









































































Text
АКАДЕМИЯ НАуК СССР
фауна
ссср
РАКООБРАЗНЫЕ
том
вып.
. 5
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА * ЛЕНИНГРАД
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 47
ФАУНА СССР
Главный редактор акад. Е. Н. ПАВЛОВСКИЙ
Редактор А. А. ШТ АКЕЛЬБЕРР
РАКООБРАЗНЫЕ
Том VII, вып. 5
Я. А. БИРШТЕЙН
ПРЕСНОВОДНЫЕ ОСЛИКИ (ASELLOTA)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1951 • ЛЕНИНГРАД
Главный редактор акад. Е. Н. Павловский.
Редактор тома А. А. Штакелъберг
ПРЕДИСЛОВИЕ
Водяные ослики широко распространены в пресноводных водоемах.
Они имеют немаловажное значение в жизни озер и прудов нашей страны
и в питании населяющих их промысловых рыб. Кроме того, водяные
ослики представляют собою удобный объект для различных эксперимен-
тальных исследований, как гидробиологических, так и морфологических
(регенерация, рост и т. д.).
Серьезное систематическое изучение этой группы началось всего
30 лет назад. Оно велось весьма интенсивно, и число известных видов се-
мейства Asellidae и других немногочисленных пресноводных Asellota
возросло в несколько десятков раз, причем были открыты не только
новые виды, но и новые роды и семейства. Полученные данные показали,
что пресноводные Asellota представляют значительный интерес прежде
всего в зоогеографическом отношении.
Изучение пресноводных Asellota нашей , страны приводит к выводу
о близости их к западноевропейским. Поэтому я счел необходимым дать
сводку данных по этой группе для всей Палеарктики, исходя также из
того, что такой сводки в мировой литературе не существует, а отдельные
специальные работы разбросаны по многочисленным, часто трудно до-
ступным изданиям. Все известные из пределов СССР виды просмотрены
мной заново и их описания и изображения оригинальны. В отношении
зарубежных видов пришлось ограничиться в большинстве случаев литера-
турными данными. Довоенная литература учтена полностью, некоторый
работы военного и послевоенного времени остались мне недоступными.
Нет никакого сомнения в том, что при дальнейшем изучении пресно-
водной фауцы СССР, в особенности фауны подземных вод, будут открыты
новые пресноводные Asellota. Я надеюсь, что эта сводка обратит на них
внимание исследователей и поможет разобраться в систематическом поло-
жении и зоогеографических отношениях вновь найденных форм.
Я очень признателен ряду лиц и, прежде всего, Е. Ф. Гурьяновой,
М. М. Кожову и покойному Г. Ю. Верещагину за передачу мне на обра-
ботку сборов водяных осликов, Л. А. Зенкевичу и Е. В. Боруцкому — за
просмотр рукописи и Н. Г. Луппо — за помощь в подготовке ее к печати.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Надсем. Aselloidea
1 Сем. Asellidae
1. Род Asellus Geoffroy St. Hilaire
1. Подрод Asellus s. str.
Стр.
1. A. (s. str.) aquaticus (L.) ......................................... 57
la. A. (s. str.) aquaticus messerianus Birst............................. 60
lb. A. (s. str.) aquaticus cavernicola Rac............................... 61
1c. A. (s. str.) aquaticus fribergensis Schneider........................ 61
Id. A. (s. str.) aquaticus abyssalis Odenwall .......................... 62
2. A. (s. str.) hilgendorfi Bovallius . . ............................. 62
2a. A. (s. str.) hilgendorfi' martynovi Birst............................ 64
2b. A. (s. str.) hilgendorfi ryukyuensis Ueno............................ 65
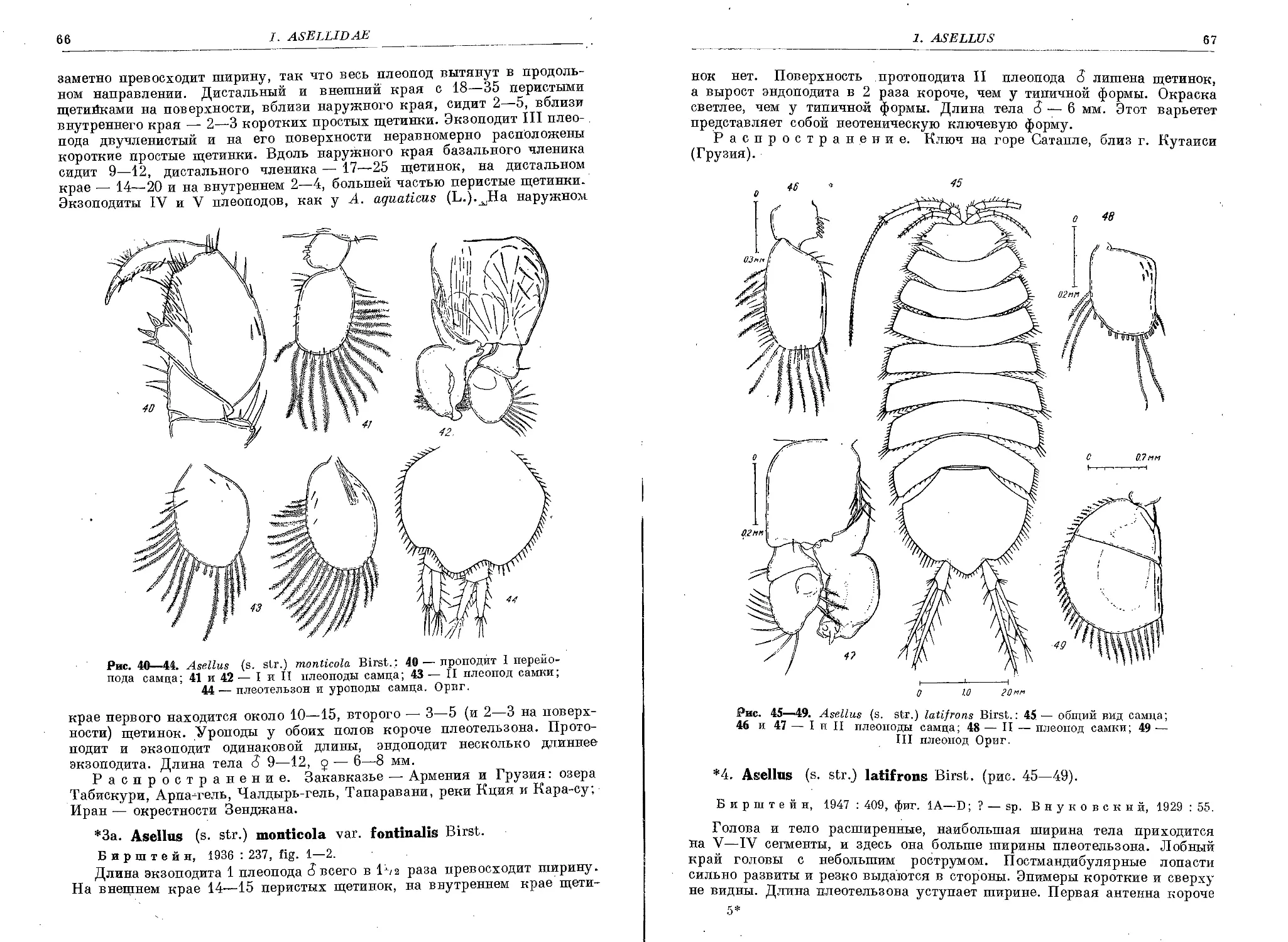
3. A. (s. str.) monticola Birst....................................... 65
3a. A. (s. str.) monticola fontinalis Birst.......................... . 66
4. A. (s. str.) latifrons Birst.......................•................. 67
5. A. (s. str.) epimeralis Birst........................................ 68
2. Подрод Mesoasellus Birst. \
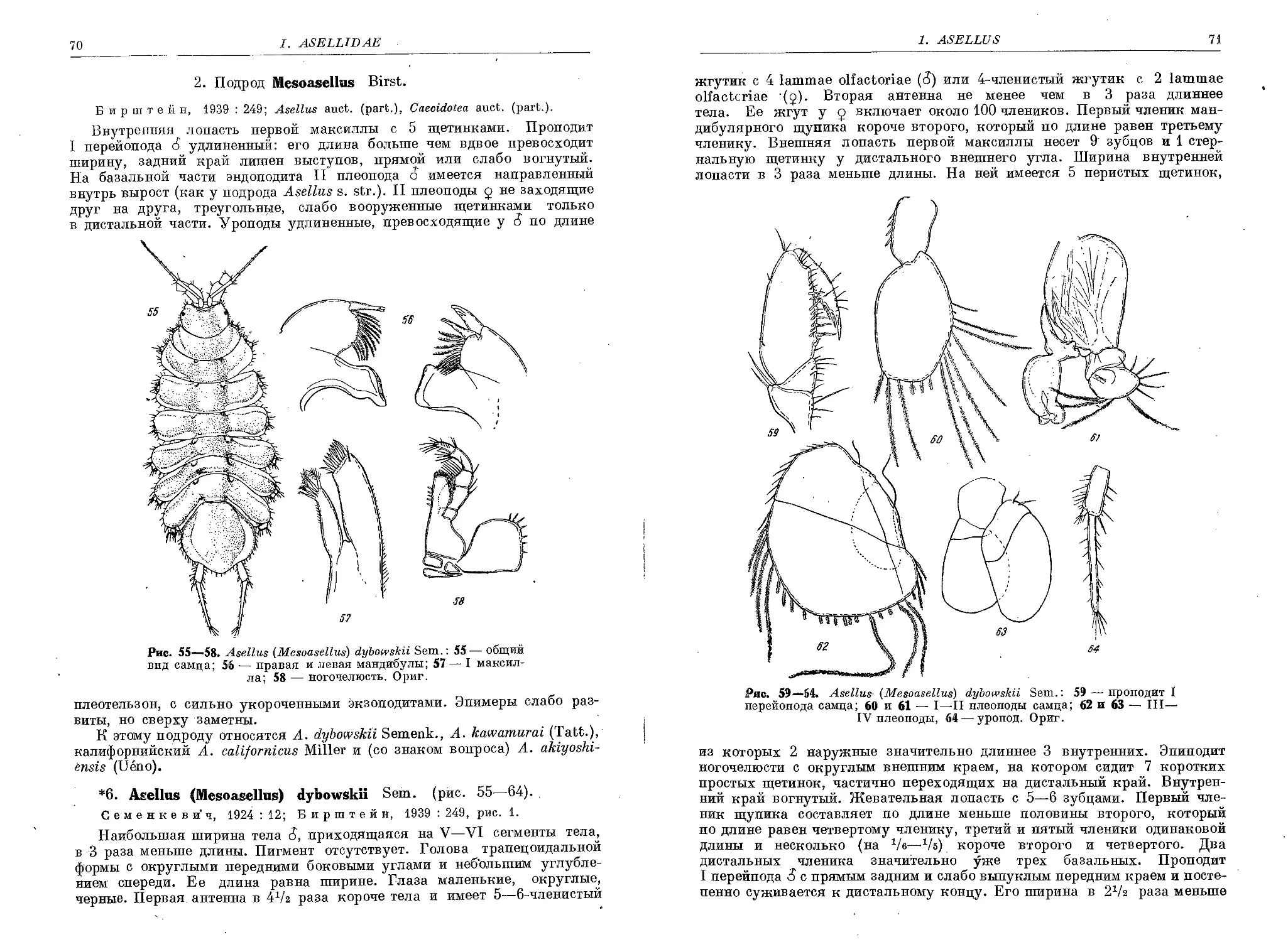
6. A. (M.) dybowskii Sem...................................................... 70
7. A. (M.) kawamurai (Tattersall)............................................ 72
8. A. (M.) akyoshiensis (Ueno) .............................................. 73
3. Подрод Baicaloasellus Stammer
9. A. (B.) baicalensis Grube................................................... 74
10. A. (B.) korotnevi Sem........................................................ 75
11. A. (B.) minutus Sem......................................................... 78
12. A. (B.) angarensis Dyb.................. . . ............................... 79
4. Подрод Proasellus Dudich.
13. A. (Pr.) moridianus Rac..................................................... 81
13a. A. (Pr.) meridianus belgicus Arc............................................ 82
14. A. (Pr.) polychaetus Dud.................................................... 83
14a. A. (Pr.) polychaetus dudichi Strouhal....................................... 84
15. A. (Pr.) remyi Monod....................................................... 84
16. A. (Pr.) infirmus Birstein ................................................ 85
17. A. (Pr.) gjorgjevici Karaman............................................... 87
17a. A. (Pr.) gjorgjevici litoralis Karaman...................................... 87
18. A. (Pr.) cavaticus Schiodte................................................ 88
18a. A. (Pr.) cavaticus foreli Blanc............................................. 89
18b. A. (Pr.) cavaticus walteri Chappuis..................................... 90
18c. A. (Pr.) cavaticus valdensis Chappuis..................... •............... 90
18d. A. (Pr.) cavaticus hercegovinensis Kar...................................... 91
18e. A. (Pr.) cavaticus montenegrinus Kar........................................ 91
19. A. (Pr.) hypogeus Rac.................................................... . 91
20. A. (Pr.) snelaeus Rac ................................................' . . 92
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
20а. A. (Pr.) spelaeus aquae-calidae Rac....................................... 93
20b. A. (Pr.) spelaeus anophthalmus Kar........................................ 93
20c. A. (Pr.) spelaeus rhausinus Remy.......................................... 94
20d. A. (Pr.) spelaeus longisetus Remy ........................................ 94
21. A. (Pr.) collingei nom. nov. ............................................ 94
22. A. (Pr.) hussoni Remy.................................................... 95
23. A. (Pr.) hermallensis Arc................................................. 96
24. A. (Pr.) gauthieri Monod................................................. 96
24a. A. (Pr.) gauthieri theodori, subsp. nov................................... 97
25. A. (Pr.) thermonyctophilus Monod ......................................... 98
26. A. (Pr.) istrianus Stammer................................................ 99
27. A.' (Pr.) amautovici Remy................................................. 100
28. A. (Pr.) monodi Strouhal............................................... 101
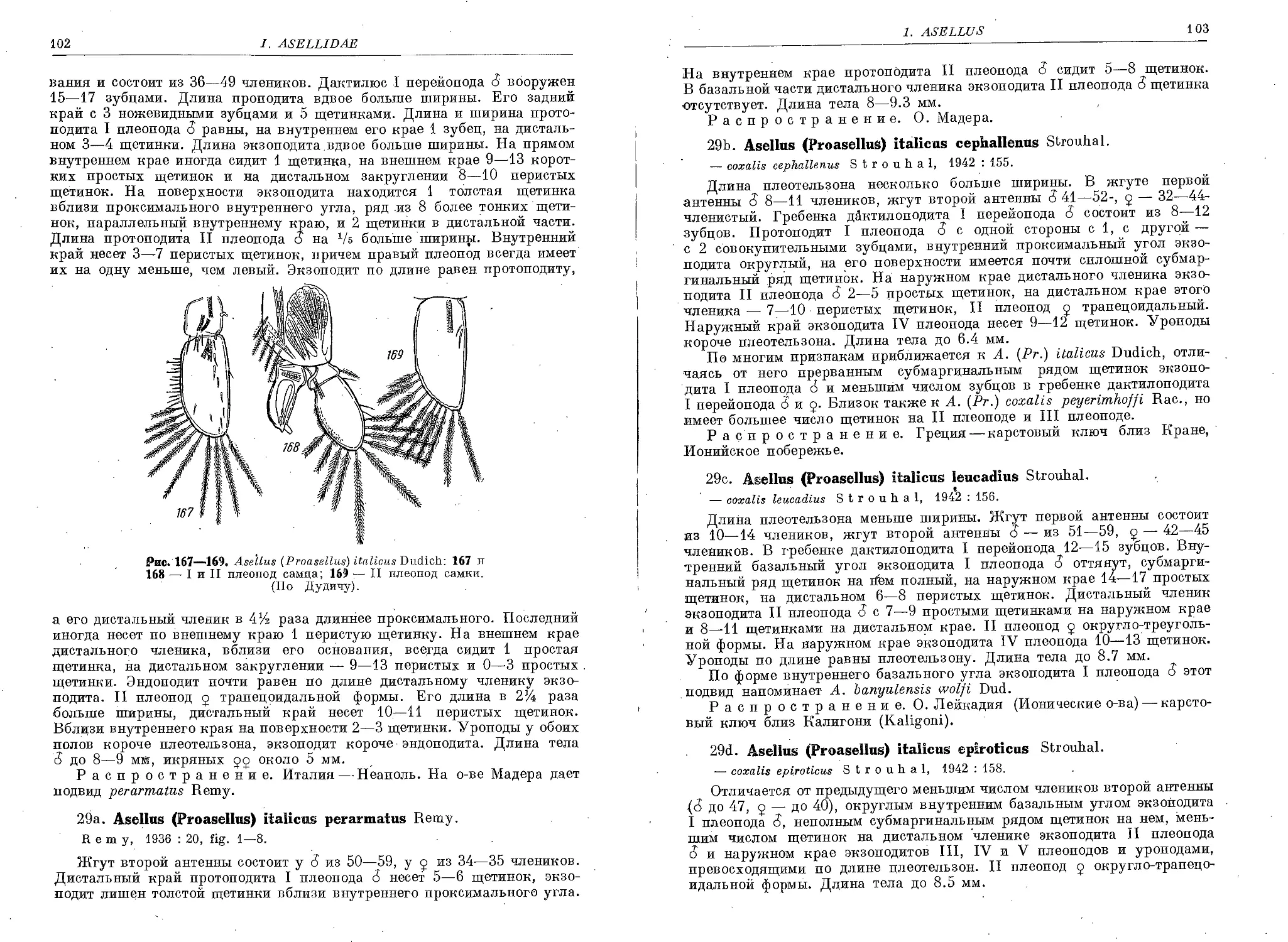
29. A. (Pr.) italicus Dud ................................................... 101
29a. A. (Pr.) italicus perarmatus Remy ....................................... 102
29b. A. (Pr.) italicus cephallenus Strouhal................................... 103
29c. A. (Pr.) italicus leucadius Strouhal..................................... 103
29d. A. (Pr.) italicus epiroticus Strouhal.................................. 103
29e. A. (Pr.) italicus corcyraeus Strouhal.................................... 104
30. A. (Pr.) banyulensis Rac................................................. 104
30a. A. (Pr.) banyulensis wolfi Dud........................................... 105
, 31. A. (Pr.) karamani Remy.................................................. 105
32. A. (Pr.) coxalis Dollfus ................................................ 106
32a. A. (Pr.) coxalis peyerimhoffi Rac. ...................................... 107
32b. A. (Pr.) coxalis cyanophilus Dud..................;......................... 108
32c. A. (Pr.) coxalis africanus Monod.......................................•. . Ю8
33. A. (Pr.) seabrai Braga...................................................... 108
34. A. (Pr.) peltatus Braga ..................................................... HO
5. Подрод Conasellus Stammer
2. Род Johanella Monod
1. J. purpurea Monod ................................'.............................. 112
I 3. Род Stenasellus Dollfus.
1. St. asiaticus Birst, et Star........................................... 115
2. St. virei Dollfus.............................................................. 117
3. St. breuili Rac ........................................................ . . H7
4. St. gjorgjeviii Rac............................................................ 118
4a. St. gjorgjevi 'i robustus, (Mehely) .......................................... 119
5. St. skopljensis Kar............................................................ 119
6. St. buchneri Stammer........................................................... 120
7. St. hungaricus (Mehely)........................................................ 120
8. St. nobrei Braga............................................................... 121
9. • St. bureschi Rac......................................................... 122
4. Род Synasellus Braga
1. S. mariae (Braga)............................................................. 124
la. S. mariae exiguus (Braga),............................................... 124
Надсем, Paraselloidea
2. Сем. Microparasellidae
1. Род Microparasellus Kar.
1. M. puteanus Kar......................................................... 127
2. Род Microcharon Kar.
i. M. stygius (Kar.)...................................................... 128
2. M. latus Kar............................................................. 129
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ 7
3. М. profundalis Kar...................................................... 129
За. М. profundalis kumanovensis Kar......................................... ISO
3b. M. profundalis kosovensis Kar...................................: . . . ISO
3c. M. profundalis beranensis Kar........................................... 131
3. Сем. Janiridae
1. Род Jaera Leach
1. Подрод Jaera s. str.
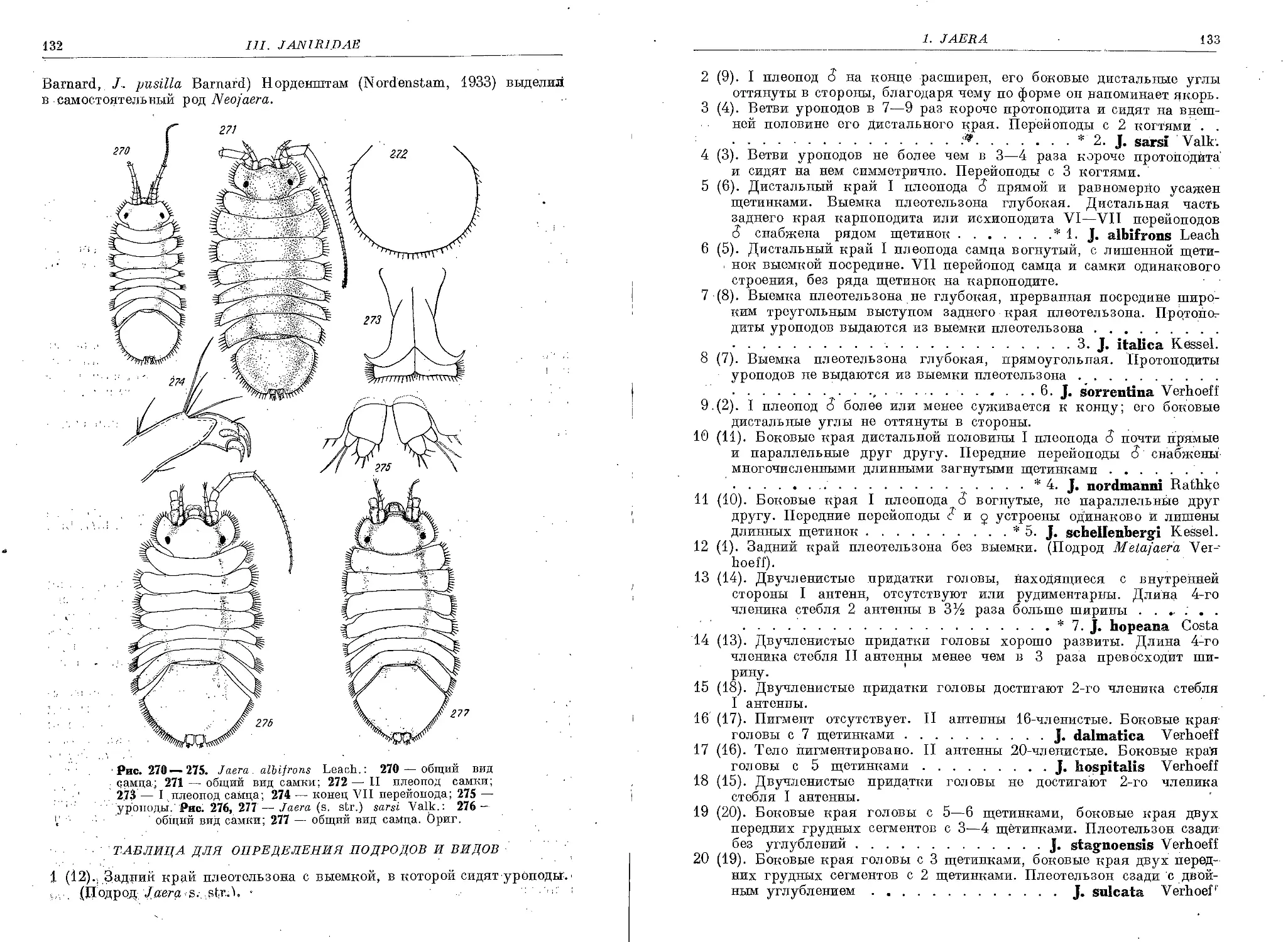
1. J. (s. str.) albifrons Leach........................................ . 134
2. J. (s. str.) sarsi Valkanov............................................ 135
2a. J. (s. str.) sarsi caspica Kesselyak................................... 136
3. J. (s. str.) italica Kesselyak......................................... 136
4. J. (s. str.) nordmanni Rathke.......................................... 137
5. J. (s. str.) schellenbergi Kesselyak................................... 137
6. J. (s . str.) sorrentina Verhoeff . ............................... . 138
2. Подрод Metajaerd Verhoeff
7. J. (M.) hopeana Costa.................................................... 139
ВВЕДЕНИЕ
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
Водяные ослики (сем. Asellidae, Janiridae и Microparasellidae) отно-
сятся к одному из подотрядов
к Asellota. Это — небольшие
рачки, не превышающие в
длину 15 мм (Asellus hil-
gendorji с о-ва Сахалина), а в
некоторых случаях не дости-
гающие и 1 мм (некоторые
Microparasellidae). Окраска
бурая, сероватая, часто с
узором из более светлых пя-
тен. Она зависит от присутст-
вия зернышек меланина, со-
средоточенных в подстилаю-
щих эпидермис звездчатых
пигментных клетках. В ме-
стах прикрепления мышеч- ,
ных пучков к наружному
скелету эти клетки отсут-
ствуют, и, таким образом,
расположение светлых пятен
непосредственно определяет-
ся мышечной системой. Спин-
ная сторона тела и конечно-
стей окрашена интенсивней •
брюшной, но освещение
брюшной стороны вызывает
ее потемнение (Needham,
1942). У форм, населяющих
подземные воды и большие
глубины, пигмент отсут-
равноногих раков (Isopoda), а именно
Рис. 1. Самка Asellus (s. str.) aquaticus (L.) e
брюшной стороны: гол. — голова; гр. —грудь;
бр. — брюшко, ai — I антенна; й'> — II ан-
тенна; I—VII — перейоподы I—VII пар; у —
уроподы. Ориг.
ствует, и животные имеют
молочно-белый или, реже,
розоватый цвет. Тело сплю-
щено в дорзовентральном
направлении, более или менее
вытянуто в длину, с параллельными или почти параллельными
боковыми краями и хитинизированными покровами. Оно состоит из.
отдельных сегментов и распадается на 3 отдела: голова, грудь (перейон)
и брюшко (плеон)’ (рис. 1).
10
ВВЕДЕНИЕ
Голова срастается с первым грудным сегментом, причем на границе
слияния часто заметен шов. Передний край головы прямой, вогнутый
-или иногда образует короткий ростральный выступ (7Йicroparasellidae).
По бокам головы обычно находится по одному постмандибулярному
выступу, обсаженному щетинками. Глаза сидячие, состоящие из неболь-
шого количества фасеток (обычно 4), расположены или по бокам головы,
или, реже, на верхней ее стороне (Л. infirmus, A. remyi, Jaera). У глубо-
ководных (Л. cavaticus foreli) и многочисленных подземных форм глаза
отсутствуют, у форм, ведущих полупбдземный образ жизни, — более
или менее редуцируются (Л. gauthieri).
Часто можно наблюдать постепенные переходы от форм с вполне раз-
витыми глазами к совершенно слепым. Для Л. aquaticus cavernicola
описаны различные типы редукции глаза, проявляющиеся в недоразви-
тии или полном отсутствии хрустального конуса и зрительного нерва.
Глаза могут быть лишены пигмента (как и все тело животного) или могут
быть окрашены в розоватый цвет.1
Редукция зрительного аппарата у Agellidae затрагивает прежде всего
•зрительные ганглии. Между тем у мокриц в первую очередь редуцируются
глаза, а зрительные ганглии часто остаются неизменными.1 2
На передней части головы сидят 2 пары антенн: первые и вторые
антенны. Первые антенны значительно короче вторых и состоят из 3-чле-
нистого основания и жгута, включающего 3—15 члеников. На члениках
основания, особенно на их дистальных частях, сидят многочисленные
щетинки, среди которых особенно заметно по одной щетинке, оперенной
на конце и служащей органом осязания. На некоторых члениках жгута
находятся колбовидные органы, так называемые «lammae olfactoriae».
-Особенно сильно развиты они у многих подземных форм; так, например,
у Мicroparasellidae их длина значительно превышает длину члеников
антенн, на которых они сидят. Функции этих органов выяснены недо-
статочно полно; описавший их впервые Лейдиг (Leidig)3 считал их орга-
нами обоняния, занимавшийся этим впоследствии Рат приписывал им,
кроме обонятельных, хеморецепторные функции. В произведенных Ра-
том опытах колбовидные органы водяного ослика, помещенного в под-
крашенную метиленовой синькой воду, «очень быстро принимали синюю
окраску, в то время как остальные части тела не меняли цвета.4 Помимо
колбовидных органов, на члениках жгута передней антенны сидят ще-
тинки, а на конце последнего членика такая же чувствительная щетинка,
как на члениках основания.
У многих видов рода Jaera между первыми антеннами располагается
пара коротких двучленистых придатков, которые, согласно Фергёф
{Verhoeff, 1943), представляют собою конечности предантеннулярного
сегмента, редуцированные у всех остальных членистоногих (рис. 297). Это
-мнение нуждается в подтверждении на эмбриологическом материале.
1 С. Kosswig und L. К о s s w i g. 1936. Ueber Augenriick- and Missbildung
bei Asellus aquaticus cavernicolus. Verb. Deutsch. Zool. Ges., 38: 274—281 (Zool. Anz.,
Suppl. 9).
2 G. de L a 11 i n. 1939. Untersuchungen an Isopodenaugen. Zool. Jahrb., Anat.,
•65, 3.
3 F. Leidig. 1860. Ueber Geruchs- iind Gehororgane der Krebse etc. Arch.
Anat., Physiol, u. Wiss. Medicin, 1860. Id. Die Hautsinnesorgane det Arthropoden.
Zool. Anz., I, 1886.
4 0. vom Rath. 1896. Zur Kenntniss der Hautsinnesorgane und des sensiblen
Nervensystems der Arthropoden. Zeitschr. wiss. Zool., 61.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
11
Основание вторых антенн состоит из 5 члеников различной длины.
У Stenasellus, М icroparasellidae и Janiridae' на третьем членике сохра-
няется экзоподит в виде чешуйки. Таким образом, морфологически только
3 первых членика основания соответствуют протоподиту. Число члени-
ков жгута колеблется в весьма значительных пределах.
Рис. 2—8. Ротовые части и конечности Asellus (s. str.) aquaticus (L.): 2 — верхняя
губа; 3 — нижняя губа; 4 — мандибула; 5 и 6 — I иП максиллы; 7 — ногочелюсть;
8 — II перейопод самца (ко — коксоподнт, б — базиподит, и — исхиоподит, м —
мероподит, ка — карпоподит, п — проподит, д — дактилюс). О риг.
Ротовые части (рис. 2—7) помещаются с нижней стороны головы.
Мандибулы, две пары максилл и ногочелюсти — парные, верхняя и ниж-
няя губа — непарные. Мандибулы находятся по бокам ротового отвер-
стия. Каждая из них состоит из тела, 3-членистого щупика, жевательной
пластинки (pars molaris) и добавочной подвижной пластинки, имеющейся
только на правой мандибуле (lacinia шоЬПщ) (рис. 4); у Synasellus и
12
ВВЕДЕНИЕ
Lirceus щупик отсутствует. Жевательная пластинка имеет цилиндриче-
скую форму и сильно хитинизирована. Добавочная пластинка несет
ряд перистых щетинок и хитинизированные зубные отростки. Длина чле-
ников щупика значительно варьирует в пределах семейства Asellidae.
Каждая первая максилла имеет 3-членистый протоподит и две лопасти —
внутреннюю и наружную. На внутренней лопасти прикрепляется 4—5
оперенных щетинок (важно для систематики), на наружно!! — несколько
простых или зазубренных на конце зубцов (рис. 5). Принципиально так
же построена и каждая из вторых максилл: она состоит из 3-членистого
протоподита и трех лопастей — внутренней и двух наружных, представ-
ляющих собой по существу одну расщепленную внешнюю лопасть. На
концах всех этих лопастей имеются многочисленные перистые щетинки
(рис. 6). По всей вероятности внешние лопасти обеих пар максилл со-
ответствуют эндоподитам, внутренние лопасти являются выростами про-
топодитов, а экзоподиты отсутствуют. Пара ногочелюстей покрывает
снизу все ротовые части. Обе ногочелюсти срастаются между собой при
основании. Каждая из них состоит из четырех частей: основания (соха),
тепа (basis), или жевательной лопасти, 5-чпенистого щупика (palpus)
и эпиподита. Основание слагается из двух члеников, жевательная лопасть
снабжена по внутреннему краю 3—5 зубцами и по дистальному краю
перистыми щетинками. К ней причленен 5-членистый щупик, членики
которого различной длины и ширины. Эпиподит всегда развит очень
хорошо, имеет вид треугольной пластинки, а у яйценосных самок значи-
тельно увеличивается в размерах и образует оостегит (рис. 7). Непар-
ные ротовые части — верхняя и нижняя губа — находятся одна спереди,
а другая сзади ротового отверстия. Верхняя губа треугольной формы,
заостренная книзу, разделена пополам поперечной линией. Базальная
часть называется эпистомом, дистальная разделена поперечной линией
на clypeus и собственно верхнюю губу (labrum) (рис. 2). Нижняя губа
продольно расщеплена узкой треугольной выемкой на две лопасти (рис. 3).
За головой следует грудь (pereion, thorax), состоящая из 7 одинаково
построенных, подвижно сочлененных сегментов. Каждый сегмент снабжен
парой перейоподов, представляющих собой одноветвистые конечности,
служащие для передвижения. Перейоподы (рис. 8) слагаются из 7 члени-
ков: коксоподита, базиподита, исхиоподита, мероподита, карпоподита,
проподита и дактилюса (dactylus). Коксоподит — наиболее короткий
членик — уплощен и срастается своим базальным краем с боковым краем
стернита сегмента груди, образуя таким образом эпимер. Его дистальный,,
покрытый щетинками, край часто видим сверху (Proasellus, Мesoasellus,
Baicaloasellus). Базиподит — наиболее длинный членик, исхиоподит
несколько короче. Оба они обычно в середине расширены. Мероподит
имеет треугольную форму. Карпоподит и проподит примерно одинако-
вой величины (у всех перейоподов, за исключением перейоподов I пары),
дактилюс значительно короче и серповидно изогнут. Края всех члеников
несут шипы и щетинки, частично чувствительные, которых особенно
много на передних краях бази- и проподита II—VII перейоподов. Задне-
нижние углы исхио- и мероподита снабжены длинными, сильно разви-
тыми шипами, передний край дактилюсов имеет по 1—6 зубцов. У икря-
ных самок коксоподиты первых четырех пар перейоподов образуют длин-
ные широкие пластинки — оостегиты, составляющие вместе выводковую
камеру (рис. 1). Оостегиты наиболее сильно развиты на IV сегменте
перейона и по направлению к голове их размеры постепенно умень-
шаются.
МОРФОЛОГИЧЕСКИЙ. ОЧЕРК
1.3
В нормальном положении живого водяного ослика оба перейопода
каждой пары S-образно изогнуты и обращены друг к другу задними
сторонами (рис. 9). Перейоподы II—VII построены совершенно одинаково
и отличаются только величиной и деталями вооружения. Наиболее корот-
кой из них является IV пара перейоподов. I пара перейоподов Asellidae
имеет несколько иное строение. Она характеризуется укороченным тре-
угольным карпоподитом и утолщенным, снабженным различными щетин-
ками и зубцами, проподитом, к которому пригибается дактилюс, так что
получается ложная клешня. Вооружение проподита I перейопода имеет
большое значение для систематики. Вдоль его заднего края расположены
щетинки и зубцы. На самом крае находится 2—4. (Asellus) или больше
(Stenasellus) ножевидных зубцов. Щетинки сидят в 3 продольных ряда:
на крае, на уровне ножевидных зубцов — субмаргинальный ростральный
ряд, и параллельно ему — субмаргинальный каудальный и продольный
каудальный ряды. На заднем крае находятся щетинки продольного тер-
гального ряда и вдоль дистального края проподита — щетинки дисталь-
ного поперечного ряда. Таково полное
вооружение проподита I перейопода
самца. Однако у целого ряда видов и у
некоторых подродов один или несколько
продольных рядов щетинок могут отсут-
ствовать или все щетинки продольных,
рядов могут сидеть беспорядочно (Asel-
lus s. str.)‘. Форма заднего края про-
подита также довольно разнообразна.
Иногда он образует один (Asellus s. str.),
два или три (Conasellus) выступа. Кроме
того, в строении I перейопода у Asellus
резко выражен половой диморфизм. У
самца проподит шире и сильнее воору-
жен, чем у самки. Первая пара пе-
рейоподов в противоположность остальным шести парам, являю-
щимся органами передвижения, служит для хватания пищи и играет
некоторую роль при спаривании.
Последний отдел тела — брюшко (абдомен, плеон) — значительно
короче груди. Брюшко состоит из 6 сегментов. Два передних сегмента
свободные, четыре задних срослись между собой в плеотельзон и покрыты
сверху единым хитиновым щитом. У рода Asellus два первых абдоминаль-
ных сегмента короткие и узкие и частично покрыты плеотельзоном. У рода
Stenasellus они почти не уступают по величине грудным сегментам, роды
Johanella и Synasellus занимают в этом отношении промежуточное поло-
жение. Соответственно-числу сегментов, брюшко имеет 6 пар конечно-
стей — 5 пар плеоподов и 1 пару уроподов. У самок первая пара плеопо-
дов, как правило, отсутствует, и счет начинается со второй пары. Первые
пары плеоподов самца служат для совокупления и поэтому трансформи-
рованы и частично редуцированы. Каждая пара плеоподов прикреплена
к середине стернита соответствующего абдоминального сегмента (рис. 10).
I плеопод самца Asellidae состоит из двух члеников — протоподита
и экзоподита. Эндоподит отсутствует. Протоподит имеет квадратную или
трапецеидальную форму и снабжен по внутреннему краю 1—4 совоку-
пительными зубцами, а по дистальному краю — обычно щетинками.
У Stenasellus эти шипы и щетинки отсутствуют. Экзоподит прямоуголь-
ной или овальной формы, его длина всегда превышает ширину, его внеш-
Рис. 9. Схематический поперечный
разрез через грудной отдел Asellus,
показывающий нормальное положе-
ние перейоподов. (По Четверикову).
14
. ВВЕДЕНИЕ
ний и дистальный края, а также иногда и внутренний несут щетинки.
У A. aquaticus на внешнем крае имеется выемка. Интересно, что, как
показал Раковица (Racovitza, 1920), эта выемка появляется только
на поздних стадиях постэмбрионального развития. Близко родственные
A. aquaticus виды — A. monticola и A. hilgendorfi — лишены ее в течение
всей жизни. Невидимому они, в особенности A. hilgendorfi, развиваясь
в том же направлении,-как и A. aquaticus, останавливаются на определен-
ной стадии, на которой и завершают свое развитие. Что же касается
A. aquaticus, то он проходит еще через стадию образования выемки на
крае экзоподита I плеопода самца. У М icroparasellidae и Janiridae экзо-
подит не вполне отделен от протоподита. Оба членика почти полностью
Рис. 16. Плеотельзон самца Asellus (Mesoasellus) dybowskii
Sem. с брюшной стороны: пен — пенис; ш — I плеопод;
пи — II плеопод; п III — III плеопод; а — анальное
отверстие; у — уроподы (эндоподит III плеопода и IV
и V плеоподы не изображены). Ориг.
лишены щетинок, только на дистальном крае экзоподита имеется не-
сколько коротких щетинок.
II плеопод <5 значительно отличается от I присутствием эндоподита,
преобразованного в совокупительный орган, а также иной формой и
строением экзоподита. Протоподит у Asellidae приблизительно всегда
квадратной формы, с простыми или оперенными щетинками по внутрен-
нему краю (у Stenasellus они отсутствуют) и иногда (у подродов Asellus
s. str., Mesoasellus и рода Stenasellus) с выемкой на внутреннем крае,
в которую может частично вкладываться эндоподит. Экзоподит двучле-
нистый. Внешний край базального членика и внешний и дистальный края
дистального членика несут щетинки. Эндоподит — копулятивный орган—
у Asellus одночленистый и имеет бутылкообразную форму, так как его
базальная часть вздута, а дистальная более узкая. На внутреннем базаль-
ном углу эндоподита подрода Asellus s. str. и М esoasellus имеется длинный
вырост, а у подрода Conasellus и многих Proasellus значительно более
короткий вырост находится с внутренней стороны базальной части эндо-
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
15-
подита. Дистальная часть имеет сложное и довольно разнообразное-
строение (рис. 11, А—Г). У подрода Proasellus легко отличить: 1) окон-
чание, имеющее обычно форму усеченного конуса (а у A. meridianus —
Рис. 11. Схема строения эндоподита II плеопода самцов подродов
Asellus'. А — Mesoasellus Birst.; В — Proasellus Dud.; В — Baica-
loasellus Stammer, Г — Asellus s. str. (ok — окончание, 6 — боко-
вая лопасть, о — отверстие, щ — щель, ц.о — цилиндрический
отросток, п — полость эндоподита, м — мускул эндоподита.
А и В — ориг., В — по Реми, Г — по Мерксу.
цилиндра), на конце или сбоку которого находится отверстие, продолжа-
ющееся далее в щель; они ведут в полость, занимающую большую часть
эндоподита; 2) боковую лопасть, расположенную над окончанием. У под-
родов Asellus s. str. и Mesoasellus боковая лопасть не выражена, а щель
очень длинная, окончание спирально закручено и края отверстия снаб-
16
ВВЕДЕНИЕ
жены зубцами. Кроме того, внутри щели находится цилиндрический
отросток, играющий роль поршня при заполнении полости эндоподита
спермой. Сходное, однако несколько более примитивное, строение свой-
ственно подроду Cinasellus. У подрода Conasellus края также снаб-
жены зубцами. У подрода Baicaloasellus также нет боковой лопасти,
окончание вытянуто в хитинизированное клювовидное образование,
щель длинная. У рода Stenasellus эндоподит состоит из двух члеников,
и его строение гораздо примитивнее, чем у рода Asellus.
Семейства Мicroparasellidae и Janiridae характеризуются иным строе-
нием II плеопода 6. Протоподиты расположены латерально, а между
ними лежат сросшиеся внутренними краями I плеоноды. Экзоподит очень
маленький, а эндоподит изогнут и суживается книзу. I и II плеоподы
$ вместе образуют крышечку для лежащих иод ними плеоподов.
I плеопод как правило, отсутствует, однако известны два случая
присутствия конечностей на стернуме I брюшного сегмента напоми-
нающих по форме II плеопод j у Stenasellus virei (Racovitza, 1920). II
плеопод j всегда имеется и у Asellidae представляет собой нерасчленен-
ную пластинку (прото- и экзоподит слились) округлой, овальной {Asel-
lus s. str.) или удлиненно-треугольной и трапецоидальной формы (Рго-
asellus) с обсаженными щетинками краями. У Мicroparasellidae и Janiridae
оба плеопода второй пары у j сливаются вместе, образуя единую кры-
шечку.
III—V плеоподы Asellidae у обоих полов имеют одинаковое строение.
Каждый плеопод состоит из небольшого протоподита и листовидных
экзо- и эндоподита. Экзоподит III пары хитинизирован, разделен швом
на две части и служит крышечкой для остальных плеоподов. Экзоподиты
IV—V пары, а также эндоподиты III—V пары почти полностью лишены
хитина и функционируют как жабры.
У A. aquaticus и A. hilgendorfi при основании и по краям экзоподи-
тов хитин сохраняется. Лишенное хитина поле средней и дистальной
части экзоподитов этих видов носит название area tschetwerikoffi. Края
и иногда поверхность экзоподита III плеопода и основные части внешних
краев экзоподитов IV—V плеоподов несут простые или перистые ще-
тинки. Нередко экзоподиты IV—V плеоподов делятся швами на 2—3
поля. Эндоподиты никогда не имеют щетинок и не разделены швами.
На поперечном разрезе через плеопод видно, что между спинным и брюш-
ным слоями гиподермы, лежащими под слоем хитина (в экзоподите III
плеопода) или эпителия, проходят кровеносные сосуды, в которых кровь
обогащается при помощи имеющихся в ней ферментов — оксидаз и пер-
оксидаз.1
Витальное окрашивание водяных осликов азотно-кислым серебром и
перманганатом калия показало, что органами дыхания служат только
эндоподиты III, IV и V плеоподов, а также часть вентральной поверх-
ности плеотельзона. У молоди, еще не покинувшей мацзупиальную сумку,
наблюдается дыхание с помощью заднего отдела кишечника. Мускула-
тура задней кишки производит 40—50 сокращений в минуту, благодаря
которым вода в задней кишке быстро сменяется.1 2
У Мicroparasellidae III—V плеоподы редуцированы и прикреплены
к II плеоподу; у Janiridae отсутствует экзоподит V плеопода.
1 3. С. Сергеева. 1930. Об органах дыхания Isopoda. ДАН СССР: 167.
2 W. Franzl. 1940. Die Atmungsorto von Asellus aquaticus L. Zool. Anz.,
132 : 44—48.
МОРФОЛОГИЧЕСКИЙ ОЧЕРК
17
Последняя пара брюшных ног — уроподы — прикрепляется к концу
плеотельзона. У рода Jaera уроподы сидят в выемке, имеющейся на зад-
ней стороне плеотельзона. Каждый уропод состоит из 3 частей — прото-
подита, экзоподита и эндоподита.. Экзоподит часто в той или иной сте-
пени редуцирован, а у Microp ar asellus отсутствует совершенно.
Постэмбриональное развитие пресноводных Asellota почти не изу-
чено. Некоторые данные имеются только о развитии двух первых пар
плеоподов самца. Раковица (Racovitza, 1920) различает 4 стадии фор-
мирования плеоподов Asellus aquaticus:
3-я стадия
4-я стадия
I плеопод
Г Короткая, треуголь-
) пая нерасчлененная
| пластинка, лишен-
I пая щетинок.
I плеопод
Пластинка разделена
на два членика, ди-
стальный членик мо-
жет быть снабжен
щетинками.
II п л е оп о д
Ветви намечены, но не от-
членены от протоподита.
На них сидят перистые
щетинки.
II плеопод
Плеопод явно дву ветви-
стый, по ветви не рас-
членены, обе ветви несут
перистые щетинки.
' Плеопод почти как у
! взрослой формы, но
число щетинок мень-
ше И видовые осо-
бенности формы пле-
опода не выражены.
Взрослая форма
Протоподит еще лишен ще-
тинок, но мускулатура
как у взрослой формы.
Щетинок на экзоподите
меньше, чем у взрослой
формы. Эндоподит начи-
нает формироваться В
копулятивный орган.
Впоследствии Нидгем (Needham, 1938, 1941) подтвердил данные Ра-
ковицы. По его мнению, Asellus aquaticus в ходе онтогенеза передних
плеоподов самца рекапитулирует стадию, соответствующую строению А.
meridianus.
Что касается II плеопода самки, то молодые экземпляры почти не отли-
чаются в этом отношении от взрослых. Различия замечаются только в ко-
личестве оперенных щетинок по краям плеоподов, которых у молодых
экземпляров меньше, чем у взрослых.
Кроме того, у молодых экземпляров почти не проявляется половой
диморфизм в строении перейоцодов и уроподов. Проподит I перейопода
взрослых самцов Asellidae значительно крупнее, чем у самок, и имеет
большее количество ножевидных зубцов, проподит IV перейопода сам-
цов также сильнее вооружен, чем у самок. Уроподы самок относительно
•короче, чем у самцов. Молодые самцы по перечисленным признакам не
отличаются от самок.
Изредка попадаются самки-интерсексы, обладающие пенисом и раз-
витыми, как у самца, I плеоподами. II плеопод обычно у таких экземпля-
ров недоразвит, обе ветви его несут щетинки и напоминают 1-ю — 2-ю
стадии развития II плеопода нормальных самцов (рис. 12). Самки-интер-
сексы A. monticola встречены в оз. Табискури. Меркс (Maercks, 1930)
находил таких самок A. aquaticus с нормальной икрой в выводковой
сумке. Тем не менее, Нидгем (Needham, 1941) указывает на возможность
толкования таких интерсексов как самцов с задержавшимися в своем
развитии передними плеоподами.
2 Фауна СССР, том VII, в. 5
18
ВВЕДЕНИЕ
Рис. 12. II илеолод интсрссксу-
альной особи Asellus (s. str.)
monticola. Birst. Ориг.
Онтогенез Jaero. albifrons изучался Форсманом (Forsman, 1944). После
откладки в марзупиум яйца имеют зеленую или бурую окраску. Через
3 дня после икрометания становится заметной сегментация зародыша,
а на 4-й день зародыш изгибается на -спинную сторону.1 Вторая стадия
эмбрионального развития начинается с отслаивания эмбриональной обо-
лочки. Через день после этого образуется бурый глазной пигмент, а через
2 дня эмбрион начинает двигаться и совершает первую линьку, после
чего переходит в последнюю стадию эмбрионального развития. В это-
время он достигает длины около 0.7 мм и постепенно выпрямляется.
Через несколько часов после линьки начинает биться сердце, на второй
дв|1ь полностью резорбируется желток. После-
второй эмбриональной линьки зародыш
представляет собою уже вполне сформиро-
вавшегося рачка, однако остается еще в
течение нескольких часов в марзупиуме-
матери.
Вышедшие из марзупиума рачки несколь-
ко отличаются от взрослых (прежде всего
отсутствием последней пары грудных конеч-
ностей) и, согласно Форсману, проходят
3 стадии манка. Первая из них характери-
зуется 8-членистыми антеннами и наличием
одной щетинки с каждой стороны головы
перед глазом. Боковые края передних груд-
ных сегментов лишены щетинок, боковые-
края остальных грудных сегментов — с од-
ной щетинкой. VII сегмент очень мал -и ли-
шен конечностей. Вторая стадия обладает
9-членистыми антеннами, 2 щетинками по
бокам головы и передних грудных сегмен-
тов и несколько более крупными размерами
VII грудного сегмента, несущего с боков
по 1 щетинке. На третьей стадии манка число-
члеников антевп превышает 9, последний
грудной сегмент почти не уступает повеличи-
не предыдущим и имеет зачатки конечностей.
После стадии манка следует стадия молоди. Самец обычно проходит-
через одну такую стадию, самка — через четыре, • но, вероятно, число-
их варьирует. На стадии молоди еще почти не проявляется половой димор-
физм. Самки нацело лишены оостегитов. Далее рачки переходят в подго-
товительную стадию, причем их гонады начинают созревать и приобре-
тают у самок зеленую окраску, у самца формируются передние плеоиоды.
Самки, обладающие вполне развитыми оостегитами, и созревшие самцы-
именуются особями в брачном наряде. Ьтметавшие икру самки теряют
оостегиты и переходят во вторую подготовительную или промежуточную
стадию, после которой снова надевают брачный наряд, т. е. развивают
оостегиты. У самцов промежуточная стадия отсутствует. О размерах
тела и продолжительности жизни всех стадий постэмбрионального раз-
вития дает представление приводимая таблица. * 11
1 Дробление яйца и органогенез Jaera albifrons подробно описаны в работе"
J. Р. Me М и г г i с h. 1895. Embryology of the Isopod Crustacea. Journ. Morphol,.
11.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 1»
Пол Стадия Длительность в днях Размеры в мм
Манка 1
2
3
Молодь I
Подготовит.
Брали, наряд 1
0.9
1.0
1.2
1.4
1.5
1.7
5
Молодь 1
» 2
» 3
» 4
Подготовит.
Врачи, наряд 1
Промежуточна
Брачи, наряд 2
5—7
4—9
5—9
10
10
10
1.4
1.0
1.8
2.1
2.3
2 4
После наступления половозрелости рачки линяют еще несколько'
раз, вырастая на 0.4—0.7 мм за каждую линьку. Минимальная величина
половозрелых самок составляет 2.4 мм, максимальная — около 5 мм,
самцов, соответственно —1.6'—3 мм.
Ливька происходит и днем, и ночью. При этом старый панцирь
лопается поперечно между IV и V грудными сегментами и после некото-
рой паузы рачек освобождается от передней половины старой шкурки.
Далее следует новый перерыв длительностью до 20 часов, и рачек сбра-
сывает заднюю половину панцыря.
Все,эти данные относятся к Jaera albifrons из морских прибрежных
вод Швеции. Возможно, что в условиях пресной воды, а также у других
видов этого рода наблюдаются существенные отличия в ходе биологи-
ческого цикла.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И РОДСТВЕННЫЕ ОТНОШЕНИЯ
Отряд Isopoda объединяет около 2000 видов, обитающих преимуще-
Втвеннб в море и на суше. Пресные воды населены весьма незначитель-
ным количеством форм равноногих раков. Однако из 7 подотрядов, на
которые в настоящее время разделяется отряд Isopoda (Asellota, Flabel-
lifera, Valvifera, Oniscoidea, Phreatoicidea, Epicaridea, Gnathiidea),
только Gnathiidea не представлены совершенно в фауне пресных вод;
с другой стороны, Phreatoicidea целиком являются пресноводными оби-
тателями, живущими главным образом в подземных и отчасти в назем-
ных водах Австралии, Новой Зеландии, Тасмании, Южной Африки.
Число представителей пресноводных Flabellifera сравнительно велико:
сем. Cymothoidae насчитывает целый ряд свободно живущих видов (Ciro-
laninae') в' пещерах Европы и Северной Америки и паразитических видов
в восточной Азии, сем. Sphaeromidae представлено несколькими видами
в водах бассейна северной половины Тихого океана и в пещерах Европы,
сем. Corallanidea — в Китае, Японии и на Зондских о-вах, сем. Ехосо-
rallanidae — одним видом в Британской Гвиане, сем. Anthuridae — одним
видом в грунтовых Ьодах Новой Зеландии.
Пресноводные Valvifera значительно уступают по количеству видов
предыдущему подотряду. Сем. Chetilidae, заключающее всего один род,
2*
20
ВВЕДЕНИЕ
известно из Южной Америки; семейство Idotheidae населяет пресные воды
бассейна Северного Ледовитого океана, Южной Америки, Индии и Но-
вой Зеландии. Oniscoidea, живущие почти исключительно на суше, за-
ходят иногда в воду, так, например, Ligidium нередко встречается
в ручьях Кавказа, Desertoniscus birsteini найден в озере Делили в Турк-
мении, пещерные Tithanetes и Spiloniscus входят в водоемы. Наконец,
некоторые Epicaridea паразитируют на тропических и субтропических
пресноводных креветках.
Но эти подотряды не характерны для пресноводной фауны СССР;
по существу только два представителя названных групп (исключая мо-
криц) обитают в пресных водах Союза — Mesidothea entomon в водоемах
Полярного бассейна и Livoneca amurensis в р. Амуре; кроме того,
несколько эвригалинных видов Idotheidae, Sphaeromidae ъ Anthuridae
встречаются в эстуариях рек, преимущественно на Дальнем Востоке.
Подавляющее большинство пресноводных Isopoda СССР, да и вообще
Палеарктики, относится к подотряду Asellota и к чисто пресноводному
надсемейству Aselloidea, заключающему одно семейство Asellidae.
Семейство Asellidae распространено только в северном полушарии;
ложной границей его распространения можно считать параллель 30°,
на севере же некоторые Asellus известны за полярным кругом — в за-
падной Гренландии и в дельте р. Лены. Основной областью распростра-
нения Asellidae является бореальная область.
К семейству Asellidae относятся 5 родов: Asellus, Stehasellus, Joha-
nella, Synasellus и Lirceus, из которых наиболее обширный ареал имеет
род Asellus, населяющий всю Европу, северную Африку, оз. Байкал,
восточное побережье Азии, восточную и западную часть Северной Аме-
рики; род Stenasellus приурочен преимущественно к юго-западной Европе
и только два из 13 входящих в него видов найдены недавно в экваториаль-
ной западной Африке (Remy, 1938; Monod, 1945) и один — в Средней Азии
(Бирштейни Старостин, 1949); монотипический род Johanella известен
только из одвого ключа в Алжире и, наконец, род Lirceus с 16 видами
занижает восточную часть Северной Америки; близкий к этому роду
монотипический род Synasellus обнаружен недавно в подземных водах
Португалии (Braga, 1942, 1944).
Таким образом, общий ареал семейства является разорванным, при-
чем разорванность ареала присуща не только всему семейству в целом,
но и отдельным его родам и подродам (рис. 13). Это обстоятельство ука-
зывает, разумеется, на значительную древность группы.
Происхождение и время вселения Asellidae в пресные воды совершенно
неясны, поскольку палеонтологические данные по Asellota отсутствуют.1
Морфологические данные также не могут помочь разрешить эти вопросы:
пресноводное надсемейство Aselloidea со своим единственным семей-
ством Asellidae резко отчленено от преимущественно морского над-
семейства Paraselloidea; промежуточные формы между обоими надсе-
мействами не известны. Из морских семейств к надсемейству Aselloidea
несколько приближается только семейство Stenetriidae, населяющее моря
побережий Южной Африки, Австралии, Индо-Китая, Вест-Индии, Юж-
ной и Средней Америки (два вида найдены в Средиземном море). Это
семейство не включается Ганзеном (Hansen, 1905) в группу Paraselloidea
(по Ганзену, семейство Parasellida,e), а выделяется в отдельное семейство,
1 V. van Straelen. 1928. Contribution а 1’etude des Isopodes Meso- et Ceno-
zoiques. Mem. Acad. Roy. Belg., Cl. sei., IX, fasc. 5 : 1—66.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
2f
равнозначное Asellidae (=Aselloidea) и Parasellidae (=Paraselloidea).
По целому ряду признаков Stenetriidae занимают среднее положение между
Рис. 13. Распространение родов и подродов сем. Asellidae: 1 — ареал подрода Asellus s. str.; 2 — ареал подрода
Proasellus Dud.; 3 — местонахождения видов подрода Mesoasellus Birst.; -4 — местонахождения видов подрода Baica-
loasellus Stamm.; 5 — ареал подрода Conasellus Stamm, и рода Lirceus Raf.; 6 — ареал Asellus tomalensis',
7 — местонахождение рода Johanella Monod; 8 — местонахождение рода Synasellus Braga. Ориг.
обоими надсемействами. Так, например, протоподиты I пары плеоподов
<5 у Stenetriidae сросшиеся, как у Paraselloidea, но их величина незначи-
22
ВВЕДЕНИИ
тельная, как у Aselloidea. I. и II пары плеоподов 5 не соединены вместе,
как у Aselloidea; строение II плеопода о среднее между Paraselloidea
и Stenasellus. Плеоподы II пары 2 сросшиеся, без следов шва, как у Pa-
raselloidea, но operculum образован III парой плеоподов, как у Asel-
loidea. IV и V пары плеоподов также построены по некоторому среднему
типу. Однако возможная близость Asellidae и Stenetriidae пока не помо-
гает разрешить вопрос о происхождении первого семейства. Тем не менее
это обстоятельство представляет значительный интерес, и оно будет рас-
смотрено несколько ниже в другом аспекте.
Вообще говоря, вселение некоторых Isopoda в пресные воды проис-
ходило в очень отдаленные времена. Уже упомянутые Plireatoicidea в три-
асе жили в пресной воде, как это доказано Чилтоном, описавшим Phrea-
loicus wianamallensis из триасовых пресноводных отложений Сиднея
(Австралия).1 Пресноводные Sphaeromidae известны из верхней юры
{('yclosphaeroma woodwardi van Straelen).1 2 Вероятно не будет ошибкой
отнести происхождение Asellidae как пресноводной группы к мезозою,
когда формировались и другие группы пресноводных Malacostraca (Deca-
*poda, Amphipoda).
Географическое распространение современных Asellidae, несмотря на
недостаточную изученность, позволяет наметить некоторые общие зако-
номерности. Весь ареал семейства разбит на 5 отдельных областей, раз-
деленных значительными пространствами суши или моря: 1) Европа
и Северная Африка, 2) оз. Байкал, 3) восточное побережье Азии с остро-
вами, 4) западная часть Северной Америки и 5) восточная часть Север-
ной Америки. Кроме того, известны изолированные местонахождения
отдельных видов в Средней Азии, Западной Сибири, Забайкалье и запад-
ной экваториальной Африке. Для европейско-североафриканского ареала
характерно присутствие рода Stenasellus (12 видов) и подрода Proasellus
(22 вида); кроме того, вся Европа населена подродом Asellus s. str.
(2 вида), отсутствующим на значительном протяжении Сибири и появляю-
щимся снова на восточных побережьях Азии (1 вид). В оз. Байкал оби-
тают 4 вида эндемичного подрода Baicaloasellus и 1 вид подрода Meso-
asellus-, на восточном побережье Азии, помимо указанного Asellus s. str.,
из Японии описано 2 вида Мesoasellus. В западной части Северной Аме-
рики известны 1 вид подрода Мesoasellus и 2 вида Asellus, систематиче-
ское положение которых не выяснено (A. lomalensis, A. adenta); восточ-
ная часть Северной Америки населена 17 видами подрода Conasellus
и ’16 видами рода Lirceus (=Мancasellus) (рис. 13).
Итак, наиболее богата видами Asellidae Палеарктика, причем пале-
арктические водяные ослики не представляют собой единой зоогеографи-
ческой группировки. Здесь можно отличить 5 основных комплексов:
1) комплекс кругосредиземноморских форм, наиболее характерным пред-
ставителем которого является подрод Proasellus', сюда’ же примыкают ,
роды Stenasellus и Johanella\ 2) комплекс евразиатских форм, к кото-
рому принадлежит подрод Asellus s. str.; 3) комплекс байкальских энде-
миков — подрод Baicaloasellus', 4) комплекс восточно-азиатских — за-
падно-американских форм — подрод Мesoasellus', 5) тяготеющий к вос-
точно-американским формам род Synasellus.
На территории Европы происходит смешение1 кругосредиземномор-
ского и евразиатского комплексов. Первый, несомненно, древнее и при-
1 Chas. Chilton. 1918. A fossil Isopod belonging to the freshwater genus Phrea-
toicus. Journ. Proc. Roy. Soc. N. S. Wales, LI : 365—388.
2 V. van Straelen, цпт. раб. : 32.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
23
митивнее второго и является исконным обитателем кругосредиземно-
морских стран. В этом отношении значительны!) интерес представляет
нахождение представителей подрода Proasellus на о-вах Додеканез
[A. coxalis по Арканджели (Arcangeli, 1929)], Сицилии [A. banyulensis
wol/i, по Дудичу (Dudich, 1925а)] и, наконец, Мадере [A. ilalicus perar-
matus, по Реми (Remy, 1936)]. Раковица (Racovitza, 1919) считал, что
родиной подрода Proasellus 1 является Малая Азия, где обитает наиболее
примитивный вид подрода — A. coxalis’, отсюда произошло заселение
северной Африки (A. coxalis peyerimhoffi), а затем, через Гибралтар,
Испании (A. banyulensis и др.) и далее южной Франции и средней Европы.
После работ Раковицы целый ряд видов Proasellus был открыт, глав-
ным образом, на Апеннинском и Балканском п-овах, что заставило Реми
(Remy, 1932) пересмотреть гипотезу Раковицы. Реми считает возмож-
ным два объяснения распространения Proasellus-. 1) Наличие Эгеиды,
существовавшей с начала третичного до начала четвертичного времени
и соединявшей Малую Азию с Балканским п-овом, причем миграция
через Эгеиду должна была осуществиться до олигоцена, так как в бассейне
оз. Охрида, совершенно изолированном с олигоцена, живет несколько
видов Proasellus. 2) В соответствии с взглядами Аргана, колос-
сальный иадвиг северного берега Гондваны на Евразию, при котором
Европа была смята в альпийскую складчатость, после чего Евразия,
покрытая в южной части Гондваной, оторвалась от Гондваны, и на месте
разрыва образовалось Средиземное море. Оказавшийся таким образом
в Европе кусок Гондваны и явился центром распространения европей-
ских Proasellus.
Следует отметить, что проблема происхождения подрода Proasellus
по существу является проблемой происхождения всей древней круго-
средиземноморской фауны и флоры в'целом. Сходное распространение
свойственно длинному ряду групп наземных моллюсков, насекомых,
.паукообразных, пресноводных Malacostraca и т. д. Можно ли припи-
сывать всем им гондванское происхождение, как это принимают для
Proasellus Раковица и Реми?
Если обратиться к числу видов подрода Proasellus, то окажется, что
в Африке известно 3 вида и 3 подвида, в Сирии и Палестине — 1 вид,
а в Европе с островами — 18 видов и 19 подвидов, т. е., что «массовый
центр ареала» (Irmscher) Proasellus лежит в Европе. Нахождение наи-
более примитивного вида — A. coxalis — в Сирии и Палестине не может
считаться надежным доказательством приуроченности центра происхо-
ждения подрода к этим странам, так как нет основания допускать одина-
ковый темп эволюции в разных частях ареала. Присутствие подвида
A. coxalis в Сицилии — A. coxalis cyanophilus — противоречит гипотезе
миграции примитивных форм через Гибралтар в Европу, предложенной
Раковицей. Широкое обобщение Реми также не представляется вполне
приемлемым. Положения Аргана, на которых основывается Реми, далеко
не доказаны, а в последнее время они подверглись серьезной критике.
Не входя в подробности этого интересного вопроса, достаточно привести
одно из возражений, собранных Мушкетовым: «Мощность пошедших на
складкообразование свит не превосходила 2000—3000 м, тогда как тол-
щина давящей глыбы (Африка) принимается до 30 000 м. Получается
явное несоответствие между, так сказать, грубым инструментом й тонкой,
1 По терминологии Раковицы, «1а lignee Asellus meridianuss.
24
ВВЕДЕНИЕ
произведенной якобы им, работой».1 Однако, если даже принять построе-
ния Аргана, то и в этом случае не получится соответствия между массой
Гондваны, якобы лежащей на Европе, и ареалом Proasellus. Нахождения
A. meridianus и A. caoaticus в Англии и Германии, A. collingei в Англии,
а также A. banyulensis и A. spelaeus в южной Франции отстоят весьма
далеко от участков Гондваны, как она изображена Арганом,1 2 и пересе-
ление туда Proasellus представляется мало вероятным. В этом отношении
интересно также нахождение единственного в СССР представителя Pro-
asellus — A. infirmus — около г. Сухуми. Кавказ, по Аргану, отделен
от Аравийского мыса иранскими и анатолийскими дугами,3 т. е. и к слу-
чаю A. infirmus объяснение Реми не подходит. Таким образом, резюмируя
сказанное, можно утверждать, что нет никаких оснований и для призна-
ния гондванского происхождения Proasellus и для привязывания его
к участкам Гондваны, лежащим на Евразии.
Советские биогеографы, сталкивавшиеся с фактами средиземномор-
ских разрывов ареалов, довольно единодушно объясняют это явление
наличием прямой континентальной связи между Закавказьем, Крымом,
Балканским полуостровом и Малой Азией, составлявших до плиоцена
и даже до начала четвертичного периода одно целое.4 В то же время вос-
точная часть Средиземного моря еще была сушей. Эти данные могут быть-
приняты и для объяснения современного распространения Proasellus.
Приуроченность ареала Proasellus к южной части Европы несомненно
стоит в связи с великим оледенением. Территория, занятая ледником,
лишена Proasellus (рис. 14). При этом у границ максимального оледене-
ния, как правило, встречаются подземные или глубоководные слепые
формы. Уже Штаммер (Stammer, 1932) для Германии показал, что
в районах распространения подземных форм Proasellus отсутствуют
наземные формы и объяснил это влиянием ледника, высказав предполо-
жение о более широком распространении Proasellus на север в доледни-
ковые времена. То же самое установил впоследствии для Бельгии
Арканджели (Arcangeli, 1935). Таким образом, Proasellus может считаться
чрезвычайно древним обитателем Европы и стран, лежащих вокруг Сре-
диземного моря, занимавшим в третичное время значительно более широ-
кий ареал, чем сейчас. На древность подрода указывает, между прочим,
нахождение некоторых его представителей на островах, в частности
на о-ве Мадере (Remy, 1936), отстоящем, как известно, на 545 км от Аф-
рики и на 800 км от Европы. Мадера вместе с о-вами Азорскими и Зеле-
ного Мыса отделилась от Старого Света до конца миоцена, и уже в это
время A. italicus существовал как вид, и на Мадере сформировался
только его подвид — A. italicus perarmatus.
Род Stenasellus также может считаться древним третичным реликтом
(Stammer, 1936). Большая часть составляющих его видов (10 из 13) оби-
тает в подземных водах южной Европы, занимая ограниченные, оторван-
ные друг от друга ареалы. Только три вида зарегистрированы вне этой
1 Д. И. Мушкетов. 1935. Региональная геотектоника.
2 Эмиль А р г а н. 1935. Тектоника Азии. ОНТИ, рис. 27.
3 А р г а н, цит. раб. : 29.
4 См. наир.: Е. В. Вульф. 1944. Историческая география растений. История
флор земного шара; И. И. Пузанов. 1927. Материалы к познанию наземных
моллюсков Крыма, ч. III. Состав, распределение и генезис крымской малако-фауны.
Бюлл. Моск. общ. исп. прир., отд, биол., XXXVI, 3—4; Я. Я. Ц е е б. 1947.
Зоогеографический очерк и история крымской гидрофауны. Учен. зап. Орловск.|пед.
инет., сер. ест. и хим., 2.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
25-
основной области распространения рассматриваемого рода — St. chap--
puisi и St. ajricanas в западной экваториальной Африке и 51. asiaticus —
в восточной Туркмении (рис. 15).
Рис. 14. Распространение ’подрода Proasellus'} Dud.; 1—граница
максимального оледенения; 2— Asellus (Proasellus} caoaticus Schiodte;
3 — другие слепые виды подрода Proasellus’, 4? — остальные виды
подрода Proasellus Dud. Ориг.
Долготный разрыв ареала (южная Европа — тропики) помимо Sten-
asellus свойствен многим подземным группам, как водным, так и сухо-
путным. Он показывает, что часть пещерной фауны Кругосредиземноморья..
имеет тропическое происхожде-
ние. Группы с таким распростра-
нением представляют собой остат-
ки тропической фауны, населяв-
шей Европу до середины третич-
ного периода. Их сохранению под
землей способствовало сходство
условий существования с одной
стороны под землей, с другой —
в тропиках, прежде всего в отно-
шении постоянства температуры,
не подвергающейся в обоих слу-
чаях суточным и сезонным коле-
баниям. По мере ухудшения кли-
Рис. 15. Распространейие рода Stenasellus
Dollf. Ориг.
мата температура подземных вод
Европы постепенно понижалась, но не подвергалась резким колебаниям,
которые могли оказаться гибельными для обитателей поверхности земли.1
Нахождение одного из видов Stenasellus в Средней Азии в отрогах.
Гиссарского хребта с несомненностью подтверждает указания на нали-
1 Я. А. Б и ррп т е й н. 1950s Пещерная фауна Западного Закавказья. Зоол.
жури., XXIX, 4.
26
ВВЕДЕНИЕ
чие в составе среднеазиатской гидрофауны средиземноморских элемен-
тов. В своем анализе фауны моллюсков бассейна Аму-дарьи Жадин1
устанавливает 6 групп видов, различающихся по происхождению и ха-
рактеру распространения, в том числе группу видов средиземноморского
и группу видов тропического происхождения. St. asiaticus можно отно-
сить с одинаковым основанием к любой из этих групп. В термальном
источнике Ходжа-койнар вместе с ним обитает моллюск Melanoid.es
tuber culatus, принадлежащий, согласно Жадину, к тропической группе
видов. В то же время подавляющее большинство видов Stenasellus (в том
числе виды, морфологически приближающиеся к St. asiaticus) населяет
подземные воды бассейна Средиземного моря. Можно предполагать, что
границы между тропическими и средиземноморскими элементами в ряде
случаев весьма неопределенны, так как многие виды со средиземномор-
-ским распространением имеют тропическое происхождение.
St. asiaticus представляет собой древнего обитателя Средней Азии.
По крайней мере по отношению к живущему вместе с этим осликом мол-
. люску Мelanoides tuberculatus палеонтологически доказано присутствие
его в Средней Азии еще в верхнем миоцене (Жадин, цит. раб.). Это под-
тверждает отмеченную выше глубокую древность рода Stenasellus, пол-
ностью гармонирующую с его примитивными морфологическими особен-
ностями.
Поэтому трудно согласиться с Арканджели (1939), который возражает
против признания Stenasellus третичным пресноводным реликтом. Этот
автор указывает на существование в середине третичного времени в Ита-
лии моря на том месте, где сейчас найден St. racovitzai, и предполагает,
что виды рода Stenasellus возникли в море, оказались «преадаптирован-
ными» к подземному образу жизни и затем вселились по подземным
щелям в систему грунтовых вод. К какому времени следует относить
процесс вселения Stenasellus из моря в пресные воды, Арканджели не
разъясняет. Шаппюи (см. Racovitza, 1950) уже указал, что в Италии не-
сомненно оставались участки суши, послужившие убежищем целому
ряду пресноводных и сухопутных третичных форм. К тому же многие
современные нахождения видов рода Stenasellus далеко отстоят от мор-
ских берегов; наиболее ярким примером такого случая можно считать
нахождение St. asiaticus в Туркмении. К этому следует добавить, что
в составе морской фауны мы совершенно не знаем форм, близких к Ste-
nasellus. Шаппюи (цит. соч.), считая Stenasellus пресноводным третичным
реликтом, склонен приписывать этому роду гондванское происхождение,
поскольку в Западной Африке найдено два далеких друг от друга вида
Stenasellus (chappuisi и africanus), что доказывает давнее обитание Sten-
asellus в Тондване. Против мнения Шаппюи говорит существование
St. asiaticus в Средней Азии. Гораздо естественней признать в Stenasellus
остаток древней тепловодной тропической фауны, захватывавшей в тре-
тичное время огромную территорию — от экваториальной Африки до
Средней Азии.
Подрод Asellus s. str. резко отличается по Своей зоогеографической
природе от только что рассмотренных. Один из входящих в него видов —
A. aquaticus — занимает всю Европу, кроме Пиренейского п-ова, Англию
и западную Гренландию; в Африке этот вид отсутствует. Восточная гра-
ница распространения A. aquaticus точно не установлена. По имеющимся
1 В. И. Жадин. 1948. Вопросы генезиса фауны и биоценозов континенталь-
.ных вод Советского Союза. Сб. «Памяти акад. С. А. Зернова», Изд. Акад. Наук СССР.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
27
сведениям и материалам, на Урале его нет. Что касается Средней Азии,
то Вальтер 1 считал отсутствие Л. aquaticus характерной особенностью
фауны ракообразных этой области. Более новые исследователи (Бир-
штейн, 1945; Бирштейн и Старостин, 1949) обнаружили в нескольких
озерах западной Туркмении особый подвид A. aquaticus, ареал которого
вероятно примыкает к нахождениям типичной формы в Иране. Кроме
того, типичная форма найдена в колодцах Мангистау. Однако восточнее
A. aquaticus не идет. Другой вид подрода, A. monlicola, живет в Закав-
казье и Иране.
На огромном протяжении Сибири и Центральной Азии представители
Asellus s. str. почти не встречаются. Исключение составляют бассейн
Оби и Иртыша, озеро Баунт в системе Ципо-Ципикаиских озер (бассейн
Витима) и дельта Лены. В каждом из этих водоемов обитает особый вид
или подвид рассматриваемого подрода (Бирштейн, 1947).
Восточная окраина Азии от Китая до побережья Охотского моря
включительно и прилегающие к ней острова (Япония, Сахалин, Риу-
Киу) заселены A. hilgendorfi. Этот вид проникает на запад до среднего
течения Амура и представлен подвидом marlynovi в дельтеЛены. В Амери-
ке подрод отсутствует (Johansen, 1920, 1926), если не считаться с сомни-
тельным указанием на нахождение A. aquaticus в западной Гренландии.
Таким образом, ареал Asellus s. str. представляется разорванным.
Между* двумя областями сплошного распространения подрода (Европа
и восточная Азия) лежит огромная Сибирь, на протяжении которой
встречаются лишь изолированные колонии Asellus.
По вопросу о происхождении Asellus s. str. высказывались разноре-
чивые мнения. Раковица (Racovitza, 1919) рассматривал A. aquaticus
в Европе как пришельца с севера, Шаппюи (1927), присоединяясь к этому
мнению, полагал, что A. aquaticus мигрировал в Европу из Сибири и от-
теснил на юг A. meridianus. Арканджели (1935) возражал против аркти-
ческого происхождения A. aquaticus, считая, что этот вид населял и
в доледниковое время среднюю и восточную Европу, а ледник заставил
его передвинуться к югу.
Все эти гипотезы нельзя считать серьезно обоснованными, поскольку
их авторам оставался неизвестным ареал Asellus s. str. в Азии. И в более
поздних работах нередко можно найти неверные указания на сплошной
палеарктический ареал A. aquaticus (Державин, 1930) или на присутствие
его на Урале, в Сибири, Китае и Японии (Remy, 1941).
Анализ имеющихся сейчас более полных и точных данных о видовом
составе и характере ареала Asellus s. str. позволяет ближе подойти к во-
просу об его происхождении.
Разрыв ареала Asellus s. str., приходящийся на Сибирь, совпадает
с разрывом ареалов многих других родов и видов растений и животных,
указанных Бергом.1 2 Согласно этому автору, объяснение происхождения
разорванных таким образом ранее сплошных ареалов следует искать
в условиях ледникового времени. '
По данным Герасимова и Маркова,3 только Западная Сибирь испы-
тала покровное оледенение. Среднесибирское плато и низины Восточной
1 A. Walter. 1889. Transkaspische Binnencrustaceen. II. Malacostraca. Zool.
Jahrb., Syst., IV.
2 Л. С. Верг. 1909. Рыбы бассейна Амура. Зап. Акад. Наук, VIII сер.,
XXII, 9.
3 И. П. Герасимов и К. К. Марков. 1939. Ледниковый период
на территории СССР. Тр. инет, географии АН СССР, XXXIII.
28
ВВЕДЕНИЕ
Сибири не имели сплошного покрова льда. Здесь шло образование нале-
дей, превращение промерзших озер в линзы льда, засыхание и погребе-
ние снежных скоплений, глубокое промерзание грунта. Сакс 1 допускает
значительно более широкое распространение ледника в Западной Сибирп
и наличие огромного по площади оледенения Среднесибирского плоско-
горья и Восточной Сибири.
Так или иначе, но суровые условия ледникового периода оказались,
очевидно, гибельными для водяных осликов, и в Сибири они почти
повсеместно исчезли. Тем интереснее обратить внимание на сохранив-
шиеся в этой области изолированные колонии представителей подрода
Asellus s. str.
Обь и Иртыш, населенные A. latifrons, в ледниковое время текли
иначе, чем сейчас, огибая наступавшие ледники и временами образуя
озеровидные водоемы. Это были независимые друг от друга реки, соеди-
нившиеся только в послеледниковое время.1 2 Поскольку их воды не были
скованы вечным льдом, можно себе представить переживание ледникового
периода водяными осликами именно в этих реках. В бассейне Лены
A. hilgendorfi martynovi встречается только в дельте; на всем остальном
протяжении Лены обнаружить водяных осликов не удалось.3 По данным
Сакса (цит. раб.), ледники с Верхоянского хребта достигали Лены, но ее
нижнее течение с дельтой всегда оставалось свободным от льда. .Озеро
Баунт, в котором обитает третий сибирский представитель Asellus s. str.—
A. epimeralis, населено и другими элементами доледниковой фауны,
в частности байкальскими, и принадлежит к системе древнего байкаль-
ского многоозерья. Замечательно, что не только по присутствию водяных
осликов, но и по наличию ряпушки Coregonus sardinella baunti Much,
намечается связь этого озера с низовьями Лены. Ряпушка живет лишь
в нижнем течении Лены, не поднимаясь выше Жиганска. По мнению Ко-
жова,4 древнее байкальское многоозерье в третичное время могло иметь
сток на северо-восток, в бассейн Лены.
Представление о сибирских водяных осликах как об остатках широко
распространенного в доледниковое время в Сибири Asellus s. str., имев-
шего тогда сплошной ареал, подтверждается и морфологическими дан-
ными. Сибирские A. epimeralis и A. latifrons обладают некоторыми общими
и притом примитивными признаками, в частности чрезвычайно крупными
по отношению к протоподиту ветвями второго плеопода <5, удлиненным
II плеоподом и прямым, лишенным выемки наружным краем экзоподита
I плеопода <5. Последний признак свойствен также дальневосточному
A. hilgendorfi и кавказско-иранскому A. monticola. У. A. aquaticus, как
было указано выше, выемка экзоподита I плеопода $ появляется только
на определенном этапе постэмбрионального развития. Соотношение вет-
вей и прртоподита II плеопода о, наблюдающееся у A. latifrons и A. epi-
meralis, также характерно для ранних стадий развития A. aquaticus.
'Таким образом, последний вид может считаться наиболее далеко зашед-
шим по пути эволюции передних плеоподов <5, т. е. наиболее молодым.
Аналогичное отклонение от примитивной формы II плеопода <5 обнару-
1 В. Н. Сакс. 1947. Четвертичное оледенение севера Сибири. Природа, № 4.
2 Н. А. Наганский. 1949. Четвертичная история долины Оби на площади
оледенения. Вопросы географии, 12.
3 Устное сообщение П. Л. Пирожникова.
4 М. М. Кожов. 1949. К истории озерных систем Забайкалья и Прибай-
калья и их фауны. Тр. Всесоюзн. гидробиол. общ., I4
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
29
живается у A. hilgendorfi и A. monticola, сохраняющих, однако, прими-
тивное строение I плеопода 6.
Все изложенное позволяет считать Сибирь центром возникновения
и распространения Asellus s. str., a A. aquaticus—северо-восточным
пришельцем в Европу, как это предполагали Раковица (Racovitza,
1919) и Шаппюи (Chappuis, 1927). Сравнительно недавнее проникно-
вение A. aquaticus в Европу согласуется с отсутствием его на Пиреней-
ском полуострове, а также в Северной Африке.
В Европе Asellus aquaticus встречается обычно и в большом коли-
честве в районах, занятых в четвертичное время ледником (север Евро-
пейской части СССР, Скандинавия). По всей вероятности, заселение
этих районов произошло после отступания ледника. Весьма интересным
в этом отношении является факт почти полного отсутствия A. aquaticus
в Арктике, особенно на арктических островах, вообще практически
лишенных настоящих пресноводных Malacostraca (Бирштейн, 1933).
Единственное исключение представляет собой Гренландия, на западном
берегу которой старыми исследователями (Fabricius, 1780; Kroyer, 1838)
был найден A. aquaticus. Присутствие здесь этого вида совершенно
необъяснимо, так как вероятность завоза его людьми очень незначительна.
Возможно, что авторами была допущена ошибка, подобная той, которая
расшифрована Иогансеном в отношении указания A. aquaticus на Лабра-
доре.1 Боваллиус (Bovallius, 1886) относит указание Кроера к Jaera.
Нахождение представителей Asellus s. str. на других островах легко
поддается объяснению. Англия и Ирландия были соединены с конти-
нентом еще в среднечетвертичное время,1 2 и ареал A. aquaticus был, оче-
видно, тогда сплошным. Точно так же и Япония и Сахалин, населенные
A. hilgendorfi, вступали в четвертичное время в соединение с мате-
риком.3
Для понимания происхождения подрода Asellus s. str. в целом важно
оценить близость его к подроду Mesoasellus. Чрезвычайное сходство
в строении эндоподита II плеопода о заставляет сближать оба подрода.
Этим признаком они резко отличаются от остальных подродов Asellus.
Другие признаки Mesoasellus напрминают подроды Proasellus, Baica-
Loasellus и Conasellus. Мesoasellus, невидимому, реликт весьма отдален-
ных времен. Три (четыре?) несомненно относящиеся к нему вида обна-
ружены по одному на больших глубинах Байкала, в пещерах Японии
и в пещерах Калифорнии. Такое распространение свойственно многим
архаическим формам, из которых для примера можно указать пиявку
Torix с близким американским родом Microbdella, Epischura, найденную
недавно на Камчатке, а также некоторых Harpacticoida.4 Все это, несо-
мненно, оттесненные реликты очень древней фауны.
Близость подродов Mesoasellus и Asellus s. str. позволяет допустить
происхождение второго от первого или, вернее, от сходных с ним форм.
Вероятно, подрод Asellus s. sjtr. зародился в Восточной Сибири, явив-
шейся очагом возникновения целого ряда пресноводных групп, как,
1 F.'Johansen. 1926. Asellus aquaticus^not found in Labrador. Canadian Field-
Nat., X (цит. no Van Name, 1936).
2 А. А. Б о p и с я к. 1934. Курс исторической геологии: 344.
3 A. II. К p и ш т о ф о в и ч. 1932. Геологический обзор стран Дальнего
Востока: 292.
4 Е. В. Б оруцкий. 1948. Материалы по фауне Copepoda Harpacticoida
Вайкала. Род Bryocamptus Chappuis. ДАН СССР, LIX, 9 : 1669—1672.
30
ВВЕДЕНИЕ
например, многих ручейников, представляющих собой для Европы также
северо-восточных пришельцев.1
Помимо одного вида Mesoasellus, в Байкале живут 4 вида эндемич-
ного подрода Baicaloasellus. Это — в высокой степени аберрантные
формы, резко отличающиеся от остальных Asellus своеобразным строе-
нием эндоподита II плеопода <5, конец которого вытянут в хитинизировэн-
ное клювовидное образование. Однако хитинизация конца эндоподита II
плеопода <5 наблюдается и у Mesoasellus, правда, в гораздо меныпей
степени. Весьма вероятно, что Baicaloasellus так же, как и Asellus s. str.,
является отдаленным потомком близких к Mesoasellus форм, причем
формирование первого подрода происходило в специфических условиях
Байкала. Такое предположение вполне соответствует взглядам Берга
на происхождение байкальской фауны,1 2 причем Baicaloasellus следует
отнести к первой группе Берга, т. е. к формам, «которые развились в са-
мом Байкале в течение его долгой геологической жизни», а Мesoasellus —
ко второй группе «остатков верхне-третичной (? Я. Б.) пресноводной
фауны Северной Азии». Сближение А. 'angarensis и ' A. baicalensis
с американскими видами, которое допускал Алпатов (1922), лишено
оснований.
Каковы яге в действительности отношения палеарктических Asel-
lidae к неарктическим? Ван-Н эм -(Van Name, 1936) полагает, что амери-
канские Asellidae произошли от форм’ Старого Света, мигрировавших
через перешеек, существовавший на месте Берингова пролива. С этим
можно согласиться только отчасти. Как указывалось выше, в Северной
Америке имеется два разъединенных Скалистыми горами ареала Asel-
lidae — западный и восточный. В западном ареале констатировано всего
два вида — A. tomalensis и A. cali/ornicus. Последний вид несомненно
относится к подроду Mesoasellus, систематическое положение первого
вида неясно из-за совершенно неудовлетворительного описания, но он
определенно не принадлежит к восточноамериканскому подроду Соп-
asellus. Присутствие A. californicus позволяет сближать фауну западно-
американских Asellidae с восточноазиатской и, поскольку большая часть
видов Мesoasellus известна из восточной Азии, можно допустить его азиат-
ское происхождение.
Фауна Asellidae восточной части Северной Америки значительно
богаче, и генетические ее отношения гораздо менее ясны. Наибольшее
количество восточноамериканских видов относится к эндемичным —
роду Lirceus (16 видов) и подроду Conasellus (17 видов). Кроме того,
известно еще 5 восточноамериканских видов (A. nickajackensis, A. adenta,
A. richardsonae, A. smithi и A. altenuatus), систематическое положение
которых остается невыясненным. До недавнего времени род Lirceus со
сросшимися с сегментами тела эпимерами и отсутствием мандибулярного
щупика стоял особняком от других Asellidae. Однако открытие Моно
(Monod, 1924) в северной Африке своеобразной Johanella изменило такое
положение. Моно указывает на чрезвычайную близость этих родов и
характеризует свою Johanella как Lirceus с мандибулярным щупи-
ком. Еще более важным с зоогеографической точки зрения фактом прихо-
дится считать нахождение в подземных водах Португалии близкого
к Lirceus рода Synasellus (Braga, 1944), что заставляет внимательнее
1 А. В. Мартынов. 1929. Экологические предпосылки для зоогеографии
пресноводных животных. Русск. зоол. журн., IX, 3.
2 Л. С. Берг. 1922. Фауна Байкала и ее происхождение. Климат и жизнь..
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ 31
отнестись к . сходству Conasellus с кругосредиземноморским Proasellus.
Сходство это довольно отдаленное, но еще Раковица (Racovitza, 1920)
отметил, что исследованный им Asellus (Conasellus) communis стоит
дальше от Asellus s. str., чем от Proasellus (по современной терминологии)»
Итак, одна из возможностей объяснения происхождения восточноамери-
канских Asellidae — это сближение их с кругосредиземноморской фау-
ной: Johanella и Synasellus — с Lirceus, a Proasellus — с Conasellus.
Попытки такого сближения для других групп пресноводных Malacos-
traca уже делались: именно так трактуют Бирштейн и Виноградов про-
исхождение восточноамериканского Cambarus.1 Если обратиться к дру-
гим пресноводным Isopoda, то окажется, что сходное распространение
свойственно преимущественно пещерным Cirolanidae-. 3 рода (Typhlo-
cirolana с 4 видами, Sphaeramides с 2 видами и Faucheria с 1 видом) на-
селяют кругосредиземноморские страны, а чрезвычайно близкие к ним
3 рода (Cirolana с 2 видами, Cirolanides и Conilera, каждый с 1 видом)
известны из Мексики, Кубы и Техаса. Раковица указывает, что среди
всех ныне живущих Cirolanidae ближе всего к Typhlocirolana стоят
восточноамериканские роды Cirolana и Conilera.1 2 Близость западноевро-
пейской и восточноамериканской древней пресноводной фауны' (Рако-
вица принимает существование Cirolanidae начиная с мелового или,
возможно, юрского периода) может быть легко объяснена связью обоих
материков посредством отрогов погрузившегося впоследствии Атланти-
ческого хребта.3 Один из них через о-ва Мадеру, Канарские и Азорские
тянулся прямо к Пиренейскому полуострову.
Таким образом, довольно вероятно двустороннее заселение Северной
Америки водяными осликами: с запада через сушу, заменявшую совре-
менный Берингов пролив, и с востока — из Западной Европы, через
отроги Атлантического хребта. При таком допущении становится понят-
ным и нахождение Asellus italicus perarmalus на острове Мадера. Реми
(Remy, 1936), описавший указанный подвид, держится именно такого
мнения. Не исключается однако и менее вероятная возможность проис-
хождения восточноамериканских Asellidae от западноамериканских;
окончательное решение вопроса, быть может, даст подробное сравни-
тельно-морфологическое изучение водяных осликов Нового Света, иссле-
дованных, к сожалению, очень поверхностно.
Обрисованная такими общими чертами картина географического рас-
пространения Asellidae может быть названа закономерной в том отно-
шении, что аналогичный характер ареала наблюдается и у других групп
древних пресноводных Malacostraca. Если взять, например, карту гео-
графического распространения пресноводных Amphipoda группы Сгап-
gonyx, составленную Шелленбергом,4 то окажется, что в северном полу-
шарии имеются те же 5 разорванных ареалов — Европа, Байкал,
Дальний Восток и западная и восточная части Северной Америки, кото-
рые были намечены выше для Asellidae. Можно проследить аналогию
и для отдельных родов и подродов. Ареал Synurella совпадает, .в общем,
1 Я. А. Бирштейн it Л. Г. Винограде в. 1934. Пресноводные Deca-
poda СССР и их географическое распространение. Зоолог, жури., XIII, 1.
2 Е. G. Racovitza. 1912. Cirolanides (premiere serie). Biospeologica, XXVII..
Arch. zool. exp. gen., V ser., X.
3 Л. С. Бер г. 1949. О предполагаемом раздвижении материков. Очерки
по физической географии : 9—36.
4 A. S chellenb erg. 1936. Die Amphipodengattungon urn Crangonyx, ihro*
Verbreitung und ihre Arten. .Vlitt. Zool. Mus. Berlin, 22, 1.
:32
ВВЕДЕНИЕ
с ареалом Asellus s. str. Совершенно тан же, нал группа Cragonyx, рас-
пространены, по Шелленбергу, бокоплавы группы Gammarus. Прибли-
зительно тот же характер распространения свойствен и Astacidae,1
с той только разницей, что речные раки не встречаются в Байкале. Нако-
нец, все эти группы древних пресноводных Malacostraca сближает между
собой отсутствие их в тропиках.1 2 В антибореальной области семейство
Astacidae замещается близкими семействами Parastacidae и Austroasla-
cidae, а группа Crangonyx представлена там несколькими, большей частью
эндемичными викарирующими родами. Формы, родственные Asellidae,
в пресных водах южных материков не известны, но преимущественно
в антибореальных морях живет сравнительно близкое семейство
Stenetriidae.
Последний список видов единственного рода этого семейства пока-
зывает, что из 25 видов рода Stenetrium 23 обитает в морях южного полу-
шария и только 2 — в Средиземном море.3
Трудно предположить, чтобы указанные совпадения оказались слу-
чайными; по всей вероятности, распространение всех древних пресно-
водных Malacostraca определялось одними, и теми же историческими
причинами. Некоторые из них, быть может, удалось подметить, но вся
проблема в целом еще очень далека от своего разрешения.
В заключение этого краткого очерка следует сказать несколько слов
и о пресноводных Paraselloidea — о родах Jaera, Microparasellus и Mic-
rocharon. Последние два рода объединяются в семейство Microparasel-
lidae, которое Караман (Кагашап, 1933) сближает с чисто морскими
семействами Мacrostylidae и Desmosomatidae. Microparasellidae до сих
пор найдены только в грунтовых водах Югославии и Румынии, но
весьма вероятно их нахождение и в других районах, в частности на Кав-
казе. Близость Microparasellidae к морским формам стоит в соответствии
с родственными отношениями других высших ракообразных, населяющих
грунтовые воды. В последнее время установлено, что значительный про-
цент подземных Malacostraca имеет близкие родственные формы не
в фауне пресных вод, а среди чисто морских видов. В первую очередь
в качестве таких форм следует назвать род Niphargus, приближаю-
щийся, как показал Шелленберг,4 к морским родам Eriopisa и Eriopi-
sella. Другой представитель подземных Amphipoda — Ingolfiella ache-
rontis из грунтовых вод Югославии — принадлежит к подотряду Ingol-
fiellidea, заключающему, кроме названного, еще 2 вида: I. abyssi, най-
денную на глубине 3000 м у входа в Дэвисов пролив, и I. littoralis, най-
денную среди кораллов в Сиамском заливе на глубине 2 м.5 Из Isopoda
замечательное семейство Microcerberidae, представленное одним видом
также в грунтовых водах Югославии, приближается по ряду признаков
к чисто морскому семейству Antharidae. Такие же связи прослежены для
некоторых пещерных Polychaeta и Archiannelida.6
1 Я. А. Бирштейн и Л. Г. Виноградов, цит. раб.
2 Единственное исключение— уже упоминавшиеся Stenasellus chappuisi и
St. africanus из экваториальной Африки.
3 A. N ordenst am. 1948. Marine Isopoda from Professor Dr. Sixten Bock’s
Pacific Expedition 1917—1918. Arkiv for Zool., 37A, 7.
4 A. Schellenberg. 1933. Niphargus-Probleme. Mitt. Zool. Mus. Ber-
lin, 19.
5L. Hertzog. 1935. Amphipoden aus dem Grundwasser von Skoplje. Zool.
.Anz., Ill, 14
6 Я. А. Бирштейн. 1940. Жизнь в пещерах. Усп. совр. биол., XIII, 3.
ГЕОГРАФИЧЕСКОЕ Р А СПРОС ГРАНЕНИЕ
33
Шелленберг 1 и Караман (1935) полагают, что известная часть фауны
грунтовых вод вселилась туда непосредственно из моря, минуя назем-
ные пресные воды. Такой переход был особенно легким для форм, кото-
рые и в море ведут «подземный» образ жизни, позволяющий им проникать
далеко в глубь грунта. Этим путем они и могли попасть из системы соле-
ных морских грунтовых вод в систему пресных. Действительно, весь-
облик Microparasellidae, Mier о cerberidae и Ingolfiellidae, имеющих вытя-
нутое червеобразное тело, при очень незначительной величине животных,
чрезвычайно характерен для представителей интерстициальной фауны,
и изложенная гипотеза по отношению к Microparasellidae может быть
принята.
Морская природа последнего рассматривающегося здесь рода —
Jeara — не вызывает уже никаких сомнений. С другой стороны, также
несомненна его тенденция к жизни в пресной воде. В настоящее время
известно 12 видов и 1 подвид Jaera, из которых 2 (7. italica и J. schel-
lenbergi) являются исключительно пресноводными, 2 живут как при
нормальной океанической солености, так и в совершенно пресной воде
(7. albifrons и J. nordmanni), 1 — преимущественно морской, перенося-
щий опреснение до 8%о (7. hopeana), 1 — солоноватоводный, не заходя-
щий в пресную воду (7. sarsi caspicd) и 1 — пресноводный, выносящий
некоторое осолонение (7. sarsi), остальные — морские. Этот род населяет
северную Атлантику, Средиземное море и Понто-Каспийский бассейн.
Вне Европы достоверно известен только вдоль североатлантического
побережья Америки. Из 12 видов 11 приурочено к бассейну Средизем-
ного моря, 1 вид (с подвидом) эндемичен Для Понто-Каспия. Такое рас-
пространение заставляет предполагать, что род Jaera сформировался
еще в третичное время в бассейне Средиземного моря и Понто-Каспийский
бассейн получил предков 7. sarsi вместе с предками других, близких
к средиземноморским ракообразных (Mysidacea, Cumacea) около сар-
матского времени, как это считал еще Совинский,1 2 принимавший, правда,
7. sarsi за 7. nordmanni. Мартынов3 оспаривал мнение Совинского, ука-
зывая на то, что «очень странно, что, несмотря на столь долгий промежу-
ток времени, она (Jaera) не отклонилась сколько-нибудь заметно от
морских (атлантических и средиземноморских) представителей этого
вида». По мнению этого автора, 7. nordmanni попала в Каспий уже в чет-
вертичное время вместе с Cardium edule. Возражение Мартынова теперь
отпадает, так как каспийская Jaera в действительности отличается от
морских представителей этого рода настолько сильно, что с полным пра-
вом выделяется в самостоятельный вид — 7. sarsi. Характер ее ареала
(Каспий с притоками, реки, предустьевые пространства и реликтовые
озера Азовско-Черноморского бассейна) с несомненностью указывает
на ее принадлежность к понто-каспийским эндемикам и заставляет вы-
черкнуть эту форму из списка четвертичных средиземноморских все-
ленцев в Каспий.
Совершенно особую проблему представляет собой вопрос о способе-
и времени вселения 7. sarsi в реки Понто-Каспийского бассейна. По
отношению к другим высшим ракообразным морского происхождения
1 A. Schellenberg. 1935. Der Niphargus des Thiiringer Waldes und die
Glacialreliktenfrage. Arch. f. Hydrobiol., XXIX. . . .
2 В. Совинский. 1904. Введение в изучение фауны Понто-Каспййско-
Араиьского морского бассейна. . . Зап. Киевск. общ. ест., XVIII.
3 А. В. Мартынов. 1924. К познанию реликтовых ракообразных бассейна
нижнего Дона, их экологии и распространения. Ежегодн. Зоол. муз. Акад Наук, XXV.
3 Фауна СССР, том VII, в. 5
34
ВВЕДЕНИЕ
этот вопрос разрешался по разному. Державин,1 Мартынов (цит. соч.)
и Бенинг 1 2 считали морских ракообразных в реках Понто-Каспийского
бассейна реликтами или псевдореликтами третичных трансгрессий, Саре, 3
Беклемишев 4 и Бирштейн5 рассматривали эти формы как активных
иммигрантов из моря в четвертичное время. За последнее время резкие
различия между этими точками зрения в значительной степени стерлись.
В своей недавней работе Державин 6 относит заселение морскими элемен-
тами каспийских рек к четвертичному времени. Вопрос о том, происхо-
дила, ли переработка морской фауны в пресноводную в самом море (Бир-
штейн) или только в его заливах (Державин), не имеет принципиального
значения и разбирать его здесь неуместно. Так или иначе, в четвертичное
время, под влиянием значительного опреснения, связанного с притоком
талых вод ледника, Jaera sarsi оказалась способной к обитанию в прес-
ной воде и попала в реки Понто-Каспийского бассейна. Каким образом
она поднялась на 1000 км вверх по Волге и на 1800 км вверх по
Дунаю — сказать трудно. С одинаковым правом можно допустить медлен-
ное активное расселение вверх по течению и пассивный перенос на обра-
станиях судов.
Штрохаль 7 обнаружил J. sarsi в одном из искусственных каналов,
'соединяющихся с Дунаем, и на этом основании считает возможным ее
активное расселение.
Хотя Державин (цит. раб.) и думает, что «вряд ли ктр-либо из сто-
ронников реликтовой теории будет считать трех понто-каспийских боко-
плавов и Corophium curvispinum, найденных в Дунае под Будапештом,
сарматскими реликтами», но именно такое мнение высказали Дудич 8
и Кессельяк (1938) по отношению к J. sarsi в Дунае (и Дудич, 1927, по
отношению к упомянутым Amphipoda). С этим мнением трудно согла-
ситься, так как сарматская фауна чрезвычайно сильно отличалась от
современной, а став на эту точку зрения, приходится допустить совер-
шенно параллельную эволюцию Jaera в разных частях ее ареала и в раз-
ных условиях, приводящую тем не менее к политопному образованию’
одного вида. Гораздо более вероятно недавнее формирование J. sarsi
в едином водоеме [последнее соединение Понта и Каспия относится
к рисс-вюрмскому (Архангельский и Шатский, 1937) или поелевюрмскому
(Борисяк, Православлев) времени] и последующее проникновение ее
в реки Понто-Каспийского бассейна. В связи с этим следует отметить
присутствие в Каспии эндемичного подвида J. sarsi caspica, приурочен-
1 А. Н. Державин. Каспийские элементы в фауне бассейна Волги. Тр.
Ихт. лаб. Упр. Касп.-Волжск. рыбн. и тюл. пром., т. 2, 1912; он же. 1924.
Пресноводные Malacostraca черноморского побережья Кавказа. Русск. гидробиол.
жури., 3.
2 А. Л. Бонин г. 1924. К изучению придонной жизни р. Волги. Саратов.
3 G. О. S а г s. 1927. Notes on the Crustacean Fauna of the Caspian Sea. — Сб.
в честь H. M. Книповича.
4 В. И. В с к л е м и ш е в. 1923. К вопросу о речных Peracarida Понто-Кас-
пийского бассейна. Русск. Гидробиолог, журн., 2.
5 Я. А. Б и р ш т е й н. 1946. К вопросу о происхождении морских ракообраз-
ных в реках Понто-Каспийского бассейна. Зоол. журн., т. XIV, 1935; о н ж е.
1946. Заметки о географическом распространении понто-каспийских бокоплавов.
Бюлл. Моск. общ. исп. прир., отд. биол., LI, 3.
6 А. Н. Державин. 1939. Мизиды Каспия. Ваку.
7 Н. Strouhal. 1939. Einige bemerkenswetre Vorkoinnisse von Wirbellosen,
•besonders Isopoden in dor Ostmark. Fostschr. Enibrik Strand, V.
8 E. Dudich. 1930. A Jaera nonnanni Rathke, egy uj viztaseka a magyer
launaban. Allat. Kozl., 27 (цит. no Kesselyak, 1937).
БИОЛОГИЯ и экология
35
ного к относительно осолоненным (12—13°/оо) частям моря и, невидимому,
не встречающегося в опресненном северном Каспии; ареал J. sarsi cas-
pica оказывается, таким образом, оторванным от ареала типичной
J. sarsi, населяющей Волгу и Урал. Можно высказать предположение,
что подвид J. sarsi caspica сформировался уже после прекращения связи
Каспийского и Азовско-Черноморского бассейна при недавнем осоло-
нении Каспия, последовавшем за послеледниковым опреснением.
Заканчивая обзор распространения Jaera, нужно указать на уже
отмеченную выше для других ракообразных особенность: этот род отсут-
ствует в тропиках, а в антибореальной области его замещает близкий,
недавно выделенный род Neojaera Nordenstam.1
БИОЛОГИЯ И ЭКОЛОГИЯ
Asellidae встречаются, главным образом, в пресных наземных и под-
земных водоемах. Наиболее обычный и широко известный вид—A. aqua-
ticus — населяет литоральную область озер, прудов и луж; В текучих
водах его количество гораздо меньше, чем в стоячих. Водяной ослик
обычно живет среди макрофитов, медленно ползая по стеблям и листьям
и изредка плавая равномерно, а не толчками, подвигаясь при этом вперед
и держа тело в горизонтальном положении. ,
‘ Боруцкий (1929) для озер Мещерской низменности сопоставляет коли-
чественное развитие A. aquaticus с типологией этих, водоемов и изобра-
жает результаты в виде схемы, приведенной на стр. 36 (звездочки обо-
значают плотность населения A. aquaticus, стрелки — пути перехода
родоемов из одного типа в другой).
Эта схема, вероятно, может быть применена и в более широком мас-
штабе, однако она далеко не универсальна. Так, например, в группе
Кончезерских озер в Карелии A. aquaticus развивается в наибольшем
количестве в озерах с явно выраженными чертами дистрофии; в Сявной
губе отмечено в среднем 600 экз. весом в 2.3 г на 1 м1 2, что составляет
15.2% общей биомассы бентоса озера. 2
Совершенно неясным остается пока факт отсутствия Asellus в водое-
мах с казалось бы вполне благоприятным для него режимом, как, напри-
мер, в оз. Севан в Армении.
Повидимому, A. aquaticus вообще представляет собой весьма эврибионт-
ную форму. Встречаясь постоянно среди гниющей растительности
и в озерах, подверженных сильным заморам, он, очевидно, способен жить
при довольно низком содержании кислорода.
Леванидов (1949) приводит данные опытов Мудрецовой-Висс, согласно
которым водяной ослик может существовать при отсутствии кислорода
от 22 часов до 2 суток при температуре 16—20°. Сходные результаты полу-
чил и сам. Леванидов. Нижним пределом содержания кислорода в воде,
достаточным для нормальной жизнедеятельности водяного ослика;
Леванидов считает 0.4—0.5 мг/л. К содержанию углекислоты и сероводо-
рода в воде водяные ослики чрезвычайно чувствительны. По сообщаемым
Леванидовым данным Мудрецовой-Висс, углекислота убивает рачков
в течение нескольких часов, а сероводород — в течение нескольких минут.
1 A. Nordenstam. 1933. Marine Isopoda of the families Serolidae, Idothei.-.
dae, Pseudidotheidao, Arcturidae, Parasellidae and Stenetriidae mainly taken from
the South Atlantic. Fourther Zool. Ros. Swed. Antarct. Exped. 1.901—1903, 3, 1.
2 H. M. Попов a. 193(1. К вопросу о продуктивности зообентоса озер Кон-,
чезорской группы в Карелии. Тр. Бородинской биол. ст., VIII, 2.
3*
36
ВВЕДЕНИЕ
р—. О лиготрофное ---------1
! I
Олиготрофное с Олиготрофное с
чертами эвтрофии ХХХХХХХ чертами дистро-
| -фии
Олиготрофно-
эвтрофное ХХХХХХ
Эвтрофное с чер-
тами олиготрофии ххххх
I
Эвтрофное XX XX
!
Ф ;
-------:----— Эвтрофное с чер- , .
тами дистрофии XXX
-------- Эвтрофно-дист-
рофное XX
ф
Дистрофное с чер-
тами эвтрофии „ у
Эвтрофное с чер-
тами дистрофии
и ясно выражен-
ной сапробиза-
цией
Олиготрофное с
различной сте-
пенью воздействия
факторов дистро-
фии
Олиготрофно-
дистрофное
"!
Дистрофное с чер-
тами олиготрофии
0-
Дистрофное <-
Олиготрофное с
чертами дистро-
фии и ясно выра-
женной сапроби-
зацией
Активная реакция среды также оказывает определенное воздействие
на водяных осликов. Они не переносят высокой кислотности и быстро
погибают, будучи помещены в воду из верховых торфяных болот.1 В опы-
тах Леванидова при рН=4.5 продолжительность жизни рачков состав-
ляла 12—24 часа, при рН=,5.0 — 3—5 суток, при pH=5^6 водяные ослики
существовали более 2 недель, но находились в угнетенном состоянии.
В природных условиях водяные ослики встречаются, как правило,
в водоемах, pH которых превышает 6.0—6.5.
Согласно: -Нидгему,1 2 нижний предел нормального существования
водяного' ослика лежит около pH=6.0, верхний предел — около рН=
=8.0. Только в этих пределах наблюдается в' условиях эксперимента
и нормальная регенерация ампутированных конечностей рачка, причем,
наиболее быстро она' происходит при рН=7.4.
Температура воды имеет меньшее значение в жизни Asellus aquaticus,
так как этот вид переносит весьма значительные ее колебания. Верхним
1 О. Harnisc h. 1924. Einige Gesichtpunkte zum Verstandnis der Fauna der
Humusgewasser. Verb. Intern. Ver. theor. ang. Limnol.
2 A. E. Needham. 1947. Sensivity of regenerating limbs o,f an aquatic Cru-
stacea to variations in the concentration of ,hydrogen and phosphate, ions in the exter-'
nal medium. Journ. exper. Zool., 106, 2,: 181—195..
БИОЛОГИЯ И ЭКОЛОГИЯ
37
температурным предел ом'существования A. aquaticus, по наблюдениям
Леванидова и Желтенковой, можно считать 28°.- При дальнейшем повы-
шении температуры рачки откочевывают вглубь водоема. Осенью, при
температуре 4°, ослики перестают двигаться и питаться, весной, при
той же температуре, питание прекращается, но подвижность сохраняется.
Для развития яиц необходима температура не менее 10—12°, для копу-
ляции — 8.5-—10°. Подземные виды водяных осликов, несомненно, весьма
стенотермны. Это относится в особенности к Stenasellus asiaticus из; тер-
мального источника в восточной Туркмении, обитающего в условиях
почти неизменной температуры (20—22°). ...
По отношению к солености A. aquaticus проявляет значительную прй-
. способляемость, так как заходит в солоноватые воды Балтийского моря
и был найден в осолоненных колодцах на п-ове Мангистау и на северном
побережье Азовского моря (соленость 0.5—1°/оо). В экспериментах Ду-
дича 1 A. aquaticus выживал и в нормальной морской воде, но в естествен-
ных условиях он никогда не был находим при солености выше З1—б^/оо.
Беляев 1 2 показал, что в условиях опыта A. aquaticus выдерживает
повышение солености до 20°/оо. Такая эвригалинность этого вида связана
с его способностью к активной осморегуляции. Осмотическое давление
его полостной жидкости (д°) в нормальных условиях обитания в прес-
ной воде составляет ;0.50—0.57°. При повышении солености окружающей
среды до 10—12°/оо осмотическое давление пблостной жидкости йодяного
ослика возрастает медленнее, чем концентрацйй солей во внешней среде;
рачек сохраняет значительную степень гипертонии. При дальнейшем
повышении- солености осмотическое давление полостной жидкости ослцка
начинает меняться параллельно изменению концентрации солей ъ окру-
жающей среде, рачек становится пойкилосмотичным, но сохраняет посто-
янную; хотя и незначительную степень гипертонии.
Совершенно так же реагируют на изменение солености -некоторые
бокоплавы оз. Байкал,3 несмотря на то, что у них функцию органа осмо-
регуляций выполняют антеннальные железы, а у водяных осликов —
максиллярные железы.4
Водяные ослики питаются растительной пищей. Согласно Виллеру,5
.излюбленной пищей рачков служат крупные диатомовые и нитчатки
обрастаний, а при их отсутствии — отмершие макрофиты и опавшие
листья деревьев. Живых макрофитов Asellus не ест. Леванидов (1949),
пользуясь «двориками» Гаевской, экспериментально показал, что «Любой
растительный материал может служить пйщей водяным осликам, но основ-
ной, предпочитаемой пищей для них являются опавйше древесные листья,
причем особенное предпочтение они отдают листьям ольхи: и вяза. Сине-
зеленые водоросли, а также листья дуба и иглы хвойных весьма неохотно
потребляются водяными осликами даже при отсутствии другой пищи.
Самый процесс переработки листьев водяные ослики осуществляют,
соскабливая поверхность мякоти, наиболее сильно затронутую разру-
1 Е. D u d i ch. 1928. A tengeviz hataza az edesvizi askakra. Magyar. Tud. Ak.
Mat. Term. Ert.,.XLV.
2 Г. M. Беляев. 1949. Осморегуляторные. способности изопод. ДАН СССР.
XVII, 6 : 1117.
3 А. Я. Вазикалова, Я. А. Бир штейн-иД. Н. Талиев. 1946.
Осморегуляторные способности бокоплавов оз. Байкал-. ДАН СССР, IX, 3 : 381.
4 А. Е. Needham. 1942. Structure and Development of Excretory Organs
of Asellus aquaticus. Joum. Microscop. Sci., 83. ; .
5 A. Wilier. 1917; N ahrungsuntersuchungen bei der Wasserassel(Asellus aqua-
ticus). Zeitschr. f. Fischerei, III, N. F. -
38
ВВЕДЕНИЕ
шением, но не разрушая даже самых тонких жилок. Постепенно вся
мякоть уничтожается, и лист превращается в кружево из жилок».
Водяные ослики наиболее многочисленны именно в тех водоемах,
в которые попадают в большом количестве опавшие древесные листья.
Леванидов по этому поводу пишет: «По данным Н. К. Дексбах (1928),
в литорали озер Мещерской низменности в зарослях Potamogeton плот-
ность населения водяных осликов составляла 120 шт. на 1 м2, а в лито-
рали лесного озера, которая была покрыта растительным аллохтонным
материалом, плотность населения доходила до 1000 шт. на 1 м2. По дан-
ным Л. Л. Россолимо, в озере Тишандра, в той его части, которая испы-
тывает сильное влияние аллохтонных органических веществ, плотность
населения равнялась 7280 особей на 1 м 2. В одном из заливов пруда
на территории Тимирязевской с.-х. академии, дно которого густо покры-
вают листья ольхи и вяза, средняя плотность, водяных осликов, опре-
деленная дночерпателем Петерсена, составила 5600 шт. на 1 м2».
За сутки взрослый водяной ослик поглощает количество пищи, состав-
ляющее несколько более 5% веса его тела. При температуре ниже 18°,
а также при пониженном содержании кислорода в воде или кислой
ее реакций, интенсивность питания падает. Пятимесячные рачки съедают
в 8 раз больше пищи, чем одномесячные. За всю свою жизнь каждый
водяной ослик потребляет 168 мг сырого веса листьев. Выяснив, кроме
того,' усвояемость пищи водяным осликом и зная продолжительность его
жизни и величину прироста за разные сезоны, Леванидов дал расчет
баланса вещества и энергии в течение жизненного цикла этого рачка:
«Водяной ослик в течение жизни получает с пищей 147 гкал. С выделе-
ниями он теряет 47 гкал; в теле водяного ослика накапливается 16.4
гкал, т. е. 11.2%. Остальная энергия в количестве 56.8% тратится на оки-
слительные процессы. Баланс органического вещества подобен энерге-
тическому: 11% вещества накапливается в теле животного, 43% теряется
с выделениями и 46% — вес продуктов окисления (СОг и НгО)».
Jaera albifrons, подобно Asellus aquaticus, также растительноядна.
Она поедает обрастания камней и водорослей, а также отмершие части
фукоидов. Даже при недостатке растительной пищи она не использует
не только живых животных, но и их трупы (Foreman, 1944).
Врагами водяных осликов являются, помимо рыб, многие хищные
цодяные насекомые — клопы, плавунцы, личинки стрекоз, а молодых
рачков уничтожают, кроме того, вислокрылки и некоторые циклопы.
В опытах Леванидова (1949) 3 окаймленных плавунца в течение 10 дней
уничтожили 500 экземпляров водяных осликов, помещенных в аквариум
размерами 86x35 см и глубиной 25 см. Jaera может становиться жерт-
вой бокоплавов (Gammarus locusta и G. duebeni), однако она хорошо защи-
щена от них своим образом жизни под камнями (Foreman, 1944).
Леванидов наблюдал массовую гибель водяных осликов в пруду под
Москвой от какой-то эпизоотии. Собранные здесь рачки погибали и будучи
помещены в аквариумы.
На водяных осликах часто поселяются простейшие — эгшбпонты —
кругоресничная инфузория Paravortieella crassicaulis Kent, сосущие
инфузории Dendrocometes paradoxus Stein, Stylocomeles digltatus Stein
и др., в марзупиальной сумке обитает симбиотическая прямокишечная
турбеллярия Castrade.llq granea (М. Вг.).
Встречаются и внутренние паразиты —-споровик Asellaria caullereyi
Poisson и личинки скребня Acanthocephalus, окончательными хозяевами
которого служат некоторые рыбы, а также лягушки.
Б ИОЛОГИЯ И ЭКОЛОГИЯ
39
Существование вне воды палеарктических Asellidae не наблюдалось;
американский A. communis способен при высыхании водоема закапы-
ваться в ил и в таком состоянии проводить несколько месяцев.
Тонкие и детальные наблюдения произведены над биологией размно-
жения A. aquaticus (Maercks, 1930). Спаривание наступает во время
линьки самки, после которой образуются оостегиты. В это время (про-
должительность 10 часов) парные половые отверстия самки, находящиеся
на брюшной стороне IV сегмента, увеличиваются в 3 раза в длину
и в 2 раза в ширину. Когда линька заканчивается, половые отверстия
закрываются оостегитами, и спаривание делается невозможным. Самец
помещается сверху самки и, удерживая ее IV перейоПодами, ожидает
линьки (рис. 16, 7). Затем, при помощи первой антенны, он просовывает
II плеоподы сбоку в щель между IV и V сегментами самки (рис. 16, III)
и вводит эндоподит одного из плеоподов в половое отверстие самки
Рис. 16. Спаривание Asellus (s. str.) aquaticus (L.). (По Мерксу).
с одной стороны на ]/3 его длины (рис. 16). При этом эндоподит и экзоподит
II плеопода перекрещиваются для того, чтобы эндоподит оказался напра-
вленным несколько в сторону (рис. 17). После того как полость эндоподита
освобождается от спермы, те же самые действия производит эндоподит
другой стороны. Парный пенис, находящийся на конце стернита VII
сегмента, в копуляции не участвует. Он только наполняет спермой
энДоподит II плеопода непосредственно перед копуляцией (рис. 16, II).
I плеопод пригибает penis, лишенный собственной мускулатуры, ко
II плеоподу. Весь процесс спаривания длится, в зависимости от тем-
пературы, От Р/г1^0 = 19°) до 6 (z° = 3.5°) часов. .
Вслед за спариванием'формируются оостегиты перейоподов иного-1
челюстей и одновременно с этим через 3 часа после спаривания, в тече-.
ние 5 минут, откладывается икра. Икра развивается в условиях постоян-
ной смены воды в выводковой сумке, что достигается ритмическими дви-
жениями оостегитов. В начале развития икры наблюдается 1 колебание
в минуту, в конце 10—15 колебаний за тот же срок. Помимо движений
оостегитов, колебательные движения совершают и отростки ногочелю-
етей, которые, по Янке,1 предназначены для очистки яиц и эмбрионов
от попадающих на них мелких частиц. Колебания отростков нОгочелю-
стей также учащаются по мере развития икры. Янке насчитывал
1 Oldwig J an ck е. 1924. Die Brutpflege einiger Malakostraken. Zool. Anz., 58.
40
ВВЕДЕНИЕ
Рис. 17. II плеопод самца
Asellus (s. str.) aquaticus
(L.) при спаривании. (По
Мерксу).
самки. Через несколько
в начале развития около 45 колебаний в течение 25 секунд, после чего
следовала пауза в 15—90 секунд, а в конце развития до 225 колебаний
в минуту. Согласно Ван-Эмдену,1 вентиляция выводковой камеры про-
изводится не за счет движения оостегитов, а за счет движений отростков
ногочелюстей. Для икры и эмбрионов Янке различает 4 стадии: 1) круг-
лые яйца; 2) яйца с листовидными отростками; 3) молодые личинки
с не вполне сформировавшимися конечностями; 4) сформировавшиеся
личинки, прошедшие первую личиночную линьку. По мере перехода
зародышей из одной стадии в другую их количество уменьшается.
Это явление стоит в связи с ростом зародышей при переходе из одной
стадии в другую, в то время как объем выводковой камеры остается
достоянным. Лишние яйца и эмбрионы выпадают наружу. Листовидные
отростки яиц второй стадии Янке рассматривает как органы, позволяю-
щие воде свободно омывать каждую икринку
благодаря тому, что между икринками остается
свободное пространство. Однако позднее Дей-
дар 1 2 показал, что эти отростки представляют
собой эмбриональные экскреторные органы.
После того как молодь покинет выводковую
сумку, самка линяет и оостегиты и отросток
ногочелюстей при этом редуцируются.
Несколько иначе происходит спаривание у
Jaera albifrons (Fbrsman, 1944). Самец распо-
лагается на спинной стороне самки, так что его
голова оказывается над плеотельзоном самки,
и удерживает ее IV и V или только V перейопо-
дами. По прошествии нескольких часов, а
иногда нескольких минут после начала спари-
вания самка линяет. Тогда самец сдвигает
несколько в сторону свой плебтельзон, опускает
вниз почти под прямым углом свои первые плео-
поды и просовывает их в щель между IV и V
грудными сегментами самки, перенося таким
образом сперму в один из семяприемников
минут самец передвигает плеотельзон на дру-
гую сторону и наполняет спермой другой семяприемник самки. По
всей вероятности, сперма из половых отверстий попадает в половые
плеоподы самца по бороздкам, проходящим по нижней стороне последних.
Через 2—10 часов после линьки самка в течение 10—25 минут
откладывает яйца. Их число колеблется от 5 до 60 и зависит от размеров
самки. Отверстия яйцеводов очень узки, и проходящие через них яйца
деформируются и принимают округлую форму только в выводковой
сумке. Вентиляция выводковой сумки осуществляется движениями оосте-
гитов (37—40 взмахов в минуту).
При спаривании Jaera italica оба партнера обращены друг к другу
брюшными сторонами, причем самец удерживает переднюю часть тела
самки 4-—5 парами задних перейоподов (Strouhal, 1942). Следует заметить,:
что из всех видов Jaera только у этого вида самцы крупнее самок.
1 Fritz van Emden. 11922. Zur Kenntniss der Brutpflege von Asellus aquaticus
nebst Bemerkungen fiber die Brutpflege anderer Isopoden. Arch, f. Naturg., 88.
2 E. D e j d a r, 1930. Die Funktion der «blattformigen Anhange» der Embrionen
von Asellus aquaticus (L.). Zeitschr. Morph. Okol. Tiere, XIX.
БИОЛОГИЯ И ЭКОЛОГИЯ
41-
Рост и размножение A. aquaticus в условиях подмосковных водоемов
исследовались в последнее время М. В. Желтенковой. Она любезно соста-
вила для настоящей сводки краткое изложение результатов своей еще
не опубликованной работы.
«Размножение начинается вскоре после освобождения водоемов
от льда, т. е. с конца апреля, и продолжается в течение всего мая, июня,
июля и большей части августа. В начале периода размножения в нем
участвуют только крупные особи (средний размер самцов 10.4 мм, самок —
8.8 мм), которые потом частично погибают, а впоследствии начинают
размножаться рачки, родившиеся весной и в начале лета (средний раз-
мер самцов в июле 4.7 мм, самок — 4.3 мм). Через короткий срок после
окончания копуляции самка линяет. Кутикула лопается поперек между
IV и V грудными сегментами. Сначала сбрасывается панцырь с задней
половины тела, причем бывают видны набитые яйцами яйцеводы, затем —
с передней половины тела, и образуется выводковая сумка, в которую
выметываются яйца. Продолжительность их развития колеблется от 14
дней в мае до 22 дней в августе, т. е. увеличивается в течение лета. Парал-
лельно с удлинением срока развития яиц уменьшается их количество,
составляющее в среднем на одну самку 98 в июне и 13 в августе, причем,
как правило, у более крупных самок насчитывается большее количество
яиц. Повторное оплодотворение самки может произойти через 5—6 дней
после выхода молоди из марзупиума. В/течение лета водяной ослик раз-
вивает самое большее 2 генерации; только оцлодотворенные до середины-
июня самки дадут потомство, способное к воспроизводству в том же году.
Число выводков каждой самки за период размножения достигает 4—5..
«Вышедшие из марзупиума молодые рачки имеют в длину 1.0—1.1 мм
при весе около 0.04 г, через 10 дней — 1.43 мм, через 21 день — 2)07 йм,
через 30 дней —’2.52 мм, через 40 дней — 3.23 мм, перёлиняв к тому-
времени 10—11 раз. Первые 25 дней,' до размера 2.4 мм, самцы и самки
растут и линяют приблизительно одинаково, а затем самки начинают
отставать в росте от самцов, благодаря более редким линькам. Сначала
линьки у обоих полов происходят через 2 дня, потом через 3—4 дня,
впоследствии через 6 дней. До наступления половозрел ости водяной
ослик должен слинять 13—15 раз. К этому времени самки достигают-
н длину 3.5—4 мм. В среднем самки становятся половозрелыми в воз-
расте 50—55 дней.
«Рост всегда связан с линькой, но не всегда линька приводит к росту..
Можно различить 4 типа линек: 1) линька молоди до наступления поло-
возрелости, сопровождающаяся интенсивным ростом, 2) линька полово-
зрелых самок, во время которой рост не происходит, 3) линька йолово-
зрелых самцов, также не растущих в период размножения, и 4) линька
после окончания периода размножения, приводящая к усиленному росту.
«Линьки половозрелых самок приводят либо к образованию марзу-
пиума, либо к его сбрасыванию. Эти линьки чередуются .одна с другой,
и промежуток между ними определяется временем инкубации икры.
Линьки половозрелых самцов не так тесно связаны с половым циклом
и происходят как до, так и после копуляций. Интервал между ними,
составляет 12—20 дней. Зимою водяные ослики не линяют и не растут.
«Учитывая все данные по плодовитости. й скорости роста и созрева-
ния водяных осликов, можно подсчитать ,их потенциальную продуктив-
ность. Среднее количество осликов на 1 м2’ в пруде в Сокольниках
в Москве — 2200 экземпляров. К концу года они могли бы дать 3 300 000,
считая, что потомство одной пары составляет 3 000 экземпляров. Если;
42
ВВЕДЕНИЕ
бы все эти ослики достигли длины 7—9 мм, т. е. веса 7.1 мг, то потен-
циальная продукция водяных осликов на 1 м* 2 дна Сокольнического
пруда равнялась бы 23.4 кг».
Цикл жизни других видов водяных осликов несомненно отличается
от описанного Желтенковой цикла жизни A. aquaticus. Так, например,
у байкальских A. angarensis и A. baicalensis размножение длится круг-
лый год.1
Образование марзупиальной сумки не связано с оплодотворением.
Она возникает у самок и в отсутствие самцов, но лишь после наступления
половозрел ости; происходит и откладка в марзупиум веоплодотворенных
и неспособных к развитию яиц. Облучение молодых осликов радием
приводит к разрушению яичников, и в таких случаях марзупиум не раз-
вивается.2
Рост Asellus происходит во время линьки. Как показал Нидгем,3
грудные сегменты Asellus aquaticus растут в длину равномерно, пропор-
ционально друг другу, для роста в ширину наблюдается более сложная
картина. Нидгем установил два градиент-поля роста в ширину — одно,
расположенное в задней части груди, другое на уровне IV грудного сев-
мента. В начале роста большее значение имеет первое поле, в конце —
второе. Голова растет значительно медленнее груди. Явление непропор-
ционального роста констатировали также Миллер и Ной4 для уроподов
Asellus californicus. В процессе постэмбрионального развития эндоподит
уроподов резко обгоняет в росте экзоподит, и разница в длине между
обеими ветвями постепенно возрастает. Аналогичное явление наблю-
дается, вероятно, и у другого представителя того же подрода — байкаль-
ского A. dybowskii.
Продолжительность жизни Asellus вероятно не превышает двух лет,
при комнатной температуре она равна одному,году. A. aquaticus при
комнатной температуре достигает половой зрелости в возрасте 3 месяцев,
при температуре 5—10° — в возрасте 13 месяцев (Maercks, 1930).
Asellus aquaticus способен к автотомии и последующей регенерации
антенн, перейоподов и уроподов (Wege, 1911; Needham, 1942). Произ-
вольное отрывание перейоподов происходит между коксо- и базиподи-
том с помощью специального мускула автотомизатора, проходящего,
в отличие от других мышц, через два членика конечности (бази- и исхио-
подит). В месте отрыва полость ноги перегорожена гиподермальной мем-
браной, а в кутикуле с вентральной стороны имеется щель, облегчающая
разлом наружного скелета.
Особенно часто наблюдается автотомия и регенерация уроподов. *Что
касается перейоподов, то только приспособленный для удержания самки
во время копуляции IV перейопод самца не способен к автотомии, так
как щель между его коксо- и базиподитом закрыта. Эта конечность авто-
томизируется, как и все остальные, у самок и у молоди, а также у самцов,
но у последних только в зимнее время, когда половой процесс прекра-
щается (Needham, 1942).
3 Г. Б. Гаврилов. 1949. К вопросу о времени размножения амфипод иизо-
под оз. Байкал. ДАН СССР, XIV, 5 : 739—742.
2 V. Haemerli-Boveri. 1926. Uber die Determination der secundaren
Geschlechtsmerkmale (Brutsackbildung) der weiblichen Wasserassel durch das Ovar.
Zeitschr. vergl. Physiol., 4, 4—5 : 668.
3 A. E. Needham. 1937. On Helative Growth in Asellus aquaticus. Proc.
Zool. Soc. London, 107, A, III.
4 A. Milton, Miller and E. A. No y. 1939. Differential Growth and
evolution in a subterranean Isopod. American Naturalist, LXXIII, 747.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ 43
Скорость регенерации VII перейопода прямо пропорциональна ско-
рости роста всего тела Asellus. Соответственно этому она происходит
у самок медленнее; чем у самцов, у крупных особей медленнее, чем
у мелких, зимою (в условиях аквариума) медленнее, чем летом.1
При регенерации перейоподов гиподерма дает гиподерму регенерата
и его внутренний и наружный скелет, базофильные амебоциты образуют
тромб, закрывающий рану, и, кроме того, фагоцитируют распадающиеся
мышцы, а мускулатура возникает за счет миобластов, мигрирующих
из старых, распадающихся мышц. Регенерат формируется под хитиновым
покровом сохранившейся части конечности, там расчленяется и появ-
ляется наружу обычно после очередной линьки, а иногда и независимо
от нее прорывает хитин.1 2 В ходе регенерации существенную роль играют
периферические нервы. Их разрушение препятствует нормальному раз-
витию процесса.3
Плеоподы не способны к автотомии, но, при их искусственном уда-
лении, регенерируют подобно остальным конечностям. Передние плео-
поды <5 при этом повторяют стадии своего нормального онтогенеза,
однако нередко наблюдаются и случаи дисгармоничного развития:
формирование левой конечности идет скорее, чем правой, или разви-
вается только дистальная часть конечности (Needham, 1941).
У Jaera albifrons наблюдалась регенерация антенны (Forsman, 1944),
у Microcharon profundalis — регенерация I. перейопода (Кагашап, 1940).
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Водяные ослики имеют существенное значение в жизни пресноводных
водоемов лесной и отчасти лесостепной полосы. Они принадлежат к тем
немногим организмам, которые используют попадающие в водоемы опав-
шие древесные листья, включая содержащиеся в них энергетические
ресурсы в круговорот энергии в водоеме и переводя их в доступную для
рыб форму. Количество опадающих листьев измеряется весьма значи-
тельными цифрами — от 2 до 5 тонн на гектар площади, т. е. 200—500 г
на 1 м2 (Леванидов, 1949).
При изобилии этого аллохтонного материала водяные ослики раз-
виваются, как указывалось выше, в огромном количестве, занимая иногда
одно из первых мест среди организмов, слагающих биомассу бентоса.
В некоторых водохранилищах, например в Рыбинском, водяные ослики
оказываются преобладающей группой донной фауны.
Водяные ослики используются в пищу многими промысловыми
рыбами. Asellus aquaticus постоянно находится в кишечнике карася,
карпа, ерша, некоторых сигов, налима. Виллер 4 характеризует этот
вид как чрезвычайно важную кормовую форму. Такое же значение
на Дальнем Востоке имеет A. hilgendorfi. В реках Сахалина желудки
налима бывают целиком набиты представителями этого вида. В Боден-
1 А. Е. Needham. 1949. Growth and regeneration in Asellus aquaticus (L.)
in relation to age, sex and season, .lourn. exper. Zool., 112, 1 : 49—78.
- Л. H. Жилкин. 1938. Исследования по сравнительной гистологии мышеч-
ной ткани. VII. Регенерация мускулатуры у ракообразных. Арх. анат., гист., эмбр.,
XIX, 3 : 402—429.
3 А. Е. Needham. 1946. Peripheral nerve and regeneration in Crustacea,
lourn. exper. Biol., 21, 134, 1945; 22 : 107.
1 A. Wilier. 1924. Die Nahrung der Fische. Handb. d. Binnenfischerei Mit-
teleuropas, 1 : 223.
ВВЕДЕНИЕ
ском озере местный сиг питается между прочим Asellus 1 (вероятно Л. cava-
ticus joreli).
Пищевая ценность A. aquaticus довольно высока,’ судя по’ данным,,
приводимым Генгом: 1 2 г'
' Живой вес 1 экз. . . . 27.17 мг
Сухое вещество .... 5.25 » ,
Вода................ 21.92 » . ,
Азот................ 0.4324 » '
Протеин............. 2.703 » ' . ’
Жир.............• - 0.2334 » •
Углеводы.............. 0.4494 » •:
Зола................ 1.865
Кальций............. 19.76 ’
Это обстоятельство, вместе с высокой плодовитостью ц крайней непри-
хотливрстью A. aquaticus, позволяет рассматривать его как возможный
объект искусственного разведения живого рыбного корма, (Леванидов).
Вместе с тем следует учитывать, что A. aqijaiicus может являться
промежуточным хозяином скребня Acanthocephalus. Некоторые виды,
в частности байкальские Asellus, благодаря своему образу жизни (под.
камнями, в щелях и выбоинах камней) остаются недоступными для рыб.
ЛИТЕРАТУРА 3
Алпатов В. В. 1922. Новый для оз. Байкал водяной ослик и сравнение байкаль-
ских видов р. Asellus с европейскими и американскими (по материалам Бай-
кальской экспедиции Московского зоологического музея). Русск. тидробиол.
жури., 11,3—4 : 1—3. .... т-
Б .и р шт е й н Я. А. 1932. Malacostraca Армении. Тр. Севанск. озерн. ст., IV,. 1—2 :
139—160. - ' ' ’ ': -
[Бирштейн Я. A.] Birstein J. А. 1932. Eine rieiie Art des Genus Asellus
Geoffroy aus Transkaukasien. Zool. Anz., 98 : 251—255.
[.Бирштейн Я. A.] Birstein J. A. 1933. Die Land-und Susswasser-Isopoden
des arktischen Gebietes. Fauna arctica, VI, 5 : 473—476. . ...‘
Бирштейн Я. A. 1936. О водяных осликах Кавказа. Сб. трудов Зоол. муз)
(при МГУ), Ш ; 235—242.
Бирштейн Я. А. 1939. О некоторых особенностях географического распростра-
нения пресноводных Malacostraca Дальнего Востока.: Зоол. журн., XVIII,
1 : 54- 69. .. ' .
Бирштейн Я. А. 1939а. Зоогеографическая характеристика водяных осликов
Байкала. ДАН СССР, XXV, 3: 248—251.
Бирштейн Я. А. 1945. Заметка о пресноводных высших ракообразных Турк-
мении и Ирана. Учен. зап. МГУ, 83 : 151—164;. ... .•
Бирштейн Я. А. 1947. О водяных осликах Сибири. ДАН СССР, LVII, 4 : 409 —
412. . . .,
Ёирштейн Я. А. и Старостин И. В. 1949. Новый для СССР род водяных
осликов. (Stenasellus) из Туркмении и его значение для зоогеографии Средней
Азии. ДАН СССР, LXIX, 5 : 691—694.
Боруцкий Е. В. 1929. Crustacea-Malacostraca водоемов Мещерской низмен-
ности (Рязанской губ.). Тр. Косинской биол. ст., 9 : 29—39.
1 G. S u г b е с к. 1916. Beobachtungen und Untersuchungen an Sandfelchen
des Bodensees. Allg. Fisch.-Zeit., 12. . ' ! '
2 H. G e n g. 1924. Der Futterwert der natiirlichen Fischnahrung. Zertschr. f.
Fischerei. 'y
3 В списке литературы приведены, главным образом, работы по систематике
палеарктических Asellidae. Из многочисленных морфолого-анатомических работ
в него внесены только важнейшие. Работы, содержащие указания на нахождение
обычных видов (главным образом Asellus aquaticus, Jaera nordmanni и J. alVijrons)r
опушены совершенно.
ЛИТЕРАТУРА
45
Внуковский В. 1929. Заметка о нахождении Asellus sp. (Isopoda, Asellidae)
в Восточной Сибири. Изв. Западносибирск. отд. Русск. геогр, общ., VI : 55—56.
Вълканов А. 1936. Бележки върху нашит бракични води. 2. Опит за тохното
хидрографской биоложко проучване. Годишн. Софийск. унив., физ.-мат.
. фак., XXXI, 3 : 209—341.
Вулканов А. 1938. Предгледъ на европейскитъ представители на рода Jaera
' ' . ' Leach 1813 (Isopoda genuina). Годишн. Софийск. унив., физ.-мат. фак.,
: ' XXXIV, 3 : 53—78.
Державин А. Н. 1930. Пресноводные Malacostraca Дальнего Востока. Русск.
гидробиол. журн., IX, 1—3 : 1—8.
К о ж о в М. 1931. К познанию фауны Байкала, ее распределения и условия обита-
ния. Изв. Биол.-геогр. н.-иссл. инет, при Гос. Иркутском унив., V, 1 : 1—170.
ЛеванидовВ. Я. 1949. Значение аллохтонного материала как пищевого ресурса
в водоеме на примере питания водяного ослика (Asellus aquaticus L.). Тр. Все-
союзн. Гидробиол. общ., I : 100—117.
.Мартынов А. В. 1932. Заметка о пресноводных Amphipoda и Isopoda Северной
Якутии. Ежегодн. Зоол. муз. Акад. Наук СССР, XXXII, 4.
Семе.нкевич IO. Н. 1924. О байкальских водяных осликах. Русск. гидробио-
логия. журн., Ill, 1—-2 : 8—13.
[Ч е т в е р и к о в С. С.) Tschetwerikoff S. 1911. Beitrage zur Anatomie
der Wasserassel (Asellus aquaticus). Bull. Soc. Imp. Natur., Moscou, N. S., 24 :
377—509.
A r c a n g e 1 i A. 1929. Ricerche faunistiche nelle isole italiane dell’Egeo. Isopodi.
Arch. zool. ital.,‘ XIII : 259—268.
Arcangeli A. 1935. Asellus delle caverhe del Belgio. Bull. Mus. roy. Hist, Nat.
Belg., XI, 37 : 1—10.
Arc.ange I i A. 1938. Etudes biospeologiques. VI. Asellus (Proasellus) hermallensis.
Bull. Mus. roy. Hist. Nat. Belg., XVI, 5 : 1—5.
Arcangeli A. 1939. Stenasellus racovitzai Razautti, Crostaceo Isopodo aquatico
cavernicolo italiano. La famiglia Stenasellidae. Boll. Mus. Zool. Anat. comp.
Torino, 46 : 17—50.
Blanc H. 1879. Faune profonde de Leman. 6 series. Bull. Soc. Vaud. Sc. Nat., XVI,
82 : 313—394. • : .
Blanc H. 1879a. Ueber den Asellus aus der Tiefenzone des Genfer Sees. Zool. Anz.,
2 : 428—431. .
Bovallius C. 1886. Notes on the Family Asellidae. Bih. K. Svensk. Vet. Akad.
Handl., XI, 5,: 1—54.
Braga J. M. 1942. Mancasellus mariae, a new blind Isopod inhabiting subterranean
waters in Portugal. Mem. Mus. zool. Univ. Coimbra, 130 : 1—12.
Braga J. M. 1942a. Un Isopode nouveau du Portugal: Stenasellus nobrei n. sp. Mem.
Mus. zool. Univ. Coimbra, 132 : 1—11.
Braga J. M. 1943. Description de 1’Asellus seabrai, isopode aveugle nouveau
des eaux souterraines du Portugal. Mem. Mus. zool. Univ. Coimbra, 145 :
1—12.
Braga J. M. 1944. Sur un Asellus nouveau des eaux superficielles du Portugal. Mem.
Mus. zool. Univ. Coimbra, 154 : 1—11.
Braga J.. M. 1944a. Note sur un genre d’Asellides nouveau du Portugal (Synasellus
h. gen.) et description d’une sousespece de ce genre. Mem. Mus. zool. Univ.
Coimbra, 158 :1—11.
Buresch J. 1929. Die Hohlenfauna Bulgarians. X Congres Intern, zool. Budapest,
II : 1427—1437. .
Carl J. 1908. Monographic der Schweizerischen Isopoden. Nouv. Mem. Soc. Helv.
Sei. nat. : 108—242.
Chappuis P. A. 1927. Die Tierwelt der unterirdischen Gewasser. Die Binnenge-
wasser, III : 1—175.
Chappuis P. A. 1942., Microcharon acherontis spec. nov. ein neuer subterraner
Isopode. Fragm. Fauna Hung., 5 : 120—122.
Chappuis P. A. 1943. Stygonectes phreaticus n. gen. n. sp. ein neuer Isopode aus
dem Grundwasser der Koros bei Baratka (Bihar). Fragm. Fauna Hung., 6 :
19—22. : .
C h.a pp u i s P. A. 1943a. Stygasellus nom. nov. fur Stygonectes Chappuis. Fragm.
Fauna Hung., 6 A: 42.
Chappuis P.. A. 1948. Copepodes, Syncarides et Isopodes des eaux phreatiques
de Suisse/Rev. Suisse zool., 55, 50 : 549—566.
Chilton Chas. 1920. Note on the Freshwater Isopods known as Asellus aquaticus.
Ahn. Mag. Nat. Hist., (9), V : 200—203.
46
ВВЕДЕНИЕ
Collinge W. Е. 1945. On the Freshwater Isopod Genus Caecidotea Packard. Ann.
Mag. Nat. Hist., (11), 84 : 815—817.
Collinge W. E. 1945a. Description of a new species of Asellus (Crustacea, Isopoda)
from the Isle of Man. Ann. Mag. Nat. Hist., (12), 91 : 497—499.
Collinge W. E. 1946. Preliminary Description of Stenasellus hazeltoni, a Caver-
nicolous Isopod new to Great Britain. Ann. Mag. Nat. Hist., (13), 98 : 140—142.
Collinge W. E. 1946a. Asellus hazeltoni, a new Freshwater Isopod. Ann. Mag.
Nat. Hist., (13), 97 : 70—72.
Collinge W. E. 1947. Description of a new Species and Variety of Asellus (Crusta-
cea, Isopoda). Ann. Mag. Nat. Hist., (13), 99 : 213—215.
Dahl F. 1916. Die Aselln oder Isopoden Deutschlands, Jena : 1—90.
Dollfus A. 1892. Note sur les Isopodes terrestres et fluviatiles de Syrie recueillife
principalement par M. 1c Dr. Th. Barrois. Rev. biol. Nord. France, 4 : 1—15.
Dollfus A. 1894. Crustaces isopodes terrestres et d’eau douce. Boll. Mus. zool.
anat, comp. Univ. Torino, 9, 177 : 3.
Dollfus A. 1897. Sur deux types nouveaux de Crustaces isopodes appartenant
a la faune souterraine de Cevennes. C. R. Acad. Sc. Paris, CXXV : 130—131.
Dollfus A. 1898, Sur deux types nouveaux de Crustaces isopodes appartenant
a la faune souterraine de Cevennes. Bull. Mus. Hist. nat. Paris, IV : 35—38.
Dollfus A. 1905. Sur quelques formes d’Isopodes appartenant a la faune souterraine
d’Europe. Ann. sc. nat., Zool., (8), XX : 365—412.
D u d i c h E. 1924. Ueber Protelsonia hungarica Mehely. Zool. Anz., LX : 151—155.
D u d i c h E. 1925. Ueber die artliche Zugehorigkeit des Asellus von Ungarn, Polen,
Dalmatian und Italien. Zool. Anz., LXIII : 1—7.
Dudich E. 1925a. Systematische Studien an italienischen Aselliden. Ann. Mus.
Nat. Hung., XXII -. 281—301.
Dybowski B. 1884. Neue Beitrage zur Kenntniss der Crustaceen-Fauna des
Baikalsees. Bull. Soc. Nat. Moscou, LX : 17—57.
F о r 1) e s S. A. 1876. List of Illinois Crustacea. Bull., № 1, Illinois Mus. Nat..
Hist.: 3—-25, figs. 1—30.
F о r s m a n B. 1944. Beobachtungen iiber Jaera albifrons Leach an der schwedische»
Westkiiste. Arkiv for zoologi, 35A, 11 : 1—33.
F r a d e, F. 1938. Un Asellus (Isopode) nouveau des eaux souterraines du Portugal.
Bull. Soc. portug. sci. nat., 13 : 17—24.
Grube Ed. 1872, Ueber einige bisher unbekannte Bewohner des Baikalsees. Jahresb.
Schles. Gesellsch., 57.
Hansen H. .J. 1905. On the morphology and classification of the Asellota-group
of Crustaceans, with descriptions of the genus Stenetrium Hasw. and its species.
Proc. Zool. Soc. London, 11 : 302—331.
II a у W. P. 1902. Observations on the Crustacean Fauna Nickajack Cave, Tenessee
and vicinity. Proc. U. S. Nat. Mus., XXV : 417—439.
Hilgendorf F. 1874. Ueber eine Siisswasserassel. Mitt. Deutsch. Gesellsch. Natur
Volkerkunde Ost-Asiens, I, 5 : 39.
Hilgendorf F. 1893. Bemerkungen uber 2 Isopoden die Japanische ’Siisswasser-
Assel (Asellus hilgendorfi Bov.) und eine neue Munna-Art. Sitzungsb. Gesellsch.
Naturf. Freunde zu Berlin, 1:1.
Karaman S. 1933. Neue Isopoden aus unterirdischen Gewasser Jugoslaviens. Zool.
Anz., 102 : 16—22.
Karaman S. 1933a. Microcerberus stygius, der dritte Isopod aus dem Grundwasser-
von Skoplje, Jugoslavien. Zool. Anz., 102 : 165—169.
Karaman S. 1933b. Ueber zwei neue Isopoden der Gruppe Asellus aus Jugoslavien.
Recueil de traveaux offert a Jivoin Georgevitch', Beograd : 103—113.
Karaman S. 1934. Beitrage zur Kenntniss der Isopoden-Familie Microparasellidae.
Mitt, uber Hohlen- und Karstforschung : 42—44.
Karaman S. 1934a. Weitere subterrane Aselliden aus Jugoslavien. Zool. Anz.,
108 : 23—26.
Karaman S. 1935. Die Fauna der unterirdischen Gewasser Jugoslaviens. Verhandl.
Intern. Ver. theor. ang. Limnologie, VII : 46—73,
Karaman S. 1936. Ein Stenasellus aus dem Grundwasser von Skoplje, St. gjorg-
jevici skopljensis n. subsp. Гласи. Скопского научи, друттва, XVII :137—141.
Karaman S. 1940. Die unterirdischen. Isopoden Siidserbiens. Гласи. Скопского
научи, друштва, XXII : 19—53.
Karaman S. 1948. Ueber das Mannchen von Stenasellus skopljensis Karaman.
Jugosl. Akad. znanosti i umjetn.
Kaulbersz G. J. 1913. Biologische Beobachtungen an Asellus aquaticus. Zool.
Jahrb., allg. Zool. n. Phys., 33 : 287—359.
ЛИТЕРАТУРА
47
Kesselyak А. 1938: Die Arten der Gattung Jaera Leach (Isopoda, Asellota). Zool.
Jahrb., Syst., LXXI, 3 : 219—252.
К u 1 c z у с к i W. 1885. Materyaly do fanny skorupiakow krayowych. Kosmos, 10 •
315—323, 405—417.
M'aercks H. H. 1930. Sexualbiologische Studien an Asellus aquaticus L. Zool.
Jahrb., allg. Zool. u. Physiol., 48 : 273—319.
Mehely L. 1924. Protelsonia hungarica nov. gen. n. sp. Ein blinder Isopod aus
Ungarn. Zool. Anz., 58 : 353—357.
M ehely L. 1927. Neue Wiirmer und Krebse aus Ungarn. Budapest : 1—19.
Miet.hc. C. 1899. Asellus cavaticus Schiodte. Ein Beitrag zur Hiihlenfauna der
Schweiz. Rev. Suis. Zool., VII : 273—319.
Miller M. A. 1933. A new blind Isopod, Asellus californi'cus and a revision of the
subterranean Asellids. Univ. California Public., Zoology, 39, 4 : 97—110.
Monod Th. 1924. Sur quelques Asellides nouveaux des eaux douces de 1’Afrique du
Nord. Bull. Soc. Hist. Nat. de 1’Afrique du Nord, 15 : 327—336.
Monod Th. 1925. Tanaidaces et Isopodes aquatiques de 1’Afrique occidentale et sep-
tentrionale. 2 partie. Bull. Soc. Sc. Nat. Maroc, 5 : 233—247.
Monod Th. 1932. Sur un Asellus aberrant (A. Remyi n. sp.) du lac d’Ohrid (Albanie).
Bull. Soc. zool. France, LVII : 206—217.
Monod Th. 1945. Un nouveau Stenasellus ouest-africain. Bull. Inst, franc: Afr.noire, 7.
Needham A. E. 1938. Abdominal appendages in the female and copulatory appen-
dages in the male Asellus. Quart. Journ. Microscop. Sci., 81, 1 : 127—150.
Needham A. E. 1941. Abdominal appendages of Asellus, II. Quart. Journ. Micro-
scop. Sci., 83, 1 : 61—89.
Needham A. E. 1942. Micro-anatomical Studies on Asellus. Quart. Journ. Micro-
scop. Sci., 84, 1 : 49—72.
Nicholls G. E.- 1929. A new species of Asellus (A. nipponensis) from Japan. Arch.
Zool. exp. gen., LXVIII, N. et R. : 33—38.
Nierstrasz H. F. und .Т.Н. S c h u r m a n s S t e к h о v e n. 1930. Isopoda
genuina. Die Tierwelt der Nord und Ostsee, XVIII : 57—133.
Odenwall E. 1927. Asellus aquaticus L. var. abyssalis nov. var. Memoranda Soc
Fauna et Flora fennica, 3 : 16.
Packard A. S. 1871. The Mammothe Cave and its habitans. On the crustaceans
and insects. Amer. Naturalist, 5 : 744—761.
P а с к a r d A. S. 1888. The cave fauna of North America with remarks on the
anatomy of the brain and origin of the blind species. Mem. Nat. Acad. Sci.
Washington, V : 1—66.
R а с о v i t z a E. G. 1919. Notes sur les Isopodes. 1. Asellus aquaticus auct. est une
erreur taxonomique. 2. Asellus aquaticus L. et A. meridianus n. sp. Arch. Zool.
exp. gen., 58, Notes et Revue ; 31—43.
R а с о v i t z a, E. G. 1919a. Notes sur les Isopodes. 3. Asellus banyulensis n. sp. 4.
A. coxalis Dollfus. 5. A. coxalis peyerimhoffi n. subsp. Arch. Zool. exp. gen.,
58, Notes et Revue : 49—П.
R а с о v i t z a E. G. 1920. Notes sur les Isopodes. 6. Asellus communis Say. 7. Les
pleopodes I et II des Asellides; morphologie et developpement. Arch. Zool. exp.
gen., 58, Notes et Revue : 79—115.
R а с о v i t z a E. G. 1922. Description de trois Asellus (Isopodes) cavernicoles nou-
veaux. Bull. Soc. Sc. Cluj, 1 : 401—410.
Racovi t za E. G. 1923. Notes sur les Isopodes. 10, Orientation de 1’Isopode et
essais de nomenclature pratique des bords et faces de son corps et de ses appen-
dices. 11. Morphologie et phylogenie des pereiopodes et de leurs phaneres. Arch.
Zool. exp. gen., 61, Notes et Revue : 75—122.
R а с о v i t z a . E. G. 1924. Diagnoses des genres Asellus et Stenasellus et description
de deux Stenasellus nouveaux. Bull. Soc. Sc. Cluj, 2 : 81—92.
R а с о v i t z a E. G. 1925, Notes sur les Isopodes. 13. Morphologie et phylogenie
des Antennes II, Arch. Zool. exp. gen., 63 : 533—622.
R а с о v i t z a E. G. 1950. Asellides. Premiere serie : Stenasellus. Arch, Zool. exp.
gen., Biospeol., 70 : 1—94.
Rathke H. 1837. Beitrag zur Fauna der Krym. Mem. Sav. Etrang. St. Petersb.,
Ill : 291—454.
Remy P. 1932. Description d’Asellus (Proasellus) Arnautovici n. sp. du lac d’Ohrid.
Bull. Soc. Hist. nat. Moselle, 33 : 73—87.
Remy P. 1934. Un Aselle nouveau du Sandjak de Novi Pazar (Jougoslavie) : Asellus
(Proasellus) Karamani n. sp. Arch. Zool. exp. gen., 76, Notes et Revue : 39—50.
Remy P. 1936. Un Aselle nouveau de Madero. «Broteria»,Ser. Cienc. Nat., V (XXXII).
fasc. 1 : 17—32.
48
ВВЕДЕНИЕ
Remy Р. 1938. Un Stenasellus en Afrique Occidentale Francaise — S. chappuisi
n. sp. de la cote d’Ivoire. Arch. zool. exp. gen., 79, Notes et Revue : 69.
R emy P. 1941. Asellotes de Jougoslavie et de Grece. Arch. zool. exp. gen., 82,
Notes et Revue, 1 : 1—25.
Remy P. 1949. Stenasellus Ruili n. sp. de la grotte de la Giraudasso a Soulatge, Aude.
Bull. mens. Soc. Linn. Lyon : 7.
Richardson H. 1905. Monograph on the Isopods of North America. Bull. U. S.
Nat. Mus., 54 : 1—727.
S a r s G. 0. 1867. Histoire naturelle des Crustaces d’eau douce de Norvege. I. Les Ma-
lacostraces. Christiania : 1—146.
S a r s G. 0. 1897. On Some Additional Crustacea from the Caspian Sea. Ежегодн.
Зоол. муз. Акад. Наук, II : 273—306.
S а г s G. О. 1899. Account of the Crustacea of Norway, 2, Isopoda. Bergen : 1—270.
Schne ide r R. 1887. Ein bleicher Asellus in den Gruben von Freiberg im Erzge-
birge (Asellus aquaticus var. Fribergensis). Sitz.-Ber. Kgl. Preuss. Akad. Wiss.
Berlin, 1887, 2 : 723—742.
S hen Chia-Jui. 1936. The fresh-water Isopods of Peiping. Bull. Fan Memorial Inst,
of Biology, VII (Zoology), 1 : 1—31.
S p a n d 1 H. 1926. Die Tierwelt der unterirdischen Gewasser. Spelaologische Mono-
graphien, XI : 1—235.
StammerH. J. 1932. Zur Kenntniss der Verbreitung und Systematik der Gattung
Asellus, inbesondere der mitteleuropaischen Arten (Isopoda). Zool. Anz., 99 :
113—131.
StammerH. J. 1936. Eine neue Hohlenwasserassel aus Spanien, Stenasellus buch-
neri und die Verbreitung der Gattung Stenasellus. Zool. Anz., 114 : 137—141.
Strouhal H. 1937. Siisswasser- und Landasseln Siiditaliens und des Monte Gargano-
Gebietes. Zool. Anz., 119 : 65—86.
Strouhal H. 1937. Asseln aus Balkanhohlen. Zool. Anz., 124 : 269—281.
Strouhal H. 1942. Vorlaufige Mitteilung fiber die von M. Beier in Nordwestgrie-
chenland gesammelten Asseln. Zool. Anz., 138 : 145—162.
Tattersail W. M. 1914. Amphipoda and Isopoda from the Lake of Tiberia. Journ.
Asiatic Soc. Bengal, 10 : 406—433.
Tatters a.11 W. M. 1920. The occurence of Asellus meridianus Rac. in Derbyshire.
Lancashire and Cheshire Naturalist, May : 273—275.
Tattersail W. M. 1921. Zoological Results of a Tour in Far East. VII. Mysidacea,
Tanaidacea and Isopoda. Mem. Asiatic Soc. Bengal, 6 : 406—433.
Tattersail W. M. 1930. Asellus cavaticus Schiodte, a blind Isopod new to the
British fauna from a well in Hampshire. Journ. Linn. Soc. London, Zool., 37 :
79-91. ' ' '
Ueno M. 1927. Notes on some subterranean isopods and amphipods of Japan. Mem.
College Sci., Kyoto Imp. Univ., Ser. B, 3 : 355—368.
Ueno M. 1933. Three noticeable freshwater Crustacea of Hokkaido. Annot. zool.
' r Japon., XIV : 115—122.
Ueno M. 1934. Subterranean Crustacea from Kwantung. Annot. zool. Japon., XIV :
445—450.
Ueno M. 1938. Asellus from the Ryukyu Islands. Trans. Natur. Hist. Soc. Formosa,
XXVIII, No 178 : 264—268.
Ueno M. 1940. Freshwater Isopoda of Manchoukuo. Rep. Limnobiol. Survey of Kwan-
tung and Manchoukuo : 309—310.
Unwin E. E. 1920. Notes upon the reproduction of Asellus aquaticus. Journ. Linn.
Soc. London, Zool., 34 : 335—343.
V a n d e 1 A. 1926. La reconnaissance sexuelle chez les Aselles. Bull. Soc. Zool.
France, 51 : 163—172.
Van Name W. G. 1936. The American Land and Fresh-Water Isopod Crustacea.
Bull. Americ. Mus. Nat. Hist., 71 : 1—535.
Ver h-o e f f K. W., 1943. Zur Morphologie, Okologie und Systematik von Sphaeroma,
Eurosphaera und Jaera. Zeitschr. Morph. Okologie der Tiere, 40, H. 2 : 276—290.
Wege W. 1911. Morphologische und experimentelle Studien an Asellus aquaticus.
Zool. Jahrb., allg. Zool. Physiol., 30 : 217—320.
-Zimmer C. 1927. Isopoda. Handbuch der Zoologie, III : 697—766.
СПЕЦИАЛЬНАЯ ЧАСТЬ
Подотряд ASELLOTA
Уроподы прикреплены к концу плеотельзона (терминально). Экзопо-
диты II или III пары плеоподов хитинизированы и образуют крышечки,
покрывающие нижележащие, служащие для дыхания плеоподы. I пара
плеоподов у 2 отсутствует. Перейоподы со свободными коксоподитами,
образующими эпимеры грудных сегментов.
Asellota могут считаться наиболее примитивным подотрядом Isopoda.
Они сохранили целый ряд архаических признаков, утерянных другими
подотрядами. Так, например, для Asellota характерны 3-членистое осно-
вание (протоподит) вторых антенн, присутствие у многих представите-
лей (Stenasellus, Microparasellidae, Janiridae,) рудимента экзоподита вто-
рых антенн, явно 3-членистый протоподит ногочелюстей, свободный кбк-
соподит перейоподов и т. д.
В пресных водах Палеарктики подотряд представлен 3 семействами —,
Asellidae, Microparasellidae п Janiridae, относящимися к 2 надсемействам,
на которые распадаются Asellota.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА НАДСЕМЕЙСТВ И СЕМЕЙСТВ
.1 (2). I (у <5) и II пара плеоподов свободные. Экзоподиты III пары плео-
подов образуют парную крышечку. (I. Надсемейство Aselloidea).
......................................... 1. Asellidae
2 (1). I и II пара плеоподов <5 соединены шипами друг с другом и обра-
зуют вместе единую парную крышечку, а у g II плеоподы совер-
шенно сливаются и образуют единую непарную крышечку.
(II. Надсемейство Par aselloidea).
3 (4). Тело вытянутое в длину, червеобразное, грудной отдел распа-
дается на два отдела — передний, состоящий из 4 сегментов, и зад-
ний, состоящий из 3 сегментов...................2. Microparasellidae
4 (3). Тело расширенно©, овальное; грудной отдел расчленен на сегменты
равномерно...........................................3. Janiridae
I. Надсемейство ASELLOIDEA
Протоподиты I пары плеоподов <5 свободные, вполне отделенные друг
от друга; экзоподиты подвижно сочленены с протоподитами и длиннее
этих последних. Протоподиты II пары плеоподов $ не соединены с I плео-
подом, не Длиннее или незначительно длиннее экзбподитов. Экзоподиты
двучленистые, приблизительно такой же длины, как эндоподиты. Плео-
поды II пары у g не срастаются между собой. Экзоподиты III пары плео-
•4 Фауна СССР, том VII, в. 5
50
I. ASELLIDAE
подов крупные, хитинизированные и служат крышечкой для эндоподитов..
а также для остальных плеоподов. IV и V пары плеоподов построены
так же, как III пара, но уступают ей по величине, и их экзоподиты почти
совершенно лишены хитина; у обоих полов их строение одинаково.
Надсемейство Aselloidea включает единственное семейство Asellidae..
1. Сем. ASELLIDAE
Тело удлиненное, равномерно расчлененное на 7 сегментов, сплющен--
ное в дорзовентральном направлении. I пара перейоподов — хвататель-
ная, остальные 6 пар — ходильные. Два первых абдоминальных сег-
мента свободные, укороченные и в той или иной мере редуцированные;.
I пара плеоподов 6 незначительной величины, с протоподитом и экзо-
подитом и обычно не срастается при основании. II пара плеоподов о
также незначительной величины, с двучленистым экзоподитом, не усту-
пающим по длине заметно эндоподиту. Плеоподы II пары g маленькие,,
не срастаются между собой. III пара плеоподов образует крышечку,;
покрывающую полностью IV и V пары плеоподов, обладающих эндо-
подитами и служащих для дыхания.
Пять относящихся к этому семейству родов близко родственны между
собой и составляют естественную группу чисто пресноводных голаркти-
ческих Asellota. Дудич (Dudich, 1924), Штаммер (Stammer, 1936) и Аркан-
джели (Arcang'eli, 1939) считают возможным выделение в качестве осо-
бого семейства рода Stenasellus на основании строения первых плеоподов.
и антенн и первых абдоминальных сегментов, однако это мнение не пред-
ставляется неоспоримым тем более, что в отношении последнего при-
знака роды Johanella и Synasellus занимают промежуточное положение
между Stenasellus и Asellus.
Коллиндж (Cbllinge, 1945) предложил разделение семейства Asellidae-
на 3 подсемейства, каждое из которых заключает по одному роду — Asel-
linae, Caecidotinae и Mancasellinae. Эти подсемейства различаются отно-
шениями длины и ширины головы и плеотельзона, а также строением
эпимеров грудных сегментов. У Asellinae ширина головы и плеотельзона
превосходит их длину, у Caecidotinae имеют место обратные соотношения,
у Mancasellinae плеотельзон «языковидный». У Asellinae и Caecidotinae
эпимеры отделены от сегментов, но у последнего подсемейства они выем-
чатые, у Mancasellinae эпимеры, сросшиеся с сегментами.
Система Коллинджа не может быть принята. Прежде всего она.
не включает 3 (т. е. 60%) рода семейства Asellidae — Stenasellus, Syn-
asellus и Johanella. Далее она преувеличивает различия между Asellus-
и Coecidotea, которые, как будет показано ниже, в действительности
настолько незначительны, что оба рода должны считаться синонимами.
Достаточно указать на существование нескольких видов Asellus, обла-
дающих удлиненными головой и плеотельзоном. При этом у большинства
видов этого рода эпимеры с выемками. Наоборот, несколько видов Сае-
cidotea имеют широкую голову. Наконец, выбранные Коллинджем внеш-
ние относительные признаки павряд ли отражают истинные родственные-
отношения между водяными осликами. Так, например, из двух сибирских
и, несомненно, родственных между собою видов подрода Asellus s. str. —
A. latifrons и A. epimeralis—у одного длина плеотельзона меньше-
ширины, у другого — наоборот.
Правильнее Считать семейство Asellidae единым и относить к нему.
5 самостоятельных родов — Asellus, Stenasellus, Johanella, Synasellus.
1. ASELLUS
51'
и Lirceus.1 Последний род представлен 16 видами из Северной Америки'
и подробно рассматриваться здесь не будет.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОБ
1 (6). Мандибула снабжена щупиком.
2 (3). Дна первых сегмента абдомена значительно короче и уже грудных
сегментов тела..............................1. Asellus G. St. Hilaire'
3 (2). Два первых сегмента абдомена мало уступают по длине и ширине
грудным сегментам тела.
4 (5). На третьем членике основания второй антенны имеется рудимен-
тарный экзоподит. Коксоподиты перейоподов свободные. Уроподы
не короче или незначительно короче плеотельзона.....................
..........................................3. Stenasellus Dollf.
5 (4). Экзоподит второй антенны отсутствует. Коксоподиты перейоподов
полностью срастаются с краями сегментов тела. Уроподы значи-
тельно короче плеотельзона.....................2. Johanella Monod.
6 (1). Мандибула лишена щупика.
7 (8). Два первых сегмента абдомена короче грудных сегментов,,
но хорошо видимы сверху. Дактилюсы II—VII перейоподов кон-
чаются одним когтем......................... 4. Synasellus Braga
8 (7). Два первых сегмента абдомена редуцированы и сверху не заметны..
Дактилюсы II—VIII перейоподов кончаются двумя когтями . . .
•...............................................(Lirceus Raf.)-
1. Род ASELLUS G. ST.-HILAIRE
G. St; - H i 1 a i г e, 1764, Hist, abregee Ins. etc., 2 : 672; R acovilz a,.
1924 : 83;—Caecidotea P a c k a r d, 1871 : 752; Van N a m e, 1936 : 465. .
. Тип -рода: Asellus aquaticus (Linne) sensu Racovitza.
Длина clypeus превышает ширину. Вторая антенна лишена рудимента-
экзоподита. Внутренняя лопасть второй максйллы вдвое (или около того)-
шире наружных лопастей. Ногочелюсти с эпиподитом, преобразованным
у яйценосных в оостегит. Перейоподы у $ и q:b большинстве случаев
различного строения, в особенности проподит Кперейопода. I и II сег-
менты плеона редуцированы, значительно более узкие, чем VII сегмент
нерейона, и в значительной своей части покрыты его задним краем..
II плеопод <5 с двучленистым экзоподитом и нерасчлененным эндоподитом,
служащим совокупительным органом. II плеоподы j овальные, округлые-
или удлиненно трапецеидальной форды, всегда довольно крупных раз-
меров. Плеоподы IV и V совершенно или частично лицгены хитина и функ-
ционируют как жабры.
Род Asellus четко отделен этими признаками от прочих родов семейства
Asellidae. Что касается рода Caecidotea Packard, то нет достаточных осно-
ваний признавать его самостоятельность.. Это мнение высказывалось
Форбсом (1871), Гейем (1876), Шаппюи (1927), Миллером (1933) и Бир-
штейном (1939). Однако, большинство американских авторов держится
1 В последнее время Габричт и Мэкин показали, что род Mancasellus Harg. пра-
вильнее именовать Lirceus Raf., и описали еще 8 новых видов и 1 новый подвид этого
рода из США (hargeri, alabamae, richardsonae, megapodus, hoppinae ozarensis, biden-
tatus, bicuspidatus, garmani и trilobus) [Hubricht L. and J. C. Mackin. 1949.
The fresh water isopods of the genus Lirceus (Asellota, Asellidae). Amer. Midland Nat.,
42, No 2 : 334—349].
.4*
52
I. ASELLIDAE
противоположного взгляда, правда, не приводя в его пользу никаких
аргументов (Richardson, 1905; Van Name, 1936). Только в недавнее
время Мэкин и Габричт 1 попытались обосновать самостоятельность
Caecidotea. Они считают, что виды этого рода) связаны между собою
более тесными родственными отношениями, чем с видами рода Asel-
lus, но никак не доказывают этого положения. Вместе с тем они допу-
скают возможность открытия в будущем более тесных родственных «меж-
родовых» отношений и установления полифилетической природы Cae-
cidotea с последующей ликвидацией этого рода. Следует при этом отметить,
что авторы в той же работе описывают один вид {adentd), стоящий,
по их мнению, ближе всего к Asellus cavaticus. Этот вид в заголовке статьи
и в подписях под рисунками именуется Caecidotea, а в тексте — то Cae-
cidotea, то Asellus. Очевидно, авторы сами не знают, к какому роду его
следует относить, что наглядно показывает нереальность границ между
Asellus и Caecidotea.
На крайней позиции стоит, как уже указывалось, Коллиндж (1945).
Не считаясь с такими общепринятыми признаками, как строение плео-
подов, ротовых частей и проподитов I перейопода, позволяющими объе-
динять водяных осликов в естественные группы, он основывается только
ла пропорциях головы и плеотельзона и на форме эпимеров. Не дав себе
труда проверить, насколько применимы выбранные им признаки к систе-
матизации всех Asellidae, он на основании просмотра весьма ограничен-
ного материала не только решительно разделяет Asellus и Caecidotea,
но и возводит их в ранг подсемейств. Посмотрим, насколько это спра-
ведливо.
Отличия Caecidotea от Asellus, согласно Пакарду (Packard, 1888),
сводятся к следующему: удлинение тела; длина плеотельзона, превос-
ходящая ширину; отсутствие глаз; длина второй антенны, равная длине
тела; незначительная ширина лопастей первой максиллы, вдвое меньшая,
чем у Asellus', ширина конечных члеников щупика ногочелюсти при
такой же; как у Asellus, ширине средних члеников; длина пятого чле-
ника основания, второй антенны, вдвое превосходящая длину четвертого
членика. Как известно, слепота и удлинение тела свойственны вообще
подземным и глубоководным членистоногим. Ряд слепых европейских
Asellus приведен ниже. С другой стороны описано два вида Caecidotea,
обладающие глазами (С. kawamurai Tatt. и С. oculata Mack, et Hubr.).
Длина плеотельзона превосходит ширину у многих Asellus (Л. cavaticus
Schiodte, A. giorgievici Kar., A. hypogeus Rac., A. spelaeus anophthalmus
Kar.). Вторая антенна не короче тела у A. aquaticus (L.), A. hilgendorfi
Bov. и превосходит длину тела у A. korotnewi Semenk., A. minutus Semenk.
и A. dybowskii Semenk. Пятый членик основания второй антенны у неко-
торых Caecidotea менее чем вдвое длиннее четвертого членика (С. ala-
bamensis Stafford, С. richardsonae Hay, C. tridentata Hungerford, C. aki-
yoshiensis Ueno). Суженные конечные членики щупика ногочелюстей
свойственны, например, A. remyi Monod и некоторым другим Asellus
и, наоборот, не наблюдаются у С. richardsonae Нау и С. smithii Ulrich,
лопасти максиллы узкие и у многих Asellus (подрод Proasellus Dud.).
Таким образом, нет никаких специфических для рода Caecidotea призна-
ков, и его следует объединить с родом Asellus.
1 J. G. Mackin and L. ilubtich t. 1940. Description of seven new spe-
cies of. Caecidotea (Isopoda, Asellidae) from Central United States. Trans. Amer. Mic-
roscop. Soc., LIX. 3 : 383—397.
1. ASELLUS
53
Если виды, входившие в род Caecidotea, разбить на группы по при-
знакам, принятым при разделении Asellus на подроды (вооружение первой
максиллы, форма проподита I перейопода <3 и двух первых пар илео-
подов), то получится следующая картина: С. stygia Pack., С. alabamensis
Staff., С. tridentcta Hungerford, C. antricola Greaser. C. dimorpha, stilo-
dactyla, packardi, spalulata, acuticarpa и oculata Mack, et Hubr. могут
быть включены в подрод Conasellus Stammer. С. kawamurai Tatt. следует
объединить с A. californicus Miller и A. dybowskii Semenk. в подрод Meso-
asellus, к которому примыкает, возможно, п С. akiyoshiensis Ueno, а систе-
матическое положение С. nickajackensis Pack., С. richardsonae Hay
и С. smithii Ulrich остается пока неясным из-за недостаточной изучен-
ности их морфологии, точно так же как и систематическое положение
еще трех американских азеллид —A. attenuatus Richardson, A. adenta
Hubr. et Mack, и A. lomalensis Harford. Весьма вероятно, что все эти
5 видов составляют особый единый подрод. Возможно также, что неко-
торые из них относятся к Mesoasellus. В нижеследующий обзор они
не включены.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ И ВИДОВ
1 (10). Внутренняя лопасть первой максиллы с 4 щетинками. (1. Подрод.
Asellus s. str.).
2 (3). На наружном крае экзоподита I плеопода <3 имеется выемка.
• • ................................... *1. A. aquaticus (L.)
3 (2). Экзоподит I плеопода $ лишен выемки на наружном крае.
4 (9). Уроподы короче плеотельзона. В жгуте I антенны не больше
7 члеников. II плеопод $ вытянут продольно и его длина больше
ширины.
5 (6). Эндоподит II плеопода <3 заметно короче протоподита. Ширина
плеотельзона равна ширине задних грудных сегментов................
..................................... *3. A. monticola Birsk
6 (5). Эндоподит II плеопода <5 превосходит по длине протоподит.
Ширина плеотельзона меньше ширины задних грудных сегментов.
7 (8). Ширина плеотельзона больше длины. Эпимеры сверху незаметны.
Голова с расширенными боковыми краями и рос!румом.................
...................................... *4. A. latifrons Birst.
8 (7). Длина плеотельзона больше ширины. Эпимеры сильно развиты
и хорошо видны сверху. Голова лишена рострума и её края не рас-
ширены ....................................*5. A. epimeralis Birst.
9 (4). Уроподы длиннее плеотельзона или равны ему. В жгуте I антенны
не менее 10 члеников. II плеопод д округлый, его длина и ширина
равны менаду собой ....... ................*2. A. hilgendorfi Bov.
10 (1). Внутренняя лопасть первой максиллы с 5 щетинками.
11 (18). Эндоподит II плеопода <5 в базальной части имеет направлен-
ный внутрь вырост (как у Asellus s. str.)..Длина проподита I перейо-
пода больше чем вдвое превышает ширину. (2. Подрод Mesoasellus
Birst.).
12 (13). Вторая антенна значительно длиннее тела. Ширина плеотельзона
не уступает его длине...................... *6. A. dybowskii Sem.
13 (12). Вторая антенна короче тела. Длина плеотельзона превышает
его ширину.'
14 (15). Глаза имеются. Передний крап головы почти прямой..........
. ....................................7. A. kawamurai (Tatt.)
-’54
I. ASELLIDAE
15 (14). Глаза отсутствуют. Передний край головы с треугольной выем-
кой.
16 (17). Внешний край II плеопода g вогнутый и снабжен щетинками.
Базальный членик экзоподита III плеопода короче дистального . .
......................................... 8. A. akiyoshiensis (Ueno).
47 (16). Внешний край II плеопода g прямой или выпуклый и лишен
щетинок. Базальный членик III плеопода длиннее дистального . .
......................................... (A. califomicus Miller ).
-18 (11). Эндоподит II плеопода <5 лишен направленного внутрь выроста.
Длина проподита I перейопода <5 превышает ширину не более чем
в 2 раза.
19 (72). Задний край проподита I перейопода <5 прямой, лишенный выро-
стов.
.20 (27). Конец эндоподита II плеопода <5 вытянут в клювовидное хити-
низированное образование. Ширина II плеопода g в дистальной
части больше, чем в базальной. (3. Подрод Baicaloasellus Stamm.).
21 (22). Уроподы, не менее чем в 3 раза, короче плеотельзона. Ширина
обоих члеников экзоподита II плеопода 3 превосходит их длину
...........................................*12. A. angarensis Dyb.
22 (21). Уроподы не короче плеотельзона. Ширина дистального членика
экзоподита II плеопода 3 меньше длины.
23 (24). Вторая антенна короче тела. Внешний край экзоподита IV плео-
пода без выемки и лишен длинных перистых щетинок. Дистальный
членик экзоподита II плеопода 8 незначительно короче базального
......................................... *9. A. baicalensis Grube
-24 (23). Вторая антенна длиннее тела. Внешний край экзоподита IV
плеопода с выемкой, позади которой сидят 4—5 длинных перистых
щетинок. Дистальный членик экзоподита II плеопода <5 по край-
ней мере в 1х/г раза длиннее базального.
-25 (26). Протоподит* II плеопода 8 менее чем в Р/граза длиннее экзо-
подита, который несет не более 10 перистых щетинок. II плеопод
2 при основании незначительно уже, чем в дистальной трети. Жгут
второй антенны более чем в 2 раза длиннее основания..................
......................................*10. A. korotnewi Sem.
26 (25). Протоподит II плеопода 8 более, чем в 1 1/г раза длиннее экзо-
подита, который несет 13—15 перистых щетинок. II плеопод g при
основании почти в 2 раза уже, чем в дистальной трети. Жгут вто-
рой антенны менее чем в 2 раза длиннее основания.....................
..........................................*11. A. minutus Sem.
27 (20). Конец эндоподита II плеопода 8 не хитинизирован и имеет
всегда окончание и боковую лопасть. II плеопод g в дистальной
части уже, чем в базальной. (4. Подрод Proasellus Dud.) ,
28 (55). Глаза есть.
29 (30). На поверхности экзоподита I плеопода 3 и II плеопода g сидят
многочисленные щетинки................................
............................ 17а. A. gjorgjevici litoralis Kar.
30 (29). На поверхности экзоподита I плеопода 3 и II плеопода g не более
5—6 щетинок.
31 (54). Правый и левый протоподиты I плеопода 3 разделены до осно-
вания. На конце эндоподита II плеопода о имеется боковая лопасть,
а окончание конической или цилиндрической формы.
32 (33). Окончание эндоподита II плеопода 3 цилиндрической формы
и значительно длиннее боковой лопасти . ‘ . . 13. A. meridianus Rac.
1. ASELLUS
55
•33 (32). Окончание эндоподита II плеопода 5 не длиннее боковой лопасти
и имеет обычно форму усеченного конуса.
34 (35). Экзоподит I плеопода <5 и II плеопод j несут всего по 4 перистых
щетинки. Дактилюсы II—VII перейоподов с 2 зубцами..................
................................... 24. A. gauthieri Monod
35 (34). Число перистых щетинок на экзоподите I плеопода 8 и на II
плеоподе <р больше 4 или все щетинки неоперенные. Число зубцов
на дактилюсах перейоподов больше двух.
- 36 (39). Глаза расположены на верхней плоскости головы. Боковые
лопасти головы и края сегментов тела расширены. Протоподит
II плеопода 8 несет с внутренней стороны простые щетинки. Про-
подит I перейопода <5 с 2 ножевидными зубцами.
- 37 (38). Вторая антенна короче половины тела и имеет в жгуте менее
30 члеников. На поверхности экзоподита I плеопода 8 и II плео-
пода $ в базальной части, вблизи внутреннего края, имеется по 1 —
2 щетинки. Протоподит I плеопода ос I совокупительным зубцом
...............................................15. A. remyi Monod
- 38 (37). Вторая антенна длиннее половины тела и имеет в жгуте более
30 члеников. Щетинки на поверхности экзопоДита I плеопода 8
и II плеопода j отсутствуют. Протоподит I плеопода <5 с 2 совокупи-
тельными зубцами............................*16. A. infirmus Birst.
• 39 (36). Глаза расположены по бокам головы. Боковые края головы
и тела почти не расширены.’ Проподит I. перейопода 8 с 3—4 ноже-
видными зубцами. Протоподит II плеопода 8 с перистыми щетин-
ками на внутреннем крае.
40 (45). Щетинки наружного и дистального краев экзоподита I плеопода
8 приблизительно одинаковой длины, не перистые. Наружный край
эндоподита II плеопода 8 вогнутый, наружный и внутренний края
дистального членика параллельны, и он имеет прямоугольную
форму.
41 (42). Экзоподит I плеопода о менее чем в 1 Щ раза длиннее прото-
подита, снабженного 1 совокупительным зубцом. II плеопод j тра-
пецеидальный с вогнутыми внутренним и наружным краями. Уро-
t поды 8 длиннее плеотельзона, уроподы j равны по длине плеотель-
зону................................... 27. A. arnautovici Remy.
42 (41). Экзоподит I плеопода 8 более чем в 1 Щ раза длиннее прото-
. подита, снабженного 2—3 совокупительными зубцами.' II плеопод
$ округло-треугольный с прямым внутренним и выпуклым наруж-
ным краем. Уроподы обоих полов короче плеотельзона.
43 (44). Дистальный членик экзоподита II плеопода 8 несколько менее
чем в 2 Чг раза длиннее проксимального и несет на дистальном
крае 6—7 перистых щетинок .... 28. A. monodi Strouhal
44 (43). Дистальный членик экзоподита II плеопода <5 более чем
в 2 Чг раза длиннее проксимального и несет на дистальном крае
8—10 перистых щетинок ...... 26. A. istrianus Stammer
45 (40). На дистальном крае экзоподита I плеопода 8 имеются перистые
. щетинки, значительно превосходящие по длине простые щетинки
наружного края. Наружный край эндоподита II плеопода' 8 выпу-
клый, наружный и внутренний края дистального членика экзо-
подита выпуклые, и он имеет овальную или яйцевидную форму.
•46 (47). Внутренний базальный угол экзоподита I плеопода 8 оттянут
назад и покрывает часть протоподита. Дистальный край прото-
подита несет 3—8 толстых щетинок. II плеопод £ трапецОидаль-
56
I. ASELLIDAE
ной формы и лишен стернальных щетинок..........................
...................................... 14. A. polychaetus Dud.
47 (46). Внутренний базальный угол экзоподита I плеопода 3 округлый
или прямоугольно-округлый и не налегает на протоподит. II плео-
под J треугольной, округло-трапецоидальной формы или трапе-
цеидальной формы, ио в последнем случае несет стернальные
щетинки.
48 (49). Ширина экзоподита I плеопода 8 менее чем вдвое уступает длине.
Длина протоподита II плеопода 8 меньше /Ширины. II плеопод
2 трапецеидальной формы и песет 1—2 стернальные щетинки
..............................................31. A. karamani Remy
49 (48). Длина экзоподита I плеопода 8 более чем вдвое превышает
ширину. Длина протоподита II плеопода 8 больше ширины. II
плеопод округло-треугольной или округло-трапецоидальной
формы.
50 (51). Вдоль внутреннего края экзоподита I плеопода 8 расположен
сплошной или прерванный субмаргинальный ряд стернальных
щетинок.......................................29. A. italicus Dub.
51 (50). Стернальные щетинки на экзоподите I плеопода 3 не образуют
правильного субмаргинального ряда.
52 (53). Внутренний край протоподита II плеопода 3 имеет не более-
4 щетинок. II плеопод £ округло-треугольной формы....................
.............................. 29. A. banyulensis Rae,.
53 (52). На внутреннем крае протоподита II плеопода 3 более 4 щети-
нок. II плеопод 2 округло-трапецоидальной формы......................
, . . ............................. 32. A. coxalis Dollfus
54 (31). Правый и левый протоподиты I плеопода 3 срастаются при осно-
вании на Vi своей длины. На эндоподите II плеопода 3 нет боко-
вой лопасти, а окончание очень широкое и короткое, с разверну7
тыми краями.................................. 34. A. peltatos Braga
55 (28). Глаза отсутствуют.
56 (67). Эндоподит уроподов длиннее экзоподита, или обе ветви при-
близительно одинаковой длины.
57 (58). Уроподы короче половины плеотельзона. На экзоподите I плео-
пода 8 и II плеопода 2 имеются многочисленные стернальные
щетинки. Края экзоподитов I и П плеоподов 8 и II плеопода 2
с многочисленными (20 и более) длинными перистыми щетинками.
. ........................... 17. A. gjorgjevici gjorgjevici Кат.
58 (57). Уроподы длиннее половины плеотельзона, равны или превос-
ходят его по длине. На экзоподите I плеопода 8 и на II плеоподе-
2 не более 2 стернальных щетинок. Края экзоподитов I и II плео-
подов 8 и II плеопода д с редкими короткими щетинками, из кото-
рых часть может быть перистыми.
.59 (64). Экзоподит I плеопода 8 несет на дистальном крае не перистые
щетинки. Внутренний базальный угол эндоподита II плеопода
8 образует отросток более длинный, чем направленное в про-
. тивоположную сторону утолщение наружного базального угла
этого членика. Эндоподит II плеопода 8 обычно длиннее экзопо-
дита.
60 (63). Уроподы не более, чем на J4 короче или длиннее плеотельзона.
Окончание эндоподита I плеопода 8 не вытянуто в трубковидное-
образование, боковая лопасть длиннее окончания, но гораздо уже-
самого эндоподита.
1. ASELLUS
57
61 (62). Длина плеотельзона превышает ширину. Уроподы длиннее
плеотельзона (или менее чем на */4 короче)........................
................................... 18. A. cavaticus Schiodte
62 (61). Длина плеотельзона меньше ширины.1 Уроподы на ]/4 или
более короче плеотельзона.....................20. A. spelaeus Rac.
63 (60). Уроподы более чем на V4 короче плеотельзона. Окончание
эндоподита вытянуто в трубковидное образование и длиннее боко-
вой лопасти. Последняя по ширине почти не уступает эндоподиту.
• • ....................................33. A. seabrai Braga
64 (59). Экзоподит I плеопода i несет на дистальном крае перистые
щетинки более длинные, чем на наружном крае. Внутренний
базальный угол эндоподита II плеопода S не образует отростка,
или этот отросток короче наружного утолщения базальной части
эндоподита. Экзоподит II плеопода <5 длиннее эндоподита.
65 (66). Протоподит I плеопода <5 с 1—3 стернальными щетинками
и I щетинкой на наружном дистальном углу, экзоподит с 28—36-
щетинками. На дистальном членике экзоподита II плеопода <5 8—
11 перистых щетинок.......................... 22. A. hussoni Remy
66 (65). Протоподит I плеопода О лишен стернальных щетинок и
щетинки на наружном дистальном углу. На дистальном членике
экзоподита II плеопода <5 7 перистых щетинок.................... ...
• ...................................... 19. A. hypogeus Rac.
67 (56). Эндоподит уроподов короче экзоподита.
68 (69). Эндоподит уроподов стилетовидный и по крайней мере в 3 раза
уже экзоподита........................ 21. A. collingei, nom. nov.
69 (68). Эндоподит уроподов не уже или незначительно уже экзоподита,
нормального строения.
70 (71). Проподит I перейопода $> с 3 ножевидными зубцами, <5 — с 9 ноже-
видными зубцами. Урон оды приблизительно вдвое короче плео-
тельзона ................................ . 23. A. hermallensis Arc.
71 (70). Проподит I перейопода j с 2 ножевидными зубцами. Уроподы
незначительно короче плеотельзона..............................._.
........................... 25. A. thennonyctophilus Monod
72 (19). Задний край проподита I перейопода S снабжен 2—3 несущими
ножевидные зубцы мощными выростами, соединяющимися осно-
ваниями (5. Подрод Conasellus Stammer).
1. Подрод Asellus s. str.
D u d i c h, 1925a : 299; Stammer, 1932 : 129; Б и p ш т е„й н, 1936 : 235-
Тип подрода: Asellus aquaticus (Linne) sensu Racovitza.
Внутренняя лопасть I максиллы с 4 щетинками. Проподит I перейо-
пода о с одним выступом на задней части. На базальной части эндопо-
дита II плеопода $ имеется направленный внутрь вырост. II плеоподы
2 округлые, слабо вытянутые вдоль, заходящие друг за друга своими
внутренними краями.
*1. Asellus (s. str.) aquaticus (L.) Rac. (рис. 18—26).
Linne, 1758, Syst; Nat., ed. X : 637 (Oniscus)', Geoffrey St.-
Hilaire, 1764, Hist. abr. Ins. : 672; G. 0. Sa r s, 1867 : 93, t. 3, f. 6—28, t. 9—
10; G. O. S a r s, 1899 : 97, t. 39; Tschetwerikof f, 1911 : 377, tt. 7—8;..
1 Исключение — A. spelaeus anophthalmus Kar.
58
I. ASELLIDAE
Racovitza, 1919 : 37, fig. 1—6; Stamm er, 1932 : 114, fig. 1—2; — vul-
garis Latreille, 1803, Hist. nat. Crust. Ins., VI : 348; — groenlandicus
Kroyer, 1838, Kong. Danske Vid. Afh., VII : 318; H a n s e n, 1888, Vid. Medd.
Nat. Foren. : 190; — goplanus Kulczycki, 1885 : 41'1, f. 22—26; — goplanus
stvitesiaha Kulczyck i, 1885 : 413, f. 27—28, 37; — aquaticus cracoviensis К u 1-
c z у с к i, 1885 : 410, f. 13—16, 18, 21; — arthrobranchialis D u d i c h, 1925 : 281,
fig. 1—6; — aquaticus gordoni, mackini et hazeltoni Col’linge, 1945 : 817.
Окраска буроватая, с узором из более светлых пятен. Наибольшая
ширина тела приходится на III и IV сегменты, которые шире плеотель-
Рис. 18. Asellus (s. str.)
• aquaticus. Самец co спин-
ной стороны. Ориг.
зона. Длина последнего больше ширины. Первая
антенна не достигает конца основания второй
антенны. Жгут первой антенны состоит из 9—
15 члеников. Вторая антенна по длине равна телу
(<5) или немного короче тела (д). Число члеников
ее жгута сильно варьирует между 40 и 75. На
выступе проподита I перейопода о сидит 3—4 но-
жевидных зубца, дактилюе с 18—20 зубцами.
Выступ проподита I перейопода j развит гораздо
слабее, чем у 6, и несет 2—3 ножевидных зубца.
Дактилюе IV перейопода S с 5 шипами. Прото-
подит I плеопода £ приблизительно квадратной
формы, ширина и длина равны. Внутренний край
протоподита с 4—5 совокупительными зубцами.
На внешнем крае протоподита может присутство-
вать 3—4 щетинки. Длина экзоподита I плеопода
S в 1 Чг раза больше ширины. На середине внеш-
него края имеется более или менее глубокая, но
всегда ясно заметная выемка. Весь экзоподит
в дистальной части шире, чем в базальной. Вдоль
базальной части внутреннего края проходит ряд
из 4—7 щетинок. На дистальном крае и дисталь-
ной части внешнего края сидит 9—20 длинных
перистых щетинок и несколько коротких простых.
На краях выемки —несколько простых щетинок.
Протоподит II плеопода S приблизительно прямо-
угольной формы — с закругленным внешним
базальным углом. На дистальной части внутрен-
него края находится 4—5 щетинок, на поверхности
протоподита, вблизи наружного края, — 4—5
щетинок и на дистальном крае, при основании
экзоподита, — 1—3 щетинки. Длина протоподита
несколько превышает ширину. Экзоподит в 2 раза
короче протоподита. Его базальный членик коро-
че и уже дистального, с 3—4 щетинками на внеш-
нем крае. Дистальныхг членик овальный, его ши-
рина превышает длину; на нем находится две
группы щетинок: на наружном крае 9—15 про-
стых щетинок, а на внутреннем 4—6 более
длинных церистых щетинок. Между этими группами сидит отдельно
1—2 простых или перистых щетинки. Эндоподит длиннее экзоподита,
суживается к концу и округло расширяется при основании. Базальная
часть имеет длинный, направленный внутрь, отросток. II плеопод j
округлый, не вытянутый в продольном направлении, по краям сидит
15—22 перистых щетинки, на поверхности может присутствовать еще
1. ASELLUS
59
до 6 коротких простых щетинок. Экзоподит III плеопода двучленистый,
причем базальный членик короче дистального. На внешнем крае базаль-
ного членика 10—15 (5) или 8—10 (j), на внешнем и дистальном краях
дистального членика 25—35 щетинок, из которых сидящие на дистальном
Рис. 19—26 Asellus (s. str.) aquaticus (L.): 19 — плеопод самца; 20 —
проподит I перейопода самца; 21 — то же самки; 22 — I плеопод самца;
23— II плеопод самки; 24 — III плеопод; 25 и 26 — IV и V плеоподы.
Ориг.
;крае и части внешнего края — перистые. Кроме того, на поверхности экзо-
подита находится большое количество коротких простых щетинок. Экзо-
подиты IV и V плеоподов разделены неясными линиями, кончающимися
у краев небольшими выемками, на три части. На внешнем крае базальной
части экзоподита IV плеопода находится 4—9 щетинок. На внешнем
60
I. ASELLIDAE
л
крае экзоподита V плеопода — 3—6 щетинок. Уроподы S длиннее плео-
тельзона, уроподы J равны по длине плеотельзону или незначительно
короче его. Протоподит в 2 раза короче эндоподита, экзоподит несколько
короче эндоподита. Длина тела S до 12 мм, j — до 9 мм.
Признаки этого вида весьма постоянны и не подвержены географи-
ческой изменчивости. Описания представителей A. aquaticus как само-
стоятельных видов и форм базировались на недоучете возрастных изме-
нений (Kulczycki, 1885). Расчлененность экзоподитов IV и V плеоподов,
послужившая Дудичу (1925) основанием для выделения особого вида,
наблюдается у всех представителей подрода Asellus s. str., а для A. aqua-
ticus была довольно точно изображена еще Сарсом (1899), Коллиндж
(1945) различает три цветовых варьетета этого вида: gordoni, с головой
и грудью, густо покрытыми волнистыми коричневатыми полосками,
и одноцветным коричневатым плеотельзоном, mackini, с телом желтова-
того цвета, и hazeltoni, с лирообразным темным лентовидным пятном
на голове, протягивающимся от одного глаза к другому. Следует отме-
тить, что расцветка A. aquaticus варьирует в широких пределах и навряд
ли отдельные отклонения от типичной для вида окраски нуждаются
в специальных наименованиях. В материале из СССР описанные Кол-
линджем варьететы обнаружить не удалось.
Распространение. Вся Европейская часть СССР, Кавказ,
Мангистау; Западная Европа — Венгрия, Югославия, Германия, Фран-
ция, Италия, Англия, Норвегия, Швеция, Дания; Западная Гренландия
(?); Иран. Восточная граница распространения не установлена, но во вся-
ком случае в Сибири этот вид отсутствует. В Америке отсутствует; ука-
зание Пакарда (1897) на нахождение на Лабрадоре относится, как пока-
зал Иогансен (1926), к наземной мокрице. Asellus aquaticus из Онтарио
есть на самом деле A. communis Say.1
Несколько форм отличаются от типичной достаточно четко и могут
считаться самостоятельными.
*la. Asellus (s. str.) aquaticus messerianus Birst, (рис. 27—29).
Бирштейн, 1945 : 158, рис. 4.
Первая антенна незначительно короче стержня второй. В ее жгуте —
8 члеников. Вторая антенна несколько длиннее тела с 45-—55-членистым
жгутом. Проподит первого перейопода <5 с хорошо выраженным выступом,
на котором сидит 3—4 ножевидных зубца, гребенка дактилоподита
из 8 зубцов. Дактилоподит IV перейопода S с 4 зубцами, проподит с
6 шипами в заднем (каудальном) и 6 щетинками в переднем ряду, карпо-
подит с 6 шипами в заднем и 2 шипами в переднем ряду. Наружный край
протоподита I плеопода 8 лишен щетинок, внутренний край с 4 сово-
купительными зубцами. Наружный край эйзоподита со слабо выражен-
ной выемкой и сплошным, не прерванным, рядом щетинок. На наружном
и дистальном краях 12—14 перистых щетинок, вдоль внутреннего края —
4—5 стернальных щетинок. Длина и ширина протоподита II плеопода z
S равны; длина экзоподита менее чем в Р/а раза уступает длине про-
топодита. Базальный членик экзоподита с 1 щетинкой на наружном крае.
Близ . наружного базального угла протоподита 2—3 стернальных
1 F. J о h а п s е п, 1920. The largerfreshwater Crustacea from Canada and Alaska,.
Part I and II. Amphipoda and Isopoda. Canadian Field-Nat., XXXIV (цпт. no van.
Name, 1936).
1. ASELLUS
61
28
Рис. 27—29 Asellus (s. str.)
'aquaticus messerianus Birst.:
27— I плеопод самца; 28 — II
плеопод самца: 29 — IV пе-
рейопод самца. Ориг.
имеется смесь нормальных
щетинки. Экзоподиты IV и V плеоподов, как у типичной формы, но всего
с 4 щетинками на наружных краях. Уроподы длиннее плеотельзона;
протоподит вдвое короче экзоподита, последний короче эндоподита.
Длина, тела — до 9 мм.
Подвид хорошо отличается от типичной формы прежде всего, неглу-
бокой выемкой наружного края экзоподита I плеопода 8 и сплошным'
рядом щетинок на этом крае, далее отношениями длины экзоподита и про-
топодита II плеопода 8, равными длиной и шириной протоподита, более
слабым вооружением IV перейопода и мень-
шим числом члеников жгута первой антенны.
Распространение. Западная
Туркмения — озера Делили (бассейн Атрека),
родник Сакка (Большие Балханы), озера
Ясха и Каратогелек в древнем русле Запад-
ного Узбоя.
*lb. Asellus (s. str.) aquaticus forma
cavermcola Rac.
R acovitza, 1925 : 573, i. 193; S lam-
in e r, 1932 : 118; Kosswig C. and Ko sswig
L,, 1936, Zool. Anz., Suppl. 9 : 274.
Эта подземная форма отличается от типич-
ной только редукцией пигмента и глаз, при-
чем удается обычно проследить все переходы
от нормально окрашенных особей с нормаль-
но развитыми глазами до совершенно ли-
шенных пигмента и глаз особей. Слепые осо-
би всегда лишены пигмента, но экземпляры,
лишенные пигмента, нередко обладают нор-
мально развитыми глазами. По мере углубле-
ния в пещеру, как показал Коссвиг, процент
слепых и депигментированных экземпляров
не возрастает, а как в пещере, так и вне ее,
типичных экземпляров A. aquaticus и экземпляров A. aquaticus cavernicola.
Распространение. Колодцы южного берега Крыма, пещеры
у Адельсберга (Италия) и близ Триеста. Целый ряд указаний A. cavati-
cus Schiodte из пещер Западной Европы, несомненно, относится к этой
форме.
1с. Asellus (s. str.) aquaticus var. fribergensis Schneider.
Schneider, 1887 : 723, t. XII,, fig. 1—10.
Помимо отсутствия пигмента отличается от типичной формы некото-
рыми мелкими структурными особенностями: выступ проподита I плео-
пода 8 развит гораздо слабее и весь проподит относительно уже, на дак-
тилюсе I перейопода 8 всего 7—9 более длинных, чем у типичной формы,
зубцов; по краям II плеопода j около 15 щетинок, и весь плеопод слабо
вытянут продольно. Глаза имеются, но фасетки явно отделены друг
от друга, так как пигмент глаз недоразвит. Длина тела не превышает
<8 мм. .
Распространение. Германия —пещера близ г. Фрейбурга;
62
I. ASELLIDAE
Id. Asellus (s. str.) aquaticus var. abyssalis Odenwall.
О d e n w a 1 1, 1927 : 16, fig. 1.
Окраска светлее, глаза меньше, чем у типичной формы. Покровы
настолько тонкие, что тело прозрачно. На краях II плеопода j всего»,
около 10 щетинок. Длина тела — 5 мм.
Распространение. Финляндия — озеро Лаппаярви, на глу-
бине 30—40 м.
*2. Asellus (s. str.) hilgendorfi Bovallius (рис. 30—35).
Bovallius, 1886 : 13; Hilgendorf, 1893 : 1; — aquaticus Tatter-
sail, 1921 : 415, t. 16; Д e p ж а в и н, 1930 : 3; Shen, 1936 : 21, t. 4; — nip-
ponensis Nicholls, 1929 : 34, fig. 1—5; U e n o, 1940 : 309, fig. 1—2.
Форма тела и окраска, как у A. aquaticus (L.). В жгуте цервой
антенны 9—14 члеников, первая антенна короче основания второй
антенны. Вторая антенна не достигает конца тела, в ее жгуте 50—65
члеников. На выступе проподита I перейопода $ 3—4 ножевидных зубца
и 1 щетинка, j — 2 ножевидных зубца. Число зубцов на дактилюсе зна-
чительно варьирует от 7 до 20. Длина протоподита I плеопода S равна
его ширине; на внутреннем его крае 2—5 совокупительных зубцов,
на поверхности протоподита, вблизи внешнего дистального угла, —
0—3 щетинки. Экзоподит в 2 раза длиннее протоподита, его внешний
край без выемки; дистальный и внешний края экзоподита несут до 20
перистых щетинок и несколько коротких простых, сидящих на поверх-
ности экзоподита. Кроме того, вблизи внутреннего базального угла, также,
на поверхности экзоподита, сидит группа из 2—5 щетинок, а иногда
имеется ряд щетинок вдоль внутреннего края. Длина протоподита
II плеопода о превышает ширину. Поверхность протоподита, в отличие
от других видов подрода, лишена щетинок; только вдоль внутреннего-
края имеется 2—4 щетинки. Длина экзоподита менее чем в 2 раза усту-
пает длине протоподита, на внешнем крае базального членика экзопо-
дита сидит 2—5 щетинок, внешняя группа щетинок дистального членика
состоит из 8—15 простых щетинок, внутренняя группа — изЗ—8 перистых
щетинок. Эндоподит незначительно короче протоподита, его строение —•
как у предыдущего вида. II плеопод j округлый, не вытянутый в про-
дольном направлении, с 18—25 перистыми щетинками по краям;
на поверхности плеопода 2—5 очень коротких щетинок. Экзоподит
III плеопода, как у предыдущего вида, но число щетинок по его краям
больше и вдоль шва, разделяющего экзоподит на два членика, имеется
ряд из 2—8 коротких щетинок. Остальные стернальные щетинки сидят
реже, чем у предыдущего вида, преимущественно вдоль внешнего края.
Расчленение экзоподитов IV и V плеоподов, как у других видов подрода,
но число щетинок в базальной части внешнего края больше: для IV плео-
иода — до 20, для V плеопода — до 10. Уроподы 3 длиннее плеотель-
зона, у J такой же длины, как плеотельзон или немного короче. Длина
тела S до 15 мм, j до 12 мм.
Дальневосточный водяной ослик был впервые указан для Японии
Гильгендорфом (Hilgendorf, 1874) как Asellus sp. Боваллиус (Bovallius,
1886) в своем обзоре Asellidae дал японской форме название A. hilgen-
dorjii, но описания не привел. Далее Гильгендорф (Hilgendorf, 1893)
снова под тем же названием отметил японского ослика, указав некоторые
признаки, недостаточно для него характерные. В 1921 г. Тетерсолл
1. ASELLUS
63:..
описал Asellus из Японии, считая его идентичным европейскому A. aqua-
ticus., причем на рисунках обозначил некоторые признаки, резко отли-
чающие восточную форму от европейской, не обратив на них внимания
в тексте. Совершенно то же самое сделал в 1936 г. Шен при описании
Asellus из Китая. Державин (1930) указал для советского Дальнего
Востока Л. aquaticus, а Бирштейн (1932). выразил сомнение в точности
Рнс. 30—35. Asellus (s. str.) hilgendorfi Bov.; 30 — проводит I перейопода самца: :
31 и 32 — I и II плеоподы самца; 33— II плеопод самки, 34и 35 — III а 29 IV плео- -
поды самца. Ориг.
этого определения. Наконец, Никольс (Nicholls, 1929) описал из Японии
новый вид A. nipponensis, игнорируя данные предыдущих авторов. Все
указанные Никольсом признаки совпадают с рисунками, приведенными'
Тетерсоллом и Шеном, и нет никакого сомнения в том, что все цитиро-
ванные авторы имели дело с одной и той* же формой, которой следует -
возвратить ее первое название — A. hilgendorfi Bov. Сравнение осликов -
из Японии, собранных Державиным на Хоккайдо, с амурскими
и сахалинскими указывает на принадлежность их к одному виду.
Распространение. Водоемы восточного побережья совет- -
ского Дальнего Востока (Амур, Суйфун, Уссури и т. д.), кроме Камчатки,.
I. ASELLIDAE
o,jsa Сахалин и Шантарские, Япония (Tattersail, 1921; Nicholls, 1929),
р^цтай (Shen, 1936), включаю Манчжурию (Ueno, 1940).
*2а. Asellus (s. str.) hilgendorfi martynovi Birst, (рис. 36—39).
Бирштейн, 1947 : 411, рис. 2; — aquaticus M a p т ы н о в, 1932 : 534;
gjrstein, 1933 : 474 (partim).
Вырост на заднем крае проподита I перейопода 3 выражен очень слабо.
jjaK у $>, и несет всегда 3 иожевидных зубца, 1 плеопод 5, как у типич-
Рис. 36—39. Asellus (s. str.) hilgendorfi martynovi Birst.: 36 — npo-
подпт I перейопода самца; 37 и 38 — I и II плеоподы самца;
. 39—II плеопод самки. Ориг.
цой формы, но экзоподит значительно шире, его длина менее чем
н Р/г раза превышает ширину. Экзоподит II плеопода $ значительно
отличается от такового типичной формы присутствием длинной перистой
детинки на дистальном крае'внешней группы щетинок дистального чле-
ника, чего не наблюдается у других форм Asellus s. str. Базальный чле-
ник экзоподита несет всего 1 щетинку на внешнем крае. Щетинки II плео-1
пода J как бы сдвинуты, по сравнению с типичной формой, к наружному
краю так, что около половины дистального края лишено щетинок;
на внутреннем крае' сидит 2—3 щетинки, на поверхности, вблизи внут-1
1. ASELLUS
65
реннего края, 3—4 более коротких щетинки. Число щетинок на поверх-
ности экзоподита III плеопода меньше, чем у типичной формы.. На
базальной части экзоподита IV плеопода — 17, V плеопода — 8 щетинок.
Длина тела <5 7 мм, g — 6 мм.
Мартынов (1932), исследовавший эту форму, не обратил внимания
на строение плеоподов и поэтому определил ее как A. aquaticus. Бир-
штейн (1933), не имея оригинальных экземпляров, привел данные Мар-
тынова. Строение первых плеоподов. <5 не оставляет никаких сомнений
в принадлежности якутского ослика к A. hilgendorfi, однако указанные
отличия от типичной формы, насколько можно судить по просмотрен-
ному материалу, постоянны и позволяют рассматривать якутскую форму
как подвид.
Распространение. Озера в дельте р. Лены.
2b. Asellus (s. str.) hilgendorfi ryukyuensis Ueno.
— nipponensis ryukyuensis U eno, 1938 : 264, fig. 1—10.
Отличается от типичной формы меньшим количеством члеников в жгу-
тах обеих антенн и более слабым вооружением проподита I перейопода
S. Жгут I антенны S состоит всего из 6, j •— из 5 члеников, жгут II
II антенны соответственно — из 40 и 32 члеников. Выступ на проподите
I перейопода 3 не выражен и имеется всего 1 или 2 ножевидных зубца.
Дистальный край проподита несет ряд мелких щетинок. Дактилюе
I перейопода всего с 6 зубцами.
Распространение. О-ва Риу-киу — наземные водоемы и грун-
товые воды.
*3. Asellus (s. str.) monticola Birst, (рис. 40—44).
В i.r s t e i n, 1932 : 251, fig. 1—5; Stammer, 1932 : 130; Бирштейн,
1936 : 236.
Окраска, как у A. aquaticus (L.). Последний и предпоследний сег-
менты тела S наиболее широкие; их ширина равна ширине плеотель-
зона. У J средние и задние сегменты и плеотельзон равной ширины.
Первая антенна короче основания второй антенны. В жгуте первой
антенны 7 члеников. Вторая антенна короче тела. Ее жгут в Р/г —
2 раза длиннее основания и состоит из 45—50 (3) или 40—45 (ф) члени-
ков. На выступе проподита I перейопода S 3 ножевидных зубца, дакти-
люс с 6—8 зубцами, j имеет соответственно 2 ножевидных зубца
на выступе проподита и 5—6 зубцов на дактилюсе. Ширина проподита
I плеопода S превышает длину, на внутреннем крае 2—5 совокупитель-
ных зубцов. Экзоподит в 2 раза длиннее протоподита. Его внутренний
кр'ай прямой, выемка отсутствует, на нем 6—10 коротких простых щети-
нок. На внешнем и дистальном краях 15—20 длинных перистых щети-
нок и несколько коротких простых (частично на поверхности). Ширина
протоподита II плеопода S равна его длине, на внутреннем крае нахо-
дится ряд из 3—5 щетинок, на поверхности, вблизи внешнего края,
сидит 2—5 щетинок; экзоподит менее чем в 2 раза короче протоподита,
базальный членик с 1—2 щетинками на внешнем крае, овальный дисталь-
ный с двумя группами щетинок — внешней, состоящей из 7—13 простых
щетинок, и внутренней, состоящей из 3—7 перистых щетинок. Эндопо-
дит построен так же, как у A. aquaticus, с направленным внутрь выростом
в базальной части, но относительно протоподита он гораздо длиннее
(он составляет около 3/а длины протоподита). Длина II плеопода $>
5 Фауна СССР, том VII, в 5
66
I. asellidae
заметно превосходит ширину, так что весь плеопод вытянут в продоль-
ном направлении. Дистальный и внешний края с 18—35 перистыми
щетийками на поверхности, вблизи наружного края, сидит 2—5, вблизи
внутреннего края — 2—3 коротких простых щетинки. Экзоподит III плео-
пода двучленистый и на его поверхности неравномерно расположены
короткие простые щетинки. Вдоль наружного края базального членика
сидит 9—12, дистального членика — 17—25 щетинок, на дистальном
крае — 14—20 и на внутреннем 2—4, большей частью перистые щетинки.
Экзоподиты IV и V плеоподов, как у A. aquaticus (Ь.).^На наружном
Рис. 40—44. Asellus (s. str.) monticola Birst.: 40 — проподит I перейо-
пода самца; 41 и 42 — I и II плеоподы самца; 43 — II плеопод самки;
44 — плеотельзон и уроподы самца. Ориг.
крае первого находится около 10—15, второго — 3—5 (и 2—3 на поверх-
ности) щетинок. Уроподы у обоих полов короче плеотельзона. Прото-
подит и экзоподит одинаковой длины, эндоподит несколько длиннее
экзоподита. Длина тела <5 9—12, j — 6—8 мм.
Распространение. Закавказье — Армения и Грузия: озера
Табискури, Арпа-гель, Чалдырь-гель, Тапаравани, реки Кция и Кара-су;
Иран — окрестности Зенджана.
*3а. Asellus (s. str.) monticola var. fontinalis Birst.
Бирштейн, 1936 : 237, fig. 1—2.
Длина экзоподита 1 плеопода S всего в Iх/2 раза превосходит ширину.
На внешнем крае 14—15 перистых щетинок, на внутреннем крае щети-
1. ASELLUS
67
нок нет. Поверхность протоподита II плеопода S лишена щетинок,
а вырост эндоподита в 2 раза короче, чем у типичной формы. Окраска
светлее, чем у типичной формы. Длина тела S — 6 мм. Этот варьетет
представляет собой неотеническую ключевую форму.
Распространение. Ключ на горе Сатапле, близ г. Кутаиси
(Грузия).
Рис. 45—49. Asellus (s. str.) latifrons Birst.: 45— общий вид самца;
46 и 47 — I и II плеоподы самца; 48 — II — плеопод самки; 49 —
III плеопод Ориг.
*4. Asellus (s. str.) latifrons Birst, (рис. 45—49).
Бирштейн, 1947 : 409, фиг. 1А—D; ? — sp. Внуковский, 1929 : 55.
Голова и тело расширенные, наибольшая ширина тела приходится
на V—IV сегменты, и здесь она больше ширины плеотельзона. Лобный
край головы с небольшим рострумом. Постмандибулярные лопасти
сильно развиты и резко выдаются в стороны. Эпимеры короткие и сверху
не видны. Длина плеотельзона уступает ширине. Первая антенна короче
68
I. ASELLIDAE
стержня второй антенны, ее жгут состоит из 6 члеников, несущих 3 (с?)
или 2 (j) чувствительных щетинки. Вторая антенна не заходит за сере-
дину длины тела; ее жгут состоит из 30—35 члеников. Проподит I перейо-
пода <5 со слабо выраженным выростом, снабженным 4 ножевидными
зубцами. Дактилюе с 8—9 зубцами. Экзоподит I плеопода S почти в 3 раза
длиннее и лишь незначительно шире протоподита, с почти параллельным
наружным и внутренним краями и всего 9—11 перистыми щетинками
на дистальном крае. Протоподит с 4—5 ножевиднымй зубцами. Длина
протоподита II плеопода 3 равна ширине. Эндоподит заметно длиннее
протоподита, с коротким и широким внутренним базальным выростом.
Дистальный членик экзоподита широкий, с 3—4 перистыми щетинками
на внутреннем крае, 9—10 простыми щетинками и 1 перистой щетинкой
на наружном крае. II плеопод $ заметно вытянут в длину и несет около
20 перистых щетинок на дистальном крае и 5—7 щетинок на поверхности
близ базального внутреннего угла. Длина экзоподита III плеопода менее
чем в Р/г раза больше ширины; его базальный членик в Р/г раза
меньше дистального. Наружный край экзоподита IV плеопода несет
около 15, V плеопода — 6—7 щетинок. Уроподы незначительно короче
плеотельзона. Длина тела S до 10 мм, $> — 7 мм. Для этого вида чрез-
вычайно характерна форма головы, с ее расширенными боковыми краями
и небольшим рострумом.
Распространение. Бассейн Оби и Иртыша, на север по край-
ней мере до 64° 55' широты. В пойме Оби водяные ослики нередко дости-
гают массового количественного развития.1 !
*5. Asellus (s. str.) epimeralis Birst, (рис. 50—54).
Бирштейн, 1947 : 410, фиг. 1 Е—Н.
Тело стройное, максимальная ширина его приходится у 3 на V—VII
сегменты, у g — на VII сегмент. Эпимеры всех сегментов очень сильно
развиты и далеко выдаются в стороны. Длина плеотельзона больше
ширины. Первая антенна достигает конца стержня второй антенны
и имеет 6—7-членистый жгут с 4 (3) или 2 (j) чувствительными щетин-
ками. Длина второй антенны составляет около 3/i длины тела. Ее жгут
состоит из 35—40 члеников. Проподит I перейопода 3 с 3 ножевидными
зубцами на нормально развитом выступе и 9—10 зубцами на дактилюсе.
Экзоподит I плеопода 3 овальной формы, значительно шире протопо-
дита и несет около 25—30 перистых щетинок на внешнем и дистальном
краях и 10—15 простых щетинок на поверхности близ внутреннего
базального угла и вдоль внутреннего края. II плеопод 3, как у Asellus
latifrons, но число щетинок на наружном крае дистального членика экзо-
подита больше, и перистая щетинка отсутствует. II плеопод j незначи-
тельно вытянут в длину, его базальный внутренний угол округло прямо-
угольный, дистальный край с 25—30 перистыми щетинками; щетинок
на поверхности нет. Длина экзоподита III плеопода почти вдвое пре-
вышает ширину; базальный членик по длине приблизительно равен
дистальному. На наружном крае экзоподита IV плеопода — более 25,
1 Е. А. Заринская-Чаликова. 1929. Предварительное сообщение
о гидробиологических работах на Куноватском Сору (р. Обь) летом 1929 года. Бюлл.
Общ. изучения края при Музее Тобольского севера, 4 : 15—21.
1. ASELLUS
69
V плеопода — 2—5 щетинок. Уроподы значительно варьируют в раз-
мерах, но всегда короче плеотельзона. Длина тела 6 до 10 мм, g — до
7 мм.
, Отличается от других видов подрода необычайно сильно развитыми
эпимерами и вытянутым в длину плеотельзоном, напоминая этими при-
знаками некоторые виды подрода Proasellus.
Рис. 50—54. Asellus (s. str.) epimeralis Birst.: 50 — общий вид самца; 51 и 52 —
I—II плеоподы самца; 53— II плеопод самки; 54 — III плеопод. Ориг.
Р аспространение. Озеро Баунт в системе Ципо-Ципикан-
ских озер, бассейн Витима. В Ципо-Ципиканских озерах найдено
несколько видов байкальского происхождения; можно думать, что эти
озера входили в состав древнего многоозерья, передавшего часть своей
фауны Байкалу. Однако нет никаких оснований сближать Asellus epi-
meralis с байкальскими водяными осликами. Вероятно, в составе фауны
Ципо-Ципиканских озер, наряду с байкальскими формами, сохранились
и древние общесибирские элементы, не перешедшие в Байкал и вымер-
шие на обширных пространствах Сибири. Именно к ним следует отно-
сить описанный вид.
70
I. ASELLIDAE
2. Подрод Mesoasellus Birst.
Бирштейн, 1939 : 249; Asellus auct. (part.), Caecidotea auct. (part.).
Внутренняя лопасть первой максиллы с 5 щетинками. Проподит
I перейопода £ удлиненный: его длина больше чем вдвое превосходит
ширину, задний край лишен выступов, прямой или слабо вогнутый.
На базальной части эндоподита II плеопода $ имеется направленный
внутрь вырост (как у подрода Asellus s. str.). II плеоподы g не заходящие
друг на друга, треугольные, слабо вооруженные щетинками только
в дистальной части. Уроподы удлиненные, превосходящие у S по длине
Рис. 55—58. Asellus (Mesoasellus) dybowskii Sem.: 55— общий
вид самца; 56 — правая и левая мандибулы; 57 — I максил-
ла; 58 — ногочелюсть. Ориг.
плеотельзон, с сильно укороченными экзоподитами. Эпимеры слабо раз-
виты, но сверху заметны.
К этому подроду относятся A. dybowskii Semenk., A. kawamurai (Tatt.),
калифорнийский A. californicus Miller и (со знаком вопроса) A. akiyoshi-
ensis (Ueno).
*6 . Asellus (Mesoasellus) dybowskii Sem. (рис. 55—64).
С e м e н к e в и ч, 1924 : 12; Бирштейн, 1939 : 249, рис. 1.
Наибольшая ширина тела S, приходящаяся на V—VI сегменты тела,
в 3 раза меньше длины. Пигмент отсутствует. Голова трапецеидальной
формы с округлыми передними боковыми углами и небольшим углубле-
нием спереди. Ее длина равна ширине. Глаза маленькие, округлые,
черные. Первая антенна в 41/г раза короче тела и имеет 5—6-членистый
1. ASELLUS
71
жгутик с 4 laminae olfactoriae (<5) или 4-членистый жгутик с. 2 lammae
olfactcriae (j). Вторая антенна не менее чем в 3 раза длиннее
тела. Ее жгут у j включает около 100 члеников. Первый членик ман-
дибулярного щупика короче второго, который по длине равен третьему
членику. Внешняя лопасть первой максиллы несет 9 зубцов и 1 стер-
нальную щетинку у дистального внешнего угла. Ширина внутренней
лопасти в 3 раза меньше длины. На ней имеется 5 перистых щетинок,
Рис. 59—S4. Asellus (Mesoasellus) dybowskii Sem.: 59 — проподит I
перейопода самца; 60 и 61 — I—II плеоподы самца; 62 и 63 — III—
IV плеоподы, 64 — уропод. Ориг.
из которых 2 наружные значительно длиннее 3 внутренних. Эпиподит
ногочелюсти с округлым внешним краем, на котором сидит 7 коротких
простых щетинок, частично переходящих на дистальный край. Внутрен-
ний край вогнутый. Жевательная лопасть с 5—6 зубцами. Первый чле-
ник щупика составляет по длине меньше половины второго, который
по длине равен четвертому членику, третий и пятый членики одинаковой
длины и несколько (на Ve—-Vs) короче второго и четвертого. Два
дистальных членика значительно уже трех базальных. Проподит
I перейпода 5 с прямым задним и слабо выпуклым передним краем и посте-
пенно суживается к дистальному концу. Его ширина в 2Чъ раза меньше
72
I. ASELLIDAE
длины. На заднем крае 4—5 ножевидных зубца и параллельный им ряд
(субмаргинальный) из 8—10 толстых щетинок. На переднем крае 1 ще-
тинка в дистальной трети и группа из 3—4 щетинок при основании дак-
тилюса. Дактилюс несет 4—6 длинных зубцов на заднем крае. Карпо-
подит узкий; его длина и ширина почти равны. Дактилюсы остальных
перейоподов удлиненные и имеют от 2 до 5 зубцов, причем число зубцов
на дактилюсах задних перейоподов больше, чем на дактилюсах передних.
Длина протоподита I плеопода $ в l3/t раза больше ширины. На вну-
треннем крае находится 4 длинных совокупительных зубца ножевидной
формы. Дистальный край лишен щетинок. Экзоподит овальной формы.
На дистальном крае — 6, на внешнем 8 длинных перистых щетинок,
за исключением крайних, почти равных по длине экзоподиту. На базаль-
ном крае и на поверхности, а также на внутреннем крае, щетинки отсут-
ствуют. Протоподит II плеопода S удлиненный. Его ширина в IV2—
13/а раза меньше длины. Две щетинки имеются только при основании
экзоподита. Экзоподит в 2l/z раза короче протоподита. Длина каждого
из его члеников значительно меньше ширины. Дистальный членик оваль-
ной формы, несколько шире и длиннее базального. Он несет 6—7 про-
стых щетинок, направленных наружу, 3—4 несколько более длинных
перистых щетинок, направленных внутрь, и 1 перистую щетинку посре-
дине, между обеими этими группами. Эндоподит всего на 1/t короче про-
топодита. В базальной его части имеется длинный, направленный внутрь
вырост. Дистальная его часть построена, в общем, как у Asellus aqua-
ticus (L.), но зубцов на лопастях больше, и они сильнее развиты. Ширина
экзоподита III плеопода всего на —Vs меньше длины. По краям его
дистального членика сидит около 25 перистых щетинок, на дистальном
крае превосходящих по длине весь членик. На внешнем крае базального
членика около 10 щетинок. Длина экзоподита IV плеопода почти в 2 раза
больше ширины. Экзоподит двучленист; базальный членик в 3V2 раза
короче дистального. На внешнем крае базального членика сидит
2 щетинки. Эндоподит на короче экзоподита. Его длина менее чем
в IV2 раза превышает ширину. V плеопод в общем походит на IV, но его
экзоподит лишен щетинок по внешнему краю. Уроподы незначительно
длиннее плеотельзона. Протоподит в 2 раза короче эндоподита. Экзо-
подит очень короткий, в 8—9 раз короче эндоподита (Семенкевич ука-
зывает отношение экзоподита к эндоподиту — Vs—Ve, так что, невиди-
мому, длина ветвей уроподов подвержена значительным индивидуальным
колебаниям, как это известно для других видов подрода Mesoasellus).
Длина тела <5 8.5—10 мм, g — 5.6 мм.
Распространение. Оз. Байкал, на больших глубинах.
7. Asellus (Mesoasellus) kawamurai (Tatt.) (рис. 65—69).
T a 11 e г s a 1 1, 1921 : 47, t. XV, fig. 11—18 (CaecidoteaY, Ueno, 1927 : 360,
fig- 3.
Тело удлиненное, его ширина в 7 раз меньше длины, с параллель-
ными краями. Все сегменты тела примерно одинаковой величины. Голова
со слабо вогнутым передним краем и маленькими, состоящими из 3 фасе-
ток, глазами на поверхности. Плеотельзон составляет около V« длины
всего тела. Его длина в IV2 раза больше ширины. Жгут цервой антенны
состоит из 13 члеников. Вторая антенна длиннее половины тела и имеет
в жгуте около 100 члеников. Проподит 1 перейопода удлиненный, с пря-
мым задним и выпуклым передним краями. На базальной части заднего
края проподита находится 2 ножевидных зубца. Протоподит 1 плеопода
1. ASELLUS
73
<5 имеет на внутреннем крае 4 совокупительных зубца. Экзоподит несет
длинные щетинки на внешнем и дистальном краях; его ширина в 2 раза
меньше длины. Длина протоподита II плеопода S незначительно пре-
восходит ширину. Базальная часть эндоподита с длинным, направленным
внутрь выростом. Длина двучленистого экзоподита III плеопода
в 2Va раза больше ши-
рины. На его дисталь-
ном крае сидят доволь-
но короткие перистые
щетинки. Редкие прос-
тые щетинки имеются
также на внешнем крае
базального членика. Эн-
доподит по длине ра-
вен 2/з экзоподита. Эк-
зоподиты IV и V плео-
подов также двучлени-
сты и шире эндоподи-
тов. Уроподы <5 значи-
тельно длиннее плео-
тельзона, их экзоподиты
по крайней мере в 2 раза
короче эндоподитов.
Длина последних силь-
но варьирует. В неко-
торых случаях экзопо-
диты двучленисты (Ue-
no, 1927). Уроподы §
короче плеотельзона.
Длина тела <5 16—17,
2—-11 мм.
Распростране-
ние. Япония — пеще-
ры близ Отсу и Киото.
8. Asellus (Mesoa-
sellus) akiyoshiensis
(Ueno) (рис. 70-71).
Рис. 65—69. Asellus (Mesoasellus) kawamurai (Tatt.):
65 — общий вид; 66 — I перейопод самца; 67 и 68 —
I и II плеоподы самца; 69—III плеопод. 70—71.
Asellus (Mesoasellus) akiyoshiensis (Ueno), самка: 70 —
общий вид; 71—II плеопод (65—69,- по Тетерсоллу;
70—71, по Уэно).
U eno, 1927 :356, fig.
1—2; 1933 : 115 (Caecidotea).
Описана только j.
Длина тела в 4 раза
больше ширины. Глаза
отсутствуют. Четыре
передних сегмента тела
одинаковой длины и ширины, три зад-
них несколько короче и уже. Ширина плеотельзона на 1lt меньше
длины. Его задний край треугольно-округлый. Первая антенна на Vs
короче основания второй антенны, ее жгут состоит из 6—10 члеников.
Вторая антенна достигает 3Д длины тела и имеет в жгуте около 70 чле-
ников. Передцдй край проподита I перейопода почти прямой, без высту-
пов, с 3 ножевидными зубцами и несколькими щетинками. На заднем
его крае 3 щетинки. Длина II плеопода $> в 1V2 раза больше ширины..
74
I. ASELLIDAE
На внешнем его крае сидит 3 щетинки и столько же на поверхности плео-
пода близ внешнего края. Длина экзоподита III плеопода в Р/г раза
больше ширины, причем первый членик короче второго. Эндоподит удли-
ненный, незначительно превышающий длину первого членика. По внеш-
нему краю экзоподита сидит около 11 щетинок, по дистальному — около
8 и по внутреннему — около 3. Уроподы варьируют по длине и часто
асимметричны. Их экзоподит незначительно короче эндоподита. Длина
тела до 13 мм.
Распространение. Япония, известковая пещера Такиана,
близ Акиоши, префектура Ямагучи; Ямабана близ Саппоро (Хоккайдо).
3. Подрод Baicaloasellus Stamm.
Stammer, 1932 : 129; Бирштейн, 1939: 249.
Тип подрода: Asellus baicalensis Grube.
Внутренняя лопасть первой максиллы с 5 щетинками. Проподит
I перейопода S лишен выступа, широкий: его ширина менее чем вдвое
уступает длине. Эндоподит II плеопода S без выроста в базальной части.
Его конец клювовидно вытянут, хитинизирован, лишен боковой лопасти.
II плеоподы § вытянутые в продольном направлении, трапецеидальной
формы, расширяются по направлению к дистальному концу и не заходят
друг за друга. Эпимеры хорошо развиты и видимы сверху.
Штаммер пишет, что помимо A. baicalensis Grube и A. angarensisDyb.,
к этому же подроду, по всей вероятности, следует отнести и A. korotnewi
Semenk., A. minutus Semenk. и A. dybowskii Semenk.
Если по отношению к первым двум видам это безусловно справедливо,
то последний вид — A. dybowskii Semenk. оказался принадлежащим
к подроду Mesoasellus Birst.
* 9. Asellus (Baicaloasellus) baicalensis Grube (рис. 72, A; 73—80).
Grube, 1872 : 57; Dybowskii, 1884 : 51, figs; Алпатов, 1922 : 1;
Семенкевич, 1924 : 9; К о ж о в, 1931 : 35 (partim); Stammer, 1932 :
123, figs. 12—14.
Ширина тела Примерно в 2 Vs (<5)—-2 (g) раза меньше длины.
Максимальная ширина приходится на V—VI сегменты. Ширина плео-
тёльзона превышает длину. Окраска головы обычно немного темнее
окраски сегментов тела. Первая антенна по длине равна 2/з основания
второй антенны и имеет в жгуте 4—5 (g) или 5—7 (<3) члеников, несущих
две lammae olfactoriae. Вторая антенна составляет обычно около 3/i
длины тела и имеет в жгуте 20—25 члеников, однако нередко встре-
чаются экземпляры как с более короткими, так и более длинными антен-
нами, соответственно чему вариирует и число члеников их жгута (10—
27). Ширина проподита I перейопода S в 2 раза меньше длины. На заднем
крае имеется 2 ножевидных зубца и небольшое (9—12) число щетинок,
на переднем крае 2 щетинки, на дистальном — 3—4 щетинки. Дакти-
люс с 4 зубцами и очень длинным когтем. Проподит I перейопода g
с параллельными задним и передним краями; с 1 ножевидным зубцом
и 3—4 щетинками на заднем крае и 2—3 зубцами на коготке. Дактилюсы
остальных перейоподов с 2—3 зубцами. Форма протоподита I плеопода
S приближается к прямоугольной. Его длина в P/s—13/< раза больше
ширины. Экзоподит овальной формы, примерно в 13/л раза более длин-
ный и широкий, чем протоподит. На дистальном крае и части внешнего
1. ASELLUS
75
СИДИТ
щетинок. Длина экзоподита III плеопода
края сидит 13—15 длинных перистых и 3—9 коротких простых щетинок.
Ширина протоподита II плеопода <5 почти в 2 раза меньше длины. Экзо-
подит значительно короче эндоподита. Его базальный членик менее чем
в 1х/г раза короче дистального. На внешнем крае базального членика
сидит 3—4 щетинки, из которых 1—2 перистые, на внешнем крае ди-
стального членика 1—2, на дистальном крае его 2 перистых щетинки.
II плеопод J с прямым внутренним и вогнутым внешним краем, сильно
расширяющийся в дистальной трети. Ширина у основания в Р/г раза
меньше ширины дистальной трети, которая несколько более чем в 2 раза
уступает длине. На дистальном крае и части внешнего края
14—19 длинных перистых ~
на Vi превышает ширину.
На внешнем крае базаль-
ного членика 3—5, на
внешнем и дистальном
краях дистального члени-
ка около 20 перистых ще-
тинок. Ширина экзоподи-
та IV плеопода в 2 раза
меньше длины. Внешний
край без выемки и пери-
стых щетинок и несет в ба-
зальной части очень тонкие
и короткие многочислен-
ные щетинки. Уроподы $
на Уз—Vi короче плебтель-
зона. Экзоподит по длине
равен протоподиту и на 1/з
короче эндоподита. Длина
тела S 3.5—5.5 мм;
2.7—3.5 мм.
Распростране-
ние. Оз. Байкал, в при-
брежной области.
Рис. 72 А — Asellus (Baicaloasellus) baicalensis
Grube, самец, Б — Asellus (Baicaloasellus) anga-
rensis Grube, самец. Ориг.
*10. Asellus (Baica-
loasellus) korotnewi Sem.
(рис. 81—86).
Семенкевич,
1924 : 10; — baicalensis Ножов, 1931 : 35 (partim).
Ширина тела в 2Уг раза меньше длины. Ширина плеотельзона пре-
восходит длину. Первая антенна S не достигает половины длины осно-
вания вторых антенн. Жгут состоит у <5 из 5, у g из 4 члеников, несет
всего одну lamma olfactoria и составляет меньше половины длины осно-
вания. Вторая антенна длиннее тела, ее жгут более чем в 2 раза превы-
шает основание и состоит у S из 30—45, у j — из 25—28 члеников. Про-
подит I перейопода 3 постепенно суживается к дистальному концу и имеет
слабо вогнутые задний и передний края. Его ширина в 2 раза меньше
длины. На заднем крае 3 ножевидных зубца и 7—11 щетинок в субмар-
гинальном ряду, на заднем крае 1 щетинка, на дистальном крае 3—4
щетинки спереди и 4—5 сзади. Дактилюе S с 5 зубцами и 4—6 щетин-
ками; дактилюе J с 3—4 зубцами. Дактилюсы остальных перейоподов
с 4 зубцами. Длина протоподита I плеопода <5 в 2 раза больше ширины.
76
I. ASELLIDAE
На середине внутреннего края сидит I совокупительный зубец. Экзо-
подит овальной формы, всего на 1/з—'/t более длинный и широкий,
чем йротоподит, и несет 8—И длинных перистых и 3—5 коротких про-
Рис. 73—80. Asellus (Baicaloasellus) baicalensis Grube: 73 — пропо-
дит I перейопода самца; 74 — то же самки; 75— II плеопод сам-
ки; 76— I плеопод самца; 77 и 78— IV и III плеоподы; 79 —
II плеопод самца; 80— IV перейопод самки. Ориг.
стых щетинок на дистальном крае и части внешнего края. Длина прото-
подита II плеопода <5 почти в 2 раза превосходит ширину. Экзоподит
равен 2/з длины протоподита. Базальный членик в Р/г—13/л раза
короче дистального и несет 2—3 щетинки на внешнем крае. Дистальный
членик имеет 3 перистых щетинки на внешнем крае и 1 перистую щетинку
1. ASELLUS
77
на конце. Эндоподит длиннее экзоподита. II плеопод j незначительно
расширяется к дистальному концу. Внешний его край слабо вогнутый.
Длина в 2г/з—2^2 раза больше ширины. На дистальном закруглении
сидит 10—12 длинных перистых щетинок. Длина экзоподита III плео-
пода на 1/з—больше ширины. На внешнем крае базального членика
4—5 щетинок, на краях дистального членика около 20 щетинок. Ширина
Рис. 81—86. Asellus (Baicaloasellus) korotnewi Sem.: 81 — проподит I
перейопода самца; 82 и 83— I и II плеоподы самца; 84— II плеопод
самки; 85 — IV плеопод; 86 — V плеопод. Ориг.
экзоподита IV плеопода в 2 раза меньше длины. На его внешнем крае
имеется глубокая выемка, от которой начинается линия, делящая экзо-
подит на 2 части. Перед этой выемкой сидит 3—4 длинных перистых
щетинки. Базальная часть внешнего края опушена тонкими короткими
щетинками. Эндоподит такой же длины, как экзоподит, но значительно
уже. Эндоподит V плеопода достигает всего т/г—2/з длины экзоподита.
Экзоподит узкий, его длина больше чем в 2 раза превышает ширину,
щетинки отсутствуют. Уроподы длиннее плеотельзона. Длина тела 3
4.5—6.5 мм; j 3—4 мм.
78
I. ASELLIDAE
распространение. Оз. Байкал, в прибрежной зоне..
*11. Asellus (Baicaloasellus) minutus Sem. (рис. 87—91).
Семенкевич, 1924:11.
Ширина тела в 2х/« раза меньше длины. Ширина плеотельзона равна
длине. Первая антенна немного длиннее V2 длины основания второй
Рис. 87—91. Asellus (Baicaloasellus) minutus Sem.: 87 — проподит
I перейопода самца; 88— II плеопод самки; 89 и 90 — I ик11
плеоподы самца; 91 — IV плеопод самца.
антенны; на двух ее члениках имеются lammae olfactoriae. Вторая
антенна незначительно длиннее тела. Ее жгут меньше чем в 2 раза длин-
нее основания, и состоит из 25—32 (5) или 18—23 (^) члеников. Пропо-
дит I перейопода б с выпуклыми передним и задним краями, слабо, сужи-
вающийся к дистальному концу. Его длина вдвое (или несколько больше)
превышает ширину. -На заднем крае 2 ножевидных зубца и 2—4 щетинки.
На дистальном крае — 2—3 щетинки, на переднем крае 1 щетинка посре-
дине и 2—3 при основании дактилюса. Дактилюс с 4 зубцами (у $ 2 зубца).,
Дактилюсы II и III перейоподов — с 3, IV — с 1, V и VII — с 2 шипами..
1. ASELLUS
79-
Протоподит I плеопода $ субтрапецоидальной формы, с 1—2 совокупи-
тельными зубцами в базальной четверти внутреннего края. Его длина
в Р/г раза больше ширины. Экзоподит овальный, несколько менее чем
в 2 раза длиннее и шире протоподита. Его ширина почти в Р/г раза
меньше длины. На дистальном закруглении 14—17 перистых щетинок.
Протоподит II плеопода <5 прямоугольной формы, со скошенным внутрен-
ним дистальным углом. Его длина в Р/г раза больше ширины. Длина
экзоподита равна ширине протоподита. Базальный членик в Р/г раза
короче дистального, с 2—3 щетинками на наружном крае. Дистальный
членик несет 3—4 коротких перистых щетинки. Эндоподит по длине
равен протоподиту. II плеопод $ значительно шире в дистальной трети,
чем при основании. Внутренний его край прямой, наружный — вогну-
тый, дистальный — округлый. На дистальном крае сидит 11—13 длин-
ных перистых щетинок.’ Максимальная ширина вдвое меньше длины.
Длина экзоподита III плеопода на 1/t' превосходит ширину. На краях
дистального членика—около 15 щетинок, на внешнем крае базального
членика — 5 щетинок. Поверхность лишена щетинок. Экзоподит IV
плеопода двучленистый, с выемкой на наружном крае, у границы
двух члеников. Недалеко от этой выемки на базальном членике
сидят 4—5 длинных перистых щетинок. Ширина экзоподита вдвое меньше
длины. Эндоподит незначительно короче экзоподита и обычно не покры-
вается им. Экзоподит V плеопода длиннее эндоподита. Уроподы S равны
по длине плеотельзону или незначительно превосходят его. Экзоподит
короче эндоподита. Длина тела 6 3.5—4.5 мм, $> 2.5—3.5 мм.
Распространение. Оз. Байкал, в прибрежной зоне.
* 12. Asellus (Bale al о asellus) angarensis Dyb. (рис. 72, Б; 92—99).
Dybowski, 1884:23; Алпатов, 1922:1; G e м e н к e в и ч, 1924 : 8;
К о ж о в, 1931 : 34.
Максимальная ширина тела, приходящаяся на VII сегмент, в 2х/г
раза меньше длины. Плеотельзон незначительно уже задних сегментов
и шире передних, % двумя глубокими выемками на заднем крае, в кото-
рых находятся уроподы. Ширина плеотельзона превосходит длину.
Окраска темная, иногда почти черная. Часто заметны более темные,
симметрично расположенные пятна. Голова, как правило, светлее тела.
Первая антенна короче основания второй антенны. Ее жгут состоит
из 4—5 члеников. Вторая антенна короче половины тела. В ее жгуте
14—15 члеников. Длина проподита I перейопода <5 в Р/г—i3/t раза
больше ширины. На его заднем крае 2 ножевидных зубца и 4—10 щети-
нок в субмаргинальном ряду. Передний край с 2 щетинками. Дактилюс
имеет на заднем крае 3—4 зубца. Задний край проподита I перейопода
о с 1 ножевидным зубцом, дактилюс с 2—3 зубцами. На дактилюсе
остальных перейоподов 2—3 зубца. Протоподит I плеопода <5 прямо-
угольной формы; его длина в Р/г раза больше ширины, на его внутрен-
нем крае сидят 2—4 совокупительных зубца. Экзоподит овальной формы,
его ширина примерно в Р/г раза меньше длины, на его дистальном
и внешнем краях 13—17 длинных» перистых щетинок. Длина протопо-
дита II плеопода <5 в Р/г раза больше ширины. Экзоподит очень корот-
кий, в 3 раза короче протоподита. Его базальный членик трапецеидаль-
ной формы, значительно более широкий, чем неправильно треугольный
дистальный членик. Ширина обоих члеников превосходит их длину.
Дистальный членик несет 4—5, базальный на внешнем крае — 3—5
80
I. ASELLIDAE
коротких перистых щетинок. Эндоподит в 2 раза длиннее экзоподита.
II плеопод j незначительно расширяется в дистальной трети, где его
ширина в 2 раза уступает длине. На дистальном закруглении сидит
И—17 длинных перистых щетинок. Длина экзоподита III плеопода
Рис. 92—99. Asellus (В aicalo asellus) angarensis Grube: 92 —
проподит I перейопода самца; 93 и 94 — I и II плеоподы
самца; 95 — I максилла; 96 и 97 — III и IV плеоподы; 98 —
II плеопод самки; 99—плеотельзон с уроподами. Ориг.
на х/4 превышает его ширину. На внешнем крае базального членика —4,
на краях дистального членика — 25—35 щетинок. Длина экзоподита
IV плеопода в 2 раза больше ширины. Экзоподит двучленистый. Эндо-
подит по длине равен экзоподиту. Длина экзоподита V плеопода также
в 2 раза больше ширины, но эндоподит заметно короче экзоподита. Уро-
поды в 3 раза короче плеотельзона; эндоподит по длине равен протопо-
диту, экзоподит незначительно короче. Длина тела <5 до 7 мм, $ до 5.5 мм.
Распространение. Оз. Байкал, в прибрежной зоне; истоки
Ангары.
1. ASELLUS
81
4. Подрод Proasellus Dud.
Dudich, 1925a : 230; Stammer, 1932 : 129; Бир ш т e й н, 1935 : 237.
Тип подрода: Asellus meridianus Racovitza.
Внутренняя лопасть первой максиллы с 5 щетинками. Проподит
I перейопода $ без выступа, эндоподит II плеопода $ лишен выроста,
его окончание более или менее вытянуто и снабжено с наружной стороны
боковой лопастью. II плеоподы j вытянутые в продольном направлении,
суживающиеся к дистальному концу, незаходящие друг за друга, тре-
угольной или трапецеидальной формы. Эпимеры хорошо развиты и видны
сверху.
Подрод имеет кругосредиземноморское происхождение. Некоторые
сомнения вызывало отнесение к Proasellus описанных Моно (Monod,
1924) из Северной Африки A. gauthieri a A. thermonyctophilus, для кото-
рых Моно отметил и изобразил всего 3 щетинки на внутренней лопасти
первой максиллы. Переисследовав по моей просьбе типы, Моно конста-
тировал, что допустил в своем описании ошибку и что на самом деле
названные виды имеют 5 щетинок на внутренней лопасти первой максиллы
{рис. 157).
Из всех подродов рода Asellus подрод Proasellus наиболее богат видами.
Некоторые из них недостаточно четко отграничены друг от друга. Это
относится в особенности к группам видов coxalis—banyulensis— italicus
и cavaticus— spelaeus. Пока были известны только типичные представители
указанных видов, их отличия между собой казались достаточно опре-
деленными. Но описания многочисленных подвидов в значительной мере
стерли межвидовые грани, и в настоящее время исследователи испы-
тывают серьезные затруднения в определении видовой принадлежности
относящихся к этим группам форм. Так, например, в своей обработке
греческих водяных осликов Штрохаль (Strouhal, 1942) лишь условно
считает описанные им новые формы подвидами coxalis, указывая в то же
время на большую близость некоторых из них к banyulensis, а других —
к italicus. Нет сомнения в том, что указанные группы видов требуют
серьезной ревизии, которая приведет, вероятно, к объединению несколь-
ких видов, но для такой ревизии требуется соответствующий коллекци-
онный материал, пока отсутствующий в наших музеях. Поэтому .здесь
приводятся все описанные виды Proasellus (за исключением португаль-'
ского lusitanicus) в принятом до сих пор толковании и лишь с неболь-
шими изменениями в отношении видовой принадлежности некоторых
подвидов.
13. Asellus (Proasellus) meridianus Rac. (рис. 100—103).
Racovitza, 1919 : 41, fig. 7—12; Stammer, 1932 : 118, fig. 3—4; —
aquaticus auctorum partim.
Окраска светлее, а светлые пятна па спинной стороне тела крупнее,
чем у A. aquaticus.1
Вторая антенна на 1/з короче тела. Ее жгут имеет менее 50 члеников.
Проподит I перейопода 5 с 3 ножевидными зубцами. Протоподит I плео-
пода 6 с 1—2 совокупительными зубцами на внутреннем крае и иногда
со щетинкой на наружном дистальном углу. На наружном и дисталь-
ном краях экзоподита 12—22 простых коротких щетинок, на поверх-
1 D. J. S с о u г f i е 1 d. Difference in coloration of the head in Asellus aquaticus
and A., meridianus. Essex Naturalist, 27, 1940 (цитир. no Needham, 1942).
6 Фауна СССР, том VII, в. 5
8
I. ASELLIDAE
ности, у проксимального внутреннего угла, 1—2 щетинки, у дисталь-
ного наружного — 1 щетинка. На внутреннем крае протоподита II плео-
пода <5 3—4 щетинки. Экзоподит на Vs длиннее протоподита, ширина
прямоугольного дистального членика в 21/,2 раза меньше длины, на его
дистальном закруглении сидит 10—15 длинных перистых щетинок. Эндо-
Рис. 106—103. Asellus (Proasellus) meridianus
Rac.: 100 — проподит I перейопода самца;
101 и 102 — I и II плеоподы самца; 103— II
плеопод самки. (По Раковине).
подит короче экзоподита и ха-
рактеризуется сильно развитым
толстым вытянутым оконча-
нием, у основания которого
находится короткая боковая
лопасть. II плеопод j трапе-
цеидальной формы и несет по
дистальному краю около 12 пе-
ристых щетинок, а вблизи ба-
зальной части внутреннего
края 1—2 простых щетинки.
Уроподы не короче плеотель-
зона.
Распространение.
Франция, Голландия, Германия
(Нижний Рейн), Англия, часто
совместно с A. aquaticus (L.).
Штаммер показал, что в некото-
рых притоках Нижнего Рейна
по направлению вверх по тече-
нию число экземпляров A. me-
ridianus постепенно умень-
шается, а число экземпляров
A. aquaticus увеличивается. Он
поставил это в связь с влиянием
загрязненных фабриками вод,
заставляющих A. meridianus
уходить в верховья рек. Однако
в данном случае не исключена
возможность вытеснения более
древнего Л. meridianus более со-
вершенным A. aquaticus. В усло-
виях аквариума живет хуже,
чем A. aquaticus, что связано,
вероятно, с относительно мень-
шей поверхностью дыхатель-
ных плеоподов (Needham, 1942).
13а. Asellus (Proasellus) meridianus belgicus Arc.
Arcangeli, 1935 : 3, fig. 11—16.
В жгуте первой антенны 7—8 члеников, на которых 2 lammae olfac-
toriae, в жгуте второй антенны — 36 (6) или 32 (j) члеников. Проподит
I перейопода <5 с 2 ножевидными зубцами. Протоподит I плеопода <5
с I совокупительным зубцом, экзоподит с 5 перистыми щетинками
на дистальном крае. II плеопод £, в общем, как у типичной формы, только-
дистальный членик экзоподита постепенно расширяется к своему концу
и несет всего 8—9 перистых щетинок. II плеопод р так же почти как
1. ASELLUS
83
у типичной формы, но посредине его на поверхности находится 1 пери-
стая щетинка.
Распространение. Бельгия — пещера близ Льежа.
14. Asellus (Proasellus) polychaetus Dudich (рис. 104—106).
D u d i c h, 1925a: 287, fig. 7—12; — Strouhal, 1937 : 65.
И—17 зубцов,
$ имеет по нижнему краю
Рис. 104—106. Asellus (Proasellus) polychaetus Dud.
104 и 105— I и II плеоподы самца; 10а— II плео.
под самки. (По Дудичу).
Окраска коричневатая, с белыми пятнами. Сегменты груди значи-
тельно шире плеотельзона. Ширина последнего равна длине (3) иди
незначительно ее превышает (j). Первая антенна короче стебля второй
антенны и имеет жгут из 10—14 (3) или 7—9 (§) члеников. Вторая
антенна короче тела. Ее жгут состоит из 52—68 (о) или 42—48 (j) чле-
ников. Дактилюе I перейопода 3 имеет по нижнему краю И—17 зубцов,
а по верхнему 7—9 щети-
нок. Проподит несет на
заднем крае 3—4 ножевид-
ных зубца, а на переднем
3—5 щетинок. Длина и
ширина протоподита I
плеопода 3 почти равны.
На внутреннем крае 1 зу-
бец, на дистальном крае
5—9 щетинок и крупная
(2/з длины протоподита)
щетинка на внешнем ди-
стальном углу. Длина эк-
зоподита вдвое больше ши-
рины. Его внутренний
проксимальный угол от-
тянут вниз и заходит на
внутренний дистальный
угол протоподита, чего не
наблюдается у других ви-
дов Asellus. Внутренний
край прямой, внешний —слабо выпуклый, с 8—15 щетинками. На
дистальном крае сидит 7—10 перистых щетинок, на поверхности — I
щетинка на удлиненном внутреннем проксимальном углу, 2 •— 4
щетинки составляют неполный ряд, параллельный внутреннему
краю, и еще 3 щетинки расположены в дистальной части. Длина
протоподита II плеопода 3 больше ширины (17 : 14). Его внутренний
дистальный угол несет 5—8 крупных перистых щетинок. Экзоподит
по длине равен протоподиту, а на внешней стороне его проксимального
членика сидит 1 перистая щетинка (иногда еще по 1 перистой щетинке
по обоим краям), на внешней стороне дистального членика — 1—2 про-
стых щетинки и на дистальном округлом крае 10—13 перистых щетинок,
из которых по 2—3 щетинки с каждого края сильно укорочены. Эндо-
подит короче экзоподита, с суженным концом. II плеопрд трапеце-
идальной формы. Его длина в 2х/г раза больше ширины. На внешнем
крае сидит 8—10 перистых щетинок и 2 щетинки на поверхности,, вблизц
внутреннего края, в проксимальной части. Экзоподит IV плеопода раз-,
делен тонкими бороздками на 4 поля и слабо хитинизирован. Уроподы
у обоих полов короче плеотельзона. Эндоподит у 3 на х/з, у } на Vi
6*
84
I. ASELLIDAE
длиннее протоподита. Экзоподит равен (<5) или немного короче ($>) эндо-
подита. Длина тела i 8—10 мм, $> 7.5—8.5 мм.
Распространение. Италия — г. Сарно, в 49 км от Неаполя,
в р. Сарно и ее притоках, совместно с A. aquaticus и Sala consilina.
14а. Asellus (Proasellus) polychaetus dudichi Strouhal.
Strouhal, 1937 : 67, fig. 1—2.
Внутренний базальный угол экзоподита I плеопода <5 гораздо меньше,
чем у типичной формы, дистальный край протоподита всего с 3—6
щетинками; на его внутреннем крае, помимо 1—2 совокупительных зуб-
цов, имеется 1—2 щетинки. На внутреннем крае протоподита II плео-
пода S — 9—10 перистых щетинок. Дистальные края II плеопода j
и экзоподита I плеопода $ зубчатые.
Распространение. Италия — Фальконара Албанезе (Falco-
nara Albanese).
15. Asellus (Proasellus) remyi Monod (рис. 107—111).
Monod, 1932 : 206, fig. 1—4,
Передний край головы вогнутый, передние боковые лопасти ее сильно
развиты.; Глаза находятся на поверхности головы сверху. Тело посте-
Рис. 1S7-—111. Asellus (Proasellus) remyi
Monod.: 107 — общий вид; 108'—проподит
I перейопода самца; 104 и ПО— I и II
плеоподы самца; 111 — II плеопод самки.
(По Моно).
пенно расширяется по направле-
нию назад, так что VII сегмент
самый широкий. Коксоподиты пе-
рейоподов развиты очень сильно.
Ширина плеотельзона больше
длины. Его задняя часть с двумя
выемками и образованной ими
треугольно-округлой задней ло-
пастью. Окраска буровато-серая,
края тела бесцветны. Первая ан-
тенна по длине почти равна осно-
ванию второй антенны и имеет
жгут, состоящий из 7 (5) или 6(g)
члеников с 3 (5) или 2 (j) lam-
inae olfactoriae. Вторая антенна
немного длиннее ‘/а тела с 21—28-
члениковым жгутом. Задний край
проподита I перейопода <5 с 2 но-
жевидными зубцами. Дактилюс
с 4 зубцами. Длина протоподита
I плеопода <5 равна ширине или
на х/з ее превосходит; на внут-
ренней стороне 1 зубец. Экзопо-
дит не больше чем в 1х/г раза
длиннее протоподита, овально-
сердцевидной формы, с 1—2 про-
ксимальными щетинками на по-
верхности и 13—18 щетинками по
дистальному и наружному краям.
Длина и ширина протоподита II
плеопода <5 равны между собой,
1. ASELLUS
85
на внутренней его стороне 3 перистых щетинки. Экзоподит
незначительно длиннее протоподита. Проксимальный членик вдвое
короче дистального, несущего 8—12 щетинок. Длина эндоподита'в 2 раза
больше ширины. Его проксимальный внутренний угол почти прямой.
II плеопод J трапецеидальной формы, со слабо вогнутым внешним краем.
На дистальном крае сидит 11 перистых щетинок, на поверхности —
в основной части, вблизи внутреннего края — 2 простых щетинки. Уро-
поды короче плеотельзона. Экзоподит по длине равен протоподиту
и немного короче эндоподита. Длина и ширина тела <5 4.5—5 X
2.1—4 мм, § 4.5 х 1-8 мм.
Распространение. Оз. Охрида, между камнями у берега.
*16. Asellus (Proasellus) infirmus Birst, (рис. 112—120).
Бир штейн, 1936 : 237, рис. 3—13.
Окраска значительно светлее, чем у A. aquaticus. Края тела. почти7
параллельны друг другу. Сегменты с развитыми коксоподитами. Ширина
плеотельзона равна длине или немного ее превосходит. Боковые лопасти
головы расширены. Маленькие глаза, состоящие всего из 3 фасеток,
расположены недалеко от краев головы, но на ее верхней поверхности.
Первая антенна короче основания второй антенны и имеет в жгуте 7 чле-
ников. Вторая антенна немного длиннее половины тела; ее жгут состоит
у S из 35, у g из 31 членика. Проподит !• перейопода S, длина которого
вдвое превосходит ширину, имеет 2 ножевидных зубца. Дактилюс с 5—6
зубцами. Дактилюс IV перейопода <5. с 2 зубцами, проподит по верхнему
краю с 3, по нижнему с 5 щетинками. Длина протоподита I плеопода
<3 незначительно превосходит его ширину. Щетинки отсутствуют.
На внутреннем крае 2 совокупительных зубца. Экзоподит в Р/г раза
длиннее протоподита, но той же ширины. Его ширина превосходит
половину длины. На дистальном крае и половине внешнего края
находится около 10 простых щетинок. Длина и ширина протоподита
II плеопода £ почти равны; внутренний его край с 2 щетинками.
Экзоподит в Р/г раза длинее протоподита. Его дистальный членик
почти равен по длине протоподиту и вдвое превосходит базальный.
На дистальном крае дистального членика сидит 9 перистых щетинок,
самые длинные из которых достигают длины всего экзоподита.
Длина эндоподита вдвое больше ширины. Окончание и боковой вырост
примерно равной длины. Длина II плеопода j вдвое больше ширины.
Форма его приближается к прямоугольно-трапецоидальной. Внешний
край с небольшим выгибом. Косой дистальный край несет 8 перистых
щетинок, длина которых равна 2/з длины всего плеопода. Длина экзо-
подита III плеопода превышает ширину всего на д/з. На внешнем крае
базального членика 7—10 коротких щетинок, на внешнем и дистальном
краях дистального членика — около 25 щетинок. На поверхности экзо-
подита всего 4 щетинки, сидящие на дистальном членике вблизи его
внутреннего края. Экзоподит IV плеопода неясными линиями разделен
на 3—4 части и несет по внешнему краю около 7 щетинок. Экзоподит V
плеопода неясной линией разделен на две части и имеет на внешнем крае
около 4 щетинок. Уроподы вдвое короче плеотельзона. Протоподит
равен по длине экзоподиту и в Р/г раза короче эндоподита. Длина тела £
6 мм, ширина 2 мм, с 5 х 1-75 мм.
: Расширенные тело, голова и плеотельзон, укороченные антенны и урр-
поды (а также строение плеоподов £) указывают на близость описанного
86
I. ASELLIDAE
вида к A. remyi. Указанные признаки, невидимому, связаны с жизнью
между камнями и появляются конвергентно у неродственных между
собой форм (Lirceus, A. communis Say, A. dngarensis Grube, Johanella
purpurea Monod). Поэтому трудно согласиться с Моно, считающим, воз-
можным выделение A. remyi Monod в особый подрод. Вооружение рото-
Рис. 112—120. Asellus (Proasellus) infirmus Birst.: 112 —
общий вид самки; 113 — проподит I перейопода самца;
114 и 115 — I и II плеоподы самца; 116 — II плеопод
самки; 117 и 118 — IV и V плеоподы; 119— I максилла;
120—III плеопод. Ориг.
вых частей, а также строение плеоподов заставляют считать как A. remyi
Monod, так и A. infirmus Birst, типичными Proasellus. ' '
Распространение. Западное Закавказье — ключ на правом
берегу реки Гумисты близ г. Сухуми.
Экология. A. infirmus найден в маленьком холодном ключе
между камнями. Повидимому, он ведет полуподземный образ
жизни.
1. ASELLUS
87
17. Asellus (Proasellus) gjorgjevici К ar. (рис. 121—124).
К а г a m a n, 1933b : 103, fig. 1—2.
121
Ka-
123
Рио. 121—124. Asellus (Proasellus)
gjorgjevici Kar.: 121 — общий вид;
122 и 123 — I и II плеоподы самца;
124 — II плеопод самки. (По
раману).
Пигмент и глаза отсутствуют. Длина плеотельзона больше ширины.
Обе пары антенн очень короткие, вторая антенна составляет около х/з
длины тела, первая вдвое короче. Жгут первой антенны короче основа-
ния и состоит из 7 члеников, из кото-
рых на 4 сидят lammae olfactoriae;
жгут второй антенны по длине равен
основанию и состоит из 16—17 члени-
ков. На нижней стороне основания вто-
рой антенны сидят длинные щетинки (их
длина равна длине конечного членика
•основания). Проподит I перейопода й
довольно широкий. Его длина в 1х/з
раза больше ширины. На заднем крае
•сидят многочисленные ножевидные зуб-
цы (около 10) и щетинки. Протоподит
I плеопода <5 с 1 зубцом по внутрен-
нему Краю и многочисленными щетин-
ками (около 10) по наружному и дис-
тальному краям. На поверхности про-
топодита также имеются 2 — 3 щетинки.
Экзоподит в 2 раза длиннее протопо-
дита. Его ширина всего в Р/г раза
меньше длины. На внешнем крае на-
ходится около 10 простых щетинок,
на дистальном закруглении — около 20
перистых щетинок. На поверхности
сидит свыше 20 щетинок. Длина про-
топодита II плеопода 6 равна ширине.
На внутреннем и наружном краях про-
топодита находится по несколько длин-
ных перистых щетинок. Иногда щетинки
сидят и на поверхности у дистального
края. Экзоподит в 1х/г раза длиннее
протоподита и несет на дистальном за-
круглении около 20 очень длинных
перистых щетинок. Эндоподит по длине
равен протоподиту. Длина II плеопода
имеет неправильно трапецеидальную форму. На дистальном крае и
дистальных частях внутреннего и наружного краев сидят свыше 20
ристых щетинок. На внутреннем и наружном краях и на поверхности
находятся многочисленные простые щетинки. Уроподы значительно (по-
чти в 3 раза) короче плеотельзона. Длина тела до 10 мм.
Распространение. Оз. Охрида, от 40 м до максимальных
глубин (286 м).
17. Asellus (Proasellus) gjorgjevici liter alls Kar.
2 вдвое больше ширины.
Он
на
пе-
К а г am an, 1933b : 108.
Отличается от типичной формы нормально развитыми глазами и зна-
чительно более укороченным и расширенным телом.
Распространение. Оз. Охрида, на глубинах меньше 40 м.
88
I. ASELLIDAE
18. Asellus (Proasellus) cavaticus Schiodte (рис. 125—128).
Schindte, 1871, in Leydig, Jahresb. Ver. Vaterb. Naturk. Wiirtb. : 79;
Fries, 1879, Zool. Anz., 2 : 129, 150; M i e t h e, 1899 : 273, t. 11—13; Carl,.
1908:125; T a t t e r s a 1 1, 1930:82, 3 fig., t. 6; Stammer, 1932:125,.
fig. 10—11; — sieboldii de Rougemont, 1875, Bull. Soc. Sc. nat. Neuchatel : 152. —
cavaticus leruthi Arcangeli, 1935, 2, fig. 1—10.
Ширина тела в 4 раза меньше длины. Эпимеры слабо развиты. Длина
плеотельзона несколько больше ширины. Глаза и пигмент отсутствуют.
Рис. 125—-128. Asellus (Proasellus)
cavaticus Schiodte: 125 — проподит
I перейопода самца; 126 и 127 — I
и II плеоподы самца; 128—II плео-
под самки. Ориг.
Первая антенна равна 3/4 длины ос-
нования второй антенны. Ее жгут ра-
вен по длине основанию и состоит из.
8—12 члеников, несущих 5—6 (<5) или
3—4 (j) lammae olfactoriae. Вторая
антенна не короче или незначительно,
короче тела и имеет в жгуте 38—44
членика. Длина проподита I перейо-
пода <5 в 2х/г —3 раза превышает его
ширину. Его задний край снабжен
4—7 ножевидными зубцами и несколь-
кими щетинками. На заднем крае — 2
щетинки. Длина и ширина протоподита
I плеопода $ равны. На его внутреннем
крае находятся 2—3 совокупительных
зубца. Ширина экзоподита вдвое мень-
ше длины. На его внешнем и дисталь-
ном краях сидит около 20—21 корот-
ких щетинок, значительно варьирую-
щих по длине, из которых несколько-
проксимальных могут быть перистыми.
На поверхности экзоподита в базальной
его трети — 3 щетинки: 2 вблизи внут-
реннего проксимального угла и 1
(иногда отсутствующая) — посредине.
Длина протоподита II плеопода <5 на
V4 превышает его ширину. Внутрен-
ний край на середине несет 2—3 пе-
ристые щетинки. Экзоподит почти в
IV2 раза короче протоподита. Его-
базальный членик шире, но короче
дистального и несет по внешнему краю
1—3 перистые щетинки. Длина дисталь-
ного членика превосходит его ширину.
На внешнем его крае сидят 7—8, на дистальном 3—4 коротких перистых ще-
тинок. Эндоподит по длине равен экзоподиту или незначительно превосхо-
дит его. В его базальной части имеется небольшой, направленный внутрь
вырост, несколько напоминающий вырост у видов подрода Asellus s. str.,
но гораздо более короткий. Окончание эндоподита хорошо выражено,
овальной формы и превосходит боковую лопасть. II плеопод $ округло-
треугольной формы. Его длина вдвое больше ширины. На дистальном крае
сидят 6—10 коротких перистых щетинок, на поверхности 2—4 простые
щетинки. Уроподы равны по длине плеотельзону или незначительно-
превышают его. Протоподит и экзоподит одинаковой длины, а эндоподит
1. ASELLUS
89-
несколько (на V5—1/J) длиннее каждого из них. Уроподы несут длин-
ные щетинки. Длина о до 8 мм.
Распространение. Южная и западная Германия, Швейца-
рия, Англия, Югославия, Бельгия.
Экология. Обитатель ключей, колодцев и пещерных водоемов.
Признаки этого вида, как следует из сравнения работ Мите, Фриза,
Тетерсолла и Штаммера и из исследования материала, довольно посто-
янны. Нет никаких оснований выделять в особый подвид бельгийского
A. cavaticus, описанного Arcangeli как A. cavaticus leruthi Arc. Этому
автору, повидимому, осталась неизвестной подробная работа Тетерсолла^
который приводит рисунки и описание A. cavaticus, ничем существенным
не отличающегося от бельгийского A. cavaticus.
Представители этого вида из альпийских озер, грунтовых и фреа-
тических вод Швейцарии, а также из некоторых районов Югославии,
значительно отклоняются от типа и выделяются в особые подвиды. Всем
им, однако, свойствен наиболее характерный признак вида — напра-
вленный внутрь вырост базальной части эндоподита II плеопода <5.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДВИДОВ
1 (2). Эндоподит II плеопода 8 более чем вдвое длиннее экзоподита.
Экзоподит лишен щетинок.................A. cavaticus walteri Chapp.
2 (1). Эндоподит II плеопода 8 менее чем вдвое длиннее или даже короче-
экзоподита. Последний всегда несет щетинки.
3 (8). Эндоподит II плеопода 6 заметно короче протоподита и короче,
равен или лишь незначительно длиннее экзоподита.
4 (7). Голова с вдавлениями с боков и спереди. Экзоподит I плеопода
8 менее чем в 1 V, раза длиннее протоподита.
5 (6). Эндоподит II плеопода 8 короче экзоподита. Вторая антенна
достигает конца тела . . . . .A. cavaticus montenegrinus Kar.
6 (5). Эндоподит II плеопода 8 равен по длине экзоподиту. Вторая
антенна доходит до 2/3 Длины тела ................................
..............................A. cavaticus hercegovinensis Kar.
7 (4). Голова без вдавлений с боков и спереди. Экзоподит I плеопода
8 вдвое длиннее протоподита .......................... . ............
.................................A. cavaticus cavaticus Schiodte-
8 (3). Эндоподит II плеопода 8 заметно длиннее протоподита и экзо-
подита.
9 (10). Экзоподит I плеопода 8 несет стернальную щетинку близ вну-
треннего базального угла. Эндоподит II плеопода 8 более чем
в 1 1/2 раза длиннее экзоподита ... A. cavaticus foreli Blanc
10 (9). Экзоподит I плеопода 8 лишен стернальной щетинки. Эндоподит
II плеопода 8 менее чем в 1 ]/2 раза длиннее экзоподита. . . . .
.................................A. cavaticus valdensis Chapp.
18а. Asellus (Proasellus) cavaticus foreli Blanc.
— foreli Blanc, 1879:377, t. 13; 1879a : 428; .Forel, 1902, Le Leman,.
Ill : 92, fig. 179; — cavaticus Hofer, 1896, Die Verbreitung der Tierwelt im Boden-
see : 14; Stammer, 1932 : 127; — cavaticus var. foreli Carl, 1908 : 127 ;— cava-
ticus subsp. foreli Chappuis, 1948 : 558, fig. 2—7.
Длина плеотельзона вдвое больше ширины. В жгуте первой антенны
4—5, а иногда до 10 члеников, из которых на 3 последних сидят Laminae
olfactoriae; число их у 8 и j одинаково. Вторая антенна достигает сере-
90
I. ASELLIDAE
дины длины тела и имеет 28—29-членистый жгут. Перейоподы как
у типичной формы. Длина протоподита I плеопода 8 в 1 2/5 раза больше
ширины; на внутреннем крае 2—4 совокупительных зубца. Экзоподит
овальной формы, его ширина вдвое меньше длины; на его дистальном
крае и дистальной половине наружного края находится 14—15 щети-
нок, близ внутреннего базального угла — 1 стернальная щетинка.
Ширина протоподита II плеопода 8 в 1 3/й раза уступает длине; на вну-
треннем крае 2 щетинки. Экзоподит вдвое короче протоподита. На наруж-
ном крае его базального членика 1—2 перистые щетинки, на наружном
й дистальном краях дистального членика 7—9 перистых щетинок. Эндо-
подит по длине более чем в 1 1/2 раза превосходит экзоподит. Уроподы
длиннее половины .плеотельзона, их эндоподит вдвое длиннее протопо-
дита, экзоподит незначительно короче. Длина тела до 5 мм, ширина —
до 1.3 мм.
Распространение. Швейцария — в озерах Женевском,
Боденском и Фирвальдштедтском; Франция — озера Аннеси и Буржэ.
Биология. Обитает на больших глубинах альпийских озер.
'Опускается до 100 м и ниже.
18b. Asellus (Proasellus) cavaticus waiter! Chappuis.
Chappuis, 1948 : 560, fig. 8—11.
Длина тела в 5 3/ю раза превышает ширину. Длина головы больше
ее ширины. Протоподит I плеопода 8 треугольной формы; его длина
в 1 2/г> раза больше ширины, на внутреннем крае всего 1 совокупитель-
ный зубец. Ширина экзоподита почти в 1% раза меньше длины,
по длине он в 17/ю раза превосходит протоподит. На наружном
и дистальном крае всего по 1 щетинке. Длина протоподита II плеопода
8 на %, больше ширины. Экзоподит втрое короче протоподита. Оба чле-
ника экзоподита одинаковой длины и лишены щетинок. Длина экзопо-
дита в 33/б‘раза превосходит его ширину и более чем вдвое длину экзо-
подита. Наружный вырост его базальной части выражен слабо. Строение
антенн, перейоподов и уроподов остается неизвестным. Длина тела 2.3 мм.
Необычайные пропорции частей II плеопода 8 и редукция щетинок
на всех исследованных конечностях могли бы позволить выделить эту
*форму в особый вид. Однако эти признаки варьируют, ив 10 км от нахо-
ждения описанной формы были пойманы водяные ослики, приближаю-
щиеся по указанным признакам к типичному cavaticus.
Распространение. Швейцария — окрестности Греллингена
•и Мюнхенштейна.
Биология. Малые размеры, удлиненные тело и голова указы-
вают на приспособление к обитанию в капиллярных ходах, заполненных
фреатическими водами. Рачки были добыты с помощью насоса.
18с. Asellus (Proasellus) cavaticus valdensis Chappuis.
Chappuis, 1948 : 563, fig. 12^-15.
Жгут первой антенны 6-членистый и несет 4 lammae olfactoriae. Про-
топодит I плеопода 8 трапецеидальной формы и имеет 1 или 2 совокупи-
тельных зубца. Длина экзоподита в 22/s раза больше ширины. На дисталь-
ном крае расположены 4 более коротких, на наружном — 13 более длин-
ных щетинок. Длина протоподита II плеопода 8 в 12/з раза больше
ширины. Длина экзоподита составляет 0.6 о.т длины протоподита. Оба
1. ASELLUS
91
членика экзоподита одинаковой длины, базальный несет 3 перистых
щетинки па наружном крае, дистальный — 5 перистых щетинок на наруж-
ном и I перистую щетинку на дистальном крае. Эндоподит на % длин-
нее экзоподита. Строение второй антенны и уроподов неизвестно. Длина
тела 3.7 мм.
Распространение. Швейцария — единственный экземпляр
был пойман под камнями в источнике Орбе.
18(1. Asellus (Proasellus) cavaticus hercegovinensis Kar.
Karaman, 1933b : 107, fig. 3.
Голова спереди и по бокам с вдавлениями. Вторая антенна равна
всего 2/з длины тела. Проподит I перейопода 5 значительно шире, чем
у типичной формы. Экзоподит I плеопода 5 лишь незначительно длиннее
протоподита, на внутренней стороне которого находится только 1 сово-
купительный зубец. На поверхности экзоподита сидит только 1 щетинка
вблизи внутреннего проксимального угла; щетинка на основном чле-
нике экзоподита II плеопода <5 отсутствует. Вырост в базальной части
эндоподита II плеопода <3 выражен слабо.
Распространение. Югославия — пещера Ветреница и дру-
гие пункты Далмации и Герцеговины.
18е. Asellus (Proasellus) cavaticus montenegrinus Kar.
Karaman, 1934a : 23, fig. 1.
Голова спереди без вдавления, а боковые вдавления значительно
менее глубокие, чем у предыдущего подвида. Вторая антенна по длине
почти равна телу, в ее жгуте до 37 члеников. Протоподит I плеопода
S почти такой же длины, как экзоподит, с 1 совокупительным зубцом
на внутренней стороне. Экзоподит лишен щетинок на поверхности.
II плеопод <5 как у типичной формы, с выступом в базальной части эндо-
подита. II плеопод J несет несколько коротких щетинок на внешнем
крае. Длина тела до 6 мм.
Распространение. Внутренняя и южная Югославия.
19. Asellus (Proasellus) hypogeus Rac. (рис. 129—130).
Racovitza, 1922 : 407, fig. 7—9.
Глаза и пигмент отсутствуют. Ширина тела в 4 раза меньше длины.
Длина плеотельзона незначительно превышает ширину. Первая антенна
на Va короче основания второй антенны, ее жгут состоит из 7 члеников
с 4 lammae olfactoriae. Вторая антенна на Ve короче тела, с 50-членистым
жгутом. На заднем крае проподита I перейопода £ 2 ножевидных зубца
и иногда третий, редуцированный. Дактилюс с 6 зубцами. Длина про-
топодита I плеопода <5 на /4 больше ширины. На прямом внутреннем
крае 1 зубец, наружный край округлый. Экзоподит овальный, на х/з
длиннее протоподита; его ширина на 37з меньше длины, на наружном
крае 12 коротких простых щетинок, на дистальном —• 6—7 довольно
коротких (вдвое более коротких, чем экзоподит) перистых щетинок,
у внутреннего проксимального угла 1 щетинка. Длина протоподита
II плеопода <5 на 14 больше ширины, на дистальной половине его вну-
треннего края сидят 4 перистых щетинки. Экзоподит немного короче
протоподита. Его проксимальный членик, на наружной стороне которого
.находится 1 простая щетинка, в 3 раза короче дистального членика,
92
II. ASELLIDAE
несущего около 7 перистых щетинок и на середине наружного края
2 простых щетинки. Эндоподит по длине равен экзоподиту. Его внешний
край с широкой и
довольно глубокой выемкой, а дистальный конец имеет
удлиненный вырост на-
ружу. Экзоподиты IV и V
плеоподов лишены щети-
нок. Эндоподит V плеопода
длиннее, но уже экзопо-
дита. Уроподы немного
длиннее плеотельзона. Их
экзоподиты на % короче
эндоподитов и незначи-
тельно короче протоподи-
тов. Длина тела S 6 мм.
Распростране-
ние. Алжир—пещера в
Itri-bou-Aman.
Rac.,
hypogeus
Раковине.
Рис. 129—139. Asellus (Proasellus)
I и II плеоподы самца. По
20. Asellus (Proasellus)
spelaeus Вас. (рис. 131 —
133).
Racovitza, 1922:403,
fig. 1—4.
Глаза и пигмент отсут-
ствуют. Ширйна тела в 4
и ширина плеотельзона равны. Первая
раза меньше длины. Длина
антенна на 1/з короче основания второй антенны. Ее жгут вдвое короче ос-
нования и имеет 7 (<5) или 5 (j) члеников, на которых находится 4 (<5)
или 2 (j) laminae olfactoriae. Вторая антенна на 1/з (<5) или вдвое (j) ко-
роче тела. В жгуте около 50 (S) или 40 (j) члеников. Проподит I перейо-
Asellus (Proasellus) spelaeus
Рис. 131—133.
Rac.: 131 и 132— I и II плеоподы самца;
133 — плеопод самки (По Р'аковице).
роче тела. В жгуте около 50 (<3)
пода S с двумя ножевидными
зубцами, дактилюс с 4 зубцами.
Длина протоподита I плеопода
S на %, больше ширины. На
внутренней стороне 1 зубец.
Экзоподит на J4 длиннее про-
топодита. Его длина на х/з боль-
ше ширины. Внутренний и ди-
стальный края несут около 13
простых коротких щетинок. На
поверхности, вблизи внутрен-
него проксимального угла на-
ходится 1 толстая щетинка.
Длина протоподита II плеопода
<5 незначительно больше ши-
рины. На внутреннем его крае
4 перистых щетинки. Экзопо-
дит на х/з короче протоподита. Его проксимальный членик, несущий на на-
ружном крае! простую щетинку, в 1Д раза короче дистального, на котором
сидит 2 простых и 2 перистых щетинки. Эндоподит несколько длиннее
и толще экзоподита, с прямым проксимальным внутренним углом.
II плеопод £ с сильно выгнутым внешним краем, несущим 3 перистых
1. ASELLUS
93
щетинки. Около середины внутреннего края на поверхности сидит 2 про-
стых щетинки. Экзоподиты IV и V плеоподов лишены щетинок. Эндо-
подит V плеопода немного шире и длиннее, чем экзоподит. Уроподы
на %. (5) или на Чз (д) короче плеотельзона. Их экзоподиты немного
короче эндоподитов и на 1/з длиннее протоподитов. Длина тела $ 5.5 мм,
g 4 мм.
Распространение. Франция —пещера Оксибар (Oxibar)
(департ. Нижних Пиренеев).
20а. Asellus (Proasellus) spelaeus aquae-calidae Rac.
Racovitza, 1922 : 406, fig. 5—6.
Жгут первой антенны состоит из 6 члеников и несет 3 lammae olfacto-
riae, жгут второй антенны из 40 члеников. Экзоподит I плеопода <5 зна-
чительно более удлиненный, чем у типичной формы, и снабжен большим
числом щетинок (16). Эндоподит и экзоподит II плеопода S по длине
и ширине равны. На наружном крае проксимального членика экзопо-
дита 1 перистая щетинка, на внешнем и дистальном краях удлиненного
дистального членика 5 перистых щетинок. Длина тела 6 — 5.5 мм (д
не известна).
Распространение. Франция—пещера 0-шод (Eaux-chaucles),
департамент Нижних Пиренеев.
20b. Asellus (Proasellus) spelaeus anophthalmus Kar.
— meridianus anophthalmus К ar am an, 1934a : 24, fig. 2.
Длина плеотельзона несколько больше ширины. Число члеников
жгута первой антенны — 4—5, второй антенны — до 25. Длина прото-
подита I плеопода <5 значительно превышает ширину. Экзоподит с округ-
лыми внешним и внутренним краями, широкий, с небольшим количе-
ством (около 10) щетинок на наружном и дистальном краях. Протопо-
дит, экзоподит и эндоподит II плеопода 6 почти одинаковой длины.
Щетинки в количестве около 3 присутствуют только на дистальном чле-
нике экзоподита. Уроподы короче плеотельзона, их протоподиты и экзо-
подиты одинаковой длины, эндоподиты несколько длиннее. Длина тела
до 2.7 мм.
Реми (Remy, 1941) правильно отметил, что эта форма не может счи-
таться подвидом A. meridianus, как думал Караман. Действительно, строе-
ние и вооружение передних плеоподов $, отсутствие глаз, длина уропо-
дов и некоторые другие признаки резко отличают anophthalmus Kar.
от A. meridianus Rac. Реми выделяет anophthalmus в особый вид. Однако
этот автор не проводит сравнения anophthalmus с другими видами Proa-
sellus. Между тем по всем основным признакам, в особенности по строе-
нию передних плеоподов <5, anophthalmus чрезвычайно близок к A. spe-
laeus Rac., отличаясь от него преимущественно короткими антеннами,
удлиненным плеотельзоном и отсутствием толстой щетинки близ вну-
треннего проксимального угла экзоподита I плеопода 8. Последний
признак свойствен описанным Реми (1941) вариететам anophthalmus.
Поэтому представляется более правильным считать anophthalmus под-
видом A. spelaeus Rac.
Распространение. Черногория — ключи Беране, Рибница,
Андриевица, Печ (Berane, Ribnica, Andrijevica, Pec).
Экология. Эта форма найдена только в ключах. '
94
I. ASELLIDAE
20c. Asellus (Proasellus) spelaeus rhausinus Remy.
Remy, 1941 : 4, fig. 1—9.
Длина плеотельзона на x/s превосходит ширину. Жгут первой антенны
27—30-членистый и достигает 2/з длины тела. Форма I плеопода <5 как
у anophthalmus, но близ наружного дистального угла протоподита и близ
внутреннего базального угла экзоподита находится по одной щетинке.
Внутренний.дистальный угол протоподита II плеопода <5, а также наруж-
ный край проксимального членика экзоподита несут по одной ще-
тинке. Наружный и дистальный края II плеопода j с 4—5 щетин-
ками, на поверхности — 2 щетинки. Длина тела <5 3.5 мм, 9 — до
4.25 мм.
Распространение. Югославия — ключ в 4—5 км к северо-
востоку от Дубровника.
20d. Asellus (Proasellus) spelaeus longisetus Remy.
Remy, 1941 : 9, fig. 10—-13.
Отличается от предыдущей формы большим количеством щетинок
(14—19) на внешнем и дистальном краях экзоподита I плеопода S. Из них
3—5 дистальных удлинены и оперены. Близ внутреннего базального
угла этого членика 2 щетинки. По краям II плеопода 9 9—12 перистых
щетинок. Длина тела 4.5—5 мм.
Распространение. Югославия — ключ близ Билека (Bileca)
и Паганьяка печина (Paganjaca pecina) в Герцеговине.
21. Asellus (Proasellus) collingei Birstein, nom. nov. (рис. 134—136).
— hazeltoni Collinge, 1946 : 72, fig. 1—4 (nomen praeoccupatum).
Длина тела в 4 раза больше ширины, которая остается постоянной
на всем протяжении тела. Голова спереди срезана прямо и имеет тра-
D4
Рис. 134—136. Asellus
(Proasellus) collingei
Birst., nom. nov.: 134 —
голова;) 135 — плеотель-
зон; *136 — уропод.
(По Коллинджу).
Распростр
Уэллсе.
пецоидальную форму, так как ее задний край зна-
чительно длиннее переднего. Глаза отсутствуют.
Первая антенна короткая. Жгут второй антенны
равен по длине стержню и достигает V грудного
сегмента. Длина плеотельзона превышает ши-
рину. Через его покровы со спинной стороны
просвечивают 2 продольных желтых лентовидных
пятна. Уроподы короткие и толстые. Их эндоподит
узкий, стилетовидный, экзоподит вдвое длиннее и
втрое или более шире эндоподита. Длина тела
<3 — 8 мм.
За год до опубликования весьма поверхност-
ного описания этого вида его автором был напе-
чатан короткий диагноз цветового вариетета обыч-
ного водяного ослика, которому было дано на-
звание A. aquaticus var. hazeltoni (см. стр. 60).
Очевидно, название вышеописанного вида должно
быть изменено, как nomen praeoccupatum, что
здесь и предлагается.
а н е н и е. Англия, пещера Dah-je-ogop в
Южном
1. ASELLUS
95-
22. Asellus (Proasellus) hussoni Remy (рис. '137—140).
Remy, 1941 : 12, fig. 14-22.
Глаза и. пигмент отсутствуют. Наружные края головы вогнуты. Длина
плеотельзона равна его ширине. Жгут первой антенны состоит из 8—9
(8) или 6—7. (о) члеников, из которых на последних 4—5 находятся lam-
inae olfactoriae. Вторая антенна длиннее
39—45 (б) или 27—34 ($>) члеников.
Проподит первого перейопода о с двумя
ножевидными зубцами, дактилюс с 6
зубцами. Длина протоподита I плео-
пода 3 приблизительно в 1% раза
больше ширины; на его наружном ди-
стальном углу 1 щетинка, вдоль внут-
реннего края 1—3 стернальных ще-
тинки, на внутреннем крае 1 совоку-
пительный зубец. Экзоподит почти
вдвое длиннее протоподита, но равен
ему по ширине и несет 28—36 частично
перистых щетинок на наружном и ди-
стальном краях и 1 —-2 стернальных
щетинки близ внутреннего базального
угла. Длина протоподита II плеопода
3 на Уз больше ширины; внутренний
его край с 2—4 перистыми щетинками.
Экзоподит короче протоподита; на на-
ружном крае его проксимального чле-
ника могут находиться 1—2 перистые
щетинки, на наружном и дистальном
краях дистального членика 8 — 1.1 пе-
ристых щетинок. Длина последнего чле-
ника больше ширины. Длина эндопо-
дита вдвое больше ширины, его внут-
ренний край выпуклый, боковая лопасть
шире и длиннее основания. II плеопод
2 трапецеидальной формы; наружный
и внутренний его края прямые, при-
чем наружный вдвое короче внутрен-
него. На дистальном крае 12 — 13 пери-
половины тела и имеет в жгуте
138 139
Рис. 137 —140 Asellus (Proasellus}
hussoni Remy: 137, 138 — I и II
плеоподы самца; 139 — II плеопод
самки; 140 — плеотельзон самца^с
уроподами. (По Реми)..
стых щетинок, близ внутреннего края 1—2 стернальных щетинки.
Длина экзоподита III плеопода на 2/з—3/з превышает ширину; прокси-
мальный членик несет 9—И гладких щетинок на наружном крае,
дистальный — 22—27 перистых щетинок на наружном крае и 2—3 глад-
ких стернальных щетинки вдоль внутреннего края. Протоподит IV
плеопода с 1 щетинкой на внутреннем крае, экзоподит, длина которого
вдвое больше ширины, с 2—4 щетинками на наружном крае прокси-
мального членика. Длина экзоподита V плеопода более, чем вдвое пре-
вышает ширину; его наружный край лишен щетинок или несет 1—2
щетинки. Уроподы по длине равны плеотельзону. Протоподит и эндо-
подит одинаковой длины, экзоподит равен каждому из них или незна-
чительно короче. Длина тела 3 8.5 мм, — 7 мм.
Распространение. Югославия, пещера’ Резница (Reznica)
в Герцеговине, в 15 км от пещеры Ветреница (Попово поле).
96
I. ASELLIDAE
Рис."141—144. Asellus (Proasellus)
hermallensis Arcangeli: 141 — ко-
нец I перейопода самца; 142 и
143 — I и II плеоподы самца;
144— II плеопод самки. (По Ар-
канджели).
Распространение,
стях Hermalle-sous-Argenteau;
25. Asellus (Proasellus) hermallensis Arc. (рис. 141—144).
Arcangeli, 1938 : 1, fig. 1—10.
Глаза и пигмент отсутствуют. Максимальная ширина тела, приходя-
щаяся на IV сегмент тела, почти в 4 раза меньше длины. Длина плео-
тельзона больше ширины. Жгут первой антенны состоит из 4—5 члени-
ков, жгут второй антенны — из 22 члеников. Длина проподита I перейо-
пода S более чем вдвое превышает ширину, его задний край с 9 ноже-
видными зубцами. Дактилюе несет 4 зубца.
Ширина протоподита I плеопода S заметно
уступает длине, на внутреннем его крае 2
совокупительных зубца. Длина экзоподита
несколько менее чем вдвое превышает ши-
рину, на дистальной части наружного края
сидят 5 длинных перистых щетинок, пере-
ходящих частично на дистальный край.
Длина протоподита II плеопода 6 незна-
чительно превосходит ширину; внутренний
край несет 3 перистых щетинки. Экзо-
подит несколько короче протоподита, его
дистальный членик почти вдвое длиннее
базального и снабжен 5 длинными пери-
стыми щетинками, сидящими на дисталь-
ном- крае, и 2 короткими щетинками на
наружном крае. Эндоподит длиннее и
толще экзоподита и имеет короткое широ-
кое окончание и короткую, но довольно
узкую боковую лопасть. Длина II плео-
пода J более чем вдвое превышает ши-
рину, наружный край выпуклый и в своей
дистальной части несет 5 длинных пери-
стых щетинок. Экзоподит III плеопода,
в отличие от других видов, лишен пери-
стых щетинок. Уроподы приблизительно
вдвое короче плеотельзона, их экзоподиты
значительно длиннее эндоподитов. Длина
тела 4.4 мм, ширина 1.2 мм.
Бельгия, грунтовые воды в окрестно-
рачек добыт с помощью насоса.
42. Asellus (Proasellus) gauthieri Monod (рис. 145—149).
. Monod, 1924 : 332, рис. 23—33.
Пигмент отсутствует. Глаза развиты слабо. Длина тела в 4 раза пре-
восходит ширину. Голова и плеотельзон уже сегментов груди. Длина
нлебтельзона больше его ширины. Жгут первой антенны состоит из 6—7
(<5) или 5 (j) члеников, несущих 4 (6) или 3 (j) lammae olfactoriae.
В жгуте второй антенны 45—49 (<3) или менее 30 (д) члеников. Пропо-
дит I перейопода S имеет на заднем крае 2 ножевидных зубца, у $ соот-
ветственно 2—3. На дактилюсах остальных перейоподов имеется только
по 2 крупных зубца. Длина протоподита 1 плеопода $ превышает его
ширину. На внутреннем крае — 1 совокупительный зубец и 1 щетинка
на дистальном крае. Длина экзоподита более чем в 1Уг раза превос-
ходит ширину. На дистальном крае 4 перистых щетинки, не достигаю-
1. ASELLUS
97
щих половины длины экзоподита, на внешнем крае 11—12 коротких
простых щетинок. Ширина протоподита II плеопода 6 значительно пре-
восходит его длину. Экзоподит по длине равен протоподиту. Сильно
расширяющийся к концу базальный членик несет на внешнем крае
короткую простую щетинку. Дистальный членик, суживающийся к концу,
почти в 3 раза длиннее базального. На дистальном крае 4—5 перистых
щетинок, на внешнем крае — 4 коротких простых щетинки. Эндоподит
незначительно короче экзоподита. Базальная его часть имеет .два неболь-
ших округлых выроста: один — направленный внутрь, один — направлен-
ный наружу. Длина эндоподита в 3 раза больше ширины, промеренной
посредине. Окончание эндоподита вытянутое, с параллельными краями,
боковая лопасть короткая и широкая. II плеопод с прямым внутрен-
ним и выпуклым внешним краем. Его длина более чем в Р/г раза больше
ширины. На дистальном крае 4 перистых щетинки, не достигающие
но длине половины длины плеопода. Длина экзоподита III плеопода
8 и J в 1Уг раза больше ширины. На внешнем крае проксимального чле-
Рйс. 145—144. Asellus (Proasellus) gauthieri Monod: 145— проподит I не-
рейопода самца; 146 и 147 — I и II плеоподы самца; 148 —II плеопод
самки; 14)— IV перейопод самца. По Моно.
ника 4 щетинки, на внешнем крае дистального — 6 щетинок, на дисталь-
ном крае — 7 щетинок. Ветви уроподов одинаковой длины, на %, длин-
нее протоподита. Длина тела 4—6 мм.
Распространение. Алжир.
Экология. Найден в ключе, близ его выхода из-под земли, между
камнями. Вероятно, ведет и подземный образ жизни (редукция глаз).
24а Asellus (Proasellus) gauthieri theodori, subsp. nov. (рис. 150—
155).
Глаза и пигмент развиты очень слабо. Максимальная ширина тела
приходится на VI грудной сегмент. Длина плеотельзона значительно пре-
восходит его ширину. В жгуте первой антенны 8 5—7 члеников с31ашшае
olfactoriae, j — 5 члеников с 2 lammae olfactoriae. Жгут второй антенны
Состоит из 35—50 (d>) или 30—35 (§) члеников. Проподит I перейопода
8 с 2 ножевидными зубцами на заднем крае. Дактилюсы всех перейопо-
дов с 2 зубцами. Длина протоподита I плеопода <5 незначительно больше
ширины, на его внутреннем крае—1—2 совокупительных зубца,
щетинки отсутствуют. Экзоподит всего в Г/г раза длиннее протоподита,
его длина несколько менее чем в Р/г раза превышает ширину, он сужи-
вается к дистальному концу и несет 4 перистые щетинки на дистальном
крае и около 10 простых щетинок на внешнем крае. Длина и ширина
7 Фауна СССР, том VII, в. 5
98
I. ASELLIDAE
протоподита II плеопода <5 равны, на внутреннем крае находится 3—4
перистых щетинки. Базальный членик экзоподита менее чем в 3 раза
короче дистального, он резко расширяется к концу и лишен щетинок.
Дистальный членик почти не суживается к концу, на его дистальном
крае сидят 4 перистых, на внешнем крае — 2—3 простых щетинки.
По длине эндоподит не уступает протоподиту, его строение, как у типич-
ной формы. Длина II плеопода £ в 2 раза превосходит ширину; внутрен-
ний его край, прямой, наружный — выпуклый. На дистальной части
наружного края и на дистальном крае находится 6—7 более длинных,,
чем у типичной формы, перистых щетинок. Базальный членик экзопо
Рис. 150—155. Asellus (Proasellus} gauthieri theodori, n. subsp.: 150-—конец
I перейопода самца; 151 и 152 — I — II плеоподы самца; 153 — II плеопод
самки; 154 — плеотельзон самца с уроподами; 155 — конец IV перейопода
самца. Ориг.
дита III плеопода несколько короче дистального. Уроподы 8 по длине,
равны плеотельзону, их экзоподиты несколько короче эндоподитов;
уроподы- £ короче плеотельзона. Длина тела — до 5 мм.
Несколько экземпляров Л. gauthieri theodori, subsp. nov. обнаружено
в коллекции североафриканских водяных осликов, присланной Моно
(Monod, Париж), который определил эту форму как новый подвид,
но не описал ее. Она отличается от типичной главным образом иными
пропорциями, отдельных частей I и II плеоподов 8 и li плеопода £,
а также укороченными уроподами.
Распространение. Ключ в гроте Рарифре (Rharifre), Алжир.
25. Asellus (Proasellus) thermonyctophilus Monod (рис. 156—159).
— gauthieri subsp. thermonyctophilus Monod, 1924 : 334, fig. 34—43; —ther-
monyctophilus Mono d, 1925 : 243, pl. LI.
1. ASELLUS
99
Известна только j. Длина тела более чем в 4 раза превосходит ширину.
Пигмент и глаза отсутствуют. Длина плеотельзона больше ширины. Жгут
первой антенны состоит из 5 члеников, несущих 2 lammae olfactoriae.
Дактилюс II—VII перейоподов с 1 крупным зубцом. Уроподы короче
плеотельзона. Экзоподит незначительно длиннее протоподита й эндо-
подита. Длина тела 4.80 мм, ширина 0.08 мм.
Распространение. Алжир.
Найден в теплом (20°) ключе, совместно
с Typhlocirolana fontis Gurney. Трогло-
бионт.
Рис. 156—159. Asellus (.Proa-
sellus) thermonyctophilus Mo-
nod, самка: 156 — общий вид;
157 — 1 максилла; 158 — I
перейопод самца; 159 — дак-
тилюс IV перейопода. 156, 158
и 159 — по Моно, 157 — ориг.
рис. Моно.
Рис. 160—163. Asellus (Proasellus) istrianus
Stammer: 160 — I плеопод самца; 161 — II
плеопод самца; 162 — II плеопод самки;
163 — I перейопод самца. (По Ш таймеру).'
26. Asellus (Proasellus) istrianus Stammer (рис. 160—163).
Stammer, 1932 : 121, fig. 6—9.
Наибольшая ширина приходится у <5 на VI сегмент, у q — на IV—V
сегменты. Ширина плеотельзона превосходит длину. Окраска серая или
серовато-бурая, со светлыми пятнами, передняя часть головы окрашена
темнее. По средней линии вдоль тела проходит темная полоса. Первая
антенна короче основания второй антенны и имеет 9—10 (<3) или 6—8
($) члеников жгута с 3 lammae olfactoriae у обоих полов. Вторая антенна
вдвое короче тела, ее жгут состоит из 35—44 (5) или 36—39 (^) члеников.
Проподит I перейопода S с 3 ножевидными зубцами, 2—3 щетинками
в верхнем ряду, 4—5 щетинками в дистальном и 2—3 в заднем ряду.
Длина и ширина протоподита I плеопода 6 равны. На внутреннем его
крае — 2 совокупительных зубца, на закругленном дистальном внеш-
нем углу — 2 щетинки. Длина экзоподита в 1/4 раза больше ширины.
На внешнем и дистальном краях сидят 21—26 простых щетинок; на
7*
100
I. ASELLIDAE
поверхности — 1 толстая щетинка у внутреннего проксимального
угла и 2—3 щетинки у внутреннего дистального угла. Длина и ширина
протоподита II плеопода <5 равны. На внутреннем его крае находится
4—6 перистых, щетинок. Дистальный членик экзоподита в 4 раза длин-
нее проксимального и несет на своем дистальном крае 8—10 длинных
перистых щетинок. На наружном крае проксимального членика —
1 перистая щетинка, иногда отсутствующая. Эндоподит короче экзопо-
дита, его ширина вдвое меньше длины. Конец эндоподита короткий.
II плеопод § треугольной формы, его ширина у основания вдвое меньше
длины, на дистальной половине внешнего края сидит 8—10 перистых
щетинок. Экзоподит IV плеопода разделен неясными линиями на 4 поля.
На дистальной части его внешнего края — 3—6 щетинок. Экзоподит
V плеопода разделен неясной линией на 2 поля и имеет на дистальной
части внешнего края 4 щетинки. Уроподы значительно короче плеотель-
зона. Протоподит короче экзоподита, который в свою очередь короче
эндоподита. Длина тела <5 5—7.5 мм, j 4.5—6 мм.
Распространение. Темная цистерна в 15 км от г. Триеста.
27. Asellus (Proasellus) amautovici Remy (рис. 164—166).
Remy, 1932 : 73, fig. 1—9.
Максимальная ширина <5 приходится на VI—VII сегменты.^ Длина
и ширина плеотельзона равны. Жгут первой антенны состоит у 6 из 9—
11 члеников, у j из 6 члеников, и несет 3—4 (<5) или 2 (j) lammae olfac-
Рис. 164—166. Asellus (Proasellus} arnautooici Remy, 164—I
плеопод самца, 165 — II плеопод самца, 166 — II плеопод
самки. (По Реми).
toriae. В жгуте второй антенны до 65 (<5) или до 40 (j) члеников. Про-
подит I перейопода 6. эллипсоидальной формы. Его длина вдвое превос-
ходит ширину. Задний край снабжен 3 ножевидными зубцами. Дакти-
люс с 10—11 зубцами. Протоподит I плеопода <5 трапецеидальной формьц
с округлыми углами. Его длина на Vs—Vs превосходит ширину. На вну-
треннем крае — 1 совокупительный зубец. Экзоподит овальный, всего
на х/з—Vs . более длинный, чем протоподит. На его дистальном крае
1. ASELLUS
101
и дистальной части внешнего края сидит 8—10 простых коротких щети-
нок. На поверхности, вблизи проксимального внутреннего угла, нахо-
дится 2 коротких перистых щетинки. Длина протоподита II плеопода
<3 незначительно превосходит ширину. На внутреннем крае 3 перистых
щетинки. Экзоподит незначительно длиннее протоподита. Базальный
членик в 4 раза короче дистального и лишен щетинок. Дистальный чле-
ник удлиненный, с параллельными краями. Его длина в 2—2% раза
больше ширины. На дистальном крае 8—10 перистых щетинок, из кото-
рых наиболее длинные почти в 1% раза превосходят длину всего чле-
ника. Эндоподит короче экзоподита. Окончание его короткое, широкое.
Боковая лопасть тоже короткая и широкая, но превосходящая по длине
окончание. II плеопод £ удлиненный, трапецеидальной формы, с вогну-
тыми внутренним и внешним краями. Длина его в ЗУг—33/< раза пре-
восходит ширину. Дистальный край несет 9 перистых щетинок, в боль-
шинстве своем равных по длине всему плеоподу. В базальной части,
вблизи внутреннего края, находится 1 щетинка. На поверхности экзо-
подита III плеопода только 2 ряда щетинок. Один, параллельный внеш-
нему краю, другой — внутреннему. Кроме того, 1—2 щетинки сидят
вблизи внутреннего дистального угла. На внешнем крае экзоподита
IV плеопода 4—5 (<3) или 2 (j) простых щетинки, на внешнем крае V
плеопода — 1—2 простых щетинки. Уроподы длиннее плеотельзона
(<3) или такой же длины (j). Экзоподит и протоподит одинаковой длины
и равны 4/s длины эндоподита. Длина тела 3 5.75 мм, ширина 2 мм;
длина тела £ 2.5 мм, ширина 1.2 мм.
Распространение. Югославия — оз. Охрида, литораль.
28. Asellus (Proasellus)' monodi Strouhal.
Strouhal, 1942 : 159.
Длина плеотельзона незначительно меньше ширины. Жгут первой
антенны <5 8—10-членистый, в жгуте второй антенны — 35—40 члени-
ков. В гребенке дактилоподита I перейопода <5 9—11 зубцов. Длина про-
топодита I плеопода $ равна ширине; внутренний край с одной стороны
тела с 2, с другой — с 3 совокупительными зубцами. Экзоподит оваль-
ный, его длина в 1% раза больше ширины и несколько более чем в 1%
раза длиннее протоподита. На его краях находится 14—23 щетинки.
Дистальный членик экзоподита II плеопода <3 с 1—2 щетинками
на наружном крае и 6—7 перистыми щетинками на дистальном крае.
Второй плеопод £ округло-треугольной формы, внутренний его край
прямой, наружный выпуклый, длина вдвое больше ширины. Наружный
край базального членика экзоподита III плеопода <5 с 8—13 щетинками,
на наружном и дистальном краях дистального членика 4—7 коротких
простых и 17—20 более длинных перистых щетинок. Уроподы у обоих
полов заметно короче плеотельзона. Длина тела до 6.4 мм.
Распространение. Греция —карстовый ключ в Катарранти,
у подножия горы Катафиги.
29. Asellus (Proasellus) italicus Dudich (рис. 167—169).
D u d i c h, 1925 : 3, fig. 1—3; 1925a : 300.
Наибольшая ширина тела <3 приходится на VI сегмент, £ — на III,
IV и VI сегменты. Ширина плеотельзона несколько больше длины. Пер-
вая антенна короче основания второй антенны и имеет в жгуте 8—11
члеников. Жгут второй антенны по крайней мере вдвое длиннее осно-
102
I. ASELLIDAE
вания и состоит из 36—49 члеников. Дактилюс I перейопода <5 вооружен
15—17 зубцами. Длина проподита вдвое больше ширины. Его задний
край с 3 ножевидными зубцами и 5 щетинками. Длина и ширина прото-
подита I плеопода <5 равны, на внутреннем его крае 1 зубец, на дисталь-
ном 3—4 щетинки. Длина экзоподита вдвое больше ширины. На прямом
внутреннем крае иногда сидит 1 щетинка, на внешнем крае 9—13 корот-
ких простых щетинок и на дистальном закруглении 8—10 перистых
щетинок. На поверхности экзоподита находится 1 толстая щетинка
вблизи проксимального внутреннего угла, ряд .из 8 более тонких щети-
нок, параллельный внутреннему краю, и 2 щетинки в дистальной части.
Длина протоподита II плеопода <5 на Vs больше ширишл. Внутренний
край несет 3—7 перистых щетинок, причем правый плеопод всегда имеет
их на одну меньше, чем левый. Экзоподит по длине равен протоподиту,
Рис. 167—169. Asellus (Proasellus) italicus Dudich: 167 п
168 — I и II плеопод самца; 169 — II плеопод самки.
(По Дудичу).
а его дистальный членик в 4Уг раза длиннее проксимального. Последний
иногда несет по внешнему краю 1 перистую щетинку. На внешнем крае
дистального членика, вблизи его основания, всегда сидит 1 простая
щетинка, на дистальном закруглении — 9—13 перистых и 0—3 простых
щетинки. Эндоподит почти равен по длине дистальному членику экзо-
подита. II плеопод j трапецеидальной формы. Его длина в 214 раза
больше ширины, дистальный край несет 10.—11 перистых щетинок.
Вблизи внутреннего края на поверхности 2—3 щетинки. Уроподы у обоих
полов короче плеотельзона, экзоподит короче эндоподита. Длина тела
3 до 8—9 мм, икряных yj около 5 мм.
Распространение. Италия — Неаполь. На о-ве Мадера дает
подвид perarmatus Remy.
29а. Asellus (Proasellus) italicus perarmatus Remy.
Remy, 1936 : 20, fig. 1—8.
Жгут второй антенны состоит у 3 из 50—59, у j из 34—35 члеников.
Дистальный край протоподита I плеопода <5 несет 5—6 щетинок, экзо-
подит лишен толстой щетинки вблизи внутреннего проксимального угла.
1. ASELLUS
103
На внутреннем крае протоподита II плеопода <5 сидит 5—8 щетинок.
В базальной части дистального членика экзоподита II плеопода <5 щетинка
отсутствует. Длина тела 8—9.3 мм.
Распространение. О. Мадера.
29b. Asellus (Proasellus) italicus cephallenus Strouhal.
— coxalis cephallenus Strouhal, 1942 : 155.
Длина плеотельзона несколько больше ширины. В жгуте первой
антенны <5 8—11 члеников, жгут второй антенны <5 41—52-, j — 32—44-
членистый. Гребенка дактилоподита I перейопода i состоит из 8—12
зубцов. Протоподит I плеопода <5 с одной стороны с 1, с другой —
с 2 совокупительными зубцами, внутренний проксимальный угол экзо-
подита округлый, на его поверхности имеется почти сплошной субмар-
гинальный ряд щетинок. На наружном крае дистального членика экзо-
подита II плеопода <5 2—5 простых щетинок, на дистальном крае этого
членика — 7—10 перистых щетинок, II плеопод j трапецеидальный.
Наружный край экзоподита IV плеопода несет 9—12 щетинок. Уроподы
короче плеотельзона. Длина тела до 6.4 мм.
По многим признакам приближается к A. (Pr.) italicus Dudich, отли-
чаясь от него прерванным субмаргинальным рядом щетинок экзопо-
дита I плеопода 6 и меньшим числом зубцов в гребенке дактилоподита
I перейопода <5 и £. Близок также к A. (Pr.) coxalis peyerimhoffi Rac., но
имеет большее число щетинок на II плеоподе и III плеоподе.
Р а сп ространение. Греция—карстовый ключ близ Кране,
Ионийское побережье.
29с. Asellus (Proasellus) italicus leucadius Strouhal.
«
— coxalis leucadius Strouhal, 1942 : 156.
Длина плеотельзона меньше ширины. Жгут первой антенны состоит
из 10—14 члеников, жгут второй антенны о — из 51—59, — 42—45
члеников. В гребенке дактилоподита I перейопода 12—15 зубцов. Вну-
тренний базальный угол экзоподита I плеопода <5 оттянут, субмарги-
нальный ряд щетинок на йем полный, на наружном крае 14—17 простых
щетинок, на дистальном 6—8 перистых щетинок. Дистальный членик
экзоподита II плеопода <5 с 7—9 простыми щетинками на наружном крае
и 8—11 щетинками на дистальном крае. II плеопод j округло-треуголь-
ной формы. На наружном крае экзоподита IV плеопода 10—13 щетинок.
Уроподы по длине равны плеотельзону. Длина тела до 8.7 мм.
По форме внутреннего базального угла экзоподита I плеопода S этот
подвид напоминает A. banyulensis wolfi Dud.
Распространение. О. Лейкадия (Ионические о-ва) — карсто-
вый ключ близ Калигони (Kaligoni).
29d. Asellus (Proasellus) italicus epiroticus Strouhal.
— coxalis epiroticus S trouhal, 1942 : 158.
Отличается от предыдущего меньшим числом члеников второй антенны
(S до 47, д — до 40), округлым внутренним базальным углом экзоподита
I плеопода <5, неполным субмаргинальным рядом щетинок на нем, мень-
шим числом щетинок на дистальном членике экзоподита II плеопода
<5 и наружном крае экзоподитов III, IV и V плеоподов и уроподами,
превосходящими по длине цлеотельзон. II плеопод 2 округло-трапецо-
идальной формы. Длина тела до 8.5 мм.
104
I. ASELLIDAE
Распространение. Греция — ключ к северо-западу от Арта
в Эпире.
29е. Asellus (Proasellus) italicus corcyraeus Strouhal.
— coxalis corcyraeus Strouhal, 1942 : 158.
Очень близок к A. (Pr.) epiroticus. В отличие от этого подвида про-
топодит I плеопода 3 с левой стороны может нести 2 совокупительных
зубца, эндоподит II плеопода S с щелевидным, а не овальным отверстием,
экзоподит IV плеопода <5 снабжен большим количеством щетинок (6—11),
чем у epiroticus (4—5). Длина тела до 8.8 мм.
Распространение. О. Корфу.
30. Asellus (Proasellus) Ibanyulensis Вас. (рис. 170—173).
Racovitza, 1919а : 49, fig. 13—20.
Окраска тела светлокоричневая. Первая антенна несколько короче
основания второй антенны. В ее жгуте 12—13 (3) или 9—10 (j) члени-
, ков с 2—4 lammae olfactoriae. Вто-
Рис. 170—173. Asell us (Proasellus) banyul-
ensis Rac.: 170 — проподит I перейопода
самца; 171 и 172— I и II плеоподы самца;
173 — II плеопод самки. (По Раковице).
рая антенна достигает длины
тела. Число члеников ее жгута,
достигает 62 (3) или 40 (j>). Про-
топодит I плеопода 5, почти квад-
ратный, с закругленными углами,
с 1 совокупительным зубцом с
внутренней стороны и 1—4 щетин-
ками на дистальном крае. Экзопо-
дит овальный, в 2)4 раза превы-
шающий длину протоподита; его'
длина в 2 раза больше ширины.
На внешнем крае находится 12—13
простых коротких щетинок, на
дистальном крае 6 — 7 перистых
щетинок; кроме того, 2 — 4 ще-
тинки на поверхности экзоподита
вблизи дистального конца и 1—2—
вблизи основания экзоподита.
Протоподит II плеопода S не-
сколько удлиненный (длина на /4
больше ширины), с 4 перистыми
щетинками на дистальной поло-
вине внутреннего края. Экзопо-
дит такой же длины как протопо-
дит, но в 13/< раза уже. Прокси-
мальный его членик несет 1 пери-
стую щетинку на внешнем крае.
Дистальный членик овальный,
продолговатый (его длина в 1%
раза больше ширины), с 8—9 пе-
ристыми щетинками по дисталь-
ному краю и 1—3 короткими ще-
тинками на внешнем крае близ,
основания. Эндоподит в 1% раза
уже и на 141 короче, чем экзо-
1. ASELLUS
105
подит. II плеопод 2 треугольный, удлиненный. Его длина в 2/4 раза
больше ширины. На дистальной половине внешнего края сидит 10 пе-
ристых щетинок, на поверхности у внутреннего края, вблизи основа-
ния— 2 коротких простых щетинки. Длина тела 3 8.5 мм, 6.5 мм.
Распространение. — Франция — Баныольс-сюр-Мер (Banyuls-
sur-Mer), Италия (Модена, Генуя, Болонья, Портофино).
Экология., Обитатель ручьев, где живет между камнями и под.
опавшими листьями.
30а. Asellus (Proasellus) banyulensis wolfi Dud.
D u d i c h, 1925a : 293, fig. 13—18.
Очень близок к предыдущему, от которого отличается главным обра-
зом иным вооружением I и II плеоподов и отчасти их формой. Проподит
I перейопода <5 вооружен 4 ножевидными зубцами. Его длина вдвое-
больше ширины. На дистальном крае экзоподита I плеопода 3 нахо-
дится 8—11 перистых щетинок. На внутреннем крае протоподита II плео-
пода 3 3 перистых щетинки. Края дистального членика экзоподита
II плеопода менее выпуклы, чем у типичной формы. Внешний край несет-
2 щетинки, а внутренний 9—12 перистых щетинок. II плеопод j лишен
щетинок вблизи внутреннего края, а на внешнем имеет только 9 перистых
щетинок. Уроподы у обоих полов короче плеотельзона. Протоподит
на % (3) или вдвое (j) короче эндоподита, который или равен (3), или,
незначительно короче ($>) экзоподита.
Распространение. Сицилия — река Кане в Сиракузах.
31. Asellus (Proasellus) karamani Remy (рис. 174—176).
Remy, 1934 : 39, fig. 1—11.
Максимальная ширина тела 3 приходится на VI сегмент, j — на III
сегмент. Ширина плеотельзона несколько больше длины. В жгуте пер-
Рис 174—176. Asellus (Proasellus) karamani Remy: 174 и 175 —
I и II плеоподы самца; 176 — II плеопод самки. (По Реми).
вой антенны — 9—12 (3) или 7—8 (j) члеников, с 3—4 lammae olfacto-
riae. Жгут второй антенны состоит из 45—50 (3) или 35—40 (j). члеников.
Проподит I перейопода 3 овальный. Его длина в • 2 раза превосходит-
106
I. ASELLIDAE
ширину. На заднем крае 3—4 ножевидных зубца. Дактилюе с 10—12 зуб-
цами. Протоподит I плеопода <5 трапецеидальной формы с 1 совокупи-
тельным зубцом на внутреннем крае и с 1 щетинкой близ середины
.дистального края. Длина и ширина протоподита равны. Экзоподит оваль-
ной формы. Его длина в 13Д раза больше ширины. Внутренний край
лишен щетинок; на дистальном и внешнем краях — 20—30 щетинок,
из которых 6—9 дистальных перистые и длинные (длиннее половины
экзоподита), а остальные короткие и простые, за исключением 2—3 про-
ксимальных, коротких, но перистых. На поверхности экзоподита, у вну-
треннего дистального угла, — 1—3 щетинки и у проксимального края —
2—4 щетинки. Длина и ширина протоподита II плеопода <5 равны.' Вну-
тренний край несет 4—5 перистых щетинок. Экзоподит по длине равен
протоподиту или несколько его превосходит. Базальный членик в 3—4
раза короче дистального и имеет на внешнем крае 1 перистую щетинку.
.Длина дистального членика несколько менее чем вдвое превосходит его
ширину. На дистальной части дистального членика сидит 11—15 пери-
стых щетинок, из которых самые длинные, находящиеся посредине
дистального края, достигают длины всего экзоподита. Эндоподит
несколько короче экзоподита. Окончание его широкое, но короткое,
а боковая лопасть очень сильно развита и равна по длине Vs—Vi всего
эндоподита. Длина II плеопода $> почтив 1Уг раза больше ширины. Форма
его трапецоидальная. Внутренний край в 2 раза длиннее наружного
и снабжен 1—5 щетинками. На дистальном крае сидят 8—11 длинных
перистых щетинок, на поверхности плеопода, в базальной его половине,
1—2 щетинки. Наружный край экзоподита IV плеопода с 1—4, наруж-
ный край экзоподита V плеопода с 2—3 короткими простыми щетинками.
Уроподы 6 по длине равны плеотельзону, у j несколько короче. Ветви
одинаковой длины и вдвое длиннее протоподита. Длина и ширина тела
5 8.5—8.8 х 2.5 мм, $ — 5-5.5 х 1.75-2 мм.
Распространение. Югославия. Найден в ключе.
32. Asellus (Proasellus) coxalis Dollf. (рис. 177—180).
Dollfus, 1892 : 12, fig. 11; Racovitza, 1919a : 55, fig. 21—35.
У j тело менее удлиненное, чем у 6, максимальная ширина приходится
на середину тела, а у о тело постепенно расширяется по направлению
к плеотельзону. Длина и ширина плеотельзона равны. Окраска светлее,
чем у A. aquaticus. Жгут первой антенны у о 8-члени(йгый, у j 7-члени-
стый, не достигающий конца основания второй антенны. Вторая антенна
составляет около 2/з длины тела и имеет у <5 свыше 50, а у j меньше 50
.(25—36) члеников. Проподит I перейопода $ овальный, его длина
вдвое больше Ширины, на заднем крае 4 ножевидных зубца и многочи-
сленные мощные щетинки. Дактилюе с 6—7 овальными зубцами по ниж-
нему краю. Протоподит I плеопода $ квадратный, его ширина равна
длине. На внутреннем крае I совокупительный зубец (иногда отсутст-
вующий на правом плеоподе), дистальный внешний край с 1—2 щетин-
ками. Экзоподит прямоугольный, удлиненный. Его длина в 2 раза
больше его ширины и в 2% раза больше длины протоподита. На округ-
лом дистальном крае сидит 5 перистых щетинок, достигающих 3/t длины
членика, на наружном крае — около 12 коротких простых щетинок,
на поверхности, у внутреннего дистального угла, — 3 коротких простых
-щетинки и 1 толстая щетинка у внутреннего базального угла. Длина
протоподита II плеопода 6 на 1/t превосходит ширину. Внутренний край
1. ASELLUS
107
несет 4—5 перистых щетинок. Экзоподит по длине равен протоподиту.
Базальный членик короткий, с 1 перистой щетинкой на наружном крае.
Дистальный членик в 3 раза длиннее базального, эллипсоидальной
формы, с 5 короткими простыми щетинками на наружном крае и 5 длин-
ными перистыми на дистальном.
Эндоподит на Пз короче экзопо-
дита; его окончание такой же
длины и ширины, как боковая
лопасть. Длина II плеопода £в
23/s раза больше ширины. На-
ружный и дистальный края не-
сут около 10 перистых щетинок,
в базальной части внутреннего
•края имеется 2 щетинки. Поверх-
ность экзоподита IV плеопода
двумя линиями делится на 3
поля. Кроме того, есть зачаток
третьей линии, начинающейся
в базальной трети наружного
края. На наружном крае около
6 простых коротких щетинок.
Экзоподит V плеопода одной
линией разделен на 2 поля и
несет на базальной части наруж-
ного края около 4 простых ще-
тиной. Уроподы у <5 такой же
длины, как плеотельзон, у $
на Vi» короче. Экзоподит и эндо-
подит одинаковой длины и
почти в 2 раза длиннее прото-
подита. Длина и ширина тела
<57x2 мм, j 5 х 1.5 мм.
Распростра некие.
Обычнейший водяной ослик Си-
рии и Палестины. Его под-
виды описаны из северной Аф-
рики и Сицилии.
Рис. 177 — 189. Asellus (Proasellus) coxalis
Dollfus 177 — проподит I перейопода самца;
178 и 179 — I и II плеоподы самца; 180-— II
плеопод самки. (По Раковице).
32а. Asellus (Proasellus) coxalis peyerimhofH Rac.
Racovitza, 1919a : 68, fig. 36—51; Monod, 1925 : 243.
В отличие от типичной формы, дистальный край протоподита I плео-
пода <5 несет 3—5 щетинок. На внешнем и дистальном краях экзоподита
около 14 перистых щетинок и около 4 простых. На поверхности экзо-
подита — продольный ряд из 4—5 щетинок. Щетинки у внутреннего
дистального угла более толстые, щетинка у внутреннего базального
угла отсутствует. Длина протоподита II плеопода 6 в 1% раза превосхо-
дит ширину. Перистая щетинка на наружной стороне базального чле-
ника экзоподита отсутствует. Дистальный членик несет 7—8 перистых
щетинок и 3 простых. Боковая лопасть эндоподита шире и вдвое длиннее
окончания. Уроподы о на Vi длиннее плеотельзона.
Распространение. Алжир.
108
I. ASELLIDAE
32b. Asellus (Proasellus) coxalis cyanophilus Dudich.
D u cl i c h, 1925a : 296, fig. 19—20.
На дистальном крае протоподита I плеопода <5 2 щетинки. На поверх-
ности экзоподита 1 щетинка у проксимального внутреннего угла, 1 —
примерно посередине экзоподита и 3 — у его окончания. На дистальном
крае 11 перистых щетинок, на наружном — 12 простых, на внутреннем —
1 простая щетинка в базальной трети. Длина протоподита II плеопода
<5 несколько больше ширины. Экзоподит и эндоподит одинаковой длины.
Базальный членик экзоподита с 1 перистой щетинкой на наружном крае,
дистальный членик несет на дистальном крае 12 перистых щетинок,
на внешнем крае — 3 простых. Лопасть и окончание эндоподита одина-
ковой длины.
Распространение. Сицилия.
32с. Asellus (Proasellus) coxalis africanus Monod.
Monod, 1924 : 330, fig. 12—22.
Очень близок к A. c. peyerimhoffi Rac., от которого отличается мень-
шей величиной (5—7 мм вместо 9—10 мм), более светлой сероватой
окраской, присутствием всего 3 ножевидных зубцов на проподите
I перейопода <3 (вместо 4 у А. с. peurimhoffi), значительно удлиненным
дистальным члеником экзоподита II плеопода 3, который у А. с. africanus
Monod в ЗУг раза длиннее базального, а у Л. с. peyerimhoffi Rac. менее
чем в 3 раза длиннее базального членика, почему у первого весь экзо-
подит почти вдвое длиннее эндоподита, укороченным экзоподитом уро-
подов, составляющим 3/а длины эндоподита против 4/а длины эндоподита
у А. с. peyerimhoffi Rac., и отсутствием щетинок на внешнем крае базаль-
ного членика 1П плеопода.
Распространение. Сев. Африка (Берберия).
33. Asellus (Proasellus) seabrai Braga (рис. 181—185).
Braga, 1943 : 1, figs. 1—10.
Глаза и пигмент отсутствуют. Тело с параллельными боковыми кра-
ями, удлиненное, его длина в 4% раза превышает ширину. Голова тра-
нец оидальной формы, ее ширина почти в 1Уг раза больше длины. Коксо-
подиты перейоподов сверху не видны. Длина плеотельзона почти в 1Уг
раза больше ширины. Первая антенна значительно короче стержня второй
антенны; ее жгут вдвое короче стержня и состоит из 5—7 члеников, несущих
5 чувствительных колб, превосходящих по длине соответствующие членики.
Вторая антенна достигает VI грудного сегмента и имеет в жгуте 34—49
(<3) или 33—43 (j) члеников. Проподит I перейопода 3 эллипсоидальной
формы с прямым задним краем, несущим 3—4 ножевидных зубца. В гре-
бенке дактилюса 4—6 коротких зубцов. Дактилюсы остальных перейо-
подов с одним дополнительным коготком, расположенным так, что
он образует с основным когтем подобие клешни. Длина протоподита
I плеопода <3 несколько превышает ширину. На прямом внутреннем крае
сидят 3 толстых совокупительных зубца, близ выпуклого наружного
края — 1—2 стернальных щетинки. Экзоподит овальный, удлиненный,
его длина в 2Уг раза превышает ширину, близ внутреннего базального
угла сидят 2—3 стернальных щетинки, на дистальной половине наруж-
ного края и на дистальном крае — 21—24 гладких щетинки. . Длина
протоподита II плеопода 3 на 1!з превышает ширину. Вдоль внутреннего
1. ASELLUS
109
Рис. 181—185. Asellus (Proasellus) seabrai Bra-
ga: 181 — проподит I перейопода самца; 182 и
18J — I и II плеоподы самца; 184— II плео-
под самки; 185 — IV плеопод. (По Брага).
края находятся 1—2 перистые щетинки и иногда '1 гладкая щетинка.
Экзоподит в 2 раза короче протоподита, его базальный членик широкий
и короткий с 1 щетинкой на наружном крае, дистальный членик более
чем в 1% раза длиннее базального, его длина превосходит ширину,
он несет 3—5 гладких направленных наружу щетинок и 3—4 более длин-
ных перистых направленных внутрь щетинок. Эндоподит несколько
длиннее экзоподита. Его базальная часть образует два выроста — напра-
вленный наружу цилиндрический вырост и направленный внутрь кони-
ческий вырост. Окончание вытянуто в длинную трубку, боковая лопасть
по ширине почти не уступает
эндоподиту и не достигает конца
окончания. Длина II плеопода
§ в 2 раза больше ширины, на-
ружный его край слабо выпук-
лый, внутренний прямой. На на-
ружном крае 1—3 гладких ще-
тинки, причем на правом плео-
поде всегда больше, чем на ле-
вом, на дистальном углу 6—7
гладких щетинок, на внутрен-
нем крае иногда 1 щетинка.
Длина экзоподита III плеопода
на % превосходит ширину.
На наружном крае базального
членика 15 гладких щетинок,
на внутреннем — 4. На краях
дистального членика 30 щети-
нок. Эндоподит почти в 1% раза
короче и в 2 раза уже экзопо-
дита. IV и V плеоподы сход-
ного строения. Длина их экзо-
подитов более чем в 1Уг раза
превышает ширину, эндоподиты
эллипсоидальной формы, на Уз
более короткие и в 2 раза более узкие, чем экзоподиты. Уроподы значи-
тельно короче плеотельзона. Экзоподит длиннее протоподита и короче
эндоподита.
Длина и ширина тела $> — 6.5 х 1-4 мм, j — 5.5 X 1.4 мм.
При описании этого вида Брага выразил сомнение в возможности
отнесения его к подроду Proasellus, учитывая оригинальное строение
II цлеодода <5. Наряду с этим он указал на близость A. seabrai к A. ther-
monyctophilus по таким признакам, как форма плеотельзона и уроподов
и вооружение дактилюсов перейоподов. Однако в устройстве II плео-
пода <5 A. seabrai нет ничего такого, что бы не встречалось у других видов
Proasellus, частично оставшихся неизвестными португальскому иссле-
дователю. Строение базальной части эндоподита чрезвычайно сходно
у форм группы A. cavaticus, трубковидное окончание эндоподита наблю-
дается у A. meridianus, а необычайная ширина боковой лопасти не может
считаться особенно существенным систематическим признаком. Таким
образом, имеются все основания без колебаний относить португальский
вид к подроду Proasellus, сближая его с группой cavaticus.
Распространение. Португалия — окрестности Порто; Лека-
да-Пальмейра (Leca da Palmeira), в колодце, -совместно с Synasellus
110
I. ASELLIDAE
yiariae Braga и ближе не определенным слепым' бокоплавом из группы
Crangonyx.
34. Asellus (Proasellus) peltatus Braga (рис. 186—190).
Braga, 1944 : 1, fig. 1—11.
Длина тела в 2Уг раза больше ширины, максимальная ширина при-
ходится у <5 на VI, у j на III сегмент. Ширина головы в 1% раза
превышает длину, постмандибулярные выросты Сильно выражены. Глаза
Рис. 186—190. Asellus {Proasellus) peltatus Braga:
186 — передние грудные сегменты снизу; 187 —
I перейопод самца; 188 и 181 — I и II плеоподы
самца; 190— II плеопод самки. (По Брага).
длины. Экзоподит овальный, вдвое длиннее
маленькие. Края грудных сег-
ментов сильно расширены и
далеко перекрывают коксо-
подиты перейоподов. Длина
плеотельзона несколько боль-
ше ширины. Первая антенна
короче основания второй; в ее
жгуте 7—8 (<3) или 5—6 (j)
члеников, с 3 (5) или 2 (5)
lammae olfactoriae на послед-
них члениках. Вторая антен-
на достигает IV грудного сег-
мента и имеет в жгуте 40—45
(<3) или 28—37 (j) члеников.
Проподит I перейопода <5
овальной формы, его длина
в 13/э раза больше ширины.
На заднем крае 3—4 ноже-
видных зубца. В гребенке
дактилюса 7—9 зубцов. Дли-
на протоподита I плеопода 3
незначительно превышаетши-
рину; наружный край вы-
пуклый, внутренний прямой,
лишен зубцов и может иметь
лишь 1 гцетинку близ дисталь-
ного угла. Правый и .левый
протоподиты срастаются вну-
тренними краями на % своей
и немного шире протоподита;
на дистальной половине его наружного края и на дистальном крае
находится 20—30 щетинок, из которых 10—14 дистальных более длин-
ные и оперены. Протоподит II плеопода о квадратный, его длина на х/&
больше ширины, прямой наружный край несет 1—4 гладких щетинки,
вдоль внутреннего края находится 4 — 6 более крупных щетинок, из
которых наиболее длинные (3—5) оперены. Экзоподит немного длиннее
половины длины протоподита, его базальный членик трапецеидальной
формы и снабжен 2 гладкими щетинками на наружном крае, дисталь-
ный членик на :,/з длиннее и значительно уже базального и несет на
наружном, дистальном и дистальной половине внутреннего края 12—18
щетинок; из них 4 — 7, находящихся на внутреннем крае, наиболее длин-
ные и перистые. Эндоподит цилиндрической формы и равен по длине
экзоподиту. При его основании находятся два выроста, направленные
внутрь и наружу, причем на первом сидит короткая щетинка. Окон-
1. ASELLUS
111.
чание эндоподита укорочено и расширено, щель широкая, боковая
лопасть отсутствует. II плеопод j треугольный, сильно вытянут в длину
и несет 1—2 гладких коротких щетинки на внутреннем крае, 1 гладкую,
щетинку на наружном крае, 3—5 длинных перистых и 5—8 гладких
дистальных щетинок и 3—5 коротких стернальных щетинок. Длина
экзоподита III плеопода в 1% раза превышает ширину: по его краям
сидят около 70 гладких щетинок. Овальный эндоподит на х/з уже и короче
экзоподита. Экзоподит IV плеопода лишен щетинок, но оперен вдоль
наружного края; эндоподит несколько короче и уже экзоподита. V плео-
под имеет сходное строение, но его наружный край гладкий. Уроподы
незначительно длиннее плеотельзона, с короткими протоподитами. Ветви
одинаковой длины, в 2% раза длиннее протоподита. Длина и ширина
тела <5 8 X 3.2 мм, j 5x2 мм.
Распространение. Португалия—в маленьком ручье, питае-
мом ключем, на дороге из Penafiel в Амаранте (Amarante).
5. Подрод Conasellus Stamm.
Stammer, 1932 : 130; Caecidotea auctorum partim.
Тип подрода: Asellus communis Say.
Внутренняя лопасть первой максиллы с 5 щетинками. Проподит
I перейопода <5 несет на задней стороне два выступа, изредка три
[A. tridentata (Hung.)], соединяющиеся своими основаниями. Эндоподит
II плеопода <5 образует в базальной части небольшой вырост, обращен-
ный в сторону экзоподита, а наружная его сторона лишена выроста.
II плёоподы q удлиненные, с немногочисленными щетинками только,
в дистальной части, не заходящие друг за друга, с наибольшей шириной
у основания.
Подрод эндемичен для Сев. Америки и поэтому подробнее здесь не рас-
сматривается. В него входят следующие виды: A. communis Say, 1818;
A. stygius (Packard), 1871; A. intermedius Forbes, 1876; A. brencauda.
Forbes, 1876; A. alabamensis (Stafford), 1911; A. tridentata (Hungerford),
1922; A. antricola (Creaser), 1931; A. incisus Van Name, 1936; A. denta-
dactylus Mackin et Hubricht, 1938; A. montanus Mackin et Hubricht, 1938;
A. hobbsi Moloney, 1939; A. stilodactylus (Mackin et Hubricht), 1940;
A. packardi (Mackin et Hubricht), 1940; A. dimorphus (Mackin et Hub-
richt), 1940; A. spatulatus (Mackin et Hubricht), 1940; A. acuticarpus (Mackin
et Hubricht), 1940 и A. oculatus (Mackin et Hubricht).* 1
Недостаточно описанные формы рода Asel-lus.
В последнее время из Англии описано еще две формы рода Asellus,,
систематическое положение которых не может быть уточнено, так как
автор этих описаний принимал во внимание только внешние признаки
и не сообщил почти никаких сведений о строении конечностей.
1 Помимо сводки Van Name (1936) см.: J. С. Mackin and Leslie Н ub г i с h t.
1938. Records of Distribution of Species of Isopods in Central and Southern United
States with Description of Four New Species of Mancasellus and Asellus (Asellota, Asel-
lidae). Amer. Midi. Natural., 19; James O. Moloney. 1939. A new cave Iso-
poda from Florida. Proc. Unit. St. Nat. Mus., 86, 3057; .L Mackin and
L. Hubricht. 1940. Description of seven new species of Caecidotea (Isopoda, Asel-
lidae) from Central United States. Trans. Amer. Microscop. Soc., LIX, 3.
112
I. ASELLIDAE
Asellus crypticus Collinge (рис. 191—193).
Collinge, 1945 : 498, figs.
Тело расширяется к заднему концу. Голова без боковых выступов
« вогнутым передним краем; глаза маленькие. Эпимеры I—IV грудных
193
191 192 ' с-—--и
Рис. 191—193. Asellus cryp-
ticus Collinge: 191—голова:
192 — эпимеры V — \ II
грудных сегментов, 193 —
млеотельзон. (По Кол-
лннджу).
сегментов на концах притуплены, эпимеры
V—VII сегментов выдаются в стороны и кон-
чаются остриями. I перейопод развит сильнее,
чем у A. aquaticus. На спинной стороне пле-
отельзона два параллельных ряда из трех слив-
шихся между собою круглых коричневых пятен,
из которых средние наиболее крупные. Экзопо-
дит уроподов длиннее эндоподита. Длина тела
8.-5 мм.
Нахождение. Англия — остров Мэн.
Рис. 194—197. Asellus quicki Col-
linge: 194 — голова; 195 —• груд-
ные сегменты; 196 — плеотель-
зон; 197 — уропод. (По Кол-
линджу).
Asellus quicki Collinge (рис. 194—197).
Collinge, 1947 : 213, figs.
Передний край головы вогнутый, боковые выступы головы отсут-
ствуют. Антенны длинные, в их жгутах 40—50 члеников. Грудные сег-
менты построены одинаково. Плеотельзон
несколько уже грудных сегментов, квад-
ратной формы, со слабо выемчатыми зад-
небоковыми краями. Протоподит уроподов
короткий, эндоподит длиннее экзоподита.
Длина тела <5 — 7 мм, д — 6 мм. От
A. meridianus и A. aquaticus отличается
более крупными глазами, менее сильными
I перейоподами, более широкими уропо-
дами и отсутствием мандибулярных высту-
пов.
Распространение. Англия —
Сксвик, Говер, Глеморган (Oxwick, Go-
wer, Glamorgan).
2. Род JOHANELLA Monod.
Mono d, 1924 : 327.
Тип рода: Johanella purpurea Monod.
Эпимеры полностью срослись с сегментами тела. Дактилюсы II—VII
перейоподов с двумя когтями почти равной величины. II плеопод д,
расширенный в базальной части и резко суживающийся к концу. Уро-
поды очень короткие, с широкими короткими протоподитами и равными
ветвями.
Род Johanella Monod чрезвычайно близок к североамериканскому
роду Lirceus Raf., от которого отличается присутствием мандибуляр-
ного щупика. По мнению Моно (Monod), Johanella есть Mancasellus
(= Lirceus) с развитым мандибулярным щупиком. Род монотипичен.
1. Johanella purpurea Monod (рис. 198—202).
М о п о d, 1924 : 328, fig. 1—11.
Окраска темнокрасная. Края головы и сегментов тела уплощены
и оттянуты наружу. Глаза отсутствуют. I и II сегменты плеона видны
2. JOHANELLA, 3. STENASELLUS
113
сверху. Они уже и короче сегментов перейона и имеют хорошо выражен-
ные, заостренные, обращенные несколько назад плевры. Длина плео-
тельзона несколько больше его ширины. Жгут первой антенны состоит
всего из 3 члеников, несущих, каждый по lamma olfactoria. Жгут второй
антенны 24-членистый. Мандибула со щупиком.^ Внутренняя лопасть
первой максиллы несет 4
перистых толстых йщтинки
и 1 простую тонкую; внеш-
няя, значительно более
широкая лопасть несет 10
щетинок, оперенных с од-
ной стороны, и одну, опе-
ренную с двух сторон.
Проподит I перейопода
овально-грушевидный, с
округлым передним краем.
На прямом заднем крае
.сидит 10 шипов, из кото-
рых. один развит значи-
тельно сильнее и оперен.
Три таких же шипа име-
ются на ' заднем крае
карпоподита и 1 на зад-
нем крае мероподита. Дак-
тилюс с 3 зубцами. Дак-
тилюсы остальных перейо-
подов с 2 когтями. II плео-
под треугольный. Ширина
его равна длине. На конце
сидит 2 коротких пери-
стых щетинки, на середине
нижнего края одна ще-
тинка. Уроподы очень ко-
Рис. 198—202. Johanella purpurea Monod.: самка:
198 — общий вид; 199 — I максилла; 200 — I пе-
рейопод; 201 — дактилюсы иерейоподов; 202 — II
плеопод. (По Моно).
роткие. Ширина протопо-
дита превышает его длину. Экзо- и эндоподит одинаковой длины, не-
сколько длиннее протоподита, конической формы, вооруженные немно-
гими щетинками. Длина тела 5.20 мм, ширина 1.92 мм. Самец неизвестен.
Распространение. Алжир — в теплом сернистом источнике
у Эль-Амана.
3. Род STENASELLUS Dollf.
D о 1 1 f u s, 1897 : 130; Racovitza., 1924 : 84; — Protelsonia M e h e 1 y,
1924 : 353.
Тип рода: Stenasellus virei Dollfus.
Глаза отсутствуют. Ширина clypeus значительно превышает длину.
Вторые антенны с коротким рудиментарным экзоподитом на третьем
членике основания. Внутренняя лопасть вторых максилл с одним рядом
щетинок и такой же ширины, как и внешние лопасти. Ногочелюсти
с эпиподитом, преобразованным у яйценосных самок в оостегит. Перейо-
поды у обоих полов одинаковые. Первый и второй сегменты плеона нор-
мально развиты и не уступают по ширине сегментам перейона. II плео-
под <5 с двучленистым эндоподитом,- значительно более длинным, чем
двучленистый экзоподит. II плеопод £ треугольный, маленький. Экзо-
8 Фауна СССР, том VII, в. 5
114
I. ASELLIDAE
подиты IV и V плеоподов хитииизированы. Уроподы обычно длиннее
или незначительно короче плеотельзона.
Подземные формы, обитающие преимущественно в южной Европе..
Кроме тог®, один вид известен из Средней Азии и два из экваториаль-
ной Африки. Из палеарктических видов St. racovitzai Razautti из север-
ной Италии (Тоскана) и St. luili Remy из южной Франции здесь не рас-
сматриваются, так как их описания остались мне недоступными.
Судя по строению первых двух сегментов плеона, ногочелюстей, вто-
рых антенн и II плеопода <5, Stenasellus сохранил целый ряд архаиче-
ских признаков и может считаться более примитивным родом, чем Asellus..
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ 1
1 (22). Эндоподиты IV—V плеоподов цельные, не расщепленные про-
дольно.
2 (19). Уроподы длиннее или лишь незначительно (менее чем на Vs)
короче плеотельзона.
3 (10). Конец эндоподита II плеопода <5 более или менее округлый..
4 (7). Экзоподит I плеопода 3 шире, чем протоподит. Конец эндопсн
дита II плеопода <5 вооружен зубцами.
5 (6). Наружный край экзоподита I плеопода 3 вогнутый. Экзоподит
II плеопода б почти в 4 раза короче протоподита ...............
............................. *1. St. asiaticus Birst, et Star.
6 (5). Наружный край экзоподита I плеопода <5 выпуклый. Экзоподит
II плеопода S в 2 раза короче протоподита ... 3. St. breuili Rac.
7 (4). Экзоподит I плеопода о не шире, чем протоподит. Конец эндо-
подита II плеопода <5 лишен крупных зубцов.
8 (9). Экзоподит I плеопода S значительно уже протоподита, его наруж-
ный край -вогнутый, внутренний выпуклый. 4. St. gjorgjevici Rac.
9 (8). Экзоподит I плеопода 3 такой же ширины, как протоподит,
и имеет прямоугольную форму..................(St. africanus Monod).
10 (3). Конец эндоподита II плеопода <5 заострен.
11 (12). Дистальная часть эндоподита II плеопода 3 вытянута в клюво-
видное образование................................St. buiii Remy
12 (И). Дистальная часть эндоподита II плеопода 3 не вытянута в клю-
вовидное образование.
13 (14). Экзоподиты IV—V плеоподов дистально расширены и полностью
покрывают эндоподиты..........................St. racovitzai Razautti
14 (13). Экзоподиты IV—V плеоподов не расширены в дистальной части..
15 (16). Экзоподиты IV—V плеоподов короче эндоподитов...............
..................................... 6. St. bucbneri Stammer
16 (15). Экзоподиты IV—V плеоподов не уступают по длине эндоподи-
там.
17 (18). Экзоподиты IV—V плеоподов заметно уж° эндоподитов . . . .
.........................................; . . 2. St. virei Dollf..
18 (17). Экзоподиты IV—V плеоподов по ширине равны эндоподитам
.............................................. 8. St. nobrei Braga.
19 (2). Уроподы на Vs короче плеотельзона.
20 (21). В жгуте второй антенны не менее 30 члеников. На конце II плео-
пода J 1 щетинка............................. 9. St. bureschi Rac.
21 (20). В жгуте второй антенны не более 20 члеников. На конце II плео-
пода 2 3 щетинки...........................7. St. hungaricus Mehely
1 Частично по Шаппюи (см. Racovitza, 1950).
3. STENASELLUS
115
22 (1). Эндоподиты IV—V плеоподов расщеплены продольно, двуло-
пастные.
23 (24). Протоподит I плеопода <5 составляет более Vs длины экзоподита.
Первый и второй членики эндоподита II плеопода <5 сросшиеся
....................................... 5. St. skopljensis Kar.
24 (23). Протоподит I плеопода <3 составляет около г/7 длины экзоподита.
Первый и второй членики эндоподита II плеопода <3 разделены . .
...........................................(St. chappuisi Remy).
*1. Stenasellus asiaticus Birstein et Starostin (рис. 203—210).
Б ирштейн и Старостин, 1949 : 691, рис.. 1.
Тело удлиненное с почти параллельными боковыми краями. Глаза
и пигмент отсутствуют. Передние углы головы почти прямые. Голова
погружена в I грудной сегмент не более чем на V« его длины. Первая
антенна незначительно превосходит длину стебля второй антенны.
Ее жгут у <3 11-членистый, причем на 6 члениках сидят чувствительные
колбочки, незначительно превосходящие по длине соответствующие чле-
ники; у о жгут первой антенны состоит из 12 члеников, а чувствитель-
ных колбочек — 9. Вторая антенна доходит до середины длины тела
и имеет в жгуте около 50 члеников. Ее рудиментарный экзоподит несет
дистально 2 щетинки. Внутренняя лопасть I максиллы вооружена
3 длинными перистыми и 1 короткой гладкой щетинками. 3-й членик
щупика ногочелюсти незначительно уже 2-го. Коксоподиты перейопо-
дов далеко перекрываются краями соответствующих сегментов (как
у St. gjorgjevici) и прикрепляются не на середине, а на I—IV сегментах
близ переднего и на V—VII сегментах близ заднего края. Проподит
I перейопода с 10—12 зубцами. Дактилюсы остальных перейоподов.
с I длинным коготком. Длина плеотельзона превышает его ширину. Экзо-
подит I плеопода <5 расширен в своей дистальной половине, его наруж-
ный край снабжен полукруглой выемкой. Внутренний край и внутрен-
няя часть дистального края несет около 20 щетинок, причем щетинки
внутреннего края перистые. Ширина экзоподита примерно в 1 % раза
меньше его длины. Длина и ширина протоподита I плеопода <3 равны
между собой, зубцы на его внутреннем крае отсутствуют. Длина
протоподита II плеопода <3 в 2 раза превосходит ширину; стер-
нальные щетинки отсутствуют. Проксимальный членик его экзоподита
с 1 длинной и 1 короткой щетинкой на наружном крае, дистальный
с 6 короткими гладкими щетинками; длина дистального членика меньше
его ширины. Длина 1-го членика эндоподита составляет около 2/з длины,
протоподита. 2-й членик несколько длиннее 1-го, расширен в средней
части и суживается дистально и проксимально. Он неясно подразделен
прямым поперечным швом на 2 части. II плеопод j грушевидной формы..
Его длина в 1% раза превышает ширину. На наружном и дистальном:
краях — 10 коротких щетинок. Кроме того, имеются 4 стернальных
щетинки. Экзоподит III плеопода двучленистый; его наружный и дисталь-
ный края округлые, внутренний край прямой. Дистальный край снаб-
жен 9—10 короткими щетинками, кроме которых имеется два ряда корот-
ких стернальных щетинок. Эндоподит в Р/г раза короче и в 2% раза
уже экзоподита. Длина экзоподита IV плеопода в 2 раза превышает
ширину; его наружный край выпуклый, а внутренний вогнутый. Округ-
лый дистальный край несет 14 щетинок, из которых большинство —.
перистые. Эндоподит на Vs короче и уже экзоподита. Экзоподит V плео-
8*
416
I. ASELLIDAE
пода овальный, его длина в 2 раза больше ширины, четкий шов разделяет
его на 2 членика, из которых дистальный в 2% раза короче базального
и покрыт вдоль внутреннего и дистального краев мелкими зубчиками.
Эндоподит по длине и ширине немного уступает экзоподиту. Уроподы
Рис. 253—210. Stenasellus asiaticus Birst, et Star.: 203 — общий вид; 204 — про-
подит I перейопода; 205 и 206 — I и II плеоподы самца; 207 — II плеопод
самки; 208 — III плеопод; 209 и 210 — IV и V плеоподы. Ориг.
короче плеотельзона. Эндоподит уроподов заметно длиннее и шире экзо-
подита. Длина тела <5 и .$> 11.5 мм.
Окраска живых рачков розоватая.
От всех остальных видов рода Stenasellus этот вид отличается прежде
всего строением I и II плеоподов 3, а именно вогнутым наружным краем
экзоподита I плеопода и вторичным расчленением эндоподита II плео-
пода. Наряду, с этим можно отметить некоторые черты сходства между
St. asiaticus Birst, et Star, и Si. breuili Racovitza, в частности значитель-
ную ширину экзоподитов I, IV и V плеоподов, отсутствие зубцов на про-
3. STENASELLUS
117
топодите I плеопода и его форму. Африканский St- chappuisi Remy также
обладает широкими экзоподитами указанных конечностей, но их форма
и вооружение совершенно иные, эндоподиты III—V плеоподов продольно
расщеплены, а строение I—II плеоподов <5 весьма своеобразно. Сравни-
тельно короткие уроподы сближают нашу форму с St. hungaricus Mehely.
Распространение. Восточная Туркмения •— термальный
источник Ходжа-кайнар у подножия горы Кара-джумалак-тау в горной
системе Кугитанг-тау (западные отроги Гиссарского хребта).
Биология. St. asiaticus обитает в подземном течении родника,
температура воды которого постоянна и колеблется между 20—22° С,
а жесткость значительна (30.9 немец-,
ких градуса). Может считаться ти-
пичным эвтроглобионтом, подобно
другим видам рода Stenasellus. Крас-
ная окраска в живом состоянии
свойственна многим подземным водя-
ным осликам.
2..S tenasellus virei Dollf. (рис.
211—214).
D о 1 1 f и s, 1897 : 130; 1898 : 36,
fig.; Racovitza, 1925 : 85, fig. 1—5;
1950 : 7, fig. 1—17.
Первая антенна с 7—12 lammae
olfactoriae, вдвое более длинными,
чем членики жгута антенны, на
которых они сидят. Вторая антенна,
достигающая IV грудного сегмента,
состоит из 35 члеников. Проподит
I перейопода несет на переднем крае
7 щетинок и 14 шипов, из которых
3 ножевидные. Дактилюсы осталь-
ных перейоподов с одним добавоч-
ным коготком. Ширина экзоподита
I плеопода <5 вдвое меньше длины.
Экзоподит несет 28 — 30 простых
щетинок. Длина протоподита II плео-
пода <5 равна ширине. На его поверх-
ности имеется несколько щетинок.
Базальный членик экзоподита с 1—2
щетинками по внешнему краю. Экзо-
подит IV плеопода несет 6—7 пе-
Рис. 211—214. Stenasellus virei Dollfus:
211 — 'II антенна; 212 и 213 — I и II
ристых щетинок на дистальном кон- плеоподы самца; 214—IV плеопод.
це. Его длина в 5 раз больше ши- 0 РаковипеГ
рины. Уроподы по длине равны плеотельзону, Их эндо- и экзоподит
одинаковой ширины. Длина тела 5 7—9 мм, 8—11 мм.
Распространение.' Пещеры Франции (в департаментах Ариеж,
Верхняя Гаронна, Верхние Пиренеи, Ло) и Испании (Уэска).
3. Stenasellus breuili Rac. (рис. 215—216).
Racovitza, 1925:86, fig. 6—3; 1950 : 61, fig. 18—23; Stammer,
1936 : 139.
Lammae olfactoriae сидят на 6-ом (<Д или 8-ом (5) члениках первой
антенны и превосходят длину членикой в 1% раза. Вторая антенна дости-
118
I. ASELLIDAE
Рис. 215—216. Stenasellus ЪтеиШ Rac., !—II пле-
оподы самца. (По Раковице).
гаёт IV грудного сегмента и ее жгут состоит из 35 члеников. На вну-
треннем крае проподита I перейопода находится 14—15 щетинок и 11—
14 зубцов, из которых 3 ножевидных. Дактилюсы остальных перейопо-
дов с 2 добавочными коготками. Длина экзоподита I плеопода $ вдвое
больше ширины; края эк-
зоподита несут 17 пери-
стых и 13 гладких щети-
нок. Ширина протоподита
II плеопода <5 незначи-
тельно больше длины; его
поверхность с2шипиками.
Базальный членик экзопо-
дита с 1 щетинкой на внеш-
нем крае, дистальный — с
17—18 перистыми щетин-
ками; конец эндоподита
вооружен 6—8 зубцами.
Ширина экзоподита IV
плеопода в 2 раза меньше
длины. На конце экзопо-
дита 10—И перистых ще-
тинок. Ширина экзоподита
V плеопода в 3 раза мень-
ше длины. Уроподы длин-
нее плеотельзона; их эндоподит шире экзоподита. Длина тела 8 мм.
Распространение. Испания — пещера Аицквири (Aitzquirri)
{близ Онате (Onate), провинция Guipuzcoa] и Акелар (Akelar) [близ Онате
(Onate), пров. Наварра].
4. Stenasellus gjorgjevici Rac. (рис. 217—221).
Racovitza, 1925 : 90, fig. 9—14; 1950 : 68, fig. 23—30.
Laminae olfactoriae сидят на 5—6-ом члениках первой антенны и пре-
восходят длину члеников в 1)4 раза. Жгут второй антенны, достигающий
Рис. 217—221. Stenasellus gjorgjevici Rac.: 217 — I перейо-
под самца; 218 и 219— I и II плеоподы самца; 220
и 221 — IV и V плеоподы. (По Раковице).
3. STENASELLUS
119
IV грудного сегмента, состоит из 40 члеников. Проподит I перейопода
усажен по внутреннему краю 11 щетинками и 14 зубцами, из которых
2 ножевидных. Дактилюсы остальных перейоподов с 1 добавочным когот-
ком. Длина экзоподита I плеопода $ в 4 раза больше ширины.
На дистальном конце экзоподита 10—11 простых щетинок. Длина про-
топодита II плеопода S в 1У2 раза больше ширины, шипы на его поверх-
ности отсутствуют. Экзоподит IV плеопода с двумя перистыми щетинками
на конце. Его длина в 4 раза больше шйрины. Длина экзподита V плео-
пода в 514 раза больше ширины. Уроподы незначительно длиннее плео-
тельзона, с ветвями равной ширины. Длина тела S 9 мм.
Распр остранение. Восточная Сербия — пещера Раваничка
печина (Ravanicka pecina) в округе Морава.
4d. Stenasellus gjorgjevici robustus (Mehely).
— hungarica robusta Mehely, 1927 : 10, fig. 7 (Protelsonia}.
Внутренний дистальный угол экзоподита I плеопода 8 с 2 длинными
щетинками, конец экзоподита с 1 толстой щетинкой. II плеопод 8 как
у типичной формы, но на внешнем крае базального членика экзоподита
имеется щетинка.
Подвид описан очень плохо, причем автор указывает, что эта форма
стоит ближе к St. gjorgjevici Rac., чем к St. hungaricus (Mehely), к кото-
рому почему-то все-таки относит Свой подвид. Судя по рисунку, этот
подвид, несомненно, относится к St. gjorgjevici Rac.
Распространение. Венгрия —пещеры близ Печ (Рве).
5. Stenasellus skopljensis Kar. (рис. 222—224).
Karaman, 1936 : 137, fig. 1—3; 1948.
223
Рис. 222—224. Stenasellus skopljensis Kar.: 222 — I
перейопод; 223— III—V плеоподы; 224— II плеопод
самки. (По Караману).
На первой антенне $> 7 laminae olfactoriae, ее жгут состоит из 11 чле-
ников. Жгут второй антенны 34-членистый. Задний край проподита
I перейопода с 12 зуб-
цами. На дактилюсах
II — VII перейоподов,
кроме одного дополни-
тельного коготка, при-
сутствует 1—2 щетинки,
©ба членика эндоподита
II плеопода 8 срос-
шиеся. Основным отли-
чием от St. gjorgjevici
gjorgjevici (и от прочих
видов Stenasellus, кроме
St. chappuisi Remy) яв-
ляется продольное рас-
щепление эндоподитов
III—V плеоподов, каж-
дый из которых состоит
из двух листовидных лопастей, причем наружная лопасть шире внут-
ренней. В остальном сходен с St. gjorgjevici gjorgjevici. Длина тела 8 мм.
Распространение. Южная Сербия —грунтовые воды близ
Скоплие.
120
I. ASELLIDAE
6. Stenasellus buchneri Stamm, (рис. 225—229).
Stammer, 1936 : 137, fig. 1—5.
Окраска розоватая. Передний край головы прямой, передние углы
почти прямые, слегка закругленные. Жгут первой антенны 5 состоит
из 12—13, § — из 9—12 члеников. Lammae olfactoriae находятся у 3
на 7—9-ом, у j на 6—8-ом члениках. Вторая антенна достигает конца
III сегмента тела и имеет в жгуте 40—47 члеников. Проподит I перейо-
пода с 7—10 односторонне зазубренными ножевидными зубцами и 4—5
простыми щетинками в маргинальном ряду, 4—5 щетинками — в суб-
маргинальном каудальном ряду и 4—5 щетинками —в дистальном ряду.
Дактилюсы II—VII перейоподов
226 22?
Рис. 225 — 229. Stenasellus buchneri
Stammer: 225 — общий вид; 226 и
227 — I и. II плеоподы самца; 228
и 229 — IV и V плеоподы.
(По Штаммеру).
с 1 зубцом. Длина экзоподита I плео-
пода 3 в 2 раза больше ширины. Ди-
стальный и внешний края — с 28—31
церистыми, внутренний край— с 7—11
простыми щетинками. Внутренний край
протоподита иногда с 1 зубцом. Ши-
рина протоподита II плеопода S незна-
чительно уступает длине, внешний
край выпуклый и на поверхности про-
топодита, близ внешнего края, сидят
1—4 щетинки. Базальный членик экзо-
подита с 1 щетинкой по внешнему
краю; длина дистального членика на
г/з больше ширины, на его внешнем
и дистальном краях — 8—10 щетинок.
Длина базального членика эндоподита
составляет 4/а длины протоподита, ди-
стальный членик по длине равен про-
топодиту и на конце заострен. II плео-
под 2 маленький, треугольный с округ-
лым внешним углом и 6—8 щетин-
ками на внешнем крае. Экзоподит III
плеопода длинный, прямоугольный, с
закругленными внешним и внутрен-
ним краями и 6—8 щетинками на внутреннем крае; экзоподит
разделен швом на 2 равные поля. Эндоподит маленький, в 2 раза
уже и на 3/а короче экзоподита. Экзоподит IV плеопода ланцетовидный,,
его длина в 3 раза больше ширины; на его дистальном крае 8—9 щетинок-
Он разделен одним заметным и одним неясным швом на 3 части. Эндо-
подит несколько шире и длиннее экзоподита. Экзоподит V плеопода
также ланцетовидный, с одним швом, отделяющим его конечную часть.
Его длина в ЗУг раза превосходит ширину, на конце и внутреннем крае
находятся мелкие треугольные зубчики. Эндоподит на % длиннее экзо-
подита, его длина в 2% раза больше ширины. Уроподы по длине равны
плеотельзону, их экзо- и эндоподиты одинаковой длины. Длина тела
3 7—8 мм, 2 6.5—7.5 мм.
Распространение. Испания — пещеры в провинции Сантандер.
7. Stenasellus hungaricus (Mehely).,
М ё h е 1 у, 1924 : 353 (Protelsonia); D u dich, 1924 : 151.
Жгут первой антенны состоит из 7—-9 члеников, несущих у взрослых
22 — 3, у взрослых 33 — 4—5 lammae olfactoriae. В жгуте второй
3. STENASELLUS
121-
антенны 17—20 члеников. Экзоподит I плеопода $ с 3 длинными щетин-
ками на внутреннем дистальном углу и длинной и несколькими мелкими
щетинками на внешнем дистальном углу. Дистальный и базальный чле-
ники экзоподита II плеопода S одинаковой длины. На внешнем крае-
базального членика 1 щетинка, на внешнем крае дистального членика
4—5 щетинок. II плеопод j треугольно-округлой формы, с 3 щетинками
на конце. Конец экзоподита IV плеопода о несет 2 (у $ — 3) длинных
щетинки, конец экзоподита V плеопода с мелкими зубчиками. Уроподы
составляют 2/з длины плеотельзона. Экзоподит короче эндоподита. Длина
тела — 7.5, ширина — 1.5 мм.
Распространение. Венгрия —пещера Абалигет.
8. Stenasellus nobrei Braga (рис. 230—235).
Braga, 1942 : 1, fig. 1—10.
Тело с параллельными краями, ширина головы несколько превышает-
длину, лобный край вогнутый. V грудной сегмент короче ^остальных,
VII значительно длиннее. Свободные брюшные сегменты несколько х уже-
Рис. 230—235. Stenasellus nobrei Braga: 230 и 231 — I и II
плеоподы самца; 232—плеотельзон с уроподом; 233
и 234— IV и V плеоподы; 235 — III плеопод. (По Брага).
грудных. Длина плеотельзона в 1% раза больше ширины. Первая антенна
по длине равна стеблю второй антенны. Ее 14—15-членистый жгут в
1% раза длиннее стебля и несет 8—10 чувствительных колбочек, постепенно-
уменьшающихся в размерах в проксимальном направлении; прокси-
мальная колбочка составляет 3/4, дистальная в 1J4 раза длиннее соот-
122
I. ASELLIDAE
ветствующего членика. II антенна заходит за IV сегмент. Ее жгут состоит
.у <5 из 51—55, у g —-из 40—43 члеников. Длина проподита I перейо-
пода вдвое больше ширины. В субмаргинальном ряду 9—11 щетинок,
в маргинальном — 2 ножевидных зубца, расположенных близ основания,
9—11 коротких односторонне гребенчатых шипов и 9—11 дистальных
щетинок, в субмаргинальном ростральном — 2—3 коротких шипа, 6—9
односторонне гребенчатых шипов и 3 щетинки. Дактилюс с 11—15 зуб-
цами. Дактилюсы остальных перейоподов с 2 дополнительными когот-
ками. Протоподит I плеопода 3 прямоугольный, его длина в 1% раза
меньше ширины, на внутреннем его крае 1 совокупительный зубец.
Экзоподит в 3% раза длиннее протоподита, его длина в 2Уч раза больше
ширины. Внутренний и дистальный края выпуклые, наружный — вогну-
тый. На 2/з внутреннего края и на дистальном крае сидит 50—52 корот-
ких гладких щетинок, более длинных на внутреннем крае. Длина прото-
подита II плеопода 3 на 'к больше ширины, его внутренний край вогну-
тый с выемкой для базального членика эндоподита, наружный выпуклый,
ближе к наружному краю — 3 стернальных шипа, на внутреннем крае
у некоторых экземпляров — 1 шип. Экзоподит значительно короче про-
топодита. Длина его базального членика в 2 раза меньше ширины,
на наружном крае 1 перистая щетинка; длина дистального членика в 1%
раза больше ширины, внутренний его край прямой, на выпуклых наруж-
ном и дистальном краях 12—14 перистых щетинок, из которых самые
длинные достигают всего 2/з длины членика. Дистальный членик эндо-
подита в 1У2 раза длиннее базального, его длина в 2Уг раза больше
ширины; на его конце 5 очень мелких зубчиков. II плеопод $ треуголь-
ной формы, с закругленными углами. Его длина почти в 1% раза пре-
вышает ширину, на дистальном крае и на дистальной части наружного
края сидит 8—12 коротких гладких щетинок, кроме того имеется 5—7
стернальных щетинок. Экзоподит III плеопода овальной формы с пря-
мым внутренним и выпуклым наружным и дистальным краями. Базаль-
ный членик с 3—4 стернальными щетинками вдоль наружного края
и 5—8 — вдоль дистального края, дистальный членик равен по длине
и ширине базальному и несет на дистальном крае 7—9 коротких щетинок
и 2 стернальные вдоль внутреннего края. Эндоподит значительно короче
и уже экзоподита; его длина в четыре раза превосходит ширину. Длина
экзоподита IV плеопода в ЗУ раза больше ширины, дистальный членик
ио длине почти равен базальному и несет на конце 8—9 перистых щети-
нок, весь наружный край опушен. Эндоподит незначительно короче,
но несколько шире экзоподита. Длина экзоподита V плеопода в 3 раза
больше ширины, базальный членик в 2% раза длиннее дистального,
щетинки отсутствуют. Эндоподит по длине и ширине равен экзоподиту.
Уроподы незначительно длиннее плеотельзона. Экзоподит короче эндо-
подита, но равен ему по ширине. Протоподит в 4 раза короче экзоподита.
Длина и ширина тела <3 11x2 мм, j — 13 х 2.5 мм. Окраска
розовая.
Распространение. Португалия—Foz do Douro в окрестно-
стях Порто. Обычен в колодцах.
9. Stenasellus bureschi Racovitza (рис. 236—241).
Racovitza, 1950:83, fig. 31—37;? — lacutnicensls Buresch, 1926 (nomen
nudum).
Известна только j. Длина тела более чем в 5 раз превышает ширину;
тело менее сплющено в спинно-брюшном направлении, чем у других
3. STENASELLUS, 4. SUNASELLUS
123
видов рода. Первая антенна несколько длиннее стержня второй; ее жгут
6-членистый. Вторая антенна достигает середины III грудного сегмента
и имеет 31-членистый жгут. Проподит I перейопода короткий и широ-
кий. II плеопод треугольной формы, его длина равна ширине, внутрен-
ний край прямой, наружный выпуклый, а в дистальной части несколько
вогнутый; одна короткая щетинка располагается дистально, одна —
на наружном крае; Длина экзоподита III плеопода в 13Д раза больше
ширины; базальный и дистальный членики равной длины, на дисталь-
ном крае последнего находится 7—8 гладких щетинок; стернальные
щетинки отсутствуют. Длина эндоподита почти равна длине экзоподита,
ширина вдвое меньше. Длина экзоподита IV плеопода в 4 раза превы-
Рис. 236—241. Stenasellus bureschi Rac.: 236 — I перейо-
под; 237—239 — III—V плеоподы; 240 — II плеопод;
241 — плеотепьзон с уроподами. (По Раковице).
шает ширину; членики его неясно отделены друг от друга, причем длина
базального членика составляет 1.75 от длины дистального, несущего
на конце 2 щетинки. Овальный эндоподит значительно превышает
по ширине и несколько — по длине экзоподит. Длина экзоподита V
плеопода в 5.5 раз больше ширины; он не расчленен и лишен щетинок.
Эндоподит овальной формы, короче экзоподита, его длина несколько
более чем вдвое превосходит ширину. Уроподы на х/з короче плеотель-
зона; ветви одинаковой длины, в 2—2.5 раза длиннее протоподита. Длина
тела — 8 мм.
Распространение. Восточная Болгария—-кантон Драгоман,
пещера Berende Isvor.
4. Род SYNASELLUS BRAGA
Braga, 1944 : 4; — Mancasellus partim. Braga, 1942 : 1.
Тип рода: Synasellus mariae (Braga).
Длина головы больше длины I грудного сегмента. Мандибула без
щупика. Дактилюсы II—VII перейоподов с одним боковым шипом,
124
I. ASELLIDAE
расположенным близ основания когтя. I—II брюшные сегменты видимы
сверху, но заметно уже последнего грудного сегмента. V плеопод пред-
ставлен одной пластинкой. Плеотельзон составляет около % длины тела,
его длина больше ширины. Уроподы длиннее плеотельзона или такой же
длины.
По строению передних брюшных сегментов и IV плеопода этот род
напоминает род Stenasellus, отсутствие, мандибулярного щупика и форма
базальной части эндоподита II плеопода 3 сближают его с Lirceus
(=Mancasellus), передние плеоподы S Synasellus не отличают его от Asel-
lus, а клювовидно вытянутая дистальная часть эндоподита II плеопода
S Synasellus очень сходна с соответственным члеником видов подрода
Baicaloasellus. В то же время наблюдающееся у Synasellus упрощенное
строение V плеопода свойственно надсемейству Paraselloidea и не типично
для Aselloidea. Повидимому, Synasellus, наряду с признаками прими-
тивной организации, ставящими его в промежуточное положение между
родами семействк Asellidae, обладает и некоторыми признаками, говоря-
щими о редукции под влиянием подземного образа жизни. Род моноти-
пичен.
1. Synasellus mariae (Braga) (рис. 242—247).
Braga, 1942 : 1 (MancasellusY, 1944 : 1, fig. 1—4, 6, 8.
Тело вытянуто в длину. Голова трапецеидальной формы, длиннее
и уже I грудного сегмента, глаза отсутствуют. Жгут первой антенны
Рис. 242—247. Synasellus mariae Braga.: 242— I
плеопод самца; 243 — II плеопод самца, 244 —
плеотельзон с уроподами; 245 — IV плеопод;
246 — конец эндоподита II плеопода самца;
247 — V плеопод. (По Браге).
состоит из 6—8 члеников, в
жгуте второй антенны 36—43
членика. Эпиподит ногоче-
люсти икряной J маленький,
треугольной формы с закруг-
ленными углами и лишен
щетинок. Проподит I перейо-
пода S с 2—3 ножевидными
зубцами и 12—16 щетинками
в субмаргинальном ряду.
Длина протоподита I плео-
пода 3 значительно превос-
ходит ширину, щетинки и
зубцы отсутствуют. Правый
и левый протоподиты срос-
шиеся в базальной четверти
своими внутренними краями.
Экзоподит в 1% раза длин-
нее, но не шире протоподита,
с округлым дистальным краем
и 13—14 гладкими щетинками
на дистальном крае и ди-
стальной части наружного
края. Протоподит II плео-
пода 3 прямоугольной фор-
мы, с прямым внутренним
и слабо вогнутым наружным краем; его длина в 1)4 раза превышает
ширину. Экзоподит более чем вдвое короче протоподита. Его ба-
зальный членик трапецеидальной формы, с 3—4. щетинками на на-
3. SYNASELLUS
125
ружном крае. Дистальный членик треугольный, несколько длин-
нее, но уже базального, с одной щетинкой на конце. Эндоподит соста-
вляет около половины длины протоподита; его -окончание вытянуто
в клювовидное образование, боковая лопасть не выражена, щель узкая.
II плеопод § треугольный, его длина в 1% раза больше ширины;
на наружном крае 1 щетинка и в. дистальной части 5—6 щетинок.
Длина экзоподита III плеопода вдвое больше ширины; на наружном
крае базального членика 5—7 щетинок, на наружном крае дистального
членика 3—5 щетинок, на внутреннем крае — 6—8 щетинок. IV плео-
под двуветвистый. Экзоподит значительно шире эндоподита, с неясным
расчленением. Длина эндоподита в 4% раза больше ширины; он дву-
членистый, причем дистальный членик вдвое длиннее базального.
V плеопод представлен одной удлиненной эллипсоидальной лопастью
(эндоподит?), разделенной на два членика; дистальный членик вдвое
длиннее и шире базального. Уроподы длиннее плеотельзона; их ветви
и протоподит приблизительно одинаковой длины. Длина тела S 6 мм,
J 6.5 мм.
Распространение. Португалия — грунтовые воды в 40 км
от Вила-де-Пальмейра (Vila de Palmeira).
la. Synasellus mariae exiguus (Braga).
Braga, 1944 : 5, fig. 5, 7, 9.
В жгуте второй антенны до 31 членика. Экзоподит I плеопода S сужен
на конце и несет всего 6—7 очень коротких щетинок. Наружный край
протоподита II плеопода $. выпуклый, на наружном крае базального
членика экзоподита 1—2 щетинки. Число щетинок на II плеоподе j
и экзоподите III плеопода меньше, чем у типичной формы. Уроподы
по длине равняются плеотельзону. Длина тела 4 мм.
Распространение. Португалия — темный колодезь в Вила-
де-Парадес (Vila de Paredes).
Недостаточно описанный представитель
семейства
Stenasellus (?) hazeltoni Collinge (рис. 248—250).
Collinge, 1946 : 141.
Тело удлиненное. Ширина головы превышает длину. Лобный край
зазубрен, поверхность гранулирована. Задн
со спинной стороны гранулированы. Жгут
антенулы 6-членистый. 5 свободных брюшных
сегментов, из них 2 передние королей уже
задних. Плеотельзон удлиненный, с выемкой
на заднем крае. Уроподы неизвестны. Окраска
светлорозовая. Длина 10 мм.
Местонахождение. Англия — пе-
щера в Чидер Клифе (Cheddar Cliffs), Сом-
мерсет.
Систематическое положение этого вида
не может считаться установленным. Кол-
линдж относит его к роду Stenasellus, осно-
вываясь на наличии свободных хорошо раз-
витых брюшных сегментов. Однако у всех
известных видов Stenasellus только 2 перед-
края грудных сегментов
Рис. 248 — 250. Stenasellus (?)
hazeltoni Collinge: 248 — го-
лова и I грудной сегмент
249 — брюшной отдел; 450—1
антенна. (По Колинджу).
126
II. MICROPARASELLIDAE
них брюшных сегмента не сливаются с остальными. К сожалению, ни
ротовые конечности, ни плеоподы вида не описаны. Весьма возможно,
что его следует выделить в особый род, но скупое и поверхностное-
описание не позволяет сделать какие-либо определенные заключения.
Можно также предполагать, как думает и Шаппюи (см. Racovitza,.
1950), что английский автор имел дело с мокрицей, а не с водяным
осликом.
II. Надсемейство PAR ASELLOIDEA
Протоподиты I пары плеоподов 5 срастаются своими внутренними
краями, их экзоподиты неподвижно сочленены с протоподитами, причем
экзоподиты значительно короче протоподитов. Протоподиты II пары
плеоподов 5 крупные, соединены при помощи шипов с 1 парой плеопо-
дов и вместе с ней образуют крышечку для III—V пар плеоподов. Эндо-
подиты и протоподиты II пары плеоподов значительно длиннее экзопо-
дитов. Плеоподы II пары g срастаются без следов шва в единую крупную
пластинку — крышечку. III—IV пары плеоподов двуветвистые, у g они
отсутствуют или рудиментарны. V плеопод у обоих полов одноветвистый.
II. Сем. MICROPARASELLIDAE
сегмент абдомена свободный. Глаза и пигмент отсутствуют. Первые
антенны значительно короче вторых. Вторые антенны короткие, не дости-
гают половины длины тела, с чешуйкой на третьем членике основания.
Все перейоподы одинаковой длины и строения. Дактилюсы с двумя когот-
ками. Формы мелкие, до 2 мм длины.
Населяют грунтовые воды Югославии и Румынии. Из трех родов
этого семейства найденный в Румынии монотипический род Stygasellus
здесь опущен.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Перейоподы прикреплены к нижней стороне сегментов. Уроподы
укороченные, одноветвистые................1. Microparasellus Kar.
2 (1). Перейоподы прикреплены к бокам сегментов. Уроподы не короче
или немного короче плеотельзона, двуветвистые.....................
..........................................2. Microcbaron Kar.
1. Род MICROPARASELLUS KAR. .
К а г a m a n, 1933 : 17 (partim); 1934 : 44; 1940 : 21.
Тип рода: Microparasellus puteanus Karaman.
Края сегментов тела с многочисленными мелкими выемками. Lammae
olfactoriae первых антенн сильно развиты и много длиннее члеников
жгута, на которых они сидят. Вторые антенны менее чем вдвое длиннее
первых. Ширина головы больше, чем длина. Имеется короткий рострум.
Мандибула со щупиком. Перейоподы прикреплены к нижней стороне
сегментов тела. У S 7 пар перейоподов, у g обыкновенно седьмая пара
отсутствует. Уроподы очень короткие, сближенные, с расширенным про-
топодитом и маленьким, сидящим на протоподите экзоподитом. Эндо-
подит отсутствует.
1. MICROPARASELLUS
12T
1. Microparasellus puteanus Kar. (рис. 251—257).
К a r a m a n, 1933 : 17, fig. 1; 1934 : 44; 1940 : 32, fig. 51—73.
Голова и сегменты тела почти одинаковой ширины. Ширина каждого
сегмента вдвое больше длины. Выемки на дистальных краях шире, чем
на проксимальных. Первые два сегмента плеона срослись в один,, ширина
которого такая же, как грудных сегментов, но длина вдвое меньшая.
Длина плеотельзона больше ширины. По его краям следы выемок. Ман-
дибула несет нормально развитый щупик. Первая максилла несет только
2—3 щетинки на дистальном конце узкой внешней лопасти. На внеш-
ней лопасти второй максиллы 8—10, на обеих внутренних по 3—4 зазуб-
Рис. 251—257. Microparasellus puteanus Kar.: 251 — плеотельзон
самца снизу; 252 — плеотельзон самки снизу; 253 — общий вид;
254 — I антенна; 255 и 256 — I и II плеоподы самца; 257 — пе-
рейопод. (По Караману).
репных шипа. Первая антенна состоит из 6—7 члеников, из которых:
два апикальных несут по одной длинной lamma olfactoria, достигающей
2/з длины всей антенны. Вторая антенна вдвое длиннее первой антенны.
Она имеет 6-членистое основание и 8-членистый жгут. На внутренней
стороне третьего членика основания сидит чешуевидный экзоподит.
Перейоподы одинакового строения и длины, прикреплены к ниж-
ним краям сегментов тела. На их дактилюсах сидит по два одинаковых
когтя. У большинства седьмая пара перейоподов отсутствует или
иногда она очень мала, нормально расчленена, но лишена когтей. У о —
7 пар перейоподов. Длина I пары плеоподов о превосходит ширину.
Протоподиты срослись как между собой, так и с ветвями плеопода.
II пара плеоподов 3 состоит из двух разделенных плеоподов. Они схо-
дятся посередине плеотельзона. На внутренней стороне каждого пахо-
128
II. MICROP ARASELLIDAE
.дится двучленистый совокупительный орган. I и II пары плеоподов
образуют вместе крышечку. Рудиментом I пары плеоподов, вероятно,
может считаться листовидный вырост II плеопода. Большинство gg
лишено I пары плеоподов. II пара представляет собой округлую пла-
стинку-крышечку с остатком III пары плеоподов. Следующие две пары
плеоподов имеют одинаковое строение у <5 и g. Первая из них имеет по про-
топодиту и по две расчлененные ветви. Вторая состоит из листовидного
выроста стернума и пальцеобразного двучленистого экзоподита. Уро-
поды построены у обоих полов одинаково. Длина тела около 1 мм.
Распространение. Югославия — грунтовые воды Скоплие
и Куманова.
Экология. Типичная форма грунтовых вод. В желудке обнару-
жен ил.
2. Род MICROCHARON KAR.
Karaman, 1934 : 42; 1940 : 21.
Тип рода: Microcharon stygius Karaman.
Края сегментов тела гладкие. Вторая антенна более чем вдвое длин-
нее первой антенны. Длина головы не меньше ширины. Перейоподы
с расширенными базиподитами и прикреплены по бокам сегментов тела.
Плеоподы как у Microparasellus. Уроподы хорошо развиты, у 3 пре-
восходят по длине плеотельзон, двуветвисты. Их эндоподиты вдвое
длиннее экзоподитов.
Известно 4 вида — три из грунтовых вод Югославии, один (М. ache-
rontis Chappuis) — из грунтовых вод Румынии. Описание этого послед-
него здесь не приводится.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (4). Протоподит уроподов не короче плеотельзона и длиннее эндопо-
дита. Длина IV—VI грудных сегментов превышает ширину.
2 (3). Длина экзоподита уроподов меньше ширины протоподита. Груд-
ные сегменты приблизительно одинаковой длины, причем длина
передних сегментов больше ширины . . . . 1. М. stygius (Kar.)
3 (2). Длина экзоподита уроподов равна ширине протоподита. Передние
грудные сегменты заметно короче задних и их длина меньше
ширины....................................3. М. profundalis Kar.
4 (1). Протоподит уроподов короче плеотельзона и эндоподита. Длина
IV—VI грудных сегментов меньше их ширины.........................
......................................... 2. М. latus Kar.
1. Microcharon stygius (Kar.) (рис. 258—260).
Karaman, 1933 : 20, fig. 2 (Microparasellus); 1934 : 44, fig. la; 1940 : 31,
fig. 28—35.
Тело сильно вытянуто в длину, у g несколько расширено. Длина
сегментов тела и головы обычно больше (у 3) ширины или (у g) равна
ей. Шесть первых грудных сегментов равной длины, седьмой часто
короче остальных. Длина плеотельзона превышает его ширину. Первая
антенна состоит из 7—8 члеников и несет у 3 3—4, у g 1—2 lammae ol-
2. MICROCHARON
129
factoriae. Вторая антенна с 8—9-членистым жгутом. Перейоподы как
у Microparasellus, но с расширенными базиподитами. Четыре первых
пары прикреплены к передним, 2—3 задних к задним частям краев сег-
ментов. Протоподит уроподов <5 широкий и превосходит длину плеотель-
зона. Длина тела до 2 мм.
Распространение.
Найден в большем количе-
стве экземпляров и в боль-
шем числе колодцев, чем
предыдущий, в Скоплие.
2. Microcharon latus Kar.
(рис. 261—262).
Karaman, 1934 : 43, fig.
lb, 2; 1940 : 21, fig. 1—27.
Тело шире, чем у преды-
дущего. Ширина сегментов
тела превосходит их длину.
Плеотельзон округлый, его
ширина и длина равны. Пе-
рейоподы и вторые антенны
развиты сильнее, чем у пре-
дыдущего вида. Протоподит
уроподов короче плеотель-
зона. Эндоподит сидит на про-
топодите терминально, а эк-
зоподит на наружном крае
сбоку. Эндоподит в 1У2 раза
длиннее протоподита и в 2
раза длиннее экзоподита.
Длина тела около 2 мм.
Р аспр остранение.
Югославия — грунтовые воды
и . ключи долины Скоплие,
грунтовые воды и ключи оз.
Охрида, в большом количе-
стве экземпляров.
Рис. 258 — 260.Microcharon stygius (Kar.): 258 —
общий вид; 259 — I антенна; 260— I перейопод.
Рис.261—262. Microcharon latus Kar.: 261 — об-
щий вид самца; 262 — общий вид самки.
(По Караману).
3. Microcharon profundalis Кат. (рис. 263—269).
Karaman, 1940 : 33, fig. 36—40.
По ширине тела занимает среднее положение между двумя предыду-
щими видами. Передние сегменты расширяются в дистальном, задние
в проксимальном направлении. Плеотельзон шире грудных сегментов,
его длина превосходит ширину. Жгут второй антенны 8-членистый.
Перейоподы как у М. latus. Конец I плеопода S расширен в стороны,
образуя 2 боковых зубца. II плеопод g часто с выемкой на середине
дистального края. Протоподит уроподов 3 и g по длине превосходит
плеотельзон; в конце он значительно уже, чем при основании. Длина
экзоподита равняется ширине протоподита, экзоподит на 1/з длиннее.
Длина тела до 1.4 мм.
9 Фауна СССР, том VII, в. 5
130
III. JANIRIDAE
Распространение. Югославия: глубокие колодцы Скоплие,
в которых встречается чаще, чем другие виды этого рода. В других рай-
онах Югославии замещается подвидами.
Рис. 263—269. Microcharon profundalis Kar.: 263—общий вид
самца; 264 — плеотельзон с уроподами снизу; 265 — 1 антенна;
206-269 — I плеопод самца М. р. profundalis, kwnanovensis, hoso-
vensis и beranensis. (По Караману).
За. Microcharon profundalis kumanovensis Kar.
Karaman, 1940 : 37. fig. 41, 46, 49.
I плеопод 3 уже, чем у типичной формы. Его конец прямой и лишен
боковых расширений. На конце каждого I плеопода <3 3 щетинки,
на наружном крае—-2 щетинки. Листовидный придаток на конце пер-
вой антенны по длине равен расстоянию от конца первой антенны до ее
основного членика, оперенный придаток на внутреннем крае 2 членика
основания первой антенны достигает ее конца.
Распространение. В колодцах г. Куманово и нескольких
окрестных сел приблизительно в 40 км на восток от Скоплие.
ЗЬ. Microcharon profundalis kosovensis Kar.
Karaman, 1940 : 37, fig. 42.
Оба первых плеопода <5 разделены щелью, расширяющейся близ
середины плеопода. На конце каждого плеопода по 2 щетинки. Листо-
видный придаток на конце первой антенны по длине равен расстоянию
от Конца первой антенны до середины ее основного членика.
1. JAERA
131
Распространение. Колодцы и грунтовые воды в Косовска
Митровича (Kosovska Mitrovica) в 110 км на северо-восток от Скоплйе.
Зс. Microcharon profundalis beranensis Kar.
Karaman, 1940 : 37, fig. 43.
Конец I плеопода £ vl его наружные края с выемками. Оперенный
придаток на внутреннем крае 2 членика основания первой антенны пре-
вышает по длине ее жгут.
Распространение. Колодцы и грунтовые воды городка
Беране [в 80 км к западу от Косовска Митровича (Kosovska Mitrovica)].
3. Сем. JANIRIDAE
Тело продолговатое, овальное. Голова с расширенными боковыми
краями; глаза, если имеются, расположены дорзально. Вторая антенна
обычно с редуцированным чешуевидным экзоподитом. Ротовые части
нормально развиты. Перейоподы с 1—2 добавочными коготками на дак-
тилюсах. Уроподы обычно двуветвистые.
Из многих чисто морских родов в пресных водах Палеарктики встре-
чается только 1 род.
1. Род JAERA LEACH
Leach, 1914, Encyclop., VII : 434; Kesselyak, 1938:226;—Jaeridina
Milne Edwards, 1840, Hist. nat. Crust.: 150; — Jaira M e i n e r t, 1877, Dan.
Nat. Tidsskr., ser. 3, II : 80.
Тип рода: Jaera albifrons Leach.
Тело овальное, широкое, очень сильно сплюснутое дорзовентрально;
края грудных сегментов расширены и уплощены. Первая антенна очень
короткая, с 2-членистым жгутом. Все перейоподы приблизительно оди-
наковой длины, короткие. Плеотельзон пресноводных представителей
на середине заднего края с вырезом, в котором сидят уроподы. Уроподы
очень короткие, двуветвистые, незначительно заходят за задний край
плеотельзона. Их ветви в несколько раз короче протоподита.
Систематика этого рода до последнего времени оставалась чрезвы-
чайно запутанной. С одной стороны некоторые виды были описаны под
разными названиями, с другой стороны новые виды описывались как
старые (J. nordmanni Sars = J. sarsi Valk.). В род Jaera включались
виды, относящиеся к другим родам (J. filicornis Grube =J. deshayesi
Lucas = Stenetrium deshayesi (Lucas). Благодаря работам Вълканова
(1938) и Кессельяка (1938), в этот вопрос в настоящее время
внесена достаточная ясность. В европейских водах насчитывается
11 видов Jaera, из которых 2 являются исключительно пресноводным^:.
1 — солоноватоводный, заходящий в пресную воду, 2 •— широко эври-
галинные, живущие и в совершенно пресной воде и в условиях нормаль-
ной океанической солености, 5 морских и 1 морской, но выносящий опрес-
нение до 8°/()|). 4 вида, в том числе и этот последний, констатированы
в пределах СССР. Антарктические виды (J. 'antarctica Pfeffer, J. s'drrata
9*
132
III. JANIRIDAE
Barnard, J. pusilia Barnard) Норденштам (Nordenstam, 1933) выделил
в самостоятельный род Neo/aera.
Рис. 270—275. Jaera. albifrons Leach.: 270 — общий вид
• <?амца; 271 — общий вид самки; 272 — II плеопод самки;
\ . 27$ — I плеопод самца; 274 — конец VII перейопода; 275 —
уроподы.'Рис. 276, 277 — Jaera (s. str.) sarsi Valk.: 276 —
V ’• общий вид самки; 277 — общий вид самца, бриг.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ И ВИДОВ
1 (12)., Задний край плеотельзона с выемкой, в которой сидят уроподы.
. (Подрод ./«era s. sir.), • \
1. JAERA 133
2 (9). I плеопод 3 на конце расширен, его боковые дистальные углы
оттянуты в стороны, благодаря чему по форме он напоминает, якорь.
3 (4). Ветви уроподов в 7—9 раз короче протоподита и сидят на внеш-
ней половине его дистального края. Перейоподы с 2 когтями . .
...............................................* 2. J. sarsi Valk.
4 (3). Ветви уроподов не более чем в 3—4 раза короче протоподита'
и сидят на нем симметрично. Перейоподы с 3 когтями.
5 (6). Дистальный край I плеопода 3 прямой и равномерно усажен
щетинками. Выемка плеотельзона глубокая. Дистальная часть
заднего края карпоподита или исхиоподита VI—VII перейоподов
3 снабжена рядом щетинок.....................* 1. J. albifrons Leach
6 (5). Дистальный край I плеопода самца вогнутый, с лишенной щети-
нок выемкой посредине. VII перейопод самца и самки одинакового
строения, без ряда щетинок на карпоподите.
7 (8). Выемка плеотельзона не глубокая, прерванная посредине широ-
ким треугольным выступом заднего края плеотельзона. Протопоп
диты уроподов выдаются из выемки плеотельзона.....................
..........................................3. J. italica Kessel.
8 (7). Выемка плеотельзона глубокая, прямоугольная. Протоподиты
уроподов не выдаются из выемки плеотельзона.......................
.................., . ... .............6. J. sorrentina Verhoeff
9 .(2). I плеопод 3 более или менее суживается к концу; его боковые
дистальные углы не оттянуты в стороны.
10 (11). Боковые края дистальной половины I плеопода 3 почти прямые
и параллельные друг другу. Передние перейоподы 3' снабжены'
многочисленными длинными загнутыми щетинками................... .
................................* 4. J. nordmanni Rathke
11 (10). Боковые края I плеопода 3 вогнутые, не параллельные друг
другу. Передние перейоподы 3 и j устроены одинаково й лишены
длинных щетинок.........................* 5. J. schellenbergi Kessel.
12 (1). Задний край плеотельзона без выемки. (Подрод Melajaera Ver-
hoeff).
13 (14). Двучленистые придатки головы, находящиеся с внутренней
стороны I антенн, отсутствуют или рудиментарны. Длина 4-го
членика стебля 2 антенны в ЗУг раза больше ширины . ......
.......................................* 7. J. hopeana Costa
14 (13). Двучленистые придатки головы хорошо развиты. Длина 4-го
членика стебля II антенны менее чем в 3 раза превосходит ши-
рину.
15 (18). Двучленистые придатки головы достигают 2-го членика стебля
I антенны.
16 (17). Пигмент отсутствует. II антенны 16-членистые. Боковые края-
головы с 7 щетинками.........................J. dalmatica Verhoeff
17 (16). Тело пигментировано. II антенны 20-членистые. Боковые края
головы с 5 щетинками.........................J. hospitalis Verhoeff
18 (15). Двучленистые придатки головы не достигают 2-го членика
стебля I антенны.
19 (20). Боковые края головы с 5—6 щетинками, боковые края двух
передних грудных сегментов с 3—4 щетинками. Плеотельзон сзади
без углублений...............................J. stagnoensis Verhoeff
20 (19). Боковые края головы с 3 щетинками, боковые края двух перед-
них грудных сегментов с 2 щетинками. Плеотельзон сзади с двой-
ным углублением................................J. sulcata VerhoeF
134
III. JANIRIDAE
1. Подрод Jaera s. str.
Verhoeff, 1943 : 289.
Боковые края грудных сегментов расширены и далеко выдаются
в стороны за места прикреплений перейоподов. Боковые края головы
тоже расширены, заострены на концах .и выдаются вперед до уровня
конца рострума. Плеотельзон сзади с выемкой.
Подрод заключает свободноживущие эвригалинные морские, соло-
новатоводные и пресноводные виды. Только J. sorrentina Verh. и J. sarsi
caspica Kessel, не были обнаружены в пресной воде.
*1. Jaera (s. str), albifrons Leach, (рис. 270—275).
Leach, 1814, Edinb. Encycl., VII : 434; Гурьянова, 1932, Опред-
фауны СССР, 4:21, фиг. 7, Вълкаиовъ, 1938 : 59, фиг. 7—9: Forsman, 1944 : 1,
fig. 1—3; — nivalis К г б у е г, 1838, К. Dansk. Vid. Selsk. Afh., 7.: 303, pl. 4>
fig. 21;— krcyeri Milne Edwards, 1840, Hist. nat. Crust., Ill : 149;—
marina G. 0. 8 a r s, 1899 : 104, t. 43; Kesselyak, 1938 : 228, fig. 4—2.
Окраска бурая, неравномерная, часто голова светлее тела. Длина
тела менее чем в 2 раза превышает ширину, у 8 шире, чем у g. II—V сег-
менты тела J одинаковой ширины, I, VI и VII — несколько уже, у 8
VI—VII сегменты такой же ширины, как и предыдущие. Ширина головы
более чем в 2 раза превышает длину. I антенна очень короткая, недостигаю-
щая конца предпоследнего членика стебля II антенны, со жгутом, состоя-
щим из 2 члеников и несущим на конце две lammae olfactoriae. II антенна
длиннее половины тела, со жгутом, в 2 раза более длинным, чем осно-
вание, состоящим из 24—40 члеников. Дактилюсы перейоподов с 3 ког-
тями, из которых один значительно меньше двух других. Карпоподит
VI—VII перейоподов 8 к концу расширяется и несет на дистальной
части заднего края ряд щетинок.
Наряду с такими особями и часто вместе с ними встречаются 88,
не отличающиеся по строению карпоподита от gg, но обладающие рядом
толстых щетинок на исхиоподите. Дистальный край дистальных чле-
ников I плеопода 8 прямой и покрыт равномерно 14—25 щетинками;
на боковых краях, перпендикулярных дистальному, щетинки сидят
гуще (на каждом крае их 15—20). Длина протоподита II плеопода 3
почти в 2 раза больше ширины, на дистальной половине его внешнего
края сидит несколько щетинок. II плеопод g округлый, его ширина
равна длине, края равномерно покрыты щетинками. Экзоподит III плео-
пода двучленистый, дистальный членик к’ороче базального; эндоподит
несколько шире экзоподита. Экзоподит IV плеопода одночленистый
и значительно уже эндоподита. V плеопод одноветвистый. Ветви уро-
подов в 3 раза короче протоподита, со щетинками на концах. Внутрен-
ний дистальный угол протоподита с I щетинкой, внешний дистальный
угол лишен щетинок. Край выемки плеотельзона прямой, без выступа
посредине. Длина тела о до 3 мм, g до 5 мм (по Kunkel — до 7 мм).
Распространение. Побережье Северного Ледовитого оке-
ана на восток до восточного берега Чешской губы, атлантическое побе-
режье Европы на юг до южной Франции, Великобритания, Исландия,
западная Гренландия, Балтийское, Средиземное, Адриатическое и Чер-
ное моря, атлантическое побережье Северной Америки от Лабрадора
до 41° N. В пресной воде в эстуариях баренцов ом орских речек, в оз.
Могильном, в южной части Кольского залива, далее в устье Эльбы,
в устьях рек Нормандии. Наиболее широко распространенный вид Jaera.
1. JAERA
135
*2. Jaera (s. str.) sarsi Valk. (рис. 276—281).
Вълкановъ, 1936 : 314; 1938 : 61, fig. 10—17; Kesselyak, 1938 : 237,
fig. 1—3, 12, 13; — nordmanni auctorum.
Окраска как у предыдущего вида. I, VI и VII сегменты тела j отно-
сительно средних сегментов значительно уже, чем у J. albifrons. Передне-
боковые углы головы развиты сильнее и более выдаются вперед. Жгут
I антенны 2-членистый, с одной lamma olfactoria на конце, жгут II
антенны — 22—36-членистый. В отличие от остальных видов, длина
Рис. 278—281. Jaera (s. str.) sarsi Valk.: 278 — I плеопод
самца; 279 — П плеопод самки; 289-— уроподы самца; 281 —
конец VII перейопода самца. Ориг. Рис. 282—285. Jaera
(s. str.) sarsi caspica Kessel.: 282—284—I плеопод самца;
285 — II плеопод самки. Ориг.
второго членика щупика ногочелюстей превосходит ширину. Дактилюсы
перейоподов снабжены всего 2 когтями. Дистальный край дистальных
члеников I плеопода <5 округлый, с выемкой посредине и двумя выем-
ками по бокам, разделяющими край на 4 доли. Две средние доли несут
по 2—5 щетинок; крайние — по 2—7 щетинок; в выемках щетинок нет.
II плеопод 5 как у J. albifrons. II плеопод j резко суживается к концу
и имеет сердцевидную форму. Остальные плеоподы как у J. albifrons.
Ветви уроподов сидят на наружной половине дистального края прото-
подита. Они в 7—9 раз короче протоподита, эндоподит более чем в 2 раза
шире экзопрдита. Внутренняя половина дистального края протоподита
удлинена так, что ее край находится на одном уровне с концами ветвей.
На ней сидят 2—3 щетинки. 1—3 щетинки находятся на внешнем
дистальном углу протоподита. Длина тела 3 до 2.6 мм, j до 8 мм.
136
III. JANIRIDAE
. Этот вид был описан Сарсом (Sars, 1897) из Каспийского моря под
названием J. nordmanni Rathke. Впоследствии многочисленные русские
(Державин, Бенинг, Мартынов, Водяницкий) и, отчасти, иностранные
(Dudich) авторы обозначали Jaera, встречающихся в реках и озерах
Понто-Каспийского бассейна, тем же именем. Только в 1936 г. Вълка-
нов показал, что из-за неудовлетворительного описания J. nordmanni,
данного Рамке, Саре допустил ошибку и что на самом деле описанная
им из Каспия форма не имеет ничего общего с J. nordmanni и должна
быть выделена в самостоятельный вид, названный J. sarsi.
Распространение. Волга (до 1000 км от устья), Урал, Дон,
Днепр, Дунай (до 1800 км от устья), Таганрогский залив, Одесско-Буг-
ский лиман, оз. Абрау, Варненское, Гебедженское (Болгария).
2а. Jaera (s. str.) sarsi caspica Kessel, (рис. 282—285).
Kessely a k, 1938 : 240, fig. 14; — nordmanni G. O. Sars, 1897 : 298,
t. XVI, fig. 2—6.
Отличается от типичной формы строением I плеопода 5. Наружные
дистальные углы I плеопода о оттянуты гораздо сильнее, чем у типичной
формы, и загнуты назад почти до уровня середины плеопода. Боковые
выемки дистального края выражены слабее, чем у типичной формы.
Конец II плеопода несколько более округлой формы. Эти признаки
подвержены известным вариациям. Изредка попадаются экземпляры,
у которых одна половина плеопода типична для J. sarsi typica, а другая —
для J. sarsi caspica (рис. 284). По величине тела разницы между подви-
дом и типичной формой не наблюдается.
Распространение. Побережья среднего и южного Каспия,
до глубины 50—60 м, при солености 11—13°/ои. В опресненном северном
Каспии не обнаружен. В Волге встречается типичная форма (в проти-
Рис. 286—288. Jaera (s. str.) italica Kessel.: 286 —
IV перейопод самца; 287 — I плеопод самца; 288 —
конец плеотельзона с уроподами. (По Кессельяку).
I
I
I
|
I
воположность мнению Кес-
сельяка).
3. Jaera (s. str.) italica
Kess. (рис. ’ 286—288).
Kessel у ak, ,1938:244,
fig. 16—19; Remy, 1941 :22,
fig. 27 — 29; Strouhal,
1942 : 150.
Максимальная ширина
тела 3 приходится на VI,
2 •— на III — IV сегменты
тела. Передне-боковые уг-
лы головы развиты слабее,
чем у других видов. Жгут
I антенны с одной lamma
olfactoria, в жгуте II ан-
тенны $ около 31, j —22
члеников. Средниечленики
трех передних перейопо-
дов 3 несут многочисленные длинные закрученные щетинки, карцопо-
дит IV перейопода S снабжен по внутреннему краю 2—3 рядами шипов
с 1—4 остриями, каждый дактилюе перейоподов с 3 изгнутыми шипами.
I плеопод i приблизительно как у J. sarsi, но он расширен не только
на конце, но и при основании, наружные дистальные углы оттянуты
1. JAERA
137
сильнее, а выпуклый дистальный край прерван всего одной, зато более-
глубокой выемкой посредине. Выемка заднего края плеотельзона неглу-
бокая и разделена на две симметричные части широким треугольным
выростом. Ширина протоподита уроподов j превосходит длину, ветви,
приблизительно вдвое короче протоподита, длина протоподита уроподов
S почти вдвое превосходит ширину.
В отличие от остальных видов $ крупнее Длина S до 3.6 мм„
ширина до 2 мм; длина j до 2.33 мм, ширина до 1.13 мм.
Распространение. Сицилия — окрестности • Сиракуз:
р. Киане и Фонте-Аретуза; Югославия — ключ Слатина Вода (Slatina
Voda) в Герцеговине; Греция — ключ около Кране (Кгапе) в Ионии.
*4. Jaera (s. str.) nordmanni Rathke (рис. 289—293).
Rathke, 1837 : 388, t. 6, fig. 1—5 (Janira); Bate and Westwood,
1878, Hist. Brit. Sessileyed Crust., 11 : 320, figs.; Вълкановъ, 1938 : 57,.
fig. 1—6; Kessely ak, 1938 : 231, fig. 6—8.
Жгут I антенны с одной lamma olfactoria, жгут II антенны состоит-
из 20—35 члеников и незначительно превосходит по длине половину
тела. Дактилюсы перейоподов во-
оружены 3 шипами. Средние чле-
ники трех передних перейоподов
5 несут длинные закрученные ще-
тинки. На внутреннем крае кар-
поподита IV перейопода S нахо-
дится 5—7 толстых шипов, снаб-
женных на конце несколькими
остриями. I плеопод 3 при осно-
вании расширен, к середине резко
суживается и внешние края ди-
стальной его половины оказы-
ваются параллельными друг другу.
Наружные дистальные углы совер-
шенно не оттянуты в стороны.
Дистальные членики заострены
к концу и несут на наружных
краях около 7 щетинок. II плео-
под J округлой формы, дисталь-
ный край посредине слегка вогну-
тый. Длина протоподита уропо-
дов в 1)4—13Л раза превосходит
ширину, эндоподит в 1)4—2 раза
длиннее экзоподита и в 3—4 раза
короче протоподита. Длина тела
до 3.5 мм.
Р аспр остранение. Чер-
ное море — побережье Крыма и
Рис. 289 — 293. Jaera (s. str.) nordmanni
Rathke: 289 и 290 — III и IV перейоподы
самца; 291-—I плейопод самца; 292—II
плеопод самки; 293 — конец плеотель-
зона с уроподами (289—292 по Кессель-
яку, 293 по Вълканову).
Болгарии, Средиземное море(?), побережья Англии, Ирландии и Франции,
Азорские о-ва. В пресной воде — в Англии и в ручье на Азорских о-вах
(«У. guernei Dollfus»).
5. Jaera (s. str.) schellenbergi Kessel, (рис. 294—295).
Kessel yak, 1938 : 242, fig. 15a, b.
Жгут I антенны с одной lamma olfactoria, жгут II антенны 24—26-
членистый. Дактилюсы передних перейоподов с 2, задних (взрослых.
138
III. JANIRIDAE
экземпляров) — с 3 шипами. Внутренний край карпоподита IV перей-
опода <5 с 10—11 зубцами, более слабыми, чему/. nordmanni', передние
перейоподы 8 без длинных закручен-
иых щетинок. I плеопод 8 расширен
Рис. 294—295. Jaera (s. str.) schellen-
bergi Kessel., 1—П плеоподы самца.
(По Кессельяку).
Распространение,
на восточном берегу Истрии, в
при основании, суживается к середине,
а к концу снова расширяется, благодаря
чему его наружные края вогнутые. Ди-
стальные членики заострены к концу,
их наружные края снабжены щетин-
• ками, выемка между ними глубже, чем
; у J.. nordmanni. На границе между
’ дистальными и базальными члениками
, на наружных краях находится по од-
ному небольшому выросту. Эндоподит
II плеопода 8 длиннее, чем у J. nord-
manni. Выемка плеотельзона разделена
на две части широким выростом. Длина
тела 8 до 2.3 мм, $> до 2.7 мм.
Карстовый ключ на горе Маджиоре,
совершенно пресной воде.
6. Jaera (s. str.) sorrentina Ver-
hoeff (рис. 296—299).
Verhoeff, 1943 : 284, fig. 7—10.
Голова с крупными выдающимися
-вперед боковыми лопастями, дохо-
дящими до основания 2 антенн. Жгут
2 антенны 27-членистый. I—III пе-
,рейоподы <5 без закрученных ще-
тинок, карпоподит IV перейопода
с рядом шипиков, заканчивающихся
одним (а не тремя-четырьмя, как
.у J. italica) острием. Плеоподы как
у J. italica Kessel. Выемка плеотепь-
зона глубокая, прямоугольная; про-
топодиты уроподов не выдаются за
ее края. Пигментизация густая и
распространяется на уроподы.
Распространение. Ита-
лия— близ Сорренто, под камнями
в полосе прибоя. Самцы вдвое мно-
гочисленнее самок (как у J. italica
Kessel.).
2. Подрод Metajaera Verhoeff.
Verhoeff, 1943 : 290.
Рис. 296—299. Jaera (s. str.) sorrentina
Verhoeff: 296 — I плеопод самца; 297 —
правая половина переднего края головы
с I антенной и дву членистым придатком;
298 — IV переопод самца; 299 — конец
„ плеотельзона с уроподами. (По Фер-
Ьоковые края грудных сегментов гёфу).
не расширены и перейоподы при-
крепляются близ краев сегментов. Боковые края головы не расширены
и не образуют заостренных спереди лопастей. Плеотельзон сзади без
выемки.
1. JAERA
139
поверхность плео-
К этому подроду относятся близкие между собою мелкие морские
виды, охарактеризованные в большинстве случаев недостаточно по-
дробно. Более детальное их описание, на которое ссылается в своем обзоре
Фергёф (1943), как на находящееся в печати, не увидело света или
осталось мне недоступным. Все перечисленные в определительной
таблице виды подрода найдены у берегов Италии и Далмации при нор-
мальной морской солености. Только один вид — J. hopeana Costa —
обитает и в Черном море, т. е. переносит некоторое опреснение.
*7. Jaera (Metajaera) hopeana Costa (рис. 300—-302).
A. Costa, 1853 in 0. G. Costa, Fauna del Regno di Nipoli. Crostacei Isopodi,
Fasc. 83 : 1, pl. VII, fig. l;KesseIyak, 1938 : 235, fig. 9—10;—albifrons Совин-
ский, 1896, Зап. Киевск.общ. ест., XV, 1 : LI (partim); — charrieri Monod, 1925 :
'238, pl. 49, fig. A—D, pl. 50, fig. A—F; Вълкановъ, 1938 : 65, fig. 18—23.
Передне-боковые углы головы развиты слабо. Задний край плеотель-
зона на конце прямой, лишен выемки. —
тельзона несет щетинки. Жгут I ан-
тенны с 2 lammae olfactoriae, II антенна
не длиннее половины тела, у $ не-
сколько короче, чем у <j. В ее жгуте
около 20 члеников. ДактилюсыII—VII
перейоподов с 3 шипами. I плеопод S
в базальной части расширен, к концу
суживается. Наружные дистальные уг-
лы образуют два длинных, тонких,
загнутых назад выроста, снабженных
только на концах 3—4 короткими ще-
тинками. II плеопод j приблизительно
полукруглой формы. Его ширина зна-
чительно превосходит длину. Длина
тела <3 1.3, о — до 2 мм.
J. hopeana — комменсалист Sphae-
roma serratum (F.), прикрепляющийся
к стернитам этого вида. На одной
Sphaeroma может находиться до трех
экземпляров У. hopeana. Сущность
взаимоотношений между Sph. serratam
и J. hopeana не выяснена. Моно (Mo-
nod, 1925) допускает возможность пара-
зитирования Jaera на Sphaeroma. В сво-
бодном состоянии/, hopeanaвстречается в
Распространение. Черное,
моря. В Черном море найден у побережий Варны, Севастополя и Херсо-
неса. Как отметил Вълканов (1938), нет никаких сомнений в том, что
Jaera, найденная Совинским 1 у Севастополя и Херсонеса на Sphaeroma
serratum и определенная последним автором как J. albifrons, на самом
деле является J. hopeana. Наименьшая соленость, при которой была
до сих пор находима J. Iwpeana, равняется 8°/00 (устье р. Омбла, впада-
ющей в Адриатическое море).
Я находил этот вид в сборах Sphaeroma serratum из Севастопольской
бухты.
1 В. К. Совинский. О некоторых новых и мало известных Isopoda’x Чер-
ного моря. Протокол 5-го очередного собрания Киевск. общ. ест., 1 октября 1894 г.
Зап. Киевск. общ. ест., XV, вып. 1: Ы—LIII, 1896.
(Metajaera)
302
Рис. 300—302. Jaera
hopeana Costa: 300— I плеопод сам-
ца; 301 — II плеопод самки; 302 —
плеотельзон с уроподами (300 по Кес-
сельяку, 301—302 по Вълканову).
исключительно редких случаях.
Адриатическое и Средиземное
АЛФАВИТНЫЙ УКАЗАТЕЛЬ НАЗВАНИЙ ЖИВОТНЫХ 1
abyssalis, Asellus aquaticus 62
abyssi, Ingolfiella 32
Acanthocephalus 38, 44
acherontis, Ingolfiella 32
acherontis, Microcharon 128
acuticarpus, Asellus 53, 111
adenta, Asellus 22, 30, 52, 53
africanus, Asellus coxalis 108
africanus, Stenasellus 25, 26, 32, 114
akyioshiensis, Asellus 53, 54, 70, 73
alabamensis, Asellus 52, 53, 111
albifrons, Jaera 18, 19, 33, 38, 40, 43.
131, 132, 133, 134, 135, 139
Amphipoda 22, 31, 32
amurensis, Livoneca 20
angarensis, Asellus 30, 42, 54, 74, 75,
79, 80, 86
anophthalmus, Asellus spelaeus 52, 93
antarctica, Neojaera 131
Anthuridae 19, 20, 32
antricola, Asellus 53, 111
aquae-calidae, Asellus spelaeus 93
aquaticus, Asellus 11, 14, 16, 17, 26, 27,
28, 29, 35, 36, 37, 38, 41, 42, 43, 44,
51, 52, 53, 57, 60, 62, 64, 65, 81, 82,
85, 112
Archiannelida 32
arnautovici, Asellus 55, 100
arthrobranchialis, Asellus 58
Asellidae 3, 9, 10, 13, 16, 17, 20, 21, 22,
30, 35, 39, 49, 50, 51, 124
Asellinae 50
Aselloidea 20, 21, 22 , 49, 124
Asellota 3, 17, 19, 20, 49, 50
Asellus 13, 14, 15, 16, 20, 27, 29, 35,
37, 42, 43, 44, 50, 51, 81, 111, 114,
124
Asellus, subgen. 13, 14, 15, 16, 21,
22, . 26, 27, 28, 29, 30, 57, 64, 70,
88
asiaticus, Stenasellus 25, 26, 114, 115,
116, 117
Astacidae 32
attenuatus, Asellus 30, 53
Austrastacidae 32
baicalensis, Asellus 30, 42, 54, 74, 75,- 76
Baicaloasellus 12, 15, 16, 21, 22, 29, 30,
74
banyulensis, Asellus 23; 24, 56, 81, 104.
baunti, Coregonus sardinella 28
belgicus, Asellus meridianus 82
beranensis, Microcharon profundalis 131
birsteini, Desertoniscus 20
breuili, Stenasellus 114, 117, 118
brevicauda, Asellus 111
buchneri, Stenasellus 114, 120
buili, Stenasellus 114
bureschi, Stenasellus 114, 122, 123
Caecidotea 50, 51, 53, 70, 72, 73, 101
Caecidotinae 50
califomicus, Asellus 30, 42, 53, 54, 70
Cambarus 31
caspica, Jaera sarsi 33, 34, 35, 134, 135,
136
cavaticus, Asellus 24, 25, 52, 57, 88, 89,
109
cavernicola, Asellus aquaticus m. 10, 61
cephallenus, Asellus italicus 103
chappuisi, Stenasellus 25, 26, 32, 115, 117
charrieri, Jaera 139
Chetilidae 19
Cirolana 31
Cirolanidae 31
Cirolanides 31
Cirolaninae 19
colleryi, Asellaria 38
collingei, Asellus 34, 57, 94
communis, Asellus 31, 39, 60, 86, 111
Conasellus 13, 14, 21, 22, 29, 30, 31,-57,
111
Conilera 31 ,
Corallanidae 19
corcyraeus, Asellus italicus 104
coxalis, Asellus 23, 56, 81, 101, 107
cracoviensis, Asellus 58
Crangonyx 31, 32, 109
crassicaulis, Paravorticella 38
crypticus, Asellus 112
Cumacea 33
1 Названия семейств и высших таксономических единиц выделены жирным
шрифтом; синонимы набраны курсивом, прочие названия—прямым шрифтом,
причем названия родов и подродов начинаются с заглавной буквы. Жирными
цифрами обозначена страница, на которой дано описание.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
141
curvispinum, Corophium 54
cyanophilus, Asellus coxalis 23, 108.
Cymothoidae 19
dalmatica, Jaera 133
Decapoda 22
dentadactylus, Asellus 111
deshayesi, Jaera 131
Desmosomatidae 31
digitatus, Sty locometer 38
dimorpha, Asellus 53, 111
dudichi, Asellus polychaetus 84
duebeni, Gammarus 38
dybowskii, Asellus 42, 52, 53, 70, 71, 74
entomon, Mcsidothea 20
Epicaridea 19
epimeralis, Asellus 28, 50, 53, 68, 69
epiroticus, Asellus italicus 103
Epischura 29
Eriopisa 32
Eriopisella 32
exiguus, Synasellus ma iae 125
Exocorallanidae 19
Faucheria 31
filicornis, Jaera 131
Plal ellifera 19
fontinalis, Asellus 65
fontis, Typhlocirolana 99
foreli, Asellus cavaticus 44, 89
fribergensis, Asellus aquaticus var. 61
Gammarus 32
gauthieri, Asellus 10, 55, 81, n6, 97
gjorgjevici, Asellus 52, 56, 87
gjorgjevici, Stenasellus 114, 118, 119
Gnathiidea 19
goplanus, Asellus 58
gordoni, Asellus aquaticus var. 58, 60
granea, Castradella 38
groenlandicus, Asellus . 58
guemei, Jaera 137
Harpacticoida 29
hazeltoni, Asellus aquaticus va . 58, 60, 94
hazeltoni, Stenasellus 125
hercegovinensis, Asellus cavaticus 89, 91
hermallensis, Asellus 57, 96
hilgendorfi, Asellus 9, 14, 16, 27, 28,
29, 43, 52, 53, 6?, 63
hobbsi, Asellus4lll
hopeana, Jaera'93, 133, 139
hospitalis, Jaera 133
hungaricus, Stenasellus 115, 117, 120
hussoni, Asellus 57, 95
hypogeus, Asellus 52, 57, 91, 92
Idotheidae 20
incisus, Asellus 111
infirmus, Asellus 10, 24, 55, 85, 86
Ingolfi cllidae 33
Ingolfiellidea 32
intermedia, Asellus 111
knnnJa 9 19 20. 22 31 32
istrianus, Asellus 55, 99
italicus, Jaera 40, 133, 136, 138
italicus, Asellus 24, 56, 81, 101, 102
Jaera-10, 17, 29, 32, 33, 34, 35, 38, 40,
131
Jaera s. str. 132, 13,
Jaeridina 131
Jaira 131
Janiridae 9, 11, 14, 16, 49, 131
Johanella 13, 20, 21. 22, 30, 31, 50, 51,
112
karamani, Asellus 56, 105
kawamurai, Asellus 52, 53, 70, 72, 73
korotnewi, Asellus 52, 54, 74, 75, 77
kosovensis, Microcharon profunadalis 130
kroyeri, Jaera 134
kumanovensis, Microcharon profunadalis
130
latifrons, Asellus 28, 50, 53, 67
latus, Microcharon 128, 129
leruthi, Asellus cavaticus 88, 89
leucadius, Asellus italicus 103
Ligidium 20
Lirceus 2, 12, 20, 30, 31, 51, 86, 124, 122
litoralis, Asellus gjorgjevici 54, 87
littoralis, Ingolfiella 32
locusta, Gammarus 38
longisetus, Asellus spelaeus 94
lusitanicus, Asellus 81
mackini, Asellus aquaticus var. 58, 60
Macrostylidae 32
Malacostraca 22, 23, 29, 31, 32
Mancasellinae 50
Mancasellus 22, 123, 124
mariae, Synasellus .109, 123, 124
marina, Jaera 134
martynovi, Asellus hilgendorfi 27, 28, 64
meridianus, Asellus 15, 17, 23, 24, 27,
54, 81, 82, 109, 112
Mesoasellus 12, 14, 15, 16, 21, 22, 29,
30, 53, 70, 72, 74
messerianus, Asellus aquaticus 60
Metajaera 133, 138
Microbdella 29
Microcerberidae 32, 33
Microcharon 32, 126, 128
Microparasellidae 9, 10, 11, 14, 16, 33,
49, 126, 132
Microparasellus 17, 32, 116, 129
minutus, Asellus 52, 54, 74, 78
montenegrinus, Asellus cavaticus 89, 91
monodi, Asellus 55, 101
montanus, Asellus 101
monticola, Asellus 14, 17, 18, 27, 2§,
29, 53, 65, 66
Mysidacea 33
Neojaera 35, 132
nickajackensis, Asellus 30, 53
Niphargus 32
nipponensis, Asellus 62', 63
nivalis. Jaera 134
142
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
nobrei, Stenasellus 114, 121
nordmanni, Jaera 33, 131, 136, 137
oculatus, Asellus 52, 53, 111 '
Oniscoidea 19, 20
Oniscus 57
packardi, Asellus 53, 111
paradoxus, Dendrocometes 38
Parasellidae 20, 21 .
Paraselloidea 20, 21, 22, 124, 126, 149
Parastacidae 32
peltatus, Asellus 56, 109, 110, 111
perarmatus, Asellus italicus 23, 24, 31,102
peyerimhoffi, Asellus. coxalis 23, 107
Phreatoicidea 19, 22
Polyehaeta 32
polychaetus, Asellus 56, 84
Proasellus 12, 14, 15, 16, 21, 22, 23, 24,
25, 29, 31, 52, 69, 81, 86
profundalis, Microcharon 43, 128, 129,
130
Protelsonia 113, 119, 120
purpurea, Johanella 86, 112, 113
pusilia, Neojaera 132
puteanus, Microparasellus 127
quicki, Asellus 112
racovitzai, Stenasellus 26, 114
rhausinus, Asellus spelaeus 94
remyi, Asellus 10, 55, 84, 86
richardsonae, Asellus 30, 52, 53
robustus, Stenasellus gjorgjevici 119
ryukyuensis, Asellus
sarsi, Jaera 33, 34, 35, 131, 132, 133,
135, 136
schellenbergi, Jaera 133, 137,- 138
seabrai, Asellus 57, 108, 109
serrata, Neojaera 131
serratum, Sphaeroma 139
sieboldi, Asellus 88
skopljensis, Stenasellus 115, 119
smithii, Asellus 30, 52, 53
sorrentina, Jaera 133, 134, 138
spathulatus, Asellus 53, 111
spelaeus, Asellus 24, 57, 92
Sphaeromidae 19, 20, 22
Sphaeromides 31
Spiloniscus 20
stagnoensis, Jaera 133
Stenasellus 11, 13, 14, 16, 20, 22, 24,'25,.
26, 50, 51, 113, 114, 116, 117
Stenetriidae 20, 21, 22, 32
Stenetrium 32
stilodactylus, Asellus 53, 111
Stygasellus 126
stygius, Asellus 53, 111
stygius, Microcharon 128
sulcata, jaera 133
switesiana, Asellus goplanus 58
Synasellus 11, 13, 20, 21, 22, 30, 31„
50, 51, 123
Synurella 31
theodori, Asellus gautie i 97, 98
thermonyctophilus, Asellus 57, 81, 98»
qq Ifiq
Titanethes 20
tomalensis, Asellus 21, 22, 30, 53
Torix 29
tridentata, Asellus 52, 53, 111
tuberculatus, Melanoides 26
Typhlocirolana 31
valdensis, Asellus cavaticus~"89, 90
Valvifera 19
virei, Stenasellus 16, 114, 117
vulgaris, Asellus 58
waiter!, Asellus cavaticus 89, 90
wianamattensis, Phreatoicus 22
wolfi, Asellus banyulensis 23, 105
woodward!, Cyclosphaeroma 22
СОДЕРЖАНИЕ
Стр.
Предисловие................................................................ 3.
Систематический указатель видов............................................. 5
Введение ................................................................... 9
Морфологический очерк.................................................. 9
Географическое распространение и родственные отношения............... 19'
Биология и экология.................................................. 35
Хозяйственное значение.............................................. 43
Литература..........................................г................. 44
Специальная часть.......................................................... 49'
1 сем. Asellidae...................................................... 50
1 род Asellus............................................ • • • 51
2 род Johanella................................................ 112'
3 род Stenasellus................................................ ИЗ
4 род Synasellus ............................................... 123
2 сем. Microparasellidae............................................ 126
1 род Microparasellus.......................................... 126
2 род Microcharon............................................... 12&
3 сем. Janiridae.................................................... 131
1 род Jaera....................................................... 131
Алфавитный указатель названий животных . . ........................... 140>
Печатается по постановлению
Редакционно-издательского совета
Академии Наук СССР
*
Редактор издательства К. А. Борисов.
Технический редактор Р. А. Ароне.
Корректоры 3. Г. Сегаль и И. И. Удимое
*
РИСО АН СССР № 4422. М-29089. Подписано
к печати 31|V 1951 г. Печ. л. 12,33. Уч-йзд.
листов 11,4. Бумага 70ХЮ8/16. Бум. листов 4,5.
Тираж 2000. Зак. № 32. Цена в перепл. 12 руб.
1-я Тип. Издательства Акад. Наук СССР.
Ленинград. В. О., 9л.,д. 12.