






























































































































































































































Author: Дубицкий А.М.
Tags: материальные основы жизни биохимия молекулярная биология биофизика энтомология комары
Year: 1970




Text
А. МДУБИЦКИЙ
НРОВОСВСУЩИЕ ном
НАЗДХСТЛНД
АКАДЕМИЯ НАУК КАЗАХСКОЙ ССР
ИНСТИТУТ зоологии
ИЗДАТЕЛЬСТВО „НАУКА” КАЗАХСКОЙ ССР
BLOODSUCKING MOSQUITOES
(DIPTERA,CULICIDAE) OF KAZAKHSTAN
A.M.DUBITZKY
ALMA-ATA 1970
КРОВОСОСУЩИЕ КОМАРЬЕ
( giptera, culicidae) КАЗАХСТАНА
А. М.ДУБИЦКИЙ
AAP/SA-ATA 1970
УДК 577.170.49
Кровососущие комары — массовые эктопаразиты, причиняющие
большой вред народному хозяйству. Многие из них являются переносчи-
ками заразных болезней.
В книге даны сведения по диагностике комаров, описана методика
сбора, хранения и изучения фаз их раззития, рассматриваются ареалы
встречающихся в Казахстане видов, биология местных популяций и за-
кономерности их распространения. Впервые для Казахстана, составлена
таблица для определения видов. Текст иллюстрируется картосхемами
расаространения комаров и фотографиями характерных мест вы плода.
Книга послужит справочным и практическим пособием для меди-
ка неких и ветеринарных работников, энтомологов, а также преподава-
телей и студентов соответствующих вузов.
Илл. 55, табл. 8, библ.: отечественная—126, иностранная—24 назв.
Ответственный редактор
доктор биологических наук, профессор А. В‘. ГУЦЕВИЧ
In this book is given information about collection, identification,
keeping and studying of bloodsucking mosquitoes. It is also introduces the
reader with the forming of the fauna mosquitoes, their spreading, the
conditions of their existence, and their distribution in different landscape
areas of Kazakhstan. The text is illustrated by the maps of the spreading
of the mosquitoes and the photographs of their places inhabit.
The book is intended for the wide circles of scientific and practical
medical and veterinarian workers, biologists, entomologists and parasi-
tologists.
Fig. tabl. 8, bibl. 150.
Editor in Chief
Doctor of Biological Sciences, Professor A. V. GUTSEVITCH
4—9—2
56—70 M.
Предисловие
С каждым годом новые и новые районы Казахстана приобре-
тают все большее значение в промышленном и сельскохозяйственном
потенциалах страны. Наряду с их освоением происходит интенсифи-
кация и расширение производства в ранее освоенных местностях.
Разумеется, что во всех случаях жизнедеятельность населения
приурочена к естественно или искусственно обводненным участкам,
зачастую являющимся местами выплода разнообразных кровососу-
щих двукрылых, из которых в большинстве районов Казахстана до-
минируют комары. Назойливость их мешает людям нормально рабо-
тать и отдыхать, снижает производительность труда и способствует
распространению различных заболеваний.
Несмотря на это, обобщающие сведения о кровососущих кома-
рах в Казахстане до последнего времени отсутствовали, а отдельные
эколого-фаунистические исследования не создавали цельного пред-
ставления об их видовом составе и биологии. Это приводило к изве-
стным трудностям в осуществлении истребительных мероприятий,
поскольку общие положения не отражали многообразия ландшафтно-
климатических условий республики. Поэтому в рекомендациях На-
учного Совета по проблеме «гнус», изложенных А. С. Мончадским.
К. А. Бреевым и А. В. Гуцевичем (1962), основной упор делается на
эколого-фаунистическое изучение отдельных регионов и на разра-
ботку локальных систем мероприятий.
Всестороннее исследование видового состава, распространения и
экологии отдельных видов, наряду с научно-теоретическими обобще-
ниями, будет иметь большое значение и для выяснения роли комаров
в передаче того или иного заболевания и его ликвидации. Нагляд-
ным примером этому служит опыт борьбы с малярией.
Помимо собственных сборов п наблюдений по различным райо-
нам Казахстана в порядке обработки материала и подтверждения
чгрс-деления просмотрены коллекции Института зоологии АН КазССР,
республиканской и некоторых областных санэпидстанций, Средне-
азиатского противочумного института и Казахского института эпиде-
миологии, микробиологии и гигиены. При изучении изменчивости
кровососущих комаров нам была предоставлена возможность просмо-
треть коллекционный материал по комарам СССР и некоторых со-
предельных территорий, хранящийся в Зоологическом институте АН
СССР.
Выполнение указанных разделов работы было бы немыслимо без
направляющей и практической помощи заведующего паразитологи-
ческим отделом ЗИН АН СССР профессора А. С. Мончадского. Во
ерем я совместных экспедиционных работ и неоднократных приездов
в Ленинград мы постоянно встречали поддержку и помощь в работе
со тороны профессора А. В. Гуцевича. Ценные замечания и пожела-
ния по оформлению рукописи были сделаны профессорами
А. А. Штакельбергом и П. П. Перфильевым.
Б процессе работы мы ощущали чуткую поддержку и вниматель-
ное стлошение со стороны нашего первого учителя академика АН
КазССР. профессора И. Г. Галузо, руководителя тематических ис-
.ледовгняй доктора биологических наук В. В. Шевченко, а также ди-
эскцпи и коллектива Института зоологии АН КазССР.
Автор выражает самую сердечную признательность и благодар-
ное всем товарищам, которые оказали то или иное содействие в
вы пр опии настоящей работы.
СИСТЕМАТИЧЕСКИЙ ПЕРЕЧЕНЬ ВИДОВ КОМАРОВ
ФАУНЫ КАЗАХСТАНА
ОТРЯД DIPT ERA
Семейство CULTCIDAE
Подсемейство CULICINAE
ТРИБА A N О Р М EL IN I
I. Род ANOPHELES MEIG.
Подрод Anopheles Meig. Стр.
1 . An.] algeriensis Theob. 39
An. iAn.) claviger Meig. 40
d An. iA77.) hurcanus Pall. 42
1 An. (AfM maculipennis Meig. . 45
An. An.f maculipennis messcae Fall. 45
Ain. к An.} maculipennis sacharovi Favre 48
Подрод M у z о m у i a Blanch.
5 Ал;. (AT.) pulcherrimus Theob. 50
6 An. (ЛЕ1 superpictus Grass! 51
ТРИБА C U 11 С I N I
II. Род URANOTAENIA L.-ARRIB.
Ur\ iinguiculata Edw. 52
HI. Род CULISETA FELT
Подрод Allotheobaldia Brol.
I Cs. (All.) longiareolata Macq. 57
Подрод С u 1 i s е t a Felt
2. Cs. (Cs.) alaskaensis Ludl. 60
3. Cs. (Cs.) annulala Schrk. 62
4. Cs. (Cs.) bergrothi Edw. 63
Подрод C u 1 i c e i 1 a Felt
5. Cs. (Cc.) morsitans Theob. 64
6. Cs. (Cc.) silvestris Sching. 64
IV. Род MANSONIA BLANCH.
Подрод CoquiHettidia Dyar
1. M. (C.) richiardii Fic. 65
V. Род AEDES MEIG.
Подрод Oehlerotatus L.-Arrib.
1. Ae. (0.) behningi Mart. 87
о Ae. (O.) beklemischevi Den. 88
3. Ae. (0.) cantans Meig. 89
4. Ae. (0.) caspius Pall. 91
4a. Ae. (O.) caspius caspius Pall. 91
4 b. Ae. (O.) caspius dorsalis Meig. 96
5. Ae. (0.) cataphylla Dyar 99
6. Ae. (O.) cyprius Ludl. 99
7. Ae. (O.) communis De Geer 102
8. Ae. (0.) detritus Hal. 103
9. Ae. (O.) diantaeus H. D. K. 104
10. Ae. (0.) excrucians WaL 106
11. Ae. (O.) flavescens Miill. 107
12. Ae. (O.) hexodontus Dyar 111
(?) Ae. (O.) hungaricus Mihalyi 112
13. Ae. (O.) impiger Walk. 112
14. Ae. (O.) intrudens Dyar 114
15. Ae. (O.) kasachstanicus Guts. 115
16. Ae. (0.) leucomelas Meig. 118
(?) Ae. (O.) marine Ed. et Et. Serg. 118
17. Ae. (0.) montchadskyi Dub. 119
18. Ae. (O.) nigrinus Eck. 120
19. Ае. (О.) nigripes Zett. 12С
20. Ае. (О.) pionips Dyar . 121
21. Ае. (О.) pulchritarsis Rond. 122
22. Ае. (О.) pullatus Coq. 12г
23. Ае. (О.) punctor Kirby 12Г
24. Ае. (О.) rempeli Vock. 125
25. Ае. (О.) riparius D. К. 126
26. Ае. (О.) simanini Guts. 127
27. Ае. (О.) sticticus Meig. 13С
28. Ае. (О.) stramineus Dub. sp. n. 132
29. Ае. (О.) subdiversus Mart. 135
Подрод Aedes Meig.
30. Ae. (Ae.) cinereus Meig. . 134
30a. Ae. (Ae.) cinereus cinereus Meig. 134
30b. Ae. (Ae.) cinereus esoensis Yam. 137
30c. Ae. (Ae.) cinereus rossicus D. G. M. 138
Подрод Aedimorphus Theob.
31. Ae. (Aed.) vexans Meig. . 138
31a. Ae. (Aed.) vexans vexans Meig. 13f
31b. Ae. (Aed.) vexans nipponii Theob. 14?
VI. Род CI LEX L.
Подрод Barraudius Edw.
1. Cx. (B.) modest us Fic. 148
2. Cx. (B.) pusUlus Macq. 152
Подрод IV e о c u 1 e x Dyar
3. Cx. (N.) hortensis Fic. . 15?
4. Cx. (N.) martinii Medsch. 155
5. Cx. (N.) territans H. D. К. 15€
Подрод C u 1 e x L.
6. Cx. (Cx.) pipiens L. 156
7. Cx. (Cx.) theileri Theob. 16C
8. Cx. (Cx.) torrentium Mart. 16C
(?) Cx. (Cx.) vagans Wied. 161
I. Особенности наружного строения
Имаго
По внешнему облику кровососущие комары напоминают многих
некровососущпх насекомых, от которых они отличаются длинным и
тонким хоботком колюще-сосущего типа. Наиболее полные сведения
по наружному и внутреннему строению кровососущих комаров мож-
но найти у А. А. Штакельберга (1937), А, В. Гуцевича (1959) и В. Н.
Беклемишева (1919). Нами приводится описание основных детален
наружной морфологии комаров, имеющих таксон омическое и гозна-
нгыельное значение.
Комары — сравнительно небольшие, стройные насекомые с длин-
ными, тонкими ногами и хорошо разграниченными головой, грудью
и брюшком. Размеры мелких форм (Uranotaenia unguiculata, Culex
modulus) едва достигают 5 мм. а крупных видов (Culiseta cllaskaen-
sis. Aedes flavescens) — приближаются к 10—12 мм. Детали их на-
ружи с го строения приводятся на рис. 1.
Овальная или круглая голова почти вплотную причленена к гру-
ди. Разделяющий их шейный склерит практически не виден. Спере-
ди на голове имеется сложный комплекс вытянутых ротовых органов,
именуемых хоботком. Его составляют верхняя губа, подглоточник.
парные верхние и нижние челюсти и нижняя губа. Последняя очень
массивна. При проколе кожи она направляет колющие части хоботка
и удерживает их. По бокам у основания хоботка находятся два ниж-
нечелюстных щупика, размеры которых могут достигать длины хо-
поъка (триба Anophelini) или быть в три-четыре раза короче (триба
Cui emi). К передней части головы причленена пара 15-члениковых
усике в, или антенн. У самцов усики опушены длинными, тонкими во-
лоекзми, отчего они кажутся лохматыми. Непосредственно за усика-
ми расположены сложные фасеточные глаза, между которыми иног-
да имеется довольно длинный хохолок чешуек. Голова самцов легко
10
отличима от головы эмок по более длинным щупикам и их пышной
опушенности.
Грудь комаров подразделяется на три отдела: передне-, средне- и
заднегрудь. Сверху почти вся грудная часть покрыта спинным скле-
ритом среднегруди. который заканчивается обособленным цельно-
краГним (триба Anopkelmi} или трехлопастным (триба Culiclnl) щит-
ком (рис. 2). Между щитком
и брюшком располагается
сферической формы образова-
ние, называемое погтнотум
или посгскутеллум На боч-
ках груди имеются той пар-
ных склерита (рис. 3*. Перед-
ние склериты называются
про-. кезс- и метаэлистерна-
ми, а задние — про-, мезо- и
метаэпимерами. На бочках
груди имеется по паре дыха-
лец: переднее между про-
эпп мерой и мезэплмерой.
а заднее — у основания
крыла.
Боковые поверхности гру-
ди у большинства видов по-
крыты чешуйками и волоска-
ми. Их наличие и расположе-
ние имеют большое таксоно-
мическое значение (рис. 4).
Присутствие или отсутствие
дыхальцевых и задыхальце-
вых щетинок является родо-
вым признаком. Дыхальце-
вые щетинки расположены
вдоль переднего края дыха-
лец» непосредственно з стиг-
мальком углублении. Их не
следует путать со щетинками,
располагающимися на про-
эпимерах и направленных
дорзально. Задыхальцевые
щетинки лежат у верхнего
края мезапистерн и также на-
правлены дорзально (эис. 3),
Рис. 1. Строение тела комара. По Штакель-
бергу. 1937. 1 — голова л ее придатки; П —
грудной отдел; III — брюшко и его (I—в)
членики.
11
Крылья комаров с жилками, более или менее покрыты волоска-
ми и чешуйками. Жилкование их специфично. Помимо костальной
и субкостальных жилок имеются четыре радиальные, две медиаль-
ные, две кубитальные и одна анальная (рис. 5). Вторая и третья ра-
диальные жилки, разветвляясь недалеко от края крыла, образуют
ны, ВМС — вентральные мезэпистерны,
ММ—мез эпимеры, ДТС—дорзальные мета-
эпистермы, ВТ С — вентральные метаэггис-
терны, ТМ — метэпимеры, СП — передняя
стигма, СЗ — задняя стигма, К — коксы.
Рис. 4. Расположение пятен чешуек на
грудных плейритах. По Сазоновой,
1958.
1 — гипостигм альное, 2 — переднее
Группы щетинок: 1—пронотальные. 2—
проэпистернальные, 3 — проэпимерные,
4 — задыха лицевые, 5 — стерноплевраль-
ные, 6 — предкрыловые, 7 — верхние мез-
эпимерные, 8 — нижние мезэпимерные.
парастигмальное, 3 — заднее парастиг-
мальное, 4 — метастигмальное, 5 —
на вентральных мезэпистернах, 3.—
на мезэпимерах, 7 — посткоксальное
пятно.
12
Рис. о. Жилкование крыла Culicini (свер-
ху) и Anophelini (снизу). По Беклемише-
ву, 1949.
Жилки: 1 — костальная, 2 — субпос-
тавка я, 3. 4 и 3— первая — третья ра-
диальные, 6* — задняя радиальная (R44-5),
7 и 8 — передняя и задняя медиальные,
9 и 10 — передняя и задняя кубиталь-
ные. 11 — анальная, 12 — радио-медиаль-
ная, или средняя поперечная, 13 — медио-
кубитальная, или задняя поперечная,
14 — плечевая.
переднюю вилку, а две медиальные образуют заднюю. Радиаль-
ную жилку с медиальной соединяет поперечная радио-медиальная, а
медиальную с кубитальной — поперечная медно-кубитальная. Ноги
комаров состоят из девяти
члеников: тазика, вертлуга,
бедра, голени и пяти члени-
ков лапки. Первые два члени-
ка очень небольшие, и прак-
тически видны только семь
последующих, начиная с бед-
ра. Последний членик лапки
снабжен парой коготков, а у
некоторых видов—двумя при-
сосками-пульвиллами (рис. 6),
или эмподием.
Брюшко комаров состоит
из восьми однотипных члени-
ков и двух, модифицирован-
ных в половые придатки.
Верхняя, склеротизованная
часть каждого сегмента, на-
зывается тергитом, а ниж-
няя — стернитом. По бокам
тергиты и стерниты соеди-
няются посредством плей-
ральной пленки, позволяю-
щей брюшку при приеме кро-
ви увеличиваться до значи-
тельных размеров. Брюшной
отдел комаров в той или иной
степени бывает покрыт че-
шуйками, своеобразное расположение которых (поперечные перевязи,
продольная линия, боковые треугольники) и цвет учитываются при
определении видов.
В значительной мере при диагностике видов используются осо-
бенности строения генитального аппарата самок и самцов. У самок
существенное значение имеет длина половых придатков — церок, а
у самцов — строение гипопигия. Устройство последнего довольно
сложно и отличается значительными родовыми и видовыми особен-
ностями. Многие виды с достоверностью могут быть определены толь-
ко по гипопигиям.
Основные отделы гипопигия самцов образованы за счет девято-
го сегмента. Причем, тергит и стернит этого сегмента имеют вид не-
13
Рис. 6. Пульвиллы на концах лапок. По Беклемишеву, 1949.
А — Culiseta; Б — Aedes; В — Culex. 1 — коготки, 2 — этеподий, 3 — пульвиллы.
широких пластинок, расположенных в основании гениталии. Зато
боковые отростки девятого сегмента достигают максимального раз-
вития и представлены двумя двучленистыми образованиями (рис. 7—
8). Наиболее массивные отделы носят названия гонококситов (кокси-
Рис. 7. Гипопнгий самцов Anopheles (вид с
дорзальной стороны). По Беклемишеву, 1949.
1 — анальный конус, 2 — гоностиль (стиль),
3 — гонококсит (коксит), 4 — конец эдеагу-
са, 5 — класпеты, 6 — шипы в основании го-
ностиля, 7 — внутренний шип на гоностиле,
8 — девятый тергит.
тов), а вершинные — гоности-
лей (стилей). На внутренних
поверхностях кокситов могут
быть 1 — 2 выпуклых образо-
вания, именуемых бородавка-
ми. В зависимости от места
нахождения они называются
базальными или вершинны-
ми. Форма, расположение и
хетотаксия бородавок имеют
большое таксономическое зна-
чение.
Внутренняя часть гипо-
пигия несет парные образова-
ния — класпеты. Проксималь-
ная часть их носит название
стволика, а верхняя — крыла.
У родов Culiseta, Uranotaenia.
Culex и Mansonia класпеты от-
сутствуют. Среднюю часть ги-
попигия занимает совокупи-
тельный орган — эдеагус, со-
стоящий из базальных пла-
стинок, парамер и фаллозо-
14
мы. Устройство последней бывает очень сложным. Так, например, у
рода Culex она распадается на 2—3 отдела, что служит важным диаг-
ностическим признаком. Анальное отверстие открывается с дорзаль-
Рис. 8. Гипопигий самцов Aedes и Culex. По Штэкельбергу, 1937.
А — Aedes: 1 — гонококсит, 2 — гоностиль, 3 — вершинная бородавка, 4 — базаль-
ная бородавка, 5 — стволик класпеты, 6 — крыло класпеты, 7 — эдеагус, 8 — ло-
пасти девятого тергита; Б — Culex: 1 — гонококсит, 2—предвершинная бородав-
ка, 3 — гоностиль, 4 — конечный придаток гоностиля, 5 — девятый тергит, 6 —
десятый стернит, 7 — базальный придаток десятого стернита, 8 — первый придаток
мезозомы, 9 — второй отдел мезозомы.
ной стороны гипопигия. С боков его окружают рудименты десятого
сегмента. Тергиты этого сегмента представляются в виде небольших
вытянутых пластинок, а стерниты (род Culex) — удлиненных обра-
зований с шипами на конце, направленными дистально.
Личинки
Наиболее полные сведения по наружному и внутреннему строе-
нию личинок можно найти у А. С. Мончадского (1951). Мы лишь
кратко осветим наиболее важные детали их морфологии, имеющие
таксономическое значение.
Внешне личинки комаров довольно хорошо отличаются от дру-
гих гидробионтов (рис. 9). Четко отграниченные голова, грудь и
15
брюшко составляют вытянутое тело. Из них только брюшная часть
имеет ясно выраженную сегментацию, подразделяясь на девять от-
дельных члеников. Грудь состоит из трех слитых воедино сегментов,
о существовании которых можно судить по наличию трех комплексов
торакальных волосков. Передняя, средняя и задняя части груди с со-
ответствующим волосяным комплексом носят название про-, мезо- и
метаторакс. Голова личинок, как правило, резко отличается от дру-
гих частей тела темной пигментацией.
Рис. 9. Личинки комаров. Общий вид со спинной стороны. По Мончадскому, 1951.
Слева—личинка Anophelini, справа—Culicini. Римскими цифрами обозначены чле-
ники брюшка, римскими цифрами в скобках — первичные членики.
16
В процессе развития личинки комаров проходят четыре стадии,
опровождающиеся сбрасыванием кутикулярной шкурки (линька).
Перед линькой личинка становится малоподвижной и внешне набу-
хает. Если такую личинку рассматривать под микроскопом, то под
отслоившейся шкуркой хорошо заметны волосяные пучки следующей
А
Рис. 10. Голова личинки. До Мончадскому, 1951, с изменениями.
А — AedeS; Б — Anopheles. ЛЩ — лобный щиток, ЛШ — лобный шов, ТШ — те-
менной шов; 1 и 2 — внутренний и наружный волоски наличника, 3 — задний во-
лосок наличника, 4, 5 и 6 — лобные волоски (внутренний, средний и наружный),
7 — теменной волосок, 8 — транссутуральный, 9 — волосок у основания усиков,
10 — глазной волосок, 11 — преклипеальные щетинки.
Б
тадии. Разрыв кутикулы начинается с головной капсулы по фрон-
тальному и теменному шву, а затем по средней линии груди. Перво-
начально у личинок освобождаются голова и грудной отдел тела.
Спустя некоторое время личинка выскальзывает из старой шкурки.
В тех случаях, когда ей не удается это сделать произвольно, она рез-
ким движением сбрасывает прикрепленную к хвостовому концу
шкурку. Слинявшие личинки светлы на вид и малоподвижны. В этот
период отмечается наиболее интенсивный рост, замедляющийся по
мере хитинизации кутикулы. Стадию личинок можно определить по
отношению ширины головы к ширине переднегруди или по абсо-
лютному размеру головы (Мончадский, 1951).
Головную капсулу образуют следующие склериты: наличник
(клипеус), лобный, затылочный (воротничок), горловой и два щеч-
ных. Хетотаксия их имеет чрезвычайно важное диагностическое зна-
чение (рис. 10). Наличник на переднем крае несет или одну (Aedes),
2—80 17
или две (Anopheles) пары небольших волосков, называемых прекли-
леальными. На этом же склерите есть еще три лары волосков —
наружные, внутренние и задние. Две первые пары имеются только у
личинок Anopheles и отсутствуют у всех других Culicinae. На лобном
склерите прикреплены три наиболее сильно развитые пары (внутрен-
ние, средние, наружные) фронтальных волосков. Ветвление и взаи-
морасположение их являются одним из существенных таксономиче-
ских признаков. Последний ряд волосков дорзальной поверхности
головы также представлен тремя парами: наружными — глазными,
средними — транссутуральными и внутренними — теменными. Все
они расположены на уровне простых глазков головы и редко дости-
гают значительного развития.
В передней части головы хорошо различимы три лопасти верх-
ней губы: непарная средняя и две боковые. Все они образованы пуч-
ками длинных, изогнутых волосков, играющих роль в доставке пи-
щи в ротовую полость. У личинок, добывающих пищу путем скари-
фикации, средняя лопасть несет ряд крупных зазубренных шипов.
С вентральной стороны передней части головы расположено ротовое
отверстие, которое образуют: сверху — надглоточник, с боков — ман-
дибулы и снизу — максиллы.
У заднего конца средней трети головы расположены две пары
глазков: простые и сложные. Простые глазки имеют вид небольших
темных пятнышек, а сложные — полулунно окружающих их удли-
ненных образований. Простые глазки существуют на протяжении
всех стадий развития личинок, а сложные — только у старших, яв-
ляясь зачатками глаз имаго.
На границе передней и средней третей головы, ближе к лате-
ральной стороне, причленены парные одночленистые усики. Поверх-
ность их может быть покрыта мелкими шипиками. Тело усиков всег-
да имеет придаток в виде пучка или одиночного волоска. Характер
его ветвления, величина и расположение имеют большое диагностиче-
ское значение.
Грудь у личинок кровососущих комаров является самой широ-
кой частью тела. Она значительно уплощена в дорзовентральном на-
правлении и имеет округло-обтекаемую форму. Хетотаксия груди
играет большую роль в диагностике комаров Anophelini и менее — у
Culicini.
Брюшко у личинок кровососущих комаров состоит из десяти чле-
ников, семь из которых однотипны, два предпоследних, слившись,
образовали сложный восьмой членик, а десятый вследствие этого
стал девятым по счету (рис. 9). Каждый членик, кроме двух послед-
них, несет до 13 пар волосков, из которых таксономическое значение
для Anophelini имеют лишь две пары: звездчатые и боковые. Пер-
18
а боковые — толь-
ь-
। a
д
во
нп
I
зл
, по
I V
I '
6Л
пл
личинок
Anopheles (схематизировано). По Мончадско-
му, 1951.
ЗЛ, БЛ, ПЛ — задняя, боковая и передняя
лопасти, ЦП — центральная пластинка, 30,
СО и ПО — ее задний, средний и передний
отделы. Ст — стигмы, СтО — стигмальные
отростки, ВП и НП — верхняя и нижняя (ос-
нование) поверхности задней лопасти, а, в, с,
d, f, 1—5 — волоски стигмальной пластинки.
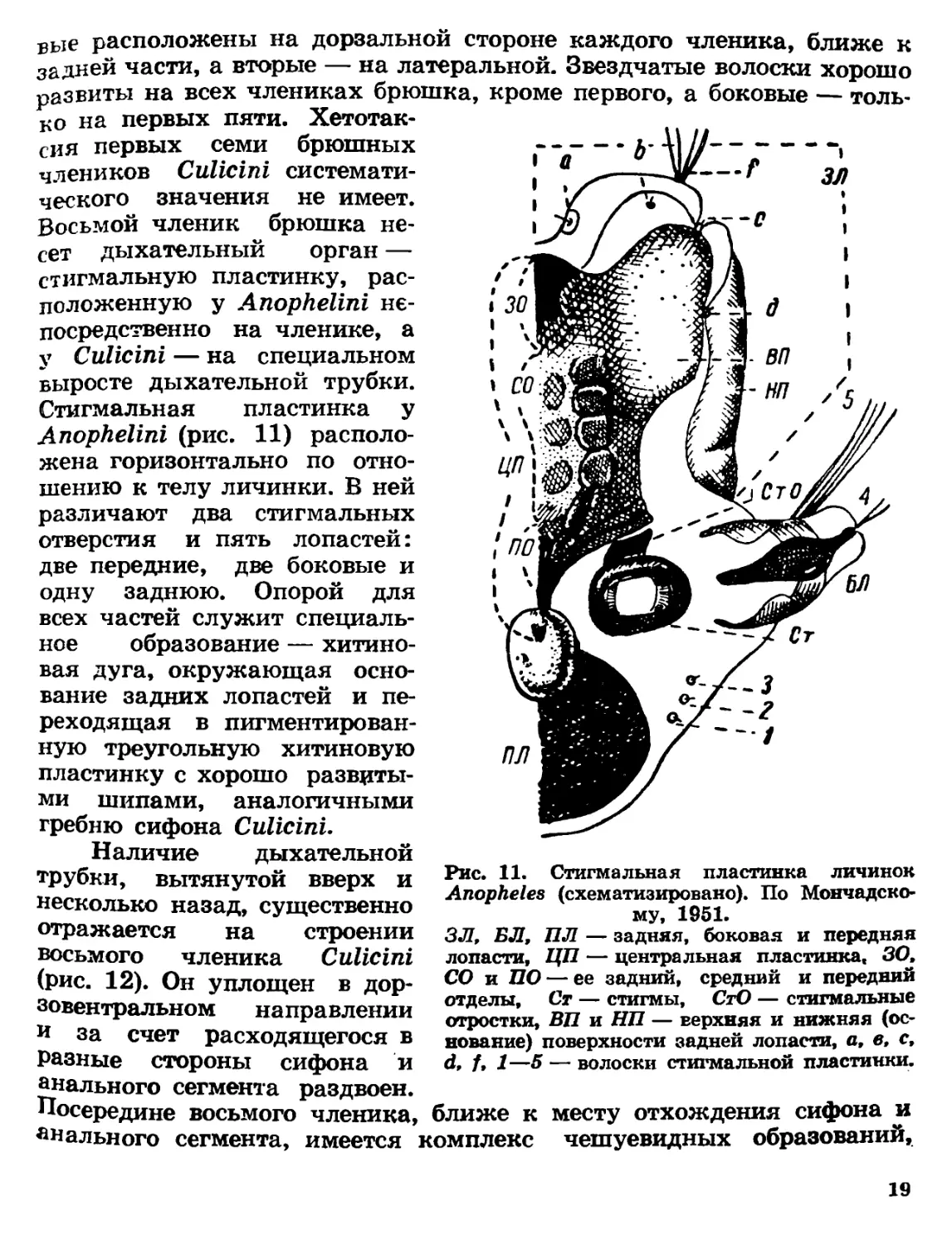
Рис. 11. Стигмальная пластинка
вые расположены на дорзальной стороне каждого членика, ближе к
задней части, а вторые — на латеральной. Звездчатые волоски хорошо
развиты на всех члениках брюшка, кроме первого,
ко на первых пяти. Хетотак-
сия первых семи брюшных
члеников Culicini системати-
ческого значения не имеет.
Восьмой членик брюшка не-
сет дыхательный орган —
стигмальную пластинку, рас-
положенную у Anophelini не-
посредственно на членике, а
у Culicini — на специальном
выросте дыхательной трубки.
Стигмальная пластинка у
Anophelini (рис. 11) располо-
жена горизонтально по отно-
шению к телу личинки. В ней
различают два стигмальных
отверстия и пять лопастей:
две передние, две боковые и
одну заднюю. Опорой для
всех частей служит специаль-
ное образование — хитино-
вая дуга, окружающая осно-
вание задних лопастей и пе-
реходящая в пигментирован-
ную треугольную хитиновую
пластинку с хорошо развиты-
ми шипами, аналогичными
гребню сифона Culicini.
Наличие дыхательной
трубки, вытянутой вверх и
несколько назад, существенно
отражается на строении
восьмого членика Culicini
(рис. 12). Он уплощен в дор-
зовентральном направлении
и за счет расходящегося в
Разные стороны сифона и
анального сегмента раздвоен.
Посередине восьмого членика,
Посередине восьмого членика, ближе к месту отхождения сифона и
анального сегмента, имеется комплекс чешуевидных образований,
19
именуемых «щеткой». Количество чешуек в щетке, их форма и рас-
положение играют большую роль при диагностике видов. С дисталь-
ной стороны щетка обрамлена пятью волосками, три из которых раз-
виты наиболее сильно и обычно состоят из нескольких ветвей. Верх-
ний пучок расположен у основания сифона, средний — на уровне
середины щетки, а нижний — у основания анального членика. В про-
межутках между средним и крайними волосками находятся простые
верхний и нижний промежуточные волоски.
Рис. 12. Строение последних члеников личинок Culicini. По Мончадскому, 1951.
А—задний конец личинки Culicini сбоку: VII—V111 — членики брюшка, 6—13 —
волоски позади щетки (6 и 13 — крайние верхний и нижний, 9 — средний, 7 и
И—промежуточные); Б — чешуйка щетки; В — зубец гребня; Г — сифон личин-
ки Culiseta сбоку. Стрелками показаны линии промеров длины сифона и ширины
его основания.
Сифон представляет собой трубковидный вырост членика. Внут-
ри него проходят парные трахейные стволы, заканчивающиеся стиг-
мальной пластинкой. Форма сифона имеет тенденцию к сужению в
вершинной части, хотя у некоторых видов он почти прямой или даже
вторично расширяющийся к вершине. Вдоль боков сифона, ближе к
20
задней стороне, тянется ряд зубцов, образующих так называемый
гребень. У некоторых видов личинок от основания сифональной труб-
ки отходят парные изогнутые отростки, носящие название «ушки»,
р зависимости от вида и рода личинок сифон может нести от одного
ДО восьми пучков или одиночных волосков. Волоски стигмальной
пластинки в это число не входят. Их может быть три пары, из кото-
рых более крупными являются волоски, расположенные на задних
клапанах. Все эти признаки, особенно сифональный индекс (отноше-
ние длины к ширине у основания), имеют большое диагностическое
значение.
Анальный членик отходит от брюшка под углом 30—45°. Сна-
ружи его полностью или частично прикрывает хитиновый склерит —
седло. На спинной стороне членика, позади седла, располагаются две
лары хвостовых волосков, а на брюшной — плавник. Последний обра-
зован сильно развитыми, ветвящимися волосками. Анальное отвер-
стие окружено двумя парами матово-белых образований, называе-
мых жабрами. Функция их не соответствует названию. Настоящая
задача анальных жабер — осморегуляция. Размер их может значи-
тельно варьировать, что во многом зависит от концентрации соли
в воде.
II. Методика сборов, хранения
и изучения комаров
Подробные сведения по вопросам сборов, хранения и изучения,
комаров можно найти у А. С. Мончадского (1952). Внеучетные сборь^
имаго осуществляются произвольными энтомологическими сачками,
эксгаустерами, морилками, а личинок — ванночкой (рис. 13), водным
сачком и специальным ковшом с ручкой. Дупла в деревьях иссле-
дуются при помощи груши с большим резервуаром и длинной вытя-
нутой трубкой, аналогичной описанному А. С. Мончадским (1952)
выкачивателю.
Учетные сборы проводятся по тщательно разработанной методи-
ке при строгом соблюдении режима исследований.
Личинки отлавливаются водным сачком (D = 15 см) один раз в
неделю в наиболее характерных местах выплода. Учет нападающих
имаго проводится эксгаустером в течение 20 мин на себе и при по-|
мощи колокола Мончадского и Радзивиловской в модификации;
К. П. Чагина при 5-минутной экспозиции. Исследования суточной ак-|
тивности осуществляются 1 раз в неделю или в десять дней за
45 мин до восхода солнца, через 1 час после восхода, в 10, 16 час, за;
1 час до захода и 45 мин после захода солнца в двух или более ха-
рактерных стациях: открытой и затененной. Место, время и дата от-
лова указываются как на прилагаемой к сбору этикетке, так и в жур-
нале сборов и наблюдений.
Влияние абиотических факторов внешней среды учитывается
при помощи психрометра Ассмана, анемометра Фюсса и люксомет-
ра, который может быть заменен фотоэкспонометром «Ленинград»
первого выпуска с таблицей для перевода в люксы. Одномоментное
определение температуры воды производится водным термометром, а
суточные колебания — максимальным и минимальным термометра-
ми. Высота обследуемых районов и атмосферное давление учиты-
ваются по показаниям высотометра или, в крайнем случае, баромет*
22
pa с нанесенной шкалой высот. Для исследования химического соста-
ва среды обитания личинок пробы воды из характернейших мест
выплода комаров отбираются в хорошо закупоренные бутылки, этике-
тируются и отправляются в химическую лабораторию, где и прово-
дится их анализ. Полученные сведения заносятся в специальный жур-
нал, а отдельные наблюдения и выводы — в полевой дневник.
Рис. 13. Отлов личинок комаров при помощи ванночки и грушевидной пипетки.
Отловленные личинки умерщвляются в подогретом 70—96° спир-
те, для чего используется спиртовка, а в полевых условиях — кусочек
ваты, смоченный в спирте. Стигмальное отверстие при этом раскры-
вается, а сегменты тела расправляются. Затем личинки расклады-
ваются по небольшим пробиркам, которые сохраняются в широкогор-
лых полиэтиленовых банках с завинчивающимися крышками. Не-
значительная перекладка пробирок ватой в сочетании с такой посу-
дой полностью гарантирует сохранность сборов даже при маршрут-
ных исследованиях. В полевых условиях массовые монотипичные
сборы большей частью определяются под бинокулярным микроско-
пом МБС-1 прямо в воде или в спирте. В тех случаях, когда у иссле-
дователя нет должного опыта, можно заключать личинок в жидкость
23
Фора, не требующую предварительного обезвоживания и просветле-
ния объекта, а затем просматривать их под микроскопом.
Умерщвление имаго производится серным эфиром или хлоро-
формом, для чего через засасывающий патрубок эксгаустера осто-
рожно до конца трубки проталкивается палочкой смоченная нарко-
тиком ватка. При замаривании сборов, отловленных сачком, его
донная часть опускается в специальную банку с предварительно зало-
Рис. 14. Схема полиэтиленовой морилки для отлова имаго (оригинал).
женной в нее ваткой с эфиром, а затем закрывается крышкой. По ис-
течении 5—10 мин донная часть сачка вынимается из банки, и
умерщвленные комары раскладываются по матрасикам. Наиболее со-
хранившаяся, непотертая часть сборов тут же накалывается на энто-
мологические булавки или на минуции, что является идеальным ус-
ловием для хранения и определения комаров. Массовые, малоценные
виды раскладываются на ватные слои. Матрасики для имаго лучше
делать размером 8X16 см, что дает возможность просматривать,
подсчитывать и определять комаров непосредственно под бинокуля-
ром МБС-1, не снимая е матрасика. В том случае, если отсутствуют
булавки, минуции или не представляется возможным наколоть насе-
комых, для сохранения редких и ценных сборов лучше использо-
вать пробирки, отделяя каждую партию из 3—4 экземпляров ватным
тампоном.
Очень хорошую сохранность отловленных комаров обеспечивает
сконструированная нами морилка (рис. 14). Сделана она из капроно-
вого тюбика из-под крема, у которого по диаметру капроновой вин-
ной пробки вырезается донная часть. В проделанное отверстие встав-
ляется капроновая пробка с круглым вырезом, диаметр которого рас-
считан на влет комара. В верхнюю часть тюбика вставляется ватный
тампон, который прижимается слоем корковой пробки с вырезанным
внутри отверстием, заклеенным мельничным газом.
Перед началом исследования ватный тампон пропитывается эфи-
ром, наливающимся через горловину, которая затем завинчивается
крышечкой, и морилка готова к работе. Севший на человека или жи-
вотное комар или какой-либо другой кровосос накрывается нижней
24
частью морилки. Комар делает попытку взлететь, попадает через от-
верстие пробки внутрь камеры, производит два-три интенсивных взма-
ха и падает, оглушенный эфиром. Выпадению комаров из морилки
препятствует конусообразное устройство пробки. Кровососы оказы-
ваются в промежутке между наружной стенкой морилки и внутрен-
ней вдающейся частью. Выделяющаяся влага адсорбируется полос-
кой фильтровальной бумаги, вложенной внутрь.
Одной заправки эфира хватает на 30—40 мин работы. Таким
образом, морилка может быть использована как для 20-минутных
учетных, так и для неучетных фаунистических сборов. За 20 мин ею
отлавливалось до 170 кровососов. При более интенсивном нападении
желательно или сократить учетное время, или подразделить 20-ми-
нутные сборы на четыре пятиминутных с удалением собранных пор-
ций, для чего вынимают пробку, а комаров высыпают в подставлен-
ную коробку с ватным слоем.
Предлагаемая модель морилки проста в изготовлении, не бьется,
не ломается и быстро заправляется эфиром. Полупрозрачные стенки
позволяют контролировать количество отловленных кровососов. Соб-
ранные таким способом комары отличались хорошей сохранностью и
не требовали последующего умерщвления.
Возможность количественного и качественного учета вредите-
лей, основанного на положительном фототаксисе многих видов насе-
комых, давно привлекает внимание энтомологов. Для кровососущих
двукрылых это имеет особое значение, поскольку среди них имеются
как зоофильные, так и антропофильные виды, и отлов на животном
и человеке не вскрывает всего многообразия видового состава. К жи-
вотным трудно применим хорошо зарекомендовавший себя при от-
лове на людях метод количественного учета, предложенный А. С. Мон-
чадским и 3. А. Радзивиловской (1947), хотя подобные исследования
широко практиковались К. А. Бреевым (1950, 1951).
В других случаях отечественные исследователи применяли для
отлова кровососущих двукрылых насекомых светоловушки, исполь-
зуемые для отлова насекомых — вредителей сельского хозяйства. И
только в последнее время В. П. Щербина (1964) описал ловушку, ка-
чественно отличавшуюся от всех предыдущих своей портативностью
и мощным источником света.
Трудность получения типовой светоловушки побудила нас про-
думать конструкцию, которую можно было бы изготовить в обычных
условиях. При ее создании мы учли недостатки ранее описанных
конструкций (одностороннее привлечение на свет, травмирование и
спрессовывание насекомых при работе аспиратора).
Предлагаемая система светоловушки представляет собой вось-
миугольную конструкцию, выполненную из прозрачного органиче-
25
ского стекла (рис. 15). Длина ее 65 см, ширина 40 см и высота 25 см.
Три боковые стенки ловушки представляют суживающиеся внутрь
четырехугольные конусы, на концах которых имеются неболь-
шие отверстия для влета насекомых (1,5X2,5 см). Длина боковых гра-
ней конусов — 27,5 см длинной и 23,5 см короткой. Четвертая сторона
ловушки занята движущейся по пазам прозрачной дверцей, которая
в случае необходимости заменяется рамкой с марлевым рукавом.
Рис. 15- Ловушка из органического стекла для отлова кровососущих
двукрылых (оригинал).
Ребра ловушки укреплены приклеенными внутри планками из тако-
го же материала, а к выходным отверстиям раструбов приклеены
пластинки с соответствующими отверстиями, которые служат одно-
временно крепежным устройством и, выступая за края раструба,
препятствуют вылету насекомых. К низу ловушки приклеены ножки
из органического стекла длиной 7 см. Размеры дверцы ловушки
(20x25 см) позволяют свободно ставить внутрь небольшую клетку с
приманочным животным (морская свинка, крольчонок, птица).
В нижнем боковом углу ловушки укрепляется мотоциклетная
фара, которая питается от аккумуляторной батареи или от аккуму-
лятора машины. От этого же источника получает энергию небольшой
электромотор, приводящий в движение лопасти инспираторного про-
пеллера. Всасывающее устройство вделано в нижнюю стенку ловуш-
ки и прикрыто сверху мельничным газом, натянутым на ободок со-
ответствующего диаметра.
В случае разрядки аккумулятора можно пользоваться обычной
переносной лампой, включаемой в сеть. Чтобы насекомые не обгора-
26
ли, лампа вставляется в специальный стеклянный колпак или в лит-
ровую стеклянную банку с прокладкой по краям.
В качестве учетного сбора используется 60-минутный отлов на-
секомых через 1 час после захода солнца. При этом учитываются
факторы внешней среды: ветер, температура и влажность воздуха.
В стационарных условиях ловушка обычно устанавливается на кры-
шу дома, сарая и т. п. на высоте 2—3 м. Во время маршрутных иссле-
дований ее помещают на кабину автомашины, а в природных биото-
пах в местах активности кровососущих двукрылых — на переверну-
тый предохранительный футляр.
По истечении срока насекомые, отловленные в ловушку, или вы-
бираются эксгаустером для опытов живыми, или умерщвляются для
хранения. В последнем случае выходные отверстия затыкаются
ватой, а внутрь ловушки ставится чашка Петри с парадихлорбен-
золом.
Особую ценность сборы описываемой ловушкой представляют в
природных биотопах, где имеются виды, отличающиеся стенофагией,
а возможно, и афагией и в обычных условиях на человека и домаш-
них животных не нападающие. Отлов на свет в совокупности с обна-
ружением личинок — чуть ли не единственный способ нахождения
таких видов.
Многие виды кровососущих двукрылых с достоверностью опре->
деляются лишь по самцам, которых обычно очень трудно обнаружить
в природе, так как они питаются соками растений. В ловушку одина-
ково хорошо залетают как самки, так и самцы. Общее соотношение
отловленных особей противоположного пола составляет примерно
2:1. В первые дни выплода больше отлавливается самцов, а в после-
дующие — самок.
Конструкция ловушки предусматривает получение материала не
только в большом количестве, но и высококачественного. С этой целью
в ловушку ставится чашка Петри с парадихлорбензолом и включает-
ся лишь источник света. Тепло от лампы способствует фумигации па-
ров яда, и залетевшие насекомые погибают в ловушке примерно с той
же скоростью, что и в морилке с парадихлорбензолом, не успев
обиться.
Для экспериментальных исследований в зависимости от харак-
тера работ имаго или выводятся из куколок, или отлавливаются в
светоловушку, или собираются эксгаустером. В любом случае насе-
комые тут же пересаживаются в обтянутый мельничным газом садок,
у которого вместо дверцы сделан марлевый рукав.
В садках всегда должны находиться чашки Петри с плавающей
на воде фильтровальной бумагой и сосуд с сахарным раствором, из
которого выступает пропитанная сиропом гигроскопическая вата.
27
В них комары живут довольно долго и по мере надобности для отдель-
ных экспериментов отлавливаются эксгаустером.
В таких садках насекомые и кормятся кровью. Для этого в садок
на некоторое время вносится специальная фиксирующая дощечка с
распластанной на ней животом вверх белой мышью или морской
свинкой. Затем напившиеся самки индивидуально отлавливаются и
после определения их видового состава (через стекло эксгаустера) по-
мещаются в отдельные садки.
Рис. 16. Обтянутый с трех сторон мельничным газом садок из органиче-
ского стекла для индивидуального содержания имаго (оригинал).
Садки для индивидуального содержания комаров представляют
собой каркас из органического стекла 10X10 см с двумя прозрач-
ными боковыми стенками. Остальные части садка обтянуты прикле-
енным к каркасу мельничным газом, который на одной из стенок
переходит в небольшой рукав с завязками (рис. 16). В такие садки
также ставятся чашки с плавающей на воде, но не закрывающей ее
полностью фильтровальной бумагой и сосуды с сахарным сиропом.
Во многих случаях самки в садках подкармливаются кровью повтор-
но, для чего к стенке прикладывается брюшная часть фиксированно-
го животного или внутрь помещается распластанная белая мышь.
Отложенные яйца подсчитываются под бинокуляром и в зави-
симости от цели опытов делятся на части, одна из которых всегда
служит контролем. Для каждого варианта опыта желательно ис-
пользовать не менее 30 яиц.
Для выявления режима, при котором возможно переживание
яиц комаров Aedes во внешней среде, отложенные кладки делятся на
28
партии, содержащиеся все время в воде, на влажной фильтровальной
бумаге и сухими (после 20-дневного пребывания во влажной среде)
как в лабораторных, так и в естественных условиях. Затем все испы-
тываемые партии яиц сохраняются в естественных условиях до сле-
дующей весны. Аналогичные варианты проводятся в лаборатории в
зимний период на других партиях яиц, промороженных в холодиль-
нике. Сухие яйцекладки заливаются водой.
Для определения условий развития личинок последние выращи-
ваются в дистиллированной, колодезной, речной, болотной и озерной
воде с различными комбинациями природного субстрата, планктона
и некоторых лемнид (ряска, лягушечник и др.). Подкормка осуществ-
ляется растертым в молоке гематогеном, растертым навозом, живыми
п сухими (растертыми) дафниями. Местами развития могут служить
аквариумные банки, кристаллизаторы, чашки Коха и полиэтилено-
вые баночки из-под плавленого сыра. Последние особенно удобны во
время маршрутных и внелабораторных работ. Вставляясь одна в дру-
гую, они занимают мало места, очень легки и не бьются.
При организации радиусных исследований местности часто бы-
вает необходимо доставить в лабораторию личинок и куколок с тем,
чтобы получить идентифицированный выплод имаго. Применив для
этой цели обычные термосы и сосуды Дьюара с пробками, можно
транспортировать личинок и куколок на большие расстояния по не-
ровной, тряской дороге и доставлять их в лабораторию живыми. В по-
следующем из них происходит нормальный выплод имаго. Таким
путем нам удавалось доставить материал с высоты 2600—2800 м
н. у. м., в то время, когда температура в водоемах не превышала 18—
20°, непосредственно в Алма-Ату, где вода в кристаллизаторах с ли-
чинками холодолюбивых видов нагревалась до 30—32° и тем не менее
происходил выплод.
Исследования мест откладки яиц самками комаров на различ-
ные участки временного усыхающего водоема и определение обилия
яйцекладок в различных местах выплода проводятся путем взятия
проб почвы размером 20x20 слс (толщина пробы 2—3 см) с различ-
ных уровней сухого водоема. Пробы почв помещаются в кюветы и за-
ливаются водой. Выплаживающиеся личинки отсаживаются в неболь-
шие чашки и доращиваются в лабораторных условиях с целью диаг-
ностики.
Определение круга прокормителей комаров проводится посред-
твом ловушек с находящимся внутри животным и липучек по мето-
дике П. А. Петрищевой (1961). Липучки размером 20X30 см,
смазанные касторовым маслом, расставляются около нор животных,
гнезд птиц, в заброшенных помещениях, провалах могил, в кроне
Деревьев, колодцах, пещерах и т. п. Основная масса материала в том
29
случае, когда насекомые полностью обволакиваются маслом, конеч-
но пропадает. Определить удается только хорошо различимые виды
и те экземпляры, у которых прилипли лишь ноги и которых удается
всосать в эксгаустер, заморить и впоследствии просмотреть в лабо-
ратории.
Для определения хемотаксисов кровососущих двукрылых было
испытано два типа ольфактометра, один из которых, так же как и
описанная хемоловушка, оригинальный. Он нашел свое отражение
в публикации (Дубицкий, 1965а).
Изучение жуков-плавунцов (Dytiscidae), их личинок, жуков-во-
долюбов (Hydrophilidae), жуков-вертячек (Cyrinus), гладышей (Noto-
nectd), плавтов (Naucoris), ручейников (Trtchoptera), личинок стре-
коз (Odonata), личинок некровососущих комаров (Chaoborinae) и дру-
гих водных обитателей как хищников проводится при изолирован-
ном содержании в банках и аквариумах их отдельных представите-
лей с определенным числом личинок, подсчитываемых через каждые
12 час. Аналогично этому исследуются личинкоядные рыбы.
Прикрепившиеся к имаго суперэктопаразиты подсчитываются под би-
нокуляром, а патогенные микроорганизмы изучаются при помощи
микроскопа.
Испытания продолжительности отпугивающего действия репел-
лентов проводятся на группах людей, состоящих из 3—6 человек.
Один из них в процессе исследования всегда служит контролем и
производит учетные сборы с определением условий внешней среды.
Испытуемые препараты в количестве 2—3 мл наносятся ватным там-
поном на кожный покров рук, шеи и лица и растираются тонким
слоем. Человек выполняет при этом ту или иную физическую нагруз-
ку в тени и на солнце. В любом случае проводится не менее двух
повторных испытаний. Критерием окончания защитного действия
репеллента считается первый укол кровососа, подтвержденный в те-
чение 5 мин вторым и третьим нападением.
Растворы инсектицидов для определения чувствительности ли-
чинок и выбора токсичных доз готовятся согласно рекомендациям
ВОЗ (серия технических докладов, № 265, 1964) в 50 мл этилового
спирта с таким расчетом, чтобы при добавлении 1 мл этого раствора
к 349 частям воды получались бы концентрации 0,0008, 0,004, 0,02,
0,1, 0,5 и 2,5 части на миллион. Для контроля берется 50 мл чистого
спирта. При манипуляциях с личинками и растворами применяются
пипетки с резиновыми баллончиками на 3 мл и серия хорошо отмы-
тых градуированных пипеток.
Для одного полного опыта в природе отлавливается 500—700
личинок с таким расчетом, чтобы среди них можно было отобрать
около 300 особей, принадлежащих к одному и тому же виду. Отлов-
30
ленные личинки (в III или в начале IV стадии) перед опытом содер-
жатся в воде из того же водоема, из которого были отловлены. Ли-
чинок с клиническими признаками заболеваний в опыт не берут.
Эксперименты начинают с того, что в 12 маленьких мензурок нали-
вают по 25 мл воды и помещают в каждую по 20—25 личинок.
В каждый из 12 стеклянных сосудов диаметром примерно 7,5—
Ю см (стеклянные банки из-под консервов) наливают 225 мл воды с
•гем, чтобы слой воды в них составлял 5—7 см. Во всех случаях не-
обходимо использовать дистиллированную или дождевую воду. Тем-
пература ее в процессе опытов должна составлять 21—25°. В каж-
дую банку при помощи пипетки вводят 1 мл соответствующего раст-
вора испытуемого инсектицида и сильно размешивают. Раствор
самой низкой концентрации готовится в первую очередь. С каждой
концентрацией ставится по 2—4 опыта. В контрольные сосуды к воде
добавляют по 1 мл этилового спирта.
Через 15—20 мин в сосуды переливают воду с личинками из
мензурок. Подсчет смертности производят через 24 час. Мертвыми
считают личинок, которые не двигаются при прикосновении иглой к
сифону или к затылочной области. Погибающие (подающие признаки
жизни) личинки учитываются совместно с мертвыми.
Ш. Повидовой эколого-
фаунистический обзор
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ТРИБ И РОДОВ
ПОДСЕМЕЙСТВА CULICINAE ПО ВЗРОСЛЫМ КОМАРАМ
(По А. А. Штакельбергу, с сокращениями и небольшими
изменениями)
1 ( 2). Щупики самок длинные, равны по длине хоботку. У сам-
цов они тоже почти равны длине хоботка, последние члени-
ки значительно утолщенные. Щиток цельнокрайний
Триба Anophelini (род Anopheles Meig.).
2 ( 1). Щупики самок короткие, во много раз короче хоботка. У сам-
цов они различной длины: или длиннее хоботка, или равны
ему; в редких случаях они короткие, как у самок (род Ura-
notaenia, подрод Aedes Meig.) Щиток трехлопастной
. . . . . . . Триба Culicini.
3 ( 4). Анальная жилка упирается в задний край крыла на уровне
ответвления общего ствола радиальных жилок (R2+R3 и
R4+R5). Микротрихии на пластинке крыла заметны лишь при
сильном увеличении . . Род Uranotaenia L.-Arrib.
В Казахстане, как и вообще в СССР, встречается один пред-
ставитель этого рода — U. unguiculata Edw.
4 ( 3). Анальная жилка упирается в задний край крыла на более
или менее значительном расстоянии от места ветвления об-
щего ствола радиальных жилок (R2+R3 и R4+R5), ближе к
вершине крыла. Микротрихии на пластинке крыла хорошо
заметны.
5 (10). Пульвиллы под коготками на лапках отсутствуют или име-
ются в рудиментарном состоянии.
6 ( 7). Дыхальцевые щетинки имеются Род Culiseta Felt.
7 ( 6). Дыхальцевые щетинки отсутствуют.
8 ( 9). Задыхальцевые щетинки отсутствуют, коготки самок без зуб-
чиков Род Mansonia Blanch.
32
В Казахстане встречается лишь один представитель этого ро-
да — М. (С.) richiardii Fic.
9(8). Задыхальцевые щетинки имеются, коготки самок, как пра-
вило, с зубчиками . . Род Aedes Meig.
Ю(5) Пульвиллы под коготками на лапках хорошо развиты . .
Род Culex L.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ТРИБ И РОДОВ ПОДСЕМЕЙСТВА
CULICINAE ПО ЛИЧИНКАМ IV СТАДИИ
(По А. С. Мончадскому, с сокращениями)
1(2) Стигмы, окруженные стигмальной пластинкой, помещаются
непосредственно на спинной стороне у заднего края восьмого
членика брюшка (триба Anophelini) . . . .
Род Anopheles Meig.
2 ( 1). Стигмы, окруженные стигмальной пластинкой, помещаются
на конце дыхательной трубки сифона, отходящей от спинной
стороны у заднего края восьмого членика брюшка (триба
Culicini).
3 ( 4). Сифон и клапаны стигмальной пластинки видоизменены в
буравящий аппарат ... . Род Mansonia Blanch.
4 ( 3). Сифон и клапаны стигмальной пластинки развиты нор-
мально.
5 (10). Сифон с одной парой пучков на задней поверхности
6 ( 7). Пучки помещаются у основания сифона..................
.............. . Род Culiseta Felt.
7 ( 6). Пучки около середины сифона или ближе к его вершине.
8(9). По бокам восьмого членика брюшка имеется по склеротизо-
ванной пластинке, от заднего края которой отходят чешуй-
ки щетки. Зубцы гребня расщеплены на концах . .
.’...................................Род Uranotaenia L.-Arrib.
9 ( 8). По бокам восьмого членика брюшка только щетка из чешуек,
расположённых в один ряд или пятном различной формы.
Зубцы гребня обычно шиповидно заострены на концах
...............................................Род Aedes Meig.
Ю ( 5). Сифон с несколькими парами пучков на задней и боковых
поверхностях...................................Род Culex L.
3-80
33
Род ANOPHELES MEIG.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА
ANOPHELES MEIG. ПО ВЗРОСЛЫМ НАСЕКОМЫМ
(По А. А. Штакельбергу, с сокращениями и небольшими
изменениями)
1 (12). Костальная жилка по переднему краю крыла имеет не более
двух светлых пятен; сюда же относятся виды, имеющие
только темные пятна на крыльях, и виды, совсем не имеющие
пятен (подрод Anopheles Meig.).
2 ( 7). Крылья с более или менее одноцветными чешуйками без
пятен; основание передней вилки крыла (R2+R3) расположе-
но явственно ближе к основанию крыла, чем основание сред-
ней вилки (медиальная передняя-!-медиальная задняя жил-
ка).
3 ( 4). Пучок белых чешуек и волосков на лбу отсутствует (имеется
слабо развитый пучок желтоватых волосков). Среднеспинка
одноцветная (без продольной полосы), с довольно длинными
темно-бурыми волосками ......................
An. (An.) algeriensis Theob.
Встречается лишь в южных районах Казахстана.
4 ( 3). Белый пучок чешуек и волосков на лбу имеется.
5 ( 6). Преобладающая окраска черно-серая со свинцовым отливом;
на среднеспинке продольная серая полоса; пучок чешуек на
переднем краю среднеспинки сильно развит и чисто белого
цвета; последний членик щупиков около % длины предыду-
щего . . . An. (An.) plumbeus Steph.
Возможен в южных и юго-восточных районах Казахстана.
6 ( 5). Преобладающая окраска желто-бурая, бурая или сероватая;
пучок чешуек на переднем крае среднеспинки слабее развит
и имеет желтоватый оттенок; продольная полоса на средне-
спинке седовато-серая; последний членик щупиков не более
половины длины предыдущего ...................................
. An. (An.) claviger Meig.
Возможен на всей территории Казахстана.
7 ( 2). Крылья с темными или светлыми пятнами; основания пе-
редней и средней вилок на крыльях расположены на одина-
ковом расстоянии от основания крыла.
8 (11). Крылья с темными пятнами.
9 (10). Общая окраска темно-бурая; среднеспинка с широкой серой
продольной полосой, занимающей почти всю ее верхнюю по-
верхность. Темные пятна на крыльях выражены явственно
34
An. (An.) maculipennis messeae Fall.
Заселяет всю территорию Казахстана севернее 46—48° с. ш.
По горным районам проникает далеко на юг.
10 ( 9). Общая окраска светло-желтовато-бурая; среднеспинка на
всем своем протяжении одноцветно светло-бурая или желтая,
иногда можно различить полосу в отделе среднеспинки, при-
легающем к щитку. Темные пятна на крыльях выражены
слабо . An. (An.) maculipennis sacharovi Favre.
Заселяет южную часть Казахстана, южнее 46—47° с. ш.
11 ( 8). Крылья на переднем крае по костальной жилке с двумя бе-
лыми пятнами . . An. (An.) hyrcanus Pall.
Распространен повсеместно, кроме Северного Казахстана (до
50° с. ш.).
12 ( !)• Костальная жилка по переднему краю крыла с четырьмя и
более светлыми пятнами (подрод Муzomy ia Blanch.).
13 (14). Брюшко и грудь сверху густо покрыты белыми чешуйками,
образующими с боков по заднему краю тергитов торчащие в
сторону пучки. Последние членики задних лапок и колечки
на передних и средних лапках белые ...............
An. (М.) pulcherrimus Theob.
Встречается лишь в южных районах Казахстана.
14(13). Брюшко с боков без пучков чешуек; последние членики зад-
них лапок темные, передние и средние лапки одноцветные,
иногда едва кольчатые ...
. . . . . . An. (М.) superpictus Grass!.
Встречается лишь в южных районах Казахстана.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА
ANOPHELES MEIG. ПО ГИПОПИГИЯМ САМЦОВ
(По А. А. Штакельбергу, с сокращениями и изменениями)
1 (Ю). Коксит при основании с внутренней стороны по большей час-
ти с двумя (редко с одной или тремя) крепкими щетинками,
из которых по меньшей мере одна расположена на сосочко-
видном бугорке (подрод Anopheles Meig.).
2 ( 3). Коксит при основании с одной щетинкой.................
...............................An. (An.) algeriensis Theob.
Горные и предгорные районы юга и юго-востока Казахстана.
. Коксит при основании с двумя-тремя щетинками.
5) . Коксит при основании с тремя крепкими щетинками, из ко-
торых две разветвлены . An. (An.) claviger Meig.
Южный и Юго-Восточный Казахстан, возможен на остальной
территории.
35
5 ( 4). Коксит при основании с двумя простыми, не разветвленными
щетинками.
6 ( 9). Шипы класпет сближены, но не слиты.
7 ( 8). Фаллозома короткая и широкая, без листовидных придат-
ков . . . An. (An.) plumbeus Steph.
Возможен в южных и юго-восточных районах Казахстана.
8 ( 7). Фаллозома длинная и узкая, на конце с листовидными при-
датками . An. (An.) maculipennis Meig.
Распространен повсеместно. В южной половине Казахстана
преобладает подвид An. т. sacharovi, а в северной — Ап. т.
messeae.
9 ( 6). По крайней мере некоторые шипы класпет слиты, образуя
подобие хитиновой пластинки. Выросты заднего края девято-
го тергита длинные и узкие .......................
. . . . . . An. (An.) hyrcanus Pall.
Распространен повсеместно, кроме северных районов Казах-
стана.
10 ( 1). Коксит с внутренней стороны при основании с 4—6 крепкими
щетинками, сидящими непосредственно на теле членика, а
не на сосочковидных выростах его (подрод Myzomyia Blanch.)
An. (М.) pulcherrimus Theob.,
...................................An. (M.) superpictus Grassi.
Встречается в южных районах Казахстана.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДВИДОВ
ANOPHELES MACULIPENNIS MEIG. ПО ЯПЦАМ
(По В. Н. Беклемишеву, с сокращениями)
1(2). Поплавки отсутствуют или рудиментарны (немногим шире,
чем оторочка, длиной не более Vs длины яйца). Верхняя по-
верхность яйца беловато-серая, без темного рисунка
. . . . An. maculipennis sacharovi Favre.
Распространен в южной части Казахстана.
2 ( 1). Воздушные камеры яйца обычно шире, чем оторочка, длина
их не менее ’/з длины яйца. В состав рисунка верхней по-
верхности яиц непременно входят поперечные полосы.
3 ( 4). За исключением черных поперечных полос и черных концов
яиц, рисунок на верхней поверхности отсутствует; фон свет-
ло-серебристый, рисунок контрастный. Если в виде исключе-.
ния встречаются темные пятна между поперечными полоса-
ми, то лишь бледные. Поплавки большие, занимают около 0,4
длины яйца An. maculipennis maculipennis Meig.
е *
36
Возможен в западных и северо-западных районах Казах-
стана.
4 ( 3). Наряду с поперечными полосами, которые иногда могут быть
разбиты на пятна, вся остальная поверхность яйца заполнена
рисунком из темных и светлых пятен или поперечных полос.
Длина поплавков больше 0,4 длины яйца . . . .
..........................An. maculipennis messeae Fall.
Распространен в северных и горных районах Казахстана.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ЛИЧИНОК IV СТАДИИ
ВИДОВ РОДА ANOPHELES MEIG.
(По А. С. Мончадскому, с сокращениями)
1 (12). Основания внутренних волосков наличника сильно сближе-
ны, расстояние между ними не больше, а обычно меньше, чем
между наружным и соответствующим ему внутренним во-
лоском.
2 ( 3). Лобные волоски короткие и простые. Боковые волоски на
четырех-шести члениках брюшка длинные, явственно перис-
тые ................................. An. (An.) plumbeus Steph.
Возможен в южных и юго-восточных районах Казахстана.
3 ( 2). Лобные волоски длинные и перистые. Боковые волоски на 4—
6 члениках брюшка неперистые, но распадаются недалеко от
основания на несколько ветвей.
4 ( 7). Наружные волоски наличника простые или слабо разветвле-
ны на концах или около середины.
5 ( 6). Волоски наличника начиная от середины или второй трети
несут тонкую вторичную перистость. Лепестки звездчатых
волосков брюшка длинные и узкие. Гребень на боках основа-
ния стигмальной пластинки в среднем из 20—25 зубцов
An. (An.) algeriensis Theob.
Встречается лишь в южных районах Казахстана.
6 ( 5). Волоски наличника гладкие, без тонкой вторичной перисто-
сти. Задние волоски наличника из двух-пяти ветвей. Лепест-
ки звездчатых волосков брюшка без вытянутой концевой ни-
ти, с гладкими или слабо зазубренными краями. Стигмы не
соединены с центральной пластинкой; без отростков. Гребень
на боках основания стигмальной пластинки из 12—16 зуб-
цов . ..... An. (An.) claviger Meig.
Возможен на всей территории Казахстана. Предпочитает гор-
ные и предгорные биотопы, на равнинных участках — род-
ники.
‘37
7 ( 4). Наружные волоски наличника древовидно разветвлены.
8 ( 9). Волосок на теле усика длинный, около половины длины по-
следнего, отходит от середины или лишь немного смещен к
основанию усика .... An. (An.) hyrcanus Pall.
Распространен повсеместно, кроме Северного Казахстана (до
50° с. ш.).
9 ( 8). Волосок на теле усика очень короткий, не превышает шири-
ны последнего и отходит недалеко от его основания.
10 (11). Наружные волоски наличника доходят до краев расправлен-
ных боковых лопастей верхней губы или заходят за них,
Стигмальная пластинка меньших размеров (длина 0,8—
0,5 мм, ширина между концами боковых лопастей 0,39—
0,52 мм), слабо пигментирована; срединная часть заднего от-
дела центральной пластинки светлая, ячейки и складки ее
переднего отдела слабо развиты ....
. . . An. (An.) maculipennis sacharovi Favre.
Заселяет южную часть Казахстана, южнее 46—47° с. ш.
11 (10). Наружные волоски наличника не доходят до краев расправ-
ленных боковых лопастей верхней губы. Стигмальная пла-
стинка больших размеров (длина 0,5—0,57 мм, ширина меж-
ду концами боковых лопастей 0,56—0,59 мм), темно пигмен-
тирована ; срединная часть заднего отдела центральной
пластинки, ячейки и складки ее переднего отдела темные, хо-
рошо развиты . . An. (An.) maculipennis messeae Fall.
Заселяет всю территорию Казахстана, севернее 46—48° с.ш.
12 ( 1). Основания внутренних волосков наличника сильно удалены
друг от друга (расстояние между ними больше, чем между
наружным волоском и соответствующим ему внутренним).
13 (14). Наружные волоски наличника тонковетвистые, с числом вет-
вей от 6 до 12—15. Задние волоски наличника 2—4-ветвис-
тые, в виде редкого исключения простые. Центральная пла-
стинка в передней части среднего отдела заметно суживается;
срединная часть центральной пластинки слабо пигменти-
рована в виде резко ограниченного светлого медальона. Пе-
редний отдел центральной пластинки имеет форму наконеч-
ника стрелы: боковые его отростки вытянуты; отростки у
стигмы отсутствуют . . . An. (М.) pulcherrimus Theob.
Встречается лишь в южных районах Казахстана.
14(13). Наружные и задние волоски наличника простые; централь-
ная пластинка широкая, не суживающаяся в передней части
среднего отдела; резко очерченный светлый медальон в сре-
динной части центральной пластинки отсутствует: передний
отдел центральной пластинки не имеет боковых отростков;
38
отростки стигм имеются у большинства особей
. . . . An. (М.) superpictus Grassi.
Встречается лишь в южных районах Казахстана.
Подрод Anopheles Meig.
An. (An.) algeriensis Theob. (карта I)
Совместно c An. claviger и An. plumbeus является одним из ти-
пичных олиготермофилов. От двух этих видов отличается сухолюби-
востью. Распространен в основном в Средиземноморской подобласти
Палеарктики.
Чрезвычайно редкий вид. В последние годы не обнаружен в Ка-
захстане никем из исследователей. Ранее указывался для Казахстана
в работах Э. Р. Геллера (1939), П. А. Вельтищева (1943), А. А. Шта-
кельберга (1937) со ссылкой на сборы А. И. Лисовой.
Различные авторы (Лисова, 1932; Петрищева, 1936; Штакель-
берг, 1937; Беклемишев, Желоховцев, 1945; Мончадский, 1951) мес-
тами выплода считают небольшие, сильно затененные водоемы и род-
никовые заболоченности, нередко с большой примесью солей. В Ка-
захстане местами его развития должны быть последние, поскольку
в опытах Ениколопова (по Беклемишеву и Желоховцеву, 1945) тем-
пература в 20° приводила к гибели основной массы личинок. Кроме
того, эти водоемы не замерзают и в них возможна зимовка личинок.
С этой точки зрения не объяснима находка П. А. Вельтищевым (1943)
личинок An. algeriensis в мочажине у корней тростника в окрестно-
стях поселка Джулек (Кзыл-Ординская область).
В Ташкентской области (Лисова, 1932) предимагинальные фазы
этого вида отлавливались с июля по октябрь в арыке с холодной со-
леной водой родникового происхождения. Личинки держались в за-
росшей прибрежной части водоема совместно с An. claviger и An. hyr-
canus. В конце сентября отмечались лишь личинки первых стадий.
В иного типа водоемах — сильно заросшем озере, подпитываемом
родниковыми водами, личинки встречались совместно с An. claviger,
An. superpictus, An. т. sacharovi и Cx. territans.
В бассейне Сыр-Дарьи этот вид отловлен П. А. Вельтищевым
(1943) в двадцатых числах июля. Причем, одновременно встречались
личинки и имаго. Подобные сведения с более ранними фенологиче-
скими сроками приводит и П. А. Петрищева (1936) для Туркмении.
Личинки и взрослые насекомые отлавливались там лишь в мае, июне
и июле. В остальное время, несмотря на круглогодичные наблюдения,
этот вид не был обнаружен.
39
По данном А. И. Лисовой (1932), в Узбекистане редко залетал в
помещения и почти не нападал на человека. В опытах заражался
лишь возбудителем трехдневной малярии, и то в незначительном ко-
личестве (4,5%). Эпидемиологическое значение невелико.
П. А. Вельтищев (1943) указывает на чрезвычайно высокую
агрессивность самок к человеку. На рогатом скоте и верблюдах, па-
сущихся в этих местах, отмечен не был.
An. (An.) claviger Meig. (карта I)
Широко распространенный олиготермофил. Обычен в Южном
Казахстане; севернее Семипалатинска не обнаружен. Возможен в
Центральном и Северном Казахстане, поскольку в Западной Сибири
этот вид доходит до 60° с. ш. (Беклемишев, Желоховцев, 1945).
Рис. 17. Местообитания личинок Anopheles claviger в горах Каратау.
Вид малочисленный, никогда и никем не отмечавшийся в Ка-
захстане в массе, хотя личинки встречаются в большом количестве.
Развитие приурочено к холодным, чистым водоемам. В Южном
и Юго-Восточном Казахстане таким условиям удовлетворяют всевоз-
можные разливы горных ручьев, родников и небольших рек (рис. 17).
Вода в них отличается слабой минерализацией (до 225 мг!л), слабо-
щелочной реакцией (pH 7,6) и малой окисляемостью (2,6 мг!л). По
данным Е. М. Глаголевой (1944), может встречаться в воде с при-
40
месью солей до 8960 мг)л и слабокислой реакцией (pH 5,8—7,6). Да-
же в жаркие летние дни температура воды в этих водоемах не под-
нимается выше 10—12°. Личинки предпочитают неглубокие (10—
15 см) выходы грунтовых вод в сильно затененных участках. В горах
Заилийского Алатау обилен на высотах 1200—1300 м н. у. м., редок
в лесолуговом поясе (1600—1900 м. н. у. м.) и отсутствует на более
высоких участках. Ниже 800—900 м н. у. м. не отмечен. В Татрах
эти комары встречались лишь на высотах 700—800 м н. у. м. и были
активны в сентябре и октябре (Trpis, 1960).
На сравнительно небольших высотах в Казахстане встречается
в ранневесенние и осенние месяцы. Жаркие летние месяцы, вероят-
но, неблагоприятны для его существования и поэтому обнаружить
личинок в это время в низинных районах практически невозможно.
Как и в Туркмении (Петрищева, 1936), в летнее время сохраняются
лишь немногочисленные имаго, укрывшиеся в густом растительном
покрове. На высотах 1200—1700 м н. у. м., отличающихся известной
прохладой, развитие An. claviger, наоборот, осуществляется лишь в
летнее время и беспрерывно. В выборе мест для откладки яиц руко-
водствуется термостимулами (Bates, 1949), выбирая водоемы с тем-
пературой ниже 20°.
Е. А. Плетневым (1943) активность имаго отмечена в Алма-Атин-
ской области с конца марта, а нами — лишь с начала мая. Осенний
подъем численности наблюдался со второй половины августа.
Необходимыми условиями для существования этого вида яв-
ляются относительно «мягкая» зима и наличие подходящих биото-
пов. Переживание неблагоприятных зимних условий происходит в
фазе личинки.
Наблюдение показало, что активность личинок, зимующих в не-
замерзающих родниковых разливах, сохранялась в течение всего
зимнего сезона. Развитие шло медленно даже у личинок, принесен-
ных в лабораторные условия. При 18—21° оно длилось с начала фев-
раля до конца марта — середины апреля. Такое замедленное развитие
Н. И. Ходукин и А. И. Лисова (1953) объясняют личиночной диапау-
зой, регулируемой длиной дня.
Л. В. Денисовой (1946) установлена возможность автогенного
развития яичников у этого вида в Алма-Атинской области.
Наблюдения по кругу его прокормителей скудны и разрозненны.
Надо полагать, что питание вида происходит на всех теплокровных,
а возможно, и пойкилотермных животных, оказавшихся в месте его
обитания. В горных условиях на человека нападает редко. Домаш-
них животных в местах его обитания мало. Остается предположить,
что он питается на различных видах птиц, которые обильны в этих
местах (кеклики, горлинки и т. п.).
41
В лабораторных условиях хорошо заражается всеми тремя вида-
ми Plasmodium (Беклемишев, 1949), но эпидемиологическое значение
его невелико.
An. (An.) hyrcanus Pall, (карта I)
Основная часть ареала этого вида находится в Ориентальной
области. В СССР в него попадают южные и восточные районы стра-
ны. Распространение этих комаров в таких местах довольно хорошо
совпадает с изотермой июля +20° (Беклемишев, Желоховцев, 1945).
Эврипластичный мезотермофил. Его экологический диапазон
ближе всего подходит к An. т. messeae, от которого An. hyrcanus от-
личается более низкой эврибионтностью. Соответственно этому его се-
верная граница проходит намного южнее, чем у An. т. messeae.
Выяснение северной границы ареала An. hyrcanus являлось, по
мнению Э. К. Мартини (1926), одной из первоочередных задач. В свое
время Э. Р. Геллер (1939) определил северную границу этого вида в
Казахстане по 48 параллели. Анализ последующих сведений позво-
ляет отодвинуть ее на два градуса севернее.
Распространение этого вида в Казахстане в пределах рассмот-
ренного ареала отличается частотой и плотностью встречаемости. Ис-
ключение представляют высокогорные районы, куда он не проникает.
Наивысшей точкой его обнаружения следует, вероятно, считать ок-
рестности г. Алма-Аты (800 м н. у. м.).
Местами выплода служат обычно постоянные и полупостоянные
водоемы с различной степенью солености. Предпочтение отдается
небольшим полупостоянным скоплениям воды, густо заросшим над-
водной растительностью (Phragmites, Typha, Carex) или находящим-
ся в затененных местах. В условиях Таджикистана личинки встре-
чаются в воде с минерализацией 140—730 мг!л и pH 7—7,6 (Глаго-
лева, 1944).
На юге Казахстана часто встречается совместно с An. pulcherri-
mus и An. т. sacharovi, а в более северных районах предпочитает
открытые, хорошо прогреваемые водоемы, нередко сопутствуя Ап. т.
messeae. Рисовые плантации и связанные с ними фильтрационные и
сбросовые разливы — одни из излюбленных мест выплода этого вида.
О возможности развития личинок в поймах горных рек сообщает
Е. И. Балкашина (1939). В Восточном Казахстане местами выпло-
да служат прибрежные разливы озера Зайсан и богатые надводной
и подводной растительностью полупостоянные водоемы поймы
Иртыша.
Первое поколение в большинстве районов Казахстана развивает-
ся в открытых, хорошо прогреваемых водоемах, а последующие — в
42
сильно затененных, с высокой тростниковой растительностью. Дале-
ко от мест дневок не улетает, что говорит о высокой гидрофильности
этого вида. Массовое размножение его связано с хорошо обводненны-
ми влажными участками, что необходимо для существования имаго,
Рис. 18. Местообитания личинок Anopheles hyrcanus, An. maculipennis
sacharovi и An. pulcherrimus в Южном Казахстане (Яны-Курган).
в то время как развитие личинок приурочено к достаточно прогре-
ваемым водоемам. Этими двумя факторами и диктуется заселяемость
An. hyrcanus тех или иных биотопов. Хорошо инсолируемые участки
основной площади Казахстана в значительной мере удовлетворяют
данным требованиям. Отсюда и отмеченное обилие его в этих
районах.
На юге Казахстана первые личинки появляются с начала мая.
Численность их очень мала, что не всегда устанавливается. В массе
личинки встречаются с начала июля и бывают обильны до конца ав-
густа. Затем количество их постепенно уменьшается и единичные
экземпляры существуют до первой декады октября.
Сезонный ход численности взрослых комаров на юге Казахста-
на соответствует фенологии личинок. Перезимовавшие особи этого
43
вида начинают отлавливаться с середины апреля. Численность их бы-
вает незначительной до середины июля, после чего отмечается зна-
чительный подъем, сохраняющийся до второй декады сентября. Ана-
логичную картину, с несколько сдвинутыми к середине сезона край-
ними сроками, показывает этот вид и в бассейне р. Или. В Гурьевской
области (Жук, 1967) численность имаго постепенно возрастает,
показывая три пика: во второй декаде июня, в первой декаде июля
и в конце августа. Эти подъемы, вероятно, соответствуют времени
прохождения циклов и выплоду нового поколения. В зависимости от
климатических условий и широты местности вид может проделывать
от 2 до 4 генераций.
Местами дневок служат густые заросли, расположенные неда-
леко от мест выплода. Отличаясь от An. maculipennis меньшей свето-
чувствительностью, этот вид активно нападает на людей и животных,
попавших в места дневок. В помещения, как правило, на дневки не
залетает. Миграция из естественных биотопов в населенные пункты —
редкое явление, о чем свидетельствует малочисленность экземпля-
ров, отловленных как непосредственно в помещениях, так и в выстав-
ляемую на ночь светоловушку. Местами дневок комаров в условиях
Гурьевской области служат заросли прибрежной растительности по
побережью Каспийского моря, где они почти круглосуточно напа-
дают на людей и животных.
Зимует в фазе имаго. Убежищами во время зимовок служат те
же заросли (тростниковые и древесно-кустарниковые), в которых осу-
ществляются дневки. Об обнаружении зимующих самок в заброшен-
ных камышитовых постройках сообщает В. Н. Беклемишев (1944), а
в складированном саксауле — Е. А. Плетнев (1943). Нами перезимо-
вавшие самки этого вида отлавливались только в природных биото-
пах. Чаще всего ими являлись прикорневые сплетения густых тугай-
ных зарослей и метелки тростника. В двух случаях отмечен вылет из
сложенных на хорошо прогреваемых участках связок тростника. Как
•показали исследования Е. Б. Виноградовой (1960), возникновение и
окончание зимней диапаузы самок регулируется световой продолжи-
тельностью дня.
В северных частях ареала (территория Восточного и Юго-Восточ-
ного Казахстана) неблагоприятные зимйие условия, вероятно, приво-
дят к гибели основной массы зимующих в открытой природе особей.
Сохраняются лишь единичные экземпляры, зимующие в достаточно
укрытых убежищах (заброшенные помещения, прикорневые заросли,
норы животных). Вследствие этого весенняя популяция крайне мало-
численна.
Суточный ход активности характеризуется одновершинной кри-
вой с пиком в вечернее время. Количество нападающих начинает
44
увеличиваться после захода солнца и бывает особенно многочислен-
ным в период наступления сумерек и через час-полтора после этого.
Так, в период массового лёта (начало сентября) в окрестностях Яны-
Кургана через 15 мин после захода солнца в течение 20 мин на об-
наженной руке было отловлено 149 комаров. В местах дневок напа-
дает круглосуточно, вылетая к добыче даже на освещенные участки.
В Казахстане отмечается его повышенная агрессивность по отно-
шению к человеку. Наряду с этим охотно нападает и на различных
животных, находящихся в зоне его активности. Единичные экземп-
ляры отловлены на липучки около гнезд вороны, сорокопута, болот-
ного луня и гнезд-нор береговой ласточки.
Возможный переносчик трехдневной малярии. Плазмодиями
тропической малярии не заражается. В природных условиях перено-
сит возбудителя туляремии (Олсуфьев, Руднев, 1960), который может
в них сохраняться 26 дней. В Зайсанской котловине от этого вида вы-
делен нейротропный вирус группы «А» (Жуматов и др., 1966).
An. (An.) maculipennis Meig.
В Казахстане достоверно известно существование двух подвидов
этого генетически и экологически расчлененного вида.
An. (An.) maculipennis messeae Fall, (карта II)
В отличие от An. т. sacharovi заселяет более северную часть
Казахстана. Если условно отнести сведения о нахождении An. macu-
lipennis к подвиду messeae, то этот подвид в северных районах Кзыл-
Ординской области может встречаться в незначительных количествах
совместно с An. т. sacharovi (Павловский, Петрищева, 1937; Плет-
нев, 1943; Вельтищев, 1943). Ориентировочной границей распростра-
нения его на юге Казахстана служит хребет Каратау, на юго-восто-
ке — Киргизский хребет, на юго-западе — Северное Приаралье. В го-
ры заходит до 1200—2000 м н. у. м., что соответствует в тех местах
поясу горных степей.
По сравнению с An. т. sacharovi — более холодолюбивый и ме-
нее солевыносливый подвид. Местами выплода служат хорошо про-
греваемые, большие по площади мелководья. В южной части ареала
таким требованиям отвечают припойменные разливы различных
рек, родников и оросительных каналов. Значительный выплод проис-
ходит на заболоченных участках, образующихся за счет фильтрации
и ввиду отсутствия коллекторной сети. В северных районах республи-
ки местами выплода в основном служат заводи, старицы и припой-
менные разливы различных рек, придорожные канавы, а также мел-
ководные участки многочисленных озер и заболоченностей (рис. 19).
45
Развитие личинок происходит в водоемах с умеренной минера-
лизацией (743—993 лгг/л на юге, 317 мг}л на севере), повышенной
щелочностью (pH 7,4—7,85) и окисляемостью (7,8—10 мг{л). В про-
цессе усыхания водоемов солевая концентрация и окисляемость мо-
гут увеличиваться. В весеннее время развиваются в незаросших во-
доемах. В дальнейшем предпочитают слабо заросшие стации.
Рис. 19. Местообитания личинок Anopheles maculipennis messeae в севе-
ро-восточных районах Казахстана.
В бассейне р. Или вылет с замовок и нападение на человека от-
мечаются с начала апреля при 19—20°. В более ранний период (сере-
дина марта) почти при такой же дневной температуре воздуха (17—
18°) вылета и нападения не отмечается. Объясняется это более значи-
тельными перепадами суточной температуры в марте, при которых
ночью ртутный столбик опускается ниже нуля. При затяжной, холод-
ной весне вылет задерживается до середины апреля.
В северных районах республики вылетает с зимовок в конце ап-
реля — первой половине мая (Геллер, 1939; Павлова, Павлов, 1963).
Частые похолодания и резкие суточные смены температур в мае за-
держивают прохождение первого гонотрофического цикла в усло-
виях Кустанайской области (Прыгунова, 1966). Подобное, с поправ-
кой на более раннее время, можно отметить и для более южных райо-
нов. Продолжительность гонотрофического цикла варьирует и в
постоянных условиях, что, вероятно, зависит от объема выпитой кро-
46
ви и стадии развития яичников. Так, в лабораторных условиях при
23—29° период переваривания крови и созревания яиц длился от 3
до 5 дней.
В бассейне р. Или первые личинки этого подвида появляются в
начале, а в северных областях — в конце мая; в Восточном Казах-
стане в начале третьей декады мая уже отлавливались личинки
III стадии.
Сезонный ход численности этого подвида в южных и северных
районах не очень различается. В обоих случаях до июля численность
комаров низкая: в южных частях ареала — в результате недостатка
мест выплода, а в северных — за счет более медленного развития
насекомых. В июле горные реки, являющиеся основными поставщи-
ками водных ресурсов для южных мест обитания этого подвида, бы-
вают особенно полноводными. Разливаясь, они создают благоприят-
ные места выплода. Температура воды в местах обитания личинок
доходит днем до 30—32°. При таком режиме период развития в лабо-
раторных условиях заканчивается в 15—18 дней. С середины июля
до конца августа отмечается максимальная численность имаго.
Уменьшение количества нападающих отмечается на севере Казахста-
на со второй половины августа, а в более южных районах — с конца
августа. На севере Казахстана первые ожиревшие самки обнаружи-
ваются на дневках в третьей декаде августа, а в сентябре уже встре-
чаются особи, ушедшие на зимовку (Прыгунова, 1966). В южных час-
тях ареала кровососы летят в зимние укрытия во второй половине
октября.
В зависимости от наличия подходящих биотопов и температур-
ной характеристики местности An. т. messeae в Казахстане проделы-
вает от 2 до 5—6 генераций. Количество их увеличивается с севера
на юг. По данным Э. Р. Геллера (1939), в лесостепной зоне этот под-
вид проделывает 3—4 генерации, в степной и полупустынной — 4—5,
а в пустынной — 5—6.
Суточный ход численности этого подвида в среднем течении
р. Или отличается от обычного двухвершинного. В летнее и жаркое
время утренняя и предвечерняя активность его стушевана. Нападение
отмечается лишь с наступлением сумерек и продолжается 1—1,5 час.
Связано это, вероятно, с тем, что комары очень легко теряют воду
(Виноградская, 1953), и поэтому активность их переносится на более
прохладное время.
Отличается повышенной агрессивностью к животным. Заметно,
что даже в тугайных зарослях в период наибольшей численности на
человека нападают лишь единичные экземпляры, тогда как окру-
жающая растительность содержит много напившихся самок. Оказа-
лось, что в этих местах, как правило, все лето пасутся лошади, круп-
47
ный и мелкий рогатый скот, которые и подвергаются нападению
комаров. Лишь ранней весной, когда животные пасутся вдали от мест
их обитания, самки очень агрессивны по отношению к человеку. Та-
кая зоофильность подвида накладывает отпечаток и на места его дне»
вок. В бассейне р. Или ими служат различные заросли, окружающие
места выплода. При наличии источников питания в естественных ус-
ловиях в населенные пункты залетают редко. Об этом говорят как
непосредственные отловы в помещениях, так и контрольные сборы
насекомых в светоловушку. В этих же природных биотопах осуществ-
ляется зимовка основной части популяции.
Иначе ведут себя комары по мере продвижения на север. Скуд-
ность природных убежищ и суровая зима вынуждают самок залетать
на зимовку в овощехранилища, подвалы, погреба, а по данным
3. Р. Геллера (1939), и в жилые помещения.
Основные переносчики трехдневной малярии в Средней Европе
и Северной Азии (Беклемишев, 1949; Foote and Cook, 1959), комары
этого подвида в Казахстане в пределах своего ареала являются пере-
носчиками Pl. vivax.
An. (An.) maculipennis sacharovi Favre (карта II)
Теплолюбивый, солевыносливый подвид. Ареал его охватывает
южную часть Казахстана, где он доминирует над другими видами
Anopheles. С севера и запада его распространение ограничено хреб-
том Каратау и Аральским морем (Беклемишев, Желоховцев, 1945).
По предгорьям Тянь-Шаня наблюдается продвижение этого вида на
северо-восток до Зайсана, где он редок, а отдельные находки нивели-
руются массовостью An. т. messeae.
Местами выплода являются многочисленные, заросшие тростни-
ком и рогозом озера, заболоченности и временные, создавшиеся за
счет фильтрации водоемы (рис. 18). В таких стациях личинки засе-
ляют прибрежные открытые участки, обильно поросшие погружен-
ной водной растительностью. Микробиотопами их служит поверх-
ностный слой воды в пространствах между переплетающимися нит-
чатыми водорослями (Spirogyrd). Подобные условия создаются на
рисовых, сбросовых и фильтрационных полях, которыми так изоби-
лует его ареал.
Как правило, места выплода представляют хорошо прогреваемые
водоемы с повышенным содержанием солей. Концентрация их может
повышаться по мере усыхания водоема (если он небольшой), и перед
выплодом содержание солей достигает такого количества, что обна-
жающиеся берега покрываются белой коркой. Меньшее количество
48
чинок наблюдается и в пресных, менее прогреваемых водоемах.
В бассейне Сыр-Дарьи они встречаются совместно с личинками
An hyrcanus и особенно An. pulcherrimus.
Первые самки этого подвида, из числа перезимовавших, появ-
ляются в бассейне Сыр-Дарьи в конце марта — начале апреля. Чис-
ленность их, как правило, незначительна, и после первоначального
подъема активности количество нападающих резко уменьшается. В
начале и в середине мая появляются особи нового поколения, чис-
ленность которых сохраняет определенную стабильность до июля,
после чего возрастает. В естественных условиях, где нет рисосеяния
и мелководных, прогреваемых участков, по количеству нападающих
на человека эти комары занимают 6—7 место. В районах с интенсив-
но развитым рисосеянием численность их намного выше, но трудно
поддается учету. В Южном Казахстане отмечается склонность к
зоофилии. Чаще и охотнее самки нападают на крупный рогатый скот,
лошадей и свиней. Поэтому численность их на дневках в животновод-
ческих помещениях бывает очень высокой. Е. Н. Павловский и П. А.
Петрищева (1937) насчитывали до 200 самок на 1 м2 коровника.
Такое обилие подвида обусловлено его высокой плодовитостью,
быстрым сроком развития и возможностью осуществления повторных
гонотрофических циклов. Кладки An. т. sacharovi, при полном на-
сыщении кровью, содержат от 250 до 439 яиц. Гонотрофический цикл
при 22—23° заканчивается в лабораторных условиях за 3—7 дней.
Столь же быстро происходит и последующее развитие. В лабора-
торных условиях при 23—29° период от-яйца до имаго осуществляет-
ся за 13—14 дней, а при 24—32° он заканчивается за 11 дней. Это
дает основание считать, что с весны до"осени возможно развитие 5—8
генераций этого политермофила. Аналогичные сведения по экспери-
ментам в природных условиях приводит И. К. Иванов (1948а).
Повышение численности этого подвида приурочено ко времени
второго летнего паводка Сыр-Дарьи, когда за счет разливов и подня-
тия воды в постоянных водоемах увеличивается площадь прогревае-
мых незатененных участков. Особенно много личинок в прибрежных
разливах постоянных водоемов.
Осеннее уменьшение численности начинается со второй полови-
ны августа. В последующее время до середины октября отмечается
постоянная незначительная миграция ожиревших самок в зимние
Убежишя.
Один из самых массовых переносчиков малярии в Средней Азии
И южных районах Казахстана. Обусловливается это его большой чис-
ленностью, количеством гонотрофических циклов и повторностью
кровососания. В период существования малярии передавал возбуди-
телей трехдневной, четырехдневной и тропической малярии. В опы-
тах процент заразившихся самок составлял 45—62% (Лисова, 1932).
80 49
Подрод Myzomyia Blanch.
An. (M.) pulcherrimus Theob. (карта III)
Теплолюбивый стенобионтный вид. Распространен на большей час-
ти территории Средней и Юго-Западной Азии.
Существование его возможно лишь при условии продолжитель-
ного и знойного лета и не слишком холодной зимы (Беклемишев, Же-
лоховцев, 1945). Соответственно этому в его ареал попадают лишь
самые южные районы Казахстана. Распространение на север ограни-
чено хребтом Каратау, а единственную находку в Джамбулской об-
ласти (Геллер, 1939) следует, вероятно, отнести за счет «пульсации»
ареала. На запад не идет дальше Кзыл-Орды. Обнаружение
П. А. Вельтищевым (1943) на станции Казалинск двух напитавших-
ся кровью самок следует, вероятно, считать случаем завоза.
В пойме Сыр-Дарьи местами выплода служат многочисленные
постоянные водоемы, озера, старицы, затоки, фильтрационные разли-
вы, плавни и т. п. В конце лета личинки встречаются совместно с
An. hyrcanus, а ранее — с An. т. sacharovi. Развитие отмечено в
пресных и солоноватых водоемах. Большую минерализацию не вы-
носят. Личинки предпочитают незатененные надводными растения-
ми участки водной площади с богатой погруженной растительностью
(рис. 18). В хлопководческих районах Чимкентской области Э. Р. Гел-
лером (1939) и Е. А. Плетневым (1943) развитие отмечено в водоемах
оросительных систем.
Для An. pulcherrimus, как и для многих других теплолюбивых
видов вблизи от северной границы их ареала, свойственна весенняя
задержка развития. Этим и определяется его сезонный ход числен-
ности.
На юге Казахстана первая генерация вследствие малочисленно-
сти обычно не обнаруживается. В заметном количестве личинки по-
являются во второй декаде июля, а в конце этого месяца уже проис-
ходит выплод имаго. В июле численность самок прогрессивно воз-
растает, достигая максимума в конце августа — начале сентября.
Единичные самки еще встречаются до конца октября. Температура
15—16° является пределом активности этого вида в осеннее время.
В более южных районах бассейна Сыр-Дарьи она на один градус вы-
ше (Беклемишев, Митрофанова, 1936).
Суточная активность характеризуется одновершинной кривой,
пик которой приходится в теплые дни на сумеречное время, а в про-
хладные — на середину дня. В. Н. Беклемишев и Ю. Г. Митрофанова
(1936) для станции Сыр-Дарьинской отмечают активность нападения
в предрассветное время. Залетает и активна нападает в помещениях.
50
По данным П. А. Петрищевой (1936), А. А. Штакельберга (1937)
д С. Мончадского (1951), зимуют личинки. Причем от суровости
зимы зависит их смертность, чем обусловлена неравномерность чис-
енности личинок в отдельные годы. В Казахстане зимующие личинки
отмечены лишь Е. А. Плетневым (1943) и Е. А. Балкашиной (1939),
Высшая точка их обнаружения 1300 м н. у. м., хотя находки в гор-
ных и предгорных районах не характерны для этого вида. Являясь
теплолюбивыми ксерофилами, эти комары заселяют, по нащим наб-
людениям, пустынные, равнинные районы, где абиотические факторы
внешней среды соответствуют их экологическому диапазону.
Комары этого вида — активные кровососы. Нападают как на лю-
ей, так и на различный скот, причем на последний более охотно. В
соответствии с этим и при выборе дневок имаго предпочитают скот-
ные постройки жилым.
An. (М.) superpictus Grassi (карта Ш)
Южный теплолюбивый вид. На юге Казахстана проходит север-
ная граница его ареала. Единичные экземпляры встречаются в пойме
Сыр-Дарьи, южнее Кзыл-Орды. По мнению В. Н. Беклемишева и
А. Н. Желоховцева (1945), на северо-западе не заходит за Кзыл-Орду,
а на северо-востоке не переходит хребет Каратау. На более широкое
распространение An. superpictus указывает Е. А. Плетнев (1943). Он
обнаруживал его в окрестностях Алма-Аты и в бассейне р. Или., По-
следующими исследователями эти находки подтверждены.не были.-
Сведения о местах выплода этого вида в Казахстане довольно
противоречивы. Е. И. Балкашина (1939) указывает на обилие его в
пойме Сыр-Дарьи, в то время как конкретные места обнаружения
приурочены к предгорным районам. Е. А. Плетнев (1943).находил
личинок в горных источниках с жесткой родниковой водой., и в мик-
роводоемах, образованных затоками горных рек.
Сезонность An. superpictus характерна как для типичного поли-
термофила. В Казахстане личинки обнаруживаются в конце июля.
Максимальной численности вид достигает в начале сентября. Послед-
ние личинки отмечались в середине, а в теплую осень — в конце-ок-
тября (Балкашина, 1939). В условиях Таджикистану (Занина, 19&7)
вылет первой генерации происходит в июне, что, вероятно* характер-
но и для Казахстана, но не прослежено. < п >
Вид в Казахстане чрезвычайно редкий, хотя в некоторые :год^т
при благоприятных условиях происходил- довольно массовый вы-
плод, что было связано с возникновением в. Чимкентском районе
Устойчивого малярийного очага (Балкашина*: Мекленбурцева^; 1'942).
Малочисленность его в обычные годы в значительной степени равй-
сит от обилия осадков. Личинкам это грозит смывом в неподходящие
для развития участки, а на имаго, как на типичных ксерофилов, не-
благоприятно действует возникающая при этом чрезвычайная влаж-
ность (Беклемишев, Желоховцев, 1945).
В отличие от другого политермофила — An. pulcherrimus — зи-
мует во взрослом состоянии. Местами зимовок служат помещения для
скота, птичники и жилые постройки, хотя последние и в меньшей
степени (Плетнев, 1943). В. Б. Дубининым (1946) в южных районах
Казахстана отмечается зимовка самок в норах (влажного типа) раз-
личных млекопитающих.
В период распространения малярии в СССР был очень опасен в
эпидемиологическом отношении. Является вторым по значимости,
после A. maculipennis, переносчиком малярийных плазмодиев, в том
числе Pl. falciparum.
Род URANOTAENIA L.-ARRIB.
Ur. unguiculata Edw. (карта IV)
Средиземноморский представитель тропической группы видов,
широко распространенный в Средней Азии и менее — в других юж-
ных районах СССР. В Казахстане редок; встречается южнее 48° с.ш.
Местами выплода служат прибрежные участки постоянных во-
доемов, питаемые грунтовыми водами (рис. 20). Состав воды харак-
теризуется повышенной минерализацией (1000—1100 мг/л), слабо-
щелочной реакцией (pH 7,6) и значительной примесью сернистых
соединений (286 мг/л), окрашивающих находящиеся в воде предметы
в сизовато-белый цвет. Запах воды сероводородный, привкус болот-
ный. Личинки заселяют богатые планктоном литоральные незарос-
шие участки таких водоемов. Е. А. Плетнев (1943) указывает на воз-
можность развития в водоемах с подпочвенной водой родникового
происхождения. Встречается совместно с Cs. alaskaensis, Сх. pipiens
и Сх. modestus.
Взрослые личинки обнаруживаются в бассейне Сыр-Дарьи со
второй, а на Или — с третьей декады августа. Предимагинальные фа-
зы существуют до конца сентября. За этот период отмечается два
пика увеличения численности личинок: в начале третьей декады ав-
густа и в начале второй декады сентября, что говорит о развитии
двух генераций. В другое время года личинок обнаружить не удава-
лось, несмотря на многолетние полисезонные исследования известных
мест выплода. Аналогичные сведения, но со сдвинутыми к зимнему
времени сроками, приводит и Киркпатрик (Kirkpatrik, 1925) для
Каира.
52
Подобно другим политермофилам, имеющим на территории Ка-
захстана северную границу своего ареала, показывает весеннюю за-
держку развития. Первые имаго в бассейнах рек Или и Сыр-Дарьи
начинают отлавливаться во время роения, кошения растительности
Рис. 20. Заросший постоянный водоем в бассейне р. Или — местообита-
ния личинок Uranolaenia unguiculata.
сачком и в светоловушку со второй половины августа. Характерно,
что даже при полисезонных наблюдениях, охватывающих крайние
сроки активности комаров в бассейнах рек Сыр-Дарьи (1964) и Или
(1965), отловить зимующих или вышедших из зимовки самок не
случалось. Подобные результаты были получены и в Туркмении, где
личинки, куколки и имаго встречались только с мая по октябрь (Пет-
рищева, 1936).
Местами дневок служат приозерные заросли. П. А. Петрищева
(1962), помимо этого, обнаруживала днюющих самок в колодцах,
шахтах, штольнях, дуплах деревьев и в гнездах птиц.
Вид, вероятно, орнитофильный. На человека и домашних живот-
ных не нападает. По мнению П. А. Петрищевой (1936), питаются
кровью птиц. В сыр-дарьинских плавнях личинки обнаружены среди
безбрежных, неглубоких (40—50 см) разливов около заросшего трост-
ником островка, заселенного колонией пеликанов.
63
Род CULISETA FELT
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CULISETA FELT
ПО ВЗРОСЛЫМ НАСЕКОМЫМ
(По А. А. Штакельбергу, с небольшими сокращениями
и дополнениями)
1 ( 2). Среднеспинка с резко выраженными продольными белыми
полосами, образующими подобие лировидного рисунка; бед-
ра и голени спереди с резко выраженными белыми пятнами.
Костальная жилка спереди на большей части покрыта белы-
ми чешуйками; щупики самцов короче хоботка (подрод А1-
lotheobaldia Brol.) ......................
.............................Cs. (All.) longiareolata Macq.
В Казахстане встречается повсеместно, но мозаично.
2 ( 1). Среднеспинка без белых продольных полос, иногда с более
или менее резко выраженными белыми пятнами; если же с
полосками, то последние не образуют лировидного рисунка;
бедра и голени спереди в одноцветных темно-бурых чешуйках
или же с более или менее значительным числом вкраплен-
ных чешуек. Костальная жилка покрыта темными чешуйка-
ми ; щупики самцов длиннее хоботка.
3 ( 8). Радио-медиальная и медио-кубитальная жилки расположены
на одной прямой (подрод Culiseta Felt).
4 ( 7). Лапки с более или менее широкими белыми колечками.
5 ( 6). Первый членик задних лапок посередине без светлого колеч-
ка ; бедра без предвершинного светлого ’ кольца. Крылья с
темными пятнами; лапки с белыми колечками
.......................................Cs. (Cs.) alaskaensis Ludl.
Повсеместно, включая предгорные, а на юге и юго-востоке и
горные районы.
6 ( 5). Первый членик задних лапок посередине со светлым колеч-
’*• ком; бедра с явственно развитым белым кольцом в вершин-
ной трети...............................Cs. (Cs.) annulata Schrk.
В пределах пустынной, полупустынной и степной зон.
7 ( 4). Лапки одноцветные, без светлых колечек.................
. . Cs. (Cs.) bergrothi Edw.
Северные и восточные районы Казахстана.
,.8.( 3). Медио-кубитальная поперечная жилка расположена явствен-
но ближе к основанию крыла, нежели радио-медиальная;
крылья без темных пятен или с едва намечающимися темны-
ми пятнами (подрод Culicella Felt).
£4
9 (Ю). Длина первого членика передних лапок самца равна или ме-
нее длины остальных члеников той же лапки, вместе взятых;
длина передней лапки самца менее длины голени той же па-
ры . . Cs. (Сс.) silvestris Sching.
10 ( 9). Длина первого членика передней лапки самцов явственно
превосходит длину остальных члеников той же лапки, вмес-
те взятых; длина передней лапки более длины голени той же
ноги. Щупики самцов длинные, основание вершинного чле-
ника щупиков лежит, как правило, явственно дистальнее
основания сосательных лопастей хоботка; покров третьего и
четвертого члеников щупиков самцов на всем протяжении
покрыт темно-бурыми чешуйками, как правило, лишь с не-
значительными включениями белых чешуек
.......................... . . Cs. (Сс.) morsitans Theob.
Восточный Казахстан, возможен в Северном.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CULISETA FELT
ПО ГИПОПИГИЯМ САМЦОВ
(По А. А. Штакелъбергу, с сокращениями и изменениями)
1 ( 2). Девятый тергит по бокам с длинными выростами: фаллозо-
ма весьма крупная (подрод Allotheobaldia Brol.)
..........................Cs. (All.) longiareolata Macq.
В Казахстане встречается повсеместно, но мозаично.
2(1). Девятый тергит простой: фаллозома умеренного размера
(подроды Culiseta Felt, и Culicella Felt)
3 ( 4). Коксит перед вершиной с внутренней стороны без сильно вы-
пуклой бородавки, но с небольшим участком густо сидящих
волосков...............................Cs. (Cs.) alaskaensis Ludl.
Повсеместно, включая предгорные, а на юге и юго-востоке и
горные районы^
4 ( 3). Коксит без предвершинной бородавки и без резко ограничен-
ного участка густо сидящих волосков.
5 ( 8). Базальная бородавка коксита, как правило, с одной-двумя
длинными и крепкими щетинками.
6 ( 7). Базальная бородавка большая, ширина ее равна или больше
высоты (лапки с широкими белыми кольцами)
. . . . . . Cs. (Cs.) annulata Schrk.
В пределах пустынной, полупустынной и степной зон Казах-
стана.
7 ( 6). Базальная бородавка небольшая, чуть выступающая (лапки
без светлых колечек) .... Cs. (Cs.) bergrothi Edw.
Северные и восточные районы Казахстана.
55
8 ( 5). Базальная бородавка коксита, как правило, с тремя-восемью
крепкими и длинными щетинками.
9 (10). Базальная бородавка коксита с тремя-четырьмя крепкими и
длинными щетинками . . . Cs. (Сс.) morsitans Theob.
Восточный Казахстан, возможен в Северном.
10 ( 9). Базальная бородавка коксита с пятью-восемью, редко с че-
тырьмя крепкими, слабо изогнутыми щетинками
....................................... Cs. (Сс.) Silvestris Sching.
Северные районы Казахстана.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ЛИЧИНОК IV СТАДИИ
РОДА CULISETA FELT
(По А. С. Мончадскому, с небольшими сокращениями
и дополнениями)
1 ( 8). Сифон относительно короткий и толстый (индекс ниже 4).
Главные стволы трахей широкой лентовидной формы. Усики
короче головы, со слабо развитым пучком.
2 ( 3). Гребень из небольшого числа хорошо развитых зубцов по
всей длине сифона. Тело усиков лишено шипиков; пучок на
усике расположен у середины, ближе к дистальному концу,
короткий, не более чем из трех ветвей (подрод Allotheobaldia
Brol.)..............................Cs. (All.) longiareolata Macq.
В Казахстане встречается повсеместно, но мозаично.
3 ( 2). Гребень сифона из зубцов, сидящих у основания, в дисталь-
ном направлении продолжается в виде ряда длинных волос-
ков. Усик покрыт редкими шипиками: пучок на нем распо-
ложен у середины, ближе к основанию, короткий, состоит
более чем из пяти ветвей (подрод Culiseta Felt).
4 (5). Усики в длину достигают половины длины головы. Средние
лобные волоски из шести-восьми ветвей. Щетка в среднем бо-
лее чем из 60 чешуек. Гребень сифона занимает 2/з его дли-
ны; зубцы в нем численно немного преобладают над волоска-
ми. Впереди плавника три-четыре пучка, из которых два
своими основаниями прободают кольцо седла
..........................................Cs. (Cs.) bergrothi Edw.
Северные и восточные районы Казахстана.
5 ( 4). Усики менее половины длины волосков. Средние лобные во-
лоски не более чем из трех ветвей. Щетка в среднем менее
чем из 50 чешуек.
6 ( 7). Сифон относительно более короткий и широкий, слабо сужен-
56
ный к вершине, отношение его длины к ширине основания
равно 2,4:3,0, а к ширине у вершины — 3,4:3,9. Чешуйки
щетки узкие, удлиненные, с параллельными боковыми края-
ми, без заметного расширения на конце основания
..............................Cs. (Cs.) alaskaensis Ludl.
Повсеместно, включая предгорные, а на юге и юго-востоке и
горные районы.
7 ( 6). Сифон относительно более длинный и тонкий, значительно
суженный к вершине, отношение его длины к ширине осно-
вания 3:4, а к ширине у вершины — 5:6. Чешуйки щетки
слегка сужены, с явственно расширенным основанием
................................ Cs. (Cs.) annulate, Schrk.
В пределах пустынной, полупустынной и степной зон Казах-
стана.
8 ( 1). Сифон относительно длинный и тонкий (индекс выше 5).
Главные стволы трахей тонкие, округлой формы. Усики длин-
нее головы, с сильно развитым пучком, расположенным за
серединой (подрод Culicella Felt).
9 (10). Внутренние лобные волоски из пяти-девяти ветвей. Чешуйки
заднего края щетки имеют на своем теле продольную, заост-
ренную к концу, более темную полосу. Волосок на задних
клапанах стигмальной пластинки крючкообразно изогнут
только на самом конце. Отношение длины пучка на сифоне к
длине последнего около 0,28. Жабры в 1,5—2 раза длиннее
седла......................... Cs. (Сс.) silvestris Sching.
Северные районы Казахстана.
10 ( 9). Внутренние лобные волоски из двух-трех ветвей. Дистальные
чешуйки щетки без темной полосы на теле. Волосок задних
клапанов стигмальной пластинки сильно крючкообразно
изогнут по всей длине. Отношение длины сифонального пуч-
ка к длине сифона 0,32 и выше. Дистальные зубцы гребня
доходят до четверти длины сифона от основания. Жабры до-
стигают половины длины последнего членика брюшка . .
..................................Cs. (Сс.) morsitans Theob.
Восточный Казахстан, возможен в Северном.
Подрод Allotheobaldia Brol.
Cs. (All.) longiareolata Macq. (карта IV)
Южный мезотермофильный вид. Значительная часть его ареала
лежит в Эфиопской области (Hopkins, 1952). В Палеарктике распрост-
ранен в основном в Средиземноморской подобласти. В Казахстане
встречается повсеместно, но чаще южнее 50° с. ш.
57
Вид, в значительной степени перешедший к синантропному об-
разу жизни. В Казахстане многими исследователями (Балкашина,
1939; Плетнев, 1943; Боженко, 1948) отмечалось развитие его в боч-
ках с водой, в различных ямах, копанках и загрязненных водоемах,
которые еще имеются в населенных пунктах. Подобные же водое-
мы, бедные или совсем лишенные травянистой растительности, слу-
жат местами развития этого вида и в природных условиях. Обычно
личинки обитают в слабо минерализованных (207 мг/л) водоемах с
довольно высокими кислотными показателями (pH 5,05) и большой
окисляемостью воды (23 мг!л). Развитие отмечается до периода за-
растания водоема элодеидами, после чего заселяются другие незарос-
шие участки.
В пустынных районах единичные личинки обнаруживались в
слабощелочной воде (pH 7,6) со значительной примесью сернистых
соединений (286 мг(л) и повышенной минерализацией (1039 мг)л).
Предпочитает пресную воду, но может развиваться в воде с соле-
ностью 1% и выше (Мончадский, 1951). Развитие личинок, достав-
ленных с высоты 2600 м н. у. м. в лабораторию (830 м н. у. м.), пре-
кращалось в воде из того же водоема, с той же растительностью
вследствие их гибели. Отрицательную роль при этом, видимо, играет
резкий переход от умеренных (15—18°) к высоким температурам
(27—33°), так как, по наблюдениям А. В. Маслова (1961а), оптималь-
ные условия выживаемости личинок находятся в пределах 30°. В лет-
нее время развитие отмечено совместно с Сх. pipiens, Сх. hortensis и
Сх. martinii, а осенью — лишь с Сх. pipiens.
На юге и юго-востоке Казахстана склонен к заселению горных
и предгорных биотопов (рис. 21), а в более северных районах — рав-
нинных. Отмеченное явление не случайно. Подобно Cs. alaskaensis у
этого вида в пустынных районах отмечается летняя депрессия в са-
мый жаркий период сезона (июнь — август). Личинки и выплажи-
вающиеся имаго обнаруживаются лишь в апреле-мае и в августе-
сентябре. Причем, у Cs. longiareolata депрессия выражена резче. Если
Cs. alaskaensis в крайние сроки теплого сезона (май и конец августа)
дает массовый выплод, то личинки Cs. longiareolata встречаются в
таких условиях лишь единично. Это говорит о более высокой олиго-
термности вида по сравнению с Cs. alaskaensis, что также подтверж-
дается вертикальным распределением этих видов в горных условиях.
Cs. alaskaensis поднимается в горы до нижних участков лесо-
лугового пояса (1600—1770 м н. у. м.), a Cs. longiareolata обнаружен
в субальпийском поясе высокогорных степей (2600—2800 м
н. у .м.).
О более высокой олиготермности этого вида говорит и то, что
личинки его в поясе горных степей в массе отлавливались в конце
58
сентября, в то время как личинок Cs. alaskaensis уже не было. Сезон-
ность Cs. longiareolata в Египте также свидетельствует о его холодо-
пюбивости. Там он обилен с декабря до июня (Kirkpatrik, 1925).
Рис. 21. Местообитания личинок Culiseta longiareolata и Cs. alaskaensis в го-
рах Заилийского Алатау.
Приведенные факты не совсем увязываются со сведениями об
экологии этих видов. По литературным данным (Маслов, 1961а), бо-
лее холодолюбивым должен быть Cs. alaskaensis, что объясняется,
вероятно, диапазоном экологической пластичности этих видов, кото-
рый у Cs. alaskaensis в- условиях Казахстана несомненно ниже.
В пустынных предгорных районах единичные личинки обнару-
живаются лишь в начале мая и конце августа, тогда как в поясе гор-
ных степей (1200—1300 м н. у. м.) в массе они отлавливаются в кон-
це сентября, а в субальпийских степях — в первой половине июля.
Во всех рассмотренных случаях максимальная температура во-
ды в местах развития не превышала 18—22°. Плотность личинок бы-
ла высокой лишь в осеннее время — более 100 экземпляров на 1 м2
водной поверхности. В этот период личинки заселяли заполненные
грунтовой водой небольшие ямки (рис. 21). В остальных случаях
59
встречались в ограниченном количестве. В Семипалатинской области
личинки обнаруживались в течение всего лета (Боженко, 1948). Ос-
новную часть личиночного питания составляет, вероятно, перифитон.
Отсутствие его увеличивает смертность личинок в 4 раза (Маслов,
19616).
Выплаживаясь в больших количествах около жилья человека,
на людей обычно не нападает. Питается, вероятно, кровью различных
птиц. П. А. Петрищевой (1936) 97% собранных экземпляров было от-
ловлено в курятниках.
Зимует в фазе имаго. Убежищами служат жилые и нежилые по-
мещения, но чаще подвалы и погреба (Плетнев, 1943). В природных
условиях зимующие имаго обнаруживались в растительных зарос-
лях, в пещерах и норах различных животных (Петрищева, 1936).
Подрод Culiseta Felt.
Cs. (Cs.) alaskaensis Ludl. (к a p f a IV)
Циркумбореальный представитель голарктической фауны. Ти-
пичен для полосы хвойных лесов (Сазонова, 1963). В Казахстане
встречается как в зоне хвойных лесов, так и во всех других древес-
ных ассоциациях. Соответственно этому тяготеет к северным, восточ-
ным и возвышенным предгорным участкам Центрального, Южного и
Юго-Восточного Казахстана. По? поймам рек может далеко проникать
на равнинные степные и пустынные участки. В горы поднимается до<
пояса горных лесов включительно (1500—2300 м н. у. м.). Массово
встречается в горных степях и лесостепях (500—1500 м н. у. м.).
Места выплода разнообразны. Предпочитает небольшие мелко-|
водные водоемы (рис. 22) типа фильтрационных разливов, копанок и
различных ям с грунтовой водой. Эти водоемы характеризуются от-
сутствием погруженной растительности, слабощелочной реакцией
(pH 7,6—8), повышенной минерализацией и значительной примесью
сернистых соединений (286—1242 мг!л). Личинки и погруженные я
такую воду предметы приобретают сизовато-белую опалесценцию.
Э. Р. Геллером (1939) обнаружен в бочке с водой. Часто встречается
совместно с Cx. pipiens, Cx. modesties и U. unguiculata. В горных райо-
нах обычен в разливах ручьев и выходах грунтовых вод, образующие
заболоченные участки (рис. 21). Вода в них отличается чистотой
прозрачностью, слабой минерализацией и нейтральной реакцией
(pH 7,2). Развитие отмечено совместно с Cx. pipiens и An. claviger. I
период максимальной численности — до 300 личинок на 1 ле2 вод
ной площади. Оптимум развития личинок находится в предела!
13—25°.
60
в бассейне среднего течения Или фенология этого вида своеоб-
разна. Ранней весной он появляется чуть раньше Сх. modestus и Ап.
messeae (конец марта — начало апреля). В это время активно на-
падает на человека и животных. Затем, на протяжении примерно ме-
Рис. 22. Заросший постоянный водоем в бассейне р. Или — местообита-
ния личинок Culiseta alaskaensis и Cs. longiareolata.
сяца (с середины апреля по вторую декаду мая), отмечается числен-
ное преобладание личинок этого вида над другими. Выплод — во вто-
рой декаде мая. Несмотря на высокую численность, нападения
выплодившихся имаго на человека или животных не наблюдается.
С конца мая до конца августа ни имаго, ни их личинки не отмечают-
ся. Не отлавливаются эти насекомые в указанный период и в свето-
ловушку.
В конце лета (вторая половина августа) первыми появляются
личинки, а позже — имаго, которые, вероятно, и уходят на зимовку.
Аналогичную фенологию этого вида приводит В. П. Боженко (1948)
Для Семипалатинской и Восточно-Казахстанской областей. Такая
Двухвершинность сезонного хода численности говорит об олиготерм-
ности этого вида. Вполне возможно, что у него имеется летняя деп-
61
рессия имаго, вызванная действием высоких суточных температур.
Подтверждением этому служит одновершинность его развития в гор-
ных условиях, где личинки встречаются в течение всего летнего се-
зона до первых чисел сентября включительно. Объясняется это нали-
чием в прохладных горных участках оптимальных условий для раз-
вития личинок и существования имаго.
В предгорных районах юго-востока Казахстана (600—800 м
н. у. м.) отмечается переход к синантропному образу жизни. В нача-
ле марта отлавливается в помещениях, подвалах, канализационных
системах и других укрытиях. Залет в эти помещения осенью проис-
ходит в течение сентября и в начале октября.
Местами дневок служат заросли тростника и кустарника, нахо-
дящиеся поблизости от мест выплода. В горных биотопах
встречается в ельниках, ущельях, а в пустынях — в тугайном лесу.
Зимует там же, иногда совместно с An. maculipennis, в фазе
имаго.
Кроме отмеченных ранневесенних нападений на человека и жи-
вотных агрессивности в дальнейшем не проявляет. В другое время,
вероятно, питается на птицах и других обитателях зарослей около
водоемов.
В бассейнах Или и Сыр-Дарьи массовый выплод отмечается в
местах обитания лысух, куликов и пастушковых.
Cs. (Cs.) annulata Schrk. (карта V)
Основная часть ареала этого вида лежит в Средиземноморской!
подобласти Палеарктики. Проникает в пограничные районы Евро-
пейско-Сибирской подобласти, доходя до Дании, Норвегии, Финлян-
дии (Штакельберг, 1937; Мончадский, 1951; Маслов, 1967).
Распространение в Казахстане ограничено в основном южными'
районами, где представлен подвидом Cs. annulata subochrea Edw. Об-
наружение в Семипалатинской области отодвигает северную границу
его ареала до 50° с. ш.
Вид чрезвычайно редкий, никогда и никем не обнаруживаемый
в Казахстане массово. Соответственно скудны сведения по его эколо-
гии. Е. Н. Павловский и П. А. Петрищева (1937) отмечали развитие
личинок в г. Туркестане в июле в сильно загрязненных, зловонных
лужах, содержащих в себе сточные воды со скотных дворов, а
Э. Р. Геллер (1939) обнаруживал личинок в бочке с водой. ’
В Туркмении личинки развиваются в мелких заболоченностях^
образующихся за счет паводков и фильтраций. Предпочитают водоем
мы, бедные растительностью, но богатые гуминовыми основаниями
(Петрищева, 1936). Развитие происходит очень быстро. Приурочен^
62
оно к весеннему и начальнелетнему периодам (с февраля до июня),
последующее время отмечаются лишь единичные личинки и ку-
колки. Как и предыдущему виду, С. annulate, subochrea, вероятно,
свойственно летнее угнетение, хотя и менее выраженное. В бассейне
Сыр-Дарьи единичные экземпляры отлавливались в конце мая — на-
чале июня в небольших, слабо заросших водоемах.
Местами дневок в Туркмении являются пещеры, щели, провалы
могил, растительные заросли и норы различных животных (Петрище-
ва, 1962). Во многих из этих укрытий и происходит зимовка вида,
обычно в фазе имаго, в то время как в некоторых водоемах всю зиму
развиваются личинки. О зимовке этих комаров при умеренном и юж-
ном климате в фазе личинки сообщают Везенберг-Лунд (Wesenberg-
Lund, 1920—1921) и А. Величкевич (1936). Нам, несмотря на край-
ние фенологические сроки исследований, ни разу не удалось обнару-
жить зимующих личинок, поскольку водоемы, где происходит их
развитие, полностью промерзают. Это дает нам основание считать,
что в резко континентальных условиях юга и юго-востока Казахста-
на комары зимуют в фазе имаго. Об этом же говорят наблюдения
Трпиша (Trpis, 1965) в Чехословакии.
Cs. (Cs.) bergrothi Edw. (карта V)
Палеарктический бореальный вид. Распространен в основном в
лесных районах, находящихся севернее 50° с. ш. В аналогичных
участках Южного Алтая обнаружен и В. П. Боженко (1948), у кото-
рого проходил под прежним наименованием — Cs. borealis. Просмотр
материала И. Г. Прыгуновой показал, что имеющиеся у нее имаго,
условно относимые к Cs. ochroptera, более подходят к Cs. bergrothi.
Обнаружены они на севере Кустанайской области.
Сведения по экологии очень скудны. Личинки встречаются в
различных стоячих водоемах: от тундровых заболоченностей до лес-
ных луж и канавок вблизи от жилья человека (Мончадский, 1951).
В лесном поясе Южного Алтая отлавливались в разобщенных плёсах
высыхающего горного ручья на высоте 1600—1800 м н. у. м. Разви-
тие отмечено в чистой прозрачной воде с каменистым дном. Водоемы
защищены от солнечных лучей окружающей кустарниковой расти-
тельностью. Плотность заселения биотопов личинками незначитель-
ная (5—7 на 1 м2). При дневной температуре 28° вода в небольших
водоемах прогревалась до 20—22°. Это совпадает с температурным
оптимумом данного вида (10—22°), выведенного А. В. Масловым
(1961а) для разных участков Дальнего Востока. При таком режиме
Развитие Cs. bergrothi заканчивается в 10—18 дней. Это обеспечи-
вает возможность существования в южных частях ареала, какими
63
являются рассматриваемые районы, двух-трех генераций. Нами в се-
редине июня отловлены, по всей видимости, личинки первого поко-
ления, а В. П. Боженко (1948) в июле-августе обнаруживал, вероят-
но, личинок второй-третьей генерации.
Эфемерность личиночных биотопов и их разреженность под-
тверждают мнение А. С. Мончадского (1961) о том, что вид зимует
в фазе имаго.
Исследования А. В. Маслова (19616) показали чрезвычайно вы-
сокую эвритрофность личинок этого вида. При наличии в водоеме
планктона они ведут себя как фильтраторы, а при перифитонном
источнике питания — как скарификаторы. Экологические условия
Восточного Казахстана предрасполагают ко второму типу пи-
тания.
Сведения по сезонной и суточной активности в Казахстане от-
сутствуют. Нами отмечено нападение лишь двух самок среди хвой-
но-лиственного леса в дневное время в середине, июня. Вполне воз-
можно, что нападала самка новой генерации, поскольку сбор прово-
дился на высоте примерно 1400 м н. у. м. (с. Масляха).
Подрод Culicella Felt
Cs. (Сс.) morsitans Theob., Cs. (Cc.) silvestris
Sching. * (к a p т a V)
Чрезвычайно редкие в Казахстане виды. Обнаружены в Восточ-
ном (С. morsitans) и Северном (С. silvestris) Казахстане. Установлены
по небольшому числу имаго. Достоверность существования этих мор-
фологически близких видов в пределах Северного и Восточного Ка-
захстана требует дальнейшего изучения. Для точной идентификации;
нужны серии из личинок, самцов и самок, чего, к сожалению, пока
не имеется. Ареалы Cs. morsitans и Cs. silvestris проходят близко от
Северного Казахстана (Пецкий, 1952 а, б).
Имаго Cs. silvestris отловлены в зарослях травы среди березовых
колков (Прыгунова, 1966). Нападений на человека не отмечено. Мес-
тами обитания личинок Cs. morsitans Матесон (Matheson, 1944) счи-
тает заполненные водой лесные ямы, но чаще лесные разливы. Био^
топы заселяет не регулярно. На человека не нападает.
♦ До недавнего времени Cs. silvestris проходил под диагнозом Cs. ochroptera-
Исследования А. В. Маслова (1967) показали, что это один и тот же вид, который
должен называться по раннему описанию.
64
Род MANSONIA BLANCH.
Подрод Coquillettidia Dyar
M. (C.) richiardii Fic. (карта VI)
Широко распространенный в Европе и Средней Азии вид. Вся
территория Казахстана, кроме горных районов, попадает в его ареал.
Несмотря на столь широкое распространение, встречается мозаично.
Рис. 23. Слабопроточная речка на юго-востоке Казахстана — местооби-
тания личинок Mansonia richiardii.
Объясняется это, вероятно, специфичностью личиночного развития,
которое осуществляется только в постоянных и полупостоянных во-
доемах с богатой погруженной высшей растительностью, из которой
личинки получают воздух для дыхания (рис. 23). Выше 600—800 м
н. у. м. не обнаружен.
Сроки появления первых особей в различных климатических зо-
нах Казахстана различны. В бассейне Сыр-Дарьи первые особи по-
являются в начале мая, в бассейне Или —в конце мая, а в Гурьев-
5—80
65
ской (Амиргазиев, 1966) и Кустанайской (Прыгунова, 1966) облас-
тях— с конца июня, что соответствует выплоду первой генерации.
Дальнейший ход численности также различен в южных и се-
верных районах Казахстана. Для бассейнов рек Сыр-Дарьи и Или он
характеризуется затяжкой массового развития до конца июля. Свя-
зано это с недостатком мест выплода, площадь которых увеличивает-
ся после летних паводков рек, наблюдаемых в начале и середине
июля. В результате разливов образуется масса мелководных, хорошо
прогреваемых водоемов с богатой рдестовой растительностью. В них
в конце июля и происходит массовый выплод второго летнего поко-
ления. В августе численность комаров нарастает за счет выплода
третьего поколения.
С наступлением первых периодов ночного похолодания (конец
августа — начало сентября) численность постепенно снижается, и в
конце сентября имаго не встречаются. Подобную сезонность М. richi-
ardii наблюдал и А. С. Мончадский (1949) в Таджикистане. Им также
отмечены развитие трех поколений и уход на зимовку личинок чет-
вертого.
В северных и западных районах республики заметное увеличе-
ние численности начиналось сразу после появления первых особей,
особенно во второй половине июля. В середине августа количество
нападающих уменьшалось, а в конце месяца прекращалось совсем
(Амиргазиев, 1966; Прыгунова, 1966). Такая фенология говорит о
развитии двух летних генераций и уходе на зимовку третьей.
Нападает в различных зарослях или вблизи от них. Залет в по-
мещения носит случайный характер. От мест выплода далеко не уле-
тает, хотя неоднократно отлавливался в светоловушку, находящуюся
в 2—3 км от этих участков.
В бассейнах рек Или и Сыр-Дарьи местами дневок служат раз-
личные тугайные и кустарниковые заросли. В этих биотопах напа-
дает в течение целого дня, хотя наибольшая активность проявляется
в вечерние часы, задолго до темноты. В Кустанайской области днем
комары прячутся в густых травянистых зарослях, откуда вылетают
с наступлением полной темноты (Прыгунова, 1966). Основная масса
нападает в лесных зарослях.
В выборе объектов нападения не проявляет заметной предпочи-
таемости. Одинаково охотно нападает на людей и на различный скот.
В пустынных районах довольно часто отлавливался на липучки око-
ло нор корсаков, ежей, сурков, сусликов, песчанок, у гнезда сороки,
у гнезда-норы береговой ласточки. Агрессивность высокая, нападения
не прекращаются даже при отмахивании.
Зимует в фазе личинки. Местами зимовки могут служить толь-
ко глубокие непромерзающие водоемы с характерной погруженной
CG
растительностью. Гибелью личинок в промерзающих водоемах мож-
но объяснить весеннюю подорванность численности популяции. Под-
тверждением этому служит указание Везенберг-Лунда (1920—1921)
о том, что в весеннее время личинки не обнаруживаются в мелковод-
ных водоемах (менее 7з метра), которые были обильно заселены
ими в летнее время. Особенно это относится к южной части Казах-
стана, где основное развитие происходит в эфемерных, летних водое-
мах. На севере республики места выплода вследствие меньшей при-
уроченности к неглубоким временным водоемам, вероятно, менее
подвержены зимнему промерзанию, и в результате этого численность
восстанавливается быстрее.
Род AEDES MEIG.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА AEDES MEIG.
ПО ВЗРОСЛЫМ НАСЕКОМЫМ
(По А. А. Штакелъбергу и О. Н. Сазоновой; с сокращениями
и дополнениями)
1 (58). Хоботок явственно длиннее передних бедер.
2 (25). Лапки со светлыми колечками.
3 ( 8). Светлые колечки лапок опоясывают вершины предыдущих и
основания последующих члеников.
4 ( 5). Брюшко сверху с более или менее ясно выраженной продоль-
ной полосой из белых чешуек (у атипичных экземпляров
брюшко может быть одноцветно-белым или с небольшими
темными пятнами на боковых участках тергитов).
а. Среднеспинка в светло-буровато-желтых или золотистых че-
шуйках с двумя узкими продольными полосами белых че-
шуек (у переходных экземпляров спинка одноцветная или с
чуть заметными полосами) . ...............
. . . . . Ае. (О.) caspius caspius Pall.
Распространен в пределах пустынной, полупустынной и степ-
ной зон Казахстана.
б. Среднеспинка с широкой темно-бурой продольной полосой;
боковые отделы среднеспинки в большинстве случаев покры-
ты серебристо-серыми чешуйками (у переходных экземпля-
ров края среднеспинки окаймлены темно-бурыми чешуйками;
светлые участки в виде широких продольных полос)
Ае. (О.) caspius dorsalis Meig.
Распространен в пределах степной и лесостепной зон Казах-
стана. По горным районам выклинивается далеко на юг.
67
5 ( 4). Брюшко без продольной полосы белых чешуек.
6 ( 7). Покровы тела светло-бурые; перевязи на брюшке и кольца
на лапках выражены неярко ...................
................................. Ае. (О.) marine Serg.
С достоверностью не установлен. Встречающиеся на побе-
режье Каспия и около солоноводных водоемов пустынной зо-
ны Казахстана популяции с такими признаками укладывают-
ся в диапазон изменчивости Ае. с. caspius.
7(6). Покровы тела черно-бурые, перевязи на брюшке и кольца на
лапках контрастные . . . Ае. (О.) pulchritarsis Rond.
Возможен в южных и юго-восточных районах Казахстана.
8 ( 3). Светлые колечки на лапках опоясывают лишь основания
члеников.
9 (10). Задние лапки при основании члеников с узкими (не более 1/<
длины среднего членика задних лапок) светлыми колечками.
а. Задние лапки при основании члеников, с очень узкими (*/б—
7в длины среднего членика задних лапок) светлыми колечка-
ми. Тергиты брюшка лишь с более или менее узкими, ровны-
ми или чуть суженными посередине белыми перевязями
.............................Ае. (Aed.) vexans vexans Meig.
По всей территории Казахстана.
б. Задние лапки при основании члеников со светлыми колечка-
ми, достигающими ’/5—’А длины среднего членика задних
лапок. Тергиты брюшка с более широкими, обычно суженны-
ми посередине перевязями и вкраплениями светлых чешуек
в средней части тергитов, образующих подобие продольной
полосы. Светлые покровы тела с желтоватым оттенком. Сред-
ней величины комары ..........................
..............Ае. (Aed.) vexans nipponi Theob.
Горные и предгорные районы юго-востока Казахстана.
10 ( 9). Задние лапки при основании члеников с более широкими
светлыми колечками, которые занимают на средних члени-
ках около половины или более длины соответствующих чле-
ников.
11 (22). Тергиты с перевязями или пятнами светлых чешуек или в
беспорядочно перемешанных темных и светлых чешуйках
(у меланистически окрашенных особей светлые чешуйки мо-
гут отсутствовать, но тогда все брюшко темноокрашенное).
12 (13). Передние тергиты брюшка со светлыми округлыми пятнами,
располагающимися на вершине предыдущего и на основании^
последующего тергита (более узкая часть пятна состоит из
1—2 рядов чешуек). Иногда светлые пятна в виде треуголь-
ников расположены вдоль средней линии брюшка. Светлые
68
колечки на средних члениках задних лапок занимают '/г
длины членика. Пятно белых чешуек на вентральных мез-
эпистернах доходит до их переднего края
................................Ае. (О.) behningi Mart.
В Казахстане распространен севернее 47° с. ш.
13 (12). Передние тергиты брюшка с одной или двумя перевязями
светлых чешуек. >
14 (17). Передние тергиты с четкими узкими перевязями белых че-
шуек при основании; перевязи светлых чешуек у их верши-
ны есть лишь на задних тергитах. Светлые колечки на
средних члениках задних лапок обычно занимают менее */г
их длины. Пятно белых чешуек на вентральных мезэпистер-
нах чуть не доходит до их переднего угла (обычно до угла
доходят лишь одиночные чешуйки, а иногда и вовсе не до-
ходят).
15(16). Передняя поверхность задних бедер пестрая; светлые и тем-
но-бурые чешуйки на ней беспорядочно перемешаны. Пе-
редние и задние парастигмальные и метастигмальные пятна
обычно разобщены. Боковые отделы среднеспинки в светлых
кремовых или беловатых чешуйках с двумя-тремя парами
бронзовых пятен, прилегающих к широкой срединной полосе
такого же цвета. Коготок на лапке круто изогнутый
Ае. (О.) cantans Meig.
Повсеместно, севернее 47° с. ш.
16(15). Передняя поверхность задних бедер светлая; темные чешуй-
ки примешиваются к светлым лишь у вершины бедра. Перед-
ние и задние парастигмальные и метастигмальные пятна
обычно слиты своими основаниями. Боковые отделы средне-
спинки в золотистых чешуйках, часто с дополнительными,
очень узкими бронзовыми полосками одного цвета, с широ-
кой срединной полосой. Коготок на лапке полого вытянутый
Ае. (О.) riparius D. К.
В Казахстане встречается мозаично, севернее 47° с. ш.
17 (14). На передних тергитах по две перевязи светлых чешуек: бо-
лее широкая при их основании и более узкая у вершины.
Жилки крыльев с более или менее значительной примесью
светлых чешуек. Светлые колечки на средних члениках зад-
них лапок обычно превышают 7г их длины. Если светлые
перевязи на тергитах неясные, то на задних голенях торча-
щие треугольные чешуйки образуют муфту.
18 (19). Светлые кольца на лапках очень широкие: занимают 5/б или
5/z длины члеников, оставляя темной лишь вершинную часть.
69
На задних голенях торчащие треугольные чешуйки образу-
ют видимое расширение — муфту...........................
...........................Ае. (О.) kasachstanicus Guts.
Юго-восток Казахстана (бассейн р. Или).
19 (18). Светлые кольца на лапках не превышают 2/3 длины члеников.
Задние голени без торчащих треугольных чешуек.
20(21). Передняя поверхность передних бедер светлая, почти сплошь
в кремовых чешуйках, к которым в небольшом числе (глав-
ным образом в вершинной трети бедра) примешаны бурые
чешуйки. Основания костальной и субкостальной жилок с
группами светлых чешуек; последние никогда не покрывают
сплошь эти жилки на значительном протяжении. Обычно
светлых чешуек на жилках мало. Пятно светлых чешуек на
вентральных мезэпистернах доходит- до переднего угла. Сред-
неспинка покрыта золотистыми или бронзовыми чешуйками
с узкими полосками из светлых чешуек на боковых участках.
Их может быть по две с каждой стороны, редко они выраже-
ны неясно. Передние тергиты с четкими светлыми полоска-
ми, границы которых не размыты. На темных участках пе-
редних тергитов светлые чешуйки вкраплены лишь единично
.......................................Ае. (О.) beklemischevi Den.
Обнаружен только севернее 50° с. ш.
21 (20). Передняя поверхность передних бедер очень пестрая, в пере-
мешанных черных и желтых чешуйках. Среди темных чешу-
ек на жилках крыла обычно вкраплено большое количество
желтых чешуек, что придает крыльям пестроту. Основание
костальной жилки сплошь покрыто светлыми чешуйками,
редко светлых чешуек на крыльях мало. Пятно светлых че-
шуек большей частью не доходит до переднего угла вент-
ральных мезэпистерн. Среднеспинка покрыта бронзово-
красноватыми чешуйками, более светлые золотистые по кра-
ям. Светлые перевязи на тергитах нечеткие, обычно с размы-
тыми границами, или же тергиты на всем протяжении пест-
рые. В южных частях ареала встречаются светлоокрашен-
ные особи с брюшком, покрытым однотонными (одноцветны-
ми) светло-желтыми чешуйками ..................................
................................. Ае. (О.) excrucians Walk.
Распространен севернее 47° с. ш.
22 (11). Тергиты без светлых перевязей или пятен. Брюшко сплошь
покрыто желтыми однотонными чешуйками. Крупные жел-
тые комары.
23 (24). Все тело комара (среднеспинка, тергиты брюшка) одного цве-
та, покрыто яркими охристо-желтыми чешуйками. Нижние
70
мезэпимерные щетинки имеются. Светлые колечки на сред-
них члениках задних лапок очень широкие, занимают около
2/з длины. Коготок на лапке большой......................
............................... Ае. (О.) cyprius Ludl.
Встречается мозаично, севернее 48° с. ш., предпочитает гор-
ные и предгорные районы Казахстана.
24 (23). Брюшко сильно отличается по цвету от среднеспинки. Терги-
ты брюшка покрыты светло-серовато-желтыми, неяркими че-
шуйками, обычно с незначительной примесью темных чешу-
ек. Среднеспинка спереди в одноцветных красновато-бронзо-
вых, а в заднем отделе у щитков — в соломенно-желтых че-
шуйках. Нижние мезэпимерные щетинки отсутствуют. Свет-
лые колечки на средних члениках задних лапок более узкие,
занимают около */г длины членика или немного более. Кого-
ток на лапке очень большой и довольно пологий
........................ . . . Ае. (О.) flavescens Mull.
Повсеместно, кроме самых южных районов Казахстана.
25 ( 2). Лапки без светлых колечек.
26 (27). Брюшко в светлых чешуйках без перевязей на тергитах и
треугольных пятен по бокам брюшка; темные покровы тела
(кроме продольной полосы на среднеспинке) светло-бурые;
чешуйки на проэпимерах широкие и прямые. Относительно
крупные комары . . . Ае. (О.) subdiversus Mart.
Распространен в Западном, Северном и прилежащих участ-
ках Центрального Казахстана.
27 (26). Брюшко с перевязями на тергитах или треугольными пятна-
ми светлых чешуек по бокам; темные покровы тела темно-
или черно-бурые; чешуйки на проэпимерах узкие и изогну-
тые. Мелкие или средней величины комары.
28 (31). Голова и среднеспинка в черных, довольно густо расположен-
ных длинных и толстых щетинках (они бросаются в глаза).
На проэпимерах черные щетинки расположены по всей зад-
ней половине склерита. Посткоксальное пятно чешуек име-
ется. Покровы тела, особенно ног (не чешуйки), черные.
29 (30). Коготок на лапке круто изогнут, дополнительный зубчик
более половины длины коготка ..................
.............. Ае. (О.) impiger Walk.
Южный Алтай.
30 (29). Коготок на лапке удлиненный, слабо изогнут, дополнитель-
ный зубчик менее трети длины коготка............................
.............. Ае. (О.) nigripes Zett.
Южный Алтай.
31 (28). Щетинки на голове и среднеспинке менее развиты. На про-
71
эпимерах щетинки расположены в один или два неправиль-
ных ряда вдоль заднего края склерита. Посткоксальное пят-
но чешуек имеется или отсутствует.
32 (45). Дорзальные мезэпистерны почти сплошь покрыты светлыми
чешуйками или светлые чешуйки образуют небольшое гипо-
стигмальное пятно непосредственно под передней стигмой.
33 (36). Почти на всех участках тела, кроме тергитов брюшка, преоб-
ладают белые чешуйки. Основные жилки крыла покрыты ши-
рокими перемешанными белыми и темными чешуйками.
34 (35). Посткоксальное пятно имеется ..................
....................................Ае. (О.) simanini Guts.
Встречается на юго-востоке Казахстана, возможен в южных
районах республики.
35 (34). Посткоксальное пятно отсутствует • ..............
......................................Ае. (О.) detritus Hal.
В пустынной и полупустынной зонах Казахстана.
36 (33). Светлых чешуек почти на всех покровах тела значительно
меньше половины. Жилки крыла покрыты узкими бурыми
чешуйками с более или менее значительной примесью свет-
лых.
37 (40). Посткоксальное пятно имеется. Дорзальные мезэпистерны
почти сплошь покрыты белыми чешуйками (свободен от них
только небольшой участок ниже обычного места гипостиг-
мального пятна).
38 (39). Хоботок в темно-коричневых чешуйках без примеси светлых
чешуек. Среднеспинка в красновато-бронзовых чешуйках, с
узкой каемкой светлых чешуек спереди, с боков и сзади у
щитка ......................
..................................Ае. (О.) cataphylla Dyar.
В равнинных районах Казахстана распространен севернее
48° с. ш.; по горным и предгорным участкам доходит до
43° с. ш.
39 (38). Хоботок с более или менее значительной примесью светлых
чешуек, особенно в средней части. Среднеспинка в золотисто-
желтых или светлых чешуйках с широкой продольной поло-
сой чешуек бронзового цвета посередине, которая у атипич-
ных экземпляров может отсутствовать............................
..................................Ае. (О.) leucomelas Meig.
В равнинных районах Казахстана распространен севернее
48° с. ш.; по горным и предгорным участкам доходит до
43° с. ш.
40 (37). Посткоксальное пятно отсутствует. Дорзальные эпистерны не
сплошь покрыты белыми чешуйками (последние образуют
72
на них лишь отдельные пятна). Непосредственно к низу под
передней стигмой имеется небольшое гипостигмальное пятно
светлых чешуек.
41 (44). Пятно белых чешуек на мезэпимерах явно доходит до ниж-
него края последних. Нижние мезэпимерные щетинки всегда
имеются и нормально развиты.
42(43). Передние бедра не пестрые. Среднеспинка с двумя узкими
продольными темными полосками или без них. Тергиты в ко-
ричневых чешуйках с треугольниками светлых чешуек по
бокам, которые часто на двух, трех и четырех передних тер-
гитах соединяются перемычкой из светлых чешуек
............................ . . . . Ае. (О.) pullatus Coq.
Горные и предгорные районы Восточного, Юго-Восточного и
Центрального Казахстана.
43 (42). Передние бедра пестрые. Среднеспинка с широкой, несколько
отличающейся по цвету от основного фона, продольной поло-
сой и двумя узкими полосками светлых чешуек, тянущими-
ся от боковых краев щитка до его середины. Светлые перевя-
зи на тергитах без резких сужений. Боковые треугольники
выражены слабо .....................
............................... Ае. (О.) montchadskyi Dub.
Юго-восток и прилежащие районы Восточного Казахстана.
44 (41). Пятно белых чешуек на мезэпимерах не доходит до их ниж-
него края. Нижние мезэпимерные щетинки обычно отсутству-
ют или единичны (1—2) ...............
...............................Ае. (О.) intrudens Dyar.
Северный, Восточный и прилежащие участки Центрального и
Юго-Восточного Казахстана.
45 (32). Дорзальные мезэпистерны на значительном протяжении го-
лые; под передней стигмой нет гипостигмального пятна свет-
лых чешуек. Светлые чешуйки образуют там только перед-
ние и задние парастигмальные пятна, которые могут сливать-
ся друг с другом.
46 (53). Пятно белых чешуек на мезэпимерах всегда доходит до их
нижнего края.
47 (50). Светлые перевязи при основании двух-пяти тергитов более
или менее резко суживаются к середине (у атипичных эк-
земпляров перевязи могут быть ровными). Посткоксальное
пятно имеется.
48 (49). Все жилки крыла сплошь в темных, почти черных, чешуй-
ках, очень редко в основании костальной жилки имеются
одна-три желтоватых (не белых) чешуйки. Среднеспинка с
боков и у щитка в золотисто-желтых чешуйках, с широкой
73
полосой буровато-бронзовых чешуек посередине. Изредка
встречаются особи с иным рисунком на среднеспинке. Перед-
няя поверхность передних бедер в темно-коричневых чешуй-
ках, светлые чешуйки единичны (их больше при основании
передних бедер). Передние и задние голени темные. Коготок
на лапке круто изогнут, основной коготь загибается почти
сразу за зубчиком ...................
............................... Ае. (О.) punctor Kirby.
Северные и восточные районы Казахстана.
49 (48). Основание костальной жилки на некотором протяжении в
светлых чешуйках, которые могут доходить до уровня осно-
вания анальной жилки. Среднеспинка сплошь в золотисто-
бронзовых или красновато-бурых чешуйках, нередко с двой-
ной срединной полосой. Передняя поверхность передних бе-
дер с большим количеством светлых чешуек среди преобла-
дающих светло-коричневых. На передних и задних голенях
светлые чешуйки снаружи образуют продольную полосу. Ко-
готок на лапке длинный и слабо изогнутый . . . .
................................. Ае. (О.) hexodontus Dyar.
Восточный Казахстан.
50 (47). Светлые перевязи на двух-пяти тергитах без всяких сужений
посередине. Посткоксальное пятно имеется или отсутствует.
51 (52). Посткоксальное пятно имеется. Жилки крыла в темных че-
шуйках; в основании костальной жилки могут быть одиноч-
ные светлые. Среднеспинка в золотисто-желтых чешуй-
ках, с одной широкой или двумя узкими средними полосами
темно-коричневых, бронзовых узких чешуек или без них. Ко-
готок на лапке вытянутый Ае. (О.) pionips Dyar.
Восточный Казахстан.
52 (51). Посткоксальное пятно отсутствует. Основание костальной и
субкостальной жилок крыла на более или менее значитель-
ном протяжении (как у Ае. hexodontus) в светлых чешуйках.
Среднеспинка обычно с двумя узкими продольными, почти
черными полосками; остальная часть среднеспинки покры-
та золотисто-желтыми или серовато-белыми чешуйками, тем-
ные полоски иногда не развиты. Коготок на лапке более кру-
то изогнут . . . Ае. (О.) communis de Geer-
В Казахстане распространен севернее 48° с. ш.
53 (46). Пятно белых чешуек на мезэпимерах явно не доходит до их
нижнего края. Остальные признаки в ином сочетании.
54 (55). Пятно белых или кремовых чешуек на вентральных мезэпи-
стернах явно доходит до их переднего края. Тергиты с ярко-
74
белыми широкими, лишь слегка суживающимися посереди-
не перевязями. Передние и средние бедра спереди пестрые, в
коричневых и ярко-белых чешуйках; задние голени снару-
жи с продольной полосой светлых чешуек. Коготок на лапке
с маленьким и тупым зубчиком ..................
............................... Ае. (О.) nigrinus Eck.
Встречается в Восточном Казахстане.
55 (54). Пятно белых или кремовых чешуек на вентральных мезэпи-
стернах или явно не доходит до их переднего угла, или близ-
ко подходит к нему, одиночные светлые чешуйки могут до-
ходить до переднего угла этого склерита. Тергиты брюшка
без широких светлых перевязей; вместо них имеются лишь
светлые треугольники по бокам тергитов, которые могут со-
единяться узкой перемычкой из светлых чешуек. Передние и
средние бедра темнее, не пестрые.
56 (57). Пятно белых чешуек на вентральных мезэпистернах явно не
доходит до их переднего угла. Задние голени темные. Голова
и боковые отделы среднеспинки в ярких золотисто-желтых че-
шуйках; посередине среднеспинки имеется широкая (не всег-
да выраженная) полоса темно-бурых чешуек с бронзовым
отливом. Коготок на лапке средней величины
. . . . . . Ае. (О.) diantaeus Н. D. К.
Встречается в Восточном Казахстане, возможен в Северном.
57 (56). Пятно белых чешуек на вентральных мезэпистернах близко
подходит к переднему углу склерита, отдельные чешуйки мо-
гут доходить до самого угла. На наружной поверхности зад-
них голеней светлые чешуйки образуют продольную полоску.
Голова и боковые отделы среднеспинки в беловатых чешуй-
ках, посередине последней имеется широкая полоса бронзо-
вых чешуек. Коготок на лапках маленький . . . .
....................... . . Ае. (О.) sticticus Meig.
Северные и восточные районы Казахстана.
58 ( 1). Длина хоботка не превышает длины передних бедер (под-
род Aedes Meig.)
а ( г). Брюшко сверху более или менее одноцветное, без светлых
перевязей.
б ( в). Голова в преобладающих светлых чешуйках, примесь их от-
мечается по бокам среднеспинки, на щитке и в проксималь-
ной части хоботка; темные чешуйки брюшка черно-бурые, с
металлическим отливом . . . . .
........................ Ае. (Ае.) cinereus rossicus D. G. M.
Встречается мозаично, чаще в северных районах Казахстана.
s( б). Голова в преобладающих бурых чешуйках; среднеспинка и
75
хоботок без примеси светлых чешуек; темные чешуйки брюш-
ка темно-бурые, часто также с металлическим отливом
................ . Ае. (Ае.) cinereus cinereus Meig.
Повсеместно, кроме высокогорных районов республики.
г ( а). Брюшко сверху с более или менее выраженными поперечны-
ми светлыми перевязями ..................
.................... Ае. (Ае.) cinereus esoensis Yam.
Встречается в предгорных районах юго-востока Казахстана,.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА AEDES МЕЮ.
ПО ГИПОПИГИЯМ САМЦОВ
(По А. А. Штакельбергу, с изменениями и дополнениями)
1 (50). Класпеты имеются (подрод Ochlerotatus L.-Arrib.).
2 ( 5). Как базальная, так и вершинная бородавка коксита слабо
развита, последняя трудно различима.
3 ( 4). Базальная бородавка усажена более или менее равномерны-
ми волосками, щетинки отсутствуют ...........................
.............. . . Ае. (О.) marine Serg.
С достоверностью не установлен. Встречающиеся на побе-
режье Каспия и около солоноводных водоемов пустынной
зоны Казахстана популяции с такими признаками уклады-
ваются в диапазон изменчивости Ае. с. caspius.
4(3). Базальная бородавка, помимо волосков, несет одну крупную
щетинку ... . Ае. (О.) pulchritarsis Rond.
Возможен в южных и юго-восточных районах республики.
5 ( 2). Коксит с развитыми вершинной и базальной бородавками
или по меньшей мере одной из них.
6 (43). Базальная бородавка с одной-тремя крепкими щетинками,
значительно превосходящими по толщине другие волоски или
щетинки. ।
7 (12). Базальный отдел коксита с внутренней стороны с тремя ще^
тинками, из которых две расположены на базальной боро-
давке.
8 ( 9). Коксит в вершинной половине своей внутренней поверхно-
сти с направленным внутрь пучком густых волосков
.................................... Ае. (О.) diantaeus Н. D. К<
Встречается в Восточном Казахстане, возможен в Северном.
9(8). Коксит в вершинной половине без направленного внутрь
пучка волосков.
10 (11). Коксит у вершины с направленным дистально пучком во-
лосков; стволик класпет близ середины с пальцевидным вы-*
76
11
12
13
14
15
16
17
18
19
20
ростом (у атипичных экземпляров последний выражен очень
слабо)............................ Ае. (О.) intrudens Dyar.
Северный, Восточный и Юго-Восточный Казахстан.
(10) . Коксит у вершины без направленного дистально пучка во-
лосков; стволик класпет без выроста.........................
. . . . . . . . Ае. (О.) pullatus Coq.
Горные и предгорные районы Восточного, Юго-Восточного и
Центрального Казахстана.
(7) . Базальный отдел коксита с внутренней стороны несет не бо-
лее двух крепких щетинок.
(14) . Базальная бородавка коксита с двумя крепкими щетинками;
вершинная бородавка слабо развита . . . . .
................ . Ае. (О.) caspius Pall.
а. Базальная бородавка сильно выпуклая; основания крупных
щетинок бородавки удалены друг от друга на половину
или более длины бородавки ................................
............................ Ае. (О.) caspius dorsalis Meig.
Распространен в степной и лесостепной зонах Казахстана. По
горным районам выклинивается далеко на юг.
б. Базальная бородавка слабо выпуклая; основания крупных
щетинок бородавки сближены ..................
. . ............. Ае. (О.) caspius caspius Pall.
Пустынная, полупустынная и степная зоны Казахстана.
(13). Базальная бородавка коксита несет не более одной щетинки.
(18). Вершинная бородавка развита слабо.
(17). Внутренняя поверхность коксита в относительно коротких
волосках (длина их меньше ширины коксита). Выпуклая
часть крыла класпеты с острым углом ........................
................. Ае. (О.) impiger Walk.
Горные участки Северного и Восточного Казахстана.
(16). Внутренняя поверхность коксита в относительно длинных
волосках (длина их превышает ширину коксита). Выпуклая
часть крыла класпеты без острого угла.......................
............. Ае. (О.) nigripes Zett.
Южный Алтай.
(15). Вершинная бородавка развита хорошо.
(24). Вершинная бородавка коксита длинная (от вершины члени-
ка она тянется в проксимальном направлении примерно до
его середины) и усажена короткими щетинистыми волосками.
(21). Длина крыла класпет примерно в три раза превосходит его
ширину; базальная бородавка не отделена от тела коксита
Ае. (О.) punctor Kirby.
. . . ..............Ае. (О.) hexodontus Dyar.
Северные и восточные районы Казахстана.
77
21 (20). Длина крыла класпет не превышает его ширины; базальная
бородавка в дистальном своем отделе явственно отделена от
тела коксита.
22 (23). Вершинная бородавка более или менее равномерно выпук-
лая; базальная бородавка короче и шире.........................
.....................................Ае. (О.) nigrinus Eck.
Встречается в Восточном Казахстане.
23 (22). Вершинная бородавка в дистальном отделе значительно более
выпуклая, нежели в проксимальном; базальная бородавка
длиннее и уже............................. Ае. (О.) sticticus Meig.
Северные и восточные районы Казахстана.
24 (19). Вершинная бородавка ограничена, как правило, вершинной
третью или четвертью коксита.
25 (36). Стволик класпет относительно короткий и прямой.
26 (33). Крыло класпет в основной трети цилиндрическое и лишь к
вершине имеет расширение.
27 (28). Базальная бородавка, помимо крепкой щетинки, усажена
очень короткими, торчащими волосками более или менее
одинаковой длины . . Ае. (О.) flavescens Miill.
Повсеместно, кроме самых южных районов Казахстана.
28 (27). Базальная бородавка, помимо крепкой щетинки, усажена
довольно длинными щетинистыми волосками.
29 (30). Ширина базальной бородавки превосходит ее высоту пример-
но в два раза. Длина крыла класпеты в 4—5 раз превышает
ширину . ...
. . Ае. (О.) montchadskyi Dub.
Юго-Восток Казахстана.
30 (29). Ширина базальной бородавки примерно равна ее высоте. Дли-
на крыла класпеты в 6—8 раз превышает ширину.
31 (32). Внутренняя поверхность коксита в относительно редких ко-
ротких волосках (противоположные концы их не заходят
друг за друга). Пластинчатое расширение крыла класпеты
небольшое и развито лишь в средней части, ближе к основа-
нию крыла. Фаллозома удлиненная, с легкой перетяжкой
.......................................... Ае. (О.) simanini Guts.
Юг и юго-восток Казахстана.
32 (31). Внутренняя поверхность коксита в относительно густых
длинных волосках (противоположные концы их заходят друг
за друга). Пластинчатое расширение крыла класпеты развито
в средней и дистальной частях на более значительном про-
тяжении. Фаллозома короткая, яйцевидная, без перетяжки
..................... . . . Ае. (О.) detritus Hal.
В пустынной и полупустынной зонах республики.
78
33 (26). Пластинчатое расширение крыла класпеты начинается близ
основания крыла.
34 (35). Крыло класпет весьма широкое, высота базальной бородав-
ки явственно (примерно вдвое) превосходит ее ширину при
основании .... Ае. (О.) cantans Meig.
Повсеместно, севернее 47° с. ш.
35 (34). Крыло класпет более узкое, ланцетовидное; высота базаль-
ной бородавки едва превосходит ее ширину при основании
. Ае. (О.) riparius D. К.
В Казахстане встречается мозаично, севернее 47° с. ш.
36 (25). Стволик класпет длинный и изогнутый.
37 (38). Коксит с внутренней стороны в коротких щетинистых волос-
ках, чуть меньше или равных его ширине..........................
. . . . . . Ае. (О.) cyprius Ludl.
Встречается мозаично, севернее 48° с. ш.
38 (37). Коксит с внутренней стороны в длинных щетинистых волос-
ках, в 1,5—2 раза превосходящих ширину коксита.
39 (40). Вершинная бородавка коксита с большим количеством щети-
нистых волосков ..................
Ае. (О.) communis De Geer.
. Ае. (О.) pionips Dyar.
В Казахстане распространен севернее 48° с. ш.
40 (39). Вершинная бородавка коксита несет лишь несколько щети-
нистых волосков . . . .
41 (42). Лопасти девятого тергита, как правило, с 6—10 короткими
прямыми щетинками . . ...
Ае. (О.) cataphylla Dyar.
В равнинных районах Казахстана распространен севернее
48° с. ш.; по горным и предгорным участкам доходит до
43° с. ш.
42(41). Лопасти девятого тергита с 8—12 направленными несколько
наружу длинными щетинками ... . .
. . . . . . Ае. (О.) leucomelas Meig.
В равнинных районах Казахстана распространен севернее
48° с. ш.; по горным и предгорным участкам доходит до
43° с. ш.
43 ( 6). Базальная бородавка без крепкой щетинки.
44 (45). Помимо базальной бородавки, усаженной плоскими ланцето-
видными щетинками, коксит имеет около нее вторую боро-
давку с двумя тонкими и длинными щетинками
. . . . .............Ае. (О.) subdiversus Mart.
Распространен в Западном, Северном и прилежащих участ-
ках Центрального Казахстана.
79
45 (44). В базальной части коксита имеется лишь одна бородавка с
длинными или короткими щетинистыми волосками.
46 (47). Базальная бородавка сильно развита, высота ее равна шири-
не или чуть больше ширины при основании . . . .
........................... . Ае. (О.) behningi Mart.
В Казахстане распространен севернее 47° с. ш. '
47 (46). Базальная бородавка плоская и широкая.
48 (49). Базальная бородавка на половину своей ширины выступает^
над поверхностью коксита и густо покрыта короткими щети-
нистыми волосками, рыжеватыми в основной трети и темны-
ми к вершине .... Ае. (О.) kasachstanicus Guts.
Обнаружен только на юго-востоке Казахстана (бассейн р. Или).1
49 (48). Базальная бородавка более широкая (до % длины коксита),
совершенно плоская или чуть выступающая, покрыта корот-
кими, очень тонкими одноцветными волосками . .
........................... Ае. (О.) excrucians Walk.
Распространен севернее 47° с. ш.
50 ( 1). Класпеты отсутствуют.
51 (52). Стиль с двумя ветвями, на конце без придатка (подрод Aedes
Meig). ................................... Ае. (Ае.) cinereus Meig.
а. Длинная ветвь стиля на вершине двураздельная
................ . . Ае. (Ае.) cinereus cinereus Meig.
Повсеместно, кроме высокогорных районов республики.
б. Медиальная ветвь стиля простая, но короткая (длина ее рав-
на примерно V3 длины боковой ветви); вырост основной час*
ти верхней поверхности коксита простой, ступенчатый, не1
двухвершинный; морщинистый участок нижней поверхно-
сти коксита удлиненной прямоугольной формы
. . . . . . Ае. (Ае.) cinereus esoensis Yam.
Встречается в предгорных районах юго-востока Казахстана,
в. Медиальная ветвь стиля простая, но длинная (длина ее рав<
на примерно половине длины боковой ветви); вырост основ
ной части верхней поверхности коксита явственно двухвер<
шинный; морщинистый участок нижней поверхности кокси
та коротко-прямоугольной формы . . . . . <
......................... Ае. (Ае.) cinereus rossicus D. G. M.
Встречается мозаично, чаще в северных районах республики.
52(51). Стиль на дистальном конце с более или менее развитым при*
датком (подрод Aedimorphus Theob.) .
............................ Ае. (Aed.) vexans Meig
По всей территории Казахстана.
80
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ЛИЧИНОК IV СТАДИИ РОДА AEDES MEIG.
(По А. С. Мончадскому, с сокращениями и дополнениями)
1 ( 2). Длина усика больше длины головы.......................
.................Ае. (О.) diantaeus Н. D. К.
Встречается в Восточном Казахстане, возможен в Северном.
2 ( 1). Длина усика меньше длины головы.
3 ( 4). Лобные волосики расположены в один дугообразный ряд:
дистальные зубцы гребня расставлены; щетка на восьмом
членике брюшка из наибольшего числа (до 20) чешуек с
остроконечным шипом; пучок на сифоне лежит за середи-
ной, ближе к концу, из трех-шести тонких коротких ветвей
(подрод Aedes Meig.).
а. типичные личинки с признаками подрода
. . Ае. (Ае.) cinereus cinereus Meig.
б. типичные личинки: на передней поверхности дыхательной
трубки, кроме предвершинных щетинок, имеется две пары
тонких, коротких, едва заметных волосков из двух-пяти вет-
вей . . . Ае. (Ае.) cinereus rossicus D. G. M.
в. типичные личинки: на боковых поверхностях дыхательной
трубки у ее вершины имеются четыре пары дополнительных
очень коротких и тонких волосков ..........................
Ае. (Ае.) cinereus esoensis Yam.
4 ( 3). Лобные волоски каждой стороны расположены треугольни-
ком : средние лежат впереди внутренних.
5 ( 8). Усики лишены шипиков. Жабры длинные, длиннее седла.
6 ( 7). Пучок на усике в виде короткого простого волоска. Зубцы
zгребня длинные, шиповидные; их длина увеличивается от
основания к вершине сифона. Щетка из 12—15 чешуек . .
. . . Ае. (F.) geniculatus 01.
Возможен в западных, южных и юго-восточных районах Ка-
захстана.
7 ( 6). Пучок на усике из двух-четырех коротких ветвей. Зубцы
гребня нормального строения. Щетка из шести-десяти че-
шуек ... . Ае. (О.) pulchritarsis Rond.
Возможен в южных и юго-восточных районах республики.
8 ( 5). Усики покрыты хотя бы редкими шипиками. Если шипики
развиты очень слабо (Ае. marine), то жабры очень короткие,
шаровидные.
9 (10). На передней поверхности сифона, кроме предвершинных ще-
тинок, имеются две-четыре пары хорошо развитых грубых
волосков, а на боковых поверхностях, около основания ди-
^—80
81
10
11
12
13
14
15
16
17
18
19
20
82
стальных зубцов гребня, находится по боковому волоску. Пу-
чок на сифоне в виде длинного простого волоска. Наиболее
дистальный из расставленных зубцов гребня заходит за осно-
вание пучка, расположен в концевой трети сифона, почти у
его вершины . . Ае. (О.) subdi versus Mart.
Распространен в Западном, Северном и прилежащих участках
Центрального Казахстана.
( 9). На передней поверхности сифона дополнительные волоски»,
кроме предвершинных щетинок, отсутствуют.
(14) . Дистальные зубцы гребня широко расставлены и заходят за
основание пучка сифона.
(13) . Седло на последнем членике брюшка не доходит до его ниж-
него края . . Ае. (О.) cataphylla Dyar.
В равнинных районах распространен севернее 48° с. ш., по
горным и предгорным участкам доходит до 43° с. ш.
(12) . Седло кольцом опоясывает последний членик . . . .
. . . Ае. (О.) nigripes Zett.
Южный Алтай.
(11). Дистальные зубцы гребня не заходят за основание пучка
сифона.
(32). Впереди плавника помещается четыре-шесть более коротких
пучков, не объединенных общим основанием. Пучок у cejJC-
дины сифона (кроме Ае. mariae) в длину не меньше ширины
трубки у места отхождения пучка.
(17). Кроме обычных волосков, вся поверхность тела покрыта гус-
тыми рядами темных мелких шипиков. Голова и длинный
сифон (индекс 4—5) охряно-желтого цвета; гребень на «сифо-
не с двумя-тремя более крупными, широко расставленными
зубцами . Ае. (О.) cyprius Ludl.
Встречается мозаично, севернее 48° с. ш.; предпочитает гор-
ные и предгорные участки.
(16). Поверхность тела голая, без мелких шипиков, только с во-
лосками.
(21). Щетка из 6—12 чешуек.
(20). Чешуйки щетки расположены в один неправильный, часто
изогнутый углом ряд. Средние и внутренние лобные волоски
из двух ветвей. Сифональный индекс 3,5—4 ....
Ае. (О.) riparius D. К.
В Казахстане встречается мозаично, севернее 47° с. ш.
(19). Чешуйки щетки расположены в два, реже в три ряда. Сред-
ние и внутренние лобные волоски простые. Сифональный ин-
декс 2........................... . Ае. (О.) nigrinus Eck-
Встречается в Восточном Казахстане.
(18). Щетка из 15—45 чешуек, расположенных по меньшей мере
* в два неправильных ряда или образующих пятно трапецие-
видной формы.
q2 (23). Шипы на усиках очень слабо развиты. Едва различимы. Сед-
ло последнего членика брюшка слабо развито, оставляет сво-
бодной его переднюю треть. Жабры очень короткие, округлой
формы. Сифональный индекс не более 2
Ае. (О.) mariae Serg.
С достоверностью не установлен. Встречающиеся на побе-
режье Каспия и около солоноводных водоемов пустынной
зоны Казахстана популяции с такими признаками уклады-
ваются в диапазон изменчивости Ае. с. caspius.
23 (22). Шипы на усиках хорошо развиты. Седло хорошо развито.
Жабры не менее половины длины седла. Сифональный ин-
декс не менее 3.
24 (27). Волоски на концах задних (самых крупных) клапанов стиг-
мальной пластинки утолщены и крючкообразно изогнуты (в
редких случаях чуть изогнуты или прямые).
25 (26). Щетка из 30—40 (чаще 32—36) чешуек
. . . . . Ае. (О.) excrucians Walk.
Распространен севернее 47° с. ш.
26 (25). Щетка из 20—28 (в среднем 22) чешуек
Ае. (О.) behningi Mart.
В Казахстане распространен севернее 47° с. ш.
27 (24). Волоски на дистальном конце задних клапанов стигмаль-
Woft пластинки не утолщены и, как правило, только слегка
изогнуты.
28 (31). Дистальные зубцы гребня расставлены.
29 (30). Наиболее дистальные из зубцов гребня не заходят за середи-
ну сифона. Пучок сифона из пяти-шести ветвей
..................... . . Ае. (О.) flavescens Mull.
Повсеместно, кроме самых южных районов Казахстана.
30 (29). Наиболее дистальные один-три зубца гребня расположены
за серединой сифона. Пучок сифона из двух-четырех ветвей
Ае. (О.) beklemischevi Den.
Обнаружен только севернее 50° с. ш.
(28). Дистальные зубцы гребня не расставлены, промежуток меж-
ду двумя последними зубцами приблизительно равен таково-
му между двумя предыдущими. Плавник из 18—19 пучков,
окруженных общим основанием, и из четырех-восьми пучков
впереди него . . Ае. (О.) cantans Meig.
Повсеместно, севернее 47° с. ш.
83
32 (15). Впереди плавника не более трех пучков, не объединенных
общим основанием. У атипичных экземпляров: Ае. caspius,
Ае. communis, Ае. pullatus, Ае. vexans, Ае. kasachstanicus
возможны четыре-пять не объединенных общим основанием
пучков плавника.
33 (44). Жабры обычно короче седла, в исключительных случаях не
более чем на '/з превышают его длину, как правило, не пиг-
ментированы.
34 (35). Все чешуйки щетки не имеют длинного, главного шипа, а на
конце и на боках несут ряд шипиков, из которых апикаль-
ные наиболее длинные. Пигментация всех покровов тела
чрезвычайно слабая; чешуйки щетки, зубцы гребня еле раз-
личимы. Жабры короткие, шаровидные..........................
............................... Ае. (О.) detritus Hal.
В пустынной и полупустынной зонах республики.
35 (34). Хотя бы некоторые чешуйки щетки с длинным главным ши
пом. Покровы тела пигментированы интенсивнее; чешуйки
щетки, зубцы гребня хорошо выделяются. Жабры овальны*
или вытянутые, более ’/з длины седла.
36(37). Ветвление пучков плавника начинается далеко от основания;
длина неразветвленного стебелька средних пучков в 1,5—3
раза больше длины боковых отростков их основания. Неко
торые из ближайших к брюшной стороне чешуек щетки <
длинным главным шипом, остальные с двумя-тремя боле
длинными шипиками у вершины и с двумя-тремя более кс
роткими шипиками по бокам ...................
Ае. (О.) leucomelas Мец
В равнинных районах Казахстана распространен севернс
48° с. ш., по горным и предгорным участкам доходит л
43° с. ш. I
37 (36). Ветвление пучков плавника начинается близко от их основ
ния; длина неразветвленного стебелька не превышает длит
отростка его основания. Нижние чешуйки щетки не отлит
ются от остальных наличием резко выраженного главное
шипа: такие чешуйки могут единично встречаться в любЛ
месте щетки.
38 (41). Средние и внутренние лобные волоски из одной-двух ветвей
Личинки средней величины.
39 (40). Сифон относительно короткий и широкий, отношение шире
ны к длине не превышает 1:3. Гребень из 15—26 зубцов I
среднем 20—22) занимает около половины длины сифон!
У части чешуек щетки главный шип не выражен . . I
Ае. (О.) caspius Pal
84
Пучки на сифоне расположены у его середины и состоят из
трех-шести ветвей. Внутренние хвостовые волоски из четы-
рех-десяти ветвей. Средний из волосков позади щетки из пя-
ти-шести ветвей . Ае. (О.) caspius dorsalis Meig.
Распространен в степной и лесостепной зонах Казахстана. По
горным районам выклинивается далеко на юг.
б. Пучки на сифоне расположены за его серединой, ближе к
вершине и состоят из пяти-десяти ветвей. Внутренние хвосто-
вые волоски из 12—15 ветвей. Средний из волосков позади
щетки из 7—14 ветвей . . . . . .
Ае. (О.) caspius caspius Pall.
Распространен в пределах пустынной, полупустынной и степ-
ной зон Казахстана.
40 (39). Сифон более стройный, индекс выше 3. Гребень из 13—15
зубцов (в среднем 14), занимает 1/з сифона. У всех чешуек
щетки главный шип выражен более или менее хорошо
Ае. (О.) simanini Guts.
Предгорные районы юго-востока Казахстана.
41 (38). Средние и внутренние лобные волоски из трех-четырех вет-
вей.
42 (43). Волоски на задних клапанах стигмальной пластинки тол-
стые. Боковые волоски на седле из двух-четырех ветвей. Ха-
рактерна хорошо развитая вторичная перистость головных и
сифональных пучков тела . . . Ае. (О.) montchadskyi Dub.
Юго-восток и прилежащие районы Восточного Казахстана.
43 (42). ^Волоски на задних клапанах стигмальной пластинки тонкие,
еле заметные. Боковой волосок на седле одинарный, как ис-
ключение, двойной. Вторичная перистость головных и сифо-
нальных пучков выражена слабо . .
. . . Ае. (О.) kasachstanicus Guts.
Юго-восток Казахстана.
44 (33). Жабры не менее чем в 1,5 раза длиннее седла, чаще всего
пигментированы.
45 (50). Число чешуек в щетке более 30; чешуйки, как правило, ли-
шены главного шипа, а по краям и на конце несут ряд ши-
пиков.
46 (47). Средние и внутренние лобные волоски простые, лишь в виде
редкого исключения из двух ветвей
. . . Ае. (О.) communis De Geer.
В Казахстане распространен севернее 48° с. ш.
47 (46). Средние и внутренние волоски из трех и более ветвей.
48 (49). Щетка обычно из 50—60 чешуек, не имеющих главного шипа.
Усики тонкие, длинные, до 2/з длины головы или немного
85
длиннее . . Ае. (О.) pionips Dyar.
Восточный Казахстан.
49 (48). Щетка из 30—40 чешуек, часть из которых с главным ши-
пом. Усики более короткие и толстые, около половины дли-
ны головы . . . . Ае. (О.) pullatus Coq.
Горные и предгорные районы Восточного, Юго-Восточного и
Центрального Казахстана.
50 (45). Число чешуек в щетке меньше 30.
51 (54). Седло опоясывает кольцом последний членик брюшка.
52(53). Число чешуек в щетке 10—20; чешуйки мелкие — 0,07—-
0,08 мм в длину . . Ае. (О.) punctor Kirby.
Северные и восточные районы Казахстана.
53(52). Число чешуек в щетке пять-девять; чешуйки более крупные,
0,11—0,13 мм в длину Ае. hexodontus Dyar.
Восточный Казахстан.
54 (51). Седло не опоясывает кольцом последний членик брюшка, в
крайнем случае доходит только до нижнего края его боков.
55 (56). Внутренние лобные волоски простые и грубые. Пучок на си-|
фоне не заходит за его середину . . . . .
Ае. (О.) impiger Walk.
Южный Алтай.
56 (55). Внутренние лобные волоски из двух-пяти ветвей. Пучок на
сифоне расположен за его серединой, ближе к вершине.
57 (60). Дистальные зубцы гребня (1—3) всегда широко расставле-
ны, заметно крупнее остальных.
58 (59). Пучок на дыхательной трубке за серединой, ближе к верши-
не всегда впереди наиболее дистального из зубцов гребня, из
трех-восьми (чаще 4—6) тонких ветвей, более коротких, чем
ширина трубки у места отхождения пучка .
. Ае. (Aed.) vexans Meig.
Распространен повсеместно.
59 (58). Пучок на дыхательной трубке у середины или немного за
ней, из трех-восьми (чаще 4—6) ветвей, равных ширине труб-;
ки у места его отхождения или длиннее. Если число расстав-
ленных зубцов гребня больше двух, пучок лежит между их
основаниями . Ае. (О.) intrudens Dyar.
Северный, Восточный и прилежащие участки Центрального
и Юго-Восточного Казахстана.
60 (57). Зубцы гребня расположены на равном расстоянии друг от
друга, как исключение, один наиболее дистальный может
быть немного удален от остальных. В щетке 16—27 (чаще
20—24) чешуек . . . Ае. (О.) sticticus Meig.
Северные, центральные (горные) и восточные районы Казах-
стана.
86
Подрод Ochlerotatus L.-Arrib.
Ae. (О.) behningi Mart, (карта VI)
Характерный для степной и лесостепной зон СССР вид (Сазонова,
1959). В Казахстане до этого не регистрировался, был известен из
пограничных районов Оренбургской (Федоров, 1951) и Саратовской
областей (Martini, 1926). Оказался одним из массивных видов на се-
вере, востоке и западе Казахстана.
По экологии и фенологии напоминает Ае. excrucians и Ае. can-
tans, совместно с которыми чаще всего и встречается. В предгорных
районах Восточного Казахстана в 1966 г. выплод произошел в конце
мая. По мере поднятия в горные районы встречались личинки более
ранних стадий. Если в степном поясе (500—700 м н. у. м.) в конце
мая отмечались куколки, то в лесостепном преобладали личинки
III—IV стадий. Окукливание последних наступило в конце перлон
декады июня, а выплод имаго произошел 4—5 дней спустя. В сильно
заросших и затененных биотопах не встречается. Предпочитает отно-
сительно глубокие (40—80 см) водоемы, заселяя в них более глубо-
кие центральные части (рис. 24).
Все места выплода с более или менее чистой водой и с прелой
прошлогодней растительностью на дне. Температура в процессе раз-
вития личинок колебалось от 15 до 27°. Большую часть времени ли-
чинку проводят в придонной части, редко выплывая для дыхания.
В лабораторных условиях доращиваются хуже, чем Ае. cantans. Вы-
плод имаго из определенных личинок (куколок) получить удавалось
редко.
В Северном Казахстане местами выплода являются всевозмож-
ные разливы и затоки воды среди лесных участков поймы Иртыша.
В Баян-Ауле места выплода те же, что и у Ае. cantans и Ае. excru-
cians (заболоченности в местах разлива горных ручьев и выходы
грунтовых вод на пониженных участках). Изучение экологии и ана-
лиз сборов дают основание считать его одним из массовых предста-
вителей группы «cantans» в северных, восточных и северо-западных
районах Казахстана.
Плотность личинок не достигает высоких пределов, но заселяе-
мая ими площадь бывает велика. За 20 мин на человека может на*
падать до 70 особей. От затененных участков далеко не улетает, пред-
почитая оставаться в зарослях. В помещениях не обнаруживался.
Роения не отмечено.
Поздневесенний моноцикличный вид с периодом активности в
первую половину летнего сезона. Зимует в фазе яйца. Нападает как
На человека, так и на домашних животных (крупный рогатый скот,
л°П1ади).
87
Рис. 24. Местообитания личинок Aedes behningi, Ае. cantans и Ле. excrucians
в Восточном Казахстане.
Ае. (О.) beklemischevi Den. (карта VI)
Вид, описанный 3. М. Денисовой (1955) под приведенным наз£
нием и О. Н. Сазоновой (1956) под названием Ае. grandilarva. I
приоритету оставляется первое название. Распространен в лесной г
не СССР. В Казахстане установлен по сборам В. П. Божен!
И. Г. Прыгуновой, С. А. Скворцовой и В. М. Деньгуба в пределах Kj
танайской, Павлодарской и Восточно-Казахстанской областей. Южн
48° с. ш. не обнаружен. Эти разрозненные сведения не создают ясно
представления об экологии Ае. beklemischevi в Казахстане. Тем :
менее можно дополнить сведения О. Н. Сазоновой (1959) тем, что <
встречается не только в лесной, но и в лесостепной зоне СССР.
Личинки отловлены в середине апреля в Семипалатинской и Па
лодарской областях (В. П. Боженко, В. М. Деньгуб) и в начале мая
Кустанайской области (И. Г. Прыгунова). Во всех случаях местаг
выплода являлись открытые участки временных водоемов. I
О. Н. Сазоновой (1959), местами выплода служат временные открыт!
водоемы на сухих и пойменных лугах, в лесных заболоченностях
припоселковых лужах, где он встречается совместно с Ае. cyprit
Ае. flavescens к Ае. excrucians.
88
Напоминает указанные виды и по фенологии. В. П. Боженко
(1948) отлавливал имаго в середине мая — начале июня. Аналогич-
ные сроки приводит О. Н. Сазонова (1959) для Рыбинского водохра-
нилища. Ею отмечено частичное отрождение особей второй осенней
генерации, но уже во временных дождевых лужах.
Судя по характеристике мест выплода и другим экологическим
особенностям, этот вид должен быть довольно широко распространен
е лесостепной и на интразональных участках степной зоны Казах-
стана. Об этом говорят просмотренные сборы из различных северных
районов республики.
Ае. (О.) cantans Meig. (карта VI)
Ареал охватывает основную часть бореальной зоны Голарктики
п вклинивается на территории СССР в Средиземноморскую под-
область (Крым, Северный Кавказ). В Казахстане распространен се-
вернее 48° с. ш. По предгорьям Тянь-Шаня выклинивается до 44° с. ш.
Поздневесенний вид, развивающийся дольше других моноцик-
личных видов (Сазонова, 1959). Объясняется это более высокой тер-
мофильностью. М. Ф. Шленова (1952) установила, что развитие на-
чинает^ при 10—11°, но протекает очень медленно. Оптимум разви-
тия при 19—25°. При 30° личинки быстро гибнут. Таким образом,
верхний и нижний пороги его развития приближаются к полициклич-
ным видам. В горных районах Восточного Казахстана, наоборот,
развивается быстрее, чем Ае. excrucians и Ае. behningi, а выплод про-
исходит раньше.
Развитие личинок в Восточном Казахстане протекало при 15—
24°. В равнинных районах выплод имаго происходит во второй поло-
вине мая, а на более возвышенных участках (1000—1200 л н. у. м.) —
в начале июня. Личинки встречаются среди припойменных зарослей
и в лесных водоемах. Предпочитают открытые или слабо затененные
участки глубиной от 20 до 40 см (рис. 24). Вода в них отличается
слабой минерализацией (107 мг!л), большой мягкостью (0,6°), незна-
чительной окисляемостью (9,9 мг!л) и слабощелочной реакцией (pH
7,7). Столь низкие показатели объясняются тем, что большинство
этих водоемов образовано талыми или паводковыми водами. В одном
случае развитие отмечено в быстро высыхающем затоке талых и
дождевых вод глубиной 10—15 см. В этом случае, несмотря на зате-
ненность водоема, развитие произошло быстрее, а личинки IV стадии
и куколки с выплаживающимися из них имаго были примерно на */з
меньше обычных.
В высокогорных условиях Южного Алтая (Рахмановские ключи)
на высоте 1900 м н. у. м. места выплода крайне ограниченны. Разви-
89
тие отмечено лишь в длительно существовавших приозерных затоках.
Спустя три года эти водоемы при постройке дороги были ликвидиро-
ваны и популяция этого вида была подорвана настолько, что, несмот-
ря на обилие водоемов других типов, ни личинки, ни имаго этого ви-
да обнаружены не были.
Все места выплода характеризуются обилием на дне опавшей
прошлогодней листвы. Большую часть времени личинки проводят на
дне, редко, но на довольно продолжительный период (37—64 сек),
всплывая к поверхности. В одном случае отмечено развитие в заточ-
ной яме глубиной 120 см. Хорошо доращиваются в лабораторных
условиях, гибель незначительная, выплод дружный.
В Павлодарской области местами выплода являются всевозмож-
ные разливы и затоки воды в лесных участках поймы Иртыша. Ана-
логичные данные приводит Э. Р. Геллер (1939), обнаруживавший этот
вид на покрытых густой растительностью островах Иртыша. В горных
участках Казахского мелкосопочника (Баян-Аул) местами выплода
являются различные низинные заболоченности — места разлива сте-
кающих с гор потоков и выходы грунтовых вод на пониженных
участках. В этих условиях его выплод происходит в менее заросших
и затененных биотопах.
Один из массовых видов в Северном и Восточном Казахстане, на
возвышенных участках Казахского мелкосопочника и в северо-запад-
ных районах республики (Иргиз, Тургай, верховья Илека, Уила).
Крайние южные точки обнаружения около 48° с. ш. (северное побе-
режье Каспия, южная часть Алаколя). Здесь отлавливаются единич-
ные экземпляры.
Плотность личинок может достигать 70 экземпляров на 1 м2 вод-
ной поверхности. За 20 мин на человека нападает до 370 особей этого
вида. В дневное время держится в зарослях, нападая в течение все-
го дня. Наибольшая активность отмечается в предвечерние и вечер-
ние часы, менее — в утренние. Далеко от зарослей не улетает, в по-
мещениях не обнаруживается. Как такового, роения не отмечено, но
зато наблюдались довольно частые перелеты самцов на высоте 1—
1,5 м, что, вероятно, связано с поисками самок.
Личинки и имаго встречались в одних биотопах с Ае. excrucians,
Ае. behningi, Ае. cinereus, Ае. communis, Ае. pullatus и Ае. punctor.
Выше 1500 м н. у. м. (нижняя кромка лесного пояса горной части Ал-
тая) не обнаружен. .
Поздневесенний моноцикличный вид, преобладающий в первой
половине теплого сезона. Зимует в фазе яйца. Самки в равной степе-
ни агрессивны по отношению как к человеку, так и к домашним жи-
вотным (отмечено нападение на коров и лошадей).
Возможный переносчик туляремии (Олсуфьев, Руднев, 1960).
90
Ae. (О.) caspius Pall.
В Казахстане встречаются оба подвида этого политипичного ви-
да. Хотя существует много суждений об их видовой самостоятельно-
сти, на основании имеющихся у нас данных мы не можем рассмат-
ривать их в качестве видовых таксонов.
Ае. (О.) caspius caspius Pall, (карта VII)
Широко распространенный в южных и средних частях Палеарк-
тики подвид. В СССР его северная граница проходит где-то около
50 параллели. Не переходит он этот рубеж и в Казахстане. Характе-
рен для пустынной, полупустынной и степной зон Казахстана, где
заметно преобладает над другими представителями рода Aedes.
Места выплода крайне разнообразны, эвритопность Ае. caspius
поразительна. Личинки могут развиваться как во временных, так и в
постоянных водоемах с колеблющимся уровнем воды, что объясняет-
ся способностью этого вида в отличие от многих других видов Aedes
откладывать яйца как непосредственно на воду, так и на влажную
почву. В лабораторных условиях 70% кладок были произведены сам-
ками на воду и 30% —на влажную фильтровальную бумагу. Такая
особенность благоприятствует беспрерывному развитию вида в самых
разнообразных биотопах, где есть вода, и вместе с тем в случае высы-
хания водоема он гарантирован от гибели ввиду сухоустойчивости
яиц, свойственной всем представителям Aedes. При отсутствии воды
кладку производит в наиболее пониженные, чаще заливаемые участ-
ки водоема.
В больших постоянных водоемах развитие личинок происходит
в прибрежной, пологонаклонной части со слабым током воды и при
отсутствии волнобоя. В таких местах их можно встретить совместно с
Сх. modestus, Сх. pipiens. An. hyrcanus, An. maculipennis и Ur. ungui-
culata. В неглубоких постоянных и полупостоянных водоемах (15—
10 см) развитие может происходить по всей площади. Из временных
водоемов местами выплода служат буквально все типы периодиче-
ских скоплений воды: фильтрационные разливы, затоки и скопления
воды в понижениях рельефа, выходы грунтовых вод, разливы родни-
ков, мелких речушек и оросительных каналов, поля фильтрации и
рисовые чеки, скопления воды около колодцев и водопроводов, заво-
ди крупных рек и особенно разливы во время паводков (рис. 25). Не-
обходимым условием является хорошая инсоляция и слабое зараста-
ние водоема. В более южных районах Казахстана, где прогреваемость
воды и без того высокая, развитие может происходить и в заросших
водоемах. В такого типа местах личинки встречаются совместно с
91
Ae. vexans и Ae. cinereus. В Бетпак-Дале выплод отмечен даже в не-
глубоком (40 см от поверхности почвы) колодце «Кара-Кудук».
Такое разнообразие мест выплода было бы невозможно для несо-
левыносливого вида, поскольку большинство водоемов в пустынной и
Рис. 25. Заболоченные участки в низовьях р. Сыр-Дарьи — места массового
выплода Aedes с. caspius.
полупустынной зонах Казахстана в той или иной степени характери-
зуется примесью солей. Галофильность его чрезвычайно высока. Раз-
витие личинок отмечено даже в водоемах с налетом соли на усыхаю-
щих участках побережья. Диапазон химического состава воды в
местах обитания очень разнообразен. Достаточно указать, что разви-
тие личинок отмечено при колебании жесткости воды от 3,8 до 66,2°,
общей минерализации от 399 до 30 000 мг!л, слабощелочной среде
и с той или иной комбинацией серных, аммиачных, углекислых, хло-
ристых, кальциевых и магниевых соединений. В воде с нижним
пределом указанных индексов и в слабо минерализованной ъстреча-
ются как исключение. Лабораторные опыты показали, что концент-
рация солей в среде обитания влияет на выплод личинок, который
происходит лишь в допустимой концентрации (1—2%). Встречаемость
же и развитие личинок в более солоноводных водоемах обусловлены,
вероятно, постепенной приспосабливаемостью личинок по отношению
к повышающемуся в результате испарения воды проценту содержа-
92
ния соли. Это говорит о том, что механизм солевыносливости подвида
наряду с адаптацией в процессе эволюции заключается и в его боль-
шей индивидуальной приспосабливаемости в период развития.
Сезонный ход численности с поправками на температурный ре-
жим местности более или менее однотипен. Первые комары появляют-
ся в конце апреля — начале мая. Они, как правило, малочисленны,
но при благоприятных для развития температурах и наличии обра-
зовавшихся после половодья пустынных рек, временных водоемов
численность их быстро возрастает. В последующем с теми или иными
перепадами, зависящими от наличия временных водоемов, обилие
их сохраняется до середины августа.
Массовая численность отмечается в середине или конце июля,
но в связи с задержкой летнего паводка может оттягиваться к середи-
ге августа.
Отдельные особи нападают до наступления заморозков, причем
последние теплые дни наблюдается довольно массовый залет их в
жилые и заброшенные помешения, где они усиленно нападают на че-
ловека. В условиях Южного Казахстана массовый залет наблюдался
во второй половине октября при 18—22°. Нападение самок было та-
ким ожесточенным, "каким оно бывает лишь в летнее время. Интерес-
но отметить, что в тугаях в это время можно встретить только еди-
ничные экземпляры, и то с пониженной агрессивностью.
Самки Ае. с. caspius менее чувствительны к влиянию температу-
ры и освещенности среды, чем многие другие виды Aedes, и поэтому
суточный ритм активности их в значительной степени растянут и
стушеван. Утреннее нападение начинается с первыми признаками
рассвета. Наиболее активно оно проявляется перед восходом солнца
и непосредственно в период восхода. Затем, постепенно снижаясь, ак-
тивность сохраняется до 7—9 час утра, а в прохладные дни — и доль-
ше. В непосредственной близости от мест укрытия комары нападают
весь день, при умеренных температурах сильнее, а при высоких
(35—37°) — слабее. Вечерняя активизация наступает после спадения
дневного зноя и бывает ощутимой за 1—1,5 час до захода солнца и
особенно сильной после захода. В светлые, лунные ночи депрессии
не отмечается, что характеризует в сумме диапазон световой актив-
ности этого подвида. Охотно летит на свет и поэтому особенно часто
и много отлавливается светоловушкой.
При определенных благоприятных условиях развитие может
идти весь летний период. Скорость развития и количество генераций
определяются температурным режимом. В Казахстане ввиду сильной
инсоляции условия очень благоприятны. За теплый период подвид
может проделывать до 5—6 генераций, из них ярко выраженных
(связанных с массовыми разливами) в пустынной зоне Казахстана
93
три: в мае, июне и июле. В Гурьевской области отмечается два пика:
в мае и июле (Жук, 1967).
Лабораторные исследования показали, что в весеннее и осеннее
время выплод личинок очень растянут. Объясняется это тем, что ко-
личество яиц в водоемах с колеблющимся уровнем воды беспрерывно
пополняется самками, как первых генераций, так и последующих.
При этом склонности яиц к диапаузированию летом не отмечается,
временное прекращение развития связано лишь с высыханием водое-
мов. В данном случае выход яиц из этого состояния отмечается сразу
же после залития их водой. Следовательно, в летнее время возможно
как непосредственное развитие Ае. с. caspius, так и чередующееся с
эмбриональным покоем. Такая двойственность говорит о чрезвычай-
но большой приспособленности этого подвида к существованию в за-
сушливых условиях, чем, по всей видимости, и определяется доми-
нирование его во многих районах пустынной и полупустынной зон
Казахстана.
Уязвимость этих комаров к воздействию неблагоприятных фак-
торов внешней среды проявляется лишь при позднелетнем или осен-
нем выплоде и наступающем затем резком похолодании. Но и в этом
случае они застрахованы от полной гибели за счет асинхронности от-
рождения личинок и преждевременного наступления диапаузы у час-
ти осенних яйцекладок. Частичное отрождение личинок Ае. с. cas-
pius в весеннее и осеннее время подтверждают данные Н. В. Хелевина
(1958а, 1959), который в опытах с близким подвидом Ае. с. dorsalis
показал, что при пониженных температурах наступает асинхрон-
ность выплода.
В аридных условиях Казахстана возможен и весенний. подрыв
численности популяции. Происходит он в тех случаях, когда при за-
тяжной, теплой, но дождливой осени многочисленные самки откла-
дывают яйца во временные небольшие затоки, которые быстро пере-
сыхают весной, вызывая массовую гибель развивающихся в них ли-
чинок.
Лабораторные опыты показали, что к задержке эмбрионального
развития способны не все яйца Ае. с. caspius, а лишь те, которые
пробыли в оптимальных условиях (влажность, температура) не ме-
нее 4—5 дней. К этому времени, как показали исследования Н. В. Хе-
левина (1946, 1958а, 1959), у многих видов Aedes происходит фор-
мирование личинок. Следовательно, до 4—5 дней яйца Aedes
подвержены губительному воздействию неблагоприятных факторов
внешней среды. Если судить по удлинению в зависимости от темпера-
туры эмбрионального периода развития, то при более низких темпе-
ратурах формирование личинок должно затягиваться, а срок уязви-
мости их яиц соответственно удлиняться.
94
Опыты с промораживанием личинок показали, что нахождение
их в течение 12—14 час в замерзшей при — 6—10° воде к гибели не
приводило. Активность их начиналась по мере оттаивания. На после-
дующем развитии это не отражалось. В опыте испытывались личин-
ки I, II и III стадий. Гибель личинок отмечена лишь в двух случаях,
когда вследствие сильного промерзания тонкий слой льда потрескал-
ся и, вероятно, травмировал личинок.
Сезонный ход численности и условия возникновения диапаузы
дают основание считать, что постепенное нарастание обилия этого ви-
да происходит в результате переживания неблагоприятных зимних
условий незначительным количеством диапаузирующих осенних кла-
док. Основная же масса популяции, развивающаяся беспрерывно, не
успевает закончить развитие до осеннего периода и гибнет при на-
ступлении похолодания. Неоднократные наблюдения в течение ряда
лет подтверждают, что даже после периода с большой численностью
восстановление популяций на следующий год происходит постепен-
но, начиная с небольших количеств. Прохождение холодового перио-
да для яиц, находящихся в диапаузе, не обязательно. Во многих
опытах выплод происходил после залития сухих яиц водой независи-
мо от сезона и условий содержания.
Количество откладываемых яиц колеблется от 95 до 233 (в сред-
нем 170). В весенний период оно, как и размер вып л вживающихся
имаго, Меньше. Самки летних генераций по размерам значительно
крупнее. Возрастает к осени и среднее количество откладывае-
мых яиц.
При развитии личинок в эфемерных, бедных питательными ве-
ществами водоемах отмечается еще одна характерная экологическая
особенность. Самки летних генераций начинают нападать через 6—
8 дней после выплода. До этого они роятся и питаются соками расте-
ний. А. С. Мончадский (1956) причину удлинения предагрессивного
периода объясняет ускоренными темпами развития личинок, в ре-
зультате чего у вылетающих имаго плазма клеток яйцевых фоллику-
лов еще лишена жировых и желточных включений, т. е. самки нахо-
дятся на стадии «N» и еще более ранней. Отмеченный период идет на
дозревание яйцевых фолликулов по крайней мере до II фазы. Наши
наблюдения полностью подтверждают это, тем более что рассматри-
ваемое явление обычно наблюдается лишь во временных, быстро пе-
ресыхающих водоемах, в которых едва успевает закончиться пред-
имагинальное развитие. В более глубоких и постоянных водоемах это
же поколение выплаживается на 4—5 дней позже, но становится аг-
рессивным на 3—4 сутки.
Продолжительность жизни составляет 1—2,5 месяца. Об этом
говорит сравнительный анализ времени выплода и степени потерто-
Р5
сти отлавливаемых самок. Кроме того, об этом можно судить по то-
му, что иногда приходится встречать самок этого подвида в местах,
где, казалось бы, совершенно отсутствуют условия для их выплода.
Так, в 1963 г., остановившись на ночлег в пустынной части Дарья-
лык-Такыра, мы подверглись в вечернее время ожесточенному напа-
дению Ае. с. caspius. Утром удалось отловить несколько экземпля-
ров имаго, укрывшихся на дневку в трещинах саксаула и в углубле-
ниях почвы. Водоемов в окрестности в радиусе 70—100 км не было.
Обследование местности показало, что 1—1,5 месяца назад до этой
территории доходила вода весеннего разлива.
Роение отмечается перед наступлением сплошной темноты. Ориен-
тирами обычно служат отдельные деревья, а в зарослях — открытые
поляны, на которых рои клубятся над какими-либо ориентирами.
Отмечено роение и в населенных пунктах, где комары клубились
прямо во дворе. Высота роения не превышает 1,5—2,5 м. Наблюдает-
ся оно только в теплые, безветренные дни. В одном случае роение
отмечено совместно с Ае. vexans, который при этом доминировал ко-
личественно.
Дневными убежищами чаще всего служат различные раститель-
ные заросли, а там, где их нет, — трещины почвы, расщелины скал,
провалы могил, колодцы, заброшенные помещения, пещеры и норы
различных животных. П. А. Пе’грищева (1962) отлавливала этих кро-
вососов в норах барсуков, волков, шакалов, лис, корсаков, песчанок,
сусликов, крыс, варана и черепахи. Мы к этому списку можем доба-
вить еще сурка, дикобраза, обыкновенную полевку, у нор которых
на липучки попадались самки этого подвида.
Из домашних животных отмечен на лошадях, коровах, ослах.
Охотно нападает на человека, нередко преследуя его на значительные
расстояния. Мигрирует в поселки, находящиеся от мест выплода в
2—3 км. Один из кровососов, залетающих для кровососания в поме-
щения, где активно нападает при электрическом освещении. Напив-
шиеся самки стремятся вылететь, а голодные нередко остаются на
дневку, но в дневное время нападают редко. Наряду с Cx. modestus
является самым массовым для южной половины Казахстана крово-
сосом.
Повинен в трансмиссии туляремийного микроба, который может
в нем сохраняться до 12 дней (Олсуфьев, Руднев, 1960). В Восточном
Казахстане от Ае. с. caspius выделен штамм нейротропного вируса
группы «А» (Жуматов и др., 1966).
Ае. (О.) caspius dorsalis Meig. (карта VII)
Подвид, морфологически и экологически близкий с Ае. с. cas-
pius. В отличие от последнего заселяет более северную часть Па-
96
леарктики. Широко распространен в северных районах США и Ка-
нады (Matheson, 1944; Carpenter, La Casse, 1955).
Основная часть ареала лежит севернее 50 параллели. Подвид,
характерный для лесной и лесостепной зон (Мончадский, 1951). В Ка-
захстане распространен в основном в лесостепной зоне. По горным,
предгорным районам и поймам равнинных рек отмечается экстразо-
нальное проникновение в степную и пустынную зоны. В результате
этого по отношению к Ае. с. caspius является симпатрическим.
Места выплода этого подвида подобно Ае. с. caspius представ-
ляют открытые, хорошо прогреваемые водоемы. В Кустанайской об-
ласти таким требованиям удовлетворяют неглубокие, лишенные рас-
тительности временные водоемы, а в Восточном Казахстане — мел-
кие ямы с талой водой и пойменные разливы. Развитие личинок
отмечено в воде со слабощелочной реакцией (pH 7,7), незначительной
общей и компонентной минерализацией (317 мг/л), но с большой
окисляемостью (21,8 мг!л), что, по всей вероятности, объясняется
обилием гниющего растительного материала.
Один из самых многочисленных в Северном и Восточном Казах-
стане кровососов. В Павлодарской области 50% нападающих кома-
ров составляют эти насекомые. Личинки последних стадий обнаружи-
ваются в начале мая, вылет имаго происходит в средних числах этого
месяца^-Выплод второй генерации приходится на конец июля, а тре-
тьей — на конец августа. Массовой численности кровососы достигают
после выплода третьей генерации. В это время на человека за 20 мин
нападает до 600 особей. Затем наступает снижение численности, и в
конце сентября комары исчезают совсем (Прыгунова, 1966). Такая фе-
нология определяется прогрессивным увеличением обилия комаров за
счет сохранения особей предыдущей генерации и выплода новой.
Вместе с тем часты и наслоения генераций, сглаживающие обыч-
ную картину. В засушливые и холодные годы численность комаров
не достигает высоких цифр, а количество генераций уменьшается
до двух.
Трехвершинный ход численности Ае. с. dorsalis характерен и
для горных (1800—3500 м н. у. м.) районов юго-востока Казахстана,
что соответствует лесостепному поясу. Как и на севере Казахстана,
первые особи появляются во второй декаде мая, а исчезают в сен-
тябре. Резкие увеличения численности отмечены в июне, июле и ав-
густе. На отдельных горных участках является доминирующим кро-
вососом. По сведениям Ж. Исимбекова (1966), роение происходит в
тихое вечернее время при 11—18° на высоте 1,5—3 м. Ориентирами
служат кустарники, различные заросли и пасущиеся животные.
Количество роящихся самцов колеблется от единиц до сотен.
7—80
97
Суточная активность не отличается от таковой Ае. с. caspius.
Наибольшее количество нападающих отмечено в утреннее и вечернее
время. В укрытиях нападает и днем. Температурный порог активно-
сти значительно ниже, чем у Ае. с. caspius. Если нападение последне-
го прекращается при температуре ниже 16—17°, то у Ае. с. dorsalis
это приближается к верхнему порогу активности. В горах Заилийско-
го Алатау оптимальной для нападения является температура в пре-
делах 12—18°.
Местами дневок служат различные естественные укрытия, но
чаще всего травяной покров и кустарниковые заросли. В лесу отлав-
ливается редко, даже в период максимальной численности, хотя на
разреженных пойменных участках Павлодарской области приурочен
к таким местам. В северных районах Казахстана отмечается своеоб-
разная концентрация имаго возле водоемов, что, вероятно, связано с
локальностью мест выплода в этом районе.
Подобно Ае. с. caspius, развитие этого подвида может осуществ-
ляться в летнее время без эмбриональной диапаузы. Тем самым жиз-
ненная схема его приближается к Anopheles и Culex, но способность
впадать в диапаузу сохраняется частично в середине и полностью в
конце теплого периода года.
Наиболее полно экология Ае. с. dorsalis освещена в серии работ
Н. В. Хелевина (1946, 1958а, 1959). Автором доказана возможность
развития личинок первой генерации при 3—5°. С повышением тем-
пературы скорость развития и процент выплода увеличиваются. По-
рог развития следующих поколений находится около 18,2—18.3°, а
температура около 30° является оптимальной.
В осенний период наступление диапаузы возможно лишь у яиц
последней летней генерации, и то по истечении 4—6 суток, необходи-
мых для формирования личинки. В летнее время задержка выплода
личинок происходит под влиянием неблагоприятных условий среды
(высыхание водоема, низкие температуры). Самки летнего поколения
способны в течение 9 суток переносить понижение температуры до
+ 1° и при последующем повышении температуры откладывать жиз-
неспособные яйца. При 8—9,5° самки, находящиеся на различных
фазах гонотрофического цикла, не только заканчивают формирова-
ние яиц, но и производят кладку. Как летним, так и осенним яйце-
кладкам свойственна асинхронность развития, что является экологи-
ческой приспособленностью к сохранению популяции в меняющихся
условиях внешней среды. Все это говорит о чрезвычайно высокой
экологической пластичности подвида.
Активный кровосос. Охотно нападает как на человека, так и на
домашних животных (лошади, крупный рогатый скот). Питания на
диких животных не отмечено, хотя массовость его на отдельных
98
участках, редко посещаемых человеком и домашними животными
(горные районы), дает основание считать, что в круг его прокормите-
лей могут входить и дикие животные.
Доказанный переносчик вируса японского энцефалита, западно-
го конского энцефаломиелита и энцефалита Сан-Луи (Kissling, 1960).
Ае. (О.) cataphylla Dyar (карта VIII)
Распространен циркумбореально в пределах лесной зоны Гол-
арктики. В РСФСР на юг, за пределы лесов, почти не выходит (Са-
зонова, 1959). В Казахстане встречается в лесостепной зоне, выкли-
ниваясь по поймам рек в степную. По лесному поясу горных хребтов,
окаймляющих Казахстан с востока, проникает в юго-восточные райо-
ны Казахстана и далее в горные районы Киргизии (Конурбаев, 1965).
Распространен на высоте 1700—2700 м н. у. м.
В равнинных районах Казахстана местами выплода служат раз-
личные естественные углубления рельефа, ежегодно заливаемые та-
лой водой. Места выплода, как правило, располагаются поблизости
от лесных участков и пойменных зарослей (рис. 26). В горных районах
выплод происходит в долинах по поймам равнинных участков гор-
ных рек. В равнинных районах Северного Казахстана личинки стар-
ших возрастов обнаруживаются в начале мая. Массовое появление
имаго происходит в середине этого месяца. Через 2—3 дня после вы-
плода начинается их нападение. Продолжительность периода актив-
ности длится около месяца. Последние особи отлавливаются в первой
декаде июня (Прыгунова, 1966). Вид моноцикличный.
Как и для других ранневесенних видов, для Ае. cataphylla ха-
рактерны утренняя и вечерняя активность и возможность дневного
нападения при приближении к местам дневок. Днюют самки в рас-
тительном покрове и всевозможных зарослях. Отличается агрессив-
ностью, залетает в поселки, расположенные в 2 км от мест выплода
(Прыгунова, 1966).
Ае. (О.) cyprius Ludl. (карта VIII)
Европейско-сибирский вид. На территории СССР обитает в лест-
ной и лесостепной зонах, интразонально вклиниваясь в степную.
В Казахстане, помимо лесостепной и степной, обнаружен в полупуст-
тынной и пустынной зонах. Южнее 45,5 параллели не отмечался.
Проникновение его в эти зоны шло, вероятно, по Иргиз-Тургайскому
междуречью и пойме Сарысу. Указание Д. И. Благовещенского (1937)
11 Е. Н. Павловского с П. А. Петрищевой (1937) о нахождении этого
вида в пойме Сыр-Дарьи касается, вероятно, светлой формы Ае. fla-
vescens.
99
Немногочисленность его в Казахстане отмечалась буквально все-
ми исследователями, что объясняется, вероятно, относительной оли-
готермнюстью этих комаров. Во всех случаях наблюдалась их при-
уроченность либо к сильно затененным, прохладным биотопам, либо
Рис. 26. Местообитания личинок Aedes cataphylla в горах Заилийского
Алатау.
к горным участкам. Причем, численность их увеличивалась по мере
поднятия на более значительную высоту. Так, например, в предгорьях
Южного Алтая (окр. Зыряновска, 470 м н. у. м.) этот вид обнаружить
не удалось. На высоте 800—1000 м н. у. м. (пос. Козлушка) встреча-
лись единичные экземпляры, а на высоте 1400—1600 м н. у. м. (Мяг-
кие ключи) он был обычным. То же самое касается Павлодарской
области. Если в пойме Иртыша около Павлодара встречались лишь
единичные экземпляры, то в Баян-Ауле (1200—1400 м н. у. м.) на-
падение было довольно массовым.
100
В Восточном Казахстане местами выплода служат сильно зарос-
шие разливы небольших горных ручьев (рис. 27). Иногда личинки
встречаются и в лесных заточных водоемах, заселяя их наиболее
глубокую и затененную часть. Независимо от типа водоема ветре-
Рис. 27. Заболоченные участки в Восточном Казахстане — местообитания
личинок Aedes cyprius.
чаются лишь единичные экземпляры. Это вполне согласуется с чис-
ленностью имаго в этих местах. Развитие личинок отмечено при
14—18°.
Наряду с Ае. excrucians в горных районах является самым позд-
но выплаживающимся видом из группы «cantans». Личинки IV ста-
дии обнаруживаются в начале второй декады июня, имаго появляют-
ся пятью-шестью днями позже. В отличие от других представителей
этой группы чаще нападает в дневное время. От мест дневок улетает
на более значительные расстояния.
Поздневесенний моноцикличный вид. Активен на всем протя-
жении большей части теплого сезона. Весьма агрессивен к человеку,
нападает и на скот.
101
Ae. (О.) communis De Geer (карта VIII)
Циркумбореальный голарктический вид. В СССР — типичный
обитатель лесной зоны. В Казахстане приурочен к лесным массивам
Южного Алтая и Казахского мелкосопочника (Баян-Аул, Каркара-
линск). На западе Казахстана обнаружен в дельте Волги.
Местами выплода в Восточном Казахстане являются различ-
ные затененные и полузатененные временные водоемы. Чаще всего
это разливы рек и затоки талых вод. Химический состав воды в мес-
тах выплода отличается слабой минерализацией (100—150 л«г/л), не-
большой жесткостью (0,6—1,2°), незначительной примесью органиче-
ских и неорганических соединений и слабощелочной реакцией (pH
7,2—7,7). В период развития личинок водоемы лишены всякой вод-
ной растительности, но дно их устлано прелой прошлогодней травой
и листвой.
Личинки III—IV стадий обнаруживаются в начале третьей де-
кады мая, а в конце месяца отмечаются первые нападающие особи.
Нападение наблюдается на 2—3 день после выплода. Развитие личи-
нок происходит при 8—20°. Довольно хорошо доращиваются в лабо-
раторных условиях при 20—22°. Это говорит о более высоком темпе-
ратурном оптимуме по сравнению о результатами наблюдений
М. Ф. Шленовой (1952) в Московской области.
Личинки в водоемах часто доминируют над остальными пред-
ставителями Aedes. С одного квадратного метра отлавливалось до
140 экземпляров. Это вполне соответствует тому количеству имаго,
которое наблюдается после выплода. В Восточном Казахстане (пос.
Козлушка) в утренние часы (7—7.20) нападало до 119 самок, в 10 и
16 час — единичные экземпляры (9—13), а в вечернее время (21—
21.20) — до 195 особей. Помимо человека, отмечено нападение на ко-
ров и лошадей.
Более чем для других видов, для Ае. communis характерен в
Восточном Казахстане массовый залет в помещения. За один вечер
при застекленных окнах и открытой для проветривания двери в се-
редине июня в комнату залетела 81 самка. В помещении нападают
круглосуточно, но в массе — во время рассвета и наступления су-
мерек.
Один из самых ранних видов. Выплод имаго происходит в конце
мая в более низкогорных районах и в начале июня в лесостепном
поясе горной зоны (900—1000 м н. у. м.). В предгорьях Южного Ал-
тая развитие Ае. communis происходит на высоте 500—2000 лен.у.м.
Выше разреженных участков лесного пояса самки не отлавливались.
В более высокогорных районах северо-восточного Алтая вид не был
найден (Желоховцев, 1938). Вполне возможно обнаружение его и в
102
равнинных участках лесостепной зоны Северного Казахстана, по-
скольку в граничащей с этими районами Омской области он обы-
чен (Нецкий, 1952). Моноцикличный, ранневесенний вид. Зимует
в фазе яйца.
От самок Ае. communis, собранных в природе, выделены вирус
лимфоцитарного хориоменингита (Глущенко, Гуцевич, Дудкина,
1957) и культура возбудителя туляремии (Олсуфьев, Руднев, 1960).
Ае. (О.) detritus Hal. (карта VIII)
Основная часть ареала в Средиземноморской подобласти Па-
леарктики с выклиниванием в более северные районы. В СССР рас-
пространен в основном в пустынной и полупустынной зонах. Такая
же закономерность характерна и для Казахстана. Крайней северной
точкой обнаружения вида являются южные предгорья Тарбагатая
(Геллер, 1939). В северных частях своего ареала распространен мо-
заично, в южных — повсеместно, но в специфичных водоемах.
Теплолюбивый, галофильный вид. По солевыносливости с ним
не может сравниться никакой другой представитель Aedes, хотя
вместе с ним встречаются иногда личинки Ае. с. caspius, Ае. flaves-
cens и Ае. montchadskyi. При этом если другие галофильные виды
развиваются в соленой воде вынужденно, то для Ае. detritus это
обычные водоемы. Выдерживает концентрацию соли до 5,2% (Kirkpat-
rick, 1925).
Местами выплода служат различные солоноватые скопления
воды с большой примесью гуминовых веществ, отчего цвет воды не-
редко становится буроватым. Сухой остаток растворенных в воде
солей может достигать 30 000 мг/л с преобладанием углекислых
(11 834 мг/л), хлористых (6985 мг!л) и сернистых (5452 мг)л) соеди-
нений.
Личинки держатся в заросших прошлогодним тростником час-
тях водоема, где питаются нитчатыми водорослями, нередко заби-
раясь в их гущу. В одном открытом заточном плёсе между барханами
развитие отмечено при совершенно обедненных кормовых условиях.
Дно водоема было покрыто лишь редкой выгоревшей прошлогодней
растительностью. В таких случаях восполнение недостатка перифи-
тона, вероятно, компенсируется за счет фильтрации гуминовых ве-
ществ.
Предпочитает незаросшие, хорошо прогреваемые мелководья.
Температура воды в них может колебаться от 8 до 22°, но может до-
стигать и 30—32°. В условиях Казахстана нам не приходилось встре-
чать места массового выплода этого вида. В бассейнах рек Или и
Сыр-Дарьи личинки начинают появляться с конца марта или в на-
103
чале апреля, после таяния снегов. В конце апреля или в первой поло-*
вине мая начинается лёт имаго. Численность ИХ никогда не бывает
высокой, хотя в бассейне Сыр-Дарьи она чуть выше, чем в бассейне
Или. Зимует в фазе яйца.
По литературным данным невозможно создать ясное представле-
ние о сезонном ходе численности этого вида. В условиях Казахстана
во всех случаях он ведет себя как моноцикличный вид. Об одной
генерации Ае. detritus говорят и сведения Л. А. Молотовой (1966) по
Туркмении. Совершенно противоположны данные П. П. Чинаева
(1964а) для многих районов Узбекистана, в том числе для верхнего
течения Сыр-Дарьи, где, по его наблюдениям, Ае. detritus встречает-
ся на протяжении почти всех летних месяцев. Аналогичные сведения
приводит и П. А. Петрищева (1936) для Туркмении. Возможно, в тех
местах этот вид и полицикличен, но в условиях Казахстана он осу-
ществляет лишь одну генерацию. Об этом можно судить как по мес-
там его выплода, которые заполняются водой лишь в весеннее время,
так и по морфо-биологической близости к комарам группы «commu-
nis», дающим лишь в отдельных случаях частичный повторный
выплод.
Местами дневок служит различный растительный покров, кото-
рый в большинстве случаев чрезвычайно обеднен у мест выплода,
и поэтому имаго рассеиваются по ближайшим стациям. Днюющие
комары отлавливаются в колодцах, оврагах, щелях, углублениях
почвы. Очень часто убежищами служат норы различных Животных.
П. А. Петрищева (1962) отлавливала самок в гнездах щурки, горного
голубя, ласточки и в норах варана. Малочисленность этого вида не
позволяет полностью определить круг его прокормителей. На челове-
ка нападает редко. Это говорит о том, что в местах обитания ему,
вероятно, приходится питаться на различных диких или пасущихся
домашних животных.
Ае. (О.) diantaeus Н. D. К. (карта IX)
Циркумбореальный голарктический вид. За пределы лесной зо-
ны не выходит, встречается редко.
В Казахстане отмечен впервые. Обнаружен в отрогах Южного
Алтая. Приводится для граничащих с Казахстаном районов Западной
Сибири (Кухарчук, 1965) и Центрального Алтая (Желоховцев, 1938).
Личинки последних стадий обнаруживаются в конце мая в раз-
личных, чуть прогреваемых солнцем, затененных водоемах. Особен-
но много их бывает в заточных ямах. Вода в них характеризуется
повышенной щелочностью (pH 7,7), увеличенной окисляемостью
(9,9 мг!л) и чрезвычайно слабой минерализацией (107 мг)л сухого
104
остатка). Дно водоемов обычно устлано прошлогодней листвой и гнию-
щими растениями. В отличие от многих других видов личинки пред-
почитают держаться в более глубокой, затененной части водоема. На
поверхность всплывают редко. Личинки хорошо различимы визуаль-
Рис. 28. Затененный водоем в Восточном Казахстане — местообитания ли-
чинок Aedes diantaeus, Ае. hexodontus, Ае. impiger, Ае. pullatus и Ае.
punctor.
но по длинным усикам, заметным в виде двух темных точек с боков
головы. Отлавливается и в других лесных водоемах, но обязательно
затененных. Плотность заселения достигает 26 личинок на 1 л«2 вод-
ной площади.
Температура воды в местах обитания личинок обычно не превы-
шает 8—10°. Места выплода чаще всего находятся на высоте 800—
1000 м н. у. м. Выше 1200 и ниже 800 м н. у. м. не отмечены. Личин-
ки встречаются совместно с Ае. communis, Ае. sticticus, Ае. punctor
и Ае. pionips.
Развитие происходит довольно долго. Имаго выплаживаются од-
ними из последних из группы ecommunis». Объясняется это более
узким диапазоном развития личинок, в результате чего они заселяют
более холодные, затененные участки водоемов. Тем не менее хорошо
выдерживает доращивание в лабораторных условиях при 19—24°.
105
Имаго появляются в начале июля. Массовое нападение отмече-
но через 2—3 дня после выплода. В зарослях нападают весь день.
В вечерние часы особенно активны. От мест выплода далеко не уле-
тают, предпочитая держаться на затененных участках. Активны на
протяжении почти всего теплого сезона — с июня по сентябрь (По-
лякова, Мирзаева, 1966).
Вид моноцикличный, дающий одну весеннюю генерацию. Как и
все виды этой группы, зимует в фазе яйца.
Предпочтения в выборе источников питания не отмечалось. Наб-
людалось нападение на человека, крупный рогатый скот и лошадей.
Очень агрессивен.
Из комаров этого вида выделен вирус лимфоцитарного менинги-
та (Глущенко, Гуцевич, Дудкина, 1957; Гуцевич, Подолян, 1959).
Ае. (О.) excrucians Walk, (карта IX)
Ареал охватывает бореальную часть Голарктики. Характерен
для лесостепной и частично степной зоны СССР (Штакельберг, 1937).
Широко распространенный и обычный для северных и восточных
частей Казахстана вид. Южнее 50° с. ш. отмечен лишь в Зайсанской
котловине и в районе Тургая.
Экология во многих чертах сходна с Ае. cantans. Местами вы-
плода служат временные лужи и полупостоянные заболоченности,
богатые гниющими растительными остатками. Обычен в весенних
пойменных водоемах. Встречается совместно с Ае. cantans, Ае. beh-
ningi, Ае. cinereus, Ае. communis, Ае. cataphylla, Ае. leucomelas,
Ае. pionips и др.
Чаще всего встречается в различных временных или полупосто-
янных водоемах, находящихся поблизости от кустарниковых или
древесных зарослей (рис. 27). Химический состав воды характеризу-
ется слабой минерализацией (100—150 мг!л), небольшой жесткостью
(0,6—1,2°), слабощелочной реакцией (pH 7,2—7,7) и незначительной
примесью отдельных солей.
Мелкие водоемы глубиной менее 15—20 см, как правило, не за-<
селяются. Предпочитает обширные разливы глубиной от 20 до 50 см,
иногда заселяет и более глубокие ямы (60—120 см) и в этом отноше-
нии приближается к Ае. cyprius. Личинки большую часть времени
проводят на дне, редко всплывая к поверхности. Только интенсивное!
взмучивание воды вызывает их массовое всплытие. Личинки IV ста-
дии отличаются медлительностью движений и слабой реакцией на
внешние раздражения. Плотность заселения до 40 особей на 1 м2.
В период развития личинок водоемы лишены какой-либо вегети-
рующей растительности. Основная масса прошлогодней растительно-
106
сТи в полупрелом виде устилает дно таких водоемов, создавая за счет
обилия перифитона прекрасные кормовые условия для развиваю-
щихся личинок. В конце развития и после выплода такие биотопы
обычно густо зарастают осоками и разнотравьем, в которых выпло-
дившиеся имаго днюют и осуществляют гонотрофический цикл и
впоследствии откладывают яйца. Густой растительный покров и со-
хранившаяся почвенная влага обеспечивают оптимальные условия
цля прохождения эмбрионального развития, а в последующем пред-
охраняют отложенные яйца от чрезмерного высушивания.
В низменных районах Восточного Казахстана выплод имаго про-
исходит в начале второй половины мая. На более возвышенных
участках (600—800 м н. у. м.) комары появлялись в конце мая, а в
лесостепном поясе (900—1000 м н. у. м.) в зависимости от степени
затененности водоема — в первой половине июня. Предельная высо-
та обнаружения в Южном Алтае около 1500—1600 м н. у. м., что со-
ответствует примерно разреженным участкам лесного пояса этой
горной системы.
В условиях Северного Казахстана личинки последних стадий
обнаруживаются во второй половине мая (Прыгунова, 1966). Выплод
имаго происходит в конце мая и затем держится более или менее на
постоянном уровне до второй половины августа. Затем численность
имаго резко уменьшается, и в середине сентября отлавливаются пос-
ледние экземпляры.
Как и Ае. flavescens, этот вид в зависимости от условий среды
обитания может давать частичный повторный выплод. Об этом со-
общают Г. И. Пецкий (1952), О. Н. Сазонова (1958), Д. Д. Нефедов
(1962 а) и И. Г. Прыгунова (1966).
Суточная активность характеризуется обычным для этой груп-
пы двухвершинным ходом с наибольшей численностью нападающих
в утренние и вечерние часы. В Восточном Казахстане с 7—7.20 на
человека нападало в середине июня 28 самок, в 10 и 16 час — от 3
до 9 и с 21 до 21.20 — 37.
Активный кровосос, нападающий на людей и домашних живот-
ных. Вблизи от убежищ агрессивен даже в дневные часы.
От отловленных в природе напившихся крови самок неоднократ-
но выделялись культуры возбудителя туляремии (Олсуфьев, Руд-
нев, 1960).
Ае. (О.) flavescens Mull. (карта IX)
Ареал охватывает северную и среднюю части Голарктической
области. Приурочен к лесным и лесостепным участкам. В Казахста-
не по предгорьям Тянь-Шаня и поймам рек доходит до 42° с. ш., ос-
новная же часть ареала лежит севернее 48-й параллели.
107
Типичный представитель лесостепных видов. Соответственно
этому обилен на севере республики и редок в Центральном и Юго-
Восточном Казахстане. Независимо от зональных условий места
выплода этого вида приурочены к поймам больших у малых рек.
Предпочитает неглубокие, хорошо прогреваемые, временные водоемы^
слабо заросшие растительностью {Phragmites, Typha, Carex). Места
выплода всецда находятся вблизи от густых кустарниковых или трост-
никовых зарослей (рис. 29).
Личинки предпочитают слабощелочную (pH 7,2—8,1), незначи-
тельно (641—742 м?1л) минерализованную воду, но могут встречать-
ся в водоемах с различной степенью минерализации (до 29 894 мг!л
включительно). В последнем случае состав воды отличался значи-
тельной щелочностью (pH 9,3), преобладанием сернистых соединенш
(5452 мг)л), углекислых (11 834 мг!л) и хлористых (6985 мг)л) солей
Экологическая пластичность Ае. flavescens позволяет ему сущест
вовать в сильно засоленной водной среде, но массовый выплод отме
чается лишь в слабо минерализованных водоемах.
Развитие личинок возможно по краям постоянных водоемов <
пологими берегами и сильно колеблющимся уровнем воды. По мер
усыхания мест выплода личинки концентрируются в центральных
более глубоководных участках водоема. В весенних потоках и ручь
ях держится среди водорослей, которые ослабляют силу тока воды
Характерной особенностью личинок является скрытность. Как пра
вило, они избегают открытых мест, прячутся среди густых зарослей
прошлогодней травы или поваленного тростника. Вполне возможно
что это вызвано трофическими связями, поскольку излюбленной сре
дой обитания личинок является вода, богатая взвешенными остатка
ми разлагающихся растений. Личинки часто встречаются совместна
с Ае. cinereus и Ае. kasachstanicus, а позже — с Ае. с. caspius.
В период усыхания водоемов могут жить и заканчивать разви
тие в межкочковых понижениях, где под слоем растительных остат
ков долгое время сохраняется вода.
Выплод личинок возможен при 7—8° (Сазонова, 1959). В даль
нейшем развитие протекает при 14—19° и длится 40—45 дней
Так, например, на юге и юго-востоке Казахстана первые личинки
обычно появляются в конце марта — начале апреля, а развитие ии
продолжается до начала мая. Выплод же происходит в первой дека!
де мая. 1
Очень часто водоемы с личинками покрываются ночью коркой
льда. Испытание в лабораторных условиях показало, что при мину(
6—10° личинки I стадии свободно переносят полное промерзание во*
ды в чашках с толщиной водного слоя от 1,5 до 6 см. В последующем
они нормально растут и развиваются. 1
108
Первые выплодившиеся особи на юге и юго-востоке Казахстана
появляются в конце апреля — начале мая. Нападать начинают с се-
редины мая; массовое нападение — в конце мая — первых числах,
июня. К концу июня численность имаго постепенно уменьшается.
На севере Казахстана первые выплодившиеся особи появляются
ро второй половине мая, массовый лёт в июне — июле и снижение
численности — в августе. В Гурьевской области немногочисленные
экземпляры этого вида отлавливались со второй половины мая до на-
чала июня. В отрогах Тянь-Шаня первые особи появляются во второй
декаде июня, массовое нападение — в июле и снижение численно-
сти — в конце августа.
При благоприятных условиях (температура, осадки, разливы)
в отдельные годы может давать частичный повторный выплод. Отно-
сительная моноцикличность этого вида описана Н. В. Хелевиным
(19586), который в различные годы отмечал от 0,6 до 30,6% повтор-
ного выплода. В июле 1964 г. подобное было отмечено нами в сред-
нем течении Или, а Ж. Исимбековым в августе — в Текесской меж-
горной долине. Температурные и гидрологические условия способст-
вовали этому. В первой половине июня шли частые дожди и наблюда-
лось затопление мест выплода. Температура воздуха не достигала
высоких показателей, что также благоприятствовало отрождению
второй генерации. Соответственно в Омской области наблюдается поч-
ти закономерное развитие двух генераций этого вида (Пецкий,
1952, а, б).
Возможность повторного выплода личинок была проверена в ла-
бораторных условиях. Из 17 полученных кладок частичное отрожде-
ние личинок отмечено лишь в двух. Часть кладок содержалась под
водой, а часть — на плавающей фильтровальной бумаге. Выплод
происходил лишь из яиц, содержащихся не менее 13—15 дней на
влажной фильтровальной бумаге до погружения их в воду. Процент
отродившихся личинок колебался от 9 до 15. Из оставшихся яиц вы-
плод не произошел даже при повторных просушиваниях и погруже-
ниях в воду.
В условиях Казахстана, где водный режим подвержен большим
колебаниям, это, вероятно, одна из форм экологической приспособ-
леннофи для сохранения вида. Благодаря ей возникает возможность
откладки яиц на различные уровни усыхающего временного водоема.
Весной кромка воды временного водоема дальше от центра, а летом —
ближе. На следующий год может не наступить полного затопления,
и тогда выплод происходит только в яйцах, отложенных ближе к
центру. Подобная картина была отмечена в 1965 г., когда незначи-
тельное половодье было лишь весной, но тем не менее часть популя-
ции все же выплодилась.
109
Недостаточность специфических биотопов у южной границы аре-
ала восполняется экологической приспособленностью к среде обита-
ния. Продолжительность жизни самок около двух месяцев. За это
время они, по данным Н. В. Волозиной (1958), успевают проделывать
до 8 гонотрофических циклов. На юго-востоке Казахстана их значи-
тельно меньше, поскольку продолжительность переваривания крови
длится от 7 до 12 дней, а период активности этого вида не превыша-
ет 1—1,5 месяца.
Чаще всего самки откладывают от 150 до 250 яиц, но иногда, в
зависимости от степени насыщения кровью и фазы развития яични-
ков, ограничиваются 30—60. Из кладок, содержавшихся в лаборато-
рии без промораживания, весной при залитии водой происходил вы-
плод личинок. Это соответствует выводам Н.В. Хелевина (1958 б) о
том, что отрождение личинок Ае. flavescens возможно и без промора-
живания яиц, находящихся в диапаузе. Вполне вероятно, что это
экологическая адаптация к более теплым условиям среды обитания,
а не просто асинхронность развития, как объясняет Н. В. Хелевин.
Кратковременное промораживание яиц в летнее время к прекраще-
нию диапаузирования не приводило, что само по себе уже говорит о
необязательности промораживания.
Подобно Ае. kasachstanicus самки этого вида не всегда довольст-
вуются однократным питанием и сосут кровь повторно. Возможно
автогенное развитие яичников ((Виноградова, 1965 а). Предпочитает
откладывать яйца на воду, а иногда и на влажную поверхность. Яй-
ца, отложенные на воду, обычно плавают, хотя некоторые сразу по-
гружаются на дно. Может быть, это связано с частичным повторным
выплодом, так как в одной и той же партии яиц отрождение личинок
произошло из яиц, находившихся на поверхности воды.
Копуляция может, вероятно, осуществляться как в роях, так и
без них. Роение на высоте 2—2,5 м над различными растениями или
крупными животными с 20.30 до 21 час наблюдал в Текесской меж-
горной долине Ж. Исимбеков (1966). Нам не удалось обнаружить рое-
ния этого вида, зато были отмечены активные поиски самцами самок
среди кустарниковых зарослей. В лабораторных условиях самки, со-
держащиеся после выплода в течение 3—5 дней с самцами, впослед-
ствии пили кровь и откладывали яйца, из которых выплаживались
личинки, что говорит о стеногамности этого вида.
Суточная активность Ае. flavescens характеризуется двухвер-
шиипостью, характерной для основной массы комаров. Утренняя ак-
тивность, наблюдаемая с рассвета до полного восхода солнца, слабее
вечерней, отмечаемой в предсумеречные часы. Вечерней активности
предшествует некоторое оживление, выражающееся в коротких пе-
релетах, концентрации в отдельных участках и т. п. Сплошная темно-
по
та действует угнетающе, и, наоборот, при нахождении вблизи от
мест дневок объекта питания наблюдается довольно интенсивное
дневное нападение. От мест выплода и дневок далеко не отлетает, в
населенные пункты не залетает. В светоловушку отлавливается чрез-
вычайно редко.
Днюют имаго в зарослях кустарника или в густом припоймен-
ном травостое. На севере Казахстана местами дневок служит траво-
стой на открытых участках лесных колок и припойменных зарослей.
Самки агрессивны как по отношению к человеку, так и к раз-
личным домашним животным (крупный рогатый скот, лошади, ос-
лы). Нападает на верхние или средние участки тела человека или жи-
вотного. За один 20-минутный сбор на человеке отлавливалось до
100 особей этого вида. В двух случаях по одной самке было отловле-
но в бассейне Сыр-Дарьи у гнезда болотного луня.
Массовый 1k широко распространенный кровосос на севере Ка-
захстана и в отдельных припойменных и высокогорных участках
Центрального и Юго-Восточного Казахстана. Один из видов, созда-
ющих весеннее неблагополучие местности по гнусу.
В опытах передавал через укус возбудителя туляремии, продол-
жительное время (8—14 дней) выделяя его с фекалиями (Олсуфьев,
Руднев, 1960).
Ае. (О.) hexodontus Dyar
До 1959 г. считался неарктическим видом. О. Н. Сазонова (1959)
отделила Ае. hexodontus от Ае. punctor и привела его для фауны
СССР, что требует дальнейшего подтверждения.
В Казахстане обнаружен лишь в отрогах Южного Алтая на вы-
соте 900—1000 м н. у. м. Личиночными стациями служили различ-
ные лесные и затененные пойменные водоемы (рис. 28). Вода в них
отличалась прозрачностью, слабощелочной реакцией (pH 7,2—7,7),
незначительной минерализацией (107—148 мг!л) и повышенной окис-
ляемостью (9,9—10,7 мг)л). Дно водоемов было устлано гниющей
прошлогодней листвой и травой. Температура воды в период разви-
тия колебалась в пределах 8—16°.
Личинки последних стадий обнаруживаются во второй половине
мая. Численность их невелика, до 5 экземпляров на 1 м2. Подобно
Ае. punctor и Ае. diantaeus они предпочитают более глубокие, зате-
ненные участки водоемов. Много общего у этих видов и в других де-
талях экологии. Окукливание наступает в последних числах мая, а
в первых числах июня уже происходит выплод имаго. В более зате-
ненных и прохладных водоемах все сроки развития удлиняются на
10—12 дней.
111
Соответствует малой численности личинок и незначительное ко-
личество выплаживающихся имаго. В основной массе более много-
численных видов присутствие Ае. hexodontus было незаметнС|. Это
затрудняло определение его суточной активности. В целом она совпа-
дала с суточным ритмом всей группы «communis*. Представлен в
сборах с человека и с крупного рогатого скота.
Ае. (О.) hungaricus (?) Mihalyi
Вид, недавно описанный по материалам из Венгрии (Mihalyi,
1955, 1961). Предположительно указывается для фауны Чехослова-
кии (Kramar, 1958).
В Восточном Казахстане обнаружены личинки IV стадии, впол-
не соответствующие видовому диагнозу этого вида. От Ае. sticticus
хорошо отличаются простыми внутренними лобными волосками, а
также формой и величиной чешуек щетки. Сравнение их с паратипа-
ми Ае. hungaricus, имеющимися в коллекциях ЗИН АН СССР, пока-
зало их довольно четкую идентичность.
Личинки были отловлены во временном лесном водоеме на вы-1
соте 900—1000 м н. у. м. (пос. Козлушка). Время обнаружения —,
третья декада мая. Сопутствующими были личинки Ае. communis,
Ае. pullatus и Ае. punctor. Эти отдельные фрагменты экологии также
соответствуют описанию Ае. hungaricus.
Единственная в СССР находка не дает полного права утверждать
с достоверностью наличие вида на территории Советского Союза. Тем
более не понятна разобщенность предполагаемого ареала. Вместе с
тем поскольку до последнего времени сведения об Ае. hungaricus в
определениях комаров фауны СССР отсутствовали, то вполне воз-
можно, что по личинкам этот вид мог относиться либо к Ае. sticticus,
либо к Ае. impiger. Нам удалось его выделить лишь благодаря ново-
му варианту таблицы для определения личинок комаров рода Aedes,
любезно присланной нам профессором А. С. Мончадским для апро-
бирования.
Ае. (О.) impiger Walk.
Арктический вид. Распространен в северной части Америки
(Matheson, 1944) и на крайнем севере Европы (Штакельберг, 1937;
Мончадский, 1951). По мнению Натвига (Natvig, 1948), характерен,
кроме того, для горных районов. В СССР его ареал охватывает зон$
тундры с выклиниванием на юг по горным районам (Румш, 1948;
Сазонова, 1959). В Неарктике также приурочен к горным районам,
доходя до 2800 м н. у. м. (Matheson, 1944). Экология изучена слабо.
112
Вследствие запутанной синонимики (Ае. parvulus, Ае. nearcticus)
ведения о распространении и биологии очень фрагментарны.
Обнаружен лишь в Восточном Казахстане и на возвышенных
лесных массивах Казахского мелкосопочника на высоте 900—
1200 м н. у. м. Среди массы других представителей группы «commu-
nism личинки этого вида встречались спорадически. Массовой кон-
центрации нигде не отмечено. Местами выплода служат сильно зате-
ненные лесные водоемы, постоянно подпитываемые тающими снега-
ми (рис. 28). Температура воды в них при суточном режиме воздуха
10—26° не поднималась выше 8—10°. Вода относительно чистая,
прозрачная, слабо минерализованная (148 мг!л), чуть щелочная
(pH 7,2), с повышенной окисляемостью (10,7), что, вероятно, опреде-
ляется гниением прошлогодней листвы, устилающей дно водоема.
Развитие происходит совместно с Ае. pullatus, Ае. punctor, Ае. hexo-
dontus и Ае. communis.
Личинки II стадии обнаружены в конце мая. Развитие их шло
долго. Окукливание наступило лишь в начале второй декады июня.
Выплод имаго произошел в середине июня одновременно с Ае. hexo-
dontus и Ае. punctor. Из-за гибели личинок лабораторное культивиро-
вание не удавалось. Нападение отмечено через 1—2 дня после вы-
плода. Вследствие крайней малочисленности вида суточную актив-
ность проследить не удалось. Наблюдалось нападение на человека.
Вид холодолюбивый. Проделывает одну генерацию в год.
Своеобразие находок в предгорьях Алтая говорит об их вероят-
ной связи с фауной центральной части этой горной системы. Судя по
холодолюбивости и предпочтению горных участков его присутствие
зозможно в других районах Алтая на более значительных высотах,
что отмечено А. Н. Желоховцевым (1938) в северо-восточной части.
Трудно судить о путях проникновения этого вида в данный регион.
Скорее всего это гляциальный реликт, поскольку сведения о его на-
хождении на сопредельных территориях Западной Сибири и Забай-
калья отсутствуют (Гуцевич, 1939; Полякова, 1963; Кухарчук, 1965;
Полякова, Мирзаева, 1966). Такая обособленность дает основание
предполагать более широкое распространение этого вида в леднико-
вый период. В то время такой прерывности ареала, вероятно, не су-
ществовало.
Ае. (О.) intrudens Dyar (карта IX)
Ареал охватывает лесную зону Северной Америки и Евразии.
Соответственно этому распространен на территории РСФСР. В Казах-
стане до последних лет считался редким. Проведенные исследования
показали, что этот вид заселяет не только всю северную половину
8—80
113
Казахстана, но и проникает по отрогам Тянь-Шаня далеко на юг
(44° с. щ.). Встречается даже в пойменных зарослях пустынной части
бассейна Или, куда, вероятно, проникает по поймам впадающий в
нее горных рек. Разобщенность находок единичных экземпляров на
северном побережье Каспия (пойма Волги) с основной частью ареала
находит свое объяснение в исследованиях Э. К. Мартини (1926), об-
наружившего Ае. intrudens в пойме Волги в окрестностях Саратова,
откуда он также мог проникнуть по пойменным лесам до дельты
Волги.
Ранневесенний моноцикличный вид, развитие которого во всех
случаях приурочено к лесным биотопам. На востоке и юго-востоке
Казахстана это лесостепные и лесные участки соответствующих поя-
сов горной зоны. В равнинных частях Казахстана выплод его отме-
чается в пойменных лесах и зарослях протекающих здесь рек. Такая
приуроченность объясняется тем, что вид имеет, по данным
М. Ф. Шленовой (1950), самый низкий из группы * communis* предел;
вылупления личинок ( + 4°), а температура 10—13° является верхним
порогом развития. В Казахстане таким условиям могут удовлетво-
рять лишь сильно затененные, слабо прогреваемые водоемы, на ко-
торых мало сказывается действие характерной для этих мест силь-
ной инсоляции.
В Восточном Казахстане местами выплода Ае. intrudens слу-
жат те же самые биотопы, в которых происходит развитие Ае. com-
munis, Ае. punctor, Ае. pullatus, Ае. hexodontus и Ае. pionips. Не по-
казывая специфической предпочитаемости, встречается как с каж-
дым в отдельности, так и в групповых биотопах. Характерной особен-
ностью этого вида является крайняя малочисленность его популяций»
Во всех случаях встречаются лишь единичные личинки. Плотность
заселения водоемов не превышает 3—8 экземпляров на 1 м2. Разви-
тие отмечено при 8—17°. Более высокие температуры приводят, ве-
роятно, к гибели предимагинальных фаз. Так, например, вследствие-
гибели личинок нам ни разу не пришлось получить в лабораторных,
условиях при 20—23° выплода Ае. intrudens, хотя удавалось это осу-
ществить у других олиготермофилов (Ае. pullatus, Ае. punctor,
Ае. communis). В Восточном Казахстане выплод имаго происходит
вслед за Ае. communis в первых числах июня.
Своеобразна фенология этого вида в горных условиях Северного
Тянь-Шаня. Вследствие отмеченной олиготермности выплод его про-
исходит в первых числах июля. В начале второй декады отлавлива-
ются лишь напившиеся крови непотертые самки. Убежищем им слу-
жит густой травостой на затененных участках кочкарниковых сазов.
Появляется раньше Ае. pullatus, но в меньшем количестве. Сезонный
ход численности подобен Ае. cataphylla и Ае. leucomelas.
114
Суточный ход активности характерен для видов, нападающих
лри низких температурах. Утренняя и вечерняя активность пониже-
на по сравнению с равнинными видами, но зато отмечается интенсив-
ное дневное нападение. Объектом нападения служит скот, пригоняе-
мый в это время на летний выпас. На человека по сравнению с Ае. ca-
taphylla, и Ае. leucomelas нападает меньше. Местами дневок служат
высокий травостой, кустарниковые заросли и ельники.
Ае. (О.) kasachsrtamcus Guts, (карта X)
Вид, описанный А. В. Гуцевичем (1962) по сборам из окрестно-
стей Или иска. Распространен в припойменных участках среднего те-
чения р. Или. До 1966 г. его личинки не были известны, что затрудня-
ло определение подродовой принадлежности вида.
Полевыми наблюдениями и лабораторными опытами установле-
но. что это поздневесенний моноцикличный вид. Первые имаго появ-
ляются в конце апреля — первой половине мая. Численность их за-
тем прогрессивно увеличивается и держится на высоком уровне весь
июнь. С начала июля замечается уменьшение количества особей, и
в последних числах этого месяца можно отловить лишь единичные,
сильно потертые (возрастной признак популяции) экземпляры.
По фенологии и экологии этот вид очень напоминает Ае. flaves-
cens, с личинками которого и происходит его совместное предимаги-
нальное развитие. Личиночными стациями служат весенние разли-
вы небольших рек и скопления талых вод в неровностях почвы с
большим количеством опавшей прошлогодней листвы, густой травы
или тростника (рис. 29). Больше всего таким условиям удовлетворя-
ют тугайные водоемы. Бода в местах развития личинок слабо мине-
рализована, до 742 мг/л, с небольшим содержанием карбонатов, суль-
фидов, хлоридов, но с увеличенным количеством аммиачных солей;
pH воды слабощелочная — 7,2—8,05. В одном случае личинки пер-
вых возрастов были обнаружены совместно с Ае. simanini в харак-
терных для этого вида значительно минерализованных водоемах.
Выплод личинок происходит во второй половине марта в теп-
лую, солнечную погоду, когда температура воздуха поднимается до
19—20°, а мелководные участки прогреваются до 12—13°. В услови-
ях ранней, но затяжной весны 1966 г. предимагинальное развитие
Длилось 35—45 дней. Первые выплодившиеся самцы появляются в
конце апреля, но в это время еще встречаются личинки IV стадии и
куколки. Последние экземпляры выплаживаются в первой декаде
мая.
115
Наблюдения за фенологией и экологией ведутся с 1963 г. За это
время отмечено ежегодное увеличение численности вида и освоение
новых биотопов.
Рис. 29. Характерный для Aedes kasachstanicus и Ае. flavescens водоем в
бассейне р. Или.
Самцы и самки появляются одновременно. Отмечается строгая
приуроченность их к растительным зарослям. Все время находятся
в тугаях и кустарниках, на удаленных от зарослей участках не напа-
дают. Характерна дневная и предвечерняя активность вида, которую
можно объяснить особенностью его копуляции. В зарослях нападение
самок отмечается в течение всего дневного времени, но наиболее ак-
тивно с 18 до 19 час, при температуре воздуха 24—26°, относительной
влажности 55—65%, освещенности 12 000—24 000 люкс и скорости
116
ветра 1,5—2 м'сек. После 19 час (t^ 20°, Н = 78°/о, освещенность
6000 люкс, штиль), хотя еще светло, активность резко обрывается, и
в сумеречное и ночное время нападения не происходит. Роения сам-
цов не отмечено, однако, как только в зарослях появляется человек
или животное, комары вылетают из убежищ и начинают кружить во-
круг него на высоте 1—1,5 м. Завидя летящую к объекту нападения
самку, самец моментально устремляется к ней и копулирует на лету.
По-видимому, дневная активность самок не что иное, как био-
логическое приспособление ^этого вида, направленное на увеличение
возможности обнаружения их самцами в течение светлого периода
времени.
Самцы, постепенно уменьшаясь в количестве, исчезают в сере-
дине июля. Лёт самок затягивается до последних чисел июля. В пос-
ледние дни, как правило, нападают самки, имеющие незначительное
количество полупереваренной крови в желудке. В начальный период
активности такого не отмечается. Лабораторные исследования пока-
зали, что приема любого количества крови достаточно для прохожде-
ния гонотрофического цикла и откладки определенного количества
яиц. Период переваривания крови при 20—25° длится 10—11 дней,
а при 23—29° — 5—7 дней. В отдельных случаях он затягивается до
13—23 дней. В садках самки пьют кровь менее охотно, чем в природе.
Количество откладываемых яиц колеблется от 14 до 115, чаще
50—70. Для яйцекладки предпочитают влажную фильтровальную бу-
магу,* хотя иногда яйца откладываются в воду у ее уреза. Некоторые
самки откладывали яйца в сироп, находившийся в садке для под-
кормки. Выплода личинок не происходило ни в партиях яиц, содер-
жавшихся 3 месяца в воде при оптимальной температуре, ни в под-
вергавшихся различной степени просушке и даже кратковременно-
му промораживанию.
В окрестностях пос. Илийска численность вида невелика, массо-
во отмечен в сборах из более верхних участков поймы р. Или (Аяк-
Калкан, Борохудзир). В последнее время обнаружен и в ниж-
нем течении р. Или. По наблюдениям четырех лет, численность напа-
дающих никогда не бывает особенно высокой. За 20 мин на человека
нападало 3—5 самок.
В обычные годы единичные экземпляры Ае. kasachstanicus до-
вольно редко встречаются среди массы комаров других видов. В усло-
виях засушливого 1965 года, когда летние генерации других видов
были крайне ничтожны, этот вид в характерном для него биотопе
являлся доминирующим. Одинаково охотно нападал на людей, ло-
Нгадей и крупный рогатый скот.
117
Ae. (О.) leucomelas Meig. (карта X)
/
Ареал охватывает Европейско-Сибирскую подобласть Палеарк-
тики. В Казахстане основная его часть лежит севернее 48° с. ш. По
горным и предгорным районам выклинивается до 43° с. ш. За преде-
лами юго-востока Казахстана известен из Иссык-Кульской котлови-j
ны (Конурбаев, 1965).
Ранневесенний моноцикличный вид. В равнинных районах Ка-
захстана развитие личинок происходит во временных водоемах, об-
разующихся за счет таяния снега и весеннего половодья. Химически!
состав воды в этих биотопах характеризуется повышенной минералц
зацией (3312 мг!л), большой жесткостью (20,6°), увеличенным содер
жанием сернистых (142 ли/л) и углекислых (732 лгг/л) соединений i
щелочной реакцией (pH 8). Водоемы открытые, хорошо прогревав
мне, хотя и приуроченные к тугайным зарослям. Дно их, как пра
вило, усеяно опавшими листьями или прошлогодней прелой расти
тельностью.
В горных районах (1400—2600 м н. у. м.) местами выплода слу
жат скопления воды, образующиеся в результате таяния льда и раз
ливов рек на равнинных плоскогорьях. Чаще всего обнаруживаются
совместно с другими представителями группы «communis* — Ае. са
taphylla, Ае. intrudens. В отличие от последних по поймам горны:
рек проникает в низинные районы бассейна Или (окр. Илийска). Вь
плаживается во второй декаде апреля и активен всего лишь в течс
ние 15—20 дней. В Кустанайской области выплод имаго проиоходи
в конце второй декады мая. Спустя 4—5 дней наблюдается массовы:
лёт комаров, который, постепенно снижаясь, прекращается в конц
июня (Прыгунова, 1966). В горных районах (2400—2600 м н. у. м.
выплод отмечен в конце второй декады июля.
Активные кровососы. Нападают на людей и домашних живоч
ных. Далеко от мест выплода не улетают. Держатся в заросших ста
циях. В горных условиях днюют в травяной растительности, напада
на пасущийся скот.
Ае. (О.) mariae (?) Ed. et Et. Serg.
Средиземноморский вид, распространенный в различных частя;
европейского, африканского и западно-азиатского побережья Среди
земного моря. В Казахстане личинки и имаго с признаками Ае. та
riae встречаются в солоноводных водоемах и у побережья морей Бал
хаш-Арало-Каспийского бассейна. Места их обнаружения соответс!
вуют экологии этого приморского стенобионта. Тем не менее мы нч
можем с достоверностью утверждать его наличие на территории Ка
118
захстана, поскольку существует вся гамма особей с переходными к
д_е. с. caspius признаками.
Ае. (О.) montchadskyi Dub. (карта X)
Недавно описанный (Дубицкий, 1968) вид. Личинки обнаруже-
ны в 1966 г. в бассейне р. Или во время поисков неизвестных тогда
Рис. 30. Характерный для Aedes montchadskyi временный водоем около
тугайных зарослей в бассейне р. Или.
предимагинальных фаз Ае. kasachstanicus и Ае. simanini. Распрост-
ранение прослежено от Илийска до Борохудзира. Обнаружен
В. Н. Ануфриевой в районе Зайсана (материал поступал для подтверж-
дения).
Вид относительно редкий. Места выплода очень ограниченны.
Единичные личинки отлавливаются во всевозможных весенних за-
токах талой воды. Чаще всего они встречаются в небольших времен-
ных водоемах, расположенных как непосредственно в тугайных за-
119
рос лях, так и около них (рис. 30). Предпочитает слабую минерализа-
цию, но подобно другим распространенным здесь видам часто встре-
чается в воде со значительной концентрацией солей. Соответственно
этому обычно встречается с личинками Ае. flavescens и Ае. kasach-
stanicus, реже с ранневесенней' генерацией Ае. с. caspius, а в одном
случае обнаружен даже с Ае. detritus. Численность личинок колеб-
лется от 1 до 12, обычно 1—3 на 1 лг2. Значительное количество личи-
нок было обнаружено лишь в затененной яме глубиной 10—40 см и
площадью 1 м2.
По фенологии Ае. montchadskyi можно отнести к группе поздне-
весенних видов. Выплод личинок отмечается в конце марта — нача-
ле апреля. Окукливание происходит в конце апреля — начале мая.
Через 5—7 дней после этого наступает выплод имаго. Нападение на
человека отмечается крайне редко, возможно, вследствие малочис-
ленности. Сезонная и суточная активность не прослежена.
Ае. (О.) nigrinus (?) Eck.
Вследствие большого сходства с Ае. sticticus распространение
не совсем выяснено. Отмечен в Южной Скандинавии и в Центральной
Европе (Natvig, 1948; Kramar, 1958). В СССР с достоверностью заре-
гистрирован в Эстонии, в Ленинградской, Волгоградской и Москов-1
ской областях (Штакельберг, 1937; Мончадский, 1951; Ремм, 1957«
Сазонова, 1959).
В Казахстане самцы этого вида обнаружены лишь в предгорьях
Южного Алтая (окр. Зыряновска). Выплод получен из куколок, со-
бранных в полузатененных лесных водоемах. Поскольку здесь же в1
значительном количестве обитает и Ае. sticticus, к определению это-
го вида мы подходим с большой осторожностью, хотя просмотренные
гипопигии по степени выраженности базальной бородавки различа-
ются заметно. Непонятно отсутствие личинок и самок Ае. nigrinus,
отличающихся от Ае. sticticus.
Учитывая большую степень изменчивости комаров, которые встре-
чаются в Восточном Казахстане, мы все же решили условно вклю-
чить этот вид в список обнаруженных видов.
Ае. (О.) nigripes Zett.
Один из наиболее холодолюбивых голарктических видов. По мне-
нию Натвига (1948), подобно Ае. pullatus отличается приуроченно-
стью к горным районам. В Казахстане единичные экземпляры обна-
ружены в лесных, горных биотопах в предгорьях Южного Алтая.
Экология местной популяции не изучена. Две самки пойманы на се-
120
бе в первых числах июня, а самец выплодился из куколки, отловлен-
дой в полностью затененном лесном водоеме. Эти единичные наход-
ки не создают ясного представления о численности и фенологии вида.
Требуются дальнейшие исследования.
Ае. (О.) pionips Dyar (карта X)
Вид до недавнегб времени считался неарктическим. Для СССР
впервые указан О. Н. Сазоновой (1959), которая считает, что за счет
имагинального сходства с Ае. communis, а личиночного — с Ае. pul-
latus этот вид проходил раньше под тем или иным диагнозом. Вы-
сказывается предположение, что сообщения многих авторов о нахож-
дении Ае. pullatus в более южных, равнинных областях относятся к
Ае. pionips. По мнению Матесона (1944), этот вид представляет лишь
вариант Ае. communis.
Экология Ае. pionips изучена слабо как у нас, так и за рубежом.
О. Н. Сазонова (1959) отмечает только, что личинки обнаружива-
лись в лесных водоемах верхнего течения Клязьмы, развитие проте-
кало долго, выплод комаров поздний и сильно растянутый.
В Восточном Казахстане обнаружена смешанная популяция всех
трех видов: Ае. pullatus, Ае. communis, Ае. pionips. Личинки III—
IV стадий отловлены в третьей декаде мая во временных заточных
водоемах. Во всех водоемах вода слабощелочная (pH 7,2—7,7), с не-
значительной минерализацией (107—148 мг)л) и повышенной за счет
гниения находящейся на дне прошлогодней листвы окисляемостью
(9,9—10,7 мг!л). Водоемы, кроме одного, затенены деревьями и кус-
тарниками. Температура воды 24—26 мая не превышала 8—9°, а в
начале июня она повысилась до 12—14°.
Для нижней границы лесного пояса (900—1000 м н. у. м.) ха-
рактерно следующее. Личинки заселяют неглубокие участки водое-
мов, не избегая солнечного освещения. Окукливание начинается в
последних числах мая, а выплод имаго отмечен в начале июня. Этот
вид появляется следом за Ае. communis. В более низменных районах
выплод имаго происходит на 6—8 дней раньше, в то время как в сред-
ней части лесного пояса и в сильно затененных водоемах он наблюда-
ется во второй половине июня. Развитие происходит совместно с
Ае. communis, Ае. pullatus, Ае. sticticus и Ае. diantaeus. Позже в этих
же водоемах выплаживаются Ае. cantans, Ае. excrucians, Ае. behnin-
gi и Ае. с. cinereus.
Один из массовых в тех местах видов. Плотность заселения водо-
емов достигает 30 личинок на 1 м2. Выплодившиеся имаго начинают
нападать на следующий день. В поисках добычи рассеиваются на
200—300 м. Совместно с некоторыми другими видами группы *сот-
121
munis» залетают для нападения в помещения. Суточная активность в
первых числах июня характеризуется двумя пиками (утренний и ве-
черний) на открытых участках и одним (дневной) в затененных мес-
тах. Самки очень агрессивны по отношению к человеку и домашним
животным.
Вид, несомненно, моноцикличный. Зимует в фазе яйца. Сведения
о переносе этим видом каких-либо заболеваний отсутствуют.
Ае. (О.) pulchritarsis Rond.
Основная часть ареала лежит в Средиземноморской подобласти
и заходит в Ориентальную область (Штакельберг, 1937). В Казахста-
не найден лишь в предгорных районах (Алма-Ата, Чимкент), гораздо
шире распространен в Среднеазиатских республиках (Узбекистан,
Туркменистан, Таджикистан).
Чрезвычайно редкий вид. Развитие личинок происходит в дуп-
лах деревьев, следовательно, места выплода крайне ограниченны. От-
сюда его редкая встречаемость и мозаичность распространения. В ус-
ловиях Туркмении чаще всего обнаруживался в дуплах ореха и кара-
гача (Петрищева, 1936). В прикорневых дуплах деревьев, располо-
женных недалеко от водного источника, вода сохраняется дольше,
поэтому наблюдалось развитие нескольких поколений. В основном
же возможность выплода была лишь в периоды с частыми осадками
(весеннее время). П. А. Петрищева (1936) отлавливала личинок, ку-
колок и имаго с апреля по июнь. За это время отмечался выплод трех
генераций. Е. А. Плетнев (1943) отлавливал единичные экземпляры
в конце июня.
В большинстве районов Казахстана естественная и декоративная
древесная растительность (сосна, ель, осина, береза, тополь и др.)
слабо подвержена кариетическим процессам с дуплообразованием.
В то же время в пустынных участках, где преобладает туранга, мож-
но встретить целые рощи деревьев с заполненными водой дуплами.
Жизненный цикл этого вида, кроме ограниченности биотопов,
ничем не отличается от других представителей Aedes. Яйца отклады-
ваются или непосредственно на воду, или на влажную органическую
подстилку дна дупла. В зависимости от наличия или отсутствия во-
ды происходит выплод личинок или задержка развития. Зимует в
фазе яйца. Помимо дупел встречается в пещерах, гротах, щелях. Об-
наружен в убежищах джейранов, кабанов, ежей и норах-гнездах не-
которых птиц, которые, вероятно, и являются прокормителями этих
комаров (Петрищева, 1962).
122
Ae. (О.) pullatus Coq. (карта XI)
Распространен в бореальной зоне Палеарктики и Неарктики. Тя-
готеет к горным и предгорным районам. В более северных районах
заселяет и низменные участки. В РСФСР характерен для северо-та-
ежной зоны и горных районов, но проникает и в полосу смешанных
лесов (Сазонова, 1959). В Казахстане за счет приуроченности к лес-
ным горным участкам вклинивается в глубь степной зоны (Баян-Аул,
Каркаралинск), а в отрогах Тянь-Шаня доходит даже до уровня пус-
тынной зоны (Заилийский Алатау). Как и в примере с Ае. punctor,
такая экстразональность говорит о реликтовом происхождении отме-
чаемой мозаичности.
В северо-восточных районах Казахстана встречается на высоте
900—1800 м н. у. м., что соответствует зоне лесостепей. Заселяет раз-
личные затененные и полузатененные водоемы, образованные талы-
ми и паводковыми водами. Чаще всего ими служат различные ямы,
впадины и затоки горных ручьев й родников, а нередко и образуемые
ими заболоченности. Вода чистая, слабо минерализованная (100—
150 лгг/л), с незначительной примесью различных органических и не-
органических соединений и слабощелочной реакцией (pH 7,2—7,7).
В период развития личинок водная растительность отсутствует. В
лабораторных условиях (20—23°) большинство личинок гибнет. Полу-
чить выплод имаго удается лишь в редких случаях.
Развитие личинок происходило при 8—18° и соответственно это-
му продолжалось до конца первой декады или середины июня. Чаще
всего они встречались совместно с Ае. communis, Ае. pionips, Ае. pun-
ctor и Ае. hexodontus. Время выплода совпадало с вылетом Ае. pun-
ctor. По численности обнаруживаемых личинок этот вид занимал
второе место после Ае. communis. С одного квадратного метра отлав-
ливалось до 100 экземпляров.
Выплодившиеся имаго не отличались большой светочувствитель-
ностью. Нападение отмечалось и в дневное время. Подобно Ае. com-
munis летели для нападения в ближайшие села. Часто залетали в по-
мещения. В числе источников питания были люди, крупный рогатый
скот и лошади.
В отличие от Восточного Казахстана в отрогах Тянь-Шаня
Ае. pullatus встречался лишь на высоте 2600—2800 м н. у. м. (пояс
субальпийских лугов). Места выплода — кочкарниковые торфяники,
находящиеся в местах разлива родниковых и талых вод (рис. 31).
Личинки встречались как в межкочковых канавках, так и в местах
сплошного разлива воды. Глубина водоемов не превышала 25—
35 см. Если в Восточном Казахстане дно водоемов было устлано бо-
гатейшим органическим детритом, то здесь имелись лишь чахлые
123
остатки прошлогодней растительности. Химический состав воды был
бы однотипным, не будь очень значительного уклонения в кислотную
сторону (pH 4,9) и увеличения окисляемости до 51,3 мг!л.
Рис. 31. Кочкарниковые «сазы» в субальпийском поясе Заилийского Ала-
тау — местообитания личинок Aedes pullatus и Ае. intrudens.
Личинки развивались при температуре воды 4—22°. Такой про-
греваемости на этой высоте способствовала значительная инсоляция.
Личинки последних стадий отлавливались лишь в середине июля.
Плотность заселения ими водоемов не превышала 17 экземпляров
на 1 м2. Доставленные в лабораторию в термосах, личинки продолжа-
ли развитие при 23—27°, и в этих условиях у них через 1—2 дня на-
чиналось окукливание, а через 5—6 дней — выплод имаго. Следова-
тельно, термическая пластичность вида чрезвычайно высокая. Во
всех случаях ограничивается одной генерацией. Зимует в фазе яйца.
Сезонный ход численности не прослежен, но если судить по вре-
мени наступления в тех местах холодов, то активность комаров долж-
на прекращаться к середине августа. За это время имаго проделыва-
ет, вероятно, не более двух гонотрофических циклов.
Местами дневок служит травяной растительный покров, на ко-
тором самки в утренние и вечерние часы сидят, не прячась от солнеч-
ных лучей. Вспугнутые — далеко не улетают, а опускаются на бли-
124
жайшие растения. Нападение отмечалось только днем, что, вероят-
но, связано с утренним и вечерним холодовым угнетением, поскольку
ночные температуры слабоположительны (4—6°). В условиях Север-
ной Америки в^д обнаруживается также на высоте 2400 м н. у. м.
(Matheson, 1944).
Недостатка в источниках питания самки не испытывают, по-
скольку период их активности совпадает с выпасом овец и лошадей.
Замечено нападение и на человека.
Ае. (О.) punctor Kirby (карта XI)
Вид с ареалом, охватывающим бореальную зону Палеарктики и
Неарктики. В Казахстане за счет приуроченности к лесным горным
участкам выклинивается в степную зону (Баян-Аул, Каркаралинек).
Местами выплода являются сильно затененные, неглубокие во-
доемы с температурой воды 8—17э. Чаще всего такие водоемы подпи-
тываются грунтовыми водами пли стекающими с гор ручьями
(рис. 28). Личинки Ае. punctcr никогда не встречались массово. Чис-
ленность их не превышала 20 особей на 1 лх2 водной поверхности.
Развитие шло совместно с Ае. sticticus, Ае. pullatus, Ае. communis
и Ае. hexodontus. Столь незначительная численность этого массово-
го во многих местах вида объясняется, вероятно, отсутствием специ-
фических мест выплода. Для Ае. punctor характерно развитие в сфаг-
новых лесных болотах (Сазонова, 1959).
В зависимости от степени затененности водоема и его глубины
выплод имаго происходит в первой половине июня. Нападение отме-
чается на второй-третий день после выплода. В лесных участках и
вблизи от зарослей активность не прекращается даже днем.
Выше 2700 м н. у. м. нами не обнаружен, тогда как А. Н. Жело-
ховцев (1938) отлавливал его в Центральном Алтае на высоте 1600—
2300 м н. у. м. В горах Южного Алтая нижней границей встречаемо-
сти являются лесные участки лесостепного пояса (900—2000 м
н. у. м.). На такой же высоте обнаружен в Баян-Ауле, а в Каркара-
линске — чуть выше (1300—1400 м н. у. м.).
Зимует в фазе яйца. Во второй половине лета может осуществ-
лять повторный выплод (Сазонова, 1956). От собранных в природе
комаров был выделен штамм возбудителя туляремии (Олсуфьев, Руд-
нев, 1960).
Ае. (О.) rempeli Vock.
При обследовании горных участков Южного Алтая в пределах
Большенарымского района Восточно-Казахстанской области были
125
обнаружены личинки, в значительной мере отличающиеся от всех
известных палеарктических видов. Согласно таблице для определе-
ния комаров Северной Америки (Carpenter, La Casse, 1955), они под-
ходят к тезе для Aedes rempeli Vock.
Личинки обнаружены на высоте 1950 м н. у. м. в водоемах, на-
ходящихся недалеко от выхода горячих (30—37°) ключей. В ука-
занном районе они обычны. В других местах Южного Алтая (окрест-
ности Усть-Каменогорска, Лениногорска, Зыряновска) не обнаруже-
ны. Личинки встречались как в сильно затененных лесных водоемах,
так и в более открытых, но предпочитали первые. Плотность заселе-
ния не превышала 16 личинок на 1 м2, обычно же приходилось 3—
8 личинок на 1 м2 водной поверхности. Развитие происходило сов-
местно с Ае. pullatus, Ае. communis, Ае. punctor и Ае. sticticus. В
конце июня отмечались личинки IV стадии и происходило их окук-
ливание. Имаго выплодились в начале июля.
По особенностям строения и экологии этот вид, несомненно, от-
носится к группе Ае. communis. От палеарктических и голарктиче-
ских видов, согласно таблице для определения личинок комаров, со-
ставленной А. С. Мончадским (1951) , он отличается как от группы
видов, имеющих в щетке более 30, так и от группы видов, имеющих
в щетке менее 30 чешуек. В первом случае описываемый вид ближе
к Ае. communis, от которого хорошо отличается строением седла,
охватывающего целиком последний членик, более стройным сифоном,
формой и количеством чешуек в щетке и многими другими особенно-
стями. Во втором случае приближается к Ае. punctor и Ае. hexodon-
tus, хорошо отличаясь от них количеством, формой и величиной че-
шуек щетки, простыми, средними и внутренними лобными волоска-
ми, сифональным индексом и многими другими особенностями.
Ае. (О.) riparius D. К. (карта XI)
Бореальный вид. Распространен в лесной зоне Палеарктики и
Неарктики. В РСФСР ареал охватывает не только лесную зону, но и
выклинивается значительно южнее (Сазонова, 1959). В Казахстане
распространен как в северных равнинных, так и в горных районах.
Имаго отловлены в Восточно-Казахстанской, Павлодарской и Актю-
бинской областях, а личинки обнаружены в Павлодарской и Куста-
найской областях. Известен из сопредельных территорий Западной
Сибири и Алтая (Желоховцев, 1938; Кухарчук, 1965).
Поздневесенний моноцикличный вид. Незначительная числен-
ность во всех районах Казахстана, кроме Баян-Аула, подтверждает
мнение О. Н. Сазоновой (1959) и Д.Д. Нефедова (1962 6) о его сравни-
тельно редкой встречаемости. В массе обнаружен лишь в Баян-Ауле
126
(Павлодарская обл.) в зарослях около заболоченностей, образован-
ных стекающими с гор ручьями, и в низменных скоплениях та-
лых вод.
В предгорно-степной зоне Восточного Казахстана самцы и сам-
ки выплаживались в середине июня. В такое же время этот вид от-
ловлен в Актюбинской и Павлодарской областях. Развитие проходи-
ло в тех же водоемах, где развивались другие виды группы «cantans».
Таким образом, фенология и экология этого вида и других представи-
телей данной группы подобны.
Из-за малочисленности Ае. riparius трудно судить о его агрес-
сивности и круге хозяев. Возможно, что в Баян-Ауле высокая чис-
ленность нападающих на человека самок объясняется недостатком
других источников питания, так как в местах наблюдения из воз-
можных прокормителей имелись только птицы.
Ае. (О.) simanini Guts, (карта XI)
С 1962 г. в окрестностях пос. Илийска Алма-Атинской области
неоднократно отлавливались особи, условно относимые нами к Aedes
leucomelas. Просмотр и сравнение этих экземпляров с коллекциями
Зоологического института АН СССР показали, что это Aedes simanini,
описанный А. В. Гуцевичем (1966) по сборам из Узбекистана. Морфо-
логическое строение и экология личинок оставались неизвестными,
что послужило основой для их детального изучения. В Казахстане
обнаружен только в бассейне р. Или. Распространение прослежено от
Илийска до границы с Китаем.
Личинки последних стадий в окрестности Илийска отмечаются в
хорошо прогреваемых биотопах со второй половины марта, а в более
затененных — в период затяжной весны, с первых чисел апреля.
Развитие отмечено в различных временных водоемах, таких как:
заточные мелководья в тугаях, непроточные канавы, скопления та-
лых вод в низинах и т. п. Чаще всего личинки обнаруживаются в за-
грязненных мусорными отбросами ямах на окраине поселка и в за-
точных лужах с прелой прошлогодней листвой (рис. 32). Вода в этих
водоемах отличалась повышенной щелочностью (pH 7,9—8,4), значи-
тельной минерализацией (до 3679 мг/л), присутствием аммиачных
соединений, карбонатов, сульфатов и хлоридов; цвет колебался от
слабо-желтого до красновато-бурого. На аллювиально-песчаных
участках, в верховьях р. Или, личинки обнаружены в слабо минера-
лизованных водоемах, образованных талыми водами.
Развитие отмечено при теплой солнечной погоде с дневными
температурами 15—20°. Вода в местах обитания личинок прогрева-
ется при таких условиях до 12—13°. Активность их снижается при
127
8—9° и почти полностью прекращается при 4—56. В таких случаях
личинки, находящиеся в придонной части водоема, неподвижны и
лишь изредка поднимаются к поверхности воды. Погода в это время
бывает неустойчивой, и при похолодании водоемы ночью промерза-
Рис. 32. Заточный водоем
около И.тийска — местообитания личинок
Ае. simanini.
ют. На личинок это, однако, не действует. При дневном оттаивании
снова проявляется их активность. Эксперименты, проведенные в ес-
тественных условиях, показали, что личинки и куколки этого вида
способны выдерживать ночные промораживания в 5—7-сантиметро-
вом слое воды при 5—7° в течение 3—4 дней. Оледенение обычно за-
хватывало куколок у поверхности воды, а личинок — в придонной
части. Личинки, перенесшие промораживание, были способны к даль-
нейшему развитию, а из куколок, как обычно, выплаживались имаго.
В одном случае самка выплодилась через 1 час после оттаивания
куколки.
В природных биотопах при суточных перепадах температуры от
—7 до +22° на прохождение всего периода развития требовалось
около 30—35 дней. В лабораторных условиях при 17—34° эти сроки
128
сокращались на 8—12 дней. Прогревание поверхности воды рефлек-
тором стимулировало выплод имаго. Вылет их в естественных усло-
виях происходил в течение 5—7 дней. В первые 1—2 дня выплажи-
вались только самцы, в средний период отмечался смешанный вылет,
и в конце появлялись только самки. В затененных и плохо прогрева-
емых водоемах все развитие затягивалось на 8—10 дней.
Численность личинок колебалась от 11 до 73 на 1 м2 водной пло-
щади, что зависело в основном от места нахождения водоема, време-
ни его заполнения, глубины и степени эфемерности. В небольших и
сильно затененных разливах личинки не обнаруживались. Не отме-
чено развития и в водоемах глубиной больше 50 см (ямы, копанки,
глубокие западины и т. п.).
Открытые минерализованные водоемы заселялись личинками
только этого вида; в затененных, с богатой, опавшей листвой разли-
вах развитие происходило совместно с Ае. leucomelas, а позже —с
Ае. flavescens и Ае. kasachstanicus. Смертность личинок как в естест-
венных, так и в лабораторных условиях весьма незначительная. Воз-
можность развития при минимальных температурах в природе и при
17—34° в помещении говорит о высокой экологической пластичности
этого вида. Личинки совершенно свободно переносили двухдневную
транспортировку в стеклянных банках на большое расстояние.
В условиях ранней, но затяжной весны 1966 г. выплод имаго
происходил в начале, а во время поздней, холодной весны 1967 г.—
лишь в конце апреля. Роящиеся самцы были замечены спустя 5—7
дней. Рои были небольшие, обычно по 15—25 самцов. Самки залета-
ли единично. К залетевшей в рой самке устремлялся один из самцов,
копуляция осуществлялась на лету. Рои обычно клубились на высо-
те 1,5—2 м на открытых участках тугайных зарослей в предвечернее
время. Ориентирами чаще всего служили небольшие, высотой 30—
40 слг, кустики.
Как показали пятилетние наблюдения, период активности этого
вида бывает непродолжительным по времени, но очень дружным.
В обычные годы основная масса самок нападает с 15 по 25 апреля.
За 20 мин около тугаев отлавливалось до 57 особей. К концу апреля
число нападающих резко уменьшалось (до 3—4 самок за учетный
сбор), и в начале мая можно было встретить лишь единичные экземп-
ляры этого вида.
Период суточной активности охватывает все дневное время, с 9
до 19 чос; наивысшая активность наблюдается с 10 до 13 час. Ре-
зультаты учетных сборов, проведенных 19 апреля 1966 г., приводятся
в табл. 1. От мест выплода имаго далеко не улетают. Залета в посе-
лок, расположенный в 200—800 м от мест выплода, не отмечено. Вы-
плодившиеся самки концентрируются в находящихся поблизости за-
9—80
129
Суточный ритм активности Ае. simanini
Таблица 1
Время наблюдения Темпера- тура, °C Влаж- ность, % Скорость ветра, м/сек Осве- щен- ность, люкс Кол-во нападаю- щих ко- маров
За 30 мин до восхода солнца 7 82 0,4 12000
Через 1 час после восхо- да солнца 11 71 0,7 16000 .—
В 10 час 19 57 1,3 48000 57
В 16 час 25 42 0,9 48000 21
За 30 мин до захода солнца 20 49 0,5 16000
Через 30 мин после за- хода солнца 14 74 0,8 1000 —
рослях, откуда при приближении человека и животных вылетают на
открытые места для нападения.
Нападению в равной степени подвергались люди, крупный рога-
тый скот, лошади, ослы. Отловлены на липучку около гнезда вороны.
Гонотрофический цикл при 19—23° длился 8—10 дней.
Количество откладываемых яиц колеблется от 73 до 161. Из яиц,
содержащихся в течение трех летних месяцев в воде и на влажной
фильтровальной бумаге, выплода личинок не произошло. Это подт-
верждает моноцикличность вида. Экология его в значительной мере
напоминает Ае. leucomelas.
Сведения, приводимые Ж. Исимбековым (1966) о наличии в бас-
сейне Или Ае. subdiversus, следует отнести к Ае. simanini.
Ае. (О.) sticticus Meig. (карта XII)
Ареал охватывает бореальную зону Америки и Евразии. В Ка-
захстане встречается лишь в северных районах Кустанайской, Восточ-
но-Казахстанской областей и в горных районах Казахского мелкосо-
почника (Баян-Аул).
Ранневесенний моноцикличный вид, приуроченный к развитию
в лесных, затененных водоемах. В основном это мелководные затоки
талой воды в лесной чаще (рис. 33). В период развития температура
в них колеблется от 8 до 17°, химический состав отличается слабой
минерализацией (148 мг/л), небольшой примесью различных соеди-
нений и слабощелочной реакцией (pH 7,2). Дно водоемов обильно
130
устлано прелой растительностью, а иногда — опавшей хвоей. Не-
смотря на ограниченность мест выплода, численность личинок в во-
доемах не бывает значительной, до 10 на 1 №. Развитие происходит
совместно с Ае. pullatus, Ае. communis и Ае. punctor, которые доми-
нируют в этих биотопах.
Рис. 33. Характерные для Aedes sticticus, Ае. cinereus и Ае. vexans лес-
ные разливы в Восточном Казахстане.
Развитие продолжается обычно дольше, чем у других видов груп-
пы «communis*. В Восточном Казахстане в зависимости от степени за-
тененности водоема выплод имаго отмечался 5—15 июня. Тогда же
И. Г. Прыгунова (1966) наблюдала появление этого вида в Кустанай-
ской области. Судя по обнаруженным в августе личинкам, куколкам
и имаго, там произошел частичный осенний выплод Ае. sticticus. Пов-
торный выплод, так же как и длительное развитие в горных усло-
виях, характеризует его как более термофильный вид из группы
«communis».
Нами и И. Г. Прыгуновой (1966) отмечено, что этот вид не выле-
тает из лесной чащи, нападая днем (чаще на людей). Другие сведе-
ния по экологии отсутствуют. Матесон (1944) сообщает о массовых
нападениях самок на людей и диких животных, а также о способно-
сти самок мигрировать на большие расстояния.
131
Ае. (О.) stramineus Dubitzky sp. n.
Многим исследователям, судя по имеющимся в Зоологическом
институте АН СССР коллекциям, неоднократно попадались крупные
соломенно-желтые комары, по ряду признаков (кольца на лапках,
строение гипопигия) напоминавшие Aedes caspius, но значительно от-
личавшиеся от них одноцветной спинкой и брюшком. Б. Н. Казанцев
(1931) образно назвал их «песочными». Анализируя встречаемость
таких экземпляров в различных районах Казахстана, мы высказали
предположение (Дубицкий, 1967), что по совокупности морфологиче*
ских и экологических признаков они не похожи ни на Ае. caspiust
ни на Ае. с. dorsalis. Обнаружение личинок этих комаров и получе-
ние идентифицированного выплода показало существенное отличив
их от всех близких голарктических таксонов в экологии, морфологии
и во внешнем облике имаго, что дает нам основание описать их в ка-
честве нового вида. (Морфологическое описание в печати).
Вид относительно редкий. Массово обнаружен лишь в пойме
верхнего течения р. Или на территории Казахстана (Панфиловский
район Талды-Курганской области). Местами выплода являлись до-
вольно большие (40—60 №), открытые, солоноводные водоемы. Вода
в них имела светловато-бурый цвет и слабощелочную реакцию (pH
8,35—8,5). На 1 л воды приходилось 12503—15896 мг различных со-
лей, в основном хлоридов и карбонатов. Общая жесткость по мере
усыхания водоемов колебалось от 193 до 382 мг-жв!л. Впоследствии
эти водоемы были покрыты белым налетом соли.
В зависимости от прогреваемости водоемов личинки появляются
в первую или вторую декаду апреля. Развитие идет в течение 30—40
дней. В условиях затяжной весны 1969 г. выплод имаго происходил
15—25 мая. Фенология всецело соответствует многочисленным Ае.
detritus и Ае. montchadskyi, совместно с которыми он чаще всего и
встречается. Характерно, что обильные в тех местах личинки Ае. с.
caspius хотя и появляются в этих же водоемах дней на 10 позже, но
выплод имаго происходит раньше выплода данного вида. За весь
теплый сезон Ае. stramineus осуществляет одну генерацию. Залитие
в сентябре проб почв, взятых с разных уровней высохшего водоема,
выплода личинок этого вида не стимулировало, тогда как личинки
Ае. с. caspius нормально развивались. Выплод личинок произошел
лишь в почвах, содержавшихся половину зимы во внешней среде. Об
одной генерации Ае. stramineus свидетельствует период лёта имаго,
который в 1969 г. длился с 15 мая по 20 июня, хотя отдельные особи
встречались в июле.
Все это говорит о том, что Ае. stramineus относится к группе
поздневесенних, моноцикличных видов и отличается, таким обра-
132
зом, от всех других представителей комплекса Ае. caspius. Анало-
гичные сведения с более ранними фенологическими сроками появле-
ния «песочной формы» приводит и Б. Н. Казанцев (1931) для Буха-
ры, a Freeborn (1926) — для Калифорнии (цит. по Б. Н. Казанцеву,
1931). Анализ сроков отлова «песочных Ае. caspius» в других райо-
нах Казахстана также показал, что в различных ландшафтно-клима-
тических условиях республики они встречаются лишь поздней весной
и ранним летом.
По морфологическим особенностям описываемый вид относится
к группе Ае. caspius. Как по внешним признакам имаго, так и по мор-
фологии гипопигия и личинок Ае. stramineus заметно отличается от
всех голарктических видов. Имаго этого вида не имеют какого-либо
рисунка на спинке или брюшке. Ае. stramineus характеризуется на-
личием на лапках светлых колечек, охватывающих вершину преды-
дущего и основание последующего членика, чего нет у представите-
лей всех других групп подрода Ochlerotatus. По строению гипопи-
гия, согласно таблице для определения комаров Северной Америки
(Carpenter, La Casse, 1955), этот вид отличается от Ае. campestris
наличием двух щетинок на базальной бородке, от Ае. melanimon —
расставленностью двух базальных щетинок и отсутствием расшире-
ния крыла класпет.
По строению генитального аппарата он ближе к двум подвидам
Ае. caspius, но от Ае. с. dorsalis хорошо отличается базальной боро-
давкой языковидной формы без резких переходов к кокситу и от-
сутствием расширения крыла класпет, а от Ае. с. caspius — вершин-
ным положением апикальной бородавки, хорошо выраженной склад-
кой, соединяющей апикальную бородавку с базальной, и расположе-
нием щетинки на внутренней поверхности волосистой части базаль-
ной бородавки. Кроме того, Ае. stramineus имеет более стройные
ветви коксита (отношение ширины к длине более 1:3), имеющие не-
большой наружный прогиб в проксимальной трети.
По личинкам описываемый вид хорошо отличается от всех рас-
смотренных представителей 4—6 ветвистыми внутренними лобными
волосками и некоторыми другими особенностями. По этому признаку
он приближается к Ае. montchadskyi (Дубицкий, 1968), отличаясь от
него слабой вторичной перистостью волосков и наличием простого
бокового волоска на седле.
Ае. (О.) subdiversus Mart, (карта XII)
Вид, считающийся редким. Описан из Саратова (Martini, 1926).
Известен из Северного и Северо-Западного Казахстана, где местами
(пойма Тургая) является массовым. В Казахстане отмечается тяготе-
ние его к полупустынной и южным районам степной зоны.
133
Ранневесенний моноцикличный вид. В Павлодарской и Куста-
найской областях личинки старших возрастов обнаруживаются во
временных водоемах, образованных талыми водами, на открытых,
хорошо прогреваемых местах во второй декаде мая. Первые окры-
ленные особи появляются с середины, а массовый лёт п роение — с
конца мая. Численность выплодившейся генерации сохраняется до
середины июня, а затем она идет на убыль. Последние экземпляры
отлавливаются в третьей декаде июня. Таким образом, продолжи-
тельность активной жизни самок не превышает 1—1,5 месяца.
Активный кровосос. Нападает как на людей, так и на животных,
причем предпочитает последних. Днюет в траве недалеко от водоемов,
особенно активен в сумеречное время, но может нападать и днем
(Прыгунова, 1966).
Подрод Aedes Meig.
Ае. (Ае.) cinereus Meig.
Широко распространенный в пределах Голарктики вид. За счет
своей эвритопности встречается в самых разнообразных ландшафтно-
климатических условиях, показывая большой диапазон морфологи-
ческой изменчивости. Последнее послужило основанием для возведе-
ния отдельных форм в видовые ранги, что не подтверждено последую-
щими исследованиями.
Ае. (Ае.) cinereus cinereus Meig. (карта XII)
В СССР встречается в таежной, лесной и лесостепной зонах. Рас-
пространение его в Казахстане не ограничивается лесостепной зоной.
По поймам равнинных и горных рек выклинивается в степную и пус-
тынную зоны. Тяготеет к северным и восточным районам Казахста-
на, редок в южных. Крайней южной точкой его массового нахожде-
ния следует считать среднее течение р. Или. Единичные экземпляры
встречаются в пойме Сыр-Дарьи. В горы поднимается до нижней гра-
ницы лесного пояса. Распространение и биотопическая приурочен-
ность напоминают Ае. flavescens, с которым он чаще всего и встре-
чается.
Поздневесенний полицикличный подвид. Развитие первого поко-
ления на юго-востоке Казахстана чаще всего происходит с Ае. fla-
vescens, а позже — с Ае. vexans и Ае. с. caspius. На востоке республи-
ки личинки встречаются совместно с Ае. cantans, Ае. behningi и Ае.
134
excrucians. Соответственно этому предпочитает лесные и густо зарос-
шие временные и постоянные водоемы с колеблющимся уровнем во-
ды (рис. 33). Местами выплода служат весенние разливы талых вод.
а также постоянные и полупостоянные водоемы с колеблющимся
уровнем воды. Встречается в эфемерных водоемах проточного харак-
тера. Предпочитает водоемы, богатые прошлогодней травянистой рас-
тительностью .
Развитие отмечено в водоемах с небольшой минерализацией
(100—650 мг)л), сравнительно мягкой водой (0,6—6,2°), слабощелоч-
ной реакцией (pH 7,2—8,1), незначительной примесью различных
органических и неорганических соединений.
В бассейне Или первые личинки обнаруживаются в конце мар-
та— начале апреля при дневной температуре воды 11—14°, близкой
к нижнему порогу развития, определенному М. Ф. Шленовой (1952)
как 13—15°. В опытах этого автора развитие при таких температу-
рах заканчивалось в 23 дня. Мы в естественных условиях при повы-
шающихся в апреле температурах воды (17—19°) отмечали более
длительное развитие. Выплод из куколок, личинки которых были об-
наружены в конце марта, происходил лишь в начале мая.
Оптимум развития личинок находится в пределах 23—24° (Шле-
нова, 1952). Такие температуры свойственны главным образом весен-
нему периоду. Как правило, развитие последующих генераций про-
исходит в сильно затененных водоемах. В жаркое, сухое лето разви-
тие летних поколений не отмечается. Объясняется это, с одной
стороны, незначительным затоплением водой мест выплода этого под-
вида, а с другой — высокой прогреваемостью воды в таких водоемах.
Как сообщила М. Ф. Шленова (1952), температуры в 26—29° приво-
дят к гибели основной массы развивающихся личинок, а при 31—33°
развития не происходит совсем. Поэтому частичное развитие и вы-
плод имаго в жаркое сухое лето наблюдаются в водоемах, питаемых
холодными грунтовыми водами. О том, что развитие этого подвида
невозможно при высоких температурах, говорит гибель личинок, вы-
плодившихся в лабораторных условиях при 22—29°. Из других кла-
док при таких же и более высоких температурах личинки вообще не
выплаживались. Частичное отрождение их отмечено лишь в марте
при температуре 18—20°.
Такая стенотермность ограничивает распространение этого под-
вида в южных районах Казахстана. Единичные находки его относят-
ся к сильно заросшим водоемам с невысокой температурой воды.
Будь этот подвид моноцикличным, он, несомненно, был бы оолее рас-
пространен в Южном Казахстане, где весенние температуры опти-
мальны для первой генерации, в то время как летние — пессималь-
ны для последующих.
135
Неблагоприятными воздействиями высоких летних темпепатур,
вероятно, можно объяснить меньшее число генераций этого подвида
на юго-востоке Казахстана по сравнению с данными И. Г. Прыгуно-
вой (1966), приводимыми для северных районов республики. Там ли»
чинки последних стадий и куколки обнаруживались в первых чис-
лах июля и в середине августа. Это, вероятно, второе и третье поко-
ления, так как первое должно выплаживаться совместно с Ае. can-
tons и Ае. excrucians примерно в начале июня, что отмечено для Пав-
лодарской и Восточно-Казахстанской областей, где личинки послед-
них стадий обнаружены в конце мая, а выплод имаго произошел в
начале июня.
Сезонный ход численности в благоприятные для развития годы
не отличается резкими взлетами и падениями. Первые экземпляры
на юго-востоке Казахстана начинают появляться с начала мая. Чис-
ленность их возрастает за счет продолжающегося выплода до начала
третьей декады этого месяца, а затем держится на определенном уров-
не до середины августа, резко уменьшаясь к его концу.
Такая картина характерна для фенологии двух сближенных ге-
нераций, выплод второй из которых приурочен ко времени старения
и гибели первой. Соответствует этому и сезонный ход численности ви-
да на севере Казахстана, где лёт имаго происходит с мая по октябрь
на протяжении всего сезона. В горных условиях юго-востока Казах-
стана развиваются два летних поколения: одно — в июне, второе — в
июле. Соответственно лёт имаго происходит с середины июня до кон-
ца августа.
Количество откладываемых яиц колеблется от 4 до 120. Данные
Н. В. Волозиной (1957) говорят о таком же непостоянстве (13—151).
Автор указывает также на большие расхождения сроков перевари-
вания самками крови (от 4 до 10 дней). Нами отмечены колебания в
пределах 6—19 дней. В отличие от М. Ф. Шленовой (1952) и Н. В. Во-
лозиной (1957), часто получавших частичный выплод личинок этого
подвида без дополнительных воздействий (высушивание, обработка
высокими или низкими температурами), мы только в одном случае
получили выплод личинок из кладок одного года, содержавшихся в
воде при 22—29°. Вероятнее всего, приспособившись к существованию
в таких термо- и ксерофильных условиях, подвид приобрел способ-
ность прекращать развитие при пессимальном режиме и продолжать
его при наступившем оптимуме. В зависимости от этого он может про-
делывать одну или две генерации, что, вероятно, дает основание го-
ворить о его моно- и полицикличности.
Особенностью яиц Ае. с. cinereus является непродолжительная
сухоустойчивость. Отрождения личинок из яиц, высушиваемых в те-
чение месяца после 11-дневного пребывания во влажной среде, не
136
происходило. Но такие яйца были способны переносить охлаждение
до 37,5° (Во лозин а, 1957). Совокупность этих факторов и определяет,
вероятно, основные черты экологии вида — более высокую числен-
ность и частоту встречаемости в северных районах и затененных
местах.
Суточная активность по сравнению с многими другими видами
Aedes четче выражена в утренние и вечерние часы. Утренний пик ак-
тивности начинается до восхода солнца и вскоре заканчивается. Ве-
черняя активность проявляется дольше, чем у других видов Aedes.
Вылет днюющих имаго отмечается незадолго до захода солнца и за-
канчивается в полных сумерках. Днем нападает только в густых за-
рослях и на затененных участках.
Объясняется это крайней влаголюбивостью подвида (Виноград-
ская, 1948). Характерно его продвижение к объекту нападения. Уви-
дев добычу, комары приближаются к ней, наподобие москитов, ко-
роткими перелетами, не вылетая надолго из зарослей травы. Напа-
дать начинают с нижних участков тела человека или животного,
продвигаясь такими же короткими перелетами к голове. Особенно на-
зойливы перед дождем.
Днюют в густых зарослях травы, тростника, тугаев и кустарни-
ков, в непосредственной близости от водоемов. Далеко от таких
участков не отлетают. В поселки в связи с этим залетают редко. В све-
толовушку, даже в период массового лёта, попадали лишь единичные
экземпляры.
Строгая приуроченность к влажным биотопам сужает круг его
прокормителей. Агрессивно нападает на человека и животных. От-
ловлен на липучки около нор барсуков и песчанок, гнезда болотного
луня, норы-гнезда береговой ласточки.
В оптимальных условиях по численности нападающих в период
своей активности занимает 3—4 место. Вдали от водоемов и зарослей
количество нападающих резко уменьшается. Как компонент гнуса
имеет значение для северных, восточных и меньшее — для юго-вос-
точных частей Казахстана; редок в южных районах республики.
Один из основных переносчиков возбудителя туляремии (Ол-
суфьев, Руднев, 1960). От отловленных в природе самок выделен ви-
рус лимфоцитарного хориоменингита (Гуцевич, Подолян, 1959).
Ае. (Ае.) cinereus esoensis Yam.
Палеархеарктический подвид. Распространен в Японии, на Даль-
нем Востоке и в Забайкалье (Штакельберг, 1937; Гуцевич, 1937, 1940,
1941; Yamaguti, La Casse, 1950). В Казахстане с достоверностью не
установлен.
137
При изучении географической изменчивости комаров Казахста-
на замечено лишь, что некоторые из личинок и самцов, отловленных
на юго-востоке и востоке Казахстана, обладают признаками, свойст-
венными Ае. с. esoensis. Обнаружение их носит хаотический харак-
тер. Какие-либо экологические закономерности в их распределении
или распространении отметить не удалось. Они всецело перекрыва-
лись экологической пластичностью и эврибионтностью Ае. с. ci-
nereus.
Ае. (Ае.) cinereus rossicus D. G. М. (к а р т а XII)
Встречается в умеренной зоне Палеарктики. В Казахстане рас-
пространен по всей территории республики. Некоторые исследователи
ввиду тождественности личинок этого подвида с Ае. с. cinereus отно-
сят его к морфам Ае. с. cinereus. Приведенные выше сведения, а так-
же наблюдения по распространению, фенологии и экологии дают
основание относить его лишь к подвидовому рангу, и то со многими
оговорками. У многих же авторов он до последнего времени проходит
как полноправный вид.
Подвид, считавшийся в Казахстане крайне редким. Исследова-
ния последних лет позволили выделить серии личинок, самцов и са-
мок из всех районов Казахстана. Характерна мозаичность распрост-
ранения, уменьшающаяся по мере продвижения на юг республики.
Отличается несоответствием морфологических признаков личинок и
выплаживающихся из них имаго. Из личинок с признаками Ае. с. ros-
sicus выплаживаются самцы и самки Ае. с. cinereus, и наоборот.
Экология, в основных чертах, кроме отдельных зональных осо-
бенностей, повторяет жизненную схему Ае. с. cinereus. Без специаль-
ных эколого-морфологических исследований не представляется воз-
можным выделить специфические для Ае. с. rossicus особенности.
Требуются дополнительные исследования этого интересного вопроса.
Экологические описания, приведенные для Ае. с. cinereus в условиях
Казахстана, в настоящее время соответствуют и Ае. с. rossicus.
Подрод Aedimorphus Theob.
Ае. (Aed.) vexans Meig.
Широко распространенный в пределах всех зон Голарктики, кро-
ме тундровой, вид. В южной части ареала заходит в Ориентальную и
Эфиопскую области. В пределах СССР распространен на север до лес-
ной зоны, хотя и вклинивается в нее по поймам рек (Мончадский,
138
1951). В Казахстане встречается повсеместно, достигая высокой чис-
ленности как на юге, так и на севере республики. Следует отметить
при этом мозаичность его распространения, обусловленную, как нам
кажется, приуроченностью к определенным условиям обитания. В
пустынной, полупустынной и степной зонах республики, характери-
зующихся значительной прогреваемостью вследствие высокой солнеч-
ной радиации личиночных биотопов, выплод приурочен к заросшим
участкам пойм протекающих здесь рек. На севере же и востоке
республики, где инсоляция меньше, его развитие приурочено к более
открытым или слабо заросшим водоемам. В Казахстане встречаются
два его подвида, хорошо различающиеся морфологически и не отличи-
мые по экологии.
Ае. (Aed.) vexans vexans Meig. (карта XIII)
Ae. vexans vexans является полицикличным, сравнительно теп-
лолюбивым видом. Места его выплода и условия развития во многом
схожи с Ае. с. caspius и Ае. с. dorsalis. Личинки развиваются во мно-
гих эфемерных, временных и полупостоянных водоемах (скопления
талых вод, пойменные разливы, затоки и фильтрационные заболо-
ченности оросительной сети, выходы грунтовых вод, копанки, ямы,
каналы, отстойники, поля фильтрации, озера и заболоченности с ко-
леблющимся уровнем воды). В воде с повышенной концентрацией
солей выплод не отмечен. Предпочитает водоемы, незначительно за-
росшие погруженной растительностью, а также находящиеся в ок-
ружении древесных и кустарниковых зарослей и частично затенен-
ные ими. На юге и юго-востоке Казахстана таким требованиям удов-
летворяют пойменные водоемы, а на севере и востоке — лесные
разливы (рис. 33). В водоемах личинки этого вида встречаются сов-
местно с Ае. sticticus.
Развитие отмечается в водоемах с минерализацией от 641 до
3515 мг/л, незначительным сдвигом реакции в щелочную сторону
(pH 7,2—7,6) и различным соотношением тех или иных соединений
в допустимых пределах. В более высоких солевых концентрациях вы-
плод не отмечен, хотя такие водоемы довольно часто попадаются в
радиусе активности этого вида.
Амплитуда температур, при которых возможно развитие, лежит
в пределах 14—34° (Шленова, 1952). При крайних ее вариантах в од-
ном случае наблюдается затяжка развития, а в другом (верхний пре-
дел) — большая смертность личинок. Оптимум развития 29—30°. Та-
ким образом, температурные критерии этого вида близки к Ае. с. cas-
pius, но развитие его и массовый выплод происходят значительно
позже. Объясняется это специфичностью требований Ае. v. vexans к
139
условиям внешней среды. Как уже отмечалось некоторыми исследо-
вателями (Шленова, 1952; Мончадский, 1950; Сазонова, 1959), раз-
витие личинок начиналось лишь после повторного заполнения водой
участков с кладками этого вида, что М. Ф. Шленова (1952) объясняет
необходимостью прохождения стадии высыхания яиц.
По нашим наблюдениям в Казахстане, выплод личинок может
осуществляться и без обязательного двукратного залития водой. Сам-
ки Ае. V. vexans гибли в садках, но яйца на воду не откладывали.
Однако на влажную фильтровальную бумагу кладки производили
охотно, причем в основном по краям ее. Во всех случаях из отложен-
ных яиц через 8—9 дней при 23—33° выплаживалось 68—79% личи-
нок. Из оставшихся яиц и кладок, содержавшихся определенное вре-
мя на влажной бумаге, а потом высушенных, выплод отмечался лишь
после повторной заливки водой. Выводы М. Ф. Шленовой (1952) так-
же подтверждают, что для нормального прохождения эмбрионально-
го развития и выплода необходим период высушивания. При этом для
последующей реактивации яиц необходимо воздействие высоких
(30—34°) температур. В условиях Казахстана, согласно эксперимен-
тальным данным, это применимо лишь для яиц, у которых по тем
или иным причинам наступила задержка эмбрионального развития.
Основная же часть их при благоприятных условиях развивается по-
следовательно .
Отмеченное явление, по-видимому, обусловлено тем, что разви-
тие основной массы личинок происходило во временных, быстро пе-
ресыхающих водоемах. К моменту начала кладки яиц выплодившей-
ся генерацией водоемы уже высыхали или были близки к этому,
что, вероятно, привело к выработке избирательных таксисов. Самки
откладывают яйца у уреза воды или на влажную поверхность участ-
ков, подвергающихся периодическому затоплению. Яйцекладки на
воду с последующим выплодом личинок являлись бы в таких случаях
биологическим тупиком. На влажной же почве к периоду полного
высыхания у них наступает эмбриональная диапауза, в процессе ко-
торой кладки подвергаются основательному воздействию сухости и
высоких летних температур, которые из-за сильного прогревания
почвы достигают больших величин (до 50°). Адаптацией к таким воз-
действиям явилась выработка ксеротермических свойств в фазе яйца.
Наследственно закрепившись, эти качества в сочетании с периодом
диапаузирования гарантируют сохранность популяции при любых
климатических отклонениях. Вместе с тем при благоприятных усло-
виях возможен непрерывный выплод личинок, что характеризует об-
щую тенденцию к увеличению биологического потенциала вида.
В природных условиях это проявляется следующим образом. В
умеренной зоне СССР, а также в аридных зонах Казахстана затяжка.
140
весеннего развития и приуроченность его к повторному затоплению
водоемов происходят за счет того, что в период между двумя затоп-
лениями перезимовавшие яйца испытывают ксеротермические воз-
действия. В более южных районах Казахстана прохождение этого пе-
риода осуществляется в теплую, сухую осень и поэтому частичное
отрождение личинок из диапаузирующих яиц наблюдается при соот-
ветствующей температуре во время первых паводков. Массовый же
выплод зависит от количества залитых кладок. Как правило, он при-
урочен не ко времени незначительных затоплений водоемов талыми
водами, а к периоду весеннего половодья различных рек и связанных
с ними водоемов. При этом уровень воды поднимается до значитель-
ных пределов и все яйцекладки заливаются.
Нами, а также К. А. Амиргазиевым (1966) и В. П. Боженко
(1948) в конце апреля обнаружено незначительное количество личи-
нок этого подвида во временных, хорошо прогреваемых водоемах.
Массовое развитие личинок на юге, юго-востоке и западе Казахстана
отмечается в первой-второй декаде мая. Связано это, как уже указы-
валось, с повторным затоплением мест выплода в более северных час-
тях и обширным весенним паводком рек в южных.
Высота паводка и соответственно площадь затопления мест вы-
плода играют для этого подвида первостепенное значение. Так, в
1962 г. весенний паводок в мае на Сыр-Дарье (100 км ниже Чардары)
был умеренным, а число выплодившихся комаров незначительным.
На следующий год уровень воды намного поднялся, площадь времен-
ных водоемов увеличилась в 7—8 раз и количество развивающихся
личинок возросло до 17 тыс. на 1 м2. Подобное явление было отмече-
но и в бассейне Или в 1963 и 1964 гг. Объясняется оно ярусностью
кладок по профилю водоема.
Первые, наиболее многочисленные кладки производятся еще в
период довольно большого уровня воды на более высокие ярусы, по-
следующие — в низменные участки высыхающего водоема и послед-
ние, незначительные по количеству, — лишь на влажное дно высох-
шего водоема. Поэтому количество выплодившихся личинок после-
дующих генераций зависит от уровня воды и площади последующего
затопления.
Поскольку непрерывный выплод вследствие откладки яиц около
уреза воды ограничен, количество последующих генераций и их чис-
ленность определяются частотой и высотой последующих разливов.
На юге, юго-востоке и западе Казахстана, как правило, бывает еще
одно поколение этого вида, связанное с летними паводками пустын-
ных рек и нагонными ветрами на северном побережье Каспия. В го-
ды с незначительными паводками численность первой генерации бы-
вает ничтожно мала, а выплод второй не отмечается совсем.
141
На севере Казахстана личинки последних стадий обнаруживают-
ся в первых числах июля в водоемах, пересохших после весеннего
снеготаяния и вторично затопленных дождевыми водами. Развитие
второй генерации отмечено в конце июля — начале августа (Прыгу-
нова, 1966).
На юге Алтая личинки последних стадий отловлены в середине
июля на высоте 1300—1400 м н. у. м. во временном открытом водое-
ме, образованном затоком талых вод. Поскольку на такой высоте это
первое затопление, то остается предположить, что выплод личинок
произошел из прошлогодней кладки, которая испытала действие
температуры и сухости прошлым летом.
Сезонный ход численности в остальных частях Казахстана соот-
ветствует фенологии личинок. На юге, юго-востоке и западе респуб-
лики небольшое число выплодившихся имаго может быть обнаруже-
но в начале мая. Основной выплод происходит в конце мая — начале
июня (в зависимости от климатических условий местности). Числен-
ность выплодившихся комаров может достигать колоссальных цифр.
21 мая 1963 г., в пойме Сыр-Дарьи за 100 взмахов энтомологического
сачка в сумеречное время отловлено 3275 экземпляров. Последую-
щий выплод, наблюдаемый обычно в конце июля — начале августа,
уже не столь многочислен и в зависимости от гидрологического ре-
жима подвержен большим колебаниям. В годы с теплым и дождли-
вым летом по численности нападающих этот вид занимает третье
место после Ае. с. caspius и Cx. modest us, а в определенные периоды
даже доминирует над ними. В такое время активность самок за счет
сближенных по времени двух выплодов держится на высоком уровне
с июня до середины августа. Последние особи отлавливаются в конце
августа — начале сентября.
На севере Казахстана первые окрыленные особи появляются в
начале июля. Численность первой генерации незначительна. Выплод
второй генерации, численно превосходивший первую в 12 раз, отме-
чался в начале августа, а максимальный лёт — в третьей декаде это-
го месяца (Прыгунова, 1966). Соответствует описанному сезонная
активность вида в горных частях бассейна Или на высоте 2000 м
н. у. м.
Продолжительность предимагинального развития превышает та-
ковую Ае. с. caspius. При 16—17° развитие затягивается до 34 дней
(Шленова, 1952). В условиях Казахстана при 23—29° оно заканчи-
вается в 15—16 дней. Такое же время затрачивалось на предимаги-
нальное развитие в естественных условиях дельты Волги (Мончад-
ский, 1956). Гонотрофический цикл при 23—29° длится 4—6 дней.
Количество откладываемых в лабораторных условиях яиц не превы-
шает 120, но чаще бывает в пределах 50—60 штук. Период эмбрио-
142
нального развития при 23—29° длится 8—9 дней. При испытании на
солевыносливость выплаживание и дальнейшее развитие личинок
происходило лишь в 1—2% растворе NaCl. Возможно, этим и опре-
деляются особенности откладки яиц около уреза воды, уровень ко-
торого всегда поднимается по мере поступления пресной воды, что
предохраняет яйца от попадания в значительную солевую концент-
рацию.
Нами роение отмечалось только в вечернее время, хотя А. С. Мон-
чадский (1951) наблюдал его и в утренние часы. Местом роения чаще
всего служат открытые участки среди зарослей, а ориентиром — про-
езжая дорога или какой-либо заметный предмет. Комары клубятся в
неисчислимом количестве на высоте 1,5—2 м. Среди основной массы
самцов и самок Ае. v. vexans встречаются единичные экземпляры
Ае. с. caspius. Как и все другие представители рода Aedes, этот вид
зимует в фазе яйца.
Суточный ход активности соответствует обычному. Основная
масса нападающих приходится на утренние и вечерние часы. Утрен-
ний пик начинается в период рассвета и заканчивается через 1 —
1,5 час после восхода солнца. Вечерний пик начинается за 45—90 мин
до захода и продолжается до глубоких сумерек. В светлые лунные
ночи активность снижается, но не прекращается совсем. Наибольшую
агрессивность самки проявляют перед восходом солнца и сразу же
после него. В дневное время нападают только в зарослях и то в не-
значительных количествах. Далеко от мест выплода и густых укры-
тий не улетают.
Местами дневок служат всевозможные растительные заросли и
естественные укрытия (пещеры, расщелины в скалах, колодцы).
П. А. Петрищева (1962) обнаруживала самок в убежищах кабанов и
джейранов, норах барсуков, волков, шакалов, степных кошек, дико-
бразов, песчанок, сусликов, хомячков, крыс, ежей, гнездах щурок,
сычей, стрижей и ласточек, мы к тому же — в гнездах ворон.
Подвид, предпочитающий диких и домашних животных, но охот-
но нападающий и на человека. Нередко залетает в поселки. Больше
всего нападает на домашних животных.
Возможный переносчик вирусов энцефалита Сан-Луи, западно-
го и восточного конских энцефаломиелитов, лимфоцитарного хорио-
менингита (Chamberlain, 1958; Глущенко, Гуцевич, Дудкина, 1957).
В условиях Казахстана Ае. vexans оказался способным передавать
возбудителя туляремии (Олсуфьев, 1938).
Ае. (Aed.) vexans nipponii Theob.
В Казахстане этот подвид встречается как самостоятельно, так
и совместно с номинальным. Распространение его охватывает юго-
143
восточные районы республики. Из других районов СССР распростра-
нен на Дальнем Востоке (Гуцевич, 1937, 1940). Основная же часть
его ареала занимает территорию Китая и Японии. На Дальний Вос-
ток проник, вероятно, диффузно из сопредельных районов Китая. Ка-
захстан отделен от Китая высочайшими отрогами Тянь-Шаня, кото-
рые для этого мезотермофила должны служить естественной прегра-
дой, однако здесь имеются два сравнительно низких перевала, веду-
щих в Джунгарию, по которым в основном осуществляется обмен
фауной (Афанасьев, 1960). Ае. vexans nipponii не только стал оби-
тать совместно с номинальным подвидом, но в некоторых случаях и
доминировать над ним, особенно в предгорных районах Джунгарского
и Заилийского Алатау.
Так же как и А. В. Гуцевичу (1937), нам не удалось отметить
каких-либо отличий в экологии этих двух подвидов.
Род CULEX L.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CULEX L.
ПО ВЗРОСЛЫМ НАСЕКОМЫМ
(По А. А. Штакелъбергу, с сокращениями и изменениями)
1 ( 4). Первый членик задних лапок явственно короче задних го-
леней (длина его равна 4/г,—6/? длины голени). Брюшко без
светлых поперечных перевязей. Мелкие темные формы (под-
род Barraudius Edw.).
2 ( 3). Брюшко с боков со светлой продольной полосой, иногда об-
разующей более или менее развитые треугольные выросты
по переднему краю тергитов
. . Сх. (В.) modest us Fic.
Повсеместно, кроме горных и лесных участков.
3 ( 2). Брюшко с боков при основании тергитов с более или менее
развитыми обособленными светлыми пятнами
. . . . . Сх. (В.) pusillus Macq.
В пределах пустынной зоны Казахстана.
4 ( 1). Первый членик задних лапок равен длине или чуть длиннее
задней голени (подроды Neoculex Dyar и Culex L.).
5 ( 6). Передние и средние бедра и голени спереди темные, со свет-
лой продольной полосой . . Сх. (Сх.) theileri Theob.
Предгорные районы в пределах пустынной зоны Казахстана.
6 ( 5). Передние и средние бедра и голени спереди без светлых про-
дольных полос.
144
7 (12). Светлые поперечные перевязи брюшка расположены на вер-
шине тергитов.
8 ( 9). Последние членики щупиков самцов голые или почти голые;
задние голени у обоих полов у вершины с белым пятном
......................Сх. (N.) hortensis Fic.
Предгорные и горные районы Южного, Юго-Восточного и Вос-
точного Казахстана. Возможен в аналогичных условиях на
всей остальной территории.
9 ( 8). Последние членики щупальцев самцов покрыты длинными
волосками.
10 (11). Общая окраска темная . . . Сх. (N.) territans Walk.
Распространен повсеместно, но мозаично.
11 (10). Общая окраска желтая . . . Сх. (N.) martini! Med.
Горные участки Южного и Юго-Восточного Казахстана.
12 ( 7). Светлые боковые пятна или поперечные перевязи брюшка
расположены при основании тергитов .............................
......................... . Сх. (Cx.) pipiens L.
Повсеместно, в равнинных и горных районах.
. . . ...............Сх. (Сх.) torrentium Mart.
В предгорных районах Южного и Юго-Восточного Казахста-
на. Возможен на остальной территории.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CULEX L.
ПО ГИПОПИГИЯМ САМЦОВ
(По А. А. Штакелъбергу, с сокращениями и небольшими
изменениями)
1 ( 4). Коксит на наружной поверхности в мелких чешуйках; пред-
вершинная бородавка расположена едва за его серединой.
2 ( 3). Стиль относительно длинный и тонкий ...
. . . . . Сх. (В.) mcdestus Fic.
Повсеместно, кроме горных и лесных районов.
3 ( 2). Стиль короткий и толстый . . . Сх. (В.) pusillus Macq.
В пределах пустынной зоны Казахстана.
4( 1). Коксит без чешуек; предвершинная бородавка расположена
явственно за его серединой.
5 (10). Предвершинная бородавка коксита на своем дистальном кон-
це без пластинчатых придатков; десятый стернит на верши-
не с одним рядом относительно крупных и длинных зубцов.
6 ( 7). Коксит на вершине с широким пальцевидным выростом, на
значительное расстояние выступающим за основание
10—80
145
Cx. (N.) hortensis Fic.
Предгорные и горные районы Южного, Юго-Восточного и Вос-
точного Казахстана. Возможен в аналогичных условиях на
всей остальной территории.
7 ( 6). Коксит на вершине без пальцевидного выроста.
8 ( 9). Стиль узкий и длинный, к вершине постепенно суживающий-
ся • • ... Сх. (N.) territans Walk.
Распространен повсеместно, но мозаично.
9 ( 8). Стиль относительно короткий, за серединой расширенный .
. . . . . . . . Сх. (2V.) martinii Med.
Горные участки Южного и Юго-Восточного Казахстана.
10 ( 5). Предвершинная бородавка коксита на своем дистальном кон-
це несет одну хитиновую пластинку, десятый стернит на вер-
шине с мелкими, расположенными в несколько рядов шипи-
ка ми.
11 (12). Десятый стернит без базального придатка или же он имеется
в рудиментарном состоянии ....
. .............................Сх. (Сх.) pipiens L.
Повсеместно в равнинных и горных районах.
12(11). Десятый стернит с явственно развитым, достигающим обык-
новенно довольно значительной величины базальным при-
датком.
13 (14). Мезозома в вершинной части разделена на две пластинки, из
которых одна образует два-три направленных латерально
зубца .... . . Сх. (Сх.) theileri Theob.
Предгорные районы в пределах пустынной зоны Казахстана.
14 (13). Мезозома в вершинной трети разделена на три резко обособ-
ленных узких пластинки, из которых одна направлена ди-
стально .... Сх. (Сх.) torrentium Mart.
В предгорных районах Южного и Юго-Восточного Казахста-
на. Возможен на остальной территории.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CULEX L.
ПО ЛИЧИНКАМ
(По А. С. Мончадскому, с сокращениями и небольшими
изменениями)
1 ( 2). Сифон в длину в среднем в 3 раза превышает ширину осно-
вания. Сифональные пучки расположены зигзагом на задней
поверхности сифона . . . Сх. (В.) pusillus Macq..
В пределах пустынной зоны Казахстана.
146
2 ( 1). Длина сифона по меньшей мере в 4 раза превышает ширину
основания.
3 ( 8). Все (или кроме одного-двух у вершины) сифональные пучки
расположены на задней поверхности сифона зигзагообразно
или парами, сильно сближенными по средней линии.
4 ( 7). Трахейные стволы лентовидные, шириной больше половины
диаметра сифона, овального сечения.
5 ( 6). Пучки на сифоне расположены пятью сближенными пара-
ми; три базальные пары иногда располагаются зигзагом.
Дистальные зубцы гребня длинные, широко расставленные,
базальные слабо развиты . . . Сх. (Сх.) theileri Theob.
Предгорные районы в пределах пустынной зоны Казахстана.
6 ( 5). Пучки на сифоне расположены зигзагообразно. Иногда вер-
шинные пучки короткие ...............
Сх. (В.) modestus Fic.
Повсеместно, кроме горных и лесных участков.
7 ( 4). Трахейные стволы узкие, меньше половины диаметра сифо-
на, округлого сечения. Ближайшие к основанию два-три пуч-
ка волосков на сифоне не менее чем в три раза длиннее его
диаметра; лежат между более широко расставленными ди-
стальными зубцами гребня . . . Сх. (N.) hortensis Fic.
Предгорные и горные районы Южного, Юго-Восточного и Вос-
точного Казахстана. Возможен в аналогичных условиях на
всей территории.
8 ( 3). Сифональные пучки расположены парами по краям задней
поверхности сифона; часть из них явственно расположена на
боковой поверхности.
9 (12). Главные трахейные стволы в сифоне тонкие, с круглыми по-
перечными сечениями.
10 (11). Сифон (индекс около 7) явственно расширен у вершины. Зуб-
цы гребня с одним-двумя дополнительными зубчиками по
нижнему краю . . . . Сх. (N.) territans Walk.
Распространен повсеместно, но мозаично.
11 (10). Сифон (индекс 7,5—11) не расширен у вершины или это рас-
ширение едва заметно. Зубцы гребня с 3—5 дополнительны-
ми зубчиками по нижнему краю ...................
Сх. (N.) martinii Med.
Горные участки Южного и Юго-Восточного Казахстана.
12 ( 9). Главные трахейные стволы в сифоне широкие, более полови-
ны диаметра сифона, с продолговато-овальным поперечным
сечением.
13(14). Сифональный индекс больше 6; из сифональных пучков
наиболее боковое положение на сифоне занимает ближайшая
147
к вершине четвертая пара ......................
.................................Сх. (Сх.) torrentium Mart.
В предгорьях Южного и Юго-Восточного Казахстана. Возмо-
жен на остальной территории.
14(13). Сифональный индекс меньше 6; из сифональных пучков
наиболее боковое положение занимает вторая к вершине
пара.
а. Сифон более длинный (индекс больше 4,5); жабры заметно
длиннее седла, обе пары одинаковой длины; задние пучки
плавника не длиннее, чем жабры.............................
.....................Сх. (Сх.) pipiens pipiens L.
Повсеместно, в равнинных и горных районах.
б. Сифон более короткий (индекс не больше 4,5); жабры не
длиннее седла. Нижняя пара их намного короче верхней;
задние пучки плавника в */2—2 раза длиннее, чем жабры
. Сх. (Сх.) pipiens molestus Forsk.
Возможен повсеместно. Развивается круглогодично.
Подрод Barraudius Edw.
Сх. (В.) modestus Fic. (карта XIII)
Широко распространенный в южной части Палеарктики вид.
В пределах СССР ареал охватывает пустынную, полупустынную и
степную зоны; во многих местах вклинивается в лесостепную. Такая
же интразональность наблюдается и в Казахстане, где он наряду с
Сх. pipiens по праву может считаться самым распространенным
видом.
Наиболее массовый в пустынной и полупустынной зонах Казах-
стана вид. Местами выплода являются различные постоянные и по-
лупостоянные озера и заболоченности с обильной надводной и под-
водной растительностью (рис. 34). Одинаково хорошо развивается в
рисовых чеках, особенно с расчлененным рельефом, затоках и фильт-
рационных разливах оросительной сети. Часто встречается совместно
с An. т. sacharovi, An. hyrcanus и An. pulcherrimus. В больших ко-
личествах выплаживается в прудах и поселковых водоемах. В период
массового развития заселяет даже эфемерные водоемы, сопутствуя
Ае. с. caspius и Ае. vexans.
На севере Казахстана развитие происходит в постоянных, откры-
тых, хорошо прогреваемых водоемах. Такого обилия, как на юге, не
достигает. В специфических условиях северного Прикаспия первое
поколение выплаживается в различных постоянных водоемах, а с
143
конца июля — в култучной зоне, мелководные участки которой после
окончания паводков быстро зарастают водной растительностью и яв-
ляются особо благоприятными для развития личинок. Поскольку в
пустынной и полупустынной зонах Казахстана многие водоемы
Рис. 34. Околопоселковый водоем в Южном Казахстане (Яны-Курган) —
место массового выплода Culex modestus и Сх. pipiens.
имеют различную степень солености, в лабораторных условиях была
проверена возможность выплода и развития личинок этого вида в
воде с различным содержанием солей. Испытаны были 1, 2, 4 и 10%
растворы поваренной соли, а в качестве контроля — дистиллирован-
ная вода. Полный выплод и дальнейшее развитие личинок происхо-
дили лишь в дистиллированной воде и в 1% растворе NaCl. Это подт-
верждается и аналитическими сведениями о составе воды в местах
выплода. Обычной являлась минерализация около 1000 мг!л, и
только в Гурьевской области концентрация солей в водоемах дости-
гала 7000 мг!л. О сравнительно небольшой солевыносливости этого
вида сообщает и П. П. Чинаев (19646), хотя П. А. Петрищева (1936)
относит его к группе солевыносливых.
В бассейне Сыр-Дарьи первые перезимовавшие особи Сх. modes-
tus начинают нападать с наступлением теплых весенних дней, когда
149
температура воздуха повышается до 16—18°, что соответствует при-
мерно середине и второй половине марта. Численность комаров в это
время невелика, а в апреле, когда основная масса перезимовавших
особей уже напивается крови, их вообще трудно обнаружить.
Выплодившиеся особи нового поколения, уже в гораздо большем
количестве, появляются в начале мая. Их потомство и создает пер-
вый массовый пик численности этого вида. Выплод происходит в
основном во временных водоемах с длительно сохраняющимся за
счет фильтрации от русла Сыр-Дарьи уровнем воды. Поэтому и сов-
падают периоды массового выплода Сх. modestus с послепаводковы-
ми периодами Сыр-Дарьи.
Если не считать резкого возрастания численности после массо-
вого выплода, то обилие этих кровососов держится на довольно высо-
ком уровне с начала июня до конца августа. Затем числен-
ность нападающих резко снижается, и к концу сентября комары
исчезают.
Обращают на себя внимание обилие комаров в осенний период и
малочисленность их весной. Объясняется это двумя причинами. Во-
первых, осенью в водоемах гибнет множество личинок и куколок, не
успевших выплодиться до наступления холодов. Так, в 1964 г. мы
наблюдали довольно значительное обилие личинок последних стадий
и куколок до 24—26 октября. Они погибли, когда водоемы стали
покрываться ледяной коркой. Во-вторых, гибнут зимующие имаго.
По нашим наблюдениям, несмотря на то, что особи этого вида в осен-
нее время встречаются во многих укрытиях (заросли тростника, кус-
тарника, норы животных и т. п.), весной перезимовавшие имаго от-
лавливаются только в густых зарослях тугаев. Резкие температурные
перепады с частыми осадками и пронизывающими холодными вет-
рами, вероятно, губительно действуют на зимующих в недостаточно
укрытых местах имаго.
Сезонный ход численности Сх. modestus и его особенности в бас-
сейне Или подобны описываемому. Первые нападающие особи появ-
ляются в начале марта, когда еще не сошел снег, но наступают теп-
лые, солнечные дни. Нападение самок отмечается лишь в зарослях
тростника или около них. В горных степях (1200—2000 м) лёт кома-
ров начинается во второй декаде мая и продолжается до первой де-
кады сентября (Исимбеков, 1966).
В северных районах Казахстана перезимовавшие самки появля-
ются в первых числах мая, массовое развитие личинок происходит во
второй декаде июня, а выплод имаго — в середине июля. Числен-
ность комаров бывает наивысшей в середине августа, после чего на-
чинает резко падать, и к середине сентября имаго исчезают (Прыгу-
нова, 1966).
150
В Гурьевской области вылет перезимовавших самок в зависимо-
сти от температурных условий года происходит с конца марта или в
апреле. Первые личинки обнаруживаются в начале мая, а выплод
имаго происходит в первых числах июня. Массовой численности ко-
мары достигают в конце июля, после чего наступает снижение, за ко-
торым следует новый небольшой подъем (середина сентября), закан-
чивающийся к концу сентября.
Количество летних генераций зависит от климатических условий
местности и суммы эффективных температур. Продолжительность
периода переваривания крови — от 5 до 15 дней. При суточных ко-
лебаниях температуры в лаборатории от 17 до 27° он продолжался
15 дней, а при 28—29° — всего 5 дней. Соответственно этому и диа-
пазон продолжительности развития от яйца до имаго зависит от тем-
пературы среды. При 24—29° развитие заканчивалось в 13 дней, а
при 21—23° оно продолжалось 16—21 день. Результаты опытов по-
казывают, что сравнительно длительные периоды весеннего развития
вида в южных и северных районах Казахстана аналогичны.
Приуроченность к постоянным водоемам, а следовательно, мень-
шая зависимость от гидрорежима в сочетании со скоростью развития
и беспрерывностью выплода создают наиболее благоприятные усло-
вия для размножения этого вида в пустынной и полупустынной зо-
нах Казахстана. Поэтому в местностях, изобилующих постоянными
и полу постоянными водоемами, он доминирует над другими видами
комаров. За теплый период подобно An. т. sacharovi может проде-
лывать 7—8 генераций. На севере республики их, вероятно, всего
две, поэтому он не выделяется количественно. Яйца откладывает на
воду склеенными в форме «лодочки». В каждой полной кладке от
118 до 179 яиц. Опыты с высушиванием кладок показали, что пребы-
вание без воды на солнце более трех часов, а в помещении — около
суток приводит к гибели яиц.
Сх. modestus, как и некоторые другие виды Culex, в местах дне-
вок или поблизости от них нападает в течение всего дневного време-
ни. Массовый пик численности (табл. 2) наблюдается в предвечерние
и сумеречные часы, более слабый — в предрассветные, до 6—7 час.
Совместно с Ае. с. caspius часто залетает в ближайшие поселки, где в
значительном количестве отлавливается в светоловушки. В отличие
от Ае. с. caspius ограничивается нападением на открытом воздухе, в
жилые помещения залетает редко.
Местами дневок служат всевозможные заросли и растительные
группировки. Залетевшие в поселок особи укрываются на день в раз-
личных щелях, под навесами и в кроне деревьев. На севере Казахста-
на местами дневок служат лесные колки и густая травянистая рас-
тительность вблизи от водоемов.
151
Таблица 2
Суточный ритм активности Сх. modestus в различные месяцы (Яны-Курган)
Время наблюдения Март Апрель Май Июнь Июль Август Сен- тябрь
t воздуха Кол-во на- падающих t воздуха Кол-во на- падающих t воздуха Кол-во на- падающих । t воздуха Кол-во на- падающих t воздуха Кол-во на- падающих t воздуха Кол-во на- падающих t воздуха Кол-во на- ' падающих
За 30 мин до вос- хода солнца 2 14 15 21,5 1 15,8 3 17,0 1 7,5
Через 1 час после восхода солнца 8 20 — 16 25,0 20,5 2 19,0 7 11 —
В 10 час 16 1 26 — 28 — 32,0 — 26,2 — 27,0 — 20 8
В 16 час 18 3 29 — 27,5 — 35,0 — 29,0 — 30,0 — 22 4
За 30 мин до за- хода солнца 13 1 23 2 22 — 28,5 184 17,8 28 26,5 98 21 19
Через 30 мин пос- ле захода солнца 8 — 20 4 19 9 27,0 199 15 58 22,0 158 19 33
Зимуют в фазе имаго в природных условиях. Укрытиями слу-
жат густые растительные заросли, буреломы, метелки тростника.
Е. А. Плетнев (1943) находил зимующих имаго в надворных построй-
ках, сложенных дровах, валежнике и, что сомнительно, в жилых по-
мещениях.
Одинаково охотно нападает на людей и различных животных,
но поскольку последних бывает больше в его биотопах, то основная
масса самок питается кровью млекопитающих и птиц. Исследования
Э. О. Конурбаевым (1965) отловленных в природе особей показали,
что 67 % самок имели в желудке кровь млекопитающих и птиц и лишь
33% содержали кровь человека. Больше и чаще других обнаруживал-
ся в норах корсаков, барсуков, ежей, сусликов, дикобразов, большой и
гребенчуковой песчанок; отмечался около гнезд сороки, вороны, луня
и сорокопута.
Возможный переносчик туляремии. В дельте Волги от этих ко-
маров выделены штаммы F. tularensis (Олсуфьев, Руднев, 1960).
Сх. (В.) pusillus Macq. (карта XIV)
Вид с ареалом, охватывающим южные районы Средиземномор-
ской подобласти. На территории СССР обнаружен1 лишь в Средней
Азии и Казахстане. Широко распространен в Туркмении (Петрище-
ва, 1936). В значительных количествах встречается в Узбекистане, в
152
низовьях Аму-Дарьи (Чинаев, 1964). Обнаружен в бассейне Сыр-
Дарьи и в окрестностях Илийска. Э. Р. Геллер (1939) приводит све-
дения о нахождении этого вида в окрестностях Талды-Кургана, что
требует дальнейшего подтверждения.
Южные районы Казахстана, по всей вероятности, являются са-
мыми северными частями его ареала. Найден в низовьях Чу (Раев-
ский, Виноградская, 1934). Редко, но встречается в окрестностях
Илийска. Все это говорит о крайней разреженности популяции Сх.
pusillus примерно около 44—45° с. ш., что свойственно обычно гра-
нице ареала видов.
Развитие личинок происходит в водоемах с повышенной кон-
центрацией солей, обычно при 0,5—1,25% солевом содержании (Kirk-
patrick, 1925).
Во всем прочем экология вида всецело напоминает Сх. modestus,
с той лишь разницей, что численность его в 8—10 раз меньше. Пере-
зимовавшие особи отлавливаются в середине мая, а готовящиеся зи-
мовать — в начале октября. Наибольшая численность отмечается в
августе и начале сентября. Единичные экземпляры отлавливаются в
первых числах октября. Аналогичные сведения по сезонности Сх. pu-
sillus в Египте приводит Киркпатрик (Kirkpatrick, 1925).
Несмотря на значительное количество выплаживающихся кома-
ров (1889 личинок за один учетный сбор), на человека нападали еди-
ничные экземпляры. То же самое отмечал П. П. Чинаев (19646) в ни-
зовьях Аму-Дарьи, где при численности личинок до 2000 на 1 м2 има-
го на человека нападали крайне редко. П. А. Петрищева (1962) отно-
сит его к группе «птичьих» видов. Чаще всего он отлавливался ею в
гнездах сизоворонок, золотистой щурки, удода, горного голубя и за-
каспийского филина. Мы обнаруживали этот вид в гнездах болотного
луня и береговой ласточки. Одна самка была отловлена в Кызыл-
Кумах около норы дикобраза, а другая — во время нападения в су-
меречное время на агаму.
Подрод N е о с u 1 е х Dyar
Сх. (N.) hortensis Fic. (карта XIV)
Ареал несколько шире Средиземноморской подобласти. В СССР
распространен в южных районах. В Казахстане по предгорным и гор-
ным районам, к которым он тяготеет, доходит до 50 параллели. При-
уроченность к предгорным районам, по всей вероятности, обусловле-
на потребностью для развития чистых, слабопроточных вод. Таким
условиям соответствуют слабо заросшие растительностью заводи гор-
153
ных рек и заполненные водой ямы и низины. Один из немногочис-
ленных видов, заселяющих высокогорные районы Тянь-Шаня. В Заи-
лийском Алатау личинки обнаружены в поясе субальпийских лугов
(2600—2800 м н. у. м.).
Рис. 35. Небольшие водоемы в русле горной реки — местообитания ли-
чинок Culex hortensis» Сх. martinii и Сх. pipiens (субальпийский пояс).
Личинки Ш—IV стадий отловлены в начале второй декады ию-
ля в слабо минерализованных (207 жг/л) водоемах с довольно высо-
ким кислотным показателем (pH 5,5) и большой окисляемостью воды
(23 мг)л). В период развития личинок водная поверхность водоема,
представлявшего заполненную водой прирусловую ямку (рис. 35), на-
половину заросла лемнидами (Lemna trisulca и L. minor). Ни ранее
(апрель), ни позднее (сентябрь) личинки в этом водоеме не встреча-
лись. Доставленные в термосе из водоемов с температурой 15—18е в
лабораторные условия (t 25—33°), они вскоре погибали.
В условиях высокогорья, вероятно, успевает проделывать две ге-
нерации, так как до начала — середины июня водоемы еще скованы
154
льдом. В сентябре, при той же температуре (15—18°), личинок уже не
было. По всей видимости, на развитие одного поколения уходит око-
ло месяца.
В Туркмении активен с апреля по ноябрь (Петрищева, 1936). В
Казахстане обнаруживается с середины апреля по сентябрь. Местами
дневок служат различные растительные укрытия. П. А. Петрищева
(1962) указывает на его дневную приуроченность к крупным пеще-
рам, гротам и норам различных животных. Зимуют имаго в скопле-
ниях древесного валежника, зарослях камыша, надворных построй-
ках и жилых помещениях (Плетнев, 1943). Возможность зимовки в
жилых помещениях требует проверки, поскольку вид экологически
приурочен к нежилым местам и не отличается агрессивностью по от-
ношению к человеку. В Южном Казахстане зимующие имаго обнару-
жены в норах грызунов (Дубинин, 1946). Об этом же, а также о
питании на этих животных ранее сообщала П. А. Петрищева
(1936, 1962).
Сх. (N.) martinii Medsch. (карта XV)
Распространен в Средиземноморской и Пустынно-Степной (по
Б. А. Кузнецову, 1950) подобластях Палеарктики (Югославия, Венг-
рия, Турция, Таджикистан, Узбекистан). Ареал этого вида охваты-
вает в основном горные страны. На их территории также отмечается
приуроченность к горным и предгорным районам.
Личинки обнаружены на субальпийских лугах (урочище Ассы,
2600 м н. у. м.) в заполненной водой грунтовой выемке (рис. 35). Во-
доем постоянного характера, подпитывается грунтовыми водами те-
кущей рядом р. Ассы. Вода характеризуется слабой минерализацией
(107 мг/л), незначительной примесью органических и неорганических
соединений и слабощелочной реакцией (pH 7,2). В момент обнаруже-
ния личинок (июль) водоем был слабо заселен Lemna minor и L. tri-
sulca. Позже (сентябрь) эти растения так разрослись, что совершенно
закрыли водную поверхность. В это время личинки уже не встре-
чались.
Амплитуда суточных колебаний температуры на такой высоте
характеризуется 4—26°. Температура воды днем при 22° доходила
до 18°. Личинки обнаруживались совместно с Сх. pipiens, Сх. horten-
sis и Cs. longiareolata, которые численно преобладали над рассматри-
ваемым видом. Личинки Culex были IV стадии, a Cs. longiareolata —
третьей. В находящемся поблизости временном разливе в это время
происходил выплод Ае. pullatus. При проверке водоемов в сентябре
личинок обнаружить не удалось, хотя личинки Сх. pipiens были.
155
Температура воды в пределах 4—15° сохраняется в этих местах
до конца августа — начала сентября, что, вероятно, дает возмож-
ность проделывать две генерации: в июле и августе.
Во время периодических посещений в течение двух лет нападе-
ния на человека не отмечено. Летом долина служит выпасом (джай-
ляу) для крупного рогатого слота, лошадей, овец, которые, по-види-
мому, и являются источником питания. Из диких животных следует
отметить обилие сурков.
Сх. (N.) territans Н. D. К. (к а р т а XIV)
Широко распространенный в Голарктике вид. Ранее прохо-
дил под диагнозом Сх. apicalis. На территории СССР встречается пов-
семестно, до таежной зоны. Характерен для многих районов Средней
Азии. В Казахстане встречается всюду мозаично.
Единичные разрозненные находки не создают представления об
экологии и фенологии этого вида в Казахстане. Отмечается приуро-
ченность его к горным и предгорным биотопам. Личинки обнаружи-
вались в горно-лесостепной зоне Алтая в середине июня. Развитие
происходит в небольших заточных лужах (рис. 36). Единичные ли-
чинки отловлены в конце мая и на юго-востоке Казахстана в болотах,
заросших тростниками, подпитываемых родниковой водой.
В Туркмении, встречаясь в предгорных районах, активен только
в июне и июле (Петрищева, 1936). В это же время встречается и в Ка-
захстане. На человека не нападает. Держится в биотопах, где обиль-
ны различные виды птиц и амфибий. О предпочтительном питании
комаров этого вида на лягушках сообщает Матесон (1944).
Возможный переносчик туляремии, сохраняет возбудителя до
24 дней (Олсуфьев, Руднев, 1960).
Подрод Culex L.
Сх. (Сх.) pipiens L. (к а р т а XV)
Чрезвычайно эврипластичный вид. Ареал охватывает все уме-
ренные и жаркие широты. В СССР распространен повсеместно, кроме
самых северных районов. Казахстан целиком попадает в его ареал.
Представлен здесь номинальной формой Сх. pipiens pipiens.
Подобно Сх. modestus наиболее многочислен в пустынной и полу-
пустынной зонах, предпосылкой чему служит обилие в этих зонах
характерных мест выплода. Предпочитает различные постоянные и
полупостоянные, хорошо прогреваемые солнцем водоемы (припой-
менные разливы, грунтовые заболоченности, речные затоки, поля
156
фильтрации, мелкие озера и т. п.). Охотно заселяет заполненные во-
дой ямы, копанки, траншеи, шурфы, бассейны, цистерны, бочки.
Характерной особенностью водоемов являются незначительная
засоленность, слабое развитие водной растительности и обилие вод-
Рис. 36. Горный ручей в Восточном Казахстане, в заводях
которого обнаружены личинки Culex territans.
ных простейших. Личинки могут встречаться в воде с общей жестко-
стью 3,8—35,2°, сухим остатком 399—7029 мг!л, окисляемостью до
23 мг!л и реакцией от щелочной (8,4) до кислой (5,05). Развитие ли-
чинок отмечено даже в заполненной дождевой водой консервной бан-
ке. Предпочитают водоемы с глубиной до 40 см. Чаще всего встреча-
ется совместно с Cs. alaskaensis и Сх. modestus, реже —с Сх. horten-
157
sis, Ur. unguiculata a Cs. longiareolata. В постоянных заросших
водоемах обитает в прибрежных открытых участках.
В Казахстане в равной степени встречаются как природные, так
и синантропные популяции этого вида. Более других приспособлен к
развитию в условиях больших городов. Заселяет фонтаны, бассейны,
фильтрационные поля, пруды, бочки с водой. Большое количество
выплаживается в разливах и затоках арычной системы.
В горных условиях встречается на прогреваемых участках всех
горных поясов до субальпийских лугов включительно. Заселяет вы-
ходы грунтовых вод, разливы ручьев и затоки горных речек (рис. 35,
36). Сопутствует An. claviger, Cs. alaskaensis, Cs. longiareolata и
Cx. hortensis.
Сезонный ход численности своеобразен. Зимующие самки от-
лавливаются в условиях Алма-Аты в январе, феврале и марте. Акти-
визации их способствуют теплые, солнечные дни с температурой 5—
12°. В такую погоду зимующие в подполье самки проникают через
щели в полу и активно летают в помещении. Исчезают с понижени-
ем наружной температуры. У самок, зимующих в канализационной
системе, активность отмечается на протяжении почти всего зимнего
периода, хотя развития не наблюдается. Температура в это время ко-
леблется в пределах 14—17°. Весной комары покидают убежища и в
течение теплого периода не появляются. В этом случае имагинальная
диапауза, вероятно, прерывается весьма рано.
В природе единичные самки начинают отлавливаться с конца
марта — первых чисел апреля. В конце апреля встречаются первые
личинки. Численность их сначала значительно уступает количеству
Cs. alaskaensis, но в конце мая превосходит его. Затем на протяжении
всего летнего периода по количеству личинок, развивающихся в по-
стоянных и полупостоянных водоемах, этот вид совместно с Сх. mo-
destus доминирует над другими. Скорость развития Сх. р. pipiens
уступает таковой Сх. modestus. В лабораторных условиях при 21—28°
развитие длится 21—26 дней. Выплод личинок из отложенных яиц
наступает на следующий день, окукливание происходит через 17—
22 дня, смертность растущих личинок незначительная.
О более высокой термофильности этого вида по сравнению с
Сх. modestus говорят и наблюдения А. С. Мончадского (1951), отме-
чавшего, что при средней температуре 14° развитие личинок резко
замедляется. Нами это неоднократно наблюдалось в осеннее время
(конец сентября) как в равнинных, так и в горных условиях. Разви-
тие последней генерации затягивалось почти до холодов. Личинки
отличались крупными размерами и развитым жировым телом. Тем-
пература воды днем колебалась от 15 до 17°, а ночью опускалась до
6—8° (вероятно, нижний предел развития). Развитие личинок Сх. то-
158
destus в естественных водоемах осенью заканчивалось при температу-
ре воды 12—15° днем и соответственно при более низкой — в ноч-
ной период. Личинок Сх. р. pipiens в этом водоеме уже не было.
О сезонности имаго можно судить только по обилию предимя^м.
нальных фаз и отлову имаго на свет, так как взрослые комары на че-
ловека нападают редко. В Южном и Юго-Восточном Казахстан? мас-
совое развитие и выплод наблюдаются в мае, июне, июле и августе.
В сентябре численность снижается. В горных условиях (1300—
2800 м н. у. м.) период активности значительно короче — с июня по
август, а в годы с теплой осенью — по сентябрь включительно. В
Гурьевской области личинки появляются в апреле, выплод первой
генерации происходит в мае, массовая численность — с июня по ав-
густ и уход на зимовку — с третьей декады августа (Жук, 1967). В
Восточном Казахстане личинки и имаго обнаруживались с середины
мая по сентябрь (Боженко, 1948). В условиях затяжной, но дружной
весны 1966 г. развитие личинок отмечено лишь с начала июня. Коли-
чество генераций может колебаться от 2—3 на севере республики до
4—6 на юге.
Местами дневок служат всевозможные растительные заросли и
укрытия, пещеры, щели, трещины скал, колодцы вблизи от мест
выплода. П. А. Петрищева (1962) сообщает о частых обнаружениях
этого вида в норах и норах-гнездах барсуков, волков, шакалов, дико-
бразов, песчанок, сусликов, ежей, крыс, сизоворонок, щурок, черепах,
ящериц. На большинстве хозяев этих убежищ Сх. pipiens питается
кровью. Столь обширный круг хозяев, который по наблюдениям в Ка-
захстане может быть дополнен воронами, болотными лунями, сакса-
ульными сойками, сорокопутами, удодами, береговыми ласточками,
свидетельствует о явной зоофильности этого вида. Тем не менее всеми
авторами отмечалось нападение комаров на человека как вблизи от
жилья, так и в природе. В лабораторных условиях нам неоднократно
удавалось наблюдать питание самок на человеке, но получить яйце-
кладки не пришлось. Они гибли на 2—3 день.
Охотно летят на свет. Чаще других отлавливаются в светоло-
вушку, выставляемую во время как стационарных, так и маршрут-
ных работ.
Зимует в фазе имаго в самых разнообразных природных стаци-
ях в южных районах. На севере Казахстана более приурочен к чело-
веческому жилью. Зимует в убежищах, свойственных An. т. messeae,
нередко вместе с ним.
В сг.язи с большой синантропностью, возможностью выплода в
самых разнообразных водоемах и наблюдаемым переходом на пита-
ние кровью человека может со временем приобрести чрезвычайно
большое значение в условиях больших городов и населенных пунктов.
159
Подобное было отмечено в Одессе (Найдич, 1962, 1964), где этот вид
в больших количествах выплаживался на полях фильтрации и стал
массово нападать на людей.
Возможный переносчик вирусов японского энцефалита, западно-
го и восточного конских энцефалитов, лимфоцитарного хориоменин-
гита, энцефалита Сан-Луи, энцефалита лошадей в СССР, а из бакте-
риальных инфекций — туляремии и бруцеллеза (Bates, 1949; Bjorn-
son, 1961; Chamberlain, 1958; Kissling, 1960; Олсуфьев, Руднев,
1960; Ременцова, 1962). В Средиземноморской зоне является перенос-
чиком Wuchereria bancrofti (ВОЗ, № 233).
Сх. (Сх.) theileri Theob. (карта XV)
Основная часть ареала этого вида находится в Эфиопской обла-
сти. В СССР он охватывает южные районы европейской части и Сред-
нюю Азию. В Казахстане распространен в южных и юго-восточных
районах. Встречается очень редко. Раньше, по данным Е. И. Балка-
шиной (1939) и Э. Р. Геллера (1939), встречался чаще. Пульсацию
ареала, видимо, вызвали климатические факторы или уменьшение
мест выплода в связи с культурным освоением местностей.
По данным Е. А. Плетнева (1943), имеет тяготение к горным рай-
онам. Э. Р. Геллер (1939), наоборот, находил его в поймах равнинных
рек. Нами обнаруживался в постоянном слабо заросшем тростником
неглубоком водоеме. Относится к числу зоофилов, вблизи от жилья
не встречается, хотя на человека нападает. Предпочитает кровь птиц
и соответственно может быть обнаружен в их гнездах (Петрищева,
1962). Зимует в фазе имаго в норах грызунов и более крупных жи-
вотных. В условиях Туркмении активен с марта по октябрь (Петри-
щева, 1936).
Возможный переносчик лихорадки Рифт (Reeves, 1962).
Сх. (Сх.) torrentium Mart, (карта XV)
Ареал охватывает Европейско-Сибирскую подобласть Палеарк-
тики. Распространен от Ленинграда до Дальнего Востока.
В Казахстане этот вид тяготеет к горным районам. В северных
районах и на прилежащей территории РСФСР не встречается. Обна-
ружен на высоте 2000 м н. у. м. Встречается совместно с Сх. р. pipi-
ens в отдаленных от жилья человека биотопах. Других сведений по
его экологии в Казахстане нет.
Очень похож на Сх. р. pipiens, поэтому к определению этого вида
следует относиться с осторожностью. С достоверностью может быть
установлен лишь по особенностям строения гипопигия.
160
Cx. (Cx.) vagans Wied. (?)
Основная часть ареала находится в Ориентальной области. На-
ряду с этим довольно широко распространен в Палеархеарктической
подобласти Палеарктики. В СССР отмечен в граничащих с нею рай-
онах Забайкалья и Дальнего Востока. В Казахстане личинки обнару-
живаются в Алма-Атинской и Восточно-Казахстанской областях.
Отсутствие в сборах имаго и четко отграниченных от Сх. pipiens
сведений по его экологии не дают основания с достоверностью утверж-
дать, что этот вид обитает на территории Казахстана.
IV. Внутривидовая изменчивость
Пространственная протяженность Казахстана и разнообразие
ландшафтно-климатических условий определяют возможность суще-
ствования на его территории самых различных видов кровососущих
комаров. У большинства из них в той или иной части республики
проходит северная или южная граница распространения. На таких
участках, как известно, условия обитания вида близки к пессималь-
ным и вид может прореагировать на них проявлением соответствую-
щих эколого-физиологических приспособительных реакций (Мончад-
ский, 1961).
В других случаях, вследствие зональной гетерогенности Казах-
стана и экологического разнообразия интразональных биотопов,
большое влияние на морфо-биологические особенности комаров долж-
ны оказать общие условия существования эвритопных видов (влия-
ние стабильных факторов, по А. С. Мончадскому, 1961).
При просмотре и анализе видов комаров, имеющих в Казахстане
ту или иную границу ареала, выяснилось, что их морфологическая
изменчивость нарастает по мере продвижения к пределам распрост-
ранения и бывает особенно значительной в крайних точках ареала.
Изменчивость проявляется в цветовых аберрациях, расплывчатости
видовых признаков и появлении морфологических отклонений (Ду-
бицкий, 1967 а).
Цветовые аберрации заключаются в основном в посветлении
окраски чешуек тела у южной границы бореальных видов (Ае. cata-
phylta, Ае. communis, Ае. intrudens, Ае. punctor, Ае. sticticus, Ае. beh-
ningi, Ае. cantans, Ае. cyprius, Ае. excrucians, Ае. flavescens) и потем-
нении южных представителей (An. т. sacharovi, Ае. с. caspius, Cs,
annulata subochrea). Аналогичная изменчивость комплексов Ае. cas-
pius, Сх. pipiens, Ае. scutellaris, по мнению Макса (Marks, 1954) и
162
Мэттингли с Найтом (Mattingly, Knight, 1956), обусловлена темпера-
турным фактором. Влияние же на окраску отдельных популяций сре-
ды обитания не меняет общего представления об изменчивости.
Расплывчатость видовых признаков проявляется у группы «com-
munis» в стирании полосатости среднеспинки, а у группы «cantans»—
в нивелировании контрастности окраски брюшка и спинки (Ае. fla-
vescens) и в затушевывании рисунка брюшка (Ае. excrucians, Ае. beh-
ningi, Ае. cantans).
Морфологические отклонения, свойственные некоторым видам
(Ае. communis, Ае. pullatus), чаще всего отмечаются у личиночной
фазы. Они выражаются в основном в уменьшении количества чешуек
щетки, их укрупнении и увеличении количества не объединенных об-
щим основанием пучков плавника.
Помимо географической изменчивости у различных видов и их
популяций наблюдаются экологические и сезонные отклонения. За-
мечено, например, что личинки одних и тех же видов (Ае. caspius,
Ае. vexans, Сх. pipiens, Сх. modestus, Cs. alaskaensis), развивающиеся
в затененных, заросших водоемах, окрашены всегда темнее, чем на
открытых местах. Имаго также имеют более темную окраску.
Большое влияние на цветовые вариации оказывает и состав во-
ды. В водоемах, богатых сернистыми соединениями, личинки кома-
ров приобретают серовато-белый оттенок, а в воде со значительным
содержанием солей железа и гуминовых веществ — красноватый.
При повышенной минерализации воды во всех случаях выпла-
живающиеся виды имеют более светлую окраску. В совокупности с
этим изменяются некоторые морфологические признаки личинок
(длина жабер, величина сифональной трубки и др.), а у имаго увели-
чивается количество светлых чешуек.
Аберративный характер носит и сезонная изменчивость популя-
ций. Как правило, первое поколение полицикличных видов имеет бо-
лее темную окраску, чем последующие. Связано это, вероятно, с по-
вышением у предимагинальных фаз содержания пигментов кутикулы,
способствующих большей поглощаемости тепловых лучей, необходи-
мых личинкам, развивающимся в относительно прохладной весенней
воде. Аналогичное явление отмечено А. С. Мончадским (1951).
Сезонная изменчивость проявляется в размерах весеннего поко-
ления полицикличных видов, зимующих в фазе имаго. Отмечено, что
у последних генераций Сх. pipiens, Сх. modestus и Cs. alaskaensis раз-
витие протекает дольше, а личинки имеют более крупные размеры
и хорошо развитое жировое тело. Как и у многих других групп
насекомых, это объясняется необходимостью создания питатель-
ных запасов для имаго, которым предстоит выдержать зимовку
в природе.
163
У географических рас (подвидов) помимо указанных на стыке
ареалов отклонений отмечается и индивидуальная наследственная
изменчивость, достаточно хорошо иллюстрирующая несостоятель-
ность их видового подразделения. Ярче всего она выражена у Ае. cas-
pius. В местах существования двух его подвидов — Ае. с. caspius и
Ае. с. dorsalis — отмечается переход признаков одного к особенно-
стям наружного строения другого.
Примером наследственной изменчивости является также симпа-
трическое отношение двух подвидов Ае. vexans. В местах соприкосно-
вения их ареалов отмечается интеграция отличающих их признаков.
У Ае. v. vexans это проявляется в расширении перевязей брюшка и
появлении единичных вкраплений светлых чешуек в центральной
части тергитов, а у Ае. v. nipponii — в сокращении ширины колец на
лапках, уменьшении ширины перевязей и их разобщенности.
Сложнее определить наследственную изменчивость некоторых
морфологически близких симпатрических видов. Описанные на ос-
новании отдельных морфологических или экологических особенно-
стей, они то относятся к подвидам, то возводятся в видовой ранг. В
Казахстане две таких пары: 1) Ае. punctor — Ае. hexodontus, 2) Ае.
communis — Ае. pionips. По имаго обе пары с достоверностью не от-
личаются, и лишь по личиночным фазам удается установить их раз-
личие (Carpenter, La Casse, 1955).
В предгорных и горных районах Южного Алтая, где все четыре
вида заселяют одни и те же биотопы, нами обнаружена вся гамма
переходных признаков одного вида в другой. У Ае. communis это вы-
ражалось в увеличении числа ветвей средних и внутренних лобных
волосков, а у Ае. pionips, наоборот, — в их редукции. Индивидуальный
выплод имаго из ранее определенных куколок (с фиксацией личиноч-
ной шкурки) не показал предполагаемых существенных различий у
самок.
Общность не только ареалов, но и биотопов, наличие переходных
признаков у личиночной фазы и отсутствие достоверных отличий у
имаго говорят против видовой самостоятельности Ае. punctor и Ае. he-
xodontus. Правда, И. А. Рубцов (1959), ссылаясь на «закон Ваттштей-
на», считает, что симпатрическое и экологическое сосуществование,
наоборот, определяет видовое различие морфологически близких
видов. В данном случае с этим нельзя согласиться, поскольку отли-
чия обоих видов основываются лишь на личиночных признаках, при-
чем переходных, и отсутствуют различия в таких таксономически
важных органах, как гениталий самцов. Нет согласованности в оцен-
ке их видовой самостоятельности и в Неарктике. Матесон (1944) счи-
тает их подвидами, а Карпентер и Ла Кассе (1955) — полноправны-
ми видами. Трудно установить ранг этих таксонов, поскольку не
164
известна причина дивергенции. Скорее всего это групповые экологиче-
ские морфы (в понятии А. Г. Шарова, 1955), вынужденные в услови-
ях границы ареала заселять общие биотопы.
В пользу большой преобразующей роли экологических факторов
говорят и другие сведения. А. С. Мончадский (1958, 1961) по степени
и времени воздействия на эволюционирующие организмы делит их
на изменяющиеся и стабильные элементы окружающей среды т На
распространенных в Казахстане видах можно проследить влияние и
тех и других.
Обитая в основном в пустынных и полупустынных районах,
такие эврипластичные виды, как Ае. с. caspius и Сх. modestus, вынуж-
дены заселять почти все экологические ниши, в том числе и солоно-
водные водоемы, которыми изобилуют указанные регионы. Это, есте-
ственно, не могло не отразиться на их морфо-экологических особен-
ностях: общем уменьшении длины сифональной трубки, сокращении
величины жабер и незначительных, главным образом цветовых, из-
менениях имаго. Экземпляры с такими признаками напоминают
Ае. mariae и Сх. pusillus, что, помимо прочего, подтверждается и их
экологией. Сх. pusillus считается галофильным видом, а Ае. mariae —
приморским стенобионтом, развивающимся в соленой воде.
Таким образом, на организм при нахождении его в иных услови-
ях обитания воздействуют стабильные элементы среды, в данном слу-
чае повышенная солевая концентрация.
Вместе с тем вполне возможно, что причиной дивергенции могут
быть и изменяющиеся элементы окружающей среды. Во многих мел-
ководных водоемах Казахстана концентрация солей повышается по
мере испарения воды. Естественно, что в процессе таких неоднократ-
но повторяющихся воздействий заселяющие водоем организмы, в
частности Ае. с. caspius и Сх. modestus, активно реагируют на них.
Наличие переходных форм и возможность изменения признаков у
одного поколения говорят о том, что это ответные реакции второго
типа (по А. С. Мончадскому, 1961). Правда, особенность яиц Ае. с.
caspius прекращать развитие при высокой солевой концентрации ука-
зывает на вероятность изменения обмена веществ, т. е. переход к
третьему, наиболее сложному типу ответных реакций. Возникнове-
ние приспособительных реакций дает возможность таким адаптан-
там существовать в водоемах с повышенной концентрацией солей.
Пример мозаичной изменчивости иллюстрируется политипично-
стью Ае. cinereus. Обладая обширным ареалом, который значительно
перекрывает территорию Казахстана, этот вид на различных участ-
ках показывает определенные пути морфоза. В восточных районах
Казахстана распространены типичные популяции этого вида. В Се-
верном Казахстане наряду с типичными встречаются имаго и личин-
165
ки, определяемые как Ае. с. rossicus, причем из куколок Ае. с. rossi-
cus выплаживались Ае. с. cinereus, и наоборот. В Западном Казах-
стане отмечаются переходные формы.
Наибольшая вариабильность наблюдается в предгорных районах
юга и юго-востока Казахстана. Этим участкам свойственны особи, по
тем или иным признакам напоминающие Ае. с. cinereus и Ае. с. ros-
sicus и определяемые как Ае. с. esoensis. Как и в Северном Казахста-
не, здесь нет соответствия между признаками личинок и выплажива-
ющихся из куколок имаго. Из личинок (куколок) с признаками Ае. с.
rossicus выплаживаются имаго с типичными признаками Ае. с. cine-
reus и гипопигиями, соответствующими Ае. с. rossicus. Как среди ли-
чинок, так и среди самцов отмечаются экземпляры Ае. с. esoensis.
Последний, будучи распространен в Японии, на территории СССР был
зарегистрирован лишь на Дальнем Востоке, причем также совместно
с Ае. с. cinereus (Гуцевич, 1937, 1940, 1941). Это подтверждает мне-
ние В. Н. Беклемишева (1960) о том, что естественные суперпопуля-
ции являются, как правило, комплексами более мелких популяций,
в которых в зависимости от степени обособленности могут протекать
противоположно направленные процессы.
Связанные целым рядом переходных признаков, эти подвиды не
показывают какой-либо географической обособленности. Здесь, по
нашему мнению, сказывается прямое влияние гетерогенности усло-
вий внешней среды на обособленные популяции этого вида. Дело в
том, что, отличаясь громадным ареалом, Ае. cinereus вследствие не-
совершенства дыхательной системы (Виноградская, 1948), не позво-
ляющей ему удерживать влагу, может существовать лишь в местах
с густым растительным покровом. А такие участки в Казахстане
приурочены лишь к определенным оазисам и поймам рек. Это соз-
дает пространственную разобщенность отдельных популяций, кото-
рые в зависимости от экоклиматических условий могут обладать оп-
ределенными вариациями, возведенными в подвидовой ранг (Ае. с.
rossicus, Ае. с. esoensis). Таким образом, еще раз подтверждается мне-
ние В. Н. Беклемишева (1960) о сложной гетерогенности суперпопу-
ляций видов, обладающих громадным ареалом.
Другим примером географического морфоза, происходящего пу-
тем изоляции популяций, может служить дивергенция Ае. pullatus.
Имея вытянутую по лесной и субальпийской зонам Тянь-Шаня часть
ареала, этот вид в значительной мере обособился от основной части
популяции. В результате в Заилийском Алатау отмечаются формы,
крайние варианты которых (среднеспинка в белых чешуйках, гипо-
стигмальное пятно из 1—3 чешуек, перевязи без сужений и др.) об-
ладают всеми признаками нового вида. И лишь исследование проме-
жуточных звеньев позволяет установить их видовую идентичность.
166
Аналогичное влияние пространственной протяженности ареала про-
слеживается и на популяциях Ае. leucomelas и Ае. intrudens.
Наличие в Казахстане различных форм изменчивости и особен-
но ее крайние варианты в значительной мере затрудняют видовую
диагностику Culicinae. Этому же препятствует излишняя раздроблен-
ность некоторых групп комаров (communis, cantans, caspius, cinereus)
на отдельные репродуктивно не изолированные таксоны и наличие
ряда переходных форм между ними.
Фауна кровососущих комаров СССР в основном изучена непло-
хо. Она отражена в сводках А. А. Штакельберга (1937), А. С. Мон-
чадского (1951), А. В. Маслова (1967) и отдельных сообщениях. Одна-
ко для более глубокого регионального исследования местных попу-
ляций с целью организации экологически обоснованных мер борьбы
с ними необходимо изучение внутривидовой систематики, которая
отражала бы политипичность видов кровососущих комаров и все про-
цессы их дивергенции.
Нам представляется необходимой единая классификация внутри-
видовых единиц, в полной мере характеризующая весь комплекс
морфо- и геновариаций. Для кровососущих комаров, как нам кажет-
ся, этому соответствовала бы система К. М. Завадского (1961), но без
предлагаемого им пол у видового ранга. Остальные шесть категорий
(подвид, экотип, местная популяция, экоэлемент, изореагент, биотип)
охватывают все наиболее важные формы естественной дифференциа-
ции видов и реально отражают обособленность внутривидовых под-
разделений.
Основной структурной единицей в этой системе признается мест-
ная популяция, что соответствует главному тезису многих определе-
ний вида (Семенов-Тян-Шанский, 1910; Станчинский, 1927; Шаров,
1955; Майр, Линсли, Юзингер, 1956; Завадский, 1961) о составе пос-
леднего из группы более или менее однородных особей. Более высшие
категории (подвид и экотип) определяют соответственно географиче-
скую и экологическую расы, у которых в последующем может про-
изойти изоляция и видообразование.
Все внутривидовые подразделения, стоящие в системе К. М. За-
вадского (1961), ниже популяции отражают совокупность воздейст-
вий факторов внешней среды в пределах внутрипопуляционной из-
менчивости.
Экоэлемент, например, представляет собой формы, уже облада-
ющие известной долей генетической самостоятельности, но в резуль-
тате постоянного скрещивания не могущие изолироваться. Это основ-
ные группы сложной популяции, являющиеся как бы зачинателями
не только экологических, как считает К. М. Завадский (1961), но и
географических рас, поскольку в основе тех и других лежит влияние
167
факторов внешней среды. В составе популяции, как нам кажется, они
будут до тех пор, пока не произойдет их экологическая или географи-
ческая изоляция, и тогда основное значение будет иметь доминирую-
щий генетический комплекс, который может привести к частичной
репродуктивной изоляции, т. е. переходу к надпопуляционным еди-
ницам.
Изореагент или морфо-биологическая группа, по определению
К. М. Завадского (1961),— совокупность организмов внутри популя-
ции, имеющая одинаковую или различную наследственную основу,
различимая морфологически и одинаково реагирующая на усло-
вия среды. У кровососущих комаров представителе!! этого подразде-
ления можно найти среди галофобов, галофилов, ксерофилов, мезо-
филов, гигрофилов, феногрупп, трофогрупп и т. п.
Самая низкая категория — биотип характеризуется как группа
организмов, имеющих тождественный генотип и отличающихся от
другой группы хотя бы одной мутацией. Это элементарная единица
генетической структуры популяции, которая в настоящее время как
бы символизирует первичную мозаичность популяции. Роль ее опре-
делится при генетических исследованиях.
Вся сложность проблемы внутривидовой изменчивости и формо-
образования диктует необходимость пересмотра многих видовых под-
разделений.
V. Экологические
особенности комаров Казахстана
1. МЕСТА ВЫПЛОДА И УСЛОВИЯ ОБИТАНИЯ
Предлагаемая типология мест выплода кровососущих комаров
в Казахстане основывается на ландшафтной характеристике всевоз-
можных биотопов, как в значительной степени отражающей диапа-
зон экологических требований того или иного вида к определенной
среде обитания (Дубицкий, Деньгуб, 1967 а).
Все естественные места выплода кровососущих комаров в Ка-
захстане относятся к четырем основным типам: лесным, степным,
пустынным, горным. Пятый тип составляют искусственные водоемы,
образуемые в результате хозяйственной деятельности человека.
Характерной чертой всех лесных водоемов являются значитель-
ная затененность, обилие на дне прелой прошлогодней растительно-
сти, слабая испаряемость и относительно низкая температура воды.
Чаще всего они образуются в результате заполнения талыми и сточ-
ными водами всех естественных понижений, оврагов, ям, балок
и т. п. Прогреваемость их зависит от степени затененности водоема,
его глубины и подпитывания грунтовыми или сточными водами. В
период развития личинок температура воды в таких водоемах колеб-
лется от 5 до 17—19° и лишь в менее затененных может достигать
21—23°.
Поскольку водоемы этого типа образуются в основном снеговы-
ми и паводковыми водами, они характеризуются слабой минерали-
зацией, незначительной примесью органических и неорганических
соединений и слабощелочной реакцией. Наличие гниющих расти-
тельных остатков, обросших перифитоном, создает обилие пищи для
личинок. Образование таких водоемов приурочено к периоду уста-
новления положительных температур (конец апреля — начало мая).
Во время развития личинок они лишены вегетирующей растительно-
сти, которая появляется позже, густо покрывая высохший или высы-
169
хающий водоем, и создает оптимальные условия для сохранения диа-
паузирующих яиц новой генерации.
Таблица 3
Типология лесных водоемов и приуроченность к ним
различных видов Culicinae
Вид комаров Небольшие заточ- ные водоемы Обширные заболоченности Водоемы озерно- го типа
пол- ностью затенен- ные частично откры- тые пол- ностью затенен- ные частично откры- тые пол- ностью затенен- ные частично откры- тые
Ае. behningi + 4- 4- 4-4-4-
Ае. с. dorsalis "1" -|" 4- I 4—
Ае. cantans —|— 4-4-4- 4- 4~
Ае. cataphylla ++ 4-
Ае. communis ++ н—Н 4~ +
Ае. cyprius 4-4-4- —1— 4—
Ае. diantaeus +++ 4-4-4-
Ае. excrucians 4- 1 1 4~ 4- 4-4-
Ае. flavescens 4—Ь 4- 4-4-4- 4-
Ае. hexodontus 4-4-4-
Ac. impiger +
Ае. intrudens + 4—Ь 4- 4—h
Ае. leucomelas 4- 4~
Ае. pionips 4- 4-4-4- 4-
Ае. pullatus 4-4-4- 4-4- 4-
Ае. punctor 4-4-4- 4—h 4—Н 4"
Ае. riparius 4- 4~ 4-
Ае. sticticus 4- 4—Ь
Ае. с. cinereus 4-4-4- 4-
Ае. с. rossicus 4- 4- 4—h
Ае. vexans 4—h +++
Cs. alaskaensis 4- 4~4~
По занимаемой площади, глубине, степени зарастания и близо-
сти грунтовых вод лесные водоемы можно подразделить на: 1) не-
большие заточные водоемы, 2) обширные заболоченности, 3) водоемы
озерного типа. Небольшие заточные водоемы (3—10 м2) обычно за-
селены представителями группы «communis» (табл. 3), реже видами
группы «cantans». Обширные, подпитываемые грунтовыми верховод-
ками, длительно не высыхающие заболоченности, наоборот, предпо-
читают1 представители группы «cantans» и реже виды группы «com-
munis». Прибрежные, хорошо прогреваемые участки больших зато-
170
ков озерного типа являются излюбленными местами выплода Ае.
сexans, Ае. cinereus и менее холодолюбивых видов — группы «сап-
tans» (Ае. behningi, Ае. cantans).
Причина такой предпочитаемости кроется в экологических осо-
бенностях этих групп. Как уже говорилось, бореальные представите-
ли группы «communis» имеют самый низкий температурный опти-
мум. Выплод и развитие их отмечаются при 5—18°. Выплаживаясь
ранней весной, они вполне успевают пройти развитие до периода вы-
сыхания водоемов. В значительной степени это, вероятно, является
также адаптацией для окончания развития до наступления периода
значительного прогревания водоемов, что, как показала М. Ф. Шле-
нова (1952), может оказаться для нпх губительным. Выбор таких
мест выплода обусловлен, как нам кажется, стремлением самок от-
кладывать яйца на влажную почву высыхающих к тому времени во-
доемов.
Обладая столь же длительным развитием, но выплаживаясь
вследствие более высокого температурного оптимума (12—25°) зна-
чительно позже, виды группы «cantans» не успевают закончить раз-
витие в небольших, быстро пересыхающих водоемах. Адаптацией их
к предохранению от откладки яиц в таких условиях является, как
нам кажется, стремление самок производить кладки на воду, которая
еще имеется в более обширных, долго не высыхающих водоемах.
Встречаемость представителей группы «communis» в небольших ко-
личествах в обширных водоемах, а видов группы «cantans» — в бо-
лее мелких, эфемерных можно объяснить откладкой яиц после пов-
торных гонотрофических циклов. К этому периоду небольшие скоп-
ления воды могут настолько пересохнуть, что самки группы
«communis» находят влажные места лишь на усыхающих участках
более обширных водоемов, и, наоборот, вследствие прошедших дож-
дей небольшие водоемы заполняются водой, и на нее откладывают
яйца самки группы «cantans». Судя по литературным данным (Шле-
нова, 1952; Сазонова, 1959), в лесной части РСФСР такого четкого
разграничения нет и надобность в нем вследствие более умеренного
климата отпадает. В Казахстане же, где рассматриваемые лесные
участки находятся в зоне высокой солнечной радиации и интенсив-
ного испарения влаги, это привело к довольно четкой экологической
адаптации. Подобное явление отмечается и у других видов у границ
их ареалов.
Лесные водоемы, как и одноименная растительная формация,
встречаются лишь в самых северных районах Казахстана, в более вы-
ровненных предгорьях Алтая и на некоторых возвышенных участках
степной зоны, где они приобретают характер экстразональных вклю-
чений. Во многих случаях водоемы такого типа отмечаются в поло-
171
гих долинах отрогов Тянь-Шаня и в поймах крупных рек и их прито-
ков (Урал, Ишим, Тобол, Ульба, Бухтарма и т. п.), находящихся се-
вернее 48—50° с. ш.
Таблица 4
Типология естественных степных водоемов и приуроченность к ним
различных видов Culicinae
Вид комаров Озера, стари- цы, рукава Весенние зато- ки и разливы Разобщенные плёсы мелких рек Выходы грун- товых вод
затенен- ные откры- тые затенен. откры- тые затенен- ные откры- тые затенен- ные откры- тые
An. hyrcanus
An. т. messeae
М. richiardii
Ае. с. caspius
Ае. с. dorsalis
Ае. cataphylla
Ае. flavescens
Ае. leucomelas
Ае. cinereus
Ае. vexans
Сх. modestus
Сх. territans
Сх. pipiens
Cs. longiareolata
Степные водоемы менее многообразны, чем лесные. Общими для
них являются слабая затененность (кустарниковая и редкая древес-
но-кустарниковая растительность), довольно хорошая прогреваемость,
сильная зарастаемость и самый разнообразный химический состав.
Большинство из них благоприятны для выплода комаров весной и
ранним летом. Затем площадь их уменьшается, открытые пространст-
ва вследствие зарастания сокращаются, а концентрация солей повы-
шается. Как видно из табл. 4, число видов Aedes, приуроченных к
этим водоемам, снижается, но зато увеличивается количество
представителей Culex, Anopheles, Mansonia. По сравнению с лесны-
ми водоемами здесь значительно сокращены число и площадь вре-
менных разливов и затоков. Однако оставшаяся часть является стой-
ким продуцентом кровососущих комаров в течение всего летнего се-
зона. Как и в предыдущем случае, наблюдается экстразональная
встречаемость степных водоемов на открытых участках лесостепной
зоны.
172
Большинство временных и постоянных водоемов восточной час-
ти степной зоны сильно засолены. Развитие личинок комаров в них
невозможно. Проведенные исследования показали, что для галофоб-
ных видов допустима концентрация до 2% солей, а для галофильных—
в пределах 6—8%. В том случае, когда концентрация солей повыша-
ется по мере усыхания водоемов, развивающиеся в них личинки мо-
гут выдерживать большее насыщение. Наши наблюдения и результа-
ты опытов говорят об относительной галофильности многих видов
Anopheles, в обычных условиях предпочитающих пресную воду.
Пустынные водоемы еще более однообразны по сравнению со
степными. В основном это более или менее заросшие участки водной
поверхности, хорошо прогреваемые солнцем, подверженные сильной
испаряемости и богатые бентосом, планктоном и нектоном. Особый
тип пустынных водоемов представляют пойменные разливы и зато-
ки, характеризующиеся слабой примесью органических и неоргани-
ческих веществ. Образуются они во время таяния снежного покрова,
но чаще в период половодья пустынных рек и в период действия на-
гонных ветров в водоемах морского порядка (Каспий, Арал, Балхаш).
Такие разливы могут тянуться на десятки километров и почти повсе-
местно заселяться личинками кровососущих комаров. Многие из
глубоких затоков в результате грунтового подпитывания от русла
реки не пересыхают совсем или пересыхают лишь к концу лета. Не
случайно большинство распространенных в этих местах видов при-
урочено к таким водоемам (табл. 5). Многие из них имеют большую
концентрацию солей и тогда заселяются галофильными видами ко-
маров.
Пустынные озера как по условиям развития, так и по приуро-
ченности заселяющих их видов во многом напоминают степные. От-
личительной их чертой являются небольшая глубина, меньшая пло-
щадь и более интенсивная степень зарастания. Концентрация соли
во многих из них превышает допустимую норму, и это препятствует
развитию в них галофобных личинок. Из галофильных видов такие
водоемы обычно заселяют Сх. pusillus, а иногда и Ае. с. caspius. Озе-
ра с незначительной концентрацией солей являются излюбленными
местами выплода комаров Culex, Anopheles, Culiseta, развивающих-
ся без эмбриональной диапаузы (табл. 5). При этом виды первых двух
родов, кроме An. hyrcanus, предпочитают открытые, хорошо прогре-
ваемые участки озер, а виды Culiseta, напротив,— прибрежную зате-
ненную часть. Вместе с ними обычно встречается и Ur. unguiculata.
На пониженных участках песчаных пустынь нередко можно
встретить естественные выходы грунтовых вод. Чаще всего они отме-
чаются в предгорных районах и вблизи от теряющихся в песках ру-
сел небольших рек. Вода в них обычно держится в первой половине
173
лета, затем пересыхает и появляется осенью. Наполненные водой,
такие водоемы охотно заселяются многими видами комаров, но осо-
бенно представителями Culiseta и холодолюбивыми видами Anophe-
les. В результате грунтового подпитывания и сильного зарастания
надводной растительностью температура воды в них никогда не до-
стигает высоких показателей. Аналогичны им по описанным услови-
ям и заселяемости личинками и родники.
Таблица 5
Типология естественных пустынных водоемов и приуроченность к ним
различных видов Culicinae
Вид комаров Озера Припойменные затоки и разливы Выходы грунто- вых вод
затенен- ные открытые затенен- ные открытые затенен- ные открытые
An. algeriensis 4- 4-
An. claviger 4-
An. hyrcanus An. т. sacharovi 4—1—F + + + 4- _| |_ 4- 4-
An. pulcherrimus An. superpictus Ur. unguiculata 4—F 4- 4- 4-
Cs. longiareolata 4~ +
Cs. alaskaensis —|— + 4-
Cs. annulata M. richiardii 4-4-4- 4-4- 4—F
Ae. c. caspius Ae. cataphylla 4- 4- 4-4-4- 4—F
Ae. detritus 4- 4—F
Ae. flavescens 4-4-
Ae. hasachstanicus +
Ae. leucomelas 4-
Ae. simanini + 4-4-
Ae. cinereus + +4- + 4-
Ae. vexans Cx. modestus + _| | Ц 4—I- 4- 4- 4-
Cx. pusillus 4~ 4-
Cx. pipiens 4~ 4- 4- 4- 4-4- 4-4-4-
Выделение горных водоемов обусловлено спецификой встречаю-
щихся в них видов и своеобразием экологии доходящих до них рав-
нинных представителей. Специфика фауны и экологии горных биото-
пов отмечается и в Центральной Европе (Trpis, 1965). На различных
высотах вода в местах выплода комаров чистая, прозрачная, с бога-
тым бентосом, а иногда и планктоном. Даже на хорошо прогревае-
174
мых участках температура ее редко достигает 20—21°. Казалось бы,
условия — идеальные лишь для олиготермных видов, но помимо них
здесь встречаются, причем на довольно большой высоте (субальпий-
ский пояс), такие эвритопные виды, как Сх. pipiens и Сх. longiareolata
(табл. 6). В этом случае они обитают там, куда не доходят многие
олиготермные представители. Кроме того, только в горных водоемах
нами обнаружены Cs. bergrothi, Сх. martinii.
Таблица 6
Типология естественных горных водоемов и приуроченность
к ним различных видов Culicinae
Вид комаров Среднегорные затоки и разливы Высоко- горные затоки (откры- тые) Высоко- горные разливы (откры- тые)
затенен- ные открытые
An. claviger +++ +
Cs. longiareolata ++ +
Cs. bergrothi + +
Cs. alaskaensis +++ -|—[_
Ae. cataphylla ++
Ae. flavescens +
Ae. intrudens + +
Ae. leucomelas +
Ae. pullatus Ч- + +++
Ae. cinereus +
Ae. vexans + 4-
Cs. territans
Cx. hortensis 4—h
Cx. martinii +
Cx. pipiens ++ 4- 4~ 4~ + +
Своеобразна экологическая приуроченность
встречающихся в
них видов. Как и в равнинных условиях, представители Aedes пред-
почитают периодически пересыхающие водоемы, но если там — зате-
ненные биотопы, то здесь, наоборот,— открытые, хорошо прогревае-
мые, что соответствует принципу вертикальной смены стаций
(Бей-Биенко, 1959). Виды Culex, Culiseta и Anopheles также чаще за-
селяют эфемерные, хорошо прогреваемые, мелкие водоемы, хотя и
избегают до осеннего периода прямого пребывания на солнце. Сильно
заросшие элодеидами водоемы не заселяются и освобождаются от ли-
чинок по мере зарастания.
В общем же вследствие своей специфичности и гипотермичности
условий в горных водоемах личинки комаров редки. Исключение
175
представляют широкие пологие долины с южной экспозицией, при-
ближающиеся по условиям к равнинным.
Одну из многочисленных групп мест выплода кровососущих ко-
маров представляют различные искусственные водоемы, связанные
с хозяйственной деятельностью человека. К ним, прежде всего, отно-
сятся фильтрационные разливы и площади сброса поливных вод,
песчаные, глинистые и щебнистые карьеры, придорожные кюветы и
углубления в местах поднятия всевозможных дамб и перемычек. Ре-
гулярно подпитываемые грунтовой и заточной водой, они являются
стойкими продуцентами комаров в течение всего летнего периода. В
зависимости от мест нахождения такие водоемы приобретают специ-
фику лесных, степных или пустынных водоемов озерного типа.
Для южных районов Казахстана немаловажное значение имеют
и околопоселковые выемки грунта в местах производства самана, раз-
личные хозяйственные ямы, копанки и приусадебные водоемы —ха-
узы. В них выплаживаются комары, обычные для грунтовых вод пус-
тынных водоемов. Некоторое значение в продуцировании отдельных
представителей Culex и Culiseta имеют городские декоративные водо-
емы, фонтаны и залитые водой бочки. Для развития и выплода ко-
маров пригодна и ирригационная сеть в местах слабого водотока.
Большую роль в поддержании очагов выплода различных видов
комаров и их массовом продуцировании играют всевозможные пру-
ды и рисовые чеки. Последние до периода вегетации, а частично и в
период вегетации риса, обладают идеальными условиями для разви-
тия и выплода Сх. modestus, Ае. с. caspius, Ап. т. sacharovi и некото-
рых других. Их продуцирующая роль повышается в том случае, если
около имеются постоянные фильтрационные или озерные водоемы,
являющиеся беспрестанным поставщиком яйцекладущих самок. В
несколько меньшем объеме, но аналогичное значение имеют неглубо-
кие, слабо заросшие пруды, которые характеризуются спецификой
озерных водоемов.
Рассмотренные типы мест выплода кровососущих комаров дале-
ко не в полной мере отражают все их многообразие. Вследствие по-
разительной экологической пластичности и при наличии на террито-
рии Казахстана благоприятных абиотических и биотических условий
(хорошая прогреваемость водоемов, обилие бентоса, планктона и нек-
тона) они заселяют буквально все временные и постоянные скопления
воды вплоть до неглубоких колодцев, корыт для водопоя скота и ду-
пел деревьев.
2. ЖИЗНЕННЫЕ ЦИКЛЫ
Среди различных экологических групп кровососущих комаров,
встречающихся в Казахстане, имеются виды, зимующие в фазе има-
176
го, яйца и личинки. Соответственно этому в процессе эволюции у них
выработались определенные жизненные схемы, характеризующиея
приспособленностью к условиям обитания.
В южных районах Казахстана преобладают виды, зимующие в
фазе имаго. Это в основном представители родов Culex и Anopheles.
Их жизненная схема отличается прямым (без эмбрионального диа-
паузирования) развитием. За теплый период года они производят
столько генераций, сколько им позволяют температурные условия
того или иного климатического района и термические флюктуации
различных лет. В лабораторных условиях эти виды развиваются от
яйца до имаго при 22—30° за 13—20 дней. Наши исследования и све-
дения, приводимые Э. Р. Геллером (1939), говорят о том, что поли- и
мезотермофильные виды Culex и Anopheles (Сх. modestus, Сх. pipiens,
Сх. pusillus, An. т. messeae, An. т. sacharovi, An. hyrcanus и др.)
могут проделывать от двух генераций на севере Казахстана до 5—
8 на юге.
Количество гонотрофических циклов, вследствие гибели большо-
го количества яйцекладущих самок, для всей популяции установить
трудно. Отдельные особи, по данным Т. С. Детиновой (1962), могут
проделывать до 9—12 циклов. Условия в южной части Казахстана
вполне благоприятствуют этому. Во время экспериментальных иссле-
дований при 22—33° гонотрофический цикл у An. т. sacharovi и Ап.
hyrcanus заканчивался за 3—7 дней, а у Сх. modestus и Сх. pipiens
он в редких случаях превышал 3—4 дня. Этим и объясняется про-
грессивное увеличение численности данных видов во второй поло-
вине лета.
Все виды, кроме синантропных популяций An. maculipennis и
Сх. pipiens, зимуют в густых зарослях тугаев, под отслоившейся корой
деревьев, в завалах тростника и в метелках, охотно используют раз-
личные заброшенные помещения и провалы могил. В условиях посел-
ков и городов An. maculipennis и Сх. pipiens в неблагоприятный зим-
ний период залетают в подвалы, овощехранилища, погреба, животно-
водческие помещения и т. п. Сх. pipiens, кроме того, приспособился
зимовать в полостях канализационной и отопительной систем.
Представители рода Culiseta отличаются холодолюбивостью. Они
распространены в основном в северных районах республики (Cs. ber-
grothi, Cs. morsitans, Cs. silvestris, Cs. alaskaensis) и частично в юж-
ных (Cs. longiareolata, Cs. annulata). На севере успевают проделать
два или три цикла. В более южных районах высокие летние темпера-
туры действуют на них угнетающе, и их развитие ограничивается
весенней и осенней генерациями. Осенний залет на зимовку в убежи-
ще, сезрнный ход численности и заселяемость эфемерных, полностью
промерзающих водоемов говорят, в противоположность существую-
12—80 177
щим мнениям, о невозможности зимовки этих видов в фазе личинки,
по крайней мере в Казахстане, что подтверждается ранневесенней ак-
тивностью имаго и появлением личинок первой генерации.
Развитие личинок этих видов, кроме представителей Anopheles,
часто затягивается до глубокой осени. Внезапные похолодания не-
редко приводят к гибели не успевших закончить развитие предимаги-
нальных фаз. В этих случаях весенние популяции бывают численно
подорванными, но сохраняются за счет асинхронного развития, в ре-
зультате чего часть их успевает выплодиться и надежно укрыться.
Близки к рассмотренной группе и виды, зимующие в фазе ли-
чинки, из них массовых, широко распространенных лишь два: Ап.
claviger и М. richiardii. Первый — представитель олиготермных ви-
дов, второй — мезотермных эврибионтов. Экологически они прямо
противоположны. An. claviger развивается и зимует в холодной, не-
замерзающей воде горных речек и родников, а М. richiardii предпо-
читает водоемы, хорошо прогреваемые летом и не промерзающие
полностью зимой. При таких условиях личинки An. claviger могут
периодически подниматься к поверхности воды, а личинки М. richi-
ardii потребляют кислород, содержащийся в подводной части расте-
ний, к которым они прикрепляются. За теплый период времени эти
виды производят по 2—3 генерации. У An. claviger их количество
лимитируется депрессирующим влиянием высоких летних темпера-
тур» а у Af. richiardii — затяжкой весеннего и осеннего развития.
Продолжительность гонотрофического цикла у обоих видов не
отличается особой растянутостью. У An. claviger, по данным
В. Н. Беклемишева (1949), она при 14—16° длилась 5—6 дней, а у
М. richiardii, в наших экспериментах, при 23—33° составляла 6—
8 дней. Эмбриональный период продолжается 4—6 дней, по истече-
нии которого происходит массовое отрождение личинок. По наблю-
дениям за естественными популяциями можно заключить, что даль-
нейшее предимагинальное развитие в летнее время осуществляется за
18—20 дней. У An. claviger такое же время затрачивается на анало-
гичный период в условиях прохладных горных биотопов, где темпе-
ратура воды не поднимается выше 18—20°, а ночью опускается до
9—12°.
Относящиеся к этой же группе An. algeriensis и An. pulcherrimus
встречаются лишь в южных районах Казахстана и требуют специ-
фических условий для своего развития.
Для An. algeriensis необходимы прохладные, влажные биотопы
и продолжительный теплый период, а для An. pulcherrimus -г- силь-
но прогреваемые водоемы.
Наиболее многочисленную по представителям составляет груп-
па видов, зимующих в фазе яйца. Все они относятся к роду -Aedes,.
178
По количеству проделываемых генераций их можно подразделить на
моно- и полицикличные. Большинство моноцикличных видов (Ае.
communis, Ае. intrudens, Ае. punctor, Ае. pullatus, Ае. diantaeus,
Ае. sticticus, Ае. simanini, Ае. montchadskyi, Ае. detritus, Ае. leuco-
melas, Ае. cataphylla, Ае. impiger, Ае. nigripes, Ае. nigrinus, Ае. he-
xodontus, Ае. rempeli, Ае. pionips) выплаживаются лишь ранней вес-
ной и встречаются в северных районах Казахстана, а также в горных
участках и экстразональных биотопах более южных районов. Разви-
тие их осуществляется при 5—18° в течение 35—40 дней. В лабора-
торных условиях они развивались при 20—25’, хотя в экспериментах
М. Ф. Шленовой (1952) на большинство личинок температура в
19—21° действовала губительно. Это указывает на региональную
адаптацию данных видов к более высоким температурам, характер-
ным для многих хорошо инсолируемых участков Казахстана. В од-
ном случае выплод Ае. pullatus, доставленного в лабораторию с высо-
ты 2600 м н. у. м., происходил при 23—31°.
Не менее велика пластичность этих комаров к противополож-
ным условиям. Промораживание Ае. simanini при 5—7° в условиях
эксперимента при одновременном наблюдении за естественными во-
доемами показало, что при дневном оттаивании личинки и куколки
способны к дальнейшему развитию. В резко континентальных райо-
нах Казахстана это имеет большое значение. Для активизации диа-
паузирующих яиц обязательно прохождение холодовой стадии, хотя
у некоторых видов отмечается повторный летний выплод. Гонотро-
фический цикл при 19—20° длится 8—10 дней. Яйцекладки осущест-
вляются на влажную почву усыхающих водоемов, где, даже в усло-
виях юго-востока Казахстана, они могут сохраняться в течение года.
Температура воздуха там в летнее время колеблется в тени от 18
до 45°, а почва на затененных участках прогревается до 18—34°.
Другая группа моноцикличных видов (Ае. cyprius, Ае. flaves-
cens, Ае. excrucians, Ае. kasachstanicus, Ае. cantans, Ае. subdiversus,
Ае. behningi, Ае. riparius) выплаживается поздней весной. Их темпе-
ратурный оптимум несколько выше. На примере Ае. flavescens его
можно определить как 12—25°, но продолжительность развития соот-
ветствует предыдущей группе и составляет 40—45 дней. У некоторых
представителей поздневесенних видов возможен повторный летний
выплод. Период развития Ае. flavescens при летнем выплоде осуще-
ствляется при 20—28° за 30—31 день. Пока не совсем ясно, какими
факторами управляется бицикличность данных видов. А. С. Мон-
чадский (1961) видит основу их в меняющихся элементах среды оби-
тания, приводящих к выработке ответных эколого-приспособительных
реакций к таким условиям. Применительно к Казахстану бициклич-
ность можно охарактеризовать как переход вследствие благоприят-
179
ных условий (температурный режим, влажность, продолжительность
теплого периода) к полицикличности. Это уже отклонение от обяза-
тельного прохождения диапаузирующими яйцами холодовой стадии.
Подобно Ае. simanini, у наиболее южного представителя — Ае.
flavescens также обнаружена способность выносить кратковремен-
ное (10—13 час) промораживание личинок.
Гонотрофический цикл у отдельных видов этой группы (Ае. fla-
vescens, Ае. kasachstanicus) при 20—30° продолжается 7—14 дней,
но чаще — 8—10. Яйцекладки в лабораторных условиях производят-
ся самками чаще непосредственно на воду и редко на влажную по-
верхность (плавающая фильтровальная бумага). Повторный выплод
личинок отмечался лишь из кладок, содержавшихся в течение 20—
32 дней на влажной поверхности, а затем залитых водой. Вполне воз-
можно, что это начало биологического расслоения данных видов на
определенные расы в результате изменившихся условий среды оби-
тания.
Основная часть полицикличных видов Aedes (Ае. caspius, Ае. ve-
xans, Ае. cinereus) в летний период развивается беспрерывно, т. е. по
типу комаров Culex, Anopheles и Culiseta. Лишь Ае. vexans своеобраз-
но реагирует на изменения условий внешней среды. Имея центром
своего географического происхождения по отношению к предыдущим
видам совершенно противоположный регион (Ориентальная область),
он отличается от других, распространенных в Казахстане, видов
Aedes способностью выходить из диапаузы не после проморажива-
ния, а после просушивания. И все же удается получить его прямое
развитие.
Ае. с. cinereus отличается большой асинхронностью развития,
затрудняющей определение факторов, которые приводят к торможе-
нию развития и его окончанию. Вероятнее всего, в летний период оно
вызывается высокими температурами, а в осенний — низкими.
Цикл развития Ае. с. caspius служит примером максимального
упрощения жизненной схемы представителей Aedes. Этот подвид
прекращает развитие при возникновении неблагоприятных условий
внешней среды и дает выплод личинок после окончания их действия.
Продолжительность жизненных циклов обоих видов зависит от
температурных условий. Так, при 23—36° развитие Ае. с. caspius
продолжается 15—16 дней, при 19—29° растягивается до 22—г
27 дней, а у Ае. с. cinereus при последнем режиме, наоборот, бывает
минимальным — 15—16 дней.
Яйцекладки у Ае. с. caspius и Ае. с. cinereus, по наблюдениям в
лабораторных условиях, обычно производятся непосредственно на
воду. Через 3—10 дней происходит выплод личинок, которые нор-
мально растут и развиваются; у Ае. с. caspius — при любых естест-
180
венных температурах, а у Ае. cinereus — лишь при температурах ни-
же 25—26°. Более высокие температуры (30—33°) выплод личинок
последнего задерживают, что полностью согласуется с наблюдения-
ми М. Ф. Шленовой (1952). Из яиц Ае. v. vexans в том случае, если
они попадают в воду, через 8—9 дней происходит выплод основной
массы (68—79%) личинок и дальнейшее развитие. Но самки во всех
случаях предпочитают откладывать яйца на урез воды. При посте-
пенно развивающейся сухости яйца впадают в диапаузу, а уже затем
для их реактивации требуется дополнительное просушивание при вы-
соком термическом режиме (30—34е), который для южной части Ка-
захстана обычен.
За теплый период времени, с учетом существующих порогов раз-
вития, Ае. с. cinereus и Ае. vexans проделывают не менее двух-трех
генераций. Ае. с. caspius в южной части Казахстана при благоприят-
ных условиях может осуществить пять-шесть поколений, а Ае. с. dor-
salis в пределах своего более северного ареала — лишь две-три гене-
рации. В отдельных случаях прохождению последовательного развития
Ае. с. caspius препятствует повышающаяся при усыхании водое-
мов концентрация солей. Это тормозит развитие, которое прекраща-
ется сразу же после опреснения воды. Как и другие виды, развиваю-
щиеся при значительных перепадах суточных температур, обуслов-
ленных резкой континентальностью климата, личинки Ае. с. caspius
способны выдерживать 8—10-часовое промораживание.
Подводя итог, можно сказать, что жизненные циклы кровососу-
щих комаров в Казахстане определяются условиями среды обитания.
В процессе эволюционного развития произошло приспособление на-
секомых к существованию в различных природных биотопах, выра-
жающееся как в усложнении жизненных циклов (появление летней
депрессии у Culiseta), так и в упрощении более сложных (переход к
беспрерывному развитию у Aedes). Процесс этот, по всей вероятности,
еще не закончен, поскольку имеется несоответствие между условия-
ми обитания и возможностями существования в них тех или иных
видов. Осуществлению его способствует влияние факторов внешней
среды в совокупности с экологической валентностью вида.
3. СЕЗОННАЯ И СУТОЧНАЯ ПЕРИОДИКА ЛЁТА
Естественно, что единого представления, а тем более изображе-
ния периодики лёта и нападения комаров для всей многообразной
территории Казахстана не может быть. В этом отношении более по-
казательны сведения по одному, наиболее характерному участку то-
го или Иного региона и их интерпретация по отношению к другим.
Критерием достоверности при этом могут обладать лишь полисезон-
181
ные наблюдения, проведенные в одном месте в течение двух и бо-
лее лет.
Исходя из этого здесь приводятся сведения по сезонной актив-
ности комаров для двух наиболее изученных противоположных ре-
гионов Казахстана (юг и север) и их экстраполяция на другие участ-
ки (Дубицкий, Дешевых, 1966).
Рис. 37. Сезонный ход численности кровососущих комаров в юж-
ных районах Казахстана.
В бассейне Сыр-Дарьи (пос. Яны-Курган, граница средней и дель-
товой части, Кзыл-Ординская область) первые комары из числа пе-
резимовавших в природе популяций начинают нападать во второй
декаде марта (рис. 37). Численность их невелика, обычно 1—5 эк-
земпляров за 20 мин. В апреле количество нападающих комаров уве-
личивается за счет выплода отдельных видов в разливах талых вод.
И даже в этот период обилие их незначительно, а следовательно, они
не причиняют людям и животным серьезного беспокойства. Так про-
должается до конца мая, после чего наступает период заметного уве-
личения массы насекомых. Происходит это после весеннего поло-
водья Сыр-Дарьи, в результате чего образуется множество водоемов
временного и полупостоянного типа. Обилие выплодившихся в них
кровососов и составляет первый массовый пик, после чего наблюда-
ется некоторое снижение численности, объясняемое уменьшением
мест выплода и периодом созревания нового поколения. Другой, не
менее значительный подъем численности наблюдается во второй по-
ловине июля. Он связан со вторым летним половодьем Сыр-Дарьи,
182
Рис. 38. Сезонный ход численности кровосо-
сущих комаров в северных районах Казах-
стана. По И. Г. Прыгуновой, 1966. 1 — в ле-
состепной, 2 — в степной зоне.
и сопутствующие им полисезонные
которое наступает в наиболее жаркий период за счет таяния высоко-
горных снегов в верховьях реки.
После каждого пика на протяжении 1—1,5 месяца создается
численно устойчивое обилие комаров. В это время их можно встре-
тить в местах, где, казалось —
бы, совершенно отсутствуют
условия для выплода, в том
числе и на редко увлажняе-
мых участках.
На севере Казахстана, по
нашим наблюдениям (Дубиц-
кий, Деньгуб, 1967 б) и све-
дениям И. Г. Прыгуновой
(1966), первые нападающие
особи из числа перезимовав-
ших самок начинают напа-
дать в конце первой декады
мая (рис. 38). Численность их
первоначально незначитель-
ная, резкое увеличение ее от-
мечается в конце мая — на-
чале июня в результате вы-
плода ранневесенних видов
{Ае. intrudens, Ае. leucomelas,
Ае. cataphylla, Ае. sticticus). В
середине июня к ним приплю-
совываются поздневесенние
(Ае. excrucians, Ае. flavescens,
Ае. behningi, Ае. cantans, Ае.
beklemischevi, Ае. subdiversus)
виды (Ае. с. dorsalis, Ае. с. caspius, Ае. с. cinereus, Ае. с. rossicus).
Умеренные летние температуры способствуют длительной жизни
выплодившихся генераций. Вместе с тем наступление наиболее теп-
лого периода года активизирует в июле второй массовый выплод по-
лицикличных видов, среди которых в этот период определенное
значение имеют An. т. messeae, Ае. vexans, Сх. modestus и особенно
М. richiardii. По-прежнему обильны Ае. с. dorsalis, Ае. с. cinereus и
некоторая часть популяций весенних видов.
Наибольшая численность комаров отмечается во второй полови-
не августа. К этому периоду выплаживается третья генерация Ае. с.
dorsalis, М. richiardii, Сх. modestus, Ае. vexans. Обилие прошедших в
первой половине июля дождей даже активизирует повторный выплод
некоторых не строго моноцикличных видов. Такой суммированный
183
выплод и обусловливает наиболее высокую численность комаров за
весь теплый период. Наступающие в конце августа резкие похолода-
ния приводят к гибели основной части массовых популяций и залету
на зимовку видов, зимующих в фазе имаго.
Приведенный ход численности характерен для лесостепной зо-
ны. Исследования, осуществленные в припойменных участках степ-
ной зоны, показали подобную же трехвершинную кривую активно-
сти комаров, но уменьшенную в 10—12 раз и с наиболее значитель-
ным пиком в весенний период.
Анализируя имеющиеся сведения о сезонной активности крово-
сосущих комаров в Казахстане, можно заметить, что периодика лёта
и нападения характеризуется в основном двумя и тремя подъемами
численности. Графически двухвершинный ход численности, с теми
или иными поправками, совпадает с двумя последними подъемами
численности трехвершинной кривой. Объясняется это просто. Пер-
вый массовый пик характерен для более северных районов Казах-
стана, где весной происходит выплод ранне- и поздневесенних моно-
цикличных видов. По мере продвижения на юг количество и числен-
ность таких видов становятся меньше. Соответственно этому сезонный
ход численности приобретает характер двухвершинного. Иногда и в
южных и юго-восточных районах Казахстана отмечается высокая
весенняя численность, что обусловливается количеством талых вод и
весенними осадками.
Следовательно, на всей многообразной территории Казахстана
первый (майский) пик численности характерен для выплода олиго-
термных моноцикличных видов, а остальные два характеризуют раз-
витие двух массовых поколений мезо- и политермных представите-
лей Culicinae, и только в отдельные годы в южных районах весеннее
увеличение численности создает Ае. с. caspius. Это ни в коей мере не
характеризует количество проделываемых генераций, которых мо-
жет быть больше, а говорит о трех массовых подъемах численности,
обусловленных гидрологическим режимом теплого времени года.
В южной части Казахстана двухвершинность сезонного хода
численности определяется, как уже указывалось, весенним и летним
половодьем пустынных рек, берущих свое начало на снежных участ-
ках Тянь-Шаня. В северной части Казахстана, лежащей севернее
50° с. ш., первый и второй пики численности определяются весенни-
ми разливами талых и паводковых вод, с той лишь разницей, что
олиготермные виды начинают развиваться еще в «ледяной воде», а
мезо- и политермные — в более поздний период, когда водоемы про-
греваются до 12° и выше. Выплод тех и других представителей следу-
ет один за другим, и это объясняет сближенность первого и второго
пиков численности. Третий массовый подъем численности отмечает-
184
ся в конце июля — начале августа после заполнения мест выплода
водой от обычных в этот период дождей.
Несколько отличается от двух описанных типов сезонный ход
численности комаров в Северном Прикаспии. Первый (майский) пик
численности, как обычно, отмечается после выплода комаров в раз-
ливах и затоках талых и паводковых вод. В дальнейшем это опреде-
ляется гидрологическим режимом Каспийского моря, повышение
уровня которого приводит к заполнению неисчислимых по площади
мест выплода. Изменения прибрежного уровня воды Каспия, в сбою
очередь, происходит во время действия штормовых юго-западных
ветров, называемых «морянами».
Определяя в общих чертах сезонную периодику лёта кровососу-
щих комаров в Казахстане, можно заключить, что помимо обычного
более или менее значительного обилия, связанного с наличием посто-
янных мест выплода, могут быть два или три необычайно высоких
подъема численности, обусловленных весенними и летними паводка-
ми и дождевыми затоками. Фенология их определяется ландшафтно-
климатическими особенностями того или иного региона, гидрологи-
ческими флюктуациями различных лет и доминированием моно- или
полицикличных видов.
Суточная активность кровососущих комаров более или менее од-
нотипна для всех регионов Казахстана. Наиболее хорошо она изуче-
на при полисезонных наблюдениях в Южном, Юго-Восточном и Се-
веро-Восточном Казахстане. Суточный ритм численности всех напа-
дающих там видов был более или менее однотипен. По времени
нападения суточную активность можно разделить на два периода:
весенне-осенний и летний. Последний характерен для всего периода
активности кровососов с нападением их в утренние и вечерние часы.
Весной и осенью ночная прохлада ограничивает нападение насеко-
мых в утренний и вечерний периоды.
В летний период утренняя активность комаров начинается до
восхода солнца и постепенно уменьшается после его восхода. Через
1 час после этого еще наблюдаются единичные нападения, а через
1,5—2 час они прекращаются совсем. Ведущее значение в этом при-
надлежит депрессивному действию светового фактора ‘(Мончадский,
1956). Наши исследования ольфакторных реакций комаров на тем-
пературный фактор показали, что на близком расстоянии большое
значение имеет выделяемое хозяином тепло (Дубицкий, 1965 а).
Теснейшая зависимость этих двух факторов ограничивает ак-
тивность кровососов в течение всего дневного времени. Понижение
температуры, порой даже незначительное, в сочетании с уменьшени-
ем освещенности вызывает их вечернюю активность. Начинается она
за 1,5—2 час до захода солнца и бывает особенно сильной в период
185
наступления темноты. Как сообщал А. С. Мончадский (1953), сплош-
ная темнота снижает активность комаров. В это время нападают осо-
би, находящиеся поблизости от человека и воспринимающие запахи
продуктов его метаболизма (Дубицкий, Якунин, 1966). В теплую по-
году при хорошем лунном освещении нападение комаров бывает до-
вольно интенсивным и в ночное время.
В противоположном случае взаимодействие температуры и осве-
щенности наиболее ярко проявляется во время необычных атмосфер-
ных явлений. Наблюдение за активностью комаров в бассейне Или
во время солнечного затмения в конце сентября 1968 г. показало,
что численность кровососов, сдерживаемая интенсивной инсоляцией
среды, стала заметно увеличиваться и была наиболее значительной в
момент максимального затмения. Исследования были проведены в
период, когда ночная прохлада уже начала ограничивать вечернюю
активность, а интенсивному дневному нападению препятствовала
высокая температура. В таких условиях уменьшение освещенности
среды привело к активизации всех комаров, находившихся на днев-
ках. Нападаемость их увеличилась в 11 раз по сравнению с дневной
активностью, в 6 раз по сравнению с предвечерней и соответствовала
обычной летней сумеречной нападаемости. Аналогичная картина от-
мечалась и А. В. Гуцевичем (1939) в Забайкалье.
Рассмотренная активность характерна для большинства откры-
тых или слабо заросших ландшафтов республики. В лесных участках
хотя и существуют два массовых периода нападения — утренний и
вечерний, но контраст между ними сглаживает обычная в тех мес-
тах дневная активность. С одной стороны, это объясняется затенен-
ностью лесных биотопов, а с другой — значительным обилием моно-
цикличных весенних видов, отличающихся слабой дневной депресси-
ей. Соответственно этому период суточной активности будет
увеличиваться в пасмурную прохладную погоду днем и в ясную теп-
лую полнолунную ночь.
В периоды, когда утренняя температура бывает ниже 14—15° в
южных районах и 8—10° в северных, нападения комаров не отмеча-
ется. Этот температурный предел ограничивает активность комаров
и в вечернее время. В такие периоды нападение комаров переносится
на более теплое дневное время. Но и в этих случаях существует об-
щий порог активности. В среднем течении Сыр-Дарьи и в бассейне
Или при температурах ниже 15—16° нападение комаров прекраща-
ется. В условиях равнинного Узбекистана такой порог, вероятно, вы-
ше. По данным Е. С. Когай (1964), первые нападения перезимовав-
ших в тех условиях имаго отмечались при 18—20°. О возможности
смещения температурных границ активности вида в зависимости от
широты местности впервые сообщил А. С. Мончадский (1956). По его
186
мнению, порог активности одних и тех же видов комаров по ее
продвижения с юга на север будет уменьшаться, и наоборот Высгк^
занное положение всецело подтверждается наблюдениями в* различ-
ных ландшафтно-климатических зонах Казахстана.
4. ПАРАЗИТЫ И ВРАГИ КОМАРОВ В КАЗАХСТАНЕ
Не будь естественных регуляторов, численность комаров в при-
роде была бы безграничной. Значительная часть каждой популяции
комаров гибнет от болезнетворного влияния различных микроорга-
низмов, от паразитов и хищников. Уменьшение численности проис-
ходит как во время личиночного развития, так и в период имагиналь-
ной жизни. В Казахстане этот вопрос никем не изучался.
Таблица 7
Результаты исследования некоторых гидробионтов как хищников
Хищник Вид подсаженных личинок Стадия личинок Кол-во опытов Поедаемость
за 12 час за 24 час
Личинки стрекоз (Odo- Ae. c. caspius IV 6 1—3 2—7
nata, Anisoptera) Ae. flavescens IV 2 — 1—2
Ручейник (Trichoptera,
Nemataulius) Ae. c. caspius IV 7 — 1—2
Гладыш (Hemiptera, No- Ae. c. caspius II 2 17—21 22—25
tonecta) Ae. c. caspius IV 3 20—33 50
Ae. flavescens IV 3 13—23 27—50
Плавт (Hemiptera, Nau- Ae. c. caspius II 2 11—21 17—27
coris) Ae. c. caspius IV 2 9—10 14—20
Ae. flavescens IV 2 15—30 16—40
Плавунец (Coleoptera, Ae. c. caspius II 2 — —
Dytiscidae) Ae. flavescens IV 3 43—45 50—53
Личинки плавунца (Dy- Ae. c. caspius IV 2 42—48 51—61
tiscidae) Ae. flavescens IV 2 24—28 30
Голец (Nemachilus dor- Ae. c. caspius II 8 46—300
salis) Ae. c. caspius III 3 147—160 183—378
На протяжении нескольких лет мы отмечали отсутствие или не-
значительное количество личинок комаров в местах обитания мелко-
водных рыб и мальков. В 1967 г. были поставлены опыты по выявле-
нию хищного значения рыб по отношению к предимагинальным фа-
зам комаров (Дубицкий, Дешевых, Деньгуб, 1967). Результаты их,
а также изучения некоторых других гидробионтов как хищников
приведены в таблице 7.
187
Наибольшей прожорливостью отличаются рыбы. За сутки один
голец съедал 183—378 личинок II—III стадий Ае. с. caspius. Как
только ему подсаживали личинок, он начинал метаться из стороны
в сторону, хватая их на различной глубине, но чаще у поверхности
воды. Это напоминало поведение гольцов в естественных мелковод-
ных разливах. Обладая, как показали наши опыты, большой прожор-
ливостью, превосходящей даже гамбузию (145 личинок за 24 час, по
В. Н. Беклемишеву, 1949), гольцы заселяют большинство медленно
текущих водоемов, речных затоков и озер многих рек пустынной зо-
ны (Берг, 1949). Предварительные результаты исследований дают ос-
нование считать серого гольца (Nemachilus dorsalis) весьма перспек-
тивным в борьбе с личинками комаров в пустынной зоне Казахстана.
По плодовитости, прожорливости, способности проникать в самые
мелководные участки (Мартехов, 1963) этот вид, при соответствующей
организации мероприятий, может сыграть такую же роль в подрыве
популяций комаров, как гамбузия в Средней Азии (Казанцев, 1959).
Из других гидробионтов большой прожорливостью в отношении
личинок комаров отличаются личинки жуков-плавунцов (Dytiscidae),
гладыши (Notonecta) и плавты (Naucoris). Частично это объясняется
тем, что они не съедают личинок, а лишь высасывают их содержи-
мое. Иногда это бывает так ювелирно сделано, что от личинок остает-
ся лишь кутикулярная оболочка с пищеварительной и трахейной си-
стемой. Им, вероятно, также принадлежит значительная роль в
истреблении личинок Culicinae, тем более, что численность их в мес-
тах выплода комаров довольно значительная.
Иначе ведут себя по отношению к личинкам комаров жуки-пла-
вунцы Dytiscidae. Привыкнув, вероятно, питаться более крупной до-
бычей, они первоначально не реагируют на личинок комаров, даже
таких крупных, как Ае. flavescens. Зато после 8—10-дневного пребы-
вания без пищи они с такой жадностью набрасываются на подсажен-
ных личинок, что за одну ночь пожирают около пятидесяти крупных
экземпляров.
Совершенно незначительная роль в уничтожении предимагиналь-
ных фаз комаров принадлежит личинкам стрекоз (Anisoptera) и ру-
чейникам (Nemataulius). Добычей им обычно служили лишь отдель-
ные ослабевшие или находящиеся в предличиночной депрессии осо-
би; здоровых и активных схватить удавалось редко. Более активно
охотились за личинками жуки-вертячки (Gyrinus) и водные скорпи-
оны (Nepidae). Отдельные представители их съедали в лабораторных
условиях за сутки от 2 до 17 особей.
В отличие от Dixinae другие представители семейства Culicidae —
личинки некровососущих комаров подсемейства Chaoborinae могут
приносить значительный вред предимагинальным фазам кровососу-
188
щих комаров. Исследованию трофических связей, экологии и вредо-
носного значения представителей этого подсемейства посвящена се-
рия работ А. С. Мончадского (1959, 1964). По его данным, представи-
тели рода Mochionyx могут съедать за сутки от 1,5 до 27 личинок
кровососущих комаров.
Мы отмечали довольно значительную заселенность личинками
Chaoborinae временных водоемов в горно-степной зоне Южного Ал-
тая. Количественное соотношение их с личинками кровососущих ко-
маров было 1:10—15. Нападению подвергались особи такого же раз-
мера и более мелкие. Личинки хаоборин обычно были неподвижны в
толще воды и при приближении личинки комара активно набрасы-
вались на нее с головной стороны. Их ротовое устройство позволяет
целиком захватывать голову личинки, по размеру чуть меньшую го-
ловы хищника. Отмечена нападаемость на личинок Ае. cantans, Ае.
communis и Ае. pullatus. Личинки хаоборин обнаруживались в зате-
ненных лесных и пойменных водоемах в мае и июне, в период разви-
тия ранневесенних видов Aedes. В начале июня численность их начи-
нает уменьшаться.
Из паразитов имаго часто отмечается поражение комаров вод-
ными клещами (Hydrachnellae), которые иногда в большом количест-
ве встречаются как на самках, так и на самцах. На одной самке
An. maculipennis мы насчитали 19 экземпляров Hydrachnellae. Чаще
всего ими поражались полицикличные виды, развивающиеся в по-
стоянных водоемах. Причем, выплаживающиеся имаго уже бывают
заражены этими суперпаразитами. Их часто можно видеть впивши-
мися в межсегментарную кутикулу брюшка с дорзальной и с вент-
ральной стороны, реже на бочках груди и около кокс. У незначитель-
но пораженных комаров патологических признаков не наблюдается.
При массовом паразитировании отмечаются вялость имаго и понижен-
ная активность. Вполне возможно, что очень пораженные экземпля-
ры далеко от мест выплода не улетают и, погибая там, способствуют
прохождению жизненного цикла этих суперпаразитов. Отпадения
клещей даже с мертвых комаров мы не отмечали, возможно, за счет
того, что сразу их замаривали. Существуют сведения, что в процессе
жизни комары от них освобождаются.
В отдельных случаях отмечается нападаемость водомерок (Ger-
ridae) на выплаживающихся из куколок имаго. Более активно, осо-
бенно в южных районах Казахстана, комаров поедают стрекозы.
Последнее можно объяснить тесной экологической сопряженностью
мест их выплода и обитания. Отлов нападающих на комаров стрекоз
показал, что все они относятся к двум родам: Aeschna и Libellula.
Нападения на комаров обычных в этих местах стрелок (Agrion idae)
не отмечено. Активность стрекоз начиналась, как правило, через 1 —
189
2 час после восхода солнца и продолжалась до глубоких сумерек.
Утром они чуть заставали конец утренней активности комаров, а
днем, летая среди зарослей, охотились за насекомыми, случайно вы-
летевшими из дневных убежищ. С этой целью в южных районах Ка-
захстана стрекозы летают вокруг животных, пасущихся среди кус-
тарниковых зарослей. Здесь им удается поймать как вспугнутых на-
секомых, так и вылетевших для нападения.
Основная активность стрекоз проявляется в вечернее время, ког-
да спадает дневная жара и комары в массе вылетают из убежищ.
Предпочитают охотиться за роящимися насекомыми, нередко приво-
дя к распаду роя. Во время нападения подлетают близко к человеку
и, кружась вокруг него, быстро разгоняют нападающих комаров.
Пойманных насекомых пожирают на лету, не переставая охотиться.
Если активность стрекоз в вечернее время отмечается на высоте
1—1,5 м над землей, то чуть выше над ними с еще большей скоро-
стью охотятся летучие мыши. Предметом их нападения служат те же
роящиеся комары, которые в это время поднимаются на высоту до
2—3 м. В этот период около мест выплода комаров наблюдается зна-
чительная концентрация летучих мышей, которая в обычное время
не отмечается. Зная прожорливость рукокрылых, не трудно предста-
вить, сколько они могут уничтожить комаров, особенно во время их
роения.
В качестве врагов комаров приводится большой перечень видов
птиц (Суитмен, 1964). Не проводя исследований пищевой массы орни-
тофауны в местных условиях, об этом судить трудно. Из активных
истребителей комаров отмечены лишь береговые ласточки (Riparia
rupestris). Активность этих птиц приходится на дневные часы. Охо-
тятся они в основном за вспугнутыми или вылетающими из дневных
убежищ для нападения на добычу комарами, поэтому в южных рай-
онах Казахстана часто сопровождают пасущийся скот и даже иду-
щего человека. Предпочитают охотиться на открытых, сильно зарос-
ших травянистой или осоковой растительностью участках. Летают
низко над землей (15—25 см над поверхностью травостоя). Высокая
активность отмечена в местах обитания комаров Ае. с. caspius и Сх.
modestus.
В настоящее время начаты более глубокие исследования по вы-
явлению естественных паразитов, хищников и возбудителей заболе-
ваний с целью использования их для интегрированной борьбы с ко-
марами. В этом плане еще предстоит выяснить их пищевые связи,
возможность лабораторного культивирования и, главное,— степень
вредоносности для других насекомых.
190
VI. Основные закономерности
распространения и генезис фауны
комаров Казахстана
Наибольшая численность комаров в Казахстане приурочена
(рис. 39) к обводненным и обильно орошаемым естественными осад-
ками равнинным участкам. В основном это северные и восточные
районы республики. Годовое количество выпадающих там осадков
колеблется от 250 до 500 мм, основная их масса приходится на теп-
лый период года. Созданию оптимальных для комаров мест выплода
в немалой степени способствует равнинный, но сильно расчлененный
рельеф местности, изобилующей мелкими и крупными озерами, за-
токами талых и паводковых вод, грунтовыми и пойменными заболо-
ченностями. Водная площадь отдельных участков занимает до 40%
территории некоторых районов Кустанайской, Северо-Казахстанской,
Кокчетавской, Целиноградской, Павлодарской и Восточно-Казахстан-
ской областей.
Соответствует описанному и климатическое разнообразие при-
родных условий Казахстана. Если на севере республики средняя тем-
пература самого холодного месяца —22, а минимальная —49, то на
юге есть районы, где температура самого холодного месяца выше ну-
ля (Пономарев, Борсук, 1927).
В пустынной и полупустынных зонах Казахстана массовый вы-
плод комаров приурочен к поймам больших и малых рек, временным
и постоянным озерам и заболоченностям. Численность комаров не
уступает наиболее неблагополучным районам Северо-Восточного Ка-
захстана. Объясняется это спецификой пустынных рек, уровень воды
в которых выше прилежащих участков. Отмечаемые ежегодно позд-
невесенний и летний паводки этих рек, наступающие в период тая-
ния горных снегов, приводят к образованию неисчислимого количе-
ства хорошо прогреваемых и богатых питательными веществами пой-
менных мелководий. Многие из них, находящиеся поблизости от
191
русла, за счет грунтовой фильтрации относятся к категории постоян-
ных водоемов.
Во многом сходны с описанным побережья Алакульских озер,
Балхаша, Аральского и Каспийского морей. Дельты впадающих в них
рек за счет низинного положения и обилия снесенных отложений
Рис. 39. Относительная плотность популяций кровососущих комаров в Казахстане:
1 — малая, 2 — средняя, 3 — высокая.
сильно расчленены и в большинстве случаев представляют питаемые
отдельными рукавами обширные заболоченности. Колеблющийся, но
не усыхающий совсем уровень воды на этих участках способствует
выплоду неисчислимых полчищ самых разнообразных видов кома-
ров. Немалую роль в этом играют ветры, действие которых проявля-
ется тем сильнее, чем больше водоем. Вода, движимая ветром боль-
шой силы, может затоплять все прибрежные понижения рельефа,
создавая благоприятные для комаров места выплода. Особенно это
относится к северному побережью Каспия, где на протяжении летне-
го времени периодически поднимающийся уровень воды в водоемах
култучной зоны способствует выплоду комаров рода Aedes.
Горные и предгорные участки Казахстана по обилию нападаю-
щих комаров не имеют первостепенного значения. Места выплода в
них строго ограничены и пригодны лишь для развития холодолюби-
вых стенобионтных видов.
192
В столь различных экологических условиях возможно существо-
вание самых разнообразных видов комаров. Общая численность их
составляет 53 вида. Среди них шесть представителей рода Anopheles,
шесть — рода Culiseta, 31 относится к роду Aedes, восемь — к роду
Culex и по одному — к родам Uranotaenia и Mansonia.
Таблица 8
Эколого-фаунистические комплексы кровососущих комаров Казахстана
Род комаров Комплекс
лесно й лесостепной | степно й пустынный
Anopheles Uranotaenia claviger mac. messeae hyrcanus pulcherrimus superpictus mac- sacharovi algeriensis unguiculata
Culiseta Mansonia bergrothi morsitans silvestris alaskaensis annulata richiardii longiareolata
Aedes communis excrucians c. dorsalis c. caspius
cataphylla intrudens flavescens cyprius cinereus stramineus detritus
Culex leucomelas nigripes pul la t us punctor sticticus diantae us impiger pionips nigrinus rempeli cantans behningi riparius beklemischevi subdiversus vexans pipiens torrentium territans pulchritarsis modestus pusillus theileri hortensis
Анализ видового состава и экологии комаров фауны Казахстана
позволил отнести слагающие ее виды к четырем эколого-фаунисти-
ческим комплексам: лесному, лесостепному, степному и пустынно-
му (табл. 8). В основу их выделения положены принципы географи-
ческой однородности, однотипности распространения и экологическо-
го сходства видов.
В достаточной степени эти группировки характеризовало бы по-
нятие «Фаунистический комплекс» в определении Г. В. Николь-
ского (1947), Хотя и оно не отражает всей сущности рассматрйваемых
группировок. Под этим термином автор подразумевает группу видов,
13—80
193
связанных общностью своего географического происхождения с опре-
деленной зоной, к условиям которой и приспособлены виды, слагаю-
щие комплекс. Мы к этому определению добавляем еще такой крите-
рий, как экологическое сходство. К кровососущим комарам не сов-
сем применим одноименный принцип зоогеографического анализа
ассоциаций видов того или иного региона, столь удачно примененный
В. В. Шевченко (1961) при изучении слепней. Объясняется это, веро-
ятно, тем, что ареалы комаров, как правило, слишком обширны, что-
бы быть специфичными для какого-либо небольшого региона, по-
скольку дивергенция у них более медленная и видовой эндемизм вы-
ражен слабо. В настоящее время подобное комплексирование
применимо для таких обширных категорий, как географическая об-
ласть. что и сделано О. Н. Сазоновой (1962).
К лесному фаунистическому комплексу относятся холодолюби-
вые, моноцикличные виды Aedes и полицикличные, олиготермные
виды Culiseta. Почти все представители этого комплекса распростра-
нены на севере СССР. На юг они выклиниваются по реликтовым ост-
ровным лесам и поймам рек до степной и полупустынной зон, что
всецело характерно для Казахстана. На его территории представите-
ли лесного комплекса довольно многочисленны в лесостепной зоне,
охватывающей северные районы республики. По мере продвижения
на юг распространение их приобретает мозаичность. Ограниченные
участки их ареала встречаются на возвышенных лесных массивах
Центрального Казахстана и в поймах крупных и малых рек в запад-
ной части. Такая зональность отмечается до 48—47° с. ш. Адаптация
их к этим условиям выражается в более раннем по сравнению с лесо-
степной зоной развитии, кратковременности периода активности и
строгой приуроченности к характерным биотопам.
По отрогам Тянь-Шаня отдельные представители этого комп-
лекса (Ае. leucomelas, Ае. cataphylla) доходят до Южного Казахста-
на и сопредельных районов Киргизии (43—42° с. ш.). В горной части
юга и юго-востока Казахстана в зависимости от высоты местности
они являются или доминантами, или субдоминантами. Доминируют
и в поясе лесов и субальпийских лугов и менее многочисленны в зоне
горных степей. Иногда по поймам холодных горных рек проника-
ют в полупустынную и даже в пустынную зону (окрестности Илий-
ска), т. е. в вертикальном распространении отмечаются те же законо-
мерности, что и в равнинной части Казахстана. Наиболее холодолю-
бивые представители этого комплекса (Ае. pullatus, Ае. intrudens)
поднимаются до 3000 м н. у. м., что соответствует поясу холодных
горных пустынь (сырты). Распространение в горных условиях видов
рода Culiseta ограничивается зоной гордых степей и нижними откры-
тыми участками лесной зоны.
194
Не совсем ясно экологическое положение трех недавно описан-
ных видов — Ае. simanini (Гуцевич, 1966), Ае. kasachstanicus (Гуце-
вич, 1962) и Ае. montchadskyi (Дубицкий, 1968). По морфологическим
и экологическим особенностям первый должен, вероятно, относиться
к лесному комплексу, а второй и третий — к лесостепному.- В на-
стоящее время решить этот вопрос не представляется возможным,
так как неизвестны ареалы этих видов за восточными пределами
Казахстана.
Представители лесостепного фаунистического комплекса холо-
долюбивы в меньшей степени, чем предыдущие виды. Диапазон их
пластичности шире, что позволяет им в равной степени существовать
в лесной и степной зонах. Некоторые из них (Ае. excrucians, Ае. fla-
vescens) проникают на север до 68—69° с. ш. (Румш, 1948). Предста-
вители этого комплекса наряду с некоторыми степными видами яв-
ляются доминирующими в одноименных зональных районах Север-
ного и Восточного Казахстана. Иногда отмечается развитие второго
(летнегэ) поколения этих в общем-то моноцикличных видов.
Отдельные представители отмечаются намного южнее, чем лес-
ные виды. Например, Ае. flavescens встречается на протяжении всего
нижнего отрезка течения Сыр-Дарьи и Или (43—44° с. ш.). Некоторые
виды (Ае. cyprius, Ае. subdiversus) немного не доходят до этого ру-
бежа, а Ае. behningi, Ае. excrucians и Ае. cantans распространены в
Казахстане примерно до 48° с. ш. Южнее всех видов заходит Ае.
claviger, но встречается он лишь в прохладных горных и предгорных
биотопах.
В горных и предгорных частях Казахстана виды этого комплек-
са распространены неравномерно. В предгорьях Алтая и Зайсанской
котловины многочисленны Ае. excrucians, Ае. cantans, Ае. cyprius и
Ае. flavescens, а в отрогах Тянь-Шаня — лишь Ае. flavescens, который
встречается здесь в поясе горных степей и нижних участках лесного
пояса. На юг он доходит до северных районов Киргизии (Конурбаев,
1965), но не достигает Таджикистана (Кешишьян, 1941).
Виды, входящие в степной комплекс, еще более пластичны, чем
лесные и лесостепные, и поэтому, как правило, отличаются большой
пространственной протяженностью ареалов. Достаточно сказать, что
в эту группу входят Ае. vexans и Ае. cinereus, ареалы которых про-
стираются от южных пустынь до таежной зоны, а Сх. pipiens за счет
своей синантропности заходит значительно севернее (Румш, 1948).
Установить зональную принадлежность этих видов можно лишь по1
предпочитаемости степных биотопов и экстразональности выклинива-
ния в пустынную и лесостепную эоны.
Если сравнить представителей этого фаунистического комплекса
с предыдущими, то бросается в глаза увеличение количества видов
195
Culex и Anopheles. Группа Aedes представлена эврибионтными поли-
цикличными видами в зависимости от климатических условий, мо-
гущих давать одно или несколько поколений. В южных частях своих
ареалов развитие и выплод всех этих видов приурочены к хорошо
прогреваемым водоемам с окружающей густой растительностью, а
на севере они проникают только до затененных и сильно заросших
водоемов. Экстразональными биотопами в первом случае служат
поймы южных рек, а во втором — открытые пространства с хорошо
прогреваемыми временными водоемами.
В горных местностях встречаются все виды этой группы, кроме
An. hyrcapus, М. richiardii, Cs. annulata. Для них, вероятно, недоста-
точно прогревание горных водоемов. Все остальные виды заселяют
пояс горных степей и нижележащие участки. Ае. с. dorsalis встреча-
ется на разреженных участках лесного пояса горной зоны, а иногда
и на хорошо прогреваемых площадях субальпийских лугов.
Представители степного фаунистического комплекса распростра-
нены в Казахстане довольно неравномерно. Пять наиболее эвриплас-
тичных видов (Сх. pipiens, Cs. annulata, М. richiardii, Ае. vexans, Ае.
cinereus) встречаются не только по всему Казахстану, но и за его
пределами. Все другие виды Culex и An. hyrcanus, охватывая южные
районы республики, доходят до 48—50° с. ш. Ареалы Ае. с. dorsalis
и Ап. т. messeae занимают северную часть Казахстана, доходя до
46—48° с. ш., а при благоприятных условиях выклиниваются за этот
рубеж. Южнее они замещаются близкими формами (Ае. с. caspius,
Ап. т. sacharovi).
Основная масса видов степного комплекса в ту или иную сторо-
ну простирается за пределы Казахстана. Проникновение An. т. mes-
seae на север до южной границы тайги большинство авторов (Бекле-
мишев, Желоховцев, 1945; Румш, 1948; Мончадский, 1951) объяс-
няют значительной синантропностью этого вида и приуроченностью
развития к незатененным, хорошо прогреваемым водоемам, а мест
зимовок — к различным помещениям. Многие более крайние точки
его обнаружения являются результатом завоза с транспортом и про-
движения за человеком во время расчистки тайги (Румш, 1948).
В пустынный фаунистический комплекс входят виды, основная
часть ареала которых лежит за южными пределами Казахстана. Как
и представители предыдущего комплекса, все они полицикличны, но
отличаются высокой сухоустойчивостью. Это позволяет им существо-
вать в сухих, жарких районах, какими является заселяемая ими
зона. Рассматриваемая группа не однородна по термофильности:
встречаются как строгие политермофилы (An. pulcherrimus, An. su-
perpictus, An. т. sacharovi, Ае. pulchritarsis, Сх. pusillus), так и эври-
пластичные представители данной категории (Сх. modestus, Cs. Ion-
196
giareolata, Ae. c. caspius, Ur. unguiculata). К этому же комплексу
относится группа из трех видов: Сх. theileri, Сх. hortensis, An. algerien-
sis, тяготеющих к развитию в прохладной воде горных и предгорных
биотопов. Такое несоответствие между стремлением к прохладным
биотопам и распространением в сухих жарких районах В. Н. Бекле-
мишев и А. Н. Желоховцев (1945) на примере An. algeriensis объяс-
няют потребностью в продолжительном теплом сезоне, который дает
возможность осуществлять в прохладных биотопах две генерации,
необходимые для сохранения вида.
Все представители пустынного фаунистического комплекса рас-
пространены южнее 48° с. ш. Лишь два вида (Cs. longiareolata и Сх.
modestus) помимо пустынной охватывают степную зону и встречают-
ся в открытых участках лесостепной. В противоположность этому
ареалы двух наиболее термофильных видов — An. pulcherrimus и
An. superpictus — заходят только в самые южные районы Казахста-
на, примерно до хребта Каратау. В горы ни один вид высоко не под-
нимается. Ае. с. caspius и Сх. modestus отмечены в самых низинных,
хорошо прогреваемых местах пояса горных степей. Ае. detritus, Сх.
pusillus и Ur. unguiculata вообще в горах не встречаются: первые
два, возможно, по причине отсутствия в горах специфических соло-
новодных водоемов, а третий на территории Казахстана, как уже гово-
рилось, отмечается только в поймах рек.
Анализируя ареалы всех рассмотренных групп, можно заметить,
что распространение на равнинных участках Казахстана основной
массы представителей лесного фаунистического комплекса не идет
южнее 50° с. ш., а представителей лесостепного — 47—48° с. ш.
Вместе с тем к 47—48° с. ш. приближается северная граница ареалов
пустынных видов, а большинство степных не проникает севернее
50° с. ш. Это говорит о наличии на территории Казахстана двух кли-
матических рубежей: около 48° и около 50° с. ш.
Отмеченное явление не случайно. Где-то около 50° с. ш. прохо-
дит большая ось материка, могущая служить границей двух основ-
ных климатических районов: северного и южного. Районы, лежащие
к югу от этой оси, отличаются сухостью и большей континенталь-
ностью климата, чем область к северу от нее. Основное значение имеет,
вероятно, сумма летних температур, которая севернее 50° с. ш. резко
убывает, что дает возможность подходить к этому рубежу с севера
олиготермофилам, а с юга — мезотермофилам.
Второй рубеж, проходящий около 47—48° с. ш., характеризует-
ся наиболее резким переходом от высоких летних положительных ба-
лансов к отрицательным зимним температурам. Теплое солнечное ле-
то создает благоприятные условия для распространения многих теп-
лолюбивых животных, в том числе и пустынных видов комаров, тогда
197
как низкие зимние температуры ограничивают возможность их пере-
живания в зимний период. Вместе с тем последний фактор позволяет,
вероятно, по экстразональным биотопам проникать до этой широты
некоторым лесостепным видам.
Экологической предпосылкой для экстразона л ьного распростра-
нения в Казахстане видов степного комплекса служит значительная
величина солнечной радиации, которая даже в северных районах рес-
публики в летний период приближается к показателям, свойственным
15° с. ш. Соответственно этому в пустынных районах республики их
развитие приурочено к затененным прохладным биотопам, а в лесо-
степной зоне, наоборот, к открытым, хорошо прогреваемым. Это впол-
не отражает основное положение принципа смены стаций (Бей-Биен-
ко, 1959).
Из всего этого следует, что основные закономерности распрост-
ранения и экологии комаров в Казахстане в значительной мере со-
ответствуют ландшафтно-климатическим особенностям различных
природных зон республики. Предпосылкой служит история форми-
рования современной фауны кровососущих комаров, в которой до-
вольно четко прослеживается наличие европейско-сибирских, среди-
земноморских, южно-азиатских и палеархеарктических предста-
вителей.
Первичная фауна кровососущих комаров Казахстана связана,
вероятно, с тропическими и субтропическими представителями ро-
да CuZex, существовавшими, по данным Эдвардса (Edwards, 1932),
уже в юре, а сформировавшимися, вероятно, в триасовом периоде.
Это подтверждается и местными палеонтологическими данными
(рис. 40). В начале третичного периода на значительной территории
Казахстана климат отличался аридностью, относимой В. М. Синици-
ным (1962) и Е. М. Лавренко (1962) к типу влажных саванн. Как и в
настоящее время, с аридными, теплыми условиями в то время бы-
ли, вероятно, связаны представители родов Culex и Anopheles, имею-
щих центром своего видообразования тропическую зону Старого Све-
та. Сфотографированный ископаемый экземпляр комара также,
вероятно, относится к роду Culex. Об этом можно судить по выпукло-
сти среднеспинки, сравнительно небольшой длине ног и наличию
перевязей на брюшке, что отличает его от рода Anopheles.
Распространению их по всей современной территории Казахста-
на способствовало отмечаемое в палеогене продвижение на север к
бореальной зоне элементов тропического ландшафта. К этому же
времени О. Л. Крыжановский (1965) относит миграцию на территорию
настоящего Казахстана некоторых родов саранчовых, чернотелок и
златок, имеющих родственные связи с представителями пустынь
южного полушария. К числу наиболее ранних представителей Culi-
198
cidae следует, по-видимому, отнести виды, проникшие в настоящее
время далеко на север (Сх. pipiens, Сх. modestus, An. т. messeae. Ап.
claviger). Продвижение других видов было, вероятно, ограничено
Рис. 40. Ископаемый комар (7 мм). Юрское озеро в отрогах
Каратау близ с. Михайловки (Джамбулская обл.). Находка
А. Г. Шарова. Фото Л. С. Соболева.
наступившим в олигоцене общим похолоданием климата и отступле-
нием тропической флоры и фауны в область южных широт. Замеще-
ние в неогене субтропических саванн их бореальным аналогом —
степями еще более ограничило жизненный оптимум этих теплолюби-
вых форм. К существованию в таких условиях могли приспособиться
лишь виды, далеко зашедшие на север и в значительной мере адапти-
ровавшиеся к похолоданию. Более теплолюбивые представители этих
групп (An. superpictus, An. algeriensis. An. pulcherrimus, Сх. pusillus)
так и не вышли за пределы современной пустынной зоны. Вероят-
ность такого же заселения умеренной зоны Северной Америки вида-
ми Culex предполагает и Росс (Ross, 1964).
199
К середине третичного периода можно, по-видимому, отнести по-
явление на территории Казахстана бореальных видов комаров, отно-
сящихся к родам Aedes (подрод Ochlerotatus) и Culiseta. Их экстра-
поляция происходила совместно со сменой саванн ландшафтом ши-
роколиственного леса. Большинство этих видов — лесные обитатели,
и ареал их повторял распространение листопадной флоры. Господст-
вовавший тогда влажный мягкий климат способствовал расселению
многих умеренных форм растительности.
Начавшееся затем наступление сухого континентального клима-
та остановило продвижение на юг бореальной флоры, а вследствие
продолжавшегося похолодания исчезли тропические и субтропиче-
ские виды растений. Опустынивание прогрессирует в неогене и в чет-
вертичном периоде (Лавренко, 1962). Леса редеют и распадаются на
отдельные массивы, разобщенные степными ассоциациями. Вместе
с лесной растительностью оттесняются бореальные виды комаров. От-
дельные их представители сохраняются лишь в экстразональных
лесных биотопах по возвышенным участкам и поймам рек. Отсюда
и отмечаемое ныне атипичное выклинивание фауны этих северных
видов в южные районы Казахстана, где они являются не чем иным,
как бореальными реликтами.
Вместе с продвижением пустынь шло дальнейшее распростране-
ние представителей Culex и Anopheles. В этом случае нынешняя фау-
на Казахстана оказалась в несколько лучшем положении, поскольку
не все представители этих родов продвинулись так далеко на север.
Среднеазиатская фауна Culex и Anopheles несколько богаче.
Таким образом, фауна комаров Казахстана формировалась за
счет северных и южных элементов. Пути их проникновения не соот-
ветствуют всецело такой полюсности, существует и зональная миг-
рация. С севера, как уже указывалось, продвигались представители
лесного фаунистического комплекса. Поэтому почти все виды кома-
ров Севера СССР встречаются в Северном и Восточном Казахстане.
В таком же направлении, но с уклоном к зональной миграции
происходило, вероятно, заселение нынешней территории Казахстана
и представителями лесостепного комплекса. В пользу этого говорит
их значительное выклинивание как в лесную, так и в аридную зону
и отсутствие некоторых из них (Ае. behningi, Ае. beklemischevi) в
восточной части лесостепной зоны.
По очаговости распространения и наличию ярко выраженного
эпицентра можно заключить, что видообразование трех представите-
лей этих двух комплексов (Ае. subdiversus, Ае. kasachstanicus, Ае,
montchadskyi) произошло, вероятно, на территории Казахстана или
на прилежащих участках Джунгарии. Об этом говорят сравнительно
высокая численность Ае. subdiversus в районе Тургайской столовой
200
страны, а Ае. kasachstanicus и Ае. montchadskyi— в бассейне Или и
отмечаемая экспансия на сопредельные территории. Единичные эк-
земпляры Ае. subdiversus встречаются в Саратовской, Оренбургской,
Гурьевской и Карагандинской областях. По поймам Урала, Иргиза,
Тургая он, вероятно, проник в полупустынную зону, а по пойме То-
бола и его притокам — в Западную Сибирь.
Очевидна и связь лесостепного комплекса с фауной соответст-
вующей зоны РСФСР, граничащей с Казахстаном. Все виды этого
комплекса встречаются в окрестностях Саратова и Оренбурга (Martini,
1926,1928), расположенных у южной границы лесостепей.
Более расчленены пути проникновения на территорию Казахста-
на представителей степного и пустынного фаунистических комплек-
сов: западный, южный и восточный. С запада, по-видимому, проник
Ае. detritus, обычный для Средиземноморья. Путь основной части
видов Culex, хотя и проходил с юго-запада из Эфиопской области, но
осуществлялся через пограничные районы Средней Азии. Там мы
встречаем не дошедший до Казахстана Сх. tritaeniorhynchus.
Аналогичным путем шло распространение и некоторых сре-
диземноморских видов Anopheles (An. superpictus, An. algeriensis).
Ориентальные виды продвигались двумя путями: восточным, по от-
ношению к Памиро-Тянь-Шаньской горной системе, и западным. В Ка-
захстане они сомкнулись, о чем можно судить по встречаемости на
юго-востоке республики двух географических рас Ае. vexans {Ае. v.
vexans, Ае. v. nipponii). Причем, смешение этих форм наблюдается
лишь в более низких, граничащих с Китаем, районах.
На основании всего изложенного можно заключить, что фауна
Казахстана, территориально объединяющего самые разнообразные
ландшафтно-климатические условия, носит составной характер.
Этим же объясняется сравнительно богатое ее видовое разнообразие.
Основными путями, по которым происходило ее формирование, слу-
жили южный — для Culex и Anopheles и северо-западный — для
Aedes и Culiseta. Процесс становления фауны, вероятно, не за-
кончен. Об этом говорят обнаружение некоторых новых для науки
видов (Ае. kasachstanicus, Ае. montchadskyi, Ае. stramineus) и их экс-
пансия. В будущем следует ожидать проникновения в Казахстан не-
которых южных (Сх. univittatus) и горных (An. lindesayi, An. marteri)
видов, уже дошедших до Таджикистана (Кешишьян, 1941).
В дальнейшем видовой состав комаров Казахстана может в не-
значительной степени пополниться как за счет обнаружения ранее не
встречаемых здесь видов, так и за счет происходящего процесса фор-
мообразования.
Заключение
Многообразие ландшафтно-климатических условий Казахстана
определяет существование на его территории самых разнообразных
таксономических и экологических групп кровососущих комаров. Об-
щая численность их достигает 53 видов, что составляет примерно две
трети всей фауны Culicinae СССР.
Соответствуют такому количеству и биологические особенности
этих кровососов. Среди них имеются виды, зимующие в фазе имаго,
яйца и личинки. В зависимости от условий среды обитания многие
из них или ограничиваются одной-двумя генерациями, или проде-
лывают их столько, сколько позволяет теплый период года. Такая
пластичность жизненных циклов дает возможность кровососущим ко-
марам заселять почти все участки Казахстана, где существует по-
стоянная или временная водная сеть. Особенно много их в северных
и восточных районах республики, где водный режим более постоя-
нен, а сумма выпадающих осадков больше. В значительной мере это-
му способствует обилие всевозможных открытых и закрытых во-
доемов.
Основные закономерности распространения и экологии комаров
в Казахстане в известной мере соответствуют ландшафтно-климати-
ческим особенностям различных природных зон республики. Пред-
посылкой этому служит история формирования современной фауны
кровососущих комаров, в которой довольно четко прослеживается на-
личие европейско-сибирских, средиземноморских и иалеархеаркти-
ческих представителей. Заполнив во времена сложного геологическо-
го прошлого нынешней территории Казахстана самые разнообразные
экологические ниши и приспособившись к определенным условиям
202
среды обитания, большинство рассмотренных представителей образо-
вали естественные группировки, охарактеризованные нами как эко-
лого-фаунистические комплексы.
Определенные закономерности вырисовываются и при анализе
внутривидовой изменчивости встречающихся в Казахстане видов.
Большинство из них представляют дивергировавшие формы, нахо-
дящиеся на той или иной фазе морфологической, экологической и
репродуктивной изоляции. Особенно ярко это прослеживается у вы-
клинившихся значительно на юг по соответствующим поясам горных
систем бореальных представителей. Испытав воздействие новых
условий среды и почти разобщившись с основной частью популяции,
многие из этих форм получили признаки новых таксонов. Все это
наряду с отмеченным пополнением видового состава и обнаружением
условноэндемичных, новых для науки видов дает основание считать,
что процесс становления фауны комаров Казахстана не завершен.
Основную массу распространенных в северных и горных райо-
нах Казахстана видов составляют моноцикличные представители ро-
да Aedes. Выплаживаясь весной, они проделывают за теплый период
лишь одну генерацию, что восполняется значительной массовостью
и большим числом гонотрофических циклов. Это, в свою очередь, со-
здает предпосылки для массового весеннего выплода личинок из
перезимовавших яиц. Вполне соответствует этому и сезонный ход
численности комаров в рассматриваемом регионе. Он характеризует-
ся массовым появлением насекомых в начале лета и их высокой ак-
тивностью на протяжении почти всего теплого периода года.
Более южную часть Казахстана заселяют в основном полицик-
личные виды, зимующие в фазе имаго. Неблагоприятные зимние
условия с частыми оттепелями и резкими похолоданиями в значи-
тельной мере подрывают популяции этих видов. Поэтому численность
их нарастает медленно и бывает массовой в самый теплый период
летнего сезона, что достигается путем последовательного осуществле-
ния нескольких летних генераций, быстрейшему прохождению кото-
рых способствуют высокие температуры и сильная инсоляция водое-
мов. И хотя по сравнению с предыдущим регионом места выплода
комаров в южной части Казахстана более ограниченны, численность
их на отдельных обводненных участках может даже превосходить та-
ковую в северных и восточных районах республики.
Разумеется, приведенные сведения значительно обобщены, по-
скольку некоторые представители моноцикличных видов проникают
на юг, а полицикличные, наоборот, заходят далеко на север. Кроме
того, отмечается и зональная приуроченность отдельных групп кро-
вососущих комаров к определенным ландшафтам, что послужило
основанием для отнесения их к различным эколого-фаунистическим
группировкам.
203
Уточнение видового состава комаров Казахстана, распростране-
ния и экологии местных популяций наряду с теоретическими обобще-
ниями отдельных вопросов позволили предложить комплекс меро-
приятий, направленных на снижение численности кровососущих ко-
маров и защиту от их нападения. Основные положения этих рекомен-
даций приведены в специальных публикациях и наставлениях.
В целом мероприятия, направленные на снижение численности
кровососущих комаров в Казахстане, вполне осуществимы и экономи-
чески выгодны. Проведение всего комплекса борьбы и защиты от ко-
маров в условиях Северного Прикаспия по разработанной системе
позволило проверить на практике обоснованность предлагаемых ре-
гиональных мероприятий и рекомендовать их для других районов
Казахстана.
ПРИЛОЖЕНИЕ
КАРТЫ РАСПРОСТРАНЕНИЯ
ОТДЕЛЬНЫХ ВИДОВ КОМАРОВ В КАЗАХСТАНЕ
Стр.
Триба Anophelini 207—208
I. Род Anopheles Подрод Anopheles Подрод Myzomyia 207—208 207 208
Триба Culicini 208—214
II. Род Uranotaenia 208
III. Род Culiseta Подрод Allotheobaldia Подрод Culiseta Подрод Culicella 208—209 208 208—209 209
IV. Род Mansonia 209
V. Род Aedes Подрод Ochlerotatus Подрод Aedes Подрод Aedimorphus 209—213 209—212 212 213
VI. Род Culex Подрод Barraudius Подрод Neoculex Подрод Culex 213—214 213 213—214 214
Карта I. 1 — Anopheles claviger; 2 — An. algeriensis; 3 — An. Hyrcanus.
Карта II. 1 — Anopheles maculipennis messeae; 2 — An. maculipennis sa-
charovi.
Карта III. 1 — Anopheles pulcherrimus; 2 — An. superpictus.
Карта IV. 1 — Uranotaenia unguiculata: 2 — Culiseta longiareolata; *3—
Cs. alaskaensis.
Карта V. 1 — Culiseta annulata; 2 — Cs. bergrothi; 3 — Cs. morsitans; 4 —
Cs. silvestris.
44_________ 60 66______________72 ____73
Карта VI. 1 — Mansonia richiardii; 2 — Aedes behningi; 3 — Ae. beklemi-
schevi; 4 — Ae. cantans.
14—80
54 60 €6 72
Карта VII. 1 — Aedes caspius caspius; 2 — Ae. caspius dorsalis.
Карта VIII. 1 — Aedes cataphylla; 2—• Ae. cyprius; 3 — Ae. communis;
4 — Ae. detritus.
Карта IX. 1 — Aedes diantaeus; 2 — Ae. excrucians; 3 — Ae. flavescens;
4 — Ae. intrudens.
Карта X. 1 — Aedes pionips; 2 — Ae< kasachstanicus; 3 — Ae. leucomelas;
4 — Ae. montchadskyi.
Карта XL 1 — Aedes pullatus; 2 — Ae. punctor; 3 — Ae. riparius; 4 — Ae.
simanini.
54 60____________66____________T2____________78
Кар т a XIL 1 — Aedes sticticus; 2 — Ae* subdiversus; 3 — Ae. cinereus cine-
reus; 4 — Ae. cinereus rossicus.
Карта XIII. 1 — Aedes vexans; 2 — Culex modestus.
Карта XIV. 1 — Culex pusillus; 2 — Cx. hortensts; 3 — Cx. territans.
Карта XV. 1 — Culex pipiens; 2 — Cx. theileri; 3 — Cx. torrentium; 4 —
Cx. martinii.
ЛИТЕРАТУРА
Амиргазиев К. А. Обзор кровососущих двукрылых северного побережья
Каспия. В сб.: «Кровососущие двукрылые (гнус) Казахстана». Алма-Ата, 1966.
Афанасьев А. В. Зоогеография Казахстана. Алма-Ата, 1960.
Балкашина Е. И. Фауна Culicinae Южно-Казахстанской области. «Мед.
паразитол. и паразитарн. болезни», 1939, т. VIII, № 5.
Балкашина Е. И., Мекленбурцева Е. Н. К эпидемиологическому
значению An. superpictus в Чимкентском районе. «Мед. паразитол. и паразитарн.
болезни», 1942, т. XI, № 1—2.
Б е й-Б и е н к о Г. Я. Принцип смены стаций и проблема начальной дивер-
генции видов. «Журн. общей биологии», 1959, т. XXX, № 5.
Беклемишев В. Н. Экология малярийного комара. М., 1944.
Беклемишев В. Н. Учебник медицинской энтомологии. М., 1949.
Беклемишев В. Н. Пространственная и функциональная структура попу-
ляций. Бюлл. Моск, об-ва испыт. природы, т. 65, № 2, 1960.
Беклемишев В. Н., Желоховцев А. Н. Ареалы некоторых видов
Anopheles СССР и причины, их обуславливающие. Бюлл. Моск, об-ва испыт. приро-
ды, т. 50, № 1—2, 1945.
Беклемишев В. Н., Митрофанова Ю. Г. К экологии взрослой сам-
ки Anopheles pulcherrimus Theo. Паразитол. сб. ЗИН АН СССР, т. VI, 1936.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. Ч. П. М. — Л.,
1949.
Благовещенский Д. И. Материалы по фауне наружных паразитов Arth-
ropoda — животных Казалинского и некоторых других районов Южного Казахста-
на. Тр. Казахст. базы АН СССР, вып. 2, 1937.
Боженко В. П. Материалы к фауне Culicidae Восточного Казахстана. «Изв.
АН КазССР, серия паразитол.», 1948, № 6.
Б р е е в К. А. О поведении кровососущих двукрылых и оводов при нападении
их на северного оленя и ответных реакциях оленей. Сообщение I. Паразитол. сб.
ЗИН АН СССР, т. XII, 1950.
Б р е е в К. А. О поведении кровососущих двукрылых и оводов при нападении
на северного оленя и ответных реакциях оленей. Сообщение II. Паразитол. сб. ЗИН
АН СССР, т. ХП1, 1951.
ВеличкевичА. К фауне и экологии комаров южного берега Крыма. Па-
разитол. сб. ЗИН АН СССР, т. VI, 1936.
ВельтищевП. А. К вопросу о распространении представителей рода Апо-
215
pheles (Diplera, Culicinae) в северной части Южного Казахстана. «Изв. АН КазФАН
СССР, серия паразитол.», 1943, № 1.
Виноградова Е. Б. Автогенное развитие яичников у кровососущих кома-
ров. «Зоол. журн.», 1965, т. XLIV, вып. 2.
Виноградская О. Н. Функциональные приспособления абдоминальных
стигм у комаров. Докл. АН СССР, т. 9, № 6, 1948.
Виноградская О. Н. Участие трахейной системы в испарении воды у
Anopheles maculipennis messeae Fall, и сезонная изменчивость дыхальцевого индек-
са у видов подсемейства Culicinae (Diptera, Culicidae). «Энтомол. обозр.», 1953,
т. ХХХШ, № 4.
ВОЗ (Всемирная Организация Здравоохранения). Серия технических докладов,
№ 233. Женева, 1963.
ВОЗ (Всемирная Организация Здравоохранения). Серия технических докладов,
№ 265. Женева, 1964.
Волозина Н. В. К фауне и экологии комаров рода Aedes в Ивановской
области. «Мед. паразитол. и паразитарн. болезни», 1958, т. XVII, № 6.
Геллер Э. Р. К вопросу о фауне кулицид КазССР. «Мед. паразитол. и пара-
зитарн. болезни», 1939, т. VIII, вып. 5.
Глаголева Е. М. Биотопы личинок Anopheles в Таджикистане и их хи-
мизм. Дисс. Душанбе, 1944.
Глущенко П. Г., ГуцевичА. В., Дудкина М. С. Исследование кома-
ров, как переносчиков вируса лимфоцитарного хориоменингита на западе Украины.
Докл. АН СССР, т. 113, № 5, 1957.
Гуцевич А. В. Материалы по изучению гнуса (кровососущие двукрылые
насекомые) на Дальнем Востоке. Тр. Воен.-Мед. акад. им. С. М. Кирова, т. VIII, 1937.
Гуцевич А. В. Материалы по изучению кровососущих двукрылых насеко-
мых (гнуса) в Забайкалье. Тр. Воен.-Мед. акад. им. С. М. Кирова, т. XIX, 1939.
Гуцевич А. В. Материалы по изучению кровососущих двукрылых (гнуса)
североуссурийской тайги. «Зоол. журн.», 1940, т. XIX, № 3.
Гуцевич А. В. Материалы по изучению кровососущих двукрылых (гнуса)
южной части Уссурийской области. Тр. Воен.-Мед. акад. им. С. М. Кирова, т. XXV,
1941.
Гуцевич А. В. Комары — семейство Culicidae. В кн.: ♦Лабораторный прак-
тикум медицинской паразитологии». М. — Л., 1959.
Гуцевич А. В. Новый вид кровососущего комара (Diptera. Culicidae) из Ка-
захстана. ♦ Энтомол. обозр.», 1962, т. ХЫ, № 4.
Гуцевич А. В. Новый вид комара из Узбекистана — Aedes (Ochlerotatus)
simanini (Culicidae). «Зоол. журн.», 1966, т. XLV, вып. 3.
ГуцевичА. В., Подолян В. Я. Изучение комаров в связи с их ролью как
переносчиков нейротропных вирусов на западе Украины. «Зоол. журн.», 1959,
т. XXXVIII, вып. 3.
Денисова 3. М. Автогенное развитие яичников An. bifurcatus Алма-Атин-
ской популяции. «Мед. паразитол. и паразитарн. болезни», 1946, т. XV, № 1.
Денисова 3. М. Новый вид Aedes (Ochlerotatus). «Мед. паразитол. и парази-
тарн. болезни», 1955, т. XXIV, № 1.
Детинова Т. С. Методы установления возрастного состава двукрылых насе-
комых, имеющих медицинское значение. ВОЗ. Женева, 1962.
Дубинин В. Б. Обитатели нор млекопитающих Южно-Казахстанской области
и их значение для человека. «Изв. АН КазССР, серия паразитол.», 1946, № 4.
ДубицкийА. М. К методике изучения ольфакторных реакций кровососу-
щих двукрылых. «Мед. паразитол. и паразитарн. болезни», 1965, т. XXXIV, № 2.
ДубицкийА. М. Об изменчивости кровососущих комаров и о внутривидо-
вых таксонах. Совещание по объему вида и внутривидовой систематике (тезисы
докл.). Л., 1967.
216
Д у б и ц к и й А. М. Новый вид комара Aedes montchadsky sp. п. ♦Паразитоло-
гия», 1968, т. II (3).
Дубицкий А. М., Деньгуб В. М. Места выплода и условия развития
комаров в Казахстане. Мат-лы науч.-теор. конф, проф.-препод, состава. Павлодар,
2967 а.
Дубицкий А. М., Деньгуб В. М. Сезонная и суточная активность напа-
дения кровососущих комаров в Казахстане. Мат-лы науч.-теор. конф, проф.-препод,
состава. Павлодар, 1967 б.
Дубицкий А. М., Дешевых Н. Д. Сезонная и суточная активность кро-
вососущих комаров в бассейне Сыр-Дарьи. Тр. Ин-та зоологии АН КазССР, т. XXV,
1966.
Дубицкий А. М., Д е ш е в ы х Н. Д., Д е н ь г у б В. М. О паразитах и вра-
гах комаров в Казахстане. Мат-лы науч.-теор. конф, проф.-препод, состава. Павло-
дар, 1967.
Дубицкий А. М., Якунин Б. М. Результаты исследования хемотаксисов
у кровососущих двукрылых. «Изв. АН КазССР, серия биол. наук», 1966, № 5.
Желоховцев А. Н. Материалы по фауне комаров (Culicidae, Diptera) Алтай-
ского заповедника. Тр. Алтайского гос. заповед., № 1, 1938.
Жук Н. С. Экологические обоснования и меры борьбы с гнусом в условиях
Северного Прикаспия. Канд. дисс. Алма-Ата, 1967.
Жуматов X. Ж., Темирбеков Ж. Т., А к б е р д и н С. У., Т а г и л ь-
ц е в А. А., Сурвилло А. В. Вирусологическое и зоопаразитологическое исследо-
вание арбовирусных инфекций в Восточном Казахстане. VII итоговая научно-практи-
ческая конференция КИЭМГ (материалы). Алма-Ата, 1966.
Завадский К. М. Учение о виде. Л., 1961.
3 а н и н а 3. Л. Биологические особенности Anopheles superpictus Grass! в усло-
виях Рушанского района Западного Памира. «Мед. паразитол. и паразитарн. болез-
ни», 1957, т. XXVI, № 6.
Иванов И. К. Водный цикл развития Anopheles maculipennis v. sacharovi в
постоянных водоемах и рисовых полях Сыр-Дарвинского района. «Изв. АН КазССР,
серия паразитол.», 1948, № 6.
Исимбеков Ж. Кровососущие комары бассейна р. Или. Канд. дисс. Алма-
Ата, 1966.
Казанцев Б. Н. Цветовые вариации бухарских Aedes caspius. Паразитол.
сб. ЗИН АН СССР, т. II, 1931.
Казанцев Б. Н. Материалы по акклиматизации гамбузии в Таджикской
ССР. Сб. работ по малярии и гельминтозам, вып. 2. Душанбе, 1959.
Кешишьян М. Н. Culicidae Таджикистана. «Мед. паразитол. и паразитарн.
болезни», 1941, т. X, № 1.
К о г а й Е. С. К экологии комаров Culex modestus Fic. рисовой зоны Ташкент-
ской области. В сб.: «Природная очаговость болезней и вопросы паразитологии», № 4.
Фрунзе, 1964.
Конурбаев Э. О. Фауна и экология гнуса Иссык-Кульской котловины Кир-
гизии. Дисс. Фрунзе, 1965.
Крыжановский О. Л. Состав и происхождение наземной фауны Средней
Азии. М. — Л., 1965.
Кузнецов Б. А. Очерк зоогеографического районирования СССР. М., 1950.
Кухарчук Л. П. Материалы по фауне и экологии кровососущих комаров
Барабинской лесостепи. В кн.: «Животный мир Барабы». Новосибирск, 1965.
Лавренко Е. М. Основные черты ботанической географии пустынь Евразии
и Северной Африки. Комаровские чтения, т. XV, 1962.
Л и с о в а А. И. Экспериментальное заражение Anopheles малярийными плаз-
модиями в кишлаке Той-тюбе. Паразитол. сб. ЗИН АН СССР, т. III, 1932.
Майр Э., Л и н с л и Э., Юзингер Р. Методы и принципы зоологической
систематики. М., 1956.
217
МартеховП. Ф. Гольцы Балхашской провинции и их биоценотическое и хо-
зяйственное значение. В сб.: «Вопросы рыбного хозяйства Казахской ССР». Алма-
Ата, 1963.
М а с л о в А. В. Материалы по экологии развития кровососущих комаров. Сооб-
щение 3. Выживаемость в ходе развития у комаров группы Culiseta. «Зоол. жури.»,
1961 а, т. 40 (4).
Маслов А. В. Материалы по экологии развития кровососущих комаров. Со-
общение 6. Условия питания и ротовые органы личинок комаров группы Culiseta.
«Зоол. журн.», 1961 б, т. XL, вып. 6.
Маслов А. В. Кровососущие комары подтрибы Culisetina мировой фауны.
Л., 1967.
Молотова Л. А. Кровососущие двукрылые насекомые курортных районов
Туркмении. Дисс. Алма-Ата, 1966.
МончадскийА. С. О типах реакции насекомых на изменение температуры
окружающей среды. «Изв. АН СССР, серия биол.», 1949, № 2.
Мончадский А. С. Нападение комаров на человека в природных условиях
субарктики и факторы, его регулирующие. Паразитол. сб. ЗИН АН СССР, т. XII, 1950.
Мончадский А. С. Личинки кровососущих комаров. М. — Л., 1951.
Мончадский А. С. Летающие кровососущие двукрылые — гнус. М. — Л.,
1952.
Мончадский А. С. Ночная активность комаров и ее эпидемиологические
особенности. «Зоол. журн.», 1953, т. XXXII, вып. 5.
Мончадский А. С. Нападение комаров на человека в приморской части
дельты Волги. Паразитол. сб. ЗИН АН СССР, т. XVI, 1956.
Мончадский А. С. О классификации факторов окружающей среды. «Зоол.
журн.», 1958, т. XXXVH, вып. 5.
МончадскийА. С. Внутривидовые отношения у хищных личинок подсемей-
ства Chaoborinae (Diptera, Culicidae). «Энтомол. обозр.», 1959, т. XXXVHI, № 3.
Мончадский А. С. Понятие о факторах в экологии. «Зоол. журн.», 1961,
т. XL, вып. 9.
Мончадский А. С. Роль личинок Chaoborinae (Diptera, Culicidae) в унич-
тожении личинок кровососущих комаров. «Зоол. журн.», 1964, т. XLIII, вып. 3.
МончадскийА. С., РадзивиловскаяЗ. А. Новый метод количест-
венного учета активности нападения кровососов. Паразитол. сб. ЗИН АН СССР, т. IX,
1947.
Н а й д и ч Н. Л. Антропофильный штамм Culex pipiens pipiens L. Тезисы док-
ладов Межвуз. науч. конф, по проблеме краевой паразитол. Одесса, 1962.
НайдичН. Л. О биологии антропофильного подвида Culex pipiens L. и опы-
ты борьбы с ним в Одессе. Тезисы докладов науч.-практ. конф, по мед. энтомол. и де-
зинсекции. Москов. гор. СЭС. М., 1964.
Нефедов Д. Д. Типы водоемов, продуцирующих гнус в Горьковской области.
«Зоол. журн.», 1962 а, т. XLI, вып. 2.
Нефедов Д. Д. К познанию фауны кровососущих двукрылых Горьковской
области в связи с их эпидемиологическим значением. «Зоол. журн.», 1962 б, т. XLI,
вып. 6.
Н е ц к и й Г. И. Наблюдения активности кровососущих комаров в природных
условиях северной лесостепи Западной Сибири. «Мед. паразитол. и паразитарн. бо-
лезни», 1952а, т. XXI, № 1.
Н е ц к и й Г. И. Кровососущие комары Омской области. «Мед. паразитол. и па-
разитарн. болезни», 1952 б, т. XXI, № 6. >
Никольский Г. В. О биологической специфике фаунистических комплексов
и значение ее анализа для зоогеографии. «Зоол. журн.», 1947, т. XXVI, вып. 3.
Олсуфьев Н. Г. Роль комаров в передаче туляремии. «Советская медици-
на», 1938, № 17—18.
213
Олсуфьев Н. Гм Руднев Г. П. Туляремия. М., 1960.
Павлова М. С., Павлов А. А. Фенология An. maculipennis и сроки прове-
дения противомалярийных мероприятий в Казахстане. В сб.: «Паразиты диких жи-
вотных Казахстана», т. 19. Алма-Ата, 1963.
Павловский Е. Н., Петрищева П. А. О распространении некоторых
кровососущих паразитов по линии железной дороги Арысь—Эмба. В сб.: «О вреди-
телях животноводства в Казахстане». Вып. 2. М.— Л., 1937.
Петрищева П. А. Фауна, экология и биология Culicidae Туркмении. Пара-
зитол. сб. ЗИН АН СССР, т. VI, 1936.
Петрищева П. А. Методы изучения и профилактика лейшманиозов и мо-
скитной лихорадки. М., 1961.
Петрищева П. А. Природные биотопы кровососущих комаров в Туркмении.
В сб.: «Вопросы краевой паразитологии ТуркмССР». Т. III. Ашхабад, 1962.
Плетнев Е. А. Кровососущие комары Южного Казахстана. «Изв. КазФАН
СССР, серия зоол.», 1943, № 2.
Полякова П. Е. К познанию фауны кровососущих комаров (Diptera, Culici-
dae) в низовьях Оби. Тр. Биол. ин-та СО АН СССР, т. X, 1963.
Полякова П. Е., Мирзаева А. Г. Изучение экологин комаров (Diptera,
Culicidae) Причулымья. «Мед. паразитол. и паразитарн. болезни», 1966, т. XXXV,
№ 1.
Пономарев Е. Д., Борсук В. Н. Климатический очерк Казахстана. Кзыл-
Орда, 1927.
Прыгунова И. Г. Кровососущие комары Северного Казахстана. Канд. дисс.
Алма-Ата, 1966.
Раевский Г. Е., Виноградская О. Н. Природные условия развития ма-
лярии и фауна сем. Culicidae в долине р. Чу Киргизской АССР. «Мед. паразитол. и
паразитарн. болезни», 1934, т. III, № 6.
Ременцова М. М. Бруцеллез диких животных. Алма-Ата, 1962.
Р е м м X. Я. Материалы по фауне и экологии комаров Эстонской ССР. «Энто-
мол. обозр.», 1957, т. XXXVI, вып. 1.
Рубцов И. А. О критериях вида у некоторых насекомых. «Зоол. журн.»,
1959, т. XXXVIII, вып. 5.
Р у м ш Л. Т. Комары Севера СССР. Паразитол. сб. ЗИН АН СССР, т. X, 1948.
Сазонова О. Н. Новый вид кровососущего комара рода Aedes Агг. (Diptera,
Culicidae) из Европейской части СССР. Паразитол. сб. ЗИН АН СССР, т. XVI, 1956.
Сазонова О. Н. Таблица для определения комаров рода Aedes Mg. (Diptera,
Culicidae) лесной зоны СССР. «Энтомол. обозр.», 1958, т. XXVII, № 3.
Сазонова О. Н. Комары рода Aedes Рыбинского водохранилища и обзор
фауны рода Aedes лесной зоны Европейской части РСФСР. Тр. Дарниновского запо-
вед., вып. V, 1959.
С е м е н о в-Т я н-Ш а н с к и й А. П. Таксономические границы вида и его под-
разделений. Зап. Имп. Акад. Наук, серия 8, т. 25, № 1, 1910.
Синицин В. М. Палеогеография Азии. М. — Л., 1962.
Станчинский В. В. Изменчивость организмов и ее значение в эволюции.
Смоленск, 1927.
СуитменХ. [Sweetman]. Биологический метод борьбы с вредными на-
секомыми и сорными растениями. М., 1964.
Федоров В. Г. Некоторые данные по фауне и фенологии комаров окрестно-
стей г. Чкалова. «Энтомол. обозр.», 1951, т. XXXI, № 3—4.
Хелевин Н. В. Яйцекладка н формирование личинки у Aedes caspius dorsa-
lis Meigen. Сб. науч. тр. Ивановского гос. мед. ин-та. Иваново, 1946.
Хелевин Н. В. Влияние внешних условий на возникновение эмбриональной
диапаузы и на количество поколений у Aedes caspius dorsalis в течение сезона. «Энто-
мол. обозр.», 1958 а, т. XXXVII, № 2.
219
Хелевин Н. В. Биология яйцевой стадии Aedes flavescens. «Мед. паразитол.
и паразитарн. болезни», 1958 б, № 1.
Хелевин Н. В. Сезонность отрождения и явление эмбриональной диапаузы
у Aedes caspius dorsalis Mg. (Diptera, Culicidae). «Энтомол. обозр.», 1959, т. XXXVIII,
№ 2.
Ходукин Н. И., Л и с о в а А. И. Влияние световой ритмики на развитие
диапаузирующнх личинок A. bifurcatus L. «Мед. паразитол. и паразитарн. болезни»,
1953, т. ХХП, № 4.
Ч и н а е в П. П. Об экзофильных комарах на целинных землях Голодной степи,
Центральной Ферганы и в дельте Аму-Дарьи. «Мед. паразитол. и паразитарн. болез-
ни», 1964а, т. XXXIII, № 5.
ЧинаевП. П. О фауне и экологии Culicidae дельты Аму-Дарьи. «Зоол. журн.»,
1964 б, т. XLIII, вып. 12.
Шаров А. Г. О путях и закономерностях видообразования. «Зоол. журн.»,
1955, т. XXXIV, вып. 3.
Шевченко В. В. О закономерностях географического распространения слеп-
ней в Казахстане. В кн.: «Природная очаговость и вопросы паразитологии». Алма-
Ата, 1961.
Шленова М. Ф. Температурные условия вылупления личинок из покоящих-
ся яиц. «Мед. паразитол. и паразитарн. болезни», 1950, т. XIX, № 6.
Шленова М. Ф. Скорость развития личинок Aedes при разных температурах
и сроки их появления в природе. «Мед. паразитол. и паразитарн. болезни», 1952,
т. XXI, № 1.
Шленова М. Ф. Продолжительность имагинальной жизни и возрастной сос-
тав популяции двух видов Aedes Подмосковья. «Мед. паразитол. и паиазитарн. болез-
ни», 1955, т. XXIV, № 4.
Штакельберг А. А. Семейство Culicidae. Фауна СССР. Насекомые двукры-
лые, т. III, вып. 4. М. — Л., 1937.
Щербина В. П. Портативная световая ловушка ПСЛ-1 для сбора кровососу-
щих летающих насекомых. «Зоол. журн.», 1964, т. XLIII, вып. 10.
BatesM. The natural history of mosquitoes. New York, 1949.
Bjornson B. F. The role of mosquitoes in the transmission of human disease.
Pest Control., 29, N 4, 1961.
Carpenters. J., La Casse W. J. Mosquitoes of North America. Universi-
ty of California Press, 1955.
Chamberlain R. W. Vector relationships of the arthropodborne encephali-
tides in North America. Ann. New York Acad. Sci., v. 70, 1958.
Edwards F. W. Diptera fam. Culicidae. Genera Insectorum (P. Wyttsmann),
194 fascicule. Bruxelles, 1932.
F о о t e R. H., С о о k D. R. Mosquitoes of Medical Importance. Agriculture Hand
Book, 152, Washington, 1959.
Hopkins G. H. E. Mosquitoes of the Ethiopien region. London, 1952.
Kirkpatrick T. W. The Mosquitoes of Egypt. Cairo, 1925.
Kissling R. К The arthropod-borne viruses of man and animals. Annual Rev.
Microbiol., 14, 1960.
Kramar J. Komari bodavi — Culicinae (Fauna CSR). Praha, 1958.
M a r k s E. N. A review of the Aedes scutellaris subgroup with a study of vari-
ation in Aedes pseudoscutellaris (Diptera, Culicidae). Bull. Brit. Mus. Nat. Hyst. Ent.,
3, 1954.
Martini К Uber die Stechmiicken der Umgebung von Saratow. Тр. Саратовско-
го об-ва естествоиспыт. и любителей естествознания, т. X, № 3, 1926.
Martini Е. Beitrage zur medizinischen Entomologie und zur Malaria-Epidemio-
logie des unteren Wolgagebiets. Hamburg, 1928.
220
Matheson R. Handbook of the mosquitoes of North America. New York, 1944.
M a 11 i n g 1 у P. F., К n i g h t K. L. The mosquitoes of Arabia. Bull. Brit. Mus.
Nat. Hist. Ent., 4, 1956.
Mihalyi F. Aedes hungaricus n. sp. (Culicidae, Diptera). Ann. Hist.-Nat.
Hung. Ser. nova, 6, 1955.
Mihalyi F. Description of the larva of Aedes (Ochlerotatus) hungaricus Miha-
lyi (Diptera, Culicidae). «Acta zool. Acad, scient. hung.», 7, N 1—2, 1961.
N a t v i g L. R. Contributions to the knowledge of the Danish and Fennoscandian
mosquitoes Culicini. Norsk Entomologisk Tidsskrift. Oslo, 1948.
Reeves W. С. Вирусы, переносимые членистоногими. ВОЗ. Женева, 1962.
Ross Н. Н. The colonization of temperate North America by mosquitoes
and man. «Mosquito News», 24, 2, 1964.
T r p i s M. Stechmiicken des Tatre-national-parks. Biologickych a lekarskych vied,
VI/6. Bratislave, 1960.
T r p i s M. Typologie der Biotope in der Ostslowakai gemass der Gesellschaften von
Larven und Imagines der Stechmiicken. <Entomol. probl.», 5, 1965.
Wesenberg-Lund C. Contributions to the biology of the Danish Culicidae.
Kobenhavn, 1290—1921.
YamagutiS., La CasseV. Mosquito Fauna of Japan and Korea. Office
of the Surgeon HQ 8th Army Apo 343, 1950.
ОГЛАВЛЕНИЕ
Предисловие 5
Систематический перечень видов комаров фауны Казахстана 7
I. Особенности наружного строения 10
II. Методика сборов, хранения и изучения комаров 22
III. Повидовой эколого-фаунистический обзор 32
Таблица для определения триб и родов подсемейства Culicinae 32
Род Anopheles Meig- 34
Таблица для определения видов 34
Повидовые очерки 39
Род Ur anotaenia L.-Arrib. 52
Род Culiseta Felt 54
Таблица для определения видов 54
Повидовые очерки 57
Род Mansonia Blanch. 65
Род Aedes Meig. 67
Таблица для определения видов 67
Повидовые очерки 87
Род Culex L. 144
Таблица для определения видов 144
Повидовые очерки 148
IV. Внутривидовая изменчивость 162
V. Экологические особенности комаров Казахстана 169
1. Места выплода и условия обитания 169
2. Жизненные циклы 176
3. Сезонная и суточная периодика лёта 181
4. Паразиты и враги комаров в Казахстане 187
VI. Основные закономерности распространения и генезис фауны комаров Ка-
захстана 191
Заключение 202
Приложение
Карты распространения отдельных видов комаров в Казахстане 206
Литература 215
Дубицкин Анатолий Михайлович
КРОВОСОСУЩИЕ КОМАРЫ КАЗАХСТАНА
Утверждено к печати Ученым советом
Института зоологии Академии наук
Казахской ССР
Редактор Э. И. Иванова
Ху дож. редактор И. Д. Сущих
Техн, редактор Л. И. Шашкова
Корректоры И. Л. Подгорная, Л. С. Нестерова
* * *
Сдано в набор 4/V 1970 г. Подписано к печати
31/VH 1970 г. Формат 70X90716- Бумага № 1.
Усл. печ. л. 16,38. Уч.-изд. л. 16. Тираж 1100.
УГ 03249. Цена 1 р. 80 к.
* * *
Типография издательства «Наука» КазССР,
г. Алма-Ата, ул. Шевченко, 28. Зак. 80.