урночки-хорошо заметен пери-
стом. У фунарии перистом двойной, внутренними наружныйт состоит из двух
лряд<й^зубцов (рис. 124, 5).
Зубцы внутреннего перистома тонкие, а наружного — пченк тпд^туе,
состоят они на_ чередующихся в поперечном направлении участков, сильно
утолщенных, имеющих вид балочек, и менее толстых — в виде полосок.
Дплйпей перистоу образуется из одного ряда перистомных клеток» рас-
положенных в ирпподтдом ^направлении в верхней части корпбошгд, нахва-
тывая урночку и ^асть крышечки.ГВтот пёрйбтшшыи ряд отделен от эпидер-
миса несколькимирядами паренхимных клеток (рис. 124, S). Стенки клеток
перистомного ряда своеобразно утолщены. Наибольшее утолщение получа-
ют наружные стенки клеток и отчасти радиальные. Последние, однако, утол-
щены лишь около наружных стенок, а далее они остаются тонкими
(рис. 124, S).
Стенки клеток перистомного ряда внутри утолщены значительно мень-
ше* Несколько ниже кольца от стенки урночки к перистому протягиваются
ряды клеток с утолщенными стенками; ими перистом прочно соединен со
стенкой урночки (рис. 124, S).
стомного ряда ряярыияются в продольном направлении — по линии нёутол-
щенных участков их поперечных стенок, вследствие -чего, перистом становит-
ся двойным.
" Пёристом фунарии также сдособствувх_ра1хеивашшх спор. У нее зуб-
цы Наружном ттериглгая ллддттота тадДг^упттг пжгд д рбрЯЯУЮТ ССТОЧКУ- Зуб-
цы внутреннего перистома в сырую поводу, разбухая, располагаются между
зубцами наружного перистома и закрывают вход в коробочку*
204
ВЫСШИЕ РАСТЕНИЯ
Рис. 124. Фуяария: 4
7 —обший вид; г — поперечный разрез стебля; поперечный разрез листа; 4 — продольный разрез ко-
робочки (п — вервстом. хр — крышечка, кол — колонка, ар — археспорий, об — стенка коробочки, вп —
воздушное пространство, ш — апофиза, н —- ножка); в — участок верхней части коробочки с хвойным пе-
ристомом (ап — внутренний перистом, мп — наружный перистом); в — протонема с почками и молодым
растением); г — прорастание спор; в—часть продольного разреза незрелой коробочки (ап — эпидермис,
тых — клетки эпидермиса, лежащие над нолечксш, мкл — толстостенные клетки, сх — клбткн стенки
коробочки, ркл — утолщенные стенки клеток переставного ряда, в — воздушная полость, рхл — клетки*
впоследствии разрушающиеся, хр — археспорий)
ОТДЕЛ — МОХООБРАЗНЫЕ
205
Рис. 125. Вегетативное размно-
жение мхов:
j — выводковые почки на листе Ulo-
ta phyllautha; 9 — выводковые поч-
ки на листе Bryum erythrocarpum;
J — отдельная выводковая почка Bry-
um erythrocarpum; 4 — кучка вывод-
ковых почек на псевдоподиуме Aula-
comnium androgynum; 5 “ стебель
Tetraphis с выводковыми почками;
6 — продольный разрез верхушки
стебля Tetraphis (видны почки)
В сухую погоду они сужаются, скручиваются и споры высеиваются на-
ружу через птверстая-мдясду зубпями обоих перистомов (рис. 124, 5Y.
Прорастающая спора дает начало ветвистой протонеме, из почек кото-
рой дадер_развиваются дтистгмугебмьптло растения (рис. 124, 6). Так закан-
чивается цикл развития фунарии.
У лиственных мхов широко представлено веготятиянде размножение.
чем и пбъ^няАтся бктгтрпд яйгеление ими огодетапй почвы и произраста-
ние дернинами. Оно осуществляется различными спдсоба^. Часто прото-
нема разрывается на отдельные участки путем образования беспветныхт
разъединяющих клеток; каждый обособившийся таким образом участок*
протонемы дает начало нескольким новым растениям,
У многих мхов-на стебле, листьяхг^ризоидах может возникать ^вторич-
ная протонема в виде нити, на которой образуются почки, а затем — нЗвйе
^асТеиИЯ. У МНПГПх видов при отмирандш^нижнёй части ищииид
ветви становятся .самостоятельными растениями, на них образуются ризо-
иды, прикрепляющие растения к почве.
У многих мхов образуются ^спепияльные выводковые почки, развиваю-
щиеся^на листьях и стеблях. Стебли иногда удлиняются, образуя так назы-
ваемый псевдоподии (ложную ножку), и несут на верхушке большое коли-
чество почёк (рйЬ. 125). Почки имеют раяличную фоЬму и состоят из _1—3
или многих клеток. Опадая, выводковые почки дают начало новым расте-
ниям. ---------—
""ТИногие мхи, особенно двудомные, размножаются только вегетативно
(половое размножение у них подавлено), например abieunum,
Neckera complanata. Вегетативное размножение осуществляется путем обо-
собления молодых побегов при отмирании материнского в наземной (Mnium)
или подземной (Climatium) части.
В природе (и в опытах) известнычГИбриды мхов (межвидовые и межро-
довые), особенно в семействе Funartaceae.
Филогенетическая система недостаточно разработана.
В распространенных ныне системах, в значительной мере искусственных,
принимается же внимание форма роста мхов и особенно строение и развитие
206
ВЫСИШВ PACT
и; ।:
перистома и его зубцов. Всего выделено 82 семейства, которые в специаль-
ных бриологических работах часто группируются в порядки (тогда Bryales
рассматривается в качестве подкласса наряду со сфагновыми и андреевыми
мхами).
По Форме
мхо
хорошо выделяются две боль:
ные (Acrocarpi) и бокоплодные (Pleurocarpt).
н».
У верхоплодных, или акрокарпных, мхов архегонии, а затем и спорот
гоны развиваются на верхушке главного стебля. Стебель прямостоячий,
иеветвистый или слабоветвистый.
У бокоплодных, или плеврокарпных, мхов архегонии и спорогоны раз-
виваются на верхушке коротких боковых ветвей. Бокоплодные мхи большей
частью обильно ветвятся; стебли их часто ползучие, распластанные на суб-
страте.
Эти два подразделения не совпадают с разделением (Bryales) на группы
цо строению и развитию перистома. По строению и развитию (онтогенезу)
перистома Bryales разделяют на две довольно естественные группы:
простой, со-
штррые о’браз^тс^^лфупп клеток;"
расположенных во внутренних (нижних) слоях крышечки. Сюда относятся
Polytrichum* Pogonatum* Tetraphis и др.
2. Arthrodonteae — членисто-зибчатые мхи. Перистом состоит _из более^.
г? 11
н:
; i; I
шечки (так называемый перистомный слой, ряд). 3убцы расположены или
в 1 ряд (простой перистом) или п 9 рстда (двойной перистом, состояиЩГиз
i ur.
I: l
клеток, лежащих перед наружным перистомом. Обычно он рано исчезает
(есть у Grimmla* Orthotrichum^ рис. 126).
Arthrodonteae разделят на две подгруппы мхов — гаплолепидные и
диплолепидные.
1. Гаплолепидные мхи (Haplolepidae). Перистом простой. Наружный
слой зубцов состоит из одного ряда клеток; внутренний из 2 рядов клеток,
соединяющихся зигзагообразной линией посредине зубца (стенками сопри-
касающихся клеток). Перистомный ряд состоит из двух слоев клеток, К га-
плолепидным мхам принадлежат некоторые акрокарпные мхи: род дик-
ранум (Dicranum)* например широко распространенные в лесах Dicranum
undulatum* D. scoparium* растущие на почве у основания деревьев; виды
родов Pottia* Tor tula* Barbulauypyrue мелкие мхи, растущие плотными
дерновинками на глинистой почве, по обочинам дорог, на старых стенах,
крышах. Зубцы перистома, расчлененные от основания на многочисленные
нитевидные доли, спирально закрученные, обычно влево (рис. 426, 2).
2. Диплолепидные мхи (Diplolepideae). Перистом двойной. Зубцы внеш-
него перистома состоят снаружи из двух рядов клеток, соединенных зигзаго-
образной линией, а с внутренней стороны — из одного ряда клеток. Перис-
томный ряд состоит из четырех слоев клеток.
Зубцы наружного перистома, так же как и зубцы простого перистома,
различны по форме, но строго постоянны у разных родов и видов — линейные,
ланцетные, реже нитевидные, цельные или расщепленные, с утолщениями
в виде поперечных перегородок, пластиночек. Наружная сторона зубцов
сильнее утолщена, чем внутренняя. Зубцы или свободные, или чаще, от
основания на некотором протяжении сросшиеся, образующие так называе-
мую основную перепонку, — темно-красные, бурые, желтые, грубоватого
строения. Внутренний перистом обычно менее развит, нежной структуры
(иногда недоразвит), бесцветный или оранжевого цвета, гладкий или папил-
лозный (с сосочками). Он свободно прилегает к наружному, нричем зубцы
его противостоят зубцам наружного перистома или чередуются с ними.
Образование гаплолепидного и диплолепидного перистома членистозуб*
ОТДЕЛ — МОХООБРАЗНЫЕ
чатых мхов ясно из схематического рисунка (рис* 126, £). К диплолепидным
мхам относятся как акрокарпные, так и плеврокарпные мхи.
Из акрокарпных можно указать Funaria hygrometrica, которая подробно
рассмотрена выше (стр* 203); род Milium, многие виды которого растут
в наших лиственных лесах на влажной почве (М. undulatum — с характер-
ными широкими волнистыми листьями, М. cuspidatum — у основания де-
ревьев, М, affine — на болотах); род Вгуит — с многочисленными видами,
растущими на болотах (В. ventricosum), по краям дорог, на старых крышах
(В. caespeiicium, В. argenteum).
К плеврокарпным диплолепидным мхам принадлежат климадии (СНта-
Нит dendroides), широко распространенный на сырых лугах, болотах,
древовидный, довольно крупный мох Neckera complanata, обитающая на
стволах лиственных деревьев; род туидиум (Thuidium): Thuidium abieti-
пит встречается по солнечным склонам, опушкам, Th. recognition на сырых
лугах, лесных полянах; род брахитециум (Brachythecium), представлен-
ный рядом видов, обитающих на пнях, стволах деревьев, болотистых лугах.
К плеврокарпным диплолепидным мхам относятся также лесные напоч-
венные (крупные)
in?
Ptilium crista castrensis,
мхи — Pleurozium Schreberi,
Рис. 126. Строение перистома мхов:
1 — иредпернстоы ортоерихумк; е—спирально еанинА шфшяом тортушс *—строение норобоянн и пе-
гастима гапжшепндных и дмплолепидных мхов (схемы) э а — гавпопйщдннвнкж б — ливлошшндные мхи
(мс — наружные слои зубцов иростого пврвстоаж пшшепнднык н зуОцов наружного перистома днпполе-
парных мхов, «с — внутренние слои, «мм — ямутрешшй перистом, кот нононт, пкор — полость норсь
<кяки)
208
ВЫСШИЕ РАСТЕНИЯ
Hylocomium splendens. Эти мхи нередко образуют сплошной ковер в хвой-
ных лесах, определяя состав и название типов леса — ельники (сосняки или
боры)-зеленомошники. Ранее их относили к роду Нурпит и называли
гипновымн мхами.
ПРОИСХОЖДЕНИЕ И РАЗВИТИЕ МОХООБРАЗНЫХ
Мохообразные — древняя группа наземных растений.
Наиболее важные материалы для суждений о происхождении и родст-
венных отношениях между группами растений дает изучение их ископае-
мых остатков (данные палеоботаники). К сожалению, при тонкости струк-
туры мохообразных и их плохой сохранности в ископаемом состоянии палео-
ботанические находки очень скупы и явно недостаточны для суждения об
их филогенезе. Поэтому в вопросе о происхождении и эволюции мохооб-
разных приходится руководствоваться в значительной мере материалами,
добытыми сравнительным изучением ныне живущих их представителей (мор-
фологии, эмбриологии, цитологии, географического распространения и др.).
Эти данные не являются бесспорными. В результате у ботаников нет единст-
ва взглядов на происхождение и эволюцию мохообразных; наоборот, в этом
вопросе существует много часто исключающих друг друга, диаметрально
противоположных точек зрения. Окончательное разрешение этих вопро-
сов — дело будущего. Бесспорные остатки вегетативных органов мохообраз-
ных известны из верхнекаменноугольных отложений. Отсюда известны как
остатки печеночников, именно талломных форм, сходных с Marchantiaceae,
так и листостебельных мхов, сходных с Polytrichaceae.
В последнее время, однако, споры мохообразных (наряду со спорами
папоротникообразных) встречены в отложениях силура и девона на терри-
тории СССР, а споры явно сухопутных растений найдены даже в кембрий-
ских и докембрийских отложениях Прибалтики.
Из пермских отложений недавно (1957) описано 4 вида зеленых мхов
и новый род Protosphagnum представители которого отличаются от сфаг-
новых мхов наличием жилки в листе. Всего из палеозойских отложений
известно 7 видов печено*
|:г,1
сов и 9 видов листостебельных мхов. Для мезо-
зоя известны 13 видов печеночников, 1 род зеленых мхов и 1 вид Sphagnum.
Обильно представлены мохообразные в кайнозое. Для третичного пери-
ода зарегистрировано 34 вида печеночников и 105 листостебельных мхов.
Они почти неотличимы от современных.
В третичное время впервые появляются листрстебельные печеночники.
Таким образом, современные палеоботанические материалы свидетельству-
ют о том, что:
1) мохообразные — очень древняя и обособленная группа растений.
Известные нам ископаемые мохообразные уже высокоорганизованные рас-
тения, и, следовательно, они прошли длинный путь эволюции (по мень-
шей мере силурийского возраста);
2) оба класса мохообразных — Hepattcopsida и Bryopsida — обособи-
«I
лись очень давно, вероятно в силуре или даже еще раньше;
3) среди печеночников слоевищные (талломные) формы появились рань-
ше листостебельных.. Последние известны лишь из неогена;
4) сфагновые мхи являются весьма древней группой растений, несом-
ненно палеозойского возраста.
J Однако непосредственных предков мохообразных палеоботаника нам
пока не открывает. Все имеющиеся данные по строению ныне живущих и
искдпаемых Bryophyta^ с несомненностью, свидетельствуют о тесных свя-
i.€
зях между отдельными порядками и классами и о единстве происхождения
всех мохообразных. Однако эти связи не настолько близки, чтобы увязы-
вать подразделения мохообразных в один ряд развития и расположить их
в линейном порядке и системе.
отдал — МОХООБРАЗНЫЕ
209
В таком крупном подразделении, как отдел, развитие шло, конечно,
в различных направлениях, имеются параллельные ряды, выработка сход-
ных признаков в разных эволюционных рядах. Учитывая все имеющиеся
данные, происхождение и эволюцию мохообразных можно представить в
следующем виде.
Мохообразные—гаплоидная (гаметофитная) линия эволюции—очень древ
няя группа высших растений. Она, как и другая, диплоидная (спорофит-
ная) линия (Pteridophyta — Gymnospermatophyta — Angiospermatophyta),
произошли от каких-то древних водорослей. Эти водоросли обладали сте-
лющимися по субстрату талломами с паренхиматическим строением, были
дорзовентральны, дихотомически ветвились. У них была ясно выражена
смена поколений, причем гаметофит и спорофит были развиты одинаково
(изоморфная смена поколений). На гаметофите развивались многоклеточ-
ные гаметангии, подобные тем, которые теперь имеются у некоторых бурых
водорослей, а на спорофите — спорангии (возможно, тетраспорангии). Та-
кие водоросли перешли к жизни на суше. Их многоклеточные гаметангии
превратились в антеридии и архегонии.
Ошгпрптвпрпидд гтядл плутттесталяться внутри яртдглшхг, й та R
как это было у —^ц^рпфтрАб^яующийся из оплодотворенной
г
I
□шврехЗДЦГК Цолупаразитическо
фита, с которым шг^шаяя;
ги-
годайт! вываливался наруж
шпрпфитл птдлл1Лгр<ГКттпряш
Это-ведол ре-
веледСТЬшГ мшпчшшшш—редукции
шмх^гдпвжях тппГювалп .ла-
слитного спорангия стерпли-
стенку лшррруода^ В дальнейшем
спорофит совершенствовался. В нижней части спорофита дифференцирова-
лась гаустория, внедряющаяся в ткань гаметофита. Вырабатывались при-
способления к наилучшему рассеиванию спор в наземных условиях (раз-
витие ножки, выносящей коробочку со спорами над растением, образова-
иг.
Ill
ние пружинок, дифференциация крышечки, возникновение и совершен-
ствование перистома и т. д.). Гаметофит, первоначально пластинчатый, тал-
ломный, развиваясь в воздушной среде, обеспечивая, помимо полового
процесса, и развитие спорогона, также совершенствовался. Он развивал боль-
шую ассимиляционную поверхность. Постепенно вырабатывалась листо-
стебельная структура мохообразных, и у наиболее сложных форм (как, на-
пример, у Poly trichum среди лиственных мхов), достигших наибольших
размеров и растущих в сравнительно духих условиях, появилось подобие
столярного строения. Однако основной фушщпей гаметофита яялпотея-обес-
печение полового процесса, который осуществляется врц пттпт
ных ^дп&рмагО^дВдов. перемещающихся в каПеЛьнб-жидкой среде1. Это опре-
деляло общие возможности гаметофита в'зашюваййи суши. Мохообразные
были ограничены более увлажненными местами, в то время как другая груп-
па высших растений, с преобладанием в цикле развития спорофита (Pteri-
dophyta — Angiospermatophyta), оказалась более приспособленной к жиз-
ни в наземных условиях и вскоре завоевала сушу.
г
1 Существует и другая точка зрения на вопрос о связи спорофита с гаметофитом у
мохообразных, развивавшаяся Д. К. 3 е р о в ы м. Он считал, что в группе зеленых водоро-
слей, кроме чередования спорофита и гаметофита, уже в водной среде существовал тип
смены поколений, при котором спорофит развивался на гаметофите и никогда не был са-
мостоятельным, морфологически подобным гаметофиту поколением. По мнению Д. К« Зе-
рова, спорофит мхов происходит от спорофита типа цистокарпия красных водорослей
[Phyllophora, Liagora), Для Pteridophyta он принимает происхождение их спорофита от
водорослей, имевших независимые спорофит и гаметофит. Однако гетероморфное чередо-
вание поколений у водорослей встречается только в тех группах, где его происхождение от
изоморфного цикла несомненно. Это относится и к карпоспорофиту красных водорослей
(см. раздел «Водоросли»),
210
выс
in;
[В РАСТЕНИЯ
Таким образом, развитие мохообразных шло от талломных форм к листо-
стебельным. Трудно представить образование листостебельного строения
из талломов водорослей, минуя талломную фазу развития высших растений»
Какие водоросли могли дать начало мохообразным? Как указывалось
формы, давно достигшие сложного морфологического Строения — типа
современных Dictyota^ Laminaria и др. Только у бурых водорослей имеются
многоклеточные гаметангии. Однако бурая окраска и иное устройство клет-
ки у бурых водорослей затрудняет выведение из них мохообразных. Поэто-
му другие ученые выводят мохообразные из зеленых __ водорослей.
Действительно, равножгутиковые зеленые водоросййГ имеют с мохо-
образными общий состав пигментов (хлорофилл, ксантофилл, каротин),
в качестве запасных веществ у них образуется крахмал. Очень сходны они
по строению клетки, составу клеточных оболочек. Наконец, ультраструкту-
ра жгутиков сперматозоидов тех и других также очень сходна. Мы не. знаем
зеленых водорослей (как среди современных, так и среди ископаемых), обла-
дающих морфологически сложным, расчлененным талломом, у которых
имелись бы многоклеточные гаметангии, характерные для бурых водорос-
лей, но К. И. Мейером еще в 1930 г. для зеленой водоросли хетонемы
(Chae tonema) были описаны многокамерные антеридии. Кроме того у харо-
вых имеются многокамерные половые органы своеобразного строения. Слож-
ное расчленение тела этих водорослей свидетельствует о том, что зеленые
водоросли как бы искали пути к образованию структуры, свойственной
наземным растениям. Сторонники происхождения мохообразных из зеле-
ных водорослей считают, что среди последних ранее были формы, обладав-
е многоклеточными гаметангиями. В этом случае мохообразные выводят
из зеленых водорослей типа Chaetophorales^ у которых наблюдается гето-
ротрихозная структура талломов. Нижняя часть их ^ела стелется по суб-
страту, а от нее вверх отходят вертикальные нити, состоящие из клеток,
HI.
1ЛГ
Гилотетичадкие предки мохообразных строятся и изображаются в виде
паренхиматической пластиночки, несущей на верхней поверхности много^
клеточныД гаметангвд^ У них, таким образом, в сравнении с хетофоровыми
развивалась стелющаяся часть, а вертикальная часть редуцировалась и
представлена только"- половыми органами. Такие гипотетические гетеро-
трихозные зеленые водоросли, выйдя на сушу, могли в дальнейшем разви-
ваться описанным выше способом (превращение гаметангиев в антеридии и
архегонни, развитие защитного слоя на талломе и пр.1).
0ба класса мохообразных — Hepaticopsida и Bryopsida отошли от об-
щего ствола развития высших растений очень рано.
Эволюция печеночников (Hepaticopsida). Больше всего примитивных
признаков сохранили гаметофиты антоцеротовых. Талломы их имеют еще
ряд водорослевых черт (хроматофоры с пиреноидами, образование выводко-
вых почек и др.). Однако спорофит антоцеротовых, наоборот, построен
очень сложно. В строении спорогона имеется ряд признаков, общих с лист-
венными мхами (строение спорангия Noiothylas яшмт&г строение его у
Sphagnum), но, как и у всех печеночников, здесь имеются споры и пружин-
ки. Спорогон обладает способностью нарастать благодаря наличию меристе-
матической ткани.
Антоцеротовые стоят несколько особняком в системе Bryophyta и долж-
ны выводиться самостоятельно из водорослей. Порядок сферокарповых
(Sphaerocarpales) характеризуется талломами простого строения. Предста-
вители этого порядка имеют признаки, характерные как для маршанциевых,
1 Некоторые ученые считают возможным вести происхождение гаплоидной я дипло-
идной линий от разных водорослей. Мохообразные в этом случае выводят от водорослей
талломного ха _ " '
вое развитие гаметофитаи спорофита), еще в водной среде'выработавших радиал
пае, что имеется у современных бурых водорослей типа Fucus, Laminaria и др.
it,
JvflEI
ОТДЕЛ — МОХООБРАЗНЫЕ
Ml
так и для анакрогинных
юнгерма
:i:rr
(стр. 186), и могут быть использованы
как звено, связующее оба эти порядка.
Сами сферокарповые упираются в сво-
ем происхождении в водорослевых
предков. От сферокарповых могли
взять начало порядки маршанциевых
и юнгерманниевых. Маршанциевые
разв ивались, усложняя внутреннее
строение при сохранении талломной
структуры. Юнгерманниевые при со-
хранении простого анатомического
строения достигли сложного внешнего
расчленения, выработав листостебель-
Рве. 127. Выработка листостебель-
ной структуры у талломного анак-
ротного печеночника:
ное строение. Среди маршанциевых
наиболее сложные формы имеют воз-
душные камеры с ассимиляторами,
устьица, простые и язычковые ризои-
ды, сложные подставки {Marchantia^
Conocephalum и ДР-)- От них развива-
лись более упрощенные формы (напри-
мер, Riccia* Ricciocarpus)^ утерявшие
воздушные камеры. В коробочках у
них развиваются только споры, пру-
жинок не образуется. Среди юнгерман-
1 ~ Aneura bogotensifi. Дихотомически раз- •
ветвленный надлом принимает вид плоских
листьев, в то время нам вижипе участки,
сгруженные в субстрат. приобретают вид
коряеиища; я—Aneiira erlocaulu- Расчле-
веявв таллома пошло дальше. Выработа-
лись стебель (главная цнпиндЕтесдаи
ось), листья (плоские ветви третьего по-
рядка), «корни* (нижние тонкие вели,
прикрепляющие таллом к субстрату).
более
простыми
анакрогинные юнгерманнии, у которых преобладают талломные растения,
имеются и переходные к листостебельным формы (A naira, Fostombronia,
рис. 127). Возникновение листьев среди анакрогинных юнгерманнии про-
исходило, вероятно, не раз и в разных эволюционных рядах. Возможно,
I: II
среди них есть и редуцированные формы, где наблюдаются и вторичные
упрощения в строении вегетативного тела. Через олиственные формы обна-
руживаются связи анакрогинных юнгерманнии с акрогинными — всегда
олиственными печеночниками, особенно интересен в этом отношении под-
; г. ।'
порядок Haplomitriales. У его представителей появляются вертикально
стоящие олиственные побеги, причем Haplamitrium является еще анакро-
гинным, a Calobryum уже акрогинным печеночникам.
Акрогинные юнгерманнии насчитывают наибольшее число видов,
они особенно распространены в природе и появились, как свидетельствуют
палеоботанические данные, позже талломных.
* Эволюция лиственных мхов (Bryopsida, Mhsci) - Родственные связи меж-
ду отдельными порядками лиственных мхов, как и связи их с печеночника-
ми, еще недостаточно ясны. Из трех порядков Sphagnalest Andreaeles и Вгуа-
les — первые два (сфагновые и андреевые мхи) более примитивны и обна-
руживают явные черты сходства с олиственными юигермат
; г,
ями (однослой-
ные, без жилки листья у Sphagnum строение и вскрывание антеридиев у
Andreaea^ наличие у обоих порядков пластинчатой протонемы и др.). Оба
порядка объединяют также неполная колонка, отсутствие перистома и
хорошо выраженной ножки коробочки.
Порядок Bryales имеет более сложное строение спорофита и гамето-
фита.
Спорогон зеленых мхов значительно более дифференцирован. Он обла-
дает прекрасно развитым перистомом, имеющим очень разнообразное стро-
ение у разных представителей. Спорогоны с хорошо развитой ножкой. Стеб-
ли их имеют проводящий пучок, у наиболее сложных форм (Polytrtchum)
напоминающий стелярное строение примитивных папоротник о образ»
ных. Вместе с тем по строению антеридиев и архегониев, как и по внешне-
му облику, Andreaeales сходны с Bryales. Все это свидетельствует о тесных
212
выс
тш
В РАСТЕНИЯ
связях между Sphagnales и Andreaeales, с одной стороны, и Bryales —
с другой. Вероятно, они произошли от общего предка. В своем происхожде-
нии лиственные мхи могут быть увязаны с юнгерманниевыми, но более при-
митивные Sphagnales и Andreaeales обнаруживают отдаленные связи и с
Anthocerotales (неполная колонка, куполообразный спорангий и некото-
рые другие признаки). Разрешение этого вопроса требует накопления но-
вых данных, в частности, по эмбриологии лиственных мхов.
В последнее время некоторые ботаники (в СССР Б. М. Козо-Полян-
ский и особенно А. Л. Т ах тадж ян) предлагают рассматривать Bryophyte
в качестве вторичной, редуцированной группы растений и выводят их из
псилофитов типа риниевых.
Развитие мохообразных они представляют шедшим в направлении от
листостебельиых форм к талломным в связи с развитием этой группы расте-
ний в условиях повышенного увлажнения. У простейших мохообразных,
с этой точки зрения, спорофиты были более совершенные, чем у ныне суще-
ствующих мхов; гаметофиты их имели радиально симметричное листосте-
бельное строение. От таких форм путем постепенной редукции образовались
все остальные мохообразные. Среди печеночников радиально постро-
енные листостебельные Haplomitriales являются более древними, а дорзовеи-
тральные — более молодыми. Затем сформировались талломные юнгермаи-
нии и, наконец, маршанциевые.
Среди Bryophyta наиболее совершенными являются талломные формы.
Эта точка зрения опирается на сходство в строении спорангиев некото-
рых ^риниевых, именно Horneophyton^ со спорогонами сфагновых и андрее-
вых мхов. У мохообразных (Bryophyta) в связи с переходом в увлажненные
местообитания редуцировались проводящая система и покровная ткань,
исчезли устьица и т. д.
С этими взглядами, однако, трудно согласиться, так как принятие в
качестве исходного для мохообразных сложноустроеиного гаметофита типа
кукушкина льна мало обосновано — неизвестно твердо, каковы были гаме-
тофиты у псилофитовых.
Чрезвычайно интересным кажется открытие среди псилофитовых тал-
ломных форм.
Невозможно принять, что целый отдел растений Bryophyta^ насчитываю-
щий почти 23
видов, является результатом регресса. Эволюция расте-
ний (как и животных) — процесс прогрессивного развития. Однако это не
исключает в отдельных звеньях регрессивного развития. Регрессивное раз-
витие имеется во всех группах растительного мира (например, паразитные
и сапрофитные семейства среди цветковых — Cuscutaceae, Rafflesiaceae
и др. — имеют сильно упрощенное морфологическое и анатомическое стро-
ение, водные высшие растения претерпевают сильную редукцию проводя-
щих, механических тканей и т. д.). Оно известно и у мохообразных. Так,
например, в различных эволюционных рядах печеночников известно раз-
витие талломных форм из о лиственных. Но это не является основным на-
правлением эволюции их, по существу прогрессивной и шедшей от таллом-
ных форм к ол ист венным.
Огромное количество видов, родов и семейств среди акрогинных —
олиственных юнгерманний — свидетельствует об их молодости. Они нахо-
дятся в процессе энергичного видообразования. Да и непосредственные
данные палеоботаники свидетельствуют о том же — мезозойские и палео-
зойские печеночники принадлежат к талломным — анакрогинным юнгер-
манииевым, а олиственные печеночники появляются позже.
В развитии спорофита мохообразных совершенно ясно обнаруживает-
ся прогрессивное направление по линии усовершенствования аппарата рас-
сеивания спор (споры и пру ж
и у печено1
ков, сложный перистом у лист-
венных мхов и др.). Такого развития спорофита в других группах высших
растений мы не знаем. Взгляд на мхи как на редуцированные формы труд-
\но принять в отношении гаметофита. Гаметофит мохообразных, несомненно,
।: г,
ОТДЕЛ — МОХООБРАЗНЫЕ
213
4
прогрессировал, постепенно вырабатывалось его сложное расчленение,
увеличение поверхности, что было необходимо для лучшего обслуживания
спорофита. Так как гаметофит мхов эволюционировал в условиях воздуш-
ной среды, он приобрел ряд признаков, характерных и для диплоидной
линии развития (листостебельное строение, зачаточную стель у наиболее
организованных форм и т. д.). Ведь именно в таком направлении шла эво-
люция спорофита в ряде Pteridophyta — Angtospermatophyta^ взявшем на-
чало от безлистных форм теломного строения псилофитов (см. ниже).
Такое конвергентное развитие — постоянная черта в эволюции выс-
ших растений (например, образование сосудов у покрытосеменных и у гне-
талиевых среди Gymnospermatophyta, образование семян у Pteropsida и
Lycopsida, и т. д.).
Система мхов до сих пор основывалась на строении спорофита (споро-
гона) и главным образом перистома. Гаметофит использовался для постро-
ения системы слабо и еще мало изучен, особенно на ранних этапах его раз-
вития. В настоящее время ботаники все в большем объеме используют эти
данные. Построение системы мохообразных, основанной на гаметофите,
нужно думать, прольет больший свет на их эволюцию и происхождение.
Г. Г,
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ МОХООБРАЗНЫХ
. П1
11:1
Г.1
Практическое значение печеночников ничтожно- Скот их не поедает.
Они имеют значение лишь как пионеры заселения почв, на лесных гарях,
на голых скалах. Лиственные мхи, особенно сфагновые, имеют важное зна-
чение для человека- Они покрывают сплошным ковром болота и по отмира-
нии дают начало залежам торфа, который широко используется в народном
хозяйстве.
Сфагновые торфы малозольны, так как образуются на бедных минераль-
ными веществами верховых болотах, водный режим которых в значительной
мере обусловлен атмосферными осадками. Сфагновый торф употребляется
прежде всего в качестве топлива. Многие электростанции целиком работают
на торфе.
По запасам торфа СССР стоит на первом месте в мире. В нашей стране
ведется широкая механизированная добыча торфа, объем которой с каждым
годом увеличивается.
Широко применяют различные способы облагораживания торфяного
топлива, увеличивающие его калорийность и рентабельность (торфобрикеты,
кокс, газы). Все шире развивается сухая перегонка торфа. При этом полу-
чают ценнейшие продукты: воск, парафин, фенолы, карболовую кислоту,
сахарин, спирт (до 120 л на 1 т сухого торфа).
Широкое применение благодаря плохой' теплопроводности торф нашел
и в строительстве, где его применяют для тепловой изоляции стен чердачных
и междуэтажных перекрытии, для изоляции холодильников и т. п.
Из торфа изготовляют фасонные изоляционные изделия, предназначен-
ные для трубопроводов и других устройств, а также бумагу и картон.
Большая влагоемкостъ обеспечивает сфагновому торфу широкое и разно-
образное применение в сельском хозяйстве. Его используют в качестве под-
стилки на животноводческих фермах, при изготовлении торфоперегнойных
горшков, мульчи, а также как консервирующий и упаковочный материал
(хранение и перевозка фруктов, овощей, мяса и т. д.).
Торф широко применяют с лечебной целью как один из видов грязе-
лечения (торфотерапия) на курортах в Кашине (Калининская область),
Миргороде (Полтавская область) и в других местах, а также в больницах,
поликлиниках и т. д. Используют торф при хронических заболеваниях
органов движения, хронических воспалительных процессах брюшной по-
лости и при других заболеваниях.
214
ВЫСШИЕ РАСТЕНИЯ
। it
Благодаря выделению сфагнумом кислот он является хорошим бактери-
цидным материалом. В период Великой Отечественной войны сфагнум ши-
роко использовался как заменитель ваты.
Многие зеленые мхи произрастают сплошными зарослями главным об-
разом на низинных болотах, где образуют залежи низинного торфа, богатого
золой, так как низинные болота обводняются грунтовыми водами, обычно
сильно минерализованными.
Низинный торф имеет огромное значение как удобрение, и его широко
используют в сельском хозяйстве. Торф вывозят на поля, где он улучшает
структуру почвы и обогащает ее органическими веществами (заменяя навоз).
Мхи имеют и отрицательное значение. Поселяясь на лугах, в лесах, они
сплошным плотным ковром одевают почву, затрудняют* аэрацию. Это ведет
к закисанию почв, что влечет за собой выпадение из травостоя ценных кор-
мовых трав, а затем и к развитию болота. На лугах приходится производить
сдирание мохового покрова.
in
ВЫСШИЕ РАСТЕНИЯ
С ПРЕОБЛАДАНИЕМ В ЦИКЛЕ
РАЗВИТИЯ СПОРОФИТА
ОТДЕЛ— ПАПОРОТНИКООБРАЗНЫЕ ( PTERIDOPHYTA)
У папоротникообразных, как и у всех высших растений, имеется смена
поколений, но, в отличие от мохообразных, у них в цикле развитияjpeogjra-
ДЗЭТ-/ШОррфдрт Г^дтофит развит значительно слабее спорофита. Называемый
заррстком^ей^представлеийу^йде слоевища (таллома), не расчлененного
настеЬбльи листья. В типичномгслучагтш плоский, пластинчатый (у боль-
шинства папоротников), реже нитевидный, но у ряда представителей пост-
роен радиально-симметрично (см. плауновые, псилотовые). Как правило,
заростки у папоротникообразных зеленые, питающиеся самостоятельно,
но у ряда представителей они бесцветны и, вступая в сожительство с гри-
бами, питаются гетеротрофно (у многих Lycopodium, у Psilotales, Ophioglos-
sales). У ископаемых представителей (у всего класса Psilophytopsida), а так-
же у вымерших представителей других классов заростки неизвестны.
На заростках развиваются половые органы — антеридии и архегонии.
Строение антеридиев сходно с таковым у мохообразных, но несколько упро-
щенное и различное у разных классов. Сперматозоиды у пацоротцикообраз-
ных подвижны, обладают жгутиками, бывают двух типов — двужгутико-
вые (у Lycopsida, кроме Isoetes) или многожгутиковые'(у Sphenopstda, Ptero-
psida). Архегония Р tert dophyta также сходны с архегониями мохообразных,
но, в отличие от них, в той или иной степени погружены брюшком в ткань
заростка и только шейка возвышается над его поверхностью.
В пределах тина можно проследить редукцию антеридиев и архегониев
в связи с общей редукцией полового поколения, особенно ясно выраженную
у разноспоровых форм (см. ниже).
х— Заростки ржщамотся р _ достаточно. увлажненных (хотя бы временно)
' местообитаниях (тенистые"леса, под землей — у плауновых, у ужовнико-
вых), так как оплодотворение возможно лишь при наличии капельно-жидкой
среды и осуществляется в дождливую погоду.
Гаметофиты (заросткц!^ папоротникообразных У боль-
шинства пред^йэдтЩшй онв-Лтаут несколько недель, и-^толью^у плаунов
и у некоторых мараттиевых. папоротников заростки существуют от двух
до нескольких лет.
Из оплодотворенной яйцеклетки развивает^я^споооФит. Сначала он
.лредставлен зя_ " _ _ _
обычно имеет Ложку (стопу), зачаточный Корешок” стебелек и первый" лист.
родышем. У современных папоротникообразных зародый£
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
215
’.HI
III
III \
:Н1 ’
При помощи ножки, внедряющейся в ткань заростка, зародыш получает
необходимые для его развития питательные вещества* Сформировавшись,
корешок зародыша внедряется в землю, на нем развиваются боковые кореш-
ки* Стебелек выпрямляется, и проросток переходит к самостоятельному
существованию. Некоторое время он, однако, еще связан с заростком
(рис. 169)* Спорофит современных папоротникообразных имеет очень слож-
ное строение* Он всегда расчленен на стебель и листья* Корень у Pleridp-
г-г^уптат? ладзамиых^щаажйр тела псилофито-
вых^.— р изо м <иья
Корней нет и не было у современных PsUotopslda. Их не было и у псило-
фитов* У Salvirda и некоторых гименофилловых из Pteropsida корни отсут-
ствуют вторично, в связи с условиями жизни.
Стебли и листья одеты ЧГТУТ™*™ с хорошо развитыми устьицами.
Стебли у большинства современных представителей травянистые, но у мно-
гих вымерших папоротникообразных и у небольшого числа видов современ-
ных древовидных папоротников они деревянистые* У большинства предста-
вителей LycopsidanSphenopsidaлистья простые, мелкие^ но у Pteropsida они
крупные и часто сложные.
Внутреннее строение Pteridophyta тень сложное. В стеблях, листьях
и корнях имеются выс^кооргадазоваадсые ^ проводящие ткани* Впервые
в растительном мире приобретают типичное строение проводящие пучки*
Они обычно концентрические, но имеются коллатеральные и др. Ксилема
у большинства форм представлена трахеидами, но у некоторых представи-
телей (в корневищах папоротника орляка, в ткани узлов стебля у хвощей)
. можно обнаружить сосуды (трахеи). У папоротникообразных имеются все
' типы стели — от протостела до сложноустроенной э у с т е л и
у папоротника одоунды (Os/nunda).
Учение о стели, включая и эволюцию стелярного строения растений,
было создано именно при изучении ископаемых и ныне живущих папоротни-
кообразных.
Флозма представлена ситовидными трубками разнообразного строения.
В пределах тишПГОЯШГ проследить эволюцию ^ксилемы и флоэмы, достигших
у ряда представителей весьма высокой организации*
ооб разных путай обычно закрытые, не
имеют камбия, растения не способны к вторичному "утолщению (камбий
имеется только у Jsoetes и некоторых ужовниковых)* Ископаемые хвощовые,
плауновые и папоротники нередко имели камбий и обладали вторичным
ростом.
Спорангии со спорами развиваются обычно на листьях на верхней их
стороне у Lycopsida и на нижней стороне листа у хвощей и папоротников*
Однако у всех ископаемых псилофитов, а также у ископаемых простейших
овых, не имевших листьев, спо-
рангии располагались на конечных разветвлен
i: Г1
II.
И i
JT
и* »’
I I
: и:
торых Pteropsida спорофиллы резко отличаются от вегетативных листьев»
называемых трофофиллами (питающими листьями), например у
алраусопера (Onoclea struthiopteris).
Спорофиллы расположены поодиночке (у большинства папоротников)
или собраны в колоски различного строения (у плаунов, хвощей). Иногда
листья состоят из спороносной и бесплодной части (у ужовниковых)* Уст-
ройство спорофиллов и их собраний (колосков) очень разнообразно у разных
групп Pteridophyta.
слой (тапетум), образованный одним или несколькими рядами клеток,
военикажнцимн из наружного слоя археспория — спорогенной ткани, запол-
няющей спорангий. Содержимое клеток тапетума расходуется на питание-
развивающихся спор.
216
nr
ВЫСШИЕ РАСТЕНИЯ
! i.
in
В молодом состо археспориальные клетки^ делясь неоднократно,
образуют материнские4? клетки спор. Эти последние делятся редукционно,
образуя каждая четверку (тетраду) гаплоидных спор.
Зрелая спора окружена ддумя^^оболочками: наружной — экзоспорием
и внутренней — эндоспорием. ДГ хвощей развивается еще внешняя оболдйка—
аттчГгРориУу кодорый-*• обретётся из так называемого периплазмо-
дия — чй^расплывшихся клеток тапетума, содержимое которых оседает
на экзоспории снаружи. При созревании спор эписпорий распадается на две
спиральные ленты — элатеры, пружинки.
Спорангии обычно возникают из группы клеток листа — эпидермальных
и нескбльких рядов подэпидермальных клеток, но _______оспорангиатйых
папоротников они возникают из одной поверхност “ етки. Более древним
является первый способ возникновения спорангия? Развитие спорангия
папоротников понятно из рисунка 168. Строение спор, их форма, величина,
характер экзоспория — его структура и рисунок — имеют важное системати-
ческое значение. Споры Ptertdophyta^ сохраняясь в ископаемом состоянии,
имеют большое значение для определения возраста древних отложений
(стратиграфия земных слоев) в геологии.
Спорангии имеют различные приспособления, обеспечивающие вскры-
вание их и рассеивание спор. Освободившаяся из спорангии спора, попав
в благоприятные условия, прорастает, образуя заросток.
Среди Pteridophyia имеются как рдиноспоровые, так и разносторонне
растения. У равноспоровых папоротникообразных все Споры одинаковые.
При прорастании спор этих растений образуются обоеполые заростки
(гаметофиты), несущие антеридии и архегонии. У предста-
вителей развитие спор в спорангии вначале идет одинаково, но затем, после
образования тетрад, в микроспорангиях^все^споры (микроспоры) достигают
окончательного развития, а в мегаспорангиях. лишь несколько спор (часто
всего 4 или даже 1) сильно разрастаются и становятся очень крупными
(мегаспоры), все же другие споры отмирают.
При прорастании спор образуются раздельнополые заростки. Микро-
споры дают начало мужским заросткам с^антеридиями, а мегаспоры — жен-
ским заросткам с архегониями.
Папоротникообразные в своей жизни тесно связаны с достаточно увлаж-
ненными территориями.
Современные Pteridophyta распространены по всей Земле и представ-
лены довольно значительным числом видов (особенно папоротники), нередко
принимают заметное участие в сложении растительного покрова, например
папоротники и плауны в наших лесах, древовидные папоротники в тропиках
и субтропиках. Однако они в этом отношении уступают голосеменным и осо-
бенно покрытосеменным растениям, господствующим на Земле в современ-
ную геологическую эпоху. Но так было не всегда. Современные папоротни-
кообразные являются жалкими остатками некогда обильной и широкораз-
витой растительности, господствовавшей на Земле до появления семенных
растений.
Расцвет папоротникообразных имел место в палеозойскую эру,, особенно
в каменноугольном периоде. Папоротникообразные* былитогда широко
распространены на доступной для заселения Земле, особенно в низинах,
болотистых лесах. Среди них преобладали древовидные формы (лепидоден-
дроны, сигиллярии из плауновых, каламиты и клинолисты из хвощовых,
крупные папоротники и т. д.). Они обладали вторичным ростом и входили
в состав каменноугольных лесов. Ископаемые остатки этих растений дали
мощные залежи каменного угля.
Папоротникообразные дали начало семенным растениям. Семена появи-
лись в результате дальнейшей эволюции разноспоровых растении. Уазно-
споровые растения имеЮТоГсредн плауновидных,1ШшротШшовндных, были
они и среди членистых. Попытку выработки семян мы встречаем у плауно-
Г.Г.
III.
>tlt
III
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
217
та
видных, но настоящие семена сформировались только у Pteropsida, Такие
семенные папоротники относятся уже к следующему отделу — голосеменных
растении. Обширный отдел Pteridophyta разделяют на 5 классов.
~ПСИЛОФИТ(^ВИДНЫЕ (FSlLttPHYtOPSlDA)
В этот класс входят исключительно ископаемые растения. Это первые
из известных нам высших наземных растений, их называют часто «первен-
цами сухопутной флоры». Псилофиты известны начиная от^нижнесо-силура
до верхнего девона- Они были открыты английскими палеоботаниками
К идет о н ом и Лангом в угольных шахтах нижнего и среднего девона
1Потландии в 1917 а. (роды рпния, хорнея и астероксилон) и были названы
псилофитами по известному еще с 1859 г. ископаемому растению Psilophyton.
которое было найдено в нижаедевонских^отложениях Канады и опиСйна
Д а у с о н о м, но не нашло места в системе растений.---
В настоящее время йсилофиты нлйдапы на всех материках и нзсчиты-
вают свыше 20 родов и большое яисло вадов. -
офиты п£ишфшШу№1к, гаметофиты их до-сих пор
и довольно разнообразны.
Известны
ле найдены.
Большинство
у них не было и корней,
обладавших дихотомия
ИХ РЫЛО пТ№лСтавленО РЯДОМ ЭЛемаптарпмт
ветвденпАу- Эти
а конечных разветвлениях теломов
развивались спорангии. Участки, соединяющие т
растение к субстрату. Они были покрыты ризоидами. Эти нижние подземные
участки псилофитов называю^ р и з_о м о и д а м и. Схема строения такого
простейшего спорофита изображена на рисунке 133, 2. Спорангии имели,
таким образом, верхушечное (терминальное) положение и не были
еще связаны с листьями.
Однако у некоторых родов уже появляются листья. Многие псилофиты
имели еще водорослевый облик, но другие напоминали уже плауны, хвощи,
папоротники.
Псилофиты были земноводными растениями, нижняя их часть была
погружена в воду или в илистый грунт, а верхушки осей развивались в воз-
душной среде и в своем строении обладали всеми признаками сухопутных
растений. Они имели проводящие пучки, спорангии, снабженные миргшитй--
ной стенкпй, споры, покрытые толстой оболочкой. Все эти признаки никогда
не встречаются у водных растений.
Систематика" класса ищо недостаточно разработана. Он представлен
одним порядком Psilophytales и несколькими семействами.
ПОРЯДОК — ПСИЛОФИТОВЫЕ (PSILOPHYTALES)
Знакомство с псилофитовыми мы начнем с рассмотрения наиболее типич-
ных и хорошо изученных родов — ринии, хорнеофитона и астероксилона.
PuHua^Rhynia). Из трех вадов ринии наиболее изучены два: риния высо-
кая major) высотой' до 50 см и диаметром до 5—6 мм и риния гвине-вогана
(Rh. gwynne-vaughanii) до 20 см высотой и 2—3 мм диаметром. Оба вида
найдены в среднедевонских отложениях Шотландии (рис. 128,7, 2). Они
росли на болотах. В почве располагалось их «корневище» (ризомоид), от
ли
которого вниз отходили пучками ризоиды, а вверх — наземные побеги.
Корней не было. Наземные побеги дихотомически ветвились, верхние участ-
ки тела заканчивались спорангиями.
Анатомическое строение ри£
г г.
высокой было чрезвычайно простым.
центре телома располагалась примитивная протостель в виде концентри-
ческого проводящего пучка. В середине его находилась ксилема* представ-
В
i г г,
218
ВЫСШИЕ РАСТЕНИЯ
Рис. 128. Риния:
J —Rhynia major (m —телом, at —весом); я—Rhynia gwynne-vaughanti; 4 — поперечный разрез «стеб-
ая» (видев евцдермвс. прерывающейся устьицами, и цровцйщвй пучок в центре); 4 — спораягий; з —
споры; « — уетьще
ленная кольчатыми трахеидами. |Ьщдему~ окружала флоэма. Она состояла
из 4—5 рядов удлиненных клеток со с^шёЕЩый^ Зденками; Далее распола-
галась мощная кора. Перицикла и эндодермы у ^ринии не было. Кора разде- >
лялась на вн^р^^ю^^Аружн^. Внутренняя кора состояла из рыхло
соединенных паренхимных клеток с межклетниками, наружная — из не-
скольких слоев плотно сдвинутых клеток. Снаружи телом одет эпидермисом.
Наружные станки его клеток утолщены. В йшдермйсе имелись устьица
нормального для Высших растении строения — с двумя замыкающими клет-
ками и устьичной щелью (рис. 128, 3).
Спорангии (до 12 мм длиной и 3—4 мм шириной) вмели многослойную >
стенку (рис. 128, 4), гочисленные споры, соединенные в тет-
рады. Таким образом*; ши. как и у всех высших растений. происхо-
__ х приспособлений к вскрыванию еяю-
ел. Споры, вероятно, ос
м;г
!«' li
сТешси сдоранхиЯГуСпоры ымвду дд«пмтп толстую оболочку. У Rh. gwynne-
Mugltmli, Шсе^шёи в^ОбщетТго же строение, наГтеломАХ находились пелу-
шаро видные паренхиматические вздутия и короткие, развивающиеся из них
боковые побеги.
Род хорнеофитон (Homeophyton), включающий один вид Я. Ugnierif
известен из среднего девона. Хорнеофитон имел дихотомически разветвлен-
ные теломы. «Корневище» было с клубневидными утолщениями, в ткани
которых находились гифы гриба. Таким образом, наблюдалась примитив-
ная микориза. Анатомическое строение было сходно с ринией^но в эпидер-
мисе стебля не было устьиц (рис. 129, 7).
Спорангии развивались на концах дихотомических разветвлений. В спо-
рангий довольно далеко вдавалась колонка, на которой располагался куполо-
образный спорангий. Колонка состояла из паренхимной ткани и была окру-
жена выстилающим слоем (тапетум). Спорангий отдаленно напоминал
строение коробочки сфагновых мхов (рис. 129, 76).
Род астероксилон (Asteroxylm) известен из среднего девона. От под-
земного (подводного?) «корневища» отходили дихотомически разветвленные
1 I V* 1
— ПАПОРОТНИКООБРАЗНЫЕ
219
оси, функционировавшие как кор-
ни. Они были безлистными (рис. 130).
Надземные участки растения (до 1 м
высотой) также дихотомически ветви-
лись и были расчленены на «стебли» и
«листья». «Листья», мелкие, шиловид-
ные, представляли собой короткие
выросты стебля (так называемые
э н а ц и и). В основание каждого
«листа» входил пучок, ответвлявший-
ся от основной стели, но он не про-
никал далеко в глубь «листа». Рао
тение обликом очень сходно с пла-
уновыми.
Стебли имели актиностелы
Центральный пучок состоял из кси-
лемы, в поперечном разрезе звезд-
чатой формы, окруженной флоэмой
(рис. 130, 2). Ксилема состояла из
кольчатых и спиральных трахеид, а
флоэма — из удлиненных, с косыми
стенками клеток без ситечек. Кора
разделялась на внешнюю и внутрен- -
нюю (рис. 130,2). Во внутренней
коре имелись воздушные камеры. В
коре видны были следы пучков, шед-
ших в листья (рис. 130, 2).
Спорангии у астероксилона раз-
вивались на концах безлистных, ди-
хотомически разветвленных осей.
Спорангии (до 1 мм длиной) имели
многослойную стенку; вскрывались
они на верхушке, споры одинаковые
(рис. 130, 3).
Из других псилофитовых отметим
род псевдоспорохнус (Pseudosporoch-
nus), известный из среднего девона
Евразии и Северной Америки. Это
было деревце до 3 м высотой с до-
вольно толстым стволом. В верхней
части ствола располагались ветви,
собранные пучками. На конечных
разветвлениях вильчатых веточек
находились спорангии, содержавшие
одинаковые споры (рис. 131. /). _
насчитываем видов. Наиболее
вестей Р. princeps, описанный Дау-
соном из среднего девона Канады.
Листьев у псилофита не было. От
горизонтального «корневища» отхо-
дили надземные оси, достигавшие
1 м высоты. Они неоднократно дихо-
томически ветвились, были голыми
или внизу несли шипы (рис. 129, 2).
Молодые верхушечные разветвления
были спирально загнутыми. Споран-
гии располагались на верхушках
Рис. 129. Хорнеофитон (Horneophyton
lignieri) к псилофитон (Psilophyton
prince ps);
1 — хорнеофитон: a — общий вид; б — продоль-
ный раярез спорангия (кол — коленка, де —
нсилема. tfut — флоеиа, пар — паренхима, и —
воневца); » — псилофитон
220
ВБ»
ЕНЕ
Е РАСТЕНИЯ
некоторых разветвлении, загнутых вниз. Анатомическое строение сходно
с ринией.
Род тениокрада (Tamiocrada) встречается в нижнедевонских отложе-
ниях Западной Европы, СССР и Китая. Тениокрада имела водорослевый
облик. Теломы ее были лентовидными, протостелическими. Спорангии разви-
вались на вертикально торчавших разветвленных осях (рис. 131, 2).
Род аостермист.. (Zosterophyllum) известен из верхнего силура как
северного, так и южного полушария. Это было водное (вторично?) растение
с корневищем протостелического строения. На возвышавшихся над водой
осях развивались спорангии, собранные в колоски (рис. 132, 7).
Урода ярравия arravia) из верхнего силура Австралии известны
спороносные участки тела. Спорангии у ярравии срастались в синангии
(рис. 132, 2).
Таким образом» в настоящее время псилофитовые известны и из силура.
Эти находки свидетельствуют о том, что известные нам псилофиты являются
уже довольно сложными специализированными растениями, проделавшими
длинный путь эволюционного развития. Открытие псилофитов пролило свет
Рис. 130. Астероксилон:
1 — общий вид ( Asteroxylon elberfeldense); 2*— поперечный разрез стебля («к — внутренняя кора, х—-
ксилема, ф — флоэма, нк — наружная кора, ле — листовой след); з — продольный разрез спорангия
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
221
Рис. 131. Псевдоспорохнус и теииокрада:
1 — псевдоспорохнус (Peeudogporochnus Krejci!); а — общий вид: б — утолщенные ветки (образование
макрофилдов через утолщение тешлюв); Д — теняскрддд (Taeniocrada deeheniana)
на вопрос о происхождении листа. Листья возникли впервые у псилофитов,
и при этом двумя путями. Микрофиллы возникли как^выросты (эвдции)
на стебле (Asteroxylon), а макрофиллы возникли в результате срастания
элементарных участков тела псилофитов — теломов (типа Rhynia), Каждый
из них имел свою стелы При срастании теломов сливались и их стели. Срос-
шиеся теломы уплощались (кладодифицировались) и превращались в ли-
стья — ветви. Такой способ образования макрофиллов можно проследить
у Pseudosporochnus*
Рис. 132. Псилофитовые:
1 — эостерофиллум; Я - нрравкя; а — Yarravia оЫавда; б — J.
2
aubsphaerlca — обе из верхнего сжа^рн
вы
н > 1 гл
РАСТВНИЯ
Рис. 133. Образование листа из талонов псилофитов и основные участки тела прими-
тивного спорофита*.
1 — сероватая листа путем появления выроста; л^оОравоваиие листа дучем ушющения срастания
тажмк»; а — основные участки тела (т — тажм. ж — несом, р — римнощ. са — спорангии)
Схема, иллюстрирующая эти оба возможных способа формирования
листа, изображена на рисунке 133.
Псилофитовые являются предками Lycopsida, Sphenopsida, Pteropsida.
г
2-й КЛАСС—ПСИЛОТОВИДНЫЕ (PSILOTOPSIDA)
m
К классу псилотовидных принадлежит один порядок Psilotales, пред-
ставленный двумя родами — псилот (Psilotum) и тмезиМтер (Tmeslpteris).
Псилотовые — небольшие растения, лишенные корней и обладающие
рядом признаков, сближающих их с пеня офитами; распространены они в
тропических и субтропических областях обоих полушарий.
Род псилот (Psilotum) представлен двумя видами — Р„ triquetrum и
Р. jlaccidum^ широко распространенными под тропиками, заходя в северном
полушарии до Японии, Кореи и Флориды (США), а в южном полушарии — до
Новой Зеландии. Р. triquetrum—прямостоячее растение, растущее на почве,
тогда как Р+ flaccidum — эпифит, растущий на стволах деревьев, особен*
III
tn
Род miiesunmep (Tmesipteris) содержит также 2 вида, распространенных
в Восточной Индии, на Филиппинских островах, в Австралии и Новой
Зеландии. Наиболее распространен эпифитныи Г. tannensis.
Оба рода имеют безлистное, дихотомически ветвящееся корневище,
скрытое в почве; выше оно Переходит в, олиственные побеги, возвышающиеся
над землей или свешивающиеся с деревьев. Надземные побеги достигают
у псилота 20—100 см длины, у тмезиптера 5—40 см. Корневища покрыты
корневыми волосками и имеют примитивное протостелическое строение.
Корней нет. Они не образуются даже в зародышевом состоянии.
Надземные стебли Р. triquetrum трехгранные, а у Р, flaccidum плоские
(рис. 134). Они, как и корневища, дихотомически ветвятся. На верхних раз-
ветвлениях расположены листья и спорофиллы со спорангиями. Листья
псилотов мелкие, чешуевидные. Жилки в листе отсутствуют, но у Р« flacci-
<&т в стебле имеется листовой след. Это {свидетельствует о том, что листья
ли
отдел—ПАПОРОТНИКООБРАЗНЫЕ
псилотов редуцированные. Функция ассимиляц
1»Г.
ется стеблем.
в основном осуществля-
Стебель имеет актиностелическое строение. Центральный пучок содержит
пяти-, десятилучевую ксилему, состоящую из лестничных трахеид, но сама
сердцевина занята склеренхимными клетками. Ксилему окружает флоэма.
Пучок снаружи окружен эндодермой. Далее к периферии стебля располо-
жена кора. Снаружи стебель одет эпидермисом. Под эпидермисом в несколь-
ко слоев клеток расположена ассимиляционная ткань (рис. 134,
Трехгнездные спорангии — синангии (сросшиеся спорангии) —
расположены на верхней стороне маленьких, вильчато-разветвленных споро-
филлов (рис. 134,1 б9 а). Спорангии закладываются каждый отдельно и лишь
потом сливаются. Они имеют многослойную стенку. В спорангии развивают-
ся многочисленные одинаковые споры. По созревании спорангии вскрывают-
ся радиальной щелью вверху и споры высыпаются наружу. Из спор разви-
ваются заростки. Заросток ведет подземный образ жизни и сожительствует
с грибом, проникающим внутрь его клеток. Заростки псилота цилиндричес-
кие, дихотомически разветвленные на концах и достигают 1,8—2 см в длину
при 1—1,25 мм в диаметре. От заростка отходят многочисленные ризоиды
(рис. 135, 2). Интересной особенностью заростка является нахождение в нем
остатков примитивной стели в виде небольшого' числа кольчатых и спираль-
ных трахеид. Это свидетельствует о том, что гаметофиты (заростки) у предков
Р silotales были хорошо развитыми и имели стелярное строение. Внешне
заросток похож на молодой спорофит. Заростки обоеполые. Антеридии
поверхностные, не погруженные в ткань заростка. В них развиваются много-
жгутиковые сперматозоиды. Архегонни брюшком погружены в ткань за-
росли} (рис. .135, 2). Образующийся в результате оплодотворения зародыш
состоит из стебля и ножки, внедряющейся в ткань заростка; ни корня,
г. г.
ни первичного листа
зародыш не имеет.
Рве. 134. Пеилотовые:
1 — некий* (PsHetum triquetrum): а—общий вид; б—веточка с трехгранншвг спорангиями (сиваяги-
ямН); « — веточка с раздвоенным на верхушке спорофиллом; я— тмемштер (Tmeaipterie tannensts): а—
общий вид: б — (шорофюпм е аинангнями: « — спорофилл с удаленными еинангнямн
НН
ВЫСШИЕ РАСТЕНИЯ
ри прорастании зародыша развивается сразу корневище с корневыми
Ьсками на нем, и далее возникают надземные стебли с листьями.
Надземные побеги тмезиптера покрыты очередными листьями. В нижней
части побега они чешуйчатые, выше же с хорошо развитой пластинкой,
зеленые, кожистые. В листе развита средняя жилка. В стебле имеется листо-
вой след. При отхождении его от стебля заметны (особенно у спорофиллов)
листовые прорывы. Стебель имеет сходное с псилотом строение, но середина
пучка занята сердцевиной.
Спорофиллы вильчатые. Они расположены в верхней части побегов
(рис. 134, 2а), где от общей цилиндрической части (черешка) отходят две
пластинки. Между ними, прикрепляясь к черешку, находится двойной
спорангий (синангий), разделенный поперечной перегородкой на два гнезда.
Каждый спорангии по созревании вскрывается продольной щелью (рис. 134,
26), через которую споры высыпаются наружу.
Споры все одинаковы; прорастая, они дают начало обоеполым заросткам.
Заростки имеют червеобразную форму, ведут подземный образ жизни,
сожительствуя с грибом.
Антеридии и архегонии рассеяны по всей поверхности заростка. В анте-
ридиях развиваются многожгутиковые сперматозоиды. Образующийся после
мкл ат эп
Рис. 135. Псил№
1 — поверенный разрез стебля (хе—неклена, ф—флсвма, в центре стеля—склеренхима, амЗ—вндрдер-
на, мкл — механические клетки, от — ассимнляцнонвзя ткань, ап — епидариис); я — варосток; а — часть
мростка (ан — антеридий, ар — архегоний. р — гифы гриба); 4 — сперматовод
ОТдаЛ — ПАПОРОТНИКООБРАЗНЫЕ
оплодотворения зародыш состоит
из стебля и внедряющейся в ткань
заростка ножки. Разрастающийся
стебель развивается в виде неболь-
шого тельца, дихотомически раз*
ветвленного с обеих сторон. Мо-
лодой спорофит долго соединен с
заростком и внешне очень на него
похож (рис. 136). При дальнейшем
ветвлении из верхней части стебля
развиваются надземные побеги, на
них возникают листья; при сствле-
нижней, остающейся под зем-
: г. ।:
лей части формируются корневи-
ща- В ископаемом состоянии
псилоты неизвестны. Отношения
Рис. 136. Развитие молодого спорофита Tmes-
ipteris tannensis;
2 — молодой спорофит; 2—с двух сторон образовались
воздушные стебла; з — гаметофит о развившимся на
нем спорофитом, образовавшим надоенные и подземные
дихотомически вемшцаесн веган (а — гаметофит, сп — г
спорофит)
их к другим папоротникообразным
неясны. Раньше их сближали с
Lycopsida или с Sphenop si da. Од-
нако от плауновых они отличают-
ся отсутствием корней, многожгу-
тиковыми сперматозоидами, нали-
чием листовых прорывов в стели.
Листья их, таким образом, не
микрофилльны. Несколько ближе
они к Sphenopsida, но резко отличаются
от них отсутствием мутовчатого расположения листьев, наличием синан-
гиев и другими признаками. Вместе с тем они имеют много общих призна- *
ков с псилофитами (отсутствие корней, примитивное стеляриое строение,
листья теломной природы и др.). Пеилотовые, вероятно, являются пря-
мыми потомками псилофитов.
cUobta+CL.
tM КЛАСЕ— ПЛАУНОВИДНЫЕ (LYCOPSffiA>—
Плауновидные —^типичные листостебельные растения. У них всегда
есть коран. Плауновидпые^араИТёрицуЮтёя ^нн<рофиддиед//У их предста-
вителей сравнительно дшлкна—лисхьяг тесно 'рямммгоженные на стебле.
Плауновипные — одна ня наиболее древних ветвей ийпорпгпйипобряяттт
Остатки их известны уже начинай с силура. Наибольшего развития
они достигали в кяменноугольнпм йприппд ттялрдпла-мя* при и были пред-
ставлены мощными деревьями (лепидодендроны. сигиллярия и др.). Ос-
татки их овраяпияли мойные залежи каменного угля.
Современные представители плауновидных — травянистые растения
(плауны, селягинеллы).
на_верхушке, с одной жилкой. Листовые прорывы стели отоун»дпутпт
Спорангии расткмтожены поодиночке нЭГ~БЩТЕ5е8~стороне спорофиллов
М—, —— —— —— ль ль От от — — rfbi —
впепеме
Древнейшие представители плауновых были травянистыми растени-
ЯМН. В своем^происхожданни пни. мод^Вмта1 пйиадеиы ид црилофитошжпя
Aeteroxylon. В дальнейшем развитии они дали, с одной стороны. пяд прапаа-
ны^ представителей, а 'В"ИВЙ,бЙ пяд травянистых Дори. Небольшое число
плауновидных (плауны, селягинеллы, полушник) дожило до наших дней.
Класс плауновидных разделяют на 6 порядков.
8-602
226
ВЫСШИЕ РАСТЕНИЯ
1-Й ПОРЯДОК — ПРОТОЛЕПИДОДЕНДРОВЫЕ (PROTO LEPIDODEND RALES)
К этому порядку принадлежат наиболее примитивные травянистые
плауновидные, известные лишь в ископаемом состоянии. Остатки их находят
в отложениях верхнего силура и нижнего и среднего девона. Из силура
Австралии описана барагванатия (Baragwanathia) — травянистое растение
до 1 м высотой, стебли её дихотомически ветвились и имели актиностели-
ческое строение. Хвоевидные листья до 4 см длиной располагались на стебле
без заметного порядка. Спорангии располагались как в пазухе листьев, так
и на стеблях между листьями; в них развивались одинаковые споры; за-
ростки неизвестны (рис- 137, 7).
У дрепанофикуса (Drepanophycus), известного из девона Евразии и
Северной Америки, спорангии были расположены уже на верхней стороне
листа. Стебли его, достигавшие 60—70 см высоты, дихотомически ветвились.
Игловидные листья очень сходны с листьями псилофитов (Asteroxylori), они
располагались на стебле также без заметного порядка (рис. 137, 2).
Рис. 137. Простейшие плауновидныв?
J--барагванатия (Baragwanattra longifoiia); 9 — лренанофикус (Drepanophycus eptnaefermls): а — об*
ший вид. о — веточка (увеличено); 9— протодедадодендрон (Protolepldodeodron scharyamim). а —об-
щий вищ С — поверхность стебля. < — спорофилл со спорангием
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
Более сложное строение имел протолепидодендрон (Protolepidodendron
scharyanum и другие виды); (рис. 137, 3). Остатки его известны из нижнего
и среднего девона Евразии. Он имел горизонтальное корневище, от которого
отходили вертикальные дихотомически ветвившиеся побеги высотой 25—40 см.
Они имели протостелическое строение. Листья дихотомически ветвились
на верхушке, располагаясь на стебле правильными спиралями. По опадении
листья оставляли на стебле «подушечки», чем напоминают лепидодендроно-
вые (стр. 235—236).
Спорангии располагались уже на верхней стороне спорофиллов, близ
их основания. Спорофиллы, как и у всех представителей этого порядка,
не отличались от вегетативных листьев и чередовались с ними. Колосков
не было.
елд-се. opsida-
ЧМЙ-ПОРЯДбк"— ПЛАУНОВЫЕ (LYCOPODIAtES)
Порядок плауновых представлен одним семейством Lycopodiaceae^
которое включает два рода — Lycopodium и Phylloglossum.
Род плаун (Lycopodium) насчитывает около 400 видов, распространен-
ных по всему свету, но большинство их встречается в .тропических, по преи-
муществу горных лесах.
Плауны — небольшие травянистые растения. Однако некоторые виды
достигают 1—1,5 лс высоты, а стебли одного из них — тропического вьюще-
гося L. volubile — достигают даже 9 лс.
В СССР встречается 14 видов плауна. Широко распространен плаун
булавовидный (L. clavatum), произрастающий в хвойных, чаще сосновых
лесах. Его гибкий, дихотомически разветвленный ётебель стелется по земле»
посылая в почву корни и образуя приподнимающиеся надземные ветви
Стебель и ветви густо покрыты маленькимй линеиноншиловидными лпгтксшп
Растение как бы плывет по земле, отсюда, вероятно, оно и названо—плавун»
пдауц (рис. 138). — ~ ‘
Анатомическое строение стебля еще примитивно, Молопые ^теблипното-
стелические. У взрослых стеблей — плектостедь. Снаружи стебель одет
эпидермисом Далее внутрь следует мощная кора. Наружные слои коры
представлены механической тканью, ‘построенной др толотоотопныт к лето"
За ней идет довольно широкаяполоса прозрачной ткани ня тонкостенных, слег-
ка вытянутых радиально клеток- яя которой снова располагается механи-
ческая ТКаНЬ, пкружяютцдд еталь (рис. 138, 5).
Центральную часть стебля занимает корпентрический 'Ирб^Цпящий
пучок (центральный цилиндр^-вдель). Ксилема в пучке расположена в виде
лЭкг^врединяющихся друг с другом. На поперечных срезах, проведенных
на рдяных уровнях стебля, кяртйра ксилемы раздприагтеледстаие того, что
по вертикали ксилемщй тяж имеет разлнчйуяГто
Между участками
юрии, расположена
[ет. вследствие чего
не образуется вторичных тканей и стебель не способен к вторичноМу утол-
щен ию. — ---------- "" ”
< -За флоэмой следует пднпглгптарый пепипикл и, наконец, эндодерма»
отграничивающая лучок от коры. Стель плауна, близкая к протостёли,
называется плектостелькъ
КсИЛеМТГсОСТОИт" ИЗ ДОВОЛЬНО пгррОК^х леСТНИЧНЫХ Тряхеид (метяКР.И*
лема), ближе к периферии встречаются кольчатые и спиральные трахеиды
(протоксилемаУ? Среди трахеид разбросаны клетки древесинной парецхрмы.
Флоэма представлена ситовидными трубками и кдетаэмп^дубя^пй парен-
ДИМЕтй'ТбЙИДНие 1 МУбКм удлиненные, ня кппдях яярстренные. лишенные
содержимого,, безъядерные, с мелкими ситечками ня продольных стенках.
Сопровоя^яютцях клеток (клетой-спУЗДиц) у ^ддаувов нет. Клетки лубяной
пяренхиМыяяполнйнм^ крахмалом.
НГ.
ВЫСШИЕ РАСТЕНИЯ
Рис, 138. Плаун (Lycopodium clavatum):
1 — общий вид; г— сосрофнлд: з — спора: 4 — проросток (молодой спорофит), еще связанный с зароет*
вок; л — поперечный разрез' стебля (кс — ксилема, ф — Флоэма, «ЙО — эндодерма, жх — механическое
кольцо, ле — листовой след, мр — кора, ап — впндермне)
и одеты эпидермисом. Устыша_ расположены на ниж-
фХНРстть-яиета. Многослойный мезофилл состоит изклеь
филловыми зернами- Ряды йЛГ^раздёлены воздухоносны-
Листья сщццп
ней и верхиетНпп
ток-^богатых хлоп ___ ___________________________________
МП"полостями (межклетниками). ' " ’ ;—
В стебле ет^донтральиого пилиппра лшео-нвверх отходят, без прорывов
стели, пучки проводящей ткани. Следуя через^кору (так называемые листо-
вые следы);1 РИМ входят в лист, где становятся единственной средней жилкой
листа. * т~ ------------——- ’ ~ ~
И. По
анатомическому строению корни вполне сходны со стеблем.
В середине лета у Z. clavatum образуются спороносные колоски
(рис. 139,1), обычно 2. Реже 1,3. Они ряс.подяглтптсгя на мдртутке ветвей
На оси ямым,. нйлегая лруг_ия друтя расположапм^^орпфилл^. Они
значительно х^ьче^вегетативных листБевГ"и рт^ичпготг^рт фпр*<д
ишраеке. СпорофиллП птипокбяйдЗвидные, длинно-заостренныё^шгяэо-
хушке и ократпецы в желтоватый
rTTnpowflffi почкови^нуюформуи прикрепляотод^к, спорофиллу Корот-
кий пмрдпй В спорангии развивается большое ипличестлп спвепптепип
одинаковы^ . мелких спор (рис. 138,"X ^)-
- Споры пкпуглп-тотрладрнческой формы и и
Наружная оболо Рры (а К зЪ 6 Ь о ~
на
етчата
окраску.
с утолщенияму
; в содерйищрм
и масла. Спо-
JDia. Сшфангий
на гранях. Внутренняя оболо
спорки идхпдитсдщгтпляАма. ggPQi пляститты. а также
рангий возникаетйз Группы поверхностных клеток спор
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
f
Рис. 139. Плаун:
1 — подземный заросток (ак — антеридий* ар — архегоний, > — зародыш, ап — зпидермис. аф — гмфы
првба, р —ркэоиды); 2 —антеридий х архегоний: а —антеридий, о — сперматозоид. г —молодой архе-
гоний, « — вскрывшийся- архегоний; з — строение и развитие зародыша: а — оплодотворенная яйцеклет-
ка (зигота) поделилась поперечной перегородкой, образовав две клетки — пив; б — клетка п больше
не делится и образует подвесок, клетка д де.штзя продольной перегородкой. • — дальнейшее депеше
клеток зародыша, > — зародыш с дифференцированными органами (п —подвесок, я — ножка» тр — точ-
ка роста, ял — первый лист, хн — корень).
имдп-ш^грдтллаДиую стенку. Mjprii внутренний слой (т а п е Т V м) обычно
почти полностью разрушается, клеток используется при
развитии спор. НодТретслойной стенкой ряст^^ложоп слдроипопми слой
спОрадаш. .(а р х е с п tf"p и йк
При делении яртясплрняльнмт клеток образуются материнские клетки
дощ. В результате ряттукцдрчипгл пяления послвтуних оораяу^тл тетрпдч
Зрелые спорангтавскрываются поперечной трещиной. К этому.лремени
ось колроска нескод^к^^^^дфрастается, орпфиллы раздвигаются
и спорыэдегко высеиваются из спорангиев наружу.
Попав на землю, спора воды уносится в углубления (трещины)
бйневГ нескольк
почвы, где^на
ет. При этом наруяс- <
кзоспорий) лопается, а содержимое ы делится,
образуя аа росток, имеющий иначала в^д комочка клеток. Вскорена за-
ростке образуются^риз че
почвы гифы гриба (рис. 139,2).
Зрелый зарос достигает 2—5 лйм в попе
нает игрушечный волчбй:
ей хл
роникают из
III
рме напоми-
остоя-
230
ВЫСШИЕ РАСТЕНИЯ
тельно питаться .да-мджет. Снаружи заросток одет эпидермисом. Снизу и с
боков заростка отходят^ризоидБь^Над нижним эпидермисом расположены
в 3—5 сл9ата иле™», ^ддрржащие гифы___гриба (эндотрофная микориз а).
За ними следует слои вытянутых вертикально (паяйсяиных) илатпк Глубже
расположена ззддоайщая тканЬдЛдслетпах которой накапливается крахмал.
Середина заростку занята Обычной паренхимнтж~кянью ------"""
Если гриб почему-либо не попадает в соприкосновение с заростком,
последний отмирает на ранних фазах развития. -- -------------
la.:*?
На верхней поверхности заростка образуются антеридии и архегонри.
. Аитеридии^ целиком погружены в^исаныааростка. В нмХ развиваютс я "morn-
численные двужгутиковые, сперматозоидыгрйс. 139, 2а). Архегонми-Дрюш-
КОМ ПОГРУ^КеЙ^ В ЗТГрбсТОК, ДО шейкиИХ нпявттптдтлтгп над егл поверхностью.
В брюшке архегония развиваются яйцеклетки „брюшная, канальцевая
клетка. Шейка ^р^дстаЮхЕНЗй^В^йг^кЗйЗльцавыьшиаштками (рис. 139, 2в).
Зрелый архегонии векрывает^я на яор^у1шго) и внутрь его проникают
сперматозоиды. Один из пих слйвается с яйцеклеткой, промавлдя пттлпттп-
твОрениеГ '
При первом делении оплодотворенной яйцеклетки образую*^ся верхняя
и рдрртрр «тогам (рисЛ39, За). Верхняя клетка называется п о д в еском
и лйьгвил. блдьтпе не делптг.я Нижняя клетка (з а р о д ы ш е в а я) мнлгл-
кратно делится и дает начало зародышу. При этом у L. clavatum, как и у
всех видов с подземными заросткамиТформируется ножка — полушаровид-
ное тельпе, внедряющееся в ткань заростк а. При ее посредстве идет всасыва-
ние зародышем питательных веществ из ткани заростка Дрив 139. 3)
У сформировавшегося зародыша хорошо1 заметен стебелек, на котором
формируется первый лпстт лишенный проводящего пучка. а_ у основания
стебелька возникает придаточньпкиорень- Прорывая ткань заростка, заро-
дыш развивается далтрте д-образует молодое, растеньипе. которое, разрас-
таясь, появляемся затем на поверхности земли (рис. 138, 4), Подземные
заростки L. clavatum и некоторых других плаунов_развиваются очень мед-
ленно. От прорастания спор до формирования назаростке"йтацдбго споро-
фита проходит 15—18 лет. ----------*
У многих других видов плауда -аэ ростки^ развиваются на поверхности
земли (например, у нашего ^Г7пцгаДд$шп) или (у некоторых тропических
М'ттовУ^на к пре деревьев В этом ^случае они" становятся нитевидными, стелю-
щимися по коре. -------’
У ряда тропических видов заростки зеленые, содержат _хллрпфилцпвые
зерна и дчта я_самостоятельно. Надземные заростки развиваются быстрее
ттлдатцтых и -жпиут обычно ОДИН C63OH. “------------------
У одного из наших^шйбн^^уча^а^на северного (L* selago) — заростки
зеленой верхущвпй^матактттт^тптдгг йддхфйщг£ностью земли. Они цилиндри-
ческие и имеюФ^радиальное г.трпенпе^ У^этпгп вида, произрастающего у нас
в сырых мшистых лесах, не образуетсяколйъкон, и изе-етпгрЬфиллы, несущие
спорангии, не отличаются^пб "внешнему^вйДу 75Т"вегетативных листьев. У
L. selago в пазухах некоторых вегетативных листьев развиваются выводко-
вые почки, служащие для вегетативного размножения.
Простейшим типом заростков плауновых являются цилиндрические,
зеленые. Из них развивались, в связи с приспособлением к различным усло-
виям жизни, нитевидные и распластанные на коре деревьев заростки эпи-
фитных плаунов и кубаревидные подземные заростки, перешедшие к сожи-
тельству с грибом.
Род филлоглоссум (Phylloqlossum), представленный одним видом
Rh. drummondii, встречается в засушливых^. .районах Юго-Западной
Австралии, в Тасмании и Новой Зеландии. Это небольшое растение, в нес-
колько сантиметров высотой. В нижней части растения развит мясистый
клубень, от которого вверх отходят шиловидные листья. Позже верхушка
клубня вытягивается и дает небольшую ось, заканчивающуюся колоском.
Ют клубня отходят также несколько корней и направляющийся вниз цилин-
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
231
дрический вырост. Углубляясь в почву, он разрастается, образуя новый
клубень. По отмирании материнского растения молодой клубень после пери-
ода покоя прорастает в новое растение.
Размножение идет, как и у Lycopodium. Заростки у филлоглоссума
зеленые.
У плауновых имеется ясно выражеппая смепа поколений. Спорофит
(само растение плауна) сменяется гаметофитом (заростком). При этом споро-
фит и гаметофит существуют раздельно, являются самостоятельными расте-
ниями. В цикле развития преобладает спорофит. Редукционное деление
происходит в спорангии при образовании спор.
Своры плаунов собираются в большом количестве и используются
в аптечном деле для пересыпки пилюль, в качестве детской присыпки.
Сбор плаунов в лесах часто проводят с декоративными целями — для
украшения зданий.
3-Й ПОРЯДОК — СЕ ЛЯГИНЕ Л ЛОВЫЕ (SELAG1NELLALES)
К порядку селягинелловых относится семейство Selaginellaceae
с одпим родом селягинелла (Selaginella), который представлен почти
700 видами, произрастающими в тропической и субтропических областях,
н лишь немногие виды встречаются в умеренной зоне — в горах Евразии
(5. helve Пса) и Северной Америки (S. apus), в северных умеренных областях,
заходя местами в Арктику (S. selaginoides). В СССР произрастают 8 видов
селягинеллы.
В ископаемом состоянии селягинелловые известны начиная с верхнего
карбона. Потомками их являются современные селягипеллы.
Селягинеллы очень разнообразны по внешнему виду. Большинство
их — небольшие, обычно ползучие травянистые растения, и лишь около
50 видов обладают прямостоячим стеблем. Некоторые тропические виды,
однако,"достигают высоты до 2 м, а вьющиеся (лианы) —
длины. С другой стороны, имеются и более мелкие виды — 5^Т0 см длиной,
напоминающие но внешнему облику мхи.
Взрослое растение (спорофит) селягинеллы по внешнему облику _очень
сходно с плауном (рис. Т4О, 7). Виды с прямостоячим стеблем..имеютедкие
листья пдипяклпого рядмерА, рдлттлложянпые „спиралью. *
У видов,.со стелющимися стеблямидистья расположены в два ряда.
Каждый^ряд состоит, в св^"очередх^лз^двух тесtio сбл ижсТйгбгу л Шаъев.
лпкЧГТЕмалспькие, а нзеркнио бОЛб^ кру1111'Ь1^Г В pu^yjibiaiezggc-
тение имеет как бы дор^ОЬТШтральлие строение Мел киевер хине
листья не^Я1сняют ниптии. й~Йбследние, таким 7ибралиМ| нормально фуйкп
циопируют. --------------- - * ’ ' --------—
К почве селягинеллы прикрепляются топкими, дихотомически ветвящи-
мися^дшрнямл—(рис. 140, 7). Корни, возникают или прямо на стебле, или
тта^оепб^у органу — j>h з о <Ь о р я х (кпрнеипстгдх). Ризофоры отходят
от стебля, возникая зкзоГётню, ка1Г отсбяи. Корпи же на-ник
эндогенно, как эТд
- EW" «'з» ——тт
ч.CTft6gj№-rццрум^г пнет одттоглойпктм яттидярмнепм. Далее следует мощная
кора, наружный слой клеток которой нередко приобретает механический
х^арактср. и, наконец, проводящие пучки,
Большинство видов селягинеллы имеет такое же строение проводящей
системы, как и плауны. У них также есть концентрические замкнутые
(без камбия) проводящие пучки. В центре их расположена ксилема, окру-
женная флоэмой. из длинных трахеид. по у двух видов
селягинеллы найдены сосуды. Флоэма состоит ия „ситовидных трубок
и лубяной пярепхммы. За флоэмой расположены пе
(рис. 140, 4).
Проводящие пучки помещаются в воздушной полости, занимавшей
центральную часть стебля (нодучков, каЗГи полостей, может быть и несколь-
рипикл и эндодерма
вы
н 111: i
РАСТЕНИЯ
Рис. 140. Селягинелла:
1 — ветка с ризофорами и оиощпцшд от них корнями; 9 — часть побега с «брюшными» к «оливными»
/ листьями; Л —конец ветки с колоском; 4 — поперечный разрез стебля (в полости расположен пучок,
тн—тоаоекуаярные нита» п —перицикл); 5 —клетка листа с хлоропластом; < —поперечный разрез
листа: * — язычок (я — язычок» тр— трахещы листа).
ко), и подвешены к клеткам внутреннего слоя коры на нитях, состоящих
из одного ряда зеленых клеток. Они называются трабекулярными^, нитями
и являются клетками эндодермы.
Листья селягинеллы имеют одну (срединную) жилку. Снаружи лист
!»; ।:
или несколько пластинчатых хвематофоров (рис. 140, 5, 6). У останца штгрд
чаются на стороне листа. Мезо^шшиобразован
prnrnp раппп^ттггппьтми псснми|1нрун^ЦЧц1Г^летЕ1>ми с крупными межклет-
ннками. В клетках мезофилла находится по нескольку пластинчатых хрома-
тофоров.
У основания листа имеется группа рыхлых клеток (так называемая
подушечка), к дртпрдй па стеблятрабекулярнаяПкань. Через
эту подушечку воздух может_ протютдт^ внутрь^стебля.
У молодых листьев, на верхней стороне (несколько ваше основания),
имеется аа^плг€то^нн^чятый вырост — К5НЧ0К. ТасширенншГЪСнованием
ип
17 _
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
if»*;
а мегаспорангии — в нижней чяётй^цддп^ря. йдиятсб
шоженыкВфаолтныхфядах^а продольных срезах колоска
йоспоровость- имеет
НГ.1.1
г*
оннезаметен (рис. 140, 7). Функция его неизвестна.
' ТэОЛЫЦОе отличие Сйлягитопттм пт тигяуттпр яй^дрЯДеТСЯ В появлении
у нее разносиоровости (га т еррспория).
У селягинеллы спорангии двух родов — микроспорангии и мегаспо-
рангии. В микроспорангии развивается большое1 количество мелких спор
(рИСГ*141,I, 2), в мегаспорангиях образуются обычно четыре крупные
мегаспоры (рис. 141,3).
Спорангии расположены на верхней стороне спорофиллов,^которые, как
и у плаунов, собраны в колллк^ "Кплгмот четырехгранные, образуются
на верхушке., веточек . ' ' ' ' ,riv:
(Спорофиллы Вйептпе стпдны ^дшгятятнппыыи jtwctkctmw. по. в птлпппге
ОТ поглгетшх^^имйют хддрптп пяатпжтый няычок ГвЧтг. 141, 1}.
Споранпш сидят на небольшой ножке и имеют плч"5нтдпу™ или ебр^
но-яйцевидную форму. Они одеты двухслойной стенкой. Выстилаю—*
СЛОЙ (тЙНёТУм), РаСПОЛОЖенНЫЙ Глубже, ВОЗНИ»яет m гттярпт.д1жплДг тжг
Спорангии образуется из группы поверхностных клеток в пазухе споро-
филла, иногда несколько выше его.
Микро- и мегаспорангии собраны большей частью одном колоске, до
расположение их бывает различно. Чаще всего микроспорангии находятся
в верхней части. , ~
те и rtfiy**1*? рас™™1нены в различи"1
справа и слева).
В эволюционном развитии выс
большое яр!
(гаметофитов) j— мужских и женских.
Микроспоры при ее прорастании обрадуют рддудпрпдяппый
(упрптДппый)_мужг-»А^ц~яярпптп1г иотпрмй не ПОКИДаОТ ОбрЛОЧКИ МИКро-
споры. При первом делении микроспоры образуются 2 клетки — маленькая.
оидальная) клетка и более крупная, —
___рис. 141, rf).
дальнейшем-^антеридиальная клетка дает начало единственному
и дню. Здесь образуются клетки стенки антеридия и 2—4 спермагенные
клетки. К этому времени микроспорангий быстро вскрывается продольной
трещиной и споры с силой выбрасываются наружу (на расстояние 1—2 см).
Дальнейшее развитие мужского гаметофита протекает на земле. Д резуль-
тате последующего деления спермагенных клеток образуется бблыпое коли-
чество двужТутикишцХ'спёрматозоидов (рис? 141, б). Ко времени iiix созре-
валяя клетки стенки антеридия и ризоидальная цветка расплываются и
сперматозоиды плавают в общей массе плазмы (рис. 141, $).
Мегаспора, прорастая, образует женский заросток. У большинства
видов мегаспоры высеиваются из мегаспорангия и развитие заростков вдет
на земле. При делении ядра мегаспоры образуется большое количество
новых ядер, междукоторыми затем врзникаютпёрегородкиГ'и в итоге форми-
руется^мдюгоклеточйый- зарос/го^^ш^чГокитрюп^^ В верхкей
ч^сти зароСткл "затем наблюдается энергичное деление клеток^ в результате
1яего п^ядотаа мегаспоры лопается и заросток вьшячиддвтся пяружу. Клетки.—
заростказеленеют, на нем развиваются рмойды, и, таким образом, женский ’
заросток может питаться самостоятельно (рис. 141, 7).
В верхней части зарОСТйа развиваются* немногочисленные архегонии.
Они погружены в ткань заростка. Шейка имеет лишь 1—3 канальцевые
клетки. Д)пдрдотворение происходит во время дощдя. Сперматозоид, плавая
в воде, достигает архегония, проникает внутрь его и сливается с яйце-
• клеткой.
Оплодотворенная яйцеклетка делится поперечной перегородкой, обра~
- зуя две клетки. Верхняя клетка вытягивается и в дальнейшем образует
подвесок. Из нижней клетки развивается зародыш, состоящий из стебелька,
листочков, ножки и ризофора (рис. 141,8).
Uli
234
ВЫСШИЕ РАСТЕНИЯ
Рис. 141. Селягинелла:
2 — жродольный разрез колоска (о — ось. лиге — микроспорангий» лсгс — мегаспорангии, я — язычок);
2 — микроспораягий с микроспорами; а — мегаснорангий с мегаспорами; 4 — прорастание микроспоры и
образование редуцированного мужского заростка (р — ризоидальная клетка, ах — клетки стенки анте-
ридия, с — снермагенвые клетки); л — клетки стенки антеридия расплылись, и видны материнские клет-
ки сперматозоидов; в — сперматозоиды; 7 — женский заросток (ар — архегоний, а — зародыши, клз —
клетки заростка, об — оболочка мегаспоры); а — зародыш (п —подвесок, вк — верхушечная клетка,
ст —стебель, ад — зародышевый лист, н —ножка, я —язычок, р —ризофор).
У многих видов Selaginella начало зародышу дает не только нижняя,
но и верхняя клетка. При помощи ножки зародыш всасывает питательные
вещества из заростка. По мере разрушения его ткани зародыш продвигается
подвеском в глубь заростка. Сформировавшийся зародыш внедряется коре-
шком в землю и переходит к самостоятельной жизни, постепенно разви-
ваясь во взрослое растение селягинеллы.
У некоторых видов селягинелла (5. opus^ 5. rupestris) развитие заростка
и оплодотворение происходят внутри мегаспорангия, в то время, когда он
ОТДЕЛ — П АППРОТН Д КОПБР ДЗНЬГЯ
235
еще находится на материнском растении, и лишь после этого мегаспорангии
опадает» Отсюда до образования семени остается лишь один шаг. У некото-
рых селягинелл проросшие микроспоры переносятся ветром на мегаспоран-
гии (опыление).
Цикл развития селягинеллы сходен с плауном. Здесь также преобла-
дает спорофит (листостебельное растение селягинеллы).
Однако имеются и очень важные отличия.
У плауна все споры одинаковые, и они дают начало хорошо развитым
обоеполым заросткам.
У селягинеллы образуются споры двух видов — микроспоры и мега-
споры. При прорастании из них формируются различные заростки — муж-
ские и женские. Заростки (особенно мужской) сильно редуцированы.
Редукция гаметофита, свойственная селягинелле, тесно связана с разно-
споровостью и всегда сопутствует последней. Сильная редукция гаметофита
является основным направлением эволюции высших растений и хорошо
прослеживается в ряду Pteridophyta — Gymnospermatophyta — Angio-
spermatophyta.
4-Й ПОРЯДОК — ЛЕПИДОДЕНДРОВЫЕ (LEPIDODENDRALES)
Лепидодендровые — полностью вымершая группа растений. Впервые
они появляются в нижнем девоне, но наилучшего развития достигают в ка-
менноугольном периоде, к концу которого большинство представителей
вымирает. Отдельные формы дожили до нижней перми, а один род (Pleuromela)
известен из триасового периода мезозойской эры.
Наиболее распространенными были роды лепидодендрон и сигиллярия^
из их остатков образовались мощные залежи каменного угля.
Порядок включает 3 семейства.
Семейство лепидодендровые (Lepidodendraceae). Род лепидодендрон
(Lepidodendron) был представлен широко распространенными деревьями
до 40 м высотой и до 2 ле в поперечнике; известно свыше 100 видов лепидо-
дендрона.
Прямостоячие стволы лепидодендронов дихотомически ветвились в верх-
ней части. На конечных разветвлениях веточки были покрыты линейными,
спирально расположенными листьями. У различных видов они были от 1 до
50 см длиной. Листья имели язычок (рис. 142).
По опадании листьев на ветвях и стволе оставались их основания, так
называемые листовые подушечки (ср. селягинеллу). Они имели ромбическую
форму, возвышались над поверхностью ствола и, разрастаясь вместе с ним,
образовывали правильные ряды (рис. 142, 2).
На листовой подушечке хорошо заметны след от опавшего язычка, след
от жилки листа и два парных следа ткани, называемой парихнос
(рис. 142, 2).
У большинства видов стебли обладали вторичным утолщением. Ствол
и крупные ветви при этом были сифоностелические, а тонкие имели прото-
стель.
Стебли обладали мощной корой, на долю которой приходилось до 90%
общей поверхности поперечника стебля.
В центре стебля находилась сердцевина. К периферии располагались
далее метаксилема и протоксилема (от первичного строения стебля) и вто-
ричная ксилема, образованная лестничными сосудами. Далее располагалось
узкое кольцо камбия, за ним — кольцо флоэмы (вторичной и первичной) и,
* наконец, перицикл. Во вторичной ксилеме имелись многочисленные узкие
сердцевинные лучи.
Кора имела сложное строение и разделялась на внутреннюю (из парен-
химных клеток) и внешнюю (главным образом из механических, толстостен-
ных элементов). Далее к периферии располагался очень толстый слой внут-
ренней перидермы, образованной феллогеном, откладывавшим кнаружи
236
ВЫСШИВ РАСТЕНИЯ
Рис. 142. Лепидодендрон:
у-~общкй вяд; 1 — ствол с листоюсжя подушечками з—схема строения листовой водушочкя? (я—язы-
чок и след от евавшего язычна, ж — жилка и след ее, пр — мрихнод и следы их); 4 — поперечный
разрез стебля (е —сердцевжяи, жкс и пке — метаксилема и протоксилема пррвичшА ехе — вто-
ричная ксилема, жжб — камбий, ф — флоема, пер—перицикл, «спи — внутренняя ^первичная нора,
мпх — наружная пцшнчван кора, фв — феллоген, наб—наружная ппцперма, мжй—внутренняя на-
ридерма).
также слой внешней перидермы. Перидерма состояла из механических кле-
ток (рис. 142, 4).
Таким образом, механическая прочность ствола обеспечивалась пери-
дермой, а не ксилемой, как у современных деревьев. Лепидодендроны назы-
вают поэтому корбвыми деревьями.
В коре и перидерме заметны листовые следы — пучки, отходящие от
центральной стели без прорывов и шедшие к листьям.
В коре берут начало и паренхимные тяжи, разветвлявшиеся в наружных
слоях стебля на две группы и входившие затем в виде двух пар паренхимных
полостей в листья (парихаос). Через эту паренхимную ткань внутренние
ткани стебля снабжались воздухом.
В нижней части ствола лепидодендронов дихотомически ветвились
и давали мощные ризофоры, которые называются стивмариямц.
(
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
На стигмариях хорошо заметны следы отходящих от них корней.
На концах верхних разветвлений стебля развивались колоски. У разных
видов они имели разную величину — от 2,5 до 30 см юяшА и от 1 до 7,5 см
диаметром, К оси колоска прикреплялись спирально расположенные споро-
филлы.
На верхней стороне спорофиллов, прикрепляясь к ним своей нижней
поверхностью, находились микроспорангии (большей частью в верхней
части колосков) и мегаспорангии. Несколько выше спорангия на спорофилле
прикреплялся язычок (рис. 143,2). У многих видов колоски были однопо-
лыми.
В микроспорангиях развивались многочисленные микроспоры, в мега-
сцорангиях — от 8 до 16 мегаспор. Мегаспоры прорастали в довольно мощ-
ный женский заросток; в верхней части заростка развивались архегонии
(рис. 143, 3),
Семейство сигилляриевие (Sigillariaceae). Главным родом семей-
ства является сигиллярия (Sigillaria). Многочисленные виды ее жили вместе
& - лепидодендронами. Сигиллярии были крупными деревьями с высокими
(до 30 м) прямыми стволами. Они обычно совсем не ветвились или л
слегка ветвились на самой верхушке (рис. 144).
Узкие и длинные листья, опадая, оставляли на стебле невысокие листо-
вые подушечки, или листовые рубцы располагались прямо на коре.
Форма и величина листовых рубцов у разных видов сигиллярии очень
различны и напоминают печать, отчего растения и получили свое название
(Sigilla в переводе означает «печать»).
На листовых подушечках или рубцах заметны следы от пучка (средняя
жилка листа) и от двух тяжей парихнос (справа и слева). Выше имеется
небольшая ямка
hi
след от язычка.
Рис. 143.
Колоски лепидодендронов и семена ископаемых плауноввдных:
1 — продольный разрез колоска лепидодендрона (жде — ютроспорангни, аис — вюгасноранпн); 2—спо-
рофилл и спорангии в разрезе; г — архегоний; 4—ископаемые семена плауновндвмхз а — лепидокаркюн
(Lepidocarpon 1отахН)^пдодольный разрез через семяпочку (жа—женский заросток, и —интегумент,
иг— варкхнос, стенка мвгасворангия. пп — проводящий пучок), б — миадесмиа (мегаспорофилл с
мегаспорангнем)
ВЫСШИЕ РАСТЕНИЯ
Рис. 144. Сигиллярия:
1 — общий вид (с — стагнации); 2 — листо-
вые рубцы двух видов сигиллярии: а — 81-
glllaria inferiors б — 8, Bardiu
Сигиллярии были растениями раз-
носпоровыми, но колоски (16—20 см
длиной) чаще однополые и содержали
или микроспорангии, или мегаспоран-
гии (последние с 6—12 мегаспорами).
Семейство плевромейевые
(Pleuromeiaceae). Единственный род
этого семейства плеврамейя (Pleurome-
ia) известен из нижнего триаса мезо-
зойской эры. Плевромейя имела неболь-
шой ствол до 2 м высотой и до 10 см
диаметром. Внешне была похожа на
сигиллярию. Внизу стебель переходил
в единственный массивный ризофо р
(рис. 147), от которого во все стороны
отходили многочисленные корни. Этим
плевромейи очень напоминают по-
лушник.
В верхней части ствола распола-
гались листья. Спадая, они оставляли
листовые рубцы. Верхушка стебля за-
канчивалась колоском. На оси колоска
располагались многочисленные споро*
филлы, с мешковидными спорангиями,
сидевшими по одному на их верхней
стороне. Плевромейевые, вероятно, дали
начало порядку — Isoetales,
\ 5-И ПОРЯДОК — ЛЕПИДОСПЕРМОВЫЕ
(LEPIDOSPERMALES)
К порядку принадлежат растения,
жившие в верхнем отделе каменно-
угольного периода и обладавшие при-
митивными семенами. Известны 2 р о-
д а — лепидокарпон и миадесмия.
Лепидокарпон (Lepidocarpori) представлен лишь женскими колосками.
Предполагают, что лепидокарпоны были крупными деревьями, сходными
с лепцдодендровыми. \
В женских колосках на верхней стороне спорофиллов развивались мега-
спорангии. В них полного развития достигала лишь одна мегаспора, кото-
рая здесь же прорастала, образуя крупный заросток (рис. 143, 4).
От основания спорофиллов поднимаете^ кольцеобразный вырост, окру-
жавший мегаспорангий. Этот вырост соответствует интегументу семенных
растений, но не гомологичен ему, так как интегумент образуется из основа-
ния семяпочки.
В таком виде эта примитивная семяпочка отпадала от материнского
растения на земле, где и шло дальнейшее развитие заростка (образование
архегониев, оплодотворение), а затем и зародыша.
Миадесмия (Mia des mi а} была представлена травянистыми растениями,
внешним обликом напоминавшими селягинеллу. На верхней стороне мега-
спорофиллов, собранных в рыхлые колоски, развивалось по одному мега-
спорангию (рис. 143, 46). В них полного развития достигала одна лишь
мегаспора, прораставшая в женский заросток. Спорофиллы имели выросты,
которыми обрастали мегаспорангии, но наверху пе срастались, образуя
микропиле. \
Архегоний и зародыши пока не найдены. \
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
239
6-Й ПОРЯДОК — ПОЛУШНИКОВЫЕ (1SOETALES)
Полушниковые представлены одним семейством — Isoetaceae.
К нему принадлежат 2 р о д а —полушник (шильник, изоэт) и нашгорсшиана.
Полушник (Isoetes) насчитывает 64 вида многолетних травянистых расте-
ний, произрастающих на всех континентах, особенно в Северной Америке.
У нас встречаются три вида полушника. Из ^вих два — _ полуш/шк бЗёрН&и
(7. lacustris) и поле/ешщ# колючий (7,. ecfeinosponz) — растут в озерах на севе-
ре и в центре европейской части СССР, произрастая под водой (рис. 145, 2).
Многие виды в Америке являются земноводными растениями, растущими
на болотах и на сыром песке, *
г~ТТнЬель Полушника, ^достигающий 8—25 см высоты, короткий, луко-
вицеобразно утолщенный; к основанию он становится двух-трехлопастным.
От участков стебля между лопастями отходят многочисленные корни^ Выше
на стебле распоДй^ны_листья. Они внизу расширены. а тспдрту с у живя ют-
ся и становятся шиловидными. Внутри их, почти по всей длине, проходят
пенаре впяпу¥пттп1.мых канала? Листья уменьшаются в размерах в няттраи-
лений^Гцеитру Стебля, Верхушка стебля состоит из листовых зачатков и за-
канчивается точкой роста. Расширенное лопастное основание стебля соот-
ветствует ризофору, а верхняя часть, от которой отходят листья, является
собственно стеблем. Во внутреннем строении стебля хорошо заметна цилин-
дрическая стель. Она расширяется книзу и образует две или три лопасти,
от которых отходят пучки, направляющиеся в корни.
От верхней части стели отходят ответвления, идущие в листья. Стель
окружена широким кольцом
коры. В паренхиме, окружаю-
щей стель, имеется прослойка
меристемы, аналогичной кам-
биюА Здесь различаются ризо-
форная и стебельная меристемы.
. Камбий откладывает кнаружи
паренхимные клетки, а к внут-
ренней”части — смешанную про-
-ноНяшую ткань, состоящую из
мелких грЯ£евд"~и ситовипных
трубок. В результате деятельно^
етя*"К1й6ия^ стебель полушника
нарастает в толщину^ z
Наружные листья поД^-
шника обычно являются мега-
спорофиллами. ДА л «д_следуют
микроспорофилзш?^ит^нашш£Ц>
внутренние пелуффдрдя^ирд-
ВаНИЫе ЛИСТКИ ^рттоштля рога,
т’ативными.
~ И& зиму спорофиллы отми-
рают,^асохраняющ^ес?г ‘вегета-
тивные^ листья на следующий
год становятсямедаспорофил-
лами. j
---внутренней сто роне
расширенного основания спо-
рофиллов, в небольшой ямке,
помещаются микро- или мега-
спорангии. Они_рщшоложены
по одному, прикрепляясь—ко
дпу ямки короткой НОЖКПЙ^-
Ткань листа по краю ямки раз-
Рис. 145. Изоет, полушник (Isoetes lacustris):
1 — обшжй вид растения: я — нижняя часть споро-
филла: а — общий аид (видна ямка со спорангием),
б — продольный разрез (тр — трабекулы, и — инду-
зий, я — язычок, «к — воздушные камеры).
240
ВЫСШИВ РАСТЕНИЯ
растается в пленчатое образование, нависающее над спорангием в виде
покрывада^ (и н д у в и ft). Над ямкой расположен язычок, луковицеоб*
л/г- Л " Л -нмеж^однослой-
М ' J' I
145, 26). Они являются как бы распорками, не позволяющими
ам спорангия спадаться. В микроспорангиях развиваются мелкие
микроспоры, в мегаспорангиях — крупные мегаспоры.
Микроспора прорастает в сильно редуцированный мужской заросток,
напоминающий заросток селягинеллы. Он состоит из одной проталлиальной
(рнаоядальной) клетки, двух спермагенных клеток и четырех клеток стенки
антеридия (рис. 146, 2). Каждая спермагенная клетка дает по одному много-
жгутиковому сперматозоиду. При прорастании мегаспора образует много-
клеточный женский заросток, не покидающий мегаспоры. На верхнем его
Рис. 146. Мужской и женский заростки изоета:
1 — л — мужской зароет (Isoetes faenstris) <р — пиаояжшввя плетка, е —клена стала антеридия,
да* — вадам типе штата, се— сяерютоесшды); в— рнэрев через жеиский заросток (Isoetes whlnoi-
рм), оцртаянпийся внутри оболочки мегескмры (ар—архегсвий); 4 — архйожЫ (л — яйцекявпа.
бк — брошам вянашювая клетка, хш — нвейноиая канальцевая клетка, ш — клетки сшит шейки):
4—женский варовток с зароштаем (се — верхушка стебля, к — иеряичный тронь, в—вошка, л — авм-
чюи, 4-нрна шта}«
отдал—ПАПОРОТНИКООБРАЗНЫЕ
Mt
Рже. 147. Натгорстиана плевромейя:
обол* пд
конце образуется несколько архего-
ниев. Из оплодотворенной яйцеклет-
ки развивается зародыш. Он состо-
ит из ножки, стебелька, листочка и
зачатка корня. Разрастаясь, зародыш
разрушает оболочку мегаспоры и
вываливается наружу (рис. 146, 5).
Укоренившись в почве, растение
переходит к самостоятельной жизни.
Натгорстиана (Nathorsttana)—
ископаемое растение, известное из
нижнемеловых отложений мезозой-
ской эры. Внешне она была похожа
на полушник, отчасти и на плевро-
меню. Прямостоячий стебель натгор-
стианы достигал 12 см высоты, в
верхней части он был покрыт ли-
стьями. Из них самые верхние были
спорофиллами. Нижняя часть стебля
состояла из ризофора, от которого
отходили корни (рис. 147,1). Натгорстиана связана в своем происхожде*
с плевромейей. Изоэт является дальнейшим развитием натгорстианы.
Полушниковые, таким образом, потомки лапидодендрових.
II
.«ills
ДгйЖЛАШ— КДШОЛИСТОВИДНЬШ^ ЧЛЕНВСГЫЕ—
(SPHENOPSIDA)
Для представителей клйнолистовидных характерно расчленение стебля
на узлы и междоузлия (почему этот класс называется также членистыми
или Articulatae) и Мутовчатое расположение листьев на'стебле.
Листья Sphenopsida мелкие, но, в отличие от плауновитгпъгг они не яв-
ляются выростами стебля (э напл я м и), а возникли из коротких боковых
веточек (т е л о м о в), ставших уплоЩётаыми, листовиднымйГ~—*-------
В настоящее время клинолистовидныё^представяейвыь-на^Земле лишь
одним родом — хвощ (Equisetum). Но современные хвощи — травянио-
। Г, I;
in;
11 IS
тые растения — являются жалкими остатками некогда широко распрост-
раненного класса высших растений с большим числом родов и видов.
Клинолистовидные появляются еще в девоне, но расцвет их наблюдает-
' ся в каменноугольном периоде палеозойской эры. Тогда многие из них были
крупными деревьями, входящими (наряду с лрпнпппанртюнями и сигилляри-
ями) в состав лесов.
Отмершие остатки их дали начало яалажям каменного утяяь. Позднее
начинается угасание,, .деградация типа ктиигодтшш^тгит Ужа в начале
трпаг.. црмирамт мошные древовидные формы их. и с тех пор они представ-
лены только хвощами, дояйвшими-до-1”1*11** дней. '
В вюем происхождения ОШшливговшии«е связаны с псилофитами типа
Pgilovhuton, простейшие их представители (Яуеп1а,'Сайш>рйр«оп) Появ-
ляются впервые в девонском периоде. Они еще не пцеди явственного
членистого строения, листья~у~них~были как мутовчатые, так и очеоедайВ^
а «ппраит—» у "™»"^1йв^ййбЛИ~Вёрхушйчное расположение.
w Клинолистовидные разделяютйа 5 порядка^
242
ВЫСШИЕ РАСТЕНИЯ
Рис. (z8. Гиениевые:
1 — гиения (Hyenia elegans): а —общий вид растения; б — два бесплодных листа с повторно дихотоми-
ческим ветвлением; в—часть верхушки стебля с двумя спорофиллами; г — каламофитон (Calamophyton
primaevum); а — общий вид растения; 6.^ два спорофилла со спорангиями; в — два бесплодных листа.
ьл.
1 Й ПОРЯДОК — ГИЕНИЕВЫЕ (HYENI^XES)
Гиениевые — целиком вымершие растения. Их остатки известны из
среднего и нижнего девона. К ним принадлежали роды гиения и каламофи-
тон, относящиеся к двум семейств/м — Hyentaceae и Calamophytaceae.
Семейство гиениевые {Нуетасеае). Сюда принадлежит род гиения
{Hyenia}. В настоящее время описансг4 вида гиении. Все они были неболь-
шими кустарничками, высотой в несколько дециметров.
Лучше других изучена Н. eUgans (рис. 148). Она имела корневище,
от которого отходили вертикальные* побеги, не имевшие ясного членистого
строения. /
Стебли гиении дихотомически ветвились и были покрыты дважды-триж-
ды дихотомически разветвленными листьями. В верхних частях ветвей листья
располагались мутовками, у нижних — спирально (рис. 148,16).
Мутовки спорофиллов гиении были собраны в рыхлые колоски, распо-
лагавшиеся на верхушках особых плодовых веточек.
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
243
На верхушке спорофиллы дихотомически ветвились. На каждом из его
разветвлений располагалось по 2—3 спорангия, свешивавшихся вниз
(рис. 148» 1в).
Род каламофитон (Calamophytori) внешне напоминал гиению, но отли-
чался ясночленистым строением стебля и однажды дихотомически развет»
вленными листьями. Листья (около 1 см длиной) внизу были очередными,
а в верхней части ветвей располагались мутовками (рис. 148, 2).
Мутовки спорофиллов, как и у гиении, были собраны в рыхлые колоски.
Спорофиллы на разветвлениях имели по два спорангия.
Гиениевые произошли от псилофитов (типа Psylophyton).
ЗЛ ПОРЯДОК — КЛИНОЛИСТНЫЕ (SPHENOPHYLLALES)
К порядку клинолистных принадлежат вымершие растения, жившие
от верхнего девона до триаса.
Клинолистные имели членистые, бороздчатые стебли, обладавшие
вторичным ростом.
На стебле мутовками располагались листья.
Листья были хорошо развиты и имели клиновидную форму, откуда
произошло название порядка, а также и всего класса.
Клинолистные представлены тремя семействами, из которых мы рас-
смотрим наиболее обширное и важное семейство Sphenophyllaceae.
Главным родом этого семейства был клинолист (Sphenophyllum), пред-
ставленный большим числом видов. Клинолисты были, вероятно, лазающи-
ми растениями. Они произрастали в болотистых лесах, образуя густые
Рис. 149. Клинолисты:
j —общий вид Sphenophytium cunelfoilum; f — листья различных видов клинолиста! « — Spb. Thonii:
б — Sph, myriophyllutn; ч — Sph majus: з — поперечный разрез центральной части стебли (пб — пуч-
ковая древесина, мб — межиучновая древесина, Ол — древесинный луч, икс — иротоксилема, мкс — ме-
таксилема)
ВЫСШИЕ PACT
^Ur-
Я
Рис. 150. Клинолисты. Строе-
ние колосков и спорофиллов:
I — колосок Sphenophyllum daweo-
nli: а — продольный рдорез; б — по-
перечный; £ — дихотомически раз-
ветвленный спорофилл Sph. majus;
Л — обе части разветвленного споро-
филла Spb. fertile несут спорангии
заросли (рис. 149). Членистые стебли клинолистов достигли 1 ж длины и
имели ребристую поверхность, причем ребра переходили из одного меж-
доузлия в другое, не чередуясь (в отличие от хвощей).
Листья располагались мутовчато. В каждой мутовке было от 6 до 18
листьев. У одних видов листья были довольно широкими, цельными, у дру-
гих — узкими, дихотомически разветвленными (рис. 149, 2),
Некоторые виды (Sph. cuneifolium и др.) были гетерофилльныии: одни
листья были широкими, клиновидными, другие — узкими (рис. 149, I).
При посредстве узких листьев растения цеплялись за окружающие деревья.
Широкие листья были ассимилирующими. На стебле, в узлах, нередко раз-
вивались придаточные корни. Стебли клинолистов имели сложное строе-
ние. В центре стебля располагалась первичная ксилема, имевшая в раз-
резе треугольное очертание.
На вершине сторон треугольника располагалась протоксилема, а I
верхушки — метаксилема с более крупными трахеидами (рис. 149, Л).
Между лучами первичной ксилемы находилась первичная флоэма.
Кнаружи от первичной ксилемы возникал камбий. В результате его дея-
тельности внутрь откладывалась вторичная ксилема, вначале занолх
шая
пространства между лопастями треугольника, а затем располагавшаяся
кольцом вокруг всей первичной ксилемы. При этом против углов треуголь-
ника откладывались более мелкие элементы.
Кнаружи камбий откладывал вторичную флоэму. Дальше шла кора,
В наружных слоях коры имелся феллоген, дававший слои пробки. Споран-
ги^с-у некоторых видов клинолистов развивалась в пазухах вегетативных
листьев, но у большинства — на верхушке веточек возникали спороносные
колоски. Они достигали 10 см в длину и 1 — 1,5 см в поперечнике. "
На оси колоска мутовками располагались спорофиллы. Каждый споро-
филл состоял из двух частей — нижней, бесплодной и верхней, спороносной.
Степень развития бесплодной и спороносной частей спорофилла, а также
форма их у разных видов Sphmophyllum были очень разнообразны,
У Sph. dawsonii бесплодные части спорофиллов срастались вместе почти
по всей длине, образуя как бы чашу (рис. 150, 26). Спороносные их части
несли по три спорангия, которые висели на ножках, загибавшихся на вер-
хушке, и напоминали спорангии гиениевых.
У Sph. majus спорофиллы дихотомически ветвились. Спороносная часть
их состояла из четырех спорангиев (рис, 150, 2). Спорофиллы резко отлича-
лись от бесплодных листьев.
ОТДВЛ — ПАПОРОТНИКООБРАЗНЫХ
У Sph. fertile обе части спорофилла были спороносными и заканчива-
лись щитками, н^нижней стороне которых развивались спорангии. Эти
спорофиллы очень похожи на спорофиллы (спорангиофоры) хво-
щей (рис. 150, 3).
Большинство клинолистов бым равноспоровыми растениями, но не-
которые из них (Sph. delectiu) обладали микро- и мегаспорангиями.
Клинолистные произошли, вероятно, от гиениевых (типа гиения).
8-И ПОРЯДОК — КАЛАМИТОВЫЕ (CALAM1TALES) \ /
< и
Каламитовые — вымершие растения. Они впервые появляются в верх-
нем девоне, наибольшего развития достигают в каменноугольном периоде
и исчезают в юрском периоде мезозойской зры.
Каламитовые — деревья. Внешним обликом они иядомпиали хвощи.
Стебли их членистые, ребристые; спорофиллы Хзпораягиофоры) щитко-
видные, они несли по 4 спорангия и были собраны в колоски разнообразной
формы ^строения. Порядок представлю двумя семействами — осте-
рокаммитотх и каламитовых.
Рис. 151. Каламиты:
1 — вегетативные органы астерокалампн) а — часть стебля; б — олиственный боковой аобеа; • — инет;
й—каламит (Calainitee carlnatu«)i а — обтай вид; б — поперечный разрез стебля (ад — вторичная дре-
весивав мкс — метаксилема, «к — наринальная волость)
246
ВЫСШИЕ РАСТЕНИЯ
Рис. 152. Строение ко-
лосков каламитов:
/ — палеостахвд; 2 — кала-
мостахкс
Семейство астерокаламитовые (Asterocalamitaceae), К нему при-
надлежат более примитивные растения порядка, жившие от верхнего девона
до нижнего карбона. Наиболее распространенными были представители
рода астерокаламит (Asterocalamites) (нижний карбон). Виды его были
деревья. В стебле ребра соседних междоузлий не чередовались. Листья
длиной до 5 слс были узкими и многократно дихотомически ветвились
(рис. 151,1в). Спороносные колоски состояли из щитовидных спорангиев,
собранных в мутовки* Последние чередовались с мутовками бесплодных
листьев.
Семейство каламитовые (Calamitaceae). К нему относятся более
сложные формы. Они очень разнообразны и объединяются в сборный род
каламиты (Catamites), насчитывающий много видов.
Каламиты появляются в нижпих отделах каменноугольного периода,
достигают наибольшего развития в среднем и верхнем его отделах. Большая
часть их вымерла в пермском периоде, но некоторые (например, Neocalami-
tes) дожили до конца триасового периода мезозойской эры.
Каламиты были мощными деревьями, росли на болотах и имели длинные
горизонтальные корневища, скрытые в болотной почве, от которых отходили
вверх надземные вертикальные побеги высотой до 30 см.
Стебли расчленялись на узлы и междоузлия. Поверхность междоузлий
была ребристой. Стебли обильно ветвились. Ветви также были членистыми.
Ребра соседних-междоузлий- чапте-чепеловались. От узлов на корневищах
и В нижней части надземных~£теблей- отходили многочисленные придатоЧ-
ные-югрни (рис."~15Т, 2), Листья в узлах располагались мутовками.
—состояли изГ8 —13 свободных или сросшихся листьев, имев-
ших в длину от 0,5 до 20 см. Листья, линейные, цельные, с одной жилкой,
содержали хлорофилл и выполняли, в отличие от листьев современных До-
тийй7 фуп«цу«^фотосинтеза. благодаря йаличиюкамбия стебли каламитов
нарастали в толщину. Они состояли из коры, стели и снаружи были покры-
ты эпидермисом. Внутри стебля развивалась сердцевина, которая у взрос-
лых растений разрушалась (рис. 151, 26). Первичные пучки состояли из
протоксилемы и метаксилемы. Последняя развивалась в направлении к
центру стебля, а протоксилема рано разрушалась, и на ее месте появлялась
пучковая (каринальная) полость. Затем возникало кольцо камбия.
Камбии откладывал внутрь вторичную ксилему, а снаружи — вторичную
флоэму. Хорошо были заметны сердцевидные лучи. Кора была мощной и
содержала перидерму с механическими клетками. Корни имели строение,
сходное со стеблем, и также обладали вторичным ростом.
Спороносные колоски были очень разнообразны и описаны под разны-
ми родовыми названиями (Calamostachys, Palaeosiachya и др-).
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
В колоске каламостахиса (Calamostachys) мутовки спорофиллов чере-
довались с мутовками бесплодных листьев, которые загибались кверху и
покрывали спорофиллы (рис. 152, 2), У палеостахии (Palaeostachya) споран-
гии располагались по одному в пазухе бесплодного листа. Это, несомненно,
вторичная группа* Об этом свидетельствует ход пучка, идущего в ножку
спорангия. Пучок сначала направляется вверх (к месту прежнего располо-
жения спорангия), а затем загибается вниз и так уже входит в спорангий
(рис. 152, /).
У других каламитов строение колосков еще более сложное.
Среди каламитов были равноспоровые и разноспоровые растения. Ка-
ламиты являются прямыми потомками гиениевых (типа каламофитов).
1 ЛА
[^4П ПОРЛДОП —-ХВОЩОВЫЕ (EQUI SET ALES).
Единственное семейство Equtsetaceae представлено в настоящее
время лишь одним родом — хвощ (Equisetum), который насчитывает
32 вида, распространенных всюду на Земле, кроме Австралии и Новой Зе-
ландии. В пашей флоре встречается 12 видов хвоща, растущих на болотах
(Е. limbsum, Е. palustre), в лесах (£* silvaticum), в кустарниках (зимнезеле-
ный — Е. hiemale), на лугах, пашнях, железнодорожных насыпях и дру-
гих местах (Е. arvense, Е. pratense).
Современные хвощи — небольшие травянистые растения около 80 —
100 см высотой и 2 — 5 мм толщиной, но некоторые виды, например тропи-
ческий южноамериканский Е. giganteum, достигают 10 — 13 м длины и явля-
Тис 153УХвощ полевой (Equfsetum arvense):
1 — летний, бесплодный побег; г — весенний спороносный побег (хд — клубеньки, кол — колоски с му-
товками спорофиллов); 3 — спорофилл (спорангиофор) со спорангиями: а — вид сбоку; б — вид снизу;
4 — споры с развернутыми элатерами; 3 — спора с завернутыми элатерами; в — схема прохождения пуч-
ков в стебле хвоща •
выс
fits
ГВ РАСТЕНИЯ
Рис. 154. Хвощ полевой:
1 — схема жсяеречното равреаа хвоща области междоузлия; в — детальное строение стебля яз участка
дс—у схемы 1 (а—внйдврмяс» ж — механическая ткань а ребрышках, ос — ассимвляцвоннвд ткань в
жмвдннках, «в “ валетсудярная волость, ва— цдовдооцкВ нучок, цп — цемтрадьшя полость стебля,
™ — вддорта, Ф—флдоде жучка, зд — певтраяьный цилиндр, жте-иетмвдави, в«с—нрвтоксжле-
шв, кор — кора, отосновная псань, у — устьице, с^цеицюжьная волость)
ются лазающими растениями. Стебель лсексияонскдео хвоща (Е. Schaffnerl)
при высоте в 2 л достигает толщины 10 сж.
Остальные виды — небольшие травянистые растения. Рассмотрим стро-
ение хвощей на примере хвоща полевого (Equisetum arvense) (рис. 153). Поле-
вой хвощ — многолетнее растение с членистым ветвистым корневищем, рас-
положенным глубоко (до 1 и) в почве. Проиярастяет бндю паровым пллям1
залежам, часто*; < ««ное «а т-ние. На корневищах образуются
^уб^тадшП^брбчёкнЫе^бйковые побеги), в которых откладывается крах-
мал. От узлов корневища отходят придаточные корни.
Надзешгые побеги хвоща полевого двух видов. Одни из них вегетатив-
ные, зеленые, мутовчато-раЗВЬТвЛенные, летние. Надземные стебли ветвятсяГ.
“ злах стеблей и выходят наружу, птюби-
вГ Летни^Тпобеги к осени от-
миракА. ^ругИТдобегй(ЬёСённие> спороносные, розовато-бурые, появля-
ютёя1 ранней весной, неветвистые. На их верхушке образуется спороносный
колосок, на оси которого расположены спорофиллы, несущие спорангии со
спорами. После спороношения весенние побеги отмирают. У других видов
хвоща побеги одинаковые. Стебли хвоща расчленены на правильно чередую-
щиеся узлы и междоузлия, ребристые. Me v ' * *. < *; ।«г. • * злы
—аадрлнены паренхимной тканью. Реб«: псддпдт wawnoy^iiff череду-
ют • i • гс"й ом. Листья хвоща чешуе^тдр™, ^урцо коричневые, лише-
;i. •«।»и < । >:. Они сроедись(за исключением верхних частей, имеющих
вид зубчиков) в трубчатое влагалище, 11рнкренлест$аиювотаИйёМ к’ узлу.
Всугедствие редукцпи дшсть^ пртктйиоеки 'органомфотосинтеза _явля-
вдся-етебэдь, И связи с * • acc i t- j sn еннйя^гкЗйС^
На поперечном разрезе, про: • < ; s •: •» асти междоузлия, стебель
Имеет следующее строение. Снаружи стебель неровный, гранистый — воз-
вышенные участки (ребрышки) чередуются по окружности с несколько вдав-
ленными (ложбинками). Однослойный эпидермис покрывает стебель снару-
жи. Оболочки клеток эпидермиса пропитаны кремнеземом, что придает
стеблям хвощей большую прочность и жесткость. Внутрь от эпидермиса распо-
лагаются кора и центральный цилиндр (степь) с кольцом мелких изолиро*
ванных пучков и с полостью в центре, которая образовалась на месте сердце-
n t и л г*
: t; Г
вая у основания влагалище сросшихся
H1S
ТШТ
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
II
вины. В молодых стеблях сердцевина имеется. Под ребрышками расположены
участки механической ткани (рис. 154), а под ложбинками — асси-
миляционная ткань. Под каждым участком ассимиляционной ткани, состоя-
щей из рыхло расположенных хлорофиллоносных клеток, находится по-
лость (коровая, или валекулярная, полость, она отсутствовала у
каламитов, рис. 154).
Под механической тканью (под ребрышками) расположены проводя-
щие пучки (рис. 154, 2).
Проводящий пучок хвощей коллатеральный. Обращенная
к центру стебля его часть занята пучковой (каринальной) полостью.
Она образовалась на месте разрушившейся протоксилемы. Иногда в пучко-
вой полости сохраняются отдельные кольчатые или спиральные трахеиды.
По каринальным полостям происходит передвижение воды.
Над полостью расположена флоэма. Сбоку от флозмы имеются две
группы метаксилемы (рис. 154, 2), представленной лестничными трахеида-
ми. У Е. arvense, как и у большинства видов, вся стель (зустель) окружена
снаружи перициклом и эндодермой (рис. 154). У некоторых видов имеется
еще внутренняя эндодерма (например, у Е. hlemale) или каждый пучок окру-
жен своей эндодермой (в стеблях Е. limosum). Последний тип эндодермы яв-
ляется наиболее совершенным.
. ч Пучки, хвощей закрытые, камбия не имеют. Вторичные ткани не обра-
зуются, и стебли не способны к вторичному утолщению, чем хвощи резко
отричаютея от каламитовых.
В эпидермисе (над ассимиляционной тканью, в ложбинках) имеются
, устьица, состоящие из двух пар клеток, расположенных друг над другом.
Верхние клетки неподвижны, так как оболочки их пропитаны кремнеземом.
Под ними расположена вторая пара клеток, являющихся, собственно,, за-
мыийишрмми ^лотками; они живые и нормально функционируют (рис. 154, Z).
Узлы стебля заполнены тканью. Здесь каждый из пучков, входящий в
узел снизу, из' междоузлия, разделяется на три веточки. Средняя из них
направляется в жилку листа. Две боковые веточки сливаются каждая с боко-
вой веточкой соседнего пучка. Они и образуют пучок, идущий в верхнее
междоузлие (рис. 153, 6). Этим и объясняется чередование пучков у двух.
соседних междоузлий.
Спороносные колоски у хвощей возникают по одному на верхушке глав-
ного побега, а иногда и боковых ветвей. У большинства видов побег, несу-.
щий колосок, зеленый, ассимилирующий. Но у полевого хвоща (Е. arvense).
спороносный побег участия в фотосинтезе не принимает (см. выше).
Колосок хвоща состоит из многочисленных спорофиллов (с п о р ан-
гио ф о р о в), собранных мутовками на его оси. Спорофиллы состоят из
ножки и расположенного на ее верхушке щитовидного диска, обычно имею-
щего шестигранную форму. На нижней стороне диска, вокруг ножки, рас-
полагается 5—13 мешковидных спорангиев (рис. 153, 8), в зрелом состоя-
нии одетыд однослойной-стенкой.
Хвощ — растение равноспоровое. В спорангии образуется большое
количество одинаковых спор. Спора, кроме двух оболочек (эндоспория
к зкзоспория), одета еще третьей — наружной оболочкой (з п и с п о-
р и е м). Наружная оболочка не сплошная, а состоит из двух спирально
закрученных лент (пружинок, элатер), расширяющихся лопатковидно на
четырех концах и прикрепляющихся к споре в одном месте. Пружинки (эла-
теры) в сухую погоду раскручиваются, что содействует распространению
спор группами, кучками (рис. 153, 4). В сырую погоду они закручены во-
круг споры. Спорофиллы в колоске плотно придвинуты друг к другу. Ко
времени созревания спорангиев ось колоска несколько разрастается (у осно-
вания всех междоузлий имеется меристематическая ткань) и мутовки споро-
филлов раздвигаются. Спорангии при этом вскрываются продольной тре-
щиной, и споры высыпаются наружу. Попав на землю, споры прорастают
в -заросток. Заросток хвоща имеет .вид зеленой, многократно рассеченной
II
2Г0
ВЫСШИЕ РАСТЕНИЯ
Рис. 155. Хвощ:
1 — заростки хвоща (Е. quisetum arvense)s в — мужской заросток (ан— антеридии); б — женский зарос-
ток (ар — архегонии); в — антеридий; г — сперматозоид; б — архегоний; е — зародыш (пл — первые
листья, н — ножка, тч —точка роста, к — корень); 2 — схема формирования стели молодого спорофита
хвоща. В сторону вынесены схемы поперечных разрезов стели (ст — стель, к — ножка, л — листья,
к — корень с корневым чехликом)
пластинки размером 0,1 — 0,9 см. Антеридии и архегонии возникают или
на одном и том ясе заростке, или на разных заростках, несмотря на то что
споры морфологически одинаковы.
У многих видов хвоща имеется физиологическая разноспоровость. Спо-
ры хвощей, соединенные в группы благодаря сцеплению пружинками, при
попадании на почву оказываются в неодинаково благоприятных условиях
освещения, снабжения водой и т. д. (например, верхние и нижние споры в
кучке). Прорастая, некоторые из, них образуют более мелкие мужские за-
ростки с антериднями, другие — более крупные заростки с архегониями
(рис. 155). Это явление можно рассматривать как известный отголосок мор-
фологической разноспоровости предков хвощей, тем более что у некоторых
видов физиологическая разноспоровость является постоянной и не связана
с условиями развития заростков.
Однако для многих видов (например, Е. arvense) экспериментально
установлено, что, поливая питательным раствором мужские заростки, на
которых начали развиваться антеридии, можно добиться увеличения их
размеров и развития на них архегониев.
Антеридии хвощей погружены в ткань заростка (рис. 155, 1а). В каж-
дом из них развивается свыше 200 многожгутиковых сперматозоидов
(рис. 155, 1г). Архегонии лишь шейкой возвышаются над заростком
(рио. 155, 16). Оплодотворение осуществляется в сырую погоду. Оплодо-
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
251
творенная яйцеклетка дает начало
зародышу. Подвеска у хвощей не
образуется. Зародыш первоначаль-
но скрыт в ткани заростка. Он со-
стоит из стебелька, двух-трех ли-
сточков и корешка (рис. 155, /е).
Пробивая ткань заростка, коре-
шок укрепляется в земле, и
растение переходит к самостоя-
тельной жизни. На одном зарост-
ке часто возникает несколько за-
родышей. Молодой стебель хвоща
имеет протостелическое строение
(рис. 155, 2).
П рактическое значение х во-
щей невелико. Прочные стебли их
употребляют для чистки метал-
лической посуды и полировки де-
рева. Укороченные клубневидные
побеги, образующиеся на корне-
вищах Е, arvense, иногда употре-
бляют в пищу.
Отрицательное значение име-
ют сорные виды — Е. arvense,
E.pratense, часто £. silvaticum. Их
корневища, разрезанные на куски
при обработке почвы, развивают
по нескольку надземных побегов
каждый. Борьба с хвощами за-
труднительна.
Рис. 156 Ископаемые хвощовые:
1 — шизоневра; 2 — филлотека
В ископаемом состоянии хвощи известны из каменноугольного периода,
например вида рода Еquisetites. Найдены стебли и ветви их без споро
ношения. Кроме того, в пермских отложениях обнаружена шизоневра
{Schizoneura) — хвощ с хорошо развитыми листьями, а в юрских — филло-
тека {Phyllotheca), листья которой срастались у основания (рис. 156).
Современные хвощи появились на Земле с мелового периода. Проис*
хождение хвощовых неясно. Они пе являются прямыми потомками известных
нам каламитов, обладавших в строении вегетативных органов более вы-
сокой организацией (вторичный рост, древовидные формы). Вероятно, хво-
щи возникли от менее специализированных форм палеозойских каламитов.
<)-H~K.TIACC —
ОБИДНЫЕ, ПАПОРОТНИКИ
В отличие от хвощовых и плауновых папоротниковидные характеризуют-
ся макрофиллией — крупнолистностью. Листья их крупные, многократно
расчлененные, реже цельные. Листья папоротниковидных произошли из
крупных ветвей (теломов и их групп) простейших наземных растений —
псилофитов. Они образовались путем срастания и дальнейшего уплощения
(кладодификации) ветвей. Листья даже современных папоротни-
ков па растают верхушкой, как стебель,__двм^подчс pkilUJB J шГЬв ня я
(осевад^дцшцода. 11рй отхождении проводящей системы листа от стебля в
иёи'ббразуется листовой прорыв (листовых прорывов нет у хвощовых и плау-
новых), нередко он очень крупный и соответствует веточному прорыву.
Стебли папоротниковидных также очень разнообразны. Большинство
их имеет сравнительно слаборазвитый стебель, представленный нередко
вы
ЙП
[ИИ РАСТЕНИЯ
корневищем, часто скрытым в земле. У многих форм стебель ползучий, тон-
кий. Но имеется и ряд папоротников с прямостоячим стеблем. Среди них
известны и древовидные формы. Исходными являются прямостоячие стебли
подудревесного типа, свойственные, простейшим ископаемым папоротнико-
видным. От них произошли древовидные и травянистые формы.
Столярное строение папоротниковидных очень разнообразно. Здесь
имеются у разных групп все тины стели (протостель, сифоностель, диктио-
стель, подициклия, эустель).
Спорангии у папоротников возникают на нижней поверхности листа
в большом количестве; реже они одиночные, чаще же спорангии собраны
в кучки — сорусы, одетые особым выростом листа — покрывалом
Однано у простейших папоротниковидных спорангии одиночные и обра-
гии расположены но -краи$ листа.
Спорангии возникают или на обычных вегетативных листьях, или
на особых спороносныХ"йиетьях^(сдо^ р о ф и л л а х). Спорофиллы могут
быть совершенно схопны с вегетатипнммиПкиг.ткомтсГу р р“ф р ф и д д а м и)
или отличаются от них величиной и формой. -------
У одних папоротниковидных спорангии образуются из группы клеток
листа как поверхностных, так и субэпидермических. В этом случае стенка
взрослых спорангиев многослойная. Такие спорангии свойственны древ-
нейшим папоротниковидным и ряду более простых современных порядков.
Их немного.
У других папоротниковидных спорангии образуются из одной поверх-
ностной клетки и во взрослом состоянии имеют однослойную стенку. Эти
папоротниковые, несомненно, более молодые. Они являются и наиболее рас-
пространенными в современную геологическую эпоху.
Зрелые спорангии вскрываются при помощи специальных образований,
развивающихся на его стенках (кольцо разного устройства, группы клеток
с неравномерно утолщенными стенками и т. д.). Простейшие представители
папоротниковидных этих приспособлений не имели.
Среди папоротниковидных встречаются как равноспоровые, так и раз-
носпоровые растения.
При прорастании спор развиваются заростки (гаметофиты).
Заростки равноспоровых папоротниковидных зеленые, питающиеся
самостоятельно. Обычно они маленькие и весьма разнообразны по форме:
пластинчатые, сердцевидные, нитевидные и т. д.
У разноспоровых папоротниковидных наблюдается сильное упрощение
(редукция) заростков, особенно мужских. Заростки их имеют микроскопи-
ческие размеры. Мужские заростки теряют способность к самостоятельной
жизни и питанию.
Папоротниковидные являются в настоящее время наиболее распро-
страненной группой высших споровых растений. Они насчитывают более
10 000 видов, распространенных по всей Земле, но наибольшее число видов
отмечено для влажных тропических областей, где встречаются и древовид-
ные папоротники. Большинство современных папоротников — травянис-
тые растения.
Впервые папоротниковидные появляются в верхнем девоне.
Класс папоротниковидные разделяют на 3 подкласса.
Mb
1-Й ПОДКЛАСС — ПЕРВОПАПОРОТНИКИ (PR1MOF1L1CES)
'л
К этому подклассу относятся исключительно ископаемые растения
очень раанообравного внешнего вида и строения. Все первопапоротники
имели спорангии с многослойной стенкой. Развивались спорангии ив группы
субэпидермических клеток листа. j
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
253
Рис, 157. Протоптеридиум:
J — Protopteridium minutum; а-»побег со спорангиями я листообразно уплощенными дихотомически
разветвленными ветвями; б—схема строения сиоравтия; в—спора; х—Р. hostimeose
Простейшие первопапоротники были весьма сходны с псилофитами.
Более организованные имели уже типичный папоротниковый облик.
Систематика Primofilices еще недостаточно разработана. Обычно под-
класс разделяют на 4 порядка.
1-И ПОРЯДОК — ПРОТОПТЕРИДИЕВЫЕ (PROMPTER 1DALES)
В этот порядок входят наиболее примитивные первопапоротники, пред-
ставители рода Protopteridium- Их остатки встречаются в среднедевонских
отложениях Европы и Азии. К роду принадлежат 5 видов. Прямостоячие
стебли их несли дихотомически разветвленные ветви. Боковые веточки были
плоскими и также вильчато-разветвленными наверху (рис. 157, 2а).
Спорангии помещались па верхушках некоторых конечных разветвле-
ний (рис. 157, /б) и были снабжены продольной полоской более мелких кле-
ток,
способствовав
х их раскрыванию. Споры одинаковые.
У некоторых видов протоптеридиума (Р. hostimense) отдельные плоские
ш
ветви напоминали уже сложноперистые листья современных папоротников
и в молодом состоянии были загнуты улиткообразно (рис. 157, 2). Анатоми-
ческое строение растений неизвестно.
2-Й ПОРЯДОК — КЛАДОКСИЛОВЫЕ (CLADOXYLALES) ,
К нему относится род кладоксилон (Cladoxylori). Он был распростра-
нен от девона до нижнего карбона и представлял собой невысокий ветвистый
кустарник, до 25 см высотой. Побеги его дихотомически ветвились и дости-
гали 4 — 6 см в диаметре. На стебле и ветвях его имелись плоские, вильча-
то-разветвленные листья-веточки (рис. 158).
В верхней части развивались группами дихотомически разветвленные
спорофиллы. Каждая веточка заканчивалась одним спорангием (рис. 158,2).
Стебель имел расчлененную актиностель (рис. 158, 5).
254
ВЫСШИЕ РАСТЕНИЯ
Рис. 158. Кладоксилон:
1 — общий вид; 2 — спорофилл; 3 — 4 — вегета-
тивные листья; S — поперечный разреа стебля
Рис. 159. Зигоптерис!
1 — общий вид листа; s — пучок спорангиев;
з — два спорангия (видна двухслойная стенка
спорангия й продольное кольцо)
I
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
З Й ПОРЯДОК — ЗИГОПТЕРИДИЕВЫЕ (ZYGOPTERIDALES) /
Сюда принадлежит несколько родов. Род зигоптерис (Zygopteris) был
представлен восемью видами, известными из каменноугольных отложений.
Зигоптерис имел уже более сходный с папоротниками облик. Стебли его
толщиной до 1,5 см были окружены основаниями «листьев», образуя вокруг
него как бы чехол.
Довольно крупные листья-ветви состояли из главного черешка (рахи-
са), на котором располагались боковые разветвления, сидевшие попарно
в два ряда (справа и слева). Из них одна веточка была более тонкой
(рис. 159). Веточки, в свою очередь, разветвлялись и на конечных разветвле-
ниях несли мелкие плоские листочки.
Черешки на поперечном разрезе имели своеобразной формы ксилему —
в виде буквы Н.
Спорангии были собраны группами, возникавшими вследствие укора-
чивания ветвей. Каждый спорангий помещался на толстой ножке, имел
Рис. 160. Первопапоротники:
1 — археоптервди а — часть бесплодного побега Archaeopterls TatlfoTtaj б — часть спороносного иобега
Arch, lati folia; в — спороносный побег Arch, hlbernfca (бесплодные «листочки» чередуются оо споро*
нотными); я — микро- и мегаспора; 2 —ракоптерис (Rhacopteris panicullfera)
258
выс
fl г,
Е РАСТЕНИЯ
обычно двухслойную стенку, снабженную вертикально расположенный
кольцом, состоявшим из клеток с утолщенными оболочками (рис. 159, 5).
У представителей рода стауроптерис (Stauropteris) — из карбона —
спорангии были еще без кольца и вскрывались отверстием на верхушке.
4-Я ПОРЯДОК — АРХЕОПТЕРИСОВЫЕ (ARCHAEOPTERIDALES) ч
К
этому порядку относятся ископаемые растения, также имев:
е уже
папоротниковый облик, напоминая ужовниковые и мараттиевые. Жили
они в девонском и каменноугольном периодах палеозойской эры.
Наиболее древний р о д— археоптерис (Archaeopteris). Он известен уже
из среднего девона. Археоптерис имел крупные двоякоперистые листья с
цельными или рассеченными сегментами. Лист разделялся на спороносную
и вегетативную части (рис. 160,1в). Спороносная часть состояла из черешоч-
ков, на которых находились продолговатые спорангии, прикреплявшиеся
к ним небольшой ножкой. Археоптерис был разноспоровым растением
(рис. 160, /г).
У других родов, например у каменноугольного Rhacopterusy рас-
членение листа на спороносную и вегетативную части было еще более рез-
ким (рис. 160, 2).
Первопапоротники ведут происхождение от псилофитовых и являются
родоначальной группой растений как для следующих классов папоротнико-
видных, так и для семенных папоротников (Pteridospermae) — простейших
голосеменных растений.
2-Й ПОДКЛАСС - ЭУСПОРАНГИАТНЫЕ ПАПОРОТНИКИ
(EUF1L1CES)
У представителей этого подкласса спорангии возникают из группы
эпидермических и субэпидермических клеток и имеют многослойную стенку.
- В настоящее время подкласс представлен небольшим числом видов. Это
древняя группа папоротников, расцвет которой имел место в прежние гео-
логические периоды. -
Подкласс разделяется на 2 порядка.
/жОВНИКОВЫЕ (OPHIOG LOSSALES)
Ужовниковые — небольшие травянистые растения. Они имеют стебель
(корневище), скрытый в земле. От корневища, растущего косо вверх,
отходят надземные листья, разделенные на две части — спороносную и зе-
леную, бесплодную, — расположенные на общем черешке. Вниз от корне-
вища, укрепляясь в почве, отходят несколько довольно толстых корней.
К этому порядку принадлежит одно семейство Ophioglossaceae
с тремя родами: ужовник, ботрихиум и гелъминтостахис.
Род ужовник (Ophioglossuiri) насчитывает 45 видов, большинство ко-
торых встречается в тропиках, где они нередко являются эпифитами.
Небольшое число видов произрастает в умеренных областях.
В северных и средних районах Европейской части СССР распростра-
нен по мшистым луговинам ужовник обыкновенный (О, vulgatum, рис. 161,7).
Как и у других видов, у ужовника обыкновенного бесплодная часть листа
имеет вид овальной цельной пластинки. Спороносная часть листа стеблевид-
ная и заканчивается колоском, на оси которого в два ряда расположены
спорангии.
Спорангии срослись между слбпй; образовав синангии.
Они несколько погружены в ткань оси колоска?*^————--------------
Корневище имеет диктиостелическое строение (рис. 162, I, 2). Отдель-
ные участки стели (меристели) представлены коллатеральными пучками-
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
Шаровидные спорангииоде-
ты многослойной—СТОШСий. В
нихг^раз§нвается мпого одина-
ковых" спор. Спорангий без
• । s в ' ; • * *» • . -
нои трещинойг—
"-"Заростки ужовника бес- \
цветные, развиваются и живут |
под землей на глубине 2 —
10 см, Ойй—достигают 5 — 6 см
в длину и 0,5 — 1,5 см в тол-
щину.
Заросток ^имеетрддиад^ьное
строение и состоит из паренхим-
ных клеток?" Ризоидов' на за-
росткО~нет, а в наружныхГсдоях
зйростка расположены__гцфы
грибаП(м и" к б' р’ и з aj.
На заростке образуются /
многочисленные антеридии и I'
архегонии (рис. 162, 3). Схщр- i
мятпяпипы мтупгржгутиковые^___;
j- Возникающий в результате
оплодотворения зародыш пер-
воначально развивается^^ спот-
зарбстка, будута " в его
ткани/ Затем ^жра^мвЯетсте-
'бельТнерехрдяприи в нижней
части в корешок, окруженный
корневым че^ликом.
""Разрывая ткань заростка
и укореняясь, растение перехо-
дит к самостоятельной жизни.
Развитие молодого растения
идет очень медленно. Для пол-
ного развития листа требуется
5 — 6 лет.
Род ботрихиум (Botrychi-
ит) насчитывает 35 видов, рас-
пространенных по всей Земле.
Он отличается от. ужовника рас-
сеченной стерильной частью
листа и ветвистой спороносной
его частью.
У нас по сырым лесным
луговинам часто встречается
В, lunaria с однаждыперистой
пластинкой (рис. 161, 2). Реже
встречается В. matricariae,
имеющий триждыперисторассе-
ченную пластинку.
Спорангии свободные, не-
сросшиеся.
В стебле Botrychium име-
ется кольцо камбия. Камбий
откладывает к центру стебля
вторичную ксилему и кнару-
жи — вторичную флоэму. Стеб-
Рис. 161. Ужовниковые:
1*~ ужовник обыкновенный (Ophioglossum vulgfe»
turn); i — гроздовник ( Botricbyum lunaria); 3 — гель-
минтостахис (Helmintoetachys zeylanlca)
9—602
выа
пгл^
РАСТЕНИЯ
ли ботрихиума, таким образом, способны ко вторичному утолщению, что
11' I'
резко отличает ужовниковых от остальных современных папоротников.
Род Helmintostachys с одним видом Я. zeylanica встречается в тропи-
ческих лесах от Шри Ланка и Индии до Северо-Восточной Австралии
(рис. 161, 3).
Ужовник овые в ископаемом состоянии известны лишь с начала третич-
ного периода. Несомненно — это очень древняя группа растений. Об этом
in
свидетельствует строение заростка, сходного с заростком псилотовых, и
наличие в стеблях вторичного утолщения, что свойственно лишь древним
группам папоротников.
Расчленение листа на спороносную и бесплодную части, как мы видели
выше, имелось у древних Archaeopteridales.
2-Й ПОРЯДОК — МАРАТТИЕВЫЕ (MARATT TALES)
Мараттиевые представлены как ныне живущими, так и ископаемыми
растение
шт
. Современные мараттиевые распространены в тропических об-
ластях Старого и Нового Света. Они насчитывают 6 родов, объединенных в
одно семейство мараттиевых {Marattiaceae).
Род ангиоптерис {Angiopteris) насчитывает около 100 видов (рис. 163),
род архангиоптерис {Archangiopteris) — 4 вида, род Macroglossum —
2 вида. Эти 3 рода встречаются в тропиках Старого Света — в Восточной
Азии и на Мадагаскаре.
Род мараттия {Marattia) имеет около 60 видов, распространенных
всюду под тропиками. Род Chrystensenia представлен одним видом из Индо-
малайской флористической области. Род данеа {Danaea) имеет 32 вида в
Южной Америке.
Мараттиевые внешне похожи на распространенные в нашей стране па-
поротники из класса Leptofilices, но являются эуспооангиатными, так как
спорангий у них развивается из группы субэпидермических клеток иидоат
многослойную стенку. Стебли мараттиевых мясистые. Они неболыйяе^у
современных форм редко достигают 1 м высоты и частично скрыты в земле;
от стебля отходят очень крупные листья с многократно рассеченной пластин-»
кой. У видов Marattia они достигают 5 — 6 л длины. Лишь у одного вида
Danaea пластинка цельная. Листья мараттиевых длинночерешковые и у
основания окружены двумя крупными прилистликя^и (я флебиями).
На прилистниках нередко образуются почки, служащие для вегетативного
размножения. Отпадая от материнского растения, они укореняются, давая
начало новой особи.
Корни «мараттиевых очень толстые, придаточные, вози икают “няттебле.
Стебель в молодом состоянии протостелический, взрослый же имеет слож-
ную полициклическую диктиостель. В стеблях, корнях и черешках листьев
характерно наличие схизогенных слизевых ходов. Эти мараттиевые сход-
ны с саговниковыми из голосеменных. Спорангии .возникают на обычных
листьях — на нижней их сторонеДрис. 164, 2). У Angiopteris, Archangiop-
teris и Macroglossum спорангии почти свободные, у остальных родов они
срослись в синангии; Синангии округлые и находятся на нижней поверх-
ности листа. Расположение спорангиев и синангиев на листе различно.
У Angiopteris спорангии расположены в два ряда вдоль жилок листа, у
Marattia — недалеко от края листа, вдоль боковых жилок (рис. 164,1а, б},
у Danaea синангии округлые и разбросаны по всей поверхности листа.
У мараттиевых нет еще типичного покрывала (индуяия)^ сорусы или
синангии большинства видов окружены у основания волосками {Angio-
pteris, Marattia, рис. 164,2), и только у Danaea синангии окружены выростом
листа.
Спорангии имеют многослойную стенку. Полость спорангии выстлана
одним или двумя слоями клеток стенки тапетума.
В спорангиях развивается большое количество одинаковых спор.
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
Рис. 162. Ужовник:
1 — провод яхцая система стебля (отмацерированнаяу # — поперечный разрез стебля. Видны
отдельные пучки (меристели) коллатерального типа (да — флоэма, кс— ксилема): 5 —подзем*
ный заросток; а — общий вид; б — верхняя часть заростка (сп — молодой спорофит с первым
корнем); 4 — поперечный разрез заростка (ан — антеридии, ар — архегонии, яг — грибные гифы)
Рис. 163. Антиоптерис (Angiopteris evecta):
1 —общий вид растения; £=— основание листа с прилистниками (афлебиями)
9*
260
выс
г I г,
[В РАСТЕНИЯ
3
Рис. 164. Мараттиевые:
1 — листья со спорангиями и синангиями;
а — ангиоптериса (Angiopteris crassipes);
б — марании (Marattia fraxinea); в —хри-
стенсении (Cbrystensehia aesculi folia); 2 —
сорус ангиоптериса: а — вид сверху; б—
разрез соруса; 8 — синангии — христенсе-
нии: а — вид сверху; б — синангий в рая-
реэе.
Рис. 165. Спорангии и заростки мараттиевых:
1—продольный разрея через спорангии ангиоптериса (жсо—многоспойная стенка спорангия, т—тапетум,
ст—спарогенная скань); в—заростки мараттинг а—молодой заросток; б 1 в- заростки с молодыми спо-
рофитами
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
261
Зрелые спорангии вскрываются продольной трещиной. Споры, прора-
стая, образуют заростки, похожие на заростки наших папоротников.
Однако они имеют несколько большие размеры (до 2 см длиной) и живут
2—3 года (рис. 165).
' От нижней стороны заростка отходят ризоиды. Заростки обоеполые.
, На них образуются антеридии, а затем и архегонии. Сперматозоиды много-
'Жгутиковые.
Из оплодотворенной яйцеклетки развивается зародыш. Сформировав-
шийся зародыш имеет стебелек и первый лист, обычно цельный, с дихотоми-
ческим жилкованием. Растущий зародыш пробивает ткань заростка. При
этом стебелек выходит над его поверхностью, а образующийся вскоре при-
даточный корень внедряется в землю (рис. 165. 2).
Мараттиевые — древняя группа папоротников. Они прежде были рас-
пространены значительно шире, особенно в каменноугольном и пермском
периодах палеозойской эры. Тогда они были представлены многочисленны-
ми родами и видами. Позднее начинается их угасание, продолжающееся ,
вплоть до наших дней. Из этих древних форм упомянем древовидные виды
Aster о theca, имевшие стебли до 10 м высоты и листья до 4—5 м jsfljsw.,
Мараттиевые произошли от первопапоротников.^
3-Й ПОДКЛАСС — ЛЕПТОСПОРАНГИАТНЫЕ ПАПОРОТНИКИ
(LEPTOF1L1CES)
ш
К подклассу принадлежит большинство папоротников, около
дов, объединяемых в 270 родов. Громадное большинство Leptofilices —
травянистые растения, и лишь немногие из них являются древовидными.
Лептоспорангиатные папоротники распространены по всей Земле, но
огромное большинство видов встречается во влажных тропических лесах,
где папоротники играют громадную роль в сложении растительного покрова.
Они произрастают как на почве, так и на деревьях в виде лиан и эпифитов.
Только здесь обитают древовидные папоротники. Много папоротников рас-
тет и во влажных субтропиках.
Меныпее значение папоротники имеют в умеренных областях, где они
также приурочены к увлажненным территориям, тенистым местам, где нет
яркого солнечного освещения, и широко распространены в лесах в глубо-
ких ущельях, на болотах.
Огромное большинство Leptofilices — равноспоровые растения, но сре-
ди них имеются и разноспоровые. Leptofilices разделяют на 3 порядка:
Порядок Filicales представлен равносноровыми. наземными рас-
тениями, а порядки Marsiliales hl Salvinuiles— разноспоровыми, водными
и водно-болотными. У представителей подкласса Leptofilices спорангии обра-
зуются из одной поверхностной клетки листа и имеют однослойную стенку.
Лептоспорангиатные папоротники в большинстве своем молодая груд-
па растений. Произошли они от первоцапоротников (PrimoHltees),
POIYPOC lOP^i’dOc
1-Й ПОРЯДОК — ТИПИЧНЫЕ ПАПОРОТНИКИ (FILICALES)
К этому порядку принадлежит подавляющее большинство современ-
ных Pteropsida — свыше 9000 видов, объединяемых в 234 рода и 11 се-
мейств.
Несмотря на большое разнообразие представителей, Filicales mseisys об-
щий план строения и цикл развития.
Ознакомимся со строением и развитием их на примере нескольких па-
поротников нашей флоры.
Одним из наиболее распространенных у нас папоротников является
мужской папоротник, щитовник (Dryopteris fillx mas), растущий в сыро-
ватых лиственных лесах, по тенистым местам. Стебель его (корневище)
262
ВЫСШИЕ РАСТЕНИЯ
1
рис. 166. Мужской папоротник (Dryopteris filix mas):
7—взрослое растение (спорофит) с молодыми улиткообразно скрученными листьями; 2—часть листа с
сорусами* одетыми покрывалами (с—сорус); 8—поперечный разрез через лист с сорусом (и—индузий.
еп—спорангии); 4—спорангий с кольцом (к—кольцо)
скрыт в земле, лишь верхняя часть корневища немного возвышается над
поверхностью почвы. Корневище довольно толстое, черно-бурого цвета.
На нем спирально расположены остатки черешков опавших листьев, отми-
рающих ежегодно, а ближе к верхушке — листья, возвышающиеся над
землей (рис. 166). Взрослые растепия имеют 5—7 хорошо развитых листьев.
Корневище заканчивается верхушечной почкой. В ней находится точка
роста, окруженная молодыми, еще не дифференцированными листьями, ко-
торые разовьются уже в следующем году (рис. 167, 7).
Листья щитовника крупные, дваждыперисторассеченпые, длинноче*
решковые. Черешки покрыты тонкими чешуйчатыми коричневатыми пленка-
ми. Молодые листья свернуты улиткообразно, растут верхушкой (как сте-
бель) и развертываются от основания к верхушке, в противоположность
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
268
семенным растениям, у которых пла-
стинка листа развивается от верхушки к
основанию (базипетально, рис. 166,1).
Листья развиваются очень медлен-
но. Они ПОЯВЛЯЮТСЯ над jiewJraft тттатттк
на третий год после заложения их на
верхушках корневища.
Стебель щитовника- снаружи одет
эпидермисом, дальше ряг.ттпдпжА.пя ко-
ра, которая ра^е^ет£^шь~внухрен-
нюю и наружную. Клетки наружной
коры имеют механический характер.
***** В ткани“стебля“^аходятся много-
численные проводящие нучкёГ"разного
размера. Более крупные пучки в не*
бортом числе расположбны"’ТТраЕИЛ,БГ-
ным кольцом вокругцентральной части
стебля и являются. стеблевыми, они
тянутся вдоль стебля.
Многочисленные мелкие пучки,
разбросанные без видимого порядка,
направляются от стебелевых пучков в
листья. Бели отмацерировать стель
от мягких паренхимных клеток, она
будет иметь вид, изображенный на ри-
сунке 167, 2. Как у большинства па-
поротников, здесь имеется диктиостелу.
Она представляет собой сетку с круп-
ными ячейкалш.Каждая~ячейка"прёд-
ставляет собой прорыв стели в связи
с отхождением ^крудвото_____ЛЖЙдрго
следа. От краев ячеек в листья отхо-
2
Рис. 167. Строение корневища папорот-
ника:
Dryopteris Ш1х mas; S—мроводящая система
стебля (ошацерировано) D. cristata: в—строе-
ние диктиостели; б—переход от иротостели и
виктаостели (пс—протостель» Яе—дикпостшь)
дит группа мелких пучков.
Все пучки (отдельные меристели)
закрытые", концентрические —имеют
биЛЦёТЙли менее округлую форму.
От коры каждый пучок отграничен хорошо выраженной эндодермой.
С птюпускнмми клетками. За эндодермой расположен одно-двухсло^ "
перйцикл. Далее следует флоэцц. Оня состоит из ситовидных трубок (
'л: 1 ? I
положена ксилема. В пеппвнпй массе это метаксилема, состоящей из широ-
ких лестничных трахеид. Среди них заметны клетки древесинной паренхи-
мы. Местами имеются группы протоксилемы, состоящей из кольчатых и
спиральных трахеид. Лист щитовника одет верхним и нижним эпидерми-
сами, в клетках которых находятся хлорофилловые зерна. В нижнем эпи-
и*
дермисе много устьиц.
Мезофилл губчатый, с большими межклетниками. В листе имеются
многочисленные жилки. Это мелкие пучки такого же строения, что и в стеб-
ле (рис. 166, 5). Черешок имеет строение, сходное со стеблем. Корень со-
держит в Центре один концентрический пучок. Кора резко разделяется на
внутреннюю (из механических клеток) и наружную, состоящую из паренхим-
ных клеток. Снаружи корень одет эпидермисом. Растущие молодые части
корня несут многочисленные корневые волоски. Спорангии щитовника соб-
раны в группу — сорусы. Сорусы возникают на нижней стороне листа, вдоль
средней жилки листочков, и одеты округло-сердцевидным покрывалом
(индузием), прикрепляющимся в центре соруса и слегка вдавленным в виде
складки от центра к его периферии.
ВЫСШИЕ PACT
ы:г*
Я
В
Рис. 168. Лептоспорангиат-
ные папоротники:
Л—спорангий с кольцом и рас-
крывание спорангия: а—зрелый
спорангий; 6 — раскрывшийся
спорангий; в — клеши кольца;
а—развитие спорангии (с—клет-
ка. дающая начало спорангию,
н—клетка, образующая ножку
спорангия» «с — выстилающий
слой» пр—археспорий)
На поперечном разрезе листа в области соруса видно, что спорангии
расположены на плаценте — массивном выросте нижней Стороны лис-
та (рнс. 166, 3). Выше от плаценты отходит, индузий. Спорангии имеет вид
двояковыпуклой чечевицы и расположен на довольно длинной ножке, на ко-
торой находятся иногда булавовидные слизевые волоски. Одет он однослой-
ной стенкой и содержит внутри споры. Расположенная по гребню спорангия
полоска клеток стенки образует так называемое кольцо. Начиная от нож-
ки, кольцо опоясывает спорангий и продольном направлении почти на 2/з
его окружности (рис. 166, 4). Клетки кольца имеют неравномерно утолщен-
ные стенки. Их внутренние и радиальные стенки утолщены, в то время как
наружные остаются неутолщенными (рис. 168, 1в).
Кольцо является аппаратом, способствующим раскрыванию спорангия
и разбрасыванию спор. При созревании спорангий подсыхает. Испарение
воды в кольце идет быстрее через неутолщенные стенки. Они вдавливаются
внутрь и становятся вогнутыми. Радиальные стенки клеток при этом сбли-
жаются и кольцо стремится выпрямиться. При дальнейшей потере воды
натяжение становится, наконец, настолько сильным, что стенка спорангия
в неутолщенной части (в так называемом устье, рис 168, 2а,б) разрывается
и кольцо резко выворачивается. Однако оно быстро занимает прежнее по-
ложение, так как прочно соед
хено с ножкой. При этом движении кольца
споры разбрасываются в стороны на расстояние до 1 ж.
Спорангий мужского папоротника развивается из одной поверхност-
ной клетки (рис. 168, 2), которая возникает в виде небольшой выпуклины
и затем отделяется перегородкой. Клетка эта делится поперечной перего-
родкой на две. Из верхней клетки развивается спорангий, а из нижней —
его ножка.
Затем в верхней клетке появляются три косо направленные перегород-
ки, в результате чего выделяется центральная клетка, окруженная тремя
клетками, которые затем делятся, постепенно формируя однослойную стен-
ку спорангия. При этом в меридиональном направлении, начиная от ножки,
один ряд клеток стенки делится более энергично. Они и образуют кольцо.
Центральная клетка также вскоре делится несколько раз, в результате
чего образуются три периферические клетки, расположенные параллельно
стенке спорангии (они становятся клетками тапетума), и одна археспори-
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
альная клетка, которая делится дальше. При этом образуется обычно 16 ма-
теринских клеток спор. Последние делятся редукционно, и каждая обра-
зует по тетраде спор. Всего в спорангии, таким образом, имеется обычно
64 споры (рис. 168, 16).
Спора одета двумя оболочками — тонкой, внутренней (эндоспо-
рой) и толстой, наружной (э к з о с п о р и й). Последняя окрашена в
бурый цвет. На земле спора прорастает, образуя заросток. При прорастании
спора образует сначала небольшую зеленую клетку, которая разрывает на-
ружную оболочку споры и вытягивается. Вскоре от нее отделяется бесцвет-
ная клетка — первый ризоид, внедряющийся в землю. Зеленая клетка
далее делится поперечной перегородкой, в результате чего формируется
зеленая нить. Верхняя клетка нити делится далее в разных направлениях.
Постепенно формируется округло-сердцевидная пластинка 0,5 —0,9 см
в диаметре (рис. 169, 1а).
Заросток у папоротников зеленый, в клетках его содержатся хлоро-
пласты. Он способен к самостоятельной жизни. Обычно заростки однослойны
по краям и многослойны в середине. Точка роста заростка находится в основа-
нии выемки. От нижней поверхности заростка отходят многочисленные
^многоклеточные ризоиды, прикрепляющие заросток к почве и всасываю-
ills
_ На нижней поверхности зрелого заростка возникают антеридии и архе-
гонии (последние ближе к выемке)^ Сначала обычно развиваются антеридии,
затем уже после выполнения антеридиями их функции созревают архегонии^
Антеридии возвышаются над поверхностью заростка, имеют округлую
форму и развивают небольшое число спирально закрученных многожгу-
тиковых сперматозоидов (рис. 169, 1г).
Рис. 169. Заросток к молодой спорофит напорот
ов:
1—Dryopterls filix mas; а—заросток; б—развитие архегонии; «—зрелый антеридий; а—антеридий в мо-
мент выхода многакгугнковых сперматозоидов; О—сперматозоид; з—Adianthum (э—з—поперечный раз-
рез заростка и зародыша. р—ризоиды» ар—неопяодопсренные архегонии. вд—первый лист зародыша —•
«сшщцоля». пи—первичный корень, н—ножка)
266
ВЫСШИЕ РАСТЕНИЯ
Рис., 170. Папоротник-орляк (Pteris aquillna):
1—общий вид; а—часть листочка с индузием; а — корневище; 4 — поперечный разрез стебля (нк — на-
ружная кора» ек—внутренняя кера, мт — полукольцо механической ткани» сп—сосудистые пучки);
Л—поперечный разрез пучка (э—эндодерма, л—перицикл» ст—ситовидные трубки» лп—лубяная парен-
хима» йа—древесинная паренхима» пхе—первичная меняема» ко—ксилема)
Архегонни обычного строения, но частично (брюшком) погружены в
ткань заростка; шейка возвышается над его поверхностью (рис. 169, 2).
Так как антеридии и архегонни находятся в контакте с влажной почвой,
оплодотворение яйцеклетки подвижным сперматозоидом в дождливую
погоду Происходит без затруднений. Из оплодотворенной яйцеклетки вскоре
формируется зародыш, первоначально развивающийся за счет заростка.
Всасывание питательных веществ осуществляется при помощи иожки заро-
дыша, внедряющейся в ткань заростка (рис. 169, 2). После образования ко-
решка, стебелька и первого листа зародыш начинает самостоятельную жизнь.
Корешок его внедряется в землю, стебелек выпрямляется, и лист разраста-
ется. За первым листом на стебле образуются новые листья, и таким обра-
зом постепенно формируется взрослое растение напоротника. Первые
листья при этом простые и. имеют дихотомическое жилкование. Последую-
щие листья постепенно приобретают папоротниковый облик.
Широко распространен у нас также папоротник орляк (Pteris aquilina),
произрастающий в светлых сосновых лесах (рис. 170).
У орляка под землей, на глубине 20 — 40 см, находится длинное гори-
зонтальное, обильно разветвленное корневище, от которого отходят пооди-
ночке крупные надземные листья, имеющие длинный черешок, заканчиваю-
щийся наверху широкой тройчато-рассеченной пластинкой. Каждая из
и г. г
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
267
ins
трех крупных долей, в свою очередь, имеет перистое расчленение (рис. 170,1).
От корневища вниз отходят многочисленные черные корни. На конце моло-
дых корней находятся корневые волоски. Корневище орляка имеет еще бо-
лее сложное, полициклическое строение. Отель его состоит из двух концент-
рических диктиостелей, как бы вставленных друг в друга.
На поперечном разрезе корневища заметны обычно два крупных про-
водящих пучка овальной формы, расположенных в центре. Это внутрен-
няя стель. За ними к периферии хорошо выражены два полукольца механи-
ческой ткани (рис. 170, 4). За механической тканью расположены много-
численные мелкие округлые пучки и обычно один овальный крупный. Они
образуют в совокупности наружную диктиостелы
Пучки погружены в паренхимную ткань. Здесь хорошо заметна внут-
ренняя и наружная кора. Снаружи стебель Покрыт эпидермисом. В коре
видны многочисленные мелкие пучки, идущие в листья. Пучки орляка за-
крытые, концентрического строения. От коры пучок отграничен эндодермой,
с пропускными клетками. Далее расположен перицикл (1 — 2-слойный).
Затем следует флоэма. В ней имеются ситовидные трубки и лубяная парен-
хима. В центре пучка расположена ксилема. Она образована в основном
метаксилемой, но в центре мелких пучков имеется одна небольшая группа
протоксилемы, а в более крупных удлиненных пучках — обычно 2 груп-
пы протоксилемы, расположенные как бы в центрах эллипса. Протоксиле-
ма состоит из кольчатых и спиральных трахеид (рис. 170, 5). Метаксилема
орляка, в отличие от других папоротников, состоит из настоящих лестнич-
ных сосудов, а не из трахеид.
У орляка сорусы возникают по краю листа. Лист при этом загибается
внутрь и тонким краем покрывает ряды сорусов, являясь их защитой.
С внутренней стороны сорусов развивается индузий, имеющий вид тонкой,
бахромчатой по краям пленки (рис. 170, 2). Строение спорангия, заростков
и зародыша сходно с таковым у щитовника.
Представители Filicales чрезвычайно разнообразны и представлены
различными жизненными формами.
Большинство папоротников произрастает во влажных областях, но
среди них имеется и небольшое число ксерофитов, растущих в засушливых
районах (в Южной Африке, на Мексиканском нагорье, в Средиземноморье
и др.). Здесь папоротники имеют густо опушенные листья, свертывающие-
ся в сухое время года. Многие из них в это время сморщиваются и становят-
ся как бы безжизненными, но при выпадении дождей они снова оживают
и продолжают рост.
—Leptofilices — обычно травянистые растения небольших размеров, обла-
дающие большей частью ползучим надземным стеблем или корневищем.
Однако среди них есть крупные древовидные папоротники, достигающие
15 — 25 ж высоты, например виды диксонии (Dicksonia), алъзофилы (Also-
phila), встречающиеся в тропических и субтропических влажных лесах
Австралии и Новой Зеландии, Африки и Южной Америки, где они иногда
образуют небольшие рощи. Наряду с этим имеются маленькие папоротни-
ки размерами всего в несколько сантиметров (и миллиметров), как, напри-
мер, виды рода Trichomanes, встречающиеся в тропических лесах. Один
вид Trichomanes найден недавно на влажных скалах в горных лесах близ
Батуми (Г. iunbridgense).
В тропических лесах нередки папоротники-лианы. Их длинные тонкие
ползучие стебли перебрасываются с дерева на дерево, обвивая их или цеп-
ляясь при помощи различных выростов-колючек на стебле или черешках
листьев.
Листья некоторых папоротников-лиан достигают 30 м длины, обладая
почти неограниченным ростом, например у Lygodium articulatum. Среди
папоротников много эпифитов, произрастающих на стволах и ветвях деревь-
ев. У некоторых эпифитов наблюдается гетерофиллия, например у видов
Plat у cerium (рис. 171, I). В этом случае некоторые листья имеют округлую
j I: i
вп
2
Рис. 171. Папоротники:
1—общий вид платащернума; 2— выводковые почки на листьях (вп —выводковые почки)? а—аеплеииума
(Asplenium viviparum); б—цистоптериса (Cystopteris bulbifera); в—хеыионитиса (Hemionitls palmata);
а—ааскслея (Onoclea struthiopteris) — сп — спорофилл
форму и нижней частью плотно прижаты к стеблю, в то время как их верх-
няя часть отгибается в сторону. В результате между листом и корой дерева
образуется кармановидное вместилище, куда падают с дерева листья и мел-
кие ветви. Перегнивая, они дают слой почвы, в который проникают прида-
точные корни папоротника.
Другие листья, крупные, многократно разветвленные, свешиваются
в виде оленьих роГов. На их окончаниях с нижней стороны образуются
сорусы спорангиев.
Листья у большинства папоротников сильно расчлененные, дважды-
триждыперистые или пальчатые, но имеются и цельные (возникшие из
расчлененных листьев), например у видов Trichomanes (рис. 173, 3).
На корневище папоротников развивается большое количество прида-
л!
точных корней. Иногда они развиты очень сильно, а у древовидных папо-
ротников, развиваясь и на надземном стебле, образуют вокруг него войлоч-
ный чехол в несколько сантиметров толщины. Чехол этот легко всасывает
и удерживает воду.
У представителей семейства гименофиллоеых, например у Tricho-
manes t корней не образуется. На стебле и листьях у них возникают много-
численные выросты — «корневые волоски» (рис. 173л 3).
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
269
Рис. 172. Осмунда (Osmunda regal is):
1—разделение листьев на бесплодные и спороносные сегменты: г—5—стадии развития спорангия; е—
зрелый вскрывшийся спорангий
Чрезвычайно разнообразно устройство стели у папоротников. Здесь
можно проследить все переходы от простейшей протостели, имеющейся у
примитивных,.древних представителей, к различным типам сифоносте-
л и (как амфифлойной, так и эктофлойной), диктиосте-
ла и, наконец, э у с т е л и.
Большинство лептоспорангиатных папоротников имеет диктиостель.
Многие же имеют полициклическое строение.
У папоротников
роко распространено вегетативное размножение.
иг,
Стебли многих папоротников легко укореняются и могут распадаться на
отдельные участки, продолжающие жить самостоятельно.
Выводковые почки нередко образуются на листьях или на особых по-
бегах (рис. 171, 2). Спадая на землю, они развиваются в новые растения.
У большинства папоротников спорангии развиваются на обычных ли-
стьях, но у некоторых родов они образуются на особых листьях — споро-
филлах. Примером последних может служить распространенный у нас по
сырым лесам вдоль ручьев страусопер (Onoclea stuthtopteris). У него весной
из корневища сначала вырастает розетка двоякоперистых зеленых вегета-
тивных листьев (трофофиллов). Они достигают 1 м длины. Летом из того же
корневища вырастают бурые спороносные листья (спорофиллы). Они зна-
чительно мельче вегетативных, однаждыперистые. Дольки листа сверну-
тые, закрывающие сорусы (рис. 171, 3).
Такое разделение листьев имеется также у видов Osmunda (рис. 172, 2).
Для систематики Filicales большое значение имеют строение кольца на
спорангии, расположение сорусов и строение индузия. Filicales обычно
270
ВЫСШИЕ РАСТЕНИЯ
разделяют на Нсемейств, для характеристики которых особенное
значение имеет устройство кольца: осмундовые (Osmundaceae), (Schizaeceae)t
(Gleicheniaceae), матониевые (Matoniaceae), гименофилловые (Hymenophy-
llaceae) локсомовые (Loxomaceae), гименофиллоподиевые (Hymenophyllo-
po diace ae, семейство, близкое к предыдущему), плагиогириевые (Plagiogy-
riaceae) , диксонйевые (Dicksoniaceae), циатиевые (Cyatheaceae), многонож-
косые (Polypodiaceae).
Большинство семейств Filicales появляется в триасе и достигает значи-
тельного развития в меловой период. Таким образом, современные папорот-
ники—ровесники цветковых и современных голосеменных растении.
Наиболее обширным является семейство многоножковые (Poly-
podiaceae), куда относятся подробно рассмотренные нами выше роды
Dryopteris и Pteris. Представители этого семейства имеют кольцо, окружа-
ющее спорангии на а/3 его окружности.
Многоножковые насчитывают около 7000 видов и свыше 170 родов.
Сюда принадлежат травянистые папоротники. Помимо указанных выше,
к этому семейству относятся наши лесные папоротники — женский папорот-
ник (Athyrium filix femina)> еще ряд видов щитовника (Dryopteris spinulosa
и др.), страусопер (Onoclea struthiopteris)* а также виды многоножки (Poly-
podium) и многие другие.
Семейство многоножковых — одно из наиболее молодых в филоге-
нетическом отношении. В последнее время это семейство рассматривают как
возникшее полифилетически. Его разделяют на 4 самостоятельных семей-
ства (Polypodiaceae^ Aspleniaceae, Aspidiaceae* Pteridaceae)t которые раз-
мещают среди других семейств.
Наиболее примитивными и древними являются семейства Osmun-
daceae, Schizaeaceae, Hymenophyllaceae и Glecheniaceae. Осмундовые (Osmun-
daceae) появляются в пермском периоде, a Schizaeaceae и Hymenophyllaceae —
в каменноугольном периоде палеозойской эры.
Семейство осмундовые (Osmundaceae) представляет большой инте-
рес, как обладающее рядом промежуточных признаков, связывающих
Leptofilices с Eufilices. К Osmundaceae принадлежит род Osmunda
с 14 видами.
Наиболее известен царский папоротник (О. геgalls), растущий на боло-
тах Кавказа. Листья его разделены на бесплодную и спороносную части.
Спорангии царского папоротника имеют однослойную стенку, но снабжены
массивной, толстой ножкой. Возникают они из нескольких клеток, среди
них, однако, вскоре выделяется одна, делением которой и осуществляется
дальне:
'ЛИ
ее формирование спорангия, как и у остальных Leptofilices. Коль-
цо весьма примитивное и сходно с зачаточным кольцом мараттиевых
(Angiopteris). Оно представлено группой клеток с утолщенными стенками,
расположенных на спинной стороне спорангия (рис. 172, 5). Вскрывается
спорангий продольной щелью на две створки.
Остатки многих видов Osmunda известны из меловых и юрских отло-
жений мезозойской эры (О, regalis — из эоцена). Близкие роды (Zalesskya
и др.) известны из пермского периода палеозойской эры.
Семейство схивейные (Schizaeaceae) представлено 160 видами тро-
пических и субтропических растений. У большинства родов имеются споро-
филлы и трофофиллы. Спорангии возникают из верхушечных клеток на
краю листа (таким образом, имеется первоначально краевое, примитивное
расположение спорангиев), но затем оказываются на нижней его стороне —
вследствие усиленного роста пластинки листа над спорангием. Спорангии
одиночные, не собраны в сорусы и расположены по краям жилок
(рис. 173, 16).
Кольцо на спорангии у схизейных верхушечное, полвое, поперечное
(рис. 173, 1в) и состоит из одного ряда клеток, но часто выражено неясно.
У ископаемых представителей из карбона кольцо состояло всего из 2 —
4 клеток и было сходно с кольцом спорангия осмундовых. К семейству отно-
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
271
Рис. 173. Схжзейные и гименофилловые:
J—схизея: а—общий вид Schizaea pusilia; б—стадии развития спорангиев (по краю листа Sch. rupestris);
•—спорангий Зек. bifida (кольцо верхушечное, с продольной трещиной); я—гименофилл (Hymenophyllum
dilatatum): я—общий вид (ксп—расположенные по краю листа спорангии); б—спорангии с полным ко-
сым кольцом; 3—трихсманес (Trichomanes reniferme)
сятся роды: схизея, лигодиум и др. Схизея (Schizaea) насчитывает около
30 видов. Она растет главным образом в тропиках, но некоторые виды (на-
пример, Sch. Pusilia) встречаются в Северной Америке, до Ньюфаундлен-
да (рис. 173, la). Лизодиум (Lygodium) представлен 40 видами тропических
и субтропических растений. Стебли имеют протостелическое строение.
Семейство гименофилловые (Hymenophyllaceae) насчитывает около
650 видов, распространенных во влажных горных лесах тропиков и субтро-
пиков. Оно представлено 2 родами — Trichomanes и Hymenophyllum.
Остатки их встречаются уже в каменноугольном периоде. Это небольшие,
часто в несколько сантиметров и даже миллиметров, растения с тонкими
прозрачными листьями, цельными или расчлененными (рис. 173, 2). Стебли
их протостелические, спорангии расположены по краям листьев (древ-
ний признак); кольцо полное, косое (рис. 173; 26).
Н ПI!IУ
PACT
Salviniales
Marsileales
Pteridaceas
Dipksornacea?
plagiogygriaceae
Dsmundaceae
Aspidiaceaa Aspleriiaceaa
Oipterutea^
Matoniaceae
Рис. 174. Взаимоотношения между главнейшими семействами Filices
Polypodiaceas
GieLcheniaceae
Семейство глейхениевые (Gleicheniaceae) насчитывает 130 видов,
распространенных в тропиках, субтропиках и умеренных областях южного
полушария. Это одно из наиболее древних семейств Filicales. Ископаемые
остатки глейхениевых известны из каменноугольного периода. Сорусы без
индузия, расположены на боковых жилках диета и состоят каждый из 2 —
8 спорангиев. Иногда спорангии одиночные. Кольцо полное, косое, раскры-
вается продольной щелью, что сближает их с Osmundaceae. Сюда принадле-
жат роды Gleichenia, Dtcranopterts и др.
Филогенетические взаимоотношения между главнейшими семействами
Filicales изображены на рисунке 174. Filicales произошли от первопапорот-
ников (Primofilices). Наиболее вероятно родство их с порядком зигоптердие-
еые (Zygopteri dales).
2-Й ПОРЯДОК — МАРСИЛИЕВЫЕ (MARSILIALES)
К этому порядку принадлежит одно семейство Marsiliaceae с тремя
родами: Marstlia (90 видов), Pilularia (6 видов) и Regnellidium (1 вид).
Из них мы рассмотрим только первый род.
Род марсилия (Marsilia) распространен на всех материках. В СССР
встречается марсилия четырехлистная (М. quadrifolia) — водно-болотное
растение, произрастающее в дельте Волги и на Северном Кавказе. Марсилия
имеет ползучее корневище, скрытое в почве. От нижней стороны корпев
отходят корни, от верхней — листья, возвышающиеся над землей (рис. 175).
Листья длинночерешковые, заканчивающиеся четырьмя листочками, тес-
но сближенными попарно. Листочки имеют маленькие сочленения и способ-
ны к движению, они складываются на ночь своими нижними поверхностя-
ми. Иногда встречаются листья с шестью листочками. Корневище имеет
соленостелическое строение (амфифлойная сифоностель).
J11
J11
'ТЯ
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
ни
В центре корневища находится стель. На поперечном разрезе корневи-
ща (рис. 175, 5) видно, что стель отграничена от коры наружной эндодер-
мой. Далее к периферии расположена внутренняя и затем наружная кора.
Внутренняя кора образована главным образом склеренхимными клетками,
имеющими утолщенные стенки. Наружная кора состоит из паренхимных
клеток и содержит воздушные полости (аэренхима). Снаружи стебель
одет эпидермисом. Стель имеет сложное строение. Под наружной эндодер-
мой, отграничивающей стель от коры, расположен однослойный перицикл.
За перициклом находятся кольцо флоэмы и кольцо ксилемы. Глубже рас-
положены внутренний перицикл и внутренняя эндодерма, отграничиваю-
щая стель от сердцевины.
Сердцевина часто состоит целиком из механических клеток, или пни
перемешаны с паренхимными клетками.
В стели имеются листовые прорывы, образующиеся при отхождении
листовых следов. Сорусы располагаются по 1 — 3 на черешках листьев в
нижней их трети, имеют сложное строение и называются спорокарпи-
Зрелый спорокарпий снаружи одет толстой, очень прочной оболочкой,
прекрасно защищающей его от высыхания. Внутри вдоль выпуклой старки
цевина)
нп
МКС пкс
внп
вэн сц
Рис. 175. Марсилия:
1—(общий вид); 2—спорокарпий;
4—раскрывание спорокарпия;
4—поперечный раэрез стебля
(апг- эпидермис, аэр—аэренхима,
х—кора, как—наружная эндодер-
ма, кп—наружный перицикл,
лскс—метаксилема, пхс—прото-
ксилема, и^л—наружная флоэма,
«эн — внутренняя эндодерма,
т
выс
Е РАСТЕНИЯ
спорокарпия расположено кольцо хрящевидной ткани (рис. 175,2—4), к ко-
торому прикреплены сорусы. Каждый сорус снаружи одет двухслой-
ным индузием. Внутри соруса имеется плацента, на вершине которой распо-
ложены в небольшом числе мелкие микроспорангии и более крупные мега-
спорангии. Спорангии имеют тонкую стенку. Кольцо отсутствует.
В микроспорангии образуется большое количество микроспор. В зре-
лом мегаспорангии находится одна мегаспора. Закладывается, однако, боль-
шее число мегаспор, но затем остальные отмирают. Зрелый спорокарпий
опадает на почву. Вода проникает внутрь спорокарпия, что вызывает на-
бухание хрящевидного кольца. Набухая, кольцо увеличивается в размерах
и оказывает сильное давление на стенки спорокарпия. В результате стенка
спорокарпия раскрывается по брюшной стороне и кольцо, разворачиваясь,
оказывается вне спорокарпия. На развернувшемся кольце в два ряда рас-
полагаются сорусы (рис. 175, 4).
Микро- и мегаспоры после разрушения индузия и стенок спорангия
попадают в воду и начинают прорастать. Полное развитие они проходят в
течение суток. Микроспора, прорастая, дает сильно редуцированный муж-
ской заросток. При прорастании образуются маленькая проталлиальная
(ризоидальная) клетка и боковая клетка, делением которой образуются две
антеридиальные клетки. Каждая из них образует по одному сильно реду-
цированному антеридию, состоящему из двух клеток стенки и одной сперма-
генной клетки, которая дает начало сперматозоидам. На этой стадии зарос-
ток состоит из семи клеток (рис. 176, I). В каждом антеридии затем образу-
ется по 6 — 8 многожгутиковых сперматозоидов.
К этому времени как клетки стенки, так и проталлиальная клетка раз-
рушаются и сперматозоиды находятся в общей слизи. Затем оболочка
микроспоры лопается и сперматозоиды выходят наружу.
Мегаспора прорастает внутри мегаспорангия, образуя женский зарос-
ток. Прорастание начинается в верхней части мегаспоры, где образуется
небольшое сосочковидное вздутие, заполненное цитоплазмой. Сюда пере-
мещается ядро мегаспоры. Остальная часть мегаспоры заполнена пита-
тельными веществами (рис. 176, 2а). Ядро мегаспоры делится, и вскоре обра-
зуется перегородка, отделяющая образовавшуюся клетку от мегаспоры.
Верхняя клетка делится дальше, в результате чего образуется единст-
венный маленький архегоний, состоящий из яйцеклетки, брюшной каналь-
цевой клетки и одной шейковой канальцевой клетки. Снаружи архегоний
одет небольшим числом клеток стенки.
Растущий архегоний разрывает оболочку мегаспоры и несколько воз-
вышается над ее поверхностью. По созревании архегоний вскрывается на
верхушке (рис. 176, 2г). Сперматозоид проникает внутрь архегония и сли-
вается с яйцеклеткой.
Оплодотворенная яйцеклетка (зигота) быстро делится. Постепенно раз-
вивается зародыш. Его развитие идет сначала за счет питательных веществ
мегаспоры, всасываемых при посредстве ножки. Образуется первичный
корень, стебелек и лист. Далее корешок внедряется в почву, стебелек вы
прямляется, развиваются новые листочки. Первые листья у марсилии
ni'.G
ловидные, затем появляются двулопастные и, наконец, листья, свойствен-
ные взрослому растению.
'Марсилиевые произошли от равноспоровых папоротников^ именно от
предков современных Schizaeaceae (рис. 174).
3-Й ПОРЯДОК — САЛЬВИНИЕВЫЕ (SALVINIALES)
К порядку сальвиниевых относятся 2 семейства разноспоровых
папоротников: Salviniaceae is. Azollaceae. Семейство салъвиниевых (Salvi*
niaceae) представлено одним родом салъвиния (Salvinia), насчитывающим
8 видов, распространенных в тропиках и субтропиках2 реже в умеренных
областях обоих полушарий.
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
Рис. 176. Марсилия. Размножение:
л—стадии прорастания микроспоры и образования жужекого заростка у Marsilia vestitai £—мегаспора и
женский заросток: а—мегаспора с ядром, переместившимся в верхнюю ее часть; б, в, « — образование
архегония (я—яйцеклетка, к—брюшная канальцевая клетка» шк—единственная шейковая канальцевая
клетка» кар—клетка стенки архегония)
выс
ИГЛУ
РАСТЕНИЯ
Рис. 177. Салыишия (Salvinia natans):
л—общий вид; Я—участок стебля с двумя плавающими листьями и одним подводным, сильно рассечен-
ным (сп—спорокарпий у основания пйдводнсго листа); а—поперечный разрез стебля (в центре—проводя-
щий пучок с трахеидами, етс—внутренний слой коры, ко—наружный слой коры, вп—воздухоносные
полости); 4—поперечный разрез листц (с—сосочки на верхней стороне листа); 5—разрез спорокарпиев;
о—микроспорокарпий; б—мегаспорокарпий (мкс—микроспорангий, мге—мегаспорангий, пл—плацента,
ин—индузии)
У нас встречается S. natans, произрастающая в озерах и заводях рек
европейской части СССР, на юге. К северу она доходит до Мещеры. Это не-
большое (до 15 см) плавающее на поверхности воды растеньице. Тонкий его
стебелек несет в узлах листья, сидящие мутовкой по три. Два листа оваль-
ные, зеленые, плавающие; третий лист подводный, сильно, рассеченный,
заменяющий растению корни. Настоящих корней у сальвинии нет (рис. 177).
В анатомическом строении стебля и листьев резко выражены черты
приспособления к водной жизни: много крупных воздушных полостей,
ничтожно развиты механические элементы, сильно редуцирована проводя-
щая ткань.
Стебель в центре имеет проводящий концентрический пучок (рис. 177, 3);
ксилема его развита слабо и состоит из 1—3 небольших лестничных или
спиральных трахеид.
Ксилему окружает флрэма, состоящая из ситовидных трубок и лубя-
ной паренхимы. Перицикл и эндодерма нерезко выражены. Дальше распо-
лагается кора, в которой развиты большие воздушные полости. Клетки
внутреннего слоя коры, примыкающие к пучку, имеют утолщенные стенки.
В листе также находятся многочисленные воздушные полости (камеры),
обычно расположенные в два зтажа. Они разделены перегородками, в клет-
ках которых находятся хлорофилловые зерна.
На верхней стороне листа развиваются особые сосочки, состоящие из
округлых клеток, стенки которых покрыты слоем воска. Сосочки эти при-
дают листу светло-зеленую (беловатую) окраску и способствуют удалению
воды с поверхности листа.
?
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
г is
nt
Sift
tri
С нижней стороны листа, как и от стебля, отходят буроватые многокле-
точные волоски.
У основания подводных листьев, на коротких боковых разветвлениях,
образуются шаровидные сорусы, называемые здесь спорокарпиями. Спорокар-
пии все одинакового размера, но содержат внутри различные спорангии.
В одних спорокарпиях развиваются в большом числе мелкие шаровид-
ные микроспорангии (м и к р о с п о р о к а р п и и),' в других — неболь-
шое число более крупных яйцевидно-овальных мегаспорангиев (м е г а с п о-
р окарпии, рис. 177, 5).
Спорангий снабжен ножкой, которой прикрепляется в плаценте, вдаю-
щейся в полость соответствующего спорокарпия. Снаружи спорокарпии
одеты двойной оболочкой (индузием). Внешний и внутренний индузии со-
единены вверху и внизу, на остальном протяжении они свободны, и между
ними образуется воздухоносная полость. Ипдузий образуется из основания
листа и постепенно обрастает спорангий в виде чаши.
Спорангии одеты однослойной стенкой. Далее расположен один ряд
клеток выстилающего слоя (тапетум). В центре спорангия развиваются
клетки археспория. Делясь, они дают начало материнским клеткам спор.
Последние делятся редукционно, образуя тетрады спор. В микроспорангии
развиваются обычно 64 микроспоры. При этом клетки тапетума расплывают-
ся, а затем содержимое их застывает, образуя твердую массу (м а с с у л а—
пенистое вещество), в которую погружены споры.
* В мегаспорангии образуется большое количество мегаспор, но полного
развития достигает лишь одна, остальные отмирают. Зрелый спорангий со-
держит лишь одну мегаспору. Мегаспора одета снаружи толстым слоем пе-
нистого вещества (периспорий), образовавшегося из расплывшихся кле-
ток тапетума. Легкий периспорий способствует удержанию мегаспорангия
на поверхности воды.
Спорокарпии осенью опадают и перезимовывают на дне водоема. Вес-
ной после сгнивания их оболочек микро- и мегаспорангии всплывают на
поверхность. Микроспора прорастает в сильно редуцированный мужской
заросток, не покидающий микроспорангия (рис. 178, I). При первых деле-
ниях микроспоры внутри нее образуются три клетки (рис. 178,16). Нижняя
из них затем делится на две клетки — мелкую ризоидальную и более круп-
ную вегетативную.
Вегетативная клетка разрастается и выдвигает верхние две клетки че-
рез разрыв оболочки микроспорангия наружу (рис. 178, 1). Обо верхние
клетки делятся, и каждая образует по одному сильно упрощенному антери-
дию. Антеридий состоит из двух клеток, стенки и одной спермагенной клет-
ки. Спермагенная клетка, делясь далее, образует четыре многожгутиковых
сперматозоида. Всего таким образом на заростке образуются два антеридия
с восемью сперматозоидами. Разрастающиеся заростки пробиБают стенку
микроспорокарпия и выходят наружу. 'Стенка спермагенной клетки рас-
плывается, сперматозоиды выплывают наружу. Плавая в воде, они направ-
ляются к женским заросткам.
Мегаспора (единственная в мега спорангии), прорастая, образует жен-
ский заросток, также не покидающий оболочки спорангия. При прораста-
мегаспоры ядро ее делится, образуя две клетки, — маленькую верху-
шечную клетку и более крупную нижнюю (базальную) клетку. Из верх-
ней клетки далее образуется заросток, нижняя же сильно разрастается,
ядро ее делится, но перегородок между ними не образуется. Сильно разрос-
шаяс^ клетка заполняется питательными веществами для зародыша.
СЗаросток состоит из зеленых клеток. В верхней части он разрывает обо-
лочку мегаспорангия и высовывается наружу в виде округло-треугольной
пластинки. На ней развиваются погруженные в ее ткань 3 — 5 архегониев
(рис. 178, 2a)^f
После оплодотворения из зиготы развивается небольшой зародыш.
Он долгое время связан с заростком?) Зародыш развивает стебелек и лис-
r< is
Рис. 178. Салъвиния;
]—микроспорашжи и мужской заростош «—иикросиорангий в проросшими микроспорами; б—прораста-
ние микроспоры и развитие заростка! I—в результате двух делений образовались три клетки; II—П1—
следующие стадии (рм—ризоидальная клетка, ей—вегетативная клетка, ак—клетки стенки антеридиев,
сив—сиермйгенные клетки, образующие восемь сперматозоидов); а—сперматозоид; з—мегасаорангий и
женским заросток! п-мегаснора с выдающимся наружу женским заростком, на нем видны архегонии;
б—мегаспора с заростком и зародышем в разрезе (об—оболочка мегасиорангия, пса—оболочка мегасноры
/периспорий/, б—заросток с хлороиластаии); зародыш» еще связанный с заростком («^-зародышевый
листок» ЛаЛяЛв—следующие за ним листья, •—заросток)
ОТДЕЛ — ПАПОРОТНИКООБРАЗНЫЕ
279
И М11
точки. Первый лист зародыша щитовидный, следующие листья округлые; да-
лее развиваются овальные листья, свойственные взрослому растению. Кор-
ня у сальвинии не образуется. Все растение покрыто волосками (рис.178, 2в),
Постепенно формируется взрослое растение.
Второе семейство — азолловые (Azollaceae) — состоит из одного
рода азоллы (Azolla), 6 видов которого произрастают в тропических об-
ластях и в умеренной области Северной Америки.
Это также плавающие водные разноспоровые растения. Размножение
и развитие зародыша у Azolla в общем сходно с Salvinia. Salvtniales про-
изошли от равноспоровых папоротников, именно от предков порядка Нуте-
nophyllales (рис. 174). Много общих черт у них с родом Trichomanes.
В ископаемом состоянии остатки Salvinia встречаются в меловых и палеоге-
новых отложениях, a Azolla — в антропогеновых (в Европе — межледни-
ковых).
У высших растении, как мы видели выше, спорофит всегда развивает-
ся из зиготы, образующейся путем слияния мужской и женской гамет в про-
цессе полового размножения. Этим объясняется большая приспособляе-
мость спорофита к окружающим условиям в сравнении с гаметофитом, ко-
торый развивается из споры. У мохообразных спорофит (спорогонии) сильно
редуцирован. Получая питательные вещества от гаметофита, с которым
он физиологически связан, спорогоний не имеет сложного расчленения.
Однако он прекрасно выполняет основную задачу — образует большое ко-
личество спор и с помощью различных приспособлений обеспечивает их
распространение.
Преобладание гаметофита и необходимость воды во время оплодотворе-
ния ограничили распространение мохообразных на суше. В большинстве
своем они связаны с сырыми или увлажненными местообитаниями.
У папоротникообразных спорофит самостоятельно существует. Он до-
стигает больших размеров, имеет стебли, листья и корни сложного внутрен-
него строения. Эти' особенности папоротникообразных позволили им более
широко распространиться на суше в сравнении с мохообразными. Папо-
ротникообразные играли и играют большую роль в сложении растительного
покрова Земли. Однако гаметофиты (заростки) папоротникообразных еще
тесно связаны с водной средой. Половое размножение у них может осущест-
вляться также лишь при наличии воды в момент оплодотворения. Это огра-
ничивает возможности папоротникообразных в завоевании суши. Они свя-
заны в своем распространении с достаточно влажными и тенистыми место-
обитаниями.
Дальнейшее завоевание суши высшими растениями могло иметь место
лишь при обеспечении полового размножения без наличия водной среды.
Такая возможность реализовалась у семенных растений. В цикле развития
семенных растений, как и у папоротникообразных, преобладает спорофит.
При этом они всегда разноспоровые растения. Здесь продолжается общее
направление эволюционного развития спорофитной линии — прогрессив-
ное развитие спорофита и дальнейшая редукция гаметофита. Наиболее слож-
ного строения спорофит достигает у покрытосеменных растений.
ин
г г.
Семенные растения
Семенные растения представлены . двумя отделами: голосеменные
(Gymnospermatophyta, Gymnospermae) и покрытосеменные (Angiospermato-
phyta, Angiospermae). Они характеризуются наличием семян, развивающих-
ся из семяпочек. Семяпочка — это видоизмененный мегаспорангий.
У всех папоротникообразных гаметофит менее развит, чем спорофит.
Он имеет небольшие размеры, простое строение, существует недолгое время.
Особенно сильно редуцированы гаметофиты у разноспоровых представите-
лей папоротникообразных. Гаметофиты у разноспоровых папоротникооб-
280
ВЫСПШВ РАСТЕНИЯ
ли
гг
разных развиваются, не покидая микро- и мегаспор, и состоят нередко из
1 — 2 вегетативных клеток (например, мужские заростки Selaginella, Salvt-
nia) или из небольшого их числа (женские заростки тех же растений).
У семенных растений гаметофиты, как мужской, так и женский, реду-
цированы еще больше, чем у папоротникообразных. Уже у селягинелйвг-
наблюдается прорастание мегаспоры в женский заросток, развитие на нем
архегония и даже формирование молодого спорофита внутри мегаспоран-
гия. У некоторых видов Selaginella формирование заростка и развитие нового
спорофита (зародыша) протекает в мегаспорангии, когда он находится еще
на материнском растении. Отсюда до образования семени — один шаг.
У семенных растений прорастание мегаспоры и образование женского
заростка, оплодотворение и развитие нового спорофита — зародыша — всег-
да происходит внутри мегаспорангия, когда он находится еще на материн-
ском растении.
Семяпочка^мегаспорангий) становится семенем. Развитие семени идет
за счет материнского растения.
Зародыш обычно содержит все основные органы взрослого растения:
стебель, лист, корень. Он снабжен также запасом питательных веществ,
прекрасно защищен кожурой семени от неблагоприятных воздействий.
Семя отделяется от материнского растения и, упав на землю, может
длительное время находиться в состоянии покоя. С наступлением благоприят-
ных условий (тепло, наличие воды и т. д.) семя прорастает. Зародыш тро-
' гается в рост, используя на первых порах запасные вещества семени. Кожу-
ра семени сбрасывается, проросток вскоре укореняется, и растение перехо-
дит к самостоятельной жизни. Сильно редуцированный мужской гаметофит
переносится тем иди иным способом на семяпрчку. В своей вегетативной
части он сведен к образованию главным образом пыльцевой трубки. Пыль-
цевая трубка проводит мужские гйметы, потерявшие у большинства семен-
ных растений подвижность, непосредственно к женской гЪмете (яйцеклет-
ке), скрытой внутри мегаспорангия. Оплодотворение здесь не связано с на-
личием волы.
Выработка семени определила огромное преимущество семенных рас-
тений перед споровыми. Развитие последних тесно связано с наличием до-
статочной влажности и тепла. Ведь громадное большинство спор, не находя
благоприятной обстановки, погибает. Споры не имеют достаточной защиты,
в них нет достаточного запаса питательных веществ. Развитие гаметофита,
процесс оплодотворения у споровых требуют также определенных условий,
которые далеко не всегда могут быть обеспечены на суше.
У семенных растений для распространения служат не споры, а семена.
Прекрасно приспособленные к современным условиям жизни на Зем-
ле, семенные растения заняли господствующее положение на суше, оттеснив
споровые растения.
Попытки выработки семени встречались неоднократно в разных груп-
пах папоротникообразных (у Miadesmia и Lepidocarpon среди Lycopsida\
может быть, это имело место и у Sphenopslda}, но лишь у Pteropsida извест-
но настоящее семя, причем здесь можно проследить и процесс его формиро-
I
ин
ли
ОТДЕЛ—ГОЛОСЕМЕННЫЕ (GYMNOSPtRMAT^J^TA,
ИЛИ ,GYMNOSPERMAE> X '
Голосеменные характеризуются наличием незащищенных семяпочек
(мегаспорангиев), расположенных открыто на мегаспорофиллах; в случае
редукции мегаспорофиллов семяпочки кажутся расположенными на верхуш-
ке побега.
Прорастая, микроспора (пыльца, пылинка) попадает непосредственно
на семяпочку; оказавшись на вершине н^целлуса, она прорастает в
I
1
I
отдал голоснмкнныв
281
пыльцевую трубку, которая, следуя через ткань нуцеллуса к женскому
даростку, доставляет мужские гаметы к яйцеклетке.
Голосеменные, таким образом, являются разноспоровыми растениями.
В цикле развития, как и у папоротникообразных, у них преобладает споро-
фит (само растение). «ок мммт—й-
лупировмы. При pTQjff щодсрцй гяметпфит не покидрет свяяи с матерднг.ким
растением—развиваясь внутри семяпочки^гпе происходит образование ме-
гаспоры и ее прорастание вженский заросток (первичный эндосперм).
Мужской гаметофит также претерпевает редукцию вплоть до полной
потери вегетативных (проталлиальных) клеток у наиболее высокооргани-
зованных представителей.
— В пределах отдела хорошо прослеживается появление и совершенство-
вание пыльцевой трубки, наиболее развитой у представителей порядка
хвойных.
I г.
«г
Для уяснения эволюционных связей семенных растений со споровыми
папоротникообразными чрезвычайно интересным и яяжным„яуяд|Вря няли-
уиецодвижных сттсрмятпяпиплк^у более древних порядков —> Cycadales и
в то время как у современных хвойных, а также у Ephedra, Gne-
tum и Welwitschia развиваются неподвижные мужские гаметы — спермин.
В пределах отдела можно проследить эволюцию семени от простейших се-
мян семенных папоротников до высокоорганизованных семян с дифферен-
цированным зародышем у современных хвойных.
Голосеменные — исключительно лрпвесные растения (деревья, кустар-
. HHKH^jggm^растений, kotISJO^
''стаНйены средд^более молодого^ и совершенного отдела — покрытосемен-
чие травянистый растений — прогрессивный признак.
Они возникли из деревянистых форйг~---- *---------- "
Голосеменные — очень древняя группа растений^ Ископаемые..^статки
их известны начиная с верхнего^^евона палеозойской зры. В своемпроис-
хождении ошгття'бвязаны (^современными Ptertdophyta и должнк быть
выведены из простейших девонских папоротникообразных и представляют,
вероятно, различные ряды развития. Среди голосеменных семенные папо-
ротники и кордаиты достигают большой сложности уже в палеозое (в ка-
менноугольном и пермском периодах). Остальные порядки были особенно
• развиты в мезозое. Мезозой называют эрой господства голосеменных.
В мезозое же наблюдается полное (беннеттитовые, кейтониевые) или чао
тичное (саговниковые, гинкговые) вымирание многих голосеменных и воз^
^никновение более новых их групп (порядок С aniferales), представленных
большим числом видов, доживших и до наших дней. Появление и развитие
этих новых, более совершенных голосоменных совпало с появлением на Зем-
ле покрытосеменных растений и современных групп папоротников (ленто-
спорангиатных).
Внутри отдела голосеменцых^продолжает^ мотрлфитглткяяя (Contfero-
psida) и макро^^льтаяЦС^4ор^4а) дадаи эволюции.
Голосеменные произошли от Pteropstda и являются по своей организа-
ции (что особенно хорошо подтверждается строением их гаметофитов) про-
межуточной ступенью между пяппрптаикообраянимц и покрытосемен|рют.
По вопросу о родственных отношениях меЗйду^классамиголосеменных в
связи с их отдельными классами Ptertdophyta у ученых нет еще единого.
ill;
1И1
Одни ученые связывают все голосеменные с Pteropstda, основываясь
на том,' что только у них имелись «настоящие» семена. Однако семена были
и у микрофилльных Lycopsida. Другие считают, что саговниколистные
связаны в происхождении с Pteropstda, а шишконосные — с Lycopsida.
В этом случае голосеменные должны рассматриваться как искусственная
группа растенийполифилетического происхождения^ Окончательное ре-
шение этого вопроса — дело будущего. Раннее появление голосеменных рас-
тений совпадает по времени сосуществованием в то время разноспоровых
г. г
282
выс
11 s I:
РАСТЕНИЯ
форм как среди Pteropslda, так и Lycopsida. Особняком стоят оболочкосемен-
ные. Отсутствие древних ископаемых остатков их и резкие отличия в строе-
нии отдельных представителей не позволяют пока установить четкое отно-
шение их к другим голосеменным растениям
Голосеменные разделяют на 3 класса.
1й КЛАСС—САГОВНИКОВИДНЫЕ (CYCADOPSIDA)
Листья крупные (макрофил л и я), черешковые или сидячие. У более
примитивных форм они перистые, сходные с листьями папоротников, реже
цельные, ланцетные.
Семяпочки (мегаспорангии) расположены на листовидных или различ-
ным образом измененных мегаспорофиллах. Мегаспорофиллы одиночные
или собранные в шишки.
В классе саговниковидных выделяют 4 порядка.
1-Й ПОРЯДОК — СЕМЕННЫЕ ПАПОРОТНИКИ (PTERIDO SPERM
этому порядку принадлежат древне:
ли
не и наиболее примитивные
голосеменные растения. Семенные папоротники — целиком вымершая груп-
па растений. Они впервые появились в верхнем девоне и наибольшего раз-
вития достигли в каменноугольном периоде. Затем наблюдается угасание
семенных папоротников, и л:
ли
ь небольшое число их дожило до конца перм-
ского периода палеозойской эры.
Семенные папоротники были древовидными растениями с прямостоячим
или лежачим стеблем и крупными сложноперистыми листьями, сходными с
листьями (вайями) папоротников. Внешне они, таким образом, напоми-
К
нали современные древовидные папоротники, но отличались от них нали-
чием семян. Семенные папоротники известны в большом числе представи-
телей. Этот порядок в настоящее время разделяют на 4 — 5 семейств, из
которых наиболее важными являются семейства лигинодендровые и
медуллозовые.
Семейство лигинодендровые (Lyginodendraceae) более примитив-
ное. Представители его были лазающими растениями. Стебель моностели-
ческий. Семяпочки одиночные. Окаменелые остатки различных частей рас-
тений (листьев, стеблей, семян) из этого семейства описывались под раз-
личными названиями и в разное время; л:
ь позднее удалось установить
их принадлежность к тем или иным родам. Наиболее хорошо изучена калим-
матотека (Calimmatotheca hoeninghaustt), Стебли ее описывались ранее
под названием Lyginodendron^ корни — под названием Kaloxylon, листья —
Sphenopteris, микроспорофиллы — Grosso theca, а семена — Lagenostoma.
Стебель налимматотеки был длинный, но тонкий, до 4 см диаметром. На
нем располагались крупные (до 50 см длины), сложные, дважды-трижды-
перистые листья. Молодые листья были улиткообразно свернутыми. Анато-
мическое строение стебля довольно сложное. В стебле имелась мощная серд-
цевина. Снаружи она была окружена кольцом коллатеральных пучков.
К древесине их'примыкала вторичная ксилема, от которой к периферии рас-
полагались камбий, затем — флоэма и эндодерма. Между пучками — широ-
кие сердцевинные лучи.
Далее располагалась довольно мощная кора с значительным развитием
в ней механических элементов. В коре имелись многочисленные листовые
следы (рис. 179). Таким образом, стебли обладали вторичным ростом. Микро-
спорофиллы были расчленены на ^стерильную и спороносную части (сегмен-
ты). В стерильной части имелись перйсторассеченные листовидные пластин-
ки. Спороносные сегменты были блюдцевидными-(в виде эполет); на нижней
стороне они несли по 6 — 7 микроспорангиев (рис. 179, 3). Микроспо-
ры крупные и в проросшем состоянии (мужской заросток) содержали боль-
«
ОТДЕЛ ГОЛОСЕМЕННЫЕ
283
Рис. 179. Калиыыатотека:
I—общий вид листа; 2 — поперечный разрез стебля (сп—сосудистые пучки, первичная древесина обра-
щена к центру, к ней примыкает вторичная древесина, лх—листовые ходы, л—луб. к—нора с механиче-
скими участками, с—сердцевина); 3—миюэоспорофилл^ (стч—стерильная часть, $пч—спороносная часть.
сп-Цшорангий); 4—семяпочка (мегаспорофилл), снаружи одета плюской; 5—продольный разрез семяпоч-
ки (н—центральная часть нуцеллуса, и—интегумент* пл—плюска, пк—пыльцевая камера)
шое ЧИСЛО КЛетоК. ПыЛЬПВВЫХ трублк ттаттрпттгтягктг—не—нмоли.
Внутри микроспоры развивались, вероятно, подвижные сперматозоиды.
Мелкие (около 5,5 мм длины) мегаспорангии (семяпочки) калимматотеки
очень напоминали семяпочки современных саговниковых. Семяпочка сна-
ружи была одета интегументом, окружавшим нуцеллус почти на всем про-
тяжении. Он плотно срастался с нуцеллусом, но в верхней части оста-
вался свободным, образуя микропиле (рис. 179, 5). Нуцеллус имел на вер-
хушке конусовидный вырост, вокруг которого находилась пыльцевая ка-
мера. Интегумент в верхней части образует свою камеру, примыкающую
снаружи к стенкам пыльцевой камеры (рис. 179, 5). В центре нуцеллуса
располагался мощный эндосперм — женский заросток. У одного вида
эндосперм был найден в хорошей сохранности, в верхней его части име-
лось 3 архегония. Снаружи семяпочка почти доверху была окружена осо-
бым покровом — лопастной плюской, которая с ней не срасталась. На
плюске были развиты головчатые железки, совершенно такого же строе-
ния, что и па вегетативных органах. В семенах калимматотеки, достигав-
ших 6—7 мм длины, как и у всех семенных папоротников, зародыша до
сих пор найти не удалось.
У ряда представителей этого семейства стели имели значительно более
простое строение. У некоторых родов, известных из нижних отделов камен-
ноугольного периода, была протосте ль.
Семейство медуллоаовые (Medullosaceae). Представители этого
семейства имели прямостоячий стебель толщиной от 2 до 20 см полистели-
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 181. Кейтошгевые:
I—сагевопгерис (лист); 2—кеАгоаня: в—ннкроспорофвлл; б—микроопора с воздушными мешками; •—
мегаспорофилл с пвристорашкшоженшлкн . «завязями*; « — продольный разрез «завязи* (с — семяпочки
с интегументом)
Семенные папоротники в своем происхождении упираются в простей-
е формы папоротников (Prtmofilices) или, возможно, ведут происхожде-
ние даже от псилофитов. В свою очередь, семенные папоротники являются
исходной группой для кейтониевых, саговниковых
lit
гл
и беннеттитовых.
очень интересная и за-
представители которых
2-Й ПОРЯДОК — КЕДТОНИЕВЫЕ (CAYTON1ALES)
К этому порядку принадлежит небольшая, но
гадочная группа ископаемых семенных растений,
жили в среднеюрское время. Они были описаны в 1925 —1926 гг. и сначала
рассматривались как покрытосеменные растения. Сюда принадлежат 2 р о-
д а — кейтония и гристорпия. Известны только листья и органы размноже-
ния этих растений. Стебли и корни до сих пор не найдены.
Кроме того, между листьями и спорофиллами долго не находили связи
и описывали их как разные роды: листья — как Antholitis, микроспоро-
филлы — как Sagenopterts, мегаспорофиллы — как Caytonia и Gristorpia.
Кейто пневые были мелкими, возможно прибрежно-водными растения
обликом напоминая водные папоротники из порядка марсилиевых. Их пе-
ристые листья состояли из 3 — 6 ланцетных листочков. Характерно нали-
чие средней жилки и сетчатого жилкования (рис. 181,1). Микроспорофиллы
имели плоскую ось, на разветвлениях которой располагались пучками по
3 — 6 синаигии, по 4 микроспорангия в каждом. Получались как бы
четырехгнездные микроспорангии (рис. 181, 2). Микроспоры были снабжены
двумя воздушными мешками. Мегаспорофиллы были также перистыми. На
боковых разветвлениях их располагались тельца, похожие на завязи. Они
формировались в результате заворачивания дольки мегаспорофилла. Здесь,
на верхушке «завязи», образуется и «рыльце» (рис; 181, 2в). Такие «завязи»
внутри содержали 6 — 30 семяпочек. Семяпочки прямые. Интегумент их
был почти до основания свободным. Внутрь «завязи» вел канал. Через него
в полость «завязи» проникали микроспоры, их часто находили в микропиле
семяпочек.
Таким образом, кентониевые были в какой-то мере покрытосеменными
растениями, но к цветковым они, конечно, никакого отношения не имели,
iHTi
286
выс
111 r> i у
РАСТЕНИЯ
да и сама «покрытосеменность» здесь иного характера, так как заворачивал-
ся краями не весь мегаспорофилл, а лишь часть его. Систематическое по-
ложение кейтониевых неясно. Большинство ботаников связывают их с се-
менными папоротниками, другие же (у нас А. Криштофович) «бли*
жают их с марсилиевыми.
3-Й ПОРЯДОК — САГОВНИКОВЫЕ (CYCADALES)
Саговниковые представлены в настоящее время одним семейством
Cj/cadaceoe, которое насчитывает 9 родов и 100 видов, встречающихся в тро-
пических и отчасти субтропических областях Восточной Азии, Австралии,
Африки и Америки.
2
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Род Cycas (16 видов) распространен от Восточной Азии до Северо-
Восточной Австралии и на острове Мадагаскар; роды Macrozamia (15 ви-
дов), Bowenia (2 вида) — в Австралии; роды Stangeria (1 вид) и Encepha*
lartos (15 видов) — в Африке; род Zamia (35 видов) — в Центральной и
Южной Америке; роды Ceratozamia (6 видов) и Dioon (3 вида) — в
Мексике; род Microcycas (1 вид) — на Кубе.
Ныне живущие саговниковые являются остатками некогда широко
распространенной группы растений. Ископаемые остатки их стеблей и ли-
стьев, а также микроспорангии и семена в большом количестве начинают
встречаться лишь в мезозойских отложениях (триас и юра)1.
В мезозое саговниковые были, пожалуй, господствующими среди дру-
гих наземных растений.
Саговниковые — деревянистые растения. Стебель ^высокий, колонно-
видный, 6—8 м высоты (у некоторых Macrozamia до 20 м) или клубневид-
ный, невысокий и даже скрытый в земле (у видов S tangeria, Bowenia). У од-
ного вида Zamia (Z. polygama), произрастающего на острове Куба, стебель
очень маленький — 3 см высоты и 2 см диаметром. Листья саговниковых
обычно простоперистые, напоминают листья пальм, и* только у рода
Bowenia листья дважды-триждыперистые (рис. 182, 3). Листья крупные
(до 2 м длины), жесткие, вечнозеленые, в молодом состоянии у большинст-
ва видов улиткообразно свернуты, как у папоротников. У многих саговни-
ковых по опадании листьев сохраняются их основания/ и тогда стебли по-
крыты как бы панцирем; у других же листья опадают целиком и тогда стеб-
ли остаются голыми. Анатомическое строение стебля весьма своеобразно/
Стебель имеет сердцевину, клетки которой набиты крахмалом. За ней к пе-
риферии расположено узкое кольцо древесины, затем следует камбий, тон-
кий слой флоэмы и кора. В коре, как и в сердцевине, характерно наличие
слизевых ходов, заполненных слизью (сходство с мараттиевыми папорот-
никами).
У некоторых саговников (Cycas, отдельные виды Encephalartos) в коре
возникают последовательно кольца камбия, отслаивающие новые ряды кси-
лемы и флоэмы. Корень саговниковых длинный. От него отходит большое
число боковых корней. В тканях последних поселяются грибы и азотофик-
сирующие бактерии, а затем сине-зеленая водоросль Anabaena. Концы кор-
ней, разрастаясь, принимают кораллообразный вид.
У большинства саговниковых спорофиллы собраны в тесные собрания—
шишки.
Саговниковые — растения двудомные. Мужские и женские шишки об-
разуются на разных особях. Урода саговник (Cycas) женских шишек нет,
мегаспорофиллы свободные, развиваются на вершине стебля. Рассмотрим
строение органов размножения у саговника. В Восточной Азии широко рас-
пространен Cycas revoluta (рис. 182, 2). Это дерево до 3 м высоты. Колон-
нообразный ствол его вверху несет крону перистых листьев до 2 м длины. ;
На вершине ствола мужских экземпляров образуются мужские шишки.
Они достигают 50—70 см рдвшл. На оси шишки располоя£ены микроспоро-1
филлы. На нижней стороне микроспорофиллов находятся многочисленные
микросорусы, каждый из 2—4 микроспорангиев.
Микроспорангий расположен на толстой ножке и одет оболочкой, клет-
ки которой имеют неравномерно утолщенные стенки. Спорангий вскрыва-
ется продольной трещиной. В нем развивается большое количество эллипти-
ческих мелких микроспор. Воздушных мешков микроспоры не имеют. Еще
находясь внутри микроспорангия, микроспоры прорастают, образуя муж-
ской заросток (рис. 183). При прорастании микроспоры в результате перво-
го деления образуются две клетки. Одна из них — проталлиальная клетка;
деление другой дает начало антеридиальной клетке, плотно прижатой к
ин
in:
: ii i
1 Остатки листьев и стеблей, сходных с саговниковыми, находят еще в верхнекамен-
ноугольных отложениях, но они, вероятно, принадлежат к семенным папоротникам.
вы
МИН 11л
РАСТЕНИЯ
2
Рис. 183. Размножение саговниковых:
1—микроспорофиллы» микроспоры и мужские заростки: а—микроспорофилл цикаса; б—микроспорофилл
замни; в—сорусы замии; г—продольный разрез микроспорангия замии; 0—микроспоры знцефаляртоса;
2—прорастание микроспоры и развитие мужского заростка: а,б,в,«»0—последовательные стадии развития
заростка диоона (пт-проталлиальная клетка» зн—генеративная клетка» вг — вегетативное ядро» спк —
спермягеввая клетка» н—клетка-ножка» сестринская клетка); е—конец пыльцевой трубки замии
(пт—проталлиальная клетка» н—клетка-ножка» сп—сперматозоиды)
проталлиальной, и вегетативной, из которой в дальнейшем развивается
гаустория (пыльцевая трубка). В таком трехклеточном состоянии микро-'
спора высеивается из вскрывающегося'микроспорангия и переносится вет-
ром (или реже насекомыми у Encephalartos) на семяпочку, где и идет даль-
нейшее развитие мужского заростка. Мегаспорофиллы у всех видов рода
Cycas располагаются в небольшом числе на верхушке стебля, чередуясь
с вегетативными листьями (сходство с папоротниковидными).
Перистые мегаспорофиллы несколько сходны с вегетативными листь-
ями, но имеют меньшие размеры и окрашены в желтый или красноватый цвет
(рис. 184). В нижней части мегаспорофилла, на его разветвлениях, располо-
жены мегаспорангии (семяпочки), обычно 6, по 3 с каждой стороны
споро филла (рис. ‘184, I).
Мегдшорангии крупные, до 5 — 6 см длины (рис. 185, I). Нуцеллус на
верхушке образует пыльцевую камеру. Снаружи нуцеллус окружен мощ-
ным интегументом, состоящим из трех слоев: наружного — мясистого, сред-
ОТДВД ГОЛОСЕМКНВЫВ
ill:
него — каменистого и_внутреннего—
снова мясистого. Интегумент почти
"на всем протяжении плотно срос:
ся с нуцеллусом. На вершине семя-
почки он вытянут в виде трубочки,
образуя микропиле (пыльцевход).
Под микропиле в нуцеллусе имеется
небольшая полость — пыльцевая ка-
мера.
В центре семяпочки находится
многоклеточная, тонкостенная
ткань — эндосперм (женский заро-
сток), в верхней части которого ббра-
Рже. 184. Мегаспорофиллы саговниковых:
1 — цккаса (Cycas clrcloalis); 2—<цикаса (Cycas
revoluta); s — цератозамня (Ceratozamta mexicana)
имеет крупную яйцеклетку и рано от-
мирающее ядро брюшной канальце-
вой клетки.
Семяпочка саговниковых, как
и всех семенных растений, является
видоизмененным мегаспорангием.
Нуцеллус является новым образо-
ванием, не встрбчаюйймён^ у» папо-
ротникообразных. .
Развитие семяпочки идет сле-
дующим образом. На мегаспорофилле
первоначально возникает нуцеллус в
виде небольшого бугорка совершенно
однородных клеток. По мере его роста из основания нуцеллуса вырастает
интегумент (он принадлежит, таким образом, мегаспорангию, а не споро-
филлу). В верхней части нуцеллуса вскоре выделяется одна, более крупная
клетка. Это материнская клетка спор. Она претерпевает редукционное де-
ление, в результате чего образуются четыре гаплоидные мегаспоры. Они
расположены друг над другом цепочкой, по вертикальной оси нуцеллуса.
Вскоре три верхние мегаспоры, расположенные ближе к микропиле, отми-
рают, а нижняя прорастает, образуя женский заросток — эндосперм. За-
росток, таким образом, находится внутри мегаспорангия.
К моменту созревания архегониев края эндосперма начинают нарас- -
тать быстрее благодаря усиленному делению краевых клеток. В результате
архегонни располагаются на дне небольшого углубления (рис. 185, 2).
В строении женского заростка (эндосперм) наблюдаются черты, сближаю-
щие его с заростком папоротника. Так, клетки эндосперма содержат лейко-
пласты. При вычленении заростка из семяпочки они зеленеют, превращаясь
в хлорофилловые зерна. ;
Микроспора, перенесенная ветром на семяпочку, через микропиле по-
падает в пыльцевую камеру нуцеллуса. Здесь она набухает и начинает про-
растать, при этом вегетативная клетка вытягивается в длинную трубочку —
гаусториют внедряющуюся в ткань нуцеллуса. При помощи гаустории муж-
ской гаметофит получает питательные вещества из нуцеллуса. Антеридиальг
ная клетка делится, образуя спермагенную клетку и клетку-ножку антери- * -
1ЩЯ^рис. 183, 2). СпермагеннИИ клетка сильно разрастается!! при последую-
дает два подвижных сперматозоида, значительно превышаю-
щих размером микроспору. Сперматозоиды имеют многочисленные жгутики,
расположенные спиральной лентой на их верхушке (рис. 183, 2е). Постепен-
но нарастая, заросток продвигается передним концом в углубление эндоспер-
ма. При этом стенки клетки-ножки и проталлиальной клетки расплывают-
ся и ядра их располагаются вместе со сперматозоидами в общей плазме.
Затем пыльцевая трубка лопается на вздувшемся нижнем конце и со-
держимое ее изливается в углубление эндосперма. Здесь сперматозоиды
10-602
Г. г.
hi:
i. i
mi. »<
290
ВЫШНИЕ РАСТЕНИЯ
Рис. 185. Семяпочка и семя саговниковых:
1 — мегаспорангий (семяпочка) Cycas revolata в нродольнож разрезе (мжс — наружный мясистый слой,
дже — внутренний мясистый слой, м — юнсроиипе» пк — пыльцевая камера» нц — нуцелпус. ар — архе-
гоний, хе —каменистый слой); 2 — верхняя часть семяпочки Cycas revoluta в продольном разрезе
, (ар — архегонии, пт — пыльцевые трубки» ап — сперматозоиды» и -* интегумент» at — микропиле);
з— продольный разрез семени Dioon edule (х —корешок. т9 — эндосперм» хс — каменистый слой»
кме — наружный мясистый слой» at — микроанле); 4 — прорастание семени
плавают некоторое время, а затем один из них направляется к архегонию и
сливается с его яйцеклеткой (рис. 185, 2)» Другой сперматозоид погибает.
Оплодотворенная яйцеклетка (зигота) делится, образуя большое коли-
чество клеток, располагающихся в нижней части архегония. Нижние клет-
ки дают начало зародышу, а верхние вытягиваются в подвесок, который
продвигает разрастающийся зародыш во все новые слои эндосперма, за счет
которого идет его развитие. В таком виде семяпочка опадает. К этому време-
ни она сильно разрастается. Наружный покров ее становится мясистым,
сочным и окрашенным в красный цвет. Средний слой становится каме-
нистым, плотным. Он надежно защищает скрытый внутри зародыш. Внут-
ренний слой постепенно разрушается, и содержимое его используется клет-
ками эндосперма (рис. 185, 3). Дозревание зародыша идет уже после опаде-
ния семени. Созревший зародыш имеет зачаточный стебелек — подсемя-
дольное колено, переходящее ниже в первичный корень; на верхушке он
одет корневым чехликом (рис. 185, 3)- В верхней части стебелька распола-
гаются обычно две семядоли и конус нарастания. Зародыш окружен эндо-
спермом, который затем используется также прорастающим растеньицем.
*
ОТДЕЛ ГОЛОСИМВн н ьТВ
291
При прорастании семени семядоли зародыша остаются внутри и служат для
всасывания питательных веществ из эндосперма. Проросток укореняется,
подсемядольное колено вытягивается, распрямляется, и на вершине его раз-
виваются листочки (рис. 185, 4).
У ДРУГИХ родов саговниковых мегаспорофиллы собраны в женские
шишки. Они имеют часто большие размеры и очень различны по внешнему
виду. Так, у Macrozamia denisonii они достигают 1 м длины при массе
40 — 42 кг, но у карликовой Zamia pigmaea они длиной всего до 3 еле. Иногда
мегаспорофиллы претерпевают редукцию: вегетативные верхушечные части
их недоразвиваются, и у некоторых родов (например, у Zamia) мегаспоро-
филлы становятся щитовидными (рис. 182, 184).
У некоторых саговниковых в пыльцевой трубке образуются не два, а
четыре (Ceratozamia) или даже больше (у Mier осуcas — 16 —22) спермато-
зоидов.
Практическое значение саговниковых сводится к получению человеком
саго из сердцевины Cycas revoluta, который часто с этой целью разводят в
Японии. Многие саговники — прекрасные декоративные растения. С этой
целью их часто разводят в оранжереях и в открытом грунте. В СССР их
культивируют на Черноморском побережье Кавказа, южнее Сочи.
tn:
4-Й ПОРЯДОК — БЕННЕТТИТОВЫЕ (BENNETTГГALES)
Беннеттитовые — ископаемые растения, жившие на Земле от триасово-
го до мелового периода мезозойской эры. Особенно широко они были рас-
пространены в юрском и меловом периодах.
Характерно, что вымирание беннеттитовых совпало с появлением и
внезапным расцветом цветковых, покрытосеменных растений. Главными
и наиболее распространенными родами были Cycadeoidea (Bennettites),
Williamsonia и Williamsoniella (рис. 186). Беннеттитовые представляли со-
бой небольшие деревья, имевшие облик саговников или папоротников. Стеб-
ли их были или высокими колонновидными (Williamsonia, рис. 186, I), или
обильно, почти до основания, ветвистыми (Williamsoniella, рис 186, 2).
Листья у большинства видов были сложноперистыми, реже простыми, цель-
ными (у Williamsoniella coronata). По строению стебля беннеттиты были
сходны с саговниками. Ствол их имел мощную сердцевину и кору. Древесина
была развита сравнительно слабо, окружая нешироким кольцом сердцеви-
ну. Кольцо камбия обеспечивало нарастание стебля в толщину. Вторичная
древесина состояла главным образом из лестничных и точечных трахеид.
Характерной особенностью беннеттитов, отличавшей их от всех осталь-
ных голосеменных, являлось наличие у большинства их обоеполой пазуш-
ной шишки (стробила). Снаружи (снизу) шишка была одета верхушечными
листьями, расположенными спирально. Далее на оси шишки располагались
(обычно по кругу, циклически) перистые (реже цельные) микроспорофиллы.
На разветвлениях микроспорофиллов, находились микросп<урангии, сра-
ставшиеся по соприкасавшимся краям в синангии (сравнить с мараттиевы-
ми папоротниками). В них развивались многочисленные микроспоры без
~ ки, заканчивая ось, располага-
лись мегаспорофиллы. Мегаспорофиллы имели длинную ножку, заканчи-
вавшуюся мегаепбрангием (семяпочкой). Они чередовались со стерильными
чешуйками, которые двухсторонне расширялись на верхушке (рис. 187).
В результате семяпочка, расположенная между двумя соседними чешуйка-
ми, была хорошо защищена, прикрыта ими. Поэтому беннеттиты называют
пгшупопрытогпмпнными растениями— *“
У некоторых беннеттитовых имелись раздельнополые (вторично) шиш-
ки, как, например, у Williamsoniella (рис. 186, 1).
Семяпочки беннеттитовых (около 6—9 мм длиной) были одеты инте-
гументом, вытянутым на верхушке в длинную трубочку с отверстиями на
конце (микропиле). Далее располагался нуцеллус. Женского заростка (эндо*
10*
ин
ин
Гни
202
ВЫСШИЕ PACT
lh; 1 s
Я
Рис. 186. Беннеттнты:
1 — вильямсонвя (Williamsonia sewardiana) i а ~ обтай вид, б — лист; £ — вкльямсониедла (William-
soniella coronata): обобщай вид б —обоеаоиая шишка: з—цжкадесидеа (Cycadeoidaa шагвЫапа)
иг,
сперма) найдено не было, но часто у беннеттитов находят семена с хоро-
шо сохранившимся зародышем. Семена были безбелковыми, зародыш имел
две семядоли (рис. 187, 3).
У разных беннеттитовых стробил находился на разных эволюционных
уровнях. Так, у Cycadeoidea микроспорофиллы имеют еще папоротниковый
облик, они сложноперистые, но у Williamsoniella они листообразные, напо-
минающие расширенные тычинки цветкового растения нимфеи (Nymphaea).
Вскоре после открытия беннеттитов их щшпку (стробил) стали рассматри-
вать как прототип цветка покрытосеменных растений. Особенно эти взгляды
были развиты английскими палеоботаниками А.р бером и Парки-
ном, создавшими стробилярную теорию происхождения цветка (см. «Цвет-
ковые растения»). Согласно этой теории, наиболее примитивными цветко-
выми являются растения из группы многоплодниковых (Polycarpicae), как,
например, Magnolia, Nymphaea, многие лютиковые и др. Действительно,
обоеполая шишка Cycadeoidea внешне сходна с цветком Magnolia. Верху-
шечные ее листья образуют как бы околоцветник. Далее следуют тычинки
и* мегаспорофиллы с семяпочками (как бы примитивные пестики). Однако у
простейших цветковых растений все части цветка расположены по спирали
и свободны (например, у Magnolia, Trollius, CaZt/гд и др.), тогда как у бен-
неттитов тычинки (микроспорофиллы) расположены по кругу (цикличе-
ил
.1111»
Рже. 187. Беннвттиты. Шишка, семяпочка и семя:
I — шишка Cycadeoidea dacotenete (продольный разрез): а — иоподМ'б — зрелая; 2—продольный раз-
рез через женскую часть шишки С. gibeoniana; 3 —семяз а — общий вид семени С. albiana. б — про-
дольный разрез семени (с — семядоли, пк — подсемядопьное колено)
ски). Покрытосемявность беннеттитов иного характера и происхождения,
чем у цветковых (у цветковых пестик образовался в результате срастания
по краю завернувшихся мегаспорофиллов).
Семена беннеттитовых безбелковые, строение их более сложное, чем
семян белковых, с эндоспермом, характерных для проста
ин
их цветковых
растений. Все это делает невозможным установить происхождение цветка
покрытосеменных из шишки известных нам беннеттитов. Возможно, будут
найдены предки покрытосеменных среди более примитивных, pai
i:r
х бен-
неттитовых. Многие ботаники считают, что покрытосеменные и известные
нам беннеттитовые произошли от просте
ли
их форм семенных папоротников.
2-й КЛАСС —
ills ни
{СНОСНЫЕ (CONIFEROPSIDA)
иг.
Листья мелкие (микрофиллия), сидячие, цельные, от ланцетных до игло-
видных и чешуевидных, реже широкие, крупные. Мегаспорофиллы сильно
видоизменены, соединены в рыхлые или, наоборот,. очень плотные шишки.
В классе шишконосных выделяют 3 порядка.
1-Й ПОРЯДОК — КОРДАИТОВЫЕ (CORDATTALBS) ъ
Кордаитовые — вымершая группа голосеменных растений. Они жили
на Земле от верхнего девона до перми. К началу триасового периода мезо-
зойской эры они вымерли. Наибольшего развития кордаитовые достигли
294
ВЫСШИЕ РАСТЕНИЯ
Рис. 188. Кордаиты*.
1“ общий вид кордаита (реконструкция); 2“вет-
ка (л“Листья» Ш“собрание шишен» вп —веге-
тативная почка)
в каменноугольном периоде, встреча-
ясь как в северном, так и в южном
полушарии, образуя леса. Остатки
кордаитовых дали начало мощным
пластам каменного угля.
Наиболее изученными являются
роды Cordaltes и Cordaianthus
(рис. 188).
Кордаиты были крупными де-
ревьями, достигавшими 30 м в вы-
соту и до 1 м в поперечнике. Их
стройные стволы ветвились лишь в
верхней части. Стебли обладали вто-
ричным утолщением. По структуре
древесины одни представители были
сходны с хвойными, другие отлича-
лись хорошо развитой сердцевиной,
чем напоминали саговниковые.
Листорасположение спиральное.
Листья кордаитов были кожистыми,
широколинейными или узколанцет-
ными; они суживались к верхушке
и основанию. У одних видов листья
были мелкие, до 3 — 4 см, у дру-
гих — крупные, до 1 м кривы и до
20 см
Hi:
рины. Жилкование парал-
лельное. Жилки слабо дихотомиче-
ски ветвились.'
Кордаиты были растениями дву-
домными. Мужские и женские шиш-
ки их располагались на разных
экземплярах.
Мужские шишки (микро-
стробилы) представляли тесные
сережковидные собрания. На оси
мужской шишки спирально распола-
гались бесплодные чешуйки и микро-
спорофиллы, последние на верхушке
несли 1 — 6 микроспорангиев (рис.
189, /, 2).
Микроспоры, размером до 0,5 мм в поперечнике, имели один воздуш-
ный мешок, располагавшийся вокруг всей микроспоры (рис. 189, 5). В про-
росших микроспорах находили большое число клеток (вероятно, протал-
лиальных, рис. 189,6). Мужской гаметофит был, таким образом, менее реду-
цированным, чем у современных голосеменных растений.
Женские шишки внешне напоминали мужские, но имели более сложное
строение. На оси женской ш
листья и вперемежку с г
щлП
: 111
ки спирально располагались чешуевидные
короткие побеги. У основания этих побегов
имелось 1 — 2 листа. На верхушке побегов располагался один мегаспоран-
гий (семяпочка, рис. 189, 3).
Семяпочки имели толстый интегумент с пыльцевходом наверху. Далее
располагался нуцеллус. В центре семяпочки находился эндосперм с архе-
гопиями (рис. 189, 4). На верхушке нуцеллуса имелась пыльцевая камера,
в которую попадали микроспоры. Оплодотворение, вероятно, осуществля-
лось подвижными сперматозоидами. Зародыши неизвестны. Возможно, раз-
витие зарод
ими
а начиналось после опадения семяпочки.
К порядку Cardattales относятся также более древние голосеменные,
как, например, роды Archaeopitys, С alii ху Ion из верхнего девона, Pitys
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 189. Строение шишек кордаитов:
1 — продольный разрез мужской шишки (Gordaianthos williamsonil); З^микроспорофилл с микроспоран-
гиями; Л— продольный разрез женской шишки того же растения; 4 — семяпочка; s — проросшая микро-
спора (мужской заросток); в —верхняя часть семяпочки, часть нуцеллуса с двумя микроспорами
(лее — микроспоры, в нижней микроспоре видны проталлиадьные клетки пк)
из нижнего карбона, объединяемые в семейство Pityaceae. Это были
крупные деревья (у Callixylon стебли достигали 2 л в поперечнике), по стро-
ению древесины (мощная сердцевина, листовые следы с прорывами стели)
II
они сближаются с семенными папоротниками, но по строению листьев на-
поминают хвойные. Однако органы размножения этих растении пока неиз-
вестны.
Родственные отношения кордаитовых не совсем ясны. Вероятно, они
являются самостоятельной ветвью развития, получившей начало, как и се-
менные папоротники, из каких-то общих предков. Кордаитовые дали на-
чало гинкговым и хвойным.
296
выапшв pact
у ;»r.
Я
I: i ?
11 I
шт.
nt
I: l ?
: 11Г.
2-Й ПОРЯДОК — ГИНКГОВЫЕ (GINKGO A LES)
Порядок гинкговые в настоящее время представлен единственным ви-
дом гинкго дв]рюпастн#ии, (Ginkgo biloba). Этот вид в диком состоя
встречается лишь в горах Юго-Восточного Китая, но с давних пор широко
культивируется (история его прослежена до XI в.) в Китае в районах с уме-
то был введен в культуру японцами, а в
1727 г. Вывезен в Западную Европу, где культивируется в ботанических са-
f дах и парках. У нас в Крыму и на Кавказе гинкго разводят в открытом грун-
те в садах и парках.
Гинкго — «живое ископаемое». В прежние геологические эпохи, осо-
бенно в мезозое, гинкговые были широко распространены на Земле по всей
Евразии и Северной Америке, а также в южном полушарии. Описано 17 ро-
дов гинкговых. Впервые типичные гинкговые появляются в отложениях
палеозойской эры, в верхнем карбоне (род Dichophyllum) и в нижней пер-
ми (р о д Sphenobaiera). Однако листья типа гинкговых известны из еще бо-
лее древних слоев —' из верхнего девона (Ginkgophyllwn). Угасание л:
гинкговых начинается с мелового периода.
Ginkgo biloba — листопадное дерево до 40 ле высотой и 4,5 м в обхвате.
Дерево обильно ветвится и образует густую пирамидальную крону. Побеги
двух родов — удлиненные и укороченные. На удлиненных побегах листья
расположены рассеянно, на укороченных собраны по 3 — 5 пучками. Листья
черешковые, с пластинкой вееровидной формы. На верхушке листья боль-
шей частью двухлопастные, но на коротких побегах более или менее цель-
ные. Жилкование листьев дихотомическое. Верной и летом листья светло-
зеленые, осенью желтоватые до карминно-красных.
Анатомическое строение стебля гинкго очень сходно с хвойными. В стеб-
ле имеется слаборазвитая сердцевина. Далее к периферии расположена
вторичная древесина, образованная трахеидами с окаймленными порами
на радиальных стенках. Во вторичной древесине имеются узкие еердтутип-
ные лучи. Далее расположены камбий и кора.
i;r.
Рис. 190. Гинкго:
1 — укороченная ветвь с мужски-
ми «соцветиями» (сережками); 2—
микроспорофпплы с микроспо-
рангиями; а—укороченная ветвь
с женскими «соцветиями»; 4 —
отдельное «соцветие» ив двух
семяпочек; б — продольный раз-
рев семяпочки; 9—врелое секи
(одна семяпочка замерла) ; 7 —
верхняя часть семяпочки (ин—•
интегумент» — нуцеллус» пи—
пыльцевая камера, а — эндосперм
(заросток). ар — архегонни)
' отдал ГОЛОСЕМЕННЫЕ
г
то — растение двудомное.
: I: i
Мужские «соцветия» имеют вид
сережек (рис. 190,7). Сережки рас-
положены на верхушке короткого
побега. На длинной оси сережки
расположены микроспорофиллы.
Каждый микроспорофилл на ниж-
ней стороне расширенной нити
имеет обычно два (иногда 3 — 7)
микппспопангая.^Микроспоры бм_
воздушных мешков. _
Микроспора прорастает в за-
<;ii^ о
_ к еще внутри микроспоран-
гия.При этом сначала отчленяется
маленькая проталлиальная клет-
ка, которая вскоре разрушается.
Затем отчленяется вторая протал-
лиальная клетка. Оставшееся яд-
ро делится еще раз, образуя две
Рис. 191. Гинкго:
1 — микроспора; £ — проросшая микросцора (он — анте*
радиальная клетка, лк — гаусториальная клетка,
пт — две проталлиальные клетки, из которых нижняя
вскоре расплывается); 3 — конец пыльцевой трубки
(пт— деградировавшая проталлиальная клетка, с—два
многожгутиковых сперматозоида); 4 — сперматозоид;
S — семя в разрезе (н* — наружная сочная часть ин-
тегумента, т — каменистая часть, пс — тонкий
пленчатый слой, а — зародыш с двумя семядолями);
в — проросток
клетки: крупную антеридиаль’ную
и вегетативную (гаусториальную),
соответствующую клетке пыльце-
вой трубки. В таком трехклеточ-
ном состоянии проросшие микро-
споры после вскрывания споран-
гия высыпаются наружу и пере-
носятся ветром на семяпочки, где
и протекает дальнейшее развитие
мужского гаметофита (рис. 191,2,2)
Семяпочки, в числе двух, развиваются на вершине дихотомически раз-
ветвленного стебелька (рис. 190, 4). Каждая семяпочка окружена у основа-
ния кольцевидным валиком (воротничком), который является редуциро-
ванным мегаспорофиллом. Такие собрания мегаспорофиллов расположе-
ны в пазухах зеленых листьев на верхушке укороченных побегов (рис. 190,3).
Семяпочка (мегаспорангий) окружена толстым интегументом, не срос-
шимся на верхушке. Здесь виден узкий канал,— микропиле. Далее распо-
ложен нуцеллус. Наверху клетки нуцеллуса расплываются и образуется
пыльцевая камера (рис. 190, 7). Несколько ниже в нуцеллусе образуется
материнская клетка мегаспор, которая затем, делясь/дает четыре гаплоид-
ные мегаспоры. Из них три верхние вскоре отмирают, а нижняя начинает
делиться. При этом образуются сначала ядра, а затем между ними возника-
ют перегородки. В результате формируется многоклеточный ч эндосперм
(женский заросток). В верхней части эндосперма образуются обычно два ар-
хегония. Архегоний имеет яйцеклетку, брюшную канальцевую клетку и клет-
ки шейки (рис. 190, 7).
Микроспора, перенесенная ветром на семяпочку, попадает через микро-
пиле в пыльцевую камеру, где оболочка ее лопается и гаусториальная клет-
ка начинает расти и внедряется в ткань нуцеллуса при помощи тонких
отростцов. При посредстве этих отростков развивающийся мужской гамето-
фит получает питательные вещества из клеток нуцеллуса. ,Айтеридиальная
клетка делится на две клетки:,{слетку-ножку и спермагенную клетку. Послед-
няя сильно увеличивается в размерах и делится, образуя два подвижных
сперматозоида, снабженных,на переднем конце венцом жгутиков (рис. 191, 4).
Гаустория (пыльцевая трубка) разрастается, и нижний конец ее направ-
ляется к архегониям. В это время в ее содержимом имеется ядро проталли-
альной клетки, ядро клетки-ножки и два сперматозоида, плавающие в об-
щей плазме. Далее конец пыльцевой трубки лопается, и содержимое попа-
дает в углубление на вершине эндосперма. Здесь сперматозоиды некоторое
i: il ;
время плавают, а затем один из них направляется к яйцеклетке архегония
и сливается с ней, производя оплодотворение.
Оплодотворенная клетка приступает к делению. Вскоре образуется
большое количество ядер, располагающихся в общей плазме. Затем между
ядрами возникают перегородки и образуется многоклеточная ткань зароды-
ша. Из нижних клеток этой ткани возникают корешок, стебелек и семядоли
зародыша, а верхние клетки, вытягиваясь, образуют подвесок.
Семя одето толстой оболочкой, образованной интегументом. Наружная
часть оболочки становится сочной; под ней расположены твердая каменис-
тая оболочка и внутренний тонкий, пленчатый слой. Зародыш в семени окру-
жен эндоспермом (рис. 191, 5).
У ископаемой Baiera (из верхнего триаса), имевшей многократно рас-
члененные листья, женские шишки несли большое число семяпочек.
Филогенетические связи гинкговых с другими голосеменными не сов-
сем ясны. Обычно их выводят из кордаитовых на основании сходства в стро-
ении древесины, которое простирается и на порядок хвойных. Но гинкговые
имеют общие черты и с семенными папоротниками (размножение, строение
семяпочки). В последнее время некоторые ботаники относят гинкговые даже
к макрофилльной линии эволюции и помещают их среди С уса do psi da.
jii
ii'i
i:r,ni
l i»I
3-Й ПОРЯДОК — ХВОЙНЫЕ (CONIFERALES)
Хвойные являются наиболее многочисленными представителями голо-
семенных растений в современной флоре. Они насчитывают дколо 600 видов,
объединяемых в 55 родов и 8 семейств.
Распространены хвойные главным образом в северном полушарии.
Здесь они встречаются в наибольшем числе видов и родов. Многие из них,
например сосны, ели, лиственницы, пихты, образуют на огромных прост-
ранствах Евразии и Северной Америки обширные хвойные леса как на рав-
нинах, так и в горных областях. Нередко хвойные леса состоят из одного .
ь немногих видов.
В южном полушарии хвойные довольно широко представлены в уме-
ренных областях — на Огненной Земле, в Патагонии и особенно в Новой
Зеландии, Тасмании. Здесь они также образуют леса.
В тропических областях на равнинах хвойные почти отсутствуют, но
встречаются в горах на известной высоте, где пониженная темпера-
тура. Здесь они также образуют леса или встречаются в виде примеси к дру-
гим растениям.
Интересно отметить, что широким распространением обладают сравни-
тельно молодые виды ироды хвойных (Picca, Pinus, Abies, Larix is. др.),
в то время как более древние их формы занимают небольшие участки земной
поверхности и приурочены в основном к бассейну Тихого океана. По запад-
ному побережью Северной Америки, в Юго-Восточной Азии, на Филиппи-
нах, в Восточной Австралии, Тасмании, в Новой Зеландии, а также в Чили
сосредоточено наибольшее число древних родов хвойных. Это секвойи
и таксодиумы в Северной Америке, криптомерия, метасеквойя в Юго-Вос-
точной Азии, агатисы в Новой Зеландии, араукарии в Австралии и Чили.
Впервые хвойные появились на Земле в каменноугольном периоде па-
леозойской эры. Их остатки в виде окаменелой древесины, шишек, семян,
микроспор, отпечатков ветвей и листьев примитивных представителе^ из
целиком вымершего семейства лебахиевых (Lebachiaceae) известны
главным образом из северного полушария. В южном полушарии они встре-
чались значительно реже. В пермский период хвойныечбыли развиты в
значительном числе видов и родов, главным образом из несколько более
сложного семейства волыпциевых (Voltziaceae). Широкого развития
хвойные достигли в юрский и особенно в меловой период мезозойской эры.
Здесь появляется большинство современных родов, причем обнаружива-
ются различия в составе флоры хвойных северного и южного полушарий е
jii
KI
ОТДЕЛ ГОЛОСЕМЕННЫЕ
in:
.11
н:г
В северном полушарии сосредоточиваются такие семейства, как сосновые,
тисовые, таксодиевые, кипарисовые, в южном полушарии — араукариевые,
подокарповые.
В палеогене и неогене хвойные были широко распространены по всей
Земле и более или менее равномерно по всей Евразии и Северной Америке.
Они росли тогда далеко к северу и югу за пределами современного распро-
странения хвойных. Хвойные леса произрастали тогда в Арктике (Шпиц-
берген, Западная Гренландия) и Антарктике (Земля Грейама).
Большой урон в составе и общем распространении хвойные понесли в
ледниковый период. Многие из них вымерли, другие же были оттеснены к
югу и там переживали неблагоприятное время. Особенно пострадали они в
Евразии благодаря широтному протяжению гор. Здесь хвойные (как и дру-
гие группы растений, в том числе и покрытосеменные), оттесненные к югу
материковыми льдами, оказались также под воздействием ледников, спус-
тившихся с гор (Альпы, Карпаты, Кавказ и т. д.).
Теплолюбивые формы почти на всем протяжении Евразии вымерли,
за исключением форм, произраставших на территории Восточной Азии, где
не было ледника и с давних пор не было мощных горообразовательных про-
цессов. Флора в этих областях развивалась без потрясений, постепенно
приспосабливаясь к ухудшавшемуся климату, поэтому здесь сохранились
до наших дней многие теплолюбивые виды хвойных.
В послеледниковое время хвойные, снова распространившиеся к севе-
ру вслед за отступавшим ледником в Европе и в большей части Азии, не
могли восстановить своей прежней роли как вследствие вымирания, так и зна-
чительного ухудшения и похолодания климата. В Северной Америке благо-
даря меридиональному протяжению гор (на западе и востоце) растения смог-
ли отступить далеко к югу, где пережили ледниковый период и сохрани-
лись в значительно большем числе видов, чем в Азии. П^сле отступления
ледников они вновь начали произрастать на прежних местах, расселяясь
вдоль тихоокеанского и атлантического берегов Северной Америки. Этим
объясняется сохранение в Северной Америке таких древних теплолюбивых
хвойных, как секвойядендрон, секвойя, таксодиум и другие, которые были
широко распространены в третичное время и в Европе, сейчас же известны
лишь в ископаемом состоянии. Хвойные представлены деревьями и реже
кустарниками. Травянистых растений среди них нет.
Ветвление у них моноподиальное. Ветви расположены спирально, но
часто спирали столь сближаются, что получается мутовчатое (ложномутов-
чатое) расположение ветвей (у араукарий, елей и др.). Верхушечный побег
всегда сохраняется. Если он отмирает, его место занимает ближайшая боко-
вая ветвь. В результате крона у хвойных имеет конусовидную форму.
У одних хвойных образуются одинаковые, однородные побеги со спи-
ральным расположением листьев разнообразной формы (например, у тисо-
вых, араукариевых); у других же развиваются побеги двух родов — удли-
ненные и укороченные. Удлиненные побеги обладают неограниченным рос-
том и покрыты многочисленными спирально расположенными игольчатыми,
чешуевидными или пленчатыми листьями. Такие побеги и обеспечивают
основной прирост дерева. Укороченные побеги развиваются в пазухах вско-
ре опадающих чешуй удлиненных побегов. Они короткие, нарастают очень
медленно. У основания эти побеги одеты чешуйчатыми листьями, а на вер-
хушке несут пучок тесно сближенных зеленых листьев — в числе 2 — 5 — 8
и до 40 (рис. 194, 3).
У некоторых видов (Larix, Pseudolarix, Cedrus) зеленые листья разви-
ваются как на укороченных, так и на удлиненных побегах.
Анатомическое строение стебля хвойных в общем однообразно. Сте-
бель ттет рдд^рразвитую сертптепину- Далее к периферии располагается
хорошо развитая-^торичная древесина, затем—кольцо камбия. Вторичная
флоэма состоит изс&ТШвдвжГТрубок с ситечками на продольных стенках.
Сопровождающих клеток нет. В коре развивается мощная пробка.
300
ВЫСШИЕ РАСТ
ни;
Древесина хвойных состоит их трахеид различной длины, причем в
процессе эволюции они укорачивались. Наиболее длинные трахеиды (до
3 мм) имеются у араукариевых, наиболее короткие — у кипарисовых.
Вторичная ксилема состоит из крупных трахеид с окаймленными пора-
ми на радиальных стенках (у Тазсиз и на тангеита л ьных). У более древних
форм, например у Aga,t№s, Araucaria, наблюдается несколько рядов окайм-
ленных пор, расположенных ближе к концу трахеиды на ее стенках.
У остальных хвойных они расположены большей частью в один ряд, и число
пор, таким образом, уменьшается. Вместе с тем наблюдается образование
торуса. У древних типов (Araucariaceae) торус отсутствует или развит
слабо. У остальных форм он развит хорошо. Наличие торуса обеспечивает
лучшее проведение воды.
11;
лее древних хвойных — араукариевых и тисовых).
Древесинная паренхима впервые появляется в стеблях юрских хвой-
ных, когда появилась на Земле смена времен года и у деревьев возникли
годичные кольца утолщения. Возникла необходимость накопления запас-
ных веществ с осени для использования их весной, при возобновлении энер-
гичного роста растения. Запасные вещества стали накапливаться в парен-
ающих кольцо прироста. Они занимали, таким
образом, терминальное (верхушечное) положение (терминальная паренхи-
ма). В последующем клетки древесинной паренхимы стали возникать в рав-
ных местах годичного прироста (диффузная паренхима). Древесина прони-
зана сердцевинными лучами; они у хвойных узкие, однорядные, реже двух-
рядные.
У большинства хвойных в коре, в древесине и часто в листьях располо-
жены иногда очень многочисленные каналы, содержание смолы,
эфирные масла (скипидарную живи^,^аЖзам^,*^меющиеЬолыаое прак-
тическое значение. Из них после перегонки получают терпентин, копал,
канифоль и другие продукты.
Смоляные каналы имеют схизогенное происхождение и изнутри вы-
стланы эпителиальными клетка£у(выстилающД слой), которыми и выделя-
ются смолы. Далее следует слой мертвых клеток и за ним — слой клеток,
богатых питательными веществами (масло, крахмал) (рис. 192). Смоляные
ходы характерны для представителей семейства сосновых. У арауцариевых
их нет совсем и смола откладывается в трахеидах, окаймляющих сердце-
винные лучи; смоляные ходы встречаются лишь в осях женских шишек.
У подокарповых смоляные ходы имеются лишь в листьях, у тисовых их нет.
Дистья^ у дромадного больщинства хвойных узкие, игольчатые, полу-
чившие название хвои, но у.рядд представителей, у более древних форм,
они ланцетные или щироколаш^етнь^^ “араукария).
У рода PhylloclddBus мелкие листочки на побеге опадают, а в пазухе
их развиваются листовидные кладодии. Наконец, у ряда хвойных листья
чешуйчатые (у кипарисовых, у многих подокарповых).
Узкие листья имеют обычно одну жилку, а широкие часто со многими,
;ш
I г. 111
UII
il'IS
более крупная, средняя жилка. (Podocarpus). Листья обычно сидячие, реже
с короткими черешками, большей частью цельные, редко выемчатые на вер-
хушке (пихта). Размер листьев сильно варьцрует^ Так, игольчатые листья
елей длиной 1 — 2 см, а у некоторых сосен (например, у Pinus palustris)
достигают 30 — 40 см яяжнп. Чешуйчатые листья часто мелкие. Широкие
пластинчатые листья некоторых араукарий достигают 17 —18 см длины
и 4— S см ширины. Листья бывают в сечении плоские, четырехгранные,
округлые. Листовые следы у хвойных двойные или простые (у кипарисовых).
При выходе из стели они образуют ясно выраженные прорывы (лакуны).
У большинства хвойных лдстья расположены спирально. Суральное
"У*™?* 1 !*.первичным: нередко наблюдается супротив-
ное иди мутовчатое (сбдиженнаястйрань) листорасположение. У все Т^этих
ОТДЕЛ ГО ЛОСЕМЕн н т>ТЕ
301
Рис. 192. Сосна:
I — смоляные ходы в древесине сосны (не — выстилающие клетки» мк — слой мертвых клеток» заполнен-
ных воздухом. <п — клетки сопровождающей паренхимы, т — трахеиды); 2— поперечный разрез листа
сосны (ап — эпидермис, екп — складчатая паренхима» сх — смоляные ходы, у — устьица, эн — эндодер-
ма. ок — кЛетки с окаймленными порами, ск* —* склеренхима, пп — проводящий пучок)
растений первые молодые побеги всегда имеют спиральное листорасполо-
жение. :У некоторых хвойных (например, у кипарисовых) наблюдается срао
такие.. листьев между собой и ~ср стеблем. ------ * s
После"1№адвния^листа в месте его отрыва образуется листовой рубец,
остается так называемая листов я я подушечка. У различных ро-
дов форма и характер листового руОца (округлые, овальные, ромбические
и т. д.) и подушечки (расширяющиеся, суживающиеся на конце и т. д.)
резко различны и широко используются в систематике хвойных.
У некоторых хвойных, например у пихты, листья опадают целиком,
i Н Г Й I ((
подушечки не выступают вперед, и ветви у них гладкие.
Внутреннее строение игольчатых листьев (хвои) рассмотрим на примере
сосны. Лист сосны снаружи одет толстостенным эпидермисом, на поверх-
ности которого развитамощная—кутикула. Устьица „(обычного и^гррения)
глубоко погруженные. Это свидетельствует о приспособленности листьев
к уменртенижргпаронрд (ксеооморфныелШЯЪЯк Лнлее-сябдует несколько
слоев клеток гиподермы (подкожиды)Г^а-яейз1Яттодожен. мезофилл, состоя-
щий, ия /^шадчятой пЙЬейУШ^ (рте. 192, В мезофилле листа обычно име-
ются смоляные-ходЫт-В центре листа расположена стель, отграниченная от
мезофилла эндодермой; Под эндодермой дм^етш. один или несколько рядов
клеток трансфузионной ткаии^Клетки ее имеют окаимлещп^н поры, через
которые рсуществляется.^вяаь пу^о!^^ мезофиллом. Ц
г.трд^-ташгга склеренхимой. располоЙ№ннНЙ~^сплошным
группами клеток ш^£окам" ПТроводяших пучков. П:____________ _
ного типа. Т1учок состоит из кСвлемы^'-флбэмы и узкой полоски камбия
между ними. У пихты, тиса мезофилл состоит из палисадной и губчатой
ткани, и тогда складчатая паренхима Не развивается. Строение этих листь-
ев сходно с таковым у двудольных растении. Сходны с ними и листья хвой-
ных с широкой пластинкой (Agathis^ Araucaria).
У большинства тталйнмх глдшплй корень. Ял не-
редко (у некоторых сосен) сильно развита система^ боковых. лю^неёТ[особен-
но на болотах и в горах).
е
ГГ
302
ВЫСШИЕ PACT
LH S11
Я
Размножение хвойных осуществляется главным* образом семенами.
Вегетативное размножение развито слабо.
Спорофиллы у большинства хвойных собраны в более или менее тесные
собрания, называемые шишками (conus). Они являются или простыми побе-
гами (колоски, стробилы), на оси которых располагаются спорофиллы (на-
пример, мужские шишки — микростробилы) большинства хвойных, или
сложными побегами, представляющими собрание укороченных и мета-
ли
П1
морфизированных простых побегов, расположенных на общей оси шишки
(женские шишки большинства хвойных, например сосны — см. ниже).
У всех хвойных шишки однополые. Растения при этом могут быть одно-
домными или двудомными. Очень редко наблюдается образование обоепо-
лых
II1,1н
ек, содержащих микро- и мегаспорофиллы (например, у некото-
рых лиственнниц, сосен).
Мужские шишки обычно расположены в пазухах листьев побега, ре-
же они верхушечные (например, у кипарисовых). Они имеют удлиненную
ось, на которой расположены микроспорофиллы. Форма и число последних
у хвойных неодинаковы.
У многих хвойных наблюдается редукция микроспорофиллов вплоть
до сохранения л:
ин
ь одного, который при этом занимает часто верхушечное
положение. Этот процесс можно наблюдать среди подокарповых (стр. 313).
Нижние микроспорофиллы при этом становятся стерильными и в виде че-
шуек окружают побег. Наиболее простыми считаются радиально-симмет-
ричные микроспорофиллы. Они имеются у древнего семейства Серка*
lotaxaceae.
Микроспорангии в большом числе, висячие, как у гинкго (рис. 190, 2).
От таких форм произошли щитовидные и пластинчатые микроспорофиллы.
Щитовидные (пельтатные) микроспорофиллы имеются у .тиса.
Верхушечная часть спорофилла здесь расширена в виде шляпки. С нижней
ее стороны, вокруг ножки, в большом числе расположены микроспоран-
гии (рис. 207, 3).
У большинства хвойных имеются пластинчатые микроспорофиллы,
происшедшие, вероятно, из щитовидных в результате исчезновения части
микроспорангиев на стороне, обращенной к оси шишки (стробила).
Зрелые микроспорофиллы содержат обычно два микроспорапгия на нижней
его стороне (у сосны). Микроспорангии развиваются из группы клеток и
появляются на микроспорофилле в виде бугорков. По мере роста в них обо-
собляется группа клеток — археспорий. Их окружает выстилающий слой
(тапетум).
Далее к периферии расположены обычно три ряда клеток. Снаружи
микроспорангий одет эпидермисом (рис. 193). Клетки археспория, делясь,
дают материнские клетки спор. Последние делятся редукционно, и каждая
из них образует тетраду микроспор. Микроспора одета двумя оболочками —
экзиной и интиной. На поверхности микроспоры имеется более или менее
сложный рисунок экзины (сетчатый, ямчатый и т. д.), строго постоянный
для каждого вида растений.
Вместе со спорами папоротникообразных и мхов, а также пыльцой раз-
личных покрытосеменных микроспоры хвойных имеют огромное значение
для различения и определения как ныне живущих, так и ископаемых рас-
тений. Оболочки спор и пыльцы, содержащие очень стойкие вещества —
спорополленины, прекрасно сохраняются в ископаемом состоянии. Изучая
последовательно различные геологические напластования (залежи торфа,
озерные отложения, песчаные и глинистые отложения) и отложенную в
них пыльцу и споры, устанавливают состав и характер растительности
прежних эпох, что позволяет судить о климатических условиях данной мест-
ности в момент отложения пыльцы и спор. Полученные при этом материалы
необходимы для стратиграфии и определения возраста изучаемых слоев
земли и имеют большое практическое значение при поисках полезных иско-
паемых; строительстве промышленных сооружений и т. д. Метод спорово-
ОТДЕЛ ГОЛОСЕМЕННЫЕ
303
Рис. 193. Микроспорангии хвойных. Образование микроспор к формирование мужского
гаметофита у сосны:
1 — продольный разрез микроспорангия Pinus laricio (an — эпидермис, вс- выстилающий слой, ж-тег
мддя среду делящихся материнских клеток спор); 2^-продольный разрез через микроспораштаи
Peeudolarix (а “ зндотеций); 3 — первое деление материнской клетки спор; 4 — второе деление; 5 — тет-
рада спор: в — микроспора; 7—М — прорастайте микроспоры сосны (рк — проталлиальные клетки,
ом__антеридиальная клетка* в« — вегетативное ядро, н клетка-ножка. спг — спермагенная клетка)
304
ВЫСШИЕ РАСТЕНИЯ
пыльцевого анализа, получивший широкое развитие в СССР, продолжает
совершенствоваться и углубляться.
Микроспоры МНОГИХ ХВОрткгх снябмкду метками ВоВ-
душнЫё‘ПШИКИ Возникают вследствие расхождения интины и экзины и рас-
тягивания последней. Образующаяся при этом полость заполнена воздухом.
Число и форма воздушных мешков у разных видов варьируют. Благодаря
наличию воздушных мешков микроспоры распространяются ветром на
.большие расстояния. У сосновых обычно имеются два воздушных мешка,
они расположены по бокам микроспоры (рис. 193, б). У подокарповых их
2—6, реже один.
ли
Наиболее примитивными являются микроспоры с одним мешком/, раз-
витым вокруг всей микроспоры. Такие микроспоры были, например, у про-
стейших хвойных — ископаемых лебахиевых, вольтциевых (рис. 199).
Микроспорангии при созревании вскрываются продольной трещиной при
посредстве особо измененных клеток эпидермиса или при помощи утолщен-
ных клеток субэпидермальных слоев, как, например, у Рзеиdolorix
(рис. 193, 2).
Количество микроспор, образующихся на одном растении, очень вели-
ко. В сосновых лесах на почве нередко весной образуется слой пыльцы
в несколько миллиметров. Попадая в озера, пруды, пыльца нередко окраши-
вает воду в желтый цвет.
Микроспоры начинают прорастать еще внутри микроспорангия. При
этом образуется мужской заросток (гаметофит). У разных хвойных он раз-
вит различно.
У сосны, например, при делении ядра микроспоры образуются две
маленькие клетки (проталлиальные, или ризоидальные, клетки) — оста-
ток вегетативной части заростка (рис. 193, 9—13), Они вскоре оттесняются
к одной из стенок микроспоры и там разрушаются. После этого ядро микро-
споры еще раз делится, в результате чего образуются еще две клетки: анте-
.и
радиальная, сдвигающаяся к стенке микроспоры, и вегетативная, дающая
позднее пыльцевую трубку.
К этому времени микроспорангий вскрывается, микроспоры высева-
ются наружу. Подхваченные ветром, они переносятся на семяпочки (см.
ниже). Там антеридиальная клетка делится, образуя генеративную (сперма-
генную) клетку и клетку-ножку антеридия (сестринскую, рис. 193, 14),
Спермагенная клетка увеличивается в размерах и затем делится, обра-
зуя два спермия (у всех хвойных мужские гаметы лишены жгутиков и на-
зываются спермиями). Стенка клетки-ножки расплывается, и ее ядро, на-
ходясь в общей плазме заростка, опускается вместе со спермиями в пыль-
цевую трубку. У более простых форм наблюдается образование значитель-
ного числа проталлиальных клеток. Так, у араукарий, у подокарпусов их
15—40. С другой стороны, например, у кипарисовых и многих таксодиевых
проталлиальных клеток совсем не образуется. Здесь в результате первого
деления ядра микроспоры образуются антеридиальная и вегетативная клетки.
Женские шишки хвойных очень разнообразны и имеют часто сложное
строение: ^1и11пгу-нек0Тд'рых хвойных (например, у тисовых) шишек не обра-
зуется. Наиболее простое строение шишки имелось у древнейших ископае-
мых хвойных — лебахиевых (см. ниже). У них собрания мегаспорофиллов
были представлены проотшпс.побегами "(женские тюлоски, мегастробилы),
у основания которых расподашшсь^аагехатшшые листья. .
Спороносные побеги развивались на удлиненных вегетативных побе-
гах, в пазухах кроющих..чешуевидных .листьев. В далые^шэдс наблюдается
редукция спороносных побегов. "Вегетяттньте листья у них постепенно ре-
дуцировались до полного исчезновения, ось побега утолщалась, и споронос-
ный побег часто сводился к нескольким (или даже одному) мегаспоранги-
ям, расположенным на его небольшой утолщенной оси. Такие шишки
ин
Llil
очень напоминали женские шишки кордаитов, чем устанавливаются не-
посредственные филогенетические связи хвойных с кордаитовыми.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
305
Рис. 194. Продольный разрез женской шишки и вегетативного
побега сосны:
Л—-молодая женская шишка Pinue bankslana (кч —-кроющяе чешуи, сч —
семенные чешуи); 2 — молодой вегетативный побег (ж — кроющие чешуи»
im — укороченные побеги); 3 — укороченный побег р« laricto <чд — чешуе-
видный лист» в пазухе которого развился побег)
in
Дальнейшее развитие женских шишек типа лебахиевых шло в двух
направлениях; 1) с сохранением длинной оси вегетативного побега и 2) с
укорочением оси и сближением редуцированных спороносных побегов в тес-
льЛ, сложные шишки. Оба направления представлены у современных хвой-
ных и хорошо прослеживаются у ископаемых форм.
Первый тип шишек имеется, например, у некоторых подокарп у-
сов (Podocarpus spicatus, Р. andinus и др., рис. 203). Здесь на главной оси
«сережки», в пазухах кроющих чешуи находятся сильно редуцированные
одиночные мегастробилы, сведенные каждый к одной семяпочке (мегаспоран-
гию). У других видов наблюдается уменьшение числа семяпочек на оси, ко-
торая часто у основания несет при этом ряд стерильных чешуи. ъ
У тиса также возникают в конце концов одиночные, сильно редуциро-
ванные спороносные побеги (мегастробил ы), состоящие из одного
мегаспорофилла с одной семяпочкой (стр. 318).
Развитие шишек по второму типу имело место уже у простейших хвой-
ных.
Среди ныне живущих хвойных примитивные шишки со сближенными,
,но еще рыхло расположенными спороносными побегами, на укороченной
общей оси имеются, например, у Cephalotaxus (рис. 206).
Дальнейшее развитие в этом направлении привело к появлению слож-
ных шишек хвойных, типа сосновых.
ха сосны состоит из длинной центральной <>си^ на которой распо-
ложены кроющие чщцуи^ в naayWSпоследних развпдду^я семенные чешуж
(редуцированные спороносные побеги — мегастробилы). На их верхней по-
верхности находятся семяпочки (м е г а с п о р а н г и и).
"ЗГТольшинства хвойных обе чешуи свободны, но у многих они срастают-
ся (у кипарисовых).
Обычно наблюдается мощное развитие семенной чешуи и редукция
кроющей, сохраняющейся в виде придатка на ее спинке. Реже разрастается
кроющая чешуя, а семенная сильно редуцируется (например, у куннинга-
: 1111
.in
Женская шишка очень напоминает строение вегетативного побега
сосны (рис. 194). Ко времени созревания семян чешуи женской шишки од-
ревесневают.
306
ins
ВЫСШИЕ РАСТЕНИЯ
Рис. 195. Строение семяпочки, оплодотворение и образование зародыша у сосны:
1 — кродрлъшЛ. разрез молодых чешуек из шишки Pinna laricio («v — кроющая чешуя, с* — семенная
чешуя, нц — нуцеллус. и — интегумент, пх — пыльцевая камера); 2 — продольный разрез семяпочки
Р. laricio (и — интегумент, нц —нуцеллус. ад — эндосперм, ар — архегоний. тип — пыльцевая трубка);
Л— верхняя часть семяпочки Р. silvestris на втором году ее развития (и — интегумент, нц — нуцеллус*
ад — вндосперм. я — яйцеклетка, бк — брюшная канальцевая клетка, кш —клетки шейки архегония*
пт—пыльцевая трубка, с —мужские гаметы (спермин), яп—ядро пыльцевой трубки, хм —остаток
клетки-ножки антеридия); 4 — яйцеклетка Р. silvestrie в момент оплодотворения (лл —ядро яйцеклет-
ки, сп — спермий); 5—10 — последовательные стадии развития зародыша (по — подвесок) I -П — тетрада
мегаспор Р. laricio
Sill
Sill
Листовидная семенная чешуя, расположенная в пазухе кроющего лис-
та (кроющей чешуи), не может быть признана за лист, так как, согласно
основной морфологической закономерности, в пазухе листа может распола-
гаться лишь побег.
Семенная чешуя — это видоизмененный побег, принявший форму
листа.
Вопрос о морфологической природе женской шишки хвойных широко
обсуждался в XIX в. среди ботаников. Наряду с мнением о гомологичности
ки хвойных цветку покрытосеменных (шишка — простой побег) выска-
зывались взгляды (особенно крупнейшими морфологами А. Брауном
И Л. Челаковским) на шишку как на сложное образование, состоя-
щее из осей различных порядков, и, следовательно, образование, негомо-
логическое мужским шишкам (там — простой побег).
Последующие работы, особенно открытие и изучение древнейших
хвойных (лебахиевых и вольтциевьп^ подтвердили справедливость взгля-
дов А. Брауна и Л. Челаковского. Семенные чешуи действительно явля-
ются сильно редуцированными спороносными побегами.
Семяпочки (мегаспорангии) голосеменных бывают прямыми (у тиса),
обращенными (у подокарпусов) и промежуточного типа. Они состоят из
нуцеллуса, интегумента и расположены на короткой семяножке (рис. 195,1).
IIISIII
SUHS»
ОТДЕЛ ГОЛОСЕМЕННЫЕ
307
Интегумент или целиком свободен от нуцеллуса (например, у агатис),
или чаще срастается с ним на большом протяжении (рис; 195, 2, 3).
Интегумент большей частью дифференцирован на три слоя. Чаще всего
средний слой становится твердым, а внешний и внутренний слои — мясис-
тыми, сочными. На верху семяпочки интегумент не срастается, образуя
пыльцевход — микропиле.
Развитие семяпочки у всех хвойных протекает по одному типу. Семя-
почки возникают в виде небольшого бугорка (нуцеллус), из основания ко-
торого вскоре образуется кольцеобразный валик (будущий интегумент,
рис. 196, 2). В начале развития семяпочки ткань нуцеллуса состоит из оди-
наковых клеток, затем в более глубоких слоях нуцеллуса выделяется более
крупная археспориальная клетка, являющаяся единственной материнс-
кой клеткой мегаспор. Она делится редукционно, в результате чего образу-
ются четыре клетки-мегаспоры, расположенные обычно цепочкой друг над
другом. Затем одна из мегаспор (обычно нижняя) сильно увеличивается в
размерах, заполняется питательными веществами и также делится, тогда
как остальные три мегаспоры оттесняются кверху и там вскоре разру-
шаются.
Мегаспора прорастает в женский заросток внутри мегаспорангия. При
делении мегаспоры возникает сначала большое количество ядер, которые
располагаются в постенном слое плазмы мегаспоры. Затем между ядрами
одновременно появляются перегородки, и таким образом возникают клетки
(так называемое свободное образование клеток). Так формируется женский
заросток — эндосперм.
На заростке, в верхней его части, образуются архегонии, большей
частью их два, но нередко бывает и больше (араукария’ агатис).
Обычно архегонии свободны, отделены друг от друга несколькими сло-
ями клеток эндосперма.
Рис. 196. Развитие семяпочки сосны. Строение и прорастание семени сосны!
л — последовательные стадии развитая семяпочка и женского заростка (кф—нуцеллус. и — интегу-
мент» ос — археспорий. мв — мегаспоры» *«0 — эндосперм. ор — архегоний); £ — строение семени Pinna
edulia (и — интегумент, с — семядоли, шр — точка роста зародыша» яс — иодоемядольное колено» эд —
ендосиерм. к — корешок, нц—нуцеллус» пд — подвесок); з — прорастание семени Р. pinea (at — семен-
ная кожура» спи — гипокотиль, с — семядоли» яг — главный корень» бк— боновые корни)
308
’ ВЫСШИЕ РАСТЕНИЯ
г»
У кипарисовых, однако, архегоний (в числе 6 — 200) плотно придвину-
ты друг к другу, образуя так называемый «комплекс архегониев». Отдель-
ные архегоний (например, у сосны, рис. 195) или их группы (комплексы,
рис. 218, 7) окружены покровом из видоизмененных клеток заростка. Стен-
ки их имеют многочисленные поры. Через эти клетки архегоний получает
питательные вещества из эндосперма.
Архегоний хвойных сильно редуцирован. Он состоит из хорошо раз-
витой^ яйцеклетки й слаборазвитых шейковых клеток и брюшной каналь-
цевой клетки (рис. 195, 3).
У большинства хвойных шейка состоит из восьми клеток, расположен-
ных в два этажа, реже всего из двух клеток (у тсуги).
Брюшная канальцевая клетка у хвойных недолговечна и обычно раз-
рушается задолго до оплодотворения.
Попавшие на семяпочку микроспоры через микропиле проникают на
вершину нуцеллуса. Этому способствует высыхание жидкости, заполняю-
щей первоначально пространство между нуцеллусом и интегументом. Мик-
роспоры втягиваются внутрь семяпочки (рис.' 195).
Развитие мужского заростка продолжается на вершине нуцеллуса.
Экзина лопается, а содержимое микроспоры, окруженное интиной, вытя-
гивается в пыльцевую трубку. Она растет через ткань нуцеллуса, достигает
архегония, разрушает его шейку и входит в соприкосновение с яйцеклет-
кой. Затем пыльцевая трубка лопается на переднем конце и ее содержимое
изливается в протоплазму яйцеклетки (обычно у хвойных в зрелой пыльце-
вой трубке имеются два спермия, ядро клетки-ножки и вегетативное ядро).
По мере продвижения спермиев к ядру яйцеклетки один из них производит
оплодотворение, сливаясь с ядром яйцеклетки, а другой отмирает (рис.195).
У ряда хвойных спермин уже в пыльцевой трубке различны по величи-
- не и строению. В этом случае (например, у тиса) оплодотворение осуществля-
ется более крупным спермием.
Сразу же после оплодотворения зиготы начинается ее деление. Ядро
ее, опускаясь на дно архегония, делйтсй Дважды, с образованием четырех
ядер, которые делятся еще раз, образуя два этажа, по четыре ядра в каж-
дом.
Клетки нижнего этажа делятся далее. При первом их делении образу-
ется восемь клеток. После этого нижние клетки делятся еще раз. Возникают
еще четыре клетки. В результате из оплодотворенной яйцеклетки формиру-
ются четыре этажа, по четыре клетки в каждом. Перед нами так называе-
мый п р ед зарод ы ш (п р о з м б р ио). Далее из нижнего этажа кле-
ток развивается зародыш. Клетки второго этажа вытягиваются, образуя
подвесок. Клетки третьего этажа остаются без изменения, а клетки четвер-
того этажа — камеры — способствуют передаче питательных веществ из
эндосперма к зародышу. Затем клетки подвеска сильно удлиняются, про-
двигают формирующийся зародыш во все новые слои эндосперма и выде-
т при этом энзимы, растворяющие содержимое клеток эндосперма
(рис. 195, Р, 10).
У сосны, тсуги и некоторых других хвойных иногда наблюдается п о-
лизмбриония — образование нескольких зародышей, обычно вследст-
вие расщепления (кливаж) основного зародыша. Однако полного разви-
тия достигает обычно лишь один заррдыш, другие же вскоре отстают в раз-
витии и отмирают.
У некоторых хвойных — у пихты, ели, тисовых, араукариевых —
наблюдается образование нескольких зародышей вследствие оплодотворе-
ния нескольких архегониев (каждый оплодотворяется спермием от отдель-
ной пыльцевой трубки). Но и в этом случае полного развития достигает
____ „ J зародыш.
Зрелый зародыш состоит из зачаточного корешка, стебелька (подсе-
мядольное колено), семядолей и почечки (рис. 196, 2). У большинства хвой-
ных сохраняется подвесок (у Pinus sibirica он спирально закручен).
• г*
IH
1Г.1
ГОЛОСВМЕННЫВ
Рис. 197. Лебахия (Lebachia
piniformis):
1 — нижняя часть ветви с пряио-
стшгами женскими шишками и ви-
сячими мужскими шишками; а—
цельные н раздвоенные на верхунпге
листья •
Число семядолей зародыша различно и изменчиво. У тисовых обычно
2 семядоли, у сосновых — 3—15, у араукариевых — 4, у кипарисовых —
2—6. Зародыш окружен эндоспермом, который затем используется при
прорастании семени.
По мере превращения семяпочки в семя происходит изменение всех ее
частей. Интегумент образует твердый (большей частью) покров семени. Из
остатков нуцеллуса ^образуется тонкая плёнка, окружающая эндосперм.
Семяпочка плотно прирастает к семенной чешуе, из ткани которой обра-
зуется около верхнего конца семени (у сосйы) или вокруг всего семени лег-
кое прозрачное крылышко, способствующее распространению семян вет-
ром.
Ко времени созревания семян женские шишки сильно увеличиваются
в размерах, при этом их чешуйки расходятся и семена освобождаются.
Зрелые семена обычно небольшие, но у некоторых хвойных, например у
подокарпусов, араукарий, они крупные, достигают 15 см длины.
После периода покоя семена прорастают. При прорастании семени пер-
вым появляется главный корень, на котором вскоре образуются боковые
корешки. Семядоли некоторое время находятся внутри семени, всасывая
из эндосперма питательные вещества. Затем благодаря разрастанию под-
семядольного колена и самих семядолей проросток освобождается от кожу-
ры семени, остатки которой постепенно сдвигаются к концу семядолей и,
наконец, сбрасываются (рис. 196, 5). У некоторых араукарий семядоли под-
земные.
О' Порядок Coniferales разделяют на 10 с е м е й с т в.
Семейство лебахиевые (Lebachtaceae). К этому семейству принад-
лежат исключительно ископаемые растения. Остатки лебахиевых в виде
формального рода вальхия (Walchia) известны еще из верхнекаменноуголь-
ных отложений, но широко представлены в нижней перми северного полу-
шария.
Лебахиевые были стройными деревьями, по облику напоминавшими
некоторые апаукариевые {Araucaria excelsa). Главные ветви располагались
на стебле мутовками и отстояли горизонтально, достигая в длину до 0,5 м
и более. Они были перисторазветвленными. Боковые веточки располагались
: двухрядно, в одной плоскости.
на к
1»
Благодаря наличию камбия деревья обладали способностью к вторично-
му утолщению. Вторичная древесина была очень примитивной и состояла
: I ’ 1:
из трахеид, на радиальных стенках которых в 1—3 ряда располагались по-
ры. Смоляных каналов не было. Ветви были густо одеты игольчатыми листь-
ями до 2,5 см длиной. Листья часто были на верхушке вильчато-разветвлен-
ными (рис 197, 2).
; Лебахиевые были однодомными растениями- Одиночные мужские шиш-
ки развивались на концах ветвей (рис. 197, 2). На оси мужской шишки спи-
рально располагались многочисленные микроспорофиллы. На нижней сто-
310
роне микроспорофилла развива-*
лось по два микроспорангия.
Микроспоры были окружены
воздушным мешком, развитым
вокруг всей микроспоры, за
исключением переднего ее кон-
ца (рис. 198, 2, 3).
Женские
КИ
лебахии
располагались также на концах
ветвей. Они были значительно
более сложными и являются
прототипом шишки современ-
ных хвойных.
На оси женской
[КИ
тесной спиралью располагались
многочисленные кроющие че-
Рис. 198. Микроспорофиллы и микроспоры ле-
бахии:
1 — мнкроспорофилл Lebachia hypnoides; s—J — мик-
роспора L. piniformis в плане в в разрезе
шуи, они часто вильчато раз-
ветвлялись на верхушке (рис.
199). В пазухах кроющих че-
шуи развивались укороченные,
уплощенные спороносные побе-
ги — мегастробилы. Они име-
ли короткую ось, на которой
I И S111
I И S111
спирально располагались сте-
рильные чешуйки (листья) и
мегаспорофиллы, заканчиваю-
щиеся обычно одной прямой
или обращенной семяпочкой
(рис. 199). В семяпочках нахо-
дили эндосперм с двумя шаро-
видными архегониями. Плоские
семена (около 0,5 см длиной)
были снабжены узким крылом.
В дальнейшей эволюции из
таких спороносных побегов,
вследствие редукции листьев и
уплощения оси, развились се-
менные чешуи современных
хвойных (стр. 321).
Наиболее распространен-
ным родом семейства явля-
лась лебахия (Lebachia). Описа-
но 14 видов этого рода. Леба-
хия имела плотные, вверх тор-
чащие женские шишки (рис.
197, 2). В пазухах кроющих
чешуй располагались споронос-
ные побеги с многочисленными
Рис. 199 Женские шишки лебахиевых:
1 — мегастробилы (Lebachia piniformis); 2 — схема
строения шишки L. hypnoides; з — мегастробшлы
Ernestiodendron (и — кроющая чешуя)
стерильными чешуями и одним
фертильным мегаспорофиллом,
занимавшим верхушечное поло-
жение, на нем развивалась боль-
шей частью одна прямая семя-
почка (рис. 199).
Род эрнестиодендрон
(Ernestiodendron) характеризу-
ется более высокой организа-
цией женских шишек. У него
ОТДКП ГОЛОСШВВВБШ
311
наблюдается редукция спороносных
колосков. На главной оси шишки,
в пазухах кроющих чешуи, распола-
гались утолщенные, дорзовентраль-
ные спороносные побеги с меньшим,
чем у лебахии, числом стерильных
чешуи или совсем без них. Мегаспо-
рофиллов 3—7, каждый с одной
прямой или обращенной семяпочкой
(рис. 199, 3).
Utt
семейство волыпциевые
(Voltziaceae). Вольтциевые являются
дальнейшим развитием лебахиевых,
отличаясь от них сросшимися сте-
рильными и семенными чешуями
женской шишки. Это были деревья,
роко распространенные от верх-
ней перми до нижней юры. На вет-
111S111
Ills
вях довольно рыхло располагались
чешуйчатые или ланцетные листья.
На оси женских шишек сидели крою-
щие чешуи, в пазухах которых рас-
полагались мегастробилы. Каждый
из них состоял из 5 — 6 сросшихся
стерильных чешуи и 4—5 также срос-
шихся мегаспорофиллов; каждый
мегаспорофилл имел одну верхушеч-
ную семяпочку.
В пределах семейства легко
проследить редукцию числа мега-
спорофиллов в спороносных побегах.
У наиболее организованных предста-
вителей имеется один мегаспорофилл.
Род Voltziopsis — мегастроби-
лы с 5—6 стерильными чешуями
и 5 плодущими мегаспорофиллами
(рис. 200, 1а).
Род Voltzia — 3—5 стерильных
чешуй. Мегаспорофиллы (обычно 3)
расположены в нижней части сте-
Рис. 200. Вольтциевые и хейролепидиевые:
1 — вольтциевые: а — вольтциопсис (Voltziopsis
afrlcana); б —вольтция (Voltzia); е — глиптопе-
пис (Glyptolepis); 2 — хейролепис (Cheirolepis
muensteri): а — вегетативный побег; б — женская
шишка; « — шишковая чешуя; г — разрез шиш но-
вой чешуи (видны сросшиеся семенная и кроющая
чешуи)
рильных чешуи (рис. 200, 16).
Род Glyptolepis — стери льных чешуй 5—6, мегаспорофилла два с
обращенными семяпочками (р ис. 200, 1в).
Род Ulmannia — стерильные чешуи плотно срослись. Мегаспорофилл
один с одной семяпочкой. Спороносный побег очень сходен с семенной че-
шуей современных хвойных.
Семейство хейролепидиевые (Cheirolepidiaceae). Это семейство явля-
ется дальнейшим развитием семейства вольтциевых. Хейролепидиевые —
ископаемые хвойные деревья кипарисового облика — жили от верхнего
триаса до верхнего мела мезозойской эры. Ветви их были покрыты спирально
расположенными чешуйчатыми листьями. Древесина со смоляными ходами.
Мужские шишки с 12—16 микроспорофиллами, на каждом из них раз-
вивалось 10—12 округлых микроспорангиев. Микроспоры без воздушных
мешков.
Женские шишки на оси несли кроющие и семенные чешуи, сросшиеся
на большем или меньшем протяжении. Семенные чешуи имели вырост (э н и-
м а т и й), нависающий над семяпочкой. Кроющие чешуи цельные или на
верхушке трех-многолопастные.
312
ВЫСШИЕ РАСТЕНИЯ
1 — сросшееся семенная и кроющая чешуи: а — об-
щий вид (я — язычок на верхней строке семенной
чешуи); б — продольный разрез; 2 — мужской заро-
сток Araucaria angustifoua (ан — антеридиальная
клетка» ее — вегетативное ядро, пт — проталли-
альные клетки* яз — крахмальные зерна)
К семейству хейролепидиевых
относятся род хейролепис (Cheirole-
pis), например Ch. muensteri с трех-
лопастной кроющей чешуей и двумя
семяпочками (рис. 200, 2), р о ддре-
панолепис (Drepanolepis) с цельной
кроющей чешуей и уже с одной се-
. мяпочкой и многие другие роды
данного семейства. Хейролепидиевые
рассматриваются в качестве предков
подокарпусовых.
\/ Семе й с т в^ араукариееые
(Araucariaceae). Араукариевые - веч-
нозеденые-высокие деревья, распро-
страненные в южном полушарии.
Стебли их обладают правильным
ростом, ежегодно образуя мутовки
крупных ветвей.
В строении вегетативных орга-
нов араукариевых имеется ряд при-
митивных признаков. В отличие от
других хвойных у араукариевых
довольно мцщная сердцевина. Трахе-
иды вторичной древесины (до 3 мм)
'г*
II
«
Рис. 202. Ветви и шишки араукарии и
аг атиса:
1 — араукария бразильская (Araucaria braellfen*
ala): а — женская шнпша; б — веточка с мужской
шишкой; 2 — агатис (Agaihia macroatacbya): d —
веточка; б — женская шишка
строению И расположению пор они
напоминают трахеиды кордаитовых и
древних хвойных (лебахиевых) из
палеозоя. Древесинная паренхима
отсутствует. Смоляных ходов в стеб-
лях нет. Листья араукариевых ча-
ще большие, широкие, почти яйце-
видные или же сравнительно мел-
кие — линейные, игольчатые. Они
густо сидят на ветвях.
Мужские шишки крупные, с
многочисленными спиральными ми-
кроспорофиллами, которые на верху-
шке расширены и с нижней стороны
несут большое число микроспоран-
гиев. В шГкроспорангиях развива-
ются многочисленные микроспоры
без воздушных мешков. При прора-
стании микроспора образует 15—25
проталлиальных клеток (рис. 201,2).
Женские шишки большие, часто
округлые, распадающиеся при созре-
вании. Отличительными особенно-
стями араукариевых являются jcpa-
стание семенной и кроющей чешуй
IH
расположённой на верхней стороне
семенной чещуи. Семяпочка пыльце-
входом обращена вниз (рис. 201, 16).
В верхней части эндосперма распо-
ложены 8—15 архегониев. Клеток
шейки обычно 12. Брюшная каналь-
цевая клетка рано исчезает.
отдал ГОЛОСЕМКННЫВ
313
Рис. 203. Сережковидные
шишки Podocarpus spica-
tus:
1 — мужская ветвь: а — общий
вид; б — мвкроспорофиллы; в —
мужская шишка; я—женская
ветвь: а — обший вид; б — часть
женской шишки
Араукариевые — очень древняя группа хвойных. В ископаемом состоя-
нии они известны из мезозойских отложений начиная с триаса, причем тог-
да они произрастали и в северном полушарии, в том числе и в Европе — до
Гренландии и на территории СССР. Их непосредственные предки (род
Araucarites) встречаются в отложениях пермского периЬда. Араукариевые
представлены двумя родами — араукария агатис.
Род араукария (Araucaria) н8бЧйтывает~15видов, распространенных
в Южной Америке (в Чили, Бразилии), а также в Австралии и на островах
Новой Каледонии.
Мужские шишки араукарий достигают 15—25 см в длину. Микроспоро-
филлы — на конце с расширенной пластинкой, несущей 6 —19 микррспо-
рангиев.
Семенные и кроющие чешуи женской шишки почти на всем протяже-
нии срослись и только на самой верхушке свободны. Семяпочка погружена
в ткань семенной чешуи. Здесь же хорошо заметен так называемый язы-
чок — придаток, вырост семенной чешуи в ее верхней части (рис. 201, 1).
Семена с кожистым крылом.
Араукарии имеют ксерофитный облик, светолюбивы и по экологии на-
поминают наши сосны. Они образуют часто обширные светлые леса.
В Южной Америке (в Андах Чили и на юго-западе Аргентины) широко
распространена араукария чилийская (А. агаисапа), образующая леса. Это
дерево до 60 м высотой и до 1,5 м диаметром. Ветви расположены по 6 в му-
товке. Листья яйцевидные, ланцетные, кожистые, плотно сидят на ветвях,
достигают 2—5ш в длину и живут 10—15 лет. Женские шишки шаровидные,
до 10 слгв диаметре.
В Южной Бразилии леса образует бразильская араукария (A. brasili-
ensis) — дерево до 50 м высотой с зонтикообразной кроной. Листья узкие,
1,5—3 см длины. Женские шишки крупные, шаровидные, до 25 еле в диа-
Г. Г.
н г,
ИГ
.111
(A. bidwillii).
тралии в состав лесов входит араукария Бидвилла
314
ВЫСШИЕ РАСТЕНИЯ
Рис. 204. Подокарпус (Podocarpus
ustus):
1 — общий вид веточки; я — женская шиш-
ка; 3 — шишка в разрезе; 4 — проросшая
микроспора (кэ — клетки заростка, н —
клетка-ксяска антеридия, ак — антериди-
алькая клетка, вг—вегетативное ядро)
На острове Норфольк произрастает
декоративная высокая араукария, (Л. ех-
celsa), напоминающая обликом ель. Часто
ее разводят в оранжереях.
Род агатис (A gat his) насчитывает
около. 20 видов, распространенных в
Индо-Малайской флористической области
и далее к югу, до Восточной Австралии
и Новой Зеландии. Это высокие вечнозе-
леные деревья с мутовчатым ветвлением.
Листья часто очень крупные, округлые,
плоские, до 17—18 еле длиной (Л. macro-
phylla, рис. 202, 2). Мужские шишки не-
большие, 4—6 см длиной. Женские ши-
шки округлые. Семенная и кроющая че-
шуи полностью срослись. Семяпочка сво-
бодная, не погружена в ткань семенной
чешуи. Новозеландский агатис (Л. aust-
ralis) образует в Новой Зеландии обшир-
ные леса. Дерево до 50 м высотой, с гу-
стыми мутовками ветвей.
Араукариевые имеют большое практи-
ческое значение. У большинства видов
араукарии отличная древесина, употреб-
ляющаяся в кораблестроении, - построй-
ке домов и идущая на многие другие це-
ли. Семена араукарий (особенно Л. агаи-
сапа, Л. angustifolia)съедобны и являются
одним из основных пищевых продуктов
населения Чили. Многие виды Agathis доставляют каури-копал, или дамма-
рову смолу, имеющую большое значение как сырье при получении лаков и
красок.
< С ем ей с tjb-jq подокарповые (Podocarpaceae). Обширное семейство
(130 видов), представители которого распространены главным образом
во внетропических областях южного полушария, где они, особенно в го-
рах, входят важнейшей частью в состав лесов. За пределами Северного тро-
ка они произрастают лишь в Индии и в Восточной Азии, где доходят до
Гималаев, на юге Китая и Японии. Подокарповые представлены ка& мощ-
ными деревьями, так и небольшими кустарниками, иногда стелющимися
hi
ин
пн
по земле.
Древесина подокарповых со слабовыраженными кольцами прироста.
Смоляных ходов нет. Листья очередные, реже супротивные или почти су-
противные, очень различные у разных родов и видов.
Некоторые виды Podocarpus, например Р. wallichianus, имеют листья до
12,5 см в длину и 3,5 см в ширину. У большинства же видов они значительно
меньших размеров, ланцетные или линейно-ланцетные и даже чешуйчатые.
У рода Phyllocladus функцию листьев выполняют филлокладии —
уплощенные побеги, принявшие листовидный облик.
н г,
Мужские ш
in
ки у подокарповых чаще одиночные, реже собраны в се-
режкбвидные соцветия (например, у Р. spicatus, рис. 203, 1) и обычно у
основания одеты бесплодными чешуями. Каждый микроспорофилл несет
с нижней стороны два микроспорангия. Микроспора у большинства видов
с двумя воздушными мешками. При прорастании микроспоры образуются
две проталлиальные клетки, которые нередко делятся и образуют четыре
и даже 6—8 (у Podocarpus) проталлиальных клеток (рис. 204, 4). Дальней-
шее развитие мужского гаметофита сходно с сосновыми. Клетка, отчле-
нившая проталлиальные клеточки, делится еще раз, образуя вегетативное
ядро и антеридиальную клетку. Последняя делится и дает клетку-ножку
ОТДЕЛ ГОЛОСЕМЕННЫЕ
315
и генеративную клетку. Генератив-
ная клетка делится, образуя два
спермия, из которых обычно один
более крупный, он и производит
оплодотворение, а другой, более мел-
кий, вскоре дегенерирует.
Женские спороносные побеги
(мегаетробилы) состоят из единст-
венного мегаспорангия (семяпочки),
который окружен видоизмененной
семенной чешуей, называемой здесь
зпиматием. Они расположены обыч-
но в пазухе кроющей чешуи, с ко-
торой иногда частично или полно-
стью срастаются. У основания спо-
роносного побега развиваются бес-
плодные чешуйки или же к ним
близко подвинуты вегетативные ли-
стья (у Podocarpus ustus, рис. 204).
Такие редуцированные спороносные
побеги располагаются поодиночке
или собраны в шишки. Однако на-
стоящих, плотных шишек, свойствен-
Пых остальным хвойным, у подокар-
повых нет.
Наиболее примитивные шишки
имеются у Podocarpus spicatus (рис.
203). У него на оси шишки, споронос-
ные побеги расположены редко, да-
леко друг от друга. От таких прими-
тивных шишек развитие шло в двух
Рис. 205. Эпиматий подокарпусовых:
1 — Podocarpus andinus; я— Р. rieillardii; J —
Phyllocladus: а — эпиматий: б — ветвь с мужски-
ми шишками
направлениях.
В одном направлении наблюдается уменьшение числа спороносных
побегов в шишке до 4, 2 и 1. В результате образовались одиночные семя-
почки, как у большинства подокарпусов (рис. 204).
В другом направлении происходило сокращение оси соцветия, в резуль-
тате чего спороносные побеги оказались тесно скученными и образовалась
сложная шишка, менее сходная с женской шишкой других хвойных. Одна-
ко женские шишки подокарповых более мелкие. Эпиматий различен у от-
дельных родов семейства. Он хорошо развит у большинства видов рода
Podocarpus^ у которых окружает целиком семяпочку и срастается с инте-
гументом в виде наружного его слоя (у Р. andinus, рис. 205, I), а нередко
и с кроющей чешуей. У других родов он развит лишь у основания семяпочки
ин
в виде придатка — ариллуса (у Phyllocladus, рис. 205, 3)
Наиболее обширным и распространенным является род подокарпус
(Podocarpus), насчитывающий свыше 100 видов. Этот род распространен в
субтропической части Австралии, Азии, Африки, Южной Америки. Здесь
имеются крупные деревья и кустарники. Листья крупные или мелкие, че-
шуйчатые (например, у Р. ustus из Новой Каледонии, рис. 204, Z. Эпиматий,
сросшийся с интегументом). Практическое значение подокарпусовых сво-
дится к использованию на различные поделки очень плотной древесины
многих видов Podocarpus и Dacrydium. В большом количестве ее добывают
в Тасмании, Австралии, Новой Зеландии. В Южной Африке подокарпусы
(Р. lattfolius, Р. elongatus) дают почти половину всей продукции древеси-
ны. ^Многие виды Podocarpus разводят как декоративные. В СССР они рас-
тут на Кавказе и в Крыму.
Семейство головчатотисовые (Cephalotaxaceae). К этому семейству
принадлежит один род Cephalotaxus с шестью видами, распространенными
316
гг
в Японии, Корее, центральной и южной частях Китая (в том числе иа остро-
ве Тайвань) и в .восточной части Индии. Это небольшие деревья или крупные
кустарники, большей частью двудомные, с почти супротивными или мутов-
чатыми ветвями. Листья очередные, линейные, кожистые, расположенные
в два ряда (рис. 206).
Мужские шишки почти шаровидной формы. Они собраны по 6—11 в
i:»
111S111
группы, располагающиеся в пазухах верхних вегетативных листьев прош-
логодних побегов. Молодая шишка скрыта среди чешуек, из которых вы-
растает ко времени созревания. Она имеет сложное строение и представляет
собрание спороносных побегов. Каждый из г
: состоит из 10—12 радиаль-
ных микроспорофиллов. Микроспорофиллы с 2—5 висячими микроспоран-
гиями. Микроспоры без воздушных мешков. Мужской заросток сильно ре-
дуцирован. При прорастании микроспор проталлиальных клеток не обра-
зуется.
Маленькие женские шишки располагаются в пазухах чешуевидных
листьев у основания прошлогодних побегов, сидят на удлиненной ножке
и головчато утолщены на верхушке. На их оси крестообразно расположено
несколько пар (3—4) чешуек. В пазухе каждой чешуйки расположены по
две прямые семяпочки, между
сводчато утолщенная наверху.
которыми находится ось второго порядка,
ка является
Таким образом, женская
I И S111
Рис. 206. Цефалотаксус (Cephalotaxus drupacea):
1 —побег с мужскими шишками; а—шишка с кроющими листьявд а—4 — микростробипы (4— верху-
шетаый); д — побег с женскими шишками; в—женская шишка (в хшвухе каждой мясистой чешуи нахо-
дится по две семяпочки); 7 — продольный раарев семени
ОТДЕЛ ГОЛОСЕМЕННЫЕ
317
'til
собранием редуцированных спороносных побегов (рис. 206, 6*). Каждая семя-
почка у основания окружена воротничковидным эпиматием, который пре-
вращается затем в мясистый покров, полностью срастающийся с интегумен-
том, приобретающим каменистое строение. Из все^ семяпочек в шишке лишь
одна, редко две доходят до созревания семени, остальные отмирают (рис. 206,7).
Зародыш с двумя семядолями. Наиболее важными видами являются
С. drupacea и С. fortuned С. drupacea представляет собой дерево до 15 м высо-
той, распространено в Японии, Китае (в центральной части и на острове Тай-
вань). У нас его разводят в Крыму и на Кавказе в декоративных целях (рис.
206). Из семян добывают жирное масло, идущее на приготовление красок,
лака и на другие цели С. fortunet — дерево до 12 ж высотой, произрастает в
центральной и южных частях Китая. В СССР разводят главным об-
разом на Черноморском побережье Кавказа и в Крыму. Имеет декоратив-
ное значение. Головчатотисовые в происхождении связаны с подокарпу-
.HI
itli
Чьу С е м ейст в о дгшювыв (Тоздееое). Женских шишек нет._ Одиночные
сНяпочки расположены на концах побега. Семя окружено частично или
полностью кровелькой (ариллусом). К этому семейству принадлежат 4 р о-
д а — Taxus, Torreya, Amenthotaxus и Austrotaxus, Наиболее распростра-
ненным и обширным является род тис (Taxus)9 наслштыпяютций 10
видов.
Обыкновенный тис (Taxus baccata) —• крупное дерево, до 15 —20 ж
высотой (рис. 207). Сплошные насаждения образует редко (кое-где на Кав-
казе и в Крыму), чаще встречается в подлеске тенистых широколиственных
(буковых) лесов повсюду в Западной Европе, в горных областях Северной
Африки я Малой Азии.
В СССР, кроме Кавказа и Крыма (Ай-Петри), встречается в западной
части Белорусской ССР (Беловежская пуща) и в западной части Украин-
ской ССР (Буковина). Другие виды распространены в Гималаях, в горах
Малайи (Г. Wallichiana), на Дальнем Востоке (в том числе на острове Са-
халин), в Японии (Г. cuspidata), в Северной Америке (Г. canadensis.
Т. floridana).
Тис обыкновенный — очень долговечное растение. Отдельные экземп-
ляры его имеют возраст до 2—3 тыс. лет, достигая при этом 20 м в высоту
и более метра в толщину. Листья тиса узкие, ланцетовидные, расположены
попарно, почти супротивно на горизонтальных ветвях. Древесина, jrnca^
желтовато-красного цвета, очень твердая, с трудом поддается гниению,jja
что тис называют «негной-деревом». Древесина высоко ценится в токарном
деле, употребляется для изготовления мебели (под красное дерево) й_^руко-
яток для инструментов. Тис ценится и как декоративное растение. Он легко
переносит обрезку, что используется для формирования различных фигур—
шаров, трапеций, пирамид и т. д.
Taxus, несомненно, древний, вымирающий род. Ареал его сокращает-
ся. В третичное время тис был широко распространен по всей Евразии и
в Северной Америке. В европейской части СССР его ископаемые остатки
встречаются под Калугой. В ледниковый период тис здесь вымер и после
отступления льдов уже не смог вернуться на прежние места. Сокращению
ареала тиса способствовал в сильной степени человек. После порубок тис
возобновляется с большим трудом.
Тис — растение однодомное. Микростробилы и меГастробилы разви-
ваютёЗГ$а разных ветвях одного дерева. ...
Мужские шишки тиса развиваются на нижней поверхности ветвей, рас-
полагаясь''поодиночке между листьями (рйс?207,‘7а). Они состоят из тесного
собрания, микроспорофиллов, расположенных на коротком стебле, несущем
ниже спорофиллов несколько бесплодных чешуек (рис. 207, 2).
Каждый микроспорофилл ' состоит из короткой ножки, оканчивающей-
ся на вершине щитовидным диском. С нижней стороны диска свешиваются
6—8 микроспорангиев, срастающихся по внутренней стороне с ножкой
'til
nr.
318
выа
III;
[В РАСТЕНИЯ
Рис. 207. Тис (Taxus):
I —ветки: а —мужская; б —женская; в — созре-
лыми семенами; а —мужская шишка; а — микро-
спорофилл; 4 — женский побег с верхушечным се-
менем; 4 — продольный разрез женской ветсякя
(тч — точка роста, сдвинутая в сторону, о — ось
шишки, ак — зачаток кровельки, нц — нуцеллус.
и — интегумент, пц — пыльцевход, же — мет
ра); в — две семяпочки (nw — точка роста между
ними); 7 — женская веточка со зрелым семенем,
окруженным кровелькой: а *- общий вид; б — раз-
рез через кровельку, в — разрез семени
(рис. 207, 5). В микроспорангиях
развивается большое количество
микроспор.
Микроспоры без воздушных
мешков. Мужской гаметофит силь-
но редуцирован. При прорастании
микроспор никаких проталлиаль-
ных клеток не образуется. Микро-
спора делится на маленькую, ге-
неративную и более крупную, ве-
гетативную клетку, дающую затем
пыльцевую трубку. Женских ши-
шек нет.
Семяпочка тиса занимает лож-
новерхушечное положение на
коротком побеге, возникающем в
пазухе игольчатого листа вегата-
тивного побега. Ниже семяпочки,
на оси побега, расположено не-
сколько пар чешуйчатых листьев.
Семяпочка закладывается в пазу-
хе одной из верхних чешуи побе-
га. Затем точка роста побега
сдвигается в сторону и семяпочка
становится верхушечной (рис. 207,
5). Иногда в пазухах двух верхних
чешуи побега развиваются две
боковые семяпочки. Тогда между
ними заметна верхушка первично-
го побега (рис. 207, 6). Наконец,
известны случаи, когда, помимо
боковых семяпочек, развивается
и верхушечная. Эти данные сви-
детельствуют о том, что перед
нами сильно редуцированный
спороносный побег, сведенный обычно к одиночной семяпочке.
Семяпочка (мегаспорангий) занимает верхушечное положение и состоит
из нуцеллуса, окруженного одним интегументом. У основания семяпочка
окружена чашевидным валиком, так называемой кровелькой (или ариллу-
сом—arillus), сильно разрастающейся после оплодотворения. Морфологи-
чески кровелька соответствует семенной чешуе остальных хвойных и зпима-
тию подокарпусовых.
Зародыш с двумя семядолями. Семя с очень твердым интегументом.
Кровелька первоначально зеленого цвета, а при созревании семени стано-
вится красной и сочной. Однако она остается свободной, не прирастает к
интегументу, а лишь плотно к нему примыкает (рис. 207, 7).
Кровелька привлекает птиц, которые способствуют распространению
семян. Все части растения, а особенно молодые листья и побеги тиса, очень
11; ।
ядовиты, но кровелька безвредна для человека и животных.
Род торрея (Torreyа) представлен крупными кустарниками или де-
ревьями и насчитывает 6 видов, распространенных в Японии (Г. nucifera),
Китае (Г. grandis и др.), в США: Калифорнии (Г. califomica — дерево до
20 — 25 ж высотой) и Флориде (Г. taxifolia). Древесина торреи прочная.
Все виды имеют декоративное значение (у нас в Крыму — Г. californica).
Мужские шишки одиночные и состоят из нескольких спорофиллов.
Ниже их, на оси шишки, расположены бесплодные чешуйки. Каждый мик-
роспорофилл несет обычно четыре микроспорангия, расположенных на
нижней их стороне.
nt
ОТДЕЛ ГОЛОСЕМЕННЫЕ
319
dll
III
111
В отличие от тиса микроспорангии у торреи дорзовентральные. Мик-
роспоры без воздушных мешков. Женских шишек нет. Семяпочки сближе-
ны попарно на ветках и расположены на коротких побегах в пазухах веге-
тативных листьев. У основания каждого побега развивается 2—3 пары су-
противных чешуек. Между семяпочками заметен недоразвитый верхушечный
побег.
Тисовые обнаруживают ясное родство с Cephalotaxaceae и имеют общее
с ними происхождение от простейших хвойных.
Существуют и другие взгляды на систематическое положение тисовых
среди голосеменных. Так, в последнее время Флорин предлагает рас-
сматривать тисовые в качестве самостоятельного порядка голосеменных
растений (порядок Taxales). По его мнению, у тисовых никогда не было
женских шишек и семяпочки всегда занимали верхушечное положение.
Интегумент у них иного происхождения. По Флорину, он возник путем
обрастания нуцеллуса супротивными верхушечными листьями, являющи-
мися видоизмененными мегаспорофиллами, в то время как у других хвой-
ных интегумент образовался из лопастей мегаспорофилла, обраставших
нуцеллус. Однако и в этом случае тисовые выводят из простейших хвой-
ных (типа лебахиевых и вольтциёвых).
Семейство сддадеде (Pinaceae). Растения, относящиеся к этому
семейству, представляют собой деревья, реже кустарники, с вечнозелеными
листьями. Только роды Larix и Pseudolartx имеют опадающую на зиму
листву.
Листья игловидные или уплощенные, называются хвоей. Листорас-
г положение спиральное. Листья или одиночные, расположенные на удли-
ненных побегах, или собраны пучками на укороченных побегах. Древесина
с хорошо выраженными годичными кольцами. Трахеиды с окаймленными
порами, расположенными на радиальных стенках обычно в один ряд, реже
в два ряда.
Сердцевинные лучи узкие, одно-двухрядные. Имеются смоляные ходы,
ки сосновых большей частью однополые. Мужские шишки часто
окрашены в желтый или красный цвет. На оси их находятся многочислен-
ные спирально расположенные микроспорофиллы. На нижней их стороне
развиваются по два микроспорангия. Микроспоры (за исключением Larix,
Pseudolarix и Tsuga) с двумя воздушными мешками. Женские шишки слож-
ного строения. На оси женской шишки попарно расположены кроющие
и семенные чешуи. Расположение их очередное, спиральное. Семенная
чешуя, находящаяся в пазухе кроющей чешуи, совершенно свободная. На
внутренней (верхней) стороне семенной чешуи находятся две семяпочки.
'.III
mi:
nt
тые, замкнутые до созревания шишки. У некоторых родов кроющие чешуи
развиты слабо. Зрелое семя у большинства сосновых снабжено пленчатым
крылышком, образовавшимся из поверхностных слоев семенной чешуи
(рис. 212, 4).
Зародыш с различным числом семядолей (от 2 до 18). Сосновые — наи-
более обширное семейство порядка хвойных. К нему относится 9 родов
и свыше 220 видов.
Распространены сосновые в северном полушарии, от субарктических
районов до субтропической зоны включительно. В южном полушарии за
пределы экватора переходит лишь несколько видов сосен (Индонезия, Ява
и Борнео, Филиппины).
Многие представители сосновых образуют леса, занимающие огромные
пространства, особенно в умеренных областях Евразии и Северной Амери-
ки. В СССР обширные леса из сосен, елей, пихты, лиственницы распростра-
нены от крайних западных до восточных границ.
Сосновые имеют огромное народнохозяйственное значение. Они достав-
ляют разного вида деловую и строительную древесину, топливо (дрова).
Древесина некоторых хвойных (пихт, елей) идет на приготовление бумаги.
in
320
ВЫСШИЕ РАСТЕНИЯ
III
11Г111
11ГЛ11
А^ногие растения содержат смолу, бальзамы, которые после перегонки дают
терпетин, канифоль и другие ценные продукты.
В ископаемом состоянии сосновые встречаются начиная с верхней юры.
Произошли они от лебахиевых и вольтциевых (рис. 200). Семейство сосновых
делят на 3 подсемейства. t
/ Подсемейство (Abietoideae), Растения только с длин-
ными побегами, укороченных побегов нет. Шишки созревают в первый год.
Род пихта (Ables). Пихты имеют плоскую мягкую хвою с двумя бело-
ватыми от воска продольными полосками на нижней стороне, в которых
находятся 3—4 ряда устьиц. Хвоя расположена спирально, но на боковых
ветвях нередко в одной плоскости. На верхушке хвоя часто выемчатая. Пих-
ты — растения однодомные.
Мужские шишки овальные или цилиндрические, с желтыми или крас-
ными микроспорофиллами. Женские шишки продолговатые, торчащие
вертикально. Кроющие чешуи чаще длиннее семенных, реже короче (у сибир-
ской пихты). Семена при созревании опадают вместе с несущими их чешуй-
ками, а стержень шишки долго остается на дереве (шишки распадающие-
ся). Семядолей у проростков от четырех до десяти.
К роду пихты (A bis) относится около 40 видов, растущих в северном
полушарии, но преимущественно в горах. В СССР встречается 9 видов. Наи-
более распространенной является пихта сибирская (A. sibirica). Она про-
израстает на северо-востоке европейской части СССР, в Западной, Средней
и Восточной Сибири, а также в Тувинской автономной области и Монголь-
ской Народной Республике, образуя темнохвойные пихтовые леса, но чаще
в виде значительной примеси в лесах с преобладанием ели. Пихта сибир-
ская достигает 30 м в высоту, при диаметре до 50 ся. Она очень теневынос-
ливая Пихтовые леса сумрачные. Нижние ветви часто укореняются и дают
начало новым деревьям. Вследствие этого сибирская тайга часто труднопро-
ходима. Растет пихта как на равнине, так и в горах, предпочитая дрениро-
стые почвы.
Пихта сибирская — очень холодостойкое дерево, приспособленное
к континентальным условиям. При разведении и более мягком климате —
в западных районах СССР и в Западной Европе — побеги ее рано трогают-
ся в рост и побиваются весенними морозами. Пихта имеет большое народ-
нохозяйственное значение. Древесина ее, мягкая, ровная, без смоляных
ходов, ценится как поделочный материал (для токарных и резных работ).
Она широко используется при изготовлении резонансных досок для роялей,
скрипок, а также для получения бумаги. Из хвои добывают эфирное масло,
идущее на изготовление лаков. В молодых ветвях содержится борнеол, ко-
торый входит в состав камфоры.
В крайних западных районах СССР (в Карпатах и изолированно в Бе-
ловежской пуще), а также в горах Средней, Южной и Западной Европы,
образуя лесной пояс на высоте от 300 — 1300 м и выше над уровнем моря,
распространена пихта европейская (A. alba), дающая «страсбургский тер-
' i:r,
В СССР на Кавказе распространена пихта кавказская (Л. tiordmanni-
та) — стройное, декоративное дерево (рис. 208). Она в смеси с буком обра-
зует леса. Нередко ее разводят на Украине, в Крыму и в Белоруссии. В го-
рах Средней Азии растет пихта Семенова (A. semenowii). Несколько видов
пихты встречается на Дальнем Востоке.
В Северной Америке на равнинах широко распространена пихта баль-
замическая (A. balsamea). Она дает канадский бальзам, применяющийся
в микроскопической технике, при изготовлении препаратов. В западных
горных районах Северной Америки встречается много эндемических видов
пихты, имеющих небольшие ареалы.
Род псевдотсуга (Pseudotsuga) насчитывает 7 видов, растущих в запад-
ной части Северной Америки, в Китае и Японии. Это высокие деревья с не-
правильно мутовчатым ветвлением и конической кроной. Хвоя плоская,
ОТДЕЛ ГОЛОСЕМЕННЫЕ
321
Рае. 208. Пихта и псевдотсуга:
л— пихта кавказская (Abies nardmanniana): а —ветвь с созревшим шишками (на одной чешуи наполо-
вину опали); б — семенная чешуя с внутренней стороны; в семенная чешуя с наружной стороны (видав
кроющая чешуя); 2 — нсевдотсуга (Pseudoteuga taxifolia); а — ветвь с женской шишкой; б — лист;
в — семенная чешуя с семенами; а — кроющие чешуи
сверху желобчатая. Женские шишки висячие. Семенные чешуи округлые,
цельнокрайние. Кроющие чешуи значительно длиннее семенных, на вер-
хушке трехлопастные (рис. 208, 2). Микроспоры без воздушных мешков.
В Северной Америке шире других распространена дуглосова пихта
(Р. taxifolia). образующая леса. Она достигает 110—115 ж в высоту. В СССР
ее культивируют в Крыму, на Кавказе и в центральных районах как деко-
ративное растение.
Род тсуга (Tsuga) — высокие вечнозеленые деревья со свисающими
тонкими побегами. Листья (хвоя) плоские, на верхушке выемчатые или
тупые, чаще расположены двухрядно. Микроспоры без воздушных мешков.
Женские шишки маленькие, свисающие, нераспадающиеся. Кроющие че-
шуи короче семенных. Род тсуга насчитывает 14 видов, растущих в Север-
ной Америке, в Гималаях, в Китае и Японии. Многочисленные остатки их
известны в ископаемом состоянии в Европе (из отложений третичного пе-
риода). В Северной Америке широко распространены тсуга канадская
(7. canadensis) и тсуга каролинская (Т. caroliniana) — обе из восточных
областей.
Род ель (Piceа), Этот род содержит около 45 видов, распространенных
в Северной Европе, в европейской и азиатской частях СССР, в Централь-
ной и Восточной Азии и по всей Северной Америке. Наибольшее число ви-
дов имеется в горах Китая. Многие виды образуют леса как в горах, так и
на равнинах» В СССР растет 10 видов ели. *
Ели — высокие вечнозеленые растения с мутовчатым расположени-
ем ветвей, образующих густую конусовидную крону. Ветвление монопо-
диальное. Верхушечная почка окружена 2—5 боковыми.
Расположение хвои спиральное, но на боковых ветвях неясно двух-
рядное. У большинства видов хвоя четырехгранная, на каждой стороне с
11—602
322
ВЫСШИЕ РАСТЕНИЯ
6
209. Ель европейская (Picea excelsa):
Рис.
1 — ветвь с мужскими шишками и молодой женской;
а — микроспорофилл: а — вид снаружи; б — вид сбоку;
a — семенная чешуя: — с внутренней стороны; б — с
наружной стороны (видна кроющая чешуя); 4 — зрелая
женская шишка; 4 — чешуйка с двумя семенами; a —
семя в разрезе (виден зародыш); 1 — зрелое семя с
крылышком
IH
белой полоской устьиц; у не-
которых видов хвоя более или
менее плоская и тогда большей
частью с двумя полосками
устьиц (едь саянская). Хвоя
держится на дереве 7—9 лет.
Кора молодых деревьев глад-
кая, серая, на старых стволах
коричневая, шелушащаяся.
Древесина со смоляными хо-
дами. Древесинной паренхимы
нет. Корневая система поверх-
ностная. Продолжительность
жизни дерева 300 — 500 лет.
Ели — теневыносливые дере-
вья; даже самые нижние ветви
сохраняются в лесах при густом
стоянии деревьев.
Мужские шишки пазушные,
развиваются па побегах про-
шлого года, желтовато-бурые.
Расположены они ниже женс-
ких шишек. Микроспоры с
двумя крупными воздушными
мешками.
Молодые женские шишки
появляются на концах прошло-
годних ветвей, фиолетово-крас-
ные или зеленоватые, торчат вертикально* Зрелые шишки повислые и со-
зревают поздно осенью в первый год. Они опадают целиком после высеива-
ния семян, достигая 10—15 еле в длину, светло- или красно-бурые. Семенные
чешуи обратнояйцевидные па верхушке, по краю выемчатые, выгрызен-
но-зубчатые (рис. 209, 3). Семя крылатое. Проросток с 4—15 семядолями.
Ископаемые остатки елей известны из отложений мелового периода. В СССР
наиболее распространенными являются ель европейская и ель сибирская.
Европейская ель (Р> excelsa, или Р. abies) встречается в Западной Европе
и в европейской части СССР — до Урала. Это дерево до 20 — 50 м высотой
с диаметром ствола до 1 л (рис. 209). Она образует на огромных прост-
ранствах своего ареала чистые или с примесью березы и сосны леса, южнее—
смешанные леса с кленом, липой, дубом. Ель — растение, малотребователь-
ное к климатическим условиям, но страдает от ранневесенних замороз-
ков.
Древесина ели беловатого цвета с желтоватым оттенком, мягкая, лег-
кая, имеет большое практическое значение. Она является основным сырь-
ем для целлюлозно-бумажной промышленности, дает строевой и поделоч-
ный материал. Кора содержит дубильные вещества. При перегонке из дре-
весины получают смолу, канифоль, вар, скипидар.
Сибирская ель (Р. obovata) произрастает на северо-востоке европей-
ской части СССР, образуя северную границу леса от Белого моря до Ура-
ла, и в Западной Сибири. Она очень близка к европейской ели, отлича-
ясь более мелкими женскими шишками (4—7 см длиной) и выпуклыми на
верхушке и по краю цельными, закругленными, семенными чешуями.
В Северной Америке широко распространены и образуют леса ель ка-
надская (Р. canadensis), встречающаяся почти повсеместно в лесной зоне,
и ель черная (Р. mariana), распространенная от Лабрадора до Аляски. Ка-
надская и черная ель идет далеко на север, образуя вместе с Larix americana
северную границу леса. Ель Энеельмана (Р. engelmannii) и ель колючая
(Р. pungens) — обе йз Скалистых гор западного побережья Северной Аме-
.in
иг.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
раки — имеют красивую, серебристую хвою. Они широко культивируются
в СССР и в Западной Европе как декоративные деревья.
Подсемейство лиственничные (Laricoideae). Это растения с
удлиненными и укороченными побегами. Листья (хвоя) одинакового стро-
ения, но на удлиненных побегах расположены рассеянно, а на укорочен-
ных — листья многочисленные, собранные в пучки. К атому подсемейству
принадлежат 3 рода. Наиболее обширен род лиственница (Larix), на-
считывающий около 20 видов, распространенных в Евразии и Северной Аме-
рике. Произрастают они как в горах, так и на равнинах, образуя обширные
леса. В СССР растет 7 видов лиственницы.
Лиственницы — деревья до 30—40 м высотой* В молодости они имеют
узкоконусовидную крону. У старых деревьев крона становится широко-
конусовидной, ветви у них расположены только высоко вверху, в то время
как нижние отмирают.
Лиственницы — светолюбивые растения. Древесина лиственниц крас-
новатая, очень богата смолой и более прочная, чем у сосен. Используется
широко на шпалы, дрова и как строительный материал. Древесина очень
стойкая к разрушению в воде, что позволяет широко использовать ее в
кораблестроении и для крепления шахт. Листья (хвоя), опадающие на зиму
(отсюда и родовое название растения), мягкие, сверху закругленные, све-
иг.
тлозеленые, на нижней стороне с беловатыми рядами устьиц. К осени хвоя
станов ится светло-желтой или бурой.
Мужские шишки, образующиеся на однолетних или на старых укоро-
ченных побегах, расположены обычно поодиночке. Микроспоры без воз-
душных мешков.
Женские шишки формируются на олиствепных коротких побегах и
созревают в тот же год, но висят на дереве 2—3 года.
Зрелые ш
.in
ки не-
распадающиеся. Кроющие чешуи обычно длиннее семенных.
В СССР встречается несколько видов лиственниц. В Карпатах про-
израстает лиственница западноевропейская (L. decidua). которая растет
также в Альпах (рис. 210,1). В восточных районах европейской части СССР
и в Западной Сибири широко распространена лиственница сибирская
(L. st birica), которая продвигается далеко на север (на севере и северо-востоке
Красноярского края и в Якутии доходит до 72° с. ш), в Восточной Сибири
и на Дальнем Востоке — лиственница даурская (L. dahurica). Последняя
прекрасно переносит суровый континентальный климат и произрастает как
на дренированных, так и на заболоченных почвах. Имеет огромное прак-
тическое значение, являясь единственным высокоствольным деревом в Вос-
точной Сибири и Якутии.
В Северной Америке наиболее распространена лиственница американ-
ская (L. americana) — от тундровой зоны до широколиственных лесов, а
также лиственница западная (L. occidentalis) — в горах западных шта-
тов США.
В ископаемом состоянии лиственницы известны с миоцена.
Род кедр (Cedrus). К этому роду относятся 4 вида, распространенных
в Гималаях и в странах Средиземноморья.
Кедры высокие, до 40—50 ле, вечнозеленые деревья с широко раскидис-
той кроной. Побеги удлиненные и укороченные. На укороченных хвоя в
пучках до 30—40 штук в каждом. Хвоя жесткая, темно-зеленая, многолет-
няя, не опадающая на зиму. Древесина кедров, очень прочная и ценная,
идет на кораблестроение, постройки и изготовление мебели. Кедры часто
выращивают в Крыму, на Черноморском побережье Кавказа и в Закавказье
и
ин
как прекрасное декоративное растение.
Мужские шишки одиночные, развиваются на концах укороченных по-
бегов. Микроспоры с двумя воздушными мешками.
Женские шишки крупные, прямостоячие, бочонкообразные (рис. 210, £).
Созревают в 2—3 года. Чешуи опадающие. Семенные чешуи крупные,
черепитчатые. Кроющие чешуи очень маленькие, снаружи незаметны.
11*
ВЫСШИЕ РАСТЕНИЯ
Ископаемые остатки Cedrus известны с мелового периода.
В Западных Гималаях и в Афганистане распространен кедр гималай-
ский (С. deodar а). Остальные виды кедра имеют следующее распростране-
ние: кедр ливанский (С. libanii) — в горах южной части Малой Азии, кедр
атласский (С, atlantica) — в Атласских горах Алжира и Марокко, кедр
короткохвойный (С. brevifolia) — на острове Кипр.
Цо леем ей ст в о сосновые (Pinotdeae). Растения с удлиненными
и укороченными побегами. Листья на них различные. На удлиненных по-
бегах развиты только пленчатые, чешуйчатые листья, расположенные в
редкой спирали. В пазухах этих чешуй развиваются укороченные побеги.
Листья на них зеленые, игольчатые или уплощенные, расположенные пуч-
ками, по 2—5 в каждом пучке.
К подсемейству относится один род сосна (Pinus)> насчитывающий
около 100 видов, распространенных в умеренных областях всего северного
полушария; в субтропиках сосны встречаются главным образом в горах.
Несколько видов сосны произрастает в тропиках. Многие сосны на всем
протяжении своего ареала образуют леса — чистые или в смеси с другими
хвойными (елью, пихтой и т. д.). Южнее сосны входят в состав смешанных
лесов (с дубом, липой и т. д.).
Сосны — вечнозеленые деревья. Молодые деревья имеют пирамидаль-
ную крону, старые — округлую или зонтиковидную (Р. pinea и др.). Дре-
весина со смоляными каналами. Хвоя на укороченных побегах, длинная,
узкая, плосковыпуклая, живет 3—6 лет.
Мужские шишки в большом числе скучены в тесные собрания у основа-
ния побегов текущего года. На оси отдельной мужской шишки спирально
Рис. 210. Лиственница (Larix decidua) и кедр (Cedrus deodara):
1 — ветвь лиственницы с женской шишкой и пучками хвоинок на укороченных побегах; 2 ** ветвь иедр»
с мужскими шишками к одной женской: а — б — микроспорофиллы; в — семенная чешуя с дамяпли»»*—
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 21 l.s Сосяа. Строение шишек:
1 — продольный разрез мужской шишки; 2 —
продольный разрез микрошюрофилла; 3 — по-
перечный разрез шкроспорофилла (jk — микро-
спорангий;; 4 — микроспора (в — воздушный
мешок, ан — антеридиальная клетка, де — веге-
тативная клетка); 4 — строение женской шиш-
ки: а — общий вид; б — продольный разрез
(жч — кроющая чешуя, сч — семенная чешуя,
ом — семяпочка)
5
Рис. 212. Сосна (Pinus silvestris):
J — ветка с шишками (хш — мужская шишка,
жш — женская шишка): а — микроспорофилл:
а — вид сбоку; б — снаружи; 3 — микроспоро-
фнлл: а — кроющая чешуя,- за ней — семенная
чешуя, за которой (внизу) видны верхушки се-
мяпочек; б — обратная сторона с двумя семя-
почками и семенной чешуей; 4 — че-
шуя зрелой шишки с двумя семенами (яр —
крыло, с — семя); 3 — продольный разрез семе-
ни (виден зародыш)
прикрепляющиеся к ней короткой нож-
располагаются микроспорофиллы,
кой. На нижней стороне микроспорофилла находятся два микроспорангия
(рис. 211, 1—3). Микроспора с двумя воздушными мешками (рис. 211, 4).
Женские шишки расположены у вершины побега текущего года, почти
конечные или боковые (рис. 212), одиночные или собраны в группы. Се-
менные чешуи зрелых ш
деревянистые, они плотно прижаты друг к
другу. На их верхнем конце, со спинной стороны развиты характерные
утолщения в виде щитка. Форма
детка различна у разных сосен. Шишки
пн
созревают на второй или третий год. Семена с длинной крылаткой, реже
бескрылые. Проростки с 4—15 семядолями. В СССР наиболее распростра-
нены два вида — обыкновенная и сибирская сосна.
Обыкновенная сосна (Р. Silvestris, рис. 212) широко распространена
в европейской части СССР, в Сибири (до 66° с. ш.) и доходит до побережья
Охотского моря (около 60° с. ш.)« В Западной Европе она встречается от
Скандинавского полуострова до Пиренеев и Балкан. Обыкновенная сосна
образует леса на песчаных де супесчаных почвах. Произрастает она и на
сфагновых болотах; на юге, за пределами сплошных лесов, растет по из-
вестковым склонам и на мелу (горные сосняки).
Обыкновенная сосна очень светолюбивое дерево с прямым высоким
стволом, достигающим 20—40 м в высоту; кора красно-бурая. Крона ее сна-
чала- конусовидная, затем широкая, округлая или даже зонтиковидная.
Хвоя на укороченных побегах собрана пучками по две штуки. Она Сизова-
326
выси
t II7
В РАСТЕНИЯ
Uil.l
ИМИ
то-зел^ная, держится обычно 3 года. В центральном цилиндре Хвоинки
заметны два проводящих пучка. Женские шишки созревают на втором го-
ду, они удлиненно-яйцевидные (3—7 см длиной и 2—3 см шириной), с плот-
ными чешуями. Щитки почти ромбические (рис. 212). Семена крылатые.
Опыление у сосны обыкновенной происходит весной, когда женские шишки
еще очень маленькие. Проросшая пылинка, попав на вершину нуцеллуса
семяпочки, целый год остается в бездеятельном состоянии. Весной следую-
щего года она прорастает в пыльцевую трубочку. Вскоре совершается опло-
дотворение. К осени созревают семена, которые высыпаются из шишек
обычно зимой.
Древесина сосны чрезвычайно ценная. Она наиболее часто употребля-
ется в жилищном строительстве. Широко используется в столярном, мебель-
ном производстве, кораблестроении, вагоностроении, в авиационной про-
мышленности. Из древесины добывают живицу, идущую на получение ка-
нифоли и скипидара. В хвое содержится много аскорбиновой кислоты (ви-
тамин С).
Сосна сибирская (Р, sibiricd) называется часто сибирской кедровой со-
сной или кедром. Она широко распространена по всей Сибири и в Монго-
лии, образуя густые леса, так называемые кедрачи. В небольшом количест-
ве она встречается также на северо-востоке европейской части СССР. Сибир-
ская сосна — высокое дерево с очень густой кроной. Листья на укорочен-
ных побегах в пучках по 5 штук, длинные, темно-зеленые, плотные. В цент-
ральном цилиндре хвоинки один пучок. Женские шишки прямостоячие,
светло-бурые, крупные (6—15 см длиной и 5—8 см шириной), созревают на
третий год; содержат бескрылые семена, так называемые кедровые орешки.
Твердая оболочка орешка является интегументом, тонкая темная пленка,
окружающая эндосперм, — остаток нуцеллуса, светлая маслянистая мас-
са — эндосперм. В нем расположен зародыш. Семена сибирской сосны
употребляются в пищу как лакомство, а также для добывания масла, имею-
щего пищевое и техническое значение.
Древесина сибирской сосны, легкая, плотная, смолистая, находит
рокое применение в народном хозяйстве. Она идет на изготовление окон-
ных рам, дверных косяков, оболочек карандашей и на другие цели. Смола
(живица) при перегонке дает скипидар и канифоль.
В Западной Европе — в Альпах и Карпатах — широко распростра-
нена европейская кедровая сосна (Р. cembra). Ее у нас часто разводят в пар-
ках как декоративное растение.
От Забайкалья до Охотского побережья, а также на Курильских остро-
вах и в Японии широко распространен кедровый стланик (Р. pumiZa), рас-
’ ~ в виде кустарника или небольшого дерева. Он образует большие
заросли, дающие приют белкам и соболям. Его семена съедобны, но более
мелки, чем у сибирской сосны.
В СССР (в горах Крыма и в Западном Забайкалье), в Малой Азии и на
Балканах произрастает палласова сосна (Р. pallasiana). образующая доволь-
но крупные леса.
На Черноморском побережье Кавказа и на Южном берегу Крыма встре-
чается реликтовая, эндемичная пицундская сосна (Р. pi thy usa). В СССР —
на мысе Пицунда — имеется роща этой сосны, у самого берега моря.
В Грузинской ССР, по течению реки Иори (приток Куры), растет так-
же эндемичная элъдарская сосна (Р, eldaricd).
В Западном Средиземноморье (Италия, Франция и др.) широко рас-
пространена и культивируется пиния (Р. pinea). У нас ее разводят в Кры-
му и в Закавказье. Дерево имеет зонтиковидную крону, крупные се-
мена пинии съедобны.
Большое число видов Pinas обитает в Северной Америке (около 30).
Преобладающая часть этих видов имеет маленькие ареалы. Среди них сосна
желтая (Р. ponderosa) дает ценную древесину. Ее разводят по всей Север-
ной Америке.
Ills
нг.г.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
В восточной и северной частях Канады число видов сосны меньше, но
они часто занимают огромные территории и образуют обширные леса. Среди
них веймутова сосна (Р. strobus). Это высокое, чрезвычайно декоративное
дерево. На укороченном побеге — пучок из 5—7 длинных сизоватых хвои-
нок. Культивируют ее в Европе и у нас в пределах лесной зоны европейской
части СССР.
В Северной Америке широко распространена также банксова сосна
(Р. banksiana) и др. Дальше на юг заходит сосна монтесумы (Р. montezu-
тае), растущая в Мексиканских Кордильерах. Благодаря крупным длин-
ным хвоинкам, расположенным по 5 штук в пучке, это сосна чрезвычайно
декоративна.
Семейство таксодиевые (Taxodiaceae), Это высокие деревья, имею-
щие игольчатые или чешуйчатые листья, расположенные спирально, но
часто сближенные так тесно, что листорасположение кажется двухрядным.
Древесина с хорошо выраженными годичными кольцами. Смоляные ходы
отсутствуют. Мужские ш
.in
ки у некоторых представителей одиночные, у
других собраны в кисть или головчатые образования. Микроспорофиллы
с коротким черешком и расширенной пластинкой, по краю которой с ниж-
ней стороны свешиваются 2—9 микроспорангиев. Микроспоры без воздуш-
ных мешков. Мужской гаметофит сильно редуцирован. Проталлиальных
клеток не образуется. Оба спермия фертильны, но один больше другого.
Женские шишки большей частью маленькие, одиночные, помещаются
на концах побегов и состоят из многочисленных спирально расположен-
ных чешуй. Кроющие чешуи маленькие, рано срастаются с семенными. Семен-
ные чешуи утолщенные, на внутренней стороне несут 2—9 семяпочек. Семе-
на свободные, окружены узким крылом или бескрылые (Sciadopitys). Про-
росток с 2—9 семяпочками.
В систематическом отношении таксодиевые занимают среднее положе-
ние между сосновыми и кипарисовыми (см. ниже). К семейству таксодиевых
относят 10 родов и 16 видов. Многие роды монотипны — представлены од-
ним видом. Таксодиевые распространены в США (Калифорнии), Мексике,
в Восточной Азии и Тасмании.
Все роды — реликты, остатки прежней, широко распространенной
в третичное время в северном полушарии флоры голосеменных. Ископае-
мые остатки их известны начиная с верхнего триаса.
Наиболее мощного развития достигают представители родов секвойя
и секвой ядендрон.
Род секвойя (Sequoja). Единственный вид — секвойя красная (S. semer-
virens) — обитает в западной части Северной Америки. От Южного Орего-
на до Калифорнии простираются секвойевые леса. Климат здесь умеренно
теплый, морозы бывают редко, годовое количество осадков составляет
1000 —2000 леи, часты туманы. Секвойя — громадное дерево, до 100 м высо-
той и 5—9 м диаметром. Отдельные деревья живут до 2500 лет. Крона деревь-
ев узкоконическая. Побеги плоские вследствие ложнодвухрядного распо-
ложения листьев. Вечнозеленые листья плоские, линейно-ланцетные,
1,5—2 см длиной (рис. 213,7). Секвойя хорошо размножается черенками и
дает поросль.
Женские шишки созревают в один сезон, очень мелкие — 2—3 см дли-
ной, почти шаровидные. Семенные чешуйки с 3—7 семяпочками.
Род секвойядендрон (Sequojadendron). К нему принадлежит лишь
один вид — секвойядендрон гигантский (S. giganteum), встречающийся
в Калифорнии и отдельными насаждениями на западном склоне Сьерры-
Невады. В настоящее время все массивы взяты на учет и более крупные
объявлены заповедниками. Дерево достигает в высоту 100 м и более, при
стволе до 10 м в диаметре. Ствол до высоты 50 м не имеет сучьев. Деревья
живут до 3500 — 4000 лет. Крона коническая. Побеги сначала голубова-
то-зеленые, затем красновато-коричневые. Вечнозеленые листья спирально*
узколанцетные, мелкие (3—6, редко до 10 мм длиной).
выс
Ш1
lE растения
,111
Рис. 213. Секвойя (Sequoja sempervirens) и секвойяден дрон (Sequojadendron giganteum):
1 — секвсйя: а — ветвь с шишками (л* — мужские шишки, ж — женские); б — иегетатиамм* побег; 2 —
секвойяденарон: а — ветвь с женскими шишками; б — вегетативный побег
ки созревают на второй год. Они яйцевидные, 5—8 см длиной.
Семенные чешуи с 3—14 семяпочками (рис. 213, 2). Древесина секвойи и
секвойядендрона высокоценная, широко используется на постройки и под-
водные сооружения, так как хорошо противостоит гниению. Представите-
ли обоих родов — прекрасные декоративные растения, разводят их в Запад-
ной Европе, в СССР — в Крыму и на Кавказе. В ископаемом состоянии
секвойя и секвойядендрон найдены во многих местах северного полушария.
В Европе и Азии они были распространены вплоть до плиоцена и вымерли
ш
в ледниковое время.
Род метасеквойя (Metasequoja). Единственный вид этого рода —
метасеквойя китайская (М. glyptostroboides) — встречается в горах централь-
ной части Китая на площади в 800 од2. Род был открыт и описан в 1944 г.
китайскими ботаниками. До этого был известен по ископаемым остаткам
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 214. Метасеквойя (Metasequoja
glyptoatroboides):
1 — ветвь с женскими ткшкаык; s —зрелые
шишки; з — ветвь с мужскими шишками
' » С
из отложения неогенового периода в Японии. В меловой период и особенно
в неогене метасеквойя в числе нескольких видов была широко распростра-
нена ио всему северному полушарию. Таким образом, это тоже «живое иско-
паемое». Растение сходно с секвойей, но имеет опадающие на зиму укорочен-
ные побеги, одетые двухрядно расположенными плоскими листьями
(рис. 214).
Род болотный кипарис (Taxodium) представлен тремя видами, встре-
чающимися в юго-восточной части Северной Америки и в Мексике. Все ви-
ды являются реликтовыми растениями, широко распространенными в Евро-
пе и Азии в неогене и вымершими здесь в ледниковую эпоху.
Болотный кипарис виргинский (Т. distichum) встречается на болотах,
по берегам рек и морей юго-восточной части Северной Америки (во Фло-
риде и по берегам Мексиканского залива). Это дерево до 40—45 м высотой
330
выс
III'
[Е РАСТЕНИЯ
Рис. 215. Болотный кипарис (Taxodium distichum):
2 — ветвь с мужскими шишками; 2— ветвь с двумя женскими шишками: J —- полузрелая женская шиш-
ка; 4 — микроспорофилл с невскрывшимися микроспорангиями; 5 — микроспорофилл с вскрывшимися
микроспорангинми; в — семенная чешуя с двумя семяпочками
и 1,5 м диаметром, с широко распростертой кроной (рис. 215). Побеги
болотного кипариса различные: удлиненные со спиральными листьями и
укороченные, развивающиеся в пазухах листьев удлиненных побегов. Ко-
роткие боковые побеги несут двухрядно расположенные зеленые листья,
к осени краснеющие. Побеги эти ежегодно опадают вместе с листвой (таким
образом, наблюдается «веткопад»). Корневая система болотного кипариса
очень мощная, хорошо удерживающая растение в илистом субстрате. Ха-
рактерны так называемые дыхательные корни (пневматофор ы).
Они растут вертикально вверх и возвышаются над поверхностью земли на
1—1,5 ли
Болотный кипарис мексиканский (Г. mucronatum) встречается в Мек-
сике, образуя обширные леса (поднимается до 1500 м над уровнем моря).
Дерево до 50 м высотой. Ствол его достигает 30 м в обхвате. В отличие от
предыдущего ветви более длинные, свисающие. Короткие побеги опадают
весной следующего года, ко времени появления новых побегов. Дыха-
тельных корней нет. Это одно из наиболее долговечных растений. Отдель-
ные экземпляры его доживают до 4000 и даже до 6000 лет. Древесина
мягкая, без смоляных ходов, с хорошими механическими качествами, ус-
тойчива против гниения. Болотный кипарис мексиканский широко рас-
пространен в культуре (у нас в Крыму и на Кавказе) и является прекрасным
декоративным деревом.
Семейство кцрариеояыя, (Cupressaceae). К семейству кипарисовых
принадлежат 20 родов и 145 видов. Все они вечнозеленые деревья или кус-
тарники. Древесина без смоляных ходов. Древесинная паренхима обиль-
ная. Трахеиды с одиночными порами.
Листорасположение мутовчатое, супротивное, реже спиральное. Листья
чешуйчатые, реже игловидные, но первые листья проростков всегда
игловидные.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
331
Рис. 216. Кипарис (Cupressus semper-
virens):
I — веточка с мужскими шишками; 2 — веточ-
ка с двумя зрелыми женскими шишками; &—
микроспорофилл: а — вид со спинной стороны;
б — в разрезе; 4 — чешуя с семяпочками
: 111
ин
Мужские шишки одиночные. Мик-
роспорофиллы супротивные или в
мутовках, щитовидные, на нижней сто-
роне шляпки несут 2 — 6 микроспоран-
гиев. Микроспоры без воздушных меш-
ков. Мужские заростки сильно реду-
цированы. При прорастании микроспор
проталлиальных клеток не образуется,
сразу формируются антеридиальная и
вегетативная клетки.
В женских шишках кроющие и
семенные чешуи срастаются на всем
протяжении. Такие цельные чешуи па
оси шишки обычно расположены супро-
тивно или мутовчато, но иногда они
одиночные и находятся на верхушке по-
бега, как, например, у можжевельни-
ка. Чешуи кожистые, деревянистые
(кипарис) или сочные (можжевельник)
прямые. Семяпочек 1-5 (до 12). Архе-
гонии соединены в группы — комплек-
сы, окруженпые покровом.
Семейство кипарисовые по устрой-
ству шишек разделяют на 3 подсе-
мейства.
Подсемейство кипарисовые
(Cupressoideae). Шишки деревянистые.
Чешуи при созревании отодвигаются
друг от друга. Сюда относится род
кипарис (Cupressus). Четырнадцать ви-
дов этого рода распространены в уме-
ренно теплых областях Европы, Азии
и Северной Америки. В СССР встречается только в культуре (Крым,
Кавказ).
Кипарисы — вечнозеленые деревья с пирамидальной или раскидистой
кроной (рис. 216). Листья их чешуйчатые, черепитчатоналегающие друг на
друга. Мужские шишки одиночные, конечные. Микроспорангиев 2—6. Жен-
ские шишки округлые, с мутовчатыми чешуями, созревают на второй год.
Семяпочек много.
В странах Средиземноморья обычен (родина Иран, Сирия) кипарис
вечнозеленый (С. sempervirens). В СССР, в Крыму и на Кавказе, разводят
чаще пирамидальную разновидность кипариса вечнозеленого, так называ-
емый пирамидальный кипарис (рис. 216). В Северной Америке (в Калифор-
нии) образует леса кипарис аризонский (С. arizonica)\ в Мексике — кипарис
лузитанский (С. lusitanica).
Древесина кипарисов, мягкая, легкая, употребляется в мебельном про-
изводстве и для разных поделок.
Подсемейство туевые (Thujoideae). Шишки деревянистые. Че-
шуи при созревании отгибаются в сторону. Сюда относятся 15 родов. Наи-
более известен род туя (Thuja), включающий 6 видов. Представители
рода туя произрастают в Северной Америке и Восточной Азии; в СССР из-
вестны только в культуре. Туи — кустарники или деревья. Побеги плоские
одеты чешуевидными супротивными листьями. Женские шишки состоят из
3—6 пар накрест расположенных кожистых чешуй, заметно налегающих
друг на друга, созревают осенью первого года.
Наиболее широко распространена в культуре (в СССР от Ленинграда
до Крыма) туя западная (Т. occidentalism Родина ее восток США. Это,
.nt
ВЫСШИЕ РАСТЕНИЯ
3
Рис. 217. Можжевельник:
1 — ветвь с одногодичными (вверху) и двухгодичными «ягодами» (шишками) Juniperue ехсе&ва; 2 — ветвь
с мужскими шишками; 9 — пудовая шишка (лис — микросяорофилл. лют — мшфоспорянгай); 4 — жен-
ская шишка; $ — верхушка женской шишки (ли — мтенорофилл* с—сомпсяиа); в —«ягода»; "—жен-
ский гаметофит J. virghriana с архегониями
дерево 12—20 м высотой и до 1 м диаметром. Древесина мягкая, легкая,
очень стойкая к гниению. Известно много садовых форм.
П о д с емейство можжевеловые (J uniperoideae). Шишки мясис-
тые. Семейная и кроющая чешуи срастаются в общую шишковую чешую,
.in
которая становится сочной, мясистой л_ содержит обычно одну (2—3) семя-
почку. Шишковые чешуи срастаются вместе и образуют мясистую шишку,
принимающую вид ягоды.
В семейство входит единственный род можжевельник (J uniperus), на-
считывающий около 70 видов, произрастающих в северном полушарии, от
Арктики до субтропиков, Несколько видов растет в горах тропической зо-
ны (на юге Китая — J. formosana, на Бермудских островах — J. barbaden-
sis). Большинство можжевельников — горные виды с небольшими ареала-
ми и л
.nt
ь несколько видов имеют очень большие ареалы, например мож-
жевельник обыкновенный (J. communis), распространенный по всему северно-
му полушарию.
Можжевельники или вечнозеленые кустарники, нередко стелющиеся,
или небольшие деревца с прямыми стволами, 10—12 м (реже до 20—30 м)
высотой. Кора серо-бурая, шелушащаяся. Ветви иля прижатые к стеблю
и направленные вверх, или отстоящие. По этой причине кроны можжевель-
ников разнообразны по форме. Листья на ветвях сидят мутовками по три.
Они или игольчатые (J. communis), или чешуйчатые (в последнем случае на
молодых растениях они также игольчатые).
Большинство представителей рода — растения двудомные. Мужские
шишки расположены поодиночке на верхушках побегов прошлого года или
в пазухах листьев группами. Они состоят из нескольких пар или му-
товок микроспорофиллов, имеющих короткую ножку и широкую плас-
тинку, на нижней стороне которой расположено по 3—6 микроспорангиев
(рис. 217, 5).
Женские шишки расположены на вершине пазушных укороченных по-
к»
бегов. Внизу они окружены бесплодными листьями и заканчиваются боль-
шей частью мутовкой из трех чешуй и трех расположенных между ними
ОТДЕЛ ГОЛОСЕМЕННЫЕ
семяпочек (рис, 217, 5). Зрелая шишка шаровидная, нераскрывающаяся;
шишковые чешуи мясистые, плотные, сомкнутые, черные или красные (так
называемые ягоды), содержат 1—10 семян (чаще 3).
Древесина идет на различные поделки, для оболочек карандашей;
«ягоды» используют в медицине, иногда в пивоварении.
В СССР наиболее распространен можжевельник обыкновенный (J. com-
munis), растущий в подлеске еловых и сосновых лесов. Листья у nefco иголь-
чатые, семена несросшиеся, свободные.
В Крыму произрастают J. oxycedrus и J. excelsa.
Можжевельник красный (J. oxycedrus) — дерево 5—10 м высотой, расте-
ние двудомное. Зрелые шишки у него красно-бурые.
Можжевельник высокий (J. excelsa) — дерево до 15 м высотой, однодом-
ное. Зрелые шишки сине-черные, с сизым налетом. Древесина идет на изго-
товление оболочек карандашей,
Можжевельник казацкий (J. sabina) — низкорослый стелющийся кус-
тарник. Побеги темно-зеленые, с неприятным запахом при растирании.
Листья чешуевидные, на нижних ветвях и на молодых растениях листья
игловидные. Чешуи по форме почти ланцетные, ромбические. Растет по скло-
нам гор, на приречных песках, холмах юго-востока европейской части СССР
и в Средней Азии.
В Средней Азии встречается еще ряд видов можжевельника, образую-
щих заросли, называемые арчевниками.
Много видов можжевельника обитает на западном побережье Северной
Америки, в Японии и Китае.
3-й КЛАСС - ОБОЛОЧКОСЕМЕННЫЕ
(CHLAMYDOSPERMATOPSIDA) I
В отличие от других голосеменных оболочкосеменные характеризуют-
ся: 1) наличием настоящих сосудов во вторичной древесине, 2) наличием
покрова («околоцветника») вокруг микроспорофиллов и мегаспорофил-
лов, 3) сильной редукцией женского заростка: у гнетума и вельвичии нё'
"образуется архегониев, 4) сильной редукцией мужского заростка. Пере-
численными признаками оболючкосеменные напоминают покрытосеменные,
цветковые растения и этим привлекают к себе особое внимание ботаников.
Однако оболочкосеменные принадлежат к голосеменным растениям. Неко-
торое сходство с цветковыми в строении их органов является результатом
параллельного развития (конвергенции).
В последнее время А. Л. Тахтаджян выводит оболочкосемениые из
беннеттитовых или общих с ними, более примитивных предков. В пользу
этого взгляда он
приводит следующие
соображения: 1. У
вельвичии
в собраниях микроспорофиллов («мужских цветках») имеются редуциро-
ванные семяпочки. Это позволяет думать, что у предков оболочкосемен-
ных были обоеполые стробилы. Среди голосеменных обоеполые стробилы
характерны только -для беннеттитовых. 2. Микропилярные трубки имелись
у беннеттитовых и у некоторых семенных папоротников. 3. Семяпочки обо-
лочкосеменных сходны с семяпочками некоторых беннеттитовых. 4. Вет-
вление осей, несущих собрания спорофиллов, у вельвичии по типу своему
сходно только с ветвлением некоторых беннеттитовых (Wielandiella).
Возможность происхождения оболочкосеменных от беннеттитовых
признавал также В. Циммерман. Однако мы не имеем никаких палео-
ботанических свидетельств в пользу древности оболочкосеменных и свя-
зей их с беннеттитовыми, так как в ископаемом состоянии оболочкосемен-
ные неизвестны.
И1
ь пыльца встречается в третичных отложениях Евра-
зии. Наиболее близки к остальным голосеменным эфедровые.
Класс включает 3 порядка.
Л
334
ВЫСШИЕ РАСТЕНИЯ
1-Й ПОРЯДОК — ЭФЕДРОВЫЕ (EPHEDRALES)
Единственное семейство эфедровые (Ephedraceae) представлено
одним родом эфедра (Ephedra). Род насчитывает 40 видов, распростра-
ненных в засушливых степных и полупустынных областях Средиземно-
морья, Индии, Китая, Северной и Южной Америки. В СССР встречается
9 видов эфедры (рис. 218).
Внешним обликом эфедра напоминает хвощи и австралийские цветковые
растения — казуарины (стр. 335, 379). Некоторые виды эфедры достигают
5—8 м в высоту, но большинство их низкорослые, сильно ветвистые кустар-
ники, как, например, встречающаяся у нас на юго-востоке европейской
части СССР, в Сибири и Средней Азии эфедра двуколосковая (Е. distachya),
достигающая 15—40 см в высоту. Ветви эфедры прутьевидные, зеленые, реб-
ристые. Листья редуцированные, мелкие, чешуевидные, расположены су-
противно или мутовками, по 3—4 листа в каждой. Листья часто буреют и
опадают. Функция фотосинтеза переходит к ветвям. Ветви снаружи одеты
эпидермисом. Устьица обычно глубоко погружены в ткань стебля. Под эпи-
дермисом, в области ребер, находятся участки механической ткани, а в
области ложбинок — ассимиляционная ткань.
Эфедра — растение двудомное. На мужских экземплярах образуются
мужские шишки, по 2—4 в узлах на тонких ветвях. На оси шишки распо-
ложены 2—8 пар чешуйчатых листьев. Нижние!—2 пары из них бесплодны,
а в пазухе верхних находятся микроспорофиллы. На верхушке микроспо-
рофилла имеется от 2 до 8 микроспорангиев. Микроспорангии двугпездные
(иногда 3—4-гнездные). Основание микроспорофилла одето двумя супро-
тивными чешуевидными листочками («околоцветник»).
В микроспорангии образуется большое количество эллипсоидальных
микроспор, которые прорастают еще внутри микроспорангия. При про-
растании микроспоры в результате первого деления ее ядра отчленяется
небольшая проталлиальная клетка .Оставшееся ядро делится еще раз, от-
членяя ядро второй проталлиальной клетки. Между ними не возникает пере-
городки и клетки, таким образом, не образуется. При последующем деле-
нии ядра микроспоры формируются вегетативное ядро (в дальнейшем ядро
пыльцевой трубки) и антеридиальиое ядро, вокруг которого дифференци-
руется цитоплазма. Антеридиальное ядро делится еще раз, и образуются
спермагенное ядро и сестринское ядро (ядро клетки-ножки антеридия,
рис. 219). В таком виде проросшая микроспора переносится ветром (у неко-
торых видов эфедры, вероятно, и насекомыми) на семяпочку.
На женских экземплярах, в пазухах верхних листьев молодых побе-
гов, образуются женские шишки (рис. 218, 2). У основания женской шишки
имеется несколько пар бесплодных чешуевидных листьев. Ось ш
'.IH
ки за-
канчивается одной семяпочкой (мегаспорангием). Семяпочка одета двумя
покровами. Наружный покров более мощный, он возникает в виде двух
листовых бугорков, которые затем срастаются и обрастают семяпочку, кро-
ме верхней ее части («околоцветник»). Семяпочка окружена интегументом.
Он вытягивается на верхушке в своеобразную микропилярпую трубку,
улавливающую микроспоры (рис. 218, 5). Середина семяпочки занята ну-
целлусом, имеющим вверху углубление — пыльцевую камеру.
В нуцеллусе оформляется материнская клетка мегаспор. При ее деле-
нии (первое деление редукционное) образуются четыре мегаспоры. Три
из них отмирают, а одна (нижн'яя) прорастает в женский заросток, в верх-
ней части которого располагаются обычно два архегопия. Архегоний имеет
шейку, состоящую из двух рядов клеток, расположенных в два зтажа.
Брюшная канальцевая клетка представлена л]
.hi
ь ядром. Ниже расположена
крупная яйцеклетка (рис. 219, 2), Микроспора, попавшая на вершину мик-
ропилярной трубки, втягивается внутрь семяпочки — в пыльцевую камеру.
Здесь продолжается развитие мужского заростка. Образующаяся пыльце-
вая трубка, в которую входит вегетативное ядро, растет через ткань нуцел-
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 218- Эфедра:
Л — ветвь с .мужскими соцветиями; г—женская ветвь со зрелыми семенами: 3 — мужскэе «соцветие»*
4 — мужской «цветок»; 4 — семяпочка в разрезе (чл —бесплодные чешуевидные листья, нц — нуцеллус,
пх — пыльцевая камера, и — интегумент, наверху вытянутый в микропилярную трубку, тщ — покрав
«цветка»); 4 — женский «цветок» (чл — бесплодные чешуи); 7 — разрез семени (а — зародыш, «итожен-
ный эндоспермом); я — проросток (с — семядоли» пс — подсемндольнзе колено, н — ножка, о — оболоч-
ка семени, к — корень)
ВЫСШИЕ РАСТЕНИЯ
8
Рве. 219. Эфедра. Размножение:
1 — мшфоснора: а—отчленилась вдоплливдодая клетка; 9—образовалась вторая прсталлиальвая клет-
ка; 4 — отделились ядро генеративной клетки и вегетативное ядро (ядро будущей пыльцевой трубки);
s — ядро генеративной клетки разделилось на ядро клеткж-иежкн я яшм» сперыагенной клетки (без
перегородки); 9 — образовались два ядра — сверили; У — сформировавшийся мужской гаметофит, в пыль-
цевой трубке которого сформировались все алеменш Оч, — врошллвальные клетки. « — ядро гене-
ративной клетки, дт —ядро пыльцевой трубки, ям — ядро клетки-ножки, дег —ядро едвриагвншЛ
клетки, лс — ядра спермиев); 8 — верхняя часть эндосперма (л—яйцеклетка, бк — брюшная канальце-
вая клетка, ш — шейка архегонии. Яйцеклетка и брюшная канальцевая клетка не разделены псрегород- '
кой)
луса к архегопию. Спермагенное ядро делится, образуя два спермин. Пос-
ле разрушения конца пыльцевой трубки спермин попадают в архегоний;
один из них сливается с яйцеклеткой, производя оплодотворение, другой
отмирает. Из оплодотворенной яйцеклетки развивается зародыш. Зрелый
зародыш имеет длинное подсемядольное колено, две семядоли, первичный
корень и ножку — вырост зародыша в области корневой шейки.
Семя снаружи окружено сонным, окрашенным в красный цвет наруж-
ным покровом («околоцветником»), далее следует твердый, одревесневший
интегумент, нуцеллус и эндосперм, в котором заключен зародыш.
Практическое значение эфедры определяется наличием в ее ветвях алка-
лоида эфедрина. Эфедрин — важнейшее лекарственное средство.
2-Й ПОРЯДОК — ГНЕТОИДНЫЕ (GNETALES)
Порядок представлен одним семейством гнетовые (Gnetaceae)
и одним родом Gnetum — гнетум.
Тод гнетум (Gnetum) насчитывает 40 видов, обитающих в тропиче-
ских влажных лесах Восточной Азии, Южной Америки, а также Африки
(2 вида).
Два вида (& gnemon и G. castatum) — небольшие деревья, остальные
мдм—с длинными стеблями.
Лист£й"гнетума супротивные, широкие, кожистые, с сетчатым жилко-
ванием, напоминают листья двудольных растений (рис. 220). Мужские и
женские шишки появляются на разных особях.
III'.
Мужские шишки сережковидные, сложные. На оси шишки мутовками
расположены собрания микроспорофиллов (иногда они сидят в 2—3 ряда
в мутовке). Каждый микроспорофилл окружен покрывалом («околоцвет-
ОТДЕЛ ГОЛОСЕМЕННЫЕ
6
Рис. 220. Гнетуи:
1 — ветвь с мужскими соцветиями; 2 — мужской цветок е двумя покровами и тычинкой, имеющей два
пыльника на равных стадиях; з — ветвь с женскими соцветиями; 4 — продольный разрез семяпочки
(нц — нупеллус, tie — интегумент внутренний, вытянутый в мвкропилярпую трубку, ин — интегумент
наружный, я —покров); 5 — женский заросток, многоклеточный — в нижней часта, в верхней вдета —
с ядрами, расположенными в постенном слое плазмы; 4 — пророст-ж и зародыш.- а — проросшее семя,
б — вычлененный проросток (ст — стебель с двумя семядолями, х — корень, н — ножка)
ником»), имеющим вид усеченной пирамиды. Покрывало образовано двумя
сросшимися листочками. Снаружи, у основания, покров окружен во-
лосками.
Микроспорофилл состоит из^_уплощенной оси («нить тычинки»), закан-
чивающейся наверху двумя ^одас^гнездными 2дшкрр^пйрйнглями («пыльце-
вые гнезда»). Таким образом формируется некоторое подобие цветка по-
крытосеменного растения. Иногда в таком «цветке» имеются два микроспо-
рофилла, каждый с одним верхушечным микроспорангием. Нити их в этом
случае срастаются.
В верхней части мужской шишки, над микроспорофиллами, иногда
развивается мутовка недоразвитых мегаспорангиев.
В микроспорангин созревают многочисленные микроспоры.
Микроспора начинает прорастать внутри микроспорангия. При про-
растании микроспоры развивается мужской заросток, редуцированный
сильнее, чем у эфедры. Первое деление ядра микроспоры дает два ядра,
соответствующие одно вегетативной клетке, другое — антеридиальпой.
Они плавают в общей плазме микроспоры. Проталлиальпых клеток или про-
таллиальных ядер не отчленяется. Затем яДрп яптцридИ^ЗГЕибй клетки де-
лится и формируются два ядра: одно клетки-ножки и другое спермагенной
клетки. В таком состоянии микроспоры переносятся (ветром, но, возможно,
и насекомыми) на вершину микропилярной трубки.
Женские шишки имеют вид удлиненной сережки. На ее оси мутовка-
ми расположены семяпочки. Каждая семяпочка {мегаспорангин) окружена
снаружи покровом («околоцветником»). Семяпочка имеет два интегумента:
наружный более короткий, внутренний наверху вытянут в микротгйлярную
трубку. Далее расположен нуцеллус. Оба интегумента и обрастающий их
снаружи «околоцветник» срастаются с нуцеллусом у его основания. В ну-
целлусе формируются четыре мегаспоры, из них три верхние отмирают,
а нижняя, прорастая, развивается в своеобразный сильно редуцированный
женский ааросток. Он не имеет тканевого строения и лишен ^архегониеди
Заросток состоит из верхней вздутой и нижней узкой части (рис. 220, 5).
338
ВЫСШИЕ РАСТЕНИЯ
В начале развития разросшаяся мегаспора содержит свободно лежа-
щие в постенном слое цитоплазмы мпогофтсленные ядра. Затем нижняя
часть заростка благодаря возникновению перегородок между ядрами по-
лучает клеточное строение. В верхней части заростка клеток не образуется,
а несколько верхних свободных ядер становятся способными к оплодотво-
рению.
Попавшая на вершину, микропилярной трубки микроспора втягивает-
ся внутрь трубки при усыхании жидкости, выступавшей на ее верхушке,
и прорастает, формируя пыльцевую трубку. В нее опускаются вегетативное
ядро и спермагенная клетка. Спермагенная клетка делится, давая нача-
ло двум спермиям. Пыльцевая трубка к этому времени врастает в верх-
нюю часть женского заростка, ее оболочка на нижнем конце растворяется,
и освободившиеся спермин сливаются с ближайшими к ним ядрами женско-
го заростка, давая начало двум зародышам, однако из двух зародышей
полного развития достигает только один.
Зрелое семя окружено оболочкой, состоящей из трех слоев: внешнего,
среднего и внутреннего. Внешний слой— сочный («околоцветник»), средний
слой — каменистый (наружный интегумент), внутренний слой — в виде
тонкой пленки (внутренний интегумент). Далее расположены перисперм
(ткань нуцеллуса) и зародыш, окруженный клетками заростка. Зародыш
к этому времени состоит из двух семядолей, подсемядольного колена, пер-
вичного корня и ножки. При прорастании пожка остается в семени и вса-
сывает питательные вещества из эндосперма и перисперма.
Наружная оболочка семени съедобна. В пищу употребляются также
молодые листья гнетума.
3-Й ПОРЯДОК — ВЕЛЬВИЧИЕВЫЕ (WE LW IT SCH TALES)
К единственному семейству порядка — Welwitschiaceae — при-
надлежит один род — вельвичия (Welwitschia), представленный видом
вельвичия удивительная (W. mirabilis). Вельвичия обитает в пустынях
Юго-Западной Африки. Внешний вид вельвичии весьма своеобразен. Она
имеет редьковидный стебель, скрытый частично в земле (рис. 221, 7). Над-
земная его часть высотой около 50 см похожа на пень. Верхушка его имеет
форму седла. От стебля отходят два супротивных лентовидных листа дли-
ной до 2—3 м. Листья постепенно нарастают у основания и разрушаются
на верхушке. Растение как бы остановилось в своем развитии на стадии про-
ростка.
Вельвичия — растение двудомное. Мужские и женские шишки обра-
зуются на разных особях.
Мужские шишки собраны группами и располагаются на верхушках
дихотомических разветвлений общей оси, выходящей из пазухи листа. На
1 оси мужской шишки парами друг над другом расположены кроющие листья.
В пазухе каждого кроющего листа находится собрание из 6 микроспорофил-
лов, сросшихся основаниями нитей в короткую трубку (рис. 221, 2), Они
окружены четырьмя чешуйками («околоцветник»). Две из них (боковые) ма-
ленькие, а другие две (медианные) более крупные1. Каждый микроспоро-
филл вверху несет 3 микроспорангия, вскрывающихся радиально. В них
созревают микроспоры. В центре собрания микроспорофиллов находится
недоразвитая семяпочка с блюдцевидно-расширенной наверху микропиляр-
ной трубкой. Микроспора начинает прорастать еще внутри микроспоран-
гия — развивается чрезвычайно редуцированный мужской заросток. При
прорастании микроспоры клеток не образуется и заросток. представлен
ядрами, свободно лежащими в цитоплазме. Первым отчленяется проталли-
альное ядро. Оставшееся ядро снова делится, формируя два ядра — веге-
1 Собрание микроспорофЕллое, окруженное четырьмя чешуйками («околоцветник»),
часто называют мужским цветком.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Рис. 221. Вельвичия (Welwitschia mirabilis):
1 — молодой цветущий женский экземпляр растения? я — часть мужского соцветия: 3 — женское соцве-
тие; 4 — мужской цветок (6 микросиорофиллов и в середине редуцированная семяпочка с вытянутым ин-
тегу ментом): 3 — продольный разрез женского цветка (п — покров, и — интегумент, нц — нуцеллус); 6—
женский гаметофит (заросток) в стадии свободных ядер; 7 — проростки: а — молодой, б — на более позд-
ней стадии развития (н — ножка, с — семядоли, к — корень, л — листья)
тативное и спермагенное. В таком трехъядерном состояния микроспора
переносится на семяпочку. Опыление осуществляется ветром или насе-
комыми.
.ТКеш^ие щищки собраны также группами на общей оси, выходящей из
пазухи листьев. В нижней части отдельной шишки, на ее оси, расположены
бесплодные чешуйки (кроющие листья), сидящие накрест супротивно (ши-
шки, таким образом, четырехгранные). В пазухе каждой кроющей чешуйки
находится семяпочка, окруженная двумя сросшимися листочками, имею-
щими на спинках По одному крыловидному выросту («околоцветник»). Эти
листочки срастаются между собой и обрастают семяпочку. Некоторыми
ботаниками они рассматриваются как наружный интегумент1.
Семяпочка (мегаспорангий) состоит из нуцеллуса, окруженного инте-
Интегумент вытянут наверху в длинную микро-
гументом (внутрег
пилярную трубку (рис. 221, 5).
В нуцеллуса обычным путем формируются ,4 мегаспоры. Из нижней
мегаспоры развивается редуцированный женский заросток. Архегониев
на нем не образуется. Заросток представлен большим числом ядер, свобод-
но расположенных в цитоплазме разросшейся мегаспоры. Ядра, первона-
и»:
чально распределенные по всему заростку, впоследствии скучиваются в
1 Семяпочку а окружающие ее «листочки» («околоцветник») называют «женским цвет-
ком»*
340
ВЫСШИЕ РАСТЕНИЯ
нижней его части по 10—12. В это время между группами ядер появляются
клеточные перегородки и ткань заростка приобретает клеточную структуру.
Затем ядра сливаются, и клетки снова становятся одноядерными.
В верхней части заростка происходит идентичный процесс, только
число ядер в каждой группе меньше (2—3). В конечном итоге они
также сливаются и возникают одноядерные клетки. Далее клетки, распо-
ложенные в верхней части заростка, вытягиваются в так называемые про-
таллиальные трубки. Проталлиальные трубки растут вверх, внедряясь в
ткань нуцеллуса. Проросшая микроспора, попав на верхнюю, расши-
ренную часть микропилярной трубки, втягивается в нее вследствие усыха-
ния сахаристой жидкости и оказывается на вершине нуцеллуса, где она
образует пыльцевую трубку, которая врастает в ткань нуцеллуса и смыка-
ется с проталлиальной трубкой. В пыльцевой трубке сперма генное ядро
делится, образуются два спермия. В месте соприкосновения пыльцевой и
проталлиальной трубок их оболочки растворяются, и один из спермиев
сливается с одним из ближайших ядер проталлиальной трубки. Оплодо-
творенное ядро опускается в нижнюю часть заростка и дает начало зароды-
шу. Клетки нижней части заростка по мере развития зародыша усиленно
делятся, образуя новые питательные клетки. Увеличивающийся зародыш
продвигается в ткань заростка, используя заключенные в его клетках пита-
тельные вещества.
Зрелый зародыш окружен эндоспермом (заростком) и имеет двесемя-
доли, подсемядольное колено, перййчный корень и ножку (рис. йЙ, 7).
Семя окружено крылатым сухим наружным покровом («околоцветником»),
далее располагаются тонкая оболочка (интегумент) и сочная "Ткань (пери-
сперм, нуцеллус). При прорастании ножка остается в семени, всасывая
питательные вещества из эндосперма и перисперма. Первым появляется
корешок, затем — подсемядольное колено с двумя семядолями. Семядоли
развертываются. Почечка, увеличиваясь в размерах, развивает два листа,
расположенные наперекрест семядолям. Семядоли вскоре отмирают, а пер-
вые два настоящих листа сохраняются в течение всей жизни растения (вель-
вичия живет до 100 лет).
ОТДЕЛ — ЦВЕТКОВЫЕ (ANTHOPHYTA),
ИЛИ ПОКРЫТОСЕМЕННЫЕ (ANGIOSPERMATOPHYTA,
ANGIOSPERMAE), ИЛИ ПЕСТИЧНЫЕ (GYNOECIATAE)
Покрытосеменные (пестичные, или цветковые) — сам&й обширный отдел
растительного мира, к которому относится больше половины всех известных
видов их.
Строепие их спорофита очень разнообразно. Среди них наряду с много-
численными деревьями, кустарниками и древесными лианами имеется много
травянистых растений, причем последние по числу видов преобладают.
Общеизвестно богатство морфологических видоизменений основных вегета-
тивных органов покрытосеменных, причем многие оригинальные структуры
обычно бывают связаны с приспособлением к существованию в крайних
условиях или к выполнению специальных функций (например, листья так
называемых насекомоядных растений). В анатомическом отношении следует
подчеркнуть широчайшую распространенность у покрытосеменных настоя-
щих сосудов (трахей); их лишены только немногие представители, по всей
вероятности, древние по происхождению.
Не менее разнообразно строение органов спороношения покрытосемен-
I ных, их цветков. В современной морфологии обычно принято считать цветок
УсвдЬобразным метаморфизированным спороносным побегом, (стробилом).
\ / Для покрытосеменных особенно характерно наличие пестика (плод-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
341
Рис. 222. Гаметофит покрытосеменных:
1 — микроспора; я— начало образования пыльцевого верна (вх — вегетативная клетка, ен — генератив-
ная клетка); J —содержимое пылинки покрытосеменных;* 4 — зародышевый мешон; 5 — двойное оплодо-
творение (я — яйцеклетка, е — синергиды. акт — антиподы, пл — полярные ядра; на рис. 5 в яйцеклетке
и рядом с полярными ядрами—спермин).
ника), образуемого одним или несколькими плодолистиками, т. е,
макро- или мегаспорофиллами, сросшимися так, что в нижней части
пестика образуется, замкнутое вместилище, называемое завязью, внутри
которого помещаются семяпочки. Не менее характерно для цветковых
растении рыльце — пыльцеулавливающий орган, который обеспечивает
прорастание пылинок. После оплодотворения завязь разрастается в плод,
а из семяпочек развиваются семена. Наличие завязи и плода дало осно-
вание для наименования — покрытосеменные (Angiospermae1), в отличие
от голосеменных, не имеющих завязи.
Микроспорофиллы покрытосеменных представлены тычинками, очень
изменчивыми в деталях, но сохраняющими общий план расчленения и стро-
ения стенок пыльцевых гнезд. В частности, характерно раскрывание пыль-
ников за счет эндотеция — субэпидермального слоя клеток, имеющих
неравномерно утолщенные оболочки.
Для цветка характерна всегда одна и та же последовательность распо-
ложения цветолистьев на цветоложе. Снизу вверх прикреплены листочки
околоцветника, тычинки, составляющие андроцей, плодолистики,
в совокупности образующие гинецей.
В разнообразии цветков покрытосеменных можно отличать варианты
высокоспециализированные (обычно в связи с приспособлениями к агентам
опыления), следовательно, более позднего происхождения, и более древние,
сохранившие примитивные черты строения; удлиненное цветоложе, неопре-
деленное число свободных, т. е. не сросшихся друг с другом, спирально
расположенных цветолистьев и др.
Не менее характерен и гаметофит покрытосеменных (рис. 222). Муж-
ской гаметофит, представленный содержимым пыльцевого зерна, у покрыто-
семенных состоит всего из двух (вегетативной и генеративной) клеток.
Начав свое развитие в пыльнике, микроскопически малый мужской заросток,
- защищенный дифференцированной оболочкой микроспоры, переносится
на рыльце пестика (опыление). Женский гаметофит сильно редуцирован.
Он формируется, как правило, из единственной мегаспоры, не покидающей
1 От греч. «аггейои» — сосуд, вместилище и «спермаа — семя.
ВЫСШИЕ РАСТЕНИЯ
II М11
1’1т
мегаспорангия, т. е. нуцеллуса семяпочки, и представлен содержимым
восьмиядерного зародышевого мешка.
Неповторимая особенность покрытосеменных — открытое в 1898 г.
русским ученым С. Г. Н а в а ш и и ы м двойное оплодотворение, суть ко-
торого в том, что одна из двух мужских гамет, образующихся из"* генератив-
ной клетки мужского заростка и доставляемых пыльцевой трубкой в зароды-
шевый мешок, оплодотворяет яйцеклетку, другая же сливается со вторичным
ядром зародышевого мешка или с одним из полярных ядер до слияния их
друг с другом. Образовавшееся триплоидное ядро дает начало многочислен-
ным ядрам клеток будущего эндосперма. Эндосперм цветковых растении
рассматривается одними исследователями как второй зародыш, не достигаю-
щий нормального расчленения, видоизмененный -соответственно функции
накопления и хранения запасных питательных веществ, которые использу-
ются впоследствии нормальным зародышем при прорастании. Другие ученые
трактуют его как орган особого рода, не имеющий к зародышу никакого
отношения. Во втором случае слияние другой мужской гаметы со вторичным
ядром зародышевого мешка не считают оплодотворением.
В целом, в цикле развития покрытосеменных вполне отчетливо разли-
чимы господствующий автотрофный (за немногими исключениями) спорофит
и слаборазвитый (редуцированный) раздельнополый гаметофит, питающийся
гетеротрофно. Это соотношение вполне соответствует тому, что типично
и для голосеменных, но с некоторыми существенными отличиями. Из них
надо отметить следующие:
1. Опыление, как необходимое условие оплодотворения, состоит у голо-
семенных в доставке пыльцевого зерна на семяпочку с последующим вовле-
чением его через микропиле на поверхность нуцеллуса; у покрытосеменных
пыльцу воспринимает не семяпочка, а рыльце пестика.
2. При развитии мужского гаметофита покрытосеменных никогда
не образуются проталлиальные клетки (остаток вегетативного тела мужского
заростка), характерные для многих голосеменных. В содержимом зародыше-
вого мешка, хотя и имеются клетки (антиподы), которые нередко принимают
за остатки женского заростка, все же последний сильно отличается от жен-
ского гаметофита голосеменных, таких, как хвойные, саговниковые, гинк-
говые, где женский заросток представлен тканевым телом.
Эти и ряд других особенностей, с одной стороны, четко отграничивают
отдел покрытосеменных от голосеменных, а с другой — свидетельствуют
об исторической связи этих двух групп. В ряду высших растений с преоблада-
нием в цикле развития спорофита (равноспоровые и разноспоровые папорот-
никообразные — голосеменные — покрытосеменные) наблюдается сначала
потеря гаметофитом его самостоятельности, а затем прогрессирующее уско-
рение созревания его — достижение половозрелости без развития вегета-
тивного тела. В этом отношении покрытосеменные выглядят как завершаю-
этап исторического процесса редукции полового поколения во всех
основных линиях эволюции наземных растений, в онтогенезе которых пре-
обладает бесполое поколение. Достигнуть крайней ступени редукции за-
ростков покрытосеменные могли, только пройдя уровень
гаметофита типа голосеменных.
Исторические связи покрытосеменных с голосеменными
и в том, что первые приобрели значение господствующей группы растений
только после середины мелового периода, оттеснив голосеменных, игравших
важнейшую роль во флоре Земли с конца пермского периода до второй поло-
вины мезозойской эры.
Вопрос о времени и конкретных предках покрытосеменных растений
остается до настоящего времени нерешенным. Ископаемые остатки покрыто-
семенных в большом разнообразии появляются на границе нижне- и верхне-
меловых отложений. Однако они встречаются и в отложениях нижнего мела,
правда в несколько меньшем разнообразии. Следовательно, возникновение
этой группы можно отнести к еще более древним временам. Это подтвержда-
ими
II г г
организации
проявляются
III
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
ется тем, что в юрских отложениях найдена пыльца, неотличимая от пыльцы
некоторых современных нимфейных. Кроме того, найдены остатки древесины,
принадлежащей либо бессосудистым покрытосеменным, родственным совре-
менным магнолиевым, либо беннеттитам. Во всяком случае, в настоящее
время нельзя отрицать вероятности появления покрытосеменных ранее
мелового периода, в юре и даже триасе. По мнению некоторых ученых,
принимающих во внимание косвенные данные, цветковые появились еще
раньше, в пермотриасе. Высказывались предположения и о еще большей
древности покрытосеменных — о зарождении их на границе каменноуголь-
ного и пермского периодов.
Не менее сложен вопрос о конкретных предках покрытосеменных.
С одной стороны, какие группы растений дали начало покрытосеменным?
С другой стороны, какие группы покрытосеменных наиболее близки к древ-
ним предковым формам?
В качестве предполагаемых предков покрытосеменных указывались
различные группы древних голосеменных растений: беннеттиты, гнетовые
(типа гнетум, эфедра), семенные папоротники, кейтониевые.
Своеобразные спороношения некоторых беннеттитов напоминали цветки
покрытосеменных тем, что на общей оси стробила в той же последова-
тельности, что и в цветке покрытосеменных, располагались видоизме-
ненные листья защитной функции, микро- и мегаспорофиллы. Такой
стробил действительно имеет сходство с цветком магнолий. Однако мега-
и микроспорофиллы беннеттитов существенно отличались от плодолистиков
и тычинок цветковых — не было у них ни рыльца, ни завязи, защита семя-
почек («покрытосемянность») обеспечивалась бесплодными чешуями, а не
самими мегаспорофиллами, не было у беннеттитов и плодов.
У кейтониевых были органы, подобные завязи и рыльцу, но у них не
было ничего сходного с цветками. Вероятно, эти растения представляют
слепую ветвь эволюции. У прочих семенных папоротников не было и «no-
крытосемянности».
Теория происхождения покрытосеменных из гнетовых предполагает,
что наиболее примитивные покрытосеменные имели однополые цветки, без
околоцветника или с невзрачным околоцветником, вроде современных бере-
зовых и близких к ним семейств. Эта теория связана со своеобразным воз-
зрением на цветок как на орган, возникший из компактного собрания («соцве-
тия») нескольких стробилов («цветков») голосеменных (псевданцие-
il о сравнительно-морфологические данные все более убеждают в том,
что цветок покрытосеменных есть простой, неразветвленный побег (э в а н-
циевая теория цветка). Цветки, имеющие удлиненное цветоложе,
неопределенное число свободных, располагающихся по спирали боковых
органов, т. е. морфологически сходные с побегом, наиболее близки стробилам
макрофилльных голосеменных. Из этой стробилярной модели цветка уда-
ется вывести большинство известных вариантов цветков покрытосеменных,
обычно не однополых, и не ветроопыляемых, как требует теория происхож-
дения покрытосеменных от гнетовых, а достаточно крупных, с ярким около-
цветником, энтомофильных.
Признание цветка покрытосеменных за орган, гомологичный стробилу
макрофилльных голосеменных, приводит к заключению, что из ныне живу-
щих представителей этой группы близка к предкам покрытосеменных группа
семейств, сходных с магнолиевыми. Именно в ней с наибольшей полнотой
реализуются примитивные признаки цветка — стробила. В пользу древ-
ности всего родства магнолиевых говорит и тот факт, что у ряда относящихся
сюда растений древесина не содержит сосудов, а имеет только трахеиды,
что можно рассматривать как признак близости с голосеменными.
Если признать, что среди современных покрытосеменных наиболее
близки к их предкам магнолиевые и родственные с ними, то более отдаленных
голосеменных предшественников цветковых растений следует искать не
выс
НГ>
:е растения
иг
среди гнетовых, а в классе Cycadopsida. Это, вероятно, были макрофилльные
растения, с невысокими стволами. Они имели стробилы, у основания которых
располагались листья, выполнявшие защитную функцию околоцветника,
а выше следовали микро- и мегаспорофиллы; эти растения еще не имели
завязей, рылец и плодов. В филогенетической системе эту гипотетическую
группу голосеменных следует поместить между семенными папоротниками^
не имевшими стробилов, с одной стороны, и саговниками и беннеттитами —
с другой.
Во второй половине мелового периода покрытосеменные стали преобла-
дающей группой в составе флоры Земли. Причины этого с полной достовер-
ностью не установлены. При разработке предположений по этому вопросу
целесообразно раздельно рассматривать причины возникновения покрыто-
семенных и причины их сначала медленного, а затем необычайно быстрого
(в геологическом масштабе) распространения — «завоевания» суши.
Образование из мегаспорофиллов завязи, предохраняющей семяпочки
от высыхания, и появление сосудов свидетельствуют о приспособленности
покрытосеменных к условиям повышенной радиации. Такие условия голо-
семенные предки цветковых могли встретить в высокогорьях, выше верхней
границы леса. Проникновение на эти территории должно было повлечь глу-
бокую перестройку системы водоснабжения и развитие средств защиты
семяпочки от высыхания.
Расселение покрытосеменных на равнинах впоследствии могло быть
обеспечено повышенной изменчивостью, дававшей материал для возникно-
вения многочисленных форм видового ранга, заселявших самые разнообраз-
ные варианты местных условий. Большое значение имело совершенствование
перекрестного опыления, выразившееся особенно ярко в сопряжённой эво-
люции цветка и насекомого. Широкое распространение энтомофилии и пере-
крестного опыления способствовало возрастанию генофонда цветковых рас-
тений, в чем собственно и заключается часто подчеркиваемая их «эволюци-
онная пластичность».
Палеогеографические изменения, связанные с горообразованием и ре-
грессиями моря, расширяли территорию с достаточно аридными условиями,
к которым легче, чем голосеменные, могли приспособиться относительно
ксерофилизованные покрытосеменные растения.
Нельзя исключать и возможность того, что в эпоху, предшествовавшую
быстрому расселению покрытосеменных и, соответственно, падению роли
во флоре Земли других групп растений, произошли планетарные перемены
условий жизни растений. Гипотезу этого рода разработал наш соотечествен-
ник профессор М. И. Голенкин (1927). Анализируя особенности строе-
ния покрытосеменных, он пришел к выводу, что они — растения солнечного
климата. Автор высказал мысль, что в середине мелового периода по каким-
то збщим космогоническим причинам по всей Земле произошло резкое воз-
растание освещенности и падения влажности воздуха. Густые облака, ранее
. постоянно окутывавшие Землю, рассеялись и дали доступ ярким солнечным
лучам к поверхности планеты, или в это время возросла теплопрозрачность
атмосферы в связи с изменением ее состава. В связи с этим резко увеличилась
сухость воздуха. Громадное большинство высших архегониальных растений
того времени, не приспособленных и не имевших возможности приспосо-
биться к яркому освещению и сухости воздуха, вымерло или резко сократило
свое распространение (кроме хвойных — наиболее ксерофильных из голо-
семенных). Покрытосеменные, имевшие до того" очень ограниченное распро-
странение и представленные еще небольшим числом форм, оказались хорошо
приспособленными к яркому солнечному свету и сухости воздуха. Это обсто-
ятельство', а также чрезвычайная эволюционная пластичность (способность
к возникновению разнообразнейших приспособлений к различным внешним
условиям) обусловили быстрое распространение покрытосеменных по всей
Земле и вытеснение ранее господствовавших групп высших архегониальных
растений.
III
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ покрытс^титстгягьпе
345
Победа' покрытосеменных повлекла за со-
бой изменения и в животном населении Зем-
ли; особенно она должна была сказаться на
темпах эволюции насекомых, млекопитающих
и птиц, питающихся насекомыми.
Гипотеза Голенкина позднее получила
подтверждение в установлении факта частич-
ной нестационарности радиационного режима
Солнца, что было использовано М. В. Сеня-
ниповой-Корчагипой (1959), развившей тео-
рию Голенкина.
Не менее, если не более, сложен вопрос о
месте возникновения покрытосеменных. При
отсутствии палеоботанических данных его при-
ходится решать, главным образом основываясь
на анализе ареалов тех семейств, которые
предположительно признаются близкими к
родоначальникам цветковых. А. Л. Тахтаджян
. (1970) считает, что первичный центр расселения
цветковых растений скорее всего находился
где-то между Индией и Полинезией, т. е. в
области, расположенной в западной части
Тихоокеанского бассейна и частично захваты-
вающей бассейн Индийского океана. По всей
Рис. 223. Зародыши покрыто-
семенных:
1 — двудольные; я—однодольные
(с — семядоля, п — почечка, е — ги-
покотиль, як — корень)
вероятности, он находился в той части Земли,
которую занимает сейчас Юго-Восточная
Азия. Автор пр
ел к выводу, что в этой же области должен был находиться
Ш1
и центр происхождения цветковых.
Было сделано несколько попыток воссоздать облик первичного цветко-
вого растения. Хотя нельзя вполне исключить возможность того, что это
были растения травянистые, большинство ботаников в настоящее время
считают, что первичные покрытосеменные были древесными растениями
с невысокими стволами, немногочисленными толстыми ветвями и моноподи-
альным ветвлением. Из них развились более крупные, симподиально ветвя-
еся деревья с многочисленными толстыми и тонкими ветвями. Из древес-
иг
ных форм в разное время в разных филогенетических линиях развивались
кустарники, полукустарники и травянистые формы, вначале многолетние,
затем, в, различных родах в связи со специфическими условиями климата
и местообитаний, двулетники и однолетники. Приспособление покрытосемен-
ных к самым различным экологическим и биотическим факторам среды
обусловило огромное разнообразие вегетативных органов (особенно листьев),*
многочисленные метаморфозы, а также бесконечное разнообразие в строении
цветков и плодов. Стали более сложными и разнообразными химический
состав и физиологические реакции.
Эволюция цветка, на строении которого главным образом базируется
систематика покрытосеменных, шла у них от цветков с удлиненным (вытяну-
тым) цветоложем, обоеполых, актиноморфных, со спиральным расположе-
нием неопределенного числа свободных (несросшихся) цветолистьев, от
цветков с верхней завязью и многочисленными семяпочками — к цветкам
циклическим, с вполне определенным числом членов, нередко сросшихся
друг с другом и располагающихся на* плоском или вогнутом цветоложе,
к цветкам зигоморфным, имеющим нижнюю завязь с одной или немногими
семяпочками. Эти изменения цветка покрытосеменных повторялись в разных
эволюционных рядах, независимо друг от друга.
Общепринятой системы покрытосеменных, как,- впрочем, и многих дру-
гих отделов растительных организмов, еще нет, несмотря на десятки пред-
ложенных систем. Разногласия между авторами систем начинаются уже
с исходного положения — какие группы растений считать более примитивными
346
выс
III
IE РАСТЕНИЯ
и принимать ли для покрытосеменных моно-*, би- или полифилетическое про-
исхождение. В конце XIX — первой четверти XX в. широким признанием
пользовались две системы—немецкого ботаника А. Энглера и австрий-
ского ботаника Р. Веттштейна. В обеих системах за наиболее прими-
тивные среди покрытосеменных принимаются семейства с однопокровными
и беспокровными, невзрачными, анемофильными, малочленными цветками
(казуариновые, перечные, ивовые, березовые, буковые и др.).
В системах, принятых в настоящее время, в начале системы ставятся
семейства с хорошо развитыми многочисленными, раздельнолепестными,
двупокровными, знтомофильными цветками, как, например, семейства магно-
лиевых, лютиковых, кувшинковых, барбарисовых и другие, так называемые
11Г.1
многоплодниковые. Семейства с однопокровными и беспокровными цветками
считаются вторично упрощенными, так как многие из них имеют ценокарп-
ный гинецей, нижнюю завязь — признаки отнюдь не примитивные. Таковы
системы ботаников Буша, Гроссгейма, Козо-Полянского,
Кузнецова, Тахтаджяна (СССР), Г а л л и р а (Голландия),
Бзсси, Пула, Шафнера (США), Гетчинсона (Англия), Ш о о
(Венгрия).
Большинство систематиков делят покрытосеменные на два класса —
двудольные и однодольные. Различия этих классов определяются целым
комплексом признаков, из которых наиболее важны и доступны для непо-
средственного наблюдения следующие:
Двудольные
Однодольные
Зародыш, как правило, имеет две семя-
доли, занимающие боковое положение от-
носительно оси. Конус нарастания нахо-
дится между семядолями, завершая ось
зародыша. Часто конус окружен зачаточ-
ными листьями и вместе с ними образует
верхушечную почечку (рис. 223).
Боковые побеги, возникающие в пазухах
листьев (иногда семядолей), имеют два пред-
листа (самые нижние, недоразвитые ли-
стья), занимающих баковое положение
в трансверсальной плоскости1 или несколь-
ко смещены относительно ее.
При наличии в пазухе листа несколь-
ких почек они располагаются сериально,
т. е. в медианной плоскости.
Проводящая система стебля сифоносте-
лическая или эустелическая; в обоих слу-
чаях, как правило, развит камбий, обес-
печивающий нарастание стебля в толщину.
Имеется ясная дифференцировка стебля
на область коры и центральный цилиндр.
Листовые пластинки обычно с сетчатым
(перистым или пальчатым) жилкованием.
Листья с небольшим числом листовых сле-
дов (1—3, реже более).
Корневая система у особей, развившихся
непосредственно из семян, сохраняет глав-
ный корень долгое время или в течение
всей жизни.
Зародыш, как правило, с одной семя-
долей, которая, вероятно в результате сме-
щения, занимает концевое положение, со-
ставляя как бы продолжение оси зароды-
ша. Конус нарастания и почечка смещены
и занимают боковое положение относи-
тельно оси зародыша.
Боковые пазушные побеги с одним пред-
листом, расположенным со стороны, обра-
щенной к материнской оси, т. е. к оси пре-
дыдущего порядка (адаксИально), и, сле-
довательно, в медианной плоскости.
В случае нескольких почек в пазухе кро-
ющего листа, они располагаются в ряд пер-
пендикулярно медианной плоскости, т. е.
колл атерально.
Проводящая система атактостелическая,
состоящая из закрытых, т. е. лишенных
камбия, пучков, рассеянных по всей по-
верхности поперечного разреза стебля.
Область коры стебля не всегда выражена.
Листовые пластинки с дугонервным или
параллельным жилкованием. Листовых сле-
дов много.
Главный корень нередко отмирает рано,
и корневая система взрослых особей сос-
тоит из придаточных корней.
1 Для ориентировки в расположении элементов пазушного вегетативного побега
и пазушного цветка удобно различать медианную и трансверсальную плоскости. Медиан-
ная плоскость совпадает с осевой линией побега, несущего кроющий лист, и проходит че-
рез срединную жилку последнего, т. е. является плоскостью его симметрии. В медианной
плоскости, очевидно, лежит осевая линия и вегетативного пазушного побега, и пазушного
цветка. Трансверсальная плоскость, перпендикулярная к медианной, тоже совпадает с
осевой линией пазушного побега. Две плоскости, делящие прямые углы между медианной
и трансверсальной плоскостями, называются диагональными.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
847
Цветки круговые, полукруговые или
ациклические. В круговых цветках число
членов каждого круга кратно 5, редко 2,
еще реже 3.
Эндосперм целлюлярный, редко гело-
биальный.
Цветки круговые, редко полукруговые.
Число членов в круге обычно кратно 3,
реже 2.
Эндосперм гелобпальный или нуклеар-
ный, редко целлюлярный.
'.III
Перечень признаков, на основании которых устанавливается различие
между двумя классами покрытосеменных, можно было бы расширить. Однако
и упомянутые здесь показывают, что выделение двух классов основывается
не на одном отличии, а на целом комплексе их. Вместе с тем ни одно из разли-
чий двух классов нельзя считать абсолютным. Так, среди двудольных извес-
тен ряд случаев, когда зародыш имеет одну семядолю в результате сраста-
ния двух (Ficaria) или редукции одной из двух (Carum bulbocastaneum.
Corydalis^ некоторые Senecto). Встречается у двудольных и рассеянное раз-
мещение проводящих пучков, лишенных к тому же камбия (некоторые люти-
ковые, нимфейные, барбарисовые). «Однодольный» характер жилкования
известен у подорожника (Plantag о), а у некоторых видов этого рода и кор-
невая система мочковатая, как у большинства однодольных. Среди дву-
дольных, особенно среди более примитивных форм, нередки случаи тример-
ности цветков (многие лавровые и др.), а иногда пятичленный и трехчленный
околоцветник можно наблюдать в пределах одного вида (ветреницы).
Словом, признаки однодольных и двудольных растений могут транс-
грессировать, налегать друг на друга. Это не раз давало повод сомневаться
в реальности однодольных как самостоятельной таксономической единицы.
Однако, в целом, различия однодольных и двудольных настолько глубоки,
что авторы современных систем обычно сохраняют деление цветковых на два
класса.
Отдел цветковых, или покрытосеменных, наиболее разнообразен среди
ныне существующих растений. Он содержит около 250 000 видов, около
10 000 родов и более 300 семейств.
Покрытосеменные населяют практически всю современную сушу, от
крайних широт Арктики до Антарктики (Aira antarctica). В Гималаях на
высоте свыше 6200 м известно гвоздичное растение (Arenaria musciforrnis).
Некоторое число видов покрытосеменных вторично перешло к водному суще-
ствованию, хотя в соленой воде растет всего лишь около 30 видов.
В современную геологическую эру покрытосеменным принадлежит ос-
новная роль в создании органического вещества суши; вместе с ними сущест-
венное значение в этом отношении имеют хвойные. Покрытосеменные почти
всюду, кроме хвойных лесов, некоторых типов болот и тундр, определяют
характер ландшафта.
Значение покрытосеменных для человека исключительно велико. Все
важнейшие культурные растения — представители этого отдела. Широкое
распространение в культуре получили около 1000 видов, а всего использу-
ется человеком примерно 30 000 видов покрытосеменных, среди них пище-
вые, лекарственные, технические, кормовые, мелиоративные, озеленитель-
ные, декоративные растения. G развитием промышленности по переработке
(особенно химической) растительного сырья вовлечение покрытосеменных
в круг хозяйственных потребностей возрастает, что настойчиво выдвигает
задачу глубокого изучения биологических особенностей покрытосеменных,
без чего невозможно введение в культуру1 ценных видов, спрос на которые
быстро растет и может превысить их природные запасы.
III
И11 г.
1-й КЛАСС—ДВУДОЛЬНЫЕ (DICOTYLEDONEAE)
В состав класса двудольных входит 292 семейства и 56 порядков, что
составляет от 3/4 до */5 всех покрытосеменных*
Вследствие большого разнообразия семейств и порядков двудольных
их традиционно делят на два подкласса: раздельнолепестные, или первич-
t нопокротые (Choripetalae, Archichlamydeae), и спайнолепестные (Sympe-
talae). Основной признак, отличающий эти подразделения, — строение
III.
Представители подкласса спайнолепестных (Sympetalae) имеют
круговые цветки с двойным (хотя бы по заложению) околоцветником и почти
всегда спайнолепестным венчиком. У видов подкласса раздельно-
лепестных (Choripetalae) венчик, как правило, состоит из несросшихся ле-
пестков, спайнолепестность наблюдается только у немногих производных
относительно высокоэволюционировавших форм.
Строение венчика коррелятивно связано еще с рядом признаков в орга-
низации андроцея, гинецея, особенно семяпочек.
Разнообразный, изменчивый по строению цветка подкласс Choripetalae
wmjipL делят на две единицы меньшего объема — Monochlamydeae (одно-
покровные) и Dialypetalae (свободнолепестные), возводя их в некоторых
случаях в ранг подкласса, т. е. увеличивая число подклассов двудольных
до трех1.
Среди раздельнолепестных двудольных растений, кроме однопокровных,
которые отличаются простым, обычно невзрачным околоцветником или его
отсутствием (что вместе с рядом других признаков позволяло многим бота-
никам рассматривать эту группу как относительно примитивную), давно
обращав на себя внимание многоплодниковые, очень разнообразные по стро-
ению цветника, по плодам и по анатомической организации вегетативного
тела.
У многоплодниковых широко распространены признаки примитивности
цветка (стр. 292 и 350), как они рисуются с позиций эволюционной теории.
Если еще учесть, что среди многоплодниковых имеются растения с бессосу-
дистой древесиной, а у некоторых наблюдается неполная закрытость завязи
и другие черты примитивности, то все это позволяет рассматривать много-
плодниковые как группу, вымершие предки которой могли дать начало не-
скольким линиям эволюционного развития двудольных растений. Такое
положение многоплодниковых и достаточно большое число относимых к ним
порядков дает некоторым ученым основание для выделения многоплоднико-
вых растений также в особый подкласс или даже в два подкласса (Тахта-
джян, 1970).
В.настоящее время достаточно выяснено, что за вычетом многоплоднико-
вых и однопокровных прочие раздельнолепестные двудольные принадлежат
по крайней мере к трем параллельным направлениям эволюции. Эти ряды
генетически обособлены, и каждый из них можно рассматривать тоже как
отдельный подкласс.
Таким образом, на современном уровне знаний в пределах прежнего
подкласса раздельнолепестных двудольных можно выделить не менее 5 под-
классов.
lll'j
11 Г.».
1 Тах, например, в «Lehrbuch der Botanik» Страссбургера (1962, 1967) Ф и р б а с
выделяет среди раздельнолепестных три группы (не называя их подклассами) — A petar
lae, или Monochlamydeae, Dialypetalae и Sympetalae. Эти же подгруппы (с названием Cho-
ripetalae вместо Dialypetalae) в ранге подклассов приняты в посмертном «Курсе систематики
высших растений» Б.М. Козо-Полянского. На то, что есть основания к выделению
Monochlamydeae в самостоятельный подкласс, указывалось и в предыдущем из-
дании этого учебника (1962, стр. 405—406, 675). Такое воззрение, в сущности,
возрождает довольно давние представления, так как указанные три группы в ранге «ко-
горт», соответствующих подклассам современных авторов, фигурировали уже в системе
Эндлихера (1836—1843).
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
Что касается спайнолепестных (Sym-
petalae), то уже давно выяснено, что
основной отличительный признак их —
спайнолепестный венчик — мог возни-
кать конвергентно в разных линиях
энтомофильной эволюции как приспо-
собление, ограничивающее потребление
нектара достаточно узким, определен-
ным кругом насекомых-опылцтелей.
Вполне возможно конвергентное про-
исхождение и других признаков спай-
нолепестных. Это приводит к выводу,
что спайнолепестные — группа поли-
филетическая.
Итак, в пределах всего класса дву-
дольных в настоящее время есть осно-
вания выделять не два (раздельноле-
пестные и спайнолепестные), а по край-
ней мере шесть (рис. 224) подклассов.
1-Й ПОДКЛАСС - МАГНОЛИИДНЫЕ
(MAGNOL11DAE)
Подкласс магнолиидных объеди-
няет растения с рядом примитивных
признаков. Цветки у них спиральные,
спирально-круглые, t реже круговые,
обычно с хорошо развитым, большей
частью ярким, простым или двойным
околоцветником, с многочисленными
тычинками, развивающимися в центро-
петальной последовательности. Харак-
терен апокарпный гинецей. Зрелые
пыльцевые зерна двухклеточные или
трехклеточные. Семяпочки двупокров-
ные, крассинуцеллятные. Растения дре-
весные или травянистые, иногда бессо-
судистые или с сосудами, членики кото-
Рис. 224. Взаимосвязь подклассов покры-
тосемянных растений (по Кронквисту):
1 — двудольные; а — однодольные
рых имеют лестничную перфорацию.
В основном ‘ этот подкласс соответствует понятию многоплодниковых
(Polycarpicae).
В составе подкласса Magnolltdae рассматриваются семь порядков.
Их можно разделить па две группы: имеющие эфиромасличные клетки в
листьях и других „вегетативных органах. (Magnollales, Piperales, Aristolo-
chiales) и лишенные их (Ranunculales, Nymphaeales, Papaverales, Nepen-
thales).
Среди порядков первой группы Magnollales практически только дре-
11; I.
Ill
весные растения: деревья, кустарники, лйаньг/травы — редчайшее исклю-
чение. В двух порядках второй группы преобладают травы и (часто лазающие)
кустарники, деревья крайне редки. ~
1-Й ПОРЯДОК — МАГНОЛИЕЦВЕТНЫЕ (MAGNOLIALES)
Магнолиецветные имеют цветки с простым венчиковидным или дифферен-
цированным на чашечку и венчик околоцветником, преимущественно пра-
вильные, спиральные, гемицикл и ческие или циклические, обычно с много-
численными тычийцами. Плодолистиков много или один. Гинецей апокар]
лишь у производных форм наблюдается срастание плодолистиков и даже
350
ВЫСШИЕ РАСТЕНИЯ
Рис. 225. Магнолия:
1 — продольный разрез цветка Magnolia precia; а —
сборный алии: 3 — диаграмма цветка М. grandiflora
погружение завязи в полое цве
толоже; завязь верхняя. Цветки»
как правило, обоеполые, энтомо-
фильные. Деревья или кустарни-
ки, реже древесные лианы.
Листья почти всегда простые;
прилистники, если они есть,
срастаются друг с другом. В дре-
весине, кроме трахеид, имеются
сосуды с лестничной или простой
перфорацией; у некоторых дре-
весина только из трахеид. У боль-
шинства представителей, особен-
но в паренхимных тканях листь-
ев и стеблей, имеются масляные
или слизевые секреторные клетки.
В порядке магнолиецветных
объединяют примерно 20 семейств
(иногда больше). Несмотря на
разнообразие в строении цветка
и другие признаки, они
ся в явно родственных
ниях.
Все разнообразие
миогоплодниковых легко
находят-
отноше-
цветков
вывести
из некоторого исходного прими-
тивного типа, каким считается цветок, сох ряды шдмй признаки стробила:
радиальную симметрию, вытянутое, как бы стеблеобразное цветоложе,
неопределенное число цветолистьев, расположенных в спиральном порядке
и не срастающихся друг с другом. Гинецей примитивного цветка много-
плодниковых или апокарпный с неопределенным числом пестиков (плодни-
ков), или из одного плодолистика.
Семейство магнолиевые (Magnoltaceae) содержит свыше 200 видов.
Это древесные и кустарниковые растения с простыми, часто имеющими при-
листники листьями; одни представители вечнозеленые, другие — с опадаю-
щей листвой. Цветки в большинстве случаев обоеполые, актиноморфныев
с удлиненным цветоложем. Листочки околоцветника (6—12 и более) располо-
жены по спирали или кругами, преимущественно трехчленными. В послед-
нем случае наружная мутовка образует чашечку. Многочисленные тычинки
размещаются спирально, как и плодолистики, образующие многочленный
апокарнный гинецей с краевым положением семяпочек в завязях; реже
плодолистики срастаются.
Типичным представителем семейства является род магнолия (Magnolia.
рис. 225), 75 видов которого частично американские (от юго-восточной при-
атлантической Северной Америки до Венесуэлы), частично восточноазиат-
ские и индо-малайские. Некоторые из магнолий культивируются в СССР
в субтропических районах (Крым, Закавказье). Среди них отметим М. grandi-
flora (родом из Флориды) — вечнозеленое дерево, достигающее 30 м высоты,
с простыми, крупными, цельнокрайними, кожистыми, блестящими листьями
на коротких черешках. Прилистники их защищают в почке молодые нераз-
вернувшиеся листья. Одиночные крупные цветки достигают в поперечнике
10 см и более. Цветоложе вытянутое, конусовидное. На нем располагаются
в трехчленных кругах 6—12 крупных белых душистых листочков около-
цветника, за которыми следуют многочисленные спирально размещенные
тычинки. Каждая тычинка имеет широкую тычиночную нить и хорошо за-
метный связник, верхушка которого выступает за пределы пыльников. Около
2/3 цветоложа в его верхней части занято гинецеем. Каждый из многочис-
ленных плодолистиков, располагающихся в спиральном порядке, образует
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
351
VlVAlt AL&AKU^tiw •
самостоятельный пестик, причем края
плодолистика, согнутого вдоль его
средней жилки, срастаются и в обра-
зовавшейся одногнездной завязи поме-
щаются две семяпочки (у некоторых
видов магнолий семяпочек до 6).
Столбика пестик не имеет, и улавли-
вающая пыльцу поверхность (рыльце)
занимает верхнюю часть краев пло-
долистика. После цветения листочки
околоцветника и тычинки опадают, а
на местах их прикрепления остаются
заметные рубцы. Цветоложе разраста-
ется. Из пестиков развиваются много-
Л1
численные сероопушенные плоды-лис-
товки, раскрывающиеся по брюшному
шву. Они, образуют крупные «шишки»,
длиной 5—7 см и более. Созревшие
семена имеют красный цвет, мясистую
семенную кожуру и свисают вниз на
длинных нитях.
Из других магнолиевых следует
Рис. 226. Тюльпанное дерево (Lirioden-
dron sinensis)
упомянуть тюльпанное дерево (Lirio-
dendron), одий из видов которого (£.
tulipifera) растет в юго-восточной части
Северной Америки, другой (L. sinen-
sis) — в Китае. Цветки этого дерева
внешне напоминают тюльпаны, хотя и не имеют яркой окраски. Листья с
четырьмя острыми на концах лопастями и крупной выемкой на вершине
(рис. 226). Тюльпанное дерево дает поделочную древесину хорошего
качества. В СССР его разводят как декоративное (например, на Черномор-
ском .побережье Кавказа).
С К семейству магнолиевых примыкает еще несколько некрупных семейств,
которые не рассматривались ранее как самостоятельные. Из них отме-
тим два.
Семейство дегенериевые (Degeneriaceae) содержит один вид
Degeneria vitiensis (рис. 227), обитающий на островах Фиджи. Это дерево
высотой до 14 м> с очередными расположенными спирально листьями без
прилистников. Сосуды с лестничной перфорацией. Особенно интересны цвет-
ки. За трехчленной чашечкой в них следуют 12—18 лепестков в 3—5 кругах,
андроцей из 20—30 тычинок в 3—4 кругах и одночленный гинецей. Тычинки
плоские. Тычиночная нить в них не дифференцирована. Микроспорангии,
вытянутые, длинные, расположены двумя парами на нижней стороне плос-
кого микроспорофилла и погружены в его ткань. Выше андроцея имеется
несколько стаминодиев. Единственный плодолистик завершает ко-
роткое, наверху несколько вдавленное цветоложе. Края плодолистика
не сомкнуты и в ранних фазах- развития ясно удалены друг от друга, но во
время цветения сближаются, однако соприкосновение достигается только
в нижней части. Края плодолистика отогнуты наружу и покрыты железис-
тыми волосками, образующими рыльцевую поверхность. В завязи находится
примерно 25 семяпочек. Плод сухой (твердый), многосемянный, длиной др
5 см, с мясистым снаружи и твердым внутри (эндокарпием) околоплодником,
семена многочисленные. Семенная кожура дифференцирована на мясистую
наружную и твердую внутреннюю часть. Имеется эндосперм. Семядо-
лей 3, реже 4.
Семейство представляет большой теоретический интерес, особенно бла-
годаря незамкнутости завязи, примитивной организации рыльца и листо-
видным тычинкам.
352
ВЫСШИЕ РАСТЕНИЯ
Рис. 227. Дегенерия (Degener&s vitiensis):
1 — цветущая ветвь; 9 — цветок; з — тычинка; 4 — поперечный разрез тычинки: 5 — пестик; в — продолъ
ный разрез пестика; 7 — поперечные разрезы пестика; 8 — плод
Семейство винтеротх (Winteraceae) представляет не меньший
интерес. Оно содержит около 100 видов, распространенных главным образом
в южном полушарии, в бассейне Тихого океана, один вид — на Мадагаскаре.
Это тоже древесные растения со спиральным расположением листьев, не
имеющих прилистников. Заслуживает внимания бессосудистая ксилема
из очень длинных и толстостенных трахеид с 2—3 рядами окаймленных пор.
В состав магнолиевых ранее включали также семейство схизанд-
pome (Schizandraceae), куда относится лиана лимонник (Schizandra chinensis).
Плоды этого растения обладают выраженным тонизирующим действием.
Лимонник распространен в Китае, Японии, Корее; в СССР — в Хабаровском
и Приморском краях, на Сахалине и Курильских островах. Семейство
иллициевые (Illiciaceae) с единственным родом Illicium ранее также отно-
сили к магнолиевым. Плоды Illicium anisatum ядовиты, а 7. (Китай)
используют как пряность («звездчатый анис»).
Близко к семейству* магнолиевых стоят довольно крупные семейства,
распространенные исключительно или главным образом в тропических
областях: аноновые, мускатниковые, лавровые.
Семейство аноноте (Аппопасеае). Это семейство содержит, как и
магнолиевые, древесные формы с простыми, без прилистников листьями,
имеющими внутренние железки. Это почти исключительно тропические рас-
тения, встречающиеся в дождевых тропических лесах Старого Света. Семей-
ство содержит 2000 — 2100 видов. Из апокарпного гинецея развиваются
сборные плоды различного строения с нежной съедобной ароматной мякотью.
Используются также древесина, масло, ароматические и пряные вещества.
Некоторые аноновые введены в культуру.
У нас на Черноморском побережье Кавказа, в Крыму культивируют
как декоративное растение листопадную внетропическую (родина—юго-восток
США) ааимину (A simina triloba) — невысокое дерево, крупные съедобные
плоды которого имеют приторно-сладкий вкус и валах земляники или ана-
наса.
В особое семейство эупомацисвых (Eupomatiaceae) иногда выделяют
род Eupomatia, виды которого распространены в Восточной Австралии
и на Новой Гвинее. Оба вида этого рода — кустарники с замечательным
строением цветка; Околоцветник отсутствует, андроцей многочленный,
спиральный, а внутренние, более близкие к гинецею тычинки превращены
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОС
1ЫВ
353
в яркие лепестковидные ста-
минодии. Таким образом име-
ется как бы внутренний венчик-
Стаминодии покрыты «пита-
тельными тельцами» трихом-
ной природы; их потребляют
опылители — жуки. Гинецей
апокарпный, спиральный, но
плодолистики сросшиеся; не-
смотря на это, каждый плодо-
листик ещё слегка открыт,
столбиков нет, рыльцевая по-
верхность низбегающая вдоль
краев плодолистиков.
Семейство мускатнико-
еые (Myristicaceae) — пантро-
пическое семейство, содержа-
щее свыше 250 видов. Родст-
венные связи не вполне выяс-
нены, его сближают с анноно-
выми или лавровыми. Из об-
ширного рода Myristica (около
100 видов) наиболее известно
растение Myristica fragrans
(Молуккские острова), семена
которого (мускатные орехи) и
сильноразвитый лопастной при-
семянник (мускатный цвет) ис-
пользуются как пряность. Де-
рево культивируют во влажных
тропиках.
V Семейство лавровые
(Lauraceae) насчитывает более
2000 видов, принимающих зна-
чительное участие в структуре
лесов тропических и субтропи-
ческих областей. Лавровые —
древесные растения {за исклю-
чением паразитного Cassytha)
с кожистыми, простыми, дель-
ными или рассеченными листья-
ми, без прилистников. Цветки
обоеНОЛЫё или однополые, из
двух-, пята-, чаще трехчленных
кругов. Цветоложе дйсковидно
'ргасширеннбё^ нлй' чашевидно-—
углубленное^ К краю его прй- ~
креплены листочки околоцвет-
ника и 3—4 круга тычинок. Ты-
чиночные нити по бокам у
основания имеют железки.
Пыльники вскрываются клапа-
нами. Андроцей лавровых, как
принято считать? состоит из
пучков, содержавших по 3, мо-
Я;ет быть, по 5 тычинок, кото- "
I Я ', . i СЬ
• • “ J*м пучке, реттупированы.
12—602
Рис. 228. Коричное дерево (Cinnamomum ze-
ylanicum):
1 — цветущая ветвь; г — продольный разрез цветка*
з— диаграмма цветка
Рис. 229. Лавр благородный (Laurus nobilis)'.
1 — пветущая ветвь; 2 — ветвь с плодами; 3 — про-
дольный разрез цветка: 4 — тычинка; 5 — тычинка с
раскрывающимися пыльниками
354
ВЫ
.Htri;
га РАСТЕНИЯ
Упомянутые железки, вероятно, представляют остатки редуцированных ты-
чинок. Пестик один, по-видимому, из одного плодолистика. Завязь одног-
нездная, соднои~с!вмяпочкой. Плод ягодэобразный или костянка.
Многие лавровые находят практическое применение. Так, йя" древесины
и листьев камфорного лавра (Cinnamomum camphor а), обитающего в Китае,
Японии, получают камфору; кора коричного дерева (С. zeylanicum), распрост-
раненного в Южнойг Индии и на Шри Ланка (рис. 228), используется как
пряность (корица); плоды груши авокадо (Persea americana), встречающейся
¥ тропической Америке, съедобны; листья благородного лавра (Laurus nobilis,
рис. 229), обычного в средиземноморских странах и часто культивируемого
как декоративное дерево (в СССР — в Крыму и Закавказье), используются
как пряность (лавровый лист).
В третичном периоде лавровые были распространены и в областях ныне
умеренного и даже холодного климата. Отпечатки листьев Cinnamomum,
Sassafras и других обнаружены в'отложениях неогена и палеогена в северном
полушарии. В неогеновом и палеогеновом янтаре Балтики найдены и цветки
Ггг.
в хорошей сохранности.
2-Й ПОРЯДОК — ЛЮТИКОЦВЕТНЫЕ (RANUNCULALES)
Цветки более разнообразны, чем у Magnoliales. Они бывают ацикличес-
кими, чаще гемициклическими или циклическими, с простым или двойным
околоцветником, иногда без него, правильные, но у высокоспециализиро-
ванных форм околоцветник может быть зигоморфным. Листочки околоцвет-
ника в неопределенно большом или фиксированном числе. Тычинки обычно
многочисленные, наружные у многих превращены в лепестковидные нектар-
ники. Число плодолистиков от многих до одного; они свободные, редко в той
или иной мере сросшиеся. Цветоложе иногда вытянутое, редко вогнутое.
Травянистые или древесные (большей частью лианы или кустарники), иногда
водные растения. Листья преимущественно простые, без прилистников.
Масляные железки, характерные для магнолиецветных, отсутствуют. Про-
водящие пучки иногда рассеянные. У большинства есть сосуды, с простой
перфорацией члеников. У некоторых же сосуды отсутствуют.
Этот порядок представляет группу, возникшую от общих с маг-
ИГ
нолиевыми предков и развивающуюся параллельно им, но с преобладанием
травянистых форм. «
Семейство барбарисовые (Berberidaceae) объединяет около 650 ви-
дов растений разнообразного внешнего облика, главным образом кустарники,
отчасти многолетние травы. Цветки одиночные или, чаще, собранные в кисте-
видные цимозные соцветия. Листочки околоцветника двух категорий: наруж-
ные лепестковидные, внутренние тоже лепестковидные, но с нектарниками.
В некоторых случаях лепестков-нектарников может и не быть. Листочки
'околоцветника (до 6, в том числе 2 круга с нектарниками), как и 4—6 тычи-
нок, расположены двух- или трехчленными кругами. Иногда нити тычинок
раздражимы — приспособление к опылению. Одногнездная верхняя завязь
единственного пестика, образованного одним плодолистиком, содержит от
одной до нескольких семяпочек, расположенных вдоль брюшного шва или
у его основания. Плод — ягода, реже другого строения (орешек, коробочка).
Типичным представителем' этого семейства является род барбарис
(Berberis). Обыкновенный барбарис (В. vulgaris* рис. 230) — кустарник с
удлиненными и укороченными побегами, причем листья первых превращены
в колючки. Декоративен, хороший медонос; промежуточный хозяин хлебной
линейной ржавчины (Puccinia graminis).
Декоративное значение имеет также другой кустарник, родом из Север-
ной Америки, — магонил (Mahonia aquifolium), имеющая темно-зеленые, к
осени краснеющие, сохраняющиеся на зиму листья.
Наряду с тримерными цветками в семействе барбарисовых встречаются
и димерные (горянка — Epimedium, рис. 230, 2).
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
и; i:
jIE
355
m RUTEMIA ALBA
У золотой печати (Ну dra-
stis canadensis) — травянистое
растение из Северной Америки—
околоцветник простой, андро-
цей многочленный, с тычинками,
расположенными спирально, ги-
нецей апокарпный. Род Hyd-
rastis занимает промежуточное
положение между барбарисовыми
и лютиковыми, и его иногда вклю-
чают в состав семейства лютико-
вых.
У североамериканского но*
голиста (Podophyllum peltatum)
околоцветник из многих листочков,
расположенных кругами: 2—3 на-
ружных образуют чашечку, про-
чие (6—9) венчик. Тычинок мно-
го, по-видимому, в круговом
расположении, гинецей одночлен-
ный. По признакам цветка ного-
лист приближается к типичным
барбарисовым, отличаясь от них
отсутствием нектарников на внут-
ренних листочках околоцветника.
Расположение частей цветка
трехчленными кругами, встречаю-
щееся у барбарисовых, как из-
вестно, характерно для однодоль-
ных растений. В связи с этим сле-
дует отметить, что у ноголиста
наблюдается рассеянное располо-
жение сосудистых пучков. Это
наблюдается и у некоторых дру-
гих лютикоцветных.
Qe м е и с т в о лютиковые
((Ranunculaceae) — это обширное
семейство, содержащее до 2000
Рис. 230. Барбарисовые:
1 — барбарис (Berberis vulgaris): а — ветвь с плодами;
б, л — цветок; г — диаграмма цветка: а — горянка: а —
цветок Epimedium rubrum; б — продольный разрез
цветка Epunedium alpinum
видов, подавляющее большинство которых травы, имеющие простые,
в большей или меньшей степени расчлененные, реже сложные
листья, без прилистников, обычно спирально расположенные. В виде исклю-
чения встречаются кустарники и лианоподобные растения. Представители
семейства имеют цветки разнообразного строения. Они могут быть актино-
морфными, как у лютика, и зигоморфными, как у аконита. Околоцветник
бывает простым и двойным. Простой околоцветник, как правило, венчико-
видный. Характерно большое и неопределенное число тычинок, расположен-
ных по спирали, однако некоторые представители имеют фиксированное чис-
ло членов андроцея, расположенных кругами. Гинецей в большинстве слу-
чаев апокарпный, но иногда наблюдается частичное срастание плодолистиков.
Разнообразны цветки лютиковых и в отношении порядка распределения
членов цветка на цветоложе.
В этом семействе встречаются растения с вполне ациклическими цвет-
ками, с цветками циклическими и гемициклическими. Несмотря на такое
разнообразие строения цветка, лютиковые представляют естественную
группу, так как крайние варианты организации цветков связываются
достаточно постепенными переходами. Плоды у одних видов лютиковых —
листовки, у других — односеменные нераскрывающиеся орешки, л:
как исключение встречаются ягоды и коробочки.
jii
12*
356
ВЫСШИЕ РАСТЕНИЯ
Рис. 231. Купальница (Trollius europaeus):
1 — важная часть растения с прикорневыми листь-
ями; 2 — верхняя часть цветоносного нобега; 3 —
нектарник; 4 — тычинка; s — гинецей; в — зрелые
листочки
Рис. 232. Зимовник (Helleborus niger):
1 — цветущее растение; 2 — разрез цветка; 3 — ней
тарник; 4 —гинецей; 5 —плоды
На основании строения гинецея и плодов в семействе лютиковых доста-
точно естественно различаются два подсемейства — зимовниковые и ветрени-
цевые1.
Для подсемейства (Helleboroideae) зимовниковые характерно на-
личие нескольких семяпочек в каждой завязи и плоды листовки (редко коро-
бочки или ягоды). В пределах этого подсемейства наблюдается большое разно-
образие нектарников. В простейшем случае нектарник представляет узкий,
плоский орган с углублением на его поверхности, выстланным железистой
тканью. Такие нектарники можно видеть в цветке купальницы (Trollius
europaeus, рис. 231). Они произошли, вероятно, путем метаморфоза тычи-
нок, в пользу чего говорит их расположение в начале ортостих спирального
(с формулой 5/13) многочленного андроцея. У купальницы азиатской (Г. asia-
iicus, огоньки) нектарники более широкие, лепестковидные. От таких
примитивных нектарников могли возникнуть лепестки типа имеющихся
у калужницы болотной (Caliha palustris) или трубчатые, как у зимовника
(Helleborus, рис. 232), возможно, двугубые, как у чернушки (Nigella),
шпорцевые, как у водосбора (Aquilegia, рис. 233) и живокости (Delphinium),
Сложное строение нектар
ков в цветках аконита (Aconitum, рис. 234) пред-
ставляет дальне
ин
ее видоизменение шпорцевых лепестков-нектарников ти-
па живокости или водосбора. Цветки зимовниковых разнообразны также по
1 К семейству лютиковых иногда относят в составе особого подсемейства роды
Hydrastis и Glaucidium*, кроме того, в виде особого подсемейства долгое время в лютиковые
включали род Patonta. Все эти растения наряду с отличиями имеют тот общий признак,
что у них наружный интегумент превосходит по длине внутренний,
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
357
Рис. 233. Водосбор (Aquilegia alpina):
7—2 — цветущее растение; з — цветок; 4 — чаше-
листик; S ’-лепесток, б — тычинка и плодолисти-
ки; 7 — диаграмма цветка
Рис. 234. Аконит (Aconitum napellus):
1 — верхняя часть побега с соцветием; 2 — цветок;
3 — разрез цветка; 4 — тычинка; 5 — гинецей; б —
продольный разрез завнэи; 7 — поперечный разрез
завязи; 8 — листовки; 9 — диаграмма цветка
расположению частей. У зимовниковых наряду с практически спиральными
цветками (купальница) есть полукруговые (калужница) и вполне круговые
(водосбор); у водосбора пять тычинок самого внутреннего круга андроцея
видоизменены в лепестковидные чешуйки (сравните с Eupomatia, стр. 352).
Если у одних зимовниковых благодаря относительно крупным размерам
и яркой окраске нектарников околоцветник кажется двойным, то у других,
где нектарники мало заметны или глубоко спрятаны в цветке, околоцветник
выглядит как простой. Интересно, что вне зависимости от того, есть ли в
цветке венчик или нет, наружные органы цветка обычно венчиковидно окра-
шены.
Наконец, надо учесть, что наряду с актиноморфными (по строению около-
цветника) цветками (калужница, водосбор) среди зимовниковых есть резко
зигоморфные (живокость, аконит).
Подсемейство ветреницевые объединяет растения, имеющие в
завязи одну (или только одну плодущую) семяпочку и плоды орешки. У од-
них родов околоцветник простой (ветреница — Anemone), иногда рано опада-
ющий (василистник — Thalictrum); у других же, кроме чашечковидных лис-
точков околоцветника, имеются лепестки, представляющие плоские расширен-
ные и ярко окрашенные нектарники, медоотделительный орган которых
находится у основания и может быть прикрыт небольшой чешуйкой (лютик —
Ranunculus). Цветки ветреницевых иногда вполне ацикличны, в других слу-
чаях гемицикличны (горицвет — Adonis), всегда актиноморфны. В случае
простого околоцветника число листочков варьирует. Так, у ветреницы (Ane-
mone) и прострела (Pulsatilla) нередко вместо 5 развивается 6 листочков,
которые в этом случае располагаются в два трехчленных круга.
358
ВЫСШИЕ РАСТЕНИЯ
Рис. 235. Горицвет весенний (Adonis ver-
nal is):
2 — общий вид; я — андроцей и гинецей; 3 — диа-
грамма цветка
Подсемейства зимовниковые и
лютиковые, вероятно, имеют общее
происхождение и развивались парал-
лельно, причем у предковой формы
должны были быть спиральные цвет-
ки и плоды-листовки, (имела боль-
шое сходство с зимовниковыми).
Для семейства в целом харак-
терно наличие алкалоидов, благода-
ря чему многие представители лю-
тиковых ядовиты. Животные обыч-
но не едят их на пастбищах. Сок
. ш
лютиков едкого (Ranunculus сосет) и
ядовитого (R. sceleratus) вызывает
раздражение кожи человека, напо-
минающее ожоги и сопровождающе-
еся образованием волдырей. В опре-
деленных дозах сильно действующие
вещества, содержащиеся в лютико-
вых, оказывают целебное действие
и поэтому нашли применение в меди-
цине. Одно из наиболее важных ле-
карственных растений — горицвет
весенний {Adonis vernalis, рис. 235).
Клубни и другие части акони-
тов содержат аконитин, который
действует в малых дозах болеутоля-
юще; большие дозы могут вызвать
опасные отравления.
Ряд представителей лютиковых
культивируют как декоративные рас-
тения : водосбор (A quilegia), живо-
кость (Delphinium), spfiBBUfiL в зелени» (Nigella), аконит (Aconitum) и др.
Некоторые лютиковые засоряют посевы: живокость полевая (Delphini-
um consolida), мышехвостник (Myosurus minimus), некоторые виды лютиков.
Другие — нежелательный элемент на пастб:
И It
(ах и лугах
3-Й ПОРЯДОК — КУВШИНКОЦВЕТНЫЕ (NYMPHAEALES)
Цветки спиральные, полукруговые или круговые с двойным или простым
околоцветником, с верхней, нижней или переходного положения завязью.
Медовые железки отсутствуют. Многолетние водные травы с атактостеличе-
скими стеблевыми органами. Сосуды- отсутствуют. В паренхимных тканях
имеются членистые млечники. Порядок представлен одним семейством.
Семейство (Nymphaeceae) включает водные и болотные травы с
плавающими, погруженными или возвышающимися над водой листьями и
одиночными, обычно крупными цветками. Цветоложе бывает различное: ко-
роткое, удлиненное или вогнутое. Цветолистья в спиральном, спирально-
циклическом и круговом расположении. Околоцветник двойной или простой,
часто многочленный. Между лепестками (нектарниками у некоторых) и ты-
чинками наблюдаются постепенные переходы. Плодолистики свободные,
иногда погруженные в разросшееся цветоложе или даже срастающееся с
ним.
Это небольшое семейство можно разделить на два подсемейства.
К подсемейству Nymphaeoideaeотносят роды: кубышка (Nu-
phar), кувшинка (Nymphaea), виктория (Victoria). Кубышка желтая (Nuphar
luteum) и кувшинка белая (Nymphaea Candida, рис 236), неправильно именуе-
мая белой водяной лилией, — широко распространены на территории СССР.
ОТДЕЦ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
359
В Северо-Восточной Африке обитают кувшинки ^египетский мтось (N, lo-
tus), одна с белыми, а другая с голубыми цветками (АГ. coerulea). Родствен-
ны кувшинкам виктории, известные крупными, до 2 ле в поперечнике, листья-
ми, плавающими на воде и способными удерживать груз до 50 кг. Цветки вик-
тории крупные, до 35 см в поперечнике. V, amazontca (У. regia) распростра-
нена в водоемах бассейна Амазонки, V. cruciana — в Парагвае. Оба вида
культивируют в бассейнах, в специальных оранжереях.
Название «лотос» обычно связывают с родом Nelumblum, выделяе-
мым в особое подсемейств, о Nelumbonoideae и. содержащим всего
два вида: один (N. lutea), распространенный в Северной Америке, а другой —
лотос индийский (N, nucifera) — растет в теплых и жарких странах Южной
и Восточной Азии, а также Европы.
Растет он^Ъ нас в дельте Волги, в низовьях Куры и на Дальнем Востоке.
В Индии, Китае и Японии его культивируют ради съедобных семян и корне-
из которых изготовляют муку. Листья и цветки индийского лотоса
возвышаются над водой. Цветки имеют удлиненное обратноконусовидное
цветоложе, в которое погружены плодолистики. Гинецей апокарпный.
ип
Плод сборный из нескольких свободных и погруженных в цветочную ось од-
носемянных орешков. Семена очень долго сохраняют всхожесть. Найденные
в плейстоценовых отложениях (около 50 000 лет) в Гоби и высеянные, они
развились в нормальные растения. Корневища, ростки, орешки лотоса
съедобны.
Кувшинковые представляют существенный теоретический интерес, так
как, с одной стороны, ряд признаков сближает эту группу с однодольными
(рассеянное расположение закрытых проводящих пучков на поперечном
разрезе стебля — атактостелия, некоторые особенности образования корне-
вых волосков), с другой стороны, привлекает внимание отсутствие у нимфейных
сосудов, что можно рассматривать, по мнению А. Л. Тахтаджяна, как выра-
жение родственных связей кувшинковых с древнейшими бессосудистыми по-
крытосеменными. О том же свидетельствуют распространенная у них апо-
карпность, наличие переходов между околоцветником и тычинками и ряд
других признаков.
Рис. 236. Белая кувшинка (Nymphaea Candida):
1—плавающий лист; а—бутон; Д-фтснустввшнйся цветок: 4—внутренняя тычинка: 4—лепесткоеидвая
тычинка; а—метик с лучистым рыльцем и спиральным равношюнием рубцов листочков околоцветника
и тычинок
360
ВЫСШИЕ РАСТЕНИЯ
4-Й ПОРЯДОК — МАКОЦВЕТНЫЕ (PAPAVERALES)
Цветки обоеполые, актиноморфные или зигоморфные, как правило, цик-
лические, в большинстве случаев димерные. Гинецеи паракарпный, число
плодолистиков варьирует от 2 до 30, завязь верхняя, одногнездная, с много-
численными семяпочками. Плоды—коробочки, орешки.
Травянистые растения, редко кустарники, даже небольшие деревья с
очередными листьями без прилистников.
Макоцветные но ряду признаков сближают с лютикоцветными.
Семейство маковые (Papaveraceae) — травянистые растения с чле-
нистыми млечными трубками или млечными клетками; если млечного сока
нет, то развиты удлиненные трубчатые клетки с особым содержимым. Цветки
радиально- или билатеральносимметричные, а иногда зигоморфные, как прави-
ло, с двумя рано опадающими чашелистиками и четырьмя лепестками, распо-
ложенными в двухчленных кругах. Тычинок 2,4 или неопределенное число.
Расположены они кругами. Пестик- из двух — нескольких плодолистиков.
Завязь верхняя, одногнездная, с многочисленными семяпочками. Плод —
коробочка или орешек, семена с маслянистым эндоспермом. Семейство мако-
вых включает около 700 видов, обитающих главным образом во внетропичес-
ких областях северного полушария.
В пределах семейства маковых различают три подсемейства, рассматри-
ваемые часто как три самостоятельных семейства (рис. 237).
Для подсемейства Papaveroideae характерно наличие млечных
трубок. Цветок с двучленной, рано опадающей чашечкой, двумя двучленны-
ми кругами лепестков, многочисленными тычинками, число которых кратно
двум, и неопределенным числом плодолистиков, паракарпным гинецеем.
Число плодолистиков разное. Двучленный гинецей имеется, например, в
цветке чистотела (Chelidonium majus), мачков (Glaucium). У представите-
лей рода мак (Ра paver, рис. 238) число плодолистиков больше двух. На-
ряду с димерным типом цветка в этом же подсемействе имеются растения
с цветком тримерным. Так, вроде пдатистемон (Platy stemori), виды
которого обитают в Калифорнии, околоцветник состоит из одного трехчлен-
ного круга чашелистиков и двух трехчленных кругов лепестков. Тычинки
многочисленные, с расширенными нитями. Плодолистиков несколько, срос-
шихся друг с другом в основании и свободных наверху, где они образуют
самостоятельные столбики с медианными рыльцами. Ко времени созревания
плодов плодолистики отделяются Друг от друга. Непрочная ценокарпность
этого растения позволяет сделать вывод о возможной связи маковых с пред-
ками, имевшими апокарпный гинецей.
Кподсемейству Hypecooideae относится только один род Нуре-
соит. Формула цветка К2С Лепестки внутреннего круга обычно
трехраздельные. Млечников нет. '
Виды подсемейства Fumarioideae также не имеют млечников, их
заменяют гомологичные секреторные клетки. Для цветков характерны две
или одна (трансверсальная) плоскость симметрии. За двумя рано спадающи-
ми чашелистиками следуют два двучленных круга лепестков. Наружные
Рис. 237. Диаграмма маковых:
/—мак; а—мачок; а—гипекоум; <—дицентра; а—хохлатка
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСТЙЦГЕННЬТВ
361
либо оба имеют мешковидные
выпячивания (дицентра, или
«пылающее сердце»,— Dicentra),
либо один из них имеет длинный
шпорец (хохлатка — Coryda-
lis}. Лепестки внутреннего кру-
га, медианные по положению, в
верхней своей части вогнутые
со стороны центра цветка и
выпуклые наружу. Эти участки
лепестков обычно прилипают
друг к другу своими краями,
смыкаясь над пыльниками и
рыльцем. Тычинки—в двух трех-
членных пучках, в трансвер-
сальной плоскости. В каждом
пучке средняя тычинка имеет
нормальный четырехкамерный
пыльник, а кроме того, боко-
вые — с двухкамерными, пред-
ставляющими как
винки нормальных. Тычиноч-
ные нити в каждом пучке бо-
лее или менее сросшиеся, но
всегда со свободными участка-
ми наверху. Краевые тычинки
каждого пучка есть основание
рассматривать как возникшие
в результате продольного рас-
щепления двух медианных ты-
чинок цветков предковых форм,
причем продукты расщепления
сблизились с трансверсальны-
бы поло-
Рис. 238. Мак снотворный (Papaver somnife-
rum):
1—цветок и бутон; 2 — яестик; -поперечный разрез
завязи; 4— коробочка; 4—семя; в—продольный разрез
семени (видан зародыш)
ми тычинками и приросли к ним.
Таким образом, формула цветка Fumarioideae, в сущности, та же, что и
Hypecoideae. Papa veroideae, характеризуемые множественностью тычинок,
ка&утся как бы ияолироваштыми. Однако большое число тычинок в цветках
представителей этого подсемейства могло возникнуть и вторично, путем
расщепления. В пользу этого говорит, например, наблюдавшиеся В а р -
м и н г о м аномальные цветки выросшего на тощей почве мака. Число ты-
чинок в них было небольшое, снижаясь иногда до четырех, расположенных
в двух двучленных кругах. Многочисленные плодолистики в цветке мака
могли также возникнуть путем полимеризации или сохранились как сви-
детельство связи маковых с предками, имевшими многочисленный с неуста-
новившимся числом плодолистиковл^М1карпныи гинецей.
Практически важное растение семейства маковых — мак снотворный
(Papaver somntferum). Он известен только в культуре и используется как
масличное и лекарственное растение. Масличные сорта содержат мало млеч-
ного сока, но накапливают в семенах до 40—55% высыхающего жирного
масла, используемого и как пищевое, и как техническое (масляные краски
для живописи). Масличный мак у нас возделывают в Поволжье, на Украине,
в Татарской и Башкирской АССР. В млечном соке мака содержится опиум.
Опиум и получаемый из него пантопон, равно как и отдельные входящие в
опиум алкалоиды (морфин, наркотин, папаверин, кодеин и др.), получили
широкое применение в медицине. Специальные опийные сорта мака разводят
вэ многих странах Азии (от Ирана на западе до Японии, Китая и Индонезии
на востоке). В СССР культура опийного мака сосредоточена в Киргизской
и отчасти Казахской ССР.
362
ВЫСШИЕ РАСТЕНИЯ
Оранжевый млечный сок чистотела (Chelidonium majus) также приме-
няют в народной медицине, в частности для лечения бородавок. Дымянка (Fur
maria) — сорное растение.
5-Я ПОРЯДОК — ПЕРЕЧНОЦВЕТНЫЕ (PIPERALES)
Порядок характеризуется мелкими обоеполыми или раздельнополыми
цветками, без околоцветника или с простым околоцветником. Цветки часто
в колосовидных соцветиях.
В относящемся сюда семействе перечных (Piperaceae) находит мес-
то перец черный (Piper nigrum) — лазающий кустарник родом из тропиков
Азии. Его незрелые плоды высушивают, при этом они чернеют. Этот продукт
используют как пряность. Листья другого вида (Р. betle) вместе с кусочками
семян пальмы Агеса и с известью на юге и юго-востоке Азии используют для
жевания (возбуждающее средство).
6-Я ПОРЯДОК — КИРКАЗОНОЦВЕТНЫЕ (AR1STOLOCH 1ALES)
Цветки круговые, большей частью с простым венчиковидным околоцвет-
ником; тычинки многочисленные, гинецей обычно цеиокарпный, но плодо-
листики почти свободные.
Из семейств этого порядка отметим семейство кирказоновых
(Aristolochiaceae)9Kyj&L относятся кирказон (Aristolochia) и копытень (Asarum).
У копытепя цветки правильные, с нижней завйзью. Копытень европейский
(Asarum еигораеит) — характерный представитель флоры широколиствен-
ных лесов.
Семейство раффлезиееые (Rafflesiaceae) объединяет паразитные
бесхлорофилльные растения, живущие на корнях и стеблях тропических де-
ревьев. Вегетативное тело этих растений редуцировано иногда до гифоподоб-
ного состояния, а цветочные побеги закладываются внутри ткани хозяина.
Среди этих паразитов знамениты такие раффлезии, как Rafjlesia аг-
noldii (Индонезия — на Суматре) с цветками до метра в поперечнике,
R. patma (Индонезия — на Яве) и др.
7-Й ПОРЯДОК — НЕПЕНТЕСОЦВЕТНЫЕ (NEPENTHALES)
Полукустарники или травы. Листья спиральные, нерасчлененные, со
своеобразными приспособлениями к ловле насекомых. Цветки правильные,
круговые или спирально-круговые, с двойным или простым околоцветником
и изменчивым числом тычинок. Гинецей ценокарпный, завязь верхняя.
Сюда относится тропическое семейство непентесовых (N epenthaceae)
с одним родом Nepenthes^ распространенным на Мадагаскаре, в Се-
верной Австралии и Новой Каледонии. Листья непентесов сильно метамор-
физированы и представляют кувшинообразные органы, служащие для ловли
и переваривания насекомых. Подобные образования наблюдаются у предста-
вителей семейства саррацениевые (Sarraceniaceae), 12 видов которого обита-
ют на болотах Северной Америки, а также в Гвиане и Венесуэле.
Приведенные сведения показывают, что в пределах подкласса Magnoliidae
есть формы как с примитивным, так и специализированным строением цветка
(семейства Papaveraceae и др.), причем имеется много представителей,
связывающих крайние типы. Учитывая постепенность переходов и некоторые
другие признаки относящихся сюда растений, можно составить представле-
ние о ходе эволюции покрытосеменных в начале их развития. По-видимому,
уже на ранних этапах эволюция покрытосеменных шла несколько параллель-
ными рядами, в каждом из которых цветок изменялся от ациклического до
кругового, а в некоторых рядах выделялись ветви, приведшие к цветку зи-
гоморфного строения, что, несомненно, было связано с совершенствованием
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
363
энтомофилии; в некоторых случаях энтомофилия была утеряна и растения
стали опыляться ветром (род Thalictrum семейства Ranunculaceae).
В Процессе дальнейшей эволюции Magnoliidae наряду с циклическими
пентамерными цветками появлялись тримерные, напоминающие цветки одно-
дольных. Сопоставляя это с тем, что у некоторых многоплодниковых наб*
людаются закрытые пучки с рассеянным их расположением, можно предполо-
жить, что однодольные в своем происхождении связаны с Magnoliidae.
В процессе эволюции в пределах подкласса, несомненно, происходило
обособление некоторых групп не только на морфологической, но и на биохи-
мической основе. Так, семейства маковых и лютиковых приобрели способ-
ность вырабатывать ядовитые вещества. Однако химизм этих веществ у обеих
групп различен.
Наиболее полный ассортимент примитивных признаков сосредоточен в
порядке Magnoliales. Но разбросанность их среди семейств и родов этого по-
рядка заставляет с осторожностью относиться к широко распространенному
мнению об этой группе как о предковой по отношению ко всему подклассу
Magnoliidae и вообще к покрытосеменным. Не следует отрицать возможность
i: । j
и того, что магнолиецветным предшествовала ныне полностью вымершая
достаточно монолитная группа первичных покрытосеменных, по отношсе
к которым Magnolia и другие порядки подкласса Magnoliidae представляют
ь конечные звенья длительного эволюционного пути, в которых как ана-
хронизмы сохранились то те, то иные признаки и свойства древнейших пок-
51Н
рытосеменных.
Родственные отношения между порядками, относимыми к подклассу
Magnoliidae, не всегда ясны. Черты примитивной организации цветка со-
хранились как у древесных Magnoliales, так и у травянистых (в основном)
Ranunculales и Nymphaeales. Поэтому не исключена возможность парал-
лельного развития этих двух групп и происхождение их от общего предка, ко-
торый связывал весь подкласс Magnoliidae с гипотетическими предками
покрытосеменных. Каждый из этих базальных порядков дал начало дру-
гим, хотя и сохранившим некоторые пр
во многом достаточно далеко ушедшим от их прародичей. Так, есть основа-
ния связывать по происхождению Piperales с их редуцированными цветками
и Arlstolochiales, приобретшими сложную приспособительную структуру спо-
рэфилльной области цветка (гиностемий), с Magnoliales. Что касается тра-
вянистой части подкласса, то порядки Ranunculales и Nymphaeales, вероятно,
прошли свою историю независимо и параллельно друг другу, но в разных
средах. С Ranunculales в качестве их потомков связываются Papaverales,
эволюция которых завершается зигоморфными дымянковыми, и Nepenthales,
развивавшиеся по пути насекомоядности. Nymphaeales если и возникли от
общего с Ranunculales предка, то от очень отдаленного, древнего покрыто-
семенного, не связанного с водной средой. Водные потомки этого древнего
предка как бы законсервировали в себе ряд структур древнего этапа исто-
рии покрытосеменных, в частности бессосудистую древесину, состоящую из
длинных спиральных и кольчатых трахеид. Наземные предки Nymphaeales
могли стать родоначальниками класса однодольных, о чем свидетельствуют
анатомическая структура стебля, развитие и строение зародыша1 а также не-
которые другие особенности нимфейных.
2-Й ПОДКЛАСС - ГАМАМЕЛИДНЫЕ (HAMAMEL1DAE)
Гамамелцдяые характеризуются всегда циклическим строением цветков,
часто однополых, большей частью анемофильных, мелких, невзрачных, со
слаборазвитым или отсутствующим околоцветником. Обычно цветки собра-
ны в соцветия. Гинецей ценокарпный. Зрелые пыльцевые зерна двухклеточ-
ные, семяпочки обычно с двойным интегументом, в большинстве случаев крас-
синуцеллятные. Большей частью древесные растения. В основном этот под-
класс соответствует однопокровным раздельнолепестным (Monochlamydeae).
I
364
ВЫСШИЕ РАСТЕНИЯ
В состав подкласса Hamamelidae в зависимости от теоретических
представлений систематики включают от 9 до 15 порядков. Ниже, из числа, *
бесспорно, принадлежащих к подклассу Hamamelldae, рассматриваются
порядки: гамамелидоцветные (Hamamelidales), орехоцветные (Juglan-
dales), букоцветные (Fagales), крапивоцветные (Urticales) и казуариноцвет-
ные (Casuar inales), упоминается также порядок троходендроцветные (Troche-
dendrales). К этому же подклассу условно причислены порядки
ивоцветные (Salicales) и протейные (Proteales)1.
1-Й ПОРЯДОК — ГАМАМЕЛИДОЦВЕТНЫЕ (НАМАМЕ LIDALES)
Гамамелидоцветные — деревья и кустарники с простыми очередными,
реже супротивными листьями, имеющими почти всегда прилистники. Цвет-
ки без околоцветника, с простым или двойным околоцветником, однополые
или двуполые.
В каждом цветке два плодолистика, редко один или более двух; гинецей
апокарпный или силка рпный; в каждом гнезде от одной до нескольких се-
мяпочек.
Семейство гамамелидовые (Hamamelidaceae) — листопадные-веч-
нозеленые деревья или кустарники с очередными, обычно простыми листьями,
почти всегда с прилистниками. Цветки в ботрических соцветиях мелкие,
различного строения; обоеполые или однополые, с двойным или простым око-
лоцветником или голые; преобладают цветки четырех- пятичленные, круго-
вые; число тычинок может быть велико. Гинецей почти всегда из двух плодо-
листиков, обычно синкарпный; завязь двугнездная, с центрально-угловой
плацентацией, нижняя, полунижняя или верхняя, с одним, реже большим
числом семязачатков в каждом гнезде. Плод— коробочка.
Около 100 видов, преимущественно в субтропических странах, главным
образом в Восточной и Юго-Восточной Азии. Заслуживает упоминания род
ликвидамбар (Liduidambar), древность которого, по палеонтологическим дан-
ным, восходит к меловому периоду. В неогене представители рода были широ-
ко распространены по всей Арктотретичной области. Из современных видов
отметим ликвидамбар восточный (L. orient alls), обитающий в Малой Азии, и
распространенный в юго-восточной приатлантической Северной Америке
ликвидамбар смолоносный (L. styraciflua) — характерное дерево, достигаю-
щее 45 м высоты; в СССР на Черноморском побережье Кавказа и Крыма ви-
ды р о д а Liduidambar разводят как декоративные растения. Декоративное
значение имеют также красиво цветущие ранней весной, до распускания
листьев, кустарники из родов гамамелис (Hamamelis), корилопсис (Cory*
lopsis) и др., разводимые у нас во влажных субтропических условиях по-
бережья Черного моря (Батуми — Сухуми). Одно из наиболее крупных де-
ревьев тропических лесов Явы (Индонезия) разамала (Altingia excelsa) — тоже
представитель семейства гамамелидовых. Это дерево достигает 80 м высоты и
2 м толщины; округлая крона его начинается иногда на высоте около 50 м.
Твердая древесина разамалы с бальзамическим запахом высоко ценится, 1 * * * * * * В
1 Порядок Salicales часто сближают с Tamaricales, а порядок Proteales — с Rosales
или Saxifragales, т. е. рассматривают их как группы, относящиеся к разным направлени-
ям эволюции двудольных. По Тахтаджяну, например, ивоцветные относятся к Duleniidae,
а протейные — к Rosidae* Вопрос о положении этих двух порядков ь системе решается
учеными по-разному. Гетчинсон (1969) видит родственную связь ивоцветных с
гамамелцдоцветными. Выводи протейные из камнеломкоцветных (Тахтаджян) или
из розоцветных (Кронквист), авторы сближают протейные с группами, непосредствен-
но примыкающими к Magnoliidae. Более радикальное мнение было высказано Галли-
ром (1912), выводившим протейные как самостоятельную ветвь, параллельную Ало-
nales из Proberbertdeae. Может быть, правильнее было бы сохранить положение ивоцвет-
ных и протейных в качестве самостоятельных линий развития (подклассов), берущих свое
начало от общих предков цветковых, как это было сделано в предыдущем издании данной
книги. Но с педагогической точки зрения увеличение числа подклассов нежелательно,
и мы присоединили оба порядка к Hamamelidae, опираясь на филему Шаффнера,
В которой они сближаются с букоцветными, крапивоцветными и другими однопокровными«
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
365
В природной флоре СССР семейство гамамелидовых представлено только
железняком (Parrotia per sic а), растущим на юго-востоке Закавказья, куда
он заходит из Ирана* Дерево имеет оригинальную внешность благодаря спо-
собности ветвей срастаться друг с другом и с ветвями соседних экземпляров;
иногда срастаются даже соседние стволы. Железняк растет медленно, дре-
весина его очень мелкослойная, плотная, тяжелая (удельный вес более 1),
прочная, красноватого цвета, употребляется в столярном и токарном деле*
Семейство платановые (Platanaceae) — деревья с очередными паль-
чатолопастными листьями, имеющими крупные прилистники, которые сраста-
ются друг с другом вокруг стебля. Цветки однополые, нередко с рудимента-
ми противоположного пола, с двойным околоцветником, невзрачные (ане-
мофилия), состоят из трех четырехчленных кругов. Гинецей апокарпный*
В завязях по одной, реже по две семяпочки* Соцветия головчатые, и каждое
содержит цветки только одного пола. Растения однодомные* Каждая цвето-
носная ветвь состоит в нижней части из коротких междоузлий и покрыта
здесь нормально развитыми листьями, верхняя ее часть образована
удлиненными междоузлиями, л:
Ulf
ена ассимилирующих листьев и несет на
себе соцветия* Плоды платановых — орешки*
К этому семейству относится только один род платан, или чинара
{Platanus), содержащий всего 8 видов. В СССР растут два вида: один — в
Закавказье {Platanus digitata), другой — на юге Средней Азии {платан
восточный — Platanus orientalist
Семейство эйкомиевые {Eucomiaceae) представлено одним видом —
эйкомией вязолистной (Encomia ulmoides), распространенной в горных райо-
нах Центрального и Западного Китая* Это дерево ценится как гуттаперче-
носное. В СССР разводят в Закавказье*
В итоге краткого описания порядка Hamamelidales следует подчеркнуть
наличие признаков примитивности в строении цветка: апокарпность гине-
цея или неполное срастание плодолистиков, низбегающие рыльца* Это сви-
детельствует о вероятной связи данного порядка с апокарпными предками
покрытосеменных* В пользу этого говорит также близость Hamamelidales
с порядком Trochodendrales, который ряд ботаников сближают с магнолие-
цветными. С другой стороны, малое число плодолистиков, простой около-
цветник или его отсутствие, а также характерная для многих гамамелидо-
цветных однополость цветков сближают этот порядок с березовыми, буковы-
ми и вообще с так называемыми «сережкоцветными» покрытосеменными*
По другому представлению, Hamamelidales тесно связан с порядком
Rosales.
2-Й ПОРЯДОК — БУКОЦВЕТНЫЕ (FAG ALES)
Цветки букоцветных невзрачные, мелкие, без околоцветника jm^jc про-
стым, слаборазвитым околоцветником;" однополые, только редко обоеполые,
в сложных, часто сережковидных или другой формы соцветиях, обычно со-
ставленных из дихазиев. В~мужских цветках 2-—4 иТюлыпе тычинок, проти-
востоящих листочкам околоцветника; в женских — пестик из двух или боль-
шего числа плодолистиков* Завязь нижняя, одно- шестигнездная, с одной-
двумя семяпочками в каждом гнезде. Плод сухой, вследствие недоразвития
других семяпочек односемянный, нераскрывающиися. Ветроопыляемые рас-
тения, иногда халазогамные или с зндотропным ростом пыльцевой трубки.
Деревья и кустарники с очередными листьями, имеющими прилистни-
ки, обычно рано опадающие.
Упомянутые в характеристике порядка дихазиальные соцветия типич-
ны для относящихся сюда растений* Дихазий букоцветных (рис* 239)
всегда занимает боковое положение по отношению к главной оси всего
сложного соцв&гия* Ось первого порядка самого дихазия выходит из
< пазухи кроющего листа (Ь) и * заканчивается первым цветком (I). Эта ось
вместе с завершающим ее цветком располагается в медианной плоскости и
1
!
ВЫСШИЕ РАСТЕНИЯ
3
Рис* 239. Схема дихазиального ветв-
ления соцветия
несет на себе ниже цветка два прицвет-
ных чешуйчатых листа. Они располага-
ются в трансверсальной плоскости или
несколько смещены в сторону кроющего
листа (Ь)* Эти листья, прицветнички пер-
вого цветка дихазия, обозначаемые в диа-
граммах аир, являются в то же время
кроющими листьями боковых разветвле-
ний дихазия, т. е. его осей второго
порядка* Последние, в свою очередь,
заканчиваются цветками (2), но под ними,
как и на оси первого порядка, располага-
ются по два прицветничка боковых цвет-
ков, обозначаемые кака1 и а1 ир1.
Из пазух этих листочков опять могут
развиться боковые оси, уже третьего
порядка, также заканчивающиеся цвет-,
ками (3). Более трех ветвлений в дихази-
ях букоцветных не наблюдается. Оси всех
порядков сильно укорочены, вследствие чего цветки дихазия образуют тесную
группу. Нередко бывает, что ось первого порядка заканчивает свой рост, не
развивая цветка, хотя соответствующие прицветнички ирх развиваются.
С другой стороны, на осях второго и третьего порядков, при наличии на них
цветков, могут оказаться редуцированными либо один, либо оба прицветнич-
ка (а1 и pz). Может оказаться редуцированным и кроющий лист дихазия (Ь).
Бывают случаи, когда ось первого порядка не ветвится, тогда дихазий стано-
вится одноцветковым*
Таким образом, дихазии отдельных представителей букоцветных могут
различаться числом цветков, количеством прицветных листьев, а также ха-
рактером последних, их величиной, формой и т. п*
Семейство березовые (Betulaceqe)* Это деревья и кустарники с прос-
тыми цельными листьями, рано опадающими прилистниками* Цветки в
сложных (из дихазиев) соцветиях, однополые, однодомные^ сложенные из
двучленных кругов. Мужские цветки иногда сросшиеся с кроющим „вдетом
^дихазия^ Околоцветник из четырех сросшихся у основаншГи ля из -двух
супротивных медианных листочков, невзрачный или совсем отсутствует.
Тычинки часто в числе листочйов околоцветника противостоят им; нити их
часто расщепленные, И каждая половинка несет свой' пыльник..Женские
цветки без околоцветника, Или-он сильно редуцирован* Пестик из двух
плодолистиков, с двумя рыльцамй и не Вполне двугнездной: завязью,^
одной семяпочкой в каждом гнезде. Плод —орех*. -
Типичные представители, семейства, "включающего более 100. видов,
роды; ольха (Alnus), береза (Betula) и лещина, или орешник (Corylus).
Мужские дихазии ольхи (Alms glut inosa, рис. 240) собраны в сережки.
Каждый дихазий имеет три цветка с четырехлистным околоцветником и че-
тырьмя тычинками, противолежащими листочками околоцветника; нити
ш*
листьев, причем. листья а, 0, 01, 0г смещены к кроющему листу j(b). Женские
цветки без околоцветника и содержат каждый по одному пестику^ несущему
по два нитевидных рыльца. Женские диха!№1^вдуцветковые"'?^веледствие
редукции первого цветка), собраны в соцветия в виде небольших шишечек.
Пять, как и в мужском соцветии, прицветных листьев срастаются в пятило-
пастную чешую, которая; древеснеет и разрастается ко времени созревания
пл о до в( ©реджи), а после рассеивания последних шишечка размером до 1,5 см
темно-бурой окраски долго сохраняется на растении.
У березы (род Betula, рис. 241) и мужские и женские соцветия в виде
сережек* В каждом мужском дихазии три цветка, причем боковые цветки
своих лрицветничков не имеют, и поэтому при цветках одного дихазия имеют-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
367
ся только три прицветных лис-
та: Ь, а и р. В состав мужского
цветка^ входят два медианно
расположенных листочка око-
лоцветника, причем задний раз*
вит меньше переднего и даже
может совсем отсутствовать.
Супротивные им тычинки рас-
щеплены вдоль и несут на каж-
дой из двух ветвей по пыльни-
ку, поэтому создается впечат-
ление, что в цветке четыре ты-
чинки. Женский дихазий тоже
трехцветковый лс тремя при-
цветными листьями, которые
срастаются в трехлопастную
чешуйку. Женский цветок jae
имеет околоцветника и состоит
только из пестика с двумя_рыль-
цами. Из завязи сплюснутой
формы развивается . орешек,
имеющий по краям крыловид-
ные пленчатые выросты.
У лещин» {род Corylus,
рис. 242) — сережчатые муж-
ские соцветия. В пазухе каж-
дого кроющего листа сидит цве-
ток £ двумя прицветниками, ко-
Рис. 240. Ольха черная (Alnus glutinosa):
1 — ыпшйшя ветвь с нлодущимв шишечками; а —
ветвь с мужскими и женскими соцветиями (ж—муж-
ские, ж—женские соцветия); а—трехцветковый диха-
зий из тычиночных цветков; 4—диаграмма мужского
дихазия;- 5—двухцветковый дихазий из пестичных
цветков; а—диаграмма женского дихазия
торые срастаются с кроющим
листом^ Боковых "ТЩСТКОИ" нет;
Таким образом, здесь дихазий
одноцветковый. Мужской цве-
ток не имеет околоцветника и
состоит из четырех двурасщеп-
ленных тычинок. ' Вследствие
иг,
fll
интеркалярного роста у ос-
нования кроющего листа дихазия. (Ь) цветок оказывается смещенным
вверх и прикрепленным к верхней поверхности трехлопастной чешуи,
образованной кроющим листом и двумя прицв&тничками. Женскцо7дахав5и‘
дещины^аходятся внутри почки, а во время пдатйипя наружу"выявляются
только рыйПЩГКрб^^ дихазиальных групп женских цветков
служат молодыетйгстья, спрэтанныё под лодочными чешуями.Х^реднйй~ цвён
ток дихазия не развивается, но его прицветнички, служащие кроющими -
листьями боковых цветков, имеются. Боковые цветки имеют по дв а своих
прицветника; Таким образом, вокруг. каждого жежжого^цветканаходятся
по три небольших прицветных листа (а, а1,01 у одного и 0 х, 0 у другого
цветка дихазия), которые срастаются вместе, разрастаются ко времени со-
зревания ореха и образуют вокруг ней) .характерный покров, называемый"
плю с к о й . Женский цветок образован пестиком ця двух плодолистиков,
причем на верхушке завязи можно найти крайне редуцированный около-
цветник (завязь, ‘следовательно, нижняя).
Среди березовых существенное хозяйственное значение имеют прежде
всего березы. Так называемая белая береза представляет сборное понятие,
обнимающее даа вида: бородавчатую (Be tula, verrucosa, lisni В, pendula)
и nyuiucmyia.^cp^y (В. pubewentyTi экономическом отношении важнее пер-
вая. Дребёсинаг березы бородавчатой высоко ценится как поделочный матери-
ал и топливо. Кора используется щжлуёлении кожи.и.при.выгонке дегтя.
Из бересты изготовляют различные предметы домашнего обихода. Цешым
368
ВЫ
И11 г.
В РАСТЕНИЯ
Рис. 241. Береза повислая, или бородавчатая
(Betula pendula):
1—ветвь с женской сережкой; а—мужские сережки;
3—трехцветковый женский дихазий: а—со стороны оси
сережки; б—со стороны кроющего листа; 4—плод;
з—крокицая чешуйка плода; 6—диаграмма мужского
дихазия; 7—диаграмма женского дихазия
поделочным материалом счита-
ется так называемый кап —
большие. _наросты„на стволах,
крупных сучьях, корнях. - Срез
нароста дает сложный й "Краси-
вый рисунок, почему его ис-
пользуют для изготовления
портсигаров, коробочек- и дру-
гих изящных изделий. Образова-
ние капа связано с обильным
ветвлением и развитием спящих
почек,. Причины, вызывающие
образование капа, остаются
пока невыясненными. Особую
форму бородавчатой березы
представляет карельская бере>-
правильным, извилистым рас-
положением годичных.слоев и
сердцевинных лучей. Она вы-
соко ценится как поделочный
материал и в мебельном про-
изводстве.
Берега белая (В, alba) —
характерное ландшафтное де-
рево, встречающееся в листвен-
ных и смешенных лееах и об-
разующее чистые насаждения.
Березовые леса в Европейской
части СССР обычно имеют вто-
ричный характер, т. е. возни-
кают на месте сведенных лесов
другого типа. Но в Предуралье
и особенно в Западной Сибири
березовые леса в виде неболь-
ших рощ (колки) разбросаны
по северной части степной зоны, создавая вместе со степными сообщест-
вами характерный естественный ландшафт сибирской березовой лесостепи.
В других районах есть свои типичные и широко распространенные
виды берез. Таковы, например, каменн$я_берега (В. ermani) — в Восточной
Сибири и на Дальнем Востоке, даурская берега (В. dahurica) — в Забайкалье
и других районах Восточной Сибири. Есть и кустарниковые виды берез,
особенно своеобразна стелющаяся.бергза карликовая (В. папа), типичная для
тундры, но встречающаяся и южнее, в лесной зоне по торфяным болотам.
Из рода ольха (Alnus) достойны внимания черная и серая ольха.
Ольху серую (Л. incana) используют в столярном и токарном деле. Древесина
черной, или клейкой, ольхи (Л. glutinosa) довольно хрупкая, но оказывается
более прочно? в воде, и поэтому ее употребляют на сваи, водопроводные
желоба, колодезныехрубы, и т. п. Кору черной ольхи иногда используют
для окраски кожи и тканей.
Лещина, или орешник (Corylus avellana, рис. 242), — характерное^! ши-
роко распространенное растение подлеска лиственных (главным образом
дубовых и липовых) и отчасти хвойных лесов. Ее плоды — лесные орехи —
съедобны. Этот кустарник распространен в европейской части СССР, а также
в Западной Европе и Малой Азии. В пределах нашей страны площадь,
занятая дикой лещиной, измеряется сотнями тысяч гектаров. В семенах
лещины содержится от 60 до 70% изболев очень ценного масла, еще не на-
шедшего у нас должного использования в пищевой промышленности.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
369
Рис. 242. Лещина (Corylus avellana):
1—опиственная ветвь; а—ветвь с мужскими и женски-
ми соцветиями (м—мужские, ж—женские соцветия);
з—кроющая чепгуя и тычиночный цветок; 4—диаграм-
ма мужского «одноцветкового дихазия»; 5 — начавший
развиваться плод, окруженный прицветными листья-
ми; в—диаграмма женского дихазия; 7—плоды
Рис. 243. Дуб черешчатый (Quercus robur):
I—ветвь с мужскими сережками; з—часть мужской
сережки; 3—тычиночный цветок; 4—пестичный цветок;
5—продольный разрез пестичного цветка (а — завязь»
ст—столбик, р—рыльце, с—семяпочки, ок—•околоцвет-
ник, о—плюска); 6—ветвь с плодами
Распространенная на Кавказе, в Малой Азии, Иране, Гималаях*. Китае
лещина древовидная (С. colurna) представляет дерево 25 "ж й более высотой
и свыше 30 см в диаметре. Под именем ломбардского (орВаг (фундук) известна
лещина, растущая то кустарником, товвиде небольшого дерева (С. maxima),
родом с Балканского полуострова и из Малой Азии. Эти три вида, а также
понтийская и американская лещины введены в культуру в ряде стран (Ита-
лия, Турция, Испания, Южная Франция; у нас — на небольшой площади
в некоторых районах Кавказа). Известно немало культурных сортов.
Один из видов рода граб — граб обыкновенный (Carpinus betulus) —
обитает в Крыму, на Кавказе, в западных районах СССР, в Иране и Западной
Европе. Он является важным лесообразующим деревом, замечателен тене-
выносливостью и отличается очень прочной древесиной, которая, однако,
вследствие кривизны стволов малопригодна для строительных целей. Но дре-
весина граба служит, ценным материалом для поделок и используют ее
особенно там, где требуется большое сопротивление трению. Кору применяют
для дубления, а внутренние ее слой — для окрашивания шерсти в желтый
цвет. Ценится граб и как топливо. В нижнем поясе гор Крыма и Кавказа,
по лесным опушкам и сухим каменистым склонам, часто растет восточный
граб (С. orientalis) — небольшое деревце или кустарник с серой корой и
370
выс
иг
В РАСТЕНИЯ
Рис. 244. Каштан (Castanea vulgaris):
1—цветущая ветвь; 2—пледы
сравнительно мелкими листья-
ми. Это чрезвычайно неприхот-
ливое растение, густо олист-
венное и обильно ветвящееся,
может быть использовано для
живых изгородей, а древесина
его — на мелкие поделки.
Семейство буковые
(Fagaceae). Это деревья с оче-
редными, простыми, _ то цель-
ными, то лопастными илц в
разной степени расчлененными
пластинками листьев, с опада-
ющими прилистниками*, вечно-
зеленые и листопадные. -Соцве-
тия сережковйдные^ли ддагпвчэ-
тые, цветки однополые, однодом-
ные; редко двуполые, распо-
ложены на главной оси -соцве-
тия „дихазиальными группами
или одиночно (вероятно, й этом
случае боковые цветки диха-
зиев не развиваются). Мужские
цветки имеют околоцветник из
трезэлешшмфухов, но иногда
число листочков околоцветника
неощйщелеиное и не кратное
трем. Тычинки . нерасщеплен-
ные, _ в числе, равном числу
листочков околоцветника или
большем. Женские цветки име-
ют надпестичный тпимерный
околоцветник и пестик из трех или большего числа плодоЙист&бИГ'Завязк
трех-шестигнездная (редко двугнездная), с двумя семяпочками в каждом
гнезде. Плод — орех, окруженный у основания особым покровом — плюской,
или два-три плода погружены вместе в общую плюску.
Семейство включает 600—900 видов, распространенных в странах умерен-
ной и отчасти тропической зоны.
Широко известен из этого семейства род дуб (Quercus), один из видов
которого — дуб черешчатый^фиегсиз robur, рис. 243) — характернейшая ле-
сообразующая ЪЬркщагйбны лиственных лесов на равнинах Восточной Евро-
пы. Дуб цветет во время распускания листьев, из пазух которых вырастают
мужские и-женские редкоцветные сережки. Мужской ^веток^ располагается
в пазухе чешуйчатого кроющего листа и имеет сростнолистный шести-семи-
- , и шесть (не всегда) тычинок. .В женском соцветии
цветки расположены..на .главной оси сережки поодиночке, но каждый
цветок окружен в основании широкобокальчатым образованием, вероятно,
осевого ^происхождения. На поверхности этого бокала имеются выросты —
—недоразвитые листья. В состав женского" цветка входят шестичленнрй,
Двукруговой, но^очёнь^слабораз витый ^адНестйчнъ& около цветник и .пестик
с дрехлопастным рыльцем, столбиком и трехгнездной завязью, содержащей
по два*“сёмязачатка иЧсаждом гнеЗДё. Плбд" дуба — желудь. В основании
желудь окружен чашевидшш^бугорчатойна наружной стороне плюской,
развивающейся из упомянутого широкобокельчатогорасширения_оси, несу-
щей женский цветок. В желуде находится только одно семя, так как опло-
дотворение осуществляется только в одной семяпочке, и поэтому другие
не образуют семян. Семд без эндосперма. Зародыш с массивными семядолями,
богатыми крахмалом. ’
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
371
Рис. 245. Диаграммы женских дихазиев буковых:
1—каштан; а—бук; 3—дуб (fr—кроющий лист первого цветка, а и р—прицветники первого цветка, а’. ₽’
«1»₽* — прицветники боковых цветков (для дуба предположительно)
Род дуб (Quercus) содержит окодр200 видов (в том числе 18 видов в
СССР), многие из которых — кру
и прочную древесину. Кора идет на дублепйё~кож;из желудей изготовляют
суррогат кофе. Перидерму пробкового дуба (Quercus suber).используют-для
выделки пробки. Пробковый дуб растет в странах Западного Средиземно-
морья; может культивироваться и у нас в Закавказье и Крыму.
Близок к дубу род лазания (Pasania), 100 видов, произрастает главным
образом в Инд
и на Зондских островах.
Отличается от указанных родов строением соцветий другой представи-
тель семейства — р а.д~хсаишиш- (Castanea). Настоящий каштан (Castanea
sativa) растет в Южной Европе и у нас в Закавказье (рис." 244); Это высокое
j I s 1:
а: I
дерево с крупными, продолговатыми, плотными, цельными листьями, круп-
нозубчатымй по краю. Мужские соцветия представляют,колрсовидные_обра-
зования, г состоящие из^ торчащей вверх главной оси и расположенных на
ней мужских цветков, размещенных трех- семицветковыми дихазиями. Муж-
ские цветки ймёют^шестичленный, двухкруговой околоцветник и несколько
(до 12) тычинок. !Г пентрёТдветка"замётен рулплитдТподтмтся. Иногда встре-
ча ю^ся-обоеполые соцветия, некоторых "на главной оси внизу располагаются
жецские, а наверху — мужские дихазии. В этих случаях в пограничной зоне
между теми и другими можно наблюдать и обоеполые цветки. Женские
цветки собраны в типичные дихазиальные трехцветковые соцветия (рис.245,1),
сидящие в пазухах крокйщх^шстьев^^б). Средний пветок. лихазия-дмеет
припветнички а и0. в~ пазухах/которых располагаются .боковыеипшгки.
Весь дихазии окружен массивной плюской, почти доверху обнимающей
цветки, котофая^при созревании плодов’расщепляется на четыре створки,
покрытые крупными оСтрыми*нагкоЯцах выростами. Положение створок плю-
ски вполне соответствует прицвётничкам (а и 0) боковых цветков, поэтому
иногда считают, что в данном случае плюска имеет листовое происхождение.
В составе женского цветка имеется шесть листочков околоцветника, за ко-
торым иногда следует шесть стаминодиев. Пестик с треугольной
в разрезе нижней завязью образован 3, 6 и более плодолистиками, в связи с
чем число рылец непостоянно. “ '
Каштан ценится как плодовое. дерево^Еголкрехи^каштаны) употребляют
в цищу в сыром, вареном и печеном виде; мука из орехов идет на изготовле-
ние суррогата кофе. Древесина прочная, ценится как строевой лес и исполь-
зуется в столярном деле. В коре много дубильных веществ.
Аналогично каштану построены женские дихазциу бука(рис.4246^-2), но
в этом случае, во-первых, нет прицветничков_среднего ^цветка; во-вторых,
дихазий двухцветковый вследствие недоразвития, среднего цветка; в-третьих,
кнаружи от каждой из четырех створок плюски располагается еще
по маленькому листочку. Некоторые морфологи считают их за выросто-
вые образования, а плюску рассматривают как орган, образованный сраста-
Рис. 246. Бук (Fagus silvatica):
I—цветущая ветвь; 2—плодоносящая ветвь
с выступающими из плюски орехами
нием прицветничков, притом не
только у каштана и бука, но и
у дуба. Другие — принимают
упомянутые добавочные листья
в дихазии бука за прицветнич-
ки, а плюску у всех буковых
истолковывают как орган стеб-
левого происхождения.
Виды рода бук (Fagus) —
обитатели северных умеренных
широт Евразии и восточной час-
ти Северной Америки (рис. 246).
Пример — бук лесной, широко
распространенный в Западной
Европе, являющийся там одной
из основных лесбобразующих по-
род, заходящий на территорию
СССР (юго-запад европейской
части и Крым). На Кавказе, а
также в Крыму обычен близ-
кий к европейскому виду бук
восточный (F. orientalis). Древеси-
на бука очень ценится, так как
дает прекрасный материал для
столярных изделий, особенно ме-
бели; используется также на
постройки и топливо, обладает
высокой теплотворной способ-
ностью. Плоды бука съедобны.
Они содержат много масла, год-
ного в пищу.
Родственный букам южный бук (род Nothojagusj, встречающийся в
южном полушарии — в Южных Андах, Новой Гвинее, в Юго-Восточной
Австралии, Тасмании, Новой Каледонии, Новой Зеландии. В Чили некото-
рые виды поднимаются высоко в горы.
У некоторых букоцветных (каштан, реже бук) развиваются обоеполые
цветки, а в мужских цветках каштана и пазании имеется рудимент пестика.
Наряду с характерным для^орядкецветроопылением у каштана наблюдается
опыление .жуками, известно также посещение насекомыми цветков дуба.
Все это заставляет думать, что у предков букоцветных были обоеполые
энтомофильные цветки и что, следовательно, сами букоцветные — формы
производные, а простота строения их цветков и ветроопыление вторичны.
Производный характер букоцветных подтверждается также наличием у них
нижней завязи. Наряду с этим при оценке положения букоцветных в системе
надо учитывать, что эти растения очень древние; ископаемые остатки буко-
вых, в частности дуба, находят в меловых отложениях Северной Америки.
О древности букоцветных говорят и некоторые особенности, которые рассмат-
риваются как архаизмы, например многоклеточный археспорий (граб, дуб,
каштан, бук, лещина), заложение нескольких зарода
чн
евых мешков, хала-
зогамия. В свете этих данных сомнительно происхождение букоцветных от
высокоспециализированных форм. Более вероятны их генетические связи с
порядком Hamameli dales.
3-Й ПОРЯДОК — ОРЕХОЦВЕТНЫЕ (JUG LAN DALES)
Цветки орехоцветных однополые, ветроопыляемые, в колосьях или более
сложных соцветиях. Околоцветник простой, чашечковидный, иногда отсутст-
вует. Мужские цветки с 2—40 тычинками. Пестик из двух плодолистиков.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
373
Завязь одногнездная, содержит одну основную семяпочку с одним интегу-
ментом. По положению она нижняя, сросшаяся с покровом, подобным плюске,
образованным верхушечными листьями.
Древесные растения с очередными перистосложными листьями, без
прилистников.
Семейство ореховые (Juglandaceae) — деревья с кру
лми непарно-
а: I
перистосложными листьями.
Главный род семейства — ореховые (Juglans). Это деревья. Листья круп-
ные непарноперистые, богатые ароматическими веществами. Прилистники
отсутствуют. Плоды у ореха — костянки; сочная наружная часть около-
плодника зеленого цвета, а внутренняя, твердая, деревянистая — желтого.
Семя одно, без эндосперма, зародыш с очень своеобразно устроенными, сильно
морщинистыми семядолями. Грецкий орех (J. regia) встречается в диком сос-
тоянии от районов Восточного Средиземноморья до Гималаев. Он высоко
ценится как плодовое дерево и культивируется в большом количестве сортов.
Из семян добывают масло, околоплодник используют для получения хорошей
бурой и черной краски. Древесина этого крупного дерева плотная, твердая,
легко полируется и имеет на срезе красивый рисунок, почему и используется
как отделочный материал. Ценится также ореховый кап. Другие виды рода
Juglans растут в Северной Америке и на Дальнем Востоке (СССР).
В мужских цветках лапин, или крылоорешников (род Pterocarya),
в большинстве случаев имеется рудимент пестика. Один из видов этого
рода обитает в Закавказье.
4-И ПОРЯДОК — КРАПИВОЦВЕТНЫЕ (URTICALES)
Цветки крапивоцветных обычно однополые, реже двуполые, в цимоз-
ных соцветиях или одиночные, с простым, четырех-, шестичленным около-
цветником; пестик из двух плодолистиков, с верхней одногнездной завязью,
содержащей семяпочку. Плод — орешек или костянка. Главным образом
анемофильные и только редко энтомофильные древесные и травянистые рас-
тения с очередными или супротивными листьями, имеющими прилистники.
Для многих крапивоцветных характерны цистолиты и нечленистые млечники.
Семейство вязовые (Ulmaceae) — ветроопыляемые древесные рас-
тения без млечного сока. Цветки однополые или двуполые, одиночные или
в цимозных соцветиях. Околоцветник простой, из 4—6 свободных листочков.
Иногда листочки в большей или меньшей мере срастаются. Тычинок обычно
столько же, сколько листочков околоцветника, и они супротивны последним.
Пестик из двух медианных плодолистиков, завязь обычно одногнездная.
Плоды — орешки или костянковид
яе. В мужских цветках часто встречается
пн
рудимент пестика.
Для рода вяз (Ulmus) характерны крылатые орешки. Вязы (Z7. laevis,
U. scabra и др., рис. 247) входят в состав наших лиственных лесов как
сопровождающие породы. Древесина их прочная.
Наиболее обширный род семейства вязовых — каркас (Celtis). Он
распространен в умеренной зоне и под тропиками северного полушария.
Плоды каркаса — костянки. В СССР растут два вида каркаса: кавказский
{С. caucasica) — на Кавказе и в Средней Азии и гладкий (С- glabrata) — в
Крыму и на Кавказе. Древесина каркаса необычайно плотная, за что ра-
стение получило название «железное дерево». К семейству вязовых относится
и дзельква (Zelkova carprinifolia) — реликт третичной флоры Закавказья.
Семейство Тутовые (Могасеае) — большое (более 1500 видов), глав-
ным образом тропическое. Древесные, в виде исключения травянистые рас-
тения с очередными простыми листьями. Прилистники сильно разрастающие-
ся. Во вторичной коре—нечленистые млечники, содержащие каучук. В клетках
эпидермиса, а иногда и в более глубоких тканях обычны цистолиты. Цветки
всегда однополые. Околоцветник мужского цветка четырехлистный (но бывает
двух-* трех-* пяти- и шестилистным). Листочки свободные или сросшиеся*
374
ВЫСШИЕ РАСТЕНИЯ
иногда отсутствуют. Каждому листочку околоцветника соответствует ты-
чинка, расположенная супротивно. Редко бывает только одна тычинка.
В женском цветке околоцветник также четырехчленный, свободно -или срост-
нолистный, иногда в той или иной степени срастающийся с завязью, которая*
таким образом, может быть верхней, нижней или полунижней. Пестик образо-
ван двумя плодолистиками, один из которых может редуцироваться настолько,
что от него остается лишь небольшой зубчик при основании столбика. Семяпоч-
ка одна. Соцветия сложные, состоят из дихазиев, отличаются разнообразием
ni;
внешнего вида: метельчатые, колосовидные, головчатые и, как результат
разрастания и срастания ветвей дихазиев, колбовидные, шаровидные, полые,
блюдцевидные и т. п.
Плоды — орешки или костянки, нередко срастающиеся друг с другом*
причем общая ось соплодия делается мясистой.
Среди тутовых много практически важных и биологически интересных
растений.
Род шелковица {Мотив, рис. 248) — деревья и кустарники с разнооб-
разными по очертанию листьями (гетерофилия). Листья белой (родом из
Китая) и черной (из Закавказья) шелковицы — почти единственный корм для
тутового шелкопряда. По этой причине шелковицу культивируют во всех
районах шелководства. Соплодия шелковицы мясистые — сладкие, съедоб-
ные, но без аромата.
Бумажное дерево {В, roueeonetia papyrifera) — родом из Восточной Азии.
Кора идет на выделку прочной бумаги, из которой изготовляют зонты,
носовые платки и даже* после промасливания* водонепроницаемые плащи.
Рис. 247. Вяз равнинный, или берест
(Ulmus campestris):
1—цветущая ветвь; 2—олиственная ветвь с плода-
ми; Л—цветок; 4-плод
Рис. 248. Шелковица
(Morus alba):
1 — ветвь с соидоднями; 2 —
мужской цветок; 3—-женский
цветок
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис. 249. Баньян (Ficus bengalensis)
Соплодия хлебных деревьев (род Artocarpus) богаты крахмалом, ве-
сят до 12 кг. Их режут на ломти, пекут и едят как хлеб или готовят сначала
тесто, из которого затем выпекают хлеб. Родина хлебных деревьев — Индо-
Малайская область, но в культуре они известны и для других мест тропичео
кой части земного шара. Выведены культурные сорта.
Castilloa elastic a, Castilloa ulei (Центральная и тропическая Южная'
Америка) и другие виды рода кастилоа используются как каучуконосы.
Упас, или анчар (Antiaris toxicaria), из Юго-Восточной Индии, содержит
; г. I;
ядовитый млечный сок, который использовался местным населением для
изготовления отравленных стрел.
Молочное, или коровье, дерево (Galactodendron utile), из Южной Америки,
имеет питательный, сладковатый на вкус млечный сок, который употребля-
ют как молоко. Добывают его, надрезая кору дерева.
Самый крупный род — фикус (Ficus) — содержит около 700 видов,
почти исключительно тропических, среди которых есть крупные деревья,
кустарники, лианы и эпифиты.
Наиболее важным является F. elastica, родом из Южной Азии; его
млечный сок содержит каучук высокого качества. Это мощное дерево с сильно
развитыми дощатыми корнями-подпорками; его очень часто культивируют
как комнатное растение.
Баньян (F. bengalensis) — растение, начинающее свое развитие из
семени на ветвях деревьев и дающее воздушные корни, которые по мере раз-
растания эпифита дастигают земли; наблюдалось исключительное явление —-
376
выс:
II Pi
[В РАСТЕНИЯ
Рис. 250. Инжир (Ficus
carica):
1 — ветвь с соплодиями; 2 — разрез
соцветия; 3 — тычиночный цветок;
4 — пестичный цветок; S — пестик
образование как бы рощи, где вместо стволов стоят столбовидные корни,
поддерживающие громадную «крону» (рис. 249), под которой может уместиться
небольшая деревня.
Смоковница (F. carica, рис. 250) растет в средиземноморских странах,
в Малой Азии, Иране и Северной Индии, у нас — в Крыму и Закавказье.
Растения разводят ради мясистых соплодий, которые имеют грушевидную
форму, мясисты и очень богаты сахаром (в сушеном виде до 75%); они из-
вестны под названием винной ягоды, инжира или фиги.
Виды рода цекропия (Cecropia), распространенные в тропической
Америке, замечательны мирмекофилией; муравьи поселяются в полых междо-
узлиях стеблей и питаются особыми волосками, находящимися у основания
листьев. Эти «квартиранты» защищают дерево от нападения опасных для
него муравьев-листорезов. Виды Cecropia дают каучук, но невысокого
качества.
Семейство коноплевые (СаппаЫпасеае), Это травянистые, прямо-
стоящие или вьющиеся растения с дланевидными листьями, имеющими не-
опадающие прилистники. Млечного сока нет. Цветки мелкие, однополые.
Мужские цветки, собранные в дихазиальные метельчатые соцветия, имеют
невзрачный околоцветник из пяти листочков и пяти расположенных супро-
тивно им тычинок. Женские цветки имеют пестик, образованный двумя пло-
долистиками и несущий два столбика. Спайнолистный пленчатый околоцвет-
ник прилегает к завязи. Они тоже образуют дихазиальные соцветия, но
более скученные, в виде шишечек или густых колосовидных образований
в верхней части стебля. Плод — орешек.
Семейство коноплевых имеет два рода: конопля (Cannabis) и хмель
(Humulus). Один из видов конопли — Cannabis sativa (рис. 251) — культи-
вируется как прядильное и масличное растение. Для конопли посев-
ной характерна двудомность. Мужские экземпляры называют посконью, жен-
ские—матеркой. Соцветия сложные, образованные пазушно расположенными
дихазиями. Конопля — очень древняя культура. Волокно ее стеблей исполь-
зуется под названием пеньки для изготовления пакли, веревок, канатов,
парусов, мешков и т. д. Семена содержат до 35% жирного масла (конопля-
ное масло), которое употребляют в пищу, идет на изготовление мыла, оли-
фы, различные технические нужды. Жмыхи служат питательным кормом
для скота. В Южной Азии культивируется разновидность конопли посевной,
которая дает материал для получения пахучего, смолистого вещества — га-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
ип
а, представляющего сильнодурманящее средство, действующее подоб-
но опиуму.
Три вида рода хмель (Humulus, рис. 252) обитают в умеренной зоне
северного полушария. Humulus lupulus находит практическое применение.
Его головчатые женские соцветия при созревании плодов превращаются в
характерные ш:
\ in
ки, образованные сильно разрастающимися пленчатыми
кроющими листьями. Последние, как и сами плоды, покрыты железками,
выделения которых используются при изготовлении пива, для придания ему
особого вкуса.
Кпорядку Urticales относится еще семейство крапивные (Ur-
ticaceae). Крапивные, как и коноплевые, главным образом травянистые рас-
тения, без млечного сока, нередко со жгучими волосками. Строение цветков
напоминает цветки тутовых: они бывают как однополые, так и двуполые.
Из распространенных у нас растении этого семейства общеизвестна крапива
двудомная (Urtica diotca, рис. 253). Это многолетнее травянистое растение
быстро размножается с помощью корнев
особенно на почвах, богатых
нитратами. Мужские и женские цветки помещаются на разных особях, хотя
иногда бывают и на одной. Мужские цветки — хороший объект для наблю-
дения приспособлений к ветроопылению. В стеблях двудомной крапивы име-
ются длинные и тонкие волокна, которые при соответствующей обработке
могут быть использованы как прядильный материал неплохого качества.
Кроме крапивы двудомной, у нас нередко встречается крапива жгучая (Ur-
tica иг ens). Так называемая китайская крапива, или рами (Boehmeria nivea)*
Рис. 251. Конопля (Cannabis sativa):
1 — женское растение; 2 — мужское растение (посконь); з — тычиночный цветок; 4 — пестичный цветок,
окруженный кроющим листом; S — пестичный цветок; 6 — диаграмма тычиночного цветка 7 — диягрям-
ма пестичного цветка
ВЫСШИЕ PACT
и: i >
Рис. 252. Хмель (Humulus lupulus):
j — ветвь с соклодняш; s — мужское соцветие;
3 — женское соцветие
Рис. 253. Крапива (Urtica dioica): —►
J— мужское растение; Я — тычиночный цветок;
3—диаграмма тычиночного цветка; 4 — диаграмма
шютжчнсго цветка
принадлежит к р о д у бемерия. Из ее волокон изготовляют хорошую ткань,
а также очень прочные канаты.
Родственные связи семейств порядка Urttcales считаются настолько
несомненными, что некоторые систематики даже соединяли все крапивоцвет-
ные в одно семейство. По ряду признаков, например по распространенности
дихазиальных соцветий, по строению цветка, по преобладанию древесных
форм, порядок Urttcales близок к букоцветным. Не исключена возможность,
что крапивоцветные произошли от общих с букоцветными предков и разви-
вались как самостоятельная, параллельная Fagales линия покрытосеменных,
с преобладанием ветроопыления.
5-Й ПОРЯДОК — КАЗУ АРИНОЦВЕТН^Е (CASUARINALES)
Цветки казуариноцветных однополые. В мужском цветке имеются два
листочка околоцветника и одна центрально расположенная тычинка. Цветки
собраны в мутовчатые колосья, за
Каждый мужской цветок помещается в Пазухе кроющего листа и сопровожда-
ется двумя прицветничками. В женских\цветкахА тоже пазушных с прицвет-
ющие концы удлиненных побегов.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
379
яичками околоцветника нет,
пестик образован двумя срос-
шимися плодолистиками и имеет
в завязи два, гнезда, из которых
одно бесплодное. Из 2—4 семя-
почек, содержащихся в плоду-
щем гнезде, Sb семя развивается,
как правило! одна. Рылец два,
они нитевидные. Женские цвет-
ки собраны в головки, распола-
гающиеся на| концах укорочен-
ных ветвей. I Плоды нераскры-
вающиеся, • односемянные, с
крыловидным; придатком на
конце. I
Деревья j и кустарники
внешне напоминают хвощи бла-
годаря редукции листьев и
мутовчатому расположению как
листьев, так й; боковых ветвей,
а также вследствие наличия
продольных бороздок на меж-
доузлиях. 1
Семейство казу-
ариновые (^asuarinaceae)—
единственное в порядке и со-
держит один род казуарина
{Casuarina, рис.: 254) с 40—
50 видами, обитающими в Авст-
ралии и отчасти Ш Индо-Малай-
ской флористической облас-
ти. Ископаемые Остатки казу-
ариновых извести^ из верхне-
меловых отложений. Казуари-
новые резко выделяются разви-
Рис. 254. Казуарина (Casuarina equiset ifolia):
1 — пучок ветвей (лс — мужские соцветия» с — сопло-
дия); 9 — две мутовки мужского соцветия; 8—диа-
грамма мутовки тычиночных цветков; 4 — пестичный
цветок; 5 — диаграмма пестичного цветка; б —сопло-
дие
тием многоклеточного археспо-
рия в семяпочке й образованием многих (до 20) мегаспор, из кото-
рых только в одной сформируется нормальный яйцевой аппарат, типично вы-
ражено явление халазогамии, впервые у казуариновых и открытое. Эти при-
знаки расцениваются иногда как признаки глубокой примитивности. Сопо-
ставление их с прост ~
в некотором отношени
повод некоторым
ния, переходные м
тиков предпочитает р
цированную. Многие указывают на вероятность связи казуариновых с Нота-
melidales. Возможно, порядок казуариноцветных отделился от древнейших
покрытосеменных как самостоятельная ветвь* не имевшая, однако* дальней- ‘
и строения цветка и с тем, что древесина казуариновых
напоминает древесину голосеменных (гнетовых), дало
тематикам рассматривать казуариновые как расте-
голосеменными и покрытосеменными. Часть система-
матривать казуариновые как группу, глубоко реду-
шего развития.
6-Й ПОРЯДОК — ИВОЦВЕТНЫЕ (SALICALES)
В этом порядке только одно семейство — ивовые.
Семейство ивовые (Salicaceae). Это кустарники и деревья с очеред-
ными простыми листьями. Прилистники иногда рано опадают. Цветки муж-
ские и женские однополые, как правило, на разных особях (двудомность).
380
ВЫСШИЕ РАСТЕНИЯ
Рис. 255. Ива козья (Salix саргеа):
1 — олиственный побег; 2 — ветвь с мужскими соцветиями; з — ветвь с женским соцветием; 4 — тычи-
ночный цветок; 3 — пестичный цветок; в — диаграмма тычиночного цветка; 7 — диаграмма пестичного
цветка; 8 — продольный разрез пестичного цветка; 9 — раскрывшийся плод; 10 — семя
Околоцветник отсутствует1. В мужских цветках от 2 до 30 и более тычинок;
в женских — один пестик из двух плодолистиков. Завязь одногнездная.
Семяпочек много. Плоды — коробочки, раскрывающиеся двумя створками.
Семена с пучком волосков у основания, без эндосперма.
Семейство ивовых объединяет только три рода: тополь (Populus),
ива (Salix) и чозения (Chosenia). Последний род представлен только одним
видом, растущим у нас на Дальнем Востоке, в Японии, на Корейском полу-
острове, в Китайской Народной Республике. Чозения крайне близка к роду
Salix (рис. 34).
В роде тополь (Populus) мужские цветки состоят из нескольких тычи-
нок, окруженных в основании особым, косоворонковидно-бокальчатым ор-
ганом — диском, представляющим или редуцированный околоцветник, или
продукт разрастания цветоложа. Женские цветки также имеют бокальчатый
диск, окружающий нижнюю часть единственного пестика, образованного
двумя паракарпно соединенными плодолистиками. В состав этого рода вхо-
дят осина (Populus tremula), тополь черный, или осокорь (Р. nigra), и др. Мно-
гие виды тополей ценятся как быстро растущие деревья с мягкой, легко ре-
1 Существует мнение, согласно которому цветки ивовых имеют простой, но глубоко
редуцированный околоцветник.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСВМИННКт
381
жущейся древесиной, которая идет на изготовление спичек, предметов обихо-
да (ложки, совки и пр.)*, используется в целлюлозной промышленности и ре-
же как строительный материал. Тополи анемофильны.
В цветках ив (р о д Salix) в некоторых случаях имеется, как и у тополя,
диск, окружающий основание цветка*, у большинства он заменен двумя
нектарниками, расположенными один со стороны кроющего листа, другой—на
стороне, обращенной к оси сережки; наконец, у некоторых остался только
один нектарник. В мужских цветках содержится от 1 до нескольких тычинок;
в женских, как и у тополей, паракарпный двучленный гинецей с двумя рыль-
цами. Цветут ивы до распускания листьев или одновременно с ним. Ивы на-
секомоопыляемые. Посещаются десятками видов насекомых, реже анемо-
фильные. Многие виды склонны к гибридизации, вследствие чего границы
видов не всегда ясны.
Ветви ив идут на плетение корзин, рыболовных снастей и пр.; в коре со-
держатся дубильные вещества. Ивы чрезвычайно легко укореняются и раз-
множаются вегетативно, поэтому их используют для закрепления песков.
Ивовые — Populus и Salix (вместе с примыкающей чозенией)— несом-
ненно, близко родственны между собой и, по всей вероятности, представляют
две самостоятельные — анемофильную (род Populus) и энтомофильную
(Salix с примыкающей чозенией) — линии эволюционного развития, воз-
никшие от общего предка. Предковая форма имела обоеполые цветки, в поль-
зу чего говорит нахождение в виде уклонений обоеполых цветков у ивы
козьей (S. саргеа, рис. 255) и у гималайского вида тополя Р. glauca. Имел
ли предок ивовых околоцветник, не выяснено. Salicaceae по происхождению
очень древнее семейство, остатки которого известны из нижнемеловых от-
ложений; в верхнем мелу Salicaceae встречаются уже очень часто; в неогене
они процветали по всей умеренной зоне северного полушария. Неогеновые
виды близки к современным тропическим ивоцветнЫм. Палеонтологические
данные, таким образом, говорят о возможности непосредственной генетиче-
ской связи ивоцветных с древней
гли
ими покрытосеменными, с их предковыми
формами. Наряду с этим существует мнение, что цветки ивовых (а следова-
тельно, и сами ивовые) возникли путем редукции от высокоспециализиро-
ванных форм покрытосеменных. Некоторые ботаники связывают это семейст-
во с семейством гребенщиковых, представители которого имеют хохолок во-
лосков на семенах, семяпочки, располагающиеся у дна завязи, и другие
признаки сходства с ивовыми. Часть ученых считает, что наиболее близкие
связи должны быть у ивовых с фиалкоцветными (постенносеменными) и раз-
вивались параллельно с гребенщиковыми от общего предка.
7-Й ПОРЯДОК — ПРОТЕЙНЫЕ (PROTEALES)
Включает одно семейство. Цветки двуполые или однополые, опыляются
насекомыми или птиц ши. Околоцветник простой, четырехчленный, тычинок
четыре, они противостоят листочкам околоцветника и срастаются с ними.
Гинецей из одного пл )долистика. Завязь верхняя, одногнездная. Древесные
растения с очередными листьями без прилистников.
Семейство протейные (Proteaceae). Растения, которые относятся
к этому обширному (ороло 1400 видов) семейству, в большинстве случаев веч-
нозеленые кустарники^ и небольшие деревья. Некоторые достигают значи-
тельной (20—30 м) высоты, как, например, Knightia excelsa, растущая в веч-
нозеленых лесах Северного острова Новой Зеландии. Травянистые формы
редки. Цветки обоеполые, актиноморфные или зигоморфные, располагаются
поодиночке или парами^ в пазухах обычных или верхушечных листьев* об-
разуя разнообразные соцветия (кисти, колосья, зонтики, головки), которые
бывают окружены оберткой из специализированных листьев. Простой около-
цветник состоит из четырех листочков, вполне свободных или несколько
сросшихся и образующих трубочку. Четыре тычинки располагаются по
одной напротив листочкоЬ околоцветника, в разной степени срастаются с ни**
382
ВЫСШИЕ РАСТЕНИЯ
ми, редко оставаясь свободными. Гинецей из одного плодолистика; пестик
с верхней одногнездовой завязью, иногда сидящей на ножке (гинофор).
Семяпочек от 1—2 до неопределенно большого числа. Плод раскрывающийся
(типа листовки и коробочки) или нераскрывающийся (орех, реже костянка).
Околоплодник обычно довольно толстый, иногда сильно одревесневающий.
Протейные в основном растения южного полушария. Более их сос-
редоточено в Южной Африке и более 1/2 (свыше 700 видов) растет в Австра-
лии. Есть протейные в Южной и Центральной Америке.
Из числа порядков подкласса Hamamelidae, вероятно, наиболее древ-
ним является порядок тпроходендроцветные (Trochodendrades), В ряде
систем покрытосеменных его включают в порядок Magnoliales, подчеркивая
несколько изолированное положение этой группы среди прочих .магнолие-
цветных. Trochodendrales объединяет всего два семейства — Тго-
chodendraceae и Tetracentraceae, содержащие по одному монотипному роду.
Троходендрон (Trochodendron arabioides — Япония до Тайваня) и тетра-
центрон (Tetracentron sinense—Китай) — деревья с мелкими цветками, собран- *
ными в соцветия. В обоих случаях гинецей синкарпный, но с явными следами
происхождения от апокарпного. Долгое время считали, что оба растения
анемофильны, но сейчас доказано, что у Trochodendron энтомофильное расте-
ние, причем выделение нектара осуществляется поверхностью плодолисти-
ков. Что же касается Tetracentron sinense, то, хотя пыльца его и разносится
ветром, пылинки имеют строение, соответствующее энтомофилии.
Наиболее близкое к Trochodendrales положение занимает порядок На-
mamelidales.Bce прочие порядки подкласса Hamamelidae, вероятно, пред-
ставляют окончания самостоятельных эволюционных ветвей, берущих нача-
ло от Hamamelidales или от их предков. Особое положение занимает порядок
Salicales, который связан с прочими Hamamelidae, вероятно, только через
общего предка.
3-Й ПОДКЛАСС — КАРИОФИЛЛИДНЫЕ (CARYOPHYLL1DAE)
Этот подкласс примыкает к Magnoliidae благодаря наличию у более
примитивных групп апокарпного гинецея. Однако преобладает ценокарп-
ный гинецей. Широко распространены согнутые (кампилотропные) семяпоч-
ки. Семена с согнутым периферическим зародышем обычно содержат пери-
сперм. Травы, редко кустарники и небольшие деревья.
К подклассу Caryophyllidae относят всего 4 порядка, из которых рас-
смотрим 3.
1-Й ПОРЯДОК — ГВОЗДИЧНОЦВЕТНЫЕ (CARYOPHYLLALES),
ИЛИ ЦЕНТРОСЕМЕННЫЕ (CENTROSPERMAE)
Цветки актиноморфные, обоеполые, реже однополые, с простым или
двойным, пента- или тетрамерным околоцветником. Андроцей круговой, с
числом тычинок, кратным числу чашелистиков, иногда тычинки многочис-
ленные. Гинецей ценокарпный или апокарпный; Завязь верхняя, полуниж-
няя или нижняя. Семязачатки согнутые. Зародыш обычно согнутый вокруг
перисперма. Эндосперма нет. В подавляющем большинстве это травы, ред-
ко кустарники или небольшие деревья с простыми, цельными листьями,
обычно не имеющими прилистников. В некоторых семействах широко пред-
ставлены листовые и стеблевые суккуленты.
Семейство фитолаккоеые (Phytolaccaceae) представляет с эволю-
ционной точки зрения особый интерес. Его можно рассматривать как пред-
ковое по отношению к прочим в том же подклассе и как связующее звено
между подклассами Caryophyllidae и Magnoliidae* У фитолакковых, отлича-
ющихся разнообразием жизненных форм (от деревьев до однолетних трав),
разнообразны и цветки. Околоцветник у них обычно простой, свободнолист-
ный, травянистый или перепончатый, реже венчиковидный пли двойной. Анд-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рве. 256. Свекла (Beta
vulgaris):
1—лршюрньвая розетка к осно-
вание корнеплода; я — верхняя
цветоносная часть растения;
8 — «клубочки»; 4 — цветок (вид
сверху); 5 — продольный разрез
цветка; в — семя; 7 — зародыш
(продольный разрез)» окружаю-
щий перисперм
г r,t
роцей варьирует в числе тычинок. Иногда они в неопределенно большом чис-
ле (вследствие расщепления) располагаются в один-два круга. Изменчив и .
гинецей, как по числу плодолистиков, так и по их взаимному расположен
Гинецеи может быть апокарпным, неполносинкарпным и даже (редко) пара-
карпным. У некоторых имеется гинофор. Плодолистиков от 5 и более (иног-
да 1); каждому плодолистику соответствуют один столбики одна базальная
согнутая семяпочка; иногда в одногнездной двух- четырехчленной завязи
находится только одна семячка. Плоды — ягоды, орехи, коробочки. Семе-
на с мучнистым периспермом и всегда согнутым зародышем. Характерные
особенности цветка и плодов этого преимущественно тропического (в основ-
ном американского) семейства можно изучить, познакомившись с лаконосом
американским (Phytolacca americana), занесенным из Северной Америки сор-
ным многолетним травянистым растением, обитающим у нас на Кавказе.
Сок из ягод этого растения употребляют для окраски шелковых тканей. За ру-
бежом, в районах виноделия, растение культивируют: используется сок для
подкрашивания вин, что небезвредно.
растения, отчасти11олукустарники, редко кустарники и небольшие деревья.
Листья простые, очередные или супротивные, без прилистников. Вследствие
или срастания листьев с междоузлиями стебля многие представи-
тели кажутся безлистными. Цветки невзрачные, в цимозных дихазиальных
собраниях с укороченными осями («к л у б о ч к и »), объединенных в коло-
совидные, метельчатые и другие сложные соцветия.
Строение цветков очень разнообразное. Они обычно правильные, обое-
полые, реже однополые. Околоцветник простой, из 5 и меньшего числа тра-
вянистых или пленчатых листочков, которые у основания в разной степени
срастаются; иногда околоцветник отсутствует. Он может сохраняться, раз-
растаясь при плодах и участвуя в их распространении. Тычинки противо-
стоят листочкам околоцветника, число их равняется числу членов окодоцвет-
384
ВЫСШИЕ РАСТЕНИЯ
ника или меньшее. Гинецей из 2, реже из 3—5 плодолистиков; пестик с таким
же числом столбиков (рылец). Завязь верхняя, редко полунижняя, одно-
гнездпая, с одной базальной или почти базальной семяпочкой. Невзрачность
мелких цветков лебедовых дает повод думать, что эти растения исключитель-
но ветроопыляемые. Фактически некоторые из них зоофильны. В связи с этим
в цветках развиваются своеобразные приспособления в виде нектарников или
поедаемых опылителями крупных, окрашенных в белый, желтый и красный
цвет придатков на конце связников. Плоды большей частью сухие, с твердым
или пленчатым околоплодником (орехи), редко раскрываются крышечкой
(коробочка); иногда плод ягодообразный. Наблюдаются многочисленные
приспособления для распространения плодов в виде волосков, шиловидных
или крыловидных разрастаний листочков околоцветника и т. п. Зарод
ii п н
резко изогнутый (охватывает перисперм) или спирально закрученный; в
последнем случае перисперм может отсутствовать.
Лебедовые содержат около 100 родов и 1500 видов и связаны в своем рас-
пространении обычно с сухими, пустынными и полупустынными областями,
являясь часто ландшафтными растениями. Многие лебедовые, например
сарсазан шишковатый (Halocnemum strobtlaceum), многие виды солянок Sal-
sola, роды Suaeda) и другие, связаны с засоленными почвами. Сравнительно
немногие виды выходят за пределы указанных географических условий,
будучи обитателями морских побережий и внутренних частей материков в
качестве сорных и рудеральных (мусорных) растений.
Экономически важный представитель семейства лебедовых — род
свекла (Beta), Предком культурной свеклы (В. vulgaris, рис. 256) считают
свеклу многолетнюю (В. perennis), дико произрастающую на Атлантическом
и Средиземноморском побережьях Западной Европы, а у нас — в Восточном
Закавказье и в Каракумах. Дикий вид так изменен, что его культурная фор-
ма вполне может считаться особым видом, созданным человеком. Для свеклы
характерно пятичленное строение цветка. Завязь полунижняя. Тычинки
спаяны внизу нитями в железистый диск. Плоды с древеснеющим околоплод-
ником, срастаются по 2—3 вследствие сближенности цветков в клубочках в
характерные морщинистые соплодия. Этим и объясняется, что при прораста-
нии свеклы образуется группа из двух-трех проростков, сидящих рядом.
Свекла — растение двухлетнее. Верхняя часть корнеплода представляет
утолщение надсемядольного колена; средняя, наиболее массивная,— раз-
росшееся подсемядольное колено; только нижняя часть с отходящими от нее
боковыми корнями образуется за счет корня. Сам корень, длинный, сравни-
тельно тонкий, продолжая нижнюю часть корнеплода, растет вглубь, до
1,5 м.
Для корнеплода свеклы характерно кольцевое анатомическое строение.
Оно объясняется тем, что в процессе утолщения последовательно происхо-
дит разрастание паренхимы, в которой закладываются кольцами новые про-
водящие пучки. Такое аномальное для двудольных утолщение осевого орга-
на распространено в семействе лебедовых.
Сахарная свекла — молодое культурное растение. Ее разведение нача-
лось в конце XVIII в. в Европе. В это время возникли первые сахарные за-
воды и производство сахара из свеклы начало быстро возрастать. В начале
XIX в. содержание сахара в сахарной свекле составляло 5—6%, в настоя-
щее время благодаря упорной селекционной работе оно значительно возросло
м достигло 18—20 %.
Около 70% мировой посевной площади сахарной свеклы приходится на
Европу, в том числе около 50% на СССР. Основным районом культуры са-
харной свеклы в СССР с давних времен была Украинская ССР; здесь и в насто-
ящее время сосредоточено более половины посевов СССР. Г pat
г, II
(Ы культуры
сахарной свеклы успешно продвигаются на восток — в результате в настоя-
щее время свеклу высевают даже на Дальнем Востоке.
Целесообразно разведение сахарной свеклы и в качестве кормового рас-
тения.
' ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
385
III
Важным овощным растением является шпинат (Sptnacia oleracea). Его
употребляют для приготовления супов, пюре, соусов, приправ. По содержа-
нию белка шпинат уступает только мясуя содержит также значительное ко-
личество витаминов: А1Э Вх, В2, С и РР; употребление его в пищу считается
полезным.
. Многие лебедовые имеют значение в качестве кормовых растений в
полупустынных и пустынных областях. Некоторые (сравнительно немногие)
поедает скот в течение всего периода вегетации или только в молодом состоя-
нии. «Сочные солянки», весьма богатые минеральными солями, делаются
съедобными глубокой осенью и зимой, когда соли выщелачиваются из рас-
тений дождем и талым снегом. Растут сочные солянки на засоленных поч-
II'Ли
Сочные солянки резко выделяются среди других растений содержанием
зольных веществ, достигающим 50% сухого веса растения. Примером таких
растений может служить солерос (Salicornia herbacea) — однолетнее растение
из подсемейства Spirolobeae* широко распространенное на солончаках юго-
востока СССР. Щелочи, содержащиеся в солянках, извлекают и используют,
например, при изготовлении мыла.
Многие солянки имеют стебли, одревесневающие в нижних частях (по-
лукустарники). Ввиду того что они образуют большие (до 70 см и более в по-
перечнике) кусты, их используют как источник топлива, что для безлесных
полупустынных и пустынных районов имеет немаловажное экономическое
значение. Особенно большую роль играют такие представители подсемейства
Spirolobeae, как древовидные лебедовые — виды саксаула (Haloxylon aphyllum
и Я. persicum — черный и белый саксаул), обычные в Средней Азии. Это
растения с корявыми стволами, достигающими 4—5 м в высоту и 50—70 см
у основания; образуют редкоствольные леса. Молодые ветви саксаула прак-
тически безлистные (вследствие глубокой редукции листьев), имеют членис-
тое строение и выполняют ассимиляционную функцию. Саксаулы имеют тя-
желую, тонущую в воде древесину, соединяющую твердость с хрупкостью.
Древесина саксаула по теплотворной способности превосходит все виды
дров и лишь немного уступает хорошим сортам каменного угля. Из золы сак-
саула добывают поташ. Серьезное хозяйственное значение имеет итсегек,
или ежовник (Anabasis salsa), содержащий, в основном в молодых зеленых
веточках, инсектицидный алкалоид анабазин.
Примером сорных лебедовых может служить белая марь (Chenopodium
album). Это очень распространенное травянистое однолетнее растение с голым
стеблем и яйцевидными выемчато-зубчатыми листьями, покрытыми, особенно
в молодом состоянии, как бы мучнистым налетом. Налет образован особыми
пузыревидными волосками, позднее опадающими. Растение цветет все лето
и производит огромное количество семян — от нескольких сотен до 20 тыс.
на один экземпляр. Семена прорастают весной, но не все в один год; всхо-
жесть сохраняют очень долго. Ко времени уборки культурного растения
большая часть семян сорняка успевает осыпаться, поэтому пахотный слой
содержит большое количество зачатков сорняка. Белая марь извлекает из
почвы много питательных веществ, в том числе азота, калия и фосфора, от-
нимая их от культурных растений. Кроме того, она дает приют вредным насе-
III
II
Белую марь, как и другие виды этого рода, нередко именуют лебедой,
но желательно относить это название к другому, близкому роду Atriplex,
«иди которого отличаются однополыми цветками. Представители рода
лебеда обычно однодомны. На глинистых обрывах по берегам рек, по пустырям
и сорным местам в средней полосе растет лебеда блестящая (A. nitens) —
крупное растение с треугольными, у основания копьевидными листьями.
и распространены по преимуществу в полупустынных и пустынных рай-
13—602
Маревые сближают с фито лайковыми, часть которых (подсемейст-
в о Micro teoideae) характеризуется простым околоцветником и одногнезд-
ной завязью, содержащей только одну семяпочку; цветки мелкие..
Семейство айвовые (Aizoaceae). Это многолетние, редко одно-двух-
летние травы, иногда полукустарники и кустарники; листья обычно мясисто-
сочные. Цветки преимущественно обоеполые в дихазиальных соцветиях,
с двойным или простым околоцветником. Чашелистики или листочки прос-
того околоцветника срастаются в трубочку. Лепестки многочисленные, об-
разовавшиеся, вероятно, из тычинок, превратившихся в лепестковидные
стаминодии. Тычинки часто сросшиеся в пучки, развиваются центрифугаль-
но. Завязь верхняя, полунижняя или нижняя, несущая свободные столбики,
двухмногогнездная, с центрально-угловой плацентой, редко, вследствие раз-
рушений перегородок между гнездами, одногнёздная. Плод — коробочка,
иногда ягода.
Семейство включает около 2500 видов. Наиболее крупный род мезембри-
антемум (Mesembryanthemum) некоторые систематики рассматривают как
сборный, объединяющий около 100 самостоятельных родов. Aizoaceae име-
ют два центра распространения — Южную Африку и Австралию, но общая
область распространения охватывает более обширные территории с засуш-
ливым климатом в тропических и субтропических районах обоих полушарий.
Аизовые отличаются красотой цветков и своеобразием вегетативных ор-
ганов. У некоторых листья напоминают камни («живые камни» — род Lit*
hops)*, у других они погружены в землю, и свет проникает внутрь листьев
через прозрачную ткань, едва выдающуюся над почвой (листья с «окошечка-
ми»). У хрустальной, или ледяной; травы тело покрыто пузыревидными
прозрачными волосками с водянистым содержимым (запас влаги), благода-
ря чему растение кажется окруженным корочкой из льдинок. Некоторые аизо-
вые введены в культуру как декоративные.
Семейство кактусовые (Cactaceae). Растения этого семейства вы-
деляются крайне своеобразным обликом. Только у немногих представителей:
роды переския (Pereskia) и перескиопсис (Pereskiopsis) — имеется нормаль-
ное расчленение надземной части тела на стебель и листья. У остальных
листья подверглись редукции; их можно наблюдать только на совсем Мо-
лодых растениях, причем они бывают у одних мясистые (веретеновидные или
плоские), у других имеют форму крайне мелких чешуек, у третьих заметны
лишь в виде зачатков близ конуса нарастания. Стебли кактусов мясистые,
оригинального внешнего вида: шаровидные, колоннообразные, простые или
бедно разветвленные. Побеги круглые в поперечном сечении или уплощен-
ные. В некоторых случаях они крыловидно разрастаются и становятся лис-
товидными (филлокладии). Листьяна мясистых побегах редуцированы.
Остаются только их разрастающиеся основания — подарии. При слия-
нии подариев друг с другом у шаровидных и колоннообразных кактусов обра-
зуются продольные ребра, соответствующие листовым ортостихам. На по-
дариях есть ареолы — бугорчатые образования, возникающие из уко-
роченных побегов — пазу
। и: ।
их почек. Листовые зачатки их развиваются не
в листья, а в колючки, щетинки или жесткие волоски различной длины, кре-
пости и формы. Они могут быть покрыты разнообразными волосками — вы-
ростами эпидермальных клеток. Такие волоски образуют (на молодых арео-
лах) спутанный, как быщерстистый покров.-В ареолах развиваются и гл о-
х и д и и — легко обламывающиеся при прикосновении ш
hi
а и щетинки.
Они
легко внедряются в кожу и не всегда легко извлекаются. Мясистость стебля
обусловлена развитием в нем мощной водозапасающей ткани. У крупных
кактусов, достигающих порой метра и более в поперечнике, скапливается
громадное количество воды (до 3000 л). Вода, запасенная в стебле,
расходуется растением крайне медленно благодаря незначительной поверх-
ности, приходящейся на единицу объема, а также очень толстой кутикуле,
восковому налету и незначительному количеству устьиц. Собирание воды
осуществляется своеобразной корневой системой, широко разрастающейся .
11:1: г, 11
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
h;i
1ЫЕ
387
иг»
г»:
почти у поверхности почвы и состо-
ящей из массы корешков, перехваты-
вающих воду атмосферных осадков
по пути ее проникновения в глубо-
кие слои субстрата. При таком стро-
ении растение может переносить
длительную засуху.
Вследствие редукции листьев
фотосинтез выполняется исключи-
тельно стеблем. Накопление ассими-
лятов совершается медленно.
Строение i вегетативного тела
кактусов соответствует условиям
их жизни — они населяют главным
образом засушливые области (полу-
пустыни и пустыни), а если и про-
никают в районы, более обеспеченные
водой, то часто поселяются на вет-
вях деревьев как энифиты, в усло-
виях, затрудняющих добывание воды.
Цветки у кактусов появляются
на стебле либо поодиночке, либо
пучками; они разного размера — от
нескольких миллиметров до 15 см
и более в поперечнике. Околоцвет-
ник состоит из ' многочисленных
листочков, располагающихся по
спирали; нижние листочки травя-
нистые, кверху (внутрь цветка) они
постепенно сменяются лепестковид-
Рис. 257. Кактус, Cereus giganteus
(Калифорния)
красный, желтый или белый цвета.
Листочки околоцветника внизу спая-
ны в трубочку. Тычинки многочис-
ленные, тоже спиральные. Завязь
нижняя из нескольких плодолисти-
ков, одногнездная с многочислен-
ными постенными семяпочками,
редко гинецей почти апокарпный.
Длинный столбик заканчивается
многолучевым рыльцем, белым или
зеленым, ярко выделяющимся на
фоне лепестков. Опыление соверша-
ется насекомыми, а в некоторых слу-
чаях, по-видимому, птицами и ру-
кокрылыми.
Плоды кактусов мясистые, яго-
дообразные, реже сухие.
Область распространения как-
тусов, которых насчитывают около
2000 видов, ограничивается в основ-
ном Южной и Центральной Амери-
кой; только один род рипсалис
(Rhipsalis) известен в Тропической
Африке и далее на восточных ост-
ровах, включая Шри Ланка.
Наибольшее разнообразие какту-
сов наблюдается в Мексике. Здесь не-
13*
Рис. 258. Кактус Echinocactus (Мексика)-
РАСТЕНИЯ
Рис. 259. Кукушкин цвет (Coronaria Поз
cuculi):
1 — прикорневые листья и нижние части стеблей;
8 — верхняя часть стебля и соцветия; з — строение
цветка (чашечка удалена, л ~ лепесток с придат-
ком» пр — придаток, т — тычинка, я — пестик,
н — нектарник); 4 — чашечка; S — поперечный раз-
рез завязи; в — продольный разрез коробочки,
окруженной чашечкой (видны семена на централь-
ном семяносце)
Рис. 260. Звездчатка дубравная (Stel-
laria nemorum):
1 — нижняя; 8 — верхняя часть цветущего
растения; з — чашечка; 4 — лепесток; 3 —
андроцей и гинецей; в —пестик
которые кактусы достигают гигантских размеров. Например, цереус гигант-
ский (Cereus giganteus, рис. 257) достигает высоты до 20 м. Высокий* колонно-
образный ребристый ствол этого кактуса образует на высоте около 1,5—2 м
немногочисленные боковые ветви, горизонтальные в основании и затем почти
под прямым углом загнутые вверх. Здесь же встречаются многочисленные
огромные, более метра в поперечнике, шаровидные формы (рис. 258), а так-
же опунции с их оригинальными стволами, состоящими из сплюснутых чле-
ников. Из Мексики кактусы отчасти проникли и в южные полупустынные
районы США. Много кактусов в кампосах восточных и внутренних районов
Бразилии. Немало своеобразных видов встречается в Андах, а отдельные
формы заходят далеко на юг, вплоть до Патагонии.
Плоды ряда кактусов съедобны. Так, в тропиках большим распростра-
нением как плодовое растение, введенное в культуру, пользуется цереус трех-
гранный (Cereus triangularly. Съедобны также плоды гигантского цереуса.
Особенно широким распространением в культуре пользуется Opuntta ficus
indica — «индийская фига», распространившаяся не только в тропиках,
но и по всему побережью Средиземного моря, где это растение одичало; те-
перь оно проникло в Азию и даже в Австралию, где стало злостным сорняком
и бичом овцеводства.
Колючие кактусы используются для живых изгородей. Кактусы хорошо
приживаются в оранжереях и комнатах и служат оригинальными декоратив-
ными растениями, иногда с очень красивыми крупными цветками.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСКМ КН НЫВ
иногда полукустарники, редко кустарники, с простыми, цельными, часто
узкими (линейными, линейно-ланцетными) листьями, преимущественно без
прилистников. Листорасположение супротивное (редко спиральное). Цвет-
ки большей частью в дихазиальных соцветиях, редко одиночные (рис. 259).
Цветки актиноморфные, у большинства представителей пятичленные, из пяти
кругов. Околоцветник дифференцирован на чашечку, которая может быть
как раздельнолистной, так и сростнолистной, и венчик, лепестки которого
на верхушке нередко выемчатые или глубокодвураздельные, у многих пред-
ставителей с хорошо выраженным ноготком. На внутренней стороне ногот-
ковых лепестков при переходе от ноготка к отгибу довольно часто наблюда-
ется образование добавочных лепестковидных выростов, образующих в сово-
купности так называемый придаточный венчик, или привенчик. Тычинки
распределены обыкновенно в два круга, часто встречается обдиплостемония.
Гинецеи по числу членов изменчив (5—2 плодолистика), синкарпный, с верх-
ней завязью и центрально-угловой плацентацией. Вследствие разрушения
перегородок между гнездами завязи может образоваться плацентарная ко-
лонка. Семяпочки многочисленные. Столбиков обычно столько же, сколько
плодолистиков.
Плоды у гвоздичных—коробочки, раскрывающиеся створками или зуб*
чиками, редко орешки или ягодообразные. Семена с периспермом и согнутым
(редко почти прямым) зародышем.
Семейство гвоздичных содержит свыше 2000 видов, довольно естествен-
но подразделяется на три подсемейства.
Подсемейство приноготковые (Paronychioideae). Листья по пре-
имуществу с прилистниками, ^источки околоцветника свободные. Часто име-
ются стаминодии, представляющие как бы зачаточные лепестки. Сюда отно-
сятся роды грыжник (Hemiaria) торица (Spergula), торичник (Spergula-
ria) и др.
Подсемейство мокричные (AlSlnoideae) характеризуется раздель-
нолистной чашечкой; лепестки венчика без ноготков (редко лепестки отсут-
ствуют). Типичным представителем этого подсемейства может служить род
. звездчатка (Stellaria, рис. 260), многие виды которого являются обычными
' нашими растениями.
Звездчатка лесная (S. holostea) — растение, образующее в наших лист-
- венных лесах заросли до нескольких квадратных метров.
Кроме лесной звездчатки, у нас встречается еще Несколько видов того
же рода. Из них отметим широко распространенную мокрицу (S. media),
часто встречающуюся на рыхлых влажных почвах как сорное расте-
ние, цветущее в течение всего вегетационного периода. Растение мокрицы,
выросшее из семян, весной достигает плодоношения настолько быстро, что
уже к осени может появиться новое поколение, которое иногда тоже успева-
ет развить семена в тот же год. Способность к быстрому размножению делает
мокрицу весьма надоедливым сорняком, требующим постоянной полки. Неж-
ная, сочная зелень мокрицы представляет хороший питательный корм
для птиц.
П о д с е м е й с т в о смолеековые (Silenoideae) объединяет гвоздичные,
у которых чашечка спайнолистная. Нередко она имеет при этом цилиндри-
ческую удлиненную форму, с чем и связано наблюдающееся у смолевковых
сильное разрастание нижней части лепестка в ноготок. У многих смолевко-
вых осевая часть цветка (цветоложе) образует выше места прикрепления
чашелистиков довольно сильно вытянутое междоузлие, на котором прочие
части цветка значительно приподнимаются над дном чашечки (а н д р о ги-
нофор). К этому подсемейству относится куколь (Agrostemma githago) —
обременительный сорняк, засоряющий яровые, а в более южных районах —
и озимые хлеба. Куколь считается одним из злостных засорителей зерна;
его семена плохо высыпаются из коробочек и таким образом попадают вместе
с урожаем культуры в обмолот, но часто семена все же,осыпаются на месте.
Г111
:i г. i> i
Семена куколя содержат ядовитый алкалоид. Вслед-
ствие этого мука, полученная при размоле зерна,
не очищенного от семян куколя, опасна для здо-
ровья уже при содержании его в количестве 0,5%;
- такая мука имеет неприятный запах и горькова-
тый йкус.
Смолевка поникающая (Silene nutans) замечатель-
на своеобразным опылением и ночным цветением.
Смолка (Viscaria viscosa, рис. 261) — обычное
растение суходольных лугов, полян и опушек —
имеет яркие малиново-красные соцветия и стебель,
покрытый темными смолистыми выделениями,
защищающими цветки от ползающих насекомых,
которые используют нектар, но не способны произ-
Рис. 261. Диаграмма вести опыление. Мыльнянка лекарственная (Sapo-
цветка смолки naria officinalis) — растение, богатое сапонином,
корни которого при растирании в воде дают пену,
напоминающую мыльную, и используются как сур-
рогат мыла. Это растение имеет довольно крупные бело-розовые цветки в
скученном соцветии. Нередко в природе можно найти экземпляры с махро-
выми цветками. Махровая форма мыльнянки культивируется как декора-
тивная.
С точки зрения декоративного садоводства существенный интерес пред-
ставляет род гвоздика (Dianthus), одной из особенностей которого являются
характерные прицветные листья, расположенные парами у основания ча-
шечки и прижатые к ней, образуя как бы вторую, наружную чашечку. Даже
некоторые дикорастущие гвоздики отличаются красотой и из^яществом своих
цветков. Такова, например, гвоздика превосходная (D. superbus) с тонко и
многократно рассеченными лепестками и приятным запахом* Из числа ви-
дов, известных в диком состоянии, материалом для получения .культурных
форм послужили гвоздима бородатая barbaiu^—горное растение За-
падной Европы, имеющее соцветия, скученные наподобие компактного щитка,
и отличающееся громадным разнообразием окраски (белая, темно-фиолето-
вая, кроваво-красная, различные пятнистые), и гвоздима садовая (D. сагуо-
phyllus) с многочисленными махровыми формами, также очень разнообраз-
ными по окраске; цветки этой гвоздики крупные, обладают прекрасным, тон-
ким ароматом.
Семейство гвоздичных распространено по всему свету, но наибольшим
разнообразием гвоздичных отличается Средиземноморье.
Родственные связи и пути филогенетического развития внутри поряд-
ка Centrospermae не могут считаться вполне выясненными, но в целом в этом
обширном порядке, по-видимому, объединено несколько ветвей, развивших-
ся независимо друг от друга^ ио произошедших от общего предка* каковым
признают семейство фитолакковых (стр. 382). Одна из этих ветвей через"
посредство небольшого тропического, преимущественно африканского се-
мейства Molugtnaceae примыкает к более примитивным фитолакковым с апо-
карпным гинецеем. К этой ветви относятся два описанных выше семейства —
айвовые и кактусовые. Оба семейства сформировались, вероятно, в пустын-
ной или полупустынной среде, о чем свидетельствует характерная для них
суккулентность. Но у кактусовых органом хранения запасов воды стал
стебель, у визовых — листья. Из этого следует, что развитие этих семейств
совершалось независимо друг от друга, что подтверждается и географически:
кактусы — растения американские, визовые — преимущественно афри-
канские и австралийские.
Вторая линия развития, берущая, возможно, начало также от фитолак-
ковых, представлена семейством гвоздичных, более примитивные представи-
тели которых или не имеют (подсемейство приноготковые) лепестков, или.
(подсемейство мокричные) обладают примитивными лепестками и чашечкой
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛ» ПОКРЫТОСЕМЕННЫЕ
391
jis свободных чашелистиков. У более высокоорганизованных гвозди*
I: I »
(подсемейство смолевковые) чашечка сростнолистная и лепестки имеют слож-
ное строение. Синкарпный, в основном с центрально-угловой плацентацией
гинецей гвоздичных не противоречит предположению об их родстве с апо-
карпными, т. е. более древними фитолакковыми.
Вероятно, несколько цную связь с последними имеет третья ветвь по ряд-
ка—гвоздичноцветные (центр о семенные), — представленная двумя, возмож-
но, параллельно развившимися семействами — лебедовые и амарантовые
(последнее здесь не рассматривается). Лебедовые с их одногнездной, хотя и
состоящей из 5—2 плодолистиков, завязью и с одной семяпочкой скорее сбли-
жаются с более высокоэволюциоиировавшими фитолакковыми, выделяемыми
в особое подсемейство (Microteoideae), характеризуемое строением гинецея.
и вообще цветка, сходным с лебедовыми.
2-И ПОРЯДОК — ГРЕЧИХОЦВЕТНЫЕ (POLYGONALES)
Цветки (рис. 262) обычно в цимозных соцветиях, большей частью обоепо-
лые, с простым околоцветником из 2—6 листочков, свободных или белее или
менее сросшихся. Число тычинок 6, но вследствие расщепления бывает 9 и
более. Пестик из 2, 3 или 4 плодолистиков, с верхней одногнездной завязью,
содержащей одну основную прямую семяпочку.
Это травы или кустарники, редко древесные растения с очередными
простыми, цельными или лопастными листьями с прилистниками, которые
обычно срастаются и охватывают основание лежащего выше междоузлия, об-
разуя так называемый раструб.
Семейство гречишные (Polygonaceae) — единственное в порядке
гречихоцветных. Оно содержит около 800 видов, в том числе в СССР произ-
растает свыше 280. Наиболее обширным у нас является род горец (Polygo-
num)', куда относятся такие распространенные растения, как спорыш, или
птичья гречиха (Р. avlculare), — обычное по дорогам, на пустырях мелкое
травянистое растение; раковые шейки (P.bistorta); гречиха земноводная (Р. ат-
phibium) — водное растение с плавающими листьями и колосьями розовых
цветков, торчащими над водой, и много других вцдов.
К роду Polygonum близок род Fagopyrum, куда относится культур-
ная гречиха (F. sagittatus) — экономически наиболее важное растение се-
мейства. Это травянистый однолетник, достигающий 70 см. Цветки в щитко-
видных метелках, сидящих на цветоносах, которые выходят из пазух листь-
ев. Цветок с простым пятираздельным, .белым или розовым околоцветником.
Тычинок 8, с ними чередуются округлые нектарники. Пестик с 3 столбиками
и 3 головчатыми рыльцами. Плод —трехгранный орешек. Семя с мучнистым
массивным эндоспермом. Семядоли зародыша листовидные, складчатые.
Из семян получают гречневую крупу и муку. Из листьев в последнее вре-
мя получают рутин (антисклеротический витамин Р). Еще больше рутина
содержит дикая гречиха (дикуша), засоряющая посевы культурной. Гречи-
ха — хороший медонос и в связи с энтомофилией гетеростильное растение
(рис. 263, 264).
Из обширного рода щавель (Rumex) используется щавель кислый (R. ace-
tosay, другие виды известны как растения, засоряющие поля и луга (щавелек—
R. acetosella, конский щавель — R. confertus и другие виды крупных ща-
велей). Щавели ветроопыляемые, некоторые виды (R. acetosa) двудомные.
Вроде ревень (Rheum), представленном крупными травами с больши-
ми, часто глубокопальчаторассеченными листьями, плоды нередко разраста-
ются но ребрам в крыловидные придатки. Некоторые ревени культивируют-
ся как лекарственные (хорошее слабительное средство) и овощные растения.
Полиморфный род джузгун (CalUgonum) широко распространен у нас
в Средней Азии, особенно на песках. Это своеобразные кустарники с реду-
цированными листьями и сероватыми стеблями. Их четырехгранные плоды
имеют по ребрам пленчатые или щетинистые выросты, благодаря которым они
выс
II Г/
Е РАСТЕНИЯ
Ряс. 262. Диаграммы цветков гречишных:
Л — ревень; 9—1*речкха; з — щавель
легко перекатываются ветром и не
засыпаются песком. В оранжереях
часто разводят Muehlenbeckia pla-
tyclada — дерево Соломоновых ост-
ровов, имеющее филлокладии.
Положение порядка гречихо-
цветных в системе покрытосемен-
ных не вполне определенное. Мно-
гие сближают его с центросеменны-
ми, с семейством маревых или с
гвоздичными. Некоторые система-
тики полагают, что гречихоцвет-
ные родственны порядку крапиво-
цветных; они основываются на
сходстве с семейством тутовых в
строев
гг
цветка,
развитии при-
Рис.. 263. Гречиха (Fagopyrum sagittatum):
j—цветущее растение; у — цветок; з —разрез
цветка (в завязи видна одна основная прямая
оемяшяка); 4 —тычинки; S — пестик; в —плод;
у — попертый разрез плода: 3 — продольный раз-
рез плода
Рис. 264. Гетеростилия у гречихи:
1 — продольный разрез дщшносголбчатого цветка:
3—продольный разрез короткостолбчатого цветка
(н — нектарник, р — рыльце, п — пыльник)
листников и срастании их в
раструб.
3-И ПОРЯДОК —
СВИНЧАТКОЦВЕТНЫЕ
(PLUMBAGINALES)
В настоящее время считают,
что порядок свинчаткоцвет-
ные (Plumbaginales) близок пред-
кам Caryophyllales. В единствен-
ном семействе этого поряд-
ка — свинчатковые (Plumbagina-
сеае) — объединены кустарники,
чаще полукустарники и травы
засушливых территорий среди-
земноморских стран, Передней и
Центральной Азии, связанные с
более или менее засоленными суб-
стратами. Цветок у свинчатко-
вых с двойным околоцветником,
пентамерный. Чашелистики часто
сросшиеся, остающиеся при пло-
дах, выполняют защитную роль
и способствуют распространению
плодов. Венчик спайнолепест-
ный, иногда лепестки срастаются
не полностью. Пять тычинок про-
тивостоят лепесткам и срастаются
с ними. Гинецей паракарпный, из
5 плодолистиков; завязь верхняя,
одногнездная, содержит одну се-
мяпочку. Плоды—орехи или ко-
робочки. Семена с эндоспермом,
но без перисперма.
В СССР распространены виды
близких родов кермеков — Sta-
tice, Goniolimon и Limonium. Не-
которые из них образуют перекати-
поле. Корни кермеков содержат
дубильные вещества. К этому по-
рядку относятся и встречающиеся
ОТДЕЛ ЦВЕТКОВЫЕ, или покрытосеменные
в Средней Азии и в Закавказье акантолимоны, образующие колючие
подушки. Некоторые виды свинчатковых декоративны.
Порядки Polygohales и Plumbaginales более специализированные, чем
Caryophyllales, и, вероятно, произошли от последних. Относительно прими-
тивные представители подкласса Caryophyllidae содержатся .в порядке Са-
ryophyllales.
Как упоминалось выше (стр. 382), более примитивные представители
подкласса входят, по всей видимости, в семейство Phytolaccaceae и через них
весь подкласс Caryophyllidae примыкает к подклассу Magnoliidae,
л
4-Й ПОДКЛАСС - ДИЛЛЕНИИДНЫЕ (D1LLEN11DAE)
В качестве характерного признака диллениидных выдвигается необыч-
ная последовательность развития тычинок, а именно: при большом числе
тычинок они развиваются центрифугально. Плацентация чаще всего угло-
вая и постенная. Семена с прямым зародышем. Некоторые группы отлича-
ются спайнолепестностью, но при этом тычинок бывает больше, чем долей
венчика, или они противолепестковые. Растения этого подкласса разнооб-
разной жизненной формы.
Из относимых к подклассу Dilleniidae 12—16 порядков ниже
рассматриваются десять. Порядки Dilleniales и Theales стоят несколько
лическое строение цветков, наличие перехода между верховыми листьями и
чашелистиками, апокарпность или неполную ценокарпность гинецея, лест-
ничную перфорацию члеников сосудов и др.
Из прочих порядков три — Ericales, Ebenales и Primulales — выде-
ляются преобладанием спайнолепестности. В тех системах покрытосеменных,
где двудольные подразделяются только на подклассы свободнолепестных и
спайнолепестных, эти порядки находят себе место среди спайнолепестных.
Порядок фиалкоцветные (Fiolales) объединяет очень разные се-
мейства. Общее для них — циклические цветки с раздельнолепестным вен-
чиком, паракарпный (или вторично синкарпный) гинецей с тремя плодолисти- *
ками и верхней завязью и ряд других признаков. Этот порядок является
узловым; от него, по-видимому, взяли начало предки порядков, специализи-
рованных в том или ином отношении, например Cucurbitales с распростра-
ненными у них усиками и ягодообразными плодами, Capparales с двучлен-
ным гинецеем и др.
Порядок малъвоцветные (Mai vales), характеризуемый, подобно
Violates, циклическим, в основном пентамерным, цветком, выделяется тен-
денцией многочисленных тычинок к срастанию в пучки или в трубку-колон-
ку, окружающую гинецей. С этим порядком часто считают связанным поря-
док Euphorbiales, выделяющийся почти исключительно однополыми цветка-
ми, с сильно редуцированным околоцветником или без него.
1-Й ПОРЯДОК — ДИЛЛЕНИЕЦВБТНЫЕ (DILLENIALES)
Околоцветник спиральный, полукруговой или круговой. Тычинки мно-
гочисленные. Гинецей апокарпный или не вполне синкарпный, каждому
плодолистику соответствует несколько семяпочек. Семена с небольшим за-
родышем и обильным эндоспермом, часто с ариллусом.
Семейство диллениёвые (Dilleniaeeae) распространено в тропиче-
ских и субтропических лесах обоих полушарий. Древесные, иногда лазаю-
щие растения, редко травы. Листья спиральные, обычно цельные, цельно-
и
крайние, без прилистников. Цветки спирально-циклические с постепенным
переходом от верховых листьев к чашелистикам. Последние в неопределенном
числе или их 5 и даже 3; они остаются после цветения и часто при этом раз-
растаются. За 5—2-м лепестками следуют многочисленные спиральные, цен-
вы
Hitt;
Л РАСТЕНИЯ
трифугально развивающиеся тычинки; вследствие редукции число их может
быть и небольшим (10 и меньше). Плодолистиков много. Как и тычинки,.
они могут редуцироваться, оставаясь в ограниченном числе (до 1). Гинецей
апокарпный. Плоды раскрываются по брюшному и спинному швам, а в слу-
чае одного семени — нераскрывающиеся. Имеются присеменник и массив»
ный эндосперм, богатый крахмалом и алейроновыми зернами.
Семейство пеоновые (Paeoniaceae) содержит только один род Раео-
nia, представленный в субтропических, отчасти в умеренных областях се-
верного, полушария (2 вида в Северной Америке) травами, нередко полу-
кустарниками с крупными тройчатыми или дваждытройчатыми листьями
без прилистников, корни клубневидно утолщены. Цветки одиночные, круп-
ные. Чашелистиков 5 или более; в одних случаях они зеленые, в других с пе-
реходами по окраске и консистенции к лепесткам или к верхушечным и че-
рез них к срединным листьям цветоносного побега. Лепестков 5—8 (до 15).
Многочисленные тычинки развиваются центрифугально; множественность
тычинок, возможно, вторичная. Гинецей апокарпный из 5 — 2 плодолис-
тиков, короткие столбики завершаются расширенными рыльцами. В осно-
вании цветка имеется нектарный диск, особенно у кустарниковых (более при-
митивных) видов; у некоторых видов опылители — жуки. Семяпочки —
в двух рядах по брюшному шву завязи; наружный интегумент превышает
внутренний. Плоды—листовки с толстыми стенками и несколькими семенами.
Семейство пеоновые имеет строение зародышевого мешка и развитие за-
родыша, неизвестные у других покрытосеменных.
Пеоны широко распространены в садовой культуре. Декоративный ин-
терес представляют и дикорастущие виды, как, например, воронец (Paeonia
tenuifolia), с тонко рассеченными листьями и темно-красными цветками, рас-
тущий в степях европейской части СССР.
Пеоновые до недавнего времени включали в семейство Ranunculaceae,
выделяя в особое подсемейство.
мт.
Я-Й ПОРЯДОК — ЧАЕЦВЕТНЫЕ (THEALES)
*. Цветки чаецветных обоеполые, обычно правильные. Околоцветник спи-
ральный или циклический, пентамерный, из свободных или почти свобод-
ных листочков. Тычинки часто многочисленные, развиваются центрифугаль-
но. Нити тычинок различным образом срастаются друг с другом. Гинецей
апокарпный или, чаще, ценокарпный (синкарпный и паракарпный). Стол-
бики в числе плодолистиков или столбик один. Завязь верхняя, иногда полу-
нижняя, с многочисленными семяпочками. Семена с крупным зародышем и
скудным эндоспермом, или последнего нет.
Главным образом деревья и кустарники, иногда лианы, редко травы.
Листья очередные, реже супротивные или мутовчатые, обычно простые, цель-
ные, с прилистниками или без них. Характерно наличие межклеточных
секреторных каналов.
Семейство чайные (Theaceae). Это деревья или кустарники, обыч-
но вечнозеленые, с простыми, спирально расположенными листьями, без
прилистников. Цветки обоеполые, полукруговые или круговые, с не вполне
определенным в пределах семейства, а нередко и в пределах одного вида
числом частей. Так, число чашелистиков 7—5, лепестков 5—4, донеопреде^
денно большого числа; листочки околоцветника свободные или у основания
сросшиеся. Тычинок большей частью много, причем они иногда распреде-
лены в 5 групп и срастаются с основаниями лепестков. Плодолистиков 6—2,
редко сросшихся только у основания, обычно они образуют единый пестик
с верхней, двух-многогнездней завязью; число столбиков соответствует
числу плодолистиков, или имеется один только, столбик, который заканчи-
вается одним или несколькими рыльцами. Плацентация центрально-угловая.
Плоды — коробочки либо орехоподобные, нераскрывающиеся. Семейство чай-
ных включает около 600 видов, растущих в тропиках и субтропиках.
иг.
’ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Наибольшее значение имеет род
камелия (Camellia). Чайный куст (Ca-
mellia sinensis, рис. 265)— вечнозеленое
растение, кустарник или небольшое де-
рево, с кожистыми листьями эллипти-
ческой формы. Довольно крупные цвет-
ки имеют 5—7 чашелистиков, сохраня-
ющихся при плодах, 5—9. белых или
розовых лепестков, многочисленные ты-
чинки и пестик с верхней трехгнездной
завязью, из которой развивается трех-
гранная коробочка с одним семенем в
каждом гнезде. Родина чайного куста —
Юго-Восточная Азия; его культиви-
руют в тропиках и субтропиках (Ин-
дия, Шри Ланка, Индонезия, Китай).
Культура чая в более удаленных от тро-
пиков районах началась сравнительно
Рис. 265. Чай (Thea sinensis). Цве-
тущая ветвь
недавно.
Первые попытки производствен-
ного разведения чайного куста у нас
. в Закавказье относятся к концу про-
шлого века. Но после Великой Октябрь-
ской социалистической революции
эта культура получила в нашей стра-
не большое промышленное значение,
заняв одно из первых мест среди важ-
нейших субтропических культур. В на-
стоящее время потребности нашей стра-
11:1:
ны в чае все более удовлетворяются за счет отечественной продукции.
Чайный куст выделяют иногда в особый род Thea (чай) с несколькими
видами, сводимыми в последнее время к разновидностям. Из них следует
упомянуть чай ассамский (Г. assamica). Имеются сведения о произрастании
чайного дерева в диком состоянии в верховьях реки Меконга.
К роду Camellia относится и красивый декоративный кустарник
Camellia japonica, родом тоже из Юго-Восточной Азии, разводимый в мас-
се сортов ради крупных белых или розовых махровых цветков. Имеются пе-
реходы между верхушечными листьями, чашелистиками, а иногда и лепестка-
ми. Тычинки у камелии многочисленные. Пестик один.
Семейство зверобойные (Hypericaceae, Gut tiferae). Деревья, кус-
тарники, травы, редко лианы. Листья простые, без прилистников, цельно-
крайние, обычно супротивные или мутовчатые. В листьях имеются просве-
чивающие точки — вместилища масел. В тканях осевых органов, в череш-
ках и жилках листьев имеются секреторные каналы. Актиноморфные цветки
с двойным околоцветником собраны в цимозные соцветия. Тычинки много-
численные, часто сросшиеся пучками. Гинецей ценокарпный, состоит из
5—3 плодолистиков. Завязь верхняя, с числом гнезд, соответствующим чис-
лу плодолистиков. Иногда завязь одногнездная. Плацентация краевая, ред-
ко постенная. Столбиков столько, сколько плодолистиков, или один. Пло-
ды сухие (коробочки) или сочные (ягодообразные). Семена без эндо-
сперма.
Зверобойные распространены в тропических и субтропических областях
обоих полушарии. В умеренных областях встречаются преимущественно в се-
верном полушарии. Наиболее обширный род — зверобой (Hypericum,
рис. 266). В нем насчитывается около 350 видов, большая часть которых
встречается в Средиземноморье. В составе этого рода имеются кустарники,
полукустарники, многолетние и однолетние травы, даже одно водное травя-
нистое растение (Я. elodes). Прочие виды — травянистые многолетники^ выде-
ВЫСППШРАСТ
Рис- 266. Зверобой:
1 — Hypericum perforatum: а — нижняя часть растения; б — соцветие; в — цветок; 9 — дваграима цвет*
на; О — поперечный разрез завязи; е — плод; 8 — Hypericum aegyptiacum — положение тычинок
ляются супротивными простыми листьями с характерными просвечивающими
железками. Прилистники отсутствуют.
К тому же (или близкому) семейству относится мангустам (Garcinia
mangostana). Плоды этого дерева 5—6 см в поперечнике, покрытые толстой
темно-красной кожурой, под которой находится нежная, белая, сладкая,
ароматная мякоть. Родина мангустана, вероятно, полуостров Малакка, но в
культуре это растение встречается по всей тропической Азии.
Многие деревья этого семейства имеют значение как источник ценной
древесины. Из других добывают смолистые вещества и камеди.
Семейство двукрылоплодниковые (Dipterocarpaceae) близкородст-
венно зверобойным. Двукрылоплодниковые—деревья с очередными простыми
листьями, имеющими прилистники. Цветки собраны в метельчатые соцветия.
Цветоложе выпуклое, плоское или даже вогнутое, иногда образующее ко-
роткий андрогинофор. Чашечка и венчик пятичленные. Чашелистики у осно-
вания сросшиеся. Андроцей из 5, 10, 15 и большего числа почти всегда сво-
бодных тычинок. Гинецей из 1—3 плодолистиков, синкарпный. Завязь трех-
гнездная с двумя — многими семяпочками в каждом гнезде, верхняя иногда
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОС
л ,т 1 и: t
1ЫВ
397
Рис. 267. Двукрылоплодниковые и маркгравиевые:
1 — саповое дерево: а — листья» соцветий плоды; б — цветок; е — продольный разрез цветня; 2 — плод
дшперокарпуса; 3 — соцветие зонтичной маркгравии (ц — кувшинчатые прицветники, ца — редуциро-
ванный цветок)
I j; I
погруженная в полое цветоложе или даже сросшаяся с ним. Плод сухой,
нераскрывающийся, односемянный (орех). Своеобразной особенностью се-
мейства является то, что 2, 3 или все 5 зубчиков чашечки, сохраняющейся
при плодах, после цветения разрастаются и, принимая крыловидную форму1
способствуют распространению плодов (рис. 267, 2, 2).
Двукрылоплодниковые — одно из наиболее типичных палеотропических
семейств, виды которого обитают преимущественно в Индостане, в Индокитае,
на островах Малайского архипелага, на Новой Гвинее и Филиппинских
островах; в Тропической Африке двукрылоплодниковые менее многочисленны.
Общее число видов в семействе достигает 400* Некоторые двукрылоплод-
никовые — важные лесообразующие породы, имеют высококачественную
древесину и содержат ценные смолы.
Dipterocarpus turbinatus— могучие деревья, нередко, как и другие виды
рода Dipterocarpus, образующие почти чистые насаждения. Из их стволов
местное население изготовляет долбленые лодки (пироги), поднимающие до
100 человек. Из того же дерева добывают так называемый ардшанский баль-
зам* Источником смолы «даммара», используемой в технике (лаки, особенно
прозрачные, линолеум и др.) и медицине (пластыри), служат виды рода
Shorea. Самвое дерево (Sh< robusta), распространенное в муссонных и других
тропических лесах Юго-Восточной Азии, способно образовывать чистые или
почти чистые насаждения; имеет ценную древесину (рис. 267,1). Dryobala-
nops aromatica — одно из величественных деревьев Суматры, Борнео и Ма-
лакки, со стволом, не имеющим ветвей до высоты около 50 ж. Крона с мно-
жеством ароматических цветков. Дерево служит источником борнейской кам-
фарыл отличающейся особым ладанным запахом.
t
вы
Hili',
В PACT
Я
Рис. 268. Актинидия ананасная Мичурина
II
иг
Семейство маркгравиевые (Marcgraviaceae) — небольшое тропи-
ческое южноамериканское семейство, состоящее из лиан и эпифитов. Заме-
чательно оно диморфизмом листьев и приспособлениями к опылению. Тене-
вые побеги маркгравиевых, лазающие с помощью придаточных корней, по-
крыты сидячими, двурядно расположенными листьями, которые почти при-
легают к субстрату, напоминая в этом отношении плющ. Световые побеги
имеют листья иного строения в несут на себе поникающие соцветия. В со-
цветиях зонтичной маркграсии (Marcgravia umbellate,) морфологически верх-
ние прицветники метаморфизированы в кувшинчатые нектарники, срастаю-
• щиеся с цветоножками недоразвитых, морфологически верхних цветков.
. Нижние цветки развиты нормально, сидят на более или менее горизонтальных
цветоножках, а их отверстия направлены к морфологической вершине со-
цветия. В целом соцветие напоминает зонтик, в центре которого торчит пучок
нектарников (рис. 267., 3). Но это соцветие поникает вниз, и поэтому во вре-
мя цветения цветки занимают физически более высокое положение, чем нек-
тарники, а отверстия цветков бывают направлены (физически) вниз и из них
свешивается многочисленные тычинки, а позднее рыльца. Вниз свисают
в это время и нектарники. Маркгравиевые отчасти энтомофильны, отчасти
орннтофильны. В последнем случае в роли опылителей выступают колиб-
ри. Маленькая птичка парит около соцветия и пьет нектар, при этом она ка-
сается пыльников или рылец. Вследствие протерандрии самоопыление исклю-
чено.
К этому порядку относится и небольшое (300 видов), преимущественно
азиатское семейство актинидиевые (Actlrridlaceae), в котором наряду
с другими находит себе место небольшой род актинидия (Actinidia), ви-
ды которого распространены главным образом в Китае и Японии, но некого-
рые виды заходят к северу, в область умеренного климата, вплоть до Амур-
ской области. В пределах СССР (на Дальнем Востоке) растут три вида акти-
нидий, все — лианы, из которых наиболее крупная — актинидия острая
* (4. atguta), стебель которой достигает 10 см в диаметре. Эта лиана при-
урочена к хвойным и смешанным лесам. Плоды актинидий ягодообразные,
съедобные. Из A. kolomikta, растущей на Дальнем Востоке, И. В. Мичурин
вывел несколько сортов культурной актинидии (Ананасная Мичурина^
рис. 268).
С примитивными чаецветными или с их диллениецветными предками
связывают еще некоторые линии эволюции, из которых здесь рассматрива-
ются 3: линия мальвоцветных — молочайных^ линия вересковых и линия
,|:гг
।: г,
I
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
3-И ПОРЯДОК — МАЛЬВОЦВЕТНЫЕ (MALVALES)
II
Цветки мальвоцветных круговые, из пятичленных кругов, актиноморф-
ные, обыкновенно обоеполые, с двойным окодоцветником, причем лепестки
в почкосложении скрученные, а чашелистики створчатые. Из двух кругов
тычинок один (большей частью наружный) обычно недоразвивается, второй
• состоит из многочисленных тычинок, возникших благодаря ветвлению (рас-
щеплению) тычиночных нитей. Нити тычинок срастаются друг с другом в
II
III л
ев шогогнездноилавяз ью.
*Д^ёвёснь1ё, ршш Травянистые растения, с простыми, редко* сложными
очередными листьями, имеющими прилистники. В анатомическоъГЪтнойении
характерны слизевые ходы, слизевые клетки, лубяные волокна в коре и
наличие ветвистых волосков.
Семейство мальвовые (Malvaceae). Травянистые растения или кус-
тарники, редко деревья с простыми, обычно падьчаторасчдененят^и листья-
ми? В основании цветка часто имеется «наружная чашечка», образованная
верхушечными листьями. Чашечка^и венчик пятичленные. Лепестки венчи-
ка свободные или в лсрпйянии српстппеся, Пять тыштпк наружного круга
редуцированы,, внутренние — многочисленные (вследствие расщепления) и
только с двумя щ^льпевыми тндяпями.иыльпевые зерна круцщде со слож-
нпскул^птуриой якяиипй. Пестик ив трех — многих плодолистиков. Стол-
бик олиниди. несколько. Завязь верхняя. Плодысухие—коробочки либо J№r
носемянйые орешки.-Семейство включает свыше 1&UU видов. .
_ TL.‘ ***** — 1 I JI- 1 ijf- г
Рис. 269. Хлопчатник:
* 1 — цветущая ветвь; 3 — цветок; з — диаграмма цветка; 4 - нераскрывшаяся коробочка; 3 — поперечный
разрез коробочки; в раскрывшаяся коробочка; 7 — семя с расчесанными волосками
4
400
ВЫСШИЕ РАСТЕНИЯ
Г-1 W *1
ь ОДИН день. В JBlqhk
.111
иг. i:Tw
иг
,111
Наиболее важный род этого семейства хлопчатник (Gossypium^
рис. 269). Виды хлопчатника — сидьцовэтвиатмй тряпм .лди^кустарннкиг
достигающие 1,5—2 лс, иногда почти древовидные. Кустарниковые формы
разводят только^тоопНКйх: внетропнческое хлопководство базируется на од-
нолетней культуре хлопчатника. У- хлопчи^шса различаются два рода вет-
вей — ростовые и плодущие; первые монрпрдиадьные, вторые симподиальные
и укорочёшые. Каждая плодущая вехвь^1аканчивается_цветкомт11избоковой
почки, дежащейниже первого цветка^^лифЗ^^^ере®цветок и т. д. Ча-
шечка србСтнолистНая,пятилопастная или усечешая, без лопастей. Пять ле-
пестков вентакйГокрашеныв светло-кремовый иди желтмй цвет. Основание
тычинонцои ^рубкисрастается сГ ледестками. В составе гинецея участвуют
3 плодолистиков; гтодКю рылктуш. Цветение
Каждого цветка, продолжается лишь один день. В ^рдьшинсхве случаев
хлопчатник самоопыляется, но наблюдается и перекрестноелшыдетие, чему
отвечает налйтаё! нектарников у основания тычинпчипй /трубкиГ^Цветение
всего куста продолжается^^течение многих недель. При этом развивается
огромное количество цветочных почек,; большаягчасть которых, как и большая
часть распустившихсяцветкбв и недозрелых коробочек, к сожалению, опа-
дает, причем доля опавших завязей может достигать 70% и более.
Мерами борьбы с опаданием являются своевременное удаление боковых
ветрей^ниже первой^цлодущей и прищипывание ростовой ветки с целью ог-
ранийтагчиилб^плодущих (так называемая чеканка хлопчатника).
Плоды хлопчатника — коробочки, раскрывающиеся 3—5 створками и
содержащие многочисленные семена, покрытые длинными волосками. Во-
лоски используются как прядильный материал (хлопок). Длина их у неко-
торых сортов хлопчатника может достигать 50 и даже 60 лсж, но обычно быва-
ет 30—40 лм*. Волокна состоят почти из чистой клетчатки, имеют скрученную
форму, хорошо красятся в разные цвета. В общем мировом потреблении
волокна для изготовления Тканей на долю хлопка приходится бо-
лее 70%.
Дикие виды хлопчатника, распространенные преимущественно в тро-
пических странах Старого и Нового Света, обитают в солнечных засушли-
вых местностях. Это кустарники, полукустарники и Небольшие деревья.
Культура хлопчатника древняя. В Старом Свете доказано хлопкопря-
дение и изготовление хлопчатобумажных тканей еще за 3000 лет до н. э.
(Синд), а в Новом Свете за 2400 лет до н. э. (Перу). В половине I тысячеле-
тия до н. э. культура хлопчатника была распространена на территории сов-
ременных среднеазиатских республик.
В культуру вошли преимущественно скороспелые однолетние сорта.
Семена хлопчатника содержат около 25% масла, пригодного для пище-
вого и технического использования. Жмых, остающийся после прессовки се-
мян, богат (около 30%) белком и годен на корм скоту. Жмыховую муку
применяют на удобрение под хлопчатник.
В настоящее время наиболее обширные площади заняты культурой хлоп-
чатника в АРЕ, Индии, США. Хлопководство в СССР наиболее северное,
главным образом поливное. Более 60% хлопка получают в Узбекской ССР.
В других республиках Средней Азии (Туркменской, Таджикской, Киргиз-
ской и южной части Казахской) получают примерно ежегодного урожая.
В Закавказье культура хлопчатников сосредоточена в основном в Азер-
байджане.
Кроме хлопчатника, в семействе мальвовых имеются еще два промыш-
ленно значимых, хотя и гораздо менее ценных, волокнистых растения — ке-
наф и канатник.
Кенаф (Hibiscus cannabinus) называют еще бомбейскойцельной, так как
центром его культуры является Индия, отчасти Иран. Иодтеблей кенафа до-
бывают светлое, мягкое и гибкое волокно, мало устуц^щрЬ волокну джута
(стр. 405), а при хорошей обработке приближающееся к льняному; исполь-
зуется для производства мешковины, а также в текстильной и химической
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
401
промышленности. В СССР ке-
наф возделывают в нехлопко-
вых районах Киргизии и в по-
ливных районах Казахстана.
Яошипкик Теофраста (А bu-
llion theophrasti,A. avicennae)—
тоже травянистое растение,
достигающее 4 м в высоту, с
кру
цельными сердце-
видными листьями. Оно встре-
чается у нас в южных райо-
нах европейской части СССР,
в Средней Азии и на юге Даль-
него Востока. Культура канат-
ника издавна известна в Китае.
Волокно, получаемое из канат-
ника, используют на изготовле-
ние мешковины, веревок,
пи
гатов, а отходы употребляют в
производстве бумаги. Семена
содержат техническое масло.
Из других мальвовых сле-
дует отметить род алтей
(Althaea), корни одного из ви-
дов которого (алтей лекарст-
венный — A. officinalis, рис.
270) употребляют при различ-
Рис. 270. Алтей (Althaea officinalis):
J — цветущее растение; S — разрез цветка; з — андро-
цей; 4 — гинецей; s—цветка (п — листки-
ни подчашия)
ных катарах как средство,
уменьшающее раздражение.
Широко известна шток-роза,
нага, лмалъва», вид близкого
рода Alcea (A. rosea); из ле-
пестков ее добывают краску для
[евых продуктов и напитков,
тах под названием китайской розы Hibiscus rosa-chinensis имеет крупные тем-
iiiii
а также для тканей. Разводимый в комна-
но-красные воронковидные цветки, часто махровые.
Семейство баобабовые (Bombacaceae) — почти исключительно тро-
пическое семейство, распространенное как в Новом, так и в Старом Свете,
представители которого (200 видов) имеют нередко ландшафтное значение.
Баобабовые — древесные растения с цветками, очень похожими на цветки
мальвовых. От этого, несомненно, близко родственного семейства баобабовые
отличаются лепестками, густо покрытыми волосками, пыльниками, в боль-
шинстве случаев четырехкамерными, и гладкой (а не шиповатой, как у маль-
вовых) пыльцой. Опыляются насекомыми и птицами.
Одним из наиболее известных представителей семейства является бао-
баб (Adansonia digitata, рис. 271), растущий в саваннах Африки. Неук-
люжее по своему габитусу, это не особенно высокое дерево достигает 18—25 м
в высоту, при диаметре ствола 4—10 м. Отметки на стволах баобаба, сделан-
ные в XV в., позволяют предположить, что некоторые деревья доживают до
5000 лет. На ветвях крупной кроны располагаются пальчатосложные листья,
по форме напоминающие листья конского каштана. Обитая в саваннах, где
климат характеризуется периодической сменой сухого и влажного времени,
баобаб сбрасывает свою листву на время засухи. К этому моменту созревают
in:
плоды, имеющие вид длинных тыкв, которые, свешиваясь с дерева, придают
ему в безлистном состоянии совершенно своеобразный вид. Сочная кисло-
ватая мякоть плодов и семена съедобны. Кроме плодов, используют толстую
кору баобаба для выделки бумаги. Древесина технически малоценная, так
как она очень рыхлая вследствие обильного развития паренхимы.
tl!
ННГ,Р
PACT
л: i;
По своеобразию внешнего облика с африканским баобабом можно со**
поставить некоторые южноамериканские баобабовые с огромными ствола**
ми-водовместилищами, как каваниллезия древовидная (Cavanillesia ar bore а),
характерное дерево сухих лесов Восточной Бразилии, и типичная для каа-
тинги Южной Америки Chorisla cripifolia. У первого из них ствол имеет вид
огромной редьки; он сравнительно тонок в основании, затем расширяется ве-.
ретенообразно и довольно быстро суживается к вершине, несущей редкую
крону.
К этому семейству относятся тропические хлопчатые, или шерстяные,
деревья (Ceiba, Bombax), в плодах которых развивается масса * волосков, по-
крывающих внутреннюю поверхность околоплодника (а не семена, как у маль-
вовых). Наиболее известно из этих деревьев Ceiba pentandra, родом из Юж-
ной Америки, разводимое по всей тропической зоне. Волоски из его плодов,
образующие вату, собирают и используют, под названием капока, для набив-
ки подушек. Вследствие ломкости волосков капок не годен для изготовления
пряжи.
К семейству баобабовых относится также бальзовое дерево (Ochroma la-
gopus), растущее в тропиках Южной Америки и на Антильских островах.
На плоту (balsa — от исп. «плот>) из этого дерева, названном «Кон-Тики»,
6 скандинавских исследователей во главе с Т. Хейердалом проплыли около
8000 км от берегов Перу до Полинезии. Плотность бальзового дерева 0,12—
0^30; оно используется для теплоизоляц
спасательных поясов и буев.
Рис. 271. Баобаб (Adanscmia digitate)
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис. 272. Шоколадное дерево (Theobroma cacao):
1 "цветущая ветвь; 3—воюдн шоколадного дерева на стволе (каулифлория): 3— цветок: а —• общн*
вял. 6 — разрез цветка; з — анаграмма цветка; 3—продольный разрез нлода
н 1411
:»и
»I '< 1
Индейцы Южной Америки в давние времена плавали на плотах из этого де-
рева по океану.
Своеобразен плодовый представитель баобабовых—дуръян (Durio zibeti-
nus), родом из Индии. Крупные, величиной с голову человека, плоды этого
дерева имеют поверхность, усаженную крупными шипами. Внутри плода
находится мякоть, которую по нежности сравнивают со сливочным кремом,
но ее запах через некоторое время становится похожим на смесь запаха тух-.
лых яиц и сильно пахнущего лука.
Семейство стперкулиевые (Sterculiaceae) — исключительно тро-
пические древесные, реже травянистые растения м обоеполыми цветками.
К этому тропическому семейству, число видов которого достигает 1000, от- -
носится шоколадное дерево (Theobroma cacao, рис. 272). Это небольшое дере-
" во из Южной и Центральной Америки сейчас возделывают в тропиках Ново-
го и Старого Света. Листья крупные, темпо-зеленые, эллиптические. Цветки
мелкие, расположенные пучками., выходящими из старых ветвей (каули-
флория). Плоды в виде ребристого огурца, желтого, оранжевого или
красного цвета. В каждом плоде содержится 20—30 и более крупных семян,
называемых какао-бобами; из них получают порошок какао-масло4 идущий
на приготовление шоколада.
Семена деревьев рода кола (Cola, Западная Африка), богатые кофеи-
ном ж теобромином, используют для приготовления соответствующих меди-
цинских препаратов.
Семейство липовые (ТШасеае). Деревья,/ кустарники, редко тра-
вянистые растения с простыми, цельными или лопастными листьями, с часто
опадающими прилистниками. В вегетативных органах обыкновенно имеют-
ся вместилища слизи. Цветки, как правило, в цимозных соцветиях, обык-
новенно двуполые, реже однополые, с чашечкой и венчиком; лепестки до-
вольно крупные, ярко окрашенные, часто с нектарниками. Андроцей очень
изменчив. Основной его план — два пятичленных круга свободных тычинок,
но в такой форме он встречается только у некоторых экзотических представив
tl!
404
ВЫСШИЕ РАСТЕНИЯ
Рве. 273. Липа (Tilia cor*
data):
1 вепь липы в цвету; ляо-
3 — paspes цветка; 4 — диа-
грамма цветка; £—тиерееццВ
разрез плода и еммин
телей этого семейства. Чаще же благодаря расщеплен
возрастает, и они образуют два круга пучков, по 5 в каждом. Иногда тычин-
ки пучков одного из кругов заменяются стаминодиями или полностью реду-
цируются, и тогда в цветке остается только один круг многотычинковых
пучков. Границы между пучками могут быть неясными, и в этом случае анд-
роцей выглядит как многочленный с неопределенным числом тычинок. Гинет
или многогнездная; столбик один, с головчатым рыльцем. Плоды сочине н ли
сухие, раскрывающиеся или нераскрывающиеся.
К этому семейству в нашей флоре относится род липа (TiZia), который
содержит около 10 видов, обитающих в умеренных широтах северного полу-
шария. Наиболее распространена в СССР липа сердцевидная (Т. cordate,
рис. 273). Это крупное, уимполиально। нарастающее дерево с богато олист-
венной кроной. Взрослые^ липы, выросшие в насаждениях, имеют правиль-
ный колоннообразный ствол. Сердцевидные листья располагаются в очеред-
ном порядке и при распускании почек сбрасывают свои тедкие пленчатые
прилистники. Липа цветет в серединулета^ покрываясь массОЙ~крушвых
щиткообразных соцветии^имеющцх_3—5 ем в поперечнике.- Общий цветонос
II
сохраняющийся при плодах и, может быть, способствующий их распростра-
нению ветром. Цветок липы имеет щятилистную чашечку, 5 белых, слегка
кремовых^лепестков и многочисленные тычинки в 5 пучках. Завязь верхняя,
пдтигдеадвая, с двумя семяпочками в каждом №езде. Столбик^заканчивается
головчашм^ры&ьцеИ' При созревании плода из 5 гнезд завязиЗ йлиКнедо-
II
с яркой окраской привйекаеРнасекомых, в частности пчел. Последние соби-
рают в цветках липы нектар, выделяемый железистыми утолщениями в ос-
новании лепестков. Прекрасный по аромату и вкусовым качествам липовый
мед высоко щенится. Высушенные цветки липы используются в медицине —
«липовый цвет». Принимают его в виде настоя как потогонное средство.
ОТДЕЛ ЦВЕТКОВЫ®, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Древесина липы мягкая, ровная, легко режущаяся. Ее используют пре*
имущественно для столярных изделий. Из коры получают лыко, идущее, в
частности, на различные плетения, и мочало.
Липа — одно из .важных лесообразующих древесных пород, местами
образует чистые или почти чистые насаждения, чаще же входит в состав дуб*
рав.
В Индии и Китае культивируют джут (Сor chorus capsularis) — одно-
летнее травянистое растение, из стеблей которого получают волокно для
.приготовления таровой ткани (джутовые мешки), веревок и каната. Культура
джута возможна в Северной Азии и Закавказье.
uii
4-Й ПОРЯДОК — МОЛОЧАЕЦВЕТНЫЕ (EUPHORBIALES)
Цветки молочаецветных большей частью однополые или с рудиментами
недостающего пола, безлепестные или вовсе без околоцветника, редко около
цветник двойной. Тычинки свободные или сросшиеся.' Гинецей обычно из 3
плодолистиков, ценокарпный, с верхней завязью, содержащей одну или две
семяпочки в каждом гнезде. Плоды—коробочки, редко ягоды или орешки.
Растения очень разнообразного внешнего облика. Главным образом
деревья, реже кустарники, вьющиеся растения, многолетние и даже одно-
летние травы.
Порядок молочаецветных многие авторитетные систематики сближают
с мальвоцветными. Как у Malvalest у молочаецветных при превращении за-
вязи в плод наблюдается ее распадение на односемянные орешки. Есть
некоторые черты сходства и в строении вегетативных органов. Наряду с этим
высказывается и другое мнение — что этот порядок следует помещать в под-
класс Rosidae.
Порядок объединяет небольшое число семейств, причем включение неко-
торых из них в состав молочаецветных оспаривается.
Семейство самшитовые (Вихасеае) содержит всего около 60 видов,
распространенных в умеренных, субтропических и тропических странах. Пре-
обладают древесные и кустарниковые формы (редко полукустарники) с цель-
ными, лишенными прилистников, супротивными (реже очередными) листьями.
По Черноморскому побережью Кавказа, а также в средиземноморских
странах и на востоке до Гималаев растет самшит (Buxus sempervirens), на-
зываемый иногда «кавказской пальмой». Это теневыносливое дерево, требо-
вательное к влажности воздуха. Оно достигает 300—400-летнего возраста*
сохраняя при этом небольшой диаметр ствола и высоту до 20—21 ж. Дре-
весина твердая, используется на ценные токарные изделия, граверные доски,
на челноки для текстильной промышленности. Часто разводится как декора-
тивный кустарник, хорошо переносящий формовку.
Семейство молочайные (Euphorbiaceae) — очень крупное семей-
ство, насчитывающее около 7500 видов. Представители молочайных встреча-
ются по всему свету, за исключением полярных стран и холодных высоко-
гории. Наибольшее разнообразие молойайных наблюдается в Юго-Восточной
Азии, Тропической Африке и тропической Южной Америке, причем в Юго-
Восточной Азии сосредоточены наиболее примитивные роды. Наряду с де-
ревьями, кустарниками и травами обычного облика в составе семейства
имеются формы оригинального строения, напоминающие, например, верес-
ковые или безлистные стеблевые суккуленты — кактусы. Листья большей
частью имеют прилистники и простые, реже сложные, разнообразные по фор-
ме пластинки, иногда редуцированные, в связи с чем наблюдается образова-
ние филлокладиев (род Phyllanthus, рис. 274). Для многих молочайных
характерны млечные трубки, клетки или мешки. Цветки в простых или слож-
ных соцветиях, причем элементарные соцветия чаще всего цимозные* у мно-
гих представителей очень оригинальное строение* напоминающее строение
отдельного цветка (циации у рода Euphorbia). Цветки с двойным около-
цветником (например* род Jatrophd)^ или в околоцветнике редуцирован вен-
л: * \
чик (род тгродеетшк—Afercurtatts), а не-
редко и чашечка, благодаря чему цветок
становится голым. Число тычинок разно-
образно, от неопределенно большого чис-
ла до одной. Гинецей ценокарпным, обыч-
но из 3 плодолистиков, с трехгнездной
верхней завязью, одним (вследствие пол-
ного срастания) или 3 столбиками и с 3
или 6 рыльцами. Семяпочки обычно име-
ют обтуратор — вырост плаценты,
прикрывающий микропиле, через который
происходит проникновение пыльцевой
трубки. Плод — большей частью коробоч-
ка, распадающаяся на три орешка, реже
мясистый, ягодообразный или в форме
костянки.
У многих молочайных, например кдо-
(jRicinus), семена снабжены к а-
рункулой (коронкой), представляю-
Рис. 274. Phyllanthus specisus (цве- щ®й Разрастание ткани интегумента в
тущая ветвь с филлокладиями) области микропиле*
Наиболее распространен у нас род
молочай (Euphorbia, рис. 275). Это тра-
вянистые растения, обычно бледно-зеленой окраски, содержащие обиль*
ный ядовитый млечный сок. В сложных соцветиях молочая бросаются в
глдза зеленовато-желтые прицветники, сидящие большей частью пара-
ни
! II
I 11
рокими очертаниям. Элементарное соцветие окружено пятью небольшими
кроющими листьями, срастающимися в бокал, который напоминает сростно-
листную чашечку. По его наружной поверхности в верхней части имеются
крупные железки. Внутри бокала помещается женский цветок, который,
занимая центральное положение, сидит на длинной* ножке и выставлен на-
ружу, и много мелких мужских цветков, сгруппированных в пять изви
пазушных по отношению к листочкам, образующим бокал. Каждый мужской
цветок расположен на короткой цветоножке и состоит только из одной ты-
чинки, нить которой составляет непосредственное продолжение цветонож-
ки. Граница между цветоножкой и тычиночной нитью бывает заметна в виде
сочленения.
Женский цветок, в свою очередь, сведен только к пестику. Такое
соцветие называется циацием, и оно биологически соответствует, а мор-.
фологически сходно с одним цветком, имеющим как бы простой сростно-
листный околоцветник (бокал), большое число тычинок (пять извилин из
мужских однотычиночных цветков) и один пестик (женский цветок). Это
толкование циация молочая подтверждается тем, что у других молочайных,
при той же степени упрощения цветка, каждый мужской однотычиночный
цветок имеет свой околоцветник; то же относится и к женским цветкам (род
AntAostema, рис. 276).
Наши молочаи довольно однообразны по внешнему облику, различа-
ясь деталями строения листьев, соцветий и пр. Однако в других областях
имеется много молочаев оригинальной внешности. Среди них заслуживают.
особого внимания африканские кактусовидные молочаи, имеющие вид кан-
деляброобразно разветвленных деревьев* У этих молочаев листья редуциро-
ваны, а стебли сочные. Суккулентный характер таких растений делает их
действительно очень сходными с кактусами, непосредственного родства с
которыми, однако, молочаи не имеют. Это хороший пример морфологической
конвергенции, возникшей в связи с засушливыми условиями, причем кактусы
эволюционировали в указанном направлении в Америке^ молочаи —
в Африке.
ОТДЕЛ ЦВЕТКОВЫЕ, или покрытосимиттуте
4(КГ
Рис* 275. Молочай:
1 — Euphorbia virgata: а — бесплодный побег, б — соцветие; 2 — Euphorbia lathyruss а — циаций,
б — циаций с удаленными бокалами; 3 — диаграмма собрания циациев; 4 — раскрывшийся плод
иг. ► <
I '<»
В наших лиственных лесах довольно обычен пролесник многолетний
(MercuriaUe perennis) — многолетнее двудомное травянистое растение, об-
разующее нередко большие куртины. На промашке однолетнем (М. annua)
и на культурном шпинате впервые в опытах Камерариуса было экс-
периментально доказано' наличие половой функции у растет ”
В практическом отношении семейство молочайных имеет большое значе-
ние. В млечном соке молочайных иногда содержится каучук высокого ка-
чества и в большом количестве. Наиболее важный каучуконос бразильская
гевея (Hevea brasiliensts, рис. 277, 2). Каучук, получаемый ив гевеи, счита-
ется наилучшим. Родом из тропической Южной Америки, это дерево рас-
пространено в настоящее время в культуре и в других тропических районах,
особенно в Индонезии, и дает до 90% всей мировой продукции каучука.
Каучук получают также из маниока (Manihot glaziovli, рис. 277, 2) из Юж-
ной Америки; другой вид того же рода — М. utlltssima — культивируется
в тропиках ]5ади крупных корневых клубней, .достигающих в длину 0,5 м и
массой более 10 кг. Из них получают особую муку (маниока, или тапиока).
В свежем виде клубни некоторых видов маниоки ядовиты, во действующее
начало легко разрушается при нагрева:
I I .
«в
1 2
t
Ряс. 276. Antkostema (цвет-
ки):
1 — тычиночный цветок; г — пе‘
стичкый цветок. Оба цветка с около-
цветником
Рве. 277. Американские кау-
чуконосы из семейства моло-
чайных:
1 — гевея (Hevea brasilieiiste):
Л—каучуковый маниок (Manihot gla*
zlcvll)
Среди молочайных много масличных растении. Иэ них особенно це-
нятся тунговые деревья (р о д Aleurites). Семена тунговых деревьев содержат
до 60% тунгового масла, которое находит применение в производстве высо-
кокачественных эмалей, лаков, красок. Тунговое масло считается незаме-.
нимым сырьем в авиастроительной, судостроительной, электротехнической,
резиновой, литографской и пищевой промышленности (лаки для покрытия
консервных банок).
Наиболее ценными считают китайское (A. fordii) и японское (A. cor data)
тунговые деревья. У нас первые опыты с разведением японского тунга был»
поставлены еще в конце прошлого столетия в Закавказье. Сейчас оба вида
культивируются в советских влажных субтропиках.
К важным масличным растениям относится и клещевина, или кастор (Hi-,
cinus communis, рис. 278). Это древесное растение восточноафриканского
происхождения культивируется во виетропических странах как однолетнее.
Листья крупные, щитовидные, на длинных черешках. В семенах клещевины,
разводимой у нас с промышленными целями на Северном Кавказе, в Закав-
казье, на юге Украины, в Волгоградской области и в Средней Азии, содер-
жится до 60% жирного масла, а также ядовитый алкалоид риципол. Касто-
ровое масло применяют в медицине (слабительное средство) и в ряде отраслей
промышленности. Значение клещевины как масличного растения в послед-
нее время сильно возросло, в виду того что касторовое масло нашло широкое
применение в технике: при изготовлении различных пластмасс, лаков и дру-
гих покрытий, в качестве низкотемпературных смазок, при изготовлении
специальных составов для реактивных двигателей и т. д.
Многие тропические виды молочайных используются как красильные,
дубильные и волокнистые растения. Некоторые дают ценную древесину.
Плоды, семена и клубни многих видов съедобны и имеют большое значение
как пищевые продукты (в тропических странах).
В тропиках Африки существенное значение имеет Oldfieldia africana
ГЛ I
дающая ценную древесину (африканское шейковое дерево). Китайское сальное
(жирное) дерево (Sapium геЬЦегит} образует в семенах белый слой, богатый
маслом, идущий на изготовление свеч и мыла.
С порядком молочаецветных сближают п о р я д о. к Thymelaeales, ку-
да относится семейство волчеягодниковые (Thymelaeaceae), представ-,
ленное в нашей флоре несколькими родами, из которых назовем волчеягодник
(Daphne), распространенный преимущественно в лесной зоне, рано цвету-
щий кустарник «волчье лыко»; растение очень ядовитое.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
409
5-Й ПОРЯДОК — ВЕРЕСКОЦВЕТНЫЕ (ERICALES)
Цветки верескоцветных, как правило, пентамерные, обоеполые, редко
однополые, обычно актиноморфные, обдиплостемонные, сростнолепестные,
иногда раздельнолепестные. Тычинки, как и лепестки, прикреплены к под**
пестичному нектарному диску. Пылинки часто в тетрадах. Гинецей цено-
карпный. Завязь верхняя, реже нижняя, с одним столбиком, трех-, много-
гнездная, с многочисленными семяпочками. Плоды разнообразные. Пре-
имущественно кустарники, полукустарники, кустарнички и
неболь
иг.
деревья, реже многолетние травы. Листья простые, цельные, почти всегда без
прилистников. Как у древесных, так и у травянистых форм широко рас-
е
пространена вечнозеленость.
Преобладающую в этом порядке спайнолепестность (присущую некото-
рым вересковым раздельнолепестность признают вторичной) нельзя считать
препятствием к сближению этого порядка с чаецветными, так как в рамках
порядка Theales лепестки тоже могут срастаться. Например, у некоторых
двукрылоплодных, чайных, маркгравиевых лепестки в основании слегка
Рис. 278. Клещевина (Ricinus communis)
410
иг,
ВЫСШИЕ РАСТЕНИЯ
Рис. 280. Черника (Vaccinium myrtillus):
1 — вепса с цветкаии; Я— разрез цветка; 3 — ты-
чинка; 4 — плод; 0 — дшвдамма цветка
Рис. 279. Грушанка круглолистная
(Pyrola rotund if olia): '
j — нижяяя часть растения; я — соцдетне;
я — продольный разрез цветка; 4 — тнчин*
на; 5 — пестик; 0 — циацтша цветка
а в семействе актинидиевых венчик иногда образует короткую
страстаются,
трубочку.
Близость чаецветных и верескоцветных основывается не только на сход-
стве организации цветка, но и на некоторых анатомических особенностях дре-
весины. Интересно, что некоторые семейства, например актинидиевые, то
включают в состав чаецветных, то предлагают перенести в порядок Ericales.
Семейство грушанковые (Pyrolaceae) ькяючвт многолетние вечно-
зеленые травы, а также бесхлорофилльные сапрофитные растения. Цветки
обычно в соцветиях, реже одиночные, с раздельнолепестным или спайноле-
пестным венчиком, иногда более или менее зигоморфные, пятй-четырех-
членные. Тычинки не связаны с лепестками. Пестик один с верхней завязью,
имеющей продольные бороздки снаружи. Гнезда в завязи не вольностью
изолированы друг от друга, вследствие чего в верхней части завязь кажется
одногнездной с центральным семяносцем. Плоды—коробочки с многочислен-
ными семенами, содержащими недоразвитый, без расчленения на вегетатив-
ные органы, зародыш.
В семействе грушанковых выделяют две группы (подсемейства). К пер-
вой из них относят зеленые растения; у них пыльца в тетрадах. Виды рода
грушшка (Pyrola, рис. 279) широко распространены преимущественно в
хвойных лесах Евразии и Северной Америки. Грушанки имеют корпев:
л.
е,
от которого возникают олиственные побеги. Семена грушанок настолько
мелкие, что масса их измеряется миллионными долями грамма; они разно-
сятся ветром. Прорастание семян наблюдалось редко; размножение осущест-
вляется преимущественно вегетативным путем. У растений второй группы —
бесхлорофилльных (сапрофитных) и не имеющих тетрад — корневища нет.
Под землей у них имеется сплетение корнеподобных органов неясной морфо?
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЯННЫЕ
411
11 Т
логической природы, на которых эндогенно закладываются надземные по-
беги, в развитом состоянии незеленые и имеющие редуцированные листья.
У . относящегося сюда подъельника обыкновенного (Monotrop a hypopity s) впер-
вые была открыта (Ф. М. Каменским в 80-х годах XIX в.) микориза,
которая оказалась не только у грушанковых, но и у многих наземных
растений.
Семейство вересковые (Ericaceae). Кустарники и полукустарники,
иногда небольшие деревья; растения большей частью вечнозеленые. Листья
простые, преимущественно ланцетные, иногда чешуйчатые, без прилистников,
нередко ксероморфного облика. Цветки правильные, реже несколько зиго-
морфные, обоеполые, обычно из пяти-четырехчленных кругов. Венчик спай-
нолепестный, реже свободнолепестный. Тычинок вдвое больше, чем лепест-
ков, или столько же. Они совсем не срастаются с лепестками или соединены
только с основаниями последних. Пыльники раскрываются дырочками и
часто имеют роговидные придатки, имеющие значение при опылении насе-
комыми. Завязь увересковы! бывает как верхней, так и нижней; она четы-
рех- или пятигнездная С центрально-угловой /плацентацией. Плоды — коро-
бочки, костянки или ягоды.
К вересковым относится около 2500 видов. Обитают они в умеренных и
холодных областях обоих полушарий, покрывая местами обширные площади
с бесплодными почвами (вересковые пустоши); в тропиках — только в горах.
Способность некоторых вересковых жить при неблагоприятных условиях
можно связать с распространенностью у них эндотрофной микоризы. Име-
ются сведения, что микоризные грибы способны к усвоению элементарного
азота воздуха. По некоторым данным, гриб пронизывает все тело растения,
входит в завязь и проникает в семенную кожуру, что, надо думать, вызыва-
ет заражение молодого растения.
Ряд видов вересковых связан с болотами, другие населяют тундровые
пространства. Вересковые, обитающие на болотах, вызывают особый инте-
рес тем, что, живя при избыточном увлажнении, они обладают рядом эколо-
гических особенностей, свойственных ксерофитам.
Семейство вересковых подразделяют на 4 подсемейства. К подсемей-
ству рододендровых (Rhododendroideae) относится род рододендрон (Rho-
dodendron), имеющий около 400 видов. В СССР рододендроны распростра-
нены главным образом на Кавказе и на востоке Сибири. Многие виды этого
рода разводят как декоративные. К этому же подсемейству относят багуль-
ник болотный (Ledum palustre) — вечнозеленый кустарник, растущий на
болотах и в сырых лесах и обладающий сильным одурманивающим запахом
(«болиголов»), и циркумполярно распространенную в Арктике луавелерию
(Loiseleuria procumbens) — низкий, плотно прижатый к земле вечнозеленый
кустарничек.
К подсемейству Arbutoideae относится земляничное дерево. Среди-
земноморский вид земляничного дерева (Arbutus unedo) входит в состав ксе-
рофильных растений субтропических лесов; у нас его разводят в садах Кры-
ма и Закавказья, где в диком состоянии встречается другой вид — A. and-
rachne. Название земляничного дерева связано с тем, что его ягодоподобные,
плоды (мясистые, многосеменные, с поверхности бородавчатые ягоды) съе-
добны и по внешности напоминают землянику. К этому же подсемейству от-
носятся характерные для моховых болот подбел обыкновенный (Andromeda
polifolia) и болотная мирта (Chamaedaphne calyculata), а также растущая в
сухих борах толокнянка, или медвежье ушко (Arctostaphylos uva-ursi). Листья
толокнянки используются в медицине как мочегонное средство. Третье под-
семейство — Ericoideae — ъкзпжя&г самый обширный род Erica, со-
держащий около 500 видов, из которых примерно 450 сосредоточены в Кап-
ской флористической области. К этому роду относят древовидный вереск
(Erica arborea), обитающий на востоке Африки и по побережью Средиземного
моря в Европе. Это растение достигает 10 лс, а на Канарских островах даже
20 м высотой. К этому подсемейству относится ирод вереск (Calluna) с един-
1'1
: I \ । \
412
ВЫ»1
nr.
Е РАСТЕНИЯ
Г Г
ственным видом С. vulgaris, весьма характерным для вересковых пустошей;
вереск растет также в сосновых борах и на болотах. Растение это имеет очень
мелкие чешуйчатые листья, густо одевающие стебли; медоносное. Наиболее
важное в практическом отношении — четвертое подсемейство yacti*
пмнфмАт вмдаляАмпе некоторыми систематиками в самостоятельное семейство.
Брусничные имеют нижнюю завяэь и плод-ягоду. Широко распространены
в хвойных лесах брусника (Vactinium vitis-idaea) — вечнозеленый кустарни-
чек— и черника (Vactinium myrtillus, рис. 280) — листопадный кустарни-
чек. Ягоды этих растений съедобны. Черника имеет медицинское применение—
отвар ягод действует закрепляюще. Сок черники используют для подкраски
вин, а с примесью квасцов им окрашивают шерсть и холст в фиолетовый
цвет. Близок к чернике гонобобель, или голубика (Vactinium uliginosum), —
кустарник до 1 ж высотой, растущий в сыроватых лесах и по болотам.
Его несколько водянистые на вкус ягоды, более крупные, чем у черники, ис-
пользует население для киселей, начинки пирогов и т. п. Очень широко ис-
пользуются ягоды клюкв» (V actinium oxycoccus и V. microcarpum) — сте-
лющихся вечнозеленых кустарничков с их очень тонкими побегами; клюква
растет на болотах. Свежие ягоды клюквы содержат витамин С.
К порядку верескоцветных относят также семейство шикшиевые
(Empetraeeae), представленное в нашей флоре вороникой (Empetrum nigrum),
растущей на верховых болотах, кустарничковых пустошах и в борах. В Юж-
ной Америке и прилежащих островах распространена вороника красная
(Empetrum rubrum).
III
6-Й ПОРЯДОК — ПЕРВОЦВЕТНЫЕ (PRIMULALES)
Цветки первоцветных правильные, пятичленные, реже четырехчленные.
Тычинки в числе лепестков, расположенные против них; кроме того, иног-
да имеются стаминодии, соответствующие по положению противочашечным
тычинкам. Гинецеи ценокарпный, обычно из пяти плодолистиков, с верхней,
реже нижней завязью. Последняя—одногнездная с одним центральным (лож-
ноосевым) семяносцем. Семяпочек много—одна. Растения разнообразного
внешнего облика: в тропиках (семейства Theophrastaceae, Myrsinaceae) —
деревья и кустарники, в странах умеренного климата—исключительно тра-
вянистые растения.
Семейство первоцветные (Primulaceae). Травянистые растения с
очередными листьями, без прилистников. Цветки правильные, редко зиго-
морфные, обоеполые, но нередко диморфные (гетеростилия).
Чашечка сростнолистная, большей частью разрастающаяся после цветения
и остающаяся при плодах. Венчик спайнолепестный, но иногда рассечен
почти до основания. Кроме одного круга противолепестковых тычинок, в не-
которых случаях имеются стаминодии. Завязь верхняя, редко полунижняя,
одногнездная, с центральной плацентой. Семяпочки большей частью много-
численные, полусогнутые, с одним интегументом. Плод — коробочка.
Около 800 видов семейства первоцветных обитают главным образом в уме-
ренных широтах северного полушария. Обширный род первоцвет, или приму*
ла (Primula), объединяет более 500 видов. Наибольшее разнообразие примул
характерно для горных флор. Многие виды примул отличаются цветками
выдающейся красоты, что дало основание для использования их в садовой,
оранжерейной и комнатной культуре. Среди декоративных примул много гиб-
ридов. В садовой культуре используются, например, Primula acaulis, Р. den*
ticulata, Р. auricula и др. В комнатах содержат Р. sinensis и Р. obconica. Выде-
ления железок, имеющихся на листьях комнатных видов, вызывают иногда
воспаление кожи (дерматит.). К широко распространенным в СССР лесным
видам относится первоцвет весенний (Primula veris, рис. 281), произрастающий
на территории от Средней Европы до Восточной Сибири. К этому же семейству
относится род сольданелла (Soldanella). Виды этого рода — горные растения,
растущие в непосредственной близости к снегам. У сольданеллЫ; как и у
tie:
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
413
Рис. 281. Первоцвет (Primula veris)!
1 — общий вид; 2 — продольный разрез длинно-
столбчатого цветка; з — продольный разрез дорог*
костолбчатого цветка; 4 плод; Д —- диаграюка
цветка
Рис. 282. Цикламен.
1 — общий вид; 2—разрез цветка
наумбургии, встречающейся у нас по сырым местам, имеются рудименты ты-*
чинок внешнего круга андроцея. Род Naumburgla близок к вербейнику
(род Lysimachia), один из видов которого—елуговой чай» (Lysimachia nummul-
laria)—широко известен как ползучее луговое растение. Вместе с тем он бли-
зок к р о д у седмичник (Trientalis)* у которого венчик семичленпыи и тычи-
нок в цветке тоже 7. Виды рода цикламен (Cyclamen, рис. 282), или дрякват
распространены в горах по берегам Средиземного, Черного и отчасти Кас-
пийского морей. Все 50—55 видов цикламена декоративные. В комнатах
под названием персидского цикламена (Cyclamen persicum) разводят гибрид-
ные формы разнообразной окраски: белые, розовые, темно-красные. Довольно
крупный надземный клубень цикламена представляет разросшееся подсе-
мядольное колено. Клубень ядовит.
Родственные связи порядка первоцветных рисуются систематиками раз-
лично. Если одни сближают их с центросеменными, то другие считают их
родственными чаецветным. Возможно^ первоцветные имели общего предка с
порядком эбеновых.
7-Й ПОРЯДОК — ЭБЕНОВЫЕ (EBENALES)
Цветки эбеновых обычно актиноморфные из пяти- или четырехчленных
кругов, обоеполые, редко однополые, как правило, со спайнолепестным
венчиком, редко со свободными лепестками. Тычинки в двух или трех кру-
гах, изомерных лепесткам, или од
I круг с удвоенным по сравнению с ле-
.414
ВЫСШИЕ РАСТЕНИЯ
111'
пестками числом тычинок. Гинецей ценокарпный, с завязью верхней, по-
лунижней или нижней, многогнездной. Плацентация центрально-угловая.
Семяпочки с двумя или одним интегументом.
Деревья и кустарники с простыми листьями без прилистников, обитаю-
щие в тропической зоне и в субтропиках.
Относящееся сюда семейство эбеновые (ЕЪепасеае) содержит около
450 тропических и отчасти субтропических видов. Древесина некоторых
видов («черное дерево») хорошо полируется, отличается твердостью, плот-
ностью (удельный вес 1,2) и темной окраской ядра с незаметными годичными
слоями и сердцевинными лучами: заболонь в виде узкого кольца. Эбеновые
деревья ценятся в мебельном и других производствах. Некоторые виды
эбеновых славятся величиной и вкусовыми качествами плодов. Так, масса
плодов Diospyros kaki (хурма) достигает иногда 0,5 кз. Они имеют вкусную,
сладкую мякоть. Известно множество сортов (в Китае до 2000). Распро-
страненная от средиземноморских стран до Китая одичавшая хурма
обыкновенная (Dyospyros lotus) растет и у нас в смешанных горных лесах
Закавказья и Средней Азии. Родина ее Китай. Плоды хурмы обыкно-
венной небольшие, около 2 см, становятся приятными на вкус лишь тро-
нутые морозами, когда они теряют танниды. Древесина хурмы обыкновен-
ной желтоватого цвета и идет на столярные и токарные изделия — из нее
изготовляют челноки для текстильной промышленности.
Ряд представителей родов семейства сапотовых (Sapotaceaejz
гуттаперчевые деревья (Palaquium gutta* ныне вовсе истребленный, и Р. ob-
longifolium), мимузопс (Mtmusops) и пайена (Рауепа) — тяючншси. гутта-
перчи. Древесина многих сапотовых отличается большой твердостью (Sir-
deroxylon faerme ж некоторые другие называют «железным деревом») и це-
нится как строительша и поделочный материал. В семенах сапотовых,
например в зародышах масляного дерева (Butyrospermum parkii, Тропическая
Африка), содержится большое количество масла* используемого в п
в технических целях.
Порядок Dilleniales был, вероятно^, родоначальной группой еще
одной или даже двух обширных ветвей эволюции двудольных. Одна из них
представлена порядком фиалкоцветных (Violates)* подобно
Theales* начало нескольким параллельным линиям развития; вторая —
порядком каперцоцветных (Capparales), развивавшимся либо парал-
411
IГГ.
8-Й ПОРЯДОК — ПОСТЕННОСЕМЕННЫЕ, ФИАЛКОЦВЕТНЫЕ (V IO LAKES)
Цветки у фиалкоцветных циклические, большей частью актиноморфные,
у некоторых зигоморфные. Двойной околоцветник, как правило, из пяти-
членных кругов; число тычинок равно числу лепестков или больше. Гинецей
обычно паракарпный, из трех плодолистиков. Этот порядок содержит боль-
шое количество семейств, главным образом тропической флоры. Сюда отно-
сятся растения разнообразного внешнего облика (деревья, кустарники*
полукустарники и травы) с очередным* реже супротивным расположением
листьев, часто тлеющих прилистники.
Из числа многочисленных семейств, относимых к этому порядку, с эво-
люционной точки зрения важное значение принадлежит обширному (1300 ви-
дов) семейству фмкуртиевых (Flacourtiaceae), характерному для тро-
пических и отчасти субтропических стран. В этом семействе древесных
и кустарниковых растений цветок изменчив по числу частей как в
околоцветнике* так в андроцее и гинецее. Столбик может быть один, или
плодолистики, образуя пестик, сохраняют свободными свои окончания
(столбики). Срастаются и другие части цветка. Так, иногда тычинки сра-
стаются в пучки* обычно свободные чашелистики могут срастаться в
короткую трубочку. Но лепестки всегда свободные или отсутствуют. Завязь
верхняя* полунижняя или нижняя* одногнездная* с постенными пла-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
415
I'll
tilt
ни
центами, которые, иногда разрастаясь по направлению внутрь, делят
единственное гнездо на несколько неполных и даже полных гнезд. Семей-
ство содержит плодовые и важные лекарственные растения, некоторые из
них используют для лечения проказы.
Семейство фиалковые (Violaceae). В нашей флоре это только тра-
вянистые растения, но в тропических и субтропических странах также кус-
тарники и небольшие деревья. Листья очередные, реже супротивные, боль-
шей частью с прилистниками. Цветки обоеполые, правильные (у экзотичес-
ких представителей) или зигоморфные, с пятилистной чашечкой и с пятью
свободными лепестками; пять тычинок чередуются с лепестками. Пестик
образован тремя (5—2) вполне сросшимися плодолистиками, он. имеет верх-
нюю. одногнездную завязь с многочисленными постенно расположенными
семяпочками и один столбик с рыльцем разнообразного строения. Плод—ко-
робочка или ягода.
Семейство фиалковых в нашей флоре представлено только видами наи-
более крупного (450 видов) рода фиалка (Viola). Наши фиалки — травянис-
тые небольшие растения, в то время как в странах Средиземноморья (Viola
arborescens — Западное Средиземноморье), а также в Северной Америке из-
вестны кустарниковые формы.
С характерными признаками фиалок можно познакомиться на любом
виде, например на фиалке трехцветной (Viola tricolor) широко распростра-
ненной в светлых лесеГх, на полянах и лугах. Очередные листья этого растения
имеют большие листовидные лировидно-перистораздельные прилистники с
крупной конечной долей. Из пяти лепестков цветка нижний имеет у основа-
орец. Тычинки с очень короткими нитями; пыльники довольно круп-
ные, окружающие пестик. Связники разрастаются над пыльниками в че-
шуйчатые придатки. Две нижние тычинки имеют по одному шпорцевидиому
нектарнику. Последние вдаются внутрь шпорца лепестка, где и собирается
нектар. На пути внутрь шпорца насекомое встречает под рыльцем неболь-
шую пластинку, отгибая которую оно пачкает ее пыльцой, принесенной с
другого цветка. При обратном движении пластинка соприкасается с воспри-
нимающей поверхностью рыльца, благодаря чему происходит перекрестное
опыление. У некоторых фиалок наряду с открытыми цветками развиваются
цветки клейстогамцые, самоопыляющиеся. Коробочки трехцветной фиалки,
как и других видов, при созревании раскрываются тремя створками. Семе-
на вследствие наличия у основания мясистых придатков (присемянник,
ариллус) растаскиваются муравьями.
Трехцветная фиалка — один из родоначальников выведенных путем
гибридизации разных сортов садовой фиалки («анютины глазкиь)Л замеча-
тельной разнообразием красивых окрасок.
Семейство ладанниковые (Cisiaceae) — кустарники, полукустар-
ники и многолетние травы, преимущественно с супротивными листьями. Цвет-
ки актиноморфные, с двойным околоцветником из трех- пятичленных кру-
гов. Тычинки многочисленные. Завязь одногнездная или ложномногогнезд-
ная, образованная 3—5 плодолистиками; плаценты постенные.
Это небольшое (менее 200 видов) семейство, представители которого
распространены в более теплых районах северного полушария, в особеннос-
ти в Средиземноморье, отчасти в Северной Америке. Виды рода ладанник
(Cistus) — кустарники, типичные для средиземноморской маккии; некото-
рые содержат ароматичную смолу — ладан. Род солнцецвет (Hellanthe-
тит) представлен полукустарниками и травами.
Семейство папаевые (Caricaceae^ или Рарауасеае). В этом небольшом,
чисто тропическом (тропическая Америка и тропическая Африка) семействе
заслуживает внимания дынное дерево (Carica papaya), с древности куль-
тивируемое в тропической Америке и Азии. Невысокий неветвистый и соч-
ный ствол дынного дерева несет на вершине пучок крупных длинночерешко-
вых пальчатых с перистыми долями листьев. Цветки обычно однополые.
Мужские цветки в больших разветвленных соцветиях, выходящих из пазух
1:1
416
ВЫСШИЕ РАСТЕНИЯ
листьев; женские соцветия на более коротких ножках и развиваются ниже
листвы. Плоды — крупные (до 8 кг) ягоды, несколько напоминающие по
форме дыню; спелые употребляют как десерт, недозрелые — как овощ; бога-
ты витаминами. Млечный сок, имеющийся во всех частях растопи я и полу-
чаемый из надрезов плодов, содержит фермент папаин, подобно пепси-
ну, расщепляющий белки. Употребление плодов дынного дерева в пищу
способствует пищеварению. В США используется в кожевенной промыш-
ленности (дубление кож), в пивоварении и как лекарственное. В настоящее
время применяется при тромбозах как антикоагулянт крови. Предполагают,
что родина дынного дерева — Вест-Индия; культивируют его во всей тропи-
ческой зоне, легко дичает.
Семейство гребенщиковые (Тamaricaceae). Кустарники и деревья
с ветвями, покрытыми мелкими чешуйчатыми или шиловидными листьями
без прилистников, что создает внешнее сходство с ветвями кипариса. Цветки
обычно мелкие, с двойным околоцветником из пяти-чатирехчлатпат кругов,
собранные в соцветия. Тычинки в числе, равном числу лепестков или вдвое
большем; иногда тычинок много; пять плодолистиков образуют пестике
одногнездной вер±ней завязью. Плоды — коробочки с семенами, снабжен-
Г|
HIS
В этом небольшом (около 100 видов) семействе наиболее крупным явля-
ется род гребенщик (Tamarix), распространенный в Северной, Северо-Вос-
точной и отчасти Западной Африке, в Южной Европе, Передней, Средней
и отчасти Центральной Азии. Это кустарники, растущие по песчаным бере-
гам рек и озер, отчасти на засоленных почвах. Солеустойчивые виды гребен- .
щиков выделяют на поверхности листьев и веточек соль. В поймах средне-
азиатских рек гребенщики часто образуют густые, труднопроходимые за-
росли.
• Семейство бегониевые (Begoniateae). Растения, входящие в это се-
мейство (свыше 800 видов), распространены почти по всему тропическому
поясу. Это травы, нередко образующие клубни, или полукустарники с оче-
редными, обычно простыми листьями с прилистниками. Цветки однополые,
правильные или зигоморфные, со свободными или частично сросшимися
листочками околоцветника. Околоцветник ярко окрашенный. Плод—ко-
робочка.
В семействе всего пять родов, из которых основной — бегония (Begonia);
он включает почти все (800) виды семейства. Бегонии часто легко образуют
придаточные почки на листьях, что и используют при вегетативном размно-
жении. Бегонии разводят как декоративные растения, хорошо перенося-
щие комнатные условия, а также высаживают на дето в грунт. Некоторые
виды и гибриды бегоний имеют очень красивые, ярко окрашенные цветки,
ценят их также за красивые пестрые листья.
Обширный порядок фиалкоцветных представляет группу если и моно-
филетическую, то состоящую из нескольких параллельных рядов развития.
111:
ия ПОРЯДОК — ТЫКВЕННОЦВЕТНЫЕ (CUCURBITALES) „
Этот порядок содержит только одно семейство тыквенные (Сисиг-
bltaceae). Однолетние или многолетние травы, редко полукустарники и кус-
тарники и даже, как редчайшее исключение, маленькие деревца (род
Dendroeicyos на острове Сокотра). Для тыквенных характерно наличие усиков,
представляющих видоизмененные побеги, в которых нижняя часть по про-
исхождению стеблевая, а разветвления, обладающие раздражимостью,— мета-
морфизированные листья. Стебли простертые по земле или лазающие. Цветки
в цимозных соцветиях иди одиночные. Цветки, как правило, однополые, но
содержащие рудимент недбСтающеро пола, правильные, из четырех кругов.
Цашечка, венчик и нижние части. .тычиночных нитей, срастаясь, образуют
1 трубку ,которая,всвокгочередь, срастается с завязью; последняя вследствие
ыг, I и
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
:н4
«7
крушые, яркие (как у тыквы), ИЛИ ЖАЙЛДКЩИЯ йалид и ааиаиамтиа-, Д ПЯТИ
лепестков. Андроцей jia пяти, трех, иногда двух тычинок. Они могут быть
аппзпта счлбпдик^дг (редко) и сро<8Дц1Имися попарноУ причем одна тычинка
остаетса,свободноиили penvnwnyfYf^0, ° срастаются все
пцть тычйок. Срастание охватывает пыльншньжж^авяфееммшштся и на
.14
.14
пыльшшйГТольноу немногих тыквешгых пыльрики прямые. вболыпинстве
случаев они изогнутые, дКогда^с^жнУ.^В^каждом пыльнике развиваются
только дв^ЙЯ^^ Паракаппный гинепей обычно из тпех (10—1)
плодолис^^ов^ДблЬСТЬ"1 МЬязи большей частью почти пеликом яанятя ряя^
роййиьшс^плахщнта^
дел итсяНй "гаезд^С^ядочек обычно много, очень редко —однд. Столбик
один с тремя* к о ми с с у рал ь и ым крыльцами. Плоды тыквенных —
. либо настоящшГ',ДДДД£^^ "‘называемые тыквины, отличающиеся
от ягод -не только размерами. но и бпле^ плотным сложейием царужныу
слоев околоплодника. Тыквины очень разнообразны"по форме и размерам.
Семена^Овз’ эндосперма.
Семейство содержит от 850 до 1000 видов. Они распространены широко,
но в основном тяготеют к сухим тропическим и субтропическим областям,
отсутствуя в холодных странах. В Европе известны только 3 дикорастущих
вида. В практическом отношении некоторые тыквенные представляют боль-
шую ценность и давно вошли в культуру как бахчевые и огородные растения.
Культура огурца (Cucumis sativus) происходит из Индии. В СССР в от-
крытом грунте она возможна до 60° с. ш., а в теплицах — и севернее. Теп-
личная культура дает возможность получать свежие огурцы на протяжении
большей части года. Плоды огурцов снимают незрелыми, в период максимума
их роста. Огурцы используются главным образом как вкусовая приправа,
собственная их пищевая ценность ничтожна.
Дыни (Melo) культивируются с древнейших (около 4000 лет до н. э.) вре-
мен в странах Азии, что, конечно, связано с накоплением в плодах большого
количества сахара. Дыни теплолюбивы* В настоящее время культура их
распространилась довольно далеко на север, в основном за счет скороспелых
сортов. Дыни наиболее ценных сортов растут в СССР в Средней Азии, где
имеются как скороспелые, так и позднеспелые сорта. Последние получили
мировую известность за вкусовые качества (лучшие среднеазиатские дыни
оцениваются знатоками выше ананаса и других тропических фруктов), за
транспортабельность и лежкость. Из дынь готовят дынный мед; особого ро-
да лакомство представляет вяленая дыня; из семян можно получить нише-
вое и техническое масло. В районах культуры дынь встречается сорная ды-
ня (Melo agrestis), засоряющая посевы и скрещивающаяся с культурными
сортами дынь, что делает сорную дыню вдвойне вредньш растением.
Небольшой (5—7 видов) род арбуз (Citrullus) характеризуется разветв-
ленными усиками. На бахчах разводят столовый арбуз (CitruUus lanatus). Он
имеет плоды, достигающие 20 кг, со съедобной сладкой (6—10% сахара)
сочной мякотью розового, красного, иногда белого цвета; из нее готовят ар-
бузный мед (нардек). Кормовой арбуз ценится в засушливой полосе как ис-
точник сочного корма на силос. Мякоть кормовых арбузов грубая, содер-
жащая мало сахара, иногда горьковатая, белого, желтого или бледно-зе-
леного цвета. Все кормовые сорта арбуза толстокорые и годны для произ-
водства цукатов. Семена арбузов содержат масло, которое можно использо-
вать в пищевой и мыловаренной промышленности. Арбузы разводят в основ-
ном в СССР; свыше 50% всей площади этой культуры сосредоточено на юге,
но можно получать хорошие урожаи и в средней полосе (Московская об-
ласть). Родиной столового и кормового арбуза обычно считают пустыни Аф-
рики. В Калахари столовый арбуз в диком состоянии встречается и сейчас.
В пустынях Северной Африки и Южной Азии растет близкий родственник
арбузов — колоцинт* или колоквинт (Citrullus colocynthis^ рис. 283) — мно-
голетнее растение с мощной корневой системой и ползучим стеблем; усики
14—682
Г
418
ВЫСШИЕ РАСТЕНИЯ
Рис. 283. Колоквинт и переступень:
1 — колоквинт: а — развернутый тычиночный цветок;
б — разрез пестичного цветка; « — две сросшиеся
тычинки; в — поперечный разрез плода; й — пере*
ступень] а — две сросшиеся тычинки; б — одна
тычинка
Рис. 284. Тыква (Cucurbitа реро):
1 — разрез тычиночного цветка; а — андроцей (срос-
шиеся тычинки); а — разрез пестичного цветка; 4 -
пестик; 5 — поперечный разрез завязи
колоцинта обладают своеобраз-
ной способностью цепляться
за почву. Плоды мелкие, горь-
кие, несъедобные. Экстракт из
мякоти используется как силь-
ное слабительное.
Род тыкеа (Cucurbita).
Происхождение культурных
тыкв связано с Центральной
Америкой. За огромный проме-
жуток времени, в течение кото-
рого разводят тыквы, создано
исключительно большое разно-
образие сортов— пищевых, кор-
мовых, декоративных (пестрой
окраски), относящихся к раз-
ным видам. В СССР большое
распространение получила тык-
ва обыкновенная (Cucurbita реро>
рис. 284) и в меньшей степени
гигантская (Cucurbita maxima).
Плоды тыквы используются в
кулинарии и в кондитерском
деле. Сортами тыквы обыкно-
венной являются кабачки и
патиссоны. В теплых странах
и у нас в южных районах куль-
тивируют тыкву горлянку
(род Lagenaria)' используя
ее плоды для изготовления
оригинальной кустарной посу-
ды: кувшинов, графинов, таба-
керок и т. п.
На юге СССР культиви-
руют также люффы — виды
цилиндрическая (Luffa cylin-
drica) и острореберная (L.
acutangula), из плодов кото-
рых освобождают прочную
сеть волокон, используемую
как грубая губка или для из-
готовления купальных туфель,
головных уборов, стелек для
обуви и т. п. Родина люффы-
тропики Старого Света.
Заслуживает также упоми-
нания переступень (род
Bryonia) — лазающий много-
летник с простыми усиками и
разной окраски (в зависимости
от вида) ягодами. Виды пе-
реступеня используются го-
меопатической медициной.
Бешеный огурец (Ecballium
elaterium) встречается на юге
европейской части СССР, в
Крыму, на Кавказе- и в Сред-
ней Азии. Он обладает интерес-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
419
ной способностью активно выбрасывать семена из плодов при отделении их
от плодоножки.
Семейство тыквенных имеет много черт сходства с некоторыми семейства-
ми порядка Violates, в частности с семейством страстоцветных (Passifloraceae),
представители которого характеризуются усиками, возникшими из побега,
как и у тыквенных. С семейством Сагусасеае тыквенные сходны спайноле-
пестностью, которая в некоторой степени наблюдается и у страстоцветных.
Указывались также черты сходства тыквенных с бегониевыми. Связи тык-
венных с Violates достаточно убедительны, и в некоторых системах это се-
мейство включается в порядок фиалкоцветных или в выделяемый из него по-
рядок Passiflorales. Однако спайнолепестность и срастание тычинок выдви-
гаются и как аргумент в пользу сближения тыквенных с такими семействами,
как сложноцветные, колокольчиковые, но наблюдаемое у этих семейств со-
единение пыльников представляет лишь склеивание их, или сцепление, осо-
in
быми волосками — крючочками; у тыквенных же имеет место настоящее
срастание вплоть до образования общего пыльцевместилища.
10-Й ПОРЯДОК — КАПЕРСОЦВЕТНЫЕ (CAPPARALES)
' Околоцветник каперсоцветных в большинстве случаев двойной, круго-
вой, чяото ИЗ димерпит мд дд^рямарнктх кругов. Тычинки к
Ъбйчотсвббодные.Гинецей паракарпный, завязь верхняя, с одним (еслёГон
есть) СТОибиКОй, обычно_содержащаямного семяпочек. 'Семёна с согну-
тым зародыш^^и^слаборазвитым эндоспермом, чаще без него. Деревья,
кустарники, травы с очередными простыми (^дкд" пальчатосложными) листь-
ями, без прилиётййКотгДляттдгих представителей характерно наличие кле-
ток с мвррзином — ферментом, расщепляющим глюкозиды, содержащие гор-
чичные маслаТЧ^етки'ЪбоепЬлыеГхфавйлыаыё, редко Несколько зигоморф-
ные. 11 - 1 ---------
Этот порядок, главным образом на основании сходства в строении цвет-
ка, часто сближают с семействами маковых и дымянковых (порядок Рара-
verales), включая их в один порядок Rhoeadales. Но и в этом случае отно-
симые к Capparales семейства иногда противополагают маковым и дымяпко-
вым как особый подпорядок. Биохимические и отчасти эмбриологические
исследования последнего времени укрепили представление о Capparales как
о группе, которая имеет происхождение, независимое от макоцветных и
генетически связана с прочими Dilleniidde.
Семейство крестоцветные (Cruciferae, Brassicaceae). Травянистые
растения, редко полукустарники с очередными листьями, лишенными *НрП-
листнцвдш.. Пластинка листа простая, цельная или; р рАмлй степени расчле-
ненная. Характерны ол^^ёто^^^^остае^^и реже мно-
^Вегетативные органы к рестлттмтных име-
ют особые клетки (и д и о б л а с ты ), содержаптие мировин. Цветки в кис-
тях, которые не^
Распускание цветков обычно начинается раньше, чем соответствующая
часть оси соцветия зшсон^итсвой рост в длинуТЪб^ТОму'В11 начэдигШЕ&гёййя
цветки вместе с бутонамикажутся рятлтур^тртиуи как faщалк&м (щит-
ковидная кис т ь ). Кроющие листья,
развиваются. Цветок имеет двойной околоцветник; чашечка _рбрдзхщана
Двумя- двучленными кругами; причём чашелистики внутреннего круга
трёйсвебсал^ныё по~пдДожению, иногда в основании имеют ^углубления,
в которых^ “ ~~
Венчик из че™рз* ДЭДУ™™ - в одном круге, занимающих промежутки
между. чашелистиками (рис. 285). Андроцей у большинства крестоцвет-
ных из шесп-шчинокГДве из них^ образующие внешний" круг,1шеют более
короткие нити и располагаются в_т р а нс в ё р с а л ьн ойпл о ско-
сти симметрии цветка. У четырех тычинок"ряутрениего кругА^ которые
; 1 >
ИГ.
расположены двумя пярями. чепануюн\имися с ‘гмуинкамм наружного круга
14*
420
ВЫСШИЕ РАСТЕНИЯ
чш
2
3
Рис. 285, Строение цветка крестоцветных:
J — диаграмма; 2 — зачаток цветка (зачагкиз
кчш — наружного чашелистика» внчш—внутреннего
чашелистика» лп— лепестка» тч—наружной тычин-
ки, вт* — пары внутренних тычинок» п — пести-
ка); 3 — цветок горчицы; 4 — андроцей и гинецей
капусты
П
внчш
состоящейиз —хршшгр-
сально расположеттих. плпдпли-
стиков, сросшихся краями. От
лящая полость завязи до верти-
кальному направлению на два
гнезда, Семяпочекобычнонесколь-
ко» и прикрёвдёщО^^
екоторые морфологи считают, что
плоДОЯЙ^тшсбв у крестоцветных
четыре, причем два из них, обра-
зующих впоследствии створки
плода, стерильныеП^не^развивают
на себе'Семяпочек), а два других
одущие. Из этих последних^»
мируется рамка» на которой натя-
нута в виде пленочки ложная
перегородка, сохраняющаяся в
плоде.
На цветоложе, выше дасжоч-
КОВ ОКОЛОЦВеТНИКа. Ц^ходстг-дитАк-
тарники, имеющие у разных
прёдС^й^ИТбДблГсамые разнообраз-
II
образуются ть«ам ддрптпм ш ш
« Ч
»:г
ной, go нередко _и околомедйанных;
сливаться в ^ёдтаое кольцо? НлбДьГ— стручки, стручочки, реже орешки.
Цветок в йределкХсйиейства крестоцветных отличается удивительным
однообразием. Изменяясь 1ИйЙелвтше~и окраске лепестков (белая, желтая,
розовая, лиловая), он сохраняет почти нейзйенныДг^йдан строения .~~Иногдя,
вследствие неодинакового развития лепестков, цветок становится зигоморф- '
ным. У немногихк^естецвеыпГх'вместо шести тычинок бывает четыре_или
две^лли, цаобоаот. больщетпрг/гуц иногда тагавджи внутреннего круга сращвцы
основаниями попарно; редко совсем отсутствуют лепестки? Попарное раедрло-
жение тычйншгвнугрениетсгкрутм, «срастание» их, наблюдаемое у некоторых
крестоцветных, а также процесс развития этих тычинок, при котором снача-
ла на месте каждой пары закладывается один, в дальнейшем раздваиваю-
щийся бугорок (рис. 285, 2), дают основание считать, что внутренний круг
андроцея у предков крестоцветных состоял из двух медианных микроспоро-
филлов, которые впоследствии расщепились вдоль. Некоторые тератологи-
ческие данные говорят о том, что и четыре лепестка крестоцветшлх есть про-
дукт расщепления двух листочков венчика. Если предположение относитель-
но расщепления лепестков и медианных тычинок верно, то теоретическую
формулу цветка крестоцветных, не выделяя в ней чашечку и венчик, можно
выразить так; Рк+зСгхгАгнгхй) £(2)Т- ^та теоретическая формула вполне
соответствует формуле цветка некоторых маковых (род Нуресоит)\ она
же повторяется у некоторых каперсовых. У крестоцветных нет особо
специализированных приспособлений к перекрестному опылению — обычна
дихогамия.
Сохраняя единый, почти н ©изменяющийся план строения цветка, крес-
тоцветные эволюционировали главным образом по линии изменений плода
(рис. 286) в следующих направлениях. Из типичного длинного стручка
у многих крестоцветных выработался короткий стручок с длиной, превы-
шающей ширину не более чем в 3—4 раза, называемый стручочком. Стручоч-
iri
1 Знак умножения показывает на удвоение членов путем расцепления»
ОТДЕЛ ЦВЕТКОВЫЕ, ИДИ ПОКРЫТОСЕМШНЫЕ
ки бывают двух родов. У одних выпуклость
створок незначительна и наибольшая ширина
стручочка равна ширине внутренней перегород-
ки его (широкоперегородчатые стручочки).
У других створки стали резко выпуклыми и
наибольшая ширина стручочка благодаря
этому совпадает с плоскостью, перпендику-
лярной перегородке (узкоперегородчатые
стручки). У многих крестоцветных стручок
потерял способность раскрываться и рассеи-
вание семян происходит благодаря разламыва-
плода на односеменные членики. У таких
крестоцветных стручки имеют членистый ха-
рактер. Наконец, у некоторых крестоцветных
число семян в плоде сократилось до одного и
плод превратился в орешек (односемянной
стручочек).
Семена крестоцветных не имеют эндоспер-
ма (рис. 287)/ЗАридыш у’ЯЙХ^Ъёгда^бгнутый^
ж»
соприкасаясь.
лей, у других^щщд^яется; вдоль новерхнос-
^тй рдоой из семядолей, а у третьих* ^прц той
же ориентировке зародышевого корешка, сами
семядоли согнута вдоль средней линии и .коре-
шок оказывается лежащим в ложбинке. Нако-
нец, четвертый тип характеризуется тем, что
семядоли спирДГьд и уЯВУчены,и потому' на
поперечном разрезе семени они пересекаются
3
286.
Видоизменения
Рис.
стручка:
стручок дикой
родоаты* стручочек нрутод
два раза. СтроенйП’ТТлбда и форма зародыша — важные признаки, на
которых обычно основ^вйётсйдёление семеиства^йгС
Родь крестоцветных в составе растиуельности южного полушария и
тропической зоны, где оци„встречаются главным образом'в горах, незначи-
тельна. В пределах северного, полушария главная йасса крестоцветных сос-
редоточена в странах Средиземноморья и Передней Азии.
Многие крестоцветные имеют существенное хозяйственное значение.
Из масличных крестоцветных наиболее важны горчицы и рапс. Под име-
нем горчиц соединяют нескдлько"раз1№ЧШо видов! трепта^уюуПчшш
(Brassica juncea), черную горчицу (В. nigra), белую ёорчицу (Stnapis albi)>
Сарептская Торчица(сизаяУ^цДнолещее^ зфааяниет^
ятной родиной стагаюУТ)го-Западную Азию. Культуррм^с^лью
получения из семян жирного масла сосредоточена в СССР в Волгоградской,
масло по вкусовым качествам й. способности к длительному хранениюсчита-
ется одйшГиз лучших пишевыхрастатольных масёлл Его>используютв~ кон-
сервной, маргариновой, парфюмернойифармацевтической промышленности,
особение-тГкондитерсКдМ и хлеоопекарном деле. Содержащееся В СбШВах в
большом количестве (от 35 до 47%)' горчичное масло получают путем хо-
лодного прессования.
Жмыхи, остающиеся после выделения масла, перемалывают в тонкий
порошок, Известный по J шшГйотбрый^ используЙТ'Для
изгот йзвестной пртфавы- с^хой горг
также fop . Испбльзованйё1 ЯЕМыхов на корм ТЖоту ДОЖйкно лишь в
пгпаттпдтгых размерах, тай как сидёржаЩийСУ "'Д^ЙмхглюкозизГ^ сини-
^грин вШовает разд ражениеорганов пищейареш^я
жмых подвергают предварятельной~Ь15£аболw :
Черную горчицу у нас возделывают мало.
III
'10
ВЫСШИЕ РАСТЕНИЯ
Рис. 287. Зародыши крестоцветных:
1 — краекорешковый зародыш CheiranthiiB cheiri (зародышевый корень расположен вдоль края плоских
семядолей); я — спиннокорешковый зародыш Brassica oleracea (зародышевый корень прилегает к по-
верхности спинкк почти плоской семядоли); 3 - вдольсвдацчатый зародыш Alliaria officinalis (семя*
цолк* согнутые вдоль средней жилки, зародышевый корень, прилегающий и поверхности одной из семя-
долей, находится в ложбинке, обрадованной продольна складкой семядоли); 4 — спиралькодольный
вароцнш Bun las ori entails (семядоли закручены спирально, что видно на продольном разрезе семени, и
благодаря этому поперечный рачрез пересекает их дважды) з я — продольный разрез семени; б — попе-
речный’ з — складчатый зародыш Heiiophila (семядоли изогнуты один раз. как к у Bunias orientalis,
а второй раз — в обратном направлении, поэтому поперечный разрез пересекает их трижды); а — про-
дольный разрез; б — поперечный
Посевы белой горчицы встречаются главным образом н^яедврцоземной
полосе^Масло, получаемое из седан-бедой горчицы, находит применение, ана-
л(П№£бе^фимененивг1кяс^7£арептской гбрчйцы.”Белая горчица считается
хорошим кормовым и медоносным растением (с 1 га посева белой горчицы
можно получить до 100 кГмеда). -----
Важным масличным , растением является рапс (Brassica oleifera).J3ro
однолетнее, растение, озимоё^^Лй 'яровое. Рапсовое масло принадлежит дс
группе техническю'1полувысыхающи'х масел й йаХбДит применение "в метал-
лургической "и ряде других областей промышленности. Рапсовый жмых яв-
ляется хорошим кормом; вредных соединений, которые снижают ценность
и
II Г'
горчичных жмыхов, в нем значительно меньше.
Из числа овощных растений, относящихся к семейству крестоцветных,
наибольшее значение имеет капуста (Brassica oleracea, рис. 288). Дикий пре-
док современных культурных форм капусты обитает в Средиземноморье, по
морским берегам. В культуру капуста введена, по-видимому, еще в доисто-
рические времена. Наиболее близкой к родительской форме является листо-
вая капуста (В, oleracea var. acephala), которая не образует кочанов; она
имеет преимущественно кормовое значение. У брюссельской капусты (В.
oleracea var. gemmifera) высокий стебель, в пазухах которого развива-
ются почки, представляющие миниатюрное повторение кочана. Кочанные
капусты известны в двух основных формах: кочанной (В. oleracea var.
capitate) и. савойской (В. oleracea var. sabauda), причем последняя отличается
более тонкими, пузырчатыми или гофрированными листьями и кочаном
меньшего размера. Цветная капуста (В, oleracea var. botrytis) представ-
ляет разновидность с сильно разросшимися ветвистыми соцветиями, _но §
недоразвитыми цветками, сидящими на белых сочных цветоножках. Из того
nr.
же вида произошла кольраби — растение с реповидно утолщенным над
землей стеблем, который используется в отваренном виде в пищу. Коль-
раби характеризуется высоким содержанием витамина CL
Наиболее широкое распространение как овощное растение повседневно-
го использования получила у нас кочанная капуста. Пищевое значение ее
III
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
определяется довольно высоким, ва-
рьирующим в зависимости от сорта,
содержанием в ее продуктивных час-
тях питательных веществ, витаминов
A, Bv В2, С, Р, К и минеральных со-
лей, особенно фосфора и кальция,
варьирующих в зависимости от сор-
та.
Так как ранние всходы капусты
погибают от мороза, а период,
необходимый для вызревания ко-
чана, велик (до 100 дней у ранних
сортов, до 160 дней у поздних), то
семена капусты высевают в парники.
Рассаду при появлении первого на-
стоящего листа
1Г1
кируют, вследст-
вие чего корневая система молодых
растеньиц ветвится, разрастается и
укрепляется. После высадки расса-
ды в грунт растение образует не-
Рис. 288. Капуста (Brassica oleracea):
1 — соцветие к плоды; s — стручок; з — раскрыв*
шийся стручок
сколько крупных листьев, распола-
гающихся розеткой. Следующие
листья благодаря более1 энергич-
ному росту их нижней (наружной)
стороны оказываются загнутыми
внутрь. И5^тих^1исйёв”й“_форййру-
ется кочан, причем, чем глубже, тем листья его моложе и мельче, вплоть
до совсем небольших ближе к конусу нарастания. Всё листья ко-
чана прикреплены к толстому" укороченному стеблю. к о -
ч е р ы г е . Из сказанного видно, что кочан, по существу, представляет
гигантскую почку. Возможность роста кочерыги не исчерпывается в первый
год. Высаженная на следующий год, она продолжает расти, ее стебель силь-
но вытягивается, развивает соцветие с большим количеством цветков (двух-
летнее растение). Капуста — растение, требовательное к минеральному пи-
танию. Поэтому ее культивируют на хорошо унавоженных почвах. Велика
потребность капусты и в увлажнении; некоторые сорта потребляют в период
формирования кочана до ведра воды на одно растение в сутки. Естественно,
для получения хорошего урожая капусты требуется обильный полив.
К роду^капуста (Brassica) относится брюква (В. тмроЬгазз1са)9_д1шо11Щя
крупные мясистые корнеплоды, содержащие до 4—5% сахара, около 1,5%
белковых веществ, а также много витаминов. Произошла она от скрещивания
листовой капусты и сурепицы; известна только в культуре. Разводится в
огородах, но, к сожалению, сравнительно мало распространена. Есть как сто-
ловые, так и кормовые сорта.
Возделываемая в Советском Союзе рта (В. гара) имеет утолщенный
корнеплод с белой или желтой мякотью.
К этому же виду относится и турнепс — растение с удлиненным корне-
плодом, достигающим нередко 16 ке. По сравнению с другими кормовыми
корнеплодами турнепс относительно беден сухим веществом и богат водой.
Возделывается преимущественно как кормовое растение.
Перечисленные овощи из рода Brassica все имеют двухлетний цикл раз-
вития.
Редька, образующая, как и представители рода капуста, корнеплод и
hi
роко распространенная в огородной культуре, относится к р о д у Raphanus.
Огородная редька (R. sativus) имеет лиловые или белые с лиловыми про-
жилками цветки и несколько вздутые, внутри губчатые стручки с неясно
намеченными внутри полостями. Корнеплод содержит большое количество
витаминов и эфирных масел, которые, по-видим ом у, обладают лекарствен-
424
ВЫСШИЕ РАСТЕНИЯ
ними свойствами. Редька подраз-
деляется на несколько культурных
разновидностей, и одной из них
является редис (R. sativus иаг.
radiculaJTcopTSFKOToporo различны
по форме клубнецх,окра€ке^1гт. д.
Редис ценится как одно из самых
ранних овощных растений". Бла-
годаря быстрому росту (от посева
до созревания проходит 30—40
дней) культура рёдиса-удается да-
же на Крайнем; Севере^лтоШмеет
большое значение при недостатке
в этих местах’ свежих овощей.
В качестве приправы, а так-
же с лечебными _ целями исполь-
зуют корни хрена обыкновенного
(Armoracia rusticana), являюще-
гося одним из лучших.. противо-
цинготных растений.
Технически ценным кресто-
цветным является вайда красила
нал (I satis tinctoria), растущая в
степной полосе. Из листьев вайды
получают синюю краску(подобную
индиго), годную для окраски сук-
на. Ради этого вайду в прежнее
время широко культивировали,
особенно во Франции иь в Герма-
нии. В настоящее время значение
этого растения упало в связи с
производством синтетических
Рис. 289. Пастушья сумка (Capsella bursa- анилиновых красок. Вайда —
past oris): растение медоносное.
1 — общий вид растения; 2 — цветок; з — раскрыв*
ШИЙСЯ плод
Целый ряд крестоцветных из-
вестен как сорные растения полей
и огородов. Таковы виды гулявни-
ков (Sisymbrium), желтушник левкойный (Erysimum cheiranthoides), сурепка
(Barbarea vulgaris), дикая редька (Raphanus raphan 1st rum) — из числа
имеющих стручки, а из стручочковых — пастушья сумка (Cdpsella bursa-
pastoris, рис. 289), ярутка полевая (Thlaspi aruense), виды рыжика (Camelina),
икотник (Berteroa incana), некоторые виды крупки (Draba) и др.
Большинство произрастающих в нашей стране сорных крестоцветных —
растения однолетние, вегетативно не размножающиеся; большая энергия их
семенного размножения затрудняет борьбу с ними. Так, пастушья сумка
приносит на один экземпляр рт2 Дё?0 тыс. семян, _а одиниз видов гулявника
(DescuraiHia sophia) — т 6 до 110 тыс. й даже (особо мощные экземпляры)
до 770 тыс. При массовом произрастании таких сорняков количестао семян,
попадающих в почву^лредставляет в сумме огромную вел'ичййу. При спо-
собности семян сорняков сохр£ня№Лсхожесть длительное время и прорастать
не сразу, а в течение ряда лёт поле всегда будет находиться под угрозой засо-
рения, если не принять предупредительных мер. Некоторые из сорных крес-
тоцветных, как та же пастушья сумка, успевают дать два семенных поколе-
ния в год. Один из особенно вредных сорняков — сурепка (Barbarea vulga-
ris) — иногда сплошь покрывает поля. Сурепка — растение двухлетнее, но
может стать многолетником*, размножающимся, корневыми отпрысками, что
вместе с большой семенной производительностью (до 10 тыс. семян на одно
растение) обеспечивает быстрое размножение этого сорняка. В то же время
отдав ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
сурепка — хороший медонос. Цветки сурепки ранее использовали в кустар-
ном производстве для окрашивания шелка в желтый цвет. Примером специ-
ального сорняка является рыжик. Некоторые виды рода рыжик (Сате-
Нпа), имеющие характерные округло-грушевидные стручочки, культивиру-
ются как масличные. По исследованиям Н. В. Цингера, масличные
формы, объединяемые под названием С. glabrata. произошли путем отбора
в процессе отвеивания семян от мелкомодного рыжика (С- microcarpa). не-
редко встречающегося как сорное растение на полях. Тем же путем возник
и сорный вид С. Itnicola. засоряющий посевы льна-долгунца в северо-запад-
ных районах.
Некоторые крестоцветные вошли в культуру как декоративные. В этом
отношении на первом месте стоит левкой (Matthiola annua), родом из Южной
Европы. Декоративны также лакфиоль (Cheiranthus cheiri), ночная красавица
(Hesperis matronalis) и некоторые другие.
Семейство каперсовые (Capparidaceae) содержит околр^ЗОО видов и,
по всей вероятности, представляет параллельную й!крестрцветными rgynny,
в которой также господствует димерйай тап цветка и наблюдается расщеп-
ление тычинок. Диаграмма цветка некоторых каперсовых вполне совпадает
с диаграммой крестоцветных. Однако у других форм вследствие ^огократ-
ного расщепления тычийок андроцей стал многочисленным. Характерна и ши-
роко распространенная особенность — образование гинофора или андро-
гинофора (рис. 290). Плоды — стручки, коробочки или ягоды. Растения это-
го семейства обитают главным образом в тропических и субтропических стра-
нах и представлены травами и кустарниками. В СССР в пределах Средней
Азии, отчасти в Крыму и на Кавказе распространены виды рода каперсы
(Capparis), один из которых (С. spinosa. рис. 291) используют (цветочные
почки) в пищу под названием каперсов. В Южной Европе этот вид культиви-
руют. ч •
Семейство резедовые (Resedaceae) сближают с каперсовыми по
наличию короткого андрогинофора, но цветок здесь зигоморфный. Замеча-
1Г1
Рис. 290. Pedicellaria
(авдрогинофор)
Рис. 291. Каперсы:
1 — цветущая ветвь; в — яплюдов плод кя гинофоре
426
ВЫСШИЕ РАСТЕНИЯ
тельно, что плодолистики ера*
стаются не полностью и завязь
(из 2—6 плодолистиков) имеет
открытое отверстие наверху.
Рыльца сидячие, небольшие.
Сюда относится садовая резеда
(Reseda odorata, рис. 292) — де-
коративное растение с ароматич-
ными цветками, родом из Север-
ной Ливии (Киренаика).
Подкласс Dillenildae через
своих наиболее богатых прими-
тивными признаками предста-
вителей, сосредоточенных в ос-
новном в порядке Dilteniates^
довольно естественно примыкает к подклассу Magnoliidae* По ряду призна-
ков (ценокарпность, редукция эндосперма и др.) близкий, но вполне легко
отличимый от DillenialeSy порядок Theales филогенетически, вероятно, связан
с Magnoliidae через посредство Dillenlales. Чаецветные (TbeaZes) по всей ве-
роятности, оказались предками нескольких самостоятельно развивавшихся
эволюционных линий цветковых. Считают вполне возможным, что этот по-
рядок был предковым по отношению к верескоцветным (Erlcales); с другой
стороны, из тех же чаецветных могли возникнуть общие предки первоцвет-
ных (Primulates) и эбеновых (Ebenates). Но наиболее мощный поток эволю-
ционных преобразований на базе диллениецвётных начинается с порядка
фиалкоцветных (Violates), оказавшегося родоначальником нескольких, во
многом своеобразных направлений эволюции.
• Так, тыквенноцветные (Cucurbitales) вместе с параллельными им
страстоцветными (Passiflor ales) формировались как группа с широким
распространением различных вариантов лазающих растений, и, вероятно,
возникла эта ветвь от каких-то Violates. Другую обособленную ветвь
составили каперсоцветные (Capparates), выделяющиеся не только димер-
ностью цветков, но и химизмом (своеобразные гликозиды, мирозин).
Наконец, совершенно особую ветвь составили мальвоцветные (Malvates)
с примыкающими к ним молочаецветными (Euphor biates). Впрочем, положе-
ние этих порядков среди диллениидных двудольных можно оспаривать.
5-Й ПОДКЛАСС - РОЗИДНЫЕ (ROS1DAE)
От Dilleniidae этот подкласс отличается как по центрипетальной после-
довательности развития андроцея, так и по плацентации, которая редко бы-
вает париетальной.
Рассмотрим 8 порядков подкласса Rosidae (из общего числа их
от 16 до 19).
1-Й ПОРЯДОК — РОЗОЦВЕТНЫЕ (ROSALES)
Цветки у большинства розоцветных обоеполые, актиноморфные или
несколько зигоморфные, с двойным (реже с простым) околоцветником,
обычно из пятичленных кругов.Число тычинок равно или чаще кратно чис-
лу лепестков, иногда же их неопределенное число; расположены они кру-
гами. Гинецей из многих, нескольких или одного плодолистика, апокарпный
иди ценокарпный, круговой или неясно спиральный. Семяпочек большей
частью две, иногда несколько в каждом гнезде завязи; они прикреплены к
краям плодолистиков. Характерна тенденция к разрастанию цветоложа, ко-
торое может быть выпуклым, дисковидно расширенным и вогнутым вплоть
до бокальчатого, в связи с чем завязь иногда становится нижней. К порядку
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
427
Рис. 293. Очиток едкий (Sedum acre):
1 — цветущее растение; г — цветок; з — анаграмма
цветка
11 Г.
относятся растения разнообразно-
го внешнего облика (деревья,
кустарники, травы, суккуленты,
редко лианы), с очередными или,
реже, супротивными и даже иног-
да мутовчатыми листьями, с при-
листниками и без них.
Причисляемые сюда семейства
находятся в тесных родственных
отношениях друг с другом и иног-
да довольно естественно смыкают-
ся с некоторыми семействами мно-
гоплодниковых.
Семе й с т в о толстянковые.
(Crassulaceae). Травянистые рас-
тения, суккулентные, многолет-
ние или однолетние, с простыми,
большей частью без прилистников,
сочными листьями. Цветки обычно
в колосовидных, кистеобразных,
щитовидных соцветиях, актино-
морфные, обоеполые, лишь редко
однополые, круговые, в большин-
стве случаев с одинаковым 4—5—6 и более числом членов околоцветника
(в обоих кругах) и гинецея, но вдвое большим числом в андроцее.я Венчик
раздельнолепестный, иногда спайнолепестный. Плодолистики свободные
или срастаются между собой в большей или меньшей степени^ Завязь
верхняя. При основании плодолистиков находятся чешуйки — нектарники.
Растений энтомофильные, некоторые орнитофильные. Цветоложе часто
блюдцевидно расширенное. Плоды — свободные или сросшиеся листовки.
Толстянковые — большое семейство, содержащее до 1400 видов, насе-
ляющих главным образом засушливые территории и горные местности.
чными признаками семейства можно ознакомиться на примере
широко распространенного в европейской части СССР, в Западной Сибири,
в Крыму, на Кавказе и других местах очитка едкого (Sedum acre) — неболь-
шого травянистого растения с мясистыми листьями (рис. 293).
Еще более типичны суккуленты рода молодим (Sempervivum). Один
из его видов — молодило побегоносный (S. soboUferum) — встречается на
песчаных почвах в средней полосе европейской части СССР. У этого растения
имеется розетка очень сочных листьев, которые плотно прилегают друг к
другу, образуя как бы сплющенный шарик. На конце пазушных столонов
образуются укореняющиеся дочерние розетки.
Способность к вегетативному размножению вообще распространена у
толстянковых широко. Вроде бриофиллюм (Bryophyllum pinnatum и дру-
гие виды) в углах краевых зубчиков листовой пластинки образуются при-
даточные выводковые почки, из которых развиваются новые растения.
У Crassula cordata (Южная Африка) выводковые почки образуются в соцве-
тиях.
Будучи очень специализированными в вегетативной сфере (суккулент-
ность), толстянковые сохранили примитивное строение цветка. Однако у
которых представителей семейства наблюдается сростнолепестность д X
Umbilicus, виды рода Sedum), что делает вероятной связь между по^клас-
сами розидных и астеридных. V. Глм
Толстянковые наиболее распространены в Южной Африке, МексиЙИзй^ ^*5*3
в Средиземноморье. /'
Многие толстянковые разводят как декоративные. Таковы, например,
эхеверия сигая из тропической Америки (Echeveria glauca) с сиЩйвато-зеле-
ными розетками; крассула (Crassula falcata) из Южной Африки с толстыми
428
ВЫСШЙЕ РАСТЕНИЯ
мясистыми клиновидными листьями, расположенными вертикально и окра-
шенными в серовато-зеленый цвет; Rochea coccinea из Южной Африки —
растение с правильно крест-накрест расположенными листьями и ярко-крас-
ными цветками, разводят иногда в комнатах.
Сам ей ст в о камнеломковые (Saxifrazaceae}. В это семейство входят
многолетние (редко однолетние) травы или кустарники и небольшие деревья
с очередными, реже супротивными листьями, обычно без прилистников.
Цветки актиноморфные (рис. 294), реже слегка зигоморфные, собранные в
соцветия, двуполые или вследствие редукции однополые. Цветоложе очень
различное: плоское, вогнутое, чашевидное, иногда выпуклое. Чашелистиков
и лепестков по пять, реже по четыре и очень редко более пяти. Тычинок
вдвое больше или столько же, сколько лепестков, редко их много. Плодоли-
стиков, почти всегда сросшихся, меньше, чем лепестков, редко, столько же.
Пестик с числом столбиков, равным числу плодолистиков./Завязь верхняя,
а иногда, в случае бокальчатого или вогнутого цветоложа*, полунижняя
или нижняя, обычно пяти-, двугнездная или одногнездная, с пятью —
двумя постенными и обычно утолщенными плацентамиА Плоды— коробочки
либо ягоды. Семена мелкие, с небольшим зародышем и хорошо развитым эн-
доспермом.
Это обширное семейство содержит 1200 видов. Его представители рас-
пространены почти по всей суше, преимущественно в умеренных широтах.
На основании различий в цветках и других признаков камнеломковые под-
разделяются на несколько подсемейств, которые нередко считают даже са-
мостоятельными семействами. Из подсемейств камнеломковых здесь упоми-
наются только некоторые.
Из подсемейства Ribestoideae следует упомянуть род смородина (Ribes).
Широко распространенная в культуре чернил, смородина (R. nigrum) в'ди-
ком состоянии встречается в европейской части СССР, в Сибири и Средней
Азии — по береговым зарослям, во влажных лесах, по окраинам болот.
Дикорастущая черная смородина считается родоначальником многочислен-
ных культурных сортов. Помимо вкусовых качеств и своеобразного приятно-
го аромата, ягоды черной смород:
я ценятся за высокое содержание в них
витамина С. Кроме того, в плодах имеются витамины А, В, Р. Другие ди-
корастущие формы, близкие к Ribes nigrum^ также имеют съедобные плоды.
Культурная красная смородина произошла, как и черная, от дикорастущей.
Одним из предков ее была смородина обыкновенная (Ribes vulgare), дико рас-
тущая в Западной Европе. Близок к смородине крыжовник (Grossularia ге-
clinata.
Второе подсемейство — Hydrangeoideae — содержит также
кустарники, но с супротивными листьями. Сюда относится род чубушник,
или садовый жасмин (Philadelphus), некоторые виды которого являются лю-
бимыми декоративными растениями у нас в садах е крупными, до 2 еле в по-
перечнике, белыми приятно пахнущими цветками. К этоИу же подсемейству
относится род гортензия (Hydrangea)^ ojqxh. из японских видов которого—
гортензия крупнолистная (Н. macrophylla) — стал родоначальником эф-
фектных декоративных гортензий. У диких форм этого вида краевые цветки
бесплодны и служат для привлечения опылителей — насекомых. У культур-
ной формы, широко распространенной в цветоводстве, соцветие в виде круп-
ного шара все целиком состоит из бесплодных цветков, у которых сильно
разрослись лепестковидные крупные чашелистики; остальные части цветка
недоразвиты. Чашелистики бывают белые, розовые или голубые, часто зеле-
новатые.
Третье подсемейство — Saxtfragoideae — занимает централь-
ное место и наиболее обширно. Сюда относится большой род камнеломка
(Saxifraga), содержащий более 350 видов травянистых растений. Камне-
ломки распространены в тундре северного полушария и в горных местнос-
тях. В связи с жизнью в условиях сурового климата камнеломки часто быва-
ют низкорослыми растениями розеточного характера или образуют дернинки
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
из коротких, торчащих или из более
длинных, лежачих побегов, густо по-
u:i
крытых мелкими листьями. Нередко
наблюдаются различные формы вегета-
тивного размножения: усами (ните-
видными побегами, заканчивающимися
почкой, которая укореняется), отделя-
ющимися придаточными или пазуш-
ными почками, возникающими иногда
в соцветиях, и т. п.
Бадан (Bergenia crassifolia) — гор-
ное растение, обитающее у нас в го-
рах Сибири и Средней Азии. Известен
как важный дубитель. Дубильные ве-
щества (около 25%) содержатся в кор-
невище и листьях. Последние имеют
широкую (30 см) пластинку, длинный
черешок и образуют прикорневую ро-
зетку. Кроме тапнидо в, это растение
служит источником гидрохинона, пиро-
галловой кислоты и др.
К этому же подсемейству относится
селезеночник (Chrysosplenium alterni-
folium) — обычное у нас по сыроватым
местам небольшое травянистое растение
с верхними листьями, сближенными
под соцветием в виде плоской розетки
и окрашенными в желтоватый цвет. В
центре розетки располагается соцветие
из четырехчленных цветков. Плоды —
коробочки.
В особое подсемейство Раг-
nassioideae выделяют род, белозор
(Pamassia). В нашей флоре он пред-
Рис. 294. Камнеломка (Saxifrage
granulata):
j, 2 — цветущее растение; з - разрез цвет-
ка; 4 — диаграмма цветка
ставлен четырьмя видами, из которых
более широко распространен белозор болотный (Parnassia palustris) — тра-
»:г
вянистое многолетнее растение с черешковыми прикорневыми и сидячими
стеблевыми листьями. На конце стеблей — одиночные крупные (более 1,5 см
в поперечнике) белые цветки, в которых за пятичленными чашечкой и венчи-
ком следует широкоовальные стаминодии, рассеченные наверху на несколь-
ко нитевидных долей, заканчивающихся железками. Тычинок пять. Растет
по сырым открытым местам. /
Семейство камнеломковых считается родственным с толстянковыми, но
отличается от них не только иным характером вегетативных органов, но от-
сутствием (почти всегда) стаминодиальных околопестичных чешуек и хоро-
шо развитой питательной тканью в семенах, которая у толстянковых развита
слабо или вовсе отсутствует.
С е м е й с т в о розовые (Rosaceae) - Это листопадные или вечнозеленые
деревья и кустарники, многолетние и (редко) однолетние травы, иногда лиа-
ны. Листья большей частью очередные, имеющие прилистники, то рано опа-
дающие, то сохраняющиеся. Цветки то одиночные, то в соцветиях, актино-
морфные (редко зигоморфные), в основном с пятичленными, реже с четы-
рех-, шестичленными кругами. Цветоложе выпуклое, плоское или вогнутое.
В последнем случае говорят о «цветочной трубке» или о «цветочном бокале»,
«осевом бокале», «гипантии» и т. п. Одни считают его целиком образовав-
шимся из вогнутого цветоложа, по мнению других — это продукт срастания
базальных частей листочков околоцветника и тычиночных нитей, наконец,
согласно третьему мнению, в образовании цветочного бокала принимают
ши
г»»
II.
430
выс
иг-
Е РАСТЕНИЯ
участие и края вогнутого цветоложа, и основания цветолистьев. Чашечка
иногда двухрядная вследствие образования второго (наружного по положе-
нию) круга — подчашия. Листочки подчашия имеют следующее происхож-
дение. Каждый Чашелистик (собственно чашечки) в этом случае с при-
листником, правый прилистник одного чашелистика и левый соседнего с
ним, срастаясь друг с другом, образуют один листочек подчашия. Такое
происхождение подчашия подтверждается тем, что нередко его листочки бы-
вают наверху не вполне сросшимися (двураздельными), а иногда на месте
одного листочка подчашия находятся два — в этом случае прилистники не
срослись,.
Венчик у розовых, как правило, ярко окрашенный, лишь редко вслед-
ствие недоразвития малозаметен или отсутствует. Андроцей всегда цикличес-
кий: число тычинок равно, 'а чаще ЬУ(вое, вчетверо и более превышает число
лепестков. Гинецей также иэменчив: число плодолистиков колеблется от
неопределенно большого до одного.. Плодолистики то свободные, то срас-
таются в один пестик, который имеет столбик, отходящий у одних форм
jiT верхушки, у других — от основания завязи (гинобазиЧеский).
Завязь одногнеэдная или у растений с ценокарпным гинецеем двух-,
^многогнездндя. Семяпочек в каждом гнезде эавязи одна, две или (реже)
несколько^По положению эавязь верхняя или вследствие срастания с
бокалом нижняя^ .
Плоды розовйхочень разнообразны — листовки, орешки, костянки и
различные формы так называемых ложных плодов (яблоня, земляника,
шиповник). Семена большей частью без эндосперма.
Это семейство содержит около 3000 видов, распространено по всей Земле,
но главным образом — во внетропических областях северного полушария.
В составе его имеются многочисленные представители со сравнительно прими-
тивным строением цветка и плода, а с другой стороны — прогрессивные,
специализированные формы. Однако специализация цветка в связи с опыле-
нием сравнительно незначительна, хотя относящиеся сюда растения энто-
мофильны. У одних представителей опылители получают только пыльцу
(Rosa), у других — обильный нектар, выделяемый железистым кольцевид-
ным диском. Диск образуется цветоложем и помещается между тычинками
и плодолистиками, в некоторых случаях лишь намечаясь в виде слабозамет-
ной полоски. Встречаются приспособления к самоопылению, вступающие
в действие в конце цветения. Так, у черемухи и рябины можно наблюдать,
что в конце цветения тычинки загибаются внутрь, по направлению к рыльцу,
а столбик, тоже изгибаясь, поворачивает рыльце навстречу пыльнику одной
из тычинок, и они соприкасаются. Интересно, что этот механизм продолжа-
ет действовать даже в том случае, если растение не нуждается в опылении и
развивает семена без оплодотворения, что можно наблюдать у манжеток
(род Alchemilla). Лишь как исключение среди розоцветных встречаются
ветроопыляемые формы (черноголовник — Poterium).
Гораздо ярче выражена специализация цветка в связи с обеспечением
распространения будущие плодов. Бокаловидные образования в основании
цветка разрастаются после цветения, часто принимая яркую окраску и стано-
вясь сочными, что, несомненно, является приспособлением к распростране-
нию зачатков животными (эндоэоохория). Это наблюдается у шиповника,
яблони, рябины и др. В иных случаях наружная поверхность бокала покры-
вается крючковидными на конце выростами, обеспечивающими прикрепление
плода к телу животных (род репяшок — Agrimonia).
Большую роль в распространении плодов животными играет разрастаю-
щееся после цветения выпуклое цветоложе (род Fragaria — земляника и
клубника). У тех видов растений этого семейства, которые имеют плод,
развивающийся без участия цветочного бокала, цветоложе все-таки может
принимать участие в рассеивании плодов. Примером может служить гравилат
речной (Geum rivals). Блюдцевидно расширенное цветоложе этого рас-
тения в центре удлинено в виде цилиндрического столбца, усаженного
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
h:i;i
431
Рис. 295. Спирея:
1 — диаграмма цветка; 2 — разрез цветка
на расширенной верхушке многочисленными пестиками. При соэревании -
плодов столбик пестика, состоящий иэ двух члеников — верхнего и нижне-
го, — разрастается. Верхний членик столбика легко отделяется, а на конце
нижнего при этом обнаруживается остроконечный крючок (прицепка). Ко
времени плодоношения цилиндрическая часть цветоложа вытягивается и вы-
носит всю группу плодов (орешков) выше границы чашечки, которая после
цветения заметно увеличивается в размерах. У вишни, сливы и других, так
называемых косточковых, по мере созревания плода ткань плодолистика ста-
новится частично мясистой (мезокарпий)^ а внутренняя часть отверде-
вает (экдокарпий)и защищает семя от разрушительного действия пи-
щеварительных соков животного, поедающего плод.
Встречаются также розовые, распространяемые ветром. Такова куро-
паточья трава, или дриада (Dry as). Виды этого рода — приземистые расте-
ния тундр и высокогорий, со стелющимся ветвистым стеблем, несущим зимую-
щие листья. Плоды дриад — семянки — сохраняют при соэревании столбик,
покрытый перисторасположенными волосками, которые обеспечивают раз-
несение их ветром. Раэнообраэие цветков и плодов и большое видовое разно-
образие розовых приводит к необходимости делить это семейство на ряд под-
семейств, которые иногда считают самостоятельными семействами.
Подсемейство Spiraeoideae. Типичные черты этого подсемейства —
вогнутое или почти плоское~цветоложе, свободные или почти только "в осно-
вании сросшиеся плодолистики, двухрядность семяпочек (от нескольких
до двух) — можно наблюдать у видов рода спирея, или таволга (Spiraea,
рис. 295). Некоторые виды спиреи известны как декоративные кустарники.
К спирее’близок род рябинник (Sorbaria), один из видов которого —
рябинник рябинолистный (5. sorbifolia) — вошел в садовую культуру как
декоративный кустарник. Листья его непарноперистосложные, соцветия иэ
белых, слегка кремоватых цветков, метельчатые, многоцветковые.
Подсемейство Rosoideae. Гинецей, как и в предыдущем подсе-
мействе, многочленный, апЪкарпный, но завязь толькойс 1—2 семяпочками.
Если семяпочек две,'то в зрелое семя развивается только одна. Это привело к
замене раскрывающегося плода (листовки спирейных) нераскрывающимся.
Подсемейство содержит много общеизвестных растений. «
Род Rubus характеризуется плодом—сложной костянкой. Объединя-
ет много видов, иэ которых наиболее важна малина обыкновенная (R. idaeus,
рис. 296) — кустарник, легко размножающийся корневыми отпрысками с
двухлетними наземными побегами — в первый год они развивают только
листья, а на второй — цветут и плодоносят. Иэ каящого цветка развивается
сложная костянка (многокостянка) в виде шапочки красного или желтого
цвета, покрывающая белое губчатое цветоложе, которое конусовидно разра-
стается ко времени плодоношения. Плоды дикой малины имеют приятный,
сладкий вкус; их используют в свежем виде, варят варенье и сушат. В суше-
ном виде они считаются хорошим потогонным средством. Известно большое
432
ВЫСШИЕ РАСТЕНИЯ
Рис. 296. Малина (Rubus idaeus):
1 — плодоносящая ветвь; £ — разрез цветка; а —разрез
вшива в разросшегося цветолока (ц — цветоложе)
число сортов культурной мали-
ны, отличающихся цветом, аро-
матом и размером плодов. Куль-
турные сорта частично получе-
ны путем гибридизации.
Под названием ежевика
объединяют большое количество
видов рода Rubusa характе-
ризующихся обычно черным
или черно-красным цветом
плодов и срастанием костянок
с цветоложем. Почти все, во
всяком случае многие, виды
ежевики ценятся как ягодные
растения; в СССР особенно ве-
лико разнообразие ежевик на
Кавказе. Известен ряд сортов
культурной ежевики, но они в
общем имеют менее широкое
распространение в садах, чем
малина.
К р о д у Rubus относится
еще ряд растений со съедобны-
ми плодами, например костяни-
ка (R. saxatilis), обитающая
главным образом в хвойных
лесах и приносящая съедобные
красные плоды, не имеющие,
однако, промышленного значе-
ния; морошка (R. chamaemorus)—
травянистое растение тундр
и сфагновых болот северной
части лесной зоны с округло-
почковидными листьями и жел-
тыми плодами, похожими на
малину, которые употребляют на варенье и в сыром виде.
Для рода земляника (Fragaria, рис. 297) характерно разрастание цве-
толожа ко времени плодоношения в красное мясистое сладкое тело, по поверх-
ности которого располагаются мелкие односемянные сухие плодики (семян-
ки). Чашечка у земляники двухрядная (с подчашием). Земляника — много-
летнее травянистое растение; она энергично размножается усами, т. е. над-
земными побегами с удлиненными междоузлиями и редуцированными листья-
ми. Возникают усы в пазухах прикорневых листьев, образующих розетку.
Укореняясь* усы дают начало новым розеткам. Опыты культуры лесной зем-
ляники (F. vesca) и клубники (F. viridis, полуница) относятся еще к XIV в.
Но из этих видов, несмотря на длительную культуру, крупноплодных сор-
тов получено не было. Лучший эффект дали завезенные из Северной Америки
земляники с более крупными плодами — виргинская (F. virginiana) и чи-
лийская (F. chiloensis). Гибриды, полученные в XVIII в., стали родоначальни-
ками культурных земляник, характеризуемых крупными плодами высоких
вкусовых достоинств и объединяемых в вид Fragaria ananassa. В настоящее
время известно много сортов культурной земляники, например Рощинская,
Коралка (Виктория), Белая ананасная, Премьер и др. Культурой этих ягод-
ных растений занята огромная площадь. Плоды земляники содержат много
железа, фосфора и витамин С.
Близки к землянике лапчатки (род Potentilla). У лапчаток цвето-
ложе выпуклое, остающееся при созревании плодов сухим. Как и у земля-
ники, есть подчашие. Окраска лепестков большей частью желтая. '
3-1
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Лапчатка прямосто-
ячаял иди калган (Р. erec-
ta), отличается четырех-
членным строением цветка.
Отвары и настойки корне-
вища этой лапчатки упо-
требляют в народной меди-
цине при желудочных
' заболеваниях.
Весьма обширен род
шиповник, или рога (Ro-
sa, рис. 298). Цветки ро-
зы имеют глубоко вогну-
тый бокал, по краю кото-
рого располагаются чаше-
листики, лепестки (по 5) и
многочисленные, в нео-
гл
пределенном числе, тычин-
ки. Пестики, тоже много-
численные, погружены
внутрь бокала* но не
срастаются с ним. Из уз-
кого отверстия бокала
наружу выступают только
рыльца, сидящие на длин-
ных столбиках. При со-
зревании плодов, цветоло-
же разрастается, прини-
мая яркую — красную,
оранжевую, иногда почти
черную — окраску* ста-
новясь при этом несколь-
ко мясистым. Шиповники
известны красотой своих
цветков (розовых, иногда
белых или желтых) и за-
мечательным ароматом.
Махровые и некоторые
обычные их формы куль-
Рис. 297. Земляника (Fragaria vesca) и сабель-
ник (Comarum palustre):
1 — цветущая земляника; 2 — плодоношение; з — разрез
цветка; 4— аестс; 5 — разрез пестика; 6 — плод; 7— диа-
грамма цветка сабельника
кой древности. Известное нам громадное число декоративных сортов пред-
ставляет результат напряженной работы цветоводов многих поколений.
Промышленное применение имеет красная казанлыкская роза (Я. da-
тазсепа /. trigintipetala), из лепестков которой добывают очень ценное аро-
матическое масло. Культивируется как эфиромасличное растение; лепестки
идут на варенье. В районе культуры этой розы распространена белая казан-
лыкская роза (R* alba), дающая масло менее высокого качества; в «плодах»
содержится от 2000 до 4000 мз витамина С на 100 г сухого вещества плодов.
Еще большим содержанием этого витамина (8750 мз на НО г сухого вещества)
выделяется R. beggeriana (Тянь-Шань, Китай* Монголия). «Плоды» ряда ви-
дов диких роз (шиповников), которых только в СССР более 60 вцдов, также
богаты витамином С. Например, в ложных плодах широко распространен-
ного шиповника коричного (R. cinnamomea) этого витамина 2000 мз на 100 г
сухого вещества; кроме того, содержатся витамины Ва, К, Р и провитамин А.
Таким образом* шиповник коричный—поливитаминное растение, что очень
важно, так как авитаминозные заболевания чаще всего вызываются недос-
татком не одного витамина* а группы их.
вы
Hili';
Ж РАСТЕНИЯ
Рис. 298. Шиповник:
1 — цветущая ветвь Rosa cinnamomea: 2 —ложный
плод R. cinnamomea; J —разрез цветка R. eptaosie-
sima; 4 — пестик из цветка Нова canina; 5 — разрез
плода R. canina;- 6 — анаграмма цветка R. tomentoea
Подсемейство Ма-
loideae отличается от двух пре-
дыдущих подсемейств как стро-
ением цветка, так и плодом.
В цветке яблоневых имеется
от 2 до 5 плодолистиков, обра-
зующих синкарпный гинецей,
причем члены его не вполне
срастаются, оставаясь свобод-
ными в верхней части. Поэто-
му число столбиков равно чис-
лу плодолистикоВя-Как и числу
гнезд в завязи. (Завязь ниж-
няя, что является наиболее
наглядным отличительным при-
знаком этого подсемейства. На
вершине завязи прикреплено
много тычинок, 5 лепестков и
5 чашелистиков.^ Последние со-
храняются на верхушке мясис-
того ягодоподобного плода —
яблока.
Род яблоня (Malus, рис.
299). Виды яблонь — деревья
средней величины, с просты-
ми овальными листьями и
опадающими прилистниками.
Крупные белые, розово-белые,
редко розовые цветки сидят в
немногочисленных пучках (зонтиках). Плоды пятигнездные, с двумя семе-
нами в каждом гнезде. Сочная мякоть яблока образуется за счет тканей
цветочного бокала и наружных тканей плодолистиков. Внутренние слои
последних становятся хрящеватыми, иногда жесткими за счет волокнистых
склереид.
Известно несколько дикорастущих видов яблони, часть которых встре-
чается в равнинных, по преимуществу лиственных лесах, а часть — в гор-
ных областях. Яблони с незапамятных времен культивируются как отлич-
ные плодовые деревья, известные в множестве сортов, произошедших от раз-
ных диких видов, главным образом выведенных путем гибридизации; они
объединяются под общим именем домашняя яблоня (Malus domestic^, В СССР
широко распространены сорта: Антоновка, Апорт, Боровинка, Коричное,
Налив, Грушовка, Кандиль, Ранет и многие другие. Наилучшие сорта про-
израстают на юге. Благодаря настойчивой творческой работе И. В. Мичу-
рина наше плодоводство располагает сейчас целым рядом сортов, хорошо
переносящих зиму и дающих плоды высокого достоинства, вполне сравни-
мые с лучшими южными сортами. Из мичуринских сортов назовем: — Пепин
шафранный, Бельфлер-китайка (рис. 300), Бельфлер красный, Славянка,
Китайка золотая, Антоновка полуторафунтовая, Ренет бергамотный, Кан-
диль-китайка. Многие мичуринские сорта введены в стандартный сор-
тимент ряда областей.
Кроме домашней яблони, нередко культивируют так называемую ки-
тайку, или китайскую яблоню (М. prunifolia), имеющую мелкие плоды, ко-
торые используют для варенья. Родиной китайки считается Северный Китай.
В Восточной Сибири и на Дальнем Востоке дико растет мелкоплодная яб-
лоня (М. pallasiana), близкая к распространенной в культуре ягодной, или
райской, яблоне (М. baccata). Последняя имеет плоды диаметром всего до
1 см, желтые, краснеющие на солнце. Она, а также дусен (М. frutescens)
часто используются как подвои для культурных сортов.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
435
Опыт разведения плодо-
вых деревьев показал, что в
суровые зимы они вымерза-
ют на уровне снега. Это на-
толкнуло на мысль выращи-
вать плодовые деревья в сте-
лющейся форме.
К роду Matus близок
род груша (Pyrus). Помимо
формы, плод груши отлича-
ется наличием в мякоти,
особенно у диких видов,
каменистых клеток, а цве-
ток — свободными от осно-
вания столбиками. Культур-
ные груши, разводимые глав-
ным образом на юге, отлича-
ются высокой сахаристостью,
неясностью плодовой мякоти
и своеобразным ароматом.
В продвижении культуры
груш в более северные мест-
ности путем создания вы-
носливых и высококачест-
венных сортов большое зна-
чение имели труды И. В. Ми-
чурина. Такие дорта, как
Бере октябрьская, Дочь
бланковой, Бере зимняя и
другие, вошли в стандарт-
ный сортимент ряда облас-
тей. При выведении сорта
Бере зимняя И. В. Мичури-
ным был использован метод
отдаленной гибридизации —
цветки дикой уссурийской груши были опылены пыльцой южного
сорта груши Бере-рояль. Путем отбора из гибридных сеянцев был получен
сорт, отличающийся исключительной зимостойкостью и устойчивостью к
вредителям при хорошей лежкости и высоких вкусовых качествах (рис. 301).
Близко к яблоне и груше в систематическом отношении стоит род ря-
бима. Обыкновенная рябима (Sorbus aucuparia) имеет оранжево-красные пло-
ды, строением напоминающие яблоко, но обычно с тремя (5—2) гнездами.
Плоды рябины используют для настоек, варенья, изготовления пастилы»
Ряд видов рябины имеет не перистые, как у обыкновенной, а простые, цель-
ные или рассеченные листья. Эти виды рябины встречаются в нашей стране
преимущественно на Кавказе.
Сходное строение имеют плоды боярышника (род Crataegus). Вследст-
вие каменистого характера эндокарпия плоды боярышника напоминают
костянки с 1—5 косточками. Боярышники используются как декоративные
растения; особенно они пригодны для живых изгородей, так как развивают
прочные крупные колючки, представляющие собой укороченные безлистные
пазушные побеги.
К этому же подсемейству относятся айва (Суdonia), мушмула (Mespilus),
японская мушмула (Eriobotrya japonica) в ряд других, имеющих съедоб-
ный плод.
Подсемейство Prunoideae. Сливовые имеют глубоко вогнутый
цветочный бокал, не срастающийся с завязью. Гинецей состоит из одного
плодолистика. Из двух семяпочек, содержащихся в завязи, только одна
ВЫСШИЕ РАСТЕНИЯ
Рис, 300. Бельфлер:
1 — Бельфлер желтый; 2 —
Кита йка; з — Бедьфлер-кж-
тайка
развивается в семя. Внутренняя часть околоплодника твердеет, окружая
единственное семя плода, а наружная становится большей частью мясистой
(костянка). К этому подсемейству относится ряд высокоценных плодовых
растений.
Миндаль (род Amygdalus) характеризуется костянкой с кожистым мезо-
карпием. Широко распространенный от Средиземноморья до Центральной
Азии, миндаль введен в культуру и за пределами распространения дикой'
родоначальной формы. Известно много сортов и разновидностей миндаля
со сладкими семенами, в то время как у диких форм преобладают горькие.
Семена миндаля употребляют в пищу, а в промышленности миндаль нахо-
дит применение как масличное растение с высоким (до 50%) содержанием
масла в семенах.
Миндальное масло применяют в парфюмерии, кондитерской промыш-
ленности и в медицине. Содержащийся в миндале глюкозид амигда;
присутствии фермента эмульсина разлагается на бензальдегид и синильную
поту. Очищенное от синильной кислоты миндальное масло идет в
iyfl но использование жмыхов для скота опасно. Обычное растение в
лесостепной полосе — степной миндаль, или бобовник (Amygdalus папа).
Это небольшой кустарник, цветущий весной. Плоды его несъедобны.
Слива (род Prunus) имеет сочную, удлиненную, с поверхности голую
костянку со сплюснутой косточкой. Слива — одна иэ древнейших плодовых
культур. Дикий предок неизвестен. Возможно происхождение культурной
формы в результате гибридизации терновника и алычи.
Культурные сливЫд объединяемые под названием Р. domestica, своими
лучшими сортами представлены на юге. Стремясь к продвижению южных
слив на север, И. В. Мичурин вывел ряд ценных сортов, среди которых сле-
дует отметить Ренклод-реформа. Широкое распространение получил также
Ренклод терновый, Ренклод колхозный и другие мичуринские сорта. Общее
число сортов сливк достигает 2000.
Терновник, или терн (Р. spinosa), — высокий кустарник с укороченны-
ми побегами, превращенными в колючки, — довольно широко распростра-
нен в СССР, особенно в лесостепной полосе, где образует густые, иногда об-
ширные заросли. Плоды его, некрупные, сизо-черные, кислые, идут на при-
готовление компотов, варенья, плодового вина и уксуса.
Алыча (Р. divaricata) — дерево или высокий кустарник с колючими ши-
рокораскидистыми ветвями и желтыми, светло-красными, розовыми или тем-
но-красными обратнояйцевидными плодами. Обитает в нашей стране на Кав-
ИЛИ
: ।: г
1 Г, I '.1
I»
ли
лиг
ОТДЕЛ цветковые, или ПОКРЫТОСЕМЕННЫЕ
437
каэё и в Средней Азии, где встречается в древесно-кустарниковых лесных за-
рослях. Дикие заросли алычи дают ежегодно несколько тысяч тонн плодов,
идущих на варенье, повидло и т. п. Местами культивируется, иногда под
названием «мирабели».
Род абрикос (Armeniaca) характеризуется сочной костянкой, е поверх-
ности опушенной, с более или менее гладкой косточкой. Этот небольшой
род распространен от Дальнего Востока до западных отрогов Тянь-Шаня.
Большинство культурных абрикосов относится к виду A. vulgaris, обитаю-
щему в горах Средней Азии (СССР) и в Китае. Более половины площадей,
занятых в нашей стране культурой абрикоса, сосредоточено в среднеази-
атских республиках, остальные приходятся главным образом на южные
области Украинской ССР и Кавказ. Экономическое значение абрикосов оп-
ределяется высоким содержанием в плодовой мякоти сахаров. В сухофрук-
тах, приготовленных из среднеазиатских абрикосов, содержание сахара в
расчете на сухую массу достигает 79%, в том числе до 49% приходится на
сахарозу.
По содержанию витамина А абрикос не уступает шпинату и яичному
желтку, но витамин С содержится в небольшом количестве. Семена аб-
рикоса содержат 30—40% жира и используются для получения пищевого
абрикосового масла. Древесина твердая и ценится для токарных работ.
Род персик (Persica) также имеет сочный, тонко и коротко опушенный
околоплодник; косточка глубокоизвилистобороэдчатая. Этот род генети-
чески близок к миндалю, причем есть переходные формы. Так, у некоторых
персиков мезокарпий почти сухой и растрескивается при созревании на
половинки, у других косточка почти гладкая. Культурные сорта персика
ценятся за сочные, нежные и сладкие плоды, разнообразные по величине,
размерам и вкусу. Культура персиков распространена главным образом
на юге.
Род вишня (Cerasus) имеет сочную костянку с шаровидной, слегка
сплюснутой или, наоборот, несколько вытянутой косточкой. Довольно круп-
ные цветки, собранные в немногоцветковые зонтики, иногда сплошь покрыва-
ют дерево.
!: I.
, Рис. 301. Бере зииняя Мичурина (справа) и ее производители: (слева)
438
выа
ll*£
СЕ РАСТЕНИЯ
Рис. 302. Вишня (Cerasus vulgaris):
1 — цветущая ветвь; 2 — продольный разрез цветка;
плоды; 4 — разрез плода; 5 — диаграмма цветка
Из дикорастущих видов ви-
шен отметим антипку (С. maha-
leb), растущую в Молдавской
ССР, Украинской ССР (Крым),
в закавказских и среднеази-
атских советских республиках.
Этот вид используется как под-
вой для культурных сортов
вишен. Из тяжелой, твердой,
Q хорошо полирующейся древе -
силы антипки изготовляют трос-
V ти, мундштуки, чубуки, извест-
П ные под названием черешневых.
Другой ВИД дикой виш-
ни — степная вишня (С- fruti-
cosa) — очень широко рас-
пространен в степной и лесосте-
пной полосе, где образует за-
росли, часто совместно с бо-
бовником и другими степными
кустарниками, или растет в
подлеске более светлых лесов
и по опушкам. Суховатые,
несколько вяжущие, кисло-
сладкие плоды ее съедобны.
Как холодостойкое растение,
она используется для гибриди-
зации и получения холодостой-
ких сортов садовой вишни.
К этому же роду относит-
ся черешня (С. avium) — дере-
во, в диком виде встречаю-
щееся на территории Украин-
ской ССР и в нижнем и среднем поясах горных лесов Кавказа. От ди-
кой черешни произошли многочисленные сорта культурной черешни.
И- В. Мичурин вывел сорта черешни, пригодные для разведения на 500—600 км
севернее основной области распространения этого плодового дерева.
Культурные сорта садовой вишни объединяют в вид обыкновенная виш-
ня (С. vulgaris, рис. 302). Есть основание считать, что культурная вишня
возникла гибридным путем от скрещивания степной вишни и черешни. В сред-
ней полосе распространены Владимирская (Родителева), Любская, Гриот
украинский и другие сорта вишни. Целый ряд замечательных сортов полу-
чен И. В. Мичуриным. Сорт Плодородная Мичурина отличается выносли-
востью к зимним морозам (переносит до —40 °C) и большой урожайностью.
По этой причине он вошел в стандарт многих областей. Этот сорт уже в кон-
це 90-х годов прошлого столетия получил заслуженное признание не только
в СССР, но и в Канаде, где он вынес крайне суровую зиму, в то время как
все другие сорта вымерзли.
Род черемуха (Padus) имеет плоды некрупные, черные, округлые, с
округлой косточкой. Цветки в кистях. Плоды нашей обыкновенной черемухи
(Р. racemosa) сушат и употребляют (в перемолотом виде) для начинки пиро-
гов (Сибирь, Урал), а также для изготовления наливок. Мелкослойная,
твердая, буро-желтая древесина идет на поделки. Кора черемухи дает зе-
леную и буро-красную краску. Черемуха — хорошее медоносное растение.
В смешанных и лиственных лесах Дальнего Востока растет особый вид чере-
мухи — Р. maackii. Это дерево интересно тем, что, скрещивая его с вишней
Идеал, И. В. Мичурин получил межродовой гибрид, названный им цера-
падусом (рис. 303).
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Род лавровишня (Laurocerasus) отличается от черемухи вечнозелены-
ми кожистыми листьями. В лесах Западного Закавказья, образуя часто непро-
ходимый подлесок, растет аптечная лавровишня (L. officinalis), из листьев
которой приготовляют лавровишневую воду. Розовато-серая древесина идет
на поделки. Все растение ядовито, так как содержит (особенно в коре и поч-
ках) синильную кислоту. Однако плоды у садовых форм, крупные и сладкие,
съедобные.
Семейство розовых, как видно из предыдущего, весьма полиморфно.
Его признаки, особенно строение цветка, очень близки к признакам семейст-
ва камнеломковых. Переходные формы с числом кругов андроцея, не превы-
шающим двух, и с семенем, имеющим эндосперм, относят к камнеломковым,
а растения с противоположными признаками — к розовым. Такое искусст-
венное отграничение двух семейств лишь подчеркивает их близость и, сле-
довательно, родственность — вероятное происхождение от общих предков.
Последние, надо думать, имели апокарпный гинецей, о чем свидетельствует
наличие вполне (у розовых) и почти (у камнеломковых, род Astilbe) апо-
карпных форм, и плоды-листовки. Камнеломковые в подавляющем боль-
шинстве перешли к ценокарпии; розовые отчасти остались апокарпными,
отчасти перешли к ценокарпии, отчасти, наконец, в результате ре-
дукции гинецей стал одночленным. В каждом из этих направлений эво-
люции можно найти признаки связи с исходными формами. Нельзя счи-
тать случайным то, что односемянные плоды многих вишневых (Prunoideae)
и розовых (Rosoideae) развиваются из завязи, содержащей две семяпочки,
особенно если принять во внимание, что среди спирейных, имеющих типич-
ную листовку, есть виды, у которых вместо многих семяпочек развиваются
только четыре и даже две (Physocarpus). Апокар:
яй гинецей и листовки,
как сказано выше, типичны и для толстянковых, по общему плану цветка то-
же близких к камнеломковым и розовым. На этом основании три названных
семейства (вместе с еще некоторыми, менее крупными) и соединяют в одном
порядке. Объединяемые общностью происхождения, три главных семейства
порядка Rosales эволюционировали каждое в своем направлении. Crassu-
laceae изменяли в основном строение вегетативных органов, приспособляясь
к жизни в засушливой среде. У розовых возникли многочисленные видоиз-
11: ।
менения цветка, отчасти в связи с опылением насекомыми, отчасти в связи
Рис. 303. Церападус и его производители:
1 — вшпия Идеал; а— квяк"<*рг>лт9цяуя черемуха Маака; л—церападус № 1
440
с обеспечением распространения семян, а большая часть камнеломковых
приспособилась к условиям сурового горного и арктического климата.
Апокарпность гинецея и плоды-листовки у предполагаемых предков по-
рядка Rosales указывают на их близость к подклассу Magnoliidae.
2-Й ПОРЯДОК — БОБОВОЦВЕТНЫЕ (FA BALES)
Цветки бобовоцветных пентамерные (реже тетрамерные), зигоморфные
или актиноморфные, с двойным околоцветником, андроцеем из пятичленных
кругов и с одночленным гинецеем. Завязь верхняя, одногнездная, плацен-
тация краевая. Особенно типичным признаком порядка считается плод-боб
(или другие формы плода, производные от боба). Деревья, лианы, кустарни-
ки и травы с очередными, обычно сложными листьями, имеющими прилист-
ники. . “ "
Весьма естественный порядок, который вследствие несомненных гене-
тических связей между относимыми сюда семействами, рассматривается не-
которыми систематиками как одно семейство, объединяющее, по разным
оценкам, от* 13 000 де 17 600 видов. Этот порядок сближают с розоцветными.
Характерный для бобовоцветных одночленный гинецей со швом, обращенным
к оси соцветия, §сть-и у некоторых розоцветных. Если у бобовоцветных цветок
преимущественно зигоморфный, то наряду Ъ этим он бывает и правильным,
что типично для розоцветных. В то же время у розоцветных встречается и ав-
томорфность. В обоих порядках листья, как правило, имеют прилистники и
сложную пластинку.
Рис. 304. Мимозовые:
2 — цветущая ветвь Acacia Senegal; 2— диаграмма цветка Acacia lophantha: л —ветвь АсаЫа тЫапо-
xylon с филлодиями; 4 — диаграмма цветка Mimosa pudica; 5 — цветов Acacia mildbredii
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
441
Рис. 305. Зонтиковидная акация (Acacia spirocarpa)
Семейство мимозовые (Mlmosaceae). Растения этого семейства пре-
имущественно древесные, с дваждыперистосложными, реже перистосложны-
ми листьями, имеющими прилистники. Цветки актиноморфные (рис. 304)
(лишь редко чашечка бывает зигоморфной), мелкие, собранные в головки или
колосовидные соцветия, привлекающие опылителей яркой — желтой, роэо-
вой или красной — окраской длинных, выдающихся иэ околоцветника тычи-
ночных нитей. Околоцветник двойной или иногда вследствие недоразвития
венчика простой. Чашелистиков 5 (3—6), сросшихся, реже свободных; иногда
они сильно редуцированы ц даже могут вовсе отсутствовать. Лепестков столь-
ко же, сколько чашелистиков; они свободные либо срастаются друг с другом.
Число тычинок различно. У одних родов их столько же, сколько чашелисти-
ков, у других — вдвое больше или неопределенно большое число. Тычиноч-
ные нити совершенно свободные, или сросшиеся в трубочку, или приросшие
к венчику. У некоторых мимозовых на верхушках пыльников образуются
сидячие, реже — на ножках, железки, ко времени цветения большей частью
опадающие. Плодолистик обычно один (редко более одного), образует пестик
с одногнеэдной верхней завязью. Плод — боб, раскрывающийся или разламы-
вающийся на односемянные членики.
Семейство мимозовых характерно для тропической и субтропической эон
обоих полушарий. Наиболее крупный род акация (Acacia) содержит прибли-
зительно 700—800 видов, более половины которых обитают в Австралии. Авст-
ралийские акации часто имеют листья с сильно или даже вполне редуцирован-
в вертикальной плоскости черешком
ной пластинкой и расширенным
(филлодий). Виды рода Acacia
.и.
роко распространены тайже в Африке,
входя там в состав тропических, листопадных лесов и участвуя в соэдаг
ландшафта саванн. Африканские акации иногда имеют характерную, как бы
сплошную крону — зонтиковидные акации (рис. 305). У некоторых акаций
прилистники превращены в колючки, которые могут быть полыми внутри и
тогда служат жилищем муравьям. Последние питаются тканью особых чечеви-
цеобразных придатков на концах листочков (питательные тельца), содержа-
III
Г, Г
Hi
щих азотистые вещества и жирное масло. Многие акации доставляют гумми;
особенно ценится в, этом отношении сенегальская акация (A. Senegal), аравий-
fits
г. г
нг
ская акация (A. arabica) и некоторые другие виды, доставляющие наиболее
ценный продукт. Гумми, получаемое иэ акаций, имеет техническое название
(отсюда название клея — гуммиарабик). В коре акаций содержится много
дубильных веществ. Иэ ядра древесины Л. catechu (тропическая и субтро-
пическая Азия) получают катеху — богатый дубильными веществами про-
дукт, используемый как краска. В СССР в качестве натурализовавшегося
растения на Черноморском побережье Кавказа — в Сочи, Сухуми, Батуми и
в др гих местах—встречается акация серебристая (Л. dealbatci) родом из Авст-
Itl'
ВЫСШИЕ PACT
h:i
Рис. 306. Кампешевое дерево (Haematoxylon campechianum):
1 — цветущая ветвь; 2— цветок; з — разрез цветка; 4 — боб
ралии. Она цветет рано весной, и ее цветущие ветви вывозят на север, в част-
ности в Москву* Ее неправильно называют мимозой*
Обширный род мимоза (Mimosa) распространен главным образом в
тропической и субтропической Америке. Сюда относится мимоза стыдливая
(М. pudica), распространившаяся по всему тропическому поясу* Ее приме-
чательная способность —быстро реагировать на раздражения, причем она скла-
дывает листочки, опуская листья; ее часто содержат в оранжереях.
Семейство д^адшшшевые (Caesalpiniaceae) включает деревья, ли-
аны, кустарники и реже травы с очередными перистыми, дваждыперистыми,
редко простыми (вследствие редукции сложных) листьями, всегда имеющими
прилистники. Цветки иногда почти актиноморфные, но обычно зигоморфные,
циклические, из пятичленных, реже четырехчленных кругов* Чашелистики,
как Правило, свободные. Лепестки в числе чашелистиков, или их меньше, не-
равные; наиболее крупный задний лепесток прикрыт краями боковых,
а последние прикрываются нижними лепестками. Десять тычинок располо-
жены в два круга; обычно ‘они свободные,но иногда срастаются своими нитями*
Нередко некоторые тычинки (например, 3 задние —1 противоленестковая и
2 противочашечные) превращены в стаминодии, чем усиливается автоморф-
ность цветка, или же некоторые тычинки вовсе редуцированы, благодаря чему
андроцей может стать пятичленным. Пестик из одного плодолистика с одно-
гнездной завязью, иногда сидящей на ножке, образованной разросшимся
цветоложем. Плод — боб, то раскрывающийся, то нераскрывающийся. Семена
у некоторых представителей с эндоспермом.
Цезальпиниевые растения почти исключительно тропические. Мно-
гие древесные формы, достигающие огромных размеров, высоко ценятся за
прочность и красивую окраску ядровой древесины. Таковы, например,
кампешевое дерево (Haematoxylon campechianum, рис. 306) из Центральной
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
443
Америки, имеющее древесину
ядра сначала ярко-красную, по-
том синеющую и, наконец, при-
обретающую фиолетово-черную
окраску (из древесины получа-
ют дубильные вещества и гема-
токсилин); красное фернамбуко-
вое, или пернамбуковое (по име-
ни порта Пернамбуку в Брази-
лии), дерево (Caesalpinia echina-
ta) из Бразилии с очень твердой
желто-красной ядровой древе-
синой, которая со временем ста-
новится темно-красной (ценит-
ся в мебельной и других про-
изводствах; содержит краску
бразилин); саппоновое дерево
(С. sappan) из стран Юго-Вос-
точной Азии (Индонезия, Вьет-
нам, Камбоджа и др.) с дре-
весиной красного цвета с жел-
тым или бурым оттенком и др.
Виды рода Cassia (рис. 307),
растущие на северо-востоке
Африки и Южной Азии, дают
так называемый александрий-
ский лист (слабительное).
Плоды многих цезальпи-
ниевых очень богаты дубиль-
ными веществами.
Очень эффектно мощное
североамериканское дерево гле-
дичия обыкновенная (Gleditschia
triacanthos), образующее огром-
ные простые или разветвленные
колючки более 10 см длиной
(рис. 308). Гледичия культи-
вируется как декоративное де-
рево в Крыму, на Кавказе и в
Средней Азии; хороший медо-
нос. Один из видов этого рода—
гледичия- каспийская (G. cas-
pica) — растет в Восточном За-
кавказье. Это невысокое дерево,
с шаровидной кроной и невет-
вистыми колючками, представ-
ляет собой реликтовое расте-
ние, сохранившееся до настоя-
щего времени на Кавказе по
крайней мере с неогена.
Семейство .мотыль-
ковые (Pabaceae^ Papilionaceae).
Травы, полукустарники, кус-
тарники или деревья с перис-
то- или пальчатосложными (ред-
ко простыми) листьями, имею-
щими прилистники. Цветки
обычно среднего размера или
Рис. 307. Цезалышниевые:
J — цветущая ветвь Cassia chamaecrista; 2 — С. flori-
bunda: a — общий вид цветка; б — разреэ цветка;
в — анаграмма цветка; 3 — плод С. ohamaecrista
Рис. 308. Колючка гледичии (Gleditschia tri-
acanthos)
ВЫСШИЕ РАСТЕНИЯ
*
п:гг
мелкие и тогда, как правило, собраны в кисти, реже головки, зонтики
или колосья. Строение цветка оригинальное (рис. 309). Пять
чашелистиков образуют сростнолистную чашечку, то актиноморфную, то зиго-
морфную, иногда двугубую. Из пяти лепестков венчика задний, называемый
парусом или флагом, выделяется как наиболее крупный; его верхняя часть
(отгиб) располагается почти под прямым углом по отношению к ноготку.
Парус прикрывает краями два ноготковых боковых лепестка, называемых
ГЛ?|]
'in
свою очередь, прикрывают два нижних, тоже асимметричных и ноготковых
лепестка, которые начиная с вершины в большей или меныпей степени срас-
таются или тесно смыкаются друг с другом, образуя так называемую лодоц-
~~~~ m ----- ь как редкад исключение встречаются мо-
' тыльковые с девятью или пятью тычинками в цветке. Наиболее часто нити
девяти тычинок срастаются вокруг пестика, образуя незамкнутую трубочку,
щель которой прикрыта несколько расширенной нитью десятой, свободной
тычинки. Реже все десять тычинок срастаются или, наоборот, все остаются
; I: I
тычинок сохраняет след происхождения из двух пятичленных кругов, что
выражается в неравенстве длины тычинок: пять длинных тычинок чередуются
с пятью более короткими. Пестик всех мотыльковых состоит из одного пло-
долистика; завязь верхняя, одногнездная. В самом общем виде цветок
мотыльковых можно охарактеризовать формулой: K5C5A5+5G1. Плоды мо-
тыльковых — бобы (рис. 310), раскрывающиеся двумя створками или четко-
видные, при созревании разламывающиеся на односемянные членики. Семе-
на без эндосперма, содержат крупный согнутый зародшп с двумя массивны-
ми семядолями.
Подавляющее большинство мотыльковых опыляется насекомыми, чем
объясняется сложное строение цветка. Все энтомофильные мотыльковые
выделяют нектар, который скапливается у основания пестика — между ним
и основанием тычиночной трубки. Незамкнутость тычиночной трубки ввиду
наличия у большинства представителей одной свободной тычинки — приспо-
собительный признак, облегчающий насекомому высасывание нектара. Обыч-
на протерандри я. Некоторые мотыльковые (например, посевной го-
рох) стали самоопыляющимися.
Семейство мотыльковых насчитывает около 12 000 видов, распростра-
ненных по всему свету, преимущественно в странах с умеренным климатом;
здесь растут главным образом травянистые и кустарниковые формы, в то
время как тропические мотыльковые в основном деревья.
В пределах семейства можно различать две группы: а) мотыльковые со
свободными тычинками (Sophera, Thermopsis) и б) мотыльковые, у которых
все десять (род Cytisus — ракитник) или только девять тычинок срослись,
а десятая остается свободной и прикрывает щель тычиночной трубки. Пер-
вые наиболее близки к Caesalpt-
niaceae, через которые мотылько-
вые связываются с
ными.
По хозяйственному значению
мотыльковые — одно из важней-
ших семейств покрытосеменных
растений, что в большой степени
определяется симбиозом мотыль-
ковых (и вообще бобовоцветных)
с бактериями,
в род
которого
розоцвет-
Рис. 309. Диаграмма цветка бобовых:
I—бобы; 2 —раюггнив (п — парус, яр —
д—лодочка)
объединяемыми
Rhizobium, в пределах
различают ряд самосто-
видов, сожительству-
определенными видами
мотыл ьк овых. Благодаря раз*
с
иг
I
445
ОТДЕЛ ЦВЕТКОВЫЙ, ИЛИ ПОКРЬНЮСВМВВНЫИ
—...- —J ..... .
Рис. 310. Плоды бобо-
вых:
1 — тшшчный боб викк; 2 —
разламывающийся боб кове-
ечника; в — спирально за-
крученный боб люцерны; 4 —
односемянный боб вснарцета;
5 — крылатый одно-двусемян-
ный боб красного сандалово-
го дерева
бактерий,
проникающих из почвы через корне-
нг.
множению клубеньковых
вые волоски в первичную кору корня, паренхима коры разрастается, образуя
наросты, называемые клубеньками. Бактерии, поселяющиеся в корне, об-
ладают замечательной способностью использовать элементарный азот ат-
мосферы, вводя его в биологический круговорот обмена веществ. Белковые
вещества, вырабатываемые клубеньковыми бактериями, используются бобо-
выми растениями. При отмирании корней и других частей бобового расте-
ния накопившиеся запасы-связанного азота поступают в почву и, разло-
женные и переработанные другими бактериями, становятся источниками
азотистого питания зеленых растений.
Роль бобовых как азотонакопителей одинаково существенна и для
естественных растительных ценозов, и для посевов культурных растений.(
Установлено, что малоплодородные почвы из-за недостатка азота дают хо-
рошие урожаи после предварительной культуры на них бобовых. Этим и
объясняется введение бобовых в севообороты. В одних случаях (например,
посевы клевера, люцерны, донника) надземная масса используется на корм,
в других — бобовое растение целиком, вместе с надземными зелеными частя-
ми, запахивается в землю (зеленое удобрение). Благодаря симбиозу с клу-
беньковыми бактериями как вегетативные части, так и семена мотыльковых
отличаются высоким содержанием белков, чем объясняются хорошие кор-
- мовые и высокие пищевые качества многих из этих растений.
Из числа пищевых растений семейства мотыльковых особенно важны в
хозяйственном отношении горох, чечевица, конские бобы, соя.
Горох посевной (Pisum sativum, рис. 311). Его семена по содержанию
белков близки к мясу (хотя белки бобовых усваиваются хуже, чем белки жи-
вотного происхождения), а кроме того, сддержат значительное количество
крахмала. У некоторых сортов используют в пищу незрелые плоды (лопат-
ки), богатые сахаром. Как корм используют и надземные вегетативные части,
содержащие значительный процент белка. Будучи сильным азотособирателем,
горох оказывает заметное положительное влияние на урожай после-
дующей культуры.
Чечевица съедобная (Lens cuUnaris, Ervum lens). Это растение использу-
ется человеком с каменного века. Зерна чечевицы содержат 30% и более
белка, а зеленые части растения служат прекрасным питательным кормом
скоту. Ценится в севообороте как хороший азотособиратель.
Фасоль обыкновенная (Phaseolus vulgaris). Известны сорта, отличающие-
ся величиной, формой и окраской семян, степенью засухоустойчивости,
скороспелостью. В семенах фасоли содержится от 11 до 29% белков. Ряд
видов фасоли разводят как декоративные растения.
11
выс
111LI Ld
РАСТЕНИЯ
8 5
Гис 311. Горох (Pisum sativum)*-
1 — ветвь цветущего растения; з — незрелые плоды; з — цветок; 4 — разрез цветка; 3 — лепестки (п —
парус, * — весла, л — лодочка); в — андроцей; 7 — вестик, 8 — строение семени (с — семядоли, ж — за-
родышевый кореш», пч — почечка) <
Бобы (Faba hispida). Плоды крупные, опушенные. Стенка их в незре-
лом состоянии мясистая. Большие семена разнообразной окраски, содер-
жат много (до 35%) белка и растворимых углеводов. Семена используются
в пищу.
ш
(евым, кормовым и техническим растением является соя (Gly-
cine hispida). У многочисленных сортов сои семена содержат от 37 до 44%
белков и 17—22% жирного масла. В сумме на долю белков и масла в семе-
нах сои приходится 54—66%. Семена сои можно непосредственно употреб-
лять в пищу, а кроме того, из них путем переработки изготовляют раститель-
ное масло, сливки, сыр, творог, муку и хлеб для диабетиков, различные пе-
ченья, суррогат какао и кофе, конфеты, бисквиты, различные соусы и т. д. —
всего до 100 различных изделий. Соевое Масло, как и подсолнечное, употреб-
ляют непосредственно в пищу или из него изготавливают твердые продукты,
подобные сливочному маслу или салу. Используется соевое масло и для
технических надобностей (лаки, масло, суррогаты каучука, клей и т. п.).
Ценны и соевые жмыхи, которые идут в корм скоту, используются в пище-
вой промышленности и технике.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
447
Соя — одна из древней-
ших культур. Имеются дан-
ные, что ее разводили уже
в V тысячелетии до н. э. Ос-
новная область возделывания
сои — страны Восточной и
Юго-Восточной Азии. Куль-
тивируют ее также в некото-
рых странах Африки, в Юж-
ной и Северной Америке, в
Европе и даже в. Австралии.
В СССР культура сои рас-
пространена на Дальнем Вос-
токе, в Западной Грузии и в
южных районах европейской
части СССР.
На Дальнем Востоке ди-
ко растет уссурийская соя —
возможный дикий предок ку-
льтурного вида.
Земляной орех, или ара-
хис (Arachis hypogea), —
травянистое растение, заме-
чательное геокарпией. Пос-
ле отцветания ножка, на
которой сидит завязь (гино-
фор), удлиняется, загибается
вниз, и благодаря этому за-
вязь углубляется в почву,
где и созревают нераскрыв-
шиеся бобы. Околоплодник
их сухой, сетчатый с поверх-
ности. В каждом бобе — не-
сколько семян. Последние
содержат до 45% жира и до
37% белка. Масло, добывае-
Рис. 312. Клевер луговой (Trifolium pretense):
1 — цветущее растение; 8 — цветок (вид сбоку): Л—
цветок (вид снизу); 4 — чашечка; S — тычинка; 6 —
пестик
мое из семян земляного оре-
растите л ьных масел, конкурирующее с
консервов (сар-
ха, — одно из лучших
прованским. Оно употребляется в пищу и при изготовлении консервов (сар-
дины). Худшие сорта масла используются в мыловарении. Жмых обладает
высокими кормовыми свойствами, а переработанный на муку используется
как примесь к муке злаков, идет на изготовление печений, прибавляется в
какао при выделке низших сортов шоколада. Из арахисной муки выпекают
кондитерские изделия для больных диабетом. Земляной орех родом, вероят-
но, из Южной Америки, но это растение широко распространилось в куль-
туре не только на родине, но также в Индии, Китайской Народной Республи-
ке, в странах Африки. В СССР культура этого растения удается в Закавказье
и Средней Азии.
Целый ряд мотыльковых используются как кормовые растения. Из них
одно из первых мест занимает клевер.
Род клевер (Trifolium), В сельскохозяйственном отношении наиболее
важен красный, или луговой, клевер (Г. pra tense, рис. 312), растущий по лу-
гам и опушкам. От него произошли культурные формы клевера, морфологи-
чески не вполне четко отличимые от диких. Клеверное сено по содержанию
белка в 1,5 раза превосходит сено из злаков, а по общей питательности одна
1Г1
ища массы клевера равна почти 0,5 единицы массы овса. Кроме красного,
культивируют также розовый, шведский, клевер (Г. hybridum), В составе
пастбищных травосмесей большое значение имеет ползучий клевер (Г. repens).
$48
ВЫ
HIM'Г'
РАСТЕНИЯ
Рис. 313. Люцерна посевная (Medicago saliva):
J— общий вид цветущего растения; S — цветок; 3 —»
плод
географических условиях, ио в основном в
Люцерна (род Medfcago),
представленная во флоре Со-
ветского Союза значительным
количеством видов, подо био
клеверу, пользуется довольно
широким распространением в
культуре как бобовый элемент
севооборота. Основными хозяй-
ственно ценными видами этого
рода считаются синецветная
посевная люцерна (М. saliva^
рис. 313) и серповидная люцер-
на (М. falcata} с желтыми
цветками. Наземные стебли лю-
церны отличаются быстрым
ростом, ио требуют для этого
достаточного количества воды.
Вместе с тем эти растения спо-
собны к развитию глубоко иду-
щего стержневого корня, до-
стигающего 2 ж, а в отдельных
случаях даже 15 м (серповидная
люцерна), что позволяет расте-
нию покрывать его потребность
в воде и в условиях засушли-
вого климата. Зеленая масса
люцерны содержит много бедка,
кроме того, установлено высо-
кое содержание в люцерне ви-
таминов. Подобно другим бо-
бовым люцерна — прекрасный
а з от осо би р а тел ь; в результате
одногодичной ее культуры в
почве накапливается связанно-
го азота до 3 ц на 1 га.
Люцерну культивируют в
нашей стране в различных
более южных районах. Особен-
но большое значение она имеет при поливном земледелии, в частности в
севооборотах с хлопчатником в Средней Азии и в Закавказье.
В СССР имеется до 36 дикорастущих видов люцерны, из которых серпо-
видная люцерна давно вошла в культуру, но, кроме того, есть еще и ряд ви-
дов, представляющих хозяйственный интерес и подлежащих испытанию в
культуре.
В некоторых районах культивируют эспарцет (преимущественно Опо-
bryclus viclaefolia). Белками эспарцет богаче клевера, хороший медонос.
Кроме того, он отличается большой засухоустойчивостью.
Люпин (Lupinus) — крупный род (свыше 400 видов), представленный
(Новый Свет, преимущественно Анды) многолетними и однолетними травами,
высокогорными кустарниковыми и подушковидными растениями.
Из числа однолетних люпинов полевой культуры, важных в сельскохо-
зяйственном отношении, отметим желтый (£. luteus), узколистный (L. angus-
tifoUue) и белый (L. albus) люпины. Они хорошие азотонакопители, так что
даже тощие, песчаные, бесплодные почвы после возделывания на них люпина
дают хорошие урожаи. В связи с этим довольно широко распространена
нг.
нг.
культура люпинов на зеленое удобрение.
Семена люпина очень богаты белком и ценны как корм, но содержат
ядовитые алкалоиды и потому требуют предварительной обработки. В насто-
1
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
нг.
।
ящее время выведены малоалкалоидные сорта из желтого, белого, узколист-
ного и других видов люпина, что открывает более широкие перспективы их
использования.
L. polyhyllus — многолетнее растение, культивирующееся как декора-
тивное с эффектными длинными кистями синих, фиолетовых или розовых
цветков.
Из других мотыльковых получили распространение в севооборотах дон-
ник и вика.
Белый донник (Meltlotus albus) дает наибольшую массу, а желтый
(М. officinalis) — хороший медонос. Кормовое значение донников несколь-
ко снижается их сильным запахом, зависящим от присутствия кумарина.
Донники отличаются высокой засухоустойчивостью и солевыносливостью,
одновременно способствуя улучшению засоленных почв. Они предохраня-
ют также почву от развеивания (пыльные бури).
Из вик у нас культивируют два вида: яровую вику (Vicia sativa) и озимую
(F. villosa)\ обе используются как кормовые растения.
Ряд представителей семейства мотыльковых нашел применение в ме-
дицинской практике. Таковы солодки и термопсис ланцетовидный. Солодки —
' 'голая и уральская (Glyctrrhtza glabra, G. uralensis) — обитают в стейях и по-
лупустынях, образуя нередко большие заросли. Порошок из корней солодки
(лакричный порошок) служит легким слабительным, а также входит в состав
пилюль от кашля (отхаркивающее средство). Иногда из сушеного корня
солодки выпаривают сироп и, сгущая его, получают так называемый лак-
ричный сахар, который находит применение в пищевой промышленности
(конфеты, напитки). Сгущенный сок лакрицы используют для загущения
: составов. В Казахской ССР местами из вымоченных стеб-
лей солодки получают волокно для производства веревки.
Ланцетовидный термопсис (Thermopsis lanceolate) — растение, нередко
встречающееся в степях Сибири и в Средней Азии. Содержит ядовитые алка-
лоиды. Из термопсиса изготовляют средство против кашля взамен импорт-
ного корня ипекакуаны.
Некоторые бобовые являются красильными растениями. Так, желтые,
расположенные кистями цветки красильного дрока (Genista tinctoria) — степ-
ного и отчасти борового кустарника — дают хорошую ярко-желтую краску,
а широко культивируемая в тропической и отчасти субтропической полосе
Indigofera tinctoria — источник прочной синей краски индиго. Камеди, до-
бываемые из некоторых среднеазиатских астрагалов (род Astragalus)
под именем траганта, находят применение в текстильной, кожевенной ^спичеч-
ной, красочной и парфюмерной промышленности, а также в кондитерском
производстве.
Наконец, мотыльковые нередко разводят как декоративные растения.
Кроме упомянутого выше люпина, часто разводят душистый горошек (Lathy-
rus odoratus, родина — Средиземноморье). Так называемая сбелая акация»
(Robiniapseudoacacia) родом из Северной Америки (дерево с большими кистями
белых цветков) и глициния (Wistaria sinensis) из Восточного Китая (высокий
вьющийся кустарник или дерево с белыми и фиолетовыми цветками) извест-
ны в культуре субтропических стран и в СССР, особенно в Крыму, на
Кавказе и в Средней Азии.
Некоторые древесные формы мотыльковых дают исключительной цен-
ности древесину; таково санталовое красное дерево (виды рода Pterocarpus)
из Азии и африканское черное дерево (род Dahlbergia). Наша вжелтая
акация» (Caragana arborescens) и близкий вид дереза (С. frutex) использу-
ются для живых изгородей.
нг.
15-602
450
выс:
Е РАСТЕНИЯ
Рис. 314. Рута (Ruta graveolens):
I — верхняя часть цветущего растения; 2 — цветок;
з — разрез цветка; 4 — диаграмма цветка
3-0 ПОРЯДОК — ТЕРПЕНТИННЫЕ
(TEREBINTH ALES)
Цветки терпентинных обое-
полые, реже раздельнополые,
правильные или зигоморфные,
четырех — шестичленные. Око-
лоцветник обычно двойной, со
свободными лепестками. Анд-
роцей из двух кругов тычинок,
обдиплостемонный или дипло-
стемонный; иногда тычинок
один круг. Плодолистиков от
2 до 5, они более или менее
сросшиеся; иногда гинецей апо-
карпный. Завязь обычно верх-
няя. Характерно наличие в
цветке хорошо выраженного
диска. Преимущественно дре-
весные растения со сложными
листьями. Вегетативные органы
характеризуются вместилищами
секреторных выделений: масел,
бальзамов, смол.
Порядок терпентинные час-
то подразделяют на два — руто-
цветные (Rutales) и сапиндоцвет-
ные (Sapindales), с нечеткими
границами между ними.
Семейство рутовую
(Rutaceae). Это ветабЗЙЯеные
деревья и кустарники, редко
травы, большей частью с очеред-
ными, реже супротивными, про-
стыми (цельными или раздельными), иногда сложными листьями. В листьях
имеются масляные железки, заметные как просвечивающие точки. Цветки
обычно актиноморфные, реже зигоморфные, обоеполые, круги большей частью
пяти- или четырехчленные (рис. 314). Часто между андроцеем и гинецеем
образуется выдающийся диск — иногда в форме колонки (гинофор). Чашеч-
ка и венчик пятичленные. Чашелистики свободные, иногда срастающиеся.
Тычинок вдвое или реже в несколько раз больше, чем лепестков; иногда они
срастаются в основании. Надлепестковые тычинки превращаются в нектар-
ники или отсутствуют. Плодолистиков 5—4, реже меньше (до одного), еще
реже плодолистиков шесть — несколько. Они срастаются, но иногда не всегда
полностью, а только в области столбика. Завйзь, как правило, многог^незд-
ная, с одной-двумя семяпочками в гнезде, верхняя. Плоды разнообразные •—
коробочки, костянки, ягоды или дробные плЬды.
Это обширное семейство (около 1600 видов) характерно, за исключением
немногих форм, для тропических и субтропических стран. Среди рутовых
имеется немало растений, используемых человеком, и среди них особенно
широкое распространение в культуре получили цитрусовые (род Citrus),
входящие в состав особого подсемейства Citroideae, характеризуе-
мого сочными ягодоподобными плодами. Родина цитрусовых в Старом Свете —
тропическая и субтропическая Азия.
Листья у цитрусовых обычно с простой, продолговатой или эллипти-
ческой пластинкой, представляющей листочек сложного листа, прочие ли-
сточки которого редуцированы. Цветки располагаются большей частью
пучками в пазухах листьев* актиноморфные, с белыми или розоватыми ле-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
451
пест ками, душистые. Тычинки, вследствие расщепления, многочисленны
(до 60) и спаяны в группы (по две-три или по нескольку). Ценокарпный
гинецей, содержащий до 20 плодолистиков, образует пестик с верхней
завязью» Плоды с сочным околоплодником, во внешних слоях которого
(экзокарпий) находятся многочисленные эфиромасляные железки; эндо-
карпий губчатый. От плодолистиков внутрь гнезда плода врастают
крупные волоски с сочпым содержимым, которые в совокупности и состав-
ляют съедобную мякоть плода. Семена нередко имеют несколько зародышей
(полиэмбриония).
Цитрусовые издавна культивируются на их родине, в Юго-Восточной
Азии (Китай, Индия), откуда они уже в очень давние времена распростра-
нены человеком на запад. В настоящее время культура цитрусовых за-*ч
нимает около 800 000 га, наибольшие площади в СИТА (Калифорния, Фло- 4
рида). В странах Средиземноморья цитрусовые настолько широко вошли
в культуру, что стали там характерной чертой растительного ландшафта.
В Южной Америке некоторые цитрусовые (апельсины) одичали и в извест-
ной степени оказывают влияние на местную растительность. В СССР цитру-
совые — одна из основных культур влажных субтропиков. Цитрусовые
приносят плоды высокой цепности и по вкусу, и по полезности. Они богаты
сахарами, лимонной кислотой и витаминами (С, А,В, Р и др.). Плоды упот-
ребляют в свежем и консервированном виде. Из околоплодников изготов-
ляют цукаты. Эфирные масла цитрусовых, добываемые из листьев, лепест-
ков и плодов, различны по составу и аромату; используются в кондитер-
ском деле и в парфюмерии.
В СССР производственное значение имеют главным образом мандарин
{Citrus reticulata), лимон (С. Итоп) и сладкий апельсин (С. sinensis). Цитру-
совое хозяйство практически создало у нас в советское время. Основные
районы культуры цитрусовых — Западная Грузия (включая Аджарскую
и Абхазскую АССР) и южные прибрежные районы Краснодарского края.
Размножаются цитрусовые прививкой. В качестве подвоя используются
выращиваемый на месте холодостойкий кустарник понцирус (Ponctrus
trifoliata) из того же подсемейства Citroideae. В небольших коли-
чествах успешно разводят лимон в комнатной культуре во многих районах
СССР (Павлово — на Оке, Нижний Тагил и др.).
В СССР (на Дальнем Востоке и Сахалине) и в Японии растет бархатное
дерево^ или бархат {Phellodendron amurense, рис. 315). Пробка, покрываю-
щая ствол, достигает на толстых деревьях 5—7 см толщины, но невысокого
качества; она только отчасти может заменить пробку пробкового дуба.
Из других Rutaceae следует отметить ясенец белый {Dictamnus albus), *
произрастающий на юге и юго-востоке европейской части СССР, — травя-
нистое растение с множеством железистых волосков, выделяющих летучие
эфирные масла» В тихую погоду от зажженной спички вокруг ясенца воз-
никает пламя, но куст при этом не повреждается. От соприкосновения с
растением возникают ожоги.
Семейство фисташковые {Anacardiaceae). Древесные растения с
очередными простыми или сложными (пальчатыми, перистыми) листьями
без прилистников; в ряде случаев прилистники опадающие. Цветки в ме-
телках, мелкие, актиноморфные, обоеполые, реже, вторично однополые.
Околоцветник двойной из пятичленных кругов. Тычинок 5, редко 10. Гине-
цей бывает апокарпный, чаще с:
карпный из 2—5 плодолистиков, иногда
мономерный. Завязь верхняя или нижняя. Плод — коробочка или костянка.
' В коре многих фисташковых часто развиваются смоляные ходы.
К этому семейству относится около 600 видов, растущих в теплых, от-
части в умеренных областях. Род фисташка (Pistacid) содержит ряд цен-
ных растений. Например, настоящая фисташка {Р. иега) родом из Азии,
растущая в горах Средней Азии, приносит съедобные плоды — фисташки.
Разводят ее в странах Средиземноморья, особенно в Италии. К этому роду
относится и кевовое дерево (Р. тиНса)Л встречающееся в Крыму и местами
51: ।
15*
вша
иг,
Е РАСТЕНИЯ
Fbc. 315. Бархатное дерево
amuxense). Ветвь с плодами
на Кавказе (близ Анапы, в Гру-
зии и Азербайджане); дает цен-
ную смолу «саккыз». В среди-
земноморских странах дико про-
израстают Р. lentiscus и Р. tere-
binthus — источники смолы «мас-
тике», из которой получают так
называемый «хиосскдй терпентин».
Cotinus coggygria, известный под
названием скумпии, встречается
ИЛ:
на Северном Кавказе, в Закав-
казье
мается
до 1500 м
над уровнем моря),
а также в
странах Средиземноморья. Хоро-
шо переносит засуху. Скумпия ре-
комендована как сопутствующая
порода при насаждении полеза-
щитных полос в степной зоне.
Вроде сумах (Rhus), виды ко-
торого обитают в Старом и Но-
вом Свете, отметим сумах дубила
ный (Rh. coriaria), растущий вмес-
те со скумпией. Оба эти растения
богаты таннидами и используют*
ся как дубильные. Древесину
сумаха используют для окраши-
вания кожи в коричневый цвет,
а из корней извлекают красную
vernicifera) и некоторые
краску. Листья и побеги Rh, cori-
(Phellodendron дают черную, плоды — крас-
ную, кора побегов — желтую и
кора корней — коричневую крас-
ку- Сумах лаконосный (Rh.
другие виды из Китая и Японии содержат в
коре сок, становящийся на солнце почти чертам; его используют при
изготовлении так называемого японского лака. Исключительной
ядовитостью отличается сумах ядовитый (Rh. toxicodendron) из Северной
Америки и Восточной Азии. Уксусное дерево (Rh. typhina) родом из Север-
ной Америки с перистыми темно-зелеными листьями, принимает осенью
ярко-красную окраску, как и соцветия ко времени плодоношения, вслед-
ствие чего это растение культивируется как декоративное.
К фисташковым относятся также некоторые из южноамериканских
деревьев — квебрахо (кэбрачо), получившие свое название за исключи-
тельную твердость древесины (от исп. tquebar» — ломать, «hacho» — топор).
Schinopsis balansae в S. lorentzii дают окрашенное квебрахо. Их древесина
красная, со временем темнеющая, плотностью около 1,2, содержит до 20%
таннидов и используется для получения дубильного экстракта* а отходы идут
* на бумагу. Заслуживает упоминания также индо-малайское манговое дере-
во (Mangifera indica), культивируемое в тропиках ради плодов манго, соч-
ных, содержащих много сахара и лимонной кислоты* но с привкусом ски-
пидара.
Семейство кленовые (Асегасеае). Листопадные, реже вечнозеленые
деревья и кустарники с супротивными простыми (пальчатолопастными,
раздельными) или перистосложными листьями без прилистников. Цветки
актиноморфные, обычно с двойным околоцветником из пяти,- четырехчлен-
ных кругов, с 10—4 (часто 8) тычинками и с пестиком из двух плодолис-
тиков. Завязь двугнездная. Характерен диск, расположенный наружу от
тычинок или между ними и пестиком. Иногда цветки однополые вследст-
ОТДЕЛ ЦВЕТКОВЫ®, ИЛИ ПОКРЫТОСЕМЕННЫЕ
вне редукции тычинок или
недоразвития пестика (лож-
нообоеполые цветки). Плоды
сухие, распадающиеся на
два односемянных нерас-
крывающихся плодика, каж-
дый с крыловидным придат-
ком (крылатка). Семейство
включает около 120 видов.
В лесной полосе евро-
пейской части СССР
нг.
роко
распространен остролист-
ный клен (Acer platanoides,
рис. 316), главным образом
как примесь в лиственных
лесах. Клен — порода дос-
таточно теневыносливая (осо-
бенно в молодости), предпо-
читающая богатые и хорошо
увлажняемые почвы. Свет-
лая желтоватая древесина
клена равномерно слоистая,
твердая, мало коробится и
хорошо режется, потому ее
ценят в столярном и токар-
ном производстве. Вместе с
остролистным кленом, пре-
имущественно в полосе лист-
венных лесов, встречаются
полевой клен (A. campestre) и
неклен, или татарский клен
(A. tataricum). Большее раз-
нообразие клена характерно
Рис. 316. Клен остролистный (Acer pl a t ano i-
des):
2 — цветущая ветвь; 2 — олвственная ветвь с плодами;
а — мужской цветок; 4 — обоеполый цветок; s — разрез
обоеполого цветка
для Кавказа и Дальнего
-Востока. Многочисленны виды кленов и в юго-восточных части США.
Здесь надо отметить сахарный клен (A. saccharum), из патоки которого
добывают сахар, а также ясенелистный клен (A. negundo), именуемый
иногда американским кленом, часто разводимый у нас с целью озеле-
нения городов. Засухоустойчивое, но недолговечное дерево. Растение дву-
домное, с перистыми листьями.
Семейство конские каштаны (Hlppocastanaceae). По. характеру
вегетативных органов (деревья с супротивными пальчатосложными листь-
ями) относящиеся сюда растения (18 видов) приближаются к кленовым,
резко отличаясь от них явно зигоморфными цветками, в которых за пяти-
членными чашечкой и ярким венчиком следует 5 тычинок и пестик из трех
плодолистиков с трехгнездной завязью. Плоды — трехсемянные коро-
бочки. Семена без эндосперма. Зародыш крупный, согнутый. Семядоли мас-
сивные.
Конский каштан (Aesculus hippocastanum) — дерево, в диком состоя-
встречающееся в странах Балканского полуострова; его часто культи-
вируют в Западной Европе и СССР в парках и скверах как декоративное.
Весной оно покрывается массой крупных, до 20 см и более, конусовидных
г г
белых соцветий, красиво выделяющихся на фоне темно-зеленых 5—7-паль-
чатосложных листьев. Крупные семена, похожие внешне на плоды настоя-
щего каштана, несъедобны, так как содержат много сапонина.
Кроме охарактеризованных семейств, к порядку Terebinthales относит-
ся еще несколько, например крупные тропические семейства Saplndaceae
и Meliaceae.
454
ВЫС
in'
Е РАСТЕНИЯ
Большой порядок Terebinthales считают родственным порядку гераней-
цветных, находя много общего в строении цветков* Близость этих двух по-
рядков настолько велика, что систематическое положение некоторых се-
мейств дискуссионно. Основное отличие Terebinthales от гераниецветных —
постоянное присутствие в цветках нектарников в форме дисков.
4-Й ПОРЯДОК — ГЕРАНИЕЦВЕТНЫЕ (GERANIALES)
В вегетативных органах гераниецветных отсутствуют вместилища мас-
лянистых, бальзамических и смолистых секретов, но часто встречаются
клетки сб слизью (особенно в эпидермисе листьев). Цветки в большинстве
случаев актиноморфные, реже зигоморфные, циклические из пятичленных
.in
111, ।
кругов, с двойным околоцветником, двумя или одним (вследствие редукции)
кругами тычинок. Тычинки часто срастаются основаниями нитей. Пять
(5—2) плодолистиков образуют ценокарпный гинецей с верхней пятигнезд-
ной завязью. Преимущественно травяцйстые, реже древесные растения
и кустарники с простыми цельными или глубоко рассеченными, реже слож-
ными листьями.
Семейство кисличные (Oxalidaceae). Многолетние, иногда одно-
летние травы, редко древесные растения с очередными, обычно сложными
черешковыми листьями, большей частью без прилистников. Цветки дву-
полые, актиноморфные, из пяти пятичленных кругов (рис. 317). Чашечка
сохраняется при плодах. Лепестки свободные, иногда несколько сросшиеся
у основания. Тычинок, как правило, десять (иногда пять из них превраще-
ны в стаминодии), в двух кругах; тычиночные нити книзу сращенные. Пять
плодолистиков, срастаясь нижн
частями, образуют пятигнездную верх-
; 1ЛI
нюю завязь с несколькими, двумя или с одной семяпочками в каждом гнезде.
Верхние части плодолистиков остаются свободными. Пять столбиков с го-
ловчатыми рыльцами. Плоды — коробочки, редко ягоды.
Кисличных насчитывают около 950 видов. Распространены они пре-
имущественно в тропических и субтропических странах южного полушария.
Самый обширный род — кислица (Oxalis)* на долю которого приходится
большая часть видов, обитающих в основном в странах Южной Африки и
Южной Америки. Здесь, наблюдается большое морфологическое разнооб-
разие вегетативных органов этих растений. Так, побеги нередко превраще-
ны в клубни или луковицы, а корни становятся водозапасающими органа-
ми. Еще более разнообразны листья, которые бывают то тройчатыми, то
пальчатыми, то парно- или непарноперистыми, а у некоторых видов листо-
вые пластинки редуцированы и ассимилируют листовые черешки — фил-
лодии. Немногие представители рода заходят далеко на север, в. страны
Северной Америки и в Евразию. В СССР очень характерна для хвойного
леса обыкновенная кислица (О. acetosella, рис. 317) — небольшое травянис-
тое растение, образующее нередко целый ярус в еловых и сосновых лесах.
У кислицы два рода цветков: хасмогамные,.в массе развивающиеся весной,
и клейстогамные, достигающие полной зрелости позднее первых. Пятигнезд-
ная коробочка раскрывается пятью щелями. Ночью и в пасмурную погоду
листочки тройчатого листа кислицы опускаются («сон» листьев). Движение
листочков вообще характерно для кисличных, а у одной тропической формы
(Biophytum) складывание листочков перистого листа происходит при меха-
ническом раздражении, как у мимозы.
Семейство леновые (Ыпасеае). Травянистые растения, полуку-
старники и деревья с простыми очередными листьями, с прилистниками
или без них. Цветки правильные, пятичленные (род Ыпшп — четырех-
членные), с двойным околоцветником, раздельнолепестным венчиком, пятью
или десятью тычинками, спаянными в основании друг с другом; иногда над-
лепестковые тычийки превращены в стаминодии. Завязь пятигнездная, при-
чем каждое гнездо может подразделяться ла два дополнительной радиаль-
ной перегородкой. Плоды — преимущественно коробочки.
ОТДЕЛ ЦВЕТКОВЫЕ, ИДИ ПОКРЫТОСЕМЕННЫЕ
455
Наиболее важный р о д се-
мейства леновых, содержащий
около 500 видов, — лен (Linum),
включающий травянистые одно-
летние и многолетние растения,
реже полукустарники. Один из
видов этого рода — лен обыкновен-
ный (L. usitatlssimum, рис. 318) —
широко культивируется как
волокнистое и как маслич-
ное растение. Культурный лен —
травянистый однолетник с оче-
редными узкими листьями без
прилистников, с простым или
(у других форм) ветвистым по-
чти от основания стеблем. Цвет-
ки собраны в рыхлые д и х а -
зиальные или — при одно-
стороннем развитии — м о н о' -
хазиальные соцветия. Цве-
тение кащдого цветка продол-
жается обычно в течение одного
дня. В большинстве случаев лен
самоопыляется, но иногда наблю-
дается перекрестное опыление,
которое в условиях эскперимента
дает наибольший эффект завязы-
вания плодов.
Культура льна известна с
древнейших времен. Уже человек
каменного века (неолит) исполь-
зовал льняное волокно и семена
Рис. 317. Кислица (Oxalis acetosella):
1 — общий вид растения; 2 — разрез цветка; J —
диаграмма цветка (а, р — прицветяички)
льна, изготовляя из них в смеси с
пшеном хлеб. В Египте возделывание льна датируется III тысячелетием
до н. з., причем посевы льна занимали большие площади. В экспортной
, торговле старой Руси, особенно Новгорода, лен имел первостепенное зна-
чение. В современном земледелии лен играет значительную роль и как
волокнистое, и как масличное растение.
Ботанический вид культурного льна (L. usttatlssimum) очень полиморф-
ный, и в нем хорошо отличимы три основные группы сортов: лен-долгуиец,
лен-кудряш и лен-межеумок (рис. 319).
Основное хозяйственное значение имеет лен-долгунец, культивируемый
на волокно. Он характеризуется длинным (70—125 см) стеблем, который
слабо ветвится, и притом только в верхней части растения. Волокна обра-
зуются в стеблях. После сбора урожая стебли мочат в воде или расстилают
под дощдем (под росу) и затем теребят (механический процесс), выделяя
таким образом пучки волокон, состоящих из отдельных волокон (ме-
ханических клеток), в среднем достигающих длины 25—30 мм, Леп-долгу-
нец наибольшее распространение имеет в'СССР и Западной Европе (глав-
ным образом во Франции и Бельгии). Лен-долгунец — одна из важнег
технических культур СССР. Его возделывают в нечерноземных северных
районах РСФСР, в ряде областей Западной и Средней Сибири и на Даль-
нем Востоке, а также в Белорусской, Литовской и Эстонской ССР.
У льна-межеумка, разводимого ради получения льняного семени и в
меньшей степени волокна, стебель ветвится сильнее. Степень развет-
вленности зависит от густоты травостоя. Лен-межеумок распространен
южнее льна-долгунца, достигая границ степной зоны.
Еще более теплолюбив лен-кудряш. Он приурочен к южным равнинам
европейской части СССР, к нагорьям Кавказа и Средней Азии. Невысокий,
. I н г.
вы
ми
ИЕ РАСТЕНИЯ
Ряс. 318. Лея (Limim usitatissimum):
j-*-верхняя часть цветущего растешн; я — цветок;
з— андроцей ж вжяяцей; 4 — нродолышй разрез ва-
вяод Л — жжеречный разрез завязи; в — раскрывшая-
ся коробочка; 7 — ярдоньный разрез семеня; й — диа-
грамма цяежка.
ветвистый почти от основания,
как мае-
кудр
используется
>; 111
личное растение.
Семена льна содержат око-
ло 35% жира. Льняное масло
считается первоклассным тех-
ническим маслом, дающим луч-
шую олифу; однако его можно
употреблять и в пищу. Жмыхи
содержат значительное коли-
чество белков и используются
как питательный корм для
скота.
Семейство гераниевые
(Geraniaceae). Это травянистые
однолетние или многолетние
растения, редко полукустар-
ники и кустарники. Листья
обычно простые, большей
частью с пальчатым жилкова-
нием, цельные или рассечен-
ные, иногда сложные, с при-
листниками; располагаются
или спирально, или супротив-
но. Цветки циклические из пя-
тичленных кругов, актиноморф-
ные, но бывают более или ме-
нее зигоморфными. Тычинок
обычно 10, реже 15 или 5. Пес-
тик из 5 или 3 плодолистиков,
с верхней завязью, содержащей
в каждом гнезде по 1—2 (ре-
же более) семяпочек. Завязь
вытянута на верхушке в длин-
ный носик. Столбик один, рыль-
це пятилучевое. Плоды—боль-
шей частью коробочки или
распадающиеся на односеменные члеь
датками. Семейство гераниевые насчитывает около 780 видов.
Один из наиболее распространенных родов этого семейства — ге-
рш (Geranium, рис. 320), виды которой есть в северном и южном полуша-
рии. Среди наших представителей этого рода широко известна луговая ге-
рань (G. pratense)* встречающаяся повсюду на лугах,. лесных полянах, по
опушкам и т. п. Опыление у большинства гераниевых осуществляется на-
Hl
нг.
секомыми, получающими нектар, который выделяют железки, находящие-
ся у основания внутренних противочашечковых тычинок. В начале цвете-
иг
ния открываются пыльники, а лучи рыльца направлены в это время кверху
и прилегают друг к другу воспринимающими пыльцу поверхностями. Они
расходятся, принимая горизонтальное положение лишь во вторую поло-
вину цветения (протерандрия). Созревший плод раскрывается пятью створ-
ками, которые быстро закручиваются на спинную сторону (наружу), при
этом'семена отбрасываются на заметное расстояние.
Кроме видов герани, у нас нередко встречается оистшик (Erodium cicu-
tarium) — небольшое однолетнее сорное растение с рассеченными листья-
ми, покрытое жесткими отстоящими волосками. Цветки несколько непра-
вильные, благодаря тому что два верхних (обращенных к оси соцветия) ле-
пестка, короче прочих. Из 10 тычинок 5 без пыльников. Пять односеменных
плодиков имеют
И!Г
рально скрученные придатки, покрытые волосками ж
от
11 L-* I
ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСВМЕННЫВ
ir
11 л
II ;ч
иг.
II
образующиеся из верхних узких частей плодолистиков. Эти придатки
служат для распространения плодов и, кроме того, скручиваясь и раскручи-
ваясь при изменении влажности, способствуют ввинчиванию плодов в
80MJ
Зигоморфность цветка, заметная у аистника, в еще большей степени
выражена вроде пеларгония (Pelargonium), центр распространения кото-
рого — Южная Африка. Некоторые пеларгонин широко известны в комнат-
ной и оранжерейной культуре под названием герани.
Пеларгония роговая (Р. гозеит) — сложный гибрид, культивируется
как эфироносное растение и ценится как таковое, — запах ее масла
напоминает очень дорогое розовое масло. В СССР это растение культиви-
руется главным образом в Закавказье, на Северном Кавказе, в Крыму,
Армении и Средней Азии.
В цветке пеларгоний под задним чашелистиком образуется углубление,
имеющее характер шпорца; оно рассматривается или как видоизменение
оси соцветия, или как впячивание чашелистика, сросшееся с цветоножкой.
Лепестки имеют часто резко неравную величину, что делает цветок зиго-
морфным.
Семейство настурциевые(Tropaeolaceae) — небольшое (80 видов)
семейство, характерное в основном для горных местностей Южной Америки.
. В СССР известна по садовому растению настурция (Tropaeolum majus,
рис. 321). Настурциевые — травы, цепляющиеся за другие растения, зави-
вающиеся листовыми черешками вокруг соседних предметов или вьющиеся.
Цветки резко зигоморфные. Чашелистики вместе с цветочной осью пр
мают участие в образовании крупного шпорца.
Семейство бальзаминовые (Balsaminaceae) объединяет травянис-
стые, обычно очень сочные растения с простыми листьями. Нередко стебли
совершенно прозрачны. В них просвечивают сосудистые пучки. Цветки
зигоморфные, с трех-, пятилопастной, часто окрашенной чашечкой, задний
листочек которой образует шпо-
рец. Лепестков пять, из них
четыре верхних часто срастают-
ся попарно. Тычинок пять.
Они плотно прикладываются
к завязи, образованной также
пятью плодолистиками.
Плод — сочная коробочка,
внезапно открывающаяся от при-
косновения к ней. Пять створок
быстро скручиваются, причем
происходит разбрасывание семе-
ни.
nt
:i:r
Рис. 319. Культурный лев:
1 — долгунец;
4 —• кудряш
Из этого семейства в
СССР встречается два вида
рода недотрога—Impatient
nolitangere и Z. parvtflora. Не-
которых представителей: баль-
заминовых разводят как деко-
ративные. Особенно часто содер-
жат в комнатах под названием
бальзамина Impatiens balsamina
родом из Южной и Юго-Восточ-
ной Азии как крайне неприхот-
ливое и красиво цветущее рас-
тение.
К этому же порядку отно-
сятся семейства парно-
листниковыв (Zygophyllaceae) в
458
ВЫСШИЕ РАСТЕНИЯ
Рис. 320. Гераниевые:
Часть ц?ету?еро ^астения’ 6 чяпь корневища и основание надземной
диаграмма цветка комнатной герани; Л — плод кровяно-крас-
шж герани; • — плод пеларгонии; 5 — плод аистника
Рис. 321. Настурция (Tropaeolum majus):
1 —* Цивгущее растение; 9 * диаграша
(ш — пшорзд)
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
459
кокаиновые
(Erythroxylaceae). Парнолистниковые в боль
ИГ.
нстве
своеу
тропические деревья, из которых экономически ценно гваяковое, алл
бакаутовое, дерево (Guafacum officinale), свойственное Южной Америке (се-
верное побережье) и Антильским островам. Оно дает очень прочную, твер-
дую и тяжелую (тяжелее воды) древесину, идущую на различные поделки.
Это дерево содержит особую, сильно пахнущую смолу, богатую гваяколом,
имеющим медицинское применение.
В небольшом семействе кокаиновые отметим кокаиновое дерево (Eryth-
roxylon coca), растущее главным образом в Перу. Листья содержат важ-
ный в медицинском отношении алкалоид — кокаин.
Зигоморфные гераниецветные, несомненно, возникли от актиноморф-
ных. Более древние представители' этого порядка по общему плану строе-
ния цветка сближаются с порядком Rosales.
5-И ПОРЯДОК — БЕРЕСКЛЕТОЦВЕТНЫЕ (CELASTRALES)
Цветки бересклетоцветных, как правило, с двойным околоцветником,
четырех- или пятичленные. Тычинки в числе лепестков и чередуются с
ними. Гинецей из 2—5 и более плодолистиков, синкарпный; завязь верхняя
или полунижняя. Характерно образование в цветке диска, который отсут-
ствует или мало выражен у примитивных семейств порядка. Деревья и кус-
тарники с простыми листьями, имеющими мелкие прилистники или без них;
листорасположение супротивное или спиральное. Вместилища выделений
в вегетативных органах чаще отсутствуют.
Семейство бересклетовые (Celastraceae). Вечнозеленые или листо-
падные деревья и кустарники, иногда лианы, обычно с супротивными, ре-
же очередными простыми листьями, с маленькими, в некоторых случаях
опадающими прилистниками. Цветки актиноморфные, обоеполые или бла-
годаря недоразвитию однополые. Околоцветник двойной из четырех-пяти-
членных кругов. Тычинок столько же, сколько лепестков, причем они при-
надлежат внешнему кругу андроцея; тычинки внутреннего круга отсутст-
вуют. Два, четыре или пять сросшихся плодолистиков образуют пестик,
обычно с верхней завязью, содержащей число гнезд, соответствующее числу
плодолистиков. Столбик большей частью короткий и с головчатым, а иногда
лопастным рыльцем. В центре почти всегда присутствует железистый диск
(нектарник). Плоды различны — коробочки, костянки, ягоды. Семена час-
то с ярко окрашенным присемянником.
Большинство родов этого семейства, насчитывающего до 850 видов,
обитает в тропических и субтропических странах западного и восточного
м г. ।
полушарий. Л
>|ц
ь немногие виды вошли во флоры умерег
областей.
Так, из рода бересклет (Euonymus), представленного преимущественно
в странах Южной Азии, в Восточной Азии, включая Дальний Восток, и на
Кавказе, два вида — спутники лиственных лесов Евразии. По наиболее
часто встречающемуся в СССР бородавчатому бересклету, или бородавоч-
нику (Е. verrucosa)^ можно познакомиться с типичными чертами се-
мейства.
lilr*.
В корнях бересклетов найдена гуттаперча в количествах, имеющих про-
мышленное значение (Е. verrucosa, Е. maackii* Е. еигораеа). Разведение бе-
ресклетов возможно путем черенкования.
К более примитивному семейству падубовых (Aquifoliaceae) отно-
сится род падуб (Ilex). Один из его видов — падуб остролистный (I, aqui-
folium) — типичный элемент вечнозеленого подлеска лесов Западной, Сред-
ней и Южной Европы. Парагвайский вид падуба — I. paraguaensis — рас-
пространен в Южной Америке; отвар его листьев («парагвайский чай») упо-
требляют как освежающий ароматный напиток.
460
ВЫСШИЕ РАСТЕНИЯ
б-И ПОРЯДОК — КРУШИНОЦВЕТНЫЕ (RHAMNALES)
Цветки крушиноцветных мелкие^ четырехкруговые вследствие выпа-
дения внешнего круга тычинок. В цветке обычно развит интрястямпняль-
ный диск. Деревья, кустарники, лианы с простыми очередными, редко су-
противными листьями.
Порядок крушиноцветных, так же как и порядок бересклетоцветных,
несомненно, родствен порядку Terebinthales, представители которого имеют
пятикруговой цветок с диском. Четырехкруговое строение цветка крушино-
цветных связано с выпаданием наружного круга тычинок, в то время как
у бересклетоцветных редуцировались тычинки внутреннего круга. Таким
образом, крушиноцветные й бересклетоцветные можно рассматривать как
две ветви, возникшие от общего терпентинного предка.
Семейство кр]дии#ные (Rhamnaceae) включает почти исклю-
чительно кустарники и деревья, нередко ко;
листьями с прилистниками. Цветки обычно мелкие, собранные в цимозные
соцветия, обоеполые, с 5—4 чашелистиками и столькими же мелкими ле-
11: *
пестками, часто суженными в ноготок. Число тычинок равно числу членов
венчика^ и расположены они против лепестков, причем каждый лепесток
нередко охватывает соответствующую тычинку. Пестик один с четырех-
одногнездной завязью, содержащей по одной семяпочке в каждом гнезде.
Цветоложе углубленное, свободно окружающее завязь или срастающееся
с ней. Плоды — костянки с одной или несколькими косточками или сухие,
Г; i: ।
распадающиеся на односемянные плодики.
Наиболее обширным родом этого семейства, содержащего около
900 видов, является крушина (Rhammu). Почтя по всей Лесной зоне СССР,
а также в лесах Кавказа, в Крыму, в Западной Сибири и отчасти в Средней
Азин произрастает ломкая крушина (R. frangula, рис. 322) — крупный кус-
' тарник с очередными обратнояйцевидными листьями. Кору и плоды ломкой
крушины употребляют как слабительное. Аналогичное использование име-
ет жостер слабительный (Д. cathartica), растущий в СССР — в евро-
пейской части, в южной части Сибири до Алтая, а также в Крыму, на Кав-
казе и в Средней Азии. Виды жостера —* хорошие красительные растения.
В Крыму, на Кавказе и в Средней Азии распространено держи-дере-
ве (Paliurus spina christf) — очень колючий кустарник с загнутыми назад
ими, представляющими метаморфизированные прилистники.
Виды рода Colletia, распространенного в Южной Америке, замечатель-
•it.ii
ны своими крепкими толстыми зелеными шнпами, представляющими изме-
ненные ветви (кладодии).
Семейство еццюера&нне (Vitaceae). Лишь некоторые представители
виноградных прямостоячие кустарники или небольшие деревья, иногда
ОТДЕЛ ЦВЕТКОВЫЙ, ИДИ ПОКРЫТОСЕМЕННЫЕ
461
Рис. 323, Виноград (Vitis vinifera):
1—сшопенная ветвь в еоцввтинмк; й— виолы: з—расврывавия цветка (сбрасывание венчика); 4—
цветом коале сбрасываний венчика: 4 — раскрывшийвц цветок: • — продольный раарея ягоды
со своеобразным клубневидно утолщенным подземным или надземным во-
дозапасающим стеблем. Большинство — лазающие кустарники или лианы
с развитыми усиками, которые либо обвиваются вокруг опоры, либо обра-
вуют на концах подушечки, с помощью которых лозы могут плотно при-
крепляться к стволам, отвесным стенам и т. п. Усики виноградных стеблево-
го происхождения и представляют видоизмененные, став
соцветия. Листья с прилистниками, располагающиеся спирально, то прос-
тые, цельные или лопастные, то пальчатосложные. Цветки мелкие, в соцве-
тиях, обоеполые, но нередко и однополые, вследствие недоразвития тычи-
нок или пестика. Чашечка, венчик и андроцей пяти-четырехчленные. Ча-
шечка в большинстве случаев слаборазвитая, представленная в форме
валика у основания цветка, с небольшими, малозаметными зубчиками. Ле-
пестки чередуются с чашелистиками и при распускании цветочных почек
либо отгибаются назад, либо, срастаясь друг с другом верхними частями,
отпадают в виде колпачка. Тычинки рамещены в одном круге против ле-
пестков. Между тычинками и завязью развит железистый диск (нектарник),
то в форме отдельных железок, то в виде сплошного, по краю лопастного
кольца. Плодолистики в числе двух (реже 3—6), вполне сросшиеся. Завязь
с числом гнезд, соответстующдм числу плодолистиков, в каждом гнезде две
семяпочки. Плоды — ягоды.
Семейство виноградные (около 700 видов) представлено почти исклю-
чительно в тропических и субтропических .местностях. Наиболее богатый
:ii
HI
it»:
fell!
г I li
nt':
462
ВЫСШИЕ РАСТЕНИЯ
Ml
видами род циссус (Cissus) характерен для тропической зоны, лишь немно-
гие представители заходят в субтропики. В практическом отношении важен
род виноград (Vitis), около 50 видов которого распространены главным
образом в субтропиках северного полушария.
С древнейших времен в Старом Свете культивируют продуктивные фор-
мы дикорастущего винограда V. silvestris, объединяемые под названием
V. vinifera (рис. 323). Эта культура распространена по всему Средиземно-
морью и далее на восток, вплоть до Средней Азии. Культура виноградной
лозы была перенесена на юг Африки, в Австралию, Новую Зеландию.
В Северной Америке разнообразен местный набор видов винограда. Индей-
цы до прихода европейцев использовали дикие плоды в сыром и сушеном
виде и изготовляли виноградное вино. Виноград Старого Света был заве-
зен в Америку в XVII в.
Основную массу урожая винограда используют для получения вино-
градных вин. Часть винограда утотребляют в сыром виде. Высушенные яго-
ды специальных сортов винограда поступают в продажу под названием изю-
ма и сабзы.
Культура винограда, распространенная, как сказано, в основном в суб-
тропических районах, значительно продвинулась и в умеренную зону. Так,
в Западной Европе виноград выращивают в долине Рейна, а у нас — в За-
кавказье, на Северном Кавказе, в Украинской и Молдавской ССР, в Сред-
ней Азии. Работы И. В. Мичурина по осеверению винограда дали возмож-
ность продвинуть эту культуру до широты Смоленска — Калинина — Ива-
нова — Казани — Уфы.
В качестве декоративного растения, обрастающего стены и заборы,
часто разводят под названием «дикого винограда», а местами под явно не-
правильным названием «плюща» Partenocissus quinquefolia — лазающий
кустарник с крупными темно-зелеными пальчатосложными листьями; ро-
дина его — Северная Америка.'
11 Г, I
'.III
7-Й ПОРЯДОК — ЗОНТИКОЦВЕТНЫЕ (UMBELLIFLORAE)
Цветки зонтикоцветных, как правило, актиноморфные, с двойным
околоцветником из пяти-четырехчленных кругов, причем чашечка сильно
редуцирована. Тычинок один круг, и они чередуются с лепестками. Цено-
карпный гинецей состоит обычно* из двух плодолистиков. Завязь, нижняя,
двугнездная, с одной однопокровной семяпочкой в каждом гнезде. На
вершине Завязи в большинстве случаев развивается железистый диск. Тра-
вы, кустарники,, небольшие деревья и лианы, чаще с очередными листь-
ями, мелкими прилистниками или расширенными влагалищными основания-
ми.
Семейство аралиевые (АгаИаееае)* Аралиевые — деревья и кус-
тарники, лишь некоторые — лианы и травы. Листья обычно крупные, оче-
редные, часто пальчатые или перистые, с мелкими прилистниками в виде
перепончатых окраин черешков или иного строения. Мелкие цветки собра-
ны в сложные соцветия, состоящие из зонтиков, головок или колосьев. Ча-
шечка представлена малоразвитой цельной или зубчатой окраиной на вер-
хушке завязи. Лепестков обычно пять, они свободные. Чередуясь с лепест-
ками, располагаются пять тычинок. Ценокарпный гинецей состоит из
5—2 плодолистиков; число гнезд нижней (лишь очень редко полунижней)
завязи соответствует числу плодолистиков. В каждом гнезде находится
одна семяпочка, развивающаяся в семя, и одна недоразвитая. На вогнутой
или, наоборот, выпуклой верхушке завязи находится железистая подушеч-
ка. Плоды у аралиевых часто сочные, могут распадаться на отдельные
ПЛ О ДИК И1
Растения семейства аралиевых (около 700 видов) в основном распро^
странены в тропиках и субтропиках. Из внетропических форм надо отме-
тить род плющ (Hedera, рис. 324). В пределах СССР виды плюща встреча-
I
Ml
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
463
юте я в прибалтийских советских республиках (на юго-западе), в Крыму,
на юге Западного Приднепровья и на Кавказе. Техническое значение имеет
китайское бумажное дерево (Tetrapanax papyrifera)^ родом с острова Тай-
вань, культивируемое в Китае и Японии. Из сердцев
u:i
и его делают так на-
зываемую китайскую «рисовую» бумагу.
На севере Китая и Кореи, а также в Приморском крае встречается
жень-шень (Panax ginseng) — многолетнее травянистое растение с мясис-
тым цилиндрическим корнем, имеющим на конце два — несколько развет-
влений, придаю:
щх ему
II г»
иногда отдаленное сходство с телом человека или
вообще причудливую форму. Китайская медицина широко использовала
жень-шень уже за 3000 лет до нашей эры. Особенно ценились корни дико-
растут:
11 г *
: растений, и притом достаточно крупные. О свойствах жень шеня
создано немало легенд. Жень-шень растет одиночными экземплярами или
небольшими группами и встречается очень редко.
В настоящее время жень-шень признан официальной медициной СССР,
США и ряда других стран. В нем найдены физиологически активные вещест-
ва. Препараты корня жень-шеня применяют у нас как тонизирующие
средства. С этой целью применяют и некоторые другие аралиевые, напри-
мео маньчжурскую аралию (Aralta mandshurica).
’Семейство зонтичные (Umbelliferae, Apiaceae). Растения этого
семейства— почти всегда травы, лишь редко полукустарники и приземистые
кустарники. Спирально расположенные листья имеют ясно выраженное,
нередко очень сильно разрастающееся влагалище. Листовая пластинка
сравнительно редко цельная, чаще же пальчато- или перисто-, одно-, дву-
или многократно расчлененная. Стебли обычно имеют заметно утолщенные
узлы; междоузлия часто бороздчатые или ребристые, нередко полые внутри
благодаря раннему разрушению сердцевины.
Цветки зонтичных мелкие, собранные в соцветия сложные зонтики,
иногда головки (синеголовник — Eryngium) или простые зонтики (подлес-
ник — Sanicula, астранция — Astrantla). У основания простых соцветий
собраны мелкие листья, кроющие по отношению к краевым цветкам; в
совокупности они образуют так называемую обертку. В случае сложного
зонтика различают обертку (у основания лучей первого порядка) и обер-
точку (у основ ания лучей второго порядка); нередко обертка, а иногда и
оберточка отсу тствуют.
Цветки зонтичных (рис. 325) в основной актиноморфные. В состав
каждого цветка входят пятилистная чашечка, пятичленный раздельноле-
пестный венчик, пять тычинок и пестик с двумя столбиками и двугнездной
нижней завязью. Редко чашелистики листовидны (рис. 326); чаще на верхуш-
ке -завязи заметны пять небольших зубчиков или чашечка представлена в
в форме выступа ющей окраины близ верхушки завязи.
464
ВЫСШИЕ РАСТ
з; i \
Рис, 325, Диаграмма цветка и цветок зонтичных
Рис. 326. Аетр&нцкя (Astrantia biebersteinii):
1— цвжн в начале иврвой
цветок в женской фаве
(чж—чашелистики)
(мужской) фазы цветения;
цвстеянл* тычинки сиали
нг
[И ИI:
: н I'
Лепестки у зонтичных белые, розовато-белые, редко интенсивно-розо-
вые, а у других — желтые или зеленовато-желтые. Форма лепестка очень
чная: от узкого основания, имеющего часто вид небольшого ноготка,
лепесток расширяется в пластинку, у многих заканчивающуюся на верхуш-
ке сужением, которое обычно загнуто внутрь цветка, благодаря чему лепес-
ток на верхушке кажется двулопастным. Лепестки прикреплены к вер-
хушке Завязи и чередуются с чашелистиками.
Чередуясь с лепестками, расположены и тычинки, причем нити их,
особенно в почкосложении, дугообразно согнуты к центру цветка и лишь
позднее отгибаются наружу. Пестик образован двумя плодолистиками. На
верхушке завязи, в основании столбиков, имеется так называемый надпе-
стичный диск в виде разделенной надвое подушечки, функционирующей
как нектарник. В каждом из двух гнезд завязи одна однопокровная вися-
чая семяпочка, но часто в процессе развития цветка закладывается по две
семяпочки в гнезде.
Скученные в соцветия, цветки зонтичных хорошо заметны насекомым.
Опыление могут осуществлять короткохоботковые мухи и другие двукры-
лые, привлекаемые типичным запахом и яркостью соцветий.
Часто наблюдается дихогамия (рис. 326). Иногда недоразвиваются ты-
чинки в одних и пестики в других цветках (вторичная однополость цветков).
После оплодотворения формируется двугнездный плод, то округлый
в поперечном сечении, то сплюснутый. При созревании он разделяется на
два односеменных, сухих, нераскрывающихся плодика (сем
продолжающих, однако, висеть на особам органе, называемом карпо-
фором (столбочком). Карпофор образуется из краевых частей плодо-
листиков и представляет как бы продолжение плодоножки, разделенное
наверху на две веточки, к которым и прикреплены семянки. Такое строе-
ние зрелого плода объясняет его название — двусемянка или вислоплодник.
В околоплоднике (рис. 328) каждой семянки по длине ее проходят
пять проводящих пучков, соответственно которым на поверхности плодика
заметны пять ребрышек: два краевых и три спинных. Они отделены друг
от друга ложбинками. Под ложбинками, также на стороне семянки, обра-
щенной к плоскости,-по которой произошло расщепление плода, в околоплод-
нике расположены эфиромасляные ходы, число и размещение которых
в деталях довольно разнообразно. Иногда на месте ложбинок вслед-
ствие разрастания ткани околоплодника возникают так называемые иго-
Гг
ричные ребрышки, которые могут выступать на поверхности семянки даже зна-
чительно больше, чем основные, первичные. Семя плотно прилегает к около-
плоднику иди срастается с ним. Кроме зародыша, в семени содержится мощ-
ный эндосперм. Последний бывает трех основных форм: 1) эндосперм по
поверхности, обращенной к плоскости расщепления плода, плоский, чуть
выпуклый или слегка вогнутый («белок прямой»); 2) на той же поверхности,
вдоль ее, проходит глубокая ложбинка, благодаря чему поперечный раз- *
рез эндосперма имеет почти серповидную форму («белок согнутый»); 3) эндо-
сперм имеет вогнутую форму как на поперечном, так и на продольном раз-
резе, т. е. образует фигуру, напоминающую часовое стекло («белок выдолб-
пег.
г i:t?
Зонтичные — одно из крупнейших семейств покрытосеменных, оно
содержит около 3000 видов.
Вследствие сравнительно незначительной изменчивости цветка деле-
ние семейства зонтичных на роды основывают главным образом на особен-
ностях строения плодов.
Зонтичные распространены по всему свету, но преимущественно в уме-
ренных областях обоих полушарий. Будучи растениями нередко крупными,
с характерным внешним видом, зонтичные придают иногда специфические
черты растительным ландшафтам. Так, на разнотравных степях европей-
ской части СССР и в Сибири виды горичника (Peucedanum) и порезника
(Libanotis) создают во время цветения своеобразные аспекты. В наших лист-
венных лесах очень часто в травяном покрове господствует сныть обыкно-
венная (Aegopodium podagraria) — корневищное растение, развивающее
главным образом прикорневые листья, с большой дважды тройчатой
пластинкой. По сырым местам в лесной полосе нередко образуются пышные
заросли дудника лесного (Angelica silvestris). У основания стеблей этого
растения развиваются огромные сложные листья, а во время цветения раз-
ветвления стебля, достигающего 2 ж в высоту, украшены большими полу-
круглыми сложными зонтиками белых цветков. Еще более эффектное вне-'
чатление производят заросли дягиля лекарственного (Archangelica officinal
lt£)\ его прикорневые треугольные в очертаниях листья с дважды перистой'
пластинкой нередко бывают очень крупными, а на огромных стеблях разви-
ваются шаровидные сложные зонтики диаметром 10—15 сж.
Подобного рода крупные зонтичные иногда характеризуют целые рас-
тительные провинции. Например, в растительности Камчатки характерны
высокотравные луга с массовым развитием медвежьего дудника (Angelica'
ursina), который входит также в травяной покров светлых редкоствольных
лесов из каменной березы. Не менее замечательны гигантские зонтичные
из рода Perula9 встречающиеся в наших среднеазиатских пустынях. В пер-
вые годы жизни они в период весенних дождей развивают розетки прикор-
невых листьев, а на 3—4-й год появляются высокие прочные. стебли, пре-
вышающие рост человека и несущие огромные сложно устроенные соцве-
тия. После плодоношения растение отмирает (монокарпические многолет-
и). В южном полушарии — в Патагонии, на Огненной Земле и в дру-
гих местностях — распространены виды рода азорелла (Azorelld). Это
низкорослые кустарники, образующие подушки до 1 ж в поперек
ветви, покрытые мелкими листьями, так плотно прилегают друг к другу,
что подушка кажется камнем, да и по твердости также приближается к
камню. Дикорастущие зонтичные с хозяйственной (кормовой) точки зрения
играют скорее отрицательную роль. Большинство зонтичных скот плохо
или совсем не поедает, а некоторые оказываются даже ядовитыми. Среди
последних особенно следует отметить цикуту 9 или вех (Clcuta virosa,
рис.329), и крапчатый болиголов (Contum maculatum^ рис. 330).
:стое растение с толстым вертикаль-
Kill I
it?
а; i;
: I; I ,
ди черешках. Встречается по бохотам, болотистым берегам рек, стариц
466
ВЫСШИЕ РАСТЕНИЯ
Рис. 327. Плоды зонтичных;
* ® Т Д®№®иянка> б — нрздрльный разрез нлода (во иедианной плоскоста). mnenan
выйрурез; г — болиголова। я _ общий вид (сбоку), б-^оиеришйкйет™ з - пегп^ка/Т^^^й
мв (сбяку), б - поперечный рырев (а - ицюдыш. м _ колой», ж» Пйслнные
?*®' ^8" Фенхель (Foeniculum vulgare). Поперечный peepee семянки (* — мягляши»
*ОДИ» я проводящие пуяки, е —андосперм, ап — карпофор)
ОТДЕЛ ЦВЕТКОВЫЕ, Или ПОКРЫТОСЕМКВ НЫд
467
Рис. 329. Вех (Cicuta virosa):
1 — соцветие; 2 —продольный разрез корневища;
3 — плод; 4 — разрез плода
Рис. 330. Болиголов (Conium maculatum):
1 — часть олиственного побега с хорошо заметной
пятнистостью стебля: 3—простой зонтик; 3—цве-
ток; 4—плод; з — разрез плод»
ПРУДОВ> озер. Корневища цикуты иногда принимают за сельдерей, из-за
чего бывают отравления. Цикуту обычно скот не поедает, но, попав случай-
но в корм, она вызывает отравления. Нередки случаи отравления цикутой
детей.
Крапчатый t болиголов — довольно высокое одно-двухлетнее травя-
нистое растение с прямым, тонкобороздчатым, голым, ветвистым стеб-
лем, с красновато-бурыми пятнами на поверхности, особенно в нижней его
части. Распространен болиголов по сырым местам, а также в кустар-
никах и по берегам рек. Все части растения, как у цикуты, ядовиты Ъ для
человека, и для скота. Также сильно ядовита собачья. петрушка (Aethusa
cynapium), встречающаяся по огородам, садам, иногда в посевах настоя-
щей петрушки, с которой внешне сходна, но отличается отсутствием специ-
фического запаха петрушки.
Положительное значение для человека имеют сравнительно немногие
зонтичные. Среди пищевых на первом месте стоит всем известная морковь
(Baucus carota, рис. 331). Это — двухлетнее растение, образующее в первый
год розетку прикорневых листьев и корнеплод. В сильно разрастающейся
коровой ткани его откладываются запасные питательные вещества (главным
образом сахара). Оранжевая окраска корнеплода моркови зависит от нали-
чия в клетках коры большого количества каротина (провитамина А). Мор-
ковь ценится не только как овощное растение, но и как сочный корм для
молочного скота и особенно для выкормки конского молодняка. На второй
год жизни растение развивает прямостоячий стебель, заканчивающийся
многолучевым сложным зонтиком, в котором хорошо развита как обертка,
так и оберточка. Соцветие моркови представляет интересный объект для
вы
н 11 г, I >
РАСТЕНИЯ
Рис. 331. Морковь (Daucus
carota):
1 — соцветие; я - цветок!
з — продольный разрез ипп^а;
4 — поперечный разрез плода
биологического наблюдения. В молодой состоянии лучи зонтика к ночи за-
гибаются и все соцветие поникает. Эти движения, насколько известно, пре-
кращаются в конце цветения, после опыления. В это время лучи зонтика
загибаются внутрь, расправляясь вновь ко времени созревания плодов при
сухой погоде. При изменении влажности такие движения лучей зонтика
у плодоносящего растения могут повторяться не один раз. Семянки морко-
ви имеют пять сравнительно слаборазвитых первичных ребрышек, покры-
тых шишками, и четыре выдающихся ’ вторичных с многочисленным»
длинными крючковидно загнутыми на конце шипами. В связи с этим, во
избежание сцепления, плодики моркови перед посевом перетирают, чтобы
обломать шипы. Известно большое количество сортов моркови. Ее крупный
корнеплод возник в результате отбора и выращивания при благоприят-
ных почвенных условиях. Дикий предок моркови встречается и сейчас как
в Европе, так и в Азии, отличаясь, однако, грубым и далеко не таким объ-
емистым корнем.
Меньшее, чем морковь, значение имеют петрушка (PetroseUnum sati-
vum), сельдерей (Apiurn graveolens), пастернак посевной (Pastinaca sativa),
укроп (Anethum graveolenfy,mxim (Canon card), анис (PtmptneUa antsum),
кервель (Anthriscus cerefolium) и кориандр (Corlandrum sativum), исполь-
зуемые главным образом как пряные приправы.
Семейства, относящиеся к порядку Umbellljlorae, несомненно, близко-
родственны друг другу, причем семейство зонтичных представляет, конеч-
но, более поздний этап в развитии этого порядка. Происхождение Umbel-
Uflorae остается не вполне ясным. Ряд признаков сближает их с ТегеЫп-
thales, Celastrales и Rhamnales: одинаковый общий план строения цветка,
сходное строение и положение семяпочек и число их, четырехкруговой
цветок, наличие вместилищ выделений и другие признаки.
С другой стороны, возможно сближение зонтичных с Rosales на основа-
выраженной тенденции у последних к образованию нижней завязи, на-
личия у некоторых камнеломковых четырехкруговых цветков и одно-
покровной семяпочки.
К этому же порядку присоединяют семейство кизиловые (Сота-
сеае), которое, по-видимому, правильнее имеете с еще рядом семейств
выделять в особый порядок Comales. К кизиловым относятся кизил
(Comas mas), имеющий съедобные плоды-костянки, и золотое дерево
(Aucuba japonica) родом из Юго-Восточной Азии с золотисто-желтоватыми
ин
гл
6ТДВЛ
8-Й ПОРЯДОК — МИРТОЦВЕТНЫЕ (MYRTALES)
Цветки миртоцветных, обоеполые, как правило, актиноморфные, в
большинстве случаев с двойным околоцветником, четырех- или пятичлен-.
ных в каждом круге. Число тычинок равно числу лепестков или в два рааа
больше, а у некоторых представителей вследствие расщепления тычинок
андроцей многочленный. Плодолистиков столько же; сколько лепестков,
реже их меньше. Гинецей ценокарпный, при атом плодолистики почти всегда
сращены доверху, в результате столбик один; рыльце цельное. Плацента
краевая. Цветоложе вогнутое, образующее трубочку, редко свободное, ча-
ще срастается с завязью, вследствие чего завязь становится нижней. Пре-
имущественно древесные растения, но в нашей флоре встречаются главным
образом травы, отчасти кустарники. Листья всегда без прилистников, обыч-
но супротивные, реже мутовчатые.
Семейство дербенниковые (Lythraceae), содержащее около 500 ви-
дов, представлено в СССР главным образом видами рода дербенник (Lytb-
rum). Дербенник иволистный, или плакун-трава (Lythrum ealicaria,
рис. 332) — травянистое многолетнее растение, растущее по берегам рек
и озер — в прибрежной полосе в воде, в зарослях ольхи или ив, а также на .
заливных лугах. Обращает на себя внимание с середины лета довольно гус-
тыми крупными соцветиями из ярко-розовых цветков. Трубчатое цвето-
ложе по внешнему виду сходно с чашечкой. Оно несет по краю 12 поочеред-
но длинных и более коротких чашелистиков (зубчиков) и 6 розовых лепест-
ков. Тычинок 12. Завязь двугнездная, свободная, с одним столбиком и го-
ловчатым рыльцем. Плод — коробочка. Цветки дербенника замечательны
тристилией — здесь различают три формы цветков с различной длиной стол-
бика и тычинок. Это приспособления к перекрестному опылению, изу-
ченные у дербенника еще Ч. Дарвином.
Родственны семейству дербенниковых два тропических семейст-
ва — зоннерациевые (Sonneratiaceae) и ризофоровые (Rhizophoraceae), пред-
1 *: I:
j; ।г.
ставители которых входят в состав мангровых зарослей, характерных для
иловатых побережий морей в приливно-отливной полосе. У многих манг-
ровых имеются пневматофоры (дыхательные корни); особенно эффектны они
у представителей рода зоннерация (Sonneratia), у которых ответвления
корней отрицательно геотропичны и поднимаются над почвой на 80 см
и выше. У ризофоровых наблюдаются ходульные корни, обеспечивающие
лучшее укрепление в илистой почве, а семена прорастают еще в плоде, и
зародыш не имеет периода покоя. Проросток развивается на материнском
дереве несколько месяцев, после чего он падает на землю и быстро укоре-
няется (живородность).
Семейство миртовые (Myrtaceoe). В это семейство входят древес-
ные растения и кустарники, большей частью с супротивными листьями
без прилистников. В тканях постоянно присутствуют лизигенные округлые
эфиромасляные железки. В цветке за четырьмя-пятью чашелистиками и
столькими же лепестками следуют многочисленные тычинки, иногда срас-
в бокальчатое цветоложе, нередко завязь нижняя. Плоды сочные (ягода
или костянка) или сухие (коробочки, семянки).
Семейство миртовых содержит до 3000 видов, подавляющее большин-
ство которых обитает в тропической зоне, где эти растения нередко прини-
. мают существенное участие в составе растительных формаций. Из форм с
сочными плодами следует отметить мирту (Myrtus communis, рис. 333),
распространенную в странах Средиземноморья и часто разводимую как
декоративное растение на юге СССР. Некоторые миртовые приносят съедоб-
ные плоды; такова, например, гуаява (Psidium guajava), свойственная тро-
пической Америке и разводимая в США (Калифорния, Флорвда) и в других
теплых странах ради плодов величиной с персик, обладающих приятным
вкусом. У гвоздичного дерева (CaryophyUus aromaticus, или Eugenia cary^
470
ВЫСШИЕ РАСТЕНИЯ
Рис. 332. Дербенник (Lythrum salicaria):
1 — цветущий побег; 2 — длинностолбчатый цветок;
з— цветок с промежуточной длиной столбика: 4 —
короткостолбчатый цветок; 6 — диаграмма цветка
phyllata,' рис, 334) плоды
также сочные, но мелкие,
несъедобные, У этого вечно-
зеленого средней величины
дерева с супротивными листь-
ями, растущего в Индонезии
(на Молуккских островах),
используются в качестве
пряности («гвоздика») цве-
точные бутоны.
Все перечисленные пред-
ставители миртовых относят-
ся к подсемейству
миртовых (Myrtoidae)' ос-
новным признаком которого
является сочный плод.
эвкалиптовых (Leptospermo-
idae) плоды сухие. Наибо-
лее важный род этого под-
семейства, а вместе с тем и
всего семейства миртовых —
эвкалипт (Еиса lyр tust рис.
335). Эвкалипты растут в
Австралии, отчасти выходя
за ее пределы. В частности,
они культивируются у нас на
Черноморском побережье
Кавказа. Это деревья и
кустарники, входящие в са-
мые разнообразные расти-
тельные формации. Их на-
считывают более 600 видов.
Некоторые виды живут при
200—250 мм осадков, в то
время как другие населяют
местности очень влажные (до
4000 мм осадков в год). Не-
которые эвкалипты являются господствующими растениями на
боль
ИГ.
пространствах. Так, например, миндалелистный эвкалипт (Е. amygdalina)
образует леса на востоке Австралии. Листья большинства эвкалиптов обычно
сизоватые от воскового налета. Распространена гетерофилия. Эвкалипты
отличаются быстрым ростом. Так, например Е. globulus^ называемый «про-
тиволихорадочным деревом», за 9 лет может достигнуть 20-метровой высоты.
В связи с этим нужно отметить, что некоторые эвкалипты достигают 100 л
и более высоты. Древесина некоторых эвкалиптовых деревьев отличается
значительной твердостью. Листья эвкалиптов содержат много эфирных ма-
сел, используемых в медицине, парфюмерии и для других целей.
Семейство гранатовые (Punicaceae) очень близко миртовым и
отличается от них прежде всего отсутствием эфиромасляных железок.
Единственный род этого семейства — гранат (Punica) — представлен
в нашей флоре гранатовым деревом (Punica granatum, рис. 336), растущим
в Закавказье и Средней Азии, а также распространенным в Иране, в странах
Малой Азии и в Афганистане. Гранат культивируют ради его съедобных
плодов. Последние развиваются из нижней завязи. Созрев
нг.
:й плод грана-
та имеет размер крупного яблока; под кожистым околоплодником находит-
ся много семян, имеющих семенную кожуру с очень сочным наружным сло-
ем красного цвета и кисловато-сладкого вкуса. Околоплодник и кора грана-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
471
та (особенно его корней) богаты
дуб ильными веществами. Кору
корней, кроме того, использу-
ют как глистогонное средство.
Семейство ослинни-
ковые {Onagraceae). Это травя-
нистые растения или кустар-
ники, большей частью лишен-
ные прилистников. Цветки
актиноморфные или несколько
асимметричные, из четырех-
или двухчленных кругов, с за-
вязью нижней или полуниж-
ней; столбик с головчатым
рыльцем у одних и четырехраз-
дельным у других представи-
телей. Плоды — коробочки,
ягоды, орехи.
К этому семейству, насчи-
тывающему 650 видов, относит-
ся ряд декоративных и инте-
ресных по своей биологии
растений.
Из числа имеющих плоды-
коробочки в СССР распрост-
ранены кипреи {Epilobium) и
очень близкий к ним иван-чай
{каперский чай — Chamaene-
rium angustifollum, рис. 337).
Это обычное растение лесных
вырубок обладает способностью
к быстрому вегетативному
размножению корневищем,
энергично растущим, особен-
но в рыхлых почвах. Расте-
Рис. 333. Мирта (Myrtus communis):
I — мирта в цвету; 2 — разрез цветка; л — цветок аосле
удаления венчика и андроцея; 4 — диаграмма цветке
ние медоносное. Высокий, в
1—2 ж, стебель покрыт ланцетными листьями и заканчивается
длинной кистью довольно крупных малиново-розовых
цветков. Вследст-
вие строго акропетального развития последних на одном соцветии можно
одновременно наблюдать все фазы цветения — от бутонов (наверху) до зре-
лых плодов (внизу). Здесь очень ярко выражена протерандрия. Среди на-
секомых-опылителей надо отметить пчел. Плоды у иван-чая — коробочки,
содержащие многочисленные семена, окруженные волосками. Листья иван-
чая в прошлом использовались как'суррогат настоящего чая, чем и объясня-
ется название растения. Нежные молодые побеги съедобны.
Спорадически, особенно на песчаных почвах, на лугах, по откосам,
встречается двухлетний ослинник {Oenothera biennis). Это двухлетнее расте-
ние американского происхождения, завезенное в Европу в XVII в. Светло-
желтые цветки раскрываются вечером и испускают при этом довольно силь-
ный аромат.
К тому же роду относится так называемая ночная свеча Ламарка
{О. lamarkiana) — основной объект исследования голландского ученого Д е
Ф р и э а, автора мутационной теории.
В биологическом отношении интересна южноамериканская Jussieua
repens, растущая в тропических болотах Южной Америки. Горизонтальное
корневище этого растения дает три рода корней. Одни внедряются в ил и
служат для минерального питания, другие — пузыреобраэные — богаты
аэренхимой и являются плавательным аппаратом, наконец, третьи по(
л:г.
472
ВЫСШИЕ РАСТЕНИЯ
Рис. 334. Гвоздичное дерево (Eugenia caryophyb
lata):
J—цветущая ветвь; а— цветок; 3— ицоцкшьвый разрез
цветка; 4 —диаграмма цветка
маются вертикально вверх
и обеспечивают снабжение
подводных частей кислоро-
дом.
У других ослинниковых
плоды — ягоды. Из этой
группы следует * отметить
род фуксия (Fuchsia), виды
которого в диком состоянии
встречаются в Центральной
и Южной Америке и в Новой
Зеландии. Фуксии — неболь-
:е кустарники или дерев-
ца, легко размножающиеся
отводками. У нас известен
в комнатной культуре ряд
сортов, фуксии с красивыми
крупными цветками.
Из числа орехоплодных
иг
г. Г?
ослинниковых отметим ро-
ды цирцею, или чаровницу
(Сircaea), и водяной орех
(Trapa па tans). Circaea lutetir
ma (колдун-трава) — не-
большое травянистое расте-
ние с супротивными зелены-
ми листьями и невзрачными
цветками — живет в лист-
венных лесах, обычно на бо-
гатых и достаточно влаж-
ных почвах.
Верхушка ее ползучего
кверху и выходит на поверхность, чтобы
корпев
Ulf
д иногда загибается
затем вновь изогнуться вниз и
войти в почву. Биологическое значение такой формы роста остается не-
выясненным. Другой вид — цирцея альпийская (С. alpina) — распростра-
нен преимущественно в хвойных лесах.
Водяной орех (или чертов орех, рогульник, — Trapa natanae, рис. 338)
встречается у нас кое-где в озерах, заводях рек и т. п. На поверхности воды
он образует розетки скученных листьев с ромбической пластинкой и пузы-
ревидно-вздутыми (вследствие развития аэренхимы) черешками, которые
служат плавательным аппаратом. Некрупные цветки с полунижней завязью
выходят из пазух листьев, причем по отцветании цветоножка загибается
вниз .и созревание плода происходит под водой. Плод типа костянки; наруж-
ная часть околоплодника зеленая, но твердая и позднее сгнивает в проти-
воположность внутренней, стойко сохраняющейся и имеющей темно-се-
рую или почти черную окраску. Своеобразна форма плода: нижняя часть
его представляет пирамиду, основанием обращенную вверх, ребристую по
граням; верхняя половина конусообразно суживается и обычно заканчи-
вается характерной коронкой. Несколько выше середины от плода отходят
четыре крепких рога, концы которых покрыты острыми зазубринами. Эти
роговидные придатки — разросшиеся чашелистики. Благодаря такому
строен
плод водяного
ореха, падая на дно, заякоривается в иле и, сохра-
няясь далее в течение всей жизни растения, поддерживает связь его розеток
с дном водоема. В плоде развивается только одно семя, оно богато крах-
малом и используется в пищу. Судя по распределению мест произрастания
1ГЛ1
.11
водяного opeta и по находкам орехов в торфах, это растение—древний эле-*
мент нашей флоры.
’ I: I.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис. 335. Эвкалипт (Eucalyptus sideroxylon)
' Л— цветочная кочка: а — общкй вад» б — ародольный разрез; 2—цветок: з —ввод
Рис. 336. Гранат (Punica granatum):
1 — цветущая ветвь; 8 — диаграмма цветка; з — продоль-
ный разрез цветка; 4 — плод; 5 — продольный разрез
плода
Рис. 337. Иван-чай (Chamaenerium
angustif olium):
1 — соцветие; 2 — разрез цветка; з — плод;
4 — семя; 3 — диаграмма цветка
Порядок Myrtales можно считать самостоятельной ветвью эволюции
покрытосеменных, берущей свое начало, подобно Fabales^ от Rosales, с ко-
торыми миртоцветные сходны общим планом строения цветка, вогнутостью
цветоложа и семенами, у большинства без эндосперма.
Порядок Rosales в подклассе Rosidae занимает положение, аналогич-
ное порядку Dilleniales в подклассе Dilleniidae, Rosales стоят на уровне
эволюционного развития более высоком, чем Magnoliidae, но не достигли
уровня, соответствующего подклассу Asteridae. Rosales в подавляющей
т
ВЫСШИЕ РАСТЕНИЯ
массе раздельнолепестные, в
ряде групп (спирейные, тол- <
стянковые и др.) с апо-
карпным гинецеем, плоды —
многосемянные листовки.
Такого типа Rosales сближают
этот порядок, а вместе с
тем и весь подкласс Rost-
daec подклассом Magnoliidae.
Порядок Rosales, вероят-
но, дал начало всем прочим
порядкам подкласса Rosldae.
Связующее звено между.
F abates и Rosales, возмож-
но, представляет небольшое
пантропическое семейство
Соппагасеае, характеризу-
емое правильным цветком.
В этом семействе как бы
предвосхищаются некоторые
признаки Fabates', у некото-
рых Соппагасеае плоды—бобы,
иногда у них срастаются ча-
шелистики, а чашечка остает-
ся при плодах, облекая их в
нижней части; свободные в.
основании лепестки могут
выше спаиваться (слипать-
ся) друг с другом; наблюда-
ется также срастание тычи-
ночных нитей в короткую
трубку.
Положение Соппагасеае
в системе не вполне опреде-
ленно, но это семейство мо-
не утратившего черты, свой-
Рис. 338. Водяной орех, чилим (Trapa natans):
1 — плавающая розетка и подводные органы; 2 — разрез
цветка; J — плод; 4 — проросток (пк — подсемядольное
колено. Ci — семядоля. са — вторая семядоля)
жёт служить моделью предка Fabales, еще
ственные Rosales.
Некоторые систематики считают семейство Соппагасеае настолько близ-
ким с Fabates, что включают его в этот порядок.
О вероятной связи порядков Rhamnales и Celastrales через общего пред-
ка сказано выше.
Самостоятельную линию эволюции представляет порядок Myrtales.
Здесь реализовалась имеющаяся у Rosales тенденция к образованию цветоч-
ной трубки, которая у менее высокостоящих представителей остается сво-
бодной, а у более удаленных от предков срастается с завязью.
Порядок Uinbelltflorae, как уже было указано, выводят или непосред-
ственно от Rosales, или через порядок Celastrales.
5-Й ПОДКЛАСС — АСТЕРИДНЫЕ (ASTER1DAE)
Астеридные объединяют в основном тетрациклические, с одним кругом
тычинок, спайнолепестные цветки. Число тычинок равно числу долей вен-
чика или, вследствие частичной редукции, меньше его; тычинки никогда
не бывают противолепестковыми. Гинецей ценокарпный из 2, иногда из 5,
реже из большего числа плодолистиков. Семязачатки обычно о одним инте-
гументом, тенуинуцеллятным (редко крассинуцеллятные). Преимуществен-
но травянистые растения.
В составе подкласса Asteridae рассматриваются 7 порядков.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
475
1-Й ПОРЯДОК — ГОРЕЧАВКОЦВЕТНЫЕ
(GENTIANALES), ИЛИ
СКРУЧЕННОЦВЕТНЫЕ (CONTORTAE)
Цветки горечавкоцветных четырехкруговые.
Чашечка и венчик из 5—4 членов. Венчик спай-
нолепестный, в почкосложении обычно скручен-
ный (рис. 339), тычинки в числе лепестков
прикреплены к трубке или к зеву венчика меж-
ду его лопастями. Гинецей обычно из 2 плодо-
листиков, ценокарпный, но в некоторых семейст-
вах сращены только верхние части плодолисти-
ков и гинецей благодаря этому как бы апокарп-
ный. Завязь верхняя, столбик один. У основания
завязи обычно развит подпестичный нектарный
диск. Растения различного облика — деревья,
Рис. '339. Скрученное почко-
сложение долей венчика
в бутоне:
1 — горечавка; г — золототысяч-
ник
лианы, травы с супротивными, реже очередны-
ми простыми листьями. Прилистники отсутствуют или мелкие, руди-
ментарные, Характерна интраксилярная флоэма.
Семейство горечавковые (Gentianaceae). Это травянистые растения,
почти всегда с простыми и цельнокрайними листьями без прилистников.
Цветки одиночные или в цимозных соцветиях, почти всегда актиноморф-
ные, из четырех кругов; чашечка, венчик и андроцей пятичленные (редко
четырехчленные), гинецей двучленный, ценокарпный, с верхней одногнезд-
ной или двугнездной завязью.
Плод — коробочка, реже ягода.
Большое семейство, содержащее около 1100 видов. Наиболее крупный
род — горечавка (Gentiana), насчитывающий более 300 видов.
В СССР много горечавок известно на Кавказе и в Средней Азии. На
равнинах и в горах (до высоты 1200 м) Европы, Западной и Восточной Си-
бири часто растет легочная горечавка (G. pneumonanthe, рис. 340) с темно-
синими крупными воронковидными цветками. Высокогорные и тундровые
горечавки выделяются очень низкими (1—3 см) стеблями, длина которых
может быть меньше длины цветка.' Горечавковые обычно содержат глюкози-
ды, на чем основано медицинское использование этих растений. Из лекарст-
венных горечавок следует отметить желтую горечавку (G. lutea) родом из
Альп, в СССР в диком состоянии встречающуюся в Карпатах и культиви-
руемую. У этого вида используются корни. Подобное применение находит
и золототысячник зонтичный {Centaurium umbellatum, Erythraea centaurium,
рис. 341) — травянистое растение с многоветвистым дихазиальным соцве-
тием из розовых цветков.
С горечавковыми сближают небольшое семейство вахтовые (Мепу-
anthacead^ представленное во флоре СССР немногими видами. Широко
распространена трифоль, или вахта (Menyanthes trifoliate часто встречаю-
щаяся на болотах в лесной зоне и хорошо отличаемая по крупным тройча-
тым листьям и очень заметным белым цветкам, собранным в довольно густые
колосовидные соцветия. Лекарственное растение.
Семейство кутровые (А росу паевое). Лазающие древеснеющие рас-
тения (лианы, часто крупные), реже прямостоячие деревья и кустарники,
еще реже травы и стеблевые суккуленты. Листья обычно супротивные,
всегда цельнокрайние; прилистники очень редки. Цветки обоеполые, пра-
вильные. Численные отношения в цветке в общем идентичны горечав-
ковым.
Завязь верхняя или полунижняя, причем плодолистики нередко вторич-
но разъединенные, и, таким образом, в цветке оказываются как бы две завя-
зи; ценокарпность гинецея при этом сохраняется благодаря наличию единого
или только внизу раздвоенного столбика (рис. 342). Пыльцевые зерна обыч-
но обособлены друг от друга и лишь редко остаются связанными в тетрады.
выс
II1 с * у
РАСТЕНИЯ
Рис. 340. Горечавка (Gentfana pneu-
monanthe)i
1 — общий вид; 2 — диаграмма цветка
Рис. 341. Золототысячник (Erythraea
centaurium):
2 — общий вид; 2 — разрез цветка
UII
Itl
Плоды или в виде двух листовок, или коробочки, иногда ягоды. Семена
крылатые, часто снабженные пучком волосков.
Кутровые — обширное семейство, содержащее около 2000 видов. Рас-
пространены они преимущественно в тропиках, лишь небольшая часть на-
селяет Внетропические страны, причем более теплые районы. Некоторые
кутровые имеют практическое значение. Из африканских (Landolphia — ли-
аны), некоторых южноамериканских (Допсоттгга) и юго-восточноазиатских
(ЖШогде&Ьуа) представителей семейства получают каучук.
Лубяные волокна кутровых отличаются большой прочностью и при
значительной длине элементарного волокна представляют практический
интерес. На этом основано использование с давних времен кендыря (род
Аросупшн, рис. 343). Корни й корневища кендыря богаты глюкозидом цима-
рином — заменителем строфантина1. Последний получают из семян афри-
канских лиан строфантов (Strophanthus hispidus и др.) и используют ядо-
витый сок строфантов для отравления стрел. Важное медицинское приме-
нение получила рауволфия (Rauvolfia) — пантропический род, отсутствую-
щий в Австралии. Некоторые виды богаты алкалоидами. Из Rauvolfia по-
лучают резерпин, применяемый при лечении гипертонической болезни.
Южноамериканское дерево квебрахо (Aspidospermum quebracho-bianco)
славится, как и квебрахо из семейства Anacardiaceae, высоким содержанием
дубильных веществ — 27% в коре и несколько меньше в желтовато-белой
древесине. В комнатной культуре распространен олеандр (Neriurn oleander,
рис. 344), растущий в средиземноморских странах вдоль ручьев и речек,
I Г. Iг. г.
1 Этот глюкозид используется для борьбы о острой и хронической сердечно-сосудис-
той недостаточностью^
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
подобно нашим ивам. Содержит глюкозид, применяемый
аналогично строфантину, и антибиотик, по силе дейст-
вия превосходящий пенициллин. Нередко культивиру-
ют барвинок (Vinca).
Семейство ластовневые, или ласгйочниковые,
(Asclepiadaceae) включает растения разнообразного внеш-
него облика: лианы, реже прямостоячие кустарники или
деревья, травы, наконец, суккуленты. Листья супро-
тивные (редко очередные, мутовчатые), простые, цель-
нокрайние, обычно без прилистников. Цветки (рис. 345)
сравнительно мелкие в цимозных соцветиях, которые
располагаются главным образом внепазушно, отходя от
стебля между двумя листьями. Численные отношения в
цветке те же, что и в двух предыдущих семействах: ча-
шелистиков, лепестков и тычинок по пять, плодолисти-
Рис. 342. Про-
дольный разрез
нижней части
ков два. Подобно - растениям из семейства кутровых, Цветка нутровых
ластовневые (рис. 346) имеют плодолистики, обособлен- столбиков~
ные в нижней части и столбик с утолщением разнооб- рилкцд)*
разной формы (рыльцевая головка). Завязь верхняя,
редко полунижняя. Строение цветка ластовневых
осложняется добавочным венчиком или короной,
образуемой
местными
разрастания
I.’
венчика
торых случаях смешанного происхождения).
или тычинок (в неко-
Наиболыпее своеобра-
зие цветкам ластовневых придает гиностегий — продукт
соедине-
1ГЛ
ния тычинок и верхней части пестика. Пыльники плотно прилегают
к рыльцевой головке пятигранной формы и даже срастаются с ней, при этом
между пыльниками остаются щели, соответствующие по положению ребрам
головки. Из затвердевающих выделений специальных железистых клеток
: на каждом ребре головки формируются особые тельца — транслят о-
р ы. Они снабжены двумя расходящимися в стороны придатками — рычаж-
гг.
«— Рис. 343. Кендырь:
1 — верхняя часть растения с соцветием;
2 —цветок; 9— плод с освобождающимися
семенами
Рис. 344. Олеандр (Nerium oleander)
478
ВЫСШИЕ РАСТЕНИЯ
Рис. 345. Диаграмма цветка ваточника (к — эле-
менты коронки, п—пыльники, пгн — пыльцевые
гнезда с по л линиями, м — трансляторы)
Рис. 346. Цветок ваточника (Asclepias corn out И):
4-общий вид цветка (вн — венчик, к — коронка); г— ко-
ронка и гиностегий (к — коронка, п — пыльник, р — рыль-
це, зав — завязь); 3 — пестик с висящими на нем палли-
ниямн; 4 — липкое тельце с поллиннями
Рис. 347. Эхиднопсис
(Echidnopsis cereiformis):
суккулентное растение
семейства ластовневых,
напоминающее кактус
ками. В каждом пыльнике только два пыльцевых гнезда, причем в каждом
из них пыльца слипается в комочек — поллиний — с клейкой ножкой,
I г. г. г.
которая и слипается с одним из рычажков транслятора. Последний пред-
ставляет как бы маленькие щипчики, цепляющиеся за ножку насекомого,
В результате насекомое уносит с собой два поллиния, принадлежащие со-
седним пыльникам. Изменение в положении поллиниев во время полета
насекомого обеспечивает попадание их на воспринимающую поверхность
рыльца1. У ластовневых, не имеющих поллиниев, пыльцевые зерна соедине-
ны в тетрады и пыльца порошковатая. В этом случае трансляторы представ-
ляют ложкообразные или бокаловидные тельца, куда высыпается пыльца
из пыльников соседних тычинок. В основании такого транслятора имеются
ножка и липкое тельце, обеспечивающее прилипание к лапке насекомого.
Плоды у ластовневых сухие, раскрывающиеся по брюшному шву. Семена
в большинстве случаев с пучком шелковистых волосков.
Семейство ластовневые, содержащее около 2000 видов, представлено
в тропической зоне и в субтропиках, однако немногие представители дости-
гают умеренных широт. Большинство (около */3) родов связано со Старым
Светом, причем центр видового разнообразия находится в Южной Африке.
Здесь широко распространены суккулентные формы (рис. 347). Особенно
1 Подлипни ластовневых представляют замечательный пример конвергенции с ор-
хидными, которые не имеют даже отдаленных родственных связей с ластовневыми.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЬЙ
479
типичен род Stapelia, около 100
видов которого обитают преиму-
щественно в Капской флористи-
ческой области. Сочные стебли
стапелий имеют выступающие
продольные ребра, или поверх-
ность их покрыта, как панци-
рем, примыкающими друг к
другу подушечками. Эти рель-
ефные образования возникают
вследствие разрастания и взаим-
ного давления друг на друга
оснований листьев.
Своеобразны южноазиатские
дисхидии (Dischidia, рис. 348).
К этому роду, объединяющему
около 50 видов, относятся эпи-
фиты с небольшими супротивными
ассимилирующими листьями или
безлистные. Они прикрепляются к
деревьям многочисленными при-
даточными корнями. Замечатель-
ны специальные метаморфизиро-
ванные листья дисхидий, имею-
щие характер кубков с узкими
входными отверстиями и с толстой
кутикулой на наружной поверх-
ности. В них собираются вода и
гумус, заносимый, вероятно, му-
равьями. Сюда направляются при-
даточные корни растения. Инте-
ресно, что эпидермис, выстилаю-
щий внутреннюю поверхность
кубка, богат устьицами, что дает
повод предполагать, что водяной
пар, выделяемый внутрь кубка,
конденсируется там и образовав-
шаяся вода может быть вновь
использована растением.
Во флоре СССР семейство
ластовневых насчитывает около
40 видов. Наиболее богат вида-
ми род ластповень (Antitoxiciun,
или Vince toxicurn,). На Кавказе
растет средиземноморская неболь-
шая лиана (вьющийся кустарник)
обвойник (Periploca graced). На
Украине и на Кавказе довольно
широко распространен как сор-
няк ваточник (Asclepias cornuti,
A. syriaca), казавшийся перспек-
тивным как каучуконосное, пря-
дильное и масличное растение,
однако не нашедший до сего вре-
мени применения.
Из семейства порядка Genti-
anales следует упомянуть еще
сравнительно небольшое тропи-
Рис. 348. Дисхидия (Dischidia rafflesiana):
1 — побег с листьями и соцветием (л — лист обыч-
ный, к —кубковидный); 2 — разрез листа-кубка
(пх — придаточшлй корень)
Рис. 349. Чилибуха (Strychnos nux-vo-
mica):
1 — цветущая ветвь: 2 — плод: 2 — разрез плода
(с —семена): 4—семи
480
Н 11 Г. I я
ВЫСШИЕ РАСТ
ческое семейство Loganiaceae, к которому относится чилибуха,
или рвотный орех (Strychnos nux-vomica, рис. 349). Семена этого расте-
ния используются в медицине, как содержащие
(сильный яд), терапевтические дозы которого возбуждают центральную нерв-
ную систему, тонизируют скелетную мускулатуру и сердце, стимулируют
процессы обмена. Более значительные дозы вызывают отравление. В корнях
чилибухи и других видов рода Strychnos содержится алкалоид курарин —
действующее начало яда кураре, который употреблялся индейцами Южной -
Америки для отравления стрел. При попадании в кровь кураре вызывает
поражение окончаний дыхательных нервов. и поперечнополосатой муску-
латуры, что влечет паралич и смерть от остановки дыхания.
алкалоид Стрихнин
2-0 ПОРЯДОК — МАСЛИНОЦВЕТНЫЕ (OLEALES, ИЛИ LIGUSTRALES)
Цветки маслиноцветных четырехкруговые. Венчик правильный, акти-
номорфный, большей частью четырехчленный, спайнолепестный, иногда
очень глубоколопастной и кажущийся раздельнолепестным. Тычинок обыч-
но две. Плодолистика два. Завязь верхняя, двугнездная, с одной, редко с
большим числом семяпочек в гнезде.
Порядок содержит одно семейство маслинных.
Семейство маслинные (Oleaceae). Это деревья и кустарники, редко
полукустарники. Листья простые, обычно цельные, реже перистосложные,
без прилистников. Чашечка в основном маленькая. Венчик спайнолепест-
ный, четырехчленный (иногда, однако, может быть и многолепестным). Ты-
чинок две, редко больше. Гинецей двучленный, ценокарпный; пестик с верх-
ней двугнездной завязью, содержащей в каждом гнезде 1—2 семяпочки,
редко более (рис. 350). Плоды — костянки, ягоды, коробочки или орехо-
видные. К маслинным относится около 400 видов, распространенных в уме-
ренных и теплых областях, преимущественно в Южной и Восточной
Азии.
Экономически наиболее важное растение маслина (Olea еигораеа,
рис. 351) — сравнительно невысокое дерево, 3—4 ж, иногда до 12 ж высоты,
достигает 2 м и даже 4 м в диаметре у основания ствола. Отличается боль-
шой долговечностью (300—400 лет, отдельные деревья до 2000 лет). В мезо-
карпии костянок содержится 25—40% (до 75%) масла. Масло высшего
качества (прованское), употребляется в пищу и получило широкое примене-
ние в консервной промышленности. Повторное прессование дает второ-
сортное пищевое и невысыхающее техническое («деревянное») масло. Пло-
ды — маслины — употребляются в
Маслина — древнее культурное растение, распространенное главным об-
разом в странах Средиземноморья, где сосредоточены наиболее крупные
плантации этой культуры.
В СССР культура маслины распространена на Кавказе и в Крыму, а
также развивается на западе Туркмении, где имеются необходимые для
нее экологические условия: обильная инсоляция
и морские бризы. Древесина маслины тяжелая
(плотность 0,8—1,1), используется для изготовле-
ния дорогих столярных изделий.
Виды рода ясень (Fraxinus) — деревья,
произрастающие в Восточной Азии, Северной
Америке и в Средиземноморье. В СССР извест-
но более 10 видов, из которых широко распро-
странен обыкновенный ясень (Fraxinus excelsior,
рис. 352) — характерное дерево широколиствен-
ных лесов европейской части СССР (до Волги).
Древесина обыкновенного ясеня ’ ценится за
прочность, легкость при обработке и красивую
текстуру. Кору используют как дубитель и как
11'1'
I гл и
или
II
ill
Рвв. 350. Диаграмма
цветка сирени
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
ОД
Рис. 351. Маслина (Olea europaea):
1 — цветущая ветка; 2 — цветок; з — развернутый венчик; 4 — продольный разрез завязи; 5 — илсд;
в — продольный разрез илода
источник черной краски. Кроме того, в коре содержатся лекарственные
начала (глистогонные средства и вещества, служащие суррогатом гиттия
Широкой известностью пользуется род сирень (Syringa) — кустар-
ники с цельными, редко с перистораздельными листьями. Разводят с деко-
ративными целями. В СССР наиболее часто встречается в культуре обыкно-
венная сирень (S. vulgaris). Реже культивируют венгерскую (S. Josikaea) и
персидскую сирень (5. persica). Декоративное значение имеет также бирючи-
на {Ligu&trum vulgare), растущая на юге европейской части СССР (в Крыму)
и на Кавказе.
К этому же семейству относится род жасмин (Jasminium), распро-
страненный преимущественно в тропических и субтропических широтах,
главным образом в Юго-Восточной Азии. Цветки жасмина обладают силь-
ным запахом1.
3-Й ПОРЯДОК — ТРУБКОЦВЕТНЫЕ (TUBIFLORAE)
Цветки трубкоцветных четырехкруговые вследствие наличия только
одного круга тычинок. Околоцветник двойной со спайнолепестным венчи-
ком, форму которого определяет в основном актиноморфность или зигомор-
фность цветков. Тычинок 5 или вследствие редукции меньше. Они прикреп-
лены к трубке венчика, иногда в ее основании, между долями венчика. Ги-
нецей в некоторых семействах из 5—3. чаще же из 2 плодолистиков. Завязь
1 Под названием жасмина часто в садах разводят чубушник (Philadelphus coronata)*
относящийся к семейству камнеломковых*
16-602
ВЫСШИЕ PACT
и: f'
Рис. 352. Ясень (Fraxinus excelsior):
цветущая ветвь; 2 — обоеполый цветок; 5 — пес-
тичный цветок; 4 — тычиночный цветок; 5—вепь о
планами и листьями
гнезда
лож-
иногда
Плоды
деревья.
порядок
для которых
направленность
лом гнезд, равным числу пло-
долистиков; у .многих
завязи подразделены
ными перегородками;
завязь одногнездная,
разнообразны: коробочки, яго-
ды, орешки. Преимуществен-
но травянистые растения, ре-
же кустарники
Листья простые, без прилист-
ников.
Этот обширный
делят на несколько (до 6) под-
порядков, возводимых не-
редко в ранг порядков. Наибо-
лее крупные из них — Convol-
vulinae, Boragininae, Verbeni-
nae> Solaninae,
характерна
микропиле. Первые два объ-
единяются преобладанием ак-
тиноморфности цветков, отли-
чаясь один от другого тем, что
у Boragininae наблюдается под-
разделение каждого из гнезд
завязи на два. В подпорядках
Verbeninae и Solaninae господ-
ствует выраженная, у многих
резкая, зигоморфность цветков.
Отличием этих двух подпоряд-
ков между собой может слу-
жить характерное для Verbeni-
j»г г.
nae подразделение каждого из
гнезд завязи на два при .ма-
лом (4) числе семяпочек (у Solaninae завязь обычно двугнездная с большим
числом семяпочек в каждом гнезде).
Следующие три семейства относятся к подпорядку Convolvuli-
пае.
Семейство вьюнковые (Convolvulaceae). Обычно травы или полу-
кустарнички и кустарнички. Стебли часто вьющиеся. Листья простые, иног-
да лапчато- или перисторасчлененные, как правило, без прилистников.
Цветки актиноморфные, обоеполые, с пятью свободными или только у осно-
вания сросшимися чашелистиками, которые большей частью остаются при
плодах. Венчик спайнолепестный, у большинства довольно крупный, во-
ранковидный, трубчатый или колокольчатый, пятилепестный, но только
слегка лопастный или ; —
ших к основанию венчика и чередующихся с лепестками. Гинецей состоит
из двух (редко из 3—5) плодолистиков. Завязь верхняя, у большинства
f ГЛ II
двугнездная, с двумя семяпочками в каждом гнезде; иногда вследствие обра-
зования ложной перегородки становится четырехгнездной. Плоды — в ос-
новном коробочки или распадающиеся на односемянные орешки.
Иэ этого обширного (около 1600 видов) семейства в СССР широко рас-
пространен сорняк — полевой вьюнок (Convolvulus arvensls, рис. 353), обиль-
но размножающийся корневыми отпрысками и семенами. Вследствие глу-
бокого проникновения корня и способности его образовывать придаточ-
ные почки и побеги даже на небольших отрезках трудноискореним. Опле-
тая стебли злаков, затрудняет их уборку. Не все вьюнки имеют вьющиеся
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЯННЫЕ
стебли. Многие из них представляют
распростертые или прямостоячие
травы или колючие полукустарнички.
Близок к вьюнку род повой
(Calystegia), один иэ видов которого
(С. septum) обычен по берегам озер и
рек в зарослях кустарников. Обра-
щает на себя внимание крупными (до
беж) воронковидными цветками белой
или розовой окраски, в основании с
двумя прицветниками, покрывающи-
ми чашечку. Вьющимися стеблями и
крупными цветками повой напомина-
ет разводимый в СССР декоративный
однолетник — ипомею (Ipomoea purpu-
rea) с синими, фиолетовыми и красными
венчиками (трубка венчика белая).
Родина ипомеи — тропическая Аме-
рика.
К р о д у Ipomoea относится важ-
ное пищевое растение — батпатп,
Рис. 353. Вьюнок полевой (Convol-
vulus arvensis):
1 — цветущая ветвь; я — продольный раз-
рез цветка; з—дваграша цветка
или
сладкий картофель (Z. batatus), — травянистое растение родом из Северной
Америки или иэ Вест-Индии. Выращивают его ради богатых углеводами
утолщенных боковых корней (клубней). Батат употребляют в пищу в пече-
ном, вареном и жареном виде. Разводят преимущественно в жарких странах,
ио культивируется также и за пределами тропических и субтропических
широт. В СССР культура батата возможна нА юге Украины, в Закавказье
и на юге Средней Азии.
Семейство повиликовые (Cuscutaceae). Сюда относится только
один род повилика (Cuscuta, рис. 354). Исключительно растения-паразиты,
с тонкими стеблями, которые обвивают побеги растения-хозяина и приса-
сываются к ним гаусториями (предположительно), метаморфизированными
придаточными корнями растения. Листья у повилики редуцированы до
мелких чешуек, а мелкие, но вполне развитые цветки собраны в тесные ди-
хазиальные соцветия. Мелкие семена, которых на одном растет
образу-
11' 111
11I и
:
ются тысячи, а иногда и десятки тысяч, могут годами сохранять всхожесть.
Проросток повилики прикрепляется корнем к почве; стебелек делает кру- *
говые движения, благодаря чему «находит» растение-хозяина и обвивает
его. Корень, лишенный корневого чехлика, рано отмирает/и связь повш
с почвой прерывается.
В большинстве случаев повилика может селиться на равных растениях*
Но некоторые виды ее довольно тесно связаны с определенными хозяевами.
Например, льняная повилика (Cuscuta epilinum) паразитирует на культур-
ном льне, с которого иногда переходит на сорняки посевов льна. Повилика
приносит большой вред. Поселяясь на молодой сахарной свекле, повилика
может снизить содержание сахара более чем на 2,5%; у льна, пораженного
-повиликой, урожай тресты снижается на 20—30%, а урожай семян может
уменьшиться даже на 70%. Наблюдались случай отравления повиликой
сельскохозяйственных животных. , t
Семейство синюховые (Polemoniaceae) близко к вьюнковым. Отли-
чается от них трехчленной и, как правило, трехгнездной завязью, а также
венчиком, не образующим складок в бутоне. Семяпочек в каждом гнезда
завязи много или одна. Относящиеся сюда растения, главным образом травы
с простыми, нередко расчлененными листьями, распространены преиму-
щественно в Северной Америке (около 300 видов).
В СССР обычна еолубая синюха (Polcmonium coenileum, рис. 355). К
этому же семейству относится род флокс (Phlox), виды которого приуроче-
ны к Северной Америке и лишь одни — Ph. slblrtca — в диком состоя
; I > г
16»
.484
1 _
выс
игл
ЕГРАСТЕНИЯ
Рис. 354. Повилика:
1 — цветущее растение па хмеле; г — цветок: з — про-
дольный разрез цветка; 4 — диаграмма цветка
Рис. 355. Синюха (Polemonium coeruleum):
1 — нижняя часть растения; г — соцветие; з—циа-
грамма цветка
встречается в СССР (Сибирь и
Дальний Восток). Многие флок-
сы ценятся как хорошие де-
коративные многолетние рас-
тения . В садовой культуре
распространен РЛ. paniculata.
Из поднорядка Boragininae
отметим одно семейство.
Семейство бурачни-
ковые (Boraginaceae). Крупно е
семейство, содержащее около
2000 видов, обитающих как во
внетропических, так и в тропи-
ческих странах. Среди тропи-
ческих бурачниковых есть
древесные растения и кустар-
ники, внетропические — глав-
ным образом травы с цилин-
дрическим стеблем и спираль-
но расположенными простыми
листьями, без прилистников.
Распространено жесткое опу-
шение. Цветки собраны в завит-
ки (рис. 356), которые улитко-
образно скручены до цвете-
ния, а по мере расцветания
постепенно распрямляются,
становясь похожими на кисть.
Нередко наблюдается двойной
завиток. Цветки актиноморф-
ные, реже зигоморфные, из
четырех-, пятичленных (редко
четырехчлепных) кругов. Чаше-
листики свободные, или, чаще
чашечка спайнолистная, но? в
большей или меньшей степени
расчлененная на доли; чашечка
сохраняется при плодах, иног-
да при этом разрастаясь. Вен-
чик то с длинной (как у меду-
ницы), то с короткой (как у
незабудки) трубкой. Форма его
довольно разнообразная, вплоть
до зигоморфной. Вход в вен-
чик нередко прикрыт распо-
ложенными в зеве чешуйча-
тыми выростами лепестков,
затрудняющими проникновение
к нектарнику «нежелательных»
насекомых. Тычинки всегда
сросшиеся с трубкой венчика
и имеют то большую, то мень-
шую, но всегда небольшую
свободную часть тычиночной
нити. Гинецей из двух плодо-
листиков, расположенных в
медианной плоскости и образу-
ющих верхнюю^ вначале дву-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис. 356. Бурачниковые:
1 — цветок медуницы (Pulmonarla obscura): а — общий вид; б — продольный разрез цветка; в —
спш; 2 —диаграмма цветка анхуэы (Anchusa officinalis); з — окопник (Symphytum officinale): а — со*
цветке; б — продольный разрез цветка; 4 — плод чернокорня (Cynoglossum officinale); 5 — плод воробей-
ника (Lithospermum officinale)
гнездную, завязь с двумя семязачатками в каждом гнезде. Впоследствии
(довольно рано) у многих бурачниковых каждое гнездо завязи
делится ложной перегородкой на два, благодаря чему завязь становится
четырехгнездной и содержит по одной семяпочке в каждом гнезде. При этом
стенки каждого из четырех гнезд завязи разрастаются, и она приобретает
четырехдольчатое (четырехлопастное) строение, а столбик вйходит из'углуб-
ления между четырьмя дольками завязи (так'называемый гино баз и-
ческий столбик). Реже завязь остается цельной и столбик занимает
верхушечное положение. Столбик несет на верхушке одно головчатое или
слегка выемчатое рыльце, иногда рылец два, еще реже — четыре. Часто
в основа]
: I: а
завязи развит подпестичный диск; При созревании плодов доль-
ки завязи отделяются друг от друга в виде четырех односемянных орешков.
У тропических форм бывают сочные, костянкообразные (нераспадающиеся)
плоды.
В СССР известно более 350 видов бурачниковых. Небольшая часть их
(около 25 видов) принадлежит подсемейству Heltotropoldeae,которое
связано в своем распространении с тропическими и субтропическими широ-
тами, как и подсемейства CordiotdeaeиEhretloldeae, отсутствующие
в СССР. У Heliotropoideae завязь остается цельной и столбик располагается
на ее верхушке. На территории Советского Союза из этого подсемейства
встречаются главным образом виды рода гелиотроп (Heliotropium), расту-
щие преимущественно в Средней Азии. Некоторые гелиотропы, особенно
Н. peruvianum (Эквадор, Перу), вошли в культуру как декоративные
ИГ
ароматические растения.
Все прочие бурачниковые нашей флоры относятся к подсемейст-
в у Boraginoideae, которое характеризуется четырехдольчатой завязью и
гинобазическим столбиком. Они занимают самые различные местообитания,
будучи иногда характерными участниками тех или иных группировок расти-
4
выс
и * *. t
РАСТЕНИЯ
Рис. 357. Медуница
(Pulmonaria obscura):
1 — цветущее растеняе;* S—
кфвяодайме лкявд; а— цве-
ток
тельности. Так, например, для широколиственных лесов характерна лсе-
Synutfa неясная (Puhnonaria obscura, рис. 357) — одно из наиболее рано
цветущих растении лесной зоны, замечательное своей способностью расти
под снегом. Цветки медуницы гетеростильные, что, как известно, обеспечи-
вает перекрестное опыление.
Характерно изменение окраски венчика у медуницы (свойственное и
другим бурачниковым) от красной при распускании цветков до голубой ко
времени отцветания.
Широкой известностью пользуются незабудки — болотная
(Myosotis pakistris) и лесная (М. silvatlca).
Некоторые виды незабудок, например альпийская (Af. alpina), культи-
вируются как декоративные растения.
j Среди бурачниковых нередки сорные и мусорные растения, например
чернокорень (Cynoglossum officinale). Он имеет грязно-темно-красные цвет-
ки и обладает неприятным, «мышиным» запахом, благодаря чему его счита-
ют (не без основания) средством против мышей. Плодики его покрыты креп-
кш якорными шипнками, что встречается часто и у других бурачнико-
вых (например, У видов липучки — Lappula) — приспособление к распро-
странению животными. Наряду с этим есть бурачниковые и с совершенно
голыйи и гладкими плодиками. Так, у лекарственного воробейника (Litho-
spermum officinale) плодики с прочным околоплодником, глянцевые и белые,
как камешки. Из. числа сорняков можно назвать еще кривоцветы (Lyco-
pets) и обыкновенный синяк (Е. vulgare), имеющий крупные синие цветки,
косоворонковидный, зигоморфный венчик без чешуек в зеве, тычинки не-
I:»
равной длины, столбик на конце двурасщепленный (у Е. rubrum — цель-
ный)* каждая веточка которого с небольшим головчатым рыльцем.
ОТДЕЛ
11; i:
ТКОЕЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Полезных растег
г г.
среди бурачниковых мало. Кроме декоративных
и медоносных, можно назвать еще алканну {Alcanna tinctoria) — растение
родом из Средиземноморья, корни которого дают буро-фиолетовую краску,
применяемую в технике и микроско
Следующие два семейства — вербеновые и губоцветные — входят
в подпорядок Verbeninae. Они настолько близкие, что некоторые под-
семейства иэ вербеновых иногда относят к губоцветным. В нашей флоре
более обычны губоцветные.
Семейство губоцветные^ или лснотковые (Labtatae, Lamiaceae).
Травы, полукустарники или кустарнички, очень редко (тропическая Южная
Америка) небольшие деревья и лазающие растения. Листья простые, без
гг
III
прилистников, цельные или в различной степени расчлененные, почти всег-
да супротивные, редко мутовчатые. Стебли четырехгранные, причем листья
прикрепляются к граням (а не к ребрам). Для губоцветных характерна
способность вырабатывать эфирные масла. Цветки губоцветных в цимоз-
ных соцветиях (рис. 358), которые представляют или трехцветковые диха-
зии, или более сложные образования — двойные завитки; в этом случае
оси второго порядка дихазия продолжают ветвиться по типу завитков. Двой-
ные завитки (как и дихазии) занимают пазушное положение относительно
измененных (прицветники) или обычных вегетативных листьев. Вследствие
супротивности каждая пара двойных завитков (или дихазиев) образует как
- бы мутовку (рис. 359). Мутовки могут быть достаточно удаленными друг от
друга, но нередко междоузлия в верхней цветоносной части стебля не вытя-
гиваются и цветочные мутовки остаются сближенными, в результате полу*
чается сложное соцветие колосовидного или головчатого характера (в за-
висимости от числа сближенности мутовок). Если цветоносная часть стеб-
ля ветвится, то это еще больше усложняет соцветие и оно становится более
или менее метельчатым.
Цветки губоцвет
: I г
(рис. 360), очень
разнообразные по величине и
окраске, почти всегда зигоморфные и имеют общий план строения, опреде-
ляемый формулой: К(5) С(5) A4G(2)*
Чашечка в большинстве случаев — пятичленная, отгиб ее имеет пять
равных или неравных зубчиков. Иногда она становится двугубой: с верхней
губой иэ трех и нижней иэ двух зубчиков. В некоторых случаях задний зуб-
м: гг
чик недоразвивается и чашечка становится четырехзубчатой. Венчик пяти-
лепестный, двугубый, обычно с двумя лепестками в верхней и с тремя
в нижней губе (%). Верхняя губа бывабт вогнутой или плоской, у некого-
соцве*
разрез
Рис. 358. Строение ложной мутовки
тия глухой крапивы (о — поперечный
главной оси соцветия; *1 и — супротивно
расположенные кроюпще листья; и я —
содержащие каждый
Рис. 359. Диаграмма цветка белой
глухой крапивы и двух завитков
около него, образующих в сово-
купности половину ложной мутовки
(один двойной завиток)
по семи цветков)
488
ВЫСШИЕ РАСТЕНИЯ
Рис. 360. Цветки губоцвет-
ных:
1 — белая глухая крапива: а —
общий вид сбоку; б — продоль-
ный разрез; в — вид спереди с
отвернутой верхней губой; 2 —
пустырник в развернутом виде;
8 — чабрец: а — общий вид: б —>
завязь с нижней частью стол-
бика; 4 — мелисса; у — живучка
ее редукция. Из трех ле-
почти полная
рых представителей
пестков нижней губы средний обычно самый крупный. Наряду с таким обыч-
ным подразделением лепестков на две губы встречаются и другие (4/v °/»)-
Иногда в результате полного слияния лепестков верхней губы венчик стано-
вится как бы четырехчленным (мята) и почти правильным. Трубка венчика
в цветке губоцветных всегда выражена. Андроцей чаще всего четырехчлен-
ный, возникший из пятичленного вследствие редукции пятой (задней) ты-
чинки, существование которой у предков губоцветных доказывается появле-
нием у некоторых видов стаминодия между лепестками верхней губы.
Тычинки обычно двусильные, причем обыкновенно две верхние бывают коро-
че нижних. У некоторых губоцветных наблюдается редукция не только зад-
ней, но и двух ближайших к ней боковых тычинок, от которых сохраняются
(но не всегда) стаминодии. Плодолис- .
тики расположены в медианой нлдбкоети цветка/' "
наблюдается
каждом гнезде. В дальнейшем так называемой ложной перегородкой, вырас-
тающей от спинки плодолистиков внутрь, к центру завязи, каждое гнездо
Подразделяется па два, благодаря чему завязь становится практически
хчетырехгнездной-О^одной семяпочкой в каждом гнезде. По мере их роста
происходит разрастайие-стенок завязи, особенно вокруг семяпочек. В итоге
вполне сформированная завязь оказывается четырехдольчатой (четырех-
лопастной), причем каждая долька содержит одну семяпочку. Основание
столбика оказывается пог ывжду дольками, завязи
til
(столбик __ . Впрочем, у некоторых губоцветных, вероятно
близких к вербеновым, столбик не вполне гинобазический и даже почти
конечный (терминальный) по положению. Верхушка столбика редко бывает
цельной, обычно она двураздельная. Воспринимающие пыльцу сосочки
покрывают лишь концевые части ветвей столбика и только с внутренней
их стороны. В основании завязи находится подпестичный диск. Созреваю-
щий плод распадается на четыре орешка (соответственно числу долек завя-
зи), остающиеся сначала лежать на дне чашечки, разрастающейся ко вре-
мени плодоношения, позднее выпадающие. Семена без эндосперма, или он
мало развит.
Семейство губоцветных — одно из крупнейших. Оно содержит око-
ло 3200—3500 видов. Губоцветные известны на всех материках, но особенно
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
489
обильны в средиземноморских странах и в горных странах субтропиков.
В СССР известно около 950 видов.
Экономическое значение губоцветных определяется главным образом
их способностью вырабатывать в большом количестве разнообразные эфир-
ные масла. Поэтому многие из них специально культивируют с целью по-
лучения сырья для парфюмерной промышленности, например розмарин
(Rosmarinus officinalis), лавандула (Lavandula spica), мускатный шалфей
(Salvia sclarea), молдавский змееголовник (Dracocephalum moldavicum)^ ко-
шачий котовник (Nepeta cataria var. citriodon),
Семена некоторых губоцветных содержат ценные жирные масла. В свя-
зи с этим введена в культуру перилла, или судза (Perilla ocimoides).
Масло периллы, содержание которого в семенах достигает почти 50%,
скоро высыхающее и применяется в лакокрасочной промышленности, при
изготовлении технических тканей и для других целей. Сходное использо-
вание получило масло грузинской ляллеманции (Lallemantia iberica). Быстро
высыхающее масло ляллеманции вдет на приготовление высококачествен-
ной олифы, для смазки частей машин и т. п.
Многие губоцветные издавна используются в народной медицине^ но
немало их признано и
шении растений мята (Mentha), В культуре распространены перечная мята
(М, piperita), колосистая (М. spicata), полевая (М. arvensis var. piperascens)
и др. Мятная настойка (мятные капли) и мятное масло широко применяют
в фармацевтической промышленности. Важное значение имеет получаемый
из мятного масла ментол, обладающий местнообезболивающим и рефлек-
торво-сосудисторасширяющим действием (входит в состав известного
сердечного средства валидола). Мятное масло используют также в ликеро-
водочном, кондитерском в мыловаренном производствах.
Источником тимола, обладающего антисептическим действием и приме-
няемого для полосканий в зубоврачебной практике и в качестве противо-
глистного средства, служит тимьян обыкновенный (Thymus vulgaris) родом
из Средиземноморья. Виды тимьянов, очень многочисленные в СССР, нахо-
дят применение как эфиромасличное сырье.
Для лечебных целей издавна употребляют листья лекарственного шал-
фея (Salvia officinalis). В последнее время вошли в лечебную практику ви-
ды пустырника (Leonurus), препараты которого понижают кровяное дав-
ление и действуют на нервную систему аналогично валерьяне. Среди губо-
цветных немало хороших медоносов, которые по этой причине культиви-
руются: мелисса (Melissa officinalis), змееголовник (Dracocephalum moldar
vicum), иссоп (Hyssopus officinalis) и др. Многие губоцветные используются
разносторонне. Например, душица (Origanum vulgare) — ароматическое
(одеколоны, мыло) и медоносное растение (нектаропрбдуктивность свыше
330 кз с 1 га), а семена содержат жирные масла. Цветки душицы окра
вают шерсть в оранжево-красный цвет, а вместе со стеблями с добавлением
нг
железного купороса дают черную окраску.
Некоторые губоцветные очень красивы благодаря крупным и приятно
окрашенным цветкам, например луговой шалфей (Salvia pratensis), широко
распространенный в северной части степной зоны (рис. 361). Есть виды шал-
фея, вошедшие в культуру как декоративные. Среди них выделяются как
очень эффектные S. cocctnea и S. splendens — первый с пунцовыми, второй
с шарлахово-красными цветками (реже иной окраски). Индийский по про-
исхождению колеус (Coleus hybridus) используют как комнатно-оранжерей-
ное горшечное растение, а также высаживают в горшках на клумбы; есть
сорта с листьями почти черными, темно-фиолетовыми, темно-красными
с желтой каймой и т. п.
Семейство вербеновые (Verbenaceae) представлено преимуществен-
но в тропических и субтропических странах. Это как Древесные, так и тра-
вянистые растения, имеющие много признаков сходства с губоцветными
а отличающиеся от них терминальным столбиком^ цельной или слегка ло~
ВЫСШИИ РАСТЕНИЯ
Рис. 361. Шалфей луговой (Salvia рга-
tensia)
паствой аавязью и плодами — хво-
мися на орешки. Ароматические
растения средн вербеновых редки. -
Вербеновых около 2600 видов.
Важное промышленное значение
имеет тиковое (тгйковое) дерево (Тес*
tona grandls), растущее в странах'
Южной и Юго-Восточной Азии; отли-
чается плотной, но хорошо подда-
ющейся обработке древесиной, стой-
кой к воздействию термитов, к исти-
ранию, к морской воде; ценится в
корабельном деле и при строитель-'
стве портовых сооружений.
Биологически интересен род
A eicennia, виды которого входят в
состав мангровых зарослей. Харак-
терно образование так называемых
«дыхательных» корней (пневма-
тофоров). Прорастание семян
происходит на материнском
нии («живородность»).
Из числа растущих вне
раете-
иг.
тропи-
ков можно назвать - лекарственную
вербену (УеяЬепа offidnelii)—травя-
нистое растение, имеющее- деко-
ративное значение.
Подпорядок Solaninae, в
отличие от предыдущих, содержит
довольно большое (до 13) число
семейств, иэ которых основное вин-
мание сосредоточим на пасленовых и норичниковых.
Семейство пасленовые (Solanaceae). Это травянистые растения,
полукустарники и кустарники (иногда лазающие), а в тропиках даже не-
большие деревья. Листья пасленовых простые, цельные или расчлененные,
без прилистников, в основном очередные. Соцветия цимозные, в большин-
стве случаев завитки. Цветки правильные или зигоморфные (рис. 363). Ча-
шечка, венчик и андроцей пятичленные. Тычинки (обычно пять) располо-
жены между лепестками. Гинецей'иэ двух плодолистиков, помещающихся
косо по отношению к медианной плоскости цветка. Завдзь верхняя, дву-
гнездная или, реже, вторично четырех-пятигнездная, с мноточисленнтш
семяпочками в каждом гнезде. Плаценты массивные, прииремлаштые к
перегородке, отделяющей гнезда. Столбик один, простой, с двулопастным
или двураздельным рыльцем. Плоды — ягоды или коробочки.
Семейство пасленовых объединяет около 2300 видов. Пасленовые
роко распространены в умеренных и тропических областях всего света,
достигая наибольшего разнообразия1 в Центральной и Южной Америке.
Пасленовые, как правило, содержат алкалоиды, вследствие чего многие
ядовиты.
Некоторые пасленовые применяются как лекарственные растения. Дру-
гие (в том числе картофель и томаты) имеют большое пищевое и техни-
ческое значение. Среди пасленовых есть целый ряд декоративных раст
Из числа пасленовых, образующих ягоду, наибольшее значение имеет
род паслен (Solanum). В нашей стране в диком состоянии встречаются чер-
ный паслен (В. nigrum) — однолетник с белыми цветками и черными ягода-
ми; сладко-горький паслен (S. dulcamara) — полукустарник с фиолетовыми
цветками и красными плодами и еще некоторые виды. Все онй не имени
и»
к I: ।:
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОС
т
Ряс. 362. Диаграммы
ков пасленовых:
1 — цурман; У — белена
пвет-
практического значения, хотя ягоды черного паслена считаются съедоб-
ными.
Исключительное народнохозяйственное значение имеет картофель, полу-
чивший широчайшее распространение в культуре главным образом в уме-
ренных широтах северного полушария. Название «картофель» ^охватывает
большую группу видов (свыше 150) рода Solanum9 входящих в состав
секции Tuberarium9 для которого характерно образование подземных
побегов (столонов) с клубнями на них. Виды этой секции обитают поч-
ти исключительно в Южной и Центральной Америке, а также на юге США.
Большинство диких видов картофеля связано с системой Анд и с Тихоокеан-
ским побережьем Южной Америки/ На этой обширной территории еще за-
долго до прихода европейцев в различных районах и от разных видов воз-
никла иультура картофеля. Родиной современного культурного картой
Рис. 363. Картофель (Solanum tuberosum):
У— общий вид; у—подземная часть растения со столонами, клубнями аорлпяс 4—uotab ваига-
fSnuudteK ссцвепем; 4—цветок; 4 — разрез юта; 4—илод (яида)
492
ВЫСШИЕ РАСТЕНИЯ
Рис. 364. Белладонна (Atropa belladonna):
1 — ветвь с цветами и плодами; а — развернутый цве-
ток.
Рис. 365. Белена (Hyoscyamus niger):
1 — цветущая ветвь; я — развернутый цветок; 8 —
пестик; 4 — плод, окруженный чашечкой; 8— вскры-
вание шода
ля (Solatium, tuberosum, рис. 363)
считается Чили (включая
остров Чилоэ), откуда это по-
лезное растение, культивиро-
вавшееся местным населением,
было ввезено в Испанию меж- *
ду 1560 и 1570 гг., а оттуда
попало в другие страны Евро-
пы и в Азию. В Россию карто-
фель был завезен Петром I око-
ло 1700 г. Как в Западной
Европе, так и в нашей стране
распространение картофеля шло
вначале медленно из-за неуме-
ния возделывать его, а также
отчасти вследствие суеверий*
Повсеместное разведение в Рос-
сии началось лишь с 40-х го-
дов прошлого века. Ценность
ка ртофеля определяется вы-
соким содержанием в его клуб-
нях крахмала (60—80% от су-
хой массы или 14 — 24% от
сырой массы) и высокой уро-
жайностью (дает на единицу
площади примерно в три раза
больше сухого вещества, чем
хлебные злаки). Пищевое и кор-
мовое достоинство определяется
наличием в клубнях, кроме
крахмала, белков с высоким
содержанием в них аминокис-
лот, необходимых для челове-
ка и животных.
В клубнях обнаружены
также витамины.
Во всех частях картофеля
содержится вредный челове-
ку глюкоалкалоид соланин;
его особенно много в ботве,
ростках, ягодах и позеленев-
ших клубнях, поэтому упот-
ребление в пищу последних не
рекомендуется. В обычных
клубнях содержание соланина
невелико и при варке он раз-
рушается. За время интенсив-
ной культуры было выведено
огромное количество (до 2000)
сортов: пищевых, кормовых,
технических. Из картофеля
изготовляют до 200 различных
блюд. Он играет большую роль
и как кормовой продукт. Про-
мышленная переработка клуб-
ней дает крахмал, декстрин,
патоку, спирт и синтетический
каучук.
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
493
Картофель выращивают из клубней. Размножение семенами имеет зна-
чение в основном при выведении новых сортов.
Открытие советскими учеными (Н. И. Вавилов, С. М. Б у к а-
с о в, С. В. Ю з е п ч у к) многочисленных дикорастущих видов картофе-
ля в Южной Америке дало возможность развернуть большую работу по
селекции высокоурожайных холодоустойчивых, фитофтороустойчивых и
ин
ракоустойчивых сортов картофеля с хорошими п
К роду Solanum относится баклажан (S. melongena) родом, вероятно,
из Индии.
Родствен паслену род Lycopersicum (томат, помидор), куда относится
наш культурный помидор (L. esculentum), имеющий большое количество
сортов. Томаты родом, вероятно, из Перу и близлежащих стран. В плодах
содержатся сахара, органические кислоты, белок, витамины. Зрелые плоды
отличаются высоким противоцинготным действием.
Красный^ или стручковый, перец (Capsicum аппиит) происходит, ве-
роятно, из тропической Америки. Возделывают множество сортов. Плоды
красного перца занимают первое место среди овощных культур по богатству
витаминами (особенно С и А).
Съедобные плоды имеют некоторые виды рода физалис (Physalis),
например «земляничный томатъ, или «перуанская вишняъ (Ph, peruviana),
из Южной Америки и североамериканский «мексиканский томат» (Ph. ixo-
сагра). Их разводят в тропиках, субтропиках и даже в умеренной зоне. Плоды
обыкновенного физалиса (Ph. alkekengi), имеющего огненно-красную чашечку
при плодах, декоративны, в некоторых местах их употребляют в пищу, хотя
они имеют невысокие вкусовые достоинства. У белладонны, vljbil красавки
Рис. 366. Табаки:
2 — виргинский табак (Nicotlana tabacum): а —цветоносная ветвь; б — продольный разрез цветка;
3— махорка (N. ruatica): а — ветвь с соцветием; б — раареа коробочки
ВЫСШИЕ РАСТЕНИЯ
Hi
I г>
(Atropa belladonna, рис. 364), плоды — тоже ягоды. Это важное лекарственное
растение, в диком состоянии встречающееся на Кавказе, в Крыму, Карпа-
тах. Разводится преимущественно на Украине. Все части белладонны ядо-
виты. Известны случаи отравления детей ягодами, внешне несколько напо-
минающими вишню. В белладонне содержится алкалоид атропин, который
оказывает сложное действие на организм человека и используется в меди-
цинской практике как противоспазматическое и болеутоляющее средство.
Источником атропина и близкого к нему гиосциамина служит также
Scopolia carniolica — растение, распространенное в Средней, Южной и
Восточной Европе, а в нашей стране — на Кавказе и в юго-западных райо-
нах европейской части СССР; белена (Hyoscyamus niger) и дурман (Datura
stramonium).
Белена черная (Hyoscyamus niger, рис. 365)— двухлетнее мусорное
растение, растущее у жилья, у дорог, реже как сорное или по заброшенным
пашням. Цветки довольно крупные с грязноватой окраской венчика, прони-
занного многочисленными пурпуровыми жилками. Характерный двугнезд-
ный плод белены («крыночка») открывается крышечкой. Плод скрыт в раз-
растающейся чашечке. Белена сильно ядовита.
Дурман (Datura stramonium) также сильно ядовит. Плод — яйцевид-
ная или почти шаровидная коробочка, покрытая крепкими крупными птица-
ми и раскрывающаяся четырьмя створками.
Род табак (Hicotiana) содержит около 50—70 видов, распространен-
ных преимущественно в Северной Америке, где и возникло табакокурение.
Практическое значение имеют настоящий табак (ЛГ. tabacum, рис. 366),
махорка (N. rustica) и душистый табак (ЛГ. affinis). Первые два разводят
дня получения курительного (реже нюхательного) табака, содержащего
весьма ядовитый алкалоид — никотин. При курении происходит возгонка
никотина, и он попадает в дыхательные пути. Действие яда двоякое: сначала
он вызывает возбуждение, а потом влияет угнетающе. Ядовитое действие
никотина при курении частично нейтрализуется тем, что, быстро всасывае-
мый, он быстро же и выводится из организма. Но постоянное курение вредно
сказывается на нервной системе человека. Нередко никотин используют
как инсектицидное средство для борьбы с некоторыми вредными насекомыми
и клещами. Путем окисления никотина получают никотиновую кислоту —
витамин РР, недостаток которого в организме человека вызывает пеллагру.
Листья махорки используют для получения лимонной кислоты. Душистый
табак (N. affinis) разводят как декоративное растение.
Часто можно встретить в цветниках и петунию (Petunia hybrida)— гиб-
рид двух южноамериканских видов. У петунии, как и у табака, плоды —
коробочки.
Корни мандрагор (род Mandragora), причудливо ветвящиеся, иногда
напоминающие фигуру человека, в древности и в средние века считали «маги-
ческими» и использовали как лечебное средство, может быть, не без основа-
ния, так как в этом растении найдены гиосциамин, атропин и другие алкало-
иды. В СССР (в Средней Азии) в диком состоянии встречается всего один
вид мандрагоры — М. turcomantca, который используется в народной
(туркменской) медицине.
Семейство норичниковые (Scrophulariaceae). Подавляющая часть
видов этого семейства — травянистые, многолетние растения или полукус-
тарники, сравнительно немногие — кустарники, единичные представители —
деревья и даже лианы. Листья норичниковых простые, без прилистников,
очередные или супротивные (причем на одном и том же растении эти две
формы листорасположения могут сменять друг друга), иногда мутовчатые.
В анатомическом отношении характерно наличие внутреннего jggg* (бико-
латеральность пучков). Цветки всегда зигоморфные, четырехкруговые,
пятичленные, довольно разнообразные вследствие редукции и срастания
некоторых органов (рис. 367). Пятичленная сростнолистная (иногда лишь
в основании) чашечка у некоторых видов рода Veronica становится
I г
г. г-
; I: I >
гг
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
495
Рис. 367. Диаграммы цветков норичниковых:
I — коровяк; г — льнянка*, з — вероника
четырехчленной благодаря недоразвитию чашелистика, обращенного к оси
соцветия. Венчик то с хорошо выраженной, то с едва заметной трубочкой,
почти правильный (Verbascum) или резко зигоморфный и двугубый (Scrophu-
laria, Antirrhinum), иногда при этом еще и шпорцевый (Linar ia). В других
случаях полное срастание двух задних лепестков, соответствующих двум
лепесткам верхней губы двугубого венчика, делает последний как бы четырех-
лепестным, при этом отгиб становится нередко плоским (рис. 368). Андроцей
только у небольшой части родов состоит из пяти тычинок, не всегда одинако-
нитей (Verbascum). Часто наблюдается редукция
вых по длине и опушен
задней тычинки, причем на ее месте может присутствовать стаминодий
(Scrophularia) или только иногда появляется рудимент недостающей тычинки
(Linaria vulgaris). У других редукция андроцея пошла дальше, так что в
цветке остаются только две тычинки, которые иногда сопровождаются двумя
стаминодиями (Gratiola officinalis), или их нет (Veronica). Гинецей норични-
ковых двучленный, с плодолистиками, расположенными в медианной плос-
кости (отличие от пасленовых). Завязь верхняя, двугнездная, с плацентами,
Рис. 368. Цветки норичниковых:
1 —коровяк; s — норичник: S — накерстяяка; S —»льввиый зев: & — общий —д: б — цветок е удалей-
вой губой; S — льнянка; б — мытнжк; 7 — очанка; s — вероника
выс
nr
Е РАСТЕНИЯ
Рис. 369. Льнянка (Linaria vulgaris):
l — верхняя часть; 2 — продольный разрез цветка;
а — продольный разрез пестика и чашечки
норичниковых
родов и около
Норичниковые
связанными в перегородкой,
разделяющей гнезда. Семяпоч-
ки многочисленные. Столбик
один. Рыльце головчатое, цель-
ное или двураздельное. У осно-
вания завязи имеется медоот-
делительный диск — кольце-
видный или односторонний.
Плоды — коробочки (ред-
ко ягоды). Семена с эндо-
спермом.
Некоторые норичниковые,
имеющие слабозигоморфный
цветок и все пять тычинок,
очень сходны 6 пасленовыми,
тем более что у последних на-
блюдается склонность к зиго-
морфии.
Семейство
включает 200
3000 видов,
распространены по всему све-
ту, главным образом в преде-
лах умеренных широт. Большо-
го практического значения не
имеют, но отдельные предста-
вители используются в меди-
цине и как декоративные.
Венчики некоторых видов
коровяка, например V. phlomo-
ides, V. thapsiforme, V. songo-
ricum, используются в медици-
не как отхаркивающее и мягчи-
тельное средство. Листья и вен-
чик медвежьего ушка (V. thap-
gus), названного так по наличию на листьях густого шерстистого опуше-
ния, используются в народной медицине.
Льнянка обыкновенная (Linaria vulgaris, рис. 369)— трудноискоренимый
корнеотпрысковый сорняк. Андроцей из четырех тычинок. Венчик резко
зигоморфный, со шпорцем на нижней губе. Две губы венчика тесно сближены,
и поэтому нектар доступен только крупным насекомым (шмелям), которые,
садясь на нижнюю губу, отодвигают ее от верхней. Такой же способ проник-
новения насекомого в цветок у львиного зева (Antirrhinum majus), средне-
европейского по происхождению декоративного растения, культивируемого
в разных цветовых вариациях. Декоративное значение имеют также гибрйды
видов южно- и центральноамериканского рода Calceolaria. Нижняя губа
цветков кальцеолярий пузыревидно вздута и имеет яркую окраску, ровную
или пеструю, сложного рисунка.
Декоративное значение имеет также род губастик (Mimulus), виды
которого с красными, оранжевыми и пестрыми цветками культивируются
как рано зацветающие летники. Как комнатное растение содержат М. moscha*
tus с ароматичной зеленью.
А вран лекарственный (Qratiola officinalis)— ядовитое растение, дейст-
вующее как сильное слабительное средство. Он сохраняет эти свойства и в
сушеном виде, поэтому, попадая в сено, может вызвать расстройство кишеч-
ника у домашних животных, приводя иногда к полному истощению их и в
смерти. Используется в народной медицине.
Существенное лечебное значение имеет род наперстянка (Digitalis)9
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕ
497
распространенный преимущест-
венно в странах Средиземно-
морья. В СССР известно 6
видов. Наперстянки — в ос-
новном многолетние травянис-
тые растения. Наибольшей
известностью пользуется пур-
пуровая наперстянка (D. pur-
purea, рис. 370) — западноев-
ропейское растение, культи-
вируемое в нашей стране как
лекарственное и, частично, как
декоративное. Препараты пур-
пурной наперстянки пользуют-
ся общим признанием сильного
сердечного средства, применяе-
мого при хронической недоста-
точности сердечной мышцы и
при острых нарушениях сер-
дечной деятельности. Теми же
качествами обладают и другие
виды этого рода.
Род Veronica*— один из
наиболее крупных в семействе—
распространен в умеренных и
холодных странах обоих по-
лушарий.
Больше всего видов в Ев-
ропе и Азии. В СССР насчиты-
вается около 100 видов. Наши
Рис. 370. Наперстянка (Digitalis purpurea) i
j — цветущая ветвь; £ — ирикорневыс листья; 3 —
продольный разрез цвели; 4 — разрез завявди а —по-
перечный; о — ирлдплыплй
вероники — травы, частью од-
нолетние, частью многолетние.
Среди новозеландских и
австралийских вероник есть
вечнозеленые кустарники и
деревья.
Примером древесного растения среди норичниковых может служить
также Paulownia tomentosa — единственный вид рода павловния*
Родина ее — горы Японии, где она достигает 10—15 м высоты. Культиви-
руется павловния как декоративное дерево в СССР (в Крыму и на Кавказе)
и в Западной Европе.
Среди норичниковых есть гетеротрофные растения, преимущественно
полупаразиты. Из них довольно обычны известный под названием иван-да-
марья марьянник дубравный (Melampyrum nemorosum) с желтыми цветками,
выделяющимися на фоне сине-фиолетовых прицветных листьев, многочис-
ленные виды очанок (Euphrasia), погремки (Alectorolophus). Последние,
поселяясь на лугах и не имея кормового значения, приносят большой вред,
угнетая кормовые травы. Настоящий паразит, лишенный зеленой окраски
листьев,— петров крест (Lathraea squamaria) — единственный из шести
видов рода Lathraea, растущий в СССР. Петров крест имеет разветвлен-
ное корневище, покрытое мясистыми, короткими и широкими чешуевидными
листьями. На поверхность земли выходит только соцветие с лилово-розовыми
кроющими чешуевидными листьями и с цветками, чашечка которых пример-
но той же окраски, а венчик красный. После отцветания вся надземная часть
растения отмирает. Обычный хозяин этого паразита — орешник (Corylus
avellana), но указывается произрастание петрова креста на ольхе, яа тополях
(белом, черном) и на осине.
Ниже рассматриваемые семейства, вероятно, родственны норичниковым.
111
sin
ВЫСШИЕ PACT
Рис. 371. Пузырчатка (Utricularia vulga-
ris):
I—цветущее растение; У — продольный разрез
цветка; а — шюд; 4 — диаграмма цветка
Рис. 372. Заразиха (Orobanche
lutea) на корнях люцерны
Семейство пузырчатковые (Lentibulariaceae) отличается от но- 1
ричниковых одногнездной завязью с центральным семяносцем.
Наиболее крупный род пузырчатка (Utricularia) распространен
почти по всему свету, за исключением наиболее холодных стран. «Листья
обыкновенной пузырчатки (U. vulgaris, рис. 371) и других видов, живущих
в водоемах нашей страны, рассечены на дольки. Дольки их частично превра-
щены в пузырьки для ловли мелких беспозвоночных. В пузырьках проис-
ходит переваривание жертвы, и продукты переваривания всасываются.
Часть тропических видов пузырчаток — наземные растения, другие,
как и наши,— растения водоемов. Некоторые тропические виды эпифитны.
Бразильская U. nelumbifolia живет в розетках листьев бромелиевых, где
скапливается вода.
Семейство зараяиховые (Orobanchaceae) отличается от норичниковых
одногнездной завязью с 2—4 постенными плацентами. Тычинок четыре.
Плод — коробочка. Семена очень мелкие с нерасчлененным зародышем,
из многочисленных клеток. Все заразиховые лишены хлорофилла, паразиты.
Их насчитывается около 150 видов.
Наиболее важен род заразиха (Or о banc he, рис. 372). Некоторые виды
заразихи, поселяясь на культурных растениях (подсолнечник, конопля,
клевер, томаты, табак и др.), приносят существенный вред. Заразихи произ-
водят очень много семян (1500—2000 в коробочке, а на целом растении число
семян исчисляется десятками тысяч, а иногда до 150 тыс.). Семена легкие
и разносятся ветром. С водой внедр
тся в почву, где могут сохраняться,
не теряя всхожести, в течение 8—10 лет. Одни заразихи поражают широкий
круг растений, другие приспособились к жизни на ограниченном числе видов.
Установлено, что прорастание
семян заразихи зависит от стиму-
лирующего действия корневых
выделений растения-хозяина.
Нитевидный проросток заразихи,
встречая корень питающего рас-
тения, проникает в него, дости-
гая проводящей системы, и раз-
растается внутри, становясь при-
соской. На поверхности заражен-
ного корня проросток образует
клубенек, из которого развива-
ются цветочные. стебли и при-
даточные корни. Придаточные
корни, в свою очередь примыкая
к корням хозяина, образуют но-
вые присоски и цветоносные побе-
ги. В нелёгкой борьбе с зарази-
хой рекомендуют уничтожать ее
наземные побеги до созревания
семян, строить севообороты так,
чтобы растение-хозяин ряд лет не
возвращалось на данное поле,
провоцировать массовое прорас-
тание семян посевом рано созре-
вающих сортов (с уборкой их
. раныпе созревания семян пара-
зита), опрыскивать гербицидами.
Особо важное средство борьбы—
выведение заразихоустойчивых
г. г.
t
Ряс. 373. Подорожник (Plantago major):
1 — общий в№ я*- цветок; а — диаграмма цветка
сортов.
В пустынной и полупустын-
ной зонах СССР встречаются не-
многочисленные виды рода Cistanche. С. flava^ паразитирующий в песча-
ных пустынях Средней Азии на корнях джузгунов (CaUigonum}, достигает
от 0,8 до 2 м высоты, т. е. может быть выше человеческого роста.
Близки к норвд
t:r
новым
семейства:
геснериевые (Gesneriaeeae},
куда относятся популярные глоксинии (род Sinningia) и еуаамбарскоя
фиалка» (Saintpaulla lonantha}*, бигнонйевые (Bignoniaceae} с родом
Jaccandra (Бразилия, Аргентина), виды которого имеют ароматичную темно-
красную или фиолетово-бурую палисандровую древесину; кунжутные
(Pedaliaceae)— из семян кунжута (Sesamum indicum} получают кунжутное
масло, идущее на изготовление халвы и других «восточных сладостей».
Семейство подорожниковые (Plantaginaceae} — небольшое, содер-
жит немногим более 250 видов травянистых растений с простыми листьями,
без прилистников. Цветки в колосовидных или головчатых соцветиях, не-
взрачные, правильные или почти правильные, из четырех четырехчленных
кругов — чашечки, венчика, андроцея и ценокарпного гинецея. Тычинки
с длинными нитями и качающимися пылытдками. Плодолистиков 2—4.
Завязь верхняя, одно-, четырехгнездная, с одной или многими семяпочками
в каждом гнезде.
Плод — коробочка, открывающаяся кольцевой поперечной трещиной,
или орешек.
Преобладающее число видов семейства сосредоточено в роде подорож-
ник (Plantago, рис, 373). Биологически подорожниковые интересны тем, что
в этом семействе можно найти переходы между насекомоопыляемыми и
ветроопыляемыми формами. Цветки обычно протерогиничны. В СССР широко
распространены подорожники — большой (Р. major} и средний (Р. media}*
500
ВЫСШИЕ РАСТЕНИЯ
4-Й ПОРЯДОК — МАРЕНОЦВЕТНЫЕ (HUBIALES)
Порядок мареноцветных резко отличается от рассмотренных выше по»
рядков спайнолепестных наличием у всех представителей нижней завязи.
Порядек содержит одно семейство.
Семейство мареновые (Rubiaceae) включает травы, кустарники
или деревья с супротивными (лишь как исключение с очередными) листья-
ми, имеющими прилистники. Прилистники располагаются или между лис-
том и стеблем, или по бокам от листового черешка, между супротивными
листьями пары. В первом случае они могут срастаться, образуя колпачок,
защищающий почки и впоследствии опадающий. Прилистники, занимаю-
щие место по бокам листа, часто бывают листовидными; они могут попарно
срастаться друг с другом или, наоборот, расщепляться до основания. Это
создает внешнюю картину мутовчатого листорасположения. Одним из дока-
зательств супротивного, а не мутовчатого листорасположения служит на-
личие почек в пазухах только двух листовидных органов мутовки. Цветки
мареновых большей частью в цимозных соцветиях. Они двуполые, актино-
морфные, реже зигоморфные. Чашечка срастается с нижней завязью, окан-
чиваясь пятью или четырьмя зубчиками, которые вследствие редукции мо-
гут и отсутствовать. Венчик из того же числа членов, что и чашечка.
Трубка венчика бывает длинной у одних видов и очень короткой у других.
Изменчива и общая форма венчика (трубчатая, воронковидная). Тычинок
большей частью столько же, сколько долей венчика, и они занимают проме-
жуточное положение между последними (рис. 374). Гинецей из двух плодо-
листиков. Завязь нижняя, дву-
Рис. 374. Ясменник душистый (Asperula
odorata):
2 — общий вид растения; 9 — пяехж в — продоль-
ный разрез цветка; 4 — диаграмма цветка
гнездная, но число гнезд, как и
число семяпочек в каждом гнезде,
в пределах семейства изменяется
от одного до нескольких. Столбик
один, цельный или двурасщеплен-
ный, рыльца головчатые. Плоды
бывают как сочные (ягоды, костян-
ки), так и сухие (коробочки, дроб-
ные плоды, распадающиеся на
орешки). Семена с эндоспермом.
Семейство мареновых — одно
из крупнейших среди спайноле-
пестных. В составе его насчиты-
вают от 6000 до 7i
видов. Ос-
новная часть их сосредоточена в
тропических странах, но есть от-
дельные группы внетропического
р аспространения.
Наибольшее экономическое
значение из мареновых имеют ко-
фейные (Африка) и хинные (Юж-
ная Америка) деревья. Кофейные
деревья — виды рода Coffea (на-
пример, С. arabica, С. liberica,
С. robusta) — небольшие вечнозе-
леные деревца или крупные ку-
старники с кожистыми темно-зеле-
ными листьями и белыми цветка-
ми. Они приносят красные или
темно-синие довольно сочные пло-
ды, содержащие по два крупных,
овальных, полувыпуклых семени
с роговым эндоспермом. Освобож-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
501
Рис. 375. Кофе (Coffea arabica J:
J — ветка с плодами; » — цветок; а — продольный разрез жлода (о — семя)
Кофейные зерна
изготовляют из-
денные от околоплодника, семена поступают в продажу,
поджаривают, перемалывают и из полученного продукта
вестный напиток — кофе. Алкалоид кофеин, содержащийся в кофе, возбуж-
дает центральную нервную систему и усиливает работу сердца. Родина
широко известного культурного С. arabica (рис. 375) — Эфиопия, но введе-
ние в культуру осуществлено в Йемене, который имел связи с Эфиопией
еще в IV в. «Вторую родину» кофе нашел себе в Южной Америке, которая
в настоящее время поставляет его на мировой рынок около 90% (в том числе
Бразилия — около 60%). Как напиток кофе проник из Аравии в Турцию,
а из нее в Европу (XVII в.).
Хинные деревья принадлежат роду Cinchona, содержащему несколько
десятков видов. В большинстве случаев это небольшие (10—15 ж), довольно
стройные, вечнозеленые деревья и крупные кустарники с кожистыми листь-
ями, пахучими белыми или розовыми цветками. Растут главным обра-
зом по восточным склонам Анд, на высотах 1000—2400 м над уровнем моря—
в полосе интенсивной конденсации водяных паров. В коре содержится хи»
нин, хинидин, цинхонин, цинхонидин и другие алкалоиды (всего до 20).
Местные жители Южной Америки еще до прихода европейцев применяли
кору хинного дерева для лечения лихорадки. Европейцам лечебные свой-
ства хинного дерева стали известны в XVII в. В связи с истощением при-
родных запасов этого ценного растения XIX в. возникла необходимость
inti
его культуры.
Сейчас основная масса коры хинного дерева поступает на мировой
рынок из Индонезии (остров Ява).
В медицина используют чистый
или смесь алкалоидов «хинет», вполне оправдавшую себя в лечении маля-
рии. Приоритет использования «хинета» принадлежит советским ученым.
В СССР, даже во влажном теплом климате Черноморского побережья
Кавказа, хинное дерево вымерзает. Зеленые черенки, заготовленные летом,
перезимовывают в парниках, а весной их высаживают на плантацию под
защитой притеняющих растений. В течение лета растения достигают ро-
ста человека, их убирают и из всего растения извлекают комплекс алкало-
идов «хинет». Препараты, изготовленные из корней маленького бразиль-
802
ВЫСШИЕ РАСТЕНИЯ
ского полукустарничка ипекакуаны — Cephaelis (Uragoga) ipecacuanha,
разводимого в Индии, используются в медицине как отхаркиваю-
щее, послабляющее и рвотное средство.
Биологический интерес представляют обитающие на юго-востоке Азии
эпифитные мирмекодии (например, Myrmecodia echinata). Клубневидные
стебли мирмекодии пронизаны ходами, где поселяются муравьи, что дает
повод считать мирмекодию мирмекофильным растением.
В .СССР произрастает 200 с лишним видов мареновых. Наиболее рас-
пространены роды подмаренник (Galium) и ясменник (Asperula), оба
представленные травами с кажущимся мутовчатым листорасположением.
Некоторые виды подмаренника могут быть использованы как красильные
растения.
Хороший краситель дает марена красильная (Rubia tinctorum)—среди-
земноморское растение, которое культивировали специально для получения
из подземных органов красной краски для хлопка и шерсти. С изобретением
химического способа производства ализарина культура марены резко
сократилась.
5-Й ПОРЯДОК — ВОРСЯНКОЦВЕТНЫЕ (DIPSACALES)
Цветки ворсянкоцветных четырехкруговые, пента- и тетрамерные,
правильные, зигоморфные или актиноморфные. Венчик в почкосложении
не бывает скрученным. Тычинок столько, сколько лепестков или меньше.
Они чередуются с долями спайнолепестного венчика, обычно срастаясь с
его трубкой. Завязь нижняя. Число плодолистиков 5—2, причем некото-
рые могут быть стерильными. Семяпочки с одним интегументом. Эндосперм •
образуется по целлюлярному типу. Древесные и травянистые растения
с супротивными листьями, пластинки которых часто рассечены. Прилист-
ников нет (исключения редки); нет также и внутренней флоэмы.
Семейство жимолостные (Caprifoliaceae) включает кустарники,
лианы, деревья с супротивными листьями, как правило, без прилистников.
Цветки актиноморфные или зигоморфные, с пятичленными чашечкой
и венчиком. Тычинок большей частью пять. Гинецей из 5—2 плодолистиков.
Завязь с таким же числом гнезд, однако некоторые плодолистики могут
недоразвиваться, и тогда, соответственно, число развитых гнезд уменьша-
ется (рис. 376). Число семяпочек в гнезде — одна или много. Плоды — яго-
ды или костянки.
Это небольшое (400 видов) семейство близко к мареновым, отличаясь
от них почти полным отсутствием прилистников (если они есть, то не бывают
листовидными) и тремя (вместо обычных двух у мареновых) плодолистика-
ми. У всех жимолостных развивается мощная сердцевина. У некоторых
она разрушается и стебель становится трубчатым. Если мареновые в основ-
иг
ном распространены в тропиках, то жимолостные — во внетропических
ротах северного полушария. В тропической зоне их можно встретить
только в горах. В южном полушарии жимолостные редки.
В СССР широкой известностью пользуется род бузина (Sambucus) —
крупные кустарники с перистосложными листьями. Метельчатую бузину
(S. racemosa), имеющую красивые красные плоды, часто разводят в садах
ни
как неприхотливый декоративный кустарник, который легко дичает. Этот
более западный вид отличается от растущей в лесах Сибири и на Дальнем
Востоке сибирской бузины (S. sibtrica). В лесах южной полосы европей-
ской части СССР (в Крыму) и Кавказа растет черная бузина (S. nigra),
отличающаяся черной окраской плодов. Один из видов бузины — бузина
травянистая (S. ebulus) — многолетнее травянистое растение, в диком
состоянии встречающееся на юго-западе европейской части СССР, на Кавка-
зе и в Средней Азии.
Род калина (Уiburnum) о&ьещаняез кустарники и небольшие деревца
с простыми листьями, иногда имеющими прилистники. Прилистт
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
503
Рис. 370. Жимолостные:
1 — диаграммы цветков: а — бузвна; б — калина; в — жимолость (двухцветковое соцветие); г — общий
вид соцветия жимолости
которых видов превращены в железки. Обыкновенная калина (У. ори1изЛ
рис. 377) растет в подлеске и по опушкам лесов от Западной Европы до За»
падной Сибири и Тянь-Шаня. Листья ее трех-, пятилопастные. Краевые
цветки щитковидных соцветий бесполые, значительно крупнее плодущих.
Из трех гнезд завязи нормально развивается только одно, и в нем одна семя»
почка. Из красных костянок готовят, несмотря на низкие вкусовые качест-
ва, кисели и начинки для пирогов. У культурных сортов обыкновенной
калины все цветки становятся бесплодными и соцветие приобретает форму
белого шара; эта декоративная форма известна под французским названием
бульденеж (boule-de-netge — снежный шар). Гордовина (У. lantand} с цель-
ными, снизу беловойлочными листьями и с черными плодами разводится
как декоративный кустарник. В диком состоянии растет на юге и юго-
востоке европейской части СССР и на Кавказе.
В СССР наиболее богат видами род жимолость (Lonleera). Виды жимо-
лости — преимущественно кустарники, имеющие зигоморфные цветки с
двугубым венчиком. Цветки расположены обычно попарно и иногда
срастаются завязями. В европейской части СССР в лесах обычна лесная
жимолость (L. xylosteum) с желтоватыми цветками и ярко-красными ядо-
витыми ягодами (так называемые «волчьи ягоды»). В степной и южных час-
тях лесной зоны Заволжья, -в Южной Сибири и в Средней Азии широко рас-
пространена татарская жимолость ((£. tatarica), вошедшая в культуру
как декоративный кустарник. Плоды дальневосточных жимолостей —
съедобной и камчатской (L, edulis, Z. kamtschatlca) — используются местным
населением в сыром виде и на варенье. Каприфоль (L. caprifolium) — лаза»
ющий кустарник, разводимый как декоративное растение.
Декоративное значение имеет и снежноягодник (Symphoricatpus race-
mosa) с ярко-белыми ягодами, созревающими осенью. Родина — Северная
Америка.
В хвойных лесах с моховым покровом встречается замечательный низ-
1 мм, с изящными беловато-розовыми пахучими цветками — Ыппаеа
borealis.
504
ВЫСШИЕ РАСТЕНИЯ
Рис. 377. Калина (Viburnum opulus):
1 — цветущая метка; 2 — иериферияесквй цветок;
4 — «редакций цветок
Семейство валерьяновые,
или мауновые (Valerianaceae). Это
травянистые растения или полу-
куста рники, редко куста рники.
Листья с глубоко расчлененной
или цельной пластинкой, без
прилистников. Цветки в соцве-
тиях, часто дихазиальных или
завиткообразных. Чашечка по
заложению пятилистная, во вре-
мя цветения
в дальнейшем
JII
хохолок,
воронча-
симмет-
почти незаметная;
она разрастается,
образуя при плодах
как у сложноцветных,
тую окраину, длинные
ричные и несимметричные зуб-
цы. Венчик трубчатый, почти
всегда с пятираздельным отгибом,
склонный к двугуб ости. На труб-
ке венчика имеется мешковидная
или трубчатая выпуклость. Ты-
чинок 4 (5—1). Гинецей по за-
ложению трехчленный, но два
плодолистика и соответствующие
им гнезда завязи недоразвива-
ются, так что фактически в за-
вязи одно плодущее гнездо с
одной семяпочкой. Столбик один,
Плод — семянка (орешек), обычно
Семейство валерьяновых сближают
с одним — тремя рыльцами,
увенчанная разросшейся чашечкой,
с жимолостными. Недоразвитие у
валерьяновых двух плодолистиков имеет место и у некоторых жимолост-
ных (род Viburnum). Тенденция к редукции части тычинок у тех же жи-
молостных получила свое дальнейшее развитие у валериановых.
Валерьяновые (около 360 видов) обитают главным образом в северном
полушарии, причем наибольшим разнообразием видов отличается Среди-
земноморье. В южном полушарии основная масса видов сосредоточена в
неотропической области, и только здесь встречаются древеснеющие
формы.
Практическое значение имеет лекарственная валерьяна (Valeriana offi-
cinalis, рис, 378) — вид очень изменчивый на обширной площади своего
распространения, и поэтому многие ботаники разделяют его на несколько
самостоятельных, Валерьяна — высокий многолетник с непарноперистыми
листьями и крупным соцветием из мелких розовых цветков со своеобраз-
ным запахом.
.Il
Ценным продуктом, нашедшим себе широчайшее применение в медици-
не, считают эфирное валерьяновое масло, которое содержится во всех ча-
стях растения, но главным образом в подземных органах. Поэтому корне-
ед и корни как дикорастущей, так и культурной валерьяны заготовля-
ют в больших количествах.
Семейство ворсянковые (Dipsacaceae). Травянистые растения,
иногда с древеснеющим у основания стеблем, с супротивными листьями,
без прилистников. Цветки обычно в компактных соцветиях, напоминаю-
щих корзинки сложноцветных как по общему виду, так и по налит
основания прицветных листьев, образующих обертку (рис. 379). От слож-
ноцветных легко отличаются порядком распускания цветков. В то время
как у сложноцветных развитие соцветия идет в акропетальной последова-
и
Г, ГЛ
тельности, у ворсянковых первыми распускаются то срединные цветки, то
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМ
одновременно вначале зацветают
1ные и нижние цветки.
Соцветие ворсянковых возник»
ло в результате агломерации
многих элементарных соцве-
тий — дихазиев, причем от каж-
дого дихазия остается только
один (конечный) цветок. Бли-
не
к нему
верховые
образовав
листья срослись,
вокруг каждого цветка обер-
точку (покрывальце), похожую
на чашечку. Поэтому ее обыч-
но и называют «внешней чашеч-
кой». Цветок имеет, кроме
того, и настоящую чашечку,
она венчает нижнюю завязь и
иногда бывает v приподнята
над ней. Чашечка, как прави-
ло, пятичленная и образует по
краю пять-четыре либо большее
число зубцов - (в некоторых
случаях зубцы вовсе не раз-
виваются) . Венчик зигоморф-
ный с пятичленным, более или
менее двугубым (2/3) отгибом,
но чаще отгиб четырехчленный
вследствие полного срастания
двух задних лепестков. Тычи-
нок не больше четырех, так
как пятая (задняя) тычинка
всегда редуцирована. Гинецей
из двух плодолистиков, но пло-
дущий только один. Завязь
одногнездная с одной семяпоч-
кой; столбик один, с цельным
или в разной степени двураз-
дельным рыльцем. Плоды —
семянки.
Семейство ворсянковых со-
держит приблизительно 250
видов Обитают они в Среди-
земноморье и далее на восток,
но только очень немногие до-
ходят до Китая и Японии. Не-
которые распространены в та-
ежной полосе. Отдельные пред-
ставители по горам Восточной
Африки достигают южной око-
нечности этого материка. В Но-
вом Свете дикорастущие ворсян-
ки отсутствуют.
В СССР обычен по суходоль-
ным лугам короставник луговой
(Knautia arvensts, рис. 380) с си-
невато-лиловыми соцветиями.
Практическое значение име-
ет посевная ворсянка (Dipsacus за-
Рис. 378. Валериана (Valeriana officinalis):
2 — соцветие; а — прикорневые листья и корни;
з — цветок; 4 — плод; 5 — диаграмма цветка
Рис. 379. Соцветие и плод скабиозы (Scabio-
sa columbaria):
1 — продольный разрез соцветия; а — плод (чш — ча-
шечка; нчш — внешняя чашечка)
soe
ВЫСШИВ PACT
ж i j
Рис. 380. Короставник (Knautia arveneis):
1 — ивжиян часть растевви; й — соцвепе; а —цветов
tlvus) — высокое двухлетнее рас-
тение с продолговатыми соцвети-
ями, достигающими 8 см в длину.
По отцветании соцветия срезают.
г.г
не дожидаясь полного созревания.
К этому времени все соцветие,
включая прицветные чешуи, за-
канчивающиеся крупными, жест-
кими и упругими остриями,
загнутыми на концах, разрас-
тается и принимает вид шишки
(ворсовальная шишка, рис. 381).
ЛИ
зуют для поднятия волосков
на поверхности сукна, драпа и
фланели. Приподнятые и ровно
затем подстриженные волоски тка-
ни образуют ворс. Культивиру-
ют D. sattoui главным образом
в Западной Европе, в СССР — в
Крыму, на Кавказе и в Сред-
ней Азии.
6-Й ПОРЯДОК —
колокольчикоцветныв
(CAMPA NULALES)
Рис. 381. Ворсовальная шишка:
1 — обтай вид; а— продольный разрев
. Цветки колокольчикоцветных
одиночные или в ботрических со-
цветиях, иногда скученные — в
головках. Большей частью они
обоеполые, актиноморфные или
зигоморфные, обычно пентамер-
ные. Чашечка сростнолистная,
венчик спайнолепестный. Тычин-
ки прикреплены или к основа-
нию венчика, или к его трубке.
Плодолистиков 5—2. Завязь ниж-
гневд, равным числу плодолисти-
ков, или одногнездная. Плоды —
коробочки,
образные.
раете-
или ягодо-
ния, редко кустарники и деревья.
Листья простые, без прилистни-
ков, в спиральном (иногда в су-
противном) расположении. У мне-
ние инулина.
Из относящихся сюда семейств
только два — колокольчико-
вые и лобелиевые — характеризу-
ются заметным видовым разно-
образием.
Семейство колокольчик
ковые (Campariulaceae). Это много-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
507
Ряс. 382. Колокольчвко-
вые:
1 — колокольчик: а — продоль-
ные разрезы молодого шюлне
распустившегося цветка; б—по-
следовательные фазы развития
пестика тычинок; • —диа-
грамма цветка; Д — лобелия
(диаграмма цветка)
летние травы, редко полукустарники. Листья очередные, цельные, не имеющие
прилистников. Соцветия обычно с конечным цветком. Цветки (рис. 382)
правильные, обоеполые, с пятичленными чашечкой, венчиком, андро-
цеем и (редко) гинецеем. Иногда число листочков околоцветника меньше
(3—4) или больше пяти (6—10). Венчик колокольчатый, воронковидный
или колесовидный. Обособленность долей венчика раскрывшегося цветка
больше, чем в цветочной почке. У некоторых венчик до основания раздель-
ный, а иногда доли его остаются соединенными наверху (виды Phyteuma).
Тычиночные нити свободные от венчика, реже сросшиеся с его основанием,
в нижней части нередко расширенные. Пыльники свободные, но бывают
соединенные (склеенные) в трубочку по всей длине или только в нижней
части.
Завязь нижняя (реже полунижняя), обычно трехгнездная. Плацен-
тация центрально-угловая, производная от постенной. Столбик с трех-
раздельным (двух-, многораздельным) рыльцем. Вокруг основания столбика
часто развит нектароотделительный орган (диск) в виде выпуклого кольца.
Плоды — коробочки, реже ягоды. Семена с эндоспермом.
Колокольчиковые энтомофильны и почти все протерандричны. Пыль-
ца освобождается иэ пыльников еще до раскрытия цветка и откладывается
ца, так как эти части пестика бывают покрыты волосками или клейкой жид-
костью. После раскрытия цветка пылы
опадают или увядают. Если
они увядают, то тычиночные нити укорачиваются или изгибаются наружу.
Насекомые таким образом получают свободный доступ к нектару, выделя-
ющемуся на дне цветка. При этом они нагружаются пыльцой. После этой
«мужской» фазы цветения наступает «женская»: столбики удлиняются, ж
лопасти рыльца расходятся, благодаря чему воспринимающая пыльцу по-
верхность становится доступной для опыления.
Если не произошло перекрестного опыления, то дальне
1.111
ее разраста-
ние и изгибание лопастей рыльца обеспечивает опыление остатками пыльцы
I; I :
того же цветка.
Семейство колокольчиковых содержит около 800 видов. Они немного-
численны в тропиках и очень разнообразны во внетропических областях
северного полушария, особенно в Средиземноморье. Из 224 видов колоколь-
чиковых, произрастающих в СССР, 150 относится к роду, колокольчик
(Campanula) — наиболее крупному в составе этого семейства. Практиче-
ское значение колокольчиковых невелико; некоторые виды ценятся как деко-
ративные; утолщенные корни колокольчика реповидного (С. rapunculoides)
508
ВЫСШИЕ РАСТЕНИЯ
кое-где используются как овощ; есть колокольчики со съедобными (как*
салат) листьями. В млечном соке колокольчиковых содержится небольшое
количество каучука.
Семейство лобелиевые (Lobeliaceae) включает травянистые расте-
ния, редко кустарнички или деревья. Соцветия большей частью без конеч-
ного цветка. Цветки в противоположность колокольчиковым зигоморфные,
обычно обоеполые. За пятилопастной чашечкой следует одно-двугубый
венчик. Пять прикрепленных к его трубке или не сросшихся с нею тычинок
расположены между лопастями венчика. Тычиночные нити в основании обособ-
лены друг от друга, а выше сросшиеся; пыльники спаяны в трубку (как у
сложноцветных).
Гинецей из двух плодолистиков. Завязь нижняя, реже цолунижняя,
обычно двугнездная или почти одногнездная, с многочисленными семяпочка-
ми. Столбик один с двумя ветвями, оканчивающимися рыльцами. Стол-
бик несет воротничок из вверх направленных волосков, соответствующих
по положению и функции выметающим волоскам сложноцветных. Плоды —
ягоды или коробочки. Семена с эндоспермом.
Лобелиевые близки к колокольчиковым и отличаются от них главным
образом зигоморфностью цветка, двугнездной завязью и срастанием тычи-
нок.
В отличие от колокольчиковых лобелиевые распространены в тропиках
и в умеренной зоне южного полушария, хотя некоторые заходят довольно
далеко на север.
Синецветную Lobelia erinus — из Капской флористической области — и
красноцветные виды того же рода (L. julgens, L. splendens^ L. cardinalis) из
Северной Америки разводят в цветниках. Два вида лобелии — L. dortmami
и L. sessilifolia встречаются в СССР в диком состоянии.
7-И ПОРЯДОК — АСТРОЦВЕТНЫЕ (ASTERALES)
Сюда относят только одно, но очень крупное семейство сложноцвет-
ных, присоединяемое нередко к предыдущему порядку.
Семейство сложноцветные (Compositae, Asteraceae). В основной
массе — это травянистые растения, преимущественно многолетние, реже
однолетние, иногда полукустарники, кустарники и даже небольшие деревья;
есть также лазающие растения и суккуленты. Листья простые, обыч-.
но без прилистников, с цельными, или расчлененными ^лястинкамн, Листо-
и»
ов, с цельными, или расчлененными пластинками. Л исто-
Рис. 383. Корзинка
же — об — листочки
трубчатые цветки)
(о — расширенная ось соцветия — общее цветоло-
ные
подсолнечника I.
обертки, л — краевые ложноязычковые цветки, т— серед
i гл:
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
509
Рис. 384. Продольные разрезы корзинок сложноцветных:
1 — лопух (Arctium lappa). Все цветки трубчатые (в середине нераспустившиеся), ось соцветия плос-
кая, покрытая пленчатыми чешуйками (кроющими листьями цветков) > листочки обертки крючковид-
ные ка ненцах; 2 — ромашка аптечная (Matricaria chamomilla). Краевые клетки ложноязычковые,
вниз отогнутые, срздинные трубчатые, ось соцветия нонусовидно вытянутая, полая» без пленчатых
чешуек (об — обертка, т — трубчатые цветки)
расположение почти всегда очередное. У многих имеются млечники во всех
вегетативных органах иллсмппст1™рупдлт Характерно образование йну-
линаТТДветки всегда в соцветиях — корзинках (рис. 383). которые, в свою
очередь, могут быть собраны в сложные соцветия* разнообразного строения
и разной стег^енц комКайтности. Основу
стеблевого происхождения, обычно (но не соответственно его природе) назы-
ваемый «общим цветоложем». На нижней части оси соцветия помещаются
сближенные верховые листья, в совокупности образующие обертку. Они
бывают зеленые (листовидные) или, наоборот, заметно отличаются от
листьев как окраской, так и строением.
Строение и взаиморасположение листочков обертки вместе с формой
нижней части оси корзинки определяют общие очертания, т. е. форму оберт-
ки в целом (рис. 384). Самые внутренние листочки обертки — кроющие листья
краевых (морфологически нижних]^ цветЕОВ-.корзинки.. Пбстальные цветки
тоже могут*' иметь”кроющие листья, но пленчатые, цельные или рассеченные.
В предельном случае рассеченности пов^шость рци «соцветия кажется прос-
то волосистой. Впрочем^ вблойШт покрывающие поверхность оси корзинки,
могут быть и трихомами, не связашЕсйи по происхождению ^.кроющими
листьями цветков. Тогда, равно как в случае голой поверхности оси корзинки,
кроющие листья некраевых цветков отсутствуют.
Поверхность оси корзинки бывает покрыта неболь-
шими ямками, соответствующими -местам прикрепле-
ния цветков, иногда она имеет сетчатое или ячеис-
тое строение. В некоторых случаях цветки распо-
лагаются на небольших бу горках, представляющих.
может быть, остатки редуцированных, .цветоножек.
Число цветков в одной корзинке может быть боль-
шим (в крупных корзинках подсолнечника — тыся-
чи) или, наоборот,, ограниченным^ „(корзинка тыся-
челистника). Известны лаже одноцветковые корзин-
ки, собрание которых, в свою очередь, образует слож-
ное соцветие в форме шаровидной головки
(мордовник — Echinops). Цветки сложноцветных
обоеполые, но бывают женскими, мужскими ц да-
:гл
Рис. 385. Диаграмма
трубчатого цветка
сложноцветных
510
ВЫСШИЕ PACT
•л; i
Рис. 386. Цветки сложноцветных:
1 — трубчатый; 2—язычковый; я — двугубый; 4 —
ложнояиычюжый; а—веронковждный; б — продоль-
ный разрез трубчатого цветка; Z—развернутая тру-
бочка из пиьвшюв
2
Рнс. 387. Формы чашечки
сложноцветных:
1 — Gymnospcrma glutinosum: f—
Rernya mariensla; a — Heliantbus
annuue; 4— Arctium lappa
nr
:e:
гнезднуд)_завязь, образованную двумя тряпякшч плодолистиками, с од-
ной^^м^очкбй?"Г^олбик один; у его основания расположёнТ^нектарный
диск, наверху Столбик всюгдад^рдддёдшый На внутренней стороне вет-
вей столбика рйп^йг1Ште1Рс(^Чкн,воспринимаю1ЦИе пыльцу,а на Наруж-
ной стороШгШй покрыты спвдщь_илй только в верхней частй вив е т а to-
щи м и вол o-O:jr а м и, названнымитак соответственно их функфш. Рас-
положение, густотами длина выметающих волосков разнообразны; иногда
они могут и отсутствовать.
Ju
венчика, свободные (лишь редко срастающиеся друг с другом).
наоборот, соединены друг с другом в трубодку ираскрымкугся
ее; редко, возможно в связи с переходом к ветроопылению^ пыльники сво-
бодные.
Общие черты венчика сложноцветных — стяйндлтАстдпсть и (первич-
ная) пявдчленность. Разнообразие венчиков в прередах семейства большое.
Венчик сложцдцйетятод как ^цгдд^рфд^^^йоэтому простей-
шей его формой можно трубчатую. сохраняющую актиноморфность
и в раз------------'*— ₽-----------*------------------* —
чиКг который может иметь не вподнед>адшае Доли, становясь в таком случае
более илв-меиее зигоморфным. Зигоморфность венчикд бывает (главным
образом у экзотических сложноцветных) и в ф<цд£е двугубости с "двучлен-
ной верхней и ттехчлеЕшои^етжней губой. Очень^астднаблюдй^тся обра-
зование одногубого венчика, ~0Т'“ трубки которого отходит один язычок,
имеющий по краю пять зубчиков, соответственно участию в его образова-
нии всех пяти лепестков. Такой венчик называют язычковым. Отличают
еще ложноязычковые венчики, которые, подобно язычковым^олногубые.
но имеют иное происхождение — их рассматривают как происшедшие иэ
UJ
i u i r,
511
двугубых (*/a) в результате редукции верхней губы. Это подтверждается
наличием на крае язычка (нижней, губы) всего трех зубчиков. Венчик при-
креплен к верхушке завязи, и нижняя часть его трубочки окружает основа-
ние столбика и нектарник. Строение чашетки сложноцветных боне. ЗЯ71
разнообразно, но никогда не вст
,,:,a
n
про-
«0
rtl
пййлимплй ИИ ЛИК, на котором формируются пять бугорка
ЩИХ, ПО. ИХ положении* ЫЯКДУ ИНЧЛТиММЦ'ЛЦПНМйбн. чдртйлиг.ти^ям в даль-
нейшем развитие заложившегося зачатка чашечки может быть очень незна-
чительным, и тог^Д й еавДИйипниитяям* ниётпо шняечдцгдшВ'о вовсе не oftfca-
. hl I и
(у бессмертника), два pJflmmopuHuo у нодеошцяишшиф-н
каждый из зубчиков пяшечки иадда- ия сипйй верхушке
причем подобные.^ щетинки могут вырастать по краям
вые видоизменения чашедди называют хохолком. Биодогическое~"значе-
ние хохолка состоит л сппаистиии
сложноцветных утрачена, и его находится в явной связи с наличшдрзберт-
ки, которая закрывает всю совокупность цветков корзинк^ ВВТвремя их
формирования и развития.
Тесное сближение мелких, но многочисленных цветков на общем ложе
ir.i'
многих сложноцветных вкащдрккорштка цпйя
е> и на краевые (зигомо
е че
значительно увели-
ёвыК цветки обычно либо ложноязыч-
ковые, как у ромашек (род Matricaria), нивяника (Leucanthemu^“ifflgd№),
чивается. У наших
1 — пыльннковая трубка в цветке с вынесенной наружу пыльцой; 8 — верхушка столбика
с двумя ршьцдмн; 4— о—рост столбика развертывание рыльца (м—вмльшпш» р —рыль-
ца)
512
ВЫСШИЕ РАСТЕНИЯ
Рис. 389. Плоды сложноцветных:
1 — подсолнечник; 2 — череда; а — цикорий; 4 — чер-
тополох; а — будни; в — одуванчик
Рис. 390. Летательный аппарат Leucocarpus
pinnatifidus (к — кроющий лист; ок— около-
плодник; с — семя)
пупавок (род Anthemis), видов
Chrysanthemum, либо _ воронко-
видные или косоворрнковидные
(василек — Centaurea). ЕсдйГ у
таких сложноцветных, как золо-
тая розга (Solidago virga-aurea),
девясилы (род Inula), краевые
ложноязычковые цветки имеют
окраску одинаковую с середин-
ными, обычно желтыми, то у дру-
гих они белые (нивяник), лиловые
(виды рода Aster), розовые (не-
которые пиретрумы — Pyreth-
rum). Контрастная расцветка
еще более усиливает привлекаю-
щее влияние корзинки.
У ряда^лищжщщвдх^х ^кра-
евые цветди бесплодны, тГе. не
имеют ни андроцея, ни гинецея.
В других слу^аях в них не разви-
вается только андроцей, и тогда
краевые цветкиойвают только
ясенские^ (нивяник, ' подсолнеч-
ник). Реже все цветки корзинки
однополые. Например, у липпь-га-
мачехи (Tussilago farjara) сере-
динные цветки _корзидки вслед-
недоразвития завязей и
се]шшбчёк' физиологически муж-
ские, по периферии же корзинки
расположены пестичные цветки.
В редких случаях наблюдается
двудомность (кошачья лапка —
Antennaria dioica). Цветки сложно-
цветных протерандричны (рис. 388). В более ранний (мужской) период
цветения оБоШоЖыХ цветков столбик еще короток и его верхушка находит-
ся примерно против основания пыльниковой трубочки. При раскрывании
пыльников пыльца высыпается внутрь трубочки, а подрастающий столбик “
своими выметающими волосками выдвигает ее к выходу.
При этом выметающие волоски, сначала прижатые к поверхности стол-
бика, отгибаются в стороны. Самоопыление в это время невозможно, так как
ветви рыльца плотно прилегают друг к другу. Если даже опылители не уне-
сут пыльцу сразу, то при обнажении рылец, которое происходит благодаря
отклонению ветвей столбика в противоположные стороны, она остается на
внешней поверхности ветвей столбика, там, где нет воспринимающих сосоч-
ков. Однако во второй (женский) период цветения, в конце его, самоопыле-
ние в качестве запасного акта возможно благодаря тому, что растущие ве-
точки столбика все сильнее изгибаются наружу, закручиваясь спирально*
и в конце концов рыльцевая ткань может войти в соприкосновение с пылин-
ками на выметающих волосках. Механизм опыления иногда усложняется тем*,
что тычиночные нити некоторых сложноцветных раздражимы. От прикосно-
вения они укорачиваются, и пыльниковая трубочка при этом опускается'
вниз, пыльца же из нее выметается. Как исключение встречаются и анемо-
фильные сложноцветные, например полыни.
Плоды сложноцветных — семянки (рис. 389). Единственное семя яя^
полняет полость плода и даже йногда срастается с околоплодником. Она
имеет очень тонкую семенную кожуру и крупный прямой зародыш. Эндо-
сперма нет.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
513
, I: I;
Распространение плодов осуществляется ветром, животными и (редко)
водой. Ветер может быть агецтом разноса только потому,
чно мелкиеддЩШЙе (полыни)501)ы»й1б ‘фЖшмвшгоя
чиважштГподъемплодиков движущ^^^Дугони || WWW i г ниju»iiilfir.
Чаще всего это разросшийся ^хохолок — лучиего принимают выгодное для
полета положение, ин7йда'1Гсвяз’и скатим слепляются
разветвленияМи7~Ьбртзуя“-ишг~б1<с1ыда
У некоторых сложноцветных, как ^одуванчиков (Taraxacum) и коало*
бородников (Tragopogon), pq ас такие верхнеичастисёйяИкй {йб&Ш) ВЫЬоко
"11 л " L~r --Ц|№ --ш'' 1 "^^йс^1ш1Д1Шхо-
ттяратдк^а и обес^ечивае^ возможность^аметного у^язтгм пупдяот мате-
ринсйой особи. Режена семянкё^развиватсУся продольные крыловидные
п^ЯДЯ^КГ^и^летательный аппарат образуется из кроющего листа, что в
очень оригинальной форме известно у Leucocarpus pinnaiifidus — кустар-
никового сложноцветного с Галапагосских островов (рис, 390). Распростра-
нению семянок животными иногда способствуют видоизменения хохолка,
как это наблюдается у череды (род Bidens), плодики которой снабжены
двумя щитинками, покрытыми жесткими выростами, направленными вниз*
У лопухов (Lappa) и дурнишников (Xanthium) к шерсти животного прикреп-
ляется вся корзинка, поскольку листочки обертки снабжены остриями,
крючочками, или же они клейкие. Плоды с сочным околоплодником у сложно-
цветных хотя и встречаются, но представляют большую редкость (род
Wulffia из Южной Америки и Индии). Специальные приспособления к рас-
пространению семянок водой у сложноцветных редки.
Сложноцветные — самое крупное семейство среди покрытосеменных
и вообще среди растений. Оно объединяет около 4000 родов и, вероятно,
более 20 (но не менее 19) тысяч видов, причем в это число не входят апд-
миктические виды из таких родов, ^как ястребинка (Hieracium) и одуван-
чик (Taraxacum), число которых, по данный ИёКФЮрШЕТЪстэматиков, близко
к той же цифре. В семействе есть исключительные по числу видов роды.
Например, один род крестовник (включая Kleinia) содержит в своем со-
ставе оКОДО 1500 видов?ТТаряду с этим есть немало родов, имеющих крайне
ограниченное число видов, Сложноцветные распространены по всей Земле
и занимают местообитания, разнообразнеиптшг
гическим услонияит Рае^ЦИЙТ^го семейства исключительно, бО£ЭЖ. жиз-
фппмямд^пхпяняя. одцяко. дп преимуществу травянистый ха-
рактер. Кустарниковые и тем более древесные формы^уак правиле, низко-
рослы, приземисты; ^Деревъяг'Бб]^ происхождения.
В составе флоры СССР известно около 2400 видов сложноцветных, не
считая почтя 800 видов ястребинок.
В противоположность исключительному .видовому разнообразию в со-
временной флоре ископаемые остатки сложноцветных немногочисленны.
По ним можно предположить, что сложноцветные жили уже в палеогене.
Семейство' сложноцветных принято делить на два подсемейства:
Tubuliflorae, или Asterotdeae, й Liguliflorae, или Ciehartoideae.
---"По~дсе~м6~йств7Г“ТиЬиИ/1огде характеризуется тем, что все цветки
корзинки трубчатые (труоЦато-воронковирные, трубцаТ^таТУ^^Дрт^ i труб-
чато=^^1Ж<ДТчатыйу? или двугубы^ или Щё при трубчатых цветках в центре
по краю корзинки расположены ложноязычйоЬИ6Г"ПдЧ¥И вив р31Г1ВИЯЯ '"без
:г,ц
К этому "подсемейству^относится большая частъ^родов* (около 740). Не-
которые воды имеют большое хозяйственное значение,
’ ~ родом из пре-
в~хут ьт; наледгтгузгийй^ь-
ПЬи—w— hiЛл1. вин ц
вали первоначально как декоративное. ион Европы в конце A VIEB.
подсолнФ8ЯйИгтагОГ^^ сначала разводили как декора-
17—602
Г
514
выа
I! I SI У
РАСТЕНИЯ
Ряс. 391. Подсолнечник (Helianthus annuus):
1 — грызовой среднерусски й — акнораступшй
тивное и приусадебное растение ограниченного применения (суррогат кофе,
корм для птицы, лакомство). О том, что семена подсолнечника содержат
масло (современные сорта — 30—35 и до 50—60%^, стало известно лишь
в начале XVIII в. Культура подсолнечника как масличного растения воз-
никла лишь в начале XIX в. в России1 2, в черноземной полосе, где и сейчас
сосредоточены основные площади, засеваемые этим растением. Сорта подсол-
нечника, разводимого сейчас в Америке,— русские по происхождению.
Подсолнечное масло используют в пищу в натуральном виде и пере-
рабатывают на маргарин. Кроме того, оно имеет техническое применение
(в мыловарении, в лакокрасочном производстве, для получения олифы и
т. п.). Обладая высоким ростом (до 2,5, иногда до 4 л), толстыми травянис-
тыми стеблями й широкими листьями, подсолнечник образует большую
вегетативную массу и используется на силос. Существенное кормовое значе-
ние имеет подсолнечный жмых. В малоснежных засушливых районах нашей
страны стебли подсолнечника, оставленные нА зиму в поле, способствуют
снегозадержанию. Их используют также на топливо, а из золы получают
поташ. Спиртовые настои язычковых цветков и листьев применяют в меди-
। г <:
е^тсак противолихорадочное средство.
1 Сорт 17 008, содержащий свыше 60% масла, выведен в 1956 г. академиком
В. С. ПустовойтОм скрещиванием Я. annuus с Я. lenticularis.
2 Прием отжима масла из семян подсолнечника принадлежит крепостному крестья-
нину Бокареву из слободы Алексеевка, Бирючинского уезда. Воронежской губернии
(1829),
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
515
' Топинамбур, или земляную грушу (Helianthus tuberosus), разводят ради
подземных клубней стеблевого происхождения- ^лубци богаты инулином
и в основноягидут на корм скоту. Зеленую массу силосуют. _Довольно экер?
гично размножаясь" клубнями.' земляная грушд легка дцчает. Разводят и
употребляют в пищу'как' О^Щу^тишок"(Супага scolymus).
Большое' экономическое значение имеет род полынь (Artemisia),
содержащий около 250 видов.ГроцадДЬа большинство полыней^»- много-
летние травы и Полукустарники, реже однолетники. Они имеют.небольшие
корзинки, собранные* вг' метельчатые соцветия. Цветки" ветроопыляемые.
Засухоустойчивые и способные переносил» засолениёГ78^бторы^Г^Ъюыйш
(Л. lercheana, A. tlahxhdnica, A. terras albae)—немаловажный экологический
компонент наших полупустынь и особенно пустынь. Массовое произ-
растание, высокая питательность, приближающаяся к питательности зла-
ков, а возможно *даже более высокая^ способстадют-распространецию полыни
в этих засушливых раЙ(Я1ах~ как хорошего и надежного (вследствие высокой
отавности) Пастбищного' Корма. Поедаемы у полыней гдавнымобразом
листья и молодые побеги. Некоторые полыни используются в ином направ-
лени^TaKfJum^r^nauutb (А. cina), растущая только на небольшой
территории в Средней Азии, ^юдержит ^шнтоцжн- -^важное глистогонное
средство. Корзинки этой полыни, особенЯоооГзЯые сантонином, известны
в фармакологии под. названием выварного, семени и щщрльзуются в тех
же “целях, что й^антонин. ^Некоторое. лекарственное применение имеет
* горькая полынъ^С. absinthium)—повсеместно, известное 4>астение горького
II»
щеё пищеварение. Кроме того, настой горькой полыни в народной медицине
полынь также в ликеро-водочном производстве.
Тархун, или зстразон^А* dracunculus), ино^дд^раародзимю^ в ртородах,
употребляется как пряная приправа (Закавказье, Средняя Азия} и для из-
готовления _ эстрагонного" уксуса.
Промышленное значение в Северной Америке имеет гваюла (Parthenium
argentatum) — растение родом та Мексики и юга США. Содержит каучук
(2—16%) в клетках паренхимы коры, сердцевины и сердцевинных лучей,
в вёТвях “и *кбрййх:-“ ~
Перемолотые высущедш^е^-Грдрвди _ далматской ^ромашки (Pyrethrum
cinerariaefolium), растущей в горахвасточногоцобереадь? А дрпйтииадктп
моря и разводимой в ряда стран, в^ом,дасле и bJCCCP» подлазванием препа-
рат пиретруйГйГ^^ хорошее инсектацидное^^ нашей
ст^ЙСПе получают этот препарат еще та кавказской (Р. roseuni) и персидской
(Р. сатеит) ромашек, которые также введены в культуру. Русскимназва-
(нивяния), Pyrethrum (пиретрум) и иногда Anthemis (пупавка). В научной
литературе название «ромашка» обычно относится к роду Matricaria.
Один из видов этого рода (М. recutita)— лекарственное растение, используе-
мое для полосканий, припарок и т. п. Ромашкой также моют волосы, отчего
онц приобретают рыжевато-золотистый оттенок. Близкий вид непахучая ро-
машка (М. inodora)— широко распространенный сорняк. Заслуживает у поми-
нания сафлор (Carthamus tinctoria)— средиземноморское растение, разво-
димое кое-где как источник желтой и красной красок, используемых в не-
больших количествах в пищевой промышленности.
Многие виды сложноцветных широко известны как декоративные рас-
та Средиземноморья, георгины (Dahlia variabilis),
разводимые в тысячах сортов, родом из Мексики, садовые астры (Callistephus
sinensis), тоже известные в тысячах сортов, цинии (Zinnia elegans)
китайско-японские по происхождении^ виды рудбекии (Rudbeckia)2 бархат-
1'1 :•
516
hi i I \ t :
РАСТЕНИЯ
Г. Г.
цев (Tagetes) американского происхождения* бессмерт»икия или имморте-
ли* — виды родов Rhodanthe* Helichrysum, Xeranthemum, у которых
сухопленчатые окрашенные листочки обертки не изменяют при высыхаг
окраски, ни формы, К декоративным относятся комнатные и оранжерейные
хризантемы (Chrysanthemum indicum* Ch. етепзе)\ цинерарии — гибриды
Senecio cruentus и S, popuUfolius с Канарских островов. Иногда на каменис-
тых участках садов и на «альпийских горках» разводят густовойлочные
эдельвейсы (Leontopodium alpinum), распространенные в горах Западной
Европы (другие виды рода Leontopodium растут на Алтае* в Тянь-Шане*
в Гималаях и других горных системах Азии).
Многие виды сложноцветных — сорняки. Один у няибп^т? —
бодяк, или розовый (m^aowS} осот (61гг$Шт де/osum)- Корневая система его
глУбиКО (на дАже Б ^У ^^ЗМЙка^этр
корни вегетативного размножения, растуддд с^нана гериотитсгтпг™ (3 ж
осоту в7тороткое вреш: захваткг«ятт> большую рл<нрздь? развивая на ней
все ттлтумирГи новый ттп^йгя гугпртгпятлштмт ттптотс ПпштятптпткгйпотаиУЬГуг
воаникнутв"шг любой Пбсге корню и.» прядила, дпряяутхря на корнях
вегетативного размножения в мрсте коленчатого изгнба. Трудность вскоре- ^
нения этого сорнякй й известной степени объясняется тем.что придаточные '
почгаГ развиваются на корнях'Йн'а'болыпойГглубине. К образованию'до
способны ттдппльтпие. ллишжтП и^тюрезку дррня. Кроме тогол бодяк про
цирует огромное количество семянок.разнос.нымх ветромблагодаря налр*
волосистогохрхрлка. -
"Бодяк широко распространен в европейской части СССР и в Сибири.
Отрицательное япячдний'пн'тйрйиу лит1 В йдяуцймии рИЙАяат. Однако
здесь его место занимают другие сорняки, в частности принадлежащий к
Illi
I \ ii
темой, сходной с бодяком, и тоже интенсивно размножающийся корневыми
отпрысками. В емйЦЯЬ отбодяка., сильным уплотнением и за-
того,— ядовитое растение. На корню, скотьего не трогает, но поедает его
в сене, отчего наблюдались массовые отравления лгшгаде^г^НемйлутоЗ^ль
как сорное растение играет синий василек (Centaurea cyanus, рис. 392) —
M Ф * *
I \ | yi
попадают в уро^д*, причем не всегда легко' бывает отастйть от них посевной
материал. Ппи правильном 'непАтгии^ОЦяйсТВ& вйСЯЯВЯ_ДЕСКЦ ДСШфЦЙЙЙСЯ.
Из других сорных следует упомянуть че~ртополохпогшкакщий(Сагаиив
nutans) и другие виды рода С.агЛп1я1 ятяищцмлавмШЦик обыкновенный
(Senecio vulgaris). Некоторые виды, не будучи специально сорными, иногда
выступают в роли сорняков, например обыкновенмпй тжяпеяисткик (Achil-
lea millefolium) jtJUUB^u-Mggexa^Tiusilago farfara). Мать-и-мачеха, селящая-
ся обычно на смытых почвах, на обнажениях по берегам оврагов (на моло-
дых субстратах), иногда сильно засоряет посевы на хорошо увлажненных
глинистых почвах. Кроме того, она широко известна как одно из наиболее
рано зацвехдлшщх растений. _ ‘ L ----------~~
П о д с е м е й с т в о Liguliflarae характеризуется тем^что-всалветки
в ирщд^кв-явынкеаые^Раететия' этого подсемейства^как правило, содержат
млечный сок. Млечные <юс
с друТАМ 'Трубок, воз!
i: l :i
я гнп п х Я8
1 Хризантемы, георгины и астры ценятся главным образом махровнми формами. Мах-
ровыми называются саожноц ветныв, у которых трубчатые ц ветки превратились в языч-
ковые.
Рис. 392. Синий василек (Centaurea cyanus): v
1—верхняя цветоносная часть; В—нижняя часть; л—атоп в — иродоцьный рмрп корм®®1» 4 —косо*
воронковицный краевой цветок; 4 -* «рубчатый цветок; Т — шин • •
Llguliflorae — группа мт™»"»™ том Тц^ц^Ногае, и, вероятно,
проивошлацот них. Сюда orgpr?*^ токпторне пищевые растения второсте-
пенного аначения.'ТДУ'салат-латук (Lactucasativai. у которого съедобны
молодые листья прикоДйШЦМ 'ВДзеткн. используемые в молодомвозбасте.
Биологический интерёё лрцдщ'шишигДругой вид того жерода — £. scariolat
у которого стеблевые листья располагаются в плоскости меридиана так, что
листовые пластинки повернуты одним краем кверху, другим — книзу.
Как салат используют также andueuH(Cichorium endivia), иногда разводимый
в огородах. Родственный^ и~д1 ибшаювенгшй цикорий (С. iMtybuif, который
растет около дорог, по' бесшшДЕлш склонам и средней и чаще черноземной
полосе, известен своими изящными корзинками о язычковыми цветками
приятной голубой окраски. Формы цикория с толстым корнем культивируют
как однолетние растения, gx ^высушенные, поджаренные и измельченные
корни придают, натуральному кофе специфический привкус и более темную
окраску, ^йог^о испбл’ьзуют иногда толстые корни
венчика (TaraxacumofficinaTeY В млечном соке некоторых одуванчиков со-
держится каучук^ иногда в больших количествах, например в казахстанском
одуванчике, или кокгсазывег (Т. kdk-saghys), и крымском одуванчике, или
крым-сагызе (Т. КТВегпгГг^/Бблътпиы содержанием каучука в млечном соке
выделяется один из видов рода козелец — тау-сагыз (Scorzonera taw-
saghys).
Среди Ltgultflorqje есть, как и среди Tubultflorae^ сорные растения.
сорное растение, размножающееся как семенами, так и корневой порослью.
По сравнению с бодяком тюлевой осот более влаголюбив и имеет корневую
систему, менее глубокую (до 0,‘5 ле). Каждый вновь образующийся, побег
iiT
развивает свою корневую систему, благодаря чему пп^рупл^та
почвы при сильном размножении долевого осота бывают пронизаны массой
его корней. Они ломкие и очень живучие. Обломки корней могут служить
для возобновления растения даже при длине 3 еле. В южных и юго-восточных
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
519
О 50 см
1 1-1—1___L—I
Рис. 393. Молокан (Mulgedium tataricum). Строение подземной части
(зарисовки с натуры А. М. Быловой)
районах СССР большую роль в засорении посевов и полезащитных лесо-
посадок играет молокан (Mulgedium tataricum, рис. 393). Подобно горчаку,
молокан успешно растет на плотных и засоленных почвах. Как и у полевого
осота, его корни хрупкие, ломкие. По залеганию, строению и глубине про-
никновения .корней молокан подобен бодяку. Сорняк очень трудно искоре-
няется.
Особо следует отметить огромный род ястребиная (Hieracium), со-
держащий более 750 видов, в большинстве случаев трудно отличимых друг
от друга и часто имеющих гибридное происхождение, благодаря чему между
видами имеются многочисленные переходные формы.
Родственные отношения в пределах подкласса Asteridae остаются не
вполне ясными. Разумно предположить, что предки подкласса имели
верхнюю завязь и что, следовательно, ближайшими к предковым формам
подкласса Asteridae надо считать порядки Gentianales, Tubiflorae и Oleales.
Последний занимает среди Asteridae непрочное положение. Что касается
Gentianales и Tubiflorae, то они, вероятно, произошли от общего пред-
ка, имевшего правильный круговой цветок с двумя кругами тычинок
и верхнюю завязь, образованную более чем двумя плодолистиками. Развитие
этих двух порядков могло идти параллельно, причем в обоих рядах был
утерян внутренний круг тычинок, а гинецей у наиболее продвинувшихся
форм стабилизировался на двух плодолистиках. Цветки эволюционировали
в направлении совершенствования энтомофилии, но осуществлялось это
на разной основе.
В порядке Gentianales, при сохранении актииоморфности цветка, глубо-
ким изменениям подверглись андроцей и гинецей, вплоть до образования
сложно устроенного гиностемия и выработки сложного аппарата, обеспечи-
вающего прикрепление поллинариев к телу опылителей.
иг,
гл
I
ВЫСШИЕ РАСТЕНИЯ
пентамерности цветков и их зиго-
В порядке Tubiflorae также совершенствовался цветок как энтомофиль-
пый орган, но происходило это на основе изменений венчика. У высших форм
он стал зигоморфным, причем уменьшалось число тычинок, а иногда наблю-
далась их глубокая перестройка.
Переходя к порядкам с преобладанием нижней завязи, отметтс, что
один из них — Rubiales*— несмотря на существенные отличия (отсутствие *
внутренней флоэмы), считается близким к Gentianales, и единственное в этом
порядке семейство Rubiaceae даже включают в состав Gentidnales. Вместе
с тем порядок Dtpsacales сближают с Rubiales вплоть до включения семейства
Rubiaceae в состав ворсянкоцветных. Из этого следует, что порядки Genti-
anales, Rubiales и Dtpsacales — члены одного эволюционного ряда. Он завер-
шается семейством ворсянковых,, для которого характерно соцветие, внешне
сходное с корзинкой сложноцветных.
Два последних порядка, характеризуемых нижней завязью (Campanula-
les is Asterales), имеют много обще
морфности, высших, пре авителей отметим тендещда _ ДЯНению
пыльников вокруг столбика. парака"5пнПЙ гинстиУЙГнижнюю завяэь. Ъ био-
логин цветка имеются^ходныв лииеШЕш ния к перекрестному опылению
и самоопылению. На ногих представителей того и другого порядка
членистых млечных трубок также говорит о сходстве этих групп. Сходство
во многих признаках сочетается и с существенными различиями, которые
препятствуют объединению этих порядков в один. Campanulales и Asterales,
вероятно, две линии развития, возникшие от общего предка, который примы-
кал к общему предку подкласса Asteridae или к древнейшим Tubiflorae.
Подкласс Asteridae в целом рисуется как возникший от раздельно-
лепестных двудольных с пентамерными раздельнолепестными цветками и
верхней завязью. Советский ученый А. Л. Тахтаджян, предложивший деле-
ние двудольных на шесть подклассов, принятое в этом руководстве, считает
родоначальником Asteridae подкласс Rosidae.
HI
ЛС
. i: i
2-й КЛАСС—ОДНОДОЛЬНЫЕ (MONOCOTYLEDONEAE)
Однодольные объединяют около 25% всех видов и семейств цветковых
растений и менее 13% порядков, т. е. малочисленное, чем двудольные. Но
в некоторых условиях (например, на лугах, в плавнях устьевых частей рек)
однодольные могут преобладать над двудольными и по числу видов, ц, глав-
ное, по числу особей. Иногда преобладание однодольных распространяется
я на очень обширные территории, соизмеримые с зонами и подзонами, как,
например, на некоторые типы степей и саванны.
Большинство однодольных — травы. Реже встречающиеся среди одно-
дольных древовидные растения отличаются от деревьев ив двудольных уже
НГ.
ские травы (бамбуки, Драцены, юкки и_даже палы№). Свойственная немно-
гим ОДНОДОЛЬНЫМ способность КО вторичному утолптению стеблей fa СТВОЛОВ-
осуществляется не ва wtwr кямОия. как вчч> ^JSubho ИЛн^ДВУД0лт.яихг а путам
обряяовявкунодрй меристемы, возникяттпей в периферической части стебля.
Меристема дает начало новым проводящим щлпмшдг няратцимд. основную
паренхиму. ' ~
время редко подвергается сомнению. Обычно принимают, что оба класса
вместе составляют естественную группу общего. происхождения. Это под-
тверждается тем, что между двумя классами пветковых растений невозможно
найти резкую, грань. Даже рпитпггие в органийщТГ" »ар<мрдпа~нё'ЙВЙ1!да вы-
держано. Известно немало двудольных растений с одной семядолей: чйсииРс
(Ranunculus ftcaria), хохлатки (Coq/dalii), некоторые зонтичные и др. Хотя
,п очень редкод встречаются однодольные с двумя семядолями (Agapanthus
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
umbellatus из лилейных, Calocasia antiquorum из аронниковых). Бывают
случаи наличия второй семядоли в редуцированном состоянии.
Крайним выражением представления о тесном единстве двух классов
покрытосеменных можно считать не раз высказывавшееся мнение, что раз-
; 1: г
ЛИНИЙ двудольных И С ЭТОЙ тошен ярртпгя однппплдрые. яипАЛяямтдйкяНпт-
дельный класс, представляют группу искусственную, конвергентную.Такое
мнение оправдано относительно спайнолепестных двудольных, но что каса-
ется однодольных, то оно не имеет убедительного подтверждения. Порядки,
включаемые в класс однодольных, тесно связаны друг с другом в достаточно
стройных эволюционных рядах, наиболее специализированные члены кото-
рых через промежуточные звенья примыкают к относительно примитивным
представителям класса, вероятно родственным исходным предкам всех одно-
211»
дольных.
Филогенетические отношения между однодольными и двудольными
могли бы быть троякого рода: 1) однодольные — потомки двудольных;
2) двудольные - потомки однодольных ДВуДольныеи^дно-
дольные ^Ипбтомки каких-то первичных форм покрытосеменшд^эМ^йб^Уо-
нировавшиевполйён^завйсймо’другот друга. Третья из этах^шояшостей
вряд ли могла иметь уеёто; во всяком ^случае, принпмяя пдйойятйГ
поставить вопросы: обладал ли этот общий гипотетический предок зароды-
шем с одной или двумя семядолями? Была ли у него эустелическая или
атактостелическая структура стебля? И т. д. Таким образом, в сущности,
остаются только две первые возможности. Выбор одной из них определяется
сравнительным анализом признаков, отличающих два класса цветковых,
в первую очередь таких, как строение зародыша, анатомия стебля и строение
цветка.
В пользу представления о вторичном, производном характере одно*
семядольного зародыша свидетельствуют эмбриологические данные. Пока-
зано, что на ранних стадиях ряяяятая вяппттыши щптолольныт и ДВУДОЛЬНЫХ
ется
похожи. На йЛбс
изложение зачата _________
правилбТдбстигают полного и более илименее одинакового развития. У одно-
дольных одиц^из зачатков растет быстрее, оттесняя при этом вбок и конус
нарастания, и вторуюТостаюшуюся пряк^ртескц исчедаюшую
семядолюГМожно предагаложить, что однодольность зародыша есть резуль-
11ГЛ
г. г,
l
й в величине двух семядолей предков однодольных.
Гетерокотилия могла иметь и иной характер. Так в отношении некото-
рых однодольных "указывалось на то, что одна семядоля их двудольных
предков превратилась тс пргян. восприндгмяюптий питательные вещества
иэ эндосперма, в то время как вторая стала развиваться в зеленый ассимили-
рующий лист-(семейство аронниковш).
Предположение о возникновении одной семядоли однодольных путем
слияния (срастания) двух не получило подтверждения.
В недавнее время (1960) советский ботаник К. И. М е й е р на основа-
результатов изучения развития зародыша у нимфейных (двудольных,
но имеющих ряд признаков, сближающих их с однодольными) высказал
мнение, что развитие зародыша у однодольных происходит по сравнех
с таковым у двудольных сокращенно. Мейер наблюдал, что у нимфейных
формирование почечки, расположенной (как это бывает обычно у двудольных
растений) между двумя семядолями, начинается с появления на верхушке
точки роста стебля двух выростов, разделенных неглубокой ложбинкой,—
зачатков двух первых листьев почечки. Получается картина, весьма напоми-
нающая образование семядолей двудольных. Из двух возникших таким
образом листовых зачатков один растет гораздо быстрее и сдвигает в сторону
второй зачаток, который отстает от первого в развитии и оттесняется к осно-
ванию первого листа. Между этими листьями возникает точка роста стебля.
Ml
ВЫСШИЕ РАСТЕНИЯ
ими
Этот процесс очень сходен с развитием зародыша однодольных. Мейер вы-
сказал предположение, что единственная семядоля однодольных соответст-
вует третьему листу зародыша двудольных, считая за два первых их се-
мядоли. Таким образом, зародыш однодольных можно рассматривать как
возникший из зародыша двудольных путем выпадения в процессе раз-
вития двух первых листьев последнего и превращения третьего листа в се-
мядолю.
В анатомическом отношении выше отмечены две черты, характерные
для однодольных: закрытые, не содержащие камбия проводящие пучки и их
II г.
II Г. I ’>
ri
Г, ifl
у ряда одцодольных, особенно молодых, находят крауцовременно действую-
; ~ камбий или следы его. Это установлено как в относительно примитивных
семействах (например, частуховые, водокрасовые), так и высокоспециализи-
рованных (злаки, орхидные). Камбий в стеблях однодольных имеет явно
остаточный характер, а наличие его показьтает, что предки однодольйЪгх
имели камбиальное утолщение стезёй Г^тсутствиейлислаббе “ ^развитие
камбия и у многих двудольных подтверждает это предположение. По этому
признаку, следовательно, однодольные — вероятные потомки двудольных.
Редукция камбия и вытекающая из этого потеря стеблем способности
к камбиальному утолщению сделали возможным размещение проводящих
пучков не только в ограниченном поясе стебля, как у двудольных, но и по
всей его толще. Интересно, в связи с этим, что рассеянное (на поперечном
срезе) расположение проводящих пучков с редуцированным или отсутствую-
щим камбием наблюдается и у двудольных (Actaea, Thalictrum. некоторые
Anemone из лютиковых, Podophyllum из барбарисовых, Peperomia из пе-
речных, виды нимфейных и др.). У однодольных размещение пучков в одном
круге, т. е. по типу двудольных,— явление редкое (семейство диоско-
рейные, а также Burmanniaceae). Очевидно, в стебле, построенном по типу
однодольных, может разместиться большее число пучков, чем в одном ряду,
как у двудольных. С этим связана замечательная черта листьев однодоль-
ных— они обычно имеют широкое основание, через которое в стебель входит
большое число проводящих пучков. Они и участвуют в анатомическом
сложении стебля. Множественность листовых следов проявляется и в жил-
ковании листовых пластинок (параллельно-нервное и дугонервное). Биоло-
гически большое число проводящих путей, связывающих листья однодоль-
ных с их стеблями, иногда объясняют повышенной продуктивностью одно-
дольных.
Преобладание тримерных кругов в циклических цветках однодольных
вместо пентамерных, характерных для двудольных, так же как и выше рас-
смотренные отличия этих классов, — признак неустойчивый. У одного и
того же вида, характеризуемого пятичленными круг^ш околоцветника,
иногда наблюдаются и трехчленные мутовки. Например, у сон-трава
{Pulsatilla patens) и близких к ней видов в типе пятичленный околоцветник
часто при развитии дополнительного листочка реконструируется в два трех-
членных круга. Элементы этих кругов довольно правильно чередуются друг
с другом. Аналогичное явление известно у дубравной ветреница {Anemone
nemorosa) и др. Поэтому не приходится удивляться, что тримерные цветки
могли стать константными среди некоторых двудольных. Это можно видеть
у некоторых магнолиевых, кувшинковых, барбарисовых, гречишных, лав-
ровых и др. Вполне вероятно, что тримерность цветков однодольных воз-
ла от их пентамерных двудольных предков.
Другие различия между однодольными и двудольными также не проти-
воречат допущению происхождения однодольных от двудольных.
В разнообразии однодольных издавна отмечали три довольно обособ-
ленные группы: 1),. водные однодольные, у многих из которых имеется апо-
карпный гинецей, и в этом можно видеть отражение признаков отдаленных
предков однодольных; 2) растёйияпв тйелкими, нередко редуцированными
цветками, объединенными в соцветия, которые сопровождаются одним или
nt:»>
1.1111
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
несколькими верховыми листьями, защищающими молодое соцветие в про-
цессе его развития (выполняют роль чашечки) или функционируют как
аппарат, привлекающий животных-опылителей (аналогично венчику); эта
группа однодольных как бы аналогична зонтичным или сложноцветным из
двудольных; 3) прочие однодольные.
В настоящее время однодольные разделяют на четыре подкласса.
1-Й ПОДКЛАСС АЛИСМИДНЫЕ (AL1SM1DAE)
У алисмидных гинецей апокарпный или примитивно ценокарпный.
Зрелые семена почти всегда без эндосперма; если же он есть, то некрахма-
листый. Пыльцевые зерна обычно трехъядерные. Травянистые водные или
полуводные растения, без сосудов или с сосудами только в корнях.
В этом подклассе различают три порядка.
1-Й ПОРЯДОК — ЧАСТУХОЦВЕТНЫЕ (ALISMALES)
Околоцветник частухоцветных дифференцирован на чашечку и венчик,
завязь верхняя, плодолистики свободные (апокарпность) или почти сво-
бодные.
* Семейство
(Butomus) с единственным видом
В. umbellatus — зонтичный сусак,
который распространен в Европе
и умеренной Азии. Это многолет-
нее травянистое прибрежное ра-
стение, несущее зонтиковидное со-
цветие с прицветным листом. Ли-
сточки околоцветника окрашен-
ные, в двух трехчленных кругах.
Тычинок 6—9 и столько же сво-
бодных плодолистиков.
* Семейство частуховые
(Alismataceae). Цветки в слож-
ных соцветиях, у большинства
обоеполые, актиноморфные с
трехчленными чашечной и венчи-
ком. Тычинок и пестиков по
шесть или по многу. Гинецей из
3, 6 и более свободных или срос-
шихся только у основания плодо-
листиков, расположенных спи-
рально илП мутовчато. Семяпоч-
ки расположены в каждом пес-
тике, по одной или по нескольку
на брюшном шве одногнездной
завязи. Плоды у большинства —
семянки, снабженные воздухо-
носной тканью и разносимые
водой. Листья у многих — сет-
чато-нервные, черешковые. В ве-
гетативных органах нередко раз-
виты млечные ходы.
Семейство Пастуховых вклю-
чает больше 70 видов в умерен-
ных и холодных областях. В
СССР — 13 видов, относящихся
к 5 родам.
сусаковые (Butomaceae)
содержит один род сусак
Рис. 394. Частуха (Alisma plantago-aqua-
tica):
1 — соцветие: 2—прикорневые листья; 2 —цве-
ток; 4 ~ плоды; а—.диаграмма цветка (тычинки
противочашечные, расщепленные и раед—яутыв
в каждой паре, тан что расошюнмны по две про-
тив лепестков)
524
выа
11 i \ I:
Частуха (Alisma plantago-aquatica) часто растет по сырым и топким
местам, имеет белые или бледно-розовые некрупные цветки в пирамидальной
метелке (рис. 394) и листья в прикорневой розетке, напоминающие листья
подорожника. Она ядовита, но печеные корневища ее, богатые крахмалом,
съедобны.
Стрелолист (Sagittaria) тоже болотное и прибрежное растение, воз-
душные листья его стреловидные, в прикорневой розетке, а погруженные —
Цветки однополые: нижние пестичные, верхние — тычи-
ночные. Осенью стрелолист образует съедобные подземные клубеньки.
\ i:i
:гл
\Г 2-Я ПОРЯДОК — ВОДОКРАСОЦВЕТНЫЕ (fiYDROCHARITALES)
Водокрасовые сходны с частухоцветными тем, что околоцветник диф-
ференцирован на чашечку и венчик, но в отличие от них имеют паракарпный
гинецей* и нижнюю завязь с неполными перегородками между гнездами.
Семейство водокрасовые (Hydrocharitaceae) от предыдущего отли-
чается нижней одногнездной завязью с 3—6 пя риетальными плацентами,
у некоторых далеко вдающимися в полость завязи. Цветки у многих одно-
полые, двудомные. В тычиночных цветках часто редуцирован пестик, а в
пестичных — стаминодии. Тычиночные цветки по нескольку, а пестичные
по одному заключены до цветения в покрывало (чехол), образо-
ванное одним или двумя кроющими листьями. Плод — ягодообразный,
разрывающийся по созреваг
Растения этого семейства плавают на поверхности воды или погружены
в нее. У последних соцветия торчат из воды или плавают на поверхности ее.
У некоторых водокрасовых цветение происходит под водой и рыльца опыля-
ются плавающей нитевидной пыльцой. Широко распространено вегетативное
размножение при помощи особых зимующих почек (так называемые ту-
ри о и ы или гибернакул ы). Водокрасовых известно около 80 —
90 видов; в СССР — 6 родов и 7 видов.
Водокрас, или лягушечник (Ну drocharis morsus-ranae), — небольшое
изящное пресноводное растение с округло-сердцевидными листьями, плава-
ющими на поверхности воды, и белыми цветками (рис. 395). Его зимующие
почки, отделяющиеся от материнского растения и снабженные слизистыми
оболочками, могут легко приклеиваться к животным и переноситься в дру-
гие водоемы.
Телорез (Stratiotes abides) образует розетки из широколииейных, по
краям колючезубчатых листьев, плавающих под водой. В огромных коли-
чествах нередко встречается в озерах, старицах, прудах и т. п. Ко времени
цветения растения телореза листья всплывают на поверхность, а затем
снова погружаются. Размножаются они преимущественно зимующими
почками, опускающимися на дно.
Элодея, или ^водяная чума» (Ebdea, или Helodea canadensis),— погру-
женное пресноводное растение с трехчленными мутовками листьев. Завезен-
ная в Европу из Северной Америки, она была впервые замечена в Ирландии
в 1836 г. Затем она обосновалась на континенте и распространилась по всей
Европе (1885 г.). В конце XIX и начале XX в. обнаружена в России — под
Коломной, в Зауралье, на Дону. Местами при массовом развитии она даже
затрудняла судоходство и получила наименование «водяной чумы» и «водяной
заразы». Завезены были только женские экземпляры, не дающие без муж- -
ских растений семян, и распространение шло путем вегетативного размно-
11i \
участками побегов.
Валлиснерия (Vallisneria spiralis) — подводное растение пресных водо-
емов юга европейской части СССР, Южной Европы и других теплых стран,
часто разводится в аквариумах. Это растение имеет очень оригинальное
' перекрестное опыление.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Виды нескольких родов (Enalus,
Thalassia, Halophila) образуют местами
обширные подводные заросли на глубине
до 10 м по берегам Индийского и Тихо-
го океанов; приспособление к подводно-
му образу жизни у них зашло так дале-
ко, что цветение и опыление происходит
под водой.
3-Я ПОРЯДОК — РДЕСТОЦВЕТНЫЕ
(POTAMOGETONALES)
Рдестоцветные характеризуются
цветками без околоцветника или с
простым околоцветником.
Семейство рдестовые (Potamogetona^
,сеае)— водные многолетние растения,
или совершенно погруженные в воду,
или с плавающими на поверхности воды
листьями. Цветение происходит или
над водой, или под водой, и тогда
пыльца нитевидная (у морской травы—
Zostera) или же цветки клейстогамные.
Цветки невзрачные, собранные в ко-
лосья, реже одиночные, обоеполые или
раздельнополый, однодомные. В цвет-
ках характерно четверное число листоч-
. ков околоцветника, тычинок и пестиков
или вследствие редукции число их еще
меньше. Плоды — орешки или костян-
ковидные. Для большинства рдестовых
характерно наличие пазушных при-
листников, вообще очень редкое у од-
нодольных. '
Семейство насчитывает около 125
видов. Наиболее крупный род —
рдест (Ро tamoge tori), широко распро-
страненный по всему свету. В СССР
растет около 40 видов рдеста (рис.
396). Анемофильные обоеполые цветки
у рдестов собраны в колосья, высовы-
вающиеся из воды. В цветках против
четырех буроватых листочков около-
цветника расположены четыре тычинки.
На листьях и стеблях многих рдестов
нередко обильно отлагается углекис-
лая известь.
Виды взморника, или морской трог
вы (Zostera), с длинными узколиней-
ными листьями, нередко образуют об-
ширные подводные луга на песчаном
или илистом дне прибрежной полосы
морей. Плоские его соцветия окружены
влагалищем верхнего листа. Околоцвет-
ника нет. Мужские цветки, состоящие
из одной тычинки, и женские, состо-
Рио. 395. Водокрас (Hydrocharis
morsus-ranae)
Рис. 396. Рдест плавающий (Pota-
mogeton natans)
ящие из одного пестика, расположены
на одинаковых уровнях^ и такую пару
826
ВЫСШИЕ РАСТЕНИЯ
иногда считают за обоеполый цветок. Zostera maritima используется
под названием «морской травы» для набивки матрацев, мебели и как упа-
ковочный материал.
В подклассе Alismidae наиболее примитивным считается поря-
док Alismales, характеризуемый апокарпным гинецеем и отсутствием
сосудов в стеблях. По этим признакам Alismales пршшмг к подклассу
Magnoliidae, в частности к порядку Nymphaeales, Но вместе с тем Alismales
обладают и признаками высокой специализац ии: отсутствием эндосперма
в зрелых семенах, наличием сосудов в корнях. Такое сочетание признаков
считается показателем того, что Alismales представляют одну из боковых
ветвей эволюции, взявшую начало* вероятно* от древнейших однодольных.
2-Й ПОДКЛАСС — ЛИЛИИДНЫЕ (L1L11DAE)
Гинецей у лилиидных ценокарпный^Семеиа с эндоспермом, содержащим
различные питательные вещества, редко крахмал. Пыльцевые зерна главным
образом двухъядерные, реже трехъядерные. Околоцветник простой, венчико-
видный, Растения наземные, редко водные, в основной массе травянистые,
редко древовидные. У большинства сосуды сосредоточены только в корнях.
Энтомофильные.
К подклассу лилиидных (Liliidae) отнесены пять порядков.
1-Й ПОРЯДОК — ЛИЛИЕЦВЕТНЫЕ (L1LIALES) у
Лилиецветные — большой порядок, в котором наиболее характерно
выражено типичное для многих однодольных строение цветков и вегетатив-
ных органов. Огромное большинство этих растений — многолетние травы
с корневищами, реже с луковицами. Цветки чаще всего из пяти трехчленных
кругов, актиноморфные. Околоцветник простой, однородный, у большинства
венчиковидный, у меньшинства чашечковидный. Пестик один из трех плодо-
листиков, синкарпный, с трехгнездной завязью и центрально-угловой пла-
центацией.
Семяпочки анатропные, у большинства горизонтальные. Плод — коро-
бочка или ягода, орешек. Семена у всех с эндоспермом.
Наиболее типичным и, вероятно, исходным для прочих семейств явля-
ется семейство лилейные (Liliaceae). От них эволюция в пределах порядка
шла в разных направлениях: уменьшение числа кругов в цветках (касатико-
вые, некоторые ситниковые), образование нижней завязи (амариллисовые,
касатиковые и др.), превращение околоцветника и соответственно переход
от энтомофилии к анемофилии, образование однополых цветков.
В филогенетическом отношении лилиецветные — древний порядок,
развившийся, вероятно, из предков, общих с предками алисмидных.
г" Семейство лилейные (Liltaceae)— центральное в порядке, с наи-
более характерно выраженными признаками его. Большинство лилейных —
многолетние травы с корневищами или луковицами. Листья чаще всего уд-
линенные, ланцеУбТОЯЯйе^или линейные', "пйр^ЛЛёльно- или дугонервные,
пейыюкрайние или зубчатые. Цветки обоеполые, актиноморфные, у боль-
шинства средних или крупных размеров, хорошо заметные. Редко они оди-
ночные, обычно собраны в кисти, колосья, метёлки, зонтиковидные и другие
соцветия. 'Околоцветник у громадного большинства венчиковидный, из двух
трехчленных кругов, разднльнолистпый или сростнолистный. Тычинок
шесть —чпгтривдвух^кругахжредко — больше.Щестик один, из трех плодо-
гжт1Ж(Й7"и6разуют^х одцу верутлпш тр^хотйятгную завяовтчк^ некоторых
три, у большинстввкже один столбик. У очень'немногих завязь полунижняя.
Плаценты центрально-угловые. Семяпочки анатропные, чаще расположен-
ные горизонтально. ХТпыление у большинства происходит при помощи на-
III'I
’ЛИ
ин
III
HI
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
(Г11Г I
hi
секомых. Плод — каробпчка1. иппгла ягода. Семенас эпдплпАрхпм^ со-
держащим белки, масл^~редсекрахмал. Зародыш окружен эндоспермом.
Семейство лилейных включает 3500—4000 видов. Большинство лилей-
ных приспособлено к жизни в сухих областях. В СССР обитает около
640 видов. Многие лилейные имеют практическое значение (овощи, лекар-
ственные растения, декоративные, пр ядильные).
Из многочисленных подсемейств рассмотрим следующие 8.
Подсемейство Melanthoideae. Оно выделяется неполным срас-
танием плодолистиков. Характерный представитель — чемерица (род Уего-
frum), виды которой (всего их около 50) распространены в Неверном полу-
шарии.
В СССР часто встречается на равнинных и горных лугах У. lobelianum —
крупное растение, с широкими овальными листьями. Содержит ядови-
тые алкалоиды, частично использующиеся в медицинских целях.
Подсемейство Asphodeloideae объединяет корневищные ли-
лейные с терминальными соцветиями, ассимилирующие листья сосредото-
чены в основаниях цветоносных стеблей. К этому подсемейству относится
род эремурус, или ширят (Eremurus). Из прикорневой розетки листьев
выходит безлистный стебель высотой 0,5 — 2 ж, заканчивающийся крупной
красивой кистью цветков (у Е. robustus в кисти около 800 цветков). На юге
нашей страны (главным образом в Средней Азии) встречается 23 вида эре-
му руса. Некоторые виды разводят как декоративные.
Род красоднев, иди лилейник (Hemerocallis), объединяет 6 видов (глав-
ным образом из Южной Европы и умеренной Азии), 5 из них обитают в СССР.
Один из видов красоднева (Я. fulva) широко распространен в садах как
неприхотливое многолетнее декоративное растение. Его крупные воронко-
видные рыжевато-оранжевые цветки с глубоко раздельным околоцветником
похожи на цветки лилий. Сюда же относят род Hosta, виды рода Hosta
разводят в садах под названием функии.
Род Hemerocallis сближают также с родом Phormium, представ-
ленным двумя видами из Новой Зеландии. Один из них — новозеландский
лен (Р. tenax)— крупное корневищное многолетнее растение’ с розетками
листьев по 1—2 м длиной. Волокно этого растения используют в веревочно-
канатном производстве, при изготовлении грубых тарных и мебельных тка-
ней, иногда для имитации шелка и изготовления бумаги. В СССР новозеланд-
ский лен культивируют в небольших количествах на Черноморском побе-
режье Кавказа.
Обособленное положение в этом подсемействе занимает большой (250 ви-
дов) род алоэ (Aloe). Алоэ — листовые суккуленты, распространенные
главным образом в Южной Африке и на Мадагаскаре. Цветки со спайно-
листным околоцветником, часто опыляемые птицами. У некоторых видов
алоэ стебли несут листья по всей длине и ветвятся, иногда встречаются
одревесневшие стебли (рис. 397). *
Некоторые виды алоэ разводят как комнатные растения,' например
A. arbarescens, очень редко цветущий (отсюда название «столетник»). Алоэ
применяют в медицине (слабительное средство, биогенные стимуляторы).
Подсемейство Wurmbaeoideae содержит клубневые и клубне-
луковичные лилейные. Интересен род безвременник (Colchicum). Некото-
рые виды, его цветут осенью. Цветки безвременника имеют спайнолистный,
с длинной трубочкой околоцветник; нижняя часть цветков остается после
цветения погруженной в почву. На следующую весну появляются листья
и созревающие плоды. Создается впечатление,! что у этого растения плоды
появляются раньше цветков.
ш
1:11
1 По типу раскрывания различают септицидные и локудицидные коробочки. У пер-
вых нарушение целостности происходит по перегородкам, отделяющим гнезда друг от
друга. У вторых продольные отверстия на стенках коробочки образуются по средней т*
нив каждого плодолистика. Септицвдное раскрывание считается более древним признаком.
ВЫСПШВ РАСТЕНИЯ
Рис. 397. Алоэ (Alofi arborescens)
Безвременники содержат
ядовитые алкалоиды —- колхи-
ПН
в медицине: колхицин применя-
ют также для получения поли-
плоидных форм.
г. г
IU
Рис. 398. Гусиный лук желтый (Gagea lu-
tea):
2— общий вик й—продольный редев цветка
подсемейство ли-
лейные (Lilioideae) характери-
зуется луковицами иногда в
сочетании с корневищами; цве-
тоносные стебли олиственные;
соцветия разнообразные (кис-
ти, зонтиковидные)^ без по-
крывала.
Гусиные луки (Gagea, рис.
398) — мелкие растения с уз-
кими листьями и некрупными,'
чаще желтыми цветками. Цве-
тут рано весной и быстро за-
канчивают вегетацию.
Тюльпан (Tulipa) с круп-
ными ярко окрашенными, у
большинства одиночными и без
запаха цветками. Луковицы
пленчатые. В СССР свыше 60
видов, большинство обитает в
Средней Азии. Тюльпаны раз-
водят как декоративные (из-
- вестно около 800 сортов).
В Западную Европу куль-
тура тюльпанов перешла из
Турции в середине XVI в. Осо-
бенно большого развития она
достигла в Голландии.
Рябчик (Fritillaria) русское
название получил от пестрого
рисунка на листочках около-
цветника многих видов; неко-
торые из них декоративны; в
СССР около 25 видов — на лу-
гах, в степях, на горных скло-
нах.
Лилия (Lllium) имеет круп-
ные ярко окрашенные цветки,
часто душистые и собранные в
кисть (реже цветки одиночные).
Луковицы лилии черепитчатые
(рис. 399). В СССР 16 видов
лилий, большинство их растет
на Кавказе и в Восточной
Азии. В Сибири и на Камчат-
ке до развития там земледелия
луковицы лилий, называемых
сарана, употребляли в
Многие виды лилий разводят
как декоративные.
I ГЛ 11
1Г1
мсковые (Scilloideae) также ха-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
529
11:11
растеризуется луковицами, по-
крытыми кожистыми чешуями;
ассимилирующие листья при-
корневые; цветки в пазухах
верховых листьев.
Пролеска, или голубой
подснежник (Scilla),— растение
с красивыми, у большинства
голубыми цветками средних
размеров. В СССР — 17 видов
в черноземной полосе и южнее;
Некоторые пролески разводят-
ся как ранневесенние декора-
тивные растения.
С пролесками сближают
род Urginea, распространен-
ный в Средиземноморье. Один
из видов этого рода известен
под названием морского лука
(U. maritima). Его крупные
луковицы содержат глюкозид
сердечного действия — сциля-
рин. Разновидность морского
лука с красными луковицами
содержит крысиный яд, при-
меняемый в элеваторах, скла-
дах. Культура возможна на по-
бережье Кавказа.
Гиацинт (Hyacinthus) име-
ет воронковидный околоцвет-
Рис. 399. Лилия (Lilium mart agon):
1 — основание стебля; г — соцветие;
ник. В роде около 30 видов
(в Средиземноморье и Африке).
В СССР — 3 вида в горах
Копетдага. Близкий род Hyacintella (в СССР он распространен в степях).
Родоначальник культурных гиацинтов с прекрасными ароматными цвет-
ками — Н. orientalis, обитаюп(ий в Иране, Малой Азии, Сирии и АРЕ. В За-
падную Европу гиацинты завезены в середине XVI в.
Подсемейство луковые (Allioideae) включает луковичные и
Iг I;
корневищно-луковичные растения, для которых типично покрывало из
1—2 листьев, расположенных в основании соцветий; покрывало скрывает
до цветения зонтиковидное или цимозное соцветие.
Большой род лук (Allium) включает около 400 видов многолетних
трав, в большинстве с резким запахом. У многих, кроме луковиц, имеется
корневище.* У некоторых луков влагалища листьев, образующие луковицу,
иг
UII
мало утолщаются, и поэтому луковицы слабо развиты. Пластинки ассимили-
рующих листьев желобчатые или трубчатые, цилиндрические. Соцветия
зонтиковидные, в молодости заключенные в чехол из кроющих листьев.
Околоцветник раздельнолистный или у основания сростнолистный (рис. 400).
У многих представителей этого рода в соцветиях развиваются луковички.
Плод — коробочка. Листья плоские, желобчатые или трубчатые, цилиндри-
ческие. Некоторые виды разводят как пряные овощи: репчатый лук (А. сера,
рас. 400, 2; 401, 2); лукгбатун, ulsol татарка (A. fistulosumY, зимующий
в грунте в центральной полосе СССР; лук-порей (A. роггит, рис. 400, 2) —
один иэ нежных луков с нетипичной удлиненной луковицей; чеснок (A. sati-
vum) со «сложной» луковицей, содержащей много луковичек-одеток»
(рис. 400, 4; 401, 2)\ шнитт-лук (скорода у или резанец,— A. schoenoprasum,
рис. 400, 3) с лилово-розовыми цветками* разводимый часто как декоратив-
ный многолетник, и др.
< г. ।
. п г
530
ВЫСШИЕ РАСТЕНИЯ
Рис. 400- Луки:
1 — лук реачашй (АШшп сера); S—лук-аорей (А. рог-
гшп); Э — шшвт-аук (А. schoenoprasum); 4 — чеснок
<А. sativum)
Рис. 401. Соцветие и строение лу-
ковиц лука и чеснока:
1 — репчатый лук: а — ссцветае; б — про- ’
дольный разрез луковицы; $ —чеснок: а —
поперечный разрез луковицы; б — дочерние
яунонкчкн (зубки)
Наиболее широко и повсеместно распространен \у нас в культуре реп-
чатый лук. Листья у него трубчатые, внутри полые. Цветонос посредине
вздутый, соцветие шаровидное; цветки мелкие, зеленовато-белые, проге-
рандрические, перекрестно опыляемые. Плод — трехгнездная коробочка
с черными семенами неправильной формы (их называют «чернушка»).
Репчатый лук имеет очень много сортов, различаемых по разным при-
знакам: вкусу, окраске наружных чешуй луковицы1, форме и величине
луковиц и т. д. Острый запах лука зависит от эфирного лукового масла.
Употребление лука, как и других пряных растений, в качестве приправы
к пище увеличивает выделение желудочного сока и этим содействует лучшей
перевариваемости пищи. Кроме того, зфирное луковичное масло действует
губительно на бактерии. В луке содержатся витамины В и С. Хотя витамина
С при этом сравнительно немного, в луке содержатся, вероятно, еще какие-то
пока не изученные вещества, которые вместе с витамином С сообщают ему
высокие антицинготные свойства1 2.
1 Наружные луковичные чешуи содержат у ряда сортов антоциан, а также крася-
щее вещество кверцетин и могут применяться для окрашивания тканей и других материа-
лов в желтый цвет.
2 Высокую противоцинготную активность имеют также другие виды луков и чеснок.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
531
гл:
лн
иг.
Луковицы репчатого лука замерзают уже при —2 °, поэтому его у нас
обычно не оставляют зимовать в грунте. Лук разводят в северных, централь-
ных и южных районах СССР как двухлетнюю, а на семена — трехлетнюю
культуру. В первый год из семян («чернушки») выращивают только мелкие
луковицы, так называемый «севок». На второй год из высаженного на гряды
севка развиваются большие луковицы — «репка». На третий год высажен-
ные отборные луковицы дают цветочный стебель и семена. На юге СССР
скороспелые, преимущественно сладкие, сорта лука выращивают на «реп-
ку» в одно лето. Севернее эти сорта тоже могут давать «репку» в одно лето,
если высаживать в грунт рассаду, выращенную предварительно в пар-
никах.
В диком состоянии репчатый лук неизвестен. В горах Ирана растет
близкий к нему А. сера var. Silvestris, Есть виды с приятно пахнущими цвет-
ками. Некоторые луки разводят как декоративные. В СССР известно около
230 видов дикорастущего лука, большинство в горах Средней Азии. Некото-
рые из этих растений местное население употребляет в пищу. Таковы
черемша, или колба (A. victorialis), которая встречается преимущественно
в Сибири; медвежий лук, или черемша (A. ursinum),— в европейской части
СССР и на Кавказе и др. На лугах и пастбищах лук нежелателен, так как
при поедании животными он придает неприятный вкус молоку.
Род Allium вместе с родом Agapanthus и некоторыми другими иногда
выделяют в особое семейство Alliaceae.
Подсемейство Asparagoideae в отличие от предыдущих подсе-
мейств имеет плоды — ягоды. Ландыш (Convallaria) — изящное растение с
белыми ароматными цветками и красными ягодами. Ландыш широко распро-
странен в лесах европейской части СССР, Кавказа, Дальнего Востока. При-
меняют его в медицине как сердечное средство. Растение ядовито, иногда раз-
водят как декоративное, хорошо выгоняется зимой.
Купена. или соломонова печать (Polygonatum), образует довольно толс-
тые корневища, развивающие ежегодно олиственные цветоносные побеги,
основания которых оставляют следы на корневищах. Ягоды темновато-сизо-
ватые.
Майник (Mafanthemum bifolium) — маленькое лесное растение, цветущее
одновременно с ландышем. Мелкие белые ароматные цветки, собранные в
кисть, построены по двучленному типу.
Вороний глаз (Paris) назван так из-за сизовато-черных ядовитых ягод,
сидящих поодиночке на вершине невысокого стебля. Цветки у него неяркие,
зеленовато-желтоватые, с 8 листочками околоцветника и 8 тычинками. В лесах
СССР встречаются 4 вида вороньего глаза.
С ландышем, майником и вороньим глазом родственна Aspidistra, один
из видов которой (A. elatior) , родом из Японии, часто, используется для озе-
ленения комнат. Это неприхотливое теневыносливое растение имеет четырех-
членные цветки. Листья ее кожистые, удлиненно-эллиптические, на длинных
черешках, темно-зеленые.
Спаржа (Asparagus) — больший род, содержащий свыше 100 видов (в
СССР — 24) трав, полукустарников, лиан (рис. 402). Функции фотосинтеза
выполняют зеленые, сильно разветвленные или линейные кладодии (видоиз-
мененные ветви), сидящие пучками в пазухах очень мелких, недоразвитых
пленчато-чешуйчатых листьев. Цветки мелкие. Обитающие в СССР виды
двудомные. Ягоды у большинства красные. Как деликатесный овощ разводят
A. officinalis var. altilis, В пищу употребляют молодые, сочные этиолирован-
ные побеги спаржи. Дикий предок ее растет на лугах, в северных степях, в
кустарниках Европы, Кавказа, Западной Сибири. Некоторые виды (А. plu-
mosus, A. scandens, A, sprengeri и др.) часто разводят в комнатах и оранжере-
ях как декоративные.
Иглица (Ruscus) — двудомные вечнозеленые полукустарники, у которых
часть побегов превращена в плоские филлокладии, а листья очень мелкие,
пленчатые. В СССР 4 вйда встречаются в лесах Кавказа и Крыма. Спаржу
г. г.
ни
S32
высх
I11ILr
РАСТЕНИЯ
Рис. 402. Спаржа (Asparagus officinalis)!
Л— кирневшце с моподаив побегами; 2—-вепса с цветками; S—развернутый тычиночный цветок;
4-* плоды
и иглицу вместе с другими родами некоторые ботаники выделяют в особое
семейство спаржевые (Asparagaceae).
Подсемейство caccana^itJieeue(Snulacoideae) имеет, как и предыдущее»
плоды — ягоды, отличается двухрядным расположением листьев с сетчатым
жилкованием листовых пластинок. Оно включает кустарники и полукустар-
:г»
ми. Мелкие цветки собраны в кисти или метелки. Плод—одно- и трехсемянная
ягода. Сюда относится сассапариль, или сарсапариль (Smilax), — деревя-
нистые лазающие лианы с черешковыми сетчато-нервными листьями. Извест-
но около 300 видов сассапариля. В СССР, в лесах Кавказа, растет 5. excelsa,
который высоко взбирается на деревья; побеги, покрытые крепкими острыми
* шипами, затрудняют передвижение по лесу. Другой вид (5. oldhami) растет
на Дальнем Востоке.
Семейство травяных деревьев (Xanthorrhoeaceae), выделенное из
лилейных, представляет своеобразную группу многолетних ксерофильных
растений с корневищем и либо тонким (более или менее разветвленным) стеб-
лем, либо (чаще) с толстым, деревянистым простым или разветвленным ство-
лом, способным к вторичному утолщению. Листья в этом случае собраны пуч-
ком на конце ствола, часто линейные. Соцветия цилиндрические или головча-
тые. Околоцветник из чешуйчатых или пленчатых, обычно неопушенных
листочков. Трехгнездная завязь с тремя свободными столбиками. Травяные де-
ревья встречаются в Новой Гвинее, Новой Каледонии, Австралии, Тасмании.
у/ Семейство агавовых (Agavaceae). Стержнекорневые или кор-
невищные растения; ствол у одних короткий, у других хорошо развитый»
имеющий способность к утолщению. Листья у короткостебельных собраны в
розетки, узкие, часто толстые, мясистые. Цветки в крупных соцветиях — ко-
лосьях и метелках; обоеполые (редко однополые), правильные или слабозиго-
морфные, с венчиковидным околоцветником, листочки которого у одних сво-
бодные, у других сросшиеся. Тычинки в двух кругах, они прирастают к
лепесткам. Завязь верхняя или нижняя, обычно трехгнездная, плацентация
центрально-угловая» семяпочек мцого или одна. Плоды — коробочки и
ягоды.
Из родов этого семейства отметим драцену (Dracaena), обитающую в Аф-
рике и Южной Азии. Пользуется известностью драконово дерево (D. draco),
растущее на Канарских островах. Оно достигает 20 л в высоту и 12—14 м в
поперечнике. Возраст его исчисляется в 5—6 тыс. лет. Из надрезов коры вы-
г г
J3
ни
иг
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
распространенный
ГЛ
гл л
текает красная смола — «драконова кровь». Этот же продукт вырабатывают и
другие виды (например, D. annabari, на острове Сокотра). Его употребляют
с древних времен для бальзамирования трупов, сейчас применяют в лако-
красочной промышленности. Драценам родственна кордилина (Cordyline)9
обитающая в Южной Азии, Полинезии, тропической Австралии. Это деревья
тоже с вторичным ростом в толщину. Виды С. australis и С. rubra иногда со-
держат в комнатах под названием драцена, а С. «лсма разводят как декоратив-
ное на Черноморском побережье Кавказа. К этим родам в систематическом
отношении тяготеет род сансевъерия (Sansevieria)
в Тропической Африке и Бирме. Виды сансевьерии культивируют как
важные волокнистые растения. У нас ее разводят как комнатное растение с
оригинальным волокнистым рисунком на мечевидных листьях.
К этому семейству относят ирод юкка (Yucca)t американский по рас-
пространению. Красиво цветущие белыми цветками У. jilarnentosa и У. glareo-
sa разводят в Южном Крыму и на Кавказе как эффектные декоративные рас-
тения.
Значительное экономическое значение имеют американские агавы (Agave\.
Около 275 видов в Мексике и прилегающих областях Северной и Южной Аме-
рики. Из листьев многих агав, разводимых и в других теплых странах (А.
sisalana, A. fourcroydes, A. cantala, A. americana), получают волокна (сизаль-
пенька, генекен и др.), идущие на канаты, шпагат, половики, грубые ткани.
Сбраживанием сахарного сока некоторых агав, выделяющегося на месте сре-
занного соцветия, в Мексике готовят алкогольные напитки («пульке»). В Юж-
ной Европе, в СССР (в Крыму и на Черноморском побережье Кавказа) агавы
разводят только как декоративные растения (главным образом А. americana).
Агавы цветут один раз в жизни, на 6—15-м году выбрасывая из розетки круп-
ных мясистых листьев высокий (до 8—12 м) канделябровидный цветонос с
тысячами цветков, после чего все наземные органы погибают.
l/С емейство амариллисовые, или нарциссовые (Amaryllidaceae). Па
строению вегетативных органов, цветков и плодов амариллисовые очень
близки к лилейным, но отличаются нижней завязью. Переход к этому встре-
чается среди некоторых лилейных, имеющих полунижнюю завязь. Листочки
околоцветника у нарциссовых нередко срастаются в трубку. Часто встреча-
ете^ привенчик (paracorolla), образованный лигулярными вы-
ростами лепестков. Привенчик помещается или наружу от тычинок, или
срастается с расширенными тычиночными нитями, образуя кольцо, подобие
бокала и т. п. Цветки собраны в цимозные соцветия, сходные с зонтиком^
окруженные у основания оберткой, состоящей из одного или более видоизме-
ненных листьев. Иногда число цветков в соцветии редуцировано до одного.
Плоды — коробочки, редко ягоды.
Семейство включает около 1000 видов, обитающих главным образом в тро-
пических и субтропических областях, преимущественно южного полушария,
с сильно выраженным эндемизмом в Южной Африке. Амариллисовые широ-
ко распространены в Средиземноморье. Отдельные группы довольно далеко
проникают в умеренные широты. Практическое значение они имеют как деко-
ративные растения. В СССР — 22 вида, все луковичные.
Подснежник (Galanthus) — ранневесеннее растение с одиночным белым
поникающим цветком (рис. 403). В СССР 7 видов, большинство растет в
лесах Кавказа; нередко разводят как декоративные G. nivalis, С. pit-
catus и др.
Нарцисс (Narcissus) имеет в зеве трубочки характерную коронку —- при- -
венчик (рис. 404). В СССР встречаются всего 2 вида дикорастущих нарциссов
на Кавказе (всего их известно около 40, главным образом на западе Средизем-
номорья). В садах часто разводят во множестве сортов южноевропейский
снежно-белый (N. poeticus, рис. 404), средиземноморско-кавказский желтый
(ЛГ. pseudonarcissus) с очень крупной коронкой; как декоративные в комнатах
и оранжереях, на юге в садах часто разводят тропические и субтропические
кринум (Crlnum)9 туберозы (Polyanthes, tuberosal из Центральной Амери-
иг.
вы
ini
РАСТЕНИЯ
Рис. 403. Подснежник белый
(Galanthus nivalis):
J —общий вид; а—продольный раз»
рез цветка
Рис. 404. Нарцисс (Narcissus pogticus):
1 —верхняя я нижняя части растения; S —
продольный разрез
1ГI
г г
ГЛ'
ки), амариллисы (виды родов Amaryllis, Hippeastrum, Nerine и др., а
также их гибриды).
Семейство касатиковые, или ирисовые (Jridaceae). Это многолетние
травы, реже полукустарники с корневищами, клубнями, клубнелуковицами,
луковицами. Лйстья**о^жкновенпо' двухрядные, узкие. Цветки в цимозных
соцйётиях, редко одиночные; правильные, зигоморфные. Шесть листочков
околоцветника лишь* редщГбывают свободнкдзд, чаще образуют трубочку,
срастаясь на большем или меньшем протяжении. Внутренний круг тычинок
отсутствует, появляясь лишь в уродливых цветках, нормально развиты три
наружв^*Тытанки7Тблько у очень немногих у^язь вертр^/r- уАпдкгпчтгт-
ва — нижняя, трехгнездная, обычно с многочисленными семяпочками при
централ ьно-угЛовой плацентаций; рОДЮэихменьше. Столбик наверху с 3 вет-
вями, которые, в свою очередь, тоже могут ветвиться; у некоторых ветви
рыльца лепестковидные. Плод — коробочка. Семена с эндоспермом, содер-
жащим запасную клетчатку. " '
Ирисовые — обширное семейство, содержащее около 1500 видов. Рас-
пространено в тропиках и субтропиках, имея два крупных центра видового
разнообразия: Капская Земля (около половины видов), тропики и субтропи-
ки Америки. Касатиковые можно рассматривать как результат дальнейшей
эволюции или амариллисовых, или лилейных.
В СССР — 4 рода и около 116 видов.
Касатиковые имеют практическое значение главным образом как деко-
ративные растения благодаря своим крупным ярко окрашенным цветкам.
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис. 405. Касатик (Iris (pseudaco-
rus):
2*—соцветие (н — наружный, в — внут-
ренний листочек околоцветника, ле —
лопасть столбика); 2 — пестик; з— диа-
грамма цветка (п — прицветник, в па-
зухе которого находится почка)
Рис, 406. Шафран (Crocus sativus);
2—общий вид; а — продольный разрез;
3—столбик с рыльцами
касатик, или ирис (Iris),
шарии. В СССР — 87 видов,
содержит около 225 видов в северном полу-
большинство на юге и в Средней Азии.
В крупных цветках его оригинален столбик с 3 большими лепестковидны-
ми лопастями, прикрывающими сверху тычинки (рис. 405). На нижней
стороне каждой лопасти столбика расположены сосочки рыльца. Многие
виды ириса и гибриды их разводят повсеместно как декоративные. Южно-
европейские виды 7. florenttna, 7. pallida, 7. germanlca разводят большими
плантациями (в СССР на юге) из-за корневищ, имеющих аромат фиа-
лок и применяемых в парфюмерии и медицине («фиалковый корень»).
У крокусов (Crocus) одиночные или парные воронковидные цветки с
очень длинной трубочкой околоцветника выходят прямо из клубнелукови-
цы, так что во время цветения завязь находится под землей, и л
позднее плод — коробочка выносится наружу. Цветут они ранней веской
или поздней осенью; многие разводят как декоративные.
Длинные красные рыльца шафрана (Crocus sativus var. autumnalis,
рис. 406), известного только в культуре (в СССР — на Апшеронском полу-
острове, в Азербайджанской ССР), применяют как пряность и изредка в
медицине. Для получения 1 кз сухих рылец нужно 40—60 тыс. цветков.
В СССР произрастает 19 видов крокусов на юге, большинство — на горных.
лугах и других открытых местах.
Гладиолус, или шпажник (Gladiolus), имеет слегка зигоморфные цвет-
ки в~рёДКом колосовидном соцветии. Насчитывают около 250 видов гладко-
in',
иг,
иг
,1ИГ
выс
III’,
РАСТЕНИЯ
л усов, боль
III.
яство находится в Юж-
ной Африке; гибриды некоторых ви-
дов во многих сортах разводятся как
декоративные; соцветия достигают
А ли В СССР имеется 9 видов, боль-
шинство произрастает на юге.
Семейство касатиковых вместе
с 2—3 мелкими экзотическими се-
мействами иногда выделяют в осо-
бый порядок Iridales.
2-Я ПОРЯДОК — СИТНИКОЦВЕТНЫЕ
(JUNCALES)
Ситникоцветные — многолетние,
редко однолетние травы, похожие
на злаки, с обоеполыми актиноморф-
ными цветками, построенными по
типу лилейных, но листочки около-
цветника пленчатые, венчик малоза-
метный (анемофилия). В типе шести-
членныйкруговой андроцей иногда
бывает трехчленным. Гинецей син-
карпный или паракарпный. Плод —
ный крахмалистый эндосперм.
Порядок этот тесно^^рищлкаеТ
к лилиецветным, б’гкрывая анемо-
фильную линию развития.
Семейство ситниковые
(Juncaceae) включает травыг с длин-
ными линейными^идн-»шиловидйьдет
нах — только в горных условиях,
оасикя (Ltuula) со многими видами
листьями, с хорошо развитыми вла-
галищами. Невзрачные цветки в
метельчатых, головчатых ядн щит-
ковидных соцветиях.
Семейство содержит около. 300
видов, распространенных в умерен-
ных и холодных странах, а в тропи-
В СССР — два рода: ситник (J uncus) и
(рис. 407). ПрактичбПЕбПГПЙрвменения
3-Я ПОРЯДОК — ОСОКОЦВЕТНЫЕ (CYPERALES)
К осокоцветным относится одно семейство осоковые (Сурегасеае). От по-
рядка злаковых, с которыми их раньше соединяли в один порядок че-
шуецветные (Glumiflorae), осокоцветные отличаются плодами — хкрашкамиг
варсшышрр*- nnpywCTMjffi аунпспепмом. ФилогеИетическц^ыводятся из лилие-
цветных через семейство ситвиковых(/ипсасеае), от которыхотДИчаднся глав-
ным образом, оуадпжг^рдной завязЬюг- * г ....... .................
Семейств о осоковые (Сурегасеае) объединяет /даогойётйПб^редко
однолетние травы1. У многолетних длинные или укороченные корнев
'ЛИ
1 Монотипный род Microdracotdes (М. tqiuiminoms — единственный вид рода) харак-
теризуется древовидным разветвленным стволом до 1 ж высотой. Растет в горах Западной
Африки. Других растений с древовидным обликом в атом семействе нет*
Рис. 408. Диаграммы цветков камыша и пу-
шицы:
Л — камыш; а—пушица
Рис. 409. Осока:
1 — диаграммы цветной: а — тычиночного; б—трех-
рыльцевого пестичного; в — двухрыльпевдго пестич-
ного; 2— схема пестичного цветка (к — кроюший лист,
«— завязь, ж — мешочеи).
тебля.
состо
в последнем случае развивающиеся из них тесно скученные надземные побе-
ги образуют более или менее плотные кусты. Стебли у больвдшсщ<г1ХШХЦЩР"
ные, без вздутых узлов, не полые (отличие от злакой). Листья расположены у
КГ яляклп)т ттревпиуптер.таетап п тттг^н^й чаСТИ
говдагялт^^о^рпхиятыяятшего стебель,
ой пластинки; язычка у болыпйнства нет, или он слабо развит.
Цветки мелкие» невзрачные. """ "От1™1™"*™) пдмылфмьтт™ Они
собраны в колосовидные соцветия'(у некоторых одноцветковые); последние.
в свою очередь, соединятгу в голпвчятыет зонтиковидные, метельчатые или в
колосовидные, сложный сопнетия- В обоеполых тетках1 окодоцветнпк я
виде шести пленочек иди чешуек, иди чаще в випе^!—6 иди многих шетинок,
или же совсем редуцирован, и цветки тогда голые; однополые цветки без
околоцветника. Из ччлпидод^отр^рнлись лутдь трп''{а^рузитгп нрут^Удщр
дяже^мен^руе, родуо бппыттРк Пыльники прикропляпы к тычиночной НИТИ СВО-
ИМ основанием (отличие от злаков). Один пестик с верхней одногнеадншкза-
nqot-тл -ррт. ряже двух плодолистиков, со столькими же столбиками (или с
одним) и рыльцами. Семяпочка одна< анатропная. Плод — орешек* зародыш в
семени окружен мучнистым эндоспермом.
Семейство осоковых включает не менее 3800 видов. Хотя большинство
видов рдоттрпоирятгдзд р наиболее массового развития осоковые
достигают в умеренных и холодных областях, где растут главным образом в
сырых и болотистых местах, на бедных почвах, малопригодных для других
растении. В СССР имеется 24 -род и около 540 видов.
Как кормовые травы (так называемые кислые злаки) осоковые в боль-
шинстве грубы* содержат мало фодфаря и кяльдпя и стоят значительно ниже
злаков. Однако в районах оленеводства* в степной зоне* на горных лугах*
особенно в песчаных и подгорных пустынях Средней Азии осоки представля-
ют ценнейший пастбищный корм.
Обоеполыецвсхкв^соколоцветником изщетинок^@мек>х^дышоах£0@£@*
(рис. 408)* без околопвейпша —-сыть. Однополые^цветки у осок (рис. 409)
м кобрезий. Из большого р о да камыш (Scirpus) в СССР широко известен
шдцщиашшнй (5. lacustris), имеющий толстыйг-томшъаеленый^щщ^беэ-
иг.
соцветиеЧрис. 410). Он широко распространен по топким берегам водоемов и
довольно глубоко заходит в воду. У пушиц (Eriophorum), растущих на сырых
1В настоящее время предложена не получившая широкого признания теория, рас- ,
сштривающая обоеполый цветок как псевданций, как соцветие из многих тычиночных
И одного пестичного цветка, прицветные листья которых послужили материалом для обра»
мвання органа* сходного с околоцветником,
ВЫШНИЕ РАСТЕНИЯ
Рис. 410. Камыш (Scirpus 1асдо
tris):
1 — общий вид; S — колосок; з — цве»
сок; 4 —мод
Рис. 411. Пушила (Eriophorum
vaginatum):
Л — общий вид растения с инодами;
S— соцветие; з —цветок; 4 — плод
лугах и болотах (рис. 411), щетинки околоцветника при плодах разрастают-
ся в крупные белые «пуховки». У сытщ или ситовника (Суperus), цветки и
кроющие их чешуи расположены двухрядно, и соцветия поэтому плоские; иэ
корневищных клубеньков чуфы, или ae^Maaggg миндаля (С. esculentus)t
P&bbqjqsmxA в теплых районах Южной Европы ив Северной Африке, получа-
ют жирное масло, применяемое как в пищу, так и в парфюмерии, а клубень-
ки ее используют как суррогат миндаля.
Папирус (Cuperus papyrus), достигающий 5 ж в высоту и образующий ог-
ромйые заросли по берегам водоемов в Тропической и Северной Африке, а так-
л - вален В древности" е полоски^ю_стеб-
дись вместо-бумаги«
, употр
лей его, склеенные по
пока не fa™* ймтймшшл ТУ я.) пергаментом. Мадагаскарский С. altenti-
folius часто разводят как декоративный в комнатных аквариумах.
Огромный род ^осош (Carex) имеет более 1
ботаников, до 2500 b2soB?ml_Q
годые. Они ообпацы в Колосов
котданпахчшцуекТРасйтеделён
:j:i
, где цветки сидят в-пазухах
рот: вверху — пёс*
—6ИШШ
однодомны; у п
стеблЯ1Лтяп(ьп(^
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
Тычиночные колосья осок простые.
Мужской слг.тшпе ия. /тух
дли трах тютттпк и располагается в па-
зухе, кроющей чешуйки. Эти чешуйки
вместе с осьиъ на которой они гидят, и
МУЖСКИМИ цветками СОСТЯ*лстт/уг муздгипй
колос (колосок) осоки (рис. 412). Пес-
тичные колосовидные соцветия сложные*
состоят из простых соцветий, похожих на
точные цветки.
В пазухе каждой кро-
ющей чешуйки сложного соцветия нахо-
дится редуцированная ветвь (побег вто-
рого порядка) с одним обращенным на-
зад (к оси всего соцветия) листом. Этот
лист кроющий. Он завертывается во-
круг сидящего в его пазухе женского
цветка и, срастаясь краями, образует
полый орган, так называемый мешо-
чек (utriculus). Сам женский цветок
состоит только из пестика и представ-
ляет в системе всего колосовидного со-
цветия ось третьего порядка (рис. 409,412).
Столбик пестика выходит наружу из вер-
ны мешочка и заканчивается у одних
1
иг
2
видов двумя, у других — тремя довольно
длинными волосистыми рыльцами1. Осо-
ки анемофильные. Пестичные цветки
становятся половозрелыми раньше тычи-
ночных (протерогиния). Плод — орешек
заключен в разрастающийся после опло-
дотворения мешочек, играющий очень
важную роль в систематике осок. Многие
виды осок растут у воды или в воде. Ме-
*3
шочки, наполненные воздухом, легко пе-
реносятся водой на значительные рассто-
:i:ti
Крупные влаголюбивые осоки быва-
ют обычно грубыми, жесткими, и их
Рис. 412. Осока:
плохо поедает скот. Мелкие осоки, рас-
тущие в полупустынях, пустынях (Carex
physodes, С. pachystilis и др.) и в горных
районах, — ценные кормовые растения.
В СССР (в Средней Азии) они на больших
1 — тычиночный цветок с кроющим лис-
том: 2 —пестичный цветок с кроющим
листом: а — продольный разрез пестично-
го цветка: 4 — общий вид осоки вздутой
(тч — тычиночные колоски)
территориях — основной корм для овец.
Род кобрезия (Kobresia) морфологически интересен тем, что у его расте-
ний кроющий лист цветка, образующий у осок мешочек, срастается своими
краями лишь в нижней части и типичного мешочка не образует. Виды коб-
резий, широко распространенные в СССР главным образом на высокогор-
ных пастбищах Алтая, Средней Азии, отчасти Кавказа (всего в СССР
14 видов), — хорошие кормовые растения.
1 Такая трактовка женских соцветий у осок возможна лишь на основании срав-
нения их с другими близкими родами: Kobresia Hemicarex и другими, а также на ос-
новании изучения ранних стадии развития мешочка, когда в нем еще иногда можно ви-
деть зачаточную ось побега второго порядка, на которой сидят ироющии лист, образу*
ющий мешочек.
540
ВЫСШИВ PACT
и: ।;
4-Й ПОРЯ ДОК — БАНАНОЦВЕТНЫЕ
(ZINGIBERALES)
Цветки бананоцветных зиго-
морфные или, Что очень редко
встречается у других растений,
асимметричные. Околоцветник у
большинства подразделен на трех-
членную чашечку и венчик. Редко
он однородный, венчиковидный.
Особенно характерен андроцей.
:ь у очень немногих банано-
Рис. 413. Диаграммы цветков банановых и
канвовых:
1 — банановые (раввиты пять тычинок» шестая реду-
цирована); * — канвовые (четыре тычинки и воловина
пятой* превращены в лепестковидные стаминодии,
вторая воловина пятой тьгшнки несет явугнеэдный
пыльник: яра —кропаний лист, ст — депестковид-
ныв стаминодии, ч — чашелистики» л—лепестки)
цветных все шесть тычинок нор-
мальные, типичные. У прочих иди
пять тычинок нормальные, а ше-
стая превращена в чешуевидный
стаминодий (семейство банано-
вых), или же (у большинства) лишь
одна тычинка (внутреннего круга)
развивает пыльцу, а прочие пре-
вращены в крупные, ярко окра-
шенные лепестковидные стамино-
\ 111
, от которых часто и зависит
яркость и заметность цветков (рис.
413).
В семействах каиновых и ма-
Рис. 414. Банан (Musa eepientum)
рантовых только одна половина
единственной тычинки развивает
двугнездный пыльник, а вторая
половина тоже превращена в ле-
пестковидный стаминодий. Завязь
нижняя, трехгнездная, реже од-
ногнездная вследствие недоразви-
тия двух других гнезд. Семяпочки
анатропные. Перекрестное опыле-
ние производится насекомыми или
у многих йтицами. Плод — коро-
бочка или сочный, ягодовидный.
Семена с эндоспермом и периспер-
мом или только с периспермом. У
многих на семенах развит арил-
лус (мясистый придаток).
К этому порядку относятся
многолетние корневищные травы.
распространенные исключительно
в тропиках и субтропиках Старено
и Нового Света. Крупные листья
их расчленены на влагалище, че-
решок и аллипсоидальную или лан-
цетовидную, цельнокрайнюю пла-
:г*
ных перистым жилкованием. Края
крупных листьев часто разрыва-
ются, и пластинки расчлен
доли, перпендикулярные главной
жилке.
Бананоцветные, вероятно,
произошли от энтомофильных пя-
отдал ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Mf
тикруговых лилиецветных с нижней завязью» причем изменения цветка осо-
бенно выразились в утрате им симметрии и в метаморфозе тычинок в лепест-
ковидные стаминодии. Бананоцветные представляют особую» не имевшую
дальнейшего развития ветвь зоофильной эволюции однодольных» параллель-
ную орхидным.
Бананоцветные объединяют свыше 2000 видов. Их делят обычно на 4
семейства: бомакоше (Musaceae) с шестью или пятью нормальными ты-
чинками; имбирные (Zingiberaceae) с одной нормальной тычинкой» с четырех-
гнездным пыльником» а прочие тычинки превращены в стаминодии; канновые
(Саппасеае) и марантовые (Marantaceae) — у двух последних семейств лишь
половина одной тычинки имеет двутнездный пыльник» вторая же половина и
прочие тычинки превращены в лепестковидные стаминодии. Отличаются два
последних семейства числом семяпочек и формой зародыша.
Наибольшее практическое значение имеют бананы (рис. 414). В мировой
торговле первое место занимает Musa pandisiaca с близким ему видом
Af. sapientium и М. acuminata с родственным ему карликовым бананом (М. са-
vendishti). Разводят более 2000 сортов бананов» употребляя в пищу их плоды.
Культура бананов распространена в тропиках всего света» причем М. acu-
minata появился в Америке» по-видимому, задолго
бананов считают Южную Азию.
до Колумба. Родиной
Рис. 415. Дерево путешествей
(Ravenala madagascariensis)
542
ВЫ
нпгло-
РАСТЕНИЯ
Рис. 416. Цветок канны
пестки, ст — стаминодии,
в — пыльник, р — рыльце)
Корневище бананов развивает наземные по-
беги, высокий ложный стебель которых образо-
ван свернутыми в трубку влагалищами листьев.
Сквозь ложный стебель прорастает пазушный цве-
точный побег, оканчивающийся соцветием — ки-
стью, нижние цветки которой пестичные, сред-
ние — обоеполые, а верхние — мужские. В кисти
может быть до 100 и больше плодов. Мучнистые
сорта бананов, богатые крахмалом, употребляют-
ся в пищу преимущественно вареными или жаре-
ными. Во фруктовых сортах значительная часть
крахмала при созревании переходит в сахар. Есть
и кормовые бананы. Эндемичный для Филиппин-
ских островов вид М. textilts разводят в тропиках
Старого Света для' получения ценного негниюще-
: г. I ;i
.1:1
го волокна, известного как «манильская пенька».
Из этого волокна изготовляют веревки, канаты,
грубые ткани, мешки.
В СССР, на Черноморском побережье Кав-
каза, нередко разводят (как декоративный) япон-
ский банан (Musa basjoo) с несъедобными плодами.
Очень своеобразно так называемое дерево путешественников (Ravenala
magadascariensis, семейство банановых, Мадагаскар). Оно достигает 3—6,
иногда 10 л в высоту (ствол настоящий) и несет на вершине огромные
двухрядно расположенные (в виде веера) длинночерешковые листья (рис. 415).
В углублениях листовых черешков, в местах их отхождения от стебля, скап-
ливается много воды, которая может быть использована путниками (отсюда
название). Дерево путешественников часто разводят в тропиках как декора-
тивное. Другой вид этого рода — R. gulanensis с очень коротким стеблем —
растет в Гвиане и северной части Бразилии, что указывает на соединение в
очень отдаленные времена (не позднее мелового периода) Мадагаскара с тро-
пической Америкой.
Из семейства Zingiberaceae интересен род имбирь (Zingiber).
Пряный, или настоящий, имбирь (Z. officinale) содержит в корневищах аро-
матическое эфирное масло, которое используется в пищевой промышленнос-
ти. Молодые побеги употребляют в пищу как овощи. Это древняя культура
Индии. Отсюда он распространился по всей тропической и субтропической
части Азии. Другие виды имбиря также богаты пряными веществами (напри-
|И
мер, некоторые виды служат источником для получения кардамона или его
заменителей).
Некоторые виды рода Curcuma содержат в корневищах много крахма-
ла, используемого в кондитерском деле и для детского питания. Из С. domes-
tica получают ценные красящие вещества.
Mar anta arundinacea из семейства Mar ant асеае служит для полу-
чения арроурута — муки из корневища, содержащего 25—27% крахмала.
Это же растение используется и как лекарственное.
Некоторые виды канны (семейство Саппасеае, рис. 416) — эффект-
ные декоративные растения в садах и оранжереях.
5-Й ПОРЯДОК — ОРХИДНЫЕ (OR
ил
DALES)
Наиболее характерным признаком этого порядка является своеобразное
строение андроцея и гинецея; из шести тычинок, характерных для однодоль-
ных, обычно развиты лишь две (редко три) или (у большинства) даже только
одна, причем нити тычинок срастаются со столбиком пестика, образуя вместе
так называемую колонку (гиностемий).
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
Рис, 417. Диаграммы цветков орхидных:
1—с двумя «ычинкамж; S—с сдвой тычинкой до сворота цветка; Л-с одной тычинкой после пово-
рота цветка
В порядке только одно большое семейство того же наименования.
Семейство орхидные^ или ятрышниковые (Orchidaceae), включает
У многих развиты ^клубни — подземные (к о р н е в ы е) или надземные
(т у б_а р тг д к н)/^таблАн«дт ^дючаюшие одно или несколько междоузлий.
Листья цростые, очередные, цельнокрайние, влагалищные, иногда стеблеобъ-
емлющие? Пветаи зигоморфные (редко околоцветник актиноморфный),
обоеполые или (очень редко) однополые. почти у всех с ярко окрашенным ле-
песткрвптртым онплоцветнцком ИЗ ДВУХ ТРехплепных кругов. У большинства
цветки, собраны в колос, похожий на кисть, вследствие того что длинная и
узкая нижняя завязь, прикрепленная основанием в пазухе кроющего листа,
бывает похожа на пветоножку. Встречаются орхидные с метельчатым соцве-
тием и с одиночными цветками. Задний (ближний к оси) лепесток внутреннего
круга пкалпцретаика jggran jgn^o отлпнаетея от остя льныт фирмой т большей
величиной и нередко окраской. Он представляет так называемую губу (рис.
417). У орхидных с непоникающими соцветиями по мере созревания цветка
завязь скручивается, цветок благодаря этому поворачивается вокруг своей
оси на 180°, и губа оказывается обращенной пе вверх, а вниз. Она служит
местом посадки опыляющего насекомого. Исходное верхнее положение губы
у таких орхидных заметно в цветочных бутонах, а на акропетально разви-
вающемся соцветии прослеживаются переходы между скрученной и нескру-
ченной завязью и, соответственно, между двумя положениями губы.
У многих орхидных (растущих не в СССР) верхняя губа вследствие
поникания соцветия оказывается в физически нижней части цветка. В этом
случае скручивания завязи не происходит. При основании губы
орхидных находится ямка с нектарником, или шпорец, па дне
скапливается нектар.
У подавляющего большинства орхидных развита лишь одна
г г.
которого
передняя1
тычинка наружного круга, у некоторых (триба башмачковых — Cypripe-
dieae) вполне развиты только две передние тычинки внутреннего круга; часть
редуцированных тычинок может сохраняться в виде стаминодиев. Л:
очень немногих индо-малайских орхидных (род Neuwiedia) оотрнвшшлъ
три тычинки (одна внешнего и две внутреннего кругов). Пестик образован
тремя плодолистиками. Завязь нижняя, почти у всех одногнездная, с тремя
париетальными плацентами, на которых расположены многочисленные (ты-
сячи, сотни тысяч и миллионы) семяпочки, развитие которых в основном про-
ходит после опыления.
При столбике, сросшемся с нитями тычинок в колонку, пыльники могут
быть обособленными, соединенными в тетрады или в комочки. У большинства
все пылинки каждой половинки пыльника склеены висцином в общий комо-
ли
; а 11
1 У цветков со скрученной завязью она кажется задней (верхней).
Рве. 418. Цветок ятрышника:
2— цветок ятрышника; я—вестик и тычинка; J—вакки поллиниев с липкини подушечками (п —пыль*
ник плк — половина ныльника. пл— поллиний. н — вакка шшлиния. л под — липкая подушечка*
с — связник, р —рыльце)
Г Г;
ГЛ
чек — поллиний, от которого вниз или вверх отходит ножка из затвер-
девшей слизи1. К свободному концу ножки приклеивается липкая подушечка
(прилипальце). Поллиний вместе с подушечкой называется поллинари-
е м. Половинки пыльника у однотычинковых разъединены связником и на-
ходятся над рыльцем (рис. 418). Из трех лопастей рыльца у однотычиноч-
ных орхидных только две бокрвые воспринимают пыльцу^, и расположены они
или обособленно, или (у большинства) слиты в одну железистую площа-
дочку, находящуюся ниже половинок пыльников. В этом случае третья
лопасть рыльца, называемая клювиком (rostellum), превращена в осо-
бый орган, вырабатывающий упомянутые выше 2 липкие подушечки (диски)
поллинариев.
Когда насекомое, сидящее на губе, всовывает головку в цветок за нек-
таром, то к ней приклеиваются липкие подушечки поллинариев. Покидая
цветок, насекомое уносит на себе поллинарии. Во время полета ножки пол-
линариев подсыхают, поллинии загибаются вниз'и вперед; при посещех
насекомым другого цветка поллинии приходятся как раз против воспринима-
ющей площадочки рыльца. Этот оригинальный способ опыления впервые
был описан немецким учителем X. К. Шпренгелем в 1793 г. Однако
интерес к опылению у орхидных появился только после опубликования в
1862 г. работ Ч. Дарвина, где приспособления к перекрестному опылению у
орхидных рассмотрены с эволюционной точки зрения. У тропических ор-
хидных с разнообразной причудливой формой цветков существует мно-
жество различных приспособлений к форме тела и повадкам опыляющих
их насекомых; у одних орхидных губа подвижна и, быстро поднимаясь
вверх, прижимает севшее на нее насекомое к колонке; у других — полл:
выбрасываются при прикосновен
Некоторые тропические орхидеи опыляются птицами. Около 200 ви-
дов относится к самоопыляющимся растениям. Наряду с этим известно
много самостерильных орхидей.
Плод орхидных — коробочка, вскрывающаяся продольными щелями.
Семена очень многочисленные (до 3—4 млн. в коробочке), чрезвычайно мел-
кие, с широкой прозрачной оболочкой, без эндосперма, С почти шаровидным
зародышем, не дифференцированным на органы. Мелкие, необычайно легкие 1 2
Г Г;
л: г.
Г Г;
1 По другим данным, ножка развивается ив стерилизованной части поллиния.
2 У орхидных с двумя и более тычинками все три лопасти рыльца воспринимают
пыльцу*
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
545
семена (масса одного семени исчисляется миллионными долями грамма) под-
хватываются даже слабыми воздушными течениями. Такой способ переноса
семян эффективен в безветренных тропических лесах, где произрастает боль-
1115
нство орхид
: и многие из них растут эпифитио. Для прорастания семян
необходимо заражение их микоризным грибом, хотя удавалось получить
прорастание и без микоризы — на субстрате, богатом органическими ве-
ществами.
При прорастании из семян обычно образуется клубневидный орган, так
называемый протокорм (возможно, соответствующий подсемядольному
колену), а уже на нем формируются побеги и корни. Развивается растение
медленно, цветение наступает только через 5—20 лет после прорастания.
У многих наших дикорастущих орхидных (ятрышников, белой «ночной фиал-
ки» и др.) под землей находятся два корневых клубня: один сморщенный (за
счет его растет стебель данного года), другой гладкий, плотный (он дает цве-
тущий побег на следующий год.)
Орхидные—второе по числу видов семейство покрытосеменных (первое—
сложноцветные), содержащее около 20 000, а по некоторым сведениям,
даже 30 000 видов.
Большинство орхидных распространено в тропиках, где они живут пре-
имущественно как эпифиты. Главные центры распространения их — Индо-
Малайская флористическая область и тропическая Америка. В Европе
растет около 250 видов, в СССР — 43 рода и 122 вида, все почвенные.
Разнообразие экологических условий местообитания орхидных и дли-
тельный процесс эволюции обусловили огромное разнообразие форм у них.
Некоторые орхидные очень мелкие, ненамного крупнее мха, a Bulbophyl-
lum minutissimum (эпифит Австралии) имеет вид клубенька в 2—3 лсж с одним
чешуйчатым листом. Однако есть и такие, как яванская лазающая лиана
галеола (Galeola), которая достигает 40 м длины. Цветки орхидных бывают от
0,2 до 25 см в диаметре. У Grammatophyllum (Зондские острова) соцветие
достигает 2—2,5 м длины; на каждом из десятков соцветий помещается 100
цветков, имеющих около 10 еле в диаметре. С другой стороны, у довольно мно-
гих орхидных цветки невзрачные, мелкие. У многих тропических эпифитных
орхидных резервуарами воды и запасных веществ, кроме клубней, бывают
сочные, мясистые листья. Характерны для эпифитов придаточные воздушные
корни, покрытые с поверхности несколькими слоями мертвых клеток (vela-
men), поглощающих воду из атмосферных осадков. У некоторых тропиче-
ских эпифитных орхидей (Phalenopsis, Taeniophyllum, Anagraecum и др.) име-
ются плоские лентовидные зеленые ассимилирующие корни.
В филогенетическом отношении орхидные представляют вершину одной
из эволюционных ветвей энтомофильных однодольных. Они, вероятно, воз-
никли от каких-то лилиецветных, где встречаются некоторые признаки, полу-
чившие у орхидных дальнейшее развитие и оригинальное оформление: зиго-
морфность цветка, нижнюю завязь, редукцию части тычинок, усложнение
процесса опыления, многочисленность зачатков семян и др.
Практическое значение орхидных очень невелико. Не вполне зрелые про-
вяленные плоды ванили (Vanilla planifolia) используют как ценную пряность.
Родина ванили — восточные районы Мексики; разводят ее во многих тропи-
ческих странах; растет как многолетняя лиана с цепляющимися придаточными
корнями. При кишечных заболеваниях применяют высушенные корневые
клубни некоторых видов ятрышника (Orchis mor io, О, militarise О» masdula
и др.), двулистной любки (Platanthera bifolia) и др. Очень многие тропические
орхидеи разводят как декоративные тепличные, оранжерейные, реже ком-
натные растения. Выведено много межродовых гибридов (даже между 3—4
родами); издаются специальные журналы, посвященные декоративным
орхидеям.
В СССР на сыроватых лугах и в лесах широко распространена белая
любка двулистная (Platanthera bifolia), или «ночная фиалка», с ду
IU5
стыми
цветками, пахнущими только вечером и ночью. Там же растет ряд видов ятп-
18—602
546
ВЫСШИЕ РАСТЕНИЯ
Рис. 420. Венерин башмачок
Рис. 419. Ятрышник
рышника (Orchis, рис. 419) и кокушника (Gymnadenia) с пурпурными или лило-
во-пурпурными цветками и др. В лесах, кустарниках изредка встречаются
виды венерина башмачка (Cypripedium), имеющие по 1—2 оригинальных круп-
ных цветка с сильно вздутой в виде башмачка губой (рис. 420), с двумя ты-
чинками (пыльца порошковатая), почти не скрученной завязью. Встречаю-
щиеся единично в лесах виды ладъяна (Corallorhiza), подбородника (Epipo-
gium), гнездовки (Neottia) лишены зеленой окраски, питаются сапрофитно и
отчасти паразитно за счет живущего в их корнях или корневищах микориз-
ного гриба.
Исходным для всего подкласса Liliidae является порядок Liliales, где
можно видеть многочисленные направления специализации, получившей
дальнейшее развитие в других порядках этого подкласса и захватившей
как отдельные органы, так и жизненную форму в целом.
Эта изменчивая группа растений дала начало двум зоофильным направ-
лениям — порядкам Zingiberales и Orchidales. Порядок Zingiberales почти не
выходит за пределы тропических стран. Их цветок изменялся на базе превра-
щения большей части тычинок в лепестковидные стаминодии. В порядке
Ordiidales, тоже в основном тропическом, но распространившемся и за пре-
делы тропиков, произошло глубокое преобразование не только андроцея,
но и гинецея, что привело к созданию .своеобразного синтетического органа—
гиностемия. Околоцветник стал не только органом привлечения опылителей,
но из него во многих случаях выработались специальные механизмы, обеспе-
чивающие точность работы опылителей. Порядок Orchidales, несомненно, одна
из вершин энтомофильной эволюции не только однодольных, но и покрыто-
семенных в целом. >
В совершенно ином направлении произошло развитие в двух других
порядках подкласса Liliidae'. Juncalesn Cyperales— тоже потомках каких-то
г г
отдал цветковый, или покрытоскмвнные
547
Г. III
Liliales, потерявших яркость околоцветника в связи с переходом к анемо-
филии. Порядок Cyperates, по-видимому, мог возникнуть от менее специали-
зированных Juncales.
Что нее касается возникновения всего подкласса Liliidae в целом, то он
произошел, вероятно, от древнейших однодольных вскоре после их обособ-
ления. Не исключено, что эти предковые формы, еще сохранившие некоторые
черты Magnoliidae, былй общими предками и Liliidae и Alysmidae.
г I.
3-Й ПОДКЛАСС — КОММЕ ЛИПИДНЫЕ (COMMELIN1DAE)
Гинецей коммелинидных ценокарпный. Семена с крахмалистым эндо-
спермом. Пыльцевые зерна трехклеточные, реже двухклеточные. Околоцвет-
ник, если он хорошо развит, двойной или редуцированный. Огромное боль-
шинство — наземные травянистые растения с сосудами во всех органах.
Энтомофильные и анемофильные.
В подклассе Commelinidae объединены пять порядков. Они довольно ес-
тественно распадаются на две группы. В порядках Commelinales^ Bromelia-
tes и Eriocaulales цветки имеют двойной околоцветник с различно окрашенны-
ми и разной консистенции чашечкой и венчиком. Здесь господствует энтомо-
филия. Вторую группу составляют порядки Restionales и Graminales^ в
обоих околоцветник простой брактеоидный, или его нет; в обоих господству-
ет анемофилия.
ГРУППА ПОРЯДКОВ ENANTIOBLASTAE1
Эта группа порядков в ряде систем рассматривается как един порядок,
но в настоящее время не без оснований ее подразделяют на четыре поряд-
ка (Commelinales, Bromeliales, Eriocaulates, Restionales). Растения входящих
сюда семейств распространены главным образом в тропиках и в южном полу-
шарии. Для них характерен мучнистый эндосперм, к которому сбоку приле-
гает зародыш, чем и объясняется название группы. То же наблюдается и
у злаков.
Enantioblastae интересны тем, что в этой группе можно наблюдать раз-
личные ступени упрощения цветка однодольных и переход от энтомофилии к
анемофилии. Но не исключена возможность, что от предковых форм этой
группы порядков возникла и энтомофильная линия, представленная поряд-
ком Bromeliales.
сг
ГЛ
Семейство коммелиношв (Commelinaceae из порядка Commeli-
nates). Многолетние или однолетние травы, часто сочные, с прямостоя-
щими или прилегающими стеблями. Листья спиральные, нередко двух-
рядные с замкнутым влагалищем. Пластинка листа часто с водозапасаю-
щей тканью. Цветки из 5 или 4 трехчленных (реже двухчленных) кругов,
правильные, редко зигоморфные. Околоцветник двойной. За трехлист-
ной (обычно) чашечкой следует трехлепестковый венчик, листочки около-
цветника свободные или у основания сросшиеся. Андроцей двухкруговой,
но один из кругов может состоять из стерильных тычинок или вовсе выпадает.
_ _ _ хя плодолистика образуют
трехгнездную завязь (одно гнездо может редуцироваться), столбик один с
головчатым или трехлопастным рыльцем. Коробочка или сухие нераскры-
вающиеся плоды.
Семейство насчитывает около 600 видов.
Наиболее известен род традесканция (Tradescantia) иэ Центральной и
Северной Америки. Многие виды его (и выделенных из него родов Zebrina,
Rhoeo ) с ползучими стеблями и довольно сочными, нередко пестрополосаты-
ми листьями часто разводят как декоративные. Волоски, покрывающие ты-
hi: »:<
1 От греч. «энантоо — напротив в «бласте» — зародыш. Группу порядков можно
было по-русски назвать «противозародшпевые».
18е
548
ВЫСШИЕ РАСТЕНИЯ
Рис. 421. Соплодие ананаса
укороченные, с розетками
оватых листьев, образующих
шш
чииочные нити Г. virginica, — удобный объект
для наблюдения струйчатого движения цито-
плазмы.
Семейство бромелиевые^ или ананас-
ные (Bromeliaceae из порядка Bromeliales).
Своеобразное семейство тропических травя-
нистых растений, живущих преимущественно
эпифитно на деревьях или на скалах. У боль-
шинства стебли
длинных
открытые кверху воронки, где скапливается
вода. Соцветия — колосья, кисти или метелки, у
многих с ярко окрашенными кроющими листь-
ями. Цветки обоеполые, актиноморфные, с око-
лоцветником, у большинства разделенным на
трехчленные чашечки и венчик. Тычинок шесть
в двух кругах, пестик один, образованный
тремя плодолистиками. Завязь трехгнездная,
причем в пределах семейства имеются все
переходы от верхней до нижней завязи. Стол-
бик один. Плод — коробочка (у родов с верхней
или полунижней завязью) или ягода (у родов с
нижней завязью).
Семейство бромелиевых очень интересно
экологически. Корни у его эпифитов слу-
прикрепления, а у некоторых (Tillandsia) сов-
воронках, образова^ых розетками листьев,
жат главным образом для
сем не развиваются. В
скапливаются вода, разлагающиеся органические остатки и даже развивается
особая фауна и флора (например, пузырчатки). Основания листьев всасыва-
ют воду и питательные вещества. У многих развиты особая водозапасающая
ткань, специальные волоски для всасывания воды и т. д.
Семейство включает свыше 1500 видов, обитающих исключительно в тро-
пической и субтропической Америке.
Как фрукты употребляют крупные ароматные кисловато-сладкие сопло-
дия ананаса (Ananas sativa), образованные плодами (большей частью бессе-
мянными), мясистой осью соцветия и разрастающимися мясистыми кроющими
листьями; из вершины соплодия выходит листоносный побег (рис. 421). Ана-
нас широко распространен в культуре и разводится вне тропиков в особых
теплицах. Из листьев ананаса при иных приемах культуры получают тонкие
шелковистые прядильные волокна.
Своеобразная Tilliandsia usneoides (так называемый «луизианский мох»
или «мексиканский мох»), широко распространенная в штате Луизиана в США,
в Мексике и др., свешивающаяся длинными (2—3 ж) прядями с деревьев,
наподобие бородатых лишайников, употребляется как набивочный материал
вместо конского волоса. Многие бромелиевые разводят как декоративные,
особенно в оранжереях.
Другая линия — анемофильная, берущая начало от семейства Cora-
melinaceae или, вероятнее, от ближайших предков его, — получила выраже-
ние в нескольких порядках и семействах того же подкласса, из которых упо-
мянем семейство канатчиковые (Restionaceae).
Семейство канатчиковые (Restionaceae из порядка Restionales}.
В большинстве канатчиковые — многолетние травы, образующие дернины,
часто сходные по облику с тростником или камышом, имеющие корневища
и ассимилирующие стебли, нередко ветвистые, двухрядно олиственные, часто
узловатые. Листья обычно с открытыми влагалищами и рудиментарными плас-
тинками, часто рано опадающие, реже — прикорневые с мечевидной пластин-
кой; частично с язычками. Цветки мелкие, собранные в колоски, а эти — в
сложные метельчатые или колосовидные соцветия. Цветки л
’.in
ь редко
обое-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
549
полые, обычно же однополые, но с рудиментами недостающих полов. Около-
цветник из чешуйчатых или сухопленчатых листочков в числе 3 + 3, 2 4- 2,
3 + 0, или его нет. Если околоцветник двухрядный, то листочки разных кру-
гов могут быть одинаковыми, явственно отличаться друг от друга. Они боль-
шей частью свободные или внутренние, в основании сросшиеся. Тычинок 3
или менее, до одной, принадлежащие внутреннему кругу. Пыльники двугнезд-
ные, прикреплены к нитям спинкой. Плодолистики в числе 3—1, завязь с
соответствующим числом гнезд, с 3—1 столбиками, с длинными перистыми
или скрученными рыльцами. Плоды — коробочки или орешки. Зародыш лин-
зовидный, прилегающий семядолей к эндосперму. Растут часто большими
скоплениями, как злаки и осоки.
Около 400 видов канатчиковых произрастает в
Австралии, на южных оконечностях Африки и Южной
Америки, на полуостровах Индокитай и Малакка. Мно-
гие систематики считают возможным из предков этого
семейства филогенетически вывести злаки, с которыми
Restionaceae действительно имеют ряд черт сходства.
5-И ПОРЯДОК — ЗЛАКОЦВЕТНЫЕ (POALES)
Этот порядок содержит только одно семейство зла-
ков (Gramineae, или Ашдоде). Подавляющее большин-
ство злаков — травы. Лишь редко стебли твердые, од-
ревесневающие (бамбуки). Стебли злаков обычно тон-
кие, цилиндрические, внутри в междоузлиях почти у
всех полые (соломина). Листья очередные», расположен-
ные двухрядно (угол расхождения 180°). Они состоят
из длинного цилиндрического влагалища, охватываю-
щего стебель, и обычно узколинейной пластинки (рис.
422). В месте отхождения пластинки от влагалища на-
ходится так называемый яды чок (ligula) в виде не-
большого пленчатого выроста ИЛй“ресничек. Некото-
рые морфологи считают язычок за два сросшихся при-
листника. Биологическая роль его, по-видимому, со-
Рис. 422. Лист злака,
охватывающий сте-
бель (уз — вздутие на
узле, ел— влагали-
ще, яа—язычок, пл-^
пластинка, ст — сте-
бель)
стоит в том, что он препятствует воде затекать внутрь
между влагалищем и стеблем. Основание влагалища
обычно слегка утолщено и образует хорошо заметный,
слегка вздутый листовой узел (рис. 423). Края влага-
лища плотно охватывают бтеббль, у большинства не
срастаются друг с другом (отличие от осок). Нижние
части междоузлий, защищаемые влагалищем, долго со-
храняют меристематический характер и в течение
значительного времени обеспечивают интеркалярный
рост стеблей.
Как у многолетних, так и у однолетних злаков вет-
вление стебля происходит почти всегда лишь в ниж-
ней его части (под землей) или, у некоторых, над по-
верхностью земли. Здесь находится так называемый
узел, точнее, %она кущения — несколько очень тесно
сближенных узлов, разделенных лишь коротк
ПЛг.
меж-
доузлиями; каждая пазушная почка из числа находя-
щихся в зоне кущения может дать начало надземному
или подземному боковому побегу; последние образуют
свои зоны кущения (рис. 424). У многолетних злаков
часто из почек зоны кущения формируются длинные
побеги, растущие горизонтально под землей, — корне-
вища. Такие злаки в луговодстве называют корне-
вищными (пырей ползучий, костер безостый и
др., рис. 424).
Рис. 423. Схемати-
ческий разрез узла
злака (пс — полость
стебля, ел — ялстп-
вое влагалище, п —
перегородка в уз-
ле, пп — проводящие
пучки)
550
ВЫСШИЕ РАСТЕНИЯ
Рис. 424. Подземное ветвление длиннокорневшцного злака зубровки душистой с вне-
влагалищными побегами (по П. А. Смирнову)
Листья на корневищах имеют вид небольших беловатых или буроватых
чешуек, образованных, собственно, только влагалищами листьев; они позд-
нее отпадают. Почти во всех узлах корневище образует придаточные корни.
На некотором расстоянии от исходной зоны кущения корневище загибается
кверху и образует надземный стебель. В подземной части этого стебля обра-
зуется новая зона кущения, дающая начало новым воздушным побегам, в
свою очередь ветвящимся, и новым корневищам... Дальнейший рост корневищ
и образование надземных побегов происходит по той же схеме.
Корневища обеспечивают вегетативное размножение злаков при доволь-
но быстром, особенно на рыхлых субстратах, захвате площади. У злаков, рас-
тущих иа песках и на илистой почве, вегетативное размножение осуществля-
гл
ется, кроме того, длинными стелющимися по земле и укореняющимися в уз-
лах побегами, так называемыми отпрысками или столонами, которые также
дают из некоторых узлов прямостоячие побеги (свинорой, мятлик обыкновен-
ный — Роа trivially иногда тростник и др.).
У так называемых кустовых злаков все наземные побеги сближены
друг с другом. В этом случае длинные горизонтальные корневища не образу-
ются, а возникающие в зонах кущения подземные побеги растут на очень
небольшом расстояЕ
г г
косо-вверх или горизонтально, ио затем быстро за-
гибаются вверх. Это относится к так называемым рыхлокустовым
з л а, к а м, примером которых могут служить тимофеевка луговая, ежа сбор-
ная, овсяница луговая и др. Если боковые побеги с самого начала растут вер-
тикально, почти параллельно материнскому стеблю (рис. 425), то образуются
плотные кусты и, соответственно, такие злаки характеризуются как плот-
нокустовые. Таковы, например, щучка, ковыли.
Дочерние побеги, развивающиеся в зонах кущения, образуются из почек,
находящ
нгг
:ся между влагалищем листа и стеблем. Эти побеги или выходят
наружу, пробивая влагалище (в невлагалищные, или экстра-
вагинальные, побеги, рис. 426), илц растут внутри влагалища (в н у т-
ривлагал ищны е, или интравагина^ьные, побеги) и в даль-
нейшем выходят из него рядом с материнским стеблем (рис. 427). Во втором
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
551
случае бывают более плотные кусты, но экстр а вагинальные побеги встреча-
ются и у плотнокустовых злаков.
Побеги у злаков монокарпические, развивающиеся один, два года или
несколько лет (моно-, ди-, полициклические побеги). Однако не все побеги
обязательно достигают цветения, они могут отмирать и ранее (побеги с не-
полным циклом развития), не достигнув цветения. У каждого побега отмирает
вся его надземная часть, остаются живыми только подземные или приземные
участки, которые входят в состав корневищной основы многолетнего злака,
построенной, как правило, по симподиальному типу. Возникающие от нее
новые поколения подземных и надземных побегов продолжают жизнь злака,
в то время как более старые участки корневищной основы отмирают. Таким
образом, особь многолетнего травянистого злака представляет систему по-
бегов, более (корневищные злаки) или менее (кустовые злаки) заметно меняю-
щую со временем положение в пространстве ассимилирующих и репродуктив-
ных частей.
Ветвление злаков не всегда концентрируется только у основания надзем-
ных побегов (в зонах кущения); бывает и так, что боковые ветви возникают
рассеянно в разных участках и подземных, и даже надземных стеблей. Из
встречающихся в нашей стране злаков ветвление побегов можно наблюдать
у тростника (Phragmites communis)^ некоторых видов вейника (Calamagros-
И$)9упырея (Agropyron repens), белой полевицы (Agrostis alba) и некоторых
других.
Описанное выше ветвление злаков всегда сопряжено с образованием в
конечном счете олиственных надземных побегов. Это приводит к ежегодному
обновлению ассимилирующих органов и увеличению синтезирующей способ-
ности за счет возрастания до известного возраста числа надземных побегов.
Если надземный побег проходит полный цикл развития, то обнаруживается
еще вторая область его ветвления — область соцветия, почти всегда у злаков
сложного. Биологическое значение ветвления соцветий заключается в воз-
можности сформировать на одном надземном побеге значительное число ко-
лосков и цветков и расположить их соответственно условиям опыления
и распространения семян.
Рис. 425. Схема кущения:
1 — рыхлохустового злака; а — шютнокуетового с виввпагалищаши побегами; А — плотнокустового е
внутрналагалшцныян побегами (з — проросшее верно. я — уровень почвы; таждому ясбегу соответст-
вует оцта нрадатееды* корень)
552
ВЫСШИЕ РАСТЕНИЯ
Рис. 426. КущениЬ злака вейника
тростниковидного с вновлагалищ-
ными побегами (по П. А. Смир-
нову)
Рис. 427. Кущение злака булаво-
носца седого с вну тривлага лишни-
ми побегами (по П. А. Смирнову)
Сложные соцветия злаков (рис. 428) состоят из простых соцветии, по-
строенных по типу колоса и называемых колосками. Колоски могут
быть собраны в сложные колосья, как, напрймёр“у ряШ, пшеницы, пырея и
др. (при этом колоски не имеют ножек, или они очень короткие), или в метел-
ку, с хорошо развитыми и повторно ветвящимися ветвями, несущими колоски
(овес, мятлик, костры и др*). У некоторых злаков сложное соцветие представ-
ляет колосовидную метелку (ложный колос, султан) с многочисленными, тес-
но сближенными короткими ветвями, несущими колоски (тимофеевка, лисо-
хвост и др.). Реже встречаются кистевидные (перловник) и головчатые слож-
ные соцветия.
Каждый колосок имеет главную ось, на которой в двухрядной последо-
вательности (как листья на вегетативном побеге) располагаются специализи-
рованные чешуевидные листья, резко отличающиеся по размерам, форме и
строению от листьев вегетативной части побега. Чешуевидные листья колос-
ков гомологичны влагалищам ассимилирующих листьев. Обычно первые (сни-
зу) два чешуевидных листа, сидящих на оси колоска, бесплодные, т. е. не
образуют в своих пазухах цветков. Эти чешуйки принято называть к п л а г-
к о в ы м и, различая по их положению нижнюю и верхнюю колос-
ковые чешуи. Колосковых чешуй иногда бывает и более двух (просо, канаре-
ечник, душистый колосок), у других — только одна (плевел) или даже их
совсем нет (белоус, леерсия). Чешуйки, располагающиеся на оси колоска вы-
ше колосковых, плодущие — с цветками в пазухах. Они называются н^и ж и и-
м и ц в е т к а » й м w (TTBeTKniiTJMTr j ттлтпму1то каждая является кроющей
по отношению к одному из цветков колоска, нижними — соответственно их
положению по отношению к соответствующему цветку). Число нижних цвет-
ковых чешуй в колоске различно, от 1 до 10 и более. Когда их много, верхних
цветков в пазухах может и не быть. Нижние цветковые чешуи у многих зла-
ков имеют более или мепее развитое продолжение — ос ть (arista), выходя-
щ ую из вершины нижней цветковой чешуи йли от ее спинки. Морфологически
ости разнообразны: они могут быть прямыми, коленчатыми, скрученными,
покрытыми волосками (иногда длинными, например у перистых ковылей)
или голыми. По происхождению ости представляют резко метаморфизирован-
ные листовые пластинки.
Занимая место в пазухе нижней цветковой чешуи, каждый цветок (по
другому мнению, цветок вместе с сопровождающими его прицветными листь-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
553
ями) представляет сильно укороченный побег второго порядка по отношению
к оси колоска. В состав этого пазушного побега входят (в последовательности
снизу вверх) верх няя цветковая чешуя, околоцветн ые
пленки, андроцей и г и н е ц е й. Из этих органов, бесспорно, к
цветку относятся только два последних — андроцей и гинецей.
Верхняя цветковая чешуя имеет обычно весьма характерное строение —
она двукилеватая (рис. 429) — с двумя сильными жилками (к и л и), но обычно
без средней "жилки. Плоская пленчатая часть этой чешуи, заключенная
между двумя килями, всегда обращена к оси колоска. Если каждый из килей
примыкает к плоской части верхней цветковой чешуи, то с другой стороны
каждого киля имеется продолжение пленчатого тела чешуи, направленное в
сторону кроющего листа (в сторону нижней цветковой чешуи). Таким образом,
на поперечном разрезе верхней цветковой чешуи получается фигура в виде
прямой скобки с боковыми сторонами, направленными в сторону нижней
цветковой чешуи и охватываемыми краями последней (рис. 430). Кроющая
чешуя рассматривается или как предлист (стр* 346), имеющий типичные
для однодольных форму и положение, или как продукт слияния двух наруж-
ных листочков околоцветника цветка предков злаков, причем третий листо-
чек редуцирован.
Выше верхней цветковой чешуи расположены околоцветные пленки —
л о д и к у л_ы (lodiculae). Они помещаются со стороны, обращенной к ниж-
нёи цветковой чешуе, т. е. к кроющему листу цветка. Околоцветные пленки
обычно небольшие, беловатые. У некоторых злаков (южноамериканский злак
Streptochaeta, бамбуки, некоторые ковыли) не две, а три околоцветные плен-
ки, причем третья обращена к верхней цветковой чешуе. Многие ученые рас-
сматривают околоцветные пленки как сильно редуцированный внутренний
круг околоцветника, трехчленный при наличии трех лодикул или потеряв-
ший один из членов, когда лодикул только две. Иногда же (душистый коло-
Рис. 428. Соцветие у злаков:
J — сложный колос; 2—8 — колосовидная метелка. или ложный колос (часть колосков уда-
лена. чтобы показать веточки соцветия); 4 —веточка колосовидной метелки с колосками; 6 — ложный
яйцевидный колос; в — сложные колосья, пальчаторасположенные; 7 — кисть иэ колосков
554
ВЫ
Hl II'1 >
РАСТЕНИЯ
тч
Рис. 429. Схема (по П. А. Смирнову) трехцветковогэ
колоска злака с недоразвитым зачатком четвертого
цветка (кчш — колосковые чешуи, нцчш — нижняя
цветковая чешуя, вцчш— верхняя цветковая чешуя,
пл — околоцветные пленки, или лодикулы, /пч — ты-
чинки, пс — пестик)
Рис. 430. Диаграммы цветков злаков:
1 — эмпирическая; 2 — теоретическая (части* шобрансевнке
пунктиром, в типичном цветке отсутствуют, нцчш — нин вяя
цветковая чешуя, вцчш — верхняя цветковая чешуя* пл — ородо-
цветные пленки, тч — тычинки, пс — пестик)
сок, лисохвост) лодикул нет вовсе. По мнению других ботаников, обычно
присутствующие две околоцветные пленки — это продукт расщепления од-
ного сильно редуцированного листового органа. Если присутствует третья
лодикула, то она соответствует следующему (но уже не расщепленному) листу
такого же типа.
Итак, существуют три взгляда на околоцветник злаков: 1) он состоит из
двух кругов, причем остатком наружного круга является верхняя цветковая
чешуя, а внутреннего — околоцветные пленки; 2) верхняя цветковая чешуя —
предлист; от околоцветника в цветке злаков сохранился только один (внутрен-
ний) его круг — околоцветные пленки; 3) верхняя цветковая чешуя и около-
цветные пленки — это сильно измененные верховые листья; цветок злаков,
таким образом, голый — совершенно лишенный околоцветника.
Андроцей у злаков в большинстве случаев трехчленный. Одна из трех
тычинок располагается между двумя околоцветными пленками, две другие
обращены к верхней цветковой чешуе. Если принимать лодикулы за редуци-
рованные листочки околоцветника, то единственный круг тычинок цветка
злаков соответствует наружному кругу андроцея в цветке лилейных, внутрен-
ний же редуцирован. Возможность существования у предков злаков двукру-
гл
гового тримерного андроцея подтверждается наличием шести тычинок, рас-
положенных в два круга в цветках риса, стрептохеты, сахарного тростника,
многих бамбуков (у последних нередко тычинок много, что представляет
результат расщепления). Тенденцию уменьшения числа тычинок можно ви-
деть в том, что у некоторых злаков (зубровка, душистый колосок) число их
снижено до двух, а иногда даже до одной (цинна). В строении тычинки надо
отметить, что тычиночная нить прикреплена не к основанию пыльника (как
у осоковых), на внутренней стороне, благодаря чему пыльник легко качает-
ся при малейшем движении воздуха (приспособление к ветроопылению).
Цветок завершается пестиком с одногнездной вер*™* йяпият.тг содержа-
щей одну семяпочку. Столбик может быть длинным или коротким и завер-
шается чаще всего двумя перистыми (следовательно, имеющими большую
поверхность) рыльцами. Наличие у некоторых злаков трех рылец (Strepto-
chaeta^ некоторые бамбуки) и трех сосудистых , пучков в стенке завязи неко-
торых бамбуковых подводит к мысли о трех плодолистиках в составе гинецея
злаков. По мнению многих специалистов, гинецей злаков образован един-
ственным плодолистиком»
отдел ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕ£
i;irr
Рис. 431. Схематический продольный разрез
зерновки пшеницы (а — зародыш, щ — щиток,
ал — алейроновый слой, амд — эндосперм)
Рис. 432. Продольный разрез зародышей зла-
ков:
1 — хорошо выражен впнбласт; 3-^3 — эпибласт редуци-
рован (э — эшголаст)
4
При цветении злаков происходит сильное набухание лодикул, вследствие
чего верхняя цветковая чешуя отодвигается от нижней, тычинки и разветвлен-
ные рыльца высовываются в образовавшиеся щели. При этом тычиночная
нить быстро удлиняется. Пыльники раскрываются продольной трещиной, и
пыльца, скапливающаяся в нижней ложкообразно изогнутой половине пыль-
ника, легко рассеивается при малейшем движении воздуха. Все злаки ветро-
опыляемые* 1, в большинстве случаев опыление перекрестное. Но есть среди
них и самоопыляющиеся с нераскрывающимися (клейстогамными) цветками.
Таковы, в частности, некоторые культурные злаки (пшеницы, ячмени, овес,
просо, рис).
Плод злаков — зерновка, характеризующаяся плотным соедине-
нием семенной кожуры с околоплодником, лишь у немногих бамбуков плод
ореховидный или ягодообразный. Ости на нижних цветковых чешуях несут
много устьиц и, испаряя много воды, содействуют лучшему притоку питатель-
ных веществ к созревающим зерновкам. У безостых видов много устьиц на
концах чешуек.
Наибольшую часть зерновки (в среднем около 74% по массе) составляет
крахмалистый эндосперм, из-за которого и возделывают хлеб-
ные злаки. Наружный слой его,называемый алейроновым, (клей-
ко в и н н ы м), состоит из толстостенных клеток, богатых белками (алей-
роновые зерна), жиром, витаминами (рис. 431).
Зародыш лежит у основания зерновки и примыкает к эндосперму щит-
ком (scutellum), который обычно считается семядолей зародыша злаков.
У многих злаков на зародыше со стороны, противоположной щитку, развива-
ется еще чешуйка, называемая эпибластом и считаемая многими морфо-
логами за рудимент второй семядоли (рис. 432)*. Осевая часть зародыша со-
стоит из почечки, покрытой колпачком (колеоптиле), и зародышевого
корня, окруженного тканью колеоризой. В промежутке между почеч-
1В тропическом южноамериканском роде Pariana, относимом к подсемейству овся-
ницевых, известно опыление насекомыми.
1 Морфологическая природа эпибласта спорна. Его считают или рудиментом второй
семядоли, или более или менее выраженным остатком влагалища единственной функцио-
нирующей семядоли щитка. Есть мнение, что эпибласт — настоящая семядоля, а щиток
представляет продукт разрастания оси зародыша и является, следовательно, органом не
листовой, а стеблевой природы.
ВЫСШИЕ РАСТЕНИЯ
nil
Рис. 433. Хлебные и крупяные злаки:
(Secale cereaU); ^ —шпешща мягкая (Triticum vulgare): в —безостая, б — остистая
durum); 4 - овво <Ате“ sativa): а - раскидистый, б - одногривый:
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
557
Рис. 434. Хлебные и крупяные елани:
1—просо (Panicum miliaceum); г— рас (Oryza sativa): а —остистый, б —безосты®; *«рволш (Sor-
ghum Japonicum); 4 — аркугара (Sorgnum cernuum); 4 — чумиза (Setaria Italia)
558
ВЫСШИЕ РАСТЕНИЯ
Hl.
кой и зародышевым корнем располагается морфологически неясно дифферен-
цированное подсемядольное колено (гипокотиль), с ко-
торым и связан щиток.
По созревании зерновок у большинства дикорастущих злаков ось колос-
ка разламывается выше колосковых чешуй (между местами прикрепления
нижних цветковых чешуй), и зерновки выпадают, окруженные цветковыми
чешуями, реже колоски опадают целиком. Неосыпаемость зерновок у куль-
турных злаков, биологически вредная для дикорастущих, — результат
многовекового отбора человеком. Также результатом отбора является и голо-
зерност ь многих хлебных злаков, у которых зрелые зерновки отделяются, не
покрытые цветковыми чешуйками.
Распространение плодов дикорастущих злаков происходит главным об-
разом посредством ветра, чему способствуют у многих различные волоски,
ости, легкие упругие цветковые или колосковые чешуйки и т. п. У некоторых
(щетинники и др.) щетинки при плодах, покрытые шипиками, крючками,
цепляются за шерсть животных и разносятся ими.
При прорастании зерновок злаков клетки щитка выделяют ферменты,
удлиняются, проникая в эндосперм, и высасывают заключенные в нем пита-
тельные вещества, передавая их начавшим расти частям зародыша. Первый
лист — колеоптиле (перышко) — выходит наружу в виде трубчатого сверну-
того, на вершине твердого и шиловидно заостренного влагалища. Он способ-
ствует прохождению прочих листьев почечки через почву.
К злакам относятся свыше 700 родов и от 8000 до 10 000 видов. Они рас-
про странены по всей Земле, вплоть до крайних пределов существования выс-
ших растений. На огромных пространствах саванн, степей, прерий, лугов,
а также на дюнах они образуют массовые и устойчивые скопления. Наиболь-
шее разнообразие видов приходится на тропические области.
В экономическом отношении злаки являются наиболее важным семейст-
вом среди всех растений.
Среди культивируемых хлебных злаков (рис. 433, 2, 3, 5; 434, 2) первые
места в мировом земледелии по площадям, занятым под культурой, и по коли-
честву урожая занимают пшеница, рис, кукуруза и ячмень. По качеству и
разнообразию доставляемых продуктов на первом месте стоит пшеница. Пятое
место после них в мировом земледелии занимает овес, шестое — рожь. В тро-
пиках и субтропиках Азии и Африки очень велика роль сорго; оно, по-види-
мому, занимает седьмое место.
Дикорастущие луговые злаки также имеют большое значение в народном
хозяйстве. По данным И. В. Л а р и н а, в среднем для СССР злаки составля-
ют около 25—30 % от всего урожая сенокосов и пастбищ. Особенно велико их
значение в степной зоне (80—90% всего урожая трав в сене и на пастбищах).
На первом месте по распространению и кормовому значению среди дико-
растущих злаков в СССР стоит род пыреи (Agropyron), на втором месте —
овсяницы (Festuca), за ними идут костры (Вготиз) и мятлики (Роа), а дальше,
в убывающем по значению порядке, — лисохвосты (Alopecurus), тимофеевки
(Phleum), полевицы (Agrostis), ежа (Dactylis), ковыли (Stipa), вейники (Са-
lamagrostis) и др. В СССР около 25—30 родов злаков играют существенную
роль в травянистом покрове и кормовых ресурсах естественных лугов и паст-
бищ.
Ископаемые злаки известны с мелового периода, но отнести эти древние
остатки к современным родам не удается. Черты предковых форм приходит-
ся восстанавливать на основании сравнительно-морфологических данных.
Наиболее близкими к предковым формам считают те, у которых сохранились
примитивные признаки в структуре цветка, поэтому едва ли не большинство
систематиков держится того мнения, что цветок злаков возник из цветка типа
лилейных путем преобразования в процессе приспособления к ветроопыле-
нию. Поэтому такие признаки, как наличие третьей околоцветной пленки или
второго трехчленного крута тычинок, считается показателем относительной
близости к предковым формам, сходным с современными Restionaceae из
отдел ЦВЕТКОВЫЙ, ИЛИ ПОКРЫТОСЕИВННЫЯ
и*;
подкласса Commelinidae. Наоборот, уменьшение числа тычинок до 2 или до 1,
отсутствие лодикул и прочее рассматриваются как отклонение от предкового
типа цветка.
Обширное семейство злаков подразделяют в разных системах на разное
количество подсемейств. Ниже рассмотрим 6 подсемейств из выделяемых 91.
Подсемейство бамбуковые (Bambusotdeae). К этому подсемейству
относится свыше 200 видов, распространенных почти исключительно в тро-
пиках и субтропиках. В пределах СССР имеются 3 вида рода саса (Заза) на
Сахалине. Стебли (рис. 435) многолетние, высокие (до 30—40 м), одревеснева-
ющие. Листовые пластинки соединены с влагалищем черешковидным суже-
нием. Соцветия метельчатые из двух-, многоцветковых колосков. Колосковых
чешуй две или больше. Столбики срастаются друг с другом. Плоды — зер-
новки, но встречаются костянки и даже ягодоподобные. В районах естествен-
ного распространения, отчасти и в культуре бамбуковые находят разносто-
роннее применение и имеют существенное экономическое значение. Из ство-
лов бамбуков строят дома, мосты, плоты, их используют на водопроводные
трубы, на изготовление мебели, музыкальных инструментов, циновок, сосу-
дов для жидкостей и сыпучих тел, молодые побеги используют как овощи.
Плоды сочноплодных бамбуков иногда достигают большого размера.
Так, например, у Melocanna bambusoldes (Индия) плоды величиной с апельсин,
с сочной съедобной мякотью. Семена этого растения без эндосперма, и заро-
дыши прорастают в плодах (в и в и п а р и я). В междоузлиях некоторых
бамбуков (Bambusa arundinacea, Dendrocalamus strictus) образуется кремнистое
кристалловидное вещество «табашир», которое' ценится в тропической Азии
как народное лекарственное средство от желтухи, дизентерии, туберкулеза и
некоторых других болезней.
К подсемейству бамбуковых некоторые систематики относят неотропи-
ческий род стрептохета (Streptochaeta), содержащий два вида и представ-
ляющий большой теоретический интерес.
Стрептохета (5. spicata) растет в лесах. Листья ее яйцевидные, с много-
нервными пластинками, на черешках. Соцветия колосовидно-кистевидные.
Колоски одноцветковые. За 4—5 небольшими колосковыми чешуями следует
одна нижняя цветковая чешуя. Верхняя цветковая чешуя почти до основания
раздвоенная. Околоцветных пленок 3, они крупные (до 1 см длиной). Тычи-
нок 6. Столбик одни с тремя рыльцами. Строение цветка стрептохеты ис-
пользуется как один из аргументов в пользу возникновения цветка злаков
из цветков типа лилейных.
П оде емейство оесяницееые (Festucoideae), или мятликовые (Poot-
deae}, объединяет более трети родов и около трети всех видов злаков. Овся-
ницевые характеризуются тем, что хлорофиллоносная паренхима заполняет
все пространство между верхним и нижним эпидермисом листовой пластинки,
поскольку оно не занято проводящими пучками и механической тканью (то
же строение листьев свойственно бамбуковым). Строение цветков и колосков
у овсяницевых в целом соответствует описанному в общей характеристике
семейства. Число цветков в колоске изменяется от нескольких до одного.
Иногда наблюдается редукция колосковых или верхней цветковой чешуи. Об-
ластью распространения этого подсемейства являются преимущественно вне-
тропические страны северного полушария.
В составе этого подсемейства упомянем ряд триб.
Триба оесяницевые (Festuceae), Колоски обычно с обоеполыми цветка-
ми, с двумя колосковыми чешуями, чаще более короткими, чем цветковые.
Нижние цветковые чешуи или без ости, или с осте видным окончанием. Ты-
чинок 3, редко 2. Сюда относятся важные кормовые растения: овсяницы (Fes-
tuca), мятлики (Роа), костры (Bromus), ежа (D act у Its glomerat а), трясунки
(Briza) и другие, используемые как на природных лугах и пастбищах, так и
в посевах.
1 По Эвглеру.
560
ВЫСШИЕ РАСТЕНИЯ
Сред* родов этой трибы выделяется плевел (Lolium), характеризуемый
одной колосковой чешуей. В отличие от большинства овсяницевых у плевела
колоски собраны в сложном колосе и прижаты к главной его оси своей узкой
стороной. Льняной плевел (L> remotum) засоряет лен. Плевел опьяняющий
(L. temulentum), растущий в посевах яровых культур, имеет ядовитые зер-
новки, в которых живет мицелий гриба. Итальянский многоукосный райграс
(L, multiformis) — прекрасный кормовой злак, ценный на пастбищах и сено-
косах.
Триба ячменевые (Hordeae) характеризуется сложным колосом, ко-
лоски одно-, многоцветковые, с типичным для злаков строением цветков,
прикрепляющиеся по одному или по нескольку на выступах главной оси со-
цветия и обращенных к ней своей широкой стороной. Ячменевые — одна из
наиболее важных в экономическом отношении групп злаков.
Злак первенствующего значения — пшеница.
К р о д у пшеницы (Triticum) относится 20 однолетних, яровых или ози-
мых, видов, из которых только 4 встречаются дикорастущими.
Колосковые чешуи у пшениц вздутые, неравнобокие, с килем. Колоски
2, 3, 5, 7-цветковые и располагаются по одному. Цветки обоеполые, обычно
самоопыляющиеся. Всходы пшеницы зеленые, у некоторых с сизым налетом.
В мировом земледелии (в том числе и в СССР) имеют значение 2 вида пше-
ницы — мягкая (Т. vulgare, рис. 433, 2) и твердая (Т. durum; рис. 433, 3).
Мягкая пшеница распространена шире, так как ее можно возделывать и в
более холодных районах. Зерновка у мягких пшениц на изломе мучнистая;
остей в колосе нет, или они по
Рис. 435. Бамбуки
длине короче колоса, и притом
сильно расходятся в стороны.
У твердой пшеницы зерновка на
изломе стекловидная; почти все
сорта имеют ости, превышаю-
щие длину колоса и параллель-
но или слаборасходящиеся.
Лучшие сорта муки-крупчатки,
макаронной муки и манной
крупы получают из твердой
пшеницы. В СССР твердую пше-
ницу выращивают по преиму-
ществу как яровую и на засуш-
ливом юге, и на юго-востоке
европейской части, в Предкав-
казье, в степной части Казах-
ской ССР, в Алтайском крае,
Новосибирской, Омской облас-
тях. В сухом континентальном
климате при небольшой влаж-
ности почвы пшеница содержит
больше белков и меньше крах-
мала. Поэтому пшеница наших
засушливых юго-восточных
районов особенно высоко це-
нится.
Как мягкая, так и твердая
пшеница имеет много разновид-
ностей и свыше 4000 сортов.
Наиболее распространен-
ные отечественные сорта
мягкой пшеницы относятся
к разновидностям
lutescens, milturum, erythrosper-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
561
тшн, ferruglneum, caesium, hostianum, albidum, alborubrum, а сорта твердой
пшеницы — к разновидностям hordeiforme, melanopus, coeru-
lessens.
Из сортов пшеницы у нас отобран ряд лучших, так называемых райони-
рованных, рекомендованных как для культуры в определенных районах
страны. Советские селекционеры создали и продолжают создавать высоко-
сортные сорта пшениц; лучшие из них не превзойдены по своим качествам во
всем мире.
За советский период посевная площадь под
ин
юницей
значительно уве-
личилась. Границы возделывания пшениц значительно продвинулись на вос-
ток и на север. Озимые пшеницы продвинулись далеко на восток за Дон и
Волгу — в Казахстан, Сибирь. Яровые пшеницы возделывают даже в Ка-
рельской АССР, Архангельской области, Якутской АССР, у Полярного кру-
га и даже севернее его. Громадные площади заняты пшеницей на целинных и
залежных землях. Культура пшеницы известна с доисторических времен.
Раньше в культуре была широко распространена малотребовательная полбо-
видная пшеница с ломким колосом и зерновками, не выпадающими из пле-
нок, — полба (Г. speltа), полуполба, или эммер (Т. spelt а), и однозернянка (Т.
monococcum). Сейчас она почти совершенно вытеснена мягкой и твердой пше-
ницей, так как полбовидная пшеница менее урожайна, дает муку с худшими
мукомольными и хлебопекарными качествами, требует дополнительных зат-
рат на отделение от зерна пленок (цветковых и колосковых чешуй).
Близок в систематическом отношении к пшенице род пырей (Agropyron).
Некоторые многолетние виды его скрещивают в СССР с пшеницей с целью
получения гибридов, соединяющих высокие качества зерна пшеницы с засухо-
устойчивостью, зимостойкостью, солевыносливостью и многодетностью пырея
(работы Н. В. Ц и ц и н а). Ряд степных видов пырея, известных под назва-
нием житняка (A. pectlntformae, A. desertorum, A. sibiricum и др.), культиви-
руют как очень засухоустойчивые многолетние кормовые травы, улучшающие
структуру почвы. Многолетний пырей ползучий (A. repens) — злостный, труд-
но искоренимый сорняк. В то же время это ценное и урожайное кормовое
растение на заливных лугах, в лиманах и на залежах. Острец (А. готовит),
растущий в солонцеватых степях и на залежах юго-востока европейской части
СССР, Сибири и Средней Азии, тоже злостный сорняк в посевах хлебных зла-
ков и одновременно ценная и урожайная кормовая трава на залежах.
Рожь (Secale cereals, рис. 433, 1) является у нас основным хлебным зла-
ком в тех районах, где культура пшеницы имеет пока еще малое распростра-
нение; на Крайнем Севере она уступает место яровому ячменю, имеющему
меньший вегетационный период. В мировом земледелии рожь занимает по
площади шестое место среди хлебных злаков, причем 50—60% всей мировой
площади, засеваемой рожью, приходится на СССР. Колосковые чешуи у ржи
узкие, шиловидные. Стержень колоска состоит из топких прямых члеников.
Колоски двухцветковые и располагаются по одному на стержне колоска. Ниж-
ние цветковые чешуи с длинной верхушечной остью. Цветки ржи обоеполые,
перекрестноопыляющиеся. Молодые всходы ржи фиолетово-коричневые или
коричневатые. Озимые сорта ржи более урожайны, раньше созревают и по-
этому меньше подвержены влиянию засухи. Яровые сорта (ярица) возделыва-
ют там, где из-за климатических условий озимая рожь не удается (например,
в Восточной Сибири). В СССР выведен ряд высокоурожайных сортов ржи,
получены ржано-пшеничные гибриды.
По содержанию белков и усвояемости, а также по разнообразию приме-
нения рожь уступает пшенице и во многих местах постепенно вытесняется ею-
Предполагают, что культурная рожь возникла из той сорной ржи, кото-
рая засоряла и теперь засоряет посевы озимой пшеницы и озимого ячменя в
горах — на Кавказе, в Средней и Малой Азии.
По мере продвижения культуры пшеницы на север и в горы пшеница
выпадала, а более выносливая рожь лучше выживала, превратившись в конеч-
ном счете в культивируемое растение.
иг
Дикорастущие виды ржи (их около 12) практического значения не имеют.
Ячмень (Hordeum, рис. 433, 5). Колосковые чешуи у ячменей узколанцет-
ные или шиловидные, постепенно переходящие в короткую ость. Нижние
цветковые чешуи, за редким исключением, с длинной остью. Колоски одно»
цветковые и сидят по три на выступах общего стержня (главной оси) колоска.
Средний колосок в каждой группе сидячий, обоеполый, плодущий; два боко-
вых — на коротких ножках, недоразвиты или только с пыльниками, бес-
плодные; лишь у культурного ZZ. vulgare все три колоска в группе плодонос-
ные и боковые колоски почти сидячие.
Культура ячменя в мировом земледелии занимает'четвертое место.
У обыкновенного, или многорядного, ячменя (И. vulgare = Н. polysti-
chum) все три колоска в группе плодоносят, а у двухрядного жменя (Я. disti-
chum) плодоносит лишь один средний колосок в группе, а два боковых реду-
цированы.
Культурный ячмень большей частью самоопыляющееся растение. Опы-
ление его происходит еще до выхолащивания. В жаркую погоду ячмень цве-
тет открыто, но и в этом случае часто самоопыляется.
Зерно ячменя используется на выработку перловой и ячневой круп, в пи-
воварении, для изготовления суррогата кофе, на корм домашнему скоту.
Триба овсовые (Aveneae) объединяет, как и Festuceae, метельчатые зла-
ки с колосками, содержащими разное число цветков, но чаще двухцветковы-
ми. Колосковые чешуи преимущественно такой же длины, как нижние цвет-
ковые. Эти последние часто с коленчато изогнутой остью, выходящей из
спинки. В этой группе центральное место надо отвести роду овес (Avena).
Овес посевной (Л. saliva, рис. 433, 4) занимает в мировом земледелии по посев-
ным площадям пятое место среди злаков (после пшеницы, ржи, кукурузы,
ячменя). Растение в основном кормовое; кроме того, из зерновок изготовля-
ют питательную крупу, муку для киселей и супов, толокно. Некоторые виды
овса (Л. fatua, овсюг и др.) — трудно искореняемые однолетние сорняки.
Высокопитательный французский, или высокий, райграс (Arrhenatherum
elatius) — ценная сенокосная трава. Щучка (Deschampia caespitosa) — плотно-
кустовой малопитательный злак, засоряющий луга, — растет преимуществен-
но на сырых местах. В этой же трибе находят место полевицы (Agrost is),
некоторые из них — важные кормовые растения; род вейник (Calamagro-
st is) — высокие многолетники с крупными метелками; метлица, или метла
(Apera spica-venii), — плодовитый сорняк озимых хлебов, образующий до
12 000 зерновок на одном растении.
Триба тростниковые (Arundineae) объединяет высокостебельные злаки
с широкометельчатыми соцветиями из многоцветковых колосков. Тростник
обыкновенный (Pharagmites communis) — почти космополитное растение, ис-
пользуемое на плетение, как дешевый строительный материал и на бумагу.
Высокая трава пампасов (Cortaderia argentea), более известная как Gynerium
argentaium, имеет очень красивые серебристые метелки. Иногда ее разводят
как декоративное растение.
Триба канареечниковые (Phalarideae) также характеризуется метельча-
тым соцветием, нередко, однако, сильно сжатым и даже колосовидным. В ко-
лоске за двумя обычными колосковыми чешуями иногда располагаются еще
две, более или менее редуцированные, без цветков в пазухах или иногда с муж-
ским цветком. Вполне развитый обоеполый цветок один с двумя околоцветными
пленками или без них и с 2, 3, 4 или 6 тычинками. К этой группе относится
оу бровка, или лядник (Hierochloe odorala, Н. australis), применяемый для изго-
товления ароматной настойки (запах кумарина). Душистый колосок (Antho-
xanthum odoratum) тоже с запахом кумарина; цветки душистого колоска с
двумя тычинками и без лодикул. Канареечник (Phalaris canariensis), родом с
Пиренейского полуострова, — источник канареечного семени, идущего на
корм канарейкам. Выделенный в последнее время из предыдущего р о д Dig-
raphis (тоже называемый канареечником или двукисточником) содержит один
вид D. arundinacea — крупный злак со стеблем, достигающим 1—2 м; рас-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
563
пространен у воды и на сырых лугах. Важными кормовыми растениями из
этой трибы являются лисохвост (Alopecurus) и тимофеевка (Phleum), имею-
щие колосовидные метелки (султан). Луговую тимофеевку часто культиви-
руют вместе с клевером.
Триба ковылевые (Stipeae) — группа злаков с одноцветковыми колос-
ками. Нижняя цветковая чешуя своими краями крепко охватывает верхнюю»
окружая и зрелый плод. Характерна ость, выходящая иа вершины или в не-
посредственном соседстве с нею, коленчато изогнутая» часто длинная и неред-
ко характерно пе р и стоо пушенная, околоцветных пленок 2, часто 3. Из боль-
шого рода ковыль (Stipa, 250 видов) в СССР известно около 60 видов, рас-
пространенных в степной зоне и в горах. Листья некоторых ковылей, особен-
но молодые, поедает скот. Остистые плоды ковылей на пастбищах нередко
внедряются в тело животных. Распространенная в Западном Средиземноморье
Stipa tenacissima (трава альфа,или экспарто) используется как источник сырья
для бумажного производства. Чий (Lasiagrostis splendens), растущий в пус-
тынно-степной полосе Заволжья, Южной Сибири и Средней Азии, — круп-
ное растение, образующее дерновины, со стеблями до 3 м высотой. Идет на
плетение (маты; циновки), может использоваться для изготовления бумаги,
а корни — для щеток. К этой же группе относится бор (Milium effusum) —
характерный злак широколиственных лесов.
Триба белоусовые (Nardeae) содержит один род с единственным видом
белоус (Nardus stricta) — многолетнее растение с односторонними метелками
из одноцветковых колосков. Имеется только одна колосковая чешуя. Лоди-
кул в цветке нет. Завязь с одним столбиком и одним рыльцем. Распространен
В Евразии И Грепляндии-
Наконец, укажем, что упомянутый выше насекомоопыляемый южноаме-
риканский злак Pariana (30 видов) также относится к подсемейству
Festucoideae и выделяется в особую трибу Parianeae.
Подсемейство полевичковые (Eragrostoideae) характеризуется на-
личием трех жилок в нижней цветковой чешуе, причем боковые приближены
к краю чешуи. Плоды помещаются между цветковыми чешуями свободно или
отделяются от растений без чешуи. У некоторых (род Sporobolus) около-
плодник ко времени созревания плода отделяется от семени и семя оказыва-
ется голым.
Из относя
к г.
хся к этому подсемейству растений упомянем род
Boute-
1оа9 виды которого характерны для некоторых типов североамериканских
прерий. Например, трава грома (В, gracilis) вместе с другим злаком подсе-
мейства — бизоновой травой (Buchloe dactiloides) — господствует в низко-
травных прериях в США. Виды рода селин (Aristida) — характерные рас-
тения пустынь. В пустынях Средней Азин некоторые виды седина закрепляют
подвижные пески. Свинорой (Cynodon dactylori) — хорошая пастб:
(ная тра-
ва, становящаяся, однако, обременительным сорняком в посевах хлопчатника
и суходольного риса.
Подсемейство рисовые (Oryzoideae) примыкает к овсяницевым по
анатомическому строению листьев. Колоски одноцветковые, причем у рисо-
вых не только нижняя, но и верхняя цветковая чешуя имеет по несколь-
ку жилок и среди них выделяется средняя (!). Обе чешуи твердые. Андро-
цей у рисовых из трех, шести и более (редко иэ 1) тычинок.
Большинство видов — водные растения, или гигрофиты. Распространены
преимущественно в тропиках и субтропиках. Рис посевной (Oryza saliva) —
яровое однолетнее растение, возделываемое на площадях, в совокупности
приближающихся к полутора миллионам гектаров. Рис — источник углевод-
ного питания для половины человечества (Китай, Япония, Юго-Восточная
Азия, Индия, Пакистан и др.). Известно несколько тысяч сортов. Возделыва-
ют на полях, покрываемых на время вегетации водой, но есть и суходольные
сорта. Кроме зерен, идущих в пищу, используют рисовый крахмал, солому
(шляпы, циновки, бумага, особенно папиросная и т. д.). Культура риса
очень др свняя, в Юго-Восточной Азии она существует более 4—5 тысячелетий.
вы
Н I > It
РАСТЕНИЯ
Представители рода Zizania — высокие водные растения Северной
Америки и Восточной Азии. Z. aquatica на востоке США образует большие
заросли. Используется под названиями «Тускарора», канадского, дикого или
водяного риса. Плоды питательные, основания стеблей употребляют как ово-
щи, травянистые частя — отличный корм для скота. Другой вид (Z. lattfolld)
распространен на Дальнем Востоке, используется как кормовое, молодые
побеги съедобны.
Подсемейство просовых (Panlcoideae). Очень крупное подсемей-
ство, содержащее не менее 1500 видов, отличающихся анатомическим строе-
нием листьев. Клетки хлоренхимы листьев располагаются вокруг проводя-
щих пучков радиально в виде двух концентрических слоев (венцовый, или
паникоидный, тип хлоренхимы). Своеобразно строение чаще двухцветковых
колосков. На оси колоска располагаются две колосковые и две нижние цвет-
ковые чешуи, но первая из цветковых чешуй не всегда содержит тычиночный
цветок.В случае его отсутствия колосок бывает одноцветковым. В пазухе вто-
рой нижней цветковой чешуи находится нормальный обоеполый цветок. Эта
вторая чешуя становится твердой и вместе с верхней цветковой чешуей, тоже
твердеющей, покрывает плод, создавая подобие орешка. Сюда относится по-
севное просо (Panicum miliaceum) — древняя засухоустойчивая культура.
Очищенные (обрушенные) от цветковых чешуй зерна проса известны под
названием пшена. Чумиза (гоми, кунак — Setaria italica) — ценная про-
довольственная (крупа, мука) и кормовая культура. Близкий к чумизе могар
(Setaria moharium) — ценное кормовое растение. Некоторые другие виды того
же рода — сорняки (Setaria glauca, S. viridis и др.), как и куриное просо
(Echinochloa crus-galli). Преимущественно нетропический род Paspalum
вместе с дикорастущими видами проса (Panicum) играет существенную роль
в составе южноамериканских саванн и пампасов, а виды Pennisetum, в
частности слоновая трава (Р. benihami), характерны для саванн Африки.
Наконец, виды рода Spinifex характерны для некоторых формаций Австра-
лии.
Подсемейство бородачевые,или сорговые (Andropogonoideae), объеди-
няет свыше 750 видов. Внутренним строением листьев они напоминают просо»
вне. Одно- или двухцветковые колоски располагаются почти всегда парами.
Первый колосок при этом на ножке, второй — сидячий. Оба колоска могут
иметь обоеполые или один сидячий колосок с обоеполыми цветками, а дру-
л гой содержит только мужской цветок. Второй колосок может вовсе редуциро-
ваться. Иногда оба колоска состоят из однополых цветков.
Распространены сорговые главным образом в жарких странах, где они
вместе с некоторыми просовыми составляют основную массу злаков (саванны,
прерии).
К большому роду
сорго (Sorghum) принадлежит ряд видов важного
экономического значения, разводимых во многих странах как
t Ш11
;евые или
кормовые, особенно в Африке и Азии (в СССР — на юге европейской части, в
Средней Азии и Дальнем Востоке): обыкновенное сорго (S. vulgare), джугара
(S. сегпиит, рис. 434, 4), гаолян (S. japonicum, рис. 434, 5), дурра (S. durra,
Африка, Индия), суданка (S. sudanense) и др. Из стеблей сахарного сорго (S.
saccharatum), содержащих 12—18% сахара, получают сладкий сироп для кон-
дитерской промышленности и выгонки спирта. Из соцветий веничного сорго
(S. technicuni) изготовляют щетки, метелки. Гу май Л или джонсонова трава
(S. halepense), — кормовое растение и вместе с тем часто злостный сорняк,
обладающий исключительной засухоустойчивостью.
Сахарный тростник (Saccharum officinarum) встречается во многих стра-
нах только в культуре и семян почти никогда не образует. Размножают его
стеблевыми черенками. Стебли его (4—5 см толщиной) содержат 13—18% са-
хара. Вегетационный период сахарного тростника очень длинный, поэтому
культура его возможна лишь в жарких странах (в Европе — в южных райо-
нах Испании и Италии — на Сицилии). В СССР сахарный тростник выращи-
вают в некоторых местах Таджикистана для получения сока, используемого
in
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
565
в производстве рома и сиропов. Родоначальником сахарного тростника раньше
считали 5. spontaneum, дико растущий в тропических и субтропических
районах Азии и Африки, а в СССР — в Туркмении и Таджикистане по бере-
гам рек. Но, вероятно, сахарный тростник является гибридом двух или не-
скольких видов, происходя
к г.
х из Индии. На юго-востоке Азии в травяном
покрове тиковых лесов с диким сахарным тростником растет аланг-аланг
(Imperata arundinacea) — высокостебельный злак, легко становящийся обре-
менительным сорняком на пашнях и на лесных расчистках. Другой вид этого
рода (7. cylindrical) — характерное растение африканских саванн. Этот вид
встречается и в СССР — на Кавказе и в Средней Азии.
Род кукуруза (Z е а) характеризуется высокими (до 3, а у южных сортов
до 6 м и более) неполыми стеблями. Цветки однополые, растение однодомное.
Мужские цветки в двухцветковых колосках, собранных в крупную метелку
на вершине стебля. Колоски в метелках парные — один с более длинной,
другой с короткой ножкой. Каждый колосок имеет 2 колосковые чешуи и две
пары цветковых чешуи. В составе цветка — две околоцветные пленки и три
тычинки. Иногда нижний цветок недоразвивается. Женские колоски собра-
ны на утолщенной боковой ветви в виде початка. В них различаются две ко-
лосковые чешуйки и тоже две пары цветковых, но цветок только один. Он
представлен завязью с длинным столбиком на вер:
II
зе, лопастным. Около-
цветных пленок нет. Второй (нижний) цветок в колоске отсутствует.
Кукуруза имеет мировое значение наряду с пшеницей и рисом. Она
культивировалась в Америке еще до открытия ее европейцами и сейчас ши-
роко распространена во множестве сортов. Ее используют на муку, крупу,
на зеленый корм и на силос, иэ зародышей получают пищевое масло. Из
кукурузы изготовляют консервное зерно, хлопья, столовый крахмал, крис-
таллическую глюкозу. СССР занимает второе место по площади, занятой
культурой кукрузы.
В подклассе Commelinidae более низкое положение занимают порядки
Commelinales и Eriocaulales. Эти энтомофильные порядки близки к предкам
подкласса. В порядке Restionales цветок претерпел упрощения в связи с пе-
реходом к анемофилии. Завершение анемофильной эволюции цветка достиг-
нуто в обширном порядке злакоцветных, который многие систематики считают
родственным порядку Restionales,
Энтомофильные Commelinales по расчленению околоцветника и другим
признакам цветка примыкают к порядку Bromeliales, Есть сходство этих двух
порядков и в строении семени. Имея это в виду, можно представить Bromelia-
les и Commelinales как группы, связанные общим предком, который, вероятно,
примыкал к порядку Liliales из подкласса Liliidae,
4-Й ПОДКЛАСС — АРЕЦИДНЫЕ (AREC1DAE)
Гинецей арецидных ценокарпный, реже апокарпный. Семена с эндоспер-
мом. Цветки мелкие, собраны в различные соцветия, обычно сопровождае-
мые особым, более или менее крупным листом (покрывалом), который защи-
щает собрание цветков во время их формирования или играет роль в привле-
чении опылителей. Многие растения древовидные, имеют широкие листья с
непараллельным жилкованием, с развитыми черешками; другие — травы.
Сосуды есть во всех частях растения.
В подклассе всего 4 порядка.
1-И ПОРЯДОК — ПАЛЬМОЦВЕТНЫЕ (ARECALES, ИЛИ PALMALES)
Порядок содержит одно крупное семейство.
Семейство прмшл (Palmae, или Palmaceae, или Агесасеае) представ-
лено деревьями с колоннообразным неветвящимся стволом (у африканских
дум-пальм из рода Hyphaene ствол ветвится) и кроной из крупных
черешчатых листьев с перисто- или пальчаторассеченной пластинкой. Ветре-
ВЫСШИЕ PACT
и: I;
Рис. 436. Палыш:
1—2— соцветия; з-4— веночки соцветий с тычиночными цветками; 5— веточка соцветия с пестичными
цветками
чаются пальмы с настолько укороченными стволами, что кажется, будто
листья, образуя розетки, выходят прямо из земли («бесстебельные» южноаме-
риканские пальмы — Astrocarium acaule и Attalea spedabilis). У многих есть
подземные побеги, и на поверхность выходят вместе несколько сближенных
стволов или розеток. Известно немало пальм-лиан с длинными тонкими стеб-
лями, на которых листья располагаются в удалении друг от друга. Такие
пальмы, прикрепляясь к опоре различными прицепками, часто перебрасыва-
ются с одного дерева-опоры на другие — это южноазиатские ротанги (род
Calamus). Длина их стеблей при толщине 3 см может достигать 300 м — это
наибольшая длина стеблей у наземных растений. В целом жизненные формы
пальм очень разнообразны. Крупные листья пальм достигают иногда рекорд-
ных размеров. Так, у амазонской Raphia taedigera черешок листа может до-
стигать 4—5 м в длину, а пластинка 19—22 м в длину при ширине 10—12 м.
Листья пальм, кроме черешка и пластинки, имеют хорошо выраженное влага-
лище. Пластинка в почке развивается как цельная, и ее перистое, пальчатое
или веерное расщепление происходит позднее.
Цветки пальм собраны в сильно разветвленные метельчатые сопветия.
одетые, по крайней мере вначале, одним или несколькими плотными кроющи-
ми листьями простой (влагалшпнпй) фирмы — пптертл»пплм (дрд/Ал) На ргуг-
вях соцветий цветки сидят, как в коллльят, или у многих, как в почяткд*
(spadix), т. е. более или менее погруженные в мясистую ость.
Оопвотия (рис. 436) ^пазушные, только у немногих верхушечные; в послед-
нем случае растения монокарпические, погибающие после цветения (саговая
пальма — Metroxylon, талипотовая пальма — Cory ph a umbraculifera). Со-
цветия пальм крупные. Наибольшего размера они достигают у талипотовой
пальмы — 14 мвысотбй и 12 м шириной; цветков в соцветии около 50000.
Цветки пальм мелкие, актиноморфные, реже слабозигоморфные, обое-
полые или у большинства вследствие редукции однополые. Растения с одно-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
567
полыми цветками однодомные, резке двудомные
(например, финиковая пальма). Околоцветник не-
взрачный, из двух трехчленных кругов, причем
листочки нарут&огоГкруга несколько мельче внут-
ренних. Тычинок у большинства шесть, они рас-
положены в двух трехчленных кругах; у некото-
рых тычинок больше — до нескольких десятков,
а у очень немногих только три. Плодолистиков
три, иногда свободных или^ уболыпинства, срос-
шихся и образующих
иг.
одну трех- или одногнезд-
ную верхнюю завязь. В каждом гнезде завя-
Рис. 437. Разрез плода
кокосовой пальмы (мп —
межплодник, внп'—
внутриплодник, а—эндо-
сперм, вн — внутренняя
полость, а — зародыш)
зи по одной семяпочке, но семя развивается
только в одном из гнезд. Опыляются пальмы
ветром, и пыльца образуется в огромных коли-
чествах. Однако встречаются и энтомофильные
пальмы. Плод — костянка, у некоторых ягода.
Плодаучщйи5Гнекрупнеё ягоды смородины,
у других могут иметь почти полуметровый попе-
речник и массу 10—25 кг (сейшельская пальма —
Lodoicea seischelariun). Это самые крупные плоды у древесных растений. Семе-
на с крупным, у некоторых очень твердым «роговым» эндоспермом, часто тес-
но срастаются с эндокарпием.
К пальмам относится около 3400 видов, растущих в разнообразных место-
обитаниях в тропических странах, особенно в бассейне Амазонки, на остро-
вах Малайского архипелага, Океании. Немногие пальмы выходят за пределы
тропиков. Так, в Европе встречается на крайнем юго-западе низкорослая
пальма хамеропс (Chamaerops hiunilis). До ледникового периода она росла и в
Средней Европе. В неогене европейских пальм было больше. Они доходили
до Гренландии, росли и на территории нашей страны. Древесные остатки
иг.
пальм известны уже с мелового периода.
По значению в жизни человека пальмы уступают только злакам, а по
разнообразию использования они занимают первое место среди всех семейств
цветковых растений. Дл£ многочисленного населения тропических областей
Азии, Африки, Южной Америки, бесчисленных островов Океании их значе-
ние исключительно велико. От многих пальм получают продукты, экспорти-
руемые и в другие страны и занимающие видное место в мировой экономике.
У таких пальм, как кокосовая, пальмира-пальма (Borassus flabelliformis) и
многие другие, все наземные части находят то или иное применение в быту.
Кокосовая пальма (Cocos nuctfera) широко распространена по побережьям
океанов и часто разводится как полезное растение. Плоды ее — костянки,
обычно неправильно называемые кокосовыми орехами (рис. 437), — достига-
ют размера 30 X 20 см и в свежем виде весят до 8 кз, содержат одно семя.
При формировании семени кокосовой пальмы ее эндосперм сначала весь
жидкий, прозрачный, затем в нем появляются капли эфирного масла, возни-
кает эмульсия молочного цвета. В дальнейшем периферические слои эндо-
сперма, близкие к нуцеллусу, отвердевают, превращаясь в белую мякоть,
напоминающую круто сваренный яичный белок. Внутренние части эндоспер-
ма долго остаются жидкими. Жидкая фаза эндосперма известна под именем
кокосового молока, она питательна и утоляет жажду. Слой отвердевшего
эндосперма толщиной до 2 см используют для получения масла, идущего в
пищу, на мыловарение и пр. Жмых, получаемый при выжимке (прессовании)
масла, идет в корм скоту, а твердый деревянистый внутриплодник —
на токарные изделия (пуговицы и т. п). Межплодник кокосового ореха про-
низан массой волокон, идущих после высыхания его (под названием кокосо-
вого волокна или копры) на циновки и щетки. Стволы употребляются как
строительный и столярный материал. Листья применяются кай кровельный
материал для жилищ, а волокно листьев идет на веревки. Очень молодые
листья употребляются как овощи. Сок, получаемый на месте срезан-
568
выс
и is
Е РАСТЕНИЯ
Рис. 438. Финиковые пальмы в Египте
ных молодых соцветий, сбраживают на вино, а при выпаривании сока обра-
зуются сладкий сироп и сахар.
Финиковая пальма (Phoenix dactylifera, рис. 438), разводимая с доистори-
ческих времен в жарких областях Африки и Азии, имеет некрупные продол-
говатые односемянные плоды-ягоды, богатые сахаром (до 54%). Во многих
местах они составляют главную пищу местного населения, а в странах им-
порта — лакомство. Известно свыше 1
ill
сортов этого
вида.
Пальма
двудомная. Разводят ее главным образом корневыми отпрысками. В СССР,
на юго-западе Туркменской ССР, ведутся опыты по акклиматизации финико-
вой пальмы. Другие виды финиковых пальм за пределами их родины разводят
как декоративные растения в комнатах, на юге — в садах (в СССР на Черно-
морском побережье Кавказа, на Южном берегу Крыма).
Из мякоти (межплодника) плодов африканской масляной пальмы
(Elaeis guineensis) получают техническое пальмовое масло, а из семян ее —
пальмоядровое пищевое масло. Из плодов некоторых южноамериканских
пальм тоже добывают ценные технические масла.
Из мягкой сердцевины стволов саговых пальм (Metroxylon* Малайский
архипелаг, Новая Гвинея) получают крахмал, идущий на изготовление на»
стоящего саго. Сахаристый сок, получаемый из стеблей на месте срезанных
молодых соцветий пальм (африканской винной —Raphia vinifera* индо-малай-
ской сахарной — Arenga saccharifera и др.), используют для приготовления
пальмового вина и пальмового сахара. Под названием пиассавы волокна из
листовых влагалищ многих пальм используются при изготовлении щеток
(для мытья улиц, лошадей, до/я картофелемоек), реже на канаты и циновки..
Мягкие волокна из листьев некоторых африканских видов рафий применя-
ются под названием мочало-рафия в садоводстве как подвязочный и обвязоч-
ный (при прививках) материал. С листьев южноамериканской карнаубской
пальмы (Copernicia cerifera) и со стеблей, тоже южноамериканской, восковой
пальмы (Ceroxylon andicola) получают воск, находящий разнообразное при-
менение. Верхушечные почки, молодые листья, сердцевину стволов многих
пальм используют в пищу («пальмовая капуста» и т. п.). Плоды многих
пальм употребляют как фрукты.
Для широко распространенного в Юго-Восточной Азии жевания бетеля
применяют вместе с перцем-бетелем (Piper betle) и жженой известью ломтики
семян арековой пальмы* или пинанга (Areca cathechu). Очень твердый
эндосперм семян южноамериканских видов Phytelephae под названием расти»
тельной слоновой кости употребляют на изготовление пуговиц и других
имитаций костяных изделий.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
569
Многие виды пальм разводят как декоративные в оранжереях и жилых
помещениях, а в более теплых странах в садах и на улицах. В СССР в откры-
том грунте выращивают около 20 видов пальм (на Черноморском побережье
Кавказа и, отчасти, на Южном берегу Крыма). Наиболее распространены
виды восточноазиатского трахикарпуса (Trachycarpus), которые способны
без существенных повреждении переносить суровые зимы (например, зиму
1949/1950 г.).
2-Й ПОРЯДОК — АРОННИКОЦВЕТНЫЕ (ARALES)
Цветки 'аронникоцветных в початках, сопровождаемых кроющим ли-
стом — покрывалом, мелкие, с околоцветником или без него; иногда, редуци-
рованные до одной тычинки и одного плодолистика, встречаются цветки,
погруженные в ось початка. Верхняя завязь с тремя — одним гнездами и с
одной—несколькими семяпочками в каждом гнезде.
Аронникоцветные — травы (часто очень крупные), лианы, эпифиты, иног-
да древовидные растения, обычно наземные, реже водные.
емейство ароидные, аронниковые (Агасеае). Наиболее характер-
ным признаком ароидпых служат их толстые, мясистые соцветия — початки,
в основании которых находится один большой кр_о_ющий"лйст (чехол, покры-
вало, крыло — spatha), нередко окружающий початок. У некоторых этот
лист зеленый (аир); у большинства ярко окрашенный и функционально игра-
ющий роль венчика, делающий все соцветие хорошо заметным для насеко-
мых-опылителей. Цветки мелкие, более или менее редуцированные, тесно
сидящие на початке. У одйих (например, у аира) околоцветник обычно типа
однодольных, у других (белокрыльник, аронник и др.) он редуцирован и
цветки голые.
Цветки обоеполые (аир, белокрыльник, антуриум) или однополые (арон-
ник, филодендрон и др.). Чаще всего тычиночные цветки расположены в
верхней части соцветия, пестичные — в нижней; редко растения двудомные.
Число тычинок у разных родов варьирует от 9 до 3—2 и даже 1; у многих
они более или менее сращены. Нередко часть тычинок превращена в стамино-
дии. Пестик один с трех-или одногнездной завязью, у некоторых погружен-
ный в цветоложе. Опыляются в большинстве насекомыми, которых привлека-
ет яркая окраска покрывала и часто неприятный запах. Плод — ягода. Се-
мена с обильным эндоспермом; распространяются они у большинства пти-
цами.
К ароидным относится около 1800 видов, растущих главным образом в
тропиках и субтропиках. Это или многолетние наземные травы с корневища-
ми, клубнями, или лианы, лазающие при помощи придаточных корней,
или эпифиты с воздушными корнями, Нередко потом укореняющимися в поч-
ве. Немало переходов от наземных форм и лиан к эпифитам. Листья у боль-
шинства крупные, с черешками и влагалищами, сетчато-нервные, часто серд-
цевидные, копьевидные, стреловидные. В анатомическом отношении харак-
терно для многих наличие млечников, смоляных ходов, групп слизистых
клеток, волосков в межклетниках.
Хозяйственное значение ароидных невелико. Толстые корневища и клуб-
ни некоторых видов хотя и содержат едкие и даже ядовитые вещества, но при
тепловой обработке освобождаются от них и становятся годными в пищу.
Особенно большое значение имеет в этом отношении таро (Colocasia esculent а)—
широко культивируемое пищевое растение исконного населения тропиков
и субтропиков Старого Света. В пищу употребляют в вареном виде крупный
подземный корневищный клубень. Многие тропические виды родов
Anihurium, С alodium, Colocasia и других часто разводят в теплицах как деко-
ративные растения с ярко окрашенными соцветиями или с красивыми, не-
редко пестрыми листьями.
В СССР растут лишь 13 видов ароидных из 7 родов. Широко распростра-
нен по берегам водоемов и на болотистых лугах аир (Acorus calamus) — круп-
570
ВЫСШИЕ РАСТЕНИЯ
Рис. 439. Белокрыльник (Galla palustris):
7— обща* гаде >—цветок; в— плоды
/
Рис. 440. Ряска (Lemna minor):
1 — ббщий вид; 2 — соцветие с двумя тычиночными
и одним (увядшим) пестичным цветком
ное растение с мечевидными листь-
ями и таким же кроющим листом
(чехлом), сдвигающим вбок поча-
ток. Уроженец Юго-Восточной
Азии, он распространился по Ев-
ропе, но цветет у нас нечасто, ни-
когда не приносит плодов вслед»
ствие недоразвития пыльцы и се-
мяпочек и размножается перено-
сом корневищ водой. Толстые
ароматические корневища его,
под названием «аирный корень»,
применяют в медицине и парфю-
мерии, а эфирное масло из них —
в ликерно-водочном и кондитер-
ском производстве.
В таких же экологических
условиях, как аир, обитает бе-
локрыльник (Сalia palustris), на-
званный так за ярко-белую ок-
раску верхней стороны крупного
яйцевидного кроющего листа
(рис. 439)« Вываренные вводе кор-
невища его съедобны, и в север-
ных районах их иногда подмешива-
ли к ржаной муке. В южных райо-
* нах растет несколько видов арон-
ников. В оранжереях и комнатах
очень часто разводится под назва-
нием монстеры или филодендро-
на (Мonstera deliciosa), с крупны-
ми перистораздельными и проды-
рявленными листьями и воздуш-
ными корнями. На родине (Запад-
ная Мексика, Гватемала), где она
растет как лазающая лиана в ле-
сах, ее ароматные крупные почат-
ки употребляют вместо фруктов.
V Семейство о я с к о-
.Ill
III
i г
(Lemnaceae).
Растения
этого мален ько го семейства на
первый взгляд не похожи на аро-
идные, однако рясковые связаны
с ними через очень упрощенный
тропический род пистия (Pistia.
из семейства Агасеае); в початках
пистии находится лишь один
женский цветок, представленный
одним пестиком, и немного мужских цветков,
состоя:
(каждый)
всего из двух сросшихся тычинок. У рясковых эта редукция пошла еще
дальше, охватила и вегетативные и генеративные органы, и ряски могут слу-
жить среди цветковых примером крайней редукции всего растения (рис. 440).
Вегетативное тело рясок представлено маленьким зеленым листовидным
стебельком, плавающим на поверхности воды или, реже, как, например,
у Lemna trisulca* погруженным в воду. У некоторых рясок (Spirodela) у осно-
вания стебля имеются два очень маленьких редуцированных пленчатых лис-
точка. От листовидного стебля отходит вниз один (у ряски — Lemna ) или
несколько (у многокоренника — Spirodela) нитевидных неветвящихся ко-
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОС
571
решков. Они, вероятно, способствуют сохранению более устойчивого положе-
ния растения на воде, а поглощение.воды и питательных веществ происходит
всей поверхностью растения, соприкасающейся с водой. У волъфии (Wolffia
arrhiza) ее очень маленькие (1—1,5 мм)9 сильно выпуклые снизу стебельки
совсем не имеют корней.
Ряски цветут чрезвычайно редко, но очень быстро размножаются веге-
тативно при помощи боковых побегов, образующихся в небольших углубле-
ниях листовидного стебля, в так называемых кармашках; новые растеньица
:ri
г. г
или остаются долго соединенными с материнскими, или скоро отделяются
от них.
Соцветие ряски — очень маленький редуцированный початок — тоже
выходит из бокового кармашкД при основании стебля и окружено одним неж-
ным пленчатым кроющим листом (чехлом). У вольфии кроющего листа нет.
В соцветии находятся один женский тдтпкт состоящий только из одного
пестика, без околоцветника, и один-два мужских цветка, тоже не имеющих
околоцветника и состоящих каждаЖиЛодиойийшщйкТДветки протероге-
ничны и, по-видимому, анемофильны. Плод одногнездный, с 1—7 семенами,
не раскрывающийся или раскрывающийся поперечно.
На зиму ряски, плавающие на воде, погружаются в воду. У многокорен-
ника развиваются особые зимующие побеги, погружающиеся на дно. Весной
в них образуются воздухоносные полости, часть крахмала растворяется, и
растение всплывает.
К рясковым принадлежат 3 рода: Lemna, Sptrodela, Wolffia — и около
25 видов. Некоторые из них широко распространены почти по всей Земле.
Вольфия растет преимущественно в тропиках. В СССР произрастают 5 ви-
дов из 3 родов.
и г.
8-Й ПОРЯДОК — ПАНДАНОЦВЕТНЫЕ (PANDANALES)
Цветки панданоцветных однополые с рудиментарным околоцветником
или вовсе без околоцветника. В мужских цветках одна — несколько тычи-
нок и иногда остатки редуцированного гинецея; в женских — одно- или мно-
гочленный гинецей. Травы, древоподобные растецця и лианы с линейными
листьями.
Семейство пондомовые (Pandmaceae) включает деревья, кустарники
или древесные лианы (Freycinetia) с длинными, спирально расположенными,
линейными, по краям плотными, по средней жилке остропильчатыми листья-
ми; у многих вцдов они собраны пучком на вершинах стеблей (рис. 441).
Цветки вторично однополые, без околоцветника, в початках или головках.
Плоды — ягоды или костянки, у многих срастающиеся в соплодия; иногда
съедобные (Pandamu edulis).
Семейство включает около 880 видов, растущих только в тропиках Старо-
го Света и объединяемых в 3 рода. Наиболее известны панданы (Pandanus) —
характерные деревья морских и речных побережий. Сравнительно тонкие,
ложнодихотомически разветвленные стволы их имеют на вершинах ветвей
пучки очень длинных листьев. Из нижней частив стволов вырастают придаточ-
ные корни, достигающие толщины руки. Сама ^нижняя часть стеблей нередко
отмирает, и деревья стоят как бы на ходулях. •,
Семейство ежеголовковые (Sparganiaceae). Прибрежно-вод-
ные растения с узкими листьями и шаровидными пазушными соцветиями —
мужскими и женскими (рис. 442). Однополые, однодомные цветки в шаровид-
ных головках, собранных в слабо разветвленные пазушные соцветия. Около-
цветник из 6—3 маленьких чешуевидных листочков. Тычинок тоже 6—3.
Пестик образован одним, редко двумя плодолистиками, завязь верхняя с од-
ной, редко двумя семяпочками. Анемофилы. Плод — костянка с губчатым
воздухоносным межплодником. Включает один род — ежеголовка (Sparger
тит) с 20 видами, обитающими в холодных и умеренных странах.
572
ВЫСШИЕ РАСТЕНИЯ
Рис. 441. Пандан
V Семейство рогозовые
(Typhaceae). Как и ежеголовко-
вые, это небольшое семейство
прибрежно-водных и болотных
растений, состоящее всего из од-
ного рода рогоз (куга, чекан —
Typha) приблизительно с 15 вида-
ми, распространенными по все-
му северному полушарию и в
Южной Африке (в СССР — 8 ви-
дов). Наиболее обычны рогоз ши-
роколистный (Т. latifolia} и рогоз
узколистный (Т. angustifolia). Это
многолетние травы с толстыми,
богатыми крахмалом корневища-
ми, высокими стеблями и длин-
ными тесьмовидными двухрядны-
ми листьями, собранными у ос-
нования стебля (рис. 443).^Цветки
мелкие, однополые, однодомные^
собртнтшё п ддирные циЛНйдри-
чша£н&початки. Мужской початок
расположен пад ясонским. Около-
цветник редуцирован в волоски.
В мужских цветкдх у болыпинст-
ва три тычинки со\росшимися на
большом протяжении нитями; в
женских —один пестик; одногнезд-
ная завязь снабжена длинной
волосистой ножкой (гинофор),
содержит одну семяпочку и несет
наверху очень длинный столбик,
f £в()1ММ^немофилия. После оплодотворе-
ния женский початок сильно раз-
растается и становится темно-бурым. Плоды — орешки с остающимся стол-
биком и сильно удлиняющимися волосками — распространяются ветром.
Рогозы, растущие нередко обширными зарослями, могут иметь разнооб-
разное применение: листья употребляются в бондарном деле, для плетения
циновок и т. п.
Растение целиком может служить для изготовления бумаги; его корне-
вища можно употреблять в пищу и для получения крахмала; волоски
при плодах — как упаковочный и набивочный материал, а вместе с
шерстью — на производство фетра для шляп.
Подкласс Arecidae в своем происхождении, вероятно, связан с
Liliidaeвернее, с их ближайшими предками. Эволюция цветков в этом под-
классе шла, по-видимому, во всех линиях развития в сторону редукции около-
цветника и перехода к однополости. Параллельно с этим эволюционировали
ин
и соцветия, но уже в двух направлениях: сложные метельчатые соцветия у
пальм и початковидные в трех прочих порядках. Эволюция вегетативного те-
ла своеобразна. В порядке пальмоцветных из травянистых стеблей предпола-
гаемых предков подкласса выработались колоннообразные одревесневающие
стволы; в трех прочих порядках одревесневающие стебли, вероятно, развива-
лись параллельно с разными формами травянистых (надземные, корневища,
клубневидные). Непосредственная генетическая связь между порядками,
относимыми к этому подклассу, через ныне живущие формы мало вероятна,
и родство их, может быть, правильнее представлять как происхождение
из общего предка.
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
573
Рис. 442. Ежеголовка (Sparganium neg-
lectum):
1 — соцветия; 2 — пестичный цветок; з — тычи-
ночный цветок; 4 — соплодие; 5 — диаграмма ты-
чиночного цветка; 0 — диаграмма пестичного
цветка
Рис. 443. Рогоз (Typha
latifolia):
1 — соцветие; 2 — тычиноч-
ный цветок; з — пестичный
цветок; 4 — диаграмма пес-
тичного цветка
ш;
ОБЗОР ЭВОЛЮЦИИ ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
Выше были рассмотрены два класса и десять подклассов покрытосемен-
ных растений, причем в большинстве случаев указывались возможные родст-
венные связи между ними. На основании этого материала можно составить
обобщенное представление об основных путях эволюции, которые привели к
современному разнообразию покрытосеменных.
Предки покрытосеменных, как указывалось ранее, остаются до настоя-
щего времени неизвестными. Но, несомненно, вся эта группа возникла от
голосеменных и обособилась от них, вероятно, еще на ранних этапах всей
истории семенных растений. Из современных покрытосеменных, соответст-
венно стробилярной, или эуанциевой, теории цветка, по-видимому, наиболее
близок к древнейшим покрытосеменным подкласс Magnoliidae. Однако Mag-
noliidae имеют уже настолько специализированный аппарат для привлечения
насекомых-опылителей, что естественно предположить существование в
прошлом еще более древней группы покрытосеменных. Такие гипотетические
предковые покрытосеменные могли быть неустойчиво энтомофильными расте-
ниями, у которых перенос пыльцы совершался и ветром, и насекомыми —
собирателями пыльцы. По всей вероятности, у них достаточно широко было
распространено и самоопыление.
574
ВЫСШИЕ РАСТЕНИЯ
Piperales
Pec. 444. Схема эволюция цветковых растений
Urticales
I Fagale
Ericales
Malvales
Umbellales
Juglandales
Celastrales
Primulales
benales
Rhamnales
Trochodendrales
. ДВУДОЛЬНЫЕ
Capparales
Polygonales
Terebinthales
Myrtales
GeranialeS;
Theales
Dilleniales
Resale
Aristolochiales
Magnoliales
ПРЕДКОВЫЕ ЦВЕТ
РАСТЕНИЯ .
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСВ
Vi; /1; i; i
Asterale
1 Dip» ecal
Solanales\
Boraginales
rbenales
Rubiatei
Graminales
Сурега
Arales
Bromeliales
Papaverales
Commeli
A recal
(Palmae)
ОДНОДОЛЬНЫЕ
Nymphaeates
es
Gentiapaie
nvolvulales
Zingiberales
Potamoget onales
Hydrocharitales
Liiiales-
; НОВЫЕ
Restipnalps
576
ВЫСШИЕ РАСТЕНИЯ
Гипотетические покрытосеменные предки современных цветковых дали
начало, вероятно, нескольким линиям эволюции (рис. 444). Главной из них.
т. е. имевшей впоследствии наибольший успех в освоении самых разнообраз-
условий жизни, была линия, в которой рано выработались Magnoliidae*
Другой была линия, представленная подклассом Hamamelidae.
В конце прошлого — начале текущего столетия было широко распрост-
ранено представление об однопокровных, которые в основном соответствуют
Hamamelidae, как о группе весьма древней, примитивной, близкой к предкам
покрытосеменных. Подтверждение этому видели в преобладании среди одно-
покровных древесных растений, в их анемофильности, в морфологической
простоте и однополостности цветков. Эти особенности присущи и голосемен-
ным, среди которых вовсе нет травянистых растении, которые, как долгое
время думали, опыляются только ветром, у которых нет и не было яркого
околоцветника, а каждое спороношение, за редкими исключениями, со-
держит только микро- или макроспорофиллы. В пользу глубокой древности
однопокровных говорят и палеонтологические данные.
Учение о происхождении покрытосеменных от голосеменных через од-
нопокровные дополнялось специально разработанной австрийским ботани-
ком Р. Веттштейном теорией происхождения цветка покрытосеменных
из «соцветия» голосеменных (типа эфедры). Эта теория получила название
теории псевданция (ложного цветка).
Научная критика концепции происхождения всего многообразия покрыто-
семенных от однопокровных вскрыла прежде всего, что морфологически
«простое» строение цветка может быть следствием упрощения, возникшего в
связи с особым направлением отбора. Это подтверждается наличием остаточ-
ного околоцветника (дуб, лещина), появлением в цветках рудиментов недо-
стающего пола и развитием в качестве исключения обоеполых цветков вместо
однополых.
Если простота цветков однопокровных вторичная, то приходится заклю-
чить, что эта группа прошла длительный путь эволюции, возникнув от пред-
ков с обоеполыми цветками. Но обоеполость цветков тесно связана с энтомо-
филией. Поэтому и распространенная у однопокровных анемофилия должна
была прийти на смену энтомофилии, что возможно в процессе приспособления
к жизни большими одновидовыми скоплениями. По-видимому, именно бла-
годаря этому от каких-то энтомофильных лилиецветных возникли анемофиль-
ные осоки и злаки; анемофильные полыни, несомненно, являются производ-
ными энтомофильных сложноцветных.
Подвергалась глубокой критике и псевданциевая теория происхождения
цветка, оказавшаяся неубедительной.
В итоге, на смену представлению о первичности и примитивности одно-
покровных сформировалось прямо противоположное воззрение — однопо-
кровных стали рассматривать как группу, прошедшую длительный путь эво-
люции и возникшую от высокоспециализированных покрытосеменных.
При этом допускалось, что различные порядки однопокровных — произ-
яе разных эволюционных линий покрытосеменных. Это означало отрица-
ние группы как единого естественного таксона.
Часть систематиков, признавая однопокровные за целостную единицу,
сближали и продолжают сближать его с порядком Hamamelidales,
Вопрос о происхождении Hamamelidales решается двояко. Одни ботаники
(из современных авторов Hutschinson, 1969) производят этот порядок
от Rosales, другие (Тахтаджян, 1959,1966,1970; Cronquist, 1968)
сближают его с Magnoliidae через предка, общего с Trochodendrales.
Trochodendrales относили и сейчас иногда относят к магнолиецветным.
Характерная для них бессосудистая древесина говорит об их древности, что
согласуется с примитивным строением цветка (отсутствие околоцветника,
неопределенное число спирально расположенных тычинок, неустойчивое
число плодолистиков и неполное их срастание). Невзрачность цветков и се-
режчатые соцветия (род Tetracentrori) соответствуют опылению ветром, но
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
577
строение пыльцы и выделение нектара, дополненное прямым наблюдением
(Trochodendrori), свидетельствуют об энтомофильности. Однако анемофилия
у этих растений, по-видимому, не исключена — существует рядом с опылени-
ем насекомыми.
Значит, порядок Trochodendrales можно рассматривать как относительно
близкий неустойчиво-энтомофильным древнейшим цветковым растениям,
а линию Trochodendrales — Hamamelidales — прочие Hamamelidae — как
параллельную линии Magnoliidae.
Древние предки покрытосеменных могли дать начало и другим линиям
развития, где неустойчивая энтомофильност'ь сохранялась долгое время, пока
не вырабатывались специализированные в отношении опыления группы.
Так могли возникнуть энтомофильные Proteales и Salicales. В пределах Sali-
cates дифференцировались энтомофильный (Salix) и анемофильный (Populus)
стволы.
Casuarinales, в схеме филогенеза покрытосеменных растений условно при-
соединенные к подклассу Hamamelidae, имеют ряд признаков сходства с
UrticalesK Hamamelidales, но своеобразие этого порядка все же столь велико,
что целесообразнее считать его самостоятельной ветвью развития (вероятно,
смыкающейся в глубине времен с предками Hamamelidales).
Подкласс Magnoliidae состоит, по-видимому, из «верхушек» разветвлений
давно вымершего «основного» ствола и, следовательно, не столько выражает ход
эволюции на относительно раннем этапе истории покрытосеменных, сколько
характеризует уровень эволюционного развития, достигнутый главным об-
разом относительно особенностей цветка. Magnoliidae* 1 сохранили относитель-
но примитивные черты организации цветка и порой малосовершенные приспо-
собления к опылению насекомыми. Эволюционное потомство Magnoliidae
было очень разнообразно. Оно, несомненно, представляет не одну, а три до-
статочно обособленный линии прогрессивной эволюции двудольных — под-
классы Caryophyllidae, Dilleniidae и Rosidae*. Спайнолепестные двудольные с
тетрациклическими цветками объединены в подклассе Asteridae, примыкаю-
щем к подклассу Rosidae. Следовательно, класс двудольных расчленяется на
6 (а если признавать за самостоятельный подкласс Ranunculidae, то на 7)
подклассов. Из них непосредственно с Magnoliidae связаны Caryophyllidae,
Dilleniidae, Residue. Кроме этих подклассов, Magnoliidae rorjh начало классу
однодольных^
Подкласс Caryophyllidae связывается с многоплодниковыми через семей-
ство Phytolaccaceae, в котором есть растения с апокарпным гинецеем. В случае
синкарпного. гинецея срастание плодолистиков у Phytolaccaceae неполное,
иногда только в основании, но, во всяком случае, число столбиков всегда со-
ответствует числу плодолистиков. Иногда, при созревании плода, плодолис-
тики отделяются друг от друга. Примитивной чертой организации цветка яв-
ляется и простой околоцветник, типичный для этого семейства, но в некото-
; и t
рых родах наблюдается образование 'второго круга околоцветника за счет
превращения наружных тычинок в лепестковидные стаминодии. И то и дру-
гое распространено также у многоплодниковых. Phytoloceaceae с одним кру-
гом тычинок, противостоящим листочкам околоцветника, и с одночлен-
ным гинецеем, вероятно, представляют вторичные формы-
Порядок Caryophyllales, наиболее обширный в составе подкласса Caryo-
phyllidae', объединяет несколько ветвей эволюционного развития, возникших,
вероятно, от общего предка. Обособление этих ветвей, некоторые из которых
достойны выделения как подпорядки (например, специфичные по экологии
стеблевые суккуленты — кактусы, листовые суккуленты — аизовые, галофи-
ты — многие лебедовые), по всей видимости, началось рано. В одних грушках
околоцветник остался простым, в других он стал двойным, причем это могло
1 В данном руководстве подкласс Magnoliidae принят в объеме А. Л. Тахта-
*» д ж я н а, 1966.
1 По Т ах т а Д ж я н у, еще Hamamelidae и Ranunculidae.
** 19—602
< г
578
выа
in;
Я РАСТЕНИЯ
осуществляться двумя разными путями — превращением части тычинок в
лепестки и образованием чашечки из верховых листьев, придвинувшихся к
цветку. В отдельных случаях проявляется тенденция к спайнолепестности
(срастание внутренних лепестков у некоторых кактусов). Спайнолепестность
как более устойчивое явление реализовалась в порядке Plumbaginales, кото-
рый примыкает к Caryophyllales по строению гинецея и по некоторым анатоми-
ческим признакам (аномальное кольцевое утолщение стебля). У Plumbagina-
les венчик спайнолепестный, раздельнолепестность имеет место лишь как
исключение. Подобно многим растениям со спайнолепестным венчиком, здесь
сохранился только один круг тычинок, которые располагаются против эле-
ментов венчика.
Особую линию эволюции Caryophyllidae представляет порядок Polygo-
nales, вероятно также возникший от Caryophyllales и развивавшийся независи-
мо от Plumbaginales.
Подкласс Dilleniidae достаточно явственно примыкает к подклассу Mag-
noliidae своими примитивными представителями из порядков Dilleniales и
Theales. От этих двух порядков берут начало основные направления разви-
тия. В одном из них эволюция завершается верескоцветными {Ericales), при-
митивные представители которых (семейство Clethraceae) имеют цветок еще
всегда раздельнолепестный. В семействе Ericaceae наряду с раздельнолепест-
ностью примитивных представителей в более прогрессивной его части спайно-
лепестность становится преобладающей. В этом же направлении изменяется
и положение завязи — от верхней к полунижней и нижней.
Вторую линию развития в пределах Dilleniidae завершают порядки Pri-
mutates и Ebenales. Их связь С раздельнолепестными не подлежит сомнению.
В семействе Primulaceae есть растения, у которых подавлено характерное для
спайнолепестных развитие трубки венчика (роды Lysimachia, Naumburgia,
Trientalis)', кроме того, известны некоторые представители с вполне раздель-
нолепестным венчиком (род Pollettera). Порядок Ebenales, вероятно, имеет
происхождение, общее с Primulales.
Третья ветвь этого подкласса наиболее обширная. Она начинается
большим порядком Violates (соответствует в основном порядку постенно-
семенных Parietales — других систем), от которого или от близких его пред-
ков, в свою очередь, взяли начало три ряда развития, существенно отличные
по своему содержанию. В одном из них широко распространены димерные
цветки. Этот ряд начинается порядком Capparales, в котором более низкое по-
ложение занимает семейство Саррагасеае, а завершается крупным семейством
Cruciferae.
иг
Второе направление характеризуется сохранением пентамерной основы
цветка. Несмотря на то что число тычинок уменьшается, наблюдается соедине-
ние пыльников, вырабатываются спайнолепестность и нижняя завязь. Веге-
тативное тело становится лежащим или благодаря усикам лазающим. К этой
группе примыкает порядок Cucurbitales. Наконец, возможно происхождение
от предков Violates и третьей ветви, центральное положение в которой зани-
мает порядок Malvales с примыкающим к нему порядком Euphorbiales. Связь
последнего с мальвоцветными принимается рядом систематиков; что же каса-
ется происхождения Мalvales, то наряду с принятым здесь есть и другое пред-
ставление, согласно которому этот порядок близок к герашдецветным.
Подкласс Rosidae может быть выведен из Magnoliales через порядок Rosa-
les, вернее, через низшие формы этого порядка, имеющие апокарпный гине-
цей и плод—листовку. От розоцветных, в свою очередь, берет начало не-
сколько филетических линий двудольных.
Одна их них объединяет семейства порядка Fabates. Начиная с форм,
имеющих правильные (Mimosaceae) или слабозигоморфные (некоторые Сае-
salpiniaceae) цветки, это направление развития завершается порядком Fabales,
выделяющимся совершенными приспособлениями к опылению насекомыми.
В этом направлении эволюционного развития от низших форм до высших со-
храняются верхняя завязь и раздельнолепестность венчика.
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
579
Вторая линия объединяет семейства порядка Myrtales и характеризует-
ся преобладанием тетрамерных цветков (у Rosales известно немало случаев
тетрамерности цветков). Другая характерная черта Myrtales — наличие
гипанция. Каково бы ни было происхождение этого органа, листочки около-
цветника и тычинки кажутся прикрепленными к его краю. Иногда гипанций
может срастаться и с завязью, которая становится нижней или полунижней.
Образование гипанция наблюдается и в некоторых группах семейства Rosa-
ceae. Myrtales имеют только один столбик (исключения редки) и, следователь*
но, синкарпия здесь полная, причем даже рыльце цельное (очень редко оно
лопастное или ветвистое). В противоположность этому при синкарПном гине-
цее в порядке розоцветных обычно верхние части плодолистиков остаются
свободными, образуя самостоятельные столбики. Следовательно, Myrtales
можно рассматривать как группу, в которой получили дальнейшее развитие
некоторые черты Rosales. Возникла она, вероятно, от более простых Rosales
и эволюционировала параллельно с ними.
Третья линия, берущая начало от Rosales, характеризуется преобладани-
ем двойного околоцветника из пятичленных кругов, пентамерным двукруго-
вым андроцеем (или, часто, производным от такового) и синкарпным гинецеем,
довольно изменчивым по числу плодолистиков.
Основное место в ней занимает порядок Terebinthales с характерным вмео
тилищем выделении в вегетативных органах и развитием медоотделительно-
го диска в цветке. У высших представителей появляются зигоморфные цвет-
ки. От предков этого порядка могли ответвиться Geraniales с изначально пра-
вильными цветками, приобретающими зигоморфность у высших форм, в
частности у Tropaeolaceae и Polygalaceae. В цветках семейства Geraniaceae час-
то развиваются нектарные железки, которые можно истолковать как стами-
нодии, но, вероятнее, это разрастания цветоложа, т. е. образования, соот-
ветствующие диску Terebinthales*, однако полного развития диск здесь не полу-
чил. Тенденцию к разрастанию цветоложа можно усмотреть и в характерной
для Geraniales обдиплостемонии, связанной с интеркалярным ростом цветоло-
жа, приводящим к смещению противочашечных тычинок к центру цветка.
Характерное для Terebinthales образование диска сохранилось и еще в одной
линии, возникшей, вероятно, от ранних Terebinthales и представленной
двумя параллельно развивавшимися порядками Rhamnales и Celastrales.
Порйдок Celastrales некоторые ученые рассматривают как близкий к предко-
вым формам подкласса Asteridae.
Как четвертую линию, берущую начало от розоцветных, можно рассмат-
ривать порядок Vmbelliflorae. Сходство с Rosales выражается в наличии ниж-
ней завязи, четырехкругового цветка с малым числом плодолистиков (семей-
ство камнеломковые) и однопокровной семяпочки.
Семейство Сотасеае сравнительно с семейством Vmbelliferae более прими-
тивное. Признавая родство зонтикоцветных и мареноцветных через Согпа-
сеае, необходимо допустить, что эти два порядка эволюционировали парал-
лельно. В конце одного из параллельных путей эволюции сформировалось
семейство зонтичных с характерными для него компактными соцветиями —
сложными зонтиками.
Вопрос о конкретных Rosidae, которые могли бы дать начало последнему
подклассу двудольных — Asteridae, нельзя считать решенным. Происхожде-
ние Asteridae возможно от предков Celastrales, так как существует мнение,
что Celastrales имеют общие черты с порядком Oleales, ррястъеншм. порядку
Gentianales,
Подкласс Asteridae в самом начале своего развития образовал две круп-
ные ветви, обе эволюционировавшие на базе пентамерного тетрациклического
цветка, возникшего вследствие редукции внутреннего круга тычинок пяти-
кругового цветка. Обе линии привели к выработке приспособлении к опыле-
нию насекомыми в строении и цветка и соцветий, что хорошо видно на при-
мере сложно устроенных, иногда густоколосовидных соцветий некоторых
губоцветных или скученных соцветий сложноцветных и высших ворсянковых.
19 •
ВЫСШИЕ PACT
и; I
ГЛ fl
Два основных направления эволюции Asteridae осуществились в порядках
Tubtjlorae и Gent inales, В обширном порядке Tubiflorae более примиптнил
семейства, объединяемые в подпорядок Contmvulineae, сохранили еще непос-
тоянство числа плодолистиков (2—5). От этого порядка возникли потомки с
практически однотипным двучленным гинецеем. Но если у одних (подпоря-
док Solanineae) кяяццхк плодолистик производит много семяпочек и, соот-
ветственно этому, плоды—коробочки или ягоды, то у других число семяза-
чатков сократилось до двух на каждый плодолистик, а плоды стали дробными,
с образованием односемянных орешков. В этом случае наряду с сохряпишпи-
ми (в основном) актицоморфность цветка (подпорядок Boraginineae^ вырабо-
тались резко зигоморфные формы со сложными (шалфей) приспособлениями к
опылению (подпорядок Verbentneae).
Параллельная ветвь — порядок Gentianales — при сохранении актино-
морфности достигла образования сложных приспособительных структур в
цветке (семейства Аросупасеае и Asclepiadaceae). Эта крупная ветвь дала на-
чало порядку Rubiales, в котором завязь стала нижней, и связанному с ним
порядку Dipsacales, в котором строение цветка усложнилось — образовалась
«наружная чашечка», появилась зигоморфность, а расположение цветков
достигло тесной скученности (конвергенция с корзинкой Сложноцветных).
От этой же ветви, начинающейся порядком Gentianales, возникла еще
группа порядков со своеобразными приспособлениями к опылению, с тен-
денцией к соединению (склеиванию, сцеплению, срастанию) тычинок вокруг
столбика. Это порядки Campanales и Asterales. В обоих наряду с актиноморф-
ными (трубчатыми или колокольчатыми) цветками достаточно широко рас-
пространены зигоморфные (язычковые и ложноязычковые цветки сложно-
цветных, неправильные цветки Lobeliaceae). Следовательно, и во второй линии
Asteridae эволюция цветка завершилась зигоморфностью, но она была до- }
стигнута после преобразования завязи из верхней в нижнюю, чего не было ;
Однодольные, как было указано выше, произошли от подкласса Magno- у
liidae. Строение цветка и распределение проводящих пучков в стебле сближа-
ют однодольные с Nymphaeales, имеющими к тому же своеобразное развитие
зародыша, проливающее свет на происхождение однодольности. Nymphaea^
tee, в свою очередь, имеют много черт сходства С Мagnoliidae. Против мнения
о связи Magnoliidae^Nymphaeales — Monocotyledoneae нередко возражают,
указывая , что Nymphaeales — группа, связанная с водной средой, и что труд-
но себе представить возникновение от нее сухопутного (в основном) класса
однодольных. Отсутствие сосудов у нимфейных в этом случае толкуют как
явление дегенерации. Это возражение, однако, устраняется, если выводить
однодольных не из самих Nymphaeales, а из их предков, которые могли быть
сухопутными растениями. У этих анцестральных форм была примитивная,
бессосудистая водопроводящая система, сохранившаяся у Nymphaeales
(А. Л. Тахтаджян), где она как бы законсервировалась вследствие отсутствия
«стимула» к замене трахей более совершенным аппаратом—сосудами. У назем-
ных потомков той же предковой группы — у однодольных — эта замена про-
изошла в порядке совершенствования приспособленности к жизни на суше.
Отделение однодольных как самостоятельного эволюционного ствола
могло совершиться в очень ранние эпохи филогенеза покрытосеменных, и,
может быть, начало однодольных надо искать не среди Мagnoliidae, а в пред-
шествовавшей им группе древнейших покрытосеменных Protoangiosperma£\
Однодольные, подобно двудольным, подразделяются А. Л. Тахтаджя-
ном на подклассы: Alismidae, Liliidae, Commelinidae и Arecidae. Все они вос-
ходят или почти восходят к древним предкам однодольных.
1 Если бы удалось подтвердить происхождение древнейших однодольных от неустой-
чиво-эвтомофильных предков покрытосеменных, было бы естественнее делить этот отдел
не ва два (двудольные и однодольные), а на три класса. Третий должен соответствовать -
современному подклассу Hamamelidae^
ОТДЕЛ ЦВЕТКОВЫЕ, ИЛИ ПОКРЫТОСЕМЕННЫЕ
581
ИГ
Подкласс Alimidae сохранял ряд признаков примитивности (апокарп-
ный гинецей, отсутствие сосудов в стеблях, листьях, а иногда и в корнях).
Из трех порядков этого подкласса два — Alternates и Poiomagetondles, вероят-
но» связаны друг с другом как предки с потомками. Третий порядок —
Hydrocharitales, по всей видимости, берет начало если не от самих Alismales,
то от их предков и представляет сильно специализированную водную
группу.
Подкласс Liliidae, вероятно, возник от тех же предков, что и Alimidae.
В отличие от последних Liliidae имеют гинецей ценокарпиый; от апокарпноо
ти сохранились следы в виде обособленных столбиков. В редчайших случа-
ях плодолистики бывают свободны почти до основания, иногда они отделя-
ются друг от друга ко времени созревания плода. В пределах Liliidae значе-
ние базовой группы принадлежит порядку Liliales, характеризуемому в
основном правильными тримерными пятикруговыми цветками. В результате
специализации появлялись группы с одним кругом тычинок, иногда даже зи-
гоморфные и с нижней завязью (касатиковые), а также такие, у которых око-
лоцветник стал спайнолистным; иногда возникали и тетрамерные цветки.
Значительного разнообразия достигли LiZiaZ^aпо жизненным формам. Диапа-
зон изменчивости в этом отношении от трав, в основном геофильных, До
древоподобных растений.
Наметившаяся в пределах Liliales тенденция к зигоморфности цветка
получила дальнейшее развитие в двух порядках — Zingiberales и Orchtda-
les, из которых второй представляет высочайшую вершину знтомофильной
эволюции покрытосеменных.
Но Liltales послужили основой и для одной из анемофильных ветвей.
В ней можно отличить две ступени, соответствующие: 1) порядку Juncales,
по морфологии цветка близко примыкающему к Liliales, и 2) порядку Сурега-
les, у которых выработались однополые ветроопыляемые цветки.
Может быть, узловое или базальное значение Liliales и шире, если счи-
тать, что два других подкласса: Commeltnidae и Arecidae— возникли от древ-
них Liliidae. Но не исключено,, что каждый из них ответвился не от
Liliidae, а возник непосредственно от неизвестных еще древнейших пред-
ков однодольных.
Древние по происхождению представители подкласса Commelinidae (по-
рядки Commelinales и Bromeliales) входят в энтомофильную группу. Если
Bromeliales специализировались главным образом как эпифитные растения и
являются, вероятно, одними из тупиков эволюции, то в пределах Commeli-
nales намечается тенденция к изменению цветка в сторону анемофильности.
Более полное выражение этой тенденции находим в порядке Bedionales, в
котором появляются особенности, свойственные злакам. Последние выделя-
ются в особый порядок, венчающий анемофильную эволюцию подкласса Сот-
melinidae.
Подкласс Arecidae был охарактеризован материалом порядков: Аге-
cales, Arales и Pandanales. Последний из них близок к Arales*, может быть,
оба имеют общее происхождение и отделились от основного ствола эволюции
Arecidae, а именно от пальмоцветных. Специально следует отметить развитие
у представителей A rales компактных соцветий и «покрывала»— ярко окрашен-
ного листа в основании початка. Роль этого «покрывала» как средства при-
влечения опылителей к компактному соцветию из мелких цветков очевидна.
Наряду с этим от порядка Arales ответвилась линия, приведшая к редукции
не только соцветий цветков, но и вегетативного тела, это направление эволю-
ции завершают ряски.
Что же касается наиболее крупного порядка рассматриваемого подклас-
са — Arecales, то здесь высокая специализация сочетается с относительно
примитивными признаками, такими, как пятикруговые цветки и апокарпный
(не у всех) гинецей. Имея это в виду, особенно апокарпность, необходимо
предположить, что Arales возникли не от Liliidae, а от более ранних однодоль-
I. г
582
ВЫСШИЕ РАСТЕНИЯ
Схематический обзор родственных связей покрытосеменных растений
позволяет сделать несколько обобщающих выводов.
Эволюционное развитие отдела Angiospermae нельзя представить себе в
виде единой цепи, которая охватывала бы все подклассы, порядки и семей-
ства, начиная с наиболее примитивных и кончая наиболее высокоорганизо-
ванными. Линейное расположение невозможно и в пределах отдельно взятых
классов и подклассов. Ход эволюционного развития осуществлялся в виде
нескольких параллельных потоков, которые соответствуют подклассам и
классам. Таким образом, система связей между покрытосеменными не может
быть уподоблена родословному дереву; она, скорее, соответствует «кустарни-
ку» на том этапе его развития, когда он уже потерял главный корень. Но в
таком случае приходится признавать, что однокоренное, монофилетическое
происхождение покрытосеменных продолжает оставаться гипотезой, хотя и
очень вероятной. Гипотетичность монофилетического происхождения покры-
тосеменных усугубляется и тем, что пока нет возможности безоговорочно
указать группу древних растений, вероятнее всего голосеменных, которая
дала начало первым цветковым. При таких обстоятельствах не приходится
удивляться, что по временам возникали и продолжают появляться гипотезы
о полифилетическом возникновении цветковых. Однако полифилетические
воззрения находятся не в лучшем положении, так как выдвигают задачу
отыскания не одного, а нескольких предков покрытосеменных, что не смяг-
чает, а обостряет трудность проблемы.
В схеме родственных отношений покрытосеменных растений достаточно
четко отражен параллелизм в развитии подклассов и даже классов. Одно
из его проявлений состоит в том, что почти в каждом подклассе имеются
л энтомофильные и анемофильные ветви. Распространенность анемофильных
приспособлении в разных эволюционных потоках свидетельствует о том, что
существуют какие-то общие условия, при которых анемофилия представляет
существенное преимущество в борьбе за существование. Одно из таких усло-
вий — способность произрастания массовыми одновидовыми скоплениями,
в которых опыление ветром, при производстве большого количества пыльцы,
перестает быть ненадежным. Но произрастание в форме массовых скоплений
возможно при условии,что внутривидовая конкуренция становится инструмен-
том саморегуляции численности, поддерживающим ее на уровне, необходи-
мом для подавления конкурентов. С этой точки зрения устойчивое ветроопы-
ление, если оно обеспечивается надежными микробиологическими особеннос-
тями цветков и соцветий, представляет высокое достижение в ходе историче-
ского развития цветковых.
Но все же более распространенным у покрытосеменных оказалось опы-
ление животными: насекомыми, птицами, рукокрылыми.
Почти в каждом подклассе можно найти растения (главным образом зоо-
фил ьные) с правильными и неправильными (зигоморфными) цветками, с
раздельнолепестным венчиком, с верхней и нижней завязью. На основании
морфогенеза, спайнолепестности, зигоморфности и нижней завязи можно с
уверенностью сказать, что эти признаки являются более поздними, вторич-
ными по сравнению с первичными: раздельнолепестностью и актиноморф-
ностью цветка, верхней завязью. Эти данные, принимая во внимание приспо-
собительное значение вторичных признаков, помогают филогенетику разме-
щать порядки и семейства в пределах эволюционных рядов соответственно
формированию во времени. Однако факт, что указанные вторичные морфобио-
логические особенности имеют место в энтомофильной эволюции во всех под-
классах, приходится рассматривать как закономерность. Нет необходимости
•видеть причину этого во «внутренних свойства т > растений, «предопределяю-
щих», как иногда думают, эволюционные изменения. Возможности изменений
органов у растений в целом очень ограничены. Сдвиги в онтогенезе цветка в
сторону более раннего наступления базального роста у оснований примордиев
лепестков, что приводит к возникновению спайнолепестности,— достаточная
основа для последующего отбора, поскольку трубка венчика ограничивает
»
ОТДЕЛ ЦВЕТКОВЫЕ. ИЛИ ПОКРЫТОСЕМЕННЫЕ
583
круг потребителей продуктов, вырабатываемых цветком, и одновременно
защищает эти продукты от посетителей, которые не могут произвести опыле-
ние. Аналогичное можно сказать и о возникновении других вторичных осо-
бенностей цветка.
Основные контуры эволюционных рядов покрытосеменных растений
были построены главным образом на основе учета разнообразия цветков и
плоДов. В настоящее время зти ряды или те, которые получены в результате
критического пересмотра ранее построенных, все чаще находят подтверждение
в особенностях вегетативного тела. В этом отношении очень важные ре-
зультаты были получены при анатомическом изучении покрытосеменных рас-
тений, в частности их водопроводящей системы. Настоящие сосуды, как из-
вестно, свойственны только покрытосеменным. Если подобные образования и
встречаются в более древних группах (голосеменные, папоротникообразные),
то это только аналоги сосудов. Но членики сосудов покрытосеменных раз-
личны. Среди них можно отличать и более типичные, и приближающиеся к
трахеидам. Кроме того, известно, что среди покрытосеменных есть и бессосу-
дистые растения. Такие группы естественно рассматривать как более древние.
В некоторых эволюционных рядах, отвечающих прогрессивному изменению
цветка, обнаруживается последовательное нарастание признаков совершен-
ствования водопроводящей аппаратуры растения. Подтверждаются эти ряды
и другими анатомическими признаками.
В течение последних десятилетий большого прогресса достигла особая
ветвь морфологии — морфология пыльцы. Ее данные, привлекаемые к по-
строению филогенетических рядов, помогают ориентироваться в высоте орга-
’ низации, а также в наличии сходств, которые можно истолковывать как пока-
затели родства.
Хотя представления о ходе эволюционного развития, показанные прила-
гаемым схематическим рисунком (рис. 444), нельзя считать окончательным,
все же в основном они правильно отражают, во-первых, взаимосвязь по край-
ней мере более крупных подразделений покрытосеменных, а во-вторых, раз-
мещение в каждом из стволов более поздних (по возникновению в пределах
данной филогенетической ветви) и более ранних таксонов. Эта схема позволяет
выделить в многообразии покрытосеменных три уровня. Первый, наиболее
низкий уровень включает примитивных покрытосеменных, их неизвестных
предков и обширный подкласс Magnoliidae. Этому уровню противоположен
другой, наивысший, характеризуемый спайнолепестностью и часто зигоморф-
ностью цветка, нижней завязью, тенденцией к мельчанию цветков и об-
разованию компактных соцветий. Этот уровень достаточно ясен у энтомо-
фильных двудольных. У высших энтомофильных однодольных соответст-
венно наблюдается зигоморфность цветков, сопровождаемая порой глубокими
перестройками андроцея и гинецея, а также появление сложных аппаратов,
обеспечивающих анемофилию. Между этими двумя уровнями находит себе
место промежуточный, в котором отразилось много частных проявлений
эволюционного процесса.
1
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ
(Цифры «о звездочкой указывают страницу, где имеется рисунок.)
Абрикос 437
Абсидия 116
Авокадо 354
Айран 496
Аврикуляриевые 146, 156
Агава 533
Агавовые 532
Агариковые 140, 142, 143,
163
Агатис 298, 300, 312, 313,
314
Аглаозония 79
Азимина 352
Азота 279
Азотовые 279
Азорелла 465
Азотобактер 22, 25, 31, 158
Азотфиксирующие бакте-
рии 25
Айвовые 386
Аир 569
Аистник 456, 458*
Айва 435
Акация 441*
Аконит 356, 357*, 358
Акрокарпные мхи 203, 207
Актинидиевые 398
Актинидия 398*
Актиномицес 28
Актиномицеты 27, 28, 29*,
31, 157
Аланг-аланг 565
Алария 82
Александрийский лист 443
Алектория 169
Алисмцдные 523
Алканна 487*
‘ Алов 527, 528*
Алтей 401*
Алыча 436
Альбуго 114*
Альзофила 267
Альтернарвя 154
Аманита 144
Амариллис 534
Амариллисовые 533
Анабена 39*, 40
Ананас 548
Ананасные 548
Анаэробные бактерии 26
Ангиоптерис 258, 259*
Андреа 198
Андреевне мхи 198*
Анис 468
Анкистродемус 47*, 49
Анововые 352
Антириум 569
Антоновка 434
Антоновка полуторафунто-
вая 434
Антоцерос 191, 193*, 194
Антоцеротовые 179, 191,
193*, 210
Аифельция 93
Анхуза 485
Анчар 375
Анютины глазки 415
Апельсин 451
Апиоцистис 46 *
Апорт 434
Аралиевые 462
Аралия 463
Араукариевые 299, 300, 309,
312, 313
Араукария 298, 300, 312,
313
362
Арахис 447
Арбуз 417
Арецидные 565
Аристояохнецветные
Ароидные 569
Аронник 569
Аронниковые 521, 569
Аронникоцветные 569
.Артишок 515
Архангионтерис 258
Археоптерис 255, 256*
Археоптерисовые 255
Архимицеты 108
Аскогимениальные 125, 131
Асколокуляриые 125, 129
Аскомицеты 117,125* 137*.
138, 155, 156, 157
, Аепергилл 126*, 127
Аспергилловые 126,
157
Асплениум 268*
Астазия 94
Астерцдные 474
Астерионелла 74*
Астерокаламит 245*, 246
Астерокаламитовые 245
Асте^оксилон 217, 218, 219,
127
Астра 515
Астрагал 449
Астранция 463, 464*.
Астроцветные 508
Афанизоменон 40
Афаномицес 112
Афиллофоровые 140 ~
Ахлия 112
Ацетабулярия 56
Аэробные бактерии 26
Багрянки 84
Багульник 411
Бадан 429
Базидиальные грибы 136
Базидиальные лишайники
163, 168
Базидиомицеты 136, 137*,
138, 156, 163
Бакаутовое дерево 459
Баклажан 493
Бактерии 21, 22*, 23—32
Бактериофаг 32, 33
Бальзамин 457
Бальзаминовые 457
Бальзовое дерево 402
Бамбук 520, 559 >
Бамбуковые 559
Банан 540, 541*
Банановые 540
Банаиоцветные 540
Бангиевые 91
Баньян 375*
Баобаб 401, 402*
Баобабовые 401
Барагванатия 226*
Барбарис 354, 355*
Барбарисовые 347, 354, 355.
522
Барвинок 477
Бархат 451
Бархатное дерево 451, 452*
Бархатцы 515
Батат 483
Батрахоспермум 87, 88*, 91
Бациллы 22*
Башмачковые 543
Бегониевые 416
Бегония 416
Беджиатоа 27*
Безвременник 527
Белан “акация 449
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ
585
Белена 491, 492% 494
Белладонна 492*, 493
Белозор 429
Белокрыльник 569, 570*
Белоус 563
Белоусовые 563
Белые мхи 194
Белый гриб 144
Бельфлер 436
Бельфлер-китайка 434, 436
Бельфлер красный 434
Бемерин 378
Беннеттитовые 281, 285,
291—293, 343
Бентосные водоросли 40,
67, 75, 93, 95, 97
Береза 366, 367, 368*
Бере зимняя 435, 437*
* Березовые 366, 367
Бере-рояль 435, 437
Бересклет 459
Бересклетовые 459
Бересклетоцветные 459
Берест 374 >
Бессмертник 511, 516
. Бешеный огурец 418
Бигнониевые 499
Биаонова трава *563
Бирючина 481
Бластокладцевые 110
Бледная поганка 144
Бледная спиротеха 27
Блязия 187*, 188
Бобовник 436
Бобовоцветные 440
Бобовые 25, 444, 445
Бобы 446
Бовения 286*
Бовиста 144, 145*
Бодяк 516 >
Бокоплодные мхи 206
Болетус 144
Болетусовые 144
Болиголов 411, 465, 466,
467*
Болотный кипарис 329
Бор 563
Бородавчатые 564
Ботридиум 67*, 97
Ботрихиум 256, 257
Боярышник 435
Брахятецнум 207
Бриопсис 55*, 56
Бриофиллум 427
Бромелиевые 548
Брусника 412
Брюква 423
Брюссельская капуста 422
Брюшнотифозные бактерии
26
Будяк 512
Бузина 502
Бук 372*
Буковые 370
* Букоц ветные 364, 365
Булавоиосец 552
Бульденеж 503
Бумажное дерево 374, 463
Бурачниковые 484, 485*
Бурея ржавчина 152
Бурые водоросли 34, 75
Бурые мхи 199
Вайда 424
Валериана 504, 505*
Валериановые 504
Валлиснерня 525
Валония 55% 56
Вальхия 309 1
Ваниль 545
Василек 512, 516, 517*
Василистник 357
Ваточник 478*, 479
Вахта 475
Вахтовые 475
Вейник 551, 552, 558. 562
Вельвцчиевые 338
Вельвичия 338, 339*
Венерин башмачок 546*
Вентурия 129, 130
Вербейник 413
Вербена 490
Вербеновые 487, 489
Вереск 411
Вересковые 409, 411
Верескоцветные 409, 426
Вероника 412, 495
Вертицнллиум 154
Верхоплодные мхи 206
Веселка 145
Ветреница 347, 357, 522
Ветренициевые 356, 357
Вех 465, 467*
Взморник 525
Вибрионы 22*
Вика 445, 449
Виктория 359
Вильямсониелла 292
Вильямсония 292
Виноград 461*, 462
Винтеровые 352
Вирусы 32, 33
Вишня 437, 438*
Водокрас 524, 525*
Водокрасовые 522, 524
Водокрасоцветные 524
Водоросли 21, 33—100
Водорослн-гетероконты 66
Водосбор 356, 357*, 358
Водяная лилия 358
Водяная сеточка 47
Водяная чума 524
Водяной орех 472, 474*
Волнушка 143
Волчеягодник 408
Волчеягодниковые 408
Вольвокс 44, 45*
Вольвоксовые 42, 43*, 61,
94
ВольтциеВые 298, 304, 311*
Вольтциопсис 311
Вольтцня 311
Вольфия 571
Воробейник 485, 486
Воронец 394
Вороний глаз 531
Ворсянка 506
Ворсянковые 504
Ворсянкоцветные 502
Вошерия 68
Высшие растения 171, 214
Вьюнковые 482
Вьюнок 482, 483*
Вяз 373, 374*
Вязовые 373
Галеона 545
Гамамелидные 363
Гамамеяидовые 364
Гамамелис 364
Гаолян 557% 564
Гаплолепидные мхи 206,207
Гапломитриевые 190
Гапломитриум 187*. 190
Гастеромицеты 144, 145*,
156
Гваюла 515
Гвоздика 390
Гвоздичное дерево 469, 472*
Гвоздичноцветные 382
Гвоздичные 389 '
Гевея 407, 408*
Гелидиум 93
Гелиотроп 485
Гелоциевые 134
Гельминтостахис 256, 257*
Георгины 515
Геранецветные 454, 459
Гераниевые 456, 458
Герань 456, 458*
Геснериевые 499
Гетерогенератные 78
Гетеромерные лишайники
164
Гетероталличные водоросли
35
Гиацинт 529
Гигартина 91*
Гидродикцион 47, 48*
Гидрурус 65, 66*
Гиениевые, порядок 242,
243
Гиениевые, семейство 242
Гиения 242*, 243.
Гименомицеты 137, 139,
156, 157, 163
Гименофидл 271*
Гимеиофилловые 215, 268,
270, 271*
Гимеиофиллоподиевые 270
Гинкго 296, 297
Гинкговые 281, 296
Гнмофяцеты 154*, 157
Гладиолус 535
Гледичия 443
Глейхениевые 272
Глеокапса 38, 39* 164
Глеоспориум 155
Глиптолепис 311
Глициния 449
Глоксиния 499
Глухая крапива 487, 488
Гнездовка 145*
Гиездовковые 145
Гнетовые 336, 343
Гнетоидные 336
Гветум 336, 337*, 343
Голенкиния 47*
Головневые 146—149, 156
Головня 146, 147*
Головчатотисовые 315, 317
Голосеменные 279, 280
Голосумчатые 119
Голубика 412
586
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
Гожеомерные лишайники
164
Гомоталличные водоросли
35
Гомфонема 72*
Гониум 44, 46
Гонобобель 412
Гордовина 503
Горец 391
Горечавка 475, 476*
Горечавковые 475
Горечавкоцветные 475
Горицвет 357, 358*
Горнчнни 465
Горлянка 418
Гормогониевые 38
Горох 445, 446*
Гортензия 428
Горчак 516
Горчица 421
Горянка 354, 355*
Граб 369
Гравилат 430
Гранат 470, 473*
Гранатовое дерево 470
Гранатовые 470
Грацилярия 93
Гребенщик 416
Гребенщиковые 416
Грецкий орех 373
Гречиха 391, 392*
Гречихоцветные 391
Гречишные 391
Грибы 21, 104, 155, 157,
160, 165
Гристорпия 285
Гроздовник 257*
Груздь 143
Груша 435
Грушанка 410*
Грушанковые 410
Грушовка 434 ,
Грыжник 389
Гуаява 469
Губастик 496
Губоцветные 487
Гулявник 424
Гукай 564
Гусиный лук 528*
J i: г. <
Дазикладус 56
Дания 92*
Данеа 258
Двудольные 346, 347
Дв^щрылоплодниковые 396
Девица в зелени 358
Девясил 512 *
Дегенериевые 351
дегенерия 352*
Делессерия 92*, 93
Дербенник 469, 470*
Дерево путешественников
541, 542*
Дереза 449
Держи-дерево 460
Десмидиевые 58, 59*, 60
Десмидиум 59*, 60
Джонсонова трава 564
Джугара 557*, 564
Джузгун 391, 499
Джут 405
Дзельква 373
Диатомеи 69
Диатомовые водоросли 69,
93, 94, 95, 97
Дикая редька 424
Дикранум 206
Диксониевые 270
Диксония 267
Диктиота 78, 79*
Диктиотовые 76, 79*
Диктиухус 112
Диллениевые 393
Дидленнецветиые 393
диллениидные 393
78
ион 65, 66*
Диоон 288
Диоскорейные 522
Диплококки 22
Диплелепидвые мхи 206,
207
Диподасковые 119
Диподаокуо 119, 1-0*
Диптерокарпус 397
Дискомицетные лишайники
168
Дискомицеты 133, 156, 163
Дисхидия 479*
Дифтерийные бактерии 26
Дицентра 361
Дождевики 144, 145
Домовый гриб 141, 142, 457
Донник 445, 449
Драконово дерево 532
Драпарналвдиопснс 50
Драпарнальдия 50, 51*, 93
Драцена 520, 532
Дренанолепис 312
Дрепанофикус 226*
Дриада 431
Дрожалковые 145, 156
Дрожжевые грибы 119, 120,
160
Дрожжи 120, 121*, 160
Дрок' 449
Дуб 369*, 370, 371
Дубовик 144
Дудник 465
Дунализлла 44
Дурман 491. 494
Дурнишник 513
Дурра 564
Дурьян 403
Дусен 434
Душистый горошек 449
Душистый колосок 562
Душица 489
Дымянковые 419
Дынное дерево 415
Дыня 417
Дюренея 89
Дягиль 465
Ежа 558, 559
Ежевика 432
Ежеголовка 571, 573*
Ёжеголовковые 571
Ежовиковые 141
Ежовник 385
Ель 319, 321, 322*
Жасмин 481
Железное дерево 373
Железняк 365
Железобактерии 25, 27
Желтая акация 449
Желто-зеленые водоросли
66
Желтушник 424
Женский папоротник 270
Жень-шень 463
Живокость 356, 358
Живучка 488
Жимолостные 502, 503*
Жимолость 503
Житняк 561
Жостер 460
Замия 286*, 288
Заразиха 498*
Зараэиховые * 498
Звездчатка 388*, 389
Зверобой 395, 396*
Зверобойные 395
Зеленые бактерии 25
Зеленые водоросли 41—61,
97, 163
Зеленые мхи 199
Земляная груша 515
Земляника 430, 432, 433*
Земляничное дерево 411
Земляничный томат 493
Земляной миндаль 538
Земляной орех 447
Зкгиема 58*
Зигнемовые 57, 58*
Зигомицеты 114, 155
Зигоптеридиевые 255, 272
Зигоптерис 254*, 255
Зимовник 356*
Зимовниковые 356
Злаки 522, 549, 550, 553
555, 556, 557
Злакоцветные 549
Змееголовник 489
Золотан печать 355
Золотан розга 512
Золотистые водоросли 65
Золотое дерево 468
Золототысячник 475, 476*
Зоннерациевые 469
Зоннерация 469
Зонтикоцветные 462
Зонтичные 463, 464, 466
Зоохлорелла 95
Зостеролист 220
Зостерофиллум 221
Зубровка 562
Ива 380, 381
Иван-да-марья 497
Иван-чай 471, 473*
Ивовые 379
Ивоцветные 364, 379
Иглица 531
Изогенератные 76
Изоет 239*, 240*
Икотник 424
Иллициевые 352
Имбирные 540
Имбирь 541
Инжир 376
Иноперкулятные
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
Ипекакуана 502
Ипомея 483
Ирис 535
Ирисовые 534
Исландский мох 162
Иссоп 489
Итсегек 385
Иудино ухо 146
Каваниллезия 402
Казуарина 379*
Казуариновые 379
Казуариноцветные 364. 378
Кактус 387*, 388
Кактусовые 386
Каламит 245, 246
Каламитовые, порядок 245
Каламитовые, семейство
245, 246
Каламостахис 246, 247
Каламофитон 242*, 243
Калган 433
Калимматотека 282, 283
Калина 502, 503, 504*
Каллитамнион 85*, 92*. 93
Калобриевые 187*
Калобриум 187*, 190, 191
Калужница 356
Кальвация 144, 145
Камелия 395
Камнеломка 428
Камнеломковые 428
Кампешевое дерево 442*
Камфорный лавр 354
Камыш 537, 538*
Канареечннковые 562
Канатник 401
Канатчиковые 548
Кандида *154
Кандиль 434
Кандиль-китайка 434
Канна 541, 542
Канвовые 540
Каперцовые 425
Кшерцоцветные 414, 419,
Каперсы 425
Каприфоль 503
Капуста 422
Кариофиллидные 382
Каркас 373
Картофель 490, 491*
Касатик 535*
Касатиковые 534
Кастилоа 375
Кастор 408
Каулерпа 55*, 56
Каштан 370, 371
Квебрахо 452, 476
Кевовое дерево 451
Кедр 323, 324*
Кейтония 285
Кейтониевые 281, 285, 343
Кендырь 476, 477*
Кервель 468
Кермек 392
Кизил 468
Кизиловые 468
Кипарис 331*
Кипарисовые, подсемейст-
во 331
Кипарисовые, семейство 299,
300, 309, 330
Кипрей 471
Кирказон 362
Киркааоновые 362
Кислица 454, 455*
Кисличные 454
Китайка 434, 436
Китайка золотая 434
Клавариевые 163
Кладоксиловые 253
Кладоксилон 253, 254*
Кладония 168, 169
Кладотрикс 26
Кладофора 54, 93
Кладофоровые 54. 61
Клевер 445. 447
Клен 453*
Кленовые 452
Клехцевина 406, 408, 409*
Климаций 207
Клинолист 243, 244*
Клинолистные 243
Клинолистовидные 241
Клостериум 58, 59*
Клубеньковые бактерии 25,
445
Клубника 430, 432
Клюква 412
Кобрезкя 539
Ковылевые 563
Ковыль 558, 563
Кодиум 55*, 56
Козелец 518 \
Козлобородник 513 '
Кокаиновое дерево 459
Кокаиновые 459
Кокки 22*
Кокосовая пальма 567
Кок-сагыз 518
Кокупшик 546
Кола 403
Колеохете 52
Колеус 489
Колоквинт 417, 418*
Колокольчик 507
Колокольчиковые 507*
Колокольчикоцветные 506
Колоцинт 417
Кольраби 422
Коммелинидные 547
Коммелиновые 547
Кониофора 141
Коноплевые 376
Конопля 376, 377*
Коноцефалум 184, 185*
Конские бобы 445
Конские каштаны, семей-
ство 453
Конский каштан 453
Конферва 67
Конъюгаты 56, 6Г
Копринус 144
Копытень 362
Кораллина 93
Кораллиновые 93
Кордаитовые 293
Кордаиты 294*, 295
Кордилина 533
Кориандр 466, 468
Коричное дерево 353*, 354
Коровье дерево 375
Коровяк 495
Короставник 506*
Корсиния 176
Космариум 58, 59*
Костер 552, 558, 559
Костяника 432
Котовник 489
Кофейное дерево 500, 501*
Кочанная капуста 422
Кошачьи лапки 512
Крапива 377, 378*
Крапивные 377
Крапивоцветные 364, 373
Красное фернамбуковое де-
рево 443
Красные водоросли 84, 85*,
86—93
Красный перец 493
Красоднев 527
Крассула 427
Кренотрикс 27
Крестовник 516
Крестоцветные 419, 420, 424
Кривоцвет 486
Кринум 533
Криптомерия 298
Криптонемиевые 89
Крокус 535
Кронарциум 153
Крупка 424
Крушина 460*
Крушинные 460
Крушиноцветные 460
Крыжовник 428
Крылоорешник 373
Крым-сагыз 518
Ксантидиум 59*
Ксантория 168
Кубышка 358
эка 358, 359*
пел
игл
HUI
Куколь 389
Кукуруза 565
Кукушкин лен 199, 200*,
202*
Кукушкин цвет 388*
Кунжут 499
Кунжутные 499
Купальница 356*
Купена 531
Куриное просо 564
Куропаточья трава 431
Кутлерия 78
Кутровые 475
Лавандула 489
Лавр 353*, 354
Лавровишня 439
Лавровые 347, 352, 353
Ладанник 415
Ладанниковые 415
Ладьян 546
Лаконос 383 .
Лактариус 143
Лакфиоль 425
Ламинариевые 78, 80*, 82
Ламинария 80*, 81, 82
Ландыш 531
Лапина 373
Лапчатка 432, 433
Ластовень 479
588
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
Ластовневые 477
Ласточниковые 477
Лебахиевые 298, 304» 309
Лебахия 308, 310
ъ1Э*
Лебедовые 383
Левкой 425
Леканора 168
Лея 455, 456*, 457*
Лён-долгунец 455
Лёи-кудряш 455
Лён-межеумок 455
Леновые 454
Лепидодендровые 235
Лепидодендрон 225, 235,
236*, 237
Лепидокарпон 237*, 238
Лепцдоспермовые 238
Лептомитус 112
Лептоспорангиатные папо-
ротники 261, 264*
Лептотрикс 27
Лессония 81
Лецидея 168
Лещина 366, 367, 369
Лигннодендровые 282
Лигодиум 271
Ликвидамбар. 364
ЛиКогалн 102
Ликопердон 144, 145*
Лилейник 527
Лилейные 521, 526, 528
Лилиецветные 526
Лилиидиые 526
Лилия 528, 529*
Лимон 451
Лимонник 352
Лннгбия 39*» 40
Липа 404*
Липовые 403
Липучка 486
Лисички 141
Лисохвост 558, 563
Лиственница 323, 324*
Лиственничные 323
Лиственные мхи 178, 194,
205
162
Листовая капуста 422
Листостебельные растения
173, 179
Литотамнион 92*, 93
Литофиллюма 93
Лишайники 21, 95
163, 166*, 168*
Лишайники корковые 162
163*
Лишайники кустистые 162
163*
Лишайники листоватые 162,
163*
Лишайники яакипвые 162,
163
Лишайниковая манна
Лобелиевые 508
Лобелия 507, 508
Локсомовые 270
Ломбардский орех 369
Лопух 509, 513
Лотос 359
Луазелерня 411
Луговой чай 413
Лук 529, 530*, 531
168
Луковые 529
Лучистые грибы 27
Лучины 61
Львиный зев 495, 496
Льнянка 495, 496*
Любка 545
Люпин 448
Лютик 357, 358
Лютиковые 347, 355, 522
Лютикоцветные 354
Люффа 418
Люцерна 445, 448*
Лягушечник 524
ЛяллеЫацпия 489
Магнолиевые 350
Магнолиепветные 349, 350
Магнолия 350
Магония 354
Манник 531
Мак 360, 361*
Маковые 360, 419
Макоцветные 360
Макроцистис 81
Мадина 431, 432*
Малломонада 65
Малломонас 66*
Мальва 401
Мальвовые 399
Мальвоцветные 393, 399,
426
Манговое дерево 452
Мангустан 396
Мандарин 451
Мандрагора 494
Манжетка 494
Маниок 407, 408*
Марантовые 540
Мараттиевые папоротники
258, 260*, 270
Мараттия 258
Маргаритка 515
Маревые 383
Марена 502
Мареновые 500
Маренопветные 500
Маркгравиевые 397*, 398
Марнгравия 397*, 398
Марсилиевые 272
Марсилия 272, 273*, 275*
Маршанциевые 179,184,211
Маршанция 179*, 180,182*,
183*, 184*
Марь 385
Марьянник 497
Масленок 142, 144, 158
Маслина 480, 481*
Маслинные 480 •
Маслнноцветные 480
Масляная пальма 568
Масляное дерево 414
Маслянокислые бактерии 26
Матониевые 270
Мать-и-мачеха 512, 516
Мауновые 504
Мачок 360
Медвежье ушко 441, 496
Медуллоза 284*
Медуллововые 282, 284
Медуница 485*, 486*
Меэембркантемум 386
Мезотениевые 60, 61
Меэотевиум 60
Мелисса 488, 489
Мелозира 70*, 75
Мелймпсора 153
Мелямпсоровые 152, 153
Мелянкониевые 154, 155
Мерулиевые 141
Метасеквойя 298, 328, 329*
Метцгерия 188
Миадесмия 237, 238
Микобадтерки 26
Микосферелла 131 ,
Микрастериас 58, 59*
Микрококки 22
Микросфера 128, 129
Микродистис 39*
Миксамебы 102
Миксобактерии 27
Миксомицеты 100,101*, 102
Мимоза 442
Мимозовые 440*, 441
Мимузопс 414
Миндаль 436
Мирмеиодия 502
Мирта 411, 469, 471*
Миртовые, подсемейство 470
Миртовые, семейство 469
Миртоцветные 469
Многоиоренник 570
Многоножка 270
Многоножковые 270
Многопиодннковые 292, 349
Можжевеловые 332
Можжевельник 332*, 333
Мокрица 389
Мокричные 389
Молодило 427
Молокан 519*
Молочаецветные 405, 426
Молочай 406, 407*
Молочайные 405, 408
Молочное дерево 375
Молочнокислые бактерии
26, 31
Монилия 134
Моиоблефаридовые 110, 155
Моноблефарис 111*
Монстера 570
Мордовник 509
Морковь 467, 468*
Морошка 432
Морскан капуста 82 *
Морская трава 525
Морской лук 529
Мортьерелла 116
Мотыльковые 443, 444
Моховик 144
Мохообразные 177, 178,
208, 209, 213
Мужоция 58*
Мужской папоротник 261,
Мукор 106, 115
Мукоровые 115, 157, 160
Мускатниковые 352, 353
Мухомор 144
Мучнеросные грибы 127,
128
Мушмула 435
Мыльнянка 390
Мытник 495
Мышехвостник .858
УКАЗАТЖЛЬ РУССКИХ НАЗВАНИЙ
58»
Мята 488, 489
Мятлик 550, 552, 558, 559
Мятликовые 559
Навикуля 70е, 73, 74е
Надбородник 546
Наперстянка 495, 496, 497е
Нарцисс 533, 534е
Нарциссовые 533
Настурциевые 457
Настурция 457, 458е
Натгорстиана 239, 241е
Недотрога 457
Незабудка 486
Нектрия 131
Немалион 90е, 91, 92е
Немалионовые 89
Ненентесовые 362
Непентесоцветные 362
Несовершенные грибы 108,
153, 154
Нечленисто-зубчатые мхи
' 206
Нивяник 511, 512, 515
Низшие растения 21
Нимфейные 347, 522
Нимфея 292
Нитрифицирующие бакте-
рии 25
Ноголист 355
Ноготки 515
Ноктилюка 64е
Норичник 495
Норичниковые 494, 495, 496
Ностой 39е, 40,1)7, 164,192
Нототилас 192, 193е
Ночесветка 64
Ночная красавица 425
Ночная свеча 471
Ночная фиалка 545
Обвойник 479
Оболочкосеменные 282, 333
Овес 552, 556*, 562
Овсовые 562
Овсяница 558, 559
Овсяницевые 559
Огурец 417
Однодольные 346, 520
Однозернянка 561
Однопокровные 348
Одуванчик 512, 513, 518
г; I
Окопник 485
Олеандр 476, 477е
Оленин мох 162
Ольпидиум 108, 109е
Ольха 366, 367*. 368
Оноилея 268е
Оомицеты 104, 155
Опенок 142, 143, 158
Оперкулятные дискомице-
ты 133
Орех 372
Ореховые 372
Орехоцветные 364, 372
Орешник 366, 368, 497
Орляк 266е
Орхидные, порядои 522, 543
Орхидные, семейство 543,
545
Осина 380
Ослинник 471
Осмунда 269е
Осмундовые 270
Осока 538, 539е
Осоковые 536
Осокорь 380
Осокоцветные 536
Осот 516, 518
_ Оспилляторкя 38, 39е
Очанка '495, 497
Очиток 427е
. Павловния 497
Падина 78, 79е
Падуб 459
Падубовые 459
Лазания 371
Пайена 414
Палеостахия 246, 247
Палочковидные бактервв 22
Пальма 520,566е
Пальмоцветные 565
Пальмы, семейство 565
Пальчатая трава 133
Пандан 571, 572*
Панлановые 571
Панданоцветные 571
* Пандорина 44е, 46
Папаеяые 415
Папирус 538
Папоротники 251, 263, 268е
Папоротниковидные 251
Папоротникообразные 214,
215
Пармелия 168
Парнолистниковые 457
Парша яблони 130
Паслен 490
Пасленовые 490
Паспалюм 133
Пастернак 468
Пастушья сумка $24*
Педиаструм 47* % *8
Пеларгония 457, ч58*
Пеллия 186*, 187
Пельтигера 168
Пениум 58
Пеницилл 106, 126* '
Пеннатные дцатомовые 69,
73, 74е
Пион 394
Пионовые 394
Первнчнопокровные 348,349
Первичносумчатые 119, 121
Первонапоротвики 252,
256е, 261, 272
Первоцвет 412, 413е
Первоцветные, порядои 412
Первоцветные, семейство
412
Перескиопсис 386
Переския 386
Переступень 418е
Перец 362
Перец стручковый 493
Перечноцветные 362
Перечные 362, 522
Перидинеи 63, 64
Перидиниевые водорослн 95
Перидиниум 64е
Перилла 489
Периспориевые 127, 129
Перистые диатомовые 69*
72, 73е
Пернамбуковое дерево 443
Пероноспора 113е
Пероноспоровые 112, 113
Персик 437
Перуанская вишня 493
Пестичные 340
Петров крест 497
Петрушка 466, 468
Петуния 494
Пецица 134
Пецицевые 134
Печеночники 176, 179* 185,
208
Печеночные мхв 179, 210
Пилоболюс 116е
Пкнанг 568
Пиннулярия 69е, 72 _ у
Пиреномицетные лишайни-
ки 168
Пиреномицеты 129, 157,462
Пиретрум 512, 515 /
Пиронема 122, 423е, * 134
Пиррофитовые водоросли
63, 64
Пистия 570
Питиум 113
Пихта 300, 319, 320, 321е
Пихтовые 320
Плагиогириевые 270
Плазмодиофора 102, 103е
Плазмодиофоровые 102
Плазмопара 113е
Плакун-трава 469
Планктонные водоросли 40,
48, 65, 67» 75, 95
Пластинниковые 143
Платав 365
Платановые 365
Платвстемон 360
Платицериум 268е
Плаун 225, 227, 228е, 229е,
230
Плауновидные 225, 226е,
237
Плауновые 227
Плевел 560
Плевраге 131
Плевроиарпные мхи 207
Плевроиокк 51е, 52
Плевромейевые 238
Плевррмейя 238, 241е
Плектасковые 126е, 127
Плодосумчатые 122
Плющ 462, 463е
Пневмококк 22
Повилика 483, 484е
Повиликовые 483
Повой 483
Погремок 497
Подбел 411
Подберезовик 144
Подлесник 463
Подмаренник 502
Подокарповые 299, 302, 314
Подоиарпус 305, 306, 314е,
315
Подорожник 499е
Подорожниковые 499
Подосиновик 144
Подснежник 529, 533, 534е
590
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
Подсолнечник 508*, ' 511.
512, 513, 514*
Подъельник 411
Покрытосеменные 177, 279,
340, 573
Полба 561
Полевица 551, 558 , 562
Полевичковые 563
Полисифония 85*, 93
Полигона 94
Полифаг 110
Полиэдер 48* >
Полушник 239
Полушниковые 239, 241
Полынь 512, 513, 515
Помвдор 493
Понцирус 451
Порезник 465
Пориевые 140
Порожистая парша карто-
феля 103
Порфира 87*, 91, 93
Постельсия 81
Постенносеменные 414
Почвенные водоросли 95,
97
Примула 412
Приноготковые 389
Пролеска 529
Пролесковые 528
Пролесник 406, 407
Просо 557, 564
Просовые 564
Простейшие 40
Прострел 357
Протейные, порядок 364,
Протейные, семейство 381
Протококковые 46, 47*, 94
Протолепидодендровые 226
Протолепидодендрон 227
Протоптеридиевые 253
Протоптеридиум 253
Протосифон 48*
Прототека 94
Псевдоспорохнус 219, 221*
Псевдотсуга 320, 321*
Псилот 222, 223*, 224
Псилотовидные 222
Псилотовые 223*
Псилофит 177, 179, 217,
219, 225
Псилофитовидные 217
Псилофитовые 217, 219,221*
Псилофитон 219*
Псилофиты 217
Птичья гречиха 391
Пузырчатка 498*
Пузырчатковые 498
.Пукциниевые 152, 153
Пукциния 149,150, 151, 152
Пупавка 512, 515
Пурпурные бактерии 25
Пустырник 488
Пушица 537, 538*
Пшеница 556, 560
Пылающее сердце 361
Пырей 551, 558, 561
Рабдонема 71*
Равножгутиковые водорос-
ли 42
Радула 189*
Разамала 364
Раздельнолепестные 348,349
Разножгутиковые водорос-
ли 66, 67, 68
Разноспоровые растения 281
Райграс 560, 562
Раковые шейки 391
Ракоптерис 256*
Рами 377
Рапс 421, 422
Раувольфия 476
Раффлезиевые 362
Раффлезия 362
Рвотный орех 480
Рдест 525*
Рдестовые 525
Рдестоцветные 525
Ревень 391
Редис 423
Редька 423
Резеда 426
Резедовые 425
Ренет 434
Ренет бергамотный 434
Ренклод-Реформа 436
Репа 423
Репяшои 430
Ржавчинники 149
Ржавчинные 149, 156
Ржавчинный гриб 149, 151,
152
Ризоклониум 54
Ризопус 116
Ризофидиум 110*
Ризофоровые 469
Ринин 217, 218*
Рипсалис 387
Рис 557, 563
Рисовые 563
Риччиевые 185
Риччиокарпус 185*
Риччия 185*
Рогатиковые 141
Рогоз 572, 573*
Рогозовые 572
Рододендрон 411
Рододендровые 411
Рожь 556*, 561
Роза 433
Розидные 426
Розмарин 489
Розовые 429, 439
Розоцветные 426
Ромашка 509, 511, 515
Ротанги 566
Роччелла 169
Рудбения 515
Рута 450*
Рутовые 450
Рутоцветные 450
Рыжик (гриб) 143
Рыжик (покрытосеменное)
424, 425
Рябина 435
Рябинник 431
Рябчик 528
Ряска 570*, 571
Рясковые 570, 571
Сабельник 433
Сагеноптерис 285
Саговая пальма 566, 568
Саговники 286*
Саговниковые 281, 285, 286,
287, 288*, 290
Саговниколистные 281, 282
Садовый жасмин 428
Саксаул 385
Салат латук 517
Саловое дерево 397*
Сальвиниевые 274
Сальвиния 274, 276*, 277*,
278
Самшит 405
Самшитовые 405
Сандаловое дерево 445
Сандаловое красное дерево
449
Сансевьера 533
Сапиндоцветные 450
Сапотовые 414
Саппановое дерево 443
Сапролегниевые 111, 157
Сапролегния 106, 111, 112*
Саргассум 82
Саррацениевые 362
Сарсазан 384
Сарсапариль 532
Саса 559
Сассапарнлевые 531
Сассапариль 531
Сафлор 515
Сахарный тростник 564
Сахаромицес 120
Сахаромицеты 119
Свекла 383*
Свинорой 550, 563 '
Свинчатковые 392
Свинчаткоцветные 392
Свободнолепестные 348
Седмичник 413
Секвойя 298, 327, 328*
Секвойядендрон 327, 328*
Селягинелла 225, 231, 232*,
233*, 234
Селягинелловые 231
Селезеночник 429
Сельдерей 468
Семенные папоротники 255,
282, 343
Семенные растения 279
Сенная палочка 23, 24*
Септория 155
Серобактерии 27
Сибиреязвенные бактерии
Сигилляриевые 237
Сигиллярия 225, 235, 237,
238*
Синеголовник 463
Синедра 74*
Сине-зеленые водоросли 36*,
37, 38, 39*. 40, 95, 97,163 ‘
Силура 65, 66*
Синхитриум 108, 109
Синюха 483, 484*
Синюховые 483
Синяк 486
Сирень 481
Ситник 536
Ситниковые 536
Ситникоцветные 536
Ситовник 538
а
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
591
Сифоновые 54, 55*, 56, 61
Скабиоза 505
Склеротиния 134
Скополия 494
Скрученноцветные 475
Скумпия 452
Слива 436
Слизевики 100
Сложноцветные 508, 509
Слоновая трава 564
Смоковница 376
Смолевиа 390
Сиолевковые 389
Смолка 390
Смородина 428
Сморчки 134
Снежноягодник 503
Сныть 465
Собачья петрушка 467
Солерос 385
Солнцецвет 415
Солодка 449
Соломонова печать 531
Сольданелла 412
Солянка 384, 385
Сон-трава 522
Сорго 564
Сордария 131
Сосна 300, 301, 302, 324,
325*, 326
Сосновые, подсемейство 324
Сосновые, семейство 299,
300, 304, 309, 319
Соя 445, 446
Спайнолепестные 348
Спаржа 531, 532*
Спаржевые 532
Спирейные 431
Спирея 431
Спириллы 22*
Спирогира 57, 61
Спиротения 60
Спиротеха 22*, 27
Спонгоспора 103
Спорынья 131, 132*, 161
Спорыш 391
Стаураструм 59*
Стауроптерис 256
Стафилококки 22*
Стеркулиевые 403
Столбнячные бактерии 26
Страстоцветные 419, 426
Страусопер 215, 269, 270
Стрелолист 524
Стрептококки 22*
Стрептохеты 559
Строфант 476
Строчки 134
Суданка 564
Судза 489
Сумах 452
Сумчатые грибы 117, 163
Сумчатые лишайники 168
Сурепка 424
Сусак 523
Сусаковые 523
Сфагновые мхи 194, 195,
208
Сфагнум 195*, 196*
Сфацелия 132
Сфацелляриевые 76
Сферелла 43*
Сферокарповые 191, 210
Соерокарпус 179, 191, 192*
Ci юропсидные 154*, 155
С|юротека 128
Сферотилюс 26
Схизандровые 352
Схизейные 270, 271*
Схизея 271*
Сценедесмус 47*, 49
Сцеплянки 56
Сыроежка 143
Сыть 537
Табак 493*, 494
Табеллярия 72, 74*
Таволга 431
Таксодиевые 299, 327
Таксодиум 298
Таро 569
Тархун 515
Тау-сагыз 518
Тафрина 121, 122
Тафриновые 121
Теиковое дерево 408, 490
Телефора 163
Телефоровые 140, 141
Телорез 524
Тениокрада 220, 221*
Термопсис 449
Терн 436
Терновник 436
Терпентинные 450
Тетраспора 46
Тиковое дерево 490
Тиллециевые 149
Тимофеевка 558, 563
Тимьнн 489
Типичные бактерии 22
Типичные папоротники 261
Тис 306, 317, 318*
Тисовые 299, 300, 309, 317,
319
Тмезиптер 222, 223*
Тмин 468
Толипотрикс 39*
Толокнянка 411
Толстянковые 427
Томат 491, 493
Топинамбур 515
Тополь 380
Торица 389
Торичник 389
Торрея 318
Торфяные мхи 194
Трава грама 563
Традесканция 547
* Трахеломонас 41*
Трахикарпус 569
Требуксия 47
Трейбия 188
Трентеполия 51*, 52, 97,
164
Трибонема 67*
Трифоль 475
Трихобактерии 26
Триходесмиум 40
Трихоманес 271*
Тростник 550, 551, 562
Тростниковые 562
- Троходецдроцветные 364,
382
Трубкоцветные 481
Трутовик 141*, 142, 157,
160
Трутовиковые 141
Трюфелевые 135
Трюфель 135, 136
Трясунка 559
Тсуга 321
Тубериулезные бактерии
26
Тубероза- 533
Туевые 331
Туидиум 207
Тунговое дерево 408
Тутовые 373
Туя 331
Тыква 418
Тыквенноцветные 416,426
Тыквенные 416
Тысячелистник 516
Тюльпан 528
Тюльпанное дерево 351*
Ужовник 256, 257*, 259*
Ужовниковые 215, 255,
257*, 258
Узамбарская фиалка 499
Укроп 468
Уксусное дерево 452
Уксуснокислые бактерии 26,
31
Улотрикс 49, 50*
Улотриксовые 49—52, 61,
164
Ульва 50, 51*
Ульвовые 50
Унцинула 128, 129
Упас 375
Уромицес 153
Уснея 169
Устилягиновые 149
Устиляго 146*
Фаг 32
Факус 41*
Фаллюс 145*
Фасоль 445
Фенхель 446*
Феотамнион 65, 66*, 82
Фиалка 415
Фиалковые 415
Фиалкоцветные 393, 414,
426
Физалис 493
Фикомицеты 104, 109, 155
Фикус 375
Филлоглоссум 230
Филлотека 251
Филлофора 93
Филодендрон 569, 570
Фильтрующиеся вирусы 32
Финиковая пальма 568*
Фисташка 451
Фисташковые 451
Фитолакковые 382
Фитофтора 113*
Флокс 483
Флориден 91
Фона 155
Фоссомброния 187*, 188
Фрагмидиум 153
Фрагмобазидиомицеты 139,
145, 156
УКАЗАТКЛЬ РУССКИХ НАЗВАНИИ
Фричиелла 52
Фузариум 154
Фуксия 472
Фукус 81, 82*. 83*
Фукусовые 76
Фулиго 102
Фунария 203, 204*
Фундук 369
Хамеропс 567
Хамесифоновые 38
Хара 62*
Харовые водоросли 61
Хвойные 298, 308
Хвощ 241, 247*, 248, 249,
250*
Хвощовые 247
Хейролепидиевые 311*
Хейролепис 311, 312
Хемионитис 268*
Хетофоровые 61
Хетоцерос 74*, 75
Хилосцнфус 189*, 190
Хинное дерево 500
Хитридиевые грибы 110*
Хламидобактерии 26
Хламидомонада 42, 43*, 61,
94
Хламидомояас 42
Хлебное дерево 374
Хлопчатник 399*, 400
Хлопчатные деревья 402
Хлорамебы 67*
Хлорелла 47*, 49
' Хлорококк <47*
Хлорококковые 46, 56, 61
Хмель 376, 377, 378*
Холобазидиомицеты 138
Хорнея 217
Хорнеофитон 217, 218, 219*
Хохлатка 361, 521
Хрен 424
Хризантемы 516
Хриэомонадовые 65, 66*,
95
Хломулина 65, 66*
Хроококк 38, 164
Хроококковые 38, 164
Хурма 414
Царский папоротник 270
Цветковые 340, 574
Цветная капуста 422
Цезалъпиниевые 442, 443*
Цекропия 376
Центрические диатомовые
69, 72, 74*
Центросеменные 382
Церамиевые 91
Церанадус 438, 439*
Йерацнум 63, 64*
ереуо 388
Цетрария 168
Цефалотаксус 316
Циатиевые 270
Цикадеоидеа 292
Цикас 286*
Цикламен 413*
Циклоспоровые 76, 81
Циклотелла 74*, 75
Цикорий 512, 517
Цикута 465
Цилиндроцнстис 60
Цимбелла 72*, 74*
Цинерария 516
Цинния 515
Знрцея 472
иссус 462
8истоаира 82
истококк 47, 163*,
Цистоптерис 268*
Зистопус 114
итрусовые 450
164
Чабрец 488
Чаецветные 394, 426
Чай 395*
Чайные 394
Чайный гриб 120
Чайный куст 395
Чаровница 472
Частуха 523*, 524
Частуховые 522, 523
Частухоцветные 523
Чемерица 527
Череда 512, 513
Черемуха 438
Черемша 531
Черешня 438
Черника 410*, 412
Черноголовник 430
Черное дерево 414, 449
Чернокорень 485, 486
Чернушка (гриб) 143
Чернушка (покрытосемен-
ное) 356
Чертополох 512, 516
Чеснок 530* .
Чечевица 445
Чешуецветные 536
Чилибуха 479*, 480
Чилим 474*
Чинара 365
Чистотел 360, 362
Чистяк 521
Членисто-зубчатые мхи 206
Членистые 241
Чозения 380
Чубушник 428
Чумиза 557», 564
Чуфа 538
Шалфей 489, 490*
Шампиньон 142, 144, 158
Шавтрансия 88
Шаровидные бактерии 22
Шафран 535*
Шелковица 374*
Шерстяные деревья 402
Шизонерра 251
Шильник 239
Шиповник 433, 434*
Ширяш 527
Шишконосные 281, 293
Шляпочные грибы 136, 161
Шоколадное дерево 403*
Шпажник 535
Шпинат 385
Щавель 391
Щитовник 261, 270
Шучка 562
Эбеновые 413, 414, 426
Эвглена 41*
Эвгленовые водоросли 40, 94
Эвдорина 44*, 46
Эверния 168*, 169
Эвкалипт 470, 473
Эвкалиптовые; подсемейст-
во 470
Эгагропила 93
Эдельвейс 516
Эдогониевые 52, 61
Эдогоииум 52, 53*
Эйкомиевые 365 '
Эйкомия 365
Экзоасковые 121
ЭизобазидиаДьные 139
Экзобазидиум 139
Эктокарпус 76, 77*, 78
Элодея 524
Эммер 561
Эмпуза 117*
Эцдивий 517
Эцдомицетовые 119
Энтомофтора 117
Энтомофторовые 115, 117
Эпихлоэ 131
Эреыаскус 118*
Эремурус 527
Эршшфе 128
Эривнфовые 127
Эрнестиодендрон 310
Эспарцет 445, 448
Эстрагон 515
Эуаструм 59*
Эубактерии 27
Эупомациевые 352
Эуспорангиатные папорот-
ники 225
Эфедра 334, 335*, 336, 343
Эфедровые, порядок 334
Эфедровые, семейство 334
Эхеверия 427
Эхинопснс 478*
%
Юкка 520, 533
Юнгерманниевые 186, 211
Юнгерманнии акрогинные
188, 211
Юнгерманнии анакрогин-
ные 186, 187*, 211
Яблоневые 434
Яблоня 343, 435*
Ярравия 220, 221*
Ярутка 424
Ясенец 451
Ясень 480, 482*
Ясменник 501, 502
Яснотковые 487
Ястребинка 513, 519
Ятрышник 544, 545, 546*
Ятрышниковые 543
Ячменевые 560
Ячмень 556*, 562
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Abies 298, 320, 321*
- Abietoideae 320
Absidia 116
Abutilon 401
Acacia 440, 441*
Acer 453*
Aceraceae 452
Acetabularia 56
Achillea 516
АсЫуа 112
Aconitum 356, 357*, 358
Acorns 569
Acrocarpi 206
Acroptuon 516
Actaea 522
Actinidia 398
Actinidiaceae 398
Actinomyces 28, 162
Actinomycetes 27
Adansonia 401, 402*
Adonis 357, 358*
Aegopodium 465
Aesuius 453
Aethusa 467
Agapanthus 520, 521, 531
Agaricaceae 142. 143
Agaricalqs 142
Agaricus 144
Agathis 300, 301, 314
Agavaceae 532
Agave 533
Aglaozonia 78
Agrimonia 430
Agropyron 551, 558, 561
Agrostemma 389
Agrostis 551, 558, 562
Annfeltia 93
Aira 347
Aizoaceae 386
Alaria 82
Albugo 114
Alcanna 487
Alchemilla 430
Alectoria 169
Alectorolophus 497
Alethopteris 284
Aleurites 408
Algae 33
Alisma 523*, 524
Alismales 523, 526
Alismataceae 523
Alismidae 523, 526, 580
Alliaceae 531 #
Allioideae 529
Allium 529, 530*, 531
Alnus 366, 368
Aloe 527, 528*
Alopecurus 558, 563"
• Alsmoidea 389
Alsophila 267
Alternaria 154
Althaea 401*
Altingia 364
Amanita 144
Amaryllidaceae 533
Amaryllis 534
Amenthotaxus 317
* Amygdalus 436
Anabaena 40, 95
Anabasis 385
Anacardiaceae 451, 476
Anacrogynales 186
* Ananas 548
-> Anchusa 485
Andreaea 194, 198
Andreaeaceae 198
Andreaeales 198, 199, 211
Andromeda 411
Andropogonoideae 564
Anemone 357, 522
* Anethum 468
. Aneura 188
Angelica 465
Angiopteris 258, 259*, 270
Angiospermae 279, 340, 341
Angiospermatophyta 235,
279, 340
Ankistrodesmus 49, 95
Anonaceae 352
Antennaria 512
'Anthemis 512, 515
Anthoceros 191, 192
Anthocerotaceae 191
Anthocerotales 191
Antholitis 285
Anthophyta 340
Anthostema 406, 408*
Anthoxanthum 562
Anthriscus 468
Anthurium 569
Antiaris 375
Antirrhinum 495, 496
Antitoxicum 479
Aphanizomenon 40
Aphanomyces 112
Aphyllophorales 140, 156
Apiaceae 463
Apiocystis 46
Apium 468
Apocynaceae 475
Apocynum 476
Aquilegia 356. 357*, 358
Aquifoliaceae 459
Araceae 569
Arachis 447
Aralia 463
Araliaceae 462
Araucaria 300, 301, 309,
313, 314
Araucariaceae 300, 312
Araucarites 313
Arbutoideae 411
Archaeopitys 294
Archaeopteridales 255, 258
Archaeopteris 255, 256*
Archangelica 465
Archangiopteris 258
Archicmamydeae 348, 349
Archimycetes 108 ,
Arcbostaphylos 411
Arctium 509, 510
Areca 568
Arecaceae 565
Arecales 565, 580
Arecidae 565, 572, 580'
Arenaria 347
Arenga 568
Arisuda 563
Aristolochia 362
Aristolochiaceae 362
Aristolochiales 349, 362, 363
Armeniaca 437
Armilariella 143
Armoracia 424
594
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Artemisia 515
Arthrodonteae 206
Articulatae 241
Artocarpus 374
Asarum 362
Asdepiadaceae 477
Asclepias 479
Ascolichenes 168
Ascomycetes 108, 117
Asimina 352
Asparagoideae 531
Asparagaceae 532
Asparagus 531, 532*
Aspergulaceae 126
Aspergillales 126
Aspergillus 126, 127
Asperula 502
Asphodeloldeae 527
Aspidiaceae 270, 272
Aspidistra 531
Aspidospermum 476
Aspleniaceae 270, 272
Asplenium 268*
Astasia 94
Aster 512
Asteraceae 508
Asterales 508, 520, 580
Asteridae 474» 519, 520»
579
Asterocalamitaceae 245
Asterocalamites 246
Asteroideae 513
Asterotheca 261
Asteroxylon 218, 225
Astragalus 449
Astrantia 463» 464*
Astrocarium 566
Athyrium 270
Atriplex 385
Atropa 492*» 494
Attalea 566
Avena 556*
Avicennia 490
Aucuba 468
Auricidaria 146
Auriculariales 146» 156
Austrotaxis 317
Azolla'279
Azollaceae 274» 279
Azorella 465
Azotobacter 25
Bacillaceae 26
Bacillariophyta 69, 170
Bacillus 23, 30, 162
Bacteriaceae 26
Bacteriophyta 28, 31, 170
Bacterium 25
Baiera 298
Balsaminaceae 457
Bambusa 559
Bambusoideae 559
Bangiephyceae 87, 91
Baragwanathia 226*
Barbarea 424
Barbula 206
Basidiolichenes 168
Basidiomycetes 108, 136
Batrachospermum 87,89, 91
Beggiatoa 27
Begonia 416
Begoniaceae 416
Bellis 515
Bennettitales 291
Bennettites 291
Berberidaceae 354
Berberis 354
Bergenia 429
Berteroa 424
Beta 383*
Betula 366» 367. 368*
Betulaceae 366
Bidens 513
Bigneniaceae 499
Biophytum 454
Blasia 188
Biastocladiales 110
Boebmeria 377
Boenia 287
Boletaceae 144
Boletus 144
Bombacaceae 401
Bombax 402
Boraginaceae 484
Boragininae 482, 484
Boraginoideae 485
Borassus 567
Borrelia 27
Botrychium 257*
Botrydium 67
Bouteloa 563
Bovista 144
Brachythecium 207
Brassica 421, 422, 423
Brassicaceae 419
Briza 559
Bromeliales 547, 548, 565
Bromus 558, 559
Broussonetia 374
Bryales 198, 499, 203, 205,
206, 211
Bryopsidales 54
Bryonia 418
Bryophyllum 427
Bryophyta 177, 178, 179,
202, 208, 210
Bryopsida 194, 208, 211
Bryopsis 56
Bryum 207
Bulbophyllum 545
Burmanniaceae 522
Butomaceae 523
Butomus 523
Butyrospermum 414
Buxaceae 405
Buxus 405
Cactaceae 386
Caesalpinia 443
Caesalpiniaceae 442
Caladium 569
Calamagrostis 551,
Calamitaceae 246
Calamitales 245
558
Calamites 246
Calamophytaceae 242
Calamophyton 241, 242*, 243
Calamostachys 246, 247
Calamus 566
Calceolaria 496
Calendula 515
Calimmatotheca 282
Calla 570
Calligonum 391, 499
Callithamnion 85, 93
Callixylon 294, 295
Calluna 411
Calobrium 190, 211
Caltha 356
Calvatia 144, 145
Calystegia 483
Camelina 424, 425
Camellia 395
Campanula 508
Campanulaceae 507
Campanulales 506, 520
Candida 154
Cannabinaceae 376
Cannabis 376, 377*
Cannaceae 540, 541
Cantharellus 141
Capparaceae 578
Capparales 393, 414, 419,
426, 578
Capparidacea 425
Capparis 425
Caprifoliaceae 502
Capsella 424*
Capsicum 493
Caragana 449
Carduus 516
Carex 538, 539
Carica 415
Caricaceae 415, 419
Carpinus 369
Carpoascomycetidae 122
Cartbanus 515
Carum 347, 468
Caryophyllaceae 389
Caryophyllales 382, 393, 577,
578
Caryophyllidae 382, 393,
577, 578
Caryophyllus 469
Cassia 443*
Cassytha 353
Castanea 370, 371
Castilloa 375
Casuprina 379*
Casuarinaceae 379
Casuarinales 364, 378
Caulerpa 55*, 56
Caytoma 285
Caytoniales 285
Cavanillesia 402
Cecropia 376
Cedrus 299, 323, 324*
Ceiba. 402
Celastraceae 459
Celastrales 459, 468, 474,
579
Celtis 373
Centaurea 512, 516, 517*
Centricae 69, 73
Centrospermae 382, 390
Cephaelis 502
Cephalotaxaceae 302, 315,
319
Cephalotaxus 305, 315, 316*
Ceramiales 91
Cerasus 437, 438*
Ceratium 63
Ceratozamia 287, 291 *
Cereus 387*, 388
Ceroxylon 568
Cetraria 168
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
595
Chaetoceros 71, 75
Chaetophorales 50
Chaetophorinae 50
Chamaedaphne 411
Chamaenerium 471, 473*
Chamaerops 567
Chamaesiponeae 38
Chantransia 88
Charophyta 61, 170
Cheiranthus 425
Cheirolepidlaceae 311
Cheirolepis 311, 312
Chelidonium 360, 362
Chenopodiaceae 383
Chenopodium 385
Chilocyphus 190
Chlamydobacteriales 26
Chlamydomonas 42, 94
Chlamydospermatopsida 333
Chloramoeba 67
Chlorella 49, 96
Chlorococcales 46
Chlorococcum 47
Chlorophyceae 42
Chlorophyta 41, 170
Choir omyces 136
Choripetalae 348, 349
Chorisia 402
Chosenia 380
Chromulina 65
Chroococceae 38
Chroococcus 38, 164
Chrysanthemum 512, 516
Chrysophyta 65, 170
Chrysospienium 429
Chrystensenia 258
Cby tridiales 110
Cicnorioideae 513
Cichorium 517
Cicuta 465, 467*
Cinchona 500
Cinnamomvm 353*, 354
Circaea 472
Cirsium 516
Cissus 462
Cistaceae 415 :
Cistanche 499
Cistus 415
Citroideae 450, 451
Citrullus 417
Citrus 450, 451
Cladonia 169
Cladophora 54
Cladothrix 26
Cladoxylales 253
Cladoxylon 253
Clavanaceae 141
Claviceps 131, 132, 133
Clavicipi tales 131
Climatium 205, 207
Closterium 58
Coccaceae 26
Cocos 567
Codium 56
Coffea 500, 501*
Cola 403
Colchicum 527
Coleochaetae 52
Coleus 489
Colletia 460
Colocasia 521, £>69
Comarum 433
Commelinaceae 547, 548
Commelinales 547, 565
Commelinidae 547, 559,
565, 580
Compositae 508
Concnocelis 87'
Conferva 67
Coniferales 281, 298, 309
Coniferopsida 281, 293
Coniophora 141
Conium 465, 467*
Conjugatophyceae 56
Connaraceae 474
Conocephalum 184, 185*
Contortae 475
Convallaria 531
Convolvulaceae 482
Convolvulinae 482, 580
Convolvulus 482, 483*
Copemicia 568
Coprinus 144
Coralline 93
Corallinaceae 93
Corallorhiza 546
Corchorus 405
Gordaianthus 294, 295
Gordaitales 293, 294
Cordaites 294
Cordioideae 485
Cordyline 533
Coriandrum 468
Coriolus 142
Cormophyta 173
Comaceae 468
Cornales 468
Comus 468
Coronaria 388*
Corsinia 176
Corydalis 347, 361, 521
Corylus 366, 367, 368, 369*,
497
Corynebacteriales 26
Corynebacterium 26, 30
Corypha 566
Cosmarium 58
Cotinus 452
Crassula 427
Crassulaceae 427
Crataegus 435
Crenothrix 25, 27
Crinum 533
Crocus 535*
Cronartium 153
Cruciferae 419, 578
Cryptonemiales 86
Cucumis 417
Cucurbita 418
Cucurbitaceae 416
Cucurbitales 393, 416, 426,
578
Cupressaceae 330
Cupressoideae 331
Cupressus 331*
Curcuma 541
Cuscuta 483
Cuscutaceae 483
Cutleria 78
Cyanophyta 170
Cyatheaceae 270, 272
Cycadaceae 286
Cycadales 286
Cycadeoidea 291
Cycadopsida 281, 282, 298
344
Cycas 287, 288, 291
Cyclamen 413
Cyclosporeae 81
Cydotella 73
Cydonia 435
Cylindrocystis 60
Cymbella 73
Cynara 515
Cynodon 563
Cynoglossum 485, 486
Cyperaceae 536
Cyperales 536, 546, 547
Cyperus 538
Cypripedieae 543
Cypripqdium 546
Cystococcus 47, 164
Cystopteris 268
Cystopus 114 •
Cystoseira 82
Dactylis 558, 559
Dahlbergia 449
Dahlia 515
Danaea 258
Daphne 408
Dasycladus 56
Datura 494
Daucus 467, 468*
Degeneria 351, 352*
Degeneriaceae 351
Delesseria 93
Delphinium 356, 358
Dendrocalamus 559
Dendrosicyos 416
Denteromycetes 153
Derbesia 56
Descurainia 424
Desmidiales 58
Desmidium 60
Dialypetalae 348
Diantnus 390
Diatomeae 69
Dicentra 361
Dichophyllum 296
1 Dicksonia 267
Dicksoniaceae 270, 272
Dicotyledoneae 347
Dicranopteris 272
Dicranum 206
Dictamnus 451
Dictyota 78
Dictyuchus 112
Digitalis 496, 497*
Diueniaceae 393
Dilleniales 393, 414,
473
Dilleniidae 393, 419,
473, 577
Dilophus 78
Dinonryon 65
Dinophyceae 63
Diospyros 414
Diplolepidae 206
Dipodascaceae 119
Dipodascus 119
Dipsacaceae 504
Dipsacales 502, 520,
Dipsacus 506
Dipterideae 272
Dipterocarpaceae 396
426,
426
580
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЯ
Dipterocarpus 397
Dischidia 479*
Discolichenes 168
Discomycetiidae 133
Draba 424
Dracena 532
Dracocephalum 489
Drapamaldia 50
Drapamaldiopate 50
Drepanolepis 312
Drepanophycus 226*
Dryas 431
Dryobalanops 397
Dryopteris 261’, 262*,
Dudresnaya 89, 91
Dunaliella 44
Durio 403
Ebenaceae 414
Ebenales 393, 413, 426, 578
EcbalUum 418
Echeveria 427
Echinochloa 564
Echinops 509
Echinopsis 478*
Ectocarpus 76, 78
EhretiOideae 485
Elaeis 568
Elodea 524
Empetraceae 412
Empetrum 412
Empusa 117
Enalus 525
Enantioblastae 547
Encephalartoe 287
Endomycetaceae 119 -
Endomycetales 119
Entomophtora 117
Entomophtorales 115, 117
Ephedra 334
Ephedraceae 334
Ephedrales 334
Epichloe 131
Epilohium 471
Epimedium 354, 355*
Epipogon 546
Equisetaceae 247
Equisetales 247
Equisetites 251
Equisetum 241. 247
Eragrostoideae 563
Eremascus 118*
Eremurus 527
Erica 411
Ericaceae 411, 578
Eric ales 393, 409, 410, 426,
578
Ericoideae 411
Eriobotrya 435
Eriocaulales 547, 565
Eriophorum 537, 538*
Ernestiodendron 310
Erodium' 456
Ervum 445
Eryngium 463
Erysimum 424
Erysiphaceae 127
Erysiphales 127, 155
Erysiphe 128
Erythraea 476* v
Erythroxylaceae 459
Erythroxylon 459
Euascomycetidae 122
Eubacteria 26
Enbacteriales 26
Eucalyptus 470, 473
Eucaryote 16
Encomia 365
Eucomiaceae 365
Eudorina 44
Eulilices 255
Eugenia 469, 472*
Euglena 41
Euglenophyta 40, 170
Euonymus 459
Euphorbia 405, 406, 407*
Euphorbiaceae 405
Eupborbiales 393, 405, 426
Euphrasia 497
Eupomatia 352
Eupomatiaceae 352
Eusclerotinia 134, 135*
Evemia 169
Exoascales 121
Exobasidiales 139
Exobasidium 139
Faba 446
Fabaceae 443
Fabrics 440, 473, 474, 578
Fagaceae 370
Fagries 364, 365, 378
Fagopyrum 3911 392*
Fagus 372*
Ferula 465
Festuca 558, 559
Pestaceae 559
Festacoideae 559, 563'
Fibuloporia 142
. Ficaria 347
Ficus 375*, 376
Filicales 261, 267, 269, 270,
272
Filices 251
Flacourtiaceae 414
Fbrideqphyceae 87, 89, 91
Foenlculum 466*
Fomes 142, 160
Fomitopsis 142
Fossombronia 188
Fragaria 430, 432, 433*
Fraxinus 480, 482*
Freyeynetia 571
Fritillaria 528
Fritschiella 52
Fucales 81
Fuchsia 472
Fuligo 102
Fumarioideae 360, 361
' Funaria 203, 207
- Funariaceae 205
Fungi 104, 108
Fungi imperfecti 153
Fusarium 154, 161
Fusicladium 130
Gagea 528*
Griactodendron 375
Galanthus 533, 534*
Grieola 545
Galium 502
Garcinia 396
Gasteromycetiidae 144
Gelidium 93
Genista 449
Gentians 475, 476*
Gentianaceae 475
Gentianales 475, 479, 619,
520, 579
Geraniaceae 456
Geraniales 454, 579
Geranium 456
Gesneriaceae 499
Geiim 430
Gigartinales 91
Ginkgo 296
Ginkgoales 296
Ginkgophyllum 296
Gladiolus 535
Glaucium 360
Gleditschia 443
Gleichenia 272
Gleicheniaceae 270, 272
Gleosporium 155
Gloeocapsa 38, 164
Glumiflorae 536
Glycine 446
Glycirrhiza 449
Glyptolepis 311
Gnetaceae 336
Gnetales 336
Gnetum 336 h
Gompbonema 731
Goniolimon 392 ’
Gonium 44
Gossypium 400
Gracilaria 93
Gramlnales 547, 549
Gramineae 549
Grammatophyllum 545 '
Gratiola 495, 496
Gristorpia 285
Grossotneca 282
Grossularia 428
Guajacum 459
Gymnadenia 546
Gynmoascomycetidae 119
Gymnocarpeae 168
Gymnosperma 510
Gymnospermae 279
Gymnospermatophyta 235,
279
Gynoeciatae 340
Gyromitra 134
" Haematoxylon 442
Halocnemum 384
Halophila 525
Haloxylon 385
Hamamelidaceae 364
Hamamelidae 363, 382, 576
Hamamelidales 363, 364.
365, 379, 382, 576, 577
Hamamelis 364
Hancornia 476
Haplolepidae 206
Haplomi triales 179,190,191,
211
Haplomitiinm 190, 211
Hedera 462, 463*
Helianthemum 415
Helianthus 510, 513, 514*.
515*
Helichrysum 516
Hellotropioideae 485
УКАЗАТКЛЪ ЛАТИНСКИХ НАЗВАНИЯ
Heliotropium 485
Helleboroideae 356
Helleborus 356
Helmintostachys 257* t 258
Helodea 524
Helotiales 134
Helvella 134, 138
Hemerocallus 527
Hemionitis 268
Hepaticae 179
Hepaticopsida 179. 208. 209
Herniaria 389
Hesperia 425
Heterochloridales 67
Heterococcales 67
Heterocontae 66
Heterogeneratae 78
Heterosiphonalee 67, 68
Heterothrichales 67
Hevea 407. 408*
Hibiscus 401
Hieracium 513. 519
Hippeastrum 534
Hippocastanaceae 453
Holobasidiomycetidae 138
Hordeae 560
Hordeum 556*. 562
Hormogoneae 38
Horneophytop 179.218. 219*
Hosts 527
Humulus 376. 377, 378*
Hyacintella 529
Hyacinthus 529
Hydnaceae 141
Hydrangea 428
Hydrangeoideae 428
Hydrastis 355
Hydrocharis 524, 525*
Hydrocharitaceae 524
Hydrocharitales 524
Hydrodictyon 47
Hydruras 65
Hyenia 241. 242
Hyeniaceae 242
Hylocomium 208
Hymenomycetiidae 139
Hymenophyllaceae 270. 271.
Hymenophyllales 279
Hymenophyllopodiaceae 270
Hymenophyllum 271
Hymenophytum 188
Hyoscyamus 492*. 494
Hypecoideae 360. 361
Hypecoum 360. 420
Hypericaceae 395
Hypericum 395. 396*
Hyphaene 565
Hypbales 154
Hyphomycetales 154
Hypnum 208
Hyssopus 489
Hex 459
Hliciaceae 352
Illicium 352
Impatiens 457 >
Imperata 565
Intugof era 449
Inonotus 142
Inula 512
Ipomoea 483
Iridaceae 534
Iridales 536
Iris 535*
Isatis 424
Isocontae 42
ISoetaceae 239
Isoetales 238
Isoetes 214. 215, 239*. 240*
Isogeneratae 76
Ixocomus 144
Jaccandra 499
Jasminum 481
Jatropha 405
Juglandaceae 373
Juglandales 364. 372
Juglans 373
Juncaceae 536
Juncales 536. 546. 547
Juncus 536
Jungennanniales 186
z Juniperoideae 332
Juniperus 332
Jussieua 471
Kaloxylon 282
Knautia 506
Knjghtia 381
Kobresia 539 t
Krombholzia 144
Labiatae 487
Lactarius 143
Lactaca 517
Lagenaria 418
Lagenostoma 282
LaUemanita 489
Lamiaceae 487
Laminaria 80*
Laminariales 78
Landolphia 476
Lappa 513
Lappula 486
Lancoideae 323
Larix 298. 299. 319, 323.
324*
Latbraea 497
Latbyrus 449
Lauraceae 353
Laurocerasue 439
Lauras 353*. 354
Lavandula 489
Lebachia 308, 310
Lebachiaceae 298. 309
Lecanora 168
Lecidea 168
Ledum 411
Lemanea 89
Lemna 570*. 571
Lemnaceae 570
Lens 445
Lentibulariaceae 498
Leontopodium 516
Lepidocarpon 237*. 238
Lepidodendraceae 235
Lepidodendrales 235
Lepidospermales 238 -
Leptofilices 258. 261, 267.
270
Leptomitus. 112
, Leptospermoidae 470
Leptothrix 25, 27
Lessonia 80*. 81
Leucantbemum 511, 515
Leucocarpus 512, 513
Liagora 89. 91
Libanotis 465
Lichenophyta 162, 170
Liguliflorae 513. 516. 517.
Ligustnim 481
Liliaceae 526
Liliales 526, 546, 547. 565
Liliidae 526, 546. 565, 572.
580
Lilium 528, 529*
Limtmium 392
Lfnaceae 454
Linaria 495. 496*
Linnaea 503 >
Linum 454, 455. 456*
Upergularia 389
Liquidambar 364
Liriodendron 351
Litbophyllum 93
Litbopsis 386
Lithospermum 485. 486
Lithotnamnium 93
Lobelia 508
Lobeliaceae 508
Lodoicea 567
boganiaceae 480
Loiseleuria 411
Lolium 560
Lonicera 503
Loxomaceae 270
Luffa 418
Lupinus 448
Luzula 536
Lycogala 102
Lycoperdon 144
Lycopersicum 493 t
Lycopodiaceae 227
Lycopodiales 227
Lycopodium 214,227. 228*
Lycopsida 177, 214. 215,
222, 225, 281, 282
Lycopsis 486
Lyginodendraceae 282
Lyginodendron 282
Lygodium 267, 271
Lyngbya 40
Lysunachla 413
Lythraceae 469
Lytbrum 469. 470*
Macrocystis 80*. 81
Macroglossum 258
Macrozamia 287. 291
Magnolia 292. 350, 363
Magnoliaceae 350*
Magnoliales 349. 354. 363»
g%2 578
Magnoliidae 349, 362. 363.
382, 426. 473. 526» 573.
576. 577. 583
Mahonia 354
Maiantbemum 531
Mallomonas 65
Maloideae 434
Mains 434, 4374
Malvaceae 39-
598
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Malvales 393, 399, 405, 426,
578
Mandragora 494
Mangifera 452
Manihot 407, 408*
Maranta 541
Marantaceae 540, 541
Marasmius 143, 158
Marattia 258
Marattiaceae 258
Marattiales 258
Marchantia 179
Marchantiaceae 184, 208
Marchantiales 179
Marcgravia 398
Marcgraviaceae 398
Marsilia 272, 274*
Marsiliaceae 272
Marsiliales 261, 272
Matoniaceae 270, 272
Matricaria 509, 511, 515
Matthiola 425
Medicago 448*
Medullosa 284*
Medullosaceae 284
Melampsoraceae 152
Melampyrum 497
Melanconiales 155
Melanthoideae 527
Meliaceae 453
Melilotus 449
Melissa 489
Melo 417
Melobesia 86
Melocanna 559
Melosira 71, 75
Mentha 489
Menyanthaceae 475
Menyanthes 475
Mercurialis 406, 407
Meruliaceae 141
Serulius 141
esembryanthemum 386
Mesotaeniales 60
Mesotaenium 60
Mespilus 535
Metasequoja 328, 329*
Metroxylon 566, 568
Metzgeria 188
Miadesmia 238
Micosphaerella 131
Micrasterias 58
Microcycas 287, 291
Microsphaera 129
MicrOteoideae 386, 391
Milium 563
Mimosa 440*, 442
Mimosaceae 441, 578
Mimulus 496
Mimusops 414
Mnium 205, 207
Moluginaceae 390
Monilia 134
Moniliales 154
Monoblepharidales 110, 114
Monoblepharis 111
Monochlamydeae 348
№2£®otyledoneae 520
giySnpa 411
Erysiphto^Q
Erythraean.
ErythroxylacOt
Erythroxylon K,
Mortierella 116 Olea 480. 481*
Morus 374* Oleaceae 480
Mougeotia 58 Oldfieldia 408
Mucor 115, 116 Oleales 480, 519, 579
Mucorales 115, 116 Olpidium 108
Muehlenbeckia 392 Onobrychis 448
Mulgedium 519* Onoclea 215, 268*, 269, 270
Musa 540, 541* Oomycetidae 114
Musaceae 540 Ophioglossaceae 256
Musci 194, 211 Ophioglossales 255
Mycetes 104 Ophioglossum 256, 257*
Mycobacteriales 26, 32 Opuntia 388
Mycobacterium 26 Orchidaceae 543
Mycophyta 104, 170 Orchidales 543, 546
Myosot is 486 Orchis 545, 546
Myosurus 358 Myristica 353 Origanum 489 Orobanchaceae 498
Myristicaceae 353 Orobanche 498*
Myrmecodia 502 . Oryza 563
Myrtaceae 469 , Oryzoideae 563
Myrtales 469, 473, 579 Oscillatoria 38
Myrtus 469, 471* Myrtoidae 470 Osmunda 269*, 270 Osmundaceae 270, 272
Myxobacteria 27 Oxalidaceae 454
Myxomycetes 100, 102 Oxalis 454. 455*
Myxophyta 100, 102, 170
Padina 78
Narcissus 533, 534* Padus 438
Nardeae 563 Paeonia 394
Nardus 563 Paeoniaceae 394
Nathorstiana 241* Paleostachya 246, 247
Naumburgia 413 Palaouium 414
Navicula 73 Pa] iurus 460
Neckera 205, 207 Pa] maceae 565
Nectria 131 PaJ males 565
Nelumbium 359 Panax 463
Nelumbanoideae 359 Pandanaceae 571
Nemalion 91 Pandanus 571
Nemalionales 91 Pandorina 44
Nematodonteae 206 Panicoideae 564
Neocalamites 246 Panicum 557, 564 \
Neottia 546 Papa ver 360, 361*
Nepenthaceae 362 Papaveraceae 360, 362
Nepenthales 349, 362, 363 Papaverales 349, 360, 419
Nepenthes 362 Papaveroideae 360, 361
Nepeta 489 Nerine 534 Papayaceae 415 Papifionaceae 443
Nerium 476, 477* Pariana 563
Neuropteris 284 Parietales 578
Neusseria 30 Parianeae 563
Nicotiana» 493*, 494 Paris 531
Nidulariaceae 145 Parmelia 168
Nigella 356, 358 Parnassia 429
Nitrobacter 25 Parnassioideae 429
Nitrosococcus 25 Paronychioideae 389
Nltrosomonas 25 Parrotia 365
Noctiluca 64 Partenocissus 462
Nostoc 40, 97, 164, 192 Parthenium 515
Nothofagus 372 Notothylaceae 191, 192, • Pasania 371
194 Paspalum 133, 564
Notothylas 192, 194 Passifloraceae 419
Nuphar 358 Passiflorales 419, 426
Nymphaea 292, 358, 359* Pastinaca 468
Nymphaeales 349, 358, 526, 580 363, Paulownia 497 Payena 414
Nymphaeaceae 358 Pedaliaceae 499
Nymphaeoideae 358 Pediastrum 48 Pedicellaria 424
Ochroma 402 Pelargonium 457
Oegoniunf 52 Pellia 187
Oenothera 471 Peltigera 168
Oenotheraceae 471 Penicillium 126, 127, 161
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
599
Решит 58
Pennatae 69
Pennisetum 564
Peperomia 522
Pereskia 386
Peresciopsis 386
Peridineae 63
Perilla 489
Peri pl oca 479
Perisporiales 127
Peronospora 114
Peronosporales 112, 114
Persea 354
Persica 437
Petroselinum 468
Petunia 494
Peucedanum 465
Peziza 134
Pezizales 134
Phacus 41
Phaeophyta 75, 82, 170
Phaeothamnion 65, 82
Phalenopsis 545
Phallus 145
Phaseolus 445
Phellodendron 451, 452*
Philadelphus 428
Phleum 558, 563
Phlox 483
Phoenix 568
Pholiota 143
Phoma 155
Phormium 527
Phragmidium 153
Phragmites 551
Phragmobasidiomycetidae
Phycomycetes 108, 109
Phycomyxineae 102
Phyllanthus 405, 406*
Phyllocladus 300, 314, 315
Phylloglossum 227, 230
Phyllophora 93
Phyllotheca 251
Physalis 493
Phytelephas 568
Phytolacca 383
Phytolaccaceae 382, 577
Phytophthora 113*, 114
Picea 298, 321, 322*
Pilobolus 116
Pilularia 272
Pimpinella 468
Pinaceae 319
Pinnularia 73
Pinoideae 324 -
Pinus 298, 300, 308, 324
Piper 362
Piperaceae 362
Piperales 349, 362, 363
Piptoponis 142
: Pirola 410*
Pirolaceae 410
Pistacia 451
Pistia 570
• Pisum 445*
Pityaceae 295
Pitys 294
Plagiogyriaceae 270, 272
Plantaginaceae 499
Plantago 347, 499*
Plasmodiophora 192
Plasmopara 114
Platanaceae 365
Platanus 365
Platanthera 545
Platoma 89
Platycerium 267
Platystemon 360
Plectascales 126
tPleurage 131
Pleurocarpi 206
• Pleurococcus 52
Pleuromeia 235, 238, 241*
Pleuromeiaceae 238
Pleurozium 207
Plumbaginaceae 392
Plumbaginales 392, 393, 578
Poa 550, 558, 559
Podocarpaceae 314
> Podocarpus 300, 305, 312.
314*, 315
Podophyllum 355, 522
Podospora 131
Polemoniaceae 483
Polemonium 483, 484*
Polyanthes 533
Polycarpicae 292, 349
. Polyeder 48
Polygonaceae 391, 579
Polygonales 391, 393, 578
> Polygonatum 531
Polygonum 391
Polyphagus 110
Polypodiaceae 270, 272
Polypodium 270
Polyporaceae 141
Polysiphonia 93
Polytoma 94
Polytrichaceae 178, 208
Polytrichum 199,
202, 206
Poncirus 451
200, 201,
Poideae 559
Populus 380, 381, 577
Poria 142 .
Poriales 140, 142 1
Porphyra 87, 91, 93
Postelsia 81
Potamogeton 525*
Potamegetenaceae 525
Potamogetonales 525
Potentilla 432
Poterium 430
Potoniea 284
Pottia 206
Primofilices 252, 253, 261,
272, 285
Primula 412, 413*
Primulaceae 412, 578
Primulales 393, 412, 578
Procaryota 16
Protascales 119
Proteaceae 381
Proteales 364, 381, 577
Protoangiospermae 589
Protoascomycetidae 119
Protococcales 46
Protolepidodendrales 226
Protolepidodendron 227
Protopteridales 253
Protopteridium 253
Protosiphon 48
Protospnagnum 208
Prototheca 94
Protozoa 40
Pninoideae 435
Prunus 436
Psalliota 144
Pseudolarix 299, 319
Pseudospariales 129
Pseudosporochnus 219, 221*
Pseudo tsuga 320, 321*
Psidium 469
Psilophytales 217
Psilophytopsida 214, 217
Psilophyton 219*, 241
Psilotales 177, 214, 222,
223
Psilotopsida 215, 222
Psilotum 222, 223*
Pteridaceae 270, 272
Pteridophyta 214, 215, 216,
217. 235, 281
Pteridospermae 255, 282
Pteris 266*, 270
Pterocarpus 449
Pterocarya 373
Pteropsida 177, 214, 215,217,
222, 251, 261, 281, 282
Ptilidium 188, 189 *
Ptilium 207
Puccinia 149, 150, 151,,152
Pucciniaceae ' 152
Pulmonaria 485*, 486*
Pulsatilla 357, 522
Punica 470, 473*
Punicaceae 470
Pyrenocarpae 168
Pyrenolichenes 168
Pyrenomycetiidae 129
Pyrethrum 512, 515
Pyronema 122, 123*, 134
Pyrus 435
Pyrrhophyta 63, 170
Pythium 113
Quercus 369*, 370, 371
Radula 188, 189
Rafflesia 362
Rafflesiaceae 362
Ranunculaceae 355, 363
Ranunculales 349, 355, 363
Ranunculus 357, 358, 520
Raphanus 423, 424
Raphia 566, 568
Rauvolfia 476
Ravenala 541, 542*
Regnellidium 272
Remya 510
Reseda 426
Resedaceae 425
Restionaceae 548, 549, 558
Restionales 547, 548, 565
Rhabdonema 71
Rhacopteris 255, 256*
Rhamnaceae 460
Rhamnales 460, 468, 474,
579
Rhamnus 460*
Rheum 391
Rhipsalis 387
Rhizobium 25, 444
Rhizoclonium 54
Rhizophidium 110
too
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
Rhizophoraceae 469
Rhizopus 116
Rhodante 516
Rhodochorton 91
Rhododendroideae 411
Rhododendron 411
Rhodophyta 84, 170
Rhoeadales 41'9
Rhoeo 547
Rhopalodia 71
Rhus 452
Rhynia 217, 218*
Ribes 428
Ribesioideae 428
Riccardia 188
Riccia 178, 185*
Ricciaceae 185
Ricciocarpus 185*
Ricinus 406, 408, 409*
Robinia 449
Roccella 169
Rochea 428
Rosa 430, 433, 434
Rosaceae 429
Rosales 426, 439, 459, 468,
473, 576, 578, 579
Rosidae 405, 426, 473, 520,
577, 578
Rosmarinus 489
Rosoideae 431, 439
Ruhia 502
Rubiaceae 500, 520
Rubiales 500, 520, 580
Rubus 431, 432
Rudbeckia 515
Rumex 391
Ruscus 531
Russula 143
Ruta 450*
Rutaceae 450, 451
Rutales 450
Saccharomyces 120, 121*
Saccharomycetaceae 119
Saccharum 564
Sagenopteris 285
Sagittaria 524
Saintpaulia 499
Salicaceae 379, 381
Salicales 364, 379, 382, 577
Salicomia 385
Salix 380*, 381, 577
Salmonella 30
Salsola 384
Salvia 489, 490*
Salvinia 215, 274, 276*, 279
Salviniaceae 274
Salviniales 261, 272, 274
Sambucus 502
Sanicula 463
Sanseviera 533 <
Sapindaceae 453
Sapinpales 450
Sadonaria 390
Sapotaceae 414
Saprolegnia 111
Saprolegniales 111, 114
Sargassum 82
Sarraceniaceae 362
Sasa 559
Sasafras 354
Saxifraga 428
Saxifragaceae 428
Saxifragoideae 428
Sea biosa 505
Scenedesmus 49, 95
Schinopsis 452
Schizaea 271*
Schizaeaceae 270, 272, 274
Schizandra 352
Schizandraceae 352
Schizoneura 251
Schizosaccharomyces 120,
121*
Sciadopitys 327
Scilla 529
Scilloideae 528
Scirpus 537, 538*
Sclerotinia 134, 138
Scopolia 494
Scorzonera 518
Scrophularia 495
Scrophulariaceae 494
Secale 556*, 561
Sedum 427*
Selaginella 231, 280
Selaginellaceae 231
Selaginellales 231
Sempervivum 427
Senecio 347, 516
Septobadasidium 146
Septoria 155
Sequoia 321, 328*
Sequoiadendron 327, 328*
Serputa 141
Sesamum 499
Setaria 557*, 564
Shorea 397
Sigillaria 237, 238*
Sigillariaceae 237
Silene 390
Silenoideae 389
Sinningia 499
Siphonales 54
Sisymbrium 424
Smilacoideae 532
Smilax 532
Solanaceae 490, 580
Solaninae 482, 490
Solanum 490, 491, 493
Soldanella 412
Solidago 512
Sonchus 518
Sonneratia 469
Sonneratiaceae 469
Sophora 444
Sorbaria 431
Sorbus 435
Sordaria 131 ’
Sorghum 557, 564
Sparganiaceae 571
Sparganium 571, 573*
Sperghlaria 389
Sphacelia 132
Sphacelotheca 147
Sphaeriales 131
Sphaerocarpaceae 191 /
Sphaerocarpales 191, 210
Sphaerocarpus 191
Sphaeropsidales 155
Sphaerotheca 128, 129
Sphaerotilus 26
Sphagnaceae 194
Sphagnales 194, 198, 109,
Sphagnum 194, 208
Sphenobaiera 296
Sphenophyllaceae 243
Sphenophyllales 243
Sphenophyllum 243, 244
Sphenopsida 177, 214, 215,
222, 225, 241
Sphenopteris 282
Spinacea 385
Spinifex 564
Spiraea 431
Spiraeoideae 431
Spirillaceae 26
Spirochaetae 27
Spirodela -570, 571
Spirogyra 57
Spirolobeae 385
. Spirotaenia 60
Spongospora 103
Stangeria 287
Stapelia 479
Staphylococcus 30
Statice 392
Stauropteris 255
Stellaria 388*, 389
Sterculiaceae 403
Stipa 558, 563
• Stipeae 563
Stratiotes 524
Streptocheta 559
Streptococcus 30
Stromatinia 134
Strophanthus 476
Strychnos 479*, 480
Suaeda 384
Sympetalae 348
Symphoricarpus 503
Symphytum 485
Synchytrium 108, 109 .
Synura 65
Syringa 481
Tabellaria 73
Taeniocrada 220, 221*
Taeniophyllum 545
Tagetes 516
Tamaricaceae 416
Tamarix 416
Taphrina 121, 122
Taphrinales 121 '
Taraxacum 513, 518
Taxaceae 317
Taxales 318
Taxodiaceae 327
Taxodium 329
Taxus 300 317, 318*
Tectona 490
Telephoraceae 140
Terebinthales 450, 453, 454,
460, 468, 579
Tetracentraceae 382
Tetracentron 382, 576
Tetrapanax 463
Tetraspora 46
Tetrasporales 46
Thalassia 525
Thalictrum 357, 363, 522
Thallophyta 21
Thea 395*
Theaceae 394
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
ем
Theales 393, 394, 409, 414,
426, 578
Theobroma 403*
Thermopsis 444, 449
Thlaspi 424
Thuidium 205, 207
Thuja331
Thyjoideae 331
Thymelaeaceae 408
Thymelaeales 408
Thymus 489
Tilia 404*
Tiliaceae 403
Tillandsia 548
Tilletia 147
Tilletiaceae 149
Tmesipteris 177, 222, 223*,
225
Torreya 317, 318
Tortula 178, 206
Trachycarpus 569
Tradescantia 547
Tragopogon 513
Traps 472, 474*
Trebouxia 47, 164
Tremellales 145, 156
Trentepohlia 52, 164
Treponema 27
Treubia 188
Tribonema 67
Trichobacteriales 26
Trichodeemium 40
Trichomanes 267, 268,271*, *
279
Trientalis 41’3
Trifolium 447*
Triticum 556*, 560
Trochodendrales 364, 365,
382, 576
Trochodendron 382 •
Trollius 356*
Tropaeolaceae 457, 579
Tropaeolum 457, 458*
Tsuga 319, 321
Tuber 135
Tuberales 135
Tuberaiium 491
Tubiflorae 481, 519, 520,
580
Tubiporus 144
Tubuliflorae 513, 517
Tulipa 528
Tussilago 512, 516
Typha 572, 573*
Typhaceae 572
Ulmaceae 373
Ulmannia 311
Ulmus 373, 374*
Ulotrichales 49
Ulothrix 49
Ulva 50
Ulvaceae 50
Umbelliferae 463
Umbelliflorae 462, 468, 579
Umbilicus 427
Uncinula 129
Uragoga 502
Uredinales 149, 156
Uredinella 146
Urginea 529
Urocystis 147
Uromyces 153
Urtica 377, 378*
Urticaceae 373, 377
Urticales 364, 373, 378, 577
Usnea 169
Ustilaginaceae 149
Ustilaginales 146, 148
Ustilago 147, 148
Utricularia 498*
Vaccinium 410*, 412
Vaccinoideae 412
Valeriana 504, 505*
Valerianaceae 504
Vallisneria 525
Valonia 56
Vanilla 545
Vaucheria 68
Venturia 129, 130
Veratrum 527
Verbascum 495
Verbena 490
Verbenaceae 489
Verbeninae 482, 487, 580
. Veronica 494, 495, 497
Verpa 134
Verticillium 154
Vibrio cholerae 30
Viburnum 502, 504*
Vicia 449
Vinca 477
Vincetoxicum 479
Viola 415
Violaceae 415
Violates 393, 414, 419, 426,
578
Viscaria 390
b Vitaceae 460 '
Vitis 461*, 462
Voltzia 311
Voltziaceae 298, 311
Voltziopsis 311
Volvocales 42
Volvox 44
Walchia 309
Welwitschia 338, 339*
Welwitschiaceae 338
Whitteleseja 284
Williamsonia 291
Williamsoniella 291, 292
Willoughbya 476
Winteraceae 352
Wistaria 449
Wolffia 571
Wulffia 513
Wurmbaeoideae 527
Xanthium 513
Xanthophyta 66» 170
Xanthoria 168
Xanthorrhoeaceae 532
Xeranthemum 516
Xerocomus 144
Yarravia 220, 221*
Yucca 533
a
Zamia 286*, 287, 291
Zea 565 ?
Zebrina 547 ;
Zelkowa 373 /
Zingiber 541
Zingiberaceae 540, 541
Zingiberales 540, 546
Zinnia 515
Zizania 564
Zoochlorella 49
Zostera 525, 526
Zosterophyllum 220
Zygnema 58
Zygnematales 57
Zygomycetidae 114
Zygophyllaceae 457
Zygopteridales 255, 272
Zygopteris 255
УКАЗАТЕЛЬ ТЕРМ
И
IOB
Автогамия 106
Автоспора 47*, 48, 67
Автотрофное питание 25, 94
Агар-агар 84
Азотобактерин 31
Акинеты 38
Актиномикозы 28, 30
Актиностель 219, 253
Аллелопаразитизм 158, 159,
167
Амебоидная структура 34
Амфигастрии 180, 188
Андрогинофор 389, 396, 425*
Андроцей 341, 353, 553
Анемохория 160
Антагонизм 161
Антеридиальная клетка 234,
287, 314, 337
Аяте^идиальные подставки
Антеридии 21, 35, 46, 62,
68*, 72*, 83*, 86, 111,
113, 156, 174, 197, 214,
250, 257, 265, 277
Антибиотики 28, 31, 127,
161, 162
Апланосиера 35, 49
Апотеций 124, 125*, 133*,
134*, 155, 165
Апофиза 201
Арбускулы 159
Ареолы 386
Ариллус 315, 318, 393, 540
Артроспора 105
Архегони альные подставк и
182
Архегонии 174*, 182, 197,
214, 230, 234, 240*, 250,
257, 265, 275*, 307
Археспориальная клетка
307
Археспории 176, 183, 215,
99Q ЧЛ9
Архикарп 122, 134*, 156
Аск 107, 117
Аскогенные гифы 118, 122,
136
Аскогон 122, 130, 156
Аскемы 125
Аскоспора 106*, 118, 123,
136, Г66
Аскостромы 125, 130
Ассимиляторы 180, 181, 200
Ауксилярные клетки 89,
90*
Ауксоспора 70, 71
Афлебии 258, 259*
Аэренхима 273
Базальная клетка 123, 124,
277
Базидиоспора 106*, 107,
136, 139, 145, 160
Базидия 106*, 107, 108,
138*, 139*, 140, 144
Бактериозы 30
Бактериофаг 32
Белая гниль 135
Бентос 35, 96
Бесполое размножение 34,
41, 42, 47, 48, 49, 52,
53, 67, 76, 105, 106, 108
Биотип 148
Боковая конъюгация 57
Брожение 25, 26, 160
Брюшная канальцевая клет-
ка 174, 230, 240, 289
Брюшные чешуйки 180
Валекулярная полость 248,
249
Вариация 14
Вегетативная клетка 163,
277, 278, 280, 337, 341
Вегетативное размножение
38, 41, 44, 57, 60, 67, 76,
86, 105, 166, 181 205
Вегетативное тело 104, 118
Ведьмины кольца 143, 157
Везикулы 159
Вид 13
Вневлагалищные побеги 550
Внутривлагалшцные побе-
ги 550
Воздушные мешки 285, 294,
304, 310
Выводковые корзиночки 181
Выводковые почки 181, 268,
269
Выметающие волоски 510
Выстилающий слой 215, 277,
302
Галофиты 385
Гамета 35, 43, 48, 49, 50,
53, 77,.99, 175
Гаметангий 21, 35, 106,
110, 116, 175, 209
Гаметангиогамия 106, НО
Гаметофит 35, 42, 50, 76,
77, 100, 176, 177, 209,
252, 279, 302, 341
Гаплоидный набор хромо-
сом 35
Гаустория у грибов 105*,
Гаустор^ия у мохообразных
Гетерогамия 35, 41, 43, 46,
49
Гетероспория 233
Гетеротрофное питание 25,
94
Гетерофиллия 267
Гетероциста 38
Гиалиновые клетки 196
Гиалодермис 196
Гимений 133, 139, 140
Гименофор 140*, 141, 143
Гинецей 341, 349, 553
Гинобазический столбик
485
Гиностегий 477
Гипокотиль 558
Гипотека 69*
Гифы 28, 105*
Глазок 34, 46, 76
Глеба 144
Глохидии 386 '
Гниение 25
Головневые споры 146, 147
' Гонидиальный слой 164, 165
Гонидии 164
Гормогоний 38, 39*, 40
Гормоцисты 38
Грибница 104
Двойное оплодотворение
341 342
Дикарион 106, 118, 122,
136, 144, 146
Диктиостель 252, 263, 269
Дипланетизм 111
Диплоидный набор хромо-
сом 35
Дистальный конец 137
Жгутики 23*, 33, 41, 42,
86, 108
Зародыш 214, 230, 277, 279,
280, 306, 318, 338, 345,
422, 555
Зародышевый мешок 341
Заросток 81, 177, 214, 223,
229, 230, 233*, 240, 250,
257, 260, 265*, 277 , 281,
283, 288, 337
Зигогамия 114, 116
Зигота 35, 41, 43, 46, 50,
57, 58, 77, 99
Зооспора 34, 42, 43, 46
47*, 48,49, 50,53, 66—68,
81, 102, 108, 110—112
Зооспорангий 34, 68, 80*,
106*
Зоохория 160
УКАЗАТЕЛЬ ТЕРМИНОВ
608
Идиобласты 419
Изидин 166
Изогамета 35, 4ЭГ -
Изогамия 35, 41, 46; 48,49
Индузий 239, 240, 252, 258,
263, 276*
Интегумент 238, 283, 288,
296*, 307, 339
Интравагииальные побеги
550
.1» и
Калиптра 183, 201
Капиллиций 102, 144
Каринальная полость 245,
246, 249
Кариогамия 106, 118, 121,
123
Карпогон 86, 89, 90*, 91
Карпоспора 87, 88, 90*
Карпоспорофит 86, 87—90,
99
Карпофор 464
Кила 102
КладодификаЦия 251
Класс 14
Клейстокарпии 124, 125*,
126*, 127, 128
Клетка-ножка 304
Клубеньки 445
. Клювик 544
Коккоидная структура 34,
67
Колеоптиль 555
Колеориза 555
Колонка 2Q1, 543
Колосковая чешуя 552
Колпачок 183
Кольцо (у грибов) 143
Кольцо (у мохообразных)
201, 203
Кольцо (у папоротникооб-
разных) 262, 264, 270,
271, 272
Конидиеносец 106*, 112,
ИЗ, 126, 154, 155
Конидия 106, 135, 146, 153
Конъюгация 35, 41, 60
Копуляция 146
Коремии 154
Корненосцы 231
Кровелька 318
Кроющая чешуя «311, 312
Крышечка 201, 203
Ламеллярная структура 36,
37
Леканоровые апотеции 165
Лестничная конъюгация 57
Лецидиевые апотеции 165
Листовая подушечка 236,
301
Листовой рубец 237, 238,
301
Литораль 96
Лодикула 553
Ложе 118, 129, 155
Ложная ножка 197 /
Макрофиллы 221
Макрофильная линия эво-
люции 281, 298
Мегаспора 233*, 234, 238,
240, 256*, 274, 277 , 307,
334, 338
Мегаспорангий 233*, 234,
238, 239, 276*, 280, 282,
287, 303*, 305, 339
Мегаспорокарпий 276*, 277
Мегаспорофилл 239, 282,
285, 287, 288, 291
Мегастробилы 304,305, 310,
317
Мезокарпий 431
Мейоспора 42, 99
Месомы 217
Мешочек 539
Микориза 144, 157, 158, 257
Микориза эктотрофная 158
Микориза зктозцдотрофная
159
Микориза эндотрофная 158,
230
Микроспора 233*, 256*, 274,
278, 281, 284, 289, 297,
304, 310, 341
Микроспорангий 233*, 234,
238, 239, 274, 276, 277,
283, 287
Микроспорокарпий 276*,
277
Микроспорофиллы 239, 282,
284, 291, 294, 309, 310,
317, 341
Микростробил 294, 317
Микрофиллы 221
Микрофильная линия эво-
люции 281
Миксотрофное питание 40.
41, 94
Митоспора 42, 99
Мицелий 28, 104, 105*,
128, 133
Монадная структура 33, 65
Моноспора 86
Морфа 14
Мутуалистический симбиоз
158
Несептированный мицелий
104
Нитчатая структура 34
Нуклеоиды 22
Нуцеллус 281, 283, 288,
339
Облигатные паразиты 33,
107, 112, 149
Обтуратор 406
Общее покрывало 142
Оидии 105, 119
Околоцветная пленка 553,
554
Ообластеуиыб йити 89, 90*,
91
Оогамия 35, 41, 43, 46, 49,
52, 61, 68
Оогоний 21, 35, 62, 68*,
82, 111, 113
Ооспора 111
Отдел 15
Пальмеллевидная стадия 43
Пальмеллоидная структура
34
Паразитизм 158
Парафизы 80*, 125, 133,
134*, 200
Паренхиматическая струк-
тура 34
Парихнос 235, 236
Партеногонидии 44
Пелликула 40
Перианций 182
Перйдермий 152
Перидии 134*, 144, 149
Перидиолы 145
Периплазма 113
Периплазмодий 216
Перипласт 40
Перистом 199, 201, 203,
206, 207*
Перитеций 124, 125*, 129,
£31, 132, 156, 165
Перифитон 96
Перихеций 182, 190
Пикнида 149, 150*, 15V,
166
Пикноспора 149, 150, 155
Пиреноид 46, 57, 58, 84,
163, 192
Плазмогамия 106, 121, 122
Плазмсдесмы 37, 85
Плазмодий 100, 101*, 103
Планктон 35, 95
Пластинчатая структура 34
Плацента у папоротникооб-
разных 264, 276*
Плектенхима 105
Плектостель 227
Плеоморфизм 107
Плодовое тело 104, 107»
124, 125*, 133, 139, 142*,
155
Пневматофоры 330, 490
Подарии 386
Подвесок у голосеменных
308
Подвесок у грибов 115, 116,
128
Подвесок у папоротникооб-
разных 230, 233*
Подвид 14
Подкласс 15
Подпорядок 15
Полициклия 252
Полиэмбриония 308
Поллинии 478, 544
Половое размножение 41,
43, 46—49, 52, 53, 56. 57,
60, 62, 86, 105, 106, 109,
118
Полушник 225
Порода 17
Порядок 14
Предзародыш 308
Предлистья 553
Предросток 178
Привенчик 533
Прилипальце 544
Прокарпий 91
Проталлиальная клетка 234,
287, 288
Протокорм 545
Протонема 178, 179, 202
Протостель 173, 215, 227,
235, 252, 263, 284
604
УКАЗАТЕЛЬ ТЕРМИНОВ
Проэмбрио 308
Пряжка 137. 138
Псевдомиксис 106, 156
Псевдопарафизы 140*
Пыльцевая трубка 280, 281,
288, 308, 338
Разновидность 14
Разноспоровость 233, 234,
250
Рак 109*, 131
Рахис 255
Редукционное деление 35,
42, 88, 100, 109, 183
Рестелия 152
Ризины 164
Ризойдальная клетка 234,
240*4277, 278
Ризоиды 62, 178, 180, 190,
199
Ризомицелий 104, 155
Ризомоид 215, 217
Рнзоморфы 105
Ризосфера 158
Ризофоры 231, 233*, 236
Родопласты 84
Сапрофиты 25, 94, 100, 107,
Сбалансированный парази-.
тизм 158
Секция 15
Семейство 14
Семенные чешуи 306, 311,
3*2
Семя 280, 290*, 293*, 296*,
307, 318
Семяпочка 280, 282, 283*,
288, 290*, 293*, 296*, 306,
339
Септированный мицелий 104
Серая гниль 135
Се^ддевкна 164, 273, 294,
Сестринская клетка 304
Сеть Гартига 159
Симбиоз 95, 140
Симбионты 157
Синангий 223*, 224, 258,
285, 291
а новая структура 34
костель 235, 252, 269
Скафидии 82, 83*
Склероций 101, 105, 131,
132*, 133
Слоевище 21, 173, 214
Смена поколений 177
Соматогамия 106, 156
Сорали 166
Соредии 166
Сорт 14
Сорус 78, 80*, 81, 109, 252,
258, 262
Спермагенвая клетка 234,
240*, 277, 304, 337
Сперматизация 124
Сперматозоиды 35, 46, 62,
83, НО, 178, 214, 234,
240*, 277, 281
Спермаций 86, 87, 90*, 106,
149, 166
Спермин 281, 308, 338
Спермогоний 149, 166
Спора 23, 24*, 28, 29*, 34,
38, Д01*, 103, 107, 176,
209, 216, 228*, 280
Спорангиеносец 115, 116*
Спорангий 86, 101, 106,
115, 116*, 176, 215, 228,
234, 241, 257, 260, 262
Спорангиоспоры 106, 109,
115
Спорангиофор 245,247*, 249
Спорогенные нита 89
Спорогон 177, 178, 193*,
197, 210
Спорокарпий 273, 277
Спороносец 28, 29*
Спорофиллы 215, 225, 228*,
229, 234, 239, 241, 244,
252, 268, 302
Спорофит 35, 42, 50, 76. 77,
99, 176, 177,л 209, 214,
262, 279, 340
Стаминодий 351, 371
Степь 173, 177, 199, 221,
248, 301
Стеригма 126
Стигма 34
Стигмарии 236
Стилоспора 155
Столоны 491, 550
Стробил 291, 292, 302, 343
Строма 118, 129, 132, 133,
155
Субгимениальный слой 143
Сублитораль 96
Сумка 107, 108/ 117, 121,
123, 133, 138*, 155
Суспензор 116
Таксон 15
Таллом 21, 51*, 86, 87,
163, 164*, 173, 180, 192,
210
Тапетум 215, 229, 302
Телейтоспора 150,151*,153*
Телом 217, 241
Тетрада спор 176, 183, 192,
216, 277
Тетраспорангий 76, 78, 85*,
87, 89 4
Тетраспорофит 87, 88, 89,
Тетраспоры 76, 78, 86, 87
Толерантный паразитизм
Торус 300
Трама 140, 143, 145
Трансляторы 477
Триба 15
Трихогана 86, 90*, 122
Трофофилл 215, 252
Туберндий 543
Уредоспора 150, 151*
Урночка 201, 203
Факультативные паразиты
107, 112
Феопласты 76
Физоды 75
Филлокладии 386
Фитобентос 96
Фитопланктон 95
Фосфобактерин 31
Фрагмобазидия 139*, 145,
146, 156
Фруктовая гниль 135*
Хемосинтез 25
Хемотаксис 111
Хламидоспора 105
Холобазидия 138, 139*
Хологамия 35, 41, 43, 61
Хроматоплазма 36
Хроматофоры 37,40—42,46,
47, 54, 57, 58, 63, 65, 67,
68, 76, 78, 84, 91, 163,
164, 192
Цветковая чешуя 552
Цветоложе 340
Ценоцит 54
Центроплазма 36
Цеома 152
Цистиды 140*
Цистокарпий 85*, 88*, 89,
90*
Частное покрывало 143
Черная ножка 108
Шейковая канальцевая
клетка 174, 230, 234, 240
Шейковые клетки 240, 308,
312
Шишка 286, 291, 292, 293*.
295, 302, 305*, 310, 313*,
319
Щитковидная кисть 419
Щиток 62, 63, 325, 555
Экзогенные споры 106
Экяоспсрий 184, 216, 228,
249, 265
Экстравагинальные побеги
550
Элатеры 179, 216, 247*
Элатофор 188
Элитораль 96
Экация 219, 241
Эндогенные споры 106
Эндокарпий 431
Эндосперм 281, 283, 288,
296*, 307, 340, 347, 555
Эндоспорий 184, 216, 228,
249, 265
Эндотеций 341
Эпибласт 555
Эпиматий 311,' 315
Эшшлазма 124
Эписпорий 216, 249
Эпитека 69*
Эпифиллы 188
Эпифиты 96, 188, 222, 261
Эпифрагма 201
Эталий 101
Эустель 215, 252, 269
Эццдий 149, 150*
Эцидиоспора 149, 150*
Язычок у злаков 54 9
Язычок у плауновидных
232, 233*, 239, 240
Яйцеклетка 35, 46, 83, 174.
230, 234, 281, 308, 334
СОДЕРЖАНИЕ
Предисловие........................5
Введение ..........................6
НИЗШИЕ РАСТЕНИЯ
*
Обща* характеристика............21
Отдел — бактерии (Bacterophyta) 22
1-й класс — типичные бактерии
(Eubacteria)...........................—
2-й класс — миксобактерии
(Myxobacteria)...................... 27
3-й класс — спирохеты (Spirochaetae) —
4-й класс — актиномицеты, или
лучистые грибы (Actinomycetes) —
Значение бактерий и актиномицетов
в природе и в жизни человека . . 28
Происхождение й эволюция бактерий 31
вирусы..........................32
ВОДОРОСЛИ Aldaee . .........
Отдел — сине-зеленые, или циано-
вые, водоросли (Cyanophyta). . . 36
Отдел — авт леновые водоросли
(Euglenophyta) 40
Отдел — зеленые водорослн (СЫо- .
rophyta)............................41,
1-й класс — равножгутиковые зеле-
ные водоросли (Chlorophyceae, или
Isocontae) ........................'42
1-й порядок — вольвоксовые
(Volvocales) . . . ,...............—
2-й порядок — хлорококковые, или
протококковые (Cblorococcales,
или Protococcalee)................46
3-й порядок — удотриксовые
(Ulothrichales)...................
4-й порядок — эдогониевые
(Oedogoniales) ...................
5-й порядок — кладофбровыо
(Cladophorales)...................
6-й порядок — сифоновые (Sipho-
nales, или, Bryopsidales) . . .
2-й класс — конъюгаты, или сцеп*
лянки (Conjugatophyceae) . . .
1-й порядок — зигнемовые (Zyg-
nematales) .......................
49
52
—«в
54
56
57
2-й порядок —' десмидиевые (Dee-
midiales) . ,.....................58
3-й порядок — мезотениевые (Ме-
sotaeniales).....................60
Филогенетические связи зеленых
водорослей..................... - —
Отдел — харовые водоросли, или
лучицы (Charophyta).................61
Отдел — пиррофитовые водоросли
(Pyrrhophyta).......................63
Отдел — хрязомонадовые, или зо-
лотистые, водоросли (Chrysophyta) 65
Отдел — разножгутиковые, или жел-
то-зеленые, водорослн — гетеро-
конты (Heterocontae, Xanthophyta) 66
Отдел^диатомовые водорослн, дна-
томен (Diatomeae, или Bacillario-
phyta) ...........................69.
1-й класс — перистые, или пеннат-
ные, диатомовые (Pennatae) 73
2-й класс — центрические диатомо-
вые (Centricae)...................
Отдел — бурые водоросли (Phaeo-
phyta) ...........................
Группа у-Йпрядко«г— язсгенерат-
ные (Isogeneratae) .... 76
Группа порядков — гетерогенерат-
ные (Heterogeneratae) ... 79
Группа порядков — циклоспоро-
вые (Cyclosporae) ...... 61
- Отдел — красные, водорослн, или
багрянки (Rhodophyta)...........34
1-й класс — бангиёвые (Bangiophy-
сеае) ........... 91
2-й класс — флоридеи (Florideophy-
сеае).............................
Экология и распространение водо-
§ослей.......................* * 93
кологнческие группировки водо- :
рослей ............................Q5
Значение водорослей в природе и в
жизни человека.....................ду
Размножение водорослей, чередо-
вание поколений и смеиа ядерных .
фаз.............................. 9|
006
СОДЕРЖАНИЕ
Отдел — миксомицеты, или слизе-
вика (Myxophyta)...............100
Отдел — грибы (Fungi, или Mycetes,
или Mycophyta) . . ..... 104
1-й класс — архимицеты (Archimy-
cetes) ...........................
2-й класс — фикомицеты (Phycomy-
cetes)............................
1-й порядок — хитридиевые (Chyt-
ridiales).......................
2-й порядок — бластокладиевые
(Blastocladiales) .......
3-й порядок — моиоблефаридовые
(Monoblepharidales) . . . . .
4-й порядок — сапродегвдевые
(Saprblegniales) . . . , .
5-й порядок — пероиосгфровые
(Perdnosporales) . . . .1 г .
6-й порядок — мукоровыеММи-
corales).................. . .
7-й порядок — энтомофторовые
(Entoipophthorales).............
108
Ю9
Ц0
ill
112
115
117
4-й порядок — ржавчинные, ржав-
чинники (U red in ales).........149
5-й (формальный) класс — несовер-
шенные грибы (Fungi Imported!.
Denteromycetes)...................153
1-й порядок — гифомицеты (Hypho-
mycetales, или Hyphales, или
Moniliales).....................154
2-й порядок — мелянкониевые
(М elan coni ales)..............155
3-й порядок — сферопсидные
• (Sph aeropsi dales) —
Филогенез грибов.................. —
Экология грибов...................157
Практическое значение грибов . .160
Антагонизм. Антибиотики...........161
Отдел — лишайники (Lichenophyta) 162
Эволюционные отношения низших
растений..........................
З-fi класс — аскомицеты, или сум-
чатые грибы (Ascomycetes) . . . . ‘ _
1-й подкласс — голосумчатые (Рго-
toascomycetidae, ил» Сушпоазсо- f
mycetidae) . . . . . .119
1-й порядок — первичносумчатые,
иди эндомицетовые. (Protascales,
или Endomycetales)................• —
2-й порядок — тафрвдовые, или
экзоасковые (Taphnnales, или
Exoaacales) . . . . ’ . . . .. 121
бщая характеристика^. .
ВЫСШИЕ РАСТЕНИЯ С ПРЕ ОБ ЛАДА-
НИЕМ В ЦИКЛЕ РАЗБИТИЯ ГАМЕТО- г
ФДГА._^. -............• -
фцщд^имаШйбраДные: (Bryophy taL> ~~
1-й класс — печетаечныеяхи,
nit
2-й подкласс — плодосумчатые
(Euascomycetiadae, шеи Сагроазсо-
mycetidae)...........7 . . . ;.
1-й порядок — плектасковые, или
аспергилловые (Plgctascales, или
Aspergillales) ...........
2-и порядок — периспориевые, или
эризифовые (Perisporiales, или Егу-
siphales) . . ..............
Группа порядков — циреномицеты
f^renomycetiidae) •
Группа порядков — ддскомицеты
(Disomycetiidae)
3-й порядок—трюфелевые (То-.
—beralw) .........................
4-й класс — базйдибЙЕщейтГЪди ба-
вндиальные грибы (Basidiomycetes)
1-й подкласс — холобАзидномн^Нш.
(Holobasidiomycetidae) .... 7^33
139
не
186
122
126
127
12$
133
135
1-й порядок — экзобазидиальные
(Exobasidiales) . . . . . . .
Группа порядков -^именоййцеты
(Hymenomycetiidae) .
2-й порядок —порирвый^ или афиш
л 4
Fl I
3-й порядок — агарикоиые (Agari-
cales)................ . . . .
Йуппа порядков — гастеромицеты
asteromycetiidae)................
2^й подкласс — фрагмобазидиоми-
цеты (Phragmobasidiomycetidae)
1-й порядок — дрожалковые
(Tremellales)
2-й порядок — аврикуляриевые
(Auriculariales).................
.3-й порядок—головневые (Ustila-
140
142
144
145
146
порядок — ю
ngermanniales). ______
водяорядотг^^нйкро’гивные
юнгерманниевые (Anacrogynales)
2-й подпорядок — акрогинные
юнгерманниевые (Acrogynales) 1
3-Й подпорядок — гапломитрие-
вые (Haplomitriales).........
порядок — сферокарповые
188
190
„‘ _ 191
ые^Хп^к
порядок — андреевые мхи (
198
поряд
и класс -г-
pajj}4fttT 1—1 __
’Практическое значение мохообраз-
ных .................................... 213
Е РАСТЕНИЯ С ПРЕОБЛАДА-
БЫС
нк
НИЕМ Б ЦИКЛЕ РАЗВИТИЯ СПОРО-
ФИТА ...................
214
Отдел папоротникообразные
(Pteridophyta) . 4 . . . ? . , _______
1-й класс — псилофитовидные (Psi-
lophytopaida)....................217
порядок — псилофитовые (Psilo- ;
t, ,р5уtales) ,
СОДЕРЖА!
;ГГ.
1-й порядок —
вне (Protpl
ядок
лепвдодендро-
овне (Ly-
л епидодёндровые
(Lepidodendrales) . . . .
5-й порядои — лепидоспермовые
(Lepidospermales) .. .
"МЬ порядокполушниковню
(JsoQtales) ..................239
^й ц^асс — клинолистовидные, чле
е (Sphenopsida).............241
ft порядо^ — гиениевые (Нуе-
' порядок
по
ИИ*
252
подкласс ’— первопапоротник]
_____ ____ "Г ♦ Г г.
1“й порядои — протоптеридиевые
(Protopterid ales) . . .
2-й порядои — иладоисиловые
(Cladoxylales)................
3-й порядок — зигоптеридиевые
(Zygopteridales)..............
4-й порядок — археоптерисовые
(АгсЬдщщ^еп^ез) . я. _v
^Йподкласс — эуспорангиатиые
’ces
па
261
подкласс — лептоспорангиатные
папоротники (Leptofilices) . . .
1-й порядок — типичные папорот-
ники (Filicales)................
порядок — марсилиевые (Маг-
siliales
272
порядок
Mniales)__j.
279
Отдел голосеменные (Gymnosperma-
tophyta, или* Gynmospermae) * . .
1-й класс — саговниковидные (Суса-
dopsida)
280
шеи {Ptend оврегтаё) . . , ~
2гй порядок — кейтонмевые (Сяу
toniales) . ._ < . . , .
к
ШИШКОВ!
о
4ий^порддок —‘ кордаитовые (Сог-
daitales).....................
2-й порядок — гннкговые (Gink-
goales) .
340
349
#
360
3-й класс — оболочкосеменные (СЫа-
mydospermatopsida)...............333
1-й порядок — эфедровые (Ephed-
rales) . . ....................334
2£й порядок " тнетоидные—(Оп»^
/•tales) ..........
^3-й порЙдЬк — яыышчшпше (Wei-
witschiales)...................
Отдел цветковые (Anthophyta), или
покрытосеменные (An^ospern^ato-
phyta, Angiospermae), или пестичные
(Gynoeciatae)...................
1-й класс — двудольные (Dicoty-
ledoneae) ......................
1-й подкласс — магнолиидные (Mag-
noliidae)............1 . v . .
1-й порядок — магнолиецветные
(Magnoliales) . ............
2-й порядои — лютикоцветные
(Ranunculales)' . . .
3-й порядои — кувшиикоцзетные
(Nymphaeales) ... ....
4-й порядои—макоцветные (Рара-
verales) .....................
5-й порядок — перечноцветные (Pi-
g erales)
-и порядои — кирказоноцвет-
ные (Anstolochiales) ..........
7-й порядои — непентесоцветные
(Nepenthales).................
2-й подкласс — гамамелидные (На-
- mamelidae).....................
1-й порядок — гамамелидоцветные
(Hamamelidales) ......
2-й порядок — букоцветные (Fa-
gales) .......................
3-й порядок — орехоцветные
(Juglandales).................
4-й порядои — ирапивоцветные
(Urticales)...............
5-й порядои — иазуариноцветиые
(Gasuannales).................
6-й порядои — ивоцветные (Sa-
licales) .....................
7-й порядок — протейные (Protea-
les)..............2 . . . .
3-й подкласс—иариофиллидные (Ca-
ryophyllidae) .................
1-й порядок—гвоздичноцветные
(CaryopnyUales), или центросе-
менные (Centrospermae) л . .
2-й ' порядои — гречихоцветные
(Polygonales) . . '...........
3-й порядок — свинчаткоцветные
(Plumbaginales) ..............
4-й подкласс—диллениидцые (Dil-
leniidae) . . '................
‘ 1-й порядок -г диллениецветные
(Dilleniales)..................
2-й порядок — чаецветные (Thea-
les) .........................
3-й порядок — мальвоцветные
(Malvales)....................
‘4-й порядон —- молочаецветные
(Euphorbiales) .............
5-й порядок — верескоцветные
(Ericales)..........\ . . .
6-й Порядок — первоцветные (Pri-
mulales)......................
7-й порядок — эбеновые (Ebena-
les)..........................
362
363
373
378*
379 ’
381
391
392
394
409
412
413
608
СОДЕРЖАНИЕ
8-й порядок — постенносеменные,
фиалкоцветные (Violates) . . .
9^й порядок — тыквенноцветные
SCucurbitales) .......
0-й порядок — каперсоцветные
(Capparales)....................
5-й подкласс—роаидные (Rosidae)
1-й порядок — розоцветные (Ro-
sales) ..................* . . .
2-й порядок — бобовоцветные
(Fabales).......................
3-й порядок — терпентинные
(Terebinthales).................
4-й порядок — гераниецветные
(Geraniales)....................
5-й порядок — бересклетоцветные
(Celastrales) ..................
6-й порядок — круппшоцветные
(Rhamnales) .........
7-й порядок — зонтикоцветные
(Umbellifjorae).................
8-й порядок' — миртоцветные
(Myrtales)..................- .
6-й подкласс —астеридные (Aste-
ridae) ..........................
1-й порядок — горечавкоцветные
(Gentianales), или скрученноцвет-
ные "(Contortae) ...............
2-й порядок — маслиноцветные
(Oleates, или Ligustrales> . . .
•,-3-й порядок — трубкоцветные
(ТпЬШогае).......................
4-й порядок — мареноцветные
414
416
419
450
459
460
462
469
474
475
480
481
500
о-и порядок — ворсянкоцветные
(Dipsacales) . . ..............
6-й порядок — колокольчикоцвет-
ные (Campanulales) ......
7-й порядок — астроцветные fAs-
terales)
502
506
508
2-й класс — однодольные (Мопосо-
tyledineae) . . ..................
1-й подкласс — адисмидные (Ali-
smidae) . . • •
1-й порядок — частучоцветные
(Alismales) . , , .’. .* .
2-й порядок — водокрацЬцветные
(Hydrocnaritales)................
3-й порядок — рдестоцветные
(Potamogetonales)................
2-й подкласс —лялиидные (Lili id ае)
1-й порядок — лилиецветные (Li-
liales)........................
2-й порядок — ситнйкоцветные
(Juncales)......................
3-й порядок — осокоцветные (Су-
perales) ........................
4-й порядок — бананоцветные
(Zingiberales) . . . .
5-й порядок — орхидные (Orchi-
dales) .......................*.
520
523
524
525
526
540^
542
3-й подкласс — коммелинндные
(Commelinidae)..................
Группа порядков Enantioblastae
5-й порядок — злакоцветные
(Poales) ......................
4-й подкласс—ареццдные (Areeidae)
1-й порядок — пальмоцветные
(Arecales, или Paimales) . . .
2-й порядок — аронникоцветные
(Arales) .......................
3-й порядок — панданоцветные
(Pandanales)...................
Обзор эволюция покрытосеменных
547
549
565
569
571
растений......................573
Указатель русских названий расте-
ний . . ......................584
Указатель латинских названий . . 593
Указатель терминов ...... 602
Нмколац Александрович Комарницкий
Леонид Васильевич Кудряшов
Алексей Александрович Уранов
БОТАНИКА
СТметежопшма растимый
Редакторы Э. С. Янчевская, М. Ф. Труевпева
Переплет художника Ю. М. Сигова
Художественный редактор В. Г. Ежков
Технический редактор Т. И. Зыкина
Корректор А; А. Рукосуева
Сдано в набор 23/VIII 1974 г. Подписа-
но к печати 1 /VII 1975 г. 70xl08l/lf.
Бумага тип. № 2. Печ. л. 38. Усл. печ.
л. 53,20. Уч.-изд. л. 58.06. Тираж 40 тыс.
экз.
Ордена Трудового Красного Знамени,
издательство «Просвещение» Государст-
венного комитета Совета Министров
РСФСР по делам издательств, полигра-
фии и книжной торговли. Москва, 3-й
проезд Марьиной рощи 41.
Ярославский >фиграфкомбннат «Союз-
полиграфпрома» при Государственном
комитете Совета Министров СССР ио де-
лам издательств, полиграфии и книж-
ной торговли. 150014. Ярославль, ул.
Свободы. 97. Зак. № 602.
Цена без переплета 1 р. 63 к., пере-
плет 35 к.