/













































































































































































































































































































Text
Проф. И. И. Полянский
(1872—1930)
Проф. И. И. ПОЛЯНСКИЙ
СЕЗОННЫЕ ЯВЛЕНИЯ
В ПРИРОДЕ
ПОСОБИЕ ДЛЯ УЧИТЕЛЕЙ
Издание 6-е
(2-е посмертное)
Под редакцией В. И. ПОЛЯНСКОГО
и Ю. И. ПОЛЯНСКОГО
ГОСУДАРСТВЕННОЕ УЧЕБНО-ПЕДАГОГИЧЕСКОЕ ИЗДАТЕЛЬСТВО
МИНИСТЕРСТВА ПРОСВЕЩЕНИЯ РСФСР
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
Ленинград 1956
Иван Иванович Полянский
СЕЗОННЫЕ ЯВЛЕНИЯ В ПРИРОДЕ
Редактор В. В. Аникиев
Худож. редактор В. Б. Михневич
Техн, редактор Л. А. Леонтьева
Корректор А. А. Морозова
Сдано в набор 13/VII 1956 г. Подписано к печати 19/Х 1956 г. М-23830. Формат бумаги 60х92!/1в
Печ. л. 18,54-0,125 л. вклейка. Уч.-изд. л. 18,7-|-0,04 л. вклейка. Тираж 20 000 экз.
Цена без переплета 5 р. 10 к.
Переплет коленкоровый 1 р. 50 к.
Ленинградское отделение Учпедгиза
Ленинград, Невский пр., 28. Заказ № 1252
Типография № 2 Управления культуры Ленгорисполкома
Ленинград, Социалистическая, 14
ОТ РЕДАКЦИИ
Первое издание «Сезонных явлений в природе» проф.
И. И. Полянского вышло в 1910 г., почти пол века назад.
Автор этой книги — видный методист-естественник и один из
пионеров в деле разработки экскурсионного метода в преподава-
нии естествознания, по книгам которого учились и учили многие
педагоги советской средней школы.
И. И. Полянский с 1902 по 1930 г. осуществлял большую
работу по подготовке учителей естествознания и по пропаганде
экскурсионного дела. Им было организовано в разных районах
СССР несколько десятков курсов для учителей-биологов средней
школы. Особенно широко эта работа развернулась после Великой
Октябрьской социалистической революции. С 1919 года И. И. По-
лянский возглавлял работу по организации экскурсионных стан-
ций вокруг Ленинграда, сыгравших важную роль в деле приоб-
щения учителей средней школы к изучению природы родного
края. В течение многих лет И. И. Полянский состоял профессо-
ром ряда высших учебных заведений Ленинграда. Его перу при-
надлежит ряд книг и статей по вопросам общей биологии и ме-
тодики естествознания.
Настоящий труд относится к числу наиболее популярных про-
изведений автора, о чем наглядно свидетельствует факт второго
посмертного издания его, выходящего в свет спустя более чем
четверть века после смерти автора (умер в 1930 г.).
Со времени выхода первого посмертного издания «Сезонных
явлений в природе» (1937 г.) был опубликован ряд популярных ра-
бот, затрагивающих ту же тему (Н. С. Щербиновский, Се-
зонные явления в природе, 3-е изд., Сельхозгиз, 1954; А. В. Ко-
жевников, Весна и осень в жизни растений, 2-е изд. Моск.
4
общ. испыт. природы, 1950; Н. В. Попов, Фенологические на-
блюдения в школе, Учпедгиз, 2-е изд., 1953, и др.). Однако эти
весьма ценные пособия преследуют несколько иные цели и от-
нюдь не лишают значения настоящую работу, весьма полно охва-
тывающую данную проблему и специально предназначенную для
учителей.
Настоящее издание просмотрено и дополнено в соответствии
с новыми научными данными, накопившимися в литературе за по-
следние годы. Подготовка к печати ботанических глав проведена
В. И. Полянским, зоологических — Ю. И. Полянским. Большую
помощь при этом оказали С. В. Герд, О. В. Заленский, Г. А. Но-
виков, С. И. Радченко, Ал. А. Федоров и Г. Э. Шульц.
Особой благодарностью редакторы обязаны Г. Э. Шульцу,
предоставившему в их распоряжение ряд материалов, особенно по
главам «Осенняя листва и листопад» и «Зимовье растений».
В книгу введен ряд новых рисунков; все прежние рисунки
перерисованы заново.
Введение
Одним из наиболее ярких явлений в окружающей нас при-
роде является смена времен года. Мы, однако, настолько привыкли
к тому, что за зимою следует весна, а затем лето и осень, что часто
не обращаем должного внимания на грандиозные изменения, ко-
торые совершаются при этом вокруг нас. Эти изменения не исчер-
пываются только колебаниями температуры, количеством осад-
ков, направлением ветров и т. п. метеорологическими явлениями.
Они затрагивают и мир живых существ. При изучении растений и
животных мы наблюдаем множество явлений, носящих сезонный
характер. Достаточно вспомнить, например, весеннее пробуждение
растений, их цветение, листопад, перелеты птиц и т. п., чтобы пред-
ставить себе, насколько значительны эти явления и как велика их
роль для человека.
Сезонные явления в природе протекают, конечно, неодинаково
в разных климатических областях земного шара. Но в той или иной
форме мы встречаемся с ними повсюду — от снежных пустынь
Арктики до девственных тропических лесов.
В данной книге имеются в виду главным образом сезонные явле-
ния, наблюдаемые в условиях лесной и степной зон Европейской
части СССР. Однако в ряде случаев приводятся примеры, касаю-
щиеся также и других районов — как более северных, так и юж-
ных. Это дает возможность лучше уяснить общее значение тех
или иных приспособлений растений и животных к меняющимся
условиям внешней среды.
Прежде чем обратиться к отдельным сторонам этих явлений,
необходимо остановиться на некоторых общих положениях и вопро-
сах, которые нужно иметь в виду при чтении последующих глав
книги.
Изучая сезонные изменения, происходящие в мире растений и
животных, мы постоянно наталкиваемся на биологически целесо-
образный, полезный для сохранения вида характер этих измене-
ний. Ниже мы найдем многие десятки примеров таких приспособле-
ний растений и животных, носящих сезонный характер.
6
Введение
Приспособленность организмов к окружающим условиям яв-
ляется их характерной чертой, и без нее они, очевидно, не могли
бы существовать и обеспечивать себя потомством. Приспособления
эти весьма разнообразны. Имея дело с сезонными явлениями, мы
сталкиваемся преимущественно с приспособлениями ритмического
характера, причем убеждаемся в большом разнообразии ритмов
развития применительно к разным климатическим областям.
Для правильного понимания изложенного ниже фактического
материала необходимо, однако, обратить внимание на то, что це-
лесообразные приспособления у организмов к условиям существо-
вания далеко не носят абсолютного характера.
Не существует целесообразности в строении и приспособлениях
организмов «вообще», а существуют приспособления к определен-
ным, конкретным условиям существования. Покровительственная
окраска животного, например, является целесообразной только до
тех пор, пока есть фон, под цвет которого животное окрашено. Если
этот фон почему-либо исчезнет или изменится, покровительствен-
ная окраска сразу перестанет быть полезным приспособлением.
На юге летом трава обычно выгорает, становится совершенно
желтой. При этом многие насекомые, окрашенные в зеленый цвет
(покровительственная окраска под цвет зеленой травы), сразу те-
ряют свое защитное приспособление. Целесообразное приспособ-
ление к определенным условиям существования в изменившейся
обстановке перестало быть таковым, и зеленые насекомые резко
бросаются в глаза на желтом или коричневом субстрате. И лишь
на растениях, сохранивших зеленый цвет, их окраска по-прежнему
является покровительственной.
Цветки красного клевера имеют ряд приспособлений, обеспе-
чивающих привлечение опыляющих их насекомых. К числу таких
приспособлений следует отнести окраску цветка, наличие нектара
в глубине венчика, форму венчика, облегчающую посадку насе-
комого. Однако все эти приспособления являются целесообразны-
ми, полезными лишь в присутствии совершенно определенных
насекомых, главным образом, шмелей, производящих опыление
красного клевера. Большинство других насекомых не может до-
стать нектара из цветка красного клевера и произвести опыление.
При отсутствии шмелей приспособления клевера к опылению
уже нельзя назвать целесообразными.
Если растение попадает в чуждую ему обстановку, то весьма
«совершенные» приспособления к опылению могут оказаться бес-
полезными. Так было, например, с яблонями, завезенными в Австра-
лию после ее открытия европейцами. В первое время они почти не
давали плодов, так как в Австралии отсутствовали пчелы, к опыле-
нию которыми главным образом приспособлены цветки яблони.
Таким образом, уже из этих примеров видно (а их можно привести
неограниченное количество), что мы говорим о целесообразности не
Введение
7
как об особом свойстве, присущем живому организму, а говорим
о полезных (целесообразных) в данной конкретной обстановке при-
способлениях. Понятие органической целесообразности нельзя мы-
слить оторванно от среды обитания, от конкретных условий суще-
ствования растения или животного.
С другой стороны, неправильно было бы думать, что приспособ-
ления организмов к данной среде всегда одинаково совершенны,
что организм всегда «идеально» приспособлен к условиям существо-
вания. Можно привести множество примеров относительного ха-
рактера целесообразности, «несовершенства» приспособлений. Они
указаны во многих местах этой книги, и здесь мы остановимся лишь
на некоторых.
Мы видели выше, что цветок красного клевера приспособлен
к опылению главным образом шмелями. Однако нектар цветка
клевера, служащий «приманкой» для шмелей, производящих опы-
ление, иногда поедается другими насекомыми (некоторыми жу-
ками), прогрызающими венчик и не производящими опыления.
В данном случае наличие нектара, являющееся одним из приспо-
соблений для опыления, не приводит к полезному для растений
биологическому эффекту.
Перелет птиц представляет собой, несомненно, полезное при-
способление к условиям сурового климата. У перелетных птиц
по сравнению с оседлыми мы находим особо мощное развитие ма-
ховых перьев в крыле, мускулатуры и т. п., облегчающее дли-
тельные странствования, которые им приходится предпринимать
дважды в году. Между тем, все эти приспособления далеко не обе-
спечивают безопасность перелета. Огромное количество птиц по-
гибает во время миграции. Особенно некоторые, как, например,
перепелки, обладающие относительно слабыми крыльями и тяже-
лым полетом, гибнут массами при перелете через моря.
Наличие приспособлений к перелету у птиц налицо, но эти
приспособления отнюдь не всегда можно считать совершенными,
идеальными. Целесообразность и здесь носит относительный ха-
рактер.
У животных, живущих в наших пресных водоемах, имеется
ряд приспособлений к перенесению зимних неблагоприятных усло-
вий подо льдом. К числу их относится понижение интенсивности
обмена веществ, зимнее оцепенение, прекращение приема пищи и
т. п. Тем не менее, иногда зимой значительная часть обитателей
водоема (в том числе и рыб) погибает. Имеющиеся приспособления
не всегда обеспечивают перенесение зимних условий.
Таким образом, у организмов имеются самые разнообразные
полезные приспособления к условиям существования,в частности,
и к сезонным изменениям. Однако эти приспособления оказыва-
ются относительными, они не носят абсолютного характера; орга-
низмы далеко не идеально приспособлены к условиям среды.
8
Введение
Естествоиспытатели XVII и XVIII столетий, считавшие виды
растений и животных неизменными, в явлениях целесообразности
видели обычно проявление общей гармонии природы, исходящей
от ее творца. Этой точке зрения, разделяемой по существу и сейчас
некоторыми идеалистами, был нанесен сокрушительный удар уче-
нием Чарлза Дарвина. В книге «Происхождение видов путем есте-
ственного отбора или сохранение благоприятствуемых пород в
борьбе за жизнь», вышедшей первым изданием в 1859 г., Дарвин
не только раскрыл основные закономерности эволюции видов, но
дал также естественное, материалистическое объяснение происхо-
ждению полезных приспособлений, т. е. органической целесооб-
разности.
В развитии и укреплении материалистических позиций в биоло-
гии теория Дарвина сыграла и сейчас играет огромную роль.
Мы не можем в этой книге, посвященной сезонным явлениям
в природе, давать подробный очерк дарвинизма. Это не входит
в нашу задачу. Изложение основных положений эволюционного
учения Дарвина можно найти в ряде книги прежде всего в класси-
ческой работе К. А. Тимирязева «Чарлз Дарвин и его учение».1
Здесь напомним только, что особо важное значение в эволюцион-
ном процессе Дарвин придавал естественному отбору, т. е. выжи-
ванию тех организмов, которые в силу изменчивости лучше приспо-
соблены к данным конкретным условиям существования. «Выра-
жаясь метафорически, — говорил Дарвин,—можно сказать, что есте-
ственный отбор ежедневно и ежечасно расследует по всему свету
мельчайшие изменения, отбрасывая дурные, сохраняя и слагая
хорошие, работая неслышно и невидимо, где бы и когда бы ни пред-
ставился к тому случай, над усовершенствованием каждого органи-
ческого существа в связи с условиями его жизни, органическими
и неорганическими».1 2
Естественный отбор протекает на основе изменчивости, дающей
материал для эволюционного процесса, и наследственности, закреп-
ляющей, фиксирующей этот материал. Без них эволюция, оче-
видно, была бы невозможна. Однако для того, чтобы попять, как
могло возникнуть в процессе эволюции все многообразие живых су-
ществ и их приспособленность к условиям жизни, обязательно нуж-
но учитывать третий основной фактор эволюционного процесса —
естественный отбор.
По этому поводу К. А. Тимирязев писал: «Если действие среды
может объяснить возникновение измененной формы, то пока один
только дарвинизм может объяснить как из этого, по существу без-
различного, материала создались организмы, т. е. формы прилажен-
ные, приспособленные к условиям своего существования».3
1 Имеется ряд изданий.
2 Чарлз Дарвин, Соч., т. III, изд. АН СССР, 1939, стр. 330.
3 К. А. Тимирязев, Соч._, т. VI, Сельхозгиз, 1939, стр. 289.
Введение
9
Жизнь в ее современных формах есть продукт длительной
истории. Разнообразные полезные, целесообразные приспособле-
ния организмов к условиям существования также возникли истори-
чески на основе естественного отбора полезных уклонений. Таким
образом, целесообразность представляет собой не изначальное, та-
инственное, присущее организмам свойство, а возникающее в про-
цессе развития органического мира приспособление к конкретным
условиям жизни в самом широком смысле этого слова. Поэтому
она может носить только относительный, а отнюдь не абсолютный
характер.
Поскольку целесообразность в строении и отправлениях орга-
низма есть исторически возникающее качество, а не изначально
присущее им свойство, очевидно, что в разные моменты эволюции
мы должны встречаться с организмами в различной степени при-
способленными к условиям существования.
Все перелетные птицы обладают хорошо развитыми крыльями.
Однако это качество развито у разных птиц в различной степени.
Стриж обладает гораздо более совершенными, приспособленными
для длительного и дальнего полета крыльями, чем, скажем, пе-
репелка.
Естественный отбор на основе наследственной изменчивости
создает новые виды, приспособленные к тем условиям, в которых
они существуют. Эти виды дадут начало новым видам, приспособлен-
ным к тем же или новым условиям существования. Целесообраз-
ность относительна на любой ступени развития органического мира.
Эту относительность, с которой мы столь часто встречаемся в при-
роде, ясно видел Дарвин. В «Происхождении видов» эти мысли
выражены им с полной четкостью: «Не следует изумляться и тому,
что все приспособления в природе, насколько мы можем судить,
не абсолютно совершенны, как, например, человеческий глаз, или
что некоторые из них не соответствуют нашему представлению о
приспособленности. Нечего удивляться и тому, что жало пчелы,
направленное против врага, причиняет смерть самой пчеле; тому,
что трутни производятся в таком большом числе ради одного един-
ственного акта, а затем умерщвляются своими бесплодными' се-
страми; той изумительной трате пыльцы, которая наблюдается у
нашей сосны; той инстинктивной ненависти, которую пчелиная
матка питает к своим собственным плодовитым дочерям; тому,
что наездники питаются живым телом гусениц, и вообще ни одному
подобному случаю. Согласно теории естественного отбора скорее
представляется удивительным то, что не открыто еще большего
числа подобных случаев отсутствия абсолютного совершенства».1
Рассматривая естественный отбор как основной фактор органи-
ческой эволюции, необходимо обратить внимание еще на одно явле:
1 Чарлз Дарвин, Соч., т. III, изд. АН СССР, 1939, стр. 653.
10
Введение
ние, значение которого для понимания происхождения видов отметил
Дарвин. Речь идет о так называемых коррелятивных изменениях.
Это понятие Дарвин в «Происхождении видов» характеризует сле-
дующими словами: «Я разумею под этим выражением тот факт,
что вся организация во время роста и развития находится в такой
тесной взаимной связи, что, когда слабые изменения проявляются
в какой-нибудь одной части и накопляются естественным отбором,
другие части также претерпевают изменения».1 Каждый раститель-
ный и животный организм представляет собой исторически сложив-
шееся органическое целое. Изменение какой-либо одной части орга-
низма влечет за собой изменение и других его частей. Характер
связей между различными органами далеко еще не всегда бывает
достаточно ясен. В некоторых случаях эта связь зависит от функ-
ционального, физиологического взаимодействия различных частей
организма. Известно, например, что большинство так называемых
вторичных половых признаков у позвоночных животных тесно свя-
зано с деятельностью и степенью развития половых желез.
Дело в том, что у многих животных самец от самки отличается
не только половыми железами, но рядом других признаков, анато-
мически не имеющих ничего общего с половыми железами и их
протоками. К числу таких признаков у птиц относятся, например,
более яркое оперение самцов по сравнению с самками, наличие у
петуха более развитого, чем у кур, гребня, наличие у самца благо-
родного оленя рогов, которые отсутствуют у самки, ит. п. Все эти
и подобные им признаки и называются вторичными половыми.
Рядом опытов установлено, что развитие их зависит от особых
веществ (так называемых гормонов), которые выделяются поло-
выми железами в кровь.
В иных случаях может иметь место и другой характер связей
между разными признаками организмов. Приведем еще несколько
примеров коррелятивной изменчивости. Дарвин обращает внима-
ние на тот факт, что у растений из семейства сложноцветных в со-
цветиях краевые цветки обладают хорошо развитым венчиком (каж-
дый может в этом убедиться, рассматривая соцветие маргаритки или
поповника). При этом усиленное развитие венчика сопровождается
частичным или полным недоразвитием тычинок и пестиков. У голу-
бей оперение ног почти всегда сопровождается развитием перепо-
нок между пальцами и др.
Явление коррелятивной изменчивости имеет большое значение
для понимания эволюции организмов. Исходя из него, мы можем
понять, каким образом на основе естественного отбора могут воз-
никать и изменяться признаки, не имеющие непосредственно по-
лезного значения для сохранения вида. Если представить себе, что
в результате естественного отбора изменяется строение какого-
1 Чарлз Дар в ин, Соч., т. III, изд. АН СССР, 1939, стр. 375.
Введение
11
либо органа в направлении, полезном для организма, то это изме-
нение в силу корреляции может повлечь за собой ряд других изме-
нений, которые не будут полезными, а в отдельных случаях даже
могут оказаться вредными для сохранения вида.
Мы встречаемся в природе на каждом шагу с различными при-
способлениями к условиям существования, сложившимися в про-
цессе эволюции организмов на основе естественного отбора. Тем
не менее, не следует думать, что каждый признак, каждая особен-
ность организма имеют непосредственно приспособительное значе-
ние. Если мы сравним между собой близкие виды, например, бабо-
чек или других насекомых, то далеко не всегда возможно бывает
установить, какое значение в смысле непосредственного приспособ-
ления к условиям существования имеют те или иные различия
между видами.
В ряде случаев это может объясняться тем, что приспособитель-
ное значение данной особенности еще не вскрыто нами, хотя в дей-
ствительности оно налицо. Но несомненно, что часто этого нет и
мы имеем дело с так называемыми «индифферентными» признаками,
не приносящими организму, обладающему ими, ни вреда, ни пользы.
На первый взгляд эти факты как будто противоречат теории
естественного отбора. На самом деле имеет место как раз обратное.
Наличие у организмов «индифферентных» признаков действитель-
но говорит против признания изначальной целесообразности орга-
низмов, которую провозглашают идеалисты. Коррелятивная из-
менчивость, на которую обратил внимание Дарвин и которая с тех
пор подвергалась обстоятельному изучению, не только делает
понятным с точки зрения теории естественного отбора появление
«индифферентных» признаков, но является вместе с тем прекрасной
иллюстрацией относительности органической целесообразности.
Говоря о наличии у организмов признаков, не имеющих непо-
средственного приспособительного значения, необходимо, наконец,
иметь в виду, что при изменении условий существования полезные
приспособления могут потерять свое значение и перейти в катего-
рию «индифферентных» или даже вредных признаков.
В этой книге, посвященной сезонным явлениям в природе, мы
будем, однако, обращать главное внимание именно на приспособле-
ния организмов к условиям существования, к тем условиям, ко-
торые создаются в природе в связи со сменою времен года.
ВЕСЕННЕЕ ПРОБУЖДЕНИЕ РАСТЕНИЙ
Снежный покров еще не успел освободить землю, а уже на пер-
вых проталинах, появляющихся в полях и на лужайках, наш взор
ласкает свежая изумрудная зелень.
Что же это за растения и когда они успели выйти из земли и
образовать зеленые листочки? Это прежде всего — злаки, т. е. ра-
стения, близкие к нашим полевым хлебным растениям (принадле-
жащие к одному с ними семейству). Злаки обладают способностью
противостоять сильным зимним холодам, по миновании ко-
торых они возобновляют свой рост и развитие. Зеленые побеги
злаков, как хлебных — на озимых полях, так равно и луговых,
вышедшие из земли еще предыдущей осенью, под влиянием морозов
лишь приостановили свой рост и теперь, весной, они снова возобно-
вили его.
На проталинах, т. е. также до окончательного удаления снеж-
ного покрова, появляются и настоящие весенние растения, не да-
вавшие всходов осенью.
Одной из существенных их особенностей является весьма быстрое
развитие цветков, часто опережающее развитие листьев. У одних
ранних растений цветки достигают полного развития, а еще нет ни
одного листочка, у других цветки и листья выходят одновременно,
но последние отстают в развитии. Откуда же берется материал для
построения столь быстро появляющихся цветков?
Переработка неорганических веществ, доставляемых растению
почвой и атмосферой, в органические вещества происходит в ли-
стьях. Как известно, этот процесс, называемый фотосинтезом,
осуществляется здесь при непосредственном участии зеленого пиг-
мента хлорофилла. В результате фотосинтеза создаются необходи-
мые для построения растений вещества: сахара, белки, жиры, крах-
мал, клетчатка и др.
Ранние весенние цветки, появляющиеся до развития молодой
листвы, очевидно, не могут пользоваться веществами, в ней при-
готовленными. Они строятся за счет органических веществ, синте-
зированных растением еще в прошлом году, т. е. в прошлогодних
Особенности ранних весенних растений
13
листьях. Эти вещества были отложены в подземных органах — кор-
невищах, луковицах, клубнях и в корнях. Содержащегося в них
запаса оказывается достаточно для построения не только цветка,
но и других частей растения. Если листья являются тем местом,
в котором происходит фотосинтез, и если до их образования органи-
ческие вещества синтези-
роваться не могут, то
ясно, что и листья до того
момента, пока они сдела-
ются способными к этому,
должны развиваться за
счет запасов органических
веществ, уже имеющихся
в растении. Так обстоит
дело у всех растений, в том
числе и у развивающихся
из семян. В семенах также
отложен запас органиче-
ских веществ, за счет кото-
рых развивается зародыш и
образуются первоначально
корень и листья со стеб-
лем. Но существенная раз-
ница заключается в следу-
ющем. В семенах—запас
небольшой. Развивающие-
ся за счет его органы —
маленькие. Дальнейший
р ост идет уже за счет орга-
нических веществ, создан-
ных самим растением, и
26/lV
1
I t11
потому—относительно мед-
ленно. В подземных же ор-
ганах запас несравненно
больший. Потому здесь
Рис. 1. Рост стебля цветоносного побега
мать-и-мачехи (Tussilago farfara)
(по Серебрякову)
и возможен более стремительный рост не только цветка, но и всего
вообще растения. Этому в большой мере способствует и то
обстоятельство, что быстро развивающиеся части формируются
в подземных органах заранее и весной имеет место преимущественно
удлинение уже заложенных в них частей побега, а не их новообра-
зование (рис. 1). Ранние весенние растения в силу этого быстро раз-
виваются.
Как показали исследования высокогорных растений Памира,
характеризующихся способностью вегетировать ранней весной, су-
ществует определенная зависимость между скоростью их развития
и осенней подготовкой зимующих почек. Ранее всего (почти одно-
14
Весеннее пробуждение растений
временно с распусканием листьев) зацветают растения, у которых
в зимующей почке имеется вполне сформировавшийся цветок. На-
против, растения, у которых цветок в зимующей почке находится
в зачаточном состоянии, хотя и начинают развиваться с самой
весны, но цветут только во второй половине лета.
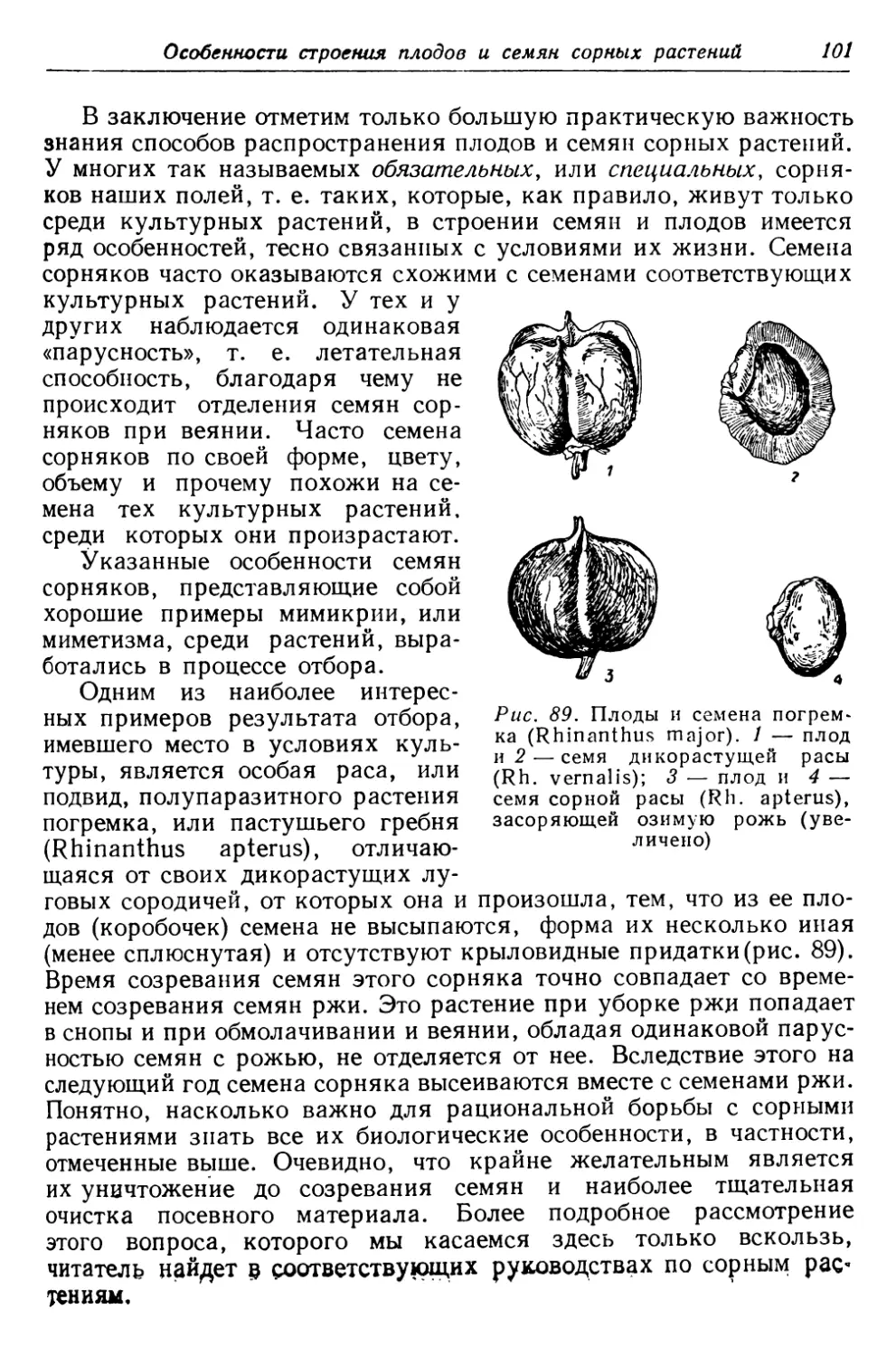
Для иллюстрации указанных особенностей ранних весенних
растений обратимся к наиболее известным примерам.
На глинистых обрывах, по канавам и т. п. повсеместно встре-
чается раннее весеннее растение мать-и-мачеха, расцветающее
еще до удаления снежного по-
крова. Цветки мать-и-мачехи
похожи на цветки одуванчика.
В обоих случаях мы имеем не
отдельный цветок, а целое со-
цветие (корзинку), т. е. собра-
ние отдельных маленьких цве-
точков. Но в то время как у
одуванчика цветочная корзинка
находится на совершенно глад-
ком стебельке, у основания ко-
торого, близ земли, имеется
целая розетка листьев, раз-
вивающихся ранее цветка, у
мать-и-мачехи мы видим дру-
гое. Здесь соцветия сидят на
стебельках, на которых вместо
листьев имеются только малень-
кие желтоватые чешуйки (рис.2).
Настоящие зеленые листья по-
являются у мать-и-мачехи поз-
же. В земле находится толстое
мясистое корневище, перепол-
ненное запасом органических
веществ, образованных прошло-
годними листьями. Это—именно
Рис. 2. Мать-и-мачеха (Tussilago
farfara)
корневище, т. е. видоизмененный подземный стебель, а не корень,
о чем говорят находящиеся на нем маленькие чешуйки. Они пред-
ставляют собой видоизмененные листья, листья же бывают только
на стеблях, но отнюдь не на корнях. Корневище с одного конца
удлиняется, с другого отмирает. Ранней весной из нескольких
почек корневища поднимаются, пробуравливая землю, цветочные
побеги. Когда эти побеги отцветут, на них образуются, подобно
одуванчику, пушистые белые головки. После их увядания из других
почек корневища вырастают новые побеги, но уже без цвет-
ков, а с большими зелеными листьями. В этих листьях будут
в течение лета синтезированы органические вещества, кото-
Отдельные представители весенней флоры
15
рые обеспечат растению возможность цветения будущей весной.
Вслед за мать-и-мачехой, главным образом в лесу, появляются
голубая (рис. 3), а вскоре за нею и белая перелеска, или ветреница
(рис.4).Ита, и другая, подобно мать-и-мачехе, обладают корневи-
щем, в котором за предыдущее
лето в значительном количестве
накопились питательные запасы.
Корневище перелесок идет не-
глубоко в земле, параллельно
поверхности, имея с нижней
стороны многочисленные корни.
Корневище понемногу растет
под землей и ветвится, но по
мере того как с одного конца
оно нарастает, старая часть
его понемногу отмирает и раз-
рушается. Образующиеся каж-
Рис. 4. Ветреница (Anemone nemorosa).
1—все растение с корневищем; 2—пе-
стики, скученные в центре цветка
(увеличено)
Рис. 3. Голубая перелеска
(Hepatica nobilis)
дой весной из нарастающего кончика корневища цветочные побеги,
очевидно, каждый год оказываются на новом месте. Следовательно,
благодаря росту корневища растение постоянно передвигается.
У белой перелески листья развиваются одновременно с цветком.
У голубой перелески рядом с цветками также видны характерные
трехлопастные листья. Но эти листья прошлогодние, перезимо-
16
Весеннее пробуждение растений
вавшие под снегом, потому они и имеют несколько поблекший
вид, с буроватыми пятнами. Молоденькие, еще невполне расправив-
шиеся листочки можно видеть на тех экземплярах, где цветки
уже отцветают. На белую ветреницу весьма походит очень часто
встречающаяся и еще более широко распространенная желтая,
или лютичная ветреница. Вскоре вслед за ветреницами расцве-
тает калужница, или куриная слепота (рис. 5). Растет она по
сырым лугам, канавам и болотам. Крупные червонно-золотые
Рис. 5. Калужница (Caltha
дуница обращает на себя
цветки калужницы нередко
почти сплошь покрывают
такие места, образуя широ-
кие золотистые полосы. У
этого растения цветки по-
являются вслед за листьями.
Запас веществ отлагается
в корневище.
Из ранних весенних рас-
тений, развивающихся за
счет запасов, отложенных
в прошлом году в корневи-
ще, назовем еще медуницу
(рис. 6), часто также попа-
дающуюся в лесу. Ее цвет-
ки распускаются вслед за
palustris) распусканием листьев, одно-
временно с ^ветреницей. Ме-
внимание тем, что цветки ее, в зависи-
мости от возраста, меняют свою окраск'7 начиная с нежной розо-
вой и кончая фиолетовой. Изменение окраски происходит вслед-
ствие изменения реакции клеточного сока, в котором растворен
пигмент антоциан (см. ниже стр. 47). В одном соцветии обычно на-
ходятся цветки на разных стадиях развития, имеющие разную
окраску. Этот факт, повышая вероятность перекрестного опыления,
приносит растению определенную пользу. На этом вопросе мы,
однако, не станем здесь останавливаться, так как он будет рас-
смотрен в следующей главе.
Одновременно с медуницей в лесах и на лугах можно найти
гусиный лук, или гусятник (рис. 7) — небольшое растение с желто-
вато-зелеными цветками, в виде шестилучевой звезды, и линейными
листьями. Как показывает самое название эт^го растения, оно имеет
луковицу, из которой и развивается. Луковица гусиного лука
имеет форму маленького яйца, обращенного вверх своим заострен-
ным' концом. Она представляет собой один видоизмененный лист,
наполненный питательными веществами. За счет их ранней весной
быстро развиваются листья со стеблем и вслед за ними —
цветок.
Отдельные представители весенней флоры
17
Из других луковичных весенних растений назовем всем изве-
стные гиацинт и тюльпан, сделавшиеся любимыми культурными
растениями. В диком состоянии они растут на юге. Если разрезать
их луковицу, то видно, что главную массу ее, как и у обыкновен-
ного лука, составляют плотно налегающие друг на друга мясистые
чешуи, сидящие основанием на сильно укороченном стебле (донце);
следовательно, вся луковица
представляет собой видоиз-
мененный стебель с листьями.
В лесах южной и юго-
восточной частей СССР ранее
других растений появляются
на проталинах и даже из-под
снега пролески (рис. 8 и 9).
Рис. 7. Гусиный лук желтый
(Gagea lutea)
Рис. 6. Медуница (Pul-
monaria officinalis)
Это тоже луковичные растения. О них еще будет речь ниже. Все
указанные луковичные растения относятся к одному семейству—
лилейных.
Из растений, развивающихся за счет запасов, отложенных в клуб-
не, ранним и распространенным представителем весенней флоры
является хохлатка, встречающаяся преимущественно в лесу (рис.
10). Она имеет красновато-фиолетовые цветки с длинными шпор-
цами, где откладывается нектар, служащий приманкой для на-
секомых; в более южной части СССР встречаются хохлатки с
18
Весеннее пробуждение растений
желтоватыми и беловатыми цветками. Цветки собраны в кисти.
Вынув подземную часть этого растения, с первого взгляда можно
Рис. 8. Молодой всход
пролески из луковицы
Рис. 9. Распустившаяся пролеска (Scilla
sibirica)
подумать, что оно имеет луковицу. Однако разрезав эту предпола-
гаемую луковицу, мы увидим, что она не состоит из мясистых, на
Рис. 10. Хохлатка (Cory-
dalis Halleri). 1 — верх-
няя часть стебля с цвет-
ками; 2 — нижняя часть
стебля с клубнем, разре-
занным вдоль
легающих друг на друга чешуй. Это—клу-
бень, также переполненный запасом ве-
ществ, образованных в прошлом году.
Клубень хохлатки, подобно известному
клубню картофеля, представляет собой
также видоизмененный подземный сте-
бель. Стеблевое происхождение клуб-
ней определяется по тому же признаку —
по нахождению на них зачаточных видоиз-
мененных листьев или почек, расположен-
ных в определенном порядке. Последнее
особенно ясно видно на клубне картофеля.
В хорошо заметных на нем ямочках (па-
зухах) помещаются почки, так называе-
мые глазки, развивающиеся в следующем
году в новые надземные побеги. На очень
молодых клубнях, около ямочек можно
заметить маленькие чешуйчатые листочки.
На сырых местах ранней весной часто
попадаются золотисто-желтые цветки чи-
стяка, или жабника (рис. И), напоми-
нающие по виду обыкновенные лютики.
Выкопав осторожно подземную часть это-
го растения, можно видеть, что, кроме
обыкновенных корней, от стебля отхо-
Отдельные представители весенней флоры
19
дит несколько продолговатых клубней. Нет надобности добавлять
что в этих клубнях откладываются запасы органических веществ
которые используются растением весной. Но по своему происхо
ждению клубни чистяка
иного типа. Это не видо-
измененные стебли, а взду-
тые отложенным питатель-
ным запасом боковые кор-
ни; такие клубни и назы-
ваются поэтому корне-
клубнями. В пазухах, т. е.
в том месте, где от стебля
отходят листья, у чистяка
развиваются маленькие
Рис. И. Чистяк (Ficaria verna). 1—стебель
с цветком и листьями; 2 — цветок (вид
снизу); 3 — корни с клубнями
грязно-желтые шишечки,
напоминающие своим про-
долговатым видом зерна
пшеницы. В этих шишеч-
ках также скопляется за-
пас органического вещества, вследствие чего они могут употреб-
ляться в пищу человеком. Они легко отпадают от стебля, и сле-
дующей весной из них также развиваются
Рис. 12. Селезеночник
(Chrysosplenium alterni-
folium), отдельно—цве-
ток (увеличен)
новые растеньица. При этом дождевая вода
иногда разносит их на значительное рас-
стояние. Шишечки эти, представляющие
собой как бы маленькие клубеньки, полу-
чили название выводковых почек.
Ветреницы, калужницы, чистяк, лю-
тики — все эти растения, ввиду сходства
в строении их цветка, относятся к одному
семейству — лютиковых. Лютиковые, а
южнее и лилейные преобладают весной.
Одним из первых весенних растений,
расцветающих вскоре после мать-и-мачехи,
является золотничка, или селезеночник
(рис. 12). Он очень распространен на сырых
местах, в особенности по канавам, ручейкам
и т. п. Растение это небольшое, с очень ма-
ленькими бледно-желтыми цветками и с
желтоватыми же верхними листьями. Бла-
годаря тому, однако, что растения растут
группами, они хорошо заметны. У селезеночника корневище тонкое,
с малым запасом органических веществ. В связи с этим находится
и малый размер этого растения, все части которого к тому же были
заложены еще в предыдущем году и перезимовали под снегом.
Даже ранее мать-и-мачехи на пригреваемых солнцем проталинах
20
Весеннее пробуждение растений
можно встретить цветущую маргаритку — каждому хорошо из-
вестное растение. Ее распростертые на земле в виде прикорневой
розетки листья и закрытые цветочные корзинки всю зиму пере-
несли под слоем снега. Таким образом, это растение правильнее счи-
тать прошлогодним. Оно мало чувствительно к холоду, и его при-
остановленное зимой развитие, подобно злакам, быстро возобно-
вляется ранней весной. Иногда морозы снова прерывают его, но
после их окончания растение опять продолжает развиваться.
Это явление приостановки развития на зиму и его возобновле-
ния весной особенно широко распространено у растений крайнего
севера и высокогорий. Быстро наступающая здесь зима с ее снегами
и морозами нередко застает их в цвету. Как показали исследова-
ния последнего времени, цветки и стебли под снегом прекращают
свою жизнедеятельность и погибают, а перезимовавшие листья
весной, по мере развития молодых листьев и цветочных побегов,
скоро отмирают. Однако роль их как хранителей запасных веществ
на первых этапах развития молодого растения очень велика. Воз-
можно также, что у некоторых видов перезимовавшие листья ран-
ней весной, до образования новых листьев, способны к фотосинтезу.
Корневища, луковицы и клубни, давая начало молодым расте-
ниям, являются, таким образом, органами размножения. Как бе-
рущие начало от зимующих подземных органов, представляющих
обычно подземные видоизменения стебля с видоизмененными ли-
стьями и с почками, ранние весенние растения, очевидно, должны
быть названы растениями многолетними. Луковичных и клубне-
вых растений особенно много в средней и южной полосе.
Ранние весенние растения в пределах СССР широко распростра-
нены в лесной, степной, полупустынной и, отчасти, пустынной зо-
нах. Народное название их — подснежники. На юге широкое рас-
пространение этой группы указывает на приспособление ее к ве-
сенним условиям, являющимся здесь наиболее благоприятным вре-
менем года для развития растений, так как лето бывает очень за-
сушливым. Хорошей переживаемости луковиц, клубней и других
подземных органов весенних растений на юге способствует то,
что почва здесь зимой промерзает незначительно, что очень важно,
так как располагаются они обычно неглубоко.
В лесной зоне растения-подснежники приурочены главным об-
разом к лиственным лесам, в которых защита от охлаждения, по
сравнению с хвойными лесами, больше. Здесь почва, находящаяся
под кронами деревьев и покрытая опавшей осенней листвой, те-
ряет (излучает) тепла менее, чем на местах открытых. Мороз в лесу
также проникает на меньшую глубину, почему погруженным в
почву луковицам и клубням в лесу гораздо меньше грозит опас-
ность вымерзания.
Однако наряду с этими положительными сторонами произраста-
ния в лиственном лесу, есть и отрицательная сторона, а именно —
Где растут ранние весенние растения 21
малое количество света, проникающего сквозь кроны деревьев по-
сле распускания листвы. Поэтому летом здесь развиваются лишь
теневыносливые формы. С этим связано быстрое развитие ранних
весенних растений до распускания листьев на деревьях. Исследо-
вания показывают, что сезонный максимум фотосинтеза у них также
падает на этот период.
Хвойные леса круглый год покрыты хвоей, в том числе и весной.
Само собой разумеется, последнее обстоятельство отражается и
на количестве весенних растений. В особенности это следует ска-
зать о лесах еловых. Сосна, будучи породой светолюбивой, много
пропускает света, а ель, хвоя и ветви которой гуще — теневыно-
слива. Света до почвы в еловом лесу доходит значительно меньше,
почему травянистая растительность как весенняя, так и летняя
под пологом елового леса весьма бедная.
Приспособленность большинства растений-подснежников лес-
ной зоны к жизни в условиях лиственного леса очень велика. В ду-
бравах и других широколиственных лесах они встречаются весьма
часто, но вне пределов лиственного леса распространяются с боль-
шим трудом. Этот факт используется для установления смены ра-
стительности. Обильное произрастание белой ветреницы, медуницы,
чистяка и других по опушкам и прогалинам современных хвойных
лесов является верным признаком того, что в былые эпохи эти
места были заняты широколиственным лесом, который впоследствии
заменился хвойным лесом.
У древесных растений цветки весной чаще распускаются раньше
листьев. Ольха и орешник расцветают приблизительно в одно время
с мать-и-мачехой и другими самыми ранними травянистыми расте-
ниями, когда ни на этих, ни на других деревьях (исключая, ко-
нечно, хвойные) листьев нет и помину. Снег еще не успел сойти
с полей, а поникнувшие над водой ветви ольхи уже покрыты длин-
ными, свешивающимися с них цветочными сережками. Вот потя-
нул легкий ветерок — и все дерево окуталось золотистой пылью.
Это пыльца из ольховых сережек. Стоит в это время потрясти
ветви орешника, и мы увидим то же самое:сережки также сильно пы-
лят. Вскоре распускаются, тоже опережая листья, сережки ив,
осины. На ветках вязов и ильмов появляются крошечные «букетики»
мелких буреньких цветочков, которые легко бросаются в глаза
только потому, что на деревьях к этому времени нет еще ни одного
листочка.
Наконец пробуждается жизнь и в листовых почках. Твердый
чешуйчатый панцырь, защищавший от холода во время зимнего
покоя нежный зачаточный побег, начинает становиться тесным для
него. Почки разбухают с часу на час. Зимний покров наконец ло-
пается, и заключенная в них пробудившаяся к жизни листва вы-
ходит на свет. У очень многих деревьев, как у березы и дуба, цве-
тение совпадает с распусканием листвы.
22
Весеннее пробуждение растений
Все деревья с опадающею на зиму листвою, как и ранняя тра-
вянистая растительность, весною также нуждаются в отложен-
ном заранее запасе органических веществ. Эти вещества нужны не
только для образования ранних цветков, появляющихся до распу-
скания зеленой листвы, но и для первоначального развития
последней. В самом деле, пока зачаточные листочки находятся под
покровом чешуй, пока они не вышли на свет, не распустились и
не позеленели, они синтезировать органические вещества иемогут;
следовательно, они не только не могут давать материал для роста
других частей растения, но и для собственного роста первоначально
нуждаются в готовом запасе. Он откладывается с осени в защищен-
ных от холода местах: в стволах и корнях под корою и в сердцевине.
В самом начале весны запас этот снова начинает подниматься
кверху — к почкам. Как же это происходит? Весной корни уси-
ленно всасывают из почвы влагу. Поднимаясь кверху по сосудам,
вода захватывает и запасные органические вещества, растворяя их.
Насыщение водой приводит в активное состояние находящиеся
в растении ферменты, которые и осуществляют с большой скоро-
стью различные химические превращения. В силу этого нераство-
римые вещества переходят в растворимые, например, крахмал
превращается в сахар.
Следует, однако, иметь в виду, что в молодых ветвях многих
деревьев, в клетках сердцевинных лучей древесины и перифери-
ческих слоев сердцевины, имеется хлорофилл (особенно зимой,
весной и летом). Есть основания полагать, что весной, еще до
распускания листьев, эти клетки способны поэтому осуществлять
фотосинтез, обеспечивающий возможность ростовых процессов.
Если ранней весной, еще до распускания листьев, просверлить
в дереве отверстие, то из него будет выделяться водянистый сок
(пасока). Явление это называется «плачем растений». Смотря по
растению и условиям, выделение сока неодинаково обильно. Наи-
более значителен плач у березы, затем у клена, бука, виноград-
ной лозы. Береза может дать до 8 и более литров сока в сутки,
причем такое выделение сока длится несколько недель, до распу-
скания листьев. С развитием последних «плач» прекращается.
На вкус сок березы, а также клена и многих других деревьев слад-
кий, так как в нем в начале весны растворено значительное коли-
чество сахара.
Без весеннего солнца растения, конечно, не пробудились бы от
зимнего покоя. Однако не в нем одном причина. В самом растении
или его зимующем органе в течение зимы, вероятно, имели место
какие-то внутренние процессы, заранее подготовлявшие то превра-
щение отложенных про запас веществ, которое происходит с про-
буждением жизни и образованием за счет этих запасов цветков,
листьев и т. д. Оттого-то жизненные процессы в пробуждающихся
весенних растениях развиваются со столь значительной энергией.
Выход молодых ростков из земли
23
Нередко можно видеть, что молодые всходы прорастают через лед
и снег. Постоянно это явление можно наблюдать на упомянутых
выше пролесках. В лесах южной и юго-восточной частей СССР
они появляются прежде всего на проталинах, рано образующихся
у подошвы деревьев. Но как только снеговая вода просочится под
соседние сугробы, прорастание начнется неудержимо и под снегом.
Листья пролески, сложенные остроконечной трубочкой (рис. 8),
в большом количестве выходят из-под снега, надвигаясь от окраины
сугроба к центру. Принимая во внимание, что весною под снегом
образуется ледяная корка, совершенно непонятно, каким образом
нежные всходы оказываются способными преодолеть такое сопроти-
вление, если не допустить, что они действуют не столько механи-
чески, сколько теплом, растопляя лед. В этом же легко можно убе-
диться па мать-и-мачехе. Если раскопать осторожно снег около
ручья, по течению которого встречается мать-и-мачеха, то под
снегом нередко можно найти всходы этого растения. При более
тщательном исследовании обнаруживается, что всходы окружены
небольшой оттаявшей полостью.
Таким образом создаются как бы естественные «парнички»,
температура в которых значительно повышена. В этих условиях
почки, заложенные еще в прошлый вегетационный период, начинают
развиваться, несмотря на сплошной снеговой покров. Откуда же
берется это тепло? Некоторая его часть вырабатывается самим ра-
стением в процессе дыхания. Как известно, при дыхании сложные
органические вещества, окисляясь, распадаются до веществ более
простых, причем имеет место выделение энергии. Однако таким пу-
тем тепла образуется сравнительно немного, и значительно боль-
шую роль играет тепловая энергия солнечных лучей. Дело в том,
что последние проникают через довольно глубокий слой снега.
Так, установлено, что снежный покров в 2—3 см толщины пропу-
скает до 50% падающей на его поверхность лучистой энергии
солнца, слой в 10 см — 2% и в 50 сл* — 1%. Под снегом, на по-
верхности почвы, световая энергия переходит в тепловую, которая
при образовании ледяной корки не может быстро излучаться. Кроме
того, и сами растущие части растения поглощают свет, также пе-
реходящий в тепло, способствующее их нагреванию. Именно в этом
и заключается главная причина образования вышеописанных
«парничков» и растапливания ледяной корки растущими растениями.
У многих лютиковых, цветущих ранней весной, а равно и в дру-
гое время в сырых местах и имеющих блестящий и вогнутый около-
цветник, как у чистяка, калужницы и др., повидимому, имеет место
собирание солнечных лучей с помощью околоцветника. Вы-
сказано предположение, что такой околоцветник играет роль
рефлектора, который отражает солнечные лучи, направляющиеся
на внутренние части цветка — тычинки и пестики — и таким об:
разом согревающие последние.
24
Весеннее пробуждение растений
Прорастающим растениям и их молодым нежным побегам при-
ходится встречаться с разными неблагоприятными условиями.
При выходе из земли эта последняя представляет уже первое
из них. В самом деле, молодые, пробудившиеся к жизни органы —
нежные, почва же — твердая, часто обладает большой плотностью
и содержит в себе острые камешки или угловатые песчинки. Ока-
зывается, молодые ростки снабжены рядом защитных приспособ-
лений, обеспечивающих им возможность выхода наружу.
Чтобы не быть раздавленными тяжестью земли, покрывающей
молодые ростки, необходимо, чтобы их листочки были крепко со-
единены друг с другом и
составляли одну плот-
ную массу; необходимо
также, чтобы с поверх-
ности, где происходит
соприкосновение с зем-
лей, эта масса имела
наиболее прочные части
и чтобы форма ростка
могла обеспечить пробу-
равливание земли. Все
это мы и находим в
дей ств ител ь ности.
Рис. 13. Выход из почвы ландыша (Conval-
laria majalis)
В одних случаях листья, свертываясь друг около друга, обра-
зуют остроконечное тело, пробуравливающее землю как бы шилом
(рис. 8 и 13). При этом у многих видов подземные листовые или
цветочные почки покрыты низовыми листьями, играющими защит-
ную роль. На верхушке этих листьев имеются группы клеток,
способных набухать и увеличиваться в размерах. Благодаря этому
кроющие листья удлиняются и прободают своим концом землю.
На этом функция низовых листьев заканчивается.
Защита нежных листочков и пробуравливание почвы нередко
осуществляются с помощью особых остроконечных чешуек, покры-
вающих отдельные листочки и слагающихся в общем таким обра-
зом, что в совокупности они имеют коническую форму. У многих
растений (у гиацинта и других луковичных, у многочисленных ор-
хидных наших лесов и лугов и пр.) нет особых кроющих листьев
и чешуек, и пробивание земли осуществляется обыкновенными ли-
стьями, т. е. такими, которые потом зеленеют и выполняют обычную
функцию листьев—образование органического вещества. В таких
случаях кончик листовой пластинки бывает превращен в настоящий
«таран». Обыкновенно он имеет вид башлыка, сидящего на сложен-
ных кончиках других листовых пластинок. Острие этого башлыка
состоит из упругих, способных набухать и увеличиваться, бес-
хлорофильных клеток. Когда конус выдвинется и листья разверз
нутся над землей, упругие доселе клетки листовой верхушки чах-
Защитные приспособления молодых цветков и листьев
25
нут и засыхают, буреют и становятся хрупкими, так что кончики
этих листьев кажутся как бы подсушенными. Наконец, необходимо
упомянуть, что у весьма многих растений в образовании остроко-
нечия и пробуравливании им земли главное участие принимает
стебель, причем листья или оказываются прижатыми к его вер-
хушке, или же последняя вовсе лишена листьев и покрыта только
чешуйками (как у спаржи).
Образование остроконечных верхушек, пробуравливающих зе-
выхода на ее поверхность у про-
млю, — не единственный спосоо
растающих растений. У многих
растений листовой черешок или
кончик стебля при этом изги-
бается как бы в колено или
крючок (рис. 14). Верхушка
этого коленчатого изгиба плот-
ная, упругая и совершенно
гладкая, как бы лощеная, т. е.
обладает такими же свойствами,
как и верхушка остроконечий,
и также прокладывает путь ме-
жду частицами почвы. Такой
способ выхода — самый распро-
страненный у растений с рас-
сеченными или мелко разделен-
ными листовыми пластинками,
которые не приспособлены для
складывания в пробуравливаю-
щие землю остроконечия. Пока
Рис. 14. Росток ветреницы, проби-
вающийся сквозь землю
изогнутый черешок или стебель
не проложит дороги через землю, листовые пластинки напра-
влены вниз. Когда же изгиб вышел из-под земли, он выпрям-
ляется, вытягивая за собой и листовую пластинку, которая
затем уже расправляется над землей.
У многих многолетних и двулетних растений выход молодых
ростков облегчается старыми листьями (у лабазника, некоторых
подорожников, деревеня, наперстянки и др.). Они образуют каналы
или, во всяком случае, пути, по которым молодые ростки сравни-
тельно свободно выходят на поверхность.
Но вот молодые почки, цветочные и листовые, освободившиеся
из-под земли или сбросившие с себя зимний чехол на древесной
ветке, распускаются. И после этого им грозит ряд неблагоприятных
внешних воздействий: последние заморозки, внезапный снег или
сильный дождь, сильно уже палящие и иссушающие в ясные дни
лучи солнца и т. п. Они не могли бы нормально развиваться, если
бы не имели разнообразных защитных приспособлений, большин-
ство которых, как только молодой орган окрепнет, исчезает. Позна-
комимся с некоторыми из этих приспособлений.
26
Весеннее пробуждение растений
Некоторое значение в качестве средства защиты от окружаю-
щей низкой температуры имеет выделение собственного тепла, ко-
торое образуется благодаря дыханию. Период цветения сопровож-
дается заметным повышением температуры: в исключительных
случаях внутри больших соцветий (например в початках некоторых
аройниковых) она достигает и даже превосходит температуру че-
ловеческого тела. Сравнительно малая величина большинства цвет-
ков, в том числе и весенних, делает затруднительным непосредствен-
ную проверку этого явления. Однако есть косвенное доказательство
его существования, именно — непрекращающееся выделение водя-
ного пара даже при значительных падениях температуры. Это легко
можно видеть в тех случаях, когда пар выделяется в более или ме-
нее замкнутой цветочной полости, на холодные стенки которой он
садится в виде капель. Такая роса образуется, например, внутри
расширенных чашечек первоцвета, под листьями обвертки закрыв-
шихся головок одуванчика, внутри свернувшихся листочков про-
растающей купены и др.
Закрывание цветков на ночь и днем при наступлении холодной
погоды, а также поникание их вниз (что наблюдается у многих
весенних растений) способствуют сохранению этого выделившегося
при дыхании тепла. Поникание цветков особенно важно как защит-
ное средство против попадания внутрь цветка ночной росы или дож-
девой воды, которая, как увидим в следующей главе, могла бы
нанести вред цветку. Это явление имеет место и у летпих растений
перед наступлением ненастной погоды или выпадением росы, но
наиболее значительно оно развито у ранних весенних растений.
На молодых нежных листочках еще недостаточно развита ко-
жица: ее внешние стенки еще не покрыты защитным слоем (кути-
кулой) и потому пропускают воду и водяные пары. Подверженная
солнечным лучам и порывам ветра зеленая ткань такого листа
тотчас высохла бы. Этого не происходит, однако, благодаря сле-
дующим приспособлениям.
Кто не наблюдал, что молодые листья не сразу развертываются
и расправляются? На их поверхности еще значительное время
остается морщинистость, складчатость и т. п. Благодаря этому по-
верхность, непосредственно подверженная солнечным лучам и на-
тиску ветра, значительно уменьшается. Уменьшается она еще и по-
тому, что листья молодых побегов более сближены друг с другом,
занимая в общем меньшее пространство и до некоторой степени за-
щищая друг друга. В плохую погоду и холодные ночи молодые
листья часто еще более завернуты, извиты, сложены. Нередко
листья принимают вертикальное положение, причем к солнцу,
равно как и к дождю, оказывается обращенным их край или ребро.
Солнечные лучи в таком случае как бы скользят по поверхности
листа, не проникая внутрь него, благодаря чему уменьшается испа-
рение.
Защитные приспособления молодых, цветков и листьев
27
Весьма часто молодые листья бывают покрыты клейкими веще-
ствами, предохраняющими их от чрезмерного испарения и засыха-
ния. Вещества эти, как только листовая пластинка разовьется и
кожица достаточно укрепится, обыкновенно исчезают; общеизве-
стный пример представляет черная ольха. У очень многих ра-
стений молодые листья покрыты мелкими волосками, как бы пуш-
ком, представляющими защитное средство и против солнца, и против
холода, и против дождя, и против иссушающего действия атмо-
сферы. Эти волоски у одних растений спадают с листьев во взрос-
лом состоянии. Так, у конского каштана листья во время развер-
тывания почки покрыты как бы густой шерстью, которая, однако,
скоро теряется, так что на взрослых листьях можно лишь кое-где
заметить сохранившиеся остатки волосков; подобное явление мож-
но наблюдать у многих ив, у серебристого тополя, у эдельвейса и пр.
У других растений от волосков освобождается лишь верхняя сто-
рона листа. Прекрасный пример представляет мать-и-мачеха:
нижняя сторона ее вполне развившихся больших листьев покрыта
густыми беловатыми волосками, а верхняя — совершенно глад-
кая. Если приложить нижнюю сторону к губам или щеке, то в срав-
нении с верхней она кажется гораздо теплее: с нижней стороны —
как родная мать пригреет, с верхней—точно мачеха холодом об-
даст; отсюда и название растения «мать-и-мачеха». Наконец, у не-
которых растений волоски остаются на обеих сторонах и во взрос-
лом состоянии. И в этом случае защита со стороны волосков бо-
лее действенна гв молодом возрасте, когда листья меньшего
размера, и следовательно, волоски на них более сближены, т. е.
более густы. Взрослые листья и цветки ранних весенних растений
уже не имеют такого количества защитных приспособлений, как мо-
лодые или как взрослые же органы, но летней и осенней флоры.
У летних растений, в зависимости от того, на влажных или су-
хих местах растут они, наблюдаются резко выраженные биологи-
ческие особенности. У растений сухих мест (на песчаной и камени-
стой почве, на обращенных к солнцу возвышенных склонах и т. п.)
встречается ряд приспособлений против чрезмерного испарения
воды. К ним в первую очередь относится меньшая поверхность ли-
стовой пластинки, что достигается, между прочим, большою рассе-
ченностью ее. В крайних случаях дело доходит до полного ее исчез-
новения (спаржа, дрок, кактусы и др.), причем функцию фотосин-
теза берут на себя снабженные хлорофиллом клетки коры стеблей.
Большое значение имеет также утолщение кутикулы, белые во-
лоски и восковые налеты, отражающие солнечные лучи, и др. Ра-
стения, у которых резко выражены такие приспособления, называ-
ются ксерофильными. У растений сырых мест указанных приспособ-
лений нет: поверхность листовой пластинки у них обычно боль-
шая, вменыней степени рассеченная, кожица нежная, почему лист
не кажется грубым, и т. д.; такие растения называются гидрофиль-
28 Весеннее пробуждение растений
ными. Среди ранних весенних растений этих двух резких противопо-
ложностей нет, так как и на тех местах, где летом бывает сухо, вес-
ной имеется влага. Листья и цветочные лепестки их имеют в связи с
этим относительно большую поверхность. В то же время они в об-
щем более сочны и нежны, обладают более тонкой кожицей. Если
мы весной сделаем букет, например из ветрениц, то букет скоро
завянет, так как нежные органы этого растения быстро испаряют
воду. Таким образом, в весенней флоре много растений, отвечающих
более гидрофильному типу. Однако вместе с тем многие весенние
растения имеют и ксерофильные признаки (опушение, утолщение
кутикулы и др.). Последнее, повидимому, связано с тем, что при
низкой температуре воздуха ранней весной поступление воды в ко-
рень оказывается замедленным.
ЦВЕТЕНИЕ РАСТЕНИЙ
В течение многих столетий строение цветков, их яркая окраска,
выделяемый ими нектар и аромат представляли загадку для мно-
гих ученых.
В самом конце XVIII столетия немецкий ученый натуралист-
любитель Шпренгель поднял, наконец, завесу над этой тайной.
В своей книге «Открытая тайна природы в строении и оплодотворе-
нии цветов» он убедительно доказал, что строение цветка приспо-
соблено к его опылению: окраска, запах, нектар — все это способ-
ствует привлечению насекомых, которые, собирая для себя на
цветках пищу, в то же время переносят пыльцу с тычинок на рыль-
ца. Для доказательства этой общей мысли Шпренгелем приведено
множество отдельных примеров приспособлений цветков к посеще-
нию насекомых.
К сожалению, современники Шпренгеля не оценили его откры-
тия. Его книга и описанные в ней наблюдения вскоре были забыты.
Только спустя более полустолетия множеством новых фактов Дар-
вин не только подтвердил мысли Шпренгеля, но еще далее пошел
в изучении этого явления. С именем Дарвина связано выяснение
значения перекрестного опыления — обстоятельства, хотя и под-
меченного, но не оцененного Шпренгелем. Длинным рядом опытов,
продолжавшихся в течение многих лет, доказана важность пере-
крестного оплодотворения, увеличивающего скорость роста, жизне-
деятельность и плодовитость растений. Дарвин сеял семена ра-
стений, полученные как в результате перекрестного опыления, так и
самоопыления, в совершенно одинаковых внешних условиях.
В большинстве случаев разница в величине и силе роста оказыва-
лась поразительной. Кроме того, растения, полученные в резуль-
тате перекрестного оплодотворения, приносили семена и лучшего
качества, и в большем количестве. Впоследствии обнаружены ра-
стения, у которых собственная пыльца, упавшая на рыльце, совсем
не производит оплодотворения; таковы, например, резеда, хохлатка
(рис. 10), льнянка, живокость, фиалка собачья и др. Найдены даже
растения, собственная пыльца которых действует на рыльце как
яд (из семейства орхидных в Бразилии).
30
Цветение растений
Шпренгель смотрел на открытые им замечательные приспо-
собления цветков к посещению насекомыми как на доказатель-
ства «премудрости творца». И только после победы эволюцион-
ной идеи все они получили совершенно иное, материалистическое
объяснение. Все эти факты, со многими из которых мы познакомимся
ниже, могут быть поняты только с точки зрения дарвиновской те-
ории естественного отбора, великолепной иллюстрацией которого
они являются.
После Дарвина ряд выдающихся исследователей работал и про-
должает работать над изучением биологии цветков. Не только про-
фессиональные ученые, но и множество ботаников-любителей увле-
каются этими работами. Таким образом накопился колоссальный
материал, полный интереснейших обобщений, и он с каждым годом
все увеличивается.
Среди цветковых растений наших стран около 1/10 опыляется
при содействии ветра.аУ большинства остальных перепое пыль-
цы с цветка на цветок, приводящий к перекрестному оплодотво-
рению, совершается при помощи насекомых.
Насекомые, посещая цветки, получают на них пищу. Что же
именно берут насекомые с цветков?
Есть немало цветков, у которых насекомые поедают цветень,
т. е. ту самую пыльцу, вырабатываемую в пыльниках, главное
значение которой заключается в опылении цветка. Пример пред-
ставляют цветки мака, розы, перелески, ветрениц, кувшинки,
паслена и др. Строение этих цветков сравнительно простое, так
как у них пет специальных приспособлений для хранения нектара.
Все такие цветки стоят вертикально. Когда раскроются, они имеют
форму чаш или раковин, направленных кверху, вследствие чего
пыльца не падает вниз (см. рис. 3,4). Сладкого сока такие цветки
обыкновенно не выделяют.
Главными потребителями цветня являются различные мелкие
жучки и мухи. Шмели и пчелы также собирают большое количе-
ство цветня, делая из него в своих гнездах и ульях запасы пищи для
личинок. Как известно, у этих насекомых сбор цветня облегчается
тем, что на их теле, главным образом на конце брюшка и го-
ленях задних ног, имеются особые волоски и щетинки. Ротовые
конечности их приспособлены для размельчения поедаемой пыльцы.
Казалось бы, что поедание насекомыми пыльцы должно было
привести к невозможности оплодотворения.
Но этого не бывает. Все цветки, лишенные нектара и привле-
кающие насекомых цветнем, имеют много тычинок, производящих
большое количество цветня. Несмотря на массовое уничтожение
его насекомыми, пыльцы хватает и на опыление рылец, тем более
что для этого требуется небольшое количество ее.
К этому нужно прибавить то существенное обстоятельство,
что пыльца цветков, опыляемых насекомыми, липкая, пристает
Что собирают насекомые на цветках
31
к их телу, следовательно, переносится ими и на рыльца других цвет-
ков. Она липкая прежде всего от выделяющегося на поверхности
пылинок клейкого вещества. Тому же содействует и форма пы-
линок. Под микроскопом видно, что они обычно покрыты многочи-
сленными ребрами, гребнями, иглами, шипиками, бородавками и
т. п. (рис. 13). Все это благоприятствует как прилипанию пыльцы
к телу насекомых, так и сцеплению пылинок между собой, следова-
тельно, захвату и переносу их насекомыми в большом количестве.
Рис. 15. Пыльца различных растений под микроско-
пом: 1 — TbiKBbi(Cucurbita); 2— гвоздики (Dianthus);
3 — горечавки (Gentiana); 4—хохлатки (Corydalis);
5 — кувшинки (Nymphaca); 6 — одуванчика (Tara-
xacum)
Несравненно более важным пищевым материалом, который
потребляется насекомыми, является сладкий сок, называемый не-
ктаром, или цветочным медом. Его выделяют до 90% растений,
посещаемых насекомыми.
Относительно наименования нектара цветочным медом следует
заметить, что он по химическому составу не тождествен с известным
нам пчелиным медом. Последний не получается пчелами из цвет-
ков совсем в готовом виде, а является уже результатом перера-
ботки нектара в теле пчелы. Нектар содержит большее или мень-
шее количество сахара и наряду с ним различные другие веще-
ства. Смотря по их содержанию, меняются консистенция, цвет и
аромат сока: то он водянист и бесцветен, то густ, тягуч и темен и
различным образом пахнет. Количество его также весьма разно-
образно.
Нектар выделяется медовыми железами, или нектарниками.
У разных растений они образуются на различных частях цветка:
на тычинках, лепестках, столбиках пестиков, реже на чашелисти-
ках, цветоложе и пр.
32
Цветение растений
В одних цветках нектар помещается открыто, в других—в за-
крытых местах на глубоком дне. Есть растения, у которых сок не
остается на месте выделения его нектарниками, а оттекает отсюда
в шпорцевидные и тому подобные сок ©вместилища (например, у
фиалки, хохлатки—см. рис. 10, льпянки, орхидей—см. рис. 34).
С местонахождением меда в цветках связано посещение их опре-
деленными насекомыми. В случаях, когда мед находится на откры-
тых частях цветка и выступает, следовательно, непосредственно
наружу, главными посетителями являются жучки и короткохобот-
ные мухи. Бабочки и шмели с длинными хоботками не могут слизы-
вать медвяного покрова. Наоборот, у тех растений, где мед спря-
тан в ямочках, трубочках, шпорцах ит. п., он является недоступ-
ным для короткохоботных насекомых. Такие цветки в наших
странах являются главным источником пищи для бабочек и шмелей.
В странах тропических к насекомым присоединяются мелкие птич-
ки — колибри (рис. 16) в Америке, нектариицы в восточном полу-
шарии и другие, достающие из цветков мед своими удлиненными
клювами и производящие опыление. По исследованиям американ-
ских ученых, число растений, опыляемых с помощью колибри,
довольно значительно. Короткохоботные насекомые иногда, в
виде исключения, достают нектар путем «взлома», разгрызания ча-
стей цветка (у глухой крапивы, ятрышника и др.). Эти случаи
представляют значительный интерес, так как они ясно указывают,
что взаимоотношения насекомых и цветков, обычно трактуемые
как классический пример обоюдной пользы, в действительности
принимают иногда характер прямого хищничества.
Каждый цветок, как правило, выделяет немного нектара. Эта
особенность является также приспособительным признаком, так как
насекомые вынуждены в силу этого производить облет большого
количества цветков, что способствует перекрестному опылению.
Иногда насекомые пользуются клеточным соком нежных, тон-
ких и сочных лепестков, прокалывая и высасывая их. Это можно на-
блюдать у зверобоя, гиацинтов, золототысячника, ракитника, на-
ших луговых орхидных и др. Случается, что пищей насекомых
служат ткани и группы клеток, отходящие от кожицы известных
частей цветка и представляющиеся невооруженному глазу в виде
сосочков, бородавочек, волосков и вздутий (у традесканции, ко-
ровяка, венерина башмачка, вербейника и др.). Сюда же должны
быть отнесены мучнистые и пыльцевидные налеты на цветках
(на губе) некоторых орхидных. По первому впечатлению эти налеты
представляют пыльцу. На самом же деле это не настоящая, а ложная
пыльца, состоящая из кучки рыхлых округлых клеток, отделяю-
щихся на поверхности лепестков.
Цветки не только доставляют насекомым пищу. Цветки, имеющие
форму больших висячих колокольчиков, служат им убежищем на
ночь и в ненастную погоду. Таковы, например, кроме всем извест-
Цветки, — убежища для насекомых
33
ных колокольчиков, цветки наперстянки (рис. 17). В полости та-
ких цветков ночью температура выше, чем в окружающем похо-
лодевшем воздухе. Само собой разумеется, что бабочки с их боль-
шими крыльями не могут скрываться внутри цветков. Их посети-
Рис. 16. Цветок Solandra (из сем. пасленовых) во время
посещения колибри
телями являются главным образом жучки, мухи и сетчатокрылые,
не имеющие постоянных жилищ, а также очень мелкие насекомые,
так называемые пузыреногие, или трипсы. С восходом солнца они
вылетают наружу. Естественно, что во многих случаях ими захваты-
вается при этом пыльца и переносится на другие цветки.
У аройников и кирказонов насекомые, попав на дно цветка,
задерживаются там довольно долго, как бы в заключении. У кир-
34
Цветение растений
казона (рис. 18) простой венчиковидный околоцветник при основа-
нии имеет шарообразно вздутую часть, где помещаются тычинки
и пестик. Это расширение переходит в довольно длинную узкую
трубку, выстланную жесткими обращенными внутрь волосками.
Трубка заканчивается листообразным расширением в виде язычка
или лопасти. Мелкие мухи и другие насекомые, садясь на эту часть,
легко проходят через трубку внутрь
цветка, но выйти обратно, вследствие
указанного расположения волосков, не
могут. Оказавшись в ловушке, насекомые
начинают бегать по стенкам расширенной
части цветка, задевая созревшие к тому
времени пыльники и обмазываясь пыльцой.
Эго происходит в течение нескольких дней,
Рис. 18. Кирказон (Aristolochia clemaiitis).
Ветвь с листьями и цветками в натуральную
величину
Рис. 17. Цветки на-
перстянки пурпурной
(Digitalis purpurea)
по прошествии которых щетинки в трубке засыхают и отпадают
и насекомые свободно выходят наружу. Попадая в следующем
цветке в такую же западню, они опыляют принесенною пыльцою
его рыльце, которое созревает здесь ранее пыльников. По удалении
насекомого наружная лопасть околоцветника, завядая, закрывает
входное отверстие, и весь цветок, прежде имевший вертикальное по-
ложение, поникает, вследствие чего насекомые не посещают опло-
дотворенных цветков.
Значение окраски цветков для привлечения насекомых
35
Наконец, в некоторых, сравнительно редких, случаях, насе-
комые пользуются цветками для отложения в них яиц. Так, не-
которые ночные бабочки кладут яйца в цветки, именно в завязь
гвоздичных. Вышедшие из яиц гусеницы питаются семяпочками,
или, вернее, молодыми незрелыми семенами, потом прогрызают
стенку завязи, выбираются наружу и окукливаются в земле. Боль-
шого вреда растению при обилии семяпочек бабочки этим,
повидимому, не приносят, пользу же приносят несомненную, про-
изводя опыление цветков, притом перекрестное. Большинство та-
ких гвоздичных, например слепец, цветет ночью (см. рис. 30, 31).
Указанные особенности цветков не могут, однако, непосред-
ственно привлекать насекомых. Необходимо еще, чтобы цветки были
заметны насекомым. Необходимы определенные внешние приметы,
или «приманки». Как известно, такой «приманкой» служат окраска
цветков и запах. Для того чтобы насекомое увидело цветок, не-
обходимо прежде всего, чтобы его окраска отличалась от преобла-
дающего тона обстановки. В связи с этим мы и видим, что на зеле-
ном фоне листвы цветки имеют другую окраску, образующую с зе-
леной резкий контраст.
Это явление представляет собой биологически целесообразный
для растения признак. Такие цветки имеют больше шансов на посе-
щение их насекомыми, а следовательно, и на перекрестное опыле-
ние. Объяснение происхождения этой, так же как и других относи-
тельно целесообразных особенностей, следует искать в естествен-
ном отборе. Более заметная для насекомых окраска цветка дает для
растений, обладающих ею, определенный перевес в борьбе за су-
ществование по сравнению с другими растениями, данной особен-
ности лишенными. Первые, очевидно, имеют больше шансов на оста-
вление потомства. Поэтому этот признак был подхвачен естествен-
ным отбором, в процессе которого в конце концов и выработалось
то контрастирование окраски цветков с окружающей их средой,
примеры которого мы можем наблюдать вокруг нас .
Ранней весной, когда новая растительность еще не развилась,
основной тон почвы не зеленый, а бледно-желтый или бурый от
прошлогодней опавшей листвы и увядшей травы. На такой почве
и контрасты цветов должны быть иными. Синие цветки на желто-
буром фоне заметны лучше, нежели на зеленом. В связи с этим, ве-
роятно, так много весенних цветков, распускающихся над засох-
шей листвой, окрашено именно в синий цвет. Вообще наблюдения
над окраской цветков в разные времена года показывают, что отно-
сительное количество разных оттенков их меняется. Так, для расти-
тельности побережья Балтийского моря было вычислено, что белая
окраска наиболее часто встречается в апреле и мае, а наименее —
поздней осенью. Желтый цвет в мае замечается в наибольшем коли-
честве, затем летом желтых цветков меньше, а к осени — опять
больше. Красных цветков весной очень мало, в течение лета они
36
Цветение растений
прибывают, а больше всего их замечается осенью. Синие и фиоле
товые цветки весной и осенью многочисленнее, чем летом. Кром(
изменяющегося тона ландшафтов, это изменение окраски цветков
вероятно, связано и с тем, что в разные времена года летают разные
насекомые. Весной летают одни насекомые, летом другие, а осенью-
опять иные. Ниже мы увидим,
что разные насекомые отдают пред-
почтение разным цветкам.
Главным носителем окраски,
привлекающей насекомых, явля-
ются лепестки, образующие вен-
чик. Когда в общежитии говорят
о цветках, то прежде всего разу-
меют их красивый венчик. Ввиду
такой функции венчика естест-
венно ожидать, что ярче окрашена
та сторона его, которая обращена
к прилетающим насекомым. И дей-
ствительно, у венчиков, имеющих
форму кувшина или колокольчика,
который свешивается вниз, так что
насекомые при пролете видят одну
только наружную сторону его, эта
именно сторона и оказывается окра-
шенной более ярко. Наоборот, у
Рис. 19. Василистник водосбо- венчиков звездчатых и чашевид-
ролистный (Thalictrum aquile- ных, обращенных к насекомым
gifolium) внутренней стороной, эта послед-
няя и более заметна. Это можно
наблюдать, например, у гусиного лука (рис. 7), у которого внутрен-
няя сторона околоцветника желтая, наружная же — зеленая, а
также у чистяка (рис. 11), птицемлечника, очного цвета и мн. др.
В некоторых случаях, когда лепестки бывают мало заметны,
в яркие цвета окрашиваются чашелистики. Пример представляют:
вереск (рис. 41), лиловая окраска цветков которого принадлежит
чашелистикам, речной гравилат, у которого чашелистики окрашены
в красно-бурый цвет, калужница (рис. 5) и купальница (рис. 51) —
с желтыми чашелистиками, борец —с синими или желтыми и др.
Иногда в яркий цвет окрашены одновременно и лепестки, и чашели-
стики, как например у живокости и др.
Реже венчиков и чашелистиков насекомые «приманиваются»
к меду тычинками, окрашенными в различные цвета. Примером
могут служить цветки василистника и ивы. У водосборолистного
василистника (рис. 19) нет и следа лепестков, да и чашечка мало
развита, зато тычинки имеют яркую лиловую окраску, утолщены
по концам и в распустившемся цветке образуют пышную шаровид-
ную кисть. В тычинковых сережках ивы (рис. 20, 2), где собрано
Носители окраски в цветках
37
множество мелких цветков, лишенных яркого околоцветника, также
заметно выделяются золотистые тычинки, отливающие золотом и
пурпуром под лучами весеннего солнца. Яркоокрашенные тычиноч-
ные нити характерны для большинства мимозовых, распространен-
ных главным образом в тропических странах.
Да и у многих цветков с развитым v ярко окрашенным венчи-
ком золотистые тычинки делают их более заметными для насеко-
Рис. 20. Ива (Salix). 1 — соцветия, начинающие распус-
каться; 2 — распустившееся соцветие с тычинками; 3 —
отдельный увеличенный цветок с тычинками; 4 — распу-
стившееся соцветие с пестиками; 5 — отдельный увели-
ченный цветок с пестиком
мых. Стоит только вспомнить о цветках наших лютиков, яблоневых
и грушевых деревьев.
Наконец, иногда случается, что не сами цветки бросаются
в глаза по своей окраске, а окружающие их листья — прицвет-
ники. Распространенный пример—Иван-да-Марья (рис. 21). Это
нежное растение тенистых мест очень привлекательно на вид, по-
тому что листья, в пазухах которых сидят желтые цветки, имеют
прелестную лиловую окраску. У селезеночника, или золотнички,
«золотят» не столько мелкие желтоватые цветки, сколько верхние
желтоватые же листья, около которых располагаются цветки
(рис. 12).
Если цветки мелки, то и при яркой окраске они в отдельности
мало заметны. Такие цветки обычно сгруппированы в разнообраз-
ные соцветия — колосья, кисти, зонтики, корзинки и пр., вслед-
ствие чего они лучше видны насекомым.
38
Цветение растений
Благодаря такому соединению мелких цветков насекомые за
то же самое время посещают большее количество их. В соцветиях
между отдельными цветками нередко наблюдается разделение функ-
ций. Так, например, цветки калины (рис. 22) расположены в одной
плоскости и образуют соцветие шиток. В то время как срединные
цветки мелки и плодовиты, краевые представляютсобой одни венчики
и, образуя вместе белый венок, привлекают насекомых к срединным
Рис. 22. Калина (Viburnum opu-
lus). 1 — срединный цветок; 2 —
краевой и срединный цветок (в
менее увеличенном виде, чем 1)
Рис. 21. Иван-да-Марья (Ме-
lampyrum nemorosum). 1,2 —
два цветка в разных положе-
ниях; 3 — плод
цветкам. Подобное разделение функций особенно распространено у
сложноцветных. Как известно, у этого громадного семейства, пред-
ставителями которого могут служить васильки, ромашки, маргарит-
ка,одуванчик, георгин и т. д., то, что на первый взгляд производит
впечатление одного крупного цветка, при ближайшем рассмотре-
нии оказывается тесным скоплением многочисленных мелких цвет-
ков, т. е. соцветием (корзинкой). Такие цветкоподобные соцветия
называются аншодиями.
Синий василек (рис. 23) имеет в середине соцветия мелкие не-
взрачные трубчатые цветки с тычинками, пестиком и медом на дне
венчика. Совсем другое представляют краевые цветки. Они окра-
шены в великолепный лазоревый цвет, имеют больший размер и
делают все соцветие хорошо заметным издали. Эти цветки бес-
плодны и их функция, очевидно, заключается в привлечении на-
секомых к срединным цветкам, способным к плодоношению. У много-
Носители окраски в цветках
39
численных сложноцветных, имеющих в середине корзинки не-
взрачные трубчатые, а по краям язычковые цветки, последние обы-
кновенно также бесплодны. Примеры подсолнечника, ромашки,
маргаритки общеизвестны. У многих зонтичных, например, у мор-
кови, сидящие по краям зонтика цветки не бесплодны, т. е. имеют
Рис. 23. Василек (Centaurea суа-
nus). 1 — краевой цветок; 2—сре-
динный цветок; 3 — плод (/, 2,
3 — увеличено)
тычинки и пестик, но у них, в
отличие от других цветков,
внешние лепестки, направлен-
ные кнаружи, значительно уве-
личены (рис. 24); благодаря это-
му весь зонтик делается за-
метнее.
Очень часто яркость и цвет-
ков и соцветий увеличивается
благодаря сочетанию несколь-
ких окрасок на разных частях
их. Бывают различно окра-
Рис. 24. Краевой цветок
моркови (Daucus carota) (уве-
личено)
шенные лепестки на одном цветке; например, у мотыльковых
часто можно заметить, что лепесток, вывернутый кнаружи и
называемый парусом, окрашен иначе, чем лодочка и весла.
Встречаются резкие контрасты в окраске между венчиками и на-
ходящимися в середине цветка тычинками и пестиком; например,
у картофеля и паслена на фиолетовой звезде вырисовывается жел-
тый конус тычинок. Бывают цветные контрасты между венчиком
и чашечкой или прицветниками; пример представляют различные
виды марьянника, как упомянутого выше Иван-да-Марья (рис. 21),
цветки которого желты, а прицветники синего, фиолетового или
красного цвета. В корзинках сложноцветных краевые цветки весьма
часто отличаются по окраске от срединных: так, у поповника,
ромашки, маргаритки краевые язычковые цветки белые, а внутрен-
ние трубчатые — желтые.
40
Цветение растений
Рис. 25. Цветок нарцисса (Narcissus poetl-
cus). Коронка в центре цветка оторочена
красной (на рисунке — черной) каймой
У множества цветков лепестки испещрены разноцветными
пятнами, линиями, узорами, оторочками и т. п. Так, трехцветная
фиалка, трехцветный вьюнок, трехцветная вика получили назва-
ние трехцветных за разрисовку их лепестков. У орхидей, губоцвет-
ных и норичниковых подобная разрисовка представляет обычное
явление. У нарцисса белая
коронка на венчике снаб-
жена красной оторочкой
(рис. 25), а у голубых
цветков незабудки вход в
короткую трубку венчика
окружен вздутым желтым
кольцом.
Как показали опыты,
иногда цветные знаки на
венчиках имеют значение
«указателей» пути к меду.
В таком случае они обык-
новенно имеют вид пятен,
точек или линий, сходя-
щихся к центру цветка,
как, например, у герани,
гвоздики и других, или
среднего светлого глазка,
как у незабудки и огуреч-
ной травы. Если оттянуть
вниз оранжевую губу жел-
того цветка льнянки обык-
новенной, то посетитель-
ница льнянки — бабочка
хоботник обыкновенный,
называемая также пав-
линьим хвостом (Масго-
glossum stellatarum из се-
мейства бражников), хотя
и подлетает к такому цветку, но попасть хоботком в отверстие
венчика не может. Предположение о существовании «медовых
дорожек» или «медовых значков» подкрепляется и тем, что это
явление наблюдается лишь у растений с запрятанным медом
и никогда не бывает у цветков, раскрывающихся только по ночам,
хотя бы мед их и был заключен в очень глубоких трубках.
Цветовой контраст часто достигается благодаря тому, что вен-
чики цветков, находящихся на одном растении, на различных ста-
диях развития меняют свою окраску. В состоянии почки они бы-
вают, например, красными, после раскрытия становятся фиолето-
выми, а потом, ко времени отмирания,—синими или малахитово:
Зрение насекомых
41
зелеными. Когда такие цветки собраны вместе, то получается пе-
строе, бросающееся в глаза сочетание красок. Особенно замеча-
тельны в этом отношении сочевники, многие растения из семейства
бурачниковых и др. Мы уже называли одно известное в этом отно-
шении раннее весеннее растение из семейства бурачниковых, имен-
но—медуницу (рис. 6), где наблюдается следующая последователь-
ность окраски: нераспустившиеся еще цветочные почки медуницы
окрашены в розовый цвет, развернувшиеся вполне цветки — в ли-
ловый или фиолетовый, отцветающие же переходят в голубой или
даже синий. Подобная же последовательность в окраске цветков
наблюдается у незабудки, относящейся также к семейству бурач-
никовых. Такое сочетание различно окрашенных венчиков на одном
растении, делая его более заметным для насекомых, имеет еще
и другое значение. На медунице, например, было замечено, что
пчелы избегали посещать те цветки ее, которые имеют голубую или
синюю окраску, т. е. которые уже оплодотворены, и, следователь-
но, лишены нектара. Растениям это также полезно, так как неопы-
ленные еще экземпляры благодаря этому обстоятельству имеют
больше посетителей и, следовательно, больше шансов на опы-
ление.
Нельзя не упомянуть также о контрасте цветков, достигаемом
близким соседством растений с различной окраской. Какое при-
хотливое сочетание красок на цветущих лугах! И эта пестрота в
общем, несомненно, делает отдельные цветки более заметными.
Когда среди голубых колокольчиков попадаются, например, крас-
ные цветки гвоздики, герани и других, они, конечно, более броса-
ются в глаза, нежели в том случае, если бы колокольчиков не было.
И наоборот — синий цвет колокольчиков благодаря присутствию
цветков другой окраски становится также заметнее.
Наблюдения заставляют признать, что насекомые, да и другие
животные, не безразлично относятся к разным окраскам. У них
есть как бы «излюбленные» и «нелюбимые» цвета, которые для
различных животных различны. Так, для пчел и шмелей «излюб-
ленными» цветами являются, повидимому, синий и фиолетовый;
желтый уже менее привлекает их; к зеленому они «равнодушны», а
красный обыкновенно обходится ими, т. е. является «нелюбимым»
цветом. Повидимому, он совсем не воспринимается ими. Джон
Лёббок произвел, между прочим, такой опыт над пчелами. Он по-
мещал капельки меда на стеклах, разложив их на кусках синей,
желтой, оранжевой, красной, зеленой, черной и белой бумаги, при-
чем бумажки неоднократно перемещались. Пчелы обыкновенно
направлялись к тому стеклу, которое лежало на синей бумаге.
Вопрос о составе цветного зрения пчелы тщательно изучил
немецкий ученый Фриш. Им был поставлен прежде всего вопрос:
какие цвета спектра различает пчела? Для разрешения этой задачи
Фриш прибегнул к методу так называемой «дрессировки». Опишем
12
Цветение растений
кратко некоторые из опытов Фриша, которые легко повторить на
любой пасеке.
Фриш брал ряд (чаще всего — 15 или 31) серых бумажных ква-
дратов различной интенсивности окраски (от белого до черного —
рис. 26) и расположенных рядом. В совокупности все эти квадра-
•синий
Рис. 26. Прибор Фриша для «дрессировки»
пчел на синий цвет. 15 квадратов серого
цвета различной интенсивности и среди
них один синий. Пчелы собрались на синем
квадрате
ты образовывали квадрат
большого размера. Среди
этих серых квадратов по-
мещался один квадрат то-
го цвета, способность раз-
личать который подверга-
лась изучению.
Первая часть опыта
сводилась к «дрессировке».
На квадраты ставились
маленькие стеклянные со-
суды (часовыестекла), при-
чем в то часовое стекло,
которое ставилось на цвет:
ной квадрат, наливался
сахарный сироп (пища).
Пчелы в большом количе-
стве собирались на цвет-
ном квадрате, где они
получали пищу. При этом
место цветного квадрата
в ряду других серых все
время менялось, чтобы
пчела не могла запом-
нить его положения от-
носительно серых квад-
ратов. Подобного рода
«дрессировка» продолжа-
лась дня два, чтобы пче-
лы могли «привыкнуть» к испытуемому цвету. После этого стек-
лышко с сиропом удалялось. Если пчелы различают цвета спект-
ра, т. е. имеют цветное зрение, то они должны в течение некоторого
времени попрежнему собираться на цветном квадрате, с цветом ко-
торого у них ассоциируется наличие пищи. Если же цветное зрение
у них отсутствует и они различают только интенсивность окраски,
то подобная картина не должна иметь места, так как в ряду окру-
жающих квадратов имеются такие, которые по интенсивности окра-
ски совпадают с испытуемым цветным. Многочисленные опыты Фри-
ша показали, что «дрессировка» легко удается на синий и желтый
цвет. На синих и желтых квадратах пчелы продолжают собираться
в большом количестве и после окончания «дрессировки», т. е. после
Условные рефлексы насекомых
43
того как сироп убран. Красный же цвет пчелами не различается.
Пчелы в отношении своего цветного зрения подобны до известной
степени людям, страдающим цветной слепотой (дальтонизмом),
т. е. неспособностью различать оттенки красного цвета. Фиолето-
вый, зеленый и другие цвета пчелы различают, повидимому, лишь
постольку, поскольку в них имеются элементы синего и желтого.
Таким образом, хотя у пчелы и имеется цветное зрение, но оно бо-
лее ограничено, чем у человека.
Используя указанный метод «дрессировки», Фриш поставил
также ряд экспериментов
для выяснения вопроса,
в какой мере пчелы спо-
собны различать форму
цветка. Для этой цели им
использовались особые
кормушки, внутрь кото-
рых помещались сосуды
с сиропом, а входные от-
верстия их имели раз-
личную форму (рис. 27).
При этом выяснилось, что
пчелы различают и форму
предметов. Однако послед-
нее имеет место, повиди-
мому, лишь на небольших
расстояниях, так что они
«близоруки».
Изложенный выше ме-
тод «дрессировки» Фри-
ша представляет собой по
Рис. 27. Установка четырех ящиков при
дрессировке на звездообразную и «генциа-
новую», имитирующую венчик горечавки,
форму отверстия
существу не что иное, как применение метода условных ре-
флексов, разработанного великим физиологом И. П. Павловым.
В этих опытах пища (сироп) представляет собой безусловный
раздражитель. Цвет квадратов, на которые ставится сироп, или
форма отверстия домика — условные раздражители. При соче-
тании безусловного и условного раздражителей у пчел и других
насекомых вырабатывается условный рефлекс (условная временная
связь), в результате которой насекомое реагирует на условный раз-
дражитель так же, как и на безусловный.
Изложенные выше опыты с полной очевидностью показывают,
что у насекомых условные рефлексы играют исключительно важную
роль в их ориентировке в окружающей среде и в отыскании пищи.
Привлечение насекомых цветками, их окраской и формой, основано
на выработке целой системы условных рефлексов.
У пчел, как у «общественных» насекомых, в посещении цветков
немаловажную роль играют также своеобразные формы «общения»
44
Цветение растений
между «рабочими» пчелами одного улья. Если какая-нибудь пчела-
сборщица возвращается в улей с богатой добычей, то она исполняет
особый «танец», выражающийся в быстром беге по кругу. Этот
«круговой танец» действует «мобилизующе» на других пчел, которые
стимулируются этим к вылету из улья за взятком. Разумеется, этот
«язык пчел» не включает в себя никаких элементов сознательного
общения, а представляет собой сложную систему рефлексов.
Указанные приемы
эксперимента подверг-
лись дальнейшей разра-
ботке со стороны Кнолля
по отношению к другим
насекомым—главным об-
разом мухе жужжало,
подражающей по внеш-
нему виду перепончато-
крылым (осам, шмелям),
и бабочке хоботнику
обыкновенному. Опыты
с разноцветными бумаж-
ками велись следующим
образом.
Вырезались квадра-
ты серой бумаги 6 X
Х6 см различной интен-
сивности окраски и
наклеивались на кар-
тон в шахматном по-
Рис. 28. Муха жужжало (Bombylius fuligi-
nosus) задерживается у квадрата синелило-
вого цвета, расположенного среди серых
квадратов разной интенсивности окраски
(стрелка обозначает направление полета)
рядке. Среди серых квадратов для опытов с мухой жужжало
наклеивались один или два синих; если —два, то различной ин-
тенсивности. Вся дощечка покрывалась сверху стеклом, чтобы
предупредить возможный от бумажек запах, и ставилась наклонно
среди цветущих растений, каковыми в Южной Далмации, где произ-
водились опыты, были гадючий лук кистецветный, образующий
кисти синелиловых цветков, посещаемых этой мухой. Жужжала
подлетали к синему квадрату, замедляли свой полет, делали иногда
над ним петлю и, наконец, улетали дальше (рис. 28). Если вклеива-
лись два синие квадрата, окрашенных с различной интенсивностью,
то мухи подлетали к ним обоим, но чаще всего к интенсивно окра-
шенному. На квадраты же серые они «никакого внимания не обра-
щали». Кроме того, вблизи цветущих растений раскладывались раз-
ноцветные бумажки — серые, черные, зеленые, красные, коричне-
вые, синие, лиловые; жужжала подлетали только к синим и лило-
вым бумажкам и иногда касались их. В 15 стеклянных трубок
90 X 15 мм были вложены различной интенсивности серые бумаж-
ки и в одну — синелиловая. Трубки были расставлены на стержнях
Условные рефлексы насекомых
45
по кругу, на расстоянии 3 см друг от друга, недалеко от цветущих
растений. Из всех трубок жужжала подлетали только к синелило-
вой. Для решения вопроса, играет ли роль в привлечении жужжал
белая каемка у входа в цветок гадючьего лука кистецветного, на
синей бумаге, вкладываемой в трубку, делались белые кружки
с черными пятнами. Около этого места насекомые замедляли свой
полет, откуда следует, что белый кант в цветке играет роль, когда
насекомое, привлеченное общей окраской цветка, садится на него,
чтобы запустить свой хоботок за пищей. Такие же и некоторые дру-
гие приемы, о которых отчасти будет упомянуто ниже, разработаны
Кноллем в применении к указанной бабочке. Для нее наиболее при-
влекательными цветами оказались желтый и синелиловый.
Некоторые бабочки привлекаются красными цветками, которые
и бывают иногда приспособлены к опылению этими насекомыми.
Более позднее появление этих цветков, сравнительно с синими,
можно поставить в связь с тем, что и бабочки выходят из куколок,
сравнительно с пчелами, позже. Колибри также охотнее других ле-
тят на кроваво-красные цветки. В связи с этим в Америке, где
водятся колибри, много и цветков такой окраски. Темнобурые
цветки охотно посещаются осами. Цветки, напоминающие по окраске
гниющее мясо, привлекают мух и жуков. Белые цветки, по-
видимому, не избегаются никакими животными. Когда темнеет,
белый цвет заметнее других, а потому белые цветки посещаются
ночными или сумеречными бабочками.
Только что появившиеся из куколки бабочки, а равно и другие
насекомые, летят на цветки определенной окраски. По наблюде-
ниям Кнолля, бабочка хоботник обыкновенный, как только она
вышла из куколки, несмотря на отсутствие опыта, посещает, как
правило, цветки двух групп — с желтыми и синелиловыми венчи-
ками. Взрослые же бабочки этого вида в средней Европе посещают
преимущественно цветки синелиловые, а на юге—желтые. Почему
со взрослой бабочкой происходит эта дифференциация? Потому
что в средней Европе желтые цветки (например, различных сложно-
цветных), в противоположность синелиловым, в большинстве или
совсем не содержат или содержат очень мало пищи для бабочек.
На юге же много желтых цветков, богатых нектаром.
Значит, в результате индивидуального опыта, у бабочек, так же
как и у пчел, вырабатываются условные рефлексы, устанавливаются
условные связи в одном случае с синелиловыми, в другом — с жел-
тыми цветками.
Этот вывод из наблюдений подтверждается опытами приуче-
ния и переучивания насекомых к различным цветкам — своего
рода «дрессировкой», проделанной Фришем над пчелами и Кноллем
над упомянутыми насекомыми, в особенности над бабочкой хобот-
ником обыкновенным. Над последней проделывались в комнате,
между прочим, следующие опыты.
46
Цветение растений
К окну подвешивались пробирки с различно окрашенными
жидкостями, причем в стенках пробирок делались углубления,
которые наполнялись сахарным сиропом. Бабочки (в Южной Дал-
мации, где они посещают главным образом желтые цветки) летели
преимущественно на пробирки желтые. Но если последние не снаб-
жались сахарным сиропом, а им, наоборот, снабжались пробирки
с синим содержимым, бабочки постепенно приучались летать на
эти последние, где и сосали сироп. Подобный же результат получался
и по отношению к пурпуровому цвету. Следовательно, дело нужно
представлять следующим образом.
При появлении на свет бабочки — а равно и другие насеко-
мые—летят на цвета, по отношению к которым у них имеется врож-
денное (инстинктивное) тяготение соответственно производимому
на орган зрения раздражению и восприятию цветов. Так как в при-
роде это оптическое воздействие идет со стороны цветущих расте-
ний, то бабочки устремляются на них и здесь находят пищу.
Опыт, который возникает с первыми же полетами, указывает, какие
именно из оптически привлекающих объектов, т. е. в данном слу-
чае цветков, содержат и какие не содержат корм, и потому число
посещаемых видов цветков постепенно сокращается до таких толь-
ко, окраска которых ассоциирована с пищей.
Чтобы покончить с окраской цветков, остановимся еще на во-
просе о том, от чего именно зависит эта окраска? Чем окраши-
ваются лепестки и т. д.? Как известно, зеленая окраска листьев за-
висит от зеленого пигмента — хлорофилла, которым пропитыва-
ются находящиеся в растительных клетках тельца — хлоропласты.
Но хлорофилл — не единственный пигмент в клетках зеленых
частей. Ему сопутствует другой пигмент — желтый (в свою очередь,
также состоящий большей частью из двух пигментов). За более
ярким зеленым пигментом — хлорофиллом — желтый обыкновен-
но не виден. Однако его можно обнаружить, например, следующим
образом.
Сделаем настой листьев (для скорости опыта — истолченных
с песком) на крепком спирту, который окрасится в изумрудно-зеле-
ный цвет от растворившегося в нем хлорофилла; отфильтро-
вав спирт, прибавим к нему бензина, несколько капель воды и
взболтаем: бензин образует верхний изумрудно-зеленый слой, а
спирт как более тяжелый опустится вниз и окрасится в желтоватый
цвет. Значит, с помощью этих двух жидкостей обнаруживается
присутствие в листьях разных по цвету пигментов. Осенью хлоро-
филл разрушается, а желтый пигмент остается, чем объясняется
осеннее пожелтение листьев. Представим теперь себе, что хлоро-
филл слабо развивается, желтый же пигмент образуется; тогда,
очевидно, и окраска органа, где это имеет место, должна быть,
подобно пожелтевшим осенним листьям, желтой. Такой именно
случай мы и имеем в желтых лепестках цветков. Смотря по коли-
Роль запаха в привлечении насекомых
47
честву образовавшихся зеленого и желтого пигментов, зеленая
окраска в лепестках легко переходит в желтую и наоборот и не-
редко они встречаются рядом; например, у гусиного лука (рис. 7)
внутренняя сторона листочков околоцветника золотисто-желтая,
наружная же — почти зеленая. Красные, синие и фиолетовые тона
зависят от присутствия особого пигмента — антоциана. Иногда он
окрашивает и листья растений, например, у красной капусты,
бегоний и др. Синие и красные тона осенней листвы также зависят
от развивающегося осенью в листьях антоциана. Он бывает то си-
ним, то красным и разных других промежуточных оттенков, в за-
висимости от того — кислый он, щелочной или нейтральный. Если
приготовить отвар из листьев лиловой капусты, лиловых, синих
или красных лепестков, то он обыкновенно имеет соответствующий
цвет. При прибавлении нескольких капель кислоты этот раствор
краснеет. При нейтрализации кислой жидкости щелочью (напри-
мер нашатырным спиртом) красный цвет снова переходит в фиоле-
товый, а при дальнейшем прибавлении щелочи — в синий, зеленый
и даже желтый. Клеточный сок чаще всего бывает кислый, редко—
нейтральный, а потому и антоциан в нем красный или фиоле-
товый. Сама же протоплазма клеток большей частью слабощелоч-
ная, и антоциан в ней поэтому окрашен в синий цвет.
Наряду с окраской внешним средством для привлечения насе-
комых является запах цветков.
Когда цветок обладает только окраской или только запахом,
тогда нетрудно подметить и решить, что именно привлекает насе-
комое. Но сложнее обстоит дело, когда у цветка имеется и то, и
другое. В таких случаях, чтобы оценить значение каждого фактора
в отдельности, нужно расчленить их влияние. Это удалось до-
стигнуть Кноллю следующими методами, примененными по отно-
шению к указанным насекомым.
Оп прикрывал цветки стеклянными трубками,закрытыми с верх-
него конца (годятся энтомологические пробирки соответствующего
диаметра или стеклянные цилиндры) или открытыми с обоих кон-
цов. При такой постановке опыта (рис. 29) цветок виден только через
стекло, запах же идет лишь из отверстий трубок. Муха жужжало
к прикрытым таким образом цветкам Muscari racemosum, располо-
женным среди неприкрытых, подлетала так же, как и к последним.
К прикрытым цветкам подлетала только на уровне цветка, задер-
живала здесь свой полет на некоторое время и летела далее (рис. 29).
Несмотря на то, что запах шел из концов трубки — из одного ниж-
него или из обоих, — насекомое ни разу не проникало в трубку
через ее отверстие. Значит, подлетая к цветку, насекомое это ре-
агировало только на определенное оптическое раздражение. Однако
и по отношению к запаху это насекомое небезучастно. Кноллвста-
вил свежие пахнущие экземпляры гадючьего лука кистецветного
рядом с экземплярами уже отцветающими и лишенными запаха
48
Цветение растений
Насекомое подлетало к тем и другим, но садилось и сосало нектар
только из пахнущих цветков. Отсюда следует, что данное насекомое
подлетает к цветку только под влиянием оптического раз-
дражения — общей окраски цветка. Сосательные же движения
хоботком производит под влиянием главным образом запаха, т. е.
химического раздражителя, и частью, быть может, дополнительного
оптического — белой каемки венчика. Подобный же опыт покры-
тия цветков трубками по отношению к бабочке хоботник обыкно-
венный показал, что она запахом не привлекается. Напротив, дру-
гая из подвергавшихся эксперименту бабочек из семейства нимфа-
лид ни на какие цвета не реагировала, но реагировала на запахи.
Рис. 29. Направление полета мухи жужжало (Bombylius fuliginosus), подле-
тающей к цветкам гадючьего лука кистецветного (Muscari racemosum) как
открытым, так и покрытым стеклянным колпачком
Для опыта служили, между прочим, искусственные цветки-кор-
мушки, сделанные из вощеной бумаги; в какие бы цвета они ни были
окрашены, бабочки не реагировали на них, если даже в них нали-
вался сахарный раствор. Прибавление же к раствору сока из сливы,
издающего запах, вызывало реакцию хоботка, независимо от цвета
кормушек. У хоботника обыкновенного реакция на те же кор-
мушки вызывалась, наоборот, их цветом, но не запахом содержи-
мого. Значение запаха Кнолль определял также при помощи ветра:
если насекомое летит па цветок только против ветра, значит, запах
играет роль в его привлечении; если же направление ветра зна-
чения не имеет, т. е. если насекомое летит на цветок и по ветру
(в этом случае запах относится от насекомого) — оно, очевидно,
руководствуется только окраской цветков.
Вопрос о роли запаха в привлечении насекомых наиболее об-
стоятельно был разработан по отношению к пчеле Фришем. Он
использовал и здесь метод «дрессировки», который применялся им
при изучении цветного зрения пчелы (см. стр. 41). Фриш устано-
вил, что острота обоняния у пчел значительна. Опа играет большую
роль при посещении цветков и сборе нектара и пыльцы. Однако
для пчелы запах является раздражителем, действующим глав-
ным образом на небольших расстояниях. В привлечении же пчел
на значительном расстоянии главную роль играет окраска цветка.
Классификация запахов
49
Лишь руководствуясь окраской и подлетая на близкое расстояние
к цветку, пчела начинает ощущать запах. Острота обоняния пчелы,
согласно Фришу, сравнима с остротой обоняния человека.
За последние годы нервная деятельность пчел и некоторых дру-
гих насекомых и в особенности возникновение у них условных ре-
флексов на цветные и другие раздражители- подверглись деталь-
ному и всестороннему изучению в Институте физиологии Академии
наук СССР, в лаборатории проф. М. Е. Лобашева. Этими исследо-
ваниями, в частности, установлено, что у пчел условные связи
возникают очень легко и быстро (через 2—14 сочетаний условного
раздражителя с безусловным) и быстро же могут исчезать и пере-
страиваться. Уже в улье, когда прилетает сборщица со взятком и
начинает свой «танец» (см. стр. 44), раздавая часть взятка окружаю-
щим ее пчелам, у последних устанавливаются условные связи меж-
ду пищевым раздражителем и запахом цветков, который приносит
пчела-сборщица. Эти возникающие еще в улье условные связи
играют большую роль в процессе «мобилизации» пчел на вылет из
улья, о чем уже говорилось выше. По этому поводу М. Е. Лобашев
пишет: «Если бы пчелы не имели в высшей степени скорой выра-
ботки и угасания условных рефлексов через контакт с другой пче-
лой, «общественная» или стадная жизнь пчел была бы невозмож-
ной. Можно думать, что форма общественной жизни у пчел и воз-
никла благодаря возможности быстрого замыкания временной
связи».1
Таким образом, из всех изложенных опытов следует, что для
одних насекомых имеет значение только окраска цветков, для
других —только запах, для третьих, каковых, повидимому, боль-
шинство, — и то, и другое, но у разных представителей в различной
степени. Вопрос этот нуждается в дальнейшей разработке на боль-
шем числе представителей. Естественно ожидать, что разнообраз-
ные насекомые на разные запахи, как и на разные цвета, реагируют
различно.
Разнообразие запахов, издаваемых различными цветками, мо-
жет поспорить с разнообразием их окрасок. Весьма многие цветки
безошибочно могут быть опознаны только по их характерному
запаху. Кто не знает аромата ландыша, гиацинта, резеды, сирени
и т. д.? Чтобы лучше разбираться во всех этих запахах, их, ко-
нечно, следует классифицировать, внести в их разнообразие опреде-
ленный порядок, т. е. собрать вместе сходные запахи и создать из
них центры, вокруг которых могли бы группироваться остальные,
подобно тому как все разнообразие музыкальных звуков группи-
руется около семи основных тонов музыки, и все разнообразие
цветовых оттенков — около семи цветов радуги. Однако класси-
1 М. Е. Лобашев, Изучение приспособления животных методом
рефлексов, Журн. общ. биол., 1955, XVI, 2.
5о
Цветение растений
фикация запахов еще не установлена, что объясняется как значи-
тельной субъективностью обонятельных ощущений, так, частью,
и тем, что химическая природа веществ, обусловливающих запахи,
еще мало изучена. Первая попытка в этом отношении сделана Кер-
нером, различающим пять групп растительных запахов. Одну груп-
пу образуют запахи,*появляющиеся при разложении белковых ве-
ществ. Запахи эти напоминают то разлагающееся мясо млекопи-
тающих, то тухлую рыбу, то навоз и т. п. Из наших растений та-
кой запах испускают во время цветения аройники, кирказоны. Из
растений других стран особенно выделяются в этом отношении ба-
ланофореи, раффлезии и некоторые южноафриканские растения;
раффлезии — паразитные растения, водящиеся на островах Борнео,
Суматре, Яве и на Филиппинах и имеющие самые большие из всех
растений цветки (до одного метра в поперечнике!). Интересно от-
метить, что цветки с гнилостным запахом обыкновенно имеют и окра-
ску разлагающегося мяса, представляющую комбинацию цветов
бледнорозового, грязно-фиолетового и т. п. Одно из веществ, обра-
зующееся при гниении белков и неприятно пахнущее, — индол,
почему Кернер и назвал эту группу запахов индолоидными.
К индолоидным запахам близки аминоидные, имеющие в основе
своей вещества амины (особенно—триметиламин), которые также
представляют продукт распада белковых веществ. Наиболее ха-
рактерные из этих запахов (обязанные триметиламину) можно срав-
нить с запахом селедочного рассола. Пример представляют запахи
цветков боярышника, таволг, калины, рябины, плюща и др.
Самая широкая группа цветочных запахов обязана аромати-
ческим веществам (производным от бензола); эти запахи названы
Кернером бензольными. Сюда относятся самые приятные ароматы.
Таковы запахи гвоздики, сирени, ландыша, резеды, жасмина, жи-
молости, белой акации, фиалки, ванили, ясминника и мн. др.
Замечательно, что многие запахи повторяются в растениях са-
мых разнообразных семейств, особенно это наблюдается по отноше-
нию к бензольным запахам. Например, запах гвоздики свойствен
не ей одной, но также многим видам заразихи, некоторым орхид-
ным, нарциссу; запах гиацинта, кроме самого гиацинта, — многим
смолевкам, пеларгониям с темными цветками; запах фиалок ощу-
щается у многих крестоцветных — у левкоев, желтофиоли и др.;
очень распространен запах ванили и т. д.
Много можно привести и обратных примеров, когда сходные
виды имеют совершенно различные запахи. Так, различные виды
волчника пахнут то ванилью, то сиренью, то фиалками, то гвозди-
кой; весьма разнообразны запахи орхидных и запах роз; запах
душистого горошка не повторяется в других горохах, запах белой
акации чужд желтой и т. д.
Четвертая группа запахов — парафиноидные (обязанные ве-
ществам, производным от углеводов парафинового ряда). Сюда
Время появления запаха
относятся запахи роз, валерианы, лип, винограда, гледичий и др.
Вероятно, сюда же относится запах свежего желтого пчелиного
воска и меда.
Наконец, последняя группа растительных запахов — терпе-
ноидные (обязаны бескислородным эфирным маслам—терпенам).
Такой запах имеют цветки померанцев, нередко он встречается у
губоцветных, например, у лаванды, тимиана.
Нередко бывает, что у одного цветка выделяются два запаха.
В таких случаях определение запаха, конечно, затрудняется. Наи-
более оно затрудняется, когда из двух ароматов получает преобла-
дание то один, то другой и когда такое соотношение изменяется
в течение дня.
Различные насекомые обнаруживают к различным запахам не-
одинаковое отношение. Одни запахи привлекают одних насекомых,
другие — других. Так, неприятные индолоидные запахи привле-
кают тех насекомых, преимущественно мух и жуков, которые обы-
кновенно водятся на падали, гниющих экскрементах и т. п. ’Как
уже сказано, цветки с таким запахом (кирказоны, раффлезии и
др.) обыкновенно и по окраске походят на падаль.
Большинство бабочек, пчел и шмели гнилостного индолоидного
запаха обыкновенно не замечают. Аминоидные запахи чаще всего
привлекают жуков, в особенности бронзовок, затем мух, перепон-
чатокрылых, но не привлекают бабочек. Запах меда привлекает
главным образом пчел,ос и дневных бабочек; насекомые же, пред-
почитающие индолоидный запах, меда совершенно не чувствуют.
Чувствительность насекомых к привлекающим их запахам
иногда достигает большой остроты. Несомненно, однако, что во-
прос об обонянии насекомых требует еще дальнейшего изучения.
Появление запаха совпадает с тем временем, когда летают на-
секомые, опыляющие данные цветки. То же следует сказать вообще
и о распускании цветков. Так, многие гвоздичные растения, посе-
щаемые и опыляемые ночными бабочками, днем совершенно не пах-
нут, и венчики их даже закрываются, с вечера же они раскрыва-
ются и начинают выделять сильный аромат. Такова, например, смо-
левка. Днем лепестки ее завернуты кончиками внутрь, закрывая
отверстие цветка, и обращены кнаружи невзрачной стороной;
аромата нет, и вообще цветок производит впечатление увядшего
(рис. 30); поэтому дневные насекомые не посещают его. На ночь
картина меняется (рис. 31). Венчик расправляется, и цветки начи-
нают сильно пахнуть. Каждый цветок распускается три ночи кряду:
в первую ночь выставляются наружу первые пять тычинок, во вто-
рую— другие пять (в цветках гвоздичных всего 10 тычинок), а
на третью ночь, когда все тычинки уже завяли, высовываются
столбики с рыльцами (в числе трех). Обращенная ночью кнаружи
сторона лепестков у слепца белая; такова вообще окраска ночных
цветков, что связано с тем, что ночью белый цвет самый заметный.
52
Цветение растений
В противоположность ночным цветкам многие дневные, посещаемые
дневными бабочками, пчелами и шмелями, с заходом солнца пере-
стают испускать запах; таковы цветки бобровника, белозора, не-
которых видов сливы и др.
У тех из опыляемых насекомыми цветков, у которых одна из
особенностей, делающих их заметными, например, окраска, недо-
статочно развита, естественно ожидать значительного развития
другой особенности—запаха.
Так оно и оказывается па
самом деле. Резеда, виноград,
плющ, гледичия и т. п. —
все это растения с невзрач-
ними, хотя и опыляемыми
насекомыми цветками. Зато
опи отличаются сильным раз-
витаем аромата. Тем же ка-
\ чеством отличаются и цветки,
fju \ растущие в тени, как у фиа-
\ > - — лок, или запРятанные в ли‘
1 стве, как у лип. Обратные
ОД примеры встречаются среди
й растений с большими, ярко
а мь окрашенными цветками, ра-
В Жд) стущих на открытых местах; у
1 них аРомат часто отсутствует,
ж Таковы тыквы, пионы, под-
1| солнечники, многие горе-
W чавки и др. Однако яркая
окраска и сильный аромат
Рис. 30. Смолевка (Silene nutans) дне^м далеко не всегда исключают
друг друга. Розы, гвоздики,
левкои, сирень, многие тропические орхидные и другие пред-
ставляют нам примеры совмещения того и другого средства привле-
чения насекомых в сильно выраженной степени. Есть растения,
у которых пахнут листья: таковы, например, мяты, ясминник и др.
Аромату ночных цветков, кроме привлечения ими ночных насеко-
мых, приписывается еще и другое значение. В ясные, тихие ночи
растения подвергаются значительному охлаждению вследствие
сильного лучеиспускания. Для цветения же нужна более или менее
высокая температура. Хотя цветки вырабатывают тепло усиленным
дыханием, но все же сильное лучеиспускание могло бы чрезмерно
понизить температуру цветка. Функцию защиты от охлаждения
и выполняют эфирные масла, придающие цветку его аромат.
Пары испаряющихся эфирных масел окружают цветок как бы про-
зрачным облаком. Пары эти перехватывают излучаемую цветками
теплоту и, таким образом, являются для них как бы стеклян-
Приспособления цветков к опылению насекомыми
53
Рис. 31. Смолевка (Silene nutans) ночью. На
один цветок садится ночная бабочка
ным колпаком, предохраняющим их от излишнего охлаждения.
Все указанные приспособления в высокой степени повышают
вероятность посещения цветков насекомыми. Однако само это по-
сещение еще не является гарантией того, что они произведут опы-
ление. В связи с этим у цветков выработался еще ряд особенностей,
способствующих их опылению с помощью насекомых. Необходимо,
чтобы насекомые могли
свободно располагаться на
цветке или вообще полу-
чить доступ к его внутрен-
ним частям и чтобы, полу-
чая на нем пищу, они в то
же время обязательно за-
хватывали пыльцу и остав-
ляли ее на рыльце другого
такого же цветка. Такие
приспособления мы дейст-
вительно и видим.
Проще всего обстоит
дело у цветков прямостоя-
щих, имеющих правильные
открытые венчики, каковы
лютики, ветреницы, лап-
чатка, маки, зонтичные,
сложноцветные и т. п. Эти
цветки доступны очень мно-
гим насекомым — жукам,
мухам, осам и др. Рас-
полагаясь на поверхности
такого цветка или соцве-
тия, насекомое обыкновен-
но нижней частью своего тела захватывает пыльцу, которую затем
переносит на рыльце другого экземпляра. Но наряду с цветками
такой простой формы существует множество других, имеющих
специальные приспособления к посещению и опылению насекомыми.
Приспособления эти чрезвычайно многочисленны и разнообразны.
Если обратиться к рассмотрению отдельных представителей флоры,
то можно найти сотни и тысячи иллюстраций этого.
Специальные приспособления наблюдаются уже в положении
цветков. Мы видим цветки, направленные вбок или вниз, причем
у многих растений такое положение цветков, наиболее удобное
для посещающих их насекомых, наблюдается только тогда, когда
цветки уже готовы к опылению. Например, у колокольчиков можно
наблюдать, что их цветоножки, первоначально отвесные, незадолго
до раскрывания венчика так сильно изгибаются, что вход в цветок
оказывается обращенным вниз, а такое положение крайне^выгодно
54
Цветение растений
для шмелей и пчел; у жимолости и ночной свечи цветок до созре-
вания торчит прямо кверху, а когда созреет для приема насеко-
мых, поворачивается в сторону; разнообразные примеры изменения
положения цветка ко времени его созревания находим среди
Рис. 32. Щмель на цветке дрока
(Spartium junceum)
орхидных, у некоторых мотыль-
ковых и др.
К числу посетителей цветков,
обращенных входным отверстием
вбок, относятся, между прочим, такие
насекомые, которые сосут мед на
лету, кружась около цветка, каковы
ночные бабочки; сюда же относятся
и птички колибри. Таким посетите-
лям не нужно точки опоры, не нужно
приспособлений, могущих служить
пристанищем: их и нет действительно
у опыляемых этими животными цвет-
ков. Однако большинство насекомых
(пчелы, шмели, осы) нуждается в та-
ких пристанищах, на которых они
располагаются, прежде чем проник-
нут к месту отложения меда. В связи
с этим на соответствующих цветках
имеются разнообразные «прилетные
площадки». Таково значение нижней
губы у губоцветных, медовой губы
у норичниковых, так называемой
лодочки и весел (крыльев) у мотыль-
ковых (рис. 32). Естественно, что
наличие таких приспособлений у
цветков, направленных вбок, делает
цветки неправильными: в большин-
стве случаев здесь лепестки неоди-
наковой формы, так как они нерав-
номерно развиты.
Нет возможности перечислить все
специальные приспособления, бла-
годаря которым расположившееся на цветке насекомое, доста-
вая мед, нагружается пыльцою и переносит ее на рыльце другого
такого же цветка. Для примера укажем на цветки шалфея и орхид-
ных. У шалфея пыльники соединены с так называемым коромыслом.
Когда насекомое, обыкновенно шмель, проникает в полость цветка,
оно неизбежно соприкасается с коромыслом. Пыльники от этого на-
клоняются, ударяют насекомое по спине и обсыпают его пыльцой.
По наблюдениям акад. Н. Г. Холодного, перенос пыльцы на рыльце
происходит следующим образом. Взлетая с цветка, шмель взды-
Взаимная зависимость растений и насекомых
55
мает облачко пыльцы со своей спинки. Парящая пыльца подхва-
тывается токами воздуха и часть ее заносится на рыльце (рис. 33).
У орхидей (рис. 34)
комочки клейкой пыль-
цы приклеиваются ко
лбу посещающих их на-
секомых в виде «рогов»
(поллинариев), которые
затем переносятся на
рыльца других экземп-
ляров того же растения.
Укажем еще, что внутри
цветков нередко наблю-
даются различные пре-
Рис. 33. Опыление шалфея (Salvia glutinosa)
(по Холодному)
грады в виде узких про-
ходов, выступов, утолщений, волосяного покрова и т. п. Значе-
ние этих преград заключается, между прочим, в том, что благо-
даря им проникающий в глубину за медом .хоботок движется
И
Рис. 34. Опыление ятрышника, или
кукушкиных слезок (Orchis maculata).
I — цветок сбоку; на губе муха с дву-
мя комочками цветня (/—2) на голове;
II — средняя часть цветка и губы
ятрышника: 1 — пыльники, 2 — рыльце,
3 — вход в шпорец; III — комочек
цветня 1 с ножкой 2 и прилипаль-
цем 3
чилось там с клевером. Он стал
семена лишь с появлением шмелей,
именно по тому пути, следуя
которому, он то покрывается
цветочной пыльцой, то остав-
ляет ее на рыльце пестика.
Приспособления цветков
к опылению их определен-
ными видами насекомых при-
водят к тому, что в случае
отсутствия этих насекомых
(например, при перенесении
растений в чужую страну)
растения оказываются почти
или совершенно бесплодными.
Яблоня, например, попав в
Австралию, первое время со-
вершенно не давала плодов,
несмотря на роскошное цве-
тение, пока одним из пересе-
ленцев не были привезены
туда пчелы. То же самое слу-
давать в большом количестве
так как ни одно из местных
насекомых не оказалось подходящим для совершения опыления.
И среди насекомых много таких, которые находятся в подоб-
ной же зависимости от посещаемых ими цветков. Это те насекомые,
для которых цветки являются единственным источником питания.
Таковы, например, почти все бабочки. Их хоботок, представляя
совершенное приспособление к сосанию меда из цветочных нектар-
56
Цветение растений
ников, другой пищи и в другой обстановке взять не может. Исчезни
цветки, —и отряд чешуекрылых (бабочек), за исключением тех от-
носительно немногих представителей, которые питаются соком,
вытекающим из коры некоторых деревьев, из плодов и т. п.,неми-
Рис. 35. Сосна (Pinus silvestris). I — ветка сосны, на которой находятся муж-
ские шишечки/, женские шишечки 2 и зеленая годовалая шишка.?; II — ши-
шка с созревшими семенами; III — мужская шишечка; IV — женская ши-
шечка; V — семенная чешуя из женской шишечки с двумя семяпочками
(III, IV, V — при разных увеличениях)
нуемо должен погибнуть.1 Следовательно, приспособление и зави-
симость между цветками и насекомыми — взаимные.
На этих примерах хорошо видна вся относительность целесо-
образности в строении цветков и насекомых. И то, и другое, взятое
само по себе, оторванно от окружающей среды, не является ни це-
лесообразным ни нецелесообразным. Целесообразность проявляется
только в отношении к опредепенной обстановке. Изменяется эта
обстановка, — и то, что было раньше целесообразным, становится
теперь вредным, ведущим организмы к вымиранию.
1 Необходимо, однако, отметить, что некоторые бабочки (например, са-
турнии) совсем не питаются в половозрелом состоянии.
Опыление с помощью ветра
57
Наряду с насекомыми другим фактором, производящим опыле-
ние цветков, является ветер. По приблизительному подсчету, ве-
тром опыляется от 10 до 12 тысяч видов, что составляет около т/10
части всех цветковых растений. Сюда относится большинство де-
ревьев наших стран: хвойные (рис. 35), ольхи (рис. 36), орешники
(обыкновенный и грецкий), дубы, буки, березы, тополи; из южных
древесных растений к ветроопыляемым принадлежат пальмы. Из
Рис. 36. Ольха (Alnus glutinosa). 1 — ветка с цветками (весной); 2 — ветка
с листьями и заложенными соцветиями (осенью); 3 — цветок с пестиками
(увеличено); 4 — группы цветков с тычинками (увеличено); 5 — шишечки
с созревшими и высыпающимися плодами
травянистых растений сюда относятся: обширное семейство злаков
и близкие к ним осоки и ситники, щавели, конопля, крапива, подо-
рожники и мн. др. Ветром опыляются также многие широко рас-
пространенные полевые культуры: рожь, кукуруза, хмель, конопля,
сахарная свекла и др. В странах экваториальных процент ветро-
опыляемых растений — наименьший, а в полярных — наиболь-
ший; очевидно, это находится в зависимости от количества насе-
комых.
Само собой разумеется, что цветки ветроопыляемых растений,
обладая теми же главными частями, по форме и особенностям строе-
ния последних существенным образом отличаются от цветков, опы-
ляемых насекомыми. У тех и у других — совершенно различные
приспособления.
Ветроопыляемые цветки не обладают средствами, привлекаю-
щими насекомых. Здесь нет ни ярко окрашенных лепестков, ни меда,
58
Цветение растений
и, в
Рис. 37. Пыльца
сосны (под микро-
скопом)
ни запаха, нет разнообразных приспособлений для приема насеко-
мых и т. д. В силу этого цветки ветроопыляемых растений весьма
невзрачны, почти не выделяются на фоне зеленой листвы и потому
мало заметны. В обиходном наречии они обыкновенно даже и не
считаются цветками. Нельзя сказать, что цветки эти совсем не по-
сещаются насекомыми. Насекомых, питающихся пыльцою, нередко
можно видеть на цветочных сережках орешника и березы, на ко-
лосьях злаков и т. д. Но здесь насекомые не могут переносить
пыльцу на рыльца, так как все строение цветков к этому непри-
частности, так как большинство их раздельно-
полые, причем, женские (пестичные) цветки, ли-
шенные пыльцы, не посещаются насекомыми.
Правда, когда насекомое садится на цветок
ветроопыляемого растения, от сотрясения может
выпадать пыльца, которую попутный ветер мо-
жет относить на рыльца. Но зато при безвет-
рии или противном ветре пыльца эта едва ли
попадет по назначению, так что в итоге посе-
щение насекомых скорее вредно, чем полезно
типичным ветроопыляемым растениям.
Пыльца этих растений вполне приспособлена
к переносу ветром. Она имеет вид пыли или
муки. Отдельные пылинки чрезвычайно мелки
и легки, их поверхности гладкие и сухие, не
склеиваются в комья и не пристают к другим
предметам. Уже легкое движение воздуха подхватывает эту пыльцу
и относит на значительное расстояние.
У многих хвойных, например у сосны, каждая отдельная пы-
линка снабжена особым летательным аппаратом: с двух сторон
у нее, между наружной и внутренней оболочкой,имеется по взду-
тию, наполненному воздухом (рис. 37); благодаря этому пылинки
приобретают подобие воздушных шаров.
Несомая ветром пыльца летит и падает на землю и на различ-
ные предметы. При таких условиях попадание пылинок на
рыльца, очевидно, является редкой случайностью. Для того чтобы
это происходило достаточно часто, пыльцы, очевидно, должно
быть большое количество. Так оно и есть на самом деле. Встрях-
ните цветущую ветвь сосны, орешника или ольхи — и полетят
целые тучи пыльцы; повторите тот же самый опыт с цветущим
кустарником смородины или крыжовника, и вы не заметите
ничего подобного: их цветки опыляются насекомыми.
Особенно много пыльцы производится сосною и другими хвой-
ными. В годы обильного цветения их в сосновых лесах при умерен-
ном ветре поднимаются громадные облака пыльцы и носятся выше
макушек деревьев; тогда не только женские (пестичные) шишечки,
цо и хвоя, сучья и ветки этих и соседних деревьев, листья и цветки
Особенности ветроопыляемых, цветков
59
Рис. 38. Тычиноч-
ная сережка ореш-
ника (Corylus avel -
Гапа) (увеличено)
трав на ближайших лугах и даже почва кажутся усыпанными жел-
той пылью, прозванной «серным дождем». Если в период цветения
хвойных пройдет дождь, который смоет пыльцу, то нередко в та-
ких случаях стекающие с пригорков дождевые воды кажутся окра-
шенными в желтый цвет и оставляют полосы и пятна желтого по-
рошка. Это дало повод встречавшемуся ранее поверью о падающем
с неба серном дожде. Число пылинок, раскидываемых одной особью
кукурузы, составляет десятки миллионов, а од-
ной особью крапивы—миллиарды. Сколько же
их на цветущей сосне?
Столь значительная продуктивность пыльцы
является целесообразным приспособлением к
опылению с помощью ветра. Однако нельзя не
отметить всю относительность этой целесооб-
разности. Гибель многих миллиардов пыльце-
вых зерен, из которых для опыления оказы-
вается использованным только ничтожно малый
процент, — вот что мы находим у всех ветро-
опыляемых растений.;
Попаданию пыльцы на рыльца цветков у
этих растений способствует также и то, что они
обыкновенно встречаются большими группами.
Каждому это более или менее хорошо известно
из собственных наблюдений не только над дре-
весными ветроопыляемыми породами, но и над
травянистыми—злаками, огоками, крапивой и
другими, образующими обыкновенно заросли.
Форма и положение цветков и соцветий
ветроопыляемых растений приспособлены к
более легкому выдуванию пыльцы ветром.
Так, тычинковые сережки, на осях которых располагаются муж-
ские цветки, как это видно на примерах ольхи (рис. 36), орешника
и многих других деревьев, легко болтаются по ветру во все стороны;
такому их раскачиванию способствует и их положение на кончиках
тонких наружных ветвей.
Понаблюдайте в начале весны за развитием соцветий ольхи
или орешника, и вы увидите такую картину: ко времени созревания'
пыльцы ось тычинкового соцветия оказывается вытянутой в длину,
так что отдельные цветки отодвигаются друг от друга (рис. 38);
пыльцевые мешочки в это время лопаются, освобождаются от
пыльцы, которая попадает на спинную сторону следующей ниже
лежащей чешуйки (прикрывающей пыльники), откуда пыльца
легко сдувается всяким порывом ветра. У многих других ветроопы-
ляемых растений пыльники выдвинуты из цветка и висят на нитях
так, что ветер при малейшем дуновении легко может раскд^
пивать их,
60
Цветение растений
Это можно наблюдать, например, у злаков (рис. 39), осо-
ковых,
у конопли, хмеля, подорожника и мн. др.
Рис. 39. Цветки злака (французского рай-
граса — Arrhenatherum elatius) с раскрывши-
мися пыльниками; внизу — при ветре (уве-
личено)
У некоторых ветро-
опыляемых растений
цветень с силой вы-
брасывается на воздух
в момент раскрывания
пыльников. Из наших
растений это особенно
хорошо можно наблю-
дать у крапивы. Кра-
пива р аз дел ьноп о л а
(рис. 40). Четыре ты-
чинки мужских цвет-
ков свернуты внутрь
так, что пыльцевые ме-
шки сидят в центре
цветка, тесно прижа-
тые друг к другу. В ро-
систое, ясное летнее
утро хорошо бывает
видно, как от стеблей
крапивы то там, то здесь
поднимаются облака
пыли. Это происходит
благодаря разрастанию
тычиночных нитей, от-
чего получается некоторое напряжение,
сыхании росы; в момент,
дела, пыльники рас-
крываются и выбрасы-
вают пыльцу на воздух.
Рыльца ветроопы-
ляемых цветков при-
способлены к улавли-
ванию цветня. Обыкно-
венно они выставлены
наружу и покрыты во-
лосками или развет-
вляются, приобретая
форму то перьев, то
кистей, то султанов и
т. п. (рис. 39). Попа-
дающие между этими
увеличивающееся при вы-
когда напряжение достигает своего пре-
Рис. 40. Цветки крапивы (Urtica dioica). 1—пе-
стичный, 2 и 3— тычиночные (до раскрыва-
ния и в момент раскрывания пыльников;
увеличено)
разветвлениями и волосками пылинки запутываются, точно мухи
в паутине,
Приспособления, препятствующие самоопылению
61
На пути цветня от тычиночных цветков к пестичным, чтобы
пылинки попадали на рыльца, не должно быть препятствий. Путь
этот не должен быть загражден, например, широкими листьями,
на которых застревала бы и погибала большая часть пыльцы. Со-
ответственно этому мы и видим, что цветки, опыляемые ветром, со-
браны в колоски, сережки или метелки, располагающиеся на по-
верхности растения, на наружной части кроны. Кроме того, боль-
шое число ветроопыляемых растений цветет ранней весной, когда
листва еще не распустилась, как у ольхи, орешника, ясеня, или
распустилась, но еще не вполне, как у березы. Весной и ветры бы-
вают более часты.
Говоря о расположении цветков на деревьях, интересно отмс-
тить, что женские цветки большей частью находятся выше муж-
ских. Значение этого факта в том, что пыльники раскрываются не
во всякую погоду, а лишь в теплые, преимущественно довольно
тихие дни. При таких условиях образуется восходящее воздушное
течение. Оно-то и поднимает пыльцу кверху, на рыльца женских
цветков.
Так как, несмотря на все указанные приспособления, громад-
ное количество пыльцы у ветроопыляемых растений пропадает
даром, то, очевидно, этот способ опыления и эта форма цветков
являются более примитивными, нежели у растений, опыляемых
насекомыми. Среди ветроопыляемых наиболее низкую ступень
представляют хвойные. У них семяпочки лежат открыто, нет при-
способленного для улавливания пыльцы рыльца. Следовательно,
шансов на попадание пыльцы по назначению здесь менее всего, а
непроизводительная трата ее более расточительна. В связи с этим
и производится ее здесь, как мы видели на примере сосны, особенно
много.
Между растениями ветроопыляемыми и насекомоопыляемыми
могут быть указаны переходные ступени. Существуют растения,
которые более приспособлены к опылению ветром, но могут также
опыляться и насекомыми; такой случай можно наблюдать, на-
пример, у подорожника. Есть и обратные случаи. Так, вереск
(рис. 41) особенно усердно посещается пчелами. Однако к концу пе-
риода цветения его упругие тычинки с пыльниками, легко выбрасы-
вающими пыльцу, нагибаются к самому краю цветка; резкий осен-
ний ветер сдувает пыльцевидный цветень и далеко разносит его во-
круг, причем он нередко попадает на торчащие из цветка рыльца
отдаленных растений.
И насекомые, и ветер, будучи в состоянии перенести пыльцу на
значительное расстояние, тем скорее могут перебросить ее на рыльце
того же самого цветка. Следовательно, чтобы опыление было только
перекрестным, необходимы приспособления, которые ограждали
бы рыльце цветка от собственной его пыльцы и таким образом ме-
шали бы самоопылению. Наблюдения показали, что такие
Цветение растений
приспособления против самоо
притом в высшей степени
Рис. 41. Вереск (Calltma vul^aHs). 1—
разрез цветка (чашечка уда. ена); 2—
отдельный цветок; 3—раскрывающаяся
коробочка (7, 2, 3 — увеличено)
опыляющий и опыляемый цветки обязательно находятся и на раз-
ных особях, у однодомных же — как на одной и той же особи, так
ления действительно имеются,
разнообразные. Познакомимся
с наиболее типичными из
них.
Прежде всего отметим
явление разделения полов
у растений.1 Как известно,
у множества растений име-
ются цветки или только с
тычинками—тычиночные, или
мужские цветки, или только
с пестиками—пестичные, или
женские цветки. У одних
однополых растений и тычи-
ночные, и пестичные цветки
находятся на одном и том же
экземпляре; это — однодом-
ные растения, пример кото*
рых представляют сосна, ель,
береза, дуб, ольха, орешник,
осока, кукуруза, тыква и др.
У других — мужские и жен-
ские цветки распределены
на разных особях; это—дву-
домные растения, каковы ива,
тополь, осина, конопля,
хмель, щавель, крапива и
мн. др. Очевидно, у однопо-
лых растений опыление цвет-
ков (пестичных) возможно
только пыльцой с другого
цветка (тычиночного). Мало
того, у двудомных растений
и на разных. Последнее обязательно для многих однодомных, ты-
1 Следует иметь в виду, что тычинки и пестики являются органами бес-
полого поколения, в которых развиваются микроспоры и макроспоры,
представленные здесь в форме пыльцевых зерен (в пыльнике) и зародыше-
вого мешка (в семяпочке). Пыльцевые зерна (микроспоры) дают начало .муж-
скому, а зародышевый мешок (макроспора) — женскому половому поко-
лению. Поэтому старое представление о цветке как об органе полового раз-
множения неправильно и должно быть оставлено. Говоря о «разделении
полов» у растений, следует помнить, что этим широко распространенным
термином, так же как и выражениями «мужской» и «женский» цветок,
можно пользоваться только условно. Подробно об этом ?см. в учебниках
ботаники для вузов.
Приспособления, препятствующие самоопылению
63
чиночные и пестичные цветки которых, находящиеся на одном
и том же растении, развиваются в разное время.
У многих двуполых растений опыление цветка собственной пыль-
цой вовсе исключается или чрезвычайно затрудняется расположе-
нием тычинок и пестиков относительно друг друга. Например,
обратим внимание на белую или желтую лилию; в цветках этих
растений столбик настолько выдается за нагруженные пыльцой
пыльники, что на рыльце эта пыльца попасть не может (рис. 42).
Если на цветок прилетает насекомое, предварительно нагруженное
пыльцой с другого цветка лилии, и опускается прямо на столбик,
то оно производит перекрестное опыление. Подобное же соотно-
шение между частями
цветка можно наблю-
дать у многих кресто-
цветных, бурачниковых,
норичниковых, вьюнко-
вых, жимолостных и
других растений.
Своеобразно соотно-
Рис. 42. Тычинки и пестик белой лилии
(Lilium candidum). Три тычинки удалены.
Рыльце на 26 мм высовывается из-за пыль-
шение между тычин- ников
ками и пестиками, на-
зываемое гетеростилией (разностолбностью). Оно заключается
в неодинаковой высоте тычинок и пестиков на разных растениях
одного и того же вида. Именно — в одних цветках высоко поднято
рыльце, сидящее на длинном столбике, пыльники же глубоко
скрыты в венчике; в других, наоборот, выглядывают пыльники,
а рыльце скрывается в глубине. Эти два сорта цветков всегда поме-
щаются на различных экземплярах растения. Пример гетеро-
стилии представляет растение первоцвет весенний (рис. 43 и 44)
и многочисленные другие первоцветные, многие бурачниковые,
например, известная нам медуница (рис. 6), некоторые горечавки
и др. Опыты показали, что у гетер остальных растений опыление
длинностолбчатого цветка пыльцою короткостолбчатого и наобо-
рот дает лучшие результаты. Другими словами, лучшие результаты
получаются при опылении рыльца пыльцой пыльников, находя-
щихся с ним на одной высоте. А к такому именно опылению посред-
ством насекомых гетеростильные цветки и приспособлены. В са-
мом деле, насекомое, вводящее свой хоботок в короткостолбчатый
цветок, коснется пыльников такою частью тела, которая придется
против рыльца длинностолбчатого цветка. Точно так же, при из-
влечении нектара из длинностолбчатого цветка, пыльца, находя-
щаяся в глубине венчика, пристанет к такой части тела, которая
придется против рыльца цветка короткостолбчатого. Почему же
длинностолбчатые цветки гетеростильных растений лучше опыля-
ются короткостолбчатыми и наоборот? Это зависит от особенностей
рыльца и пыльцы в том и другом случае. Наблюдения Дарвина
64
Цветение растений
показали, что в цветках длинностолбчатых рыльце снабжено гораздо
более длинными сосочками, а пылинки, наоборот, заметно мельче,
чем в цветках с короткими столбиками. Длинные сосочки на рыльце
первой формы легко задерживают более крупные пылинки второй
формы, и наоборот, более короткие сосочки второй соответствуют
_ более мелкой пыльце первой
Рис. 43. Первоцвет весенний
(Primula veris). 1 — отдельный
распустившийся цветок; 2 —
лопнувший плод (коробочка)
формы.
Самое распространенное при-
способление, мешающее самоопыле-
нию, заключается в неодновремен-
ном развитии мужских и женских
органов каждого отдельного цвет-
ка. Явление это носит название
дихогамии. Ранее описанные при-
способления можно назвать разъ-
единением полов в пространстве,
а дихогамию — разъединением их
Рис. 44. Два цветка первоцвета
в продольном разрезе: справа —
короткостолбчатый, слева—длин-
ностолбчатый. 1 — рыльце; 2 —
столбик; 3 — венчик; 4 — пыль-
ники; 5 — чашечка; 6 — завязь
во времени. Дихогамия может быть двух видов: у одних растений ты-
чинки развиваются раньше пестика, и пыльца высыпается из пыль-
ников, когда рыльце еще не готово к ее восприятию; у других, на-
оборот, готовое к восприятию рыльце выставляется из цветка в то
время, когда его пыльца еще не созрела. Первый случай называется
протерандриею, второй — протерогинией). Возможность опыления
в том и другом случае легко понять, если иметь в виду, что цветки
никогда не распускаются все одновременно, а одни раньше, другие
позже. За исключением самого начала и самого конца периода цве-
тения, всегда будет готовая пыльца в одних и готовое рыльце в дру-
Приспособления, препятствующие самоопылению
65
гих цветках того же вида. Насекомое, перелетая с цветка на цветок,
в одном коснется раскрывшихся пыльников, в другом — созрев-
шего рыльца и таким образом произведет перекрестное опыление.
Протерандрия встречается в природе чаше протерогинии. Есть це-
лые большие семейства, у представителей которых пыльники со-
зревают ранее рыльца; таковы почти все сложноцветные, колоколь-
чиковые, зонтичные, гераниевые (рис. 45), многие лютиковые, гвоз-
дичные (рис. 46) и др. Протерогинию можно наблюдать у подорожни-
ков, многих злаков, у водных рас-
Рис. 45. Герань (Geranium Рис. 46. Протер.андричные цветки гвоз-
silvaticum) с протерандрич- дики (Dianthus)
ними цветками
Весьма часто приспособления, направленные к разъединению
полов в пространстве, соединяются с дихогамией, т. е. разъедине-
нием их во времени. Это сочетание двоякого рода приспособлений
можно наблюдать среди цветков гетеростильных, однополых и т. п.
Так, у весьма многих однодомных растений пестичные и тычиноч-
ные цветки, находящиеся на одном растении, созревают не одно-
временно— первые ранее последних (протерогиния).
В результате опыление производится пыльцой не только с дру-
гого цветка, но и с другого растения. Это наблюдается, например,
у однодомных древесных растений — ольхи, орешника, березы,
дуба, вяза и др. У них рыльца делаются способными к восприятию
пыльцы на несколько дней ранее созревания последней в тычиноч-
ных цветках того же растения.
Изучение приспособлений, служащих для скрещивания, и тех
выгодных последствий, которые связаны с перекрестным опыле-
нием, привело некоторых исследователей к крайним увлечениям.
Некоторыми из ближайших последователей Дарвина, продолжав-
66
Цветение растений
ших его изыскания в области биологии цветения, было высказано
мнение, что самооплодотворение или совсем не существует в расти-
тельном царстве, или встречается очень редко. Позднейшие наблю-
дения обнаружили, однако, что такое мнение ошибочно. Оказы-
вается, что в природе имеется большое число самых разнообразных
растений, размножающихся обыкновенно самоопылением. Сюда от-
носится множество травянистых растений с мелкими невзрачными
цветками. Для примера можно указать на крестовник, звездчатку
обыкновенную, осот, торицу, некоторые лютики, кипреи и др.
Рис. 47. Цветок куколя (Agrostemma githago).
I — продольный разрез цветка; II и III — ты-
чинки и пестики после удаления покровов. 1—внеш-
ние, 2 — внутренние тычинки, 3— рыльца, 4 — за-
вязь. На трех изображениях показано последова-
тельное развитие тычинок и пестиков
Сверх того, самоопыление встречается и среди тех растений, где
преобладает перекрестное опыление. В большинстве случаев само-
опыление устранено или затруднено у них лишь в начале цветения.
Под конец же в цветке, близком к увяданию, иногда даже создаются
условия, облегчающие самоопыление.
Познакомимся с одним из разительных примеров самоопыления,
который можно наблюдать у общеизвестного растения — куколя
(рис. 47),растущего среди хлебных посевов. У него два круга ты-
чинок — наружный и внутренний. Сначала выбрасывает пыльцу
наружный круг тычинок; затем развертываются столбики (проте-
рандрия); к концу периода цветения, наконец, начинают подни-
маться тычинки внутреннего круга и выносят свои пыльники; стол-
бики к этому времени настолько высовываются наружу, что могут
получить от этих тычинок пыльцу.
Вообще говоря, средства, которыми достигается самоопыление,
также весьма разнообразны: то тычиночные нити под конец цвете-
ния начинают усиленно расти, благодаря чему пыльники подни-
маются до уровня рыльца, с которым и соприкасаются, как это мы
видели сейчас у куколя; то удлиняются лепестки и поднимают при-
росшие к ним тычинки; то, наоборот, удлиняется столбик или за-
Самоопыление
67
вязь, и кверху поднимается рыльце, дорастая до стоявших выше него
тычинок; то изгибаются или цветоножка, изменяя прежнее поло-
жение цветка так, что на рыльце начинает сыпаться собственная
пыльца, или столбик, или тычиночные нити, вследствие чего пыль-
ники и рыльца входят в соприкосновение (рис. 48); то венчик су
живается и также сближает пыльники с рыльцем; то венчик вместе
с приросшими к нему тычинками опа-
дает, и таким образом рыльце про-
пускается сквозь кружок пыльников
и т. д. Словом, происходит удлине-
ние, укорачивание или изменение
относительного положения отдельных
частей цветка, в результате чего он
под конец цветения опыляется собст-
венной пыльцой.
Дихогамия — один из указанных
выше способов предохранения от
самоопыления — чаще всего встре-
чается в виде неполной дихогамии:
несовпадение в созревании тычинок
и пестиков наблюдается в начале
периода цветения, а потом, прежде
чем опередившие в своем развитии
органы (тычинки —у протерандрич-
ных цветков и пестики — у протеро-
гиничных) успеют завянуть, созре-
вают и органы отставшие. В этих
случаях вначале возможно только
перекрестное опыление, а после де-
лается возможным и самоопыление.
Там, где самоопыление преобла-
дает, сближение пыльников с рыль-
Рис. 48. Цирцея (Circaea alpi-
na). 1 — недавно распустив-
шийся цветок (передний лепе-
сток удален); 2 и 3 — цветок
на позднейших стадиях разви-
тия; 4 — плод (/, 2, 3, 4 —
увеличено)
цветения; там можно
вокруг
цами происходит не только под конец
видеть, например, что первые все время расположены
вторых и созревание тычинок и пестиков наступает одновременно
(обыкновенная звездчатка, торица, некоторые лютики, кипреи).
Весьма интересен способ самоопыления (автогамии), известный
под названием клейстогамии. У некоторых растений на одном и том
же экземпляре образуются два совершенно различных сорта цвет-
ков. Одни цветки обыкновенные: они сравнительно крупны, бро-
саются в глаза и доступны насекомым, которыми и опыляются.
Другие наоборот, — мелкие, невзрачные и совсем не раскрыва-
ются, производя впечатление бутонов. Внутри этих последних под
прикрытием слаборазвитых покровов развиваются также тычинки,
пестик и происходит самоопыление. Образование таких закры-
тых самоопыляющихся цветков называется клейстогамией, а
68
Цветение растений
самые цветки — клейстогамными. Они встречаются у многих из
наших обыкновенных растений, например, у некоторых фиалок
(рис. 49), у недотроги, кислицы и др. У растений, обнаруживаю-
щих клейстогамию, таким образом, имеет место разделение функ-
ций между различными цветками; в одних происходит перекрестное
Рис. 49. Фиалка (Viola hirta). Слева—
целое растение: 1 — клейстогамные
цветки, 2 — плоды, 3 — засохший ве-
сенний цветок; I — увеличенный клей-
стогамный цветок, на нем: 1 — вен-
чик, 2 — чашечка; II — тот же цветок
с удаленной чашечкой
опыление, в других — толь-
ко самоопыление. У некото-
рых растений, например, у
обыкновенного льна, если
ко времени цветения упорно
стоит неблагоприятная пого-
да, цветки совершенно не
раскрываются, и внутри зам-
кнутых венчиков происходит
самоопыление. Такие случаи
представляют как бы переход
к клейстогамии.
Если перекрестное опыле-
ние, где оно существует, ве-
дет к образованию более здо-
ровых семян, каково же зна-
чение имеющего место наряду
с ним самоопыления? У тех
растений, где перекрестное
опыление является преобла-
дающим, а приспособления
к самоопылению развиваются
лишь под конец цветения,
этот последний способ имеет
значение, так сказать, «за-
пасного». Да и сами при-
способления к самоопыле-
нию у таких растений чаще
всего проявляются в полном
объеме в том случае, когда
предварительно не соверша-
лось перекрестного опыления. На цветки клейстогамные, су-
ществующие у растений вместе с раскрывающимися цветками, сле-
дует смотреть тоже как на запасные, дополнительные. Такие слу-
чаи, очевидно, нисколько не противоречат положению, что пере-
крестное опыление выгодно для растения. Но как помирить с этим
положением существование растений, у которых, как мы видели,
имеет место только самоопыление или оно, во всяком случае, пре-
обладает над перекрестным опылением? Для ответа на этот вопрос
наука в настоящее время еще не располагает достаточными данными.
Еще не выяснено с желательной полнотой и точностью, почему на-
Защита цветков от намокания
69
ходящиеся в завязи, в семяпочках, яйцеклетки только тогда начи-
нают усиленно питаться, размножаться и превращаются в конце
концов в зародыши семени, когда с ними слились мужские половые
клетки, образующиеся в пыльцевых трубках.1 Неизвестно еще,
что же такое привносят последние в яйцеклетку?
Пока можно высказаться только в общей форме, что, вероятно,
в них есть такие составные части, которые, попав в яйцеклетку,
выводят ее из состояния покоя или равновесия, заставляя усиленно
усваивать питательный материал и т. д. Другими словами, в хими-
ческом составе мужской половой клетки, по сравнению с яйцеклет-
кой, есть некоторые вещества, которые играют роль как бы воз-
будителей последующих энергичных физиологических процессов.
Если так, то естественно, что эти особенности или различия в со-
ставе сильнее выражены между половыми клетками с различных
экземпляров растения. С другой стороны, и близость в составе ме-
жду этими элементами, взятыми с одного цветка, не у всех расте-
ний одинакова. Где эта близость более, там мужская половая клетка
не может быть достаточным возбудителем для яйцеклетки, вслед-
ствие чего и самоопыление не дает удовлетворительных результа-
тов. Наоборот, где эта близость менее и вообще соотношение хими-
ческого состава более благоприятное, там может происходить само-
опыление, в связи с чем и не развились приспособления для скрещи-
вания. У многих самоопыляющихся растений роль дополнительного
возбудителя для яйцеклетки играют, быть может, внешние условия.
Дело в том, что большинство этих растений обладает способностью
к большому распространению. Попадая из одной местности в дру-
гую с отличными от первой условиями, они испытывают влияние
последних как новых возбудителей. Последняя мысль в свое время
была высказана еще Уоллесом.
Образование приспособленного к опылению цветка вызывает
необходимость его защиты от различных неблагоприятных усло-
вий. Прежде всего цветкам нужна защита от воды, падающей на
них в виде дождя или росы. Вода пагубно действует на нветень.
Пылинки быстро впитывают ее, набухая настолько, что нередко
1 Женская половая клетка (яйцеклетка) находится внутри семяпочки.
Последние располагаются внутри завязи в том или ином количестве (у раз-
ных растений от одной до нескольких тысяч). После оплодотворения семя-
почка развивается в семя, а вся завязь целиком—в плод. Из оплодотво-
ренной яйцеклетки образуется зародыш семени. Процесс оплодотворения
заключается в слиянии мужской и женской половых клеток. Мужская по-
ловая клетка (спермий) у семенных растений (у всех покрытосеменных и у
большинства голосеменных) устроена весьма просто и представляет собой
ядро, окруженное тонким слоем протоплазмы (генеративная клетка). Гене-
ративные клетки образуются в пыльце при ее прорастании в пыльцевую
трубку на рыльце. При наличии внутри завязи несколько семяпочек опло-
дотворение внутри каждой из них происходит независимо от других. Под-
робности об оплодотворении у растений см. в учебниках ботаники.
70
Цветение растений
даже лопаются. Словом, после смачивания пылинки, за самыми
немногими исключениями, делаются уже негодными, погибают.
Да и нектар нуждается в защите от воды, которая может вымывать
его. В странах тропических, где цветение начинается по окончании
периода дождей, этой опасности не существует. Не то в наших ши-
ротах. Однако здесь у растений в процессе естественного отбора
выработался ряд разнообразных приспособлений, защищающих вну-
тренние части цветка от смачивания.
У очень многих растений над цветками или их внутренними ча-
стями имеются постоянные покровы. Так, у липы цветки защищены
особым прицветным ли-
Рис. 50. Цветки липы (Tilia tomentosa)
стом, или прицветни-
ком, под которым они
расположены (рис. 50),
у недотроги (рис. 61)
и некоторых других
растений непосредствен-
но над каждым цвет-
ком находится по ли-
сту: какой бы сильный
дождь ни шел, дожде-
вые капли отскакивают
от этих листьев. У мно-
гих аройниковых, цвет-
ки которых собраны в
початок, этот послед-
ний /во время раскры-
вания пыльников почти
весь окутан большим, общим для всех цветков кроющим листом,
так называемым чехлом. Чаще роль защитных покровов, оберегаю-
щих пыльцу, исполняют части самого цветка. Например, в обшир-
ном семействе мотыльковых, известном каждому по душистому
горошку, тычинки вместе с пестиком совершенно скрыты в так на-
зываемой лодочке цветка. Насекомое, садясь на цветок, тяжестью
своего тела может отогнуть лодочку вниз и обнажить тычинки, но
от дождя они вполне защищены. У весьма также обширного и обще-
известного семейства губоцветных роль покрова выполняет верхняя
губа венчика (рис. 32); чаще всего она имеет вид шлема, под выпук-
лым сводом которого тычинки оказываются скрытыми от дождя.
В лесах и на лугах довольно часто встречается растение, известное
под именем купальницы, а также называемое в народе бубенчиками
(рис. 51). Довольно крупные шарообразные цветки его действи-
тельно походят на бубенчики. Цветок этот имеет многочисленные
золотистые чашелистики, которые прикрывают друг друга так, что
совершенно не видно скрытых внутри цветка многочисленных ты-
чинок и плодников. Дождевые капли неминуемо скатываются с его
Защита цветков от намокания
71
поверхности, а насекомые, именно сильные насекомые — пчелы —
проникают внутрь него только после усилий, затраченных для рас-
крытия его лепестковид-
ных чашелистиков.
В цветках касатиков
тычинки скрываются под
широкими лентовидными
рыльцами и т. п.
Есть немало таких
растений, которые не име-
ют никаких специальных
защитных покровов, но
их положение всегда тако-
во, что вода не попадает
внутрь. Это растения,
цветки которых постоян-
но обращены отверстием
вниз: их покров образует
Рис. 51. Цветки купальницы (Trollius euro-
paeus). 1 — целый цветок; 2 — цветок
в продольном разрезе
как бы естественную кры-
шу, по которой скатываются дождевые капли, не смачивая внут-
ренних органов. Хороший общеизвестный пример такого положе-
ния представляют цветки ландыша
(рис. 52); то же можно видеть у
вереска (рис. 41), брусники и др.
У некоторых, преимущественно
альпийских, растений (например,
у некоторых первоцветов, насе-
ляющих склоны высоких гор) устье
цветков не имеет никакой покрыш-
ки и всегда обращено кверху, бу-
дучи открытым для дождя и росы.
Однако и здесь смачивания пыльцы
не происходит. Возможность его
устранена тем, что венчик этих
цветков переходит в трубку, ко-
торая внизу сужена и даже как
бы перетянута. В этой суженной
части и находятся тычинки с пе-
стиком. Насекомые с тонким хо-
ботком без труда проникают сюда,
Рис, 52, Ландыш (Cbnvallaria ma- Доставая на дне венчика нектар
jalis) и захватывая попутно пыльцу.
Водяные же капли не могут туда
проникнуть, задерживаясь на верхней части венчика как бы на
тарелке, откуда скатываются ветром.
Во всех описанных случаях цветок хорошо защищен от сма-
72
Цветение растений
чивания благодаря своему устройству или постоянному положе-
нию. Но еще более распространенный способ защиты заключается
в изменении, при наличии неблагоприятных условий, или относи-
тельного положения отдельных частей цветка или положения
всего цветка, или же того и другого вместе.
Очень многие цветки и даже целые соцветия с открытыми и
обращенными кверху венчиками перед наступлением ненастной
погоды и даже на ночь закрываются. Таково большинство предста-
Рис. 53. Цветочные корзинки чертополоха (Carlina acaulis).
Налево — во время дождя и ночью, направо — днем в ясную
погоду
вителей громадного семейства сложноцветных, куда относятся наши
ромашки, маргаритки, георгины, васильки, ястребинки, чертопо-
лохи (рис. 53), а также многие колокольчики с поднятыми кверху
цветками, розы, пионы, опунции, кувшинки, шафраны (рис. 54)
и мн. др. Суточное открывание и закрывание цветков у некоторых
растений довольно точно приурочено к определенному времени.
Это дало возможность знаменитому шведскому естествоиспыта-
телю XVIII столетия Линнею установить «цветочные часы» по
времени открывания и закрывания определенных растений в опре-
деленные часы дня.
У другой, не меньшей группы растений изгибаются цветоножки,
и цветки из бокальчатых и чашевидных превращаются в свеши-
вающиеся вниз колокольчики (рис. 55). У одних этот изгиб совер-
шается на все то время, когда в цветке есть созревшая пыльца, нуж-
дающаяся в защите; таковы многие представители семейств па-
сленовых, норичниковых, лилейных, первоцветных, бурачниковых
Защита цветков от намокания
73
и др.; когда оплодотворение совершится, время цветения минует,
цветоножки часто снова выпрямляются. У других цветоножки по-
никают вниз периодически—в плохую погоду или на ночь; приме-
ром могут служить многие виды кислицы,мака,горицвета, лютика,
ветреницы, лапчатки, звездчатки, ясколки, паслена, тюльпана.
Подобно цветоножкам, у некоторых растений изгибаются целые
оси соцвгтий, так что вся кисть или колос и т. п. свешивается
Рис. 54. Цветки шафрана (Crocus multifidus). Налево —
в ненастье и ночью, направо — в ясный день
вниз. Часто одновременно наблюдаются два или даже три способа
защиты. Например, у многих ветрениц и лютиков, у перелески и
кислицы не только лепестки складываются над тычинками, но и
цветоножки наклоняются и свешиваются; в цветочной корзинке
маргаритки, осота и многих других сложноцветных вечерем и в сы-
рую погоду не только краевые язычковые цветки образуют крышу
над пыльниками срединных, но и стебелек, несущий соцветие,
также наклоняется, и корзинка поникает.
Чтобы нагляднее видеть, в какой мере распространены указан-
ные средства защиты цветков от неблагоприятных-условий, лучше
всего обратить внимание на резкую перемену общей картины цве-
тущего луга в зависимости от погоды и времени суток.
В теплые летние дни, при ясной погоде и безветрии зелень лу-
гов испещряется многочисленными раскрытыми цветками. Боль-
шинство из них обращено к солнцу; есть даже такие (например,
подсолнечник), которые прямо следуют за солнцем: утром обра-
74
Цветение растений
щены на восток, днем на юг, вечером на запад. Но вот наступает ве-
чер, солнце прячется за горизонт, подул прохладный ветерок, по-
явилась роса, насекомые спрятались. Нераздельно с этим и физио-
номия луга, пестревшего днем яркими цветами, также меняется:
лепестки складываются, цветочные головки закрываются, цветки
и соцветия наклоняются к земле. Такая же перемена, но в еще
Рис. 55. Защита пыльцы. Налево цветки герани (Geranium Ro-
bertianum)—/, короставника (Scabiosa lucida) — 2, колоколь-
чика (Campanula patula) — 3 днем в ясную погоду. Направо
те же цветки в ненастье и ночью
более резкой степени, совершается при наступлении ненастной
погоды. Только у сравнительно немногих растений наших лугов,
например, у подорожника, цветки остаются открытыми и в ненастье.
Но в этих цветках в росистые ночи и во время дождя замыкаются
треснувшие в сухую погоду пыльники, вследствие чего пыльца
также оказывается скрытой.
Как же объяснить все эти целесообразные, полезные для расте-
ний закрывания лепестков, загибания цветоножек и т. д.? Ближай-
шая причина этих явлений лежит в изменении напряжения тканей
у производящих движение органов. Это же изменение, в свою оче-
редь,‘вызывается изменениями температуры, освещения и влаж-
ности воздуха. Установлено, что в некоторых случаях освещение
растения ночью и затенение днем приводит к тому, что обычный ход
процесса меняется на обратный. Однако такая прямая зависимость
Защита цветков от ползающих насекомых
75
между освещением и положением частей растения имеет место не
всегда. Иногда движения в цветках связаны с явлениями
роста.
Такое объяснение, однако, ничего не говорит о том, почему и
как появилась у растений эта столь выгодная для них способность
к подобного рода реакциям на внешние воздействия. Очевидно,
что она, так же как и другие целесообразные особенности орга-
низмов, исторически, в течение многих поколений развилась у них
в процессе естественного от-
бора, т. е. выживания наиболее
приспособленных в борьбе за
существование.
Среди насекомых растения
имеют не только величайших
друзей, содействующих опыле-
нию, но также и врагов. Тако-
выми являются прежде всего
ползающие насекомые—муравьи
и всевозможные гусеницы и
пр. Они только поедают содер-
жимое цветка, его части и даже
листья растения, не принося
ему никакой пользы. В самом
деле, если даже какое-либо
ползущее насекомое, посетив
цветок, нагрузится пыльцою—
мало шансов, чтобы эта пыльца
была доставлена на рыльце дру-
гого цветка того же вида. Насе-
Рис. 56. Стебель смолки (Viscaria
vulgaris). Под листьями видно кольцо
смолистого вещества, в котором за-
вязли насекомые
комое должно для этого спуститься с одного растения, проползти
по почве и подняться на другой цветок. Проползая между
листьями, сучьями, частичками почвы и т. п., насекомое трется о
них, и пыльца удаляется с него. Известны жуки цветоеды,
поедающие лепестки, но не способствующие опылению. Среди
летающих насекомых нежелательными, приносящими ущерб посе-
тителями являются насекомые чересчур мелкие для цветка. Если
такое насекомое, проскользнув внутрь цветка, не задевает ни за
пыльники, ни за рыльце, то от него нет никакой пользы, но в то
же время, похитив мед, оно лишит цветок приманки для привлече-
ния полезных ему насекомых. Против подобных насекомых у ра-
стений выработался ряд защитных приспособлений.
От ползающих насекомых наиболее обыкновенным средством за-
щиты являются выделяемые растением клейкие вещества. Хо-
рошо известный пример представляет смолка — одно из самых
обыкновенных луговых растений, встречающееся на более сухих
местах; красные цветки этого растения по внешнему виду и строе-
76
Цветение растений
нию походят на цветки гвоздики, и относится оно к семейству гвоз-
дичных. Смолка получила свое название оттого, что на поверхности
Рис. 57. Линнея (Linnaea
borealis). Вверху—чашечка
стебля этого растения его кожицей
выделяется широкими кольцами лип-
кое смолистое вещество. Мелкие пол-
зающие насекомые, а иногда мелкие
мушки и другие крылатые насекомые
завязают в этой липкой смоле и поги-
бают (рис. 56). У многих растений клей-
кое вещество выделяется не прямо ко-
жицей стебля, а особыми железистыми
волосками. Местом выделения клей-
и прицветные листочки,
усаженные железистыми во-
лосками; внизу — цветок
(менее увеличено, чем верх-
ний рисунок)
кого вещества является или стебель,
или чашечка цветка (рис. 57), или
листья и пр. Интересно, что к такому
же средству защиты, имеющему место
в природе, прибегают и садовники:
когда они хотят оградить плодовые деревья от вредных гусе-
ниц, то наносят на ствол дерева ловчие кольца.
Есть растения, у которых подобная
же защита достигается с помощью не
клейкого, но скользкого вещества,
именно воска. Например, есть ивы, цве-
тоносные ветви которых покрыты вос-
ковым налетом, словно инеем. Попав
на такое место, ползущее насекомое
начинает скользить и, несмотря на
усилия, падает на землю.
У некоторых растений цветки за-
щищены от ползающих насекомых с
помощью воды. У таких растений су-
противно расположенные листья сра-
стаются своими основаниями, образуя
воронки, где скопляется и держится
дождевая вода. Стоящий среди такой
воронки стебель представляет как бы
остров, окруженный со всех сторон во-
дой и поэтому недоступный для пол-
зающих насекомых. Хороший пример
такого растения представляет встречаю-
Рис. 58. Ворсянка (Dipsa-
cus laciniatus)
щаяся в южной и западной части СССР
ворсянка (рис. 58). Подобное средство
также применяют садовники, когда ста-
вят горшки с растениями в плошки с водой. У произрастающей
в Средней Азии ферулы насекомые часто попадают в чашевидно-
раздутое влагалище листа.
Защита цветков от улиток
77
Для защиты от мелких насекомых служат разнообразной
формы складки, бородавки, бугорки, волосяные преграды и т. п.,
встречающиеся у входа в цветок. Большие, сильные насекомые пре-
одолевают эти препятствия. Для мелких же они в большинстве слу-
чаев делают внутренние части цветка недоступными.
Защитную роль, очевидно, играют и наблюдаемые у некоторых
растений (например, у хлопчатника) внецветковые нектарники,
видимо, отвлекающие нежелательных посетителей цветка своими
сахаристыми выделениями.
В числе врагов, опасных для цветков, должны быть названы еще
улитки, поедающие их лепестки. Какая же защита от них? Клей-
кие вещества не могут составить преграды для улиток. Поэтому
против них мы видим у многих растений другую защиту: шипы и
щетинистые волоски на стеблях, направленные остриями вниз;
пример представляет известная нам медуница. Чем ближе к цветку,
тем эти образования обыкновенно жестче и тверже. Если улитка
начнет ползти по такому растению, она натыкается на торчащие
навстречу острия волосков и ранит об них свое мягкое тело. Кроме
улиток, такие волоски защищают и от других мягких животных,
например,от гусениц и тлей. Скопление плотных, густо посаженных
друг против друга чешуй, покрывающих соцветие (корзинку) черто-
полоха и многих других сложноцветных, несомненно, также имеет
защитное значение.
РАСПРОСТРАНЕНИЕ ПЛОДОВ И СЕМЯН
По мере того как лето близится к концу, цветков становится все
меньше и меньше. Начинают увядать и самые растения. Но в течение
лета из завязей опыленных цветков образовались плоды, а в пло-
дах — семена. Из семян же на следующий год появляются новые
растения.
Однако плоды и семена служат не только для размножения, но
и для распространения растений по поверхности земли, для расши-
рения занимаемой ими территории. Если бы все семена падали около
самого растения, на котором они образовались, они не могли бы все,
проросши, развиться во взрослые растения, так как им было бы
тесно. Для этого необходимо, чтобы семена рассеивались на воз-
можно большие расстояния. Это и происходит на самом деле благо-
даря разнообразным приспособлениям к распространению.
Не у всех растений распространение совершается с помощью
отдельных семян или плодов. Иногда, как мы увидим ниже, эту
функцию несет целое соплодие. На примере чистяка (см. стр. 19)
мы уже встречались с распространением с помощью выводковых
почек. Как известно, некоторые растения распространяются посред-
ством побегов. Такова, например, водяная чума, или элодея, за-
везенная в Европу свыше ста лет назад, а в настоящее время явля-
ющаяся самым обычным обитателем наших водоемов, несмотря на
то, что плодоношения в Европе ни разу не наблюдалось.1 Наконец,
в дальнейшем мы встретимся и со случаями распространения по-
средством целого растения. Таким образом, «единицы для распро-
странения» у разных растений могут быть весьма различны. В на-
стоящее время «единицы распространения растений» — чем бы они
ни являлись — называются диаспорами. Диаспорой, следователь-
но, может быть отдельное семя, плод, соплодие, побег, выводковая
почка, целое растение и т. п.
1 В Европе встречаются почти исключительно только женские экзем-
пляры элодеи.
Распространение семян самими растениями
79
Главнейшие приспособления плодов и семян, обеспечивающие
растениям возможность широкого расселения по поверхности земли,
вполне доступны непосредственному наблюдению в природе, глав-
ным образом во вторую половину лета и осенью.
Разбрасывание семян самими растениями. На это явление обра-
тил внимание, между прочим, еще Гёте — поэт и в то же время
страстный ботаник, обогативший науку чрезвычайно важной исти-
ной, что все части цветка суть видоизмененные листья. Вот какой
случай описывает он: «Я принес домой несколько семенных коро-
бочек мягкого аканта (Acanthus mollis) и положил их в открытый
ящик. Затем однажды ночью я услышал треск и вскоре после того
прыганье по потолку и по стенам как бы мелких телец. Я не мог
себе объяснить этого, но заметил после, что мои стручки раскрылись
и семена были рассеяны вокруг. Сухость комнаты довела в течение
немногих дней созревание до такой упругости».
Существует обширная группа плодов — бобы, стручки, коро-
бочки и другие, — семена из которых разбрасываются подобным
образом. Сущность механизма заключается в том, что во время
созревания этих плодов в их стенках, окружающих семена (около-
плодник), возникает большое напряжение. Следствием его является
разъединение ткани в определенных местах, не выдержавших на-
пряжения. Образовавшиеся свободные края стенок, подобно осво-
бодившейся пружине, которая перед тем была чрезмерно закру-
чена или растянута, свертываются, или загибаются, или произво-
дят сильное сжатие и т. п. Находящиеся на стенках семена при этом
разбрасываются в окружающее материнское растение пространство.
В подробностях этого механизма у отдельных плодов наблюдает-
ся большое разнообразие. Познакомимся с несколькими наиболее
характерными примерами.
Если в жаркий день поздним летом или раннею осенью идти
по садовой аллее, обсаженной желтой акацией (чилигою), а также
по местности, поросшей дроком или заборным горохом, то можно
слышать постоянно повторяющийся легкий треск. Это раскрыва-
ются двустворчатые, высохшие плодики растений — бобы. Створки
быстро закручиваются, и при этом выбрасывают семена на значи-
тельное расстояние. Закручивание обязано находящемуся на вну-
тренней стороне створок слою деревянистых длинных клеточек.
При высыхании эти клетки или волокна укорачиваются. Когда их
напряжение превзойдет крепость соединения створок, последние
разъединяются и закручиваются. Так как волокна расположены
наискось по отношению к продольной оси плода, то закручивание
происходит винтообразно. Подобным же образом закручиваются
бобы и у других растений. Если плод короток, створки делают
один или пол винтового оборота; если же он длинен, то—два или
даже три винтовых оборота, и створки опорожненного плода при-
нимают вид завитого локона (рис. 59 и 60).
80
Распространение плодов и семян
В низкой лесистой местности, по берегам ручьев и в иных сы-
рых местах целыми зарослями встречается сочное, нежное светло-
зеленое растение с желтыми цветками, висящими на тонких цвето-
ножках и напоминающими по форме музыкальную трубу. Это
недотрога, или «не тронь меня». Что означает такое название? Ра-
стение не жжется и не колется. Но как только мы дотронемся паль-
цем до его созревшего плода — стручковидной, состоящей из пяти
частей коробочки, как коробочка эта с силой разбрасывает вокруг
себя семена (рис. 61). Пять ее ство-
рок, или плодолистиков, при этом
спирально закручиваются внутрь.
Рис. 60. То же, что и на
рис. 59. Линия аб показы-
вает направление деревяни-
стых волокон
Рис. 59. Растрески-
вание бобов мотыль-
кового растения
Причина закручивания в следующем. Створки плода состоят каждая
из трех слоев клеток. Один слой, лежащий непосредственно под
кожицей, сильно набухает и, будучи окружен другими слоями,
сдерживающими его, сильно напрягается. Ко времени созревания
плода связь между отдельными створками ослабевает и легко раз-
рывается. Для большей наглядности створки можно сравнить с
пружинами, натянутыми вдоль столбика с помощью тонких нитей.
При малейшем сотрясении нежные нити, удерживавшие пружины,
разрываются и освободившиеся пружины — а в нашем плоде
створки — сразу и с силой закручиваются. Если ветер качает эти
растения, если сквозь чащу недотроги проходит человек или
животные, — плоды лопаются и семена разбрасываются далеко
вокруг. Таким путем происходит распространение в новые места
поселения, тем более что животные, на которых попали семена
недотроги, могут в своей шерсти занести их на более значительное
расстояние. Благодаря такому способу рассеивания семян, другой,
Разбрасывание семян самими растениями
81
близкий вид этого рода (Impatiens parviflora) в настоящее время
широко распространился по ботаническим садам Европы.
В тенистых сырых лесах обыкновенным обитателем является
кислица — небольшое нежное растение с тройчатыми листьями,
напоминающими листья клевера, расцветающее раннею весною бе-
лыми с красными жилками цветками. От присутствия в тканях ки-
слой на вкус ядовитой щавелевокислой соли (являющейся защитой
Рис. 61. Недотрога (Impatiens noli tangere). 1 — расте-
ние с цветками; 2 — плод
против животных) кислица и получила свое название. Из созревшей
коробочки этого растения семена выбрасываются также благодаря
слою набухающей ткани, находящемуся здесь только не в створках
плода, а в кожуре самих семян (рис. 62). Первоначально растре-
скиваются высохшие стенки самой коробочки, образуя продольные
щели, и семена обнажаются. Находящийся под наружным слоем их
кожицы слой набухающей ткани пропитывается водой и поэтому
увеличивается в размерах. Наружный слой кожицы не выдерживает
этого давления, и кожица разрывается вдоль семени. Оба образо-
вавшихся края ее моментально закручиваются, выворачиваясь
наизнанку. Семя получает от этого сильный толчок и выбрасывается
через отверстие коробочки. Если подавить немного более или менее
спелый плод кислицы, то семена выскочат с большой силой. У со-
вершенно же зрелого плода это происходит само собой, как описано.
Кто не знает цветков фиалки? Большинство видов этого расте-
ния обладает следующим весьма своеобразным механизмом для
рассеивания семян.
Плод — коробочка, раскрывающаяся тремя створками. Стенки
каждой створки, отклонившись в сторону, изгибаются таким
82
Распространение плодов и семян
образом, что створка принимает форму ладьи. Между краями ладьи-
семена. Края с силой сближаются и выдавливают семена (рис. 63).
Последние выбрасываются на несколько метров, чему способствуют
также их форма и гладкая поверхность. Выбрасывание это со-
вершается подобно тому, как «стреляют» плодовыми косточками и
семенами, сжимая их между большим и указательным пальцами.
Сближение краев створок вызывается
Рис. 62. Кислица (Oxalis acetosella)
неравномерным высыханием
различных слоев ткани.
Очевидно, на внутренней
стороне высыхание боль-
шее, почему в эту сторону
и происходит изгибание и
сжатие створок.
На лугах и между ку-
старниками довольно обык-
новенным растением яв-
ляется луговая герань
(рис. 64), а на сырых ме-
стах, по откосам канав —
болотная герань. Первое
растение с крупными
фиолетово-синими, а вто-
рое—с пурпуровыми цвет-
ками. Иначе герани назы-
ваются журавельниками.
Такое название дано им
за форму плода, похожего
на голову журавля или
аиста. Эта оригинальная
форма плода связана с ори-
гинальным способом раз-
брасывания семян. В середине плода имеется пятигранная длинная
колонка. Вдоль нее натянуто пять плодолистиков, или створок.
У основания колонки, между нею и каждым плодолистиком, поме-
щается по одному семени. Благодаря этому здесь образуется полу-
шаровидное вздутие — как бы голова, а остальная длинная часть
колонки с прилегающими к ней плодолистиками представляет как
бы длинный клюв. Когда семена созреют, плодолистики высыхают,
но высыхают, как это мы видели у фиалки, неравномерно. Наруж-
ный слой высыхает скорее, чем внутренний. Вследствие этого пло-
долистики закручиваются; нижний край их отделяется от колонки,
и они спирально свертываются снизу вверх, где остаются соединен-
ными с колонкой. Нежная высохшая ткань, которою плодолистики
до этого времени были связаны, не представляет препятствия та-
кому свертыванию. Что же происходит с семенами? При быстром
загибании нижнего края плодолистика, где в углублении, как бы
Разбрасывание семян самими растениями
83
в ладони, лежит семя, это последнее отбрасывается, описывая боль-
шую дугу. В жаркие дни высыхание и закручивание происходят
быстрее, и семена отбрасываются дальше. Оставшийся столбик с пя-
тью закрученными у верхнего конца его плодолистиками произво-
дит впечатление канделябра.
Еще более замечательный случай разбрасывания семян пред-
ставляет бешеный огурец — растение, часто встречающееся на юге
Европы и разводимое в некоторых местах для медицинских целей.
Плод этого растения — не-
большой огурец. Когда семе- <э
на созреют, окружающая их
ткань, наполняющая всю
внутренность плода, превра-
щается в слизистую массу.
В стенках его развивается
слой слизистых клеток, рас-
ширяющийся и производящий
поэтому давление на внутрен-
нюю массу. Связь плодоножки
с огурцом (основание плодо-
ножки проникает внутрь)
вследствие ослизнения внут-
реннего содержимого ослаб-
ляется. Огурец, наконец,отва-
ливается. В то же мгновение
через образовавшееся отвер-
Рис. 63. Фиалка (Viola elatior)
стие на том месте, где в огурец
входило основание плодовой
ножки, под напором слизи-
стого слоя стенки внутреннее водянистое содержимое вместе с се-
менами стремительно выбрасывается наружу на расстояние иногда
до 6 метров и более (рис. 65). Легкое прикосновение к созревшему
огурцу человека или проходящего животного вызывает такой эф-
фект. Выбрасываемая масса может при этом обдать лицо. Попадая
на животное, семена в шерсти его относятся на более значительные
расстояния.
Наконец, остановимся еще на одном приспособлении для раз-
брасывания семян, связанном с деятельностью ветра.
Это упругие стебли и плодоножки, играющие роль как бы
«метательных орудий». Наклонение таких стеблей ветром вызывает
то, что в силу упругости они стремительно возвращаются в нормаль-
ное положение, причем далеко отбрасывают сидящие на них плоды
и семена. Хороший пример этого представляет мак. Всмотревшись
в созревшую коробочку обыкновенного мака, или, как ее называют,
«маковую головку», легко заметить наверху, под лапчатыми краями
рыльца, ряд маленьких отверстий (рис. 66). При раскачивании
84
Распространение плодов и семян
стебля, на котором находится коробочка, через эти отверстия,
как из перечницы, вылетают семена. Рагппост^анеалый по полям
Рис. 64. Луговая герань (Geranium рга-
tense). Отдельно справа — раскрыв-
шийся плод
Рис. 65. Бешеный огурец (Ес-
ballium elaterium). Веточка с
листом и плод, выбрасывающий
семена
средней и особенно южной Европы мак-самосеика за такой способ
рассеивания семян и получил свое название. У большей части других
Рис. 66. Коро- Рис. 67. Раскры-
бочка мака (Ра- тая коробочка
paver) тюльпана (Ти-
lipa)
растений, образующих коробоч-
ки, рассеивание семян происхо-
дит подобным же образом (тюль-
паны — рис. 67, гвоздичные —•
рис. 68, колокольчики — рис. 69
и т. д.).
Описанные примеры знако-
мят только с наиболее типич-
ными способами разбрасывания
семян самими растениями. Не-
смотря на обилие этих способов,
такой путь распространения рас-
тений далеко не может быть по-
ставлен на первом месте. Он на-
блюдается только у раскрываю-
щихся плодов, да и то не у всех. Большее значение имеют дру-
гие факторы: частью—вода, а в особенности—ветер и животные.
Распространение плодов и семян водой
85
Этими путями обеспечивается распространение плодов и семян
на значительно большие расстояния.
Распространение плодов и семян водою. Когда от проливного
дождя образуются потоки, быстро стекающие по почве, то они смы-
вают не только песок и землю. Они увлекают и упавшие се-
мена, которые откладываются затем уже на новом, более отдален-
ном месте, Семена и плоды уносятся еще далее, если тем же дож-
девым потоком или ветром они приносятся в текущую воду ручьев
и рек. На песчаных отмелях горных ручьев, на берегах рек, после
Рис. 69. Коробочки колоколь-
чика (Campanula), имеющие по
одному отверстию сбоку
Рис. 68. Раскрытые ко-
робочки гвоздичного
(Silcnc)
того как вода спала, постоянно находится много отложенных пло-
дов и семян самых различных растений. Многие из них, правда,
не имеют тут будущности и погибают за отсутствием условий для
дальнейшего развития или потому, что утратили во время переноса
свою всхожесть; но некоторые прорастают, а иные развиваются
даже роскошно.
Водой могут быть захвачены и отнесены на более или менее зна-
чительное расстояние всякие семена, не обладающие какими-либо
специальными приспособлениями. Но есть растения — правда,
немногие, — плоды которых приспособлены к вымыванию из них
семян дождевой водой. Так, есть несколько видов вероники —од-
ного из очень распространенных у пас травянистых растений, —
плодовые коробочки которых в сухую погоду замкнуты, а намок-
нув во время дождя, широко раскрываются (рис. 70). Дождь вы-
мывает из коробочек семена и уносит их. Таким приспособлением
обладают, между прочим, виды вероники, обитающие по берегам
текучих и стоячих вод. Выгода такого способа распростране-
ния семян у этих видов ясна. Ветер мог бы занести их на сухие
86
Распространение плодов и семян
места, на которых данные растения произрастать не могут. Дожде-
вая же вода сносит вымытые семена на влажную землю болота или
в неглубокую воду ручья или лужи, представляющих самые благо-
приятные места обитания для этого вида вероники.
Из встречающихся у нас растений, приспособленных к такому
способу распространения семян, назовем еще очиток едкий. Это
маленькое мясистое растение обитает среди
камней, по скалам, на сухих каменистых
склонах, в расщелинах старых стен и на
бесплодном песке. Его пять плодиков,
соединенных основаниями, представляют
как бы пятилучевую звезду. В сухую
погоду они закрыты, в дождливую же
раскрываются так широко, что мелкие
семена легко вымываются дождевыми кап-
лями (рис. 71). В этом легко убедиться,
Рис. 70. Вероника, или дуб-
ровка (Veronica chamaed-
rys). 1—отдельный цветок;
2—плод (1, 2—увеличено)
Рис. 71. Очиток едкий (Sedum acre). 1—за-
крытый плод; 2 — открытый плод (увели-
чено)
если положить плод очитка в воду. Вместе с водой семена очитка
попадают в трещины земли, в расщелины камней, стен и т. п. Этим
объясняется появление очитка даже на совершенно отвесных стенах.
Плоды и семена наземных растений, попавшие благодаря ветру
или рекам в море, могут далеко уноситься морскими течениями.
В особенности это нужно сказать про плоды легкие, не тонущие.
Попав к новым, далеким берегам, они там во время волнений мо-
гут быть выброшены на сушу. Но могут ли их семена, долгое время
Распространение плодов и семян ветром
87
пробывшие в соленой морской воде, прорастать? Чтобы ответить на
этот вопрос, неоднократно производились специальные исследова-
ния, которыми в свое время занимался еще Чарлз Дарвин.
Опыты показали, что некоторые семена не утрачивают всхоже-
сти, пролежав в морской воде более года. Наличие двух условий—
легкости плода и продолжительной выносливости семян по отноше-
нию к морской воде — свойственно немногим растениям. Но там,
где оба эти условия имеются, возможно распространение растений
по морю на очень большие расстояния. Хороший пример представ-
ляет кокосовая пальма. Под кожистым, непроницаемым для воды
слоем ткани в кожуре кокосового ореха содержится воздухо-
носный волокнистый слой, благодаря которому орех плавает. На-
ходящиеся внутри семена долго сохраняют свою всхожесть. Пере-
несенный на очень далекое расстояние и выброшенный на берег
кокосовый орех, если не препятствуют почвенные и климатические
условия, прорастает. Известно, что кокосовая пальма — одно из
первых растений, появляющихся на коралловых островах без вся-
кого содействия человека. К берегам Норвегии нередко прибива-
ются плоды одного бобового растения, произрастающего в тропи-
ческой Америке. Как показали опыты в ботанических садах, се-
мена эти не теряют всхожести от столь далекого путешествия.
На берегу Норвегии они, конечно, погибают от неподходящего
для них климата.
Нет нужды останавливаться на том, что плоды водных расте-
ний распространяются при посредстве воды. Впрочем, в стоячих
водах, где нет течений, на помощь является еще ветер. При этом
в плодах, например, у болотных осок, кувшинок, частух и других,
развиваются особые приспособления в виде воздухоносных покро-
вов и тому подобных образований. Благодаря им плоды плавают
и распространяются дующими вдоль поверхности ветрами.
Распространение плодов и семян ветром. На последних приме-
рах, а также выше (стр. 83) мы уже видели, что распространение
семян некоторых растений усиливается благодаря ветру. Однако
этот фактор имеет часто первостепенное значение. Хорошо известно,
что в воздушной пыли содержатся не только мертвые минеральные
и землистые частицы, но наряду с ними — зародыши разных ми-
кроорганизмов. Споры многих грибов и бактерий благодаря их
микроскопическим размерам могут долгое время находиться во
взвешенном состоянии в воздухе и воздушными течениями разно-
ситься всюду. Плесневые грибы, например, встречаются по всей
земле; столь широким распространением они, несомненно, обязаны
ветру.
Среди семян также имеются чрезвычайно легкие, пылеобразные,
способные долгое время находиться в воздухе. Сюда прежде всего
следует отнести семена орхидей; у одного вида последних средний вес
семени составляет только 0,000002 г. И некоторые другие растения,
88
Распространение плодов и семян
Рис. 72. Крыла-
тые плоды березы
(Betula)
Рис. 73. Кры-
латый плод
вяза (Ulmus)
(увеличено)
особенно живущие в глубоком перегное, имеют чрезвычайно легкие
семена, от 0,0001 до 0,000003 г; таковы, например, семена грушан-
ки одноцветной (0,000004 г), белозора болотного (0,00003 г) и др.
Долгому «парению» в воздухе этих семян
способствуют также их более или менее сплю-
щенная форма и такое расположение центра
тяжести, вследствие которого они
обращаются к земле своею широ-
кой стороной.
Но и более крупные семена
могут носиться по воздуху благо-
даря разного рода летательным
приспособлениям: крыловидным
пластинкам, каемкам, пучкам
длинных волосков и т. п., играю-
щим роль как бы крыльев, пару-
сов и парашютов. При помощи
всех подобных придатков площадь
сопротивления воздуху при падении и ветру значительно увели-
чивается при ничтожном в то же время увеличении веса. Ветер
поэтому может уносить такие семена на большое расстояние.
Рис. 74. Плоды клена (Acer)
У наших деревьев можно наблюдать
все главнейшие типы этого рода приспо-
соблений. Так, у плодов березы (рис. 72)
и вяза (рис. 73) имеется
перепончатая каемка,
образующая как бы
крылья с двух сторон;
у клена (рис. 74), со-
сны, ели (рис. 75), ясе-
ня—пластичный крыло-
видный придаток; у ив
(рис. 76), осины, топо-
лей — пучки волосков;
лапина (рис. 77) обра-
зует крылатые плоды
(орешки). Когда плоды
тополей созревают, по-
Ги 7 Слева—
сехченная чешуя
из шишки ели
(Picea) с двумя
семенами; спра-
ва—семя с кры-
лом
луоткрываются и семена из них высеиваются, то в местности,
засаженной этими деревьями, иногда можно наблюдать нечто
вроде снега. Интересную особенность представляют плоды клена:
при падении они быстро вращаются в горизонтальной пло. когти,
благодаря чему долго остаются в воздухе. У липы при плодах
остается большой прицветник (см. рис. 50), играющий роль паруса.
У белой акации ту же самую роль играют плоские створки бобов,
которые отделяются от растения вместе с семенами.
Распространение плодов и семян ветром
89
Волосистые придатки или хохолки разнообразной формы осо-
бенно распространены в семействе сложноцветных. Здесь они обла-
дают способностью складываться под влиянием сырости и раскры-
ваться при высыхании. Примером может служить хорошо всем из-
вестный одуванчик (рис. 78). Когда плодики его созревают, то
обвертка в сухую погоду раскрывается, и волоски хохолков расто-
пыриваются, принимая вид парашютов, имеющих большую по-
верхность; таким образом формируются шаровидные головки со-
плодий. Плодйки легко отделяются от плодоложа. Достаточно
Рис. 76. Лопнувшие коробочки
ивы (Salix), из которых выпадают
семена с волосками
Рис. 77. Крыла-
тые плоды лапины
(Pterocarya ptcro-
сагра)
даже небольшого порыва ветра, чтобы они разлетелись па значи-
тельное расстояние.
Снабженный парашютом плодик медленно опускается на
землю, а пока опустится, может совершить большое путешествие.
В вечернем влажном воздухе, а тем более в сырую погоду, гигроско-
пические парашюты снова складываются и закрываются обверткой
до следующих теплых лучей солнца. Такую же картину можно ви-
деть у латука (рис. 79) и других сложноцветных. Приспособлением
к распространению плодов воздушными течениями является и пу-
зыревидное вздутие их оболочки, что можно наблюдать у многих
растений Закавказья и Средней Азии (например, у астрагалов).
Распространение семян при помощи летательных приспособ-
лений обыкновенно совершается в горизонтальном направлении.
Но в то же время в местностях гористых оно может происходить
и вертикально. При восходящем токе воздуха такие семена могут
подниматься очень высоко, благодаря чему ими постепенно засе-
ляются края и карнизы высоких крутых утесов.
У некоторых семян и плодов крылья и парашюты соединены
с ними только во время полета. Так, когда крылатое семя сосны
где-либо осядет, то пленчатое крыло его сейчас же отделяется и
90
Распространение плодов и семян
семя теряет способность лететь далее. Еще резче это явление выра-
жено у плодиков татарников. Поддерживаемые парашютом срав-
нительно крупные семянки этого растения спокойно плавают по
воздуху. Но как только на пути встретится препятствие, происхо-
Рис. 78. Одуванчик (Taraxacum officinale).
1 — развитие плода; 2 — отдельный цветок
(увеличен)
дит столкновение и се-
мянка мгновенно отде-
ляется и падает на
землю. Этим и объяс-
няется частое произра-
стание татарников вдоль
стен и заборов, на ко-
торые наталкиваются
летящие плоды.
В других случаях
летательные приспособ-
ления остаются все
время соединенными с
плодами или семенами
и часто содействуют
Рис. 79. Плодовая го-
ловка латука (Lactuca)
прицеплению семени к месту прорастания или его зарыванию в
землю. Прекрасный пример последнего представляет наш степной
злак — ковыль. Семянка ковыля снабжена длинною, изогнутою
под прямым углом перистою остью (рис. 80). Нижнее колено ее
обладает способностью скручиваться по винтовой спирали при вы-
сыхании и раскручиваться при действии влажности. При этом верх-
нее колено двигается наподобие часовой стрелки, но если оно встре-
тит при своем движении препятствие в каком-нибудь неподвиж-
ном предмете, например, в почве, то вращаться вокруг своей про-
Распространение плодов и семян ветром
91
дольной оси начинает сама семянка. Так как конец ее заострен, то
при вращении он, наподобие бурава, постепенно углубляется в
землю, причем обратному движению мешают волоски, обращенные
концами кверху. В результате изменений состояния влажности се-
мянка зарывается в почву на большую или меньшую глубину.
Подобным же механизмом самозарывания обладают плодики аист-
ника, или грабелек (рис. 80)—растения, часто встречающегося у нас
на полях, возле дорог и на
выгонах и относящегося к од-
ному семейству с упомяну-
тыми ранее журавельниками.
Существует и много дру-
гих так называемых само
зарывающихся плодов и се-
мян с разнообразными при-
способлениями, служащими
главным образом для зарыва-
ния семени в почву, а частью
для передвижения. Примером
последних могут служить
многие виды овса, например,
очень распространенный у
пас в южной части СССР ди-
кий овес, или овсюг (рис. 80).
Здесь мы также видим отхо-
дящие от семян длинные ко-
ленчатые ости, но эффект
вращения их другой. Дело в
том, что у овсюга отпадают
от материнского растения не
отдельные зерновки, а целые
колоски, имеющие по две зер-
новки и по две ости. Понятно,
Рис. 80. 1 — плод ковыля (Stipa реп-
nata); перистая часть ости изображена
не вся; 2 — плод аистника (Erodium
cicutarium); 3 — колосок овсюга (Avena
fatua)
зарывание в землю целого колоска,
наподобие бурава, невозможно: колосок слишком велик, чтобы
свободно вращаться, и вращается поэтому горизонтальное колено
ости. Концы остей при вращении упираются в землю и сообщают
колоску неправильное переваливающееся движение.
В местностях с длинным периодом засухи у многих растений
развиваются плоды и соплодия, перекатываемые ветром по ровной
поверхности земли. Весьма богаты такими представителями страны
около Средиземного моря и степные области. Отличительной осо-
бенностью таких плодов и соплодий, очевидно, должен быть легкий
вес и более или менее круглая форма. Так, многие зонтичные, свой-
ственные горным степям Востока, имеют эллипсоидные гладкие
плоды величиной с лесной орех. Если человеку, закрывшему глаза,
положить на ладонь несколько таких плодов, то он не ощущает
92
Распространение плодов и семян
тяжести, не замечает, что ему положены плоды. Поразительно
малый вес их зависит от того, что в них развился слой, напоминаю-
щий бузинную сердцевину. Понятно, такие плоды, отпав, перекаты-
ваются по степи ветром, пока не попадут куда-нибудь в трещину
высохшей глинистой почвы или в щель скалы. Будучи смочены
дождем, они раскрываются, и семена высыпаются наружу.
Некоторые растения, не отделяя плодов, целиком вырываются
ветром с корнем или отрываются у основания корня и затем катятся
Гис. 81. Оторвавшееся соцветие кермека (Goniolinion tataricuin)
в виде шаров или клубков. Такие растения, называемые «перекати-
поле», в большом количестве встречаются на открытых равнинах,
в степях и пустынях СССР. Это не один какой-либо определенный
вид, а растения, относящиеся к разнообразным группам, с большими
и маленькими плодоносящими кустами. Таковы, например, кермек
(рис. 81), качим, зопник и др. Таким образом, схожая форма неза-
висимо выработалась в процессе естественного отбора в различных
семействах.
Большой интерес представляет вопрос о том, каким образом
эти растения при созревании семян отделяются от земли. Однолет-
ние растения обычно выдергиваются с корнями, а многолетние —
отламываются близ основания стебля вследствие того, что стебель
здесь становится хрупким и переламывается в сухую погоду даже
от незначительного толчка (ветер, проходящее животное). Реже
отделение многолетних растений происходит благодаря перегни-
ванию стебля при основании. Последнее наблюдается у растений
с очень твердым стеблем (например, у зонтичных) и имеет место
Распространение плодов и семян ветром
93
поздней осенью или даже зимой. Что касается шаровидной формы,
характерной для «перекати-поле», то она достигается обильным
ветвлением, загибанием ветвей и т. д. и образуется или в ранних
фазах развития растения или значительно позже, иногда после
цветения. Часто случается, что несколько многоветвистых кустов
при перекатывании переплетаются и образуют шары, величиною
с воз сена. Наблюдалось, что при вихрях такие шары поднимаются
с почвы и большими скачками носятся по степи.
У некоторых «перекати-поле» выработались специальные при-
способления, обеспечивающие рассеивание семян на большую пло-
щадь. К ним относятся: волоски, развивающиеся около плодиков
Рис. 82. Иерихонская роза (Anastatica hierochuntica)
и затрудняющие высыпание семян; всегда загнутые внутрь зубцы
коробочки, сильно суживающие отверстия, через которые высы-
паются наружу семена. В результате растение во время перека-
тывания разбрасывает семена не сразу, а постепенно. Интересно,
что у близких видов, не образующих «перекати-поле», эти осо-
бенности отсутствуют.
В степях Египта, Аравии и Сирии распространено одно (кре-
стоцветное) растение, которое еще в средние века привозилось
участниками крестовых походов, под названием «иерихонской
розы». Когда это растение созреет, небольшие вэтви его дугообразно
загибаются внутрь, и таким образом сидящие на концах развет-
влений замкнутые грушевидные стручковые плоды как бы окру-
жаются крепкой решеткой (рис. 82). Растение теперь имеет вид
клубка или закрытой «розы». Ветер катает его, как мяч. Как только
влага смочит его, что бывает обыкновенно с наступлением зимних
дождей, ветви расходятся и выпрямляются. Плодики тогда также
вскрываются,и семена могут падающим дождем вымываться из ство-
рок плода. Есть еще другая «иерихонская роза» (из сложноцвет-
ных), произрастающая в пределах от северной Сахары до Пале-
стины и особенно часто встречающаяся в окрестностях Иерихона.
У этого растения не ветви, а расположенные розеткой листочки
обвертки смыкаются над созревшею плодоносною головкой и
У4
Распространение плодов и семян
остаются в закрытом состоянии до тех пор, пока не наступят
зимние дожди, раскрывающие «розу» и уносящие плодики.
Распространение плодов и семян животными и человеком.
Этот способ распространения почти столь же разнообразен, как и
распространение воздушными течениями.
Во многих случаях прорастают семена, прошедшие через ки-
шечный канал животного и выброшенные вместе с испражнениями.
С целью точнее установить возможность распространения растений
таким путем, проф. Кернер фон-Марилаун произвел ряд следующих
опытов. Он кормил различных животных плодами и семенами от
25 различных видов растений и помещал для прордстания, с одной
стороны, испражнения этих животных, а с другой, для сравнения,
семена тех же растений, но не употребленные для кормления. Почти
все плоды и семена, принятые в пищу млекопитающими животными,
были разрушены — или уже в самом начале, или при жвачке. Из
испражнений коровы проросли весьма немногие избежавшие из-
мельчения при жвачке зерна проса; из лошадиного навоза — еди-
ничные семена чечеви1Ы и плоды овса; точно так же были лишь
единичные случаи прорастания семян из помета свиньи.
Взятых для опытов птиц Кернер, по результатам опытов, раз-
делил на три группы. Одну группу составляют такие птицы, кото-
рые размельчают все, даже самые твердые плоды и семена, в своем
мускулистом, снабженном терками и, обыкновенно, наполненном
песком и мелкими камешками желудке; сюда отнесены: индейка,
курица, голубь, клест, снегирь, щегол, чиж, синица, сойка и утка.
Впрочем, последующие опыты показали, что эта характеристика
требует известных оговорок, так как, оказывается, даже птицы
с сильным мускулистым желудком (куры), в котором разрушаются
твердые семена, не могут повредить семян омелы с их сильно раз-
витой мякотью и слизистой оболочкой. К другой группе отнесены
вороны и галки, у которых косточки и твердокожие семена приня-
тых в пищу мясистых плодов без вреда проходят через кишечный
канал, но мягкокожие семена и плоды сплошь разрушаются. Между
прочим, в испражнениях этих птиц после кормления находились
вишневые косточки 15мм в поперечнике, которые все были всхожи.
Третью группу из взятых для опыта птиц составляли: овсянка,
певчий дрозд, каменный дрозд и малиновка. Из плодов и семян,
прошедших через их кишечный канал, проросло: у овсянки 75%,
у певчего дрозда 85%,у каменного дрозда 88% и у малиновки 80%.
Сравнительно с однородными плодами и семенами, не употребляв-
шимися для кормления и высеянными только для контроля, прора-
стание семян, прошедших через кишечный канал, большей частью
было замедлено, но у некоторых таких семян происходило раньше
и ростки развивались пышнее.
Этими опытами и другими наблюдениями с несомненностью уста-
новлено, что распространение семян, принятых в пище птицами,
Распространение плодов и семян животными и человеком
$5
главным образом певчими, совершается в большом количестве.
Наибольшее значение в этом отношении имеют дрозды, распростра-
няющие около 30 видов ягодных растений, но также и многие дру-
гие птицы. О масштабах данного явления свидетельствуют, напри-
мер, наблюдения, проведенные в сосновых посадках Савальского
лесничества (Воронежская область), согласно которым птицы за-
несли сюда 18 видов кустарников: красную и черную бузину, кру-
шину, калину, черемуху, рябину, боярышник, черную смородину,
два вида шиповника и др. В лесной даче Сельскохозяйственной
академии им. К. А. Тимирязева с помощью птиц расселились:
рябина, красная бузина, мушмула, городовина, магония, кизиль-
ник обыкновенный. Преимущественно таким путем распространя-
ются растения с мясистыми плодами, в которых находятся семена,
защищенные твердой оболочкой от самых энергичных агентов,
включая сюда и желудочный сок. У таких плодов наблюдается ряд
свойств, являющихся приспособлением к распространению при по-
мощи животных.
Такое значение имеет прежде всего образование сочного или мя-
систого околоплодника. Предоставленные сами себе, эти плоды,
в конце концов, загнивают, так что зародыши семян не могут вос-
пользоваться веществами, содержащимися в мякоти плода. При-
способлением является и то, что такие плоды в сущности содержат
весьма мало питательных веществ. Птицы, питающиеся ягодами,
должны поедать их огромные количества, чтобы собрать достаточно
годного материала, служащего для построения и восстановления
тканей их тела. А раз поедается много плодов, то, следовательно,
проходит через кишечный канал и рассеивается много и семян.
Однако с другой стороны, нельзя не признать, что большое коли-
чество несъедаемых птицами и сгнивающих плодов приводит все же
к непроизводительной трате значительного количества веществ,
за счет которых был построен плод.
Большую роль птицы играют, в частности, в распространении
омелы (Viscum album L.) — паразитного растения, встречающе-
гося на разнообразных деревьях и кустарниках в большей части
Европы (от Португалии до Черного моря, от Сицилии до Осло).
В связи с паразитическим образом жизни омела лишена корня и
получает воду с растворенными в ней веществами от растения-
хозяина, с которым плотно срастается. Ее ягодообразные плоды
(в зрелом состоянии — белого цвета) не обладают летательным
аппаратом и не могут разноситься ветром. Уже римляне знали, что
семена омелы распространяются дроздами. Последние охотно по-
едают плоды этого растения и вместе с испражнениями выделяют
семена, прилипающие к стволу или ветвям деревьев или кустар-
ников, посещаемых птицами. Долгое время считалось, что прора-
стать могут только те семена, которые прошли через пищевари-
тельный канал птиц, но специальные опыты показали ошибочность
96
Распространение плодов и семян
этого мнения. Кроме дроздов, семена омелы распространяются и
некоторыми другими птицами.
Яркие цвета, запах и соответствующий вкус плодов заметны и
привлекательны для животных. Но указанные качества появляются
лишь ко времени зрелости семян. До этого времени плоды, наобо-
рот, обыкновенно скрыты среди листвы материнского растения,
окрашены в зеленый схожий с листвою цвет и лишены аромата.
Но как только зрелость приблизится, их покровы приобретают яр-
кую окраску, и притом хорошо заметную на данном фоне. На зе-
лени четко выделяется красный цвет. И у растений вечнозеленых
(например, брусники, толокнянки и др.), а равно и у тех, листва
которых хотя и опадает на зиму, но ко времени созревания плодов
еще не принимает осенней окраски (земляника, малина, смородина,
рябина, вишня, красная бузина и др.), —плоды красного цвета.
Если же ко времени созревания плодов листва принимает уже осен-
нюю красную или желтую окраску, то у таких растений плоды ча-
сто окрашены уже в другие, наиболее контрастирующие с ней
цвета, напримэр синий. Окраска плодов обязана известному нам
пигменту, окрашивающему лепестки цветков и осеннюю листву, —
антоциану (см. стр. 47 и 104).
Ягоды, как известно, ‘ созревают преимущественно во вторую
половину лета и осенью. Это также имеет большое значение для рас-
тений, так как на это время падают перекочевывание и перелет
птиц. Понятно, что поедание плодов в это время приводит к широ-
кому распространению растений, что не имело бы места в более ран-
нее время года, когда птицы живут каждая в ограниченном районе.
Многие сочные плоды ядовиты, что, казалось, должно было бы
служить препятствием к их распространению при помощи живот-
ных; однако в действительности этого нет. Наблюдениями обна-
ружено, что ядовиты они обыкновенно только для человека и жвач-
ных животных, но не для птиц. Птицы, в отличие от жвачных, не
разрушают семян, и яды, содержащиеся в сочных плодах, для них
не опасны. Наблюдалось, что, например, певчий дрозд (вневоле)
с жадностью съедал множество ягод белладонны, содержащих
атропин, без всякого вреда для себя. Точно так же семена белены
и наперстянки и ягоды бузины не оказывали на него никакого вред-
ного действия.
Установлено, что деятельность птиц по распространению семян
рябины и других растений в известной мере сокращается лесными
полевками. Последние в большом количестве поедают лежащие
на земле ягоды, утерянные птицами, а также семена, содержащиеся
в экскрементах птиц, после того как экскременты размываются
дождями.
Еще большее количество плодов и семян распространяется бла-
годаря животным другим способом, а именно — прицепляясь или
приклеиваясь к ним при помощи разнообразных приспособлений.
Распространение плодов и семян животными и человеком
97
Эти последние можно разделить на две категории: разнообразные
цепкие придатки и клейкие выделения или другие склеивающие
вещества.
Колючие, служащие для прицепления к животным придатки
чрезвычайно разнообразны. Приблизительно десятая часть семен-
ных растений имеет плоды
и семена, распространяемые
посредством когтевидных или
крючковатых отростков. Как
только млекопитающее или
птица коснется их, они при-
цепляются к волосам, щетине
Рис. 83. Подмаренник цепкий Рис. 84. Городской гравилат (Geum urba-
(Galium aparine). Налево ввер- num). 1 — цветок в продольном разрезе;
ху — плод (увеличен) 2 — плоды; 3 — отдельный плод, на-
право — в разрезе (увеличено)
или перьям. Животное потом старается освободиться от неприят-
ных придатков, и семена попадают в почву. Таковы, например,
плоды подмаренника (рис. 83), гравилата (рис. 84), моркови
(рис. 85), череды (рис. 86) и мн. др. Есть немало растений, у ко-
торых цепляются не отдельные плоды, а целые соплодия; общеиз-
вестные примеры представляют лопушник (рис. 87) и репейник
(рис. 88). У наших растений колючки на плодах сравнительно не-
большие, но в других местах встречаются плоды с громадными
колючками, причиняющими бедствия животным. Так, в Южной
Африке на плодах одного растения (Harpagophyton procumbens)
вырастают крючковатые шипы величиной с вороньи лапы. Они
составляют бедствие для домашних и диких жвачных животных.
Когда живущие в Трансваале и у Оранжевой реки козы нечаянно
98
Распространение плодов и семян
наступят на них, то копыта животных обхватываются острыми
шипами, и несчастные животные бегут от боли как бешеные.
Часто проходит несколько дней, пока коробочка треснет и отпа-
дет. У некоторых растений отходящие от плодов колючки пря-
мые и впиваются в ноги наступивших на них животных.
Менее распространенный, но все же частый случай—приклеи-
вание плодов и семян к животным. У некоторых растений, напри-
мер, у разных видов льна, семена покрыты слоями клеток, наруж-
ные стенки которых
ос л из н я ются, стано-
вятся липкими и мо-
гут приклеиваться к
животным, которые
прикоснутся к ним.
В других случаях
Рис. 85. Плод мор-
кови (Daucus carota)
(увеличен)
семена приклеиваются мякотью раздавленных плодов, напри-
мер, у омелы, паслена. У полыни плодики становятся липкими по-
сле дождя: однажды была поймана сова, которая только что зале-
тела в заросль полыни, гоняясь за полевой мышью, — она вся
оказалась покрытой этими плодиками. Семена, не липкие сами по
себе, часто пристают к животным вместе с размокшей землей,
илом и т. п. Дарвин, исследуя ил, приставший к водным птицам,
в 63/4 унциях его нашел множество семян, из которых 537 проросли,
а из 6V2 унций твердой земли, снятой с лапы красной куропатки,
проросло не менее 82 растений.1
Береговые болотные растения и многие сорные травы, расту-
щие на полях и по краям дороги, распространяются именно таким
образом.
Известно, что некоторые животные, например, белка, хомяк,
кедровка устраивают в расщелинах скал, в земляных норах и дру-
1 Английская унция равняется 28,35 г.
Распространение плодов и семян животными и человеком
9()
гих укромных местах запасы семян и плодов. Случается, что жи-
вотное не приходит за таким сделанным им запасом, или забывая
место кладовой, или же, что вероятнее, сделавшись добычей хищ-
ной птицы. Как бы там ни было, но оставленные семена прорастают
на новом месте.
Некоторые семена, например, чистотела, душистой фиалки и
других, разносятся муравьями. На таких семенах обыкновенно
имеются мясистые придатки, служащие пи-
щей муравьям. Из-за них муравьи и несут
семена к себе в жилище. Унесенные мура-
вьями под землю или в щели стен, семена
прорастают там на следующий год, хотя при-
датки у них и откушены. Случается, что еди-
ничные семена с откушенным придатком ле-
жат по дорогам, по которым двигались
муравьи. Этим и объясняется то, что неко-
торые растения обыкновенно произрастают
на муравьиных дорогах; например, в вен-
ском ботаническом саду постоянным спутни-
Рис. 88. Репейник,
или репяшок (Agri-
monia eupatoria). От-
дельно — плод (уве-
личен)
ком муравьиных дорог яв-
ляется чистотел.
В числе семян, прино-
симых муравьями в их жи-
лища, часто встречаются
семена лугового марьянни-
ка, которые не идут в пищу.
Что же привлекает к этим
семенам? Привлекает, воз-
можно, их «обманчивая на-
ружность». Они чрезвы-
чайно сильно — и цветом,
Рис. 87. Соплодие
лопушника (Arc-
tium lappa)
и размером, и формой — похожи на муравьиные коконы, содержа-
щие куколок и называемые в просторечии «муравьиными яйцами».
Насекомые закапывают эти семена так, как они это делают с соб-
ственными коконами. Вот почему луговой марьянник часто воз-
вышается над муравьиными ходами. Известная выгода от этого для
муравьев заключается в том, что они поедают сладкий сок, выделяю-
щийся на определенных местах листьев растения.
Такое полезное сходство растений и животных или их отдель-
ных органов с другими растениями, животными или их орга-
нами называется миметизмом или мимикрией. Сходство это, раз
приобретенное, давая преимущество обладателям его над другими
организмами, не имеющими этого сходства, естественно закреп-
ляется в ряде поколений. Так, в нашем примере марьянник с се-
менами, сходными с муравьиными яйцами, имеет благодаря му-
равьям преимущество в распространении и, естественно, в течение
100
Распространение плодов и семян
многих поколений мог вытеснить других сородичей, которые этого
сходства не имели.
Указываются и другие случаи миметизма у растений, хотя
этот вопрос еще мало исследован. Есть семена, похожие по виду на
гусеницу или червя (Scorpiurus vermiculata), многоножку (Bis-
serula pelecinus), жуков (клещевина, ятрофа), пауков и др.
Может быть, польза для растений от такого «подражания» за-
ключается в том, что птицы, обманутые этим сходством, прежде
чем бросить схваченные ими семена, уносят их на некоторое рас-
стояние, способствуя тем самым распространению растений.
Сходство с насекомыми может служить защитой и в том случае,
если поедание данных семян зерноядными птицами оказывается
невыгодным для растения.
Подмечено несколько случаев миметизма и среди цветков:
именно — есть цветки, не имеющие нектара, но по внешнему виду
похожие на цветки, которыми вырабатывается нектар. Первые
из этих цветков, принимаемые насекомыми за вторые, посещаются
ими, вследствие чего опыляются как те, так и другие.
Наконец, обратим еще внимание на распространение растений
человеком. Кто пробирался в начале лета через заросли тополей
и ив или в конце лета — через заросли репейников, тот знает,
как много пристает к платью цепких и липких плодов и семян.
При сношениях между отдаленными странами, сознательно или
бессознательно, растения данной страны заносятся в новые, отдален-
ные местности. Известно, что много растений Старого Света куль-
тивировано в Новом. Вместе с семенами культурных растений пе-
ревозится много диких видов — сорных трав, которые постепенно
захватывают большие районы. Многие экзотические виды переплыли
моря на наших судах, и в наших портах ими основываются настоя-
щие «колонии». Несомненно, что роль океанского и морского транс-
порта в обогащении флоры чуждыми ей растениями чрезвычайно
велика. Но и постройка железной дороги, устройство канала спо-
собствуют появлению в данной местности новых видов, и в наше
время нередко приходится наблюдать быстрое распространение
благодаря подобным причинам форм, неизвестных в данной мест-
ности.
Особенно большое значение в этом отношении имеют железные
дороги. Так, например, установлено, что по линии Октябрьской
ж. д. на территорию Калининской области проник ряд южных
растений, завезенных сюда с фуражом, зерном и т. п. Интересно,
что многие из них являются на своей родине обитателями песков,
распространяющимися по железнодорожной насыпи далеко на север.
Конечно, особо выдающееся значение имеет деятельность чело-
века в распространении плодов и семян культурных растений.Од-
нако вопрос этот выходит из рамок настоящей главы, посвященной
лишь дикой флоре.
Особенности строения плодов и семян сорных растений
101
В заключение отметим только большую практическую важность
знания способов распространения плодов и семян сорных растений.
У многих так называемых обязательных, или специальных, сорня-
ков наших полей, т. е. таких, которые, как правило, живут только
среди культурных растений, в строении семян и плодов имеется
ряд особенностей, тесно связанных с условиями их жизни. Семена
сорняков часто оказываются схожими с семенами соответствующих
культурных растений. У тех и у
других наблюдается одинаковая
«парусность», т. е. летательная
способность, благодаря чему не
происходит отделения семян сор-
няков при веянии. Часто семена
сорняков по своей форме, цвету,
объему и прочему похожи на се-
мена тех культурных растений,
среди которых они произрастают.
Указанные особенности семян
сорняков, представляющие собой
хорошие примеры мимикрии, или
миметизма, среди растений, выра-
ботались в процессе отбора.
Одним из наиболее интерес-
ных примеров результата отбора,
имевшего место в условиях куль-
туры, является особая раса, или
подвид, полупаразитного растения
погремка, или пастушьего гребня
(Rhinanthus apterus), отличаю-
щаяся от своих дикорастущих лу-
говых сородичей, от которых она и произошла, тем, что из ее пло-
дов (коробочек) семена не высыпаются, форма их несколько иная
(менее сплюснутая) и отсутствуют крыловидные придатки (рис. 89).
Время созревания семян этого сорняка точно совпадает со време-
нем созревания семян ржи. Это растение при уборке ржи попадает
в снопы и при обмолачивании и веянии, обладая одинаковой парус-
ностью семян с рожью, не отделяется от нее. Вследствие этого на
следующий год семена сорняка высеиваются вместе с семенами ржи.
Понятно, насколько важно для рациональной борьбы с сорными
растениями знать все их биологические особенности, в частности,
отмеченные выше. Очевидно, что крайне желательным является
их уничтожение до созревания семян и наиболее тщательная
очистка посевного материала. Более подробное рассмотрение
этого вопроса, которого мы касаемся здесь только вскользь,
читатель найдет в соответствующих руководствах по сорным рас’
тениям.
Рис. 89. Плоды и семена погрем*
ка (Rhinanthus major). 1 — плод
и 2 — семя дикорастущей расы
(Rh. vernalis); 3 — плод и 4 —
семя сорной расы (Rh. apterus),
засоряющей озимую рожь (уве-
личено)
100
Распространение плодов и семян
многих поколений мог вытеснить других сородичей, которые этого
сходства не имели.
Указываются и другие случаи миметизма у растений, хотя
этот вопрос еще мало исследован. Есть семена, похожие по виду на
гусеницу или червя (Scorpiurus vermiculata), многоножку (Bis-
serula pelecinus), жуков (клещевина, ятрофа), пауков и др.
Может быть, польза для растений от такого «подражания» за-
ключается в том, что птицы, обманутые этим сходством, прежде
чем бросить схваченные ими семена, уносят их на некоторое рас-
стояние, способствуя тем самым распространению растений.
Сходство с насекомыми может служить защитой и в том случае,
если поедание данных семян зерноядными птицами оказывается
невыгодным для растения.
Подмечено несколько случаев миметизма и среди цветков:
именно — есть цветки, не имеющие нектара, но по внешнему виду
похожие на цветки, которыми вырабатывается нектар. Первые
из этих цветков, принимаемые насекомыми за вторые, посещаются
ими, вследствие чего опыляются как те, так и другие.
Наконец, обратим еще внимание на распространение растений
человеком. Кто пробирался в начале лета через заросли тополей
и ив или в конце лета — через заросли репейников, тот знает,
как много пристает к платью цепких и липких плодов и семян.
При сношениях между отдаленными странами, сознательно или
бессознательно, растения данной страны заносятся в новые, отдален-
ные местности. Известно, что много растений Старого Света куль-
тивировано в Новом. Вместе с семенами культурных растений пе-
ревозится много диких видов — сорных трав, которые постепенно
захватывают большие районы. Многие экзотические виды переплыли
моря на наших судах, и в наших портах ими основываются настоя-
щие «колонии». Несомненно, что роль океанского и морского транс-
порта в обогащении флоры чуждыми ей растениями чрезвычайно
велика. Но и постройка железной дороги, устройство канала спо-
собствуют появлению в данной местности новых видов, и в наше
время нередко приходится наблюдать быстрое распространение
благодаря подобным причинам форм, неизвестных в данной мест-
ности.
Особенно большое значение в этом отношении имеют железные
дороги. Так, например, установлено, что по линии Октябрьской
ж. д. на территорию Калининской области проник ряд южных
растений, завезенных сюда с фуражом, зерном и т. п. Интересно,
что многие из них являются на своей родине обитателями песков,
распространяющимися пожелезнодорожной насыпи далеко на север.
Конечно, особо выдающееся значение имеет деятельность чело-
века в распространении плодов и семян культурных растений. Од-
нако вопрос этот выходит из рамок настоящей главы, посвященной
лишь дикой флоре.
Особенности строения плодов и семян сорных растений
101
Рис. 89. Плоды и семена погрем*
ка (Rhinanthus major). 1 — плод
и 2 — семя дикорастущей расы
(Rh. vernalis); 3 — плод и 4 —
семя сорной расы (Rh. apterus),
засоряющей озимую рожь (уве-
личено)
В заключение отметим только большую практическую важность
знания способов распространения плодов и семян сорных растений.
У многих так называемых обязательных, или специальных, сорня-
ков наших полей, т. е. таких, которые, как правило, живут только
среди культурных растений, в строении семян и плодов имеется
ряд особенностей, тесно связанных с условиями их жизни. Семена
сорняков часто оказываются схожими с семенами соответствующих
культурных растений. У тех и у
других наблюдается одинаковая
«парусность», т. е. летательная
способность, благодаря чему не
происходит отделения семян сор-
няков при веянии. Часто семена
сорняков по своей форме, цвету,
объему и прочему похожи на се-
мена тех культурных растений,
среди которых они произрастают.
Указанные особенности семян
сорняков, представляющие собой
хорошие примеры мимикрии, или
миметизма, среди растений, выра-
ботались в процессе отбора.
Одним из наиболее интерес-
ных примеров результата отбора,
имевшего место в условиях куль-
туры, является особая раса, или
подвид, полупаразитного растения
погремка, или пастушьего гребня
(Rhinanthus apterus), отличаю-
щаяся от своих дикорастущих лу-
говых сородичей, от которых она и произошла, тем, что из ее пло-
дов (коробочек) семена не высыпаются, форма их несколько иная
(менее сплюснутая) и отсутствуют крыловидные придатки (рис. 89).
Время созревания семян этого сорняка точно совпадает со време-
нем созревания семян ржи. Это растение при уборке ржи попадает
в снопы и при обмолачивании и веянии, обладая одинаковой парус-
ностью семян с рожью, не отделяется от нее. Вследствие этого на
следующий год семена сорняка высеиваются вместе с семенами ржи.
Понятно, насколько важно для рациональной борьбы с сорными
растениями знать все их биологические особенности, в частности,
отмеченные выше. Очевидно, что крайне желательным является
их уничтожение до созревания семян и наиболее тщательная
очистка посевного материала. Более подробное рассмотрение
этого вопроса, которого мы касаемся здесь только вскользь,
читатель найдет в соответствующих руководствах по сорным рас^
тениям.
102
Распространение плодов и семян
Все вышеизложенное дает представление о том, насколько раз-
нообразны приспособления плодов и семян к распространению их
по поверхности земли.
В заключение обратим еще внимание на то, что образование
того или другого плода данным растением находится в связи со
строением его и с условиями местообитания. Так, если бы, напри-
мер, крылатка, служащая для полета семян, развилась на низких
растениях, то она была бы бесполезна; но на травах и кустарниках
она и не встречается, а только на высоких деревьях — клене,
грабе, сосне и др. Наоборот, цепкие плоды на высоких ветвях от-
сутствуют. Оказывается, что европейские растения, образующие
подобные плоды, обычно не превышают 2 м. Не образуют их, с дру-
гой стороны, и очень низкорослые виды, а равно и водные рас-
тения.
Целесообразность этого станет вполне понятной, если мы вспо-
мним, что эти плоды переносятся млекопитающими животными.
Все это можно особенно наглядно наблюдать в лесу, где со-
вместно произрастают растения, резко отличающиеся по своей вы-
соте. У деревьев, составляющих верхний ярус леса, семена и плоды
в большинстве случаев разносятся ветром. В так называемом под-
леске, образованном более низкими древесными растениями (рябина,
калина, шиповник, крушина, кизильник и др.), мы встречаем глав-
ным образом плоды-ягоды, поедаемые птицами, способствующими
распространению этих растений. Ягоды вообще свойственны почти
исключительно только древесной растительности, среди которой
и живут птицы, питающиеся ими. Несомненно, что распространение
семян этих растений с помощью ветра не могло бы быть достаточно
эффективным, так как лиственный полог больших деревьев в зна-
чительной мере защищает их от ветра. Среди травянистых растений
леса у многих из наиболее высоких трав плоды и семена разносятся
ветром. Внутри леса ветер — наибольшей силы, хотя по сравнению
с местами вне леса он, конечно, незначителен. Но чем ниже мы
будем спускаться, тем количество подобных растений будет стано-
виться все меньше и меньше. Среди наиболее низких растений
леса преобладают такие, которые характеризуются приспособле-
ниями к самостоятельному разбрасыванию семян, а также расте-
ния, семена которых разносятся муравьями и другими мелкими
животными.
Все описанные в этой главе факты представляют собой яркую
иллюстрацию деятельности естественного отбора, в процессе ко-
торого в течение длинных геологических периодов времени, на про-
тяжении многих тысяч и миллионов поколений и развились все
эти целесообразные особенности. Этот вопрос был уже рассмотрен
нами в «Введении», почему мы и не будем здесь к нему возвра-
щаться.
ОСЕННЯЯ ЛИСТВА И ЛИСТОПАД
Осенью наши лиственные леса и сады меняют цвет своего
наряда. На место монотонной летней окраски выступает большое
разнообразие весьма ярких тонов.
Листья грабов, кленов и берез становятся светло-желтыми,
дубов—буровато-желтыми, вишни, рябины и барбариса — пун-
цово-красными, черемухи — пурпуровыми, бирючины и берескле-
та — фиолетовыми, осины — оранжевыми, ольхи — мутного бу-
ро-зеленого оттенка, а листья буков представляют все переходы
от желтовато-красного к буро-красному цвету и т. п. Особенно эф-
фектно все это разнообразие цветов, когда они сочетаются в одном
общем ландшафте. Такие картины наблюдаются в смешанных лесах
северного умеренного пояса.
Осенняя перемена окраски листьев не ограничивается, однако,
только деревьями и кустарниками, но простирается и на низко-
рослые травы. В лесистых местностях они очень мало обращают на
себя внимание, скрываясь за крупными древесными породами, и
лишь изредка придают осеннему пейзажу характерную черту. Но
иное дело там, где высокоствольных деревьев совсем нет и первен-
ствующую роль играют именно низкорослые травы: таковы области
полярной флоры, а в особенности высокие горы, поднимающиеся
выше границы лесов. Листва небольших трав и полукустарников
и в особенности карликовых кустарников, образующих мохнатые
ковры, приобретает красные, фиолетовые и желтые тона со всеми
переходными оттенками, не уступающие в яркости живым цветам.
В то время как листья голубицы принимают фиолетовый оттенок,
черника одевается в яркокрасный цвет, альпийская толокнянка
приобретает поразительно яркую пурпуровую окраску и пр. С по-
лучившими осеннюю окраску растениями перемешаны вечнозеле-
ные породы, преимущественно хвойные, столь обильно встречаю-
щиеся в арктических и горных странах. Наконец, к этому
ландшафту присоединяются белые и серые лишайники, например,
так называемый олений и исландский мхи, кажущиеся осенью бе-
лыми полосами и пятнами на красном, фиолетовом и желтом фоне.
104
Осенняя листва и листопад
Где же кроется причина осенней перемены в окраске листвы,
и имеет ли она какое-либо значение для растений?
Что касается происхождения осенних цветов листвы, то оно
объясняется глубокими изменениями жизнедеятельности листо-
вых тканей с приближением неблагоприятного зимнего времени.
Весной и летом хлоропласты более или менее равномерно распре-
деляются в постенном слое протоплазмы. Это обусловливает ярко-
зеленую окраску листьев. С наступлением же осенних холодов
хлоропласты скучиваются в компактные глыбки, причем, по мне-
нию некоторых ученых, происходит обособление протоплазмы от
клеточных стенок. Это приводит к изменению ярко-зеленой окраски
листьев в темную и тусклую. Такие сезонные изменения окраски
хвои отчетливо наблюдаются у наших вечнозеленых хвойных по-
род: ели, сосны, можжевельника и др.
У подавляющего большинства деревьев и кустарников холодной
и умеренной зоны приспособление к зиме с ее морозами пошло в на-
правлении образования листопадных форм, сбрасывающих листву
на зиму. Осенняя расцветка листопадных пород есть следствие
связанного с этим сезоном отмирания листвы. Мы уже знаем (см.
стр. 46), что в листьях, наряду с зеленым пигментом — хлорофил-
лом, всегда присутствуют еще желтые пигменты — ксантофилл,
каротин и другие, которые за хлорофиллом, как обладающим боль-
шею яркостью, бывают невидимы. Осенью у листопадных пород
в процессе подготовки листвы к опадению хлорофилл разрушается,
а желтые, замаскированные ранее хлорофиллом, пигменты высту-
пают во всей своей красе, и лист принимает золотисто-желтую
окраску. При этом желтые пигменты химически несколько видоиз-
меняются.
Иначе обстоит дело с красным, синим и другими осенними цве-
тами листвы. Здесь разрушение хлорофилла идет обычным путем,
но сюда присоединяется еще образование нового красящего пиг-
мента, также нам уже известного, именно антоциана. Доказать
это легко. Если взять, например, красный лист и прокипятить его
в воде, то он сделается зеленым или желтым, смотря по тому, как
далеко ушел процесс разрушения зеленого пигмента. Горячая вода
убьет клеточки листа; красный пигмент, растворенный в клеточном
соке, перейдет вследствие этого в воду; хлорофилл же, не маски-
руемый более ничем, а если он уже разрушен, то желтый пигмент,
сообщает теперь свою окраску листу, так как, в отличие от анто-
циана, они в воде не растворяются. Как мы уже видели, говоря об
окраске цветочных лепестков (стр. 47), различные оттенки анто-
циана зависят от того, в кислой ли, щелочной или нейтраль-
ной среде он находится. Если антоциан растворен в кислом клеточ-
ном соке, то, смотря по крепости раствора, он имеет малиновый
или розовый цвет; в нейтральном соке антоциан фиолетовый; на-
ходясь в щелочной среде, он приобретает уже синий оттенок, Вот
Причины появления антоциана
105
эти-то разнообразные оттенки антоциана и сочетание его с другими
постоянными пигментами листа и придают разнообразие осеннему
расцвечиванию листвы.
Откуда берется антоциан и почему он образуется в громадном
количестве именно осенью? На этот вопрос дают ответ весьма инте-
ресные исследования Овертона, в значительной мере нашедшие
затем подтверждение и в исследованиях других ученых. Овертон
во время путешествия на Альпы обратил внимание на удивительное
обилие и разнообразие красной листвы на горах по сравнению с до-
линами. В то же время, исследуя содержимое клеток в красных ли-
стьях, он нашел в них значительные количества сахара. Это обстоя-
тельство побудило его по возвращении предпринять ряд опытов для
решения следующих вопросов: 1) находится ли появление красного
пигмента в какой-либо связи с обилием сахара в окрашенных ли-
стьях; 2) какое участие принимает свет в образовании красного пиг-
мента и 3) влияет ли на образование его температура. На два послед-
них условия — свет и температуру — было обращено внимание
потому, что яркий свет и низкая температура (особенно по ночам)
представляют наиболее существенные отличия условий произра-
стания в горах. Опыты производились в большом количестве и над
разнообразными растениями. На основании их Овертон пришел
к следующим выводам: у большого числа растительных видов,
принадлежащих к самым различным семействам, появление
красного пигмента находится в определенной зависимости от коли-
чества сахара в клеточном соке; свет составляет существенное
условие образования красного пигмента: в опытах, произведенных
в совершенной темноте, несмотря на присутствие сахара, красный
пигмент не появлялся; температура, независимо от времени года
и стадии развития растения, также оказывает большое влияние
на образование антоциана, этому благоприятствует температура
невысокая, приближающаяся к 0°; при прочих равных внеш-
них условиях красная окраска появляется тем реже и тем менее
интенсивна, чем температура окружающей среды выше. Осенью
температура воздуха понижается, а равно происходит в значитель-
ном размере образование в листьях сахаров, передвигающихся
затем в другие части растения.
Последующие исследования, впрочем, показали, что из этих трех
факторов, влияющих на образование антоциана, наличие света
не является обязательным. У многих растений антоциан образуется
и в полной темноте. Повидимому, разные растения оказываются
в этом отношении существенно неодинаковыми.
Значение сахаров для образования антоцианов объясняется та-
ким образом. Антонианы представляют продукт взаимодействия
сахаров с органическими веществами — флавонами. Флавоны в
свою очередь также — производные сахаров. Флавоны в тканях
растений появляются в тех случаях, когда расход сахаров на про-
106
Осенняя листва и листопад
цессы нормального обмена задерживается. Низкие, близкие к нулю
температуры тормозят синтез белков и других сложных органиче-
ских веществ. В результате этого в тканях растений сахара накап-
ливаются в избытке. Это создает условия для синтеза флавонов и,
в конечном счете, — антоцианов. Нарушение нормального обмена
веществ в растениях может происходить не только при низких
температурах. И в этих случаях часто образуются антоцианы.
Механически поврежденные или пораженные грибными парази-
тами ткани окрашиваются нередко в фиолетовый или красный
цвет. Вокруг уколов сосущих насекомых часто образуются харак-
терные окрашенные пятна или кольца. Иногда и при нормальном
обмене веществ, когда в больших количествах накапливаются са-
хара, образуются антоцианы. Многие плоды (крыжовник, слива,
вишня, смородина, земляника и т. д.), по мере созревания, как
известно, зеленый цвет меняют на красный или фиолетовый; в тоже
время с созреванием крахмал превращается в них в сахар.
Какое же значение для растений имеет их осеннее расцвечива-
ние? И на этот вопрос окончательного ответа еще не дано.
Осенняя окраска проявляется так ярко, приводит в осенние
месяцы в лесах и садах к такой богатой гамме различных оттен-
ков, цветовы-х контрастов, что издавна привлекала к себе внима-
ние человека. И в наши дни специалисты-озеленители подбирают
в декоративных насаждениях ассортимент пород, обеспечивающий
возможно более красочное осеннее расцвечивание. А приносит ли
эта расцветка пользу самим растениям? Не есть ли это просто одно
из проявлений отмирания листвы перед листопадом? Повидимому,
первоначальное появление антоциана в растительных тканях было
простым следствием обмена веществ, избыточного накопления саха-
ров и само по себе не было для растений ни вредным, ни полезным.
Но мы знаем немало примеров, когда естественный отбор превращал
безразличные свойства организмов в полезные приспособления. Так,
в процессе приспособления к животному миру у растений появи-
лись ярко окрашенные антоцианом и другими пигментами около-
цветники — органы приманки животных-опылителей (см. стр. 46).
Яркая окраска многих плодов также стала приманкой животных,
распространяющих семена (см. стр. 96). Правдоподобно не раз
высказанное предположение, что антоциан, окрашивающий в
темные тона молодые побеги и листочки растений-подснежников,
служит как бы экраном и способствует таким образом превраще-
нию части поглощенной тканями лучистой энергии солнца в тепло-
вую. Повышение же температуры в холодную погоду содействует
более энергичному протеканию ряда важных жизненных процессов
в теле растения (превращение веществ, их передвижение, испарение
воды и пр.). Значение антоциана как фактора, способствующего
согреванию листа, доказано опытным путем. Установлено, что
участки листа, окрашенные антоцианом, при действии солнечных
Значение листопада
107
лучей нагреваются на два градуса, а то и выше, нежели участки
зеленые. Именно поэтому окрашенные антоцианом растения осо-
бенно широко распространены на крайнем севере, в высокогор-
ных условиях, а в более умеренном климате — рано весной, т. е.
во всех случаях, когда растения испытывают недостаток в тепле.
Однако это объяснение биологической роли антоциановой
окраски трудно приложить к осеннему изменению цвета листьев.
Полезность его для растений остается неустановленной. Но не
исключена возможность, что наука вскроет такие случаи, когда
естественный отбор поставил на службу растению и яркую осеннюю
расцветку его листьев.
За изменением окраски листьев следует их опадение — осенний
листопад.
На грустные размышления наводит картина листопада. Глядя
на нее, кажется, что стихийные силы мертвой природы побеждают
живой растительный мир. Ближайшее знакомство с делом показы-
вает, однако, что этот вывод неверен и что листопад представляет
собой одно из важнейших приспособлений против неблагоприятных
для растения зимних условий.
Опадение листьев свойственно всем деревьями кустарникам и вы-
текает из особенностей роста этой группы растений. Старые листья
по мере разрастания кроны все сильнее затеняются. Возможность
их ассимиляции все более и более падает. Старые листья постепенно
отмирают и опадают. Во влажном тропическом климате эта смена
листьев происходит постепенно, не будучи приурочена к определен-
ному времени года. Каждый лист нередко способен жить и ассими-
лировать несколько лет. Деревья и кустарники влажных тропи-
ков, как правило, вечнозелены. В нашем северном климате деревья
живут и развиваются при ежегодной смене лета и суровой зимы.
Естественный отбор выработал в этих условиях строгую сезонную
периодичность в сроках листопада, с ежегодным сбрасыванием всей
листвы однажды в году — осенью. Таким образом возникла листо-
падность.
Основное значение осеннего листопада состоит в том, что, теряя
листья, растения тем самым спасаются от засыхания, которое по-
вело бы к неминуемой гибели. Листья представляют громадную по-
верхность испарения для содержащейся в растении влаги. В теплое
время года эта убыль влаги пополняется равномерно притоком ее
из почвы, откуда она всасывается корнями. Но с охлаждением почвы
всасывательная деятельность корневых волосков падает; она по-
нижается настолько, что хотя испарение влаги листьями вследствие
низкой температуры тоже уменьшается, тем не менее потеря воды
растением теперь не может уже возмещаться. Это, несомненно,
доказывается следующими опытами.
Если охладить до немногих градусов выше нуля почву, в кото-
рой посажены растения с листьями, усиленно испаряющими влагу,
110
Осенняя листва и листопад
Можно признать несомненной зависимость листопада от темпе-
ратуры почвы, именно от охлаждения ее, в связи с количеством
испаряемой листьями влаги. Один и тот же вид гораздо ранее ли-
шается листвы на горах, нежели в долинах и низменностях. При-
чина этого, очевидно, в том, что на больших возвышенностях
охлаждение почвы наступает ранее, чем внизу. Холодные ветры на
горах быстро отнимают у почвы теплоту, что влечет за собой пре-
кращение всасывательной деятельности корней и раннее наступле-
ние листопада. К тому же ведет охлаждение почвы осенью и для
наших лиственных деревьев. Как только всасывание корнями влаги
падает, так что продолжающееся испарение ее через листья не воз-
мещается поступлением из почвы, это пагубно отражается на
листве, — она опадает.
При таком объяснении естественно ожидать, что у тех деревьев,
у которых листья способны к большему испарению, у которых,
следовательно, скорее нарушается равновесие между количеством
воды испаряемой и всасываемой корнями, и листопад наступает ра-
нее. Так оно и оказывается на самом деле. У деревьев, листья ко-
торых испаряют менее влаги, осенний листопад или наступает
позже, как, например, у дуба, или же совсем отсутствует, как у хвой-
ных. Особенность листьев последних заключается в том, что они
покрыты более толстым слоем кутикулы, т. е. пленки, пропитан-
ной особым веществом, сообщающим ей значительную непроницае-
мость. Благодаря ей наши сосны и ели расходуют сравнительно
небольшое количество воды. У дуба кутикула развита менее, чем
у хвойных, но более, чем у других наших лиственных деревьев.
Если энергию испарения воды листьями у различных деревьев
выразить в цифрах и принять ее для липы за 100, то она в среднем
выразится: для бука — 71, клена—64, ели—19 и сосны — 16.
Зависимость листопада от всасываемой и испаряемой растением вла-
ги делает понятным и тот факт, что, при прочих равных условиях,
листва долее остается там, где почва и воздух более влажны: из
влажной почвы корни могут более всасывать воды, а влажный
воздух уменьшает испарение ее через листья. В тенистых, сырых
лесных ущельях и оврагах остаются зелеными не только папорот-
ники, но даже листья берез, буков и ясеней, в то время как здесь
же рядом, на сухом открытом холме побуревшая листва уже давно
опала с подобных деревьев на засохшие вайи папоротников.
Вот и все, что в настоящее время можно сказать о ближайших
внешних причинах утраты деревьями листьев с наступлением осени.
Но этим вопрос о причинах листопада далеко еще не исчерпывается.
Следующие факты говорят, что к влиянию одних внешних условий
эти причины нельзя свести. Известно много примеров культивиро-
вания наших лиственных деревьев в таких местах земного шара, где
ни температура воздуха, ни количество осадков не изменяются
существенно с наступлением зимнего времени. Однакоже и там мно-
Летний листопад
111
гие из наших зеленых соотечественников стоят по зимам голыми
среди чуждых им вечнозеленых деревьев и кустарников. На острове
Мадере уже много лет культивируются наши дубы и другие деревья,
и, несмотря на то, что температура самого холодного месяца
(января) здесь достигает+15,4°, листья у них на зиму сбрасыва-
ются. Сеянцы, выращенные в оранжереях при постоянных благо-
приятных условиях, лишаются листьев приблизительно в то же
время, когда происходит листопад у деревьев, растущих на воле.
Из подобных наблюдений ясно следует, что главные побудитель-
ные причины к наступлению листопада кроются не во внешних
условиях. Здесь действует закон наследственности — передачи от
одного поколения другому того, что выработалось в течение дол-
гого времени в процессе естественного отбора.
Сроки же листопада для каждого вида в различных географиче-
ских районах определяются наследственными свойствами вида и се-
зонным ходом изменений во внешней среде. Молодые листья приобре-
тают способность нормально расцвечиваться и опадать не сразу.
Невызревшие в должной мере листья в случае заморозка или рез-
кой засухи «убиваются», засыхают, но нормальное пожелтение и
листопад у них никогда не наблюдается. Обычные среднерусские
породы (береза бородавчатая, клен остролистный, липа мелколист-
ная и др.), будучи перенесены в Мурманскую область, не успевают
к концу короткого субарктического лета вызревать, и листва у них
осенними заморозками побивается еще зеленой. В климате средней
полосы СССР листья этих же пород за 4—5 месяцев успевают вызреть
и листопад совершается нормально. Интересно, что при культуре
в Средней Азии среднерусские древесные породы желтеют и сбра-
сывают листву после 7—8 месяцев вегетации. Следовательно, сроки
листопада определяются не только возрастом листьев, по и наступле-
нием метеорологической осени.
Мы рассматривали до сих пор листопад как приспособление ра-
стений к морозной зиме. Существует, однако, и другое направле-
ние приспособлений, приведшее к листопадности,—приспособле-
ние к перенесению знойно-засушливого времени года. Наивысшего
развития этот тип листопадности приобретает в тропиках — в са-
ваннах. Но и в пределах СССР в зонах пустынь и полупустынь
большое значение имеет летний листопад в начале знойно-засушли-
вого периода. Среднеазиатские миндали, при выращивании их на
богаре, с наступлением летней засухи теряют значительную часть,
а иногда и всю листву, и в это время закладывают почки возобновле-
ния. Летний листопад наблюдается и у многих полукустарников,
например, у полыней и ряда солянок. Осенью, при наличии дождей
листообразование у этих растений возобновляется. Биологическое
значение летнего листопада тоже, что и осеннего — предохранение
растения от засыхания.
Самый механизм листопада заключается в следующем.
112
Осенняя листва и листопад
Перед опадением листьев в основании их черешков появляются
слои особых тонкостенных клеточек. Это так называемые раздели-
Рис. 90. Молодой раздели-
тельный слой (аб) в осно-
вании листового черешка
Plectranthus fruticosus; вг—
сосудистоволокнистый пу-
чок (под микроскопом)
тельные слои. Их можно хорошо рассмо-
треть только под микроскопом (рис. 90).
Вследствие быстрого размножения этих
клеток снаружи, против разделитель-
ного слоя появляется вздутие, отличаю-
щееся от грубых старых тканей более
светлым цветом и некоторой прозрач-
ностью. Когда разделительные слои до-
стигнут соответствующей толщины, их
тонкостенные клетки отделяются одна
от другой, причем оболочки нигде не
разрываются и не повреждаются. По
всей вероятности, соединяющее их меж-
клеточное вещество растворяется орга-
ническими кислотами, благодаря чему
связь между клетками нарушается и
листья опадают. Это происходит даже
само собой, при отсутствии внешних
побудительных причин. А когда послед-
ние имеются, листопад совершается
с особенной легкостью. Каждый порыв
ветра срывает множество листьев. За-
мерзание и последующее оттаивание клеточного сока, связанное
с изменением его объема, также сильно способствует их отделению.
Рис. 91. Сложные листья конского каштана
(Aesculus hippocastanum) во время листопада
Вот почему и явилось
предположение, будто
морозы служат непо-
средственной причиной
листопада. Разделитель-
ный слой иногда обра-
зуется не в нижней ча-'
сти черешка, а распола-
гается так, что от че-
решка остается малень-
кий чешуеобразный %
остаток, который слу-
жит защитой для разви-
вающейся в его пазухе
почки, например, у жас
мина,
ных
слой, кроме основания
главного черешка, возникает еще ниже каждого листочка (рис. 91).
Вследствие этого такие листья, составленные из многих листочков,
У листьев слож-
разделительный
Механизм листопада
113
рассыпаются осенью при каждом порыве ветра, как карточные
домики. Поверхность на месте отделения черешка затягивается
пробковым слоем и всегда бывает гладкой и определенной формы для
каждого вида растения; здесь нет раны, в которой могло бы на-
чаться гниение. Иногда пробка образуется и раньше, способствуя
отпадению листа.
Для размножения клеток, образующих разделительный слой,
требуется известная температура внешней среды. Рано и внезапно
наступающие в некоторые годы морозы могут воспрепятствовать
появлению разделительных слоев, и листья тогда замерзают, не
успев отпасть. В такие годы на многих деревьях в течение всей
зимы остаются сухие, побуревшие листья.
Время появления разделительного слоя оказывается зависящим
от длины периода дневного освещения: чем он короче, тем скорее
появляется разделительный слой. Прямые опыты показали, что
затемнение растений на некоторое количество утренних и вечерних
часов вызывает более ускоренный листопад. Таким образом, уко-
рачивание дня к осени является одним из факторов, стимулирующих
сбрасывание листьев.
Посредством изменения в тканях, подобных описанному, отде-
ляются также иногда лепестки цветков, тычинки, самые цветки,
оставшиеся неопыленными, спелые плоды, листовые черешки,
если оторвать от них пластинки и т. д. Следовательно, листопад
представляет только частный случай из ряда однородных явлений.
Продолжительность листопада различных деревьев неодина-
кова. Так, у гинкго листопад продолжается всего несколько
дней, а у грабов и дубов — несколько недель, причем осе-
нью у этих деревьев опадает лишь часть листьев, а остальные опа-
дают только в конце зимы. Существует разница еще в следующем
отношении. У одних деревьев от листьев начинают обнажаться
крайние ветви, и отсюда уже листопад постепенно доходит до
основания; у других же он имеет обратное направление. Примером
первого порядка служат ясени, лещины и буки, а второго — липы,
ивы, тополя, груши.
ЗИМОВЬЕ РАСТЕНИЙ
Зимой растительный мир производит впечатление совершенно
мертвого. Ни в покрытых снежным покровом растениях, ни у от-
крыто стоящих обнаженных от листвы деревьев, подверженных
действию трескучих морозов, незаметны какие-либо жизненные
проявления. На самом деле, растения, за исключением однолетних
трав, не погибают зимой. В них, на первый взгляд, прекращаются
жизненные процессы; однако это не смерть, а глубокий покой.
Достаточно появиться первым лучам весеннего солнца, чтобы в ра-
стении началось движение воды и органических веществ, пробуди-
лись почки, затем развернулись листья и т. д.
Оказывается, видимое прекращение жизненных отправлений
у зимующих растений неполное. Это вытекает из следующих каж-
дому доступных опытов и наблюдений.
В октябре или ноябре, до наступления морозов, срежем ветки
нескольких деревьев, например: ольхи, орешника, ивы, тополя,
липы, клена. В теплом помещении поставим эти ветки нижним кон-
цом в сосуд с водой и будем наблюдать — не разовьются ли их почки.
Результат окажется отрицательный: ветки или совсем не распу-
стятся и засохнут, или же распустятся через несколько недель, а то
и месяцев. Если этот опыт проделать несколько позже — после не-
скольких морозов, — результат будет более благоприятный. А
в конце зимы — в феврале, марте — срезанные ветви в большинстве
случаев через несколько дней дадут молодые побеги. Значит, у
деревьев осенью и зимой, помимо внешних неблагоприятных усло-
вий, есть какой-то внутренний тормоз роста. К весне, после перене-
сенных холодов этот тормоз прекращается. Другими словами,
зимой внутри дерева совершаются какие-то благоприятные для
роста внутренние изменения—какие-то жизненные процессы.
После того как эти изменения произошли, причиной продолжаю-
щегося в природе покоя растений остаются уже только внешние
неблагоприятные условия — не окончившиеся еще холода.
Интересно, что для некоторых древесных пород умеренной зоны
зимние холода являются необходимым условием не только для
Растения во время зимнего покоя
115
Рис. 92. Бак для теплых ванн,
снизу подогреваемый радиатором,
а сверху закрываемый войлоком
нормального возобновления роста побегов весной, но и для нормаль-
ного развития цветков. Так, установлено, что у персика, сливы,
черешни и некоторых других плодовых деревьев образование цве-
точных почек идет гладко только при температурах около 0°. Если
же температура зимой (как это в отдельные годы бывает на юге
СССР) держится все время значительно выше 0°, цветочные почки
этих пород далее не развиваются и весной засыхают и осыпаются.
В такие годы урожай плодовых насаждений заметно снижается.
Изменения, имеющие следствием
устранение тормоза к росту,
можно вызвать в дереве посред-
ством некоторых искусственных
мер, которые в практике цветово-
дов применяются для выгонки ра-
стений зимой. Так, под влиянием
эфиризации можно заставить цве-
сти сирень в декабре и даже ра-
нее. Для этой цели кустики сирени
помещают в плотно закрываемый
ящик при температуре 17—19°
и рядом с ними ставят чашку с
серным эфиром (30—40 г эфира на
гектолитр воздуха), который бы-
стро испаряется, и, таким образом,
ящик наполняется его парами.
Через двое суток пребывания в та-
кой атмосфере сирень в теплом и влажном помещении скоро расцве-
тает. После приема эфиризации разработан (Молишем) более про-
стой метод ускоренной выгонки — метод теплых ванн. Он состоит
в помещении сирени или других растений и ветвей их в теплую
воду около 30—35°. Если растение в горшке, — последний остается
над водою, а вода закрывается, чтобы не остыла (рис. 92). Сирень
хорошо выгоняется после 9—12-часового пребывания в теплой
ванне. Так же хорошо выгоняется ландыш и некоторые другие ра-
стения. Подобные опыты приводят к заключению, что пары эфира
и теплая вода,действуя как раздражители,более ускоренным обра-
зом развивают в почках те способствующие выгонке процессы, ко-
торые совершаются в природе в период зимнего покоя в течение
большего времени.
Что же именно происходит в деревьях в период покоя? Исследо-
вание состава синтезированных в течение летнего вегетационного
периода запасных органических веществ показывает, что зимой
происходят превращения этих веществ. Летом клетки паренхимы
(мякоти) дерева наполняются крахмалом. Весной следующего года
этот крахмал превращается в сахар, который, будучи в отличие от
крахмала растворимым, уносится восходящим водяным током вверх
116
Зимовье растений
и дает материал для развития почек в побеги. Напрашивается пред-
положение, что зимой крахмал остается в дереве. Однако оказы-
вается, что уже поздно осенью, еще до наступления сильных холо-
дов, запасной крахмал в древесных тканях почти полностью исче-
зает, превращаясь главным образом в сахар или жир. Веспой же,
перед пробуждением растения, крахмал снова появляется. Ниже
мы увидим, что благодаря сахару и жиру увеличивается холодо-
стой к ость расте н и я.
Исследования показывают, что в почках наших древесных по-
род зимой во многих случаях не приостанавливается и процесс
роста. Согласно наблюдениям проф. Н. И. Железнова, произведен-
ным свыше 100 лет назад (1851 г.), почки деревьев в условиях
Москвы продолжают расти в течение января—февраля и временно
прекращают видимый рост лишь при падении температуры ниже
— 10°. Позднее было установлено, что при этом происходит
деление клеток и дифференциация зачатков листьев и цветков.
Возможно, что у многих, а может быть и у большинства наших зи-
мующих растений, втом числе и травянистых,приостановка роста на
зимний период не является полной.
Почему же открытые, предоставленные действию морозов части
растений не погибают зимой, мало того — в них еще совершаются
некоторые жизненные процессы?
Оказывается, у них имеется ряд приспособлений, дающих им
возможность переносить зимние холода. Такое значение, как мы
уже знаем, имеет прежде всего осенний листопад. Мы видели, что
опадение листьев, делая невозможным дальнейшее испарение на-
ходящейся в растениях влаги, предохраняет их от засыхания.
Теперь прибавим, что прекращение испарения влаги опавшими
листьями имеет еще и другое, побочное значение для зимующих
растений. Испарение воды, переход ее из жидкого состояния в газо-
образное, сопровождается большим поглощением тепла и потому
производит значительное охлаждение. Это зависит от того, что ча-
стицы газообразных веществ гораздо более подвижны, чем веществ
жидких. Приобретение такой подвижности частицами жидких ве-
ществ при переходе в газообразное состояние совершается за счет
тепла, отнимаемого от окружающей среды. Если бы такое отнятие
тепла от растений происходило во время зимы, то этим увеличива-
лось бы действие на них морозов. Осенний листопад предохраняет
их от этого.
Листья не могут перенести морозов и погибают, но для всего
растения в целом их гибель является фактом положительным.
Оставшиеся же ветви и толстые древесные стволы не боятся моро-
зов. Прежде всего в них, в отличие от листьев, развивается под
кожицей слой особой ткани, играющей защитную роль. Это — проб-
ковая ткань. Весной и в начале лета молодые растущие древесные
ветки похожи на травянистые стебли: они так же сочны, мягки
Приспособления растений к зимним условиям
117
зеленого цвета. Но уже среди лета у большинства деревьев почки
с поверхности начинают сереть или буреть, так как под кожицей
вместо содержащей хлорофильные зерна мякоти (паренхимы) об-
разуется иная ткань — пробковая. Она состоит из нескольких ря-
дов клеток, у которых протоплазма отмирает, а стенки пропиты-
ваются пробковым веществом, обладающим, как это известно по
бутылочным пробкам, непроницаемостью для жидкостей и газов.
С каждым годОхМ число слоев пробковой ткани увеличивается.
Внутренняя полость клеток пробковой ткани вместо протоплазмы
наполнена воздухом, чем объясняется легкость пробки. На стволах
и крупных ветвях березы клетки пробковой ткани, образующей
слои бересты, заполняются белым порошкообразным веществом
(бетулином), сообщающим березе характерный белый цвет.
Благодаря непроницаемости для жидкостей, пробковая ткань
предохраняет дерево от того же, от чего и листопад — от
потери воды и засыхания. Если бы не пробка — вода испарялась
бы зимой через стебель. Хотя это испарение было бы несравненно
меньшим, чем через листья, но и оно при прекратившейся, вследствие
охлаждения, всасывательной деятельности корня могло бы пове-
сти к пагубным последствиям. Правда, и пробка, не пропускающая
через свою поверхность воды, не останавливает все-таки совершенно
испарения' ее деревом: на поверхности ветвей и стволов остаются
«продушины», служащие для дыхательного обмена воздуха; это
гак называемые чечевички, соответствующие устьицам листьев;
через них вместе с газовым обменом происходит испарение, кото-
рое удается обнаружить и в зимний период. Этот процесс при не-
благоприятных условиях водного режима может даже, повидимому,
в отдельных случаях привести молодые проростки деревьев к ги-
бели от зимнего высыхания. Но более взрослым деревьям такая
опасность едва ли угрожает ввиду того, что пробка у них развита
сильнее, а запасы влаги в стволах и корнях достаточно велики.
В указанном, т. е. в предохранении дерева от засыхания, за-
ключается функция пробки, а не в том, будто она, как часто
говорят, защищает, благодаря плохой теплопроводности заклю-
ченного в ней воздуха, от охлаждения и, следовательно, играет
роль как бы «теплого платья». Платье человека и волосяные
покровы животных только потому согревают организм, что он бес-
престанно вырабатывает тепло. Платье и покровы умеряют тепло-
отдачу, по не прекращают ее. То же нужно сказать и относительно
пробки. Тонкие слои пробковой ткани на ветвях в состоянии задер-
жать проникновение мороза едва ли на час, а на стволах, где проб-
ковые слои толще, быть может, на несколько часов; морозы же
длятся зимой неделями или месяцами. Некоторое значение плохой
теплопроводности пробковой ткани, быть может, в том, что некото-
рое замедление в отдаче растением тепла охладевшему воздуху уме-
ряет быстроту и резкость колебаний температуры внутри растения.
118
Зимовье растений
Сказанное о значении пробкового слоя на ветвях и ство-
лах должно быть отнесено и на почечные чешуи. Наружные крою-
щие чешуи пробковеют, т. е. приобретают свойства пробковой
ткани, не пропускающей воду. Не от охлаждения, а от засыхания
защищаются благодаря этим чешуям миниатюрные нежные зачатки
листьев и цветков будущего года, скрытые в глубоких почках. Это-
му, помимо свойств самих кроющих чешуй, содействует еще то,
что промежутки между почечными чешуями, через которые все же
могли бы проникнуть водяные пары, часто наглухо заливаются смо-
листыми или клейкими веществами. Не опадающие на зиму листья,
как иглы наших хвойных, имеют кожицу, покрытую толстой плен-
кой непроницаемой для воды кутикулы, почему листья эти жестки
на ощупь. Устьица их на зиму в значительной степени закрываются.
Вследствие этого испарение воды через эти листья зимой сокра-
щается до незначительного минимума. И такое испарение является,
однако, причиной меньшего содержания воды в древесине деревьев
зимой, почему лесопромышленники ценят зимнюю древесину боль-
ше летней.
Другие приспособления деревьев и вообще зимующих растений
против замерзания имеют свое основание прежде всего в постоян-
ных физических и химических свойствах растительных тканей.
Главная масса древесины и внутренняя часть коры (луб) состоят
из элементов (в древесине — так называемые сосуды и трахеиды,
в лубе — так называемые ситовидные трубки), представляющих
собой чрезвычайно узкие, т. е. не что иное как капиллярные, труб-
ки. В этих же последних вода замерзает при более низкой темпера-
туре против обычной нормы. В трубках, имеющих диаметр от 0,3
до 0,4 мм, она переходит в лед лишь после охлаждения до —7,
—10°. В живых клетках их внутреннее содержимое, растягивая
упругую оболочку, вызывает со стороны последней большое давле-
ние, часто достигающее нескольких атмосфер. Это обстоятельство
также понижает температуру замерзания клеточного содержимого.
Если из растения выжать сок, то оказывается, что вне растения
он замерзает ранее, т. е. при более высокой температуре, чем внутри
растения. Выжатый сок успеет замерзнуть, а внутри растения он
продолжает оставаться в жидком виде, т. е. находится в переохлаж-
денном состоянии, и требует для своего замерзания дальнейшего
понижения температуры.
Растительные клетки содержат вещества, температура замерза-
ния которых значительно ниже температуры замерзания воды. Та-
ковы растворы различных веществ —минеральных солей, сахари-
стых веществ и других, — входящих в состав клеточного сока.
Опытами замораживания растворов солей установлено, что неко-
торые из них могут быть охлаждены до —5° и даже иногда до — 10°
и ниже, не замерзая. Равным образом понижается температура за-
мерзания воды и в том случае, если она, не растворяя в себе другого
Приспособления растений к зимним условиям
119
тела, хорошо пропитывает его, располагаясь в мельчайших про-
межутках между его частицами, как это видим в так называемых
коллоидах, например, в клейстере, желатине, курином белке. Таким
именно образом пропитывает вода, содержащая притом в растворе
различные вещества, и живые клетки.
Наблюдениями и опытами установлено, что низкая температура
сама по себе не убивает растения. Состояние переохлаждения не
является для него пагубным. Смерть наступает вследствие обез-
воживания протоплазмы, являющегося результатом выморажива-
ния воды, или повреждения протоплазмы кристалликами образую-
щегося в ней льда. Замерзание даже незначительных количеств
воды внутри протоплазмы легко может вызвать гибель клетки.
Если медленно охлаждать растение, так чтобы оно не образовы-
вало льда (явление переохлаждения), растение может перенести
значительное понижение температуры. Если лед быстро образует-
ся, температура, как известно, поднимется. Однако теперь, в усло-
виях льдообразования, дальнейшее ее понижение окажется уже
гибельным — растение погибнет при температуре, при которой
в условиях переохлаждения оно сохраняло жизненность. Понятно
поэтому, что все указанные физико-химические причины, обуслов-
ливающие понижение точки замерзания воды в растениях, явля-
ются вместе с тем защитным средством и против их смерти от холода.
А так как количество воды в растении зимой убывает, то концен-
трация (крепость) находящихся в нем растворов увеличивается.
Следовательно, зимой повышается и химическая защита против
замерзания.
Если температура понизилась до такой степени, что в данном
растворе или веществе, представляющем подобие клейстера, начи-
нается процесс замерзания, то прежде всего замерзает, как оказы-
вается, не раствор или не клейстер целиком, а выделяющаяся из
них чистая вода. То же самое происходит и в растениях. Когда в них
начнется замерзание, то замерзает отнюдь не все содержимое кле-
ток сразу, а только часть воды, выделяемой из растворов. Притом
вода эта выходит за пределы клеток и попадает в межклеточные
пространства, где и начинается образование льда. На наружных
поверхностях клеточных оболочек появляются мелкие кристаллы,
которые затем могут разрастаться в довольно крупные кристалли-
ческие скопления льда, разрывающие ткани растения. В боль-
шинстве случаев для такого образования ледяных кристаллов
вне растительных клеток требуется охлаждение до —2, —3°.
Известно, что появление в переохлажденных жидкостях хотя
бы маленького кристаллика вызывает быстрый рост кристалличе-
ской массы, т. е. быстрое замерзание. Однако появляющиеся в меж-
клетниках ледяные кристаллы по отношению к внутреннему содер-
жимому клеток такой роли не играют, так как они все время оста-
ются отгороженными от клеточного сока. Перегородкой для них
120
Зимовье растений
являются клеточная оболочка и поверхностный, более плотный,
полупроницаемый слой протоплазмы. Эта плазматическая перепонка,
так называемый кожистый слой протоплазмы, пропускает сквозь
себя воду, но медленно.
Многочисленными наблюдениями установлено, что описанное
выделение воды в межклетники и начавшееся замерзание ее здесь
еще не ведут за собой смерти живых клеток. В чем же тогда при-
чина гибели растений от мороза?
Немало исследований было произведено различными учеными
над выяснением сущности и ближайших причин вымерзания ра-
стений. Весьма обстоятельные работы в этой области советских
ботаников — акад. Н. А. Максимова и проф. И. И. Туманова,
привели к следующим заключениям.
Наиболее чувствительным к холоду оказывается наружный,
кожистый слой протоплазмы. Разрушительное действие мороза
на него заключается в изменении коллоидальных веществ прото-
плазмы — в их необратимой коагуляции (свертывании). Ближай-
шей причиной коагуляции является отнятие воды растущими ле-
дяными кристаллами, т. е. как бы высушивание вещества прото-
плазмы, а частью — и механическое давление льда на нее. После
оттаивания такая протоплазма оказывается уже мертвой.
Установлено, что сопротивляемость протоплазмы разрушитель-
ному действию мороза увеличивается от присутствия в растворе
некоторых веществ, главным образом сахара, масла, а затем раз-
личных солей, являющихся, таким образом, защитными веще-
ствами. Сахар, не препятствуя замерзанию растений, смягчает
вредные последствия его — предохраняет в большей или меньшей
степени белковые вещества протоплазмы от свертывания вследствие
замерзания. Это следует из таких опытов.
Если заморозить раствор куриного белка, то после оттаивания
часть его оказывается свернувшейся, выпавшей в осадок, который
более не растворяется (необратимая коагуляция). В присутствии
же сахара замерзнувший раствор белка после оттаивания ока-
зывается прозрачным — осадка в нем не образуется. Подобный же
результат получается в опытах замораживания в присутствии и
отсутствии сахара, выжатого из растений, и профильтрованного
сока. Такое же действие, очевидно, должен оказывать сахар и на
белки протоплазмы. После этого понятно, какое значение имеет
превращение в растениях зимой крахмала в сахар. Вместе с повы-
шением концентрации растворов сахар представляет химическое
средство защиты против замерзания — средство, которым расте-
ние оказывается вооруженным зимой. С наступлением весны,
когда сахар опять начинает превращаться в крахмал, палает и хо-
лодостойкость растения: известно, что возвращающиеся после ве-
сенних оттепелей хотя бы не особенно сильные морозы производят
несравненно более жестокие опустошения среди растений, чем
Приспособления растений к зимним условиям
121
самые суровые зимние холода. Подобное явление можно вызвать
и искусственно. Возьмем весной веточку кедра, тол ько-что вынесшую
суровые зимние морозы, поместим ее в сосуд, окруженный замора-
живающей смесью (лед или снег с солью), и дадим температуре
упасть до —8° или —10° — ветка будет убита морозом.
Наоборот, если растения, выросшие в тепле, выдерживать не-
которое время при температуре лишь незначительно выше 0°, —
их выносливость по отношению к морозу повышается. При этом
количество сахара в их клетках оказывается увеличенным. Расте-
ние, таким образом, как бы «закаляется» против мороза.
Это накопление сахаров у озимых растений, повидимому, свя-
зано с замедлением роста при низких температурах. В этих усло-
виях скорость образования сахаров в процессе фотосинтеза хотя
и снижается, но меньше, чем скорость их потребления на рост и
развитие. К тому же при пониженной температуре задерживается
крахмалообразование и главными продуктами фотосинтеза оказы-
ваются сахара. У древесных растений большое значение в процессе
закаливания имеет то, что они накапливают большое количество
запасного крахмала, который при соответствующих условиях
осенью превращается в защитные вещества.
Закаливание растений представляет собой, по современным воз-
зрениям, сложный и длительный процесс, в котором можно разли-
чить две фазы. Первая фаза проходит при температурах приблизи-
тельно от 0 до -|-6° и характеризуется увеличением общего содер-
жания сахаров. Под влиянием прохождения этой фазы температура
вымерзания, например, озимых растений, понижается с —7 и —9°
до —10 и —12°. Она протекает с разной быстротой и при благопри-
ятных условиях может быть пройдена за 5 дней и даже меньше.
Совершенно иной характер носит вторая фаза закаливания. В от-
личие от первой фазы, она имеет место при слабых морозах (прибли-
зительно от —2 до —5°) в замерзших растениях, при условии обез-
воживания клеток, вследствие перехода части воды в лед. Такое обез-
воживание снижает количество льда, которое может образоваться
в клетках.
Вероятно, при этом происходят также какие-то детальнее еще
не изученные изменения в самой протоплазме, делающие ее более
выносливой к вымораживанию. В течение второй фазы устойчивость
растений к холоду значительно возрастает. Так, в одном опыте
у озимо1** пшеницы Лютесценс 1060/10 после прохождения первой
фазы закаливания при температуре—13° выжило 17% растений, а
после прохождения второй фазы —96%. Вторая фаза протекает
сравнительно быстро, за несколько дней, но только после предва-
рительного прохождения первой фазы. Таким образом, обезвожи-
вание и низкая температура способствуют закаливанию растений
только при наличии в них защитных веществ, накопившихся в те-
чение первой фазы.
122
Зимовье растений
У различных солончаковых растений их высокая холодостой-
кость, повидимому, должна быть поставлена в связь с находящи-
мися в них в растворе солями. Различие в количестве содержащих-
ся в клетках веществ влияет на различную степень их холодо-
стойкости.
Неблагоприятно для холодостойкости большое количество воды
в клетках. Увеличение содержания ее, особенно резко проявляю-
щееся при прорастании семян или при распускании почек, должно
оказываться вредным, с точки зрения холодостойкости, в двоя-
ком отношении. С одной стороны, при этом разбавляется раствор
содержащихся в клетке защитных веществ. С другой, если в клеточ-
ном соке много воды, то уже при сравнительно незначительном мо-
розе образуется большое количество льда, а он своим давлением
оказывает вредное механическое влияние на протоплазму, содей-
ствуя ее коагуляции. Из сказанного понятно, почему растения
сочные в общем обладают меньшей холодостойкостью, чем расте-
ния ксерофильные, приспособившиеся к существованию при не-
большом количестве влаги. У последних — меньшее содержание
воды и в клеточном соке более крепкий раствор защитных веществ.
Сильные морозы, не убивая благодаря указанным защитным
приспособлениям открыто стоящих деревьев, однако, нередко вы-
зывают в них так называемые морозобоины, т. е. продольные тре-
щины в стволах. Они образуются как результат сильного сжатия на-
ружных слоев древесины от сильного охлаждения. Сжатие иногда
достигает такой степени, что связность слоев древесины оказывается
недостаточной, чтобы противостоять ему: она вдруг нарушается и де-
рево с треском лопается. Морозобоины наиболее часты на листвен-
ных породах—их можно найти в каждом саду и парке. Очень часто
по краям морозобоин дерево омертвевает, и здесь образуются на-
плывы. Последние иногда вновь поражаются морозами и отмиракэт.
На этом месте появляются паразитные грибы, нередко ведущие
к гибельным для плодовых деревьев или отдельных их ветвей ра-
ковым заболеваниям. По морозобоинам внутрь дерева иногда про-
никает вода с зародышами микроорганизмов, что, в конце концов,
может повести к образованию большого дупла, лишающего дерево
стойкости против бурь и сильных ветров. Исследованиями
И. В. Мичурина, развитыми далее проф. Б. С. Мошковым и другими
учеными, установлена интересная зависимость зимостойкости дре-
весных растений от длины периода дневного освещения. В преды-
дущей главе мы видели (см. стр. ИЗ), как влияет этот фактор на
время листопада. Оказывается, однако, что влияние это не ограни-
чивается одним только листопадом, но распространяется и на дру-
гие процессы, связанные с зимним покоем растений, как, напри-
мер: развитие пробки, остановку роста, накопление в коре и в дре-
весине запасных питательных веществ и т. п. Ускорение всех про-
цессов, характерных для зимнего покоя, вызываемое укорочением
Защита зимующих низкорослых растений
123
периода дневного освещения, связано и с повышением зимостой-
кости. Искусственно укорачивая длину дня (т. е. затемняя расте-
ние), И. В. Мичурину удалось в Тамбовской области без укрытия
на зиму благополучно выращивать деревца южных персиков и
абрикосов, а Б. С. Мошков добился того, что южные породы, обыч-
но не выносящие ленинградской зимы (например, белая акация,
абрикос и др.), перезимовывали здесь без всякого укрытия. Другие
исследователи, применив искусственное укорочение летнего дня
на Кольском полуострове, достигли того, что здесь, за полярным
кругом, благополучно перезимовал в открытом грунте ряд средне-
русских деревьев и кустарников: тополей, берез, кленов, чернъй
и красной смородины, желтой акации и др. Замечательна перези-
мовка в этих условиях сеянцев белой акации.
Таким образом, в этих исследованиях удалось, активно вмешав-
шись в цикл развития растения, управлять наступлением периода
покоя и зимостойкостью, увеличив ее по желанию эксперимента-
тора.
Выше рассмотрены защитные приспособления против холода —
против засыхания и замерзания зимой—для частей растений,
открыто стоящих на воздухе, т. е. для стволов и ветвей деревьев.
Для более низких растений и для частей растений и органов, скры-
тых в земле и под снежным покровом, роль важнейшей защиты
играет этот последний, так как он плохой проводник тепла; дру-
гие приспособления развиваются здесь в меньшей степени. В част-
ности, снежным покровом защищены: находящиеся в земле корни,
корневища, луковицы, клубни; лежащие на земле семена и споры;
наконец, многие травянистые растения, у которых перезимовывают
и их надземные органы, и кустарники. Мощный снежный слой
в состоянии защищать почву и покрытые им растения от чрезмер-
ного охлаждения даже и во время тридцатиградусных морозов,
которые нередки для наших широт. Опыт показал, что в северном
умеренном поясе в малоснежные зимы погибает масса растений,
между тем, как в случае обильного снега они хорошо переносят
сильнейшие морозы. Многие виды кустарников и низкорослых
деревьев часто оказываются вымерзшими как раз по то самое ме-
сто, до которого достигал снежный покров. Садовники отлично
знают плохую теплопроводность снега и потому часто пользуются
им для защиты наиболее чувствительных к холоду растений, при-
гибая их к земле и набрасывая на них кучи снега. В лиственных
лесах защитным средством против охлаждения является также ли-
ства, опадающая с деревьев осенью и покрывающая почву и низ-
кие растения.
Зависимость формы и строения растения от снежного покрова,
а также и от направления движения снега особенно наглядно вы-
ступает у представителей флоры севера и альпийского пояса найгих
горных систем, где снежный покров, как известно, достигает
124
Зимовье растений
значительной мощности, а в горах бывают снежные лавины. Осо-
бенности строения этих растений являются хорошим приспособ-
лением к условиям их обитания и представляют собой яркий
пример деятельности естественного отбора.
Весьма характерную форму имеют некоторые северные и горные
гольцовые и альпийские кустарниковые растения, примером ко-
торых может служить широко распространенный восточнее Бай-
кала кедровый стланец (рис. 93). Его ветви, в отличие от сибирского
Рис. 93. Кедровый стланец (Pinus pumila)
кедра, в типичных случаях не растут кверху, но от основания
ствола стелются по почве, соприкасаясь с ней, а дальше изогнуто
приподнимаются. В своих обычных местообитаниях кедровый стла-
нец достигает высоты в 2—4 ли сильно ветвится. У верхней гра-
ницы своего распространения в горах он представлен стелю-
щимися формами, не превышающими 40—50 см высоты при длине
ствола Р/г—21/г м. Стелющиеся ветви стланца образуют придаточ-
ные корни, позволяющие егду произрастать в самых неблагоприят-
ных условиях (на почвах с высоким уровнем вечной мерзлоты, боло-
тах, горных осыпях). Стланец разрастается очень густо, и ство-
лики его сильно ветвятся. Во взрослых зарослях ветви до того
скучены, что человеку пробраться через них местами невозможно.
Помимо направления роста, приспособление к тяжелым снежным
покровам заключается еще в большой упругости ветвей. Если
влезть на такое растение, то оно сильно нагибается, а по освобож-
Приспособления горных растений к снежному покрову
125
дении от тяжести опять выпрямляется. Точно так же упругие ветви
наклоняются под давлением снежной массы, причем их верхушки
касаются даже земли, и иглистые ветви прижимаются к почве.
Зимой кедровый стланец часто заносится снегом и пригибается
к земле. Места, где он летом образовывал густые чащи, кажутся
тогда лишенной растительности заснеженной равниной. Опыты по-
казали, что закрытые снегом ветви перезимовывают благополучно,
а возвышающиеся над снегом погибают от мороза. Когда снег
начинает таять, то освободившиеся от тяжести сучья и ветви сами
собой постепенно поднимаются и наконец принимают свое прежнее
положение.
Подобно кедровому стланцу стелются еще многие другие тун-
дровые и горные кустарники, например, северные карликовые
березы (ерники),можжевельники,горная сосна и другие, вследствие
чего все они низкорослы.
У всех стланцевых форм деревьев и кустарников главный по-
бег рано отстает в росте и меняет свое направление из вертикального
в наклонное или горизонтальное. Многочисленные боковые побеги
обгоняют в росте главный побег и растут также наклонно или го-
ризонтально.
Впрочем, одним только приспособлением к снежному покрову
нельзя объяснить все случаи низкорослости альпийских растений
и связанной с нею способности прижиматься к земле. В той области,
где снежный покров достигает наибольшей мощи, еще высятся
кедры, лиственницы и ели, обладающие упругими ветвями, не об-
ламывающимися от снега. Наоборот, на таких больших высотах
над уровнем моря, где массы снега уже убывают, большие деревья
не встречаются, а есть только низкорослые. В этом обстоятельстве
сказывается еще другого рода приспособление против холода.
Почва зимой бывает теплее атмосферы. Помимо защитного
действия снежного покрова это объясняется еще тем, что почва
поглощает больше тепловых лучей, чем атмосфера, и сама по себе
не так легко отдает тепло, как воздух. Чем выше подниматься
над уровнем моря, тем разница в температуре почвы и воздуха бо-
лее и более будет увеличиваться. Здесь воздух разрежен, он легче
пропускает лучистую энергию солнца, почему усиливает нагре-
вание почвы, но сам не нагревается. Измерения показали, что на
вершине Монблана сила солнечных лучей на 26% больше, чем на
уровне Парижа, что их химическая сила на высоте 2600 м над уров-
нем моря на 11% значительнее, чем у самого моря. На Гималай-
ских горах, на высоте 300 м над уровнем моря термометр зачернен-
ный (благодаря чему им поглощаются солнечные лучи) показывает
иногда на солнце на 40—50° больше, чем в тени. Итак, на возвышен-
ных местах наземные части растений окружены относительно более
холодным воздухом, чем на низменностях; наоборот, в почве на
высотах скрыто гораздо более тепла. При столь большом термине-
126
Зимовье растений
ском контрасте почвы и атмосферы низкорослость является целесо-
образной особенностью.
В этом, вероятно, кроется объяснение происхождения встре-
чающихся на возвышенностях карликовых растений среди таких
видов, которые на местах низких представляются крупными фор-
мами. По той же причине листья подобных растений отличаются
малыми размерами, цельнокрайней или округлой формой и соби-
раются в розетки. Это предохраняет их от излишнего излучения
тепла в пространство.
Наличие наиболее благоприятных температурных условий в при-
почвенном слое воздуха привело еще к одному своеобразному
приспособлению. При понижении температуры, особенно ниже
0°, боковые ветви большинства наших древесных пород несколько
опускают свои вершины, изгибаясь дугой книзу. При повышении
температуры имеет место обратное движение. Это — следствие
эксцентричности анатомического строения боковых ветвей, благо-
даря которой верхняя и нижняя их половины при перемене темпе-
ратуры изменяют свой объем неравномерно. Последнее и обуслов-
ливает их изгибание. Этот процесс особенно сильно выражен у
стланцевых древесных растений. При наступлении холодов они
пригибаются к земле, располагаясь на неблагоприятное время года
в слое воздуха, непосредственно прилегающем к почве. Отмечены
случаи, когда кедровый стланец, летом поднимавшийся над землей
на 100—140 см, после наступления морозов, еще до выпадения снега,
настолько пригибался к земле, что возвышался над ней всего лишь
на 30—40 см.
Такое изменение ориентации растения в пространстве произво-
дит впечатление активной его реакции на внешние воздействия,
причем реакции целесообразной, помогающей перенести неблаго-
приятное время года. Поэтому поводу следует иметь в виду, что дан-
ная полезная особенность, как и все другие, исторически разви-
лась в процессе естественного отбора и ее происхождение не может
быть понято вне отбора.
Стланцевые формы кустарников превосходно приспособлены
к суровому климату северной части Дальнего Востока. Здесь за-
росли кедрового стланца нередко простираются на многие десятки
километров, чередуясь с группами травянистых многолетников,
кустарников и редколесий, и определяют направление хозяйствен-
ного использования значительных территорий.
Высокая приспособленность стланцевых форм деревьев и ку-
старников к суровому климату привела известного сибирского
садовода А. Д. Кизюрина к мысли культивировать плодовые де-
ревья близ северных пределов их распространения в припочвенном
слое (не свыше 30—35 см над поверхностью почвы), размещая здесь
все дерево с его стволом и ветвями. Так были созданы стелющиеся
культуры плодовых (рис. 94). А. Д. Кизюрин рекомендует сажать
Зимовье травянистых растении
127
молодые деревца-однолетки в наклонном положении, под углом
30—40°. Для удержания их в наклонном положении применяются
деревянные приколки, крючки и т. п. Стелющиеся плодовые куль-
туры Кизюрина и его последователей с успехом переносят сибир-
ские зимние стужи и обеспечивают хорошее плодоношение.
В советских среднеазиатских республиках и на Кавказе, где
достигают своего северного предела культуры инжира, граната
Рис. 94. Стелющаяся яблоня (г. Омск)
и маслины, широко применяется пригибание этих растений на зиму
к земле.
Интересный способ зимовки представляют травянистые дву-
летние и многолетние растения. Их наземные органы весьма
нежны и потому у большинства погибают. Однако жизнь на этом
не прекращается. Части, дающие начало новым листьям и стеблям
у этих растений, как уже не раз говорилось, помещаются в почве
в виде корневищ, луковиц и клубней, где и перезимовывают. Эти
органы в течение лета переполняются органическими веществами,
приготовляемыми в листьях и затем проводимыми вниз. На них
образуются чешуйчатые низовые листья и зачатки новых листьев
и цветков, которым только в следующем году суждено будет увидеть
свет.Зимой они защищены почвой и снегом от холода, а с наступле-
нием весеннего тепла из них выходят на поверхность новые стебли
и листья, которые зеленеют, цветут, приносят плоды и снова обра-
12В
Зимовье растений
зуют на солнечном свете и в тепле органические вещества для новых
подземных луковиц, клубней и корневищ.
Замечательно то, что названные подземные образования, пред-
ставляющие собой не что иное, как видоизменение стебля с ли-
стьями и зачинающимися побегами, тем глубже сидят в почве, чем
сильнее данное место подвержено опасностям мороза и лучеиспу-
сканию. Глубина нахождения в почве, смотря по условиям, различ-
на не только для различных видов растений, но в некоторых слу-
чаях даже для различных экземпляров одного и того же вида.
Хороший пример этого представляет известная уже нам (см. рис.
10) хохлатка, растущая в различных местностях — ив лиственных
лесах, и среди кустарников, и на открытых лужайках. В первом и
втором случае, т. е. под защитой деревьев и кустарников и их опав-
шей листвы, плохо проводящей тепло, клубни хохлатки находятся
на расстоянии лишь нескольких сантиметров от поверхности почвы.
Наоборот, на лужайках, где снег часто сдувается сильными ветрами
и незащищенная почва промерзает на значительную глубину, клубни
оказываются значительно далее от поверхности. У многих расте-
ний, имеющих клубни (например, из семейства орхидных), глубина
их нахождения и глубина промерзания почвы настолько тесно
соответствуют друг другу, что, определив первое, можно с точно-
стью предсказать и второе, так как клубни обыкновенно распола-
гаются недалеко от границы промерзающего слоя.
Каким же способом происходит углубление названных подзем-
ных образований в землю и как объяснить это явление? Как пока-
зывают наблюдения, корневища сначала растут отвесно вниз, и это
направление роста сменяется горизонтальным лишь по достижении
глубины, уже безопасной в отношении промерзания.
Клубни и луковицы или с самого начала залегают на определен-
ной глубине или же втягиваются туда благодаря сокращению кор-
ней, вызываемому действием холода. При сокращении конечные
разветвления корня, т. е. его нижние части, будучи прикреплены
к почве многочисленными корневыми волосками, остаются на своем
месте, верхние же части, а с ними вместе клубни и луковицы, вслед-
ствие этого, перемещаются вглубь. Сократительной способностью
отличается только центральная часть корня; кора же, окружающая
ее, не сокращается и потому образует на сокращенном корне склад-
ки и морщины. Это хорошо можно наблюдать на корнях куль иви-
руемых всюду гиацинтов и на корневищах различных видов наших
касатиков.
Сокращение корней под воздействием холода обязано их боль-
шой чувствительности ко всякого рода влияниям. Достаточно лег-
кого прикосновения к молодому корешку, чтобы заставить его пе-
ременить направление роста и таким образом избежать опасности
поранения. Если па кончик корня действуют два слоя почвы —
сырой и сухой, то он изгибается в сторону сырого слоя. Равным
Зимовье травянистых растений
129
образом, если в почве неравномерно распределены питательные
элементы, то корень всегда растет в том направлении, где их больше.
Если в почве находятся вредные вещества, то корень растет в сто-
рону от них. Подобные изменения направления роста под влиянием
одностороннего внешнего раздражения носят название пгропизмов.
Но если корни растений так чувствительны к внешним раздра-
жениям, причиняемым различными препятствиями, к поранениям,
влаге и пище, то после этого ничего удивительного не представляет
и чувствительность к холоду, заставляющая корни сокращаться
и втягивать вглубь луковицы и т. д. Об этой чувствительности,
между прочим, говорит еще следующий факт. На далеком севере
почва на известной глубине замерзает на целый год. Летом оттаи-
вают только верхние слои, глубже которых остается постоянный лед.
На оттаивающих слоях появляется растительность, в том числе
хвойные деревья. И вот у этих-то последних корни растут сначала
прямо вниз, а по достижении промерзлого слоя изгибаются в сто-
рону и принимают горизонтальное направление, продолжая свой
рост в оттаявших слоях. Здесь чувствительность к холоду берет
перевес над силой земного притяжения, по причине которой корень
нормально растет в вертикальном направлении — сверху вниз.
У некоторых многолетних растений, как о том говорилось уже
в главе «Весеннее пробуждение растений», перезимовывают и
листья — часто в виде розеток, прижатых к земле, или на стелю-
щихся тоже прижатых к земле стеблях. Таковы, например, из наи-
более известных растений: маргаритка, первоцвет, медуница, ман-
жетка, подорожник, одуванчик, чистотел, коровяк, сурепка, зем-
ляника, купырь, Черноголовка, глухая крапива, яснотка пур-
пуровая и др. Перезимовывают наземные молодые всходы у озимых
однолетников, каковы, например, озимые рожь и пшеница.
Не обладая развитыми защитными приспособлениями против
вымерзания в такой мере, как деревья, и потому вымерзая в боль-
шинстве в бесснежные зимы, зимующие наземные органы указан-
ных и других травянистых растений охлаждению все же под-
лежат, и потому в соответствующей степени защитные приспо-
собления (сахар и пр.) для борьбы с холодом у них к зиме выра-
батываться должны. Это следует, например, из таких наблюдений.
Если мы будем выращивать указанные, хорошо перезимовываю-
щие растения не под открытым небом, где они постепенно переходят
от лета к осени и зиме, а в теплой оранжерее и затем сразу выста-
вим на мороз, они неминуемо погибнут. Более того, если выкопать
их зимою из грунта, выдержать некоторое время в земле и снова
вынести на мороз, они также погибнут. В природных же условиях
не редкость, что во время осенних заморозков озими хрустят под
ногой, и тем не менее это промерзание не наносит им вреда. Это
объясняется уже знакомым нам (стр. 121) явлением закаливания
растений против холода.
130
Зимовье растений
Однолетние растения, за исключением озимых, погибают зи-
мою. Однако эта гибель всех индивидов каждого рода не ведет
к истреблению видов. Они сохраняются благодаря семенам и спо-
рам, которые лучше, чем сами растения, переносят стужу и со-
храняют способность к прорастанию, т. е. пребывают в состоянии
покоя, не умирая, более продолжительное время. Производились
опыты охлаждения семян ячменя, пшеницы, тыквы, гороха и др.
до —250, —259° (между прочим, опусканием их в жидкий водород),
и семена после этого не теряли своей всхожести. Это такая выно-
сливость, перед которой наши самые сильные морозы совершенно
бледнеют и в которой вряд ли когда может встретиться необхо-
димость. Она, несомненно, связана с малым содержанием воды в се-
менах — с их сухостью. Если разбухшие в воде семена вынести на
мороз, то они потеряют свою всхожесть. Разбухшие же и высохшие
затем в теплом помещении семена всхожести не теряют.
Выносливость спор у папоротников, хвощей, мхов, грибов, во-
дорослей и бактерий приблизительно такая же, как и у семян.
Здесь находим ту интересную особенность, что у многих растений
перед зимовкой образуются особые зимние споры, сильно отличаю-
щиеся от тех, которые образовывались летом. Эти случаи нередки
среди паразитных грибов — тех большей частью микроскопически
мелких организмов, которые массами поселяются на различных
частях высших растений.
Весьма распространенным примером являются паразитирующие
на многих хлебных и плодовых растениях ржавчинные грибы. При
помощи микроскопа видно, что они, подобно другим грибам, со-
стоят из сплетения мелких нитей (гиф), которые проникают в ткань
листьев и стеблей. Гифы поглощают питательные соки растения-
хозяина. На их концах отшнуровываются маленькие одноклеточ-
ные крупинки — споры, скопления которых в виде ржавых оран-
жево-бурых пятен во второй половине лета можно наблюдать на
стеблях наших культурных злаков. Эти легкие споры разносятся
ветром и, попадая на листья других экземпляров злаков, выра-
стаю'! в грибные нити, проникающие внутрь растения и, в свою
очередь, также образующие споры; таким образом, паразит распро-
страняется в течение лета.
В начале осени образуются уже другие споры — зимние, отли-
чающиеся более толстой оболочкой и состоящие из двух клеток
(рис. 95); кучки этих спор представляются на поверхности листав
виде темнобурых точек. Будучи покрыты толстой оболочкой и со-
держа минимальные количества воды, зимние споры легко переносят
холода, по миновании которых они и начинают прорастать. Другие
виды паразитных грибов зимуют в виде особых, весьма плотных
образований, так называемых склероциев, которые, благодаря ма-
лому содержанию в них воды, отлично переносят действие низких
температур. Примером таких грибов является спорынья, склеро-
Зимовье низших растений
131
ции которой, известные под названием «рожков», высовываются
из колосьев злаков в конце лета. Опавшие на землю и зимующие под
снегом рожки прорастают весной, образуя особые головчатые гри-
бочки на длинных ножках; внутри головок образуются споры, ко-
торые и заражают завязи ржи (рис. 96).
У многих шляпочных грибов перезимовывают подземные части,
так называемая грибница, или мицелий, представляющая собой
сплетение гиф, имеющих вид
ветвящихся нитей, состоящих
из одного ряда однообразных
клеток. На ней в следующем
году вырастают новые плодо-
вые тела — то, что в просто-
Рис. 96. Спорынья (Claviceps pur-
purea). 1 — колос ржи с рожками
(склероциями) спорыньи; 2—скле-
роций с развившимися на нем вес-
ной плодоношениями
Рис. 95. Линейная ржав-
4HHa(Puccinia graminis).
Кучка летних спор, ме-
жду которыми — одна
зимняя спора (под ми-
кроскопом)
речьи и называют «грибом», тогда как в действительности это лишь
часть растения-гриба, находящегося в виде грибницы в почве.
В случае отсутствия снежного покрова и сильных морозов часть
грибницы может, конечно, погибнуть, но часть ее сохраняется
в более глубоких непромерзших слоях почвы и весной начнет
разрастаться. Достаточно сохраниться нескольким нитям, чтобы
они, с наступлением благоприятных условий, разрослись в новую,
густо разветвленную грибницу, от которой пошли бы надземные
плодовые тела. Плодовые тела, образующие споры, сами потому
обыкновенно погибают, что в большинстве случаев они содержат
в себе очень большое количество воды (до 90%). Есть, однако, и
исключения. Так, на стволах лиственных деревьев, под корочкой
льда иногда попадаются неповрежденные шляпки гриба «бархат-
132
Зимовье растений
ной ножки» (рис. 97), с желтой плоско-выпуклой шляпкой и кашта-
новой ножкой, у основания черноватой, с бархатистой кожицей.
Есть еще и другие виды шляпочных грибов, попадающихся зимой
на стволах деревьев. Большая или меньшая выносливость их про-
тив мороза связана с малым содержанием в них воды. Известно так-
же, что на стволах деревьев хорошо
перезимовывают многие трутовики,
имеющие обычно вид полукруглых
шляпок, плотно прикрепляющихся к
стволу сбоку, откуда они заимствуют
пищу посредством разветвляющейся
внутри ткани дерева грибницы. Ткань
зимующего трутовика — его плодового
тела — очень твердая и сухая. Ясно
заметные на поверхности трутовика
Рис. 98. Зимующая ризо-
морфа гриба «бархатная
пожка» (Collydia veluti-
pes) (уменьшено в 20 раз)
Рис. 97. Зимующий на деревьях гриб
«бархатная ножка» (Collydia velutipes)
кольца говорят о числе лет его, так как ежегодно от края при-
бавляется новое кольцо — подобно тому, как в древесине дерева.
У некоторых грибов выработалось чрезвычайно оригинальное
приспособление для сохранения жизненности мицелия — так на-
зываемые ризоморфы. Лучше всего их можно наблюдать и иссле-
довать у всем хорошо известного опенка, обычно образующего
свои плодовые тела на старых пнях, по нередко являющегося
весьма опасным паразитом наших лесных пород. Пораженное опен-
ком дерево довольно скоро отмирает, и тогда, отламывая куски коры
с мертвых стволов и гнилых пней, легко обнаружить между корой
и древесиной длинные, ветвящиеся черновато-бурые шнуры и ленты,
образующие в общей сложности сеть, очень похожую на корневую
Зимовье низших растений
133
систему какого-нибудь высшего растения. Эти довольно твердые
шнуры и ленты достигают иногда нескольких метров длины. Они
имеют роговидную консистенцию и в сухом состоянии очень ломки,
но разрываются с трудом, обнаруживая в месте разрыва белую
ткань. Исследование показало, что это образование состоит из плот-
ного сплетения грибных нитей, хорошо перезимовывающих в та-
ком состоянии. Потом на концах таких шнуров возникают бугорки,
превращающиеся в пло-
довые тела опенок. Не-
которые другие шля-
почные грибы тоже об-
разуют ризоморфы (рис.
98). У некоторых гри-
бов зимующий мицелий
принимает вид склеро-
циев, о чем мы уже
говорили выше.
У большинства па-
поротников остаются на
зиму скрытые в земле
многолетние корневи-
ща. Под защитой снеж-
ного покрова перези-
мовывают плауны, мхи,
многие лишайники.
Большинство лишайни-
ков растет у нас на
деревьях, т. е. зимой
они не защищены сне-
гом. Но и у них, как
и у некоторых из жи-
вущих на земле лишай-
Рис. 99. Воздушные водоросли. 1 — 2—Зеле-
ные водоросли с коры деревьев: 1 — плевро-
кокк обыкновенный (Pleurococcus vulgaris),
2 — трентеполия теневая (Trentepohlia um-
brina); 3—4—Синезеленые водоросли с влаж-
ных скал: 3—глеокапса скальная (Gloeocapsa
rupestris); 4 — сцитонема глазчатая (Scyto-
ncma occllatum) (под микроскопохм)
ников (например, так называемые олений мох и исландский мох),
выработалась способность, благодаря отдаче в засушливое время
года почти всей влаги, оставаться жизнеспособными. Вследствие
этого они переносят зиму, будучи открытыми. То же нужно ска-
зать и о некоторых мхах. Громадной выносливостью отличаются
бактерии и особенно споры их. Как и с семенами, делались опыты
охлаждения некоторых бактерий сих спорами до температуры около
—252°. Будучи помещены после этого в питательные среды, споры
развивались в бактерий.
Большой интерес представляет вопрос о зимовье так называ-
емых «воздушных водорослей». Так называют водоросли, обитающие
на различных возвышающихся над почвой твердых субстратах —
на коре деревьев, на заборах, на скалах и т. д. Здесь встречаются
некоторые представители зеленых, синезеленых и диатомовых
134
Зимовье растений
водорослей, развивающихся нередко в массовых количествах
(рис. 99). Условия их жизни характеризуются частой и резкой сме-
ной температуры и влажности. И если в засушливые периоды года
многие из них высыхают настолько, что могут быть растерты в по-
рошок, то зимой они промерзают и, тем не менее, сохраняют свою
жизнеспособность. Опыты показывают, что их выносливость к низ-
ким температурам весьма велика. Так, например, относящаяся
к этой группе зеленая водоросль стихококк палочковидный
(рис. 100) после выдерживания ее в течение 8 часов при температуре
—69, —75° частично сохраняет свою жизнеспособность и, будучи
перенесена в благоприятные условия, при нормальной темпера-
туре обильно размножается.
Рис. 100. Стихококк палочковидный
(Stichococcus bacillaris) (под микро-
скопом)
Еще более выносливы к низ-
ким температурам некоторые
диатомовые водоросли, оста-
вавшиеся живыми после воз-
действия на них температу-
ры —200°. Замечательно, что
наиболее типичные воздуш-
ные водоросли не образуют
каких-либо специальных по-
коящихся стадий, и приспособления к перенесению низких темпе-
ратур и других неблагоприятных условий заключаются у них во
внутренних физиологических особенностях клетки: в повышении
вязкости протоплазмы, в накоплении в клетках масла и т. п. Это
дает им возможность быстро переходить от покоя к вегетации и
наоборот, что и необходимо в условиях частой смены благоприят-
ных и неблагоприятных воздействий, характерной для многих их
местообитаний.
Нижний предел температуры, переносимый взрослыми цветко-
выми растениями, для различных видов различен. Так, рябина,
сирень, липа, береза, ольха, орешник, смородина, крыжовник и
многие другие выдерживают морозы в —21° и ниже без малейшего
вреда. Дуб и бук вымерзают при —25°; сливовые и вишневые де-
ревья — при —31°; яблони и груши—при—33°. Многие растения
гибнут уже при —10°, —12°, а другие и при еще более высоких тем-
пературах: миндальное дерево — при —9°, —10°, земляничное
дерево — при —6°, —7°, дрок цветущий — при —2°, —3°, чайное
дерево — при —1°, —2°. Иногда случается, что растения, относя-
щиеся к одному роду, обладают резко отличающеюся выносли-
востью. Так, например, клен обыкновенный легко переносит холода
в —20°, а клен вечнозеленый гибнет при —6°, —7°. Каштан дикий
выносит ту же температуру, что и обыкновенный клен, а красный
каштан гибнет при —10°, —12°. Это различие зависит, конечно,
от индивидуальных особенностей растений и прежде всего от сте-
пени развития тех или других приспособлений к перенесению хо-
Стадийное развитие растений
135
лода, начиная с указанного в начале этой главы развития пробко-
вой ткани.
У многих растений зимний покой является необходимым условием
для дальнейшего развития. Мы уже видели это на примере веток
наших древесных пород, которые, будучи срезаны осенью или зи-
мою, развиваются в теплом помещении обычно только после того,
как они подверглись действию мороза (см. стр. 114). Точно так же
семена многих растений не прорастают без предварительного охлаж-
дения.
Как известно, ряд сортов культурных растений (например,
хлебных злаков) при*весеннем посеве до самой осени не плодоносит.
Для того чтобы у этих так называемых озимых сортов вызвать
образование колоса, их высевают с осени, и в этих случаях, после
зимнего покоя, развитие протекает нормально и доходит до коло-
шения. Согласно исследованиям акад. Т. Д. Лысенко, для опреде-
ленного момента в развитии озимых растений необходим некоторый
комплекс внешних условий — в первую очередь понижение темпе-
ратуры (лучше до 0° или +2°) в течение того или иного срока,
различного для разных сортов. При посеве осенью озимые сорта
находят эти условия в естественной обстановке в поле, после чего
их развитие в дальнейшем доходит до конца. Однако, как пока-
зал Т. Д. Лысенко, обычно весьма длинный вегетационный период
этих растений может быть искусственно сокращен методом так
называемой яровизации. Яровизированные семена озимых растений,
будучи посеяны весной, плодоносят в течение одного лета, т. е. ве-
дут себя как яровые. Метод яровизации сводится к воздействию на
только-что начавшие прорастать семена определенных внешних
влияний и заключается в подвергании семян некоторым определен-
ным манипуляциям (понижение температуры и др.).
В основе метода яровизации лежит разработанная Т. Д. Лы-
сенко теория стадийного развития растений. Сущность этой те-
ории сводится к тому, что весь процесс развития однолетнего се-
менного растения состоит из отдельных физиологических этапов —
стадий развития, протекающих в строгой последовательности, так
что последующая стадия не может быть пройдена, пока не закончена
предыдущая. При этом каждая стадия развития требует для своего
протекания различных внешних условий. Одной из этих стадий
и является та, которую Т. Д. Лысенко называет стадией яровиза-
ции. Нормально озимые сорта проходят эту стадию в поле, уже в зе-
леном раскустившемся виде. При искусственной же яровизации
необходимый для нее комплекс внешних условий, среди которых
ведущим является понижение температуры, растение получает еще
до посева, находясь в виде едва начавших прорастать семян.
Однако поскольку данная стадия оказывается пройденной, расте-
ние может вступить в следующую стадию своего раз&ития, которую
Т. Д. Лысенко называет световой стадией. Для нее понижение
136
Зимовье растений
температуры уже не нужно, но зато необходимо достаточно продол-
жительное (не менее 10—12 часов) ежедневное освещение. Только,
после прохождения обеих этих стадий растение, развиваясь далее,,
сможет перейти к выколашиванию.
Не останавливаясь более на этом вопросе, далеко выходящем:
из рамок данной главы, возвращаемся к нашей теме.
Итак, и у деревьев, и у других зимующих растений зимний по-
кой их наземных и подземных органов не абсолютный. Хотя види-
мые процессы питания и т. д. в это время не совершаются, тем не:
менее внутренние процессы превращения веществ не прекращаются.
Это напоминает зимнюю спячку животных, у которых, как мы узнаем
далее, физиологические отправления в это время до крайней сте-
пени понижаются, но не прекращаются.
На западе СССР, в полосе океанического климата, в январе
и феврале иногда устанавливается такая теплая погода, что на
южных склонах почва оттаивает и на несколько дней принимает
температуру, при которой может происходить прорастание семян,,
а у деревьев и кустарников — и почек. Если бы и действительно,
семена, клубни и почки начали при таких условиях прорастать,,
то последующие морозы могли бы погубить молодые ростки. Однако,
такие факты обычно не наблюдаются, так как в условиях океани-
ческого климата у растений в процессе естественного отбора выра-
боталась способность не выходить из состояния покоя в случае-
наступления преждевременных оттепелей. Но в континентальном:
климате Сибири, где длительные зимние оттепели не наблюдаются
и тепло устанавливается только при переходе к устойчивой весне,,
местные растения таким приспособлением не обладают. Они тро-
гаются в рост с первым же теплом и теряют до следующей зимьп
свою зимостойкость. Вот почему сибирская пихта и некоторые дру-
гие подобные ей деревья у себя на родине без вреда переносят в.
состоянии зимнего покоя морозы свыше —40°, а будучи переса-
жены в условия Западной Европы, гибнут там, тронувшись во время
ранних оттепелей в рост, от последующих небольших мо-
розов.
Наши многолетние растения настолько приспособлены к зим-
нему покою, прочно вошедшему в их жизненный цикл, что он еже-
годно повторяется у них и в том случае, если они перемещаются
в теплые местности. Мы уже знаем, что деревья и кустарники наших
широт даже в теплых странах ежегодно теряют листву там, где мест-
ные растения круглый год зеленеют, цветут и приносят плоды. По-
сле же листопада они пребывают некоторое, хотя и менее продол-
жительное, время в состоянии покоя. Некоторые деревья, правда,
постепенно утрачивают эту способность, но только спустя очень
продолжительное время. Так, вишня и персик постепенно превра-
тились на острове Цейлоне в вечнозеленые деревья, но эта перемена
совершилась в высшей степени медленно.
Биологические типы растений
137
В иных условиях зимуют водные растения, но об этом будет
речь в главе «Жизнь подо льдом».
Способ зимовки, другими словами — система приспособлений
к зимовке, а в засушливых областях — система приспособлений
к засушливому времени года, кладет, как мы видели, настолько
сильный отпечаток на растения, что их по сходству приспособле-
ний можно распределить по группам, получившим название биоло-
гических типов или жизненных форм. Датский ботаник Раункиер,
главным образом по расположению на растении почек возобновле-
ния и степени их защищенности от зимних невзгод, различает ряд
основных жизненных форм, которые характеризуются следующим
образом.
1. Мегафанерофиты и мезофанерофиты (условно обозначаются
буквами ММ), куда относятся крупные деревья, высотой от 8 до
30 м (мезофанерофиты) или свыше 30 м (мегафанерофиты). Почки
их в нашем климате прикрыты защитными чешуйками, а у тропи-
ческих деревьев ничем не прикрыты. Наши крупные деревья: ель,
сосна, дуб, береза и т. д., представляют примеры мезофанерофитов.
2. Микрофанерофиты (М) — кустарники и небольшие деревья,
высотою от 2 до 8 м\ на такой же высоте находятся и почки их.
Примеры: ива, крушина, яблоня, рябина, сирень, лещина, свидина.
3. Нанофанерофиты (N) — кустарники высотой от 0,3 до 2 м.
Примеры: смородина, можжевельник, волчник.
4. Хамефиты (Ch) — растения, у которых почки находятся на
коротких вертикальных или распластанных побегах не выше
0,3 м над землей и с начала зимы укрыты в толще снегового по-
крова. Это имеет место или у мелких кустарничков (например,
черника, брусника, багульник), или у полукустарников, у которых
верхняя часть побегов к зиме отмирает (например, многие виды
полыней и многолетних солянок), или, наконец, у травянистых ра-
стений, у которых побеги к зиме не погибают, а зимуют, несколько
приподнимаясь над поверхностью почвы (например, копытень, лу-
говой чай, барвинок и др.).
5. Гемикриптофиты (Н) —растения, почки возобновления ко-
торых лежат на одном уровне с поверхностью земли, а поднимаю-
щиеся над землею части их отмирают на зиму. К этому биологи-
ческому типу относится не менее половины видов наших цветковых
растений. Таковы многие растения, примеры которых приведены
выше, образующие на зиму прикорневые розетки.
6. Геофиты (G) — растения, почки возобновления которых ле-
жат под поверхностью земли. В эту группу входят растения, имею-
щие корневища, луковицы, клубни, примеры которых мы видели
в первой главе среди представителей весенней флоры. Сюда отно-
сятся также некоторые трудноискоренимые корневищные и корне-
отпрысковые сорняки, например, пырей, бодяк, осот, а на юге —
горчак и гумай.
138
Зимовье растений
7. Гелофиты и гидрофиты (НН) — имеющие почки возобновле-
ния под поверхностью воды. Сюда относятся растения болотные
(гелофиты) и водные (гидрофиты).
8. Терофиты (Th) — однолетние растения, почек возобновле-
ния не образуют. Неблагоприятное время года переживают в виде
семян. Сюда относится, между прочим, большинство сорняков.
9. Эпифиты (Е) — растения, живущие на поверхности других
растений, но не паразитирующие на них. Таковы живущие на на-
Рис. 101. Схематическое изображение биологических типов растений (по
Раункиеру). Покоящиеся побеги, их части и почки возобновления — черные,
те же части, которые при начале неблагоприятного времени года отмирают,
обозначены только контурами. При сравнении изображений видно, что чем
дальше направо идет наблюдатель, тем растения сильнее защищены от небла-
гоприятного времени года: почки возобновления лежат все глубже (у 2 —
близко над землей, у 3—на поверхности земли, у 4—8—в земле или на дне
бассейна). 1 — фанерофит; 2 — хамефит; 3 — гемикриптофит; 4 и 5 — гео-
фиты; 6 — гелофит; 7 и 8 — гидрофиты
ших деревьях лишайники и некоторые мхи, среди же цветковых
растений представителей этого типа в нашей флоре не имеется.
10. Суккуленты (S) — растения, имеющие сочные стебли, как-
то: кактусы, молочаи и т. п.
Геофиты, гелофиты и гидрофиты объединяются в группу крипто-
фитов.
Помещенное здесь схематическое изображение дает более на-
глядное представление о разных биологических типах наших ра-
стений, установленных Раункиером по способу зимовки (рис. 101).
Система жизненных форм Раункиера, наряду с рядом бесспор-
ных достоинств, обеспечивших ей широкое признание, не свободна
и от недостатков. В ней отсутствуют наземные низшие растения (во-
доросли, грибы, лишайники), а также мхи, играющие на значитель-
ных пространствах нашей Родины существенную роль в раститель-
ном покрове. Подразделение фанерофитов на подгруппы по росту
(мега-, мезо-, микро- и нанофанерофиты) искусственно. Следует
Биологический спектр флоры
139
также иметь в виду, что одна только высота расположения почек
над поверхностью земли еще не определяет зимостойкости расте-
ний, зависящей от ряда причин, среди которых особенно важны
свойства самой протоплазмы.
В различных типах растительности преобладающая роль при-
надлежит разным жизненным формам. Так, в зоне тундр из цветко-
вых растений наибольшую роль играют хамефиты (кустарнички),
в лесной зоне—фанерофиты и гемикриптофиты, в степной — ге-
микриптофиты, в пустынной — хамефиты (полукустарники) и те-
рофиты и т. д.
Если дать характеристику всем растениям данной местности,
распределив их по жизненным формам, выразить в процентах число
представителей каждой отдельной жизненной формы и располо-
жить полученные результаты в виде таблицы, то мы будем иметь
то, что Раункиер назвал «биологическим спектром» флоры этой
местности. Последний сравнивается с так называемым «нормальным
биологическим спектром», который был составлен Раункиером пу-
тем обработки двух независимых, взятых без особого выбора, спис-
сков видов цветковых растений земного шара, давших совпадаю-
щие результаты.
В нижеследующей таблице приводится «нормальный биологи-
ческий спектр», с которым сопоставляются биологические спектры
цветковых растений Костромской области (по А. Е. Жидовскому)
и Гиссарской долины в Таджикской ССР (по Г. Э. Шульцу1).
Местность
Костромская
область
Гиссарская до-
лина
Нормальный
спектр
Чис- ло видов Фанерофиты Хамефи- ты (Ch) Геми- крипто- фиты (Н) Криптофиты Терсфи- ты (Th)^
всего из них всего из них
S Е ММ М N G НН
790 7,2 — — 1,4 з,з 2,5 4 51,6 19,б[12,9 6,7 17,6
750 5,2 0,5 — 0,7 1,5 2,5 5,6 20 27,3 20 7,3 41,9
1000 46 2 3 8 18 15 9 26 6 4 2 13
Нормальный спектр в известной степени отражает количествен-
ные соотношения разных жизненных форм флористически наибо-
лее богатых областей мира — тропиков и субтропиков. Оба при-
веденных в таблице советских района расположены в умеренной
зоне, и в них, по сравнению с нормальным спектром, в несколько
раз падает удельный вес фанерофитов и возрастает удельный вес
1 Биологический спектр флоры цветковых растений Гиссарской долины
публикуется здесь впервые.
140
Зимовье растений
криптофитов. В этом — приспособление растительности к зиме
с ее морозами, а в Средней Азии — также к знойно-засушливому
лету. С другой стороны, биологические спектры Костромской
области и Гиссарской долины (крайний юг СССР) обладают
своими особенностями, отражающими различия в условиях
среды. Это тем более важно, что биологический спектр флоры
Костромской области дает в сущности представление о количествен-
ном соотношении жизненных форм почти во всей нашей средней,
а также и западной лесной полосе. Видовой состав этих лесов
беден. Число древесных и кустарниковых видов составляет всего
лишь около 7% от всего состава флоры. Резко преобладают геми-
криптофиты, т. е. многолетние травы с отмирающими на зиму над-
земными частями и с почками возобновления, расположенными под
защитой снега на поверхности почвы. Затем следуют криптофиты,
с почками возобновления, скрытыми в почве. Далее идут терофиты,
т. е. однолетники, в большинстве случаев сорняки.
Биологический спектр флоры Гиссарской долины несет на
себе печать присущего этому району знойно-засушливого лета.
Скудная древесная растительность встречается главным образом по
берегам рекили на поливных участках. Основная масса раститель-
ности летом выгорает. Число видов деревьев (ММ) и кустарни-
ков (М) падает, по сравнению с Костромской областью, почти в два
раза. На раскаленной летом почве не находят себе благоприятных
условий и гемикриптофиты, которых здесь в два с лишним раза
меньше, чем в Костромской области. Лучше всех приспособлены
к этим условиям представители двух жизненных форм — геофитов,
скрывающих свои почки в почве, и терофитов, переживающих не-
благоприятные периоды года в виде семян. Как видно из таблицы,
число терофитов составляет здесь свыше 40%, а число терофитов
и криптофитов вместе — почти 70% всего видового состава флоры.
Так, сравнивая биологические спектры флор разных областей,
можно судить о климате последних.
СЕЗОННЫЕ ПЕРЕМЕНЫ ПОКРОВОВ
И ОКРАСКИ У животных
Сравнение животного населения разных стран с несомненно-
стью убеждает, что свойства покровов животных находятся в тес-
ной связи с условиями жизни и окружающей обстановкой.Степень
развития покровов птиц и млекопитающих находится в тесной за-
висимости от климатических условий, а окраска покровов в боль-
шинстве случаев более или менее соответствует окраске окружаю-
щих животное предметов внешней среды.
Так, большинство тропических млекопитающих обыкновенно
имеет редкую, жесткую и короткую шерсть, некоторые иногда бы-
вают голы, а животные холодных стран, наоборот, имеют густой,
длинный и мягкий волосяной покров. Норвежские и сибирские ло-
шади имеют шерсть более густую и длинную, нежели лошади более
теплых стран.
У животных полярных стран часто наблюдается белая
окраска — под цвет снега и льдов; пример белого медведя всем хо-
рошо известен. У животных пустынь обычна желтая или бурая
окраска; лев, верблюд, различные антилопы, змеи, ящерицы,птицы,
насекомые и т. д. — все они в пустынях окрашены под цвет песка.
В тропических странах обитающие высоко в горах обезьяны
обладают значительно более густой шерстью, чем обезьяны, живу-
щие у подножья гор. В вечнозеленых тропических лесах часто на-
блюдается зеленая и зеленовато-бурая окраска животного насе-
ления.
Такая соответствующая окружающей обстановке окраска, на-
зываемая покровительственной, или охранительной, выгодна для
животных. Она выгодна и для хищников, делая их незаметными
и таким образом помогая подкрадываться к добыче; выгодна и для
преследуемых видов, помогая им скрываться от врагов. Так/белый
медведь мог бы погибнуть от голода, если бы он был бурого или чер-
ного цвета, так как тюлени, которых он преследует, уже издали
замечали бы его приближение на белом фоне полярных снегов и
льдов и скрывались бы. Насекомые, преследуемые птицами, если
142
Сезонные перемены покровов и окраски у животных
бы они не приобрели окраски под цвет зеленой травы или древес-
ной коры, смотря по тому, где живут, могли быть истреблены пер-
натыми и т. д. У некоторых видов, наряду с покровительственной
окраской, выработалось сходство с окружающими предметами и
по форме; например, среди насекомых есть похожие на древесные
листья, сучья, плоды и т. п. По мнению Уоллеса, покровительствен-
ная окраска в ее различных видоизменениях свойственна, вероятно,
половине всех животных, населяющих земной шар.
Развитие покровительственной окраски и формы, как и других
приспособительных признаков растений и животных, обязано
действию естественного отбора. Сохранение полезных уклонений
в длинном ряде следующих друг за другом поколений привело
к тем совершенным формам приспособления к среде в отношении
окраски животных, с которыми мы встречаемся в окружающей
нас природе.
Во всех странах, где теплое время года чередуется с холодной
снежной зимой, в покровах животных не было бы полного соответ-
ствия с климатическими условиями, если бы они всегда оставались
одинаковыми. Теплый покров, защищавший от зимних морозов,
был бы обременителен в летнюю жару. Окраска, гармонирующая
с летними тонами, не подходила бы под белоснежный пейзаж зимы,
на котором она уже не укрывала бы, а наоборот, выдавала живот-
ное. Соответственно такой изменчивости условий существования
изменяются периодически и покровы животных, подвергаясь линьке.
Линька, т. е. сбрасывание наружных покровов и замена их но-
выми, всюду существует в животном мире, но в странах с холод-
ными зимами она имеет резко выраженный сезонный характер.
В то время как в тропическом и прилегающих к нему поясах смена
шерсти у млекопитающих и перьев у птиц совершается постепенно,
в течение всего года, в странах холодных эта смена происходит
в определенные сроки, между летним и зимним сезонами. У тропи-
ческих животных летние и зимние покровы не различаются между
собой; у зимующих же в холодных странах млекопитающих и птиц
зимние покровы более теплые, нежели летние, а у некоторых ви-
дов, кроме того, и окраска их меняется под цвет зимнего пейзажа.
И чем далее на север, тем эти качества зимних покровов все более
и более усиливаются.
У млекопитающих различают более длинные и жесткие волосы—
ость — и короткие мягкие волосы—подшерсток. Для сохранения
тепла главное значение имеет подшерсток. Он особенно обильно
развивается у многих животных холодного климата, у обитателей
же тропиков часто отсутствует совершенно.
У всех полярных млекопитающих зимой значительно больше
волос, чем летом, и волосы много длиннее. Получается пышный,
густой мех, который защищает не только спину и грудь, но и все
остальное тело до самых пальцев. Например, у мускусного быка,
Покровы млекопитающих
143
живущего па крайнем севере Америки и в Гренландии, по обеим
сторонам тела почти до самой земли свешивается длинная косма-
тая шерсть, наподобие покрывала, каким защищает человек
домашних животных; шея и грудь защищены, кроме того, силь-
но разросшейся гривой. У песцов (рис. 102, 103), зайцев, лем-
мингов (мелких грызунов) длинные шелковистые волосы и мяг-
кий волнистый подшерсток покрывают все тело и густо опушают
даже пальцы. Хвост песца необыкновенно пушист; свертываясь,
животное прячет в него свою морду.
Рис. 102. Песец (Vulpes lagopus) в летнем одеянии
Шерсть северного оленя, по признанию жителей далекого се-
вера и полярных путешественников, доставляет наилучшую защиту
от ветра и непогоды полярных стран.
Для человека гораздо большую ценность представляют меха,
добытые от северных млекопитающих зимой, когда их мех особенно
густой и теплый.
Свойства меха неодинаковы у животных мелких и крупных.
На дальнем севере мелкие животные часто защищаются от ветра и
снежных бурь тем, что заносятся снегом, под которым находятся
во время непогоды. Вообще эти животные менее подвержены дей-
ствию ветра. Мех их поэтому должен защищать только от низкой
температуры, в связи с чем он должен быть густым и состоять из вол-
нистых, шелковистых и длинных волос. Более крупные животные,
кроме мороза подвержены действию ветра и пурги. Поэтому они
нуждаются в «наряде» из грубых и толстых волос. У северного
оленя, например, такие волосы, сильно суживающиеся или за-
остряющиеся у основания, сидят на коже так тесно, что са-
мый сильный ток воздуха, направляясь даже против шерсти, по
144
Сезонные перемены покровов и окраски у животных
словам акад. Миддендорфа, в состоянии произвести в покрове не
более, как узкий конусообразный промежуток, который достигает
поверхности кожи разве своей острой, как игла, вершиной.
Если длину волоса сравнить с величиной тела, то окажется, что
у мелких видов длина волос относительно большая и покровы их
относительно теплее. Так, у северного оленя длина каждого волоса
составляет небольшую часть диаметра тела, а у лемминга она почти
равна диаметру тела. Это тесно связано с причинами, можно ска-
зать, «геометрическими». Дело в том, что количество тепла, выраба-
Рис. 103. Песец в зимнем одеянии
тываемого организмом, зависит в значительной степени от объема
тела, а количество тепла, излучаемого в пространство,—от по-
верхности его; соотношение же между величиной объема и поверх-
ности у больших и малых тел неодинаково. Если объем увеличи-
вается или уменьшается, то и поверхность при сохранении прежней
формы тела тоже увеличивается или уменьшается, но в меньшее
количество раз. Так, если мы имеем куб, каждое ребро которого
равно 1 м, то вся поверхность этого куба будет равна 6 кв. м, так
как каждая из шести плоскостей его равна 1 кв. м. Объем этого куба
равен 1 куб. м. Если представим себе, что куб этот вырос настолько,
что каждое ребро его стало равно 2 лг, то каждая плоскость его
в таком случае равна 4 кв.м., а поверхность — 24 кв. м., т. е. уве-
личивается сравнительно с первым кубом в 4 раза; объем же этого
нового куба будет равен 8 куб. м, т. е. возрастет в 8 раз. Вообще
увеличение поверхности будет равно квадрату того числа, пропор-
ционально которому возросло каждое ребро, а увеличение объема —
кубу. И так будет всегда, какие бы увеличения тела, при какой бы
форме его мы ни брали: увеличение поверхности будет отставать
Сезонное изменение окраски
145
от увеличения объема сообразно указанному соотношению. Таким
образом, у крупных животных при большом объеме — поверхности
сравнительно маленькие, а у маленьких, при малом объеме —
поверхности сравнительно велики. Стало быть, мелкие животные,
по сравнению с крупными, вырабатывают теплоты относительно
мало, но излучают ее в пространство сравнительно много.
Усиленное развитие у них покровов компенсирует усиленную
теплоотдачу, так как сохраняет значительное количество тепла.
Перемена окраски при линьке представляет распространенное
явление среди тех млекопитающих холодных стран, которым зимой
приходится много бегать по снежной поверхности. Примером могут
служить ласка, горностай, песец, заяц-беляк.
Заяц-беляк (Lepus timidus), широко распространенный на се-
вере Европы и Азии (в Европейской части СССР южная граница
его распространения проходит примерно по линии, идущей че-
рез Чернигов, Тамбов, Саратов, Чкалов), а также встречающийся
в Альпах и Пиренеях, зимой бывает бел, как чистейший снег.
Только кончики ушей его остаются черными. Эту особенность, может
быть, следует рассматривать как своеобразную «метку», обусловли-
вающую возможность нахождения друг друга особями одного вида.
Весной, когда начинает пригревать солнце и сходит снежный по-
кров, окраска зайца-беляка начинает меняться. Это изменение
окраски связано со сменой волосяного покрова — линькой. Сна-
чала сереет спина, и на боках к белым волоскам примешивается
все более и более серых. Вскоре заяц становится неравномерно пе-
гим или крапчатым и, наконец, приобретает характерную рыжевато-
бурую окраску. Время, в течение которого заяц-беляк носит свой
«зимний наряд», зависит от продолжительности зимы. У северной
границы его распространения это — 9—10 месяцев, у южной гра-
ницы ареала — 4—5 месяцев. Такое же соответствие продолжи-
тельности зимы и времени ношения зимнего наряда наблюдается
и у большинства других животных, имеющих сезонную смену
шерсти или оперения.
Очень резко выражены различия в летней и зимней окраске
у песцов, широко распространенных во всей зоне тундр в Европе,
Азии и Северной Америке. Летняя окраска песцов землисто-бурая
с голубоватым оттенком; зимою — чисто белая (рис. 102, 103).
Столь же резки сезонные различия окраски и у некоторых других
мелких хищников — ласок и горностаев.
Ласки, одетые летом в каштановые шкурки, на зиму облекаются
в белоснежные теплые «шубы», и притом чем дальше на север, тем
полнее совершается эта смена и тем чище становится ее белый цвет.
Ласки широко распространены по всей территории СССР и на юг
спускаются даже в Малую и Центральную Азию и Северную
Африку. В южных частях их ареала зимней окраски ласки совсем
не приобретают.
146
Сезонные перемены покровов и окраски у животных
Очень отчетливо выражено сезонное изменение окраски у гор-
ностая. У этого кровожадного маленького хищника летняя окраска
Рис. 104. Ласка (Mustela nivalis) наверху и горностай (М. erminea) внизу
в летнем одеянии
Рис. 105. Ласка (налево) и горностаи (направо) в зимнем одеянии
со спины светло-коричневая, с брюшной стороны—лимонно-жел-
тая, зимой же он чисто белый, за исключением самого кончика
хвоста, который остается черным (рис. 104, 105).
У других животных разница в цвете между их летней и зим-
ней одеждой менее заметна; примером могут служить волк,
Сезонное изменение окраски
147
заяц-русак, белка. У волка весеннее линяние — в апреле, осен-
нее— с конца сентября до половины ноября. Зимняя шерсть
пушистее, ее ости с черными концами; подшерсток густой, желто-
вато-белый. Летом подшерсток реже, короче и желтее; черных кон-
чиков и остей меньше. В общем, летний мех имеет желтовато-бу-
рый оттенок, переходящий в желто-рыжий, а зимний—серый, но
последний гораздо чище и светлее.
У зайца-русака (Lepus europaeus), распространенного по всей
территории Советского Союза примерно от 64° с.ш. и южнее, в юж-
ных частях его ареала рыжеватая с темными пестринами окраска
сохраняется в течение всего года. Ближе к северной границе рас-
пространения русака его окраска зимой несколько меняется, он
становится более светлым, сероватым. Однако белой окраски, ха-
рактерной для зайца-беляка, русак никогда не приобретает.
Типичные обитатели леса — белки, в Европейской части СССР
и в Западной Сибири летом окрашены в рыжий цвет, зимой они
заметно светлеют, становятся серыми, но обычно с некоторой при-
месью бурых и рыжих тонов.В Восточной Сибири и на Дальнем Во-
стоке летняя окраска белок бурая, иногда почти черная, зимой она
несколько светлеет до темно-серой. У белки, проводящей большую
часть жизни на деревьях, где она строит гнездо (в дуплах или на
ветвях), ее серая зимняя окраска является, вероятно, покровитель-
ственной и имеет защитное значение.
Есть, наконец, млекопитающие, у которых цвет шерсти при
линьке совсем или почти совсем не изменяется, что также, однако,
связано с условиями их жизни. Сюда относятся все животные се-
вера, а также степей или высоких гор, проводящие зиму в спячке
в норах, как сурки,суслики и др. Им не приходится бегать по снегу,
а потому и белый цвет шерсти для них не имеет защитного значе-
ния. Упомянутый ранее мускусный бык зимой сохраняет свою
бурую окраску, делающую его весьма заметным на снежной
равнине. Но это животное стадное, и его безопасность обусловли-
вается именно тем, что животные собираются в маленькие стада,
для которых враг не страшен. Для него, повидимому, гораздо по-
лезнее быть окрашенным так, чтобы иметь возможность распозна-
вать животных своего вида на далеком расстоянии, чем быть скры-
тым окраской от врагов, против которых имеется другое средство
борьбы. Соболь также сохраняет свою бурую окраску в течение
всей сибирской зимы. Нужно при этом иметь в виду, что соболь
является опасным хищником, у которого среди хищных животных
и птиц мало врагов. Соболь охотится не только днем, но
и ночью.
Наоборот, белый медведь, почти всегда окруженный снегом или
льдом, всегда бел; не только цвет его не изменяется, но и вообще
разница между зимним и летним мехом выражена слабее, чем у дру-
гих животных.
MS
Сезонные перемены покровов и окраски у животных
Отдельно остановимся на линьке птиц, имеющей много свое-
образного и интересного. Для птиц линька имеет, пожалуй, даже еще
большее значение, так как перья служат для них не только покро-
вом, защищающим их тело, но вместе с тем необходимы для полета.
Необходимость линьки животных, помимо приспособления к раз-
личным сезонным условиям существования, вызывается и тем,
что покровы изнашиваются. У птиц это последнее особенно бывает
заметно. Именно — в сухом континентальном климате их перья
превращаются во что-то жалкое, обтрепанное. Бородка и стержень
пера на значительной части своей длины расщепляются и потом
обламываются; окраска становится грязно-бурой, только закрытые
части пера еще сохраняют в известной степени свою первоначаль-
ную окраску. В умеренном и влажном климате перо хотя и не в та-
кой степени, но все же изнашивается. У птиц же перелетных, к ко-
торым относится большинство наших птиц, их продолжительное пу-
тешествие с севера на юг и обратно, сопряженное с энергичными
движениями крыльев и пребыванием в странах с различными кли-
матическими условиями, должно, конечно, также вредно отра-
жаться на состоянии перьев и вызывать их смену.
У перелетных птиц их новое оперение должно быть готово ко
времени отлета. Поэтому естественно, что у огромного большин-
ства птиц северного полушария линька приурочена к концу лета
и главным образом к началу осени. К этому времени прекращается
воспитание птенцов и перья изнашиваются. При этой линьке вы-
падают как мелкие перья, так и крупные, т. е. маховые (на
крыльях) и рулевые (на хвосте).
Обыкновенно линька продолжается не особенно долго. Исклю-
чение составляют крупные орлы, у которых она очень продолжи-
тельна, с чем связано и их гнездованье — не ежегодно, а через
год. У этих птиц линька прекращается совершенно только на
время вывода птенцов, а затем все время идет постепенная смена
перьев, которые поэтому оказываются смешанными; рядом с совер-
шенно новыми перьями есть совсем или отчасти износившиеся.
• Во время линьки огромное большинство птиц не утрачивает
способности летать, так как маховые перья выпадают не одновре-
менно все. Полет несколько ослабевает, но правильность движений
не нарушается. Однако в этом отношении есть исключение, которое
представляют утки, гуси и лебеди; у них маховые перья выпадают
все сразу, после чего они уже не в состоянии летать, пока эти перья
не вырастут снова. На крайнем севере, в тундрах, гуси не успе-
вают еще приобрести маховые перья, как наступает уже время пе-
релета на зимние «квартиры». Гуси собираются в стаи для перелета,
не имея еще возможности лететь и, как указывает Мензбир, частично
начинают миграцию «пешком». Эта своеобразная ускоренная линь-
ка находится в соответствии с малой продолжительностью
полярного лета.
Линька птиц
149
У большинства птиц бывает одна линька в год, хотя у некоторых
опа наблюдается по два и у немногих даже по три раза. Вторая и
третья линьки обыкновенно бывают частичными: сменяются только
мелкие перья — все или часть их, а маховых линька не затраги-
вает. Исключение представляют только птицы,совершающие очень
большие перелеты, у которых маховые перья изнашиваются после
путешествия в один конец. У таких далеких странников, напри-
мер, у ржанок, пролетающих в один конец до 15 ОООклг, маховые
Рис. 106. Белая куропатка (Lagopus albus) в летнем оперении
перья сменяются дважды в год: осенью — перед отлетом на зимовку,
и весною — перед возвращением на родину. Пример частичной
линьки представляют и те самцы, у которых в брачный период
развиваются различные украшения в виде гривок, воротников и
т. п.; все эти перья, появляющиеся во время брачного периода,
по миновании его теряются птицей.
У многих птиц, особенно у зимующих, происходит и перемена
окраски. Например широко распространенная у нас оседлая птица
белая куропатка,1 встречающаяся в тундре, а также во всей лесной
зоне и даже в лесостепи, летом, как про нее говорят, «пестра от го-
ловы до пят», а зимою — «белее снега» (рис. 106, 107). У нее летнее
1 Белая куропатка вполне оседлая птица в лесной и лесостепной зоне,
из тундры же она зимой откочевывает на юг к северной границе леса.
150
Сезонные перемены покровов и окраски у животных
перо бурое, с пестринами, осенью оно отчасти заменяется бу-
рым же, но гораздо более темным, с редкими белыми перышками.
К зиме белые перья окончательно вытесняют собой бурые; весной —
наоборот. Насколько бурая окраска защищает куропатку летом,
настолько белая полезна при жизни на снегу. Помимо цвета, зим-
нее оперение белой куропатки отличается от летнего и другими ка-
чествами. Летнее перо ее рыхлое, сравнительно плохо защищаю-
щее тело птицы; зимнее, напротив — жесткое, служащее надеж-
ной защитой от суровых морозов. Сквозь бородки летнего пера
Рис. 107. Белая куропатка в зимнем оперении
можно ясно видеть предметы, у зимнего же бородки гораздо теснее
сплочены. Пуха и мелких перьев зимой больше. Эти особенности
зимнего оперения, делающие его более теплым, свойственны вообще
всем зимующим на севере птицам.
Немало есть птиц, у которых весенние яркие цвета к осени
пропадают, заменяясь более скромными. И это вполне целесо-
образно. Весной и летом зеленые листья деревьев и трав с разно-
образными яркими цветками дают надежный приют птичке, как бы
ярко она ни была окрашена. Другое дело — осенью. Побуреют
травы, пожелтеют и осыплются листья с деревьев, голо станет
в степи и в лесу. На таком открытом и однородном по тону ланд-
шафте трудно скрыться пестро одетой птичке от зорких глаз вра-
гов. Скромный наряд, подходящий под цвет побуревших листьев,
обнажившихся ветвей и почвы, делает ее менее заметной.
У некоторых птиц летняя окраска появляется на смену зимней
без смены перьев, а только благодаря уничтожению светлых краев
перьев их зимнего наряда. Лучшим примером такого изменения
Линька птиц
151
оперения без линьки может служить наш домовый воробей. Осе-
нью, когда самец покрывается зимним оперением, он гораздо свет-
лее, чем летом: о черном пятне на шее нет и помина, остается только
небольшое пятно на подбородке. Весной опять восстанавливается бо-
лее пестрый наряд, а между тем ни одного пера воробей не сбрасы-
вает. По мере приближения весны светлая кайма на перьях, благо-
даря обтрепыванию их по краям, постепенно суживается; кончики
перьев спадают, и окраска нижних перьев выходит наружу. Таким
образом, черные перья хорошо видны у воробья на шее летом—те
самые, которые приобретены им предшествующей осенью во время
линьки; зимой они кажутся бурыми или серыми благодаря тем
светлым каймам, под которыми скрыта летняя окраска. То же са-
мое наблюдается у горихвосток, некоторых овсянок, зябликов, по-
дорожников и других; стоит только приподнять у них перья зим-
него покрова и там можно увидеть летний покров, скрытый под
светлыми каймами зимнего.
Наконец, немало есть птиц, у которых изменение окраски про-
исходит и без уничтожения светлых краев перьев. Такие птицы
просто «перекрашиваются», т. е. одно красящее вещество (пигмент)
в перьях заменяется другим; вместе с тем может измениться и ри-
сунок. Такая именно «перекраска» происходит обыкновенно весной
у самцов, приобретающих иногда без всякой линьки очень краси-
вый брачный наряд. Хороший пример такой перекраски предста-
вляет луговая пиголица, или чибис, резко разнящаяся в зимнем и
брачном наряде. Усиление окраски весной иногда объясняется
и тем, что перо стирается, от чего находящееся в нем красящее ве-
щество выступает сильнее.
Линька захватывает не только перья, но и другие роговые обра-
зования, каковы оболочки клюва и когти некоторых птиц. Так,
у чечеток роговой чехол клюва на зиму удлиняется, а на лето уко-
рачивается. У некоторых чистиков линяют роговые оболочки клю-
ва, наросты около глаз и в углах рта, и эта линька сопровождается
даже переменой окраски линяющих частей, что совершенно изме-
няет облик птицы. У белых куропаток зимой когти отрастают
до очень большой длины, на лето же укорачиваются.
Перо птицы сложнее и массивнее волоса, и процесс новообразо-
вания перьев связан с более резким изменением питания кожи,
нежели развитие новых волос. Отсюда понятно, что для птицы
линька не проходит с такой легкостью, как для млекопитающих.
Лишь в редких случаях, например, у орлов, где линька растяги-
вается на огромный промежуток времени, она протекает безболез-
ненно. Обычно же линька ослабляет птицу. Это доказывается
увеличивающейся среди линяющих птиц смертностью, наблюдае-
мой даже у птиц домашних. Становясь более беззащитными, птицы
во время линьки пугливы и обычно прячутся в чаще травы, среди
опавшей листвы и т. п.
152
Сезонные перемены покровов и окраски у животных
Во время линьки птицы страдают от преследования врагов.
Здесь перед нами еще один пример относительности целесообразности
в природе. Явление линьки, несомненно, целесообразно, однако
в процессе линьки птица претерпевает ряд неблагоприятных, бо-
лезненных изменений.
Чем же объяснить изменение качества покровов, сообразное
температуре и цвету окружающей обстановки? Каким образом
установилось это соответствие, столь полезное для животных?
Появление у животных холодных стран более теплой «одежды»
по сравнению с южными видами и усиление этого качества на зиму
благодаря линьке в значительной мере зависит прежде всего от
самой температуры. Наблюдения показывают, что если животное,
меняя место жительства, попадает в иные климатические условия,
то и покровы его изменяют свои свойства. Так, в тропической Африке
овцы совершенно не имеют волны: они покрыты там тонкими и ред-
кими волосами. Тамошние собаки почти голы. Но если этих собак
перевезти в Европу, они через некоторое время приобретают во-
лосяной покров.
В Сибири, в бассейне Оби и Иртыша живут белки, обладающие
особенно пушистой ценной шерстью. Эти белки называются теле-
утками. В конце 20-х и 30-х годов были осуществлены широкие ме-
роприятия по акклиматизации телеуток в Крыму и на Кавказе.
Они здесь хорошо прижились и размножились. Однако шерсть их
в этих новых условиях стала гораздо менее пушистой и длинной.
Очевидно, это зависит от более теплого климата, а главное, от от-
сутствия холодной, суровой зимы. Вероятно, холод действует как
раздражитель на кожу и вызывает более усиленный рост волос.
Однако подобного рода изменения шерстного покрова под
непосредственным воздействием температуры все же носят ограни-
ченный характер. Более мощное развитие шерсти у животных хо-
лодных стран представляет собой наследственный признак, раз-
вившийся как полезное приспособление в процессе эволюции на
основе естественного отбора.
Нужно полагать, что животные, окраска которых оказывалась
более соответствующей господствующему цвету среды, скрываясь
от хищников или легче отыскивая добычу, выживали в большем
количестве, чем те, которые этих признаков не имели. Из их потом-
ков таким же образом переживали те, у которых это полезное ка-
чество усиливалось, и т. д. Таким образом, в природе совершался
на основе изменчивости естественный отбор особей, обладающих
данным полезным для них качеством. В странах с холодной зи-
мой такой отбор совершался в двояком направлении: летом истреб-
лялись более всего животные, оставшиеся белыми, а зимой — остав-
шиеся при линьке бурыми. Приобретенные таким путем полезные
признаки закрепились благодаря наследственности. Влиянию глав-
ным образом естественного отбора следует приписать и регулиро-
Сезонный диморфизм
15.3
Рис. 108. Бабочка Vanessa prorsa-le-
vana. Слева—весеннее поколение, спра-
ва — осеннее
ванне продолжительности наличия летнего и зимнего меха. По-
добным же образом объясняется происхождение и множества дру-
гих целесообразных приспособлений как в животном мире, так
и в растительном; примеры последних мы видели в предыдущих
.главах.
Линька совершается также у холоднокровных позвоночных и
беспозвоночных животных. Но здесь она имеет иной характер. Эти
животные не остаются на зиму в бодрствующем состоянии: зары-
ваясь в землю, под кору или опавшую листву деревьев и т. п.,
они погружаются в глубокое оцепенение. Их линька не носит по-
этому ясно выраженного сезонного характера, а совершается
главным образом вследствие
того, что покровы изнашива-
ются или при росте живот-
ного становятся ему тесными.
Говоря о сезонных из-
менениях окраски у живот-
ного, следует упомянуть о
сезонном диморфизме — су-
ществовании у одного вида
двух сменяющих друг друга,
в зависимости от времени года, поколений, отличающихся по
окраске или по другим признакам. Явление это наиболее известно
у бабочек. У некоторых крапивниц, брюквенниц и других бабо-
чек зимнее поколение, вылупляющееся из яиц, отложенных на
зиму, окрашено светлее, чем летнее, вылупляющееся летом из
яиц, отложенных весенним поколением. В частности, наиболее
обычный пример сезонного диморфизма представляет одна из кра-
пивниц (Vanessa или Araschnia prorsa-levana), весеннее поколение
которой, выходящее из перезимовывающих куколок, отличается
более светлой рыжей окраской (V. levana), а поколение осеннее,
развивающееся из яиц и куколок летних, отличается темно-бурой
окраской (V. prorsa) (рис. 108). В средней Европе, где лето про-
должительнее, темноокрашенных поколений у этой бабочки бы-
вает два и три, а кроме того, наблюдается третья, промежуточ-
ная форма. Экспериментально установлено, что причиной такого
изменения окраски является влияние температуры: подвергая
действию холода куколок летнего поколения этой бабочки, по-
лучали особей с рыжей окраской, а из перезимовывавших куко-
лок под влиянием тепла получали особей более темных. Покро-
вительственного значения эта перемена окраски, повидимому,
не имеет. Явление сезонного диморфизма наблюдалось также
у некоторых червей (турбеллярий), коловраток, ракообразных
(дафний) и некоторых других беспозвоночных животных.
ПЕРЕЛЕТЫ ПТИЦ
Теплое, благотворное дыхание весны и осенние холода одина-
ково чувствуются как растительным, так и животным миром, и
там, и здесь давая о себе знать целым рядом заметных явлений. Наи-
более бросающимися в глаза и в то же время наиболее доступными
наблюдению из весенних и осенних явлений в животном мире, по
справедливости, следует признать весенний и осенний перелеты
птиц. Явление это имеет и глубокий научный интерес. Насколько
оно интересует ученый мир, можно судить уже по тому, что в Европе
и Америке открыта целая сеть станций для наблюдения за переле-
тами птиц. Орнитологические станции имеются и в Советском Союзе.
Это явление замечательно уже с его внешней, так сказать, описа-
тельной стороны, но еще больший интерес представляет выяснение
тех причин, которые заставляют большинство представителей на-
шего пернатого царства лететь осенью за тридевять земель и вес-
ной возвращаться обратно, притом лететь в оба конца, за редкими
исключениями, не сбиваясь с пути, по определенным дорогам и в
определенном порядке.
В текущем столетии в изучении перелета птиц много дал метод
кольцевания. За последнее время его использование на многих
орнитологических станциях сделалось массовым и приобрело зна-
чение систематически применяемого научного метода. Сущность
его состоит в том, что пойманных на месте гнездования или на про-
летных путях птиц метят, именно надевают на одну ногу кольцо,
на котором указано название организации, проводящей кольцева-
ние, город, страна и порядковый номер кольца. Под соответствую-
щим номером в книгах учреждения, осуществляющего кольцева-
ние, записываются возможно более подробные сведения об усло-
виях, при которых осуществлялось кольцевание: вид птицы, дата,
примерный возраст (птенец, взрослая), условия, при которых околь-
цована (в гнезде, на пролете), и т. п. Кольца делаются алюминие-
вые, легкие, не причиняющие неудобства птице (рис. 109, ПО).
Ясно, что сопоставляя место и время поимки окольцованной
птицы с местом и временем окольцевания, можно установить пути
Кольцевание птиц
155
перелета, быстроту его, выяснить отношение птиц к старым местам
гнездовья и гнездам и разрешить много других вопросов биологии
птиц (продолжительность жизни и др.).
Впервые метод кольцевания был применен датским ученым Мар-
тенсеном в 1899 г., когда им было окольцовано 102 экземпляра пе-
релетных птиц (аистов, скворцов, уток). После первого опыта Мар-
Рис. 109. Цевка утки
с кольцом бюро коль-
цевания и кольцо в
расправленном виде
НШЮТН HORPONT
BUDAPEST 19716
MUS. TOOL
HELSINFORS
FINLAND
MUSEUM
GOTEBDRS
SWEDEN
VOGELWARTE
ROSS! T TEN
GERMANIA
D
22023
Рис. ПО. Марки колец
некоторых европейских
станций
тенсона, оказавшегося весьма удачным, метод кольцевания полу-
чает широкое распространение во всем мире. С 1904 г. в больших
масштабах кольцевание птиц проводилось Немецким орнитологи-
ческим обществом на Росситенской станции (на берегу Балтийского
моря близ устья Немана). До 1930 г. кольцами этой орнитологиче-
ской станции было окольцовано свыше 300 тыс. птиц. К 1938 г. было
получено обратно свыше 15 000 колец из самых различных стран.
Очень широко осуществлялось кольцевание птиц на орнитологи-
ческой станции на о. Гельголанде, через который проходят пролет-
ные пути многих видов птиц. Эта работа развертывалась все шире
и шире, и к 1930 г. в Европе было уже свыше 40 пунктов, где систе-
матически осуществлялось кольцевание птиц.
В Северной Америке метод кольцевания стал применяться поз-
же (1909 г.), но в очень широких масштабах. Работа эта прово-
дилась Американской ассоциацией по кольцеванию птиц. За 18 лет
(с 1920 по 1938 г.) в США было окольцовано свыше 27 400 тыс.
156
Перелеты птиц
экземпляров, из них 152 964 кольца вернулись обратно. В Америке
в работу по кольцеванию широко вовлечены орнитологи-любители
(более 2 тыс.), что обеспечивает очень широкий масштаб проводи-
мых мероприятий.
В дореволюционной России кольцевание птиц в небольших
масштабах началось впервые в 1913 г. Русским орнитологическим
комитетом, но работа эта была прервана начавшейся войной.
Более широкие масштабы эта работа приобрела уже после Ок-
тябрьской революции, начиная с 1924 г. Сначала она была сосредо-
точена на биостанции юных натуралистов в Сокольниках (Москва).
К ней были широко привлечены юннаты, работавшие под руковод-
ством Н. И. Дергунова. Биостанцией был установлен свой собст-
венный тип кольца с надписью БЮН Moskwa.
Биостанцией юных натуралистов было окольцовано около
100 тыс. птиц. С 1932 г. в СССР решением конференции по изуче-
нию фауны СССР дело кольцевания было централизовано и руко-
водство им возложено на Центральное бюро кольцевания птиц при
Управлении по заповедникам, которое организует и объединяет
всю эту работу (рис. 109).1 В настоящее время в Советском Союзе
кольцевание проводится по единому плану в различных районах
государственными заповедниками, научно-исследовательскими уч-
реждениями, юннатскими, краеведческими и охотничьими организа-
циями и т. п. Особенно широко эта работа проводится в Прибал-
тийских республиках, по территории которых проходят многие
основные пролетные пути. Например, организованной в 1946 г.
Латвийской орнитологической станцией за период с 1946 по 1950 г.
окольцовано 97 299 птиц, до 1 января 1951 г. станцией получены
сведения о нахождении 2393 птиц. Столь же широко ведутся орни-
тологические работы с применением метода кольцевания в Ли-
товской и Эстонской ССР.
Наряду с кольцеванием большую помощь в изучении переле-
тов птиц оказали наблюдения, проводившиеся с самолетов, благо-
даря которым удалось уточнить, в частности, вопрос о высоте по-
лета птиц.
Далеко не все еще выяснено в этом грандиозном явлении при-
роды, но область загадочного и неясного постепенно все сокра-
щается. Изложим кратко важнейшие научные достижения в этой
области.
Какие птицы перелетные? Из птиц умеренных стран северного
полушария только немногие остаются в течение всего года в той
области, где они родились. Сюда обычно относили воробьев, во-
рон, галок, сорок, тетеревов, глухарей, белых и серых куропа-
ток, рябчиков и некоторых других. Это — оседлые птицы. Сюда
1 Адрес его: Москва, 139, Орликов пер., д. 1/11, Центральное бюро
кольцевания.
Оседлые, кочевые, перелетные птицы
157
же следует отнести птиц кочевых, которые по окончании вскарм-
ливания молоди более или менее значительными стаями бродят из
одного места в другое в поисках пищи. Они не следуют при этом
какими-либо определенными путями и не удаляются слишком да-
леко от своего места рождения, куда раннею весной спешат возвра-
титься. Таковы, например, дятлы, клесты, коноплянки, щеглы,
разные синицы.
Огромное же большинство птиц умеренного и холодного поясов
северного полушария принадлежит к перелетным и совершают еже-
годно два больших перелета: осенью — с севера на юг, иногда
с отклонениями к юго-западу и юго-востоку, а весною — в обрат-
ном направлении. Они, таким образом, имеют два постоянных ме-
стожительства: одно — в более холодных странах и другое — в
более теплых, куда отправляются на зиму. Чем далее на север,
тем все более и более увеличивается процент перелетных птиц.
На Таймыре, самом северном полуострове Азии, по наблюдениям
академика Миддендорфа, нет уже ни одной оседлой птицы. По на-
правлению же к югу картина меняется в обратном смысле: число
видов перелетных птиц убывает, пока, наконец, при приближении
к тропическим странам большие регулярные перелеты не исчезают
совершенно.
Резкой границы между птицами перелетными и оседлыми, ра-
зумеется, нельзя провести. Кочующие птицы представляют пере-
ходную группу от птиц оседлых к птицам с наиболее короткими
перелетами. Один и тот же вид, в зависимости от климатических
условий, для одной местности может быть перелетным, а для дру-
гой — оседлым или кочевым. Например, грачи для Северной Ев-
ропы являются птицами перелетными, а в южной части СССР есть
оседлые колонии грачей, живущие там целый год. Перепелка для
нашей фауны — странствующая птица, а в Южной Африке она жи-
вет оседло. Применение метода кольцевания сокращает количество
птиц, относимых к группе оседлых. Например, такая птица, как
серая ворона, живущая у нас и летом и зимою и потому в своей
оседлости не возбуждавшая ни у кого сомнения, благодаря приме-
нению к ней метода кольцевания должна быть причислена у нас
к группе кочующих. Оказывается, что ленинградские вороны по-
падаются зимой в Германии и средней Франции, куда они, очевидно,
улетают на зимовку. У нас же зимуют индивиды серой вороны, при-
летающие из более северных районов. Повидимому, и галка яв-
ляется перелетной птицей, а также некоторые другие птицы, счи-
тавшиеся прежде оседлыми.
Зависимость перелетов от места обитания птиц очень наглядно
иллюстрируется примером утки кряквы. Общеизвестно, что в СССР
и на большей части территории Европы кряквы являются птицами
перелетными. Осенью они улетают в юго-западном направлении.
В Англии же, где зима в силу морского климата является более
158
Перелеты птиц
мягкой и водоемы не замерзают, кряква оказывается оседлой
птицей и никуда не улетает (это доказано точными наблюдениями
с применением метода кольцевания).
У некоторых перелетных птиц область зимовки и область гнез-
дования не резко отделены друг от друга и непосредственно сопри-
касаются (например, овсянка, зеленушка). Другие же, и послед-
них огромное большинство, имеют два ареала. Один из этих
ареалов представляет собою область гнездования; другой — об-
ласть зимовки. В местности, разделяющей эти оба ареала, птицы
бывают лишь при пролете.
Куда же именно летят от нас пернатые странники? Где же их
зимовки? Птицы, прилетающие на лето в Европу, зимуют в Аф-
рике, в Малой Азии и в Европе — на южных берегах ее; только
немногие из них удаляются в Среднюю и Южную Азию. Птицы,
гнездящиеся в Северной Азии, летят на зиму в Аравию, Иран,
Индию, Китай, даже в Австралию и Новую Зеландию. Каждый вид
имеет свое место, где он проводит зиму. Так, многие из наших се-
верных водных птиц зимуют на южном берегу Каспийского и Чер-
ного морей, на озерах Средней Европы, например, в Швейцарии, в
южной части Англии, по берегам Франции, Италии. Кулики, бе-
касы, грачи, аисты зимуют в Африке по берегам реки Нила. Пев-
чие птицы—соловьи, пеночки, малиновки и т. п.— улетают в
Среднюю Африку. Ласточки, кукушки забираются дальше других
вглубь Африки, до самого экватора и южнее, и проводят зиму в
наиболее жаркой части света. Большая часть наших орлов зимует
в Египте, журавли добираются до Средней Африки и т. п.
Наблюдения показывают, что во многих случаях, чем севернее
летит птица летом, тем южнее она удаляется на зиму. Поэтому даль-
ность перелета для различных птиц сильно варьирует. Для самых
северных видов расстояние между летним и зимним местожительст-
вом достигает более 10 000 км. Например, длиннохвостая крачка
гнездится в Арктике, зиму же она проводит в Антарктике (где
в это время лето) (рис. 111). Многие северные виды куликов, ле-
туя в области арктической тундры, на зиму улетают в умеренную
зону южного полушария (южной Австралии, Новой Зеландии,
южной Африки).
Примером коротких перелетов могут служить горные жаворонки,
подорожники, свиристели, улетающие на зиму с севера на юг Ев-
ропы, причем некоторые из них не спускаются далее пределов юж-
ной части Украины. Относительно недалеко улетают также гнез-
дящиеся у нас летом кряквы (рис. 112).
И наши широты являются местом зимовья некоторых птиц,
прилетающих сюда из более северных стран. Когда закончится пе-
релет от нас на юг местных перелетных птиц, когда опустеют леса,
поля и парки, появляются с севера зимние пернатые гости: снегири,
свиристели, чечотки. Они проводят лето в более северных лесах
Поведение птиц на местах зимовья
159
или в тундре и спускаются к нам с наступлением холодов. Зимою
мы видим, сверх того, вокруг себя — в садах, на деревьях вблизи
жилищ, на дворах — разных синиц, овсянок. Но эти птицы и лето
проводят в наших лесах, зимой же они перемещаются ближе к жи-
Рис. 111. Распространение и миграции амери-
канских особей длиннохвостой крачки (Sterna
paradisaea). Черные точки — места гнездова-
ния; треугольники — места зимовок; стрелки—
направления и пути пролета
лищу человека, где можно легче найти пищу и быть в большей без-
опасности от хищников.
На местах зимовья птицы ведут себя совершенно иначе, нежели
в северных странах. Там они гнезд не вьют, потомства не выводят
и не поют. Это относится ко всем перелетным птицам на всех местах
зимовья, в том числе и на отдаленном, благодатном по климату
юге.
160
Перелеты птиц
Как и во время перелета, птицы на юге все время держатся ста-
ями. Особенно велики бывают стаи водных птиц на озерах близ
Суэцкого канала.
Время прилета к нам и отлета от нас птиц в значительной сте-
пени постоянно для одного и того же вида, но для различных ви-
дов различно. Осенний прилет северных птиц у нас начинается в
Рис. 112. Направления перелета крякв (Anas platyrhynchos)
августе или даже в конце июля и длится в течение сентября и ок-
тября. Прилет птиц с юга начинается с половины февраля и длится
до половины мая. Для каждого вида, взятого в отдельности, при-
лет занимает в большинстве случаев всего несколько дней, тогда как
отлет продолжается неделями, а для некоторых видов даже меся-
цами. Состояние погоды оказывает значительное влияние на время
отлета, хотя не для всех птиц, прилет же от погоды зависит менее и
отличается большим постоянством. Он редко колеблется в пределах
более 10 дней для каждого данного вида. А есть птицы, у которых
это колебание составляет только 2—3 дня. Так, тупики появляются
в некоторых местах их гнездовья ежегодно день-в-день, как будто
Время прилета и отлета
161
их кто созывает каким-либо сигналом, и совери ей ю независимо
от того — тепло или холодно, тихо или ветрено. Время прилета
морских птиц к некоторым из их гнездовий настолько точно, что
составляет дату в календаре лиц, заинтересованных этим собы-
тием.
Находящаяся близ Москвы (в Сокольниках) Биостанция юных
натуралистов имени К. А. Тимирязева, где тщательно велись фе-
нологические наблюдения, получила следующие данные о посто-
янстве времени прилета некоторых птиц: горихвостка в 1919 г.
прилетела в окрестности биостанции 15 апреля, в 1920 г. — 15-го,
в 1921 г.—17-го, в 1922 г.—17-го, в 1923 г.—17-го и в 1924 г.—20 ап-
реля; мухоловка-пеструшка в 1920 г.— 19 апреля, в 1921 г.—
19-го, в 1922 г.— 17-го, в 1923 г.— 16-го и в 1924 г.— 20 апреля;
кукушка в 1919 г.— 28 апреля, в 1920 г.— 27-го, в 1921 г.— 21-го,
в 1922 г.— 26-го, в 1923 г.— 28-го и в 1924 г.— 18 апреля; славка-
черноголовка в 1920 г.— 3 мая, в 1921 г.— 5-го, в 1922 г.— 4-го,
в 1923 г.— 7-го и в 1924 г.— 8 мая; таким же постоянством во вре-
мени прилета отличаются и почти все другие славки. В Германии
кукушка ожидается между 17 и 19 апреля. На Майне .говорят о
ней: 17-го она уже может прилететь, 18-го должна бы была поя-
виться, 19-го же должна быть на месте во что бы то ни стало. Стрижи
прилетают 28 апреля. Стрижи, первыми покидающие наши широты
осенью, могут служить примером и точного отлета, сообразующе-
гося также более с известными числами месяца, нежели с состоя-
нием погоды. Прибытие перелетных птиц на места гнездовья, вообще
говоря, согласуется с нашим календарем точнее, нежели весеннее
цветение растений. Это обстоятельство заслуживает большого ин-
тереса, если учесть дальность пути и всевозможные случайности
и невзгоды, которым птицы подвергаются во время перелетов.
Прилет птиц совершается определенными фазами или «волнами»,
совпадающими с основными фенологическими явлениями в ходе
весны и начала лета.
Первую «волну» составляют у нас грачи, которые появляются
вместе с первыми проталинами. Время их прилета в Московской
области — 19 марта (среднее за 35 лет).
Вторая «волна» прилета—это жаворонки, скворцы, зяблики,
появляющиеся в самых последних числах марта до 1—2 апреля.
Третья «волна», обычно совпадающая с ледоходом и началом
второго периода весны, включает большое число видов птиц (белые
трясогузки, чибисы, зорянки, дрозды, некоторые хищники—пус-
тельга, черный коршун; прилет водоплавающей и болотной дичи:
вальдшнепов, бекасов, гусей, уток). Для Московской области—это
первая декада апреля.
Четвертая «волна» (половина апреля) характеризуется приле-
том многих видов мелких воробьиных птиц (варакушки, пеночки-
теньковки, мухоловки-пеструшки).
162
Перелеты птиц
В конце апреля наблюдается пятая «волна» прилета, совпадаю-
щая обычно со значительным потеплением. В это время прилетают
кукушки, вертишейки, деревенские ласточки.
Вначале мая—шестая «волна»; это—серые мухоловки, со-
ловьи, пеночки-пересмешки, стрижи.
Наконец, в конце мая и первых числах июня прилетают еще не-
которые птицы (иволга, сорокопут-жулан, чечевица, камышевки,
овсянка-дубровник). Их прилет можно рассматривать, как послед-
нюю, седьмую, очень растянутую «волну».
Существует определенная зависимость между сроком прилета
и общей длиной пролетного пути. Те птицы, которые прилетают
в начале весны, зимуют недалеко. Напротив, позднее других при-
летают птицы, область зимовки которых отстоит на большом рас-
стоянии от мест гнездовья (иволга, стриж, кукушка зимуют в сред-
ней и южной Африке, за 5—6 тыс. километров от мест гнездовья).
Сроки прилета птиц, зимующих недалеко, в гораздо большей
степени зависят от условий погоды, чем птиц, прилетающих изда-
лека. Это и понятно, так как погодные условия на местах гнездовья
никак не сказываются на отдаленных местах зимовок и отлет птиц
определяется не метеорологическими, а иными факторами, о чем
речь будет идти ниже.
Интересно, что, несмотря на дальние путешествия, возвращаю-
щиеся к нам птицы находят свои прошлогодние места жительства,
а некоторые из года в год поселяются даже в прежних гнездах.
Этот факт достоверно доказан по отношению к аистам, ласточкам,
скворцам и другим гнездящимся вблизи домов птицам. Именно —
еще до систематического применения метода кольцевания птиц
метили по-разному: например, надевали кольца на ноги или
привязывали ленточки, и на следующую весну видели у себя
тех же самых птиц. Массовое и систематическое применение ме-
тода кольцеваний подтверждает прежние наблюдения. Так, околь-
цованные одним венгерским орнитологом ласточки возвращались
в свое родное гнездо до шести лет подряд. Многие птицы, напри-
мер грачи и стрижи, прилетев на прежнее место, вьют новые гнезда
где-либо по соседству со старыми. Таким образом в определенный
район ежегодно возвращаются одни и те же птицы или их потомство,
которое может не смешиваться с птицами того же вида из соседнего
района: кулики возвращаются на свои родные болота, перепела —
на свои поля, соловьи — в свои рощи и т. д. Этим объясняется тот
факт, что и среди перелетных птиц складываются местные формы.
Перелетные птицы какого-нибудь небольшого района приобретают
иногда, в связи с местными условиями, какие-нибудь особенности,
которые не передаются птицам того же вида из соседнего района.
Так складываются, например, разные напевы соловьев по районам;
различают напевы курских, бердичевских, московских и других
соловьев.
Порядок прилета и отлета
163
Формирование местных форм, отличающихся по характеру пе-
сни, подробно изучил А. Н. Промптов (1930) у обычнейшей птицы
наших лесов — зяблика.
Не нужно, однако, думать, что возврат на прежнее место гнез-
довья представляет собой нечто совершенно неизбежное и «фаталь-
ное». Напротив, у некоторых птиц наблюдалось явление противо-
положного характера, когда места гнездовья птиц после прилета
резко меняются. Установить это удалось применением метода коль-
цевания. Например, точно установлены случаи, когда утка кряква,
окольцованная утенком в Шотландии, была найдена с выводком
птенцов в Дании, а выведшиеся в Англии чирки-свистунки были
обнаружены на гнездовье в Германии. Это явление разлета, наз-
ванное английским орнитологом Томпсоном термином «абмигра-
ция», касается преимущественно молодых особей. Однако оно пред-
ставляет собой гораздо менее распространенное явление, чем воз-
врат птиц на прежние места гнездования.
Ранее всех улетают на юг птицы, питающиеся насекомыми:
стрижи, щурки, иволги, кукушки, соловьи, варакушки, гори-
хвостки, пеночки, славки, ласточки и др. Такой порядок вполне по-
нятен, так как этот птичий корм исчезает ранее другого. К концу
лета насекомых бывает уже значительно менее. Одни из них поги-
бают, отложив яички, другие прячутся в укромные места, где и про-
водят зиму в спячке. Многие гусеницы зарываются в землю.
Птицы, частично или полностью питающиеся растительной пи-
щей, могут находить корм и позднее; с этим связан более поздний
отлет таких птиц, как дрозды, перепела, куропатки.
Самые ранние весенние гости у нас — грачи, скворцы и поле-
вые жаворонки, самые поздние —ласточки, стрижи и щурки, ра-
нее других улетающие в конце лета. Если соединить линиями точки
одновременного прилета особей одного и того же вида в разных
пунктах страны, то получатся линии, названные (Миддендорфом)
изопиптезами. Они в общем соответствуют изотермам, т. е. линиям,
соединяющим местности с одинаковой средней температурой, и
следовательно, указывают на соответствие времени прилета с об-
щим ходом весны. У нас изопиптезы обстоятельно разработаны по
отношению к кукушке и некоторым другим видам птиц проф.
Д. Н. Кайгородовым. Изопиптезы, как и изотермы, в европейской
части СССР спускаются с запада на восток, т. е. чем западнее район
в Европе, тем прилет раньше, так как климат теплее (рис. 113).
В соотношении между временем прилета и отлета разных видов
птиц наблюдается следующая закономерность: птицы, ранее уле-
тающие на юг, позже возвращаются, и наоборот. Такое соотноше-
ние понятно: птицы, для которых оказываются неблагоприятными
климатические условия начинающейся осени, точно так же не могут
найти подходящих условий и в начале весны, когда и насекомых
еще мало и недостаточно тепло. Климатическими условиями объяс-
164
Перелеты птиц
няется и то, что птицы более северные, по сравнению с более юж-
ными, ранее улетают на места зимовья и позже возвращаются: на
севере теплое время года короче. Так, например, ласточки-касатки,
выводящиеся в Южной Европе, покидают юг Африки, где они зи-
муют, в половине февраля, но те из них, котррые выводятся в север-
ной части СССР, остаются в Африке до половины апреля.
Рис. ИЗ. Изопиптезы первого кукования кукушки (по Кайгородову)
На время отлета птиц как с севера на юг, так и с юга на север
оказывает влияние и их размножение. Первыми у каждого вида под-
нимаются от нас такие птицы, которым не удалось вывести птенцов
или которые случайно потеряли яйца или молодых птенцов. Они не
испытывают связывающего влияния со стороны инстинкта заботы
о потомстве, заставляющего не оставлять места гнездовья, прежде
чем не окрепнут все птенцы и не сделаются также способными к
перелету. У таких птиц инстинкт перелета становится преждевре-
менно настолько сильным, что они покидают свои летние жилища
иногда до полного окончания линьки. За ними и под их руко-
водством летят молодые, но уже окрепшие птенцы, влекомые врож-
Проявление стадности при перелетах
165
денным побуждением к перелету. Через некоторое время, напри-
мер, через неделю после молодых, летят взрослые самцы. Самки
летят уже после них, так как они задерживаются «материнскими
обязанностями» по отношению к более слабым и молодым птенцам,
которых и захватывают вместе с собой.
Весной порядок возвращения отчасти обратный. Первыми яв-
ляются самцы, за ними самки, потом годовалые птицы и нако-
нец — наиболее слабые и больные особи. Так как срок весеннего
Рис. 114. Аисты (Ciconia alba) перед отлетом
прилета для каждого вида, по сравнению с осенним отлетом, очень
короткий, то указанный для весны порядок путешествия птиц не
так резко бросается в глаза, как порядок осенний. И тот и другой
порядок имеют, конечно, исключения. Есть некоторые птицы, на-
пример скворцы, у которых молодые индивиды улетают на юг ра-
нее взрослых, совершенно одни, т. е. без всякого «руководства».
Отлету птиц обыкновенно предшествуют «стаенье», собирание
их в большие стаи, и кочевки из одного места в другое, вызывае-
мые сокращением корма в местах гнездовья (рис. 114). Кочевки стай
у некоторых видов незаметно переходят в отлет, у других он начи-
нается резко. Однако кукушка, выросшая в чужом гнезде вне об-
щения с другими кукушками, удод и некоторые другие отлетают в
одиночку.
Жизнь стаей во время перелета имеет для птиц очень большое
биологическое значение. Так, пока одни птицы кормятся или отды-
хают, другие их сторожат. «Сторожа», в роли которых выступают
166
Перелеты птиц
наиболее опытные и старые птицы, всегда предупредят об опасно-
сти.1 Вот, например, как описывает ночлег прилетающих диких
гусей С. Т. Аксаков.
«Гуси завертывают голову под крыло, ложатся или, лучше ска-
зать, опускаются на хлупь2 и брюхо и засыпают. Но старики со-
ставляют ночную стражу и не спят поочередно или так чутко дрем-
лют, что ничто не ускользает от их внимательного слуха. При вся-
ком шорохе сторожевой гусь тревожно загогочет, и все откликаются,
встают, выправляются, вытягивают шеи и готовы лететь; но шум
замолк, сторожевой гусь гогочет совсем другим голосом—тихо,
успокоительно, а вся стая, отвечая ему такими же звуками, снова
усаживается и засыпает. Так бывает не один раз в ночь, особенно
уже в довольно длинные сентябрьские ночи. Если же тревога была
не пустая, если, точно, человек или зверь приблизился к стае, —
быстро поднимаются старики и стремглав бросаются за ними моло-
дые, оглашая зыбучий берег и спящие в тумане воды и всю окрест-
ность таким пронзительным криком, что можно услышать его за
версту и более . . . Когда же ночь проходит благополучно, то сто-
рожевой гусь, едва забелеет заря на востоке, разбудит звонким кри-
ком всю стаю, и она снова, вслед за стариком, полетит уже в зна-
комое поле и точно тем же порядком примется за ранний завтрак,
какой наблюдался недавно за поздним ужином».
В стаях многих птиц во время перелета есть вожаки, которые
летят впереди, выбирают места остановок, подают пример к про-
должению пути после отдыха и т. п. В стаях иногда соединяются
по нескольку различных видов; так образуются большие сборища
различных ласточек, уток и др. Есть птицы, путешествующие в
одиночку, но их немного, например, кукушка, козодой.
Проявлением общения птиц во время перелетов являются их
голоса. Некоторые виды испускают положительно целый хор зву-
ков. Их можно слышать на перелетных путях по ночам — и вес-
ной, и осенью, когда стая за стаей пролетает по стемневшим небе-
сам. По всей вероятности, эти звуки служат главным образом для
собирания перелетающих стай и показывают верное направление
пути для молодых и неопытных. Птицы же, молчащие в этом слу-
чае, могут воспользоваться криком шумливых и следовать за ними.
Вот как пишет об этом Г. Гэтке, посвятивший наблюдению над
перелетами птиц на о. Гельголанде (в Немецком море) пятьдесят
лет жизни. * У
1 Разумеется, та «взаимопомощь» и «разделение труда», с которыми мы
встречаемся у птиц, основаны на инстинктивной деятельности. В этом отно-
шении они резко отличны от того, что имеет место в человеческом обществе.
У животных разные формы так называемой «общественной жизни», как и
другие полезные приспособления, развились на основе естественного от-
бора. Они представляют собой наследственно закрепленные инстинкты.
' Хлупь — древнерусское слово, означает кончик крестца у птицы.
Расположение птиц в стаях
167
«Картина природы, на фоне которой разыгрывается такое бога-
тое проявление животной жизни, сама по себе представляет уже
нечто необыкновенно привлекательное. Ровная, тихая, темная ночь
без луны, без звезд, совсем слабый юго-восточный ветерок —та-
ковы условия для наиболее величественного развития явлений
перелета птиц; если атмосфера при этом еще значительно напитана
влажностью, то явления эти приобретают особенную силу. Повсюду
вокруг глубокий мрак, среди которого огромное светлое тело маяка
как бы парит в воздухе,— широкие лучи, которые расходятся от
него во все стороны и простираются, кажется, до бесконечности в
мутном воздухе,— сознание близости громадного моря вокруг нас
и полное безмолвие всей природы,— все это наполняет душу серь-
езным, почти торжественным чувством. И вот среди этой абсолют-
ной тишины внезапно раздается сначала одиночное «сцып» певчего
дрозда, кое-когда также высокие призывные звуки жаворонка,
затем опять минутная или двухминутная пауза, вдруг прерываемая
далеко слышным «гигик» черного дрозда и разнообразным «тир-р-р»
пролетающей стайки песочников; наконец, начинаешь слышать
зов большого количества жаворонков, приближающихся и удаляю-
щихся стайками различной численности, хриплое «эр» бекасов, яс-
ное «тют» золотистой ржанки, громкое звучное «крю-ю» тулеса,
далеко разносящийся призыв большого кроншнепа, многосложное
«так-так-так» рябинника, протяжное «цип» дрозда-белобровика,
звуки спешно проносящейся, сильно растянувшейся стаи исланд-
ских песочников, легко узнаваемых по их разнообразному, быстро
испускаемому «тютт-тютт-тютт-тютт», и бесчисленные свистящие,
скрипящие и трескучие голоса, незнакомые здешним охотникам
и птицеловам, напоминающие своей мелодией скрипящую телегу,
но частью принадлежащие, очевидно, серой цапле и ее сородичам,
испускающим очень громкие и грубые звуки. Весь небесный свод
теперь наполнен хаосом сотен тысяч голосов, раздающихся вблизи
и издали».
Так как в местах зимовья перелетные птицы все время держатся
стаями, то в особом стаденье перед отлетом здесь нет нужды. Но
здесь перед отлетом обращает на себя внимание другое — перемена
в поведении птиц, сделавшихся более беспокойными и возбужден-
ными.
Интересно расположение птиц в стаях во время перелетов. Для
различных птиц оно весьма разнообразно. Утки, например, в боль-
шинстве случаев летят рядом; гуси — друг за другом, причем каж-
дый последующий находится не прямо позади предыдущего, а не-
сколько в стороне от него; ибисы — образуют изгибающуюся ли-
нию; гагары—совершенно прямую; журавли, а также и гуси рас-
полагаются клином или углом, одна сторона которого обыкновенно
бывает длиннее другой (рис. 115 и 116). В последнем расположе-
нии многими усматривается та выгода, что клин легче рассекает
168
Перелеты птиц
воздух, нежели беспорядочная стая. Более всех работы в этом
случае приходится на долю передней птицы, поэтому она через не-
Рис. 115. Расположение разных
видов птиц при перелете: 1—утки,
2 — гуси, 3 — ибисы, 4 — журав-
ли, 5—гагары, 6—кулики
которые промежутки времени сме-
няется. С этим объяснением, од-
нако, не все соглашаются. Дело в
том, что при клинообразном, как
и при всяком другом расположе-
нии, птицы следуют не непосредст-
венно друг за другом, а через про-
межутки. Поэтому и воздух рас-
секается не клином, как целым,
а каждой птицей в отдельности.
Как бы то ни было, благодаря
определенному расположению от-
дельных видов перелетающих птиц,
опытный глаз в большинстве слу-
чаев уже по одному общему виду
стаи определит, какие птицы ее
составляют, хотя бы они летели
'настолько высоко, что отдельную
птицу нельзя было бы узнать.
Как высоко и как быстро летят
птицы?
Еще относительно недавно мно-
гие орнитологи утверждали, что
высота полета птиц очень значительна. Так, например, Гэтке опре-
делял высоту полета некоторых птиц в 5000—6000м. Несомненно,
Рис. 116. Пролетная стая гусей
что высокий полет представляет для птиц значительные преиму-
щества. С большой высоты легче ориентироваться в окружающей
местности, птицы меньше подвергаются нападениям хищников и
Высота и скорость перелетов
169
т. п. Повидимому, над морем птицы летят на большей высоте, чем
над сушей.
Известный русский орнитолог акад. Мензбир, однако, уже давно
предостерегал против переоценки высоты полета птиц.
Предостережения Мензбира оказались справедливыми. Иссле-
дования на самолетах и воздушных шарах за последнее время за-
ставили значительно снизить предполагавшуюся ранее высоту
птичьего полета во время перелетов. Прежде всего наблюдения на
различных воздушных аппаратах установили, что на высоте 7000 м
температура воздуха равняется —33° и давление равно лишь 298 мм
ртутного столба; на высоте 5000 м температура воздуха равняется
—20° и давление равно V2 атмосферы. Уже a priori трудно допу-
стить, чтобы птицы, улетающие от зимних холодов, поднимались
в слои атмосферы, где всегда жестокий холод и давление наполо-
вину разрежено. В Германии было предпринято 100 полетов воз-
душных шаров специально для изучения высоты, на какой птицы
совершают свои перелеты, и во время этих полетов были замечены
как исключение только одна ласточка на высоте 450 м, два аиста
и один сарыч — на 900 м и один жаворонок — на 1900 м. Таким об-
разом, подъем на высоту свыше 1000 м представляет среди птиц
редкое явление.
Глазомерные определения высоты полета птиц Гэтке, очевидно,
ошибочны: в вертикальном направлении на фоне ясного неба, осле-
пляющего наши глаза солнечным светом, определять расстояния
значительно труднее, чем в горизонтальном. Для проверки вычис-
лений Гэтке Луканус в Германии подвешивал чучела птиц к аэро-
статам *и при подъеме последних наблюдал, когда чучела переста-
вали быть видимыми. Оказалось, что, например, ястреб-перепелят-
ник становится невидимым уже на высоте 850 м. Между тем, Гэтке
видел его летящим на высоте . . . 3000 м\ В общем на основании всех
последних данных установлено, что птицы совершают свои пере-
леты обычно на высоте от 100 до 400 м.
Относительно быстроты полета птиц наблюдения последнего
времени также расходятся с прежними вычислениями,повидимому,
тоже преувеличенными. Для разных видов птиц быстрота перелета
прежде определялась свыше 100, а в некоторых случаях свыше 200
и даже 250 км в час. На орнитологической станции Росситен (в Гер-
мании) эта быстрота довольно точно определялась путем наблюде-
ний за перелетающими птицами с двух пунктов, расположенных
один от другого на расстоянии Ч2 км. Таким способом высчитано,
что серая ворона летит со скоростью 780 м в минуту, или около
50 км в час, ястреб-перепелятник — 720 м в минуту, или около
45 км в час, скворец — 1236 м в минуту, или около 75 км в час.
В прежнее время очень высокие цифры быстроты перелета получа-
лись относительно некоторых птиц, которых во время весеннего пе-
релета никому не удалось наблюдать в промежуточны* местностях.
170
Перелеты птиц
Таковы, например, черноголовая ржанка, варакушка. Получа-
лось впечатление, будто весь свой перелет из Северной Африки
до севера Европы они совершают в одну ночь, т. е. со скоростью
300 км в час. Последние наблюдения подтверждают, однако, спра-
ведливость другого объяснения того, почему эти птицы невидимы
на путях весеннего пролета: их трудно заметить, так как они делают
остановки в чрезвычайно болотистых и малодоступных для наблю-
дения местностях.
Не следует, однако, думать, что птицы все время летят с полным
возможным для них напряжением. Это наблюдается при перелете
разве только через моря. Над сушей же полной быстроты полета
птицы обычно не развивают и, кроме того, делают остановки для
сбора пищи и отдыха. За немногими исключениями птицы летят
или только днем, или только ночью. Значительное большинство со-
вершает свой перелет только ночью. В особенности предпочи-
таются лунные ночи. Грандиозные массовые ночные перелеты
птиц через моря можно наблюдать вблизи маяков, яркий свет ко-
торых привлекает птичьи стаи.
Некоторые плохо летающие птицы совершают свои переселения
пешком, перелетая на значительном расстоянии главным образом
только реки, озера и моря, а некоторые морские птицы совершают
свои переселения вплавь. К числу сухопутных путешественников
в нашей фауне принадлежат коростели, или дергачи,— плохо лета-
ющие, по превосходно бегающие птицы,— а также перепела. Вот
интересные подробности путешествия перепелов.
По-видимому, эти птицы пускаются в путь поодиночке
и собираются в стада по дороге. Пешком эти стада отправляются
на юг и собираются в южных оконечностях полуостровов Среди-
земного и по берегам Черного морей, через которые потом летят
в Африку и Малую Азию. Плохие летуны — перепела — летят над
самой поверхностью воды. Много их при этом гибнет, особенно если
во время перелета разыгрывается буря. Прибыв на берег, пере-
пела быстро оправляются и бегут далее. Из Северной Африки часть
их добирается до Судана и даже до мыса Доброй Надежды.
Интересно отметить, какое значение имеет для летящих птиц
направление ветра. Прежде предполагали, что наиболее благо-
приятный для перелетов ветер — боковой, дующий косо к направ-
лению перелета, или же слабый встречный. Сильный противный
ветер замедляет скорость полета, а попутный ставит перья птиц
«дыбом» и поэтому является самым опасным. Однако за последнее
время собраны многочисленные данные, заставляющие пересмот-
реть этот вопрос. Оказывается, что птицы предпочитают слабый
попутный ветер, который является наиболее благоприятным. Силь-
ные ветры гибельны для птиц. После сильных северных ветров осе-
нью, во время перелета перепелов через Черное море в Малую Азию,
ца поверхности моря находят тысячи погибших птиц этого вида.
Пролетные пути
171
В зависимости от продолжительности остановок, от дальности
расстояния и быстроты полета находится продолжительность пе-
релетов. Осенний перелет в этом отношении существенно отличается
от весеннего. На юг птицы летят не торопясь, с остановками, иногда
довольно длинными. Для большинства видов продолжительность
путешествия в этот конец занимает от одного до двух месяцев. Об-
ратное путешествие протекает значительно быстрее. В Европе боль-
шинство возвращающихся весной птиц достигает своей родины в
течение одного безостановочного перелета. Редкое исключение пред-
ставляет вальдшнеп. К радости охотников, он подвигается на се-
вер медленно, по педелям останавливаясь на подходящих для пего,
изобилующих пищею местах.
Направляясь к конечной цели перелета, птицы в то же время
избирают определенные дороги или. как говорят, пролетные пуши,
а не летят через любую местность, лишь бы было ближе. Есть из-
любленные местности для пролета: по берегам морей и океанов,
вдоль рек и озер, а также гор, долин и лесов. Берега материков
представляют удобозапоминаемую дорогу, на которой легко ориен-
тироваться. При этом птицы с могучими крыльями, следующие по
этим дорогам, отступают от береговой линии там, где она извилиста,
перелетая заливы и мысы; птицы же более слабые следуют изгибам
суши. При перелете через континенты самым излюбленным путем
являются реки. Помимо того, что они служат показателями пути,
воды рек и произрастающая на их берегах растительность заклю-
чают в себе самую разнообразную пищу. Поэтому вдоль рек мы
видим птиц всех родов: сухопутных и водяных, насекомоядных и
зерноядных, воробьиных, ржанковых, и даже океанических чаек,
или крачек, и т. д. Тем не менее, существует определенная связь
между характером пролетных путей того или иного вида птиц и
его биологией. Водоплавающие птицы преимущественно летят по
водным артериям — рекам и вдоль морских побережий. Лесные
птицы придерживаются лесных ландшафтов, хотя иногда и следуют
вдоль течения рек.
При слиянии рек, вдоль которых следуют птицы, соединяются
и стаи последних. В конце концов к устьям больших рек образуются
громадные стаи. Горы представляют менее распространенные пути
перелета, но и они для некоторых видов играют весьма важную
роль, служа для них прекрасными путеводителями. Если по на-
правлению пути встречаются горные долины, ущелья и проходы,
птицы летят по ним. Птицы лесные и при перелете, где есть возмож-
ность, держатся опушек лесов, рощ и других древесных насаждений.
Изучение пролетных путей птиц, проходящих над современной
территорией СССР, было начато еще в середине XIX в. В 50-х го-
дах прошлого столетия обширные наблюдения над перелетами птиц
опубликовал известный русский орнитолог К. Ф. Кесслер. Он от-
рицал наличие определенных отграниченных пролетных путей и
172
Перелеты птиц
считал, что птицы летят «широким фронтом». В противоположность
этому гельсингфорский орнитолог Пальмен в обширной работе
(1876 г.) устанавливает наличие определенных пролетных путей
для водоплавающих и болотных птиц Европы и Азии. Но особенно
большое значение имел капитальный труд выдающегося москов-
ского орнитолога акад. М. А. Менз 1ира «Пролетные пути птиц в
Европейской России», вышедший в 1886 г., в котором устанавли-
ваются основные пролетные пути, а также высказываются инте-
ресные мысли о происхождении явления перелетов. Этот труд и до
настоящего времени не утратил своего значения.
Мензбир основные пролетные пути Европейской части СССР ха-
рактеризует следующим образом (цитируем по его позднейшей книге
«Миграции птиц», 1934 г.): «1) Норвежский путь, который, начи-
наясь от берегов Сибири, идет берегом Ледовитого океана и далее
берегом Норвегии; 2) Балтийский путь, начинающийся с северо-
востока в Ледовитом океане, разбивающийся на несколько рука-
вов и через Финский залив и Балтийское море направляющийся
на запад; 3) Понтийский, или Черноморский путь, начинающийся
на северо-востоке и отчасти востоке, целым рядом разветвлений
пересекает Восточную Европу, идя с северо-востока на юго-запад,
спускается течениями рек к Черному морю и далее идет к Босфору;
4) Каспийский путь идет с севера на юг, отчасти придерживаясь
течения Волги, а далее па восток продолжается многими рукавами,
пересекая Уральский хребет и Мугоджары в направлении с северо-
востока на юго-запад, огибает Каспийское море и ведет к берегам
Ирана. Нельзя не видеть, что общее направление этих пролетных
путей было определено еще Миддендорфом, затем точнее они были
вычерчены Пальменом и с еще большими подробностями нанесены
на карты мною» (рис. 117).
В исследовании пролетных путей птиц в Средней Азии особенно
большое значение имели работы акад. П. П. Сушкина («Птицы сред-
ней Киргизской степи», 1908 г.), установившего 12 пролетных пу-
тей (главным образом по долинам рек Урала, Эмбы, Иргиза и по
цепи озер).
В дальнейшей разработке и уточнении вопроса о пролетных
путях птиц особенно большую роль сыграл метод кольцевания,
о чем мы уже подробно говорили выше.
С учетом всех этих данных А. Я- Тугаринов в 1937 г. рисует сле-
дующую общую картину пролетных путей на территории Совет-
ского Союза, которых он насчитывает восемь (рис. 118).
Атлантическое направление (I), по которому
летят почти все птицы, населяющие полярное побережье до Тай-
мыра на восток. Большинство этих птиц зимует в Западной Европе
и по берегам Средиземного моря.
Средиземноморско-африканское направ-
ление (II), ведущее из Европейской части СССР и Западной Си-
Пролетные пути
173
Рис. 117. Карта пролетных путей Европейской части СССР (по Мензбиру)
Рис. 118. Схема главнейших направлений пролетных путей (по Тугаринову)
Перелеты птиц
Пролетные пути
175
бири к европейским и африканским берегам Средиземного моря.
По этому направлению летит множество видов птиц: воробьиные
(например, ласточка, рис. 119) кулики, хищники, цапли, аисты и др.
Каспийско-Иранское направление (III) —
из Западной Сибири к Каспийскому морю и вдоль его берегов в
Иран. Зимовка птиц от берегов Каспия до Персидского залива.
Этим путем летят многие птицы (казарки, дупеля, гаршнепы, не-
которые камышевки и др.).
Индийское направление (IV) собирает птиц из
южной части западносибирских степей и Казахстана и ведет в
Индию вплоть до низовьев больших индийских рек. По этому на-
правлению летят птицы самых различных отрядов.
Хинганское направление (V) ведет в Китай,
Вьетнам, на Филиппинские и Зондские острова. На него собираются
птицы преимущественно из тайги Средней и Восточной Сибири.
Сначала путь этот в своей южной части идет почти прямо на восток,
а затем круто поворачивает на юг. По этому пути летят главным об-
разом лесные воробьиные птицы Сибири.
Приморское направление (VI) проходит вдоль
восточного побережья Азии в юго-восточную Азию. По нему летят
птицы Приамурья, северной части Якутии (лесные утки, казарки,
кулики, хищники и др.).
Камчатское, или островное, направление
(VII) — с Камчатки вдоль Курильской гряды в Японию и далее на
юг. По этому пути летят не только водяные птицы, но многие «су-
хопутные» виды. Это — обитатели восточной окраины Азии, Кам-
чатки, Анадырского края.
Американское направление (VIII) — с северо-
востока Азии через Берингов пролив на Аляску и далее на юг вдоль
берегов Сев. Америки. Этим путем следуют некоторые птицы, гнез-
дящиеся на северо-востоке (белый гусь, канадский журавль, неко-
торые кулики).
Для западноевропейских, особенно для германских, видов птиц
Луканус намечает три основных пути перелета: 1) Западный бере-
говой — вдоль побережья Балтийского и Немецкого морей, далее
по северному и западному берегам Франции на Пиренейский полу-
остров и, наконец, через Гибралтар в Африку; этот путь, служащий
преимущественно для водяных птиц, совпадает с указанным выше
Балтийским путем; 2) Адриатическо-тунисский путь, по которому
из Венгрии и Австрии и далее к востоку лежащих областей многие
птицы устремляются через Карпаты к берегам Адриатического моря
и далее через Сицилию в Тунис. Этим путем летят многие певчие
птицы, цапли и др.; 3) Итальянско-испанский путь—также из
Австрии и Венгрии в обход Альп, через северную Италию на Кор-
сику и Сардинию или вдоль южного побережья Франции в Испа-
нию.
176
Перелеты птиц
Чтобы иметь понятие о том, как много птиц летит во время пе-
релета по наиболее торным, так сказать, частям пути, посмотрим,
что делается у устьев Волги осенью. М. Н. Богданов описывает та-
кую картину:
«Крики птиц заглушают человеческие голоса, так что вы не
услышите вашего товарища, ежели он будет в ста шагах от вас.
Каких, каких птиц тут нет! Стаи белоснежных лебедей, уток, гу-
сей, нырков покрывают все озера и притоки реки. Между ними
чинно плавают стада
Рис. 119. Стая ласточек над морем
пеликанов, ныряют чер-
ные бакланы. Над по-
верхностью воды реют
разные чайки. Берега
усеяны пестрыми груп-
пами журавлей, кол-
пиц, куликов, цапель,
караваек. Все это на-
перерыв ловит рыбу,
с криком, шумом, дра-
ками. Дунет северный
ветер, хлынет дождь
с изморозью — и вся
эта пестрая толпа птиц
с шумом поднимется на
воздух, разобьется на
стаи по породам и по-
летит . . .». Применение
метода кольцевания дало возможность выяснить много частных
вопросов касательно путей перелета птиц. Так, оказалось, что
особи одного и того же вида, обитающие в сравнительно неболь-
шой области, иногда летят разными пролетными путями. В За-
падной Европе это установлено, например, относительно чаек,
вальдшнепа, аиста и других, причем точно прослежены их пути.
К этому нужно прибавить известный и ранее факт, что пути осен-
него и весеннего пролета у некоторых видов не совпадают. Так,
соловей западный пролетает над Гельголандом в апреле и мае,
осенью же его там ни разу не замечали; кулик-тулес почти никогда
или очень редко наблюдался весной на Мальте, между тем осенью
его там видят постоянно; обыкновенная горлица в мае и июне ле-
тит над Гельголандом в громадном числе, а осенью в гораздо мень-
шем; красный веретенник летит на север главным образом через
восточную часть СССР, а возвращается на юг через Западную Ев-
ропу и т. п.
Метод кольцеваний обнаружил следующее на первый взгляд
парадоксальное явление. Некоторые птицы летят вначале не в на-
правлении конечной цели пути, а значительно в сторону, иногда
Пролетные пути
177
Рис. 120. Схема пролетных путей речных
чаек в Европе (по Луканусу)
даже прямо противоположную, т. е. на север. Это объясняется, оче-
видно, тем, что таким путем они попадают на пролетный путь, так
сказать, на столбовую дорогу. Например, часть речных чаек из
Южной Германии и Богемии летит осенью сначала почти на север,
придерживаясь рек Эльбы и Рейна и устремляясь к побережью
Немецкого моря, где на пролетном пути присоединяется к другим
особям того же вида (рис. 120).
Все ли птицы летят по пролетным путям? Нужно заметить, что
рядом с большинством орнитологов, признававших определенные
пролетные пути для
птиц, существовали и та-
кие, которые если и не
отрицали совершенно
пролетных путей, то
сильно ограничивали их
значение и выдвинули
принцип «широкого
фронта», согласно кото-
рому птицы перелетают
широкой волной по
всему пространству, ог-
раничиваемому линия-
ми, соединяющими край-
ние западные и восточ-
ные пункты областей
гнездования и зимовья.
Защитником учения
о «широком фронте» в
России был, как мы
видели, Кесслер, эту же точку зрения защищал известный немецкий
орнитолог Гэтке. Метод кольцевания дал основания для примире-
ния этих точек зрения. Для многих видов птиц, без сомнения, су-
ществуют определенные, прочно установившиеся пути перелета.
Это следует сказать прежде всего про водных и болотных птиц, ко-
торые главным образом и летят по указанным выше пролетным пу-
тям. С сухопутными же птицами дело обстоит иначе. Многие из них
также имеют свои пролетные пути, но со значительными осложне-
ниями: пути эти перекрещиваются под разными углами, расхо-
дятся и сходятся. А некоторые птицы летят по типу «широкого
фронта». Благодаря значительным вариациям первого типа, оба они,
т. е. перелет сухопутных птиц по пролетным путям и широким фрон-
том, связаны между собой промежуточными ступенями и таким об-
разом постепенно переходят один в другой.
Обычно на начальных этапах перелета птицы движутся «широ-
ким фронтом». По мере передвижения их к югу все более и более
определенно складываются пролетные пути. Акад. Сушкин в своей
178
Перелеты птиц
работе «Птицы средней Киргизской степи» по этому поводу пи-
сал: «Пролетные пути очень трудно определимы у северной окраины
области, но чем дальше к югу, тем яснее они обрисовываются. Про-
летный путь тем яснее выражен, чем дальше отстоит пункт наблю-
дения от области гнездовья данного вида или данной стаи. Лишь
постепенно, из кочевок за покормкой, слагается пролет, и для пол-
ного развития его, несомненно, нужно и известное время, а иногда
и значительное пространство».
Относительно пролетных путей в заключение приходится ска-
зать, что все они — как указанные выше, так и другие определяе-
мые орнитологами пути
Рис. 121. Крылья оседлой птицы нырка
(слева) и перелетной камнешарки (справа)
перелета птиц для раз-
личных районов — име-
ют значение лишь об-
щих схем, от которых
различными птицами де-
лается не мало частных
отступлений.
Естественно, что у
перелетных птиц, в от-
личие от оседлых, раз-
вились в процессе эво-
люции особые приспо-
собления к столь дале-
ким путешествиям. Эти
приспособления более
всего заметны в крыль-
ях и оперении. Крылья большинства перелетных птиц длинны и
остроконечны, т. е. имеют форму, наиболее всего приспособленную
к длинному и продолжительному полету (рис. 121). В особенности
резко выражена эта форма у птиц, совершающих самые длинные
перелеты, например, у камнешарки, пролетающей более 10 000 км,
у ласточки и др. Этот вопрос был подвергнут специальному изуче-
нию на большом материале Н. А. Гладковым (1935 г.). Его иссле-
дования в общем вполне подтвердили наличие особенностей раз-
вития крыла у перелетных птиц по сравнению с оседлыми.
Оперение перелетных птиц характеризуется прилеганием перьев
к телу. Неплотный пушистый покров можно встретить только у
плохо летающих видов. В этой особенности перелетных птиц так-
же заключается важное приспособление к летанию, так как торча-
щие и развевающиеся перья оказывают сопротивление при быст-
ром движении по воздуху. Линька перелетных птиц, как уже указы-
валось, предшествует времени перелета. Она совершается или два
раза в год — весною и осенью, или же только о,гин раз—осенью.
Несмотря на все приспособления, на все природные способности
к перелету, птицы встречаются во время длинных путешествий со
Гибель птиц во время перелетов
179
многими опасностями и в большом количестве погибают. Очевид-
ным доказательством огромной смертности среди перелетающих
птиц служит тот факт, что из всей массы их, улетающей от нас на
юг, весной возвращается сравнительно немного. Приспособления
к перелетам далеко не гарантируют птицам безопасность в пути.
Осенью, незадолго до перелета, можно наблюдать огромные сборища
стрижей и деревенских, городских и береговых ласточек, но никто
не видел подобного количества этих птиц весной. Многие из них
при отлете еще молоды, не имеют достаточно окрепших крыльев,
а между тем, в пути их ждет много бедствий. Утомление, бури,
хищники, проливные дожди — все это губит много тысяч птиц.
Как это мы уже видели на примере перепелок, переутомлению птицы
более всего подвергаются при перелете через моря, где и гибнет
их особенно много. Известно много случаев, когда море выбрасы-
вало на берег массы разных утонувших птиц, особенно после бури.
Нередко летящие над морем птицы вследствие утомления садятся
на встречающиеся на пути корабли, а долетевшие до берега до того
переутомлены, что их можно ловить руками.
Из хищников самыми большими врагами перелетных птиц яв-
ляются различные ястребы, соколы: они постоянно носятся в воз-
духе вблизи стай более мелких птиц. Большие соколы следуют на
огромные расстояния, между прочим, за перелетающими утками
и безнаказанно пожирают их; ястребы охотятся за славками, дроз-
дами, вьюрками и различными воробьиными птицами. Каждая
местность, через которую приходится следовать перелетным птицам,
имеет своих хищников, выжидающих свою добычу. Многие из хищ-
ников сами принадлежат к перелетным птицам, причем они летят
более или менее одновременно с птицами, являющимися их добычей.
Так, прилет сапсана всегда совпадает с прилетом в эти местности
уток; чеглок появляется только поздней весной, когда в лесах
множество мелких птиц, и т. д.
Во время пролета много птиц уничтожает и человек.
Случаются годы, когда во время весеннего прилета птиц, напри-
мер, в апреле, возвращаются зимние холода с большими морозами
и метелями. Это также нередко бывает причиною гибели многих
птиц, так как обратно в таких случаях они обыкновенно не возвра-
щаются. Неблагоприятные метеорологические условия, как силь-
ные боковые ветры и бури, могут сбивать птиц с пути, угонять их
на сотни миль в сторону от прямого направления и пригонять в
страны, которых они при нормальных условиях перелета никогда
не посещают. Непривычные условия также бывают причиною бед-
ствий и гибели птиц.
Иногда массовая гибель перелетных птиц может происходить
и в местах их зимовок. На южном берегу Каспия зимует большое
количество различных, преимущественно водоплавающих, птиц.
Обычные мягкие зимы в этом районе благоприятствуют зимовке.
180
Перелеты птиц
Но изредка на южном Каспии бывают необычайно холодные зимы,
сопровождающиеся обильным снегопадом и замерзанием водоемов
(особенно суровыми и многоснежными были здесь зимы 1924—1925
и 1949—1950 гг.). При этом наблюдалась массовая гибель птиц.
По данным бюро кольцевания (Михеев, 1953), в год, следующий
за суровой зимой на южном Каспии, резко сокращается прилет
водоплавающей дичи в различные районы Европейской части СССР.
Мы подошли к самым важным и трудным вопросам: что застав-
ляет птиц, несмотря на все невзгоды длинных путешествий, совер-
шать ежегодные регулярные перелеты, и как объяснить то, что они
летят по строго определенным путям, с которых при нормаль-
ных условиях, за редкими исключениями, не сбиваются?
Что касается причин перелетов, то на первый взгляд может по-
казаться, что осенью птиц гонит от нас холод, а весною привлекает
возвратившееся тепло. Такое объяснение, предлагавшееся прежде,
несостоятельно. Во-первых, оперение птиц перелетных в большин-
стве случаев не хуже, чем у оседлых, так что перенесение холода
почти одинаково возможно и теми и другими. Во-вторых, многие
птицы, как мы уже видели, улетают и прилетают в определенные
числа месяца, совершенно не сообразуясь с состоянием погоды. Так,
например, полевой жаворонок возвращается к нам уже в марте,
когда все еще покрыто снегом и сковано льдом, с другой стороны —
кукушка улетает уже в конце июля или в августе, когда продол-
жает стоять самое теплое время. Все это показывает, что холод не
является основной причиной перелетов. Должны быть другие, бо-
лее существенные причины.
Если принять во внимание, что кочевья птиц из одной местности
в другую совершаются вследствие недостатка корма и что осенние
отлеты для большинства тоже начинаются предварительным коче-
вьем, то справедливее существенную причину отлета видеть в умень-
шении пищи. В самом деле, с наступлением осенних холодов плоды
и даже листья растений опадают, насекомые постепенно скрываются
и т. д.; снежным же покровом и все оставшееся на поверхности за-
крывается от глаз. Ясно, что все встречающиеся у нас пернатые про-
кормиться в зимнее время здесь не могут. Для птиц водных пригод-
ный род пищи совершенно закрывается под ледяной поверхностью,
почему они и улетают от нас на зиму в полном составе. То же нужно
сказать про насекомоядных, которые, как мы видели, первыми на-
чинают отлеты. В странах южных с приближением лета усло-
вия питания также изменяются в неблагоприятную сторону. От
большой жары и сухости здесь в это время частично выгорает
растительность и уменьшается количество насекомых.
Сверх того побудительной причиной прилета к нам птиц из
стран южных на летнее время является потребность воспроизве-
дения, неудержимое стремление выводить и выращивать птенцов.
Доказательство этому усматривается в том, что взрослые, уже
Причины перелетов
181
окрепшие птицы, у которых инстинкт к продолжению рода
вполне развит, первыми приступают к перелету. Однолетние же
птицы, не вполне созревшие в половом отношении, прилетают
позже и часто даже не долетают до мест гнездовья своего вида;
иные даже в течение всего лета остаются вблизи своих зимовок.
Над некоторыми перелетными птицами сделано наблюдение, что
они, будучи посажены в клетку, к весне, когда созревают их по-
ловые продукты, начинают петь и обнаруживают признаки бес-
покойства, свойственного птицам на пролете.
Однако одним недостатком пищи, вместе с изменением темпера-
туры и стремлением к воспроизведению причины перелетов не ис-
черпываются. Первые прилетающие весною птицы появляются у
нас тогда, когда снежный покров еще не сошел и они находят
менее пищи, чем на юге. Осенью очень многие птицы улетают от
нас не тогда, когда уже есть нечего, а ранее,— когда пищи еще до-
вольно много. И летят они не исхудалыми, а наоборот, отъевши-
мися, жирными. Значит, не чувство голода заставляет их пус-
каться в далекое путешествие, а какая-то внутренняя, более глу-
бокая причина. Еще более заставляют признать это следующие
обстоятельства. Молодь, ни разу не бывшая за пределами родины,
иногда улетает прежде старых птиц. Самки некоторых птиц, на-
пример, ласточек, часто улетают на юг, оставляя на произвол
судьбы своих неокрепших еще птенцов: настолько сильно охваты-
вает их вдруг стремление к перелету. Подобное беспокойство прояв-
ляют даже такие птицы, которые посажены в клетки птенцами
и выкормлены там, которые ни разу не участвовали в перелетах
и незнакомы ни с холодом, ни с предшествующим голодом. Пре-
пятствий для выведения птенцов в местах зимовья, за исклю-
чением разве только континентальных тропических стран, где солн-
цем выжигается растительность и наступают неблагоприятные ус-
ловия для жизни, тоже нет. Все это показывает, что, независимо от
внешних обстоятельств, в самой природе птиц есть что-то такое,
что заставляет их с наступлением осени и весны предпринимать
длинные путешествия, что у них есть безотчетное, неудержимое,
инстинктивное стремление к перелетам, унаследованное от предков
и передаваемое последующим поколениям. Выше упоминалось о том,
что возвращающимся к нам ранней весной птицам иногда прихо-
дится терпеть от поздних морозов и снега. В таких случаях птицы
могут приостановить свое движение вперед, но назад в большинстве
случаев не поворачивают, хотя бы им пришлось мерзнуть, голодать
и гибнуть массами. Это также доказывает, что птицы руководству-
ются главным образом бессознательным, врожденным инстинктом.
В сезонных изменениях поведения птиц и, в частности, в воз-
никновении у них «стремления» к перелету, в качестве внутренних
физиологических причин большую роль играет деятельность же-
лез внутренней секреции. К числу таковых, как известно, относятся
182
Перелеты птиц
и половые железы, Во время пребывания птиц на зимовке половые
железы их (как семенники, так и яичники) слабо развиты. К весне,
когда птицы продолжают еще находиться на местах зимовок, на-
чинается процесс развития половых желез. При этом в кровь вы-
деляются гормоны, которые и оказывают на нервную систему, а
через нее и на все поведение птиц активизирующее влияние, сказы-
вающееся в пробуждении «инстинкта перелета». Развитие половых
желез в свою очередь находится под контролем других желез внут-
ренней секреции и, в особенности, придатка мозга—гипофиза. Вы-
деляемые в кровь продукты секреции гипофиза стимулируют раз-
витие половых желез,.а гормоны последних, попадая в кровь, из-
меняют через нервную систему весь характер поведения птицы.
Но развитие половых желез в свою очередь зависит от условий
окружающей среды. В настоящее время накоплен значительный
экспериментальный фактический материал, говорящий о том, что
на развитие половых желез существенное влияние оказывает про-
должительность дневного освещения, а именно постепенное увели-
чение продолжительности дня, которое в природе имеет место вес-
ной.
Приведем в качестве примера описание одного из опытов, до-
казывающих влияние света на развитие половых желез у птиц.
Канадский биолог Родан провел следующий опыт (1924—1927 гг).
Осенью во время отлета он поймал 200 самцов перелетной птички
овсянки-юнко (Junco hiemalis). Пойманных птиц он разделил на
две равные партии. Обе они были помещены в двух одинаковых
вольерах, расположенных на открытом воздухе. Птицы содержа-
лись на одинаковом пищевом рационе. Начиная с ноября, один из
вольеров после захода солнца дополнительно освещался электри-
ческим светом (1050 ватт). Продолжительность этого дополнитель-
ного освещения ежедневно увеличивалась на несколько минут соот-
ветственно с тем, как это происходит в природе после зимнего рав-
ноденствия. Второй вольер дополнительно не освещался. В резуль-
тате, у опытной группы птиц развитие семенников началось значи-
тельно раньше, и уже в декабре семенники достигли таких разме-
ров, которые обычно бывают весной перед началом перелета.
В контрольной же группе семенники продолжали оставаться не-
развитыми. Поскольку все остальные условия в опыте были
совершенно одинаковыми, усиленное развитие семенников можно
отнести только за счет дополнительного освещения. Аналогичные
результаты были получены и на других ’видах птиц.
Было бы, однако, ошибочно думать, что свет является единствен-
ным фактором, прямо действующим на развитие половых желез.
Наряду со светом, безусловно, имеют значение и иные факторы
внешней среды, о которых упоминалось уже выше, в частности,
температура, характер пищи и др.
Однако влияние факторов внешней среды, а также связанных с
Причины перелетов
183
ними внутренних физиологических причин недостаточно для того,
чтобы объяснить самый факт возникновения перелетов у птиц.
Откуда же взялся «инстинкт перелета», почему у одних он есть, а
у других — оседлых — его нет? Чтобы ответить на этот вопрос,
нужно обратиться к факторам исторического порядка, постараться
выяснить, каким образом перелеты возникли в процессе эволюции
птиц, как они связаны с изменениями, происходившими на земле
в предшествующие геологические эпохи.
Многие ученые возникновение перелетов ставят в связь с суще-
ствованием в северном полушарии ледниковой эпохи.
Известно, что в геологический период, непосредственно пред-
шествовавший современному, на земле госпедствовали сильные хо-
лода. Громадный ледник, спускавшийся со скандинавских и фин-
ских высот, покрывал большую часть Европы. Насколько климат
был холоден, например, во Франции, доказывает нахождение
в этой стране остатков северного оленя, лемминга и других поляр-
ных животных. В Европейской части СССР ледник спускался до
Киева и Кременчуга.
До ледниковой эпохи на севере господствовал климат умеренно-
теплый, т. е. значительно более теплый, чем в настоящее время.
Птицы спокойно жили на своих местах, надобности в перелетах
не было. По мере же охлаждения климата для большинства перна-
тых эта надобность наступила. Часть их, не улетая далеко, приспо-
собилась к новым условиям существования, большая же часть, по
мере наступления ледника, оттеснялась все далее и далее к югу.
Вследствие образовавшегося здесь перенаселения птицы с наступ-
лением летних оттепелей опять, насколько позволял климат, под-
нимались к северу. Сначала эти передвижения на север были ко-
роткие, но когда климатические условия стали изменяться в благо-
приятную сторону и ледник начал отступать, открывая для суще-
ствования все большие и большие пространства, перелеты стано-
вились все длиннее и длиннее.
Так, в течение длиннейшего ряда поколений, на протяжении
многих веков, постепенно создалась и укрепилась способность к пе-
релетам, которая закрепилась благодаря естественному отбору. Пе-
редаваясь благодаря наследственности из одного поколения в дру-
гое, она и дала в результате то врожденное бессознательное стрем-
ление, которое мы называем перелетным инстинктом. Перелеты,
благодаря инстинкту, продолжают из года в год повторяться на
прежние расстояния. Из этого объяснения происхождения переле-
тов ясно, что место летнего пребывания птицы, являющееся местом
гнездовья и вывода птенцов, представляет настоящую родину птицы.
Косвенное подтверждение справедливости этого объяснения
можно видеть в фактах зарождающегося перелета, наблюдаемого
в настоящее время у некоторых птиц в связи с климатическими пе-
ременами; таковы: кайры, некоторые виды чаек, гага обыкновенная,
184
Перелеты, птиц
гага гребенушка. Все эти виды по ареалу своего распространения
принадлежат или к полярным, или к арктическим странам. Они
не совершают периодически правильных больших перелетов. Тем
не менее, чрезвычайно суровые зимы побуждают их к весьма не-
правильным перелетам в области, лежащие южнее их местожи-
тельства. Здесь они не остаются долго и удаляются обратно, когда
наступает несколько более мягкая погода, так что движения их хотя
и имеют характер перелета, но перелета неправильного и неуверен-
ного. Вполне естественно, что если климатические условия станут
здесь суровее, то эти зарождающиеся перелеты перейдут в правиль-
ные и систематические путешествия, какие совершают современные
нам перелетные птицы. •
А пролетные пути? Их тоже ставят в связь с передвижением или
расселением птиц из южных областей в северные, вызванным глав-
ным образом отступлением ледника на север. Пути перелета при-
близительно повторяют собой пути расселения вида. Косвенным
подтверждением такого взгляда на происхождение пролетных пу-
тей может служить следующее на первый взгляд непонятное обстоя-
тельство. Птицы во время перелетов иногда без всякой видимой
причины и надобности меняют направление, делают большие крюки
и т. п. Например, в Закаспийской области замечено, что птицы,
летящие на север, вниз по реке Мургабу и долетевшие до выхода
этой реки из гор в степи, направляются не прямо на север, а круто
поворачивают на запад и летят между степями и горами Каспий-
ского моря, откуда уже следуют на север. Таким образом, они
делают большой крюк в несколько сот километров. Этот факт нельзя
объяснить иначе, как только предположением, что пролетный путь
здесь сложился ранее, когда геологические условия местности были
иные, чем в настоящее время.
Известно, что степь, примыкающая к Каспийскому морю, была
некогда покрыта морем, которое соединяло Каспийское море с
Аральским. Южный берег этого моря находился там, где теперь
степь граничит с горами и где теперь летят птицы. Значит, ранее
это направление избрано потому, что здесь была береговая линия,
а теперь птицами повторяется тот путь, которым летели их предки.
Иногда наблюдается, что птицы без видимой надобности отступают
от береговой линии и летят над морем, удлиняя опасную часть пути.
Подобные случаи обычно объясняются иным распределением суши
и моря в -предшествующие геологические эпохи. Предполагается,
что в силу образовавшейся привычки птицы летят вдоль исчезнув-
шей береговой линии когда-то существовавших островов и мате-
риков. В отдельных случаях это подтверждено уже данными гео-
логии и сравнительным изучением морских глубин. Сопоставляя
эти данные с пролетными путями птиц, в настоящее время со зна-
чительной вероятностью можно предполагать о существовании в
отдаленные прошлые времена суши и берега между Новой Кале-
Причины перелетов
185
донией и Новой Зеландией, след чего остался в виде острова Нор-
фолька между Аляской и Полинезией, между Индийскими полу-
островами и Мадагаскаром и др. По всем этим направлениям в на-
стоящее время перелетают разные птицы.
Связь возникновения пути перелетов с расселением вида осо-
бенно отчетливо выступает у тех птиц, которые расселяются и в
настоящее время, расширяя ареал своего обитания. Хорошим при-
мером служит дубровник (Emberiza aureola). Эта птичка широко
распространена в Сибири. Оттуда началось ее расселение в Восточ-
ную Европу по широким заливным лугам больших рек, оно про-
исходит буквально у нас на глазах. В настоящее время дубровник
обычно встречается в поймах Северной Двины, Камы, верхней и
средней Волги. Прилетает он с Востока очень поздно (в самом конце
мая или начале июня). Сразу после выведения птенцов, в конце
июля начинается его отлет на Восток за Урал по тому же пути, по
которому он проник в Европу. Из Сибири дубровник улетает на
зиму в Индию. Таким образом, гнездящиеся в Восточной Европе
популяции дубровника, следуя по путям своего расселения, про-
делывают длинный путь, измеряемый тысячами километров. Дуб-
ровники «не знают», что совсем недалеко от мест их гнездовья к
югу имеются места вполне подходящие для зимовки. В противопо-
ложность дубровнику, птицы, проникшие в Сибирь в процессе своего
расселения из Европы, на зиму улетают на Запад. Например, ду-
пель (Capella media), гнездящийся в Сибири вплоть до восточных
ее районов, на зиму летит через Европу и зимует в южной
Африке.
Большинство существующих пролетных путей более или менее
совпадает с путями расселения вида. Однако в ряде случаев возни-
кают вторичные изменения пролетных путей, обычно связанные с
сокращением их протяженности. В качестве примера можно ука-
зать на пролетные пути белого аиста. Его распространение в Европу
шло с юго-востока по восточному берегу Средиземного моря из
Африки. Этот же путь сохраняется и теперь в качестве пролетного.
Но кроме того, возник еще второй путь, ведущий из западной ча-
сти Европы через Гибралтар в Африку. Этот путь является вто-
ричным, он не связан с первоначальным путем расселения вида.
К приведенному объяснению происхождения перелета птиц и
перелетных путей в настоящее время можно сделать следующие
дополнения. Относительно некоторых видов наших перелетных птиц
установлено, что они, несомненно, южного происхождения. Таковы,
например, стрижи, иволги, кукушки, сизоворонки, удоды и др. Зна-
чит, происхождение их перелета в связь с надвиганием и отступанием
ледника ставить нельзя. Чем же объяснить его? Это, очевидно,
объясняется постепенным расширением области гнездования, что
обеспечивало молодое поколение достаточным количеством корма.
Так как северные области изобилуют летом нужной для них пи-
jgg Перелеты птиц
щей, то эти птицы и начали для гнездований, т. е. на летний период,
улетать все далее и далее на север. Пути, по которым шло это рас-
селение их, отвечают их пролетным путям,— в соответствии с
тем, как образовались пролетные пути и для птиц северного про-
исхождения. Для южных птиц характерно то, что они осенью уле-
тают на юг раньше других птиц,— часто как только выведут птен-
цов, когда еще в окружающей природе достаточно для них и корма,
и тепла. Так, мы уже видели, что стрижи и иволги прилетают вес-
ной позже всех других птиц, а улетают на юг первыми.
Как же объяснить способность птиц не сбиваться на длинном
пути и с удивительной точностью находить прежнее место, часто
включительно до гнезд? Некоторые орнитологи, поражаясь этой
способностью птиц, допускали, что у птиц врожденным является
не только стремление улетать со своей родины и обратно, но и зна-
ние дороги. Такое предположение, странное по существу, не согла-
суется с некоторыми фактами. Если бы молодые птицы обладали
врожденным знанием путей перелета, то им не нужно было бы ле-
теть под руководством старых птиц; между тем, они обыкновенно
имеют руководителей при перелете. Что же касается тех случаев,
когда молодые птенцы летят одни, то данных утверждать, что они,
помимо общего направления на юг, следуют все время определен-
ным путям — поскольку последние не подсказываются самой при-
родой,— не имеется. Кроме того, бывают случаи, с молодыми пти-
цами в особенности, что они сбиваются или отклоняются с пути,
которым следует большинство.
Авторитетными орнитологами в настоящее время признается,
что у птиц является врожденным не знание путей перелета, а чув-
ство направления, способность ориентироваться относительно стран
света. Известно, что почтовые голуби, выпущенные из трюма ко-
рабля, удалившегося от берега настолько, что последнего не видно,
будучи выпущены, сразу определяют правильное направление для
полета. Все вообще птицы, улетающие на значительное расстояние
в поисках пищи, безошибочно находят гнезда, расположенные где-
нибудь в чаще леса. Подобные случаи мы объясняем не врожденным
знанием всевозможных путей, а именно способностью ориентиро-
ваться в направлении, в связи с остротой зрения и хорошо разви-
той «памятью» птиц.
В Германии было произведено массовое окольцовывание аистов,
давшее возможность установить, что они имеют два пути перелета —
юго-западный и юго-восточный. Однако, несмотря на это, два мо-
лодых окольцованных аиста, в одиночку выпущенные из Германии
и Австрии, отправились не по этим путям, а полетели прямо на юг
и были убиты в Италии. Значит, им было свойственно лишь общее
ориентировочное направление на юг.
Относительно роли острого зрения птиц следует заметить, что
его значение не нужно преувеличивать для птиц, которые летят
Перелеты в вертикальном направлении
187
преимущественно ночью. Они им пользуются при определении пути,
очевидно, не все время, а главным образом в частных моментах пе-
релета: для поисков корма, выбора подходящего места для отдыха
и для ближайшей ориентировки в некоторых случаях. Птицы, ле-
тящие днем, повидимому, в большей мере опираются на зрение,
активно определяя дорогу. Это подтверждается следующим об-
стоятельством: стоит только туману закрыть землю, как невообра-
зимая сумятица поднимается в рядах птиц, они взлетают, кружатся
на одном месте, испуская тревожные крики. Что касается памяти
птиц, то она у них довольно хорошая.
Очень большую роль во всем поведении птиц, в том числе и раз-
витии способности их к перелету, играет обучение их родителями и
развитие целой сложной системы условных рефлексов. Выходящие
из гнезда птенцы сначала весьма беспомощны. В процессе их «обу-
чения», в связи с высокой степенью развития у птиц нервной системы
и органов чувств, быстро вырабатывается система условных реф-
лексов, способствующая ориентировке птиц в природе, нахождению
ими пищи и т. п. В частности и «запоминание» пути в пролете так
же связано с выработкой ряда условных рефлексов, с «регистра-
цией» в мозгу разнообразных внешних впечатлений, исходящих
от окружающего птицу внешнего мира.
Наряду с перелетами с севера на юг и обратно, в странах нагор-
ных имеют место периодические перемещения птиц с возвышенно-
стей в места низменные и наоборот. Это — перелеты или кочевки
в вертикальном направлении. Они преобладают в странах жарких,
хотя встречаются и в таких умеренных по климату местностях,
как Великобритания и Скандинавия. Все гнездящиеся на Альпах
птицы, как-то: белая куропатка, альпийская ворона, горный вью-
рок, альпийская завирушка и прочие, осенью, при выпадении снега,
спускаются в долины; за ними же спускаются и хищные птицы. На
Кавказе также, например, альпийская завирушка и горный сне-
гирь гнездятся на высоте до 2800 и над уровнем моря, между верхней
границей лесной растительности и нижней границей вечных снегов,
а зимуют в долинах, много ниже пояса лесной растительности.
Но вертикальный перелет выражен особенно ясно в Индии. Мно-
гочисленные виды переселяются для вывода птенцов из долин на
склоны Гималайских гор, поднимаясь на несколько тысяч метров
над уровнем моря и возвращаясь с наступлением холодов в низ-
менные местности; вальдшнеп выводится здесь на высоте 2800 м9
а зимует в равнинах. Передвижения таких птиц так же правильны,
как и передвижения птиц, перелетающих на тысячи миль с севера
на юг и обратно.
Весенний и осенний перелеты птиц существуют и в Северной
Америке. Мало того, здесь эти явления, пожалуй, грандиознее
даже, чем в Старом Свете. Причина в том, что переходы от одного
времени года к другому в Северной Америке резче и, кроме того,
188
Перелеты птиц
птицы не встречают на своем пути горных цепей, преграждающих
им дорогу. Здесь уже самими физико-географическими условиями
страны пернатым странникам, можно сказать, предначертано три
тракта: на востоке и западе — вдоль морских побережий, а по-
середине материка — по бассейну «отца всех рек» — Миссисипи.
В нижнем течении Миссисипи, среди субтропической природы
южных штатов, многие из перелетных птиц находят уже вполне
подходящий для них климат, достаточное количество пищи и
вообще хорошие условия для зимовки.
В южном полушарии крупных периодических перелетов птиц
нет, так как там совершенно другие условия. Распределение мате-
риков и морей там иное, чем в северном полушарии, представляя
последнему почти полную противоположность. В южном полуша-
рии мы видим преобладание площади моря над площадью суши
и иное распределение суши, вследствие чего здесь создаются со-
вершенно особые климатические условия. Климат Южной Африки
и Южной Америки не настолько суров, чтобы в них могли гнез-
диться полярные формы. Антарктический же материк лежит так
близко к южному полюсу, что здесь даже полярные формы не на-
ходят для себя подходящих условий. Поэтому в южном полушарии
имеют место только небольшие переселения, но недлинные регуляр-
ные перелеты. Кроме того, в южном полушарии не было леднико-
вого периода — по крайней мере, в таком масштабе, как в се-
верном,— явившегося главнейшей причиной возникновения длин-
ных перелетов птиц в северном полушарии.
ПЕРЕСЕЛЕНИЯ ДРУГИХ ЖИВОТНЫХ
Не одни только птицы в зависимости от времени года меняют
места своего обитания, улетая осенью на юг, а весною возвращаясь
обратно. Подобные переселения имеют место и среди других живот-
ных. Они совершаются в меньшем масштабе, с меньшей правиль-
ностью и менее известны, но представляют также весьма интерес-
ное биологическое явление. Заслуживают внимания прежде всего
переселения (миграции) млекопитающих животных.
Основная причина этих переселений, как и перелета птиц,—
смена времен года и связанное с этим наступление неблагоприят-
ных условий жизни. У нас таким неблагоприятным сезоном яв-
ляется зима, а в странах тропических—лето.
В умеренных широтах особенно большую роль в возникнове-
нии сезонных миграций млекопитающих играет снежный покров.
Его значение в жизни зверей, наряду с температурой, светом и
влажностью, чрезвычайно велико. Снег затрудняет движение жи-
вотных; для многих, особенно для травоядных, частично или пол-
ностью прекращает доступ к пище. С другой стороны, для некото-
рых мелких зверьков, живущих на поверхности почвы и в норах,
он является важным фактором сохранения тепла, защитой от сви-
репых морозов. К годичным сезонным переменам жизненных ус-
ловий присоединяются резкие колебания их в ту или другую —
благоприятную или неблагоприятную—сторону, совершающиеся
в разных странах через большее или меньшее количество лет. Бы-
вают из ряда вон выходящие засухи, приводящие к неурожаю, или
чрезмерные зимние холода и т п.; в таких случаях переселения
животных, естественно, совершаются в большем против обычного
масштабе.
К этому присоединяется еще весьма важная причина — размно-
жение животных, вызывающее захват новых мест обитания. Если
теплый урожайный год вызвал усиленное размножение, а в особен-
ности если вслед за тем произошло резкое изменение климатических
условий в неблагоприятную сторону, то образовавшийся избыток
животного населения часто переселяется в новые места обитания.
]lJ0 Переселения других животных
Характер переселений очень разнообразен. Наиболее обыч-
ным для наших стран является простое кочеванье. Животные пере-
ходят из одной местности в другую, в особенности с наступлением
зимы — в поисках пищи. Усиленное размножение и наступление
чрезвычайно неблагоприятных условий вызывают спорадические
массовые переселения. При этом одни выселившиеся животные на-
зад не возвращаются, другие — возвращаются. В первом случае
мы имеем только странствования, во втором — переселения в соб-
ственном смысле слова. Эти переселения, как происходящие под
влиянием непостоянных причин, носят характер случайных. Но
есть животные, совершающие и правильные сезонные переселения,
подобные перелетам птиц. Впрочем, провести резкую границу меж-
ду всеми этими явлениями — кочеваньями или странствованиями,
случайными и правильными периодическими переселениями — не-
возможно: они связаны рядом переходов. И у птиц есть подобные
постепенные переходы от кочеванья к правильным перелетам, с
той разницей, что последние преобладают.
Все сказанное о причинах и характере переселений животных
лучше выяснится при знакомстве с самими фактами. К ним и перей-
дем, причем остановимся на более известных и типичных примерах.
Пример кочеванья представляет наш обыкновенный хищник —
волк. Волчица устраивает логово и рожает детенышей в самом на-
чале весны. Волчата (в числе 4—6, а иногда и больше) родятся бес-
помощными, слепыми и 4—5 недель выкармливаются молоком.
С полуторамесячного возраста они переходят на мясную пищу.
К осени волчата вырастают настолько, что уже могут само-
стоятельно охотиться. Однако они не отделяются от родителей и
продолжают охотиться вместе с ними. Осенью и зимой происходит
довольно беспорядочная кочевка отдельных выводков или небольших
волчьих стай. Рассказы о стаях волков в сотни голов являются вы-
мыслом. Направление этих кочевок в основном связано с отысканием
пищи, которая у волков разнообразна: от крупных копытных жи-
вотных (лоси, рогатый скот, овцы, олени) до мелких зверьков (мы-
ши, полевки) и даже лягушек и крупных насекомых.
Обычно районы этих кочевок не очень велики (десятки кило-
метров), но в некоторых случаях в поисках корма (в особенности
в зоне тундр) волки могут проходить большое расстояние, измеряе-
мое сотнями километров.
Подобным образом кочуют многие травоядные животные, на-
пример, лоси — обитатели наших лесов. Лоси живут небольшими
стадами, состоящими из 2—3 самок с телятами и одного, редко двух
самцов (рис. 122). Старые самцы ведут одинокую жизнь.
Питаются лоси в основном ветками деревьев и кустарников,
древесными лишайниками, обгрызают кору молодых деревьев. Во
многих местностях, где высоты снежного покрова не превышают
60—70 см, лоси не предпринимают зимних миграций и живут бо-
Миграции млекопитающих
191
лее или менее оседло. В тех же районах, где высота снежного по-
крова выше указанной, лоси зимой мигрируют в места с менее вы-
соким снежным покровом. В данном случае снег не препятствует
лосям в добыче пищи, так как корм у них надснежный, но снежный
покров затрудняет их движения. Например, большинство лосей
с западных склонов Уральского хребта, где снег очень высок,
зимой мигрируют на восточные, где высота снежного покрова
Рис. 122. Лось (Alces alces)
значительно меньше, а весною возвращаются вновь на западные
склоны. Закономерные зимние миграции лосей наблюдаются в Си-
бири в Обь-Иртышском бассейне и в др.
Подобного рода зимние миграции, непосредственно связанные
с высотой снежного покрова, имеют место и у другого более мел-
кого, чем лось, представителя семейства оленьих—у косули. С за-
падных склонов Урала они также переходят зимой на восточные.
В тех же местах, где высота снежного покрова невелика (например,
Тувинская Автономная область), косули круглый год ведут осед-
лый образ жизни, не предпринимая сколько-нибудь значительных
миграций.
В годы сильных засух, когда солнце выжигает траву и насту-
пает неурожайное время, в разных местах наблюдаются массовые
переселения и таких животных, которые в обычных условиях не
192
Переселения других животных
предпринимают сколько-нибудь значительных странствований. Из
постигнутой неурожаем местности переселяются и травоядные, и
хищные, и другие. В наибольшем масштабе наблюдаются пересе-
ления грызунов — особенно в тех случаях, когда предшествующее
благоприятное время (урожайное лето, теплая зима) вызвало уси-
ленное размножение их: грызуны—мыши, белки и т. д.— отли-
чаются, как известно, громадной плодовитостью.
Еще в более грандиозном размере совершаются подобные пере-
селения леммингов, или пеструшек.
К леммингам относится несколько родственных полевкам видов
грызунов, живущих в тундре и северной лесной зоне. Из них пест-
рушка, или обыкновенный лемминг (Lemmus lemmus), населяет
тундры Кольского полуострова. Шубка у него пестрая, неправильно
разрисованная большей частью в бурые, желтые, серые и черные
тона, мех очень хорошо развит. По размерам крупнее домашней
мыши, но несколько мельче крысы.
Лемминги питаются растительной пищей (стеблями и корнями
трав, листьями ив, мхами). Зимой они не впадают в спячку, а жи-
вут под снегом в норах, изредка выходя на поверхность снежного
покрова. Лемминги очень плодовиты, они размножаются несколько
раз в год, в каждом помете 5—10 детенышей.
При благоприятных условиях количество леммингов быстро
возрастает. Их массовое размножение сопровождается поеданием
пищи, следствием чего являются массовые перекочевки. Грызуны
передвигаются несметными количествами, преодолевая на своем
пути разнообразные, в том числе и водные, преграды. Значительная
часть их в результате перекочевок погибает. Вслед за леммингами
передвигаются и хищники: песцы, поляр! ые совы.
Неблагоприятные условия питания вызывают иногда массовые
перекочевки и у белок. Белка — лесной зверек, но при массовых
переселениях она может преодолевать безлесные пространства и
даже переплывать реки.
Вызванные неблагоприятными условиями массовые перекочевки
иногда приводят к тому, что данный вид животного оказывается
в местности, лежащей за пределами ареала его распространения.
В отдельных случаях это может привести к расширению ареала
распространения вида. Так было, например, с белками. В 1920 г.
из лесов Восточной Сибири, преодолев тундру, белки проникли на
Камчатку, где они раньше не водились, а в настоящее время живут
там и размножаются.
Говоря о млекопитающих, совершающих регулярные сезонные
переселения, наиболее напоминающие перелет птиц, следует отме-
тить прежде всего летучих мышей, похожих на птиц своей способ-
ностью летать. Некоторые виды этих своеобразных млекопитающих,
например, обыкновенный кожан, к концу лета пробираются далеко
на север. На севере СССР они появляются между 60 и 65° с. ш.
Миграции млекопитающих
193
обыкновенно в августе. Крайняя граница их распространения здесь
лежит около 68—70°. Но неизвестно ни одного случая нахождения
летучих мышей в указанных пределах в состоянии зимней спячки.
Значит, они здесь — временные гостьи, залетающие сюда не более,
чем на шесть недель, так как с октября здесь начинается уже суро-
вое время года. Детенышей своих они здесь не выводят, а выводят их
главным образом между 54—60° с. ш.— месте обыкновенного пре-
бывания европейских летучих мышей. Точно так же в Северной
Америке известны два вида перелетных летучих мышей, летом жи-
вущих в Канаде, а зимой переселяющихся в южные штаты Север-
Рис. 123. Северный олень (Rangifer tarandus)
пой Америки. В жарких странах, где летучие мыши встречаются
в значительно большем количестве, переселения их еще заметнее.
В период засухи многие из них перелетают в горы, другие же виды
отправляются в более отдаленные местности, возвращаясь в оби-
таемую ими область лишь по прошествии известного времени. Часть
видов направляется в более холодные месяцы ближе к экватору,
а другая, наоборот, в теплые месяцы ищет пристанища в более хо-
лодных местностях или поднимается в горы. В некоторых случаях
причина этих перемещений летучих мышей может быть объяснена
климатическими условиями, но обыкновенно эти животные следуют
за насекомыми или летят туда, где созревают поедаемые ими плоды.
Правильные сезонные переселения совершает северный олень
в Сибири (рис. 123). К концу мая он покидает леса вдоль южной
окраины тундры. Находясь здесь зимою, он был более защищен от
холода, и здесь ему легче было доставать из-под снега мох и лишай-
ники, покрывающие тундру. Летом же олени большими стадами
направляются на открытую тундру к северу частью потому, что
находят тут на мшистых равнинах лучшую пищу, частью также
194
Переселения других животных
избавляясь от мух и комаров, которые с наступлением весны по-
являются в воздухе южной тундры бесчисленными тучами. Олени
идут постоянно по одним и тем же путям, протоптанным с незапа-
мятных времен и ведущим к самым удобным переходам и перепра-
вам через реки. На таких местах оленей поджидает человек. Вес-
ной, когда олени очень исхудали, а шкуры их не имеют цены вслед-
ствие дыр, проделанных личинками оводов, охотники оставляют
их в покое и дают им свободный доступ к хорошим пастбищам. Зато
на возвратном пути осенью они подстерегают откормленных, по-
крытых новой шерстью оленей на местах их переходов через реки
и убивают здесь животных целыми массами.
Одна из основных причин закономерных сезонных миграций
северных оленей между тундрою и лесом заключается в большом
несоответствии между запасами летних кормов и тех кормов, ко-
торые доступны в тундре для использования зимой. Например,
по расчетам И. В. Друри (1949) в тундре зимой может прокормиться
в 4—5 раз меньше оленей, чем на той же площади летом. За послед-
ние 50—100 лет поголовье дикого северного оленя значительно
уменьшилось, произошла замена его домашним, прирученным оле-
нем. Тем не менее, миграции дикого оленя на огромных простран-
ствах тундр Азии являются и теперь одним из поразительных по
своему масштабу явлений в жизни животных. На прилагаемой карте
(рис. 124) показаны основные пути сезонных перекочевок дикого
северного оленя в настоящее время. Протяженность этих путей до-
стигает 500 — 750 км. На побережье Ледовитого океана олени
ранней весной переходят по льду даже на острова, откуда в на-
чале зимы после ледостава уходят. Регулярно кочуют олени
и на Новой Земле с ее северной оконечности к южной.. Что же ка-
сается европейской тундры и, в частности, Кольского полуострова,
то в недавнем историческом прошлом здесь также осуществлялись
обширные сезонные миграции оленя. В настоящее время картина
резко изменилась. Настоящего дикого северного оленя здесь почти
уже нет. Многочисленные же стада прирученного оленя перекоче-
вывают и меняют пастбища по воле человека.
Олени идут всегда в определенном порядке: самки и телята
впереди, самцы следуют за ними. Во главе стада обыкновенно идет
крупных размеров самка. Отдельные стада состоят каждое из
200—300 голов, но стада эти находятся на весьма близком рас-
стоянии друг от друга. Иногда таким образом составляются целые
полчища; рога их образуют как бы странствующий лес, лишенный
листьев. Так же точно северные олени переселяются в Северной
Америке. Ежегодно направляются они из Гренландии на материк
и обратно, переходя по замерзшим проливам с одного острова на
другой.
Подобные правильные передвижения в прежние времена, при-
мерно до половины прошлого столетия, совершали по степям Се-
Осенние кочевки диких северных оленей
Рис, 124. Основные направления осенних кочевок диких северных оленей в Арктике и Субарктике
к местам зимовок (показаны стрелками) (по А. А. Насимович, 1955)
196
Переселения других животных
верной Америки бизоны — крупнейшие, почти вымершие теперь
жвачные (рис. 125). Осенью, к началу зимы, они двигались на юг,
спасаясь от северных холодов, метелей и зимней бескормицы. Вес-
ной переселялись на север, уходя от южного зноя, из выжигаемых
солнцем мест, в которых оставалось уже мало пищи. Переселения
происходили неторопливо, с остановками на лучших пастбищах, из
года в год по одним и тем же путям и в строго определенном порядке.
Во главе стада стоял сильный старый бизон. В случае нападения
врага телята становились внутри стада, их окружали самки, кото-
Рис. 125. Стадо бизонов (Bison americanus)
рые располагались рогами наружу, а внешнее кольцо составляли
самцы: низко опустив лбы, они представляли благодаря своим ро-
гам живую крепость. Индейцы следовали за стадами бизонов со
всем своим домашним скарбом. Тянулись также за ними стаи аме-
риканских луговых волков (койотов), которые нападали на слабых,
больных или раненых животных, отставших от стада.
В степях Южной Африки массовые переселения животных, пре-
имущественно копытных, вызываются сильными жарами. Здесь
живет много диких, близких к лошадям полосатых зебр, В изо-
билии водятся многие виды антилоп — стройных животных раз-
ной величины, с различными прямыми и закрученными рогами,
с раздвоенными копытами, напоминающих отчасти наших коз.
Стадами бродят эти животные по степям, питаясь обильной вы-
сокой травой. Привольно живется им, пока не наступают летние
жары, совершенно изменяющие картину. Жары эти чрезвычайно
сильны. Трава желтеет, выгорает, и прежде пышная степь
превращается почти в пустыню. Вода в водоемах, служащих водо-
Миграции рыб
197
поями степным животным, почти ежегодно высыхает. От прежних
речек кое-где остаются ямы, наполненные теплой, почти непро-
точной водой. Около пересыхающих водоемов собираются прежде
всего табуны зебр. К ним присоединяются антилопы и страусы.
Стадо охватывает неудержимое стремление к перемещению, и начи-
нается движение в поисках воды и свежей, еще не выгоревшей травы
(рис. 126). К одному стаду присоединяются другие. Присутствие
страусов в стаде зебр полезно для тех и других: высокие страусы
Рис. 126. Смешанное стадо зебр (Hippotigris zebra) и страусов (Struthio),
спасающихся бегством
издали видят подкрадывающегося врага и своим беспокойством вы-
дают его зебрам, а эти последние легче могут отбиться от мелких
хищников и, защищая себя, вместе с тем защищают от них и страу-
сов. Особенно большие стада составляются иногда из мелких ан-
тилоп — газелей-прыгунов. Их стихийное движение не могут ос-
тановить никакие преграды. Движутся животные на более возвы-
шенные места или к югу, подальше от экватора. С наступлением
более мягкого времени года, с приближением периода дождей, т. е.
тамошней зимы, начинается обратное движение тех же животных.
Подобно травоядным африканских степей, живут стада особой
породы диких ослов — куланов — в Средней Азии и в Монголии.
Куланы, или джигетаи, похожи на лошадь и на осла. Грива, хвост
и копыта у них такие, как у осла. Уши длиннее лошадиных, но
короче ослиных. Ростом куланы невелики, красивы, стройны, очень
легки на бегу. Весной они пасутся небольшими табунами, которые
летом постепенно увеличиваются. Увеличение табунов начинается
198
Переселения других животных
с погони самцов за самками. Это приводит в движение отдель-
ные небольшие табуны, которые, носясь по степи, встречаются один
с другим, соединяются вместе и достигают в конце концов тысячи
и более голов. Драки между самцами мало-помалу затихают, и
неправильный бег по разным направлениям постепенно переходит
в правильное странствование. Из средних частей степи стада
направляются к ее окраинам, где она переходит в горы. Там больше
воды и травы. Весной происходит переселение в обратную сторону.
В настоящее время количество куланов невелико. Они близки
к полному уничтожению.
В горах переселение животных происходит не с севера на юг
и обратно, а сверху вниз — осенью и снизу вверх — весной. Эти
переселения короче, но они наблюдаются, можно сказать, у всех
горных животных. Серна, каменный козел, благородный олень,
кабаны и другие горные травоядные животные весной поднимаются
в горы на альпийские пастбища, а осенью снова спускаются в гор-
ные долины. Вслед за копытными и грызунами совершают сезон-
ные вертикальные миграции в горах и хищники: волки, рыси,
барсы.
Правильные сезонные переселения наблюдаются у некоторых
рыб. Они известны под именем хода рыб.
Икра многих морских рыб может развиваться только в пресной
воде. Эти рыбы ежегодно перед наступлением времени метания икры
массами отправляются в реки. Рыбы, принадлежащие к одному виду,
собираются во время этого путешествия в большие стаи, или косяки,
и направляются к устьям рек. К идущим стаям присоединяются все
новые и новые группы. Прежде чем двинуться в путь по реке, рыбы
обыкновенно останавливаются у устья. Почему? Полагают, что
рыбы постепенно привыкают здесь к пресной воде. Переход из
соленой воды сразу в пресную был бы резким, у устьев же рек
морская вода опресняется постепенно.
Чтобы дать наглядное представление о тех громадных массах,
какими движутся рыбы по рекам, приведем несколько примеров.
В реках Дальнего Востока в больших количествах ловится кета.
Эта рыба входит в наши дальневосточные реки такими мас-
сами, что буквально запружает всю реку. Вот как описывал
такую картину акад. Миддендорф.
«С неудержимым стремлением мчится вверх по течению в горную
реку соединенная в миллионы штук кета. Положительно кипит вода,
принимающая рыбный вкус, весла бессильны сделать что-либо в
этой массе рыб. Напротив, если воткнуть весло в эту живую массу,
то оно долгое время движется стоймя вместе с плывущей рыбой.
Если же лодка идет вдоль отлогого берега, то крайние ряды рыб
вытесняются на берег, где они и погибают. Все-таки основная масса
стремится вперед, напрягает свои силы в борьбе с мчащимся пото-
ком, в борьбе с порогами, поднимаясь все выше,> все в гору, до тех
Миграции рыб и икрометание
199
пор, пока не станет некуда идти. Из воды уже высовываются спин-
ные плавники, вылезают спины, а кета еще идет вверх . .»
По наблюдениям В. К. Солдатова (1912), ход кеты в Амуре пред-
ставляет грандиозное явление.
«Обычно валовому или полному ходу рыбы предшествует сначала
появление единичных экземпляров кеты; количество ее начинает
увеличиваться иногда довольно правильно, а иногда резко и быстро;
иногда рыба начинает идти в промысловом уже количестве, и про-
мышленники ждут полного хода, как ход опять начинает умепь-
Рис. 127 Ход горбуши (Oncorhy nchus gorbuscha)
шаться, и опять ловятся лишь единичные экземпляры. Полный ход
представляет действительно грандиозное зрелище, поражающее
даже человека, видавшего богатые ловы рыбы.
Десятки, сотни тысяч рыб, стая за стаей, побуждаемые каким-то
могучим внутренним импульсом, устремляются в Амур; встретив
препятствие в виде стенки заездка, рыба останавливается у нее в
громадных количествах и, делая тщетные попытки преодолеть эту
преграду, выпрыгивает поминутно из воды, производя такой шум
и всплески, что их можно слышать издали. Все пространство у
заездка наполняется сплошь рыбой, а все еще приваливают новые
стаи ее Всюду видны головы, спинные плавники кеты, повсюду
вода пенится, как в котле, от массы сталкивающейся между собой
и выпрыгивающей из воды рыбы . Простым крючком можно в
несколько минут наполнить лодку рыбой. В способе и садке заездка
200
Переселения других животных
творится что-то невообразимое. Сотни рыб ежеминутно перегоняются
из способа в садок, но на смену являются все новые стаи рыбы . . .
Люди изнемогают от усталости и не успевают вычерпывать рыбу
из садка. Нередко рыбы идет столько, что заполняется и способ и
садок, и за неимением места лов ее прекращается».
Во всех северных морях обитают лососи, которые массами под-
нимаются для метания икры во все реки, впадающие в умеренную
и северную части Атлантического, Тихого, а также Северного Ле-
довитого океанов. В реки Камчатки лососевые (кета, горбуша,
красная, чавыча) заходят такими массами, что даже затрудняют
движение лодки, случайно попавшей в их стаю, а их спины и плав-
ники положительно рябят зеркало воды (рис. 127). Подобными же
массами идет сельдь по Волге из Каспийского моря. Достаточно
сказать, что во время одного хода сетями вылавливается иногда
до 35 млн. штук сельдей. Такими же массами входит вобла в южные
рукава Терека и т. д.
Значительное большинство рыб совершает свой ход для метания
икры весной; только у некоторых, например у лососевых, у неко-
торых миног метание икры падает на осень. При этом каждый вид
мечет икру в свое определенное время, подлежащее некоторым ко-
лебаниям, в зависимости от погоды, главным образом — от времени
вскрытия рек и температуры воды. Отдельные виды следуют друг
за другом в определенном порядке. Так, на нижней Волге наблю-
дается следующая последовательность весеннего хода: как только
вскроется лед, начинает идти белуга; за ней идут севрюга, вобла,
судак (появляющийся иногда даже ранее воблы), лещ, сельдь, сом
и осетр.
Различные виды рыб входят в разные реки. Так, из Каспийского
моря белорыбица и сельдь идут почти исключительно в Волгу, от-
части встречаясь только еще в Урале. Терек далее отстоит от Куры,
нежели от Волги, тем не менее по входящим в него рыбам он имеет
более общего с Курой, чем с Волгой. Так, в Терек и Куру входят
шемая и каспийский лосось, из которых первая почти не встре-
чается в Волге, а второй бывает здесь лишь случайным гостем. Внеш-
ние условия, особенно сильный ветер, течения, до некоторой степени
могут оказать влияние на ход рыбы. Известно, например, что в
северной части Каспийского моря рыба, обычно направляющаяся
главной своей массой к устью Волги, при сильном северо-запад-
ном ветре сворачивает со своего обычного пути на восток к устью
Урала, в который и входит для икрометания. Бывает и наоборот:
при северо-восточном ветре рыба, направляющаяся в Урал, свора-
чивает к западу и входит в Волгу. Подобные примеры наблюдаются
и в других местах.
Во время хода рыбы проходят иногда сотни, а по длинным ре-
кам, как Волга и сибирские реки, тысячи километров. В большинстве
случаев рыбы даже не питаются на всем длинном пути. Они потреб-
Миграции рыб и икрометание
201
ляют жировые и белковые запасы своего тела и к месту метания
икры приходят сильно истощенными. Многие из лососевых рыб
даже погибают после икрометания, совершая, таким образом, по
выражению Миддендорфа, «кочевание до смерти». Относительно
кеты многие американские, японские и русские авторы указывают,
что и та кета, которая после икрометания скатывается вниз по реке
до моря, обречена на гибель. В. К. Солдатов говорит по этому по-
воду следующее.
Рис. 128. Места нереста европейского (Anguilla fluviatilis)
и американского угрей (Anguilla rostrata)
«Как-то трудно поверить, чтобы мириады лососей, посещающих
ежегодно реки Тихоокеанского побережья от берегов Японии до
берегов Аляски и Северной Америки, обречены были все на неиз-
бежную гибель. Еще более поразительной является эта гибель по-
тому, что она постигает и молодых, и старых рыб, и самцов, и са-
мок, не связываясь, таким образом, ни с полом, ни с возрастом . . .
В стаях кеты встречаются рыбы нескольких поколений, и если
мы должны допустить поголовную гибель кеты, то тем самым мы
допускаем и тот в высшей степени удивительный факт, что лососи
Тихоокеанского побережья нерестуют лишь раз в жизни и притом
потребность нереста пробуждается у них в различное время их
жизни».
Далеко не у всех проходных рыб имеет место «кочевание
до смерти». Большинство видов после нереста возвращается или,
202
Переселения других животных
как говорят, «скатывается» сильно истощенными в море, где вновь
начинает энергично питаться.
Если по пути встречаются пороги и т. п., рыбы обыкновенно
употребляют все усилия, чтобы преодолеть их. В плоти-
нах шлюзованных рек иногда устраивают так называемые «рыб-
ные лестницы», имеющие целью облегчить рыбам проход через
высокую плотину, иногда около 2 м высотой. «Изумительно,—
говорит очевидец-наблюдатель подъема лососей по таким лестни-
цам, — то упорство, с которым лососи стараются перепрыгнуть
плотину, потому что, сколько бы они ни производили безуспешных
прыжков, они до тех пор не успокоятся, пока не достигнут цели».
Огромное большинство проходных рыб, как мы видели выше,
входит для икрометания из моря в реки. Подобные миграции носят
название анадромальных. Однако у угря направление миграции
является противоположным. Он для икрометания из рек спускается
в море. Это так называемая катадромалъная миграция. Угри, оби-
тающие в европейских реках, по достижении определенного воз-
раста (обычно в конце лета) спускаются в море и направляются
в Атлантический океан к берегам Северной Америки (рис. 128).
Здесь, повидимому, на больших глубинах, происходит икромета-
ние, после которого угри погибают. Из икры выходят небольшие,
почти совсем прозрачные рыбки, совершенно непохожие на взрос-
лого угря (так называемый Leptocephalus, описанный ранее как
самостоятельный вид, рис. 129). Эти личинки угря направляются
к берегам Европы, которых они достигают только через 3 года. Их
миграция является в значительной мере пассивной, так как они
несутся струями теплого течения Гольфстрема. К моменту прибли-
жения к берегам Европы Leptocephalus претерпевают метаморфоз
и превращаются в небольших так называемых «стеклянных угрей»,
которые и входят в реки. В пресных водах угорь живет от 5 до 20
лет, после чего спускается в море.
Эта замечательная история жизни европейского угря выяснена
относительно недавно работами главным образом датских и нор-
вежских ученых.
Американский угорь, являющийся по отношению к евро-
пейскому другим видом, предпринимает также катадромальную
миграцию в Атлантический океан. Но его метаморфоз проте-
кает более быстро и Leptocephalus американского угря пре-
вращается в «стеклянных угрей», входящих в реки Америки,
в течение одного года.
Выше говорилось о длинных путешествиях морских рыб в реки
для метания икры. Рыбы, совершающие такие миграции, назы-
ваются проходными. Различают еще рыб полупроходных. Это —
те, которые водятся в опресняемых устьями больших рек участках
морей и поднимаются вверх по рекам недалеко от устья. Есть мор-
ские рыбы, которые мечут икру у морских берегов. Так, треска и
Миграции рыб и икрометание
203
живущая в океанах сельдь большую часть года проводят вдали от
берега. Перед нерестом же они предпринимают далекие миграции;
например, треска, живущая в Баренцевом море, в период нереста
собирается в большие косяки и в апреле—мае направляется к Ло-
фотенским островам, расположенным у северо-западного берега
Норвегии, где и происходит икрометание. После этого взрослая
треска вновь возвращается в Баренцево море, характеризующееся
высокой кормностью. Икра трески является пелагической,т. е. пла-
вающей в толще морской воды. Выметанная у Лофотенских остро-
Рис. 129. Личинки угря (Leptocephalus)
вов, икра пассивно несется
теплым Нордкапским тече-
нием (Гольфстрем) к Мур-
манским берегам. Во время
этого пассивного дрейфа
икры постепенно происхо-
дит ее развитие. В конце
июня — начале июля из
икринок выходят мальки.
К этому времени икра
оказывается в большин-
стве своем уже у Мурман-
ских берегов. Большин-
ство мальков трески в те-
чение первого года жизни
находится около Мурман-
ского берега и питается в
прибрежной зоне. Затем,
по мере дальнейшего роста,
треска отходит от берегов
и начинает питаться в открытом море. По достижении половозре-
лости (в возрасте 5—6 лет) она предпринимает весной описанную
выше нерестовую миграцию к Лофотенским островам, а затем
вновь возвращается в Баренцово море для нагула. В течение своей
жизни треска совершает нерестовые миграции несколько раз.
Мурманская атлантическая сельдь широко распространена в
северной части Атлантического океана и в Баренцовом море, где
она находит обильную пищу. Половая зрелость наступает в воз-
расте 5—6—7 лет. Половозрелая сельдь предпринимает весной об-
ширные миграции к Лофотенскому мелководью в районе Лофотен-
ских островов, где и происходит нерест. Пелагическая икра и раз-
вивающиеся из нее мальки, так же как и у трески, дрейфуют на се-
веро-восток по струям Нордкапского течения.
Периодические сезонные миграции наблюдаются и у рыб, всю
жизнь проводящих в реках.
Почти все наши речные рыбы для метания икры поднимаются
вверх по течению на большие или меньшие расстояния, смотря по
204
Переселения других животных
видам. В большинстве случаев эти передвижения совершаются во
время половодья, почему наблюдать их затруднительно. Так же,
как и у проходных рыб, здесь наблюдается определенный порядок,
в котором отдельные виды следуют друг за другом. Так, по наблю-
дениям на Майне, первою начинает путь обыкновенно уклейка, за
нею следуют пескарь, плотва, красноперка, лещ, окунь, щука, ерш,
карп, карась, линь, усач, голавль, елец, подуст, вьюн, судак, бы-
Рис. 130. Выметанная икра окуня (Perea fluviatilis)
чок, речная минога. Рыбы собираются большими стаями и не
останавливаются перед препятствиями, если пришло время выме-
тать икру, для которой нужно найти подходящее место. Щука,
например, перед выметыванием икры, подпускает рыбака чуть
не вплотную и нередко остается на том же самом месте, где сорва-
лась со снасти; ее нетрудно в этом случае поймать тут же вторично.
Зато, выметав икру, рыба становится весьма пугливой.
Само икрометание состоит в том, что самка выпускает икру, ко-
торую самец тут же поливает молоками.
Этот процесс называется нерестом, а места, где происходит
икрометание,— нерестилищами, У различных рыб эти места весьма
разнообразны. Так, одни рыбы, например карповые, выбрасывают
икру среди растений, главным образом в местах разлива вод, на
растениях же путается икра окуня, имеющая вид слизистых шнуров
(рис. 130). Другие, как осетровые, нерестятся на каменистом дне.
Лососевые откладывают икру среди мелких камней в верховьях
рек.
Как ни разнообразны нерестилища, все они имеют достаточное
количество воздуха и тепла, необходимых для развития икры. Там,
Миграции рыб и икрометание
205
где быстрое течение, вода освежается и пополняется воздухом благо-
даря течению. Где есть растения,— вода обогащается кислородом,
выделяемым растениями в процессе фотосинтеза. В мелких местах
воздуха вообще больше растворено в воде, чем в глубинах. Тепла для
икры одних рыб требуется больше, для других — меньше. На
мелких местах вода скорее прогревается солнцем, и поэтому теплее.
Количество икры, именно отдельных икринок, выметываемых
рыбами, чрезвычайно большое. Щука дает около 100 000 отдель-
ных икринок, окунь — 300 000, камбала — больше 3 млн., треска
свыше 9 млн.; сом, осетр и белуга мечут икру также миллионами.
Моря, вероятно, было бы недостаточно, если бы все икринки, вы-
метанные рыбами в один год, развились и все вышедшие из них
рыбки достигли величины своих родителей. Это обилие икры стоит в
связи с тем, что, оставленная выметавшей ее рыбой на произвол
судьбы, она гибнет в громадном количестве. Из миллионов икри-
нок некоторая часть остается неоплодотворенной; миллионы ее вы-
брасываются волнами на берег и засыхают; много также попадает
в слишком глубокую воду, где икра не развивается; начавшие же
развиваться икринки подстерегаются бесчисленными полчищами
врагов и прежде всего — самими же рыбами. При таких условиях
только необыкновенно большое количество икры может гаранти-
ровать, что не вся она погибнет и размножение рыб не прекратится.
Есть некоторые исключения, именно — некоторые рыбы не ос-
тавляют икру на произвол. Так, форели и лососи откладывают свою
икру в вырываемые ими на дне ямки и затем, по оплодотворении,
засыпают ее песком и галькой. Эти рыбы и икры дают сравнительно
мало — около 25 000 икринок. У тех рыб, которые выметывают
относительно немного икры, икринки обычно крупнее, чем у рыб,
обладающих высокой плодовитостью. Например, у лососевых ик-
ринки довольно большие, у карповых рыб они мельче, наконец,
у тресковых, которые отличаются особенно высокой плодовитостью,
икринки очень мелки.
Колюшка обнаруживает заботу о потомстве: она приготовляет
настоящее гнездо. У трехиглой колюшки местом для него является
ямка на дне (рис. 131). У девятииглой — гнездо устраивается среди
растений и является настоящим архитектурным сооружением,
имеющим вид округлой муфточки обыкновенно с двумя отвер-
стиями на противоположных концах; материалом для гнезда слу-
жат травинки и корешки. В нем самки откладывают свои немного-
численные (не более 100) икринки, где они и оплодотворяются.
После нереста истощенная рыба (у тех видов, у которых не про-
исходит поголовной гибели отнерестовавшей рыбы) обыкновенно
с жадностью набрасывается на корм и направляется в обратный путь.
Теперь отдельные особи уже не собираются стаями, а плывут каж-
дая сама по себе. Поэтому обратный ход рыбы менее заметен.
Рыба, приступающая к нерестовой миграции, находится в осо-
206
Переселения других животных
бом физиологическом состоянии, характеризующемся определен-
ной степенью созревания половых продуктов (различным для раз-
ных видов) и пробуждением «миграционного инстинкта». При этом
существенную роль играют, как это установлено рядом исследо-
ваний, железы внутренней секреции (эндокринные железы). Вы-
деляемые ими секреты (гормоны) поступают в кровь и, действуя
через нервную систему, меняют все поведение рыбы и ее отношение
Рис. 131. Гнездо трехиглой колюшки (Ga-
sterosteus aculeatus)
к внешним раздражителям.
Из описания хода рыб
ясно, что это явление имеет
много общего с перелетом
птиц: та же отчетливая
сезонная периодичность, та
же привязанность к опре-
деленным путям. Мы да-
леко не всегда можем ска-
зать, какие факторы внеш-
него и внутреннего поряд-
ка вызывают ход рыбы.
Чем объясняется ориенти-
ровка рыб? Простая ссылка
на инстинкт, который, не-
сомненно, играет здесь
роль, не может служить
исчерпывающим объясне-
нием. Привязанность к
определенным речным бассейнам частью, вероятно, объясняется
характером рек —быстротой течения, которую предпочитает дан-
ная рыба, и т. п. Но кроме того, здесь, повидимому, играет роль
и передаваемая из поколения в поколение привычка — инстинкт,
закрепленный естественным отбором. Высказывается предположе-
ние, что часть проходных рыб ранее обитала в пресной воде. Затем
они начали из устья постепенно спускаться в море за пищей — сна-
чала близко, потом все далее и далее, и таким образом посте-
пенно приспособились к жизни в морской воде. Однако икра и мо-
лодь попрежнему требуют для своего развития пресной воды. По-
этому рыбы и продолжают возвращаться для метания икры в то
место, где они ранее жили. Таким образом, можно думать, и соз-
дался этот замечательный инстинкт периодических странствований
у многих рыб. Рыбы, морские по происхождению, приплывая к
берегам для метания икры и наталкиваясь на речное русло, могли
заходить в него. Найдя в нем благоприятные условия для размно-
жения, рыбы с каждым годом, с каждым поколением могли все
дальше и дальше захватывать для нереста пресные воды и, таким
образом, мало-помалу приобрести инстинкты настоящих проходных
рыб.
Миграции беспозвоночных животных
2(П
Рис. 132. Вредная черепаш-
ка (Eurygaster integriceps)
Захождение в пресные воды для икрометания и кормления было
одной из форм расширения ареала обитания. Последнее, несомненно,
представляло признак, полезный в борьбе за существование. Ве-
роятно, поэтому инстинкты миграций у рыб и закрепились через
естественный отбор.
Ход рыбы имеет большое значение для человека, пользующе-
гося этим ходом для ловли рыбы.
Ряд особенно важных в промысловом отношении рыб в СССР
относится к проходным или полупроходным рыбам (лососе-
вые, большинство видов каспийской сельди, вобла, осетровые и
мн. др.). Основные моменты их промысла
тесно связаны с массовым вхождением
рыбы в реки, когда и происходит ее вы-
лов. В силу этого изучение миграций
рыб и факторов, их определяющих, пред-
ставляет не только теоретический, но и
огромный практический интерес. Поэ-
тому понятно, что ряд научных учреж-
дений Советского Союза изучает биоло-
гию рыб и, в частности, их миграции.
Явления массовых сезонных мигра-
ций имеют место и у некоторых беспоз-
воночных животных. Среди насекомых
закономерные массовые сезонные пере-
селения совершает, например, широко
распространенный в степной зоне вре-
дитель зерновых культур—клоп вредная черепашка (рис. 132).
Весной и в первую половину лета черепашки живут преимуще-
ственно на хлебных полях. Здесь они питаются и размножаются.
Во второй половине лета, после уборки хлебов, начинается массо-
вый отлет черепашки с полей в места с древесной и кустарниковой
растительностью. Они могут пролетать много километров и иногда
при этом поднимаются в горы в пояс древесной растительности.
В лесной зоне черепашки зимуют, забираясь при наступлении холо-
дов под слой опавших листьев, в дернину. Весной, как только по-
вышается температура воздуха, черепашки вновь перелетают на
поля. Такой же характер носят сезонные переселения у некоторых
других насекомых, например, у некоторых видов жуков из семей-
ства божьих коровок (Coccinellidae). В Средней Азии божьи ко-
ровки осенью поднимаются в горы, где зимуют группами в лесной
подстилке, под пнями и т. п. На лето они спускаются на поля.
В отличие от черепашек, большинство божьих коровок является
хищниками: они питаются другими насекомыми, преимущественно
тлями.
Менее регулярный и более спорадический характер носят пе-
релеты азиатской саранчи (Locusta migratoria) и других саранче-
208
Переселения других животных
вых. Этот вредитель культурных растений постоянно живет и раз-
множается в южной полосе Европейской части СССР, по берегам
нижнего течения больших рек (Волга, Дон, Днепр). В годы, особенно
благоприятные для массового размножения саранчи, она появляется
в огромных количествах и совершает миграции в различных направ-
лениях от мест выплода. Переселение саранчи начинается обычно
летом в стадии личинки, когда она не имеет крыльев и движется
«пешком». После последней линьки насекомые приобретают крылья,
и тогда миграция совершается по воздуху. Стаи перелетной саранчи
могут достигать огромных размеров. Когда такая «туча» сядет на
землю, то она уничтожает всю зелень до последней былинки, всю
листву на деревьях.
В отличие от вредной черепашки, массовые миграции саранчи
не носят, однако, закономерного ежегодного характера и зависят
от условий размножения насекомого в разные годы.
Регулярные сезонные миграции совершают и некоторые морские
беспозвоночные животные. Особенно замечателен в этом отношении
камчатский краб (Paralithodes kamtschatica) — важное промысло-
вое животное советских дальневосточных морей (Японского и Охот-
ского). Зимуют крабы на больших глубинах. Весной они мигри-
руют к берегам на мелкие места, где происходит их усиленное пи-
тание и размножение. Поздно осенью крабы опять уходят на глу-
бины. Одной из важных причин, обусловливающих миграции кра-
бов, является, повидимому, температурный режим. Поздней осенью
происходит резкое охлаждение прибрежных вод, температура же
глубинных вод является более высокой. Крабы направляются в
более теплую воду. Весной же, когда происходит прогревание при-
брежных вод, они направляются к берегу. Изучение миграций этого
важного в промысловом отношении животного имело большое зна-
чение в организации его промысла.
ГНЕЗДОВЬЕ ПТИЦ И УХОД ЗА ПТЕНЦАМИ
Весной, как только пернатые путешественники вернутся из
своего далекого странствования, они сейчас же принимаются за
постройку гнезд, а там — за кладку яиц, вывод молодых птенцов
и уход за ними. Разумеется, забота о потомстве у птиц носит совер-
шенно другой характер, чем у человека. Она является результатом
не сознательной, а инстинктивной деятельности. Постройка
гнезда — вот первое, в чем проявляется забота о потомстве у птиц.
Гнезда птиц весьма разнообразны: от самых незатейливых по-
строек можно видеть постепенные переходы к таким, которые в
состоянии удовлетворить строгим требованиям архитектурного
искусства.
Есть, впрочем, сравнительно немногочисленные птицы, которые
кладут яйца прямо на землю. Так, например, козодой, выбрав в
бору совершенно сухое место, откладывает свои 2 яйца на почву,
большей частью под прикрытием нависшей древесной ветви или
среди оленьего мха, а иногда — на гнилом пне. Подобным же об-
разом поступают некоторые куриные птицы, откладывающие яйца
в каком-нибудь углублении в почве, найденном или вырытом са-
мою птицей. В таких случаях мы не имеем гнезда в строгом смысле
слова, так как здесь нет даже подстилки, вообще нет «строительной
работы» со стороны птицы. Такая же кладка яиц встречается у
некоторых прибрежных птиц, кладущих яйца прямо на берег, а
также у некоторых экзотических птиц, например: страус кладет
свои яйца в песок, насиживая их только ночью, а днем предостав-
ляя согревающему действию солнца.
На крайнем севере на побережье Варенцова моря (Новая Земля,
Мурманский берег Кольского полуострова) некоторые морские птицы
образуют огромные поселения на скалах, круто спускающихся к
воде. Это так называемые «птичьи базары» (рис. 133). В состав та-?
кого «базара» входят тысячи, а иногда и десятки тысяч птиц. «Ба-
зарными птицами» являются кайры (рис. 134), чайки-моевки, чи-
стики, тупики и некоторые другие. «Базары» представляют собой
временные объединения, «колонии» птиц на период их размножения
Рис, 133. «Птичий базар» на Восточном Мурмане (фото Л. О. Белопольского)
210 Гнездовье птиц и уход за птенцами
Птичьи базары
211
«Базары» могут образоваться несколькими видами птиц или же
только одним видом. Особенно большие базары на Мурмане и
Новой Земле образуют кайры. Большинство видов этих птиц ни-
Рис. 134. Кайра (Urla troile) (фото Л. О. Белопольского)
каких гнезд не строят. Они откладывают яйца непосредственно на
уступы скал и здесь же их и высиживают. Интересно отметить, что
в связи с таким своеобразным способом откладки и высиживания
яиц меняется форма последних. Например, у кайры яйцо груше-
видное (рис. 135). Такие яйца очень плохо катаются, так как у них
212
Гнездовье птиц и уход за птенцами
сильно смещен центр тяжести и при толчке они только крутятся
на месте. Это является очень важным биологическим приспособле-
нием, препятствующим скатыва-
Рис. 135. Яйцо кайры. Кружком
показан центр тяжести, стрел-
кой—направление силы тяжести;
п — воздушная камера (ориг.
схема Л. О. Белопольского)
нию яиц с узких скалистых усту-
пов и карнизов в море.
Однако большинство птиц вы-
водит птенцов в более или менее
сложно устроенных гнездах.
Наиболее простой формой гнез-
да являются ямки, в которых
птицами сделана подстилка. Та-
кие гнезда бывают у тетеревиных
птиц, они представляют неглубо-
кие ямки в земле, выстланные сте-
бельками, сухими листьями. То
же видим у жаворонка (рис. 136),
выстилающего ямку стебельками травы, у чибиса, пользующегося
для подстилки также сухой травой. Трясогузки и некоторые
овсянки из того же мате-
риала свивают уже чашевид-
ные гнезда.
Есть птицы, которые роют
для гнезда норы в земле.
Наиболее известный пример
представляют береговые ла-
сточки (рис. 137) (неправиль-
но называемые иногда бере-
говыми стрижами). Труднд
подумать, чтобы эта неболь-
шая птичка с маленьким клю-
вом могла прорывать ходы в
твердом песчанике, а это так.
Береговые ласточки выры-
вают норы в берегах рек и
обрывов, хотя бы они были
очень тверды. Приступая к
работе, птица прежде всего
систематически пробует клю-
вом в разных местах, пока не
найдет наиболее подходящего
местечка: она не будет рыть в
рыхлом грунте, который мо-
Рис. 136. Полевой жаворонок (Alauda
arvensis) и его гнездо
жет оседать, но не будет останавливаться и на очень твер-
дом, если есть грунт помягче. Облюбовав место, птичка начинает
усиленно работать, при этсм она действует клювом, ногами и
крыльями. В постройке гнезда в одинаковой мере участвуют самки
Различные типы гнезд
213
и самцы. Таким образом вырывается ход, достигающий в длину у
некоторых птиц до 2х/2 м. Обычно норка идет слегка вверх, благо-
даря чему в ней не может скопляться дождевая вода. Направление
норки, если на пути не оказалось какой-либо преграды, вроде камня
или корня,— прямое. На конце нора расширяется. Здесь поме-
щается собственно гнездо. Оно весьма простого устройства, пред-
Рис, 137. Береговая ласточка (Riparia riparia) и ее гнезда
ставляет собой кучу сухой травы и мягких перышек, сдавленных
тяжестью тела птички. В подобных же норах и гнездах живут
зимородки. В норах же гнездятся красные утки и пеганки, в нор-
ках или щелях между камнями — щурки и розовые скворцы.
Многие птицы устраивают гнезда на большей или меньшей вы-
соте над землею, преимущественно на деревьях и кустарниках.
В параллель с норками могут быть поставлены дупла, в кото-
рых, как известно, устраиваются гнезда дятлами (рис. 138 и 1-39).
В большинстве случаев дятел выдалбливает дупло своим креп-
ким клювом. Сначала направление выдалбливаемого хода горизон-
тальное, а затем он поворачивает вертикально вниз и расширяется
на конце, образуя собственно гнездо. Оно весьма просто, так как
214
Гнездовье птиц и уход за птенцами
подстилкой его является та самая древесная труха, которая полу-
чается при выдалбливании дупла. Опасными для леса дятлов, не-
смотря на их, повидимому, разрушительную работу,считать нельзя,
так как они пользуются большей частью деревьями, начавшими
гнить. Из наших деревьев осина более всех других подвержена
сердцевинной гнили. Поэтому там, где есть осина, чаще можно уви-
деть дятлов. Другие птицы, гнездящиеся в дуплах, как-то: синицы,
вертишейки, поползни, пе-
стрые мухоловки, гори-
хвостки — не в состоянии
сами выдалбливать дупло,
Рис. 138. Большой пестрый дятел
(Dryobates major)
Рис. 139. Гнездо дятла
а потому принуждены строить гнезда в дуплах, образовавшихся
естественным путем или покинутых дятлами.
Пищухи устраивают гнезда в щелях за отставшей от ствола
корой.
Птицы, не пользующиеся для гнезда почвой, ямками, норками
или дуплами,— а таких птиц большинство,— строят гнезда более
сложные, так чтобы они могли служить достаточною опорой как для
самих птиц, так и для их яиц и птенцов. Вот тут-то мы и видим глав-
ное разнообразие в отношении как формы гнезд, так и тех материа-
лов, из которых они сооружаются.
Хищные птицы устраивают довольно сходные между собой
гнезда из грубого материала. Так, у орла мы видим довольно мас-
сивную постройку из сучьев (рис. 140)—внизу более толстых,
а кверху постепенно все более и более тонких. Наверху гнезда на-
Различные типы гнезд
215
Рис. 140. Орел (Aquila) и его гнездо
с птенцами
ходится неглубокое вдавление — гнездовой лоточек. Гнездо имеет
широкий край, так называемый приполок. На нем складывается
принесенный для птенцов корм. Сюда же птенцы, когда подрастут,
выходят из гнезда и ожидают возвращения родителей, отправив-
шихся на охоту. Подобным же образом устроено и гнездо ястреба,
отличающееся меньшим размером. По сравнению с гнездами, о ко-
торых будет речь ниже, гнезда
хищников, менее совершенны.
У больших хищников они
массивны и заметны издали.
Часто хищники устраивают
свои гнезда на высоких вы-
ступах скал, прикрытых свер-
ху нависшими карнизами,
иногда—на высоких деревьях
и притом в глуши, вдали от
человеческого жилья.
С виду похожие на гнез-
да хищников, но более проч-
ные, при меньшем размере
их, устраивают на верхуш-
ках деревьев грачи (рис. 141)
и вороны. Эти гнезда иногда
сохраняются целые годы.
Ими пользуются и мелкие
хищники.
Чаще всего для постройки
гнезд применяется материал
мягкий, состоящий из травя-
нистых стеблей, листьев,
волос, пуха и т. п.; таким
материалом пользуются почти
все наши певчие птицы. Наиболее обычная форма устраиваемых
из него гнезд — чашковидная, с более или менее приподнятыми
краями.
Особенно красивы гнезда зяблика (рис. 142) и щегла.
Зяблики обычно гнездятся на деревьях, чаще всего у места от-
хода крупных ветвей от ствола, на высоте нескольких метров. Осо-
бенно охотно строятся гнезда на деревьях, обильно обросших мхом
и лишайниками. Гораздо реже находим гнезда зяблика в крупном
кустарнике.
Главным материалом для гнезда являются растительные во-
локна, которые зяблик добывает, между прочим, расщипывая во-
локнистую кору растений; вплетаются также нежные мхи, пау-
тина, растительный пух и лишайники. Мхи и в особенности
лишайники очень искусно укрепляются на наружной поверхности
216
Гнездовье птиц и уход за птенцами
гнезда, придавая ему вид естественного нароста на дереве. Внутри
гнездо тщательно выстилается растительным пухом. Получается
глубокая чашка для яиц—
Рис. 141. Гнезда грачей (Corvus frugi
legus)
Рис. 142. Зяблики (Fringilla coelebs)
у гнезда
глубина ее превосходит
ширину. Благодаря свой-
ству материалов гнездо
очень крепко и упруго,
так что принимает свою
прежнюю форму даже по-
сле сильного сжатия.
На гнездо зяблика по-
хоже гнездо щегла.
В гнезде иволги та осо-
бенность, что оно почти
висячее (рис. 143): оно по-
мещается близко к оконеч-
ностям ветвей, обыкно-
венно в развилинах их.
Материалом для постройки
служат полувысохшие ли-
стья, трава, березовая ко-
ра, различные волокна,
шерсть и паутина; внутри
гнездо выстлано тонкими
травинками, шерстью и
перьями.
Многие мелкие птички
вьют гнезда из одной тра-
вы, например, сорокопут
жулан и славка Черного-
ловка. Свыше ста довольно
цепких травинок нужно
сорокопуту на постройку
внешней части гнезда.
К этой стенке он присоеди-
няет вторую—из мха, пере-
плетенного с более мелки-
ми травинками; самые тон-
кие волокна идут на внут-
реннюю отделку. Гнездо
Черноголовки имеет форму
неглубокой чашечки из
очень мелких травяни-
стых стеблей. Сюда отно-
сится и гнездо соловья, состоящее обыкновенно снаружи из сухих
прошлогодних листьев, внутри же—из тонких, сухих травинок.
Различные типы гнезд
217
Некоторые птицы при пост-
ройке гнезда к растительным
материалам добавляют жирную,
влажную землю, которая, вы-
сыхая, играет роль цементирую-
щего вещества, например, у чер-
ного дрозда.
Большинство болотных птиц
строит гнезда из травы и ка-
мыша у берега в тростниках.
Устройство этих гнезд хотя до-
вольно грубое, но интересно в
виду преодоления тех трудно-
стей, с которыми встречается
птица.
Водяная курочка строит гнез-
до из сухих листьев и стеблей
растений обычно на горизонталь-
ных ветвях прибрежной ивы,
крупных стеблях тростника или
других растений.
Дроздовидная камышевка,
встречающаяся в южной полови-
не Европейской части СССР, под-
вешивает свое гнездо (рис. 144) и
укрепляет его, чтобы оно могло
держать птицу с яйцами и птен-
цами и чтобы эти последние не
выпали из него. Одной тростинки
для такой тяжести недостаточно:
нужно найти, по меньшей мере,
три, отстоящих на равном рас-
стоянии друг от друга. Так как
при этом еще требуется, чтобы
стебли на определенной высоте
над водой обладали достаточной
твердостью и в этом отношении
соответствовали друг другу, то
такую комбинацию не так легко
найти. Затем, чтобы положить
первоначальную основу гнезда,
Рис, 143. Гнездо иволги (Oriolus
oriolus)
птичке даже не за что как еле- Рис. 144. Гнездо дроздовидпой ка-
дует уцепиться: приходится лап- мышевки (Acrocephalus arundinaceus)
ками держаться за гибкий сте-
бель тростника. Птица набирает сухих листьев камыша, вод-
ных растений длиною от 20 до 35 см, мочит и соединяет их, делая
218
Гнездовье птиц и уход за птенцами
из них плотную тесемку, которую укрепляет на надлежащей высоте
в отгибе листа и обматывает вокруг тростника; потом направляет
тесемку к другой тростинке, которую обматывает так же, и т. д.,
повторяя эту операцию несколько раз, пока не удастся прочно
соединить тростинки между собой.
Рис. 145. Деревенские ласточки (Hirundo rustica) и их гнездо
Расположенные друг над другом в горизонтальном направле-
нии круги из такой тесемки переплетаются снизу вверх так, как
корзинщик плетет корзину. Так как камыш качается ветром, то,
чтобы яйца или молодые птенцы не могли при этом выпасть из гнезда,
оно делается глубоким. Внутри гнездо устилается тонкими травами
и шелковистыми метелками камыша. Если поблизости тростни-
ков нет, го камышевка вьет гнездо на прибрежных кустах.
Есть птицы, которые строят свои гнезда целиком из минераль-
ных материалов, лишь изнутри выстилая их мягкой подстилкой.
Таковы, например, наши ласточки. Деревенская ласточка (рис. 145)
строит свое гнездо почти всегда в жилых местах: под карнизами
домов, в конюшнях, сараях, па чердаках и т. п. Каково бы ни было
место, ласточка строит гнездо так, чтобы оно находилось под защи-
Различные типы гнезд
219
той от дождя, который не мог бы превратить его в бесформенную
массу грязи. Гнездо делается из жирной земли, которую птица со-
бирает клювом и перемешивает со слюной. Так как рот ласточки
мал, то ей приходится совершить иногда до 500 путешествий для
постройки одного домика. Слюна, засыхая, придает земле большую
твердость. Ласточка перемешивает ее, кроме того, с шерстью, во-
лосками, травянистыми
стеблями. Внутри гнез-
до выстилается шер-
стью, перьями, тонки-
ми травами. Подобным
же образом строится
гнездо и городской ла-
сточки. Разница в том,
что у деревенской ла-
сточки гнездо — откры-
тое сверху, у город-
ской же — закрыто со
всех сторон, имеет
только небольшое от-
верстие сбоку для вле-
та и вылета.
Китайский стриж
саланган лепит кро-
шечное гнездо исклю-
чительно из своей слю-
ны, прикрепляя его пре-
имущественно к стен-
кам скалистых утесов.
Слюна эта выделяется
в виде густой, липкой
массы и застывает. Суп
из гнезд саланган считается в Китае лакомством, почему эти
гнезда и служат предметом торговли.
Гнезда закрытые, обладающие только отверстием для влета и
вылета, представляют наиболее совершенные формы. Если гнездо
городской ласточки, построенное из земли, не может помещаться
открыто, потому что дождь размочил бы его, то другие закрытые
гнезда, сделанные из более прочного материала, не нуждаются
в посторонней защите. Из наших птиц такие гнезда строят крапив-
ники, пеночки и водящиеся на юге СССР ремезы. Гнездо крапив-
ника (рис. 146) представляет шар, сплетенный из мха и стебельков,
с одним входным боковым отверстием.
Еще совершеннее гнездо ремеза (рис. 147). Оно имеет вид ко-
шелька, сплетенного из тонких растительных волокон, с входною
боковой трубкой, наподобие горлышка бутыли; на дне гнезда —
Рис. 146. Гнездо крапивника (Troglodytes
troglodytes)
220
Гнездовье птиц и уход за птенцами
довольно толстый слой из растительного пуха. Это гнездо прибли-
жается по своему устройству к еще более совершенным гнездам
тропических птиц — ткачиков и портних; последние делают гнезда
между листьями, сшивая их при помощи растительных волокон
и накладывая внутрь в
(Remiz penduli-
Рис. 147. Гнездо ремеза
ntis)
качестве гнездовой выстилки раститель-
ный пух (рис. 148).
Как видно уже бы-
ло из описаний
ных гнезд, их
ные формы в
приспособлены к тем
условиям, в которых
живут различные виды
птиц. Степень этой при-
способленности, однако,
у разных птиц различ-
на. Выражением при-
способленности являет-
ся и то, что обыкно-
венно гнезда бывают
удачно скрыты: в осо-
бенности это следует
сказать про гнезда мел-
’ких птиц и расположен-
ные невысоко. Найти
их не легко, так как
они спрятаны или по-
хожи на окружающую
обстановку. То впле-
тенные в гнездо пленки
бересты делают его не-
заметным на березе, то
оно маскируется вклю-
— теми же, самыми, ко-
отдель-
различ-
общем
лишаиниками
ченными в него мхом и
торые покрывают соседние ветви, и т. д.
Способность строить гнезда известного типа, несомненно, яв-
ляется врожденной и, следовательно, принадлежит к числу ин-
стинктов. Молодые птицы, еще ни разу не упражнявшиеся в гнездо-
строительстве, ко времени кладки яиц строят такие именно гнезда,
какие свойственны их виду. Это они делают, не учась у других птиц.
Обыкновенно птицы очень рано покидают гнезда, в которых они
вывелись, вырастают на свободе, а когда наступает время самим
приняться за постройку, энергично берутся за дело. Если у кана-
реек вынуть из клетки гнездо, сооруженное ими самими, и заменить
его войлочным, то молодые канарейки, выведшиеся в таком гнезде
и получившие потом такое же готовое гнездо, когда придет время
Различные типы гнезд
221
кладки яиц, всегда начинают сами строить гнезда из положенных
в клетку мха и волос. Нельзя совсем отрицать значение индивиду-
ального опыта птиц в смысле приобретения навыка строить гнезда,
но роль его, во всяком случае, второстепенная.
В связи с конкретными условиями местообитания гнезда птиц,
при общем соответствии определенному типу постройки, подлежат
Рис. 148. Гнездо длиннохвостой портнихи (Ortho-
domus longicauda)
видоизменениям в деталях. Наблюдается некоторая изменчивость
в строении гнезд в пределах данного вида птиц. Городская
ласточка представляет очень хороший пример такой приспособ-
ляемости. Исследователь гнезд и жизни этих птиц В. А. Ваг-
нер пишет: «Обыкновенно эти ласточки выбирают для своих по-
строек такие места, в которых гнездо могло бы прикасаться к сте-
нам двумя или тремя плоскостями. Всего чаще поэтому мы встре-
чаем их или на карнизах домов или в углах окон. Но иногда ока-
зывается невозможным найти такое место, и птицы вынуждены за-
нимать места, не соответствующие их обычным требованиям . . .
Получаются гнезда с резко измененной формой вследствие того,
что местом для их устройства избраны дугообразные углубления
222
Гнездовье птиц и уход за птенцами
под окнами ... В тех случаях, когда гнездо приходится строить
на одной плоскости, оно имеет форму правильного полушара» и
т. д. Сокол сапсан гнездится не только на деревьях или скалах,
но и на земле, причем или делает гнездо из хвороста, или же кла-
дет яйца прямо на землю. Славки делают гнезда обыкновенно из
Рис. 149. Кукующая кукушка (Cuculus canorus)
растительных стебельков, но иногда они строят их даже из сбро-
шенных змеиных шкурок.
Совершенно своеобразна форма заботы о потомстве у кукушек.
Существует около 40 видов кукушек, встречающихся в различных
преимущественно тропических странах. У нас встречается кукушка
обыкновенная (Cuculus canorus), распространенная почти по всей
территории СССР, преимущественно в лесной зоне. Кукушка
(рис. 149) не строит гнезда, а подкладывает яйца в гнезда других
Ридов, преимущественно мелких воробьиных птиц. Зарегистриро-
Забота о потомстве у кукушек
223
вано, что кукушка откладывает свои яйца в гнезда около 150 видов
птиц, но особенно сильно страдают от нее приблизительно 20 ви-
дов. Диапазон «паразитизма» кукушек очень широк — от таких
довольно крупных птиц, как галки и сороки, до маленького кра-
пивника. Яйца кукушки, сообразно ее росту, небольшие. Они
весят около 3 г, тогда как яйца дрозда, размеры которого прибли-
зительно соответствуют кукушке, весят 7—8 г.
Откладка кукушкой яиц в чужие гнезда связана с рядом весьма
своеобразных приспособлений. Маленькие птички инстинктивно
чувствуют в кукушке врага и всячески отгоняют ее от своего гнезда
с отложенными яйцами. Неред-
ко кукушка-самец отвлекает «хо- Гкд
зяев» гнезда, и в этот момент X
кукушка-самка проскальзывает к С /.
гнезду и откладывает свое яйцо.
При этом она выбрасывает из гнезда ' \б
одно «хозяйское яйцо», так что \/
общее число яиц не изменяется. \ х 1
Обычно яйцо кукушки по размеру /
и окраске напоминает яйца того J
вида птиц, которому оно «подки-
дывается». Повидимому, каждая Рис- Птенец кукушки вы-
самка кукушки откладывает яйца брасывает яйцо из гнезда
определенной расцветки, имити-
рующие таковые определенного вида птиц. «Владельцы» гнезда
обычно не замечают замены и высиживают яйцо кукушки наряду
со своими собственными. Кукушонок после выхода из яйца первые
часы ведет себя спокойно. Но вскоре у него проявляется своеобраз-
ный инстинкт, под влиянием которого он выбрасывает все яйца или
птенцов (если они уже вывелись) из гнезда. Это выбрасывание со-
вершается следующим образом. Особыми движениями кукушонок
захватывает яйцо (или птенца) на спину, придерживая его при
этом крыльями. Сам он в это время приближается к краю гнезда и
резким толчком выбрасывает свою ношу за его пределы (рис. 150).
В результате кукушонок остается в гнезде один. Его «приемные
родители» не замечают происшедшей перемены и усиленно выкарм-
ливают «убийцу» собственных птенцов. На этом примере видно, что
забота о потомстве у птиц далека от сознательной деятельности и
представляет в основе своей систему врожденных инстинктов, за-
крепленных естественным отбором.
Выкидывание кукушонком из гнезда всех птенцов имеет для
кукушки важное биологическое значение. Если бы все птенцы ос-
тавались на месте, то родители не были бы в состоянии прокормить
как собственное потомство, так и прожорливого «приемыша».
После окончания постройки гнезд птицы приступают к кладке
яиц. Число яиц у разных видов различно. В общем наблюдается,
224
Гнездовье птиц и уход за птенцами
что птицы сильные или хорошо защищенные откладывают меньше
яиц, нежели птицы слабые или гнездящиеся открыто. Так, круп-
ные хищники, как грифы, орлы, коршуны, кладут обыкновенно по
1—2 яйца; птицы эти, при большой силе и прекрасном вооружении
острым клювом и когтями, гнездятся высоко на скалах или верши-
нах деревьев и потому не подвержены нападениям со стороны вра-
гов. Мелкие же воробьиные птицы кладут большей частью по 4—б
яиц, а некоторые, например синицы, даже по 10—12 и более. Много
яиц кладут также куриные птицы, так как они хотя и обладают боль-
шим размером, но слабо вооружены, и гнезда их лежат на земле,
а потому доступны хищникам; в частности, глухарка кладет до 12
яиц, тетерка до 16, серая куропатка до 24, фазан до 12 яиц и т. д.;
яйца откладываются одно за другим в течение немногих дней. Уси-
ленное размножение птиц слабых и обладающих плохо защищен-
ными гнездами является одним из средств сохранения их вида про-
тив истребления со стороны хищников.
В окраске яиц также наблюдаются защитные приспособления.
У всех птиц, яйца которых кладутся в закрытые места, будут ли
это дупла деревьев, норки в земле или простые гнезда, яйца или
белые, или очень бледные, однообразного цвета. То же наблюдается
и в тех случаях, если птица, как козодой, при высиживании почти
не сходит с яиц, а сама окрашена под цвет окружающей обстановки,
или —если птица, сходя с яиц, прикрывает их чем-нибудь, что
видим у серой куропатки, у кряквы и др. Так как яичная скорлупа
состоит главным образом из углекислой извести — вещества белого
цвета,— то Уоллес полагает поэтому, что белый цвет птичьих
яиц — их основной, первоначальный цвет. Разнообразная окраска
и пятнистость яиц — явление позднейшее, развившееся у таких
птиц, которые гнездятся открыто и часто оставляют свое гнездо.
Значение окраски здесь покровительственное. Она делает яйца
мало заметными на фоне окружающей обстановки.
Хорошим примером покровительственной окраски могут слу-
жить яйца различных куликов, откладываемые в углублениях при-
брежной почвы и окрашенные под цвет глины или песка, смешан-
ных с темными камешками; яйца тетеревов вполне соответствуют
цвету лесной подстилки; яйца прибрежных птиц, откладываемые
на морском берегу, сливаются с окружающим их песком или галь-
кой (рис. 151); яйца полевого жаворонка также настолько мало
заметны, что на них легко наступить, не увидев их, и т. д. Большин-
ство яиц различных птиц испещрено бурыми и черными пятныш-
ками и черточками по тому или другому фону, так что, находясь
в тени гнезда, при сложной игре света и тенистых пятен от листьев
и т. д., какая бывает при весеннем солнце, они совсем незаметны.
Эта окраска яиц у птиц под цвет субстрата представляет собой один
из ярких примеров целесообразного приспособления, выработав-
шегося на основе естественного отбора. Окраска скорлупы яиц
Насиживание яиц
225
изменчива. Из поколения в поколение преимущественно сохраня-
лись те яйца, которые по своей окраске наиболее соответствовали
цвету субстрата. В результате длительного действия отбора и вы-
работалось это замечательное соответствие между окраской скор-
лупы и фоном, на котором лежат яйца.
Если у птицы взять 1 или 2 яйца, она обыкновенно вскоре по-
полняет недостаток. Если взять все яйца, птица обыкновенно кла-
Рис. 151. Яйца ржанки (Charadrius pluvialis), отложенные среди
камней
дет их вновь, для чего обычно строится новое гнездо. Повиди-
мому, вторая кладка действует на самку ослабляющим образом,
так как яиц в этом случае кладется обыкновенно меньше и они
имеют более бледную окраску.
Чтобы находящийся в яйце зародыш мог развиваться, необхо-
дима определенная температура. Вот потому-то отложенные в гнез-
дах яйца подвергаются насиживанию, т. е. согреванию теплотой
тела высиживающей птицы. В огромном большинстве случаев вы-
сиживает самка. Редко самец чередуется с самкой, например, у
страусов, гусей, лебедей. Еще того реже яйца высиживаются ис-
ключительно самцом, например, у плавунчиков. При высижива-
нии у птицы образуются так называемые «наседные пятна», т. е.
на брюхе выпадают или выщипываются самой птицей перья в об-
226
Гнездовье птиц и уход за птенцами
ласти разветвления брюшной кожной вены. Эти места, богатые
кровью, особенно способствуют согреванию яиц. Высиживающая
птица сидит на яйцах обыкновенно весьма прилежно и нередко
остается в гнезде, не обращая внимания на явную опасность.
Интересные приспособления, связанные с высиживанием яиц,
имеют место у тропической птицы носорога (рис. 152). Птицы
эти водятся в лесах Индии и Африки. Гнездо делается в дупле,
Рис. 152. Птица носорог (Bucorvus) у гнезда
высиживает яйца самка. Самец же на все время высиживания прочно
замуровывает дупло, оставляя лишь небольшое отверстие только
для просовывания клюва самки. Строительным материалом для
замуровывания служит глина, смешанная с собственным пометом
и особыми выделениями желудка. Самец регулярно приносит пищу
и кормит самку. В период насиживания самка носорога претерпе-
вает линьку, которая у других видов птиц обычно бывает после
окончания высиживания и выкормки птенцов. Лишь после выклева
птенцов самец освобождает самку из «темницы». Все это вместе взя-
тое следует рассматривать как приспособление защитного порядка*
«Замуровывание» защищает самку от проникновения в гнездо вся-
ких врагов, весьма многочисленных в условиях тропического леса.
Еще более поразительные приспособления к насиживанию на-
Насиживание яиц
227
Рис. 153. Насиживающий
королевский пингвин (Apte-
nodytes patachonica); яйцо
находится в<карманчике»—
складке кожи над лапами
блюдаются у некоторых антарктических пингвинов, например у
королевского пингвина. Эти птицы, живущие в вечных льдах у
южного полюса, не могут насиживать яйца на земле (как делают
это некоторые другие виды пингвинов) или строить гнездо, так как
вокруг них только лед. У этих птиц выработалось очень своеобраз-
ное приспособление к высиживанию яиц «на ходу» (рис. 153). У
самок и у самцов этого вида на брюшной стороне у основания ног
имеются особые складки кожи—карманы, куда и закладываются
яйца. Стоящая на льду птица держит яйца в кармане и таким об-
разом «насиживает» их. В то время как,скажем,самка держит яйцо,,
самец плавает в море и охотится за рыбой. Затем происходит смена.
Самец приближается вплотную к самке
и особым движением с помощью клюва
яйцо перекладывается в нему в «кар-
маны», самка же приступает к питанию
в море. Карман на брюхе королевского
пингвина несколько напоминает анало-
гичные образования у австралийских
сумчатых млекопитающих.
У некоторых арктических птиц
имеются интересные физиологические
приспособления к развитию птенца
в яйце. Наблюдения Л. О. Белополь-
ского показали, что весной, в момент
откладки яиц кайрами на «птичьих
базарах» (см. выше стр. 211), на кар-
низах скал еще частично сохраня-
ются снег и лед. Кайры не строят
гнезд, а непосредственно откладывают
яйца на субстрат. В результате по-
лучается крайне неравномерное нагревание яиц. Та сторона яйца,
которая обращена к телу насиживающей птицы и где располагается
зародыш, нагревается до температуры близкой к 40°; противопо-
ложная сторона яйца, которой оно соприкасается с субстратом,
обладает гораздо более низкой температурой, приближающейся
к 0°. Этими условиями развитие яиц кайр резко отличается от дру-
гих птиц, у огромного большинства которых имеет место более или
менее равномерный прогрев всего яйца. Интересно, что специаль-
ные опыты, проведенные Л. О. Белопольским, показали, что яйца
кайр потеряли способность развиваться при условии равномерного
прогрева. Если их поместить в инкубатор, то они погибают. Таким
образом, различие температур стало, повидимому, для кайр необ-
ходимым условием развития яйца.
Совершенно иными путями, чем у большинства птиц, дости-
гается повышение температуры, необходимое для развития заро-
дыша в яйце, у австралийских сорных кур. Эти птицы зарывают
228
Гнездовье птиц и уход за птенцами
свои яйца в кучи гниющих листьев, которые, разлагаясь, разви-
вают теплоту, заменяющую высиживание.
Продолжительность высиживания у мелких птиц вообще меньше,
чем у крупных. У самых мелких птиц оно длится 10—11 дней, у
большинства мелких пташек — 13—15 дней, у более крупных
птиц — 18—19 дней; у куриных, водяных и хищников средней
величины период насиживания—24 дня, у буревестников—35 дней,
у лебедя — от 35 до 40, у кондора —даже до 54 дней, у австра-
лийского эму — восемь недель. Вообще говоря, более длительным
является период насиживания у выводковых птиц, более ко-
ротким — у птенцовых (см. ниже).
Рис. 154. Однодневные птенцы: 1 — серой куропатки (выводко-
вая); 2 — полевого конька (птенцовая); 3 — орлана-белохвоста
(переходный тип)
Вышедшие из яиц птенцы часто также бывают окрашены под
цвет окружающей обстановки.
Выведшихся птенцов нужно воспитать, пока они не сделаются
способными к самостоятельной жизни. Этот период протекает не у
всех птиц одинаково. У одних—птенцы выходят из яиц уже
довольно окрепшими: их глаза раскрыты, тельце густо покрыто
пухом, а ножки так сильно развиты, что маленькая птичка,
только-что выйдя на свет, или очень скоро после вылупления, уже
начинает бегать; притом ее тело еще снабжено довольно значитель-
ным запасом питательного желтка, что и дает цыпленку возмож-
ность некоторое время просуществовать без корма. Птицы, выво-
дящие таких птенцов, называются выводковыми; к ним относятся
куры, тетерева, утки, гуси, чайки, чибисы, бекасы и др. У других
птиц, называемых птенцовыми, птенцы покидают яичную скор-
лупку в беспомощном состоянии —слепыми, слабыми и большей
частью с голым тельцем; питательный желток у этих птенцов
весь потреблен, так что с первого же часа после выхода на свет они
уже должны получать пищу от родителей. Примером птенцовых
Забота о потомстве
229
птиц являются воробьиные, голуби, дятлы, хищные птицы и
др. (рис. 154). Вся жизнь родителей во время гнездования уходит
на заботы о прокормлении потомства, на поддержание чистоты в
гнезде, на предохранение голых телец от холода и на защиту гнезда
от всяких опасностей. Сами родители, питаясь впроголодь, сильно
худеют и изнуряются. Пища птенцам выбирается более нежная;
так, зерноядные в большинстве случаев выкармливают своих птен-
цов сначала насекомыми, а затем уже зернами, размягченными в
зобу. У большинства птиц самка или самец, возвратившись с кор-
мом, дают его по очереди каждому птенчику.
Если внимательно присмотреться к росту и развитию птенцов
и заботливому уходу за ними родителей, то увидим, что у птенцов
проявляются прирожденные инстинкты, но многому они научаются
от родителей путем подражания.
У птенцовых птиц молодые, только что выклюнувшиеся,птенцы,
едва способные вытянуть шею, высоко поднимают голову и раскры-
вают рты, как только мать-самка садится на гнездо или наблюдатель
осторожно прикоснется к краю гнезда пальцем: очевидно, в этом
случае проявляется унаследованный от рождения инстинкт. Затем
даже совсем еще недоразвившиеся птенцы без всякого :научения по-
нимают звуки родителей и отвечают им. Охотники отлично знают,
что тревожные звуки, издаваемые какою-либо птицею, «понятны» и
другим, отличным от нее птицам. Это — природные звуки, и значе-
ние их схватывается инстинктивно.
Чему же «учатся» птенцы у своих родителей? Несомненно, обуче-
ние имеет место при приобретении навыка находить пищу и уменья
летать.
Очень важно, чтобы птица научилась находить пищу. Цыплята
наших домашних кур, например, уже вскоре после своего рожде-
ния начинают клевать все мелкие предметы, привлекающие их вни-
мание, но отличать среди них полезное для себя они научаются,
как то доказано точными исследованиями, лишь путем опыта. На-
личие индивидуального «опыта» особенно важно у птенцов хищ-
ных птиц. Они должны уметь схватить налету быструю птицу или
броситься с вышины вниз на крохотную мышку ит. п. Неудивительно
поэтому, что молодые птицы, еще не научившиеся правильно опре-
делять расстояние, в начале большею частью упускают свою до-
бычу. Но родители целыми неделями обучают их, и, в конце кон-
цов, птенцы овладевают этими навыками.
Этот процесс «обучения» следует представлять себе как образо-
вание у молодой птицы целой системы условных рефлексов, лежа-
щих в основе ее поведения в природе. Наряду с прирожденными
инстинктами, эти вновь возникающие условные связи играют ис-
ключительно большое значение во всей жизни птиц, в их приспо-
соблении к условиям внешней среды.
В обучении полету не нуждаются только некоторые из лучших
230
Гнездовье птиц и уход ва птеяцаяги
летунов, например, некоторые ласточки и стрижи. Большинство
же птиц проходит большую или меньшую «школу летания». Роди-
тели приучают птенцов к полету сначала на более короткие рас-
стояния, например, с дерева на землю и обратно, а затем на более
длинные. При этом родители иногда подталкивают птенцов, не-
уклонно следуют за ними, готовые при неудаче оказать помощь,
показывают пример летания и т. д.
К числу существенных родительских обязанностей птиц от-
носится также охрана от врагов как яиц, так и молодых птенцов.
Мелкие воробьиные птицы, в случае приближения какого-либо
врага к гнезду, летают вокруг и выражают свое беспокойство жа-
лобными криками. Крупные и сильные птицы вступают в откры-
тую борьбу с врагом; например, лебеди наносят ему удары силь-
ными крыльями. Мелкие пти1Хы иногда стаями отгоняют хищника,
подкравшегося к их гнезду, преследуя его и провожая громкими
криками. Применяется и такой прием защиты: птица «притворяется
раненой», не способной летать, благодаря чему отвлекает пресле-
дователя от гнезда, а потом сама улетает.
Интенсивность материнского инстинкта у птицы, выражаю-
щегося в насиживании яиц и уходе за птенцами, изменяется, сначала
повышаясь, а потом понижаясь. В первые дни насиживания птицы
несравненно легче бросают свои гнезда, чем в последующие. Выс-
шей степени привязанность к гнезду достигает ко времени вылуп-
ления птенцов из яиц. В это время поймать их на гнезде совсем
нетрудно. Косари нередко скашивают косой голову сидящей на
яйцах перепелки, которая не слезает с гнезда, несмотря на шум
медленно приближающегося косаря. По выходе детенышей из яйца
дело мало-помалу меняется: материнский инстинкт начинает идти
на убыль, повторяя в своем регрессивном развитии те же ступени,
только в обратном порядке.
Говоря о «материнском чувстве» или «долге» у птиц, нужно,
однако, иметь в виду, что эти выражения здесь — образные, не
имеющие того смысла, как в применении к человеку. Насколько
слепа эта потребность, не освещающаяся сознанием,—доказывается
например, такими фактами. Много раз наблюдали, что птицы, у
которых ломали и отнимали яйца, продолжали сидеть па разбитых
яйцах, садились на камни, на железные предметы и сидели на них
по всем «правилам» высиживания. Когда проходило определенное
время высиживания, потребность эта ослабевала, и птица прекра-
щала высиживание. Наблюдалось, что если у птицы заменить яйца,
уже насиженные ею, чужими свежими, то она будет сидеть на них,
но вывода птенцов может и не дождаться, а слетит с гнезда, когда
окончится свойственная ее виду продолжительность высиживания.
Очевидно, здесь только удовлетворяется инстинктивная потреб-
ность сидения при отсутствии элемента сознания. То же следует
сказать и о кормлении птенцов.
Забота о потомстве
231
Если разыскать гнездо с молодыми птенцами и положить его
в клетку, то родители будут прилетать и доставлять корм птенцам
через решетки клетки. Они будут делать это даже и в том случае,
если клетку повесить у человеческого жилья, — преодолеют ин-
стинкт самосохранения. Но когда птенцы подрастут, родители по-
степенно перестанут прилетать к ним, хотя бы последние неистово
кричали в клетке и им угрожала голодная смерть.
Родители перестают кормить узников к тому самому времени,
когда они перестали бы это делать на свободе. Почему так? Да по-
тому, очевидно, что к исполнению и этих «родительских обязанно-
стей» они побуждаются не сознательной любовью, а внутренним без-
отчетным, инстинктивным влечением, которое проявляется тоже
только в течение определенного времени. Если у птицы отнять
птенцов, она ищет, где бы удовлетворить свою потребность в их
кормлении. Если она услышит характерный писк чужих
птенцов, летит кормить их, не останавливаясь даже перед опас-
ностью быть наказанной со стороны их настоящих родителей. Но
когда пройдет положенное время, потребность кормить проходит,
и «благотворительность» птицы прекращается. Насколько безот-
четны эти родительские стремления, следует и из того, что сидящая
на гнезде птица не различает своих яиц от чужих, которые ей мо-
гут быть положены, не различает даже и птенцов. На этом основано
то, что кукушка, не вьющая своего гнезда, кладет яйца для выси-
живания в чужие гнезда, обыкновенно — в гнезда мелких насе-
комоядных птиц.
Забота о потомстве, гнездостроение и т. п. представляют собой
полезные для сохранения вида инстинкты. Так же, как и другие
целесообразные признаки в строении и поведении животных, они
выработались постепенно в процессе эволюции на основе естест-
венного отбора.
ЗИМОВЬЕ животных
Зима с ее морозами и снегом является главным «врагом» всего
живущего в умеренных и арктических странах. Она угрожает и хо-
лодом и голодом. У организмов выработался ряд приспособлений
для перенесения этих неблагоприятных условий.
Самым замечательным из этих приспособлений является зимняя
спячка. Часть млекопитающих и животные позвоночные и беспо-
звоночные с непостоянной температурой тела погружаются зи-
мой в глубокий сон или оцепенение, но не умирают, а при появле-
нии теплых лучей весеннего солнца снова пробуждаются к жизни.
Другие млекопитающие и остающиеся у нас зимовать птицы бодр-
ствуют зимою, ведут открытый образ жизни и разыскивают скуд-
ную пищу. От холода они защищены теплыми покровами — шер-
стью и перьями, предохраняющими тело от потери тепла; свойства
этих покровов, т. е. их густота и длина, находятся в зависимости
от климата и времени года. Некоторые млекопитающие обеспечи-
вают себя на зиму тем, что заранее заготовляют запасы пищи, ко-
торые и потребляют затем постепенно. Обратимся к рассмотрению
отдельных примеров зимовья животных, сосредоточив главное вни-
мание на интереснейшем из относящихся сюда явлений — зимней
спячке.
Из наших млекопитающих не впадают в спячку все копытные
и ластоногие. Среди же других отрядов имеется большее или мень-
шее количество представителей, подверженных зимней спячке.
Из хищных впадают в спячку медведь и барсук. Медведи перед
наступлением зимы приготовляют себе берлогу в естественных пе-
щерах или углублениях, которые вырывают сами; часто также в
темной чаще леса они устраивают себе настоящее жилище, вроде
шалаша, из хвороста и листьев; они наносят туда травы, мху и т. п.
и таким образом приготовляют себе мягкую подстилку. Впрочем,
некоторые медведи ложатся прямо на открытом месте, и уже пада-
ющий снег образует над ними покров. Медведица обнаруживает
большую «взыскательность» в отношении своей берлоги, так как
последняя служит ей в то же время колыбелью для детенышей.
Зимовье млекопитающих
233Г
Медведица залегает в берлогу ранее медведя, который бродит до
тех пор, пока находит пищу, и выходит оттуда иногда при отте-
пелях.
В лесах Дальнего Востока встречается черный медведь, гораздо
более мелкий, чем наш бурый медведь. Он устраивает обычно зим-
нюю берлогу в больших дуплах деревьев.
По сравнению с другими животными зимний сон медведя не от-
личается крепостью. Большим шумом, например, выстрелами из
ружей около берлоги, его можно разбудить. Такой разбуженный
медведь уже не залегает в спячку в ту же зиму и весьма опасен.
В поисках добычи он забирается на скотные дворы, режет коров,
нападает на людей и т. п.
В зоологических садах, где медведей регулярно кормят в тече-
ние всей зимы, они не удаляются в логовище даже во время силь-
ных морозов. Отсюда видно, что их заставляет засыпать не столько
холод сам по себе, сколько трудность добывания пищи зимой.
Существует поверие, будто медведь во время зимней спячки выса-
сывает из своей лапы жир, которым и питается. Нелепость этого
предположения вне всякого сомнения. Привычка сосать лапу свой-
ственна медведям во всякое время года, причем никакого «высасы-
вания» никогда не наблюдается.
Барсук вырывает себе на зиму нору в земле. Обычно в одной
и той же норе он зимует несколько лет подряд. Каждый год нора
все усложняется, и в конце концов образуются довольно сложные
подземные сооружения с несколькими выходами. Норы роются
обычно на холмах с южной их стороны в легком песчаном грунте/
Внутри норы барсук постоянно поддерживает чистоту. Спит
барсук зимой в своей норе в течение 5—7 месяцев. Временами он
на короткие сроки просыпается.
Не подвергаются спячке из хищников лисицы и волки, усиленно
рыскающие, как известно, зимой в поисках пищи; хорьки, выдры.
Среди белых медведей делают берлогу во льдах и впадают в
спячку, повидимому, лишь беременные самки. В берлоге же у них
родятся детеныши. Остальные особи белых медведей бодрствуют
круглый год.
Настоящей глубокой зимней спячке подвержены почти все ле-
тучие мыши. Как только становится холодно, они удаляются в;
укромные места, соединяясь при этом большими группами. Не-
редко летучие мыши какой-либо местности собираются в одно
убежище, которым пользуются из года в год. Таким убежищем слу-
жат для них чердаки зданий, дупла старых деревьев, а в гористых
местностях — пещеры, гроты и т. п. Зацепившись когтями задних
ног за какую-либо поверхность, свесившись головой вниз, обвер-
нувшись при этом крыльями, как плащом, они тесно прижимаются
друг к другу, сохраняя таким образом тепло, и погружаются в глу-
бокий сон, пока весеннее тепло снова не выманит из зимних убе--
234
Зимовье жизотяых
жищ насекомых, служащих им пищей. В очень холодные зимы часть
их погибает.
Из насекомоядных в глубокую зимнюю спячку погружается еж.
Уже с осени он при помощи своих игл натаскивает в густом кустар-
нике целую кучу из мха, листвы и т.п. Снаружи эта куча растрепана,
бесформенна, внутри же, где помещается животное на зимовку с
наступлением первых зимних холодов, тщательно и мягко выстлана.
Из других насекомоядных крот бодрствует всю зиму, следуя за
червями в глубь земли.
Землеройки, находясь зимой
в защищенных местах, точно
так же бодрствуют там и на-
ходят пищу в достаточном
количестве. Не подвергается
спячке и выхухоль.
Бодрствующих зимой
среди грызунов значитель-
но больше, чем среди дру-
гих отрядов; у них мы нахо-
дим также разнообразные
случаи собирания запасов
пищи.
Заяц ведет зимой откры-
тый образ жизни, питаясь
преимущественно корой де-
ревьев. Дикий кролик тоже
не собирает запасов и не
впадает в спячку, хотя живет
в норах под землей. Некото-
рые из мышей делают за-
Рис. 155. Хомяк (Cricetus cricetus)
пасы на зиму, другие — нет, но все бодрствуют в холодное время
и находят себе корм даже под снегом; для человека они стано-
вятся особенно тягостными именно зимой. Наши белки собирают
на зиму запасы пищи в дуплах, за отставшей корой и в тому
подобных укромных местах. При неблагоприятной зимней погоде
белки часто целыми днями не покидают своего убежища, но зим-
ней спячке не подвергаются.
Обильными запасами снабжает себя на зиму полевка-экономка,
получившая от этого и свое название. Она наносит в свою нору так
много кореньев, что вызывает удивление, как может набрать эту
массу такой маленький зверек в местности, бедной растительно-
стью. Все корни разгрызаны на куски.
В степях Заволжья и в Сибири обитает’другой грызун —пищуха,
или сеноставец, запасами которого пользуются местные жители.
Он заготовляет на зиму сено не в норке, слишком тесной для этой
цели, а у входа в норку. Зверьки складывают его в настоящие
Зимовье млекопитающих
235
стожки, имеющие в диаметре от 15 до 30 см и высоту 12—18 cjw, а для
защиты от дождя прикрывают его широкими листьями. Зимой
пищухи прокладывают ходы от нор к этим стожкам и могут обхо-
диться таким образом без зимней спячки. Когда начинает чув-
ствоваться недостаток подножного корма для скота, монголы гонят
свои стада в местности, где много пищу-х, и предоставляют овцам
отрывать стожки из-под снега. Во время бескормицы местные
жители сами собирают такое сено и кормят им своих лошадей.
Рис. 156. Суслики (Citellus citellus)
Хомяки (рис. 155), суслики (рис. 156), сурки, байбаки (рис. 157)
представляют пример грызунов, впадающих зимою в спячку и в то
же время собирающих запасы пищи. Естественно поэтому, что мы
находим у них норы с более сложным устройством. Так, жилище хо-
мяка состоит из двух камер: в одной он помещается сам, а другая
служит кладовою. В жилую камеру открывается несколько ходов.
Один из них ведет наружу и во время спячки обычно заваливается
землей. Другой тоже ведет наружу, но не доходит до поверхности
земли и кончается слепо. Через него хомяк по окончании спячки
выходит наружу, проделывая отверстие. Наконец, специальный
ход соединяет жилую камеру с помещением, где собраны запасы
пищи. Жилая камера выстилается сухой травой. Самка приступает
к заготовке зимнего продовольствия лишь после того, как отгонит
от себя последних детенышей: она хватает поэтому то, что найдет,
и помещает запасы кое-как. Наоборот, старые самцы занимаются
этим делом продолжительное время и обыкновенно сносят в нору
зерна более, чем в состоянии сами съесть в самую продолжитель-
236
Зимовье животных
ную зиму. Различные сорта хлеба у них тщательно рассортированы,
что является результатом, вероятно, разновременного поспевания
разных хлебов. Несмотря на богатые запасы, хомяки засыпают
зимой, по крайней мере на самые холодные месяцы, пробуждаясь
только на время и пользуясь при этом своими «сокровищами». Глав-
ную массу запасов они потребляют весной, уже после пробуждения,
пока держат вход в нору еще запертым. Количество зерна, заготов-
ляемого хомяками, очень значительно. Оно достигает иногда 6—7 кг.
Рис. 157. Байбак, или степной сурок (Marmota bob ас)
Подобно хомякам строят норы суслики (рис. 158). С наступле-
нием заморозков суслик совершенно заделывает наружный проход
в норку и затем начинает рыть другой ход, который, однако, про-
кладывает не насквозь, а только до поверхности: весной же, после
пробуждения от зимнего сна, животное проделывает отверстие, ко-
торое и служит ему для сообщения с внешним миром (рис. 159).
Горные, или альпийские, сурки (рис. 160) впадают зимой в глу-
бокий и непрерывный сон, причем они запасают себе сено. Летом
и зимой сурки живут в различных норах. На теплое время года
каждая пара вырывает небольшую норку, в которой находит кров
и защиту со своими детенышами. Осенью подготовляется зимнее
помещение — более низко на горе, чем летнее, глубже и большего
размера, в которое сурки залегают для зимовки целою семьей или
даже соединяются по нескольку семей. Оно тщательно выстилается
сеном. Впрочем,до сих пор считается недоказанным, едят ли сурки
при пробуждении сено, которое они сносят в свою нору, или оно
служит им исключительно подстилкой. Во всяком случае, весной
сурок выходит из норы очень исхудалым. А близкий родственник
Зимовье млекопитающих
237
сурка — байбак, живущий в степях целыми колониями, выхо-
дит из норы весной почти таким же жирным, каким залег
осенью в спячку. Очевидно, это животное питается зимой сеном,
обильный запас которого делается осенью. В связи с тем, что сурки
Рис. 158. Суслик (Citellus colombianus) в «жилой камере»
во время спячки
и другие грызуны заделывают на время вход в нору, температура
в «жилой камере» бывает значительно выше, чем температура окру-
жающего воздуха. Специальные измерения показали, что она обычно
не падает ниже 0°.
Уместно обратить
здесь внимание на то,
что у некоторых жи-
вотных, делающих за-
пасы пищи, имеются
специальные приспо-
собления , облегчающие
им переноску пищи в
нору; с каждой сто-
роны ротовой полости
у них образуются более
или менее обширные
Рис. 159. Схема строения гнезда суслика
складки, так называемые защечные мешки. Благодаря такому при-
способлению зверек может издалека переносить в нору значитель-
ное количество зерна и других пищевых веществ. Большее или
меньшее развитие защечных мешков наблюдается, вероятно, у всех
животных, собирающих зерно. У хомяка они настолько велики,
что он тащит в пору иногда сразу до 50 г зерна.
238
Зимовье животных
Змеи прячутся па зиму в глубокие норы, где впадают в оцепе-
нение. Некоторые из них при этом собираются группами. Так зи-
муют, между прочим, наши гадюки (рис. 161). Их иногда находят
штук по 15—20 вместе под корнями можжевельника, под старыми,
полуистлевшими пнями ольхи или березы и т. п., куда они соби-
раются при наступлении морозов и где остаются до весны. Лесо-
рубы при выкапывании старых пней нередко наталкиваются на
Рис. 160. Альпийский сурок (Marmota marmota)
подобные находки и убивают змей. Зимний сон гадюки не очень
крепок. Если ее потревожить, она поднимает голову, медленно
ползет и высовывает язык.
Подобно змеям цепенеют на зиму и ящерицы, укрываясь в при-
родные углубления — трещины и ямки, иногда довольно глубокие.
Земноводные .зимой также впадают в спячку. Лягушки обычно
зимуют в водоемах подо льдом,куда они забираются поздней осенью.
Относительно молодых лягушек, не достигших половозрел ости,
имеются данные о зимовке их на суше, где они зарываются в мох,
залезают под кору пней и т. п. Точно так же на суше зимуют широко
распространенные в наших пресноводных водоемах тритоны.
Некоторые рыбы, как указывается в следующей главе (стр. 257),
зимой также впадают в состояние оцепенения.К числу их относятся,
например, караси, карпы, сомы.
Зимовье насекомых
239
Из беспозвоночных животных зимуют насекомые — одни в виде
яиц, другие в виде личинок, третьи в виде куколок, четвертые в
виде совершенно развитых насекомых. Для зимовки насекомые
скрываются под камни, мох, в кучу старых листьев, под кору де-
ревьев, в щели зданий и т. п. Зимующие яйца и куколки в коконах
где-либо на растениях и т. п. на
часто остаются прикреш
открытом воздухе. Мно-
гие насекомые в совер-
шенно взрослом состоя-
нии также подвержены
зимнему оцепенению.
Чтобы убедиться в
этом, достаточно про-
извести следующий
опыт: в холодную осень
или бесснежную зиму
отправиться в лес, на-
брать горсть сухих ли-
стьев и затем в теп-
лой комнате просеять
через проволочное ре-
шето на лист бумаги;
тогда можно увидеть,
что это не простая тру-
ха из сухих листьев, а
вместе с нею и живой
мир насекомых, оцепе-
невших и примерзших
к листьям. Еще лучше
порцию лесной подстил-
ки поместить в банку,
закрытую марлей, по-
ставить в теплой ком-
нате. Из нее поползут
Рис. 161. Зимовка гадюк (Vibera perils)
различные насекомые. Равным образом много насекомых можно
видеть на предметах, уносимых водой во время весеннего ледо-
хода,— кусочках дерева, тростнике, семенах растений и т. п. Все
эти организмы перезимовали в развитом состоянии.
Из бабочек в стадии взрослых насекомых зимуют траурница,
лимонница. Из жуков — некоторые жужелицы, листоеды. Среди
двукрылых в стадии взрослых насекомых проводят зиму некото-
рые комары и, в частности, малярийный комар. При этом самцы ма-
лярийного комара погибают осенью, на зиму остаются лишь опло-
дотворенные самки. Пробудившись весной, они откладывают яйца.
В стадии куколки проводит зиму широко распространенная у
нас бабочка капустница. Ее куколки прикреплены к различным
240
Зимовье животных
предметам (заборы, стволы деревьев и т. п.) и мало защищены про-
тив зимних холодов. Куколки капустницы способны переносить
сильное охлаждение. Однако в суровые зимы большая часть их
погибает.
Самый обыкновенный из наших жуков — майский жук — три
года зимует в виде личинки, глубоко зарывающейся в землю. К
концу четвертого лета личинка, достигшая уже полного своего раз-
вития, опять зарывается в землю и превращается в жука еще до
наступления зимы; следовательно, четвертый раз майский жук зи-
мует в форме развитого насекомого, которое на следующее лето
после кладки яиц умирает. Подобно майским жукам зимуют хлеб-
ные жуки, с той разницей, что их развитие совершается не в три,
а в два года. В стадии личинки зимуют также гусеницы некоторых
бабочек, в том числе гусеницы одного из важных вредителей зерно-
вых культур — озимой совки. При этом они закапываются в почву
на глубину 15—20 см. Те гусеницы, которые почему-либо остаются
ближе к поверхности почвы, погибают.
Многие жуки имеют однолетний период развития: отложив
яички, они умирают, яички же перезимовывают, и из них затем
выводятся новые особи. То же наблюдается и у многих других на-
секомых, например, саранчи. Осенью она откладывает яйца, в числе
30—60, которые заключаются в оболочку, опускаются в круглые
ямочки в земле и закрываются землею; после этого саранча отмирает.
Насекомые, ведущие так называемый «общественный» образ
жизни, именно муравьи и пчелы, отличаются в отношении зимовки
тем, что муравьи не всегда подвергаются спячке, а пчелы—никогда.
Когда температура воздуха не очень низка, в глубине муравейника
жизнедеятельность для его населения оказывается возможной.
Только при температуре —2° в глубине гнезда, что может быть
лишь при больших морозах, муравьи впадают в оцепенение. Сред-
ством для поддержания их существования зимой являются тли —
мелкие насекомые, выделениями которых муравьи питаются. Тля
впадает в спячку и пробуждается при такой же температуре, как и
муравьи. Те из муравьев, которые не обладают инстинктом держать
у себя в муравейнике этих полезных им насекомых, переползают
в первые заморозки и поздней зимой при благоприятной погоде к
подножью деревьев, где находится тля. В это время тля дает очень
мало выделений, но и муравьям нужно для поддержания жизни
зимой тоже немного. Когда холод усиливается, муравьи собираются
в кучу тысячами особей. Такое скопление их, конечно, способствует
сохранению тепла.
Пчела принадлежит к немногочисленным насекомым, которые,
подобно птицам и млекопитающим, могут быть названы, с извест-
ным правом, «теплокровными» животными. При низкой внешней
температуре существование пчелы зависит от собственной внутрен-
ней теплоты, предохраняющей ее от окоченения. Под влиянием тем-
Зимовье насекомых
241
пературы ниже 5° пчела, изолированная от других пчел, быстро
погибает.
Специальное исследование Н. И. Калабухова (1933) показало,
что пчелы обладают очень малой холодоустойчивостью. Среди пчел,
взятых из ульев и помещенных при температуре от 0 до 1°, через
13—24 часа переживает 33%, при —2—3° лишь 16,5%, а при
—4—6°—5,8%. Такая малая холодоустойчивость пчелы зави-
сит, повидимому, от того, что у нее в теле почти не откладывается
запасных питательных веществ, которые имеются в большом коли-
честве у впадающих в спячку насекомых. Кроме того, у пчел при
низкой температуре нарушается всасывающая функция кишеч-
ника, так что даже если в кишечнике и имеются питательные веще-
ства, то они на холоду не усваиваются пчелой.
Таким образом, пчела, которая при низкой температуре не пи-
тается, погибает главным образом из-за истощения. Но в улье, где
сосредоточено свыше 20 тысяч особей, близко прижимающихся
друг к другу, охлаждение уменьшается от уменьшения охлаждаю-
щейся поверхности. Теплота, производимая пчелами, продолжаю-
щими зимой потреблять пищу, запасенную летом, теряется в не-
значительном количестве. В улье среди пчел бывает тепло даже
при больших морозах. Именно, при исследованиях оказалось,
что в центре собравшихся в кучу пчел температура доходит до 12—
15°, у периферии этой кучки достигает только 7—8°, между тем,
как в углах того же улья образовывался толстый слой льда и тем-
пература понижалась до 2—5° ниже нуля.
Главными условиями сбережения пчел зимой, как следует из
сказанного, является: большое число пчел в улье и достаточное ко-
личество пищи, которая, превращаясь в соки организма, дает ма-
териал для окисления.
Перезимовывают у пчел только матки и рабочие пчелы, а
трутни, выгнанные из ульев, умирают поздней осенью. У родст-
венных пчелам ос, шмелей и шершней перезимовывают только одни
матки, закапывающиеся в землю или забирающиеся под поросшие
мхом камни и защищенные трещины деревьев.
Несмотря на то, что у насекомых и других беспозвоночных
имеется ряд приспособлений для перенесения зимних неблагопри-
ятных условий, значительная часть их все же погибает. К сожале-
нию, имеется лишь очень небольшое количество исследований, ко-
торые бы освещали вопрос о проценте переживания зимующих на-
секомых. Так, у картофельного жука в окрестностях Чикаго (Се-
верная Америка), по данным Тоуэра, в условиях зимовки погибает
около 20% особей. Очень обстоятельные проводившиеся в течение
15 лет исследования другого американского зоолога Гейниса над
хлопковым долгоносиком показали, что у этого жука процент пе-
реживающих зиму индивидов очень невелик, причем он зависит
от минимальной температуры зимы. Так, при минимальной темпе-
242
Зимовье животных
ратуре —7° пережило зиму 6,14°/0 особей, при температуре —13° —
лишь 0,31%, а при температуре —17°—лишь 0,01 %. Имеются
также данные для некоторых клопов, у которых переживает зиму
лишь около 5?6. Известно также, что число гибнущих за зиму осо-
бей очень велико у капустницы, которая, как мы видели, зимует в
стадии куколки.
Все это заставляет думать, что смертность среди зимующих на-
секомых очень велика, причем она зависит от условий зимовки и,
в первую очередь, от минимальной температуры зимы.
Большинство пауков устраивает себе на зиму из шелковистых
паутинных нитей плотные логовища, которые помещаются под ко-
рою деревьев, в трещинах земли и т. д. В. А. Вагнер, между про-
чим, следующими опытами доказал выносливость наземных пауков
(крестовиков и др.), в отличие от водяных, по отношению к низкой
температуре. Он оставлял тех и других на зиму в пеотопляемом по-
мещении, в котором температура спускалась до —20°. Наземные
пауки оставались живыми и обнаруживали свою деятельность вся-
кий раз, как приносились в теплую комнату. Впрочем, здесь они
гибли, а оставшиеся на холоде до весны благополучно пережили
зиму и с наступлением теплых дней разбрелись. Подобные наблю-
дения дают основание полагать, что все пауки, подвергающиеся
спячке при низких температурах, не перезимовывают в тепле. Во-
дяные же пауки, оставленные на холоду, все погибали. Естественно,
что у них не выработалась способность к перенесению низких тем-
ператур, так как в воде, где они зимуют, такой температуры не
бывает.
Другая обстановка зимовки у пауков, живущих в норах, каковы
тарантулы. На зиму тарантул копает нору, превосходящую по
своей глубине летние норы. Он докапывается до слоя земли, не про-
мерзающего даже в суровые зимы. Здесь он устраивает небольшое
расширение, в котором делает из шелковистых нитей паутины плот-
ный мешок и устраивается, как в гнездышке. Раз тарантулу не при-
ходится сталкиваться с морозами, то понятно, что он и не приспо-
соблен к перенесению их. Будучи извлечен зимой из норы на мо-
роз, он быстро погибает.
Наземные моллюски (улитки, слизни) зимою также впадают
в спячку и могут выносить^овольно низкие температуры. При этом
улитки выделяют особую известковую крышечку, закрывающую
устье раковины.
Как показали опыты, моллюски, живущие в воде, являются
менее стойкими к низким температурам, чем наземные.
После знакомства с отдельными примерами зимней спячки на
разнообразных представителях животного царства воспроизведем
общую картину этого явления и обратим внимание на те внутренние
физиологические процессы, которыми оно сопровождается.
Прежде всего нужно отметить тесную зависимость этого явле-
Зависимость спячки от внешних условий
243
ния от условий, в которых животные находятся. Животные, под^-
вергающиеся в наших широтах оцепенению на свободе, могут бодр-
ствовать круглый год, если их содержать в иных условиях, чем
в открытой природе. В зоологических садах можно видеть много
подобных примеров, из которых пример медведя нам уже из-
вестен.
Сравнивая различные местности, мы видим, что чем далее на
юг, чем короче и мягче зимы, тем больше видов, которые не впадают
в настоящую спячку, а просто укрываются где-нибудь на самое
холодное время.
Говоря о влиянии севера и юга на количество подвергающихся
спячке животных, нужно, однако, сделать оговорку, что на край-
нем севере, в арктическом поясе, процент впадающих зимой в оце-
пенение млекопитающих животных не наибольший, как может по-
казаться с первого взгляда и как думали ранее, а наоборот, таковые
здесь почти отсутствуют. Затишье, наблюдаемое в природе с наступ-
лением многомесячной полярной ночи, объясняли прежде именно
тем, будто в спячку впадает все оседлое население полярной страны.
Это оказалось неверным.
С одной стороны, полярные млекопитающие относятся, за ис-
ключением белого медведя, к таким семействам, представители
которых не подвержены зимней спячке и в умеренном климате.
С другой — весьма вероятно и то, что подобное летаргическое
состояние довело бы большинство арктических зверей до смерти,
так как едва ли было бы им достаточно теплоты собственного тела,
даже при самом экономном расходовании ее, для борьбы с низкой
температурой окружающего воздуха в течение продолжительного
времени, тем более, что земля, в которой роются норы, тоже мер-
злая. В связи с этим арктические млекопитающие проводят боль-
шую часть полярной ночи в бодрствовании. Даже белый мед-
ведь — сородич бурого, спящего зимой в берлоге, — и тот бодр-
ствует большую часть арктической зимы. Полярные волк и лисица,
лемминг (грызун) и заяц деятельны в течение всей зимы; мелкие
животные, как лемминги, проделывают себе ходы под снегом.
Северные олени зимою в бочьние морозы для защиты от холода
держатся более тесными стадами, чем ранее, плотно прижимаясь
друг к другу.
Животные же с непостоянной температурой тела в арктиче-
ском поясе подвергаются оцепенению на более продолжительное
время, чем в умеренном.
Время наступления и продолжительность спячкиt зависят как
от продолжительности зимы, так и от особенностей каждою (Вида
животных. В средней полосе СССР животные начинают укрываться
на зиму около половины сентября, а пробуждаются с начала
или половины апреля. В отношении некоторых грызунов (сурки,
суслики) установлено, что начало спячки зависит не только от тем-
244
Зимовье животных
пературы, но и от качества пищи. Так, если лето сухое, влага от-
сутствует, пища тоже является сухой, — сурки впадают в спячку
еще в июле. Получается явление так называемой «летней спячки»,
которая переходит в зимнюю. Если влаги достаточно, то сурки засы-
пают лишь в сентябре. Из приведенных описаний спячки отдельных
животных уже видно, что сравнительно мало времени пребывает
в спячке медведь, залегающий в зимнее логово не ранее половины
октября и пробуждающийся около половины марта, а при оттепе-
лях даже в январе от времени до времени оставляющий свою бер-
логу.
Температура, при которой наступает оцепенение, неодинакова
для различных видов, да и для одного и того же вида иногда раз-
ными наблюдателями указывалась различная. Так, температура
оцепенения сурков указывалась между +5° и +8° Ц, темпе-
ратура оцепенения летучих мышей и ежей — между +5° и +7°,
оцепенение сусликов наблюдалось при 0°, при +10° и +13°, сони —
между 0° и +18°. В Московском зоологическом саду барсуки не
принимали пищи и лежали неподвижно в устроенной в клетке норе
только при температуре ниже —20°. Такое понижение температуры
наступления спячки, очевидно, находится в зависимости от полу-
чения барсуком пищи. Время залегания в спячку зависит также
от пола и возраста. В отношении сурков, например, известно, что
раньше других залегают взрослые самцы, после них — самки и
позже всех — молодые сурки.
Зимняя спячка наступает не сразу. Сначала животное начинает
меньше есть и двигаться, но больше спать. Затем принятие пищи
и движения совершенно прекращаются. Животное большей частью
свертывается клубком (еж, суслик, сурок и др.), что уменьшает
поверхность теплоизлучения, и кажется как бы мертвым. В сере-
дине периода спячки сон бывает более глубок, чем в начале и конце.
Для большинства он непрерывен, но некоторые животные во время
спячки просыпаются и снова засыпают.
Пробуждение от спячки также совершается не вдруг, а более
или менее постепенно.
Сурки, суслики и хомяки в течение первых 10—12 часов после
прекращения спячки бывают вялы и неподвижны, производя впе-
чатление больных. Это зависит от того, что нормальные отправле-
ния организма восстанавливаются не сразу.
Состояние зимней спячки теплокровных животных характери-
зуется тем, что все физиологические отправления их в это время
сильно понижены. Кровообращение совершается значительно мед-
леннее, чем в активном состоянии. Представление о том, насколько
замедляется кровообращение, можно получить путем изучения
числа сердечных сокращений. В следующей таблице (которую мы
заимствуем из. книги Н. И. Калабухова) приведены соответствую-
щие цифры.
Физиологическое состояние во время спячки
245
Число сердцебиений
в спячке и в активном
у млекопитающих
состоянии
в ИЙ Состояние
Су рок f Активное . . ( Спячка. . .
Суслик ( Активное . . ( Спячка . . .
Хомяк ( Активное . . [ Спячка . . .
Летучая мышь f Активное . . 1 Спячка . . .
Температура тела Число сердцебиений в минуту
33—37° 88—140
3—11° 3— 15
35—37,8° 100—350
5— 9Э 5— 19
36—38° 150—200
5— 7° 12— 15
37,3° 420
4—1° 16
Иногда сердцебиение даже прекращается на несколько минут.
Интересное наблюдение сделано Дюбуа над сурком: сердце
этого животного, убитого во время зимней спячки, продол-
жает биться в течение Зх/2 часов и долее после смерти, тогда как у
сурка, убитого при нормальных условиях жизни, сердце перестает
биться очень скоро. Следовательно, в состоянии зимнего оцепене-
ния теплокровное животное делается в этом отношении похожим
на «холоднокровное»; например, у лягушки, черепахи и других
сердце, вынутое из организма, продолжает долгое время сокра-
щаться.
Вследствие редкости и слабости сердечных сокращений движе-
ние крови совершается медленно, что непосредственно можно на-
блюдать под микроскопом в капиллярах летательной перепонки
летучей мыши. Пульсирование заметно только в артериях шеи.
Давление крови в сосудах слабое. При перерезке их кровь выте-
кает медленно.
Количество крови во время спячки уменьшается, причем она
сосредоточивается по преимуществу во внутренних органах и боль-
ших грудных сосудах. Поверхностные части тела, а равно мозг
и его оболочки в это время весьма бедны кровью, несколько ме-
няется даже и состав ее. Количество белых кровяных телец сильно
уменьшается не только абсолютно, но и относительно.
Точно так же в состоянии зимнего оцепенения в значительной
степени понижается потребность в дыхании и газовый обмен в ор-
ганизме. Так, спящие суслики дышат 1—4 раза в минуту, между
тем как бодрствующие — больше 100 раз; спящий сурок дышит
7—8 раз, еж—4—5, летучая мышь—5—6, соня—9—10 раз.
У сурка дыхание иногда совершенно прекращается на несколько
минут. Вдыхание вообще происходит медленнее выдыхания; у сус-
лика первое в четыре раза продолжительнее второго. Раздражения
погруженных в зимнюю спячку животных, даже незначительные —
легкий шум, слабое прикосновение, подергивание за хвост, разре-
246
Зимовье животных
жение воздуха и т. п.»— вызывают глубокий вздох и учащение ды-
хания; равным образом раздражения увеличивают и число сердеч-
ных сокращений. Количество потребляемого во время спячки кис-
лорода падает до или 1/4Q того, что потребляется животным во
время бодрствования. Отношение между количеством выделяемой
углекислоты и потребляемого кислорода также становится иным,
именно выделение углекислоты падает более, чем уменьшается
потребление кислорода. Так, у спящего сурка потребление кисло-
рода уменьшается в 41 раз по сравнению с бодрствующим, а выде-
ление углекислоты — в 75 раз. Так как во время зимней спячки
питание совсем или почти совсем прекращается, то понижение ды-
хания и связанное с этим уменьшение окисления является необхо-
димым для сохранения жизни.
Ослабление процессов кровообращения и дыхания ведет к силь-
ному понижению температуры тела. Оно тем значительнее, чем хо-
лоднее окружающий воздух. Замечено, например, что соня при
температуре воздуха 3° имела температуру З1^0, при 4° воздуха—
10°, при 15° воздуха—14°. Подобная же изменчивость температуры
тела в зависимости от температуры окружающего воздуха конста-
тирована и у других впадающих в зимнюю спячку животных (см.
выше таблицу на стр. 245). Таким образом спящие теплокровные
животные до известной степени делаются подобными животным,
температура тела-которых и в бодрственном состоянии изменяется
в зависимости от температуры окружающей среды, обыкновенно
лишь немного превышая ее.
Так как во время спячки животные в большинстве случаев со-
всем не принимают пищи или принимают, но очень мало и через
долгие промежутки времени, то, соответственно этому, у них за-
медляется и процесс пищеварения. У животных, подвергающихся
непрерывной спячке, он почти- совсем прекращается, хотя выделе-
ние соков пищеварительными железами не приостанавливается
окончательно. Пробовали доставать у этих животных пищевари-
тельные соки из разных мест пищеварительного канала и действо-
вать ими на пищевые вещества. В результате или переваривания
совсем не происходило или, если и обнаруживалось действие фер-
ментов, то весьма слабое.
Наблюдения над общим обменом веществ в организме во время
спячки привели к следующим выводам.
Животные, недавно впавшие в оцепенение, теряют в весе в 5—7
раз меньше, чем при голодании в бодрственном состоянии в течение
одинаковых промежутков времени. При глубокой спячке те же жи-
вотные теряют уже в 25 раз меньше, чем при норме. Незадолго до
начала пробуждения, когда сон менее глубок, потери в весе воз-
растают в несколько раз, но все-таки они далеко не достигают тех
размеров, какие имеют место в бодрственном состоянии. Организм
спящего сурка за 160 дней зимовки приблизительно теряет столько,
Физиологическое состояние во время спячки
247
сколько голодающий, но бодрствующий сурок в течение 12 дней.
Потеря веса во время спячки может быть еще большей, если живот-
ное прикасается к какому-либо хорошему проводнику тепла и если
у него острижена шерсть. И то и другое вызывает усиленную потерю
теплоты, а через это — более усиленное окисление, что пополняет
эту потерю.
Нужно иметь в виду, что во время спячки происходит не только
количественное изменение обмена веществ в сторону его пониже-
ния, но и глубокие качественные изменения. В частности, наряду
с процессами окисления, приводящими к образованию таких про-
дуктов, как углекислый газ и вода, во время спячки имеют место
процессы неполного окисления. К числу этих процессов относится
частичное окисление жира, приводящее к образованию гликогена
(животного крахмала), углекислого газа и воды. Благодаря этому
во время спячки происходит уменьшение количества жира/а гли-
коген (животный крахмал) не только не уменьшается, но даже уве-
личивается в количестве.
Из сказанного следует, что во время спячки животные сильно
худеют. Так, барсук уходит в свою нору на зимовку хорошо отъев-
шимся животным, а весной появляется совершенно исхудавшим.
Главным материалом, запасаемым на зиму и употребляемым во время
спячки, является жир. У всех млекопитающих, впадающих зимою
в спячку, к моменту спячки накопляется большое количество жира.
Так, например, у сусликов перед залеганием в спячку вес увели-
чивается почти вдвое, причем это увеличение идет за счет накоп-
ления жира. За время спячки почти весь жир расходуется. Но
кроме жировой уменьшаются в весе и другие ткани и целые органы.
Кишечный канал во время спячки суживается и укорачивается.
Объем легких и печени также уменьшается.
Еще в 1857 г. Валентин сравнил вес различных органов и тка-
ней сурка до и после спячки. Им получены были следующие цифры
уменьшения первоначального веса:
Жировая ткань теряет...................99,3 %
«Железа спячки» 1......................68,7 %
Печень.................................58,7 %
Диафрагма..............................45,6 %
Легкие.................................44,5 %
Кожа с подкожной клетчаткой ...........35,3 %
Мышцы................................ 30,3 %
Сердце................................. 27,48%
Скелет ................................11,69%
Иногда животные, впадающие в спячку, не успевают израсхо-
довать за зиму весь жир, и значительное количество его остается
до весны.
1 «Железой спячки» называется особое скопление жировой ткани в
грудной области.
248
Зимовье животных
Обнаружены также изменения в микроскопическом строении
некоторых тканей и органов, хотя этот вопрос исследован еще мало.
Так, во время спячки изменяется форма некоторых клеток в сли-
зистой оболочке желудка, и железы его укорачиваются — неко-
торые на х/3 по сравнению с железами бодрствующих животных.
Интересно отметить, что в половых железах у некоторых млеко-
питающих во время спячки наблюдается подготовка к периоду
размножения. Так, у самцов сурков перед спячкой половые железы
уменьшены в объеме, и образования в них сперматозоидов не про-
исходит. За время спячки объем семенников увеличивается, и к
моменту пробуждения в них происходит формирование спермато-
зоидов. Пробудившиеся от спячки сурки благодаря этому сразу
оказываются способными к размножению. Однако у ежей во время
спячки, в противоположность сурку, образования семенных клеток
не происходит.
Раз понижены процессы обмена веществ, то неизбежным резуль-
татом является угнетение мышечной и нервной систем. Степень
угнетения стоит в связи, с одной стороны, с продолжительностью
оцепенения, а с другой — с индивидуальными особенностями жи-
вотного. Еж в период глубокой спячки совершенно не чувствителен
к уколам и другим сильным раздражениям. Сурка в это время можно
катать, как шар; если даже бросить его с высоты 1—Р/г м, и тогда
он не пробуждается. Более сильные раздражения сурков, напри-
мер, нанесение порезов и т. п., сначала тоже не вызывают пробуж-
дения, но спустя известное время после операции животное про-
сыпается.
Сравнительно более чувствительными во время зимней спячки
остаются, например, соня и летучая мышь; первая издает крик при
легком раздражении ее, а у летучих мышей легкие уколы вызы-
вают более или менее сильное рефлекторное движение. Функцио-
нирование органов чувств во время спячки весьма слабо выражено.
У сурков наблюдалось расширение зрачков. Лучи света, направ-
ленные на их глаза, не вызывают пробуждения. Шум и даже писто-
летный или ружейный выстрел не оказывают на них никакого дей-
ствия. Пахучие вещества, приложенные к носу во время спячки,
вызывают пробуждение только после долгого действия.
Ближайшей причиной спячки является, как мы уже знаем,
холод. В дополнение к сказанному по этому поводу интересно при-
вести еще наблюдение проф. Г. В. Никольского над погружением
в спячку тушканчиков в Средней Азии. В степях Туркестана раннею
весной, когда по ночам еще были морозы, днем и даже вечером
было достаточно тепло, так что тушканчики повыскакивали из
своих нор. С вечера проф. Никольский ставил на них капканы, а
рано утром почти в каждом из них находил тушканчика. Обыкно-
венно они попадались за заднюю ногу или хвост, так что не полу-
чали существенных повреждений. Между тем, на первый раз туш-
Причины спячки
249
канчики были приняты за мертвых, так как они были неподвижны,
тело их было холодно и даже слегка окоченело. Проф. Никольский
собрался уже было снимать с одного из них кожу, но, когда внес
его в юрту и отогрел у костра, тушканчик ожил. Таким образом,
холод ночи заставил тушканчика заснуть, несмотря на неудобство
положения и боль, причиненную капканом. Помещение сурков или
сусликов летом в ящик, где температура —10° Ц, вызывает через
24 часа быстрое понижение температуры тела до +5° и затем, че-
рез некоторое время, засыпание.
Однако холод нельзя считать единственной причиной спячки.
На примерах медведя и барсука мы уже видели, какое влияние в
этом случае оказывает пища. Медведь не залегает в зо-
ологических садах в спячку, если его регулярно кормить, даже и
при больших морозах, а барсук при таких условиях выдерживает
температуру до —20°, когда только и оцепеневает. Значит, нельзя
сказать, что эти животные, погружающиеся на свободе в спячку
около половины октября, не могли выносить температуру этого
времени года, стоящую на несколько градусов выше нуля, и засы-
пали исключительно под ее непосредственным действием. Очевидно,
здесь играют роль также уменьшение добычи пищи и зимняя голо-
довка, заставляющие их заблаговременно отправиться на «зимние
квартиры».^
У других упоминавшихся теплокровных животных влияние
пищи на спячку не так заметно, но во всяком случае и здесь нельзя
отрицать его. Нельзя сказать, чтобы, например, хомяки, байбаки,
суслики да и те же тушканчики и т. д., оцепеневающие при нор-
мальных условиях до наступления морозов, не могли, при их по-
кровах, выносить такую температуру. Вероятнее всего, и здесь
сентябрьская температура действует на них, не только охлаждая,
но в то же время и являясь предвестником предстоящего неблаго-
приятного времени года, когда нельзя будет прокормиться. Зим-
няя спячка выработалась как приспособление не только к холод-
ному, но в то же время и к голодному времени года. Если
бы все животные в течение зимы продолжали вести обычную жизнь,
пребывая в бодрственном состоянии, то, как уже упоминалось,
для них нехватало бы пищи и большинство неминуемо должно
было бы погибнуть. Кроме того, как отмечено выше (стр. 243), на
наступление спячки влияет также качество пищи. Сухой корм
у сусликов вызывает спячку даже в июле.
Существенной причиной понижения всех жизненных отправле-
ний организма является, повидимому, накопление в нем углекис-
лоты. Мы уже видели, что уменьшение количества выделяемого
углекислого газа у погруженного в спячку животного, по сравне-
нию с бодрствующим, значительнее уменьшения потребляемого им
кислорода. Выделяясь в меньшем количестве, чем потребляется
кислорода, углекислый газ насыщает кровь. По всей вероятности,
250
Зимовье животных
он как бы наркотизирует организм. Это подтверждается таким опы-
том Дюбуа. Заставляя сурка дышать воздухорл с излишним содер-
жанием углекислоты, он вызывал у этого животного сон со всеми
признаками зимней спячки, т. е. с падением температуры, замедле-
нием дыхания и т. п.
Большой интерес представляет изучение тех внутренних изме-
нений, которые происходят в теле животных с непостоянной темпе-
ратурой тела в период зимнего оцепенения. Выше мы уже видели,
насколько глубокими являются те различия в процессе обмена ве-
ществ, которые отличают млекопитающих животных, находящихся
в состоянии зимней спячки, от бодрствующих. Между тем, многие
так называемые «хладнокровные» позвоночные и беспозвоночные
зимою впадают в еще более глубокое оцепенение, сопровождаю-
щееся полным прекращением всех видимых жизненных отправле-
ний и значительным понижением температуры тела. Происходит
ли при этом замерзание соков тела животного? Как долго они мо-
гут находиться в подобном сост§янии? Прекращаются ли при этом
процессы обмена веществ? Все эти вопросы представляют глубокий
теоретический и практический интерес. Однако далеко не все в
этой области явлений уже достаточно выяснено. Остановимся кратко
на изложении некоторых главнейших результатов, полученных ра-
ботами различных исследователей.
Начало исследований в области изучения влияния низких тем-
ператур на насекомых и других животных связано с именем извест-
ного русского ученого проф. П. И. Бахметьева, впервые разрабо-
тавшего теорию так называемого анабиоза при замерзании. Под
анабиозом понимают особое состояние «скрытой жизни», при кото-
ром имеет место более или менее полное прекращение жизненных
процессов, производящее впечатление смерти. Однако при восста-
новлении благоприятных внешних условий (например, повышение
температуры) животные, находившиеся в состоянии анабиоза, вновь
«оживают».
П. И. Бахметьев при помощи особых приспособлений (термо-
электрического термометра) изучал изменение температуры тела
насекомых (бабочек и др.) при замерзании. Он показал, что соки
насекомого, замерзающие вне его тела при —1° или —1,5°, внутри
тела могут переохлаждаться, не замерзая, до —10° и даже до —20°,
подобно тому как переохлаждается вода, покрытая слоем масла,
до —20°, не замерзая при этом. Подобная способность переохлаж-
дения в теле насекомого связана с тем, что здесь имеются много-
численные капиллярные пространства между и внутри органов.
Жидкости же в капиллярных пространствах способны достигать
значительной степени переохлаждения. В состоянии переохлажде-
ния насекомое окоченевает, перестает двигаться, но, будучи вынуто
из прибора, в котором производится охлаждение, легко оживает.
При дальнейшем охлаждении наступает замерзание соков насеко-
Анабиоз насекомых при замерзании
251
мого. При этом происходит температурный скачок, и температура
повышается до —1° или —1,5°. Эю повышение температуры объяс-
няется выделением теплоты, которое всегда имеет место при замер-
зании переохлажденных жидкостей. При дальнейшем действии хо-
лода температура замерзшего уже насекомого снова падает. И если
падение Достигает той границы, до которой повышалась темпера-
тура соков насекомого при переохлаждении и при которой нача-
лось замерзание (критическая температура), т. е. до —10°, —20°,
то тогда уже насекомое погибает. При этом Бахметьев считал, что
после температурного скачка и последующего охлаждения при-
близительно до —4,5° все соки тела насекомого превращаются в
лед. Вот это состояние замерзшего, но вместе с тем не погибшего
окончательно животного Бахметьев и называл состоянием анабиоза.
Проф. Бахметьев показал, что чем гуще соки насекомого, тем тем-
пература замерзания их ниже, тем большая степень переохлажде-
ния может быть достигнута.
Однако последующие исследования заставили многое дополнить
и изменить в первоначальных взглядах Бахметьева на анабиоз у
насекомых. Было показано (главным образом работами Пэйн,
1926—1929), что большая или меньшая выносливость насекомых
к холоду обусловливается содержанием воды в теле, которое умень-
шается к холодному времени года. Так, у одного и того же вида
личинок дровосеков температура переохлаждения в январе равня-
лась —24°, температура замерзания соков тела —14°, в августе
же соответственно —2° и —1,2°. Таким образом один и тот же
вид, но в разное время года обнаруживает весьма различную хо-
лодоустойчивость. При этом содержание воды в теле личинки в
августе оказывалось значительно выше, чем в январе, с чем, веро-
ятно, и связано столь значительное различие в отношении темпе-
ратуры замерзания.
Исследования Н. Л. Сахарова (1928) ставят под сомнение утвер-
ждение Бахметьева о том, что после температурного скачка про-
исходит вскоре полное замерзание соков в теле насекомого. На-
против, образование льда после температурного скачка происхо-
дит постепенно. Например, в опыте с зимующими гусеницами зла-
тогузки образование льда началось лишь при —11°, а при —17,35°
лишь 15,22% содержавшейся в теле воды перешло в лед. Другие
опыты Сахарова показали, подобно исследованиям Пэйн, зависи-
мость точки замерзания от количества воды, а также жира. Чем
меньше воды, тем ниже температура переохлаждения. Повидимому,
окончательная гибель насекомого происходит задолго до того, как
все соки его превратятся в лед. Вероятно, в природных условиях
большинство зимующих насекомых находится в состоянии пере-
охлаждения соков тела.
В состоянии переохлаждения у насекомых физиологические про-
цессы полностью не приостанавливаются, хотя и протекают очень
252
Зимовье животных
медленно. Того состояния «скрытой жизни» («анабиоза» в узком
смысле слова), во время которого полностью прекращаются все
физиологические процессы, у насекомых, повидимому, не суще-
ствует.
В этом отношении показательны, например, исследования
И. В. Кожанчикова (1935) над гусеницами кукурузного и лугового
мотыльков. Этот ученый наблюдал у них в состоянии переохлаж-
дения (при —6°, —12°) поглощение кислорода, т. е. процессы ды-
хания, которые хотя и были очень замедлены, но не прекратились
окончательно.
В связи с этим интересно отметить, что у зимующих насекомых
с осени, подобно млекопитающим, накопляется в теле значитель-
ное количество жира. Этот жир, повидимому, и расходуется посте-
пенно во время зимнего оцепенения.
За последнее время, однако, появились исследования, показы-
вающие, что некоторые насекомые способны переносить настолько
низкие температуры, при которых трудно допустить сохранение
в тканях организма воды в жидком состоянии. Например, Л. К. Ло-
зина-Лозинский у гусениц кукурузного мотылька проводил охла-
ждение до —79° (в твердой углекислоте) продолжительностью до
24 часов. После оттаивания часть гусениц обнаруживала двигатель-
ные реакции и реагировала на раздражение. При таких низких
температурах гусеницы становятся совершенно твердыми и раз-
биваются на куски при падении. Было высказано предположение
(Алпатов), что и при этих условиях внутри клеток сохраняются
небольшие количества незамерзшей воды. Вопрос этот нельзя счи-
тать еще окончательно решенным. Во всяком случае такая холо-
достойкость, которая наблюдается у гусениц кукурузного мотылька,
является среди насекомых довольно редким исключением.
Влияние низких температур на «хладнокровных» позвоночных
за последние годы также подверглось довольно обстоятельному изу-
чению (главное значение имеют исследования Бородина (1932) в
Америке и Калабухова и Никольского (1934, 1935) в* СССР).
Прежде существовало убеждение, что рыбы, лягушки, змеи и яще-
рицы могут, замерзая, превращаться в куски льда, а затем, оттаи-
вая, вновь оживать. Указанные весьма тщательные исследования
не подтверждают этих данных. У рыб и лягушек сколько-нибудь
значительного переохлаждения соков тела и температурного скачка
вовсе не наблюдается. Как только замерзают соки тела, животное
безвозвратно погибает. У ящериц и черепах наблюдается неболь-
шое переохлаждение и температурный скачок, после которого, при
продолжающемся охлаждении, начинается образование льда и бы-
страя гибель животного.
«Таким образом,— пишет Калабухов (1935),— следовало придти
к выводу о том, что длительное переживание организма при темпе-
ратуре ниже 0° возможно лишь в состоянии переохлаждениям
Анабиоз у млекопитающих при замерзании
253
Бахметьев незадолго до своей смерти (1912) предпринял иссле-
дование по анабиозу у млекопитающих, а именно у летучих мышей.
Он наблюдал у них явление переохлаждения и температурный
скачок, после которого, несмотря на замерзание соков, мышь ожи-
вала.
Однако последующие работы Калабухова (1934, 1935), про-
веденные с очень точной методикой, не подтверждают этих данных.
Летучие мыши могут претерпевать значительное охлаждение до
0° и даже ниже. При этом у взрослых летучих мышей переохлажде-
ние соков не наблюдается. Образование льда в их теле начинается
при температуре около —1°, после чего животное погибает. У мо-
лодых же летучих мышей наблюдается небольшое переохлажде-
ние и температурный скачок. Но как только произойдет образова-
ние льда, животное оказывается безвозвратно погибшим.
Интересно отметить что, согласно исследованию Калабухова
(1935), переход к состоянию переохлаждения зависит также от ско-
рости и равномерности понижения температуры. Чем медленнее и
равномернее охлаждение, тем легче достигается состояние пере-
охлаждения. Эти условия легче достигаются у мелких животных,
чем у более крупных.
Самый, так сказать, «механизм» гибели животного от замерза-
ния, так же как и у растений (см. стр. 118), имеет много общего с
высыханием. При образовании в тканях кристалликов льда проис-
ходит своеобразное «высушивание» протоплазмы клеток, а также
механическое разрушение клеток образующимися кристалликами
льда.
Проф. П. Ю. Шмидт дает следующее определение анабиоза:
«...я объединяю в одну категорию явлений анабиоза все биоло-
гические явления, связанные с более или менее значительным угнете-
нием и остановкой жизни и сопровождаемые «утратой воды жи-
вым веществом».
Если мы вспомним, что зимой огромное количество животных
нашей фауны, в том числе и насекомых — вредителей сельского
хозяйства, впадает в подобное состояние, то ясным станет большое
значение дальнейшего изучения этих явлений.
жизнь подо льдом
Когда ледяной покров сковывает водную поверхность, невольно
рождается подозрение, что жизнь в охлажденных водах вымерла.
Никаких признаков ее незаметно, если даже воду обнажить ото
льда. Исчезли водные растения,плававшие на поверхности, не видно
ни рыб, ни прыгающих в воду лягушек, никаких других живых
существ. Вода кажется мрачной, безжизненной, как безжизненно
на ее покрытых снегом берегах, где летом было неисчислимое ко-
личество разнообразных живых существ.
В действительности дело обстоит далеко не так, как предста-
вляется с первого взгляда. Уж если наземные живые существа, ко-
торых не видно зимой, в большинстве случаев не погибают, а только
погружаются в зимнюю спячку, то тем менее вероятно ожидать
гибели водной фауны и флоры. Менее — потому, что вода пред-
ставляет собой благоприятные условия для зимовья, в силу ее ис-
ключительных физических свойств.
В отличие от других тел природы, вода, как известно, обладает
той замечательной особенностью, что при охлаждении ее ниже
+4° Ц она не сжимается, а начинает расширяться. Наиболее плот-
ная, т. е. наиболее тяжелая вода при Н-4°. Особенно значитель-
ное расширение происходит при самом замерзании воды. Лед при
0° имеет объем на 9% больший, чем объем воды при 0°. Значит,
настолько лед и легче воды.
Не будь этой особенности, подчиняйся вода без отступления об-
щему закону сжимания тел при охлаждении, тогда лед, как имею-
щий температуру более низкую, чем вода, и следовательно, боль-
ший удельный вес, должен был бы опускаться на дно. Вновь обра-
зовавшийся ледяной покров опять опустился бы и т. д., пока вода
не промерзла бы до дна. Но благодаря указанной особенности этого
не происходит — лед остается наверху. А так как он плохой про-
водник тепла, то охлаждение сквозь него идет медленно. Медленно
поэтому утолщается и ледяная кора. Толщина льда в 1 м является
редкостью и для наших широт достигается только в особо суровые
зимы. На дно же должна опускаться вода, имеющая наибольшую
Зимовье водных животных
253
Рис. 162. Жук вер-
тячка (Gyrinus)
(увеличено)
плотность, т. е. с температурой +4°. Обыкновенно такая темпера*
тура при плохой теплопроводности льда и самой воды господствует
уже на небольшом расстоянии ото льда, и во всяком случае на дне
даже неглубоких бассейнов эта температура держится зимой по-
стоянно.
Естественно, что на дно погружается перед наступлением зимы
и большинство обитателей воды, чтобы с наступлением весны снова
подняться оттуда. Многие из них впадают в состояние покоя, или
зимней спячки, в связи с чем большей частью зарываются в тину
или в ил. Обратимся к рассмотрению отдельных
примеров.
Лягушки осенью возвращаются в водоемы.
При их мягкой гладкой коже и отсутствии
защитных покровов на теле они почти неиз-
бежно замерзли бы, если бы остались на суше.
На дне же они зарываются в незамерзающую
тину и спят там в течение всей зимы. При этом
они собираются целыми кучами, так что неко-
торые участки ила на середине пруда чуть не
целиком состоят из живой массы лягушек и
других спящих животных. Как же не задыха-
ются лягушки, раз органом дыхания у них слу-
жат легкие, которыми в воде дышать невоз-
можно? Не задыхаются потому, что в состоя-
нии глубокой спячки физиологическиё процес-
сы, не прекращаясь совсем, совершаются в
весьма незначительных размерах. Для лягушек
достаточно и того небольшого количества кислорода, какое полу-
чают они через свою кожу из воздуха, растворенного в воде. В не-
которых случаях, однако, зимой наблюдалась массовая гибель ля-
гушек, обусловленная чаще всего недостатком кислорода. Пони-
жение физиологической активности не всегда является для лягушки
достаточной «гарантией» для перенесения зимних условий. Это при-
способление лишь относительно целесообразно.
Из других земноводных остаются на зиму в воде саламандры,
зарываясь, подобно лягушкам, в ил или тину, а тритоны еще в се-
редине лета выходят из воды на сушу.
К числу самых обыкновенных обитателей стоячих вод принад-
лежат жуки—плавунцы, водолюбы, вертячки. Особенно обращает
на себя внимание последний. Это маленький жучок, величиной
с небольшую горошину (рис. 162). Когда он начинает кружиться
на поверхности воды при ярком солнечном свете, он кажется оде-
тым в блестящий панцирь, сверкающий разными цветами. Его дви-
жения быстрые, извилистые, круговые. По большей части эти жуч-
ки держатся целыми группами, собираясь на поверхности какого-
либо тихого заливчика. Скользя по его поверхности во всевозмож-
256
Жизнь подо льдом
ных направлениях, описывая удивительные кривые линии, они ни-
когда не сталкиваются и не мешают друг другу. Быстрота и лов-
кость их движений обязаны особому устройству их ног, представ-
ляющих собой как бы весла. Когда вертячки ныряют в воду, они
уносят туда с собой пузырек воздуха для дыхания.
Рис. 163. Большой прудовик (Limnaea stagnalis) летом
С приближением зимы поведение этих проворных животных ме-
няется. Движения замедляются, и, наконец, они удаляются на дно
пруда, где зарываются в тину. Всю зиму они проводят в глубокой
Рис. 164. Большой прудовик зимою
спячке, а с наступлением
весны поднимаются на по-
верхность.
Подобным же образом
погружаются на дно водо-
ема и другие водные жу-
ки, водные клопы, много-
численные куколки и ли-
чинки насекомых, моллю-
ски и др.
Обратим внимание на
самого обыкновенного мол-
люска, именно на прудо-
вика. Эта улитка обладает
красивой завитой остро-
конечной раковиною. Она
хотя и живет в воде, но
органом дыхания имеет не
жабры, а легкие и потому нуждается для дыхания в атмосфер-
ном воздухе. Моллюск часто всплывает на поверхность воды,
открывая свой легочный мешок, чтобы набрать в него воздуха
(рис. 163).
Зимой прудовик погружается на дно, где и проводит в спокой-
ной спячке холодное время года, зарывшись в ил (рис. 164). Как
холоднокровное и в физиологическом отношении относительно мало
деятельное животное, улитка, погрузившись в глубокий сон, об-
ходится зимой без притока свежего воздуха.
Зимовье водных животных
257
Как проводят зиму рыбы? У некоторых из них также наблю-
дается зимний сон. Явление это известно для рыб карповых, осет-
ровых и сомов. В холодное зимнее время, под ледяным покровом,
эти рыбы сбиваются массами на дне глубоких ям и там лежат
неподвижно рядами, окутанные толстым слоем слизи. В это
время рыбы действительно засыпают, становятся вялыми, слабо
реагирующими на внешние раздражения. Только после известного
промежутка времени к такой рыбе, если ее потревожить, возвра-
щается обычная восприимчивость, и она вполне просыпается от
зимнего сна. Естественным порядком такое пробуждение проис-
ходит при наступлении весеннего тепла, а иногда даже во время
случайных зимних оттепелей. Места, где залегает рыба на зиму,
в районе Каспийского бассейна, известны под названием «ятовей».
Они наблюдаются и в море, и в реке. Особенно славятся ятови с
красной рыбой 1 в реке Урале, где производился особый вид рыбо-
ловства — багренье. Подобные же залежи красной рыбы в зимнее
время известны на Иртыше, где они также создали особый тип про-
мысла под названием «юрового». У большинства рыб зимней спячки
не наблюдается. Наиболее общим явлением оказывается пониже-
ние жизнедеятельности, не доходящее, однако, до состояния сна.
В это время рыбы принимают пищу в малых количествах.
Нужно заметить, что условия зимовья далеко неодинаковы для
животных в бассейнах глубоких и мелких, в бассейнах, имеющих
приток свежей воды, в текучих и в совершенно замкнутых, каковы
пруды. Зимой, когда водная поверхность скована льдом, отделя-
ющим ее от атмосферы, почти прекращается поступление воздуха
в воду. Прекращается и другой источник обогащения ее кислоро-
дом— жизнедеятельность водных растений, выделяющих кисло-
род. При таких условиях находящимся в воде животным прихо-
дится довольствоваться зимой для дыхания преимущественно тем
кислородом, который был растворен в воде до покрытия ее льдом.
Очевидно, если бассейн глубокий,— запас кислорода в нем значи-
тельный и условия зимовья более благоприятные. В мелких же
бассейнах, не имеющих притока свежей воды, животные подо льдом
могут задыхаться. Известно, что в мелких прудах зимой иногда
погибают все рыбы и раки, а также многие лягушки. Главная при-
чина подобных катастроф — недостаток кислорода.
К этому присоединяется еще и то, что в каждом пруде вслед-
ствие гнилостных процессов, протекающих в условиях недостатка
кислорода, появляются сероводород и аммиак, а оба эти газа ядо-
виты для многих водных животных, в особенности для рыб. Летом
эти газы могут диффундировать и окисляться в присутствии сво-
бодного кислорода. Зимой же прекращается поступление кисло-
рода и вода обогащается этими вредными веществами, из которых
1 «Красной рыбой» называют ценные породы осетровых рыб.
258
Жизнь подо льдом
сероводород, будучи вредным сам по себе, отнимает еще от воды
кислород.
Эти совершающиеся зимой в непроточных водоемах процессы
приводят иногда к так называемым «заморам»— массовой гибели
рыбы. «Заморы» могут приносить большой экономический ущерб.
Для спасения рыб в прудах во льду пробиваются проруби. Ча-
сто можно видеть, как рыбы тяжело дышат около них, что служит
признаком недостатка кислорода. Иногда, если проруби не помо-
гают, в пруд накачивают воздух.
Наряду с погружающимися в спячку есть ряд и таких живот-
ных, в изобилии встречаемых летом в лужах и прудах, которые
зимой совершенно исчезают, независимо от условий зимовья.
Это организмы однолетние. Так, зимой не бывает большинства даф-
ний, или водяных блох,— мелких ракообразных; почти нет коло-
враток, нет мшанок; не найдем также зимой губок. Все эти живот-
ные, погибая сами, образуют на зиму покоящиеся стадии. Дафнии
откладывают особые зимние яйца, окруженные плотной оболочкой;
пресноводные губки — бадяги — осенью распадаются на большое
число мелких почечек (геммул) и т. п. Такие покоящиеся зимой за-
родыши продолжают свое развитие ближайшей весной, когда ус-
ловия существования снова сделаются благоприятными. В данном
случае, следовательно, имеет место гибель индивидуумов, но не ги-
бель вида. Гибель индивидуумов здесь неразрывно связана с обра-
зованием зимующих стадий.
С другой стороны, есть животные, которые, не погибая и не по-
гружаясь в спячку, продолжают бодрствовать подо льдом даже
и в мелких водоемах. Если мы прорубим лед и под ним проведем
сачком из тонкой сетки, то в большинстве случаев мы поймаем мел-
ких веслоногих — низших ракообразных (например, циклопов).
У них зимний покой не наблюдается. Мало того, яйцевые
мешки, которые носят самки, показывают нам, что на зиму падает
время размножения этих животных. Найдено много и других мел-
ких животных, не впадающих зимой подо льдом в спячку; видное
место среди них занимают простейшие: амёбы, жгутиковые и инфу-
зории. Однако в тех водоемах (преимущественно небольших пру-
дах с илистым дном), где развивается зимой сероводород, исчезают
и эти организмы.
Некоторые животные подвергаются или не подвергаются спячке
в воде зимой, в зависимости от условий. Так, некоторые насекомые,
погружающиеся в оцепенение где-нибудь в прудах, покрытых льдом,
ползают и плавают в местах, обнажаемых ото льда, например,
у притоков или оттоков озер. Недостаток кислорода под сплошным
ледяным покровом и другие неблагоприятные условия могут, та-
ким образом, служить одной из дополнительных причин спячки
и влиять на то, чтобы понижение жизнедеятельности организма и
сон были более глубокими.
Зимовье водных растений
259
Что делается с животными, если неглубокий бассейн промер-
зает до дна? Какова степень устойчивости пресноводных организ-
мов к отрицательным температурам?
Исследования в этом направлении производились многими уче-
ными, среди них особенно полными, с применением точных методов
исследования являются недавние работы Граевского (1948). Ему
удалось показать, что большинство пресноводных животных об-
ладает очень малой холодостойкостью и не выносит замерзания.
Для пресноводных моллюсков, пиявок и других беспозвоночных
температуры — 1°, —2° являются уже смертельными. В этом от-
ношении многие наземные беспозвоночные, как мы видели в преды-
дущей главе, являются гораздо более стойкими. Имеются, однако,
некоторые наблюдения, показывающие, что в условиях Арктики,
где промерзание пресноводных водоемов до дна является обычным
ежегодно повторяющимся явлением, некоторые живущие там бес-
позвоночные могут переносить зимнее промерзание и «оживать»
весной. Так, по наблюдениям Грезе, Таймырское озеро (на Тай-
мырском полуострове), являющееся довольно мелководным, ме-
стами промерзает до дна. Тем не менее, в нем сохраняются живыми
некоторые личинки комаров-толкунов и круглые черви, которые
после оттаивания не обнаруживают никаких признаков поврежде-
ния. Вопрос этот требует еще дальнейшей разработки.
Ряд исследований показал, что в водоемах в лед вмерзает зна-
чительное количество организмов, причем большинство их при от-
таивании оживает. Это объясняется тем, что температура льда ни-
когда не бывает очень низкой, и поэтому промерзания соков вмерз-
ших в лед организмов не происходит. Они находятся в состоянии
некоторого переохлаждения соков тела и при оттаивании возвра-
щаются к активной жизни. Масса организмов, вмерзающих в толщу
льда, получила название «пагон». Исследования показали, что в
состав пагона водоемов окрестностей Москвы входит более 100 ви-
дов растений и животных. Это—моллюски, личинки некоторых на-
секомых (стрекоз, комаров-толкунов), пиявки, водоросли и др.
Теперь обратимся к растениям. Замечательно, что и у них наи-
более распространенный способ зимовья подо льдом такой же,
какой у животных, именно опускание на дно. В связи с этим об-
разуются особые зимующие почки или отпрыски (побеги). По вы-
ражению Кернера, в стоячих водах прудов и болот при наступле-
нии морозов происходит настоящее «бегство вглубь», напомина-
ющее то, что мы видели с луковицами, клубнями и корневищами
наземных растений. Рассмотрим наиболее известные примеры.
Всем известно в изобилии плавающее на поверхности стоячих
вод маленькое растение — ряска. Скопление рясок, имевшее ле-
том прекрасный вид, осенью представляет другую картину; боль-
шая часть растеньиц погибла, листья обесцветились. Но под ними
находятся другие растеньица, более толстые и темнее окрашенные.
260
Жизнь подо льдом
Это их потомки—отпрыски, образовавшиеся вегетативным путем.
Они непрочно соединены со старыми растениями и благодаря
легкому толчку отделяются от них. Отделившись, темные отпрыски
погружаются на дно. Погружаются по той причине, что наполнены
запасом питательных веществ (крахмалом) и лишены воздухонос-
Рис. 165. Рдест курчавый (Potamogeton cEispus). Развитие зимних отпрысков
и погружение их на дно
ных полостей. Такие полости имеются у старых экземпляров,
от которых произошли эти отпрыски, а равно и у молодых
отпрысков, образующихся летом. Погрузившись на дно, осенние
отпрыски дают молодое поколение будущего года.
Как же весной они попадают снова на поверхность воды? Они
развивают новые, наполненные газом летние побеги, которые и
выносят их наверх.
Обыкновенным водным растением является также курчавый
рдест, отличающийся курчавыми краями своих листьев от других
Зимовье водных растений
261
плосколистных видов этого рода. Хотя у этого растения, в отличие
от ряски, нижняя часть укреплена на дне, где всегда защищена от
холода, тем не менее, мы
и здесь видим подоб-
ный же способ зимовья.
Осенью на верхушках
отдельных растений
курчавого рдеста обра-
зуются молодые побеги с
мелкими, тесно сидящи-
ми листьями (рис. 165).
Это зимние отпрыс-
ки, от которых зависит
будущее существование
растения. У их основа-
ния образуется легко
разрывающийся разде-
лительный слой. Напол-
ненные питательными
веществами, образовав-
шимися в процессе
фотосинтеза, и не имея
воздухоносных поло-
стей, отпрыски эти,
подобно зимующим от-
прыскам ряски, оказы-
ваются тяжелее стебля,
на котором они образо-
вались, и тяжелее воды.
Поэтому они отделяются
от стебля и погружают-
ся на дно, где благодаря
своим острым шилооб-
разным концам укреп-
ляются в тине (рис. 166).
Теперь они являются,
в сущности, просто от-
делившимися почками
или отводками, совер-
шенно подобными тем,
которые садовники от-
резают и отсаживают
в наших садах. С на-
Рис. 166. Зимующие на дне отпрыски кур-
чавого рдеста
Рис. 167. Отпрыски курчавого рдеста, начи-
нающие расти весной
ступлением весны от-
прыски начинают развиваться и превращаются во взрослые ра-
стения с длинными листьями и стеблями (рис. 167), на конце
262
Жизнь подо льдом
которых, над самой поверхностью воды образуются головки или
колосья маленьких зеленых цветочков.
У других водных растений, например, у водокраса, роголист-
ника, пузырчатки, образуются зимующие почки. Подобно побегам
курчавого рдеста они опускаются на дно и весной начинают разви-
ваться (рис. 168 и 169).
Рис. 168. Водокрас (Hydrocharis morsus гапае). Образование зимних почек
и погружение их на дно
Телорез — растение, своими шиповатыми листьями несколько
напоминающее алоэ, весной и ранним летом плавает на поверхно-
сти воды. Здесь оно производит новые мечевидные листья и цветки,
высовывающиеся наружу. Отцветши, растение погружается вглубь,
там у него зреют семена и образуются почки новых дочерних особей.
Около конца августа растение снова всплывает на водную поверх-
ность. Проросшие за это время дочерние растеньица во всем, кроме
величины, сходны с материнскими; они образовались из почек, си-
девших на концах удлиненных побегов, выросших между листьями
розетки, и окружают материнское растение. И вот осенью плети,
связывавшие материнское растение с дочерними, отмирают, и все
изолированные розетки опускаются теперь вместе с материнскою
особью на дно. Там они перезимовывают и весной снова появляются
ца поверхности.
Зимовье водных растений
263
У всем известных кувшинок погибают их круглые листья; на
дне же перезимовывает их толстое, чешуйчатое корневище. Так же
зимуют сусак, частуха, стрелолист. У водяного лютика плавающая
на поверхности часть растения погибает, а сохраняется только
невзрачный, тонкий покрытый редкими щетинковидными листьями
стебель, укоренившийся в илистом дне. У живучей элодеи перези-
мовывает большая ее часть, и даже оторванные льдинами кусочки
стебля образуют новые побеги в других местах озера.
Рис. 169. Зимние почки водокраса, поднимающиеся весною
Заслуживает внимания зимовье живущих в воде водорослей.
Подо льдом, при температуре от 0 до 4°, наряду с микроскопиче-
скими животными, можно найти и некоторые микроскопические во-
доросли — диатомовые, сине-зеленые и даже зеленые: планктон
зимой не всегда исчезает, хотя и сильно сокращается как в коли-
чественном, так и в качественном отношении.
Каким образом подо льдом, покрытым снегом, через который
свет может проникать только в чрезвычайно малом количестве,
могут жить хлорофиллоносные организмы, для нормального разви-
тия которых необходим свет? Разгадка в том, что простейшие во-
доросли в большинстве являются миксотрофами, т. е. могут пи-
таться не только продуктами фотосинтеза — усвоения углерода
на свету,— но и готовыми органическими веществами, растворен-
ными в воде.
Большинство зеленых дерновинно-кустистых водорослей, об-
разующих летом зеленые дерновинки в воде, отмирает к зиме, обес-
печивая будущие поколения с помощью спор; таковы нитчатки из
264
Жизнь подо льдом
группы сцеплянок (например, спирогира, зигнема). Однако есть
среди водорослей и перезимовывающие на дне — они весной и
летом всплывают на поверхность воды большими ватообразными
зелеными подушками. Причиной всплывания являются скопления
пузырьков кислорода, который начинает усиленно выделяться во-
дорослями в процессе фотосинтеза. Стойкими против холода ока-
зываются из зеленых водорослей вошерии, лучицы, кладофоры.
Из сине-зеленых водорослей осциллатории и близкие к ним формы
совершенно не образуют спор или каких-либо других приспособ-
лений против неблагоприятных условий существования, а между
тем, они всюду распространены, не исключая мелких водоемов.
Это объясняется тем, что они без вреда для себя могут переносить
низкие температуры.
снежный ПОКРОВ
Снежный покров представляет крупнейшее сезонное явление
в странах с умеренным климатом. Зима с ее снегом и лето с его теп-
лом — вот, можно сказать, два крайних, противоположных полюса,
между которыми вращается изменение внешних условий в окру-
жающей нас природе в течение года. Снежный покров одевает
территорию Советского Союза зимою почти на всем его протя-
жении, поэтому ближайшее знакомство с ним особенно важно.
Мы уже видели, какое влияние оказывает снег с сопутствующими
ему холодами на растения и животных. Теперь познакомимся ближе
со снежным покровом и его разносторонним влиянием на окружа-
ющую природу.
Изучение снежного покрова обязано главным образом наблю-
дениям на метеорологических станциях. Таких станций больше
всего на юго-западе и в центре Европейской части СССР. В азиат-
ской части СССР наибольшее количество станций приходится на
южную часть Сибири; много их также вдоль сибирской железно-
дорожной магистрали.
Исключительное разнообразие климата на территории СССР
обусловливает разнообразие режима снежного покрова. Как из-
вестно, большая часть СССР расположена в умеренном поясе и ха-
рактеризуется временным снежным покровом. Но на крайнем се-
вере (острова Новой Земли, архипелаг островов Земли Франца
Иосифа и Северной Земли) снег выпадает во все месяцы, не ис-
ключая и летних. Однако на июль—август здесь в среднем прихо-
дится всего лишь по 6 дней со снегопадом, тогда как общее число
дней со снегопадом в году превышает 135. В этих северных райо-
нах, а также в горах, снежный покров лежит, не стаивая, в течение
10—11 месяцев, а местами и круглый год. С другой стороны, на во-
сточном побережье Каспийского моря снег выпадает не каждый год,
и в среднем за много лет число дней в году со снегопадом равно 4.
На юге и западе СССР снег не образует устойчивого покрова.
Ниже приводятся некоторые данные, касающиеся фактической
стороны вопроса: размеров снежного покрова в различных местах
СССР и его распределения во времени.
266
Снежный покров
На большем протяжении Европейской части СССР в течение
года выпадает более 100 мм осадков в виде снега, а на вос-
токе, в средней части СССР —более 150 мм. На средней Волге
толщина снежного покрова превосходит 175 мм, а на средней Каме
достигает почти 200 мм. На западе и на крайнем севере толщина
снежного покрова не достигает 100 мм. По направлению к югу
она еще значительнее уменьшается, не достигая устьев Волги
и Урала и по берегу Черного моря 50 мм.
Что касается времени появления первого снежного покрова,
то оно для разных районов СССР весьма различно. В высоких ши-
ротах Арктики установить эту дату крайне трудно, потому что,
как мы уже указывали, отдельные снегопады наблюдаются здесь
во все летние месяцы. В Европейской части СССР среднее время
появления снежного покрова изменяется от начала октября на се-
веро-востоке до начала января (на юге Крыма и Черноморском
побережье Кавказа). Последняя дата относится также к Средней
Азии. Это наиболее позднее среднее время появления снежного
покрова в пределах СССР. Ранее всего (конец августа) снежный
покров в Азиатской части СССР устанавливается на Новосибирских
островах. Интересно, что более раннее появление снежного покрова
обычно имеет место на возвышенностях (Средне-Русская, Приволж-
ская и др.). Поданным многолетних наблюдений для Европейской
части СССР разница между сроками наиболее раннего и наиболее
позднего начала снежного покрова колеблется между 35 и 85 днями.
Общая продолжительность залегания снежного покрова (снеж-
ный период) также весьма различна для разных районов СССР,
как это видно на прилагаемой карте (рис. 170). Для Европейской
части СССР продолжительность залегания снежного покрова в
общем увеличивается с юго-запада на северо-восток, а для Азиат-
ской части СССР — с юга на север.
В Западной Европе снег представляет обычное явление зимой
на Скандинавском полуострове и в Германии. В Париже со снегом
бывает до 14 дней в году; в Марселе — до 3. Как сравнительно
редкое явление снег наблюдается в Южной Испании, Южной Ита-
лии и Алжире. В Азии он еще не редкость в Иерусалиме и Багдаде;
в Северной Америке наблюдается на берегах Мексиканского залива,
а в Южной — известен уже в Бразилии. Само собой разумеется,
здесь идет речь о местностях, сравнительно невысоко поднима-
ющихся над уровнем моря. На высоких горах имеются области веч-
ных снегов даже и под тропиками.
От количества выпадающего снега зависит толщина (высота)
снежного покрова. Последнюю легко измерить с помощью рейки —
деревянного шеста в х/г—2 м длиной, с нижнего конца заострен-
ного и с нанесенными делениями (в сантиметрах), начиная снизу.
Неровности рельефа и разные преграды оказывают влияние на
распределение снега, на толщину снежного покрова. Известно,
Рис. 170. Продолжительность снежного периода (в днях) (по Рихтеру)
Продолжительность снежного периода 267
268
Снежный покров
например, что около изгородей и других предметов, оказывающих
препятствие движению снега во время метелей, снег скопляется
в виде сугробов. При этом, если изгородь проницаемая,— снег скоп-
ляется не только перед изгородью, но и позади нее, т. е. не с повет-
ренной только стороны, но и с подветренной, и на последней сугроб
может быть даже значительнее. Это объясняется тем, что сквозь
проницаемую преграду ветер проходит, но его сила,т. е. скорость,
при этом уменьшается, вследствие чего проникшие через преграду
вместе с воздухом снежинки уже не в состоянии быть взвешенными
в последнем и падают позади преграды, образуя здесь сугроб.
Чтобы наглядно показать влияние на толщину снежного по-
крова окружающих условий, следует произвести съемку. Для этого
нужно выбрать участок, приблизительно* в километр длиною,
на котором бы встречались поле, кустарник, лесок, канавы и т. п.,
и несколько раз в зиму сделать промер снежного покрова по этому
направлению; на ровных местах делать промеры реже, а на не-
ровностях и при переходе от одного типа участка к другому,— по
возможности чаще. Результат промера нужно нанести на график,
причем вертикальный масштаб для наглядности нужно взять зна-
чительно крупнее горизонтального. Промеры по выбранному на-
правлению желательно делать хоть раз в месяц; после же метели
необходимо сделать дополнительный промер, который наглядно
покажет влияние тех или других препятствий на распределение
снежного покрова.
Средняя многолетняя толщина снежного покрова в СССР пока-
зана на прилагаемой карте (рис. 171). Из ее рассмотрения виднр,
что наибольшая толщина наблюдается в трех районах: на запад-
ных склонах Северного Урала и в его западных предгорьях, в за-
падной части Средне-Сибирского плоскогорья и в примыкающих
к нему с запада частях Западно-Сибирской низменности, и на даль-
невосточном побережье (в районах Камчатки, Сахалина и низовьях
Амура). Наименьшая толщина снежного покрова характерна для
юга Европейской части СССР, равнин Средней Азии, южного Забай-
калья и южных районов дальневосточного Приморья.
На большей части СССР максимальная толщина снежного по-
крова падает на март, в северных районах — на апрель, а в юж-
ных — на февраль и даже январь. Средняя толщина в 10 см (соот-
ветствующая началу санного пути), на крайнем северо-востоке Ев-
ропейской части СССР устанавливается обычно уже в конце ок-
тября, а на юге и западе — в середине или конце февраля.
Количество воды, заключающееся в снежном покрове, зависит,
с одной стороны, от его толщины, а с другой—от плотности.
Вообще плотность снега небольшая -—он рыхлый. Средняя плот-
ность свежевыпавшего снега равна 0,1 плотности воды, т. е. при
гаянии он дает объем воды в десять раз меньший. В местах открытых
плотность снега благодаря действию ветра бывает больше, а в
Рис. 171. Средняя многолетняя толщина снежного покрова (в см} (по Рихтеру)
Толщина снежного покрова 269
270
Снежный покрод
огражденных, например, между строениями или в лесу,— меньше.
Плотность снега зависит также от формы снежинок, изменяющейся
в зависимости от температуры (см. ниже). При больших холодах
объем снега в 15—20 раз более объема получающейся при таянии
воды, между тем, при температуре, близкой к 0°, когда снег падает
в виде хлопьев, объем его только в 3—5 раз более объема воды, при
оттепели же — он еще меньше. По мере лежания снежный покров
уплотняется, так что глубокие слои его плотнее лежащих выше.
Понятно также, что плотность изменяется в зависимости от времени
года, достигая в большинстве районов СССР своего максимума в
апреле.
Плотность снега имеет первостепенное практическое значение,
так как определяет большинство физико-механических свойств
снежного покрова. Она влияет на проходимость местности в зимних
условиях, от нее в значительной мере зависит объем снегоуборочных
работ, с ней приходится считаться при вычислении снежных нагру-
зок на крышах зданий и т. п.
Плотность снега легко определить следующим способом. Взяв
небольшой цилиндрический сосуд (например, жестянку от консер-
вов), погрузить его в снег до наполнения, не сдавливая при этом
снега. При помощи дощечки или лопаточки приподнять сосуд с
наполняющим его до краев снегом, внести в теплое помещение и
дать снегу растаять. Потом измерить (при помощи градуирован-
ного цилиндра) объем полученной из снега воды и сравнить его с
объемом всего цилиндра, т. е. воды, наполняющей его до краев.
Отношение между этими объемами и будет указывать на плотность
снега. Можно поступить еще таким образом. Определить вес: от-
дельно сосуда, сосуда со снегом и сосуда с водою; вычтя из двух
последних весов вес сосуда, определим веса наполняющих его снега
и воды, отношение между которыми и будет соответствовать плот-
ности снега.
Разнообразие снежного покрова в разных районах СССР и его
большая роль в природе и народном хозяйстве делают необходи-
мым районирование территории СССР по характеру снежного по-
крова. Такое районирование у нас было впервые произведено проф.
Г. Д. Рихтером (1945). В основу его положены максимальная тол-
щина снежного покрова и продолжительность многоснежного пе-
риода. За максимальную толщину снежного покрова принята наи-
большая среднедекадная его толщина, взятая из многолетних сред-
них величин. Под многоснежным периодом понимается период,
в течение которого среднедекадная толщина снежного покрова,
также выведенная из многолетних средних данных, превышает
30 см. Последняя цифра взята потому, что до толщины в 30 см снеж-
ный покров обычно не препятствует передвижению, тогда как при
большей его толщине условия для транспорта резко ухудшаются
и приходится прибегать к защите от снежных заносов, к расчистке
Значение снежного покрова в природе 271
снега и т. п. Кроме этих двух основных показателей, при райони-
ровании приняты во внимание условия залегания, характер нако-
пления и физико-механические свойства снежного покрова.
По максимальной среднедекадной толщине снежного покрова на
территории СССР различают 5 основных зон: I — свыше 70 см\
II — от 50 до 70 см\ III — от 30 до 50 см\ IV — от 10 до 30 см\ V —
менее 10 см. Сверх того выделены еще арктическая (VI) и горная
(VII) зоны. В арктической зоне максимальная среднедекадная тол-
щина снежного покрова колеблется от 20 до 50 см, так что она в
этом отношении отвечает III и IV зонам, резко отличаясь от них,
однако, по условиям снегонакопления, физико-механическим свой-
ствам снега и продолжительности его залегания. Горная зона ха-
рактеризуется неравномерностью залегания снега (в связи с раз-
нообразием рельефа) и различной продолжительностью залегания
(в зависимости от абсолютной высоты). В пределах зон установлен
ряд районов, различающихся между собой по продолжительности
залегания снега, по условиям залегания и по свойствам снежного
покрова (рис. 172).
Каково значение снежного покрова в природе?
На первый взгляд кажется, что снег представляет побочное зим-
нее явление, сопутствующее низкой температуре, что все другие
зимние перемены, в том числе и спячка растений и животных, за-
висят также только от холода, но отнюдь не от снега. На самом
деле—далеко не так. Будучи порождением известных климати-
ческих причин,снег в то же время является и важнейшим климати-
ческим фактором, простирающим свое влияние на окружающую
природу не только зимой, но весной и летом, т. е. даже тогда, когда
земля уже обнажена от снега.
Появляясь вследствие холода, снег в то же время и сам понижает
температуру окружающего воздуха. Это происходит от несколь-
ких причин. Во-первых, от того, что снег обладает способностью
отражать лучи. Достаточно сказать, что снежная поверхность от-
ражает от 80 до 94% падающей на нее лучистой энергии, тогда как
почва, лишенная снега, всего лишь 10—30%.
Наглядным доказательством этой способности являются по-
крытые вечным снегом горы тропических стран. Несмотря на оби-
лие тепла, посылаемого сюда солнцем, снег здесь не тает, так как
лучи почти целиком отражаются в пространство. В ясные, тихие
дни и на равнине, особенно перед началом весны, когда солнце на-
чинает озарять землю лучами, можно наблюдать, что снег не под-
дается им. Таяние снега начинается тогда, когда вблизи его или
на его поверхности имеются предметы, поглощающие солнечные
лучи и затем нагревающие снег, или когда приносится теплый воз-
дух со стороны. Весной при наступлении оттепелей можно наблю-
дать, что около деревьев, столбов и т. п. образуются воронки вслед-
ствие таяния снега и что грязный снег, т. е. покрытый частичками
Рис. 172. Районы снежного покрова (по Рихтеру)
272 Снежный покров
Охлаждающее влияние снега
273
инородных тел, поглощающих солнечные лучи, тает быстрее. Для
наглядности произведем следующий опыт. Приготовим две метал-
лических пластинки (например, из жести), размером в 10-42 кв. см.
Одну из них окрасим белой, другую черной краской. В ясный,
солнечный день ранней весны поместим эти пластинки на ровном
месте на снег. Черная пластинка скоро погрузится в снег, потому
что она нагреется солнечными лучами, отчего снег под нею будет
таять. Белая же пластинка не погрузится, потому что благодаря
белому цвету она будет отражать солнечные лучи; аналогично этому
отражает лучи и белый снег. Подобно черным пластинкам, весной
можно видеть погрузившимися, как бы провалившимися в снег
различные упавшие на него темные предметы—листья, ветки
и т. п.: они поглощают солнечные лучи, нагреваются, и снег под
ними тает. Но главным образом таяние связано с теплым ветром,
дующим с морей или с суши, свободной от снега. Вот почему в ме-
стах, защищенных от ветра, например, на лесных полянах, со всех
сторон окруженных деревьями, снег долго не тает.
Вторая причина охлаждающего влияния снега на атмосферу
заключается в том, что он обладает весьма большой способностью
излучать тепло в пространство. Нужно заметить, что вообще твер-
дые тела обладают способностью излучения тем большею, чем рых-
лее строение их поверхности. Крайними в этом отношении будут та-
кие тела, как порошки, пыль, хлопья, губка, войлок, мех или пух.
Сюда же относится и снег, в особенности свежевыпавший, рыхлый.
Значение этого обстоятельства, т. е. охлаждающего влияния снега
вследствие излучения, заметно в особенности бывает в ясные ночи,
когда притока тепла от солнца нет, а между тем излучение его снеж-
ной поверхностью происходит; облака препятствуют большому
излучению. Вследствие указанных причин поверхность снега обычно
бывает холоднее поверхности голой почвы, причем разница темпе-
ратур там и здесь в холодные дни достигает 3—8°. Чтобы констати-
ровать охлаждающее влияние снега вследствие излучения тепла,
произведем следующий простой опыт. Во время мороза, при полном
отсутствии ветра, положим на открытом месте один термометр на
снег и другой на почву, обнаженную от снега, чтобы изолировать
от охлаждающего влияния последнего. Удобно это сделать во время
весенних утренних заморозков, когда можно воспользоваться об-
разовавшимися проталинами на поверхности почвы и нет надоб-
ности специально расчищать почву от снега. Термометры должны
быть положены таким образом, чтобы их резервуары не касались
ни снега, ни почвы, чтобы показания термометров относились бы
к температуре воздуха над снегом и почвой. Температура над
снегом окажется более низкой. Наблюдение следует проводить
ночью или утром перед восходом солнца, когда имеется только
редкий рассеянный свет, так как днем будет иметь место погло-
щение солнечных лучей почвой и вследствие этого нагревание
214
Снежный покров
над нею воздуха. Наблюдения утром при слабом свете показали,
что, когда температура воздуха над снегом была, например, —8°,
над обнаженной от снега почвой в то же время —6,5°. Охлажда-
ющее влияние снега на воздух более всего касается, конечно, ниж-
них слоев последнего, которые поэтому бывают холоднее слоев,
лежащих выше. На основании сделанных наблюдений можно пред-
полагать, что такое распределение температуры над снегом, т. е.
понижение ее по мере приближения к снежной поверхности, про-
стирается вверх на несколько десятков метров.
Наконец, следует упомянуть еще об одной, хотя не постоянно
и не везде одинаково действующей, но все же весьма важной при-
чине охлаждающего влияния снега. Она заключается в том, что
снег препятствует наступлению оттепелей, вызываемых приноси-
мым со стороны теплым воздухом. Неспособность снега нагре-
ваться выше 0° обусловливает то, что теплый воздух легко охлаж-
дается. Поэтому зимние оттепели там, где существует обширный
снежный покров, очень непродолжительны, температура подни-
мается немного выше 0°, да и то только при очень сильном ветре.
Как только он утихнет и небо прояснится, температура быстро па-
дает. Для местностей, недоступных влиянию теплых ветров, эта
причина не имеет значения, но она играет видную роль там, где
такие ветры дуют зимою.
Из сказанного следует, что в присутствии снега температура
воздуха зимой, при прочих равных условиях, бывает ниже, чем
в отсутствии его. Таким образом, снег — не только побочное явле-
ние, сопутствующее зимнему холоду, но вместе с тем и причина,
усиливающая суровость зимы.
Снег оказывает также значительное влияние на влажность воз-
духа. Хотя снег и лед представляют собой воду в твердом состоянии,
тем не менее, они испаряются. Особенно значительно испаряется
снег, потому что он шероховатый и, следовательно, имеет боль-
шую свободную поверхность. Однако вследствие низкой темпе-
ратуры поверхности снега, уступающей температуре воздуха,
испарение часто сменяется конденсацией и снежный покров
тогда сушит воздух. Поэтому абсолютная влажность воздуха над
снегом не особенно велика. Однако влажность относительная,
т. е. оцениваемая с точки зрения того количества влаги, какое при
данной температуре может содержаться в воздухе,— значительна,
и превышает таковую над бесснежными пространствами. По мне-
нию проф. А. И. Воейкова и других, она приблизительно равняется
относительной влажности атмосферы над пресноводными озерами,
т. е. там и здесь воздух близок к насыщению парами. Если бы
снег не был холоднее прилегающего к нему воздуха, а особенно
если бы он был теплее, тогда испарение его было бы еще значи-
тельнее.
Большого внимания заслуживает очищающее влияние снега на
Влияние снега на чистоту воздуха, его движение и состав
27,5
воздух. Как бы ни казалась нам чистой атмосфера, она на самом
деле бывает переполнена бесчисленным множеством разнообраз-
ных пылинок, среди которых встречаются и заразные микробы.
Эта пыль отчасти очищается дождем, но гораздо лучше снегом, бла-
годаря тому, что снежные хлопья при своем падении кружатся в
воздухе, и, покрыв землю, снег препятствует поднятию с ее поверх-
ности новой пыли. Благодаря этому в больших городах, где вес-
ной, летом и осенью воздух бывает очень пыльным, зимой гигиени-
ческие условия улучшаются. Скопление пылевых частиц загрязняет
снег, и известны факты, когда загрязнение это было столь зна-
чительным, что придавало снегу ту или иную окраску. В большин-
стве подобных случаев пыль, выпадавшая вместе со снегом, оказы-
валась принесенной издалека, из районов, лишенных снежного по-
крова. Так, например, в марте 1907 г. на территории б. Самарской
и Саратовской губерний выпал желтый снег, анализ которого по-
казал наличие в нем большого количества лессовых частиц. По-
следние, очевидно, были занесены из Арало-Каспийских степей
юго-восточным ветром, сопровождавшим снегопад.
Падающие со снежными хлопьями частички пыли при таянии
снега соединяются между собой все теснее и теснее и, в конце кон-
цов, образуют заметный черный слой. Поглощая солнечные лучи,
этот слой содействует дальнейшему таянию снега, а соединившись
с почвой, удобряет ее.
Снежный покров ослабляет движение воздуха и, следовательно,
умеряет силу ветра. Такая роль его объясняется следующим об-
разом.
Нижний слой воздуха вследствие охлаждающего влияния снега
делается более плотным, более тяжелым и, следовательно, менее
подвижным. Это обстоятельство в связи с тем, что температура воз-
духа над снежным покровом не увеличивается с высотой, а умень-
шается, является причиной того, что нижний слой воздуха не во-
влекается в круговорот верхними течениями. Наши зимние метели
обычно не сопровождаются бурями. Последние бывают значительно
сильнее осенью. С другой стороны, не следует забывать, что снеж-
ный покров сглаживает поверхность, устраняя тем самым препятст-
вия на пути движения воздуха. Это, казалось бы, должно способ-
ствовать увеличению скорости ветра. Однако в силу преоблада-
ющего влияния вышеуказанных обстоятельств она при снежном
покрове обычно меньше. К тому же энергия ветра в случае рыхлого
снегового покрова частично затрачивается на перенос снега, при-
чем, повидимому, создаются вихревые токи. Все это также способ-
ствует ослаблению ветра.
Снег оказывает определенное влияние на состав слоев воздуха
над снежным покровом. Это связано с тем, что он, обладая плохой
газопроницаемостью, нарушает нормальный газообмен между ними
и поверхностными слоями почвы. Между тем, этот газообмен в
276
Снежный покров
значительной мере определяет состав приземных частей воздуха.
Важно также, что почва, не покрытая снегом, промерзает зимой
на большую глубину, что приводит к ослаблению или даже полной
приостановке протекающих в ней биохимических процессов.
Поэтому можно сказать, что снежный покров, предохраняя почву
от промерзания, способствует тому, что биохимические процессы
в ней продолжаются и в зимнее время.
Прямые наблюдения показывают, что при снежном покрове ко-
личество углекислоты и озона в воздухе увеличивается. Напротив,
количество аммиака зимой в воздухе оказывается уменьшенным:
по некоторым данным оно почти вдвое меньше, чем летом. Послед-
нее, повидимому, связано со способностью снега поглощать неко-
торые газы из атмосферы. Если принять во внимание огромную
общую поверхность снегового покрова, то роль его в этом отноше-
нии должна быть весьма значительной. Весной при таянии снега
эти поглощенные зимой из воздуха вещества попадают в почву.
Значение снежного покрова не ограничивается одними только
зимними месяцами. Его влияние простирается и на последующее
время, когда снег уже отсутствует, причем оказывается еще более
важным по своим последствиям.
Весной и летом не было бы такой, как теперь, растительности,
если бы не существовало снежного покрова. Мы уже знаем, что
снег — плохой проводник тепла и потому явлйется как бы «теплым
одеялом» для зимующей флоры и предохраняет большую часть ее
от вымерзания. Вследствие этого в северном умеренном поясе в
малоснежные зимы погибает множество растений, между тем как
в случае обильного снега они без вреда для себя переносят силь-
нейший мороз.
Снежный покров, нарушая нормальный теплообмен между поч-
вой и атмосферой, играет роль теплоизоляционной прослойки, «раз-
рывающей» круговорот тепла между ними. В силу этого в темпера-
турном отношении начинается как бы «самостоятельная жизнь»
почвы и воздуха. И этот разрыв в теплообороте оказывается тем
большим, чем толще снежный ’покров.
О том, насколько мала теплопроводность снега и насколько пре-
дохраняет он землю и находящиеся на ней растения от промерза-
ния, наглядно говорят, например, следующие наблюдения. Зимой
1892—1893 гг., когда около бывш. С.-Петербурга морозы дохо-
дили до —38° и даже до —40°, в окрестностях города почва, по-
крытая снегом, даже на небольших глубинах оказалась не про-
мерзшею. В среднем, за январь 1893 г. на глубине 20 см под снегом
было 4-0,23°, между тем, на той же глубине в почве, очищенной
от снега, средняя температура за январь оказалась —14,06°. В от-
дельных случаях разность температур между покрытой и непо-
крытой снегом почвой достигала за эту зиму еще больших размеров;
например, 15 января 1893 г. температура на поверхности снега
Теплопроводность снега
277
была —39,3°, на поверхности почвы под снегом —3,0°, в той же
почве на глубине 20 см была —0,9°, а в почве, лишенной снега,
на той же глубине —23,9°. Сопоставление результатов подобных
наблюдений приводит к заключению, что обнаженная почва даже
на глубине 40—80 см обнаруживает в главных чертах те самые
температурные колебания, какие наблюдаются на ее поверхности,
хотя и в смягченном виде и с опозданием. Между тем, почва, при
достаточной толщине снежного слоя, даже на поверхности своей,
непосредственно под снегом, показывает лишь сравнительно малые
колебания, весьма редко превышающие 1—3°. А на глубине 20 см
температура в покрытой снегом почве почти совсем не колеблется
и держится большей частью выше 0°. Подобные наблюдения очень
легко провести, имея в течение зимы площадку, обнаженную от
снега, т. е. очищаемую всякий раз по выпадении его, и сравнивая
глубину промерзания почвы на этой площадке и под снегом. Если
два одинаковых небольших сосуда (например, баночки, энтомо-
логические пробирки) наполнить водой и в морозный день один
из них .закопать в снег, а другой оставить на воздухе, то в по-
следнем сосуде вода замерзнет скорее. Этой-то защите со стороны
снега от охлаждения и замерзания, вследствие его плохой тепло-
проводности, и обязано спасительное влияние снежного покрова
па растительность.
Степень теплопроводности снега зависит от его рыхлости. Ска-
занное выше о значении рыхлости для излучения тепла приложимо'
и к теплопроводности, только в обратном смысле: чем большая рых-
лость, тем большая способность излучения и тем меньшая тепло-
проводность. В порошкообразной форме даже металлы плохо про-
водят тепло и хорошо его излучают.
Рекомендую провести следующий опыт, показывающий влия-
ние рыхлости снега на его теплопроводность. В морозный день на-
полнить снегом на открытом воздухе две чашки или просторных
банки, измерив температуру снега. В одной чашке снег, если он
сильно рыхлый, уплотняется немного (встряхиванием или посту-
киванием чашки), а в другой—уплотняется сильно, насколько
возможно (давлением на него). Затем чашки со снегом вносятся в
теплое помещение, и в них одновременно погружаются на одина-
ковую глубину (около 2—3 см) термометры. Сейчас же по погру-
жении в снег в обоих термометрах начнется понижение ртути, но
в чашке с сильно уплотненным снегом значительно быстрее; в конце
концов, в обоих термометрах ртуть упадет до температуры снега
на открытом воздухе, что в чашке с рыхлым снегом произойдет
медленнее (например, до температуры —15° — минут через десять).
Спустя некоторое время начнется поднятие ртути в термометрах,
причем и этот процесс быстрее пойдет в уплотненном снегу. Если
обогревшиеся чашки со снегом опять вынести на мороз, то нач-
нется медленное опускание ртути в термометрах, причем в плотном
278
Снежный покров
снегу опять сравнительно более быстрое, чем в рыхлом. Вы-
воды из этих наблюдений ясны. Медленность прогревания снега,
принесенного с мороза в теплое помещение, и медленность охлажде-
ния прогретого снега, вынесенного на мороз, говорит о его плохой
теплопроводности. Более медленное опускание ртути в термометре,
опущенном в рыхлый снег, по внесении его в теплое помещение,
затем более медленное поднятие ртути здесь и опять более медлен-
ное опускание ее на морозе, по сравнению с опущенным в плотный
снег, —все это показатель меньшей теплопроводности и, очевидно,
связано с рыхлостью, т. е. с большим количеством воздуха в рых-
лом снегу, и следовательно, говорит о том, что плохая теплопро-
водность снега есть результат находящегося в нем воздуха.
В природе был проведен длинный ряд наблюдений и изме-
рений по влиянию рыхлости снега на его теплопроводность, на-
пример, Г. Абельсом, вблизи Свердловска. В результате он пришел
к заключению, что теплопроводность снега прямо пропорциональна
квадрату его плотности. Чем рыхлее снег, тем он лучше защищает
почву и находящуюся на ней растительность от охлаждения. Теп-
лопроводность снега зависит также от его влажности: мокрый снег
обладает большей теплопроводностью.
Если во время морозов снег защищает почву от потери тепла,
то, наоборот, при температуре выше 0° он ее охлаждает. В начале
весны значительная часть энергии, посылаемой солнцем, тратится
на таяние снега. Температура тающего снега равна 0°; такой же
остается и температура почвы до окончательного удаления снеж-
ного покрова, так как она пропитывается снеговой водой. Ассман
сделал попытку вычисления того количества тепловой энергии,
какое требуется для удаления снежного покрова. Он нашел, что
для превращения в воду 240 000 млн. куб. см снега, выпавшего в
Германии с 19 по 22 декабря 1886 г., необходимо 960 биллионов
калорий тепла, действие которых равно 172 млн. лошадиных сил.
Если принять во внимание, какие громадные пространства покры-
ваются зимой снегом и иметь в виду всю толщину снежного по-
крова, то можно приблизительно представить и все то громадное
количество тепла, какое поглощается весною снегом.
Это охлаждающее действие снежного покрова не только не вре-
дит растительности, но, наоборот, не менее важно для нее, чем со-
гревающее действие его зимой. Весной тепло не сразу становится
непрерывным. Обычно оно многократно прерывается заморозками,
заставляющими замерзать воду и поверхность почвы. Если бы от
первых весенних лучей растительность начала оживляться, то эти
заморозки оказались бы убийственными для нее. Начавшие прора-
стать семена, нежные всходы травянистых растений и молодые по-
беги кустарников и деревьев не выносят даже самого легкого мо-
роза. Тающий снежный покров оказывает и в данном случае спа-
сительную услугу: поддерживая почвенную температуру на одном
Искусственное задержание снега
279’
уровне, неблагоприятном для прорастания, но и не гибельном для
зимующих растений, он не позволяет им пробудиться от зимнего
сна, прежде чем весна не установится окончательно и прорастание
не сделается совершенно безопасным.
Если снег и согревает (почву) и охлаждает (воздух и почву),
то естественно спросить: какое же влияние его значительнее?
Сопоставляя то и другое, проф. А. И. Воейков пришел к заключе-
нию, что согревающее значение снега значительнее охлаждающего.
Весеннее охлаждение почвы сравнительно с зимним согреванием
кратковременно и незначительно. То же следует сказать и об
охлаждении воздуха, потому что не только охлаждаемая снегом, но
даже общая масса его весьма мала по сравнению с тою частью зем-
ной коры, на которую простирается согревающее влияние снега.
Если бы зимой не было снега, то охлаждение земного шара подви-
нулось бы, несомненно, далее, чем теперь. Благодаря снежному
покрову даже в холодные зимы, при шестимесячном лежании снега,
вероятно, уже на глубине одного метра температура в самом холод-
ном месяце не ниже средней годовой температуры на поверхности
суши и в нижнем слое воздуха.
Снежный покров имеет для растительности еще то весьма важ-
ное значение, что является для нее источником влаги. Весенний
период прорастания — самый важный в жизни растений. Поэтому
прорастающее растение не должно испытывать недостатка во влаге,
который легко может оказаться пагубным. В нашем континенталь-
ном климате при редко падающих дождях это всегда случалось
бы, если бы почва не запасалась с весны влагой от тающего снега,
о чем мы подробнее узнаем из следующей главы.
В виду важности снежного покрова для растительности боль-
шое значение для сельского хозяйства имеют мероприятия, способ-
ствующие искусственному задержанию снега на почве, особенно
там, где его мало выпадает или где он легко сдувается ветром. За-
держание и накопление снега на полях обеспечивает увеличение
запасов влаги в почве и предохраняет ее от промерзания, что в
свою очередь благоприятствует более полному поглощению почвой
талых вод. Однако правильная и рациональная организация работ
по снегозадержанию возможна только на обширных площадях.
В дореволюционной России, при раздробленном частном земле-
пользовании соответствующие меры не могли получить сколько-
нибудь широкого распространения, и только в условиях СССР,
после победы колхозно-совхозного строя, положение в этом отно-
шении резко изменилось. Достаточно указать, что в 1948 г. снего-
задержание в СССР было проведено уже на площади в 14 млн. га.
Комплекс мероприятий, имеющих своей целью улучшение тепло-
вого режима и увлажнение почвы путем воздействия на снежный по-
кров, может быть разнообразен, причем те или иные меры должны
применяться в соответствии с местными условиями и с обязательным
:280
Снежный покров
учетом того, какую задачу нужно решить в первую очередь.
Для регулирования температурного режима почвы под снегом
рекомендуется создавать рыхлый и равномерный слой снега. Извест-
ное значение в этом отношении имеет осенняя распашка полей без
последующего боронования и бороздовой посев озимых культур.
Накапливающийся между бороздами снег способствует утеплению
почвы. Для увеличения накопления снега полезно также покры-
вать озимые всходы соломой, разбрасывая ее около 2/3 tn на 1 га,
или хворостом. В тех случаях, когда поля уже покрылись доста-
точно толстым слоем снега, рекомендуется втыкать в него ветки или
длинные стебли (подсолнечника, кукурузы и других растений).
Иногда на пашнях вместе с озимыми культурами узкими полосками
сеют быстро растущие растения (сурепицу, белую горчицу и др.),
образующие к наступлению снегопадов своего рода «кулисы», удер-
живающие снег. В условиях континентального климата (напри-
мер, в Сибири) известное значение для накопления снега имеет по-
сев озимых по неубранному жнивью (стерне) яровых и пропашных
культур. Большой положительный эффект дают кустарниковые и
древесные полезащитные полосы.
Иногда бывает нужным не сохранить тепло в почве, а наоборот,
охладить ее. Такая необходимость может возникнуть при обиль-
ном снегопаде на талую почву, что может привести к выпреванию
или вымоканию озимых. В этих случаях снег уплотняют, укаты-
вая его деревянными катками. Уплотненный снег, как мы уже
знаем, имеет большую теплопроводность, чем рыхлый.
Все указанные выше меры регулирования температурного ре-
жима почвы под снегом способствуют вместе с тем накоплению зим-
них запасов влаги. В тех случаях, когда необходимо решить только
эту последнюю задачу, надобность в равномерности снежного по-
крова, очевидно, отпадает и можно с успехом применять перенос*
ные щиты или заборы, около которых снег накапливается в виде
сугробов. Во второй половине зимы с этой целью искусственно
устраивают также снежные валы, кучи и снежные заборы или
используют специальные снегопахи. Понятно, что и укатывание
снега будет способствовать его накоплению.
Нередко возникает необходимость в сельскохозяйственных це-
лях регулировать в ту или иную сторону время залегания снежного
покрова. Для того чтобы задержать таяние снега, его уплотняют
и покрывают торфом, опилками и другими теплоизолирующими
материалами. Для ускорения таяния снег посыпается темными по-
рошками, поглощающими солнечные лучи (золой, сажей, землей
и т. п.).
Выше было уже указано, что на плотность снежного покрова,
а следовательно, и на его теплопроводность влияет форма снежных
кристаллов. Ближайшее изучение их показало, что они весьма раз-
нообразны, но в основе их лежит одна кристаллографическая си-
Форма снежинок
281
стема — гексагональная (шестиугольная). Отдельные снежинки
большей частью представляют сложные фигуры, в которых преоб-
ладает число шесть. Это — шестиконечные звездочки (рис. 173),
шестиконечные пластинки, шестигранные призмы и разнообраз-
ные сложные комбинации этих фигур; самые простые формы сне-
жинок — иглы. Для рассматривания форм снежинок удобно при-
менить следующий простой прием. Взять небольшую доску или
Рис.
173. Разные формы снежинок (увеличено)
картон, обтянуть черным сукном и выставить на открытый воздух,
когда падает редкий снег. На темном фоне отдельные снежинки,
когда их немного, рельефно выступают. Рассматривать их, чтобы
лучше видеть форму, следует через лупу. Можно исследовать сне-
жинки и просто на рукаве пальто.
Размеры снежинок зависят от температуры воздуха: чем ниже
температура, тем они мельче. Вместе с тем меняется и их форма.
При очень низких температурах чаще всего выпадают мелкие пла-
стинки или иглы, при температуре близкой к 0° —звездочки, иногда
слипающиеся в хлопья; при положительных температурах нередко
падает ледяная «крупа», т. е. непрозрачные шарики разных разме-
ров, образовавшиеся благодаря намерзанию на снежинку воды.
Установлено также, что существует определенная зависимость ме-
жду формой снежинок и типом облаков, дающих снег. Так, низкие
разорванные дождевые облака образуют крупные разветвленные
282
Снежный покров
пластинчатые снежинки, высокие перистые и перистослоистые —
пластинчатые и стержневидные снежинки и т. д.
Высчитано, что средний вес снежной звездочки составляет около
0,1 мг, пластинки —0,007 мг, а иглы —еще в несколько раз ме-
нее. На вес серебряного гривенника звездочек придется 15—20
тысяч, а пластинок и призмочек — по 200—300 тысяч. Эти цифры
соответствуют только кристаллам средней величины; наибольшие
же и наименьшие кристаллы каждой формы отличаются друг от
друга по размерам в сотни и тысячи раз.
По приведенным числам легко рассчитать, что если землю по-
крыл довольно плотный снежный покров в метр толщиной, то на
квадратный метр выпало от нескольких миллиардов до нескольких
десятков миллиардов снежинок, т. е. в несколько раз больше, чем
живет людей на земном шаре!
Водные пространства в странах с холодной зимой покрываются-
льдом, который в свою очередь покрывается снегом. Лед, как из-
вестно, тем отличается от снега, что в то время как снег состоит из
отдельных кристалликов, между которыми помещается воздух, лед
представляет сплошную твердую массу. Из этого отличия происте-
кает большая плотность и, следовательно, большая теплопровод-
ность льда, а также и его цвет. Лед сохраняет бесцветность и
прозрачность воды, снег же имеет белый цвет, который обязан на-
ходящемуся между кристалликами воздуху. В последнем легко убе-
диться: если лед превратить в порошок (толчением или скоблением),
он приобретает вид белой снегообразной массы. Если снег напитать
водой, то последняя вытеснит воздух и снег потеряет белизну.
Находящийся на поверхности льда слой снега является причиной
сильного понижения теплопроводности. Это доказывается уже тем,
что ледяной покров, обнаженный от снега, сильно утолщается.
Поэтому-то предназначенный для колки лед в течение зимы обыкно-
венно очищают от снега.
Принято думать, что на реках, озерах и т. д. лед образуется
только сверху и что под поверхностным ледяным покровом темпе-
ратура воды до самого дна выше 0°. Исследованиями, однако, уста-
новлено, что это положение неверно. В реках, вообще в текучих
водах, вследствие течения происходит смешение верхних слоев
воды с нижними. То же происходит и в морях и больших озерах,
особенно у берегов и на местах мелких, благодаря волнению. Вслед-
ствие же смешения верхних и нижних слоев воды сравнивается по
всей глубине и температура. Охлаждаемая наверху до температуры
замерзания вода спускается вниз, где также происходит образо-
вание льда.
Этот процесс представляется в таком виде. После понижения
температуры воды до точки замерзания при продолжающемся мо-
розе происходит легкое переохлаждение всей массы воды ниже
точки замерзания. Тогда начинается образование ледяных кристал-
Донный лед
283
лов на разных глубинах и на поверхности охлаждаемых от сопри-
косновения с водой до той же температуры твердых предметов.
Особенно благоприятное место для образования ледяных кристал-
лов представляет само дно благодаря наличию прилегающего к
поверхности дна неподвижного слоя воды. Вследствие замерзания
воды происходит выделение тепла, но в то же время при продолжа-
ющемся морозе происходит и теплоотдача всеми слоями воды, смеши-
ваемой течением или волнением. Сталкиваясь на течении и смер-
заясь между собой, кристаллы льда находятся во всех слоях воды
в форме «ледяного сала» или «шуги» и, примерзая к грунту, водорос-
лям на дне ит. п., образуют губчатые массы рыхлого донного льда.
Отделяясь от дна и всплывая, донный лед при морозах крепнет с
поверхности и образует местный речной ледоход, предшествующий
замерзанию поверхности реки. В закрытых поверхностным льдом
частях реки образование донного льда прекращается, но прибыва-
ющий с верхних частей реки донный лед, цепляясь за неровные под-
водные выступы льдин, образует иногда весьма значительные за-
громождения русла реки, т. е. «заторы», иногда вызывающие мест-
ные наводнения. Образование донного льда представляет также
опасную помеху для работы гидростанций.
ПОЛОВОДЬЕ
Явления природы, регулярно повторяющиеся из года в год,
менее приковывают к себе наше внимание, нежели то, что совер-
шается неожиданно. Это, в частности, следует сказать про весеннее
половодье. Мы слишком привыкли к этому величественному явле-
нию и недостаточно оцениваем его. О каком-нибудь сильном летнем
ливне значительно больше разговора и его дольше помнят, нежели
о сильном весеннем разливе реки, хотя никакой ливень ни по ко-
личеству воды, ни по значению в природе не может сравниться даже
и со слабым сравнительно половодьем.
Весеннее половодье, сопровождающееся большим разливом рек,
и потому, между прочим, заслуживает нашего внимания к себе, что
это наше местное явление, в большом масштабе свойственное рекам
СССР. Ширина многих, сравнительно узких рек увеличивается при
этом в 10—50 раз. Разливы Дона, Волги и других крупных рав-
нинных рек превышают 10—20 км. В восточной Германии поло-
водье уже значительно слабее, а далее на запад постепенно сходит
почти на нет. Так, в то время как на Оке (в верхнем ее течении)
наибольшая амплитуда колебания уровня (с одной стороны — в
половодье, с другой — летом и осенью) достигает 17—18 ж, а в ниж-
нем течении Волги — 14—17 м, для Рейна (в Эммерихе) эта раз-
ница равняется всего только 1 м. В Сибири весенние разливы рек,
по мере движения на восток, также постепенно падают и исчезают.
Здесь это зависит от того, что сибирские реки — в меньшей степени
реки равнин, чем реки Европейской части СССР. Уже Обь получает
много воды с Алтая — от таяния снега и от летних дождей. К во-
стоку от Енисея вся местность гориста, и потому горные снега дают
все более и более воды, в долинах же зима становится все беднее
и беднее снегом. На горах снег тает позже, чем на равнинах. По-
этому, по мере движения на восток Сибири, взамен весеннего раз-
лива рек все большее и большее значение приобретают летние па-
водки, зависящие от таяния снега и от дождей на горах. Однако
масштаб этого явления здесь не меньше, а даже больше, чем в Евро-
пейской части СССР. Так, на Оби, Лене и других больших си-
Количество весенних вод
285
бирских реках амплитуда колебания уровня воды достигает 15—
20 ;и, а на Нижней Тунгуске — даже 30 м. Разливы Оби и Лены так-
же превышают 10—20 км. К востоку от Байкала снега выпадает
уже очень мало, за исключением некоторых прибрежных местно-
стей и горных склонов, летние же дожди становятся все обильнее,
так что в бассейне Амура от них главным образом и зависят раз-
ливы рек, достигающие очень крупных размеров. На Амуре при
амплитуде колебания уровня в 12—14 м разливы бывают на 10—
30 км. В Северной Америке, особенно в Канаде, снега выпадает
много более, чем в бассейне Волги. Но там в то же время много
озер, которые настолько регулируют течение рек, что наводнения
бывают только от ледяных заторов. Материки южного полушария
имеют настолько умеренную зиму, что на них не накопляется боль-
ших масс снега: он тает вскоре после того как выпал.
Количество весенних вод, получающихся во время нашего поло-
водья от таяния снега, находится в прямой зависимости от массы
этого последнего. В пределах Европейской части СССР снеговое
питание рек составляет от 50 до 100%, а количество снега —1/4
всех осадков; на востоке и севере оно доходит до 1/3, на западе умень-
шается до 1/5, уменьшается также и на юге. Сами по себе эти цифры,
пожалуй, еще не очень внушительны. На основании их одних при-
шлось бы признать, что поверхность нашей страны — ее почва и
водные бассейны — в значительно большей зависимости находится
от дождей, падающих в теплое время года и составляющих около
3/4 всех осадков.
В действительности картина оказывается иной, если принять
во внимание еще другие данные — количество испаряющейся влаги.
Наблюдения на метеорологических станциях, расположенных в
разных местах СССР, показывают, что в теплое время года, с апреля
по октябрь, испарение всегда превышает осадки; в холодные же
месяцы, с ноября по март, осадков больше, нежели испарения.
Следовательно, на развитие растительности расходуется главным
образом вода, скопленная в почве поздней осенью, зимой и ранней
весной, т. е. запас атмосферных осадков, выпадающих преимущест-
венно в виде снега. Ко времени пробуждения растительности они
глубоко пропитывают почву, образуя грунтовые воды, и надолго
служат источником питания для корней. В то же время, перепол-
няя реки весною и поступая в них потом из грунтовых вод, эти
осадки служат главным источником питания и для речных бассей-
нов. Летние же осадки идут преимущественно на покрытие потерь,
происходящих от испарения. Мелкие дожди, падающие на про-
сохшую почву, едва успевают смочить тонкий слой последней, как
уже принесенная ими влага снова испаряется.
Что касается сроков весеннего половодья, то они зависят от
климатических условий и связанного с ними времени снеготая-
ния. В СССР на реках юго-западной части Восточно-Европейской
286
Половодье
низменности половодье наблюдается в среднем в марте—апреле,
что является наиболее ранним сроком. Самое позднее половодье
отмечено для рек северной части Западно-Сибирской низменности
(июнь—июль).
Чтобы дать более наглядное представление о весеннем расходе
вод и, в частности, о разливе рек, приведем некоторые результаты
13-летних наблюдений (с 1884 по 1897) над половодьем в верховье
Оки, от истоков ее до города Орла, собранных бывш. Гидротехни-
ческим отделом.
Площадь, которую охватили эти наблюдения, составляет
4300 кв, км, длина реки на этом расстоянии равняется 100 км. Ве-
сеннее половодье продолжается здесь в среднем 56 дней — от конца
февраля до конца апреля (ст. ст.). За это время рекою протекает
3/4 всего годового количества воды: полный годовой расход ее здесь
определен в 450 млн. куб, м, из которых 340 млн. приходится на
время весеннего половодья, а остальные составляют расход дру-
гого времени года в 309 дней. Средний расход воды в одну секунду
за время половодья определен в 72 куб. м, а за остальное время —
в 4 куб, м. Увеличение расхода воды, в среднем за 13 лет,
начинается с половины февраля, так что средний секундный рас-
ход, равнявшийся в январе 4 куб, м, в среднем за февраль равен
уже 7,2 куб, м, средний расход в марте равен 88 куб, м, в апреле —
40 куб, м, а в начале мая половодье прекращается, и средний май-
ский секундный расход падает до 4 куб, м.
Исчисление всего количества снега на площади бассейна вер-
ховьев Оки и отношение этого количества к расходу вод во время
весеннего половодья дало возможность определить, что только
45% зимних осадков стекает весной непосредственно в реку. Ос-
тальная же часть главным образом пополняет подземные запасы
вод и питает ключи, так как испарение воды в это время при отсут-
ствии растительности и низкой температуре небольшое. Произве-
денные в то же время наблюдения над дождем дали возможность
определить, что после больших ливней только 12% воды стекает
в реку, а из всего количества дождя, выпадающего в период от окон-
чания половодья до покрытия реки льдом, только 7% поступают
в реку как поверхностно, так и подземным стоком. Если принять,
что общее количество осадков, выпадающих в виде дождя, втрое
превышает количество снега, и следовательно, 7% этих осадков рав-
няются приблизительно 21 % всей массы снега, то и в таком случае
выходит, что в меженные месяцы, т. е. между двумя половодьями,
верховья Оки питаются главным образом за счет зимних запасов
влаги, поступающих из подземных ключей.Это видно,между прочим,
и из того, что в те годы, когда количество снега было большое, а
половодье по каким-либо причинам сравнительно слабое,— рас-
ход воды в меженные месяцы бывал наибольший сравнительно с
другими годами. Наоборот, в те годы, когда при малом количестве
Факторы, определяющие размер половодья
281
снега был большой весенний разлив реки, меженный расход
воды в ней бывал очень мал.
Так как Ока представляет одну из типичных рек Европейской
части СССР, то изложенные выше результаты наблюдений над раз-
ливом верховьев Оки рисуют довольно общую картину половодья
для всех почти наших рек. Отклонения в ту или другую сторону в
отношении продолжительности, а следовательно, и силы половодья
зависят главным образом оттого, в каком направлении течет река —
широтном или меридиональном. Если река течет в направлении
географической широты местности, т. е. с запада на восток или на-
оборот, то в таком случае таяние снега по ее длине, ввиду близо-
сти климатических условий и одинаковых температур, совер-
шается одновременно и разлив реки — дружный. Если же река
течет в меридиональном направлении, т. е. с севера на юг или нао-
борот, тогд^ таяние снега на ее протяжении, а следовательно, и
половодье растягивается, ввиду постепенного продвижения тепла
с юга на север, на больший промежуток времени.
Приведенные результаты, как уже указано, представляют сред-
ний вывод из наблюдений за 13 лет. Но если взять разливы отдель-
ных лет, то они друг от друга отличаются иногда очень сильно.
Каждый из собственных наблюдений знает, что как время наступ-
ления половодья, так и его продолжительность и сила разлива под-
лежат колебаниям. Иногда весенние разливы рек производят боль-
шое опустошение и причиняют жителям большие убытки. Таков,
например, бывший в 1908 г. чрезмерный разлив реки Москвы, за-
лившей более низкие части города и причинившей жителям ее
убыток на сумму около 200 млн. рублей.
От каких причин зависит сила разливов? Нельзя ли эти причины
заранее учесть и на этом основании предвидеть размер половодья,
что было бы чрезвычайно важно для принятия, в случае необходи-
мости, мер предосторожности?
Пока вопрос о предвидении (прогнозах) наводнений не получил
еще надлежащей разработки. Тем не менее, известный специалист
в этой области, Н. С. Нестеров, предсказавший московское навод-
нение 1908 г., находит возможным ответить на него утвердительно.
Главнейшие факторы, определяющие размер разлива реки, он счи-
тает подлежащими заблаговременному более или менее точному
учету.
Эти факторы следующие: 1) количество осадков, выпадающих
за зиму в речном бассейне; 2) расход этих осадков в течение зимы;
3) продолжительность периода таяния снега весной и 4) коэффи-
циент стока воды по поверхности речного бассейна.
Количество осадков определяется по толщине или глубине и
плотности снежного покрова посредством реек на большинстве ме-
теорологических станций СССР; на некоторых станциях ведутся
наблюдения и за плотностью снега.
288
Половодье
Зимний расход осадков имеет также большое влияние на силу
разлива. Расход этот зависит главным образом от оттепелей, во
время которых талая вода отчасти стекает, отчасти просачивается
в почву. Точный учет этого расхода можно производить путем си-
стематических наблюдений над дебитом воды (т. е. количеством
воды в единице времени) в ключах или родниках, наблюдениями
над уровнем грунтовой воды и уровнем реки перед ледоставом и
в течение зимы. Колебания этого уровня выражают собой в общих
чертах расход снеговых и грунтовых вод в продолжение зимы в
речном бассейне. Представим себе два случая: один — когда уро-
вень воды в реке за зиму упал чрезвычайно сильно, и другой —
когда вследствие бывших зимой оттепелей уровень воды перед на-
ступлением половодья поднялся высоко. Очевидно, что при прочих
равных условиях (при одинаковой толщине снежного покрова
перед половодьем и т. д.) во втором случае весенний разлив должен
быть сильнее,так как в первом — значительное количество снеговой
воды уйдет на пополнение сильно упавшего уровня реки. Ясно,
что определение расхода зимних осадков имеет большое значение
в предвидении наводнений.
Третий из указанных факторов, определяющих размеры весен-
них разливов рек, — продолжительность периода таяния снега —
в свою очередь зависит от следующих причин: 1) от характера
растительного покрова (лес или травянистые растения); 2) от усло-
вий погоды во время весеннего таяния снегов и 3) от плотности
снега. Значение леса как фактора, увеличивающего продолжитель-
ность таяния снега, будет указано ниже; здесь отметим лишь, что
этот фактор поддается точному учету. К сожалению, нельзя того
4же сказать относительно погоды, что, конечно, вносит известную
условность в предсказание наводнений. Высокая температура,
дожди и сильные теплые ветры значительно ускоряют таяние снеж-
ного покрова. Большим противовесом при этом могут служить ноч-
ные заморозки, сковывающие талые воды и тем задерживающие
сток их и удлиняющие период снеготаяния. Что касается плотно-
сти снега, то рыхлый снег тает значительно быстрее. Частые снеж-
ные заносы зимой — наглядное свидетельство наличия рыхлого
снежного покрова. Если зима изобиловала заносами и если за ме-
телями не было больших оттепелей, во время которых снег, уплот-
няясь, садится, — всегда надо опасаться быстрого разлива
воды.
Последний из названных факторов, обусловливающих размер
разлива рек, именно коэффициент стока талых вод, помимо топо-
графии почвы (ровная или наклонная и в какой степени) и со-
става ее (быстро или медленно пропускает воду),в большей степени
зависит от состояния почвы — мерзлая или мягкая почва нахо-
дится под снежным покровом. По поверхности мерзлой земли сток
сильно увеличивается. Еще более увеличивается он, когда промерз-
Значение лесов
289
шая земля покрылась ледяной корой, образующейся в том слу-
чае, если после оттепели следуют морозы.
Итак, из приведенного краткого обзора главнейших факторов,
обусловливающих высоту весеннего разлива вод, видно, что все
они, за исключением погоды при таянии снегов, могут заранее под-
лежать точному учету, на основании которого возможно заранее
правильно определить размер разлива данной реки. Наблюдения
над состоянием снегового покрова и другими факторами, обуслов-
ливающими половодье, в настоящее время ведутся в ряде пунктов
СССР. Руководство гидрологическими прогнозами сосредоточено
в Главном управлении гидрометеорологической службы СССР.
На все указанные факторы и на общую картину половодья силь-
ное влияние оказывает лес. Он влияет прежде всего на толщину
снегового покрова, так как часть снега задерживается кронами
деревьев. Особенно много осадков задерживается в хвойных насаж-
дениях, где листва (хвоя) не опадает на зиму. По измерениям
Н. С. Нестерова, в лесной даче Московского сельскохозяйственного
института в березовом насаждении 70—75-летнего возраста задер-
живается зимних осадков 4—5%, а в хвойных насаждениях —до
50—55%. С изменением состава, возраста и сомкнутости древес-
ного насаждения эти цифры сейчас же меняются.
Благодаря защите древесного полога от нагревания солнечными
лучами и от лучеиспускания, колебание температуры воздуха в
лесу значительно меньше, чем вне леса. Циркуляция воздуха там
также меньше. Вследствие этого там не бывает такого таяния снега
зимой от оттепелей, какое наблюдается на открытых местах.
Благодаря той же защите древесного полога от солнца и ветра
и задерживанию дождевых осадков, таяние снега в лесу наступает
позднее и идет гораздо медленнее, продолжаясь на 3—5 недель
дольше, чем на открытом месте. За 10 лет наблюдений в Петровско-
Разумовском, около Москвы, период весеннего таяния снегов и
стока талых вод в лесу продолжался от 26 до 27 дней, тогда как в
поле процесс оканчивается иногда в 5—6 дней. Влияние состава
леса на таяние снега видно из следующих приводимых Н. С. Не-
стеровым данных об обнажении земли от снега в 1908 г. На полях,
вырубках и вообще открытых местах снег сошел к 22 апреля, в
молодняках, несомкнувшихся — к 24 апреля, в редколесье юж-
ного склона — к 26 апреля, в сосняках — к 7 мая и в ельниках —
к 14 мая. Такая последовательность в таянии снега замечается
всегда и вовсе не зависит от условий погоды.
Чем дольше продолжается период таяния снега, т. е. чем на боль-
ший промежуток времени растягивается сток воды, тем он менее
интенсивен. В то же время, чем дольше талые воды омывают поверх-
ность почвы, тем они дольше, а следовательно, и в большем коли-
честве поглощаются ею, что в свою очередь также оказывает влия-
ние на силу разлива.
290
Половодье
Влияние леса на количество стекающей воды весьма сильно
еще и по следующим причинам.
В лесу почва не так промерзает, как на открытых местах. Дре-
весный полог защищает лесную почву от лучеиспускания. Лесная
подстилка, состоящая из опавших листьев и сучьев, будучи пло-
хим проводником тепла, играет для почвы роль как бы «одеяла»;
такое же значение имеет и снежный покров, покрывающий почву
в лесу более равномерным слоем. В том же направлении действуют
и происходящие в лесной подстилке процессы гниения и брожения,
сопровождающиеся выделением теплоты. Благодаря этой слож-
ной защите и слабому движению воздуха в лесу, лесная почва или
совсем не замерзает зимой, или же если и замерзает, то гораздо
позднее и на значительно меньшую глубину, чем на открытых ме-
стах, причем она рано оттаивает, находясь еще под толщей снега.
Поэтому впитывание воды лесной почвой значительно больше,
чем в поле.
Вместе с тем на склонах, хотя бы и на крутых, лесная почва с
рыхлым слоем подстилки играет роль гигантской губки: она погло-
щает воду медленно тающего снега и по насыщении отдает влагу
нижележащим слоям. Если, несмотря на это, и окажется избыток
осадков, он стекает медленно, постоянно задерживаясь стволами,
бесчисленными сплетениями корней и т. п., причем на каждом шагу
стекающая вода может попасть в ненасыщенное еще место.
Благодаря всем указанным условиям лес, ослабляя весенние
разливы рек, накопляет большой запас Цочвенной влаги, которая
идет затем на питание грунтовых вод и ключей и самих же рек
в меженные месяцы, уменьшая их летнее и осеннее мелководье.
В этом великая роль леса как могучего регулятора вод в эконо-
мии природы.
Так как в лесу и на открытых местах таяние и образование мак-
симального количества талых вод происходит не одновременно, то
поэтому и наиболее сильный сток воды в лесистых местностях про-
исходит не сразу, а по частям. Такое значение леса давно уже под-
мечено русским народом. В лесистых местностях северных и сред-
них районов СССР население различает в половодье три воды, из-
вестные под разными местными названиями. Так, в районе Костромы
различаются полевая, лесная и коренная воды, в Горьком —сне-
жица, лесная и подошвенная. Первая вода стекает с поверхности
открытых мест (полей, лугов и пр.), вторая, появляющаяся позд-
нее первой,— тоже верховая вода, но стекающая с лесных площа-
дей; третья — грунтовая вода, в большом количестве поступающая
в реки в лесистых местностях, но уже много времени спустя (через
1—Р/г—2 месяца) после окончания стаяния снегов.
Вместе с тем замечено также, что с вырубкой лесов все три воды
«сходятся» в одно время. Это вполне понятно: запасы грунтовых
вод с уничтожением леса уменьшаются, лесной воды нет, и посту-
Образование оврагов
291
пает в повышенном размере одна лишь «полевая» вода. Другими
словами, неизбежным последствием уничтожения лесов являются
наиболее сильные разливы рек весной и последующее мелководье,
ухудшающее сплавоспособность и судоходность рек.
Из сказанного ясно, что наилучшим средством для борьбы с
опустошительными весенними разливами рек и для рационального
использования в возможно большем количестве снеговых вод яв-
ляется сбережение лесов, правильное лесное хозяйство и разведе-
ние леса там, где он вырублен, особенно по склонам долин, берегов,
ложбин и т. п. Хвойный лес, в котором таяние снега задерживается
долее всего, должен быть поставлен в этом отношении на первое
место.
В местностях безлесных весенние разливы являются причиной
образования оврагов. В средней и особенно южной части СССР ов-
рагов чрезвычайно много, и они обязаны своим возникновением и
развитием частью большим ливням, а более всего — весенним во-
дам. На склоны оврагов устремляется талая вода с окружающего
поля или степи. Мутные потоки ее, скатываясь со склонов вниз,
производят огромную механическую работу. Они размывают и раз-
рушают почву, промывают глубокие промоины, увлекают целые
глыбы пород, словом, вызывают усиленное разрастание оврагов и
образование новых. Небольшое углубление, появившееся на от-
логом месте, например, сделанная человеком канава или бороздка,
если сюда по наклону устремляется поток воды, может через ряд
лет превратиться в большой овраг. Если по берегу образовавшегося
оврага человек уничтожает лес, а особенно, если подпахивает и
травянистый покров, который сплетением многочисленных корней
все же скрепляет почву,— то этим оказывается большое содействие
быстрому росту оврага. К сожалению,такое содействие оврагообра-
зованию со стороны человека имело и теперь иногда еще имеет
место.
Являясь главной причиной образования оврагов, весенние
разливы оказывают сильное влияние на окружающую местность,
вредное для интересов человека.
На равнине или на пологих склонах вода, стекая медленно,
более или менее равномерно пропитывает почву и увеличивает за-
пасы грунтовых вод. В местности же, изрезанной оврагами и рыт-
винами, вода, быстро скатываясь вниз по этим естественным жело-
бам, сбегая в реки и речки и, следовательно, увеличивая весенний
разлив их, почти не увлажняет почвенного слоя. По наблюдениям,
почва вблизи оврагов всегда суше, нежели вдали от них. Это зави-
сит еще от того обстоятельства, что в овраги зимой сдувается боль-
шое количество снега, а открытая местность около них в соответст-
вующей мере оголяется. Кроме того, оврагами увеличивается испа-
ряющая поверхность почвы. Из всего этого ясно, что овраги оказы-
вают влияние, обратное тому, какое оказывает лес, регулирующий
292
Половодье
и задерживающий быстрое стекание полой воды, а через это
увеличивающий запасы грунтовых вод и увлажнение почвы.
Это еще далеко не все. Оврагами сокращается площадь годной
для обработки земли. С одной стороны, в них весенними водами,
а равно и ливнями в большом количестве сносятся с окружающего
поля наиболее тонкие и плодородные частицы почвы, с другой —
на скатах самих оврагов обнажаются глины, пески и другие породы,
не имеющие хозяйственного значения. Затем огромное количество
продуктов разрушения пород и почв, выносимых из оврагов, отла-
гаясь в реках, озерах и прудах, способствует их обмелению.
Хорошо известно, что заливные луга, как правило, дают
обильные травы. Вешние воды несут много мелких илистых частиц,
которые, осаждаясь в долине, удобряют ее почву. Однако в области
развития оврагов мы видим противоположную картину. Обильное
количество глины, песка и т. п., выносимое здесь водой, засоряет
луг, может сделать его совершенно бесплодным и даже превратить
в болото.
Чтобы предупредить эту вредную деятельность весенних вод,
необходимо, как уже сказано, бережное обращение человека с ле-
сом и вообще с растительностью, в особенности на склонах уже
образовавшихся оврагов. Сверх того необходимы и более активные
меры борьбы. К этим мерам относятся: обсаживание оврага, в осо-
бенности вершины его, растительностью и выстилание хворостом,
устройство поперек оврага и над вершинами его запруд и т. п. Кол-
хозы черноземной полосы СССР особенно энергично проводят борьбу
с оврагами, развитие которых наносит большой вред сельскому хо-
зяйству.
С целью большей утилизации полой воды на юге СССР в неко-
торых хозяйствах начинают применять так называемое лиманное
орошение, т. е. устраивать запруды для получения заливных лугов.
На полях весьма целесообразным средством для этой же цели яв-
ляется так называемый «черный пар», т. е. вспахивание парового
поля не весной, а предшествующей осенью. Между бороздами бо-
лее удерживается снега, через вспаханную землю талая вода лучше
просачивается, кроме того, за счет борозд увеличивается и всасы-
вающая поверхность почвы.
Л ИТЕРАТУРА
Алехин В. В. Растительность СССР в основных зонах, 2-е изд., «Со-
ветская наука», М., 1951.
Вузовский курс ботанической географии СССР, содержащий из-
ложение главнейших закономерностей распространения растительного
покрова и характеристику основных ботанических зон СССР. Много
внимания уделено биологии отдельных видов.
Беляева Л. Т. Ботанические экскурсии в природу, Учпедгиз, М., 1955.
Пособие для учителей, освещающее вопросы организации ботани-
ческих экскурсий в природу.
Васильев И. М. Зимовка растений, изд. АН СССР, М., 1956.
Сводка современных представлений о зимовке растений в связи
с практическими задачами растениеводства.
Голлербах М. М. Споровые растения как объект школьных экскур-
сий, Лен. гор. ин-т усоверш. учителей, Л., 1945.
Содержит интересный материал по биологии водорослей, грибов,
лишайников, мхов и папоротникообразных.
Дарвин Чарлз. Происхождение видов путем естественного отбора
или сохранение благоприятствуемых пород в борьбе за жизнь. Ряд из-
даний.
Основной классический труд Дарвина.
Доппельмаир Г. Г., Мальчевский А. С., Новиков Г. А.
и Фалькенштейн Б. Ю. Биология лесных зверей и птиц, Гос-
лесбумиздат, М. — Л., 1951.
Учебник, рассчитанный на студентов лесотехнических вузов. Со-
держит много материала по биологии отдельных видов лесных живот-
ных.
Калабухов Н. И. Спячка животных, 2-е изд., «Советская наука»,
М., 1946.
Книга, весьма полно освещающая вопросы распространения явле-
ния снячки в природе, физиологических изменений при спячке и влия-
ния внешних факторов на возникновение и течение спячки.
Кожевников А. В. Весна и осень в жизни растений, 2-е изд. Моск,
об-ва испытателей природы, М., 1950.
Научно-популярная книга, содержащая большой материал по био-
логии наших древесных и травянистых растений, изложенный приме-
нительно ко временам года: весне и осени.
Комаров В. Л. Введение в ботанику, «Советская наука», М., 1949.
Вузовский курс ботаники, содержащий большой и разнообразный
материал по биологии растений.
Любименко В. Н. иИльин М. М. Сорные растения наших полей
и огородов, Гос. Учебно-пед. изд-во, М. — Л., 1931.
Общая биологическая характеристика сорняков и описание от-
дельных видов сорных растений.
294
Литература
Максимов Н. А. Краткий курс физиологии растений, 8-е изд., Сель-
хозгиз, М., 1948.
Вузовский курс физиологии растений. Специальный раздел посвя-
щен устойчивости растений против неблагоприятных внешних воздей-
ствий.
Мальцев А. И. Сорная растительность СССР и меры борьбы с нею,
3-е изд., Сельхозгиз, М. — Л., 1936.
Вузовский курс. Большое внимание уделено биологическим осо-
бенностям сорных растений.
Н а с и м о в и ч А. А. Роль режима снежного покрова в жизни копытных
животных на территории СССР, изд. АН СССР, М., 1955.
Монографическая работа, подробно освещающая роль снежного
покрова в жизни копытных; основана на собственных многолетних ис-
следованиях автора.
Наумов Н. П. Экология животных, «Советская наука», М., 1955.
Вузовский курс, затрагивающий в ряде других проблем также и
сезонные явления в мире животных.
Новиков Г. А. Роль позвоночных животных в жизни леса, «Животный
мир СССР», т. 4, изд. АН СССР, М. — Л., 1953.
Работа содержит богатый материал, характеризующий многооб-
разное значение позвоночных животных в жизни леса.
Огнев С. И. Жизнь леса, 5-е изд. Моск, об-ва испытателей природы,
М., 1950.
Описание жизни животных и растений леса по сезонам.
Полянский И. И. Ботанические экскурсии, 2-е изд., Учпедгиз, Л. —
М., 1950.
Книга содержит описание ботанических экскурсий (в лес, на луг,
на болото для изучения водной растительности, сорной раститель-
ности и т. п.) и отдельных растений нашей флоры, с обращением осо-
бого внимания на их биологию.
Попов Н. В. Фенологические наблюдения в школе, 2-е изд., Учпедгиз,
М., 1953.
Методическое пособие для учителей, освещающее вопросы органи-
зации фенологических наблюдений в школе, значение их в учебно-вос-
питательном процессе и в практической деятельности.
ПромптовА. Н. Сезонные миграции птиц, изд. АН СССР, М. — Л.,
1941.
Изложение современного положения вопроса о перелетах птиц.
Включает большой материал по биологии птиц.
Райков Б. Е. и Римский-Корсаков М. Н. Зоологические
экскурсии, 6-е изд., Учпедгиз, Л., 1956.
Богатое биологическим материалом руководство по зоологическим
экскурсиям в природу.
Рихтер Г. Д. Снежный покров, его формирование и свойства,
изд. АН СССР, М. — Л., 1945.
Научно-популярная книга. Отдельная глава посвящена снежному
покрову СССР.
Рихтер Г. Д. Роль снежного покрова в физико-географическом про-
цессе, изд. АН СССР, М. — Л., 1948.
Основной труд по данному вопросу, освещающий влияние
снежного покрова на природные условия: климат, почву, расти-
тельный и животный мир. Специальный раздел посвящен характери-
стике снежного покрова на территории СССР, в частности, его райони-
рованию.
Соколов А. А. Гидрография СССР (воды суши), Гидрометеоиздат, Л., 1952.
В книге излагаются закономерности географического распростра-
нения вод, основные особенности водного и ледового режима рек и озер
и кратко описываются главнейшие реки и озера СССР.
Литература
295
Тимирязев К. А. Чарлз Дарвин и его учение. Ряд изданий.
Классическое общедоступное изложение учения Ч. Дарвина.
Туманов И. И. Физиологические основы зимостойкости культурных
растений, Сельхозгиз, Л., 1940.
Основной труд по физиологическим основам зимостойкости важ-
нейших сельскохозяйственных культур.
Туманов И. И. Основные достижения советской науки в изучении мо-
розостойкости растений, изд. АН СССР, М., 1951.
Небольшая книга, характеризующая современное положение проб-
лемы морозостойкости растений и значение трудов советских исследо-
вателей в ее разработке.
Туров С. С. Жизнь птиц, 2-е изд. Моск, об-ва испытателей природы,
М., 1951.
Богатая фактическим материалом, популярно написанная книга,
освещающая основные вопросы биологии птиц, а также их распреде-
ление по ландшафтным зонам СССР.
Формозов А. Н. Снежный покров, как фактор среды, его значение
в жизни млекопитающих и птиц СССР, изд. Моск, об-ва испытателей
природы, М., 1946.
Подробная характеристика значения снежного покрова в разных
зонах СССР для жизни млекопитающих и птиц.
Формозов А. Н. Шесть дней в лесах, 6-е изд., Детгиз, М., 1950.
Популярная книга, излагающая наблюдения юных натуралистов
над жизнью млекопитающих и птиц.
Фриш К. Из жизни пчел, Биомедгиз, М. — Л., 1935.
Популярное изложение работ автора над поведением пчел.
X е й н р о т Оскар. Из жизни птиц, Гос. изд. иностр, литературы, М.,
1947.
Научно-популярный очерк, затрагивающий различные стороны
биологии птиц: их размножение, окраску, питание, движение и нерв-
ную деятельность.
ШенниковА. П.Экология растений, изд-во «Советская наука», М., 1950.
Вузовский курс экологии растений. В одной из глав специально
рассматривается вопрос о фенологических наблюдениях.
Шиголев А. А. иШиманюк А. П. Сезонное развитие природы Ев-
ропейской части СССР, Госиздат, М., 1949.
Книга содержит методические указания к производству феноло-
гических наблюдений и обширный календарь природы для Европей-
ской части СССР (за 15 лет).
Шмидт П. Ю. Миграции рыб, 2-е изд. АН СССР, М. — Л., 1947.
Сводка, освещающая основные проблемы миграций рыб; рассчи-
тана на подготовленного читателя.
Шмидт П. Ю. Анабиоз, 4-е изд. АН СССР, М. — Л., 1955.
Полная сводка вопросов, связанных с влиянием низких темпера-
тур на организм. В 4-м, посмертном, издании дополнена статьей проф.
Л. К. Лозина-Лозинского, критически излагающей содержание иссле-
дований в области анабиоза за последние годы.
Шульц Г. Э. и Шам раевский В. В. Фенологические наблюде-
ния. Практическое руководство к производству наблюдений над сезон-
ными явлениями живой природы, ГЭНИИ, Л., 1941.
Описание ряда работ по фенологии.
Щербиновский Н. С. Сезонные явления в природе, 3-е изд , Сель-
хозгиз, М., 1954.
Научно-популярная книга, описывающая сезонный ход развития
природы в разных географических условиях — от Арктики до тропиков.
Специальная глава посвящена методике фенологических наблюдений.
СОДЕРЖАНИЕ
От редакции ................................................ 3
Введение ................................................... 5
Весеннее пробуждение растений ..............................12
Цветение растений ..........................................29
Распространение плодов и семян .............................78
Осенняя листва и листопад .................................103
Зимовье растений ..........................................114
Сезонные перемены покровов и окраски у животных ...........141
Перелеты птиц .............................................154
Переселения других животных ...............................189
Гнездовье птиц и уход за птенцами .........................209
Зимовье животных ..........................................232
Жизнь подо льдом ..........................................254
Снежный покров ............................................265
Половодье .................................................284
Литература ................................................293