























































































































































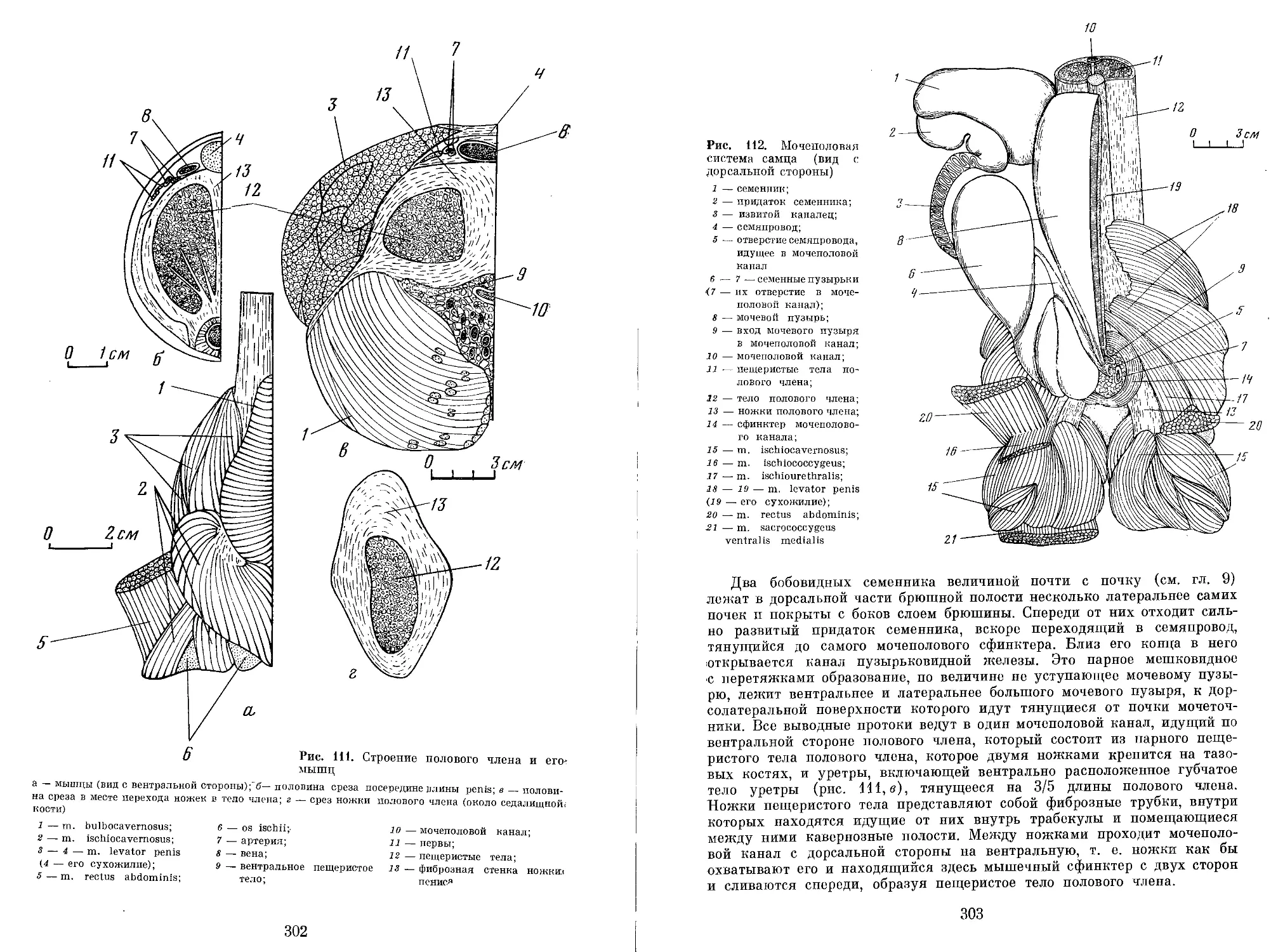




















































Author: Яблоков А.В. Воробьева Э.И.
Tags: mammalia млекопитающие биология морские млекопитающие
Year: 1986




Text
ЛАМАНТИН
Морфологические
адаптации
-Наука-
АКАДЕМИЯ НАУК СССР
Институт эволюционной морфологии
и экологии животных им. А. Н. Северцова
ЛАМАНТИН
Морфологические
адаптации
Ответственный редактор
академик В. Е. СОКОЛОВ
МОСКВА
«НАУКА»
1986
УДК 599
ЛАМАНТИН. Морфологические адаптации. М.: Наука, 1986.
В коллективной монографии на оригинальном материале и с привлече-
нием обширной библиографии рассматриваются вопросы эволюции сирен
и место среди них ламантинов, морфология и адаптивные особенности
кожного покрова, опорно-двигательной системы, пищеварительного
тракта, сосудистой системы и т. п. американского ламантина. Особое вни-
мание уделено специфичности адаптаций ламантина и всех сирен в ходе
их эволюции в условиях водной среды.
Табл. 16. Ил. 155. Библиогр. 9 с.
Рецензенты:
доктор биологических наук Э. И. ВОРОБЬЕВА,
доктор биологических наук А. В. ЯБЛОКОВ
и2005000000-213
042 (02)-86
277-86-11
© Издательство «Наука», 1986
ПРЕДИСЛОВИЕ
Изучение сирен — небольшого отряда водных млекопитающих лежит в дав-
них традициях отечественной зоологии. Мы по праву гордимся классиче-
ской монографией первого директора Зоологического института император-
ской Санкт-Петербургской Академии наук академика Ф. Ф. Брандта,
во многом заложившей основы науки о сиренах — сиренологии. Именно на
территории нашей страны, на Командорских островах экспедицией Бе-
ринга были обнаружены последние живые морские коровы, остатки особой
северной ветви сирен, некогда обитавших по всему побережью Северной
Пацифики. В наших музеях хранятся обильные материалы по этим уни-
кальным животным. Заметное повышение интереса к этому отряду млеко-
питающихся, выражающееся в росте числа публикаций, связанных с сире-
нами, в том числе и работ обобщающего характера, проведении особых
международных совещаний по сиренам, создании специального интерна-
ционального центра по изучению ламантинов, вызвало к жизни предлагае-
мую ниже работу, посвященную морфологии и адаптивным особенностям
одного из представителей современных сирен — ламантина. Особый упор
в ней сделан на выяснение специфики сирен, и в частности ламантина,
развившейся в ходе приспособления их к исключительно водному образу
жизни.
Непосредственным поводом к постановке настоящего исследования
было разрешение правительства республики Куба па отлов одного круп-
ного самца ламантина. В связи с этим по приглашению президента Кубин-
ской академии наук Кубу посетила группа советских зоологов во главе
с академиком В. Е. Соколовым, в которую также входили доктора биоло-
гических наук П. П. Гамбарян и В. Б. Суханов и кандидаты биологиче-
ских наук Л. М. Мухаметов и О. Ф. Чернова. Все они вместе с кубинскими
товарищами участвовали в отлове ламантина в водах залива Батабано
у берегов полуострова Сапата в бухте Матанамбре в сентябре 1980 г.
и в дальнейшей перевозке его судном в Сурхидеро-де-Батабано, а затем
автотранспортом в Гавану, где он был помещен в небольшой бассейн на
территории Института зоологии АН Кубы. Здесь был проведен ряд нейро-
физиологических экспериментов (см. гл. 12), а затем после усыпления
животного смертельной дозой дитилина на четвертые сутки после отлова
оно было зафиксировано 10%-ным раствором формалина, накаченным в
спинную аорту под давлением (с применением садового опрыскивателя).
Одновременно был взят ряд гистологических и электронномикроскопиче-
ских проб кожного покрова, кишечника, брюшной мышцы, печени. Консер-
3
вированный таким образом ламантин был самолетом переправлен в СССР
в Зоологический институт АН СССР, на базе которого проводилась даль-
нейшая морфологическая его обработка. Скелет животного поступил в От-
дел млекопитающих ЗИН АН СССР (№ 31795).
Естественно, дать полное анатомическое описание всех систем органов
ламантина по одному экземпляру было невозможно, поэтому основное вни-
мание было уделено кожному покрову, скелету и мышечной системе, пи-
щеварительному тракту, некоторым деталям сосудистой системы, от изу-
чения которых в первую очередь можно было ожидать ответа на вопрос
о сущности морфологических адаптаций ламантина. В ряде случаев были
привлечены дополнительные материалы из зоологических хранилищ Мо-
сквы, Ленинграда, Киева, Харькова, Львова. Учитывая общую направлен-
ность монографии, было решено предпослать ей обзор современных пред-
ставлений об эволюции и системе сирен, роли морфологических адаптаций
в их эволюции и месте ламантинов в системе как своеобразную точку
отсчета при обсуждении предлагаемого оригинального материала.
К работе были привлечены сотрудники Института эволюционной мор-
фологии и экологии животных АН СССР, Зоологического института
АН СССР, Ленинградского государственного университета, Московского
государственного университета, Института зоологии АН УССР, Института
эволюционной физиологии и биохимии АН СССР. В оформлении моногра-
фии большую помощь оказали сотрудники ИЭМЭЖ АН СССР Е. Г. По-
тапова и И. Н. Попова. Особую благодарность необходимо вынести нашим
кубинским товарищам за великолепную организацию той части работы,,
что была проведена на Кубе.
В. Е. Соколов:
ВВЕДЕНИЕ
Современные сирены (Mammalia, Sirenia) представляют небольшую груп-
пу морских исключительно растительноядных млекопитающих, носящую
явно реликтовый характер. Никогда не покидая водной среды, они зани-
мают в ней важную экологическую нишу, будучи единственными из круп-
ных животных консументами первого порядка в тропических водах. Рас-
цвет сирен — в прошлом, хотя они никогда не были очень разнообразными.
В настоящее время сохранились лишь два рода сирен: ламантины Triclie-
chus Linnaeus, 1758 с тремя видами: Т. manatus L., 1758; Т. senegalensis
Link, 1759; Т. inunguis (Natterer, 1883) и дюгони Dugong Lacepede 1799;
с единственным видом D. dugon (Muller, 1775). Ламантины обитают в при-
брежных водах Мексиканского залива, Карибского моря и северо-восточ-
ного побережья Южной Америки (Т. manatus — ламантин‘), в реках бас-
сейна Амазонки и Ориноко (Т. inunguis — бразильский ламантин), в реках
и у побережья Западной Африки (Т. senegalensis — африканский ламан-
тин), дюгони в Индопацифике. В недавнее историческое время существо-
вали на Командорских островах в северной части Тихого океана гигант-
ские морские коровы Hydrodamalis gigas (Zimmermann, 1780), истреб-
ленные человеком к 1741 г.
1 Русские названия сирен в целом, отдельных родов и видов все еще не устоялись,
и в литературе существует значительный разнобой. С одной стороны, это вызвано
использованием русифицированных форм латинских (сирены, дюгонь) или мест-
ных названий (ламантин) без учета первичного объема этих понятий. Так, ламан-
тинами (слово французского происхождения) за рубежом называют чаще лишь
африканского ламантина (Mohr, 1957), а американские формы именуются манати
(название идет от одного из карибских диалектов, из которого взято имя Mana-
tus — младший синоним Trichechus). У нас общепринято ламантинами (в множе-
ственном числе) называть всех представителей рода Trichechus. Кроме того,
иногда ламантином (в единственном числе) называется Т. manatus (Соколов, 1979),
который в нашей литературе часто обозначается и как американский ламантин
(Томилин, 1971), и как обыкновенный ламантин (Огнев, 1949). В зарубежной лите-
ратуре его часто называют американский манати (Murie, 1872а). С другой сторо-
ны, во многих языках мира существует понятие «морские коровы». Видимо, впер-
вые оно сложилось у ассирийцев (Simpson, 1932) применительно к дюгоню. Неда-
ром Симпсон закрепляет это название в качестве местного или народного за этим
родом (Simpson, 1945). В дальнейшем, однако, морскими коровами стали почти
повсеместно называть всех сирен (Mohr, 1957; Howell, 1930; Domning et ah, 1982;
Гептнер, 1967), отмечая этим их способность преобразовывать растительную мас-
су пищи в мясо и молоко, подобно настоящим коровам (Anon., 1974). В то же вре-
мя в русской литературе сложилась устойчивая традиция закреплять это назва-
ние только за одной уже исчезнувшей группой сирен (Hydrodamalidae одних авто-
ров, Hydrodamalinae других) (Томилин, 1971; Соколов, 1979), причем то же назва-
ние без изменения фиксируется и за родом (Hydrodamalis) и в единственном чис-
ле— морская корова — за видом (Н. gigas) (Соколов, 1979). Чаще, однако, это
животное называют стеллерова корова (Walker, 1975; Наумов, Карташев, 1979) или
стеллерова морская корова (Brandt, 1866; Томилин, 1971; Domning, 1972). Все это
надо иметь в виду, встречая разночтения в отечественной литературе.
В настоящем издании приняты названия, рекомендованные В. Е. Соколовым
(1979).
5
Реликтовый характер сирен не только вызывает особый интерес у ис-
следователей, но и заставляет спешить с их изучением. Опасность их ско-
рого исчезновения была ясна уже в прошлом веке (Murie, 1872). Само
изучение сирен имеет давнюю историю. Литература по ним насчитывает
более тысячи названий. Полная библиография приведена в ряде специаль-
ных изданий (Allen, 1881; Freund, 1950; Whitfield, Farrington, 1975; Hu-
sar, 1975; Ronald et al., 1978; Leutenegger, 1982). Первые известия о сире-
нах можно найти уже у Плиния Старшего, хотя у древних авторов они
всегда выступают в мифологическом облике (например, сирены и Одиссей
у Гомера). Действительно, облик этих животных удивителен: объемистое
туловище, лишенное задних конечностей, с хвостом, вооруженным мощ-
ным плавником, и неожиданно с очень подвижными передними конечно-
стями, преврщенными в ласты; на сравнительно небольшой голове, си-
дящей на короткой шее, очень маленькие глаза, ушных отверстий не вид-
но, верхняя губа превращена в своеобразный короткий хобот, или, как его
называют, губной диск. Таковы ламантины на уже ставшей классической
и неоднократно воспроизводившейся в печати литографии (рис. 1), заим-
ствованной из работы Мьюри (Murie, 1980). Дань изучению сирен отдали
многие классики мировой зоологии: Ж. Кювье, Р. Оуэн, О. Абель,
Дж. Г. Симпсон. Лучшую монографию по сиренам опубликовал русский
академик Ф. Ф. Брандт (Brandt, 1876 (1974), 1861—1868). В своей работе
с поистине символическим названием «Simbolae Sirenologie» он не только
подытожил свои многолетние исследования остеологии знаменитой мор-
ской коровы (или, как он сам ее называл, Steller’s Seekuh), взятой в срав-
нении с другими сиренами, но и собрал воедино все, что было известно
к этому времени по морфологии, палеонтологии и биологии сирен. Непре-
ходящее значение этой работы до наших дней отмечают все серьезные
исследователи сирен (Simpson, 19322).
В целом если в изучении сирен в прошлом веке преобладали сугубо
морфологические работы большей частью описательного характера
а в первой половине нашего века — работы по палеонтологии сирен (и при
этом обобщающих исследований было мало), то за последние годы появил-
ся ряд крайне важных публикаций, затрагивающих уже более широко
естественную историю сирен (их эволюцию, морфологию, поведение, эко-
логию, охрану и т. п.). Так, была издана первая монография по этологии
ламантина (Hartman, 1979), в основе которой лежат наблюдения над не-
большой популяцией этих животных, обитающих в относительно чистых
водах Кристал-Ривер во Флориде, что позволило описать их пищевое, по-
ловое, социальное и т. д. поведение, что было невозможно в других частях
ареала, так как ламантины обычно живут в замутненной воде. Появились
два морфологических описания мышечной системы дюгоня и бразильского
ламантина (Domning, 1977b, 1978а) с рядом интересных соображений по
локомоторным приспособлениям и работе их челюстного аппарата. Подго-
товленные целой серией исследований в области палеонтологии сирен
(Simpson, 1932; Sickenberg, 1934; Reinchart, 1959, 1976), появились ра-
боты того же Д. П. Домнинга (Domning, 1977а, 1978b, 1982; Domning et
al., 1982), в которых эволюция отдельных групп сирен уже рассматрива-
ется как смена последовательных адаптаций в связи с изменением при-
родной обстановки в местах их обитаний. Естественно, предложенные
автором «модели» — схемы эволюционных преобразований в семействе Tri-
chechidae и подсемействе Hydrodamalinae во многом гипотетичны, их под-
2 В библиотеке Зоологического института АН СССР (г. Ленинград) хранится экземп-
ляр этой монографии с большими рукописными вставками на латинском языке,
сделанными карандашом, возможно, рукой самого акад. Ф. Ф. Брандта.
6
Рис. 1. Различные позы ламантина (Trichechus manatus) по зарисовкам с животных,
содержащихся в аквариуме (по: (Murie, 1980))
тверждение или опровержение требует более глубоких знаний по спе-
цифике приспособлений современных сирен, привлечения новых фактов и
групп фактов, новых их интерпретаций. Важно подчеркнуть методологи-
ческое значение подобного подхода к изучению сирен — чтение хода эво-
люции через вскрытие частных адаптаций. На необходимость изучения
приспособительных сторон морфологии сирен уже указывалось в превос-
ходной книге А. Б. Хауэлла «Морские млекопитающие. Их адаптации
к жизни в воде» (Howell, 1930). Между строк в ней можно выделить ка-
жущуюся нам очень современной мысль, что изучение адаптаций должно
вестись через специфику приспособительной эволюции каждой группы
животных (вспомним, что во времена Хауэлла речь шла об определении
адаптивного смысла отдельных структур), и этот подход более плодотво-
рен, чем сравнительное исследование, скажем, сирен и китообразных как
групп, равно порвавших с сушей, в ходе которого преимущественное вни-
мание уделяется лишь различиям в уровне приспособления к водной среде,
что затушевывает различие путей приспособления сирен, с одной стороны,
и китообразных — с другой, и то, что сирены овладели единственной в сво-
ем роде экологической нишей, в которую и не пытались проникнуть киты
и дельфины.
В результате в последние годы меняются даже сами представления
о сиренах. Если раньше их история полагалась простой, лишенной внешне
крупных событий и именно это представлялось главной сложностью в их
изучении (Repenning, 1977), то теперь было показано, какие глубокие из-
7
менения произошли в ходе самой эволюции сирен. Достаточно сказать,
что у них реализовались два противоположных по результатам принципа
приспособления к увеличившемуся стиранию зубов: одно — путем развития
гипсодонтных корней у дюгоней, другое — путем выработки горизонталь-
ного замещения коренных зубов у ламантинов при увеличении числа од-
новременно функционирующих зубов. Это приводит к необходимости пе-
рестать считать историю сирен историей утраты ими плезиоморфных
признаков (Savage, 1977) или сводить характеристику сирен и отдельных
их групп, кочующую из одной сводки в другую (Jones, Johnson, 1967;
Гептнер, 1967; Соколов, 1979), к перечню черт только современных их
представителей.
Можно отметить также, что в силу своей специфики сирены облада-
ют рядом преимуществ перед другими животными в качестве модельной
группы при изучении таких форм эволюции, как «квантовая эволюция»
по Симпсону (Domning, 1978с) и некоторые другие.
Необходимость всестороннего и глубокого изучения всех аспектов есте-
ственной истории сирен вызывается также резким сокращением их числен-
ности, что требует незамедлительных природоохранительных мероприятий,
основанных на конкретных знаниях. |3а историческое время число сирен
сократилось до предела. Все их виды занесены в Красную книгу, одни как
находящиеся в угрожаемом положении, другие — как стоящие на грани
вымирания (Ronald et al., 1978). Так, еще в конце прошлого века в неко-
торых прибрежных районах Северного Квинсленда (Австралия) отмеча-
лись тысячные стада дюгоней. Одно такое стадо занимало площадь до 5 км
длиной и 300 м шириной (Bertram, 1981). Это, конечно, исключительный
случай, но и сейчас еще в водах Австралии по неизвестным причинам со-
средоточена большая часть дюгоней, тогда как в других частях их огром-
ного ареала их численность, видимо, всегда была незначительна. В лите-
ратуре можно найти сведения и о количестве уничтоженных за определен-
ный срок ламантинов. Так, называется цифра в 8000 экземпляров убитых
ламантинов за два года в начале нашего столетия (Anon., 1974). Можно
назвать две основные причины современного катастрофического падения
численности сирен: разрушение их природных местообитаний — закрытых
спокойных мелководий в тропических и субтропических зонах земного
шара, в заводях рек, речных старицах, прибрежных зонах моря, и осо-
бенно уничтожение их человеком, единственным их врагом. Разительный
пример — уничтожение человеком всего за 27 лет довольно многочислен-
ных морских коров, обитавших у берегов Командорских островов и откры-
тых в 1741 г. экспедицией Витуса Беринга. Других природных врагов си-
рены не имеют, хотя и были отмечены случаи нападения на них акул,
крокодилов, косаток (Anderson, 1981b).
Человек может использовать до 80% туши ламантина (Anon., 1974).
От него получают мясо с очень высокими пищевыми качествами (вкус,
высокая сохранность в присоленной воде или в собственном жиру), от-
личный, приятный на вкус и цвет жир, шкуру, идущую в обработку, кость,
используемую на замену слоновой. Уже это объясняет, почему со времени
открытия ламантинов экспедицией Колумба на них шла непрерывная
охота испанскими колонистами, буканьерами, владельцами сахарных план-
таций (на корм для рабов). В наше время охрану сирен, а они практиче-
ски во всех странах, куда заходит их ареал, защищены законами, затруд-
няет то, что большинство этих стран относится к категории развивающих-
ся, где экономические проблемы стоят очень остро и действие законов
подчас оказывается малоэффективным. Правительству Австралии прихо-
дится даже разрешать определенным группам населения (аборигенам,
8
жителям островов Торресова пролива) официально охотиться на дюгоней,
учитывая только потребность их в источниках питания, но и важную
роль, какую играет охота на дюгоней в социальной жизни этих народов.
Таким образом, как это иронично ни звучит, судьба сирен находится в
руках их единственного врага — человека (Ronald et al., 1978) и их охра-
на нуждается в более действенных средствах.
Озабоченность человека судьбой сирен выразилась в ряде международ-
ных мероприятий, приходящихся на 70-е годы нашего века. В 1974 г.
в Джорджтауне (Гайяна) был созван семинар, собравший участников из
восьми стран, на котором был обсужден вопрос о необходимости и возмож-
ностях создания Международного центра по изучению ламантинов и под-
готовке рекомендаций к составлению программы исследований такого
центра (Anon., 1974). Были сформулированы следующие цели, которые
должны стоять перед специалистами, изучающими ламантинов: 1) реше-
ние фундаментальных задач биологии ламантинов с особым упором на ос-
новные вопросы физиологии, эндокринологии, анатомии, биологии размно-
жения, биохимии, этологии, экологии и смежных дисциплин; 2) даль-
нейшее изучение проблем, связанных с использованием ламантинов в
борьбе с зарастанием водоемов и ирригационных сооружений — страшным
бичом тропических вод. Расчет здесь строится на прожорливость ламан-
тинов, потребляющих за день растительной продукции до 20% собствен-
ного веса, неприхотливость их вкуса. Успешные опыты такого рода были
проведены в Гайяне, где некоторые каналы уже десяток лет поддержива-
ются в чистом виде за счет обитающих в них ламантинов. Из этого выте-
кает 3-я задача: сохранение популяций ламантинов, изучение возможно-
стей повышения их численности, включая одомашнивание и искусственное
разведение, а также восстановление разрушенных местообитаний. В ка-
честве 4-й задачи ставится поощрение науки в развивающихся странах,
общее руководство и координация исследований по ламантинам. В каче-
стве местоположения Центра была предложена Гайяна, в водах которой
обитает еще довольно многочисленная популяция ламантинов (от 1000
до 10 000), самая массовая из всех известных. Объединение усилий уче-
ных разных стран уже приносит свои первые плоды. Так, опубликован
первый обширный обзор (Biological Synopsis) по всем вопросам, связан-
ным с ламантинами: номенклатуре и классификации, эволюции и геогра-
фическому распространению, морфологии и некоторым деталям физиоло-
гии и биохимии, образу жизни, популяционной экологии, охране и т. п.
(Ronald et al., 1978). Один список литературы, прилагаемый к обзору,
насчитывает более 800 названий.
Не менее важным следует считать Первый Международный семинар
по биологии и охране дюгоней, состоявшийся в г. Таунсвилл (Австралия,
Северный Квинсленд) в Университете Джемса Кука в 1979 г. и собравший
участников из семи стран и представителей местного населения. На семи-
наре было заслушано большое количество сообщений о современном со-
стоянии, предполагаемой численности популяций дюгоней, их экологии,
поведении, морфологии, роли в морских экосистемах. Особенно были под-
черкнуты важность сохранения дюгоней как единственного строго мор-
ского растительноядного млекопитающего (Heinsohn, 1981) и моральная
ответственность за это Австралии как страны, в водах которой обитают
две наиболее обширные и многочисленные популяции дюгоней из пяти
существующих (Nishiwaki et al., 1981). По результатам работы семинара
был опубликован большой том трудов (см.: (The Dugong..., 1981)) с при-
ложением методики стандартных измерений и взятия материала для ис-
следований.
9
Все сказанное выше говорит о происходящем в настоящее время рас-
ширении фронта исследований сирен. И в этих условиях мы надеемся,
что предлагаемая вниманию читателя новая работа, посвященная ламан-
тину, послужит нашим вкладом в общее дело.
В заключение необходимо отметить, что основным объектом исследо-
вания послужил крупный по современным понятиям экземпляр ламанти-
на, самца живым весом в 402 кг с длиной (измеренной на лежащем
брюхом вверх животном от конца морды до конца хвостового плавника,
следуя по изгибам тела) 321 см. Его экстерьер характеризуют также
некоторые дополнительные параметры. Так, расстояния, взятые от конца
морды, составляют: до пупка 123 см, до половой щели 132 см, до аналь-
ного отверстия 210 см; длина хвоста от анального отверстия до конца
82 см, максимальная ширина хвостового плавника 84 см; расстояние от
переднего конца рыла до уровня переднего края ласта, принимаемое
за длину головы и шеи, составляет 54 см, длина ласта от подмышки до
дистального конца (так называемая задняя длина ласта) — 50 см, макси-
мальная ширина ласта — 21 см.
Глава первая
ОБЩАЯ ХАРАКТЕРИСТИКА И ЭВОЛЮЦИЯ СИРЕН
И МЕСТО ЛАМАНТИНОВ В ИХ СИСТЕМЕ
(Mammalia: Sirenia)
Под морфологическими адаптациями подразумевается отражение в строе-
нии животных процесса приспособления к среде в широком смысле
(физико-географической, биологической) и, следовательно, это — катего-
рия, имеющая определенную эволюционную направленность (вектор-
ность). Такой подход требует изучения морфологических адаптаций
с точки зрения не только расшифровки их смысла (адаптивного значе-
ния), но и их становления в эволюции определенных групп животных.
Следовательно, их изучение является предметом равно как функциональ-
ной и экологической морфологии, так и палеонтологии, значение которой
в изучении адаптаций сирен особенно возросло в последнее время. Отсюда
вытекает целесообразность предпослать разбору собственного морфологи-
ческого материала изложение современных представлений о систематике
и эволюции всех сирен, ископаемых и современных, адаптивных тенден-
циях, отмечаемых у них, а также попытаться дать некоторое представле-
ние о самой адаптивной сущности сирен в целом и отдельных их групп.
Усиление внимания к этим вопросам систематики и эволюции сирен, поч-
ти полностью сосредоточенных сейчас в руках палеонтологов, составляет
характерную черту современного этапа науки о сиренах, начало которого
следует вести с известной работы Дж. Г. Симпсона «Ископаемые сирены
Флориды и эволюция Sirenia» (Simpson, 1932). В настоящее время это
направление достигло расцвета в серии работ Д. П. Домнинга, которые
дают целостное представление о разных этапах развития сирен и направ-
лениях их эволюции. Этот автор предложил крайне интересные экологи-
ческие модели развития таких специализированных групп сирен, как
ламантины (Trichechidae) с одной стороны, и Hydrodamalinae, тихоокеан-
ская ветвь сирен —с другой (Domning, 1977а, 1978с, 1982). На основе
этих работ, как и ряда других, и составлен предлагаемый очерк. Некото-
рые стороны адаптивной природы сирен здесь будут рассмотрены лишь
вкратце, поскольку к их разбору необходимо будет возвращаться еще раз
в последующих главах.
Сирены представляют собой морфологически единую группу млекопи-
тающих, рассматриваемую обычно в ранге отряда, вся специфика которой
связана с питанием водной растительностью рек и прибрежных частей
морей и океанов. Это единственные растительноядные млекопитающие,
ставшие облигатными обитателями водной среды, полностью порвавшими
связь с сушей. Именно приспособление к этой узкой экологической нише,
где они практически не имеют в настоящее время конкурентов, определи-
ло их относительно малое разнообразие, их географическое распростра-
нение, направления морфологических преобразований в отдельных
линиях развития. Лишь сейчас начинает выясняться вся глубина этих
преобразований в пределах отряда, затрагивающая челюстной аппарат,
околоушную и носовую области черепа, весь локомоторный аппарат. В то
же время мы еще очень мало знаем о биологии, и в частности физиологии
сирен, о характере требований, которые предъявляют к их организму
специфическая среда, сам образ жизни, особенности поведения. Все это
заставляет внимательно относиться к морфологии сирен как источнику
11
сведений, позволяющих в какой-то мере реконструировать и предсказы-
вать еще неизвестные особенности их экологии, двигательной активности,
деятельности сенсорных и других систем. В этом плане крайне важны
тенденции морфологических преобразований, выявляющиеся в эволюции
сирен и помогающие понять действительную экологическую специфику
ламантинов и дюгоней, такую неявную, если о ней судить только по тому,
что мы знаем в настоящее время об их биологии.
В системе млекопитающих сирены занимают весьма обособленное
положение, усугубляющееся тем, что прямые предки их почти неизвестны.
Долгое время господствовало мнение, что сирены — это лишь стадия по
пути к другим постоянным обитателям морей и океанов — китообразным,
с которыми их и помещали в одну группу (Ж. Кювье, Р. Оуэн, Э. Коп),
хотя уже было отмечено их отличие от всех других водных и полуводных
млекопитающих и определенное сходство с хоботными (Blainville, 1816).
Была и компромиссная точка зрения: сирены промежуточны между хо-
ботными и китообразными. Обзор всех этих и ряда других представлений
можно найти в литературе (Brandt, 1846, 1861/1868; Simpson, 1932).
Однако все же следует отметить, что принципиальный вывод об отсутствии
связи сирен с китообразными был сделан в результате изучения иско-
паемой сирены Halitherium schinzi и при этом были подчеркнуты черты,
сближающие их с примитивными копытными типа тапира (Lepsius,
1881), вернее, как мы это воспринимаем сейчас, с таким адаптивным ти-
пом, что представлен тапиром. Окончательно мнение о близости сирен
и хоботных утвердилось после открытия в одних и тех же эоценовых
отложениях Египта примитивных представителей обеих этих групп, сходст-
во которых по ряду важных признаков уже было бесспорно (Andrews,
1906). Это привело к тому, что Sirenia и Proboscidea были включены
вместе с Hyracotheria (даманы) и вымершими Embritopoda в группу
Subungulata в ранге отряда (Weber, 1928) или только с даманами в над-
отряд Paenungulata (Simpson, 1945; и многие за ним). В последней пред-
ложенной системе млекопитающих (McKenna, 1975) хоботные и сирены
включены в особую группу (mirorder) Thethytheria, в которую входят
также вымершие бегемотообразные Desmostylia, раньше рассматриваемые
в качестве подотряда самих сирен (Simpson, 1932), а теперь в ранге
самостоятельного отряда (Reinhart, 1953). Даманы в этой системе сбли-
жены с непарнокопытными и помещены вместе с ними и наиболее прими-
тивными первичными копытными Condylarthra в миротряд Phenacodonta.
Во всех случаях принимается, что общих предков сирен, хоботных и тех
или иных названных выше более мелких групп вымерших млекопитающих
следует искать среди кондиляртр в доэоценовое время (Domning et al.,
1982).
Все эти изменения в системе и в рангах отдельных групп вызваны
в основном различием в подходах исследователей к построению системы,
эволюционным у Симпсона и откровенно кладистским у Маккенны.
Исследование же истинных взаимоотношений отдельных групп примитив-
ных копытных, или «субунгулят», на основе новых и переописания
старых материалов все еще является насущной задачей териологии
(Simpson, 1932). Пока же система остается неустойчивой. Так, Moeri-
therium, примитивное хоботное из эоцена Северной Африки, некоторыми
авторами сближается с сиренами даже больше, чем с хоботными (Tobien,
1971), а другими выделяется в самостоятельный отряд Moeritherioidea
(Copens, Beden, 1978). Есть идея, что именно от них могли произойти
десмостилии (Domning, цит. по: Coppens, Beden, 1978). Другие авторы,
указывая на большую близость черепа меритерия к таковому сирен, чем
настоящих хоботных Elephantoidea, подчеркивают, что это сходство
12
основано на симплезиоморфных признаках или вызвано конвергенцией
и общим сходством сирен и хоботных и, следовательно, не может быть
основанием для отрыва меритерия от более поздних хоботных (Tassy,
1979). Все это, однако, не изменяет общего представления, что какие-то
кондиляртры Старого Света (место происхождения не оспаривается
никем'), возможно обитавшие на границе воды и суши, дали начало как
сухопутной линии развития, приведшей к хоботным, так и амфибиотиче-
ской (меритерии и особенно десмостилии) и отдельно сугубо водной
(сирены) (Reinhart, 1959).
Среди общих признаков меритериев и примитивных сирен один (нали-
чие билофодонтных зубов) имеет апоморфный характер, что сближает
их еще с одной небольшой группой вымерших млекопитающих
Barytheria, возможно также имеющей с ними общих предков (Savage,
1977). Главная и практически единственная аутапоморфная черта сирен,
отделяющая их от других млекопитающих, включая меритерия,— это раз-
витие у них пахиостоза, т. е. утолщения и уплотнения костей, в том числе
и длинных трубчатых, за счет утери гаверсовых каналов и полостей для
костного мозга, что, как правило, увязывается всеми с переходом к вод-
ному образу жизни. При этом существует, мягко говоря, тенденция у ряда
современных исследователей (Savage, 1977) оставлять в стороне адап-
тивные черты сирен, более четко отличающие их от наземных родичей
(начавшаяся редукция задних и модификация передних конечностей,
смещение вверх ноздрей и т. д.), поскольку аналогичные изменения
происходят и у других водных млекопитающих. Односторонность такого
подхода кладистов, очевидно, должна быть преодолена путем изучения
собственно сиреновой специфики этих процессов и осознания принци-
пиального равноправия апоморфных признаков, с одной стороны, и так
называемых адаптивных — с другой, в систематике. Собственно, развитие
пахиостоза тоже вызвано приспособлением к среде. Задача же исследова-
теля — как раз установить, на каком уровне и в силу каких причин при-
обретение того или иного признака было ведущим в эволюции изучаемой
группы животных. Причем в разных ситуациях одно и то же морфологи-
ческое изменение может определять различия на высоком таксономиче-
ском уровне и на уровне родов, видов. Это, видимо, относится и к вент-
ральному отклонению ростральной части черепа и симфиза нижней
челюсти и соотносительному развитию носовых и лобных костей и т. д.,
т. е. к признакам, по традиции используемым в систематике сирен.
Пахиостоз у сирен захватывает в первую очередь длинные кости
конечностей, ребра, отдельные кости черепа, особенно скуловую дугу,
надглазничный отросток, предчелюстные кости (см. рентгенограммы
в Kaiser, 1974). Как критерий, помогающий установить даже по фраг-
ментам наличие сирен среди ископаемых остатков, он очень полезен.
Однако его абсолютизация в какой-то мере препятствует пониманию ста-
новления сирен как водных животных, механически исключая формы без
пахиостоза из числа потенциальных переходных форм, снимая остроту
вопроса о становлении самого пахиостоза.
В настоящее время существует лишь два рода сирен: Trichechus
с тремя видами, находившийся, видимо, в стадии биологического про-
гресса до встречи с человеком, и Dugong с одним видом (Bertram, Bert-
ram, 1973). Еще один род Hydrodamalis с одним видом был истреблен
J В последнее время с нахождением достоверных остатков сирен эоценового возра-
ста, с одной стороны, на Яве, а с другой — в Северной Америке (Флорида, Алаба-
ма) заставляют некоторых авторов (Reinhardt, 1976) склониться к мнению, что
нет позитивных доказательств средиземноморского происхождения сирен.
13
человеком к середине XVIII столетия. Все это специализированные
формы, представляющие крайние ветви филогенетического дерева сирен.
В ископаемом состоянии известно еще более двух десятков родов (разные
авторы насчитывают от 26 до 32 родов и до 60 видов2), большей частые
принадлежащих к обширному семейству Dugongidae, представители кото-
рого доминировали в прибрежных участках морей и океанов со среднего
эоцена до плейстоцена. Расцвета сирены достигли в миоцене, когда число
их родов насчитывало около 10—12 (Domning, 1978b).
Система сирен в настоящее время выглядит так:
ОТРЯД SIRENIA ILLIGER, 1811
СЕМ. PRORASTOMIDAE СОРЕ, 1889. С. эоцен, Большие Антильские острова
Prorastomus Owen, 1855 (типовой вид: Р. sirenoides Owen, 1855).
Р. sirenoides Owen, 1855; с. эоцен (лютеций), Ямайка.
СЕМ. PROTOSIRENIDAE SICKENBERG, 1934. Н. эоцен, Ю. Америка;
с. эоцен, Ю. Азия, С. Америка; с. и в. эоцен, С. Африка, Европа.
Protosiren Abel, 1907 (типовой вид: Р. fraasi Abel, 1907); н.-в. эоцен, Ю. и С. Аме-
рика, Европа, С. Африка, Ю. Азия.
Р. fraasi Abel, 1907; н. эоцен, Аргентина; с. эоцен, Венгрия, Индия; с. и в_
эоцен, Египет.
? Р. minima (Desmarest, 1822: Hyppopotamus); с.-в. эоцен, Франция.
Остатки Protosiren sp. описаны (Domning et al., 1982) из среднего эоцена США
(Флорида, Сев. Каролина).
СЕМ. DUGONGIDAE GRAY, 1821. С. эоцен —ныне; С. Африка, Европа,.
Азия, Австралия, С. Америка.
ПОДСЕМ. HALITHERIINAE CARUS, 1868 sensu Abel, 1913. С. эоцен — плиоцен, Евро-
па; с. эоцен — и. миоцен, С. Африка; с. олигоцен, С. Америка.
Halitherium Капр, 1838 (включая Manatherium Hartlaub, 1886). ? эоцен, Европа;
олигоцен, Европа, ? Ц. Америка; н. миоцен, Европа, С. Америка (типовой вид:.
Hippopotamus dubius Cuvier, 1824).
Н. dubius Cuvier (1824: Hippopotamus), включая H. cuvieri Delfortrie, 1872;.
олигоцен — миоцен, Франция.
Н. schinzi Каир, 1838; н. олигоцен, Бельгия; с. олигоцен, ФРГ.
Н. schinzi f. typica Каир, 1838; с. олигоцен, ФРГ, Швейцария.
Н. schinzi f. delheidi (тип Manatherium delheidi Hartlaub. 1886; включая Meta-
xytherium pergense Toula, 1899; M. guettardi Blainville, 1843; H. uytterhoeveni
Lefevre, 1889); н. олигоцен, Бельгия, Франция.
?H. antillense Matthew, 1916; с. олигоцен, Пуэрто-Рико.
Н. alleni Simpson, 1932 (включая Н. antiquum Allen, 1925=Manatus inornatus
Leidy, 1873 и M. antiquus Leidy, 1860); миоцен, США (Ю. Каролина).
Н. canhami Flower, 1874; олигоцен, Англия.
Н. fossile (Cuvier, 1823: Manatus); миоцен, Франция.
Н. christoli Fitzinger, 1842; с. олигоцен (стампий), Австрия, ФРГ.
Н. bellunense Zigno, 1875; н. миоцен (бурдигал), Италия.
Н. olseni Reinhart, 1976; с.-в. миоцен, США (Флорида).
Из с.-в. эоцена Испании описан «Н» sp. (см. Domning et al., 1982), из эоцена-
Венгрии несколько зубов, отнесенных к «Halitherium subappeninum» (Sickenberg,
1934); остатки сирен, возможно близких к этому роду (cf. Halitherium), отмечены
в районе Индопацифики (Domning, 1978b).
Eotheroides Palmer, 1899 (включая Eosiren Andrews, 1902; Archaeosiren Abel,.
1913; Masrisiren Kretzoi, 1941). С.-в. эоцен, С. Африка, Европа (типовой видг.
Eotherium aegyptiacum Owen, 1875).
2 Не все эти роды основаны на хорошем материале, возможно, часть их в дальней-
шем будет признана невалидными. Сравнение их представляет обычные трудности:
ввиду фрагментарности и, следовательно, частой несопоставимости остатков, от-
сутствия серийного материала. Большинство родов монотипичны, другие с боль-
шим числом видов,— скорее всего, являются сборными.
14
E. aegyptiacum (Owen, 1875: Eotherium; включая «Manatus» coulombi Filhol,
1878); с. эоцен, Египет.
E. abeli Sickenberg, 1934; в. эоцен, Египет.
E. libycum (Andrews, 1902: тип рода Eosiren); в. эоцен, Египет.
Е. stromeri (Abel, 1913: тип рода Archaeosiren); в. эоцен, Египет.
Е. majus (Zdansky, 1938: Eotherium); с. эоцен, Египет.
В отложениях с. и в. эоцена Румынии, Венгрии, Франции описаны остатки,
обозначенные как Е. sp. (см.: (Domning et al., 1982)); в с. эоцене (лютеций) Ливии
обнаружены несколько черепов и части посткраниального скелета, возможно, ново-
го рода, близкого к Eotheroides (см.: (Savage, 1977)).
Prototherium Zigno, 1887 (включая Mesosiren Abel, 1906; Paraliosiren Abel, 1906);
в. олигоцен, Европа, ? Африка (типовой вид: Halitherium veronense Zigno, 1875).
P. veronense (Zigno, 1875: Halitherium); в. эоцен, Италия (включая Protosiren
dolloi Abel, 1904, тип рода Mesosiren Abel, 1906; Paraliosiren suessi Abel, 1906).
Фрагменты челюстей с зубами, сходными с Prototherium, найдены в отложениях
возможно эоценового возраста в Сомали (Savage, 1977).
Metaxytherium Christol, 1840 (включая Felsinotherium Capellini, 1871; Halysiren
Kretzoi, 1941); н. миоцен — плиоцен, Европа; с. миоцен — плиоцен С. Америка;
? н. миоцен — плиоцен, Сев. Африка (типовой вид: Hippopotamus medius Desma-
rest, 1822), миоцен, Ю. Азия.
М. medius (Desmarest, 1822: Hippopotamus;=M. cuvieri Christol, 1840); с. мио-
цен (виндобон), Франция, Швейцария.
М. krahuletzi Deperet, 1895; н. миоцен (бурдигал), Австрия, Швейцария.
М. lovisati Capellini, 1886; в. миоцен, Бельгия.
М. meyeri Abel, 1904; н. миоцен (бурдигал), Австрия.
М. beaumonti Christol, 1840; н. миоцен (бурдигал), Франция.
М. calvertense Kellogg, 1966; с. миоцен, США (Мэриленд, Виргиния).
М. riveroi Varona, 1972; с. миоцен, Куба.
М. forestii Capellini, 1865 (тип рода Felsinotherium Capellini, 1871); плиоцен,
Италия.
М. serresi (Gervais, 1859: Halitherium); н. плиоцен (плезаний), Франция; плио-
цен, Марокко (М. cf. serresi).
М. floridanum Hay, 1922; плиоцен, США (Флорида).
М. ossivallense (Simpson, 1932: Felsinotherium); плиоцен, США (Флорида).
М. subappeninum (Bruno, 1839: Chirotherium); плиоцен, Италия.
М. gastaldi (Zigno, 1878: Felsinotherium); плиоцен, Италия.
Остатки, неопределимые далее как М. sp., отмечены в н. миоцене Ливии и плио-
цене Марокко (см.: (Domning, 1978b), в миоцене Индии (Savage et al., 1975), в с.-в.
миоцене Аргентины (Reinhart, 1976); все плиоценовые формы рода часто выделя-
лись в род Felsinotherium (Simpson, 1932, и др.)).
Halianassa Studer, 1887 ex Meyer, 1838; н. миоцен, 3. Европа (тип рода: Н. stu-
deri Studer, 1887 ex Meyer, 1838: Manatus studeri, nomen nudum).
H. studeri Meyer, 1838; н. миоцен, Австрия, ФРГ, Швейцария.
Н. collinii Meyer, 1838; олигоцен, ФРГ, Швейцария.
Thalattosiren Sickenberg, 1928; с. миоцен, Европа (типовой вид: Metaxytherium
petersi Abel, 1904).
Th. petersi (Abel, 1904); с. миоцен (тортон), Австрия.
Hesperosiren Simpson, 1932; с. миоцен, С. Америка (типовой вид: H. crataegensis
Simpson, 1923).
H. crataegensis Simpson, 1932; с. миоцен, США (Флорида).
Dioplotherium Cope, 1883; ?н.— с. миоцен, С. Америка (типовой вид: D. manigaul-
ti Соре, 1883).
D. manigaulti Соре, 1883; н. или с. миоцен, США (Ю. Каролина).
D. allisoni Kilmer, 1965: Halianassa (?); с. миоцен, США (Калифорния).
Caribosiren Reinhart, 1959; с. олигоцен, Б. Антильские о-ва (типовой вид: С. tur-
ineri Reinhart, 1959).
15
C. turner! Reinhart, 1959; с. олигоцен, Пуэрто-Рико.
Prohalicore Flot, 1886; с. миоцен, Европа (типовой вид: Р. dubaleni Flot, 1886).
Р. dubaleni Flot, 1886; с. миоцен, Франция. ч
Rytiodus Lartet, 1866 (тип подсемейства Rytiodinae Abel, 1919); н. миоцен, Евро- |
па, С. Америка (типовой вид: R. capgrandi Lartet, 1866). й
R. capgrandi Lartet, 1866; в. олигоцен (н. миоцен: аквитан), Франция. *
Предполагается, что из нижнемиоценовых отложений Сев. Африки (Ливия) бу-
дет описан новый вид (см.: (Savage, 1977)).
Anomotherium Siegfried, 1965; в. олигоцен, Европа (типовой вид: A. langewiesc-
hi Siegfried, 1965).
A. langewieschi Siegfried, 1965; в. олигоцен, ФРГ.
ПОДСЕМ. MIOSIRENINAE ABEL, 1919; плиоцен, Европа.
Miosiren Dollo, 1890; и. плиоцен, Европа (типовой вид: М. kocki Dollo, 1890).
М. kocki Dollo, 1890; н. плиоцен (понт). Бельгия.
ПОДСЕМ. DUGONGINAE GRAY, 1821 sensu Simpson, 1932; голоцен — ныне; Индопа-
цифика.
Dugong Lacepede, 1799 (=Halicore liliger, 1811); современный, Индопацифика
(типовой вид: Trichechus dugon Miiller, 1775).
D. dugon (Muller, 1775: Trichechus); современный, Индопацифика.
В ископаемом состоянии известны лишь фрагменты из голоцена Индопацифи-
ки (Domning, 1978b).
ПОДСЕМ. HYDRODAMALINAE PALMER, 1895 sensu Simpson, 1932. С. миоцен — ныне
(уничтожены в XVIII в.); побережья северной части Тихого океана.
Dusisiren Domning, 1978; с. миоцен — и. плиоцен, С. Америка (типовой вид: Ме-
taxytherium jordani Kellogg, 1925).
D. jordani (Kellogg, 1925); в. миоцен—н. плиоцен, CBIA (Калифорния) (вклю-
чает Halianassa vanderhoefi Reinhart, 1959).
D. reinharti Domning, 1978; с. миоцен, США (Калифорния).
В миоценовых отложениях Калифорнии указаны еще два неадекватно извест-
ных вида, обозначенные как Dusisiren sp. В и sp. D. (см.: (Domning, 1978с)).
Hydrodamalis Retzius, 1794 (=Rytina или Rhytina liliger, 1811); в. плиоцен —
современный, С. Америка, С.-В. Азия (типовой вид: Manati gigas Zimmermann,
1780).
Н. cuestae Domning, 1978; с.-в. плиоцен, США (Калифорния).
Н. gigas (Zimmermann, 1780: Rytina) (=Rytina stelleri Retzius, 1794); поздний
плейстоцен — современный (уничтожен), Командорские острова, Алеутские
острова.
В Японии описан Н. sp. (Shikama, Domning, 1970) из верхнего плиоцена. Очевид-
но, в это время Hydrodamalis был распространен по всему побережью северной час-
ти Тихого океана от Калифорнии до Японии.
DUGONGIDAE INCERTAE SEDIS.
Paralitherium Kordos, 1977; в. эоцен, Европа (типовой вид: Р. tarkanyense Kordos,
1977).
Р. tarkanyense Kordos, 1977; в. эоцен, Венгрия.
Anisosiren Kordos, 1979; с. эоцен, Европа (типовой вид: A. pannonica Kordos,
1977).
A. pannonica Kordos, 1977; с. эоцен, Венгрии.
Sirenavus Kretzoi, 1941; с. эоцен, Европа (типовой вид: S. hungaricus Kretzoi,
1941).
S. hungaricus Kretzoi, 1941; с. эоцен, Венгрия.
СЕМ. TRI CHE CHI D AE GILL, 1872; миоцен — ныне; Ю. Америка, побережья
Мексиканского залива и Карибского моря, 3. Африка.
Sirenotherium Paula Couto, 1967 (условно отнесен к этому семейству); н. мио-
цен, IO. Америка (типовой вид: S. pirabensis Paula Couto, 1967).
S. pirabensis Paula Couto, 1967: н. миоцен, Бразилия.
Potamosiren Reinhart, 1951; с. миоцен, Ю. Америка (типовой вид: Р. magd'alen-
sis Reinhart, 1951).
16
P. magdalensis Reinhart, 1951; с. миоцен, Колумбия.
Ribodon Ameghino, 1883; в. миоцен — н. плиоцен, Ю. и С. Америка (типовой'
вид; R. limbatus Ameghino, 1883).
R. limbatus Ameghino, 1883; в. миоцен, Аргентина.
В нижнеплиоценовых отложениях США (Сев. Каролина) отмечен R. sp. (Dom-
ning, 1982).
Trichechus Linnaeus, 1758 (=Manatus Briinnish, 1772); плиоцен — ныне, Ю. и С.
Америка, 3. Африка (типовой вид: Т. manatus Linnaeus, 1758).
Т. m. manatus Linnaeus, 1758; в. плиоцен — плейстоцен, США (Флорида, Сев. Ка-
ролина), ныне, побережья Мексиканского залива, Карибского моря, северо-
востока Ю. Америки.
Т. manatus L., 1758; Карибское море, побережье северо-восточной части.
Ю. Америки 3.
Т. m. latirostris (Harlan, 1824: Manatus latirostris); США (Флорида).
Т. senegalensis Link, 1795; ныне, побережье и реки 3. Африки от Сенегала до*
Анголы.
Т. inunguis (Natterer, 1883: Manatus); ныне, реки Ю. Америки (бассейн Ама-
зонки и Ориноко).
SIRENIA INCERTAE SEDIS.
Ishatherium Sahni et Kumar, 1980; ?н. эоцен (с. эоцен), Ю. Азия (типовой вид:
I. subathuensis Sahni et Kumar, 1980).
I. subathuensis Sahni et Kumar, 1980; ?н. эоцен (с. эоцен), Индия.
История ранних сирен была связана с Тетисом — морем, простиравшим-
ся в мезозое и начале кайнозоя в широтном направлении от западных по-
бережий Атлантического океана через Южную Европу и Северную Афри-
ку до Юго-Восточной Азии. В палеогене он представлял самый центр мор-
ских тропиков того времени. В его прибрежных мелководьях с хорошей
инсоляцией еще с мела произрастали морские покрытосеменные (морские
травы, или «seagrasses») из семейств водокрасовых (Hydrocharitaceae) и
рдестовых (Potamogetonaceae). Приспособление к питанию этими растени-
ями (отнюдь не водорослями, как иногда указывается в литературе) в
настоящее время рассматривается как исходное в становлении сирен как
группы (Domning, 1977, 1978 Ь, с, 1982; Domning et al., 1982). По сущест-
ву, современные сирены остались верными этой экологической нише, а их
нынешнее географическое распространение также не выходит за пределы
древнего Тетиса. Не изменили они и тропикам. Тем не менее эта внешняя
неизменность условий существования не помешала сиренам пройти дли-
тельный путь эволюционного развития, дифференциации их в разных на-
правлениях, выработки ряда приспособлений, смысл которых не всегда-
еще ясен.
Самые древние сирены отмечены в нижнеэоценовых отложениях. Их
фрагментарные остатки по сути ничего не говорят ни об облике этих жи-
вотных, ни о степени приспособления к водной среде. Хотя по находкам в
Венгрии (Kretzoi, 1953) можно думать, что их ребра были охвачены пахи-
остозом, т. е. эти сирены уже могли быть облигатными гидробионтами,,
однако в нижнеэоценовых (может быть среднеэоценовых) отложениях
Индии открыт Ishatherium (Sahni, Kumar, 1980), относимый к сиренам,,
кости еще недостаточно плотны, а строение посткраниального скелета
якобы (доказательства не приведены) свидетельствует о том, что это было
четвероногое животное, которое равно могло жить на суше и в воде. Все-
это вызывает сомнения, как и само отнесение ишатерия к сиренам (Dom-
ning et al., 1982).
Первые достаточно полные и не вызывающие сомнений находки сирен
3 Существование подвидов Trichechus manatus вызывает сомнение (Moore, 1951а;
Hartman, 1971).
17
датированы средним эоценом, причем они говорят уже и о значительном
их разнообразии, и о немалой предшествующей истории их развития. Пер-
вая генерация ранних сирен представлена пока лишь одной среднеэоцено-
вой формой (Prorastomus sirenoides), которая выделена в особое семейство.
Предполагается, что нахождение ее (Ямайка) вдали от предполагаемого
центра происхождения сирен в районе современного Средиземноморья го-
ворит уже о закате прорастомид, оттеснении их последних представителей
на окраину ареала более прогрессивной группой сирен протосиренид. К со-
жалению, прорастомус известен лишь по черепу и одиночному атланту и,
следовательно, нельзя сказать, порвал ли он полностью с сушей или был
амфибионтом на манер карликового бегемота или тапира. В то же время он
имеет ряд очень примитивных признаков, особенно в слуховой области че-
репа и в строении атланта, которые вызывают мысль о связях его с конди-
ляртрами (Savage, 1977). Как строение зубной системы (полная эутерие-
вая зубная формула з~{ 5~3 ' J мелкие, одинаковые по размеру резцы,
простые брадиодонтные коренные с двумя поперечными лофами; макси-
мально возможное число функционирующих предкоренных), так и прямые
челюсти — все говорит об отсутствии какой-либо специализации в характе-
ре питания сравнительно с ближайшими сухопутными родичами.
В свое время к прорастомидам условно была отнесена еще одна форма
ранних сирен Sirenavus hungaricus из среднего эоцена Венгрии (Kretzoi,
1941). Однако переизучение его черепа (Kordos, 1982) показало, что сим-
физарная часть его нижней челюсти, видимо, была уже в какой-то мере
отогнута вниз. По мнению автора, это говорит о специализации челюстного
аппарата сиренавуса и невозможности, следовательно, помещать его в одно
семейство с прорастомусом. Хотя Кордош и не предлагает конкретных ре-
комендаций относительно сиренавуса, само открытие эоценовых сирен с
возможной донной специализацией представляется важным, поскольку от-
сутствие их при наличии подходящих местообитаний уже вызвало удивле-
ние (Domning et al., 1982).
Вторую генерацию эоценовых сирен представляют Protosirenidae (см.
филогенетическую схему на рис. 2). При этом один и тот же род Protosi-
ren обнаружен начиная с раннего эоцена в Аргентине (Sereno, 1982) по
поздний эоцен Франции (Sickenberg, 1934) и Египта (Andrews, 1906; Abel,
1907). Первыми среди сирен протосирениды достигли пантетисного рас-
пространения. Их остатки также обнаружены в среднеэоценовых отложе-
ниях Флориды и Северной Каролины (Domning et al., 1982) и Индии (Sah-
ni, Mishra, 1975). Есть мнение, что позднемиоценовый Miosiren из Бельгии
был последним представителем этого семейства (Domning et al., 1982),
хотя обычно его рассматривают в составе особого подсемейства в составе
другого семейства дюгонид (Simpson, 1932; 1945; Reinhart, 1959). Протоси-
рениды уже приобрели дефинитивный сиреновый облик, за исключением,
может быть, сохранения еще функционирующих в качестве ластов задних
конечностей. Во всяком случае, их таз (рис. 3) хотя и уменьшен в разме-
рах, но еще мало отличается по форме от таза наземных млекопитающих,
у него сохраняется глубокая ацетабулярная яма, так же как и сам скелет
свободной конечности. Однако в черепе уже произошло увеличение слож-
ной по составу (барабанная и каменистая кости и сосцевидная часть че-
шуйчатой кости), но сросшейся в единое образование околоушной кости
(os perioticum), которая потеряла полностью твердое сочленение с окру-
жающими костями черепа; крыло-клиновидный канал утрачен. В то же
время протосирены сохранили примитивную полную зубную систему.
Лишь первые верхние резцы слегка увеличены, а ростральная часть чере-
па слегка отогнута вниз.
18
о
о
&
В ходе своей весьма умеренной радиации, не выходящей за рамки одно-
го рода, протосирениды, видимо, дали начало двум другим семействам сирен,
с одной стороны, путем изоляции во внутренних водах Южной Америки
Trichechidae, а с другой — оставшимся в открытых водах Тетиса Dugon-
gidae. Именно последние полностью вытеснили к концу эоцена протосире-
нид и заняли доминирующее положение в типичной сиреновой экологиче-
ской нише — богатых водной растительностью морских прибрежных
мелководьях, дав третью генерацию сирен, которая не только сохранила
пантетисное распространение, но и проникла в акваторию Тихого океана.
Примитивные представители дюгонид, такие, как Eotheroides и Proto-
therium, из среднего и позднего эоцена Северной Африки и Европы еще
сохраняют черты, сближающие их с прорастомидами (раньше эотероидеса
относили к ним, см. Simpson, 1932; Reinhart, 1959) и протосиренидами.
В частности, у них всех полная эутериевая зубная система с тремя резца-
ми, клыком, пятью предкоренными и тремя коренными (морфологическую
характеристику Eotheroides см. у Domning, 1978). У всех последующих
послеэоценовых дюгонид число зубов начинает сокращаться, исчезают все
или часть резцов, клыки и предкоренные, в некоторых случаях меняется
даже структура коренных. Однако для многих форм дюгонид характерны
бивнеобразно увеличенные первые резцы (у дюгоня они хорошо выраже-
ны у самцов, а у самок маленькие, не прорезающиеся через десну). Этот
Рис. 3. Два пути редукции таза у сирен (вид слева)
а — Protosiren, с. эоцен; б — Eotheroides, с. эоцен; в — Halitherium,"’миоцен; г — Halianassa;
плиоцен; д — Dugong dugon, современный; е — Hydrodamalis gigas, современный; ж — Trichechus
manatus, современный (а, в, г, ж по; (Abel, 1904); б — по: (Abel, 1929); д, е — по фотографии в:
(Kaiser, 1974))
20
процесс увеличения резцов у ряда сирен иногда сравнивают с появлением
бивней у хоботных, говоря о сходстве в этом отношении обеих групп. Одна-
ко у хоботных бивни образуются из второго верхнего резца или, как счи-
тают некоторые авторы, даже из третьего, а у сирен — из первого, что хоро-
шо видно у примитивных дюгонид, у которых, как у Eotheroides, при со-
хранении всех резцов i1 уже слегка увеличен. Степень утраты задних
конечностей у примитивных дюгонид не ясна. Таз и у прототерия, и у
эотероидеса мало отличается от протосиренового, и обычно в диагнозах
(Simpson, 1932; Reinhart, 1959) указывают, что у этих форм задние конеч-
ности могли функционировать как ласты. Однако изучение бедренной ко-
сти Eotheroides показало, что скорее всего рудиментарные кости свободной
конечности у него лежали в толще мускулатуры и, таким образом, снару-
жи задние конечности не были видны (Siegfried, 1967).
Семейство Dugonidae включает подавляющее большинство послеэоце-
йовых родов сирен. Несмотря на неоднократные попытки упорядочить их
систематику (Abel, 1919, 1928; Simpson, 1932, 1945; Kretzoi, 1941; Rein-
hart, 1959), она все еще остается весьма условной. Поэтому ряд авторов
(Domning, 1978с) подчеркнуто придерживаются максимально возможной
в этих условиях консервативной точки зрения, аргументированной еще
Дж. Г. Симпсоном, что в настоящее время следует большинство форм дюго-
нид без ясно выраженных черт специализации рассматривать в составе од-
ного подсемейства Halitheriinae (см. схему филогении у Dechaseaux, 1958,
с. 359), существовавшего со среднего эоцена по плиоцен. И хотя уже сам
Симпсон говорил, что это — сборная группа, тем не менее выделить в ней
какие-либо четкие линии развития пока не удается. Так, серьезные возра-
жения (Domning, 1978с) вызвали попытки выделить из халитериин само-
стоятельное подсемейство Halianassiinae на основании соотносительного
развития носовых и лобных костей (Reinhart, 1959). В дальнейшем и сам
этот автор отказался от своего предложения (Reinhart, 1976) и даже при-
знал род Halianassa синонимом Metaxytherium.
Структурно Eotheroides может рассматриваться как генерализованная
форма, которая могла дать начало всем другим дюгонидам (Domning,
1978b). Как уже отмечалось, у него сохраняется максимально полная зуб-
ная система с пятью предкоренными зубами, из которых dp5, кажется, не
имел заместителя (Sickenberg, 1934), коронки зубов типичные для палео-
геновых дюгонид (на верхних коренных два поперечных гребня: прото-
лоф, состоящий из протокона, протоконула и паракона, и металоф из гипо-
кона, метаконула и метакона, разделенных свободной долинкой). Много
примитивных черт и в черепе: умеренно отогнутый вниз ростр, расширен-
ные надглазничные отростки лобных костей, хорошо развитые носовые и
слезные кости, скуловая кость расширена позади орбиты, имеется хорошо
развитый ламбдовидный гребень на чешуйчатой кости, латеро-дорсальные
фланги боковых затылочных костей не расширены, тело нижней челюсти
узкое, впереди от большого foramen mentale имеется несколько дополни-
тельных отверстий.
В ходе эволюции халитериины претерпевают ряд прогрессирующих
морфологических изменений, уровень развития которых последовательно
отражают такие обширные рода, как Halitherium (олигоцен — н. миоцен) и
Metaxytherium (миоцен — плиоцен), с максимально возможным широким
географическим распространением каждый. Среди этих изменений можно
назвать (Abel, 1904; Simpson, 1932): исчезновение всех или большей части
резцов и сокращение числа коренных зубов (от 4pm+3m у Halitherium, до
lpm+3m у Metaxytherium); постепенное превращение билофодонтных
зубов в бунодонтные, бугорчатые в основном за счет смещения вперед ме-
таконула и появления добавочных бугорков (у Halitherium этот процесс
21
только начинается); увеличение i1 в виде своеобразного бивня (tusk); рас-
ширение крыши черепа, так что височные гребни становятся более парал-
лельными; укорочение черепа и расширение переднего носового отвер-
стия; редукция носовых и слезных костей; уменьшение надглазнично-
го отростка (у Halitherium еще широкий, как у Eotheroides, а у Metaxy-
therium уже узкий); смещение вперед вентрального расширения скуловой
кости; увеличивается и все более отгибается вниз (придонная специали-
зация) ростр; верхнезатылочная кость все больше начинает вклиниваться
вентрально между боковыми затылочными костями, приближаясь к боль-
шому затылочному отверстию; укорачивается и становится выше нижняя
челюсть, при этом все более расширяется ее симфиз. Намечается и ряд тен-
денций в изменении посткраниального скелета: уменьшается число груд-
ных позвонков, обладающих полуфасетками для сочленения с головками
ребер, занимающими, таким образом, интерцентральное (между телами
двух смежных позвонков) положение (у Halitherium 12 позвонков с полу-
фасетками, 3 с двумя фасетками и 1 с одной; у Metaxytherium лишь 9 по-
звонков с полуфасетками); расширяется предостная ямка лопатки, стано-
вится выше ее ость, а акромиальный отросток становится более выступаю-
щим: расширяется гленоидная впадина; на плечевой кости малый бугорок
становится меньше, а большой увеличивается, при этом их оси более дивер-
гируют; увеличивается дельтовидный гребень плечевой кости; локтевая и
лучевая кости все более перемещаются в одну плоскость; удлиняется IV
melacarpale относительно длины лучевой кости; все более редуцируется
лобковая кость, а седалищная удлиняется и становится уже, теряется аце-
табулярная яма (см. рис. 3). Все эти тенденции по-разному реализуются
у разных форм халитериин. Так, у Rytiodus развиваются большие упло-
щенные бивнеобразные резцы, а у Thalattosiren и Garibosiren они вообще
утрачены. Сильно варьирует и степень вентрального отклонения ростра и
симфиза нижней челюсти. Возможно, что эта черта вообще очень пластич-
на и сама по себе не может говорить о тесном или, наоборот, удаленном
систематическом сходстве (Domning, 1982).
Единственным представителем аберрантного подсемейства дюгонид
Miosireninae является Miosiren kocki с шестью коренными зубами, из ко-
торых три первые одинаковы по размерам и просты по форме, а последую-
щие (т1-3) лишены добавочных бугорков, но имеют передний и задний
шельфы, причем последний т3 редуцирован до простого вертикального
столбика. Полагают, что миосирен был моллюскоядом (Domning, 1978b).
Его корни неясны. Это потомок или самых примитивных халитериин или,
может быть, даже протосиренид (Abel, 1919).
Последние Halitheriinae датированы плиоценом. Их вымирание, как
сейчас представляют (Domning, 1982) по крайней мере в западной части
ареала (современный Карийский бассейн), было вызвано их неконкуренто-
способностыо по сравнению с вышедшими из изоляции сиренами другого
типа (Trichechidae). Эвстатическое понижение уровня мирового океана
в плиоцене вызвало значительное увеличение площади суши и восстанов-
ление системы стоков, что повлекло за собой увеличение эрозии. Огромное
количество взвешенных частиц ила и песка хлынуло в океан. Питающиеся
придонной растительностью брахиодонтные дюгониды при этом, возмож-
но, испытали селективное давление на выработку более износоустойчивых
зубов. Потенциально поздние халитериины, видимо, были способны выра-
ботать гипсодонтию. Так, у Dioplotherium allisoni из миоцена Калифор-
нии на уже крайне стертом т3 все еще не был закрыт задний корень (Dom-
ning, 1978с). Однако проникновение в зону их обитания более эврибионг-
ных ламантинов с уже сложившейся системой «функциональной гипсо-
донтии» (см. ниже) поставил поздних халитериин в явно невыгодное nt>
22
ложение: им нужно было догонять конкурентов по уровню приспособления
к новым условиям. Стройность этой гипотезы, однако, нарушает вымира-
ние халитериин в центральной части их ареала, приходившейся на терри-
торию Средиземноморья, Южную Европу и Северную Африку, где не
было конкурентов в лице ламантинов.
Лишь в относительно изолированных водах Индопацифики (восточная
часть ареала, см. рис. 2) особая ветвь дюгонид, представленная собственно
дюгонем, развила настоящую гипсодонтию: на сохраняющихся обычно двух
коренных зубах (т2-3) корни на каждом зубе сливаются в один большой,
широко открытый, что обусловливает их постоянный рост.
Современный Dugong неизвестен в ископаемом состоянии, если не
считать фрагментов из голоцена. Тем не менее ясно, что по крайней мере
с позднего эоцена в Индопацифике обитали какие-то сирены. Их остатки
(Reinhart, 1976) не имеют точной систематической принадлежности, даже
те, что получили свои названия (Indosiren javanensis Koenigswald, 1952 из
позднего миоцена Явы и Chronozoon australe de Vis, 1883, возможно, из
плиоцена Австралии). Однако, видимо, какие-то халитериины, близкие
к Halitherium, были здесь в раннем миоцене (Domning, 1978b), специали-
зируясь в питании на обширных «морских лугах» (seagrass meadows).
Естественно полагать, что дюгони — потомки халитериин, достигших
структурного уровня Halitherium или даже Metaxytherium и оказавшихся
в условиях изоляции в восточной части ареала позднетретичных дюгонид.
При этом дюгони, что несомненно свидетельствует о тесном их родстве
с халитериинами, продолжили дальше те морфологические преобразования,
основные тенденции которых характеризовали эволюцию последних
(Simpson, 1932). Так, еще более уменьшенным оказалось число зубов
(кроме бивнеобразного i *, сохраняются еще только два-три заднекорен-
ных) ; переднечелюстные кости еще более вздулись и вытянулись
назад и ростр стал еще больше загнутым вниз — отклонение до 70°
(соответственно переднее носовое отверстие оказалось смотрящим прямо
вверх); носовые кости или полностью исчезли, или сохраняются в виде
небольших рудиментов; череп в значительной степени укоротился; верх-
няя затылочная кость вошла в край большого затылочного отверстия;
нижняя челюсть стала очень короткой и высокой; желоб двуглавой мышцы
плечевой кости расширился и стал глубже; прошла дальнейшая редукция
таза, седалищная кость стала еще более длинной и узкой, палочковидной
(см. рис. 3), ацетабулярная яма исчезла; осталось всего семь грудных
позвонков с полуфасетками (у восьми позвонков имеются две фасетки,
одна для головки и вторая для бугорка, а у четырех — одна, т. е. головка
и бугорок соответствующего ребра слились).
В то же время современного дюгоня характеризует ряд черт собствен-
ной специализации, что позволяет выделять его в отдельное подсемейство
дюгонид — Dugonginae. В первую очередь следует назвать утерю им эмали
на коренных зубах (она еще имеется на непрорезавшихся через десну
зубах, см. Fernand, 1953), что приводит при первом же стирании к утрате
системы бугорков на зубе, весьма напоминающей таковую третичных
дюгонид. Это явление большинство авторов определяет как дегенерацию
зубов, отсутствующую у халитериин (Simpson, 1932). Однако у них раз-
виваются открытые корни и достигается постоянный рост зубов, что
трудно назвать дегенерацией. Как уже отмечалось, это связано с увели-
чением выноса в океан твердых частиц и увеличением стирания. При
этом подчеркивают, что состав пищи современных тропических и суб-
тропических сирен, скорее всего, оставался неизменным практически
с эоцена. Флора морских покрытосеменных (seagrass flora) существовала
уже в раннем мелу, и ее ареал был близок к современному. И хотя
23
эволюция этих растений почти неизвестна, однако можно думать, что
в ней не было ничего, что могло бы отразиться на сиренах (Domning, 1982;
Domning et al., 1982).
Среди других признаков, характерных для дюгоней, называют еще
сильный наклон внутрь processus retroversus чешуйчатой кости (Dom-
ning, 1978b), смещение вперед под орбиту расширения скуловой кости,,
укорочение лобной кости и обонятельной камеры, определенная дегене-
рация турбиналий, ведущая, например, к исчезновению эктотурбиналий,.
облегчение ушных костей, в частности менее вздутые tegmen tympani и
pars mastoideus; изогнутость лучевой кости, что привело к расширению
межкостного пространства в предплечье (Simpson, 1932); относительную
тонкость ребер, значительно менее охваченных пахиостозом (в них даже
есть губчатая кость), чем это свойственно всем другим, в том числе и
более древним примитивным сиренам. Одни авторы склонны не придавать,
этому большого значения (Simpson, 1932), другие, наоборот, считают, что
это дает основание вести линию развития дюгоней прямо с раннего эоцена
(Abel, 1904, 1919). Однако возможно, что такие ребра появились у дюго-
ней вторично и их предки имели плотные толстые ребра, как у всех
позднетретичных дюгонид.
Дюгони пережили эпоху оледенения, влияние которой в области индо-
пацифики, видимо, было минимальным. Однако и другая группа сирен
(Hydrodamalinae), часто называемая за рубежом тихоокеанскими или
северными морскими коровами, также сохранилась до современности
(уничтожена человеком), перенеся все события, связанные с оледенением,
в несравненно более суровых условиях севера Тихого океана. Более того,
вся их эволюция была подчинена приспособлению к этим условиям. Их
ареал, прежде (в позднем плиоцене) занимавший в виде узкой полосы
все побережье северной части Тихого океана от Калифорнии до Аляски
и Командорских островов и далее до Японии, сократился уже после оледе-
нения, что, несомненно, не было связано с ухудшением климата.
Во всяком случае, всего 20 тысяч лет назад морские коровы рода Hydro-
damalis жили в Калифорнии. Тем более сохранились они к концу своего-
существования в самой северной части своего ареала. История Hydroda-
malinae представляет одну из удивительных страниц в жизни всех сирен,
которые всегда были и остались до нашего времени, за этим единственным
исключением, теплолюбивыми животными, крайне болезненно отзываю-
щимися на понижение температуры воды.
Предками гидродамалин были позднетретичные сирены из подсемейства
Halitheriinae, которые сохраняли еще ряд черт, исходных для Dugonginae
и для Hydrodamalinae и в какой-то мере сохранившихся у последних.
Сюда относится относительно (по сравнению с дюгонем) небольшое рас-
пространение переднечелюстных костей назад, более длинное и узкое
переднее носовое отверстие, наличие еще носовых костей, более длинная
лобная кость, более выступающие и изолированные птеригоидные отрост-
ки, более крупные и расширенные восходящие отростки нёбных костей,
более удлиненная обонятельная камера и менее дегенерированные турби-
налии, более толстые ушные кости (os periotica) и широко охваченный
пахиостозом посткраниальный скелет. Все эти черты имелись у относи-
тельно поздних форм халитериин типа Metaxytherium (Simpson, 1932).
Таким образом, истоки и дюгонин и гидродамалин восходят примерно1
к одному структурному типу, но дальнейшие пути их морфологических
преобразований разошлись. У Hydrodamalinae развилась менее отогнутая
вниз ростральная часть черепа, более удлиненное и низкое тело нижней
челюсти с удлиненным симфизом, отклонение которого вниз не превышает
50°; носовые кости еще сохраняются, хотя и в рудиментарном виде, они
24
прикреплены с медиальной стороны передних отростков лобных костей;
затылочная часть черепа расширена в поперечном направлении, более
округла по очертанию (нет выемок в местах контакта верхней и боковых
затылочных костей), ламбдовидный гребень почти сглажен, foramen
magnum вытянуто поперек; хоаны расширены; полностью отсутствуют
функционирующие зубы. Таковы сирены рода Hydrodamalis (Simpson,
1932). Специфичен для этой группы и ряд других особенностей: чрезвы-
чайное утолщение эпидермиса и слоя подкожного жира, практически от-
сутствующего у тропических сирен, высокая модификация передней конеч-
ности, увеличение размеров тела и т. п. Все это заставляет придать им
статус самостоятельного подсемейства в пределах Dugongidae, но в то же
время значительно большая их близость к дюгоням (происхождение от
общих предков), чем к ламантинам, категорически отрицает возможность
рассматривать их на правах семейства (Weber, 1928; Гептнер, 1967; То-
милин, 1971; Соколов, 1979) без соответствующего повышения ранга
трихехид и всех сирен в целом.
Появлению Hydrodamalinae предшествовало (и предопределило)
важное событие в истории сирен. Сразу две их формы, одна — типичный
представитель поздних халитериин Dioplotherium и другая, рассматрива-
емая в настоящее время уже как первый гидродамалид Dusisiren rein-
harti, через центральноамериканский морской проход проникли примерно
18 миллионов лет назад (в раннем миоцене) в бассейн Тихого океана,
в район современной Калифорнии, очутившись там в определенной изоля-
ции (Domning, 1977а, 1978с); обратный обмен членов с карибскими сире-
нами был затруднен; дальнейшее продвижение на юг было ограничено так
называемым «мексиканским барьером», на север — климатическими фак-
торами. В эту эпоху на западном побережье Северной Америки господ-
ствовал тропический климат, изрезанная береговая линия способствовала
существованию множества мелководных заливов и лагун, относительно
обособленных от открытого моря, хорошо инсолированных,— идеального
места для роста seagrasses, обычной пищи сирен по всему их ареалу. Оче-
видно, эти морские покрытосеменные были достаточно разнообразны: одни
были сравнительно небольшими, прижатыми ко дну, другие более высоки-
ми, поднимающимися ближе к поверхности. В определенном количестве
юбитали здесь и бурые (типа ламинарий), и красные водоросли. Однако в
целом водоросли предпочитали воды у открытой береговой линии, более
каменистый грунт, а бурые водоросли еще и более низкие температуры.
Один из пришельцев — диоплотерий, достигавший около 3 м длины,
был приспособлен к питанию придонной растительностью. У него была
сильно отогнутая (до 70°) вниз ростральная часть черепа, вооруженная
парой несколько уплощенных бивнеобразных резцов, покрытых эмалью
с одной стороны, что обеспечивало постоянное самозатачивание их при
стирании (у атлантических диоплотериев эмали на резцах вообще не
было). Как у всех позднетретичных халитериин, у него было сокращенное
число зубов: от молочной генерации в нижней челюсти йрцгц-з во взрос-
лом состоянии сохранялись только заднекоренные. Иной тип адаптации
характеризовал вторую форму тихоокеанских раннемиоценовых сирен
Dusisiren reinharti, также несомненного потомка атлантических халите-
риин. У него отсутствовали резцы, а зубная формула была сокращена
/ 2_5 1_3
(г/р 2—5'm 1—3 с сохранением у взрослых лишь m 1—3), но главное,
у него был значительно более прямой ростр (около 45° вентрального
отклонения), что позволяло дузисирену использовать в пищу растения,
доходящие до поверхности воды, крупные seagrasses и, очевидно, попутно
бурые водоросли, которые хотя и росли на значительной глубине, но
25
посылали свои листовые пластинки вверх к поверхности. Именно этот
тип адаптации к питанию в более поверхностных слоях воды дузисирена
предопределил его дальнейшую столь необычную для сирен судьбу.
Пока условия обитания в новом месте мало чем отличались от при-
вычных для сирен еще по Карибскому бассейну, вполне было возможно
существование в одном регионе двух форм сирен, разведенных к тому
же в разные экологические ниши. При этом диоплотерий еще делил свою
нишу с другими крупными растительноядными млекопитающими — десмо-
стилиями, также питающимися на дне. Последние пришли к берегам Кали-
форнии из Азии, пройдя длинный путь от Японии через Камчатку и Аляс-
ку. Их самые ранние находки в Калифорнии датированы тем же ранним
миоценом. Резко изменилась ситуация примерно 12 миллионов лет назад.
Тектонические причины вызвали начавшееся похолодание и регрессию
моря, сформировалась открытая береговая линия, сократилась площадь
мелководий в бассейнах закрытого типа, с понижением температуры ба-
ланс между seagrasses и водорослями был нарушен в пользу последних.
Естественно, это обострило конкурентные отношения между диоплоте-
риями и более приспособленными к холоду десмостилиями. В результа-
те диоплотерии исчезли к позднему миоцену, а дузисирен оказался в
какой-то степени преадаптирован к новым условиям и в связи с обыч-
ной для сирен неприхотливостью к пище был меньше затронут измене-
ниями в пищевой базе, а на дальнейшее нарастание неблагоприятных по
отношению к обычным сиренам условий он ответил рядом эволюцион-
ных преобразований в одном без филетического ветвления ряду, пред-
ставленном двумя родами и шестью видами Hydrodamalinae, иллюстри-
руя, таким образом, «quantum evolution» Симпсона.
Главное направление эволюции гидродамалин — приспособление к
обитанию в условиях бурных (течения, прибой) холодных вод при откры-
той береговой линии, к питанию почти исключительно у поверхности
воды за счет произраставших в мелководьях водорослей. Полностью это
приспособление было реализовано у рода Hydrodamalis, достигшего рас-
цвета к позднему плиоцену и распространившегося в это время по всем
побережьям северной части Тихого океана. В ходе этого приспособления
у гидродамалин последовательно происходил ряд морфологических пре-
образований, коррелированных с изменениями внешних условий и
предъявляемыми ими к животным требованиями: началось (у Dusisiren
sp. В) и затем продолжилось дальше разрастание латеро-дорсальных
флангов боковых затылочных костей, знаменуя увеличение латераль-
ных поворотов головы: затем (у D. jordani) исчезла на этих костях fossa
condylaris, что уменьшило дорсовентральные ракурсы в атланто-заты-
лочном суставе; последующая (D. sp. D.) утрата фаланг в передней ко-
нечности, видимо, свидетельствует об окончательном освоении местооби-
таний и образа жизни, аналогичных тем, что наблюдал Г. Стеллер (Stel-
1ег, 1751) у морских коров на Командорских островах; и, наконец, пол-
ная утрата во взрослом состоянии зубов у Hydrodamalis cuestae (у юве-
нильных животных имевшиеся зубы были сдвинуты в заднем направле-
нии и их гомология не ясна), что совпало с резким (почти вдвое) уве-
личением размеров тела. Один из описанных экземпляров Н. cuestae до-
стигал 9 м длины, возможно, они были и больше. В сравнении с ним
Н. gigas с Командорских островов был, видимо, меньше (максимальный
отмеченный экземпляр имел длину 7,88 м). Сирены этого рода приобре-
ли необыкновенную плавучесть (в отличие от наземных млекопитающих
вес скелета морских с увеличением общих размеров возрастает в мень-
шей степени, чем объем тела) и ряд других приспособлений.
Домнинг (Domning, 1978с) рассматривает увеличение тела как при-
26
способление к холоду через изменение соотношения поверхность/объем.
В этом же направлении работает возросшая толщина эпидермиса и гипо-
дермы; уменьшает теплоотдачу и сухая спина, удерживаемая большей
частью над уровнем воды, и сокращение поверхности ластов с утратой
фаланг. В то же время увеличенная плавучесть облегчает пребывание
на мелководье, удержание тела в направлении движения воды, а толстая
кожа защищает от интенсивного стирания о камни и травм при ударах
о скалы в волноприбойной зоне.
Наиболее важным представляется вопрос о смысле утраты зубов. Тот
же автор (Domning, 1977а, 1978с) связывает это по времени с моментом
изменения в адаптивной зоне, выразившимся в полном переходе к пи-
танию от требующих пережевывания более волокнистых морских покры-
тосеменных к водорослям, листовые пластинки которых необходимо
лишь разделять на части (не исключено и проглатывание их в целом
виде). С последней задачей вполне справляются роговые пластинки в
передней части рострального выроста переднечелюстных костей и на
удлиненном симфизе нижней челюсти. Особо подчеркивается, что эти
роговые pads не занимают у Hydrodamalis места зубов (они имеются у
всех сирен в этом же месте и лишь слегка увеличена их площадь у этого
рода), а зубы исчезают в связи с отсутствием необходимости в перети-
рающих (поперечных) движениях челюстей. Таким образом, стирания
зубов у морских коров не должно было быть. В соответствии с этим у
Hydrodamalis ослаблены альвеолярные части верхней и нижней челюстей,
а также мышцы, отвечающие за их поперечные движения. Так, у Hydro-
damalis по сравнению с Dusisiren, в рационе которого еще были sea-
grasses, произошли изменения в скуловой дуге, крыловидных отростках,
височном гребне и т. п. Наоборот, структуры, связанные с продольными
движениями, или остались неизменными, или получили усиленное раз-
витие. Нельзя также забывать, что тенденция к сокращению числа зубов
вообще характерна для всего семейства Dugongidae, и Hydrodamalinae
могли продолжить эту тенденцию дальше, как это происходит у них с
уменьшением носовых костей, надглазничного отростка лобных костей,
проникновением верхнезатылочной кости к foramen magnum, уменьше-
нием числа позвонков с двумя полуфасетками до семи (восемь позвон-
ков с двумя фасетками, четыре с одной), увеличением sulcus bicipitalis
плечевой кости, расположением локтевой и лучевой кости без пересече-
ния в одной плоскости, слиянием запястных костей, дальнейшей редук-
цией таза (см. рис. 3).
Ламантины (Trichechus) среди современных и вымерших сирен зани-
мают особое положение, сочетая в своем строении много примитивных
черт с более специализированными, уклоняющимися от типичных для
большинства сирен (Simpson, 1932). Морфологические отличия ламанти-
нов от других современных родов (Dugong, Hydrodamalis) описывались
или суммировались неоднократно (Brandt, 1846, 1861/1868; Weber, 1928;
Simpson, 1932; Petit, 1967; Kaiser, 1974), как и различия между тремя
видами ламантинов (Petit, 1967; Ronald et al, 1978). Однако причины
появления этих признаков и их смысл, как правило, остаются неизвестны-
ми. Отсюда вытекает пристальное внимание к предложенной недавно
гипотетической модели эволюции Trichechidae (Domning, 1982), которая
не только показывает, как могла идти эта эволюция, но и вскрывает
адаптивное значение развития ряда характернейших признаков ламанти-
нов и этапы этого развития. Принципиальным является также показ
того, что ряд черт собственно ламантинов, обычно распространяемых на
всех потенциальных представителей Trichechidae, сформировались доволь-
но поздно в эволюции семейства (мио-плиоцен), начало которой было
27
положено еще в эоцене в условиях изоляции (экологической, генетиче-
ской), продолжавшейся до плейстоцена.
В совокупности признаков, определяющих современных ламантинов,
можно различить три категории. Одни показывают, что ламантины имеют
сходство главным образом с эоценовыми сиренами (протосиренидами или
самыми ранними дюгонидами) и более позднее их ответвление от общего
ствола сирен невозможно. Так, у ламантинов относительно мало развиты
предчелюстные кости (рис. 4, а, б), особенно их лобные отростки, которые
Рис. 4. Череп Trichechus manatus, современный, Карибское море, Куба; экз. ИЗ-5945-
а — сбоку; б — сбоку с удаленными скуловой, височной и околоушной костями; в — сверху; г —
спереди; д — сзади; е — снизу (а, б, г —е — вид при горизонтальном положении альвеолярного
края верхних челюстей, в — вид в плоскости лобных костей); ж — нижняя челюсть сбоку; з — ниж-
няя челюсть сзади; и — л — околоушная кость (и — вид с латеральной стороны и немного снизу,.
к — вид с каудальной стороны, л — вид с дорзомедиальной стороны).
1 — ala vomeris; 46 — lamina lateralis proces- 82— ramus mandibulae;
2 — alisphenoid (большое кры- sus pterygoidei; 83 — sulcus postzygomaticus;
ло); 47 — lamina medialls proces- 84 — sulcus septalis;
3 — angulus mandibulae; sus pterygoidei; 85 — sulcus tympanicus;
4 — angulus praemaxillare; 48 — lamina perpendicularis 86 — supraoccipitale;
5 — apertura nasi ossea; ossis ethmoidalls; 87 — sutura coronalis;
6 — basioccipltale; 49 — linea temporalis; 88 — sutura lambdoidea;
7 — basis mandibulae; 50 — maxilla; 89 — sutura maxillojugularis;
8 — basisphenoid; 51 — nasale; 90—sutura occipitomastoidea;
9 — caput mandibulae; 52 — orbita; 91 — sutura parietoocclpitalis;
10 — choanae; 53 — orbitosphenoid (малое gz — sutura parietotemporalis;.
11 — collum mandibulae; крыло); - 93 — sutura sagittalis;
12 — condylus occipitalis; 54 — palatinum; 94 — sutura squamosa;
13 — corpus mandibulae; 55 — parietale; 95 — sutura sphenosquamosa
14 — crista frontalis; 56 — pars basilaris ossis sphe- 96 — symphysis mandibulae;
15 — crista lambdoidea; noidalis; 97 — tegmen tympani;
16 — crista occipitalis externa; 57 — pars mastoidea ossis pe- 93 — temporale;
17 — crista parietalis; riotici; 99 — tubermentale;
18 — crista temporalis; 58 — pars petrosa ossis perio- 100 — tuberculum articulate;
19 — ethmoideum; tici; 101 — tuberculum pharyngeum;.
20 __ ethmoturbinalia; 59 — pars squamosa ossis tem- 102 — vomer;
21 — exocclpitale; poralis; юз — 104 — вход и выход
22 — facies insertiones appara- 60 — pars tympanica ossis ре- в канал, соединяющий но-
tus hyoidei; riotici; совую капсулу с под?
23 —• fenestra rotunda; 61 — periotica; глазничным каналом;
24 — foramen endolymphaticum; 62 — praemaxilla; 105 — желоб для евстахиевой-
25 — for. incisiva; 63 — presphenoid; трубы;
26 — for. infraorbitale; 64 — processus ascendes ossis 106 — канал, ведущий из кры-
27 — for. magnum; jugularis; лонебной ямы в хоану;
28 — for. mandibulae; 65 — pr. ascendes ossis maxil- 107 — канал, ведущий из хоаны
29 — for. mastoideum; laris; вперед в глазничную об-
30 — for. mentalis; 66 — рг. condylaris; ласть;
31 — for. opticum; 67 — pr. coronoideus; 108 — 109 — костная капсула
32 — for. ovale; 68 — pr. fonticularis; последнего верхнего ко-
33 — for. sphenopalatina; 69 — pr. frontalis praemaxillae; репного (108) и нижнего
34 — fossa digastrica; 70 — рг. mastoideus; коренного (109) зубов;.
35 — fossa mandibularis gle- 71 — pr. orbitalis ossis frontalis; 110 — отверстие для г. alveola-
noidalis; 72 — pr. palatinus maxillae; ris medii n. maxillaris
36 — fossa masse terlca; 73 — pr. paroccipitalis (V2);
37 — fossa pars mastoidei; 74 — pr. pterygoideus; ill — отверстие для r. alveola-
38 — fossa pterygoidea; 75 — pr. retroversus; в ris rostralis n. maxillaris;.
39 — fossa temporalis; 76 — pr. supraorbltalis; 112 — отверстие канала для вы -
40 — frontale; 77 — рг. ventralis ossis jugalis; хода n. trochlearis (VI) 1
41 — incisura intercondylaris; 78 — pr. zygomaticus maxillae; 113 — отверстие, соединяющее
42 — incisura mandibulae; 79 — pr. zygomaticus ossis tem- слезную область с под-
43 interparietale; poralis; глазничным каналом;
44 — jugale; 80 — protuberantia mentalis; 114 — ямка для прикрепления
45 lacrimale; 81 — protuberantia occipitalis m. semispinalls capitis
externa;
28
29
Cd
32
Рис. 4е
2 Ламантин
33
Рис. 4 ж, з
34
Рис. 4 и, к, л
не налегают на надглазничные от-
ростки лобных костей, как у дюго-
ня, а ростральная часть черепа от-
носительно мало отклонена вент-
рально: у Trichechus senegalensis
наклон (в град.) достигает 15—40
при ж=25,8 (М=20), у Т. inun-
guis 25—36 при х = 30,4 (iV=35),
у Т. manatus 24—52 при ж=38,2
(ZV=72), тогда как у дюгоня он
равен 67—72 (Domning, 1978с,
1982). Носовые кости у ламанти-
нов обычно (не всегда) имеются,
хотя и очень небольшие, срастаю-
щиеся с лобными костями, буду-
чи как бы погруженными в них
(рис. 4, б). Надглазничные от-
ростки лобных костей хорошо
развиты и расширены за орбитой.
Череп в целом и обонятельная
камера относительно длинные;
верхнезатылочная кость отделена
от foramen magnum (рис. 4, б);
угол, образованный нижними по-
верхностями основной затылоч-
ной и основной клиновидной
костей (около 215°), такой же,
как у олигоценовых дюгонид,
но меньше, чем у дюгоня и Hyd-
rodamalis (230°) (Simpson, 1932).
Скуловая кость сильно расши-
рена в посторбитальной обла-
сти (у ламантинов здесь находит-
ся сильный вентральный отросток
jugale, см. рис. 4, а), тогда как
у современных Dugong и Hydro-
damalis это расширение смещено
под орбиту. В то же время в от-
личие от последних и по крайней
мере от позднетретичных дюгонид
типа Metaxytherium у ламантинов
нет контакта скуловой и предче-
люстной костей (рис. 4, г, е), что,
несомненно, является примитив-
ным признаком (уже отмечалось,
что у эоценового Prorastomus ску-
ловая кость даже не была вклю-
чена в переднюю стенку орбиты).
Очевидно, примитивно и широкое
окно между основными клиновид-
ной и затылочной, височной и бо-
ковой затылочной костями, в ко-
тором помещаются слуховые, кос-
ти, слившиеся вместе (рис. 4, в),
так как и у дюгоня и у Hydroda-
35
2*
malis сосцевидный и парокципптальный отростки более направлены впе-
ред, что ведет к укорочению постгленоидной области черепа. Коренные
зубы ламантинов с раздельными замкнутыми корнями и низкой корон-
кой, покрытой эмалью (а не цементом, как у дюгоня), билофодонтные
с острыми бугорками, организованными по примитивной схеме, близкой
к эоценовым дюгонидам. Тело нижней челюсти низкое (рис. 4, ж). В за-
пястье сохраняются шесть-семь отдельных костей, тогда как у олигоце-
нового Halitherium их было меньше. Седалищная кость относительно
мало модифицирована по форме и соответственно не может быть выве-
дена из сильно измененного узкого ischium постэоценовых дюгонид.
Вторая категория признаков включает те, что отсутствуют у ранне-
третичных дюгонид, но развиваются у них позже. Сюда входят редук-
ция носовых и слезных костей с утратой слезного канала, редукция
резцов, клыков и переднекоренных зубов, эпифизов позвонков, утрата
задних конечностей и редукция лобковых костей таза. Очевидно, все эти
преобразования идут у ламантинов и поздних дюгонид параллельно. Воз-
можно, они адаптивны (Simpson, 1932), но смысл их не ясен.
Основной интерес вызывают черты, специфичные для ламантинов.
В первую очередь это относится к сильному увеличению в числе заднеко-
ренных зубов (рис. 4, в), вызванному приобретением способности к не-
ограниченной смене их в горизонтальном направлении, когда более пе-
редние стертые зубы выталкиваются более задними и выпадают, а сзади
идет непрерывное появление новых «вторичных» моляров. В результате
одновременно функционируют семь-восемь зубов в каждой половине че-
люсти, а всего во время жизни через челюсти животного может проходить
до 80 зубов (Savage, 1977; Domning, 1978b; Domning, Hayek, 1984). Сама
горизонтальная смена зубов в той или иной степени встречается и у дру-
гих млекопитающих, но неограниченность ее — черта, присущая только
ламантинам. В связи с увеличением длины зубного ряда у них, видимо,
венечный отросток нижней челюсти (рис. 4, ж) начинает расти не прямо
вверх, как у всех дюгонид, а вверх и вперед (Domning, 1982). Другим от-
личием ламантинов является практически замкнутая сзади орбита, отгра-
ниченная от височной ямы сильно сближенным, уже отмеченным надглаз-
ничным отростком лобной кости и особым восходящим отростком jugale
(рис. 4,а), подкрепленным с внутренней стороны выступом вверх задне-
го наружного угла processus zygomalicus maxillae (рис. 4, в, е).
У дюгонид же орбита всегда широко открыта назад. Специфично для
ламантинов заметное вздутие tegmen tympani и pars mastoidea околоуш-
ной кости и развитие особого сочленения между нижней челюстью и кры-
ловидными отростками клиновидной области основания черепа (Domning,
1978а), а также расширение скулового отростка височной кости, что, воз-
можно, связано со звуковой рецепцией (Bullock et al., 1980). Большие
изменения затрагивают посткраниальный скелет ламантинов. Так умень-
шается число позвонков как во всем туловище, так и в шейном (до 6 против
7 у всех дюгонид и подавляющего большинства других млекопитаю-
щих) и спинном отделах (до 15—17, тогда как у Protosiren их было 20,
см. Sickenberg, 1934, а у большинства дюгонид, включая самого дюгоня,
и Hydrodamalis обычно 19). В лопатке ламантинов сильно удлиняется
акромиальный отросток (он растет и в онтогенезе), а в humerus редукции
подвергается большой бугорок, а малый смещается вперед дальше от го-
ловки плеча и соединяется с большим бугорком, что приводит к исчезно-
вению биципитального желоба, характерного как для дюгонид, так и для
большинства млекопитающих. Очень своеобразна редукция таза у ламан-
тинов (см. рис. 3, ж). В отличие от дюгонид здесь исчезает не только
лобковая кость, но и подвздошная, а седалищная сохраняется в виде суб-
треугольной широкой пластинки.
36
В целом эволюция Trichechidae в настоящее время представляется сле-
дующим образом (Domning, 1978). Какие-то представители протосиренид,
имевших в эоцене пантетисное распространение, достигли в раннем или
среднем эоцене Южной Америки, где в условиях генетической изоляции
от родительской группы и дали начало трихехидам. Нельзя исключить,
что предками ламантиновых могли быть и самые ранние дюгониды, стоя-
щие на уровне Eotheroides. Однако их остатков в Южной Америке, дати-
рованных эоценом, нет, а протосирениды обнаружены (Sereno, 1982).
До миоцена трихехиды не появлялись на жизненной арене. В то же время
самые ранние их представители известны лишь по разрозненным остаткам
отдельных зубов, фрагментам челюстей и, следовательно, трудно говорить
об уровне развития у них собственно трихехидных черт. Ясно лишь, что,
видимо, до среднего миоцена селективное давление на челюст-
ной аппарат трихехид и дюгонид было одинаково, что привело у них к
параллельному сокращению коренных зубов до m 1—3 или у взрослых
даже до ш 2—3. Предполагается, что ранние трихехиды, как и современ-
ные ламантины, обитали в водах прибрежных рек и эстуариев, тогда как
дюгониды, в том числе миоценовый Metaxytherium, предпочитали морские
воды, т. е. обе группы были разведены в разные экологические ниши.
К позднему миоцену Trichechidae уже были широко распространены по
побережью Южной Америки в районе Карибского моря и тропической
части Атлантического океана. Во всяком случае, их остатки обнаружены
и на севере в бассейне р. Магдалены (Potamosiren), и на востоке близ
устья теперешней Амазонки (Sirenotherium), и на юге у Ла-Платы
(Ribodon). Но у первых двух более ранних по возрасту, еще не было го-
ризонтальной смены зубов, а их коренные были крупными (условная пло-
щадь одного моляра типового экземпляра Potamosiren magdalenensis опре-
делена в 420 мм2, другого экземпляра, возможно, Р. sp., более чем в
600 мм2), явно несменяемыми. Не был, видимо, направлен вперед и ве-
нечный отросток потамосирена.
В позднем миоцене на эволюции сирен стали сказываться последствия
начавшегося в миоцене цикла Андского орогенеза, приведшего к увели-
чению системы стоков и соответственно эрозии почв. В ответ на появление
более абразивной пищи, возможное появление в диете водных злаков с по-
вышенным содержанием кремнезема впервые у Ribodon появилась гори-
зонтальная смена зубов; сами зубы стали мельче (361 мм2 у наибольшего
моляра), что вызывало увеличение на единицу площади зоны прикуса дли-
ны эмалевых гребней, что равносильно усложнению коронки и дает боль-
шее сопротивление стиранию, а необходимость поддержания общей пло-
щади прикуса относительно величины животного потребовала увеличения
числа зубов (до 6) сравнительно с 2—3 у Potamosiren. Ламантины (Tri-
chechus) в дальнейшем развили дальше эти тенденции, уменьшив в ряду
Т. senegalensis—Т. manatus—Т. inunguis вдвое условную площадь наиболь-
шего по размеру моляра (286,6 мм2—263,2 мм2—187,3 мм2), увеличив чис-
ло зубов до 7 (редко 8) у первых двух видов и до 8 у последнего. Кроме
того, у Т. inunguis произошло еще и дополнительное усложнение коронки.
Таким образом, этот вид по многим показателям должен рассматривать-
ся как более прогрессивный (и в отношении зубного аппарата, и в при-
способлении к наиболее жесткой пище, которую Т. inunguis берет пре-
имущественно у поверхности воды среди так называемых «floating
meadows», или плавающих лугов, главную массу которых составляют
Graminaea, что потребовало перестройки движения в атланто-затылочном
суставе и, в частности, более вертикального затылка4, и в отношении
4 У Т. manatus верхнезатылочная кость составляет с теменной угол около 120° (см.
рис. 4, а), а у Т. inunguis около 90°.
37
ластов, которые у этого вида исключительно длинные). Наоборот, Т. ma-
natus и близкий к нему Т. senegalensis относительно более примитивны.
Видимо, их общий предок остался обитателем северо-восточного побе-
режья Южной Америки, откуда одни его потомки проникли в Карибское
море, а другие, видимо пользуясь морскими течениями, в Западную Аф-
рику. Однако эта экспансия должна была происходить позже, уже в
плейстоцене.
Более сложной должна была быть история происхождения Т. inunguis.
В среднем или позднем миоцене какие-то ламантины проникли во внут-
ренний водный бассейн, существовавший в Южной Америке на месте
теперешних верховий Амазонки. Возможны две версии: это могли быть
потомки Ribodon, которые из бассейна р. Магдалены проникли через
расположенный рядом центральноамериканский морской проход в Тихий
океан и далее в западную Амазонию через проход, связывающий океан
с внутренним водоемом и существовавший до начала позднего миоцена.
Это могли быть и прямые потомки ламантинов из прибрежных популя-
ций, попавшие в Амазонию в плиоцене уже после закрытия тихоокеан-
ского прохода, что было связано с поднятием Анд, через невысокие водо-
разделы, отделявшие внутренний бассейн от бассейнов Ориноко на севере,
Ла-Платы на юге и низовьев Амазонки на востоке (возможно, через
перехват течений). Этот первично солоноватоводный внутренний бассейн
и был, очевидно, местом формирования Т. inunguis из форм близких по
уровню развития к Т. manatus (остатки таких форм обнаружены в плио-
плейстоценовых отложениях Западной Бразилии). Изолированный харак-
тер этого бассейна, не имеющего постоянных внешних стоков, обеспечи-
вал генетическую изоляцию попавших сюда ламантинов от прибрежных
популяций. Повышенное плодородие его вод, получающих питательные
вещества с Анд, привело к преимущественному развитию здесь злаковых
растений, тогда как у прибрежных популяций основу диеты по-прежнему
могли составлять seagrasses в сочетании со злаками. Соответственно се-
лективное давление на амазонскую популяцию могло быть сильнее, что и
потребовало дальнейшей эволюции жевательного аппарата Т. inunguis,
развития наиболее мелких и сложных моляров, большего их числа. В свою
очередь прибрежные популяции, распространяясь в Карибское море и
пользуясь преимуществами своей «функциональной гипсодонтии», вытес-
нили отсюда последних дюгонид и вторично оказались в прибрежно-мор-
ских местообитаниях, исключительно богатых seagrasses, где злаки уже
могли не играть важной роли в их питании. Это приостановило дальней-
шее развитие их зубной системы в направлении указанных тенденций.
Даже больше, заставило обратить внимание на бесхозную придонную
морскую экологическую нишу, занимаемую ранее дюгонидами. В резуль-
тате именно у этих популяций Т. manatus наиболее сильно выражено
вентральное отклонение ростральной части черепа (среди 35 экземпляров
флоридской популяции ламантинов 40,1°, а в выборке из 15 экземпляров
из Южной Америки 33,8°). Африканский ламантин появился в Аф-
рике лишь недавно и не имел еще времени на значительное обособление
от предковой группы ламантинов с побережья Южной Америки. В неко-
торых особенностях он даже примитивнее, чем представители карибской
популяции Т. manatus. В свою очередь, Т. inunguis появился в низовьях
Амазонки, что привело к разрыву ареала прибрежной популяции Т. ma-
natus близ устья Амазонки, лишь в плио-плейстоцене, когда был размыт
водораздел между западной и восточной частями Амазонии, и в Южной
Америке установилась современная дренажная система.
Такова общая картина эволюции сирен, составленная на основе учета
как апоморфных, так и адаптивных их черт. В некоторой степени к близ-
38
ким результатам подошел Р. Саваж (Savage, 1977), анализируя только
первые из них. На рис. 5 представлена его кладограмма, долженствующая
отражать филогению сирен, но скорее она иллюстрирует последователь-
ность некоторых морфологических преобразований и в ряде случаев их
параллельность. Этот путь изучения эволюции не дает понимания причин
развития и, как пишет сам автор, даже не позволяет ставить вопрос о
какой-то естественной иерархизации сирен. Естественно обратить внима-
ние на большую продуктивность первого пути, по которому пошел Дом-
нинг в серии своих работ (Domning, 1977а, 1978b, с, 1982). Однако труд-
ности формального определения отдельных групп сирен существуют. Клю-
чевым событием в происхождении той или иной ветви по современным
взглядам часто считается, например, начальная географическая изоляция,
как в случае Trichechidae (Domning, 1982), а не последующее приобре-
тение ими тех или иных производных признаков, хотя и наиболее харак-
терных для данной группы в лице их ныне живущих представителей.
Отсюда трудность распознавания ранних членов групп и необходимость
определять тот или иной таксон совокупностью тенденций с указанием
начального и конечного состояния. Сама характеристика групп поневоле
становится очень лаконичной. Тем не менее кажется целесообразным
такие характеристики привести.
f
§
§
§
§
i
в
I
I
А;
fy/M маю
Щ/77
'2
Т/аса/са рейшир/rfaz
/ее/; слезрае а/пдере-
тие утра vena
^аз/арсРсн/п/зал зуде/ап сас/пглза
Рис. 5. Кладограмма взаимоотношений основных родов сирен отр. Sirenia
(по: (Savage, 1977))
Цифрами обозначены точки ветвления «филогенетического» древа
л»
у.
See
afffiev
у/перя/м
fiape/m/e jyfie/
4,2
4/
1^
fasalia ।
pefiyaapafiame । §
//икмаеслал
смела ayfiafi
SI7/
si
рудал/е/гтгрм/
fie twee ve/и 4 jm/w-
Wm: зуда, тсш/с
’разделеле/ лсРлз/ма rw/ш
'Мшм а/псря////; п/ю/е/глмгаРа/е jyfo умен/а/е////
. vuc//e /ле далее ve/и /' А/даеадразлма; mrxrtia
реЛ/царала/////; рас/пру/и реала апп/ыу/п елиз
Раструм дтаглу/п fie/аз, г/ажпае резае/ рудалзел/т/аал//:
/'слегка ydeaavea; лареалз/е й/падааараРа///гт/е ’
fij/yaafian afinacme апеуаапазараЛ/па
Рсге/гса; Радлз/й а/раз з/сазг/и; рерулцал зад/ш aeaevaacmea; РазлилаеРе//ае //а/аастма:
' //аалу/па с /еаздреи
§
39
Prorastomidae Cope, 1889
Ростральная часть черепа и симфиз нижней челюсти удлиненные, пря-
мые; скуловая кость исключена из переднего края орбиты; носовые кости
большие, достигающие сзади орбит; слезные кости и канал есть; передний
край орбиты расположен на уровне р4; череп за орбитами сильно сужен;
околоушная кость неподвижно соединена спереди и сзади с окружающи-
ми костями; tegmen tympani еще не расширено в виде почки, как у дру-
гих сирен; алисфеноидный канал есть; на нижней челюсти до пяти под-
бородочных отверстий, из которых два передних большие; зубная форму-
3 15 3
ла 'т-'а- ; резцы мелкие, равного размера, коренные простые, субквад-
ратные, брадиодонтные, с двумя поперечными лофами; атлант в отличие
от других сирен с широкими плоскими крыльями и выступающим назад
отростком на вентральной дуге.
Protosirenidae Sickenberg, 1934
Сирены небольшого размера (до 2,5 м длины); ростральная часть черепа
и симфиз нижней челюсти слабо отогнуты вниз; скуловая кость входит
в передний край орбиты, передний край которой находится на уровне
промежутка между р1 и р2; орбита широко открыта в височную яму; над-
глазничные отростки лобных костей широкие; носовые кости соприкаса-
ются по средней линии, но не достигают орбит; околоушная кость увели-
чена и представляет единое образование, подвижно соединенное с окру-
жающими костями черепа; tegmen tympani расширено, почкообразно;
алисфеноидный канал, как у большинства сирен, утрачен; верхнезаты-
лочная кость не входит в край foramen magnum; одно большое и малень-
_ г 3.1.5.3
кое, дополнительное, подбородочное отверстие; зуоная формула g-j 5 д- ;
первые верхние резцы слегка увеличены; таз мало редуцирован, имеется
хорошо развитый ацетабулюм; задние конечности, возможно, могли функ-
ционировать как ласты.
Dugongidae Gray, 1821
Сирены среднего, крупного и очень крупного (до 9 м длины) размера;
предчелюстные кости обычно сильно развиты и имеют более или менее
сильный наклон ростральной части вниз; передний край орбиты на уров-
не середины ряда предкоренных; скуловая кость по крайней мере у позд-
нетретичных и современных форм контактирует с предчелюстной; алис-
феноидный канал утрачен; подбородочное отверстие обычно одно; полная
эутериевая зубная формула у ранних форм или сокращенная до m 2—3
у более поздних; первые верхние резцы, как правило, увеличены, но мо-
гут и отсутствовать вообще; коренные бунобилофодонтные (в крайних
случаях сильно модифицированы или утрачены); туловище длинное,
в шейном отделе 7, в спинном 19—20 позвонков; задние конечности реду-
цированы; в тазе прогрессивно редуцируются лобковые кости, а седалищ-
ная и подвздошные становятся длинными, узкими; в humerus имеется хо-
рошо развитый биципитальный желоб, запястные кости склонны к слия-
нию и уменьшению в числе.
Halitheriinae Abel, 1913
Дюгониды обычно сравнительно небольшого (до 2,5—3 м) размера, редко
большего (до 5 м); ростральная часть черепа обычно отогнута вниз; не-
большие слезные кости соприкасаются по средней линии или у более
40
поздних форм разделены лобными костями; слезные кости более или ме-
нее редуцированы, слезный канал отсутствует лишь у некоторых поздних
форм; обонятельная часть черепа относительно длинная; верхнезатылоч-
ная кость обычно не достигает края foramen magnum; зубная формула от
ЗЛ.5.3. 1.0.1-2.3 0.0.1-2 3 '
ЗЛ.бХ у эоЦеновых форм до - 0 0 л 3 или оол 3 у более
поздних; первые верхние резцы обычно расширены, реже утеряны;
коренные брахиодонтные с эмалью и закрытыми корнями; форма корон-
ки от билофодонтной до бунодонтной; т3 нередуцированный, со сложной
коронкой.
Miosireninae Abel, 1919
Дюгониды до 3,5—3,75 м длины; затылок широкий; череп округлый верх-
незатылочная кость достигает foramen magnum; слезный канал есть; ниж-
няя челюсть короткая, высокая; зубная формула i‘c0p4m3; первые верх-
ние резцы увеличены, т3 сильно редуцирован в виде цилиндрического
столбика, коронка упрощенного строения; первые три премоляра равной
величины, просты по строению, р4 увеличен и сильно моляризован; корни
коренных замкнутые, эмаль есть.
Dugonginae Gray, 1821
Дюгониды большого размера (до 5—6 м длины, но обычно около 3 м);
ростральная часть черепа сильно (до 70) отогнута вниз; носовые кости
рудиментарны, широко разделены лобными; обонятельная часть черепа
укорочена; затылок узкий, верхнезатылочная кость входит в край fora-
men magnum; processus retroversus чешуйчатой кости сильно отогнут
внутрь; зубная формула q 0 0 2__-- J верхний резец большой, бивнеобраз-
ный у самцов, у самок маленький; коренные зубы без эмали, покрыты
цементом, корни слитые, незакрытые; ребра относительно тонкие, внутри
частично губчатые.
Hydrodamalinae Palmer, 1895
Дюгониды большого размера от 3 м у ранних форм до 9 м у более позд-
них; ростральная часть черепа удлиненная, умеренно отклоненная вниз
(не более 45°); носовые кости рудиментарные, разделенные лобными
костями или отсутствуют; обонятельная часть черепа относительно длин-
ная; processus retroversus слабо выражен, не отогнут внутрь; тело нижней
челюсти низкое, длинное; функционирующие увеличенные резцы отсут-
ствуют; зубная формула ранних форм во взрослом состоянии m 1—3,
у поздних форм функционирующие зубы вообще отсутствуют; фаланги
на передних конечностях у поздних форм утрачены.
Trichechidae Gill, 1872
Сирены среднего размера (до 4,5 м длины); предчелюстные кости срав-
нительно небольшие; ростральная часть черепа относительно мало от-
клонена вниз; носовые кости рудиментарны, маленькие, сращенные с
лобными; скуловые кости входят в передний край орбиты, но не контак-
тируют с предчелюстными, сильно расширены позади орбиты; орбита
более или менее обособлена от височной ямы; затылок широкий; нижняя
челюсть низкая, венечный отросток, по крайней мере у более поздних
форм, наклонен вперед, а нижняя челюсть имеет дополнительный кон-
такт с основанием черепа через крыловидные отростки; функционирую-
41
щие резцы отсутствуют; зубная формула у ранних форм сведена
к m 1—3 или даже m 2—3, у поздних за счет появления дополнитель-
ных моляров их число возрастает до 6—8, смена которых идет сзади
наперед всю жизнь; туловище несколько укорочено, в шейном отделе
всегда 6, в спинном 15—17 позвонков; биципитальный желоб в humerus
утрачен, запястные кости мало слиты; лобковые и подвздошные кости
редуцированы, седалищная в виде широкой в основании субтреугольной
пластинки.
Подводя итоги вышесказанному, необходимо суммировать, что же нам
известно о морфологических адаптациях сирен в целом и отдельных их
представителей. Учитывая, что ряд сторон их адаптаций, особенно в от-
ношении ламантинов, будет еще рассмотрен в последующих главах, от-
метим здесь лишь основные, которые должны учитываться при дальней-
шем анализе собственного материала.
Возникновение сирен было связано с развитием у них уникальной
способности менять свой удельный вес, что при среднем удельном весе,
близком к таковому воды, позволяет подниматься и погружаться в воду
без участия локомоторных органов при минимуме мышечных усилий
(гидростатическая адаптация по Domning, 1978b, с). Именно с этой спо-
собностью связано развитие пахиостоза скелета и ряд приспособлений
для поддержания тела в горизонтальном положении (горизонтальная
диафрагма, удлиненные легкие, моноподиальное ветвление бронхиально-
го ствола, см.: (Pick, 1907)), целесообразность которого для сирен была
подчеркнута особым постулатом (Domning, 1977а). Сирены — облигатно-
водные растительноядные млекопитающие, первичным в жизни которых
было питание морскими покрытосеменными (seagrasses). Отсюда — оби-
тание в мелководьях у побережий морей тропических и субтропических
зон, умеренная подвижность, определенное ослабление дистантных сен-
сорных систем (ввиду отсутствия в этих местообитаниях естественных
врагов при обилии корма). Отсюда их общий внешний облик: укорочен-
ная шея, ластовидные передние конечности, отсутствие задних конеч-
ностей, горизонтально расширенный хвостовой плавник, как главный
орган локомоции. Из сказанного выше вытекает и изменение наклона
ростральной части черепа и симфиза нижней челюсти в зависимости от
способности брать пищу преимущественно с одного уровня (со дна, у по-
верхности или в толще воды). В этом направлении развивались морфо-
лЬгические адаптации дюгонид.
Общим для всех сирен оказалась необходимость в мио-плиоцене раз-
лития специальных приспособлений, противодействующих усилившемуся
стиранию зубов. С одной стороны, стирание увеличилось в связи с воз-
росшим попаданием абразивных частиц вместе с пищей. На это дюгони
(ответили развитием гипсодонтных коренных зубов. С другой стороны, на
это влияние у ламантинов было наложено широкое проникновение в прес-
ные воды рек и опресненных эстуариев злаков с повышенным содержа-
нием кремнезема в самих растениях (дюгонид с их морским образом
жизни это практически не коснулось). На аналогичные изменения, про-
изошедшие на суше и приведшие к широкому развитию степей с их рас-
тительным преимущественно злаковым покровом, наземные копытные от-
ветили появлением настоящих гипсодонтных зубов с высокой коронкой
и открытыми корнями. Ламантины пошли иным путем, развив «функцио-
нальную гипсодонтию», которая обеспечила им преимущество в конку-
рентной борьбе с позднетретичными дюгонидами и возможность вторично-
го проникновения в прибрежно-морские биотопы с возвратом к питанию
большей частью морскими покрытосеменными.
42
Специфичными оказались адаптации Hydrodamalinae, которые пере-
шли с питания морскими покрытосеменными на водорослевую диету
(бурые, красные водоросли). С одной стороны, это позволило им отка-
заться от пережевывания пищи (зубы стали излишни), а с другой —
послужило предпосылкой для их приспособления к условиям начавше-
гося ухудшения климата, в частности похолодания, вызвавшего сокраще-
ние seagrasses и, наоборот, расцвет водорослей. При этом на первый план
у гидродамалин выдвинулись терморегуляционные адаптации (увеличе-
ние объема тела, утолщение эпидермиса и слоя подкожного жира и т. п.).
Важную роль в этой линии развития сирен играло приспособление
к добыче корма у поверхности воды (модификация передних конеч-
ностей, возрастание защитной роли эпидермиса, вторичное увеличение в
определенной степени подвижности шеи).
Глава вторая
СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ
КОЖНОГО ПОКРОВА ЛАМАНТИНА
Кожный покров млекопитающих как пограничная с внешней средой сис-
тема органов в первую очередь подвергается ее действию, которое во
многом определяет свойства и особенности строения кожного покрова.
Как известно, эволюция покровов позвоночных протекала в направлении
образования поверхностных многослойных пластов клеток и разделения
функций между двумя его компартментами: эпидермисом и дермой.
Специализация эпидермиса шла по пути усовершенствования в первую
очередь барьерных свойств, в то время как соединительнотканные компо-
ненты принимали на себя механические и терморегуляторные функции.
Для вспомогательных и специальных целей у млекопитающих развился
целый ряд производных эпидермиса: волосы, вибриссы, железы и т. д.
У вторичноводных млекопитающих, к которым относятся и сирены,
кожный покров показывает черты адаптации к обитанию в воде. При
сравнительно-анатомическом методе исследования наиболее очевидные
различия оказались в дермальном слое и эпидермальных производных
(Соколов, 1973; Harrison, Thurley, 1974; и др.), в то время как в эпи-
дермисе, по мнению некоторых исследователей, особенности адаптации не
улавливаются (Ling, 1970). Последнее объясняется универсальностью
принципа организации эпидермиса млекопитающих (на основе процесса
кератинизации) и недостаточностью наших знаний о тонких механизмах,
благодаря которым осуществляются его функции. Очевидно, что выявле-
ние модификаций признаков, свойственных эпидермису животных, зани-
мающих разные экологические ниши, требует более специальных и тон-
ких методов исследования.
Для изучения кожного покрова ламантина и выявления адаптивных
характеристик наряду с традиционными анатомическими и гистологиче-
скими методами нами было проведено электронно-микроскопическое ис-
следование эпидермиса, что позволило обсудить особенности процесса
кератинизации.
Подкожная жировая клетчатка служит связующим звеном между
кожей и подкожной мускулатурой и играет большую роль в термоизоля-
ции у водных млекопитающих, а также в запасании питательных
веществ. Обитание ламантина в теплых экваториальных водах, безуслов-
но, должно наложить отпечаток на строение и степень развития под-
кожной клетчатки. Мы предприняли попытку описать и обсудить эти
особенности в сравнительном и функциональном аспектах.
Проведено также специальное исследование вибриссного аппарата
ламантина в сравнении с таковым у китообразных, ластоногих и назем-
ных млекопитающих.
Кожный покров сирен давно привлекает внимание исследователей.
Первые описания касаются внешнего вида кожи, толщины ее слоев, осо-
бенностей сочленения эпидермиса и дермы, расположения и размеров
волос и щетин у морской коровы (Steller, 1749; Brandt, 1871), амери-
канского ламантина (Murie, 1872; 1872а).
О закономерностях изменений толщины кожи у молодых особей
дюгоня и ламантина позволяют судить графики, приведенные в ряде
работ (Matthes, 1929; Domning, 1977).
44
Основные гистологические исследования кожи сирен до сих пор про-
водили на эмбриональном материале (в лучшем случае у новорожден-
ных или молодых особей) или на топографически неопределенных про-
бах кожи. Все это зачастую делает несопоставимыми полученные разны-
ми авторами данные. Так, в распоряжении Доша (Dosch, 1915), который
первым описал гистологическое строение кожи африканского ламантина
(Trichechus senegalensis) и дюгоня (Dugong dugon), был материал,
предоставленный в его распоряжение Кюкенталем: пробы кожи спины
разновозрастных эмбрионов и новорожденных особей. Это позволило опре-
делить закономерности формирования эпидермиса, дермальных сосочков
и вибрисс, а также сравнить особенности строения кожи у этих двух
видов. Кроме того, Дош представил доказательства закладки обычных
волос и последующей их дегенерации у эмбриона дюгоня. Кожа ново-
рожденного дюгоня и бразильского ламантина (Т. inungius) исследована
В. Е. Соколовым (1973). Им же дана общая характеристика кожи для
отряда сирен, в целом сходная с таковой, приводимой другими авторами
(Brandt, 1871; Weber, 1928; Petit, 1955; Ling, 1974). Вибриссы дюгоня
(их размеры, расположение, гистологическое строение) изучены наиболее
полно (Bryden et aL, 1978; Kamiya, Yamasaki, 1981).
Подробное гистологическое исследование кожи, приписываемой мор-
ской корове (Rhytina stelleri), выполнено лишь в 1957 г. по образцам,
возраст которых определен в 188 лет (Haffner, 1957). Ранее Г. Стеллером
(Steller, 1751) и А. Брандтом (Brandt, 1871) была описана короподобная
поверхность кожи, благодаря которой этот зверь получил название «Вог-
kentier» ’, рассмотрены слои кожи и характер сочленения эпидермиса
и дермы. Хаффнер всестороннее изучил образцы кожи морской коровы:
внешнюю морфологию, отдельные слои эпидермиса и дермы, провел экс-
периментальные исследования ее физических свойств.
Большинство исследователей обсуждают вопросы анатомического
сходства кожи различных представителей отряда сирен, а также кожи
сирен и хоботных (филогенетически близких отрядов), сирен и китооб-
разных (двух отрядов настоящих водных млекопитающих). Мнения по
этому вопросу противоречивы. Так, некоторые исследователи находят,
что кожа морской коровы сходна с таковой у ныне живущих сирен
(Brandt, 1871; Dosch, 1915), в то время как другие авторы (Haffner,
1957) придерживаются противоположного мнения. Маттес (Matthes, 1929)
отмечает сходство кожи у ламантинов и дюгоней и некоторое ее отличие
от кожи морской коровы. Выявлено большое сходство между кожей мор-
ской коровы и кожей слонов, что считается одним из подтверждений
родства этих отрядов (Haffner, 1957). Сходное предположение было сде-
лано и ранее (Weber, 1928). Предприняты попытки сравнения толщины
кожи сирен, бегемота и различных китов (Matthes, 1929).
Гистологическое описание строения кожи ламантина (Trichechus ma-
natus) в литературе до сих пор отсутствовало. Мы детально изучили
гистологическое строение кожи взрослого самца. Пробы кожи (52 пробы
с различных участков тела (рис. 6, табл. 1)) фиксировали 10%-ным нейт-
ральным формалином, заливали через анилин в парафин; срезы окраши-
вали гематоксилин-эозином. Эластиновые волокна выявляли орсеином.
Промеры толщины кожи и подкожной клетчатки проводили на фикси-
рованном материале. Основные промеры препаратов выполнены по опре-
1 По Хаффнеру (Haffner, 1957), короподобная поверхность кожи морской коровы —
результат деятельности кожных паразитов. Однако есть мнение, что все сохранив-
шиеся фрагменты кожи могли принадлежать гладким китам (Balaenidae) (Dom-
ning, 1978с).
45
Таблица 1. Толщина кожи взрослого самца американского ламантина
Проба, Место взятия пробы Кожа (без подкожной жировой клетчатки), мм Эпидермис Роговой слой Дерма, мм Высота дермаль- ных сосочков Подсосочковый слой Подкож- ная жиро- вая клет- чатка, мм
ММ в процен- тах к тол- щине кожи мкм в процен- тах к тол- щине эпи дермиса
мкм в процен- тах к тол- щине эпи дермиса мм в процен- тах к тол- щине кожи
1 Между ноздрями 10,0 1,0 10 113 и 9,7 752 75 9,0 4,0 3,0 7,1 18,0 7,0 11,0 9,0 15,0 90
2 Вокруг глаза 4,6 0,6 13 113 19 4,4 397 66 87
о 4 5 Вокруг слухового отверстия Середина верхней губы Верхняя губа с вибриссами 4,5 7,6 18,9 1,5 0,5 0,9 33 7 5 113 45 34 8 9 4 4,3 7,4 18,7 1274 342 697 85 68 77 67 93 —
6 7 Угол рта и нижняя губа Внутренний край нижней губы 7,4 14,8 0,4 3,8 5 26 90 350 23 9 7,2 14,1 197 3056 49 80 95 74 -
8 Середина морды между глазами 9,7 0,7 7 113 16 9,4 367 52 93
9 10 Середина головы между слуховы- ми отверстиями 15,7 0,7 4 67 10 15,5 520 74 96 1,0
Середина спины между лопат- ками 21,7 0,7 3 113 16 21,4 368 53 21,0 97 3,0
11 12 107 см от края морды 147 см 195 см 229 см Дорсальная сторона хвостового 21,7 0,7 3 113 16 21,5 474 68 21,0 97 2,0
22,8 0,8 4 ИЗ 14 22,6 574 72 22,0 97
14 15 18,6 0,6 3 113 19 18,4 374 62 18,0 97 5,2
18,4 11,5 0,4 0,5 2 4 ИЗ 113 28 23 18,2 11,3 174 274 44 55 18,0 11,0 98 96 6,0
плавника, 254 см от края морды Середина дорсальной стороны
16 9,5 0,5 5 ИЗ 23 9,3 297 59 9,0 95 5,1
. хвостового плавника
17 Край хвостового плавника, 15 см от средней линии 3,8 0,8 21 ИЗ 14 3,4 597 75 3,0 79 -
18 Середина горла, 25 см от края нижней челюсти 5,4 0,4 7 68 17 5,2 219 55 5,0 93 3,0
19 Середина горла 3,5 0,5 14 ИЗ 23 3,3 297 59 3,0 86
20 Середина груди между плавни- 5,5 0,5 9 79 16 5,3 337 67 5,0 91 2,5
21 ками
То же 7,5 0,5 7 90 18 7,3 297 59 7,0 6,0 93
22 Середина груди с вибриссой 6,6 0,6 9 45 8 6,4 442 74 91
23 24 Середина брюха, 134 см от аналь- ного отверстия 4,5 0,5 И 56 И 4,4 388 78 4,0 89 2,0
То же, 97 см 9,6 0,6 6 ИЗ 19 9,4 373 62 9,0 94 7,7
25 Наружная кожа препуция 11,5 0,5 4 ИЗ 23 11,3 274 55 11,0 96
26 Внутренняя кожа препуция 4,3 0,3 7 ИЗ 38 4,1 120 40 4,0 93 —
Таблица 1 (окончание)
Проба, № Место взятия пробы Кожа (без подкожной жировой клетчатки), мм Эпидермис Роговой слой Дерма, мм Высота дермаль- ных сосочков Подсосочковый слой Подкож- ная жиро- вая клет- ча!ка, мм
ММ в процен- тах к тол- щине кожи мкм в процен- тах к тол- щине эпи дермиса мкм в процен- тах к тол- щине эпи- дермиса мм в процен- тах к тол- щине кожи
27 Перинеальная область 9,4 0,4 4 из 28 9,2 174 44 9,0 11,0 96
28 Середина брюха, 50 см от аналь- ного отверстия 11,9 0,9 8 из 13 11,7 674 75 92 7,0
29 Середина брюха, 19 см 8,5 0,5 6 113 23 8,3 274 55 8,0 10,0 8,0 94 7,0
30 Циркуманальная область 11,2 1,2 11 из 9 11,0 974 81 89
31 Постанальная область, 10 см от анального отверстия 8,6 0,6 7 из 19 8,4 431 72 93 7,2
32 Середина постанальной области 8,6 0,6 7 из 19 8,4 397 66 8,0 93 10,0
33 Вентральная сторона основания хвоста, 59 см от анального отвер- стия 11,7 0,7 6 169 24 11,4 361 52 11,0 94
34 Вентральная поверхность основа- ния хвоста 6,6 0,6 9 из 19 6,3 318 53 6,0 91 -
35 Вентральная поверхность хвоста, 78 см от анального отверстия 5,7 0,7 12 из 16 5,5 474 68 5,0 88 -
36 Каудальный край хвоста 5,8 0,3 14 90 11 5,5 541 68 5,0 86
37 Бок шеи, 30 см от края морды 8,5 0,5 6 ИЗ 23 8,3 291 58 8,0 94
38 Плечо, 46 см от края морды 6,6 0,6 9 56 9 6,5 488 81 6,0 91 1,0
39 Бок тела, 73 см от края морды То же, 126 см 25,6 0,6 2 113 19 28,4 374 62 25,0 98
40 30,6 0,6 2 ИЗ 19 30,4 374 62 30,0 98
41 Бок тела, 164 см от конца морды То же, 204 см 32,5 0,5 2 ИЗ 23 32,3 274 55 32,0 99
42 19,5 0,5 2 34 7 19,4 353 71 19,0 97 1,5
43 То же, 227 см 29,7 0,7 2 113 16 29,5 474 68 29,0 98 16,1
44 Край хвостового плавника, 46 см от средней линии 14,8 0,8 5 ИЗ 14 14,6 574 72 1<0 95
45 Плечевой отдел грудного ласта 4,4 0,4 9 ИЗ 28 4,2 5,3 174 44 4,0 91
46 Середина наружной стороны ласта 5,6 0,6 И ИЗ 19 318 53 5,0 89
47 Наружная сторона грудного ласта 4,8 0,8 17 113 14 4,6 568 71 4,0 83
48 Передний край грудного ласта 8,7 0,7 8 79 И 8,6 601 86 8,0 92
49 Задний край грудного ласта 4,5 0,5 И 68 14 4,4 398 80 4,0 89
50 Середина внутренней стороны ласта 3,6 0,6 17 169 28 3,2 205 34 3,0 83 —
51 Внутренний край угла рта 11,5 0,5 4 ч 23 5 11,4 387 77 11,0 96
52 Роговая пластина губ 6,4 2,4 38 700 29 4,7 700 29 4,0 63 -
ца ламантина
деленной схеме (рис. 7) (максимальные значения промеров представле-
ны в табл. 1). Составлена схема распределения вибрисс; измерены
вибриссы и расстояние между ними. Проведено электронно-микроскопи-
ческое изучение пробы кожи с середины брюха. Материал фиксировали
в 2,5 %-ном глютаральдегиде на фосфатном буфере с постфиксацией
в 1%-ной OsO4. Образцы заливали в эпон 812, срезы окрашивали уранил-
ацетатом и цитратом свинца и исследовали в электронном микроскопе
JEM-100C.
ПОВЕРХНОСТЬ И ТОЛЩИНА КОЖИ
Окраска кожи изученной особи ровная, темно-серая, без пятен. Цвет
кожи зависит от ее состояния (Murie, 1872): высохшая кожа ламантина
имеет серую окраску, мокрая — черную. В отличие от описанных Мыори
особей у изученного нами взрослого самца отсутствуют тускло-желтые
участки кожи на морде. Известны индивидуальные вариации окраски ла-
мантина: с возрастом кожа светлеет, ее окраска варьирует у разных
особей от грязно-серой до голубова-
то-серой (Ronald et al., 1978). Встре-
чаются белые особи, по-видимому,
альбиносы. Южноамериканские ры-
баки по окраске различают до 4 раз-
новидностей ламантина. Поврежде-
ния кожи (от столкновений с мотор-
ными лодками, от гарпунов и пр.)
оставляют заметные темные пятна
на коже. Кожа новорожденных ла-
мантинов мягкая, гладкая и темпо-
серая, но уже через несколько дней
она грубеет (Ronald et aL, 1978), а у
взрослых покрывается бороздами и
Рис. 7. Схема промеров слоев кожи на
вертикальном срезе
1 — толщина кожи;
2 — толщина эпидермиса;
3 — толщина рогового слоя эпидермиса;
4 — толщина дермы;
5 — высота дермального сосочка;
6 — толщина подсосочкового слоя;
7 — толщина подкожной жировой клетчатки
а — а — линия горизонтального среза на рис. 91
48
1 — эпидермис; в — поверхностная борозда; 12 — пучок коллагеновых во
2 — дермальный сосочек; 7 — вибрисса; локон’
з — подсосочковый слой; 8 — роговой слой; 13 — кровеносные сосуды;
4 — подкожная жировая клет- 9 — клетки шиповатого слоя; 14 — эластиновое волокно;
чатка; ю — клетки базального слоя; 15 — жировая клетка
5 — поверхностное углубле- 11 — меланоцит;
ние;
49
Рис. 9. Эпидермис ламантина: поверхность кожи спины (проба № 10) с углубления-
ми в эпидермисе (а, б — под сканирующим электронным микроскопом)
« — ув. х 50;
б — ув. х 150;
в — вертикальный срез кожи
хвостового плавника (про-
ба № 15), ув. X 100;
г — горизонтальный срез кожи
хвостового плавника (про-
ба № 16) на уровне дер-
мальных сосочков (по ли-
нии а — а на рис. 7),
ув. X 25
1 — роговой слой;
2 — эпидермальная перегород-
ка;
з — дермальный сосочек;
4 — углубление («ямка») в эпи-
дермисе
50
складками (Dosch, 1915). У морской коровы окраска также была ровной
без пятен; цвет кожи изменялся от темно- до серо-коричневого (Steller,
1751). У изученного нами самца на коже обнаружены рачки рода Bala-
nus. Интересно, что на образцах кожи морской коровы также присутству-
ют ракообразные (Mohr, 1957) и другие кожные паразиты (Haffner,
1957).
Кожа ламантина образует крупные складки на морде, в подмышках,
в основании хвоста, описанные в литературе (Murie, 1872). У изучен-
ного нами самца складки кожи расположены сходно. Общий план строе-
ния кожи ламантина представлен на рис. 8.
Одна из специфических особенностей строения поверхности кожи
сирен, отличающих их от других млекопитающих (и в частности, от хо-
ботных и китообразных) — наличие многочисленных углублений —
«ямок» 2. Они имеют округлую форму (рис. 8, 9, а, б), выстланы рого-
вым слоем толщиной до 210 мкм и, сужаясь, глубоко проникают в эпи-
дермальные перегородки (рис. 9, в). Размеры «ямок» небольшие: на спи-
не, боках и хвосте их диаметр 0,1—0,5 мм, а на внутренней поверхности
ласта — 1,0—2,1 мм. Расстояние между «ямками» на ластах максимально
3,1 мм; на других участках тела колеблется от 0,7 до 1,0 мм. «Ямки»
соединяются неглубокими бороздами. В коже спины эти борозды идут
перпендикулярно длинной оси тела. В других участках (бок туловища,
хвостовой плавник, грудные ласты) борозды образуют сетчатый рисунок
(см. рис. 8). Ширина борозд на спине достигает 525 мкм, а на внутрен-
ней поверхности грудного ласта не превышает 35 мкм. «Ямки», кони-
чески сужаясь, вдаются в дерму, их внутренние стенки покрыты пятью-
шестью радиально расходящимися складками. С внутренней поверх-
ности эпидермиса складки «ямок» и борозды образуют звездчатую систему
гребней. Вся поверхность эпидермиса (в том числе и «ямки») покрыта
мелкими (диаметром до 339 мкм) округлыми бугорками, которые с внут-
ренней стороны эпидермиса выглядят как округлые отверстия — ячеи.
В них входят дермальные сосочки.
«Ямки» — углубления в коже морды, остающиеся у взрослых особей
после дегенерации ювенильных вибрисс, имеются у дельфинов (Palmer,
Weddell, 1964). Некоторые исследователи полагают, что у взрослых
животных фолликулы вибрисс превращаются в специализированные так-
тильные органы, реагирующие на изменения давления воды и на низко-
частотные колебания.
Интересно провести специальные исследования по онтогенезу кожи
ламантина, что, возможно, позволило бы выяснить происхождение кож-
ных «ямок». Как и у других современных сирен, кожа ламантина по-
крыта редкими вибриссами. Их описание приводится ниже.
Толщина кожи изученного самца достигает значительной величины
и колеблется от 3,6 мм (проба 50) до 32,5 мм (проба 41) (см. табл. 1),
что превышает пределы изменений толщины кожи, определенные ранее
для сирен (Barrett, 1935). Характер изменения толщины кожи (без учета
подкожной жировой клетчатки) различен на разных сторонах туловища
(рис. 10). Вдоль средней линии спинной поверхности толщина кожи рез-
ко увеличивается между лопатками (проба 10), достигает максимума в
середине спины (проба 13), затем резко уменьшается в области основа-
ния хвоста. Вдоль средней линии брюшной поверхности толщина кожи
2 В литературе этот вопрос специально не изучался. Судя по рисунку поверхности
кожи эмбриона дюгоня (Weber, 1928), поверхность кожи у этого вида также имеет
«ямки». Упоминания об углублениях в коже ламантинов и дюгоней также имеют-
ся в литературе (Dosch, 1915).
51
Г I L- .1_____L_
ч в 9 10 11
। । । ± и____
1819 ™ 22
I I I Z7 1_
2 3 37 39
t Плавники
Верхняя
губа.
29 27 29 31 33 35
25 28; 30 32 । 39
90 91 92
Рис. 10. Изменение толщины
кожи вдоль туловища взрос-
лого самца ламантина (без
учета подкожной клетчатки)
1 — по середине спины;
2 — по середине брюха;
з — по середине бока
93 Провы кажи
Анус
Основание
хвостового
плавника
колеблется с небольшой
амплитудой и достигает
максимальных значений в
середине брюха, в пост-
анальной области и в ос-
новании хвоста (проба
33). Вдоль середины бока
кожа резко утолщается в
средней и задней частях
туловища (пробы 39—41)
и резко уменьшается к ос-
нованию хвоста. Таким об-
разом, на туловище кожа
наиболее утолщена в сре-
динной части, что соответ-
ствует литературным дан-
ным по дюгоню (Domning,
1977) и африканскому ламантину (Matthes, 1929). Вместе с тем имеются
и отличия. Например, у дюгоня в отличие от ламантинов обнаружено
заметное утолщение кожи на дорсальной поверхности основания хвосто-
вого плавника; у ламантина в коже брюха не происходит плавного изме-
нения толщины кожи, как у дюгоня и африканского ламантина. У ламан-
тина и дюгоня толщина кожи брюха меньше, чем на спине и боках ту-
ловища. Однако у ламантинов кожа боков туловища толще, чем на спине,
а у дюгоня наоборот. У ранее исследованных взрослых ламантинов тол-
щина кожи сходна с таковой у изученного нами самца (Quiring, Harlan,
1953).
Сравнение абсолютных значений толщины кожи у разных сирен
и других морских млекопитающих по литературным сведениям не имеет
смысла из-за различий в размерах, возрасте, поле исследованных особей.
Вычислить относительную толщину кожи, сопоставив ее с каким-либо
поперечным промером, например диаметром туловища, невозможно ввиду
пренебрежения к этому параметру большинства исследователей. Поэтому
мы сделали попытку пересчитать собственные и литературные данные по
отношению к длине туловища, понимая при этом условность такого дей-
ствия. У нашего ламантина толщина кожи на спине (среднее по про-
бам 10—14) составляет 0,7%, на середине бока (39—41) — 0,93%,
брюха (23—24, 28—29) — 0,3%. Аналогичные цифры получаются по дру-
гим взрослым ламантинам: на спине 0,7%, на брюхе — 0,5% (Quiring,
Harlan, 1953). У эмбриона африканского ламантина (Matthes, 1929)
на спине и брюхе (по 6 пробам, взятым позади ластов) было всего 0,3%,
а на боку —0,5%. Возможно, с возрастом толщина кожи на спине уве-
личивается. У дюгоней (Domning, 1977) толщина кожи составляет соот-
ветственно 1,1; 0,7; 0,6%, это подтверждает вывод о том, что у дюгоня
кожа толще, чем у ламантинов, и максимально утолщена не на боку,
а на спине.
У китообразных кожа считается наиболее толстой, но в отношении
52
к длине тела (Соколов, 1973) она составляет 0,3—1,5%, что вполне сопо-
ставимо с относительной толщиной кожи сирен. Так, у близких по раз-
мерам к нашему ламантину короткоголового (самка) и полосатого
(самец) дельфинов толщина кожи составляет на спине 0,6 и 0,5%,
на боку 0,5 и 0,3%, на груди 0,5 и 0,4%, т. е. у дельфинов кожа на
спине и боках тела несколько толще, чем у ламантина, а на груди толще
(у ламантина по пробам 20—22 на груди всего 0,2%). В отличие от
сирен у китов толщина кожи на протяжении туловища более или менее
постоянна (Соколов, 1973).
ЭПИДЕРМИС
Эпидермис толстый, состоит из четырех слоев: базального, шиповатого,
зернистого (этот слой различим на электронно-микроскопическом уров-
не), рогового (рис. 11). Толщина эпидермиса в различных участках тела
неодинакова и изменяется в значительных пределах (см. табл. 1, рис. 12).
На спинной поверхности эпидермис более тонок на верхней губе
(проба 4), заметно толще и более или менее одинаков на передней поло-
вине туловища, но несколько утолщается к середине (12), а затем почти
вдвое становится тоньше, чтобы увеличиться к хвостовому плавнику.
Относительная же его толщина (в процентах к толщине кожи) больше
па переднем конце морды и в области основания хвоста. Толщина эпи-
дермиса на брюхе меньше на середине горла, далее назад она возрастает,
но неравномерно с локальными пиками (пробы 22, 28, 30) в местах, где
возрастает и толщина кожи.
Ближе к хвосту эпидермис не-
сколько толще, чем впереди у
ластов. Относительная толщи-
на по брюху меняется очень
неравномерно (имеются места с
заметным увеличением толщи-
ны эпидермиса, как у проб 19,
23, 30, или, наоборот, с утонь-
шением — пробы 24, 31, 32), и
лишь к хвосту наращивание
толщины эпидермиса становит-
ся закономерным. В целом эпи-
дермис на брюхе относительно
толще, чем на спине и боках
тела.
Толщина эпидермиса на бо-
ках более или менее одинакова,
за исключением таких спе-
цифических мест, как около
слуховых отверстий, где он рез-
ко утолщен. Наблюдается неко-
Рис. И. Вертикальный срез кожи
спины (проба № 10) ламантина,
ув. Х50
1 — роговой слой эпидермиса;
2 — шиповатый слой эпидермиса;
з — базальный слой эпидермиса;
4 — эпидермальная перегородка;
5 — дермальный сосочек;
•6 — подеосочковый слой
53
2 3 37 39 40 41 42 43 Пробы коти
Рис. 12. Изменение толщины эпидермиса вдоль туловища ламантина
z — по середине спины; 2 — по середине брюха; з — по середине бока
торое возрастание толщины к хвосту, как это было замечено и на других:
участках тела. Однако относительная толщина эпидермиса на боковых
частях головы значительно больше (более чем в пять раз), чем на самом
туловище, где она поддерживается на более или менее постоянном уров-
не (около 2% от толщины кожи) до самого хвостового плавника. Таким
образом, как правило, более толстая кожа покрыта более тонким эпидер-
мисом, а дерма соответственно утолщается. В коже туловища само соот-
ношение эпидермиса и дермы можно определить следующим образом:
вдоль средней линии спины (пробы 10—14) 1 : 30, груди (20—22) 1 : 10,.
брюха (23, 24, 28, 29) 1 : 13, бока (39—43) 1 : 47.
Обратную зависимость между степенью развития эпидермиса и дермы
можно объяснить образованием более мощного защитного эпидермально-
го слоя при слабом развитии дермы, как это было высказано при обсужт
дении строения кожи морской коровы (Matthes, 1929). Действительно,,
вокруг туловища тонкий эпидермис имеется в самой толстой боковой
коже и самый толстый эпидермис — в тонкой коже брюха. Сходное со-
отношение этих слоев кожи найдено на хвостовом плавнике: на каудаль-
ном краю плавника, где толщина кожи незначительна (проба 17, 36),
эпидермис составляет 1/4—1/7 толщины дермы; на вентральной поверх-
ности (34, 35)—1/10—1/8 при толщине кожи 6,6—5,7 мм, в то время
как на дорсальной стороне кожа толще (15, 16), а эпидермис тоньше —
1/22—1/19. На грудном плавнике эта закономерность также подтверж-
дается: на вентральной стороне ласта (50) эпидермис составляет 1/5 дер-
мы при толщине кожи 3,6 мм, а на утолщенном переднем краю
(48; 8,7 мм) — 1/12. В то же время показано, что у китообразных тол-
щина эпидермиса увеличивается на участках кожи, подвергающихся
большей механической нагрузке (передние края плавников, передняя
часть головы) (Соколов, 1973).
У ламантина эпидермис утолщен на голове и органах движения:
на каудальном краю хвостового плавника (принимающего активное учас-
тие в передвижении этого зверя), на наружной и внутренней поверх-
ностях ласта (см. табл. 1). Утолщенный эпидермис защищает ушные от-
верстия, ноздри, глаза, губы, анальное отверстие и препуций.
По литературным данным, толщина эпидермиса у ламантина (проба
кожи не указана) составляет 1.16 толщины дермы (Murie, 1872), что
сходно с относительными промерами эпидермиса груди и брюха изучен-
ной нами особи. Но ни в одной из проб кожи этого экземпляра толщина
эпидермиса туловища не достигает абсолютного значения, указанного»
Л4
ранее (Murie, 1872) — 1,6 мм, как и значений, приводимых для родов
Trichechus и Dugong — 2,5 мм (Matthes, 1929). Промеры толщины эпи-
дермиса исследованного самца более сходны с данными В. Е. Соколова
(1973) для других сирен: толщина эпидермиса в коже спины дюгоня
равна 0,6, а у бразильского ламантина — 1,0 мм. Отношение эпидермиса
и дермы (в данной пробе кожи) сходно с нашими данными по ламантину
п равно: у дюгоня на спине — 1 : 30, на груди — 1 : 15; у бразильского
ламантина соответственно 1 : 22 и 1 : 20 (подсчитано по данным Соколо-
ва (1973), табл. 69, с. 330).
Среди сирен эпидермис достигал максимальной толщины (25,5 мм)
у морской коровы и в отличие от эпидермиса ламантинов и дюгоня он
был в пять раз толще дермы (5 мм) (Steller, 1749). По данным Хаффне-
ра (Haffner, 1957), толщина эпидермиса в исследованных им образцах
кожи морской коровы достигла 7,5 мм, а толщина дермы — 5—10 мм.
Высказано предположение, что у этого вида отсутствовала четкая грани-
ца между дермой и подкожной жировой клетчаткой (как это имеет место
у китов (Соколов, 1973) и поэтому ее толщина — относительно мини-
мальная среди сирен — занижена (Matthes, 1929). Возможно, нижняя
часть дермы была срезана при взятии проб. Вероятное объяснение силь-
ному развитию эпидермиса, грубой бугорчатой поверхности кожи этого
вида — необходимость усиления защитных и теплоизолирующих свойств
кожи при обитании в прибрежных мелководных участках холодного
моря, а также во льдах. Неудивительно, что для эпидермиса морской
коровы были характерны эластичность, упругость, отслаивание толстых
поверхностных слоев эпидермиса (десквамация) (Haffner, 1957).
Толщина рогового слоя изменяется в значительных пределах
(см. табл. 1). Его абсолютная толщина максимальна на губах (про-
ба 7, 52) и минимальна на боку туловища (41). Относительная толщина
рогового слоя3 (в процентах к толщине кожи) не превышает 29%,
за исключением внутренней кожи препуция (39%). В некоторых участ-
ках кожи наблюдается обратная зависимость между толщиной эпидермиса
и толщиной рогового слоя. Это объсняется одной из основных функций
рогового слоя — механической защитой кожи. Роговой слой максимально
развит на губных пластинах, у углов рта, на плавниках, на препуции
/см. табл. 1). Как у ламантина и китообразных, у дюгоня роговой слой
толще на участках кожного покрова, подвергающихся большой механи-
ческой нагрузке (Соколов, 1973).
Внутренняя поверхность эпидермиса имеет сложную ячеистую струк-
туру. Наружные отделы эпидермиса образуют тонкий сплошной слой,
от которого в глубь дермы идут толстые эпидермальные перегородки,
делящиеся на более мелкие вторичные перегородки. Вторичные пере-
городки заканчиваются сосочковидными врастаниями в дерму (см. рис. 8).
Поверхностный сплошной слой состоит из рогового, трех—шести рядов
клеток базального, шиповатого и зернистого слоев, и его толщина, как
правило, не превышает ИЗ мкм. Число рядов клеток в эпидермальных
перегородках (в направлении, перпендикулярном к поверхности кожи)
колеблется от 12 (при толщине эпидермиса 0,7 мм) в середине морды
между глазами до 91 на краю нижней губы (при толщине эпидерми-
са 0,8 мм). Число и форма эпидермальных перегородок на 1 мм вер-
тикального среза неодинаковы в разных участках кожи; число перегоро-
док максимально (85) в углах рта и на нижней губе и минимально (19)
3 Толщина рогового слоя в среднем составляет 1/7 толщины эпидермиса, но в неко-
торых пробах (14, 16, 19, 25, 33, 36, 37, 45, 51, 52) она равна 1/2 —1/4 его толщины,
что сходно с относительным развитием рогового слоя у морской коровы (Haffner,
1957).
55
на середине горла — проба 19. В коже спинной поверхности оно не пре-
вышает 41, на груди и брюхе — 25, на боку — 32. На грудном ласте пере-
городки расположены чаще (до 46 на 1 мм среза), чем в коже туловища.
Перегородки направлены не только вертикально в глубь дермы, но и
разветвляются во всех трех плоскостях. Форма поперечника эпидермаль-
ных ячей и соответственно дермальных сосочков изменяется на разных
уровнях от округлой до звездчатой. Звездчатое строение эпидермальных
ячей американского ламантина не только увеличивает площадь соедине-
ния эпидермиса и дермы, а следовательно, и укрепляет это соединение,
но и увеличивает поверхность продуцирующего слоя эпидермиса
Последнее способствует быстрому нарастанию эпидермиса и его рогового
защитного слоя, что важно для восстановления рогового слоя при по-
стоянном его слущивании в плотной водной среде.
ДЕРМА
Дерма четко отделена от эпидермиса и образована дермальными сосоч-
ками и подсосочковым слоем4 5. Толщина дермы изменяется в разных
участках кожного покрова и составляет 52 (проба 52) — 95 % (41) тол-
щины кожи. На морде дерма достигает максимальной толщины на верх-
ней губе, покрытой вибриссами; вдоль середины спины ее толщина
колеблется незначительно; дерма гораздо тоньше на брюшной стороне и
на ластах (см. табл. 1). Высота дермальных сосочков у ламантина колеб-
лется в значительных пределах: она составляет от 29 (проба 52) до
(85%) (3) толщины эпидермиса (см. табл. 1). Высота дермальных сосоч-
ков отличается на обеих сторонах грудного ласта: они короче на его
внутренней поверхности. На хвостовом плавнике дермальные сосочки
хорошо развиты по краям плавника и на его вентральной поверхности.
На дорсальной поверхности хвоста высота сосочков занимает немного
более половины толщины эпидермиса. Таким образом, дермальные сосоч-
ки у американского ламантина, как и у других сирен (Dosch, 1915;
Haffner, 1957; Соколов, 1973), подходят близко к поверхности кожи;
в некоторых участках они почти достигают рогового слоя. Вероятно, это
повышает чувствительность кожи.
Наши данные не подтверждают зависимости высоты дермальных со-
сочков от величины механической нагрузки, выявленной у китообразных
(Соколов, 1973). Видимо, демпфирующие свойства кожи у сирен зависят
не от высоты дермальных сосочков, а от их распределения по телу
и формы. У нашего ламантина, как и у бразильского (Соколов, 1973),
не обнаружены дермальные валики, имеющиеся у китов.
Надо отметить, что сосочковидное сочленение эпидермиса и дермы
характерно не только для настоящих водных (сирены, киты) и полувод-
ных (морж, бегемот) млекопитающих. Такое сочленение имеется в утол-
щенных и обычно лишенных волос участках кожи наземных млекопитаю-
4 Мы попытались измерить с помощью курвиметра длину наружного и внутрен-
него слоев эпидермиса на рисунках, изготовленных на рисовальном аппарате с
вертикальных срезов. На вертикальных срезах длина базального слоя в различных
участках кожного покрова увеличивается за счет перегородок в 1,2—4 раза по
сравнению с более гладкой поверхностью кожи, т. е. предположительно площадь
эпидермиса уменьшается от внутренней его стороны к наружной, а площадь дер-
мы, напротив, увеличивается к границе с эпидермисом и уменьшается к подкож-
ной клетчатке. Максимальное увеличение поверхности базального слоя отмечено
на ластах и хвостовом плавнике, что подтверждает усиление продуцирующей спо-
собности базального слоя в участках кожи, подвергающихся большей механиче-
ской нагрузке.
5 Эти слои выделяются нами условно. Их сопоставление с сосочковым и сетчатым
слоями дермы наземных млекопитающих находятся вне интересов монографии.
56
1цих: на подошвах лап, на носу, губах. По-видимому, такое сочленение,
помимо увеличения прочности и продуцирующей способности, придает
коже упругость, что доказано в экспериментах (Haffner, 1957). По дан-
ным Хаффнера, эластичность мокрой кожи морской коровы соответствует
таковой автомобильной резины. У морской коровы, как и у ныне живу-
щих сирен, эпидермис и дерма соединяются взаимопроникающими выро-
стами. Своеобразие сочленения эпидермиса и дермы у этого вида заклю-
чается в том, что дермальные сосочки имеют простую цилиндрическую
форму, небольшой диаметр, сильно вытянуты по высоте и их число на
единицу площади очень велико. Эти особенности затушевывают сходство
общего плана строения кожи у морской коровы и ныне живущих сирен,
и неудивительно, что некоторые авторы (Haffner, 1957) вообще отрицают
это сходство.
Дермальные сосочки содержат рыхлые тонкие (толщиной 11 — 17 мкм)
пучки коллагеновых волокон, многочисленные кровеносные сосуды, ред-
кие эластиновые волокна. Пучки коллагеновых волокон идут параллель-
но длинной оси сосочка (перпендикулярно поверхности кожи). Диаметр
кровеносных сосудов в дермальных сосочках равен 5—10 мкм, т. е. со-
суды преимущественно капиллярного типа. Их стенки образованы эндо-
телиальными клетками. Последние имеют веретеновидную форму: утол-
щены в области расположения ядра, а к периферии уплощены. На по-
перечном срезе сосуд содержит 5—7 клеток. Капилляры в дермальных
сосочках идут вдоль пучков коллагеновых волокон и имеют вид петель
или мелкоячеистых сетей. В основании дермальных сосочков капилляры
образуют разветвленную капиллярную сеть, простирающуюся параллель-
но поверхности кожи.
Длина капиллярной сети, заключенной в 1 мм3 сосочков, неодинакова
в различных участках кожного покрова и варьирует от 29,4 до 74,5 мм
Место взятия пробы (п = 5) Длина капиллярной сети в 1 мм3 (в мм) Поверхность капилляр- ной сети в 1 мм3 (в кв. мм)
Губа 31,7 1035,1
Ухо 51,4 1675,6
Грудь 29,4 958,4
Плечо 44,7 1457,2
Спина 27,5 896,5
Бок 33,0 1075,8
Брюхо 34,0 1108,4
Грудной ласт: Дорсальная сторона 54,0 1760,4
Вентральная сторона 34,4 1121,4
Хвостовой плавник 74,5 2448,7
Примечание. Поверхность капиллярной сети вычисляли по формуле 8=лЯ2Л,
где 8 — поверхность капиллярной сети; Я—радиус капилляра; h — длина капил-
ляра в 1 мм3.
Определение плотности капилляров проводили на гистологических препаратах
по методу G. М. Блинкова и Г. Д. Моисеева (1961), с использованием формулы
для капиллярных сетей: L = 2пс, пс = Nc/S, где L — длина капилляра в 1 мм3
подэпидермальной зоны, пс — плотность (число) концов капилляров на 1 мм2,
Nc — число концов капилляров, приходящихся на сетку окулярмикрометра, 8 —
площадь в 1 мм2 сетки окулярмикрометра при данном увеличении. Подсчет про-
водили по пяти полям зрения каждого образца.
Различная плотность капиллярной сети свидетельствует о разном
уровне метаболизма в коже рассматриваемых участков тела американ-
ского ламантина. Чем больше длина капиллярной сети в единице объема.
57
тем выше уровень метаоолизма, т. е. перенос продуктов оомена через
стенку капилляра. Этот уровень зависит не только от длины капилляр-
ной сети, но и от величины ее поверхности, через которую осуществляет-
ся транспорт веществ. Как известно, величина диаметра капилляра свя-
зана с физиологическим состоянием организма, температурой окру-
жающей среды. Полученные нами данные о величине поверхности
капиллярной сети в коже ламантина свидетельствуют, что в участках
тела, обладающих наибольшей подвижностью (хвостовой плавник, груд-
ной ласт, губы), поверхность капиллярной сети, заключенная в единице
объема кожи, наибольшая (т. е. уровень метаболизма здесь высок),
а в коже туловища (спина, грудь) ее величина снижается более чем
в два раза.
Диаметр и строение капилляров в коже ламантина изменяется в раз-
ных участках тела. На границе сосочков и подсосочного слоя длина
капилляров резко уменьшается, а диаметр капилляров увеличивается
до 23 мкм. В этих сосудах, проходящих через подсосочковый слой, рас-
полагаются многочисленные артерио-венозные анастомозы, достигающие
в наружном диаметре за счет резкого увеличения толщины стенок 113 мкм.
Под эндотелием этих анастомозов находятся специфические эпителиоид-
ные клетки, которые хорошо выявляются при окраске гематоксилин-
эозином. Артерио-венозные анастомозы имеют форму прямых или изви-
тых трубочек разной длины и диаметра. В одном участке кожи встре-
чаются различные артерио-венозные анастомозы: с зияющей полостью,
частично или полностью закрытой полостью. Это свидетельствует об их
различном функциональном состоянии. Анастомозы, имеющие закрытую
полость, не могут пропускать кровоток из артерий в вену, и кровь пол-
ностью направляется в капиллярную сеть, лежащую под эпидермисом.
Открытые анастомозы, напротив, дают возможность перехода кровотока
из артериальных сосудов кожи в венозные, минуя капиллярную сеть,
располагающуюся под эпидермисом. Принимая во внимание способность
артерио-венозных анастомозов регулировать ток крови, направляя его к
поверхности кожи или в глубь организма, следует заметить, что такая
ангиоархитектоника кровеносных сосудов кожи способствует выполнению
терморегулирующей функции. В тех случаях когда возникает необходи-
мость отдать избыток тепла организма во внешнюю среду, артерио-веноз-
ные анастомозы закрываются и весь кровоток поступает в терминальные
подэпидермальные капиллярные сети. Если, наоборот, необходимо сохра-
нить тепло, то анастомозы раскрываются и пропускают ток крови в ве-
нозную систему, минуя поэпидермальную сеть. Возможно, проникновение
сосудов в дермальные сосочки ближе к поверхности тела облегчает
терморегуляторные процессы.
Толщина пучков коллагеновых волокон различна в поверхностных и
глубоких слоях подсосочного слоя. Тонкие (толщиной 17—23 мкм) по-
верхностные пучки коллагеновых волокон в нижних частях дермы утол-
щаются до 56—79 мкм. Переплетение пучков различно в разных участ-
ках кожного покрова. При беспорядочном «хаотичном» переплетении
толщина пучков незначительна (не более 17 мкм), при ромбовидном или
взаимно перпендикулярном пучки имеют большую толщину.
Характер переплетения пучков коллагеновых волокон в подсосочко-
вом слое является одной из адаптаций кожного покрова к преодолению
механических нагрузок. У наземных млекопитающих «механический
слой» (Браун, 1945) (сетчатый слой кожи) различен на спинной и брюш-
ной сторонах тела. На брюхе кожа более тонкая и эластичная, тогда как
на спине, где она испытывает большую силу натяжения, кожа, толстая
и состоит из мощных пучков коллагеновых волокон, переплетающихся во>
58
взаимно перпендикулярном направлениях. У водных зверей структура
подсосочкового слоя определяется однотипностью натяжения. У китооб-
разных и тюленей пучки коллагеновых волокон располагаются диаго-
нально к длинной оси тела. Такая ориентация волокон наиболее выгодна
при плавательных движениях (Браун, 1945). Сходное направление воло-
кон описано в коже новорожденного ламантина (Dosch, 1915). У изучен-
ного нами ламантина кожа частей туловища (бок, спина), на которые
воздействует постоянный силовой вектор определенного направления,
имеет взаимно перпендикулярную или ромбовидную вязь грубых толстых
пучков коллагеновых волокон (рис. 13, а), что отмечается и у других
представителей отряда (Haffner, 1957; Соколов, 1973). На границе с под-
кожной жировой клетчаткой толстые тяжи коллагеновых волокон тянут-
ся параллельно поверхности кожи в отличие от китов, у которых пучки
коллагеновых волокон проходят так лишь на границе подкожной жиро-
вой клетчатки и подкожной мускулатуры. По-видимому, такое располо-
Рис. 13. Вязь пучков коллагеновых волокон в подсосочковом слое
« — толстые пучки коллагеновых волокон переплетаются во взаимно перпендикулярных направле-
ниях (проба № 39); б — тонкие пучки коллагеновых волокон переплетаются в различных направле-
ниях под разными углами (проба № 46), х 100
59
жение пучков коллагеновых волокон в коже туловища характерно в
целом для ламантинов (Brandt, 1871). Кожа активно двигающихся
частей (хвостовой плавник, ласты, препуций, голова), подверженных
механическому воздействию, меняющему свое направление, имеет петли-
стое «хаотичное» (иногда ромбовидное) переплетение тонких пучков кол-
лагеновых волокон (рис. 13, в).
Эластиновые волокна немногочисленны в подсосочковом слое, про-
ходят вдоль пучков коллагеновых волокон. Их толщина составляет не
более 2 мкм в коже плеча и 6 мкм в коже брюха. Эластиновые волокна
многочисленнее в коже брюха, чем на спине. В коже углов рта они
могут образовывать неплотное сетевидное переплетение, что способствует
эластичности губ ламантина.
ПОДКОЖНАЯ ЖИРОВАЯ КЛЕТЧАТКА
На взятых образцах кожи ламантина подкожная жировая клетчатка раз-
вита неравномерно (см. табл. 1). Ее толщина изменяется в значительных
пределах. Жировые клетки лежат сплошным слоем, не разделены на
группы, имеют крупные (до 113X113 мкм) размеры. Пучки коллагено-
вых волокон направлены под острым углом к поверхности кожи (на вер-
тикальных срезах). Толщина многочисленных одиночных эластиновых
волокон не превышает 6 мкм. Они не образуют сетевидного переплете-
ния, а, извиваясь, идут вдоль пучков коллагеновых волокон.
Неравномерное развитие подкожной жировой клетчатки, по-видимо-
му, характерно для сирен, оно варьирует и в зависимости от места и от
индивидуальных особенностей животного и, в частности, зависит от упи-
танности зверя. Так, она «отсутствует» 6 у неполовозрелой самки дюгоня
(Domning, 1977b), в то время как у новорожденного дюгоня подкожная
жировая клетчатка имеется в коже спины и груди (Соколов, 1973).
У молодого бразильского ламантина подкожная жировая клетчатка до-
стигает толщины 12,3 мм на спине и 9,1 мм на груди (Соколов, 1973).
У ламантина жир в подкожной клетчатке развит по-разному у разных
особей. У взрослого самца он имеется лишь в отдельных местах (наши
данные); а у молодого самца отсутствовал. В то же время самка (Murie,.
1872, р. 134) «была жирная как свинья, у нее сразу под кожей лежал
слой необыкновенно плотного жира, который обертывал все тело».
На спине толщина жира достигала 2,5—5,0 см. На брюхе она Не превы-
шала 2,5 см. Вес этого жира составлял ‘/в веса тела самки.
Судя по литературным описаниям, подкожная жировая клетчатка
была очень сильно развита у морской коровы. Она была «толщиной
с ладонь» (Steller, 1751), что равно примерно 10 см. Возможное объяс-
нение этого явления — обитание морской коровы в холодных водах.
По-видимому, у этого вида подкожная жировая клетчатка (как и у ки-
тов, у которых она сильно развита, а жировые клетки в зимнее время
имеются во всех слоях дермы (Соколов, 1973)) служила термоизоля-
ционным слоем.
У нас нет данных по изменчивости подкожной жировой клетчатки в
течение года; сезонные изменения степени ее развития могут оказаться
значительными.
6 Подкожная клетчатка как комплекс соединительнотканных фасций, обеспечиваю-
щих связь кожи с подкожной мускулатурой, не может отсутствовать, но малое
количество жировых клеток говорит о ее отсутствии.
60
ВИБРИССЫ
Сплошной волосяной покров и специфические кожные железы (за исклю-
чением желез век и молочных желез самок), а также обычные потовые
железы отсутствуют.
Редкие на туловище и густые на морде волоски представляют собой
вибриссы. Вибриссы располагаются по всему телу, но наиболее много-
численны на морде (рис. 14). Соответственно расстояние между вибрис-
сами на морде минимально, а максимально на середине спины. Наиболее
постоянно расстояние между вибриссами на голове: возле глаз, на верх-
ней и пижней губах, на горле, груди; максимальный разброс в расстоя-
нии между вибриссами наблюдается на середине спины, на внутренней
поверхности ласта, на затылке. Основные примеры, касающиеся самих
вибрисс, приведены ниже.
У дюгоня (исследованы две взрослые самки) вибриссы реже распо-
ложены по бокам туловища, на вентро-латеральной поверхности и на
хвосте (Kamiya, Yamasaki,1981), что вполне сходно с расположением
вибрисс у ламантина.
Редкие бесцветные вибриссы описаны у ламантина (Murie, 1872).
В отличие от изученного нами ламантина, у которого длина вибрисс
изменяется от 5 до 17 мм, у исследованной Мьюри самки (около 160 см
длины) длина вибрисс туловища равна примерно 38 мм (1,5 дюйма),
но может достигать и 50 мм (2 дюйма), т. е. длина вибрисс ее в не-
сколько раз превышает промеры вибрисс исследованного нами самца.
Самые короткие вибриссы найдены на нижней и верхней губах и в уг-
лах рта, а самые длинные на загривке, боку, в основании хвоста. Ни
вентральной поверхности тела и на ластах вибриссы короче, чем на дор-
сальной поверхности.
Рис. 14. Размещение вибрисс на теле (а)
и на морде (б) взрослого самца ламантина
Кружком с точкой обозначены толстые вибриссы
61
Участок тела, тип вибрисс Толстые вибриссы Длина наружной части вибриссы (w = 5, lim, мм) Расстояние меж- ду вибриссами (n = 5, lim, мм)
Верхняя губа, нижняя губа, угол рта Промежуточные вибриссы 5-7 5-15
Нижняя губа 5-10 4-13
Тонкие вибриссы Вокруг глаза 10-12 20-25
Затылок 10-12 15-30
Загривок 15-17 21-45
Середина спины 10-13 34-59
Дорсальная поверхность основания хвостового плавника 12-15 25-30
Середина бока 10-15 35-40
Горло 10-11 16-18
ГРУДЬ 9-10 30-32
Брюхо 8-9 40-45
Ано-генитальная область 9-10 21-35
Дорсальная поверхность хвостового плавника 8-9 34-36
Вентральная поверхность хвостового плавника 7-9 29-39
Наружная поверхность грудного ласта 8-12 15-25
Внутренняя поверхность грудного ласта 7-9 10-35
По толщине и строению вибриссы делятся на две основные группы.
Первую составляют тонкие вибриссы (диаметр стержня 0,05—0,06 мм),
располагающиеся по всему телу. На верхней и нижней губах и в углах
рта имеются 84 толстые и короткие вибриссы диаметром до 1,5 мм, от-
носящиеся ко второй группе. Кроме этих двух групп, на верхней и
нижней губах имеется еще промежуточный тип — вибриссы диаметром
0,9 мм. На морде вибриссы располагаются беспорядочно, концентрируясь
в углах рта (см. рис. 14). На роговых пластинках губ также найдены
тонкие вибриссы.
У дюгоня, как и у ламантина, вибриссы делятся на две основные
(длинные тонкие и толстые короткие) и промежуточную (длинные
утолщенные) группы. На верхней губе длина тонких вибрисс составляет
15—20 мм, а толщина 1—3 мм; длина толстых вибрисс 5—7 мм, что
сходно с размерами губных вибрисс американского ламантина (Kamiya.
Yamasaki, 1981).
Строение вибриссной сумки ламантина в целом типично для млеко-
питающих (Andres, 1966). Однако имеются и отличия. Вибриссная кап-
сула изученного нами ламантина имеет мощные коллагеновые стенки,
образующие своеобразный коллагеновый мешок, в котором помещается
стержень вибриссы (рис. 15). Детали строения и размеры составных
элементов фолликула отличаются у тонких и толстых вибрисс.
Фолликулы тонких вибрисс устроены наиболее просто. Толщина
стенки коллагенового мешка и диаметр кровеносных лакун незначитель-
ны (табл. 2). Кольцевой валик и кольцевой синус имеются. Около каж-
дого фолликула располагаются четыре-пять голокриновых желез удли-
ненно-мешковидной формы (рис. 15, а). Они лежат на глубине 700 мкм
от поверхности кожи, имеют небольшие размеры, и их выводные прото-
ки впадают в вибриссную сумку. Проток каждой железы широкий
(диаметром 79 мкм) и довольно длинный (146 мкм). Секреторный эпи-
телий типичный голокриновый: герминативный слой однорядный, его
62
клетки плоские с темным ядром удлиненной формы. Длина выбриссной
сумки неодинакова в разных участках кожного покрова (см. табл. 2).
У тонких вибрисс кожи брюха найдены мышцы, поднимающие их.
Мышцы прикрепляются к середине вибриссной сумки и имеют толщину
525 мкм не вертикальном срезе. Рудиментарные мышцы, поднимающие
вибриссы, найдены также у эмбриона дюгоня (Petit, 1955). У ламанти-
на нерв диаметром 56 мкм пронизывает коллагеновый мешок на уровне
нижнего кавернозного синуса. Иннервация вибрисс морды изучена у
дюгоня (Kamiya, Yamasaki, 1981). Как и у нашего ламантина, толстый
нервный тяж (ответвление тройничного нерва) пронизывает коллагено-
вую капсулу сбоку в нижней трети. Затем нерв резко поворачивает вверх
вдоль вибриссы и дает терминальные окончания в области сосочка луко-
вицы и на стекловидной мембране. Иннервация туловищных и головных
вибрисс различна (Kamiya, Yamasaki, 1981) и требует специального ис-
следования.
У тонких вибрисс сердцевина может отсутствовать на всем протяже
нии или быть развитой только в нижней части стержня (71% толщины
стержня). Поперечные срезы вибрисс имеют округлую форму. Корковый
слой пористый.
Фолликулы толстых вибрисс углов рта имеют сравнительно толсто-
стенную коллагеновую оболочку и хорошо развитые кавернозные сину-
Рис. 15. Строение фолликула тонкой вибриссы на вертикальном срезе кожи груди
(а) и на горизонтальном срезе кожи угла рта (б) взрослого самца ламантина, Х43
1 — эпидермис; 2 — дерма; 3 — коллагеновая капсула вибриссы; 4 — кольцевой кровеносный синус;
5 — кольцевой валик; в — голокриновые железы; 7 — стержень вибриссы
63
Таблица 2. Промеры основных структур вибриссной сумки (в мм; п=5)
Проба кожи, тип вибрисс Вибриссная сумка 1 Толщина Размеры го- локриновых желез Диаметр вибриссы
длина диаметр коллаге- нового мешка кровенос- ного си- нуса
Толстые вибриссы Верхняя губа 13,2 5,2 1,о 1,о — 1,5
Промежуточные вибриссы Нижняя губа — 1,9 0,4 0,3 — 0,9
Тонкие вибриссы Середина брюха — 2,5 0,5 0,6 — 0,2
Грудь 1,1 1,0 0,1 0,2 0,02X0,01 0,05
Хвостовой плавник 2,2 1,4 0,01 0,5 0,02X0,005 0,06
Грудной ласт - 0,9 0,03 0,3 - 0,06
сы (см. табл. 2). Их строение типичное: хорошо выражен кольцевой со-
единительнотканный валик (не найденный у вибрисс дюгоня (Kamiya,
Yamasaki, 1981)), кольцевой синус и стекловидная мембрана. Различи-
мы меркелевы клетки. Голокриновые железы не обнаружены, что отли-
чает этот тип вибрисс от тонких вибрисс туловища. Особенность строе-
ния фолликулов толстых вибрисс состоит также в симметричном враста-
нии продольных (вдоль вибриссной сумки на вертикальном срезе)
эпидермальных перегородок в коллагеновую капсулу. Для апикальной
части волосяного влагалища характерно наличие эпидермальных перего-
родок и дермальных сосочков.
Толстые вибриссы имеют примерно одинаковую толщину на всем про-
тяжении, лишь их апикальные концы сильно стерты и закруглены (что
подтверждает большую механическую нагрузку на концевые отделы
вибрисс). Толстые вибриссы светлые и грубые, в поперечнике имеют
округлую форму. В отличие от тонких вибрисс у толстых сердцевина
всегда имеется, она занимает 56% основания вибриссы и сужается к ее
вершине. Корковый слой плотный, с неровной внутренней поверхностью.
Кутикула образована крупными и мелкими чешуйками различной фор-
мы. Свободные края чешуек сильно изломаны.
Разделение вибрисс на толстые губные и тонкие туловищные харак-
терно и для других представителей отряда. Так, у морской коровы длина
толстых («толщиной с гусиное перо») белых вибрисс губ достигала
38 мм, что значительно превышает длину вибрисс губ ламантина, а объ-
ясняется, по-видимому, не только более крупными размерами морской
коровы, но и более активным участием вибрисс в пищевом поведении
у этого зверя. По литературным описаниям, с помощью межгубных ще-
тин (вибрисс?) морская корова очищала и срезала стебли растений
(цит. по: Гептнер, 1967). У ламантина толстые прочные с мощной ме-
ханической структурой вибриссы губ также участвуют в пищедобыва-
тельной деятельности (аналогично вибриссам моржа) (Hartman, 1979).
Вибриссы губ дюгоня, как и вибриссы ламантинов, участвуют в отборе
пищи (Kamiya, Yamasaki, 1981).
В отличие от ныне живущих сирен у морской коровы вибриссы были
многочисленнее на вентральной поверхности запястья и пясти: их длина
«оставляла здесь 12—13 мм. Эти вибриссы были жесткими и упругими и
64
служили своеобразной «метелкой», функция которой недостаточно ясна:
возможно, она способствовала передвижению и удержанию на дне или
помогала зверю вырывать водоросли (Steller, 1751).
Распределение вибрисс по телу сирен значительно отличается от та-
ковой у других водных и полуводных млекопитающих — китообразных и
ластоногих, у которых вибриссы имеются только на голове. У усатых
китов вибриссы сконцентрированы на нижней челюсти, на дорсальной
поверхности верхней челюсти, а у тюленей (кроме моржа) — у глаз,
у ноздрей и на верхней губе (Яблоков, Клевезаль, 1964).
Сравнение с вибриссами китообразных и ластоногих показало, что по
абсолютной толщине вибриссы углов рта и губ сходны у моржа, хохла-
ча, гренландского тюленя (Яблоков, Клевезаль, 1964) и ламантина.
Используя предложенный А. В. Яблоковым и Г. А. Клевезаль (1964)
коэффициент погружения вибриссы (отношение корня вибриссы к ее
наружной части) можно определить, что утолщенные вибриссы верхней
и нижней губ и углов рта ламантина лежат так же глубоко в дермальном
слое (коэффициент равен 1,76), как и вибриссы усатых китов (коэффи-
циент изменяется у разных видов от 1,16 до 2,06 (Яблоков, Клевезаль,
1964)). Тонкие вибриссы туловища ламантина залегают сравнительно
неглубоко — коэффициент погружения у вибрисс груди составляет
0,11, у вибрисс хвостового плавника — 0,25.
Сравнение строения вибрисс ламантина с таковыми у ластоногих и
китообразных показывает определенные черты сходства и различия. Как
известно, сумки вибрисс китов тесно связаны с окружающей дермой с
помощью соединительнотканных тяжей и неподвижны, а вибриссные
сумки ластоногих снабжены мышцами и являются подвижными образо-
ваниями (Яблоков, Клевезаль, 1964). Разница в строении тонких и тол-
стых вибрисс ламантина предполагает, что вибриссная сумка первых
подвижна: она снабжена мышцей, а вибриссная сумка вторых относи-
тельно неподвижна в дерме, хотя и не имеет соединительнотканных
тяжей, закрепляющих ее в дермальном слое, подобно вибриссам китов.
Большая неподвижность толстых вибрисс губ ламантина по сравнению
с вибриссами морды ластоногих объясняется характером питания этого
зверя: вибриссы не травмируются столь сильно, как у моржа или рыбо-
ядных ластоногих, и поэтому нет необходимости в их амортизации.
Тонкие подвижные вибриссы туловища, конечностей и хвоста, по-види-
мому, контролируют направление потоков воды (действуют как гидро-
локаторы) и регистрируют прикосновение водорослей при передвижении
зверя. Тактильная функция вибрисс способствует ориентации ламантинов
в воде и относительно друг друга (Hartmon, 1979). Возможно, именно
особенности обитания ламантинов на мелководье, часто в мутной воде
(в отличие от китообразных — за редкими исключениями обитателей
открытых водных пространств) обусловили наличие вибрисс на туловище,
конечностях, хвостовом плавнике.
Сравнение деталей строения вибрисс ламантина с таковыми у других
водных млекопитающих показало, что степень развития соединительно-
тканной капсулы корня вибриссы сходна у ламантина и у ластоногих
(Яблоков, Клевезаль, 1964). В то же время система кровеносных сину-
сов напоминает таковую у китообразных. По-видимому, у настоящих
водных млекопитающих происходит утолщение соединительнотканной
капсулы корня вибриссы, уменьшение синусов, стабилизация корня виб-
риссы в дермальном слое.
У ламантина железы сохраняются только у тонких вибрисс тулови-
ща. У эмбриона дюгоня (длиной 51 см) также обнаружены сальные же-
лезы вибрисс туловища (Dosch, 1915), которые с возрастом, видимо,
3 Ламантин
65
исчезают. Во всяком случае, у новорожденного дюгоня длиной ИЗ см»
они не найдены (Соколов, 1973). Обитание в водной среде делает неце-
лесообразным и невозможным выполнение железами защитносмазочной
и терморегуляторной функций. Слабое развитие сальных желез, отсутст-
вие потовых и специфических желез свидетельствуют о неучастии желе-
зистого аппарата в осуществлении химической коммуникации у ламан-
тинов. Вместе с тем известно, что ламантины имеют хорошо развитый
обонятельный нерв и обонятельные луковицы; в неволе они обнаружива-
ют пищу, брошенную в бассейн. Хотя под водой их ноздри закрываются
клапанами, в литературе имеется предположение о восприятии химиче-
ских стимулов через обонятельный эпителий с током воды, через рот и
хоаны (Ronald et al., 1978).
На эмбриональном материале (африканский ламантин, дюгонь) пока-
зано, что в их коже имеются закладки обычных волос, но по мере раз-
вития эмбриона они исчезают (Dosch, 1915). Это служит доказательством
редукции волосяного покрова у сирен. В то же время формирование
вибрисс у сирен предшествует закладкам обычных волос, что вполне со-
гласуется с данными по развитию волосяного покрова у других млеко-
питающих (Соколов и др., 1984). Сопоставление сроков онтогенетиче-
ского развития обычных волос и вибрисс представляет интерес для об-
суждения вопросов последовательности филогенетического развития этих
структур.
ЖЕЛЕЗА КОЖИ ВЕК
В коже век ламантина Мейбомиевые сальные железы, характерные для
большинства млекопитающих, отсутствуют. В литературе имеется упо-
минание о присутствии в коже век ламантина специальных желез
(Dosch, 1915). Действительно, у самца ламантина на внутренней поверх-
ности угла глаза открывается выводной проток железы. Эта железа по
строению сложная альвеолярно-трубчатая: ее выводной проток ветвится»
секреторный отдел распадается на трубки, заканчивающиеся гроздью'
альвеол (рис. 16). Выводной проток диаметром ИЗ мкм открывается на
конъюнктиве верхнего века. Его стенки многослойны, состоят из мелких
кубических клеток с темным центрально расположенным ядром. Цито-
плазма этих клеток почти бесцветна, не окрашивается красителями. Диа-
метр вторичных протоков, впадающих в выводной проток, колеблется от
23 до 34 мкм. Вставочные отделы неширокие (диаметром 23 мкм), из-
витые, выстланы плоским однослойным кубическим эпителием. Темное
мелкое ядро располагается в базальной части цитоплазмы. Последняя —
слабо базофильная. Секреторные альвеолы диаметром 28 мкм имеют
необычный вид: лишь некоторые из них полностью заполнены живыми
клетками. Эти клетки полигональной формы с центрально расположен-
ным ядром и слабо базофильной цитоплазмой. Основная масса секретор-
ных клеток содержит слоистый бесформенный секрет. Живые клетки
располагаются по периферии таких альвеол. По-видимому, в альвеолах
происходит секреторный процесс, сходный с голокриновым. Имеются
миоэпителиальные клетки. Они лежат вокруг секреторных трубок и вы-
водных протоков. Судя по слабой базофилии секреторного эпителия, же-
лезы выделяют белково-слизистый секрет. Наблюдения за живым зверем
показали, что отделение секрета происходит на суше интенсивно:
удалось собрать примерно 1 мл секрета. Биохимический анализ, прове-
денный Б. М. Граевской, выявил в секрете присутствие белка в количе-
стве 0,6 мг на 100 мл, pH 6,3. Это говорит о некотором сходстве по со-
ставу секрета железы века ламантина со слюной.
66
ПРОЦЕССЫ КЕРАТИНИЗАЦИИ В ЭПИДЕРМИСЕ
Наиболее важным механизмом в осуществлении барьерной функции
кожи всех позвоночных является способность эпидермиса к постоянно-
му обновлению. Это качество проявляется уже у низших позвоночных и
становится высоко запрограммированным универсальным процессом у
млекопитающих. Упорядоченное движение клеток от базального слоя
вверх к конечной стадии дифференцировки, в результате которой клетка
гибнет и слущивается, является фундаментальным свойством многослой-
ных покровных эпителиев. В эпидермисе всех наземных млекопитающих
описанный процесс заканчивается не просто десквамацией клеток, а об-
разованием рогового слоя, состоящего из корнеоцитов и межклеточного
цемента с особыми барьерными свойствами.
Поскольку кератинизирующиеся эпителии представляют собой дина-
мическую систему, то особенности их изучения в сравнительном аспекте
заключаются в оптимальном выборе элементов, которые могут отражать
адаптивные изменения.
При рассмотрении процесса дифференцировки клеток эпидермиса
можно выделить несколько таких основных характеристик, служащих
одновременно и морфологическими маркерами программы кератинизации.
К ним относятся:
Рис. 16. Строение железы века взрослого самца ламантина:
а — ув. X 40; б — ув. X 100
2 — секреторная альвеола; 2 — вторичный проток; 3 — секреторная трубка; 4 — выводной проток
67 Э*
1. Кератиновые филаменты (промежуточные филаменты 7—10 нм в
диаметре), которые образуются из белковых субъединиц разного моле-
кулярного веса. В терминальной стадии дифференцировки они подверга-
ются посттрансляционным модификациям и стабилизируются благодаря
появлению внутри- и межмолекулярных S—S-связей.
2. Десмосомы и полудесмосомы — специализированные клеточные
контакты.
3. Внутриклеточные гранулы Одланда — специфические для много-
слойных эпителиев органеллы, ответственные за образование межклеточ-
ного цемента.
4. Межклеточный цемент — комплекс веществ, обеспечивающий коге-
зию клеток и барьер проницаемости рогового слоя.
5. Кератогиалиновые гранулы — депо белка филаггрина, необходимо-
го для стабилизации кератиновых филаментов, и источник свободных
аминокислот в корнеоцитах.
6. «Поперечно-сшитая», или роговая, оболочка корнеоцита, замещаю-
щая его плазмалемму, придает клетке повышенную устойчивость к дей-
ствию некоторых агентов.
7. Исчезновение органелл из клеток верхних слоев эпидермиса —
показатель полного процесса кератинизации (Prunieras, Delescluse,
1984).
Перечисленные признаки, с одной стороны, отражают основные эта-
пы дифференцировки, а с другой — изменяются под влиянием эндоген-
ных и экзогенных факторов. При обсуждении строения эпидермиса кожи
ламантина они и будут в центре внимания.
Эпидермис ламантина представляет собой мощный многослойный
пласт клеток, находящихся на разных стадиях дифференцировки. Ба-
зальный слой состоит из клеток, плотно примыкающих друг к другу и
представляющих собой неоднородную популяцию. Среди кератиноцитов
можно выделить по крайней мере три их разновидности. Клетки первого
типа цилиндрической формы, вытянутые перпендикулярно поверхности
кожи. Они имеют длинные цитоплазматические выросты, уходящие в.
дерму. Ядро овальной формы с хорошо выраженной оболочкой, снабжен-
ной порами. В ядре хорошо различимы одно или два ядрышка. Цито-
плазма содержит многочисленные свободные рибосомы, несколько профи-
лей гранулярного эндоплазматического ретикулума и многочисленные
пучки тонофиламентов. Умеренное количество митохондрий распределе-
но по всей цитоплазме. Митохондрии округлые или овальные по форме,
имеют электронноплотный матрикс и хорошо различимые кристы. Не-
большой аппарат Гольджи представлен несколькими короткими цистер-
нами и немногочисленными мелкими везикулами. Он располагается в
близком соседстве над ядром. В клетках редко обнаруживается несколько'
гранул меланина, чаще его нет, хотя отростки меланоцитов находятся
с ними в тесном контакте. Иногда можно наблюдать аппарат Гольджи в
виде мультисистемы. Такой аппарат занимает несколько участков цито-
плазмы, но группируется все равно вокруг ядра.
Второй тип клеток отличается от первых отсутствием цитоплазмати-
ческих выростов, более умеренным содержанием рибосом и тонофиламен-
тов. Пигментных гранул в таких клетках значительно больше, и распо-
лагаются они над ядром. Латеральная плазмалемма образует выражен-
ную складчатость.
Третий тип, редко встречающихся клеток, имеет электронноплотную
цитоплазму и ядро. Они более узкие и удлиненные, чем кератиноциты,
описанные выше. Их цитоплазма содержит те же органеллы и включе-
ния, что и клетки первого типа.
68
Между соседними клетками образуются немногочисленные десмосомы,
а к базальной мембране клетки прикреплены полудесмосомами. Послед-
ние представлены большим числом, и их размеры достигают 600 нм в
Длину.
Из специализированных клеток эпидермиса в базальном слое распо-
лагаются клетки Лангерганса. Встречаются они очень редко и выделяют-
ся светлой цитоплазмой, дольчатым ядром и отсутствием десмосом на их
плазмалемме. Клетки содержат митохондрии с плохо различимыми кри-
стами и немногочисленные везикулы. Цитоплазма полностью лишена
филаментозного материала, что также отличает эти клетки от кератино-
цитов.
Меланоциты представляют собой второй тип специализированных
клеток эпидермиса. Они имеют крупное ядро, иногда с хорошо выражен-
ными порами на ядерной оболочке. Цитоплазма содержит многочислен-
ные гранулы меланина и меланосомы разной степени зрелости. Прост-
ранство между гранулами занято свободными рибосомами, цистернами
гранулярного эндоплазматического ретикулума и некрупными митохонд-
риями.
Шиповатый слой состоит из нескольких рядов кератиноцитов,
структура которых постепенно меняется по мере продвижения их
вверх. Уплощается форма клеток, и в верхних слоях длинная ось кера-
тиноцита ориентируется параллельно поверхности кожи. Как только
клетка покидает базальный слой, на их плазмалеммах строятся много-
численные десмосомы, которые на препарате выглядят хаотично распо-
ложенными. Это впечатление создается потому, что демосомы распола-
гаются на многочисленных клеточных выростах. В верхних слоях шипо-
ватых клеток, где складчатость плазмалеммы расправляется, десмосомы
образуют четкие ряды вертикально стоящих электронноплотных полос.
Количество филаментов в клетках возрастает, и их массы постепенно
заполняют больший объем цитоплазмы. Если в нижних шиповатых клет-
ках можно различить органеллы, описанные для базальных кератиноци-
тов, то в верхних это удается сделать с трудом, прежде всего из-за
обилия филаментозного материала. В средних слоях шиповатых клеток
тонофиламенты теряют сродство к красителям, в них не определяются
отдельные филаменты, так что они выглядят почти аморфными массами
средней электронной плотности. На этой стадии дифференцировки в ке-
ратиноцитах появляются гранулы Одланда. Они представляют собой
округлые образования диаметром 100 нм, окруженные мембраной. Их
содержимое средней электронной плотности, без или с более плотной
сердцевиной. Они располагаются по всей цитоплазме или сконцентриро-
ваны в околоядерной зоне (рис. 17, а). Количество этих органелл быстро
возрастает.
Два-три слоя клеток, расположенных непосредственно под роговым
слоем, следует рассматривать как зернистый слой. Несмотря на то что
типичных кератогиалиновых гранул в эпидермисе ламантина не образу-
ется, в указанных клетках наблюдаются некоторые особенности, характе-
ризующие их как стадию зернистого слоя. Нижние клетки содержат
максимальное количество гранул Одланда, а затем оно резко сокращает-
ся. Уменьшается количество десмосом, хотя структура их сохраняется.
Деформируется и уменьшается ядро клеток.
Переход в следующую стадию дифференцировки — роговой слой, про-
исходит резко, без промежуточных форм. Роговой слой состоит из
25—26 рядов клеток сильно уплощенных, полностью лишенных орга-
нелл. Они заполнены топофиламентами. Тонофиламенты располагаются
рыхло в нижних слоях, выше они ориентируются вдоль клетки, а в
69
верхних— содержимое корнеоцитов гомогенное. В отдельных клетках
встречаются пигментные гранулы, расположенные поодиночке или груп-
пами по пять-шесть гранул. Иногда обнаруживаются вакуоли разного
размера. В корнеоцитах всех слоев хорошо выражена утолщенная плаз-
малемма (рис. 17, б). Между собой клетки соединены взаимопроникаю-
щими выростами. В некоторых местах сохраняются остатки десмосом.
Внутренняя ламеллярная структура утрачена, а центральная часть дес-
мосомального промежутка заполнена гомогенным контрастным веществом.
Межклеточное пространство представляет собой электронно-полупрозрач-
ное содержимое.
Из представленного описания следует, что процесс кератинизации в
эпидермисе ламантина протекает в соответствии с полной универсальной
программой, исключающей лишь образование кератогиалиновых гранул.
Это означает, что дифференцировка клеток завершается образованием
полноценного рогового слоя, состоящего из клеток, лишенных органелл и
окруженных оболочкой, и межклеточного цемента. Следует отметить еще
две особенности в программе кератинизации эпидермиса ламантина:
обилие гранул Одланда и появление их в средних (а не в верхних, как
это описано для других животных) слоях шиповатых клеток, а также
повышенное образование тонофиламентов и изменение их окраски в тех
же шиповатых клетках на ранних стадиях дифференцировки.
Значительное развитие тонофиламентов, наблюдаемое в эпидермисе
ламантина, характерно вообще для толстых эпидермисов. Это отмечено
у китообразных (Соколов и др., 1971), у приматов, включая отдельные
Рис. 17. Клетки шиповатого (а) и рогового (б) слоя эпидермиса, ув. Х4000
7—.пучки тонофиламентов; g— гранулы Одланда; 3— десмосомы; 4—корнеоцит; S — орого-
вевшая оболочка
70
участки кожи человека (Breathnach, 1971). В тонком эпидермисе (на-
пример, лабораторных мышей и крыс) тонофиламенты представлены
довольно тонкой сетью и не образуют мощных пучков.
Обычно тонофибриллярному аппарату клетки, являющемуся ее ци-
тоскелетом, приписывают опорную и амортизационно-защитные функции
(Михайлов, 1979). Это вполне согласуется с тем, что у морских млеко-
питающих, и у ламантина в частности, мощно развиты тонофиламенты.
Кожа этих животных испытывает большие физические нагрузки. Но оче-
видно, что в процессе дифференцировки тонофиламенты играют и какую-
то другую роль, поскольку имеют место не только количественные, но и
качественные изменения этих структур, происходящие на молекулярном
уровне (Birchmeier, 1984), а также перераспределение и изменение их
ориентации в клетке.
Большое количество филаментов идет на образование десмосом, коли-
чество которых в эпидермисе морских млекопитающих огромно. У кито-
образных их насчитывается до нескольких сот на клетку (Harrison,
Thurley, 1974). В эпидермисе ламантина они, вероятно, не менее много-
численны, поскольку плотность их расположения на срезах кожи анало-
гична таковой у дельфина.
Как правило, кератиновые филаменты представляют собой электрон-
ноплотные образования, которые становятся более светлыми лишь в ро-
говом слое, где они выделяются на фоне темного матрикса (Brody,
1959). Это изменение окраски совпадает с появлением S—S-связей в мо-
лекулах кератина. В эпидермисе ламантина филаменты меняют сродство
к красителям уже в средней части шиповатого слоя. Изменение окраски
в данном случае может свидетельствовать о каких-то биохимических про-
цессах, происходящих в них, но какова их природа сказать трудно. Воз-
можно, что это также связано со стабилизацией (образованием межмоле-
кулярных S—S-связей) филаментов, но не исключается возможность и
появления артефакта во время подготовки образца, что требует про-
верки.
Способность образовывать кератин клетками покровного эпителия
проявляется в филогенезе еще на ранних этапах эволюции. Синтез этого
белка и образование из него филаментов служит стержневым элементом
программы дифференцировки клеток эпидермиса, сохраняющимся даже в
самых экстремальных условиях. Хотя их образование в кератиноцитах
обязательно, может меняться относительная пропорция высоко и низко-
молекулярных, связанных и не связанных дисульфидными мостиками
кератинов. И как следствие, будут меняться свойства корнеоцитов.
Например, большая резистентность к воде связана с большим содержа-
нием цистина в молекуле кератина, а от количества дисульфидных мо-
стиков в нем зависят механические свойства кератиновых филаментов
(Robbins, Fernee, 1983).
Полная программа дифференцировки кератиноцитов включает и
образование кератогиалиновых гранул, которые не обнаруживаются
в эпидермисе ламантина. Эти структуры, присутствующие в подавляю-
щем большинстве кератинизирующихся эпителиев позвоночных, давно
привлекали внимание исследователей, но их функции, равно как и хи-
мический состав, оставались неизвестными. Только в последние годы
было показано, что кератогиалиновые гранулы состоят из филаггрина,
белка, который синтезируется в зернистом слое. С его помощью осуще-
ствляется стабилизация молекул кератина, которые благодаря этому про-
цессу, происходящему в роговом слое, становятся нерастворимыми и
более устойчивыми к механическим воздействиям (Scott, Harding,
1981а; Green et al., 1982). Поскольку филаггрин является короткоживт-
71
щим белком, то в процессе терминальной дифференцировки корнеоцита
он распадается на отдельные аминокислоты. Назначение этих аминокис-
лот и их превращения в клетках рогового слоя еще до конца не ясно.
Однако полагают, что некоторые из них, являясь осмолитами, обеспечи-
вают осмотическое давление в клетке, другие могут участвовать в защи-
те организма от ультрафиолетового облучения (Scott, Harding, 1981).
На основании этих данных представляется, что филаггрин — очень
важный и необходимый компонент дифференцировки. Однако имеются
данные, что кератогиалиновые гранулы в эпидермисе некоторых млеко-
питающих встречаются не во всех клетках зернистого слоя (Соколов,
1973). Сообщается о появлении и исчезновении гранул в зависимости от
сезона (Ling, 1974). Наконец, у некоторых наземных и у изученных
представителей морских млекопитающих кератогиалиновые гранулы не
обнаруживаются (Соколов и др., 1971; Соколов, 1973; Harrison, Thurley,
1974). Не известно, однако, означает ли отсутствие кератогиалиновых
гранул отсутствие филаггрина. Белок может синтезироваться в клетках
и выполнять свои функции, не накапливаясь в виде гранул. Решение
этого требует применения специальных методов исследования, например
иммуноцитохимических. Если окажется, что кератинизация в эпидерми-
се протекает без участия филаггрина или его аналога, то возникает
вопрос, что происходит с молекулами кератина и какими свойствами
обладают корнеоциты в этом случае. По морфологическим характери-
стикам корнеоциты эпидермиса ламантина и человека, например, анало-
гичны. Но, как известно, морфологическое сходство не всегда означает
функциональную идентичность.
Одной из наиболее ярких черт эпидермиса ламантина является оби-
лие гранул Одланда. Причем увеличено не только их содержание в
клетке, но и эпидермальных слоев, в которых они встречаются значи-
тельна больше, чем у наземных млекопитающих. У ламантина они начи-
нают образовываться в средней части шиповатого слоя, в то время как
у наземных они обнаруживаются только в верхних шиповатых клетках
и в зернистом слое.
Гранулы Одланда — специализированные органеллы многослойных
эпителиев также не являются приобретением млекопитающих. Они встре-
чаются уже у амфибий (Hayward, 1979; Odland, 1981) и постоянно при-
сутствуют в коже наземных позвоночных. Некоторыми авторами было
уже замечено, что количество этих органелл увеличено в неороговеваю-
щих эпителиях млекопитающих (Hashimoto et al., 1966; Squier, 1977).
Повышенное содержание кератиносом (синоним гранул Одланда) на-
блюдали и в коже дельфинов (Соколов и др., 1971; Harrison, Thurley,
1974), однако авторы лишь констатировали факт, но никак его не ком-
ментировали, поскольку в то время об этих органеллах было мало что
известно.
Только недавно удалось выяснить их роль в связи с изучением мак-
ромолекулярных основ барьера проницаемости кожи млекопитающих.
Было показано, что барьер этот локализован в роговом слое эпидерми-
са (Eriksson, Lamke, 1971), а обеспечивается межкорнеоцитным прост-
ранством, заполненным цементирующим веществом (Elias, 1981, Elias
et al., 1977). Оно состоит из различных свободных жирных кислот, по-
лярных и нейтральных липидов и гликолипидов (Elias et al., 1981;
Werts, Downing, 1982). Источником всех этих веществ и служат грану-
лы Одланда (Elias et al., 1979).
Небезынтересно отметить один факт из описания экспериментов Хар-
рисона и Турлея (Harrison, Thurley, 1974). Авторы изучали действие
пресной воды на кожу морских дельфинов (Tursiops truncatus). 72-ча-
72
совое пребывание зверей в пресной воде вызывало усиленное отделение
верхних слоев эпидермиса, что приводило к удалению сформированного
защитного пласта. В оголившихся слоях клеток многочисленные гранулы
Одланда выбрасывали свое содержимое в межклеточное пространство, что
в норме не наблюдалось. Описанную реакцию кератиносом можно рас-
сматривать как компенсаторную в ответ на экстремально изменившиеся
условия.
Если гранулы Одланда действительно ответственны за барьер прони-
цаемости, то их увеличенное образование в эпителиях, соприкасавшихся
с водой, вполне объяснимо. Вода, являясь универсальным растворите-
лем,— среда агрессивная. В процессе эволюции и вырабатывается меха-
низм, который обеспечивает барьер для проникновения этого растворите-
ля внутрь организма.
Если наземные животные контактируют с водой лишь эпизодически
(во время дождя, непродолжительного плавания, водопоя), то водная
среда обитания требует более надежного механизма, с запасом прочно-
сти. С этим и связано увеличение содержания кератиносом в коже мор-
ских млекопитающих.
Одной из морфологических особенностей гранул Одланда в эпидерми-
се ламантина является отсутствие внутренних ламеллярных структур.
В коже наземных млекопитающих и у дельфина (у единственно изучен-
ного в этом отношении представителя морских млекопитающих) гранулы
имеют ламеллярное строение (Соколов и др., 1971; Breathnach, 1971;
Harrison, Thurley, 1974). Однако морфология кератиносом ламантина не
уникальна. В неороговевающих эпителиях млекопитающих и в коже че-
ловека при некоторых заболеваниях эти органеллы тоже не имеют ла-
меллярного содержимого (Squier, 1977; Hayward, 1979). Ответить на
вопрос, с чем это связано и как отражается на свойствах барьера пока
трудно. По крайней мере показано, что неороговевающие эпителии также
непроницаемы для пероксидазы хрена (низкомолекулярный белок, ис-
пользуемый в качестве метки для изучения, в частности, барьера прони-
цаемости), как и эпидермис (Squier, 1973). Изучение химического соста-
ва кератиносом только начинается, и пока не проведено сравнительного
анализа этих органелл из разных эпителиев. Не исключено поэтому, что
состав содержимого гранул Одланда ламантина, а в связи с этим и
свойства межклеточного цемента окажутся при детальном изучении не-
сколько иными, чем у наземных млекопитающих или дельфина.
Переход к наземному образу жизни индуцировал появление в клет-
ках эпидермиса позвоночного уникального образования: «поперечно-сши-
той» оболочки. Эта структура была выявлена только после введения
электронно-микроскопических методов исследования, а представления о
механизме ее образования и некоторых функциях сформировались лишь
в самое последнее время. Адаптационное значение «поперечно-сшитой»
оболочки пока не получило должной оценки, несмотря на то, что ее
появление, видимо, имело решающее значение в эволюции эпидермиса.
Клеточная адаптация к осмотическому стрессу является кардинальным
биологическим процессом, который защищает организм от летальных
эффектов обезвоживания.
Биохимические исследования последних лет показали, что оболочка
состоит из белка кератолинина, который синтезируется клетками зерни-
стого слоя и откладывается под плазмалеммой корнеоцита. Под действием
трансглутаминазы в белке образуются е-у (глутамил)-лизиновые сшивки,
которые придают ему свойства прочности и нерастворимости даже в
сильных реагентах (Rice, Green, 1977; Buxman et al., 1980). Элиминация
органелл и образование оболочки — процессы, вероятно, взаимосвязан-
73
ные, о чем можно судить по данным кинетических исследований, а также
по отсутствию оболочки в неороговевающих эпителиях. В ороговевшей
клетке, лишенной органелл, плазмалемма функционировать уже не мо-
жет и замещается оболочкой. Она придает корнеоциту механические
свойства и предохраняет его от потери воды, выполняя функции осмоти-
ческой мембраны. Полностью ороговевший слои клеток — это не только
повышенная (по сравнению с неполностью ороговевающим эпидермисом)
механическая защита, но и возможность выдерживать осмотические
стрессы и изменение солености воды в больших пределах, вплоть до пол-
ностью пресной, а также длительное пребывание организма на границе
двух сред вода—воздух.
В связи с вышесказанным интересно сравнить некоторые свойства
эпидермиса ламантина, живущего в прибрежной зоне, и дельфина, оби-
тающего в открытом море. Эти животные имеют приблизительно одина-
ковую толщину эпидермиса и рогового слоя (Соколов, 1973; Harrison,
Thurley, 1974). Однако str. corneum у дельфина в противоположность
полностью ороговевшему эпидермису ламантина образован неполностью
ороговевшими клетками, сохранившими органеллы (Соколов и др., 1971;
Speerman, 1973; Harrison, Thurley, 1974). Вероятно, корнеоциты в этом
случае лишены «поперечно-сшитой» оболочки или она плохо выражена
(ни у одного из авторов нет указания на утолщение плазмалеммы кор-
неоцитов). Неполное ороговение обусловливает и внешние признаки
кожи. У дельфина она мягкая, гладкая, легко ранимая, подвергается
быстрому высыханию на воздухе7. Такой эпидермис в большей степени
подвержен действию не только физических и химических факторов, но и
заражению микроорганизмами. Одним из возможных защитных механиз-
мов от перечисленных воздействий и является известная способность кле-
ток эпидермиса дельфинов к быстрой и обильной десквамации. Однако
такая расточительная потеря клеток возможна только при условии столь
же обильного восполнения их из базального слоя. И этот аппарат у ки-
тообразных имеется. По данным Брауна и др. (Brown, 1983), пролифе-
ративный индекс в эпидермисе дельфина в 1,3 раза превышает этот по-
казатель у человека. Глубокая и сложная складчатость внутренней сто-
роны эпидермиса китообразных (Соколов, 1962; Giocometti, 1967) созда-
ет огромную площадь пролиферирующего слоя, которая оказалась в
8,7 раза выше (на единицу поверхности), чем у человека, а отсюда сле-
дует, что и пролиферативная способность эпидермиса китообразных в
11,3 раза выше (Brown et al., 1983).
Пребывание морских китообразных в пресной воде, как известно, но-
сит эпизодический характер. Эксперименты с переносом дельфинов в
пресную воду (о них уже упоминалось выше) показали, что последняя
вызывает в эпидермисе значительные необратимые изменения поверхно-
стных его слоев, приводящие к отслаиванию пластов большой площади.
При этом клетки подвергаются баллонной дистрофии, т. е. набухают
(Simpson, Gardner, 1972). Это явление вполне объяснимо, если принять
во внимание, что неполностью ороговевшие корнеоциты эпидермиса
7 Благодаря полностью ороговевшему str. corneum кожа ламантина кажется более
грубой, чем у дельфина, что усугубляется его поверхностным рисунком. «Ямки» и
борозды (см. выше) на эпидермисе точно повторяют углубления на поверхности
дермы, так же как это имеет место в коже человека. Дермальные сосочки у ламан-
.. тина располагаются между «ямками» и на стенках «ямок». Самые высокие сосочки
дают небольшую выпуклость на поверхности кожи, как бы приподнимая эпидер-
мис. У дельфина дермо-эпидермальное соединение таково, что ни поверхность дер-
мы, которая почти ровная, ни одинаковой высоты дермальные сосочки, располо-
женные четкими рядами, не отражаются на поверхности эпидермиса.
74
дельфина лишены «поперечно-сшитой» оболочки. Такие клетки, приспо-
собленные к определенным условиям солености, оказавшись в среде с
низкой осмолярностью, набухают, что обусловлено осмотическим всасы-
ванием воды. Мы не располагаем подробными данными о тонком строе-
нии эпидермиса речных дельфинов, но из краткого описания Харрисона
и Турлея (Harrison, Thurley, 1974) можно сделать вывод о том, что
эпидермис обитателей рек отличается от эпидермиса морских дельфинов,
а роговой слой его больше напоминает таковой у наземных животных.
Из сказанного выше следует, что описанный своеобразный механизм,
обеспечивающий барьерную функцию эпидермиса китообразных, являет-
ся адекватным для жизни в открытых морских просторах с незначитель-
ными колебаниями осмотического давления, однако он оказывается не-
достаточным для жизни в пресной воде. Обитание млекопитающих в
эстуариях выдвигает несколько другие требования к барьерным свойст-
вам кожи. Неполностью ороговевающий эпидермис не может в этих ус-
ловиях осуществлять надежную защиту организма. Гомеостаз сохраняет-
ся лишь благодаря полностью ороговевшему слою клеток, имеющих
вместо плазмалеммы «поперечно-сшитую» оболочку. Следует учитывать,
что механизм осморегуляции значительно сложнее, чем простое обеспе-
чение непроницаемости клетки оболочкой (которая не абсолютно, а лишь
относительно непроницаема).
Само понимание барьерной функции кожи не ограничивается обсуж-
даемыми в этой статье качествами эпидермиса млекопитающих. Оно
включает в себя более сложные многогранные свойства кожи, связанные
не только с функцией кератиноцитов. Специализированные клетки Лан-
герганса и меланоциты вносят свой вклад в кожный барьер, определяя
его иммунологические и химические свойства. Однако это требует спе-
циального исследования и обсуждения.
Рис. 18. Поверхность дермы (под сканирующим электронным микроскопом)
1 — «ямка»; 2 — дермальные сосочки; 3 — коллагеновые волокна
75
Таким образом, проведенный анализ полученных данных позволяет
считать, что эпидермис ламантина имеет сходное с эпидермисом других
млекопитающих строение. Однако в нем обнаруживаются некоторые осо-
бенности, выявляемые на ультраструктурном уровне.
Во-первых, в эпидермисе сохраняются черты, свойственные наземным
млекопитающим, и, очевидно, предкам ламантина. Дифференцировка
кератиноцитов заканчивается образованием рогового слоя, состоящего из
полностью ороговевших клеток с «поперечно-сшитой» оболочкой и меж-
клеточным цементом. Во-вторых, эпидермис приобретает черты, свойст-
венные эпидермису водных млекопитающих,— увеличенное по сравнению
с наземными формами содержание гранул Одланда.
Сказанное выше дает возможность предположить, что универсальная
программа дифференцировки кератиноцитов, выработанная на ранних
этапах прогрессивной эволюции позвоночных, решается у млекопитаю-
щих также неоднозначно. Она реализуется в той степени, в какой это
определяется условиями обитания животного. Модификации, появившие-
ся на основе уже имеющихся признаков и выработанные в процессе
адаптации, закрепляются генетически.
Глава третья
МОРФОЛОГИЯ
И ОСНОВНЫЕ НАПРАВЛЕНИЯ СПЕЦИАЛИЗАЦИИ
ОСЕВОГО СКЕЛЕТА СИРЕН
Осевой скелет давно уже не привлекал внимание исследователей. Клас-
сическое непревзойденное по полноте описание его у Hydrodamalis gigas
в сравнении с ламантином и дюгонем было дано Ф. Ф. Брандтом (Brandt,
1846; 1861 — 1868). Однако по условиям того времени у Брандта отсутст-
вовали как эволюционный подход, так и стремление видеть в осевом ске-
лете механическую конструкцию, особенности которой определяются спе-
цификой его функционирования. В большинстве последующих работ,
в которых упоминается осевой скелет сирен, приводятся по нему лишь
самые общие сведения, как правило не выходящие за рамки сводки
Брандта. Следует отметить лишь работу Мьюри (Murie, 1972а), который
подробно рассмотрел соотношение осевого скелета с окружающими его
мягкими тканями. С другой стороны, внимания заслуживают отдельные
описания остатков ископаемых сирен, в том числе и фрагментов осевого
скелета (Lepsius, 1881—1892; Abel, 1904; Simpson, 1932; Sickenberg,
1934; Kellogg, 1966; Domning, 1978c; и др.), дающие материал для по-
нимания структурных преобразований посткраниального скелета, привед-
ших к возникновению специфических его особенностей у трех современ-
ных форм сирен. Однако обобщение этих данных еще не проведено,
и одной из причин этого является отсутствие как четкого понимания
морфологической и адаптационной специфики осевого скелета ламантина,
дюгоня и стеллеровой морской коровы, так и хорошей его иконографии.
Даже в атласе фотографий по отдельным частям скелета сирен (Kaiser,
1974) основное внимание уделено черепу и его элементам, а посткрани-
альный скелет фактически не затронут.
Основным материалом для настоящей главы послужили результаты
изучения двух скелетов взрослых ламантинов Trichechus m. manatus
(один в Институте зоологии АН УССР, обозначенный как ИЗ-5945,
и второй в Зоологическом институте АН СССР — ЗИН-31795 *). Оба
экземпляра были добыты в одном и том же месте в Карибском море,
первый в 1964 г. В. И. Таращуком, второй в 1980 г. (см. Предисло-
вие). Дополнительно были использованы один монтированный скелет
ламантина меньших размеров: Т. т. (?) latirostris (как Т. latirostris
этот экземпляр был изучен Брандтом, однако место его происхождения —
Суринам позволяет усомниться в правильности определения) в ЗИН
АН СССР (ЗИН-677), другой неполный разборный (ЗИН-6835) также из
‘Суринама и Т. manatus неизвестного происхождения в Музее природы
1 На основании предварительного изучения последнего в печати появилась работа,
касающаяся осевого скелета ламантина (Manzij, Piliptshuk, 1984), содержащая ряд
неточностей и крупных ошибок в интерпретациях имеющихся фактов и в частно-
сти пропагандирующая представление о якобы большей близости ламантинов и
морских коров (S. 283). Не спасает дело и внесенное позже в корректуру резюме
обратный тезис о близости дюгоней и морских коров (S. 257, 293), противоречащий
тексту. Настоящим мы объявляем фактические данные, включенные в указанную
статью, несоответствующими действительности. Для предлагаемого ниже исследо-
вания весь материал был переописан заново.
77
Таблица 3. Абсолютная и относительная длина скелета и его отделов у сирен
Вид, экземпляр । Общая длина (по прямой), мм Кондилобазальная длина черепа, мм (% от общей длины) Длина, мм (% от общей длины)
1 позвоночного столба 1 шейного отдела грудного отдела постторакаль- ного отдела
Trichechus m. manatus 2550 360 2190 135 1015 1040
(ИЗ-5945; Куба) (14,1) (85,9) (5,3) (39,8) (40,8)
Trichechus m. manatus 2723 370 2359 133 1055 1165
(d, ЗИН-31795; Куба) (13,6) (86,4) (4,9) (38,7) (42,8)
Т. т. (?) latirostris 1895 295 1600 90* 720 790
(ЗИН-677; Суринам) (15,6) (84,4) (4,7) (38,0) (41,7)
Т. manatus 2413 324 2089 89* 940 1060
(Х-175, место?) (13,4) (86,6) (3,7) (39,0) (43,9)
Dugong dugon 2099 350 1750 90 699 960
(ЗИН-3220; Красное море) (16,7) (83,3) (4,3) (33,3) (45,7)
D. dugon 1810 295 1515 80 560 870
(ЗИН-679; Красное море) (16,3) (83,7) (4,7) (30,9) (48,1)
Hydrodamalis gigas 6230 580 5650 290 2080 3280
(3M-3464; Командорские острова) (9,3) (90,7) (4,6) (33,4) (52,7)
* Цифры, возможно, занижены из-за трудности измерения длины шейного
отдела на монтиро-
ванных скелетах.
Харьковского государственного университета им. А. М. Горького
(Х-175). Со сравнительными целями были исследованы два скелета Dugong
dugon из Зоологического института АН СССР, один разобранный (ЗИН-
3220) 2 и другой монтированный, но частично разобранный при изучении
(ЗИН-679); оба от экземпляров с Красного моря, и наиболее полный и
хорошо сохранившийся в естественном сочетании частей скелет Hydro-
damalis gigas, хранящийся в Зоологическом музее Львовского государст-
венного университета им. И. Франко (3M-3464), и полностью доступный
для тщательного изучения (отдельные элементы в монтированном скеле-
те легко снимаются). Дополнительно просмотрены остатки морских ко-
ров в Зоологическом институте АН СССР (один монтированный скелет
был описан Брандтом), в Зоологическом музее МГУ и Палеонтологиче-
ском музее Института зоологии АН УССР. Все морские коровы проис-
ходят с Командорских островов. Характеристика основных материалов
приведена в табл. 3.
Изучение материала включало анатомическое описание и изображе-
ние наиболее важных элементов скелета. Все рисунки по скелету в этой
и других главах выполнены с натуры художниками Т. Д. Раковой,
К. И. Улановой и К. П. Мешковым под руководством одного из авто-
ров (В. Б. Суханова). Важное место в изучении занимало определение
следующих параметров, касающихся осевого скелета: 1) вес отдельных
позвонков, всего скелета и его отделов, определение частного центра тя-
жести позвоночного столба; 2) объем грудных позвонков, их невральных
2 У этого скелета, видимо, при разборке пропал блок, включающий все шейные и
первые три грудные позвонка вместе с их ребрами. Отсюда вытекала необходи-
мость вычисления некоторых параметров по отсутствующим частям путем экстра-
поляции на основе второго скелета с учетом разницы в размерах.
78
дуг и тел; 3) длина тела позвонка, абсолютная и относительная —
в процентах к длине позвоночного столба; 4) общая высота позвонка, вы-
сота тела (по задней сочленовной поверхности) и невральной дуги (аб-
солютная и относительная); 5) ширина позвонков в поперечных (диапо-
физы) или поперечнореберных отростках (абсолютная и относительная —
в процентах к высоте позвонка), тела позвонка (по задней сочленовной
поверхности и в месте срединного сужения — минимальная ширина те-
ла) ; 6) момент сопротивления сил напряжения позвоночного столба к
внешним силам, действующим в вертикальной плоскости, который опре-
деляется по «формуле Слайпера» (Slijper, 1946): bh2, где b — ширина
тела, a h — высота тела позвонка; в горизонтальной плоскости — b2h',
7) общие размеры спинномозгового канала, высота и ширина позвоноч-
ного отверстия; 8) высота и ширина остистого отростка (абсолютная и
относительная), угол его наклона; 9) ширина позвонка по центрам зад-
них суставных отростков (постзигапофизов, абсолютная и относитель-
ная — в процентах к ширине тела; 10) угол наклона суставной поверх-
ности (в поперечной проекции) постзигапофизов; 11) положение центров
фасеток для сочленения с головкой и бугорком ребра относительно вы-
соты позвонка (по вертикали) и переднего края тела позвонка (по го-
ризонтали), расстояние между ними; 12) ширина диапофиза, абсолютная
и относительная в процентах к длине тела позвонка; 13) положение диа-
пофиза в горизонтальной и вертикальной проекции (углы наклона);
14) вес ребер, расстояние между их концами (хорда), длина по на-
ружной поверхности, максимальная высота изгиба по перпендикуляру к
хорде до точки на внутренней поверхности, расстояние по хорде от го-
ловки до этого перпендикуляра, абсолютная и относительная — в про-
центах к длине хорды. Все горизонтальные промеры взяты вдоль длин-
ной оси позвоночного столба, совпадающей в каждом позвонке с ходом
спинномозгового канала, все вертикальные — по перпендикуляру к этой оси.
Все полученные данные были сведены в цифровые таблицы, на осно-
ве которых составлены графики, отражающие ходом своих кривых ха-
рактер изменения отдельных параметров по длине позвоночного столба.
Тем самым они отражают изменения в возможностях функционирования
отдельных его участков. Нашу задачу мы видели не столько в том,
чтобы понять, как действуют те или иные элементы скелета (это требует
дополнительных исследований), сколько в том, чтобы выявить путем
сравнения специфику в функционировании позвоночного столба и ребер
у изучаемых форм современных сирен. В этом плане меньше помогают
конкретные цифры, пределы вариаций которых могут быть велики, а ог-
раниченность материала не дает возможности дать им оценку, чем срав-
нение кривых на графиках, как правило вскрывающих наиболее харак-
терные особенности изучаемых объектов. Это подтверждает почти полная
идентичность кривых, построенных на основе изучения осевого скелета
двух экземпляров ламантина. Объем главы, к сожалению, не позволяет
привести весь полученный материал, и мы ограничимся лишь обсужде-
нием основных результатов исследования. Сравнением с ископаемым ма-
териалом мы преследуем цель определить насколько возможно, на каком
уровне возникают те или иные особенности, специфичные для разных
форм современных сирен, и выявить возможные параллелизмы в их раз-
витии.
В большинстве случаев мы придерживались международной ветери-
нарной анатомической номенклатуры. Однако в отношении некоторых
отростков позвонков удобнее было применить номенклатуру, разработан-
ную первично Р. Оуэном, дополненную другими авторами и суммирован-
ную А. Ремане (Remane, 1936). Термины в этой системе не только лако-
79
ничнее, поэтому мы их используем в тексте, но и позволяют разобраться
в генезисе таких сложных образований, как поперечные, поперечно-
реберные и реберные отростки, и в особенностях соединения позвонков и
ребер. Вся терминология приведена в подписях к рисункам. Там же дано
сопоставление в терминах обеих систем. В дальнейшем русские термины
«поперечный» или «поперечно-реберный» отростки будут использоваться
в тексте только в своем обыденном значении, если оно не вызывает сом-
нения и речь не идет о его происхождении. В других случаях предпоч-
тительнее будет использоваться термин «диапофиз» (отросток, несущий
фасетку для сочленения с бугорком ребра), как и другие в этой систе-
ме (парапофиз как выступ, несущий фасетку для сочленения с головкой
ребра; синапофиз, если произошло слияние диапозифа с парапофизом;
плеврапофиз, если к диапофизу и пар апофизу присоединяется рудимент
ребра; мерапофиз, если обособившаяся от остальной части ребра головка
прирастает к парапофизу, как у некоторых китов; пре- и постзигапо-
физы; метапофиз как синоним сосцевидного отростка и анапофиз как
синоним processus accessorius).
Осевой скелет сирен, состоящий из позвоночного столба, ребер и
грудной кости, очень специфичен в связи с тем, что фактически единст-
венным локомоторным органом стал хвост, тогда как у китов в локомо-
ции участвует все туловище; грудной отдел скелета ламантина очень
обособлен и не выполняет ни локомоторной, ни активной дыхательной
функции, скорее его роль в создании постоянного горизонтального гра-
диента, а основные его морфологические особенности объяснимы участи-
ем его в гидростатических приспособлениях и в опоре для скелетной
мускулатуры.
ОБЩАЯ ХАРАКТЕРИСТИКА ПОЗВОНОЧНОГО СТОЛБА
Межпозвоночные диски, эпифизы тел позвонков. Позвоночный столб си-
рен слагается из позвонков и межпозвоночных дисков, причем и те и
другие очень необычны. У всех современных сирен тела позвонков пол-
ностью лишены костных эпифизов, характерных почти для всех млеко-
питающих. Исключение составляют лишь сирены и однопроходные
(Flower, 1885), у которых их нет даже на ранних стадиях развития.,
Однако у раннетретичных и позднетретичных сирен они еще есть3.
Так, тонкую костную пластинку, выстилающую тело позвонка спереди и
сзади, отмечают на некоторых грудных позвонках относительно молодого
экземпляра Metaxytherium calvertense (Kellogg, 1966), при этом предпола-
гается, что с возрастом она могла превратиться в настоящий эпифиз.
Очевидно, что утеря эпифизов у сирен шла параллельно в разных лини-
ях их развития. Межпозвоночные диски сирен в отличие от других вод-
ных млекопитающих, особенно китообразных, развиты крайне слабо. Это
отмечают все анатомы, имевшие дело с препаровкой ламантинов или
дюгоней. Так, Мьюри (Murie, 1872а) указывает, что у самки Trichechus
manatus длиной около 160 см толщина дисков не превышала 2,5 мм
(0,1 дюйма). Исходя из положения суставных фасеток на зигапофизах,
промежуток между телами позвонков не мог превышать 5—7 мм в за-
висимости от отдела, где этот промежуток определялся. К сожалению,
была упущена возможность точного определения толщины межпозво-
ночных дисков у взрослого ламантина, фиксированного формалином.
Опубликованные данные по этому экземпляру (Manzij, Piliptshuk, 1984,
3 Иногда они есть и у ламантина в качестве индивидуальной вариации (Albrecht,
1883).
80
Tab. 6) приводят совершенно фантастические цифры якобы, например,
в грудном отделе толщина диска могла достигать 13—14 мм, а во всем
позвоночном столбе на них отводится около 20% длины, что, конечно,
нелепо: суставные поверхности пре- и постзигапофизов при этом вообще
не касались бы друг друга.
Таким образом, следует заключить, что малое развитие межпозвоноч-
ных дисков, как и ряд других особенностей в осевом скелете, говорит
о повышенной жесткости позвоночного столба, особенно в грудном от-
деле, в ущерб его эластичности.
Отделы осевого скелета. Позвоночный столб сирен можно подразде-
лить всего на три отдела: шейный, грудной, все позвонки которого несут
ребра, и, наконец, последний — послегрудной включающий поясничный,
крестцовый и хвостовой отделы наземных млекопитающих. Однако в ли-
тературе часто можно встретить упоминания о наличии у сирен и этих
отделов как самостоятельных структурных подразделений. Как точка
отсчета при этом используется позвонок, к которому связкой прикрепле-
на рудиментарная тазовая кость (крестцовый позвонок), или первый
позвонок, несущий шевронные кости (он считается первым хвостовым).
Тем не менее нет никакой уверенности, что упомянутая выше связка
крепится именно к тому позвонку, к которому должны были быть при-
членены подвздошные кости, и, следовательно, выделение на этой основе
крестцового позвонка крайне условно, а противопоставление пояснич-
ного и хвостового отделов теряет смысл. Наличие шевронов как крите-
риев тоже сомнительно. Число их варьирует индивидуально. У экз. ИЗ-
5945 они впервые появляются между III и IV послегрудными позвонка-
ми, а у экземпляра ЗИН-31795 — между IV и V, тогда как связка от
таза у последнего идет к I. Однако известно, что у ламантинов она мо-
жет крепиться на первом позвонке, снабженном шевроном, т. е. на I
«хвостовом», который, таким образом, одновременно является и «крест-
цовым» (Brandt, 1861—1868). В то же время у наземных млекопитаю-
щих отмечены случаи, когда шевроны соединены с истинными пояснич-
ными позвонками (насекомоядные).
Таким образом, в дальнейшем мы будем говорить почти всегда только
о шейном, грудном и послегрудном отделах позвоночного столба, причем
только граница между первым и вторым четко выражена появлением
первого ребра, головка которого сочленена с телами последнего шейно-
го и первого грудного позвонков, а бугорок — с диапофизом последнего
из названных позвонков. Напротив, граница между грудным и послегруд-
ными отделами иногда замаскирована. У экз. ИЗ-5945 она легко опре-
деляется, как проходящая между последним позвонком, имеющим ребра
(XVI th) 4, и первым, лишенным их. У экз. ЗИН-677 XVII th имеет боль-
шие ребра, а следующий позвонок, рассматриваемый как I pth, на одной
стороне имеет сравнительно короткий, короче, чем на II pth, поперечный
отросток со следом прирастания очень маленького ребра, а на другой это
ребро сохраняет свою самостоятельность. У экз. ЗИН-31 795 после XVII th
с большими ребрами следует еще один с ребрами, несколько укорочен-
ными, но не сросшимися с поперечными отростками и вступающими в
контакт с образованием суставов с ребрами предшествующей пары (см.
дальше). Хотя этот позвонок с полным правом мог бы рассматриваться
как грудной, мы его определяем как I pth, тем более что таз крепится
4 В дальнейшем для обозначения в тексте и на рисунках позвонков будут даваться
римскими цифрами их порядковый номер в отделе, а сами отделы — буквами: с —
шейный, th — грудной, pth — послегрудной, иногда 1 — поясничный, s — крестцо-
вый, cd — хвостовой.
81
у этого экземпляра связкой именно к нему. Таким образом, заднюю гра-
ницу грудного отдела целесообразно проводить за последним позвонком,
несущим большие ребра и ясно участвующим в образовании грудной
клетки, функционируя вместе с другими позвонками, тогда как следую-
щий позвонок, имеющий или нет свободные ребра, имеет определенно
другую функцию: или особую, как у экз. ЗИН-31 759, или сходную с
остальными передними послегрудными («поясничными») позвонками.
Число позвонков. В состав позвоночного столба у ламантинов входят:
у экз. ИЗ-5945 — 6с + 16th+ 24pth (возможно, 1—2 последних хвостовых
отсутствуют) =46 (? +1—2); у экз. ЗИН-31 795 — 6с +17th + 26pth = 49;
у экз. ЗИН-677 — 6c+17th+25pth = 48; у экз. Х-175 — 6c+17th+22pth
(последние cd видимо, отсутствуют) >45; у дюгоней: у экз. ЗИН-3220 —
7c + 18th + 27pth (последние позвонки утрачены) >52; у экз. ЗИН-679 —
7с +18th + 25pth (3—6 последних cd утрачены, заменены муляжом) =53
или 56; у морской коровы 3M-3464 — 7c + 19th + 35pth (часть, возможно,
отсутствует) >61; у монтированного скелета в Зоологическом музее
АН СССР — 7c+19th+30pth = 60. Привлекая литературные данные
(Brandt, 1861—1868; Murie, 1872а; и др.), можно уверенно говорить, что
у ламантинов шесть шейных позвонков; в грудном отделе число позвон-
ков варьирует от 15, что является правилом для Trichechus inunguis
(Petit, 1955; Domning, 1978a, 1982) до 16—17 (редко 18) у других ла-
мантинов, в послегрудном от 25 до 27. Таким образом, общее число их
позвонков варьирует от 46 (у Т. inunguis) до 49 (редко 50—51, см.:
(Murie, 1972а); у T.m. latirostris, см.: (Ronald et al., 1978). Вариации
объясняются индивидуальной изменчивостью, может быть, иногда возра-
стом, но не полом. Ошибочные указания ранних авторов на наличие
якобы у некоторых ламантинов семи шейных позвонков были опроверг-
нуты (Murie, 1972а) на тех же самых материалах, на которые эти ав-
торы ссылались, причины появления ошибок как в отношении ламанти-
нов, так и других сирен, были обсуждены Брандтом (Brandt, 1861—
1968) и другими (Murie, 1972а; Petit, 1928).
Тенденция уменьшения числа позвонков от более примитивных форм
ламантинов (Т. manatus, Т. senegalensis см.: (Hatt, 1934)) к более
специализированному Т. inunguis четко выражена. Эта тенденция про-
должает ту же тенденцию, что была связана с происхождением ламанти-
нов от эоценовых предков типа Protosiren, имевшего 7c+20th+31+ls+
неопределенное число cd, и ранних дюгонид, подобных Eotheroides с та-
кой же формулой позвоночного столба (Sickenberg, 1934). Фактически,
у ламантинов уменьшено количество позвонков во всех отделах: в шей-
ном, грудном и послегрудном, что видно при сравнении их с дюгонида-
ми, вымершими и ныне живущими.
Общее число позвонков у дюгоня определяется как 55—62 (Brandt,
1861—1868), у морской коровы от 60 до 61 (может даже более),
у Dusisiren jordani — 64 (Domning, 1978с). При этом у всех дюгонид,
включая Hydrodamalis gigas, имеются 7 шейных позвонков. Утвержде-
ния ранних авторов (Steller, 1751; и др.) всего о 6 позвонках оказа-
лись ошибочными и были документально опровергнуты еще в прошлом
веке (Brandt, 1861—1868; Woodward, 1885). Возрождение в наши дни
этих ошибок основано на пренебрежении фактическим материалом и
поверхностном знании специальной литературы (Гептнер, 1967; Manzij,
Piliptshuk, 1984). У экземпляра 3M-3464 и в монтированном скелете
морской коровы в Зоологическом музее АН СССР по 7 шейных позвон-
ков; в скелете, принадлежащем МГУ,— 6, но один позвонок был утерян
при передаче его в Москву (его видел Брандт). При монтировке двух
скелетов в Киеве (в Палеонтологическом музее и музее КГУ) в шейный
82
отдел было поставлено 6 позвонков (оба скелета сборные;. Окончательно
решен вопрос о количестве шейных позвонков у гидродамалин нахожде-
нием у непосредственных предков стеллеровой морской коровы Dusisiren
jordani и Hydrodamalis cuestae тех же 7 шейных позвонков (Domning,
1978с). Грудной отдел дюгонид включает от 18 (наши материалы) до
19 (Abel, 1904; Simpson, 1932), реже 20 (Eotheroides, Miosiren) или даже
21 (Dusisiren) позвонок. Точное количество послегрудных позвонков у
дюгонид установить трудно в связи с обычным отсутствием в музейных
материалах какого-либо количества последних хвостовых позвонков
(с возрастом их число также увеличивается за счет продолжающегося
окостенения в конце хвоста). Для дюгоня, очевидно, характерно 30—36
позвонков в этом отделе (Brandt, 1861—1868), для морской коровы и
Dusisiren 35.
Таким образом, у всех дюгонид больше позвонков в шее, в грудном
отделе и особенно в послегрудном, чем у ламантинов. В то же время у
последних минимально количество позвонков в «поясничной» части после-
грудного отдела, всего 2, тогда как у дюгоня и примитивной морской
коровы Dusisiren их 3, а у Hydrodamalis gigas 6 или даже 8 (Kleins-
mith, 1951). Кроме того, у дюгонид имеется еще так называемый «крест-
цовый» позвонок, а у ламантинов его нет, так как тазовые кости соеди-
нены связками с первым позвонком, снабженным шевронами, т. е. с
«хвостовым». Из этого следует также заметная тенденция специфиче-
ского удлинения «поясничного» отдела у морской коровы за счет собст-
венно хвоста.
Пропорции осевого скелета. При взгляде сбоку на скелеты современ-
ных сирен (рис. 49), приведенных к одному размеру, сразу бросаются в
глаза различия в пропорциях отделов у разных форм. Резко изменяется
размер черепа, его величина максимальна у дюгоня, уступает ему череп
ламантина и относительно маленькая голова у морской коровы (см.
табл. 1). Сходные цифры по последней имеются в литературе: 9—10%
(Brandt, 1846, 1861—1868), 9,2% (Kaiser, 1974). Шейный отдел кажется
большим у ламантина и морской коровы, а грудной ламантина явно уве-
личен. Если привести к одному размеру длину черепа (рис. 20, а),
то оказывается, что у дюгоня минимальна относительная длина тулови-
ща, у ламантина она имеет промежуточное значение, у морской коро-
вы — максимальное. Кроме того, четко выявляются различия в длине шеи
относительно черепа, что имеет большое функциональное значение: у дю-
гоня шея составляет всего */4 от длины черепа, у ламантина около
‘/з, а у морской коровы почти половину. При приведении к одному раз-
меру позвоночного столба (рис. 20, б) становится очень заметно, что у
дюгонид грудной отдел сокращен и почти одинаков у обеих форм, а у ла-
мантина он увеличен, несмотря на то, что число позвонков у последнего
здесь меньше. Вообще у ламантинов длина грудного отдела почти равна
длине послегрудного. Сходные данные имеются относительно Т. senega-
lensis (Slijper, 1936). В то же время у дюгоня после грудной отдел пре-
вышает грудной на 15—20%, а у морской коровы на 21%. Значительное
увеличение количества позвонков в «поясничном» отделе морской коровы
сравнительно с дюгонем сказывается на линейных размерах этой части
позвоночного столба меньше, чем можно было бы ожидать, что связано
с относительно большей длиной тел первых послегрудных позвонков
дюгоня.
Таким образом, намечаются два пути специализации позвоночного
столба: ламантиновый и дюгонидный — с расхождением по ряду дета-
лей дюгоня и морской коровы в пределах общего типа.
Позвоночный столб ламантина слабо изогнут, максимум выгиба вверх
83
Рис. 19. Общий вид монтированных скелетов морской коровы Hydroda-
malis gigas (а), дюгоня Dugong dugon (б) и ламантина Trichechus ma-
natus (в), приведенных к одному размеру
Рис. по фотографиям, сделанным в Национальном музее естественной истории Смит-
соновского института, США (см.: (Kaiser, 1974))
84
приходится на уровень V th. Судя по скосу тел I—III th назад верхней
частью, в этой области позвоночный столб относительно почти горизон-
тального шейного отдела поднимается вверх, но высота выгиба не превы-
шает 6,2% длины грудного отдела (длина вычислена по проекции отдела
на прямую, а высота по нижним поверхностям тел позвонков). За V th
начинается слабый и постепенный наклон длинной оси позвоночного стол-
ба вниз (тела позвонков слегка скошены вперед), и она пересекает го-
ризонтальную плоскость, проходящую по шейному отделу за три позвон-
ка до окончания грудного отдела. Все последующие позвонки, видимо,
лежат ниже этой горизонтальной плоскости. У дюгоня отмечается значи-
тельно больший прогиб вверх спинного отдела (Kaiser, 1974), но никаких
подробностей не приводится. Интересно, что у морской коровы (экз.
3M-3464, очевидно, один из немногих скелетов в музеях мира, большинст-
во позвонков которого, по крайней мере в шейном и грудном отделах,
принадлежат одному индивиду) практически все позвонки имеют не-
большой скос вперед, т. е. можно полагать, что вопреки обычным утвер-
ждениям, что у морских коров позвоночный столб занимал горизонталь-
ное положение, на самом деле он шел назад и слегка вниз относительно
шейного отдела. Возможно, это соотносится с питанием этих животных
над уровнем моря или у его поверхности.
Масса скелета. Для таких животных, как сирены, ведущих всецело
водный образ жизни, масса скелета как части, удельный вес которой
превышает таковой воды, является одной из важнейших характеристик.
Масса всего скелета ламантина равна 29 293 г. (экз. ИЗ-5945), из кото-
рых на череп приходится 10,2%, на конечности — 11,6%, на осевой ске-
лет — 78,2% (24% позвоночный столб и 54,2% ребра). У другого экзем-
пляра (ЗИН-31795) масса всего скелета определена в 35 755 г, в томчисле
череп 14,3%, конечности 11,2%, осевой скелет 74,5% (позвонки 25% и
ребра 49,5%), т. е. скелет от общей массы животного составляет около
9,0%, что в общем не превышает относительной массы скелета у круп-
ных наземных млекопитающих (для крупного рогатого скота указыва-
ется 9—13%, причем половина из них падает на скелет конечностей,
шестая часть на череп и лишь треть на осевой скелет). Необычным для
осевого скелета, однако, будет очень большая масса ребер, которая почти
в два раза превышает массу позвоночного столба. У дюгоня ребра менее
массивны (как и сам скелет имеет больше губчатого вещества), и их
масса лишь в 1,3 раза превышает массу позвоночного столба. На череп
у дюгоня (ЗИН-3220) приходится 25,6% массы, на конечности 7,6%, на
весь осевой скелет 66,8%. Таким образом, череп дюгоня в соответствии
со своими большими размерами более чем вдвое массивнее, чем у ламан-
тина, а конечности имеют менее развитый скелет (в 1,5 раза меньше по
массе). Меньшую массу имеет и сам осевой скелет дюгоня.
Распределение масс вдоль длинной оси животного закономерно меня-
ется, и характер этого изменения сильно отличается у ламантина и дю-
гоня (рис. 21). Спереди, за более тяжелыми первыми двумя шейными
позвонками следуют самые легкие III и IV шейные. Масса каждого
позвонка постепенно растет в каудальном направлении и достигает
своего максимума примерно к V грудному позвонку у дюгоня и VIII у
ламантина, но дальше у ламантинов она начинает сокращаться к концу
грудного отдела, а у дюгоней держится на одном уровне. Первые три
послегрудные позвонка у ламантина заметно наращивают свою массу
частично в связи с вхождением в их состав рудиментов ребер, а далее
назад масса позвонков этого отдела постепенно уменьшается. У дюгоня
самым тяжелым позвонком является I послегрудной, последующие, как
и у ламантина, становятся легче, причем относительная масса после-
85
Рис. 20. Относительные размеры
отделов осевого скелета при
приведении к одному размеру
длины черепа (а) и длины по-
звоночного столба (б)
Г — Trichechus manatus (экз. ИЗ-5945);
2 — Т. manatus (экз. ЗИН-31795);
3 — Dugong dugon (экз. ЗИН-3220);
4 — D. dugon (экз. ЗИН-679);
5 — Hydrodamalis gigas (экз. 3M-3464>
Рис. 21. Посегментное распреде-
ление масс вдоль позвоночного,
столба у ламантина по экз. ИЗ-
5945 (I) и дюгоня по экз. ЗИН-
3220 и ЗИН-679 (2)
По оси абсцисс дан порядковый номер
позвонка (сегмента), по оси ординат —
вес в процентах к общему весу позво-
ночного столба. Сплошной линией по-
казан вес позвонков, пунктирной —
вес сегмента (позвонок + оба ребра).
Сплошная стрелка указывает границы
грудного отдела у ламантина, штрихо-
вая — у дюгоня
грудных позвонков (в процентах от массы позвоночного столба)
в 1,5—2 раза больше таковой ламантина. Таким образом, если рассмат-
ривать только позвоночный столб, то у дюгоня большая его масса будет
сосредоточена в задней половине туловища, а у ламантина посередине.
Иначе обстоит дело с ребрами. У ламантина опять самые массивные реб-
ра приходятся на середину грудного отдела (IX—XII th), но у дюгоня
на переднюю часть (IV—VI), компенсируя тем самым повышенную тя-
жесть постгрудного отдела. Это различие хорошо видно на кривых, от-
ражающих изменение массы всего сегмента (позвонок с относящимися
к нему ребрами, причем, как правило, в каждом сегменте основная мас-
са представлена ребрами). Обращает внимание, что у дюгоня кривая
массы сегмента плавно переходит в кривую массы позвонков послегруд-
ного отдела. Это может говорить о том, что увеличение массы этих по-
86
звонков в основном идет за счет включения в состав их поперечно-ре-
берных отростков рудиментов ребер. У ламантинов же мы видим иную
картину: в месте соединения обеих кривых имеется глубокий прогиб
вниз. Природа его становится яснее при обращении к другому экземпля-
ру ламантина (ЗИН-31 795), у которого имеется не только дополнитель-
ный грудной позвонок, но и позвонок переходного характера (I pth)
с крошечными, но не приросшими к позвонку ребрами. И если предполо-
жить, что дальше ребра не будут уменьшаться в размерах, то все равно
их масса в сумме с массой последующего позвонка будет недостаточна,
чтобы объяснить значительную прибавку массы первых послегрудных
позвонков. Именно постепенное возрастание их собственной массы при-
водит к тому, что самым массивным у ламантина является не первый
послегрудной, а последующие позвонки. Таким образом, кажется, что
поперечно-реберные отростки первых послегрудных позвонков своей дли-
ной обязаны в первую очередь не рудименту ребра, что может быть у
дюгоня, а увеличению самого отростка (синапофиза).
Следует отметить еще две особенности ребер ламантина, касающиеся
их массы и отличающие их от ребер дюгоня. Во-первых, последние ребра
ламантина, кроме уже отмеченных рудиментов у I pth экз. ЗИН-31 795,
тяжелее соответствующих позвонков, и лишь одно ребро последнего сег-
мента равно ему по массе, тогда как у дюгоня равно по массе позвонку
лишь пятое от конца ребро, а последние четыре легче соответствующих
позвонков. Во-вторых, передние ребра ламантина наиболее массивны в
их позвоночном конце, каудально это место смещается дистально по
ребру, оказываясь около середины его на XI—XII ребрах и в самом
дистальном конце у последних ребер. У дюгоня все ребра массивнее в
своем позвоночном конце. В то же время удельный вес позвонков в мас-
се сегмента у ламантинов выдерживается на большей части грудного
отдела позвоночного столба (с III по XIV th) примерно на одном уровне
(около 17—20%), будучи больше в самой передней части груди (26% в
I сегменте) и в самом конце ее (32% в XVI сегменте). У дюгоня же
практически на всем протяжении позвоночного столба с III th (20%)
идет нарастание относительной массы позвонков до 50% в XVIII сегменте.
Интересно, что, несмотря на все перечисленные различия ламантинов
и дюгоня в отношении распределения масс по длине позвоночного стол-
ба, у них обоих частный центр тяжести скелета (с учетом черепа и
скелета конечностей, а также принимая во внимание общую длину ске-
лета и длину каждого из позвонков в абсолютных единицах, а не в ус-
ловных, по числу сегментов) оказывается в одном месте — в области
примерно IX грудного позвонка.
Длина тел позвонков (рис. 22) ламантинов, начиная с самых корот-
ких в шейном отделе (IV с), прогрессивно нарастает в каудальном на-
правлении, достигая максимума в начале второй половины грудного от-
дела (X—XI th), а затем опять начинает уменьшаться вплоть до конца
позвоночного столба. У дюгоня характер изменений длин позвонков за-
метно отличается. Главное отличие (отличия, относящиеся к шейному
отделу, будут оговорены при его рассмотрении) — в сдвиге максимума
кривой длин позвонков в начало послегрудного отдела. То же самое на-
блюдается у морской коровы, но у нее нет определенного пика с макси-
мумом длины, а после достижения определенной величины она на боль-
шом протяжении позвоночного столба (с XI th по XI pth, а может,
и дальше) остается более или менее на одном уровне, и лишь центр
этого интервала падает на начало послегрудного отдела. Важно подчерк-
нуть, что относительная длина всех грудных позвонков ламантинов на-
много превосходит таковую дюгоней и морской коровы (так, у X th она
87
Рис. 22. Изменение относительной длины тел позвонков у ламантина (1, 2), дюгоня
(3) и морской коровы (4)
1 — экз. ИЗ-5945; 2 — экз. ЗИН-31795; 3 — экз. ЗИН-3220 с IV шейного до конца и ЗИН-679 для
I—IV шейных; 4 — экз. 3M-3464
По оси абсцисс дан порядковый номер позвонков, по оси ординат — их длина (в процентах к длине
позвоночного столба). Стрелки указывают границы грудного отдела
составляет 48% от высоты позвонка у экз. ИЗ-5945 и 58% У экз. ЗИН-
31795, а у того же позвонка дюгоня всего 30%, у морской коровы 34%),
т. е. в грудном отделе ламантина при его большей относительной длине
за счет увеличения длины позвонков могло сократиться их число, что
ведет к уменьшению общей гибкости отдела, так как сокращает коли-
чество межпозвоночных дисков. В то же время важно подчеркнуть, что
у таких дюгонид, как Eotheroides, Halitherium, Metaxytherium, Miosiren,
а также у Protosiren отношение длины тела позвонка к высоте всего
позвонка колеблется около значений, свойственных для дюгоня и морской
коровы. Следовательно, удлинение тел позвонков у ламантина должно
рассматриваться как апоморфный признак.
Высота позвонков, их тел и невральной дуги. В целом позвонки ла-
мантина производят впечатление более низких, чем у дюгоня, и не
только из-за их относительного удлинения. Так, у дюгоня (ЗИН-3220)
с меньшей общей длиной позвоночного столба (80% от такового ламан-
тина ИЗ-5945) длина тела X грудного позвонка составляет 56% от тако-
вой ламантина, тогда как высота уступает высоте позвонка ламантина
очень немного, составляя 88% ее.
Изменение высоты позвонков по ходу позвоночного столба (рис. 23)
у ламантина аналогично изменению длины. За относительно высокими
первыми двумя шейными позвонками следуют низкие III—VI с. Быст-
рый рост в высоту начинается с I грудного позвонка, максимума он
достигает в области X th, затем идет постепенное уменьшение высоты,
достигающее в конце отдела уровня, соответствующего IV—V th, и про-
должающееся дальше в хвост. При этом выясняется, что если в области
первых трех грудных позвонков позвонки в первую очередь набирали
высоту за счет невральной дуги, то далее шло преобладающее увеличе-
ние вертикального диаметра тела позвонка. В уменьшении высоты по-
звонков во второй половине грудного отдела большую роль играет со-
кращение невральной дуги, тело же меняет свою высоту более медлен-
но. В целом высота невральной дуги у ламантина в передней части
88
туловища превышает высоту тела позвонка; они сравниваются близ кон-
ца грудного отдела (XV th); каудальнее дуга становится ниже тела. Ис-
чезает невральная дуга у ламантинов рано: ее уже нет на XV послегруд-
ном позвонке, и дальнейшее уменьшение хвостовых позвонков связано
уже, естественно, только с изменением самих тел позвонков.
Ход изменения высоты всего позвонка у морской коровы внешне
очень сходен с ламантиновым. Отчетливо выраженный пик приходится
на заднюю половину грудного отдела, и быстрое падение высоты начи-
нается уже в конце его. Однако анализ кривых убеждает, что сходство
здесь достигается разными способами. В самом начале грудного отдела
резко возрастает величина дуги, а тело в своем росте опаздывает. Од-
нако к VIII th начинается быстрый рост и тела, которое достигает
максимальной высоты в области XI th. В дальнейшем на протяжении
двух десятков позвонков тело остается почти неизменным, и даже
после уменьшение его незначительно. Таким образом, отвечает за изме-
нение высоты позвонка почти исключительно невральная дуга, кроме
короткого интервала (4—5 позвонков в первой трети грудного отдела).
89
Резко отличается характер изменения высоты позвонка у дюгоня.
IV грудной позвонок достигает уже почти максимальной высоты (хотя
о максимуме можно говорить скорее в районе XI—XV th), а дальше, на
протяжении почти двух десятков позвонков, высота остается более или
менее постоянной, да и последующее уменьшение ее до XII послегруд-
ного позвонка идет очень медленно. В то же время само тело позвонка
изменяется очень необычно: нарастание его высоты очень медленное,
и максимума она достигает лишь на V—VIII послегрудных позвонках,
когда высота всего позвонка уже уменьшается. Почти на всем протяже-
нии позвоночного столба (до XIII—XIV pth) невральная дуга выше тела,
и исчезает она у дюгоня только к XXI послегрудному позвонку. Таким
образом, характер изменения тела позвонка почти сходен у дюгоня
и морской коровы, отличаясь заметно от наблюдаемого у ламантина,
но по высоте всего позвонка морская корова больше похожа на послед-
него.
Относительная высота невральной дуги (в процентах к высоте позвон-
ка) в области шейного и самых первых позвонков грудного отделов бо-
лее или менее близка у всех трех форм, но у ламантина она, после мак-
симума у II—III грудных позвонков, начинает уменьшаться более или
менее равномерно до VII pth, а затем несколько более ускоренным тем-
пом, тогда как у дюгоня и морской коровы, достигнув максимальной вы-
соты в районе III—IV th, большей, чем у ламантина, в дальнейшем на-
чинает сокращаться более медленно, чем у последнего, в результате чего
разница в относительной высоте дуги у них все более и более нарастает,
достигая в конце грудного отдела 17%, У X pth —30% и т. д. Следует
указать, что у морской коровы невральная дуга исчезает не раньше, чем
на XXX pth.
Тела позвонков и моменты сопротивления. В работе позвоночного стол-
ба большую роль играют форма и пропорции тел позвонков, которые раз-
личны у всех трех изучаемых форм сирен. На форму тел влияет как
соотношение вертикального и поперечного диаметров, так и наличие на
теле тех или иных отростков (парапофизы, вентральный гребень, степень
развития ножек дуги и т. п.). У ламантина сочленовная поверхность тел
шейных и самых передних грудных позвонков напоминает овал, вытяну-
тый в поперечном направлении, следующие за ним позвонки приобре-
тают сперва почковидную форму с впадиной сверху для спинномозгового
канала, а затем по мере увеличения вертикального диаметра и осо-
бенно с появлением в задней части тела V грудного позвонка
(у экз. ЗИН-31795 — IV th) вентрального гребня приобретает отчетли-
вую сердцевидную форму. Лишь последний грудной и первые после-
грудные позвонки опять становятся почковидными, затем последователь-
но шестигранными (две верхние грани связаны с ножками невральной
дуги, две нижние — с прикреплением шевронных костей, две боковые —
с основанием поперечно-реберных отростков), пятигранными (с исчезно-
вением невральной дуги две верхние грани как бы сходятся в одну),
ромбовидными (со сближением правых и левых половин шевронов две
нижние грани объединяются) сильно уплощенными дорсовентрально.
Последнее изменение формы связано с более ранним исчезновением
невральных дуг, чем шевронных костей.
Соответственно в шейных и первых грудных позвонках ширина тела
значительно превышает его высоту. Каудально и то и другое возрастает,
но из-за того, что высота тела растет быстрее, сами тела становятся от-
носительно выше и выше, а относительная ширина уменьшается.
На VII—XIV грудных позвонках ширина лишь немного превышает
высоту. Таким образом, самые широкие позвонки у ламантина лежат в
90
шее и передней части грудного отдела, а также ближе к концу позво-
ночного столба, а самые высокие в задней половине грудного отдела.
У дюгоня (ЗИН-679) шейные и I грудной позвонки имеют почти
прямоугольную форму (высота опять меньше ширины), которая сменяет-
ся через переходную трапециевидную с более длинным нижним основа-
нием (III th) на овальную (IV—V) и затем сердцевидную форму, но
значительно более низкую и широкую, чем у ламантинов. Эта уплощен-
ность возрастает к концу грудного отдела, и IV pth опять становится
овальным. Далее назад с уменьшением относительной ширины тела
позвонков становятся шестигранными, а в области XIII—XV послегруд-
ных позвонков даже восьмигранными (боковые грани как бы подразде-
ляются на две). Самые последние позвонки хвоста опять сильно упло-
щены. Отличительной чертой формы тел позвонков морской коровы
является их относительная ширина. У шейных и первых двух грудных
позвонков сочленовые поверхности овальные, далее назад они почковид-
ные, и эта форма сохраняется до XI послегрудного позвонка, после чего
она приобретает вид лежащей на боку восьмерки, а к XXV pth она ста-
новится пятиугольной, а в самом конце хвоста почти круглой. Таким
образом, и у дюгоня, и у морской коровы главное отличие, касающееся
относительной ширины тела позвонка (в процентах к ее высоте), заклю-
чается в том, что у них после самых широких II послегрудного позвонка
у первого и IV у последней происходит почти до самого конца хвоста
постепенное увеличение высоты тела при уменьшении ширины. В то же
время у морской коровы постгрудные позвонки имеют большую относи-
тельную ширину, чем у дюгоня. У ламантина, наоборот, более или менее
ровная относительная ширина тела первых 12—13 послегрудных позвон-
ков сменяется их дальнейшим уплощением, и лишь последние четыре
хвостовых позвонка становятся несколько выше. Заметно больше также
относительная ширина последних шейных позвонков у дюгоня и особенно
морской коровы, чем у ламантина.
Отмечая недостаточность того или иного параметра для оценки позво-
ночного столба как механической конструкции, поскольку ни один из них
сам по себе не говорит о силах, возникающих в нем, Е. Слайпер
(Slijper, 1946) предложил использовать формулу, используемую в науке
о сопротивлении материалов для расчета момента сопротивления, разви-
ваемого прямоугольной балкой в отношении силы, действующей на ее
сгибание: W'=*/e bh2, где W — момент сопротивления, Ъ — ширина
и h — высота балки (при силе, действующей на балку в вертикальной
плоскости). Учитывая разницу в форме сечения прямоугольной балки и
позвоночного столба, автор указывает, что правильнее было бы исполь-
зовать формулу W—Jle (/ — момент инерции, е — расстояние между
центром тяжести и наиболее высокой точкой непрямоугольного сечения),
но использование ее затруднительно, особенно на монтированных скеле-
тах в музеях. Проведенное Слайпером сравнение кривых, построенных на
цифрах, полученных обоими способами на скелете одного животного
(лев), показало почти полную их идентичность. Следовательно, ис-
пользование величины bh2 вполне допустимо для определения характера
изменения момента сопротивления по ходу позвоночного столба. Таким
образом, в формуле Слайпера — bh2, как ее стали называть последую-
щие исследователи, увязываются вместе сразу два параметра — ширина
и высота тела позвонка (его сочленовная поверхность). Абсолютная
величина момента сопротивления не имеет большого значения, опреде-
ляясь в основном размерами животного, но положение пика на кривой
указывает то место позвоночного столба, на которое действуют максималь-
ные внешние силы. Соответственно минимальные значения моментов со-
91
противления, отмечаемые на графике падением кривой, указывают на
область позвоночного столба, нагрузка на которую наименьшая. Слайпер
(Slijper, 1946) при этом оговаривает роль дополнительных факторов
(развитие остистых отростков, сочленения в зигапофизах, сила связок
позвоночного столба), которые могут вносить дополнительный вклад в вы-
полнение главной статической функции позвоночного столба — сопро-
тивления изгибанию в вентральном направлении (выгиб направлен
вверх) и, таким образом, компенсировать недостаточную силу конструк-
ции, даваемую одними соединениями тел позвонков.
Нами были рассчитаны моменты сопротивления позвоночных столбов
обоих экземпляров ламантинов, дюгоня, морской коровы, а также, ис-
пользуя данные параметров тел позвонков, приведенные в литературе,
Halitherium schinzi (Sickenberg, 1934), Metaxytherium calvertense (Kel-
logg, 1966) и Dusisiren jordani (Domning, 1978c), что позволяет оценить
глубину специфичности наблюдаемых различий. До сих пор кривая мо-
ментов сопротивления была опубликована лишь для дюгоня (Cozzi,
1981). Бразильского ламантина исследовал Слайпер но, видимо, не сочтя
его сильно отличающимся в этом отношении от морской свиньи (Phocae-
na phocaena), не поместил соответствующего графика в своей работе
(Slijper, 1946, р. 42).
Анализируя полученные материалы (рис. 24), следует отметить, что
ламантин принципиально отличен от дюгоня и морской коровы. За не-
большим пиком в конце шейного отдела и некоторым уменьшением мо-
мента сопротивления в самом начале грудного (I—II th) следует стре-
мительный подъем кривой к XI грудному позвонку, и за ним такой же
стремительный спуск к концу отдела и дальше более полого в хвост.
У дюгоня нет никакого локального подъема в шейном отделе, а наращи-
вание величины bh2 в грудном отделе идет не с самого его начала
(лишь с VI—VII th) и совершается более или менее постепенно до
самого конца его, а продолжается до максимума в районе V послегруд-
ного позвонка (или первого-второго позвонка, снабженного шевроном).
За довольно четко выраженным пиком следует падение кривой, более
быстрое, чем подъем в груди. У морской коровы имеется подъем кривой
в области задних шейных позвонков, более выраженный, чем у ламан-
тина, затем падение в начале грудного отдела и замедленное поднятие
на первых восьми грудных позвонках (аналогия с дюгонем), сменяемое
стремительным подъемом до максимума в пределах всего трех позвонков.
Таким образом, максимум достигается уже на XI th (как у ламантина),
но за ним не следует падение кривой. Более или менее одинаковые зна-
чения моментов сопротивления сохраняются до X—XI послегрудного
позвонка, и лишь после этого кривая опускается к концу хвоста. Можно
сказать, что середина этого плато на вершине кривой приходится опять
же, как и у дюгоня, на начало послегрудного отдела, т. е. кривую, харак-
терную для морской коровы, исключая шейный пик, вполне можно срав-
нить с дюгоневой, у которой вершина основного пика срезана горизон-
тальной линией. А обе они вместе отличаются от кривой ламантина
сдвигом основного пика назад, в начало послегрудного отдела.
В этом смысле характер изменения моментов сопротивления по ходу
позвоночного столба, свойственный дюгонидам, значительно ближе к
тому, что наблюдается у большинства китообразных, где вершина пика,
или середина более растянутого максимума, почти всегда приходится на
конец поясничного отдела (Cozzi, 1981). Однако это сходство следует
признать поверхностным, как и сделал Б. Коцци, выделивший дюгоня
в особую группу по характеру кривой, отличную от трех других, свой-
ственных изученным им китообразным (три типа, один из которых с тре-
92
Bh2
Рис. 24. Изменение по ходу позвоночного столба «момента сопротивления» (bh2)>
силе, действующей на сгибание в вертикальной плоскости
1 — Trichechus manatus, экз. ИЗ-5945; 4 — Halitherium schinzi;
2 — Dugong dugon,экз. ЗИН-679 и ЗИН-3220; 5 — Dusisiren Jordani
3 — Hydrodamalis gigas, экз. ЗМ-3464; Стрелками указаны границы грудного отдела'
мя подтипами). У всех китообразных уменьшен собственно грудной
отдел, но увеличен поясничный. Подъем кривой, таким образом, не огра-
ничен первым из них, а продолжается в пояснице в меньшей (полосати-
ки, крупные киты, крупные дельфины, речные дельфины) или равной
(нарвалы, кашалоты, клюворылы) степени. Лишь в случае небольших
длиннотелых дельфинов, питающихся рыбой в поверхностных слоях
воды, отмечено, что максимум достигается уже к концу груди, но здесь
этот максимум растянут на всю поясницу и начало хвостового отдела.
Таким образом, эта кривая приобретает вид, напоминающий таковую
морской коровы.
У сирен поясничный отдел сильно сокращен (см. выше). Сравнение
с китами заставляет думать, что он включен в грудной отдел, а не просто
редуцирован, увеличение же «поясничной» части послегрудного отдела
у морской коровы, очевидно, является вторичным и идет за счет хвоста.
Вследствие этого сокращения поясницы сравнительно с наземными пред-
ками и иной организации двигательного аппарата, чем у китообразных,
пик кривой у дюгонид помещается в начале собственно хвостового отде-
ла, напоминая этим кривую, характерную для кашалота.
Сопоставление кривых дюгоня и Halitherium в соответствии с пред-
ставлениями о их родственных связях (см. гл. 1) доказывает их прин-
93
ципиальное сходство: положение главного пика, слегка срезанного у ха-
литерия, и растянутость подъема, и большая крутизна опускания,
и меньшее значение bh2 в начале грудного отдела. В противоположность
этому кривые морской коровы и Dusisiren почти идентичны (увеличен-
ный пик в шее, падение в начале грудного отдела, стремительный рост
в середине груди, плато на вершине, растянутое на всю заднюю полови-
ну грудного и начало послегрудного отделов). Таким образом, гидродама-
линовый характер строения позвоночного столба Dusisiren подтверж-
дается. В небольшой срезанности вершины Halitherium можно усмотреть
начало тенденции, более явственно выраженной у более позднего по вре-
мени Dusisiren, или же это общая черта примитивных дюгонид, которая
в дальнейшем у дюгоня была замещена появлением характерного дюго-
невого пика на кривой, а у гидродамалин развилась дальше.
Особое положение ламантинов в отношении характера изменения
bh2 интересна тем, что он не поддается сравнению с какими-либо дру-
гими животными, ни наземными, ни водными. Сдвиг пика на его кривой
вперед в грудной отдел, очевидно, можно объяснить крайней необходи-
мостью уменьшить сопротивление вентральному сгибанию хвостового от-
дела, так характерному для него, но не для других сирен. Соответствен-
но, видимо, надо признать, что, хотя нам и неизвестно, как осуществ-
ляется локомоция дюгоня в действительности, тем не менее конструктивно
она организована иначе, чем у ламантинов.
Момент сопротивления поперечным силам, действующим на позвоноч-
ный столб (bh2), изменяется на протяжении позвоночного так же, как
момент сопротивления вертикальным силам. Это говорит о том, что уси-
ление конструкции позвоночного столба сирен идет синхронно и в высо-
ту, и в ширину, несмотря на то, что горизонтальные силы в их жизни,
как у большинства млекопитающих, играют значительно меньшую роль,
а амплитуды боковых изгибов туловища крайне ограничены. Можно
лишь указать, что в связи с постоянным превалированием поперечного
диаметра позвонков над вертикальным, значения b2h больше, чем bh2.
Невральная дуга через свои отростки также определяет конструктив-
ные особенности позвоночного столба. Некоторые параметры, связанные
с ними, имеет смысл в общем виде рассмотреть здесь, до обсуждения
особенностей позвонков различных отделов позвоночника.
Максимальная ширина позвонков определяется в шейном отделе сте-
пенью развития плеврапофизов, в грудном — диапофизов, а в послегруд-
ном — поперечнореберных отростков. Поскольку различна природа этих
отростков и их функции и они соответственно прямо не сопоставимы, то
само понятие «максимальная ширина» становится формальным. На пер-
вое место здесь выступает длина этих отростков, дающих опору мышцам
и ребрам (что целесообразно обсудить для каждого отдела отдельно),
а не увеличение за их счет собственной ширины позвонка. Тем не менее
укажем, что у всех сирен максимальная ширина позвонка изменяется
более или менее одинаково. За широким атлантом следует узкий эпистро-
фей. Последующие шейные позвонки увеличиваются в ширину, а груд-
ные опять сужаются. Переход грудного отдела в послегрудной ознамено-
ван расширением I pth у дюгоня или более растянутым (на два-три по-
звонка) расширением у морской коровы и ламантина, за которым следует
постепенное сужение позвонков, кроме самых последних у дюгоня, опять
несколько расширяющихся. При сопоставлении относительной макси-
мальной ширины позвонка при общем сходстве обнаруживаются и раз-
личия в постторакальной области (рис. 25), связанные с уже отмечен-
ной выше разницей в высоте ее позвонков.
94
Рис. 25. Относительная ширина позвонка (в диапофизах, или поперечнореберных
отростках) у ламантина (7 — экз. ИЗ-5945), дюгоня (2 —экз. ЗИН-3220 и ЗИН-679)
и морской коровы (3 — экз. 3M-3464)
По оси абсцисс дан порядковый номер позвонка, по оси ординат — ширина позвонка (в процентах
от высоты позвонка). Стрелками отмечены границы грудного отдела
Рис. 26. Ширина позвонка между центрами суставных поверхностей постзигапофи-
зов ламантина (7) и дюгоня (2)
а — ширина в мм; б — относительная ширина в процентах к ширине задней сочленовной поверхно-
сти тела позвонка.
Л/п/п
1 — экз. ИЗ-5945; 2 — экз. ЗИН-3220 и ЗИН-679.
Стрелками отмечены границы грудного отдела
95
Ширина позвонков в постзигапофизах также в целом изменяется
у ламантина и дюгоня одинаково (рис. 26, а): к концу шейного отдела
она возрастает, в начале грудного резко уменьшается почти в два раза
к III—IV th. У дюгоней за этим следует медленное и постепенное
уменьшение расстояния между постзпгапофизами одного позвонка вплоть
до их утери на V th. У ламантина за сужением в начале грудного
отдела небольшое расширение к XIII th и лишь после идет окончатель-
ное постепенное сужение к II pth, за которым постзиганофизы уже от-
сутствуют. Различия между ламантином и дюгонем выступают более от-
четливо при обращении к относительной ширине (рис. 26,6).
Положение фасеток постзигапофизов у сирен сильно меняется на
протяжении позвоночного столба, но характер этого изменения у всех
более или менее одинаков (рис. 27). Наибольший наклон (оценка шла
Рис. 27. Изменение положения постзига-
пофизов ламантина (1 — экз. ИЗ-5945) и
дюгоня (2 — экз. ЗИН-3220 и частично
ЗИН-679)
По оси абсцисс дан порядковый номер позвонков,
по оси ординат — угол наклона (а) плоскости сус-
тавной поверхности постзигапофиза (в градусах)
относительно сагиттальной плоскости (в попереч-
ной проекции). Стрелки указывают границы груд-
ного отдела
только в трансверзальной проекции) имеют постзиганофизы эпистрофея.
Далее назад наклон уменьшается, в начале трудного отдела даже стано-
вится почти горизонтальным у ламантина и чуть ли не выворачивается
слегка внутрь у дюгоня. Но за этим плоскость суставных поверхностей
опять постепенно принимает все более наклонное положение к XI th
у ламантина и XII у дюгоня, чтобы к концу грудного отдела опять слег-
ка уменьшить наклон. К сожалению, у морской коровы не были на месте
замерены углы наклона фасеток зигапофизов и лишь сравнены из поло-
жения на IV и X th всех трех форм. В первую очередь в их отноше-
нии бросаются в глаза отличия по величине и месту относительно
невральной дуги. Наклон же, изменяясь в сходном направлении у всех
сирен, характеризуется разными углами на одном и том же по порядку
позвонке, но у разных форм, что опять же будет оговорено в дальнейшем.
Спинномозговой канал. Позвоночные отверстия на позвонках сирен
сильно различаются по форме (см. дальше) и относительной величине.
Спинномозговой канал у изученных форм меняется также на протяже-
нии позвоночного столба, то сужаясь, то расширяясь в том или ином на-
правлении, что связано не с изменением поперечного сечения мозга,
который занимает в канале относительно небольшое место, а с развитием
вдоль него более или менее развитых пучков чудесной сети. У ламантина
наибольшие по общим размерам позвоночные отверстия находятся в
самом конце шеи (самое большое), в середине и в самом конце груд-
ного и начале послегрудного отделов, наименьшие в области III—IV груд-
ных позвонков и в хвостовой части позвоночного столба. Вертикальный
и поперечный диаметр отверстий меняются независимо друг от друга,
часто в разных направлениях, так что спинномозговой канал расширяет-
ся то в высотном направлении, то в широтном, ширина достигает макси-
96
мума в I и VI шейных и XVI грудном позвонках, высота — у II шейно
го и у V—X грудных, более или менее равными оба диаметра являются
лишь у II шейного, IV и XIII грудных позвонков; высота превышает
ширину лишь в области между двумя последними (IV и XIII), у осталь-
ных позвонков, кроме уже названных, ширина превалирует над высотой.
У дюгоня ширина на всем протяжении позвоночного столба превы-
шает высоту. Максимальным по поперечному сечению спинномозговой
канал является в области последних шейных и первых грудных позвон-
ков. Никакого расширения близ границы грудного и поясничного отде-
лов нет. Лишь у второго шейного, как у ламантина, у VIII грудного
п позади VII послегрудного позвонков высота приближается к значе-
ниям, близким к ширине. Максимально широкие и низкие позвоночные
отверстия у IV—V шейных и самых последних грудных позвонков.
В целом позвоночные отверстия дюгоня значительно крупнее, чем
у ламантина.
ШЕЙНЫЙ ОТДЕЛ
Укороченная шея сирен во многом сходна с другими водными млекопи-
тающими, далеко ушедшими вперед в своей специализации. Как основ-
ную причину укорочения шеи именно сирен А. Б. Хауэлл (Howell, 1930)
называет адаптацию по питанию, когда нет необходимости в большой
подвижности шеи, а само животное плавает относительно медленно и во-
просы обтекаемости, лобового сопротивления перед ним не встают. Пита-
ние же массовой водной растительностью вполне обеспечивается поворо-
тами головы вместе с телом.
Шейные позвонки ламантина и дюгоня (рис. 28) были подробно
описаны в специальной статье (Petit, 1928), а морской коровы в моно-
графии (Brandt, 1861—1868), что делает излишним полное повторение
этих описаний здесь. Ж. Пти особое внимание уделил индивидуальным
вариациям в форме позвонков, невральной дуги и особенно в области
«поперечного комплекса», как он называет плеврапофиз шейных позвон-
ков (сам термин плеврапофиз этот автор употребляет более узко, чем
это принято, прилагая его лишь к рудименту ребра, вошедшему в состав
плеврапофиза в обычном смысле, как правило, замыкающему латерально
или латерально и вентрально поперечное отверстие). Анализ этих вариа-
ций позволил гомологизировать разные части поперечного комплекса.
Так, было показано, что диапофиз последних шейных позвонков соответ-
ствует таковому грудных, но к нему прирос бугорок шейного ребра,
«парапофиз» представляет слияние головки ребра с собственно парапо-
фпзом (выступом, или просто участком, на теле позвонка, несущим фа-
сетку для головки ребра) и шейку ребра, тогда как пластинка, замыкаю-
щая поперечное отверстие латерально, является остатком тела шейного
ребра. На основе этого анализа автор аргументировал положение, выска-
занное еще Ф. Ф. Брандтом (Brandt, 1863), что последний (VII) шей-
ный позвонок, имеющийся почти у всех сирен, вошел у ламантинов в со-
став грудного отдела в качестве I грудного, вопреки существовавшим
ранее представлениям, что уменьшение числа позвонков в шейном отделе
ламантина вызвано исчезновением предпоследнего (VI) позвонка (Blain-
ville, 1839; Flower, 1864; и др.) или III позвонка (Murie, 1872а). Послед-
ний автор на пользу своей гипотезе привлекал тот факт, что III шейный
позвонок часто срастается с эпистрофеем (и следовательно, он функцио-
нально мало значим), а также открытие им рудиментарной головки
ш. scalenus, предположительно принадлежавшей III позвонку, но с исчез-
новением последнего сместившейся на эпистрофей дополнительно к уже
4 Ламантин
97
Рис. 28. Шейные и первый грудной позвонки ламантина (а, в, д, экз. ИЗ-5945) и дюгоня (б, г, е, экз. ЗИН-679)
а, б — вид сбоку; в, г — вид сверху (позвонки несколько раздвинуты); д, е — вид снизу
1 — arcus dorsalis, дорсальная дуга;
2 — arcus ventralis, нижняя дуга;
3 — corpus vertebrae, тело позвонка;
4 — crista dorsalis, дорсальный гребень;
5 — crista epistrophei, гребень эпистрофея;
6 — dens зуо;
1 — facies articularis cranialis, задняя су-
ставная поверхность;
8 — facies articularis ventralis, нижняя су-
ставная поверхность;
9 — foramen intervertebrale, межпозвоно ч-
ное отверстие;
10 — for. transversariuni, поперечное отвер-
стие;
1 — fossa atlantis, ямка атлант а;
12 — fovea articularis atlantis, краниальная
суставная ямка (для затылочных мы-
щелков);
13 — fovea costalis caudalis, задняя реберная
ямка;
14 — fovea costalis cranialis, передняя ребер-
ная ямка;
15 — fovea costalis transversalis, реберная ям-
ка поперечного отростка;
16 — pleurapophysis, плеврапофиз;
11—processus articularis caudalis (=post-
zygapophysis), задний суставной отро-
сток (постзигапофиз);
18 — рг. articularis cranialisl(=praezygapop-
hisis), передний суставной отросток (пре"
зигапофиз);
19—pr. spinosus, остистый отросток;
20 — рг. transversus dorsalis (=diapophysis),
верхний поперечный отросток (диапо-
физ);
21 — pr. transversus ventralis (=parapophysis),
нижний поперечный отросток (парапо-
физ);
22 — spatium interarcuale, междуговое про-
странство;
23 — sulcus dorsalis atlantis, дорсальная бо-
розда атланта;
24 — tuberculum dorsale, дорсальный бугорок;
25 — tuberculum ventrale, вентральный бу-
горок
CD
00
6 25 В
Рис. 28 д.
100
существующей на нем собственной сильно развитой головке лестничной
мышцы.
Продолжая несколько дальше мысли Ж. Пти, можно сделать вывод,
что предпосылкой превращения последнего шейного позвонка в грудной
у ламантина была меньшая дегенерация последних шейных ребер.
Именно этим объясняется подмеченная раньше (Petit, 1928) особенность
парапофиза VI позвонка, сильно выгибающегося в своем основании
вперед (см. рис. 28, д), что приближает его к задне-боковым углам тела
предшествующего позвонка, напоминая тем самым о бывшем межвертеб-
ральном положении головки шейного ребра VI шейного позвонка.
Первые два шейных позвонка сирен, как у всех млекопитающих,
крайне специфичны: они обеспечивают максимум движений головы
в вертикальной плоскости и относительно длинной оси (вращение).
Атлант сирен, как обычно, представляет костное кольцо, состоящее из
двух боковых масс, соединенных сверху и снизу более тонкими дугами.
Спереди это кольцо несет пару больших суставных ямок для затылоч-
ных мыщелков, а сзади пару ямок для сочленения с эпистрофеем
(рис. 29). В то же время он не только сильно отличается от атланта
других млекопитающих, в частности отсутствием крыльев атланта (рас-
ширенных, более или менее горизонтально поставленных пластинок по
бокам позвонка), которые заменены у сирен боковыми выступами, впол-
не сопоставимыми с настоящими поперечными отростками, но и заметно
варьирует у разных форм сирен. В целом атлант ламантина относительно
выше, чем у дюгоня и морской коровы. У последней ширина явно пре-
валирует над высотой.
У ламантина верхняя дуга намного массивнее нижней: она сильно
вздута и ее толщина (25—30% от высоты позвонка) почти не уступает
высоте остистого отростка эпистрофея (фактически, при измерениях под
высотой остистого отростка понимается не только высота отростка, но и
высота у средней линии самой пластинки дуги позвонка). В передне-зад-
нем направлении верхняя дуга также расширена. В сагиттальном сече-
нии она субтреугольна: одна вершина треугольника направлена вперед,
вторая, образующая tuberculum dorsale, вверх и назад, третья — назад.
Каудальная поверхность дуги уплощена и несет треугольную площадку,
служащую упором для очень массивного остистого отростка эпистрофея.
У дюгоня и морской коровы верхняя дуга значительно тоньше (около 15%
от высоты позвонка), не вздута в середине, где она лишь незначительно
толще более латеральных частей дуги, и по массивности не превышает
нижнюю дугу. Дорсальный бугорок развит слабо, и вперед от него идет
невысокий гребень. У морской коровы на дорсальной поверхности дуги
вместо бугорка имеется шероховатость, на которую опирается нависаю-
щий сверху передний выступ эпистрофея. Еще одно отличие, вызванное
тем же утолщением дуги ламантина, имеет верхняя дуга дюгоня и мор-
ской коровы: при виде спереди она имеет вид настоящей дуги окруж-
ности, тогда как у ламантина обе ее ветви сходятся к средней линии под
углом около 90°, напоминая двухскатную крышу.
Поперечные отростки атланта у ламантина выступают в виде мощных
выступов, более толстых сверху (часть, соответствующая диапофизу)
и тонких снизу (парапофиз). Нижняя часть более короткая, верхняя
удлинена и отогнута вверх и назад. Дорсально дистальные концы отрост-
ков поднимаются на уровень верхнего края позвоночного отверстия.
Таким образом, весь отросток имеет вид толстой балки, несколько упло-
щенной, поставленной косо сверху вниз и вперед (широкая задняя по-
верхность с небольшой fossa atlantis обращена назад и немного вниз).
У экз. ЗИН-31 795 с правой стороны отросток прободен поперечным от-
101
14
Рис. 29. Атлант ламантина
(а—д, экз. ИЗ-5945) и дюгоня
(3, экз. ЗИН-679)
а — спереди;
б — сзади;
в, д — сбоку;
г — снизу
1 — arcus dorsalis;
2 — arcus ventralis;
3 — fossa atlantis;
4 — fovea articularis atlantis;
5 — fovea articularis caudalis,
каудальная’ суставная
ямка;
6 — fovea dentis, ямка зуба;
7 — lamina arcus vertebrae,
пластинка дуги позвонка;
8 — massa lateralis, боковая
масса;
9 — pediculus arcus vertebrae,
ножки дуги позвонка;
10 — pleurapophysis, плеврапо-
физ;
11 — pr. transversus dorsalis (dia-
pophysls);
is — pr. transversus ventralis
(parapophysis);
13 — sulcus dorsalis atlantis;
14 — tuberculum dorsale;
IS — tuberculum ventrale
102
Показаны взаимоотношения
затылочного мыщелка (J)
с атлантом (2), изображен-
ным в положении с
наибольшим совпадением
суставных поверхностей
(сплошная линия), в наи-
более верхнем (штриховая)
или нижнем (штрих-пунк-
тир) положении. Вид спра-
ва. Альвеолярный край зуб-
ного ряда горизонтален
Рис. 30. Схема атланто-затылочного сустава ламантина (а, экз. ИЗ-5945), дюгоня
(б, экз. ЗИН-3220) и морской коровы (в, экз. 3M-3464)
верстием для прохождения a. vertebralis. У дюгоня поперечные отростки
очень сильно отличаются от ламантиновых: они имеют вид округлых в
сечении, удлиненных и несколько сужающихся к концу выступов боко-
вых сторон позвонка, направленных почти горизонтально (на уровне
середины высоты позвонка) и менее отклоненных назад, чем у ламанти-
на. У морской коровы поперечные отростки напоминают таковые ламан-
тина (четырехугольное сечение, почти вертикальная постановка макси-
мальной ширины, толщина), но они менее отклонены вверх (не доходят
до уровня верхнего края позвоночного отверстия) и назад.
Краниальные суставные ямки атланта варьируют индивидуально по
форме. Так, у ламантина экз. ЗИН-31 795 (см.: (Manzij, Piliptshuk, 1984),
A.bb. 7В) они имеют почковидную форму, заметно отличающуюся от
экз. ЗИН-5945 (рис. 22, а). Более важная их особенность — вертикаль-
ная протяженность суставной поверхности, которая у ламантина, как у
тюгоня, не поднимается выше дорсального края позвоночного отверстия,
а у морской коровы заходит несколько выше. Каудальные суставные
ямки ламантина и морской коровы имеют большую вертикальную протя-
женность, дорсально они идут выше бугорков для крепления lig. trans-
versum atlantis, а у дюгоня только до этого уровня. Это определяет
большую амплитуду вращательных движений в атлантоосевом суставе
у двух первых.
Свобода движений в атланто-затылочном суставе в первую очередь
определяется формой затылочного мыщелка. У всех сирен его кривизна
(рис. 30) не одинакова на всем протяжении поверхности. У ламантина
форма мыщелка наиболее приближается к сферической, хотя и здесь
кривизна больше в верхней части, меньше в нижней. Сопоставление кри-
визны мыщелка и fovea articularis atlantis показывает, что наиболее кон-
груентны их поверхности в средней части мыщелка. Как правило, это
означает, что в таком положении на сустав падает наибольшее рабочее
напряжение. Как в более верхнем, так и нижнем положении (рис. 23, а)
конгруентность уменьшается, что не запрещает таких положений, но
делает их менее эффективными. Учитывая большую протяженность рабо-
чей поверхности мыщелка и меньшую в соответствующих суставных по-
верхностях атланта, можно оценить амплитуду движения в вертикаль-
ной плоскости в атланто-затылочном суставе примерно в 65—70° с рав-
ным отхождением от горизонтальной оси зубного ряда вверх и вниз.
Наиболее нагруженное положение в суставе соответствует горизонталь-
ному или слегка наклоненному вперед положению зубного ряда.
Длина окружности затылочного мыщелка дюгоня и морской коровы
значительно короче, чем у ламантина, меньше и кривизна, а высота су-
103
ставных поверхностей атланта больше. Все это связано с меньшей ампли-
тудой движений в суставе. Так, в линии морских коров (Dusisiren rein-
harti—D. jordani—Hydrodamalis) показаны сокращение вертикальной
длины поверхности затылочных мыщелков и утеря fossa condilaris dor-
salis, в которую упирается и край атланта при гиперэкстензии головы
(Domning, 1978 с), что, безусловно, говорит об уменьшении ракурсов
головы в вертикальной плоскости у морских коров, происшедшем неза-
висимо от дюгоней. В то же время у позднетретичных халитериин, среди
которых были предки и гидродамалин и дюгонин, амплитуда движений
головы в вертикальной плоскости была большая. В то же время у мор-
ских коров расширяются затылочные мыщелки, что в сочетании с уве-
личением флангов боковых затылочных костей говорит об увеличении
латеральных движений в атланто-затылочном суставе. Это сопровождает-
ся сужением суставных поверхностей на атланте (Brandt, 1861—1868)
по сравнению с ламантином. Кривизна в верхней части мыщелка дюгоня
меньше, чем в нижней, что обратно ламантину и морской корове. Наи-
большая конгруентность достигается здесь в более верхнем положении,
но сам мыщелок смещен относительно направления зубного ряда вниз.
У морской коровы, наоборот, при более вентральном положении мыщелка
по сравнению с ламантином конгруентность наибольшая в нижнем поло-
жении атланта, т. е. допускается, что самым рабочим положением для
нее является наклоненная вниз голова, тогда как у дюгоня голова
в атланто-затылочном суставе больше поднята (отсюда необходимость
при взятии корма со дна большого вентрального отклонения ростральной
части черепа), а у ламантина занимает более горизонтальное положение.
Таким образом, у обеих изученных форм дюгонид мы видим, что измене-
ния в атланто-затылочном суставе у них равно направлены на расшире-
ние именно тех возможностей движения головы, которые смягчают их
узкую специализацию в отношении аппарата захватывания пищи
(см. гл. 1), но при этом они диаметрально противоположны соответствен-
но своей пищевой специализации. Ламантины же пошли по пути макси-
мального расширения возможностей движения головы в вертикальной
плоскости.
Атлант Protosiren (Domning et al., 1982, fig. 25) во всех отношениях
больше похож на таковой ламантина: массивность верхней дуги, схожде-
ние обеих ветвей на манер двухскатной крыши, большая тонкость ниж-
ней дуги, форма и крепость поперечных отростков. Однако атлант прото-
сирена значительно шире (более 170% от высоты позвонка против 140%
у ламантина, но ближе к дюгоню и морской корове), угол, образованный
скатами, несколько больше — около 100°, поперечные отростки меньше
подняты вверх. Сходство атланта ламантина и протосирена не противо-
речит представлениям, что именно протосирениды дали начало трихехи-
дам (гл. 1), однако лишь форма и увеличенная толща верхней дуги
специфичны для них обоих, все остальные отмеченные признаки свой-
ственны большинству дюгонид. Так, у эоценового Eotheroides (Sickenberg,
1934: pl. 4, fig. 8), миоценового Metaxytherium (Abel, 1904: Taf. 6,
fig. 1) и плиоценового Dusisiren (Domning, 1978c: pl. 8) форма попереч-
ных отростков также близка к таковым протосирена. Следовательно, она
была свойственна большинству древних сирен и сохранилась в таковом
виде у морской коровы, и лишь у дюгоня она претерпела сильную моди-
фикацию. Но в то же время у всех дюгонид верхняя дуга закруглена
и не утолщена, что скорее всего говорит об общем для них характере
взаимоотношения невральных дуг атланта и эпистрофея.
Эпистрофей (рис. 31) ламантина характеризуется прямой при виде
сбоку невральной дугой, ось которой слегка отклонена от вертикали
104
Рис. 31. Эпистрофей ламантина
а, г — сбоку; б — спереди; в —
1 — arcus vertebrae, дуга
позвонка;
2 — corpus vertebrae;
3 — crista epistrophei;
4 — dens;
S — extremitas caudalis, зад-
ний конец, или ямка
позвонка (задняя сочле-
(а—в, экз. ИЗ-5945)
сзади
новная поверхность тела
позвонка);
6 — facies articularis caudalis,
задняя суставная поверх-
ность;
7 — facies articularis cranialis;
8 — facies articularis ventra-
lis;
и дюгоня (г, экз. ЗИН-679)
9 — for. vertebrale, позвоноч-
ное отверстие;
10—pr. articularis caudalis
(=postzygapophysis);
11 — pr. splnosus;
12 — pr. transversus dorsalis
(—diapophysis);
13 — pr. transversus ventral is
(=parapophysls)
105
назад. Дорсально дуга очень массивна, и ее остистый отросток представ-
ляет собой сильное вздутие дуги с клиновидным выступом, направлен-
ным назад и, нависая над невральной дугой III шейного позвонка, упи-
рающимся в нее. Фиксация этого упора подтверждается срастанием
обоих позвонков у экз. ЗИН-31795 (рис. 32). Невральная дуга эпистро-
фея дюгоня построена иначе: ножка ее сохраняет то же направление,
что и у ламантина, но дорсальнее, сразу за постзигапофизом, дуга резко
меняет направление на более переднее. В результате ее верхушка
(остистый отросток) своей самой массивной частью образует упор для
дорсального бугорка атланта, налегая на него сзади и немного сверху.
У морской коровы невральная дуга прямая, как у ламантина, но обра-
зует мощный, треугольный в поперечном сечении и сужающийся вперед
выступ, на нижней поверхности которого имеется уплощенная площадка,
налегающая на верхнюю дугу атланта, т. е. тот же принцип ограниче-
ния движения атланта вверх относительно эпистрофея путем создания
своеобразного замка, защелкивающегося сзади и сверху, что и у дюгоня,
здесь развит дальше.. Таким образом, дюгони и морские коровы, с одной
стороны, и ламантин — с другой, решили одну и ту же задачу «замка»
Рис. 32. Сросшиеся эпистро-
фей и третий шейный позво-
нок ламантина (экз. ЗИН-
31795)
а — сбоку; б — сзади;
14 — for. intervertebrale;
IS — for. transversarium;
IS — pleurapophysis;
17 — pr. articularis cranialis
(=praezygapophysis);
18 — рудимент spatium interarcu-
ale;
остальные обозначения, как на
рис. 31
Рис. 33. Последний шейный
позвонок ламантина (VI с)
(а, б, экз. ИЗ-5945) и дюгоня
(VII с) (в, г, экз. ЗИН-679)
а, в — сбоку; б, г — сзади;
19 — fovea costalis caudalis;
20 — lamina arcus vertebrae;
21 — tuberculum dorsale;
22 — tuberculum ventrale;
остальные обозначения, как на
pud. :• 1 и 32
106
диаметрально противоположными способами: у ламантинов упор верх-
ней дуги атланта направлен назад и передается дальше через остистый
отросток эпистрофея на невральную дугу III шейного позвонка
(т. е. замок смещен каудально), а у дюгонид его влияние ограничено
эпистрофеем и не передается на последующие позвонки. В результате
у ламантина в передней части шейного отдела произошла значительная
иммобилизация, что и подтверждается наиболее частыми у них случаями
сращения II и III шейных позвонков (Petit, 1928). У дюгонид сращения
каких-либо шейных позвонков наблюдаются реже. Пути, по которым по-
шли дюгониды и трихехиды, разошлись, видимо, еще в эоцене. Во вся-
ком случае такой ранний представитель дюгонид, как Eotheroides, уже
имел направленный вперед выступ остистого отростка эпистрофея (Si-
ckenberg, 1934, pl. 5, fig. 4). Близкую форму имела дуга эпистрофея и
у Protosiren (Там же: р. 77, fig. 13).
Судя по суставным фасеткам и общей конфигурации атланта и эпи-
строфея, вращательные движения в атлантоосевом суставе сирен очень
незначительны по сравнению с большинством других млекопитающих.
107
Остальные шейные позвонки более сходны между собой, кроме по-
следнего, готовящего условия к переходу позвоночного столба от шейного
отдела к грудному (рис. 33). Характерная особенность последнего шей-
ного позвонка сирен — наличие на его теле в каудолатеральном его углу
особой ямки, обращенной назад и немного вбок, для головки ребра, зани-
мающей интервертебральное положение (Brandt, 1863). Степень разви-
тия этой ямки варьирует. Обычно, как у ламантина ИЗ-5945, она хорошо
развита, но у экз. ЗИН-31 795 эти фасетки практически отсутствуют в
связи с необычным строением головки ребра, которая утеряла переднюю
фасетку, сохранив на ее месте острый выступ, упирающийся в заднюю
поверхность парапофизной части плеврапофиза латеральнее от тела
позвонка,' где имеется небольшая вдавленность, совсем отличная от обыч-
ной суставной фасетки. Зато у ребра неожиданно образовался второй
бугорок с настоящей суставной поверхностью (см. дальше), для которой
в дистальной части задней поверхности плеврапофиза VI шейного
позвонка развилась настоящая фасетка. По всем шейным позвонкам этой
части отдела идет направленное спереди назад изменение ряда парамет-
ров. Так, плеврапофизы меняют свое положение относительно продоль-
ной оси шеи, опускаясь от эпистрофея к V позвонку, а затем вновь под-
нимаясь вверх (рис. 28, а, б). Если соединить их дистальные концы
линией, то она образует волнообразную кривую с нижним пиком в
V позвонке и постепенным подъемом к концу шеи, продолжающимся в
начале грудного отдела диапофизами грудных позвонков. Важно подчерк-
нуть, что наиболее низко опущенный плеврапофиз принадлежит одному
и тому же V позвонку всех сирен, хотя у дюгоня и морской коровы он
занимает более вентральное положение (ниже тела позвонка), чем у ла-
мантина (немного выше или на уровне вентрального края тела). Все это
говорит, что последовательность шейных позвонков у последнего не была
нарушена, т. е. выступает против выпадения III позвонка.
При взгляде снизу на шейный отдел (рис. 28, д, е) в первую очередь
бросается в глаза короткость тел III—VII позвонков у дюгоня. Фактиче-
ски это широкая и невысокая субпрямоугольная тонкая пластинка
(рис. 33, в, г), стоящая вертикально и в большей или меньшей степени
прогнутая спереди назад. Ножки дуги также напоминают тонкие пластин-
ки, объединенные с плеврапофизами в единое образование, отходящее от
тела вбок и слегка вперед. Латерально эти пластины несколько утолща-
ются, формируя диапофизы, которые направлены перпендикулярно к
длинной оси шеи. В результате весь позвонок кажется вдавленным спе-
реди и выпуклым сзади. Дорсально стоящая поперек пластина ножки
дуги отгибается назад (линия сгиба стоит косо относительно вертикаль-
ной плоскости: при виде сзади она идет сверху вниз и медиально, при
виде сверху — назад и медиально). Таким образом, формируются две ле-
жащие почти друг над другом плоскости: одна, обращенная вверх и не-
сколько вперед и медиально, используется как передняя суставная
поверхность, вторая, смотрящая вниз назад и латерально,—как задняя
суставная поверхность. Таким образом, зигапофизы как выступающие
вперед и назад отростки здесь отсутствуют. Каудально за этими фасет-
ками невральная дуга, дугообразно выгибаясь, идет назад и вверх. В ре-
зультате получается сложный боковой профиль у всех этих шейных поз-
вонков (см. рис. 33, в), которые, сочленяясь, как бы вкладываются друг
в друга (передние в более задние), а их суставные отделы оказываются
наложенными друг на друга, подобно черепице (см. рис. 28, б). Все это
приводится к сокращению междуговых пространств в шейном отделе дю-
гоня, а вместе с укорочением тел позвонков и к короткости всего отдела
сравнительно с ламантином (см. рис. 28, в). В целом у морской коровы
108
наблюдается то же, что и у дюгоня (короткие тела, уплощенность всех
позвонков, их вкладывание друг в друга). Наоборот, у ламантина тела
позвонков относительно длиннее, междуговое пространство широкое, нож-
ки дуги толще, а зигапофизы развиты хорошо (презигапофизиы, как вы-
ступы вперед ножки дуги, латерально укрепленные массивными диапо-
физами, а постзиганофизы как выросты назад и вбок пластинки дуги,
в результате суставная фасетка последнего располагается не под тако-
вой первого, как это было у дюгоня, а позади) и при виде сверху от-
делены друг от друга латеральной вырезкой.
Формирование дюгонидного облика III—VII шейных позвонков нача-
лось очень рано в эволюции. Во всяком случае уже у эоценового эотеро-
идеса (Sickenberg, 1934; pl. 4, fig. 1, 9; pl. 5, fig. 1) видно, что тела
позвонков укорочены, в сочлененных позвонках наблюдается вкладывание
позвонка в позвонок, презигапофизы развиты и более обособлены от
ллеврапофизов, невральная дуга не образует коленообразного изгиба
назад. Как примитивный признак, очевидно, следует рассматривать нали-
чие длинного остистого отростка, по крайней мере на VI позвонке. Не-
которое обособление зигапофизов сохраняется еще у миоценового Hespe-
rosiren (Simpson, 1932) и у морской коровы, но окончательно исчезает
у дюгоня, т. е. сохраняется у халитериин и гидродамалин, но не у дюго-
нин. У протосирена (Sickenberg, 1934: р. 78, fig. 14) зигапофизы также
обособлены, а невральная дуга прямая, возможно, позвонки не налега-
ли друг на друга, но тело позвонка короче, чем у ламантина. У всех
третичных дюгонид, как и у современных, плеврапофизы последних шей-
ных позвонков (особенно V) опускаются ниже тела позвонка, что можно
рассматривать как плезиоморфный признак, утрачивающийся только у
ламантинов, тогда как уплощение тела позвонков и невральых дуг,
ведущее к налеганию позвонка на позвонок, следует рассматривать как
апоморфный признак дюгонид (вернее, укороченность тел позвонков —
общая черта всех сирен, но у ламантинов оно остановилось рано в эво-
люции, а у дюгонид, особенно у дюгонин, пошло дальше).
Следует также отметить, что у ламантина тела последних шейных по-
звонков прогрессивно увеличиваются в длину (см. рис. 22), и эта тенден-
ция продолжается в передней части грудного отдела, тогда как у дюго-
ней резкое наращивание длины тела начинается только с I грудного по-
звонка. Это указывает на определенную торакализацию оставшихся зад-
них шейных позвонков. Наоборот, у дюгоней можно говорить о церви-
кализации I грудного позвонка, несущего сходную по форме с последними
шейными позвонками невральную дугу, а также имеющего еще относи-
тельно плоское тело и участвующего в оговоренной выше системе вкла-
дывания позвонка в позвонок. У морской коровы характер изменения дли-
ны тел шейных позвонков промежуточен между дюгонем и ламантином,
что может быть отражает более примитивное состояние, унаследованное
еще от халитериин, тогда как дюгони отклонились от общего направления
эволюции дюгонид в одну сторону, а ламантины, предки которых разде-
лили общие с халитериинами признаки,— в другую.
Заметно отличаются ламантины и дюгони по степени расставленности
зигапофизов одного позвонка. У ламантина расстояние между центрами
постзигапофизов (см. рис. 26) шейных позвонков значительно меньше,
чем у дюгоня, причем эта разница нарастает в каудальном направлении
(на 30% от ширины тела эпистрофея до 55% у VI позвонка), поскольку
это расстояние на IV—VI шейных и I грудном позвонках ламантина
практически неизменно, а у дюгоня непрерывно нарастает. У обеих форм
имеется позвонок, презигапофизы которого максимально широко расстав-
лены по сравнению с предыдущими позвонками, а постзиганофизы начи-
109
нают сближаться (тенденция, характеризующая самые первые грудные
позвонки). У дюгоня это — последний (VII) шейный позвонок, а у ла-
мантина лишь первый грудной, что опять говорит о поглощении у него
последнего шейного позвонка грудным отделом. В то же время ширина
шейных позвонков в дистальных концах плеврапофизов значительно боль-
ше у ламантина (см. рис. 25), т. е. длина их плеврапофизов относи-
тельно ширины тела позвонка и расстояния между зигапофизами больше,
чем у дюгоней. И опять сокращение этой ширины, характерное для груд-
ного отдела (ширина в фасетках диапофизов), у ламантинов начинается
только со II грудного позвонка, а у дюгоня с I грудного, свидетельствуя
о сдвиге на один сегмент границы между шейным и грудным отделами,
произошедшем у ламантинов. Это должно увеличивать рычаг приложения
лестничных мышц спины к шейным позвонкам, а поскольку те практи-
чески не имеют латеральной подвижности, то, видимо, должны служить,
для удержания передней части тела в фиксированном положении.
ГРУДНОЙ ОТДЕЛ
Грудной отдел осевого скелета, пожалуй, наиболее специфичен у сирен
среди других млекопитающих. Если у китообразных приспособление к
постоянному обитанию в воде и активному плаванию было связано с
уменьшением грудного и сильным увеличением поясничного отделов,
а также у большинства с усилением гибкости почти всего туловища, то
у сирен адаптация к сходным условиям в сочетании с отсутствием необ-
ходимости в активном плавании, далеких миграциях и т. п. пошла диа-
метрально противоположным путем: грудной отдел необычайно увеличил-
ся в результате возрастания числа позвонков, несущих большие ребра,
а поясничный отдел соответственно уменьшился; подвижность в грудном
отделе резко уменьшилась, и он практически оказался исключенным из
активного участия в локомоции или в дыхании, превратившись в стер-
жень, вокруг которого организована работа более подвижных частей тела:
активных манипуляторов — передних конечностей (ластов), шеи и основ-
ного локомоторного органа — хвоста. Необходимо разобраться, в какой
мере эта характеристика относима ко всем сиренам и не страдает ли она
односторонностью.
Несмотря на сходство, возможно кажущееся в форме тела, образе жиз-
ни и повадках, конструкция грудного отдела ламантина принципиально1
иная, чем у дюгоня, и все их позвонки (рис. 34—39) отличаются друг
от друга. Кроме того, они сильно меняют свою форму, величину по ходу
позвоночного столба, что, безусловно, отражается и в возможностях их к
движению в том или ином направлении. Меняется и характер связи по-
звонков с ребрами. Нам кажется целесообразным дать сначала описание
двух типичных позвонков из передней (IV th) и задней (XI th) половин
грудного отдела ламантина и дюгоня, а затем проследить изменение ос-
новных параметров по длине отдела. Первый и последний позвонки отде-
ла требуют отдельного рассмотрения.
Четвертый грудной позвонок ламантина (рис. 36, а, в, д) имеет отно*-
сительно длинное тело, составляющее около 40% от высоты позвонка,
более узкое спереди и широкое сзади за счет большего развития высту-
пов — задних парапофизов. Посередине длины тело подразделено силь-
ной перетяжкой (минимальная ширина тела) на переднюю половину с
очень короткими передними парапофизами, несущими глубокие реберные
ямки, поверхность которых обращена наружу и вперед (рис. 40, а, б).
Их вентральный диаметр больше горизонтального, а форма напоминает
правильный овал. В задней части тела задние парапофизы имеют также*
110
Рис. 34. Первый грудной позвонок ламантина (экз. ИЗ-5945)
•а — спереди; б — сверху; в — сбоку
1 — arcus vertebrae;
2 —[corpus vertebrae;
-3 — crista dorsalis;
4 — extremitas cranialis, передний конец поз-
вонка (головка позвонка, передняя со-
членовная поверхность тела);
Л — extremitas caudalis;
6 — facies articularis caudalis;
7 — facies articularis cranialis;
8 — for. vertebrae;
9 — fovea costalis caudalis;
JO —fovea costalis cranialis;
11 — fovea costalis transversalis, реберная
ямка поперечного отростка;
12 — incisura vertebralis caudalis, задняя по-
звоночная вырезка;
13 — incisura vertebralis cranialis, передняя
позвоночная вырезка;
14 — lamina arcus vertebrae;
15 — margo caudalis process! spinosi, задний
край остистого отростка;
16 — margo cranialis processi spinosi, перед-
ний край остистого отростка;
17 — pediculus arcus vertebrae;
18 — pr. articularis caudalis (= postzygapophysis);
19 — pr.articularis cranialis (=praezygapophysis)
20 — pr. spinosus;
21 — pr. transversus dorsalis (= diapophysis);
22—pr. transversus ventralis (= parapophysis
111
20
Рис. 35. Первый грудной позвонок дюгоня (экз. ЗИН-679)
а — спереди; б — сверху; в — сбоку
Обозначения как на рис. 34
глубокие задние реберные ямки овальной формы, обращенные назад и
вбок. Их длинные диаметры расположены косо спереди вниз и назад.
Вместе с передними ямками последующего позвонка они образуют единую
суставную ямку для головки ребра, очертания которой вписываются в
широкий овал с почти горизонтальным положением длинного поперечни-
ка. При этом передняя часть этой общей реберной ямки (задняя ребер-
ная ямка) больше выступает латерально, чем задняя, и вследствие этого
общая поверхность в целом обращена вбок и несколько назад. Вентраль-
но, в месте пережима, тело округлено и не имеет киля. Соответственно
передняя и задняя сочленовные поверхности имеют почковидную форму с
небольшой выемкой сверху, являющейся частью спинномозгового канала.
Невральная дуга соединена с телом очень толстыми ножками с суб-
прямоугольным сечением во фронтальной плоскости. Длинная ось этого
прямоугольника расположена косо спереди назад и внутрь, но при взгля-
де сбоку и спереди кажется, что ножка имеет равную толщину в любом
направлении. От верхней части ножки латерально, вверх и вперед
Рис. 36. Четвертый грудной позвонок ламантина (а, в, д, экз. ИЗ-5945) и дюгоня
(б, г, е, экз. ЗИН-3220)
а, б — спереди; в, г — сзади; д, е — сбоку
3 — insertio ligamenti flavi, место прикрепления lig. flavum, желтой связки; остальные обозначе-
ния, как на рис. 34
112
из
отходят очень сильные диапофи-
зы, дистальные концы которых
подняты выше уровня позвоноч-
ного отверстия. Форма сечения
диапофиза четырехугольная, углы
более или менее сглажены. Наи-
большая сторона четырехуголь-
ника обращена вверх, наимень-
шая — вперед, две другие вниз и
вперед (нижняя) и вниз и назад
(задняя). Передняя и нижняя по-
верхности разграничены переги-
бом переднего края диапофиза,
идущим косо снизу от верхнего
края передней реберной ямки
вверх и латерально к верхнепе-
реднему углу дистального конца
диапофиза. Второй, менее выра-
женный перегиб, идущий от того
же места латерально, вверх и на-
зад к задненижнему углу диапо-
физа, делит нижнюю и задние
поверхности последнего. Верхняя
и задняя поверхности у самого
основания диапофиза сходятся
под острым углом, но ближе к се-
редине угол возрастает и стано-
вится более сглаженным. Перед-
няя поверхность с верхней обра-
зуют угол, близкий к прямому,
с нижней — чуть больший. Все это
приводит к тому, что каждая по-
верхность имеет свою форму:
верхняя почти прямоугольная,
слегка расширяющаяся вместе с
диапофизом дистально; передняя
и задняя субтреугольны с верши-
нами, обращенными к концу диа-
пофиза; нижняя также треуголь-
на, но основание треугольника ле-
жит у конца диапофиза.
Нижняя поверхность диапофи-
за в своем заднелатеральном углу
несет овальную, почти плоскую
фасетку — реберную ямку попе-
речного отростка для сочленения
Рис. 37. Одиннадцатый грудной позво-
нок ламантина (а, в, г, е, экз. ИЗ-5945)
и дюгоня (б, д, экз. ЗИН-3220)
а, б — спереди; в — сзади; г,д — сбоку;
« — сверху
24 — crista ventralis, вентральный гребень;
25 — pr. mamillaris (= metapophysis), сосце-
видный отросток (метапофиз);
остальные обозначения, как на рис. 34 и 36
114
115
Рис. 38. Последний (XVI) грудной
позвонок ламантина (экз. ИЗ-5945)
а — сбоку; б — вид спереди и слегка свер-
ху и сбоку с ребром
2в — caput costae, головка ребра;
27 — fovea costalls, реберная ямка;
28 — tuberculum costae, бугорок ребра; осталь-
ные обозначения, как на рис. 34, 36 и 37
с бугорком ребра. Она имеет
овальную форму с длинным по-
перечником, расположенным на-
клонно снизу вверх и назад. По-
верхность ее обращена латерально
и несколько вниз и вперед. Центр
фасетки смещен назад относи-
тельно вертикали, проходящей че-
рез центр передней реберной
ямки. Выше фасетки дистальный
конец диапофиза, входящий в
капсулу реберно-поперечного су-
става, крайне неровен. Если
смотреть прямо на конец диапо-
физа, то видно, что передний край
диапофиза образует выступающий
вперед клиновидный фланец, н<
суставом. Самая передняя точка этого фланца выступает в виде тупого
бугра (рис. 40, а), связанного с отхождением m. semispinalis cervicis.
На задней стороне диапофиза, примерно на расстоянии */3 длины от его
конца, расположена бугорчатость для отхождения начальных медиаль-
ных сухожильных ножек их. longissimus dorsi.
В верхней части ножек дуги, на уровне их соединения с пластинкой
дуги, латеральнее позвоночного отверстия (округлого, с почти равными
вертикальным и поперечным диаметрами) вперед выступают сильные,
длинные, заходящие за уровень краниального края тела отростки — пре-
зигапофизы. Они явственно отграничены от диапофизов глубокими вы-
емками в переднем крае послед-
них. Передние суставные поверх-
ности на этом позвонке лежат
почти в горизонтальной плоскости.
Позади них в пластинке дуги и
смежной части диапофиза распо-
ложены особые стопорные ямки,
ограничивающие скольжение фа-
сеток постзигапофизов впередиле-
жащего позвонка назад, т. е. пере-
разгибание позвоночного столба.
Постзигапофизы еще более длин-
Рис. 39. Первый послегрудной позво-
нок ламантина (а, б, экз. ЗИН-31795)
с переходным характером между груд-
ными и поясничными позвонками и
последний грудной (XVIII) позвонок
дюгоня (экз. ЗИН-3220)
а — спереди; б, в — сбоку
— fovea costalis, реберная ямка;
.2 — synapophysis, синапофиз
116
20
ные, чем презигапофизы (передняя вырезка позвонка очень небольшая,
задняя очень глубокая). В сочленении с презигапофизами последующего
позвонка они отграничивают сверху длинное и низкое межпозвоночное
отверстие, длинная ось которого занимает горизонтальное положение
(рис. 40, в).
Пластинка дуги короткая, так как спереди она глубоко врезана между
обоими презигапофизами, причем эта вырезка доходит назад до уровня
середины ножек дуги. Дорсально пластинка несет относительно неболь-
шой остистый отросток, основание которого (примерно */з его высоты)
почти перпендикулярно к плоскости пластинки и горизонтальной пло-
скости, а дистальные две трети отогнуты примерно под 45° назад. Осно-
117
Рис. 40. Одиннадцатый грудной позвонок (а, б) и взаимное положение двух смеж-
ных грудных (IV и V) позвонков (в—е) у ламантина (а, в, д) и дюгоня (б, г, е)
а, б — снизу; в, г — сбоку; д, е —
1 — crista ventralis; 9
2 — facies articularis cranialis;
3 — foramen intervertebrale; 10
4 ,3 — processus transversus
ventralis (s.parapophysis); 11
4 — передний парапофиз,
5 — задний парапофиз;
в — spatium interarcuale;
7 — tuberculum m. longissimus
dorsi;
3 — tub. m. semispinalis dorsi;
tuberositas lig. costotrans.
versa rium;
бугорок медиальный на 13
заднем краю диапофиза; 74
бугорок на вентральной
поверхности диапофиза и
гребень, идущий от бугор-
ка к заднему краю по-
следнего;
— гребень, идущий от осно-
вания остистого отростка
к переднему краю диапо-
физа;
- синовиальный желобок;
• стопорная ямка;
римскими цифрами обоз-
начен порядковый номер-
позвонка в отделе, а стрел-
ками — место фасетки»
диапофиза
118
вание отростка как бы срезано вертикально спереди и сзади с образова-
нием здесь особых треугольных площадок, на которых крепится эластич-
ная желтая связка — ligamentum flava. Снаружи эти площадки
ограничены выступающими гребнями, которые идут сверху от остистого
отростка вниз, расходясь в стороны. Сзади эти гребни переходят в ребра
жесткости на дорсальной стороне постзигапофизов, а спереди продолжа-
ются вперед по медиальным краям презигапофизов, ограничивая изнутри
упомянутые выше стопорные ямки, но до передних суставных поверхно-
стей не доходят. Сами площадки для lig. flava, таким образом, представ-
ляют правильные треугольные углубления с характерной системой двух
взаимно перпендикулярных (горизонтальный и средний вертикальный)
гребней. Сам остистый отросток имеет в сечении вид очень высокого,
с узким основанием треугольника. Передний его край острый, задний сре-
зан и имеет срединный гребень, идущий вертикально от загнутого назад
заднедорсального угла остистого отростка (характерная черта именно
этого позвонка) вниз к такому же гребню по центру задней площадки
для lig. flava. На середине заднего края остистого отростка находятся
парные бугорки для крепления сухожильного отхождения m. inter spinalis.
Дорсально остистый отросток сзади утончается, и его сечение приобрета-
ет вид тонкой двояковыпуклой линзы.
IV грудной позвонок дюгоня (см. рис. 36, б, г, е) имеет намного бо-
лее короткое тело, составляющее 23% от его высоты. Дуга несравненно
больше тела. Позвоночное отверстие просто огромное, субпятиугольное.
Тело снизу округлое, никакого пережима посередине его длины нет, нет
и киля. Передняя реберная ямка лежит выше по телу, наполовину уже
на ножке дуги. Фасетка в виде вытянутого длинного эллипса, располо-
женного почти вертикально (сверху вниз и слегка назад). Ее поверхность
обращена латерально и вниз. Парапофизарного выступа в этом месте нет,
и фасетка сидит в глубокой ямке на теле и ножке дуги. Задний пара-
пофиз практически тоже отсутствует. Задняя реберная ямка меньше
передней. Ее вытянутая косо снаружи вниз и медиально фасетка обра-
щена преимущественно назад и в меньшей степени латерально и вниз.
Ножка дуги тоньше, чем у ламантина, в сечении овальная, длинная
ось этого овала поставлена косо изнутри вперед и наружу. По передне-
латеральному краю ее проходит отчетливый гребень, тянущийся от дор-
сального края передней реберной ямки латерально по нижнепереднему
краю диапофиза и отделяющий заднюю поверхность последнего и самой
ножки от более глубокой передней, представляющей желоб, в котором
идет из спинномозгового канала широкий плоский пучок чудесной сети,
переходящий дистальнее в желоб ребра (sulcus costae) предшествующего
позвонка. У ламантина аналогичный пучок чудесной сети выходит скорее
в передней части межпозвоночного отверстия и прижимается к задней,
а не передней поверхности ножки позвонка. Диапофизы у дюгоня зани-
мают более вентральное положение, примерно на уровне середины позво-
ночного отверстия, а не в верхней его части и лежат почти в горизон-
тальной плоскости и лишь немного вперед. В сечении они похожи па
треугольник с округлой верхней стороной, более прямой передней и под-
ходящей к ней под острым углом задненижней. Задняя вершина Этого
треугольника тупая, широко закругленная. Дистальный конец диапофиза
расширен во всех направлениях, округлен. Реберная ямка поперечного
отростка почти круглая, слегка смещена вниз и назад от центра диапофи-
за, а ее центр расположен почти на одном уровне с центром общей ям-
ки для головки ребра. По передней стороне диапофиза намечается тупой
гребень, разделяющий верхнюю и передненижнюю его поверхности. На
середине гребня, как и у ламантина, помещается бугорок для m. semi-
spinalis servicis.
119
Положение и форма презигапофизов отличны от таковых ламантина.
Это — широкие, короткие, лопастевидные выступы пластинки дуги, рас-
положенные частью над позвоночным отверстием, частью над ножками
дуги. Фактически они представляют медиальное продолжение только что
упомянутого гребня на передней стороне диапофиза, от которого их отде-
ляют лишь очень небольшие выемки. Суставные поверхности презигапо-
физов широкие, большие, расположенные здесь почти горизонтально.
За ними, как у ламантина, имеются стопорные ямки, но они значитель-
но шире. Скорее можно сказать, что в передний край пластинки дуги и
диапофиза врезана горизонтальная, слегка выдвинутая вперед, полукруг-
лая площадка, задний край которой и образует стопорный уступ. Эти
площадки медиально ограничены гребнями, идущими от медиальных кра-
ев презигапофизов назад и вверх по основанию остистого отростка и схо-
дящимися дорзально, выделяя небольшую площадку для lig. flava. В от-
личие от ламантина, где она намного больше, у дюгоня ее плоскость еще
и не вертикальна, а обращена вперед и вверх. Медиальная вырезка между
презигапофизами в связи с общим укорочением позвонка у дюгоня менее
глубокая. Постзиганофизы тоже короче, их каудальные концы не дохо-
дят до уровня конца остистого отростка, а у ламантина заходили даль-
ше назад. Несмотря на короткость, и презигапофизы и постзиганофизы
выступают за соответствующие края тела позвонка. Медиально постзига-
пофизы отделены друг от друга очень небольшой вырезкой, сверху еще
прикрытой сильно наклоненными назад остистым отростком. Таким об-
разом, они вместе образуют как бы козырек, нависающий над презигапо-
физами последующего позвонка, что ведет к крайнему сужению междуго-
вого пространства (рис. 40, е). Межпозвоночное отверстие имеет в свою
очередь вид узкой щели, наклоненной спереди вниз и слегка назад
(рис. 40).
Остистый отросток длинный, его наклон назад больше, чем у ламан-
тина, и при малейшем переразгибании спины налегает на передний край
остистого отростка последующего позвонка. Таким образом, у дюгоня
чрезвычайно сокращено пространство между остистыми отростками смеж-
ных позвонков (см. рис. 40, г), в котором должны помещаться mm. in-
terspinales. В сечении остистый отросток дюгоня треугольный, даже в бо-
лее суженной дорсальной части. Спереди отросток также имеет вид вы-
сокого треугольника, боковые стороны которого от самого основания более
или менее прямо сходятся к концу отростка, тогда как у ламантина лишь
основание отростка сравнимо с равносторонним треугольником, над кото-
рым возвышается стоящая вертикально пластинка, слегка утончающаяся
к концу.
Подводя итоги, следует отметить, что многие параметры, по которым
IV грудные позвонки ламантина и дюгоня различаются, связаны с из-
менением длины позвонка у первого. Это — длина тела, ширина диапофи-
зов и остистых отростков, расстояние между концами пре- и зигапофи-
зов. Из конструктивных особенностей различия в первую очередь связа-
ны с положением и формой презигапофизов, направлением и местом
диапофизов, расстоянием между остистыми отростками и вообще невраль-
ными дугами смежных позвонков и т. д. Отличается развитие lig. flava,
передняя площадка которого у дюгоня маленькая, Д-образная, а задняя
имеет вид глубокой треугольной выемки в задней поверхности пластинки
дуги и основания остистого отростка между обоими постзигапофизами и
обращена назад и вниз.
У морской коровы IV грудной позвонок во всех основных деталях схо-
ден с дюгоневыми: презигапофизы в виде широких горизонтальных ло-
пастей, мало ограниченных от несколько коротких диапофизов; диапофизы
120
помещаются более вентрально и занимают положение более близкое к
горизонтальной плоскости, хотя и чуть больше поднимаются вверх, но
не в такой степени, как у ламантина; позвоночное отверстие крупное,
тела позвонков короче, чем у ламантина; крепление lig. flava организо-
вано, как у дюгоня: переднее крепление помещается в глубокой выемке
задней стороны невральной дуги, что препятствует разрастанию этой эла-
стичной связки дорсально, сама эта поверхность очень низкая; у обеих
форм презигапофизы расставлены шире относительно тела (около 100%
против 60% у ламантина); остистый отросток сильно наклонен назад,
налегая, подобно черепице, на остистый отросток последующего позвонка,
междуговое пространство маленькое, межпозвоночное отверстие укоро-
чено, и его вертикальный диаметр больше горизонтального. Самым замет-
ным отличием IV th морской коровы от дюгоня является низкое и ши-
рокое тело позвонка и более короткие диапофизы.
XI грудной позвонок (см. рис. 37) ламантина несет те же специфич-
ные для рода черты: длинное тело, пережим в его середине, длинные и
крепкие зигапофизы, вертикальные площадки для lig. flava, поднятые
вверх диапофизы, невысокие остистые отростки, как бы переломленные
на уровне своей нижней трети, глубокие вырезки между зигапофизами
и соответственно большое междуговое пространство, овальные, вытянутые
спереди назад и низкие межпозвоночные отверстия, большое расстояние
между остистыми отростками смежных позвонков. Однако имеются и
большие различия, связанные с положением этого позвонка в задней по-
ловине грудного отдела.
Тело относительно дуги стало очень большим (длина около 50%,
а ширина более 55% от высоты позвонка), а форма его сочленовных по-
верхностей стала сердцевидной. Вентрально на теле появился большой
крепкий гребень. Передняя реберная ямка несколько сместилась назад от
переднего края тела, ее поверхность обращена латерально. Задняя ребер-
ная ямка уменьшилась, и ее поверхность больше обращена назад, а зад-
ние парапофизы выступают больше наружу и вверх, вынося задние ребер-
ные ямки частично выше верхнего края задней сочленовной поверхности
тела позвонка. Ножки дуги изменили свою ориентацию на более продоль-
ную. Позвоночное отверстие стало несколько выше, изменило форму на
узкую шестиугольную, слегка более широкую сверху, чем снизу. Шири-
на диапофизов почти не изменилась, несмотря на увеличение позвонка,
но относительно длины позвонка они стали уже. Их направление измени-
лось: не вверх и вперед, а вверх и назад. Реберная ямка поперечного
отростка (фасетка диапофиза) стала крупнее и имеет теперь вид овала
с горизонтальным положением длинного поперечника. Выступающий
вперед фланец диапофиза исчез. В сечении диапофиз стал эллипсовидным,
с округлым передним и задним краями без каких-либо отчетливых греб-
необразных перегибов, разделяющих его поверхности. На заднем его крае
по-прежнему имеется бугорок для сухожильной ножки m. longissimus
dorsi. Презигапофизы стали очень мощными, более толстыми, чем узкие
суживающиеся назад постзиганофизы. Большие передние суставные по-
верхности обращены внутрь и вверх, т. е. из горизонтальных стали на-
клонными. На их наружных, поднимающихся вверх фланцах имеется от-
четливое шероховатое возвышение — метапофиз, или сосцевидный отро-
сток. В результате этих изменений при виде спереди презигапофизы
стали похожи на широкие, открытые вверх и внутрь желоба. Задние су-
ставные поверхности стали узкими, но их длина совпадает с длиной пе-
редних суставных поверхностей последующего позвонка, говоря об огра-
ничении сгибательных движений, но определенной возможности двигаться
121
вокруг длинной оси (вращение), что было исключено в передней части
грудного отдела, и о невозможности боковых движений.
XI грудной позвонок дюгоня, хотя и увеличился в размерах, но про-
порции его тела и невральной дуги изменились мало. Тело относительно
короткое, широкое. Пережима как такового нет, но вентрально тело не-
сколько суживается в середине своей длины, как бы становясь килеобраз-
ным. Вентрального гребня, как у ламантина, нет. Форма тела слегка
сердцевидна, но по своим пропорциям и угловатости сильно отличается
от таковой ламантина. Передняя реберная ямка отодвинулась от крани-
ального края тела и расположилась прямо под центром фасетки на диа-
пофизе. Форма ее и ориентация передней оси остались неизменными.
Задняя реберная ямка исчезла полностью. Ножки дуги стали массивнее,
увеличились спереди назад и также изменили положение на более про-
дольное. Позвоночное отверстие уменьшилось, сохранив более или менее
прежнюю форму, ширина его превышает высоту. Диапофизы перемести-
лись ниже по ножке дуги и помещаются теперь не в верхней части по-
звоночного отверстия, а на уровне его середины. Их длина уменьшилась.
При виде сбоку бросается в глаза опущенный вниз фланец диапофиза.
Реберная ямка поперечного отверстия маленькая, эллипсовидная, вытя-
нутая в горизонтальном положении, т. е. сохранила свою ориентацию.
Ее поверхность почти плоская, обращена латерально, немного назад и
вниз. На заднем крае диапофиза расположены два бугорка, меньший ди-
стальнее и более крупный медиальнее примерно на уровне центра ножки
Дуги.
Презигапофизы потеряли лопастевидную форму, увеличились в дли-
ну, представляя теперь мощные, треугольные при виде сверху отростки
с широким основанием. Передние суставные поверхности обращены
внутрь и вверх и по форме не похожи на желоба, как было у ламанти-
на. Слабее развиты и боковые их фланцы, а бугристость метапофиза на
этом позвонке выражена слабо. В этом суставе (X—XI th) допустимы не-
большие сгибательные и вращательные движения. По наружной поверх-
ности презигапофизов косо вниз и назад к передненижнему углу конца
диапофиза идет округлый гребень. Постзигапофизы остались короткими,
они не достигают каудального края тела и остистого отростка. Суставные
их поверхности меньше, чем на презигапофизах последующего позвонка,
т. е. здесь вращательные движения больше, чем в предыдущем суставе.
В связи с удлинением презигапофизов увеличилось междуговое прост-
ранство, но межпозвоночное отверстие по-прежнему осталось узким с ко-
сой ориентацией сверху вниз и назад. Остистые отростки более длинные,
чем у ламантина, и сильно расширены в переднезаднем направлении.
В основании они почти перпендикулярны к длинной оси позвонка, но дор-
зальная половина их немного отогнута назад. Из-за короткости тел и
ширины остистых отростков расстояние между последними у двух смеж-
ных позвонков остается небольшим, но налегания одного остистого
отростка на другой, как это было в передней части грудного отдела, здесь
быть не может. Передняя площадка для lig. flava увеличена, стала сход-
ной с тем, что наблюдается у ламантина, но задняя, хотя и тоже стала
больше, осталась во впадине на задней поверхности остистого отростка
и пластинки дуги.
Таким образом, уже путем сравнения двух грудных позвонков ла-
мантина и дюгоня обрисовываются характер изменений, совершающихся
в пределах грудного отдела сирен, и его различия у разных форм, на чем
необходимо остановиться подробнее.
В первую очередь обращает на себя внимание массивность и величи-
на тел грудных позвонков ламантина, их явное преобладание над нев-
122
Рис. 41. Изменение общего объема (а), т/1
•объемов невральной дуги (в) и тела
позвонка (б) на протяжении шейного 220
и грудного отделов позвоночного стол-
ба ламантина (1 — экз. ИЗ-5945) и
двух грудных позвонков дюгоня (2 — 1В0
экз. ЗИН-3220)
Стрелкой указаны границы грудного отдела
140
ральной дугой. Измерения общего 100
объема позвонков и отдельно их
дуг и тел показали (рис. 41), что gg
у ламантина идет непрерывное
нарастание объема позвонка с I
по X th в 4,4 раза, и это связано
главным образом с увеличением
его тела в 7,3 раза. В начале груд-
ного отдела равно увеличивается объ
•более или менее равны, но с V th дальнейшее возрастание объема дуги
останавливается, а рост тела продолжается дальше. За XII грудным
позвонком общий объем позвонков начинает сокращаться и опять сперва
за счет преимущественно тела, а через пару сегментов уменьшаться
начинает и дуга. Замеры объемов двух грудных позвонков дюгоня пока-
зывают, что у него в передней части грудного отдела объем дуги больше
объема тела, а у X th их объемы сравниваются. Дальше идет относи-
тельное увеличение объема тела, связанное с некоторым уменьшением
дуги, главным образом за счет редукции диапофизов, и продолжением
увеличения тела. Общий объем позвонков у дюгоня растет до конца груд-
ного отдела.
При рассмотрении общей характеристики позвоночного столба уже
было показано, что у всех современных сирен в каудальном направлении
идет нарастание массы с I по VIII th в 2,7 раза (см. рис. 21), увели-
чение длины (см. рис. 22), высоты (см. рис. 23) и ширины (см. рис. 25)
позвонков и значения показателя Слайпера (bh2) (см. рис. 24). Теперь
то же показано и для объема позвонков. Все это легко объяснимо, по-
скольку все эти параметры связаны с линейными размерами позвонков
или являются их производными. Однако из этого следует два важных вы-
вода. Во-первых, пик нарастания всех этих параметров у ламантина при-
ходится на заднюю половину грудного отдела, а к концу его уже проис-
ходит значительное снижение всех показателей (масса, объем в 1,6 раза),
а у дюгоня (и с некоторыми особенностями у морской коровы) — на са-
мый конец грудного и начало послегрудного отделов. И если характери-
стика моментов сопротивления у дюгоня сопоставима с таковой некоторых
китообразных, то ламантин не имеет никаких аналогий ни с какими-либо
морскими, ни с наземными млекопитающими. Во-вторых, хотя изменение
значений различных линейных параметров происходит в одном направ-
лении, синхронно, то темпы и степень этих изменений не совпадают ни
у самих параметров (например, высота и длина, длина и ширина, объем
дуги и тела), ни у ламантина и дюгоня. Следовательно, они специфичны
для разных форм сирен.
Форма тела, вентральные гребни, диафрагма. Различия в форме тел
позвонков у ламантина и дюгоня зависят от большего развития у перво-
го передних и задних парапофизных выступов, что вместе с удлинением
тел приводит к формированию отчетливых срединных пережимов на всех
грудных позвонках, делающих их похожими при взгляде снизу на пе-
123
сочные часы (см. рис. 40, а). Величина минимальной ширины тела в
этом пережиме почти на всем протяжении грудного отдела (IV—XV th)
остается неизменной, лишь на I и XVI th (рис. 28, д') она заметно боль-
ше соответственно в пределах II—III th идет сокращение ширины до
обычного уровня. Таким образом, изменение других широтных парамет-
ров на телах позвонков ламантина (ширина сочленовной поверхности,
ширина в парапофизах) не связано с изменением минимальной шири-
ны тела. У дюгоня с его укороченными телами позвонков и более низким
положением диапофизов, а следовательно, с иной организацией реберно-
позвоночных суставов, таких пережимов нет вообще, парапофизы как са-
мостоятельные выступы на теле практически не развиты. Имеется лишь
небольшое вентральное сужение тела в середине позвонка (см. рис. 40, б).
Такое же сужение наблюдается у морской коровы и ряда других иско-
паемых дюгонид. Таким образом, формирование пережима на теле груд-
ных позвонков специфично для ламантина. Не наблюдалось его и у его
возможных предков — протосиренид (см.: (Domning et al., 1982, fig. 27)).
Очевидно, возникновение пережима было связано с необычным удлинени-
ем тел позвонков.
Подчеркнутая сердцевидность тел позвонков ламантина в определен-
ной степени вызвана необходимостью поддерживать мощный вентраль-
ный гребень (см. рис. 37, а). Его несут большинство грудных позвонков:
V—XVI у экз. ИЗ-5945; V—XVII у экз. ЗИН-677; IV—XVII у экз.
ЗИН-31 795 и Х-175. Таким образом, они имеются на протяжении 11—14
позвонков, включая, как правило, последний грудной (см. рис. 38, а).
В ходе анатомирования экз. ЗИН-31 795 выяснилось (см. гл. 5), что
именно на этих гребнях крепятся сухожильные тяжи, входящие в со-
став срединной сухожильной части диафрагмы, занимающей в результа-
те, как это принято писать в литературе, горизонтальное положение на
всем протяжении длинного грудного отдела позвоночного столба. Из это-
го следует важное положение — по наличию таких гребней или структур,
их замещающих, можно судить о протяженности этой горизонтальной ча-
сти диафрагмы у других, в частности и ископаемых, сирен, а также о сте-
пени наклона передней более вертикальной (поперечной) части диафраг-
мы. Поскольку у ламантина наиболее краниальное крепление диафрагмы
(IV или V th) близко по положению к отхождению начальных мышеч-
ных ножек диафрагмы (дистальный конец III ребра) и заднему краю
грудины, на котором диафрагма также укреплена, то ясно, что здесь по-
зади перикарда диафрагма занимает почти вертикальное положение, от-
деляя полость, занятую сердцем, от брюшной полости, где помещаются
желудок и прочие внутренности. Правое и левое же легкие помещаются
целиком в обособленных друг от друга длинных горизонтальных карма-
нах, тянущихся до конца грудного отдела.
Как у дюгоня, так и у Hydrodamalis gigas количество позвонков, снаб-
женных вентральными гребнями, сильно уменьшается (до 4—6 позвон-
ков): они имеются на XII—XV th у одного дюгоня (ЗИН-3220) и на
XII—XVII th у другого (ЗИН-679), на XI—XVI th у морской коровы
(3M-3464). Причем у последней четко выраженные гребни сидят только'
на XIII и XIV позвонках, а на остальных имеются в этом месте шеро-
ховатости, возможно, тоже связанные с прикреплением диафрагмы. О вы-
пячивании некоторых грудных отростков в брюшную полость в области
желудка и печени писал еще Г. Стеллер (Steller, 1751), отмечая также,
что тела остальных позвонков остаются круглыми вентрально. Следова-
тельно, у дюгоня и морской коровы горизонтальные карманы грудной по-
лости не доходят до конца грудного отдела; сильно укорочены они и спе-
реди (может быть, у этих форм укорочены легкие?), а передняя часть
124
грудобрюшной преграды занимает более косое положение, поскольку
спереди и латерально она начинается от грудины и дистальных концов
III—XVII ребер (Domning, 1977), а медиально крепится значительно кау-
дальнее (XII грудной позвонок). Более косое положение диафрагмы на-
блюдается у многих водных млекопитающих, в том числе и у китообраз-
ных и ластоногих (Howell, 1930), но она никогда не заходит так далеко
назад, как у сирен,— до уровня конца брюшной полости, а крепится где-
то вблизи конца грудного отдела. Учитывая, что у тех же китообразных
очень длинный поясничный отдел, становится ясным, что в этой ситуации
при любом косом положении диафрагмы дорсокаудальный конец грудной
полости не может достигать конца брюшной полости. Таким образом,
ясно, что возникновение горизонтальных карманов грудной полости,
связанное с горизонтальным положением большей или меньшей части
диафрагмы, является спецификой только сирен. В то же время кажется,
что в разных линиях сирен — в дюгонидной и трихехидной она развива-
ется самостоятельно.
Судя по опубликованным материалам, ни у эоценовых (Eotheroides,
Protosiren), ни у олигоценовых (Halitherium) форм вентральных гребней
не было, что, возможно, говорит об отсутствии у них диафрагмы гори-
зонтального типа или, во всяком случае, об отсутствии ее крепления на
телах позвонков. Нет гребней и у более позднего Miosiren, аберрантной
формы с древними корнями. Впервые в палеонтологической летописи
формы с вентральными килями появляются в миоцене, например
Hesperosiren, у которого гребни отмечены на XII—XIV th. Есть они у
плиоценовых видов Metaxytherium. Так, на серии из X—XIV грудных
позвонков был обнаружен гребень на XIV th и явно его не было на
X th (Simpson, 1932). Можно, следовательно, считать достоверным, что
более поздние Halitheriinae уже имели вентральные гребни, но лишь на
более задних грудных позвонках и на небольшом их числе, т. е. по дюго-
невому типу. На XI—XVII грудных позвонках имеются гребни и у от-
носительно примитивного представителя гидродамалин Dusisiren jordani
(Domning, 1978 с). Можно заключить, что гребни и сама косая диафраг-
ма появились у дюгонид лишь на поздних стадиях эволюции, но еще до
расхождения дюгонин и гидродамалин. Отсюда следует, что у ламанти-
нов, корни которых нужно искать среди эоценовых сирен (см. гл. 1),
горизонтальная диафрагма развилась независимо и на другой конструк-
тивной основе, предусматривающей полный отказ от реберного дыхания
(см. дальше) и позволившей большую протяженность горизонтальных
карманов грудной полости.
Парапофизы, реберные ямки. На телах грудных позвонков в зависи-
мости от их положения в отделе имеются две или всего одна реберные
ямки для сочленения с головкой ребра. Они сидят или прямо на теле
или даже в его углублении, как на большинстве позвонков дюгоня, или
на особых выступах, носящих разную форму (отдельного отростка, про-
сто утолщения на теле и т. п.),— парапофизах. В передней части груд-
ного отдела ребра имеют двураздельную головку, сочленяющуюся с тела-
ми сразу двух позвонков, у которых для этого в задней части тела пред-
шествующего позвонка имеется задняя реберная ямка (задняя
полуфасетка), а в передней последующего — передняя реберная (перед-
няя полуфасетка). Обе они входят в один сустав головки ребра. По ходу
позвоночного столба варьируют степень развития того или иного пара-
пофиза, регистрируемая по ширине позвонка, измеряемой по наружным
краям полуфасеток; положение парапофиза на теле относительно его
переднего и заднего краев, величина полуфасеток, их форма, направление
их поверхности.
125
У ламантина на первых двух грудных позвонках (см. рис. 34) обе
нолуфасетки или равны, или передняя больше задней. На последующих,
наоборот, задняя больше передней по вертикальной протяженности
(см. рис. 36, д, 37, г). Форма полуфасеток обусловлена во многом ха-
рактером связи с краями тела позвонка и межпозвоночными дисками.
У I—III th обе полуфасетки, а в IV th одна задняя — полностью отгра-
ничены острым гребнем от окружающей поверхности тела. Эти фасетки
имеют форму, близкую к кругу или широкому овалу. Начиная с IV th,
передняя полуфасетка, а с V th обе полуфасетки открываются более или
менее широко в область межпозвоночных дисков, как бы разрывая на-
ружу краевые гребни. В этом случае они приобретают ушковидную фор-
му. Задняя полуфасетка остается открытой до самого своего исчезновения
(ее нет уже на XIII th), а передняя постоянно с X th, а в качестве
индивидуальной вариации и с VIII th, оказывается опять строго ограни-
ченной кольцевым гребнем. Форма ее становится грушевидной с узким
концом, обращенным вниз и вперед, а на XIII—XIV th — вверх. С ис-
чезновением задней полуфасетки на теле остается всего одна, бывшая
передняя фасетка, которая, начиная с XIII th, постепенно сдвигается
назад ближе к середине тела позвонка (до этого она располагалась вбли-
зи переднего края тела). Форма этой единственной парапофизной фасет-
ки обычно почковидная с выемкой, обращенной вверх и назад
(см. рис. 38, а). На всем протяжении грудного отдела ламантина перед-
няя полуфасетка своим верхним краем достигает уровня верхнего края
тела, а задняя полуфасетка, начиная с IV th и до своего исчезновения,
за счет задних парапофизов выступает частично за верхний край тела
(см. рис. 37, в). Почти во всех грудных позвонках центр сустава головки
ребра располагается краниальнее центра ребернопоперечного сустава
(фасетка на диапофизе), и лишь у первых двух — прямо под ним.
У дюгоня значительно меньшее количество ребер имеют двураздель-
ную головку. Соответственно уже на VII (а не XIII th) и всех после-
дующих позвонках отсутствует задняя полуфасетка (рис. 37, д). Центр
сустава реберной головки только первых грудных позвонков располагает-
ся впереди от фасетки диапофиза, а у всех остальных он смещается под
нее. Форма суставных реберных полуфасеток всегда узкая, овальная, вы-
тянутая в вертикальном направлении, что сообразуется с короткими те-
лами. Похоже, что передняя полуфасетка всегда обособлена от межпоз-
воночного диска и лишь задняя связана с ним. Ни на одной
.полуфасетке дюгоня не наблюдалось кольцевидного гребня, как это было
у ламантинов. Наоборот, характер окружающей поверхности говорит, что
сама капсула сустава отодвинута здесь от краев суставной поверхности,
позволяя ребрам большую свободу движений, тогда как у ламантина она
плотно облегает суставные поверхности. Как будет показано дальше, есть
основания полагать, что у дюгоня капсула головки ребра сообщается под
диапофизом с капсулой поперечно-реберного сустава (с V th и дальше
назад). Парапофизы, как выступы на теле позвонка, у дюгоня развиты
очень слабо (и лишь в задних грудных позвонках). Передние реберные
поверхности сидят практически в яме на теле, причем, как это было от-
мечено при описании IV грудного позвонка (см. рис. 36), частично на
теле, частично выше его уже на ножке дуги. Это положение сохраняется
до X th, а затем диапофиз, смещаясь вниз, оттесняет эту фасетку ниже,
заставляет ее сперва частично залезать на его вентральную поверхность
(см. рис. 37. д') и менять соответственно свою плоскость на более вент-
ральную, а затем укорачивать ее в дорсовентральном направлении. Так,
на XIII th диапофиз уже спустился до уровня верхнего края сочленов-
ной поверхности тела позвонка, в результате фасетка полностью оттес-
126
нена на тело. При этом диапофиз укорачивается, что ведет к сближению
фасеток диапофиза и парапофиза, причем первая становится все меньше
и меньше. На XVII th диапофиз уже так мал, что представляет всего
небольшой козырек над овальной фасеткой парапофиза, с длинной осью,,
лежащей в горизонтальной плоскости, а сама фасетка диапофиза пол-
ностью исчезла. Ребро стало одноглавым (бугорок исчез) и сочленяется
с единственной фасеткой, сидящей на вершине короткого отростка —
синапофиза, а состав которого вошел парапофиз и рудимент диапофиза.
На XVIII, последнем грудном позвонке (см. рис. 39, в) исчез этот диа-
пофизарный козырек, а синапофиз стал больше выступать над поверх-
ностью тела позвонка.
Положение и судьба обеих парапофизарных полуфасеток у морской
коровы аналогичны тому, что наблюдается у дюгоня: передняя полуфа-
сетка сидит частично на теле, частично на ножке; центр сустава ребер-
ной головки в большинстве случаев лежит на одном уровне по вертикали
с фасеткой диапофиза; задняя полуфасетка отсутствует, начиная с
IX th; в конце грудного отдела диапофиз опускается ниже и на XVIII th
фасетки диапофиза и парапофиза практически слиты и сидят на едином
отростке — синапофизе. Близкая к этому картина описана у Metaxythe-
rium calvertense (Kellogg, 1966). На последних грудных позвонках
(XVIII—XXI th) Dusisiren jordani имеется синапофиз с единственной
реберной головкой (Domning, 1978 с). Таким образом, на всех этих по-
звонках ребра становятся настоящими одно головыми (бугорок исчез или
слился с головкой). Как уже отмечалось в литературе (Simpson, 1932;
см. гл. 1), у всех позднетретичных дюгонид происходит сокращение чис-
ла позвонков, имеющих две полуфасетки на теле и одну фасетку на
диапофизе (позвонки с тремя фасетками), и увеличение числа позвонков
с двумя фасетками (исчезла задняя полуфасетка): у дюгоня и морской
коровы их становится соответственно 8 (7) и 9 (8), против 9 и 6 у мио-
ценовых и плиоценовых Metaxytherium, Hesperosiren, 12—6 (в среднем)
у олигоценового Halitherium (здесь цифры приводятся разные: 10—13
и 5—8 (Sickenberg, 1934); 12 и 3 (Simpson, 1932). У эоценовых халите-
риин, таких, как Eotheroides, отмечено 12 позвонков с тремя фасетками и
7 с двумя. У большинства дюгонид последний грудной или два последних
имеют одну единственную фасетку, результат слияния фасеток парапофи-
за и диапофиза (выпадение данных Симпсона из общего ряда связано,
возможно, с неправильным увеличением числа однофасеточных позвон-
ков до 4, что при совпадении у всех авторов числа грудных позвонков
у той или иной формы механически ведет к уменьшению числа позвон-
ков с двумя фасетками). Таким образом, состояние, описанное выше для
ламантина, соответствует таковому раннетретичных дюгонид, исключая
то, что у ламантина вообще в пределах грудного отдела не происходит
слияния парапофизной и диапофизной фасеток, даже на последнем по-
звонке. Его можно видеть только на I послегрудной позвонке
(см. рис. 39, б). В то же время считаемый ближайшим предком трихе-
хид Protosiren, судя по описанию (Sickenberg, 1934), если оно правиль-
но, имеет необычную форму сочленения ребер с позвонками. Первые 14
грудных позвонков у него имеют по две полуфасетки, но лишь 5 первых
из них несут фасетки на диапофизе, т. е. все последующие ребра, начи-
ная с VI, сочленены только с телом позвонка двухраздельными головка-
ми спереди, а сзади (с XV ребра) лишь головками с одной фасеткой на
них. Любопытную аналогию с ламантином составляет единственный дру-
гой представитель сирен, не имеющий в грудном отделе синапофизов и,
следовательно, истинных одноглавых ребер, Miosiren, у которого, может
быть, даже большее число позвонков, чем у ламантина, может нести три
127
фасетки (две полуфасетки и фасетку): по данным Л. Долло (Dollo,
1889), 17 из общего числа в 20 грудных позвонков, по данным О. Зи-
кенберга (Sickenberg, 1934), от 14 до 17.
Невральная дуга сирен, как и у всех млекопитающих, имеет сложный
рельеф, который удобнее анализировать по отдельным деталям: ножка
дуги, диапофиз (его наклон, сечение, степень развития, рельеф, фасетка),
зигапофизы (степень развития, размер и наклон суставных поверхно-
стей), метапофизы, площадки для крепления lig. flava остистый отросток
(величина, форма, наклон).
Ножка дуги I грудного позвонка в сечении имеет вид длинного узкого
прямоугольника с длинной осью, направленной примерно под 60° к
продольной оси позвонка. До V th включительно она помещается на
передней части тела позвонка. Каудальнее начинается постепенное сме-
щение ее назад ближе к середине длины позвонка. При этом ориентировка
длинной оси ее сечения также постепенно изменяется на более про-
дольное, сама она становится толще, а ее сечение к концу почти оваль-
ным. Здесь ножка уже полностью располагается по длинной оси поз-
вонка.
У дюгоня происходит аналогичное изменение положения длинной
оси ножки с почти перпендикулярного в I th до продольного в задних
грудных позвонках. В последних грудных позвонках дюгоня ножка пред-
ставляет тонкую широкую пластинку, ориентированную спереди назад.
Диапофиз. Верхние поперечные отростки, или диапофизы, на I груд-
ном позвонке ламантина помещаются в нижней половине ножки дуги
(см. рис. 34, а) и направлены почти горизонтально вбок (при виде спе-
реди) и немного вперед (при виде сверху). Таким образом, нижний край
диапофиза здесь находится на уровне верхнего края тела позвонка, а ре-
берная ямка поперечного отростка (фасетка диапофиза) своим краем
даже немного заходит вентральнее этого уровня. В этом можно видеть
переходный характер I th, поскольку плеврапофизы шейных позвонков,
как сказано выше, больше наклонены вниз, и подъем начинается в самом
конце шеи (VI с). Каудальнее диапофиз на грудных позвонках несколь-
ко поднимается по ножке выше середины позвоночного отверстия.
Можно сказать, что центр основания диапофиза всегда находится близ
области максимальной ширины позвоночного отверстия в данном по-
звонке. Эта область, как и соответственно диапофиз, в каудальном на-
правлении смещается в верхнюю половину позвоночного отверстия.
Наклон диапофиза по ходу грудного отдела меняется как в вертикаль-
ной, так и в горизонтальной плоскостях (рис. 42). Подъем дистальногоя>
конца диапофиза вверх достигает максимума у IX th. Дальше идет срав-
нительно незначительное понижение к последнему грудному позвонку.
Таким образом, практически на всем протяжении грудного отдела диапо-
физы подняты так, что пространство между ними и остистым отростком
значительно уменьшается (см. рис. 37, а), что, как будет показано в
гл. 5, связано с относительно слабым развитием у ламантинов эпакси-
альной мускулатуры. В то же время это приводит к необычному для си-
рен поднятию диапофизных фасеток, увеличению расстояния между
ними и фасетками парапофиза, что отражает своеобразие прикрепления
ребер у ламантина. У дюгоня диапофиз I грудного позвонка (см. рис. 36)
находится в более низком положении по сравнению с ламантином, а сам
процесс подъема совершается быстрее: уже на II th диапофиз принимает
близкое к горизонтали положение и до самой своей редукции на задних
грудных позвонках остается таким. При этом и сидит он на ножке дуги
почти все время не выше середины позвоночного отверстия. Таким обра-
зом, пространство для эпаксиальной мускулатуры, учитывая еще и дли-
128
Рис. 42. Изменение направления диапофизов и поперечно-реберных отростков по-
звонков относительно горизонтальной (а) и вертикальной (б) плоскости у ламанти-
на (/ — экз. ИЗ-5945) и дюгоня (2 — экз. ЗИН-3220 и ЗИН-679)
Стрелки указывают границы грудного отдела
а — отклонение от горизонтальной плоскости, подъем кривой «а» соответствует изменению направ-
ления на более переднее;
< |3 — отклонение от вертикальной плоскости, подъем кривой «б» соответствует изменению направ-
ления на более переднее
ну остистых отростков, у дюгоня значительно больше, чем у ламантина
(см рис. 37, б). Больше оно и у морской коровы.
Небольшое отклонение диапофиза I th ламантина вперед постепенно
уменьшается, и на VI позвонке диапофиз направлен прямо вбок (при
виде сверху). Изменение его направления продолжается и к предпослед-
нему позвонку (XIV th) достигает максимума, что, как уже было пока-
зано, отражается на относительном положении центров фасетки диапофи-
за и парапофиза. На последних двух грудных позвонках заднее отклоне-
ние начинает уменьшаться, кульминируя уже в послегрудной области
(II pth). У дюгоня идет почти сходное изменение направления диапофи-
за в горизонтальной плоскости, но диапофиз I th гораздо сильнее обра-
щен вперед; наклон в заднем направлении у них меньше, а максимум
его достигается раньше по длине отдела (на уровне конца средней трети,
т. е. за 6—7 позвонков до конца отдела).
5 Ламантин
129
Сечение диапофиза по середине его длины меняется. У ламантина
диапофизы передних грудных позвонков широкие, уплощенные. Форма
сечения напоминает неправильный четырехугольник, но передняя сторона
быстро сокращается в размерах, и к VII th сечение принимает вид ши-
рокого низкого треугольника с вершиной, обращенной вниз и смещенной
назад за уровень середины основания. Внешне передневерхний угол:
этого треугольника выглядит широким, сужающимся вперед и вперед и
наружу фланцем впереди и немного выше фасетки диапофиза. Этот фла-
нец уже хорошо был виден на IV th (см. рис. 36, д), максимально ши-
рок он на VI th, но далее назад быстро начинает сокращаться в разме-
рах и на VIII уже почти не виден. Сечение диапофиза принимает вид
вытянутого в длину по оси позвонка овала, который становится каудаль-
но все короче и короче. Ширина диапофиза (в переднезаднем направле-
нии) сначала растет до VII грудного позвонка, потом более или менее
равномерно начинает сокращаться, и диапофиз последнего грудного поз-
вонка имеет меньший диаметр, чем I th.
У дюгоня сечение диапофиза I грудного позвонка в виде очень ши-
рокого, но низкого треугольника с основанием, обращенным назад, са-
мым острым углом вниз, вершиной, сдвинутой вверх вперед. Каудальнее-
происходит расширение верхней короткой стороны, небольшая переориен-
тация передней поверхности и нижнего угла, смещающегося намного
вперед. Таким образом, диапофиз расширяется в переднезаднем направ-
лении, а в высоту относительно уменьшается. Все эти изменения, проис-
ходящие в пределах первых шести грудных позвонков, приводят к тому,,
что сечение диапофиза теперь напоминает полуовал (овал, срезанный
снизу и немного наискось спереди и снизу — вверх и назад). Характер-
ной чертой диапофиза дюгоня VI и еще в большей степени последующих
позвонков становится направленный вниз (вперед и вниз) передний
фланец, заходящий ниже диапофизной фасетки, как бы прикрывая ее
спереди. Ширина диапофизов дюгоня, как и у ламантинов, увеличивает-
ся к VI—VII th, но далее почти до самого конца грудного отдела шири-
на остается неизменной. Лишь после XIV th диапофиз несколько сужа-
ется, но даже последний XVIII грудной позвонок имеет более широкий
диапофиз, чем I th. Все это связано в первую очередь с необычной ко-
роткостью первых грудных позвонков дюгоня: они почти плоские и этим
мало отличаются от шейных позвонков. Можно говорить, что если у ла-
мантина последний шейный позвонок был подвергнут торакализации и
стал грудным (I th), то у дюгоня наблюдается противоположный процесс
цервикализации, уподобления первых грудных позвонков шейным.
Ширина диапофиза дюгоня в большинстве случаев превышает высоту
в два-три раза, а у ламантина лишь в полтора раза. Относительная ши-
рина диапофиза (в процентах к длине тела) в передних грудных позвон-
ках и ламантина и дюгоня почти одинакова (около 60%), но у ламанти-
на потом она возрастает на 15%, а у дюгоня на 25%. Это приводит к
тому, что диапофизы дюгоня относительно длины тела шире, чем у ла-
мантина, и в связи с общей короткостью тела расстояния между
диапофизами смежных позвонков у дюгоня несравнимо меньше, чем у
ламантина. Соответственно и ребра у дюгоня более сближены. У обеих
форм самые длинные диапофизы (максимальная ширина позвонка по
диапофизам) имеются у I th (вернее у дюгоня на VII шейном позвон-
ке, что еще раз свидетельствует о цервикальном происхождении I th ла-
мантина), а каудально идет уменьшение их длины (см. рис. 25) до ми-
нимума у последнего грудного позвонка.
Рельеф диапофизов сильно варьирует индивидуально, и для оценки
его истинного значения необходим серийный метериал. Так, на ламанти-
130
не экз. ИЗ-5945 может быть заметная шероховатость или бугор, а на экз.
ЗИН-31 795 их нет. Все это затрудняет описание рельефа, которое мы
ведем, основываясь главным образом на первом из этих экземпляров.
На переднем крае диапофиза I th ламантина на середине перегиба,
отмечающего границу между верхней и передней поверхностями диапо-
физа, помещается отчетливый богорок для m. seniispinalis cervicis. На
заднем крае диапофиза имеется такой же бугорок для отхождения сухо-
жильной ножки ш. longissimus dorsi. Фактически, оба они отмечают на
этом позвонке два медиальных угла (передний и задний) более или
менее видной шероховатости на латеральной трети дорсальной поверх-
ности диапофиза, служащей для крепления поперечно-реберной связки.
Передни!! бугорок прослеживается до IV th включительно в соответст-
вии с количеством ножек m. semispinalis cervicis (см. гл. 5). Далее на-
зад его замещает небольшой бугорок, смещенный медиальнее к самому
основанию презигапофиза на VII th, а еще каудальнее переходящий на
латеральную поверхность бокового фланца презигапофиза. Очевидно, этот
бугорок соответствует метапофизу других млекопитающих, на котором
крепятся мышечные пучки mm. miltifidus, глубокой порции m. semispina-
lis dorsi, но здесь они не развиты в настоящие отростки.
Задний бугорок на II th (и особенно на последующих) обособляется
ют шероховатости для связки и перемещается на задний край диапофи-
за. Иногда он очень слабо выражен. Вообще у ламантина бугорки на
диапофизе развиты хуже, чем у дюгоня. Не прослеживается какой-либо
рельеф и на нижней поверхности диапофиза.
У дюгоня от основания остистого отростка косо к передненижнему
углу дистального конца диапофиза идет гребень, наиболее выступающая
вперед часть которого и образует бугорок для m. semispinalis cervicis.
Этот гребень был отмечен при описании IV грудного позвонка
(см. рис. 36, а). Вперед гребень прослеживается до I th, далее назад
постепенно суживается, а бугорок приближается к презигапофизу. Как
самостоятельный, он виден еще на VI th, но на VII уже слит с прези-
гапофизом, резко увеличивая латеральное протяжение последнего. Это
показывает метапофизную природу этого бугорка на передних грудных
позвонках. На более каудально расположенных грудных позвонках мета-
пофиз выступает в виде более или менее утолщенного бокового фланца
на презигапофизах. На задней поверхности диапофиза имеются два ту-
пых, скорее прощупывающихся, чем ясно видных, бугорка. Один из них
лежит медиальнее, фактически уже на ножке дуги ниже и латеральнее
постзигапофиза, второй ближе к дистальному концу у задневерхнего края
нижней шероховатости, служащей для крепления, очевидно, капсулы
поперечно-реберного сустава. На V грудном позвонке к этому бугорку
подходит гребень, отходящий от середины основания диапофиза на его
вентральной поверхности, идущей вперед на треть длины диапофиза к
отчетливому бугру у задневерхнего угла передней реберной ямки, а за-
тем поворачивающий назад и наружу к заднему краю диапофиза, где и
находится второй из двух бугорков на заднем крае его. Этот бугорок
служит местом отхождения сухожильных ножек m. longissimus dorsi. При-
рода более медиального бугорка неясна. Единый на IV th (и III; у более
передних позвонков его не видно), на V th он явно подразделяется на два
бугорка, один в обычном месте, другой несколько латеральнее (слева рас-
стояние между ними 5 мм, а справа 10 мм). Далее назад эти отдельные
бугорки сближаются и сливаются опять в один на VII—VIII позвонках.
Очень заметен этот бугор на XII—XIII th. С утончением ножки дуги на
последующих позвонках этот бугорок оказывается на самом заднем крае
(ребре) ножки. На XVIII th от него остается совсем маленький выступ.
131
5*
Еще один бугорок на диапофизе дюгоня расположен на дистальном конце
отчетливого гребня, идущего от верхнего края передней реберной ямы к
передненижнему углу дистального конца диапофиза, к вершине передне-
го, направленного вниз фланца, уже упомянутого выше. Этот гребень от-
деляет переднюю, гладкую поверхность дпапофиза и ножки дуги, обра-
зующую нечто вроде широкого желоба, открытого вперед от вентральной
поверхности, более шероховатой и ограниченной сзади уже описанным
гребнем на вентральной поверхности диапофиза. Позади последнего
гребня поверхность диапофиза опять гладкая. Таким образом, складыва-
ется впечатление, что эта шероховатая вентральная поверхность постав-
лена в иные условия, чем передняя и задняя. Возможно, она входит в
капсулу единого реберно-позвоночного сустава, которую поддерживает
система названных гребней и бугорков на вентральной поверхности
диапофиза в большом отступе от краев собственно сочленовной фасетки
парапофиза. Уже говорилось, что у ламантина ничего похожего нет,,
и у него сустав головки ребра не сообщается с поперечно-реберным су-
ставом. Важно отметить, что на IV грудном позвонке гладкие передняя
и задняя поверхности смыкаются на направленной вниз вершине тре-
угольника, образованного сечением диапофиза (см. выше). Это относится
и ко всем впереди лежащим грудным позвонкам. И лишь начиная с V
грудного позвонка, что совпадает с изменением формы сечения диапофи-
за, появляется шероховатая вентральная поверхность, возможно, говоря-
щая о формировании единой капсулы реберно-позвоночного сустава (на
эти детали не обращали внимания исследователи, имевшие возможность
анатомировать дюгоня; связочный аппарат сирен почти неизвестен). Мо-
жет быть, именно это событие открыло для дюгоня возможность значи-
тельного сближения сочленовных фасеток диапофиза и парапофиза,
а следовательно, головки и бугорка на ребрах, на более каудально рас-
положенных грудных позвонках.
В соответствии с положением диапофиза относительно всего позвонка
у ламантина центр реберной ямки поперечного отростка (фасетка дпапо-
физа) помещается в начале грудного отдела ближе к переднему кракт
тела (по отвесу), несколько впереди центра передней полуфасетки па-
рапофиза. С IV грудного позвонка центр начинает смещаться за уровень
этой полуфасетки и в области XIII—XIV th находится посередине тела,
а у XV и XVI th даже несколько позади его. На I грудном позвонке
фасетка имеет вид широкого овала с горизонтальной длинной осью. Ка-
удальнее по ходу позвоночного столба фасетка начинает постепенно ме-
нять свое положение (поворот против часовой стрелки при взгляде сле-
ва), сперва на наклонное вперед (при этом фасетка уменьшается в ши-
рину, овал становится уже), затем на VI th почти вертикальное
(фасетка укорачивается), за ним ось продолжает вращаться еще дальше
в том же направлении и к X th опять принимает горизонтальное поло-
жение. При этом длина фасетки опять становится больше (ширина оста-
ется неизменной), и лишь на XV th она уменьшается в размерах, ста-
новится почти круглой, а на XVI фасетка уже совсем крошечная, и в нее
упирается не фасетка бугорка ребра, а острый выступ его (см. рис. 38, б).
На I th фасетка глубокая, полусферическая, на остальных позвонках
почти плоская. Плоскость фасеток обращена на I th вбок, вперед и вниз,
на последующих преимущественно вбок и вниз, на XIV—XV th вбок,
вниз и назад. По отношению к торцевой поверхности диапофиза фасетка
занимает всю ее на I th, нижние 2/з на II—III th, смещается в нижне-
задний угол на IV—V th. С сокращением переднего фланца диапофяза
фасетка опять оказывается по центру диапофиза, а на XI—XII th даже'
несколько вперед от него. С уменьшением ширины диапофиза у задних
132
грудных позвонков фасетка опять начинает занимать всю торцевую его
поверхность. С поверхности фасетки диапофиза идут отчетливые желоб-
ки, выпускники для синовии. С I по XV th такой желобок имеется свер-
ху или обычно в передневерхнем углу фасетки. С XII th дополнительно
появляется выпускник снизу, связанный на этом и XIII th с маленькой,
но глубокой ямкой в нижней части суставной поверхности, имеющейся на
VIII—XIII грудных позвонках. Постоянное присутствие верхнего вы-
пускника и его большая величина на большинстве позвонков свидетель-
ствуют, что капсула реберно-поперечного сустава отодвинута у ламанти-
на от самой суставной поверхности сверху значительно дальше, чем
снизу.
Фасетки диапофиза дюгоня очень отличаются от ламантиновых.
Главное — их длинная ось не меняет своего положения, оставаясь на
всем протяжении позвоночного столба горизонтальной. На I—IV th фа-
сетки почти круглые, на I фасетка занимает весь торец, на II переднюю
его часть, на III середину, на IV—V th заднюю часть. На последующих
позвонках фасетка имеет овальную форму и помещается по середине
торца. На сухом скелете, имевшемся в нашем распоряжении, система
выпускников у дюгоня почти не развита и кажется, что суставная по-
верхность широко открыта вниз в капсулу единого реберно-позвоночного
сустава.
У морской коровы на I—II th фасетка субтреугольна, верхнее ее ос-
нование широко округлено, вершина обращена вниз и слегка вперед. На
IV th вершина смотрит слегка назад. Далее назад она сглаживается,
и фасетка становится овальной с горизонтальной длинной осью. Фасетка
относительно невелика по размеру по сравнению с торцевой поверхно-
стью диапофиза. На VII—X th она лежит в середине ее внизу, а далее
назад (особенно хорошо видно на XIII th) смещается в задненижний угол
торца. Важно отметить, что у дюгоня и у морской коровы центры фасет-
ки диапофиза помещаются значительно ниже (на уровне начала верхней
трети позвоночного отверствия), чем у ламантина (примерно на треть
высоты этого отверстия, выше его верхнего края).
Зигапофизы. Презигапофизы у ламантина на I грудном позвонке еще
очень короткие (см. рис. 34), и передние суставные поверхности еще на-
половину или даже на 2/3 своей длины лежат позади переднего края
диапофизов. Тем не менее в отличие от дюгоня (см. рис. 35) презигапо-
физы хорошо обособлены от диапофизов отчетливыми, хотя и неглубоки-
ми еще выемками. Расстояние между центрами передних суставных по-
верхностей у I th максимальное для всего грудного отдела, и медиальная
вырезка между презигапофизами представляется широкой дугой окруж-
ности, радиус которой вдвое превышает глубину вырезки. С постепенным
сдвигом назад диапофизов и увеличением длины самих презигапофизов
последние все больше и больше начинают выступать вперед. У IV—V th
передняя суставная поверхность уже целиком впереди от каудального
края выемки, отделяющей презигапофиз от диапофиза (см. рис. 40, д').
Презигапофизы быстро сближаются, и минимальное расстояние между
ними достигается уже на V грудном позвонке. Соответственно меняется
глубина и форма вырезки между ними: на II th она еще дугообразна
(радиус примерно равен ее глубине), на III th трапециевидна, почти
субквадратна, на последующих позвонках имеет вид более или менее
длинного прямоугольника. К XI th (см. рис. 37, е) задний край перед-
ней суставной поверхности уже лежит на середине длины презигапофи-
зов (на большинстве грудных позвонков этот край находится примерно
на одном уровне с передним краем тела позвонка, а на средних X—XI th
даже впереди от него, а сам презигапофиз выступает вперед на 2/э
133
своей длины). Стопорные ямки в задней части дорсальной поверхности
презигапофизов прослеживаются до VI грудного позвонка. Дальше, с на-
чалом постепенного наклона суставных поверхностей внутрь, они исче-
зают, теряя свой смысл. Слегка наклонное положение передних сустав-
ных поверхностей I и II th сменяется горизонтальным уже на III груд-
ном позвонке, но вскоре опять начинает изменяться на наклонное с
VII th. Наклон уже хороню заметен на VIII и IX th, а с X и дальше
(см. рис. 37, а) до конца отдела наклон достигает своего максимума,
а презигапофизы за счет слияния с метапофизами получают боковые,
завернутые вверх фланцы, придающие им при виде спереди желобообраз-
ную форму. Таким образом, в районе VII—VIII th у ламантина проис-
ходит изменение характера движений, осуществляемых в дугоотростча-
тых соединениях; возможные в передней части грудного отдела неболь-
шие сгибательные (разгибание ограничено стопорными ямками) и
боковые движения сменяются заметными вращательными (относительно
длинной оси позвоночного столба). Сгибательные движения в этой обла-
сти, скорее всего, ограничены сильно увеличенными сочленовными по-
верхностями тел позвонков. Наличие в задней части вращательных дви-
жений подчеркивается наличием метапофизов и сохранением самих
презигапофизов в послегрудном отделе, когда контакт между зигапофи-
зами смежных позвонков уже нарушен, постзигапофизы полностью ре-
дуцируются, а на презигапофизах исчезают передние суставные поверх-
ности. Таким образом, на метапофизах крепление коротких пучков
системы m. transverso—spinalis, отвечающих за вращательные движения
(mm. multifidus et submultifidus), ведет к сохранению презигапофизных
отростков, на которых метапофизы помещаются в каудальной части
грудного отдела и в послегрудном отделе.
Необходимо, однако, отметить, что зигапофизы относятся к очень
пластичным структурам и легко изменяют свой наклон в зависимости от
наклонностей того или иного индивидуума. Вышеприведенное описание
было сделано по экз. ИЗ-5945, но у экз. ЗИН-31 795 значительно большее
число передних грудных позвонков имеют горизонтальные или почти го-
ризонтальные передние суставные поверхности, например X th. Тем не
менее оба экземпляра имеют общую тенденцию к изменению положения
презигапофизов в задней части грудного отдела.
У дюгоня, как уже отмечалось при описании IV грудного позвонка
(см. рис. 40, е), презигапофизы как самостоятельные отростки практиче-
ски отсутствуют, а передние суставные поверхности сидят прямо на
горизонтальной пластинке дуги, практически не выступают за передний
край диапофизов и не обособлены от них особыми выемками. Лишь по-
степенно презигапофизы увеличиваются: они уже хорошо заметны на
VIII th и на XI th достигают максимальной длины, более или менее
сохраняющейся до конца грудного отдела. Если у ламантина с его тол-
стыми и широкими ножками дуги и узкими позвоночными отверстиями
презигапофизы помещаются посередине ножки, то у дюгоня они сидят
частично над большими позвоночными отверстиями. Однако если соотне-
сти расстояние между центрами передних суставных поверхностей с
максимальной шириной тела позвонка, то оказывается, что у дюгоня
презигапофизы расставлены шире, чем у ламантина. Так, на IV th это
расстояние у дюгоня определяется примерно в 100%, то у ламантина
всего 60%, на X th 80 и 50% соответственно. Передние суставные по-
верхности первых грудных позвонков близки к горизонтальной плоскости,
наклон становится заметнее к середине отдела. Однако боковые фланцы
презигапофизов у дюгоня не выступают так заметно вверх, как у ламан-
тина, и презигапофизы не имеют желобовидного характера (рис. 37, а п
37,6).
134
Характерная особенность презигапофизов морской коровы — их ши-
рокий лопастеобразный вид, напоминающий передние грудные позвонки
дюгоня, но имеющийся практически на всем протяжении грудного отдела.
Лишь на XVI и дальнейших позвонках презигапофизы несколько обо-
собляются от диапофизов. Практически на всех грудных позвонках, кроме
самых последних (XV—XIX), передние суставные поверхности почти
горизонтальные, чем морская корова резко отличается от ламантина и от
дюгоня. Очевидно, вращательные движения в позвоночном столбе ей не
свойственны, а латеральные более обычны.
Постзигапофизы на I грудном позвонке ламантина выступают мало,
скорее здесь они представляют часть пластинки дуги, на заднелатераль-
ных углах которой лежат с вентральной стороны задние суставные по-
верхности (см. рис. 34). Фактически задние края постзигапофизов и са-
мой невральной дуги на этом позвонке находятся на одном уровне.
Центры постзигапофизов значительно сближены сравнительно с таковы-
ми презигапофизов того же позвонка. Суставные поверхности больше,
их ширина больше длины. По площади они почти равны передним су-
ставным поверхностям следующего позвонка. Как и последние, они име-
ют небольшой наклон (наружу). Начиная со II th, постзигапофизы на-
чинают удлиняться, уже на IV th задние суставные поверхности лежат
за пределами заднего края тела позвонка. Между постзигапофизами об-
разуется глубокая вырезка, входящая вместе с аналогичной вырезкой
между презигапофизами последующего позвонка в междуговое простран-
ство, имеющее вид длинного прямоугольника и открытое сверху
(см. рис 40, д). Если на первых грудных позвонках (I—III) ширина и
длина задних суставных поверхностей более или менее равны между со-
бой и равны таковым передних суставных поверхностей соответствующе-
го позадилежащего позвонка, то далее назад они становятся значительно
уже, как сужаются и сами постзигапофизы, не имеющие других функ-
ций, кроме как нести соответствующую суставную поверхность, и все
более и более противопоставляющиеся мощным толстым презигапофизам.
Длина задних суставных поверхностей в задней части грудного отдела
оказывается немного меньшей (примерно на ‘Д), чем длина соответст-
вующей передней суставной поверхности, например в дугоотростчатом
соединении XII и XIII th, но ширина вдвое уступает таковой на прези-
гапофизе XIII th. Это опять говорит, что вращательные движения здесь
имеют сравнительно большую амплитуду, а сгибательные относительно
невелики. Это состояние продолжается вплоть до сочленения последую-
щего грудного позвонка с I pth. Наклон постзигапофизов изменяется в
том же направлении, что и презигапофизов, со смещением на один сег-
мент вперед. Четко выраженный диафрагмальный позвонок (Slijper,
1946), т. е. позвонок, у которого презигапофизы имеют горизонтальное
(тангенциальное), а постзигапофизы наклонное (радиальное) положение,
у сирен отсутствует. У ламантина ближе всего к нему VI th.
Постзигапофизы дюгоня всегда короткие, не выступающие за задний
край тела позвонка, кроме самых передних грудных позвонков, где это
происходит в результате большего наклона назад всей невральной дуги
(относительно перпендикуляра к продольной оси тела позвонка). Ника-
кой вырезки между ними нет, и фактически почти до IV th задние края
постзигапофизов лежат в одной линии с задним краем пластинки дуги,
которая, таким образом, вся в виде козырька или черепицы налегает на
презигапофизы последующего позвонка и прикрывает сверху медиальную
вырезку между последними. Это приводит к практически полному отсут-
ствию междугового пространства между первыми четырьмя грудными
позвонками. Оно мало даже между IV и V th (см. рис. 40, е), хотя на
135
IV th уже имеются слегка выступающие постзигапофизы, и к тому же
прикрыто сверху сильно наклоненным назад остистым отростком. Такое
черепитчатое перекрытие, существующее между передними грудными
позвонками, продолжает таковое, описанное ранее для шейного отдела.
Фактически, видимо, он вместе с передними грудными позвонками
функционирует как единое целое, и здесь исключены, несмотря на почти
горизонтальное положение суставных поверхностей, движения в горизон-
тальной плоскости. Разгибательные движения, в свою очередь, исключа-
ются эффективно действующей стопорной системой за счет указанного
перекрытия и почти полного налегания остистого отростка впередилежа-
щего позвонка на таковой последующего. Очевидно, дюгоню необходима
фиксация этой части позвоночного столба в неподвижном состоянии при
подъеме головы, причем ось этого подъема сдвинута далеко назад в пре-
делы начала грудного отдела. Отмеченная выше система стопорных ямок
на передних .грудных позвонках ламантина, несомненно, выполняет ог-
раниченную функцию препятствования переразгибанию, но не объединя-
ет шейный и переднюю часть грудного отдела в одно целое. У морской
коровы остистые отростки первых четырех грудных позвонков, как и у
дюгоня, налегают друг на друга. Сходное состояние изображено для
Dusisiren jordani (Domning, 1978, е, fig. 6). Судя по облику передних
грудных позвонков, налегание остистых отростков было и у Metaxythe-
rium (Kellogg, 1966, pl. 37). Однако междуговое пространство у морской
коровы несколько больше (см.: Brandt, 1861—1868, Tab. 5, fig. 1—2).
Относительное расстояние между пре- и постзигапофизами у морской
коровы близко к дюгоневому и сильно отличается от ламантинового.
Так, на IV th расстояние между презигапофизами составляет 104% от
ширины тела, а между постзигапофизами около 90%.
Дюгоневый тип соединения невральных дуг передних грудных по-
звонков прослеживается к самым истокам семейства. Налегающие друг
на друга первые три грудных позвонка показаны для эоценового Eothe-
roides (Sickenberg, 1934, pl. 4, 5). Сходны с дюгонем и презигапофизы
первых грудных позвонков. Возможное функциональное значение фик-
сации неподвижного положения в начале грудного отдела подчеркивается
здесь случаем срастания I и II th. Естественно, что более поздние дюго-
ниды, такие, как Halitherium и Metaxytherium, сохраняют этот принцип
в неизменном состоянии. Уже отмечалось, что и IV th Protosiren больше
схож с таковым дюгоня, чем ламантина. Таким образом, формирование
презигапофизов, значительно выступающих вперед уже в самом начале
грудного отдела, как и более длинных постзигапофизов, ведет к увели-
чению расстояния между смежными остистыми отростками и междугово-
го пространства и является специфической чертой ламантинов, возможно,
с самого начала трихехидной линии развития сирен. Легко видеть, что
это коррелировано с удлинением тел позвонков ламантина: одно не воз-
можно без другого. Сохранение же первичной относительной короткости
тел дюгонид не потребовало и перестройки дугоотростчатых соединений
и усиленного развития пре- и постзигапофизов.
Остистый отросток на I грудном позвонке ламантина или вообще от-
сутствует (экз. ЗИН-31 795), или развит очень слабо (ИЗ-5945), что
опять напоминает о его цервикальном происхождении. На II th ос-
тистый отросток уже, значительно выше (у ИЗ-5945 почти вдвое по срав-
нению с I th) и шире. У экз. ЗИН-31 795 отросток имеет острый задне-
верхний угол и весь задний край отростка более длинный, чем передний.
У экз. ИЗ-5945 оба края практически одинаковы, а верхняя часть от-
ростка расширена в передне-заднем направлении. Наиболее высокий
(абсолютно и относительно) остистый отросток у III th (чуть больше
136
50% высоты позвонка). У обоих экземляров он имеет сильно оттянутый
назад в виде отростка верхне-задний угол. Последующие остисгые отро-
стки ламантина постепенно абсолютно и относительно уменьшаются,
причем абсолютная высота особенно заметно падает у последних пяти
грудных позвонков, и это продолжается и дальше в послегрудном отделе.
Относительная высота, которая уменьшалась с III по XII—XIV th на
15%, к концу отдела опять несколько увеличивается, а окончательное ее
уменьшение начинается только после III послегрудного позвонка и до-
стигает всего 12% у XI pth.
В противоположность непрерывному уменьшению высоты остистого
отростка после III th у ламантина у дюгоня максимальная высота от-
ростка, достигнутая к IV th (чуть меньше 50% высоты позвонка), далее
практически не изменяется до VII послегрудного позвонка, да и за ним
уменьшение идет очень медленно, так что к XII—XVI pth высота состав-
ляет 40—30%. Таким образом, у дюгоня остистый отросток по сравнению
с ламантином развит намного больше, особенно в задней половине груд-
ного и постторакальном отделе, что можно связать с ослаблением эпак-
сиальной мускулатуры у ламантина. У морской коровы остистые отрост-
ки достигают максимума к IV—V th и, несмотря на некоторое дальней-
шее уменьшение, все же к XXIII pth составляют 30% от высоты позвон-
ка. Таким образом, относительно высокие остистые отростки — общий
признак для современных дюгонид.
У дюгоня остистый отросток I грудного позвонка очень сходен с та-
ковым VII шейного. Да и весь этот позвонок по многим параметрам мог
бы быть шейным, исключая то, что он несет свободное ребро. Форма
остистых отростков передних грудных позвонков более или менее одина-
кова (кроме, как уже сказано, I th и несколько сужающегося дорсально
отростка II th): это широкие, с почти параллельными передним и задним
краями, наклоненные назад пластинки с сечением равностороннего тре-
угольника на всем своем протяжении. Обращенная назад сторона этого
треугольника шире у основания, уже у дистального конца отростка. На-
чиная с VII th, наклон остистых отростков у дюгоня уменьшается и они
становятся шире. С появлением передней вертикальной площадки для
lig. flavum основание остистых отростков становится более перпендику-
лярным к продольной оси позвонка и более узким, чем несколько расши-
ренная дорсальная лопасть отростка с широко округлым верхним краем.
Ширина в передне-заднем направлении основания и тем более расширен-
ной лопасти остистых отростков дюгоня в отношении к длине тела позвон-
ка несравненно больше у дюгоня (ширина почти не уступает длине тела),
чем у ламантина (около 50%). В результате этого не только в передней
части грудного отдела, где остистые отростки практически налегают друг
на друга, но и далее назад промежутки между отростками смежных поз-
вонков остаются у дюгоня небольшими (в */4 ширины расширенной
дорсальной части), а у ламантина несравненно большими (соизмеримы-
ми с шириной всего отростка). Соответственно, у дюгоня практически не
остается места для развития mm. interspinales, а у ламантина они разви-
ты (см. гл. 5).
Наклон назад остистых отростков характерен и для ламантина, и для
дюгоня, но степень этого наклона меняется по ходу позвоночного столба
по-разному у обеих форм (рис. 43). У ламантина минимален наклон (по
верхней части отростка выше lig. flava) на I th, на V он становится зна-
чительно больше. На VI th наклон становится меньше, но затем на про-
тяжении семи позвонков остается неизменным, но к концу грудного от-
дела опять начинает расти с новым, еще большим, чем на V th, макси-
мумом в области II послегрудного позвонка. У дюгоня в начале груди
137
(грай}
140 г
120 -
100 -
80 -
60 -
40 -
20 -
Рис. 43. Изменение наклона
остистого отростка у ламанти-
на (1 — экз. ИЗ-5945), дюгоня
(2 — экз. ЗИН-3220 и ЗИН-
679) и морской коровы (3 —
экз. 3M-3464)
Стрелками указаны границы груд-
ного отдела
а — способ измерения угла накло-
на
идут сходные с ламантином изменения, но за V th начинается уменьше-
ние наклона до I pth, где остистый отросток принимает почти вертикаль-
ное положение. Далее он сперва опять наклоняется назад до 30° у
VII pth, потом несколько угол уменьшается и, наконец, опять увеличи-
вается. Таким образом, за V грудным позвонком изменение наклона
остистых отростков в грудном отделе у ламантина и дюгоня идет в про-
тивоположных направлениях: у первого он растет, у второго уменьшает-
ся. В то же время следует отметить, что у обеих форм нет антиклиналь-
ных позвонков.
Выше при описании IV и XI th уже было отмечено, что характерная
особенность грудных позвонков ламантина — более или менее вертикаль-
ное положение обеих площадок для прикрепления lig. flava, тогда как у
дюгоня и морской коровы задняя площадка выступает в виде глубокой
ямы на задней поверхности пластинки дуги и основания остистого от-
ростка. У ламантина сходная яма наблюдается только на II грудном по-
звонке. Начиная с III th, у него резко возрастает высота задней, а с IV
и передней площадки, что говорит об усилении lig. flava. К VIII груд-
ному позвонку высота обеих площадок у ламантина почти сравнивается
и остается таковой до конца грудного отдела. Наиболее мощной, если
судить по высоте и ширине площадок для прикрепления в их основании,
эта связка должна быть в районе X—XIII грудных позвонков. Далее
назад площадь обеих площадок уменьшается. Длина каждого lig. flavum
обусловлена расстоянием между основаниями остистых отростков двух
смежных позвонков и, как говорилось выше, оно значительно больше у
ламантина. В то же время удлинение связки дает возможность и боль-
шего ее растяжения, а следовательно, и сгибания позвоночного столба.
У дюгоня с его короткими расстояниями между позвонками развитие ямы
на месте задней площадки можно рассматривать как способ увеличить
длину lig. flavum.
Заканчивая обсуждение грудного отдела позвоночного столба, необ-
ходимо отметить, что грудные позвонки обоих экземпляров, сохраняя все
присущие им отмеченные выше специфические черты, заметно отлича-
ются друг от друга по некоторым количественным показателям. Так,
тела позвонков у экз. ЗИН-31 795 менее массивны, несколько короче,
138
а невральная дуга относительно лучше развита, чем у экз. ИЗ-5945. Не
исключено, что эти различия связаны с полом, к сожалению, пол по-
следнего экземпляра остался неизвестным.
ПОСЛЕГРУДНОЙ ОТДЕЛ
Рассматриваемый отдел позвоночного столба сирен резко противопостав-
лен своей подвижностью всей остальной его части. По существу, эта
часть туловища служит единственным их движителем, развивающим про-
пульсивную силу путем изгибания в вертикальной плоскости. Внешне,
благодаря отсутствию задних конечностей и соответственно дифференциа-
ции крестцового отдела, послегрудной отдел сирен напоминает таковой
китообразных, хотя о их соответствии говорить не приходится. Если у
сирен в этот отдел включено очень мало или, может быть, даже вообще
не включены истинные поясничные позвонки, ушедшие в ходе процесса
торакализации в грудной отдел 5, то у китообразных, наоборот, развива-
ется, как правило, огромный поясничный отдел в какой-то мере за счет
уменьшения числа собственно грудных позвонков (люмбализация). В со-
ставе послегрудного отдела китообразных, таким образом, находятся по-
ясничная его часть, включающая 1) собственно поясничные позвонки,
2) несколько бывших крестцовых позвонков, место которых отмечено вы-
ходом спинномозговых нервов, формирующих «plexus pudendus» (Slijper,
1936) и 3) некоторое число настоящих передних по положению хвосто-
вых (послекрестцовых) позвонков, не имеющих шевронных костей (та-
ких позвонков еще не было у древних китов — археоцетов, нет их и у
современного Hyperoodon, но у косаток, карликового кашалота и полоса-
тиков их уже 4, а у настоящих дельфинов до 6—7), и хвостовая часть,
позвонки которой снабжены шевронами. В результате послегрудной отдел
китообразных, как правило, составляет 70—80% длины позвоночного
столба (некоторые археоцеты, настоящие дельфины, нарваловые, каша-
лотовые, клюворылые, почти все усатые киты). Из этого ряда выделяют-
ся лишь южноамериканские речные дельфины Inia и Pontoporia (59 и
64%), укорочение послегрудного отдела которых могло быть вторичным
(Slijper, 1936, 1946), и особенно карликовый гладкий кит Сарегеа шаг-
ginata (50%), по некоторым параметрам очень сходный с сиренами. Ис-
тинное положение крестцовых позвонков у сирен остается неизвестным:
в литературе нет сведений о положении у них «plexus pudendus», не были
отпрепарированы соответствующие спинномозговые нервы и на нашем
материале, а о сомнительности использования связи рудиментарного таза
с позвоночным столбом через связку в качестве критерия выделения
крестцового позвонка уже говорилось в начале главы (это особенно отно-
сится к ламантинам). Первые лишенные шевронов позвонки послегруд-
ного отдела сирен (2 у ламантинов, 3+1 «крестцовый» у древних сирен
Protosiren, Eotheroides, Halitherium, как и у дюгоня и Dusisiren, 6—8 у
Hydrodamalis gigas), таким образом, могут быть как действительно по-
ясничными и крестцовым, так и послекрестцовыми по аналогии с более
5 В связи с этим полезно напомнить, что у млекопитающих (см. Flower, 1885) обыч-
но в туловищном отделе позвоночного столба (грудь + поясница) бывает около
19—21 позвонков (сумчатые, хищные, парнокопытные, мозоленогие, грызуны, боль-
шинство насекомоядных) и лишь у немногих их число увеличено (даманы — 28—
30; слоны — 23—24; непарнокопытные — 22—25; и особенно — до 36 — китообраз-
ные). Уменьшение этой обычной «нормы», видимо, должно рассматриваться как
вторичное явление (у южноамериканских речных дельфинов их 16—18). Весь этот
лимит израсходован более древними сиренами и современными дюгонидами на
один грудной отдел.
139
прогрессивными формами китообразных. Скорее, всего, именно последнее
относится к случаю с Hydrodamalis gigas, где произошла люмбализация
первых хвостовых позвонков, вызвавшая увеличение у этого рода «пояс-
ничных» позвонков сравнительно с непосредственными его предками, по-
добными Dusisiren. Все это влечет за собой заключение о несопоставимо-
сти по составу послегрудного отдела сирен, с одной стороны, и китооб-
разных — с другой.
Относительная величина послегрудного отдела сирен так же сильно
отличается, составляя у ламантина 47—51% длины позвоночного столба,
у дюгоня 55—58%, у Hydrodamalis gigas около 58%. У эоценовых, более
примитивных сирен Protosiren и Eotheroides, близких к предкам как ла-
мантинов, так и всех дюгонид, послегрудной отдел не превышал 54—
56% (расчет по данным Sickenberg, 1934), что, возможно, следует рас-
сматривать как состояние, исходное для более поздних сирен, сохранив-
шееся у халитериин и гидродамалин, но изменившееся в сторону умень-
шения у ламантинов. Это привело к характерному для них почти
полному равенству грудного и послегрудного отделов по относительной
длине. У упомянутых выше речных дельфинов разница в этих отделах
определяется в 27% у Inia и 35% У Pontoporia, т. е. она значительно
больше, чем у всех дюгонид. В то же время у карликового кита после-
грудной отдел имеет сходные с ламантином пропорции, хотя соотноше-
ние поясничной и собственно хвостовой его частей совершенно различно.
У ламантина минимум занимают «поясничные» позвонки: их всего два и
вместе они составляют всего 6—7% длины позвоночного столба. У Саре-
геа поясничная область занимает 15% позвоночного столба и состоит из
4 позвонков, но собственно хвост у этой формы представлен всего 14 поз-
вонками, что составляет 35% длины позвоночного столба. У ламантина
«хвостовых» (с шевронами) позвонков — 22—24 на примерно те же 35%
длины позвоночного столба, что говорит не только о большей короткости
у ламантина этих позвонков, но и главным образом о принципиально
другой механической конструкции их движителя, включающего значи-
тельно большее число центров вращения и, возможно, таким образом
призванного решать иные функциональные задачи, чем у карликового
кита. К сожалению, биология последнего, его экологическая ниша, осо-
бенности движения, питания и т. п. практически неизвестны и сопостав-
ление его с ламантином мало помогает понять их общие и специфиче-
ские черты. Мало изучены и речные дельфины, известные как формы,
обитающие в реках, что требует относительно небольшой подвижности,
малой скорости движения (увеличение ее могло бы даже стать опасным
при ограниченных пределах передвижения) и минимальной способности
к погружению (Cozzi, 1981). В какой-то мере эти черты свойственны и
сиренам, но в морфологических особенностях осевого скелета последних
это никак не реализуется, кроме уже упомянутого уменьшения длины
послегрудного отдела некоторых речных дельфинов, приближающегося
в какой-то степени к уровню, близкому у дюгонид. Однако у этих дель-
финов сокращение отдела, как и у Сарегеа, идет за счет уменьшения
числа хвостовых позвонков: 16 у Inia и 19 у Pontoporia и сохранения
примитивного небольшого числа (5—6 соответственно) поясничных поз-
вонков при типичном для китообразных укороченном грудном отделе
(13 и 10 позвонков). В результате общее число позвонков у них равно
всего 41—42. Это минимальное число, отмеченное у китообразных (лишь
у Сарегеа тоже 42 позвонка), достигнутое ими в ходе своей собственной
специализации, хотя известно, что у более древних или более примитив-
ных форм китообразных позвоночный столб состоял или состоит из мень-
шего числа позвонков, чем у более прогрессивных групп, таких, как на-
140
стоящие дельфины: касатки (50—61), Delphininae (50—69), Pliocaeninae
(60-99) (Slijper, 1936, 1946).
Из вышесказанного вытекает, что сирены по конструкции послегруд-
ного отдела, как и грудного, не имеют морфологического аналога среди
китообразных, как нет среди них и экологических аналогов.
При рассмотрении осевого скелета в целом уже обсуждался ряд па-
раметров, характеризующих послегрудные позвонки ламантина, дюгоня
и стеллеровой морской коровы, в том числе масса позвонков, длина, шири-
на и форма тел, высота тел и невральных дуг, остистых отростков. По-
этому дальше в нашем описании эти стороны морфологии послегрудных
позвонков будут затронуты лишь коротко для полноты морфологической
характеристики.
Типичные передние послегрудные позвонки ламантина (рис, 44, 45)
представлены массивным телом со слегка сердцевидной или почковидной
формой сочленовных поверхностей, ширина которых превышает их дли-
ну, несущим относительно слабо развитую невральную дугу с короткими
остистыми отростками и длинные поперечно-реберные отростки, по на-
личию которых эти позвонки легко отличаются от грудных. Именно силь-
ное развитие этих отростков отвечает за резкое изменение в сторону уве-
личения массы передних pth (см. рис. 21) сравнительно с последними
грудными позвонками. Уже отмечалось, что у дюгоней это происходит, ви-
димо, главным образом за счет включения в состав отростка рудиментов
ребер, у ламантинов же нельзя исключить и разрастания самих попе-
речных отростков, к которым прирастают названные рудименты. Такой
увеличенный поперечный отросток виден на I pth ламантина ЗИН-
31 795 (см. рис. 39, а). Бороздой, отмечающей на правой стороне место
его слияния с ребром, виден подобный отросток и у второго экземпляра
(см. рис. 44). У дюгоня же, судя по последнему грудному позвонку, это
увеличение поперечных отростков выражено в меньшей степени.
Тела первых pth ламантинов продолжают укорачиваться (см. рис. 22),
продолжая процесс, начавшийся еще в задней трети грудного отдела и
идущий до конца хвоста. У дюгоней, наоборот, наиболее длинным в по-
звоночном столбе является IV pth, т. е. здесь продолжается начавшееся
еще в конце шейного — начале грудного отдела постепенное удлинение
тел позвонков и лишь за этим позвонком сменившееся их укорочением.
Примерно так же изменяется длина тел позвонков у морской коровы,
разве что у последних грудных и первых pth длина сохраняется пример-
но на одном уровне. Особенности размерных характеристик передних
pth дюгонид, сказывающиеся в одинаковом изменении по ходу позвоноч-
ного столба длины, высоты (см. рис. 23) и ширины тел, показателя
bh2 (см. рис. 24) и так сильно отличающие их от ламантинов тем не
менее не приводят к сглаживанию одного из основных морфологических
различий между ламантинами с их длинными относительно высоты по-
звонка телами, с одной стороны, и дюгонидами, у которых длина тела
всегда относительно невелика — с другой.
Изменение формы тел послегрудных позвонков уже было описано
(с. 90—91) и нет нужды повторять его здесь. Однако необходимо отме-
тить, что у ламантина относительная ширина тел послегрудных позвон-
ков (в % к высоте тела) в первой трети отдела сохраняется на одном
уровне (между 130 и 140%, что значительно меньше абсолютного мак-
симума в передней части грудного — 170% и шейного — 184% отделов,
но больше, чем в задней части грудного), за XI pth начинается быстрый
ее рост до максимума у XX pth (185%). В последних послегрудных
позвонках относительная высота опять уменьшается до 140%. Иная кар-
тина у дюгонид: за максимумом в передней части отдела (у II pth дю-
141
2
7
Рис. 44. Первые два послегрудных позвонка ламантина
1 — Corpus vertebrae;
2 — facies articularis cranialis;
3 — insertio ligament! flavi;
4 — pr. articularis caudalis
(= postzygapophysis);
5 — pr. articularis cranialis
(= praezygapopliysis);
в — pr. costotransversarius
( — pleurapopliysis);
поперечно-реберный
отросток;
(экз. ИЗ-5945) сверху
7 — pr.mamillaris (=metapo-
physis);
8 — pr. spinosus;
9 —[synapophysis;
10 — рудимент ребра, прирос-
ший к синапофизу
б
Рис. 45. Первый (а) и второй (б) послегрудные (поясничные) позвонки ламантина,
(экз. ИЗ-5945) спереди
Обозначения, как на рис. 40
142
гоня—172% и IV pth морской коровы 200%) начинается постепенное
уменьшение высоты к 105% на XVI pth дюгоня и 110% на XXX pth
морской коровы.
Поперечно-реберные отростки сирен очень специфичны, и характер
их развития сильно отличается от наблюдающегося у китообразных. Само
по себе удлинение этих отростков не представляет что-либо особенное
для млекопитающих, в том числе и наземных. Происходит это обычно в
поясничной области и связано с необходимостью дать большую опору и
место для прикрепления ряду важных эпаксиальных и гипаксиальных
мышц, отвечающих за подвижность туловища в вертикальной и меньше
в горизонтальной плоскостях, особенно за движения задней половины
туловища и хвоста относительно передней, что так важно для млекопи-
тающих с появлением у них принципиально новой асимметричной локо-
моции. У водных млекопитающих, всецело порвавших с сушей,— сирен и
китообразных, эти движения приобрели особое значение как важнейшие,
обеспечивающие создание сил пропульсии. Соответственно у обеих этих
групп поперечно-реберные отростки послегрудных позвонков сильно уве-
личиваются как по длине, так и по ширине. У китообразных это проис-
ходит в первую очередь в поясничном отделе, само удлинение которого в
какой-то степени, видимо, связано с необходимостью умножить число
позвонков с увеличенными поперечными (и остистыми) отростками. Од-
нако, кроме того, этот процесс захватывает у китообразных и заднюю
часть грудного отдела, последние позвонки которого также имеют увели-
ченные поперечные отростки (см. рис. 30, в работе: Яблоков и др.,
1972). Происходит как бы люмбализация этой части позвоночного столба
(кроме увеличения поперечных отростков, здесь теряется связь позвонков
между собой через суставные отростки, последние ребра становятся одно-
головчатыми, соединенными лишь с концами поперечных отростков
и т. п.). Люмбализация, выражаемая в увеличении поперечных отрост-
ков, захватывает и переднюю часть хвостового отдела (5—10 позвонков в
зависимости от его длины и числа в нем позвонков).
У сирен, напротив, люмбализации грудного отдела противостоит тора-
кализация передней части поясничного отдела, ведущая к сокращению
числа позвонков, которые можно было бы, хотя бы условно, отнести к
поясничным. Длина поперечных отростков резко возрастает, начиная с
I pth, причем у дюгоней именно на этом позвонке они имеют наиболь-
шую длину, у морских коров на II—III pth (экз. 3M-3464) или IV pth
(экземпляр, монтированный в Зоологическом музее АН СССР, см. Tab.
VII в работе: Brandt, 1851—1868), у ламантинов на III (экз. ИЗ-5945;
ЗИН-677) или IV (ЗИН-31 795). Каудальнее поперечно-реберные отростки
уменьшаются в длину вплоть до XXII pth у ламантинов, а у дюгонид до
самого последнего хвостового позвонка (см. рис. 25). Причем у дюгоней
отмечается вновь некоторое их увеличение в самом конце хвоста (см.:
Brandt, 1861—1868). То же отмечено у примитивной морской коровы
Dusisiren (Domning, 1978, fig. 6). Несомненно, такое расширение должно
давать опору хвостовому плавнику. Таким образом, люмбализация хво-
стовой части послегрудного отдела позвоночного столба у сирен идет до
самого его конца, а не ограничивается передней частью (половиной или
менее), как у китообразных. Таким образом, в локомоторные движения у
сирен включен лишь укороченный послегрудной отдел, фактически со-
стоящий в основном из хвостовых позвонков, а у китообразных в них
принимает участие значительно большая часть туловища, включая даже
часть его грудного отдела. Такое смещение локомоторных функций лишь
в заднюю половину тела следует признать специфичным для сирен.
По своему генезису поперечно-реберные отростки послегрудных по-
143
звонков сирен представляют продукт слияния синапофизов (диапофиз-1-
парапофиз) с рудиментами ребер, т. е. являются плеврапофизами (на I
pth ламантина экз. ЗИН-31 795 поперечный отросток соответствует одно-
му синапофизу). Следует отметить, что такая же природа поперечно-
реберных (в послегрудной) или поперечных (в грудном отделе) отрост-
ков древних китов — археоцетов, части примитивных зубатых китов из:
сем. Squalodontidae и некоторых форм речных дельфинов (например,
Inia) (Slijper, 1936). У большинства же китообразных эти отростки ведут
свое начало только от диапофизов, путем их соединения с рудиментами
ребер, а у кашалотовых — от парапофизов.
Соответственно своему происхождению за счет слияния диа- и пара-
пофизов поперечный отросток на позвонке переходного типа (см.
рис. 39, а) у ламантина ЗИН-31 795 сидит в месте соединения ножки дуги
и тела позвонка. На II pth или I у экз. ИЗ-5945 (см. рис. 45) он сме-
щается в верхнюю часть тела, а на следующем позвонке на середину его
высоты — положение, сохраняющееся до конца позвоночного столба.
У дюгоня даже на I pth отростки занимают более низкое положение, что
связано с более низким формированием синапофиза на последнем груд-
ном позвонке (см. рис. 39, в) и более низким сравнительно с ламантином
положением диапофизов в грудном отделе.
Направление поперечно-реберных отростков меняется по ходу позво-
ночного столба как в вертикальной (при виде спереди), так и горизон-
тальной (сверху) плоскости, причем особенности этого изменения разнят-
ся у ламантина и дюгоня.
При взгляде спереди поперечно-реберные отростки I pth (ИЗ-5945)
(см. рис. 45, а) или II pth (ЗИН-31 795) ламантинов занимают почти го-
ризонтальное положение. Однако на следующем позвонке их дистальные
концы несколько опускаются вниз на уровень вентральной четверти тела
(рис. 45,6). Наклон достигает максимума на III (IV) pth, т. е. позвон-
ке с максимально длинными отростками. В результате их концы опуска-
ются ниже уровня вентрального края тела почти на полную его высоту.
Важно подчеркнуть, что именно в этом месте (между II и III pth) на-
ступает разрыв связи между смежными позвонками, осуществляемой су-
ставными отростками, и возникает возможность сильного сгибания хвоста.
Таким образом, увеличение поперечно-реберных отростков именно на
III pth и их крайнее отгибание вниз имеет явный функциональный
смысл — обеспечить более эффективное действие мышц-сгибателей хвоста,
способствующее столь характерному для ламантинов вентральному заги-
банию хвостового плавника (рис. 1). Далее назад угол вентрального на-
клона поперечно-реберных отростков начинает постепенно уменьшаться,
что соответствует подъему кривой на графике (см. рис. 42, б). К VII—
VIII pth отростки принимают опять горизонтальное положение и далее
назад продолжают несколько подниматься вверх, что продолжается до
самого конца хвоста. Поскольку для дюгонид такое сгибание хвоста не
характерно, неудивительно, что их поперечно-реберные отростки в перед-
ней части послегрудного отдела не отклоняются так сильно вниз. К тому
же наибольший наклон, например у дюгоня, наблюдается на позвонке
vV pth), поперечно-реберные отростки которого уже начали укорачивать-
ся (ср. рис. 25 и 42, б). У морской коровы поперечно-реберные отростки
первых четырех послегрудных позвонков слегка отогнуты вверх, макси-
мально на IV pth — их концы находятся на уровне верхнего края тела
позвонка. Каудальнее они сначала принимают горизонтальное положение,
а к IX—X pth отгибаются слабо вниз (на 1—1,5 см ниже тела позвонка).
Таким образом, у дюгонид почти на всем протяжении грудного и после-
грудного отделов диапофизы и поперечно-реберные отростки занимают
144
большей частью положение, близкое к горизонтальному, тогда как у ла-
мантинов они то получают очень сильный наклон вверх (в середине
th), то вниз (в начале pth), то опять несколько вверх (ближе к концу
хвоста).
У изученных нами сирен поперечно-реберные отростки I pth при виде
сверху несколько отклонены назад (см. рис. 44). Следующий позвонок у
ламантинов несет или почти перпендикулярные, или слегка отклоненные
вперед отростки, а, начиная с III pth, последние все более и более откло-
няются назад (см. рис. 42, а), достигая максимума на IX pth. Затем этот
наклон постепенно немного уменьшается. У дюгоня (Brandt, 1861—
1868, Tab. VIII, fig. 4) отростки II и особенно III pth обращены не-
сколько вперед, как бы поддерживая I pth. IV pth имеет почти перпен-
дикулярные отростки, пять последующих позвонков — опять с небольшим
наклоном вперед, за чем следует быстрое прогрессирующее изменение на-
клона в другую сторону — назад, достигающее максимума на XV pth.
В дальнейшем угол этого отклонения постепенно к XXII pth приближа-
ется опять к 90°. Аналогичная картина наблюдается, видимо, у Dusisi-
ren (Domning, 1978, fig. 6) и Hydrodamalis gigas (Brandt, 1861—1868,
Tab. VIII, fig. 2), только у изображенного Брандтом экземпляра нет вы-
прямления поперечно-реберных отростков в конце хвоста. Таким образом,
основное отличие ламантина и дюгонид заключается в том, что у послед-
них в передней части послегрудного отдела имеется целый ряд позвонков
(до 9) с поперечно-реберными отростками почти перпендикулярными к
оси позвоночного столба или даже обращенными немного вперед, тогда
как у ламантина таков только один позвонок, у всех остальных в той или
иной степени отростки отогнуты назад. Конечно, это тоже говорит о раз-
личиях в функционировании послегрудных отделов позвоночного столба
ламантинов, с одной стороны, и дюгонид — с другой.
Для ламантинов характерно образование своеобразного органа жест-
кости путем контакта или более тесного соединения концов поперечно-
реберных отростков первых двух, реже трех, послегрудных позвонков с
последним ребром. Детали этой конструкции варьируют у разных экземп-
ляров ламантинов, хотя в топ или иной форме они имеются у всех.
У экз. ИЗ-5945 I и II pth с помощью своих поперечно-реберных от-
ростков образуют упор для ребер последней пары (XVI th). Сами эти
ребра заметно отличаются своей большой тонкостью и округлым сечени-
ем от ребер предыдущей (XV) пары, широких, уплощенных и более-
длинных. Характерно положение XVI ребер: от своего позвоночного кон-
ца они направляются немного вбок, но больше назад (при виде сверху
под 45° к оси позвоночного столба) и так мало вниз, что занимают близ-
кое к горизонтали положение (угол около 20°), если смотреть сбоку.
Благодаря этому задний край этих ребер сближается с концами соответ-
ствующих поперечно-реберных отростков и вступает в контакт с имею-
щимися на них хрящевыми «подушками» (на вываренном скелете на
концах поперечно-реберных отростков видны характерные рубцы, подоб-
ные имеющимся на костных ребрах в месте их соединения с хрящевой
частью ребра). Таким образом, объясняется, почему у ламантинов длина
поперечно-реберного отростка II pth больше, чем у I pth. Форма попе-
речно-реберных отростков, несмотря на наличие определенных вариаций
(можно сравнить правый и левый отростки I и II pth на рис. 44 или
сам рис. 44 с рис. 100), имеет характерную форму: будучи широкими
уже в основании, они еще немного расширяются к середине своей длины,
а затем сужаются в виде клина с более или менее закругленным оконча-
нием или же, наоборот, округлым расширением. У данного образца об-
ращает внимание еще бугор на переднем крае левого поперечно-реберного-
145
отростка II pth. Видимо здесь также был хрящ, в который упирался зад-
ний край предшествующего отростка. Такие непостоянные, варьирующие
индивидуально связи между поперечно-реберными отростками I и II
pth были отмечены и у дюгоня (см.: (Slijper, 1936, Abb. 137)).
У ламантина ЗИН-31 795 эта система упоров усложнена как наличием
вставочного элемента — I pth переходного типа с рудиментарными, но
не сросшимися с синапофизами ребрами, так и созданием своеобразных
суставов в местах контакта с поперечно-реберными отростками II pth.
Эти ребра представляют собою небольшой согнутый под углом в 130°
костный элемент (схематично он показан на рис. 100), на задней поверх-
ности которого как раз на сгибе имеется настоящая суставная поверх-
ность, в которую входит хрящевой выступ на переднем углу конца
поперечно-реберного отростка II pth (см.: (Manzij, Piliptshuk, 1984,
Abb. 14) изображен II, а не I pth). В этих местах развивается даже на-
стоящая суставная капсула. Дистальный более узкий конец этого ребра-
рудимента через продолжающий его хрящ вступает в контакт с концом
поперечно-реберного отростка следующего III pth. Таким образом, в виде
этого ребра, лежащего полностью в горизонтальной плоскости, и поддер-
живающих его поперечно-реберных отростков двух следующих позвонков
формируется прочная основа, в которую упирается последнее длинное
ребро XVII грудного позвонка, аналогичное по строению (округлое в се-
чении) с XVI ребром экз. ИЗ-5945. Еще более длинное предпоследнее
грудное ребро, уже широкое и плоское, в свою очередь опирается на эту
конструкцию, и именно оно образует мощный рычаг, через которой пе-
ребрасываются некоторые брюшные мышцы, эффект которых как сгиба-
телей хвоста увеличивается за счет изменения направления действия сил
(см. гл. 5).
Короткое ребро, еще не сросшееся с I pth, имеется и у третьего эк-
земпляра ламантина — ЗИН-677 (см.: (Brandt, 1861—1868, Tab. VIII,
fig. 3)), но оно имеет контакт с поперечно-реберным отростком только
одного II pth.
Таким образом, несмотря на индивидуальные вариации, у всех ламан-
тинов имеется этот сложноустроенный орган жесткости, включающий
первые 2—3 послегрудных позвонка с их поперечно-реберными отростка-
ми (или рудиментарным свободным ребром на I pth) и последние два
ребра грудного отдела. Его смысл — обеспечивать эффективную работу
мышц, управляющих хвостовым плавником, а дополнительное следст-
вие — утрата подвижности в горизонтальной плоскости в самой передней
части послегрудного отдела. У дюгонид, видимо, также, хотя и в значи-
тельно меньшей степени, имеется связь последнего ребра с поперечно-
реберными отростками I pth (Slijper, 1936, Abb. 137), второй же после-
грудной позвонок не завязан в этот узел, как и предпоследнее грудное
ребро. Важно подчеркнуть, что первым действительно свободным после-
грудным позвонком у ламантина оказывается III (или IV) pth: он не
только участвует в описанной конструкции, но и не имеет связи с пре-
дыдущим позвонком через суставные отростки и у него же самые длин-
ные и самые вентрально отогнутые поперечно-реберные отростки, что
создает наиболее благоприятные условия действующим на него мышцам-
сгибателям. У этого же позвонка появляется такой мощный рычаг для
приложения сил мышц, как шевронная кость. Таким образом, именно
здесь, на контакте II и III (III и IV) pth у ламантина находится важ-
нейший центр двигательной активности послегрудного отдела. У дюгоня
такой точки нет: самые длинные поперечно-реберные отростки имеются
на I pth, наиболее отогнутые вентрально — на V pth, утрата контакта
суставных отростков приходится на границу между III и IV pth у
146
экз. ЗИН-679 и между IV и V у экз. ЗИН-3220. Шевроны имеются с
VI pth. У морской коровы наиболее длинными поперечно-реберными от-
ростками обладает III pth, а разрыв связи через зигапофизы происходит
только между IX и X pth, шевроны появляются с X pth. Это все обус-
ловливает принципиально разную подвижность послегрудного отдела у
ламантинов и дюгонид.
Увеличение поперечно-реберных отростков в послегрудном отделе си-
рен влечет за собой и значительное уменьшение боковых движений по-
звоночного столба в этой области. Практически исключены они, видимо,
у ламантинов, причем если в самом начале это вызывалось уже отме-
ченным органом жесткости, то, начиная с VII pth, этому препятствуют
жесткие лопасти хвостового плавника, имеющего вид округлой или ло-
патообразной пластинки, лежащей в горизонтальной плоскости, что, та-
ким образом, допускает лишь вертикальные движения и, в определенной
степени, вращательные. У дюгонид хвостовой плавник не только имеет
другую форму, но он еще и сдвинут каудальнее по позвоночному столбу
в область, где длина поперечно-реберных отростков уже не является по-
мехой боковым движениям позвоночного столба (у дюгоня стебель хво-
стового плавника приурочен к началу последней трети послегрудного-
отдела, а у ламантина — к концу первой трети).
Максимальная ширина поперечно-реберных отростков у ламантинов
растет от I pth до II—III (от II до IV у экз. ЗИН-31 795), у дюгоня
этот процесс более растянут, достигая максимума у VI pth. Каудальнее
у тех и других ширина отростков постепенно сокращается.
Невральная дуга, включая остистый отросток, I pth составляет око-
ло половины высоты позвонка ламантина (48% У экз. ИЗ-5945 и 51% у
экз. ЗИН-31 795, остистый отросток соответственно — 38 и 37%), что
значительно меньше, чем в области III—IV th (74 и 55% у экз. ИЗ-
5945, 68 и 50% у экз. ЗИН-31 795). Далее назад вместе с постепенным
уменьшением позвонков послегрудного отдела уменьшается абсолютная
высота дуги, а так как ее уменьшение идет опережающим темпом по
сравнению с высотой тела (см. рис. 23), то уменьшается также ее от-
носительная высота. На XV pth (экз. ИЗ-5945) или XVI (ЗИН-31 795)
невральная дуга полностью редуцируется. Остистые отростки исчезают
еще раньше (на XI—XIII pth). В пределах первых 10 послегрудных
позвонков сокращение дуги идет фактически за счет одного остистого от-
ростка. Так, в этих пределах отросток сокращается с 38 до 15% высоты
позвонка, а высота собственно дуги (вернее ножки дуги) сохраняется
близкой к 9—11%. Относительно малая величина остистых отростков ла-
мантина в «поясничной» области послегрудного отдела является важным
показателем слабого развития у него эпаксиальной мускулатуры, управ-
ляющей движениями хвоста.
У дюгоня невральная дуга I pth достигает около 63% высоты позвон-
ка, из них 47% приходится на остистый отросток. Таким образом, здесь,
как и в грудном отделе, дуга значительно превалирует над телом позвон-
ка. У морской коровы дуга этого позвонка несколько ниже (57 и 41%
занимает остистый отросток). При этом у обеих форм в связи с более
низким положением поперечно-реберного отростка на теле позвонка объ-
ем, который может занять эпаксиальная мускулатура, заметно больше,
чем у ламантина. Кроме того, у дюгоня и морской коровы, т. е. у всех
современных представителей дюгонид, сокращение невральных дуг и ос-
тистых отростков по ходу позвоночного столба идет более медленным тем-
пом, что приводит к тому, что невральная дуга у дюгоня редуцируется
только на XX pth (остистый отросток исчезает уже на XVII), а у мор-
ской коровы даже к XXXIV pth (отросток на XXV). Все это говорит о
147
большем развитии эпаксиальной мускулатуры дюгонид по сравнению с
ламантинами.
В то же время у дюгонид относительная величина невральных дуг,
включая остистые отростки, заметно уступает таковой многих китообраз-
ных. Так, у Phocaena phocaena она составляет около 78% (расчет по
графикам, приведенным в работе: (Slijper, 1936, Abb. 219), у Ziphius—
около 72%. На этом фоне еще больше выделяется ламантин, с его особо
короткими невральными дугами и остистыми отростками. Лишь у немно-
гих китообразных невральные дуги и остистые отростки сравнительно не-
велики. Это те же речные дельфины (65 и 39% У Platanista и 64 и
41% у Lipotes) и карликовый гладкий кит.
Характер изменения высоты остистых отростков по ходу позвоночного
столба ламантина, отмеченный уже выше при рассмотрении грудного
отдела, напоминает таковой древних китов — археоцетов, у которых ос-
тистые отростки наиболее высоки в начале или середине грудного отдела,
а затем идет сильное их уменьшение на протяжении задней части груди,
всей поясницы и далее в хвост. У всех других китообразных в противо-
положность этому остистые отростки, как правило, максимальны в пояс-
ничном отделе, обычно в его передней части. У дюгоня величина остистых
отростков достигает максимума к середине — началу задней половины
грудного отдела и сохраняется более или менее неизменной по IV pth
и лишь затем начинается постепенное их уменьшение. Ни у кого из из-
вестных по литературе современных китообразных это не отмечено:
у всех, кроме археоцетов, в грудном отделе остистые отростки увеличи-
ваются в высоту каудально, но максимум достигается только в пояснич-
ном отделе (может быть, это — указание на торакализацию поясничного
отдела у дюгонид, приводящую к тому, что позвонки с самыми высокими
остистыми отростками, будучи первично поясничными, включены в груд-
ной отдел?).
Таким образом, для сирен характерно сочетание в передних позвонках
послегрудного отдела сильно развитых длинных поперечно-реберных от-
ростков с относительно низкими остистыми отростками, тогда как для
китообразных отмечены совсем иные типы: короткие поперечно-реберные
и длинные остистые отростки (у археоцетов, кашалотовых и клювоголо-
вых), длинные поперечно-реберные и длинные остистые отростки (с низ-
ким положением метапофизов — усатые киты, и с высоким положением
метапофизов — морские свиньи и другие настоящие дельфины) (Slijper,
1936, 1946). Исключение составляет Caperea marginata с низкими ости-
стыми отростками в поясничном отделе (максимум приходится на VII—
XIV th) и относительно длинными поперечно-реберными отростками.
Характер наклона остистых отростков послегрудного отдела у ламан-
тина и его отличие от такового дюгоня был уже описан выше (см.
рис. 43). Ширина остистых отростков в их основании продолжает в после-
грудном отделе также постепенно уменьшаться (после максимума в зад-
ней трети грудного отдела). У дюгоня, наоборот, отростки самые широ-
кие в первых трех послегрудных позвонках (максимум у II pth) и лишь
затем начинают медленно сокращаться к XII pth, далее этот процесс
убыстряется.
Презигапофизы I pth ламантина практически не отличаются от тех,
что имеются на последнем грудном позвонке: это — крепкие расширяю-
щиеся вперед отростки, сидящие на границе пластинки и ножек дуги,
с широкими желобообразными суставными поверхностями, плоскость кото-
рых наклонена под углом около 45° к вертикальной плоскости. На на-
ружном верхнем крае презигапофизов имеется вздутие, соответствующее
метапофизу. Каудальнее по позвоночному столбу презигапофизы быстро
148
•сокращаются как по своей толщине в поперечном сечении, так и ширине
суставных поверхностей. Так, если последние на I pth имели ширину,
равную у экз. ИЗ-5945 16 мм, то на II pth они уже сокращаются до
II мм, а на III pth вообще исчезают, так как в связи с редукцией пост-
зигапофизов на II pth присутствие суставных поверхностей теряет смысл.
С III pth сохранение презигапофизов обусловлено уже лишь прикрепле-
нием соответствующих мышечных сегментов к метапофизам, т. е. факти-
чески презигапофизы начинают играть роль метапофизов. Презигапофи-
зы далее продолжают сокращаться в длине и толщине. К VIII pth они
представляют уже небольшие острые выступы, исчезающие полностью на
IX pth. Параллельно с редукцией презигапофизов идет изменение поло-
жения бугорка метапофиза. На IV pth он еще занимает положение на
наружной поверхности короткого презигапофиза, на V pth он уже пере-
мещается несколько назад, частично на ножку дуги. Далее бугорок по-
степенно спускается по наружной поверхности ножки к телу позвонка
(к VIII pth). При этом он становится все менее и менее заметным. Пол-
ностью исчезает след метапофизарного бугорка на X pth.
Постзигапофизы сильно редуцированы уже на I pth: их суставная
поверхность имеет размеры всего 7 мм в длину и 4 мм в ширину. Одна-
ко сами отростки, хоть и тонкие, еще выступают за уровень заднего края
тела позвонка. На II pth экз. ИЗ-5945 правый постзигапофиз уже практи-
чески отсутствует (имеется лишь небольшой выступ на его месте), а ле-
вый, хотя и достигает уровня края тела позвонка, но уже утерял сустав-
ную поверхность. На III pth постзигапофизы уже полностью отсутству-
ют у обоих экземпляров ламантинов.
У дюгоня постзигапофизы полностью редуцируются на V pth. Однако
контакт зигапофизов, как уже отмечалось, был утерян у экз. ЗИН-679
между III и IV pth, а у экз. ЗИН-3220 между IV и V. У морской коро-
вы постзигапофизов нет, начиная с IX pth.
Шевронные кости начинаются, как уже отмечалось, у ламантина с
III pth, у дюгоня с VI, морской коровы с X. Общее число шевронов у
ламантинов значительно меньше (12—13), чем у дюгонид (17—19), что,
видимо, связано с укорочением послегрудного отдела ламантинов и
уменьшением числа позвонков в нем. Форма шевронов и существующие
в них различия у современных сирен подробно описаны Брандтом
(Brandt, 1861-1868).
РЕБРА И ГРУДНАЯ КЛЕТКА
Грудная клетка сирен очень своеобразна и не имеет других аналогов у
других млекопитающих. Это связано с ее длиной (увеличением числа
грудных позвонков и соответственно ребер), укорочением грудины и со-
кращением числа истинных ребер, увеличением числа флуктуирующих
ребер, величиной, формой, положением ребер и т. п. Изменения в осевом
скелете коррелированы с необычным изменением положения внутренних
органов и вмещающих их полостей. Лишь самая передняя часть грудной
клетки сирен оказывается замкнутой со всех сторон. Здесь помещается
только сердце и лежащая позади него самая передняя часть легких.
Сами легкие вытягиваются в длину до конца грудной клетки в особых
парных карманах, образованных диафрагмой, занимающей вентромеди-
альное по отношению к каждому легкому положению, и телами позвон-
ков и ребрами с дорсолатеральной стороны. Таким образом, грудная
полость не занимает всей полости грудной клетки, а лишь ее незначи-
тельную переднюю и самую дорсальную часть. Брюшная полость, вме-
щающая печень, желудок, кишечник, почки и т. п., наоборот, распростра-
149
няется вперед, практически занимая пространство внутри грудной клетки,
широко открытой за грудиной вентрально, и помещаясь, таким образом,
вентральнее, а не каудальнее грудной полости. Ребра при этом не участ-
вуют в прямом поддерживании внутренних органов, вся тяжесть которых
ложится на брюшную мускулатуру, фасции живота. Все это ведет к от-
казу от реберного дыхания, минимальной подвижности ребер. Грудная
клетка, помимо обычной защитной функции для легких и сердца, прини-
мает на себя ряд новых, дополнительных функций, в частности обеспечи-
вает более эффективное действие мышц, связанных с движением хвоста,
туловища. Особую роль она и составляющие ее структуры играют в гид-
ростатике животного, способствуя такому распределению масс по длинной
оси животного, которое позволяет сохранять им в водной среде постоян-
ное горизонтальное положение тела и устойчивость к крену на бок. Это
создается путем как помещения центра тяжести скелета в центре груд-
ной клетки, так и увеличением длины и массы самих ребер, широко рас-
ставленных в стороны, что распределяет массу на более широкую пло-
щадь, способствуя особой устойчивости. Определенную роль в этом игра-
ет и положение легких (катамаранный эффект).
Исходя из величины, формы, характера сочленений ребер с позвоноч-
ным столбом ламантина (ИЗ-5945), была сделана графическая рекон-
струкция его грудной клетки в двух проекциях (сверху и сбоку), на ко-
торой отмечалось положение в пространстве тел позвонков и реберно-
поперечных суставов, дистальных концов ребер (их удаление от оси
позвоночного столба и смещение каудально относительно собственного’
сегмента), что позволило определить в некотором приближении есте-
ственное положение ребер в пространстве и общие очертания грудной
клетки, ее ширину и высоту. Оказалось, что в ее полость включаются
также первые два послегрудные позвонка (из-за очень косого положе-
ния последних ребер), что ведет к еще большему удлинению грудной
клетки. Максимальная ширина в области X—XIII th составляет около
45—48% ее длины. Вперед грудная клетка постепенно сужается до 30%
на уровне VI th и всего 25% в области передней апертуры (т. е. почти
вдвое). Сужение идет и каудально (примерно до 40%). При такой зна-
чительной ширине высота грудной клетки (в сводах правой и левой
половин) крайне незначительна (всего 22—26 см при максимальной
ширине около 52 см, т. е. относительную высоту можно оценить
в 42—50%). В результате сбоку такая грудная клетка выглядит совсем
плоской (см. рис. 19). Абсолютная высота грудной клетки практически
остается неизменной, заметные отклонения наблюдаются лишь в районе
первого ребра и самого последнего (около 19 см). В какой-то мере это
связано с тем, что дистальные концы большинства ребер находятся на
одном горизонтальном уровне, а ширина и высота грудной клетки обес-
печиваются за счет разной длины и кривизны самих ребер. Надо еще
отметить, что даже при такой незначительной высоте грудной клетки ее
внутренняя полость сильно сокращается из-за сильно вдающихся внутрь
тел позвонков (по центру высота в области XI th не превышает 30%).
Близкое значение (45%) относительной высоты грудной клетки в обла-
сти VI th указано для африканского ламантина (Slijper, 1936). Там же
приведены цифры, характеризующие многих китообразных (от 70
до 110%; лишь у немногих, как у речных дельфинов, белухи, сейвала,,
южного кита, этот показатель опускается до 60), ластоногих (80 у обык-
новенного тюленя), некоторых наземных млекопитающих (110 у красного'
волка, 70 у тигра, 120 у лошади) 6.
6 V всех указанных форм измерялась не максимальная высота грудной клетки,,
а только до тела позвонка.
150
Ребра, как правило, занимают косое положение — сверху от реберно-
позвоночного сустава латерально, вниз и в значительной степени кау-
дально. В результате, например, дистальный конец X ребра оказывается
на вертикальном уровне задней половины тела XII th (на 2 сегмента
каудальнее), конец XIV заходит еще дальше, до уровня центра I pth
(на 3 сегмента). Иначе говоря, последние ребра занимают более близкое
к горизонтальной плоскости положение, чем передние (IV ребро дости-
гает уровня задней половины V th). Еще более вертикальное положение
относительно продольной оси соответствующего позвонка занимают
самые первые ребра, но из-за выгиба здесь позвоночника вверх они все
равно имеют направленное назад положение по отношению к длинной
оси туловища. У дюгоня, возможно, грудная клетка уже, так как ребра
своими дистальными концами больше опущены вентрально. Конструкция
их реберно-позвоночных суставов заметно отличается от имеющейся у
ламантина и позволяет более свободную монтировку скелета. В резуль-
тате у всех выставленных в музеях скелетов (см. рис. 17) грудная
клетка смонтирована относительно высокой. Определенно более высокая
грудная клетка у морских коров, ребра которых мало отклонены назад
(см.: (Brandt, 1861—1868, Tab. VIII, fig. 1)). Правда, это тоже может
быть результатом неправильного монтажа.
Ребра сирен характеризуются большой величиной, массивностью, так
как в отличие от других млекопитающих, в том числе и китообразных,
они полностью состоят из плотной костной ткани, лишенной каких-либо
полостей и каналов и сравнимой со слоновой костью. Лишь у дюгоней,
скорее всего вторично, ребра имеют в своем составе определенное коли-
чество губчатой кости, а сами они тоньше и уже, чем у других совре-
менных и ископаемых сирен. Число ребер, как всегда, соответствует
числу грудных позвонков: у ламантинов 15—17 (редко 18), у дюгоней
18—19, у морской коровы 19 (но могут быть вариации), у ископаемых
дюгонид до 20—21. Из них у ламантинов только 2—3 истинные (одно
из них может быть ложным), у дюгоня — 4, у Hydrodamalis gigas воз-
можно 5. Таким образом, характерная черта всех сирен — большое коли-
чество флюктуирующих ребер при малом числе ребер, связанных с гру-
диной.
Ребра ламантинов наиболее специфичны из всех сирен и наиболее
сложно моделированы. Каждое ребро отличается от других ребер дли-
ной, шириной, толщиной, формой сечения на разных уровнях, кривизной
и искрученностью, положением бугорков и шероховатостей, связанных
с прикреплением мышц и связок, не говоря уже о различиях в развитии
головок и бугорков ребра. Длина ребер, начиная с короткого I, посте-
пенно увеличивается в 2,2 раза при измерении по наружной поверхности
(по кривизне) и в 1,6 раза — по прямой (по хорде), достигая максимума
у IX—XIII. В этой области длина ребер (по хорде) составляет около 35%
длины грудного' отдела (у дюгоня и морской коровы 45%; такое разли-
чие обусловливается, видимо, относительным удлинением отдела за счет
тел позвонков). Последующие ребра опять уменьшаются. Однако послед-
нее длиннее II (по кривизне) или III (по хорде). Все ребра сильно
выгнуты, причем у I выпуклой стороной является морфологически зад-
няя, а у большинства других, более задних по положению,— внешняя.
Индекс кривизны (отношение длины по кривизне к длине хорды) изме-
няется от 1,32 у I ребра до 1,69 у IX. У последнего ребра (XVI) кри-
визна опять меньше: 1,25. Помимо этого, большинство ребер, кроме
самых передних и последнего, скручены вдоль своей длинной оси (про-
тив часовой стрелки, если смотреть с дистального конца на левое ребро).
Сочетание этого скручивания и изгиба дает ребру форму, которую мож-
151
но сравнить с отрезком спирали, накрученной на полость грудной клет-
ки. Именно это способствует возможности удлинять ребра (гидростати-
ческое значение) при малой высоте грудной клетки и занимать им более
косое положение. Последнее обеспечивает, в свою очередь, увеличение
межреберных промежутков, а следовательно, и возможность ребер к рас-
ширению, и возможность увеличения поверхности ребер как основания
для туловищных мышц. В результате в любую плоскость поперечного
сечения туловища ламантина всегда попадают сразу несколько ребер
(Slijper, 1936, Abb. 54с). При этом функционировать эта система может
лишь при практической неподвижности ребер, что и обеспечивают ребер-
но-позвоночные суставы ламантинов. Иначе обстоит дело у дюгонид, где
скрученность ребер мало выражена. Здесь удлинение ребер вызывает
увеличение высоты грудной клетки. Ограничение подвижности, таким
образом, теряет смысл.
Ребра ламантина поражают шириной (см. рис. 19). По оценке Бранд-
та, у морской коровы они уже в 1,2—1,4 раза для средних ребер (расчет
сделан по максимальной ширине и при приведении животного к одному
размеру), а у дюгоня даже в 1,5—1,8 раза. Однако если у морской коро-
вы ребра широки почти на всем протяжении и даже в дистальном конце,
то у ламантина всегда имеется область максимальной ширины, от кото-
рой ребро сужается в оба конца. Характерно, что все флюктуирующие
ребра на вентральном конце сужаются на конус, на самой вершине кото-
рого имеется небольшой рубец для реберного хряща, имеющего у этих
ребер вид небольшой хрящевой нашлепки. Максимальная ширина ребер
ламантина нарастает с I по IX—XIV, где ширина в 2,2 раза больше
толщины (у 1 в 1,7 раза). XV ребро уже начинает сужаться. Самое
узкое XVI ребро, у которого ширина практически равна толщине. Наи-
более широкое место меняет свое положение в ряду ребер, спускаясь от
границы между первой и второй 'h длины I ребра дистальнее к сере-
дине ее (в области IX—X) и далее в дистальную половину. Толщина
ребер меняется незначительно: I — самое тонкое, IV — самое толстое
(в проксимальной трети), последующие постепенно утончаются, дости-
гая у последнего ребра величин, близких к I. Наибольшая толщина
также смещается дистальнее по ряду последовательных ребер. У дюгоня
максимальной ширины достигают I—III ребра, последующие IV—XIV за-
метно уже (примерно в 1,3 раза). Дальше ширина быстро уменьшается
и последние два ребра уже I почти в два раза (ЗИН-679). У морской
коровы самое широкое ребро I, а последующие два — самые узкие.
Остальные ребра опять расширяются до уровня, почти достигающего
ширины I. Ширина IV—X ребер держится более или менее на одном
уровне, а затем начинает постепенно уменьшаться. Таким образом, особо
широкое I ребро — черта, специфичная для дюгонид.
Морфология ребер в целом и каждого в отдельности подробно описа-
ны и сравнены у всех трех современных форм сирен (Brandt, 1861 —
1868) и поэтому многие детали из-за нехватки места мы вынуждены
опускать. Первые 4 ребра, и особенно I, сильно отличаются от осталь-
ных (рис. 46—48). Это частью связано со значительно большим числом
крепящихся здесь мышц (на 1 — 12, на IV—XV —6). На каждом ребре
имеется много бугорков и шероховатостей не всегда ясной природы.
Однако судьбу их можно проследить от ребра к ребру. Особенно заметны
среди них: 1) косо идущая по наружной поверхности ребер шерохова-
тость (на I—IV ребрах маленький бугорок немного дистальнее шерохо-
ватости для реберно-поперечной связки; на последующих ребрах шерохо-
ватость постепенно отодвигается от бугорка ребра примерно на */3 реб-
ра), связанная с местом отхождения начального сухожилия или самой
152
латеральной части начального апоневроза m. longissimus dorsi и прикреп-
ления мышечных зубцов нескольких сегментов m. iliocostalis; 2) более
или менее выраженный бугорок по заднему краю ребер, связанный с на-
чальным сухожилием мышечного сегмента m. iliocostalis (гомолог угла
ребра других млекопитающих), и на I—III ребрах расположенный почти
на одном уровне с только что описанным бугорком, но далее все более
и более спускающийся дистальнее по ребру, так что расстояние между
ним и упомянутыми бугорками (шероховатостями) возрастает (т. е. рас-
Рис. 46. Первое левое ребро ламантина
а—в— экз. ИЗ-5945; г — экз. ЗИН-31795: а — сзади, б — спереди, в, г — проксимальный конец
-сверху
1 —caput costae; 7 — facies articularis tuberculi dorsi, бугорок длинней-
2 — collum costae, шейка costae, суставная поверх- шей мышцы спины;
ребра; ность бугорка ребра; 12 — tuberculum m. longus capi-
3 — corpus costae, тело ребра; 8 — sulcus costae, борозда tis, бугорок длинной мыш-
4 — crista capitis costae, гре- ребра; цы головы;
бень головки ребра; 9 — tuberculum costae, буго- 13 — tuberositas m. scaleni pri-
5—crista costae, гребень ребра; рок ребра;) mae costae, шероховатость
€ — facies articularis capitis io — tuberculi m. intercostalis лестничной мышцы перво-
costae, суставная поверх- interni, бугорки внутрен- го ребра;
ность головки ребра; них межреберных мышц; 14— tuberositas m. serratus ven-
72 — tuberculum m. longissimus tralis, шероховатость вен-
тральной зубчатой мышцы
153
16
7
Рис. 47. Второе левое ребро ламантина (экз. ИЗ-5945)
а — спереди; б — сзади; в — сверху
IS — tuberculum m. iliocostalis, бугорок подвздошно-реберной мышцы; 16— tuberositas m. inter-
costalis extemus, шероховатость наружной межреберной мышцы; 17 — ямка для lig. costotransver-
sarius; остальные обозначения, как на рис. 46
154
О Зсм
I__1__I__।
Рис. 49. Грудина ламантина (экз. ИЗ-5945). Вид снизу
Рис. 48. Десятое левое ребро ла-
мантина (а — экз. ИЗ-5945) и дюго-
ня (6 — экз. ЗИН-3220). Вид сверху
18 — место прикрепления m. iliocostalis;
остальные обозначения, как на
рис. 46 и 47
5 — incisura jugularis, яремная вырезка;
6 — manubrium sterni, рукоятка грудины;
7 — рг. xiphoideus, мечевидный отросток
J — angulus sterni, угол грудины;
2 — corpus sterni, тело грудины;
3 — crista sterni, гребень грудины;
4 — incisura costalis, реберная вырезка;
тет поле, занятое m. iliocostalis); в результате этого бугорок смещается
по XII ребру дистальнее середины его длины; затем он опять несколько
приближается к шероховатости m. longissimus dorsi. Последнее ребро
также выпадает из общего ряда, будучи тонким и узким с почти округ-
лым, а не более или менее вытянутым овальным сечением. Бугорок этого
ребра фактически редуцировался, на его месте сохранился лишь острый
выступ, связанный, видимо, с диапофизом XVI th лишь связкой
(см. рис. 38, б). Практически это ребро стало одноглавым.
Остальные ребра ламантина — двуглавые, т. е. имеют головку и буго-
рок. На I—XIII головка двураздельная, состоящая из двух фасеток,
передней и задней, разделенных гребнем головки ребра. Каждая фасетка
ограничена острым краем, что, возможно, свидетельствует о раздельности
их капсул. Обе фасетки различаются величиной (в ряду ребер передняя
уменьшается, задняя увеличивается), формой; их длинные оси и плоско-
сти не совпадают друг с другом. Таким образом, эти ребра, связанные
с позвоночным столбом сразу в трех точках, практически лишены
подвижности, хотя в реберно-поперечном суставе первых трех ребер
определенное скользящее движение возможно из-за разной величины
фасеток на диапофизе и бугорке. Это, возможно, допускает передне-зад-
нее смещение дистальных концов ребер в пределах 5—6 см (дуга
около 15°). На XIV ребре передняя фасетка исчезает, но ие сливается
155
с задней: ее место отделено от последней рудиментом гребня головки.
Лишь с сужением самих головок на XV-—XVI ребрах одна оставшаяся
фасетка занимает весь позвоночный конец ребра. Таким образом, у ла-
мантина лишь три (4 у ЗИН-31 795) последних ребра имеют однофасе-
точную головку и связаны с позвонком только в двух точках. Как ано-
малию следует расценивать появление на I ребре экз. ЗИН-31 795 доба-
вочного бугорка (рис. 46, г), сочленяющегося с новообразованной фасет-
кой на задней стороне плеврапофиза VI шейного позвонка. Таким образом,
здесь ребро становится настоящим трехглавым. Однако у него редуци-
руется передняя фасетка головки. Следует отметить, что термин «двух-
главые и одноглавые ребра» в литературе часто используется ошибочно.
Так иногда (Slijper, 1936, Tab. 13) называют ребра, имеющие всего
лишь двураздельную или однофасеточную головки. В отличие от ламан-
тинов у дюгоня и морской коровы резко сокращено число ребер с дву-
раздельными головками (до 7), соответственно возросло количество ребер
с однофасеточными головками. Причем кажется, что передняя фасетка
на I—VII ребрах, последовательно уменьшающаяся, все же не исчезает
полностью, а, начиная с VIII ребра, входит в состав единой фасетки.
Последнее ребро дюгоня, как и у ламантина, одноглавое, но похоже, что
бугорок ребра здесь слился с головкой. Вообще, у ламантинов бугорок
ребра, начиная с I, постепенно отодвигается от головки и лишь за
X—XI ребрами начинает опять сближаться с нею. Тем не менее на
XVI ребре между головкой и упомянутым выше выступом сохраняется
расстояние в 35 мм. У дюгоня больше всего отстоит от головки бугорок
IV ребра, а дальше все время происходит их постепенное сближение.
Следует еще отметить разницу в положении бугорка у ламантина и дю-
гоня. У первого он всегда расположен близ заднего края ребра (рис. 48),
а у дюгоня с его более узкими ребрами он всегда лежит по центру
наружной поверхности. В результате у ламантина реберно-поперечный
сустав (например на X th) лежит каудальнее сустава головки ребра
(линия, соединяющая их занимает косое положение), а у дюгоня они
лежат в одной вертикальной плоскости. Все это говорит о глубокой раз-
нице в конструкции грудной клетки ламантина и дюгоня.
Грудина сирен (рис. 49) уже неоднократно описывалась в литература
и нет нужды в повторении.
Глава четвертая
СПЕЦИФИКА СКЕЛЕТА
ПЕРЕДНЕЙ КОНЕЧНОСТИ ЛАМАНТИНА
Скелет конечностей современных и вымерших сирен описывался неодно-
кратно (Brandt, 1861—1868; Krauss, 1858, 1862, 1870; Woodward, 1885;
Abel, 1904; Freund, 1904; Annandale, 1907; Simpson, 1932; Bahrdt, 1933;
Kellogg, 1966; Domning, 1978c; и др.). Тем не менее ряд особенностей:
скелета конечностей до сих пор не получил своего функционального объ-
яснения. Это объясняется как плохой изученностью работы конечностей
у современных сирен, так и общими трудностями в оценке связей между
структурой и функцией скелета. Работа конечностей у ламантина описа-
на в литературе (Hartman, 1979), но о работе ластов дюгоня и особенно
морских коров практически ничего неизвестно (Steller, 1751; Anderson,.
1981В). При этих условиях естественно и несовершенство попыток рас-
крыть пути филогенеза сирен как последовательного адаптивного про-
цесса. Совершенно недостаточны и попытки освещения истории станов-
ления ламантинов как особого эволюционного пути среди остальных
сирен. Для оценки причин и путей преобразования скелета у сирен за-
частую приходится прибегать к морфологической аргументации без спе-
циальных знаний о работе соответствующих структур. Поэтому в этой
главе в основном будут разбираться те особенности скелета конечностей,
которые лучше всего можно было трактовать как связанные с опреде-
ленными функциями. Собственно анатомическое описание опущено и
заменено подробными рисунками.
Scapula (рис. 50—52). Наиболее бросающиеся в глаза различия свя-
заны с величиною и положением ости лопатки и ее бугра, строением
акромиального отростка, бугра лопатки, коракоидного отростка и гленоид-
ной впадины лопатки. У эоценовых Protosiren, Eotheroides, Prototherium
(Abel, 1904, Taf. II; Sickenberg, 1934, PLIII) и олигоценового Halithe-
rium (Lepsius, 1882) лопатка узкая, длинная, саблевидно изогнутая со
скошенным назад коротким позвоночным краем. Ость лопатки простира-
лась почти от позвоночного края к шейке лопатки или даже ближе к
вентральному углу лопатки. Бугор ости лопатки выражен незначитель-
но. Акромиальный отросток развит слабо, выступая всего лишь как под-
нятый над поверхностью лопатки вентральный конец ости. Предостная
и заостная ямки узкие, первая обычно шире. Бугор лопатки едва наме-
чен на краниальном крае лопатки близ края гленоида. Особый коракоид-
ный отросток на нем не выражен. На каудальном крае, несколько
отступя от каудального угла, имеется небольшой выступ, связанный с на-
чалом ш. teres major. Гленоидная впадина овальная, ее длинная ось-
лежит в плоскости лопатки. Такой тип лопатки в какой-то мере близок
к имеющемуся у современных наземных копытных. Поэтому можно
думать, что и у предков сирен движения конечностей были довольно
однообразными и проходили почти исключительно в парасагиттальной
плоскости.
У дюгоня изменения сравнительно незначительны (рис. 51), что, ви-
димо, связано с ограниченной подвижностью ластов. Ость лопатки отсту-
пает от позвоночного края более чем на треть длины лопатки, но бугор
ости лопатки по-прежнему не развит. Более обособляется акромион, вы-
157
Рис. 50. Левая лопатка ламантина (экз. ИЗ-5945)
•а — с латеральной стороны;
суставной впадины
1 — acromion (= pr. hamatusj
акромион;
2 — angulus caudalis, каудаль-
ный угол;
3 — angulus cranialis, крани-
альный угол;
4 — cavitas glenoidalis, сус-
тавная впадина;
5 — collum scapulae, шейка
лопатки:
6 — facies m. triceps brachii
caput longum, поверх-
медиальной стороны;
в — спереди; г — сзади; д — со стороны
ность для крепления длин-
ной головки трехглавой
мышцы;
7 — fossa infraspinata, заост-
ная ямка;
8 — fossa subscapularis, под-
лопаточная ямка;
9 — fossa supraspinata предост-
ная ямка;
10 — incisura collum scapulae,
вырезка шейки лопатки;
11 — linea muscularis, мышеч-
ная линия;
12 — pr. coracoideus, коракоид-
ный отросток;
13 — spina scapulae, лопаточная
ость;
14 — tuber scapulae, бугор ло-
патки;
15 — tuber spinae scapulae,
бугор ости лопатки;
16 — место начала m. teres
major
б — с
ступая уже как отросток, направленный наружу, слегка вентрально и
несколько более каудально и имеющий расширенную шероховатую вер-
лиину. Разрастается каудальный угол лопатки, в результате чего расши-
ряется заостная часть лопатки, как и вся она. Это связано с увеличе-
нием, видимо, большой круглой мышцы, выступ для которой смещается
вентрально по каудальному краю на треть его длины, что увеличивает
158
площадку для отхождения этой мышцы. Все эти изменения намечаются
уже у поздних халитериин (Metaxytherium, Thalattosiren), но в меньшей
степени. М. teres major дюгоня, судя по окончанию, может лишь сгибать
плечо. Очевидно, основная осуществляемая в плечевом суставе дюгоня
функция — сгибание и разгибание. Специфичным для лопатки дюгоня
среди других дюгонид является изменение положения длинной оси гле-
ноидной впадины передним концом внутрь. Отклонение от плоскости
лопатки составляет около 30°. Сама впадина становится более вытянутой,
сужающейся вперед. Это изменение связано, по сути дела, со скручива-
нием всей гленоидной части лопатки в ее шейке против часовой стрел-
ки (при виде с вентрального угла левой лопатки). Бугор лопатки увели-
159
Рис. 51. Левая лопатка
дюгоня (экз. ЗИН-3220)
а — с латеральной стороны
б — спереди;
в — со стороны суставной
впадины
Обозначения, как на рис. 50
160
Рис. 52. Левая лопатка морской коровы Hydrodamalis gigas (экз. ЗИН-17170/1)
а — вид с латеральной стороны; б — спереди; в — сзади; г — со стороны суставной впадины
Обозначения, как на рис. 50
6 Ламантин
161
чивается в размерах, становится более протяженным по краниальному
краю лопатки и продолжается в длинный немного сужающийся кора-
коидный отросток, тупо кончающийся поверхностью, от которой начи-
нается m. coraco-brachialis. Коракоидный отросток отогнут сильно внутрь,
под углом в 70° к плоскости лопатки. Слаборазвитый коракоидный от-
росток, отогнутый слегка внутрь, наблюдался еще у олигоценового
Halitherium и миоценового Metaxytherium krahuletzi. Так что в этом
дюгонь лишь довел до крайности тенденцию, существовавшую уже у
поздних халитериин. Но у них гленоидная впадина не меняет еще своей
ориентации, а именно это изменение у дюгоня в совокупности с удлине-
нием коракоидного отростка привело к такому сильному загибанию по-
следнего внутрь.
В линии развития морских коров их ранний представитель Dusisiren
еще имел черты, во многом сходные с поздними халитериинами: ость
лопатки отодвинута от позвоночного края и вентрально доходит до шей-
ки лопатки, площадка для большой круглой мышцы увеличена по срав-
нению с эоценовыми формами, бугор лопатки близок к краю гленоида,.
представляя простое вздутие на краниальном крае лопатки без особого
выделения коракоидного отростка, длинная ось гленоида лежит в плоско-
сти лопатки. У Hydrodamalis лопатка заметно видоизменяется
(см. рис. 52). Резко укорачивается ость лопатки, отодвигаясь от ее вент-
рального угла на А/4 длины. Соответственно слабо выраженный, как у
халитериин, акромион тоже сдвигается дорсальнее (сдвигается место от-
хождения дельтовидной мышцы, уменьшается ее значение как супинато-
ра плеча). Наружная поверхность ости расширяется, превращая практи-
чески всю ость лопатки в гигантский бугор ости, прямо переходящий в
акромион (возможно, увеличился m. spinotrapezius, а т. acromiotrapezius
сократился). У Dusisiren бугор ости, отсутствующий у халитериин и
дюгоня, лишь начинает выделяться как небольшое шероховатое утолще-
ние кости. Площадка для m. teres major у Hydrodamalis продолжает уве-
личиваться, а выступ для этой мышцы, развитый, правда, слабее, чем
у предков, сдвигается по каудальному краю лопатки вентральнее, почти
к его середине. Интересно, что у морской коровы внутренняя поверх-
ность лопатки не образует желоба, как у большинства сирен, а стано-
вится плоской или даже выпуклой. Возможно, это вызвано ослаблением
m. subscapularis, главного пронатора плеча. Таким образом, у морской
коровы ротационные движения сведены к минимуму, а флексорные под-
черкнуты усиленным развитием большой круглой мышцы и превраще-
нием дельтовидной мышцы, по существу, в сгибатель плеча в связи со
смещением точек его прикрепления. Бугор лопатки у морской коровы
также увеличен в размерах, но коракоидный отросток на нем не выра-
жен. Длинная ось гленоидной впадины изменяет свое направление, буду-
чи теперь повернута передним концом наружу примерно на 15° к плос-
кости лопатки. Таким образом, ось смещена в противоположном направ-
лении по сравнению с дюгонем. В результате при разгибании в плечевом
суставе ласт морской коровы отводится несколько в сторону. То же
самое делает ласт у дюгоня, но, учитывая иное направление гленоида,
это достигается лишь сильной модификацией головки плечевой кости.
У ламантина (см. рис. 50) направление изменений лопатки совершен-
но иное. От исходного состояния у них сохраняется лишь ориентация
гленоидной впадины, близость вентрального края ости лопатки к вент-
ральному углу и, может быть, отодвигание ее от позвоночного края.
На ости сформировался мощный бугор ости, расширенный и нависаю-
щий над заостной ямкой. По форме он не имеет ничего общего с бугром
ости морской коровы, хотя причины, вызвавшие его к жизни, могли
162
быть общие — усиление задней части трапециевидной мышцы, но в от-
личие от Hydrodamalis у ламантина увеличивается и передняя ее часть,
так как ость продолжается вентрально и вперед в огромный акромиаль-
ный отросток, выходящий за передний край гленоида и выносящий его
конец далеко латерально и вперед по отношению к плечевому суставу.
Это обеспечивает большую площадь отхождения дельтовидной мышце и
усиление ее супинаторной, абдукторной и протракторной (экстензорной)
функции, что прямо противоположно общей тенденции, наблюдаемой у
всех дюгонид. Сильно развитый акромиальный отросток характерен для
млекопитающих, обладающих большой и разнообразной подвижностью в
плечевом суставе. В то же время усиление обеих частей трапециевид-
ной мышцы ламантина может означать увеличение подвижности плече-
вого пояса относительно туловища. Лопатка ламантина сильно расши-
ряется (увеличивается позвоночный край, предостная и заостная ямки),
сложной становится ее конфигурация и искривленность самой лопасти,
что, видимо, в первую очередь связано с необычно мощным m. subsca-
pularis, из-за модификации малого бугорка плечевой кости превративше-
гося в очень сильный пронатор. Опять это прямо противоположно на-
блюдаемому у дюгонид и развивается только в линии развития сирен,
ведущей к ламантинам с их усиленной манипуляционной и пропульсив-
ной деятельностью ластов (см. гл. 5). Особенностью лопатки ламантина
является также особое развитие бугра лопатки и его отодвигание от края
гленоида на уровень шейки лопатки и сильное выступание вперед.
Однако, как и у эоценовых сирен, коракоидный отросток как самостоя-
тельное образование отсутствует. Близ гленоида по каудальному краю
лопатки сформирована обширная треугольная площадка, связанная с на-
чалом очень мощной длинной головки m. triceps. У дюгоня и халитериин
площадки нет, но она появляется у морской коровы, возможно, также
в связи с усилением этой мышцы, играющей роль не только разгибателя
локтевого сустава, но в какой-то, видимо, важной и для ламантинов и
для Hydrodamalis мере сгибателя плечевого сустава. Гленоидная впадина
у ламантинов широкоовальная (отношение короткой оси к длинной 0,8).
Кривизна в передне-заднем направлении имеет радиус в 30 мм
(по экз. ЗИН-31795), в медиолатеральном — 35 мм. У дюгоня гленоид-
ная впадина более вытянута (индекс 0,7), по форме субтреугольна.
Humerus (рис. 53—57). Плечевая кость ламантинов подверглась дале-
коидущим преобразованиям, связанным с перекомпановкой большого и
малого бугорков и их гребней, что делает ее уникальной среди всех мле-
копитающих, а не только сирен. В то же время при сохранении общего
плана строения, свойственного млекопитающим, плечевая кость дюгоня
также сильно видоизменилась, но в направлении совершенно ином, чем
у ламантинов. Humerus морских коров, как и дюгоня, легко выводится
из общего предкового типа. Основные различия плечевой кости ламан-
тинов и дюгонид кроются в ориентации и степени обособления головки
плеча, степени развития и взаимоположения бугорков плеча, а следова-
тельно, и в положении мест прикрепления ряда мышц, действующих на
плечевой сустав, в строении стержня плеча, отражающем изменения,
происшедшие в проксимальном эпифизе, положении и развитии медиаль-
ного надмыщелка, взаимной ориентации осей движения в плечевом и
локтевом суставах. Все эти изменения имеют яркий адаптивный характер
и связаны с различиями в использовании ластов тремя современными
формами сирен. У наземных млекопитающих большой бугорок лежит в
парасагиттальной плоскости латеральнее (и спереди) головки, а малый —
медиальнее. Обычно длинная ось головки ориентирована спереди назад,
т. е. тоже в парасагиттальной плоскости, в которой лежит и стержень
163
6*
Рис. 53. Левая плечевая кость ламантина (экз. ИЗ-5945)
Вид с латеральной (а), дорсальной (б), медиальной (в) и вентральной (г) сторон и с проксимального
(б) и дистального (е) концов
1 — caput humeri, головка пле-
чевой кости;
2 — collum humeri, шейка пле-
чевой кости;
3 — condylus humeri, мыще-
лок плечевой кости;
4 — corpus humeri, тело плеча
5 — crista tuberculi majoris;'
гребень большого бугорка.
6 — crista tuberculi minoris,
гребень малого бугорка;
7 — epicondylus lateralis, ла-
теральный надмыщелок;
8 — epicondylus medialis, ме-
диальный надмыщелок;
9 —fossa coronoidea, венеч-
ная ямка;
10 — fossa olecrani, локтевая
ямка;
11 — labium laterale trochleae,
латеральная губа блока
плечевой кости;
12 — labium mediate trochleae,
медиальная губа блока
плечевой кости;
13 — sulcus intertubercularis,
межбугорковый желоб;
14 — sulcus spiralis (= s. m.
brachialis), спиральный
желоб;
15 — trochlea humeri, блок пле-
чевой кости;
16 — tuberculum ma jus, боль-
шой бугорок;
17 — tuberculum minus, малый
бугорок;
18 — tuberositas deltoidea, дель-
товидная шероховатость;
19 — tuberositas m. clavodeltoi-
deus, шероховатость для
прикрепления m, clavo-
deltoideus;
20 — tuberositas m. pectoralis
superficialis, поверхность
для прикрепления
m. pect. superficialis;
21 — tuberositas m. subscapula-
ris, шероховатость для
прикрепления подлопаточ-
ной мышцы;
22 — tuberositas mm. teres et
latissimus dorsi;
23 — рудимент sulcus intertu-
bercularis
23
165
166
Рис. 54. Левая плечевая кость дюгоня
Фото с экз. 1981—153 Национального музея естественной истории в Париже (по любезности
J. Р. Gasc) Вид с латеральной (а), дорсальной (б), медиальной (в), вентральной (г) сторон и с прокси-
мального (9) и дистального (е) концов. Обозначения, как на рис. 53
167
IB
Рис. 55. Левая плечевая кость морской коровы (экз. ЗИН-14576)
Вид с латеральной (а), дорсальной (б), медиальной (в), вентральной (г) сторон и с проксимального
(Э) и дистального (е) концов. Обозначения, как на рис. 53
плеча. Именно в этой плоскости производятся основные сгибательно-раз-
гибательные движения. Отведение же, приведение и ротация, если не
учитывать лазающие формы, в плечевом суставе второстепенны. Между
большим и малым бугорком у наземных млекопитающих расположен
межбугорковый желоб, в котором скользит сухожилие двуглавой мышцы
плеча. При этом сильно выступающая проксимально за головку плеча кра-
ниальная часть большого бугорка поворачивает медиально, нависая над
желобом (оси обоих бугорков как бы конвергируют медиально, стремясь
сомкнуться над желобом). Соответственно желоб у наземных млекопи-
тающих узкий. От бугорков дистальнее по стержню humerus могут идти
более или менее развитые их гребни. Их протяжение и выраженность
определяют поперечное сечение на разном уровне длины стержня.
В целом оно имеет форму слегка вытянутого овала или неправильного
с округлыми углами четырехугольника с суженным и более или менее
168
приостренным, направленным вперед концом, отмечающим положение
гребня большого бугорка (рис. 57, е). Примерно такое же строение
проксимального эпифиза наблюдается у эоценовых Protosiren и Eotheroi-
des, у которых оси обоих бугорков (при виде на эпифиз с проксималь-
ного конца) сходятся к головке плеча под углом около 35—40° (Sicken-
169
Рис. 56. Головка правой пле-
чевой кости ламантина (экз.
ЗИН-31785)
а — с проксимального конца;
б — с латеральной стороны;
в — сзади;
г — с медиальной стороны (по пре-
парату, фиксированному в фор-
малине):
1 — caput humeri;
2 — tuberculum majus;
з — tuberculum minor
Рис. 57. Блок плечевой кости
ламантина (а—в), дюгоня (г),
морской коровы (д) и лося
(е)
а—б — внешний вид поверхности
(правая кость). По препарату, фик-
сированному в формалине (экз.
ЗЙН-81795):
а — с дистального конца,
б — спереди;
в—е — взаиморасположение длин-
ной оси блока левой плечевой
кости (1), сечения диафиза (2)
и длинной оси головки (3) пле-
чевой кости
berg, 1934; Domning et al., 1982), что говорит об узости межбугоркового
желоба. У дюгонид резко разрастаются большой и малый (каудальная
часть) бугорки, расширяется и углубляется межбугорковая вырезка.
Угол, под которым сходятся оси бугорков, постепенно возрастает от более
ранних к более поздним халитериинам (до 80° у Thalattosiren, поздне-
миоценовых Metaxytherium). Наибольший угол —у дюгоня (до 95°).
Этот процесс уже давно отмечен в литературе (Abel, 1904; Sickenberg,
1934). Однако функциональные последствия такого преобразования не
привлекали должного внимания. Прогрессивное увеличение большого бу-
горка вело к удалению точек прикрепления предостной и заостной мышц
от центра вращения в плечевом суставе. Разрастание краниальной части
большого бугра у халитериин (это проявляется даже в онтогенезе дюго-
ня), кроме того, смещает прикрепление m. supraspinatus медиальнее,
ближе к парасагиттальной плоскости. Это дает увеличение плеча рычага
и уменьшает отводящий и вращающий эффект мышцы. Увеличение боль-
шого бугорка сопровождается сперва усилением, а затем и гигантским
170
ростом у дюгоня гребня большого бугорка, что сильно увеличивает плечо
действия дельтовидной мышцы. В результате стержень плеча приобре-
тает субтреугольное сечение, причем передний угол сильно вытянут и
заострен (рис. 57, г). Таким образом, стержень сильно сужается в медио-
латеральном направлении сравнительно с переднезадним. Гребень у дюго-
ня почти достигает дистальной четверти плечевой кости и выходит к ме-
диальным краям лучевой ямки и блока плечевой кости, т. е. идет косо
по передней поверхности с латеральной стороны на медиальную. По нему
у дюгоня дистально смещаются прикрепления m. brachiocephalicus и
m. pectoralis superficialis (Domning, 1977b), что усиливает их действие
как протрактора у первого, как ретрактора (сгибателя) и пронатора у вто-
рого. Увеличение гребня большого бугра и переднезаднего диаметра
стержня ведет к появлению у дюгоня огромного спирального желоба,
вместилища m. brachialis. В то же время уменьшена переднемедиальная
поверхность плечевой кости, связанная с отхождением медиальной голов-
ки трехглавой мышцы и зажатая между гребнем малого бугорка с ме-
диальной стороны, дистально переходящим в гребень медиального над-
мыщелка, и гладким перегибом стержня плеча с латеральной стороны,
продолжающим проксимально неясно выраженный гребень латерально-
го надмыщелка и подходящим к середине головки плечевой кости.
Третья более плоская сторона треугольного в сечении стержня плеча
продолжает дистально межбугорковый желоб. В целом, таким образом,
дюгоню свойственно, можно сказать, утрирование черт, наблюдающихся
у наземных млекопитающих. Только в связи с увеличением медиального
надмыщелка и его разрастанием в сторону от блока, гребень малого
бугорка отодвигается от гребня большого бугорка медиальнее. Изменяет-
ся и положение длинной оси головки плечевой кости, которая теперь при-
нимает косое положение по отношению к оси стержня плеча (произошло
вращение по часовой стрелке, если смотреть на головку левого humerus
на угол около 35° из вертикального положения).
Проксимальный эпифиз humerus морской коровы построен на тех же
принципах, что и у дюгоня. Межбугорковый желоб большой, широкий.
Оси обоих бугорков сходятся под углом около 85—90°, хотя это нараста-
ние очевидно шло параллельно с дюгонем, так как у непосредственного
предка морской коровы Dusisiren угол был меньше (75—80°), т. е. соот-
ветствовал уровню, достигнутому поздними халитериинами. Плечевая
кость морской коровы имеет более толстый стержень, плоскости которо-
го, хотя и соответствуют таковым дюгоня (сечение также треугольное),
несколько переориентированы (рис. 57, д): наибольшая ширина стержня
здесь расположена медиолатерально. В связи с развитием большого лате-
рального надмыщелка и изменением хода его гребня в сторону головки
плеча (соответствующий перегиб стержня теперь идет не косо по задней
поверхности humerus, а латеральнее к месту стыка головки и каудальной
части большого бугорка) сокращается ширина и общий объем спираль-
ного желоба. Большой бугорок, как у дюгоня, сильно развит. Однако он
и его гребень утолщен и расширен медиолатерально. На гребне большого
бугра нет такого гребня для m. deltoideus, как у дюгоня; видимо, дельто-
видная шероховатость смещена дистальнее на уровень середины длины
плеча. Малый бугорок развит значительно сильнее, чем у дюгоня. Таким
образом, крепление подлопаточной мышцы дальше отодвинуто от центра
вращения в плечевом суставе. Головка субсферическая, ее длинная ось,
лишь немного превышающая короткую, ориентирована почти так же, как
у наземных млекопитающих. Однако возможно, что это достигнуто вто-
рично, поскольку у возможных предков морских коров среди халитериин
ось головки была несколько ближе к наблюдаемому у дюгоня, в связи
171
с приобретением подчеркнуто парасагиттального движения конечностей,
необходимого для свойственного им образа жизни.
У ламантинов принципиально иное строение проксимального эпифиза.
Головка также почти сферическая (см. рис. 56), но намечаемая длинная
ось почти перпендикулярна к оси плечевой кости (вращение против
часовой стрелки на 55° от вертикали). Качественно новое — обособление
хорошо выраженной суженной шейки, что придает плечевой кости облик,
напоминающий femur. Главное изменение связано с бугорками. Очень
сильно развитый и далеко отстоящий от головки малый бугорок пол-
ностью слился с краниальной частью большого бугорка. Каудальная
часть последнего, наоборот, развита очень слабо и несколько обособлена
от краниальной части. В результате слияния бугорков межбугорковый
желоб как самостоятельное образование исчез полностью, хотя место
прохождения начального сухожилия двуглавой мышцы угадывается по
небольшой выемке между шероховатостями для прикрепления подлопа-
точной и глубокой грудной мышц. Степень развития этой выемки
(она больше развита у Trichechus inunguis (Domning, 1982) не меняет
существа дела. Сухожилие бицепса у ламантинов перебрасывается через
блок, отнесенный далеко вперед от центра вращения плечевого сустава,
и меняет свое направление на 90° (см. рис. 106, в). Сдвиг малого бугра
и места прикрепления подлопаточной мышцы во много раз усиливает
пронаторную аддукторную функцию последней. Предостная и заостная
мышцы более эффективно теперь действуют как протракторы и супина-
торы плеча, тогда как у дюгоня первая скорее работала как абдуктор,
а вторая — как супинатор и слабый ретрактор. Соответственно с измене-
нием положения и степени развития бугорков меняются и их гребни.
Практически гребень большого бугра не развит, имеется лишь неболь-
шой перегиб на поверхности стержня humerus, занимающий обычное по-
ложение, связывая каудальную часть большого бугорка с дистальным
эпифизом в области медиального края блока. Во второй четверти по ходу
этого перегиба имеется отчетливая шероховатость дельтовидной мышцы.
Гребень малого бугорка в своем проксимальном конце также смещен
вперед и идет дистально в направлении того же блока, не доходя до ко-
торого фактически соединяется с перегибом, соответствующим гребню
большого бугорка. В свою очередь, гребень латерального надмыщелка
идет дистально, подходя к головке значительно медиальнее, чем у дюго-
ня. Все это приводит к изменению профиля диапофиза плечевой кости
(рис. 57, в), в которое становится округло-четырехугольным. При этом
угол, связанный с гребнем большого бугорка, практически отсутствует и
занимает более латеральное положение, чем у дюгоня и морской коровы:
угол, соответствующий гребню малого бугорка, сместился на переднеме-
диальный край диафиза. Спиральный желоб, как таковой, перестал
быть желобом, каким он наблюдался у всех других сирен, и m. brachia-
lis покоится теперь на сравнительно ограниченной выпуклой поверхности
стержня плечевой кости.
Дистальный эпифиз сирен представлен блоком плечевой кости, сильно
развитым медиальным и слабым латеральным надмыщелками, относи-
тельно неглубокой локтевой и едва намеченной лучевой ямками.
Медиальный надмыщелок ламантина очень большой, выступающий ди-
стально за уровень медиального края блока. Он сильно отогнут назад
под углом 60°, ограничивая локтевую ямку медиально. Для наземных
млекопитающих, таких, как копытные, характерен еще больший (за 90°)
загиб надмыщелка назад, почти прижимающегося к локтевому отростку
ulna. Близкое к ламантину положение надмыщелка наблюдается у эоце-
новых Protosiren и Eotheroides (Domning et al., 1982; Sickenberg, 1934),
172
т. е. это состояние следует рассматривать как исходное для сирен. Уже
у олигоценовых халитериин надмыщелок сокращается в размерах (у всех
дюгонид он не достигает края блока), уменьшается отклонение его назад
(у Halitherium 40°). Такое положение сохраняется и у гидродамалин
(у морской коровы 35°), но у дюгонин идет дальнейшее приближение
надмыщелка к осп блока (у дюгоня 20°). У ламантина на месте гребня
медиального надмыщелка имеется толстое округлое по краю как бы его
продолжение, идущее в направлении головки плечевой кости, но доходя-
щее только до середины диафиза, оставляя медиальнее глубокие шерохо-
ватые ямы для прикрепления m. pectoralis superficialis, latissimus dorsi
et teres major. У дюгоня имеется слабо выраженный гребень медиаль-
ного надмыщелка, переходящий, как уже отмечалось, в гребень малого
бугорка. Латеральный надмыщелок у всех сирен развит слабо и дисталь-
но далеко не доходит до блока. Он связан с отхождением группы экстен-
зоров запястья и пальцев. У ламантина на нем более четко обособляются
два выступа, один на передней стороне дает место отхождения ш. Ьга-
chioradialis, второй на задней — для начального сухожилия локтевого
разгибателя запястья. Блок плечевой кости занимает по отношению к ее
диафизу более или менее косое положение, близкое к перпендикулярно-
му у дюгоня и морской коровы (80—85°), что напоминает состояние,
свойственное копытным (так у косули 85°), но значительно более на-
клонное у ламантина (около 65°). В результате плоскость, в которой
двигается предплечье, пересекается с продольной осью плеча под значи-
тельным углом, что ведет при разгибании к отведению ласта от сагит-
тальной плоскости. Суставная поверхность блока гладкая, без синовиаль-
ных ямок, канавок, разделенная слегка винтообразноидущим желобом на
две неравные части, большую латеральную и меньшую медиальную
(рис. 57, а, б). Эта винтообразность имеет следствием появление враща-
тельного момента при сгибательно-разгибательных движениях в лок-
тевом суставе (при сгибе пронация). Протяженность поверхности у
экз. ЗИН-31 795 по желобу 70 мм, в поперечном направлении по средней
части латерального мыщелка 20—22 мм, радиус кривизны последнего
16—18 мм. Толщина слоя суставного хряща, измеренная в семи точках
по всей поверхности, составляет 2,9±0,1 мм. Латеральный мыщелок
блока у дюгоня также больше медиального, но разница между ними зна-
чительно больше. Минимальная она у морской коровы. Превалирующее
развитие одного из мыщелков наблюдается и у наземных млекопитаю-
щих, особенно у тех, чьи конечности становятся исключительно локомо-
торными, например у копытных, слонов. У первых более развит медиаль-
ный мыщелок в связи с тем, что давление через плечо передается преиму-
щественно на лучевую кость, у слонов — латеральный (нагрузка на
локтевую кость). Величина суставных поверхностей обычно адекватна
действующим силам (Клыков, 1978). Следует, следовательно, заключить,
что у ламантина большая сила приходится на латеральный мыщелок и
что у дюгоня эта сила еще больше. В то же время возможно, что исход-
ным для сирен было состояние, когда медиальный мыщелок был больше
латерального. Видимо, таким он был у Protosiren (Domning et al., 1982,
fig. 31). Состояние, наблюдаемое у морской коровы, однако, может быть,
является таким вторично в связи с более парасагиттальным движением
ластов в условиях большей нагрузки (заякоривание и подтаскивание
тела вперед).
Большие различия, существующие у сирен в развитии и положении
суставных поверхностей головки и блока, их главных осей, прямо связаны
с различиями в использовании ими передних конечностей и требуют
более внимательного рассмотрения. У ламантина головка почти круглая,
173
но, как уже отмечалось, с выделяющейся длинной осью. Поверхность
головки занимает несколько больше полуокружности по длинной оси и
чуть меньше — по короткой. Такая величина обусловлена хорошо разви-
той шейкой. Радиус кривизны около 32 мм. Толщина слоя хряща, изме-
ренная в 10 точках, составляет 4,8±0,2 мм, причем больше она каудола-
терально и меньше краниомедиально. Амплитуда основных движений,
рассчитанная по совпадению крайних точек суставных поверхностей, со-
ставляет для пронации-супинации 70—80°, сгибания-разгибания 80—85°,
приведения-отведения 90—95°. Обращает внимание высокая степень раз-
вития капсулы плечевого сустава, прикрепление которой к плечевой
кости далеко отступает от самой головки (см. рис. 107). Это, безусловно,
обеспечивает большую амплитуду движений по всем направлениям, оче-
видно заметно превышающим приведенные выше цифры. Особенно вели-
ки должны быть круговые (скручивающие) движения. Тем не менее
ведущими движениями в суставе следует считать те, что происходят по
длинной оси головки плечевой кости. У ламантина это фактически приво-
дит к вращению плеча относительно его длинной оси, причем каудаль-
ному положению соответствует максимум пронации, а краниальному —
супинации. При этом плечо, занимающее почти горизонтальное положе-
ние (дистальный конец несколько опущен), двигается по дуге с радиусом,
определяемым расстоянием от центра вращения в суставе по продольной
оси диафиза плечевой кости, т. е. с очень незначительным перемещением
вперед назад. Приведение-отведение определяется движением по корот-
кой оси головки. Собственно сгибание и разгибание в привычном смысле
(как уменьшение или увеличение угла между лопаткой и плечом в пара-
сагиттальной плоскости) возможно только в крайне приведенном состоя-
нии. При нормальном (наиболее нагруженном) положении, когда совпа-
дают геометрические центры суставных поверхностей головки и гленоида,
а длинная ось головки совпадает с таковой гленоида (уже говорилось,
что в этом положении плечо принимает почти горизонтальное положе-
ние), протракторно-ретракторные движения дистального конца плеча
возможны лишь при круговых движениях в плечевом суставе (что озна-
чает изменение положения длинной оси головки относительно длинной
оси гленоида), обеспечиваемых, в частности, значительно большим радиу-
сом кривизны гленоидной впадины по сравнению с головкой. У дюгоня
с его менее обособленной головкой ее поверхность значительно меньше,
она скорее напоминает отсек поверхности эллипсоида, а не сферы, как
у ламантина. Соответственно амплитуды всех движений ограничены и
неравноценны в разных направлениях. Наибольшую амплитуду имеют
движения по длинной оси головки. Их можно назвать сгибательно-разги-
бательными и супинаторно-протракторными, причем краниальному поло-
жению (разгибание), в отличие от ламантина, соответствует пронация,
а не супинация. При нормальном положении плечо дюгоня стоит косо
по отношению к горизонтальной плоскости (примерно под 45°). Движе-
ния, связанные с отведением и приведением, наиболее ограничены, как
и круговые движения, обеспечивающие пронацию и супинацию. В целом
плечевой сустав говорит о значительно меньшем разнообразии движений
ластов дюгоней.
Глубокие различия в работе конечностей ламантинов, дюгоней и мор-
ских коров особенно четко выявляются при сопоставлении положения
длинных (наиболее рабочих) осей головки плеча с положением длинной
оси блока плечевой кости. На схемах (рис. 57, в—е) стрелками показаны
одноименные направления по этим осям (медиально по оси блока и кау-
дально по оси головки). Угол между этими двумя векторами принят за
показатель пространственной взаимосвязи между обеими осями. При этом
174
юбнаружепо, что у ламантина он составляет около 60°, у морской коро-
вы — 85°, у дюгоня — 150°. У наземных млекопитающих он ближе к пря-
мому (у косули 80°). Отсюда вытекает, что от исходного положения,
очевидно близкого к наземным млекопитающим, изменение длинной оси
головки идет у ламантина и дюгоня в прямо противоположных направле-
ниях: против часовой стрелки (если смотреть на головку со стороны ее
сочленовной поверхности) у ламантина и наоборот — у дюгоня. Очень
близко к исходном}7 состояние у морской коровы, но оно может быть вто-
ричным. У ископаемых сирен на положение оси головки внимание не
обращалось, и в литературе нет фотографий или рисунков в необходимой
для его установления проекции. Условно мы можем считать, что общая
тенденция, ведущая к дюгоню, характерна и для халитериин, которые,
однако, возможно, не пошли так далеко, как дюгонины. У Dusisiren опи-
сывается «медиодистальное» направление длинной оси головки (Dom-
ning, 1978с). В этих терминах у ламантина направление латеродисталь-
ное. Это может говорить о вторичности состояния, наблюдаемого у мор-
ской коровы.
Реальные движения ластов ламантинов были прослежены Хартманом
(Hartman, 1979). Среди огромного их разнообразия, связанного с добы-
ванием и манипуляциями с кормом, с ухаживанием, комфортным поведе-
нием, с передвижением по дну за счет попеременных толчков конечно-
стями, особое внимание привлекает активная пропульсивная работа лас-
тов, совершаемая двумя способами. В одном случае ласт выполняет гребок
назад (forward stroke), называемый нами пропульсивным гребком. В под-
готовительный период вся конечность выносится вперед (протракция),
при этом ласт двигается вперед своим узким радиальным краем, а пло-
скость ласта находится в более отведенном от туловища положении.
В конце периода лопасть ласта пронируется и переводится в поперечную
плоскость пальмарной поверхностью назад, сам ласт приводится ближе
к туловищу. В собственно пропульсивный период ласт ретрацируется,
удерживаясь в поперечной плоскости, и делает мощный гребок. Очевидно,
это и есть наиболее нагруженное движение ласта, и в нем надо искать
объяснение специфическим чертам строения плечевой кости ламантина.
Следует отметить, что ввиду ограничения движения в локтевом суставе од-
ной плоскостью, которое, однако, сопровождается обязательно вращением
лопасти сустава, весь комплекс движений, необходимых для такого рода
гребка, выполняется за счет плечевого сустава. В результате начальный
этап подготовительного периода ласт начинает из положения, когда плечо
максимально ретрацировано, ось блока развернута поперек, так что ра-
диальный край ласта обращен вперед. Затем плечо отводится (по корот-
кой оси головки) и выносится дистальным концом вперед (круговое дви-
жение в сочетании с вращением плеча в направлении длинной оси голов-
ки краниально). Чтобы удержать радиальный край ласта в переднем
положении, ласт сгибается в локтевом суставе, что вызывает супинацию,
компенсирующую изменение ориентации оси блока. Все это движения ма-
лонагружены, так как они не встречают большого сопротивления воды.
В конце периода плечо опять приводится, переводя ласт ниже под пле-
чевой сустав. В этом положении блок принимает более продольное поло-
жение, плоскость ласта ставится почти поперек и плечо пронируется
(по длинной оси головки), что и обеспечивает мощную пропульсивную
силу этого гребка. При этом ласт опять разгибается и при этом про-
нируется, удерживаясь в поперечной плоскости.
Второй тип гребка (backward stroke, или тормозящий гребок) описы-
вается Хартманом так же, за исключением того, что ласт движется спер-
ва назад ульнарным краем вперед и в конце подготовительного периода
175
разворачивается (супинация) поперек, так что пальмарная поверхность
становится обращенной вперед. В результате последующего сильного дви-
жения ласта вперед развивается сила, толкающая животное назад. Воз-
можность такого толчка не вызывает сомнения. Однако учитывая, что ко-
сти предплечья прочно сращены и не могут вращаться друг относитель-
но друга в своем дистальном конце, а также что ось блока плеча при
любом положении не может менять свое положение на 180°, трудно пред-
ставить, как может ласт развернуться своей пальмарной поверхностью
вперед. Скорее всего это движение осуществляется при нормальном по-
ложении ласта, т. е. в конце подготовительного периода осуществляется
не супинация, а пронация, а сам тормозящий гребок менее нагружен,
хотя бы из-за более выгнутой поверхности тыльной (экстензорной) сторо-
ны ласта. Как работают передние конечности дюгоня, практически неиз-
вестно. Возможно, что они не могут помогать животному передвигаться
по дну. Во всяком случае при добывании корма они оставляют на грун-
те след от ротового диска, но не от ластов. Есть сведения, что дюгонь
ими может пододвигать пищу ко рту. Общепринято, что ласты использу-
ются дюгонем для изменения направления движения, осуществляемого
за счет работы хвостового плавника (рули поворотов), или его тормо-
жения (Anderson, 1981). Сами ласты дюгоня относительно короче, чем у
ламантина, поскольку они почти до самого локтевого сустава включены в
кожный мешок туловища, что не может не ограничивать их движения.
При таких скудных сведениях остается надеяться, что анализ строения
плечевого сустава и самой плечевой кости может внести некоторую яс-
ность, какие же движения ластов доступны дюгоню и какие из них яв-
ляются главными, совершающимися по основным осям суставов. Резкие
отличия дюгоня от ламантина, касающиеся взаимоотношений длинной
оси гленоидной впадины и плоскости лопатки, длинной оси головки, про-
дольной оси плеча и оси блока, отмеченные выше, приводят к принципи-
ально иным движениям в плечевом суставе и соответственно иначе рабо-
тают их ласты. Поскольку длинная ось головки лопатки занимает близ-
кое к поперечному положение, как и ось гленоида, и в то же время близ-
ка к положению в пространстве оси блока, движение, происходящее в
плечевом суставе, в большей степени является приводящим (в краниаль-
ном направлении) — отводящим и супинаторно-пронаторным. В резуль-
тате дистальный конец плеча в небольшой степени смещается в передне-
заднем направлении, а, главное, ось блока из горизонтального положения
в крайне приведенном состоянии переводится в более вертикальное поло-
жение. При этом она все время остается в поперечной плоскости. Таким
образом, учитывая отсутствие на блоке винтообразного желоба, лопасть-
ласта при таком движении все время остается обращенной радиальным
краем вперед; пальмарная поверхность обращена к туловищу при при-
ведении, а при отведении — вниз, т. е. плоскость ласта занимает горизон-
тальное положение. Соответственно, ласты при этом несут функции ру-
лей глубины и несущих плоскостей. Это объясняет развитие гребня боль-
шого бугра в латеральном направлении, относящего далеко в сторону
прикрепление дельтовидной мышцы, выполняющей функцию супинатора,,
а не более проксимально, что вело бы к усилению разгибательных движе-
ний. В результате движения в суставе по главной оси, таким образом,
никакого активного гребка не производится. При движении в плечевом
суставе по малым осям дистальный конец плеча также совершает опре-
деленные передне-задние перемещения при небольших абдукторно-ад-
дукторных, но при этом ось блока не меняет своей ориентации в простран-
стве. Ласт при этом выдвигается вперед или назад, сохраняя радиальный
край направленным вперед и помогая удерживать заданное направление
176
Рис. 58. Левые кости предплечья ламантина
Латеральная (а) и медиальная (б) поверхности
1 — caput, radii, головка луче- 8
вой кости;
2 — collum radii, шейка;
3 — corpus radii, тело луче- 9
вой кости;
4 — facies articularis carpea
radii, лучевая часть запяст- ю
ной суставной поверх- и
ности;
5 — facies articularis carpea 12
ulnae, локтевая часть за-
пястной суставной поверх-
ности; 13
6 — fossa capitis radii, ямка
головки лучевой кости;
7 — incisura trochlearis (=inc. 14
semilunaria), суставная 15
вырезка;
— margo anterior, передний
край (= преаксиальный
край);
— margo posterior, задний
край (_ постаксиальный
край);
— olecranon, олекранон;
— рг. anconeus, крючковид-
ный отросток;
— рг. coronoideus lateralis,
латеральный венечный от-
росток;
— рг. coronoideus radii; ве-
нечный отросток лучевой
кости;
— radius, лучевая кость;
— spatium interosseum, меж-
костное пространство;
16 — sulcus tendi m. abductoris
et extensoris pollicis longus;
17 — sulcus tendi m. extensoris
carpi radialis et brachio-
radialis;
18 — sulcus tendi m. extensoris
digitalis communis;
19 — sulcus tendi m. extensoris
digitalis quinti;
20 — sulcus vascularis, сосуди-
стый желобок;
21 — tuber olecrani, локтевой,
бугор;
22 — tuberculum laterale, свя-
зочный бугор;
23 — ulna, локтевая кость;
24 — крепление кольцевой связ~
к и
движения всего животного вперед. И опять это движение не встречает
большого сопротивления воды. Активное пропульсивное (гребущее) дви-
жение ласта возможно у дюгоня только при круговом движении в плече-
вом суставе, когда длинная ось головки устанавливается в положении ко-
роткой оси гленоида. Лишь в этом случае ось блока переводится в па-
расагиттальную плоскость и лопасть ласта принимает поперечное
положение с пальмарной поверхностью назад. Судя по строению передне-
го края гленоидной части лопатки, это осуществимо у дюгоня только в
177
крайне приведенном состоянии плеча и при малых амплитудах сгибатель-
но-разгибательных движений. Пропульсивный гребок при этом требует
пронации, выполняемой подлопаточной мышцей, и сгибания в суставе,
осуществляемого этой же и большой круглой мышцей. Возможно, что
именно работа при таком положении осей в плечевом суставе отвечает
за сильно отогнутый внутрь коракоидный отросток лопатки, что позволя-
ет сухожилию бицепса не выходить из межбугоркового желоба. В то же
время ясно, что большой роли в жизни дюгоня такой гребок играть не
может.
Судя по положению длинной оси головки плеча у морской коровы, ей
свойственны преимущественно сгибательно-разгибательные движения в
парасагиттальной плоскости.
Ulna и radius (рис. 58—60). Различия костей предплечья сирен хоро-
шо известны (Abel, 1904; Simpson, 1932). Они касаются величины меж-
костного пространства, степени выгнутости, взаимного положения, но
функциональный смысл их не обсуждался. В эволюции сирен постепенно
ослабляется локтевая кость, уменьшается олекранон, растет степень сра-
щения локтевой и лучевой костей, исчезает их перекрест. В то же время
каждая линия развития современных сирен имеет свои отклонения от
Г
17
медиальной стороны
10
Зсм
Рис. 59. Скелет левой передней конечности молодо-
го дюгоня (экз. ЗИН-679) с
1 — caput humeri;
2 — carpalia distalia
(cl+2+3+4), дистальный
запястный ряд;
з — epicondylus lateralis;
4 — epicondylus medialis;
S — epiphysis radii, эпифиз
лучевой кости;
6 — epiphysis ulnae, эпифиз
локтевой кости;
7 — humerus;
s — metacarpalia, пястные
косточки;
9 — pr. coracoideus;
jo — radius;
11 — radiale + intermedium
12 — scapula;
13 — tuber olecrani;
14 — tuberculum majus;
13 — tuberculum minor;
16 — ulna;
17 — ulnare + pisif orme -f-
+ carpale distale 5
178
этих общих тенденций. У эоценовых Protosiren (Domning et al., 1982,
fig. 32) и Eotheroides (Sickenberg, 1934, Pl. IV) локтевая кость больше
лучевой, а локтевой отросток большой и крепкий, особенно у протоси-
рена. Возможно это состояние близко к исходному для сирен. У назем-
ных млекопитающих усовершенствование локомоции связано с изменени-
ем передних конечностей, которое может идти двояким путем. У хищных
и копытных происходит прогрессивное уменьшение локтевой кости при
усилении лучевой вплоть до превращения у парно- и непарнокопытных,
мозоленогих первой из них в тонкий придаток олекранона, простираю-
щийся в виде гребня у задней поверхности мощной лучевой кости. У хо-
ботных, наоборот, происходит переразвитие локтевой кости, а лучевая
становится все слабее и слабее, особенно ее проксимальный конец. Од-
нако во всех случаях олекранон остается большим, будучи рычагом при-
крепления разгибателей локтевого сустава, играющих огромную роль в
179
обеспечении пропульсии. Хотя усиление лучевой кости можно считать
генеральной линией в эволюции локомоторного аппарата наземных мле-
копитающих, более частные приспособления часто нарушают эту тенден-
цию. Так, в связи с приспособлением к рытью у свиней обе кости урав-
ниваются в своем развитии. Видимо, растительноядным полуводным мле-
копитающим, а именно такими были, очевидно, предки сирен, часто при-
ходится совмещать движения роющего характера с упиранием обеими
передними конечностями в субстрат, чтобы выдернуть то или иное вод-
ное растение. При этом большая нагрузка падает на локтевую кость. Со-
ответственно, она больше развита у вымершего полуводного хоботного
Platybelodon и у представителя целиком исчезнувшей группы десмости-
лий, близких, возможно, к предкам сирен, Paleoparadoxia (Shikama,
1968). Именно такому состоянию близки кости предплечья эоценовых си-
рен. При приспособлении к постоянной жизни в воде и утрате конечно-
стями основной роли в пропульсии, а значит и в ослаблении сгиба-
тельно-разгибательных движений в локтевом суставе, олекранон теряет
свое значение и начинает уменьшаться. Передняя конечность сирен пре-
вращается в ласты, причем у дюгоня в ласт включаются кисть и пред-
плечье, а у ламантина еще и дистальная часть плеча, выступающая на-
ружу. Соответственно у ламантина расширение ласта начинается прокси-
мальнее и обеспечивается выгибом вперед лучевой кости, расширением
межкостного пространства, увеличением локтевого сгибателя кисти. У дю-
гоня ласт расширяется более дистально и лучевая кость остается более
или менее прямой. При работе ластов как рулей поворотов, при активном
гребке основная нагрузка падает на радиальный край предплечья, что и
ведет к относительному усилению лучевой кости. Сама лопасть ласта
у дюгоня делает невозможными вращательные движения предплечья от-
носительно плеча, что обычно связано с перекрестом лучевой и локтевой
костей и их относительной подвижностью. У ламантина такая возможность
обеспечивается винтообразным блоком плечевой кости и иным строением
запястного сустава и не требует подвижности локтевой и лучевой костей,
которые прочно срастаются в проксимальном и дистальном концах. У дю-
гоня подвижность тоже теряется, поскольку дистально обе кости пред-
плечья сочленяются с единой, образовавшейся в результате слияния, про-
ксимальной костью запястья. В связи с этим у обеих форм теряется
перекрест лучевой и локтевой костей, долгое время сохранявшийся в
эволюции сирен. Так, им обладали и олигоценовый Halitherium и миоце-
новый Metaxytherium (Abel, 1904, Taf. V; Sickenberg, 1934, Pl. IX).
В результате и у ламантина и у дюгоня обе кости предплечья находятся
в одной плоскости, лучевая кость впереди. Однако перекрест сохраняется
в линии развития гидродамалин, хотя у Hydrodamalis он несет совсем
другую функцию по сравнению с тем, что было у Dusisiren, где лопасти
еще работали в основном как рули (Domning, 1978с). Три суставные по-
верхности для костей запястья у морской коровы смещены на медиаль-
ную сторону сросшихся костей предплечья. Это говорит о том, что руди-
ментарная кисть ее постоянно была отогнута медиально, образуя крючок.
Но чтобы он функционировал как своеобразный багор, необходимо его
хоть частично развернуть поперек, что и достигается сохранением пере-
крещивания локтевой и лучевой костей (Kaiser, 1974, pl. 62А). В то
же время для морской коровы необходимо укрепление костей предплечья
(они утолщаются и срастаются, межкостное пространство практически
исчезает). Возможно, вновь усиливается локтевая кость, огромным ста-
новится локтевой отросток (необходимое условие для новой функции ко-
нечностей) .
Суставная поверхность локтевой и лучевой костей ламантина, входя-
180
Рис. 61. Кости кисти и дистальная часть предплечья ламантина (экз. ЗИН-31795)
а — с медиальной стороны; б —
1 — 3 — carpale distale (1 —с^;
2 — с3; з — с4);
4 — facies articularis carpea
radii;
5 — facies articularis carpea
ulnae;
постаксиальной (задней)
6 — intermedium;
7 — metacarpalia;
8 — radiale:
9 — radius;
стороны
10 — sulcus tend! m. extensori
digitalis guinti;
11 — ulna;
12 — ulnare 4- pisiforme +
4- carpale distale 5
тцая в локтевой сустав, имеет сложную топологию. Большой латеральный
участок — двояковогнутый, его форма служит как бы отпечатком лате-
рального мыщелка плечевой кости. С медиальной стороны целостность по-
верхности нарушается синовиальным рукавом. Характерные радиусы
кривизны основного латерального участка в кранио-каудальном направ-
лении составляют 16—18 мм. Большой радиус имеет кривизну в области
локтевого отростка. Вариации в толщине суставного хряща очень малы.
Среднее значение толщины 3 мм. Сустав допускает одно, ограниченное
амплитудой в 60—65°, сгибательно-разгибательное движение. Его моделью
может служить цилиндрическая поверхность с винтообразным желобом,
расположенным на ней по ходу относительного движения звеньев. При
сгибании локтевого сустава полностью теряется контакт суставной
поверхности олекранона с блоком плечевой кости, так же как при раз-
181
V —IV—III — II —I
Рис. 62. Схема контактов косточек в запястье (а) ламантина (экз. ЗИН-31795) и
(6) дюгоня (экз. ЗИН-679)
гибании лучевая кость перестает касаться блока. Таким образом, в этом
суставе нет полной конгруентности поверхности, которая является харак-
терной особенностью сустава наземных млекопитающих (копытные,,
хищники). Запястный суставной рельеф костей предплечья (рис. 61 —
63) представлен двумя раздельными участками: седлообразной поверхно-
стью локтевой кости и разделенной в середине выпуклой поверхностью'
лучевой кости. Обе поверхности винтообразные, имеют характерные ра-
диусы кривизны вдоль дуги движения, равные 11 — 14 мм. Большие зна-
чения радиусов имеют место в переднем участке лучевой кости, мень-
шие — в желобе седла локтевой кости. Толщина слоя хряща 3,1±0,1 мм.
Кисть. Кости запястья у ламантина представлены 2—3 проксималь-
ными и 3 дистальными. Их генезис отражен в подписи к рис. 61. Все
запястье вместе с прилегающими костями предплечья представляет слож-
ное кинематическое соединение из 12—13 костных элементов, соединен-
ных 22 более или менее простыми подвижными сочленениями — сустав-
ными контактами в один общий сустав. Часть смежных костей при этом
не имеет суставных контактов (рис. 62, а). Сросшаяся с гороховидной и
V дистальной локтевая кость запястья имеет 4 суставных фасетки: боль-
шую проксимальную седлообразной формы (рис. 64) для сочленения с
ulna; две дистальных, выпуклую для сочленения с вогнутой суставной
поверхностью V metacarpale (тс) и плоскую для IV carpale distale (cd);
малую боковую для intermedium. Последнее имеет 5 фасеток: проксималь-
ную вогнутую для заднего края суставной поверхности лучевой кости, две
дистальные плоские для III и IV cd, две боковых — переднюю для слив-
шихся I и II cd и заднюю для ulnare. Сочленения промежуточной и
лучевой костей запястья в виде сустава нет: оно замещено хрящевой
прослойкой (ЗИН-31795), полностью обызвествленной у экз. ИЗ-5945..
Radiale, кроме сочленения с лучевой костью, имеет суставное соединение
только со слившимися I и II cd и несет, таким образом, только 2 фа-
сетки. IV cd сочленяется с IV и III cd, ulnare и intermedium, но не с
V cd; III cd — только с промежуточной костью и двумя II и III пястны-
ми и, наконец, слившиеся I и II cd — с radiale, intermedium, I и I me.
Таким образом обращает внимание, что все 3 дистальные кости запястья
не имеют суставных контактов между собой. Пястные кости соединяются
с костями запястья проксимальными фасетками и, кроме того, между
собой небольшими боковыми фасетками, образуя последовательную кине-
матическую цепь. Самое длинное и широкое у ламантина — V тс
(у дюгоня IV). Флексорная поверхность метакарпалий несколько упло-
щена, экстензорная выпукла. Все пястнофаланговые суставы, кроме I,
блоковидные. Каждая суставная головка пястных костей несет на себе
участок цилиндрической поверхности с незначительным пологим желобом
в середине, идущим в направлении движения. Основания проксимальных
фаланг имеют выпукло-вогнутые суставные фасетки, которые по форме
182
отвечают определенному участку поверхностей соответствующих пястных
костей. Суставы допускают движение в сторону дорсальной флексии
кисти на угол 30—35° и обратно. Пологий желоб на суставной поверх-
ности пястных костей вместе с гребнями на проксимальных фалангах
ограничивают боковые смещения фаланг относительно пястных костей.
Единственная фаланга I пальца соединяется с пястной костью небольшой
плоской фасеткой, образуя при этом малоподвижный сустав. Движения
в нем связаны с упругой деформацией суставной сумки и не имеют
строго связанных с суставной поверхностью ограничений. Средние
фаланги II —IV пальцев подвижно соединяются с первыми фа-
лангами слегка седлообразными, мало различающимися по величине су-
ставными поверхностями. При этом дистальный конец первой фаланги
имеет выпуклую на вертикальном сечении и вогнутую на горизонтальном
сечении поверхность, а проксимальный конец второй фаланги вогнутую
на вертикальном и выпуклую на горизонтальном сечениях, при этом
выпуклая — всегда по радиусу кривизны больше вогнутой. Выпуклость
и вогнутость суставных фасеток столь незначительны, что практически
можно считать эти суставы плоскими и движения в них разрешаются по
•суставным поверхностям не более 10°.
В целом запястье ламантина допускает сгибание-разгибание с ампли-
тудой около 90° (вычислено по суставным фасеткам), обеспеченное на
78% соединением костей предплечья и запястья и на 22% соединением
костей проксимального ряда с дистальным и V тс, и супинацию-прона-
цию кисти с амплитудой около 80°, когда конечность в предплечезапяст-
ном суставе согнута, и около 25° при его разогнутом состоянии. Обеспе-
чивается ротация всем множеством суставных контактов. Однако основ-
ная нагрузка приходится на сочленение с V и I шс. Поэтому сочленение
ulnare, сросшегося в V cd и pisiforme, с V шс выпуклое, также как со-
членение I + H cd с I шс, что позволяет относительно больший размах
движения, а суставные сумки с латеральной и медиальной сторон более
податливы на растяжение. Плотная соединительная ткань, в которую
включены кости кисти, относительно толстый кожный покров, лишь не-
много отступающий от поверхности костей, могут заметно ограничивать
размах движения и, видимо, расчетные амплитуды сильно завышены.
В целом можно сказать, что запястный сустав ламантина допускает за-
висимые друг от друга сгибательно-разгибательные и супинаторно-прона-
торные движения (сгибание связано с пронацией).
Запястье дюгоня существенно отличается от ламантинового. Кроме
слившихся intermedium и radiale, в одну кость объединяются все 4 ди-
стальные кости запястья. В результате в общем суставе участвуют всего
10 костей (рис. 59), связи между которыми отражены на рис. 62, б. За-
метно уменьшен сгибательный участок суставной поверхности локтевой
кости, вследствие чего дуга относительных движений в предплечезапяст-
ном суставе уменьшена. Фасетки запястных костей более уплощены, что
приводит к еще большему ограничению движения. Таким образом, раз-
нообразие движений у дюгоня ограничено наличием более жестких свя-
зей между костями. Вероятно частично эти движения осуществляются за
счет упругой деформации связей между всеми элементами запястья. За-
пястье морской коровы резко отличалось по своей структуре от запястья
ламантина и дюгоня. Стеллер (Steller, 1751) писал, что у Hydrodamalis
gigas отсутствовали фаланги пальцев, а весь дистальный конец конеч-
ности оканчивался тупо, подобно ампутированной конечности, и был за-
гнут, образуя своеобразный крюк. Это подтверждается наличием на ди-
стальном конце предплечья 2, иногда 3, почти плоских сравнительно не-
больших фасеток, смещенных на медиальную сторону (рис. 60), что
183
О 2см
I____I----1
Рис. 63. Рельеф запястной суставной
поверхности правого предплечья ла-
мантина (экз. ЗИН-31795)
а, б — вид с дистального конца; в — вид с ме-
диальной стороны; б, в — по препарату, фик-
сированному в формалине
1 — facies articularis carpea radii;
2 — facies articularis carpea ulnae
Рис. 64. Рельеф суставных поверхностей проксимального (а, б) и дистального (в, г)
рядов костей правого запястья ламантина (экз. ЗИН-31795)
а — предплечье-запястный сустав (проксимальная поверхность); б — в — среднезапястный сустав
б — дистальная поверхность проксимального ряда,
в — проксимальная поверхность дистального ряда); г — запястно-пястный сустав (дистальная
поверхность дистального ряда)
На рисунке медиальная поверхность внизу
1—3 — carpalia distalia (1 — С,+2; 2 — С3; 3 — С4); 4 — intermedium; 5 — radiale; 6 — ulnare -J-
4~ pisiforme + carpale distale 5
говорит о крайне небольших движениях в предплечезапястном суставе,
фактически о фиксации постоянно согнутого положения кисти. Сами ко-
сти запястья и пясти стеллеровой морской коровы неизвестны. Предпола-
гаемая пястная кость (Kaiser, 1974, fig. 63), очевидно, не имеет отно-
шения к пясти (Domning, 1978с). Однако последний автор описал
несколько запястных и одну пястную кость позднеплиоценовой калифор-
нийской морской коровы Hydrodamalis cuestae. Пястная кость толстая
короткая и действительно не имеет на дистальном конце суставной по-
184
верхпости, а сам этот конец расширен и закруглен наподобие копытца и
уплощен дорсовентрально. У предкового Dusisiren еще имелись фаланги
пальцев, а метакарпалии были несколько длиннее.
В заключение глав по костной системе сирен полезно привести неко-
торые данные, характеризующие костную ткань, слагающую длинные
кости конечностей ламантина. Они не содержат медулярных костно-моз-
говых полостей и целиком заполнены костным веществом. Средний
Рис. 65. Поперечный срез плечевой кости в середине диафиза
Поверхностный (а) и глубокий <б) слои
185
показатель отношения массы к объему составляет 1,5 г/см3. Измерения
удельного веса плечевой кости дали более высокое значение 1,9 г/см3.
Анализ поперечных срезов, полученных из средних участков диафизов
плечевой и локтевой костей и IV тс показывает: внутри кость имеет вид
мелкоячеистой губки, около 50% площади занимают костные полости и
сосуды, ширина которых варьирует от 0,02 до 0,5 мм. К периферии губ-
чатое вещество центральной области плавно переходит в настоящую' ком-
Рис. 66. Поперечный срез в середине диафиза локтевой кости (а) и четвертой пяст-
ной (mt IV) кости (б)
186
Рис. 67. Относительная частота распределения значений площадей гаверсовых кана-
лов в поперечном сечении компакты в середине диафиза плечевой (а) и локтевой
(б) костей и IV пястной кости (в) ламантина
Л' — число подсчитанных каналов остеонов
пактную ткань, почти полностью состоящую из остеонов (рис. 65, 66).
Слой периостальной кости развит слабо. Так, в плечевой кости он дости-
гает 0,7 мм толщины у каудальной поверхности (рис. 65, а) и почти ис-
чезает у краниальной. Среднее число остеонов на 1 мм2 в компакте равно
27 и мало варьирует при переходе от одного звена к другому. Более ва-
риабельна площадь каналов остеонов: в плечевой кости каналы зани-
мают 5% площади компакты, в локтевой 7%, в IV тс 10%. Относитель-
ные частоты распределения площадей гаверсовых каналов для компакты
костей ламантина приведены на рис. 67. Предельная сжимающая аб-
солютная нагрузка (сила, производящая полное разрушение общего об-
разца из середины диафиза при направлении нагрузки вдоль длинной оси
кости) для плечевой кости составляет 13,5 тс, локтевой 4,2 тс и IV тс
0,9 тс. Те же показатели, отнесенные к плоскости поперечного сечения
диафиза, в среднем составляет 12,5 кг/мм2 с отклонением в пределах ко-
нечности ±4%. Аналогичные измерения у хоботных дают 13,6, копытных
17,3 с вариациями ±15% и ±25% соответственно. Для образцов из диа-
физов костей ламантина характерно хрупкое разрушение, которому пред-
шествует появление на их поверхности множества косых линий, будущих
расколов образца. Как правило, они идут под углом 34—35° к продоль-
ной оси образца.
1 лава пятая
СТРОЕНИЕ, ФУНКЦИИ
И АДАПТИВНЫЕ ОСОБЕННОСТИ
СКЕЛЕТНОЙ МУСКУЛАТУРЫ ЛАМАНТИНА
К изучению мышечной системы сирен обращались многие исследователи,
особенно в первой половине XIX века. Однако почти всегда объектами
служили очень молодые неполовозрелые особи или даже эмбрионы. Един-
ственным всеобъемлющим описанием мышц Trichechus manatus (по двум
неполовозрелым экземплярам: ? — 160 см, & — 120 см) до сих пор слу-
жит монография Дж. Мьюри, посвященная общей его анатомии (Murie,
1872а). Впоследствии ее положения были дополнены этим же автором по
более крупному (около 230 см) экземпляру (Murie, 1880). Учитывая
время выполнения, работа Мьюри поражает богатством своего содержания
и точностью фактов. Однако не всем современным требованиям эти опи-
сания удовлетворяют, поскольку они не преследовали целей уяснить спе-
цифику адаптации ламантина или дать какое-либо функциональное объ-
яснение описываемым особенностям. Соответственно ряд структур описан
довольно поверхностно. Большинство последующих авторов обращались
преимущественно к отдельным узлам в мышечной системе на разных ви-
дах ламантинов (см. сводную таблицу у Domning, 1978а). Среди них мож-
но выделить лишь обстоятельный разбор мышц в связи с изучением раз-
вития передних конечностей (Bahrdt, 1933), лицевых мышц (Saban,
1975). К сожалению, не были опубликованы результаты исследования му-
скулатуры ламантина и дюгоня, проведенного крупнейшим знатоком в
области осевого скелета и мускулатуры Э. Слайпером (Slijper, 1936,
1946) с целью сравнения с китообразными или другими водными и на-
земными животными. В его монографиях эти сведения лишь вкраплены в
описание другого материала.
Новый этап в изучении мышечной системы сирен связан с именем
Д. Домнинга, не только опубликовавшего две монографии с описанием
миологии: одна —дюгоня (Domning, 1977b), другая — бразильского
ламантина (Domning, 19786), но и попытавшегося приложить эти данные
к анализу развития гидродамалиновой линии развития сирен (Domning,
1978с). Сделана им и попытка рассмотреть морфологию мышечной систе-
мы ламантина и дюгоня с функциональной точки зрения, как приспособ-
ление к плаванию, первичной переработке пищи и т. п. Впервые
у Домнинга появляются и данные по весовому анализу ряда мышц. К со-
жалению, он сделал это только по бразильскому ламантину, но не по
дюгоню, а именно последнее могло бы, как нам кажется, привести к очень
интересным результатам при сравнении с ламантином.
В нашем распоряжении находился экземпляр взрослого крупного
ламантина Т. manatus. Задачи исследования стояли так: детальное опи-
сание всей скелетной мускулатуры с особым упором на мышцы ясного
функционального назначения, изображение их в ходе последо-
вательной препаровки, сравнение их с литературными данными по дюго-
ню, бразильскому ламантину и Т. manatus и на основе сравнительного
изучения остеологии современных сирен (см. гл. 3 и 4), проведение ве-
сового и функционального анализа всей системы в целом и отдельных
188
наиболее важных ее узлов с целью попять развитие специфики ламапти-
новой линии эволюции сирен.
Для удобства сравнения мы принимаем последовательность изложе-
ния, предложенную Домнингом, и, как правило, его номенклатуру, если
у нас нет более удобного термина. Часть предлагаемых иллюстраций схе-
матизирована (мышцы головы, туловища, полового аппарата), другая
часть (мышцы передней конечности) максимально приближена к реаль-
ности. На них даже сделана попытка отразить характер перистости мышц,
как он виден на их поверхности. Большое внимание во всех случаях уде-
лено выявлению внутренней структуры мышц, его сухожильного скелета.
ЛИЦЕВЫЕ И ПОВЕРХНОСТНЫЕ МЫШЦЫ ШЕИ
Сразу по удалении кожи обнажаются поверхностные мышцы туловища,
шеи (рис. 68) и головы.
М. sphincter colli superficialis — поверхностный сжиматель шеи
(рис. 69, 71) — часть подкожной мышцы, занимающая вентральное поло-
жение в области шеи, передней части головы и груди. В краниальной
своей половине ее отдельные пучки начинаются на средней линии голо-
вы и шеи и идут назад до уровня рукоятки грудины. При этом происхо-
дит переплетение пучков правой и левой сторон, как бы врастающих друг
в друга. Далее назад начало мышцы отступает от средней линии. Отдель-
ные более или менее обособленные пучки сухожильных волокон отходят
от поверхности наружной фасции m. cutaneus trunci и m. trapezius,
р. mandibularis и следуют в переднебоковом направлении к сухожильно-
му тяжу, волокна которого идут более или менее поперек им (от средней
линии назад и вбок). Этот тяж также закреплен по всему своему протя-
жению на фасции m. cutaneus trunci. Мышечные волокна задних пучков
поверхностного сжимателя шеи начинаются на этом сухожильном тяже.
Сами пучки идут вперед и дорсально. Благодаря разному направлению
мышечных пучков, а также различию в их величине они образуют слож-
ное переплетение также и на каждой половине шеи, что вообще харак-
терно для этой мышцы. Оканчивается она на самой вентральной части
наружной поверхности фасции platysma myoides р. jugularis и поверхност-
ной фасции предплечья. Отдельные сухожильные волокна, являющиеся
продолжением поверхностного сжимателя шеи, формируют особый сухо-
жильный тяж, идущий в составе поверхностной фасции предплечья на
его латеральной (разгибательной) поверхности до уровня пястных костей.
Однако прикрепления на I пястной кости, описанного в свое время у ла-
мантина (Murie, 1827а), нами не обнаружено. Не было отмечено оно и
у бразильского ламантина (Domning, 1978а). У дюгоня вообще нет какого
либо сухожилия поверхностного сжимателя шеи, простирающегося в пе-
реднюю конечность (Domning, 1977b).
Platysma myoides (рис. 68—72) продолжает вперед подкожную мыш-
цу туловища, простираясь к голове по дорсальной поверхности шеи.
Большая ее pars jugularis начинается от переднего края сухожильного
тяжа, занимающего вертикальное положение в области лопатки и закреп-
ленного своим внутренним краем на поверхности фасции глубжележащих
мышц. Поскольку от заднего края этого тяжа назад идет m. cutaneus
trunci, то сам тяж можно рассматривать как сухожильную перемычку в
едином пласте подкожной мускулатуры, включающей обе мышцы. Конча-
ется pars jugularis на поверхностной фасции m. temporalis, образуя на
ней фиброзное утолщение, переходящее по надглазничному отростку на
скуловую дугу (см. рис. 76). Вентральные мышечные волокна заходят
дальше вперед и внедряются в переплетение мышечных волокон, т. па-
189
solabialis и m. maxillonasolabialis,
участвуя, таким образом, в дви-
жениях верхней губы. Будучи
прикрепленной с другой стороны
на фасции височной мышцы и
скуловой дуге, а с вентральной —
натягиваемой поверхностным сжи-
мателем шеи, эта часть платизмы
обеспечивает, таким образом, опо-
ру на черепе служащему основ-
ным сгибателем хвоста in. cuta-
neus trunci. Вероятно, эта часть
платизмы соответствует pars Ьга-
chialis многих млекопитающих:
сумчатые, насекомоядные, грызу-
ны (Huber, 1930; Meinertz, 1941,
1978).
Вторая часть платизмы (р. buc-
calis) представлена пятью-шестью
отдельными мышечными пучка-
ми, отходящими от внутренней
стороны фасции, окутывающей
поверхностный сжиматель шеи.
Эти пучки конвергируют вперед
к верхней губе. Судя по описанию
и рисунку, Домнинг (Domning,
1978а, fig. 4) эту часть пла-
тизмы описывает как m. sphincter
colli profundus р. oris, хотя она
лежит поверх платизмы (р. jugu-
laris), между нею и поверхност-
ным сжимателем шеи, и, таким
образом, к глубокому сжимателю
шеи отношения не имеет. Ника-
ких особых мышечных пучков
платизмы, связанных с ушным
отверстием п слуховым проходом,
как у Т. inunguis, а также р. tem-
poralis и р. scapularis, описанных
у дюгоня (Domning, 1977b), нами
не обнаружено.
Более краниально и глубже
от двух описанных мышц лежит
сложный комплекс мышц, произ-
водных глубокого сжимателя шеи
(в широком смысле), состоящий
из собственно глубокого сжимате-
ля и предглазничных лицевых
мышц.
М. sphincter colli profundus
s. str., собственно глубокий сжи-
матель шеи (рис. 69, 71), сохра-
няет в своем начале тесную связь
с поверхностным сгибателем шеи.
Начинается он по средней линии
191
Рис. 69. Мышцы шеи и передней части туловища (вид снизу)
•На правой стороне рисунка снят m. sphincter colli superficialis и удален ласт
1 — ш. acromiodeltoideus;
2 — m. acromiotrapezius;
3 — m. clavodeltoideus;
4 — in. clavotrapezius;
5 — 6 — m. cutaneus trunci
<5 — pars dorsalis,
6 — p. ventralis);
7 — m. digastricus;
8 — m. masseter lateralis;
9 — 10 — platysma myoides
(9 — p. buccalis,
10 — p. jugularis);
11 — 15 — m. sphincter colli
(11 — profundus;
12 — 15 — superficialis;
13 — начальный апоневроз,
14 — 15 —конечный апоневроз,
крепящийся на предпле-
чье — 14 и кисти — 15)
16 — m. spinodeltoideus;
17 — т. trapezius р. mandibu-
laris
в пространстве между правой и левой нижними челюстями вместе с мыш-
цей другой стороны и более краниальными пучками поверхностного сжи-
мателя шеи, лежащими снаружи. Дальше от начала оба сжимателя раз-
деляются мышечными волокнами m. platysma myoides р. buccalis. У на-
шего ламантина собственно глубокий сжиматель шеи представлен только
192
ог st-
7 Ламантин
193
5
Рис. 71. Лицевые и подъязычные мышцы (вид снизу)
Слева — поверхностный слой (platysma myoides р. jugularis и m. sphincter colli superficialis перере-
заны и отогнуты); справа — более глубокий слой
194
Рис. 72. Глубокие лицевые и жевательные мышцы (вид сбоку)
Удалены platysma myoides р. buccalis, m. sphincter colli superficialis и части m. levator nasolabialis
1 — m. buccinatorius;
2 — m. digastricus;
.5 — m. lateralis nasi;
4 — m. levator nasolabialis;
S — m. mandibularis;
6 — 7 — m. masseter
(6 — lateralis;
7 — medialis);
8 — m. maxillonasolabialis;
9 — m. orbicularis oris;
10 — platysma myoides p. jugu-
laris;
11 — m. retractor labiorum;
12—m. temporalis;
13 — m. trapezius p. mandibula-
ris
двумя пучками, один из которых оканчивается ниже глаза и может со-
поставляться с р. palpebralis, второй идет к углу рта ( р. oris). У млеко-
питающих обычно различают четыре части этой мышцы: р. auris, р. in-
termedia, р. palpebralis и р. oris (Meinertz, 1941). Первая из них
отсутствует у ламантина, поскольку описанная под таким названием
Домнингом (1977b, 1978а) мышца на самом деле является частью трапе-
циевидной мышцы, поскольку она иннервируется XI, а не VII, как вся
лицевая мускулатура, нервом. Очевидно, именно поэтому и в описании
лицевой мускулатуры африканского ламантина (Saban, 1975) отсутствует
упоминание о р. auris глубокого сжимателя шеи, что так удивляет Дом-
1 — m. buccinatorius;
2 — т. cleidomastoideus;
з — т. digastricus;
4 — т. genlohyoideus;
S — т. lateralis nasi;
в — т. levator nasolabialis;
7 — т. mandibularis;
8 — 9 — т. masseter
(8 — lateralis,
9 — medialis);
10 — m. maxillonasolabialis;
11 — 12 — m. mentalis
(12 — пучки, оканчивающиеся
на коже подбородка);
13 — т. mylohyoideus;
14 — 1S — т. orbicularis oris
(IS — p. labialis superior);
16 — 17 — platysma myo ides
(16—p. buccalis,
17 — p. jugularis);
18 — m. pterygoideus;
19 — 20 — m. sphincter colli
(19 — profundus,
20 — superficialis);
21 — m. sternohyoideus;
22 — m. stemomastoideus; .
23 — m. stemothyroideus;
24 — m. styloglossus;
25 — m. stylohyoideus;
26 — m. stylopharyngeus;
27 — m. thyrohyoideus;
28 — m. trapezius p. mandibula-
ris
195
7*
Рис. 73. Мышцы головы
и шеи (вид сверху)
На правой стороне рисунка
часть мышц удалена
1 — m. acromiotrapezius;
2 — m. clavotrapezius;
3 — m. cleidomastoideus;
4 — m. lateralis nasi;
5 — m. levator nasolahia-
lis;
в — m. masseter media Ils;
7 — maxillonasolabialis;
8 — 9 — m. obliquus capi'
tis
(S — caudalis;
9 — cranialis);
10 — 12 — m. rectus capitis
dorsalis
(10 — major,
11 — intermedius,
12 — minor);
13 — m. rectus capitis late-
ralis;
14 — 15 — m. semispinalis
(14 — capitis,
15 — cervicis);
16 — m. splenius;
17 — m. sternomastoideus;
18 — m. temporalis
нинга, считающего, что она имеется у бразильского ламантина и у дю-
гоня. Собственно, глубокий сжиматель шеи представляет, как сказано
выше, две последние части, а р. intermedia Домнинга рассматривается
нами в виде двух отдельных мышц.
М. orbicularis oculi, круговая мышца глаза (см. рис. 70), первая из
них, представлена лишь небольшим количеством мышечных волокон в
составе соединительнотканного кольца вокруг глаза, сокращение которых
вызывает его закрывание. Вторая мышца (m. levator nasolabialis) вхо-
дит уже в комплекс предглазничных мышц, все из которых, кроме носо-
губного поднимателя, Домнинг рассматривает как р. oris глубокого сжи-
мателя шеи. Ничего подобного m. frontalis, рассматриваемого Домнингом
в качестве составного компонента р. intermedia, у нашего ламантина не
наблюдалось.
Схематично устройство предглазничных лицевых мышц можно пред-
ставить у ламантина следующим образом. Более дорсально, в области рас-
196
Рис. 74. Лицевые и жевательные мышцы
1 — m. buccinatorius;
2 — т. digastricus;
3 — т. dilatator nasi;
4 — т. lateralis nasi;
5 — m. levator nasolabialis;
6 — m, mandibularis;
(наиболее
7 — 8 — m. masseter
(7 — lateralis,
8 — medialis);
9 — m. maxillonasolabialis;
10 — m. mentalis;
11 — 12 —- m. orbicularis oris
глубокий слой, вид сбоку)
{11 — р. labialis inferior,
12 — р. labialis superior);
13 — m. retractor labiorum
14 — m. stylomyoideus;
15 —m. temporalis
ширенной верхней губы, помещаются три мышцы: сзади вперед и вниз
с дивергирующими волокнами идет m. levator nasolabialis; снизу вперед
и вверх, дивергируя, идет m. maxillonasolabialis; прямо снизу вверх идет
m. lateralis nasi. Все три мышцы тесно переплетены в своих окончаниях
в толще верхней губы, образующей так называемый губной диск, своеоб-
разный короткий хобот сирен, и управляют его движениями. Вентральнее,
в области щек, располагаются две идущие сзади наперед мышцы (m. man-
dibularis и т. retractor labiorum) и одна, идущая поперек с почти верти-
кальными волокнами m. buccinatorius s. str., которая помещается в самой
толще щеки. Кроме того, в этот комплекс входят m. orbicularis oris, рас-
положенный вокруг рта, и связанный с кожей подбородка и лежащий
впереди m. mentalis. Все они являются хорошо дифференцированными
дериватами глубокого сжимателя шеи и помещаются впереди m. sphin-
cter colli profundus s. str. Идентификация 'отдельных частей этого комп-
лекса иногда вызывает споры. Так, Домнинг (Domning, 1978а) включает
в него в качестве наиболее заднего компонента щечную часть платизмы.
М. levator nasolabialis, носогубной подниматель (рис. 70—74), начина-
ется хорошо выраженным сухожилием от дорзальной поверхности глаз-
ничного отростка лобной кости (рис. 75, а). Его мышечные волокна идут
вперед и вентрально, расходясь веерообразно к латеральной и передней
части верхней губы, причем на своем пути они дифференцируются на бо-
лее поверхностные и более глубокие, между которыми вторгается иду-
щий сзади и снизу m. maxillonasolabialis. Часть волокон platysma myoides
р. jugularis, как уже было отмечено, также пронзает мышечную массу
носогубного поднимателя, переплетаясь с его волокнами. Концевые волокна
197
Рис. 75. Площади при-
крепления мышц на че-
репе
а — вид сверху;
б — вид снизу
7 — m. acromiotrapezius;
2 — т. buccinatorius;
з — т. clavotrapezius;
4 — т. digastricus;
5 — т. lateralis nasi;
в — т. levator nasolabialis;
7 — т. longus capitis;
8 — 9 — m. masseter
(8 — lateralis,
9 — medialis);
10 — m. maxillonasolabia-
lis;
11 —m. obliquus capitis cra-
nialis;
12 — m. platysma p. jugula-
ris;
13 — 14 — m. ptcrygoideus
(13 — lateralis,
14 — medialis);
15 — m. pterygopharyngeus;
16 — 17 — m. rectus capitis
(16 — dorsalis major,
17 — ventralis);
18 — m. semispinalis capitis;
19 — m. splenius;
20 — m. sternocleidomastoi-
deus;
21 — m. stylohyoideus;
22 — m. temporalis;
23 — m. trapezius p. mandi-
bularis
последнего также переплетаются с волокнами идущего снизу m. latera-
lis nasi, часть волокон которого прободает m. levator nasolabialis. Наибо-
лее дорсальные пучки носогубного поднимателя располагаются по дор-
сальной поверхности ноздрей, и их сокращение способствует открыванию
их при входе.
М. maxillonasolabialis (см. рис. 71—74) начинается на вентральной
поверхности горизонтальной пластинки скулового отростка верхнечелюст-
ной кости (см. рис. 75, б) прямо под орбитой и немного позади нее. Прой-
дя вперед за орбиту, мышца веерообразно расширяется, отдавая волокна
как прямо вперед, так и вперед вверх, разделяя на своем пути поверх-
ностные и глубокие волокна носогубного поднимателя, с которыми они
смешиваются, образуя мелкие пучочки, прободающие общую мышечную
массу, занимающую губной диск.
М. lateralis nasi, латеральная мышца носа (см. рис. 71—74), начина-
ется от передней и латеральной поверхностей мощного хрящевидного
образования, одевающего снизу предчелюстные и самые передние кон-
цы верхнечелюстных костей (см. рис. 75, а, 77). Отсюда мышечные волок-
на направляются прямо вверх к мягкому краю верхней губы, на боковую
часть губного диска, где отдельные пучки этой мышцы переплетаются с
198
23
1
Рис. 76. Площади прикрепления мышц на черепе (вид сбоку)
1 — ш. acromiotrapezius;
2 — m. buccinatorius;
3 — m. clavotrapezius;
4 — m. digastricus;
5 — m. lateralis nasi;
в — m. levator nasolabialis;
7 — a — m- masseter
(7 — lateralis,
a — medialis);
9 — m. obliquus capitis cra-
nialis;
10 — platysma myoides p. jugu-
laris;
11 — 12 — m. pterygoideus
(11 — lateralis,
12 — medialis);
13 — m. pterygopharyngeus;
14 — 16 — m. rectus capitis dor-
salis
(14 — major,
15 — intermedius,
16 — minor);
17 — m. rectus capitis lateralis;
18 —m. semispinalis capitis;
19 — m. splenius;
20 — m. sternocleidomastoideus;
21 — m. stylohyoideus;
22 — m. stylopharyngeus;
23 — m. temporalis;
24 — m. trapezius p. mandibu-
laris;
25 — 26 — твердое небо
(25 — губчатая ткань,
26 — роговое покрытие);
27 — фиброзная перегородка
носа
таковыми носогубного поднимателя и m. maxillonasolabialis. Основная
масса волокон кончается у вентрального края ноздри и глубже по нижней
стороне носового прохода.
Остальные, лежащие более вентрально, мышцы этого комплекса часто
рассматриваются как дериваты одного m. buccinatorius (в широком смыс-
ле) . Однако они хорошо обособлены друг от друга, и их удобнее рассмат-
ривать как отдельные m. mentalis, m. buccinatorius (s. str.), m. mandi-
bularis, m. retractor labiorum, m. orbicularis oris.
M. mentalis, подбородочная мышца (см. рис. 70, 71), занимает наибо-
лее переднее вентральное положение. Она начинается от узкой полоски
вдоль передневерхнего края симфизарной части нижней челюсти (см.
рис. 77, а) впереди от foramen mentale. Волокна более дорсальной части
мышцы идут почти вдоль нижней губы, а более вентральные направле-
ны как вниз и вперед, так и вперед и латерально к коже в области
подбородка.
М. buccinatorius (s. str.), собственно, щечная мышца (см. рис. 71, 72,
74), помещается позади подбородочной мышцы. Эта самая глубокая мыш-
ца комплекса. Ее волокна простираются в толще щеки вертикально от
верхнечелюстной кости у самого основания зубов (рис. 75, б) к нижней
челюсти, а также к альвеолярному ее краю (см. рис. 77, а). Сверху эта
мышца в своей вентральной части прикрыта двумя дугами, идущими го-
ризонтально.
М. mandibularis, нижнечелюстная мышца (см. рис. 72, 74), является
более мощной из них и занимающей более вентральное и поверхностное
199
Рис. 77. Площади при-
крепления мышц на
нижней челюсти
а — вид с ла теральной сто-
роны;
б — с медиальной стороны-
Зачернено прикрепление
апоневрозов.
7 — ш. buccinatorius;
2 — ш. digastricus;
3 — m. genioglossus;
4 — m. geniohyoldeus;
5 — m. mandibularis;
в — 7 — m. masseter
(в — lateralis,
7 — medialis);
8 — m. mentalis;
9 — m. myloglossus;
10 — m. mylohyoideus;
11 — 12 — m. pterygoideus
(21 — lateralis,
12 — medialis);
13 — m. retractor labiorum;
14 — m. temporalis;
15 — губчатая ткань;
2 в — роговое покр ытие
положение. Она начинается от самого края венечного отростка нижней че-
люсти (см. рис. 77, б) на его расширенном и обращенном вперед и
внутрь переднем крае латеральнее наиболее заднего в зубном ряду зуба,
медиальнее linea obliqua на венечном отростке и сразу же каудальнее за
ш. buccinatorius. Часть волокон начинается и от верхней челюсти, сразу
позади волокон последней мышцы. У бразильского ламантина все волок-
на брали начало только от нижней челюсти. Плоское брюшко мышцы от
своего начала идет вперед и на уровне угла рта переходит в плоское
широкое сухожилие, волокна которого теряются в мышечных пучках под-
бородочной мышцы.
М. retractor labiorum, оттягиватель угла рта (см. рис. 74), представля-
ет более слабую мышцу и более глубокую в своем начале, чем преды-
дущая, рядом с которой она начинается от того же основания венечного
отростка, но чуть дорсальнее (рис. 77, 5). Отсюда эта мышца идет вперед
к углу рта, где ее волокна сливаются с волокнами следующей мышцы.
Оттягиватель угла рта был описан у дюгоня (Domning, 1977b), но не
обнаружен у бразильского ламантина (Domning, 1978а).
М. orbicularis oris, круговая мышца рта (см. рис. 72), занимает наи-
более поверхностное положение в комплексе. Ее волокна идут под кожей
вдоль краев верхней и нижней губ, огибая сзади угол рта, где эта мышца
почти не обособлена от собственно щечной мышцы.
Ротовой и носовой аппарат сирен необычен в связи с наклоном вниз
ростральной части черепа (предчелюстных и передней части верхнечелю-
стных костей) и удлиненной симфизной части нижних челюстей, несущих
200
роговые пластинки, и с необычайным увеличением наружного носового
отверстия черепа, обращенного вверх и вперед и сильно вытянутого на-
зад в результате редукции носовых костей. Мягкие стенки крупной но-
совой полости покоятся на лобных отростках предчелюстных и переднем
крае лобных костей и переходят по бокам верхней челюсти в верхние
губы, сильно нависающие сверху и с боков над нижними губами. Все это
вместе образует губной диск, напоминающий хобот сайгаков. Носовые от-
верстия обращены вверх и помещаются в передневерхних углах губного
диска. Лишь при опускании передней части верхней губы носовые от-
верстия могут открываться вперед. Перед вдохом ноздри вытягиваются
вверх над уровнем губного диска, выступают на поверхность воды и
лишь затем открываются. После одного или нескольких вдохов ноздри
закрываются, как бы втягиваются внутрь губного диска, и животное ока-
зывается готовым к погружению.
Мясистые губы ламантина, снабженные жесткими короткими вибрис-
сами и образующие отдельные подвижные выпячивания на латеровент-
ральной поверхности верхних губ, играют существенную роль в захва-
тывании пищи. Независимость в движениях отдельных участков верхней
губы и наличие на них вибрисс способствует ощупыванию пищи, ее
захвату и перемещению в сторону рта. Роговые пластинки в передней
части рта, на переднечелюстных костях и симфизе нижней челюсти слу-
жат фиксации растительных кормов, разрыванию отдельных растений,
а губы помогают перемещению пищи дальше в ротовую полость.
Несмотря на разнообразие и сложность указанных выше движений:
поднимание и опускание передней части морды (губного диска), замы-
кание и открывание ноздрей, движения верхних и нижних губ и их от-
дельных частей,— все они выполняются лицевыми мышцами, устроенны-
ми по значительно более упрощенному плану сравнительно с наземными
млекопитающими, хотя следовало бы ожидать, наоборот, их большей
дифференциации и усложнения.
Поднятие передней части области морды перед вдохом осуществляет-
ся сокращением дорзальных волокон носо-губного поднимателя и
m. maxillolabialis, который принимает на себя функцию особого
m. dilatator nasi других млекопитающих. Затем в работу включаются
волокна латеральной мышцы носа, оттягивающие дно носовых проходов
вниз, что ведет к раскрыванию ноздрей. Расслабление m. lateralis
nasi имеет следствием замыкание ноздрей. При каждом акте дыхания,
даже если животное извлечено из воды, этот процесс повторяется:
раскрыванию ноздрей всегда предшествует поднятие губного диска над
уровнем твердого нёба.
Сложное манипулирование латеровентральной части верхней губы
осуществляется за счет вентролатеральных пучков латеральной мышцы
носа, разных порций m. buccinator и, возможно, продольных пучков
платизмы и m. maxillolabialis. Зафиксированное Хартманом (Hartman,
1979, fig. 28) втягивание губного диска внутрь носовой капсулы, приво-
дящее к тому, что нижняя сторона его обращается вперед, осуществляет-
ся носо-губным поднимателем и m. maxillonasolabialis. При этом перед-
ний край латеральной мышцы носа оказывается перемещенным так, что'
ее тяга может способствовать выдвижению вперед верхней губы. При
работе наружных продольных пучков m. maxillonasolabialis латеровент-
ральный угол верхней губы может оттягиваться наружу, а более глубо-
ких пучков — внутрь. М. retractor labiorum оттягивает внутреннюю по-
верхность верхней губы, а при одновременном действии мышц, подходя-
щих к углу рта (щечная часть платизмы), верхняя губа активно
опускается вниз и нависает сверху и сбоку относительно нижней губы.
201
Таблица 4. Лицевые мышцы,
управляющие движением губ
Мышцы Масса
г %
ш. levator nasolabialis 344 18,1
m. maxillonasolabialis 576 30,3
m. lateralis nasi 490 25,8
m. mentalis 30 1,6
m. retractor labiorum 10 0,5
m. buccinatorius s. str. 450 23,7
Всего 1900 100,0
Мышцы обеих сторон 3800 0,46 *
* В % к массе тела.
Манипулирование нижней губой
осуществляется за счет действия
подбородочной и нижнечелюстной
мышц, волокна которых, как было
сказано выше, образуют впереди
сложное переплетение.
Общая масса лицевых мышц, со-
ставляет 0,46% от массы тела, что
значительно больше, чем всех мышц
подъязычного аппарата и собственно
языка (0,2%), и больше, чем это
обычно бывает у млекопитающих.
Если же учесть, что в работе лице-
вых мышц принимают также уча-
стие щечная часть платизмы, волок-
на ее скуловой части, оканчиваю-
щиеся на верхней губе, а также ряд
пучков поверхностного сфинктера
шеи, то становится ясным, что у сирен значение лицевой мускулатуры
резко возросло. Наиболее мощными из всей группы мышц (табл. 4) явля-
ются in. maxillonasolabialis и m. lateralis nasi, совместное действие кото-
рых обеспечивает открывание и закрывание ноздрей и сложные манипу-
ляции верхней губы. Самые слабые из них — m. mentalis и m. retractor
labiorum.
ЖЕВАТЕЛЬНАЯ МУСКУЛАТУРА
Этот комплекс (рис. 72—78) включает у ламантина две жевательные,
две крыловидные, височную и двубрюшную мышцы.
М. masseter lateralis (m. masseter, по Домнингу), латеральная жева-
тельная мышца (см. рис. 71, 72, 74), начинается мощным сухожилием
от вентрального отростка скуловой кости (см. рис. 75, ty, 76) и, посте-
пенно расширяясь, крепится к латеральной поверхности угла нижней че-
люсти (см. рис. 77, а) несколькими (до 9) внутренними сухожильными
апоневрозами (рис. 78, з), наличие которых придает мышце сложнопе-
ристую структуру.
М. masseter medialis (m. zygomaticomandibularis, по Домнингу),
медиальная жевательная мышца (см. рис. 72, 74) начинается от вент-
рального края скуловой кости сразу каудальнее отхождения предыдущей
мышцы, вентральной поверхности скулового отростка височной кости
впереди от суставного бугорка (см. рис. 75, б, 76) и дорзальной поверх-
ности основания того же отростка латеральнее отхождения височной
мышцы (см. рис. 75, а). Почти на всей площади своего отхождения ме-
диальная жевательная мышца начинается мышечными волокнами, лишь
на латеральном крае скуловой дуги имеется сухожильный апоневроз,
и другой апоневроз находится у границы с височной мышцей. Прикреп-
ляется мышца к наружной поверхности венечного отростка (см. рис. 77,
а) как мышечпо, так и несколькими внутренними апоневрозами (рис. 78,
ж), места окончания которых видны на кости как более или менее вы-
раженные гребни, пересекающие венечный отросток. Один из этих апо-
неврозов идет в середине толщи мышцы, второй по наружной поверх-
ности. Судя по месту своего отхождения, эта мышца соответствует
медиальной жевательной мышце других млекопитающих, лишь часть во-
локон которой, идущих от дорсальной поверхности основания скулового
отростка, может сопоставляться с m. zygomaticomandibularis. Однако
строение скуловой дуги сирен не позволяет им идти в направлении,
202
Рис. 78. Поперечные
грезы жевательных мышц
2 — m. pterygoideus medi-
alis
(7 — середина мышцы,
2 — ее начало);
3 , 4 — та. pterygoideus late-
ralis
(3 — середина мышцы,
4 — ее начало);
5 — m. temporalis на уров-
не дорсального края
венечного отростка;
6 — m. digastricus, сере-
дина мышцы;
7 — т. masseter medialis
на уровне дорсально-
го края венечного от-
ростка;
3 — m. masseter lateralis
на уровне 1/4 длины
(от начала)
О Зсм
I I I------1
свойственном настоящему m. zygomaticomandibularis. Поэтому мы счи-
таем необходимым закрепить за этой мышцей название медиальной же-
вательной, поскольку большая ее часть, если не вся она полностью, го-
мологична именно этой мышце млекопитающих.
М. temporalis, височная мышца (см. рис. 72—74) начинается от всей
чешуи височной кости сразу выше linea temporalis, спереди от crista
temporalis и латеральной поверхности теменных и лобных костей от
cristae parietalis et frontalis вентральнее вплоть до скулового отростка
(см. рис. 75, а, 76). По гребням начало осуществляется апоневрозом,
а по остальной поверхности мышечно. Прикрепляется мышца на меди-
альной поверхности венечного отростка мышечно и мощным внутрен-
ним апоневрозом, крепящимся на особых гребешках- на этой поверхности
ближе к ее дорзальному краю, а также апоневрозом, идущим по внут-
ренней стороне мышцы к бугру у основания венечного отростка
(рис. 78, б). Каудальнее венечного отростка мышечные волокна височ-
ной мышцы практически не обособлены от таковых m. masseter medialis.
М. pterygoideus lateralis (т. pterygoideus externus, по Домнингу),
латеральная крыловидная мышца начинается мышечносухожильно от
наружной поверхности крыловидного отростка основной клиновидной ко-
сти почти от суставного бугорка сверху, а вентрокаудально — до отхож-
дения медиальной крыловидной мышцы (рис. 76, 79). Мышечные волок-
на идут к мыщелковому отростку нижней челюсти, на медиальной по-
верхности которого и кончаются. Сильный сухожильный апоневроз идет
внутри мышцы (рис. 78, в, г) и крепится на гребне, пересекающем мы-
щелковый отросток сверху от особого отростка на заднем его крае вниз
и вперед.
М. pterygoideus medialis (m. pterygoideus internus, по Домнингу),
медиальная крыловидная мышца (см. рис. 71, 80, 81), начинается на
заднем крае крыловидного отростка близ его вентрального конца
(см. рис. 76, 79), где она граничит с крылоглоточной мышцей. Начало
осуществляется мышечносухожильно. Наиболее плотный апоневроз идет
203
Рис. 79. Площади прикрепления мышц
на черепе (вид сзади)
1 — m. acromiotrapezlus;
г — in. clavotrapezius;
3 — m. digastricus;
4 — m. masseter medialis;
5 — m. obliquus capitis cranialis;
в — 7 — m. pterygoideus
(6 — lateralis,
7 — medialis);
8 — m. pterygopharyngeus;
9 — 11 — m. rectus capitis dorsalis
(9 — major,
10 — intermedius;
11 — minor);
12 — m. rectus capitis lateralis;
13 — m. semispinalis capitis;
14 — m. splenlus;
15 — m. stylohyoideus;
16 — m. stylopharyngeus;
17—m. temporalis;
18 — nt. trapezius p. mandibularis
по переднему краю мышцы. 1олща мышцы пронизана несколькими апо-
неврозами (рис. 78, а, б), число которых к концу возрастает до 8—9.
Крепится мышца на медиальной поверхности угла нижней челюсти
(рис. 77, б), причем для конечных апоневрозов здесь имеются неясно
выраженные, короткие, радиально расположенные гребни.
М. digastricus двубрюшная мышца (см. рис. 71, 80, 81), начинается
на заднем крае тимпаногиоида близ его прикрепления к парокципиталь-
ному отростку боковой затылочной кости и частично на стилогиоиде
(см. рис. 76, 79). Мышечные волокна образуют единственное веретено-
видное брюшко, идущее к самому вентральному краю угла нижней че-
люсти (см. рис. 77). Латерально крепление осуществляется прямо мы-
шечными волокнами, а медиально в концевой части мышцы виден ряд
апоневрозов (см. рис. 78, е), образующих со своими мышечными волок-
нами как бы налегающие друг на друга лепестки.
Работа жевательных мышц была обстоятельно рассмотрена Домнин-
гом (Domning, 1978а), и нет нужды здесь повторять этот разбор.
Отметим лишь, что на открывание
рта работает лишь двубрюшная
мышца и отчасти некоторые пучки
латеральной крыловидной мышцы.
Все остальные мышцы комплекса
закрывают рот и обеспечивают сжи-
мание челюстей, как и возможные
боковые движения, которые, однако,
ограниченны из-за сильного развития
крыловидных отростков. Выяснение
способов первичной обработки пищи
в ротовой полости нуждается в даль-
нейшем изучении, в частности не
совсем ясны особенности смыкания
зубов, их стирания и соответственно
их участия в пережевывании корма.
Необходимы исследования челюстно-
го сустава и той дополнительной
Таблица 5. Мышцы жевательного
аппарата
Мышцы Масса
Г %
m. temporalis 235,0 36,1
т. masseter lateralis 128,0 19,7
т. masseter medialis 157.8 24,2
т. pterygoideus lateralis 28,8 4,4
m. pterygoideus medialis 59,3 9,1
m. digastricus 42,0 6,5
Всего 650,9 100,0
Мышцы обеих сторон 1301,8 0,33*
* В % к массе тела.
204
Рис. 80. Мышцы подъ-
язычного аппарата (вид
снизу)
Слева — удалены: m. genio-
hyoideus, m. mylohyoideus et
myloglossus; pr. lingualis
и нижняя челюсть
1 — m. cricopharyngeus;
2 — m. cricothyroideus;
3 — m. digastricus;
4 — m. genioglossus;
5 — m. geniohyoideus;
6 — 7 — та- hyoglossus
(6 — lateralis,
7 — medialis);
8 — m. hyopharyngeus;
s — m. masseter lateralis;
J.0 — m. myloglossus;
JJ — m. mylohyoideus;
12 — m. pterygoideus media-
lis;
J.3 — m. pterygopharyngeus;
14 — m. sternohyoideus;
15 —m. sternothyroideus;
IS — 18 — m. styloglossus
ifj fi — intermedius,
17 — lateralis,
18 — medialis);
19 — m. stylohyoideus;
20 — m. stylopharyngeus;
21 — 22 — m. thyrohyoid',
us
<(22 — minor);
.23 — чудесная сеть нижней
челюсти
'опоры, которую, по Домнингу, получает нижняя челюсть при контакте
в некоторых положениях с крыловидным отростком. Дополнительно мож-
но отметить, что распределение масс жевательных мышц (табл. 5) точно
-совпадает с размерами рычага приложения сил: наибольшая масса у ви-
сочной мышцы соответствует и наибольшему рычагу (8—10 см), а наи-
меньшая — у латеральной крыловидной — минимальной длине рычага
(1—2 см). Рычаги у остальных мышц имеют следующую длину: медиаль-
ная жевательная мышца — 7—9 см, латеральная жевательная — 5—7 см,
медиальная крыловидная — 3—5 см, двубрюшная — 4 см. Необходимо так-
же напомнить, что сильное отклонение венечного отростка вперед у ла-
тмантина сравнительно с дюгонем и морской коровой в значительной сте-
:пени увеличивает длину рычагов основных жевательных мышц.
205
Рис. 81. Мышцы подъязычного аппарата
лена)
1 — cart, cricoidea;
2 — cart, thyroidea;
з — 4 — cart, hyoidea
{3 —. corpus,
4 — pr. Hngualis);
5 — cart, stylohyoidea;
6 — lingua.
Мышцы:
7 — m. cricoarytaenoideus dor-
salis;
8 — m. crlcopharyngeus;
(вид сбоку,
9 — m. cricothyroldeus;
10 — m. digastricus;
11 — m. genioglosus;
12 — m. geniohyoideus;
13 — 14 — m. hyoglossus
{13 — lateralis,
14 — medialis);
15 — m. hyopharyngeus;
16 — m. myloglossus;
17 — m. mylohyoideus;
18 — m. pterygoideus medialis;
правая нижняя челюсть уда-
19—m. pterygopharyngeus;
20 — 22 — m. styloglossus
{20 — intermedius;
21 — lateralis;
22 — medialis);
23 — m. stylohyoideus;
24 — m. stylopharyngeus;
25 — 26 — m. thyrohyoideus-
(26 — minor);
27 — m. thyropharyngeus
МЫШЦЫ ЯЗЫКА И ПОДЪЯЗЫЧНОГО АППАРАТА
Скелетную основу языка и всего подъязычного аппарата образует подъ-
язычная кость (рис. 82—85), которая у взрослого экземпляра Trichec-
hus manatus, имевшегося в нашем распоряжении, заметно отличается от
таковой Т. inunguis (судя по описанию молодых экземпляров (Домнинг,
1978а)) и тем более дюгоня (Domning, 1977b). Единый непарный эле-
мент — базигиоид, или тело (corpus), имеет вид расправленной бабочки
с широкими крыльями и суженной в середине частью (рис. 83, б). В се-
редине он окостеневший, но с поверхности и особенно по краям хряще-
вой. Всем каудальным краем этот элемент прижат к щитовидному хря-
щу, так что передний выступ последнего вклинивается в заднюю сре-
динную выемку тела. Никаких следов обособленных задних рогов, или
тиреогиоидов, нет в отличие от бразильского ламантина и дюгоня, у ко-
торых близкий по форме непарный элемент подъязычной кости отделен
от щитовидного хряща значительным расстоянием и небольшими парны-
ми хрящами, сочлененными с заднебоковыми углами задних расширен-
ных лопастей тела. Спереди к телу подвижно прикреплены парные вы-
тянутые вперед широкие хрящи, связанные друг с другом по средней
линии фиброзной тканью. Их можно условно сопоставить с язычным от-
ростком (processus lingualis) ряда млекопитающих, хотя он всегда бы-
вает непарным и обычно не отделен от тела. У бразильского ламантина!
206
Рис. 82. Мышцы глотки
и гортани (вид с дор-
сальной стороны)
Справа удалены глоточные
мышцы и пищевод
Г — 9 — хрящи и кости:
1—cart, arytaenoidea;
2 — cart, cricoidea;
3 — cart, ctmeiformae;
4 — cart, epigloticus;
5 — cart, thyroidea;
в — cart, tracheae;
7 — corpus hyoidea;
8 — pr. pterygoideus;
9 — stylohyoid;
10 — 21 — мышцы:
10 — m. arytenoideus trans-
versus;
11 — 12 — m. cricoarytenoi-
deus
(It — dorsalis;
12 — lateralis);
13 — m. cricopharyngeus;
14 — m. hyoepiglotticus;
IS — m. hyopharyngeus;
16 — m. pterygoideus media-
lis;
17 — m. pterygopharyngeus;
20 — m. thyrohyoideus mi-
nor;
21 — m. thyropharyngeus;
22 — пищевод
и дюгоня этот отросток вообще не отмечен. Большие, длинные, уплощен-
ные и слегка расширенные вентрокраниально парные костные элементы,
причлененные к боковым крыльям тела подъязычной кости и направлен-
ные назад и вверх к черепу, можно сопоставить с малыми рогами обыч-
ной подъязычной кости млекопитающих. Их задние концы прочно сое-
динены с относительно короткими хрящами, сращенными своими расши-
ренными концами с парокципитальными отростками боковых затылочных
костей. Судя по топографическому положению, эти хращевые элементы
-следует определить как tympanohyoideum, а длинные костные элементы
как stylohyoidewn, так как именно с ними связан m. styloglossus
(см. рис. 85). Однако при описании подъязычной мускулатуры бразиль-
ского ламантина Домнинг (1978а) этот удлиненный элемент называет
эпигиоидом, а 1 укороченный хрящевой — стилогиоидом, считая, видимо,
что тимпаногиоид отсутствует. Кроме того, им выделен небольшой чле-
ник — кератогиоид, соединяющий тело с эпигиоидом. Аналогичное со-
стояние указано им и для дюгоня (Domning, 1977b), лишь кератогиоид
здесь значительно расширен. Однако следует отметить, что обычно у
млекопитающих эпигиоиды значительно меньше стилогиоидов, а не на-
оборот. Все это заставляет сомневаться в правильности интерпретаций
207
Рис. 83. Площади при-
а,
1 — cart, arytenoideus;
2 — cart, cricoldeus;
3 — cart, cuneiformes;
4 — cart, epigloticus;
5 — 6 — cart, hyoidea
(5 — corpus,
в — pr. lingualis);
7 — cart, thyroideus;
8 — cart, tracheae;
9 — stylohyoid;
10 — tympanohyoid.
Мышц ы:
11 — m. arytenoideus transver-
sus;
12 — 13 — m. cricoarytenoideus
(12 — dorsalis;
13 — lateralis);
14 — m. crlcopharyngeus;
15 —m. cricothyroideus;
16 — m. geniohyoldeus;'
17 — m. hyoepiglotticus;
18 — is — m. hyoglossus
18 — lateralis,
19 — medialis);
20 — m. hyopharyngeus;
21 — m. myloglossus;
22 —m. mylohyoideus;
23 — m. sternohyoideus;
24 — m.~ sternothyroideus-;.
25 — 27 — m.styloglossus
(25 — intermedius,
26 — lateralis,
27 — medialis);
28 — m. stylohyoideus;
29 — m. stytopharyngeus;’”'™'
30 — m. thyroarytenoideus vent-
rails (s. m. ventricularis)
31 — 32 — m. thyrohyoldeus
(32 — minor);
33 — m. thyropharyngeus
208
3 — cart, epiglotticus:
4 — 5 — cart, hyoideus
(4 — corpus;
5 — pr. lingualis);
в — cart, tliyroideus;
7 — cart, tracheae;
8 — stylohyoid;
9 — tympanohyoid.
sus;
11 — 12 — m. cricoarytenoideus
(11 — dorsalis,
12 — lateralis);
13 — m. cricopharyngeus;
14 — m. cricothyroideus;
15 — m. geniohyoideus;
(16 — 17 — m.hyoglossus
16 — lateralis,
21 — m. sternohyoldeus;
22 — m. sternothyroldeus;
23 — 25 — m.styloglossus
(23 — intermedius,
24 — lateralis,
25 — medialis);
26 — m. stylohyoideus;
27 — m. stylopharyngeus;
28 — 29 — m.thyrohyoldeus
17 — medialis);
18 — m. hyopharyngeus;
2 9 — minor);
30 — m. thyropharyngeus
Домнинга. Таким образом, мы считаем, что у нашего экземпляра ламан-
тина эпигиоиды или исчезли, или слились с образованием единого эле-
мента со стилогиоидом (этот элемент здесь будет называться stylohyoide-
шп) кератогиоиды или исчезли, или слились с телом подъязычной ко-
сти, а элементы, названные Домнингом стилогиоидами, являются
тимпаногиоидами.
Как у всех млекопитающих, толща языка ламантина пронизана мы-
шечными волокнами, идущими во взаимно перпендикулярных направле-
ниях. Они образуют ш. lingualis proprius и позволяют языку сжиматься
с боков, становясь более высоким, уплощаться или укорачиваться. Три
другие мышцы (шилоязычная, подъязычноязычная и подбородочноязыч-
ная) обеспечивают собственно подвижность языка.
М. styloglossus, шилоязычная мышца (см. рис. 80, 81), дифференци-
рована на три части, различающиеся как по своему положению, так и
местом отхождения. Их удобно обозначить по положению их окончания
в толще языка —более медиальному, промежуточному и латеральному.
Pars medialis начинается мышечно от обширной площади латеральной
поверхности стилогиоида (от 2/s его проксимальной длины) и небольшой
части тимпаногиоида (рис. 84) сразу вентральное и несколько медиальнее
отхождения двубрюшной мышцы, а кончается в корне языка близ его
209
дорсальной поверхности. Эта часть соответствует первой части
in. styloglossus бразильского ламантина и дюгоня (Domning, 1977b,
1978). Pars intermedia в своем начале полностью обособлена от предшест-
вующей, отходя мышечными волокнами от небольшого участка латераль-
ной поверхности стилогиоида у его вентрокраниального конца, сочленен-
ного с телом подъязычной кости (см. рис. 84). Волокна этой части также
идут в корень языка, но несколько латеральнее предыдущей части. Оче-
видно, эта часть соответствует второй части дюгоня, описанной Домнин-
гом, но у бразильского ламантина отсутствующей. Третья р. lateralis —
наиболее длинная порция шилоязычной мышцы. Она начинается длин-
ным сухожилием от шероховатой линии на латеральной поверхности
стилогиоида точно позади отхождения медиальной части, начиная с са-
мого ближнего к черепу (проксимального) конца его (см. рис. 83, 84).
Начальное сухожилие (см. рис. 81) имеет вид сильно вытянутого тре-
угольника, одна длинная сторона которого закреплена на кости, а ко-
роткое основание продолжается уже мышечными волокнами, идущими
вперед вдоль всей вентральной поверхности языка вплоть до его верши-
ны. Эта часть соответствует третьей части Домнинга. Однако и у дюго-
ня, и у бразильского ламантина отхождение ее сдвинуто дистально бли-
же к телу подъязычной кости.
М. hyoglossus, подъязычная мышца (см. рис. 80, 81), по своему строе-
нию отличается от описанного у дюгоня и бразильского ламантина Дом-
нингом. Фактически здесь имеются две мышцы, не имеющие связи друг
с другом и разделенные прохождением третьей мышцы (m. genioglossus),
но они обе связаны своим отхождением с гиоидом, а окончанием с язы-
ком, что и позволяет определять их как подъязычноязычные. М. hyog-
lossus lateralis начинается от дорсальной поверхности язычного отростка
соответствующей стороны (см. рис. 83, а). Сзади мышечные волокна идут
от него вверх, а далее вперед вверх и вперед, веерообразно расходясь и
образуя тонкую вентрально поставленную пластинку мышечного брюш-
ка (она образует второй от края валик на вентральной поверхности язы-
ка; первый отмечает прохождение m. styloglossus р. lateralis). Отсутствие
язычковых отростков у бразильского ламантина и дюгоня приводит к
тому, что у первого mm. hyoglossi обеих сторон (они, видимо, соответст-
вуют нашему m. hyoglossus lateralis) в своем начале срастаются друг с
другом по средней линии, где они в свою очередь закреплены на сухо-
жильных концевых частях mm. mylohyoideus et geniohyoideus, т. e. здесь
нет закрепления на подъязычной кости. У дюгоня отхождение m. hyog-
lossus сдвинуто значительно каудальнее (как и прикрепления mm. mylo-
hyoideus et geniohyoideus) и помещается на вентральной поверхности
тела подъязычной кости и частично на кератогиоиде. М. hyoglossus me-
dialis у нашего ламантина начинается мясисто от дорсальной поверхно-
сти язычного отростка (см. рис. 83, а), медиальное отхождения латераль-
ной мышцы и от передней части тела подъязычной кости. Мышечные
волокна образуют округлое в сечении брюшко, постепенно сужающееся
вперед к вершине языка. Оно заметно на вентральной поверхности язы-
ка в виде широкого валика в самой медиальной ее части. Эта часть
Домнингом вообще не описывается. Можно предполагать, однако, что он
не смог ее дифференцировать от m. genioglossus, часть волокон которого
у дюгойя, судя по его описанию, вопреки обыкновению Достигает дор-
сальной поверхности тела подъязычной кости и кератогиоида.
М. genioglossus, подбородочноязычная мышца (см. рис. 81), ня чиня -
ется в специальном углублении аборальной поверхности симфиза ниж-
ней челюсти (см. рис. 78, б) медиальнее отхождения m. geniohyoideus
крепким, идущим назад и постепенно утончающимся сухожилием и ча-
210
стично мышечными волокнами. Последние отходят также от всей дорсаль-
ной поверхности начального сухожилия и идут вверх и назад, образуя
вертикально поставленную тонкую мышечную пластинку, уходящую
в глубь языка (ее вентральный край вместе с сухожилием образует на
нижней поверхности языка валик, заметный между соответствующими
валиками m. hyoglossus medialis медиально и т. hyoglossus lateralis
латерально). Лишь в корне языка конпевые волокна т. genioglossus
сливаются с волокнами медиальной части подъязычноязычной мышцы.
В движении языка, помимо описанных выше мышц, принимает
участие еще ряд мышц, управляющих положением самой подъязычной
кости. При этом мышцы, соединяющие ее с грудиной и щитовидным
хрящем, а также мышца, связывающая грудину с щитовидным хрящем,
отвечают за оттягивание всего подъязычного аппарата назад и вниз,
а мышцы, идущие от нижней челюсти к гиоиду, восстанавливают его по-
ложение, что облегчает проникновение пищи в пищевод (при этом хря-
щевой тимпаногиоид сгибается). В составе этого комплекса насчитыва-
ется семь следующих мышц.
М. sternohyoideus, грудинноподъязычная мышца (см. рис. 71,80), на-
чинается мышечно от узкой поперечной полоски передней части внут-
ренней (дорсальной) поверхности рукоятки грудины сразу позади отхож-
дения m. sternothyroideus. Волокна мышцы, направляясь вперед, обра-
зуют плоское ремневидное брюшко. Крепятся мышцы на особой сухо-
жильной перемычке, образованной непосредственно вентральнее язычных
отростков в области срастания по средней линии правой и левой подъ-
язычночелюстных мышц (см. рис. 83, б, 84, 85). У одного из экземпля-
ров бразильского ламантина эта мышца отсутствует (Domning, 1978а),
то же отмечается для Trichechus manatus (Murie, 1872а). Очевидно, это
индивидуальная вариация у ламантинов, а возможно, и у дюгоня, у ко-
торого мышца также не наблюдалась (исследовано по одному экземпля-
ру, см.: (Domning, 1977b)).
М. sternothyroideus, грудиннощитовидная мышца (см. рис. 71, 80),
также начинается от внутренней поверхности рукоятки грудины сразу
впереди от отхождения предыдущей мышцы (рис. 86) и образует плос-
кое брюшко с параллельным направлением волокон (см. рис. 71), при-
крытое с вентральной стороны более широкой грудинноподъязычной
мышцей, причем ближе к своему окончанию грудиннощитовидная мыш-
ца помещается немного медиальнее грудинноподъязычной, идущей даль-
ше вперед. Оканчивается m. sternothyroideus на латероаборальном крае
щитовидного хряща гортани позади отхождения m. thyrohyoideus
(см. рис. 83, Ь, 84). Несмотря на плотное прилегание обеих мышц,
срастания их не наблюдается. Отмеченное Домнингом (1978а) отхожде-
ние m. sternothyroideus не только от грудины, но и от смежной хряще-
вой части первого ребра вызывает сомнение, так как сочленение ребра
с грудиной осуществляется позади рукоятки далеко от начала мышцы.
В то же время у дюгоня m. sternothyroideus вообще не связан с груди-
ной, а начинается от I ребра и его хряща (Domning, 1977b).
М. thyrohyoideus, щитовидноподъязычная мышца (см. рис. 71, 80, 86),
начинается мышечно в тесной связи с окончанием m. sternothyroideus
(отделен только сухожильной перемычкой) (см. рис. 83 б, 84) на вентро-
латеральной поверхности щитовидного храща. Брюшко мышцы — пло-
ское, широкое. Оно прикрыто с вентральной стороны передней частью
ш. sternohyoideus. Кончается щитовидноподъязычная мышца на уже
упомянутой сухожильной перемычке m. mylohyoideus глубже прикреп-
ления т. sternohyoideus (см. рис. 83, б, 84). Домнинг (1978а) выража-
ет сомнение в гомологичности данной мышцы с одноименной у болыпин-
211
Рис. 85. Мышцы гортани и подъязычного аппарата
а — детали прикрепления мышц на черпаловидном хряще (вид с латеральной стороны); б — срез по
средней линии (вид с медиальной стороны)
1 — cart, arytenoideus;
2 — cart, cricoideus;
3 — cart, epiglotticus;
4 — S — cart, hyoideus
(4 — corpus;
•5 — , pr. lingualis);
6 — cart, thyroideus;
7 — cart, tracheae:
a — stylohyoid;
9 — tympanohyoid.
Мышцы:
10 — m. arytenoideus transver-
sus;
11 — 13 — m. cricoarytenoideus
(11 — dorsalis,
12 — lateralis,
13 — ventralis (s. vocalls);
14 — m. cricothyroideus;
15 — m. geniohyoideus;
16 —m. hyoepiglotticus;
17 — 18 — m. hyoglossus
(17 — lateralis;
18 — medialis);
19 — m. myloglossus;
2 0 — m. mylohyoideus;
21 — m. stemohyoldeus;
22 — m. stylohyoideus;
23 — m. stylopharyngeus;
24 —in. thyroarytenoideus (s.m.
ventricularis);
25 — m. thyrohyoideus;
26 — слизистая гортани
ства млекопитающих. Действительно, ее прикрепление у американского
ламантина оказалось смещенным вперед к области язычных отростков и
не связано с задними рогами (тиреогиоидами) подъязычной кости и даже
с самим ее телом. Однако обращает на себя внимание сдвиг вперед у
ламантина и прикрепления m. sternohyoideus, а именно связь в оконча-
нии щитовидноподъязычной и грудиноподъязычной мышц характерна
для млекопитающих. У дюгоня прикрепление m. thyrohyoideus занимает
более типичное для млекопитающих положение, будучи связано с бази-
гиоидом.
М. thyrohyoideus minor, малая щитовидноподъязычная мышца (см.
рис. 80),— небольшая веерообразная мышца, связывающая переднелате-
ральный край щитовидного хряща с аборальным крылом тела подъязыч-
ной кости (см. рис. 83, б, 84). У бразильского ламантина она кончается
на тиреогиоиде, а у дюгоня как самостоятельная отсутствует (Domning,
1977b, 1978а). Таким образом, ее появление, как и положение мышц,
описанных ранее, обусловлено перестройкой подъязычного аппарата, на-
блюдающейся у нашего экземпляра ламантина и большей специализаци-
ей этой области ламантинов относительно дюгоней.
212
Рис. 86. Мышцы шеи и передней части туловища
(вид снизу). Глубокий слой
1 — gl. parothyroidea;
2 — gl. parotis;
3 — gl. thyroidea;
4 — m. acromlodeltoideus;
4 — m. clavodeltoideus;
6 — m. cutaneus trunci p. dor-
salis;
7 — m. intercostalis internus p.
cranialis;
a — m. masseter lateralis;
9 — m. pectoralis abdomlnalis;
10 — m. pectoralis profundus;
11 — m. pectoralis superficia-
lis;
12 — platysma myoides p. Jugu-
laris;
13 — m. rectus abdominis;
14 — m. spinodeltoideus;
15 — m. sternohyoideus;
16 — m. sternomastoideus;
17 — m. sternothyroldeus;
18 — m. thyrohyoideus;
19 — rn. transversus thoracitis;
20 — in. trapezius p. mandibu-
laris;
21 — thymus
M. stylohyoideus, шилоподъязычная мышца (см. рис. 71, 80), начи-
нается мышечно от задней стороны тимпаногиоида (см. рис. 79, 83—
85). Мышца имеет веретеновидное брюшко, идущее вперед и медиально,
которое переходит в длинное сухожилие, делящееся на два конца. Более
латеральный и поверхностный из них крепится на боковой поверхности
213
язычного отростка, а более глубокий простирается до переднего края
тела подъязычной кости (см. рис. 83, б, 84).
У бразильского ламантина окончание связано с кератогиоидом (уже
отмечалось, что у нашего экземпляра он мог слиться с телом подъязыч-
ной кости), а у дюгоня с задней частью вентральной поверхности бази-
гиоида.
М. mylohyoideus, подъязычночелюстная мышца (см. рис. 71, 80, 83—
85), лежит под языком в межчелюстном пространстве. Она начинается:
от узкой полоски медиальной поверхности нижней челюсти (linea mylo-
hyoideum), идущей под ее альвеолярным краем от уровня заднего края
второго зуба на 3—4 см далее конца зубного ряда (см. рис. 77, б)..
Мышечные волокна направляются медиально, каудально и несколько
вверх, дифференцируясь на две порции, разделенные пучком чудесной
сети, проходящей в foramen mandibulae. Более передняя по положению,
сопоставимая с in. myogloss us других млекопитающих, образует мышеч-
ный слой, волокна которого срастаются с мышцей противоположной сто-
роны непосредственно под язычными отростками подъязычного аппарата.
Волокна второй, более задней порции, частично перекрывающей сна-
ружи (с вентральной стороны) переднюю часть, связаны с уже упоми-
навшейся выше сухожильной перемычкой, лежащей перед телом подъ-
язычной кости. Она имеет треугольные очертания (см. рис. 71): одна
сторона этого треугольника образует средний шов с аналогичным сухо-
жильным апоневрозом противоположной стороны, задняя сторона связа-
на с окончанием m. sternohyoideus и т. thyrohyoideus, а третья, обращен-
ная вперед и латерально,—с задней порцией m. mylohyoideus. Как и у
бразильского ламантина, m. mylohyoideus нашего ламантина в отличие-
от дюгоня (Domning, 1977b, 1978а) не имеет прикрепления с вентраль-
ной поверхности базигиоида.
М. geniohyoideus, подбородочноподъязычная мышца (см. рис. 80, 81) т
начинается апоневрозом от края специального углубления на аборальной
поверхности симфиза нижней челюсти и по медиальному краю ветви
нижней челюсти каудальнее отхождения m. genioglossus (см. рис. 77).
Появляющиеся скоро в апоневрозе мышечные волокна образуют плоское
брюшко мышцы, которое идет назад, слегка расширяясь, и прикрепляет-
ся на переднем крае язычного отростка подъязычной кости (см. рис. 83—
85), уходя, если смотреть снизу, под m. mylohyoideus. В отличие от опи-
санного у бразильского ламантина и дюгоня m. geniohyoideus в нашем
случае не был связан с сухожильной перемычкой т. mylohyoideus. В то-
же время не было и крепления на теле подъязычной кости, что отмеча-
лось для Trichechus manatus (Murie, 1972а).
Mm. omohyoideus, jugulohyoideus et keratohyoideus у сирен не обна-
ружены.
МЫШЦЫ ГЛОТКИ И ГОРТАНИ
Глоточные мышцы связывают глотку с черепом, подъязычной костью и
хрящами гортани, образующими скелетную основу переднего конца тра-
хеи.
Гортань ламантина расположена непосредственно позади тела подъ-
язычной кости, которое своим задним краем тесно прилежит к передне-
му краю cartilago thyroidea щитовидного хряща (см. рис. 83—85).
Последний представляет собой две сильно расширенные аборально лопа-
сти, соединенные вентрально узкой перемычкой, вытянутой вперед в
виде клина, входящего в заднюю срединную выемку базигиоида. Сам
щитовидный хрящ образует как бы вентральное полукольцо, охватываю-
щее просвет трахеи, и подвижно соединен дорсально с вентральными ло-
214
пастями cart, arytenoidea черпаловидного хряща, представляющего дор-
сальное полукольцо, так что они могут несколько вдвигаться друг в дру-
га, меняя ширину просвета. На дорсальной поверхности щитовидного
хряща лежит закрепленный крайне подвижно связками сильно расши-
ренный и корытообразно выемчатый сверху cart, epiglottica надгортан-
ный хрящ, функция которого сводится в основном к замыканию просве-
та трахеи во избежание попадания туда пищи. Сзади к щитовидному и
черпаловидному хрящам примыкает cart, cricoidea, кольцевидный хрящ,
образующий полное, очень широкое кольцо, расширенное, видимо, еще
за счет прирастания сзади передних хрящевых колец самой трахеи.
Дорсально и вентрально кольцо сильно уплощено, и там сверху на коль-
цевидном хряще имеются срединные гребни, в дорсолатеральных углах
ближе к переднему краю помещаются бугры, служащие для мышечных
прикреплений. Первое хрящевое кольцо трахеи несколько вдвинуто
внутрь в кольцевидный хрящ. Дорсально оно имеет расширенную упло-
щенную площадку, а вентрально — разрыв в хряще. Второе кольцо силь-
но сращено вентрально с первым. Разрыв в третьем кольце приходится
на середину дорсальной стороны, как и у пятого, а у четвертого лежит
с правой стороны.
Перечисленные хрящи, входящие в состав гортани, служат местом
прикрепления не только глоточных мышц, но и собственно мышц горта-
ни. Число последних варьирует (от 5 до 10) у разных животных.
У ламантина их число несколько сокращено.
Пять мышц образуют собственно мускулатуру глотки.
М. hyopharyngeus, подъязычноглоточная мышца (см. рис. 82), начи-
нается мышечно от дорсальной поверхности бокового крыла тела
подъязычной кости (см. рис. 83, а, 84) и идет постеро-дорсально к глот-
ке, где, соединяясь с мышцей противоположной стороны, участвует в
образовании боковой и частью дорсальной ее стенки. У бразильского ла-
мантина и дюгоня описано отхождение от стилогиоида, а не от базигио-
ида.
М. stylopharyngeus, шилоглоточная мышца (см. рис. 82), начинается
от большей части медиальной поверхности стилогиоида и смежной части
тимпаногиоида (см. рис. 84, 85). Мышечные волокна идут широким вее-
ром вперед до уровня крыловидных отростков черепа и несколько вент-
рально и, соединяясь с мышцей противоположной стороны, формируют
дорсальную часть стенки носоглотки. Большая часть их начинается не-
посредственно от кости. Однако на дорсомедиальной поверхности брюш-
ка видно треугольное сухожильное зеркало, волокна которого начинают-
ся от самого проксимального конца стилогиоида.
М. pterygopharyngeus, крылоглоточная мышца (см. рис. 82), начина-
ется мясисто от аборального края крыловидных отростков (см. рис. 79).
Его волокна идут веером назад, оканчиваясь в дорсальной стенке глот-
ки, где они смешиваются с волокнами m. stylopharyngeus, имеющими
встречное направление, и m. hyopharyngeus с поперечным направлением.
М. thyropharyngeus, щитовидноглоточная мышца (см. рис. 82), начи-
нается мышечно от щитовидного хряща дорсальнее прикрепления
mm. sternothyroideus et thyrohyoideus. Место его отхождения вытянуто в
виде узкой полоски на дорсальной поверхности тироида и его каудаль-
ных рогов (см. рис. 83, б, 84). Мышечные волокна идут дорсально в
стенке глотки, встречаясь с встречными волокнами мышцы противопо-
ложной стороны.
М. cricopharyngeus, кольцевидноглоточная мышца (см. рис. 82), обра-
зует наиболее аборальные пучки мышечных волокон глотки. Его волок-
на начинаются на боковых мышечных буграх кольцевидного хряща
215
(см. рис. 83, б, 84) и идут антеродорсально в стенку глотки, навстречу
волокнам мышцы противоположной стороны.
У ламантина имеются восемь мышц, связанных исключительно с гор-
танью.
М. cricothyroideus, кольцевиднощитовидная мышца (см. рис. 80, 81),.
начинается мышечно на вентральном срединном гребне кольцевидного
хряща, но в каудальном направлении несколько отходит от гребня
(см. рис. 83, б, 84). Волокна идут вперед, расходясь веером вверх и
крепятся на заднебоковой стороне щитовидного хряща, вентрального'
края его каудальных рогов, вентральнее отхождения m. thyropharyngeus.
и позади прекрепления m. sternothyroideus (см. рис. 83, б, 84).
Кольцевидный хрящ с черпаловидным соединяют сразу три мышцы,
хотя обычно имеются лишь две.
М. cricoarytenoideus dorsalis, дорсальная кольцевиднощитовидная
мышца (см. рис. 80—83), большей частью прикрыта сверху кольцевид-
ноглоточной мышцей. Она начинается мясисто от дорсального срединно-
го гребня кольцевидного хряща и его каудального края (см. рис. 83, а,
84). Волокна идут вперед сбоку от пищевода и крепятся на особом мы-
шечном бугре черпаловидного хряща (см. рис. 83, а). В оральной по-
ловине мышечного брюшка на ее наружной поверхности имеется сухо-
жильное зеркало, волокна которого также конвергируют и прикрепляют-
ся на том же бугре черпаловидного хряща. Небольшая порция
мышечных волокон m. cricoarytenoideus dorsalis несколько обособлена
от орально расположенного мышечного бугра в дорсолатеральном углу
черпаловидного хряща. Волокна этой порции идут в том же направлении
и имеют общее прикрепление со всей мышцей.
М. cricoarytenoideus lateralis, латеральная кольцевиднощитовидная
мышца (см. рис. 82), большей частью прикрыта снаружи кольцевидно-
глоточной мышцей. Она начинается большей частью мясисто от утол-
щенного дорсолатерального участка заднего края кольцевидного хряща
(см. рис. 83, а). Волокна идут вперед, конвергируя к прикреплению на
особом мышечном бугре черпаловидного хряща непосредственно вент-
ральнее предыдущей мышцы (см. рис. 84).
Домнинг (Domning, 1978а) описал у бразильского ламантина еще
три мышцы, начинающиеся от щитовидного хряща и кончающиеся на
черпаловидном хряще. Это m. thyroarytenoideus, т. (?) vocalis и т. (?)
ventricularis. У дюгоня (Domning, 1977b) отмечен только первый из них.
Из всех трех два первые кончаются на мускульном отростке черпаловид-
ного хряща или m. vocalis сдвинут ближе к основанию рожкового от-
ростка, a m. ventricularis плоским сухожилием уходит в крышу глотки.
На нашем экземпляре ламантина ситуация была иная. Действительно,
на мускульном отростке крепились две мышцы, но отхождения их были
сдвинуты каудальнее и с щитовидным хрящом ни одна из них связи не'
имела. Гомология их с описанными Домнингом мышцами не ясна.
«М. thyroarytenoideus» (см. рис. 85) начинается вместе с мышцей дру-
гой стороны от медиальной связки, тянущейся от каудального края щи-
товидного хряща к краниальному краю кольцевидного (см. рис. 83, б).
(Поскольку ни у бразильского ламантина, ни у дюгоня не было такогг
глубокой выемки вентрально в заднем крае щитовидного хряща, то мож-
но предположить перенесение у нашего ламантина места отхождения на
связку, находящуюся на том же месте, где у первых двух форм был
хрящ. В то же время по своему переднему положению относительно вто-
рой мышцы «т. thyroarytenoideus» следовало бы сопоставлять с m. vo-
calis (голосовая мышца) большинства млекопитающих, у которых в этом
месте также имеется всего две мышцы (Grau, 1945), однако начинаю-
216
Таблица 6. Мышцы гортани, глотки и языка
Мышцы Масса Мышцы Масса
Г % Г %
in. cricopharyngeus m. ihyropharyngeus m. hyopharyngeus m. stylopharyngeus m. pterygopharyngeus styloglossus lateralis styloglossus medialis styloglossus intermedins genioglossus hyoglossus lateralis hyoglossus medialis stylohyoideus geniohyoideus mylohyoideus cranialis mylohyoideus caudalis 7,3 10,1 3,6 29,0 11,2 2,0 10,0 1,0 7,0 5,0 15,0 5,0 8,0 5,7 16,3 1,8 2,5 0,9 7,2 2,8 0,5 2,5 0,3 1,7 1,2 3,7 1,2 2,0 1,4 4,1 thyrohyoideus s. str. sternothyroideus sternohyoideus thyrohyoideus minor hyoepigloticus cricothyroideus cricoarytaenoideus dorsalis cricoarytaenoideus lateralis cricoarytaenoideus ventralis seu vocalis thyroarytaenoidens seu ventricu- laris arytaenoideus transversus мышцы собственно языка (1/2) Всего Мышцы обеих сторон 14,0 67,0 85,0 3,0 0,6 6,2 5,6 0,4 2,3 2,4 1,4 78,0 402,1 804,2 3,5 16,7 21,1 0,7 0,2 1,5 1,4 0,1 0,6 0,6 0,4 19,4 100,0 0,2*
* В % к массе тела.
щихся от щитовидного хряща. Возможно, это и есть «т. vocalis» Дом-
нинга. Кончается мышца на мускульном отростке черпаловидного хряща.
М. cricoarytenoideus ventralis, вентральная кольцевидночерпаловид-
ная мышца (см. рис. 85), лежит точно позади предыдущей и фактиче-
ски неотделима от нее, но ее начало лежит на внутренней поверхности
кольцевидного хряща, а оканчивается на мышечном бугре черпаловид-
ного хряща. Опять только по положению эта мышца сопоставима с
ш. ventricularis (желудочковой мышцей) других млекопитающих, но с
условием, что ее начало сместилось далеко назад на черпаловидный
хрящ. В то же время эта мышца больше схожа по описанию с
ш. thyroarytenoideus Домнинга. М. ventricularis Домнинга, таким обра-
зом, вообще не имеет аналогов у нашего ламантина. Возможно, глубо-
кая перестройка этих мышц у Т. manatus связана с отсутствием, во вся-
ком случае на нашем экземпляре, голосовой щели и голосовых связок,
а также и перестройкой самой гортани.
М. arytenoideus transversus, поперечная черпаловидная мышца
(см. рис. 82), начинается дорсальнее окончания двух предыдущих и
mm. cricoarytenoideus dorsalis et lateralis от мышечного бугра черпало-
видного хряща (см. рис. 83, а, 85), идет дорсально и, сужаясь, переки-
дывается через его вырезку, направляясь медиально к соединению с
мышцей противоположной стороны по срединному апоневрозу.
М. hyoepiglotticus, подъязычнонадгортанная мышца (см. рис. 82),
представляет небольшой пучок мышечных волокон, простирающихся от
основания надгортанного хряща к внутренней поверхности тела подъ-
язычной кости у ее переднего края (см. рис. 83, а, 85). У бразильского
ламантина описано отхождение от кератогиоида (Domning, 1978а).
М. thyrohyoideus minor (см. рис. 83—85) практически представляет
'Собой продолжение m. thyropharyngeus, который закрепляется, с одной
стороны, на щитовидном хряще, а с другой — на теле подъязычной
кости, однако слабо выраженная сухожильная перемычка в месте связи
217
его волокон с m. thyropharyngeus заставляет считать возможным описа-
ние его как особой мышцы.
Обычно в мягком нёбе млекопитающих имеются m. tensor veli pala-
tini (напрягатель нёбной занавески) и m. palatopharyngeus (нёбногло-
точная) . Обе эти мышцы описаны у бразильского ламантина и у дюгоня
Домнингом (1977b, 1978а), но при этом отмечается их слабое развитие,
особенно у ламантина, что было увязано с молодостью исследованного
экземпляра. Однако у нашего взрослого американского ламантина в.
тканях мягкого нёба вообще не было обнаружено мышечных волокон.
ТУЛОВИЩНАЯ МУСКУЛАТУРА
Все мышцы туловища можно подразделить на три главных функцио-
нальных комплекса: 1) мышцы, обеспечивающие потребности животного-
в поступательном движении, в основном вызываемом движениями хвоста
в вертикальной плоскости (мышцы спины и хвоста); 2) мышцы, управ-
ляющие движениями головы и короткой шеи (собственно шейные)
3) мышцы, обслуживающие потребности дыхания (брюшной пресс и
диафрагма). Не все мышцы, входящие в 'одну группу, имеют единое про-
исхождение с морфологической точки зрения и даже не всегда близки
по топографическому положению. Специфика сирен среди других млеко-
питающих требует подчеркнуть именно эти главные функциональные
потребности, так как во многом мышцы подчинили свои собственные,
типичные для млекопитающих функции этим основным. В связи с этпм
последовательность описания мышц туловища, принятая нами, несколь-
ко отличается от традиционной. Следует отметить, что ряд других
мышц, топографически относящихся к туловищу и связанных своим
происхождением с осевой мускулатурой, включен в два других само-
стоятельных комплекса: это мышцы, управляющие передней конечностью,
и мышцы, связанные с генитальной областью.
Мышцы туловища — спинные, грудные, хвостовые
В эту группу объединены мышцы разного происхождения (например,
сюда включена основная подкожная мышца m. cutaneus trunci) и то-
пографической принадлежности, обычно никогда не рассматриваемые
вместе. Уже говорилось, что локомоторные способности сирен обеспечи-
ваются движениями хвоста, начиная с его корня; собственно туловище
остается неподвижным в отличие от того, что происходит у большинства
китообразных. Однако на хвост начинают работать многие мышцы, ра-
нее в его деятельности не участвовавшие (это и подкожная мышца ту-
ловища, и мышцы брюшного пресса и т. д.). Кроме того, мышцы груд-
ной клетки также работают теперь в основном на обеспечение условий
для более эффективной работы хвоста. Все это, вместе взятое, и позволя-
ет нам проводить описание этих мышц вместе.
М. cutaneus trunci (panniculus carnosus), подкожная мышца тулови-
ща (см. рис. 68, 87), гигантская мышца, состоящая из двух — латераль-
ной и вентральной — порций, начинающихся самостоятельно. Их волокна
идут спереди назад и за ластом объединяются в единый мускул, прости-
рающийся к основанию хвоста.
Латеральная порция берет начало от сухожильного тяжа, подвижно
закрепленного изнутри в области лопатки по поверхности фасции, при-
крывающей мышцы пояса. Спереди к этому тяжу подходит из области
головы платизма. Таким образом, этот тяж является фактически пере-
мычкой в едином подкожном мышечном пласте, тянущемся от головы до
218
Рис. 87. Мышцы переднего
ща (вид сверху)
Слева подкожные мышцы
удалены
1 — ш. acromiotrapezius;
2 — m. clavotrapezius;
з — m. cutaneus trunci;
4 — m. infraspinatus;
3 — m. latissimus dorsi;
пояса и подкожные мышцы передней
« — m. levator nasolabialis;
7 — m. masseter medialis;
a — platysma myoides p. jugu-
laris;
9 — m. spinodeltoideus;
10 — m. spinotrapezius; '
11 — m. temporalis;
половины тулови-
12 — m. teres major;
13 — m. trapezius p. mandibula-
ris;
it — 13 — поверхностная фас-
ция
(it — наружный слой,
15 — внутренний)
219
хвоста и иногда даже рассматриваемом как одна мышца (Murie,.
1872а). Более вентральные пучки латеральной порции короче дорсаль-
ных. Они идут вниз и назад и закрепляются на вентральной порции.
Более дорсальные продолжаются до хвоста. Фасция m. cutaneus trunci
дорсально и несколько внутри закрепляется на особых бугорках VII—
XV ребер. Причем спереди это прикрепление расположено дорсальнее
(в области верхней трети ребра, у границы с т. iliocostalis), а сзади
вентральнее (в нижней четверти ребра, далеко от подвздошнореберной
мышцы). В области каждого прикрепления с ребром фасция уплотняет-
ся, образуя как бы стягивающую дорсальный край латеральной порции
m. cutaneus trunci поперечную ленту. Следует отметить, что на этих бу-
горках ребер крепится аналогичное утолщение наружной фасции m. in-
220
tercostalis externus, образующее как бы поперечную ленту, соединяющую
ребра между собой.
Вентральная порция в своем начале подразделяется на две части,
латеральная начинается апоневрозом (см. рис. 69), формирующимся за
счет соединительнотканных волокон, идущих от наружной поверхности
заднего края m. spinodeltoideus от сухожильной перемычки в m. brachi-
ocephalicus (отделяющей m. clavotrapezius и m. cleidomastoideus от
m. clavodeltoideus) и от наружной фасции m. latissimus dorsi.
В медиальной части более глубокие мышечные волокна берут начало от
вентральной поверхности реберных хрящей первых трех ребер, в то вре-
мя как поверхностные волокна являются непосредственным продолжени-
ем лежащего впереди m. trapezius р. mandibularis (глубокие волокна по-
следнего крепятся на рукоятке грудины по соседству с отхождением глу-
боких волокон медиальной части вентральной порции m. cutaneus trunci
и волокон т. pectoralis superficialis).
После объединения латеральной и вентральной порций m. cutaneus
trunci идет назад, заметно утолщаясь (рис. 88). Ближе к хвосту лате-
ральная часть мышцы своей внутренней поверхностью упирается в кон-
цы последних пяти-шести ребер, особенно в XVII ребро, занимающее
особое от других положение (оно отодвинуто латеральнее своим дисталь-
ным концом, образуя, таким образом, упор для XVI ребра и, кроме того,
укреплено сзади коротким ребром последующего позвонка, см. дальше),
а также в седалищную кость. В результате волокна мышцы меняют на-
правление при подходе к хвосту, за счет чего возрастает эффективность
ее действия. Кончается m. cutaneus trunci на наружной фасции сгибате-
лей хвоста у его основания. Таким образом, эта мышца вместе с платиз-
мой и частично с m. trapezius р. mandibularis становится самым эффек-
тивным сгибателем хвоста, что оправдывает ее большую силу: масса обе-
их, правой и левой, мышц в сумме достигает 5% массы тела ламантина,
значительно превышая массу остальных мышц, взятых по отдельности.
От внутренней поверхности m. cutaneus trunci в подмышечной обла-
сти отделяется особый пучок (см. рис. 86), направленный вперед. Он
Рис. 88. Поперечные срезы некоторых мышц туловища и пояса передних конечно-
стей (затемнены апоневрозы)
1 — 5 — ш. cutaneus trunci
(7 — начало, около platysma
р. jugularis,
2 — в месте перехода в m. tra-
pezius р. mandibularis;
3 — р. thoracalis,
4 — на месте объединения
р. lateralis с р. ventralis,
5 — на уровне anus);
6 — m. trapezius р. mandibu-
laris на уровне переднего
края грудины;
7 —• m. sphincter colli superfi-
cialis, середина;
3 — т. sternomastoideus, око-
ло грудины;
9 — 10 — т. rectus abdominis
(9 — 1/10 от начала,
10 — 1/10 от окончания);
11 — т. obliquus abdominis ex-
ternus, пучки, идущие к
сгибателям хвоста, сере-
дина;
12 — 14 — m. longissimus dorsi
(12 — краниальная треть,
13 — середина,
14 — каудальная треть);
15 — 17 —• m. sacrococcygeus
dorsalis lateralis
(15—краниальная треть,
16 — середина,
17 — каудальная треть);
18 — 19 — m. semispinalis dor-
si, середина
(19—т. sacrococcygeus dorsa-
lis medialis, середина;
20 — 21 — т. iliocostalis, сере-
дина
(20 — dorsi;
21 — caudae);
22 — 23 -— т. sacrococcygeus
ventralis
(22 — на уровне anus,
23 — середина);
24 — т. ischiococcygeus, сере-
дина;
25 — 26 — т. longus capitis
(25 — середина,
26 — на уровне затылочных мы-
щелков);
27 — т. splenius, середина;
28 — т. scalenus, середина;
29 — т. aciomlotrapezius, сере-
дина;
30 — in. clavotrapezius; сере-
дина ;
31 — т. spinotrapezius, середи-
на;
32 — 33 — т. pectoralis super-
ficialis
(32 — р. cranialis,
33 — р. caudalis);
34 — т. pectoralis profundus,,
середина;
35 — т. pectoralis abdominalis,..
середина;
36 — т. serratus ventralis, се-
редина;
37 — т. latissimus dorsi, сере-
дина;
38 — т. rhomboideus; середина»
221
переходит в сухожилие, теряющееся на внутренней поверхности т. рес-
toralis profundus. Этот пучок, очевидно, следует рассматривать как само-
стоятельную мышцу, внедряющуюся в m. cutaneus trunci изнутри и го-
мологичную с m. pectoralis abdominalis, имеющуюся у многих сумчатых,
насекомоядных, грызунов, хищных (Гамбарян, 1960). Эта мышца служит
оттяжкой, которая уменьшает продольное смещение m. cutaneus trunci,
особенно при переднем положении ластов.
Строение m. cutaneus trunci не только сильно отличается у дюгоня,
но и проливает определенный свет на саму перестройку его у ламантина.
В исходном состоянии у сумчатых, насекомоядных, хищников подкожная
мышца туловища начинается на плечевой кости и веером расходится по
всему туловищу к остистым отросткам и средней линии живота, образуя
несколько разных порций. У дюгоня (Domning, 1977b) большая часть
мышцы начинается от края нижнечелюстной части трапециевидной
мышцы. Перемещение m. cutaneus trunci вперед по вентральной поверх-
ности туловища не необычно для млекопитающих. Это было, например,
отмечено у тенреков, землероек (Dobson, 1883). Вторая часть подкожной
мышцы туловища меньше по размеру. Она берет начало от фасции в
подмышечной области, покрывая мышцу, называемую Домнингом
«ш. latissimus dorsi», которая, с одной стороны, крепится на внутрен-
ней поверхности m. cutaneus trunci, а с другой — на плечевой кости
совместно с окончанием наружной грудной мышцы. Для m. latissimus
dorsi млекопитающих вообще характерно отхождение от грудных позвон-
ков и ребер, а прикрепление вместе с большой круглой мышцей и лишь
иногда второй ножкой вместе с грудными мышцами (Гамбарян, 1960).
Таким образом, «т. latissimus dorsi» Домнинга у дюгоня не имеет спе-
цифических черт этой мышцы, что заставляет предполагать полное ис-
чезновение ее, в то время как имеющаяся мышца представляет собой
на самом деле порцию подкожной мышцы туловища, связанную в своем
отхождении с плечевой костью. Как сказано выше, это свойственно прими-
тивным млекопитающим. У ламантина уже полностью потеряна связь
m. cutaneus trunci с плечевой костью, а самой мощной частью его ста-
новится порция, связанная в своем начале с платизмой. Более развитым
становится и сращение с нижнечелюстной частью трапециевидной
мышцы.
Окончание m. cutaneus trunci дюгоня более широкое, чем у ламанти-
на. Мышца, очевидно, может и сгибать хвост, и отгибать вбок, и даже
поднимать его. Для ламантина же одного характерно перемещение его
прикрепления на вентральную сторону и превращение его в основной
сгибатель хвоста. Подводя итоги, можно сказать, что вопреки утвержде-
нию Домнинга (1977b, р. 16), что в отношении m. cutaneus trunci
«дюгонь более отклонился от исходного состояния, свойственного копыт-
ным, чем ламантин», дело обстоит как раз наоборот: ламантин ушел
далеко вперед в своей специализации, а дюгонь сравнительно близок к
типичному для млекопитающих состоянию. У дюгоня платизма значи-
тельно шире, чем у ламантина, и она, как обычно, у млекопитающих в
основном работает на движения губ. Однако у него есть еще и особая
часть платизмы, кончающаяся на лопатке, что уникально для млекопи-
тающих. В то же время скуловая ее часть распространяется на область
плеча и даже дальше, почти до передних пучков m. cutaneus trunci,
подготавливая тем самым возможное их соединение при разрастании
подкожной мышцы туловища в направлении головы. Именно это и про-
изошло у ламантинов. Платизма при этом свела до минимума число
мышечных пучков, связанных с губами (pars buccalis), и в основном ста-
ла работать на обеспечение фактической фиксации единого мышечного
222
пласта, включающего m. cutaneus trunci и platysma myoides на скуловой
кости.
М. trapezius р. mandibularis (in. sphincter colli profundus p. auris,
см.: (Domning, 1977b, 1978a)), нижнечелюстная часть трапециевидной
мышцы (см. рис. 69, 70), начинается хорошо выраженным апоневрозом
от наружной поверхности сухожилия наружной жевательной мышцы и
по нему переходит на скуловую кость, распространяясь по последней до
ее заглазничного отростка (см. рис. 72, 75, 76). Апоневроз переходит в
мощное, в поперечном сечении почти треугольной формы брюшко
(см. рис. 88), более дорсальные волокна которого крепятся на рукоят-
ке грудины, а вентральные без всяких границ вливаются в вентральную
часть подкожной мышцы.
Нижнечелюстная часть трапециевидной мышцы развита у многих
млекопитающих. Обычно она начинается на вентральном углу нижней
челюсти, но нередко перемещается по наружной поверхности жеватель-
ной мышцы на скуловую кость (Гамбарян, 1960), поэтому ее начало на
скуловой кости у ламантина не представляет каких-либо принципиаль-
ных различий от других млекопитающих. Однако, ее неполный переход
в подкожную мышцу у дюгоня и полный, даже без сухожильной пере-
мычки, переход у ламантина достаточно уникальны.
Во всех предшествующих описаниях мышечной системы сирен отме-
чается слияние всей эпаксиальной мускулатуры в единый мышечный
пласт, среди которого вполне дифференцирована лишь более вентральная
его часть, так называемая подвздошнореберная система. В то же время
все мышцы, лежащие между остистыми отростками позвонков и рядом
с ними на поперечных отростках (transversospinalis system Домнинга),
теснейшим образом соединены как друг с другом, так и с мышцами,
входящими в longissimus system и расположенными латеральнее над
проксимальной частью ребер и реберно-позвоночными суставами. Обе
системы тем не менее описываются раздельно.
Mm. interspinales, spinalis et semispinalis dorsi, межостистые, остистая
и полуостистая мышцы спины (см. рис. 88—93). В типе у млекопитаю-
щих отчетливо дифференцированы короткие мышцы, связывающие ос-
тистые отроски смежных позвонков, и, лежащая вдоль них, длинная
остистая мышца, которая закреплена на боковых сторонах остистых от-
ростков и латерально граничит с m. longissimus dorsi. Однако, спереди,
обычно в области шеи и передней части груди, между ними вклинивает-
ся полуостистая мышца головы, отходящая от сосцевидных отростков
(метапофизов) соответствующих позвонков и тянущаяся к голове. При
дальнейшем разрастании назад часть мышечных волокон, отходящих
от сосцевидных отростков грудных и поясничных позвонков, крепится
уже на остистых ртростках шейных и грудных позвонков. При такой
ситуации уже говорят о полуостистой мышце шеи (передняя часть) и
спины. Эти предварительные замечания необходимы, чтобы разобраться
в сложной структуре мышц этого комплекса.
Остистые отростки соседних позвонков соединены сложной системой
межостистых связок, располагающихся в несколько слоев. Основания
остистых отростков соединены мощным эластичным тяжем (ligaments
flava), впервые появляющимся между II и III грудными позвонками и
далее назад до VII позвонка утолщающимся и распространяющимся
вверх. В таком состоянии он идет до послегрудных позвонков и опять
начинает сокращаться, будучи представлен между IX и X pth уже тон-
ким округлым шнуром в толще имеющихся там фиброзных связок (см.
рис. 94). В целом ligg. flava представляет собой мощную упругую систе-
му, развитую значительно сильнее, чем у каких-либо других исследован-
223
Рис. 89. Детали строения
1 — г т. iliocostalis
(1 — caudae,
2 — dorsi);
3 — 4 — m. intercostalis
(3 — externus,
мышц в области перехода
4 — interims);
5 — m. interspinalis;
6 — m. longissimus dorsi;
7 — m. sacrococcygeus dors
lateralis;
К хвосту (вид сбоку)
8 — m. sacrococcygeus dorsalis
medialis;
9 — m. semispinalis dorsi
ных млекопитающих. Дорсальный приостренный край каждого из компо-
нентов, слагающих эту связку, связан с более тонкой частью межостис-
тых связок. Последние слагаются из тонкой пластинки, которая образо-
вана фиброзными волокнами, идущими от переднего края остистого
отростка позвонка, расходясь веерообразно вперед и вниз к заднему
краю остистого отростка предыдущего позвонка по обеим сторонам от
срединной соединительнотканной пластинки. Таким образом, формируется
трехслойная межостистая связка (рис. 90, б), занимающая промежутки
между остистыми отростками от IV грудного до X—XII послегрудных
позвонков, причем, начиная с V послегрудного позвонка, эти фиброзные
связки резко утолщаются, хотя их высота соответственно укорочению
остистых отростков хвостовых позвонков сокращается (см. рис. 94).
С исчезновением последних сходит на нет и сама связка. Связки меж-
ду остистыми отростками II и III, III и IV грудных позвонков представ-
лены однослойной фиброзной пластинкой. Дорсальный край этих связок
соединен с наружными апоневрозами эпаксиальных мышц спины, подхо-
дящих к нему с обеих сторон. Эти апоневрозы связаны также с вер-
хушками остистых отростков и именно здесь они утолщены и обогащены
фиброзными волокнами. Эти утолщения и соединяющие их более тонкие
участки в межостистом пространстве образуют сухожильную горизонталь-
ную пластинку — надостистую связку, достигающую толщины в 1 мм,
а в хвосте и в 2 мм. Далее вперед (между II шейным и I грудным
позвонками) в связи с укорочением остистых отростков межостистая
связка представлена уже горизонтальной фиброзной пластинкой, растя-
нутой между невральными дугами позвонков (см. рис. 94).
Непосредственно к межостистым связкам в области, начиная с IV
грудного и кончая примерно XII хвостовым позвонком, прилегают с
обеих сторон мышечные пучки, которые можно идентифицировать как
mm. interspinales (рис. 90, б). Они образованы системой вставленных
друг в друга треугольных образований, одни из которых отходят от зад-
него края остистого отростка предшествующего позвонка и идут к перед-
нему краю остистого отростка последующего позвонка, другие наоборот.
224
8 Ламантии
225
Рис. 91. Площади прикрепления мышц на позвонках шеи
а — вид сбоку; б — вид сверху; в — эпистрофей, вид сзади; г — атлант, вид снизу
1 — m. illocostalis;
г — 4 — ш. longissimus
(2 — atlantis,
3 — cervicis,
4 — dorsi);
5 — m. longus colli;
6 — 7 — m. obliquus capitis
(« — cranialis,
7 — caudalis);
8 — 12 — in. rectus capitis
(« — lateralis,
9 — ventralis,
10 — 12 — dorsalis);
10 — major, 11 — Intermedius,
12 — minor);
13 — m. scalenus;
14 — 16 — m. semispinalis
(14 — capitis,
IS , 16 — cervicis,
is — p. medialis,
16 — p. lateralis);
17 — m. serratus ventralis;
18 — m. spinalis cervicis
226
'С каждой стороны участвуют по три-четыре таких зубца (между II
и III грудными позвонками один спереди, два сзади). В основе такой
элементарной структурной единицы лежит начальное сухожилие, отходя-
щее от одного остистого отростка, и идущие от него веерообразно мышеч-
ные волокна, формирующие двуперистое брюшко и кончающиеся на
другом остистом отростке, причем сухожильные начала смещены меди-
альнее мышечных прикреплений. Наиболее дорсальные из этих единиц
берут начало сухожилием и от основания надостистой связки.
От передней части каждого остистого отростка шейных (начиная
с III) и грудных позвонков и смежной части надостистой связки отхо-
дят сухожилия, вокруг которых формируются мышечные волокна, со-
ставляющие перистое брюшко, идущее назад и отделяющее со своей ме-
диальной стороны мышечный зубец, который перекидывается через
один сегмент и сухожилием прикрепляется на заднем краю остистого
отростка соответствующего позвонка (например, с VIII идет к X груд-
ному позвонку). Затем отделяется второй зубец, проходящий уже через
два сегмента (к XI позвонку), третий (к XII позвонку, т. е. пропускаю-
щий три сегмента) и, наконец, четвертый (см. рис. 90, 93). В шейном
отделе у мышц, отходящий от III и IV шейных позвонков, имеются
лишь два зубца, далее назад их не менее трех, в области середины спины
четыре. Причем вся мышца здесь, следовательно, начинается на остистых
отростках и на них же кончается. Начиная с V грудного позвонка, кроме
уже описанных мышечных зубцов, идущих к остистым отросткам соот-
ветствующих позвонков, имеется и еще один, последний зубец, наиболее
мощный, который перебрасывается сразу через 4—7 сегментов (видимо,
чем дальше назад, тем больше) и кончается на связке, соединяющей
постзигапофиз предшествующего позвонка с диапофизом последующего.
Из таких мышечных единиц и формируется основная масса мышц опи-
сываемого нами комплекса. При этом мышечные пучки, имеющие и на-
чало и прикрепление только на остистых отростках, можно рассматри-
вать как производные m. spinalis cervicis (в области шеи) et dorsi
(в области собственно туловища), которые фактически продолжают один
другой. В то же время пучки, связывающие остистые отростки, а сзади
связки между постзигапофизами и поперечными отростками, должны от-
носиться к ш. semispinalis dorsi.
При переходе в хвост (эту часть у наземных млекопитающих обычно
рассматривают как самостоятельный m. sacrococcygeus dorsalis medialis)
строение мышечных единиц не меняется, лишь наблюдается прогрессив-
ное увеличение толщины (а следовательно, и мощности) межостистых
мышц, достигающих XX послегрудного позвонка. Видимо, и связи с этим
сухожильные отхождения мышечных единиц вытесняются с остистых
отростков на суставные, а их концевые мышечные зубцы перебрасыва-
ются по-прежнему через два-три сегмента. Конечные их сухожилия про-
стираются до последнего (XXV) послегрудного позвонка. В целом весь этот
комплекс покрыт снаружи плотной фасцией, отделяющей его от смежного
комплекса m. latissimus dorsi, волокна которого частично начинаются от
этого футляра.
Ни Домнинг, ни другие предшествующие авторы не описывают де-
тально внутреннюю структуру мышечной массы этого комплекса. Дей-
ствительно, отдельные ее части, определяемые нами как mm. interspina-
les, m. spinalis cervicis et dorsi, m. semispinalis dorsi практически не
отделимы друг от друга. Однако, разрывая их на части и прослеживая
судьбу отдельных порций мышечных волокон, все же можно, как это
сделано выше, представить общую картину строения этого комплекса.
В связи с этим кажется удивительным, что Слайпер (Slijper, 1946),
227
8*
228
а вслед за ним и Домнинг (Domning, 1977b, 1978а) отрицают присутствие
т. spinalis и у дюгоня, и у ламантинов, тогда как в нашем случае со-
ответствующие ему мышечные порции развиты достаточно сильно. По
Домнингу (1977b), у дюгоня вся масса transversospinalis system, кроме
m. seinispinalis capitis, состоит из mm. multifidus и еще более коротких
пучков. Слайпер у дюгоня тоже нашел mm. multifidus. Не исключено,
что в нашем m. semispinalis dorsi также были более глубокие и корот-
кие пучки, идущие от остистых отростков через два-три сегмента к ме-
тапофизам более каудально расположенных сегментов (в задней половине
грудного и начале послегрудного отделов).
М. longissimus dorsi, длиннейшая мышца спины (см. рис. 88, 93—
96), лежит латеральнее предшествующего комплекса и состоит, как и
последний, из отдельных мышечных единиц, теснейшим образом пере-
плетенных друг с другом. В типе у млекопитающих мышечная единица
длиннейшей мышцы спины состоит из нескольких зубцов, отходящих от
метапофизов и диапофизов нескольких смежных позвонков и, объединя-
ясь вместе и проходя через несколько сегментов назад, прикрепляющих-
ся единым окончанием к остистому отростку соответствующего позвонка
(в задней части грудного и в послегрудном отделах). Таким образом,
имеется принципиальное отличие ее от таковой m. semispinalis dorsi,
где, начинаясь от впереди лежащего остистого отростка единым сухожи-
лием, прикрепление осуществляется уже отдельными ножками на мета-
пофизах позади лежащих позвонков (т. е. в m. longissimus dorsi общее
направление действия сил: спереди и снаружи — назад и внутрь, меди-
ально, aym. semispinalis dorsi: спереди и изнутри — назад и латераль-
но). Имея в виду эту схему, легче понять особенности строения m. lon-
gissimus dorsi ламантина сравнительно с таковой других млекопитающих.
В целом весь этот комплекс простирается от IV шейного до XVII
хвостового позвонка. Обычно у млекопитающих заднюю часть его, свя-
занную с хвостом, выделяют в самостоятельную мышцу m. sacrococcyge-
us dorsalis lateralis, которая, как и m. longissimus dorsi, начинается от
метапофизов поясничных и крестцовых позвонков, однако окончание его
смещено с остистых отростков хвостовых позвонков к их суставным и
поперечно-реберным отросткам. У ламантина нет никакой дифференциа-
ции между обеими мышцами. Более того, как будет показано дальше,
черты, свойственные хвостовой части разбираемой системы обычных
млекопитающих, можно теперь обнаружить почти на всем протяжении
m. longissimus dorsi.
Мышечный сегмент (элементарная единица) m. longissimus dorsi
представлен несколькими мышечными зубцами, отходящими от разных
сегментов осевого скелета. Все они идут назад, перекидываясь через
определенное число сегментов (для каждого зубца это число различно),
соединяясь вместе, и кончаются на наружной фасции мышц, входящих в
трансверзоспинальную систему (можно сказать — в фасцию m. semispi-
nalis dorsi, поскольку именно он образует наружную часть комплекса).
В этой фасции, начиная с VII грудного позвонка, формируются особые
(10 — 11 —m. obliquus capitis 15 — m. rectus capitis lateralis;- 21 — m. serratus ventralis;
(10 — caudalis. 16 — 17 — m. scalenus 22 — m. splenius;
11 — cranialis); (16 — primae costae dorsalis, 23 — m. sternohyoideus;
12 — 14 — m. rectus capitis dor- 17 — thoracalis); 24 — m. sternomastoidtus;
salis 18 — 20 — m. semispinalis; 25 — m. sternothyroideus;
(12 — major, (18 — capitis; 26 — m. transversus abdominis;
13 — intermedius, 19 — cervicis, 27 — m. trapezius p. mandibu-
14 — minor); 20 — dorsi); laris
229
Рис. 93. Мышцы шеи и передней части грудной клетки (вид сверху)
а — слева удалены подкожные мышцы и акромиальная часть трапециевидной, перерезаны спинная
часть трапециевидной мышцы и широчайшая мышца спины; справа —удалена пластыревидная мыш-
ца и мышцы переднего пояса; б — полусхема прикрепления глубоких мышц
1 — m. Iliocostalis;
2 — т. latissimus dorsi;
3 — 4 — т. longissimus
(3 — atlantis et cervicis,
4 — dorsi);
5 — 6 — m. obliquus capitis
(5 — caudalis,
6 — cranialis);
7 — m. rhomboideus;
a — 12 — m. semispinalis
(a — capitis,
9 — dorsi,
10 — 12 — cervicis;
10 — p. caudalis,
11 — p. lateralis,
12 — p. medialis);
13—m. serratus ventralis;
14— m. spinalis dorsi;
13 — m. spinotrapezius;
16 — m. splenius
утолщения, косо идущие от остистых отростков назад и несколько вен-
трально через некоторое количество сегментов к сосцевидным или попе-
речным отросткам позадилежащих позвонков. Эти утолщения обнаружи-
вают сухожильные волокна, которые дальше назад становятся все более
и более явственными, формирующими почти самостоятельные сухожи-
лия, идущие в хвост. Именно на этих утолщениях-сухожилиях и крепит-
ся мышечный сегмент m. longissimus dorsi на значительном удалении от
соответствующего остистого отростка. В этом мы видим определенную
аналогию со смещением прикреплений m. sacrococcygeus dorsalis latera-
lis большинства млекопитающих относительно латеральнее мест, где кре-
пятся окончания отдельных сегментов самого m. longissimus dorsi, хотя
это, безусловно, производные одной системы.
Начальные зубцы m. longissimus dorsi спереди начинаются длинными
тонкими сухожилиями от поперечно-реберных отростков IV—VI шейных
позвонков (см. рис. 93, б). Далее назад отхождение аналогичных зубцов
(их можно рассматривать как основную слагающую каждого мышечного
231
Рис. 94. Поперечные срезы межостистых связок и межостистой мышцы
Римскими цифрами обозначено месторасположение связок I—XVIIth — грудные позвонки;
I — X cd — хвостовые позвонки
1 — фиброзная ткань; г — пластическая ткань; з — мышечная ткань
сегмента) располагается латеральнее, уже на наружной поверхности ре-
бер сразу дистальнее поперечно-реберных связок. На первых трех груд-
ных сегментах они по-прежнему в виде тонких сухожилий. Начиная с
IV ребра, эти зубцы начинаются уже широкими, относительно короткими
(чем дальше назад, тем больше) сухожильными апоневрозами, простира-
ющимися от поперечно-реберных связок по поверхности соответствующе-
го ребра до особых бугорков на них (в области реберного угла), места
прикрепления головок — m. iliocostalis. Ширина этих начальных апоне-
врозов постепенно возрастает от 81—85 мм на V ребре до 166 мм у
XVII ребра. Апоневроз от IV ребра частично смещен вперед на межре-
берное пространство, затянутое фасцией.
В грудном отделе появляется серия более медиально расположенных
зубцов, которые начинаются на заднем крае диапофизов, причем от
первых четырех грудных позвонков с помощью длинных тонких сухожи-
лий. Мышечные волокна, слагающие эти зубцы, появляются только че-
рез два-три сегмента. Зубец от V грудного позвонка уже с самого нача-
ла мышечносухожильный.
Примерно с IV грудного позвонка наиболее поверхностная и латераль-
ная порция волокон обособляется от основного мышечного зубца, от-
ходящего от ребра, и формирует особый мышечный зубец, перебрасы-
вающийся через большое количество сегментов (в задней части грудного
отдела через 7—8) и входящий, следовательно, в состав другого более
заднего по положению мышечного сегмента. Уже в задней половине
грудного отдела появляются мышечные пучки, также входящие в состав
соответствующего мышечного сегмента, волокна которых начинаются от
уже описанных выше сухожильных утолщений наружной фасции m. se-
232
12
5
Рис. 95. Поперечный разрез ламантина
Справа — при вдохе, слева —
1 — costae;
2 — cutis;
3 — diaphragma;
4 — pulmones;
5 — vertebrae.
при выдохе
Мышц ы:
6 — та. cutaneus trunci;
7 — та. iliocostalis;
8 — m. longissimus dorsi;
9 — 10 — m. obliquus abdo
(9 — extemus,
10 — internus);
11 —m. rectus abdominis;
12 — m. semispinalis dorsi;
13 — m. transversus abdominis
mispinalis dorsi, причем по ним они могут дотягиваться даже до остис-
тых отростков. Получается парадоксальное положение. Не только кау-
дальные окончания мышечных сегментов ламантина смещены латераль-
нее (сравнительно с обычным для млекопитающих состоянием), но и в
состав более задних сегментов входят мышечные зубцы, начало которых
смещено медиальнее (к остистым отросткам), чем это положено для ти
пичного m. longissimus dorsi. Все это, возможно, говорит об изменении
обычного направления приложения сил, развиваемых длиннейшей мыш-
цей спины, и, следовательно, переключении ее действий с собственно
спины на хвост.
В самом хвосте мышечные волокна в m. longissimus dorsi прослежи-
ваются лишь до уровня XIII—XIV послегрудного позвонка, далее вплоть
до XVII позвонка окончания этой мышцы исключительно сухожильные.
Начальные апоневрозы основных зубцов мышечных сегментов в области
хвоста перенесены на поперечно-реберные отростки, а зубцы от попереч-
ных отростков теперь связаны с суставными отростками.
М. iliocostalis, подвздошнореберная мышца (см. рис. 88—93, 95—97),
представляет собой наиболее латеровентральный компонент эпаксиальной
233
Рис. 96. Расположение m.
intercostalis externus (вид
сверху)
1 — lig. costotransversarium
dorsalis;
2 — lig, m. longissimi dorsi et
m. iliocostali;
3 — 4 — m. intercostalis ex-
ternus
(4 — пучки, соответствующие
m. levator costarum);
5 — m. longissimus dorsi.
Римскими цифрами обозначены
сегменты грудных позвонков
мускулатуры. С редукцией задних конечностей и таза она, естественно,
теряет связь с подвздошной костью и превращается в мышечную более
или менее широкую ленту, простирающуюся от V шейного позвонка
спереди до корня хвоста сзади (примерно на уровне V—VI послегрудных
позвонков) и состоящую из отдельных мышечных сегментов, вложенных
друг в друга, с общим дорсокаудальным направлением их. Самые кра-
ниальные сегменты начинаются самостоятельными сухожильными зубца-
ми от концов плеврапофизов V—VI шейных позвонков и наружной по-
верхности первых двух ребер (на I на расстоянии 19 мм от поперечно-
реберного сочленения, на II—43 мм) (см. рис. 93). Мышечные волокна,
формирующиеся вокруг каждого из этих начальных сухожилий, образуют
мышечный сегмент, отдельные пучки которого перекидываются через не-
сколько ребер и крепятся последовательно к IV, V и VI ребрам близ
начальных апоневрозов основных (реберных) зубцов m. longissimus dor-
si. Таким образом, как и у выше рассмотренных мышц, мышечный сег-
мент имеет единое начало на одном каком-либо сегменте туловища (осе-
вого скелета), а кончается сразу на нескольких, лежащих позади, при-
чем места прикрепления расположены дорзальнее (и медиальнее) мест
отхождения.
В области груди, начиная с 111 ребра, начальные зубцы становятся
сухожильномышечными и сильно расширяются. Проксимально по ребру
они граничат с широким начальным апоневрозом основных зубцов
ш. longissimus dorsi, а вентрально — с прикреплением к ребрам апо-
невроза m. cutaneus trunci. Самый латеральный (вентральный) край
этого отхождения образован сухожилием (2—3 мм), связанным с особым
бугорком на ребре, далее медиально сухожилие продолжается в тонкий
апоневроз, еще медиальнее от ребра начинаются уже непосредственно
мышечные волокна. Это широкое начало образует как бы основание вы-
тянутого треугольника, с которым можно сравнить мышечный сегмент
234
11
Рис. 97. Детали прикрепления мышц в области первых грудных ребер (вид сбоку)
о — перерезан m. scalenus thoracalis; б — часть мышцы отогнута. Римские цифры — порядковый но-
мер ребер
1 — 2 — ш. iliocostalis
(1 — cervicis,
2 — thoracitis);
з — 5 — щ. intercostalis
(3 — externus,
4 — internus,
5 — internus p. primae costae);
6 — 9 — m. scalenus
(6 — primae costae dorsalis,
t — primae costae ventralis,
8 — secundae costae dorsalis,
9 — thoracalis);
10 — 11 — m. serratus ventralb
(10 — p. cervicalis,
11 — p. thoracalis);
12 — m. transversus abdominis
235
m. iliocostalis. Вершина его образована длинным конечным сухожилием,
в которое переходят наиболее латеральные мышечные волокна, сегмен-
та, образуя его латеральную сторону. Оно крепится на другом, более
проксимальном, бугорке ребра, на котором также закреплено сухожилие,
образующее латеральный край начального апоневроза основного зубца
ш. longissimus dorsi. Самые медиальные волокна сегмента m. iliocostalis
кончаются коротким сухожилием на переднем крае следующего (через
одно) ребра, средние перебрасываются через два ребра и прикрепляются
уже на третьем, наиболее латеральные же проходят через четыре сег-
мента на пятое (см. рис. 90, а). В целом все эти окончания формируют
медиальную сторону треугольника, граничащую с m. longissimus dorsi.
Общее направление тяги, создаваемой таким треугольным сегментом,—
спереди назад и дорсомедиально. В своем начале каждый предыдущий
сегмент несколько прикрывает последующий. Следует подчеркнуть сдвиг
отхождений сегментов, входящих в состав m. iliocostalis вентральнее от-
носительно позвоночного столба, что связано с приобретением подвздош-
нореберной мышцей ламантина новых функций. Обычно же у млекопи-
тающих вся масса m. iliocostalis лежит не ниже уровня, занимаемого
телами позвонков.
Задние сегменты m. iliocostalis ламантина кончаются уже не на реб-
рах, а на утолщенной фасции m. longissimus dorsi (сегмент, связанный
своим началом с XV ребром) и на наружной фасции m. iliocostalis caudae
(сегменты от XVI и XVII ребер). Аналогичное явление обнаружено у бра-
зильского ламантина (Domning, 1978а). Интересно, что у дюгоня этой
порции вообще нет (Domning, 1977b) и iliocostalis (m. iliocostalis thora-
cis Домнинга) не заходит назад за последнее ребро. В то же время
внутренняя структура мышцы дюгоня, если она точно описана и изобра-
жена, отличается от описанной для ламантина: массу m. iliocostalis дю-
гоня составляют перистые пучки волокон, формирующиеся вокруг ко-
ротких идущих постеродорсально сухожилий и связывающие, если судить
по рисунку (Domning, 1977 b, fig. 23), смежные, может быть, через один
сегменты.
Краниально отхождения зубцов m. iliocostalis варьируют. Так, у аме-
риканского ламантина было найдено начало на эпистрофее (Murie,
1872а). Слайпер (Slijper, 1946) утверждал, что он тянется даже к ат-
ланту. У бразильского ламантина у одного экземпляра был зубец от
VI шейного позвонка, а у другого от III. У дюгоня самый передний зу-
бец был связан с VII шейным позвонком. Возможно, все эти различия не
являются существенными, будучи отражением индивидуальных вариаций.
М. iliocostalis caudae — хвостовая подвздошно-реберная мышца
(см. рис. 68, 80, 89), несомненно принадлежит системе подвздошно-ре-
берных мышц, как ее наиболее каудальное, но несколько обособлен-
ное продолжение. Ее следует рассматривать как новообразование у си-
рен (она есть и у ламантинов и у дюгоней), связанное с изменением
функций ш. iliocostalis.
Мышца имеет мощное веретеновидное брюшко в латеровентральной
области задней части туловища. Начинается она мышечными волокнами
от каудального края XVII ребра и поперечной связки (см. рис. 89),
которая простирается от тела I послегрудного и гемальных отростков
II и III послегрудных позвонков вдоль внутренней поверхности XVI реб-
ра к его дистальному концу, а также отдельным сухожильным концом к
концу XVII ребра (рис. 98, а). Кроме того, m. iliocostalis caudae имеет
отдельные мышечносухожильные зубцы от концов поперечно-реберных
отростков III и IV послегрудных позвонков и наружной фасции более
глубоко лежащего в своем начале m. sacrococcygeus ventralis lateralis.
236
1 — lig. costaevertebralis;
.2 — m. iliocostalis caudae;
3 — m. intertransversarius cau-
dae;
4— m. ischiocavernosus;
5 — m. ischiococcygeus;
6 — ni. rectus abdominis;
7 — 9 — m. sacrococcygeus
ventralis
(7 — lateralis,
8 — medialis,
9 — конечные сухожилия)
10 — чудесная сеть
Начальные сухожилия этих зубцов проникают в общее тело мышцы,
обеспечивая ее перистость. Кончается мышца мощным круглым в сече-
нии сухожилием, идущим к округлой расширенной лопасти хвоста, где
оно дифференцируется на отдельные более мелкие сухожилия, расходя-
щиеся в соединительной ткани плавника, причем часть их идет и к его
наружному краю. Эта соединительная ткань перевита мощными фиброз-
ными тяжами и пластинками, в ячеях которых расположены плотно при-
легающие друг к другу жировые клетки, способствующие постоянному
напряжению самой фиброзной ткани. Дорсально в краниальной половине
мышцы на ее наружной фасции крепятся последние мышечные сегменты
собственно ш. iliocostalis (от XVI и XVII ребер), не имеющие другого
окончания. Таким образом, их действие фактически усиливает эффектив-
ность ш. iliocostalis caudae, а последний функционирует как сгибатель
237
хвоста, поскольку линия действия его силы проходит ниже оси позвоноч-
ного столба.
Считая m. iliocostalis caudae непосредственным продолжением назад,
m. iliocostalis, мы все же не выделяем его под особым названием как
самостоятельный, хотя с таким же основанием к нему можно было бы
отнести и задние сегменты собственно m. iliocostalis, не имеющие окон-
чаний на ребрах. Также поступил и Мьюри (Murie, 1872а), предложив
назвать его не совсем удачно m. lumbocaudalis. Считая его также про-
изводным m. iliocostalis (его т. sacrolumbalis), Мьюри прямо рассмат-
ривает эту мышцу среди прочих сгибателей хвоста. Однако у его эк-
земпляра ламантина он обнаружил начало только от первых четырех:
послегрудных позвонков.
Домнинг при описании довольно сходной ситуации у бразильского,
ламантина (начало от последнего ребра близ места прикрепления к нему
ш. iliocostalis широким сухожилием и мышечными волокнами от попереч-
ных отростков I—VIII послегрудных позвонков) определяет эту мышцу
как ш. intertransversarius coccygeus, что может ввести лишь в заблужде-
ние, так как под таким названием у млекопитающих описываются осо-
бые мышечные пучки, соединяющие поперечные отростки смежных по-
звонков. Ни у дюгоня, ни у ламантина Домнинг их не обнаружил, хотя
они есть. В свое время Слайпер (Slijper, 1946), рассматривая m. ilio-
costalis млекопитающих, писал о ш. iliocostalis dorsi и m. iliocostalis;
lumborum, понимая под последним часть единой мышцы, связываю-
щую последние ребра и поперечные отростки поясничных позвонков
с таковыми более сзади лежащих и в основном с подвздошной костью.
С редукцией таза у китообразных и сирен подвздошнореберная мышца
утеряла связь с ilium и термин «т. iliocostalis lumborum» стал прилагать-
ся просто к той части общей подвздошнореберной мышцы, что лежит по-
зади ребер. При этом у китообразных, изученных значительно лучше,
чем сирены, отмечается, что m. iliocostalis тянется у них от атланта,
иногда даже от головы, до самого хвостового плавника (Slijper, 1936),
но при этом его каудальная часть не перемещается вентрально и мышца
как и у наземных позвоночных, служит разгибателем хвоста. Если же-
происходит расширение брюшка мышцы вентрально, как у морског
свиньи, то оно происходит спереди, что ведет к усилению первичной функ
ции. У сиреновых произошло обратное. Можно сказать, что каудальная-
часть подвздошнореберной системы приобрела особое значение, вопреки-
своему эпаксиальному происхождению, став сгибателем хвоста. Этим и
можно объяснить большую дифференциацию задних сегментов m. ilio-
costalis, которые приобрели вид отдельных веретеновидных головок, при-
чем задняя из них, которую мы определяем как m. iliocostalis caudae,.
вряд ли сопоставима с m. iliocostalis lumborum китообразных. Послед-
нее название, видимо, лучше оставить за последними сегментами m. ili-
ocostalis, крепящимися на фасции т. iliocostalis caudae (т. е. отсутствую-
щими, по Домнингу, у дюгоня, но имеющимися у бразильского ламанти-
на, а также у американского (по данным Мьюри (Murie, 1872а), и на-
шим). Предпосылкой приобретения задней частью m. iliocostalis сиреш
(в противоположность китообразным) новых функций явилось отсутст-
вие у них поясничного отдела и соответственно возможность использо-
вать мощные задние ребра как упоры, меняющие направление действия-
си л.
Mm. intercostales externi et levatores costarum, наружные межребер-
ные мышцы и подниматели ребер (см. рис. 89, 90, 96, 99), которые-
обычно у млекопитающих хорошо обособлены, у ламантинов представля-
ют единое целое в связи с сокращением возможности поднимать ребра-
238
’Собственно наружные межреберные машцы, имеющие, как правило, про-
стое строение, сильно модифицированы, так как изменились условия их
деятельности: межреберное пространство уменьшилось вследствие расши-
рения ребер в средней части, появилась потребность жесткой фиксации
ребер, особенно в задней области. В частности, произошла дифференциа-
ция отдельных порций mm. intercostales externi по направлению мышеч-
ных волокон, и эта дифференциация все более возрастает каудально
(рис. 99, а). Следует, видимо, различать пучки собственно наружной меж-
реберноп мышцы (mm. intercostales externus s. str.), сохраняющие ти-
пичное вентрокаудальное (больше каудальное, чем вентральное) направ-
ление и идущие от заднего края предыдущего ребра к переднему краю
последующего, и производные от нее более поверхностные порции, появ-
ляющиеся начиная с VII ребра. Одна из них, косая поверхностная (pars
obliquus superficialis), занимает более срединное по ребру положение.
Как правило, она прикрывает предшествующую часть и крепится по
наружной поверхности смежных по положению ребер, забираясь вперед
даже к переднему краю предшествующего ребра. Волокна этой порции
принимают более косое положение (больше вентрально, чем каудально).
В задней части грудного отдела наблюдается даже несколько отдельных
пучков волокон с таким направлением, один выше по ребру, другой ниже.
Интересно, что если взять сразу несколько сегментов, то прослеживается
расположение таких пучков смежных сегментов по одной косой линии
(в более краниальных сегментах они лежат выше), совпадающей с ли-
нией действия этих пучков (она пересекает позвоночный столб на два-
три сегмента впереди). Последняя, продольная поверхностная часть
(р. longitudinalis superficialis) расположена вентральнее остальных.
Частично она прикрывает предшествующую. Как и у остальных, ее во-
локна соединяют ребра двух сегментов. Однако неожиданно было обна-
ружено перекидывание одной головки через сегмент (от VII ребра к
IX), причем на обеих сторонах (рис. 99, а).
Все три порции по весу относятся друг к другу как 4:1:2. Впереди
•от VII ребра имеются только собственно наружные межреберные мыш-
цы, причем в самых передних сегментах ее наиболее вентральные волок-
на имеют более косое направление, но они обособлены от основной части,
плавно переходят в нее, а последняя так же часто распространяется на
наружную поверхность позади лежащего ребра. Вентральные волокна
этой порции не доходят до дистального конца ребра. Далее назад вент-
ральная граница ее, видимо, все дальше отодвигается от конца, а сами
волокна могут подразделиться на отдельные пучки, разделенные проме-
жутком. Возможно, что именно в такой промежуток вросли волокна глуб-
же лежащей внутренней межреберной мышцы, расположенной между
XV и XVI ребрами (рис. 99, а), и образовали своеобразную поверхностную
порцию, частично прикрывающую снаружи собственно наружную межре-
берную мышцу.
Волокна собственно наружных межреберных мышц, наиболее близкие
к позвоночному столбу, начинаются не только на заднем крае соответ-
ствующего ребра, но частично и от смежной части диапофиза и от имею-
щегося в этом месте поверхностного апоневроза (см. рис. 96). Они
соответствуют mm. levatores costarum. Поскольку они совсем не обособ-
лены от mm. intercostales externi, вряд ли имеет смысл выделять их
отдельно в этом случае, как это делает Домнинг (1978а), тем более что
соответствующих функций они выполнять не могут.
Дифференциацию отдельных частей mm. intercostales externi следует
связать с особенностями локомоции ламантина. Для успешного выполне-
ния новоприобретенных функций в качестве сгибателей хвоста mm. cuta-
239
240
neus trunci, obliqui abdomini externus et internus необходима жесткая
фиксация задних ребер, служащих им упором. Продольная поверхностная
порция наружных межреберных мышц способствует сближению дисталь-
ных концов ребер, т. е. в определенной степени сгибанию спины, но
сдвигу их назад они могут препятствовать только при фиксации грудины
впереди лежащими мышцами. Косая поверхностная часть в то же время
развивает значительный горизонтальный компонент сил, имея при этом
опору на позвоночный столб, т. е. фактически они работают на сгибание
хвоста.
Ни у бразильского ламантина, ни у дюгоня Домнинг не отметил каких-
либо особенностей mm. intercostales externi. Указывается лишь, что у ла-
мантина в последних четырех-шести межреберных промежутках их во-
локна отсутствуют в дистальных частях.
Mm. intercostales interni, внутренние межреберные мышцы (см. рис. 99),.
связывают задний край впереди лежащего рёбра с передним краем сле-
дующего за ним. Направление волокон дорсокаудальное, более крутое,,
чем у волокон mm. intercostales externi s. str. Спереди описываемые мыш-
цы занимают все межреберное пространство, в том числе и пространство’
между I—IV реберными хрящами. Волокна, связанные с ребрами и хря-
щами, полностью не отделимы и, безусловно, представляют один и тот
же комплекс. Поэтому выделение особых mm. intercartilaginei externi
(Domning, 1978a) вызывает возражения. В то же время у нашего ламан-
тина не наблюдается никаких связей этих пучков с mm. intercostales
externi. Нам не кажется также целесообразным включать в состав внут-
ренних межреберных мышц мышечные волокна (m. sternocostalis Дом-
нинга), соединяющие передние края I ребра и его реберного хряща с.
краем рукоятки грудины.
Большей частью mm. intercostales interni начинаются и кончаются у
нашего экземпляра ламантина прямо мышечными волокнами. Далее на-
зад протяженность мышц в межреберном пространстве сокращается. Они
все более и более ограничиваются его дистальной частью, а самые прок-
симальные волокна, начинают закрепляться на ребрах апоневрозом. Уже
отмечалось, что в пространстве между XV и XVI ребрами сформировался
особый веерообразный пучок волокон с узким началом на впереди ле-
жащем ребре и широким окончанием на наружной поверхности заднего
ребра. Интересно, что такое же образование было найдено у бразильско-
го ламантина и тоже между предпоследним и предшествующим ребрами
(XIV—XIII). У нашего ламантина был еще пучок волокон от XVII ребра
к фасции опускателей хвоста. Нет сомнений, что он также принадлежит
к системе mm. intercostales interni.
М. transversus thoracis, поперечная мышца груди (см. рис. 86), на-
чинается от выступа реберного хряща второго ребра, идет вперед, при-
крывая снизу реберный хрящ I ребра, и оканчивается на краниолате-
ральном углу грудины.
М. scalenus, лестничная мышца (см. рис. 97), состоит из отдельных
порций, различающихся по своему началу. Самая мощная из них на-
Рис. 99. Мышцы грудной области (вид сбоку)
а— удалены подкожные мышцы, разгибатели спины и мышцы переднего пояса; б — удалены также
и наружные межреберные мышцы'*
1 — 2 — m. intercostalis (3 — lateralis, 6 — primae costae ventralis,
(1 — externus, 4 — medialis); 7 — thoracalis);
2 — internus); 5 — 7 — m. scalenus « — m. transversus abdominis.
3 — 4 — m. sacrococcygeus ven- (5 — primae costae dorsalis,
trails
241
пинается примерно от средней 1/4 или 1/5 наружной поверхности III
ребра, и ее веретеновидное брюшко по своему ходу располагается между
двумя зубцами вентральной зубчатой мышцы. Мышечные волокна этой
части идут примерно до середины шеи, конвергируя в сухожилие, окан-
чивающееся на диапофизе эпистрофея (см. рис. 91). Мышечное начало
части мышцы, идущей от II ребра, занимает значительно большую
площадь, чем предыдущей. И ее брюшко образует широкую мышечную
пластинку, выступающую при виде сбоку как сверху, так и снизу из-
под прикрывающей его части, идущей от III ребра (см. рис. 97, а).
Однако эта пластинка очень тонкая и особенно утончена в средней части
под брюшком первой порции. От I ребра идут сразу две порции, имею-
щие свои собственные названия (см. рис. 97, б). Выше расположена
р. primae costae dorsalis, идущая от дорсальной части передней и наруж-
ной поверхности I ребра к плеврапофизам V и VI шейных позвонков.
Вторая порция (р. primae costae ventralis) своим началом занимает
дистальную половину краниолатерального края ребра, почти доходя вен-
трально до реберного хряща. Ее волокна оканчиваются на плеврапофи-
зах III—V шейных позвонков, по пути смешиваясь с волокнами, идущи-
ми в составе второй порции от II ребра. По-видимому, этот мышечный
пласт соединяется и с волокнами порции от III ребра. Таким образом,
фактически все части этой мышцы представляют одно целое — одну об-
щую мышцу, имеющую более широкое раздельное от трех первых ребер
начало и оканчивающуюся зубцами на плеврапофизах всех, кроме атлан-
та, шейных позвонков.
Последний комплекс мышц, входящий в рассматриваемую нами груп-
пу туловищной мускулатуры, включает вентральные мышцы, связанные
с хвостом. Однако их описанию целесообразно предпослать краткое опи-
сание тазовой кости, поскольку некоторые детали ее строения необходимо
знать в связи с прикреплением на ней одной из мышц хвоста и рас-
сматриваемых позже мышц генитального аппарата.
Тазовая кость (см. рис. ИЗ) представляет собой уплощенную плас-
тинку неправильной формы с несколькими выступающими отростками.
Более крепкий из них с субквадратным сечением соответствует морфоло-
гически дорзальной ацетабулярной части седалищной кости, в состав ко-
торой вошел рудимент подвздошной кости (Abel, 1904). Углы этого квад-
рата в сечении соответствуют наружным граням (гребням), делящим
поверхность отростка на почти равные продольные части. Этот отросток
занимает более латеральное положение и дальше будет обозначаться как
латеро-дорсальный отросток. Второй более тонкий и сужающийся медио-
вентральный отросток соответствует каудальному краю седалищной
кости. Морфологически передний край его представлен тупо срезанным
передним выступом с небольшой выемкой посередине. Вентральный край
кости разделен невысоким, выступающим в виде тупого угла отростком
на две более или менее равные части, более прямую переднюю, к кото-
рой крепится ножка полового члена, и слегка вогнутую заднюю, остаю-
щуюся свободной.
М. ischiococcygeus, седалищнохвостовая мышца (см. рис. 98, а), начи-
нается двумя головками, более мощной (m. ischiococcygeus Домнинга)
от вентральной и отчасти дорсальной поверхностей медиовентрального
отростка седалищной кости (см. рис. ИЗ) и более слабой (m. retractor
ischii Домнинга), тонким апоневрозом закрепленной на вентральной
фасции прямой мышцы живота, и двумя сухожильными ножками на
верхушке латеро-дорсального отростка и основании тела седалищной
кости. Обе головки за тазовой костью сливаются и крепятся на гемаль-
кых отростках IV—XI послегрудных позвонков. Близ окончания наруж-
242
пая фасция седалищнохвостовой мышцы срастается с сухожильным
зеркалом, формирующимся на m. sacrococcygeus ventralis medialis. У бра-
зильского ламантина и дюгоня эти головки, видимо, были более диффе-
ренцированы, что послужило основанием для выделения их в само-
стоятельные мышцы Домнингом.
М. sacrococcygeus ventralis lateralis, латеральная вентральная мышца
хвоста (см. рис. 98), является наиболее поверхностной на вентральной
стороне хвоста. Она начинается мышечно от каудального края средней
трети XVII ребра, более латеральной части поперечной связки, отмечен-
ной выше (см. m. iliocostalis caudae) и вентральной поверхности попе-
речно-реберных отростков передних хвостовых позвонков. Ее волокна
идут каудально и медиально. Отдельные пучки перебрасываются через
три сегмента па четвертый. Кончается m. sacrococcygeus ventralis late-
ralis тонкими сухожильными тяжами, сливающимися близ средней линии,
в единый апоневротический листок, закрепленный на гемальных отрост-
ках хвостовых позвонков. Мышечные волокна не дотягиваются до конца
позвоночного столба, и в области последних хвостовых позвонков имеют-
ся лишь концевые сухожилия. Общее направление мышечных волокон по
ходу хвоста несколько меняется, будучи спереди более продольным,
а сзади более поперечным, что связано с прогрессивным укорочением тел
позвонков. У бразильского ламантина не отмечено отхождения этой
мышцы от последнего ребра, а только от поперечно-реберных отростков.
Ill—XX послегрудных позвонков.
М. sacrococcygeus ventralis medialis, медиальная вентральная мышца
хвоста (см. рис. 91), лежит более глубоко по отношению к предыдущей
и подразделяется на две части, более краниальную и более каудальную.
Первая начинается несколькими мышечными пучками. Самый передний
из них берет начало на заднем крае тела XVII грудного позвонка и кре-
пится уже на теле III послегрудного позвонка (через два сегмента на
третий). Следующий пучок отходит двумя мышечными зубцами от тел
I и II послегрудных позвонков, а кончается общим сухожилием на по-
верхностной фасции более каудальной части мышцы на уровне примерно
IV—V послегрудных позвонков. Эти передние по своему положению пуч-
ки имеются и у Т. manatus, и у бразильского ламантина (у последнего
доходят до последнего ребра и тела предшествующего позвонка), а также
у дюгоня (от последних трех ребер (XVII—XIX) и вентральной части
XVI и последующих грудных), однако по неизвестным причинам
Домнинг описывает здесь эти пучки в составе m. sacrococcygeus ventralis
lateralis. Вполне логично эту часть сопоставлять с m. quadratus lumbo-
rum наземных млекопитающих, как это уже делалось ранее (Murie,
1872 а).
С появлением начиная с III послегрудного позвонка гемальных от-
ростков отхождения мышечных пучков смещаются на их основания.
Именно эти пучки формируют каудальную часть m. sacrococcygeus vent-
ralis medialis. От начала они идут каудолатерально, затем вентрально
и, наконец, медиально, как бы закручиваясь штопором. При этом они пе-
ребрасываются через три сегмента на четвертый. Кончаются они сухо-
жилиями, входящими в состав широкого медиального апоневроза, за-
крепленного на верхушках гемальных отростков хвостовых позвонков.
Латеральная и медиальная вентральные мышцы хвоста тесно связа-
ны в своем начале. Значительно лучше они дифференцированы в сред-
ней части хвоста, где имеется определенная степень подвижности по-
верхностного слоя относительно глубокого. Далее назад относительная
подвижность уменьшается, и с XVI—XVII хвостовых позвонков обе мыш-
цы опять практически неразделимы.
243
Mm. intertran&versarii, межпоперечные мышцы (рис. 98 б), у нашего
экземпляра ламантина обнаружены в постторакальной области до уров-
ня XIV позвонка. Все они относятся к гипаксиальной мускулатуре, так
как лежат вентральное фасции, соединяющей смежные поперечные от-
ростки. Две передние межпоперечные мышцы лежат собственно уже в
грудной области. Мышечные волокна первой из них отходят от заднего
края медиальной трети последнего (XVII) крупного ребра и прикрепля-
ются по всему переднему краю вентральной поверхности короткого
XVIII ребра, соответствующего I послегрудному позвонку (см. гл. 3).
Второй, несколько меньший пучок связывает XVIII ребро с вентральной
стороной относительно короткого поперечно-реберного отростка II после-
грудного позвонка. Следующая мышца, самая большая из всей серии, на-
чинается от вышеупомянутого отростка и, расширяясь каудально, конча-
ется на вентральной поверхности почти всего переднего края сильно
увеличенного в длину поперечно-реберного отростка III pth. Каждая
последующая мышца становится все уже и уже, держась ближе к концу
соответствующих отростков, которые далее назад постепенно укорачива-
ются. Кроме основной порции мышцы с продольным направлением мы-
шечных волокон, в II—XIII межпоперечно-реберных промежутках диф-
ференцируются особые, меньшие по ширине пучки, занимающие более
дорсальное положение. Волокна их имеют более косое направление (на-
зад и латерально) и в своем начале смещены по заднему краю впереди
лежащего поперечно-реберного отростка медиальнее отхождения основной
порции, начало которой помещается в задней части его вентральной по-
верхности. Эти пучки латерально отходят от концов поперечно-ребер-
ных отростков дальше, чем основные порции mm. intertransversarii.
Работа мышц туловища
Полностью достоверный функциональный анализ скелетномышечной си-
стемы ламантина невозможен ввиду недостаточного знания, как же в
действительности двигается животное. Для этого необходимы квалифи-
цированные ускоренные съемки под водой, применение специальных ме-
тодов для фиксации тех или иных кинематических параметров. И хотя в
настоящее время о движениях ламантина (локомоторных, поведенческих
и т. п.), в основном благодаря исследованиям Д. Хартмана (Hartman,
1979), известно намного больше, чем, скажем, о дюгоне, тем не менее и
этого слишком мало. Поэтому при оценке двигательных особенностей ла-
мантина приходится прибегать к отдельным фотографиям, рисункам их
поз в аквариумах и т. п. В этих условиях анализ, идущий от морфологии,
может много помочь как в интерпретации этих скудных фактов, так и в
дальнейшем изучении самой двигательной активности сирен.
Основной локомоторный орган всех сирен — хвостовой плавник, имею-
щий разную форму у ламантинов, с одной стороны, и дюгонид — с дру-
гой (Murie, 1872; Howell, 1932; см. также гл. 6). Работа его у ламан-
тина по своему темпу и размаху находится в прямой связи с режимом
плавания. Для ламантина отмечено три режима: плавание медленное при
скорости в 1—3 км/ч, крейсерское — 3—7 км/ч и скоростное —18—
25 км/ч (Hartman, 1979), причем последнее крайне непродолжительно и
совершается только на коротких отрезках пути. При медленном плавании
животное совершает несколько движений хвостом, затем следует пауза,
во время которой плавание осуществляется за счет инерции. Крейсер-
ская скорость прямо коррелирует с частотой ударов и размахом движе-
ния хвостового плавника. В то же время у сирен и, видимо, особенно у
ламантина характерно, что грудная часть туловища не включается в ра-
244
боту хвоста, как это бывает у китообразных. Не происходит ни сгибания,
ни разгибания ее. Число ударов хвостового плавника в минуту достигает
24—36 при крейсерской скорости и 45—50 при быстром плавании, при-
чем у молодых животных при равной скорости частота ударов должна
быть больше, чем у взрослых (36—44 взмаха при крейсерской скорости).
Общая амплитуда движения хвостового плавника в вертикальной плоско-
сти достигает 130° (для дельфинов отмечена дуга в 200°). Возможно,
что вниз плавник опускается при движении несколько ниже относитель-
но горизонтальной плоскости, проходящей через длинную ось тела, чем
поднимается вверх (Hartman, 1979, fig. 16), причем при подъеме точка
опоры сдвинута значительно краниальнее, чем при опускании. В то же
время подъем занимает вдвое меньшее время, чем обратное движение
(см. рис. с кадров киносъемки: Hartman, 1979, fig. 17). Учитывая разни-
цу в форме дорсальной и вентральной поверхности хвостового плавника
(см. гл. 6), можно сделать вывод, что вентральное движение хвоста тре-
бует большей силы, чем обратное. Следовательно, у ламантина должны
быть механизмы, обеспечивающие скоростное движение вверх и силовое
вниз.
Говоря о работе хвостового плавника, необходимо указать еще на от-
меченную Хартманом способность ламантина вращать лопасть плавника
относительно длинной оси тела и использовать его, таким образом, в ка-
честве руля. Плывущие на крейсерской скорости животные могут направ-
лять движение, наклоняться и вращаться относительно длинной оси с
помощью одного хвостового плавника. В то же время допускаются зна-
чительные его движения в горизонтальной плоскости (по 45° вправо и
столько же влево).
Обсуждая поведение ламантина, связанное с дыханием, покоем, пита-
нием, размножением и комфортной активностью, Хартман неоднократно
обращает внимание на сопутствующие движения хвоста, туловища, пе-
редных конечностей. Так, им отмечается относительно большая гибкость
туловища, хотя и в значительно меньшей степени, чем у китообразных.
Спинной отдел при этом может несколько разгибаться (выгибаться с вы-
пуклостью вниз), значительно больше сгибаться, опускать или подгибать
под себя хвост. Надо сказать, что последнее действие является чуть ли
не наиболее характерной чертой ламантина (см. рис. 1), давно уже обра-
тившей на себя внимание исследователей (Murie, 1880). Ничего подобно-
го никто не отмечал относительно дюгоня и тем более морской коровы.
В этом случае совершается совершенно необычное по амплитуде сгиба-
ние хвоста в начале послегрудного отдела, в конце концов ведущее сово-
купно со сгибанием самого хвоста к его повороту на 180°. В литературе
отмечается, что в такой позе, положив дорсальную поверхность плавника
на дно, животное может питаться, объедая растительность вокруг, стоя
как бы на якоре. Это опять подчеркивает специфичность именно вент-
рального движения хвостового плавника ламантина.
Поднятие ламантина к поверхности для дыхания и последующее по-
гружение или заныривание совершается по-разному. Покоящееся на дне
животное небольшим подъемом хвоста разворачивается мордой вперед и
вверх (возможна, видимо, помощь передних конечностей в отталкивании
'от дна). Сам подъем совершается без заметного пропеллирующего
движения хвостом при теле, направленном примерно под углом 30—
45° к поверхности воды. Лишь при самом вдохе уже у поверхности со-
вершается слабое движение хвостом вверх, способствующее еще больше-
му поднятию переднего конца морды и удержанию ноздрей над водой.
Опускание его вниз начинается еще при видных над поверхностью воды
ноздрях. Очевидно, оно ведет к переориентации длинной оси тела ламан-
245
тина в более горизонтальное положение. Затем животное было способно
в таком положении опускаться спокойно на дно, обычно на то же место,
откуда оно поднималось вверх, что опять говорит об отсутствии все это-
время каких-либо серьезных пропеллирующих движений хвоста. В то же
время ламантин может и круто нырнуть вниз. Возможно, в этом случае
ему необходимо резкое и сильное движение хвостом вниз. При крейсеро-
вании поднятие на поверхность для дыхания совершается постепенно,,
а затем опускание сопровождается резким движением хвоста вниз, ве-
дущим к тому, что спина и хвост показываются над поверхностью, рас-
секая воду.
Как уже говорилось, о плавании дюгоней известно очень мало, еще
меньше — об их двигательной активности. Максимальная зарегистриро-
ванная скорость движения дюгоня — 18 км/ч, что, возможно, и занижено.
Однако убеждение, что дюгонь более скоростное животное, чем ламантин,
базирующееся в основном на более, кажется, скоростном типе их хвосто-
вого плавника (форма похожа на плавник кита), нельзя абсолютизиро-
вать. В отношении всех сирен без исключения говорится как о медлитель-
ных животных. Но дюгони живут в море, совершают большие по продол-
жительности миграции (Anderson, 1981b), а ламантины, обитающие'
преимущественно в закрытых местах у побережья морей, в эстуариях рек
и т. п., не нуждаются в дальних перемещениях. Дюгони свободно пасутся
на глубинах до 18 м, а ламантины избегают глубин более 4 м, даже если
там достаточно подходящей пищи. Это заставляет думать, что форма хво-
стового плавника дюгоня связана скорее не с большей скоростью движе-
ния, а с меньшей утомляемостью или большей способностью к пырянию.
Специфика работы хвостового плавника ламантина оказалась такой,
что потребовала глубокой перестройки туловищной мускулатуры и скеле-
та, затрагивающей общее развитие мышц, работающих на движение
хвоста; соотношение эпаксиальной и гипаксиальной мускулатуры; рыча-
ги действия сил и углы их приложения; создание специфических приспо-
соблений, обеспечивающих более эффективную работу мышц путем из-
менения направления их действия; организация мышц в комплексы,
действующие по одной линии, или фиксации определенных участков ске-
лета как опоры для действия мышц, развитие особых морфологических
структур, обеспечивающих автоматизм ряда движений и экономящих
мышечную энергию за счет использования сил упругости и деформации.
На работу хвостового плавника влияет большинство мышц туловища.
Суммарная их масса достигает более 25% веса тела (табл. 7). Однако
среди них целесообразно различать мышцы, активно участвующие в са-
мих движениях (собственно опускатели или подниматели хвоста),
и мышцы, улучшающие условия опускания хвоста.
Среди мышц, непосредственно участвующих в сгибании хвоста
(табл. 7, 8), находятся подкожная мышца туловища, через скуловую-
часть платизмы зафиксированная спереди на черепе, подвздошноребер-
ная мышца хвоста, гипаксиальные мышцы самого хвоста (латеральная и
медиальная вентральные мышцы хвоста, седалищнохвостовая и межпопе-
речные мышцы), также те части наружной и внутренней косых мышц
живота, которые кончаются на наружной фасции опускателей хвоста
(эти мышцы будут описаны ниже).
Среди мышц, обеспечивающих эффективную работу опускателей
хвоста, т. е. фактически функционирующих вместе с ними с общей
задачей — сгибать хвостовую часть туловища, следует назвать прямую
мышцу живота (она будет описана позже), наружные и внутренние-
межреберные мышцы, подвздошнореберную мышцу и нижнечелюстную
часть трапециевидной мышцы, фиксированную на челюстной дуге чере-
246
Таблица 7. Мышцы, влияющие на движение хвоста
Масса
обеих сторон
Мышцы, группы мышц Г %
m. cutaneus trunci (включая platysma i-is) р. jugula- 26 170 24,04
т. ischiococcygeus 2520 2,32
т. sacrococcygeus ventralis lateralis 8420 7,73
m. sacrococcygeus ventralis medialis p. cranialis 1570 1,44
m. sacrococcygeus ventralis medialis p. caudalis 11250 10,33 1 Часть
m. obliquus abdominis externus 2690 2,47 мышц,
m. obliquus abdominis internus 280 0,26 1 оканчивающих- 1 ся иа опуска-
m. iliocostalis caudae 710 0,65 - телях хвоста.
m. intertransversarius 360 0,32
Всего собственно опускатели хвоста 53 970 49,57
m. rectus abdominis 5600 5,14
m. intercostalis externus 7800 7,16
m. intercostalis internus 2530 2,32
m. iliocostalis 3470 3,19
m. trapezius p. mandibularis 2890 2,65
Всего вспомогательные опускатели хвоста 22 290 20,47
m. longissimus dorsi 13 300 12,22
m. semispinalis dorsi 3720 3,42
m. interspinalis dorsi 40 0,04
m. sacrococcygeus dorsalis lateralis И 190 10,28
m. sacrococcygeus dorsalis medialis 4360 4,00
Всего подниматели хвоста 32 610 29,95
Всего 108 870 100,00
Мышцы обеих сторон 27,22 *
В % к массе тела.
па. Все вместе мышцы — собственно сгибатели хвоста, и мышцы, обес-
печивающие их работу (вспомогательные по отношению к этой функции,
что не исключает их участие в выполнении самостоятельных функций),
имеют массу, достигающую более 70% от массы всех мышц, участвую-
щих так или иначе в работе хвоста, и лишь около 30% приходится на
долю собственно поднимателей хвоста, среди которых следует назвать
межостистую мышцу спины и системы mm. semispinalis dorsi et longis-
simus dorsi с их естественным продолжением в хвост в виде mm. sacro-
coccygeus dorsalis medialis et lateralis (их масса, указанная в табл. 7,
определялась просто отрезанием от единой системы хвостовой ее части),
причем масса длиннейшей мышцы спины в целом (вместе с m. sacro-
coccygeus dorsalis lateralis) составляет более 23% от массы мышц, ра-
ботающих на хвост. Таким образом, следует заключить, что у ламантина
на опускание хвоста работает не только большая часть мышц, но и сум-
марная сила их намного больше, чем мышц, поднимающих хвостовой
плавник (соответствующие изменения в осевом скелете, связанные с
этим, были уже отмечены в гл. 3). Интересно, что у китообразных иные
соотношения эпаксиальной и гипаксиальной мускулатуры. Так, у морской
•свиньи (Phocaena phocaena) и финвала (Balaenoptera physalus) на пер-
247
Таблица 8. Сравнение масс мышц, влияющих на движение хвоста
Trichechus inunguis и Т. manatus
Мышцы, группы мышц Т. inunguis (по: (Domning, 1978)) Т. manatus
масса масса
г % г %
m. rectus abdominis 79,2 4,2 5600 5,2
т. iliocostalis 50,0 2,7 4180 3,8
т. т. obliquus abdominis externus+internus 73,4 3,9 7890 7,1
собственно опускатели хвоста 355,9 19,1 23 760 21,3
m. cutaneus trunci 523,0 28,1 24 200 21,7
т. т. intercostalis externus+internus 175,0 9,4 10330 9,3
т. trapezius р. mandibularis 115,0 6,2 2890 2,6
Всего опускатели хвоста 1371,5 73,6 78 850 70,8
m. longissimus dorsi 353,0 18,9 24 400 21,9
т. semispinalis dorsi 139,8 7,5 8120 7,3
Всего подниматели хвоста 492,8 26,4 32 520 29,2
Всего 1864,3 100,0 111 370 100,0
Мышцы обеих сторон 26,9 27,7*
* В % к массе тела.
вую приходилось от 11 до 14% от массы тела, а на вторую немного бо-
лее 7% (Раггу, 1949), тогда как эпаксиальная мускулатура нашего ла-
мантина не превышала 8%, и при этом она была так модифицирована,
что не всей своей массой участвовала в поднятии хвоста. Следует также
вспомнить, что m. cutaneus trunci, превращенный у ламантина в мощней-
ший сгибатель хвоста, иначе построен не только у китообразных, но и у
дюгоня (Domning, 1977 Ъ), у которого при такой структуре функции
сгибателя сведены до минимума. Судя по ряду особенностей осевого ске-
лета (гл. 3), у дюгоня и морской коровы также можно ожидать относи-
тельного, сравнительно с ламантинами, усиления эпаксиальной мускула-
туры в ущерб гипаксиальной.
Отнесенный выше к группе поднимателей хвоста m. longissimus dorsi
у ламантина в грудной области имеет начало на ребрах, смещенное в не-
которой степени, учитывая положение ребер ламантина в пространстве,
латерально (примерно на i/3 ребра) от позвоночного его конца. Тем са-
мым он скорее участвует в сближении ребер, хотя оно и ограниченно,
чем в разгибании грудного отдела, что может вести к движениям позво-
ночного столба в горизонтальной плоскости. Функция поднимания хвоста,
однако, у этой мышцы усиливается с последней трети туловища по мере
увеличения связи концевых сухожилий m. longissimus dorsi с остистыми
отростками через сухожильное зеркало на m. semispinalis dorsi. В связи
с этим вся система m. longissimus и была включена в число поднимате-
лей хвоста, хотя исключительно таковым он является лишь в самом хво-
сте. Другая эпаксиальная мышцы (m. iliocostalis), служащая обычно у
млекопитающих разгибателем спины, сильно смещается в своем начале
вентрально по ребрам, что изменяет направление действия ее сил, пре-
вращает ее если не прямо в сгибатель хвоста, каким стала ее каудаль-
ная порция, превратившись в m. iliocostalis caudae, то, во всяком случае,
в мышцу, способствующую работе прочих сгибателей.
248
Мышцы, включенные в число вспомогательных, работают по-разному,
и их роль в сгибании хвоста неравноценна. Так, мощная прямая мышца
живота (табл. 7) наиболее удалена от оси сгибания позвоночного столба
и имеет соответственно большой рычаг действия и угол приложения сил,
направленных на сгибание туловища. Благодаря прикреплению на тазо-
вой кости она через связку, соединяющую последнюю с I послегрудным
позвонком, служит прямым опускателем хвоста. Однако в связи с тем
что основная функция этой мышцы — работа на брюшной пресс, мы
включаем ее в число вспомогательных мышц.
Совсем особый смысл имеет система межреберных мышц и функцио-
нально близкой к ним грудной части m. iliocostalis. Подвижность ребер
у ламантина крайне ограниченна. Активное отведение (поднятие) ребер
в сторону практически исключено (редуцированы мышцы, подниматели
ребер и дорсальные зубчатые мышцы). Ширина ребер в середине и очень
маленькие промежутки между ними допускают лишь небольшое сближе-
ние и удаление их вентральных концов, что может вести лишь к сгиба-
нию и разгибанию грудного отдела, причем последнее ограничено преде-
лом растяжения коротких межреберных мышц. В то же время возможные
движения ребер не влияют на работу диафрагмы в связи с ее особым
строением. Соответственно, межреберные мышцы меняют свою основную
функцию мышц аппарата дыхания на систему фиксации грудной клетки
для облегчения опускания хвоста. Это привело к резким структурным
перестройкам их, что особенно хорошо выражено в каудальной части
грудной клетки. В наружных межгрудных мышцах дифференцируются
продольные пучки волокон, перекидывающиеся с ребра на ребро, а иног-
да и через ребро и идущие вперед почти до уровня заднего конца груди-
ны. В более краниальных межреберных промежутках такое же направ-
ление волокон имеют близ вентральных концов ребер и внутренние
межреберные мышцы. Другие пучки волокон наружных межреберных
мышц в области между VIII и XV ребрами занимают более косое поло-
жение, причем отдельные пучки, принадлежащие разным сегментам, вы-
страиваются в одну линию, образуя как бы несколько многосегментных
полос, идущих снизу вверх и вперед. Эти продольные и косые пучки
несут разную функцию. Первые из них, расположенные у вентральных
концов ребер, имеют наиболее выгодное направление тяги, чтобы препят-
ствовать сдвигу этих концов ребер назад. Однако это возможно лишь при
фиксации всей системы спереди, что и достигается с помощью нижнече-
люстной части m. trapezius и лестничной мышцы. Первая жестко фик-
сирована на черепе и грудной кости. Лестничная же мышца, которая,
будучи закреплена на плеврапофизах шейных позвонков и первых
трех ребрах, у наземных млекопитающих является одним из важнейших
двигателей шеи, у ламантина с его короткой малоподвижной шеей фак-
тически стала фиксировать передние ребра и связанную с ними грудину,
представляющие, таким образом, жесткую опору для заднего компонента
системы, собственно сгибателей хвоста.
Функция межреберных мышц по фиксации ребер становится особен-
но ответственной в конце грудного отдела, где через вентральные концы
ребер перекидываются основные опускатели хвоста. Здесь формируется
даже специальный орган жесткости позвоночного столба (рис. 100).
У нашего экземпляра ламантина он представлен тремя последними сво-
бодными ребрами (двумя XVI и XVII большими и одним маленьким у
I послегрудного позвонка) и поперечно-реберным отростком II послегруд-
ного позвонка. При этом последние два больших ребра, сильно, откло-
ненные, если смотреть сверху назад, упираются друг в друга, поскольку
XVII ребро несколько отодвинуто в сторону, как бы пересекая при
249
гает спереди расширенная часть XVII, а
Рис. 100. Скелетная основа жест-
костной системы каудальной час-
ти грудной клетки
Римские цифры — порядковый номер
позвонков соответствующей области
1 — cartilage costalis;
2 — costae;
3 — pr. costotransversarius;
4 — 5 — vertebrae
(4 — thoracalis,
5 — postthoracalis)
взгляде сзади предшествую-
щее ребро. В свою очередь,
эти ребра вступают в кон-
такт с хрящевой частью ко-
роткого XVIII ребра и осо-
бым расширением дисталь-
ного конца поперечно-ребер-
ного отростка II (настояще-
го I) послегрудного позвон-
ка. Здесь даже появляется
гиалиновый хрящ, образую-
щий вместе со связками
нечто подобное суставу меж-
ду коротким ребром и кон-
цом поперечно-реберного от-
ростка. К этой опоре прпле-
еще дальше тоже расширенная
часть XVI ребер. Таким образом, последние ребра оказываются жестко
фиксированными и дают прочную опору перекидывающемуся через них
основному сгибателю хвоста — пт. cutaneus trunci и части мышц живота.
Это приводит к изменению направления действия сил, большому увели-
чению угла приложения сил и соответственно увеличению их рычага.
Если принять во внимание, что эти сгибатели составляют по массе почти
четверть всех мышц, действующих на хвост, то будет понятно значение
этой сложной системы фиксации ребер. К этому нужно добавить, что
в результате рычаг становится очень большим, поскольку мышцы ото-
двинуты от оси движения на длину ребер, тогда как для поднимателей
хвоста рычагом служат остистые отростки, более короткие у ламантина,
чем у дюгоня. По этому же принципу организовано строение и такого
важного опускателя хвоста, как m. sacrococcygeus ventralis medialis, ко-
торый разделяется на более поверхностные латеральные и более глубо-
кие медиальные пучки. И те и другие крепятся через три сегмента на
четвертый от начала, но наружные пучки оказываются опирающимися
на внутренние, что увеличивает как угол приложения сил, так и плечо
их рычага. Одновременно снизу на телах хвостовых позвонков появляются
гемальные дуги (шевронные кости), по высоте не уступающие невраль-
ным дугам с их остистым отростком, что также увеличивает длину ры-
чагов.
Усиление ligg. flava у ламантина, сравнительно с другими млекопи-
тающими и даже, видимо, с дюгонем (см. гл. 3), превращение этой связ-
ки в мощнейший эластический тяж с сечением 3,5 X 2,3 см в задней по-
ловине грудного отдела, но более тонкий в начале груди, свидетельствует
о повышении у него необходимости перевести режим работы хвостового
плавника частью в более автоматический режим за счет сил упругости.
При этом исчезает необходимость иметь сильные эпаксиальные мышцы и
250
они ослабляются у ламантина. Появляется возможность к укорочению
остистых отростков, именно топ их части, что помещается выше площа-
док для ligg. flava, а следовательно, допускается и более значительное
сгибание туловища, чему могли бы препятствовать короткие mm. inter-
spinales. Наоборот, эластичная связка допускает максимальное сгибание
с автоматическим возвращением позвоночного столба в исходное состоя-
ние. Как не требующее мышечных усилий, осуществляемое за счет сил
упругости и, возможно, упругой деформации межпозвоночных дисков,
площадь которых (а главное вертикальная протяженность) сильно уве-
личена у ламантина сравнительно с дюгонем, возвращение хвостового
плавника в горизонтальное положение стало более быстрым. Лишь даль-
нейший его подъем требует уже мышечных усилий. Возможно, именно с
этим и связано сокращение времени, затрачиваемого на общий подъем
хвостового плавника из максимально низкого состояния в максимально
высокое при крейсерском плавании, чем на движение плавника вниз,
требующее больших усилий и в силу гидродинамических особенностей
плавника, и в силу необходимости растягивать ligg. flava. Следует при
этом отметить, что даже при естественном (более или менее горизонталь-
ном) положении позвоночного столба жесткие связки уже оказываются
в сильно напряженном состоянии.
Из более слабого развития ligg. flava у дюгоня и морских коров,
о чем можно судить по строению площадок для их прикрепления (см.
гл. 3), можно сделать предположительный вывод, что в дюгонидной ли-
нии развития сирен использование сил упругости не получило большого
применения. И здесь можно ожидать более равного развития эпаксиаль-
пой и гипаксиальной мускулатуры.
Кроме основных функций поднятия и опускания хвоста, мышцы туло-
вища выполняют и ряд дополнительных функций. По-видимому, основа
этих дополнительных движений заключена в одностороннем включении
мышц туловища. При одностороннем включении полуостистой мышцы
спины, кроме разгибательного момента, возникает и вращательный мо-
мент вокруг оси позвоночного столба. Этот момент в конечном счете поз-
воляет ставить хвостовой плавник под некоторым углом к фронтальной
плоскости туловища. По-видимому, последнее движение особенно хорошо
развито у дюгоня. Дело в том, что вращательный момент тем более пре-
валирует над разгибательным, чем более косо направлены мышцы. А это
значит, что он возрастает как за счет сокращения длины позвонков, так
и за счет меньшего количества сегментов, через которые перекидываются
волокна от начала до конца соответствующего пучка. У дюгоня тела по-
звонков короче, чем у ламантина, и, кроме того, его трансверзоспиналь-
ная система состоит практически полностью из мышечных сегментов,
перебрасывающихся через один-два сегмента на второй-третий, тогда как
у ламантинов они распространяются на четыре и более сегментов, т. е.
если у дюгоня имеются m. multifidus и еще более короткие пучки этой
системы (Slijper, 1946; Domning, 1977b), то у ламантина преимущест-
венно развита система m. semispinalis.
Одностороннее сокращение длиннейшей мышцы спины и крестцово-
хвостовых дорсальных и вентральных мышц приводит к отведению хво-
стового плавника в сторону при сохранении его фронтального положения.
В этом случае подъем и опускание хвоста приводят одновременно к раз-
вороту животного.
251
МЫШЦЫ ШЕИ
Поверхностные мышцы шеи, топографически принадлежащие этой обла-
сти, описаны выше в составе лицевой мускулатуры головы (m. sphinc-
ter colli superficialis, platysma myoides, m. sphincter colli profundus). Это
все производные подкожной мускулатуры. Ниже приведенное описание
будет связано с так называемыми глубокими мышцами шеи.
М. splenius пластыревидная мышца (см. рис. 92, 93 а), начинается
широким апоневрозом, в своем отхождении тесно связанным с наружной
фасцией подлежащего m. semispinalis capitis (в интервале от III—IV до
VI—VII грудных позвонков). Вопреки описаниям бразильского ламанти-
на и дюгоня (Domning, 1977b, 1978а) начальный апоневроз не достигает
средней линии, что более согласуется с тем, что отмечалось ранее у
Trichechus manatus (Murie, 1872а). После выхода из-под m. rhomboideus
в апоневрозе появляются мышечные волокна, которые направляются впе-
ред и вбок и формируют брюшко, в начале широкое плоское, а ближе к
прикреплению оно становится треугольным в сечении (см. рис. 88). Кон-
чается пластыревидная мышца на височной кости сразу позади височно-
го гребня и на фасции, затягивающей большое мастоидное отверстие на
стыке верхней и боковой затылочных и височных костей (см. рис. 75, а,
76, 79). В отличие от многих млекопитающих у сирен m. splenius не
прикрепляется ни к шейным позвонкам, ни к выйной связке, которая у
них вообще отсутствует.
М. semispinalis capitis, полуостистая мышца головы (см. рис. 70, 93),
начинается отдельными мышечными зубцами, сливающимися в единую
мышечную массу, идущую вперед к голове. Наиболее медиальный и дор-
сальный из них начинается сухожильной фасцией от фасциальной вы-
стилки длиннейшей и полуостистой мышц спины в месте их контакта на
уровне VIII—IX грудных позвонков (см. рис. 93). Более задние латеро-
вентральные зубцы отходят от особых бугорков первых шести ребер,
а более передние от латеральных краев дорзальной стороны постзига-
пофизов II—VI шейных позвонков (см. рис. 91). У бразильского ламан-
тина и дюгоня задние зубцы начинаются от дорзальной поверхности диа-
пофизов I—VII грудных позвонков, т. е. у нашего экземпляра отхождение
смещено латеральнее, что может быть связано с большим развитием
ш. semispinalis dorsi, занимающим все пространство над поперечными
отростками. Общее мышечное брюшко полуостистой мышцы несегменти-
ровано. Волокна его идут, конвергируя, к прикреплению (частью сухо-
жильно, частью мышечно) на верхней части верхней затылочной кости
позади затылочного гребня черепа (см. рис. 75, а, 76, 79). Своей внут-
ренней поверхностью m. semispinalis capitis сильно сращен с т. rectus
capitis dorsalis major. Прикрепление у бразильского ламантина в отличие
от нашего представляет узкую полоску, заходящую немного на чешую
височной кости и боковую затылочную кость. Необычное состояние дан-
ной мышцы у дюгоня (очень широкое прикрепление на черепе и соот-
ветственно широкое мышечное брюшко), описанное Домнингом,—несом-
ненно результат недоразумения: наиболее передневентральные волокна,
идущие от дорсальной стороны поперечного отростка атланта и, возмож-
но, постзигапофиза эпистрофея к боковой затылочной кости, представля-
ют m. obliquus capitis cranialis (см. дальше), сходный с таковым типич-
ных млекопитающих. Описанная же под, этим названием мышца дюгоня
в действительности представляет m. rectus capitis ventralis.
М. semispinalis cervicis, полуостистая мышца шеи (см. рис. 93, б),
начинается хорошо выраженными сухожильными ножками от особых бу-
горков на переднем крае диапофизов II—VI грудных позвонков (р. la-
252
teralis), а также от связки, соединяющей постзигапофиз V с диапофизом;
VI грудных позвонков (р. medialis). Мышечные волокна формируют бо-
лее или менее дифференцированные брюшки, которые объединяются в
единую мышцу, лежащую над остистыми отростками шейных и первых
грудных позвонков. Лишь брюшко от названной связки сохраняет практи-
чески свою самостоятельность вплоть до эпистрофея. Кончается m. semi-
spinalis cervicis мышечными волокнами на заднем крае гребня эпистрофея
(см. рис. 91). Кроме уже упомянутых сухожильных ножек, от V и VI
грудных позвонков имеются еще пучки мышечных волокон (р. caudalis),
которые начинаются сразу медиальнее последних и идут вперед, причем
пучок от VI позвонка крепится на III грудном позвонке сразу каудаль-
нее отхождения III сухожильной ножки, а от V позвонка — на II позвон-
ке. Эти пучки можно назвать медиальными ножками в отличие от опи-
санных ранее латеральных (сухожильных). М. semispinalis cervicis впол-
не обособлен от in. spinalis et semispinalis dorsi, который тянется далеко
вперед, достигая III шейного позвонка, и располагается спереди вент-
ральнее полуостистой мышцы шеи, а далее назад оттесняет ее от средней
линии. Домнинг (1977b, 1978а) не выделяет этой мышцы из единого
ш. semispinalis (включающего также наш m. spinalis et semispinalis
dorsi).
M. longissimus cervicis et atlantis, длиннейшая мышца шеи и атланта
(см. рис. 93), объединяет две обычно раздельные мышцы, связывающие
передние грудные и задние шейные позвонки одна с атлантом, другая с
передними шейными. В нашем случае отчетливо дифференцируются на-
чальные сухожильные ножки от специальных бугорков первых двух ре-
бер (от них начинаются мышечные волокна, идущие вперед) и мощное
сухожильное окончание мышцы на диапофизе атланта. Сухожильные-
ножки отходят также от дистальных концов поперечных отростков
остальных шейных позвонков (II—VI). Задние из них являются, видимо,
начальными, а передние, возможно, уже надо рассматривать как конце-
вые, принадлежащие собственно m. longissimus cervicis. Домнинг не вы-
деляет эту мышцу из общего комплекса m. longissimus dorsi.
М. rectus capitis dorsalis (см. рис. 70, 73) у нашего ламантина факти-
чески состоит из трех мышц, как это обычно и бывает у млекопитаю-
щих. М. rectus capitis dorsalis major, большая дорсальная мышца головы,
наиболее поверхностная и мощная из них. Она начинается мясисто от
большей части гребня эпистрофея (см. рис. 91) и, расширяясь, крепится
на особой площадке затылочной кости, занимая обширное место непо-
средственно под ш. semispinalis capitis (см. рис. 79), с которым, как уже
отмечалось, она частично срастается по своей поверхности. М. rectus ca-
pitis dorsalis intermedins, средняя дорсальная прямая мышца головы, зна-
чительно меньше предыдущей и не полностью от нее обособлена, хотя
срастания между ними в дистальной части нет. Начинается от переднего
края и части латеральной поверхности гребня эпистрофея впереди и вент-
ральнее отхождения m. rectus capitis dorsalis major (см. рис. 91). При-
крепляется на затылочной кости вентральнее и несколько латеральное
последней (см. рис. 79). М. rectus capitis dorsalis minor, малая дорсаль-
ная прямая мышца головы, наиболее глубокая и короткая из всех, на-
чинается от верхней дуги атланта (см. рис. 91) и идет прямо вперед к
прикреплению на затылочной кости близ средней линии ниже окончания
большой и медиальнее окончания средней дорсальных прямых мышц го-
ловы (см. рис. 79). У бразильского ламантина наличествует сходное со-
стояние, хотя особая средняя мышца и не выделяется. Однако Домнинг
(1978а), очевидно, в силу ошибочной интерпретации данных мышц, вы-
сказанной им ранее при описании мускулатуры дюгоня (1977b), боль-
253
штую мышцу считает отсутствующей у его ламантина, а имеющуюся хо-
рошо развитую называет средней. Дело в том, что у дюгоня им была
описана очень маленькая наиболее дорсально расположенная мышца, ко-
торую он сопоставлял с m. rectus capitis dorsalis major, а имеющуюся под
ней обширную мышцу с типичным для большой дорсальной прямой мыш-
цы головы прикреплением — средней. Логичнее, однако, было бы гово-
рить о выделении из обычного m. rectus capitis dorsalis major небольшой
дорсальной порции, не свойственной большинству млекопитающих, и от-
сутствии средней мышцы, что также является обычным. Соответственно
у ламантина, не найдя еще одной мышцы, которую можно было бы на-
звать собственно малой дорсальной мышцей, Домнинг и пришел к выво-
ду об отсутствии у него m. rectus capitis dorsalis major, поскольку термин
«средняя дорсальная прямая мышца» он отнес к верхней, большей из
двух имеющихся здесь мышц.
М. obiiquus capitis caudalis, каудальная косая мышца головы (см.
рис. 73), начинается мясисто от заднебокового угла остистого отростка
эпистрофея вентральнее отхождения m. rectus capitis dorsalis major (см.
рис. 91). Мышечные волокна направляются вперед, вниз, главным обра-
зом латерально к прикреплению на задней поверхности поперечного от-
ростка атланта.
М. obiiquus capitis cranialis, краниальная косая мышца головы (см.
рис. 70, 73, 93), одна из наиболее мощных и сложно построенных шейных
мышц. Начинается мышечными волокнами и сухожильными тяжами от
обширной площади на дорсальной поверхности верхней дуги и диапофиза
атланта, впереди от прикрепления т. obiiquus capitis caudalis, а также
отдельными более слабыми зубцами от постзигапофизов II —IV шейных
позвонков (явление, крайне необычное для млекопитающих) (см.
рис. 91). Обширное мышечное брюшко с общим косым направлением
(вперед и вверх) имеет многоперистое строение, будучи пронизано внут-
ренними плоскими сухожилиями, частью связанными с началом мышцы,
частью с ее окончанием. Всего в брюшке можно выделить до 10 отдель-
ных структурных единиц (перьев). Кончается краниальная косая мышца
головы на большей части латерального крыла боковой затылочной кости
(см. рис. 79), т. е. вентральнее прикрепления m. splenius, медиальнее
m. clavotrapezius и m. clavomastoideus, дорсальнее т. rectus capitis la-
teralis и латеральнее группы прямых дорсальных мышц головы. Мышцу,
описанную Домнингом у дюгоня как т. obiiquus capitis cranialis, скорее
всего, надо рассматривать в качестве m. rectus capitis ventralis. За это
говорит ее слабость, отхождение от вентральной стороны поперечного от-
ростка атланта, прикрепление на вентральной части боковой затылочной
кости между затылочным мыщелком и прикреплением m. brachiocepha-
licus, т. е. все, что характерно для прямой латеральной мышцы головы
млекопитающих. Наоборот, мышца, описанная Домнингом в качестве
последней у бразильского ламантина, имеет все признаки, свойственные
ш. obiiquus capitis cranialis.
М. rectus capitis lateralis, латеральная прямая мышца головы (см.
рис. 73, 92), начинается мышечно от вентральной части передней по-
верхности поперечного отростка атланта (см. рис. 91, а). Ее волокна идут
прямо вперед, формируя небольшое брюшко. Кончаются они у основания
парокципитального отростка боковой затылочной кости (см. рис. 79).
У бразильского ламантина сходная по всем параметрам мышца описана
Домнингом под названием «т. obiiquus capitis cranialis», что, несомненно,
ошибочно. В свою очередь, «т. rectus capitis lateralis» ламантина Дом-
нинга представляет m. obiiquus capitis cranialis. Одноименная мышца дю-
гоня тем не менее определена точно. Все это снимает удивляющее Дом-
254
нинга несходство m. rectus capitis lateralis бразильского ламантина от-
Trichechus manatus и дюгоня.
М. rectus capitis ventralis, вентральная прямая мышца головы,—не-
большой пучок волокон, отходящий от нижней дуги атланта впереди и
вентральнее поперечного отростка (см. рис. 91, а). Она направляется впе-
ред и крепится непосредственно спереди от затылочного мыщелка (см.
рис. 75, б). У бразильского ламантина отмечено прикрепление далее впе-
ред к tuberculum pharyngeum сразу позади окончания m. longus capitis.
У дюгоня.мышцы под таким названием вообще не описано. Однако оче-
видно, что небольшая мышца, обозначенная как m. obiiquus capitis cra-
nialis и отходящая от антеровентральной стороны поперечного отростка
атланта, полностью соответствует вышеприведенному описанию. Только
ее окончание смещено несколько латеральнее от затылочного мыщелка,
ближе к прикреплению m. brachiocephalicus.
Таким образом, вся группа прямых и косых мышц головы у сирен
значительно ближе к таковым обычных млекопитающих, чем это можно'
понять из описания Домнинга. Источником ошибок явилось включение
им у дюгоня волокон, принадлежащих m. obiiquus capitis cranialis, в со-
став m. semispinalis capitis, который никогда у млекопитающих не на-
чинается от атланта. Соответственно термин «т. obiiquus capitis crania-
lis» пришлось приложить к мышце, принадлежащей топографически к
другому региону, и вследствие этого для названия «т. rectus capitis;
ventralis» уже не нашлось мышц. Впоследствии при описании бразиль-
ского ламантина, где соответствующая мышца в силу своей большей свя-
зи с m. longus capitis уже никак не могла быть идентифицирована иначе,
чем как вентральная прямая головы, Домнингу пришлось приложить,
название «т. obiiquus capitis cranialis» к лежащей более латерально
m. rectus capitis lateralis, поскольку по своему развитию эта мышца зна-
чительно ближе к мышце, определенной у дюгоня как краниальная косая-
голова. 'А так как у ламантина уже не было тесной связи m. semispi-
nalis capitis с настоящим m. obiiquus capitis cranialis и последний пред-
ставлял вполне обособленную мышцу, то, чтобы найти выход из создав-
шегося положения, Домнингу пришлось говорить о неестественном
увеличении «т. rectus capitis lateralis», так якобы отличающем бразиль-
ского ламантина от американского и от дюгоня.
Таким образом, мышцы, описанные в данной работе у ламантина
Trichechus manatus, гомологизируются с мышцами бразильского ламан-
тина и дюгоня (в терминах, данных Домнингом) следующим образом::
Trichechus manatus
m. semispinalis capitis
m. semispinalis cervicis
m. longissimus cervicis
et atlantis
m. rectus capitis dorsalis
major
Trichechus inunguis
(Domning, 1978a)
m. semispinalis capitis
m. semispinalis (pt)
m. longissimus dorsi (pt)
Dugong dugon (Domning,
1977b)
m. semispinalis capitis
(pt) (волокна, идущие к
верхнезатылочной кости)
m. semispinalis capitis
(pt) или часть общей
transversospinal system
m. longissimus dorsi (pt)
m. rectus capitis dorsalis
(?) intermedius
C m. rectus capitis dorsalis
I (?) major
| m. rectus capitis dorsalis;
1 (?) intermedius
m. rectus capitis dorsalis
intermedius
m. rectus capitis dorsalis
minor
m. rectus capitis dorsalis
minor
m. rectus capitis dorsalis
minor
255
m. obliquus capitis cra-
nialis
m. rectus capitis lateralis
m. rectus capitis ventralis
m. rectus capitis lateralis
m, obliquus capitis cra-
nialis
m. rectus capitis ventralis
m. semispinalis capitis
(pt) (волокна, идущие
от атланта и эпистрофея
к боковой затылочной
кости)
m. rectus capitis lateralis
m. obliquus capitis cra-
nialis
Наше толкование остальных мышц описываемого комплекса не отли-
вается от такового Домнинга.
М. longus capitis, длинная мышца головы (рис. 101), начинается су-
хожильными ножками от задних краев вентральной поверхности тел
IV и III грудных позвонков, которые сливаются в одно широкое сухо-
жилие, затем тонкими сухожилиями от специальных мышечных бугорков
на вентральной поверхности II и I ребер близ их головок. Отходящие от
этих сухожилий мышечные волокна вливаются в общее мышечное брюш-
ко, которое также пополняют волокна, начинающиеся от фасции, разде-
ляющей mm. longus capitis et cervicis. На противоположной стороне вме-
сто длинных сухожильных ножек от IV и VI грудных позвонков имеется
практически мышечное начало от III и II позвонков. На уровне III—
IV шейных позвонков на наружной поверхности мышцы появляется су-
хожильное зеркало. Ближе к голове от его наружной поверхности начи-
наются мышечные волокна, которые формируют более поверхностную и
латеральную часть общего мышечного брюшка m. longus capitis. Это при-
водит к тому, что зеркало уходит в глубь мышцы, образуя в ней широкую
продольную перегородку (см. рис. 101), отделяющую более поверхност-
ную и латеральную часть волокон, которые с него начинаются, от более
глубоких и медиальных волокон, часть которых прикрепляется к
его внутренней (дорсальной) поверхности. Общее прикрепление
(см. рис. 75, б) на tuberculum pharyngeum черепа осуществляется как
мышечно (наиболее глубокие и длинные волокна медиодорсальной части
мышцы и более короткие поверхностные волокна), так и посредством от-
меченного выше плоского сухожилия.
У бразильского ламантина отмечено отхождение мышцы только от
тела I грудного позвонка и от головки I ребра, а у одного из экземпля-
ров имелся еще сухожильный зубец от плеврапофиза V шейного поз-
вонка.
У американского ламантина (Murie, 1872а, 1880) было описано со-
стояние как близкое к бразильскому ламантину, так и приближающееся
к нашему описанию (в частности, более каудально перемещено начало —
к II ребру и III грудному позвонку). Очевидно, отхождение m. longus
capitis варьирует индивидуально.
У дюгоня отмечено начало на I ребре и поперечных отростках V и
VI шейных позвонков.
М. longus colli, длинная мышца шеи (см. рис. 101), представляет
собой сложную систему слабо дифференцированных мышечных пучков,
связывающих первые грудные позвонки с шейными и атлантом и послед-
ние шейные с атлантом. Вся масса этой мышцы варьирует даже у одно-
го экземпляра, что, видимо, связано со слабой функциональной ее зна-
чимостью вследствие почти полной неподвижности шейных позвонков
друг относительно друга. Вариации захватывают и характер отхождения
той или иной порции мышцы: оно может быть сухожильным, смешанным
или чисто мышечным. В схеме строение мышцы можно представить сле-
дующим образом. Более каудальная и медиальная группа мышечных
256
Рис. 101. Длинные мышцы шеи
и головы (вид снизу). Удалена
грудина
Римские цифры — номера сегментов
1 — 2 — lig. capitis costae radiatum
(J — p. caudalis,
2 — p. cranialis);
3 — 4 — m. longus
(3 — capitis,
4 — colli);
S — m, scalenus;
6 — межреберная чудесная сеть
пучков, входящих в состав
мышцы, начинается от тел
первых трех грудных позвон-
ков и мускульных бугорков
на вентральной стороне пер-
вых двух ребер. Наиболее
медиальная и задняя порция
начинается широким сухо-
жилием, продолжающимся по
ее вентральной поверхности
на 2/3 ее длины, а кончается
мясисто на медиальной части
вентральной дуги атланта и
отдельными мышечными пуч-
ками, отходящими от ее
внутренней (дорсальной) поверхности к телам II—V шейных позвонков.
Часть мышечных волокон этой порции начинается от фасции, разделяю-
щей эту мышцу и m. longus capitis. Более латеральные порции могут
начинаться как сухожильно, так и мышечно. Кончаются они на осно-
ваниях поперечнореберных отростков IV—V шейных позвонков. Таким
образом, общее направление этой части мышцы сзади наперед и медиаль-
но, причем латеральные порции короче медиальной. Вторая группа мы-
шечных пучков начинается от задних шейных позвонков и имеет общее
направление вперед и медиально. Здесь можно выделить три порции.
Наиболее латеральная с широким мышечным началом от вентральной
поверхности VI шейного позвонка идет к атланту на задневентральную
поверхность его поперечного отростка. Средняя порция начинается сухо-
жильными зубцами от поперечно-реберных отростков VI—III шейных
позвонков, а крепится на вентральной дуге атланта медиальнее прикреп-
ления предыдущей порции. Наиболее медиальная порция отходит широ-
ким сухожильным апоневрозом от оснований поперечно-реберных отрост-
ков VI—V шейных позвонков и прикрепляется на вентральной дуге
атланта медиальнее предшествующей порции, будучи оттесненной от
средней линии лишь наиболее медиальной и длинной порцией задней
группы пучков пт. longus colli.
У бразильского ламантина Домнинг выделяет всего три неясно диф-
ференцированные порции: наиболее латеральную от тел II—VI шейных
позвонков; среднюю, более глубокую, от III—V шейных позвонков и
медиальную, наиболее обширную, от VI шейного и I—II грудных по-
звонков. Первые две порции крепятся только на атланте, а последние —
также на II—V шейных позвонках.
9 Ламантин
257
На шейных позвонках оканчивается также m. scalenus, начинающий-
ся на передних ребрах. Однако кажется, что его основная функция —
не управлять движениями шеи, а фиксировать положение ребер, т. е.
способствовать более эффективной работе мышц, связанных с движением
хвоста, поэтому он был рассмотрен раньше. Аналогично участвующие в.
движениях головы m. trapezius р. mandibularis и m. sternomastoideus
описаны в соответствующих местах.
РАБОТА КОМПЛЕКСА ШЕЙНЫХ МЫШЦ
Шея сирен (как было показано в гл. 3) малоподвижна. Единственным
значительным движением является поднимание и опускание головы,,
амплитуда его у ламантина равна 65—70°, а у дюгоня и морской коро-
вы, по-видимому, меньше. При этом известно (Steller, 1751), что у мор-
ской коровы голова при кормлении почти все время наклонена вниз.
В то же время такой наклон совершенно излишен у дюгоня с его силь-
ным наклоном ростральной части черепа вниз. Наоборот, у него должна
усиливаться функция поднятия головы (см. гл. 3). Боковые и враща-
тельные движения головы у Т. manatus весьма ограниченны, хотя, воз-
можно, и в меньшей степени, чем у дюгоня. Ограничение подвижности
шеи привело к редукции ряда коротких мышц шеи, а именно многораз-
дельной мышцы шеи, межпоперечных и межостистых мышц. Суммарная
масса всех мышц, влияющих на движение шеи и головы, по отношению
к туловищу составляет 2% массы тела. При этом в эту группу (табл. 9)
внесены также мышцы, функция которых далеко не полностью связана
с движением головы и шеи. Так, одна из наиболее мощных мышц,
включенных в этот комплекс,— нижнечелюстная часть трапециевидной
мышцы, которая, как уже говорилось ранее, практически в меньшей мере
Таблица 9. Мышцы, действующие на движение головы
Мышца, группа мышц Масса Мышца, группа мышц Масса
Г. % Г. %
m. rectus capitis dorsalis major m. rectus capitis dorsalis intermedius m. rectus capitis dorsalis minor in. obliquus capitis cra- nialis in. obliquus capitis cau- dalis m. rectus capitis lateralis m. longissimus cervicis+at- lantis m. semispinalis capitis m. semispinalis dorsi до IX th. от эпистрофея m. semispinalis cervicis m. splenius Всего поднимателп головы 249 60 56 296 36 34 73 <662 1110 351 800 4727 3,03 0,73 0,68 3,61 0,44 0,41 0,89 20,25 13,52 4,28 9,75 57,59 m. scalenus т. rectus capitis ventralis т. longus colli m. longus capitis m. trapezius p. mandibu- laris m. sternomastoideus Всего опускатели головы Всего Мышцы обеих сторон 638 32 92 96 2500 122 3480 8207 7,80 0,39 1,12 1,16 30,45 1,49 42,41 100,0 2,06*
* В % к массе тела.
258
Таблица 10. Сравнение масс мышц шеи у Trichechus manatus и Т. inunguis
(no (Domning, 1978))
Мышцы, группы мышц т. inunguis (Domning, 1978) Т. manatus
масса масса
г % г %
2 m. recti capitis dorsalis 16,8 7,4 365 5,6
т. obliquus capitis cranialis 7,0 3,0 296 4,5
m. semispinalis capitis 50,3 21,1 1662 25,7
m. splenius 19,2 8,6 800 12,4
Всего поднимателп головы 93,3 40,1 3123 48,2
in. rectus capitis ventralis 1,0 0,4 32 0,5
m. scalenus 8,3 3,6 638 9,8
in. longus colli 2,3 1,0 92 1,4
m. longus capitis 6,8 3,0 96 1,5
m. trapezius p. mandibularis 115,5 51,9 2500 38,6
Всего опускатели головы 133,9 59,9 3358 51,8
Всего 227,2 100,0 6481 100,0
Мышцы обеих сторон 1,64 1,62*
* В % к массе тела.
опускатель головы, чем опускатель хвоста, так как поверхностные ее во-
локна прямо переходят в подкожную мышцу, являющуюся у ламантинов
основным опускателей хвоста. Истинными поднимателями головы у си-
рен можно считать: полуостистую мышцу головы, составляющую 20,25%
веса всей группы мышц, действующих на голову и шею, и прямые дор-
сальные мышцы головы, в сумме составляющие 4,44% массы всей груп-
пы (См. табл. 9). Интересно отметить развитие двух косых мышц головы.
Обычно у млекопитающих из них значительно сильнее развита каудаль-
ная косая мышца головы, которая является основной мышцей, вращаю-
щей голову на зубовидном отростке эпистрофея. У ламантина она очень
слаба, и только латеральная прямая мышца несколько слабее ее. И это
неудивительно, так как вращение вокруг зубовидного отростка у ламан-
тина крайне ограниченно. Зато краниальная косая мышца головы, кото-
рая у ламантина становится в основном поднимателем, а не ротатором
головы, крайне усиливается: возрастает ее масса и одновременно она
превращается в многоперистую мышцу (см. рис. 70). При одностороннем
действии она может в небольшой степени (по-видимому, не более 10°)
поворачивать голову в атлантозатылочном суставе. Интересно отметить,
что она усиливается также и за счет значительного увеличения площади
«е отхождения, которое у наземных млекопитающих ограничивается ат-
лантом, а у ламантина захватывает и последующие три шейных позвонка.
У бразильского ламантина (Domning, 1978а) заметно сильнее разви-
ты прямые мышцы головы и слабее краниальная косая мышца головы
(табл. 10). Аналогично ослабляется у него и пластыревидная мышца.
Поскольку она, как и передняя косая мышца головы, кроме поднятия
головы, вызывает еще и ее повороты и вращения, то складывается впечат-
ление, что, по-видимому, у бразильского ламантина эти движения разви-
ты еще меньше, чем у Т. manatus. Интересно, что у дюгоня и в мень-
259
9*
ятей мере у бразильского ламантина начало га. splenius распространяется
вперед по шее почти до затылка, тем самым его волокна принимают
(особенно в передней своей части) более вертикальное направление, тог-
да как у нашего ламантина расположение волокон значительно более го-
ризонтально, что приводит к большему моменту на поворот, чем у дюгоня
и бразильского ламантина, но к меньшей — на подъем головы. Одновре-
менно эта мышца и усиливается у нашего ламантина, составляя 12,4%
массы группы при 8,4% у бразильского ламантина. Это опять свидетель-
ствует о меньшей боковой и вращательной подвижности головы у бра-
зильского ламантина.
Таким образом, у бразильского ламантина, как и у дюгоня, повышено
значение мышц — поднимателей головы. Уже говорилось о важности
подъема головы у дюгоня и о том, что наиболее нагруженным у него
является такое состояние атланто-затылочного сустава, какое ведет к
поднятию головы (см. гл. 3). Особенности питания бразильского ламан-
тина сравнительно с Т. manatus (Domning, 1978а) объясняют, почему и
у него к поднятию головы приложено больше силы. Если американские
ламантины питаются растительностью по всей толще воды, в том числе
и растущей на дне, вырывание которой требует как движения головы
вверх и в какой-то мере вбок, то бразильские ламантины используют в
основном плавающую растительность, находящуюся у поверхности воды.
Таким образом, им необходимо удержание головы в приподнятом состоя-
нии (ростральная часть черепа у них также более прямая, чем у боль-
шинства Т. manatus), но нет необходимости в резких рывках.
В заключение следует повторить, что у всех сирен самостоятельная
подвижность шеи практически отсутствует. Главными стали движения в
атланто-затылочном суставе по поднятию и опусканию головы. Исходя
из этого понятно, почему у сирен усилены мышцы, работающие именно
на поднятие головы. Очень малы вращательные движения в атлантоосе-
вом суставе. Соответственно сильно редуцирована каудальная косая
мышца головы. Вся шея фактически превращена в неподвижный стер-
жень, на котором сидит подвижно сочлененная с ним голова. Причем эта
неподвижность особенно подчеркнута у дюгоня и морской коровы с их
черепитчатым положением зигапофизов и остистых отростков (см. гл. 3%
исключающим разгибание в шейных и передних грудных суставах. Шея,
возможно, может несколько подниматься вся в целом вместе с головой,
причем ось этого поднятия у дюгонид сдвинута дальше на грудной отдел.
У ламантина нет этой системы фиксации шейных суставов, вернее, она
устроена иначе и, возможно, менее эффективно. Соответственно у них
следует ожидать и несколько большей подвижности в самой шее. Из-за
необходимости увеличения определенных движений в атланто-затылочном
суставе в связи с различиями в добывании корма выявляется определен-
ная специфика в развитии ответственных за эти движения мышц у двух
видов ламантинов, у ламантинов и дюгоней. В литературе отмечается
возможное усиление латеральной подвижности шеи у морских коров
(Domning, 1978с, см. также гл. 3). Весь ход рассуждений Домнинга по
этому поводу в целом верен, хотя очевидно, что развитие у морских коров
латеральных фланцев боковых затылочных костей связано с усилением
коротких мышц, способствующих боковым движениям, а именно кра-
ниальной косой мышцы головы, а не приданием этих функций длинной
полуостистой мышце головы, которая якобы распространилась в своем
прикреплении на эти фланцы (см. выше).
260
МЫШЦЫ БРЮШНОГО ПРЕССА И ДИАФРАГМЫ
М. obiiquus abdominis externus, наружная косая мышца живота
(см. рис. 68), начинается мышечными зубцами от наружной поверхно-
сти III—XVII ребер по косой линии, так что передний зубец отходит на
уровне конца близ прикрепления реберного хряща. При этом происходит
постепенное возрастание мощности мышечных зубцов спереди назад.
Объединяясь, мышечные зубы образуют единый мускульный пласт, во-
локна которого имеют общее направление снаружи назад и медиально.
Передняя часть мышцы оканчивается серией сухожильных зубцов, во-
локна которых прослеживаются вплоть до белой линии живота. В то же
время своей дорсальной поверхностью эти зубцы сращены в промежутках
между аналогичными зубцами m. obiiquus abdominis internus с наружной
фасцией прямой мышцы живота серией сухожильных пучков. Последние
7—8 мышечных зубцов, слагающих заднюю часть мышцы, в своем ходе
перекидываются через дистальные концы ребер, меняя, таким образом,
направление приложения сил, и кончаются на вентральной фасции
мышц, опускателей хвоста (m. ischiococcygeus lateralis et medialis, sacro-
coccygeus ventralis lateralis et medialis, m. intertransversarius). У бразиль-
ского ламантина было обнаружено отхождение m. obiiquus abdominis
externus лишь от III—XIV ребер, а у американского (Murie, 1872а) от
всех ребер. У дюгоня отхождение еще обширнее, захватывая апоневроз
между XIX ребром и поясничными позвонками.
М. obiiquus abdominis internus, внутренняя косая мышца живота
(рис. 102), начинается мышечными зубцами от внутренней поверхности
дистальных концов V—XVII ребер близ прикрепления к ним реберных
хрящей, а также от наружной фасции латерального сгибателя хвоста
(m. sacrococcygeus ventralis lateralis). Общее направление их волокон
вперед и медиально, почти перпендикулярно к направлению волокон
m. obiiquus abdominis externus. При этом самый задний зубец (от фас-
ции) перебрасывается через дистальный конец XVII ребра, а зубец от
этого ребра через конец XVI ребра. Все мышечные зубцы соединяются
в единый пласт, который медиально переходит в широкий апоневроз,
при подходе к m. rectus abdominis расслаивающийся на два листка —
внутренний и наружный, охватывающий его с обеих сторон. Каждый из
них представляет систему сухожильных тяжей, не образующих единого
слоя, которые сращены с фасцией прямой мышцы живота. Их волокна
прослеживаются до белой линии живота. Описываемая мышца бразиль-
ского ламантина, как и предыдущая, имеет менее протяженное назад
отхождение — только от IV—XV ребер. Обнаруженные Домнингом от-
дельные пучки, связывающие дистальные концы XII и XIII, XIII и
XIV, XIV и XV ребер и отнесенные им к m. obiiquus abdominis internus,
несомненно, принадлежат к системе mm. intercostales interni.
М. rectus abdominis, прямая мышца живота (см. рис. 68, 98), начина-
ется частью сухожильно, частью мышечно отдельными зубцами
(см. рис. 86) от заднего края I и от вентральной поверхности II и III
реберных хрящей, от латерального края грудины у места сочленения с
III реберным хрящом, от переднего и заднего краев дистального конца
II ребра и от свободного конца хряща IV ребра и идущей от него сухо-
жильной перемычки к телу грудной кости. После соединения всех этих
зубцов образуется единый! мышечный пласт, волокна которого направля-
ются назад вдоль всего брюха. В области анального отверстия мышечное
брюшко сужается и острым мышечным зубцом закрепляется на краниаль-
ном крае тазовой кости, по латеральному краю ножки полового члена,
проникая между m. ischiocavernosus и s. ischiouretralis к его основанию.
261
Рис. 102. Мышцы грудной области (вид снизу)
Справа — удалена прямая мышца живота, слева — удалены все брюшные мышцы
J — diaphragma;
2 — lamina fibrosa diaphrag-
matica;
3 — 5 — m. intercostalis
(.? — externus,
4 — Internus,
5 — internus p. cranialis);
6 — in. latissimus dorsi;
7 — m. obliquus abdominis
internus;
8 — m. rectus abdominis;
9 — m. scalenus p. thoracalls;
10 — m. transversus abdominis;
j j — магистральная артерия
чудесной сети;
магистральные вены чу-
десной сети, расположен-
ные под прямой мышцей
живота на внутреннем
апоневрозе т. obliquus
abdominis internus
262
Непосредственно под m. rectus abdominis (дорсальнее его) располагает-
ся чудесная сеть живота (см. рис. 102), представленная на каждой сто-
роне продольной артерией с двумя расположенными по ее бокам венами
и многочисленными боковыми разветвлениями и анастомозами.
М. transversus abdominis, поперечная мышца живота (см. рис. 102),
образует широкий мышечный пласт, закрепленный с одной стороны (на-
чало) на всех ребрах, а с другой (окончание) переходит в апоневроз, до-
тягивающийся до белой линии живота и прирастающий вместе с анало-
гичным апоневрозом внутренней косой мышцы живота к внутренней
фасции прямой мышцы живота. Начинается поперечная мышца живота
от внутренней поверхности первых трех ребер (на первом — на уровне
проксимальной трети, на II и III — в дистальной половине) и области
соединения реберных хрящей с дистальными концами всех остальных
(до XVII) ребер, на которых их отхождение вытеснено вентральнее мы-
шечными ножками диафрагмы. Если более передняя часть мышцы, как
уже сказано, оканчивается на прямой мышце живота, то наиболее ка-
удальные ее пучки (от XIV—XVII ребер) закреплены на наружной фас-
ции опускателей хвоста и буграх тазовой кости. У бразильского ламан-
тина поперечная мышца живота не заходит каудальнее XV ребра,
а у дюгоня, наоборот, продолжается за последнее ребро, отходя от внут-
ренней поверхности апоневроза, натянутого между ним и тазовыми ко-
стями, и от самих подвздошной и седалищной костей, что связано с
меньшей редукцией таза у дюгоня и иным его положением, чем у ла-
мантинов.
Diaphragma, диафрагма (см. рис. 102), как уже неоднократно отме-
чалось в литературе, у сирен принимает необычное горизонтальное поло-
жение (см.: (Murie, 1872а, fig. 39)). Мускульная ее часть начинается
14 мощными мышечными ножками от IV по XVII ребро (на XVII нача-
ло сухожилием). На IV ребре начало находится только на заднем крае
реберного хряща, на последующих ребрах — на передних и задних (на
XVII только переднем) краях их дистальных концов близ прикрепления
реберных хрящей. Все мышечные ножки направляются медиально и
вверх к сухожильной части диафрагмы, которая закреплена утолщенны-
ми сухожильными порциями на особых вентральных гребнях тел IV—
XVII грудных позвонков. Каудальнее после каждого такого прикрепления
сухожильная часть диафрагмы утончается и у задних краев тел позвон-
ков оказывается прободенной сосудистыми пучками, проходящими с
вентральной стороны вверх и латерально к чудесной сети, тянущейся по
заднему краю проксимальной трети ребер.
У Т. manatus ранее (Murie, 1872а) отмечалось отхождение диафраг-
мы от всех ребер, в том числе первых трех. Как уже отмечено, у нашего
экземпляра этого нет, более того, это место занято передней частью
m. transversus abdominis. Домнинг, описывая диафрагму бразильского
ламантина, обошел вопрос о положении ее переднего края, а поперечная
мышца живота у него не заходит вперед далее III ребра. У дюгоня тот
же Домнинг описывает отхождение диафрагмы от III—XVII ребер.
Спереди сухожильная часть диафрагмы связана с перикардом, закреп-
ленным на грудине и реберных хрящах.
Ламантины относятся к тем млекопитающим, у которых сохраняется
только диафрагмальное дыхание, а ребра имеют ограниченную подвиж-
ность и имеют малые возможности участвовать в дыхании. Однако осо-
бенности строения диафрагмы, прикрепление ее сухожильной части к
почти всем грудным позвонкам и приобретение в связи с этим горизон-
тального положения говорит, что у сирен диафрагмальное дыхание особо
эффективно. Легкие ламантина вытянуты почти до уровня анального от-
263
Таблица 11. Сравнение масс мышц
брюшного пресса у Trichechus manatus
и Т. inunguis ((по: (Domning, 1978))
Мышцы Т. inunguis Т. manatus
г. % г. %
ш. m. obliquus abdominis ех- ternus+internus 73,4 22,4 7890 36,0
т. rectus abdo- minis 79,2 24,4 5600 25,6
т. transversus 82,6 25,5 5650 25,8
abdominis diaph^agma 89,7 27,6 2820 12,9
Всего Мышцы обеих сторон * В % к массе тел 324,9 а. 100.0 4,1 21960 100,0 5,47*
Таблица 12. Мышцы
брюшного пресса
Мышца, группа мышц Масса обеих сторон
г %
m. obliquus abdomi- nis externus 3400 18,0*
т. obliquus abdomi- nis internus 1520 8,0*
m. transversus abdominis 5650 29,9
m. rectus abdominis 5600 29,6
diaphragma 2820 14,9
Всего 18 990 100,0
Масса мышц обеих сторон 47**
* Только грудной части без пучков,
оканчивающихся на хвосте.
** В процентах от массы тела.
верстия и заключены в полость между круто выгнутыми ребрами (свер-
ху и латерально) и диафрагмой (снизу и медиально) (см. рис. 95). Од-
нако объем их относительно веса тела не увеличен и равен таковому
взрослого человека и тюленя (около 5 л на 100 кг веса) (Irving, 1969:
цит. по: Яблоков, Белькович, Борисов, 1972). Это неудивительно, так как
они представляют из себя плоские однодольные мешки, толщина кото-
рых у взрослого животного достигает всего 7 см в передней части, 8,9 см
в середине и 3,8 см у заднего окончания (Murie, 1872а). Для сравнения
следует указать, что у некоторых дельфинов (афалина, морская свинья)
объем легких значительно больше (7 л на 100 кг веса). В то же время
объем одноразового вдыхаемого воздуха у ламантина в три раза больше,
чем у человека, но вполовину меньше, чем у морской свиньи, при этом
у ламантина обновляется 50% воздуха в легких против 10—20% У чело-
века и 80—90% у морской свиньи (Sholander, Irving, 1941). Своеобразие
сирен состоит, следовательно, в том, что при небольшом относительном
объеме легких они способны вдохнуть большой объем воздуха и обновить
50% воздуха, содержащегося в легких. Именно это достигается таким
необычным строением диафрагмы. Большую эффективность дыхания
дельфинов следует искать в увеличении объема легких при косом поло-
жении диафрагмы (некоторое приближение к сиренам): это обеспечивает
большой объем вдыхания воздуха. В то же время резко отличный от
ламантинов характер локомоции, которая осуществляется за счет дви-
жения всего туловища, а не только хвоста, потребовал у них увеличе-
ния постторакального отдела позвоночного столба при уменьшенном
грудном отделе (уменьшена грудина, число стернальных ребер). Это соз-
дало условия для особо эффективного действия брюшного пресса, чем,
видимо, и объясняется большой процент обновленного воздуха. Сдвиг
легких у сирен назад (целиком за сердце) также можно рассматри-
вать как предпосылку эффективного действия у них мышц брюшного
пресса.
В схеме дыхательная деятельность мышц этого комплекса сводится к
напряжению диафрагмы, что создает отрицательное давление в дорсаль-
264
ном отделении грудной полости, в результате чего в легкие насасывается
воздух. Расслабление диафрагмы и напряжение мышц брюшного пресса
приводит к выдавливанию воздуха из легких. Ребра в этом процессе
участия практически не принимают.
Общая масса мышц брюшного пресса и диафрагмы (табл. 11) со-
ставляет около 5,5 °/о от массы тела. Однако если учесть, что значитель-
ная доля m. obliquus abdominis externus (больше половины) и т. obli-
quus abdominis internus (около */s) работают в основном на сгибание
хвоста, то этот процент следует понизить до 4,7% (табл. 12). В то же
время одна из самых мощных мышц комплекса — m. rectus abdominis в
равной степени работает на собственно брюшной пресс и на сгибание
хвоста.
МЫШЦЫ, СОЕДИНЯЮЩИЕ ПЛЕЧЕВОЙ ПОЯС И ПЛЕЧО
С ТУЛОВИЩЕМ
В состав этой группы входят мышцы разного происхождения: висце-
ральные — трапециевидная и ключичнососцевидная, иннервируемые XI
нервом, производные осевой мускулатуры — ромбовидная и вентральная
зубчатая (дорсальные ветви шейных и грудных спинномозговых нервов
и n. thoracicus longus) и вторично вступившие в связь с туловищем и
развивающиеся из почки конечности — широчайшая спины (грудные
нервы, идущие от плечевого сплетения).
М. trapezius, трапециевидная мышца (рис. 87, 103), очень обширна и
мощна. Она подразделяется на 4 более или менее обособленные части с
разными функциями: m. trapezius р. mandibularis, mm. clavotrapezius,
acromiotrapezius et spinotrapezius \ которые целесообразно рассматри-
вать как отдельные мышцы. Первая из них уже была описана среди ту-
ловищных мышц, вторая обычно включается в особый функциональный
комплекс m. brachiocephalicus (см. табл. 13). М. spinotrapezius представ-
ляет дорсокаудальную часть комплекса, начинающуюся апоневрозом на
общей наружной фасции разгибателей спины (длиннейшей мышцы спи-
ны и подвздошнореберной) (рис. 93, а). Сзади он достигает уровня
VII—VIII ребер, немного не доходя до средней линии спины, а вперед
идет, постепенно приближаясь к ней, до II грудного позвонка. Мышеч-
ные волокна направляются вниз и вперед, конвергируя и прикрепляясь к
сухожильному апоневрозу на внутренней поверхности мышцы, который
кончается на краниальной части наружной поверхности бугра ости ло-
патки (рис. 103, б). Этот апоневроз составлен из отдельных сухожиль-
ных лент, сходящихся к прикреплению на манер жесткого каркаса веера
(рис. 103, г) и формирующих конечное широкое сухожилие. Аналогично
прикрепляется мышца у бразильского ламантина, но у дюгоня ее окон-
чание обширнее и простирается по наружной фасции заостноп мышцы
от бугра ости до большой круглой мышцы. М. acromiotrapezius продол-
жает дальше вперед предыдущую мышцу. Его волокна начинаются от
срединного срастания мышц правой и левой сторон и наружной фасции
ромбовидной мышцы, переходя краниальнее на ламбдоидный гребень
черепа (рис. 75, 76, 79) и идя по нему почти до самого его вентрального
края. У бразильского ламантина эта мышца вообще не начинается от
черепа, не доходя до него на 1—2 см. Мощное мышечное начало на че-
1 Эти названия прочно вошли в сравнительную анатомию и удобнее терминов, пред-
лагаемых международной номенклатурой (m. cleidocephalicus, m. trapezius
р. cervicalis et p. thoracica для mm. clavotrapezius, acromiotrapezius et spinotrape-
zius), так как они отражают общность происхождения и топографические различия.
265
266
1 — m. abductor + exten-
sor pollicis longus;
2 — 3 — hi. acromiotra-
pezius
( 3 — ее пучок к m. del-
toideus);
4 — m. brachialis;
S — m. brachioradialis;
6 — m. clavotrapezius;
7 — m. cleidomastoideus;
8 — m. deltoideus;
9 — m. extensor carpi ul-
naris;
10 — m. ext. digitorum com-
munis;
11 — m. flexor carpi ulna-
ris;
12 — m. infraspinatus;
13 — in. latissimus dorsi;
14 — in. pectoralis profundus,
15 — in. rhomboideus;
16 — m. serratus ventralis;
17 — in. spinotrapezius;
18 — m. subscapularis;
19 — m. supraspinatus;
20 — m. teres major;
21 — 23 — m. triceps brachii
(21 — caput accessorius,
22 — cap. longum,
23 — cap. mediate)
Рис. 103. Мышцы плечевого пояса, плеча и частично предплечья
Вид плечевого пояса латерально, часть ласта развернута так, что видна дорзолатеральная поверх-
ность
а — поверхностный слой (mm. trapezius, rhomboideus, serratus ventralis, latissimus dorsi, brachiocep-
halicus) отрезаны и показаны только их части, идущие к лопатке и плечу;
б — глубокий слой после снятия mm. trapezius, deltoideus, serratus ventralis и удаления части m.
teres major и m. latissimus dorsi;
e — глубокие мышцы плеча после удаления части m. triceps brachii с. longus, mm. supraspinatus
et infraspinatus сняты и показаны мышечные прикрепления на лопатке;
г — внутренняя поверхность m. spino-, acromiotrapezius
267
репе было отмечено и у других экземпляров американских ламантинов
(Murie, 1872 а), что, очевидно, специфично для этого вида. От своего на-
чала волокна мышцы идут вниз и назад почти параллельно и крепятся
вдоль всего протяжения ости лопатки от ее бугра до дистального конца
акромиона (рис. 103, б), но лишь на последнем он осуществляется прямо
мышечными волокнами к кости. Более каудальные волокна кончаются
на части наружной фасции дельтовидной мышцы и апоневрозе, укреплен-
ном в особой неглубокой, но широкой вырезке на краниальном крае ости.
У бразильского ламантина мышца крепится не мышечно, а сухожильно.
Наиболее передняя небольшая часть волокон m. acromiotrapezius не-
сколько обособляется (рис. 103, а; 104, а) и включается в m. c.Iavodeltoi-
deus, идя вместе с ним, видимо, до плечевой кости. Такое явление об-
наружено впервые и не известно ни у кого из млекопитающих. У дюго-
ня начинается сухожилием от височного гребня черепа лишь небольшая
обособленная передняя порция; основная часть мышцы также ослаблена,
мышечные ее волокна далеко не доходят до средней линии спины. При-
крепляется мышца также сухожильно. Таким образом, у ламантина
m. acromiotrapezius несравненно более мощный, чем у дюгоня, но, глав-
ное, его начало по височному гребню и окончание по очень длинному
акромиальному отростку, чего нет у дюгоня, распространяется значитель-
но вентральнее. Это усиливает вращательный момент, действующий на
лопатку, относительно ее заднего угла, что следует связать с большей
подвижностью передних конечностей ламантинов.
М. brachiocephalicus, плечеголовая мышца, образует единофункциони-
рующий комплекс мышц разного происхождения, разделяемых попереч-
ной сухожильной перемычкой на месте исчезнувшей у сирен, как и у
ряда других млекопитающих, ключицы. От черепа к ней подходят
m. clavotrapezius и т. cleidomastoideus, а от нее далее идет m. clavodel-
toideus, являющийся их фактическим продолжением к плечевой кости
(последний безусловно является производным дельтовидной мышцы,
иннервируемым n. axillaris). М. clavotrapezius (cephalohumeralis Дом-
нинга) (рис. 69, 104, а, 105, а) начинается мышечно от самого вентраль-
ного конца височного гребня и сосцевидного отростка височной кости
(рис. 75, 76, 79) и образует плоское ремневидное брюшко с параллель-
ными волокнами, поверх которого идет более узкая ключичнососцевидная
мышца. Кончается на упомянутой выше сухожильной перемычке при-
мерно на 2/3—3Л ее протяжения с латеральной стороны. М. cleidomas-
toideus (часть brachiocephalicus Домнинга), ключичнососцевидная мышца
(рис. 104, а, 105, а) начинается совместно с уже грудинососцевидной
мышцей широким сухожилием от сосцевидного отростка височной кости
непосредственно краниальнее отхождения m. clavotrapezius (рис. 76).
Обычно обе мышцы объединены в своем начале в единую грудиноклю-
чичнососцевидную мышцу, но у сирен они сразу же обособляются друг
от друга. Брюшко ключичнососцевидной мышцы узкое ремневидное с
параллельными волокнами. В сечении у своего прикрепления оно имеет
25X8 мм. Кончается на сухожильной перемычке, но, поскольку послед-
няя идет косо (способ увеличения площади для прикрепления мышц), то
более наружные волокна оказываются более длинными. М. clavodeltoideus
(другая часть brachiocephalicus Домнинга) (рис. 104—106) начинается
от той же перемычки и образует мощное вздутое брюшко, конвергирую-
щее дистально и ближе к прикреплению подразделяющееся на две само-
стоятельно прикрепляющиеся части (рис. 105, б): более поверхностные и
латеральные пучки волокон, несколько обособляющиеся от подлежащей
большей массы более медиальных волокон, переходят в сухожильный ши-
рокий апоневроз, в свою очередь переходящий в наружную фасцию m. bra-
268
Рис. 104. Мышцы плечевого пояса, плеча и частично предплечья, вид снизу: ласт
развернут так, что преимущественно видна его латеральная (экстензорная) поверх-
ность
а — поверхностный слой (mm. trapezius, pectorales superficialis et profundus, clavotrapezius, cleido-
mastoideus обрезаны); б — глубокий слой после снятия mm. clavodeltoideus, deltoideus и остатков
mm. trapezius, pectorales superficialis et profundus
1—m. abductor 4-ext. pol- 10 — m. coracobrachialis; (19 — profundus,
licis longus; 11 —m. deltoideus; 20— superficialis);
2 — 3 — m. acromiotrapezius 12 — m. extensor carpi radialis; 21 — m. subscapularis;
(3 — его пучок к m. deltoide-
us); 1
4 — m. biceps brachii;
5 — m. brachialis;
6 — m. brachioradialis;
7 — m. clavodeltoideus;
8 — m. clavotrapezius;
.9 — m. cleidomastoideus
13 — m. ext. carpi ulnaris;
14 — m. ext. digit! quinti;
15 — m. ext. digitorum
communis;
16 — m. flexor carpi ulnaris;
17 — m. infraspinatus;
18 — m. latissimus dorsi;
19 — 20 — m. pectoralis
22 — m. supraspinatus;
23 — m. teres major;
24 — 26 — m. triceps brachii
(24 — caput accessorius,
25 — cap. longum,
26 — cap. mediate)
269
270
chialis. Остальные волокна проникают между плечевой мышцей и собст-
венно дельтовидной, оставляя их латерально, и двуглавой с другой
стороны и прикрепляются на передней поверхности плечевой кости па
уровне середины ее диафиза (рис. 107, г). М. brachiocephalicus выдвига-
ет вперед вентральный угол лопатки и является протрактором плеча,
участвуя в разгибании плечевого сустава. Однако при фиксации лопат-
ки и конечности широчайшей мышцей спины и грудными мышцами
одностороннее сокращение плечеголовой мышцы вызовет поворот головы
вместе с шеей вбок (в главе 3 уже говорилось, что это движение крайне
ограничено), а двустороннее — опускание головы. Можно представить
себе, что это происходит, например, при касании ластами субстрата и со-
бирании корма на дне. Тем не менее, учитывая разницу в массе головы
и шеи и самого ласта, предпочтительнее считать, что основная роль
мышцы — управление конечностями.
М. sternomastoideus, грудинососцевидная мышца (рис. 86) начинает-
ся, как сказано выше, вместе с ключичнососцевидной, но ее длинное су-
хожильное начало сразу же обособляется и идет назад, медиально и вниз,
переходя в небольшое веретеновидное брюшко, волокна которого конча-
ются на рукоятке грудины. Мышца фиксирует грудину при неподвижной
голове или способствует опусканию последней. Возможно и небольшое
подтягивание грудины вперед, движение в некоторой степени допустимое
за счет слабой подвижности первых трех ребер (см. гл. 3).
М. rhomboideus, ромбовидная мышца (рис. 93, а, 103, а, 105, а), начи-
нается апоневрозом от наружной фасции пластыревидной мышцы, при-
мерно в 17 см от головы, затем переходит на фасцию полуостистой мыш-
цы головы, простираясь назад до уровня VI ребра на расстоянии 16 см
от средней линии спины. Апоневроз переходит в широкое плоское брюшко
с почти параллельным направлением волокон, идущих каудально и вниз
к прикреплению на каудальных 2/3 вертебрального края и узкой полоски
внутренней поверхности лопатки (рис. 105, б). Фактически можно разли-
чать две части ромбовидной мышцы, более заднюю грудную с короткими
волокнами, идущими вентрально к позвоночному краю лопатки, и более
переднюю, волокна которой идут почти в продольном направлении и об-
разуют основную массу мышцы, прикрывая с внутренней стороны дор-
сальную часть прикрепления вентральной зубчатой мышцы. Образован-
ное ими плоское, в виде очень длинного треугольника в сечении с ко-
роткой стороной вверх брюшко расслаивается. Меньшая часть передних
и занимающих более внутреннее положение волокон образует камбало-
видной формы брюшко, имеющее на своей внутренней поверхности
Рис. 105. Мышцы плечевого пояса, плеча и частично предплечья.
Вид с медиальной стороны: ласт развернут так, что преимущественно видна вентромедиальная по-
верхность плеча и медиальная (флексорная) поверхность предплечья
а — поверхностный слой; у mm. trapezius, rhomboideus, serratus ventralis, latissimus dorsi, pectora-
les major et minor показаны лишь части, идущие к лопатке и плечу; б — глубокий спой после сня-
тия mm. rhomboideus et serratus ventralis,.части m. teres major и укорочения оставшихся частей mm,
latissimus dorsi, pectorales superficialis et profundus, brachiocephalus.
1 — 2 — m. acromiotrapezius
(2 — его пучок к m. deltoide-
us);
3 — m. biceps brachii;
•4 — m. brachialis;
3 — m. brachioradlalis;
в — m. clavodeltoideus;
7 — m. clavotrapezius;
8 — m. cleidomastoideus;
9 — m. coracobrachialis;
1 0—m. deltoideus;
11 — 12 — m. flexor carpi
(11 — radialis,
12 — ulnaris);
13 — m. fl. digitorum;
14 — m. infraspinatus;
15 — m. latissimus dorsi;
16 — 17 — m. pectoralis
(16 — profundus,
17 — superficialis);
18 — m. pronator teres;
19 — m. rhomboideus;
20 — m. serratus ventralis:
21 — m. subscapularls;
22 — m. supraspinatus;
23 — m. teres major
271
ближе к концу апоневроз, к кото-
рому и крепятся мышечные во-
локна. Апоневроз сужается и
переходит в длинное сухожилие,
оканчивающееся на вертебраль-
ном крае лопатки в точке, отстоя-
щей от краниального угла на ‘/з
длины края. Остальная часть во-
локон, подстилающая это брюш-
ко, если смотреть изнутри, идет
каудальнее и тоже крепится на
сухожильном апоневрозе по своей
внутренней поверхности, который,
сужаясь, также переходит в сухо-
жилие, оканчивающееся непо-
средственно в районе каудально-
го угла лопатки. Ромбовидная
мышца оттягивает позвоночный
край лопатки вперед и вверх, но
ввиду короткости большей части
волокон амплитуда этого движе-
ния невелика.
М. serratus ventralis (serratus
niagnus Домнинга), вентральная
зубчатая мышца (рис. 92, 105, а)
состоит из двух частей, разделен-
ных только в своем начале. Шей-
ная часть начинается от плевра-
пофизов II—VI шейных позвонков
(рис. 91) и передней поверхности
I ребра на протяжении около
50 мм, начиная от бугорка. Груд-
ная часть берет начало от II до
VII ребер и отделена от первой
дорсальными пучками наружной
межреберной мышцы, связываю-
щей I и II ребра (рис. 97,6).
Места отхождения мышцы посте-
пенно отодвигаются все дальше
от бугорка ребра. Так, на II ребре
расстояние между ними достигало
83 мм, а на IV и V оно уже рав-
нялось 250 и 320 мм. Обычно на
каждом ребре прикрепление вы-
тянуто косо поперек (каудодор-
сально) по наружной поверхности.
Вентральная зубчатая мышца —
одна из самых мощных среди всех
мышц пояса и конечностей ла-
мантина (см. табл. 13). Ее волок-
на образуют толстое и широкое
брюшко, которое мышечно кре-
пится на внутренней поверхности
лопатки вентральнее прикрепле-
ния ромбовидной и дорсальнее
272
Рис. 106. Глубокие мышцы
плечевого пояса и плеча
а — вид спереди; сняты все мышцы,
идущие с наружной поверхности ло-
патки, где показаны места их от-
хождений, к плечу, и ш. clavodel-
toideus;
б — вид с вентромедиальной сто-
роны; m. brachialis перерезан,
небольшая его проксимальная пор-
ция удалена, чтобы показать место
ее отхождения от плеча, а дисталь-
ная часть отогнута;
в — топография m. biceps bracbii,
вид с медиальной стороны у дюгоня
(Сверху) и ламантина (снизу)
1 — m. biceps brachii;
2 — т. brachialis;
з — т. brachioradialis;
4 — in. clavodeltoideus;
5 — m. coracobrachialis;
6 — m. deltoideus;
7 — m. extensor carpi radialis;
8 — m. ext. digitorum communis;
9 — m. flexor carpi radialis;
10 — m. fl. digitorum;
11 — m. infraspinatus;
12 — 13 — m. pectoralis
(12 — profundus,
13 — superficialis);
14 — m. serratus ventralis;
15 — m. spinotrapezius;
16 — m. subscapularis;
17 — m. supraspinatus;
18 — m. teres major
отхождения подлопаточной мышц (рис. 105, б). Наиболее каудальные во-
локна заходят на наружную поверхность самого каудального угла лопат-
ки (рис. 103, а, б). Основная функция мышцы — фиксация лопатки отно-
сительно грудной клетки. У дюгоня вентральная зубчатая мышца более
отчетливо подразделена на переднюю и заднюю части, различающиеся
не только началом, но и прикреплением на лопатке, что заставило по-
дозревать большую возможность к переднезаднему сдвигу лопатки (Dom-
ning, 1977b).
М. latissimus dorsi, широчайшая мышца спины (рис. 87, 93, а, 103—
107), начинается каудально слабым обособленным мышечным зубцом от
фасции в промежутке между VIII и IX ребрами и все более увеличива-
ющимися зубцами от VIII по IV ребра далее вперед (рис. 92). Перед-
ние зубцы расположены сразу вентральное отхождения от ребер вентраль-
ной зубчатой мышцы и занимают по каждому ребру по 1/6 его протя-
жения, а более задние — несколько дорсальнее той же мышцы, и в этой
области часть мышечных волокон берет начало от наружной .фасции,
распространяясь от IX ребра до начального апоневроза трапециевидной
мышцы. У ламантина широчайшая мышца спины — одна из самых мощ-
ных. Ее волокна образуют широкое плоское брюшко (рис. 88) и, подходя
к плечу, конвергируют, прикрепляясь изнутри к широкому сужающему-
ся дистально сухожильному апоневрозу. Часть мышечных волокон в этой
области может сливаться с волокнами m. teres major. Поперечное сече-
ние, проведенное на этом уровне через обе мышцы (рис. 108), показыва-
ет, что брюшко широчайшей мышцы спины имеет здесь вид удлиненного
треугольника, короткой передней стороной прилегающего к задней по-
273
верхности большей по объему большой круглой мышцы. Конечный сухо-
жильный апоневроз широчайшей мышцы спины утолщен в своей задней
части и здесь как бы образует угол с основной частью апоневроза, иду-
щей по латеральной поверхности брюшка m. latissimus dorsi. Дисталь-
нее, ближе к плечевой кости, этот апоневроз вливается в аналогичный
апоневроз, образующий сухожильное зеркало на латерокаудальной по-
верхности дистальной части большой круглой мышцы, и вместе с ним
кончается на медиальной поверхности плечевой кости немного прокси-
мальнее ее середины на особой шероховатости (рис. 107, в). Широчайшая
мышца спины обычно оканчивается вместе с большой круглой мышцей,
с которой они связаны общим происхождением в онтогенезе, также у
большинства млекопитающих. Однако у ряда более примитивных форм
часть мышечных пучков, связанных в своем начале с основной массой
волокон m. latissimus dorsi, обособляется и, перебрасываясь через двугла-
вую и коракоидноплечевую мышцы, кончается вместе с поверхностной
грудной и подкожной мышцей туловища на гребне большого бугорка
плечевой кости (Гамбарян, 1960). Обычно эти пучки рассматриваются
как часть m. latissimus dorsi. У ламантина их . нет. У дюгоня, наоборот,
нет никакой мышцы, оканчивающейся вместе с большой круглой, но есть
слабая мышца (m. latissimus dorsi Домнинга, 1977b), прикрепленная на
большом бугорке в общем сухожилии с т. pectoralis profundus, а в своем
начале связанная с т. cutaneus trunci. Все это заставляет считать, что
настоящая широчайшая мышца спины у дюгоня исчезла, а называемая
так мышца является производным подкожной мышцы туловища, лишь
вторично вступающим в связь с широчайшей мышцей спины у ряда
млекопитающих. В то же время это говорит о глубокой перестройке,
которой подверглась мускулатура передней конечности дюгоня и, наобо-
рот, о сохранении ею у ламантина некоторых исходных соотношений.
Широчайшая мышца спины у ламантина является флексором и пронато-
ром плеча.
М. pectoralis superficialis (pectoralis major Домнинга, ectopectoralis
других авторов), поверхностная грудная мышца (рис. 86, 104, а, 105)
начинается двумя почти самостоятельными частями. Краниальная часть
начинается почти на всем расширении рукоятки грудины, будучи оттес-
нена лишь от средней линии прикреплением m. trapezius р. mandibula-
ris, и на сухожильной пластинке, простирающейся от латерального края
рукоятки к I реберному хрящу. Каудальная часть отходит от сухожиль-
ной части грудины, ее мечевидного отростка и наружной фасции прямой
мышцы живота. Собираясь с этого обширного начала, мышечные волокна
образуют плоское, но толстое брюшко (рис. 88) и конвергируют ближе
к своему прикреплению. При этом более передние волокна идут поверх-
ностнее и дистальнее по плечу, а более задние глубже и проксимальнее,
проникая, таким образом, под первые и образуя нечто вроде перекреста.
На внутренней (дорсальной) и более каудальной части поверхности мы-
шечного брюшка формируется сухожильное зеркало, к которому и кре-
пятся мышечные волокна, а само оно, быстро утолщаясь, превращается
в мощное короткое конечное сухожилие, прикрепленное к особой шеро-
ховатости на медиальной поверхности плечевой кости сразу дистальнее
прикрепления большой круглой и широчайшей мышцы спины, от кото-
рого его отделяет лишь узкая полоска окончания m. coracobrachialis
(рис. 107, в). Близ самого прикрепления от конечного сухожилия отходят
в дистальном направлении две длинные сухожильные оттяжки
(рис. 105, б), из которых более задняя по положению идет к медиально-
му надмыщелку плечевой кости, а вторая, более передняя, проникает под
.длинное конечное сухожилие двуглавой мышцы и идет далее дистально
274
1 — m. brachialis;
2 — m. brachioradialis;
3 — m. clavodeltoideus;
4 — m. coracobrachialis;
5 — m. deltoideus;
6 — 7 — m. extensor carpi
(6 — radialis;
7 — ulnaris);
8 — m. ext. digiti quint!;
9 — m. ext. digitorum com-
munis;
10 — 11 — m. flexor carpi
(10 — radialis,
Jl — ulnaris);
12 — m. fl. digitorum;
13 — m. infraspinatus;
14 — m. latissimus dorsi;
13 — 16 — m. pectoralis
(15 — profundus,
16 — superficialis);
17 — m. pronator teres;
18 — m. subscapularis;
19 — m. supraspinatus;
20 — m. teres major;
20 — 21 — m. triceps brac-
hii
(21 — caput accessorius,
22 — cap. mediale);
23 — капсула плечевого
сустава
Рис. 107. Площади прикрепления мышц на плечевой кости
а — вид с латеральной стороны; б — вид сзади; в — вид с медиальной стороны; г —. вид спереди;
д — вид с проксимального конца
275
параллельно с последним к прикреплению на лучевой кости. Эта послед-
няя оттяжка препятствует яереразгибанию предплечья, а первая вместе
с основным сухожильным окончанием поверхностной грудной мышцы об-
разует как бы шпильку, фиксирующую конечное сухожилие бицепса у
поверхности плечевой кости, что не только препятствует его смещению,
но и образует блок, позволяющий ему менять направление. У бразиль-
ского ламантина вместо узкого сухожилия, кончающегося на radius,
описывается апоневроз, идущий к фасции предплечья, но не крепящийся
на лучевой кости. У дюгоня окончание поверхностной грудной мышцы
находится в самой дистальной части гребня большого бугорка. У ламан-
тина мышца обеспечивает в первую очередь аддукцию плеча, в меньшей
степени она пронирует его. Возможно последняя функция несколько
увеличена у дюгоня.
М. pectoralis profundus (pectoralis minor Домнинга, endopectoralis
других авторов), глубокая грудная мышца (рис. 86, 105) начинается от-
дельными (4—5) мышечными пучками от края I ребра, от рукоятки гру-
дины и смежной части II хрящевого ребра и от фасции прямой мышцы
живота, формирующими уплощенное в сечении (рис. 88) камбаловидное
брюшко, идущее краниально и несколько латерально. Кончается частью
мышечно, частью сухожилием (за счет сухожильного зеркала по вент-
ральному краю дистальной части брюшка) на специальной шероховато-
сти краниальной части большого бугорка плечевой кости непосредственно
проксимальнее рудиментарного межбугоркового желоба и медиальнее
прикрепления к бугорку предостной мышцы (рис. 107, в, д'). Работает
глубокая грудная мышца как протрактор (экстензор) плеча и, в некото-
рой степени, его аддуктор. У дюгоня прикрепление спускается значитель-
но дистальнее по особой шероховатой линии на медиальной поверхности
гребня большого бугорка ближе к окончанию поверхностной грудной
мышцы и осуществляется плоским сухожилием, сливающимся с конеч-
ным сухожилием «т. latissimus dorsi» дюгоня (Domning, 1977в, см. вы-
ше). Соответственно мышца у него теряет протракторную функцию,
превращаясь, наоборот в ретрактор и пронатор плеча.
М. pectoralis abdominalis (рис. 86) — небольшой пучок волокон, от-
делившихся от внутренней поверхности подкожной мышцы туловища и
формирующих длинное узкое ремневидное брюшко (рис. 88), которое
проникает под поверхностную грудную мышцу и переходит в сухожилие,
срастающееся у своего прикрепления с конечным сухожилием глубокой
грудной мышцы. Именно эта мышца описана Домнингом как широчайшая
спины у дюгоня. У ламантина она до сих пор была не отмечена (Murie,
1872а; Domning, 1978а).
МЫШЦЫ, ДЕЙСТВУЮЩИЕ НА ПЛЕЧЕВОЙ II ЛОКТЕВОЙ СУСТАВЫ
Топографически в эту группу включены мышцы, соединяющие лопатку
с плечевой костью, и мышцы, помещающиеся в плече.
М. deltoideus, дельтовидная мышца (рис. 103, а, 104—106), начинает-
ся по ости лопатки вентральнее бугра ости лопатки, включая его ниж-
нюю половину, и задней и прилегающей к ней смежной части наружной
поверхности акромиона. Часть волокон отходит также от фасции, натя-
нутой в выемке переднего края ости, с которой также связаны снаружи
волокна трапециевидной мышцы, а изнутри предостной, и от небольшого
участка сухожилия, идущего по заднему краю m. spinotrapezius к бугру
ости лопатки. Мощное брюшко мышцы имеет снаружи треугольную фло-
му, его сечение по середине длины представлено на рис. 108, 1. Ближе
к прикреплению брюшко становится уже и в сечении больше напоминает
276
Тис. 108. Поперечные срезы мышц конечностей (в скобках — уровень среза от прок-
симального конца к дистальному)
1 — 2 — m. deltoideus (5/6);
3 — m. infraspinatus (7/s);
4 — т. supraspinatus (а/4);
5— в—т. subscapularis (5—'А, 6—Vs);
7 — 8 — т. triceps brachii caput longum
(7—>/2>
9—m. cleidomastoideus (5/6);
10 — ni. clavotrapezius (5/e);
11 — m. triceps brachii caput accessorius (1/2);
12— m. triceps brachii caput mediale (Vz);
параллелограмм (рис. 108, 2). Брюшко снаружи подразделено на две
части: более передние волокна идут к прикреплению поверхностнее и
более каудально, более задние, значительно сильнее конвергирующие,
наоборот, глубже и несколько вперед. Именно на внутренней, медиаль-
ной поверхности задней более утолщенной части появляется сухожиль-
ный апоневроз, переходящий в широкое плоское концевое сухожилие
мышцы длиной около 50 мм, на котором кончаются все ее волокна,
в том числе и передней части, кроме небольшой их порции, переходящей
в апоневроз, вливающийся в поверхностную фасцию плеча над плечевой
мышцей (рис. 104, б). Прикрепляется сухожилие дельтовидной мышцы
к специальной шероховатости на латеральной поверхности плечевой кос-
ти (рис. 107, а). Уже отмечалось, что небольшая часть волокон т. ас-
romiotrapezius непосредственно вливается в m. deltoideus, проходя ме-
277
Рис. 108 (продолжение)
13 — 14 — m. biceps brachii (13—>/2, 14—’/,)
15 — m. coracobrachialis (V2);
16 — 22 — m. pronator teres (16—*/,), 17—</„
18 — '/e, 19—>/„ 20 — >/2, 21— »/„ 22-»/,),
23 — 26 — m. brachioradialis (23—»/,, 24—Vi,
25—>/„ 26—•/,);
27 — 29 — m. teres major + m. latissimus dorsi
(27—V,. 28—41, 29—a/l0);
30 — 33 — m. extensor carpi radialis (30—Ч2,
31—32—7„, 33—»/„);
34 — m. extensor digitorum communis (l/2);
диальнее самого конца акромиона. Необходимо отметить, что у ламантина
от бугра ости лопатки идет каудально над заостной мышцей к гребню
для ш. teres major фасция, волокна которой идут почти под прямым уг-
лом к направлению волокон дельтовидной мышцы (небольшая их часть
278
чо
Рис. 108 (продолжение)
35 — 37 — m. extensor carpi ulnaris (35—Vi),
36—V2, -з?—’/4);
38 — 42 — m. flexor digitorum (38—3/8, 39—Чг,
40—ъ/а, 41 — 3/4, 42—7s);
43 — 50 — m. flexor carpi radialis (43—V„)
44—Vs, 45—V4, 46— 3/„ 47—Vs, 43—5/8,
49—3/4, 50—Vs),
51—52 — m. flexor digiti quinti (51— V8,)
52—7s);
.даже начинается с внутренней поверхности этой фасции). Вентральнее
эта фасция продолжается в наружную фасциальную выстилку дельто-
видной мышцы, хотя явно не является ее производным. У дюгоня от
этой поперечной фасции начинается задняя часть дельтовидной мышцы
279
S3 .
54
Рис. 108 (окончание)
53 — 56 — m. adductor digiti quinti (53-‘/8>) 57 — 60 — m. adductor digiti quart! (I—>/,;
54—71, 55—V2, 56—2/3); 2—1/1, 3— >/2, 4—»/4);
61 — m. adductor digiti tertius (72)
и к ней же подходят волокна трапециевидной мышцы. Прикрепляется
m. deltoideus у дюгоня на специальном бугре на гребне большого бугорка
далеко от продольной оси плечевой кости, но в то же время несколько
проксимальнее, чем у ламантина. В результате, если у последнего дель-
товидная мышца работает как протрактор, абдуктор и супинатор, то у
дюгоня она становится сильнейшим супинатором. Этим и объясняется
как смещенное более каудальпо начало дельтовидной мышцы, так и раз-
растание гребня большого бугра у дюгоня. Изменение положения длин-
ной оси головки плечевой кости у ламантина и, следовательно, самого
положения плеча в норме, потребовало у него максимального удлинения
акромиона, иначе дельтовидная мышца не смогла бы эффективно работать.
Но в этих условиях становится излишним и высокий гребень большого
бугорка. У морской коровы предполагается относительное уменьшение
дельтовидной мышцы (Domning, 1978с).
М. supraspinatus, предостная мышца (рис. 103, б, 104, б, 105), на-
чинается мышечно от всей предостной ямки лопатки, от части наружной
фасции, выступающей за краниальный край лопатки подлопаточной
мышцы, передней ости лопатки медиальной стороны акромиального от-
ростка, части шейки лопатки и задней поверхности ее ости (рис. 103, в,
106, а). Однако площадь, занятая началом, несколько варьирует даже на
разных сторонах. Отхождения от части внутренней поверхности лопатки,
описанного у бразильского ламантина (Domning, 1978а), нет. Брюшко
мышцы объемистое, со слабой перистой структурой, основу которой со-
ставляет появляющееся в его толще на уровне бугра лопатки широкое
280
(до 50 мм) плоское сухожилие (рис. 108, 4). Задние волокна, начинаю-
щиеся от ости и акромиона, идут поверхностно вперед и вниз и большей
частью кончаются на наружной стороне внутреннего сухожилия и лишь
частично крепятся непосредственно к большому бугорку плечевой кости.
Более передние волокна от фасции подлопаточной мышцы и предостной
ямки направляются более вентрально и глубже и идут назад и вниз,
оканчиваясь целиком на медиальной поверхности внутреннего сухожи-
лия, к которой также подходят самые глубокие и короткие волокна от
предостной ямки. Кончается предостная мышца указанным выше сухо-
жилием и частично мышечно на специальной шероховатости большого
бугорка плечевой кости (рис. 106, 107, д). Мышца у ламантина разги-
бает плечевой сустав и немного супинирует плечо. У дюгоня из-за боль-
шего разрастания большого бугорка возможно усиливается разгибатель-
ная функция.
М. infraspinatus, заостная мышца (рис. 103, 104, б, 105), начинается
от почти всей поверхности заостной ямки (лишь вентрально она потес-
нена предостной мышцей) и задней поверхности ости лопатки. Неболь-
шое количество волокон идет также от внутренней поверхности попереч-
ной фасции (см. описание дельтовидной мышцы). В дистальной 1/3
объемистого брюшка на его наружной поверхности ближе к заднему
краю появляется сухожильный апоневроз, проникающий вперед в толщу
брюшка до его середины (рис. 108, 5). Он постепенно утолщается, осо-
бенно сзади, и формирует широкое (у конца около 40 мм) плоское,
но толстое конечное сухожилие, прикрепляющееся на специальной шеро-
ховатости каудальной части большого бугорка плечевой кости
(рис. 107, а, д'). Интересно, что на внутренней стороне этого сухожилия,
прилегающего к капсуле плечевого сустава, имеется особая округлая
впадина, что доказывает частое выступание головки вперед за пределы
гленоидной впадины (как это показано на рис. 106, а), требующее фор-
мирования специального ложа для нее. Мышечные волокна, отходящие
от заостной ямки, крепятся на внутренней поверхности конечного сухо-
жилия, основы перистой структуры мышцы. Однако дополнительно к
этому они организованы еще в своеобразные лепестки, имеющие по од-
ному краю тонкое сухожилие, а по другому округлую лопасть из мы-
шечных волокон. Эти лопасти налегают друг на друга подобно листам
бумаги, а сухожилия сходятся дистально к основному конечному сухо-
жилию. Все это создает крайне сложную внутреннюю структуру мышц.
Волокна от ости лопатки занимают более поверхностное положение, идут
в вентральном направлении и, крепясь на наружной поверхности того
же сухожилия, достигают почти самого его конца. У бразильского ла-
мантина отмечено наличие двух концевых сухожилий, идущих в толще
мышцы параллельно друг другу. У ламантина заостная мышца супини-
рует плечо и разгибает плечевой сустав, у дюгоня супинирует и отводит
плечо.
М. subscapularis, подлопаточная мышца (рис. 105, 106), начинается
от большей части внутренней поверхности лопатки вентральнее при-
крепления вентральной зубчатой мышцы, небольшой части наружной по-
верхности лопатки близ ее краниалыюго края и даже от фасции, разде-
ляющей подлопаточную и предостные мышцы, выступающие вперед за
этот край. Брюшко мышцы крайне объемистое, толстое, выступающее за
края лопатки и спереди и сзади (рис. 103, в). Оно слагается из 4 по-
верхностно разделенных, сложно построенных, перистых мышечных го-
ловок, которые объединяет их внутренний сухожильный скелет. Основу
его представляет широкое, но толстое сухожилие, появляющееся в толще
мышцы чуть дистальнее середины ее длины и ориентированное примерно
281
параллельно плоскости лопатки (рис. 108, 6). Это сухожилие выходит
на наружную (медиальную) поверхность мышцы в 40 мм от ее прикреп-
ления. В глубь мышцы от этого сухожилия отходят продольные сухо-
жильные же выступы, вокруг которых и формируется перистость отдель-
ных головок. Кончается мышца сухожилием на особой шероховатости
малого бугорка плечевой кости (рис. 107, в). Учитывая огромную массу
подлопаточной мышцы и сдвиг малого бугорка вперед и его разрастание,,
следует признать, что она является очень мощным пронатором и частич-
но аддуктором плеча. У дюгоня мышца несравненно слабее.
М. teres major, большая круглая мышца (рис. 103, а, 105, а), начина-
ется мышечно от каудального края лопатки, чуть отступя от самого угла
и далее вентрально до выступа, связываемого с началом большой круглой
мышцы, и от смежной части наружной поверхности лопатки, образую-
щей треугольник, ограниченный спереди отчетливым перегибом (рис.
103, в). Волокна от наружной поверхности лопатки образуют более тон-
кую лопасть, частично прикрывающую сзади заостную мышцу. Волокна
от края лопатки формируют более утолщенную заднюю часть брюшка
(рис. 108). Дистальнее, в 8—9 см от прикрепления, на наружной по-
верхности брюшка появляется сухожильное зеркало, переходящее в кон-
цевое сухожилие, с которым срастается концевое сухожилие широчай-
шей мышцы спины. Мышечные волокна прикрепляются к внутренней
поверхности этого сухожилия и лишь самые глубокие идут дистальнее,
не достигая кости на 1 — 1,5 см. Общее конечное сухожилие обеих мышц,
крепится на специальной шероховатости на медиальной поверхности
плечевой кости (рис. 107, в) проксимальнее прикрепления поверхностной
грудной мышцы. Такое соотношение достаточно необычно для млекопи-
тающих, у которых прикрепление m. teres major связано с гребнем ма-
лого бугра, а ш. pectoralis superficialis с гребнем большого. Именно так
крепятся обе мышцы у дюгоня. У ламантина большая круглая мышца
сгибает плечевой сустав и пронирует плечо, у дюгоня это — преиму-
щественно флексор.
М. coracobrachialis, коракоидноплечевая мышца (рис. 105, 106), на-
чинается сильным широким (10 мм) сухожилием от особого рубца на
краниолатеральной поверхности бугра лопатки. Сухожилие идет медио-
каудально, оставляя снаружи (с латеральной стороны) сухожильное зер-
кало самой дистальной части подлопаточной мышцы. От этого сухожилия;
под углом отходят дорсально и вентрально мышечные волокна, форми-
рующие веретеновидное, вздутое посередине (рис. 108, 15) брюшко и
кончающиеся на втором, уже концевом широком сухожилии, лежащем
на медиальной, обращенной к двуглавой мышце поверхности брюшка.
Это обеспечивает перистость m. coracobrachialis. Концевое сухожилие
идет между большой круглой мышцей латерально и бицепсом медиально
к прикреплению на медиальной поверхности плечевой кости (рис. 107, в)
дистальнее места окончания m. teres major + m. latissimus dorsi и про-
ксимальнее прикрепления m. pectoralis superfisialis. У дюгоня прикреп-
ление смещено дистальнее к медиальному надмыщелку. Иные и условия
его работы, что связано с сильным развитием и отгибанием внутрь ко-
ракоидного отростка, от самого конца которого m. coracobrachialis начи-
нается у дюгоня. У него эта мышца приводит плечо и участвует немного
в разгибании плеча. Ее сила действует почти по прямой от начала к
прикреплению. У ламантина мышца в своем ходе делает резкий поворот
вокруг малого бугорка и прикрепляющейся к ней подлопаточной мышцы
в качестве блока. В этих условиях она обеспечивает пронацию плеча и
его приведение.
М. biceps brachii, двуглавая мышца плеча (рис. 105 ,106), начинается
282
длинным (7—8 см) слегка уплощенным (5X2,5 мм) сухожилием от пе-
редней поверхности суставной сумки плечевого сустава без непосредст-
венной связи с самой лопаткой. Далее оно идет косо вниз, будучи за-
крепленным на капсуле особым влагалищем, и, проходя через рудимен-
тарный межбугорковый желоб, перегибается вокруг слившихся малого и
большого бугорков плечевой кости и направляется вдоль последней
(рис. 106, в). Здесь мышца образует небольшое веретеновидное уплощен-
ное (в сечении овальное, см. рис. 108, 13—14) брюшко с перистой струк-
турой, поскольку его волокна отходят от начального сухожилия, продол-
жающегося до середины брюшка в виде сухожильного зеркала на его
.латеральной, обращенной к коракоидноплечевой мышце, поверхности,
а кончаются на апоневрозе на медиальной поверхности, который перехо-
дит также в длинное (6 см) тонкое, овальное в сечении сухожилие,
которое, будучи фиксировано между концевыми сухожилиями m. pecto-
ralis superficialis (см. выше), опять делает крутой поворот и по особо-
му ложу в наружной фасции плечевой мышцы направляется к пред-
плечью, где и кончается почти на середине лучевой кости (см. рис. ПО, г)
.дистальнее прикрепления m. brachialis. Сухожильной оттяжки, идущей
от плечевой кости дистально к конечному сухожилию m. biceps и опи-
санной у бразильского ламантина (Domning, 1978а), нет. У ламантина
мышца очень слабая. Возможно, она участвует в сгибании локтевого сус-
тава. У дюгоня двуглавая мышца развита сильнее (Bahrdt, 1933; Dom-
ning, 1977в, 1978а). Она начинается от основания бугра лопатки на его
наружной поверхности близ края гленоида сразу мышечными волокнами,
образующими веретенообразное брюшко. Без всякого перегиба мышца
проходит в межбугорковом желобе плечевой кости (рис. 106, в), идет
вдоль нее и кончается длинным сухожилием на венечном отростке лок-
тевой и смежной части, но несколько дистальнее, лучевой кости. Здесь
ш. biceps также сгибатель локтевого сустава, но нельзя исключить и
участие его в разгибании плечевого сустава. Скорее всего, редукция дву-
главой мышцы плеча и ее перестройка являются следствием изменений
в проксимальном эпифизе плечевой кости, вызванных потребностями дру-
гих мышц, а не особенностями функционирования самого бицепса.
М. triceps brachii, трехглавая мышца плеча (рис. 103, 104), мощный
комплекс из трех головок, объединенных только у своего прикрепления
на локтевом отростке ulna. Caput longum длинная головка берет начало
от треугольной площадки на каудальном краю лопатки вентральнее ее
угла (рис. 103, в), причем от наружной стороны этой площадки отходит
сухожильный апоневроз, образующий зеркало на небольшой части лате-
ральной поверхности брюшка мышцы. Последнее очень толстое, треу-
гольное в сечении, но в дистальной трети дорсовентрально уплощается
(рис. 108, 7—8). На его вентральной, прилегающей к остальным голов-
кам трицепса, поверхности появляется ближе к медиальному краю сухо-
жильное зеркало (проксимально оно немного проходит в толще самой
мышцы), формирующее плоское широкое (15 мм) конечное сухожилие,
прикрепляющееся к олекранону. Caput mediate, медиальная головка, на-
чинается мышечно от всей задней поверхности плечевой кости
(рис. 107, б) и частично от капсулы плечевого сустава. Дистально при-
крепление может достигать локтевой ямки и латерального надмыщелка
(на одной конечности) или не доходить до дистального конца плечевой
кости на 5 см. На дорсальной поверхности дистальных 2/5 плоского в
сечении, но широкого брюшка (рис. 108, 12) помещается сухожильный
апоневроз, переходящий в конечное плотное и широкое сухожилие, кон-
чающееся на олекраноне. В латеральную часть этого сухожилия влива-
ется сухожилие длинной головки. Часть более глубоких мышечных воло-
283
кон медиальной головки кончается непосредственно на локтевом отростке.
Caput accessorium (с. laterale Домнинга), добавочная головка, очень
небольшая по размерам, начинается преимущественно от капсулы плечево-
го сустава и от смежной части задней поверхности плечевой кости в узком
участке, зажатом между проксимальными частями отхождения плечевой
мышцы с латеральной стороны и медиальной головки трехглавой мышцы
с медиальной. Уплощенное (рис. 108) с почти параллельными волокнами
Рис. 109. Мышцы лате-
ральной (экстензорной)
поверхности ласта (пред-
плечье и кисть)
а — наружный слой;
б — глубокий слой после
снятия mm. extensor carpi
ulnaris, ext. digiti quinti et
ext. digitorum communis;
e — крепление мышц на
костях предплечья;
г — крепление мышц на ко-
стях кисти;
д — m. ext. digitorum com-
munis (особенности струк-
туры мышц)
1 — щ, abductor + ext.
pollicis longus;
2 — m. adductor digiti te-
rt ius;
3 — щ. brachialis;
4 — ni. brachioradialis;
5 — в — m. extensor carpi
(5 — radialis,
6 — ulnaris);
7 — m. ext. digiti quinti;
8 — m. ext. digitorum
communis;
9 — m. flexor carpi ulnaris;
10 — m. interossei lateralis;
11 — m. triceps caput lon-
gum;
12 — кольцевая связка
284
брюшко этой головки лежит поверх медиальной головки. Примерно на
середине длины в медиальной половине брюшка по наружной поверхно-
сти появляется сухожильный апоневроз, формирующий конечное широкое
сухожилие, сливающееся с таковым медиальной головки. Как место от-
хождения, так и соотношения этой части трехглавой мышцы с другими
головками и плечевой мышцей — все говорит, что она не может быть
гомологом латеральной головки трицепса, которая в своем отхождении
лежит латеральное плечевой мышцы. Очевидно, у всех сирен латеральная
головка, исчезла, но сохранилась добавочная, имеющаяся у низших на-
секомоядных, сумчатых, грызунов, хищных. Трехглавая мышца плеча в
целом является разгибателем локтевого сустава. Длинная головка при-
нимает участие также в сгибании плечевого сустава.
М. brachialis, плечевая мышца (рис. 103—106), начинается мышечно
от латеральной и дистально от части передней поверхности плечевой
кости (рис. 107, а, г), простираясь от капсулы плечевого сустава до ла-
терального надмыщелка. Брюшко мышцы толстое, округлое в сечении
(рис. 106, б), имеет на медиальной стороне своей дистальной части су-
хожильное зеркало, крепящееся на особой шероховатой линии, идущей
косо по передней поверхности лучевой кости от медиопроксимального
285
угла radius в дистолатеральном направлении примерно до середины ко-
сти. Кроме этого сухожилия плечевая мышца оканчивается мышечными
волокнами на проксимальной половине передней поверхности лучевой
кости (рис. 109, в, 110, г). Площадка, занятая прикреплением, имеет вид
треугольника, коротким основанием прилегающего к капсуле локтевого
сустава. Его медиальную сторону образует упомянутая выше косая шеро-
ховатость. Плечевая мышца является основным сгибателем локтевого
сустава. У дюгоня мышца развита несравненно сильнее в связи с расши-
рением спирального желоба и, соответственно, площади ее отхождения.
Помимо того, кроме прикрепления в проксимальной части лучевой кости,
m. brachialis имеет еще сухожильное окончание на медиальной стороне
дистального конца radius (Domning, 1977b).
МЫШЦЫ ПРЕДПЛЕЧЬЯ И КИСТИ
М. brachioradialis, плечелучевая мышца (рис. 109), начинается мышечно
(более поверхностные волокна) и сухожильно от латерального надмы-
щелка плечевой кости (рис. 107, а) проксимальнее всех остальных экс-
тензоров предплечья. Брюшко мышцы сжато с боков и в сечении имеет
вид треугольника с коротким основанием, выступающим наружу, и вер-
Рис. 109 г, д
286
шиной, обращенной вглубь (рис. 108, 23—26). Начальное сухожилие идет
по этому внутреннему краю брюшка, отдавая на его медиальную, обра-
щенную к плечевой мышце, и латеральную поверхности сухожильные,
постепенно расширяющиеся и утончающиеся тяжи, формирующие сухо-
жильные зеркала. Обычно латеральное зеркало, обращенное к лучевому
разгибателю запястья, шире и длиннее медиального, но в целом протя-
жение обоих может варьировать. Примерно по середине длины внутри
брюшка появляется широкое плоское сухожилие (рис. 108, 25), за счет
которого формируется конечное сухожилие, узкое (3X4 мм в сечении)
и длинное (до 80 мм). Дистально оно расширяется до 15 мм и, сливаясь
своим задним краем с сухожилием m. extensor carpi radialis, прикрепля-
ется вместе с ним на летеральной поверхности слившихся I и II сагра-
lia distale (рис. 109, б, г). Такое строение сухожильного скелета мышцы
обеспечивает ее перистость. На одной из сторон имеется небольшой со-
единительный пучок мышечных волокон, идущий от брюшка плечелуче-
вой мышцы к лучевому разгибателю запястья (рис. 109, б). Мышца раз-
гибает кисть и немного супинирует ее и предплечье. У дюгоня m. Ьга-
chioradialis кончается на переднем крае слившихся radiale и intermedium
и на I metacarpale.
М. extensor carpi radialis, лучевой разгибатель запястья (рис. 109),
начинается, как и предыдущая мышца, от латерального надмыщелка
(рис. 107, а) сразу дистальнее ее и мышечными волокнами, и сухожиль-
но. Сухожилие, сразу подразделяющееся на три самостоятельных плоских
тяжа, частично надвигающихся друг на друга вентральными краями,
образует узкий вентральный край треугольного сечения брюшка в самом
его начале (рис. 108, 33) и всю его заднюю, обращенную к m.ext. dig.
communis, поверхность. Дистальнее самый вентральный тяж формирует
сухожильное зеркало по передней поверхности брюшка, обращенной к
m. brachioradialis, которое едва достигает середины длины мышцы; са-
мый дорсальный — такое же зеркало по задней поверхности, а средний
проникает частично в глубь мышцы и способствует обособлению более
глубоких волокон в особую головку, образуя сухожильное зеркало на ее
задней поверхности. Дистальнее эта головка, занимающая более глубокое
и переднее положение, образует плоское брюшко, подстилающее плечелу-
чевую мышцу. Более поверхностные и задние по положению волокна
формируют основную головку лучевого разгибателя запястья, которая,
собственно, и выходит на поверхность предплечья. Еще до раздела мыш-
цы на две эти головки в ее толще появляется внутреннее сухожилие,
имеющее в сечении вид скобки, выпуклой стороной обращенной наружу
(рис. 108, 30). В дальнейшем оно разделится на три части: более мощ-
ная задняя послужит источником формирования широкого, толстого
(9X2—3 мм) конечного сухожилия основной головки мышцы, идущего
к основаниям II и III пястных костей. Значительно более тонкое сухо-
жилие идет вдоль его переднего края, но, подходя к запястью, отщепля-
ется и соединяется, предварительно делясь на два зубца, с конечным
сухожилием более глубокой головки и m. brachioradialis. Эта последняя
головка также образует на своей наружной поверхности широкое тонкое
сухожилие, дистально расширяющееся и своим передним краем срастаю-
щееся с конечным сухожилием m. brachioradialis до прикрепления на
I + II cd. К этому сухожилию подходят спереди волокна небольшого пуч-
ка от последней мышцы, а сзади мышечные волокна, обособляющиеся от
основной головки лучевого разгибателя запястья и формирующие допол-
нительную маленькую головку (рис. 109, б). Таким образом, мышца име-
ет сложную перистую структуру и множественное прикрепление. Как и
предыдущая, она разгибает кисть и немного супинирует ее. Перед под-
287
Рис. 110. Мышцы ме-
диальной (флексорной)
поверхности ласта (пред-
плечья и кисти)
а — наружный слои;
б — глубокий слой после
снятия mm. flexor
carpi radialfs, fl. digi-
torum superficialis et
profundus, fl. carpi
ulnaris, abductor digi-
ti quinti;
в — mm. interossei;
г — крепления мышц на
костях предплечья;
д — крепление мышц на
костях кисти
1 — 2 — m. abductor digit!
(I— quart!,
2 — quinti);
3 — m. abductoH-ext. pol-
licis longus;
4 — 6 — m. adductor digit!
(4 — quart!,
5 — quinti,
6 — tertius);
7 — m. brachialis;
3 — m. biceps brachii;
9 — 10 — m. flexor carpi
(9 — radialis,
10 — ulnaris);
11 — m. fl. digit! quinti;
12 — m. fl. digitorum;
13 — 17 — m. interossei
digit!
(13 — quart!,
14 — quinti,
15 — 16 — secunda;
15 — lateralis, 16— medialis,
17 — tertius);
18 — m. pronator teres;
19 — кольцевая связна
ходом к запястью концевые сухожилия обеих мышц фиксированы коль-
цевой связкой (рис. 109, а) в особом желобе на антеролатеральной по-
верхности дистального конца лучевой кости. У бразильского ламантина
лучевой разгибатель запястья описан очень кратко (Domning, 1978а), что
не дает представления о его сложном строении. У дюгоня отмечено при-
крепление только на II тс и специально указано, что мышца не подраз-
делена на головки (Domning, 1977b). Для многих млекопитающих (сум-
чатые, насекомоядные, грызуны, приматы, хищные) характерно наличие
длинного и короткого лучевых разгибателей запястья, самостоятельных
мышц с раздельным прикреплением на II и III тс. У хоботных обе
мышцы сливаются в одну с общим прикреплением на этих же пястных
костях (Гамбарян, Рухкян, 1974), т. е. они сходны с основной головкой
288
мышцы, описанной выше у ла-
мантина. Формирование же осо-
бой глубокой головки уникаль-
но для млекопитающих.
М. extensor digitorum com-
munis, общий разгибатель паль-
цев (рис. 109, а, д), сложно по-
строенная мышца, состоящая
из трех головок, причем две из
них также подразделены на
две головки каждая. Начинает-
ся мышца мышечно от постеро-
латеральной поверхности луче-
вой кости (рис. 109, в) и час-
тично наружной фасции m. ex-
tensor digit! quinti, а также
сухожилием, более слабо выра-
женным на левой конечности,
от латерального надмыщелка
(рис. 107, «). Все головки у от-
хождения фактически неразли-
чимы, но дистально хорошо
обособляются. Каждая из них
выходит на поверхность лишь
небольшой своей стороной, по-
скольку их брюшки сжаты с
боков. Соотношение этих голо-
вок отражено на поперечном
сечении (рис. 108, 34), причем
самая передняя головка (I)
показана снизу, над ней еще
нераздельная II, а выше два
брюшка III головки (справа
сторона, обращенная к лучевой
кости). Передняя и самая глу-
бокая I головка занимают
большую, более дистальную
часть отхождения от лучевой
кости. В проксимальной части
ее брюшка имеется сухожиль-
ное зеркало, продолжающее
сухожильное начало от плече-
вой кости. На задней поверх-
ности брюшка в дистальной
его половине появляется также
сухожильное зеркало, перехо-
дящее в длинное концевое су-
хожилие, идущее ко II пальцу,
где оно крепится последова-
тельно к основаниям двух по-
следних фаланг (рис. 109, г).
II головка начинается от узкой
полоски лучевой кости позади
и слегка проксимальнее отхож-
дения I головки и образует два
10 Ламантин
289
Рис. 110 г, д
плоских лепестковидных брюш-
ка, сложенных вместе своими
уплощенными сторонами. По
задней поверхности каждого
брюшка формируется сухо-
жильное зеркало, переходящее
дистальнее в узкое длинное
сухожилие. Оба сухожилия ди-
стально, уже в пределах за-
пястья, объединяются в одно,
идущее к III пальцу и крепя-
щееся к основаниям всех трех
его фаланг. III головка, лежа-
щая ближе к локтевому краю
предплечья и более глубокая
по положению, чем II, поверх-
ностно делится на две: более
мощную переднюю, своим окон-
чанием связанную с IV паль-
цем, и более слабую, начинаю-
щуюся преимущественно не от
кости, а от фасции m. ext. di-
giti quinti, и идущую к V паль-
цу. По задним поверхностям
обеих имеются сухожильные
зеркала, за счет которых фор-
мируются длинные конечные
сухожилия, закрепленные на
основаниях фаланг соответст-
вующих пальцев. Конечные су-
хожилия общего разгибателя
пальцев проходят в области
кисти в специальных влагали-
щах, погруженных в плотную
соединительную ткань, вплот-
ную прилегающую к костям,
а у дистального конца пред-
плечья фиксированы кольцевой
связкой в широком, обращен-
ном латерально желобе, обра-
зованном сросшимися дисталь-
ными концами лучевой и лок-
тевой костей. Для бразильско-
го ламантина отмечено форми-
рование двух основных сухо-
жилий, каждое из которых
делится на два по одному к
каждому пальцу. У африкан-
ского ламантина (Bahrdt, 1933)
не отмечено отхождение от ra-
dius, а только от надмыщелка,
но возможно, это лишь дефект
описания, хотя вообще отхож-
дение общего разгибателя паль-
цев от лучевой кости необычно
290
для млекопитающих. Поскольку у дюгоня мышца начинается так же,
как у ламантина, следует признать такое смещение ее дистально общей
чертой всех сирен.
М. extensor digiti quifiiti, разгибатель V пальцев (рис. 109, а), начина-
ется мышечно от латеральной поверхности костей предплечья в месте
слияния их проксимальных эпифизов (рис. 109, в) и частично от фасции,
разделяющей эту мышцу от предыдущей. Кроме того на одной конечно-
сти было сильное (до 5 мм в поперечнике) сухожилие, начинающееся
на латеральном надмыщелке (рис. 107, а), которое дистально шло по
переднему краю мышечного брюшка. На другой конечности сухожилия
как такового не было, лишь слабые сухожильные волокна прослежива-
лись от капсулы локтевого сустава. Брюшко мышцы в сечении представ-
ляет равносторонний треугольник с основанием наружу и вершиной
внутрь. Уже в проксимальной его половине появляется Т-образное внут-
реннее сухожилие, обеспечивающее перистую структуру. Дистально
каждое плечо этого сухожилия превращается в самостоятельное сильное
длинное сухожилие. Три таких сухожилия дистально на подходе к коль-
цевой связке объединяются в одно (5 мм), которое фиксируется на ди-
стальном конце локтевой кости в особом желобке и поворачивает почти
под прямым углом каудально, проходя, расширяясь до 9 мм, к оконча-
нию на проксимальном конце V тс, сразу дистальнее места прикрепле-
ния локтевого разгибателя запястья (рис. 109, г). У бразильского ла-
мантина также нет сухожильной связи мышцы с надмыщелком, только
с капсулой сустава. У дюгоня такое сухожилие имеется. У относительно
примитивных млекопитающих (сумчатые, насекомоядные, грызуны, хищ-
ные) разгибатель V пальца начинается только на латеральном надмы-
щелке, а кончается отдельными сухожилиями на II—V пальцах (Гам-
барян, 1960). У копытных появляется дополнительное начало от костей
предплечья, количество конечных сухожилий уменьшается (у свиней к
II и III пальцам, у полорогих к IV, у лошадей к III, см. Grau,
1943). Лишь у сирен его окончание приурочено только к V пальцу
(.у дюгоня кроме V тс, еще к проксимальной фаланге).
М. extensor carpi ulnaris, локтевой разгибатель запястья (рис. 109, а),
начинается широким (8 мм), длинным (55 мм) сухожилием от специаль-
ного выступа в дистокаудальном конце латерального надмыщелка
(рис. 107, в, б), которое дистально продолжается в сухожильное зеркало
на внутренней поверхности брюшка. В сечении брюшко имеет овальную
форму (рис. 108, 35—37). Уже к середине длины в его толще появляет-
ся широкое внутреннее сухожилие (отсюда перистость), которое выходит
на наружную поверхность брюшка в его дистальной 1/4. Конечное сухо-
жилие, плоское и очень широкое, прикрепляется к основанию V шс,
а на одной стороне продолжается и к IV шс. Мышечные волокна в свою
очередь могут достигать капсулы запястного сустава и на одной стороне
даже V шс. У бразильского ламантина отмечено прикрепление к ulnare
и V тс, а начало частично от фасции, окутывающей локтевой отросток.
Последнее в еще более выраженной форме (самостоятельное сухожилие
от латеральной поверхности олекранона) имеется и у дюгоня. Вообще,
для млекопитающих характернее отхождение двойное: от надмыщелка и
от олекранона,—хотя иногда (медведь, косуля) бывает и только от пле-
чевой кости.
М. abductor et extensor pollicis longus, абдуктор и экстензор I пальца
(рис. 109), начинается мышечно от латеральной и частично медиальной
поверхности проксимальной половины локтевой кости (рис. 109, в, НО, г)
и немного от фасции в межкостном пространстве. Брюшко мышцы очень
плоское широкое. По его наружной поверхности уже в 30—40 мм от на-
291
10*
чала формируется сухожильное зеркало, дистально переходящее в широ-
кое (15 мм) конечное сухожилие, потом сужающееся до 5 мм, но еще
дистальнее уже за кольцевой связкой, опять расширяющееся и дающее
два отдельных сухожилия: одно к радиальному концу основания II тс,
а второе к 1 тс, причем от заднего края последнего ответвляется более
тонкое сухожилие, идущее к прикреплению на основании концевой фа-
ланги этого же пальца (рис. 109, а, г). Мышечные волокна от медиаль-
ной поверхности ulna несколько обособлены от остальной массы мышцы
и формируют маленькую головку с самостоятельным конечным сухожи-
лием, вливающимся в основное сухожилие мышцы еще на уровне ди-
стальной половины предплечья (рис. 109, б). Возможно, что эта головка
гомологична m. ext. pollicis longus других млекопитающих, а основная
часть мышцы — m. abductor pollicis longus. Сопоставление одной из этих
частей с m. ext. pollicis bevis вряд ли целесообразно, поскольку послед-
ний скорее всего позднее приобретение высших приматов. У дюгоня, судя
по описанию (Domning, 1977b), обе мышцы вполне самостоятельны,
но начало абдуктора занимает большую площадь, включая и каудолате-
ральную поверхность лучевой кости.
М. pronator teres, круглый пронатор (рис. 110, а) начинается мощным
плоским (15—16 мм X 2—3 мм) сухожилием 30 мм длины от наиболее
проксимального конца гребня медиального надмыщелка (рис. 107, в),
продолжающимся дистально вдоль всей задней, обращенной к лучевому
сгибателю запястья стороны брюшка мышцы, достигая его дистальной
трети (на другой стороне даже до прикрепления мышцы на лучевой
кости, т. е. мышца подверглась сухожильному перерождению, фактиче-
ски превращаясь в связку). Кончается мышца мышечно на медиальной
стороне лучевой кости дистальнее и каудальнее прикрепления двуглавой
мышцы плеча. Брюшко пронизано рядом продольных сухожильных тя-
жей, число которых указывает на начавшееся сухожильное перерожде-
ние (рис. 108, 16—22). Превращение мышцы в связку характерно и для
ряда других млекопитающих (копытные, хоботные). У бразильского ла-
мантина круглый пронатор не был обнаружен (Domning, 1978а), у аф-
риканского он был хорошо развит (Bahrdt, 1933). Имеется он и у дюго-
ня, но практически превращенный в связку (Domning, 1977В). Очевидно’
с утратой подвижности костей предплечья мышца теряет свою функцию
и соответственно свою необходимость, что и позволяет ей исчезнуть,
слиться с m. fl. carpi radialis (Murie, 1880), превратиться в связку.
М. flexor carpi radialis, лучевой сгибатель запястья (рис. 110, а),
начинается от медиального надмыщелка проксимальнее остальных сгиба-
телей (рис. 107, в) частью мышечно, но главным образом широким пло-
ским сухожилием, делящимся на более тонкую переднюю часть, образую-
щую сухожильное зеркало на внутренней поверхности проксимальной
части главной, более передней части мышечного брюшка, и лежащую по-
зади сильно утолщенную часть, в дальнейшем расслаивающуюся на два
широких плоских сухожилия в толще брюшка. Одно из них образует
сухожильное зеркало на внутренней, обращенной к главной части брюш-
ка поверхности дополнительной плоской части. Обе части формируют об-
щее веретеновидное брюшко, грубо овальное в сечении (рис. 108,
43—50). На наружной поверхности дистальной половины брюшка также
формируется сухожильное зеркало, начало которому дают внутренние
сухожильные тяжи, тянущиеся в толще мышцы уже с проксимальной
его 1/5—1/4. Дополнительная часть брюшка также имеет на своей внут-
ренней, обращенной к главной поверхности зеркало, переходящее в сухо-
жилие, сливающееся с общим конечным очень широким и толстым сухо-
жилием (12X3—4 мм), прикрепляющимся к слившимся I и II cd и ос-
292
нованию I тс (рис. 110, д'). Все это создает сложный внутренний ске-
лет мышцы и сложную систему перистости. У некоторых экземпляров
бразильского ламантина было отмечено прикрепление только к пястным
костям. Наоборот, у африканского ламантина была связь с radiale и слив-
шимися carpalia distalia, но не с пястными костями. Очевидно, прикреп-
ление мышцы сильно варьирует, в том числе и индивидуально. В отли-
чие от дюгоня у ламантинов нет отхождения от лучевой кости.
М. flexor digitorum (superficialis + profundus), сгибатель пальцев
(рис. НО, а), начинается сухожильно и частично мышечно от дисталь-
ного конца медиального надмыщелка спереди от начала локтевого сгиба-
теля запястья. Имеет одно неподразделенное на головки брюшко, про-
ксимально лежащее на предплечье глубже лучевого сгибателя запястья.
Мышца многоперистая со сложным сухожильным скелетом как снаружи,
так и внутри. В сечении брюшко проксимальнее треугольно (широкое
основание прилежит к ulna, более короткая передняя сторона к m. fl. с. ra-
dialis, более длинная задняя обращена наружу и к m. fl. с. ulnaris),
дистально овально (рис. 108, 38—42). По наружной поверхности брюшка
помещается тонкое сухожильное зеркало, связанное с началом мышцы,
дистально достигающее середины брюшка, а на внутренней стороне
мышцы — очень сильное сухожилие, также идущее от медиального над-
мыщелка и достигающее дистальной 1/4 брюшка. Уже на уровне сере-
дины мышцы в ее толще обнаруживаются два широких, дистально еще
более расширяющихся сухожильных тяжа, которые частично налегают
друг на друга. Более задний из них вместе с начальным апоневрозом
образует скелет более поверхностной и задней порции мышцы. Он ди-
стально формирует мощное конечное сухожилие, которое сперва идет бок
о бок с таковым основной части мышцы, но потом уже в области пясти
обособляется от него и, подразделившись на два сухожильных конца,
кончающихся на антеропроксимальном и каудопроксимальном углах
основания проксимальной фаланги IV пальца, пропускает между ними
конечное сухожилие m. interosseus IV. Большая и более передняя и глу-
бокая масса мышцы связана, с одной стороны, с передним внутренним
сухожильным тяжем, за счет которого формируется конечное сухожилие
этой порции, и начальным сухожилием, с другой. Конечное сухожилие
уже к концу мышечного брюшка подразделено на два, идущих рядом
друг с другом и с сухожилием первой порции в общей фасции. Переднее
из них кончается на антеропроксимальном углу основания проксималь-
ной фаланги III пальца, более заднее и более широкое своей внутрен-
ней поверхностью закрепляется к середине того же основания и
идет дальше, закрепляясь на основаниях двух последующих фаланг
(рис. 110,5). Такое устройство сгибателей пальцев очень необычно для
млекопитающих, у которых он, как правило, подразделен на поверхност-
ный, связанный в начале с надмыщелком, а в конце двумя концами с
вторыми фалангами каждого из II—V пальцев, и глубокий, отходящий
несколькими головками как от надмыщелка, так и от ulna и radius,
а кончающийся на концевых фалангах тех же пальцев, проходя при этом
между обоими концами поверхностного сгибателя к каждому пальцу.
Видно, что у ламантина нет ничего похожего на эту схему, да и снаб-
жает эта мышца всего два пальца. Однако очевидно, что это состояние
у них может сильно варьировать: у бразильского и африканского ламан-
тинов отмечались сухожилия ко II—V пальцам, у Т. manatus ко II —
IV пальцам (Murie, 1872а), причем они прикреплялись к основаниям
всех фаланг. У дюгоня поверхностный и глубокий сгибатели выступают
как самостоятельные мышцы. М. palmaris longus, описанный у бразиль-
ского ламантина (Domning, 1978а), у нашего экземпляра отсутствует.
293
M. flexor carpi ulnaris, локтевой сгибатель запястья (рис. 110, а),
наиболее обширная мышца предплечья, лежащая по его заднему краю.
Начинается коротким широким сухожилием от медиального надмыщелка
(рис. 107, в), образующим сухожильное зеркало по переднему, обращен-
ному к m. fl. digitorum краю брюшка. Мышечные волокна начинаются от
этого сухожилия и идут дистально почти параллельно друг другу. Более
задние из них оканчиваются в толще соединительной ткани заднего края
ласта на уровне дистального конца V тс, а более передние — на заднем
крае его (рис. 110, д). Очевидно прикрепление может варьировать. Так,
у экземпляра, описанного Мьюри, оно ограничено проксимальной поло-
виной V тс. У бразильского ламантина выделено сухожилие, кончаю-
щееся на дистальном конце V тс. У дюгоня мышца построена более
сложно: имеется более передняя головка, связанная с предыдущей мыш-
цей, и задняя, соответствующая локтевому сгибателю запястья ламан-
тина. У млекопитающих обычно имеются две головки, начинающиеся и
от медиального надмыщелка и от олекранона и кончающиеся на горохо-
видной косточке. В этом случае мышца работает как сгибатель запястья,
у сирен же она скорее отводит V палец.
Короткие мышцы кисти сирен (рис. 110, б) крайне своеобразны, и их
гомология часто вызывает сомнения. Наиболее достоверна гомология сги-
бателя и абдуктора V пальца. Бесспорно наличие четырех межкостных
мышц, но медиальная межкостная мышца II пальца уже выпадает из
нормы. Происхождение мышц, обычно называемых червячными, не ясно.
У млекопитающих червячные мышцы начинаются от сухожилия глубо-
кого сгибателя пальцев и кончаются на проксимальных их фалангах.
У сирен они связаны с запястным апоневрозом и дистальными концами
III—V тс, что исключает их действие как сгибателя пальцев. Можно
было бы представить себе, что их начало сместилось на этот апоневроз
в связи с редукцией глубокого сгибателя пальцев и вообще уменьшением
числа конечных сухожилий сгибателя пальцев, как у нашего ламантина.
Но этот переход произошел у бразильского ламантина, где имелись сухо-
жилия к II—V пальцам. Мы предпочитаем определять их как абдукторы
и аддукторы соответствующих пальцев.
М. flexor digiti quinti, сгибатель V пальца, начинается от фасции
запястного сустава на уровне intermedium плоским сухожилием, образую-
щим сухожильное зеркало на наружной поверхности проксимальной
четверти брюшка, и частично более глубокими мышечными волокнами.
Остальные волокна берут начало уже от сухожильного зеркала отдель-
ными более или менее поверхностно обособленными пучками. В дисталь-
ной половине брюшка на его внутренней поверхности формируется сухо-
жильное зеркало, переходящее в конечное сухожилие, идущее по
радиальному краю мышцы. Место его прикрепления занимает узкую
полоску на дистальной части V тс и основании проксимальной фаланги
(рис. 110,6). Часть более задних волокон крепится непосредственно
к V тс. Брюшко мышцы в сечении овальное (рис. 108,51, 52) и имеет
сложный внутренний скелет.
М. abductor digiti quinti, абдуктор V пальца (рис. 110, а), начинается
на одной конечности сухожилием от внутренней поверхности retinaculum
flexorium, у места прикрепления его к ulna или от внутренней поверх-
ности локтевого сгибателя запястья. Идет в виде неправильного округло-
го в сечении (рис. 108, 54—56) брюшка к дистальной части V тс, где
она и прикрепляется сухожилием проксимальнее предыдущей мышцы и
краниодистальнее локтевого сгибателя запястья (рис. 110,5).
Группа абдукторов и аддукторов III—V пальцев (mm. lumbricales
Домнинга) состоит из отдельных маленьких мышц с перистым строе-
294
нием. Всего можно различить 4—5 таких мышц (рис. 110,6). Три из них
(ш. lumbricalis V Домнинга) начинаются вместе на фасции запястья в
области проксимальнее оснований IV и V пальцев. Более задняя мышца
(m. adductor digiti quinti) кончается на радиальном крае дистального
конца V шс; две передние (mm. abductores digiti quinti; на одной стороне
на их месте имеется только одна мышца) — на середине и на ульнарном
крае дистального конца IV шс (рис. 110,6). Четвертая мышца (m. ad-
ductor digiti quarti = m. lumbricalis IV) начинается ближе к радиальному
краю по фасции запястья по сравнению с предыдущими, а кончается на
радиальном крае дистальной части IV шс. Последняя мышца (m. adduc-
tor digiti tertius = m. lumbricalis III) от запястной фасции идет к ра-
диальному краю дистального конца III тс. Большие вариации среди этой
группы мышц говорят о малой роли, какую они играют в работе ластов
сирен.
М. interossei, межкостные мышцы (рис. НО, в), существуют у нашего
экземпляра ламантина в числе пяти; по одной к III—V пальцам и две,
медиальная и латеральная, ко II пальцу. М. interosseus medialis II начи-
нается мышечно от смежных частей оснований I и II тс и слившихся
I и II cd (рис. 110,6) и сухожильно от капсулы запястного сустава.
Начальное сухожилие продолжается по передней радиальной стороне
мышечного брюшка, а находящееся внутри него сухожилие формирует
конечное широкое тонкое сухожилие, прикрепляющееся к середине и ди-
стальному концу проксимальной фаланги II пальца. М. interosseus late-
ralis II начинается мышечно от заднего (ульнарного) края II тс и
сухожильно от капсулы запястного сустава. Брюшко мышцы узкое, за-
жатое между предыдущей мышцей и аддуктором III пальца. Кончается
широким (8—9 мм) конечным сухожилием, идущим рядом (позади)
с таковым медиальной межкостной мышцы II пальца и кончающимся на
дистальном конце проксимальной фаланги, основании и дистальном кон-
це следующей фаланги II пальца. М. interosseus III начинается от боль-
шей части III тс мышечно и образует высокое треугольное в сечении
брюшко, зажатое между лежащими несколько поверхностнее аддукто-
ром III и аддуктором IV пальцев. На его боковых сторонах находятся
плоские сухожилия, еще одно проходит в толще мышцы. Соединяясь Ди-
стально вместе, они формируют общее конечное сухожилие, кончающееся
на ульнарном крае проксимального конца I фаланги III пальца. М. inte-
rosseus IV начинается мышечно от большей части IV тс и образует
широкое уплощенное брюшко, облегающее эту пястную кость с трех сто-
рон. Оно фактически состоит из трех тесно прилегающих друг к другу и
сращенных перистых головок: средняя и более задняя из них имеют
внутри по сухожильному тяжу, выходящему более дистально на поверх-
ность, к которому с обеих сторон подходят мышечные волокна, начинаю-
щиеся от пястной кости. Более передняя головка связана, с одной сто-
роны, с идущим на поверхности сухожильным тяжем, закрепленным
проксимально на IV тс, а с другой, с широким плоским сухожилием,
сформированным на внутренней поверхности мышцы. Именно за счет
него образуется путем слияния с сухожилиями двух первых головок длин-
ное и широкое общее конечное сухожилие, проникающее между сухо-
жильными концами ш. flexor digitorum кончающимися по обеим сторо-
нам основания проксимальной фаланги IV пальца. Оканчивается это
сухожилие на дистальном конце предпоследней фаланги этого пальца.
М. interosseus V начинается мышечно от большей части V тс и неболь-
шого участка каудопроксимального угла IV шс. В дистальной половине
его плоского широкого брюшка ближе к переднему краю появляется
сухожильное зеркало, переходящее в широкое длинное конечное сухожи-
295
лие, которое кончается на основании концевой фаланги V пальца.
Описанное выше окончание большей части межкостных мышц крайне
необычно. У млекопитающих, как правило, имеется по две межкостные
мышцы к каждому, кроме I, пальцу, и их окончания связаны с пястно-
фаланговыми их суставами. Продолжение их к более дистальным фалан-
гам уникально и связано, видимо, у нашего экземпляра ламантина с ре-
дукцией конечных сухожилий m. fl. digitorum. Одно из них (к III паль-
цу) сохраняется, и лишь на этом пальце имеется нормальная межкостная
мышца. Однако такое состояние может быть индивидуальной вариацией,
поскольку на других экземплярах (Murie, 1872а, 1880) имелись обычные
короткие межкостные мышцы по две к II—V пальцам. У бразильского
ламантина отмечено три обособленных брюшка ко II пальцу, а к осталь-
ным по одной мышце (Domning, 1978а).
РАБОТА МЫШЦ КОНЕЧНОСТЕЙ
В разных группах сирен роль передних конечностей неоднозначна. Более
или менее подробно известно только, как они функционируют у ламантина
(Hartman, 1979). По поводу дюгоня фактически имеются только догадки
и отрывочные сообщения (Anderson, 1981b). Сведения, касающиеся мор-
ской коровы, ограничены наблюдениями ее первооткрывателя Г. Стелле-
ра (Steller, 1751) и относятся, видимо, только к использованию конеч-
ностей в условиях крайнего мелководья. Тем не менее определенно мож-
но сказать, что конечности ламантинов играют огромную и разнообразную
роль в их жизни, значительно большую, чем у дюгоней. Это — не только
хорошие манипуляторы, помогающие активно обрабатывать пищу (отря-
хивание песка и ила с вырванных из грунта растений, подтягивание и за-
талкивание их в рот) и участвующие в комфортном (почесывание, очи-
щение ротового аппарата и губного диска), половом и прочем поведении,
но и сильные локомоторные органы, за счет которых животное активно
поворачивается, кувыркается в воде, вращается вокруг своей длинной
оси, принимает любое положение (Хартман называет ламантинов даже
водными акробатами). При поочередном отталкивании ластами ламантин
может перемещаться по дну или отталкиваться от него. Сила передних
конечностей такова, что, например, вытащенный на палубу ламантин мог
приподнимать переднюю половину тела и некоторое время «стоять» на
кончиках ластов. Все это объясняет необычное строение скелета и ко-
нечностей и сильное развитие определенных групп мышц. Следует вспом-
нить уже упоминаемое ранее большее выступание ластов ламантина за
пределы туловища в связи с более проксимальным прикреплением мышц,
связывающих плечо с туловищем, почти сферическую головку плечевой
кости, со всей несомненностью свидетельствующую о крайне разнообраз-
ных движениях в плечевом суставе, причем движениях с большой ампли-
тудой. Так, в литературе есть сведения о том, что ламантин может пово-
рачивать плоскость своего ласта на 180° (Davilliers, цит. по: Hartman,
1979). Движение ласта пальмарной стороной назад при активных forward
stroke и вперед при backward stroke описывает и сам Хартман. Однако,
учитывая относительно ограниченную ротацию ласта относительно пле-
ча, следует отнести этот поворот за счет почти исключительно плечевого
сустава. Поэтому именно в строении головки плечевой кости и осталь-
ных структур проксимального ее эпифиза, как и в строении мышц, свя-
занных с этими структурами, мы видим целый ряд специфических для
ламантинов изменений, корни которых нужно искать в потребности жи-
вотного совершать силовые, максимально нагруженные движения среди
всего доступного их разнообразия. Для ламантина таковым будет про-
296
пульсивный гребок (см. гл. 3), специфичный именно для него. Суть про-
пульсивного гребка в активном гребущем движении назад ласта, развер-
нутого в поперечной плоскости, обеспечивающем мощный толчок вперед
(основной, или рабочий, период) и в последующем выносе ласта вперед
с плоскостью, ориентированной парасагиттально, готовящем новый гре-
бок (подготовительный период). Из них первый, встречающий сопротив-
ление воды, требует максимального усилия, второй — значительно мень-
шего. Непосредственно с пропульсией связана лопасть ласта, включаю-
щая предплечье п кисть. Плечо же представляет тот стержень, вокруг
которого и за счет которого организуется вращение плоскости ласта и
перемещение его в переднезаднем направлении. Ориентация ласта зави-
сит от положения оси блока плечевой кости в пространстве и собствен-
ного вращения вокруг своей длинной оси. Ясно, что первое обусловлено
положением блока относительно длинной оси плечевой кости, положением
головки ее относительно той же оси и положением рабочей оси самой
головки, что в свою очередь определяет положение плеча в пространстве.
Как показано в главе 3, все это вместе взятое обеспечивает плечу косое
положение относительно сагиттальной плоскости, сохраняющееся на всем
протяжении движения с совпадающими длинными осями головки плеча
и гленоида. В результате ориентация оси блока плечевой кости при про-
тракторно-ретракторных движениях остается неизменной: она переме-
щается в пространстве более или менее параллельно сама себе. Отсюда
вытекает для ламантина необходимость обеспечивать поворот плоскости
ласта в конце рабочего—начале восстановительного периодов, ставящий
его радиальным краем вперед (супинация), за счет движений в локтевом
и запястном суставах, что и достигается винтообразностью суставных
поверхностей блока и предплечезапястного сустава. Надо подчеркнуть,
что речь идет не о произвольной ротации ласта, а о сопряженной. Супи-
нация совершается только при разгибании в обоих суставах. Таким обра-
зом, в начале подготовительного периода ласт должен разогнуться, что
и обеспечивает ему необходимую для этого периода ориентацию. Пере-
нос ласта вперед осуществляется уже за счет протракции плеча, вклю-
чающей одновременно его супинацию, аддукцию и экстензию в плечевом
суставе, возможно с долей кругового движения в последнем, а также
смещение вентрального угла лопатки вперед. В конце подготовительно-
го — начале рабочего периодов за счет продолжающегося кругового дви-
жения (по часовой стрелке, если смотреть сверху на головку левого
humerus), а также некоторого сгибания локтя и кисти, лопасть ласта
пронируется и ставится в рабочее положение. Само силовое движейие
требует максимальной пронации плечевой кости, что и обеспечивается
почти поперечным положением длинной оси головки, при сгибании в пле-
чевом суставе и сохранении плеча в приведенном состоянии. При этом
вентральный угол лопатки оттягивается назад. Для самой лопасти ласта
силовым будет удержание его в несколько согнутом положении, препят-
ствующем выведению его из рабочего поперечного положения, т. е. на-
грузка будет ложиться на флексоры локтевого и запястного суставов.
Остается добавить, что вынос ласта вперед происходит в более отведен-
ном (ближе к горизонтальному) положении, его ретракцию, наоборот,
в более приведенном. Тормозящий гребок вряд ли может быть очень силь-
ным. Можно себе представить, что он может происходить при двояком
положении лопасти ласта. Если ласт повернут пальмарной стороной
назад и нагрузка, таким образом, ложится не на нее, а на более выпук-
лую тыльную сторону, то движения, совершаемые в суставах, просто
обратны таковым при пропульсивном гребке. Труднее представить под-
черкиваемое Хартманом движение с пальмарной поверхностью вперед.
297
Оно возможно только при почти на 180° круговом движении в плечевом
суставе (может быть, для этого и существует такая необычная капсула
этого сустава) и развороте локтя внутрь и наружу.
Представив себе, что же происходит с ластом ламантина во время
пропульсивного гребка, можно теперь оценить роль тех или иных мышц
в его выполнении. В суммированном виде функции каждой мышцы при
ее действии на плечевой сустав, исходя из ее топографии и т. п., вы-
глядят нижеследующим образом, при этом главная функция названа пер-
вой (равные по значению соединены знаком плюс):
Мышцы Ламантин Дюгонь
Плечеголовная Протракция (экстензия) Супинация Протракция
Широчайшая спины Флексия + пронация
Поверхностная грудная Ретракция (флексия) + ад- дукция Пронация Аддукция + пронация
Глубокая грудная Протракция (экстензия) Ретракция (флексия)
Дельтовидная Супинация Абдукция Протракция (экстензия) Супинация
Предостная Протракция (экстензия) Супинация Протракция Абдукция
Заостная Протракция (экстензия) + супинация Абдукция Супинация + абдукция
Подлопаточная Пронация Аддукция Ретракция (флексия) Аддукция Пронация
Большая круглая Ретракция (флексия) + +пронация Ретракция
Коракоидноплечевая Пронация Аддукция Аддукция Протракция
Двуглавая плеча Аддукция Протракция
Трехглавая плеча (длинная головка) Флексия Флексия
Уже сопоставление этих данных наводит на мысль, что у ламантина
увеличена роль ротаторов плеча. Их больше (9), чем у дюгоня (6), что
связано с формированием хорошо выраженной и более длинной шейки
головки плечевой кости, смещением и увеличение малого бугорка, в ре-
зультате чего функциональная ось плеча, линия, соединяющая геометри-
ческие центры головки и блока, оказывается на значительно большем
расстоянии от продольной оси плечевой кости, что увеличивает ротацион-
ный эффект многих мышц. Особенно развиты у ламантина оказываются
пронаторы, а именно пронация, как было сказано выше, обеспечивает в
первую очередь пропульсию при активном гребке. Так, основной прона-
тор ламантина — подлопаточная мышца составляет 14,8% массы всех
мышц конечности (табл. 13), что намного больше, чем у наземных мле-
копитающих (обычно 2-3%, реже 4-5%, см. Гамбарян, 1972), а по-
скольку именно она прикрепляется на малом бугорке, то становится
ясным смысл глубокой перестройки проксимального эпифиза плечевой
кости у ламантина, приведшей к увеличению рычага этой мышцы по
крайней мере вдвое по сравнению с наземными млекопитающими и дюго-
нем. Значительно увеличена у ламантина и большая круглая мышца
\5,5% против 0,9—2,3% у копытных и 1,7—4,0 у.хищных), прикрепле-
298
Таблица 13. Мышцы передних конечностей и их пояса
Мышцы по экз. ЗИН-31795 по экз. INPA-PB-6 (Domning, 1978а)
Масса мыш1 обеих сторон
Г % г %
m. brachiocephalicus 1108 4,5 — —
т. trapezius (acromio- + spino—) 1251 5,1 36,6 10,65
т. rhomboideus 676 2,8 6,9 2,0
т. latissimus dorsi 2581 10,6 22,6 6,6
m. sternomastoideus 121 0,5 — —
m. ectopectoralis (pect. major) 2612 10,7 35,2 10,3
m. endopectoralis (minor) 1078 4,4 12,6 3,7
m. pectoralis abdominalis 220 0,9 — —
m. serratus ventralis 2288 9,4 20,7 6,0
m. deltoideus 894 3,7 16,1 4,7
m. supraspinatus 1970 8,1 28,3 8,2
m. infraspinatus 1040 4,3 23,0 6,7
m. snbscapularis 3603 14,8 52,0 15,1
m. teres major 1355 5,5 19,9 5,8
m. coracobrachialis 173 0,7 6,2 1,8
m. anconeus longus 576 2,4 9,0 2,6
m. anconeus lateralis 93 0,4 4,1 1,2
m. anconeus medialis 265 1,1 3,5 1,0
m. biceps brachii 65 0,3 0,7 0,2
m. brachialis 838 3,4 9,6 2,8
m. brachioradialis 38 0,1 2,4 0,7
m. pronator teres 27 0,1 — —
m. ext. carpi radialis 72 0,3 1,9 0,6
m. ext. digit, communis ИЗ 0,5 2,8 0,8
m. ext. digiti quinti 48 0,2 1,7 0,5 .
m. ext. pollicis longus + brevis 49 0,2 1,8 0,5
m. ext. carpi ulnaris 37 0,1 1,5 0,4
m. fl. carpi ulnaris 402 1,5 5,7 1,7
m. fl. carpi radialis 96 0,4 2,3 0,7
m. abd. digiti quinti 27 0,1 1,8 0,5
in. fl. digiti quinti 41 0,2 0,7 0,2
m. fl. digitorum 323 1,3 6,2 1,8
m. interossei II 42 0,2 1,2 0,4
III 40 0,2 1,1 0,3
IV 44 0,2 1,1 0,3
V 20 0,1 0,4 0,1
m. lumbricalis III 22 0,8 0,7 0,2
IV 60 0,2 1,3 0,4
V 92 0,4 1,8 0,5
Всего 24 400 100,0 343,4 100,0
Масса всей группы мышц 6,8* 3,2*
* В процентах к массе тела.
299
ние которой также смещено вперед по диафизу плечевой кости, что
обеспечивает возможность пронировать ее, в отличие от дюгоня, где эта
мышца выполняет функцию флексора плеча. Участвует в пронации и по-
верхностная грудная мышца, значительно увеличившая свою мощь
(10,7% против 2,9—5,9 у копытных и 3,7—7,2 у хищных). Таким обра-
зом, с пронацией у ламантина связаны почти все мышцы, выделяющиеся
своей массой. Интересно, что глубокая грудная мышца, обычно у млеко-
питающих чуть ли не вдвое более сильная, чем наружная (7,9—14,2%
у копытных, 7,1 — 10,6 у хищных), у ламантина слабее последней (4,4%),
что возможно связано с сужением ее функций: она участвует в протрак-
цип плеча, осуществляемой в основном за счет работы других сильных
мышц (плечеголовная, предостная и заостная, дельтовидная). Аддуктор-
ную же функцию, характерную для наземных млекопитающих, она у
сирен утеряла.
Как уже указывалось, подготовительный период пропульсивного греб-
ка менее нагружен. Он требует супинации и протракции плеча, что и
выполняется одними и теми же мышцами: плечеголовной, предостной,
заостной, дельтовидной, и, видимо, не случайно все протраторы являются
у ламантина и супинаторами. В то же время предостная и заостная
мышцы у наземных млекопитающих супинаторами не являются, как не
является заостная мышца и протрактором. Сильное развитие у ламанти-
на акромиального отростка и соответственно смещение крепления ш. ас-
romiotrapezius и m. deltoideus латерально и вперед заметно изменяет
условия работы мышц. Обе они заметно увеличиваются. Так, у копытных
трапециевидная мышца составляет всего 1,8—3,6%, у хищных 2,0—3,4%,
дельтовидная — 0,9—1,8%. Трапециевидная мышца вместе с m. brachio-
cephalicus в подготовительный период тянут вентральный угол лопатки
вместе со всем плечевым суставом вперед, обеспечивая большее движе-
ние ласта вперед. Следует отметить, что изменение положения дельто-
видной мышцы позволяет ей выполнять значительно более разнообраз-
ные функции, чем у дюгоня, где она является лишь супинатором.
В рабочий период для успешного выполнения главной задачи, связан-
ной с пронацией, необходима также аддукция плеча, что делают те же
подлопаточная и поверхностная грудная мышцы. Последняя также уча-
ствует в ретракции плеча (флексия плечевого сустава) (первая в мень-
шей степени) вместе с большой круглой мышцей. И опять оказывается,
что главные пронаторы плеча оказываются одновременно и ретракторами
и в большей или меньшей степени аддукторами. Широчайшая мышца
спины, прикрепляющаяся вместе с большой круглой благодаря смеще-
нию прикрепления дистальнее, чем у наземных млекопитающих, усили-
вает свое действие как ретрактор плеча. В то же время она обеспечивает
оттягивание плечевого сустава и лопатки назад в конце рабочего периода.
Как уже было сказано, при пропульсивном гребке на кисть падает
громадная нагрузка: эта часть ласта наиболее расширена, и двигается
он с большей скоростью. В то же время ее перемещение в простран-
стве обусловлено движением плеча, а не самой кисти. Наоборот, мышцы
предплечья и кисти должны удерживать кисть в заданном положении,
чтобы воспрепятствовать ее разгибанию давлением воды. Самого сустава
для этого оказалось недостаточно, и у ламантинов возникает уникальный
механизм, препятствующий отведению кисти в медиальную сторону и раз-
гибанию, основным компонентом которого становится локтевой сгибатель
запястья, который необыкновенно увеличивается (1,6% от массы мышц
конечностей и 44% веса всех мышц предплечья и кисти, вместо 0,3—
0,9% и 10% соответственно — копытных; у хищных не более 16% мышц
предплечья) и из сгибателя кисти превращается в мышцу, ее отводящую
300
латерально. Меняется ее внутренняя структура, так что она служит
теперь мышцей-связкой, которая обычно у млекопитающих слаба, а здесь,
наоборот, необыкновенно усилена. Перенос ласта вперед в подготовитель-
ный период также встречается с сопротивлением воды, хотя и меньшим.
В этом случае стабильность ласта обусловливается деятельностью луче-
вых сгибателя и разгибателя запястья, плечелучевой мышцы, сместив-
шейся в своем прикреплении с предплечья на запястье, экстензора I паль-
да, коротких мышц кисти. Последние, несмотря на видимость невозмож-
ности функционировать из-за заключения всех пальцев в плотный
соединительнотканный футляр, хорошо развиты и достаточно мощны
(большинство их перистые), что можно понять как следствие потреб-
ности противостоять давлению воды. Видимо, аддукторы и абдукторы
пальцев управляют не соответствующими пальцами, а всей кистью
в целом.
Как уже отмечалось, у дюгоней ласты несут более однообразную
функцию, они глубже погружены в кожный туловищный мешок (почти
по самый локоть), двигаются в иной плоскости, чем у ламантина; крайне
необычно направление длинной оси головки плечевой кости, и, соответ-
ственно, у них иные движения являются рабочими. Плечо у дюгоней
двигается больше в парасагиттальной плоскости, хотя лопасть ласта
занимает преимущественно более горизонтальное положение. Вообще
ласты играют не активную пропульсивную роль, как это было у ламан-
тина, а пассивную. Они служат рулями поворота и глубины при про-
пульсивных силах, развиваемых работой хвоста. Соответственно движе-
ния их являются менее силовыми, они осуществляются, как правило,
тонким краем ласта вперед. Даже простое сопоставление функций кон-
кретных мышц показывает, что структурные изменения в проксимальном
эпифизе плеча, как и смещение прикреплений ряда мышц (дистально
большой круглой и плечеголовной, латерально дельтовидной), вызывают
соответствующие различия в функциях одной и той же мышцы у дюгоня
и ламантина. Ротация плеча не потеряла своего значения, но рабочим
движением его, как показано в главе 3, стала супинация, а не пронация,
выводящая лопасть ласта из положения прижатого медиальной поверх-
ностью к туловищу в отведенное в сторону и помещенное плашмя
(в горизонтальной плоскости). Основным супинатором стала дельтовид-
ная мышца (и это по сути дела ее единственная функция), эффектив-
ность работы которой обеспечена разрастанием гребня большого бугра.
Пронация потеряла свое значение, так как обратному возвращению ласта
способствует и само сопротивление воды. В результате даже исчезла ши-
рочайшая мышца спины. Видимо, уменьшились и переднезадние переме-
щения вентрального угла лопатки в связи с утратой (или, наоборот, не
приобретением) необходимости в активных гребущих движениях. С этим
связано также слабое развитие m. acromiotrapezius. В то же время
протракция плеча все еще необходима и предостная мышца получает
более эффективный рычаг за счет разрастания большого бугорка плече-
вой кости. К сожалению, мы не имеем сопоставимых данных по массе
мышц дюгоня, которые могли бы помочь понять различия в характере
приспособительных изменений мышц конечностей у дюгоня и ламантина.
МЫШЦЫ ГЕНИТАЛЬНОЙ ОБЛАСТИ
В ходе анатомирования взрослого самца Trichechus manatus был открыт
его мочеполовой аппарат. Описание его не входит в нашу задачу, отме-
тим лишь самые общие его черты (рис. 111, 112), чтобы было понятнее
положение мышц этой области.
301
мышц
а — мышцы (вид с вентральной стороны) f <5— половина среза посередине длины penis; в — полови-
на среза в месте перехода ножек в тело члена; г — срез ножки полового члена (около седалищной*
кости)
1 — m. bulbocavernosus;
2 — т. ischiocavernosus;
з — 4 — т. levator penis
(4 — его сухожилие);
5 — т. rectus abdominis;
6 — os ischii;
7 — артерия;
8 — вена;
9 — вентральное пещеристое
тело;
10 — мочеполовой канал;
11 — нервы;
12 —пещеристые тела;
13 — фиброзная стенка ножки?
пенисл
302
Рис. 112. Мочеполовая
система самца (вид с
дорсальной стороны)
1 — семенник;
2 — придаток семенника;
3 — извитой каналец;
4 — семяпровод;
5 — отверстие семяпровода,
идущее в мочеполовой
канал
6 — 7 — семенные пузырьки
<7 — их отверстие в моче-
половой канал);
8 — мочевой пузырь;
9 — вход мочевого пузыря
в мочеполовой канал;
10 — мочеполовой канал;
11 — пещеристые тела по-
лового члена;
12 — тело полового члена;
13 — ножки полового члена;
14 — сфинктер мочеполово-
го канала;
15 — m. ischiocavernosus;
16 — т. ischiococcygeus;
17 — т. ischiourethralis;
18 — 19 — т. levator penis
(19 — его сухожилие);
20 — т. rectus abdominis;
21 — т. sacrococcygeus
ventralis medialis
10
Два бобовидных семенника величиной почти с почку (см. гл. 9)
лежат в дорсальной части брюшной полости несколько латеральнее самих
почек п покрыты с боков слоем брюшины. Спереди от них отходит силь-
но развитый придаток семенника, вскоре переходящий в семяпровод,
тянущийся до самого мочеполового сфинктера. Близ его конца в него
•открывается канал пузырьковидной железы. Это парное мешковидное
с перетяжками образование, по величине не уступающее мочевому пузы-
рю, лежит нейтральнее и латеральнее большого мочевого пузыря, к дор-
солатеральной поверхности которого идут тянущиеся от почки мочеточ-
ники. Все выводные протоки ведут в один мочеполовой канал, идущий по
вентральной стороне полового члена, который состоит из парного пеще-
ристого тела полового члена, которое двумя ножками крепится на тазо-
вых костях, и уретры, включающей вентрально расположенное губчатое
тело уретры (рис. 111, в), тянущееся на 3/5 длины полового члена.
Ножки пещеристого тела представляют собой фиброзные трубки, внутри
которых находятся идущие от них внутрь трабекулы и помещающиеся
между ними кавернозные полости. Между ножками проходит мочеполо-
вой канал с дорсальной стороны на вентральную, т. е. ножки как бы
охватывают его и находящийся здесь мышечный сфинктер с двух сторон
и сливаются спереди, образуя пещеристое тело полового члена.
303
Рис. 113. Площади при-
крепления мышц на се-
далищной кости и пе-
нисе
а — вид с вентральной
стороны;
б — вид с дорсальной
стороны;
1 — m. bulbocavernosus;
2 — т. ischiocavernosus;
3 — т. ischiococcygeus;
4 — т. ischiourethralis;
5 — т. levator penis;
6 — т. rectus abdominis;
7 — т. retractor ischii;
8 — pedunculi penis;
9 — penis;
10 — место расположения
вентрального кавер-
нозного тела
В состав уретры входит и первая из четырех имеющихся у ламантина
мышц генитальной области.
М. bulbospongiosus (m. bulbocavernosus по старой терминологии и
Домнингу), луковично-губчатая мышца (рис. 111,а, в), начинается на
медиовентральной поверхности ножек пениса (рис. ИЗ, б) и от средней
фиброзной пластинки на месте соединения волокон мышц обеих сторон.
Мышечные волокна идут от начала прямо латерально и кончаются на
латеральном продолжении фиброзной оболочки пещеристого тела. Основ-
ная масса луковично-губчатой мышцы сосредоточена ближе к основанию
полового члена, дистальнее она становится тоньше, хотя и доходит до
конца губчатого тела уретры.
М. ischiocavernosus, седалищнопещеристая мышца (см. рис. 111,
112),—наиболее мощная из всего комплекса (табл. 14). Это сложный
многоперистый мускул с обширным началом как от вентральной так и
дорсальной сторон тазовой кости, занимающим на ней почти всю поверх-
ность (рис. ИЗ) . Оканчивается мыш-
Таблица 14. Мышцы полового
члена
Масса
Мышцы
Г %
m. ischiocavernosus 774 62,8
m. bulbospongiosus 226 18,3
m. levator penis 220 17,9
m. ischiourethralis 12 1,0
Всего 1232 100,0
Мышцы обеих сторон 2464 0,62*
* В % к массе тела.
ца на фиброзной оболочке ножек пе-
ниса, охватывая их с вентральной,
латеральной и немного дорсальной
сторон. По-видимому, дорсальные и
вентральные части этой мышцы мо-
гут работать независимо, так как,
судя по направлению их волокон,
их действия противоположны.
М. levator penis, мышца, подни-
мающая половой член (см. рис. 111,
112), начинается мышечно от части
дорсальной поверхности тазовой ко-
сти и сухожилием от ее краниально-
го края. Начало захватывает также
часть латеральной поверхности нож-
ки пениса. Мышечные волокна от
304
своего начала идут дорсомедиально и кончаются на широком срединном
сухожилии между мышцами обеих сторон, которое постепенно сужается
и идет в виде овального в сечении тяжа по средней линии вдоль дорсаль-
ной поверхности полового члена вплоть до его головки, где он соединя-
ется с фиброзной оболочкой пещеристого тела, доходя до когтеобразного,
покрытого роговым веществом возвышения на головке.
М. ischiourethralis (см. рис. 112) начинается тонким апоневрозом
вентральнее мышцы по вентрокраниальному краю тазовой кости у осно-
вания ножки пениса и оканчивается коротким сухожилием, срастающим-
ся с таковым мышцы противоположной стороны и лежащим в рыхлой
соединительной ткани у вентральной поверхности мочевого пузыря и над
дорсальной стороной основания полового члена. Это самая слабая мышца
комплекса (табл. 14).
М. retractor ischii (см. рис. 112) фактически не принадлежит к этому
комплексу, хотя топографически и связан с ним. Он начинается от кон-
цов передних шевронных костей и идет вперед и латерально, прикреп-
ляясь к дорсокраниальному краю тазовой кости рядом с прикреплением
прямой мышцы живота.
I
Глава шестая
ДВИЖИТЕЛЬНЫЙ КОМПЛЕКС ЛАМАНТИНА
И ЕГО ОСОБЕННОСТИ
Ламантины, как и все сиреновые и китообразные, используют для своего
передвижения в воде и создания силы тяги волновые колебания тела,
и главным образом машущие колебания мощного хвостового плавника.
Однако движительный комплекс ламантина заведомо уступает по своим
гидродинамическим качествам не только генетически отдаленным дель-
финам, но даже и представителям того же рода сиреновых дюгоням.
Тем не менее краткое рассмотрение движительного комплекса ламантина
представляет как самостоятельный интерес, так и в сравнительном аспек-
те в плане гидробионики. Известно, что хвостовые плавники взрослых
ламантина и дюгоня аналогичны по форме нескольким стадиям эмбрио-
нального развития хвостового плавника китообразных (Першин, 1969).
Кроме этого, интересно выявить, имеются ли в хвостовом плавнике
ламантина характерные для плавников китообразных так называемые
комплексные артериовенозные сосуды, которые являются структурами
системы саморегуляции упругости лопастей плавников и органами термо-
регуляции у китообразных (Першин и др., 1970).
Следует отметить, что несмотря на довольно обширную биологическую
литературу по сиреновым, сведения по движительному комплексу их
весьма ограниченны. Нам удалось воспользоваться лишь некоторыми ка-
чественными данными и иллюстрациями из ряда работ (Howell, 1930;
Anon., 1974; Heinsohn. 1976). Авторами был исследован хвостовой плав-
ник одного экземпляра карибского ламантина общей длиной 3,2 м, мас-
сой 402 кг, который был добыт у берегов Кубы. Изучена форма плавника
в плане и в профиле, выполнены гистологические исследования попереч-
ных срезов тканей и кровеносных сосудов хвостового плавника.
Ламантины как вторичноводные растительноядные млекопитающие
могут съедать морских водорослей в день до 20% массы тела. Однако
процессы обмена веществ у них весьма замедлены и низки. В местах
обитания ламантинов в прибрежных теплых водах океанов растительная
пища имеется в изобилии. Поэтому ламантины не нуждаются в большой
скорости плавания. Хотя к настоящему времени имеется мало данных по
скорости плавания сирен, все же можно привести следующие оценки.
Крейсерская скорость плавания карибского ламантина составляет от 4
до 10 км/ч. Кратковременная спринтерская скорость его, возможно, до-
стигает 25 км/ч, но на очень ограниченном расстоянии, порядка 20—30 м
и не более 100 м. При этом число ударов в минуту хвостового плавника
ламантина составляет от 20 до 30—40 на крейсерской скорости и до 50
на максимальной.
Поскольку позвоночный столб у ламантина сильно изогнут, его тело
и хвостовой плавник в плавании имеют несимметричную амплитуду коле-
баний относительно горизонтальной плоскости, большую при ударе хво-
стом вниз, нежели при взмахе вверх, о чем можно судить по фотографии
(рис. 114). Подобному строению движительного комплекса ламантина
отвечает и привычное для него плавание не по прямой траектории,
а с заныриванием до дна водоема и последующим всплытием ближе к
поверхности воды. Каждые 3—5 мин ламантин нуждается в появлении
над поверхностью воды на 3—5 с для смены воздуха.
306
Сопоставляя между собой плавание ламантина и обыкновенного дель-
фина, можно заметить грузное тело первого и стройное, прогонистое тело
второго. Очевидно, что форма тела ламантина значительно хуже обтекае-
мая, чем у дельфина, как вследствие тупорылой передней части, так и
по характеру выступающих частей — грудных плавников. Форма тела и
хвостовой плавник дюгоня занимают промежуточное положение между
ламантином и дельфином.
Известно, что водные животные, адаптированные к постоянному пла-
ванию в водной среде, имеют симметричную форму тела относительно
срединной сагиттальной плоскости их тела. Поэтому наиболее удобно,
сравнивать форму тела и плавников ламантина, дюгоня и дельфина по
Рис. 114. Подводные фото с натуры
а — бразильский ламантин Trichechus inunguis, б — дюгонь Dugong dugon, в — обыкновенный
дельфин Delphinus delphis
307
контурам в горизонтальной плоскости (рис. 115). В связи с этим отметим,
что среди ламантинов встречаются формы с различным в плане хвосто-
вым плавником. У карибского ламантина (рис. 115 а) хвостовой плавник
имеет более вытянутую и узкую форму, нежели у флоридского
(рис. 115,6); последний напоминает в плане греческую букву дельта, но
со скругленными углами. Обе эти формы в плане хвостового плавника
ламантинов напоминают таковые у небольших, малоскоростных рыб, на-
пример бычка-кругляка, что отвечает форме для водных животных, пла-
вающих с небольшими скоростями. Однако у массивного ламантина хво-
стовой плавник толстый, мощный, а у небольшой рыбы бычка он пред-
ставляет собой тонкую и гибкую упругую пластину. Это отвечает
различию их чисел Рейнольдса, которое для ламантина составляет на
крейсерской скорости порядка ReL=l-107.
Характерным отличием ламантина от дюгоня по внешнему виду их
тела является наличие у ламантина одной или двух крупных попереч-
ных складок в месте соединения корпуса тела с хвостовым плавником
(см. рис. 115). Несомненно, что эти складки облегчают возможность осу-
ществления больших амплитуд колебаний хвостовым плавником у ламан-
тина. Однако такие складки несколько ухудшают обтекаемость всего
животного водным потоком. Дюгонь же имеет более узкую и плавную
форму перехода тела в плавник и у него поперечные складки отсут-
ствуют. Очевидны более обтекаемые и изящные формы тела и плавников
у обыкновенного дельфина, скорость плавания которого выше в два-три
раза сравнительно с ламантином и дюгонем.
Исследованный авторами хвостовой плавник карибского ламантина
общей длиной L = 3,2 м имеет дельтовидную форму в плане. Длина плав-
ника вдоль оси тела h = 1,05 м, поперечный размах 1 =0,8 м, площадь
несущей поверхности S =0,5 м2, относительное удлинение плавника со-
ставляет Х= 12/S = 1,33, что втрое меньше такового у дельфинов. Уже
одно это обстоятельство свидетельствует о более низких гидродинамиче-
ских качествах хвостового плавника ламантина.
Рис. 115. Контуры животных в плане
а — карибский ламантин Trichechus m. manatus; б — флоридский ламантин Т. m. latirostris,
в —дюгонь Dugong dugon, г — обыкновенный дельфин Delphinus delphis (по: Howell, 1930)
Рис. 116. Продольные профиль (а) и разрез (б) хвостового плавника карибского ла-
мантина длиной 3,2 м
а — на расстоянии 1/3 полуразмаха от продольной оси животного; б — ближе к внешней границе
1 — эпидермис; г — сухожильный слой; 3 — гиподерма; 4 — кровеносные сосуды
308
Продольные профиль и разрез
хвостового плавника карибского
ламантина представлены на
рис. 116. Профиль сагиттального
сечения плавника (параллельного
срединной плоскости) имеет вид
с криволинейным выпуклым очер-
танием сверху и плоским на боль-
шей части длины снизу, близкий
профилю хвостового оперения
планеров. Это соответствует числу
Рейнольдса данного профиля, вы-
раженного через длину его хорды,
порядка Re (1-^2) 10е. При длине
хорды профиля 0=0,55 м, наи-
большая толщина профиля t=9%,
которая находится в расстоянии
;го=15—2О% хорды от начала за-
кругленного носика профиля.
Изучение продольного разреза
плавника показало, что эпидермис
имеет толщину 2—5 мм, выделя-
ется слой мягких сухожильных
тяжей, с пересекающимися по
диагональным направлениям во-
локнами, общей толщиной 3—
7 мм. В гиподерме содержится
большое число жировых клеток,
которые оставляют жирный след
на бумаге, приложенной к разре-
зу хвостового плавника. В средин-
ной части профиля расположены
довольно многочисленные крове-
носные сосуды.
Характерной особенностью
внешней поверхности кожи хво-
стового плавника, как и всей по-
верхности тела ламантина, явля-
ется грубая шероховатость ее
(рис. 117). Она выражена попе-
речными складками верхнего слоя
кожи, которые чередуются через
каждые 2—3 мм по длине живот-
ного и хорошо ощущаются, если
провести рукой вдоль тела. Не-
сомненно, что эта шероховатость
кожи существенно повышает гид-
родинамическое сопротивление
плаванию ламантина, однако при
достаточно мощной мышечной
системе и энергетике его двигате-
ля и небольшой потребной скоро-
сти плавания подобная шерохова-
тость, по-видимому, не представ-
ляется большой помехой для пла-
вания животного.
309
Между тем шероховатость, складчатость поверхности кожи ламанти-
на, надо полагать, играет большую роль в терморегуляции тела. Темпе-
ратура тела ламантина около 37° С, а обитает он в теплых прибрежных
водах океанических шельфов с температурой воды до 25° С. Поэтому
терморегуляция тела ламантина при непрерывном плавании даже с не-
большими скоростями составляет важную проблему, которая разрешается
в значительной мере при условии развитой, шероховатой поверхности
кожи. Это подтверждается и анализом кровеносной системы хвостового
плавника ламантина.
Кровеносные сосуды, расположенные в срединной части профиля
плавника, состоят как из отдельных артерий и вен, так и, главным обра-
зом, из сопутствующих друг другу артерий и вен, заключенных в общую,
обособленную от тканей плавника оболочку — мембрану (рис. 118).
Сопутствующие артерии и вены обычно состоят из одной мышечной тол-
Рпс. 117. Поверхность кожи
хвостового плавника ка-
рибского ламантина. Вид-
на складчатость поверхно-
сти, ориентированная по-
перек обтекающего потока
Рис. 118. Микрофото по-
перечного среза тканей в
средней части хвостового
плавника карибского ла-
мантина
1 — эпидермис;
2 — срединный слой дермы;
3 — жир;
4 — щелевидная артерия:
5 — сопутствующие артерии и
вены;
6 — т <ани сетчатого строения
310
стостенной артерии, которая имеет чечевицеобразный (щелевой) про-
свет, и располагающихся вдоль артерии двух-трех тонкостенных вен,
значительно более крупных и круглого поперечного сечения. По-видимо-
му, наличие таких кровеносных сосудов может иметь некоторое термо-
регуляционное значение для ламантина. Однако сопутствующие артерии
и вены в хвостовом плавнике ламантина, несомненно, не имеют никакого
значения для регулирования упругости тканей, в чем ламантин при его
ограниченных возможностях плавания и не нуждается. Эти особые кро-
веносные сосуды у ламантина имеют лишь самое отдаленное сходство с
комплексными артериовенозными сосудами в плавниках дельфинов и от-
личаются как морфологически, так и особенно по своему возможному
функциональному назначению.
Таким образом, сравнительно с такими быстрыми и эффективными
пловцами, как дельфины, медленно плавающий ламантин представляется
несколько худшим пловцом. Об этом говорят его плохообтекаемая форма
тела, большая шероховатость поверхности кожи, упрощенная форма и
малое относительное удлинение хвостового плавника, простейший тип
профиля последнего и отсутствие в плавнике настоящих комплексных
артериально-венозных сосудов, которые так характерны для плавников
дельфинов.
Глава седьмая
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ
ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
В многочисленной группе растительноядных млекопитающих сирены за-
нимают особое положение — это единственные в ней представители вод-
ных форм. Именно это обстоятельство и позволило в свое время оценить-
уникальные морфологические образования пищеварительной системы
сирен как двойную адаптацию: к жизни в воде и питанию раститель-
ностью (Lemire, 1968). Такая слишком упрощенная и, пожалуй, фор-
мальная трактовка вопроса не раскрывает связи важнейших адаптив-
ных проявлений, неизбежно возникающих с переходом к водному образу
жизни (касающихся не только питания, но и особенностей теплообмена,
размеров тела, двигательной активности, энергетических потребностей"
и т. п.), с морфологическими преобразованиями пищеварительной систе-
мы сирен.
Хотя морфология пищеварительной системы сирен описывалась не-
однократно (см. обзор: Ronald et aL, 1978; Reynolds, Krause, 1982), все
же результаты исследований, охватывающих разные уровни структурной:
организации, далеко не исчерпывающи, а иногда и противоречивы. Что
касается соображений о функциональной роли своеобразных структур, то*
они базируются преимущественно на предположениях, часто недостаточ-
но обоснованных.
Отсутствие в желудке зоны ороговевшего эпителия (а значит, и мор-
фологической базы для преджелудочной ферментации) и большая отно-
сительная длина кишечника (с преобладающим развитием толстого отде-
ла), превышающая длину тела в 12—15 раз (Kenchington, 1972; Spain,.
Heinsohn, 1975; Ronald et al., 1978), послужили основанием для причис-
ления представителей отряда сирен к группе «нежвачных» млекопитаю-
щих с так называемой постгастричной (кишечной) ферментацией, ана-
логичной происходящей, например, в пищеварительном тракте лошади
(Lemire, 1968; Reynolds, 1979). Это мнение особенно укрепилось после
исследования содержания жирных кислот в пищеварительном тракте-
дюгоня (Murray et al., 1977). Необычно высокая концентрация летучих
жирных кислот, обнаруженная в слепой и толстой кишках дюгоня, пока-
зала, что в этих отделах кишечника происходит более интенсивная фер-
ментация волокнистой, пищи, чем у наземных млекопитающих.
Несмотря на то что по типу пищеварения ламантины и дюгони ока-
зались в одной группе с нежвачными сухопутными млекопитающими,
все же они стоят в ней особняком. Детальное морфологическое иссле-
дование, проведенное в настоящей работе, и морфофункциональный ана-
лиз полученных, а также опубликованных ранее данных склоняют к
мнению, что представители отряда сирен очень далеки от остальных
растительноядных видов не только по строению, но и по функциони-
рованию пищеварительной системы, что подтверждается и нетипично*
высокой концентрацией летучих жирных кислот в заднем отделе кишеч-
ника, и соотношением конечных продуктов ферментации.
312
СПЕЦИФИКА ПИТАНИЯ
Функциональные исследования пищеварительной системы неизбежно свя-
заны с изучением питания; при этом особенно важны сведения о каче-
ственном составе пищи. В изучении питания сирен основное внимание
уделялось экологическим аспектам: видовому составу поедаемых расте-
ний и особенностям пищевого поведения (например, Reynolds, 1981;
Anderson, 1981а).
Исследования на многих наземных видах млекопитающих показали,
насколько может повыситься питательная ценность корма при наличии
пищевой избирательности. У ламантинов и дюгоней предпочитаемость
кормов неодинаково проявляется в разных местах обитания: одни попу-
ляции кормятся доминирующими растениями и неразборчивы в пище,
другие выбирают растения по пищевой ценности и переваримости (Hein-
sohn, Birch, 1972; Hartman, 1979; Marsh et al., 1982). Заслуживает ин-
тереса наблюдение (Hartman, 1979), что ламантины избегают грубой,
богатой клетчаткой пищи, например Cocos nucifera. В результате кормле-
ния такими растениями погиб молодой ламантин в майамском океана-
риуме. Тогда как дюгони могут потреблять грубоволокнистые стебли и
ризомы (Marsh, 1981в).
В целом водные растения, в том числе и служащие пищей сиренам,
не так богаты клетчаткой и другими структурными элементами, как на-
земные. В соответствии с этим в пище, например, жвачных, содержится
50% нейтрального детергента клетчатки, а в пище дюгоня — 30% (Mur-
ray, 1979). Кроме того, вероятно, сказывается и избирательность кор-
мов сиренами по калорийности и пищевой ценности; не исключено также,
что растительные корма сирен обогащаются за счет белка беспозвоноч-
ных, захваченных с основным кормом. Качество потребляемых кормов,
в свою очередь, определяет пищевую ценность содержимого желудка,
которая у дюгоня оказалась выше, чем у наземных растительноядных
млекопитающих: соотношение сырого белка и сырой клетчатки у дюго-
ня составляло 19 и 36% соответственно (Murray et al., 1977), тогда как,
например, у слона (по результатам аналогичного анализа) было обна-
ружено 12,3% сырого белка и 43,9% сырой клетчатки (Clemens, Ма-
loiy, 1982).
Специального внимания требует оценка отношения сирен к пище жи-
вотного происхождения, которое в известной степени служит показате-
лем специализации вида к питанию низкокалорийными кормами. Высоко
специализированные к питанию клетчатковыми кормами млекопитающие
не стоят в зависимости от внешних источников пищевого белка, содер-
жащего многие незаменимые аминокислоты (Наумова, 1974, 1976,1981).
Из сирен, по крайней мере, ламантины, по-видимому, испытывают по-
требность в животной пище. Вместе с водной растительностью они захва-
тывают большое количество живущих на ней беспозвоночных, которые
служат важным источником белка (Hartman, 1979). При содержании в
океанариумах ламантины с охотой едят рыбу, хлеб, мясо; наблюдалось
систематическое поедание рыбы и ламантинами, живущими на воле
(Povel, 1978).
'МАКРОМОРФОЛОГИЧЕСКОЕ СТРОЕНИЕ
ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
По анатомии желудок и слепая кишка исследованного экземпляра ла-
мантина (см. Введение) существенно не отличаются от описанных ра-
нее (Murie, 1872а; Harrison, King, 1965; Lemire, 1968); в макрорельефе
слизистой этих органов отмечены некоторые новые, не упоминавшиеся
в предшествующих исследованиях структуры.
313
1 — пищевод;
2 — кардиальный дивертикул;
3 — пилорический дивертикул;
4 — ампула двенадцатиперстной кишки;
5 — двенадцатиперстная кишка;
в — куполообразный клапан;
7 — полость пилорического дивертикула;
8 — отверстие, соединяющее полость ди-
вертикула с полостью желудка;
9 — пилорический сфинктер
Рис. 119. Желудок ламантина
а — схематическое изображение внешнего вида; б —продольный разрез через пилорический дивер*
тикул; в — рельеф слизистой пилорической части желудка
Желудок. Желудок ламантина (рис. 119, а) состоит из единственной
подковообразно изогнутой камеры, полностью выстланной железистым
эпителием. При визуальном осмотре желудок кажется как бы состоящим
из двух камер, разделенных перехватом. Второе расширение ранними
исследователями обозначалось как «пилорический желудок». В дальней-
шем, однако, было показано, что «пилорическая камера» имеет кишеч-
ное строение (Harrison, King, 1965; Lamire, 1968). Рассмотрение сли-
зистой желудка и дальнейший микроскопический анализ, проведенные
в настоящей работе, подтвердили, что каудальное расширение — не что
иное, как ампула двенадцатиперстной кишки, расширенная до диаметра
желудка.
Вход пищевода в желудок расположен в изгибе малой кривизны же-
лудка. Кардиальная часть желудка расширена и на вершине свода не-
сет уникальное для млекопитающих образование — кардиальный дивер-
тикул, лишенный своей собственной полости и состоящий сплошь из
железистой ткани. Между складками слизистой, заполняющей диверти-
кул, расположены небольшие микрополости, открывающиеся в централь-
ный канал. В основании дивертикула микрополости увеличиваются в
размерах, образуя своего рода «лакуны», способные вместить заметное
количество секрета. Кардиальный дивертикул сообщается с полостью
желудка тремя небольшими округлыми отверстиями.
По макроструктуре железистого слоя и степени концентрации желез
кардиальный дивертикул ламантина напоминает большую желудочную же-
лезу бобра и представителя сумчатых — коала. Но топография и форма
314
этого аналогичного ооразования у перечисленных млекопитающих раз-
личны. Целесообразность подобного обособления железистой ткани мо-
жет заключаться как в необходимости ее защиты от механического по-
вреждения крупными частицами содержимого, так и в обеспечении
периодичной секреции. Функциональная особенность железистой ткани
такого строения заключается в способности выбрасывать сразу большое
количество секрета; однако качество самого секрета и специфика его
действия может быть различной.
Среди растительноядных млекопитающих развитие дивертикула в вер-
шине свода желудка характерно также для Suidae и Bradypodidae —
таксонов, не стоящих друг с другом даже в отдаленном родстве.
Иногда дивертикулом называют ороговевшую часть желудка у Probos-
cidea. Однако аналогия этого образования у перечисленных групп млеко-
питающих носит лишь внешний, топографический характер; структура же
их (в данном случае и функция) существенно отличается от структуры
кардиального дивертикула ламантина. В огромном желудке хоботных так
называемый «дивертикул» имеет собственную обширную полость, выст-
ланную ороговевшим эпителием и служит преджелудочной бродильной
камерой. У свиней и ленивцев это, как и у ламантина, обособленная
железистая ткань, но изливающая свой секрет в бродильные камеры,
выстланные ороговевшим эпителием.
На границе между антральной частью желудка и ампулой двенад-
цатиперстной кишки имеется еще одно уникальное с морфологической
точки зрения образование — парный пилорический (или дуоденальный)
дивертикул. Это два симметрично расположенных слегка изогнутых от-
ростка, напоминающие по форме кардиальный дивертикул, но более
крупных размеров, тонкостенные, имеющие свои собственные полости,
открывающиеся в ампулу двенадцатиперстной кишки. У Trichechus se-
negalensis полости этих органов свободно сообщаются с полостью
двенадцатиперстной кишки; только продольная складка слизистой захо-
дит в начальный отдел слепого отростка (Lemire, 1968). У исследован-
ного нами ламантина полости пилорических дивертикулов полностью изо-
лированы от полости ампулы двенадцатиперстной кишки с помощью
мускульных сфинктеров и полусферических клапанов, имеющих форму
куполов, направленных вершиной в полость дивертикула (рис. 119 б).
Клапаны отделяют полости отростков от полости кишки таким образом,
что они сообщаются лишь посредством плотно закрывающихся щелевид-
ных отверстий, расположенных по одной линии и соединенных глубокой
бороздой слизистой.
На продольном разрезе через пилорический дивертикул видно, что
куполообразный клапан образован глубокой складкой слизистой с учас-
тием подслизистой оболочки и внутреннего слоя мускульной оболочки
(рис. 119, в). Эта складка развивается главным образом со стороны
«большой кривизны» пилорического дивертикула; со стороны «малой
кривизны» она незначительно вдается в полость дивертикула.
Желудок отделен от двенадцатиперстной кишки мощным мускульным
сфинктером (см. рис. 119, б).
Среди млекопитающих парный желудочный дивертикул развивается
только у гиппопотама (кстати, питающегося также водной и полувод-
ной растительностью), однако он не сравним с парным дивертикулом у
ламантина ни по структуре, ни по топографии. У гиппопотама парный
дивертикул выстлан ороговевшим эпителием, открывается в кардиаль-
ную часть желудка и несет функцию рубца. У ламантина внешне он
расположен в каудальной части желудка и выстлан железистым эпите-
лием.
315
Таким образом, все три дивертикула, расположенные в передней
части пищеварительного тракта, изолированы от полостей желудка и
кишечника. Кардиальный дивертикул, хотя и не имеет обширной по-
лости, а только небольшие пазухи, расположенные между железистой
тканью, сообщается с полостью желудка тремя небольшими отверстия-
ми; тонкостенные пилорические дивертикулы, обладающие емкими по-
лостями, изолированы с помощью куполовидных клапанов. Это позволяет
считать эти структуры не дополнительными образованиями, способными
усиливать железистую секрецию, как полагали исследователи, а спе-
циальными, предназначенными для сбора и хранения больших количеств
пищеварительных секретов и обеспечения периодичности секреции.
Слепая кишка. У растительноядных млекопитающих с постгастрич-
ным пищеварением размеры слепой кишки отражают роль происходя-
щих здесь ферментативных процессов и служат показателем степени их
клетчаткоядности. С функциональной точки зрения это объясняется тем,
что в слепую кишку попадает пища, обедненная легкоусвояемыми компо-
нентами, которые утилизируются в желудке и тонком отделе кишечника,
и относительно обогащенная клетчаткой. Поэтому чем больше трудно-
перевариваемых углеводов содержится в пище, тем более емкой должна
быть слепая кишка и тем длиннее толстый отдел кишечника. У млеко-
питающих с развитым преджелудочным пищеварением роль слепой
кишки ослабевает (компенсация функций) и она не достигает такого
развития, как у растительноядных млекопитающих с простым желуд-
ком; в отдельных случаях слепая кишка отсутствует.
Для млекопитающего с постгастричным пищеварением у ламантина
слепая кишка маленькая — по размерам уступает даже желудку. Ее
длина у исследованного экземпляра от основания дивертикула, включая
ампулу ободочной кишки, составляет всего лишь 20,5 см. И, хотя ее
диаметр почти равен длине (19 см), размеры слепой кишки у ламанти-
на гораздо меньше, чем у других млекопитающих с постгастричным
пищеварением. Например, у лошади слепая кишка в длину достигает
100 см, а ее емкость составляет 33 л (Бергнер, Кетц, 1973).
Морфология слепой кишки ламантина очень специфична. Свободный
конец слепой кишки несет два симметричных отростка — парный аппен-
дикс (рис. 120, а). Аппендикс имеет полость, которая изолирована от
полости тела слепой кишки мускульным сфинктером. Область илеоце-
кального соединения со стороны слизистой (рис. 120, б) носит довольно
примитивный характер. Илеоцекальное отверстие имеет вид небольшой
поперечной щели, замкнутой с помощью илеоцекального сфинктера, изо-
лирующего полость тонкого кишечника от полости слепой кишки. Парал-
лельно ему и дистальнее (относительно основания слепой кишки) рас-
положены щелевидные отверстия дивертикулов. Таким образом, тело
слепой кишки очень небольшое и короткое — 3,5 см; по другим данным
(Murie, 1872а) — 2,5 см. Илеоцекальный сфинктер служит условной гра-
ницей между слепой кишкой и толстым кишечником, а расширенный
участок, примыкающий к основанию слепой кишки, представляет собой
ампулу ободочной кишки, функционально, по-видимому, равнозначную
слепой кишке.
Ампула слепой кишки снабжена тремя карманами: двумя симметрич-
ными, расположенными с мезентериальной стороны (рис. 120, а),
и центральным, расположенным чуть каудальнее с антимезентериальной
стороны. Даже при визуальном осмотре слепой кишки ламантина замет-
но, что она снабжена сильной мускулатурой, что нетипично для назем-
ных растительноядных млекопитающих, для которых характерна тонко-
стенная и объемистая слепая кишка, не сохраняющая форму после
316
5 — отверстие, соединяющее полость пар-
ного аппендикса с полостью слепой
кишки;
6 — илеоцекальное отверстие
a — схематическое изображение внешнего вида;
б — рельеф слизистой илеоцекального соедине-
ния
1 — подвздошная кишка;
2 — парный аппендикс;
3 — карманы ампулы ободочной кишки;
4 — ободочная кишка;
фиксации. Слепая кишка ламантина не вместительна, а поверхность ее
слизистой не усложнена дополнительными складками.
Среди млекопитающих парная слепая кишка характерна лишь для
даманов — одной из наиболее близких сиренам групп млекопитающих.
Однако едва ли эти структуры можно рассматривать как признак фило-
генетической связи, так как, с одной стороны, ближайшие родственни-
ки ламантина — дюгони — имеют единственный непарный дивертикул в
слепой кишке, а, с другой — даманы, помимо парной слепой кишки,
расположенной дистально (относительно желудка), имеют и непарную
проксимальную слепую кишку типичного для млекопитающих строения.
МИКРОСКОПИЧЕСКОЕ ИССЛЕДОВАНИЕ
Методика. 30 образцов из разных морфологически дифференцирован-
ных структур желудка и кишечника было проведено через спирт и за-
лито в парафин. Срезы толщиной 7 мкм были окрашены гематоксили-
ном-эозином, альциановым синим по Стидмену и реактивом ШИК по
Хочкиссу.
Пищевод. В каудальной части пищевода его стенка достигает значи-
тельной толщины (до 20 мм) за счет сильного развития мускульной
оболочки, что естественно для зверя таких крупных габаритов, как ла-
мантин. Мускульная оболочка состоит из продольного и кругового слоев
гладких мускульных волокон, которые собраны в пучки, расположенные
не строго параллельно друг другу, а переплетающиеся или волнообраз-
но изогнутые. Хорошо развита подслизистая основа, состоящая из плот-
ной соединительной ткани, богатой ретикулярными волокнами и отдель-
ными небольшими пучками мускульных волокон. Кроме того, подслизи-
стая основа очень богато васкуляризована.
Слизистая пищевода (рис. 121, а) представлена преимущественно эпи-
телиальным слоем. Собственно слизистая и мускульная пластинка сли-
зистой не дифференцированы. Покровный эпителий имеет типичный для
растительноядных млекопитающих вид; он состоит из ростковой зоны
(базальной), где четко очерчены ядра и хорошо заметны границы кле-
ток, и ороговевшей (поверхностной) зоны, состоящей из пластов керати-
низированных эпителиальных клеток. В клетках базальной зоны выяв-
317
ляется мелкая ШИК-положительная зернистость, В базальную зону эпите-
лия повсюду глубоко врастают соединительнотканные сосочки (рис. 121, а).
Кардиальная часть желудка. Переход от ороговевающего многослой-
ного эпителия пищевода к железистому желудочному эпителию осущест-
вляется без образования промежуточных форм (как это наблюдается, на-
пример, при переходе от донных желез к пилорическим) и без образова-
ния пограничной складки, характерной для многих млекопитающих с же-
лезистым желудком. В области желудочно-пищеводного соединения рядом
с ороговевшим эпителием пищевода располагаются кардиальные железы,
развивающиеся далеко не у всех растительноядных млекопитающих и
залегающие у ламантина мощным слоем, который становится несколько
тоньше по мере удаления от пищевода. Кардиальные железы (рис. 122, а)
представляют собой разветвленные железистые трубочки, открывающиеся
в хорошо выраженные желудочные ямки. Они состоят из морфологи-
чески однородных клеток мукозного типа с расположенным базально
сплющенным ядром и вакуолизированной цитоплазмой. Кардиальные же-
лезы имеют хорошо выраженные шейки, построенные из высоких цилин-
дрических эпителиоцитов с базально расположенным ядром, вытянутым
по оси клетки или округлым. Желудочная поверхность в области распро-
странения кардиальных желез выстлана эпителиоцитами такого же строе-
ния. Железистые трубочки разделены хорошо развитыми широкими меж-
железистыми перегородками, богатыми ретикулярными и мускульными
волокнами и в изобилии содержащими лимфоидные элементы.
Рис. 121. Слизистая пищевода (а) и фундальные железы кардиального дивертику-
ла (б). Об. 12,5, ок. 8
а — гематоксилин-эозин; б — ШИК, гематоксилин.
1 — главные клетки; 2 — обкладочные клетки; 3 — поверхностные эпителиоциты;
318
Клетки, образующие тела кардиальных желез и апикальные концы
поверхностных эпителиоцитов, содержат заметное количество кислых
гликопротеинов и показывают очень интенсивную реакцию на нейтраль-
ные гликопротеины (рис. 122, б). Плотная пленка гликокаликса, покры-
вающего поверхностный эпителий, также состоит в основном из нейт-
ральных гликопротеинов.
Кардиальные железы распространены в желудке ламантина, по край-
ней мере, до основания кардиального дивертикула, где они имеют не-
сколько иное строение. Железистые трубочки здесь короче, хотя и
остаются извитыми, а в мукоцитах почти не выявляются кислые глико-
протеины; нейтральные гликопротеины содержатся по-прежнему в
изобилии.
Кардиальный дивертикул. Мускульная оболочка кардиального дивер-
тикула относительно тонкая, гораздо тоньше, чем в теле желудка; основ-
ная же масса этого лишенного полости органа состоит из железистой
ткани, образующей сложные складки, которые создают иллюзию дольча-
того строения дивертикула. Складки образованы только слизистой обо-
лочкой с участием мускульной пластинки слизистой, которая и очерчи-
вает «доли» желизистой ткани (см. 121, б).
Эпителиальный слой слизистой кардиального дивертикула представ-
лен фундальными (донными) железами однотипного, но достаточно спе-
Рис. 122. Кардиальные железы (а) и нейтральные МПС в них (б)
а — гематоксилин. Об. 25, ок. 6; б — ШИК, гематоксилин. Об. 12,5, ок. 6.
1 — мукоциты; 2 — межжелезистые перегородки
319
цифичного строения. Железистые трубочки длинные, большая часть
их образует тела желез; шейки короткие; хорошо выражены желудоч-
ные ямки. Базальные отделы желез, составляющие до 2/з длины тела
железы, построены преимущественно из главных клеток (см. рис. 121, б).
Обкладочные клетки встречаются здесь изредка. Их количество резко
увеличивается в шейках желез, где они оказываются преобладающим
клеточным элементом, чередуясь с заметным количеством добавочных
клеток. Зона распространения обкладочных клеток колеблется в преде-
лах от ‘/4 до ‘/г длины тела железы. Таким образом, главные клетки
сконцентрированы на значительном протяжении базальных отделов же-
лезистых трубочек, а обкладочные локализованы преимущественно в
привершинных частях. Такое соотношение и топография клеток, обра-
зующих фундальные железы, не часто встречаются у млекопитающих.
Железы аналогичного строения описаны у зайцеобразных и распростра-
нены у них лишь на небольшом участке желудочной поверхности (Нау-
мова, 1981).
Основные клеточные элементы фундальных желез у ламантина име-
ют типичное для млекопитающих строение. Главные клетки мелкие с
темной оксифильной цитоплазмой и крупным ядром, занимающим боль-
шую часть объема клетки. В основаниях желез, где локализованы глав-
ные клетки, железистые протоки расширены, в связи с чем и сами ба-
зальные части желез выглядят утолщенными. Обкладочные клетки
крупные, неправильной формы, с крупным темным ядром и светлой
цитоплазмой, показывающей слабую ШИК-позитивную реакцию. Доба-
вочные клетки, расположенные среди обкладочных, имеют светлое ядро,
а их цитоплазма содержит яркие тяжи ШИК-положительного материала,
который выявляется и в протоках желез. Поверхность фундальных же-
лез выстлана высокими цилиндрическими эпителиоцитами со смещенным
в основание вытянутым ядром. Апикальная цитоплазма поверхностных
эпителиоцитов ярко окрашивается на нейтральные гликопротеины.
Фундальная часть желудка. Согласно предшествующим исследовани-
ям (Lemire, 1968), вся кардиальная половина желудка вплоть до пере-
гиба занята донными железами. Такую картину мы наблюдаем и у
большинства других млекопитающих с железистым желудком. Однако
в теле желудка у ламантина типичных фундальных желез, подобных
сконцентрированным в кардиальном дивертикуле, не обнаружено. Желе-
зы, расположенные на большой кривизне желудка каудальнее кардиаль-
ного дивертикула, содержат в основании большое количество клеток
мукоидного типа (рис. 123, а), что ставит под сомнение их классифика-
цию как фундальных. Хотя обкладочные клетки и встречаются в кле-
точном составе этих желез, они немногочисленны, а часто единичны и
сосердоточены в основном в поверхностных частях железистых трубо-
чек; главные клетки не идентифицированы.
Хотя вопрос о классификации этих желез не обсуждается в цитиро-
ванной выше работе, ее автором, судя по приведенной фотографии, по-
лучены сходные результаты. По-видимому, эта область желудка анало-
гична области перехода фундальных желез к пилорическим, расположен-
ной у других млекопитающих с железистым желудком на большой
кривизне против угла серповидной складки, тогда как настоящие фун-
дальные железы сконцентрированы в кардиальном дивертикуле. Железы,
выстилающие тело желудка, состоящие преимущественно из клеток
мукоидного типа, в которых синтезируется огромное количество ШИК-
положительного материала и единичных обкладочных клеток, следует
скорее отнести к пилорическим, чем к фундальным.
320
Пилорическая часть желудка. Пилорические железы, расположенные
в расширенном привратнике желудка, представляют собой извитые же-
лезистые трубочки, тела которых построены исключительно из клеток
мукоидного типа. Клетки имеют округлые базально расположенные ядра
и сильно вакуолизированную цитоплазму; синтезируют преимуществен-
но нейтральные гликопротеины. Межжелезистые перегородки широкие и
содержат много лимфоидных элементов. Фиксация, к сожалению, не
обеспечила сохранность покровного эпителия как в теле желудка, так
и в его пилорической части.
Пилорический дивертикул. Этот парный орган, расположенный на гра-
нице пилорической части желудка и двенадцатиперстной кишки, имеет
кишечное строение и представляет собой дериват двенадцитиперстной
кишки. Поэтому термин «дуоденальный дивертикул», примененный ранее
к этому образованию (Lemire, 1968), вполне оправдан, так же как и
термин «пилорический дивертикул», характеризующий его топографи-
чески.
Слизистая пилорического дивертикула образует тонкие нитевидные
ворсинки, выстланные цилиндрическими эпителиоцитами со всасывающей
каемкой. Ядро эпителиоцитов овальное, базально расположенное. Хорошо
развит криптальный слой, содержащий большое количество бокаловид-
ных эпителиоцитов (рис. 123,6). Единичные бокаловидные эпителиоциты
Рис. 123. Нейтральные МПС в пилорических железах (а) и слизистая куполообраз-
ной складки пилорического дивертикула (б). ШИК, гематоксилин
а — об. 25, ок. 6; б — об. 12,5, ок. 6.
1 — мукоциты; 2 — единичные обкладочные клетки; з — ворсинки; 4 — кишечные крипты;
5 —дуоденальные железы
1/211 Ламантин 321
входят в состав покровного эпителия ворсинок. Строма ворсинок содер-
жит мускульные волокна и лимфоидные элементы. В подслизистой от-
дельными группами расположены дуоденальные железы, не достигающие
здесь сильного развития; доли дуоденальных желез оплетены мускуль-
ными волокнами. Дуоденальные железы состоят из клеток мукоидного
типа и секретируют огромное количество нейтральных гликопротеинов.
Бокаловидные эпителиоциты крипт и поверхностного эпителия ворсинок
интенсивно окрашиваются как на нейтральные, так и на кислые глико-
протеины.
Подслизистая дивертикула изобилует лимфатическими полостями и
кровеносными сосудами. Мускульная оболочка не так сильно развита,
как в желудке: она едва достигает толщины слизистой дивертикула.
Куполообразный клапан дивертикула выстлан слизистой такого же
строения, как и сам дивертикул. Клапан образован впячиванием в по-
лость дивертикула слизистой и подслизистой оболочек и внутреннего
(кругового) слоя мускульной оболочки. Таким образом, клапан с обеих
сторон выстлан слизистой, а его сердцевину составляет удвоенный и
сращенный круговой слой мускульной оболочки. Слизистая клапана об-
разует более короткие ворсинки, чем слизистая, выстилающая полость
дивертикула.
Рис. 124. Слизистая двенадцатиперстной кишки (а) и ее ампулы (б). ШИК, гема-
токсилин. Об. 12,5, ок. 6
1 — ворсинки; 2 — кишечные крипты; 3 — дуоденальные железы
322
Двенадцатиперстная кишка. В ампуле двенадцатиперстной кишки
слизистая образует высокие плотно расположенные пальцевидные вор-
синки с округлыми вершинами (рис. 124, б). Кишечные крипты глубже,
чем в пилорических дивертикулах, и содержат больше бокаловидных
эпителиоцитов. Слой дуоденальных желез здесь увеличивается. Бокало-
видные эпителиоциты обильно секретируют кислые и нейтральные глико-
протеины, а клетки дуоденальных желез — только нейтральные. Под-
Рис. 125. Слизистая подвздошной кишки (а) и кислые МПС в слизистой аппендик-
са (б). Об. 12,5, ок. 6
а — ШИК, гематоксилин; б — альциановый синий, гематоксилин
1 — кишечные крипты; 2 — бокаловидные эпителиоциты; з — уплощенные поверхностные эпите-
лиоциты; 4 — головка лимфоидной бляшки
323
и*
слизистая основа необычайно богато выскуляризована. В целом же
ампула двенадцатиперстной кишки и пилорические дивертикулы имеют
аналогичное строение.
В собственно двенадцатиперстной кишке по сравнению с ее ампулой
резко изменяется соотношение железистых элементов. Ворсинки здесь
короткие и широкие; в покровном эпителии и криптах значительно уве-
личивается количество бокаловидных эпителиоцитов, в изобилии секрети-
рующих кислые и нейтральные гликопротеины. В слизистой заметно воз-
растает количество лимфоидных элементов; в подслизистой основе
встречаются пейеровы бляшки, врастающие своими вершинами в Сли-
зистую оболочку. Слой дуоденальных желез плотный (рис. 124, а) и по
толщине в четыре-пять раз превосходит аналогичный слой в ампуле
двенадцатиперстной кишки и пилорическом дивертикуле. Количество лим-
фатических полостей и кровеносных сосудов, залегающих в подслизистой
основе, напротив, сведено к минимуму. Клетки дуоденальных желез по-
казывают интенсинвую реакцию на нейтральные гликопротеины.
Подвздошная кишка. Слизистая подвздошной кишки в каудальном
отделе образует короткие плотно расположенные пальцевидные ворсин-
ки, покрытые невысокими цилиндрическими эпителиоцитами со всасы-
вающей каемкой. Эпителиоциты имеют округлые базально расположен-
ные ядра. Среди эпителиоцитов со всасывающей каемкой изредка встре-
чаются бокаловидные эпителиоциты, секретирующие кислые и нейтральные
гликопротеины и сосредоточенные главным образом в неглубоких кишеч-
ных криптах (рис. 125, а). Эпителиоциты на вершине ворсинок утрачи-
вают цилиндрическую форму и сильно уплощаются. Эти плоские эпи-
телиоциты с удлиненными ядрами образуют на кишечной поверхности
слой, сглаживающий рельеф ворсинок и подвергающийся постепенному
слущиванию. Благодаря таким образом видоизмененному поверхностно-
му эпителию слизистая подвздошной кишки в каудальном отделе функ-
ционально имеет криптальное, а не ворсинчатое строение, что характер-
но для толстого отдела кишечника.
Слепая кишка. Слизистая слепой кишки по составу и топографии кле-
точных элементов очень напоминает каудальный отдел подвздошной киш-
ки. Только слой слизистой в слепой кишке примерно вдвое тоньше и
уплощенные эпителиоциты создают на поверхности не такой мощный
слой. Бокаловидные эпителиоциты сосредоточены в глубине крипт и
секретируют, как и повсюду, кислые и нейтральные МПС. Подслизис-
тая основа в отличие от аналогичного слоя в подвздошной кишке бо-
гато снабжена лимфатическими и кровеносными сосудами.
Дивертикул слепой кишки. Парный дивертикул слепой кишки ламан-
тина — пожалуй, единственный орган, имеющий весьма типичное для
млекопитающих строение для толстого отдела кишечника. Глубокие ки-
шечные крипты дивертикула состоят преимущественно из бокаловидных
эпителиоцитов (рис. 125, б), секретирующих в изобили кислые и нейт-
ральные гликопротеины. В подслизистой основе расположено большое ко-
личество пейеровых бляшек, открывающихся вершинами в просвет киш-
ки, что характерно для аппендикса и у других млекопитающих. В меж-
железистой ткани повсюду встречается множество лимфоидных элементов.
TRICHECHIDAE И DUGONGIDAE:
СТРУКТУРНЫЕ ОСОБЕННОСТИ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
В исследованиях пищеварительной системы ламантинов и дюгоней
часто отмечалось значительное сходство между представителями этих
двух семейств сирен. Это позволяло с легкостью экстраполировать не-
достающие данные по одному виду, используя результаты, полученные
324
3
Рис. 126. Строение желудка и слепой киш-
ки дюгоня
а — внешний вид желудка (по: (Kenchington,
1972));
б — строение желудочной камеры (по: (Kenching-
ton, 1972));
в — слепая кишка (по: (Lemire, 1968))
1 — кардиальный дивертикул;
2 — пилорический дивертикул;
3 — кольцевидный гребень;
4 — пилорический сфинктер;
5 — аппендикс
при исследовании другого вида. Однако у ламантинов и дюгоней наряду
со сходным общим планом строения пищеварительной системы имеется
и ряд отличий.
Кардиальная железа у дюгоня морфологически не дифференцирова-
на, а представляет собой конусообразное возвышение свода желудка
(рис. 126, а). Она состоит из типичных донных желез и имеет неболь-
шую собирательную полость (Kenchington, 1972; Lemire, 1968). Донные
железы, расположенные в кардиальной железе у дюгоня имеют такое же
строение, как у ламантина, но их общая масса меньше, а собиратель-
ная полость более емкая, чем у ламантина.
Полость желудка у дюгоня, внешне состоящего из одной камеры
(рис. 126, б), подразделена кольцевидной складкой (гребнем), образован-
ной слизистой и подслизистой оболочками (Kenchington, 1972), на две
полости. Складка содержит большое число гладких мускульных волокон.
Слепая кишка у дюгоня (рис. 126, в) в отличие от ламантина име-
ет непарный дивертикул (Lemire, 1968). По длине, составляющей в
среднем 19—23 см (Spain, Heinsohn, 1975) она, по-видимому, близка к
слепой кишке ламантина, однако тело слепой кишки у дюгоня больше,
а ампула слепой кишки менее дифференцирована и не достигает таких
размеров, как у ламантина.
Общая длина кишечника у дюгоня превышает длину тела в 13—15
раз, тогда как у ламантина — в 10—12. Толстый отдел кишечника у
дюгоня по относительной длине значительно превышает тонкий отдел:
его длина составляет 63—65% от общей длины кишечника (рассчитано
по данным (Spain, Heinsohn, 1975)). У ламантина, напротив, преобла-
дающее развитие получает тонкий отдел кишечника (Ronald et al.,
1978). У исследованного нами экземпляра длина тонкого кишечника со-
ставила 56% всей длины кишечника. Относительная длина слепой киш-
ки у ламантинов и дюгоней ничтожна — менее 1 %.
Судя по описаниям цитированных выше авторов и по результатам
настоящего исследования, не исключено, что плоский многослойный эпи-
телий, выстилающий толстый отдел кишечника, у дюгоня развит в боль-
шей степени, чем у ламантина.
И* Ламантин 325
Перечисленные выше особенности в строении пищеварительного трак-
та дюгоня и ламантина слишком существенны, чтобы ими можно было
пренебречь при оценке пищевой специализации. Различия подобного-
масштаба у сухопутных растительноядных млекопитающих, как правило,
отражают неодинаковую роль клетчатковых кормов в питании животно-
го. Однако морфология пищеварительного тракта рассматриваемых мор-
ских млекопитающих настолько своеобразна, что вряд ли можно прово-
дить аналогию между ними и наземными видами, не имея дополнитель-
ных функционально-морфологических данных.
МОРФОЛОГИЧЕСКАЯ СПЕЦИФИКА
ПИЩЕВАРИТЕЛЬНОГО ТРАКТА [СИРЕН
Рассмотрение морфологических модификаций желудка и кишечника у
ламантинов и дюгоней позволяет выделить основные черты в строении
этих органов у сирен, отличающие их от наземных растительноядных
млекопитающих.
Желудок железистого типа распространен только у части мелких
сухопутных млекопитающих, основной пищей которых служит раститель-
ность. Эти виды в большинстве либо потребляют мало клетчатки, либо-
дополняют свою диету животными кормами. Среди крупных растительно-
ядных млекопитающих, сравнимых по размерам с сиренами, нет обла-
дателей железистого желудка. Все крупные виды с постгастричным
пищеварением независимо от таксономической принадлежности имеют в
желудке зону ороговевшего эпителия.
Помимо отсутствия в желудке ороговевшего эпителия, его специфику
составляют железистые дивертикулы: кардиальный, в котором сконцен-
трированы фундальные железы, и парный пилорический — дериват
двенадцатиперстной кишки. Хотя кардиальный дивертикул морфологи-
чески уникален, его функциональные аналоги встречаются у многих
сухопутных млекопитающих.
Слепая кишка сирен выделяется неестественно малыми для расти-
тельноядных млекопитающих размерами. Она характеризуется также
необычным для этого органа сильным развитием мускульной оболочки,
способной к мощным перистальтическим движениям. Аппендикс (дивер-
тикул) слепой кишки, редко развивающийся у растительноядных млеко-
питающих, у сирен непропорционально велик (относительно размеров-
слепой кишки): у ламантина (парный) составляет около 2/з длины
слепой кишки, у дюгоня (непарный) — около ‘/з. Аппендикс богат лим-
фоидной тканью.
Многослойный плоский покровный эпителий, обнаруженный у сирен
в заднем отделе кишечника вместо обычного однослойного цилиндричес-
кого со всасывающей каемкой, не встречается у других млекопитающих
в аналогичных отделах кишечника. Этот эпителий распространяется и
в тонкий кишечник, что привело к редукции ворсинчатой структуры сли-
зистой.
ПИТАНИЕ И РАЗМЕРЫ
ОРГАНОВ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Большая относительная длина кишечника, особенно толстого отдела,,
характерна для сирен и часто рассматривается как следствие питания
растительной пищей и как показатель постгастричного пищеварения.
На самом деле это не совсем так. И здесь в соотношение длины тела
и кишечника вмешивается абсолютная величина животного. У ламанти-
нов и дюгоней длина кишечника превышает длину тела в 12—15 раз,,
у стеллеровой коровы — в 20,5 раз. Отражается ли в этих цифрах раз-
326
личие в качественном составе корма у сирен? Другие примеры показы-
вают, что связь между длинами тела и кишечника неоднозначна. Так
если у мелких клетчаткоядных грызунов кишечник длиннее тела в 6—
7 раз, то у крупных тюленей, несмотря на плотоядность, длина кишеч-
ника превышает длину тела в 28 раз (Gorgas, 1967). Очевидно, что
абсолютные размеры тела животного маскируют зависимость между дие-
той и длиной кишечника. Причинная подоплека этих закономерностей
пока не ясна.
Качественный состав пищи в большей степени сказывается на соот-
носительном развитии отделов кишечника. У дюгоня (и у стеллеровой
коровы) толстый отдел кишечника по длине значительно превышает тон-
кий, составляя до 65% всей длины кишечника; у ламантина он не до-
стигает и половины длины всего кишечника (44%). Возможно, в этом
проявляется специфика пищевой избирательности ламантина — меньшей
по сравнению с дюгонем и стеллеровой коровой терпимостью к содер-
жанию в растениях клетчатки. Не исключено также, что в преоблада-
нии длины тонкого отдела кишечника у ламантинов находит отражение
роль в их питании животных кормов, в которых ламантины, по-види-
мому, испытывают большую потребность, чем дюгони. О тяготении дю-
гоней к животной пище упоминаний не встречается; в содержимом
желудков дюгоней животные корма не обнаружены (Heinsohn, Birch,
1972).
Недостаток данных по биохимическому составу кормов, по физиоло-
гии пищеварения и морфологии пищеварительного тракта не позволяет
предложить более обоснованную функциональную трактовку особенностей
в соотносительном развитии отделов кишечника у ламантинов и дюго-
ней. В дальнейшем может оказаться, что именно меньшая относитель-
ная длина толстого отдела кишечника у ламантина по сравнению с дю-
гонем служит причиной меньшей эффективности пищеварения (до 70% —
у ламантина и около 83%—у дюгоня (Best, 1981)), либо, что у
ламантина есть какие-либо морфологические структуры, компенсирую-
щие относительно меньшее развитие у него толстого отдела кишечника.
Размеры желудка и слепой кишки у ламантинов и дюгоней малы от-
носительно количества ежедневно съедаемой пищи. У ламантина, съедаю-
щего до 50 кг растительности, больший диаметр желудка и слепой киш-
ки, по известным данным, составляет около 25 см. У дюгоня вес же-
лудка с содержимым составляет 5% от веса тела (Murray et al., 1977).
По сравнению с общей длиной кишечника длина слепой кишки у ла-
мантинов и дюгоней ничтожна (менее 1%). Из этого следует, что де-
понирующая роль желудка и ферментирующая роль слепой кишки у
сирен снижены и перешли к другим органам. Такими органами могут
быть только тонкий и толстый кишечник. Подтверждением этого слу-
жит большая емкость этих отделов, особенно толстого: у ламантинов
диаметр тонкого кишечника достигает 7 см, а толстого —11 см (Ronald
et al., 1978).
Биологический смысл сохранения относительно малых размеров же-
лудка и слепой кишки у таких крупных животных, как сирены, заклю-
чается, по-видимому, в оптимизации соотношения контактной поверхно-
сти и объема органа, что в определенной степени способствует и дости-
жению высокой эффективности пищеварения. У стеллеровой коровы
колоссальные абсолютные размеры желудка (длина его достигала 1,8 м)
должны были отразиться и на эффективности пищеварения. В какой-то
степени невыгодное соотношение объема желудка и его секретирующей
поверхности могло компенсироваться меньшей скоростью прохождения
пищи через пищеварительный тракт.
327
И**
ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ ЖЕЛУДКА
И ДВЕНАДЦАТИПЕРСТНОЙ КИШКИ
Обособление фундальных желез от основной желудочной полости в том
или ином морфологическом варианте довольно часто встречается у ра-
стительноядных млекопитающих и обычно бывает связано с необходи-
мостью периодичной секреции фундальных желез и с механической за-
щитой слизистой от жестких частиц пищи. У наземных млекопитающих
периодичная секреция чаще всего обусловлена резкой сменой качествен-
ного состава корма, вызванной сезонностью в питании или физиологиче-
ской копрофагией. У сирен она, безусловно, связана с водным образом
жизни, который во время кормления вынуждает поглощать большее или
меньшее количество воды, что невыгодно разбавляет пищеварительные
соки. Поэтому и целесообразна резервация секрета фундальных желез в
лакунах кардиального дивертикула, способного совершать мощную хи-
мическую атаку желудочного содержимого после всасывания избытка
воды.
В то же время изоляция фундальных желез предохраняет их от засо-
рения песком и повреждения жесткими пищевыми частицами (напри-
мер, осколками раковин моллюсков). Защитную функцию несет и мо-
дифицированный железистый слой, выстилающий основную желудочную
камеру. Обилие в нем клеток мукоидного типа обеспечивает интенсивный
синтез слизистого секрета, защищающего внутреннюю поверхность же-
лудка от механического повреждения грубым содержимым, а также от
действия соляной кислоты, продуцируемой кардиальным дивертикулом.
На основании морфологического строения желудка можно предполо-
жить, что в основной желудочной камере поддерживается высокая кис-
лотность. Это согласуется и с экспериментальными данными, показавши-
ми, что в желудке дюгоня кислотность желудочного содержимого соот-
ветствует pH 2,5 (Kenchington, 1972). Однако трудно объяснить низкое
содержание соляной кислоты в кардиальном дивертикуле (pH 5,8), где,
казалось бы, благодаря сосредоточению основной массы обкладочных кле-
ток должна преобладать наивысшая кислотность. Связано ли это с ошиб-
кой определения или с периодичностью секреции (что мало вероятно для
этого органа) должны показать повторные исследования.
Высокая кислотность, поддерживаемая в основной желудочной камере,
в большей степени, чем соленость воды (Lemire, 1968), угнетает разви-
тие микрофлоры в желудке сирен. Соляная кислота в высокой концент-
рации оказывает мацерирующее действие на растительные ткани (для
освобождения легкопереваримых компонентов растительной пищи), а так-
же декальцинирующее на раковины захватываемых с растениями мол-
люсков.
Необычное для других млекопитающих развитие ампулы двенадцати-
перстной кишки и обособление пилорических дивертикулов, имеющих ки-
шечное строение и существенно увеличивающих функциональную
поверхность двенадцатиперстной кишки, позволяет предположить, что
здесь происходит не только нейтрализация кислого желудочного содер-
жимого. В этот отдел кишечника поступает поджелудочный сок и желчь.
По-видимому, именно на этом участке сосредоточены основные процессы
пристеночного пищеварения и насыщение содержимого пищеварительны-
ми соками для дальнейшего полостного пищеварения. Это подтверждает-
ся и тем, что только в переднем отделе кишечника нормально развит
ворсинчатый слой; в остальном тонком кишечнике ворсинки редуцирова-
ны, а сам тонкий кишечник в значительной мере несет депонирующую
функцию.
328
РОЛЬ ТОЛСТОГО ОТДЕЛА КИШЕЧНИКА
В ПИЩЕВАРЕНИИ СИРЕН
Из приведенного выше описания следует, что морфологические предпо-
сылки, служащие показателем ведущей роли в пищеварении толстого от-
дела кишечника, у сирен менее выражены, чем у наземных млекопитаю-
щих. Слепая кишка, которая служит обычно наиболее важным органом
в постгастричном пищеварении, у ламантинов и дюгоней имеет неболь-
шие размеры и не может вместить достаточное количество пищи для ин-
тенсивного развития симбионтного пищеварения. Однако морфологиче-
ским данным противоречат экспериментальные. В слепой кишке у дюгоня
насыщенность содержимого летучими жирными кислотами, достигающая
183 ммолей/л (Murray et al., 1977), выше, чем у каких-либо сухопутных
млекопитающих: например, у слона этот показатель составляет
137,6 ммолей/л (Clemens, Maloiy, 1982). Более того, если у наземных
млекопитающих концентрация летучих жирных кислот в толстой кишке,
как правило, уменьшается, у дюгоня она, напротив, возрастает до
236 ммолей/л. Такого высокого содержания продуктов ферментации у
других млекопитающих не было обнаружено ни в одном отделе пищева-
рительного тракта. Даже допущение методической ошибки в получении
этих данных (Murray, 1979) не может существенно умалить роль толсто-
го отдела кишечника у дюгоня.
Чем же объясняется необычно высокая эффективность постгастрично-
го пищеварения у дюгоня? Служит ли она проявлением чисто биохими-
ческой адаптации, недоступной расшифровке при помощи морфологиче-
ских методов исследования, или существуют коррелятивные связи био-
химических процессов со структурной характеристикой толстого отдела
кишечника?
Путем сравнительного анализа в строении толстого отдела кишечника
у сирен были выделены специфичные для них важные морфологические
особенности: необычное для других млекопитающих сильное развитие
мускульной оболочки кишечника и уникальное преобразование однослой-
ного цилиндрического покровного эпителия в многослойный плоский.
Эти признаки можно рассматривать как выражение морфологической
адаптации к интенсивному перемешиванию содержимого кишечника,
обеспечивающему максимально полный контакт с эпителиальной поверх-
ностью. При такой ситуации уплощение покровного эпителия, возможно,
связано с необходимостью защиты слизистой от повреждения жесткими
частицами, содержащимися в химусе, которое может возникнуть при
усилении перистальтики. Однако неизвестно, только ли благодаря энер-
гичной перистальтике так существенно повышается интенсивность фер-
ментации.
Судя по количеству продуцируемых летучих жирных кислот, основ-
ным бродильным чаном у дюгоня служит толстый кишечник. Это под-
тверждается и преобладающим развитием у дюгоня толстого отдела ки-
шечника над тонким. Более надежным доказательством ведущей роли
толстого кишечника в пищеварении дюгоня могут служить специальные
расчеты, показавшие, что с изменением размеров животного длина тол-
стого отдела кишечника возрастает с положительной аллометрией, тогда
как длина тонкого отдела и слепой кишки изменяются изометрично
(Spain, Heinsohn, 1975).
У ламантина толстый отдел кишечника по относительной длине усту-
пает тонкому. Однако морфология этой части кишечника у ламантина
изучена недостаточно полно. Только в одной из самых ранних работ
(Murie, 1872а) у ламантина описаны два расширения в толстом отделе
329
кишечника, расположенные каудальнее слепой кишки. Первое — с хоро-
шо развитой, образующей складки слизистой, второе — тонкостенное,
сходное по структуре с остальным толстым кишечником. G морфологиче-
ской точки зрения это очень важные образования, которые могут слу-
жить функциональным аналогом слепой кишки и компенсировать малые
размеры этого органа. Одновременно они могут компенсировать меньшую
у ламантина по сравнению с дюгонем относительную длину толстого от-
дела кишечника, способствуя интенсивному протеканию ферментативных
процессов.
Изложенные материалы, построенные в значительной степени на
сравнительном анализе, показали морфологическую и функциональную
специфику пищеварительной системы сирен. Несмотря на широту мето-
дик, примененных в исследованиях этой системы органов у сирен, начи-
ная от сравнительной морфологии и кончая электронной микроскопией и
биохимией пищеварения, все же ощущается недостаток данных, не поз-
воляющий более обоснованно и достоверно интерпретировать адаптивные
и функциональные особенности органов пищеварения этих своеобразных
млекопитающих. Экстраполяция же особенностей пищеварения сухопут-
ных млекопитающих на сирен рискованна и может быть чревата ошибка-
ми, а для надежной функциональной трактовки структурных особенно-
стей пищеварительной системы требуются дополнительные специальные
исследования всех ее звеньев, так же как и многих вопросов биологии,
связанных с эволюцией пищеварительной системы.
Глава восьмая
О НЕКОТОРЫХ
ФУНКЦИОНАЛЬНО-МОРФОЛОГИЧЕСКИХ
АДАПТАЦИЯХ ВЕНОЗНОЙ СИСТЕМЫ ЛАМАНТИНА
В сложном комплексе морфофункциональных адаптаций сердечно-сосу-
дистой системы водных и полуводных млекопитающих значительная роль
отводится приспособительным изменениям строения и функции венозной
системы большого круга кровообращения (Harrison et al., 1954; Соколов,
1954; Barnett et al., 1958; Яблоков и др., 1972; Галанцев, 1977; Агарков
и др., 1974; Иванова, 1975; и др.). Этот отдел сосудистой системы, от-
ветственный за выполнение функции депонирования крови при нырянии,
в силу особой его пластичности у всех исследованных видов ныряющих
млекопитающих перестраивался наиболее значительно и в целом в сход-
ном направлении, а именно, по линии увеличения емкости венозного рус-
ла, образования своеобразных венозных депо в виде расширения отдель-
ных участков венозных магистралей и формирования обширных веноз-
ных сплетений. Сердечно-сосудистая система сиреновых до сих пор
изучена весьма фрагментарно, а сообщений об адаптивных особенностях
строения ее венозного отдела практически вообще не было (Murie, 1871,
1872а; Fausett, 1942; Жеденов, 1961; Slijper, 1961; Иванова, 1975; и др.).
Между тем изучение приспособительных особенностей строения венозной
системы этих высокоспециализированных гидробионтов совершенно необ-
ходимо для выяснения основных путей эволюционного формирования и
направленности адаптивных преобразований сосудистой системы млеко-
питающих в процессе освоения ими водной среды обитания.
Исследование выполнено на половозрелом самце ламантина (см. Вве-
дение) . Для изучения строения и топографии венозного русла отдельные
участки последнего инъекцировались рентгенконтрастной массой, изго-
товленной на основе оксида свинца. Инъекцированные рентгенконтраст-
ной массой сосуды и отдельные органы препарировались с целью изго-
товления сосудистых препаратов для дальнейшей тотальной рентгено-
графии с помощью рентгеновского аппарата 7Л2. Диаметр просвета
отдельных сосудов определялся на рентгенограммах с точностью до 0,1 мм.
Препаровка интраорганных сосудов осуществлялась под контролем би-
нокулярного микроскопа МБС-2.
Наиболее существенные изменения венозной системы ламантина в
связи с водным образом жизни обнаружены в системе каудальной полой
вены (v. cava caudalis). Эта вена формируется двумя самостоятельными
стволами, берущими начало в тазово-поясничном венозном сплетении.
В преренальном отделе обе полые вены имеют одинаковый просвет
(у каудального края почек их диаметр равен 16,3 мм). В области почки
правый ствол каудальной полой вены, лежащей справа от аорты, резко
ампулообразно расширяется и диаметр его увеличивается более чем в
два раза (рис. 127, а). В области максимального расширения диаметр это-
го сосуда составляет 36,8 мм. Этот венозный коллектор принимает три
коротких венозных ствола от дольчатой почки. Краниальнее правой почки
просвет этого ствола полой вены постепенно уменьшается и перед слия-
нием со стволом левой полой вены диаметр его становится равным
26,6 мм. Интересно, что левый ствол каудальной полой вены значитель-
331
Рис. 127. Каудальная
полая вена в области
почек (а) и печени (б)
j — левая и правая почка;
2 — почечные вены;
з — расширение правого
ствола каудальной по-
лой вены в области
почки;
4 — печень;
5 — основной ствол вены;
6 — печеночные вены
но уже и в почечной зоне увеличивается в диаметре весьма незначитель-
но (до 22,8 мм). В эту вену от левой почки также впадают три коротких
венозных сосуда (рис. 127, а). На почках ламантина венозных (звездча-
тых) сплетений, как это имеет место, например у ластоногих, не форми-
руется. Объединение обоих стволов полой вены происходит в первой тре-
ти на пути их от почек к печени. Перед слиянием полых вен диаметр
левого ствола несколько уменьшается (до 20,3 мм). Общий ствол полой
вены (диаметром 30,0 мм) входит в правую большую долю печени, где
принимает в себя почечные вены (vv. heraticae). Никаких венозных си-
нусов, как, например, у ластоногих и некоторых других полуводных мле-
копитающих (Галанцев, 1977), в печени ламантина не формируется
(рис. 127, б). Характерной особенностью почечного участка полой вены
является лишь заметное увеличение его просвета у краниального края
печени. В этом месте диаметр полой вены возрастает в два раза по срав-
нению с участком ее в преддверии ворот печени и становится равным
60 мм. Пройдя через обширный сухожильный центр диафрагмы, каудаль-
ная полая вена продолжает резко воронкообразно увеличиваться в диа-
метре. Очень короткий и широкий грудной участок полой вены, объеди-
няясь с передней полохг веной, формирует перед правым предсердием
мощный толстостенный венозный синус. Грудной участок каудальной по-
лой вены, таким образом, существенно уклоняется от общего плана
строения его у большинства других млекопитающих, в том числе водных
и полуводных.
В брюшной полости ламантина основным венозным депо крови явля-
ется не печеночный синус, как у ластоногих, а правый ствол полой вены
в почечной области. Следует отметить, что образование значительного ве-
нозного коллектора каудальной полой вены в преренальном отделе ее
наблюдается и у некоторых других млекопитающих, например у выхухо-
ли (Иванова, 1964; Черняев, 1964; Галанцев, 1977).
Диафрагмального сфинктера каудальной полой вены, типичного для
ряда видов водных млекопитающих, у ламантина не обнаружено. Полая
вена этого вида проходит через мощный центр диафрагмы, а ее наруж-
ные стенки даже не контактируют с мышечными участками диафрагмы
и никаких мышечных элементов на своей поверхности не имеют.
Краниальная полая вена (v. cava cranialis) ламантина относительно
короткая и, объединяясь с каудальной полой веной, перед впадением в
правое предсердие формирует очень мощный венозный синус. Этой осо-
бенностью ламантин существенно отличается от других ныряющих мле-
копитающих, у которых подобных синсуообразных расширений в преде-
лах грудной полости обычно не наблюдается. Видимо, в этом синусе
может депонироваться значительная часть крови в период гемодинами-
332
ческих перестроек во время ныряния. Образование объемистого венозного
синуса в грудной полости ламантина в процессе адаптациогенеза, вероят-
но, было возможно в силу своеобразной топографии сердечно-легочного
комплекса и особого горизонтального расположения диафрагмы. Мешко-
образные, уплощенные, недольчатые и очень объемистые легкие ламанти-
на лежат значительно каудальнее сердца и заходят в поясничную об-
ласть. Отделены они от желудочно-кишечного тракта горизонтально
ориентированной диафрагмой с широким сухожильным центром. С серд-
цем легкие контактируют лишь своей верхушкой. В связи с этим легкий
не мешают наполнению крови в предсердном синусе, а депонирование
крови в нем, в свою очередь, не приводит к сжатию легочной ткани и не
препятствует нормальному течению газообмена в них.
Рис. 128. Схема притока и от-
тока крови от межреберных
чудесных сетей
1 — аорта;
2 — приносящие артерии чудес-
ной сети;
3 — вены чудесной сети;
4 — чудесная сеть
Наружная яремная вена (v. jugularis externa) ламантина не имеет
никаких коллекторообразных расширений и в этом отношении сущест-
венно отличается от таковой у многих других ныряющих млекопитаю-
щих (Галанцев, 1977). Это довольно узкий сосуд, ио своему просвету
приближающийся к таковому внутренней яремной вены (v. jugularis
interna), с которой он соединяется в области первого ребра. Внутренняя
яремная вена начинается от большого венозного сплетения в шейно-за-
тылочной области и лежит с внутренней стороны сонной артерии
(a. carotis). Яремные и подключичные вены (vv. subclavia). объединя-
ясь, дают начало безымянной вене (v. anonima). Интересно, что некото-
рые авторы отмечают отсутствие клапанного аппарата в этих венах (Mu-
rie, 1872а).
Таким образом, для венозных магистралей ламантина, как и для мно-
гих ныряющих млекопитающих, характерно расширение отдельных
участков венозных сосудов и образование особых синусов, что увеличи-
вает общую емкость венозного русла. Локализация их, однако, сущест-
венным образом отличается от локализации подобных образований у
других водных и полуводных видов. Увеличение емкости венозного русла
может происходить и за счет обширных венозных сплетений, которые
у ламантина наиболее значительны в тазовой и поясничной областях.
Одним из интереснейших и своеобразных сосудистых образований ла-
мантина являются так называемые чудесные сети (rete mirabile). Они
представляют собой скопление густопереплетенных, плотноупакованных
и сильноизвитых сосудов равного или различного калибра. Подобные со-
судистые образования встречаются еще только у китообразных (Howell,
1930; Slijper, 1936, 1962; Яблоков и др., 1972; Агарков и др., 1974; Ива-
нова, 1975; Галанцев, 1977; Мельников, 1981; и мн. др.). Чудесные сети
могут быть артериальными и артерио-венозными. У ламантина нами об-
наружены участки только артерио-венозных, т. е. смешанных чудесных
сетей. Эти сосудистые участки обычно локализованы в различных облас-
333
5см
5 см
Рис. 129. Рентгенограмма
артериальной части цент-
рального участка пакета
и участка артерио-ве-
нозной (б) чудесной сети
1 — магистральные венозные
сосуды;
с — магистральный артериаль-
ный сосуд
тях грудной полости ламантина. Как и у дельфинов, у ламантина хорошо
развита система межреберной чудесной сети, артериальная кровь в кото-
рую поступает непосредственно из аорты (по одному сосуду) и выносит-
ся по двум венам во внутрипозвоночную сосудистую сеть (рис. 128).
Хорошо выражены такие же сосудистые пучки в подмышечной области
ламантина; мощная чудесная сеть лежит и под на. rectus abdominis и в
других областях.
Методом последовательной рентгенографии было установлено, что
единственный артериальный ствол этих сосудистых пучков отдает от себя
густоветвящиеся ветви, образующие плотное строго ограниченное артери-
альное сплетение (рис. 129, а). Артериальный ствол сопровождают два
параллельных выносящих венозных сосуда, в которые собирается кровь
из густопереплетенных, мелких, венозных сосудов. В результате перепле-
тения артериальных и венозных стволов и образуется сосудистый плот-
ноупакованный пакет (рис. 129, б). Чудесные сети не имеют анастомозов
с сосудами окружающих тканей, от которых они отделены соединитель-
нотканной оболочкой. Основные артериальные и венозные сосуды этих
сетей тянутся на значительные расстояния, практически не изменяя
своего просвета, лишь в терминальной части сосудистого пучка они пере-
ходят в мощную сеть мелких артерий и вен.
По поводу функциональной роли чудесных сетей было высказано мно-
го предложений, однако до сих пор окончательного экспериментального
доказательства значения их не сделано. Имеются лишь данные об уча-
стии этих сосудистых образований в кровоснабжении головного мозга
китообразных и материалы, свидетельствующие о сглаживании чудесной
сетью пульсовых волн (McFarland et al., 1966; Nagel et al., 1968; Rid-
gway, 1972; Мельников, 1981).
334
1 — аорта;
2 — расширение аорты;
3 — нисходящая часть дуги
аорты;
4 — безымянная артерия;
5 — левая сонная артерия;
6 — левая подключичная артерия;
7 — легочная артерия;
8 — расширение легочной арте-
рии;
9 — левый и правый стволы ле-
гочной артерии;
10 — артериальная связка
Рис. 130. Схема бульбообразного расширения восходящей части дуги аорты (а) и
начального участка легочной артерии (б)
Своеобразное устройство смешанной чудесной сети ламантина позво-
ляет предположить, что она создает условия в период брадикардии и сни-
жения периферического кровотока под водой для поступления из нее ве-
нозной крови, не отдавшей кислород окружающим тканям обратно в си-
стему общего кровотока. Не исключена, естественно, и демпфирующая
роль чудесных сетей. Во всяком случае, поскольку чудесные сети прису-
щи только высокоспециализированным водным млекопитающим — кито-
образным и сиренам, ясно, что они имеют весьма определенное значение
в общем комплексе адаптационных механизмов и окончательное выясне-
ние функциональной значимости их имеет, безусловно, первостепенное
значение.
В венозной системе малого круга кровообращения ламантина коллек-
торообразных образований не имеется. Слияние двух легочных вен (vv.
pulmonales) происходит практически у границы левого предсердия, и про-
тяженность общего ствола легочной вены не превышает 10—12 мм. Из
других сосудистых преобразований необходимо указать на развитие мощ-
ного бульбообразного расширения восходящей дуги аорты (рис. 130, а).
Начальный участок восходящей аорты у сердца имеет диаметр 28,1 мм,
а затем просвет его резко увеличивается до 60,5 мм в диаметре. Нисхо-
дящий участок дуги аорты уменьшается в диаметре до 34,2 мм. От вер-
хушки дуги аорты отходят последовательно три сосуда — безымянная ар-
терия (a. anonima), левая сонная (a. carotis sinistra) и левая подключич-
ная (a. subclavia sinistra).
Необходимо подчеркнуть, что подобное бульбообразное расширение
аорты обнаружено у многих видов ныряющих млекопитающих из различ-
ных отрядов (Иванова, 1975; Elsner, 1976; Галанцев, 1977; Мельников,
1981; и мн. др.). Конвергентное сходство в изменении восходящей час-
ти дуги аорты у многих водных млекопитающих различной систематиче-
ской принадлежности указывает на общность функциональной значимо-
сти этого образования для животных, объединяемых известной общ-
ностью условий существования и образа жизни. Несомненно, что у всех
ныряющих животных это расширение дуги аорты служит для выполне-
ния одной и той же функциональной роли. Вероятно, бульбообразное
расширение аорты предназначено для дополнительного обеспечения под-
держания энергии диастолического давления в периоды брадикардии и
тахикардии, развивающейся в связи с нырянием за счет расширения его
стенок.
Следует также отметить, что подобное же расширение, начального
участка артериального русла обнаружено и в малом кругу кровообраще-
ния. Легочная артерия ламантина, начинающаяся одним стволом от ар-
335
териального конуса правого желудочка, сразу же бульбообразно расши-
ряется (до 75,8 мм в диаметре). Левая и правая легочные артерии для
каждого легкого (диаметром 30,2 мм) берут начало от этого объемистого
сосудистого расширения (рис. 130, б). Бульбообразное расширение легоч-
ной артерии соединяется с аортой артериальной связкой (lig. arteriosum),
являющейся облитерированным боталловым протоком (ductus Botalli).
Амортизирующая роль подобных образований при перенаполнении кровью
и резких изменениях величин сердечного выброса, по-видимому, вполне
очевидна. Локализуется это бульбообразное расширение легочной арте-
рии над продольной бороздой двухвершинного сердца между левым и
правым предсердиями (рис. 131).
Резюмируя все сказанное, можно заключить, что сердечно-сосудистая
система ламантина несет ряд хорошо выраженных морфологических из-
менений, имеющих несомненное приспособительное значение. Естествен-
но, что не все еще морфологические преобразования этой системы у ла-
Рис. 131. Рентгенограмма сердца ламантина
1—2 — желудочки (1 — правый, 2 — левый); 3—4 — предсердия (3 — правое, 4 — левое);
•> — аорта; 6 — бульбообразное расширение легочной артерии
336
мантина описаны, а функциональное значение многих из них еще требует
экспериментального физиологического обоснования. Тем не менее стано-
вится вполне очевидным, что в процессе приспособительной эволюции
адаптивные перестройки сосудистого русла этого вида во многом разви-
вались в общем для всех млекопитающих, осваивающих водную среду,
направлении, что, видимо, в значительной мере было детерминировано
известной общностью взаимодействия этих животных с факторами внеш-
ней среды. Достаточно указать на перестройку дуги аорты и легочной ар-
терии, развитие чудесных сетей, венозных сплетений и коллекторов, ко-
торые в той или иной мере развиты у различных видов водных и полу-
водных млекопитающих. Интенсивность выраженности этих сосудистых
образований во многом зависит от уровня специализации животных, а чу-
десные сети представлены только у исключительно водных форм — кито-
образных и сиреновых. Естественно, что развитие венозных коллекторов
и других сосудистых образований у ламантина отличается по ряду дета-
лей от других ныряющих млекопитающих, в частности, по месту их лока-
лизации и форме. Несмотря на известные различия в форме, размерах и
локализации венозных коллекторов у разных видов полуводных и водных
млекопитающих, что, видимо, связано с их генотипическими особенностя-
ми, этим сосудистым образованиям, несомненно, присуща единая функ-
циональная роль — депонирование крови в период гемодинамических пе-
рестроек, вызванных нырянием.
Глава девятая
АНАТОМИЯ ПОЧКИ ЛАМАНТИНА
Морские млекопитающие в отличие от остальных морских позвоночных
не нуждаются в экстраренальных органах осморегуляции. Морские кости-
стые рыбы, морские рептилии и птицы для получения пресной воды пьют
морскую воду и (поскольку почка их не способна выводить избытки со-
лей) опресняют ее жабрами либо солевыми железами. Морские млекопи-
тающие не пьют морскую воду (Pilson, 1970), однако многие из них пи-
таются морскими беспозвоночными либо растениями, которые находятся
в осмотическом равновесии с морской водой. При этом единственным ор-
ганом осморегуляции у морских млекопитающих являются почки, кото-
рые выводят избыток солей из организма и обеспечивают у этих живот-
ных продолжительный водный баланс. Однако при этом у различных
отрядов морских млекопитающих (киты, тюлени и сирены) в анатомиче-
ском строении почек наблюдаются существенные различия. У китов поч-
ки дольчатые (Sperber, 1942; Cave, Aumonier, 1912). Количество долек у
дельфинов составляет несколько сотен, у крупных усатых китов 1,5—
3 тысячи (Яблоков, 1961). Каждая долька представляет собой отдельную-
почку («почечку»), расположенную вместе с другими на общем стволе —
мочеточнике наподобие виноградной грозди (Oliver, 1968). Почки тюле-
ней также дольчатые, однако разделение на дольки поверхностное и не-
проходит в глубь коркового слоя почки. Мочеточник внутри почки раз-
деляется сначала на два отдела — краниальный и каудальный, которые-
в свою очередь многократно ветвятся, образуя узкие протоки, заканчи-
вающиеся небольшой чашечкой, в которую опускаются сосочки почки;
(Gerhardt, 1914). При этом количество сосочков может не совпадать с ко-
личеством поверхностных почечных долек (Шуров, 1975). Почки дюгоней
в отличие от китообразных и тюленей с наружной стороны гладкие, раз-
деление на дольки у них полностью отсутствует, они сильно вытянуты в
кранио-каудальном направлении. Почечный сосочек один, слегка высту-
пает в полость лоханки, которая тянется двумя протоками к обоим кон-
цам почки (Gerhardt, 1914; Petit, 1924b, 1925). Общий вид и разрез поч-
ки дюгоня изображен у Герхардта (Gerhardt, 1914).
В своем описании морской коровы Стеллер (Steller, 1751) указывает
на то, что почки у нее дольчатые, однако он не приводит данных по ко-
личеству и размерам долек и не дает какого-либо изображения.
Некоторые разногласия в литературе имеются по анатомии почек ла-
мантина, которые касаются в основном разделения почки на дольки
(Murie, 1872а; Chapman, 1875; Beddard, 1897). По данным этих авто-
ров, у различных особей ламантина почки могут иметь либо гладкую на-
ружную поверхность, либо четко выраженное разделение на доли. Наи-
более полное исследование почек ламантина провел Ж. Пти (Petit,
1924а, Ь, 1925). Им описаны почки как взрослых особей, так и эмбрио-
нов Trichechus senegalensis. Почки эмбриона и молодого ламантина были
13 и 125 мм длины соответственно и имели гладкую наружную поверх-
ность. У взрослых особей почки, достигающие около 230 мм длины, име-
ют четко выраженное наружное деление на доли. Описывается определен-
ная закономерность увеличения степени дольчатости почки в зависимости
от возраста животного. Однако при этом нет указаний на то, с какого-
338
возраста животного или с каких размеров почки появляется дольчатость.
Предыдущие авторы описывали лишь внешнее строение почек. Крайне
схематичное изображение приведено лишь у Дж. Спербера (Sperberr
1942). Описывая срезы почки, Пти (Petit, 1924а) указывает, что у моло-
дых особей и эмбрионов ламантина наблюдается внутреннее разделение
почки на доли. Таким образом, принципиальных изменений в строении
почек ламантина в процессе онтогенеза не происходит. Пти (Petit, 1924b,
1925) также проводит сравнение строения почек ламантина, дюгоня и
морской коровы, причем данные по последней взяты у Стеллера (Steller,
1751). Почки дюгоня как по внешним анатомическим признакам, так и
по внутреннему строению существенно отличаются от почек ламантина,
будучи как внешне, так и изнутри не поделенными на доли.
В нашем распоряжении имелась правая почка взрослого самца ла-
мантина (см. Введение). Вес почки 1050 г, длина 226, ширина 143, тол-
щина 64 мм. Поверхность почки гладкая, разделяется неглубокими бо-
роздами на шесть долей (рис. 132, а). В наиболее крупную краниаль-
ную долю впадает отдельный кровеносный сосуд, возможно связанный
с почечной артерией, и в нее же открывается проток лежащего отдельно
от почки надпочечника (рис. 132, б). Перед впадением в почку этот
проток раздваивается. Каждая доля представляет собой как бы отдель-
ную почку, окруженную корковым веществом, внутри которого находятся
наружная и внутренние зоны мозгового вещества почки, и имеющую не-
посредственный выход в почечную лоханку (рис. 132, в), и снабжающую-
ся веточками артерий, отходящих от почечной артерии (рис. 133). По-
чечная артерия входит в почку в воротах и сразу же разветвляется на
четыре крупных ствола, снабжающих кровью центральный и каудальные
отделы почки (см. рис. 133). Наиболее крупная краниальная доля, веро-
ятно, снабжается артериальной кровью отдельно (рис. 132, б) через со-
суд, видимо, отделяющийся от общего ствола почечной артерии до ее
вхождения в почку. Во всяком случае, при заливке общего ствола почеч-
ной артерии рентгенконтрастное вещество не попадает в краниальную
долю. В каждой из почечных долей соотношение — кора почки : наруж-
ная зона мозгового вещества: внутренняя зона мозгового вещества —
составляет 0,6 : 1,6 : 0,8. Подобные соотношения различных зон почки
наблюдаются также у китов, дельфинов и тюленей. У них также мозго-
вое вещество почки толще, чем корковое (Sperber, 1942). Однако в отли-
чие от них у ламантина очень слабо развиты сосочки. В то же время у
дюгоня, как отмечено выше, имеется один сосочек, но, судя по его по-
ложению (см. рис. в Gerhardt, 1914), можно предположить, что нефроны
с длинными петлями Генле имеются здесь только в центральной части
почки, что необычно для почек млекопитающих. Сведения о соотношении
различных зон почек у дюгоней в литературе отсутствуют.
Поскольку почка была уже фиксирована в формалине, нам не удалось
провести полного микродиссекционного исследования структуры нефро-
на. Тем не менее можно установить, что клубочки в почке ламантина
располагаются равномерно по всему слою коркового вещества. Диаметр
клубочков довольно стабильный 0,265±0,005 мм (га = 20) и не зависит от
положения их в толще коркового слоя. По месторасположению клубоч-
ков нефроны не удается разделить на суперфициальные, интракортикаль-
ные и юкстамедулярные. Следующий за клубочком проксимальный изви-
той каналец образует многочисленные петли. Почти у всех нефронов
одна из петель проксимального извитого канальца доходит до поверхно-
сти почки. Лишь у наиболее глубоких нефронов, клубочки которых рас-
полагаются ближе всего к границе мозгового вещества, петли прокси-
мального извитого канальца не доходят до поверхности почки. Удалось
339
выделить полностью один проксимальный извитой каналец, клубочек ко-
торого находился приблизительно по середине коркового слоя. Длина
этого отдела нефрона составляла 15,3 мм. За проксимальным извитым ка-
нальцем следует проксимальный прямой отдел нефрона, спускающийся в
мозговой слой почки и составляющий нисходящий участок петли Генле.
Далее наблюдаются различия в строении у отдельных нефронов, две
крайние стадии которых выражаются в следующем: у нефронов с наибо-
лее коротким проксимальным прямым отделом отсутствует тонкий отдел
петли Генле, а у нефронов с наиболее длинным проксимальным прямым
отделом (длина его приблизительно в полтора раза больше, чем у преды-
дущих) имеется длинный тонкий отдел петли Генле.
Соответственно выявляется определенная закономерность между дли-
ной проксимального прямого отдела и тонким сегментом петли Генле,
Рис. 132. Почка ламантина
а — латеральная сторона;
1 — краниальная доля;
2 — 5 — почечные доли;
в — ворота почки;
7 — почечная артерия;
8 — почечная вена;
— медиальная сторона; в — рг
9 — артерия, снабжающая
краниальную долю;
10 — 11— надпочечник
(11 — его проток);
12 — кора почки;
13 — 14 — мозговое вещество
(13 — наружное.
14 — внутреннее);
15 — почечная лоханка;
16 — выводные протоки от
почечных долей
Рис. 133. Рентгенограмма почечных артерий в почке ламантина
1 — почечная артерия; 2—з — краниальная доля почки (з — снабжающий ее кровью сосуд).
340
однако нам не удалось обнаружить нефроны, у которых имеется тонкий
восходящий участок петли Генле. Дальнейшую структуру нефрона про-
следить не удалось. При фиксации почки хорошо сохранились заполнен-
ные кровью капилляры мозгового слоя почки. Располагаются они строго
параллельно почечным канальцам. Такая структура мозгового слоя почки
характерна для животных, обитающих в условиях недостатка пресной
воды (Гетманова, 1976; Rouffignac, 1981), т. е. у пустынных млекопитаю-
щих. Физиологически сходные условия функционирования почек должны
наблюдаться у морских млекопитающих, поскольку они живут в услови-
ях дефицита пресной воды. К сожалению, однако, данных по микроана-
томии почек китообразных и ластоногих в литературе нет. У ламантина
же, помимо сходного с пустынными млекопитающими соотношения корко-
вой и мозговой зон почки и расположения кровеносных сосудов в мозго-
вом веществе, наблюдаются отличия, выраженные в слабом развитии со-
сочков, отсутствии восходящего участка тонкого отдела петли Генле.
В то же время у них имеются как нефроны вообще без тонкого отдела
петли Генле, что считается характерным для млекопитающих, обитаю-
щих в пресных водах, так и нефроны с длинным тонким нисходящим
отделом. Такое противоречивое микроанатомическое строение почки ла-
мантина, вероятно, связано с возможностью его обитать не только в мор-
ской, но и в пресной воде.
Таким образом, следует отметить, что почки ламантина по внешним
анатомическим признакам резко отличаются от остальных морских мле-
копитающих и наибольшее сходство у них наблюдается с почками слонов
(Sperber, 1942). Однако, если сравнивать соотношение зоны коркового и
мозгового вещества, можно заметить, что здесь, наоборот, имеется сход-
ство как с остальными морскими, так и с пустынными млекопитающими,
у которых это соотношение составляет приблизительно 1 : 2 или 1 : 3,
тогда как у африканского слона оно составляет приблизительно 1 : 1
(Sperber, 1942). Таким образом, соотношение коркового и мозгового ве-
щества в первую очередь отражает приспособление к жизни в условиях
дефицита пресной воды (у всех морских и пустынных млекопитающих
мозговое вещество почки толще коркового), тогда как внешнее строение
почки больше говорит о генетическом родстве сирен с хоботными.
Глава десятая
ОБ ОРГАНЕ ЗРЕНИЯ ЛАМАНТИНА
И ДРУГИХ СИРЕН
В последние годы возрос интерес к биологии сирен, их особенностям пи-
тания и поведения. По ряду биологических особенностей все сирены пред-
ставляют «собой в высшей степени резко характеризованную группу»
(Гептнер, 1967, с. 22). Они — узко специализированные настоящие гид-
робионты, связанные с мелководьями прибрежной полосы моря, лагуна-
ми, эстуариями, устьями рек с богатой растительностью; в пелагиали не
держатся (Anderson, 1981а). Характер их питания указан в гл. 1 и 7.
При этом они пасутся не только на водных растениях, но и иногда за-
хватывают наземную растительность, свисающую над водой, высовывая
для этого голову (Best, 1981; Reynolds, 1981). При максимуме пищевой
активности в утренние и вечерние часы строгой суточной ритмики у си-
рен все же не отмечено (Hartman, 1979; Montgomery et al., 1981). Можно
говорить, следовательно, о полифазном типе и круглосуточной форме ак-
тивности (Соколов, Кузнецов, 1978) сирен. Так, в неволе можно легко
изменять форму активности с суточной на дневную и наоборот, приручая
животных брать траву из рук (Томилин, 1977).
Сирены практически не имеют врагов, исключая человека, хотя и
могут стать случайной добычей акул, крокодилов и косаток. Это — от-
носительно медленно плавающие (обычная скорость 2—3 км/ч) кажущие-
ся неуклюжими животные, погружающиеся на глубину до 20 м (Ander-
son, 1981а). При обилии пищи, минимальном давлении со стороны
окружающей среды и отсутствии врагов социальная организация этих жи-
вотных находится на «рудиментарной стадии» (Hartman, 1979). Тем не
менее у сирен хорошо развиты слух и звуковая коммуникация, тактиль-
ное и термическое восприятие; сведения о хеморецепции очень скудны и
противоречивы (Томилин, 1977; Ronald et al., 1978).
Орган зрения и вспомогательные части глаза сирен недостаточно хо-
рошо изучены, несмотря на старые данные (Putter, 1903; Walls, 1942;
Rochon-Duvigneaud, 1943) и работы, появившиеся совсем недавно (Ronald
et al., 1978; Андреев, 1982; Cohen et al., 1982). Остается открытым воп-
рос о роли зрения в пространственно-коммуникативном анализе. Боль-
шинство выше перечисленных авторов склоняются к мысли о незначи-
тельной роли зрительного анализатора в этом процессе, основываясь как
на морфологических данных, так и на экологических и поведенческих.
Отмечено (Ronald et al., 1978), что глазное яблоко невелико, оптический
нерв тонок, латеральное коленчатое тело и передние бугры четверохол-
мия очень малы по сравнению с другими морскими млекопитающими.
Г. Уолс (Walls, 1942), оценивая орган зрения сирен, отмечает, что по
сравнению с ластоногими и китообразными глаза этих животных пло-
хо приспособлены к зрению под водой. Он связывает это с характером
питания и особенностями локомоции. Однако Д. Хартман (Hartman,
1979), изучая экологию и поведение ламантинов во Флориде, приходит
к выводу о «широком использовании зрения» (с. 139). Таким образом,
разница во мнениях о роли зрения дает повод для дальнейших исследо-
ваний глаза этих животных.
Бспомогательные органы морских млекопитающих, в том числе си-
342
рен, имеют ряд специфических свойств. На наружной поверхности век
большинства млекопитающих имеются ресницы. У китов, ластоногих и
сирен они отсутствуют. Свободный край века у млекопитающих обычно
снабжен выводными протоками тарзальных, или мейбомовых желез. Они
являются модифицированными сальными железами, выделяющими глаз-
ную смазку. Однако у морских животных, обитающих в пресных и соле-
ных водоемах, тарзальные железы отсутствуют. Помимо исчезновения
этого типа желез, нет и слезных. Компенсацией тому, видимо, является
сильное развитие у этих млекопитающих гардеровой железы. У ламанти-
на она окружает большую часть глазного яблока с вентральной, меди-
альной и дорсальной сторон. По своим свойствам секрет гардеровой желе-
зы — белковая слизь, коэффициент преломления которой равен таковому
воды. Это качество секрета позволяет животным видеть предметы без
искажений в водной среде. Слизь из железы поступает в медиальный угол
глаза, заполняет конъюнктивальный мешок, покрывает роговицу. Защит-
ная и увлажняющая роль секрета этой железы очевидна и вполне ясна.
Причем выделение его очень обильно у животных, извлеченных из воды
на сушу. Отсюда, как отмечает В. Г. Гептнер (1967), легенда о «плачу-
щих сиренах» и само название ламантин (от французского lamanter —
плакать). Такая особенность «плача» сирен, дельфинов, китов и тюленей
связана со строением черепа. У наземных млекопитающих секрет желе-
зы поступает в слезно-носовой канал. Однако у морских млекопитающих
этот канал отсутствует даже в эмбриональном состоянии (Walls, 1942),
поэтому излишки секрета обильно выводятся наружу через глазную
щель.
Внутренняя поверхность века покрыта сильно васкуляризованной
конъюнктивой. Ее склеральный отдел образован многослойным плоским
эпителием, продолжающимся до границы лимба (место соединения скле-
ры и роговицы). Здесь конъюнктива покрыта слоем кератина, который
переходит на роговицу. Столь характерный для большинства млекопи-
тающих хрящевой слой в веках полностью утрачен у сирен и китов и
слабо выражен у ластоногих. Поэтому основную массу внутренней (глу-
бинной) части век составляют соединительная, жировая и мышечная тка-
ни. Из мышц особенно развит круговой мускул век. Он принимает ак-
тивное участие в акте сжимания век, способствует защите глазного ябло-
ка от механических повреждений. Палпебральная часть мускульного
слоя, осуществляющая мигание, развита гораздо хуже.
Глазодвигательные мышцы ламантина в числе семь (4 прямые, 2 ко-
сые и протрактор глазного яблока) развиты вполне удовлетворительно.
Обращает внимание мощное развитие оттягивателя глаза, который обес-
печивает передвижение глазного яблока вдоль зрительной оси. Работа
всей Глазодвигательной мускулатуры изменяет пространственное направ-
ление зрительных осей, что, видимо, позволяет животным существенно
увеличивать размеры угла общего поля зрения.
Глаза ламантина и дюгоня расположены по бокам головы, а зритель-
ные оси направлены вперед и вниз. Такое положение глаз на голове,
характерное для китообразных, слонов, некоторых копытных и мозоле-
ногих, позволяет животным видеть объекты, находящиеся не только сбо-
ку, но впереди и под ними. Вероятно, сирены могут рассматривать пред-
меты бинокулярно. Так отмечается, что ламантины, приближаясь к объ-
екту не двигают, не сканируют головой (Hartman, 1979). Полагают, что
у ламантина перекрываются монокулярные поля, т. е. присутствует угол
бинокулярного поля зрения, размер которого равен 15° (Ronald et al.,
1978). В целом же для сирен характерно панорамное зрение при незна-
чительной роли бинокулярного восприятия.
343
Таблица 15. Размеры глазного яблока и хрусталика (в мм)
Вид Диаметр глаза Ось глаза Хрусталик Источник
горизон- тальный верти- кальный диа- метр ОСЬ
Trichechus manatus 19 18,5 19 6,5 4 Наши данные
Trichechus senegalensis I 10,5 — 8,4 — — Putter, 1903
« II 15 — 16 — — (Rochon—
Dugong dugon I 22,5 — 21 7 4 JDuvigneaud, 1943
« II 25 25 23 6,9 4,4 Walls, 1942
Глазное яблоко сирен не велико; оно мельче, чем у таких маленьких
китообразных, как обыкновенный дельфин (30 мм) (табл. 15).
Г. Стеллер указывает, что «глаза (морской коровы, капустника) ле-
жат точно посредине между концом хобота и ушами, на одинаковой вы-
соте с верхней частью носа, но лишь еще немного выше; относительно
столь крупного тела они чрезвычайно малы и не больше овечьих» (цит.
по: Гептнер 1967, с. 32), т. е. около 30 мм. По данным табл. 1 глаза
сирен относятся к сферическому типу. У этих животных нет уплощенно-
сти глазного яблока, которая очень характерна для ластоногих и кито-
образных (Андреев, 1978а). То же относится к роговице, которая у ла-
мантина довольно выпуклая, в противоположность плоской роговице всех
водных позвоночных (рыб, пингвинов, морских млекопитающих). Наруж-
ный радиус кривизны роговицы больше внутреннего, поскольку толщина
ее на периферии (0,9 мм) больше таковой в центре (0,6 мм). Форма
роговицы у ламантина — горизонтальный овал (drop 7,7 мм, dIi(.PT 6 мм),
у дюгоня ее размеры равны 11X7,5 мм. Растр не очень велик (110° по
горизонтали и 80° по вертикали), что свойственно животным с дневной
или круглосуточной формой активности. Толщина склеры везде одинако-
ва (0,95 мм), чудесной сети нет, тогда как обычно у морских млекопи-
тающих (особенно у китов и дельфинов) в районе заднего полюса глаз-
ного яблока имеется значительное утолщение склеры, в котором распо-
ложена rete mirabile.
Сосудистая оболочка ламантина тонкая даже в районе заднего полю-
са (0,2 мм). Данные о тапетуме (отражательной оболочке) у сирен
противоречивы. Одни отрицают его присутствие у дюгоня и ламантинов
(Walls, 1942), другие (Rochon-Duvigneaud, 1943) отмечают его у дюго-
ня, но отрицают у бразильского ламантина. У Trichechus manatus тапе-
тум в темноте отбрасывает свет, и глаза этого животного блестят розо-
вым цветом (Hartman, 1979). Нам удалось обнаружить на гистологиче-
ских препаратах клетки тапетума у ламантина (рис. 134). Они располо-
жены в центральной части глазного дна и занимают незначительную по
размерам площадь. Клетки лежат между слоем сосудов крупного и мел-
кого калибра и хориокапиллярным слоем. Отражательная оболочка гла-
за ламантина развита плохо, поскольку образована всего 4—5 слоями
волокнистых клеток. В то же время у китообразных и ластоногих на-
считывают до 35 слоев. Видимо, по толщине тапетума можно судить о
его отражательной способности.
Ресничное тело глаза ламантина развито не очень хорошо, составляя
только пятую часть длины оси глаза. Ресничные отростки невысокие
(0,5 мм), булавовидной формы. Между основными отростками имеются
промежуточные и небольшое количество маленьких добавочных. Цин-
новы связки образуют слабо выраженный перекрест. Довольно рыхлый
344
Рис. 134. Участок попе-
речного среза сосуди-
стой оболочки п тапету-
ма
1 — слой мелких сосудов и
хориоидальный пигмент
2 — слой клеток отражатель-
ной оболочки:
3 — клетки пигментного
эпителия сетчатки
трабекулярный остов занимает незначительное пространство между рес-
личным телом и склерой. Имеется мускулатура ресничного тела, нали-
чие которой отрицал А. Рошон-Дювиньо (Rochon-Duvigneaud, 1943). Од-
нако она развита слабо, поскольку не образует единой мышечной мас-
сы: между отдельными тяжами находятся соединительнотканные
прослойки с большим количеством хроматофоров. Указанные особенности
строения ресничного тела дают основание полагать, что в глазе ламан-
тина может осуществляться аккомодация при помощи изменения кри-
визны хрусталика.
Радужина темно-коричневого цвета. Ее передняя поверхность, обра-
щенная к роговице, гладкая, без характерного для радужины млекопи-
тающих рисунка, который бывает образован циркулярными и радиаль-
ными сосудами. Корень радужины, ее часть, примыкающая к ресничной
зоне, толще зрачкового края. Сфинктер и дилятатор радужины хорошо
развиты, поскольку организованы в единые мышечные тяжи; между от-
дельными волокнами соединительной ткани мало или нет совсем. У ла-
мантина и дюгоня зрачок круглый (Rocho-Duvigneaud, 1943). По данным
Г. Уолса (Walls, 1942, с. 410), «...зрачок большой, быстро меняющий
размеры. Он горизонтальноовальный у дюгоня и круглый у ламантина.
У живых ламантинов зрачок круглый, а у мертвых — горизонтально-
овальный». На нашем морфологическом материале зрачок ламантина был
круглый. Важной особенностью радужины ламантина является ее асим-
метрия, при которой назовентральная часть уже дорсотемпоральной на
1 мм. При этом центр зрачка смещен немного вперед и вниз. Подобным
свойством обладает радужина хоботных и некоторых копытных. Асим-
метрия зрачка способствует увеличению размеров угла монокулярного
поля зрения в его назовентральном секторе, что, в свою очередь, увели-
чивает перекрывание полей зрения правого и левого глаза.
Особенностью хрусталика ламантина является желтоватая окраска,
которая играет роль цветового фильтра. «Как и другие желтые фильтры
(желтый хрусталик у миноги, змей, сусликов и обезьян, желтая рого-
вица у щуки и окуня), они могут служить для повышения остроты зре-
ния за счет уменьшения хроматической аберрации и снятия голубой
дымки, возникающей в связи с рассеянием света в мутных средах»
(Орлов, 1972, с. 86). Целесообразность приобретения желтой окраски
хрусталика ламантина очевидна в связи с обитанием в довольно мутной
воде. Хрусталик сирен относительно глазного яблока невелик и сравни-
тельно с другими морскими млекопитающими уплощен. У ластоногих и
китообразных отношение оси хрусталика к диаметру близко к единице
12 Ламантин 34ц
(0,85—0,98). Индекс уплощения хрусталика сирен (см. табл. 1) колеб-
лется от 0,57 до 0,63, что характерно для дневных наземных млекопи-
тающих. Шаровидная форма хрусталика свойственна многим водным
позвоночным: рыбам, личинкам бесхвостых и хвостатым амфибиям, китам,
тюленям: т. е. такая форма — важнейшая адаптивная черта оптической
системы, необходимая для восприятия в водной среде. А исходя из ана-
томо-оптической характеристики системы «роговица—хрусталик», следу-
ет полагать, что светопроводящие структуры глаза сирен плохо приспо-
соблены к зрению в воде. Причем орган зрения этих животных в водной
среде характеризуется гиперметропией, а в воздушной — миопией. Это
подтверждено экспериментальными данными. При воздействии атропина,
парализующего ресничную мышцу, у ламантина выявили очень не-
значительный объем аккомодации и некоторую гиперметропию (Ronald
et al., 1978). У дюгоня определили гиперметропию в воде и наличие
миопии в воздухе, которая равнялась 5Д (Walls, 1942).
Изучение световоспринимающей системы глаза ламантина показало
следующее. Сетчатка, выстилая дно глазного яблока, занимает большую
часть сферы, примерно 265°. Толщина ее невелика в центре (150 мкм)
и на периферии (100 мкм). В сетчатке четко выделяются все слоит
(рис. 135, а). Самым широким является слой фоторецепторов и их пери-
Рис. 135. Поперечный срез сетчат-
ки в центре глазного дна (а, б) при
разном увеличении и на перифе-
рии (в)
1 — отростки фоторецепторов
(1а — палочковые, 16 — колбочковые);
2 — наружный ядерный слой
(2в — перикарионы палочек,
2г — перикарионы колбочек);
з — наружный плексиформенный слой;
4 — внутренний ядерный слой;
5 — внутренний плексиформенный слой;
6 — ганглиозный слой
(6д — гигантские ганглиозны е клетки,
бе'— ганглиозные клетки)
346
карионов, а самым узким — внутренний ядерный. Пигментный эпителий
сетчатки интимно граничит со стекловидной пластинкой сосудистой обо-
лочки. Он представлен однородными уплощенными шестигранными приз-
матическими клетками. Гранулы темно-коричневого пигмента располо-
жены по внутренним граням, т. е. граням, примыкающим к наружным
членикам фоторецепторов. Однако в том районе дна глазного яблока,
где расположен тапетум, в эпителии пигмент отсутствует. Такая законо-
мерность в региональном расположении тапетума и пигмента в пигмент-
ном эпителии сетчатки характерна для других млекопитающих (Андреев,
1973-1979).
В фоторецепторном слое можно выделить два типа световоспринима-
ющих клеток (рис. 135, б). Обращает внимание форма сегментов фото-
рецепторных отростков при изучении их в световом микроскопе. Обычно
у млекопитающих внутренний членик рецептора, содержащий эллипсоид
или миоид, несколько толще или равен наружному. У ламантина же внут-
ренний членик диаметром всего 1,2 мкм, а наружный — 2,5 мкм. Длина
такой фоторецепторной клетки, которая, вероятнее всего, является па-
лочкой, равняется 19 мкм. Следует заметить, что эти клетки сетчатки
глаза ламантина сходны по форме с «зелеными» палочками бесхвостых
амфибий. Так же как у палочковых фоторецепторов, наружный сегмент
колбочек толще (4,9 мкм) внутреннего (2 мкм). Колбочки имеют
длинные внутренние и короткие наружные сегменты. Длина фоторецеп-
торов этого типа — 13 мкм. Исследование фоторецепторов сетчатки глаза
ламантина (Cohen et al., 1982), проведенное с помощью световой и элект-
ронной микроскопии, также указывает на наличие палочковых и колбоч-
ковых рецепторов. Оно определило, что у одного животного в дорсотем-
поральной области сетчатки отношение палочек и колбочек составляет
2,81, а в вентральной — 3,35, у второго животного соответственно —
2,88 и 2,55. Таким образом, можно говорить об относительно высоком
содержании колбочек в сетчатке глаза ламантина. Однако специфической,
региональной концентрации колбочек в каком-либо районе сетчатки не
имеется. Лишь на крайней периферии они полностью отсутствуют.
В соответствии с различием отростков фоторецепторов в наружном
ядерном слое удается выявить два размерных варианта перикарионов:
одни почти круглые (2,5X3 мкм), другие овальные, более крупные
(3X5 мкм). Последние, по нашим подсчетам, составляют около 30% от
общего числа перикарионов наружного ядерного слоя. Хроматиновая
сеть перикарионов первого типа характеризуется интенсивной базофили-
ей и высокой концентрацией хроматина. У клеток второго типа концен-
трация хроматина в ядре сравнительно невелика; на препаратах, окра-
шенных по методике Гендейгайна, они выглядят значительно бледнее.
Причем перикарионы этого типа могут располагаться в любом месте в
пределах наружного ядерного слоя. Указанные особенности строения
фоторецепторов и их перикарионов дают право говорить о наличии двух
типов фоторецепторов.
Плотность распределения фоторецепторов очень низка как в центре
(76 тыс/мм2), так и на периферии (58 тыс/мм2); перикарионы располо-
жены всего в 3—4 ряда. Наружный плексиформенный слой узок, с боль-
шим количеством смещенных элементов наружного и внутреннего ядер-
ных слоев. Внутренний ядерный слой тоже узкий; он образован всего
одним-двумя рядами клеток. Плотность распределения элементов этого
слоя также крайне низка (18,6 тыс/мм2 в центре и 13,5 тыс/мм2 на пе-
риферии). Внутренний плексиформенный слой, образованный аксонами
и дентритами биполярных, амакриновых и ганглиозных клеток, в три
раза шире (30 мкм) наружного плексиформенного и очень хорошо выде-
347
12*
Рис. 136. Участок тан-
генциального среза сет
чаткп в слое ганглиоз-
ных клеток
Обозначения, как на
рис. 135
ляется. В нем изредка встречаются смещенные элементы из внутреннего'
ядерного слоя и ганглиозные клетки (рис. 135, в).
Ганглиозный слой во всех районах сетчатки ламантина на попереч-
ных срезах состоит из одного ряда клеток. Так же как у дельфинов и
ластоногих, ганглиозные клетки в сетчатке ламантина отделены значи-
тельными межнейронными промежутками, причем группы, состоящие из
2—4 клеток, не встречаются. Плотность распределения ганглиозных кле-
ток мала как в центральных (500 шт/мм2), так и периферических
(170 шт/мм2) районах сетчатки. В центральной части сетчатки несколько'
вентральнее места выхода зрительного нерва была обнаружена область
с повышенной плотностью ганглиозных клеток небольшого (10—15 мкм)
размера, в которой насчитывается 11—14 клеток на 100 мкм радиального
среза сетчатки (Cohen et al., 1982). Помимо мелких клеток, в ганглиоз-
ном слое присутствуют гигантские ганглиозные клетки диаметром 35—
50 мкм (рис. 136). Эти клетки расположены не только в вентральной об-
ласти, как указывают Ж. Коэн с соавторами, но и в других районах.
Мнение этих авторов о наличии клеток диаметром 90 мкм, вероятно,
ошибочно, поскольку ширина сетчатки глаза ламантина всего 100—
150 мкм.
Количественные соотношения нейронов сетчатки и фоторецепторов по-
казывают, что в центре на одну ганглиозную клетку приходится около
40 клеток внутреннего ядерного слоя и 150 рецепторов, а на периферии
соответственно — 80 и 340. Данные по плотности распределения нейрон-
ных элементов и их отношениям указывают на то, что сетчатка глаза
ламантина очень бедна нервными элементами и фоторецепторами ПО'
сравнению с ластоногими и китообразными. Она не содержит специали-
зированных участков типа area и тем более fovea. Исходя из структур-
ных данных, которые косвенно позволяют судить о функциональных
возможностях, следует полагать, что орган зрения ламантина характери-
зуется малой остротой зрения (очень незначительно число ганглиозных
клеток) и невысокой светочувствительностью (невелика плотность фото-
рецепторов) . Помимо небольшой плотности элементов сетчатки, на ее сла-
бое развитие, несколько упрощенное строение и низкий уровень метабо-
лизма указывают: малое количество ретинальных сосудов, тонкость
внутренних сегментов фоторецепторов, слабое развитие сосудистой обо-
лочки.
На основании вышеизложенного попытаемся в целом охарактеризо-
вать орган зрения ламантина и других сирен. Отмеченные выше некото-
рые черты биологии этих животных и особенности строения глаза позво-
ляют предполагать крайне неэффективное использование зрения в про-
странственно-коммуникативном анализе и то, что орган зрения сирен не1
348
может быть ведущим анализатором. У ведущих относительно пассивный
образ жизни животных, не испытывающих пресс хищников, с низким
уровнем социальной организации, зрение обычно развито плохо.
Л. Хартман (Hartman, 1979), напротив, утверждает, что зрение ламанти-
на играет важную роль в биологии этого вида. Однако он не приводит
ни физиологических, ни морфологических данных в поддержку своей ги-
потезы. Напротив, описывая поведение животных в Кинг-Бэй, он указы-
вает на частые травмы головы о цементные блоки и другие объекты при
плавании в канале. По нашему мнению, плохая способность различать
предметы связана с дальнозоркостью животных, а не только со степенью
прозрачности воды. Высказывание Л. Хартмана о том, что у ламантинов
зрение ночью не слабее дневного, требует экспериментальной проверки.
Имеющиеся морфологические данные противоречат этому и свидетельст-
вуют о низкой световой чувствительности.
Несмотря на водный образ жизни и связанные с этим преобразова-
ния в других органах, ни в световоспринимающей, ни в светопередающей
системах глаза ламантинов и других сирен не удается обнаружить адап-
тивные черты, связанные со зрением в водной среде. У, водных животных
хрусталик имеет близкую к шару форму, а у сирен он в значительной
мере уплощен. Кривизна роговицы рыб, личинок амфибий, морских чере-
пах, крокодилов, пингвинов и морских млекопитающих очень мала, т. е.
роговица плоская, у сирен же она выпуклая, как у наземных животных.
Мы видим, что у первично и вторично водных позвоночных имеются ярко
выраженные черты специализации, диктуемые оптикой воды, которые у
сирен отсутствуют.
Световоспринимающая система, сетчатка морских млекопитающих
характеризуется, как правило, очень высокой морфологической степенью
суммации нейронных элементов сетчатки, которая определяет высокую
светочувствительность и небольшую остроту зрения. У ламантина и све-
точувствительность, и острота, вероятно, невелики. У морских млекопи-
тающих обычно наблюдается хорошее развитие сосудистой оболочки и
тапетума. И опять-таки этот отдел глаза у сирен развит значительно
хуже, чем у китообразных и ластоногих.
Орган зрения сирен представляет собой уникальный пример отсутст-
вия черт специализации к среде обитания. Даже у всех ластоногих,
проводящих в воде меньше времени, глубина адаптаций всех систем
глаза несравненно больше. Причем имеется; в виду именно орган зрения,
глазное яблоко. Во вспомогательных же органах глаза сирен наблюдают-
ся адаптивные к водной среде свойства, подобные таковым китообразных:
строение железистого аппарата (отсутствие мейбомовых желез, мощное
развитие гардеровой, редукция слезной железы), сильное развитие круго-
вого мускула век и протрактора глазного яблока и др. Причем степень
преобразований во вспомогательных органах у сирен глубже, чем у лас-
тоногих. Это, видимо, связано с более ранним переходом в водную среду
предков сирен.
Вообще выделяют четыре типа приспособления глаза и его вспомога-
тельных органов к водной среде у морских млекопитающих: термиче-
ские (наличие чудесной сети), химические (изменения окологлазничных
желез), гидростатические (мощное развитие склеры и мускулатуры гла-
за и век), оптические (изменение формы роговицы и хрусталика, повы-
шение светочувствительности сетчатки) (Putter, 1903). Видно, что у си-
рен лишь «химические» адаптации выражены в полной мере и отчасти
«гидростатические»: химизм воды требует изменения железистого аппа-
рата, а неглубокие погружения — некоторого усиления мышц век и глазо-
двигательной мускулатуры. Отсутствие глубоководного ныряния приво-
349
дит к тому, что сирены не испытывают резких температурных, световых
и барометрических изменений.
Узкая, довольно однообразная среда обитания привела сирен к та-
кому состоянию, когда в самых разных органах наряду с глубокими пре-
образованиями отмечаются черты, свойственные их наземным предкам.
Как уже отмечалось в предварительном сообщении (Андреев, 1982), из
всех наземных млекопитающих наибольшее сходство глаза ламантинов,
особенно его сосудистого отдела, наблюдается с органом зрения хобот-
ных. Это тем более интересно, что сирены и слоны объединены в над-
отряд слонообразных.
Орган зрения, с одной стороны, весьма консервативная система,
а с другой — необычайно пластичная (Андреев, 19786). Отсутствие адап-
тивных черт в строении глаза сирен подтверждает предположение о кон-
сервативности этой системы. Когда же зрение становится одним из ве-
дущих анализаторов, то оно эволюционно очень быстро приспосаблива-
ется к среде, выявляя свою необычайную пластичность, особенно в оп-
тической и световоспринимающей системах. Примером тому может быть
орган зрения ластоногих.
К сожалению, нет данных о строении глазного яблока морской коро-
вы, однако имеется краткое описание вспомогательных органов животно-
го, сделанное Г. Стеллером. Сходство этих органов у морской коровы и
других сирен несомненно. Вероятно, не будет большой ошибкой полагать,
что и орган зрения во многом схож с глазами рецентных сирен, посколь-
ку это вытекает из их экологического сходства.
Весьма соблазнительно провести параллели в степени развития и ха-
рактере использования органа зрения сирен и речных дельфинов, по-
скольку в указанных группах есть чисто пресноводные (палатанисты и
бразильский ламантин), смешанные (амазонская иния, карибский и аф-
риканский ламантины) и морские (лаплатский дельфин, дюгонь и мор-
ская корова) виды. Однако ни в литературе, ни у нас нет пока доста-
точных данных для подобного сравнения. Можно лишь предположить,
что наличие регрессивных изменений глаза некоторых речных дельфинов
(Pilleri, 1977), упрощение строения и отсутствие черт специализации
глаза сирен обусловлено местами обитания, главным образом малой сте-
пенью прозрачности воды, характером питания и типом локомоции.
Глава одиннадцатая
ОСОБЕННОСТИ МИКРО- И УЛЬТРАСТРУКТУРЫ
И ГИСТОХИМИИ
НЕКОТОРЫХ ОРГАНОВ ЛАМАНТИНА
В этой главе мы рассмотрим результаты изучения содержания липидов
в клетках скелетных мышц одного экземпляра ламантина (см. Введение)
в сравнении с некоторыми другими водными млекопитающими и резуль-
таты электронномикроскопического изучения мышечных клеток и клеток
печени этого же животного.
ГИСТОХИМИЧЕСКОЕ ИЗУЧЕНИЕ ЛИПИДОВ
В СКЕЛЕТНЫХ МЫШЦАХ ЛАМАНТИНА
И ДРУГИХ МОРСКИХ МЛЕКОПИТАЮЩИХ 1
Гистохимическая структура мышц морских млекопитающих исследована
слабо. В литературе имеются отрывочные сведения по гистохимии мышц
гренландского тюленя (Pagophilus groenlandica), калифорнийского морско-
го льва (Zalophus californianus), калана (Enhydra lutris), тихоокеанского
короткоголового дельфина (Lagenorhynchus obliquidens), морской свиньи
(Phocaena phocaena) и афалины (Tursiops truncatus) (George et al., 1971;
George, Ronald, 1973, 1975; Маньковская, 1975; Tulsi, 1975; Родионов,
1978, 1979; Соколов, Родионов, 1978; Ponganis, Pierce, 1978; Blix et al.,
1979). Микроскопическая структура мышц сирен ранее не исследовалась.
В настоящей работе изложены результаты гистохимического изуче-
ния проб мышц взрослого самца ламантина (Trichechus manatus). Для
сравнения были исследованы также мышцы гренландского тюленя
(Pagophilus groenlandica), афалины (Tursiops truncatus), серого кита
(Eschrichtius gibbosus) и белого медведя (Ursus maritimus). Выявле-
ние липидов проводили на поперечных срезах мышц с помощью окраши-
вания Суданом черным Б. За диаметр мышечного волокна принимали
среднюю от измерения его максимального и минимального диаметров.
Соотношение волокон разных типов в процентах общего числа волокон
определяли по нескольким участкам мышц, насчитывавшим, как прави-
ло, более 400 мышечных волокон. Для того чтобы оценить, какая часть
мышц по массе приходится на тот или иной тип мышечных волокон,
с помощью полученных данных рассчитывали содержание разных мы-
шечных волокон в процентах площади поперечного сечения всех воло-
кон. При этом площадь поперечного сечения мышечного волокна прини-
малась равной площади круга с таким же диаметром, а количество
представленных в мышце волокон разных типов — их процентному со-
отношению.
Проведенное исследование показало, что у морских млекопитающих
все мышцы были неоднородны по составу и содержали мышечные во-
локна, различавшиеся по концентрации в них липидов. При глазомерной
1 Пользуясь случаем, автор выражает искреннюю благодарность В. Е. Соколову,
П. П. Гамбаряну, В. Б. Суханову и Ф. В. Андрееву, предоставившим материал для
исследования.
351
оценке интенсивности окраски можно было выделить три типа волокон:
с высоким, средним (промежуточным) и низким содержанием жиров.
Все три типа мышечных волокон были хорошо различимы в большинстве
мышц китообразных, в то время как у ламантина, гренландского тюленя
и белого медведя были выделены только два типа: светло- и темноокра-
шенные. Сопоставление диаметров темных и светлых волокон, а также
интенсивности окраски темных мышечных волокон этих зверей с таковой
китообразных позволило установить, что темноокрашенные волокна
должны быть отнесены к категории мышечных волокон не с высоким,
а с умеренным содержанием жира. Однако необходимо отметить, что со-
держание жира в мышечных волокнах этого типа несколько варьирова-
ло, что связано, вероятно, с неодинаковым расходованием его в разных
мышечных клетках в период, предшествовавший смерти животного. Во
всяком случае средние диаметры мышечных волокон, немного различав-
шихся по содержанию липидов, были одинаковыми.
У ламантина средние диаметры мышечных волокон с умеренным и
низким содержанием жиров были сходны в одних мышцах, мало отли-
чались в других и достаточно резко отличались в третьих (табл. 16).
При этом светлые мышечные волокна имели, как правило, больший диа-
метр. Исключением в этом плане была височная мышца (m. temporalis),
в которой немногочисленные светлые волокна были почти вдвое тоньше
темных мышечных волокон. В других мышцах ламантина средние диа-
метры волокон с умеренным содержанием жира варьировали от 39,2 мкм
в длиннейшей мышце спины (m. longissimus dorsi) до 68,5 мкм в зубча-
той вентральной мышце (m. serratus ventralis), т. е. менялись в 1,8 раза.
Аналогичным образом изменялись диаметры мышечных волокон с низ-
ким содержанием жиров: от 47,7 мкм в наружной межреберной мышце
(m. intercostalis externus) до 82,0 мкм в зубчатой вентральной мышце,
т. е. в 1,7 раза.
Обращает на себя внимание тот факт, что мышцы ламантина содер-
жат очень много светлых, бедных жиром волокон. За исключением ви-
сочной, состоящей почти исключительно из темных волокон, и межребер-
ной мышц, во всех остальных мышцах на долю светлых волокон прихо-
дилось не менее 63,4% их общего количества в мышце. Пересчет же на
площадь поперечного сечения показал, что содержание светлой мускула-
туры в мышцах ламантина очень велико: от 68,7% в поверхностной
грудной мышце (m. pectoralis superficialis) до 98,1% в длиннейшей мыш-
це спины.
Интересно отметить, что обычно выполняющие дыхательную функ-
цию наружная и внутренняя межреберные мышцы резко отличаются
друг от друга по гистохимической структуре. Если средние диаметры и
соотношение мышечных волокон разных типов во внутренней межребер-
ной мышце были сходными с таковыми у диафрагмы, также несущей
дыхательную функцию, то диаметры волокон в наружной межреберной
мышце были примерно в 1,5 раза, а содержание светлых волокон — в
два раза меньше, чем у внутренней или диафрагмальной мышц. Эти
особенности структуры двух межреберных мышц свидетельствуют о су-
ществовании каких-то серьезных различий в их функционировании.
У афалины и серого кита средние диаметры мышечных волокон с
высоким содержанием жира (темных) были в 1,5—2 раза меньше, чем
у волокон, содержавших мало жира (светлых), а диаметр промежуточ-
ных волокон был больше, чем у темных, но меньше, чем у светлых
мышечных волокон (см. табл. 16). Из приведенных данных видно, что у
китообразных, несмотря на различия в диаметрах волокон у разных ви-
дов и у разных особей одного вида, сохраняется соотношение диаметров
352
я
Illi II
Illi II
Й g Е g 8
И
£ £ £ £
Е Е-5
Е Е Е g
8 Е
а а a а
I
353
мышечных волокон, отличающихся по содержанию жира. Интересно от-
метить, что, несмотря на очень значительные различия в абсолютных
размерах тела, диаметры мышечных волокон соответствующих типов бы-
ли у серого кита всего в 2—2,5 раза больше, чем у афалины. Средние
диаметры мышечных волокон у двух других крупных китов — кашалота
(Physeter catodon) и сейвала (Balaenoptera borealis) — также были срав-
нительно невелики —108 и 60,5 мкм соответственно (Тиняков и др.,
1976). Достаточно резкие отличия обнаружены в количественных соот-
ношениях разных мышечных волокон в мышцах серого кита и дельфинов.
У афалины на долю темных волокон приходилось от 30 до 59%, а у
кита — только 11—26% от всех подсчитанных в мышце волокон. У афа-
лины содержание светлых мышечных волокон составляло в разных
мышцах 28—4.4%, а у серого кита значительно больше — от 49 до 65%.
В двух исследованных мышцах гренландского тюленя диаметры тем-
ных и светлых волокон отличались незначительно, в количественное со-
отношение волокон двух типов было примерно равным (см. табл. 15).
Мышцы белого медведя также состояли из двух типов волокон (рис.
137, ж—д), незначительно отличавшихся по диаметрам (см. табл. 15).
Содержание светлых волокон в разных мышцах медведя менялось от
36 до 69%, т. е. почти вдвое. Следует отметить только, что, судя по
гистохимической реакции, темноокрашенные мышечные волокна медведя
содержали липидов больше, чем подобные волокна в мышцах тюленя и
ламантина.
Сопоставление площадей, занимаемых мышечными волокнами разных
типов на поперечных срезах мышц (по существу, сопоставление весо-
вых соотношений разных волокон в мышцах), показало, что ламантин
и кит резко отличаются от других исследованных морских млекопитаю-
щих тем, что их мышцы на 3/4 и более состоят из не содержащих
жира мышечных волокон, т. е. более чем на 3/4 состоят из так назы-
ваемой «белой» мускулатуры.
Поскольку основными источниками энергии в мышце являются жир
и гликоген, можно предположить, что не содержащие жира мышечные
волокна должны использовать альтернативный путь получения энергии —
за счет утилизации гликогена. Гистологическое и элетронномикроскопи-
ческое изучение ламантина (Умнова, Новоселова, 1982; и второй раздел
этой главы) гренландского (George et al., 1971) и каспийского тюленей
(Гетманова и др., 1982) показало, что их мышцы содержат мало жира,
зато богаты гликогеном, который и является, вероятно, основным энер-
гетическим субстратом в их мускулатуре.
Известно, что мышечные волокна получают необходимую им энергию
либо за счет окислительных процессов, проходящих с участием кислоро-
да, либо за счет анаэробной фазы гликолиза, протекающей при недостат-
ке или отсутствии кислорода в работающей мышце. Проведенные ранее
исследования показали, что тонкие, богатые жиром мышечные волокна
афалин обладают высокой активностью окислительного фермента сукци-
натдегидрогеназы (СДГ), богаты митохондриями и миоглобином, в то
время как толстые волокна характеризуются низкой активностью СДГ,
низкпм содержанием жира, митохондрий и миоглобина (Tulsi, 1957;
Соколов, Родионов, 1978). Косвенным свидетельством того, что мышцы
ламантина обладают достаточно низкой окислительной способностью,
может служить тот факт, что содержание в их мышечных волокнах ос-
новных энергетических органелл — митохондрий — было невелико (Ум-
нова, Новоселова, 1982).
В многочисленных опытах на млекопитающих и птицах было уста-
новлено, что мышечные волокна, получающие энергию за счет процес-
354
сов гликолиза, способны к мощной, но непродолжительной работе, в то
время как устойчивые к утомлению волокна используют, кроме гликоли-
за, еще и окислительные процессы, причем основным субстратом окисле-
ния у них, как правило, является жир. Поэтому можно предположить,
что наиболее длительно и интенсивно работающие мышцы должны со-
держать больше богатых жиром мышечных волокон с окислительным
типом метаболизма.
Рие. 137. Гистохимическое выявление липидов Суданом черным Б на поперечных
срезах мышц морских млекопитающих
а — в — ламантин Trichechus manatus: а — m. pectoralis superficialis б — m. intercostalis externus
b — m. temporalis; г — 0 — серый кит Escherichtius gihbosus; a — m. rectus abdominis, d — m. in-
tercostalis externus; e — афалина Tursiops truncatus: m. obiiquus abdominis externus; ж—з — белый
медведь Ursus maritimus: ж — m. biceps femoris, a — m. sartorius
355
Из изученных морских млекопитающих наименее подвижный образ
жизни ведут ламантины, плавающие и ныряющие много хуже тюленей
и дельфинов. Отсюда и очень низкая концентрация миоглобина в их дви-
гательных мышцах (Blessing, 1972) и, как показало настоящее исследо-
вание, малое количество мышечных волокон с высоким содержанием
жира. Напротив, височная мышца, выполняющая жевательную функцию,
состоит у ламантина почти целиком из жирсодержащих волокон (см.
табл. 15; рис. 137, в). Это объясняется, вероятно, тем, что ламантины
питаются растительной пищей и вынуждены в силу этого подолгу рабо-
тать челюстями, обрабатывая ее.
Из исследованных нами видов морских млекопитающих только в мыш-
цах китообразных были обнаружены резко различавшиеся по диаметру
и содержанию жира мышечные волокна с характерным рисунком их
взаимного расположения. По имеющимся у нас данным такая структура
мышц присуща еще двум видам дельфинов — морской свинье (Phocaena
phocaena) и дельфину-белобочке (Delphinus delphis). Все это свидетель-
ствует об определенной схожести, специфичности структуры мышц у
китообразных. Однако имеются также и глубокие различия в строении
мышц кита и дельфинов, выражающиеся в том, что мышцы кита содер-
жат значительно меньше богатых жиром и больше бедных жиром мы-
шечных волокон, чем мышцы дельфина. Большая часть мышц ламанти-
на также состоит из волокон, не содержащих жира. Эта особенность
строения мышц кита и ламантина может объясниться двумя основными
причинами.
Во-первых, понижением тонической, позной, антигравитационной на-
грузки, которая ложится на мышцы китообразных и сирен вследствие
того, что их тело в течение всей жизни находится в воде во взвешен-
ном состоянии. Поскольку выполнение этой функции всегда связывают с
красной мускулатурой, то ясно, что снижение тонической нагрузки долж-
но вести к уменьшению содержания мускулатуры этого типа в их мыш-
цах.
Во-вторых, снижением терморегуляционной нагрузки на мышцы ки-
та и ламантина по сравнению с мышцами других морских млекопитаю-
щих: у кита — вследствие крупных размеров тела и очень выгодного
с терморегуляционной точки зрения соотношения массы и поверхности
тела, а у ламантина — вследствие обитания в тропиках в мелководных,
хорошо прогреваемых водоемах. Этот пункт требует небольшого поясне-
ния.
Общеизвестна ведущая роль скелетной мускулатуры в реакциях хи-
мической терморегуляции. Было показано, что при холодовом воздейст-
вии на млекопитающих их мышцы увеличивают свою электрическую
активность (т. е. увеличивают теплопродукцию) неодинаково: одни в
большей, другие в меньшей степени, и что в неравномерном подключе-
нии разных групп мышц к увеличению теплопродукции проявляется
экологическая специфика вида (Иванов, 1965; Баженов, 1969; Давыдов,
Кузьмо, 1969; и др.). Более того, и внутри мышцы волокна нагружены
неравномерно. В опытах на крысах (Туманова, 1972; Слоним, Туманова,
1973) было обнаружено, что мышечные волокна с окислительным типом
метаболизма (так называемые «красные» волокна) имели в несколько раз
большую электрическую активность, чем мышечные волокна с гликоли-
тическим типом обмена («белые»). Следовательно, теплопродуцирующая
роль скелетных мышц связана главным образом с красной мускулатурой,
и понижение терморегуляционной нагрузки также должно вести к сни-
жению содержания в мышцах млекопитающих мышечных волокон с
окислительным типом метаболизма. Таким образом, внутренняя структу-
356
ра мышцы у млекопитающих, ее состав и соотношение в ней мышечных
волокон разных типов должны определяться не только объемом и харак-
тером выполняемой физической работы, но и участием данной мышцы в
терморегуляции (Родионов, 1980).
Ранее было показано, что в мышце блювала активность таких окис-
лительных ферментов как цитохромоксидаза, сукцинатоксидаза и сукци-
натдегидрогеназа была значительно ниже, чем в мышцах лошади, быка,
слона и других наземных млекопитающих (Lawrie, 1953). Очевидно, что
и в мышцах серого кита активность окислительных ферментов должна
быть пониженной, поскольку обитают эти киты примерно в одинаковых
условиях. К сожалению, активность ферментов в мышцах ламантина не
исследовалась. Однако с большой долей вероятности можно утверждать,
что окислительная способность его мышц также будет невелика. В поль-
зу этого говорит то, что в его мышечных волокнах содержится мало ми-
тохондрий, жира и миоглобина, а косвенно — общий низкий уровень ме-
таболизма: уровень потребления кислорода у ламантина в покое был в
5—10 раз меньше, чем у человека (Scholander, Irving, 1941).
Таким образом, несмотря на определенные различия в гистохимиче-
ской структуре мышц у таких далеких в систематическом отношении ви-
дов, как серый кит и ламантин, мышцы этих двух видов характеризуют-
ся очень важным общим для них признаком: высоким содержанием
белой мускулатуры. Основными факторами, определившими такое сход-
ное изменение состава мышц у кита и ламантина (при сохранении дру-
гих особенностей структуры), следует считать понижение тонической и
терморегуляторной нагрузок на их скелетные мышцы.
МИКРОСКОПИЧЕСКОЕ И УЛЬТРАСТРУКТУРНОЕ ИССЛЕДОВАНИЕ
СКЕЛЕТНОЙ МЫШЦЫ ЛАМАНТИНА2
Мышечная система, осуществляющая основной механизм двигательной
функции в организме, составляет у млекопитающих около 40% объема
тела. В существующих немногочисленных исследованиях, посвященных
изучению соматической мускулатуры ламантинов, приводится в основном
анатомическое описание разных групп мышц (см. обзор: Ronald et al.,
1978).
Настоящая работа посвящена выяснению гисто- и ультраструктуры
одной из скелетных мышц ламантина (m. obliquus abdominis externus).
Для световой микроскопии отдельные фрагменты мышцы фиксировали в
жидкости Буэна и заливали в парафин. Гистологические препараты ок-
рашивали железным гематоксилином с докраской по Маллори. Для выяв-
ления гликогена на срезах проводили иодную реакцию и окрашивали
препараты квасцовым кармином по методу Драйсенса (Роскин, Левинсон,
1957).
Для электронно-микроскопического изучения небольшие кусочки
мышцы фиксировали в 2,5 %-ном растворе глутаральдегида с последую-
щей дофиксацией в 1%-ном растворе четырехокиси осмия. После обезво-
живания в спиртах и ацетоне материал был залит в эпон. Ультратонкие
срезы мышц, помещенные на бленды, контрастировали уранилацетатом,
цитратом свинца, просматривали и фотографировали в электронном ми-
кроскопе Jem=100C.
2 Авторы выражают глубокую благодарность О. Ф. Черновой, Л. В. Степановой,
Г. В. Еляковой, А. Н. Студитскому, М. М. Калашниковой, Н. И. Казанской за по-
мощь в получении и обработке материала, а также обсуждения полученных резуль-
татов.
357
Рис. 138. Цепочки ядер (ЯМВ) в мышечных волокнах (МВ). Железный гематокси-
лин по Рего с докраской по Маллори. Ув. X 320 (об. 40, ок. 8)
Рис. 139. Нервный стволик (НС) и жировые клетки (ЖК) в соединительной тка-
ни мышцы. Железный гематоксилин с докраской по Маллори. Ув. X 320 (об. 4(Х.
ок. 8)
358
Косая мышца живота состоит из толстых и единичных тонких мы-
шечных волокон с отчетливо выраженной поперечной исчерченностью. На
гистологических препаратах в мышечных волокнах видны немногочислен-
ные ядра, иногда образующие короткие цепочки по периферии волокна
(рис. 138). Мышечные волокна разделены эндомизием, в котором часто
встречаются кровеносные капилляры. Пучки мышечных волокон разгра-
ничены широкими прослойками перимизия, представленного волокнистой
соединительной тканью, в которой расположены артериолы, венулы и бо-
лее крупные кровеносные сосуды, а также нервные стволики. При гисто-
химическом исследовании в мышце ламантина выявлено большое коли-
чество гликогена, диффузно расположенного в мышечных волокнах.
Исследованная мышца содержит небольшое количество жира: в соеди-
нительной ткани встречаются отдельные жировые клетки (рис. 139).
Электронно-микроскопическое изучение косой мышцы живота ламан-
тина показало, что сократительный аппарат мышечных волокон пред-
ставлен миофибриллами разной толщины с хорошо выраженными сарко-
мерами, разграниченными Z-линиями. Миофибриллы состоят из плотно-
упакованных протофибрилл (миофиламентов), наблюдаются картины
расщепления толстых миофибрилл на более тонкие. Миофибриллы отде-
лены друг от друга участками саркоплазмы — то более узкими, то доволь-
но широкими. Многие мышечные волокна содержат значительное коли-
чество саркоплазмы, в которой располагаются цитоплазматические орга-
неллы. Исследованная мышца ламантина отличается содержанием
небольшого числа полиморфных митохондрий удлиненной, шаровидной
или разветвленной формы. Митохондрии распределяются в саркоплазме
между миофибриллами без видимой периодичности. Небольшие скопления
крупных митохондрий под плазмолеммой обнаруживаются лишь в единич-
ных мышечных волокнах. Встречаются митохондрии с плотным располо-
жением крист, в то время как другие содержат немногочисленные кристы.
В митохондриях кристы ориентированы в продольном или поперечном
направлении относительно длинной оси данных органелл (рис. 140).
Саркоплазматическая сеть в исследуемой мышце представлена неболь-
шими канальцами, пузырьками, цистернами, полостями. Терминальные
цистерны саркоплазматического ретикулума и поперечные трубочки
Т-системы образуют триады, расположенные на границе А и И дисков.
На электронных микрофотографиях в саркоплазме мышечных воло-
кон, так же как и на гистохимических препаратах, при гистохимическом
исследовании выявляется большое количество гликогена. Гликоген пред-
ставлен гранулами, расположенными в основном между миофибриллами;
единичные гранулы гликогена обнаруживаются среди протофибрилл.
Величина гранул гликогена варьирует в разных волокнах. В одних мы-
шечных волокнах гранулы гликогена мелкие, а в других, нередко рядом
расположенных, они значительно крупнее (рис. 141) и могут образовы-
вать конгломераты.
Главным элементом трофического аппарата поперчнополосатых мы-
шечных волокон являются ядра (Студитский, 1977). Они имеют эллипсо-
идную форму, занимают периферическое положение, располагаясь в сар-
коплазме среди миофибрилл. Длина ядер соответствует размеру 4—8 сар-
комеров. Ядра преимущественно электроннопрозрачные, небольшие
сгущения хроматина видны по периферии ядер. В некоторых ядрах хро-
матин образует более плотные скопления в виде широких полос или глы-
бок у внутренней ядерной мембраны. Контуры ядер нередко извилисты,
иногда наружная ядерпая мембрана образует глубокие впячивания внутрь
ядра, так что в центре ядра может быть заключен участок саркоплазмы.
Ядро содержит чаще всего одно ядрышко, но встречаются ядра и с дву-
359
мя ядрышками. Ядрышки имеют плотную гранулярную структуру и лишь
некоторые — более рыхлое сетевидное строение.
В мышце ламантина выявлены также клетки-сателлиты (миосателли-
тоциты). Они располагаются между плазматической и базальной мембра-
нами мышечных волокон. В зависимости от плоскости прохождения среза
эти клетки имеют округлую форму или вытянуты вдоль мышечных воло-
кон. Крупное ядро, занимающее почти весь объем клетки, ограничено уз-
кой зоной цитоплазмы. Ядра, соответственно контуру (профилю) клетки,
имеют округлую или овальную форму и отличаются более плотной струк-
Рис. 140. Фрагмент мышечного волокна. Ув. X 32 500
MX — митохондрии; ЯМВ — ядро мышечного волокна
360
турой по сравнению с ядрами мышечных волокон (рис. 142, а). Хроматин
в виде крупных глыбок (гетерохроматина) конденсируется не только на
внутренней ядерной мембране, но обнаруживается и в центре ядер. В яд-
рах миосателлитоцитов ядрышек не обнаружено. Наружная мембрана в
некоторых участках усеяна рибосомами, видны ядерные поры (рис. 142, в).
Иногда заметен переход наружной ядерной мембраны в канальцы грану-
лярной эндоплазматической сети (рис. 142, б). Эндоплазматическая сеть
представлена наряду с канальцами колбовидными образованиями со свет-
лым содержимым, контактирующими с плазмолеммой. По наружной по-
Рис. 141. Фрагмент мышечного волокна (МВ), содержащего значительное количест-
во саркоплазмы и гликогена (Г). Ув. X 8 400
361
верхности клетки, обращенной к базальной мембране, видны микропино-
цитозные пузырьки. В одной из клеток обнаружено липидное включение
и образование волокнистого характера, напоминающее фрагмент большой
миофибриллы. Эта клетка имеет цитоплазматические отростки (рис. 143,
б). В участке мышечного волокна, граничащего с миосателлитоцитом,
нередко видны щели и полости, а также скопления мембранных струк-
тур. Иногда эти структуры имеют вид клубков или слоистых образований,
внутри них могут быть заключены участки саркоплазмы, содержащей
гликоген (см. рис. 142, в). Агрегаты мембранных структур лежат в не-
посредственной близости к плазмолемме мышечного волокна. В отдельных
Тис. 142. Миосателлитоцит под базальной мембраной мышечного волокна
а — общий вид. Ув. х21 450; б — фрагмент: отделение гранулярной эндоплазматической сети от
наружной ядерной мембраны. Ув. Х48 ООО; в — фрагмент: ядерные поры. Ув. Х60 ООО
Г — гранулы гликогена; ГЭС — гранулярная эндоплазматическая сеть; МС — мембранные струк-
туры; Мс — миосателлитоцит; МФ — фрагмент муофибриллы; MX — митохондрия; ЯМс — ядро
миосателлитоцита; ЯП — ядерная пора
362
случаях мебранные структуры в миосателлитоците и мышечном волок-
не располагаются симметрично по отношению друг к другу и разграни-
чены только плазматическими мембранами (см. рис. 143, а).
В саркоплазме косой мышцы живота обнаружены структуры типа
миелиновых фигур. Скопления концентрически расположенных мембран
можно видеть между ядрами в периферической саркоплазме волокна.
Рис. 142 б
363
Миелиноподобные тельца имеют разные размеры и плотность. Встреча-
ются аутофагосомы, сходные с крупными вакуолями, содержащими плот-
ные тельца, имеющие структуру миелиновых фигур (рис. 144).
Эндомизий, окружающий мышечные волокна, представлен бесструк-
турным войлокоподобным аморфным веществом, тонкими коллагеновы-
ми волокнами и клеточными элементами. Это клетки веретеновидной фор-
Рис. 142 в
364
мы с крупным ядром и большим количеством органелл в цитоплазме.
Характерной особенностью фибробласта является развитая гранулярная
эндоплазматическая сеть, полости которой заполнены аморфным вещест-
вом..
В эндомизии встречаются также и шванновские клетки. Эти клетки
имеют большое ядро довольно плотной структуры и содержат, кроме обыч-
ного набора органелл, миелин, представляющий собой выросты клеточной
оболочки, формирующей кольцевидные структуры из плотнорасположен-
ных мембран.
В рыхлой соединительной ткани проходят кровеносные сосуды, осо-
бенно многочисленны капилляры. Эндотелиальные клетки (эндотелиоци-
ты), окружающие стенки капилляров, имеют электронноплотное ядро.
В некоторых ядрах заметно ядрышко плотной структуры. В цитоплазме
эпдотелиоцитов располагаются рибосомы, полисомы, митохондрии разных
размеров, цистерны и канальцы гранулярной эндоплазматической сети,
комплекс Гольжди, гранулы гликогена. Характерной особенностью эндо-
телиоцитов является наличие большого числа пиноцитозных пузырьков.
Ряды пузырьков примыкают как к люминальной, так и к базальной повер-
хности клеток. Пиноцитозные пузырьки подходят вплотную к плазмолем-
ме и видны картины включения мембраны пузырька в оболочку клетки.
В эндотелиоцитах встречаются структуры типа миелиновых фигур, неред-
ко обнаруживаются липидные капли разных размеров (рис. 145). На лю-
минальной поверхности эндотелиоцитов видны микроворсинки. Они могут
быть короткими и длинными. В некоторых микроворсинках встречаются
липидные включения. Тонкий эндоплазматический вырост эндотелиоцита
иногда образует замкнутую спиралевидную структуру — «бляшку», плот-
ноприлегающую к стенке сосуда. Снаружи капилляры окружены пери-
цитами.
Таким образом, косая мышца живота ламантина имеет структурную
организацию, свойственную поперечнополосатым мышцам других позво-
ночных животных. Она состоит в основном из мышечных волокон, со-
держащих мало митохондрий, обилие саркоплазмы и значительное коли-
чество гликогена. По этим признакам такие волокна можно отнести к
белым мышечным волокнам (Шмерлинг и др., 1981). Вместе с тем в
данной мышце есть мышечные волокна с плотноупакованными миофиб-
риллами, небольшими скоплениями митохондрий под плазмолеммой. Это —
мышечные волокна переходного типа или, возможно, красные. Следова-
тельно, косая мышца живота ламантина, как и некоторые другие мышцы
данного животного, на основе гистохимического исследования (Родио-
нов, 1982) является смешанной.
Картины расщепления миофибрилл мы считаем признаком физиоло-
гической регенерации мышечных волокон.
В наружной косой мышце живота ламантина, как и в мышцах других
позвоночных животных, обнаружены миосателлитоциты. Между мышеч-
ными волкнами и миосателлитоцитом видны щели, полости, скопления
мембранных структур. Эти особенности рассматриваются иногда как одно
из доказательств концепции происхождения миосателлитоцитов из мы-
шечных волокон (Reznik, 1971; Елякова, 1972; Клингов, 1984). В одной
клетке было найдено образование типа фрагмента большой миофибриллы.
Миофибриллы лишь изредка встречаются в более зрелых миогенных клет-
ках-миобластах (Muir et., 1965). Вероятно, и данная клетка пред-
ставляет собой дальнейшую стадию развития миосателлитоцита — миоб-
ласт. Ядра миосателлитоцитов имеют более плотную структуру, чем мы-
шечные ядра. Исследователи, занимающиеся изучением интактной и ре-
генерирующей мышечных тканей, считают это показателем слабой
365
Рис. 143. Миосателлитоцит под базальной мембраной мышечного волокна
а — форма веретеновидная. Симметричное расположение мембранных структур в клетке и волокне.
Ув. Х27 390; б — на значительном протяжении клетка отделена от волокна широкой щелью.
Ув. х41 500
Л — липидное включение; МВ — мышечное волокно; остальные обозначения, как на рис. 142
366
функциональной активности данных клеток (Студитский, 1977; Клишов,
1984). Наличие миосателлитоцитов в мышце свидетельствует о способ-
ности исследованной мышцы к физиологической и репаративной регене-
рации, а сами клетки, возможно, служат структурной основой этих процес-
сов.
Появление миосателлитоцитов в мышечной ткани носит реактивный
характер и вызывается предшествующими дегенеративными процессами
(Студитский, 1977). Встречающиеся в интактной мышце ламантина мие-
линовые фигуры, аутофагосомы можно рассматривать как проявление
деструктивного процесса, результатом срабатывания структурных элемен-
тов и активизации лизосомальной системы, ответственной за перестройку
367
Рис. 144. Аутофагосома (АФ) и скопление гранул гликогена (Г) в мышечном волок-
не. Ув. X 25 000
ЯМВ — ядро мышечного волокна
отработанных компонентов мышечных волокон (Покровский, Тутельян,
1974).
Одним из структурных компонентов исследованной мышцы являются
капилляры. Наличие большого числа пиноцитозных пузырьков в цито-
плазме эндотелиоцитов свидетельствует о метаболической активности дан-
ных клеток, об активном переносе веществ из крови в окружающие тка-
368
Рис. 145. Продольный срез капилляра в мышце. Пиноцитозные пузырьки (НП)
и липидное включение (Л) в цитоплазме эндотелиоцита. Ув.Х24900
ПрС — просвет сосуда
ни и обратно. Липидные включения в цитоплазме эндотелиоцитов, веро-
ятно, связаны с интенсивным жировым обменом у данного вида живот-
ных. Известно, что сирены обладают большим запасом нежного жира
(Дозье, 1980). В то же время нельзя также исключить возможность от-
ложения липидных капель в стенке сосудов, связанного с возрастом жи-
вотного. Об этом также свидетельствует наличие «бляшек» на стенках ка-
пилляров.
369
Рис. 146. Участок печеночной дольки ламантина
Клетки печени с небольшим количеством темного пигмента. Ув. X 200 (об. 8, ок. 25)
Рис. 147. Желчный капилляр (ЖК), образованный на стыке трех гепатоцитов.
Ув. X 15 000
370
СВЕТООПТИЧЕСКОЕ И ЭЛЕКТРОННО-МИКРОСКОПИЧЕСКОЕ
ИССЛЕДОВАНИЕ КЛЕТОК ПЕЧЕНИ ЛАМАНТИНА
Ламантин является представителем водных млекопитающих, использую-
щих в пищу разные водные растения, предпочтительно покрыто- и го-
лосеменные (Hartman, 1979). Из литературы известно, что печень ламан-
тина имеет три основные доли и уплощена по отношению к диафрагме.
У особи длиной 1,8 м она имеет размеры 35,6 см от края правой доли.
У самца Trichechus manatus длиной 2,96 м, весом 424 кг печень весит
5,5 кг, а у самки длиной 3,25 м, весом 557 кг печень весит 6,24 кг
(Ronald et al., 1978).
Сведений по структуре печени ламантина в литературе нами не об-
наружено, однако известно (Caldwell et al., 1969), что печень Dugong
dugon имеет такое же долчатое строение, как у всех млекопитающих.
Печень Trichechus manatus была зафиксирована для световой микроскопии
в фиксаторе Буэна, залита в парафин, срезы окрашены крезил-виолетом
с докраской эозином, для электронной микроскопии — в 2 %-ном глута-
ральдегиде на 114М8-коллидиновом буфере при pH-7,2—7,4, с дофикса-
цией OsO4 на том же буфере, кусочки ткани залиты в эпон; ультратон-
кпе срезы, контрастированные по Рейнольдсу, просматривали в электрон-
ном мискроскопе Jem-100C.
Светооптическое исследование показало, что перенхима печени раз-
делена на дольки. В дольке гепатоциты располагаются в виде компакт-
ных переплетающихся тяжей по радиусам от периферии к центру (рис.
146). Клетки имеют полигональную форму, в цитоплазме находится одно
пли два ядра, а в каждом ядре довольно крупное ядрышко. Ядра в ге-
патоцитах располагаются в непосредственной близости от синусоидов, что
необычно для большинства животных и отмечено нами лишь у белого
медведя. В цитоплазме большинства клеток можно видеть включения чер-
по-корпчневого цвета, а светлые вакуоли встречаются в клетках на пери-
ферии долек. Синусоиды многочисленны, умеренно расширены, выстланы
эндотелиальными клетками с ядрами удлиненной формы. В просвете си-
нусоида можно видеть очень часто эритроциты и лейкоциты. Ядра эндо-
телиальных клеток располагаются довольно редко, так что можно думать,
что эндотелиальные клетки либо образуют очень длинные отростки, либо
располагаются прерывистым слоем (Джонсон, 1982).
При электронно-микроскопическом исследовании видно, что в гепато-
цитах ядерная оболочка имеет ровный, либо фестончатый край, нередко
можно впдеть широкие поры (75—150 нм). В ядре располагается крупное
широкопетлистое ядрышко. Хроматин имеет негомогенное строение. Зоны
гетерохроматина находятся вдоль ядерной оболочки и разбросаны по всей
кариоплазме. Иногда в клетке два ядра лежат так близко (на расстояния
180 нм), что можно думать о недавно происшедшем делении путем ами-
тоза или эндомитоза. Довольно часто ядра гепатоцитов располагаются
так близко от синусоида, что их разделяет полоска цитоплазмы шириной
не более 200 им.
В цитоплазме находится огромное количество митохондрий (от 65 на
срезе клетки до 100 и более). Их матрикс гомогенный, число крист неве-
лико, они короткие, располагаются беспорядочно (рис. 147, а). Диаметр
митохондрий варьирует от 0,65 до 1,5 мкм. Очень часто можно видеть
митохондии, делящиеся по кристам. Иногда митохондрии образуют це-
почки, располагаясь «конец в конец», что тоже, вероятно, является след-
ствием их деления.
Эндоплазматическая сеть представлена чрезвычайно богато, однако
преобладающей формой является не гранулярная эндоплазматическая
сеть (ГЭС), как у всеядных животных, а агранулярная эндоплазматиче-
371
Рис. 148. Синусоидальный полюс гепатоцита ламантина
а — включения первого вида (I), находящиеся на различных стадиях созревания. Ув. ХЗО ООО',
б — пино- и фагоцитоз из синусоида (С)
ЭК — эндотелиальные клетки
372
ская сеть (АЭС). АЭС представлена огромным количеством мелких, корот-
ких, уплощенных профилей и пузырьков, располагающихся очень плот-
но относительно друг друга (рис. 150). Обширные зоны коротких профи-
лей АЭС можно видеть на синусоидальном полюсе, причем иногда видно,
как эти профили выходят из гепатоцитов прямо в пространство Диссе,
которое в печени ламантина довольно широкое. Профили АЭС в свобод-
ном виде встречаются и в сосудистом русле.
Цистерны ГЭС уплощены, короткие, очень часто имеют кольцевидную
форму, располагаются в тесном топографическом контакте с митохондрия-
ми. Рибосомы не всегда располагаются на них равномерно.
Комплекс Гольджи трудно различим из-за обилия АЭС. Располагается
он либо в непосредственной близости от ядра, либо в зоне желчного ка-
пилляра, представлен преимущественно короткими плоскими цистернами,
располагающимися параллельно друг другу, или вакуолями среднего раз-
мера и мелкими пузырьками. Когда комплекс Гольжди локализован око-
ло желчного капилляра, в его зоне довольно часто можно видеть мульти-
везикулярные тела.
Гликоген в гепатоцитах встречается очень редко и в крайне незначи-
тельных количествах, липидные капли отсутствуют. Особенность гепа-
тоцитов ламантина является наличие нескольких видов включений.
К включениям первого вида мы относим гранулы размером от 0,5 до
1,4 мкм, содержащие грубозернистый электронноплотный материал. Эти
гранулы располагаются либо на синусоидальном полюсе гепатоцитов, либо
в зоне желчного капилляра. На синусоидальном полюсе эти гранулы бо-
лее мелкие, содержат меньше электронноплотного гранулярного материа-
ла (см. рис. 148, а), окружены, как правило, кольцевидной цистерной
ГЭС. Так как между ворсинками гепатоцита на синусоидальном полюсе
видны электронноплотные гранулы, лежащие на мембране гепатоцита
(рис. 148, б), можно думать, что этот электронноплотный материал путем
пиноцитоза поступает в цитоплазму гепатоцита и таким образом оказы-
вается во включениях первого вида. Когда гранулы первого вида распола-
гаются в зоне желчного капилляра, они тоже имеют различные размеры,
но всегда содержат огромное количество электронноплотного материала.
К включениям второго вида мы относим вакуоли, имеющие различные
размеры (от 0,8—5,5 мкм), нередко достигающие размера ядра
(рис. 149). Они локализуютяся на синусоидальном полюсе гепатоцитов и
в этом случае окружены огромным количеством профилей АЭС и мито-
хондрий, а также по всей цитоплазме. В этих вакуолях находятся хло-
пьевидное вещество и структуры типа миелиновых фигур. Последние всег-
да располагаются по периферии вакуоли, около ее оболочки. Следует
отметить, что не во всех вакуолях есть миелиновые фигуры. В одних их
нет, а в других —мало, в третьих — много, причем они разного размера.
Интересно отметить, что часто встречающиеся в вакуоле миелиновые
фигуры располагаются в зоне контакта данной вакуоли с митохондрией.
Создается впечатление, что эти структуры образуются после фермента-
тивного взаимодействия митохондрии с содержимым вакуоли. Встречают-
ся еще гранулы, окруженные оболочкой и содержащие гомогенный ма-
териал слабой электронной плотности. По структуре и плотности они на-
поминают микротельца. Их можно видеть в тесном топографическом кон-
такте с включениями первого и второго типа.
Желчные капилляры часто спавшиеся, с большим количеством микро-
ворсинок. В том случае, когда они открыты, в их просвете можно видеть
мелкозернистый материал и структуры типа миелиновых фигур. При этом
количество микроворсинок небольшое, многие из них набухшие, так что
создается впечатление недавно происшедшего выделения продуктов жел-
373
Рис. 149. Включения второго вида (77) в цитоплазме гепатоцита. Ув. X 30 000
Видны тесные топографические контакты митохондрий (MI) с включениями II вида
чи в желчный капилляр (Калашникова, 1981). По включениям первого
вида, всегда располагающихся в большом количестве около желчного
капилляра (см. рис. 150), можно судить о локализации желчных капил-
ляров как на электроннограммах, так и на гистологических препаратах
(см. рис.146).
В гепатоцитах вдоль синусоидов наблюдается различное расположение
374
органелл. В одних гепатоцитах непосредственно около синусоидальной
мембраны можно видеть ядро с фестончатыми краями, обилие митохонд-
рий и цистерны ГЭС. В других — митохондрии, цистерны ГЭС и гигант-
ские (величиной с ядро и меньше) вакуоли с гомогенным содержимым и
структурами типа миелиновых фигур. Наконец, в третьих гепатоцитах
вся цитоплазма на синусоидальном полюсе заполнена везикулярной АЭС
и митохондриями, так что создается впечатление, что идет активный фа-
Рис. 150. Включения первого (!) и второго (II) вида и обилие агранулярной эндо-
плазматической сети в цитоплазме гепатоцитов ондатры. Ув. X 45 000
375
го- и пиноцитоз. Кроме того, в этой зоне располагаются вакуоли с хло-
пьевидным содержимым, в котором можно видеть мелкие точки черных
включений типа гемосидерина. Различная ультраструктура гепатоцитов
на синусоидальном полюсе свидетельствует о том, что разные гепатоциты
в один и гот же момент выполняют различную функцию — либо погло-
щают сахарозу, либо, поглощая железо, синтезируют ферритин.
Несомненный интерес представляют синусоиды в печени ламантина.
Как мы уже отмечали, они довольно широкие, имеют широкое простран-
ство Диссе, иногда в нем можно видеть коллагеновые волокна, но редко.
Синусоидальные клетки имеют крупные размеры, в их цитоплазме рас-
полагаются митохондрии, цистерны ГЭС, комплекс Гольджи и довольно
крупные фагосомы. Нередко можно видеть в цитоплазме синусоидальных
клеток фагоцитированные эритроциты. Содержимое синусоидов очень
грубо структировано (в них содержится большое количество хлопьевид-
ного материала), а также в просвете синусоида можно видеть вакуоли
и пузырьки АЭС.
Проведенное исследование показывает, что в отличие от всеядных мле-
копитающих, в гепатоцитах ламантина, являющегося растительноядным
животным, содержатся огромные количества цистерн и пузырьков АЭС,
которая, как известно, участвует в метаболизме углеводов (Cardell, 1977;
De Man et al., 1966). Включения второго вида — крупные вакуоли, рас-
полагающиеся среди профилей АЭС, вероятно, являются морфологиче-
ским выражением накопления в печени сахарозы (Tribant-Vercruyssen
et al., 1980). Тесный контакт митохондрий с вакуолями, накопление в ва-
куолях структур типа миелиновых фигур, возможно, свидетельствует о
трансформации углеводов в фосфолипиды. Содержимое включений перво-
го вида, расположение этих включений и на синусоидальном полюсе ге-
патоцита и на билиарном полюсе заставляет нас думать, что в клетку
поступают из крови вещества высокой электронной плотности, вероятно
металлы (железо и др.), которые в довольно большом количестве содер-
жатся в поедаемых ламантином растениях. Тот факт, что эти включения
окружены цистернами ГЭС, вероятно, неслучаен. Можно думать, что со-
держимое этих включений обогащается в гепатоците белком, который син-
тезируется на мембранах ГЭС. Локализация включений первого и второго
вида в зоне желчных капилляров свидетельствует о том, что они выводят-
ся из клетки вместе с желчью.
Крупные ядрышки в ядрах гепатоцитов, широкие и многочисленные
поры в ядерной мембране, обилие митохондрий, профилей в эндоплазма-
тической сети — все это свидетельствует об активном функционировании
печени и отражает участие печени ламантина в метаболизме поступаю-
щих с пищей углеводов п минеральных веществ.
С целью выяснения, какие же специфические черты печеночной клет-
ки характеризуют растительноядное животное, была изучена ультраст-
руктура клеток печени ондатры. Оказалось, что, как и у ламантина, в ге-
патоцитах ондатры преобладающей формой эндоплазматической сети яв-
ляется АЭС и так же, как и у ламантина, присутствуют два вида совер-
шенно идентичных включений (рис. 150), которые также располагаются
в зоне желчных капилляров.
В изученных нами гепатоцитах зубра также преобладает АЭС, однако
характер включений совершенно другой.
Проведенное сравнение позволяет нам прийти к выводу, что для кле-
ток печени растительноядных животных характерно обилие АЭС, которая,
как уже говорилось выше, участвует в метаболизме углеводов, и наличие
разного вида включений, характер которых, по-видимому, определяется
особенностями химизма поедаемой пищи.
Глава двенадцатая
ИССЛЕДОВАНИЕ НЕКОТОРЫХ
ФИЗИОЛОГИЧЕСКИХ ОСОБЕННОСТЕЙ
ЛАМАНТИНА
В настоящее время физиологическое изучение сирен находится в началь-
ной стадии (Ronald et al., 1978). В частности, практически нет работ,
в которых были бы использованы электрофизиологические методы. Нам
не известны исследования, в которых была бы записана электроэнцефа-
лограмма ламантинов и дюгоней. Но имеются две работы, в которых бы-
ла зарегистрирована электрокардиограмма. У одного плавающего флорид-
ского ламантина обнаружено, что частота сердечного ритма замедляется
при нырянии, как и у других ныряющих животных (Scholander, Irving,
1941). У другого флоридского ламантина, находившегося на суше, реги-
стрировалась электрокардиограмма с целью выяснить, отражается ли в
морфологии зубцов элекрокардиограммы частичная раздвоенность желу-
дочков сердца у этих животных (Teeney, Hanover, 1958). Кроме того,
недавно были опубликованы две электрофизиологические работы, в кото-
рых слуховая система амазонского и флоридского ламантинов изучалась
методом вызванных потенциалов (Bullock et al., 1980; Bullock et al.,
1982).
Наша работа представляет собой фрагмент комплексного морфофизио-
логического исследования одного карибского ламантина. Впервые была
предпринята попытка зарегистрировать на плавающем представителе от-
ряда сирен электрофизиологическую полиграмму, состоящую из электри-
ческой активности головного мозга, сердца и мышц. Возможности экспе-
риментов были весьма ограничены сроками и методическими сложно-
стями.
Подопытный ламантин был половозрелым самцом (см. Введение).
Транспортировка его на корабле и автоплатформе от места отлова до
бассейна в Гаване заняла в общей сложности 7 ч. В период работы с
ним, в течение трех суток после отлова, ламантин содержался в пресной
воде в бассейне размером 11X7 м и глубиной 1,5—2,5 м. Адаптация к
условиям экспериментов была минимальной — одни сутки. В бассейне
плавали водные гиацинты, но не было замечено, чтобы ламантин питал-
ся. Хотя ламантин был малоподвижным и внешне спокойным, нельзя ис-
ключить, что у него весь этот период сохранялось состояние послеотлов-
ного стресса.
Были проведены два сеанса (с 21 до 7 и с 13 до 7 ч) непрерывной
полиграфической регистрации с помощью вживленных электродов раз-
личных физиологических показателей — электроэнцефалограммы (ЭЭГ)
двух полушарий, электрокардиограммы (ЭКГ), электромиограммы (ЭМГ)
шейных мышц, частоты дыхания. В период полиграфической регистрации
ламантин располагал акваторией 7 X 6 м при глубине 0,8 м.
Конструкция электродов и способ их вживления были такими же, ка-
кие использовались ранее на дельфинах афалинах (Мухаметов, Супин,
1975). На время операции ламантина извлекли из воды, местное обезбо-
ливание производили новокаином. Кожу и мягкие ткани над дорзальной
поверхностью черепа протыкали стальной трубкой с внешним диаметром
4 мм до упора ее в кость. Через трубку просверливали отверстие в кости
13 Ламантин 377
1
I (,4liU<T«W^ nim.wMR>^F
*ZI*H4**+*
и**и*м¥*у^*Ии^ J
5
Рис. 151. Электрополиграмма бодрствующего ламантина во время дыхательного акта
1 — ЭЭГ правого полушария; 2 — ЭЭГ левого полушария; 3 — ЭКГ; 4 — ЭМГ шейных мыппЦ
5 — отметка времени (1 с); подъем линии — отметка открывания ноздрей во время дыхательного,
акта. Калибровка амплитуды — 200 мкв
черепа и вводили электроды для регистрации ЭЭГ двух полушарий голов-
ного мозга. Электродами служили нихромовые проволочки в лаковой изо-
ляции диаметром 0,3 мм. Кроме того, такие же электроды были вживле-
ны в шейные мышцы для регистрации их фоновой ЭМГ и дыхательных
движений головы. Для регистрации частоты сердечного ритма (ЧСР) вы-
бирали ту пару электродов, расположенных на голове, которая давала
максимальную амплитуду зубцов ЭКГ. Период открывания ноздрей реги-
стрировали визуально и отмечали на электрополиграмме. Пример элект-
рополиграммы бодрствующего ламантина во время дыхательного акта по-
казан на рис. 151.
Отводящие антивибрационные кабели шестиметровой длины фикси-
ровали к стальному винту диаметром 6 мм, ввинченному в затылочное
утолщение черепа, и затем присоединяли ко входу четырехканального
электроэнцефалографа ЭЭГП4-02. Коммутационные кабели не мешали
ламантину передвигаться в отведенной акватории. Во время регистрации
ламантин большую часть времени находился под водой, лежа на дне, был
малоподвижен и только изредка плавал по кругу.
Обработка полученных электрополиграмм позволила дать количествен-
ную характеристику дыхания ламантина и частоты его сердечного рит-
ма; кроме того, оказалось возможным сравнить его ЭЭГ во время сна и
бодрствования.
Внешнее дыхание является наиболее доступным для наблюдателей
элементом поведения ламантинов, поэтому оно уже неоднократно было
описано как на свободных ламантинах в их естественной среде (Мооге,
1951в; Hartman, 1979), так и при содержании их в условиях неволи в бас-
сейнах и вольерах (Parker, 1922; Scholander, Irving, 1941). Для сравне-
ния с уже имеющимися литературными данными мы произвели количе-
ственную обработку внешнего дыхания ламантина на протяжении многих
часов непрерывной регистрации.
Во время каждого дыхательного акта ламантин медленно поднимал
голову к поверхности воды, что сопровождалось постепенным нарастанием
шейной ЭМГ (см. рис. 151). Обычно в воздух выставлялись только нозд-
ри, которые открывались для дыхательного акта. Выдох-вдох в отличие
от такового китообразных происходил бесшумно. После закрывания нозд-
рей голова резко опускалась вниз, что сопровождалось в ЭМГ шеи бы-
378
стрым падением амплитуды. Для внешнего дыхания ламантина, так же
как и ластоногих и китообразных, свойственно чередование кратковремен-
ной активной вентиляции легких с длительными дыхательными пауза-
ми. Период апноэ сопряжен, как правило, с пребыванием под водой.
Внешнее дыхание ламантина довольно редкое и колеблется от 1: до
5 дыхательных актов за 10 мин, в среднем это было 2,7±0,8 дыхатель-
ных актов за 10 мин, что было подсчитано по 39 десятиминутным перио-
дам. У трех хорошо адаптированных к неволе ламантинов (Parker, 1922)
среднее число дыхательных актов за 10 мин было 3,1 у самого мелкого,
2,4 у среднего и 2,2 у самого крупного ламантина, т. е. не отличалось
принципиально от наших цифр. Близкую частоту дыхания наблюдали у
взрослых диких животных в естественных условиях их обитания (Moore,
1951в; Hartman, 1979). При этом частота дыхания у детенышей была
значительно выше, чем у взрослых. В опытах Шоландера и Ирвинга
(Scholander, Irving, 1941) частота дыхания двух ламантинов составила
8 и 6 дыхательных актов за 10 мин, но при этом животные были им-
мобилизированы для изучения их газообмена.
Обычно ламантин за один дыхательный акт менял воздух в легких
один раз. При этом продолжительность открывания ноздрей была 5,6±
±1,8 с, при колебаниях этого показателя от 4 до 10 с. Длительность в*ы-
доха-вдоха взрослых спокойных животных в естественных условиях
была короче — 3,8 с и укорачивалась еще больше, если повышалась
двигательная активность (Hartman, 1979).
Следующие за периодом активной вентиляций легких дыхательные
паузы колебались от И до 465 с. Их средняя продолжительность была
218,4±97,4 с при п=126. У взрослых спокойных свободных ламантинов
средняя продолжительность дыхательных пауз была 232 с (Hartman,
1979). Дыхательные паузы длительностью до 150 с составили 20,6% от
общего их числа, длительностью от 151 до 300 с —62,8% и от
301 до 500 с —16,6% (рис. 152). Таким образом, животное в наших эк-
спериментальных условиях задерживалось под водой чаще всего на срок
в 2,5—5 мин и в редких случаях до 7,5 мип. По литературным данным,
сиреновые могут погружаться в воду и на большие сроки — до 8 мин
при естественных заныриваниях в вольере и до 12—15 мин при прину-
дительных погружениях в воду в эксперименте (Scholander, Irving, 1941),
до И мин 46 с (Hartman, 1979), до 16 мин 20 с (Parker, 1922).
Никакой закономерности в чередовании длительности дыхательных
пауз не наблюдалось (рис.153). Относительная продолжительность актив-
ной вентиляции легких, выраженная в процентах ко времени последую-
щих дыхательных пауз, у ламантина колеблется в пределах от 1,1 до
57,1 % и составляет в среднем 4,0%±3,5%.
Вышеописанный рисунок дыхания ламантина был основным. В то
же время во втором сеансе наблюдался период длительностью примерно
в 4 ч, когда дыхание ламантина явно изменилось. Во время дыхательно-
го акта ноздри ламантина оставались открытыми значительно дольше —
8—45 с, в среднем — 17,1±7,7 с. При этом, судя по экскурсиям грудной
клетки, ламантин менял воздух в легких во время дыхательного акта
несколько раз — от 2 до 6, в среднем — 3,0±1,1 (п=63). Дыхательные пау-
зы при этом укоротились до 128,6±69,9 с У свободных животных в ес-
тественных условиях обитания (Moore, 1951в; Hartman, 1979) и у хоро-
шо адаптированных к неволе животных (Parker, 1922) тоже могут
встречаться оба рисунка дыхания — и одиночные дыхательные акты с
коротким выставлением ноздрей в воздух, и серии выдохов-вдохов с от-
носительно продолжительным зависанием ламантинов у поверхности
воды.
379
13*
Рис. 152. Гистограмма распределе-
ния длительностей дыхательных
пауз
По оси абсцисс — длительность пауз
в секундах, по оси ординат — количество-
пауз в процентах от их общего количества
(126 пауз)
Рис. 153. Распределение во времени!
длительности последовательных ды-
хательных пауз
По оси ординат — длительность пауз,
в секундах
Взаимодействие дыхательного и сердечного ритмов у млекопитающих
является хорошо известным и давно установленным фактом. Особенно
ярко выражена зависимость частоты сердечного ритма от интенсивности,
респираторной деятельности и фазы дыхательного цикла у морских мле-
копитающих — китообразных и ластоногих, а также у многих полуводных
видов из отряда насекомоядных, грызунов и хищных (Галанцев, 1977;
Колчинская и др., 1980). Большинству из них при включении внешнего
дыхания во время ныряния или при задержке дыхания на суше свойст-
венно проявление ярко выраженной рефлекторной брадикардии. Интен-
сификация внешнего дыхания, наоборот, сопровождается развитием поло-
жительной хронотропной реакции сердца. О взаимосвязи ЧСР с дыхани-
ем у ламантинов упоминается только в одной работе (Scholander,.
Irving, 1941). По наблюдениям этих авторов ЧСР одного флоридского ла-
мантина, плавающего в вольере, была 60—50 ударов в мин во время ды-
хания и постепенно снижалась до 30 ударов в мин при заныривании. Нам
удалось провести более детальный анализ динамики сердечного ритма ла-
мантина в зависимости от фазы дыхательного цикла.
380
Рис. 154. ЭКГ ламантина в пределах одной дыхательной паузы
Каждая запись является непосредственным продолжением вышерасположенной записи. Горизон-
тальные черточки показывают время открывания ноздрей. Калибровка времени — 1с, амплитуды
200 мкв
n(R~R)
70
SO
SO
00
30
20
10
Рис. 155. Вариационные кардиоинтервалограм-
мы ламантина (для 5 различных дыхательных
пауз)
Заштрихованный вариант получен при эмоциональном
воздействии. По оси абсцисс — длительность кардиоин-
тервалов, по оси ординат — процент интервалов от об-
щего их числа во время каждой из дыхательных паув
127 159 191 2,27 2,S5 2,87 3,19 3,51 3,83 4,25
’ t(R-R)
381
Средний суточный уровень ЧСР ламантина в состоянии относительно-
го покоя составляет 27,6±1,1 удара в минуту при колебаниях этого по-
казателя от 18 до 48 мин. Таким образом, ЧСР у нашего ламантина ока-
залась ниже, чем в проведенных ранее опытах (Scholander, Irving, 1941;
Tenney, Hanover, 1958). Последние авторы, работавшие на ламантине,
извлеченном из воды, приводят только одну частоту сердечного ритма —
75 ударов в минуту.
Как уже отмечалось, ламантин остается на поверхности обычно в те-
чение нескольких секунд на период активной вентиляции легких. В этот
кратковременный период ЧСР становится заметно выше среднего уров-
ня и колеблется в разное время от 30 до 48 ударов в минуту. В среднем
же ЧСР во время дыхательных актов составляет 34,0±2,3, что
на 23,2% достоверно выше суточного уровня (при р<0,05). Ритм серд-
ца в этот период выраженной тахикардии характеризуется относительно
высокой стабильностью продолжительности сердечных циклов.
При погружении в воду в начале дыхательных пауз частота сокра-
щений сердца у ламантина довольно высока, а ритм относительно ста-
билен (рис. 154). В первые 10—20 с после погружения ЧСР колеблется
от 30 до 46 ударов в минуту и составляет в среднем 36,1 ±1,2. По срав-
нению со средним уровнем ЧСР в этот начальный период дыхательных
пауз выше на 30,8% (при р<0,01). В процессе дальнейшего пребывания
под водой у ламантина постепенно происходит увеличение длительности
отдельных кардиоциклов, особенно к концу дыхательных пауз перед оче-
редным выдохом-вдохом. В самом конце дыхательных пауз за 10—20 с
до очередного, периода активной вентиляции легких ЧСР урежается до
21—28 ударов в минуту и составляет в среднем 24,6±8,0, что на 10,9%
ниже общего среднего уровня ЧСР и на 31,9% ниже уровня ЧСР в пе-
риод тахикардической фазы после завершения активной вентиляции лег-
ких. Брадикардия во время погружения сопровождается усилением арит-
мии сердца. В отдельных случаях может наблюдаться спонтанное значи-
тельное увеличение длительности 1—2 сердечных циклов (до 3,34—
4,42 с).
Весьма показательным для характеристики сердечного ритма и дина-
мики сердечных циклов при нырянии ламантина могут быть вариацион-
ные пульсограммы, построенные на основании ранжированных рядов
кардиоинтервалов в пределах одной дыхательной паузы (рис. 155). Из
рисунка видно, что наиболее часто встречаемые значения длительности
сердечных циклов укладываются в диапазон от 1,59 до 2,23 с. Длитель-
ные кардиоинтервалы встречаются относительно редко. Например, в при-
веденных на рисунке случаях, кардиоинтервалы продолжительностью
2,87—4,25 с составляют всего 1—2%. Короткие сердечные циклы дли-
тельностью до 1,59 с также немногочисленны (от 4 до 21%) и наблю-
даются в основном только сразу после погружения животного в воду.
Максимальное значение времени сердечного цикла в состоянии относи-
тельного покоя животного, находящегося под водой, было 4,42 с, а мини-
мальное — 1,25 с. Размах вариации длительности сердечных циклов в
пределах отдельных дыхательных пауз колеблется от 0,52 до 2,56 с (в
среднем 2,15+0,2 с).
Таким образом, ритм сердца ламантина характеризуется не только
низкой частотой, но и значительной изменчивостью во времени, хотя и
менее выраженной, чем например, у ластоногих. Коэффициент аритмич-
/tz R — R макс-100 4Пп\ -
ности пульса у ламантина (Ка = —R_R мин--------101) )колеолется от 78
до 220%, в среднем составляя 142,5+13,9%.
382
При анализе особенностей сердечного ритма ламантина нельзя не от-
метить и реакцию сердца на эмоциональное воздействие. При спровоци-
рованном внезапном испуге ламантина, находящегося под водой, разви-
вается особенно сильная отрицательная хронотропная реакция. В этом
случае вариационная пульсограмма (см. рис. 155, заштрихованный ва-
риант) смещается вправо и приобретает многовершинную конфигурацию.
Такая форма вариационной пульсограммы не только отражает увеличе-
ние длительности кардиоциклов и соответственно урежение ЧСР, но и
указывает на усиливающуюся аритмию сокращений сердца.
Представленный материал показывает, что у такого высокоадаптиро-
ванного к водной среде вида, как ламантин, при произвольном погруже-
нии в воду даже на 3—7 мин в относительно спокойном состоянии бра-
дикардия выражена в значительно меньшей мере, чем у ластоногих или
полуводных насекомоядных, грызунов и хищных, у которых ЧСР при
нырянии замедляется в 4—10 и более раз (Галанцев, 1977; Галанцев,
Маминов, 1979). Само погружение в воду, которое является ведущим
стимулом в развитии рефлекторной, коротколатентной брадикардии у
большинства ныряющих млекопитающих и других водных амниот, у ла-
мантина не приводит к резкому замедлению ЧСР. В этом отношении при-
способительная реакция сердца ламантина в ответ на ныряние в значи-
тельной мере сходна с таковой у других гидробионтов — дельфинов
(Колчинская и др., 1980). При нырянии у дельфинов, которые погружа-
ются в воду на вдохе, как и ламантины, ритм сердца в течение несколь-
ких секунд остается на относительном высоком уровне. Тахикардическая
стадия сердечного ритма дельфинов характеризуется высокой стабиль-
ностью времени сердечных циклов, лишь через 3—12 с после погруже-
ния у дельфинов развивается брадиоаритмия. Сходство в изменениях сер-
дечного ритма при нырянии у сиреновых и китообразных и существенное
отличие реакций их сердца на ныряние от таковых реакций у полувод-
ных млекопитающих свидетельствует о различных путях формирования
адаптаций к нырянию со стороны сердечно-сосудистой системы у гидро-
бионтов и амфибионтов в процессе их приспособительной эволюции.
Регистрация электроэнцефалограммы ламантина была предпринята с
целью начать исследование сна этих животных, так как идентификация
сна млекопитающих невозможна без регистрации электрической актив-
ности их головного мозга.
Ранее у дельфинов афалин (Мухаметов и др., 1976) и дельфинов
азовок (Мухаметов, Полякова, 1981) было обнаружено явление одно-
полушарного медленноволнового сна. Оно состоит в том, что медленно-
волновая синхронизация электроэнцефалограммы возникает у дельфинов
только в одном полушарии — поочередно в правом и левом. Было вы-
сказано предположние, что его функциональным назначением является
обеспечение во время сна дыхательной активности, сложно организован-
ной у морских млекопитающих. Чтобы ответить на вопрос, является ли
однополушарный медленноволновый сон обязательным спутником пол-
ностью водного образа жизни млекопитающих, необходимо изучить в
этом отношении представителей отряда сирен, так как только китооб-
разные и сиреновые являются полностью водными млекопитающими.
Наша работа представляет собой первую попытку электроэнцефалографи-
ческого изучения сна одного из представителей отряда сирен.
Как показал морфоконтроль, ЭЭГ регистрировалась мононолярно от
передних отделов дорзальной поверхности коры двух полушарий. В ЭЭГ
ламантина были обнаружены обычные для млекопитающих картины де-
синхронизации, соответствующие бодрствованию, и синхронизации, соот-
ветствующие медленноволновому сну. Медленные волны имели пре-
383
имущественную частоту 1—3 в секунду. Во всех случаях медленные
волны развивались и исчезали одновременно в двух полушариях. Мед-
ленноволновая синхронизация регистрировалась преимущественно в ноч-
ное время, возможно, из-за меньшего количества пробуждающих факто-
ров. В общей сложности за время наблюдения медленные волны регист-
рировались около 3 ч. Во всех случаях медленные волны исчезали перед
дыхательным актом, заменяясь десинхронизацией, а после дыхательного
акта возникали снова, постепенно нарастая ио амплитуде.
Таким образом, в организации медленноволнового сна ламантина
и дельфинов имеется существенное различие. Основной формой сна дель-
финов является однополушарный медленноволновой сон (Мухаметов
и др., 1976; Мухаметов, Полякова, 1981). При этом дельтаволны одновре-
менно в двух полушариях не возникают. У ламантина в первых же двух
ночных сеансах медленные волны, в том числе дельтаволны, регистриро-
вались билатерально, т. е. так же, как у всех изученных к настоящему
времени наземных млекопитающих. Следовательно, уже сейчас можно
предполагать, что однополушарный медленноволновый сон является осо-
бенностью дельфинов и представляет собой лишь одну из возможных
форм приспособления полностью водных млекопитающих к дыханию во
время сна. Ламантины же, по-видимому, однополушарного сна не имеют
(это может быть доказано только в дальнейшем при увеличении числа
обследованных животных и удлинении сеансов полиграфической регист-
рации) и реализуют другую возможность — сон урывками во время дыха-
тельных пауз с пробуждениями на время дыхательных актов. Именно
такую структуру сна имеют каспийские тюлени — представители ласто-
ногих (Мухаметов и др., 1982).
Ламантин и дельфины оказались различными не только по организа-
ции сна, но и по чувствительности к общему наркозу. Хорошо известна
чрезвычайно высокая чувствительность дыхания дельфинов к различным
наркотическим веществам. По мере углубления, например, барбитурового
наркоза автономное дыхание дельфинов выключается раньше достиже-
ния хирургической стадии наркоза (Мухаметов, Супин, 1982).
Развитие наркотического состояния у ламантина было прослежено по
ходу его эвтаназии, для которой был использован нембутал. Внутримы-
шечно (в шейные мышцы) было введено в общей сложности 37,5мг/кг
нембутала пятью инъекциями по 7,5 мг/кг с интервалами в 8, 8, 13 и
11 мин. После последней инъекции ламантин был вытащен из воды на
сушу, он был полностью неподвижным и совершенно не реагировал на
разнообразные сильные раздражители и на явную боль. В то же время
он самостоятельно дышал. В сердечном ритме постепенно по мере разви-
тия наркоза исчезала дыхательная аритмия, ЧСР стабилизировалась на
уровне 25—30 ударов в минуту. Примерно через час после последней
инъекции нембутала пульс и дыхание ослабли, стали урежаться и прек-
ратились примерно за 5 мин.
Таким образом, у подопытного ламантина удалось дробными внутри-
мышечными инъекциями нембутала воспроизвести продолжительное со-
стояние наркоза хирургической глубины с полной нечувствительностью к
сенсорным и болевым раздражителям при сохранении самостоятельного
дыхания. Следовательно, дельфины и ламантины различаются и степенью
автономности их дыхания.
Глава тринадцатая
СПЕЦИФИКА АДАПТАЦИЙ СИРЕН
(вместо заключения)
Изучение сирен представляет огромный интерес и не только из-за релик-
товости и крайней экзотичности этих легендарных животных. Это —
хороший модельный объект для изучения эволюции и роли в ней адап-
таций. Приспосообительный характер многих черт сирен выявляется до-
статочно ясно, и этому способствуют относительно простые их связи со
средой, отсутствие у них конкурентов и врагов в лице крупных хищни-
ков, узкая экологическая специализация — растительноядность в услови-
ях мелководий с хорошей инсоляцией и практически неограниченными
кормовыми ресурсами. Все это способствовало при сохранении у них в
общем примитивной природы развитию сложных компенсаторных приспо-
соблений, затрагивающих многие системы органов. Палеозоогеографиче-
ская история сирен классически проста: с их древним ареалом, связан-
ным с палеогеновым морем Тетисом, лежащим в пределах тропической
и субтропической зон; с формированием по окраинам этого ареала трех
самостоятельных линий развития сирен, приведших к появлению трех
современных родов; с исчезновением примитивных групп сирен в центре
самого ареала и поздним выходом трихехид за пределы своего узкого
изолированного центра образования и их широкой экспансией в пределы
древнего ареала с частичным возвратом к питанию исходным для сирен
кормом — морскими травами (seagrasses), но сохранением потребности в
пресной воде; с возникновением особой группы дюгонин, связанных с
обитанием в исключительно морской воде и возможной повышенной
способностью к длительному плаванию и более глубокому погружению;
с необычной попыткой сирен (гидродамалины), перейдя на новый источ-
ник питания — морские водоросли, выйти за рамки тропиков и субтропи-
ков и овладеть местообитаниями в условиях пониженных температур.
К этому нужно добавить, что история сирен относительно хорошо доку-
ментирована в палеонтологической летописи, что не только позволяет го-
ворить о нескольких уровнях, достигнутых ими в ходе своего развития,
по и в ряде случаев проследить время появления тех или иных особен-
ностей, свойственных современным формам, путем сопоставления послед-
них с ископаемыми сиренами.
Изучение сирен до сих пор чаще шло под знаком представлений, что
они являются как бы стадией на пути овладения млекопитающими вод-
ной средой, в конце которого находятся китообразные (они и более ти-
хоходны, и не могут нырять на такие глубины, и гидродинамические их
характеристики явно уступают последним). Это, однако, мирно ужива-
лось, несмотря на противоречия, с признанием, что сирены, как и кито-
образные, также полностью порвали с сушей, что привело к утрате зад-
них конечностей, что они высокоспециализированы и хорошо приспособ-
лены к условиям водной среды. В результате обычаем было при сравне-
нии сирен и китообразных в первую очередь рассматривать черты, общие
для обеих групп, как результат приспособления к общим условиям сре-
ды, и различные — в качестве доказательства меньшей приспособленно-
сти сирен. При этом водная среда представлялась чем-то однородным,
равно выступающим перед лицом осваивающих ее животных, т. е. фак-
385
тически недооценивалась как специфичность разных водных экологиче-
ских ниш, так и специфичность самих путей овладения этими нишами,
накладываемая на различие в систематической принадлежности эволю-
ционирующих в этих условиях животных. На самом деле, китообразные
и сирены шли в своем развитии совершенно различными, даже не парал-
лельными путями, и совершалось это развитие в разных, а не одинако-
вых, условиях среды. Такая постановка вопроса в качестве методологи-
ческой предпосылки позволила в настоящем исследовании выявить
многочисленные, не обращавшие ранее на себя внимание специфические
особенности сирен в целом и по отдельности, в первую очередь касаю-
щиеся скелета и мышечной системы, позволяющие дать более углублен-
ную характеристику морфологических преобразований в каждой линии
развития сирен, вскрыть их адаптивный смысл (или направленность на
обеспечение определенных функций). При этом, конечно, не исключается,
что водная среда представляет к ее обитателям и ряд общих требований,
ответом на которые может быть появление в чем-то сходных морфологи-
ческих черт. Важно лишь подчеркнуть, что мы все еще не умеем доста-
точно полно и дифференцированно оценивать эти требования и их дав-
ление (через отбор) на ту или иную систему органов. Основное внима-
ние, уделенное в монографии костномышечной системе, объясняется тем,
что она равно участвует в тех изменениях, которые необходимы при пере-
ходе к обитанию в водной среде, будь то китообразные или сирены,—
изменениях, связанных с питанием (ротовой аппарат, желудочно-кишеч-
ный тракт), дыханием (верхние дыхательные пути, легкие, диафрагма,
грудная клетка, дыхательная мускулатура, сосудистая система) и локо-
моцией (осевой скелет и скелет конечностей, их мускулатура). Остальные
системы органов, в соответствии с имевшимся материалом и уровнем, до-
стигнутым в их изучении, рассмотрены более фрагментарно и, хотя при
их описании также выявлено много нового, они значительно меньше
дают для решения основной задачи монографии — выявления смысла
морфологической специфики сирен, а не просто выделения их диагности-
ческих черт.
На основе полученных данных и пересмотренного в новом свете ли-
тературного материала можно следующим образом определить адаптив-
ную специфику сирен в целом и в основных линиях их развития, при-
чем под адаптацией понимается вся цепочка, обеспечивающая приспо-
собление животного к среде: морфологическая структура — ее функции —
условия функционирования — адаптивное значение — среда, а не только
последние два звена. Естественно, даваемая нами характеристика не мо-
жет считаться всеобъемлющей и не все звенья этой цепочки ясны в рав-
ной мере. Главное внимание уделяется основному объекту изучения в
монографии — ламантину.
Всем сиренам присуще появление губного диска, своеобразного корот-
кого хобота, сформированного за счет увеличенной верхней губы и мяг-
ких тканей, окружающих носовой проход. Подвижность губного диска и
отдельных его частей друг относительно друга обеспечивает захват пищи
и перемещение ее в ротовую полость, где она перетирается совместным
действием роговых пластинок на увеличенной ростральной части предче-
люстных и челюстных костей (небная пластинка) и на симфизе нижней
челюсти (нижнечелюстная пластинка) и коренных зубов, расположенных
позади них. Образование губного диска и роговых пластинок вполне мож-
но расценивать как аутапоморфную черту сирен. В связи с развитием
подвижного губного диска сильно возрастает общая масса предглазнич-
ных лицевых мышц, управляющих движением верхней губы и клапана-
ми, отрывающими (путем оттягивания дна носовых проходов) и закры-
386
вающими наружные ноздри, помещающиеся на дорсальной стороне губ-
ного диска. При этом лицевая мускулатура не только не усложняется,
но, наоборот, упрощается, теряя ряд отдельных мышц или их порций.
В связи с отсутствием наружных слуховых органов исчезают все мышцы,
управляющие ушной раковиной; из всех дифференцированных у млеко-
питающих окологлазничных мышц остается одна круговая мышца глаза.
Изменения зубного аппарата у сирен и их значение подробно разбира-
лись в главе 1, и нет смысла их резюмировать здесь (это относится к
степени вентрального отгибания ростральной части черепа как показа-
теля уровня, на котором берется пища; храктеру смены зубов; развитию
функциональной гипсодонтности; приобретению постоянного роста корен-
ных зубов или полной их потери). Желудочно-кишечный тракт сирен
(см. гл. 7) имеет ряд специфических черт (малая величина слепой кишки,
что необычно для животных с постгастричным пищеварением, какими
являются сирены, объем которой, однако, увеличен за счет появления
особых, парных у ламантина и непарного у дюгоня, дивертикулов; уни-
кальное для млекопитающих формирование кардинального и парного
пилорического дивертикулов, последний увеличивает функционирующую
поверхность двенадцатиперстной кишки). Однако невозможность в на-
стоящее время исследовать биохимию пищеварения сирен ограничивает
интерпретацию этих данных, и в их числе различие ламантина и дюгоня
по относительному развитию тонкого (он преобладает у ламантина) и
толстого отделов кишечника. Высказывается лишь мысль, цто, возможно,
ламантины зависят в какой-то степени от животного белка, что они, в от-
личие от дюгоней, избегают грубой, богатой клетчаткой пищи.
В целом для сирен характерно развитие пахиостоза, захватывающе-
го кости черепа, конечностей, осевого скелета, в том числе и ребра.
При этом относительная масса скелета в сравнении с массой всего тела
сохраняется примерно на том же уровне, что и у наземных млекопита-
ющих. Однако для водного животного увеличение абсолютной массы ске-
лета приобретает важное значение: только он имеет удельный вес, пре-
вышающий таковой воды (расчет показывает, что за счет этого превы-
шения скелет компенсирует до 2/3 объема воды, вытесняемой
наполненными воздухом легкими), что позволяет животному, активно
изменяя объем своего тела за счет деятельности мышц брюшного пресса
и диафрагмы, менять свою плавучесть. Это ведет к уникальной способно-
сти сирен погружаться и подниматься на поверхность воды без актив-
ного участия локомоторных органов прямо вертикально без изменения
горизонтального положения тела, и в то же время минимализировать за-
траты энергии не только на ныряние, но и на саму локомоцию, поскольку
остается лишь необходимость преодолевать сопротивление воды, не счи-
таясь с силой тяжести или же чрезмерной плавучестью. Ничего подобно-
го нет у китообразных. Увеличение массы скелета и сосредоточение ее в
основном в осевой его части имеет и еще одно следствие, играющее важ-
ную гидростатическую роль: теперь масса ввиду сегментарности осевого
скелета имеет возможность так распределиться по длинной оси тела,
а также за счет ребер в стороны, чтобы способствовать удержанию ее в
преимущественно горизонтальном положении, важность чего для сирен
подчеркнута уже была ранее (Domning, 1977а). На это же направлено
вытягивание легких вдоль всего удлиненного грудного отдела и превра-
щение их в два длинных широко расставленных поплавка, увеличиваю-
щее остойчивость животного (катамаранный эффект), изменение относи-
тельного положения грудной и брюшной полостей, причем последняя
оказывается под первой.
Освоение новой экологической ниши вызвало у сирен развитие специ-
387
фического способа передвижения, лишь внешне напоминающего то, что
имеют китообразные, причем это касается как самой организации двига-
тельного акта, так и конструктивных особенностей локомоторного ап-
парата. Если пропульсия у китообразных вызывается прохождением по
всему туловищу бегущей волны изгиба в вертикальной плоскости, ам-
плитуда которого лишь уменьшена в передней части туловища в связи с
ее большей массой, то у сирен имеется четкое противопоставление не-
подвижного, или очень мало подвижного, длинного грудного отдела, не
участвующего непосредственно в создании пропульсии, и очень подвиж-
ного послегрудного, вооруженного хвостовым плавником, за счет которого
животное и двигается. При этом подвижный у наземных позвоночных
поясничный отдел оказался включенным фактически в грудной отдел
(маловероятной альтернативой кажется возможность почти полного его
исчезновения при одновременном значительном увеличении числа груд-
ных позвонков), а хвостовой отдел подвергся видоизменению, приобретя
на большем своем протяжении черты, обычно свойственные именно по-
ясничному отделу — увеличенные поперечнореберные отростки позвон-
ков, т. е. можно говорить о люмбализации хвостового отдела в противо-
положность торакализации поясничного отдела. Для китообразных ха-
рактерна обратная тенденция: не только заметное увеличение пояснично-
го отдела, вовлеченного в создание пропульсивной волны, и уменьшение
грудного, но и люмбализация задней части последнего. Соответственно,
у сирен глубокой перестройке подверглись осевой скелет и мускулатура,
и в частности ослабла эпаксиальная ее часть, а гипаксиальная, наоборот,
усилена (у китообразных соотношение обратное). Так, хвостовая под-
вздошнореберная мышца эпаксиального происхождения превратилась в
сгибатель хвоста, тогда как у китообразных соответствующая часть под-
вздошнореберной мышцы является его разгибателем. Для сирен вообще
характерно смещение таких эпаксиальных мышц, как длиннейшая
спины и подвздошнореберная, дальше от позвоночного столба по реб-
рам, что также свидетельствует об изменении их функций. Не исключено,
что активное, за счет действия мышц, разгибание спины у китообразных
у сирен в какой-то мере происходит в более автоматическом режиме в
результате использования сил упругости (у них сильно развиты ligg.
flava, хотя и несколько по-разному у ламантинов и дюгоней) и деформа-
ции, что и позволило ослабить эпаксиальный мышечный комплекс и пере-
ключить его на новые для него функции. Это лучше изучено у ламанти-
на, хотя возможно, что некоторые выявленные у него особенности, каса-
ющиеся туловищной мускулатуры, свойственны всем сиренам. Однако
пока это утверждать нельзя из-за неадекватности описания ее у дюгоня.
Как у всех китообразных, шейный отдел сирен укорочен (позвонки
уплощены спереди назад), подвижность его крайне мала (имеется систе-
ма фиксации, препятствующая разгибанию шеи; практически отсутству-
ет ротация в атлантоосевом суставе; атлант сильно отличается от того,
что имеется у наземных позвоночных) и, очевидно, шея двигается как
единое целое, что не могло не сказаться на шейной мускулатуре, в част-
ности лестничная мышца вообще потеряла связь с шейными позвонками.
Главными движениями в этой области у сирен стало поднимание и опу-
скание головы в атлантозатылочном суставе, что так необходимо им в
связи с добычей корма на разных уровнях в толще воды и с дыханием,
требующим поднятия конца морды над поверхностью. В то же время
выяснилось, что отличие шейных мышц сирен от других млекопитающих
не так велико, как предполагалось (Domning, 1977b, 1978а), о чем гово-
рит предлагаемая здесь новая система их гомологии (см. гл. 5). Боковая
подвижность шеи и головы также крайне ограничена, возможно, чтобы
388
рыскание головы из стороны в сторону не препятствовало локомоции.
Кстати, эта же причина может отвечать в какой-то мере и за само уко-
рочение шеи, обычно приписываемое необходимости достичь большей
обтекаемости, что вряд ли при свойственной сиренам скорости плавания
может приниматься во внимание.
Вопреки обычному утверждению, удлинение легких не влечет за со-
бой увеличения их объема, поскольку они сильно уплощены. Относитель-
ный их объем (5 л на 100 кг массы у ламантина) не превышает таковой
человека, ластоногих (5 л), многих наземных млекопитающих (6 л у
кошки, летучей мыши, 4 л у домовой мыши), но значительно уступает
ряду китообразных (7 л у афалины, морской свиньи, но 2,5—3 л у бу-
тылконоса, финвала, т. е. китов, способных к глубокому нырянию,
см. Яблоков и др., 1972). Выше уже было сказано, что возможно удли-
нение легких сирен связано с гидростатикой, а не самим дыханием, ко-
торое у них отличается не большим количеством вдыхаемого воздуха,
а большим процентом обмениваемого за один вдох. Может быть, именно
это обеспечивает наряду с другими функциями необычная конструкция
грудной клетки. Дыхание у сирен почти исключительно диафрагмальное.
Положение легких в широкой дуге, образованной ребром и замкнутой по
хорде диафрагмой, тянущейся на протяжении всей грудной клетки,
и само положение ребер относительно позвоночного столба (гл. 3) исклю-
чают какое-либо влияние подъема ребер на объем грудной полости.
Соответственно механизмы реберного дыхания, характерные для назем-
ных млекопитающих исчезают (так отсутствуют дифференцированные
мышцы — подниматели ребер), а межреберные мышцы принимают на
себя функцию фиксации положения ребер и тем самым увеличения же-
сткости грудной клетки (у наружных межреберных мышц даже меняет-
ся гистохимическая характеристика, см. гл. 11). В тесной связи с дыха-
нием в условиях погружения в воду стоит система запасников, накапли-
вающих кровь, имеющихся в той или иной мере у многих водных
млекопитающих, и особенно сирен и китообразных (чудесные сети, ве-
нозные сплетения и коллекторы). Однако развитие их у сирен имеет свою
специфику. Так, в брюшной полости главным депо служит правый ствол
полой вены в области почек, тогда как венозных синусов в печени нет;
в грудной полости развивается уникальный венозный синус перед правым
предсердием, чему способствует смещение легких назад за сердце, так
что они не мешают кровенаполнению этого синуса (гл. 8). Значение этих
структур как депо крови не вызывает сомнений, однако, неясно, почему
у сирен их развитие пошло своим путем.
Уже отмечалось (гл. 1), что ламантины, дюгони и морские коровы
представляют три главные пути развития сирен, причем ламантины очень
рано уклонились в сторону от пути, взятого дюгонидами, а дюгони и
морские коровы как представители разных подсемейств разошлись срав-
нительно поздно, причем произошло это на разных концах древнего ареа-
.ла, занимаемого примитивными дюгонидами. Ниже мы попытаемся оха-
рактеризовать своеобразие ламантиновой линии развития путем сопо-
ставления ее с дюгонидной, или с дюгоневой и гидродамалиновой в
отдельности. В главах 3—5 выявлено более 120 отличительных черт
разного ранга, касающихся строения осевого скелета, скелета передних
конечностей и всей мышечной системы. Естественно здесь можно будет
коснуться лишь некоторых из них. В первую очередь следует отметить
различия в гидростатических приспособлениях, направленных на удержа-
ние общего для сирен горизонтального положения тела и сохранение
центра тяжести скелета на середине грудного отдела. У ламантинов при
их относительно небольшом черепе и облегченном послегрудной отделе
389
увеличенная масса передних конечностей уравновешивается таким же
увеличением массы задних ребер. У дюгоня с большим массивным чере-
пом, но ослабленными передними конечностями смещение массы позво-
ночного столба к началу послегрудного отдела коменсируется увеличе-
нием массы передних ребер. В то же время возрастание массы ребер у
ламантина относительно массы самого позвоночного столба при их изме-
ненном положении обеспечивает распределение ее на большую площадь,
увеличивая остойчивость животного (у дюгоней ребра уже и менее мас-
сивны, поскольку содержат несколько больше губчатой ткани). О суще-
ственных различиях в биомеханике позвоночного столба ламантинов и
дюгонид', об адаптивном значении которых мы пока можем лишь дога-
дываться, говорят изменения размерных параметров позвонков по длин-
ной оси животного, которые заметно отличаются у обеих групп. Это ка-
сается темпов относительного увеличения и уменьшения высоты тела и
дуг позвонков, ширины и их длины, объема и массы позвонков, величи-
ны позвоночного отверстия, его пропорций и формы и т. п. В результате
при уменьшении у ламантинов числа позвонков во всех отделах позво-
ночного столба сравнительно с примитивными сиренами и всеми дюгони-
дами тела грудных позвонков у них оказываются значительно длиннее
(48—58% высоты позвонка), чем у дюгоня (30%) и морской коровы
(34%), что ведет к относительному возрастанию длины грудного отдела,,
но в то же время к уменьшению его гибкости, поскольку сокращается
число межпозвоночных дисков. Соответственно изменяются и другие ча-
сти позвонков: удлиняются суставные отростки, увеличиваются проме-
жутки между остистыми отростками и диапофизами, а следовательно,
и ребрами, что позволяет последним сильно расширяться, усиливаться
ligg. flava и межостистым мышцам, увеличиваться в размерах межпозво-
ночному отверстию и междуговому пространству и т. п. Все это приводит
к качественному изменению функций позвоночного столба ламантинов.
Так, изменения показателя bh2, отражающего момент сопротивления по-
звоночного столба, рассматриваемого как упругая балка, внешним силам,
действующим вертикально, и идущего параллельно изменениям других
размерных показателей позвонков, имеет у ламантина уникальный для
млекопитающих, наземных и водных, характер, достигая максимума в на-
чале второй половины грудного отдела, а не в начале послегрудного, как
у дюгонид и ряда китообразных, что, следовательно, можно оценить как
аутапоморфную черту ламантинов. Очевидно, такой сдвиг максимума всех
этих параметров вперед в пределы грудного отдела связан с необычной
способностью ламантинов подгибать хвост под себя, используя его то в
качестве опоры, то, возможно, иными, еще неизвестными способами. Это
ведет к глубочайшим перестройкам как в скелете животного, так и в его
мышечной системе, накладываемым на отмеченное выше свойственное
всем сиренам относительное возрастание роли гипаксиальных мышц и
уменьшение эпаксиальных. Достаточно сказать, что в мощнейший сгиба-
тель хвоста превратилась подкожная мышца туловища (также аутапо-
морфный признак), которая не только изменила свою ориентацию на
более вентральную сзади, но и путем установления тесной связи с пла-
тизмой и нижнечелюстной частью трапециевидной мышцы получила
спереди точку опоры на черепе, вовлекая, таким образом, в эту функцию1
мышцы, которые даже трудно было бы заподозрить во влиянии на хвост.
Мощными сгибателями хвоста служат теперь и косые и прямая мышцы-
живота, причем для усиления сгибательной мощи подкожной мышцы
туловища и косых мышц живота служат блоком, изменяющим направ-
ление приложения сил, вентральные концы последних ребер, положение1
которых фиксировано как с помощью особого органа жесткости, включа-
390
тощего ребра и первые послегрудные позвонки, так и специально диффе-
ренцированных наружных межреберных мышц. В фиксации грудной
клетки, необходимом условии приложения максимальных усилий мышц
именно на сгибание хвостовой части позвоночного столба, оказываются
замешанными и целый ряд других мышц. Сильному сгибанию хвоста
сразу же с начала послегрудного отдела способствует разрыв связи через
суставные отростки уже между II и III послегрудными позвонками, силь-
ный наклон поперечнореберных отростков III pth вниз, что увеличивает
угол приложения сил, действующих на сгиб, быстрое уменьшение высоты
остистых отростков (у ламантина они на XI pth составляют всего 12%
высоты позвонка, а у дюгоня на XII—XVI — 30—40%, столько же у
морской коровы на XXIII pth), что снижает сопротивление изгибу меж-
остистой связки. В целом можно сказать, что именно ламантины в наи-
большей степени решили задачу противопоставления жесткого грудного
отдела, неучаствующего в локомоции, подвижному послегрудному отделу.
В самом грудном отделе ламантинов можно найти массу отличий, гово-
рящих о разных условиях его функционирования у них и у дюгоней,
собственно адаптивный смысл которых еще неясен. Можно назвать и
большее сближение у ламантина суставных отростков позвонков, и более
высокое положение диапофизов, и изменение ориентации осей их фасеток
по ходу позвоночного столба, чего нет у дюгоней, и иная система крепле-
ния желтых связок, обеспечивающая большее их развитие и т. д. Ясно
только, что у ламантина в передней половине грудного отдела допусти-
мы боковые движения, полностью исключенные в задней части его и на
протяжении всего послегрудного отдела. В то же время сгибательные
движения возможны и на всем протяжении позвоночного столба (есть
система жесткой фиксации, препятствующая чрезмерному переразгиба-
нию), а с середины грудного отдела к ним присоединяются и вращатель-
ные движения. У морских коров на протяжении всего грудного отдела
допустимы преимущественно боковые движения, видимо, и в суженном
основании хвостового плавника, но вращательные должны быть сильно
ограничены. У дюгоня возможности, очевидно, промежуточные между
ламантином и морской коровой, хотя большее развитие у них, судя по
описаниям, многораздельных мышц, связанных обычно именно с враща-
тельными движениями, требует особого внимания. Послегрудной отдел
ламантинов и дюгонид также значительно различаются, что связано, ви-
димо, с иной организацией локомоторной волны (достаточно вспомнить
смещение наибольшего момента сопротивления позвоночного столба и
ряда размерных характеристик позвонков у дюгонид именно на начало
этого отдела), о чем говорит и разная форма хвостового плавника. Нали-
чие у дюгонид хвостового плавника, по форме напоминающего китовый,
вряд ли объясняется их более скоростными данными (в литературе нет
сведений о более быстром движении дюгоней сравнительно с ламантина-
ми, пока есть лишь обратные). Скорее это связано с большей продолжи-
тельностью плавания, меньшей утомляемостью, погружением на боль-
шую глубину. В то же время ясно, вопреки мнению, высказанному ранее
(гл. 6), что хвостовой плавник ламантинов с его уникальной формой и
большими размерами (он включает около 18 позвонков, что составляет
около четверти длины всего позвоночного столба) никак не должен рас-
сматриваться как промежуточная стадия к развитию плавника дюго-
ниевого типа на основе поверхностного сравнения со стадиями развития
плавника у китообразных, у которых в него включено значительно
меньше позвонков, чем у сирен (не более 12% длины позвоночного
столба, см. Slijper, 1936). Появление особой формы хвостового плавника
у ламантина вероятнее всего следует рассматривать как новоприобрете-
391
ние в связи с приспособлением его не только к участию в локомоции,
но и в функции опоры или чем-то подобном. Не случайно, что очень
многие особенности скелета и мускулатуры ламантина объяснимы лишь
как приспособления к увеличенному сгибанию хвоста.
Особая, уникальная для млекопитающих, черта ламантинов — сокра-
щение числа позвонков в шее до шести в связи с включением последнего
шейного позвонка в грудной отдел. Более того, даже VI с ламантина по
некоторым своим особенностям уподобляется грудным позвонкам (тора-
кализация). У дюгонид — противоположная тенденция: первые грудные
позвонки как бы цервикализируются и включаются вместе с шейными
позвонками в единую систему жесткой фиксации путем черепицеобраз-
ного налегания невральных дуг (передних на задние), черта, заставляю-
щая вспомнить некоторых китообразных. У ламантина, наоборот, послед-
ние (IV—VI) шейные позвонки более самостоятельны, а фиксация первых
трех осуществляется совсем иначе, чем у дюгонид (гл. 3). Таким обра-
зом, отмеченные выше общие для всех сирен особенности шейного отде-
ла позвоночного столба достигаются у ламантинов и дюгонид разными
средствами. Естественно ожидать, что такие различия должны быть
функционально обусловлены. Учитывая раннее появление «черепицы» в
эволюции сирен, следует признать, что путь, принятый ламантинами в-
своем развитии в отношении шейного отдела, является явным уклонени-
ем от общей линии эволюции сирен. Отмеченные в главе 3 различия в
строении атланто-затылочного сустава и связанные с приспособлением к
добыванию корма на разных уровнях скорее характеризуют адаптивную
специфику на видовом уровне. Важно лишь подчеркнуть, что эта адап-
тация направлена на смягчение жесткого приспособления к питанию на
дне или в толще воды, вовлекающего изменение наклона ростральной
части черепа и симфиза нижней челюсти.
Общий для сирен переход на диафрагмальное дыхание тем не менее1
сочетается с различиями в положении диафрагмы у ламантина, с одной
стороны, и дюгонид, с другой. Если у первого на всем своем протяжении
она горизонтальна, будучи закреплена на особых гребнях тел большого
числа (11—14) грудных позвонков, то для дюгонид, видимо, следует до-
пустить более косое положение передней части диафрагмы, закрепленной
у них лишь на последних 4—6 грудных позвонках. Возможно, это сви-
детельствует о несколько большей роли у них реберного дыхания. Более
подвижны у дюгонид и ребра. Достаточно сказать, что большая часть их
ребер имеет однораздельную головку и сочленяется с позвоночным стол-
бом, следовательно, лишь в двух точках, что определяет фиксацию одно-
го направления возможных движений ребер. В свое время было показа-
но, что именно это является условием участия ребер в дыхании (Slij-
per, 1936). У ламантинов, наоборот, большая часть ребер (первые 12)
имеет двураздельные головки и, таким образом, сочленяется с позвоноч-
ником сразу в трех точках, что безусловно ограничивает любое направ-
ление движения ребра. В этом же плане действуют капсулы реберно-
позвоночных суставов, видимо, очень жесткие у ламантина и более сво-
бодные у дюгоня. Последние, в частности, допускают, возможно, не толь-
ко большую подвижность, но и сближение по ходу позвоночного столба
головок и бугорков ребер, что приводит к появлению у дюгонид в конце
грудного отдела 1—4 одноглавых ребер. Но, как уже говорилось выше,
любые движения ребер сирен не увеличивают объем грудной полости и,
следовательно, мало могут способствовать повышению эффективности
дыхания. Таким образом, причины большей подвижности ребер у дюгоня
пока неясны, хотя они могут быть связаны с большей подвижностью их
туловища. Пожалуй легче понять, почему ребра ламантинов закреплены
392
более жестко. Возможно, это связано с доведенной у ламантинов до пре-
дела общей тенденцией сирен к противопоставлению жесткого грудного и
подвижного послегрудного отделов, когда ребра служат лишь местом
опоры или прямого прикрепления многих мышц туловища, участвующих
в сгибательных движениях хвоста (отсюда расширение ребер и их силь-
ное отклонение назад). В этой ситуации ребра могут работать и как
рычаги, обеспечивающие сгибание грудного отдела туловища. В целом
как морфология скелета и туловищной мускулатуры ламантина, с одной
стороны, и дюгоня и морской коровы, с другой, так и обусловленные ею
двигательные их потенции, очень сильно различаются. Дюгони и морские
коровы в этом отношении явно представляют единый тип адаптации
(речь идет о локомоции и дыхании, движениях головы и шеи). Их спе-
цифические пути развития были связаны уже с другим — с разной ролью
передних конечностей в их жизнедеятельности. Ламантины, со своей
стороны, избрали вновь отличный путь эволюции. У них резко повышена
активная роль передних конечностей, превратившихся не только в искус-
ные манипуляторы, но и в мощные пропульсивные органы, обеспечива-
ющие крайнюю их маневренность, резкие повороты (вспомним их акро-
батические способности), движения по грунту и в воде, что, видимо,
совсем не свойственно дюгоням (гл. 4, 5). Причем связанные с этим у
ламантинов конструктивные изменения в скелете и мускулатуре конечно-
стей носят явно вторичный характер, а не обусловлены сохранением черт
наземных предков с более развитыми конечностями. Достаточно назвать
уникальные для млекопитающих преобразования проксимального эпифи-
за плечевой кости и сам факт превращения в основное силовое движе-
ние в плечевом суставе пронации плеча, а не обычной ретракции (сги-
бания), что вызвало соответствующую перестройку целого комплекса
мышц. У дюгоней, как, видимо, и у всех примитивных дюгонид, перед-
ние конечности слабее развиты и работают преимущественно как рули
глубины при пропульсии, вызываемой работой хвоста, и у них развива-
ются приспособления, обеспечивающие, в первую очередь, супинацион-
ные движения плеча, переводящие ласт в рабочее положение. Уникаль-
ное использование морскими коровами передних конечностей как своеоб-
разных багров при движении в условиях мелководий и бурного волнения?
воды из-за прибоя и открытости береговой линии обусловило их пере-
стройку, обеспечивающую парасагиттальное положение их конечностей,
усиление ретракции, превращение кисти в особые крючки и т. п. Вряд
ли могут быть сомнения во вторичности этих приспособлений. Вместе со
всеми дюгонидами предки морских коров прошли длинный путь разви-
тия, связанный, в частности, с частичной редукцией передних конеч-
ностей.
В заключение хочется отметить, что настоящее исследование во мно-
гом носит предварительный характер. Не все выявленные отличия совре-
менных сирен могут еще быть объяснены как в отношении их адаптив-
ного значения, так и роли в эволюции. Необходимо продолжать работу
в этом направлении с привлечением новых материалов. В то же время и
не все, что было обнаружено нового, в ходе исследования удалось отра-
зить в этом кратком заключении.
ЛИТЕРАТУРА
Агарков Г. В., Хоменко В. Г., Хаджин-
ский В. Г. Морфология дельфинов.
Киев: Наук, думка, 1974. 166 с.
Андреев Ф. В. К структуре глаза белого
медведя,— В кн.: Экология и морфоло-
гия белого медведя. М.: Наука, 1973,
с. 99—107.
Андреев Ф. В. Структурно-функциональ-
ные особенности глаза дельфинов и
ластоногих,— В кн.: Новое в изучении
китообразных и ластоногих. М.: Наука,
1978а, с. 116—133.
Андреев Ф. В. Сравнительный анализ ор-
гана зрения наземных и водных млеко-
питающих.— Тез. докл. II съезда Все-
союз. териол.о-ва. М.: Наука, 19786,
с. 42—43.
Андреев Ф. В. Строение глаза зубра и би-
зона.— В кн.: Зубр.: Морфология, систе-
матика, эволюция, экология. М.: Наука,
1979, с. 389—398.
Андреев Ф. В. О строении глаза ламан-
тина.— В кн.: Тез. докл. VIII Всесоюз.
совещ. по изучению, охране и рацио-
нальному использованию морских мле-
копитающих. Астрахань, 1982, с. 11—13.
Баженов Ю. И. О терморегуляции при мы-
шечной работе в холоде.— В кн.: Фи-
зиологические адаптации к теплу и хо-
лоду. Л.: Наука, 1969, с. 135—140.
Бергнер X., Бетц Х.-А. Научные основы
питания сельскохозяйственных живот-
ных. М.: Колос. 597 с.
Блинков С. М., Моисеев Г. Д. Определение
плотности капиллярной сети в органах
и тканях человека и животных незави-
симо от толщины микротомного среза.—
Докл. АН СССР, 1961, т. 440, № 2,
с. 465—468.
Браун А. А. Слои соединительнотканной
части кожи позвоночных и их номен-
клатура,—Докл. АН СССР, 1945, т. 46,
№ 2, с. 80—83.
.Галанцев В. И. Эволюция адаптации ны-
ряющих животных. Л.: Наука, 1977,
191 с.
Галанцев В. П., Маминов М. И. Различия
функции сердечного автоматизма у де-
тенышей курильского тюленя и морско-
го котика,—Зоол. журн., 1979, т. 58,
№ 10, с. 1565—1573.
Гамбарян П. П. Приспособительные осо-
бенности органов движения роющих
млекопитающих. Ереван: Изд-во АН
АрмССР, 1960. 195 с.
Гамбарян П. П. Бег млекопитающих:
Приспособительные особенности орга-
нов движения: Л.: Наука, 1972. 334 с.
Гамбарян П. П., Рухкян Р. Г. Морфофунк-
циональный анализ мышц конечностей
слонов.— В кн.: Функциональная мор-
фология млекопитающих. Л.: Наука,
1974, с. 190—265. Тр. Зоол. ин-та АН
СССР; Т. 54.
Гептнер В. Г. Отряд морских коров или
сирен. Ordo Sirenia liliger, 1811.— В кн.:
Млекопитающие Советского Союза. Т. 2.
Ч. 1. Морские коровы и хищные. М.:
Высш, шк., 1967, с. 13—46.
Гетманова Т. И. Сравнительная ангиоар-
хитектоника почек некоторых грызу-
нов.— Журн. эволюц. биохимии и фи-
зиологии, 1967, т. 12, № 2, с. 128—133.
Гетманова Т. И., Филюшина Е. Е., Шмер-
линг М. Д. Некоторые морфологические
особенности скелетных мышечных во-
локон байкальской нерпы.— В кн.: Тез.
докл. VIII Всесоюз. совещ. по изуче-
нию, охране и рациональному использо-
ванию морских млекопитающих. Астра-
хань, 1982, с. 86—87.
Давыдов А. Ф., Кузьма Р. И. Терморегу-
ляция у нутрии и ондатры в связи со
специализацией к полуводному образу
жизни.— В кн.: Физиологические адап-
тации к теплу и холоду. Л.: Наука, 1969,
с. 78—90.
Джонсон П. Периферическое кровообра-
щение. М.: Медицина, 1982. 440 с.
Дозье Т. Киты и другие морские млеко-
питающие. М.: Мир, 1980. 129 с.
Елякова Г. В. Электронно-микроскопиче-
ское исследование образования миобла-
стов в регенерирующей мышечной
ткани.—Докл. АН СССР, 1972, т. 202,
№ 5, с. 1196—1198.
Жеденов В. И. Легкие и сердце животных
и человека. М.: Высш, шк., 1961. 474 с.
Иванов К. П. Мышечная система и хими-
ческая терморегуляция. М.; Л., Наука,
1965. 127 с.
Иванова Е. И. Адаптивные особенности
морфологии кровеносной и дыхательной
систем русской выхухоли,— В кн.: Мор-
фологические особенности водных мле-
копитающих. М.: Наука, 1964, с. 82—127.
Иванова Е. И. Функциональная морфоло-
гия сосудистой системы млекопитаю-
щих. М.: Наука, 1975. 55 с.
Калашникова М. М. Ультраструктура ге-
патоцита при накоплении и выделении
продуктов желчи,— Бюл. эксперим.
394
биологии и медицины, 1981, № 5, с. 620—
623.
Климов А. А. Гистогенез и регенерация
тканей. Л.: Медицина, 1984. 232 с.
Клыков В. И. Морфофункциональный ана-
лиз локтевого сустава некоторых ко-
пытных: Автореф. дис. ... канд. биол.
наук. Киев, 1978. 20 с.
Колчинская А. 3., Манъковская И. Н., Ми-
сюра А. Г. Дыхание и кислородные ре-
жимы организма дельфинов. Киев:
Наук, думка, 1980. 331 с.
Манъковская И. Н. Содержание и распре-
деление миоглобина в мышечной ткани
черноморских дельфинов.— Журн. эво-
люц. биохимии и физиологии, 1975, т. И,
№ 3, с. 263—267.
Мельник К. П. Морфология и биомехани-
ческие свойства скелета млекопитаю-
щих в связи с особенностями их локо-
моции: Автореф. дис. ... д-ра биол. наук.
Киев, 1978. 52 с.
Мельников В. В. Кровеносное русло голо-
вы, шеи и спиногрудного отдела туло-
вища китообразных (Cetacea): Автореф.
дис. ... капд. биол. наук. Владивосток,
1981. 23 с.
Михайлов И. Н. Структура и функция
эпидермиса. М.: Наука, 1979. 238 с.
Мухаметов Л. М., Полякова И. Г. Электро-
энцефалографическое исследование сна
у дельфинов азовок.— Журн. высш,
нерв, деятельности им. И. П. Павлова,
1981, т. 31, № 2, с. 333—339.
Мухаметов Л. М., Супин А. Я. ЭЭГ-иссле-
дование различных поведенческих сос-
тояний свободноподвижных дельфи-
нов.— Журн. высш. нерв, деятельности,
1975, т. 25, № 2, с. 396—401.
Мухаметов Л. М., Супин А. Я. Электро-
энцефалографическое исследование бар-
битурового наркоза у дельфинов афа-
лин.— Журн. эволюц. биохимии и физи-
ологии, 1982, т. 18, № 2, с. 191—194.
Мухаметов Л. М., Супин А. Я., Поляко-
ва И. Г. Электроэнцефалографическое
исследование сна у каспийских тюле-
ней— Докл. АН СССР, 1982, т. 266, № 3,
с. 752—755.
Мухаметов Л. М., Супин А. Я., Строко-
ва И. Г. Межполушарная ассиметрия
функциональных состояний мозга во
время сна у дельфинов.— Докл. АН
СССР, 1976, т. 229, № 3, с. 767—770.
Наумов Н. П., Карташев Н. Н. Зоология
позвоночных. Ч. 2. Пресмыкающиеся,
птицы, млекопитающие. М.: Высш, шк.,
1979. 272 с.
Наумова Е. И. Копрофагия у зайцеобраз-
ных и грызунов в свете микроскопиче-
ского и гистохимического строения пи-
щеварительного тракта.— Зоол. журн.,
1974, т. 53, вып. 12, с. 1848—1855.
Наумова Е. И. Пути специализации пище-
варительной системы потребляющих
клетчатку млекопитающих (Rodentia,
Lagomorpha) к обеспечению организма
пищевым белком.— Журн. общ. биоло-
гии, 1976, т. 37, № 2, с. 276—285.
Наумова Е. И. Функциональная морфоло-
гия пищеварительной системы грызу-
нов и зайцеобразных. М.: Наука, 1981.
262 с.
Огнев С. Н. Зоология позвоночных. М.:
Сов. наука, 1945. 519 с.
Орлов О. ТО. Об эволюции зрения у позво-
ночных.— В кн.: Проблемы эволюции.
Новосибирск: Наука, 1972, с. 69—94.
Ларин В. В., Баевский Р. М., Волков Ю. Н.,
Газенко О. Г. Космическая кардиоло-
гия. Л.: Медицина, 1967. 206 с.
Першин С. В. Оптимизация кормового
плавникового движителя в природе на
примере китообразных.— В кн.: Биони-
ка. Киев: Наук, думка, 1969, вып. 3,
с. 26—34.
Першин С. В., Соколов А. С., Томи-
лин А. Г. О регулируемой специаль-
ными сосудистыми органами упругости
плавников дельфина.— Докл. АН СССР,
1970, т. 190, № 3, с. 709—712.
Покровский А. Л., Ташев Т. А., Тутель-
ян В. А. и др. Изучение механизма об-
разования вторичных лизосом, морфо-
логические и биохимические исследова-
ния на экспериментальной модели ин-
дуцированного глюкагоном клеточного
аутофагоцита.— Цитология, 1977, т. 19,
№ 10, с. 1171—1175.
Покровский А. Л., Тутельян В. А. Лизо-
сомы. М.: Наука, 1976. 382 с.
Родионов В. А. О строении, функции и
происхождении кожной мускулатуры у
китообразных.— В кн.: Морские млеко-
питающие. М.: Наука, 1978, с. 17—27.
Родионов В. А. Гистохимическое изучение
распределения миоглобина в мышцах
некоторых млекопитающих.— В кн.:
Дыхательные белки некоторых групп
современных животных. М.: Наука,.
1979, с. 38—41.
Родионов В. А. Гистохимическое изучение-
мускулатуры грызунов.— В кн.: Грызу-
ны. Материалы V Всесоюз. совещ. Са-
ратов, 3—5 дек., 1980 г. М.: Наука, 1980,
с. 104—106.
Родионов В. А. Гистохимическая структу-
ра мышц у морских млекопитающих.—
В кн.: Тез. докл. VIII Всесоюз. совещ.
по изучению, охране и использованию
морских млекопитающих. Астрахань,
1982а, с. 313—315.
Родионов В. А. Основные особенности
анатомического строения мускулатуры
у дельфинов в связи с приспособлени-
ем к водному образу жизни.— В кн.:
Тез. докл. VIII Всесоюз. совещ. по изу-
чению, охране и использованию мор-
ских млекопитающих. Астрахань, 19826,
с. 315—316.
Роскин Г. И., Левинсон Л. Б. Микроско-
пическая техника. М.: Сов. наука, 1957,
с. 248—249.
Слоним А. Д., Туманова Н. М. К вопросу
о терморегуляторной электрической ак-
тивности мышечных волокон разных
типов у белых крыс, адаптированных к
различным температурам среды.— Фи-
395
зиол. журн. СССР, 1973, т. 59, № 3,
с. 590—595.
•Смит Д. Скелетная, сердечная и гладкая
мышечная ткань.— В кн.: Электронно-
микроскопическая анатомия. М.: Мир,
1967, с. 141—155.
Соколов А. С. Некоторые функционально-
морфологические и возрастные особен-
ности ладожского тюленя в связи с вод-
ным образом жизни: Автореф. дис. ...
канд. биол. наук. Л., 1954. 22 с.
Соколов В. Е. Структура кожного покро-
ва некоторых китообразных. Сообщение
II.— Науч. докл. высш. шк. Биол. на-
уки, 1962, № 3, с. 45—55.
Соколов В. Е. Кожный покров млекопи-
тающих. М.: Наука, 1973. 485 с.
•Соколов В. Е. Систематика млекопитаю-
щих. Отряды: китообразных, хищных,
ластоногих, трубкозубых, хоботных, да-
манов, сирен, парнокопытных, мозоле-
ногих, непарнокопытных. М.: Высш, шк.,
1979. 528 с.
Соколов В. Е., Калашникова М. М., Ро-
дионов В. А. Микро- и ультраструктура
кожи морской свиньи.— В кн.: Морфо-
логия и экология морских млекопитаю-
щих. М.: Наука, 1971, с. 26—46.
Соколов В. Е., Кузнецов Г. В. Суточные
ритмы активности млекопитающих. М.:
Наука, 1978. 264 с.
Соколов В. Е., Родионов В. А. Гистохими-
ческое изучение сукцинатдегидрогена-
зы в мышцах дельфина афалины Tur-
siops truncatus (Delphinidae, Cetacea).—
Докл. АН СССР, 1978, т. 238, № 5,
с. 1252—1255.
Соколов В. Е., Чернова О. Ф., Сэйл Дж.
Кожный покров тропических копытных.
М.: Наука, 1984. 166 с.
•Соловьев В. А. Морфогистохимическая ха-
рактеристика мионов жевательной
мышцы млекопитающих и человека.—
Арх. анатомии, гистологии и эмбриоло-
гии, 1982, т. 83, № 10, с. 65—71.
Студитский А. Н. Трансплантация мышц
у животных. М.: Медицина, 1977. 248 с.
Тиняков Г. Г., Чумакова В. П., Севастья-
нов Б. А., Макаев В. М. О структуре
мышечных волокон у китов,— Зоол.
журн., 1976, т. 55, № 2, с. 311—315.
Томилин А. Г. Отряд Сирены (Sirenia).—
В кн.: Жизнь животных. Т. 6. Млекопи-
тающие, или звери. М.: Просвещение,
1971, с. 406—410.
Томилин А. Г. Снова в воду. М.: Знание,
1977. 144 с.
Туманова Н. М. Участие быстрых и мед-
ленных волокон в изменениях электри-
ческой активности скелетных мышц
крыс при остром и хроническом дей-
ствии холода.— Физиол. журн., 1972,
т. 58, № 11, с. 1765—1768.
Умнова М. М., Новоселова И. Л. Гието- и
ультраструктура скелетной мышцы ла-
мантина.— В кн.: Тез. докл. VIII Все-
союз. совещ. по изучению, охране и ра-
циональному использованию морских
млекопитающих. Астрахань, 1982,
с. 378—380.
Черняев Э. Г. Особенности строения зад-
ней полой вены некоторых насекомояд-
ных.— Докл. АН СССР, 1964, т. 156, № 6,
с. 1483—1485.
Шмерлинг М. Д., Филюшина Е. Е., Бу-
зуева И. И. Изменения ультраструкту-
ры скелетных мышечных волокон под
влиянием острого физиологического на-
пряжения.— Арх. анатомии, гистологии
и эмбриологии, 1981, № 2, с. 43—48.
Шуров В. А. К морфологии почек мор-
ских млекопитающих.— В кн.: Морские
млекопитающие. Киев, 1975, выл. 2,
с. 182—183.
Яблоков А. В. Некоторые особенности мо-
чеполовой системы китообразных.—
Бюл. Моск, о-ва испытателей природы,
1961, т. 66, вып. 2, с. 33—38.
Яблоков А. В., Белькович В. М., Бори-
сов В. И. Киты и дельфины. М.: Наука,
1972. 472 с.
Яблоков А. В., Клевезаль Г. А. Вибриссы
китообразных и ластоногих, их распре-
деление, строение и значение.— В кн.:
Морфологические особенности водных
млекопитающих. М.: Наука, 1964,
с. 48—82.
Abel О. Die Sirenen der mediterranen Ter-
tiarbildungen Oesterreichs.— Abh. Geol.
Reichsanst., 1904, Bd. 19, H. 2, S. 1—223.
Abel O. Die Stammesgeschichte der Meeres-
saugetiere. B., 1907. 914 S.
Abel O. Die Stamme der Wirbeltiere. Ber-
lin; Leipzig, 1919. 423 S.
Abel O. Vorgeschichte der Sirenia.— In:
Weber M. Die Saugetiere. 2. Aufl. Jena,
1928, Bd. 2, S. 496—504.
Albrecht P. Note sur la presence d’epiphy-
ses terminales sur les corps des vertebres
d’un exemplaire de Manatus americanus
Desm.— Bull. Mus. hist, natur. Beige,
1883, t. 2, p. 35—38.
Allen J. A. Preliminary list of works and
papers relating to the mammalian orders
Cete and Sirenia.— Bull. US Geol. Surv.
Territories, 1881, vol. 6, N 3, p. 399—562.
Anderson P. K. The behavior of the dugong
in relation to conservation and mana-
gement.— Bull. Mar. Sci., 1981a, vol. 31.
N 3, p. 604—647.
Anderson P. K. Dugong behaviour: Obser-
vations, extrapolations and speculati-
ons.— In: The dugong: Proc, of a semi-
nar/workshop hold at James Cook Univ,
of North Queensland, 8—13 May 1979/Ed.
H. Marsh. Townsville, 1981b, p. 91—111.
Andres К. H. Uber die Feinstruktur der
Rezeptoren an Sinushaaren.— Ztschr.
Zellforsch., 1966, Bd. 75, S. 339—365.
Andrews C. W. A descriptive catalogue of
the Tertiary vertebrate of the Fayum.
Egypt, based on the collection of the
Egyptian Government in the Geological
Museum, Cairo, and on the collection in
the British Museum (Natural History), L.,
1906. 324 p.
396
Annandale N. The appendicular skeleton
of the dugong (Halicore dugong)Rec.
Ind. Mus. Calcutta, 1907, vol. 1, p. 79.
Anon. An International Centre for Manatee
Research: Rep. of a workshop held 7—
13 Fehr. 1974, Georgetown, Guyana, South
America. Georgetown: Nat. Sci. Res.
Council, 1974. 34 p.
Bahrdt H. J. Beitriige zur Entwicklungsges-
chichte der Sirenenflosse.— Jen. Ztschr.
Naturwiss., 1933, Bd. 68, H. 1, S. 193—276.
Barnett С. H., Harrison R. J., Tomlin-
son L. D. W. Variations in the venous
systems of mammals.— Biol. Rev., 1958,
vol. 33, N 4, p. 442—487.
Barrett 0. W. Notes concerning manatees
and dugongs.— J. Mammal., 1935, vol. 16,
p. 216—220.
Beddard F. F. Notes on the anatomy a ma-
natee (T. inunguis) lately living in the
Society’s gardens.— Proc. Zool. Soc. Lon-
don, 1897, p. 47—53.
Bertram G. C. L. Dugong number in retros-
pect and prospect.— In: The dugong:
Proc, of a seminar/workshop hold at Ja-
mes Cook Univ, of North Queensland,
8—11 May 1979/Ed. H. Marsh. Townsville,
1981, p. 1—7.
Bertram G. C. L., Bertram С. K. R. The mo-
dern Sirenia: Their distribution and sta-
tus.— Biol. J. Linn. Soc., 1973, vol. 5,
N 4, p. 297—338.
Best R. Foods and feeding habits of wild
and captive Sirenia.— Mammal. Rev.,
1981, vol. 11, N 1, p. 3—29.
Birchmeier W. Cytoskeleton structure and
function.— Trends Biochem. Sci., 1984,
vol. 8, N 4, p. 192—195.
Blaniville H. M. D. de. Osteographie ou des-
cription iconographique comparee due
squelette et du systeme dentaire des
mammiferes recents et fossiles pour ser-
vir de base a la zoologie et a la geolo-
gie. P., 1839, liv. 15. 140 p.
Blessing M. H. Studies on the concentra-
tion of myoglobin in the sea-cow and por-
poise.— Comp. Biochem. and Physiol. A,
1972, vol. 41, N 3, p. 475—480.
Blix A. S., Grav H. G., Ronald K. Some as-
pects of temperature regulation in new-
born harp seal pups.— Amer. J. Physiol.,
1979, vol. 236, N 3, p. 188—197.
Brandt A. Uber die Haut der nordischen
Seekuh (Rhytina borealis Ill.).— Mem.
Acad. imp. sci. St. Petersbourg. Ser. 7,
1871, t. 17, N 7, p. 1—28.
Brandt J. F. Symbolae sirenologicae quibus
praecipue Rhytinae historia naturalis il-
lustratur. Pertropoli: Typus Academiae
Scientiarum, 1846, 160 p.; Engl, transl.
Contributions to sirenology, being princi-
pally an illustrated natural history of
Rhytina. Belgrad, 1974. 122 p.
Brandt J. F. Bemerkungen die Zahl der
Halswirbel iiber der Sirenen.— Bull.
Acad. imp. sci. St. Petersbourg, 1863, t. 5,
p. 7—10.
Brandt J. F. Symbolae sirenologica. Fasc.
II, III. Sireniorum, Pachydermatum,
Zeuglodontum et Cetaceorum ordinis os-
teologia comparata, nec non Sireniorum
generum monographiae.— Mem. Acad,
imp. sci. St. Petersbourg. Ser. 7,1861/1868,
t. 12, N 1, p. 1—383.
Breathnach A. S. An atlas of the ultra-
structure of human skin. L., 1971. 398 p.
Brody S. The keratinization of epidermal
cells of normal guinea pig skin as re-
vealed by electron microscopy.— J. Ultra-
struct. Res., 1959, vol. 2, p. 482—511.
Brown W. R., Geraci J. R., Hicks B. D.,
Aubin D. J. Epidermal cell proliferation
in the bottlenose dolphin (Tursiops
truncatus).— Canad. J. Zook, 1983, vol. 61,
p. 1587—1590.
Bryden M. M., Marsh H., Macdonald В. M.
The skin and hair of the dugong, Dugong
dugon.— J. Anat., 1978, vol. 126, p. 637—
638.
Bukman M. M., Lobitz C. J., Wuepper L. D.
Epidermal transglutaminase identificati-
on and purification of a soluble substra-
te with studies of in vitro cross-linking.—
J. Biol. Chem., 1980, vol. 255,
p. 1200—1203.
Bullock T. H., Domning D. P., Best R. Avo-
ked brain potentials demonstrate hearing
m a manatee (Sirenia: Trichechus inun-
guis) .— J. Mammal., 1980, vol. 61, N 1,
p. 130—133.
Bullok T. H., O’Shea T. J., McClune M. C.
Auditore evoked potentials in the West
Indian manatee (Sirenia: Trichechus ma-
natus) .— J. Comp. Physiol., 1982, vol. 148,
p. 547—554.
Caldwell F., Sherman E., Levitsky K. The
composition of bladder bile and histolo-
gical pattern of the gallbladder and li-
ver of the sea cow.— Comp. Biochem. and
Physiol., 1969, vol. 28, Nl, p. 437—441.
Cardell R. R. Smooth endoplasmik reticu-
lum in rat hepatocytes during glycogen
deposition and depletion.— Intern. Rev.
Cytol., 1977, vol. 48, p. 221—227.
Cave A. J., Aumonier F. J. Morphology of
the cetacean reniculus.— Nature, 1962,
vol. 193, N 4817, p. 799-800.
Chapman H. C. Observations on the structu-
re of the manatee.— Proc. Acad. Natur.
Sci. Phila., 1875, p. 187.
Clemens E. T., Maloly G. M. O. The digesti-
ve physiology of three East African her-
bivores: The elephant, rhinoceros and
hippopotamus.— J. Zool., 1982, vol. 198,
N 2, p. 141—156.
Cohen J. L., Tucker G. S., Odell D. K. The
photoreceptors of the West Indian mana-
tee.—J. Morphol., 1982, vol. 173, N 2,
p. 197—202.
Coppens Y., Beden M. Moeritherioidea.—
In: Evolution of African mammals/Ed.
V. J. Maglio, H. B. S. Cooke. Cambridge;
London, 1978, p. 333—335.
Cozzi B. Osservazioni sulla morfologia del-
la colonna vertebrale nei Cetacei.— Atti
Soc. ital. sci. natur., 1981, vol. 122, N 3/4,
p. 225—235.
Dechaseaux C. Sirenia.— In: Traite de pa-
397
leontologie. P., 1958, pt 6. Mammiferes.
Evolution, vol. 2, p. 333—361.
Domning D. P. An ecological model for
Late Tertiary sirenian evolution in the
North Pacific Ocean.— Syst. ZooL, 1977a,
vol. 25, N 4, p. 352—362.
Domning D. P. Observations on the myolo-
gy of Dugong dugon (Miiller).—Smith-
sonian Contribs Zool., 1977b, N 226,
p. 1—57.
Domning D. P. The myology of the Amazo-
nian manatee. Trichechus inunguis (Nat-
terer) (Mammalia: Sirenia).— Acta Ama-
zonica, 1978a, vol. 8, N 2, p. 1—81.
Domning D. P. Sirenia.— In: Evolution of
African mammals/Ed. V. J. Maglio,
H. B. S. Cooke. Cambridge; London, 1978b,
p. 573—581.
Domning D. P. Sirenian evolution in the
North Pacific Ocean.— Univ. Cal. Pubis
Geol. Sci., 1978c, vol. 118, p. 1—176.
Domning D. P. Evolution of manatees:
A speculative history.— J. Paleontol.,
1982, vol. 56, N 3, p. 599—619.
Domning D. P., Hayek L.-A. C. Horizontal
tooth replacement in the Amazonian ma-
natee (Trichechus inunguis).—Mamma-
lia, 1984, vol. 48, N 1, p. 105—127.
Domning D. P., Morgan G. S., Ray С. E.
North American Eocene Sea cows (Mam-
malia: Sirenia).— Smithsonian Contribs
PaleobioL, 1982, N 52, p. 1—69.
Dosch F. Bau und Entwicklung des Inte-
guments der Sirenen.— Jen. Ztschr. Na-
turwiss., 1915, Bd. 53, S. 805—854.
Elias P. M. Lipids and the epidermal per-
miability barrier.— Arch. Dermatol. Res.,
1981, vol. 270, p. 95-117.
Elias P. M., Brown В. E., Fritsch P. et al.
Localization and composition of lipids in
neonatal mouse stratum granulosum and
stratum corneum.— J. Invest. Dermatol.,
1979, vol. 73, p. 339—348.
Elias P. M., McNutt N. S., Friend D. S.
Membrane alterations during cornificati-
on of mammalian squamous epithelia:
A freeze-fracture, tracer and thin-section
study.— Anat. Rec., 1977, vol. 189,
p. 577—594.
Elsner R. Diving adaptations in marine
mammals.— In: Diving medicine /Ed.
R. M. Strauss/. Grunc. a. Stratton, 1976,
p. 257—268.
Eriksson G., Lamke L. Regeneration of hu-
man epidermal surface and water barri-
er function after stripping.— Acta derma-
to-venereol., 1971, vol. 51, p. 169—178.
Fawsett D. V. A comparative study of blo-
od vascular bundles in the Florida mana-
tee (Trichechus latirostris) and certain
Cetaceans and Edentates.— J. Morphol.,
1942, vol. 75, N 1, p. 105—117.
Fernand V. S. V. The teeth of the dugong.—
Ceylon J. Sci. B, 1953, vol. 25, N 2,
p. 139—147.
Flower W. H. Note on the number of cer-
vical vertebrae in the Sirenia.— Natur.
Hist. Rev., 1864, p. 259—264.
Flower W. H. An introduction to the osteo-
logy of the Mammalia.— 3rd ed. L., 1885..
382 p.
Freund L. Die Osteologie der Halicoreflos-
se.— Ztschr. wiss. Zool., 1904, Bd. 77,
S. 363—397.
Freund L. A bibliography of the mammali-
an order Sirenia.— Vestn. Ceskosl. spo-
lec. zool., 1950, sv. 14, s. 161—181.
Functional anatomy of marine mammals/
Ed. R. J. Harrison. N. Y. etc., 1974. Vol. 2.
366 p.
Gabbott P. A. Physiological and biochemi-
cal adaptation of marine organisms.—
Bol. fisiol. anim., 1982, N 6, p. 37—56.
George J. C., Rant'd K. The harp seal, Pa-
gophilus groenlandicus (Erxleben, 1777).
XV. Ultrastructure and metabolic adapta-
tion of skeletal muscle.— Canad. J. Zool.,
1973, vol. 51, N 8, p. 833—839.
George J. C., Ronald K. The harp seal, Pa-
gophilus groenlandicus (Erxleben, 1777).
XVII. Structure and metabolic adaptation
of the caval sphincter muscles with some
observations on the diaphragm.— Acta
anat., 1975, vol. 93, N 1, p. 88—99.
George J. C., Vallyathan N. V., Ronald K.
The harp seal, Pagophilus groenlandicus
(Erxleben, 1777). VII. A histophysiologi-
cal study of certain skeletal muscles.—
Canad. J. Zool., 1971, vol. 49, N 1, p. 25—
30.
Gerhardt U. Das Urogenitalsystem der Sau-
getiere.— In: Dr. H. G. Bronn’s Klassen
und Ordnungen des Tierreichs wissens-
chaftlich dargestellt in Wort und Bild.
Leipzig, 1914, Bd. 2, T. 5, Lfg. 1/4. Mam-
malia, S. 1—143.
Giacometti L. The skin of the whale (Ba-
laenoptera physalus).—Anat. Rec., 1967,
vol. 159, p. 69—75.
Gorgas M. Vergleichend-anatomische Unter-
suchungen am Magen-Darm Kanal der
Sciuromorpha, Hystricomorpha und Ca-
viomorpha (Rodentia): Eine Studie liber
den Einfluss von Phylogenie, Spezialisa-
tion und funktioneller Adaptation auf den
Saugetierdarm.— Ztschr. wiss. Zool., 1967,
Bd. 175, N 314, S. 237—404.
Grau H. Das Muskelsystem: Der aktive
Bewegungsapparat.— In: Ellenberger-
Baum Handbuch der vergleichenden
Anatomie der Haustiere. 18. Aufl. B.,
1943, S. 194—330.
Green H., Fuchs E., Watt F. Differentiated
structural components of the keratinocy-
te.— Cold Spring Harbor Symp. Quant.
Biol., 1982, vol. 46, p. 293—301.
Haffner K. Bau, Eigenschaften und ehema-
lige Verwendung der Haut der Seit 1768
ausgerotteten Steller’schen Seekuh (Rhy-
tina stelleri Retz.).— Mitt Hamburg,
zool. Mus. und Inst., 1957, Bd. 55,
S. 107—136.
Harrison R. J., King J. E. Marine mammals.
L., 1965. 192 p.
Harrison R. J., Thurley K. W. Structure of
the epidermis in Tursiops, Orcinus, Delp-
hinus and Phocaena.— In: Functional
398
anatomy of marine mammals/Ed.
R. J. Harrison. N. Y. etc., 1974, vol. 2,
p. 45—71.
Harrison R. J., Tomlinson I. D. W., Berns-
tein Z. The caval sphincter in Phoca vi-
tulina.— Nature, 1954, vol. 173, N 4993,
p. 86—89.
Hartman D. S. Ecology and behavior of the
manatee (Trichechus manatus) in Flori-
da. 1979. 153 p. (Spec. Publ. Amer. Soc.
Mammal.; N 5).
Hashimoto K., Dibella R. J., Shklar G.
Electron microscopic studies of the nor-
mal human buccal mucose.— J. Invest.
Dermatol., 1966, vol. 47, p. 512—525.
Hatt R. T. A manatee collected by the Ame-
rican Museum Congo Expedition, with
observations on the recent manatees.—
Bull. Amer. Mus. Natur. Hist., 1934,
vol. 66, art. 4, p. 533—566.
Hayward A. F. Membrane-coating granu-
les.— Intern. Rev. Cytol., 1977, vol. 59,
p. 97—127.
Heinsohn G. E. Sirenians-draft report.— In:
Scientific consultation on marine mam-
mals, Bergen, Norway, 31 Aug.— 9 Sept.
1976. Bergen: Advis. comm, of mar. reso-
ur. res., 1976, p. 10. (AGMRR/MM/SG/
WG —4-1).
Heinsohn G. E. The dugong in the seagrass
ecosystem.— In: The dugong: Proc, semi-
nar/workshop hold at James Cook Univ,
of North Queensland, 8—13 May 1979/Ed.
H. Marsh. Townsville, 1981, p. 162—163.
Heinsohn G. E., Birch W. R. Food and fe-
eding habits of the dugong, Dugong du-
gon (Erxleben), in Northern Queensland,
Australia.— Mammalia, 1972, t. 36, N 3,
p. 414—422.
Howell A. B. Aquatic mammals: Their adap-
tations to life in the water. Springfield;
Baltimore, 1930. 338 p.
Huber E. Evolution of facial musculature
aud cutaneous field of trigeminus.—
Quart. Rev. Biog., 1930, vol. 5, p. 389—437.
Husar L. L. A review of the literature of
the dugong (Dugong dugon).—Wildlife
Res. Rep. US Fish and Wildlife Serv.,
1975, N 4, p. 1—30.
Jones J. K., jun., Johnson R. R. Sirenians.—
In: Recent mammals of the world: A sy-
nopsis of families/Ed. S. Anderson,
J. K. Jones, jun. N. Y., 1967, p. 366—373.
Kaiser H. E. Morphology of the Sirenia:
A macroscopic and X-ray atlas of the os-
teology of recent species. Basel, 1974.
76 p.
Kamiya T., Yamasaki F. A morphological
note on the sinus hair of the dugong.—
In: The dugong: Proc, seminar/workshop
held at James Cook Univ, of North Que-
ensland, 8—13 May 1979/Ed. H. Marsh.
Townsville, 1981, p. 193—197.
Kellogg R. Fossil marine mammals from
the Miocene Calvert formation of Mary-
land and Virginia. 3. New species of ex-
tinct Miocene Sirenia.— Bull. US Nat.
Mus., 1966, N 247, pt 3/4, p. 65—98.
Kenchington R. A. Observation on the di-
gestive system of the dugong, Dugong
dugon (Erxleben).— J. Mammal., 1972,
vol. 53, N 4, p. 884—887.
Kleinsmith A. Uber ein Skelet und eine
Rekonstruktion des ausseren Habits der
Riesenseekiih, Rhytina gigas Zimmerman,
1780,— Zool. Anz., 1951, Bd. 146,
S. 292—314.
Kordos L. Some complements to the know-
ledge of a Middle Eocene sirenian, Sire-
navus hungaricus.— Fram. Miner. Pala-
eontol., 1981 (1982), t. 10, p. 75—78.
Krauss F. Beitriige zur Osteologie des suri-
namischen Manatus.— Muller’s Arch.
Anat. und Pbysiol., 1858, S. 390—425.
Krauss F. Beitrage zur Osteologie des su-
rinamisches Manatus.— Arch. Anat.,
Physiol, und Wiss. Med., 1862, S. 415—427.
Krauss F. Beitrage zur Osteologie von Ha-
licore.— Arch. Anat. und Physiol., 1870,
p. 525—613.
Kretzoi M. Sirenavus hungaricus n. g.,
n. sp., ein neuer Prorastomide aus dem
Mitteleozan (Lutetium) von Felsogalla in
Ungarn.— Ann. Mus. Nat. Hung., Miner.
Geol. Palaontol., 1941, S. 146—156.
Kretzoi M. A legidosebb magyar osemlos-
Cetet — Foldt. kozl., 1953, k. 83, N 7/9,
old. 273—277.
Lawrie R. A. The activity of the cytochro-
me system in muscle and its relation to
myoglobin.— Biochem. J., 1953, vol. 55,
N 2, p. 298—305.
Lemire M. Paticularites de 1’estomac dula-
mantin Trichechus senegalensis Link (Si-
reniens, Trichechides).— Mammalia, 1968,
vol. 32, N 3, p. 475—524.
Lepsius R. Halitherium schinzi, die fossile
Sirene des Mainzer Beckens.— Abh. Mil-
telrheinisch. Geol. Vereins. Darmstadt,
1881—1882, Bd. 1, S. 1—200.
Leutenegger M. Bibliographic der Sirenen
(Trichechidae).— Zool. Gart. N. F., 1982,
Bd. 52, N 2, S. 81—105.
Ling J. K. Pelage and molting in wild
mammals with special reference to aqua-
tic forms.— Quart. Rev. Biol., 1970,
vol. 45, p. 16—54.
Ling J. K. The integument of marine mam-
mals.— Functional anatomy of marine
mammals/Ed. R. J. Harrison. N. Y. etc.,
1974, vol. 2, p. 1—43.
Ling J. K. Vibrissae of marine mammals.—
In: Functional anatomy of marine mam-
mals/Ed. R. J. Harrison. N. Y. etc., 1977,
vol. 3, p. 387—415.
Man Y. de, Blok A. R. Relationship betwe-
en glycogen and agranular endoplasmic
reticulum in rat hepatic cells.— J. Histo-
chem. and Cytochem., 1966, vol. 14, N 2,
p. 135-148.
Manzij S. F., Piliptshuk О. У. Morphologic
und Funktionsanalyse des Axialskeletes
des amerikanischen Lamantins Triche-
chus manatus Lin., 1758, Mammalia, Si-
renia.— Zool. Jb. Abt. Anat., 1984, Bd. Ill,
S. 257—295.
Marsh H. The food of the dugong.— In: The
399
dugong: Proc, seminar/workshop held at
James Cook Univ, of North Queensland,
8—13 May 1979/Ed. H. Marsh. Townsvil-
le, 1981b, p. 164—165.
Marsh H., Channelles P. W., Heinsohn G. E.,
Morrissey J. Analysis of stomach contens
of dugongs from Queensland.— Austral.
Wildlife Res., 1982, vol. 9, N 1, p. 55-
67.
Mattes E. Die Dickenverhaltnisse der Haut
bei den Mammalia in allgemeinen, den
Sirenia im besonderen.— Ztschr. wiss.
Zool., 1929, Bd. 134, N 2/3, S. 345—537.
Mauro A. Satellite cell of skeletal muscle
fibers.— J. Biophys. and Biochem. Cytol.,
vol. 9, N 2, p. 493—495.
McFarland W. L., Morgane P. J. Neurolo-
gical, cardiovascular and respiratory
adaptations in the dolphin, Tursiops trun-
catus.— Proc. 74th Annu. Conv. Amer.
Psychol. Assoc., 1966, p. 167—168.
McKenna M. C. Toward a phylogenetic
classification of the Mammalia.— In:
Phylogeny of the primates/Ed.
W. P. Luckett, F. S. Szalay. N. Y., 1975,
p. 21—46.
Meinertz T. Das oberflachliche Facialis-
gebiet der Nager.— Zool. Jb. Abt. Anat.
u. Ont., 1941, Bd. 67, H. 2, S. 119—270.
Meinertz T. Die facialsmuskulatur der In-
sektenfresser (Insectivora). 2. Pachyura
indica und Sorex araneus.— Jb. Morphol.
und mikrosk. Anat. Abt. 1, 1978, Bd. 124,
N 4, S. 58-594.
Mohr E. Sirenen ober Seekiihne. Witten-
berg; Lutherstadt, 1957. 61 S. (Die neue
Brehm-Biicherei; H. 197).
Montgomery G., Best R., Yamakoshi M.
A radio-traching study of the Amasoni-
an manatee.— Biotropica, 1981, vol. 13,
N 2, p. 81—85.
Moore I. C. Range of the Florida mana-
tee.— Quart. J. Fla Acad. Sci., 1951a,
vol. 14, p. 1—19.
Moore J. C. The status of the manatee in
the Everglades National Park, with notes
in its natural history.— J. Mammal.,
1951b, vol. 32, N 1, p. 22—36.
Muir A. R., Kanji A. H., Allbrook D. B. The
structure of the satellite cells in scele-
tal muscle.— J. Anat., 1965, vol. 99, N 3,
p. 435—444.
Murie J. Researches upon the anatomy of
the Pinnipedia. 1. On the Walrus (Tri-
chechus rosmarus, Linn.).— Trans. Zool.
Soc. London, 1871, vol. 7, p. 411—464.
Murie J. On the form and structure of the
manatee (Manatus americanus).— Trans.
Zool. Soc. London, 1872a, vol. 8, pt 3,
p. 127-202.
Murie J. On the skin etc., of the Rhytina,
suggested by a recent paper of Dr.
A. Brandt.— Ann. and Mag. Natur. Hist.
Ser. 4, 1872b, vol. 4, p. 306—313.
Murie J. Further observations on the ma-
natee.— Trans. Zool. Soc. London, 1880,
vol. 11, pt 2, N 1, p. 19—48.
Murray R. M. The importance of VFA in
dugong nutrition.— In: The dugong:
Proc, seminar/workshop held at James
Cook Univ, of North Queensland, 8—13
May 1979/Ed. H. Marsh. Townsville, 1981,
p. 166—168.
Murray R. M., Marsh H., Heinsohn G. E.,
Spain A. V. The role of the midgut cae-
cum and large intestine in the digestion
of sea grasses by the dugong (Mamma-
lia: Sirenia).— Comp. Biochem. and Phy-
siol. A, 1977, vol. 56, N 1, p. 7-10.
Nagel E. J., Morgane P. J., McFarland
W. L., Galliano R. E. Rete mirabile of
dolfin: Its pressupe-damping effects on
the cerebral circulation.— Science, 1968,
vol. 161, N 3844, p. 898—900.
Nishiwaki M., Kasuya T., Tobayama T. et
al. Distribution of the dugong in the
world.— In: The dugong: Proc, of a se-
minar/workshop held at James Cook
Univ, of North Queensland, 8—13 May
1979/Ed. H. Marsh. Townsville, 1981,
p. 8—9.
Odland G. F., Holbrook K. The lamellar
granules of epidermis.— Curr. Probl. Der-
matol., 1981, vol. 9, p. 29—49.
Oliver J. Nephrons and kidneys. N. Y.,
1968. 116 p.
Palmer E., Waddel G. The relationship
between structure, innervation and func-
tion of the skin of the bottle nose dolp-
hin (Tursiops truncatus).— Proc. Zool.
Soc. London, 1964, vol. 143, pt 4, p. 553—
568.
Parker G. H. The breathing of the Florida
manatee (Trichechus manatus.).—
J. Mammal., 1922, vol. 3, p. 127.
Parry D. A. The anatomical basis of swim-
ming in whales.— Proc. Zool. London,.
1949, vol. 119, pt 1, p. 49—60.
Petit G. Remarques sur la lobulation du
rein des lamantins.—C. r. Acad. sci. Ser.
C, Paris, 1924a, t. 178, p. 244—246.
Petit G. Sur la morphogenie du rein des
Sireniens.— C. r. Acad. sci. Ser. C, Paris,
1924b, t. 178, p. 2197—2200.
Petit G. Recherches anatomiques sur Tap-
pareil genitourinaire male des Sireni-
ens.— Arch, morphol. gen. exp., 1925,.
t. 23, p. 181—121.
Petit G. Les vertebres cervicales des si-
reniens actuels.— Arch. Mus. nat. hist,
natur., Paris, 1928, t. 6, p. 243—299.
Petit G. Ordre des Sireniens.— In: Traite
de zoologie. P., 1955, vol. 17. Mammife-
res. Les ordres: anatomie, ethologie, sy-
stematique, fasc. 1, p. 918—993.
Pick P. Zur feineren Anatomie der Lunge
von Halicore dugong.— Arch. Natur-
gesch., 1907, Bd. 73, N 1, S. 245-272.
Pilleri G. The eye Pontoporia blainvillei
and Inia bolivensis and some remarks
on the problem of regressive evolution
of the eye in Platanistoidea.— In: In-
vestigations on Cetacea. Bern, 1977,
vol. 8, p. 149—159.
Pilson M. E. Water balance in sea leons.—
Physiol. Zool., 1970, vol. 43, p. 257—269.
Ponganis P. J., Pierce R. W. Muscle me-
tabolic profiles and fibertype compositi-
400
on in some marine mammals.— Comp.
Biochem. and Physiol., 1978, vol. 17,
N 1, p. 169—191.
Powell J. A. Evidence of carnivory in ma-
natees (Trichechus manatus).—J. Mam-
mal., vol. 59, N 2, p. 442.
Prunieras M., Delescluse C. Epidermal cell
culture systems in skin pharmacology.—
Brit. J. Dermatol., 1984, vol. 27, p. 43—
57.
Putter A. Die Augen der Wassersaugetie-
re.— Zool. Jb. Abt. Anat. und Ontog. Je-
na, 1903, Bd. 17, S. 99—402.
Quiring D. P., Harlan C. F. On the ana-
tomy of the manatee.— J. Mammal., 1953,
vol. 34, N 2, p. 192—203.
Reinhart R. H. Diagnosis of the new mam-
malian order Desmostylia.— J. Geol.,
1953, vol. 61, N 2, p. 187.
Reinhart R. H. A review of the Sirenia and
Desmostylia.— Univ. Cal. Pubis Geol.
Sci., 1959, vol. 36, N 1, p. 1—146.
Reinhart R. H. Fossil sirenians and desmo-
stylids from Florida and elsewhere.—
Bull. Fla State Mus. Biol. Sci., 1976,
vol. 20, N 4, p. 187—300.
Remane A. Wirbelsaule und ihre Abko-
mlinge.— In: Handbuch der vergleichen-
den Anatomie der Wirbeltiere/L. Bolk,
E. Goppert, E. Kallius, W. Lubosch. Ber-
lin: Wien. 1936, Bd. 4, S. 1—206.
Repenning Ch. A. Symposium: Advances
in systematics of marine mammals.-—
Syst. Zool., 1976 (1977), vol. 25, N 4,
p. 301—303.
Reynolds J. E., Internal and external mor-
phology of the manatee (sea cow).—
Anat. Rec., 1979, vol. 193, N 3, p. 663.
Reynolds J. E. Behaviour patterns in the
West Indian manatee, with emphasis on
feeding and diving.— Fla Sci., 1981,
vol. 44, N 4, p. 233—242,
Reynolds J. E., Krause W. J. E. A note on
the duodenum of the West Indian ma-
natee (Trichechus manatus), with em-
phasis on the duodenal glands.— Acta
anat., 1982, vol. 114, N 1, p. 33—40.
Reznik M. La regeneration du muscle strie
squelettique: Etude de morphologie ex-
perimentale. Liege, 1971. 200 p.
Rice R. H., Green H. The cornified envelo-
pe of terminally differentiated the man
epidermal keratinocytes consists of cross-
linked protein.— Cell, 1977, vol. 11,
p. 417-422.
Ridgway S. H. Mammals of the sea: Biolo-
gy and medicine. Illinois, 1972. 765 p.
Robbins C. R., Fernee R. M. Some obser-
vations on the swelling of human epi-
dermal membrane.— J. Soc. Cosmet.
Chem., 1983, vol. 34, p. 21—34.
Rochon-Duvigneaud A. Les yeux et la vi-
sion des vertebres. P., 1943. 719 p.
Ronald K., Selley L. J., Amoroso E. C. Bio-
logical synopsis of the manatee. Ottawa:
Intern.'Develop. Rec. Cent., 1978. 112 p.
Rouffignac C. de, Bankir L., Roinel N. Re-
nal function and concentrating ability
desert rodent: The gundi (Ctenodactylus
vali).— Pfliigeers Arch., 1981, vol. 390,
N 2, p. 138—144.
Saban R. La musculature peauciere de la
tete chez un jeune lamantin du Senegal
(Trichechus senegalensis Link 1795;
Mammifere, Sirenien).— Zentr-Bl. Ve-
terinarmed. C, 1975, Bd. 4, N 3, S. 232—
248.
Sahni A., Kumar K. Lower Eocene Sire-
nia, Ishatherium subathuensis, gen. et
sp. nov. from the Type Area, Subathu
Formation, Subathu, Simla Himalayas,
H. Р,— J. Palaeontol. Soc. India, 1978
(1979), N 23/24, p. 132-135.
Sahni A., Mishra V. P. Lower Tertiary ver-
tebrates from Western India.— Monogr.
Palaeontol. Soc. India, 1975, N 3, p. 1—
48.
Savage R. J. G. Review of early Sirenia.—
Syst. Zool., 1977, vol. 25, N 4, p. 344—
351.
Savage R. J. G., Tewari B. S. A new sire-
nian from Kutch, India.— J. Palaeontol.
Soc. India, 1975(1977), vol. 20, p. 216—
218.
Scholander P. F., Irving L. Experimental
investigations on the respiration and
diving of the Florida manata.— J. Cell,
and Comp. Physiol., 1941, vol. 17, N 2,
p. 169—191.
Scott I., Harding C. R. Histidine-rich pro-
teins: Synthesis and posttranslational
modification during epidermal differen-
tiation.— Biochem. Soc. Trans., 1981a,
vol. 9, p. 389—390.
Scott I., Harding C. R. Studies on the synt-
hesis and degradation of a high molecu-
lar weight histidine-rich phosphoprotein
from mammalian epidermis.— Biochim.
et biophys. acta, 1981b, vol. 669, p. 65—
78.
Sereno P. An early Eocene sirenian from
Patagonia (Mammalia, Sirenia).— Amer.
Mus. Novit., 1982, N 2729, p. 1—10.
Shikama T. Additional notes on the po-
stcranial sceletons of Japanese Desmo-
stylia.— Sci. Rep. Yokohama Nat. Univ.
Sect. 2, 1968, N 14, p. 21—26.
Sickenberg О. Beitrago zur Kenntnis ter-
tiarer Sirenen.— Mem. Mus. toy. hist, na-
tur. Belg., 1934, N 62, p. 1—352.
Siegfried P. Das Femur von Eotherioides
libyca (Owen) (Sirenia).— Palaontol.
Ztschr., 1967, Bd. 41, N 3/4, S. 165—172.
Simpson G. G. Fossil Sirenia of Florida
and the evolution of the Sirenia.— Bull.
Amer. Mus. Natur. Hist., 1932, vol. 59,
art. 8, p. 419—503.
Simpson G. G. The principles of classifi-
cation and a classification of mammals.—
Bull. Amer. Mus. Natur. Hist, 1945,
vol. 85, p. 1—350.
Simpson J. G., Gardner M. B. Comparati-
ve microscopic anatomy of selected ma-
rine mammals.— In: Ridgway S. H.
Mammals of the sea: Biology and medi-
cine. 1972, p. 298—418.
Slijper E. I. Die Cetacean vergleichend-
anatomisch und sistematisch: Ein Beit-
401
rag zur vergleichenden Anatomie des
Blutgefass-, Nerven- und Muskelsystems,
sowie des Rumpfskelettes der Saugetiere,
mit Studien fiber die Theorie des Aus-
sterbens und der Foetalisation.— In:
Capita zoologica. Hague, 1936, vol. 7,
pt. 1/2, p. 1—590.
Slijper E. J. Comparative biologic-anatomi-
cal investigations on the vertebral co-
lumn and spinal musculature of mam-
mals.— Verh. Knkl. nederl. akad. we-
tensch. Afd. natuurk. R. II, 1946, bd. 42,
N 5, biz. 1—128.
Slijper E. J. Foramen ovale and ductus
arteriosus botalli in the aquatic mam-
mals.— Mammalia, 1961, vol. 25, p. 528—
561.
Slijper E. J. Whales. N. Y.: Basicbooks,
1962. 475 p.
Spain A. V., Heinsohn G. E. Size and weight
allometry in a North Queensland popu-
lation of Dugong dugon (Muller) (Mam-
malia: Sirenia).— Austral. J. Zool., 1975,
vol. 23, N 2, p. 159—168.
Speerman R. T. C. The epidermal stratum
corneum of the whale.— J. Anat., 1972,
vol. 113, p. 373—381.
Sperber J. Studies on the mammalian kid-
ney.— Zool. bidr. Uppsala, 1942, bd. 22,
s. 249-432.
Squier C. A. The permiability of keratini-
zed and non-keratinized oral epithelium
to horseradish peroxidase.— J. Ultra-
struct. Res., 1973, vol. 43, p. 160—177.
Sqaier C. A. Membrane-coating granule in
non-keratinized oral epithelium.— J. Ult-
rastruct. Res., 1977, vol. 60, p. 212—220.
Steller G. W. De bestiis marinis.— Nov.
Comm. Acad. sci. Petropolitanae, 1751,
t. 2, p. 289—398; Англ, nep.: The beasts
of the sea.— In: Jordan D. S. The fur
seals and fur-seal islands of the North
Pacific Ocean. Wash., 1889, pt 3, art. 8,
p. 179-218.
Tassy P. Relations phylogenetiques du
genre Moeritherium Andrews. 1901
(Mammalia).—C. r. Acad. sci. D., 1979,
t. 289, p. 85—87.
Tenney S. M., Hanover N. H. Correlative-
observations on the electrocardiograms
and morphology of the heart of the Flo-
rida manatee.— Amer. Heart J., 1958,
vol. 56, N 6, p. 933—938.
The dugong: Proc, of a seminar/workshop
held at James Cook Univ, of North Queen-
sland, 8—13 May 1979/Ed. H. Marsh.
Townsville, 1981a. 400 p.
Thibant-Vercruyseen R., Ronvcaux-Dupal
M. F., Shirion Y., Wattiaux R. Morpholo-
gical changes of the rat liver cell after
sucrose injection.— In: Proc. 7th Europ.
congr. on electron microscopy, Hague,
Netherl., Aug. 24—29. Hague, 1980, vol. 2,
p. 12.
Tobien H. Moeritherium, Palaeomastodon,
Phiomia aus dem Palaogen Nordafrikas
und die Abstammung der Mastodonten
(Proboscidea, Mammalia).— Mitt. geol.
Inst, techn. Univ. Hannover, 1971, Bd. 10,
S. 141—163.
Tulsi R. S. Observations on the structure
of the dorsal muscle in the bottle-nose
dolphins (Tursiops truncatus).— J. Anat.,
1975, vol. 119, N 1, p. 39—48.
Walker E. P. Mammals of the world. 3rd
ed. Baltimore; London, 1975, vol. 2,
p. 647—1500.
Walls G. The vertebrate eye and its adap-
tive radiation. Blomfield Hills (Mich.),,
1942. 785 p.
Weber M. Die Saugetiere. Jena, 1928.
Bd. 2. 898 S.
Wertz P. W., Downing D. T. Glycolipids
in mammalian epidermis: Structure and
function in the water barrier.— Science,
1982, vol. 217, N 4564, p. 1261—1262.
Whitfield W. K., Jun., Farrington S. L. An-
annotated bibliography of Sirenia.— Fla
Mar. Res. Pubis, 1975, N 7, p. 1—44.
Woodward H. On an almost perfect scele-
ton of Rhytina gigas (Rhytina Stelleri,
«Steller’s sea-cow»), obtained by Mr. Ro-
bert Damon, F. G. S., from the Pleisto-
cene peat-deposits on Bering’s Island.—
Quart. J. Geol. Soc. London, 1885, vol. 41„
p. 457-472.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ ............................................ 3
^ВВЕДЕНИЕ (В. Е. Соколов, Фернандо М. Гонсалес Бермудес) . . 5
Глава первая
ОБЩАЯ ХАРАКТЕРИСТИКА И ЭВОЛЮЦИЯ СИРЕН (MAMMA-
LIA: SIRENIA) И МЕСТО ЛАМАНТИНОВ В ИХ СИСТЕМЕ
(В. Б. Суханов)..........................................Н
Глава вторая
СРАВНИТЕЛЬНАЯ МОРФОЛОГИЯ КОЖНОГО ПОКРОВА
ЛАМАНТИНА (В. Е. Соколов, О. Ф. Чернова, Л. В. Степанова,
Б. И. Петрищев).........................................44
Поверхность и толщина кожи.........................
Эпидермис...........................................53
Дерма...............................................56
Подкожная жировая клетчатка.........................60
Вибриссы............................................61
Железа кожи век.....................................66
Процессы кератинизации в эпидермисе.................67
Глава третья
МОРФОЛОГИЯ И ОСНОВНЫЕ НАПРАВЛЕНИЯ СПЕЦИАЛИЗА-
ЦИИ ОСЕВОГО СКЕЛЕТА СИРЕН (В. Б. Суханов, С. Ф. Манзий) 77
Общая характеристика позвоночного столба .......... 80
Шейный отдел........................................97
Грудной отдел.......................................ИО
Послегрудной отдел..................................139
Ребра и грудная клетка..............................149
Глава четвертая
СПЕЦИФИКА СКЕЛЕТА ПЕРЕДНЕЙ КОНЕЧНОСТИ ЛАМАН-
ТИНА (В. Б. Суханов, П. П. Гамбарян, В. И. Клыков).....157
Глава пятая
СТРОЕНИЕ, ФУНКЦИИ И АДАПТИВНЫЕ ОСОБЕННОСТИ
СКЕЛЕТНОЙ МУСКУЛАТУРЫ ЛАМАНТИНА (Л. П. Гамбарян,
В. Б. Суханов).........................................188
Лицевые и поверхностные мышцы шеи..................189
Жевательная мускулатура............................202
403
Мышцы языка и подъязычного аппарата.................... 206-
Мышцы глотки и гортани...................................214
Туловищная мускулатура..................................218-
Мышцы туловища (спинные, грудные, хвостовые) .... 218
Работа мышц туловища...................................244
Мышцы шеи................................................252
Работа комплекса шейных мышц.............................258
Мышцы брюшного пресса и диафрагмы........................261
Мышцы, соединяющие плечевой пояс и плечо с туловищем . . 265
Мышцы, действующие на плечевой и локтевой суставы . . . 276
Мышцы предплечья и кисти.................................286
Работа мышц конечностей..................................296
Мышцы генитальной области................................301
Глава шестая
ДВИЖИТЕЛЬНЫЙ КОМПЛЕКС ЛАМАНТИНА И ЕГО ОСОБЕН-
НОСТИ {В. Е. Соколов, С. В. Першин, А, С. Соколов. А. Г. Томилин) 306
Глава седьмая
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ ПИЩЕВАРИТЕЛЬНОГО
ТРАКТА (Е. И. Наумова).................................312
Специфика питания..................................313
Макроморфологическое строение пищеварительного тракта . . 313
Микроскопическое исследование......................317
Trichechidae и Dugongidae: структурные особенности пищевари-
тельного тракта....................................324
Морфологическая специфика пищеварительного тракта сирен . 326
Питание и размеры органов пищеварительного тракта . . . 326
Функциональные особенности желудка и двенадцатиперстной
кишки..............................................328
Роль толстого отдела кишечника в пищеварении сирен.....329
Глава восьмая
О НЕКОТОРЫХ ФУНКЦИОНАЛЬНО-МОРФОЛОГИЧЕСКИХ
АДАПТАЦИЯХ ВЕНОЗНОЙ СИСТЕМЫ ЛАМАНТИНА (В. П. Га-
ланцев) ...............................................331
Глава девятая
АНАТОМИЯ ПОЧКИ ЛАМАНТИНА (С. П. Гамбарян) .... 338
Глава десятая
ОБ ОРГАНЕ ЗРЕНИЯ ЛАМАНТИНА И ДРУГИХ СИРЕН
(Ф. В. Андреев)........................................342
404
Глава одиннадцатая
ОСОБЕННОСТИ МИКРО- И УЛЬТРАСТРУКТУРЫ И ГИСТОХИ-
МИИ НЕКОТОРЫХ ОРГАНОВ ЛАМАНТИНА........................351
Гистохимическое изучение липидов в скелетных мышцах ламан-
тина и других морских млекопитающих (В. А. Родионов) . . 351
Микроскопическое и ультраструктурное исследование скелетной
мышцы ламантина (М. М. Умнова, И. Л. Новоселова) . . . 357
Светооптическое и электронно-микроскопическое исследование
клеток печени ламантина (М. М. Калашникова, Н. И. Казанская) 371
Глава двенадцатая
ИССЛЕДОВАНИЕ НЕКОТОРЫХ ФИЗИОЛОГИЧЕСКИХ ОСОБЕН-
НОСТЕЙ ЛАМАНТИНА (Л. М, Мухаметов, В. П. Галанцев) . . 377
Глава тринадцатая
СПЕЦИФИКА АДАПТАЦИЙ СИРЕН (вместо заключения)
(В. Б. Суханов)........................................385
ЛИТЕРАТУРА.............................................394
ЛАМАНТИН
Морфологические адаптации
Утверждено к печати
Институтом эволюционной морфологии
и экологии животных им. А. Н. Северцова
Академии наук СССР
Редактор издательства Г. М. Орлова
Художник Н. Н. Власик
Художественный редактор Н. А. Фильчагина
Технический редактор И. Н. Жмуркина
Корректоры Г. Н. Джиоева, К. П. Лосева
ИБ № 31654
Сдано в набор 05.11.85. Подписано к печати 26.03.86
Т-00265. Формат 70Х1001/,,. Бумага типографская № 1
Гарнитура обыкновенная. Печать высокая
Усл. печ. л. 33,15. Усл. кр.-отт. 33,15. Уч.-изд. л. 36,7
Тираж 1000 экз. Тип. зак. 2118
Цена 5 р. 80 к.
Ордена Трудового Красного Знамени
издательство «Наука».
117864 ГСП-7, Москва В-485, Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинекий пер., 6