
























































Author: Павлова В.Н. Тихомирова В.Н.
Tags: география растений фитогеография флора географическое распространение растений растения биология ботаника растениеводство
ISBN: 5-211-02853-8
Year: 1993




БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ
ОБЛАСТИ
Часть!
Издательство Московского университета
1993
БИОЛОГИЧЕСКАЯ ФЛОРА МОСКОВСКОЙ ОБЛАСТИ
Под редакцией
профессора В.Н.Павлова, чл.-корр. РАН В.Н.Тихомирова
Выпуск девятый
Часть первая
Издательство Московского университета
1993
УДК 581.9 (471.311)
Биологическая флора Московской области. Вып. 9, ч.1 /Под
ред. В.Н.Павлова, В.Н.Тихомирова. - М.: Изд-вс МГУ, 1993 . - 112 с.
ISBN 5-211-02853-8(4.1).
ЖВМ 5-2II-02534-2.
В книге обобщены итоги оригинальных наблюдений, эксперимен-
тальных работ и литературные данные по систематике, географичес-
кому распространению, морфологии, онтогенезу, экологии, коноор-
тивным связям, фитоценологии, хозяйственному значению видов рас-
тений, произрастающих в Московской области. Охарактеризованы как
широко распространенные раотения - лютик ползучий, мышехвоотник
маленький, виды рода василистник, виды рода сердечник, клюква
четырехлепестная, кровохлебка лекарственная, так и охраняемые на
территории области - мякотница однолистная, кокушник комарнико-
вый, лилия кудреватая.
Сборник представляет ообой продолжение издания "Вюлсгичес-
кая флора Московской области", вып. 9 в двух чаотях.
Для широкого круга специалистов-биологов, работников сельс-
кого и лесного хозяйства, деятелей охраны природы.
Рецензенты: кандидат биологических наук Ю.Е.Алексеев,
кандидат биологических наук А.В.Чичев
Редактор выпуска: М.Г.Вахрамеева
Печатается по постановлению
Редакционно-издательского совета
Московского университета
077(02) - 93 - заказное Qc) Московский государственный
ISBN 5-2II-02853-8 (4,l) университет, 1993
ISBN 5-2II-02534-2
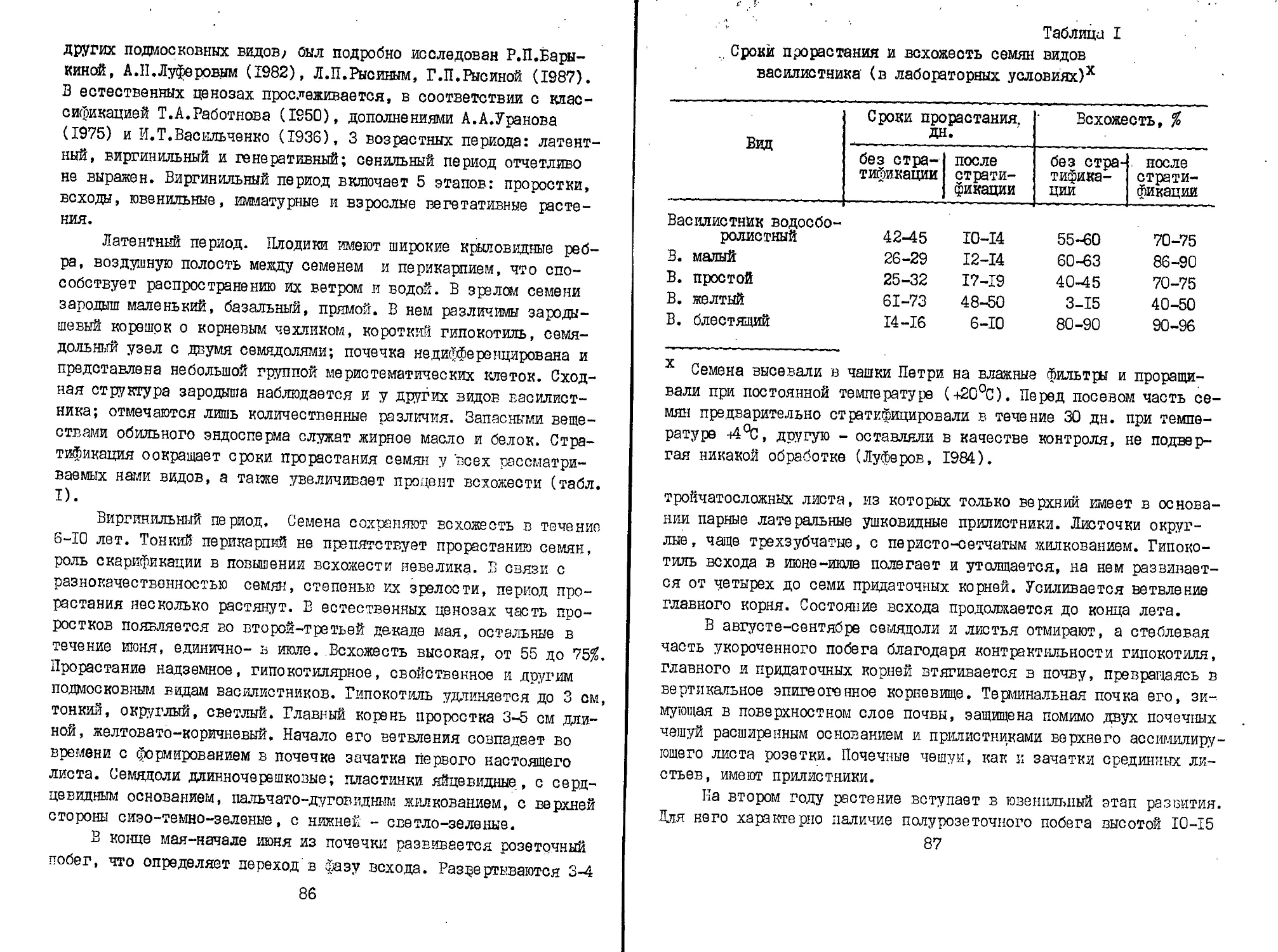
ПРЕДИСЛОВИЕ
Начиная с 1974 г. опубликовано 8 выпусков "Биологической
флоры Московской области", в которых было охарактеризовано свы-
ше 120 видов растений. В настоящем выпуске, как и в прошлом,
принимали участие сотрудники ряда вузов и научных учреждений:
Московского государственного университета, Мооковсксго гооудар-
ственного педагогического университета , I Московского медицин-
ского института, Московской сельскохозяйственной академии им.
К.А.Тимирязева и других учреждений. Характеристика растений
дана в традиционном для настоящего издания плане: морфология,
географическое распространение, онтогенез, ритм сезонного раз-
вития, экология, фитоценология, хозяйственное значение. Раздел
"Номенклатура" написан чл.-корр. РАН В.Н.Тихомировым по несколь-
ко измененному плану: форма цитации приближена к принятой в
"Новостях систематики высших растений" и "Флоре европейской
части СССР".
Географические названия, приводимые в настоящем выпуске
соответствуют состоянию на конец 1991 г.; по техническим при-
чинам было невозможно изменить характеристики распространения
отдельных видов, используя новейшие наименования ( СНГ, Прибал-
тийские страны, Украина, Кыргызстан и др.).
Основную работу по комплектованию выпуска и его редактиро-
ванию осуществили сотрудники кафедры геоботаники МТУ проф. В.Н.
Павлов, ст.н.с.М.Г.Вахрамеева, доц.Ю.Е.Алексеев. Существенную
помощь в подготовке рукописи к печати оказали научные сотруд-
ники кафедры геоботаники Т.В.Багдасарова, Т.П.Баландина, Г.В.
Мусина.
3
М.В.Нехлюдова, В.Р.Филин
СТРАУСНИК ОБЫКНОВЕННЫЙ
..оченклатура, систематическое сложение, внутривидовая систематика
Matteuccia struthiopteris (L.) Todaro, 1866, Giorn.Sci. Nat,
’cob, Paurcrto, 1:235; Голицын, i954, в Маевский, Фл.,изд.8:52;
'awal-ёе, 1964, Fl. Europ. I 19; Пояркова, 1964, в Маевский,
Фл. ,изд.9: 5; Ворошилов, 1966, Спред.раст.Моск.обл.:45; А.Бобров,
974, Фл.:вроп.части СССР, 1:74: Черепанов, 1981, Сосуд.раст.
СССР:305; В.Тихомиров, 1986,0прсд.раст.Мещеры, 1:51. - osmunda
struthiopteris L., 1753, Sp, Fl. Юбб. - Struthiopteris filica-
strum А.1., 1785, Fl.Pedem, :285; Фомин, 1934, Фл.СССР, 1:29;
Назаров, 940,,в Маевский, Фл.изд.7;40. - Onoclea etruthtopte-
ris (1. Hoffm. 1795, Deutschl. Fl. 2:12; Шмальгаузен, 1897,
Фл.Средн, и Южн.России, 2:695; Сырейщиков, I9C6, Илл.фл.Моск.губ.
.:45; он е, 1914, цит.соч.4:3; он же, 1927; Опред.рарт.Моск,
губ.:39. - Strythiopteris germanica Willd. 1809. Enum. Pl. Horti
Bervi, 1071; Кауфман,1866, Моск.фл., изд.1:626; Цингер, Т.885, Сбор
Сборник свед.:507. - Страусник обыкновенный (сем. Onooleaceae -
оноклеевне).
Страусник принадлежит к группе оноклеоидных папоротников,
которою длительное время, следуя системе, принятой в " Die natur-
lichen F^lanzenf amilieji" ( Diels, 1902), ОТНОСИЛИ К подтрибв
Onocleir.ae подсемейства Woodaieae семейства Polypodiaceae.
P.Ллойд (L’oyd, 1971) рассматривает ее в качестве подсемейства
Onoclecideae сем. Aspidiceae , которое ОН сближает C Athyria-
сеае и Dryopteridaceae. Современные птеридологи-систематики
рассматривают оноклеевне в качестве самостоятельного семейства
Onoclea ае (Pichi Sermolli, 1970) или включают их в подсемей-
ство ;.hyrioideae сем. Aspleniaceae ( Crabbe, Jermy, Mickel,
1975; Mice:, 1982).
К роду Matteuccia , согласно P.Ллойду, кооме М, struthiop-
teris •.ринадлежИТ еще M.orientalis (Hook.) Trev. M.interme-
dia ' . ’-г. в последнее врнмя установлено, что ближайшим родст-
венни ом м.struthiopteris является обитающий в Мексике.и в Гва-
темал; ВИД Onocleopsis hintonii'(F.Eellard, 1945), a M. orien-
talis no своему строению более сходна с Onoclea sensibilis,
1., а он с М. struthiopteris ( Kato, "Sahashi, 1980). Что же ка-
4
сается м. intermedia, то не исключено, что это описанной и» Се-
веро-Восточного Китая и, возможно, встречающееся также в Гимала-
ях растение ( Christensen, 1913) представляет собой, как пред-
полагает Р.Ллойд, аллополиплоид, возникший при скрещивании М.
struthiopteris и M.orientalis. Японские исследователи рассмат-
ривают оноклйевые как родственную Athyria'ceae группу, состоящую
ИЗ двух ВИДОВ Onoclea - 0.sensibilis и O.orientalis (Hook.)
Hook., двух ВИДОВ Matteuccia - М.struthiopteris и M.iiintonii
(P.Bellard)M.Kato и предположительно межродового гибрида, за ко-
торым пока сохраняется название м.intermedia. Исходя из анализа
ареалов и учитывая морфологическое сходство видов, предполагают,
что оноклеевые возникли где-то в Центральной или Северной Амери-
ке, а м.struthiopteris, ведущая начало от M.hintonii- подобно-
го предка в Северной Америке , впоследствии широко расселилась•в
Северном полушарии ( Kato, Iwatsuki, 1983).
Род Matteuccia назван по фамилии итальянского физиолога и
политического деятеля Карло Маттеуччи (I8II-I868), видовой эпи-
тет дан по сходству листа с пером страуса и происходит от греч.
struthion - страусенок и pteris - папоротник (Каден, Терентьева,
1975). В старой русской ботанической литературе растение называ-
ли "разнолистник" или папоротник черный (Анненков, 1878), что,
конечно же, хорошо отражало такие признаки растения, как димор-
физм листьев и черный цвет стволика. Позже стали употреблять на-
звание "страусопер", которое, к счастью, не прижилось и теперь
заменено на страусник.
Распространенную на востоке Северной Америки популяцию
страусника некоторые американские флористы рассматривают как са-
мостоятельный вид M.pensylvanica Willd. (Haymond, 1950), но
большинство не считает возможным рассматривать ее даже в ранге
подвида, хотя иногда и допускают, что эта популяция представляет
собой особую расу (Hulten, 1962). В пределах вида M.strutniop-
teris описано несколько разновидностей и форм (например,folia-
сеа, furcata, pubescens,serrata)(Luerssen, 1889; Hegi, 1965),
но лишь немногие из них являются "хорошими" (Prange, Aderkas,
1985). Исходя из особенностей поведения верхушечной меристемы
страусника (Klekowski, 1984), можно предполагать, что многие из
этих разновидностей и форм являются мутантами.
Географическое распространение
Почти пиркумбореалышй голарктический вид, приуроченный,
Ix-I04I 5
главным образом, к лесной зоне и к горно-лесному поясу ( Hulten,
1962; Keusel ot al., 1965; Atlas Fl.bur., TS*?2; Kato, Iwat-
suki, 1983; Frange, Aderkas, 1985).
За пределами СССР в Европе встречается от Скандинав;® на
севере до Северной Италии и Хорватии на юге (в прошлом веке вид
был собран в Сицилии на склонах Этны) и от Бельгии на западе до
границ СССР на востоке. Исходя из данных нумерического анализа
ареалов растений в Европе, вид относят К Lycopodium annotinum
.Флористическому элементу ( Birks, 1976). Редкость местонахожде-
ний страусника в Центральной Европе иногда объясняют тем, что
будучи оттеснен в плейстоцене к юго-востоку Европы вид не успел
еще захватить освободившуюся впоследствии территорию (кавЪясИ
ot al., 1976). Это объяснение не согласуется с тем Фактом, что
страусник часто встречается на Скандинавском п-ове, территория
которого была покрыта почти сплошным ледниковым щитом, над по-
верхностью которого лишь кое-где возвышались нунатаки.
В зарубежной Азии вид встречается в Северном Иране, Северо-
Восточном Китае, Корее, Японии.-Не исключено, что приводимые в
литературе ( iiult<Sn, 1962) местонахождения в Юго-Восточном Китае
относятся К Onoclea orientalis.
В Северной Америке вад распространен от 40°с.ш. до примерно
62°о.ш. Большая часть местонахождений приурочена к востоку мате-
рика, где ввд встречается от Ньюфаундленда на севере до оз.Эри
на юге. На Южной Аляске отмечены единичные местонахождения ( чи1~
tdn, 19б8; Prange, Aderkas, 1985).
В европейской части СССР северная и южная границы сплошного
ареала в основном совпадают о границами лесной зоны. Разрознен-
ные местонахождения в лесостепной и на севере степной зон в УССР,
Воронежской и Саратовской обл. приурочены к долинам рек (Флора
Юго-Востока, 1927;Atlas Fl.Sur., 1972; Фл.евр.части СССР,1974;
Раменская, Андреева, 1982; Устименко, Андр1енко, 1982; Олред.
высш.раот.Украины, 1987; Цвелев, 1988). Изолированные местонахож-
дения в равнинной части УССР некоторые ботаники рассматривают
как реликтовые, оставшиеся со времени рисс-вюрмского межледни-
ковья (Харкевич, 1953). На юге встречается в Предкавказье, на
Кавказе, в Закавказье и в Талыше, где ограничен лесным поясом и
поднимается до высоты 1600 м над уровнем моря (Соколов, 19366;
Гроссгейм, 1939).
Сплошной ареал страусника из европейской части СССР через
Урал продолжается в Западной Сибири, где большая часть местона-
6
хоадений приурочена к югу леоной зоны и к Алтаю; по долине Оби
вид доходит до 61°с.ш. (Крылов, 1927). Изолированные местонахож-
дения в Тургайской впадине приурочены к островным лесам по бере-
гам озер (Павлов, 1928; Пугачав, 1972). Далее к востоку от Алтая
и Тувы южная граница ареала в значительной мере совпадает о Го-
сударственной границей СССР, а северная, поднявшись по долине
Енисея до 63°с.ш., резке спускается к югу в Центральной Сибири,
где страусник встречен в Саянах, на юге'Средне-Сибирского плато,
в Патомском нагорье и Витимском плоскогорье (Флора Центр.Сибири,
1979; Иванова, Чепурнов, 1983; Чечеткин, 1989). В Якутии вид со-
бирался в долине Лены около 60°о.ш. и в долине Алдана (Караваев,
1958; Опред.раст.Якутии, 1974). На Дальнем Востоке страусник
распространен в Приморье, на Сахалине, Курильских с-вах и Кам-
чатке (Ворошилов, 1982). По Охотскому побережью идет до Гижигин-
ской губы, но в последние года там не собирался (Хохряков, 1985).
В Московской обл. встречен во всех флористических районах
(Ворошилов и др., 1966).
Морфологическое описание
Столонообразующий гемикриптофит-хамефит с коротким верти-
кальным стволиком, от подземной части стволика отходят корни и
столоны, а на верхушке его располагается крупная почка, окружен-
ная воронковидной кроной из спирально расположенных летнезеле-
ных стерильных и зимующих спороносных листьев. Стволик, обычно
называемый корневищем ( Luerssen, 1989), достигает 10 см в диа-
метре И 15-20 СМ, редко 60 см ( Prange, Aderkas, 1985) в высоту
и состоит из сравнительно тонкого (дс 2 см в диаметре) стебля,
окруженного плотным чахлом из тонких (диаметром 0,5-2 мм) при-
даточных корней и кос оприкрепле иных, длинных (до 10 см), широ-
ких (до 20 мм) и толстых (до 5 мм), обратноланцетных оснований
листьев с прилегающими участками стержня листа. Отмирая, осно-
вания листьев постепенно разрушаются, в связи с чем стволик
имеет вид заостренного книзу конуса, верхушка которого обычно
переходит в горизонтальный столон, давший начало растению.
Стебель на поперечном сечении многолопастной, покрыт не-
резко выраженным эпидермисом, подстилаемым склеренхимой гипо-
дермы. Внутренняя зона коры и сердцевина состоят из паренхимы,
содержащей крахмал и дубильные вещества. Проводящая система
представлена диктиостелой из небольшого числа меристел ( тег-
7
letzki, 1884; Gwynne-Vaughan, 1904; Bower, 1928). ЛИОТОВОЙ
след ,двупучковый, однолакунный. В нижней части листового пролива
от стелы отходят 1-3 корневых следа, в верхней - след, идущий к
остаточной меристеме или к возникшему из нее столону ( Kato,
Sahashi, 1980). На поперечном срезе стебля в сердцевине и коре
видны схизогенные полости, выстланные чешуйками и волосками и .
ограниченные склеренхимной обкладкой. Расположенные рядом на
одном радиусе сердцевинная и коровая полости, сливаясь между со-
бой в области листового прорыва, образуют вытянутый, расположен-
ный наискось по отношению продольной оси стебля канал, который
может сообщаться и с внешней средой; этот канал иногда называют
пазушным карманом ( Campbell, 1887; Mickel, Х938).
Поверхность корней в зоне проведения покрыта толстостенны-
ми клетками первичной коры с опробковевшими почти черными обо-
лочками. Стела диархная, поэтому боковые корни располагаются
двурядно.
Столоны на большей части своего протяжения горизонтальные,
от I см до 150 см редко до 3 м ( Prange, Aderkas, 1985) в длину,
с осью диаметром 2-10 мм, со спирально расположенными (по фор-
муле 1/3), обычно редко расставленными низовыми листьями, или
катафиллами и отходящими при их основании редкими придаточными
корнями. У катафиллов пластинка не развивается, а дуговидно
изогнутое выпуклое на спинке основание достигает 1-4 см в дайну
и I см в ширину. По своей анатомии ось столона сходна с осью
стволика.
Стерильные листья, или трофофиллы достигают 150 см, редко
300 см ( Prange, Aderkas, 1985) в длину и 25-35 (60) см в шири-
ну. Черешок короткий, 4-12 см в дайну и до I см в ширину, темно-
бурый, на ’поперечном сечении в основании узкий и ромбический, с
двумя проводящими пучками, выше - расширенный и треугольный с
адаксиальным желобком, переходящим в желобок рахиса, или стерж-
ня листа. При основании черешка долго сохраняются чешуи, при-
крывавшие зачаток листа. Рахис внизу бурый, обычно голый, в
верхней части листа - зеленый с мелкими волосками и редкими уз-
кими; чешуйками на абаксиальной стороне. Пластинка листа мягкая,
желтовато- или светло-зеленая, дважды перистая, в очертании об-
ратно ланцетная, постепенно суживающаяся к основанию и внезапно
заостренная на верхушке. Сегменты первого порядка, или перья в
числе до 70-90 пар сидят на стержне очередно или почти супро-
8
тивно. Нижние сегменты расставленные, отогнутые к основанию лис-
та, короткие, выше расположенные сегменты соприкасающиеся или
даже черепитчато налегающие"один на другой, длинные, отходящие
от рахиса почти под прямым углом или наискось, с акроскопически
загнутыми верхушками. Средние сегменты линейнс-ланцетные, посте-
пенно заостренные, до 20 см в длину и до 2 см в ширину, в осно-
вании и середине до 3/4 ширины полуплаотинки надрезаны на сопри-
касающиеся, косояйцевццные или усеченные, цельнокрайние или у
верхушки слегка зубчатые доли, снизу, особенно по жилкам, корот-
ко опушенные. Жилкование долей простопериотсе. Устьица в числе
до 20 на I мм^ располагаются на нижней стороне листа; изредка
одиночные устьица есть и на верхней стороне ( Campbell, 1887).
Спороносные листья, или спорофиллы, располагающиеся на дне
воронки из вегетативных листьев, почти прямостоячие, жесткие, к
концу лета тускло-зеленоватые, позже буреющие, до 60 см в длину
и до 7 см в ширину. Черешок длинный, равный примерно трети дли-
ны пластинки, сверху желобчатый, в основании с чешуями. Пластин-
ка однаждыперистая. Спороносные сегменты ланцетно-линейные.поч-
ти цилиндрические, четко видно перетянутые, до 7 см в длину и
8 мм в ширину, косо вверх торчащие и загнутые на верхнюю сторо-
ну пластинки так, что верхушка сегмента.нередко располагается
на противоположной от места его прикрепления стороне рахиса, а
нижерасположеиные оегменты перекрывают вышерасположенные. Плас-
тинка сегмента завернута на нижнюю сторону почти до средней жил-
ки и полностью покрывает сорусы своим однослойным краем.
Изредка у растений без всякой видимой причины или же после
удаления молодых трофофиллов образуются листья, у которых ниж-
ние сегменты стерильные, а верхние спороносные, или лиотья, у
которых основание сегмента спороносное, а верхушка стерильная
(Goebel, 1887).
Верхушечная почка надземного побега к осени обычно прикры-
та катафиллами, очень крупная - до 10 см в высоту и 5 см в диа-
метре. У взрослых растений почка содержит несколько десятков
разновозрастных зачатков листьев, самые внутренние из которых
в нормальных условиях превратятся в зрелые листья на 4-й год, а
самые внешние - в начале лета следующего года. Зачатки листьев
густо покрыты чешуйками и волосками, большая часть которых опа-
дает по окончании процесса развертывания листа. Волоски корот-
кие, простые, с конечной клеткой, вырабатывающей слизь. Чешуи
9
при созревании буреющие, от линейных до продолговато-ланцетных,
до 15 мм в длину и 3 мм в ширину, изогнутые, в основании слегка
сердцевидные, обычно заканчивающиеся волоском.
Сорусы градатные ( Bower, 1928), крупные, располагающиеся
по жилкам до 5 на каждую дольку, а потому сомкнутые настолько,
что кажутся одним сорусом. Покрывальце в очертании полукруглое,
зубчатое по краю, рано исчезавшее. Спорангии полиподпоидного ти-
па с почти вертикальным кольцом из 25-40 клеток, содержащие
обычно 64, реже меньшее число спор.
Споры светло-коричневые, билатеральные, однолучевые, анизо-
полярные, в экваториальном положении слегка напоминают семя фа-
соли (с выпуклой дистальной и слегка вогнутой до слегка выпук-
лой проксимальной стороной), в полярном положении овальные, дли-
ной 32-73 мам ( Sorsa, 1964; Lloyd, 1971; Кутафьева, 1972;
Kato, Sahaahi, 1980; Ferrarini et al., 1’986). Перина тонкая,
с мелкими (до I мкм в диаметре) отверстиями и с немногочисленны-
ми, анастомозирующими (особенно на дистальной стороне) и образу-
ющими редкую сеточду высокими гребнями.
Данные по хромосомному числу противоречивы; одни авторы
считают, ЧТО 2п =78 ( Fabbri, Kenicanti, 1970; Kurita, 1976;
L5ve, Love, 1976; Love et al., 1977) другие, что 2n =80 (Fri-
ebel, 1933; Okuno, 1936; Sorsa, I°;2; Kitui, 1965).
Обоеполые или женские заростки обычно сердцевидные, с вер-
хушечной меристемой, расположенной в выемке, голые (если не
считать ризоидов), мужские заростки нитевидные или пластинчатые,
без меристемы. Гаметангии обычного для полиподиоидных папорот-
ников строения ( Campbell, I887;liottier, 1910; V/uist, 1910;
Karpowicz, 1927,1972; Pietrykowska, 1962b;Orth, 1936; Como-
se, 1958). Сперматозоид до 65 иж длиной, спирально скрученный,
с крупным пузырьком и многочисленными жгутиками ( Bao et al.,
1988).
Онтогенез
В качестве запасного питательного вещества споры содержат
белки-глобулины (15-20%), жиры (11-27%) и фитиновую кислоту,
которые, соответственно, являются источникам азота, энергии и
фосфора (De Maggio, stetler, 1985; Стеценко, Шевченко, 1988;
Templeman et al., 1988). В отличие от спор большинства папо-
ротников споры отраусника содержат хлоропласты и в темноте по
10
большей части не прорастают. Попав во влажную среду, на свету и
при соответствующей температуре споры быстро набухают, и уже че-
рез несколько часов отмечается высокая активность многих фермен-
тов и образование крахмала в хлоропластах; при этом у страусника
обнаружен необычный для хлоропластов способ деления (инвагинация
внутренней мембраны) и своеобразные пики поглощения света
(Gantt, Arnott, 1565; Olsen, Gullvag, L97I; Jarwis, Wilkins,1973;
Cohen, De Laggio, 193b).
Дальнейшее развитие заростков прослежено в культуре при по-
севе спор в дистиллированную воду, на почву, на жидкие и агари-
зированные питательные среды ( Campbell, 1887; Lagerberg, 1908;
Lottier, 1910; Waist, 1910; и др.). На 3-6-й день после посева
появляется ризоид, а затем и хлоронема. Пластинчатые заростки
развиваются по типу Adianthiun(Uayar, Kaur,1971).
Как в культуре, так и в природе (Aderkas, 1983), в популя-
ции заростков можно встретить заростки нескольких типов: облада-
ющие верхушечной меристемой женские, мужские и обоеполые, амери-
стематические мужские и так называемые нейтральные, у которых к
моменту наблюдения гаметангии могли еще не образоваться. Амери- -
стематические мужские заростки имеют вид простой или разветвлен-
ной с антервдиями на концах ветвей нити или продолговатых прос-
тых или многолопастных пластиночек, по краю и на брюшной сторо-
не которых располагаются антеридии. Заростки с верхушечной мери-
стемой сердцевидные, шириной 1,5-3 мм, с утолщенной под меристе-
мой областью (подушкой) и однослойными крыльями. Архегонии воз-
никают на подушке, антеридии в задней однослойной части заростка
среди ризоидов или по краю крыльев, реже антеридии появляются на
верхней стороне заростка или на подушке. В культуре первые гаме-
тангии появляются на заростках спустя 2-6 недель после посева
спор. Обычно первыми появляются антеридии, а спустя некоторое
время архегонии. В старых культурах образуются крупные (до 6 мм
в длину) меристематические сердцевидные женские заростки, в зад-
ней части таких заростков могут появиться нити или лопасти, на
которых затем возникнут антеридии.
Развитие и строение гаметангиев, зародыша и проростка под-
робно описано Д.Кэмпбеллом (Campbell, 1887). Первый лист про-
ростка длинночерешковый, обычно с 4-8-лопастной пластинкой с
дихотомическим жилкованием; листовой след простой. Расчленен-
ность пластинки у последующих листьев постепенно возрастает, и,
II
наконец, появляется лист с округлым в сечении черешком и перис-
той пластинкой с зубчатыми дольками. У последующих листьев плас-
тинка постепенно приобретает очертания, свойственные листьям
взрослого растения; изменяется и филлотаксис (от 1/3 к 2/5 и
3/8). На протяжении первых 1-2 мм стебля услож.няется и структу-
ра стелы: от протостелы через соленостелу она переходит в дик-
тиостелу ( Campbell, 1887; Мейер, 1949). Завершается этап эпидо-
генеза, и растение вступает в этап менетогенеза ( Eggert, 1961).
У выращенного в лаборатории на среде Кнопа спорофита через 5 мес.
после оплодотворения было 5 листьев ( Kottier, 1910).
Начинающий развиваться из остаточной меристемы на главном
стебле столон растет первоначально косо вверх, протискиваясь
между плотно сомкнутыми и жесткими основаниями листьев. В это
время столон очень тонкий. По выходе за пределы покрывающего
главный стебель чехла из оснований листьев и придаточных корней
столон постепенно утолщается и, если он вышел на достаточной
глубине, переходит к плагиотропному росту. Если столон вышел за
пределы чехла над землей или близ поверхности земли, он первона-
чально углубляется в почву и лишь затем растет горизонтально.Со-
гласно Н.И.Шориной и Н.М.Державиной (1981), столон растет 2 го-
да. К концу второго сезона роста, находящаяся обычно на глубине
5-10 см зеленоватая верхушка столона слегка утолщается и начина-
ет изгибаться кверху. Междоузлия при этом сильно укорачиваются,
а филлотаксис от 1/3 переходит к следующим ступеням. К осени под
землей у самой ее поверхности возникает крупная, защищенная ка-
тафиллами почка с многочисленными (около 10 хорошо различимых)
зачатками листьев. Весной следующего года из внешних зачатков
над поверхностью земли развернутся 1-5 зеленых листьев дайной
до 15-50 см, а почка к осени будет содержать уже более 10 хорошо
различимых невооруженным глазом зачатков листьев.
Первые 1-2 листа молодого растения, возникшего на конце
столона, по своей морфологии (округлый черешок, расставленные
перья с зубчатыми долями) напоминают листья возниипего из зиготы
проростка, что свидетельствует о глубоком омоложении (в смысле
Н.П.Кренке, 1940) вегетативного потомства. Постепенно стволик
растет вверх, а нижняя его часть в естественных условиях зано-
сится аллювием или перекрывается растительным опадом, так что
даже у старого растения почка обычно невысоко возвышается над
поверхностью земли.Филлотаксис доходит до 5/13, а, возможно, и
более высоких ступеней. Диаметр стебля достигает своего макси-
12
мума (около 2 см).
В литературе нет сведений о том, на какой год начинает спо-
роносить растение, вознимпее из заготы. Что же касается растений,
образовавшихся из столонов, то, по нашим наблюдениям в культуре
и литературным данным ( Prange,Aderkas, 1985), спорофиллы могут
появиться на второй-третий год, после того как над поверхностью
земли поднимутся первые трофофиллы. В благоприятных условиях
культуры спорофиллы затем появляются ежегодно в числе до 15. В
природных популяциях спорофиллы, по-видимому, появляются позже,
они образуются в небольшом числе (1-7), далеко не у всех расте-
ний и не ежегодно.
У культивируемых в саду в Московской обл. экземпляров, об-
разующих в год до 25-30 листьев (считая трофофиллы, спорофиллы
и катафиллы), стволик ежегодно нарастает примерно на 1,5 см, в
Канаде ( Prange, Aderkas,1985) - на 2 см. Остающиеся на стебле
основания листьев в течение многих лет остаются живыми и служат
местом запасания питательных веществ (прежде всего крахмала), в
связи с чем их предлагают называть трофоподами (от греч.трофе -
питание, подос - нога) ( Wagner, Johnson, 1983). Верхушка осно-
ваний постепенно отмирает, они укорачиваются, и стволик приоб-
ретает редьковидную форму.
В.П.Воротников (1985), наблюдая страуоник в Волжско-Кам-
ском заповеднике, попытался выделить возрастные состояния рас-
тения по наличию спорофиллов и числу трофофиллов. Подсчитав
число имеющихся на стебле оснований листьев и число листьев в
кроне растения он определил относительный возраст растения,раз-
делив первое число на второе. Согласно В.П.Воротникову, вирги-
нильные особи (их возраст до 20 лет) образуют в среднем ежегод-
но примерно от 4 до 5 листьев, генеративные (их возраст 21-45
лет) — от 5 до 6 листьев, сенильные (возраст 46-70 лет) - от 2
до 4 лиотьев. Но, как мы уже отмечали, спорофиллы могут поя-
виться уже у двухлетних особей, и, напротив, оооби весьма пре-
клонного возраста, но находящиеся в неблагоприятных для роста
условиях, могут не спороносить до момента своей гибели.
Что же касается приведенных В.П.Воротниковым данных об от-
носительном возрасте, то следует иметь в виду, что использован-
ный им и применявшийся уже при расчете возраста щитовника муж-
ского способ ^Погорелова, Работнов, 1978; Науялис, Филин,1983),
строго говоря, приложим, во-первых, к растениям, вступившим в
фазу менетогенеза (Филин, 1990), когда объем верхушечной мери-
стемы, а, следовательно, и строение первичного тела (в том чис-
ле и такой показатель, как число листовых зачатков) в течение
многих лет меняются слабо. А, во-вторых, этот способ вряд ли
целесообразно использовать по отношению к страуснику, так как
для точного подсчета числа катафиллов, образовавшихся в текущем
году, обычно необходимо повредить верхушечную почку, данные же
по одним лишь трофофиллам и спорофиллам могут послужить причи-
ной серьезных ошибок, так как их число не остается более или
менее постоянным даже у растения, находящегося в фазе менетоге-
неза. Так, по нашим наблюдениям в 1982-1983 гг. в Звенигород-
ском р-не Московской обл. из 35 растений, у которых в 1983 г.
развернулось 5 и более трофофиллов, .лишь у одного три года под-
ряд их число оставалось неизменным (8-8-8). У 10 растений одно
и то же число трофофиллов появлялось в течение двух последующих
сезонов (напр., 5-5-6, 6-8-8, 6-6-5, I5-II-II). У остальных 24
побегов число трофофиллов за три года или постепенно возрастало,
или уменьшалось, или колебалось (например, 7-9-13, I3-I2-II,
II-I5-8, I4-I2-I3, I4-I7-I6). У растений, листья которых появи-
лись над землей на второй год наблюдений, число их на третий
год обычно увеличивалось (например, 2-7, 3-10), но иногда и па-
дало (3-2). Очевидно, если учитывать не только трофофиллы, но и
катафиллы, общее число листьев у молодого растения будет посте-
пенно увеличиваться, а у растения, находящегося в фазе менетоге-
неза, оставаться примерно на одном уровне (включая, конечно,
сюда и число спорофиллов, если они образуются). При расчете при-
мерного возраста нельзя не учитывать и еще одного обстоятельства.
По нашим наблюдениям в Московской обл. и в Мордовской АССР неко-
торые растения страусника в течение 1-3 лет не образуют листьев, I
находясь в состоянии вторичного покоя, причины которого пока не
выяснены.
Самые крупные из наблюдаемых нами в Тверской обл. и в Мор-
довской АССР экземпляров страусника имели стебли в 20-30 см дли- .
ной (не очитая участок, входящий в верхушечку® почку). На таких i
стеблях было от 300 до 450 листовы,. оснований, а при основании
стеблей еще сохранялся столон, давший начало раотению. Исходя ;
из числа листьев воех категорий, образовавшихся в последний год,
и внося поправку на этап эпидогенеза, можно предполагать, что
возраст данных экземпляров около 30-40 лет, а если учесть, что в
природе встречаются отоль же крупные стволики, в основании кото-
14
рых нет и следов столона, можно думать, что растение на данном
месте появилось и более 40 лет тому назад. Хотя рост у страусни-
ка, как и у других папоротников, детерминирован, в Московской
обл. (и, видимо, вообще в средней полосе ев]5опейской части
СССР) экземпляры страусника вряд ли доживают до. того времени,
когда деятельность верхушечной меристемы прекращается в резуль-
тате естественного старения апикальной клетки; обычно растения
погибают в результате повреждения верхушечной почки.
Сезонное развитие
Если ранней осенью перенести из природы в теплицу столон,
то его почка сразу тронется в рост, но если перенести в теплицу
надземный побег, новые листья не развернутся. Для того чтобы
почка надземного побега вышла из состояния органического покоя,
необходим определенный минимум низких температур. Растения, взя-
тые из природы в Новой Шотландии (Канада) и перенесенные в теп-
ло, пробуждались в том случае, если их выкопали после 18 ноября;
к этому времени температура почвы на глубине 5 см была 5,8°С.
100%-ное пробуждение выкопанных осенью растений отмечено при вы-
держивании их в течение 7-8 недель при 0-I°C (Prange, 1985).
После получения необходимо минимума низких температур в
природе растение пребывает уже в состоянии вынужденного покоя
до весны, когда почва и воздух достаточно прогреваются. В лабо-
раторных условиях при 6°0 пробуждается 8% растений, находящихся
в состоянии вынужденного покоя; с увеличением температуры до
18-24°C количество пробудившихся растений увеличивается до 88%,
при дальнейшем повышении температуры количество пробудившихся
растений падает до 67% ( prange, 1985).
В природе пробуждение наступает, когда почва прогреется
примерно до Ю°С. В Московской обл. листья начинают разворачи-
ваться, когда зацветает черемуха; обычно - это первая половина
мая. В Канаде за 14-20 дн. приступают к разворачиванию все го-
товые к этому листовые зачатки, при этом каждый из них, разво-
рачиваясь за 16-20 дн., дает начало трофофиллу. На скорость рас-
кручивания зачатка, точнее на деление и рост его клеток, влияют
не только температура воздуха и почвы, но такке освещенность,
обеспеченность растения водой и другие факторы, например при
недостатке влаги и сильной освещенности формируются более ко-
роткие листья.
тн
В целом, весна - наиболее критический период в жизни стра-
усника. В почке взрослого растения число имеющихся зачатков ли-
стьев примерно в четыре раза превышает число листьев, которые
образуются каждый год, т.е. в нормальных условиях обычно реали-
зуется один "набор" зачатков и по мере "выбывания" из состава
почки внешних зачатков на конусе нарастания формируются новые.
Когда напочвенные заморозки повреждают начавшие раскручиваться
"улитки", начинают разворачиваться, образуя трофофиллы, зачатки,
которые в норме развернулись бы позже в.текущем году, дав нача-
ло спорофиллам и (или) катафиллам, или даже зачатки "набора"
следующего года. К такому же результату ведет и обрывание "ули-
ток" при заготовке страусника да!я пищевых целей. Поэтому реко-
мендуют обрывать не более трех "улиток", в противном случае в
рост тронутся зачатки "набора" следующего года, что приведет к
истощению растения. Поведение растения при повреждении или уда-
лении зачатков свидетельствует о том, что развитие имеющихся в
почке зачатков жестко не детерминировано и во многом зависит от
окружающих условий.
Спорофиллы, по-видимому, начинают развиваться довольно ра-
но, но становятся хорошо заметными в центре воронки из трофофил-
лов спустя 1,5-2 мес. после того, как почка тронется в рост, т.е.
вскоре после того, как полностью развернутся вое трофофиллы. В
Московской обл. спорофиллы хорошо заметны уже к концу июня-нача-
лу июля. Они образуются у достаточно крупных экземпляров, рас-
тущих на сильно увлажненной, но не перенасыщенной влагой почве,
богатой доступными растению ионами азота, фосфора, калия. Не-
обходимо такие, чтобы сила света была достаточной для достиже-
ния точки компенсации на кривой фотосинтеза ( Prange, 1980). В
таких условиях активно образует достаточное для образования
спорофиллов количество угяеводов. Если оборвать сильно развер-
нувшиеся трофофиллы в конце мая, то вместо спорофиллов с реду-
цированней пластинкой могут появитьс,- спорофиллы, перья которых
частично стерильны ( Goebel, 1887). Роль углеводов и высокой
температуря в дифференцировке спорофиллов .’ гдтверждана и опыта-
ми по культивированию зачатков листьев родственника страусника
- оноклеи чувствительной ( Harvey,Caponetti, 1972).
.Последние из готовых к дифференцировке в текущем сезоне
зачатков внешнего "набора" образуют нередко листья с хорошо
развили? основанием и редуцированной, а то и вовсе абортирован-
16
ной пластинкой.Эти катафиллы несколько отличаются величиной и
очертанием от катафиллов столона. Иногда катафиллы не образуют-
ся, а иногда 1-2 катафилла формируются до того, как возникнут
спорофиллы. В Новой Шотландии (Канада) из 340 обследованных в
широколиственном лесу растений катафиллы образовались у 94% эк-
земпляров, а из 2 314 листьев этих растений большая часть (79%)
была трофофиллами, 20% - катафиллами и лишь 1% - спорофиллами
( Aderkas, Green, 1986).
Если у страусника удалить листья на шестой неделе с начала
пробуждения почки, то могут появиться катафиллы или новые ли-
стья вовсе не появятся. Это свидетельствует о тем, что верхушеч-
ная почка рано, впадает в состояние органического покоя ( Ader-
kas, Green, 1986). Все изложенные выше факты позволяют предпо-
лагать, что внешние условия оказывают сильное влияние на диффе-
ренцировку листовых зачатков. Очевидно, что формированию трофо-
филлов способствуют более низкие, а формированию спорофиллов —
более высокие температуры. Определенную роль играет и длина дня.
В опытах с оноклеей чувствительной показано, что развитие спо-
рангиев протекает лучше в темноте, а длинный день ингибирует их
образование ( Harvey, Caponstti, 1972). Возможно, что и у стра-
усника длинный день способствует дифференцировке трофофиллов, а
короткий - спорофиллов и катафиллов.
Когда спорофиллы только разворачиваются, на плаценте уже
возникают спорангии и идет мейоз. Наблюдаются определенные изме-
нения в морфологии развивающихся спор ( Kuramoto, 1970). К кон-
цу августа пластинка спорофилла начинает буреть, споры созревают
и могут прорасти при посеве, но края сегментов спорофиллов еще
полностью прикрывают спорангии, предохраняя их от высыхания и
вскрывания. Первые заморозки ведут к отмиранию трофофиллов, они
буреют и ложатся, прикрывая в густых зарослях страусника споро-
филлы и верхушечные почки. На покров из отмирающих трофофиллов
ложатся опадающие листья деревьев. Дополнительную защиту почке
от высыхания создает слизь, выделяемая волосками на зачатках
листьев. Растения страусника заносятся снегом. К середине зимы,
когда отмершие трофофиллы съеживаются, спорофиллы освобождают-
ся, и по мере оседания снега их концы начинают показываться
над его поверхностью, подвергаясь иссушающему действию ветра и
мороза. Чзсдъ спор и спорангиев начинает осыпаться по мере раз-
вертывания пластинок сегментов уже в декабре ( Farrar, 1976),
17
а массовое спороношение, когда края сегментов полностью отогнут-
ся, освободив спорангии начинается весной;в Московской обл. в.
конце марте-начале апреля. Спорангии и споры подхватываются вет-
ром и ложатся на поверхность тающего снега и обнажившуюся места-
ми почву, часть из них может быть затем перенесена в иное место
талыми водами.
Способы размножения и распространения
Как и другим папоротникам, страуснику свойственны бесполый
и вегетативный способы размножения. Выше отмечено, что спорофил-
лы появляются не у всех растений и в небольшом числе, но это
компенсируется тем, что каждый спорофилл образует от 100 тыс. до
I млн опор ( Farrar, 1976). Разнесенные ветром и талыми водами
не все споры попадают в условия, благоприятные для прорастания.
Уже давно была отмечена гибель значительной части спор при посе-
ве (dottier, 1910). Впоследствии было установлено, что у стра-
усника, как и у других многолетних растений, обладающих одной
инициальной клеткой в апексе стебля, очень высока вероятность
сохранения в апексе вредных мутаций ( Klekowski, Kazarinova-Fuk-
shansky, 1984 а,ь). Поэтому с возрастом побега у страусника
нарастает величина мутационного генетического груза, в результа-
те чего многие из образовавшихся особей вегетативного происхож-
дения, а потом и целые клоны будут представлять собой генетичес-
кие химеры. Генетический груз у этих химер отрицательно сказыва-
ется на спорообразовании, и значительная доля спор оказывается
нежизнеспособной (Klekowski, 1934,1985,1988).
При выращивании заростков в лаборатории отмечено, что при
посеве умеренной плотности заростки растут лучше, чем в условиях
акоенической культуры, когда их рассаживают по одному. В первом
случае уже через 4 недели после посева 23% заростков были муж-
скими, 62% - женскими, а у 15% пол еще не определился. Через 8
недель после посева в популяции было обнаружено 29% мужских за-
ростков, 42% женских и 19% обоеполых. В акоенической культуре
через 8 недель после посева пол не определился у 63% заростков,
9% заростков были мужскими, 25% - женскими и 3% - обоеполыми
( Aderkas, 1983). В популяции заростков, обнаруженной в условиях
сильной затененности и влажной атмосферы, под пологом зрелых спо-
рофитов ТО июня все заростки были еще бесполыми, 6 июля появи-
лись мужские заростки (1%) и женские (0,3%; 2 заростка из 720),
18
30 июля 55% заростков были мужскими, а 45% - бесполыми. Женских
и обоеполых заростков до II сентября найдено не было, хотя всего
за сезон наблюдений было просмотрено I 299 заростков ( Aderkas,
1983). Даже учитывая, что часть заростков каждый раз в момент -
наблюдения изымалась из популяции, что не могло не сказаться на
общих результатах, очевидно, что условия местообитания под поло-
гом зрелых спорофитов не способствуют росту и развитию заростков,
хотя некоторые исследователи ( dottier, 1910) и полагают, что пол
заростка предопределен в споре и не зависит от условий произрас-
тания гаметофита. При прорастании густо расположенных спор (на-
пример, внутри опавшего целиком спорангия) образуются нитевидные
мужские заростки ( Campbell, 1887).
Хотя заростки страусника и откликаются на антеридиоген, вы-
деляемый заростками орляка, и формируют антеридии, в субстрате
под заростками самого страусника подобного феромона пока не най-
дено ( D6pp, 1950,1959; Naf, 1956). Отмеченные выше особенности
полового созревания заростков страусника увеличивают шансы меж-
заросткового спаривания, велики также шансы и внутризаросткового
самооплодотворения ( Klekowski, 1969). Не исключено, что отмечен-
ная в культуре способность заростков к ветвлению ( Campbell,1887;
Waist, 1910 ;Mottier, 19Ю; Aderkas, 1983 ), в природных усло-
виях, после разрушения части ткани заростка, может привести к об-
разованию клона, состоящего из потомков одного гаметофита.
В литературе нет сообщений о нахождении в природе спорофи-
тов, возникших из зиготы. Это, видимо, не случайно, так как отме-
ченный у страусника наивысший среди папоротников мутационный ге-
нетический груз сказывается не только на спорообразовании. До
95% спор несет в себе спорофитные летали, генетический груз ска-
зывается не только на процессе оплодотворения, но и на развитии
зародаша и молодого опорофита ( Klekowski, 1988). В то же время
характер местообитаний некоторых изолированных популяций страус-
ника позволяет предполагать, что изредка в природе заростки и
молодые спорофиты возникают из спор. Основной же способ размноже-
ния и расселения страусника - вегетативное размножение с помощью
столонов. В стерильной культуре каждый из очагов остаточной мери-
стемы в основании листа может дать начало столону ( Wardlaw,1943,
1946; Aderkas, 1986).Высаженные на плантации куски разрезанного
на части стволика также, приживаясь, дают начало растению из од-
ного очага остаточной меристемы ( Prange, 1985).
19
'аднако в природе столонов образуется сравнительно немного,
к' образуются они обычно у- молодых растений на краю популяции.
Когда верхушка столона переходит к ортотропному росту, обычно у
основания будущего стволика появляется новый столон. В первые
года жизни у молодого растения может ежегодно появляться до 5
столонов, но по мере старения прямостоячего побега столоны на
нем образуются все реже и реже или вовсе не появляются. Верху-
шечная почка по мере роста растения постепенно слегка поднимает-
ся над. поверхностью: земли, и это, .видимо, не способствует про-
буждению очагов остаточной меристемы, находящихся над уровнем
почвы,, хотя изредка и в природе и в культуре мы наблюдали столо-
ны, возникшие над поверхностью почвы и углубляющиеся в нее.
Что же касается очагов, находящихся под землей, то следует
учесть, что к атому времени растение окружено другими экземпля-
рами страусника, все свободные ниши заняты, и растение это ка-
ким-то. образом чувствует. Однако в культуре при ежегодном удале-
нии молодых растений,.находящихся рядом с материнской особью,
последняя ежегодно активно образовывала новые столоны, во всяком
случае на протяжении 10 лет, когда шли наблюдения. Перед отмира-
нием стволика иногда отмечается как бы второй период активного
формирования столонов. Во всяком случае, у раскопанного в стра-
усниковом оероольйанике стволика с мертвой верхушечной почкой у
его верхнего конца на протяжении 7 см обнаружено 9 столонов дли-
ной от 2 до 12 см, из них три еще не перешли к ортотропному рос-
ту, а остальные дали начало 4-м однолетни:.! и 2-м двухлетним ор-
тотропным побегам.
Подобно тону, как удаление верхушки ортотропного побега ве-
дет к пробуждению очага остаточной меристемы в основании листа,
так и повреждение верхушки столона ведет к его ветвлению. При
посадке в почву отдаленных в июле от материнского растения моло-
дых столонов, еде не перешедших к ортотропному росту, мы через
год на 7 отолонах обнаружили от I до 4 боковых ветвей о 1-2 ка-
тафиллами. Очевидно, что процессы ветвления и образования столо-
нов находятся под контролем верхушечных мериотем. Поскольку
столоны иногда достигают значительной длины (до I м), отрауоник
быстро может захватывать новые территории. Видимо, именно таким
способом, "шаг за шагом" страусник после отступания ледника про-
двигался по долинам рек и их притокам, расширяя свой ареал.хотя
и не исключено., что начало новым популяциям могли положить и
20
перенесенные во время половодья в новые места выпаханные из поч-
вы стволики со столонами либо возниилие на заростках спорофиты.
Отмирание или разрыв столонов ведет к образованию клонов, кото-
рые, опять же благодаря столонам, могут длительное время удержи-
вать захваченную территорию, производя с их помощью перераспре-
деление ресурсов между особями разного возраста и моментально
закрывая "окна" от выпавших экземпляров. Можно сказать, что бла-
годаря столонам популяции страусника обретает как бы бессмертие.
При культивировании заростков на искусственной питательной
среде обнаружено явление факультативной апогамии, в результате
чего могут возникнуть пилоплоидные спорофиты ( Aderkas, 1984в).
Для получения чистых линий спорофитов в будущей селекционной ра-
боте можно использовать методы выращивания in vitro очагов ос-
таточной меристемы или листовых примордиев ( Wardlaw, 1943,1946;
Aderkas, Hicks, 1985; Aderkas, 1986). При таком выращивании
недавно обнаружено явление факультативной апоспории - образова-
ние гаметофитных выростов на ювенильных листьях и остаточной ме-
ристеме ( Aderkas, 1986). Интересно, что Onoclea orientalis
(Matteuccia orientalis) на материке (в Азии) представлена дип-
лоидной, а на Японских островах - аутотетраплоидной циторасой, у
которой танке обнаружено явление факультативной апогамии (Lloyd,
1971,1973). Не исключено, что и у страусника обыкновенного в пре-
делах его обширного ареала есть появившаяся в результате апога-
мии (или, что менее вероятно, апоспории) и последующей сингамии
аутотетраплоидная раса.
Экология
По сравнению с содержащими хлоропласты спорами других папо-
ротников (например, Onoclea ), споры страусника более устойчивы
к воздействию неблагоприятных факторов внешней среды. Лиофилизи-
рованные споры всходят и спустя три месяца после замораживания
( Barker, White, 1964). В воздушно-сухом состоянии при хранении
в темноте при 3-5°С споры сохраняют жизнеспособность до двух
лет, но при хранении при комнатной температуре всхожесть быстро
падает ( Gantt, Arnott,1965; Lloyd, Klekowski, 1970; Jarvis,
Wilkins, 1973 ; Стеценко, 1985).
Сообщают, что часть спор может прорастать и в темноте (Jar-
vis, Wilkins, 1973), но не исключено, что такие споры уже успе-
ли получить необходимый для прорастания световой импульс. В це-
ЙХ-Ю41
21
лом же для активного и массового прорастания требуется свет вы-
сокой интенсивности, и чем дольше длится действие света, тем
больше спор прорастает ( Klebs, 1916; Pietrykowaka, 1962а; Mohr,
Hill, I964-; Jarvia,Wilkins, 1973). Прорастание идет в довольно
широком диапазоне температур: от 5 до 30°С ( Kleba, 1917; Hill,
Wagner, 1974; Warne, Lloyd, 1980). В глубокой тени заростки
растут и развиваются хуже, чем при умеренном освещении (Aderkaa,
1983); они не выносят подсушивания ( Pickett, 1914). Сообщают,
что содержащие хлоропласты споры лучше прорастают на минераль-
ном, а не на органическом субстрате (Hill, Wagner, 1974), а по-
скольку в спорах страусника содержится много запасного N и Р,
на первых порах гаметофиты мало зависят от субстрата и быстро
развиваются. Некоторые исследователи ( Warne, Lloyd, 1980) пред-
полагают, что температура свыше 20°С ингибирует образование ар-
хегониев, но многочисленные наблюдения в лаборатории и в поле не
подтверждают этого.
Присущие страуснику морфолого-анатомические и физиолого-
биохимические особенности позволяют ему расти в очень разнооб-
разных условиях. Экотопы, в которых встречен страусник, объеди-
няют в две группы - экотопы равнинных и экотопы горных ландшаф-
тов (Порфирьев, 1975). К первой отнесены экотопы: I) логовых
депрессий, 2) возвышенных плато и пологих склонов водоразделов
и верхних террас речных долин, 3) низменных плато с подпочвенны-
ми грунтами постоянной влажности, 4) пойменных террас и цент-
ральных пойм. К экотопам горных ландшафтов отнесены экотопы до-
лин горных рек и склонов разной крутизны и экспозиции. Не удиви-
тельно поэтому, что страусник можно встретить на почвах разного
механического состава и генезиса: на мелко- и крупнозернистом
гумусированном аллювии, дерново-подзолистых супесях и суглинках.,
на глинах и илах, и даже на торфяно-болотных почвах с содержа-
нием органики до 60%. В Евразии страусник чаще приурочен к тем-
ноцветным, темно-серым и бурым почвам (Порфирьев, 1975); приуро-
ченность страусника к какому-либо специальному типу почв не от-
мечена и в Северной Америке (Aderkaa, Bird, 1983).
При всем разнообразии почв общей чертой почти всех экотопов
страусника является то, что он приурочен к местам с хорошим дре-
нажем, с перемещающимися по водоупорным горизонтам проточными
почвенно-грунтовыми водами, богатыми кислородом и минеральными
солями. Страусник можно считать индикатором таких вод, хотя из-
22
редка его можно встретить и в местах с временно застойным водным
режимом. В пору развертывания листьев страусник очень чувствите-
лен к недостатку воды ( Prange, 1985), но в это время воды в по-
чве достаточно, летом же зеркало грунтовых вод в местах произра-
стания страусника опускается сравнительно неглубоко, поэтому
страусник может успешно переносить и довольно продолжительные
летние засухи.
По содержанию минеральных элементов почвы, на которых про-
израстает папоротник, богаты К, Р и К ( Aderkas, Bird, 1983). В
культуре растение хорошо откликается на подкормку нитратным или
нитратным и аммонийным азотом, но подкормка только аммонийным
азотом нарушает режимы водообмена, дыхания и фотосинтеза и ведет
к некрозу листьев (prange, Ormrod, 1982). В Европе и в Северной
Америке страусник часто приурочен к известковым почвам, что дало
основание многим исследователям считать его кальцефилом. Однако
анализ разных типов почв и опыты в культуре свидетельствуют о
том, что папоротник может расти в широком диапазоне pH - от 7,5
до 4,5. Но поскольку очень низкие или очень высокие значения pH
изменяют доступность многих элементов, одни из которых необходи-
мы, а другие в больших концентрациях вредны для растения, лучши-
ми для произрастания страусника будут нейтральные или близкие к
ним субстраты, залегающие на содержащих известь материнских по-
родах ( Prange, 1980; Prange, Ormrod, 1982; Aderkas, Bird, 1983;
Prange, 1985; Prange, Aderkas, 1985).
Изучение фотосинтеза и откликов страусника на водный дефи-
цит позволяет охарактеризовать его, как типичное теневыносливое
лесное растение (Prange, Ormrod, 1983;Prange et al., 1983,
1984; Prange, 1985; Aderkas, 1986), хотя у северной границы
ареала или южнее, но в местах с очень высокой влажностью воздуха
страусник можно встретить и на совершенно открытых местах. В
культуре при освещенности, равной 70$ от полной, растение обра-
зует более крупные листья (Prange, Aderkas, 1985) и, по нашим
наблюдениям, спороносит ежегодно. В густых лесах спороносящие
экземпляры встречаются лишь там, где в древостое есть "окна’’.По
числу устьиц (около 20 на I мм^) страусник та же близок к гигро-
фитам тенистых местообитаний.
В Северной Америке прослеживается четкая связь между север-
ной границей ареала страусника и температурным режимом года
(prange, Aderkas, 1985). Эта граница совпадает со среднегодовой
23
изотермой -3,9°С, она пролегает там, где вегетационный сезон со
средней дневной температурой 5,6°С начинается не позже 20 мая и
оканчивается перед I октября, среднегодовая температура почвы
лежит в пределах от 2,2 до 8,3°0, а среднелетняя температура - в
пределах от 8,3 до 15°С. Определенно нет корреляции между север-
ной границей распространения вида и продолжительностью безмороз-
ного периода, началом морозного периода, мощностью снегового по-
крова, количеством осадков, максимальными или минимальными днев-
ными температурами. К сожалению, пока нельзя сказать, существует
ли какая-либо связь между климатом и южной границей ареала.
Для местообитаний, к которым приурочен страусник, обычно
характерна небольшая изменчивость температуры воздуха и почвы
в течение длительных периодов летом и почвы - зимой. По сравне-
нию с другими летнезелеными папоротниками, растущими под пологом
леса, страусник более морозоустойчив. Хотя его листья и погибают
при понижении температуры ниже -3°С, стволики, охлажденные в ла-
боратории в течение двух суток до -5°С, безболезненно переносят
потом охлаждение до -20°С в течение 12 ч ( Sato, 1982). Но в при-
роде стволики, и, прежде всего, важнейшая их часть - верхушечная
почка вряд ли длительное время испытывают действие очень низких
температур. Уже отмечалось, что почка очень рано впадает в сос-
тояние органического покоя. Крупные размеры почки сами по себе
снижают риск повреждения конуса нарастания низкими температура-
ми. Кроме того, вымерзанию и иссушению почки в холодный период
препятствуют: I) покров из катафиллов, 2) слизь, выделяемая во-
лосками на зачатках листьев, 3) дубильные вещества в клетках,
4) сохраняющиеся и намного возвышающиеся над почкой основания
листьев, 5) скапливающаяся при листопаде на дне "воронки" из
трофофиллов листва деревьев, 6) поникающие при заморозках листья
рядом стоящих экземпляров страусника, 7) снеговой покров. Следу-
ет также иметь в виду, что после отмирания пластинки листа обо-
лочки клеток торчащих концов листовых оснований опробковевают,
тем самым возможность испарения в этих местах сводится до мини-
мума ( Kittinger et al., 1987).
Особенно благоприятные для перезимовки условия складывают-
ся в долинах рек, там, где периодически откладывается аллювий.
Аллювий заносит стволики папоротника, поэтому верхушечная почка,
приподнимающаяся над поверхностью почвы при росте побега, вновь
оказывается на уровне земли. При пересадке страусника из леса на
открытые участки отмечено ( Prange, 1985), что спустя 3-4 года
24
листья у пересаженных растений по сравнению с листьями растений
той же популяции, но оставшихся в лесу, начинают весной развора-
чиваться позже, и таким образом папоротник как бы предусмотри-
тельно избегает напочвенных заморозков, вероятность которых на
открытом месте выше, чем в лесу. Имея в виду относительно высо-
кую морозоустойчивость страусника и то, что температуры выше
24°C несколько тормозят развертывание трофофиллов, вполне оправ-
дано отнесение страусника по температурной шкале к умеренным ви-
дам ( Bllenberg, 1978).
В черешках и рахисах листьев страусника хорошо развита
склеренхима, но ее надежность в качестве пружины, предохраняющей
листья от излома при ударе, невелика (Раздорский, 1939). Правда,
и механические нагрузки такого типа растению большей частью ис-
пытывать не приходится, так как обычно оно обитает в защищенных
от ветра местах.
Фитоценология
Наблюдений над природными популяциями гаметофитов отраусни-
ка практически нет, но, исходя из особенностей его спор и эколо-
гии спорофита, можно предполагать, что заростки ведут себя как
экспле ренты, находя оптимальные для своего существования места
на обнаженных минеральных, обогащенных органикой субстратах в
полу затененных местах.
Спорофиты страусника выступают часто в качестве доминанта
или субдоминанта третьего яруса, или яруса средних трав (Быков,
I960) в ассоциациях, где доминантами древесного яруса являются
разнообразные хвойные (пихта, ель, сибирский кедр), широколист-
венные (бук, дуб, вяз, липа, клен, яоень, каштан) и мелколист-
венные (ольха, береза, черемуха, тополь) породы. Б СССР страус-
ник можно встретить в ельниках и в производных от них липняках и
осинниках, в черневых пихтовых, пихтово-еловых и пихтово-липовых
лесах, в кедрово-пихтовых и в кедрово-широколиственных лесах, в
дубравах, вязовниках и в буковых лесах, в ольшаниках, тополевни-
ках и пр. (Быков, I960; Порфирьев, 1975; Растительность европей-
ской части СССР, 1980). Изредка страусник встречается в сообщест-
вах, где нет деревьев, например на болотах со сфагновыми мхами
или с осоками ( Aderkas, Bird, 1983). Наиболее мощные заросли
страусника можно встретить, пожалуй, в долинах равнинных и гор-
ных рек. Примечательно, что на Кавказе, этом убежище третичных
25
флор, страусник выступает в ольховых, пихтовых и буковых лесах
в долинах ручьев на горных склонах как доминант выдающейся силы,
совершенно подавляющий возобновление древесных пород (Соколов,
1936а), в том числе и такой теневыносливой породы, как бук.
Представляется вероятным, что именно ?в таких убежищах страусник
сохранялся в плейстоцене, расселяясь на равнинах по долинам рек
в межледниковья и в голоцене.
Популяции страусника могут насчитывать от нескольких десят-
ков до тысяч надземных побегов, возраст, мощность и характер
расположения которых зависят от конкретных условий. Б долинах
рек, в уремах, на территории, занятой популяцией страусника, не-
редко можно выделить различающиеся по численности страусника и
флористическому богатству травостоя участки. В сомкнутых надзем-
но и подземно зарослях страусника, где на 10 может прйходится
до 30-50 надземных побегов папоротника, преобладают по численно-
сти и массе старые экземпляры, а молодые появляются лишь в "ок-
нах”, возникающих при гибели других побегов. На таких участках
страусник создает свой микроклимат.
Боронковидные кроны из крупных трофофиллов со слегка пере-
крученными вдоль жилок и сомкнутыми и даже иногда черепитчато
налегающими один на другой сегментами эффективно перехватывают
свет и осадки, а при отмирании полностью покрывают почву. По-
этому летом под пологом страусника в таких местах фактически об-
разуется "мертвая" для других зеленых растений зона. Лишь кое-
где можно встретить одиночные побеги теневыносливых цветковых
растений, а на кусочках валежа и на некоторых стволиках самого
папоротника - зеленые мхи. Напротив, по краю популяции, где эк-
земпляров папоротника немного и они не столь мощны, по массе и
численности преобладают представители крупнотравья из цветковых-
нитрофилов. Но и здесь страусник прочно удерживает свои позиции,
а при возможности и быстро захватывает освободившиеся ниши. По-
этому вряд ли правильно относить страусник только к экоплерентам
(Разумовский, Киселева, 1979), он может быть и виолентом, и па-
тиентом; это лишний раз свидетельствует о том, что три типа
стратегий растений, в чистом виде встречаются редко.
Приводимые в литературе (Воротников, 1985; Гуреева, 1987а)
данные по возрастному спектру популяций следует оценивать с уче-
том трудностей, возникающих при определении возрастных групп
страусника.
26
Анализ флористических списков ассоциаций с участием страус-
ника позволяет утверждать, что этот папоротник не связан посто-
янно с каким-либо видом или группой видов растений, и его нали-
чие в том или ином месте обусловлено в первую очередь эдафичес-
кими факторами ( Aderkas, Bird, 1983). Высокая вегетатихвная под-
вижность страусника, присущая ему способность к перераспределе-
нию ресурсов с помощью столонов и к.развертыванию новых листьев
взамен поврежденных обусловливают стабильность синузий папорот-
ника - они сохраняются на одном месте даже при смене доминантов
древесного яруса (Порфирьев, 1961; Kisteneich, 1988). В то же
время нарушение гидрологического рента территории при мелиора-
ции, а тайке выпас скота могут быстро привести к гибели популя-
ции.
Консортивные связи
В корнях спорофита страусника обнаружена везикулярно-арбус-
кулярная микориза ( Boullard, 1957; Крюгер, Попонина, 1979;
Berch, Kendrik, 1982), но симбиоз с грибом не является обяза-
тельным условием жизни папоротника, так как он хорошо растет и в
стерильной культуре. Страусник является хозяином ржавчинного гри-
ба Uredinopsis struthiopteridis Stormer ex Diet., На ПЭПОроТНИ-
ке тайке обнаружены различные аско- и базидиомицеты: Taphrina
struthiopteris Nishida,Phoma exigua Desm.,Leptothyrium litigio-
sum(Desin. )Sacc. ,Sphaerothyrium filicinum Bub.,Sclerotium decidu-
um Davis, Ceratobasidium anceps(Bres.et Saco.)Jackson,Cyphella
capula(Holmskj.)Fr., Solenia filicina Pk.,Herpobasidium struthi-
opteris(Rostr.)Lindl.,Dothidella osmundae (Pk.et Clint.)Sacc.,
Dasyscyphus carestianus(Rabh.)Sacc. (Gregor,1938; Stevenson,1945;
Index, I960; Connors, 19.67; Gourley, 1983; Aderkas,Brewer,1983).
Изредка на страуснике паразитирует повилика европейская
( Gams, 1938).
На подземной части стволика в плотном чехле из корней и ос-
нований листьев живут дождевые черви, которые находят здесь не
только хорошее убежище от хищников, но и пищу, на надземной - мо-
гут поселяться различные зеленые мхи и даже цветковые растения.
Не подлежит сомнению, что спогн страусника используют в пи-
щу те же почвенные беспозвоночные, которые питаются спорами, на-
пример кочедыжника женского ( Schneller, 1979).
Одно время полагали, что слабая повреждаемость папоротников
27
насекомыми обусловлена наличием у папоротников каких-то веществ,
отпугивающих насекомых. В 1967 г. в листьях страусника был обна-
ружен один из экдизонов (фитоэкдизон птерозин) - гормонов, на-
рушающих цикл полного превращения насекомых ( Takemoto a.oth.,
1967), но впоследствии было установлено, что экдизоны 'папоротни-
ков не влияют на насекомых, поскольку они с трудом проникают в
организм животного, а проникнув, быстро связываются, теряя свои
свойства ( Gerson, 1979).'Поэтому не удивительно, что список на-
секомых, использующих папоротники в пищу, постоянно растет. К
настоящему времени обнаружено, что страусником питаются: из рав-
нокрылых - тли ( Amphorophora ampullata Buckton, A.laigi Ma-
son), из чешуекрылых - тонкопряды ( Stenopis auratus Grote),
листовертки ( Olethreutes anrleapitana Clemens, Spargonotis
reticulatana Clemens) и медведицы ( Papipema sp.), из перепон-
чатокрылых - ПИЛИЛЬЩИКИ ( Strombocerus delicatulus Fallen,
Strongylogaster lineata Christ., S.struthiopteridis Forsins,
Thrinax struthiopteridis Mai., Blastiootoma filiceti Klug ),
ИЗ двукрылых - цветочницы ( Chirosia betuleti Ringdahl, Ch.hy-
strioina Rondani), галлицы ( Chinodiplosis sp.) и злаковые
мухи ( Slachiptera costata Loew.,Trieimba melancholica Becker)
(Ермоленко, 1972,1975; Balick et al., 1978; Aderkas, Peterson,
1987; McCabe, Wagner, 1989). Но если, например, цветочные мухи
и галлипэ используют уже готовые галлы предшественников, то упо-
мянутые пилильщики (некоторые из них относят к примитившейшей
группе) питаются исключительно папоротниками. На страуснике в
зимнее время обычно покоятся слепняки, представители полужест-
кокрылых (Lygus rubroclarus Knight), а прямо торчащие до вес-’
НЫ спорофиллы используют ДЛЯ окукливания коконопряды (Malacos-
ma disstria Hubner)(Aderkas, Peterson, 1987).
Листья страусника на Дальнем Востоке иногда поедает пят-
нистый олень (Рябова, Саверкин, 1937), их едят овцы и коровы
( Prange, Aderkas, 1985).
Биологическая продуктивность
Вес листьев у о,иного растения страусника увеличивается по
мере их развертывания в течение первых четырех недель, после
чего практически не меняется ( Aderkas, 19866). По семилетним
наблюдениям в Татарской АССР (Райфа) на- верхней террасе Волги,
где в западинках микрорельефа в липняке страусниково-прслесни-
28
ковом-на среднеоподзоденнах супесях, страусник образует.’синузию’’
сомкнутых куртин, запиглающих до 40% площади, сухой вес надаем- о
пой фитомасс»’ (очевидно,, только листьев) варьировал ’от 18,3 г/м*
до 58,6 г/м*, вес фитомассн о,иного растения - от 3,3. г до 18,7 г .
(Порфирьев, 1261).
По наблюдениям на востоке Кг-нады в широколиственном лесу из
вяза, ясеня н'клена в популяций, насчитывающей много сотен ство-
ликов; несущих-от 2-х до 13-ти (в среднем 6,75+2,3) листьев,
дайна которых варьировала от 33 до 152 см (средняя -.85 см), су-
хой вес листьев у одного растения варьировал.; от 1,6 до 72,3 г
(средняя - 72,3 г) ( Prange, Aderkas,.-1985).
• Спороносящие-растения обычно много мощнее- неспороносящих и
образуют еще. большую.фитомассу. Дапржер, 'у отцюг’о из эдаемпад- •
ров страусника, выращиваемого в- саду Дос коде кой обл. и сохранив-
шему- на стебле- 175 старых оснований- листьев» сырой вес образовав-
шихся к осени 1990 г. 14' трофо^шов .и 10 спорофиллов равнялся,
соответственно, 450 и 200 г, сухой вое,, соответственно, 122,5 и
39,5 Г. У второго, еще. более мощного экземпляра, на стебле кото-
рого насчитано 218 старых оснований листьев, образовалось в
1990 г. 16 трофофиллов и 15 спорофиллов, сырой вес которых рав-
нялся1, соответственно,/53С и- 320 г, а сухой - 148 и 6G г. Пели
учесть, что листья для взвешивания срезали не у самого основания
и что имеются еще катафиллы* а зачатки листьев будащего года
сильно подросли, произведенная растениями надземная фитомасса
была в действительности выше указанной. Сырой вес выкопанных,
очищенных от почвы-и большей части корневой системы стволиков у
этих двух растений был равен 620 я 870 г, а сухой, соответствен-
но, - 217 я 301 г.
В Волкеко-Камском заповеднике у экземпляров, образующих
ежегодно по 5-6 листьев и отнесенных к генеративным, сухой вес
"корневищ" был равен 17-28 г (Воротников, 1985).
Хозяйственное значение
Страусник обыкновенный - один из немногих папоротников, ис-
пользуемых человеком в качестве пищевого продакта в больших ко-
личествах. Коренное население Дальнего Востока Евразии и востока
Северной Америки издавна использовало начинающиеся разворачи-
ваться листья ("улитки"), а то и развернувшиеся листья как весен-
нюю овощьв сыром виде или сушило впрок; в Норвегии его использо-
29
вали при приготовлении пива (Лучник, 1938; Gams, 1938; Fernaid,
Kindsley, 1943; Aderkas, 1984a, 1985). В последние года попу-
лярность страусника-на востоке США и Канады как ценного диетиче-
ского продукта возросла настолько, что возникли компании, зани-
мающиеся заготовкой страусника. В провинции Нью Брансуик за ко-
роткий весенний оезон иногда заготавливают 50-100 т "улиток", в
штате Мэн - 40-50 т, а в целом на востоке Северной Америки - око-
ло 225 т ( Brooks, 1979; Prange, 1980; Aderkas, Bird, 1983;
Aderkas, 1984a). Собирают папоротник по лицензиям, с одного ра-
стения рекомендуют брать не более 2-3-х "улиток". Оптимальными
условиями для сохранения питательных и товарных качеств листьев
перед замораживанием является хранение их при 0°С при 100$ влаж-
ности ( Lidster a.oth., 1989).
Существует много рецептов приготовления разных блюд из стра-
усника ( Brooks, 1979). По вкусовым свойствам отваренные "улитки"
сравнивают со спаржей. Они богаты витаминами, минеральными соля-
ми, белками, причем белки полноценны и легко усвояемы и в этом
отношении сравнимы с белком коровьего молока. Б них есть отсут-
ствующие у семенных растений ненасыщенные жирные кислоты, необ-
ходимые животному организму. Они содержат много К и мало На , а
поэтому их рекомендуют в качестве диетического продукта при бес-
солевой. диете. Данные по химическому составу листьев приведены
во многих работах (Цапалова, Плотникова, 1981; Bushway et al.,
1982; Стеценко, Табачний, I984;Prange, Aderkas, I985;Soeder,
1985). Листья страусника в больших количествах накапливают крем-
незем, но он распределен в тканях беспорядочно и не образует ка-
ких-либо частиц, а потому, в отличие от листьев орляка, листья
страусника не обладают канцерогенными свойствами ( Newberne,l976;
Hohne, Richter, .1981; Aderkas et al., 1986).
Хотя потенциально лишь в провинции Пью Брансуик от дикорас-
тущих растений можно за сезон собрать до 1000 т "улиток" ( Ader-
eas, 1984а), труднодоступность многих мест заготовки весной и
угроза подрыва жизнеспособности тех популяций, где сбор произво-
дят регулярно, заставили начать работы по внедрению страусника в
культуру на востоке Канады и США. Изучают способы размножения,вы-
ращивания, селекции наиболее урожайных клонов (Cumming, 1985;
Prange, Aderkas, 1985). Урожайность растения на заложенных пло-
щадках сравнима с урожайностью страусника в благоприятных для его
роста природных местообитаниях. В СССР листья страусника заготав-
30
ливают на Дальнем Востоке обычно впрок (сушат или маринуют).
В народной медицине страусник попользовали как средство от
лихорадки, падучей болезни, худобы, а чаше (.видимо, по аналогии
с щитовником мужским) как глистогонное средство (Анненков,1878).
Проведенные впоследствии исследования не подтвердили пригодность
страусника в качестве глистогонного средства, но при этом было
обнаружено, что содержащиеся в страуснике вещества оказывают
спазмолитическое, седативное и противовоспалительное действие,а
потому растение можно считать перспективным лекарственным сырь-
ем (Шретер, Карнитина, 1975).
Отвар "корневищ" использовали как инсектицидное средство,а
сами побеги - в корм рогатому скоту (Кормовые растения, 1950;
Полезные растения, 1951). Листья использовали для упаковки соч-
ных плодов (Роллов, 1908).
Страусник - одно из прекраснейших декоративных растений от-
крытого грунта, его крона сохраняет декоративные качества в те-
чение всего вегетационного сезона. Во многих районах СССР в ре-
зультате хозяйственного использования территории заросли стра-
усника исчезли или находятся на грани исчезновения, в связи с
чем страусник включают в списки редких и охраняемых растений.
З.П.НемченКо
ЗИШЯ КУДРЕВАТАЯ
Номенклатура и систематическое положение
Lilium martagon L., 1753, Sp.Pl. :303; Цингер, 1885, Сбор-
ник свед.:431; Кауфман, 1889, Ыоск.фя.,дед.2: 512; Шмальгаузен,
1897, Фл.Средн. и Южн.России, 2: 509; Четунников, 1901, Крит.об-
зор, 3:68; Сырейщиков, 1906, Илл.фл.Моск.губ. 1:238; он же,1927,
Опред.паст.Моск.губ.:90; Комаров, 1935, Фл.СССР, 4:288; Победи-
мова, 1964, в Маевский, Фл., изд.9;662; Скворцов, 1966, Опред.
раст.Моск.обл.:±15; Баранова, 1979, Фл.европ.части СССР,4:239;
liatthews, ±980, pl.iurop. 5:34; Черепанов', 1981, Сосуд.раот.
СССР:290, - L.pilosiusculum (Freyn) Kiscz., 1910, Trav.Kus.Acad.
Sci.Petereb..,3ot. 8:192; Победимова,1964.цит.ооч. :663; Черепа-
нов,1981; цит.соч.:290. - Лилия кудреватая (саранка, царские ку-
дри) (семейство Liliaceae -лилейные).
В. Московской области - один вид Lilium martagon L. Встре-
чается в лесных оврагах, старых парках; в парках, возможно, оди-
чало (Ворошилов и др., 1966; Кадастр ботанических объектов...,
1978).
Тип: "Hungaria, Helvetia, Siberia, Lipaiae.” (L.) (Бара-
нова, 1990).
Название вида турецкое, дано по форме цветка, напоминающей
особую Фо пл у чалмы (Баранова, 1977).
Внутривидовая систематика
Вид очень изменчив. Б природе встречаются формы с различны-
ми окраской и числом цветков, размерами растений, форлой и раз-
мерами листовых пластинок, сроками цветения, наличием и степенью
опушенноети стебля, прицветников и бутонов.
Со времени описания Lilium martagon к. Линнеем было описа-
но еще множество форм, разновидностей и подвидов ( Woodcock,
Steam, 1950; Priszter, 1967; Песауле, Орехов, 1973; Баранова,
1977). Некоторые исследователи (Мшценко, I9II; Гроссгейм, 1940;
Врищ, ±972; и др.) выделяли из вида Lilium martagon самостоя-
тельные виды: L.pilosiueculum (Freyn) Kiscz., L.caucasicum
(l.iiscz. exGrossh.) Orossh..L.cattaniae (Vis.) Vis. Исследова-
тель рода Lilium Ч.В.Еаранова считает, что признаки растений
32
выделяемых видов не выходят за пределы изменчивости вида Lili-
um martagon и в пределах этого вида выделяет четыре подвида и
несколько форм.
Lilium martagon subsp. martagon L. Ареал его охватывает
почти всю европейскую часть ареала вида до р.Волги и южные рай-
оны Сибири. Подвид характеризуется широколанцетными листьями,
собранными по 6-8 в мутовки, сиреневой окраской цветков.
В пределах L.martagon subsp. martagon выделены несколько
форм, которые встречаются по всему ареалу: f.albiflorum (Vuk.)
G.Beck - доли околоцветника белые с легким красным креплением
у основания, алой срединной полосой снаружи, устойчива против
серой гнили луковичных, встречается в Германии и Югославии;
f.album (Weston) G.Beck - цветки белые, без пятен, пыльники и
пыльца желтые, зацветает на неделю позже основного вида, растет
В Западной Европе и Сибири; f.Daugava Lacis (sin.var.koknese ) -
- околоцветник светло-сиреневый, цветет несколько раньше типово-
го подвида, семена дают всходы в год посева, встречается в Лат-
вии в окрестностях г.Гробина у пос.Кокнесе, растет на моренных
отложениях ледникового периода и, возможно, сохранилась с после-
ледникового времени (Несауле, Орехов, 1973).
L.martagon subsp. pilosiusculum (Preyn) Iljin ex B.?edtsch.
отличается от основного подвида паутинистой опушенностью бутонов
и прицветников, более узкими листьями (листовая пластинка лан-
цетной формы, 8-12 см длины и 1,5-3 см ширины), произрастает в
Западной и Восточной Сибири, на Урале, в европейской части нашей
страны (бассейны рек Еятки и Камы), за пределами СССР - в север-
ных районах Монголии.
L.martagon subsp. caucasicum Kiscz. sx Grossh. отличается
от типового подвида суженными к основанию и коротко заостренными
на верхушке листьями до 18 см дайной и до 5-6 см шириной, соб-
ранны:,ш обычно-в двух мутовках. Подвид распространен на Кавказе.
L.martagon subsp. cattaniae (Vis.) Degen - ЛИСТОЧКИ ОКОЛО-
цветннка винно-красного цвета, без пятен; характерны вюровиддае
густо-белоопушеннне бутоны, листья с блеском. Произрастает в
Албании, Югославии и северных районах Греции.
Морфологическое описание
Многолетнее луковичное растение. Луковица до ТО см в диа-
метре, яйцевидной Форш, не очень плотная, состоит из донца ко-
:-1641 23
нической формы и спирально расположенных на нем сочных чешуй. Че-
шуи золотисто-желтого цвета, узко-ланцетной формы, заостренные
на конце, цельные или с перетяжками, в количестве 25-30.
Стебель 80-150 см высоты, круглый, зеленый или неровно буро-
ватый, часто в верхней части белооиушенный, олиственный.
Листья короткочерешковые или сидячие, в средней части стеб-
ля собраны по 10-15 в 2-4 мутовки (иногда мутовки выражены нечет-
ко) , в верхней части стебля очередные. Листья в мутовках продол-
говато-овальной или обратно-яйцевидной формы, 12-13 см длины,
3,7-4,5 см ширины; в верхней части стебля ланцетные, 5-7 см дли-
ны, 1-2 см ширины (Немченко, Новиков, 1978). Форма и размеры ли-
стовых пластинок варьируют в зависимости от места произрастания.
(Priszter, 1967; ВриЩ, 1972).
У лилии кудреватой, как почти у всех видов лилий, различают
два типа корней: луковичные - растущие от донца луковицы, и стеб-
левые - развивающиеся на подземной части стебля. Луковичные кор-
ни растут с весны до поздней осени, функционируют 1-1,5 года..
Среди них есть.сильно утолщенные, поперечно морщинистые втягива-
ющие (контрактильные) корни. С помощью их луковица углубляется в
почву. Стеблевые корни начинают расти поздно весной и отмирают
осенью вместе со стеблем (Трофимов, 1963; Шиповская и др., 1972;
Баранова, 1980). Кроме функции поглощения питательных веществ
они способствуют закреплении стебля в почве.
Соцветие кистевидной формы из 3-х - 17-ти (и более) цветков.
Цветки поникшие, обычно сиреневые или розово-сиреневые (известны
формы с белыми, желтыми, винно-красными цветками), с темно-пур-
пурными пятнышками (или без них), слабо ароматные. Околоцветник
чалмовидной формы, листочки его 3-4 см длины, 1-1,3 см ширины,
4-4,2 см в диаметре, сильно отогнуты назад. Тычинок 6, тычиноч-
ные нити и отолбик голые, пыльца коричнево-красная, рыльце трех-
лопастное , темно-пурпурное.
Плод - обратно-яйцевидная шестигранная трехгнездная коро-
бочка, усеченная сверху, с заостренными ребрышками, высотой 2,5-
3 см и шириной 1,5-1,7 см.
Цветет в июне-июле.
Диплоидный набор хромосом 24 ( Stewart, 1947), 24 + I-3B
(Хромосомные числа...,1969).
Географическое распространение
Лилия кудреватая имеет самый обширный ареал сведи видов
34
рода. Западная граница ее ареала проходит в Португалии (8° з.д.),
восточная - вдоль нижнего и среднего течения р. Вилюй (124°в.д.)
и верхнего течения р.Лена; на севере доходит до низовья р.Енисей,
достигая 68°о.ш.; на юге захватывает в Европе Италию,Грецию , в
Азии - северную часть Монголии и Малую Азию (Баранова,1966).
Изолированно от основного ареала, этот вид произрастает в севе-
ро-западном районе Кавказа и на северо-западе полуострова Малая
Азия (Баранова,1990).
На территории нашей страны нреизрастает в Литве, Латвии,
Средней России, Сибири (Врищ,1972; Несауле и др.,1973; Рада-
ускене, 1976; Редкие и исчезающие растения Сибири, 1980).
В Московской обл. - найдена в Серпуховском р-не в деревне
Жерновка и деревне Зыбинка, в старом липовом парке среди зарос-
лей ежевики; в Можайском р-не в оеле Поречье, в старом парке
бывшего имения в травостое из кислицы, майника, звездчатки, па-
поротников, печеночницы; в окрестностях г. Рузы (Кадастр ботани-
ческих объектов...,,1978). В местах бывшего культивирования может
сохраняться неопределенно долго. Даже после суровых зим, например,
зимы 1978-1979 гг., эта лилия сохранилась в нашей области, обра-
зуя в последующие оезоны цветущие и плодоносящие побеги. В этом
смысле при сохранении старых парков можно говорить, что лилия
кудреватая здесь натурализовалась.
Онтогенез
Семена неправильной треугольной формы с округлым краем,
7,5-9,5 мм длины, 6,5-8,5 мм ширины, 0,5-1,0 мм толщины, с
эндоспермом, светло-буро-коричневые, тусклые. Зародыш недиффе-
ренцированный, поэтому его лучше называть предзародышем (про-
эмбрио). Последний по отношению к эндосперму занимает централь-
ное положение, он линейный или слабо согнутый, сплюснутый,
молочно-белого цвета, 5-6 мм длины, 0,5-0,6 мм ширины, 0,3 мм
толщины (Трофимов, 1963).
В лабораторных условиях семена лилии кудреватой прорастают на
9-10-Й день при температуре 20°С. Предварительное охлаждение
семян до +5°С в течение 30 дней вызывает, по-видимому, переход
в состояние глубокого физиологического покоя, исключившего рост
не только в период ее непосредственного действия, но и после парен
са семян в условия температуры +20°С (Седова,1978).
Прорастание семян подземное: проросток ведет в первый год
35
подземный образ жизни. При осеннем посеве оемян при самосеве в
течение следующего лета в почве образуются небольшие проростки
о почечкой и корнем. При прорастании в узкой части семени
около рубчика через микропиле вытягивается наружу нижняя часть
проэмбрио, достигающая 8,5-15 мм и вынооящая зачаточный корешок
и точку роста. Верхняя часть семядоли остается в семени и в даль-
нейшем выполняет гаусториальную функцию, перенося питательные
вещества из эндосперма к развивающемуся проростку.
В июне-июле нижняя часть черешка семядоли заметно утолщается,
образуя влагалище семядоли с находящейся в ней точкой роста. В
пазухе влагалища семядоли формируется почечка, в которой можно
различить несколько чешуек. В дальнейшем она увеличивается и
значительно удлиняется корень. В конце лета почечка состоит
из 2-3-х утолщенных чешуй и зачаточного лиоточка с точкой роста
в его основании. Есть и раздвоенные почечки с сильно утолщенными
чешуями и зачаточным листом . К осени формируется один корешок
длиной от 4 до 7,5 ом с 2-3 боковыми разветвлениями в нижней
части. Более утолщенные корешки с поперечными морщинами выполняют
Функцию втягивающих корней. На следующий год весной на поверхность
почвы выходит один единственный лист.
В дальнейшем при нормальном развитии молодые растения лилии
кудреватой с каждым годом увеличиваются в размерах, частично
изменяя свою структуру. Лист на втором году надземной жизни
приобретает уже овально-заостренную или яйцевидную форму, с
черешком достигает 8-10 см длины и 2-3 см ширины. На третьем
году жизни ( у некоторых более сильных - на втором) над почечкой
появляются два листочка. В трехлетием возрасте формируется сте-
бель высотой 12-25 см, и растения приобретают облик взрослых
особей. У них в верхней части стебля обычно развивается один круп-
ный лист, под ним формируется мутовка листьев (5-8 листьев), а
при двухъярусном расположении мутовки,число листьев в мутовке до-
ходит до 14 (Трофимов, 1963; Баранова, 1974).
Цветение и плодоношение начинается в 4-5-летнем возрасте
(Трофимов,1963; Баранова, 1977) и даже 7-летнем возрасте (Неса-
уле, Орехов, 1973).
По описаниям некоторых исследователей высота взрослых расте-
нии составляет от 30 до 120 см, а число цветков в соцветии равно
3-10 (Комаров,1935; Баранова, 1966). По нашим наблюдениям в Бо-
36
таническом саду МГУ растения бывают значительно выше, листья
крупней, соцветие несет.15-20 цветков.
Соответственно изменению надземных органов усложняется
структура и размеры луковиц: увеличивается количество чешуек и
корешков. У мелких луковиц бывает 2-3 втягивающих корня, у круп-
ник - до 10. Кроме того, от донца луковицы отходят более тонкие
корни с разветвленной системой боковых корешков. У взрослых осо-
бей на стебле чуть выше луковицы образуются придаточные корни.
Размеры луковиц из года в год увеличиваются в соответствии с уве-
личением количества чешуй или их размеров (при неизменном коли-
честве) .
Сезонное развитие
Ежегодно-растение развивается из главной почки вовобновле--
ния, которая закладывается в материнской, луковице. Она формиру-
ется на вершине донца, в пазухе верхней чешуи, у основания цве-
тоносного стебля. Почка возобновления закладывается в августе,
побег из нее начинает развиваться почти через два года (Еаранова,
1971). Нередко образуется 2 и даже 3 почки возобновления, в ре-
зультате чего через несколько лет образуется гнездо луковиц.
3 течение первых 3-4 месяцев после заложения почки в ней
формируются зачатки низовых чешуй луковицы, которые позже станут
запасающими. Вслед за ними (будущей весной, за год до цветения)
начинают дифференцироваться стеблевые листья. Зачатки низовых
чешуй и листьев сходны в это время по ферме и строению. Диффе-
ренциация стеблевых листьев в почке продолжается 5-6 мес. и за-
канчивается в июле-августе, т.е. через 12 мес, после ее заложе-
ния и почти за год до цветения.
В почке возобновления через 10-12 дней после окончания за-
ложения листьев междоузлие, расположенное над последней низовой
чешуей, начинает интенсивно разрастаться. Зачаток цветоносного
стебля хорошо выделяется среди низовых чешуй. Вскоре у его осно-
вания появляется бугорок - зачаток новой почки возобновления.
Одновременно на конусе нарастания цветоносного стебля дифферен-
цируются цветки. Это происходит в августе предшествующего цвете-
нию года. Таким образом, конец лета - наиболее важное время в
развитии лилии кудреватой.
Нормирование цветков длится 2-2,5 мес. и заканчивается
обычно в октябре. Зимой рост почки возобновления будущего года
х 37
3 -1041
в л?/ко вице практически прекращается, а весной начинается снов;:.
Поскольку вегетативные и генеративные органы сформированы с осе-
ни, растение быстро проходит все фенофазы: почти сразу за нача-
лом роста следует бутонизация, а затем цветение и плодоношение.
Вегетация завершается в августе. Таким образом, цикл развития
длится 24 мес.j из них 20 мес. падает на дифференциацию органов
внутри луковицы й только 4 мес. продолжается надземное развитие
цветоносного стебля (Баранова, 1980).
Цветение начинается в третьей декаде июня и продолжается
7-10 дней. Тычинки и рыльце расположены на одном уровне.
Лилия кудреватая - геофит. На зиму все надземные органы от-
мирают. Почки возобновления находятся в почве в пазухе верхней
чешуи луковицы.
Способы размножения и распространения
Лилия кудреватая - растение самоопыляющееся. Размножение се
и расселение осуществляется в природе в основном семенами. Раз-
множается также луковичками-детками, которые формируются на стеб-
левых корнях под землей.
В естественной среде обитания всхожесть семян составляет
44-29$, деток практически не образуется (Ивашкина, 1985).
Экология
Лилия кудреватая - типичный мезофит. Растет на- лесных опу ;-
ках, полянах, лугах. Произрастает на почвах достаточно увлажнен-
ных (четвертая ступень увлажнения по шкале Г.Элленберга ( silen-
berg, 1974).
По отношению к свету этот вид можно считать полутеневыноо-
ливым (по Элленбергу получает более 10, но в большинстве случаев
менее 100$ от полной освещенности). Он приурочен к лесным райо-
нам. Растет среди кустарников, в густом травостое. Б Московской
обл. найден в зарослях ежевики под пологом нескольких видов дре-
весных растений.
Лилия кудреватая сравнительно нетребовательна к теплу. Па
некоторых этапах развития прорестнов температура является регу-
лирующим прорастание фактором. Снижение ее до ч-5°С вызывает пере-
ход в состояние покоя. Семена могут находиться в состоянии покоя
до наступления более благоприятных, эффективных для роста темпе-
ратур (Седова, 1978). Луковицы зимуют в промерзшей почве.
38
Эта ления в природе произрастает на почвах,по составу близ-
ки н известковым,рыхлых,богатых (Priszter, 1967;11есауле .Орехов,
1973) Г. Эллен бе рг указывает на приуроченность ее к почвам преиму-
щественно нейтральным (седьмая ступень шкалы),но может расти на
слабокпслых и нейтральных почвах., от не очень бедных-до. .богатых
(по отношению к азоту).
Е Московской обл,'встречается в местах,где почва карбонатная,
подстилаемая известняками, относительно богатая (Кадастр ботани-
ческих объектов..., 1978).
Фитоценология
Лилия кудреватая в Европе - характерный мезофильный элемент
буковых и дубово-буковых лесов и играет там значительную роль.
Б Московской обл. найдена в сообществе старых лип, одичавших
яблонь, рябин, черемухи, ежевики.
Консортивные связи
в культуре этот вид поражается фузариозом (Fusarium oxyspo-
rum Sciii.). В годы мало благоприятные (дождливые.холодные) пора-
жается грибами серой гнили луковичных ( Botrytia elliptica(Berk.)
Cooks).
Кз вредителей ее повреждают клещ луковый и корневой,журчалки. •
Хозяйственное использование и охрана
Лилия кудреватая введена в культуру более 1000 лет назад как
декоративное растение,особенно краоива белоцветковая форма, Этот
гид о.-цж из самых распространенных в культуре (благодаря своей зи-
мостойкости) .Используется в открытом грунте для оформления раба-
ток, куртин,в миксбордере.В садовой культуре лилию кудреватую-раз-
множают луковичками-детками,делением гнезда луковиц,чешуями,семе-
нами.
Луковицы линии издавна использовали в пищу в вареном и пече-
ном виде,сок луковиц - в народной медицине как ранозаживляющее,
настой - от зубной боли. Их используют та же в красильной промыш-
ленности: с солями железа окрашивают ткани в черный цвет (Губанов,,
и др., 1976).
Ио наблюдениям в Предуралье численность особей в популяциях
ежегодно резко сокращается: в 1979 г. - 296 растений, в 1981 г.-
87, в 1984 г. - 54. В семи регионах страны лилия- кудреватая вне-
сена в список редких и исчезающих видов (Ивашкина, 1985).
39
М.Г.Вахрамеева, Т.М.Быченко, И.В.Татаренко, М.В.Экзерцева
МЯКОТНИЦА ОДНОЛИСТНАЯ
Номенклатура, систематическое положение и
внутривидовая систематика
Kalaxis monophyllosxAb.) Swartz, 1800, Kongl.Sv.Vet.Akad.
Handl. 21:234; Кауфман, 1866, Моск.фл.,изд. 1:480; Голицын,
1964, в Маевский, Фл.,изд.9:681; Ворошилов,1966, Опред.раст.
Моск.обл.:123; Смольянинова,1976, Фл.европ.части СССР, 2:18;
Черепанов,1981, Сосуд.раст.СССР: 210; Новиков,1986, Опред.раст.
Мещера, 1:132. - Ophrys monophyllos L., 1753, Sp.Pl.: 947. -
biicrostylls monophyllos (L.) Lindl. 1830, Gen Sp.Orch. Pl. :I9;
Цингер,1885, Сборник свед.:4I5; Шмальгаузен, 1897, Фл.Средн. и
Юкн.России, 2:446; Сырейщиков,1906, Илл.фл.Моск.губ. 1:270; он
же, 1914, цит.соч. 4:50; он же, 1927, Опред.раст.Моск.губ.:97;
Невский, 1935, Фл.СССР, 4:601; Назаров, 1940, в Маевский, Фл.,
изд.7:242; Голицын,1954, в Маевский, Фл.,8:694. - Мякотница одно-
листная (семейство Orchidaceae - орхидные).
Б роде Malax!s насчитывают около 70 (пс другим данным до
300) видов, значительная часть которых распространена в тропиче-
ских районах Азии и Африки, в СССР - I вид.
Мякотница однолистная - генетически однородный вид, измен-
чивость его невелика. В Чехо-Словакии, например, указывают всего
две формы этого вида -f. monophyllos (растения до 10 см высотой,
в соцветии менее 30 цветков) и f.robusta (выше 10 см, в соцве-
тии более 30 цветков) ( Frochazka.Velisek, 1983). Кроме того,
практически повсеместно на протяжении ареала, помимо типичной
однолистной изредка встречается двулистная форма. Это послужило
основанием для ряда авторов выделить самостоятельный вид Mala-
xis diphy 1.1а Cham. (Ificrostylis diphyllos Lindl.) (СыреЙЩИКОВ,
1906; Hegi, 1936; Schumacher et al., 1986; и др.). В опытах
Фюллера (Fuller, 1966) показано, что одна и та же особь в раз-
ные годы может иметь то один, то два листа. Большинство исследо-
вателей , на наш взгляд, справедливо считает двулистную мякотницу
лишь формой М.monophyllos (Флора СССР, 1935; Флора европейской
части СССР, 1976; Черепанов, 1981 и др.).
х/ Saepe "Xonophylla"
40
Географическое распространение
Ареал вида голарктический бореальный (Протопопова, 1987)
охватывает Европу, значительную часть Азии, северо-запад Север-
ной Америки. В Европе встречается преимущественно в средней и
восточной части, отсутствует на Британских островах, на север до-
ходит до Финляндии (65°50’) и Норвегии (62°с.ш.), на юг распро-
страняется до Северной Италии и Югославии, но не отмечен южнее
Альп, отсутствует на Балканском полуострове. Мякотница встреча-
ется в Малой Азии, а танке на территории от Урала до Тихого оке-
ана, Монголии, Китая и Японии (Флора СССР, 1935; Hegi, 1936;
Ziegenspeck, 1936; Ptiller, I966;Plora Europaea, 1980; Pro-
chazka, Velisek, 1983).
В пределах Советского Союза ареал мякотницы однолистной ох-
ватывает всю лесную территорию до степной зоны, в европейской
части - от Прибалтики до Карпат, от Кольского полуострова и дс
Средне-Днепровского и Средне-Волжского районов, далее она пере-
ходит через Урал и в азиатской части страны через Западную и Во-
сточную Сибирь доходит до тихоокеанского побережья, встречается
на Камчатке, Сахалине, Курильских островах (Флора СССР, 1935;
Попов, 1957; Горовой, 1966; Воробьев, 1974; Ворошилов, 1982;
Протопопова, 1987).
В Московской обл. мякотница однолистная распространена в
Северном, Клинеко-Дмитровском, Западном, Восточном районах, реже
в южной части области, но почти всегда в незначительном количе-
стве (возможно, в ряде случаев просматривается). В прошлом веке
и начале нынешнего указывалась в ближайшем Подмосковье, в част-
ности и на территории современной Москвы - Воробьевы горы, Лоси-
ный остров, Кунцево, Горенки (Кауфман, 1866; Сырейщиков, I9C6,
1927).
Морфологическое описание
Многолетнее травянистое поликарпическое растение с кратко-
временно существующим (1-3 года) корневищем и одним утолщенным
междоузлием побега (надземным зеленым клубнем), одетым влагали-
щем зеленого листа и низовыми чешуевидными и влагалищными листь-
ями. Эту структуру., свойственную многим тропическим эпифитным
орхидеям ( irmisch, 1863), называют псевдобульбой, бульбой,псев-
доклубнем, клубнелуковицей и пр. (Тихонова, 1981; Dressier,
1981; Смирнова, 1984 и др.). Многие черты эпифитизма в морфоло-
41
гии и экологической приуроченности мякотйицы однолистной, веро-
ятно, связаны с тропическим происхождением рода.
От основания клубня отходят тонкие немногочисленные корни,
густо покрытые корневыми волосками. Длина корневищного участка
между старым и молодым клубнями варьирует в зависимости от ха-
рактера субстрата, от 0,5-1,5 см у растений на относительно
плотных субстратах дс 5-7 см у особей на,моховых болотах. Однако
число чешуевидных (2) и низовых влагалищных (I) листьев, распо-
ложенных на корневище, остается неизменным.
Зеленый лист один (редко 2, еще реже 3) - эллиптический или
яйцевидный, продолговатый, туповато-заостренный, 3-10(11) ом
длиной, 1,2-5 см шириной, с многочисленными продольными жилками,
при основании суженный в черешок, объемлющий цветонос. Второй
лист, если он присутствует, значительно меньше.
Цветоносный стебель треугольный в поперечном сечении, не-
редко с изгибами, 8-30 (40) см высотой. Кисть тонкая рыхлая узко-
цилиндрическая многоцветковая 3-15 см длиной с 30-60 (до 115)
цветками. Прицветники желтовато-зеленые, ланцетные, заостренные,
равные или короче завязи. Цветки мелкие, зеленоватые, желтовато-
зеленоватые или беловато-зеленоватые. Околоцветник зигоморфный,
из 6 свободных листочков. Лиоточки наружного круга яйцевидно-
ланцетные или копьевидные, прямостоячие, с I-й жилкой, длиной
2,7-3 мм; средний листочек обращен вниз. Листочки внутреннего
круга узкие, линейные, одинаковой длины с наружными листочками,
но в 3-4 раза уже, направлены книзу и немного повернуты назад.
Третий внутренний лизточек-губа немного короче прочих листочков
околоцветника, толстоватая, при основании широкояйцевидная, к
верхушке суженная в линейно-ланцетное заострение, несколько во-
гнутая, обращена вверх (вследствие скручивания цветоножки). По
краям близ основания края губы заворачиваются в виде узких скла-
док, заканчивающихся зубчиками, ниже которых они вплоть до конеч-
ного заострения несколько утолщены и густо усажены очень мелкими
бугорками.
Андроцей, как и у других видов орхидных, сильно редуцирован
- сохранилась лишь одна плодущая тычинка. Ее нить срослась со
столбиком пестика, образовав колонку (гиностемий) короткую и
толстоватую. На вершине колонки расположены 2 пыльника, соединен-
ные связником, который, разрастаясь, образует как бы свод над
пыльниками. Пыльца склеена в два воскообразных комочка клиновид-
. 42
ной формы - подлинна, от основания.которых отходит ножка, закан-
чивающаяся клейкой подушечксй-прилипалъцем. Обе половинки (гнез-
да) пыльника расположены близко друг к другу, после цветения они
не спадают. Гинецей устроен также своеобразно. О столбике сказа-
но выше. Рыльце трехлопастное, располагается на внутренней сторо-
не колонки под пыльником. Воспринимают пыльцу только две лопасти,
средняя превращена в треугольный клювик, на котором помещены при-
липальца поллинариев. Завязь нижняя на скрученной цветоножке.
Плод - округлая коробочка, содержащая большее число желтовато-
коричневых оемян (Кауфман, 1889; Сырейщиков, 1906; Флора СССР, .
1935; Zlegenapeck, 1936; Ftiller, 1966; Prcchazka, Veliaek, 1983).
Диплоидное числе хромосом 30 (Аверьянов и др., 1981).
Онтогенез и способы размножения
Маленькие округлые плоды - коробочки (2-3 мм длиной и около
I мм шириной) содержат в условиях Московской обл. от 1500 до
4000 оемян. В Приморском крае в плодах мякотницы в среднем .
насчитывают 1473+150 семян и 11782+633 семян на одну особь.
Несмотря на большое число цветков, плодов обычно завязывается
немного - от 5(2) до 22 (33)%. Х.Цигеншпек (Ziegenapeok J936)
отмечает, что на севере ареала от 3 до 30% цветков мякотницы дают
плоды, на юге - дс 65%. По нашим.наблюдениям, интенсивность
завязывания плодов колеблется в пределах одного фитоценоза даже
одной особи в зависимости от погодных условий очень резко.
Так, в Одинцовском районе Московской обл. на щучково-таволгово-
разнстравном лугу в 1987 г. завязывание плодов произошло у 31%
цветков, в 1988 г. - у 16%, в 1990 г. вследствие дождливой пого-
ды в момент цветения плоды на этом участке не завязались совсем.
Этому,возможно, также способствовало необычно мощное развитие
некоторых из окружающих трав, скрывающих ооцветия мякотницы от
опылителей.
За 6 лет наблюдений нами было выявлено, что максимальный
срок цветения одной оооби мякотницы - 4 года, перерывы в цвете-
нии, как правило, не наблюдаются ( в отличие от многих наших ор-
хидей), после этого в большинстве случаев сразу (реже - через
I год) растение отмирает (лишь в исключительных случаях вегети-
рует еще 2-3 года). Обычно наибольшее число цветков образуется
на второй год цветения, в первый и последний год числе цветков
43
заметно меньше. Так, например, на этом же лугу в Одинцовском
районе число цветков у одной генеративной особи изменялось сле-
дующим образом: в первый год - 40, во второй - 51, в третий -
28, четвертый - 22, у другой особи соответственно - 40,55,50,43,
Число плодов не соответствует этой закономерности, а зависит от
погодных условий. Максимальное число плодов на наблюдаемой тер-
ритории у всех ценопопуляций мякотници было отмечено в 1987 г.,
минимальное - в 1990 г.
Семена мякотницы мелкие 0,4-0,7 мм, с недифференцированным
зародышем 0,2-0,3 мм, прорастают только в присутствии гриба. По
Х.Цигеншпеку ( ziegenspeck, 1936) семена созревают во вторую по-
ловину лета, с наступлением сырой погоды сразу прорастают, обра-
зуя маленький протокорм - беловатое, полупрозрачное округлое об-
разование размером около 1x2 мм. Протокорм с помощью микоризного
гриба, снабжающего растение необходимыми веществами выполняет
функцию запасания питательных веществ. В юнце второго вегетаци-
онного сезона у протокорма появляется второй членик и листовая
чешуйка - образуется так называемый микоризом, содержащий много-
численные гифы микоризного гриба ( Ziegenspeck, 1936; FrocHazka,
Velisek, 1983). На следующий год формируются еще один членик
микоризома и листовая чешуйка. Лишь после этого в конце лета
третьего года жизни растения в пазухе листовой чешуйки заклады-
вается клубневидный орган. На четвертый год под этим клубневидным
органом развивается междоузлие, неоущее чешуйчатые листья, и лишь
на 5-й год появляется первый надземный зеленый лист и растение
переходит в ювенильное состояние, которое .длится от 2-х до 4-х
лет (в неблагоприятных условиях и дольше). Зеле'ный лист еще очень
мал - от 0,2 до 1,7 (2) см длины, 0,2-0,7 ом ширины, число жилок
на нем 2-10, клубень зеленовато-беловатый, 0,2-0,4 см длиной,
О,2-0,3 см шириной. Вначале микоризом сохраняется, придаточные
корни отсутствуют, к концу ювенильного периода микоризом отмира-
ет, появляются I, реже 2 корня. Очень чаото семена прорастают в
толще отмерших оснований листьев, окружающих генеративное расте-
ние. Проростки и ювенильные растения так и живут в этом рыхлом и
влажном субстрате, предохраняющем их от высыхания и содержащем
микоризные грибы. Образуются плотные группы разновозрастных осо-
бей. Позже взрослые и ювенильные растения постепенно "уходят"
друг от друга.
Имматурное состояние характеризуется следующими морфологиче-
скими параметрам: лист имеет длину 1,8-3,0 (3,2) см, ширину 0,8-
44
2 см, число жилок 10-14, клубень дайной 0,5-0,8 см, шириной 0,4-
0,6 см, число придаточных корней 2 (редко 3). В этом состоянии
растение пребывает от I до 5 лет, чаще (во всяком случае в Под-
московье .) - 3 года.
Взрослое виргинильное растение имеет более крупный лист
длиной 3,5-5,5 (7) см, шириной 1,7-2 (2,5) см о многочисленными
жилками (16-22), клубень дайной 0,8-0,9 (1,1), шириной 0,5-0,8
(0,9) см, число корней 3-4 (5). В этом состоянии растение нахо-
дится от I до 3 лет (иногда и дольше). Генеративное состояние,
таким образом, в лучшем случае наступает в природных условиях
не ранее, чем через I0-II лет после прорастания оемени. Размеры
вегетативных органов обычно несколько больше, чем у предыдущей
группы, но незначительно. Длина листа 5-8 (9) ом, ширина 2,2-4,5
(5) ом, число жилок 22-30. Второй лист мы наблюдали крайне редко
и только у генеративных растений, причем он всегда был значитель-
но меньше, чем первый. Клубень генеративного растения 1,0-1,5 ом
длиной, 0,8-1,2 см шириной, число корней 5-6 (7). Длина соцветия
до 10 см. Цветет мякотница обычно без перерыва в течение 3-4-х
лет. В сенильное состояние переходят не все особи, большинство
отмирает после последнего цветения.
Таким образом, большой жизненный цикл мякотницы однолистной
в условиях Подмосковья составляет примерно 15-20 лет.
Возрастная структура ценопопуляций мякотницы свидетельству-
ет о достаточно интенсивном процессе возобновления - ювенильные
растения обычно составляют 2СМ0% от состава популяции, что выше
численности ювенильных особей у большинства других подмосковных
орхидей. Популяции - нормальные полночленные, с высоким процен-
том генеративных особей, что указывает на вполне благоприятные
условия произрастания вида. Сходный возрастной состав популяций
наблюдали мы и в Приморском крае (таблица).
Мякотницу однолистную можно отнести к группе вегетативных
малолетников (Высоцкий, 1915), так как продолжительность жизни
надземных клубней не превышает 2 (реже 3) лет. Обычно у оооби об-
разуется I молодой клубень. Па старых надземных клубнях могут об-
разовываться придаточные почки. Чаще всего они развиваются у ве-
гетативных растений из апикальной меристемы, что в ряде случаев
может привести к вегетативному размножению растений ( ziegenspeck,
1’936). Вегетативное размножение наблюдается крайне редко лишь
при очень неблагоприятных условиях или при нарушении целостности
игубня. Так, на территории Иркутской обл., в окрестностях Гай-
Таблица
Возрастная структура ценопопуляций мякотницы однолистной
г разных частях ареала
Возрастная группа Московская обл. низинный луг Московская обл. елово-березо- вый лес Приморс кий край пихтово- широколистный лес
Ювенильные 32 42 39
Имматурные 18 24 . 16
Взрослые веге- тативные 20 13 21
Генеративные . 30 21 24
Общая числен- ность ценопо- пуляции 65 170 106 особей
кальского целлюлозно-бумажного комбината в крайне нарушенных .ус-
ловиях на берегу отстойников шлам-лигнина были найдены несколько
генеративных особей мякотницы, имеющих одновременно по два моло-
дых надземных клубня.
В ненарушенных условиях семенное размножение можно считать
фактически единственным способом размножения мякотницы, что под-
тверждается И другими исследователями (Goebel, 1901; Ziegenspeck,
1936).
Сезонное развитие и консортивные связи
Начало вегетации и цветения мякотницы однолистной наступает
примерно на две недели позже, чем у большинства наших орхидей, в
частности любки двулистной и ятрышника Фукса. Единственный зеле-
ный лист мякотницы в Подмосковье разворачивается в начале июня,
зацветает в конце июня-иачале июля. Цветение длится долго - око-
ло месяца; иногда до .двух месяцев. Нередко в сере .дине августа
можно застать цветущие особи наряду с плодоносящими. Плоды хоро-
шо заметны уже в начале августа, в конце августа семена высыпа-
ются из плодов. Листья начинают желтеть в конце августа, в нача-
ле сентября листья быстро отмирают, так что к середине сентября
следов вегетирующих растений найти не удается, но соцветия у ге-
неративных растений часто сохраняются более года.
К этому времени (середине сентября) клубень уже полностью
46
сформирован, достигает своих предельных размеров. Сквозь белова-
тые чешуи просвечивает зеленая почка, из которой разовьется по-
бег будущего года. В конце августа в условиях Подмосковья в поч-
ке генеративных особей хорошо различимо соцветие, прилистники,
отдельные цветки.
Любопытный опыт по выращиванию мякотницы в горшке был прове-
ден Ф.Фюллером ( Fuller, 1966). Растения были высажены в горшок
27 июня I960 г. В конце марта следующего года самые сильные, и в
начале апреля все остальные дали новый побег. Соцветия появились
13 мая, новые клубни - в середине августа.
Консортивные связи изучены недостаточно. Известно, что мя-
котница однолистная перекрестноопыляемое растение, автогамия у
нее отсутствует (чему способствует устройство цвета). Опылители
- различные мелкие насекомые, в том числе и комары ( puller,
1966; Schumacher et al., 1986).
Ряд авторов считает, что мякотница тесно связана с грибами
с момента прорастания семян ( ziegenspeck, 1936; puller, 1968).
Иаши исследования в Приморском крае показали, что микоризные гри-
бы отсутствуют в корнях особей всех автотрофных возрастных состо-
яний. Лишь в старых корнях некоторых генеративных особей встреча-
лись отдельных слабозакрученные гиФы эндомикоризных грибов. Мно-
гочисленные гифы грибов ризосферы, обнаруженные на корнях вместе
с обильными длинными корневыми полосками, видимо, обеспечивают
достаточное питание растений.
Экология
Мякотница однолистная растет в условиях частичного затене-
ния, но не менее 10% от полной освещенности (третья ступень шкалы
Ландольта). В условиях Московской обл, встречается обычно на слег-
ка затененных опушках, по обочинам заросших лесных дорог и троп,
в "окнах", на лесных полянах, на полностью открытых участках -
редко.
Обычно растет на влажных почвах, однако избегает слишком сы-
рые и слишком сухие местообитания, индикатор влажных местообита-
ний (четвертая ступень шкалы увлажнения Ландольта). В условиях
Одинцовского района Московской обл. встречается на почвах с влаж-
ностью верхнего горизонта от 60 до 80%. Предпочитает почвы бога-
тые гумусом (пятая ступень шкалы Ландольта), обычна на почвах
более или менее хорошо вентилируемых, часто на супесчаных, песча-
47
но-глеевых (четвертая степень шкали Ландольта), нередко подсти-
лаемых известняками ( Mirek, Piekos-Kirkowa, 1984). Встречается
я на бедных минеральным азотом, индикатор бедных почв (пятая
ступень шкалы богатства почв Ландольта). Обитает как на кислых
(pH 3,5-5,5), так и на нейтральных и щелочных почвах. М.Л.Рамен-
ская (1983) относит его к кальцефилам. Прохазка и Величек ( Рго-
chazka, Veiisek, 1983) отмечают их на почвах с pH 6,7 до 8,1. Б
Подмосковье мякотница была нами изучена на участках таволгово-
гравилатово-разнотравного луга с jK 6,2 и на таволгово-манжет-
ковом лугу с {Я подстилки 6,4 и pH почвы 6,1, на гераниево-щуч-
ково-разнотравном лугу на перегнойно-глеевой почве с pH подстил-
ки 6,1 и pH почвы 5,7.
Не отмечено особой чувствительности к холоду, встречается в
районах с континентальным климатом, но все же предпочитает место-
обитания с теплым летом и с глубоким снежным покровом зимой.
Встречается как на равнинах, так и в горах, где поднимается
до 1200-1660 м ( Ziegenspeck, ,1936),.
Вид не отличается строгой оитоценотической приуроченностью
(ziegenspeck, 1936; Fuller, 1966; Jaros, 1972). В пределах
таежной зоны он встречается как в лесах, так и на лугах и боло-
тах, в зарослях кустарников. В лесах разнообразного состава -
хвойных (чаще мшистых еловых, травяных сосняках), широколиствен-
ных (дубовых, грабовых, буковых), мелколиственных (березовых,чер-
нсольховых), смешанных хвойно-мелколиственных, хвойно-широколист-
венных (Ziegenspeck, 1936; Hegi, 1936; Sundermann, 1962; Fil-
ler, 1966; Flora Europaea, 1980; Флора СССР, 1935). На Дальнем
Востоке (Приморский и Хабаровский края), в Сибири чаше в мшистых
кедрово-елово-широколиственных, кедрово-пихтовых (Попов, 1957;
Ворошилов, 1966,1982). В лесах предпочитает разреженные достаточ-
но освещенные участки - окна, лесные поляны и луговины с не очень
густым травяным покровом (общее проективное покрытие травяного
покрова не более 60%). В Чехо-Словакии вид указывается в сообще-
ствах союза Caricetalia fuscae, Violion caninae Galio-Carpineturn
(Jaros, 1972).
Растет на лугах разнообразного состава, но обычно на сырых,
как на равнине, так и в горах, встречается и на субальпийских лу-
гах, часто вместе С другими орхВДНЫМИ - Gymnadenia conopsea, Her-
minium monorchia, Coeloglossum viride, Dactylorhiza maculata и
др. ( Hegi, 1936; Sundermann, 1962; Prochazka, Veiisek, 1'983;
48
Протопопова, 1987). На Сахалине и Курильских островах (Воробьев,
1974) растет на лугах близ выхода ключей, особенно часто на мор-
ских террасах.
Можно мякотницу встретить и на окраинах болот, преимущест-
венно низинных, но тайке и на сфагновых (Сырейщиков, 1927; Hegi,
1936; culler, 1966; Флора Сибири, 1987).
В средней полосе европейской части СССР мякотница указыва-
ется С.В.Голицыным (1954) как довольно обычное растение, южнее -
отсутствует. ’ . ’ >
В Московской обл. (Ворошилов и др., 1966) растет в сырова-
тых хвойных лесах, хвойно-мелколиственных и мелколиственных, на •
окраинах моховых болот.
Нами встречена на территории Московской обл. как в хвойных
и смешанных лесах, так и на лугах оырых, преимущественно низин-
ных.
Мякотница однолистная - вид в конкурентном отношении слабый,
что подчеркивается рядом исследователей (Ziegenspeck, 1936; Рго-
chazka, Veiisek, 1983; Раменская, 1983). Он предпочитает расти
е условиях более или менее разреженного травостоя, избегая близ-
кого соседства растений с крупными надземными частями (например,
папоротников). Чаще растет одиночными экземплярами или неболыпи- :
'ми группами, хотя иногда попадаются и значительные скопления. На-
пример, на олуговелсй поляне среди березово-еловсго леса была
встречена ценопопуляция мякотницы, насчитывающая около 200 особей
на 100 м? на низинном таволговс-манжетково-разнотравном лугу на
площади 10 м^ ценопопуляция из 60 особей. И в том и другом случае
ценопопуляции - полночленные, с большим числом ювенильных и гене-
ративных оообей. Общая численность ценопопуляций из года в год
колеблется (при сохранении существующих условий) незначительно -
в пределах 20-10%. Однако при изменении условий, например усиле-
нии заболачивания, из 60 особей через 5 лет осталось лишь 7 взро-
слых, ювенильные растения исчезли все. В условиях интенсивной
рекреации, сопровождающейся заметным вытаптыванием, мякотница так4
же быстро исчезает.
Особый интерес представляет популяция мякотницы однолистной
в сильно нарушенных условиях - на берегу прудов-накопителей шлам ।
- лигнина Байкальского целлюлозно-бумажного комбината. Этот вид ,
местами даже доминировал в травостое, имел необычно крупные разме-
ры, мощные соцветия, высокую плотность популяции (39,1 особь на
4-I04I
I м2), отличаясь заметно, как от подмосковных, так и местных ир-
кутских растений.
Хозяйственное значение и охрана вида
Сведений о хозяйственном значении мякотницы однолистной нет.
В связи с малой декоративностью вид не подвергается прямому унич-
тожению, однако численность его сокращается в результате наруше-
ния и уничтожения его местонахождений, изменения условий место-
обитания под влиянием деятельности человека. В Чехо-Словакий,на-
пример, 50% местонахождений вида исчезли, и он отнесен к катего-
рии видов, находящихся в опасности ( Prochazka.Velisek, 1983)‘.В
Норвегии его относят к видам, находящимся в угрожаемом положении,
из 53 известных местонахождений сохранились около 30, два место-
нахождения в настоящее время охраняются государством ( Schuma-
cher et al., 1986). Охраняется мякотница однолистая и в ряде ре-
гионов нашей страны. На Украине она занесена в республиканскую
Красную книгу, на территории Московской обл. она внесена в список
растений, охраняемых постановлением Мособлисполкома и Мосгорис-
полкома от 19.I.1984 г. в числе других видов семейства орхидных.
«I.Г. Вахрамеева, Pl.0.Виноградова, п.В.Татаренко,О,В.Цепляева
КОКУШНИК КОМАЕНИКОВЫЙ
Gymnadenia conopeeax/,(L.) R.Br.,I8I3, in Alton, Hortus
Kew.,ed.2, 5:191; Кауйман, 1861, Фл.Меск.губ.,изд.1:471; Дин-
гер, 1885, Сборник свед.:418; Снрейщиков, 1906, Илл.фп.Моок.губ.
1:257; он же, 1914, цит.соч. 4:47; он же, 1927, Опред.раот.Моск.
губ.:96; Невский, 1935, Фл.СССР, 4:668; Назаров, 1940, в Маев-
ский, Фл.,изд.7:247; Голицын,1954, в Маевский, Фл.,изд.8:697; он
же, 1964, цит.ооч.,изд.9:684; Ворешилов,1966, Опред.раот.Моск.
обл.:125; Смольянинова, 1974, Фл.европ.части СССР, 2:34; Moore,
1980, Fl.Europ. 5:332; Черепанов,1981, Сосуд.раст.СССР: 310;
Новиков, 1986, Опред.раст.Мещеры, 1:135. - Orchis conopsea L.,
1753, Sp.Pl. :942. - Habenaria conopsea (L.) Benth.,1880, Journ.
Linn.Soo. 18:354; Шмальгаузен, 1897, Фл.Средн.и Юан.России,
2:461. - Кокушаик комарниковый (семейство Orchidaceae - орхид-
ные).
Изменчивость вида довольно велика, в связи с чем выделяют
РЯД внутривидовых таксонов: помимо типичной формы subsp. conop-
sea имеются еще два. подвида - subsp.alpina (Turcz. ex Reichenb.
fil.) Janch. ex Soo и subsp. densiflora (Wahlenb.) K.Richt
(Черепанов, 1981; Flora Europaea, 1980; Цвелев, 1969; Perring,
1962). Прохазка и Велипек ( Prochazka, Velisek, 1981) вместо
subsp.alpina указывают subsp. montana Bisse. Все три подвида
различаются размерами как вегетативных, так и генеративных орга-
нов, а танке условиями местообитания. Приводятся следующие разме-
ры высоты цветоноса и ширины листа для этих таксонов: subsp. co-
nopsea - высота 20-40 (50) см, ширина листа 3(5) - 10(22) мм;
subsp. montana 40-70 (80) см, 15-25 (40) мм и subsp. densiflora
- 50-100 см и 15-30 мм соответственно, последний подвид отлича-
ется очень густыми соцветиями ( Prochazka, velisek, 1983). Ф .Пер-
ринг ( Perring, 1962) показывает, что на Британских островах
subsp. densiflora помимо более крупных размеров, обладает цвет-
ками ярко-розового цвета с запахом клевера и растет на лугах на
почвах, богатых основаниями, subsp. conopsea имеет танке душис-
тые цветки, но с иным запахом и растет обычно на заболоченных
участках. Различия настолько велики, что ряд авторов считает эти
х/ Saepe "сопореа'
51
подвида самостоятельными видами. Например Л.В.Аверьянов (1981),
отмечая высокую изменчивость рода Gymnadenia на северной грани-
це ареала, проявляющуюся в морфологическом облике растений, по-
лагает, что'возникновение самостоятельного вида Gymnadenia den-
siflora связано с переходом к облигатной кальцефильноети, по-
влекшим за собой изменение морфологических и биологических (сро-
ков цветения) признаков. Н.Н.Цвелев (1969) указывает еще два
подвида — subsp. conigera С.Koch ц subsp. sibirica Turcz. ex
Lindl.
Многие исследователи выделяют ряд более мелких таксонов -
разновидностей И форм ( Schulze, 1894;Beauverd, 1935/36; Hegi,.
1936; Sipkes, 1954; Rose, 1963; Ohwi, 1969; Черепанов, 1981;
Шмейдт, Яагомяэ, 1987; и др.). Наиболее распространены следующие
разновидности и формы: var.ornitis-Beck. (чисто белые цветки),
van. ecalcarata М.Schulze (шпорец отсутствует),var.crenulata
Beck, (боковые лопасти губы более широкие, чем у типичной формы,
и зубчатые), van. intermedia peterm. , (соцветие удлиненное с
цветками по размеру в 2 раза мельче, чем у типичной формы), f.
clavata M.Schulze (внутренние листочки околоцветника короткие,
булавовидные), f. viridiflora L.Keller, H. Fleischmann
(мелкие зеленоватые цветки, губа широкая, трехлопастная, до 2,5
мм длины и 3 мм ширины).
Для кокушника комарникового так же, как и для многих других
наших орхидей характерно образование многочисленных гибридных
форм как внутриродовых, так И межродовых (Arnell, 1911; Fuchs,
1919;Alleizeit, I950;Peitz, 1972; Klimpel, 1978; Prochazka,
Veiisek, 1983;Rothmaler, 1976).
Прохазка и Велишек ( Prochazka, veiisek, 1983) отмечают на-
личие гибридов с восемью родами (Anacamptis, Dactylorhiza, Cha-
morchis, Leucorchis, Orchis, Traunsteinera, Platanthera, Coelo-
-glossum ). Е.Пейтц (Peitz, 1972) установил около 30 гибридных
сКррм у кокушника комарникового. В.Ротмалер (Rothmaler, 1976)
указывает на существование в Германии гибридов Gymnadenia сопор-
seh с Platanthera bifolia (Gymnaplatanthera chodati (Ledrd.)
Lamp., Coeloglossum viride (Gyronaglossurn jacksonii (Druce) Rol-
fe), Nigritella nigra (Gyinnigritella suaveolens S.-G.), Dacty-
lorhiza cruenta (Dactylodenia raetica Paroz. и Rein ). Известны
гибрида также с другими видами рода Dactylorhiza (D.sambucina,
D.traunsteineri, D.maculate, D.fuchsii (Alleizetta, 1950; Euchs,
1919).
Морфологическое описание
Многолетнее травянистое растение. По И.Г .Серебрякову (1962,
1964) его следует относить к классу наземных травянистых поли-
карпических растений с ассимилирующими побегами песуккулентного
типа, подклассу клубнеобразующих травянистых многолетников,груп-
пе корне клубневых многолетников. Корневая система по Г.П'.Голу-
беву (1984) - кисте корневая, образованная придаточными корнями,
отходящими от основания побега. Клубни корневого происхождения,
сжатые с боков, 4-6 (10) - пальчато-раздельные, расположены на
глубине 3-7 (8) см от поверхности почвы. Обычно одновременно .
можно видеть два клубня - старый, слегка сморщенный, функциони-
рующий в текущем году и.молодой, плотный, беловатый, расположен-
ный несколько выше старого, в верхней части'молодого клубня мож-
но видеть остроконечную почку возобновления побега будущего го-
да. Иногда в первой половине лета можно заметить короткий столон
(0,5-1,5 см), на котором располагается молодой клубень. Чаще
столон практически не виден. С возрастом форма клубня, размеры,
степень рассеченности меняются.
У ювенильных растений клубни округлые, обратно-яйцевидные,
шириной 0,2-0,5 (0,8) см, длиной 0,3-0,7 (0,8) см и гаирпюй 0,2-
0,32 см со шнуровидным окончанием (несколько напоминает клубень
молодых особей видов из рода Platanthera). Придаточных корней
один, редко два, длина их 0,5-2,5 см, глубина проникновения 0,6-
3,8 см. В имматурном состоянии клубень становится более плоским,
число шнуровидных (пальчатых) окончаний увеличивается до 2-3.
Длина клубня (нерасчлененной его части) 0,9-1,59 (1,7) см, щири-
на 0,5-0,76 см, число придаточных корней увеличивается до 3-4,
глубина их проникновения к почву от 3 до 5,5 (6) см. По взрослом
виргинильном состоянии нерасчлененная часть клубня становится
еще более уплощенной и широкой (длиной 1,6-1,8 см, шириной 0,9-
1,5 (1,9) см), число шнуровидных окончаний 4-5 (6), число прида-
точных корней 4-8, глубина их проникновения 4-6,5 (9) см, Генера-
тивные растения имеют примерно те же размеры клубня (длина 1,7-
1,8 (2,2), ширина 1,5-2,2 см), чизло шнуровцдных окончаний 5-8
(редко до 15), число придаточных корней 5-9 (II), глубина проник-
новения их 7,5-10 (11,5) см. Корни покрыты короткими, но доволь-
но густыми корневыми волосками. Направление роста корней разно-
образно в зависимости от их функций и особенностей почвы - часть
корней растет горизонтально, другие - под углом вверх, третьи -
4х-®041 53
вертикально вниз. Корни отличаются контрактильностью, что позво-
ляет постепенно заглублять клубень (у ювенильных растений он ра-
сполагается чаще на глубине 0,5-2 см, у генеративных - 6-8 см,
как было отмечено нами на душистоколосково-калганово-разнстрав-
ном лугу в Подмосковье).
В.Н.Голубев (1984) относит кокушник комарниковый к полуро-
зеточным растениям. Основание стебля взрослых растений окружено
1-2 чешуевидными и 3 влагалищными пленчатыми листьями, выпе по
стеблю 4-7 листьев с хорошо развитыми зелеными пластинками, ли-
нейные или линейно-ланцетные, матовые, гладкие, быстро сужающие-
ся, на верхушке более или менее колпачковидностянутые, образуя
"башлычок" на конце листа (что является хорошим признаком вида),
несколько вдоль сложенные и оттого снизу килеватые, 7-15 (25) см
дайной, 0,5-2 (2,5) см шириной. Верхние листья в числе 1-5, более
мелкие, прицветникообразные ("брактеи"). У вегетирующих особей
листья обычно сближены, а у генеративных - расставлены. Верхние
листья примерно в 3 раза короче нижних. 0 возрастом число зеле-
ных листьев увеличивается: у ювенильных - один (с 2 жилками), у
имматурных - 2-3 (с 6-8 жилками), у взрослых виргинильных - 4-5
листьев(с 8-10 жилками), генеративных - тоже 4-5 (с 10-16 жилка-
ми) .
Стебель (цветонос) имеет различную высоту в завизимости от
условий местообитания и таксономической принадлежности: от 10-20
до 60-80 (100) см. В Московской и прилегающих областях высота
обычно колеблется в пределах от 50 до 75 см. Соцветие - колос,
густой, цилиндрический, дайной 6-15 (25) см и шириной до 2 см. В
Подмосковье чаще дайна соцветия от 10 до 15 см. Прицветники зеле-
ные , овальше, длиннозаостренные, эллиптически-ланцетныа» неясно
трехнервные, изогнутые, в 1,5 раза длиннее завязи. Нижние при-
цветники длиннее или равны цветкам, верхние - короче. Число цвет-
ков в соцветии колеблется от 20 до 87, в Московской и прилегаю-
щих областях от 25 до 70. Окраска цветков весьма изменчива.Даже
в пределах одной популяции можно встретить цветки от лилово-ро-
зовых, красновато-розовых до светло-лилово-пурпурных, светло-ро-
зовых, почти белых. О.М.Шмейдт и Т.Яагомяэ (1987) отмечают в Эс-
тонии форму кокушника с члс.то белыми цветками. Мы наблюдали бело-
цветковую форму кокушника в Северной Осетии.
Цветки неправильные, с простым венчиковидным околоцветником,
состоящим из двух трехчленных кругов. Листочки наружнего круга -
54
продолговато-ланцетные, тупые, с тремя жилками, 0,4-0,5 ом дли-
ной, 0,2-0,3 см шириной. Боковые наружные листочки эллиптичес-
кие, отогнутые, неравнобокие, вдоль свернутые и оттого кажутся
уже, отклонены несколько в стороны и вниз. Средний листочек на-
ружного круга продолговато-яйцевидный, с 3-5 жилками, тупой, на
конце с "колпачком". Листочки наружного круга немного длиннее
листочков внутреннего круга. Боковые листочки внутреннего круга
неравнобокие, широкояйцевидные с одной стороны и ланцетные с
другой, постепенно суженные, тупые, с тремя жилками. Средний
листочек наружного круга и боковые листочки внутреннего круга
обращены вверх и сложены в виде шлема. Средний листочек внутрен-
него круга - губа, длиннее остальных листочков (до 0,5 см дли-
ны), ромбовидная, трехлопастная, с тупыми почти яйцевидными ло-
пастями, городчатыми по краю, по краям в верхней чаоти они пок-
рытыми волосками. Средняя лопасть немного длиннее боковых, с не-
сколькими ветвящимися жилками. Губа с длинным (до 1,8 см) сер-
повидно-изогнутым шпорцем, в 1,5-2 раза длиннее завязи, на са-
мом конце с небольшим утолщением. Андроцей сильно редуцирован -
сохранилась лишь одна плодущая тычинка с двухгнездным пыльником,
две другие тычинки обычно присутствуют в виде маленьких стериль-
ных стаминодиев, расположенных по обеим сторонам колонки (стол-
бика, сросшегося с нитями тычинок). Пыльца склеена в поллинии,
имеющие продолговато-округлое прилипальцэ. Клеющее вещество прп-
липальца сходно по составу с клеющим веществом у рода llctan-
thera, что подтверждает их близость ( FUller, 1978). Рыльце
почкообразное, расположено ниже пыльников. Завязь сидячая скру-
ченная, около 0,8 см длиной. Плод - удлиненная коробочка (до 1,3
и более см), раскрывается узкими щелями, содержит огромное коли-
чество мельчайших семян (Флора СССР, 1935; Ziegenspeck, 1936;
Гроссгейм, 1949; Sumrierhayes, 1951; Pliller, 1978; talsson,
1979; и др.).
Диплоидное число хромосом 40 (Цвелев, 1969; Иосковцева,
1980), однако указывают тайке на формы с 2n=80, 100 и 120
( Flora Ьпгораеа, 1380; Prochazka, Velisek, 1983).
Географическое распространение
Ареал вида - бореальный евразиатский палеарктический (
mirfiayes, 1951; Протопопова, 1987; Голубев, 1984). Кокушник ко-
марниковый обитает по всей Европе от северной Норвегии и Карелии
55
до Италии, Балканского полуострова, Крыма на юге, на западе
достигает Великобритании, Исландии, растет на Шетландских ост-
ровах. В Азии встречается на значительной территории от Запад-
ной Сибири до Монголии, Северного Китая, Кореи, Японии, а так-
же в Малой Азии, на Кавказе, в Иране (Флора СССР, 1935? zie-
genspeck, 1';3б; Flora Suropaea, 1980; FUller, 1978).
В пределах СССР встречается практически во всех районах
европейской части, в Крыму, на Кавказе, Сибири и на Дальнем Во-
стоке .
Е западной части ареала кокушник - одна из наиболее рас-
пространенных орхидей (в Великобритании, Германии, западных и
центральных районах европейской части СССР),в восточной части
(Хабаровский и Приморский края СССР) - это редкий и малочислен-
ный вид.
В Московской обл. кокушник комарниковый встречается во
всех районах (реже в Юго-восточном и Заокском районах), но обыч-
но в небольшом количестве.
Онтогенез и особенности размножения
Размножается кокушник комарниковый преимущественно семенным
путем. Семенная продукция его велика: в среднем одна генератив-
ная особь дает от 30 до 90 тысяч семян. Образование плодов про-
исходит достаточно интенсивно - от 40 до 90$ цветков завязывают
ПЛОДЫ ( Ziegensps.ck, 1936; Salisbury, 1942; Summerhayes,
1951). По нашим данным, в Московской и Тверской областях от 35
до 85$ цветков дают плоды, в других частях ареала примерно те
же цифры: Тебердинзкий заповедник 60-80%, Северо-Осетинокий за-
поведник 50-88%, "Кедровая падь" 48-60%)• В Примере ком крае
70,1% цветков дает плоды, среднее число плодов на особь 44+5,1,
число семян на особь 97173+512, число оемян в одном плоде 1675+
+205. Число семян в плодах колеблется от Т400 до 6900, в среднем
около 3000 (3150£329,4) в одной коробочке. Среднее число плодов
на одну особь в Подмосковье 17+4,6, т.е. в этих условиях одна
особь может дать около 50 тысяч семян. В Северо-Осетинском запо-
веднике на субальпийских лугах семенная продуктивность еще вше
- до 90 тыс. на генеративную особь и примерно 230 тыс. семян на
I м2. Сходные цифры приводятся для Великобритании - среднее чис-
ло плодов в соцветии 15,4+0,43, число семян в плоде от 911 до
3960 (2044+11), число семян на особь от 23 до 40 тыс. (31699+
56
+8800) ( Salisbury, 1942). Масса семени очень мала - 0,0008 мг,
оно может лететь со скоростью 0,31 м/с ( Lidller-Schneider,
1977), по нашим данным масса семени около 0,002 мт. Размеры се-
мени очень незначительны - 0,7 х 0,2 мм ( Fuller, 1978), оно
состоит всего из нескольких десятков клеток и почти не содержит
питательных веществ, зародыш в них надифференцирован.
Прорастание оемени происходит под землей и только с помо-
щью гриба-микоризообразователя, снабжающего прорастающее семя
необходимыми питательными веществами. Вначале образуется так
называемый протокорм (прокаулом) - округлое полупрозрачное тело
размером 1-2 мм с одним чешуевидным листом. Позднее формируются
еще 1-2 членика первичного корневища, внутри которых находятся
гифы гриба, все это образование вместе нередко называют микори-
30М0М - ” mykorhizom" (Ziegenspeck, 1936; Summerhayes, 1951).
Проросток обычно в точение двух лет живет под землей ( Summer-
hayes, 1951), лишь к осени второго года жизни эндогенно обра-
зуются два маленьких придаточных корня, один из которых преоб-
разуется в клубень, первичное корневище - микоризом к этому
времени отмирает ( Ziegenspeck, 1936).
На 3-й год (иногда на 2-й) появляется первый зеленый лист
- растение переходит к надземному существованию. В этом (юве-
нильном) возрастном состоянии клубень ещз не рассечен, имеет
одно шнуровидное окончание, от основания побега отходят 1-2 при-
даточных корня. На 4-й год образуется 3-4 придаточных корня,
клубень имеет две лопасти и соответственно 2 шнуровидных оконча-
ния, два зеленых листа - растение переходит в имматурное состоя-
ние. В последующие годы происходит увеличение числа листьев и
придаточных корней, размеров клубня. В генеративное состояние
кокушник переходит в лучшем случае на 6-7-й год, но чаще позд-
нее ( Ziegenspeck, 1936; Summerhayes, 1951). Судя По тому, ЧТО
в возрастном составе популяций значительную часть (от 40 до 70%)
составляют взрослые вегетативные и генеративные особи, кокушник,
видимо, во взрослом состоянии пребывает значительно дольше, чем
в ювенильном и имматурном. Однако, как и у многих других расте-
ний, в экстремальных условиях (на северной границе ареала, на
верхней границе распространения в горах) происходит задержка в
развитии особей, вследствие чего в составе популяций преобладают
молодые особи, что нам и приходилось наблюдать на Кольском полу-
острове и на Кавказе.
57
Кокушник комарниковый часто после первого цветения, образо-
вав большое число семян, отмирает, т.е. ведет себя как монокар-
пическое растение ( summerhayes, 1951). Однако в благоприятных
условиях одна и та же особь может цвести по крайней мере 2-3 ра-
за подряд, что мы неоднократно наблюдали в Московской и Тверской
областях. Видимо, в связи с монокарпичностью находится и склон-
ность популяций этого вида к значительным флюктуациям (от не-
скольких тысяч до 1-2 десятков особей), что нередко отмечается
исследователями.
Размножение кокушника, как и большинства других корнеклуб-
невых орхидей, происходит почти исключительно семенным путем. Их
клубни образуются по одному в год (так называемые замещающие) и
служат лишь для поддержания особи, а не для размножения. На сле-
дующий год у растения сохраняется лишь небольшой участок основа-
ния стебля, молодой клубень и молодые придаточные корни, поэтому
его относят к так называемым вегетативным малолетникам по терми-
нологии Г. Н. Высоцкого (1915). Лишь в исключительных случаях, на-
пример, при повреждении клубня или В условиях особо благоприят-
ного питания может образоваться более одного клубня. Каждый клу-
бень несет почку, из которой формируется надземный побег, в осно-
вании которого в свою очередь формируется клубень. Вновь образо-
вавшиеся особи вначале не разделяются, а растут плотной группой.
Полное разделение особей в пространстве происходит лишь через не-
сколько лет ( summerhayes, 1951). Отдельными исследователями уда-
валось найти в природе экземпляры кокушника комарникового с дву-
мя и более клубнями, из которых в следующем году развивались са-
мостоятельные особи (Лукс, 1970).
Наличие у этого вида спящих почек такке свидетельствует о
возможности вегетативного размножения ( ziegenspeck, 1936). Одна-
ко нам в природных условиях в различных частях ареала пока не
удалось найти убедительные доказательства вегетативного размноже-
ния этого вида. При культивировании кокушник комарниковый прояв-
ляет способность к вегетативному размножению. В.Г.Собно (1980) в
Центрально-республиканском ботаническом саду (Киев) отмечал обра-
зование одновременно двух новых клубней у кокушника, особенно при
повреждении растения. Он использовал это свойство и, делая 2-3
надреза на клубне, через 5-7 дней отмечал появление на месте над-
реза I-2-х молодых клубней. Это явление известно у растений дав-
но. Н.П.Кренке (1928) назвал его процессом реституции. Вновь по-
58
явившиеся (вторичные) клубни раотут очень быстро. Если их уда-
лить, то через 15-20 дней можно получить третий клубень, а затем
таким же способом и четвертый. Первый вторичный клубень - круп-
ный, по возрастному состоянию соответствует материнскому расте-
нию и может даже цвести на следующий год, а последние - мелкие,
формирующие на следующий год растения ювенильного облика.
Сезонное развитие и консортивные связи
В условиях Подмосковья (так же, как и вообще в равнинных
условиях Европы ( Summerhayes, 1951; Eichner, 1974) кокушник
комарниковый зацветает в первой половине июня. Однако на сильно
заболоченных участках он зацветает значительно позже - на 3-4
недели. Плодоносит кокушник в августе-сентябре. Близ северной
границы ареала (Мурманская и Архангельская области), а тайке в
горах (Северная Осетия, Теберда) зацветает не ранее июля, а близ
верхней границы распространения на субальпийских лугах - в конце
июля, августе. В.Н.Голубев (1984) по ритму сезонного развития
относит кокушник к поздневесенним-среднелетним растениям. Начало
вегетации в Подмосковье - в конце апреля-начале мая. Заложение
подземных органов происходит довольно рано, молодой клубень (еще
почти не рассеченный) становится заметным в начале июня, шнуро-
видные окончания и молодые корни появляются значительно позже -
ближе к осени, обычно не ранее второй половины июля - в августе.
К началу сентября подземные органы кокушника принимают вид, со-
ответствующий возрастному состоянию особи. К этому моменту жел-
теют и отмирают листья.
В.Г.Собко (1983) исследовал цикл развития побега нескольких
видов орхидных, в том числе и кокушника комарникового в условиях
Киева (Центральный республиканский ботанический сад). В первой
половине августа на сформированном молодом подземном побеге за-
кладывается в виде бугорка почка возобновления, содержащая две
точки роста. Из верхней точки роста в дальнейшем развивается но-
вый побег, из нижней - новый клубень. В течение осени, зимы и
ранней весны в точке роста побега каких-либо заметных изменений
не наблюдается. Новый клубень формируется в течение веоны и пер-
вой половины лета. К этому времени (к июлю) надземная зеленая
часть растения почти полностью отмирает и наблюдается начало
дифференциации конуса нарастания. В течение июля-октября форми-
руются зачатки листьев. Листья свернуты в плотный конус, вершина
59
которого вплотную подходит к поверхности почвы. В апреле-мае
почка появляется на поверхности почвы и из нее развертывается
розетка листьев и поднимается стрелка цветоноса. Во второй дека-
де мая цветонос достигает необходимого размера, раскрываются
цветки. В первой половине июня формируются плоды, после чего по-
бег начинает отмирать. Таким образом, малый жизненный цикл побе-
га у кокушника комарникового составляет, по данным В.Г.Собко,
23-24 мес. Сходную длительность развития побега кокушника указы-
вает для условий Германии Моллер ( М811ег, 1987) - 24-27 мес.
Консортивные связи
Цветки кокушника комарникового содержат нектар, обладают
приятным запахом и потому весьма привлекательны для насекомых-
опылителей. Он опыляется как дневными, так и ночными (к ночи за-
пах цветков усиливается) опылителями - бабочками, мухами, пчела-
ми, мотыльками, молями. Ресупинация (вращение) завязи позволяет
расположить цветок несколько наклонно и таким образом, что губа
становится удобной посадочной площадкой для насекомых - опылите-
лей. Опыляются от 40 до 90% цветков, самоопыление наблюдается
очень редко ( Ziegenspeck, 1936; Sommerhayes, 1951; НШеоп,
1979). Цигеншпек ( ziegenspeck, 1936) отмечает, что основные чер-
ты процесса опыления кокушника во многом сходны с опылением ви-
дов рода Platanthera (Ziegenspeck, 1936).
В течение всей жизни, а особенно в молодом возрасте, кокуш-
ник тесно связан с микоризным грибом. В корнях этого растения
найден гриб Rhizoctonia repentitis (Godfery, 1933). Наши иссле-
дования микоризы в подземных органах кокушника позволили устано-
вить, что гифы гриба содержатся в основном в придаточных корнях,
есть они и в шнуровидных окончаниях клубня, в самих клубнях гриб
отсутствует совсем.
Максимальное содержание микоризного гриба отмечено в корнях
ювенильных растений, однако и все другие возрастные группы также
тлеют микоризу. В течение года интенсивность микоризной инфекции
изменяется, как правило, в начале вегетационного периода она вы-
ше, снижаясь к середине лета, затем вновь возрастает, что связа-
но, вероятно, с повторной инвазией грибов в течение вегетацион-
ного периода (Харли, 1959).
Кокушник довольно устойчив к грибным заболеваниям, редко
поедается насекомыми. Однако, по данным чешских ученых, он может
60
поражаться грибами Melanospora repentis Plowr., Puccinia orchi-
dearum - pholaridis Kob. (Prochazka, Veiisek, 1583).
Экология и фитоценология
Кокушник комарниковый отличается довольно широкой экологи-
ческой амплитудой. К освещению не очень требователен ( Landolt,
1974), хотя предпочитает все же хорошо освещенные места, выдер-
живает затенение до 50?£ от полного (четвертая ступень шкалы ос-
вещенности Ландольта) ( Landolt, 1977).
Растет в условиях от сухо- до сыролугового (58-80 ступени
шкалы Л.Г.Раменского, 1956). В.Н.Голубев (1984) относит этот
вид к мезоксерофитам. Действительно, кокушник встречается как
на довольно сухих лугах, так и на влажных, может расти даже, на
заболоченных участках, однако предпочитает средние условия ув-
лажнения, где наиболее обилен ( Hegi, 1936). По мнению Ландоль-
та (1977), кокушник - индикатор условий средней сухости (третья
ступень шкалы влажности), избегающий как крайне сухие, так и
очень мокрые участки. По Элленбергу (1974), обычно растет на
влажных не пересыхающих почвах (седьмая ступень шкалы увлажне-
ния). В зависимости ст влажности почвы находится направление
роста и положение в почве корней и клубней. На сырых субстратах
корни кокушника растут, изгибаясь кверху, что позволяет им ос-
таваться в верхних горизонтах почвы, богатых кислородом. На су-
хиХ/Почвах корни растут прямо вниз, а клубень закладаваясь вер-
тикально, "пробуравливает" себе отверстие и облегчает проникно-
вение в более влажные слои почвы клубням, возникшим в последую-
щие ГОДЫ ( Ziegenspeck, 1936; Summerhayes, 1951). Кроме ТОГО,
расположиться в "нужном" горизонте почвы клубню помогает и кон-
трактильная деятельность корней. По мнению Цигеншпека (1936),
разветвление клубня наступает лишь тогда, когда он достигает
"правильного" горизонта. Им были встречены взрослые экземпляры,
клубни которых были расположены слишком близко к поверхности
почвы и не были разделены на шнуровидные окончания.
К богатству почвы не требователен, может расти на довольно
бедных почвах (8-II ступени богатства почвы по Л.Г.Раменскому).
По Е.Ландольту может расти на почвах, бедных минеральным азотон
(вторая ступень шкалы), так как на богатых, по его мнению, не
выдерживает конкуренции. Однако предпочитает почвы богатые гу-
мусом, являясь индикатором гумусных почв (четвертая ступень
61
шкалы Ландольта). Кокушник предпочитает щелочные и нейтральные
почвы (pH 5,5-8,5), может расти на слабокислых, никогда не
вотречаетоя на сильнокислых (четвертая ступень шкалы Ландольта,
восьмая ступень шкалы Элленберга). Большинство исследователей
отмечает, что кокушник предпочитает почвы, богатые известью,на
которых иногда образует большие скопления. Он встречается на
разных типах почв - дерновых, дерново-подзолистых, торфянисто-
болотных, горно-луговых, горно-лесных, на мелах и известняках,
предпочитает почвы более тонкой структуры - глинистые, торфя-
нистые, избегает пеочаные, обычно растет на плохо вентилируемых
почвах (пятая ступень шкалы Ландольта, 1977).
К температуре нетребователен, может расти в районах с су-
ровой зимой, в горах встречается на высоте до 2300-2800 м и
нередко растет близ края ледника (например, в Северной Осетии),
на склонах разной экспозиции и крутизны (до 15-20°).
Кокушник комарниковый - вид с довольно широким фитоценоти-
ческим ареалом, растет на лугах низинных, суходольных, поймен-
ных, альварных, субальпийских (различных по флористическому со-
ставу - душистоколосковых, краоноовсяницевых, белоусовых, мис-
кантуоовых и пр.) встречается на осоково-сфагновых болотах, в
разреженных лесах и на опушках хвойных (сосновых, еловых, лист-
венничных) , смешанных, мелколиственных и широколиственных (ду-
бовых, дубовс-грабовых), в зарослях кустарников (Флора СССР,
1935; Summerhayes, 1951; hichner, 1974; Флора европейской ча-
сти СССР, 1976; Флора северо-востока европейской части СССР,
1976; Fuller, 1978; Nilsson, 1979; Somsak, 1982). В фитоце-
нозах кокушник редко играет заметную роль, обычно встречаясь
вместе с другими орхидеями, например bistera ovata, coelogios-
sun viride, Herminium monorchis, Epipactis helleborine, E.pa-
lustris, ВИДаМИ рода Dactylorhiza (Summerhayes, 1951; FUller,
1978). Однако иногда образует значительные скопления и является
одним из доминантов травяного яруоа. Так, указывается, что ко-
кушник встречается в огромном числе .на меловых холмах юга Ан-
глии, где СН растет вместе С Ophrys apifera, Orchis ustulata
и др., а танке на райграсовых дугах, где растет вместе о Arrhe-
natherum elation, Dactylorhiza fuchsii, Anacamptis pyramidalis
( Summerhayes, 1951). Сомзак ( Soinsak, 1982) приводит пример
доминирования кокушника комарникового наряду с другими видами
в белоусниках нижнего пояса (550-800 м) Словацкого нагорья (За-
62
падные Карпаты), где выделяет ассоциацию Gymnadenio-Nardetum,
которая, по мнению автора, вообще характерца для этого района.
Нам приходилось наблюдать в Московской и Тверской областях дот
вольно значительные скопления Gyumadenla сопораеа на сырых
лугах (преимущественно душистоколосковых), а также на белоусо-
вых лугах в Советских Карпатах, где он играл роль оодоминанта
и даже доминанта. Но, к сожалению, такие сообщества встречаются
редко. Следует, видимо, признать, что кокушник комарнйковый все
же довольно устойчивый в конкурентном отношении вид, в отличие
ст большинства других наших орхидей. Об этом можно судить не
только по его достаточно широкому фитоценстическому и географи-
ческому ареалам, но и присутствию в гуотых травостоях с большим
видовым разнообразием и высоким общим проективным покрытием, не-
редко достигающие 80-90$. Известно, что большинство орхидных
предпочитает учаотки с низким проективным покрытием травяного
покрова.
Плотность популяций кокушника комарникового иногда танке
бывает весьма высокой. Например, в Московской обл. на душисто-
колосково-гераниево-раэнотравном лугу средняя плотность кокуш-
ника составляла 5,7 особей на м2, максимальная 21 особь на м2,
в Тверской обл. средняя плотность 5,5, максимальная 14 оообей
на м2, на белоусово-разнотравном лугу на Карпатах (Ивановс-Фран-
ковокая обл.) средняя плотность 8,7, максимальная 48 особей на
м2.
Сильное антропогенное влияние кокушник, как и другие орхи-
деи, переносит плохо - он отрицательно реагирует на сильный вы-
пао, интенсивное вытаптывание и сбор на букеты при рекреации.
Внесение больших количеств удобрений, особенно азотных, также
отрицательно сказывается на состоянии кокушника, так как усили-
вает конкуренцию со стороны других видов, особенно злаков ( Mi-
ler, 1987). Слабый выпас и сенокошение переносит удовлетвори-
тельно. Более того, полное отсутствие традиционного хозяйствен-
ного использования лугов (сенокошения, пастьбы) обычно оказыва-
ется отрицательно на состоянии популяций кокушника ( MBiler,
1987).
Хозяйственное значение и охрана вида
Клубни кокушника комарникового содержат лекарственные веще-
ства и под названием "салеп" применяются в официальной медицине
63
D качестве средства, употребляемого при лечении кишечных забо-
леваний, особенно у детей, такке при отравлении ядами, кроме то-
го, рекомендуется ослабленным больным как питательное средство
(Гаммерман и др., 1963; Телятьев, 1987). Издавна клубни кокуш-
ника попользовались и в народной медицине в качестве тонизирую-
щего и противовоспалительного средства. Клубни кокушника комар-
никового содержат до 50% слизи, около 30% крахмала, 13% декст-
рина, пентозаны, сахарозу и др., в надземных органах найдены
кумарины, флавоноиды, полисахарид маннан (Телятьев, 1987).
Кокушник комарниковый успешно выращивается в ботанических
садах (Лукс, 1961; Кукк, 1980; Собко, 1980). Есть опыт его ве-
гетативного размножения в культуре (Собко, 1980). Предполагают,
что кокушник можно выращивать в условиях городских насаждений
(Лукс, 1961).
Однако несмотря на широкое распространение, высокую числен-
ность в отдельных, преимущественно мало нарушенных районах, в
целом численность кокушника комарникового сокращается под влия-
нием различных антропогенных факторов, в том числе прямого унич-
тожения его местообитаний. Во многих регионах нашей страны он
относится к охраняемым видам. Кокушник включен в список видов,
охраняемых на территории Московской обл. и никоим образом не.
должен испсльзоватьоя в качестве лекарственного растения.
Г.В.Мусина, Р.И.Барякина, Н.Б.Чубатова
МЫШЕХВОСТНИК МАЛЕНЬКИЙ
l yosurus Kiininus L., 1753, Зр.В1.:284; Кауфман, 1866,
Моск.фл., изд.1:24; Цингер,1885, Сборник свед.:35; Шмальгаузен,
1895, Фл.Среда.и Южн.России, 1:12; Сырейщиков,1908, Илл.фи.
Моск.губ. 2:151; он же, 1927, Опред.раст.Моск.губ. :133; Краше-
нинников, 1937, Фл.СССР, 7:323; Невский,1940, в Маевский, Фл.
изд.7:364; Голицын,1954, в Маевский, Фл.,изд.8:77; (Putin, 1964,
.’1. ,urop. 1:238; Маценко,1954, в Маевский, Фл. ,изд.9:79; Воро-
шилов,1966, Опред.раст.Моск.обл. :167; Черепанов,1981, Сосуд,
раст.СССР: 422; Новиков, 1986, Опред.раст.Мещера, 1:179 - Мыше-
хвостник маленький (семейство Hanunculaceae - лютиковые).
Географическое распространение
Ареал мышехвостника маленького охватывает европейскую часть
СССР (за исключением севера), горы Средней Азии, Кавказ, Крым,
Западную Европу, Скандинавию, Северную Африку, Австралию. Вне
основного ареала отдельные местонахождения известны в окрестно-
стях Воркуты, на р.Юньяхе, в среднем и нижнем течении р.Вычег-
ды, в Сибири в окрестностях Томска, Тобольска, в Еарабинской
степи. Произрастает местами обильно в лесной зоне на залежах,
вдоль дорог, на склонах оврагов, на выгонах, в канавах, в степ-
ной и полустепной зонах - на лугах в мочажинах, где весной за-
стаивается вода, нередко на солонцеватых участках (Крылов,1931;
Крашенинников, 1937). В Московской обл. встречается на полях,
залежах, часто как сорное растение входит в состав культурных
фитоценозов (Смирнов, 1958; Ворошилов, 1966).
Морфология
Мышехвостник маленький - кис те корневой, розеточный одно-
летник, 3-15 см высотой, с терминальными цветками на главном и
пазушных побегах. Корневая система мочковатая, проникающая на
глубину до 8 см. Главный корень хотя и сохраняется, но теряется
среди придаточных корней, число которых колеблется от 3 до 18.
Листья все прикорневые, с узким основанием, коротким череш-
ком и линейной мясистой пластинкой 0,5-8,5 см данной и 1-1,5 мм
шириной.
65
Гокопыо побеги розеточные с двумя сближенными преданотьям»:
л основании ( Troll, 1964), благодаря чему прикорневая розетка
роптания становится более модной и может включать до 45 листьев.
У одной особи обычно развивается по 2-3, а в отдельных случаях
до ТЗ цветоносных побегов. Цветоносы цилиндрические, тонкие,
кверху слегка расширенные, особенно во время оозревания плодов,
2-5 см дайной. Цветки одиночные, мелкие, зеленоватые, актино-
морфные, с двойным околоцветником. Чашечка из 5 линейнс-продол-
говотых листочков 2,5-3,5 мм длиной и 1-2 мм шириной, при осно-
вании переходящих в короткий (1,5-2 мм дайной) постепенно заос-
тренный шпорец. Лепестков 5 (2-2,5 мм данной), пластинка их
продолговато-яйцевидная, при основании с медоносной ямкой, в
2-2,5 раза короче линейного ноготка. Тычинки в числе 5-10 не
превышают по длине лепестки. Многочисленные пестики сидят на
,длинном, выдающемся из околоцветника цветоложе, которое по мере
оозревания плодиков значительно увеличивается в дайну (до 7 см),
.'/длинеиное цветоложе обеспечивает высокую семенную продуктив-
ность растения. Плод апокарпный из многочисленных односеменных
орешковвдных плодиков, при созревании отдаляющихся от цветоложа,
иногда вентрицидно вскрывающихся. Плодики мелкие .(длиной до. 1,5
мм), трехгранные, слабо опушенные, бурые, с маленьким отходящим
от основания прямым стилодаем (Иванова, 1966). Йемена эллипсо-
идальные, сплющенные с боков, светло-коричневые, блестящие.глад-
кие, 0,8 мм дайной. 2п =16 (28) (Хромосомные числа, 1969).
Онтогенез
Онтогенез мышехвостника маленького подробно изучен Т.А.Гу-
..нян (1976). Е жизненна/, цикле четко прослеживаются три перио-
да: латентный, виргинилышй, генеративный. В зрелых семенах его
хорошо различим зародыш, отчетливо дифференцированный на 2 семя-
доли, почечку, гипокотиль и корень с корневым чехликом, окружен-
ный обильным эндоспермом. Абсолютная величина зародыша очень ма-
ла, всего 0,22 мм, что составляет 27,4% от длины эндосперма
(табл.1).
В природе семена прорастают в апреле при наличии в почве
обильной влаги. Прорастание надземное, гипокотилярное. Первым
появляется зародышевый корешок, в основании снабженный венцом
всасывающих волосков. Затем разрастается гипокотиль, вначале
бесцветный, но со временем зеленеющий, дайной от 0,5 до I см.
66
Таблица I
Количественные показатели семян мышехвостника маленького
(Иванова, 1966)
>с семени, .г Длина, мм
семени эндосперма зародыша оемядолей
6,07 0,88 0,82 0,22 0,08
Черешки семядолей срастаются на небольшом протяжении, образуя
короткую трубку. Пластинки семядолей овальные, с единственной
средней жилкой. Главный корень тонкий, бесцветный, 4-5 мм дли-
ной, с длинными корневыми волосками. Через 2-2,5 недели после
прорастания почечка образует первый ассимилирующий лист, а
вскоре второй, третий и остальные листья в очередном порядке.
С развертыванием первых листьев появляются придаточные коржи на
базальных участках гипокотиля и по одному на каждом узле укоро-
ченного побега.
Корневая система у ювенильных форм мочковатая, смешанная;
главный корень не выделяется среди 3-18 придаточных, слабо вет-
вящихся, микоризных корней, проникающих на глубину 5 см. Такой
характер подземной сферы сохраняется на протяжении последующих
фаз онтогенеза. Спустя 1,5-2 месяца после прорастания растение
приобретает облик дефинитивного. У наиболее крупных экземпля-
ров разрывается до 27 листьев в розетке. На третьем месяце рас-
тение вступает в репродуктивную фазу развития. Одновременно на-
чинают отмирать семядоли и нижние листья. Генеративными стано-
вятся терминальная и большинство пазушных почек верхних розе-
точных листьев. Массовое цветение продолжается в зависимости
от погодных условий года от 10 дней до 1,5 месяцев, цветение
дружное, хотя отдельные цветки нередко появляются и в конце ве-
гетации. Поме диссеминации растение полностью отмирает.
Сезонное развитие
Мышехвостник маленький - весенний эфемер. По нашим много-
летним наблюдениям в Московской обл. в годы с относительно су-
хим весенним сезоном период вегетации мышехвостника маленького
длился 1,5 месяца. Так, в 1981 г. цветение началось в середине
мая, плоды созрели в конце мая, а уже 31 мая наблюдалось отми-
рание всех частей растения. Во влажные годы, например в 1982 г.
вегетация у мышехвостника началась 24 апреля, цветение длилось
с середины мая до середины июля, диссеминация наступила лишь в
конце июля, т.е. весь жизненный цикл продолжался около 3 меся-
цев, что отмечала ранее и Т.А.Гуланян (1976).
Таким образом, эфемерность у мышехвостника не жестко де-
терминирована и степень выраженности ее контролируется внешними
условиями. Вместе с тем высокая лабильность ритма развития обес-
печивает возможность его произрастания на обширной территории.
Способы размножения и распространения
Мышехвостник маленький размножается семенами. Семенная про-
дуктивность мышехвостника чрезвычайно высока (табл.2). Наиболее
мощные экземпляры дают более 3000 семян.
Семена лучше прорастают на свету. В лаборатории при 20°С
свежесобранные семена прорастают через 1,5 месяца. Из семян,по-
сеянных после 3-4-месячного хранения, проростки появляются быс-
трее, через неделю. Низкие температуры обычно задерживают про-
растание семян. Всхожесть в наших опытах не превышала 50%, со-
гласно же Т.А.Работнову (1951), она может достигать 99%; такое
расхождение данных, видимо, связано с явлением гетерокарпии и
гетероспермии.
Таблица 2
Семенная продуктивность мышехвостника маленького
(Одинцовский район Московской обл.)
Число цветков на рас- тении Число осо- бей Число плодиков в плоде Семенная продуктивность одного растения
макс. мин. среднее макс. мин. среднее
I 25 190 10 74 190 10 74
2 22 250 5 86 350 40 173
3 3 380 20 171 740 300 513
4 5 340 0 176 830 570 704
5 2 290 10 194 1160 780 970
6 4 400 20 2П 1970 960 1270
7 I 400 40 181 1270
8 I 400 40 162 1300
13 I 430 20 244 3180
Плодики и семена мелкие, поэтом?/ их распространению могут
способствовать ветер и вода.
68
Фитоценология, экология, биологическая
продуктивность и консортивные связи
Мышехвостник маленький относится к весенней флоре. В Мос-
ковской обл. входит в состав культурных фитоценозов (Смирнов,
1958; Ворошилов, 1966). В Одинцовском, Солнечногорском и Серпу-
ховском районах Московской обл.,где проводились наши наблюде-
ния, растет по распаханным окраинам сырых ложбин, на пашнях и
полевых дорогах. Сравнение численности и морфологических данных
показали, что белее благоприятные условия для развития мышехво-
стник находит в более южных районах Московской обл.
По экологической классификации мышехвостник маленький све-
товой мезофит или мезогигрофит, обильно произрастающий при пус-
тынно-полупустынном увлажнении (17-22 ступени шкалы Раменского),
умеренно и единично встречающийся при сухостепном увлажнении
(12-31 ступени шкалы Раменского).
Биологическая продуктивность мышехвостника маленького нич-
тожно мала независимо от местообитания растения (табл.З). По
нашим данным, на окраинах посевов, где зеленая масса культурных
злаков (овса, ячменя, пшеницы) составляла лишь 50%, на долю мы-
шехвостника (с плотностью - 37 особей на I м^) приходилось вое-
го 5% от общей фитомасоы сорных трав (пастушья сумка, мятлик
однолетний, лютик ползучий и др.).
Таблица 3
Биологическая продуктивность мышехвостника
маленького в разных условиях местообитания
Место- обита- ние Сроки наб- люде- ния Влаж- ность поч- вы,% Сырой вео, г/м^ Воздушно-юухой вес, г/м^
под- зем- • ные орга- ны надземные под- • зем- ные надземные
ли- стья стеб- ли листья стебли
Окраина посевов овсяно- го поля ЗО.У 1981 - 0,24 0,6 1,3 - . 0,2 0,3
Окраина ржаного поля 29.У 1982 21 1,0 1,0 2,4 0,8 0,2 0,5
1.Л 1982 43 0,8 2,4 4,8 0,2 0,8 1,2
5х-1041
69
Мышехвостник маленький - микотроф. Опыляется самыми мел-
кими насекомыми, главным образом двукрылыми, а тайке наездни-
ками и жуками (Нейштадт, 1957). В литературе есть указание, что
весной при нехватке зеленого корма он иногда поедается крупным
рогатым скотом, козами, лошадьми (Работнов, 1951).
В.И.Полуянова
ЛЮТИК ПОЛЗУЧИЙ
Ranunculus repens ,L., 1753, Sp. Pl. ;554; Кауфман, 1866,
Моск.фл., изд.1:30; Цингер,1885, Сборник свед.:42; Шмальгаузен,
1895, Фл.Средн, и Юкн.России, 1:21; Сырейщиков,1908, Илл.фл.Моск,
губ. 2:162; он же, 1914, цит.соч.:581; он же, 1927, Опред.раст.'
Моск.губ.:Г36; Овчинников, 1937, Фл.СССР, 7:423; Невский,1940,
Маевский, Фл., изд.7:369; Голицын, 1954, в Маевский, Фл.изд.8:
82;Tutin, 1964, Fl.Europ. 1:227; Маценко, 1964, в Маевский, Фл.,
изд.9:81; Ворошилов, 1966, Опред.раст.Моск.обл.:169; Черепанов,
1981, Сосуд.раст.СССР:426; Новиков,1986, Опред.раст.Мещеры,I:182.
- Лютик ползучий (семейство Ranunculaceae - лютиковые).
Географическое распространение
Лютик ползучий широко распространен по всем континентам,
особенно в северной внетропической зоне.
В СССР распространен всюду, за исключением Крайнего севера.
Растет преимущественно'в равнинных районах, но поднимается в го-
ры, где не доходит до границы леса. Ареал лютика ползучего про-
ходит от Двино-Печорского бассейна до Обской 'губы; от Карелии,до
Крыма и Кавказа на юге; через Западную и Восточную Сибирь до Кам-
чатки; встречается и на Тянь-Шане (Флора СССР,1937).
Морфологическое описание
Лютик ползучий - кистекорневое многолетнее наземно-ползучее
травянистое растение с дициклическими ортотропными, полурозеточ-
ными монокарпическими побегами и плагиотропными наземно-ползучи-
ми побегами, служащими для вегетативного размножения. Корневая
система поверхностная, максимальная глубина укоренения 20-30 см.
Листья однажды-дважды тройчатые, очередные, за исключением са-
мых верхних, черешковые. Черешки 3-15 см длиной, голые или воло-
систые. Волоски одинаковой формы. Листовая пластинка 3-7 см дли-
ной, рассеченная на надрезанные или надрезанно-зубчатые доли.
Цветки обоеполые, актиноморфные, на бороздчатых цветоножках,жел-
тые, 15-30 мм в диаметре. Околоцветник двойной. Чашечка состоит
из пяти оттопыренных чашелистиков, венчик из пяти лепестков,кото-
рые вдвое длиннее чашелистиков. Лепестки тычиночного происхсжде-
71
нил и в отличие от чашелистиков снабжены лишь одним листовым
следом. Тычинок много, расположение спиральное. Пыльники вскры-
ваются продольно. Пыльцевые зерна трехбороздчатые. Гинецей
апокарпный. Столбик хорошо развит. Семязачатки гемитропные.
Плод - многоорешек, расположен на выпуклом цветоложе. Плодовая
головка шаровидная, цветоложе немного волосистое, плодики ок-
ругло-яйцевидные , ожатые, окаймленные, с почти прямым довольно
длинным носиком. Семена защищены от намокания плотным слоем
эндокарпия. Диплоидное число хромосом 32.
Лютик ползучий - растение сильно, варьирующее по размерам
(длина, ширина, рассечение листьев, число ползучих побегов).
Как отмечает Коулс ( coles, 1977), форма роста у особей лютика
ползучего очень пластична. Например, степень рассечения листо-
вой плаотинки варьирует в зависимости от положения на растении
и времени года. Наиболее рассеченные листовые пластинки на ро-
зетках листьев, которые уходят под зиму: листья на ползучих
побегах и дочерних укоренившихся розетках менее рассечены .
Степень рассечения листовой пластинки следует тайке клинально-
му изменению: растения с наиболее широкой поверхностью листа
встречаются на юго-востоке Европы, а с наименьшей - на северо-
западе. Такое же изменение наблюдается в опушении черешков ли-
стьев: на юго-востоке и северо-востоке Европы растения с не-
опушенными черешками, а в Западной Европе - с опушенными.Цвет-
ки и семена также изменяются в клинальном направлении через
всю Европу. Эти изменения более значительны в западных океани-
ческих районах, чем на востоке и северо-востоке Европы.
Онтогенез, сезонное развитие и способы размножения
Семена лютика ползучего в большом количестве встречаются
в луговых почвах. При подсчетах на одном лугу на площади в I га
было найдено свыше 250 млн.семян (Работнов, 1951). После осы-
пания некоторые семена всходят сразу, но большей частью - на
следующий год. Всхожесть оемян до 83%. Прорастание затягивает-
ся на ряд лет, благодаря чему в почве обычно содержится боль-
шое число всхожих оемян (Работнов, 1951). В лаборатории семена
всходят медленно. Первые проростки появляются на 20-30-й день.
Вохожеоть низкая (15-23%) из-за недоразвитости зародыша и его
низкой ферментативной активности (Благовещенский, 1966; Ива-
72
нова, 1966).
Как показали исследования Р.П.Барыкиной и В.И.Пустовойтр-
вой (1973), при надземном прорастании семян первым трогается
в рост зародышевый корешок, затем вытягивается гипокотиль, вы-
носящий семядоли на поверхности почвы. Пластинки семядолей
развертываются на 30-35-й день с момента посева оемян; отмече-
ны трехсемядольные зародыши. Семядоли яйцевидные или овальные,
суживающиеся черешки срастаются друг с другом при основании;
жилкование дуговидное. Поверхностный рост семядолей продолжа-
ется в течение 15-20 дней, максимальная их длина I см, ширина
7-8 мм. Первый лист развертывается на 42 день, второй на 50.
Первые листья простые, округлые, трехлопастные на длинном че-
решке; жилкование пальчато-краевое. Семядоли отмирают с появле-
нием второго ассимилирующего листа. Ювенильные растения харак-
теризуются розеточной формой роста с даинночерешковыми листьями
в числе 5-7. Затем развертываются трехраздельные листья с над-
резанно-зубчатыми долями. Почки без почечного покрова формиру-
ются в пазухах листьев. На ранних стадиях развития проростка
четко выражен главный корень, а с появлением первых листьев
образуются придаточные корни на гипокотиле и на узлах укорочен-
ного побега. В основании первичной розетки формируется мочкова-
тая корневая система (Голубев,1956,1962). С наступлением осени
однолетний побег принимает форму ортотропного укороченного кор-
невища, защищенного основаниями листьев, в пазухах которых
скрыты печки возобновления.
В последующие годы розеточная форма роста сохраняется, а
прирост главного побега происходит из терминальной почки. Нача-
ло боковым розеточным побегам дают почки возобновления, которые
находятся в базальной части главной оси. Таким образом формиру-
ется разветвленное, короткочленное корневище (Серебряков, Сере-
брякова, 1965). Корневище сохраняется живым до 4-5 лет. В при-
родных условиях ювенильный период длится 2-3 года. Первые рас-
тения зацветают на 4 год (Серебряков, 1952). В культуре цве-
тут на втором году жизни (Корсмо, 1933; Голубев, 1958), образуя
при этом цветоносный побег с розеткой длинночерешковых ассими-
лирующих листьев (Любарский, Полуянова, Маркова, 1984). После
плодоношения удлиненная часть монокарпического побега полностью
отмирает. А в пазухах розеточных листьев формируются почки во-
73
зобновления, из которых в течение лета образуются дочерние боко-
вые укороченные побеги с ассимилирующими листьями. В терминаль-
ной точке роста у них закладывается цветоносный побег будущего
года, а осенью полностью сформирована лишь его вегетативная
часть (Серебряков, 1952; Серебрякова, 1956).
Лютик ползучий зимует в виде розетки 2-3 см в диаметре с
2-4-мя листьями. Весной (в конце апреля) формируются новые ли-
стья на длинных черешках. В конце мая образуются ползучие побе-
ги. Рост и развитие растений достигает максимума в середине
июля и заканчивается в середине октября. Цветение начинается в
середине мая, а пик цветения - в конце июня. Иногда отдельные
экземпляры цветут в октябре ( Harper, 1957; Sarukhan, Harper,
1973). Семена лютика ползучего начинают созревать в конце июня
и на одном и том же растении имеют разные сроки созревания. В
период вегетации постоянно происходит смена листьев материнских
растений. Старее отмирают, появляются новые. Монокарпический
побег у лютика ползучего полурозеточный и развивается по дицик-
лическому типу: почка - розеточный вегетативный побег - удлинен-
ный олиственный генеративный побег. Возобновление симподиальное.
Почки возобновления в пазухах нижних розеточных листьев материн-
ской оси формируются как вегетативные; из них весной будущего
года развиваются надземные ползучие побеги с удлиненным междо-
узлиями и ассимилирующими листьями. Они служат для вегетативно-
го размножения растения. Растения, выросшие из семян, обычно со
второго года жизни начинают размножаться с помощью ползучих по-
бегов, но в некоторых случаях отдельные экземпляры уже к концу
августа первого года образуют моноподиальные нарастающие олист-
венные побеги (Любарский, Полуянова, Маркова, 1984). Почки, ко-
торые закладываются в пазухах листьев этих побегов, В’ июне об-
разуют боковые розеточные побеги с 2-5-ью листьями. В почках
эндогенно закладываются зачатки корней, но потом они меняют на-
правление роста с горизонтального на вертикальное и под узлом
выходят наружу. Верхушка плагиотропного побега к концу вегета-
ционного периода принимает ортотропное положение и заканчивает
развитие образованием терминальной розетки. К осени дочерние
розетки укореняются, обособляются и сохраняются до следующего
года, давая начало новым особям.
Как отмечает В.Н.Голубев (I960), розеточная структура ве-
74
гетативногс побега о приземными почками возобновления, защищен-
ными сохраняющимися основаниями листьев, - это приспособление к
перенесению холодного зимнего периода. Укоренившиеся дочерние
розетки в вегетативном состоянии погут быть от 1-го до 3-х лет.
Длина и степень ветвления ползучих побегов уменьшается с пере-
ходом от ювенильной в генеративную фазу развития (Барыкина, Пус-
товойтова, 1973). Иногда ползучие побеги могут развиваться из
пазушных почек нижних стеблевых листьев цветоносного побега.Это
наблюдается в конце цветения при наличии благоприятных условий,
например,малой плотности популяции или при недоразвитии генера-
тивного побега из-за повреждения растения весенними заморозками
или насекомыми. Такие ползучие побеги длинные, тонкие, красно-
ватые, обильно ветвящиеся с многочисленными, но плохо укореняю-
щимися розетками.
Интенсивность вегетативного размножения особей в популяции
лютика ползучего существенно различается в зависимости от плот-
ности популяции - она усиливается при малой исходной плотности,
а при высокой исходной плотности ослабевает, резко дифференци-
руясь по особям. При этом в популяциях, где преобладает генера-
тивное размножение, в плоде формируется 25-30 семян, а там, где
только около 50$ растений образуют генеративные побеги, - 15-16
семян. И в естественных популяциях лютика ползучего, и на опыт-
ных площадках в течение всего вегетационного сезона ( с июня до
конца августа) встречаются всходы семенного происхождения. Мы
наблюдали, что в сформировавшихся популяциях в течение сезона
одни проростки погибали, их место занимали новые и тоже гибли -
и так до осени, А на вновь занятых территориях и в отсутствии
конкуренции проростки развивались быстро и в августе у некоторых
появились ползучие побеги. Очевидно, сложившаяся популяция люти-
ка ползучего закрыта для семенного возобновления и семенное раз-
множение здесь обеспечивает "банк семян" в почве (Любарский, По-
луянова, Маркова, 1984).
Экология, фитоценология и консортивные связи
Лютик ползучий - мезогигрофит. Он приурочен к разнотравно-
злаковым, канареечниково-мятликово-изящноосоково-разнотравным
ассоциациям (Барыкина, Пустовойтова, 1973). Лютик ползучий не
требователен к почве, но предпочитает богатые и слабоаллювиаль-
75
ные; встречается на подзолистых почвах, на щебнистых субстратах,
где не цветет и не дает вегетативных побегов. Растение здесь
заметно опушено. При перенесении в благоприятные условия дает
вегетативные побеги, розетки которых теряют опушенность. В эк-
стремальных условиях растения приобретают ксероморфный облик.
Летом - это растения щебнистых и сухих субстратов. Осенью,когда
почва уже физиологически сухая, новые листья тоже опушены, хотя
материнские растения не имели опушения. Семена, вызревшие в су-
хое, жаркое лето, тоже имеют войлочно-белое опушение. Растения^
развившиеся из этих семян в благоприятных условиях, опушения не
имеют. Лютик ползучий хорошо переносит умеренный выпас и массо-
во встречается на сенокосных лугах. Но в луговых фитоценозах он
редко образует чистую заросль: только по краям лужиц, канавок,
где застаивается вода. Он не образует чистой заросли и на лесных
возвышенных участках. Лютик ползучий положительно реагирует на
внесение в почву аммиачной селитры и аммофоса. Лучшими условия-
ми его развития являются повышенная влажность и затенение. Как
типичный эксплерент, он при таких условиях и в отсутствии кон-
куренции с другими видами способен образовывать монодоминантные,
продуктивные (20 г на 10 м^) группировки. Но его урожай может
резко меняться, например, под влиянием подсева дягиля (7 г на
10 м^)(Работнов, 1974).
Согласно шкале Раменского (1956) увлажнение в его местооби-
тании варьирует от сыролугового (в пределах от 80 до 89 ступени)
до сухолугсвого (в пределах от 59 до 99 ступени) в лесной зоне
и от влажнолугового (в пределах от 72 до 92 ступени) до болотно-
го (61-99 ступень) в степной зоне. Нс чаще он встречается в сы-
ролуговом и влажнолуговом местообитании, при сильно переменном
увлажнении; на лугах, в местах вымочек, где произошло массовое
отмирание или резкое угнетение преобладающих.здесь злаков (лисо-
хвоста лугового, канареечника тростниковидного). Например в пой-
ме реки Оки в I95I-I953 гг. в результате длительного весеннего
застоя воды на поверхности лугов произошло массовое отмирание
лисохвоста и канареечника, что привело к резкому улучшению усло-
вий произрастания низкорослых растений (в том числе и лютика
ползучего), размножающихся надземными укореняющимися побегами
(Работнсв, 1974). Большое значение для быстрого разрастания лю-
тика имеет значительный запас семян в почве лисохвсстовых и ка-
76
нареечниковых лугов, в результате большие площади лугов были пок-
рыты ползучелютиковыми и частично полввицевыми травостоями. Рас-
тительность центральной поймы стала более однообразней. Флорис-
тический состав же почти не изменился, нс произошли изменения в
количественных соотношениях компонентов, в составе ценопопуля-
ций, а также структуре травостоев. Вместо несомкнутого призем-
ного биогеоценогоризонта сформировался сомкнутый (проективное
покрытие 80-100$) горизонт, высотой 30-40 и даже 50 см, образо-
ванный лютиком ползучим. Преобладание лютика ползучего длилось
три года, а потом создались благоприятные условия для злаков,
что привело к резкому увеличению их участия в сложении травосто-
ев и соответственно к снижению участия лютика ползучего. По уче-
ту на 10-ти площадках в I м^ продуктивность лютика ползучего из-
менялась (в относительных цифрах) следующим образом: 1953 г. -
100$, 1954 - 72, 1955 - 75, 1956 - 1$. Внесение фосфора и калия
благоприятно влияет на лютик и оказывает слабое влияние на зла-
ки. При применении гербицидов, когда участие лютика в травостое
снизилось, резко возросла продуктивность злаков. Следовательно,
когда лютик хорошо развит он снижает, эффективность использования
удобрений злаками. Полного возврата лугового сообщества к исход-
ному состоянию после отмирания лютика не произошло.
Лютик ползучий будучи вегетативным малолетником и обладая
способностью к быстрому семенною/ и вегетативному размножению,
является очень удобным объектом для различных экспериментальных
популяционных исследований. С целью изучения динамики морфологи-
ческой структуры популяции лютика ползучего мн в 1970 г. в тече-
ние двух месяцев наблюдали за 146 экземплярами лютика вместе с
их вегетативным потомством. Растения были выделены в монодоми-
нантной группировке лютика ползучего, находящейся в липняке раз-
нотравном с примесью березы и осины в Сараловском лесничестве
Волжско-Камского заповедника. В результате наблюдений выяснилось,
что из 146 растений, только 85 образовали ползучие побеги. Эта
производная выборка была нами рассмотрена, и ет выяснили, что
постепенно, в течение вегетационного сезона уменьшается число
"материнских кустов", так как они отмирают вместе с системой
своих ползучих побегов. Л успевшие укорениться "дочерние кусты"
лишь частично остаются в живых. Из табл. I видно, что в период
наблюдения постепенно увеличивались средние показатели рассмат-
77
ё
а
Таблица I
tr
4
У
n
к
§
1
5?
»к
(П
см
U0
? т ч
Ё
|Н
м» ю ш
§
ю «н<
ю ю
о ш
00
s
СО
ю
ю
IK
и
К
°? т ч
Й
о со
со со
я
ю о
ю
IK
00
к
ф ю
со
-f t
8
со со
ю
С\> 02 Ю
сч
риваемых нами признаков, заметно усиливался и размах варьирова-
ния. Но коэффициент варьирования остался в основном на уровне
40-60%, проявляя в течение периода наблюдений некоторую тенден-
цию к снижению.
Интересно сопоставить приведенные выше результате с.данными
эксперимента по пересадке особей лютика ползучего на делянку
в условиях отсутствия конкуренции (Любарский, Полуянова,1975).
В условиях прогрессирующей засухи наблюдалось нарастание варьи-
рования всех показателей и постепенное снижение первоначально
высотах положительных корреляций между признаками. Однако когда
из выборки исключались отмершие и отмирающие экземпляры, для
оставшихся более жизнеспособных растений корреляция вновь была
достаточно высокой. По-видимому, падение потенциальной жизнеспо-
собности особей в ценопопуляции лютика получего свидетельству-
ет об ослаблении функциональных связей и зависимостей у расте-
ния. Креме того, проведенные нами опыты по изучению процесса
приведения экспериментальной популяции лютика ползучего к опти-
мальной плотности позволяет сделать определенные выводы о суще-
ствовании в популяции сложного механизма, который регулирует
поддержание оптимальной плотности в популяции путем внутрипопу-
ляционной регуляции размножения и выживаемости особей.
Однако, установившаяся некоторая оптимальная плотность популя-
ции,во-первых,всегда оценивается оптимальными пределами плот-
ности, вс-вторых, реализуется подвижней оптимальной плотностной
неравномерностью территории занятой популяцией, в-третьих, про-
являет характерное свойство сезонной пульсации. Последнее
подтверждается нашими многолетними экспериментами на серии пло-
щадок пс I кв.м, проведенными на биостанции Казанского универси-
ета. Сложившиеся экспериментальные популяции лютика ползучего
поддерживают (в условиях постоянного удаления растений всех про-
чих видов) достаточно близкую характерную оптимальную плотность
.по годам наблюдений и по отдельным площадкам), ранней весной
или поздней осенью. Однако, летом наблюдается значительное вре-
менное увеличение плотности (иногда в несколько раз) разное в
зависимости от условий года. Та же закономерность наблюдается и дл>
других лет. Летняя вспышка общей плотности популяции обеспечива-
79
гтся каждый раз появлением новых укореняющихся розеток (резуль-
тат вегетативного возобновления и размножения) и появлением
исходов семенного происхождения.
Биологический смысл такой временной пульсации плотности
популяции - в повышении надежности сохранения оптимальной плот-
ности популяции за счет создания временной избыточности, препят-
ствующей проникновению на занятую популяцией территорию других
видов растений и обеспечивающей многократно завышенный механизм
обновления взрослых особей вегетативного малолетника лютика пол-
зучего. Сезонная пульсация плотности реализуется таким образом,
благодаря своевременному развитию у особей ценопопуляции доста-
точного количества ползучих и генеративных побегов, обеспечива-
ющих соответственно вегетативное и семенное размножение в опти-
мальных для механизма регуляции плотности соотношениях и свое-
временно постав.’ ищих необходимое количество укореняющихся веге-
тативно-дочерних розеток и необходимое пополнение "банка семян"
в почве. В случае малой исходной плотности это достигается,
главным образом, вегетативным размножением, в случае высокой ис-
ходной плотности - генеративным размножением. При оптимальной
плотности роль вегетативного размножения становитоя второстепен-
ной по сравнению с размножением генеративным (табл.2). Мы наблю-
дали, что в условиях сформировавшейся популяции лютика ползучего
всходы семенного возобновления в течение лета все погибли, в ус-
ловиях же отсутствия конкуренции всходы развивались быстро и уже
в августе у некоторых появились ползучие побеги, но ни одно рас-
тение в первый год не цвело. Среди них постепенно также стала
намечаться дифференциация по высоте и количеству листьев.
При этом скорость освоения территории и достижение популя-
цией оптимальной плотности с помощью вегетативного размножения
при малой исходной плотности зависит и от конкретных экологичес-
ких условий (табл.З) и от длительности жизни органов вегетатив-
ного размножения, так как связь "дочерних" особей с материнским
растением обеспечивает им конкурентную способность значительно
выше, чем у всходов, возникиих из семян. Это определяет меньшее
их отмирание в естественных условиях, по сравнению со всходами.Об
этом можно судить по наблюдениям, которые проводились в Англии
над лютиком ползучим на пастбищном лугу. Ь течение 10 мес. на
I м^ появились 14 всходов (отмерло из них 9) и возникло 179 осо-
80
бей вегетативного происхождения (отмерло 26). За этот срок из
139 взрослых особей, отмеченных в начале наблюдений, отмерло
107, но из-за энергичного вегетативного размножения к концу на-
блюдения общее число особей достигло 184. Таким образом, в дан-
ных условиях лютик ползучий размножается в основном вегетатив-
ным путем, но длительность жизни возникпих при этом особей,оче-
видно, невелика ( Sarukhan, 1970).
Таблица 2
Динамика соотношения вегетативного и генеративного размноже-
ния в экспериментальных популяциях лютика ползучего
Показа- тели Номер площа- док Исход- ное чи- сло осо- бей Годы Примечания
Т980 I98T 1982
Число 1,2 4 13 6 2 ср.из двух площ.
ползучих 3,4 9 17 13 3 ср.из двух площ.
побегов 5 25 14 14 7
6 100 30 35 II
Число Т.2 4 5 18 у- . ср.из двух площ.
генера- 3,4 9 9 35 ИГ’ ср.из двух площ.
тивных 5 25 10 46 52
побегов 6 100 70 91 124
Среднее 1,2 4 0 25 60 ср.из двух площ.
число 3,4 9 0 30 60 ср.иэ двух площ.
семян в 5 25 25 49 59
плоде 6 100 30 50 59
Таблица 3
Влияние некоторых экологических факторов на среднесуточный
прирост вегетативных побегов лютика ползучего (см)
Погода Отсутствие конкурен- ции Дополнитель- ное увлажне- ние Аммиачная селитра Аммофос
Засушливая 2,8 5,2 3,6 3,7
Дождливая 4,5 8,3 9,0 11,5
6-I04I
81
Лютик ползучий - насекомоопыляемое растение. При раскрыва-
нии цветка тычинки изогнуты внутрь и закрывают плодолистики. Со-
зревание пыльников начинается с тычинок наружного круга и посте-
пенно достигает тычинок, прилагающих к плодолистикам. После это-
го становится возможным попадание пыльцы на рыльце с помощью на-
секомых. Лютик ползучий поедается бобрами (Хлебович,1933), а его
семена поедаются глухарем и тетеревом (Соколов,1949). Корни пер-
вого и третьего порядков имеют эндотрофную или эктоэнтотрофную
микоризу. Он является эккрисотрофом и частично сапротрофом (Ра-
ботнов, 1974).
Хозяйственное значение
Известно, что лютик ползучий поедается мелким рогатым ско-
том, но в Московской обл. отмечено поедание его и крупным рога-
тым скотом (Работнов, 1940). В вареном ввде может скармливаться
индюшатам (Корневен, 1912). На сенокосах не представляет никакой
ценности, так как дает лишь малое количество поедаемого корма.
Поэтому лютик ползучий относят к луговым сорнякам и ведут с ним
борьбу: опрыскивают 0,1-0,2^-ным раствором гербицидов в фазе при-
корневых листьев, вносят удобрения для создания густых высоких
травостоев, которые подавляют лютик ползучий, так как он прекра-
щает развитие ползучих побегов при выоокой конкуренции со злака-
ми; с этой же целью регулируют водный режим, потому что при не-
благоприятных условиях увлажнения лютик ползучий даже при отсут-
ствии конкуренции не способен удержать за собой площадь. В силу
слабо развитой корневой системы он не оказывает кумулятивного
действия на среду (Любарский, 1969).
Лютик ползучий - ядовитое растение. Ядовитым началом счита-
ется наличие в растении анемонола, который обладает сильным мест-
ным действием на кожу и слизистые оболочки. Отравление лютиком
происходит на выпасах и при кормлении травой с большим содержа-
нием этого растения. В народной медицине используется как крово-
останавливающее средство (Петров, 1912).
Т.В.Еагдасарова, Р.П.Барыкина, А.Н.Луферов
Род ВАСИШЮТНИК
Род василистник - Thalictrum L. (семейство Ranunculaceae
Juaa.) включает около 150 видов, распространенных преимущест-
венно во внетропических областях обоих полушарий; лишь немногие
вида произрастают в Африке, Центральной и Южной Америке ( Lecc- •
yer, 1885; Boivin, 1944; Tamura, 1968). Наибольшее видовое ?
разнообразие отмечается в Восточной Азии.
Латинское название рода происходит от греческого "thaiic-
tron" - названия растения, относящегося, вероятно, к Thalict-
rum minus I,. (Backer, 1936; Gilbert-Carter, 1964; Каден, Те-
рентьева, 1975). Не исключено, что исходным словом могло быть
*thal£o" - "цвести, зеленеть", что связывалось с яркой окрас-
кой растения ( Beckmann, 1801).
Во флоре СССР около 25 видов василиотника. Большинство из
них - многолетние травянистые мезофиты, входящие в состав лес-
ных, луговых, степных фитоценозов. Подземная часть растений
представлена короткими или длинными корневищами и многочисленны-
ми придаточными корнями. Надземные фотофильные побеги обычно
прямостоящие, простые или разветвленные, с очередным листораспо-
ложением. Листья сложные, с прилистниками; у некоторых видов раз-
виваютоя и прилистнички. Соцветие сильно разветвленное, метель-
чатое, реже кистевидное. Околоцветник простой, чашечковидный, из
4-5 листочков, которые обычно короче тычинок. Нектарников нет.
Гинецей апокарпный. Пестики обычно многочисленные, с короткими
прямыми или немного изогнутыми стилодиями.
Плод - многоорешек; перикарпии кожистые, нередко очень тон-
кие, перепончатые, с продольными ребрами.
В Московской области 5 видов василиотника.
Ключ для определения подмосковных видов
I. Плодики на длинных ножках, повислые, с крыловидными реб-
рами. Тычиночные нити булавовидно утолщенные. Листья широкотре-
угольные, с плотными, кожистыми прилистниками и прилистничками,
расположенными при разветвлениях черешков ....................
........................................ Th.aquilegifolium I.
- Плодики сидячие, по ребрам не крылатые. Тычиночные нити
тонкие. Прилистничков нет или они тонкие, пленчатые ........ 2
83
2. Листочки узколанцетные и линейные, до 0,5 (0,8) мм шир.,
цельнокрайние, реже с 1-2 зубцами, сверху темно-зеленые, блес-
тящие, снизу сизовато-зеленые ................ lucidum L.
- Листочки иной формы, более широкие, 3-5-зубчатые, иногда
без зубцов................................................. 3
3. Метелка узкопирамидальная. Листья узкотреугольные, при-
жатые (кроме самых нижних) к отеблю,дважды тройчато- или дважды-
перистосложные ............................... Th. simplex L.
- Метелка обычно щитовидная или широкопирамидальная.Листья
широкотреугольные, отклоненные от стебля, расчлененные до (2)
3-4 порядка ............................................... 4
4. Цветоножки в начале цветения прямостоячие. Цветки бледно-
желтые. Листочки продолговато-обратнояйцевидные или эллиптичес-
кие. Черешки полые. Корневища и корни желтые ..... Th.flavum L.
- Цветоножки поникшие, цветки зеленоватые, белые или светло-
розовые. Листочки округлые, широкоэллиптические или обратнояйце-
видные. Черешки плотные. Корневища и корни сероватые или темно-
бурые ............................................ Th. minus L.
I. ВАСИЛИСТНИК ВОДОСБОРОЛИСТНЫЙ
Номенклатура и географическое распространение
Thalictrum aquilegifolium L. 1753, Sp.pl; 1:547; КауфМ.,
1866, Моск.фл.,изд. 1:17, 1889, изд. 2:4; Илл.фга. 2:144; он же,
1927, Опр. рас т. Моск, губ.: 132; Невский, 1937, Фл.СССР 7:512; он
же, 1940, в Маевский, Фл.,изд. 7:372; Голицын, 1954, там же, изд.
8: 85; Маценко, 1964, там же, изд. 9: 87; Tutin, 1964, Pl.Eur.
1:241; Ворошилов, 1966, Опр.раст.Моск.обл.: 170; Черепанов,1981,
Сосуд.раст.СССР : 427; Новиков, 1986, Опр.раст.Мещеры I: 183;
Василистник водосборолистный (семейство Ranunculaceae - лютико-
вые ).
Василистник водосборолистный относится к неморальному средне-
европейскому элементу флоры: распространен в лесной и лесостеп-
ной зонах. Граница ареала проходит по югу Швеции и Финляндии, до-
стигает южного побережья Белого моря у низовий р.Онега и Северной
Двины (г.Архангельск), затем простирается на юг, до среднего те-
чения р.Вычегды, Камы и далее к г.Саратову, Днепропетровску и
Одессе. В зарубежной Европе южная граница ареала проходит через
84
среднее течение Дуная, северную Италию и Испанию. Вид широко
распространен в странах Средней и Южной Европы, а тайке встре-
чается на западе Малой Азии (Турция) ( Meusel et al., 1965;
Atlas Fl. Ear., 1989). В Московской обл. обычное растение
(Ворошилов и др., 1966).
Морфологическое описание
Короткокорневищный кистекорневой вегетативно малоподвижный
геофит с безрозеточными моноциклическими, иногда озимыми, моно-
карпическими побегами и диморфными придаточными корнями (тол-
стыми запасающими и тонкими поглощающими). Корневище вертикаль-
ное, эпигергенное. Высота надземной части достигает 150 (180)
см. Побеги голые, полиметамерные, несущие до соцветия 3-5 че-
шуевидных Ц4-7 срединных очередных листьев. Листья дважды,
трижды перистосложные, длинночерешковые, с парными прилистника-
ми при основании и прилистничками в местах разветвления рахиса.
Пластинки в очертании широкотреугольные. Листочки длиной 2-5
см, шириной 0,5-4,5 см, обратнояйцевидные и округло-эллиптичес-
кие, трехлопастные или надрезанно-городчатые, свержу темно-зе-
леные , снизу сизоватые на черешочках.Соцветие - закрытая или
монотелическая ( Troll, 1964,1969), сильно разветвленная много-
цветковая фрондулезно-брактеозная метелка или, согласно А.А.
Фёдорову, З.Т.Артюшенко (1979), - кистевидный тирс. Количество
цветков в соцветии колеблется от 160 до 400. Цветки мелкие, на
прямых или слабо изогнутых цветоножках, актиноморфные. Около-
цветник простой, зеленовато-белый из 4 > лепестковидных листоч-
ков, которые короче тычинок в 2-3 раза. Тычиночные нити свержу
расширенные, бледно-лиловые, белые или розовые. Пестиков 6-14.
Плодики-орешки длиной 12-20 мм, шириной около 4 мм, продолгова-
то-обратногрушевидные с 3-мя неравными крыловидными ребрами,по-
вислые на длинных (4-7 мм) ножках. Перикарпий тонкий, хрупкий,
легко опадающий (Рыоина, 1973). Семена эллиптические, до 2 мм
длиной, 1,5 мм шириной, треугольно-округлые в поперечнике, тем-
но-коричневые, блестящие. Маоса I тыс.оемян составляет в сред-
нем 2,5-3,0г. Диплоидный набор хромосом 14 (Жукова, 1961; Бо-
чанцева, 1972), 14,28 (Хромосомные числа .... 1969).
Онтогенез
Большой жизненный цикл василистника водосборолистного (и
бМои 85
других подмосковных видов? сил подробно исследован Р.П.Бары-
киной, А.П.Луферрвым (1982), Л.П.Рысиным, Г.П.Рысиной (1987).
В естественных ценозах прослеживается, в соответствии с клас-
сификацией Т.А.Работнова (I960), дополнениями А.А.Уранова
(1975) и И.Т.Васильченко (1936), 3 возрастных периода: латент-
ный, виргинильный и генеративный; сенильный период отчетливо
не выражен. Виргинильный период включает 5 этапов: проростки,
всходы, ювенильные, ижатурные и взрослые вегетативные расте-
ния.
Латентный период. Плодики имеют широкие крыловидные реб-
ра, воздушную полость между семенем и перикарпием, что спо-
собствует распространению их ветром и водой. В зрелом семени
зародыш маленький, базальный, прямой. В нем различимы зароды-
шевый корешок о корневым чехликом, короткий гипокотиль, семя-
дольный узел с двумя семядолями; почечка недифференцирована и
представлена небольшой группой меристематических клеток. Сход-
ная структура зародыша наблюдается и у других видов василист-
ника; отмечаются лишь количественные различия. Запасными веще-
ствами обильного эндосперма служат жирное масло и белок. Стра-
тификация сокращает сроки прорастания семян у всех рассматри-
ваемых нами видов, а также увеличивает процент всхожести (табл.
I).
Виргинильный период. Семена сохраняют всхожесть в течение
6-10 лет. Тонкий перикарпий не препятствует прорастанию семян,
роль скарификации в повышении всхожести невелика. В связи с
разнокачествонностью семян, степенью их зрелости, период про-
растания несколько растянут. В естественных ценозах часть про-
ростков появляется во второй-третьей декаде мая, остальные в
течение июня, единично- в икоте. Всхожесть высокая, от 55 до 75$.
Прорастание надземное, гипокотилярное, свойственное и другим
подмосковным видам василистников. Гипокотиль удлиняется до 3 см,
тонкий, округлый, светлый. Главный корень проростка 3-5 см дли-
ной, желтовато-коричневый. Начало его ветвления совпадает во
времени с формированием в почечке зачатка первого настоящего
листа. Семядоли длинночерешковые; пластинки яйцевидные, с серд-
це видным основанием, пальчато-дуговидным жилкованием, с верхней
стороны сиэо-темно-зеленые, с нижней - светло-зеленые.
S конце мая-начале июня из почечки развивается розеточный
побег, чт'о определяет переход в фазу всхода. Развертываются 3-4
86
Таблица I
Сроки прорастания и всхожесть семян видов
василиотника (в лабораторных условиях)*
Вид Сроки прорастания, да. Всхожесть, %
без стра- тификации после страти- фикации без стра- тифика- ции после страти- фикации
Василистник водосбо- ролистный 42-45 10-14 55-60 70-75
В. малый 26-29 12-14 60-63 86-90
В. простой 25-32 17-19 40-45 70-75
В. желтый 61-73 48-50 3-15 40-50
В. блестящий 14-16 6-10 80-90 90-96
х Семена высевали в чашки Петри на влажные фильтры и проращи-
вали при постоянной температуре (+20°С). Перед посевом часть се-
мян предварительно стратифицировали в течение 30 дн. при темпе-
ратуре -4 °C, другую - оставляли в качестве контроля, не подвер-
гая никакой обработке (Луферов, 1984).
тройчатосложных листа, из которых только верхний имеет в основа-
нии парные латеральные ушковидные прилистники. Листочки округ-
лые, чаще трехзубчатые, с перист о-сетчатым жилкованием. Гипоко-
тиль всхода в июне-июле полегает и утолщается, на нем развивает-
ся от четырех до семи придаточных корней. Усиливается ветвление
главного корня. Состояние всхода продолжается до конца лета.
В августе-сентябре семядоли и листья отмирают, а стеблевая
часть укороченного побега благодаря контрактильности гипокотиля,
главного и придаточных корней втягивается в почву, превращаясь в
вертикальное эпигеогенное корневище. Терминальная почка его, зи-
мующая в поверхностном слое почвы, защищена помимо двух почечных
чешуй расширенным основанием и прилистниками верхнего ассимилиру-
ющего листа розетки. Почечные чешуи, как и зачатки срединных ли-
стьев, имеют прилистники.
На втором году растение вступает в ювенильный этап развития.
Для него характерно наличие полу розе точного побега высотой 10-15
87
ом о дважды тройчатосложными листьями, из которых 2-3 распола- ,
гпютсп на укороченной части побега и лишь один - на удлиненной.
Листочки обратнояйцевидные или округло-обратнояйцевидные, иног-
да трохлопастные, с сердцевидным или округлым основанием, по
краю зубчатые или городчатые. Наряду с прилистниками тлеются и
прилистнички. Последние развиваются по два (с абаксиальной сто-
роны) в местах разветвления общего черешка и черешков последую-
щего порядка. Прилистники и прилистнички ушковидные, цельнокрай-
ние, светло-коричневые, кожистые или пленчатые.
У ювенильных растений отчетливо выражена дифференциация при-
даточных корней на более длинные (10-20 см длины), толстые (2-4
мм в диаметре), ветвящиеся до 2-4-х порядков запасающие и более
короткие (4-8 см длиной), тонкие (1-2 мм в диаметре), олабо вет-
вящиеся поглощающие. Первые живут два вегетационных периода,
последние - один. Диморфизм корней сохраняется и в последующие
годы онтогенеза особи. В течение лета пазушная почка одного из
чешуевидных лиотьев (почечных чешуй) сильнее других увеличивает-
ся в размерах, ветвится и становится покоящейся почкой возобнов-
ления. Осенью главный корень и надземная удлиненная часть побега
отмирают, а укороченные междоузлия последнего формируют новый
годичный прирост эпигеогенного корневища.
Весной третьего года жизни растение переходит в имматурное
состояние. Моноподиальное нарастание главной оси, характерное
для ювенильных особей, сменяется симподиальным. Из перезимовавшей
почки возобновления в мае развивается вертикальный розеточный фо-
тофильный побег высотой 20-30 см. Одновременно с материнской тро-
гается в рост и одна из ее дочерних почек, образуя короткие (2-3
см), укореняющееся в узлах, подземное горизонтальное гипогеоген-
ное корневище с тремя-пятью чешуевидными листьями. Соотношение
числа ассимилирующих листьев на укороченном и удлиненном участках
надземного годичного побега у имматурных растений в сравнении с
ювенильными меняется на обратное: прикорневой лист всегда один,
последующие 2-3 листа отделены друг от друга удлиненными междо-
узлиями. Листья трехлетних особей, как и более взрослых, имеют
короткое влагалище, прилистники и прилистнички, выполняющие за-
щитную Функцию ( Roth, 1957; Guedea, 1968; Луферов, 1981). При
первом-втором нижних разветвлениях общего черешка число прилист-
ничков увеличивается до четырех (два более крупных располагаются
88
; абаксиальной, два других - с адаксиальной стороны листа). Прн-
листнички часто срастаются в основании. Примечательно, что на
протяжении трех первых лет число ярусов ветвления черешка сохра-
няется более или менее стабильным в пределах особи и соответст-
вует ее возрасту (Барыкина, Луферов, 1982). Эпигеогенное корне-
вище у трехлетних растений в середине лета отмирает. Следующей
почкой возобновления становится верхушечная почка гипогеоген кого
корнечица. У четырехлетних особей она выходит на дневную поверх-
ность, образуя удлиненный ортотропный побег, который характерен
для взрослых вегетативных форм. В этом возрастном состоянии рас-
тение достигает 40-80 см высоты, емкость почек возобновления
увеличивается до 6-9 метамеров, кроме 3-4 почечных подуй за-
кладывается от 3 до 5 ассимилирующих листьев (дважды или
трижды тройчато- и перистосложных)’; отмечается рассеянное ветв-
ление надземных побегов. Из пазушных почек корневища могут раз-
виться (в числе 1-3) силлептические ортотропные олиственные (по-
лурозеточные или безрозеточные) побеги обогащения. Гипогеоген-
ное корневище сохраняется живым до июля будущего года и, пере-
дав питательные вещества последующему поколения побегов, отми-
рает. Спмподиальное нарастание корневища идет по типу монохазия
или дихазия, в последнем случае наряду с возобновлением происхо-
дит, хотя и слабое, вегетативное размножение.
Генеративный период и сезонное развитие
Первое цветение василистника водосборолистного в природе
наступает примерно на седьмом году жизни (Барыкина, Луферов,
1982), а г культуре - на втором (Скворцов, Трулевич, Алферова,
1979). Почки возобновления перезимовывают в почве на глубине
1-6 см. Осенью в них генеративный, побег будущего года сформиро-
ван полностью, включая соцветие и отдельные цветки (Серебряков,
1952).
В условиях Подмосковья вегетации начинается после таяния
снега с конца апреля-в начале мая (Серебряков, 1947). Разверты-
вающиеся листья достигают окончательных размеров к концу мая.
Прикорневые листья сохраняются зелеными до глубокой осени. За-
цветает василистник 15-25 июня, отцветает к 20-25 июля. Сухие
орешки опадают в августе-ноябре.
Иногда в случае теплой и влажной осени почки возобновления
89
могут i; пигуетв-оонтябре тронуться в рост, развертывая 1-2асси-
млиируюпшх листа; развитие побегов по типу озимых наблюдается
«ок у п*, io руинных, так и взрослых вегетативных растений (Бары-
ни , .'ivlopOB, 1982).
Способы размножения и распространения
Размножение и расселение осуществляется в основном семенами
(Рысина, 1973). Вегетативная подвижность растения очень слабая.
Ваоилистник водосборолистный - типичный энтомофил. Он хорошо при-
способлен к опылению насекомыми, имеет крупные соцветия, хорошо
заметные под пологом леса. Цветки многочисленные с сильным запа-
хом, окрашенные в серо-сиреневый цвет, реже белые,- тычинки не
только продуцируют огромное количество пыльцы, но и выполняют
роль атрактантов. Семенная продуктивность в естественных ценозах
высокая и составляет от 60 до 400 семян на генеративный побег.
Число завязавшихся орешков определяется световым режимом, погод-
ными условиями, обилием опылителей. Так, в условиях Подмосковья
в смешанном елово-березовом лесу с папоротником орляком при от-
носительной освещенности 38% в среднем на генеративном побеге из
400 завязей сформировалось только 60 плодиков, что составило 15%.
На поляне в сосновом лесу при освещенности 75% из 395 орешков
только 175 оказались зрелыми, т.е. 40% от общего числа. На просе-
ке же в сосновом лесу с относительной освещенностью 66% из 160
орешков лишь 60 были зрелыми, остальные 70% щуплыми, недоразви-
тыми. Отрицательно влияют на семенную продуктивность также дожди
в период цветения. Некоторые плодики повреждаются личинками на-
секомых.
Экология
Василистник водосборолистный произрастает на среднесухих и
влажных почвах (третья ступень шкапы Ландольта). В условиях Под-
московья относительная влажность в горизонтах AoL.AoF составля-
ет от 50 до 80%, в горизонтах Aj - от 20 до 25%. Это полутеневое
растение (пятая ступень шкалы Элленберга), но может цвести и
плодоносить в условиях значительного затенения (при относительной
освещенности 15-20%). В Подмосковье при освещенности 60-70% ва-
силистник достигает высоты 180-200 см.
Растет на почвах слабокислых, иногда нейтральных или слабо-
90'
щелочных с pH - 4,5-5,5 (третья ступень шкалы Ландольта). В усло-
виях Подмосковья распространен на почвах с pH (водным) в горизон-
тах AoL, AoF и Аон от 4,8 до 6,2; с pH (солевым) в тех же го-
ризонтах от 4,5 до 5,6. Василистник водосборолистный является
индикаторе»! высокого содержания азота в почве (восьмая ступень
шкалы Эппенберга). В Московской обл. встречается на дерново-под-
золистых или бурых лесных с примесью карбонатов почвах различно-
го механического состава: суглинистых, супесчаных и песчаных.
Фитоценология
Василистник водосборолистный - опушечно-лесной вид. В Мос-
ковской обл. он произрастает в различных типах еловых, сосново-
еловых, смешанных и лиственных лесов, на водораздельных плато,
склонах террас и оврагов; нередко встречается на лесных вырубках,
полянах, опушках, в зарослях кустарников. Его участие в фитоцено-
зе обычно невелико (до 10-15% от общей площади): растения произ-
растают небольшими куртинами или одиночно. На территории Звени-
городской биостанции Московского университета встречается в сос-
няках малиново-разнотравных, в березняках орляково-разнотравных,
в ельниках липово-кленово-снытьево-зеленчуковых, в ельниках сос-
ново-разнотравных и других.
В табл.2 приведена возрастная структура популяций в различ-
ных типах леса на площадках 100 м^.
Наибольшая плотность особей наблюдалась нами на поляне в бе-
резняке орляко-разнотравном. Во всех типах леса василистник водо-
сборолистный произрастает совместно со следующими растениями:
Anthrisous aylvestris, Angelica sylvestris, Geranium sylvaticum,
Oalamagroatia arundinacea, Ajuga reptans, Potentilla erecta,
Aegopodium podagraria, Rubua aaxatilis, Hypericum perforatum И др.
Консортивные связи
Василистник водосборолистный симбиотически связан о фикоми-
цетными эндофитными грибами, образующими эндотрофную микоризу.
Эндомикориза приурочена к самым тонким корням (2-4-го порядка),
сохраняющим первичное строение.
Цветки василистника водооборолистного охотно посещаются раз-
нообразными насекомыми-опылителями (Daumann, 1969). В зависимос-
ти от подвижности всех опылителей можно разделить на две группы:
91
Таблица 2
Возрастной состав пенопопуляций василиотника
водосборолистного в различных фитоценозах
Ten леса Число особей различного0возраст- ного состояния на 100 м2 общее число осо- бей плотность растений на м2
всходы юве- ниль- ные шма- ту р- ные взрос- лые ве- гета- тивные гене- оатив ные
Еерезняк опляково- разнотрав- ннй 9 15 16 20 95 155 1,5
Сосняк малиново- разнотоав- Н1Й — 7 . 15 18 30 '' 70 0,7
Ельник вейниковый 45 28 23 ТО 30 136 т.з
Ельник липово-кле- ново-сныте- во-зелен- чуковый 8 ТО 2 8 5 33 0,3
очень подвижных и медлительных. К первой группе относятся жур-
чалки, шмели, роющие осы, пчелы, ко второй - жуки-усачи, брон-
зовки, слоники, мягкотелки, горбатки, клопы.
Опылители василиотника водосборолистного
.Двукрылые:
семейство Empididae - Empia teaaelata; семейство Syrphidae -
Syrphus groaaularia, Syrphus cinotelluii, Melanostoma mellinum,
Volucella pelluaoens, Xylota lenta, Temnoatoma ар.; семейство
Muscidae - Tricops ap., Phaonia baaalis .
Куки:
семейство Cerambicidae - Leptura inexpactata, L.viridis, Gauro-
tes virginea, Strangalia mslarura, S.arcuata, Pachyta quadripun-
ctata.Rhagion inquisitar, Monohamus; семейство Scarabaeidae -
Cetonia aurata; семейство Curculionidae; Cantharididae; Mor-
dellidae.
92
Перепончатокрылые:
Apis mellifera, Crabro cribjBarius, Bombas sp.
Клопы:
Lygocoria sp.; Colocoris sexguttatus.
Время опыления одним насекомым может быть различным. Напри-
мер, Phaonia baaalia находится на соцветии от нескольких секунд
до одиннадцати минут. Чаще всего насекомые посещают соцветие че-
рез 35-40 с. Наблюдалось, что за 10 МИН Phaonia baaalia посе-
тила одно соцветие 54 раза. Так что во время своего цветения
остаться неопыленным василистник водосборолистный практически
не может.
Основным опылителем василиотника, переносящим 53,6$ пыльцы,
является муха Phaonia basalis. На втором месте стоят жуки-усачи
(43,1$), среди которых доминирует Pachyta quadripunctata
(13,2$), Gaurotea virginea (11,2$), Leptura inexpectata
(10,4$). Все эти виды в имагинальной стадии являются актофагами,
т.е. питаются исключительно пыльцой.
Биологическая продуктивность
В зависимости от условий и местообитания биомасса надземных
частей колеблется от 60 до 83 г (сырой вес) на I 1?. В табл.З
отражена биологическая продуктивность василиотника в Одинцовском
районе Московской обл.
Таблипр 3
Биологическая продуктивность василиотника водосборо-
листного в различных фитоценозах
Тип леса Вес сырой_массы, г на Вес воздушно-сухой массы, г на I г
надземных органов подземных органов надземных органов подземных органов
Березняк орляково-раз- нотравный 235 65 45 21
Сооняк малиново-раз- нотравный 260 76 50 25
Ельник вейниковый 215 60 41 21
Ельник липово- кленово-снытвво зеленчуковый 240 83 50 21
93
Хозяйственное значение
Василис тник водосборолистный издавна используется в народ-
ной мед:щине в качестве гипотензивного, успокаивающего (Комен-
дар, K7I; Шретер, 1975), мочегонного и слабительного (Левчук,
1929) средства. Известны такке его антибактериальные свойства
(Сацыперова, 1952). Представляет интерес и как декоративное
растение (Мерло, 1965; Полетико, Мищенкова, 1967; Карписонова,
1985), а такие - медоносное (Головкин, 1986). Кормовое значение
относительно небольшое, надземные побеги иногда поедаются мел-
ким и крупным рогатым скотом, лошадьми, оленями (Работнов,
1951).
2. ВАСИЛИСТНИК МАЛЫЙ
Номенклатура и географическое распространение
Thalictrum minus !>., 1753, Sp. pl. 1:546; КауфМ., 1866,
Моск.фп. ,иэд. 1:17; 1889, изд. 2:5; Цингер, 1886, Сб.свед.: 29;
Шмальг., 1895, Фл. 1:5; Сырейщ., 1907, Илл.фл. 2:144; он же
1927; Опр.раст.Моск.губ. : 132; Невский, 1937, Фл.СССР 7:524 ;
он же, 1940, в Маевский, Фл., изд. 7:373; Голицына-1954, там же,
изд. 8:87; Маценко, 1964, там же, изд. 9:88; Tutin, 1964, Fl.
Eur. I: 241; Ворошилов, 1966, Опр.раст.Моск.обл.: 171; Новиков,
1986, Опр.раст. Мещеры Г:183. - Василистник малый (семейство
Kanunculaceae _ лютиковые).
Василистник малый - голарктический вид (Гроссгейм, 1950).
Ареал охватывает всю Европу, Малую, Переднюю и Центральную Азию,
Тибет, Китай, Корею, Японию, встречается также в Северной Амери-
ке. В СССР произрастает во всех районах, за исключением Крайнего
Севера, пустынь и полупустынь юго-востока европейской чаоти
СССР, Казахстана и Средней Азии. В Московской обл. встречается
повсеместно.
Морфологическое описание
Короткокорневишный геофит с бе~ розеточными моноциклическими
монокарпическими побегами и диморфными шауровидными придаточными
корнями; в некоторых цо:?ляциях встречают- - и длиннокорневищные
особи. Горизонтальные корневища длиной от 3-5 до 30 см, с много-
численными придаточными корнями. Стебли прямые, иногда коленчато-
94
изогнутые, высотой 60-180 (200) см, округлые, ребристые.Листья
очередные, черешковые, в очертании широкотреугольные, длиной
7-30 см и шириной 30 см, трижды, четырежды тройчато- или триж-
ды, четырежды перистосложные. Листочки длиной 1-3 (4) см, ши-
риной 0,5-3 см, округлые, обратнояйцевидные, широкоэллиптичес-
кие, трехлопастные или цельные, 3-9-зубчатые, в основании ок-
руглые, реже клиновидные, снизу более светлые. Соцветие - широ-
копирамидальная или щитковидная раскидистая многоцветковая ме-
телка. ’(ветки поникшие, мелкие, зеленоватые, белые, розовые,
разных оттопков, с 4-мя, реже 5-ью листочками простого около-
цветника, длиной 2-4 мм и многочисленными длинными висячими
тычинками. Пестиков 5-12 (15)...Орешки яйцевидные, яйцевидно-
эллиптические или продолговато-эллиптические, ребристые, длиной
2-4 мм, шириной 1-2 мм, с прямым или немного согнутым отилоди-
ем длиной С,5-1 мм.
Очень полиморфный вид, распадавдийся на ряд полиплоидных
рас и локальных форм, различающихся по величине надземных и
подземных побегов, размерам листьев, листочков, соцветий, чис- '
лу тычинок, пестиков, некоторым другим признакам ( Lecoyer,
1885; Boivin, 1944; Emura, 1972). Внутривидовая систематика.. .
разработана еще недостаточно полно и нуждается в дальнейших "
специальных исследованиях.
Диплоидный набор хромосом: 2п = 14,40,42,48,70,84; у боль-
шинства Йорм 2п -- 40,42,70 ( Kuzmanov, Dutschewsca, 1982),
реже 2п = 28 ( OvertonJ909) .
Онтогенез
Латентный период. Зародыш василистника малого, по своей
структуре сходен с зародышем василистика водосборолистного, но
отличается меньшей величиной (длина 315 + 23 мкм, ширина 112 +
+ 14 мм).
. Б естественных ценозах семена прорастают в мае-июне следу-
ющего, после диссеминации года, и лишь небольшая их часть (1-5%)
- в июле. При хранении семян в лаборатории всхожесть сохраняет-
ся в течение 5-8 лет. Стратификации приводит к сокращению сроков
прорастания, семян, а также к возрастанию процента их всхожести
(тасл.1).
Зиргинильный период. Проростки' имеют главный корень дли-
ной 3-6 см, желтовато-коричневый, с многочисленными корневыми
95
полосками . Боковые корни образуются с появлением первого лис-
тового примордая. Гипокотиль немного короче главного корня,
слегка полегающий при основании. Семядоли располагаются супро-
тивно; их пластинки яйцевидные или округло-яйцевидные, с тремя
тонкими жилками, зеленые.
Почечка трогается в рост через 6-10 дней после прораста-
ния, давая начало укороченному побегу с 1-3 тройчатосложными
листьями. Листочки округлые, трехзубчатые, тонкие, с перисто-
сетчатым жилкованием. Гипокотиль в конце мая полегает, утолща-
ется и постепенно втягивается в почву; в результате контрак-
тильности его поверхность становится поперечно-морщинистой. В
семядольном узле и на гипокотиле образуются 4-8 придаточных
корней. Главный корень ветвится и удлиняется до 6-8 см. В авгу-
сте-октябре надземная часть фотофильного побега отмирает, а его
укороченная стеблевая часть превращается в эпигеогенное корне-
вище, которое погружается в почву. Наряду с гипокотилем этому
способствуют главный и придаточные корни, такке обладающие кон-
трактильной способностью.
Ювенильные растения - двулетние; у них четко прослеживает-
ся дифференциация придаточных корней на запасающие (длина их
8-15 см, толщина 2-3 мм), ветвящиеся до 3-4-го порядка и живу-
щие 2-4 года, а такке поглощающие корни (длиной 3-7 см, толщи-
ной 1-2 мм), неветвящиеся и функционирующие один вегетационный
сезон. Надземный побег полурозеточный, высота его 5-15 см. Ли-
стья дважды тройчато- или дважды перистосложные, с коротким
влагалищем и прилистниками. При разветвлениях черешков иногда
развиваются мелкие ушковидные или неправильной формы прилист-
нички. Лиоточки округлые, округло-обратнояйцевидные, более
крупные, чем у всходов. Осенью главный корень и надземная часть
ассимилирующего побега отмирают.
Имматурные растения (трехлетние) характеризуются сменой
моноподиального нарастания главной оси симподиальным в резуль-
тате отмирания верхушечной почки и развитием короткого гипогео-
генного корневища из пазушной почки элитеогеиного корневища.
Гипокотиль в июне-июле отмирает. Надземный побег достигает ID-
35 см. Листья трижды тройчато- и трижды перистосложные. Почка
возобновления корневища в течение вегетационного периода замет-
но увеличивается в ра'змерах и, будучи защищенная 3-5 чешуями,
зимует на глубине 0,5-3 ом.
96
Взрослые вегетативные (4-7-летние) растения характеризу-
ются наличием системы оимподиально нарастающих (по типу моноха-
зия или дихазия, реже плейохазия) коротких корневищ. В некото-
рых популяциях довольно обычны особи с длинными корневищами,го-
дичные приросты которых достигают длины 5-20 см при диаметре
2-5 мм. Длиннокорневищные растения отмечались также В.Н.Голубе-
вым (1958), Г.П.Денисовой (1960,1965), С.Н.Зиман (1981). Над-;
земные побеги полурозеточные, реже безрозеточные, высота их
20-70 (120) см.
Генеративный период и сезонное развитие
Первое цветение у особей дикорастущих популяций наблюдает-
ся в Москве (Главный ботанический сад АН СССР) на 6-7 м. году
онтогенеза (Луферов, 1984) при интродукции на 2-м году (Вороши-
лов и др., 1961). Формирование соцветий будущего года происхо-
дит в августе-сентябре (Серебрякова, 1956), иногда в марте-ап-
реле . .
Ритм сезонного развития василистника малого в естественных
ценозах Подмосковья изучался Н.Н.Каденом (1950) и Т.И.Серебря-
ковой (1956). Отрастание надземных побегов обычно происходит в
конце апреля-начале мая. К концу июня рост стеблей и листьев в
основном завершается. Наряду с моноциклическими монокарпически-
ми побегами нередко развиваются также удлиненные вегетативные
(Серебряков, 1959). Нарастание корневища продолжается до авгус-
та. Число его годичных приростов, по наблюдениям разных авто-
ров, составляет 2-4 (Серебряков, 1954а; Зозулин, 1959), в уело-
виях европейского Севера - от 4-6 и даже до 40 (Бахматова, 1984),
в Подмосковье, по нашим данным, от 2-3 до 10. В результате вет-
вления и отмирания наиболее старых участков корневищ образуется
клон. Многочисленные придаточные корни расходятся горизонталь-
но, реже отвестно вниз. По наблюдениям Л.П.Рысина и Г.П.Рыси-
ной (1987), последние достигают глубины I м и ветвятся до кор-
ней 4 порядка.
Цветение происходит с третьей, иногда со второй декады
июня до конца июля-начала августа; плодоношение - во второй по-
ловине августа-октябре; отмирание надземных органов - в сентяб-
ре-октябре.
Размножение и распрог панение осуществляется семенами, а
7-I04I 97
также вегетативно посредством ди- или плейохазиально ветвящих-
ся корневищ, особенно при произрастании на рыхлых, хорошо аэри-
руемых почвах.
Экология и фитоценология
Василистник малый предпочитает сухие почвы (2-я ступень
шкалы Ландольта), избегает очень сухие, а также переувлажненные,
заболоченные. Встречается на открытых, хорошо освелвнных место-
обитаниях, но обычен и при небольшом затенении - при освещенно-
сти 50-60% (третья ступень шкалы Ландольта). Растения, развива-
ющиеся под пологом леса (освещенность 10-40%), отличаются мень-
шей биомассой; количество цветков и плодов уменьшается у них в
2-5 раз в сравнении с особями луговых ценозов.
Василистник малый произрастает на слабокислых, нейтральных
и слабощелочных почвах: pH 4,5-8,0 (третья ступень шкалы Лан-
дольта) , со средним содержанием гумуса (третья ступень шкалы
Ландольта); встречается такке и на бедных азотом песчаных, хо-
рошо аэрируемых почвах (вторая ступень шкалы Ландольта) и, по
нашим наблюдениям (Барыкина, Луферов, 1987), нередко выступает
в качестве кальцефила, произрастания на выходах известняка. Эк-
спериментальные последования, проведенные К.Эмонтсом (Emonta,
1956), показали,однако, что слабокислые почвы (рй 5,0-6,0) ока-
зывали более благоприятное воздействие на развитие растений,чем
нейтральные.
В Московской обл. василистник малый произрастает на сухо-
дольных лугах, приречных террасах среди кустарников, изредка на
заливаемой части пойменных лугов, обычен на лесных опушках, по-
лянах, в разреженных мелколиственных и смешанных лесах, на дер-
ново-подзолистых или луговых с погребенным подзолистым горизон-
том почвах, иногда при сильном задервении, а также на мелкозе-
мах, песчаных и глинистых склонах оврагов. Входит преимуществен-
но в состав разнотравно-злаковых и бобово-разнотравно-злаковых
ассоциаций, в которых нередко играет роль эдификатора.
Консортивные связи
Василистник малый опыляется преимущественно ветром, реже
насекомыми ( Datunann, 1969). В корнях обнаружена эндотрофная
микориза (Крюгер, 1961).
98
| Биологическая продуктивность
Б разных фитоценозах Подмосковья продуктивность надземной
• и подземной фитомассы василиотника малого в зависимости от эко-
' лого-фитоцэнотических условий среди различается довольно суще-
>' ственно (табл, у ). Наибольшей продуктивностью отличаются попу-
i- ляции опушечно-лесных и луговых местообитаний с достаточным ув-
лажнением и хорошей инсоляцией (участки № 1,2,3), При сильном
затенении (участок $ 4) и на пастбищах, используемых для выпаса
। крупного рогатого скота (участок № -5) фитомасса очень мала. При
i сушки надземных побегов в качестве лекарственного сырья их вес
* уменьшается в 3-4 раза (табл, 4 ).
j Таблица 4
Биологическая продуктивность василистика малого
' в различных фитоценозах
Но- мер уча- стка Название фитоценоза Вес сырой массы, г на I г Вес воздушно-сухой массы, г на I и2
надзем- ных органов подзем- ММу органов надзем- ных органов подземных органов
I Березняк орляково-раз- нотравный 370 82 95 27 2 Пойменный луг (1-я терраса) 314 78 87 25 3 Разнотравный суходольный луг 287 71 69 22 4 Широколиствен- ный лес с пре- обладанием ясеня и клена 107 34 28 12 5 ЗЛаково-разно- травный сухо- дольный луг - пастбище круп- ного рогатого скота 74 29 21 9 / / Хозяйственное значение /' Растение широко используется в народной медицине как бак-
99
торицидное (Дроботько, Айзенман, Швайгер и др.,1958; Гусева,
1961), успокаивающее, мочегонное, слабительное, раноэаживляющее
средство (Гром, 1965; Кучеров, Гуфранова, 1968; Макаров, 1974).
Клинические исследования показали возможность использования
препаратов из василистника малого для лечения сердечно-сооудис-
тых заболеваний (Овсепян, 1954), а такке в акушерстве и гинеко-
логии (Баипеев, Медведева, Бернацкий и др.,1966), в онкологиче-
ской практике (Евстатиева, 1982). Трава (надземные побеги с
цветками и зелеными плодами) входит в состав сбора Здренко,при-
меняемого для лечения папилломатоза мочевого пузыря и анацидных
гастритов. Сырье собирают в фазу бутонизации - цветения. В этот
период, по данным Т.Н.Кагарлицкой и В.А.Чижевсксй (1982), на-
капливается максимальное количество алкалоидов - основных био-
логически активных веществ василистника малого. Изучение дина-
мики восстановления биомассы его надземных побегов при разном
режиме заготовок показало, что на каждые 10 м2 необходимо ос-
тавлять несрезанными 1-2 хорошо развитых растения для обсемене-
ния. Заготовки на одних и тех же массивах можно проводить не
чаще одного раза в 3 года. Верхнюю часть цветоносных побегов
длиной 30-40 (до 60) см и все зеленые листья срезают серпом,
секатором или ножом (Луферов, 1987; Луферов, Барыкина, Акопов
и др., 1988).
Василистник малый хорошо поедается сельскохозяйственными и
промысловыми животными (Работнов, 1951). После сенокошения цве-
тоносных побегов не образуется, а формируется небольшая розетка
листьев, в связи с пробуждением почек возобновления, а также
боковых почек при основании надземного побега (Лихачев, 1959).
В культуру василиотник малый предложено ввести как декоративное
растение (Полетико, Мищенкова, 1967).
3. ВАСИЛИСТНИК ПРОСТОЙ
Номенклатура и географическое распространение
Thalictrum simplex L., 1767, Mant. 1:78; Кауфм., 1866,
Моск.фл.,изд. 1:19; 1889, изд. 2:6; Цингер, 1886, Сб.свед.: 31;
Шмальг., 1895, Фл. 1:6; Сырейщ., 1907, Илл.фи. 2:145; он же,
1927, Опр.раст.Моск.губ.: 132; Невский, 1937, Фл.СССР 7:526; он
же, 1940, в Маевский, Фл.,изд. 7:373; Голицын, 1954, там же,изд.
ТОО
8:86; Маценко, 1964, там же, изд. 9:88; Tutin, 1964, Fl. Ear.
1:242; Ворошилов, 1966, Опр.раст.Моск.обл.; 170; Новиков,1986;
Опр.раст.Метеры 1:183. - Василистник простой (семейство Ranun-
ctilaceae - лютиковые).
Василистник простой - евразиатский вид, распространенный
в Средней и Южной Европе, на юге Скандинавии и Финляндии, на
западе Малой Азии, в Китае, Монголии. На территории СССР север-
ная граница ареала проходит по югу Мурманской обл., берегам
Белого моря, достигая 67°с.ш.; восточнее - до низовий рек Ме-
зень, Цильма (г.Усть-Цильма), заходит в Заполярье (г.Нарьян-
Мар, верхнее течение р.Колвы, г.Воркута), достигает низовий
Оби и простирается по линии Новый Уренгой - Туруханск - долина
р.Нижняя Тунгуска - р.Вилюй - г.Якутск - г.Магадан (на Дальнем
Востоке редок). Затем граница ареала резко поворачивает на югб-
запад и приблизительно совпадает с восточной границей Якутии и
Читинской обл. Вид довольно обычен в горах Южной Сибири, встре-
чается в лесном и лесостепном поясе гор Киргизии и Узбекистана.
В Казахстане произрастает в северных и восточных (преимущест-
венно в горах) областях. Южная граница ареала проходит по линии
Караганда - Актюбинск - Астрахань, затем огибает сухие степи
Калмыкии и поворачивает на юг до Ставрополья. Вид встречается
на Северном Кавказе (западнее г.Орджоникидзе), в Грузии, Арме-
нии, на западе Азербайджана. На европейской части СССР южная
граница ареала проход от Ростова-на-Дону до г.Днепропетровска
и Кишинева, достигая Румынии.
Морфологическое описание
Василистник простой - вегетативно подвижный длиннокорне-
вищный геофит с полурозеточными моноциклическими монокарпичес-
кими побегами и диморфными шнуровидными придаточными корнями.
Стебли прямые, высотой 60-150 см, ребристые с дважды и трижды
тройчатосложными листьями, верхние из которых прижаты к отеблю.
Листочки продолговато-обратнояйцезидные, с округлым или клино-
видным основанием, рассеченные на 3 цельнокрайние или зубчатые
доли, реже цельные, ланцетные, у верхних листьев линейные. Со-
цветие - продолговатая метелка с направленными вверх ветвями.
Цветки преимущественно повислые. Листочки околоцветника бледно-
желтые, белые, зеленовато-розовые. Пестиков 5-10. Еыльце стрело-
7х-1041 101
видное, отреловйдно-широкотреугольное. Орешки сидячие, длиной
2-3 мм, шириной около I мм, яйцевидные, эллиптические, с прямым
или слабо изогнутым стилодием длиной до 0,5 мм.
2п » 56 ( Langlet, 1927; Бочанцева, 1972), 56, 70 ( Kuhn,
1928).
Онтогенез
Латентный период. Семена прорастают на следующий год после
диссеминации, обычно в мае-июне, иногда в июле. В'лаборатории их
всхожесть сохраняется в течение 4-8 лет. Зародыш по своей вели-
чине близок к зародышу предыдущего вида.
Виргинильный период. Проростки и воходы очень сходны с
растениями этих же возрастных состояний васдаистника малого.
У ювенильных ( двулетних) растений василистника простого
развивается полурозеточный побег высотой 5-15 см, имеющий наряду
о низовыми чешуевидными 2-5 тройчатосложных листьев (один-три из
них - прикорневые). В основании ассимилирующих листьев формиру-
ется короткое влагалище и прилистники. Листочки обратнояйпевид-
ные, продолговато-обратнояйцевидные или эллиптические, 3-5-эуб-
чатые, в основании округлые или клиновидные, с верхней отороны
блестящие. В базальной части надземного побега образуются много-
численные придаточные корни. Одни из них запасающие, дайной 8-15
см, толщиной 2-4 мм, слабо ребристые, лимонно-желтые, разветвлен-
ные до 3-го порядка, живущие около 2 лет; другие - поглощающие,
более короткие (5-10 см) и тонкие (1-2 мм толщ), о круглые, белые,
позднее светло-коричневые, неветвящиеся или слабо ветвящиеся,
функционирующие в течение одного вегетационного периода.
К концу лета второго года жизни укороченная часть надземно-
го побега погружается в почву, становясь годичным приростом эпи-
геогеиного корневища. Одна из его пазушных почек, находясь на
глубине 1-3 см, заметно увеличивается в размерах и становится
почкой возобновления. Главный корень, большая часть гипокотиля,
а также надземный побег осенью отмирают.
Имматурные (3-4-летние) растения характеризуются симподиаль-
ным возобновлением побега, наличием короткого (1-2 ом) гипогео-
генного корневища, формирующегося на смену зпигесгенному. Надзем-
ный побег полурозеточный, высотой 10-30 ом, с 3-5 тройчатосложны-
ми, дважды тройчато- или перистосложными листьями, из них 1-2
102
причгрневых. Зпигеогенное- корневище у 3-летних особей осенью
отмирс
Взрослым вегетативным (5-7-летним) растениям василиотника
простого свойственны длинные гипогеогенные корневвда, ветвящие-
ся моно-, ди- или плейохазиально. Годичные приросты корневища
длиной 5-25 см, толщиной 0,3-0,4 см; в некоторых популяциях
длиной до 3 см, толщиной 0,5-1,2 см. Фотофильные побеги полуро-
зеточные, высотой 15-60 см, с 4-6 дважды перистосложными ли-
стьями. Из них прикорневые (1-2) уже в июлв-августе отмирают.
Пазушине почки (1-4), формирующиеся в базальной части побега,в
течение вегетационного периода дают начало новым гипогеогенным
корневищем. Терминальные почки последних ветвятся до 2-3 поряд-
ка и зимуют на глубине 1-7 см. На следующий год каждая из них
развивается в ортотропный фотофильный побег, а их дочерние поч-
ки образуют очередные приросты разветвленного корневища; внуча-
тые же почки увеличиваются в размерах и только на третий год
после заложения развиваются в надземные побеги.
Генеративный период и сезонное развитие
Первое цветение в естественных ценозах наблюдается обычно
на 6-7-м году развития (Луферов, 1984), в культуре - на 2-м го-
ду (Ворошилов и др.,1961). Заложение соцветий в почках возобно-
вления происходит в сентябре-октябре или в апреле-мае следующе-
го года. Цветоносные побеги полиметамерные, до соцветия несут
5-II фотофильных листьев. Вегетативная часть надземных побегов
обычно неразветвленная. В области соцветия ветвление достигает
2-5 порядков. Годичные приросты гипогеогенных корневищ (длиной
5-50 см, толщиной 0,3-0,5 см) сохраняются живыми 3-6 лет, ино-
гда до 10. Почки возобновления зимуют на глубине 2-15 см. Те
из них, которые располагаются близко к поверхности почвы, дают
начало более коротким корневищам. Наряду с длиннокорневищными
растениями могут встретиться короткокорневищные. Последние от-
мечались нами на плотных почвах с низкой влажностью, на пастби-
щах. Размножение растений осуществляется семенами и частями
корневищ.
Экология и фитоценология
Василистник простой - мезофит. Произрастает обычно по сы-
103
ронатым лугам и кустарникам, у болот, реж^ по сухим открытым
мостам. В сырых тенистых местах это - бледно-зеленое растение
о тонкими листьями и желтоватыми цветками в ‘соцветиях, в сухих
ярко-зеленое с красновато-зеленоватыми цветками. Успешно разви-
вается на открытых хорошо освещенных участках и в полутени, но !
в целом характеризуется как светолюбивый вид. На территорий
Звенигородской биостанции встречается при относительной освещен-;
ности 65-80%. Приурочен к дерново-перегнойно-глеевым легкосуг- ;
линистым и дерново-слабоподзолистым супесчаным, хорошо аэрируе-
мым почвам, бедных минеральным азотом. Однако Наиболее мощные
экземпляры развиваются на почвах богатых гумусом. Встречается
на слабокислых (pH 4,5-7,5), иногда кислых почвах.
На территории Звенигородской' биостанции тяготеет к южным
склонам оврагов, с дерново-слабооглеенными легкосуглинистыми
почвами, с pH в гор. Ао - 5,5; в rop.Aj - 6,6. Влажность в го-
ризонтах Ао - 18%; Aj - 82%;'количество катионов Са^+ и Mg+
в гор. Ао - Са2+ - 14,4, Mg+- 1,8, в гор. Aj - Са^+ - 1,8,
Mg+ 0,5 мг/экв на 100 г почвы.
В Подмосковье василистник простой характерен для лугов,
опушек, березово-разнотравных ельников, елово-березово-разно-
травных сосняков, нередок на склонах оврагов. По нашим данным
на I встречается от 80 до 120 особей различных возрастных
групп.
В табл.5 показаны возрастные группы на 4-х участках пло-
щадью в юго-восточного о клона Мелеевского оврага, ориенти-
рованных поперек от бровки (участок I) к днищу (участок 4).
Таблица 5
Возрастной состав ценопопуляций василиотника простого на юго-во-
сточном склоне Мелеевского оврага (Звенигородская биостанция)
Возрастные группы растений Участок Я I Участок № 2 Участок № 3 Участок № 4
Ювенильные 29 2"! 15 4
Имматурные 27 26 33 21
Взрослые веге- тативные 29 24 19 52
Генеративные 30 20 15 4
104
Василистник простой в фитоценозах чаще всего произрастает
со следующими ввдами растений: купырем лесным, геранью лесной,
таволгой вязолистной, зверобоем продырявленным, снытью обыкно-
венной, ежой сборной, овсянницей луговой и др.
Биологическая продуктивность
Биомасса василиотника простого с различных участков на скло
не оврага на территории Звенигородской биостанции приведена в
табл.6.
Таблица 6
Биологическая продуктивность василистика простого на
юго-восточном склоне Мелеевского оврага
Номер уча- стка Вес сырой массы, г на I мг Вес воздушно-сухой массы, г на I Плотность особей 9 на I м^
надземных частей подземных частей надземных частей подземных частей
I 1450 350 550 182 11,5
2 651 186 340 60 8,1
3 499 134 240 49 6,4
4 864 240 460 85 9,0
5 694 208 340 78 <Г 8,2
Как видно из таблицы, величина надземной и подземной био-
массы василиотника простого различается незначительно, -исключе-
ние составляет 1-й участок. Отмеченная здесь высокая продуктив-
ность связана с лучшей освещенностью и хорошей аэрацией почвы.
Консортивные связи
Василистник простой - энтомофил, хотя возможно и его ветро-
опыление ( Daumann, 1969). Опылителями являются мухи (свм. Musi-
dae ), жуки (сем. Scarabaeidae, Mordellidae, Curculiodae, Cera-
mbicidae).
В корнях обнаружена эндотрофная микориза (Крюгер, 1961).
Хозяйственное значение
Василиотник простой применяется в народной медицине при
кожных и желудочно-кишечных заболеваниях, как ранозаживляюшре
105
И антибактериальное средство (Сацыперова, 1952; Макаров, 1974;
и др.).
. 4,. ВАСИЛИСТНИК КВЛТЫЙ
Номенклатура и географическое распространение
Thalictrum flavtun J,., 1753; Sp. pl. 1:546; Кауфм., 1866,
Моск.фл.,изд. 1:20; 1889, изд. 2:8; Цингер, 1886, Сборн.свед.:32;
Шмальг., 1895, Фл. 1:6; Сырейщ., 1907, Илл.фл. 2:146; он же,
1927, Опр.раст.Моск.губ.: 132; Невский, 1937, Фл.СССР 7:527; он
же, 1940, в Маевский, Фл., изд. 7;373; Голицын, 1954, там же,
изд. 8:86; Маценко, 1964, там же, изд. 9:87; Tirtin, 1964, Fl.
Eur. I: 242; Ворошилов,. 1966, Опр.раст.Моск.обл. :170; Новиков,
1986, Опр.раст.Мещеры, 1:183. - Василистник желтый (семейство Ra-
nunculaceae — лютиковые)..
Ареал вида - европейско-западносибирский, охватывает Южную
и Среднюю Европу, юг Фенйоскандии. На территории СССР северная
граница распространения василистника желтого проходит по югу
Мурманской обл., берегам Белого моря до низовий Печоры и г.Вор-
куты. Восточная граница ареала простирается по долине р.Обь до
г.Омска, затем круто поворачивает на запад и достигает Магнито-
горска, Саратова, Волгограда, Днепропетровска, Кишинева и грани-
цы с Румынией.
Морфологическое описание
Длиннокорневищный геофит с полурозеточными моноцикличесни-
ми монокарпическими побегами и диморфными шнуровидными придаточ-
ными корнями. Корневища длиной 15-40 (60) см, с многочисленными
придаточными корнями желтого цвета. Стебли прямые, выоотой 60-
150 ( 200) см бороздачатые. Листья широкотреугольные, дайной 10-
20 см, шириной 7-18 см, дважды, трижды тройчатосложные или два-
жды, триады перистосложные. Листочки длиной 2-5 см, шириной
1-3 см, продолговато-обратнояйцевидные, эллиптические, сверху
зеленые, матовые, снизу светлее. Метелка щитковидная или широко-
пирамцдальная, многоцветковая. Цветоножки короткие, прямые или
слегка изогнутые. Околоцветник и тычиночные нити бледно-желтые.
Тычинки прямостоячие. Орешки яйцевидные, длиной 2-3 мм, шириной
1-1,5 мм, с почти прямым стилодием. Диплоидный набор хромосом
2п = 28 ( Gregory, IS4I; Langlet, 1927; Бочанцева, 1972).
106
Онтогенез, сезонное развитие, спосооы размножения
Большой жизненный цикл и сезонный ритм вегетации василист-
ника желтого протекают сходно 'с ранее описанными у василистника
простого. Отмечаются лишь незначительные количественные видовые
различия. Размножение и расселение осуществляется семенами
(табл.Г), а такке частями корневищ с почками возобновления.
Экология и фитоценология
Василистник желтый - полутеневое растение, произрастающее
при относительной освещенности более 10% (третья ступень шкалы
Ландольта). Встречается на влажных и сырых почвах (четвертая
ступень шкалы Ландольта), среднебедных и среднебогатых (третья
ступень шкалы Ландольта), щелочных (pH от 5,5 до 8,0 - четвертая
ступень шкалы Ландольта), богатых гумусом, но корни не достигают
минерального грунта (четвертая ступень шкалы Ландольта). Предпо-
читает глинистые, торфянистые, обычно водонепроницаемые, плохо
аэрируемые почвы (пятая ступень шкалы Ландольта). Характеризует-
ся широкой экологической амплитудой. Встречается в зарослях кус-
тарников, по берегам рек и речек, на заболоченных или сырых силь-
нозадернованных лугах.
В Московской обл. произрастает у низинных болот на опушках
заболоченных лесов, в ольшаниках, в зарослях кустарников, на
сильнозадернованных влажных лугах, на сильнооглеенных суглинис-
тых почвах с pH 5,7-7,8.
Хозяйственное значение
Василистник желтый применяется в народной медицине анало-
гично василистнику простому, может быть использован для корма
сельскохозяйственных животных (Работнов, 1951) представляет ин-
терес как декоративное (Полетико, Мищенкова, 1967) и медоносное .
растение (Пельменев, Рудянская, 1967).
5. ВАСИЛИСТНИК БЛЕСТЯЩИЙ
Номенклатура и географическое распространение
Thalictrum lucidum X., 1753, sp.pl. 1:546; Голицын, 1954,
в Маевский, Фл.,иэд. 8:86; Маценко, 1964, там же, изд. 9:87;
Tutin, 1964; Fl. Eur. 1:242; Ворошилов, 1966; Опр.раст.Моск.
107
обл.; 170; Новиков, 1986, Опр.раст.Мещеры, 1:183. - Th. angusti-
follum auot. fl. Ross., non L.; Кауфм., 1866, Моск.фл.изд.1:19;
1889, изд. 2:7; Дингер, 1885, Сб.свед.: 31; Шмальг., 1895, Фл.
1:6; Сырейщ.,1907, Илл.фл. 2:146; он же, 1927, Опр.раст.Моск.
губ.: 132; Невский, 1937, Фл.СССР 7:527, он же, 1940, в Маев-
ский, Фл.., изд. 7:373. - Василистник блестящий (оемейство Ra-
nunculaceae — ЛЮТИКОВЫе).
Василистник блестящий - бореальный европейский вид (Рысина,
1973), произрастающий преимущественно на территории Средней Ев-
ропы. На юге Европы и в.южной части Фенноскандии встречается
очень редко, возможно, как заносное раотение. В СССР граница
ареала проходит по линии: Петрозаводск - Котлас - Пермь - Ка-
зань - Саратов - Волгоград - Днепропетровск - Кишинев. Как за-
носное обнаружен северо-восточнее Архангельска (в бассейне р.Ку-
лой), на юге Урала, в Магадане и в Крыму.
Морфологическое списание
Короткокорневищный геофит с безрозеточными моноциклически-
ми монокарпическими побегами и диморфными шнуроввдными прида-
точными корнями. Корневища длиной 2-4 (6) см, слабоветвящиеся.
Стебли высотой 50-200 (250) см, прямые, ребристые. Листья тре-
угольные, 2-4-перистослажные или 2-4-тройчатосложные. Листочки
длиной 2-4 ом, шириной до 0,5 (0,8) см, сверху темно-зеленые,
блестящие, снизу светло-зеленые, матовые, о завернутыми вниз
краями. Метелка пирамидальная, щитковидная, нередко раскидис-
тая, многоцветковая. Листочки околоцветника и тычиночные нити
бледно-желтые. Орешки дайной 1-2 (2,5) мм, шириной 1-1,3 мм, с
коротким, почти прямым стилодием.
Диплоидный набор хромосом 2n=28 ( Langlet, 1927; Kuhn,
1928; Бочанцева, 1972).
Онтогенез
Латентный период. После опадения плодиков с материнского
растения семена находятся в состоянии покоя до мая следующего
года. Иногда отмечается появление единичных проростков уже в
год диссеминации (Луферов, 1984), однако они очень слабые и с
наступлением заморозков обычно отмирают. В лабораторных услови-
ях семена василистника блестящего прорастают заметно раньше,
чем семяна других видов, и обладают довольно высокой всхожестью
108
(табл.I); в естественных ценозах она может достигать 90%, но в
отдельные года - всего 20-30%. По наблюдениям Г.П.Рысиной
(1973), из общего количества опавших семян весной будущего года
проросло 30-40%, на втором году - большая их часть, на третьем
- прорастание было единичным.
Зародыш ваоилистника блестящего в отличии от других видов
заметно меньшей величины (дайна его 150-170 мкм, ширина 75-95
мим).
Виргинильный период. Прорастание семян и развитие проро-
стков происходит, как и у других видов ваоилистника. К числу
его видовых отличий следует отнести образование относительно
мелких семядолей с овальными пластинками дайной 3-4 мм, шириной
2,5-3 мм, формирование надземного побега начинается на 7-10-й
день после прорастания и уже у всходов он становится полурозе-
точным, достигает длины 12 см, несет 2-4 тройчатосложных ли-
стьев, из них 1-3 - в прикорневой розетке. Листочки округлые
или широкосбратнояйцевидные, трехзубчатые. Семядоли в июне-
августе желтеют и отмирают. Гипокотиль в течение лета втягива-
ется в почву, утолщается (до 3 мм в диаметре), меняет свою ок-
раску от лимонно-желтой до коричневой, становится поперечно-
морщинистым; на нем образуются 4-8 слабо разветвленных прида-
точных корней.
Ювенильный и имматурный этапы развития у ваоилистника бле-
стящего не выражены, уже двулетние особи принимают облик взрос-
лых вегетативных растений. Надземный побег, развивающийся на
2-м году из почки семядольного узла, безрозеточный, с 3-6 чешу-
ями в основании и 3-5 дважды или трижды тройчато-, дважды или
триады перистосложными листьями. Листочки нижнего ассимилирующе-
го листа обратнояйцевидные или округлые, 3-5-зубчатые, в осно-
вании округло-клиновидные; у остальных листьев они продолгова-
то-яйцевидные, узкоэллиптические, линейные, сверху блестящие,с
несколько завернутыми на нижнюю сторону краями. В разветвлени-
ях черешка иногда развиваются ушковидные или неправильной фор-
мы прилистнички. Одновременно с ростом надземного побега одна-
две его базальных пазушных почек образуют короткое (дайной 1-4
см, шириной 0,4-1 см) гипогеогенное корневище. Терминальная
почка последнего на следующий год дает начало надземному побе-
гу. Главный корень отмирает на 2-м году онтогенеза в июле-авгу-
109
сто, гипокотиль - весной 2-го года. Придаточные корни мпогочж-
леш1ыо,д11мор$ные, морфологически сходные с таковыми у васалист-
ника простого.
Генеративный период и сезонное развитие
Генеративный период в остоствекных ценозах наступает на
4-7 году онтогенеза (Луферов, 1984), в культуре - на 2-м
(Скворцов, Трулевич, Алферова и др., 1979). У молодых генератив-
ных особей образуются обычно 1-2 цветоноса, у взрослых генера-
тивных их число может достигать 4 (7). Весеннее отрастание фото-
фильннх побегов начинается в конце апреля. Начало цветения при-
ходится на конец мая-начало июня, а завершается оно в конце июня
-начале июля. Плодоношение - в августе-сентябре. Отмирание над-
земной массы растений наблюдается в конце сентября-октябре. За-
ложение соцветий в почках возобновления происходит и сентябре -
октябре; в ото время они содержат 6-12 метамеров :: располагают-
ся под землей на глубине 1,5-8 ом; иногда "внутрипочечная" фаза
формирования соцветий отмечается только в апреле-мае следующего
года. Размножение и расселение осуществляется семенами и частя-
ми корневищ с почками возобновления.
Экология и фитоценология
Василистник блестящий произрастает при относительной ос-
вещенности более 10% (третья ступень шкалы Ландольта). Естроча-
ется на глинистых, торфянистых, обычно водонепроницаемых, плохо
аэрируемых почвах (пятая ступень шкалы Ландольта), от средне-
бедных до среднебогатых (третья ступень шкалы Ландольта), щелоч-
ных с pH 5,5-8,0 (четвертая ступень шкалы Ландольта). Б Москов-
ской обл. предпочитает задернованные слабооглеонные почвы сухо-
долов и пойменных лугов. Также распространен в заболоченных мел-
колиственных лесах, на заболоченных полянах, на сыроватых лугах.
Консортивные связи
Б корнях василиотника блестящего обнаружена эндотрофная
микориза арбускулярно-визикулярного типа. Опыляется василистник
блестящий как насекомыми, так и ветром. Цветки посещаются теми
же видами насекомых, что и у василиотника водосборолистного.
ИО
Еиологичеекая продуктивность
О
Б Подмосковье на суходольном лугу на I м произрастает до
9 разновозрастных особей. Фитомасса (сырой вес) надземных орга-
нов достигает 885 г/м2, подземных - 640 г/м2.
Хозяйственное значение
Василистник блестящий представляет интерес как лекарствен-
ное растение: его надземные побеги содержат вещества, обладаю-
щие бактерицидными и фунгицидный! свойствами (Сацыперова, 1952,
1953), а так же как декоративное в зеленом строительстве (Гей-
деман И др., 1962; Killer, 1989).
СОДЕРЖАНИЕ
Предисловие ..............................................
Страусник обыкновенный.М.В.Нехлюдова, В.Р.Филин...........
Лилия кудреватая.Э.П.Немченко ........................... .3;
Мякотница однолиотная:М.Г.Вахрамеева, Т.М.Ейченко, И.В.Та-
таренко, М.В.Экзерцева .................................... Ф
Кокушник комарниковый. М.Г.Вахрамеева, И.0.Виноградова,
И.В Татар ;нко, О.В.Цепляева............................. 3'1
Мышетарсттяк маленький. Х.В.Муоина, Р.П.Барыкина,
Н.В.Чубатова .................................;........... 6!
Лютик ползучий. В.И.Полуянова ............................ 7^
Род Василистник. А.Н.Луферов, Р.П.Барыкина,Т.В.Багдасарова. в:
Научное издание
Биологическая флора Московской области
Выл. 9. Часть I.
зав. редакцией Н.М. Глазкова
Редактор Э.И. Кране
Художественный редактор Ю.М. Добрянская
Техн.редактор Н.И. Смирнова
Н/К
Подписано в печать26.II.92.
Формат 60x90 I/I6 . Бумага тип. №2.
Офсетная печать.
Усл. печ. л. 7,0. Уч.-изд. л. 5,86.
Тираж 1000 экз. Заказное. Изд. № 2186.
Ордена "Знак Почета" издательство. Московского университета.
103009, Москва, ул. Герцена, 5/7.
Типография ордена "Знак Почета" изд-ва МГУ.
II9899, Москва, Ленинские горы