/













































































































































































































































































































































































































Author: Кузнецов Б.А. Чернов А.З.
Tags: общая зоология зоология ветеринария
ISBN: 5—10—000460—6
Year: 1989




Text
УЧЕБНИКИ И УЧЕБНЫЕ ПОСОБИЯ
ДЛЯ СТУДЕНТОВ ВЫСШИХ УЧЕБНЫХ ЗАВЕДЕНИЙ
(курс зоологий)
Б. А. КУЗНЕЦОВ
А. 3. ЧЕРНОВ
Л.Н.КАТОНОВА
ИЗДАНИЕ ЧЕТВЕРТОЕ,
ПЕРЕРАБОТАННОЕ
И ДОПОЛНЕННОЕ
Допущено Управлением высшего и среднего специаль-
ного образования Государственного агропромышлен-
ного комитета СССР в качестве учебника для сту-
дентов высших учебных заведений по специальности
«Защита растений»
МОСКВА ВО -АГРОПРОМИЗДАТ- 1989
ББК 28.6
К89
УДК 591 (075.8)
Ред а к т о р Е. В. Мухортова
Рецензент - доктор биологических наук, профессор И. Т. Покозий
Кузнецов Б. А.- и ftp.
К 89 Курозоологи^/Б.'А. Кузнецов, А. 3'. Чернов, Л. Н. Като-
новй.-—"4*е ИЗД'., ГЬер^рШ. и доп.—/М.: Агропромиздат,
1989.— 399 с.: ил,— (Учебники и учеб, пособия для студен-
тов высш. учеб, заведений).
ISBN 5—10—000460—6
В общей части учебника рассматриваются строение и развитие жи-
вотных, эволюция животного мира; в систематическом разделе опи-
сание животных. В четвертое издание (третье вышло в 1978 г.) внесены
дополнения и уточнения как в темы общей части курса, так и в описание
отдельных систематических категорий животных.
Для студентов по специальности «Защита растений».
1907000000-008 ПОС( оо
------------— 238—88
035(01)—89
ISBN 5-10—000460-6
К
ББК 28.6
© Издательство «Высшая школа», 1978
© ВО «Агропромиздат», 1989,
с изменениями
ПРЕДИСЛОВИЕ
К ЧЕТВЕРТОМУ ИЗДАНИЮ
Изучение зоологии имеет большое значение для подготовки вы-
сококвалифицированных специалистов сельского хозяйства. По-
знание животного мира важно для общебиологического образова-
ния и формирования материалистического мировоззрения. В то
же время зоология предоставляет будущим специалистам комп-
лекс научных знаний, полезных для понимания ряда задач сель-
скохозяйственного производства.
Авторы стремились создать учебник по сельскохозяйствен-
ной зоологии, в котором были бы, с одной стороны, даны осно-
вные теоретические положения зоологической науки, с другой —
изложены нужные работникам сельского хозяйства сведения о
животных.
При подготовке четвертого издания учебника внесены допол-
нения во введение и общую часть. Расширена информация о нау-
ках — отраслях зоологии. Обращено внимание на углубление све-
дений, имеющих отношение к профессиональным интересам, обу-
чающихся в сельскохозяйственных вузах. Уточнена систематика.
Главы «Экологиями хозяйственное значение птиц», «Экология
и хозяйственное значение млекопитающих», «Охрана и рациональ-
ное использование животного- мира в СССР» написаны заново
доцентом кафедры зоологии ТСХА Л. Н. Катоновой.
ВВЕДЕНИЕ
Зоология в сельскохозяйственных вузах. Зоология (греч. zoon —
животное, logos — учение) — наука о животных, часть биологии,
изучающая многообразие животного мира (строение и жизне-
деятельность, распространение, связь со средой обитания и т. д.).
Зоология способствовала обоснованию общебиологической теории
развития органической природы. На протяжении становления на-
уки о животных направления исследований и методы менялись.
В давние времена преобладал описательный метод. Но и тогда
описание животных дополнялось изучением процессов, протека-
ющих в организмах, с постановкой экспериментов. Опыты в приро-
де и лабораториях способствовали углубленному познанию живот-
ного мира, объяснению особенностей строения разных животных
и их взаимоотношений с живой и мертвой природой. В наши дни
успехи зоологии и других наук о природе сделали возможным
изыскания путей воздействия на биологические процессы в ор-
ганизмах в расчете познать способы управления их жизнедеятель-
ностью.
Теоретические изыскания расширили использование научных
сведений для удовлетворения потребностей людей и народного
хозяйства. Постепенно стали зарождаться производственные от-
расли зоологической науки и каждая со своими теоретическими
и практическими задачами. Среди них — сельскохозяйственная
зоология, накопившая материалы большого значения для разных
направлений сельскохозяйственного производства. Об этом свиде-
тельствуют приводимые примеры.
Утверждение эволюции животных, открытие факторов эволю-
ционного процесса способствовали разработке проблемы про-
исхождения домашних животных и стали основой селекционной
работы в животноводстве.
Сложны и разнообразны взаимоотношения животных с миром
растений. Большие работы проведены по проблеме взаимозависи-
мости двух царств органической природы. Обращено внимание на
почвенную фауну: описаны многие виды почвенных животных, их
экология. В почвах, не протравленных ядами, обитают животные
разных типов, особенно многочисленны беспозвоночные. Так, од-
4
ноклеточные встречаются в 1 г почвы до 1,2 млн особей, что в пере-
счете на площадь в 1 м2 составляет более 100 млрд. На
площади того же размера (1 м2) обнаружены миллионы червей —
коловраток, десятки миллионов нематод, сотни дождевых червей,
десятки и сотни тысяч клещей и первичнобескрылых насеко-
мых. Суммарная масса животных, населяющих почвенный слой
Земли, па 1 га может достигать 4 и более тонн. Выяснена ре-
шающая роль этой живой силы в образовании почвенного слоя
Земли. Почвенные животные наряду с бактериями влияют на
физико-химические процессы, протекающие в почве, на ускорение
переработки растительного опада, трупов животных, их экскремен-
тов, а в целом — па плодородие почвы. В почве животные
находятся в контакте с подземными органами растений и, сле-
довательно, представляют собой важный биотический фактор сре-
ды жизни растений.
Изучение биологии животных углубило понимание экологиче-
ских связей в органической природе и открыло путь биологичес-
кому методу борьбы с животными, приносящими вред, путем
использования их естественных врагов. Например, в борьбе с сов-
ками, вредной черепашкой и рядом других насекомых-вредителей
разводят и в нужное время выпускают па поля паразитических,
перепончатокрылых насекомых — наездников и яйцеедов.
Преимущество биологического метода над применением хими-
ческих препаратов бесспорно, так как ядохимикаты губят пеутоль-
ко вредных, по и многих полезных животных.
Но не только биологические способы борьбы с вредителя-
ми, а и ряд химических, основываются на знании строения и фи-
зиологии животных, их жизненных повадок, среды обитания, к
которой они приспособлены. Многие химические препараты приме-
няют с учетом особенностей питания или дыхания животных, с
которыми ведется борьба.
Агротехнические приемы (например, лущение стерпи) также
базируются на осведомленности о биологии животных. Учитывает-
ся распределение вредителей в слоях почвы в разные периоды
жизни.
Около 700 видов паразитических червей известны как воз-
будители заболеваний домашних животных и сельскохозяйствен-
ных растений. Данные о биологии паразитов и их хозяев —
необходимая предпосылка при выработке мер профилактики, нре-
д у и р е ж д ei i и я з а бол е в а и и й.
Прямое отношение к вопросам сельского хозяйства имеет
знание участия животных в круговороте веществ, наглядно про-
слеживаемого в периоды миграций животных, больших и малых.
В. И. Вернадский (1863—1945) —основатель науки биогеохи-
мии— обосновал роль животных в круговороте химических эле-
ментов на нашей планете, их транспортировке по лику Земли, учас-
тие в формировании земной коры и почвы. Перемещение животных
происходит повсеместно и всюду. Оно особенно обращает па себя
внимание во время массовой миграции, например, саранчи. В своей
книге «Биосфера» (1936) В. И. Вернадский приводит наблюде-
ние натуралиста Карутерса, описавшего грандиозный перелет са-
ранчи в 1889 г. с берегов Северной Африки в Аравию. Размер
лишь одной из туч натуралист определил в 5967,3 км2, массу
которой можно принять за 4,4-107 т. Такое количество саранчи,
выраженное в химических элементах и метрических тоннах, пишет
ученый, может считаться аналогичным горной породе или, вернее,
движущейся горной породе, одаренной свободной энергией. Вся
эта масса углерода, кислорода, кальция и прочих элементов,
входящих в состав живой материи, будет изъята в местах выпло-
да этих насекомых и где-то отложена после смерти мигрирую-
щих животных. Вопросы, поставленные перед зоологией сель-
скохозяйственной наукой и практикой, многообразны. Изучение
животных служит более углубленному усвоению учащимися вуза
ряда других дисциплин учебного плана. Хотя зоология имеет
непосредственное отношение к теории и практике растениеводства
и животноводства, ее изучение не может подменять другие дис-
циплины, однако будущие специалисты сельского хозяйства имеют
возможность в каждой главе курса почерпнуть комплекс сведений,
имеющих профессиональный интерес.
История зоологии. С глубокой древности люди накапливали
опыт использования естественных ресурсов, среди которых важ-
ную роль играли различные животные. Добывая их, они постепен-
но познавали жизнь и строение животных. Начало зоологии как
науке положил знаменитый древнегреческий ученый и философ
Аристотель (IV в. до н. э.). В своих работах «История животных»,
«О частях животных», «О возникновении животных» и др. он дал
описание 452 различных известных в то время животных. Значи-
тельный вклад Аристотель внес в изучение строения животных,
рассматривая их части тела во взаимосвязи.
Походы римлян в далекие страны значительно обогатили на-
уку знаниями о животных Северной Африки, Передней Азии и
Европы. Древнеримский ученый Плиний Старший (23—79 гг. н.э.)
в своей многотомной «Естественной истории» дал описание всех
известных в то время животных.
В эпоху феодализма, когда Европа была разобщена на мно-
жество мелких владений феодалов, а господствующая над об-
ществом религия тормозила развитие науки, изучение животных
испытало период длительного застоя.
Великая эпоха Возрождения (XV—XVI вв.) была временем но-
вого расцвета науки. Путешествия великих землепроходцев того
времени — Колумба, Марко Поло, Магеллана и многих других —
намного обогатили знания человечества о животном мире разных
континентов.
Накопленный к концу XVI в. обширный материал о фауне ра-
зличных частей Земли потребовал их систематизации и обобщения.
Из подобных обобщающих-зоологических работ наибольшую цен-
6
ность представляет многотомная сводка швейцарского ученого
К- Геспера (1516—1565) «История животных»подлинная
энциклопедия для того времени данных о животном мире.
В XVII в. был создан микроскоп, который открыл перед зо-
ологами обширный и удивительный мир мельчайших животных
и позволил приступить к изучению тончайших структур органов
многоклеточных животных. Из первых зоологических исследо-
ваний с помощью микроскопа надо прежде всего отметить работы
голландского натуралиста А. Левенгука (1632—1723), который
опубликовал 4-томпый труд «Тайны природы, открытые при
помощи микроскопа». Им были открыты инфузории, описаны эрит-
роциты, мышечная ткань вышних животных и многое другое.
Итальянский ученый М. Мальпиги (1628—1694) описал капил-
ляры в кровеносной системе позвоночных животных, сделал цен-
ные открытия в области микроскопического строения выдели-
тельных органов и покровов различных животных.
В физиологии много сделали М. Сервет (1511 —1543) и осо-
бенно У. Гарвей (1578—1657), описавшие кровообращение у чело-
века. В XVII—XVIII вв. зародились современная систематика
животных и палеонтология. С именем Ж. Кювье (1769 —1832)
связана разработка принципа корреляции, согласно которому все
части и органы животного организма находятся в неразрывной
связи друг с другом, и изменение одного из них влечет изменение
остальных органов тела (об этом в общих чертах ранее писал
Аристотель). Этим положением пользуются палеонтологи, восста-
навливая целое животное па основе его останков, найденных
в окаменелом состоянии. Из важнейших работ Ж. Кювье отмстим
«Царство животных» в 5 томах, «Иконография животного царст-
ва» с 450 таблицами и 6200 рисунками, многие из которых исполь-
зуются и в современной научной и учебной литературе, «Рас-
суждения о переворотах на поверхности земного шара и об
изменениях, какие они произвели», «Исследования об ископаемых
костях» (первое издание в 4 томах, четвертое -- в 10 томах).
В «Рассуждениях...» излагаются теория катастроф, распределение
ископаемых но слоям Земли и вместе с тем отказ признать смену
фаун как результат эволюции.
XIX в. знаменуется утверждением идеи эволюции органиче-
ского мира, постепенного развития всей живой природы от более
простых форм к более сложным. Развитию идеи эволюции спо-
собствовало и создание в 30-х годах XIX в. теории клеточного
строения животных и растений (Т. Шванн, М. Шлейдеп),
заложившей фундамент представления о единстве животного и
растительного мира.
Большие заслуги в развитии идей эволюции животных прина-
длежат знаменитому французскому естествоиспытателю Ж. Ла-
марку (1744—1829). Он развил и усовершенствовал систематику
животных, предложенную К- Линнеем, проделал большую работу
по изучению беспозвоночных. Но особенно ценна работа Ламарка
7
«Философия зоологии» (1809), в которой он выступает против
метафизических воззрений большинства биологов того времени о
неизменяемости видов животных и излагает первую целостную
теорию эволюции живой природы. Ламарк утверждал, что все
растения и животные постоянно изменяются и превращаются в
новые формы под воздействием внешних условий и внутреннего,
заложенного в каждом организме стремления к совершенство-
ванию. К- А. Тимирязев считал основной труд Ламарка «Филосо-
фия зоологии» произведением, в котором впервые вопрос о проис-
хождении организмов обсуждается не мимоходом, а со всей
необходимой широтой охвата, во всеоружии научных знаний того
времени. Но, создав теорию эволюции животного мира, Ламарк
дал ошибочное толкование причины этого процесса.
Окончательно победили идеи эволюции в биологии после опу-
бликования Ч. Дарвином (1809—1882) его основного труда «Про-
исхождение видов путем естественного отбора, или сохранение
благоприятствуемых пород в борьбе за жизнь» (1859). В этой
замечательной работе Ч. Дарвин не только доказал существова-
ние изменчивости видов и эволюции всего органического мира,
но и вскрыл причины этого процесса. Он объяснил целесообраз-
ность организации и приспособленность живых существ как ре-
зультат действия длительного естественного или искусственного
отбора — важнейшего фактора эволюции. Теорию эволюции Дар-
вина высоко оценил В. И. Ленин, указавший, что Дарвин впервые
поставил биологию на вполне научную почву, установив из-
меняемость видов и преемственность между ними.
Значение теории эволюции органического мира Ч. Дарвина
для естествознания и, в частности, для зоологии огромно: бы-
ло дано научное материалистическое объяснение строению и яв-
лениям жизни животных. Нет такой отрасли зоологических зна-
ний, в которой утверждение эволюционного учения не вызвало
бы коренных изменений во взглядах ученых. Победа теории эво-
люции в биологии послужила мощным стимулом к развитию всех
отраслей зоологии.
Быстрое развитие зоологических исследований в конце XIX в.
и особенно в XX в. было тесно связано с ростом животновод-
ства, рыбного и охотничьего промыслов и других отраслей
сельского хозяйства, использующих данные зоологии. Развитие
зоологической науки во многом способствовало росту и совер-
шенствованию сельского хозяйства, охране здоровья человека.
Накопление огромного фактического материала и теоретических
построений о животных и их жизни повлекло разделение зоологии
в XIX в. и начале XX в. на ряд отраслей — зоология стала
комплексной наукой.
В нашей стране зоология имеет длительную и славную ис-
торию. Еще в первых русских книгах («Русская правда» и др.)
есть упоминания о многих животных, обитавших в Древней Руси.
Но широкое развитие зоологические исследования получили в Рос-
8
сии в XVIII в., когда Академия наук организовала серию далеких
экспедиций для изучения природы различных районов страны.
Академик П. Паллас (1741 —1811) совершил путешествие в По-
волжье, Сибирь, Казахстан и на Урал, С. Стеллер (1709—1746) —
на Дальний Восток, С. Гмелин (1745—1774) — на юг Европейской
России, И. Гюльденштедт (1745—1781) — на Кавказ, И. Лепехин
(1740—1802) — по центральным и северным областям страны.
Ими были собраны большие зоологические коллекции и проведено
много наблюдений за животными посещенных районов. На основе
этих материалов П. Паллас создал капитальный труд «Русско-
Азиатская зоография», в котором дал описание всех известных
в то время позвоночных животных русской фауны.
Изучение животного мира России продолжалось и в первой
половине XIX в., когда многие ученые совершили ряд длитель-
ных путешествий в разные отдаленные районы страны. Особенно
плодотворной была трехлетняя поездка академика А. Ф. Мидден-
дорфа (1815—1894), объехавшего почти всю Сибирь и в полном
смысле слова «научно открывшего» ее для естествоиспытателей.
Большое значение для развития русской зоологической науки
имели труды профессора Московского университета К- Рулье
(1814 — 1858), в которых он развивал идеи единства организма
животного и окружающей среды, стремился показать, что измене-
ния условий жизни вызывают изменения животных. К- Рулье вы-
ступал против метафизических взглядов Ж- Кювье и других уче-
ных, отстаивающих теорию неизменности видов.
Ученик К- Рулье Н. А. Северцов (1827—1885) создал ряд
замечательных работ по экологии и зоогеографии. В них он
всегда подчеркивал необходимость изучения животных в связи
со средой обитания. С риском для жизни Н. А. Северцов проник
в горы и пустыни Средней Азии и дал ' подробное описание
фауны этой замечательной страны.
Большой вклад в зоологическую науку внес выдающийся рус-
ский исследователь академик К- М. Бэр (1792—1876). Его заслу-
женно считают одним из основоположников науки о развитии жи-
вотных — эмбриологии. Огромное значение для развития рыбного
промысла имели экспедиции К- М. Бэра на Каспийское и Азов-
ское моря.
Особенно быстро стала развиваться русская зоологическая
наука во второй половине XIX в. после опубликования Ч. Дарви-
ном теории эволюции органической природы. Передовые ученые
России — ботаник К. А. Тимирязев (1843—1920), зоологи
А. О. Ковалевский (1840—1901), И. И. Мечников (1845—1916),
В. О. Ковалевский (1842—1883) и др.— не только популяризиро-
вали и распространяли учение Дарвина, но и обогащали его
своими исследованиями.
Во второй половине XIX и начале XX в. продолжалось экспе-
диционное изучение фауны нашей страны и соседних с ней
территорий. Таковы экспедиции Н. М. Пржевальского (1839—
9
1888) и его учеников в Центральную Азию, Н. М. Книповича
(1862—1939) по морям России. Эти поездки значительно обогати-
ли знания о фауне России.
Особенно широко развернулись зоологические исследования в
нашей стране после Великой Октябрьской социалистической
революции. Коренным образом изменились содержание и объем
зоологических работ: вместо разрозненных исследований отдель-
ных ученых началось планомерное комплексное изучение фауны
страны, направленное на лучшее использование животных ресур-
сов в социалистическом строительстве, охрану и обогащение жи-
вотного мира СССР. Выдающихся успехов достигли школы за-
мечательных советских ученых: А. Н. Северцова (эволюционная
морфология), Н. К. Кольцова (генетика), Е. Н. Павлов-
ского (паразитология), К- И. Скрябина (гельминтология),
М. А. Мензбира (зоогеография), В. А. Догеля (протозоология),
В. Н. Беклемишева (сравнительная морфология беспозвоночных)
и многих других зоологов. Созданы научно-исследовательские
институты, разрабатывающие вопросы теоретической и приклад-
ной зоологии. Ежегодно десятки зоологических экспедиций на-
правляются в различные районы Советского Союза. Все эти рабо-
ты позволили создать ряд обширных монографических описаний
отдельных групп животных фауны СССР. Академия наук СССР
выпускает многотомное издание «Фауна СССР». По степени изу-
ченности своей фауны Советский Союз занимает одно из первых
мест среди стран мира.
Зоология — комплексная наука. Развитие наук вместе с накоп-
лением знаний приводит к их расчленению. В настоящее время
зоология представляет систему наук, каждая из которых имеет
свои задачи и в исследованиях использует свою методику. Они
не обособлены одна от другой, поскольку у всех общий объект
познания — животный мир.
Морфология (от греч. morphe — форма) — одна из основ-
ных и древнейших биологических наук. Ее предметом являются
изучение и объяснение внешнего и внутреннего строения организ-
ма. Описание морфологии животных — отправной пункт всякого
зоологического исследования.
Методом сравнительной морфологии более четко выявляются
характерное в строении животных и степень сложности их ор-
ганизации. Этим методом пользуются и при установлении
исторических связей между группами животных, и в экологичес-
ких исследованиях, когда анализируют приспособительные черты
у животных к среде обитания и в связи с их образом жизни.
К достижениям сравнительной эволюционной и экологической
морфологии принадлежит разработка учения об органах аналогич-
ных и гомологичных. Аналогичные органы те, что выполняют схо-
дную функцию, но строение имеют различное. Примером могут
служить крылья насекомых и птиц. У них аналогичная функция,
с помощью крыльев совершается полет, но нет сходства в их строе-
10
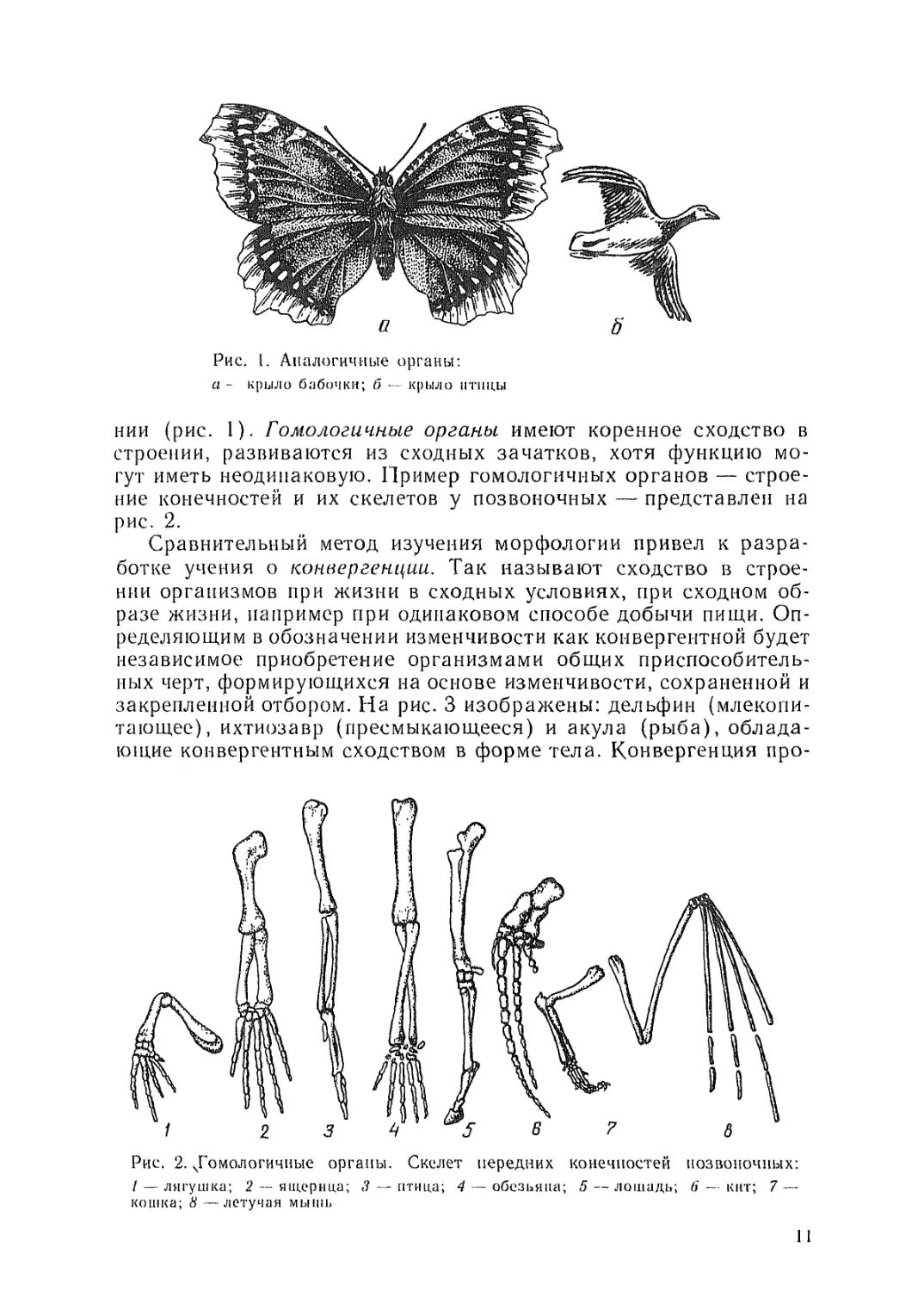
Рис. 1. Аналогичные органы:
а - крыло бабочки; б — крыло птицы
нии (рис. 1). Гомологичные органы имеют коренное сходство в
строении, развиваются из сходных зачатков, хотя функцию мо-
гут иметь неодинаковую. Пример гомологичных органов — строе-
ние конечностей и их скелетов у позвоночных — представлен на
рис. 2.
Сравнительный метод изучения морфологии привел к разра-
ботке учения о конвергенции. Так называют сходство в строе-
нии организмов при жизни в сходных условиях, при сходном об-
разе жизни, например при одинаковом способе добычи пищи. Оп-
ределяющим в обозначении изменчивости как конвергентной будет
независимое приобретение организмами общих приспособитель-
ных черт, формирующихся на основе изменчивости, сохраненной и
закрепленной отбором. На рис. 3 изображены: дельфин (млекопи-
тающее), ихтиозавр (пресмыкающееся) и акула (рыба), облада-
ющие конвергентным сходством в форме тела. Конвергенция про-
Рис. 2. хГ°МОЛОГИЧНЬ1е органы. Скелет передних конечностей позвоночных:
/ — лягушка; 2 — ящерица; 3 — птица; 4 — обезьяна; 5 — лошадь; б — кит; 7 —
кошка; 8 —летучая мышь
1 1
Рис. 3. Конвергенция:
I — акула; 2 — ихтиозавр; 3 — дельфин
является и в утрате органов, например в редукции глаз у пещер-
ных животных.
С другой стороны, обычная картина — разнообразие форм у
животных, живущих в одной и той же среде, свидетельствует о
многообразии связей с факторами среды обитания. Например,
жизнь в воде тогда сопровождается конвергентным сходством,
когда имеется общее в жизненных связях с данной средой. Рыбы,
по-разному добывающие пищу, отличаются по форме тела, строе-
нию рта и т. д.
Физиология (от греч. fazis — природа) — наука о жизне-
деятельности организма и его органов, изучающая одновремен-
но и их строение. Закон о единстве формы и функции находит
полное подтверждение в морфологии и физиологии. К теорети-
ческим достижениям в физиологии относится учение И. П. Павло-
ва о нервной системе, интегрирующей организм в единое целое,
а также работы в области эндокринологии о роли желез
внутренней секреции, о выделяемых ими гормонах, участвующих в
регуляции функций различных органов и организма в целом.
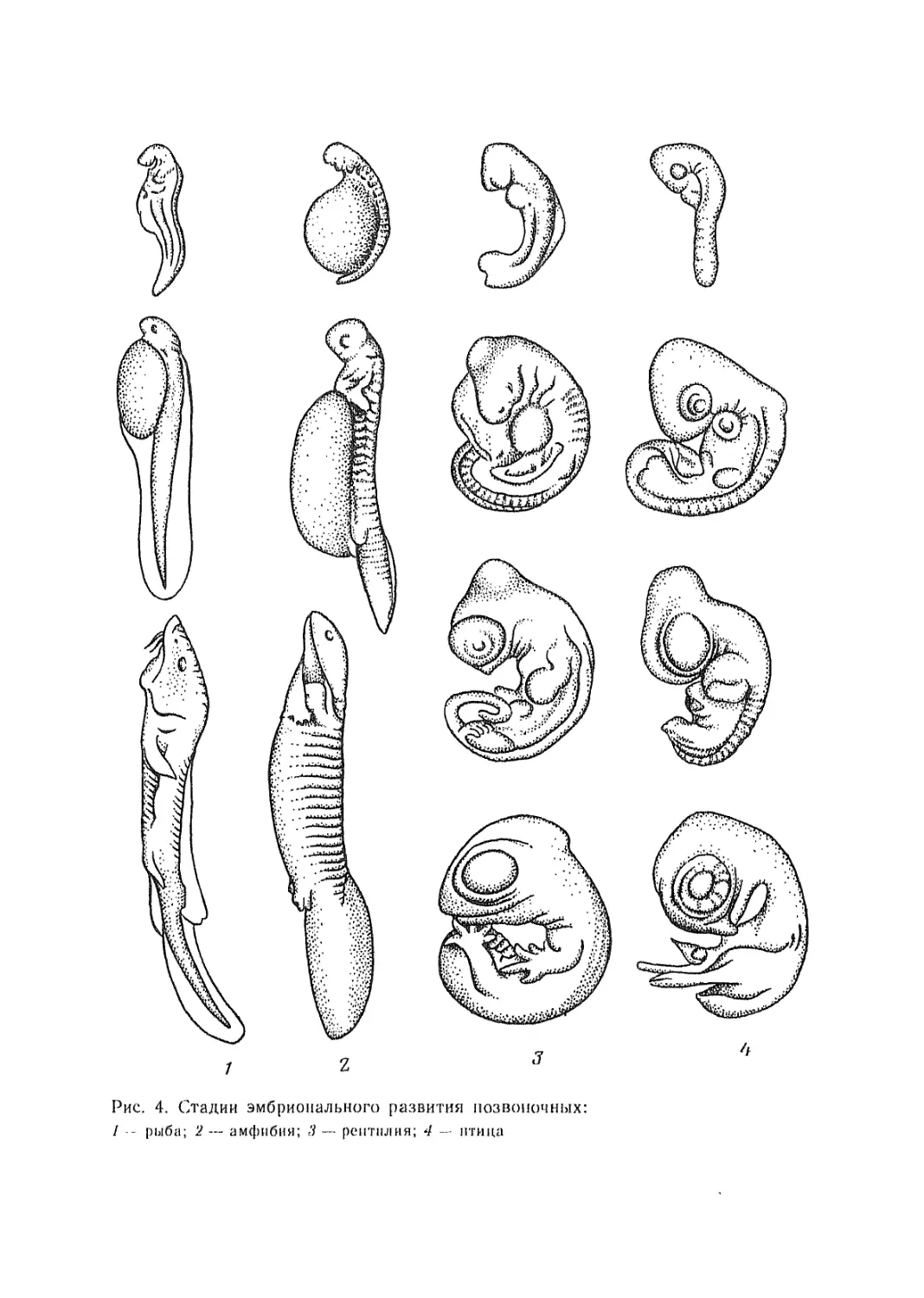
Эмбриология (от греч. embryon — зародыш) изучает
рост и развитие животных в зародышевом периоде. Материалы
сравнительной эмбриологии позволяют понять законо-
мерности в развитии эмбрионов, взаимосвязь онтогенеза (индиви-
дуального развития) с филогенезом (развитием историческим).
Пройденные этапы в эволюционном развитии животных не про-
ходят бесследно, а в какой-то мере проявляются у современных
видов на эмбриональных стадиях. Наибольшее сходство эмбрионов
животных разных типов наблюдается в раннем периоде их раз-
вития. На более поздних стадиях это сходство становится меньше
(рис. 4). Такая закономерность была сформулирована К- Бэром
12
Рис. 4. (Лидии эмбрионального развития позвоночных:
I — рыба; 2 — амфибия; 3 — рептилия; 4 — птица
как закон зародышевого сходства: в эмбриональном периоде
развития раньше других закладываются признаки типа, к которо-
му принадлежит животное, затем — признаки класса, еще по-
зднее — признаки отряда. Видовые признаки оформляются по-
следними. Закономерность повторения в онтогенезе стадий истори-
ческого развития была подмечена рядом ученых и обобщена Э. Гек-
келем в форме биогенетического закона: онтогенез есть краткое
повторение филогенеза. Для его формулировки послужили работы
Ф. Мюллера, А. О. Ковалевского, И. И. Мечникова и др. Ученые
достигли выдающихся результатов в описании зародышевого раз-
вития многих беспозвоночных животных и низших хордо-
вых и считаются основоположниками эволюционной эмбрио-
логии.
Использование биогенетического закона для выявления пре-
дков современных животных встречает трудности. Известно, что
организмы изменчивы на всех стадиях жизни, а не только во
взрослом состоянии. Выживание зависит от приспособленности к
существованию во все периоды развития. Исследования о соотно-
шении онтогенеза и филогенеза, их взаимопроникновения про-
ведены А. Н. Северцовым — автором теории филэмбриогенеза,
изучаемой в курсе дарвинизма.
Экология (от греч. oikos — жилище, местообитание)
изучает животных в связи с местом и условиями их жизни, зако-
номерности во взаимосвязях организмов со средой обитания. Од-
ной из главных проблем экологии будет исследование приспосо-
бительных черт в строении животных, в их жизненных от-
правлениях, в поведении. Последнее стало предметом изучения
особой науки — этологии (от греч. ethos — характер, нрав).
Данные экологии ценны при разработке теоретических проблем об-
щей биологии, но имеют большое значение и для практики. Так,
мероприятия по акклиматизации животных включают тщательное
изучение биологии самих животных, а также экологических фак-
торов на территории, куда решено их внедрить. Сведения по
экологии домашних животных помогают проведению мероприятий
по районированию пород, рациональному использованию пастбищ
и т. д. В растениеводстве важна осведомленность об экологии вре-
дителей возделываемых культур и животных, которых можно
использовать в борьбе с ними.
Пути экологических исследований: изучение биологии отдель-
ных видов и сообществ животных, изучение факторов среды
их обитания, изучение воздействия факторов среды.
Факторы среды. К абиотическим факторам (физико-
химическим) относятся географические, климатические, эдафичес-
кие (греч. edafos — почва) и др. При изучении водных животных
большое внимание уделяется таким факторам, как прозрачность
воды, ее температура, растворенные в воде газы, соли, показатель
активной реакции среды (pH); соединениям азота, фосфора,
железа.
14
Биотические факторы: взаимовлияние организмов одного и
разных видов друг на друга (хищник и жертва, хозяин и его
паразиты, растения и животные и т. д.).
Антропогенные факторы — воздействие человека на природу.
Оно бывает и положительным, но чаще хозяйственная деятель-
ность наносит ущерб, который бывает очень значительным.
Имеются специальные постановления об охране природы, созданы
общества охраны природы.
Все факторы взаимосвязаны. Так, количество растворенно-
го в воде кислорода зависит от температуры воды, влияющей на
растворимость этого газа, от организмов, населяющих водоем.
Одни из них обогащают воду кислородом, другие потребляют его.
Интенсивность процесса связана с температурой, освещенностью,
количеством тех или других организмов и с другими обстоятель-
ствами.
Место обитания организмов со всеми его особенностями на-
зывается биотопом (греч. bios — жизнь, topos — место), а со-
общество организмов животной и растительной природы в био-
топе — биоценозом (греч. bios жизнь, koinos — общий). Круп-
ные биотопы подразделяют на более мелкие. Так, например,
в озере есть биотоп открытой толщи воды — пелагиаль, его
биоценоз — планктон; биотоп дна — бенталь, его биоценоз —
бентос; биотоп прибрежной зоны — литораль. Каждый из биото-
пов отличается особенностями абиотических и биотических фак-
торов, к которым приспособлены организмы биоценозов.
Экология отдельных групп животных рассматривается в гла-
вах систематической зоологии.
Зоогеография изучает географическое распространение
животных, закономерности этого распространения. Зоогеогра-
фические материалы вносят вклад в решение общебиологичес-
ких проблем и, в частности, в теорию эволюции животного мира.
Изучение ландшафтных зон областей с характерным для каж-
дой из них комплексом животных и растений и физико-географи-
ческими условиями жизни; изучение конкретных связей населяю-
щих их организмов со средой -- их экологии — создаст предпо-
сылки к практическому использованию сведений, например, для
обоснования мер по реконструкции фауны.
В работу зоогеографа входят установление географических
границ распространения видов животных — их ареалов, выявле-
ние причин наблюдаемого распространения, что связано с про-
ведением разносторонних изысканий. Для своих сообщений зооге-
ографы обращаются к данным исторической геологии, палеонто-
логии и других наук. Например, известно, что в Южной Америке
живут ленивцы, муравьеды и броненосцы. Их отсутствие на со-
седнем материке объясняется геологической историей разоб-
щенностью материков. Географическая изоляция Северной Амери-
ки от Южной в прошлом и изменчивость животных привели к раз-
витию новых родов и видов, происшедших от предков, живших на
15
том же континенте в минувшие геологические эпохи. А палеонто-
логические находки тех же неполнозубых на о-ве Куба свиде-
тельствуют о связи о-ва Куба в геологическом прошлом с Южной
Америкой. Так, данные зоогеографии, исторической геологии и
палеонтологии, взаимно дополняя, придают выводам научную
достоверность.
Виды животных, свойственные только данной местности, на-
зываются эндемичными (греч. eudetnos — местный). Много энде-
миков, например, в оз. Байкал. Исключительное своеобразие
этого уникального водоема отражено в специфичности его фауны
и микрофлоры. В озере множество не только эндемичных видов,
но и родов и даже семейств. Эндемики нередки в горной местности.
При обнаружении эндемиков во всех случаях отмечаются своеоб-
разие условий жизни и наличие каких-либо факторов изоляции
(географических или иных). Большое значение имеет фактор
времени — длительность жизни при особых обстоятельствах.
Зоогеографические изыскания привели к находкам реликтов —
остатков фауны, населявшей территорию в минувшие геологиче-
ские времена. Есть реликты — свидетели ледниковой эпохи, есть
морские реликты, сохранившиеся в водоемах от времени морской
трансгрессии (наступления моря), вызванной понижением суши и
последующим отступлением моря при повышении суши.
Успехи зоогеографии послужили основанием к разделению
суши и Мирового океана на зоогеографические области по приз-
наку наличия ряда видов животных, присущих только им.
Территория СССР принадлежит к одной области — Палеар-
ктической. В нее входят также Западная Европа и северная
часть Африки. Ландшафтные зоны СССР представлены полосой
тундр на Севере, за которой к югу располагается лесная полоса,
затем —лесостепь и пустыни. Отдельные зоны составляют Кавказ,
горный Крым, низменная часть Средней Азии, горный Туркестан,
а на востоке — смешанные леса Дальнего Востока (см. главу
«Животный мир Советского Союза»).
Неоарктическая область включает Северную Америку и Грен-
ландию. По составу видов близка к Палеарктической. Эти обе
северные области объединяют в одну Голарктическую область.
Африка, кроме ее северной полосы, составляет Эфиопскую
область. Неотропическая область включает Центральную и Юж-
ную Америку и Вест-Индские острова. Восточная, или Индо-Ма-
лайская, область распространяется на Индию, Южный Китай с
прилегающими островами, включая о. Цейлон. Смежна с Восточ-
ной Австралийская область. Граница между ними — острова Ин-
донезии Ломбок и Бали. В нее входят также о-ва Тасмания,
Новая Зеландия, Новая Гвинея, многие острова Полинезии.
Материалы по зоогеографии представлены в темах систе-
матического отдела курса.
Палеонтология (от греч. palaios — древний) — наука о
вымерших животных (палеозоология) и растениях (палеоботани-
16
ка), занимается всесторонним изучением вымерших организмов
минувших геологических эпох, опираясь в своих исследованиях
на другие биологические науки. Так, палеоморфологи восстанав-
ливают по окаменелостям облик целых животных, палеоэколо-
ги — условия жизни в те времена, среду биотическую и абиоти-
ческую и приспособленность к ней организмов и т. д.
Большой след оставил в палеонтологии Ж- Кювье. Он создал
научное построение распределения окаменелостей по слоям оса-
дочных пластов Земли разного геологического возраста, но дал
ошибочное толкование наблюдаемому с позиций противника эво-
люции. Согласно Ж. Кювье своеобразие палеофауны в разных
пластах обязано периодической гибели животных в результате
грандиозных катастроф. Новый состав животных, причем выше ор-
ганизованных, в следующем более молодом слое (слое отложений)
объяснялся приходом их «откуда-то». Ученики Кювье выдвинули
фантастическую гипотезу о повторных актах творения после каж-
дой катастрофы, доведя к половине XIX в. число катастроф и по-
вторных актов творения до многих десятков.
В прогрессивном направлении палеонтологии видное место за-
няли работы В. О. Ковалевского. По Ковалевскому, палеонто-
лог должен полагать своей задачей не простую констатацию от-
личия в фаунах разных эпох, а направление усилия на поиски
среди ископаемых останков сходства между животными смежных
напластований. Автор представил по данным палеонтологии до-
казательство эволюционного развития современных однопалых ло-
шадей от многопалых предков и дал новый экологический анализ
материалов. Переход от 4- к 3-палому животному и затем к од-
нопалой лошади происходил во взаимосвязи со сменой условий
жизни и, в частности, с появлением злаков на обширных степных
пространствах. Ковалевский учитывал ряд факторов: смену ха-
рактера питания, появление новых кормовых растений, их распро-
странение и способ добычи пищи. Экологический путь анализа
привел к созданию эволюционной палеоэкологии.
Палеонтологическая летопись бедна по причине трудности
сохранения останков, подвергающихся разрушению под действием
внешних сил. Сохраняются, как правило, животные с наружным
или внутренним скелетом. Более благоприятны возможности
захоронения водных животных и тех наземных, трупы которых
сносятся водой в понижения рельефа местности и там погребаются
под выносами вод. Некоторые насекомые сохранились погребен-
ными в смоле.
В практическом отношении палеонтология предоставляет ис-
торической геологии сведения о «руководящих» ископаемых того
или иного геологического возраста. Эти данные используются гео-
логами при поисках полезных ископаемых.
Филогенетика (греч. phylon — племя, род, genesis —
происхождение) — раскрытие исторических связей в мире живот-
ных и растений. Наука возникла вместе с утверждением тео-
17
рии развития органической природы. В результате исследования
родственных отношений животных стало возможным построение
родословного древа животного мира. Филогенетические изыска-
ния стремятся проводить параллельно в трех направлениях:
сравнительно-морфологическом, эмбриологическом и палеонто-
логическом. Если выводы исследования, проведенного тремя пу-
тями, совпадут, то достоверность вызовет меньше сомнения. Фи-
логения типов животных рассматривается в разделах система-
тической части курса.
Систематика (от греч. systema — целое, составленное из
частей) разрабатывает на основе данных других зоологических
наук классификацию животных и естественную систему животного
мира, которая отражает родственные связи различных групп жи-
вотных.
Время возникновения первых систем затеряно в глубине ве-
ков. Известно, что их отличала практическая направленность
в содержании. Животные издавна подразделялись на диких и до-
машних, полезных и вредных. Из ранних зоологических клас-
сификаций, в основу которых положены сходство и отличие осо-
бей между собой, выдающееся значение имела классификация
Аристотеля — знаменитого философа и естествоиспытателя ан-
тичной Греции. Примечательно подразделение Аристотелем
животных на две группы: имеющих кровь и без крови. Первая
группа соответствует в современной классификации позвоноч-
ным, а составляющие ее отделы — в основном классам позвоноч-
ных.
Вторая группа получила в XIX в. в работах Ламарка наимено-
вание беспозвоночных. В древности применялась такая номен-
клатура, как вид и род. Под видом Аристотель подразумевал
группы организмов, сходных между собой и способных к скрещи-
ванию. Содержание термина «род» было менее определенным.
В XVII—XIX вв. вид был признан основной систематической
категорией.
Много сделал для систематики шведский врач и естество-
испытатель К. Линней (1707—1778). Им описаны тысячи видов
животных и растений. Линней утвердил в систематике ряд катего-
рий: вид, род, отряд, класс — и уточнил их содержание. Он ввел
в широкую практику двойное наименование вида — бинарную
номенклатуру вида, в которой первое слово, имя существительное,
обозначает название рода, а второе, прилагательное,— вида. Наз-
вание рода пишется всегда с прописной буквы, а видовое — со
строчной. Например, собака домашняя — Canis faniiliaris L.
После наименования вида и других таксонов ставится фамилия
автора, впервые его описавшего, лишь фамилия Линнея обозна-
чается одной буквой L.
Позже, в XIX в., в систематику были введены систематиче-
ские категории: семейство и тип.
Главные работы К. Линнея: Система природы (1735), Виды
18
Рис. <5. Половой диморфизм у бабочки (зимней пяденицы):
а - самец; б - самка
Рис. 6. Полиморфизм у муравьев (Pheidole instabilis):
1 -крупный муравей; 2-- рабочий средних размеров; 3— самка, сбро-
сившая крылья; 4 - самец
растений (1753), Философия ботаники (1751). Описания К. Лин-
неем животных и растений отличаются точностью и глубиной по-
нимания природы организмов. Он ввел название «млекопитаю-
щие», отнес человека к группе приматов и т. д. В современной клас-
сификации сохранена схема подразделения на систематические ка-
тегории, но принцип построения отличен. Систему Линнея называ-
ют искусственной. Ее автор ставил целью преимущественно ката-
логизацию органической природы. Естественная система исходит
из признания развития организмов. Она призвана отразить ге-
неалогические связи организмов. Для объединения животных в ту
или иную категорию основанием служит степень сходства как
отражение родства. К одному виду относятся животные, наиболее
близко родственные, и этим объясняются их морфологическое
сходство, физиологическое (генетическое) свободное скрещивание
с получением плодовитого потомства и географическое распрост-
ранение (вид имеет свой ареал).
В результате внутривидовой изменчивости образуются под-
виды и разновидности. Внутривидовые изменения об-ратимы. Из-
вестна неодинаковость строения в размерах, окраске и пр. у живот-
ных одного вида, но проживающих в разных географических
зонах (географическая изменчивость, экологическая и пр.). На-
секомые и моллюски отличаются особенно большой пластич-
ностью.
Многообразная внутривидовая изменчивость побудила пер-
вых эволюционистов, включая Ламарка, сделать вывод, что вид
как систематическая категория понятие условное, что в природе
нет четких видовых отличий. Популярностью пользовался тезис
Лейбница: природа не делает скачков. Отказ от признания чет-
ких видовых отличий привел к порождению множества псевдо-
видов.
Описывающий новые формы мог классифицировать по собст-
венному усмотрению как новые виды, располагая нередко малым
количеством особей или даже единичными экземплярами. Ревизия
в систематике привела к «закрытию» многих псевдовидов, оказав-
шихся разновидностями.
Видовые признаки стойкие, они отличаются исключительной
консервативностью.
Свои отличия имеет сезонная внутривидовая изменчивость.
Например, смена шерстного покрова млекопитающих. Их зимний
мех, как правило, светлой окраски, более густой. Сезонная из-
менчивость закономерна.
Морфологические отличительные особенности животных раз-
ного пола одного и того же вида получили название полового
диморфизма (рис. 5).
Многообразие форм особей одного вида называется полимор-
физмом.
Полиморфизм также характерен для пчел, муравьев, термитов
(рис. 6).
20
Основная схема современной номенклатуры в систематике:
Царство (regnum)
П од ц а р ств о (su h regn и in)
Тип (phylum)
Подтип (subphytum)
Класс (clussis)
Подкласс (хubclassis)
Отряд (огdo)
Подотряд (subordo)
Семейство (familia)
Род (genus)
Вид (species)
Подвид (sub s ре ci es)
Наименование семейства оканчивается на -idae (например,
бычьи — Bovidae), наименование подсемейства — па -inae (на-
пример, овцекозы — Caproovinae).
ОБЩАЯ ЧАСТЬ
ОСНОВНЫЕ ОТЛИЧИТЕЛЬНЫЕ ЧЕРТЫ
ЖИВОТНОГО ОРГАНИЗМА
Жизнь на Земле зародилась очень давно — около 4 млрд лет на-
зад. За этот огромный промежуток времени органическая природа
прошла сложный путь развития — от первичного живого вещества
до современных сложнейших организмов. Эволюция живой приро-
ды на нашей планете привела к образованию двух основных
групп организмов: животных и растений.
В чем же главные различия между этими группами живых
существ? Важное отличие между ними заключается в характере
обмена веществ. Животные—в основном гетеротрофные
организмы, т. е. организмы, питающиеся органическими вещест-
вами, созданными другими животными и растениями. Напротив,
подавляющее большинство растений — автотрофные орга-
низмы, способные при помощи фотосинтеза и других физико-
химических процессов создавать органические вещества своего те-
ла из неорганических веществ внешней среды. Но эта важная
черта отличия животных от растений неабсолютна. Среди расте-
ний есть некоторые группы (например, грибы), которые не могут
синтезировать органические вещества из неорганических и питают-
ся гетеротрофно. С другой стороны, у некоторых низших живот-
ных, например у обычных обитателей наших водоемов — эвглен, в
теле имеются хлорофилловые зерна, что позволяет им на свету
питаться подобно растениям. Но помещенные в темноту эвглены нс
погибают, а начинают поглощать органические вещества подобно
другим животным.
Подавляющее большинство животных обладают подвиж-
ностью, могут перемещаться или совершать различные движения.
Растения же, как правило, неподвижны. Но у некоторых высших
растений (мимоза, росянка и др.) наблюдается движение отдель-
ных частей (например, листьев), а многие низшие одноклеточные
растения могут передвигаться в пространстве. Клетки тела живот-
ных обычно не имеют целлюлозных оболочек и не содержат вакуо-
лей клеточного сока, весьма характерных для клеток растений.
Но и эти отличия в строении клеток присущи не всем животным
и растениям.
22
Таким образом, провести резкую границу между животными
и растениями невозможно. Если высшие, сложно организованные
животные и растения резко отличны, то низшие их формы обна-
руживают черты сходства. На общность указывает и клеточное
строение тела.
КЛЕТКА
Изобретение микроскопа открыло путь к познанию микрострук-
туры тела животных и растений, их строения. Клеточная теория —
учение об общих чертах строения животных и растений, о клетке
как элементарной структуре, о тканях, образованных клетками.
Использование электронного микроскопа, микроманипу-
ляторов, физико-химических методов и т. д. углубило познание
общего в строении клеток и особенного. Выяснено, что в деталях
клетки имеют специфическое строение не только у организ-
мов, далеко отстоящих в системе, но и у особей одного вида,
и у клеток разных тканей в одном организме. Установлено также,
что непрерывное физиологическое взаимодействие клеток в
организме происходит через их поверхностную мембрану.
Отошло в прошлое суждение о многоклеточном организме, как
о «государстве клеток» (механистическая теория анатома Вир-
хова). Клетки свое начало берут при дроблении яйца и в даль-
нейшем, численно возрастая путем деления, продолжают сохра-
нять взаимосвязь.
Жизненные отправления клеток обусловлены целым орга-
низмом, закономерностями его роста и развития. В клетках про-
исходит обмен веществ — процессы ассимиляции и диссимиля-
ции. Новым стало и понимание физиологической роли меж-
клеточных веществ.
В теле многоклеточных животных клетки дифференцированы
в зависимости от их функций. Они различаются по размерам,
форме и строению.
Большинство из клеток микроскопически малы, но некоторые
достигают относительно большой величины. Например, отрост-
ки некоторых нервных клеток мозга крупных млекопитающих
достигают длины свыше метра. Формы клеток (рис. 7) обуслов-
лены их функцией и положением в организме. Встречаются клетки
округлой, овальной, кубической, призматической, веретено-
видной, звездчатой и других форм. Некоторые клетки не имеют
постоянной формы, она изменяется путем образования временных
выпячиваний.
Строение клеток животных отличается большой сложностью.
Обычно в них можно различить наружную мембрану, цитоплаз-
му, клеточное ядро (или ядра) и различные органоиды
(рис. 8).
23
Рис. 7. Различные формы клеток животного;
1 — нервная клетка; 2—4 — клетки эпителия; 5 — клетка соединитель-
ной ткани; 6 — яйцеклетка; 7 — мышечная клетка
Рис. 8. Схема строения клетки жи-
вотного:
/ — ядро; 2 -- ядрышко; 3 — ядерная
мембрана; 4 — цитоплазма; 5 — аппа-
рат Гольджи; 6 - митохондрии; 7 —
лизосома; 8 — эндоплазматическая
сеть; 9 — рибосомы; 10 - клеточная
мембрана
Наружная мембрана кле-
ток животных, как правило,
очень тонка (толщина око-
ло 10“ь см). Она состоит из
трех слоев: наружного, сред-
него и внутреннего. Наруж-
ный и внутренний слои об-
разованы белками, сред-
ний — липоидами. На внут-
ренней поверхности мембра-
на образует складки и раз-
ветвления, переходящие в эндоплазматическую сеть цитоплазмы.
Мембрана служит защитной оболочкой клетки и активно
участвует в регуляции обмена веществ между клеткой и окру-
жающей средой.
Цитоплазма занимает большую часть тела клетки. Цитоплаз-
ма— сложная коллоидная система. В ее состав входят бел-
ки, часть которых соединена с липидами, различные соли, фер-
менты и большое количество воды. Общими для всех клеток
структурными образованиями цитоплазмы являются эндоплазма-
тическая сеть, аппарат Гольджи, рибосомы и митохондрии.
Эндоплазматическая сеть пронизывает всю цитоплазму клет-
ки. Опа представляет собой систему тончайших мембран. Около
ядра мембраны переходят в мембраны ядерной оболочки.
Эндоплазматическая сеть выполняет функцию остова клетки,
а по ее канальцам и синусам происходит внутриклеточный
обмен веществ, синтезируемых в различных частях клетки.
Аппарат Гольджи имеет структуру, близкую к структурным
образованиям эндоплазматической сети. Он образован мембрана-
ми, ограничивающими более крупные вакуоли и мелкие пузырьки.
Функции аппарата Гольджи еще недостаточно выяснены, но, ве-
роятно, он служит для временного накапливания некоторых про-
дуктов внутриклеточного синтеза, главным образом ферментов
и гормонов. При определенном состоянии организма эти вещества
могут быть выведены из клетки через эндоплазматическую сеть
и вовлечены в обменные процессы всего организма.
Митохондрии являются энергетическими центрами клетки и
оказывают влияние на ее многие жизненные отправления. Это
мелкие удлиненные тельца длиной 0,2—5 мкм. Они покрыты двух-
25
слойной оболочкой. Из внутреннего слоя оболочки в полость мито-
хондрии направлены многочисленные гребни. Они резко увеличи-
вают внутреннюю поверхность митохондрий, что важно для ускоре-
ния биохимических реакций накопления и превращения энергии
в клетке.
Рибосомы — мельчайшие зерна диаметром около 0,015 мкм,
расположенные преимущественно на поверхности мембран эндо-
плазматической сети. Они имеются также и в ядре клетки. Функция
рибосом заключается в синтезе белков, которые затем по каналам
эндоплазматической сети разносятся по всей клетке.
Центрозома присуща почти всем клеткам животного. Обычно
она имеет вид светлого поля, в Котором размещены 1—2, реже
больше мелких зернышек — центриол. В некоторых клетках,
особенно находящихся в состоянии деления, от центрозомы ради-
ально расходятся тончайшие лучи, образующие лучистую сферу.
Центриоли принимают важное участие в расхождении хромосом
при сложном делении клеток.
Нередко в цитоплазме клеток животных можно обнаружить
различные тончайшие нити и волоконца. Одни из них служат как
бы опорным каркасом клеток (тонофибриллы), другие обладают
способностью сокращаться (например, миофибриллы мышечных
клеток). В нервных клетках нити цитоплазмы (нейрофибриллы)
участвуют в проведении нервных импульсов.
В цитоплазме клеток тела животных периодически наблюда-
ются различные временные включения (капельки жира, зерна
и глыбки запасных белков и др.). Они возникают и исчезают в за-
висимости от баланса обмена веществ. Железистые клетки содер-
жат капли секрета, позднее выводимого из них. К клеточным
включениям относятся и зернышки различных пигментов — крася-
щих веществ, придающих клеткам ту или иную окраску. Нередко
наружный слой цитоплазмы (эктоплазма) отличается от ее внут-
ренних частей (эндоплазмы) более плотной консистенцией и от-
сутствием включенных пузырьков и зерен.
Ядро присуще почти всем клеткам животных. Лишь некото-
рые специализированные клетки (например, красные кровяные
клетки высших позвоночных) в процессе своего формирования ут-
рачивают ядро. Ядра клеток животных разнообразны по величи-
не и форме. Снаружи ядро клетки одето двухслойной ядерной мем-
браной. В ней имеются многочисленные поры, через которые осу-
ществляется обмен веществ между ядром и цитоплазмой. Описана'
также система тончайших канальцев, которые связывают ядро со
структурными элементами цитоплазмы. Кариоплазма — вещество
ядра, содержит белки, липоиды, ферменты, минеральные вещест-
ва, нуклеиновые кислоты. В ядре расположены хромосомы и яд-
рышко. Хромосомы — носители наследственной информации.
Число и форма их постоянны для данного вида животного. Они
видны в период деления ядра. Ядрышко — мелкое тельце округ-
лой формы, хорошо различимое в неделящихся клетках.
26
ТКАНИ
Клетки, за исключением половых, находятся в составе тканей.
Ткани — сложившиеся в процессе исторического развития много-
клеточных организмов структуры, образованные клетками. Они со-
держат также межклеточное вещество. Ткани входят в состав
органов и участвуют в выполняемой ими функции. Строение
той или иной ткани соответствует деятельности, которую она
осуществляет. Многообразие функций тела животного отражено в
строении органов и соответственно тканей. Различают четыре типа
тканей: эпителиальную, соединительную, мышечную и нервную. В
каждом типе тканей встречается разнообразие в строении соот-
ветственно особенностям отправлений. Так, функции кожного эпи-
телия и выстилающего кишечник неодинаковы. В этой связи в ги-
стологии (учении о тканях) принято понятие «система тканей»:
система эпителиальных тканей, система соединительных тканей и
т. д. Специфическое в структуре тканей выявляется при рассмот-
рении их места в организме и функции, выполняемой органами.
Эпителиальная ткань. Ее клетки тесно лежат одна к другой,
межклеточного вещества очень мало, иногда клетки связаны меж-
ду собой протоплазматическими мостиками. В однослойном эпите-
лии клетки лежат в один ряд, в многослойном — в несколько
рядов, скрепляемых тонофибриллами (имеющими вид переплетен-
ных дуг, опирающихся на базальную мембрану) (рис. 9). Другая
отличительная черта эпителия — неодинаковость в строении внеш-
ней части клеток и их базальной части, где расположено ядро.
Базальная мембрана клеток эпителия связывает эпителиаль-
ную ткань с находящейся под нею соединительной тканью. Для
эпителия характерна способность к регенерации — к восстановле-
нию эпителиального пласта по мере отмирания клеток, срок
деятельности которых бывает коротким, как, например, в кише-
чнике.
На долю кожного эпителия приходится контакт с внешней
средой, и его строение свидетельствует о приспособленности жи-
вотного к характерной для него среде обитания. Так, кожа рыб
отличается обилием слизистых желез, а у наземных животных —
защитных образований, предохраняющих тело от высыхания и
травм. У насекомых эпителий с хитиновой оболочкой, а в много-
слойном эпителии наземных позвоночных происходит ороговение
клеток наружных слоев (рис. 10). У эндодермального эпителия
кишечника иная функция и свои особенности в строении. Клетки
близки к цилиндрической форме, лежат в один ряд. Наружный
край с ресничками (у многих беспозвоночных) или, как у поз-
воночных, с каймой, имеющей палочковидную исчерчеиность сло-
жного строения. Физиологическая роль этого эпителия также
сложна. Через него происходит в известной мере избиратель-
ное всасывание переваренной пищи (рис. 11). На рис. 12 изоб-
ражены клетки эпителия слизистой оболочки полости носа. Они
27
Рис. 9. Многослойный эпителий:
А — плоский многослойный эпите-
лий роговицы глаза кошки; Б --
схема расположения тонофибрилл
Рис. 10. Однослойный эпителий
насекомого (поперечный разрез
через спинной щиток брюшка
личинки плавунца):
1 — эпикутикула; 2 -- экзокутнку-
ла; <? —эндокутикула; 4 — клетки
гиподермы; 5 — ядро; б - базаль-
ная мембрана
Рис. 11. Однослойный цилинд-
рический каемчатый эпителий
тонкой кишки млекопитающего;
/ — клетки эпителия; 2 - клетки
в состоянии митоза
Рис.
12. Многорядный эпителий слизистой
оболочки полости носа:
1 - - мерцательные реснички; ‘2 -- бокаловидные
клетки; 4 - базальная мембрана
клетки; 3 --- замещающие
высокие, на стороне, обращенной в просвет трахеи, с ресничка-
ми. Мерцанием ресничек обусловливается вынос из органов дыха-
ния слизи с осевшей на ней пылью и пр. В этом проявляется
защитная функция мерцательного эпителия трахеи; в легких же
эпителий плоский и несет дыхательную функцию. В железах
клетки эпителия продуцируют секрет (железистый эпителий).
В многослойном ороговевающем эпителии живыми будут клетки
базального ряда и отчасти лежащие над ним. Здесь происходит
размножение клеток делением. По мере оттеснения клеток наружу
они становятся все более уплощенными, а самые наружные слои
состоят из плоских роговых чешуек. При шелушении на их место
поступают перерождающиеся клетки глубинных слоев.
Соединительная ткань (иначе, система тканей внутренней сре-
ды). Ее морфологическая особенность — преобладание межкле-
точного вещества над клетками. В крови оно жидкое, в кости
плотное. Основные функции соединительной ткани: трофическая,
связанная с питанием организма, опорная, защитная и др. Виды
соединительной ткани: мезенхима, ретикулярная, кровь, волок-
нистая, хрящевая, костная, жировая, пигментная.
Ретикулярная ткань представляет собой рыхлое скоп-
ление звездчатых клеток, соединенных в синцитий. Из этой ткани,
например, в основном состоит селезенка, имеющая губчатое строе-
ние. Пространство ячей заполнено образующимися в селезенке бе-
29
Рис. 13. Ретикулярная ткань лимфатического узла:
1 — ретикулярная клетка; 2 — ретикулиновые волокна
лыми кровяными клетками — лимфоцитами. Способность рети-
кулярного синцития к фагоцитозу (к захвату и поглощению
инородных тел, включая бактерий) определяет защитную функцию
этой ткани (рис. 13).
Рыхлая волокнистая соединительная ткань
(рис. 14). Промежуточное вещество жидкое, с многочисленными
волокнами: относительно толстыми коллагеновыми (клейдаю-
щими) и тонкими эластиновыми. Среди клеток выделяют фибро-
бласты — клетки, порождающие волокна, а также пигментные
клетки, ретикулярные и др. У позвоночных ткань входит в состав
органов, в подкожную клетчатку.
Плотная волокнистая соединительная ткань
в основном состоит из плотно переплетенных волокон. Если при-
обладают коллагеновые волокна, ткань называют фиброзной,
если эластиновые—эластиновой. Эластиновая ткань образует
связки, придавая им растяжимость. Коллагеновые же волокна
отличаются прочностью при слабой растяжимости, выдерживают
30
нагрузку до 6 кг на 1 м2. У млекопитающих фиброзная
ткань образует, например, нижний слой кожи, идущий па выделку
кожевенных изделий.
Хрящевая ткань состоит из клеток, лежащих в капсулах,
вкрапленных в плотное основное вещество. Хрящ входит в сос-
тав скелета позвоночных и ряда беспозвоночных (у немногих
аннелид и головоногих моллюсков). Рост хряща происходит за
счет надхрящницы. В гиалиновом хряще (рис. 15) промежуточное
вещество выглядит на срезах однородным, благодаря тому что
коллагеновые волокна имеют одинаковое лучепреломление с
аморфным веществом. Покрывает суставные поверхности костей.
В эластиновом хряще эластиновые волокна преобладают над кол-
лагеновыми. Встречается, например, в ушных раковинах. Воло-
книстый хрящ состоит из плотно лежащих коллагеновых волокон
и округлых клеток. У позвоночных им образованы межпозвоноч-
ные диски.
Костная ткань образует кости. Ее промежуточное вещест-
во состоит из коллагеновых волокон и аморфной массы, пропитан-
Рис. 14. Рыхлая волокнистая соединительная ткань:
/ — пучки коллагеновых волокон; 2 -- эластиновые волокна; ,3 - фибробласты;
4 — их ядра; 5 — гистиоциты (блуждающие)
31
Рис. 15. Гиалиновый хрящ:
/ — надхрящница; 2 -•
основное вещество; 3 — хря-
щевые клетки внутри кап-
сул
ной солями. Есть кости трубчатые, пластинчатые, чешуйчатые и
другой формы и разного строения. Кости развиваются в мезенхиме,
богатой коллагеновыми волокнами. Из клеток образуются остеоб-
ласты — костеобразователи, деятельностью которых накапливает-
ся бесструктурная масса. Следующий этап —- отложение солей и
преобразование остеобластов в костные клетки — остеоциты (рис.
16), имеющие многочисленные отростки, которыми клетки, замуро-
ванные в основном веществе, сообщаются между собой. Остеоциты
располагаются концентрически вокруг Гаверсовых каналов, в ко-
торых находятся кровеносные сосуды и нервы. Кость — живая
ткань. Большая часть принадлежит крови, в частности, в достав-
ке солей и извлечении их из кости. Кости — солевой резерв,
из которого животные могут почерпнуть при необходимости мине-
ральные вещества. В костях содержится около 50 % воды, 15,7 %
жира, 12,45 % органического вещества и 21,85 % солей (но
Заварзину и Щелканову).
Кость может развиваться и на месте хряща. В этом случае
продуцирование костного вещества происходит на периферии хря-
ща при одновременном изменении его самого. Хрящ разрушается
остеобластами, в измененную ткань проникают кровеносные сосу-
ды и соединительнотканные клетки; остеобласты в дальнейшем
становятся остеоцитами.
Кровь. В состав крови входят: жидкая ее часть — плазма и
форменные элементы — кровяные клетки. Форменные элементы у
животных разных типов имеют отличия. У беспозвоночных (рис.
17) они часто бесцветны и способны к амебоидному движению.
У позвоночных (рис. 18) они подразделяются на эритроциты
(красные кровяные клетки), лейкоциты (белые кровяные клетки)
и тромбоциты (у млекопитающих представлены пластинками).
Главный орган кроветворения наземных позвоночных — костный
мозг. Лишь часть белых кровяных клеток образуется в селезенке
и лимфатических узлах. У водных позвоночных кровообразование
происходит во многих местах.
Эритроциты большинства позвоночных имеют ядро, у млеко-
питающих они безъядерные. Длительность их жизни различна.
Роль эритроцитов в дыхании заключается в поглощении ими
кислорода органами дыхания и транспортировке его к тканям.
Рис. 17. Кровяные клетки пчелы в разных стадиях развития:
/ ядро; 2 хроматиновые зерна; 3 вакуоли
2 Б. А. Кузнецов и др.
33
Лейкоциты различны по форме и выполняемым функциям.
Наиболее они разнообразны у позвоночных. Все формы лейкоцитов
имеют ядро. Многие способны к амебоидному движению. Известна
защитная функция фагоцитов (клеток-пожирателей). В наиболь-
шей мере ее выполняют нейтрофильные лейкоциты. При инфекци-
онных заболеваниях их число возрастает в 5—10 раз. Есть сре-
ди лейкоцитов ферментообразователи, что свидетельствует об
участии лейкоцитов в обмене веществ. У ряда лейкоцитов защит-
ная функция проявляется в нейтрализации ядовитых веществ.
Длительность существования разных форм лейкоцитов от нес-
кольких дней до нескольких месяцев.
Тромбоциты (образующие тромб). Участвуют в свертывании
крови, в заживлении ран.
Кровяная плазма — вязкая жидкость сложного химического
состава. Содержит белки, аминокислоты, углеводы, жиры, мине-
ральные вещества, гормоны, газы, продукты обмена. Состав плаз-
мы отражает многостороннее участие ее в жизнедеятельности ор-
ганизма. Среди функций плазмы: дыхательная — транспортиров-
ка кислорода и других газов; трофическая — транспортировка пи-
тательных веществ; регуляторная — перенос гормонов; терморегу-
ляторная — благодаря большой теплоемкости воды поддержание
водного баланса тканей путем обмена жидкостью через стенки кро-
веносных капилляров; защитная — содержание антител, антито-
ксинов.
Лимфа у низших позвоночных представлена межтканевой
жидкостью с клеточными включениями. У животных, имеющих
Рис. 18. Кровь крупного рогатого скота:
/ — эритроциты; 2 — лимфоцит; 3 — мо-
ноцит; 4с, 4п, 4ю — сегментоядерпый,
палочкоядерный и юный нейтрофилы;
5 — эозинофил; 6 — базофил; 7 — кровя-
ные пластинки
Рис. 19. Формы мышечной ткани:
а — гладкие мышечные клетки; б -
поперечнополосатые мышечные во-
локна
34
кровь, лимфатическая система развита наряду с кровеносной.
В наиболее полном виде она состоит из лимфатических сосудов
и лимфатических узлов. Ее клеточные элементы — лимфоциты, об-
ладают амебоидным движением и способны выселяться из русла
сосудов (блуждающие клетки).
Мышечная ткань. Различают мышцы гладкие и поперечнополо-
сатые. В тех и других мышцах, их клетках и волокнах, находят-
ся тончайшие волоконца — миофибриллы. Сократительные воло-
конца имеют и высшие одноклеточные, но мышечная ткань
характерна для многоклеточных животных. Гладкие мышцы — на-
иболее древняя структура мышечной ткани, развивается из мезен-
химы. Гладкие мышцы состоят из пучков мышечных клеток,
тесно прилежащих одна к другой (рис. 19, а). Клетки верете-
нообразной формы с одним продолговатым ядром. Миофибриллы
гладкие. Коллагеновое вещество связывает клетки и придает всему
пучку монолитность. Для этих мышц характерна плавность сокра-
щения и расслабления. У низших беспозвоночных и кольчатых
червей такие мышцы находятся во внутренних органах и образуют
кожную мускулатуру. Членистоногие и позвоночные, имеющие ске-
лет, кроме гладкой мускулатуры во внутренних органах, имеют
скелетные мышцы поперечнополосатые.
Поперечнополосатые мышцы (рис. 19,6) состоят из много-
ядерных волокон со сложным строением миофибрилл, способны
совершать быстрые сокращения и выносить большую нагрузку.
Скорость их сокращения в отличие от мышц гладких варьирует
от очень медленного до крайне быстрого (пчелы и многие дру-
гие насекомые делают несколько сот взмахов крыльями в секунду).
Они во всех случаях имеют прочную опору. Поперечнополосатые
мышцы обычно опираются на скелет наружный или внутренний
и относятся к мышцам произвольного сокращения. Каждое мышеч-
ное волокно покрыто сарколеммой, под которой расположено
несколько продолговатых ядер. В миофибриллах с поперечной
исчерченностыо светлые диски чередуются с темными. Каждая
миофибрилла по длине неоднородна, и это в совокупности про-
является в исчерченности всего волокна (рис. 20). Длина волокон
в отдельных мышцах достигает нескольких сантиметров. Эти
мышцы отличаются от гладких и физиологическим процессом сок-
ращения волокна.
Рис. 20. Схема строения мио-
фибрилл;
/ -- светлые диски; 2 — темные
диски; 3 — перемычки (инофраг-
мы) светлых дисков; 4 - пере-
мычки темных дисков
2*
35
Нервная ткань воспринимает и передает раздражения, посту-
пающие из внешней среды и возникающие в самом организме.
Раздражимость — одно из свойств, характеризующих живую ма-
терию.
Тканевые элементы нервной системы представлены клетками
и межклеточным веществом. Различают нервные клетки — нейро-
ны и одевающие их (сопровождающие) клетки — нейроглии
(рис. 21). В цитоплазме нервной клетки — нейроплазме — нахо-
дятся нейрофибриллы, а также своеобразное тигроидное вещест-
во — глыбки Нисселя, об участии их в деятельности нейрона мож-
но судить по изменению числа глыбок, концентрация которых при
раздражении уменьшается. По функции различают нейроны чувст-
вительные и двигательные. Нейрон имеет отростки: короткие —
дендриты и длинный — нейрит (аксон). Концевые разветвления
нейрита чувствующей клетки воспринимают раздражение и назы-
ваются рецепторами (рис. 22). Они многочисленны на поверхности
тела и во внутренних органах. От рецептора возбуждение пере-
дается по нейриту в тело клетки, а затем через дендриты —
дендритам двигательной клетки, вызывая ту или иную реакцию
Рис. 21. Мультиполярный нейрон:
/ - тело клетки с нейрофибрилламп и
глыбкамн тигроидпого вещества; 2--
дендриты; 3 — нейрит (аксон) в оболочке,
образованной клетками глин (олигоденд-
роглии); 4 — нервное окончание в мышце;
5 — мышечное волокно
Рис. 22. Рецепторы в коже и их спе-
цифическая функция:
1 колбочки Краузе (на холод); 2—
эпидермис; 3— дерма; 4 — нервные спле-
тения волосяного фолликула; 5 — тельце
Руффини (на тепло); 6 —свободные нерв-
ные окончания (на болевые стимулы);
7 — диски Меркеля и 8 — тельца Мейсне-
ра (на прикосновение); 9 - тельце Начи-
ни (на более сильное давление)
36
органа: в мышце двигательную, в железе выделение секрета и т. д.
Разветвление двигательного нейрита в органе называется эффек-
тором. Так идет реакция возбуждения по рефлекторной дуге.
Например, если в спинном мозге позвоночных, кроме двух
нейронов, есть еще промежуточный, то рефлекторная дуга будет
образована тремя нервными клетками. Нейроны одноядерные.
Установлено, что у высших позвоночных они не размножаются,
но их нейтриты при ранениях регенерируют. Нейроглия состоит из
клеток различного строения и выполняет ряд функций: защитную,
опорную, секреторную. Слой глии покрывает нейриты, у позвоноч-
ных глия выстилает мозговую полость, входит в состав голов-
ного и спинного мозга. Присутствие в ряде мест промежуточного
вещества глии в виде волокон обусловливает опорную функцию
нейроглии.
ОРГАНЫ
Жизненные процессы многоклеточных животных осуществляют:
покровы, органы движения, пищеварения, дыхания, кровообраще-
ния, выделения, нервной системы с органами чувств, внутренней
секреции, размножения. Отправления любого из органов совер-
шаются в полной взаимосвязи и при взаимодействии со всеми
другими органами, свидетельствуя о целостности организма. По-
добное же прослеживается и в жизнедеятельности структур
одноклеточного животного, особенно выраженное у высших прото-
зоа — инфузорий.
Каждый орган функционирует как неразрывная часть едино-
го организма. Например, пищеварительная функция невозможна
без воздействия нервной системы, органов дыхания и пр. Прием
пищи, ее механическая обработка, переваривание, всасывание,
дефекация — все эти" сложные процессы протекают с непремен-
ным участием других органов. Следовательно, в данном при-
мере мы говорим не об органе пищеварения, а о системе органов
пищеварения как о взаимодействующих структурах.
Органы изучаются во всех разделах систематической части
зоологии.
РАЗМНОЖЕНИЕ ЖИВОТНЫХ
Размножение — свойство живых организмов воспроизводить себе
подобные особи.
Животные размножаются бесполым и половым путем. Бесполое
размножение свойственно преимущественно низшим животным.
При бесполом размножении от материнской особи отделяется
часть ее тела или вся особь делится на две или большее число
частей, каждая из которых развивается в самостоятельное живот-
37
Рис. 23. Бесполое размножение протозоа:
1—деление амебы; 2 — почкование инфузории (сувойки); 3 —
шизогония малярийного плазмодия
Рис. 24. Формы сперматозоидов:
1 — барана; 2 — рака; 3 — круглого червя; 4 — жука
ное. Различают несколько способов бесполого размножения жи-
вотных: деление, почкование, шизогонию (рис. 23).
Деление заключается в перетяжке материнской особи на
две более или менее равные части, каждая из которых становится
новым организмом.
Почкование — такое бесполое размножение, когда на те-
ле материнской особи образуется вырост (почка), постепенно при-
обретающей форму и строение взрослого животного. После от-
деления (отпочкования) от материнского организма новая особь
начинает вести самостоятельную жизнь.
Множественное деление (шизогония) наблюдается
у ряда паразитических одноклеточных животных. Сначала ядро
материнского организма многократно делится, что ведет к об-
разованию многоядерного шизонта. Затем вокруг каждого ядра
обособляется участок цитоплазмы и шизонт распадается на
многочисленные мелкие дочерние особи (мерозоиты), что влечет
за собой быстрое накопление паразитов в теле хозяина.
Половое размножение присуще всем типам животных. При по-
ловом размножении новый организм развивается из зиготы, об-
разующейся в результате оплодотворения яйца сперматозоидом.
Женская половая клетка — яйцо (яйцеклетка) обычно имеет
округлую или овальную форму и состоит из цитоплазмы и ядра.
В цитоплазме содержится то или иное количество питательного
вещества. Яйцо, как правило, покрыто оболочками различного
строения.
Мужские половые клетки — с п е р м а т о з о и д ы имеют
разнообразную форму (рис. 24), но обычно состоят из головки,
соединительной части и хвоста, который служит для движения
в жидкой среде. Большую часть головки занимает ядро.
У одних многоклеточных животных (как правило, обитающих
в воде) наблюдается наружное оплодотворение: как
яйца, так и сперматозоиды выделяются в воду, где и происходит
их слияние. Другим животным свойственно внутреннее
оплодотворен и е: сперма вводится в половые пути самки,
и там совершается акт оплодотворения яйца сперматозоидом.
Иногда животное развивается из иеоплодотворенного яйца;
такое размножение называется девственным — парте-
ногенетическим.
НАЧАЛЬНЫЕ ЭТАПЫ РАЗВИТИЯ
МНОГОКЛЕТОЧНЫХ ЖИВОТНЫХ
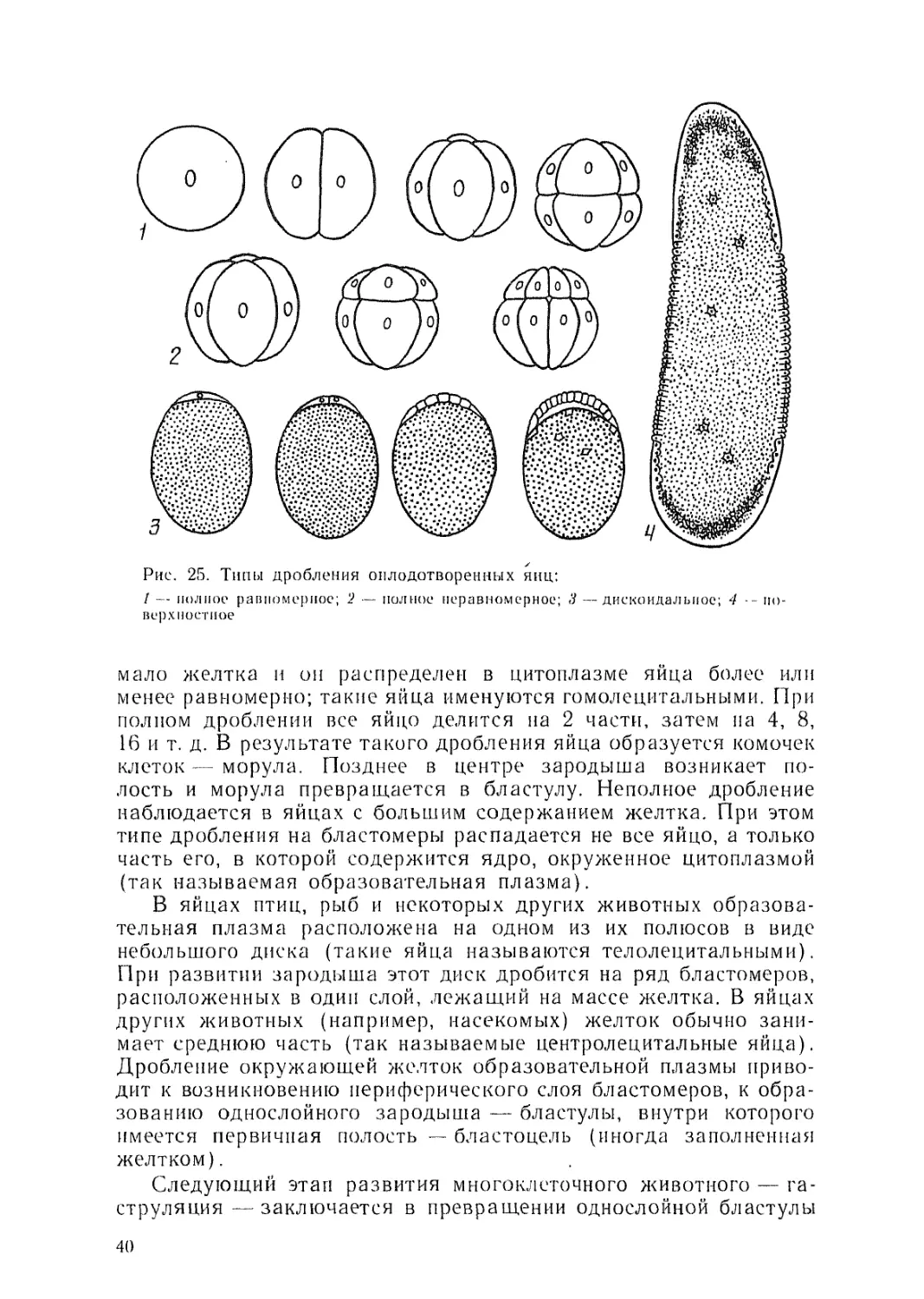
Развитие оплодотворенного яйца начинается с его дробления
на ряд клеток — бластомеров. В зависимости от количества
желтка в яйце этот процесс протекает у разных животных раз-
лично.'Дробление яйца бывает полным и неполным (рис. 25). Пол-
ное дробление яйца происходит в тех случаях, когда в нем
39
Рис. 25. Типы дробления оплодотворенных яиц:
1 — полное равномерное; 2 — полное неравномерное; 3 — дискоидалыюе; 4 -- по-
верхностное
мало желтка и он распределен в цитоплазме яйца более или
менее равномерно; такие яйца именуются гомолецитальными. При
полном дроблении все яйцо делится на 2 части, затем на 4, 8,
16 и т. д. В результате такого дробления яйца образуется комочек
клеток — морула. Позднее в центре зародыша возникает по-
лость и морула превращается в бластулу. Неполное дробление
наблюдается в яйцах с большим содержанием желтка. При этом
типе дробления на бластомеры распадается не все яйцо, а только
часть его, в которой содержится ядро, окруженное цитоплазмой
(так называемая образовательная плазма).
В яйцах птиц, рыб и некоторых других животных образова-
тельная плазма расположена на одном из их полюсов в виде
небольшого диска (такие яйца называются телолецитальными).
При развитии зародыша этот диск дробится на ряд бластомеров,
расположенных в один слой, лежащий на массе желтка. В яйцах
других животных (например, насекомых) желток обычно зани-
мает среднюю часть (так называемые центролецитальные яйца).
Дробление окружающей желток образовательной плазмы приво-
дит к возникновению периферического слоя бластомеров, к обра-
зованию однослойного зародыша — бластулы, внутри которого
имеется первичная полость — бластоцель (иногда заполненная
желтком).
Следующий этап развития многоклеточного животного — га-
струляция —заключается в превращении однослойной бластулы
40
в двухслойную гаструлу. Гаструляция у разных животных проте-
кает неодинаково (рис. 26). У форм, яйца которых проходят
полное'и равномерное дробление, бластула имеет вид пузырька
с клеточной стенкой примерно одинаковой толщины. В этом случае
гаструляция идет путем впячивания нижней части бластулы.
В результате образуется зародыш с двумя зародышевыми лист-
ками: наружными — эктодермой и внутренним -- эндодермой.
У животных, яйца которых проходят полное, но неравномер-
ное дробление, верхняя часть бластулы сложена мелкими, а ниж-
няя — более крупными бластомерами. В этом случае образование
двухслойного зародыша (гаструлы) происходит путем обрастания
нижних крупных бластомеров более быстро делящимися верх-
ними мелкими. Внутренний слой гаструлы у некоторых животных
образуется вселением (иммиграцией) отдельных клеток нижней
части бластулы в ее полость (бластоцель).
Как бы ни шел процесс гаструляции, он приводит к образо-
ванию зародыша, состоящего из двух слоев клеток: эктодермы
и эндодермы. Пространство между ними остается первичной полос-
тью тела, а гастроцель (кишечная полость гаструлы) в дальней-
шем становится полостью кишечника взрослого животного. Отвер-
стие гастроцели - бластопор (первичный рот) у большинства бес-
позвоночных животных сохраняется и становится ртом взрослых
особей. Такие животные называются первичноротыми. У другой
группы животных (иглокожих, хордовых) бластопор зарастает или
становится заднепроходным отверстием, рот же впоследствии об-
разуется вновь (вторичпоротыс).
Для низших многоклеточных животных -- губок и кишечнопо-
лостных — характерно двухслойное строение тела. Но у более вы-
сокоор га и изова н ных ж и вот-
иых между эктодермой и эн-
додермой образуется сред-
ний зародышевый листок —
мезодерма. Ее образование
происходит различно. Низ-
шие трехслойные животные
в течение всей жизни сохра-
няют первичную полость
тела, и их именуют первич-
нополостпыми. В развитии
ряда хордовых мезодерма за-
кладывается в виде парных
карманообразных выпячива-
Рис. 26. Типы гаструляции:
/--•путем иммиграции; 2 путем пи-
ваги наци и
41
г Д % £ Ж
Рис. 27. Образование вторичной полости тела у кольчатых червей (/) и лан-
цетника (//):
А — начальная стадия; Б — формирование целомических мешков; В — сформировав-
шийся целом; Г — обособление нервной пластинки в эктодерме, в эндодерме начинают
выпячиваться сомиты; Д — образовался зачаток нервной трубки, сомиты обособляются;
Е — сомиты с полостью внутри (целомом) обособились от эндодермы, начинает заклады-
ваться хорда; Ж — сомиты с целомом внутри разрослись и заняли пространство первич-
ной полости тела, нервная трубка и хорда сформировались;
/—кишка; 2 — зачаток мезодермы; 3 — первичная полость; 4 — полость целоми-
ческого мешка (целом); 5 — внутренняя стенка целомического мешка; б — наружная
часть стенки целомического мешка; 7 — спинной кровеносный сосуд; 8 ~ брюшной кро-
веносный сосуд; 9 — брюшные нервные стволы; 10 — эктодерма; // — эндодерма (киш-
ка); 12—мезодерма; 13— нервная пластинка; /'/ — нервная трубка; /5 — невроцель;
16 — хорда
ний стенки кишечника зародыша (рис. 27). Они отшнуровываются
от кишки, располагаясь по бокам от нее. Внутри каждого
мезодермального мешочка — сомита имеется полость, называемая
вторичной полостью тела — целомом. Сомиты располагаются
между экто- и эндодермой в первичной полости тела и, раз-
растаясь, вытесняют ее.
Наружные стенки сомитов соприкасаются с эктодермой, а вну-
тренние — со стенкой кишки. Когда перегородки между сомитами
рассасываются, полости сомитов сливаются. Животные, имеющие
целом, называются вторичнополостными (целомата). Остатки пер-
вичной полости сохраняются у целомата в виде каналов, щелей,
лакун. Часть их становится полостью кровеносных сосудов, при-
обретающих у большинства собственные стенки. Первичнополост-
ные лишены кровеносных сосудов, у них функцию крови выполняет
полостная жидкость, омывающая внутренние органы. В развитии
ряда целомата (например, моллюсков, членистоногих) происходит
частичное слияние первичной полости с целомом. Такую полость
42
называют смешанной, а кровеносную систему их — незамкнутой:
кровь течет то в сосудах, то разливается по лакунам.
В процессе развития эмбриона зародышевые листки путем
дифференцировки образуют ткани и органы. В формировании эпи-
телиальной ткани участвуют все три зародышевых листка. Из
эктодермы развиваются кожный эпителий и его железы, а у мно-
гих — передний и задний отделы кишечника, органы дыхания, мо-
чевые протоки; из эндодермы — эпителий среднего (пищевари-
тельного) отдела кишечника, нервная система; из мезодермы —
скелет, мышцы, кровь.
СИММЕТРИЯ ТЕЛА ЖИВОТНЫХ
У подавляющего большинства животных части тела расположены
симметрично. Лишь у немногих (например, у некоторых губок)
тело имеет неправильную, лишенную симметрии форму (рис. 28, / ).
Рис. 28. Формы симметрии:
1 — асимметричная (амеба); 2 ра-
диальная (морская звезда); ,3 - - дву-
сторонняя (речной рак)
43
Различают два основных типа симметрии животных: радиальную
и двустороннюю.
Радиальная симметрия (рис. 28, 2) характеризуется тем, что
одинаковые части тела и органы располагаются по радиусам
от срединной продольной оси животного. Тело с радиальной сим-
метрией может быть разделено на равные части несколькими
плоскостями, проходящими через эту ось. Радиальная симметрия
тела свойственна преимущественно животным, ведущим сидячий
или малоподвижный образ жизни или пассивно плавающим в воде.
Примером подобных животных могут служить гидры, медузы, мор-
ские звезды и пр.
Двусторонняя симметрия (рис. 28,3) тела животного отличает-
ся тем, что только одна плоскость может разделить его на две
равные половины. У двустороннесимметричных животных можно
различить левую и правую половины, передний и задний концы
тела. Данная симметрия присуща преимущественно животным,
способным к самостоятельным передвижениям. Как правило,
она бывает неполной, относительной. Обычно организм двустороп-
несимметричного животного снаружи более или менее правильно
симметричен (левая и правая половина его почти одинаковы),
но в расположении многих внутренних органов наблюдается яв-
ная асимметрия. Так, например, у человека сердце лежит в ле-
вой части грудной клетки, у птиц яичник расположен в левой
части таза и т. д.
Характер симметрии тела служит важным систематическим
признаком различных типов животных. Например, у иглокожих
наблюдается смена симметрии тела в течение жизни (морские
ежи и др.). Их плавающие личинки имеют двустороннюю симмет-
рию, а взрослые — радиальную.
СИМБИОЗ В ЖИВОТНОМ МИРЕ
Отношения животных, обитающих в той или иной среде, харак-
теризуются взаимовлиянием прямым, непосредственным, или ко-
свенным. Сожительство организмов разных видоее получило наз-
вание симбиоз.
Форма симбиоза по своему существу имеет для сожитель-
ствующих животных весьма различное биологическое значение:
сожительства враждебного характера; сожительства с взаимной
пользой — м утуализм (от лат. muluus — взаимный); сотрапе-
зничество (совместное использование корма) — комменса-
лизм (от лат. сот — вместе, mensa трапеза). Но комменса-
лизмом называют и те случаи, когда видимую пользу получают не
оба организма; а один из них при пребывании на другом орга-
низме или близ него, или внутри него. Последнее позволяет
предполагать, что определенные обстоятельства (прежде всего не-
достаток пищи для меньшего и слабого комменсала-нахлебника)
44
могут вызвать смену характера сожительства с переходом ком-
менсализма во временный или постоянный паразитизм.
Бесчисленное разнообразие оттенков в формах симбиоза зат-
рудняет четко очертить границы каждого из них. Это относится
и к определению понятия паразитизм (греч. parusitos -
нахлебник). Признак вредности дает особенно ясное представ-
ление о паразитическом сожительстве. В отличие от хищника
паразит, питаясь за счет другого — хозяина, как правило, не
убивает его. Однако, подтачивая здоровье, может способствова ть
его гибели. Паразит, за единичными исключениями, меньше хозяи-
на и в своей организации имеет в сравнении со свободпоживущн-
ми сородичами резко выраженные черты регрессивного метамор-
фоза — упрощения в строении, особенно заметные в нервной систе-
ме, органах движения. В процессе эволюции вместе с переходом
к паразитизму появляются и новоприобретения, имеющие при-
способительное значение.
В фауне нашей планеты животных, ведущих паразитический
образ жизни, описано свыше 80 тыс., из которых половину сос-
тавляют перепончатокрылые насекомые — наездники, на втором
месте по численности стоят нематоды и плоские черви, на треть-
ем— протозоа. В типе хордовых паразиты единичны. Паразиты
оказывают большое влияние на существование свободножпвущнх,
вызывая временами эпизоотии.
Те или иные виды паразитов локализуются во всех системах
органов животных, преобладая в пищеварительных. Так, в желуд-
ке и кишечнике обитают: ряд видов жгутиковых, инфузорий,
несколько десятков видов амеб, среди них дизентерийная амеба,
споровики (кокцидии), микроспоридии (ноземы), из плоских чер-
вей — ряд трематод, цепней, широкие лентецы, а из нематод
аскариды, острицы, трихинеллы (в период роста до ноловоз-
релости), из насекомых —личинки желудочного овода; в печени
кокцидии, фасциолы, ланцетовидный сосальщик, финны эхино-
кокка и пр.; в кровеносной системе — малярийные плазмодии,
пироплазмы, трипаносомы, мигрирующие личинки аскарид, трихи-
нелл и других нематод; в мышцах — споровики, личинки три-
хинеллы, финны цепней и др.; в сухожилиях нематоды оихоцер-
ки (у лошадей и рогатого скота); в подкожной клетчатке
ришта, микрофилярии, личинки кожного овода; в трубчатых
костях — финны эхинококка и др.; в органах дыхания - финны
эхинококка, мигрирующие через легкие личинки ряда нематод
(аскарид и др.), взрослые нематоды; в почках — очень длинная
нематода (до метра), диоктофиме; в половой системе жгути-
ковые, трематоды и пр.; в мозге — финны овечьего мозговика;
в органах чувств — трематоды (в хрусталике глаза рыб), не-
матода сингамус (в среднем ухе кошек), личинки оводов
(в носовой полости). И у растений животные-паразиты (в
большинстве нематоды) локализуются также во всех системах ор-
ганов, преобладая в корневой системе.
45
Обстоятельства жизни экто- и эндопаразитов в разных уча-
стках организма хозяина у животных и растений разных система-
тических категорий, разного возраста, живущих в разных гео-
графических широтах, крайне своеобразны, и столь же различны
приспособления в морфологии и жизненных отправлениях у пара-
зитов. Организм хозяина для паразита — среда жизни. Внешняя
среда воспринимается паразитом, иногда очень тонко, через из-
менения во внутренней среде организма хозяина, что подтверж-
дается экспериментально. Так, например, изменение человеком,
носителем бычьего цепня, распорядка работы днем или ночью
отражается на выходе из кишечника зрелых проглоттид. При бодр-
ствовании днем они выходят в вечернее время, а с переходом
на ночную работу — утром. Подобную же чувствительность к со-
стоянию организма хозяина показывают и эктопаразиты. Моноге-
нетическая трематода многоустка развивается на жабрах голо-
вастика лягушки одновременно с созреванием хозяина. Вместе с
метаморфозом головастика дозревает и паразит, переходя потом
в мочевой пузырь лягушки.
Искусственная задержка развития головастика вызывает за-
держку развития личинки многоустки.
Неодинакова у паразитов способность выживания в разных
условиях обитания. Одни приспособлены к жизни в определен-
ном органе узкого круга хозяев, другие могут паразитировать в
определенном органе, но у многих видов животных, как, на-
пример, печеночная фасциола или трихинелла. Есть среди па-
разитов и способные существовать в самых различных органах
хозяина. Так, личинки широкого лентеца плероцеркоиды встре-
чаются у щук в мышцах, в стенке желудка, кишечника, в поч-
ках, селезенке, в брюшной полости (Гнездилов).
Паразиты подобно свободноживущим организмам, обитая
в одной и той же среде, показывают разные приспособительные
черты в строении, отражающие неодинаковый характер питания,
способов добычи и обработки пищи, дыхания и пр. Например,
паразитирующие в кишечнике хозяина цестоды питаются осмоти-
чески и лишены кишечника. Их тело плоское (большая повер-
хность всасывания). Нематоды же, паразитирующие в кишечном
тракте, имеют кишечник. Их тело округлое. У некоторых пара-
зитов рот вооружен зубовидными придатками, у других — со-
сущий, у третьих функции сосания выполняет пищевод. У фито-
нематод свои особенности и разнообразие в приспособлениях при
жизни в хозяине в одинаковой среде.
Многие факты свидетельствуют о глубокой связи эндопара-
зитов с эктопаразитами и свободноживущими организмами. Ли-
чинки нематоды вешерерии, возбудителя в тропиках слоновой бо-
лезни, живущие в крови человека, появляются в периферичес-
ких сосудах больного в вечерне-ночное время, когда особенно
активны комары, распространяющие паразита в пространстве. Вы-
работка подобного способа распространения вешерерии, надо по-
46
лагать, заняла весьма длительное время, как и другие способы рас-
пространения эндопаразитов.
На общем фоне установления паразитических форм симбио-
за выделяются нематоды, показывающие примеры временного
перехода от свободного существования к эндопаразитизму. Во-
обще нематоды отличаются исключительно высокой приспо-
собляемостью. Своеобразие их морфологии, как и физиологии,
все еще не получило объяснения.
На установление взаимосвязи паразита и хозяина особое
влияние оказывают пространственные контакты. Фасциола —
паразит травоядных проходит личиночное развитие в промежу-
точном хозяине в одном из воздушнодышащих брюхоногих мол-
люсков, обитающих в зонах прибрежных растений, водоемов и на
заливных лугах, т. е. в местах, посещаемых травоядными. Завер-
шается личиночное развитие фасдиол инцистированием в воде и
на траве, что способствует попаданию молодого червя в хозяи-
на вместе с кормом. Заражение хозяина происходит по пищевой
цепи — наиболее обычным путем в распространении паразитов. У
трематод — паразитов плотоядных это обеспечивается включе-
нием второго (дополнительного) хозяина — рыбы, ракообразного
и других, являющихся пищей дефинитивного. Промежуточные хо-
зяева, если их несколько, тоже связаны как пространственно,
так и пищевыми отношениями.
О роли контактов в образовании взаимосвязи паразита и
хозяина свидетельствует и тот факт, что у человека и собаки
(первой по давности одомашнивания) общих гельминтов 36 видов
(Петров).
Общее число гельминтов, встречающихся в ветеринарной прак-
тике, свыше 1 тыс. видов (Скрябин), а в медицинской —
около 150. Распространению паразитов способствуют резервуар-
ные хозяева, попадая в которых паразиты не развиваются, но нака-
пливаются, сохраняя жизнеспособность. Такие резервуарные хо-
зяева известны для многих паразитов из разных классов.
Борьба с паразитами проводится в двух направлениях:
в предупреждении заражения, в принятии профилактических мер и
полном уничтожении паразитов, в полном оздоровлении
территории. Первый, оборонительный метод Скрябин называет
п р с з е р в а ц и е й, а второй — д с в а с т а ц и ей- В нашей оте-
чественной практике девастация поставлена главной .целью науч.-,
ной и практической паразитологии. Среди достижений в этом
трудном деле ликвидация очагов малярийных плазмодиев, ришты
и некоторых других.
РАЗВИТИЕ ЖИВОТНОГО МИРА
Палеонтологическая летопись показывает общую картину разви-
тия органической природы. Датировать точно возникновение
тех или иных групп животных трудно. Палеонтология рисует
47
эволюцию жизненных форм лишь крупными мазками. Геологи-
ческая история насчитывает около 4 млрд лет. Ее подразделяют
на эры: архейскую, протерозойскую, палеозойскую, мезозойскую,
кайнозойскую. Эра отражает крупные этапы эволюции животных
и растений. Эры подразделяются на периоды, так в палеозойской
эре различают периоды: кембрийский, ордовик, силурийский,
девонский, каменноугольный, пермский; в мезозойской — триас-
ский, юрский, меловой; в кайнозойской —третичный и четвертич-
ный (современный). Появление жизни на Земле приурочивается
к архейской эре. В отложениях протерозойской эры обнаружены
бактерии, водоросли и ряд низших беспозвоночных — радиолярии,
губки. В палеозойскую эру животный мир достиг уже большого
разнообразия. В начале эры жизнь была сосредоточена в море.
Животные были представлены беспозвоночными, а растения —
преимущественно водорослями. Первые наземные растения обна-
ружены в кембрии, в ордовике — первые позвоночные (рыбы).
В девоне найдены костистые рыбы и амфибии, в каменноугольном
периоде — стегоцефалы (из древних амфибий), хрящевые рыбы.
Произошло разделение амфибий на хвостатых и бесхвостых.
Существовали в ту поры и насекомые, и пауки. Мезозойскую эру
называют веком пресмыкающихся. Первые рептилии найдены
в каменноугольном периоде палеозойской эры, но массовое раз-
витие их произошло в мезозое. Поражают разнообразие форм и
нередко гигантские их размеры. Одни доминировали на суше,
другие жили в водоемах, третьи были способны к полету. В мезо-
зойскую эру появились первые птицы и млекопитающие, цвет-
ковые растения, тогда как плауновые, хвощи и папоротники
постепенно теряли господствующее среди растений положение.
В кайнозойскую эру флора и фауна Земли постепенно при-
обрели современный состав. В растительном мире все больше
преобладали покрытосеменные. Вымерли характерные для мезозоя
головоногие моллюски — аммониты и белемниты. Пресмыкаю-
щиеся потеряли господствующее положение среди позвоночных,
уступив место млекопитающим и птицам, которые дали большое
разнообразие форм. В эту эру (2—3 млн лет назад) появился
на Земле человек разумный — Homo sapiens.
Современная систематика выделяет большое число типов
животных. В настоящем учебнике дается описание только тех
типов животных, которые либо имеют важное значение для по-
знания эволюции животного мира, либо содержат большое коли-
чество форм, представляющих практический интерес: домашние
и промысловые виды, вредители или паразиты разводимых живот-
ных или возделываемых растений.
СИСТЕМАТИЧЕСКИЙ
ОБЗОР животных
ПОДЦАРСТВО ОДНОКЛЕТОЧНЫЕ
(PROTOZOA)
Общая характеристика. В подцарство объединены животные, но
размерам микроскопические, а по общим чертам строения в боль-
шинстве одноклеточные. Исключение составляют колониальные,
у которых тело образуют несколько клеток. Так, у вольвокса
(рис. 29) число клеток в теле исчисляется тысячами, а их диффе-
ренцировка на вегетативные и воспроизводящие приближает- т аких
колониальных животных к многоклеточным. Тело протозоа обра-
зовано протоплазмой с одним или несколькими ядрами, органел -
лами и органами временными или постоянными (движения,
пищеварения, выделения). Размножаются как бесполым, так и
половым путем. Но образу жизни — свободпожнвушне и пара -
зиты. Обитают в водоемах, почве, различных жидких и влажных
средах. Могут переносить высыхание благодаря выделению
защитных оболочек и в таком инцистировапном состоянии времен-
но сохраняться. Протозоа именуют п р о с т е й ш и м и, допуская
условную трактовку наименования типа (греч. prolos - первый).
В прошлом при малой осведомленности об этих существах они
казались простыми в сравнении с многоклеточными. В настоящее
время называть протозоа простейшими можно, лишь отдавая
дань истории.
Выяснено, что протозоа имеют сложнейшую микроскопическую
(и ультрамикроскопическую) организацию, обеспечивающую все
жизненные отправления. Обоснование в 30-х годах XIX в. кле-
точной теории строения животных и растений породило другое
вольное толкование наименования типа, как объединяющего
животных одноклето ч и ы х. Термин «одноклеточные» не охва-
тывает всего разнообразия форм, но удачен напоминанием о
предках многоклеточных животных.
Протозоа — целостные организмы подобно тому, как целост-
ными являются животные многоклеточные. Одноклеточность в
строении их тела есть своеобразная форма организации.
В основании родословного древа животных располагаются
гипотетические примитивные первоначально безъядерные формы,
которые в дальнейшем эволюционировали в клеточную форму
с характерным для клетки признаком — наличием ядра - - важ-
49
J
1
Рис. 29. Вольвокс:
А — Volvox globalor — участок колонии с половыми клетками; Б — Volvox
aureus — колония в процессе бесполого размножения;
1 — макрогаметы; 2 — мнкрогаметы; 3 — дочерние особи
кого регулятора жизненных отправлений и носителя наследствен-
ной информации. В процессе эволюции одноклеточные дали
начало животным многоклеточным. Их пути разошлись в очень
давние времена.
Протозоа эволюционировали при сохранении родоначальной
одноклеточной формы тела и микроскопических его размеров.
Функции их органоидов, жизненные отправления в основном
аналогичны жизненным отправлениям у многоклеточных. Послед-
нее определяет возможность широкого использования протозоа
для исследования многих биологических проблем, касающихся
всего животного мира. Они стали объектами изучения ультра-
структуры живой материи и ее основных жизненных свойств,
явления анабиоза (скрытой жизни) и потенциальных возможно-
стей сохранения жизни при крайних параметрах физико-химиче-
ских факторов среды, изменчивости, экологии, разработки
проблемы эволюции форм полового процесса, биологического
анализа форм бесполого и полового размножения и пр.
Кажутся безграничными тончайшие морфофизиологические
приспособления у протозоа к обстоятельствам жизни в разных
экологических условиях, при разном образе жизни и поведении.
Аналогичное известно и в мире многоклеточных. Сравнение общего
и особенного у протозоа и метазоа в строении и жизненных
отправлениях позволяет классифицировать тех и других подцар-
ствами в царстве животных.
Подцарство Protozoa подразделяется на ряд типов, число
которых продолжает уточняться систематиками. В данном учеб-
нике рассматриваются следующие типы: Sarcoinastigopliora,
Sporozoa, Cnidosporidia, Microsporidia, Ciliata.
Широкое распространение протозоа, своеобразие органи-
50
зации и практическое значение вызывали к ним большой интерес
и повлекли накопление обширного материала. Со временем
утвердились отрасли протозоологии: сельскохозяйственная, поч-
венная, ветеринарная, медицинская и другие, разрабатывающие
теоретические проблемы и практическое использование полезных
видов и меры ведения борьбы с вредными. Обитатели толщи воды
океанов: фораминиферы (с известковыми раковинами), радио-
лярии (с кремневым скелетом), кокколиты (из жгутиковых,
с известковым панцирем) — по отмирании образуют на дне мощ-
ные отложения известковых и кремниевых пород, входящих
в состав земной коры. Мел на 90—98 % состоит из мельчайших
панцирей кокколитофорид. Эти жгутиковые в живом виде содер-
жатся в 1 м3 до 800 тыс. особей в Тихом океане, до 3 млрд.—
в Атлантическом (Шумейко, 1978). Глобигериновый ил занимает
в Мировом океане площадь около 120 млн. км2, имея толщину
(местами) в несколько сот метров. Ископаемые протозоа ис-
пользуются в установлении стратиграфии осадочных пород как
руководящие формы при определении возраста отложений, что
важно в разведке полезных ископаемых и для исторической гео-
логии. Установлено участие протозоа в почвообразовании, их
санитарная роль в водоемах — как потребителей вредных бак-
терий, как извлекающих из воды ряд вредных веществ, посту-
пающих в водоем с водосборной площади и со сточными водами.
Медицинская и ветеринарная паразитология изучают биологию
паразитов человека, домашних животных и возделываемых
растений, их распространение, взаимосвязь с хозяином и другие
вопросы общей и частной паразитологии. Выяснена возможность
использования протозоа для испытания действия фитонцидов.
Получен положительный результат выращивания культуры ин-
фузорий для кормления мальков рыб в садках и прудах.
Строение и жизненные отправления. Размеры тела одно-
клеточных в большинстве микроскопические, измеряемые в микро-
нах и миллиметрах. Исключение составляют лишь немногие,
например морские корненожки — фораминиферы. Они тоже малых
размеров, но отдельные виды современных достигают 3 см, а
поперечник тела некоторых ископаемых превышает 10 см. Наи-
меньшие размеры имеют те, что паразитируют в теле самих про-
тозоа и в клетках различных органов многоклеточных животных.
Из свободноживущих малые размеры характерны для почвенных
обитателей, жизнь которых проходит в тончайшей пленке воды,
окружающей твердые частицы земли. Они значительно меньше
особей тех же видов из водоемов.
Форма тела весьма разнообразна (рис. 30), как это обыч-
но наблюдается у животных очень крупной и богатой видами
систематической категории. Причина тому — широкое распростра-
нение видов и их жизнь в разных экологических условиях.
У хороших пловцов тело удлиненное, веретеновидное, обте-
каемое, а у «парящих» в толще воды — с причудливыми выро-
51
Рис. 30. Различные формы тела протозоа:
/--амеба; 2— диффлугия; // — эвглена (£. oxyuris); 4 — ламблия; 5-
стентор; 6 — грегарина
стами, увеличивающими поверхность тела, что наряду с включе-
ниями в протоплазме (жировыми каплями, газовыми пузырьками)
способствует пассивному плаванию. Обитающие на дне водоемов
отличаются относительно большой массивностью. У многих тело
заключено в раковину, которая у одних органической природы,
у других, как у диффлугии, построена из песчинок, сцементиро-
ванных выделениями тела.
Свои особенности в строении эктопаразитов и эндопаразитов.
Так, у трипанозом — паразитов крови тело веретеновидное,
по его краю простирается протоплазматический парус, способ-
ствующий плаванию в вязкой плазме крови. Приспособительные
черты в строении других паразитов крови выражены по-иному.
Кишечные лямблии имеют тело уплощенное, с присасывательной
площадкой на нижней стороне. Присоска удерживает паразита
в кишечнике. Грегарины же (тоже кишечные паразиты) закреп-
ляются выростами переднего отдела тела (крючьями, щупальце-
образными придатками). Тело их вытянуто и подразделено на
2—3 отдела. Некоторые из грегарин наружной расчлененностью
напоминают в миниатюре плоских членистых червей. Прикрепи-
тельным аппаратом обладают и эктопаразиты.
52
Рассмотренные черты морфологии протозоа, живущих в той
или иной сходной среде, можно считать типичными лишь относи-
тельно по причине многообразия приспособлений, обусловленных
отличиями в биологии — активностью поведения, различием в
способах добывания пищи и т. д.
П о к р о в ы. На наружную пограничную зону тела приходятся
такие важные для существования организма функции, как вос-
приятие всевозможных воздействий извне, защита от механиче-
ских и химических агентов внешней среды, поглощение кислорода,
а при осмотическом питании — также поглощение пищи, удаление
продуктов обмена веществ. Выполнение всех этих функций нельзя
считать как зависящими только от структуры наружного покрова
протоплазмы. В избирательной проницаемости покровов, как
правило, участвует все тело одноклеточного животного. Выпол-
нение функций определяется жизнедеятельностью всего организма
и являет собою не механический, а физиологический процесс.
Тело корненожек, ряда эндопаразитов и многих других про-
тозоа покрыто тончайшей мембраной. Большинство жгутиковых,
инфузорий и саркодовых имеют покровы в виде тонкой, обычно
эластичной пелликулы. Благодаря присутствию в пелликуле
эластичных фибрилл она придает животному характерную для
него форму, вместе с тем допуская временное ее изменение,
и не препятствует совершать изгибы тела при движении. У ряда
инфузорий пелликула достигает значительной толщины и плот-
ности, приобретая свойства кутикулы или настоящего панциря
(если пропитана солями). Вообще покровы протозоа отли-
чаются сложностью и разнообразием строения. Как мембраны,
так и пелликула составляют с протоплазмой одно целое и у
некоторых видов в течение жизни или в отдельные ее периоды
переходят одна в другую.
Под мембраной и пелликулой нередко отчетливо виден в
световой микроскоп слой гомогенной желатинизированной про-
топлазмы— эктоплазмы. Глубже ее в теле находится жидкая
и обычно зернистая эндоплазма. Она содержит ядро или ядерный
комплекс из двух и более ядер, а также различные структуры,
обычные для клеток и специфические, свойственные протозоа как
организмам. Опорные фибриллы — один из видов скелетных об-
разований. У некоторых на поверхности тела плотные, нерастя-
жимые волоконца и пластинки. Такие покровы приобретают
свойства наружного скелета, выполняющего также защитную
роль. Немногие протозоа имеют внутренний скелет. Как наружный,
так и внутренний скелеты служат опорой для волокон (мионем),
обладающих сократимостью. По функции они подобны миофиб-
риллам — сократимым волокнам мышечных клеток метазоа. Ске-
лет разных одноклеточных отличается и по строению, и по хими-
ческому составу. У корненожек он в виде домика, раковины.
В строении раковин можно обнаружить весьма сложную струк-
туру, как, например, у пресноводной раковинной амебы арцеллы.
53
Твердая основа ее раковины — кремний — органической природы,
а входящая в нее эктоплазма имеет вид сети из призматических
альвеол соответственно призматической структуре всего скелета.
Электронный микроскоп позволяет отметить еще одну тонкую
важную деталь ультраструктуры раковины — наличие сети нор,
связующих эндоплазму с внешней средой.
Протозоа, ведущие жизнь в прикрепленном состоянии, снаб-
жены скелетом, открытым на переднем конце, через который тело
может выступать наружу. Этот участок тела служит для улав-
ливания и приема пищи. Скелет плавающих в толще воды отли-
чается тонкостью раковины и легкостью.
Трихоцисты — специфические образования у инфузорий, рас-
положенные под пелликулой. Они придают эктоплазме вид тонкой
исчерченности. При раздражении инфузории кислотами, щело-
чами содержимое трихоцист вытягивается в длинные нити, смо-
ченные веществом, вероятно, защитного действия. В электронный
микроскоп видно, что нить имеет несколько утолщенную основ-
ную часть и острый конец. Трихоцисты обычны для инфузорий,
но изредка встречаются у жгутиковых. Их рассматривают как
средство защиты и нападения. Трихоцисты встречаются в теле
животных то равномерно расположенными по всей периферии,
как у инфузорий парамеций (рис. 31), то сконцентрированными
в окружении рта. У некоторых они собраны в батареи, размещен-
ные на определенных участках тела.
В соответствии со строением и функ-
цией среди трихоцист различают: пек-
тицисты и токсицисты — орудие напа-
дения хищных инфузорий, мукоцисты —
слизистые и др. Трихоцисты служат
также для закрепления на субстрате
во время питания.
Восстановление стрекательного ап-
парата после использования происхо-
дит путем новообразования внутри тела,
на что указывает встречаемость три-
хоцист в эндоплазме.
Паразитические протозоа типа кни-
доспоридий обладают стрекательным
аппаратом, отличающимся большой
сложностью и по строению напоминаю-
Рис. 31. Инфузория туфелька (Paramecium саи-
datum):
1 — реснички; 2 — пищеварительные вакуоли; 3 —
микронуклеус; 4 — ротовое отверстие; 5 —глотка;
6 — порошица; 7 — сократительная вакуоль (резер-
вуар и радиальные приводящие каналы); 8 — макро-
нуклеус; 9 — трихоцисты
54
щим стрекательные капсулы низших многоклеточных животных
(гидры и др.). Стрекательные капсулы ряда споровиков содержат
спирально скрученную нить. У одних нить имеет трубчатое строе-
ние, у других устроена по-иному. При выстреливании одни нити
своим острым концом внедряются в тело жертвы, другие служат
для заякоривания на теле или в кишечнике хозяина.
Аналогия в строении стрекательного аппарата споровиков
и метазоа прослеживается и в том, что в образовании стрека-
тельных капсул и у тех и других важное участие принимает аппа-
рат Гольджи.
Раздражимость — восприятие факторов среды и реакция
на их воздействие. Известна способность живой материи отвечать
на разного рода воздействия характерным образом. Реакция
живых существ на раздражение представляет собой не механи-
ческий акт, а сложный биологический процесс. В живой природе
ответная реакция на раздражение всегда специфична. На один
и тот же фактор организм реагирует по-разному в зависимости
от физиологического состояния в данный момент и от природных
особенностей, Сытое одноклеточное животное, как и многоклеточ-
ное, иначе отвечает на пищевой раздражитель, чем голодное.
Организмы, близко родственные, но живущие в разных средах,
неодинаково реагируют на один и тот же раздражитель. Так,
зеленые жгутиковые скапливаются в освещенной зоне водоема,
а близкие к ним виды, но обитающие в затененных местах, лишен-
ные хлорофилла, уходят от света.
Раздражимость у протозоа проявляется вне зависимости от
высоты их организации, специфичности. Во всех случаях ответ-
ная реакция — функция всего организма.
Ответная реакция на раздражение движением называется
таксисом, на свет — фототаксисом, на электричество — гальвано-
таксисом (инфузории парамеции, сувойки и др. плывут в элект-
рическом поле к катоду), на температуру — термотаксисом, на
механический раздражитель — тигмотаксисом, на химический
хемотаксисом. Таксисы различают положительные (движение
к раздражителю) и отрицательные (уход от раздражителя). На-
пример, у амеб хемотаксис отрицательный к слабым щелочам,
положительный термотаксис к оптимальной температуре.
Среди протозоа отличаются сложным поведением инфузории,
имеющие и наиболее сложное строение тела. У ряда видов только
в связи с питанием отмечено несколько форм движения. Стал-
киваясь с резко неблагоприятными условиями, они могут дви-
гаться задним концом и уходить от опасности.
Восприятие раздражений извне, рецепторная функция, осу-
ществляется покровами и их производными: ресничками, жгути-
ками и пр., а также специальными органоидами, имеющимися
у ряда видов. Так, органы зрения имеются у жгутиковых.
Глазок (стигма) образован скоплением оранжево-красных
капель, за которым располагается слой особо дифференциро-
ванной протоплазмы, а перед пигментным экраном — прелом-
55
ляющее свет тельце. В схеме такой глазок близок к схеме строения
глаз у метазоа. У инфузорий описаны особые пузырьки с кри-
сталликом внутри. Их называют статоцистами, а по функции
принимают за органы равновесия. С изменением положения тела
находящийся в пузырьке кристаллик, перемещаясь, давит на
стенку пузыря в ином месте, что вызывает ответную реакцию
восстановления положения тела в пространстве. Подобное же
можно отметить в схеме строения органа равновесия много-
клеточных. Находки статоцистов у инфузорий почти единичны,
и оценка их функции требует дополнительных исследований.
Изучение поведения одноклеточных животных показало его
сложность, неодинаковость по отношению к воздействующим
факторам, привыканию к раздражителю (световому и др.).
Движение. Функцию движения у протозоа выполняют
временные образования — псевдоподии и постоянные — жгутики
и реснички.
Решающее значение иногда приобретают изгибы тела, как
это свойственно ряду жгутиковых и др. Многие жгутиковые и ин-
фузории из плавающих в толще воды поступательное движение
совершают преимущественно за счет винтообразного вращения
тела, направляемого движением жгутика или ресничек. В рамках
каждого из отмеченных способов движения имеется значитель-
ное разнообразие с большим числом вариаций.
Движение у одноклеточных животных, как и у многоклеточ-
ных, связано с наличием особых контрактильных (сократитель-
ных) белков, отличающихся крупными молекулами волокнистой
структуры. Такие белки (актиновые и мизиновые), ранее выяв-
ленные в миофибриллах мышечных клеток, обнаружены и в
Рис. 32. Схема ультраструк-
туры реснички
Рис. 33. Схема ультраструктуры поверхно-
стного слоя инфузории туфельки:
/-—пелликула; 2 — реснички; - базальные
тельца ресничек; 4 -- поверхностные пеллнку-
лярпые фибриллы; 5 — трихоцисты
56
Рис. 34. Стилоиихия
протоплазме протозоа, и в их органоидах движения -- жгутиках
и ресничках. Таким образом, в природе двигательной функции
у животных того и другого подцарства прослеживаются анало-
гичные черты.
Ультраструктура жгутиков и ресничек протозоа, как и жгу-
тиков и ресничек метазоа (в мерцательном эпителии, у сперма-
тозоидов и пр.), показывает сходные черты в фибриллярном
строении — наличие в каждой ресничке и в каждом жгутике
11 волоконец (нитей), расположенных продольно. Из них девять
находятся на периферии и два занимают центральное положение
(рис. 32).
Во всех случаях основание жгутиков и ресничек в пелли-
куле имеет вид гранулы, называемой базальным тельцем. У жгу-
тиковых базальное тельце часто называют блефаропластом, а у
инфузорий — кипетосомой. С базальным тельцем связаны особые
фибриллы — корневые нити, которые у жгутиковых простираются
в глубь тела, иногда достигая ядра.
Двигательный аппарат ряда высших протозоа (инфузорий)
показывает сложность расположения фибрилл (рис. 33).
. Реснички отличаются от жгутиков многочисленностью на теле
и малой длиной. Усиление функций ресничек связано со слиянием
их, что приводит к образованию толстых протоплазматических
придатков наподобие ног (как у бегающих стилонихий, рис. 34),
щупалец, мембран специального назначения, например в пред-
ротовом углублении и т. д. Иногда подобное наблюдается и у
жгутиковых.
П и т а н и е, д о б ы ч а пищи, п и щ е в а р е п и е. В типе про-
тозоа встречаются всевозможные способы питания. Широко
представлен древнейший способ — поглощение растворенных в
воде веществ поверхностью тела. Такое питание осмотическим
путем в мире животных называется сапрозойным. Оно распро-
странено среди животных, обитающих в воде, богатой растворен-
ными органическими и минеральными веществами, а также харак-
терно для эндопаразитов. Столь же широко среди одноклеточных
распространен собственно животный тип питания — голозойный,
питание оформленной пищей, поглощение которой происходит
с помощью специальных образований.
Растительный способ питания встречается преимущественно
в классе жгутиковых у тех из них, которые имеют хроматофоры,
57
Рис. 35. Сосущие инфузории
nium, напавшие на инфузорию
Didi-
содержащие пигменты: зеле-
ный хлорофилл, желтый, сине-
ватый и др. Их присутствие
обеспечивает аутотрофам фо-
тосинтез, продуктами которого
обычно являются из углево-
дов крахмал и парамил. Ряд
определенных обстоятельствах
способны переходить к осмотическому и гетеротрофному пита-
нию.
Материалы о зеленых жгутиковых свидетельствуют, с одной
стороны, о наличии в подцарстве протозоа организмов, сочетаю-
щих в себе морфофизиологические черты двух царств органиче-
ской природы: животных и растений, с другой — показывают
своеобразие их организации и жизненных отправлений.
Органами добычи пищи и транспортировки ее внутрь тела
служат: у корненожек — псевдоподии, у жгутиковых — жгутики
и иногда близ его основания рот, у инфузорий — ресничный
аппарат в предротовой области и реснички всего тела. Это общая
схема. Инфузории в добыче пищи используют стрекательные
нити, а сосущие инфузории — сосущий аппарат (рис. 35).
Добыча пищи с помощью псевдоподий признается наиболее
примитивной (древней). Способность образовывать псевдоподии
как временное явление сохранили некоторые одноклеточные из
других классов. Амебы и другие корненожки захватывают пищу
псевдоподиями (рис. 36) по-разному. Так, амебы могут обтекать
пищевой комочек, окружать его своими псевдоподиями, могут
активно втягивать его внутрь.
В пищевой рацион амеб входят водоросли иногда больших
размеров, чем амеба. Электронный микроскоп позволил выяснить
Рис. 36. Захват амебами пищевых частиц:
1—4 — амеба, захватывающая пищевую частицу; 5—7-амеба, захватывающая
нитчатую водоросль посредством «втягивания» ее внутрь тела
58
детали поглощения крупной добычи. Амеба, накрыв длинную
водоросль своим телом, образует псевдоподии, так что водоросль
оказывается в образованном ими канале. В стенках канала
появляются микропсевдоподии, назначение которых — удержи-
вать пищевой объект. Одновременно происходит втягивание нит-
чатой водоросли с закручиванием ее внутри тела амебы. Пред-
полагается, что этому способствуют как сокращение самой про-
топлазмы, так и находящиеся в ней сократимые волоконца.
Прикрепленные к субстрату протозоа питаются преимущест-
венно подвижной пищей, а плавающие — взвешенными в воде
частичками детрита и живыми организмами. Скрепленные с суб-
стратом сосущие инфузории способны улавливать своими щупаль-
цами несколько проплывающих мимо инфузорий и поедать их
(высасывать). Но бывает й так, когда плавающие хищные инфу-
зории группой нападают на одну жертву и сообща высасывают
ее. Среди протозоа есть монофаги и полифаги. Подтверждены
экспериментально наличие у инфузорий выбора пищи и способ-
ность отличать на основе хеморецепции пищевые частицы —
бактерий, дрожжей — от непищевых. В пищевой рацион некото-
рых протозоа входят и микроскопические животные из много-
клеточных.
К дополнительным способам питания можно отнести погло-
щение продуктов фотосинтеза, продуцируемых симбиотическими
хлореллами. Сожительство с ними широко представлено у прото-
зоа. Их присутствие в теле одноклеточных сопровождается
значительным повышением положительного фототаксиса (при
условии оптимального освещения).
Пища, поступившая внутрь тела, обычно переваривается в
пищеварительных вакуолях, которые возникают вместе с приемом
пищи. Лишь у некоторых инфузорий (из панцирных) имеется
внутри полость, внешне несколько напоминающая пищеваритель-
ный тракт метазоа. В случае осмотического питания вакуоли
не образуются. Пищеварительные вакуоли содержат ферменты,
воздействующие на белки (протеазы), на углеводы (карбогидра-
зы) и пр., по функции аналогичные пищеварительным ферментам
метазоа. Наблюдения за инфузориями и опыты с применением
микрохимических реакций позволили установить железистый ха-
рактер участков протоплазмы, прилегающих к вакуолям. В общем
процесс пищеварения у протозоа аналогичен в самых основных
чертах известному для метазоа, но многое остается невыясненным.
Длительность переваривания зависит от особенностей пищи
и от ее размеров. Амеба, например, переваривает крупную нит-
чатую водоросль за 3—5 дней, а мелкую пищу — за несколько
часов. Инфузория туфелька, питающаяся в основном бактериями,
переваривает содержимое вакуолей примерно за час. Неперева-
ренные остатки у той же инфузории накапливаются в опреде-
ленном месте перед порошицей. При дефекации одновременно
выбрасываются скопившиеся остатки из 4—5 вакуолей. У корне-
59
ножек при отсутствии пелликулы и вообще более простой орга-
низации тела дефекация происходит в разных его участках.
Дыхание обеспечивает получение энергии путем поглоще-
ния кислорода для окисления органических веществ: углеводов,
жиров и белков. Дыхание совершается с участием дыхательных
ферментов (оксидазы, цитохрома и др.), т. е. подобно тому, как
у многоклеточных. При распаде белков конечными продуктами
будут СО2, вода, азотистые вещества, в частности мочевина
и мочевая кислота. Процесс дыхания (диссимиляция) в сочетании
с ассимиляцией — основное свойство жизнедеятельности всякой
живой материи. У некоторых диссимиляция осуществляется
брожением, при котором происходит расщепление органических
веществ с освобождением энергии.
Протозоа имеют неодинаковую потребность в кислороде. У ин-
фузорий с их сложной организацией и поведением интенсив-
ность газообмена значительно выше, чем у амеб. Кроме того,
данные газообмена показывают приспособленность отдельных ви-
дов животных к существованию в условиях разной концентрации
кислорода. Многие протозоа способны жить при ничтожном содер-
жании его в среде. К таким относятся донные обитатели илистых
грунтов в озерах и прудах, где и в летний сезон недостаток
кислорода — обычное явление. К категории невзыскательных к
кислороду животных надо причислить ряд паразитов, которые
подобно дизентерийной амебе живут в кишечнике хозяина почти
в анаэробных условиях. Примечательна способность протозоа
водоемов временно переносить анаэробные условия в активном
состоянии или впадая в анабиоз.
Выделение, или экскреция, служит освобождению
организма от продуктов обмена веществ и избытка воды. Это
происходит через поверхность тела и с помощью специальных
периодически образующихся выделительных органов, которые у
протозоа называются сократительными вакуолями или пульси-
рующими по причине ритмичного наполнения их с последующим
выбрасыванием жидкости наружу. Вакуолей бывает одна или
несколько. Сложность строения выделительной системы неодина-
кова. Амебы имеют только вакуоль, возникающую то тут, то там,
а инфузории — вакуоль (или вакуоли) с системой приводящих
каналов, как бы дренирующих участки тела во всех направлениях.
Вакуоли занимают в инфузориях определенное место, и их опорож-
нение происходит также в определенном участке. Микрохимиче-
скими реакциями установлен железистый характер протоплазмы,
прилегающей к приводящим каналам, свидетельствуя об экс-
креторном процессе выделения воды и продуктов распада.
(Экскрецией называют выработку веществ, подлежащих удале-
нию из тела, в отличие от секреции — процесса продуцирования
веществ, нужных для жизнедеятельности организма.)
Сократительные вакуоли протозоа выполняют ряд функций.
С их помощью выводится из организма избыток воды и тем под-
60
держивается осмотическое давление в протоплазме на опреде-
ленном уровне. Эта функция характеризует выделительную систе-
му как оеморегуляторпую. Жизнь пресноводных животных прохо-
дит в гипотонической среде, и аппарат осморегуляции для них
необходим. Протоплазма содержит большее количество солей,
чем пресная вода, и в силу разницы осмотического давления про-
исходит непрерывное поступление воды внутрь тела через полу-
проницаемую оболочку покровов. Морские протозоа, ряд эндо-
паразитов и пр. из обитающих в изотопической среде сократи-
тельных вакуолей обычно не имеют. Наличие или отсутствие
вакуолей определяется также регуляторными особенностями
самих покровов. Частота пульсации значительно колеблется. Так,
у амебы интервалы пульсации равны 1—5 мин, а у инфузорий —
10 20 с. Кроме осморегуляции, на экскреторные вакуоли возла-
гается удаление продуктов обмена веществ -- углекислоты, моче-
вины, и пр. В том случае, когда снята функция вывода воды,
удаление продуктов диссимиляции и белкового обмена происходит
через покровы. О значении сократительных вакуолей как осмо-
регуляторов показывают опыты культивирования пресноводных
протозоа в соленой воде. Постепенное наращивание солености вле-
чет утрату амебами сократительных вакуолей, они их перестают
образовывать.
Р а з м н о ж е и и е протозоа происходит по-разному, и в осо-
бенностях его прослеживаются приспособительные черты. Нара-
стание числа особей у одноклеточных организмов происходит
путем бесполого размножения (рис. 37). Наиболее обычной для
свободиоживуших будет та его форма, при которой материнская
особь разделяется па две дочерние (рис. 37, /). Ядро при этом
делится кариокниетпческим путем.
Многие споровики и ряд паразитических протозоа других
классов совершают бесполое размножение с той особенностью,
что при делении материнской особи образуется несколько дочерних
(множественное деление) (см. рис. 39). Высокая плодовитость —
приспособительная черта биологии паразитов в связи с гибелью
особей на пути распространения. При почковании после деления
ядра следует неравномерное разделение протоплазмы и дочерний
организм имеет вид почки на материнском теле (рис. 37, //).
Среди инфузорий есть виды с внутренним почкованием. При этом
дочерние особи образуются внутри камеры в теле матери, что
напоминает живородность (рис. 37, ///). Половое размножение
метазоа происходит путем слияния двух половых клеток-гамет,
а у протозоа гаметами становятся особи, проходя этап созре-
вания. У одних копулируют партнеры одинакового размера
(изогамия), у других неодинакового (анизогамия) (рис. 38).
У таких, как кокцидии, плазмодии, одни гаметы подобны яйцам,
крупные, неподвижные, другие, как сперматозоиды метазоа,
мелкие, подвижные. У вольвоксов подобно метазоа только часть
клеток тела дают гаметы: мужские и женские, и есть виды раз-
дельнополые и гермафродиты.
Своеобразен процесс педогамии и аутогамии. При педогамии
копулируют дочерние особи, претерпев созревание. При аутога-
мии копулируют ядра одной и той же особи. После разделения
ядра каждая часть проходит созревание (редукционное деле-
ние) .
Половой процесс инфузорий — конъюгация. При конъюгации
происходит временное сближение двух особей с взаимным опло-
дотворением ядрами, которые выполняют функцию гамет. Ядра
в теле инфузорий двух категорий: одно крупное — макронуклеус,
другое малое—микронуклеус. Первое называют вегетативным,
второе — половым. В ходе конъюгации макронуклеус распадается,
а микронуклеус проходит сложный процесс созревания, в резуль-
тате которого в каждом из конъюгантов после двукратного де-
62
Рис. 38. Половое размножение жгу-
тикого (пзогамное)
Рис. 39. Конъюгация инфузорий:
1 -- мнкропуклеус; 2 -- макроиуклеуе
ления микронуклеуса получаются четыре ядра, из которых три раз-
рушаются, а четвертое делится, образуя два ядра (разных по
величине). Более крупное остается на месте, а меньшее мигрирует
в партнера и оплодотворяет оставшееся там ядро. Этим взаим-
ным оплодотворением завершается конъюгация, и инфузории
расходятся. Вскоре в каждой восстанавливается ядерпый аппарат
(рис. 39).
Среди инфузорий есть примеры, показывающие возможное
сближение процессов копуляции и конъюгации, когда один из
партнеров при конъюгации уменьшается в размерах и поглощается
другим. В размножении ряда прикрепленных колониальных ин-
фузорий видны приспособительные черты, выраженные в образо-
вании на колонии особых веточек с конъюгантами. Нечто подоб-
ное известно у колониальных гидроидов в типе кишечнополостных.
И и ц и с т и р о в а н и е — выделение на поверхности тела за-
щитной оболочки — цисты. Заключение тела в цисту происходит
по разным причинам: обычно с наступлением ухудшения условий
жизни, иногда перед половым процессом или после приема пищи.
У эндопаразитов иицистирование происходит обязательно перед
выходом из хозяина во внешнюю среду. Оно способствует не
только выживанию, но и распространению в пространстве (вет-
ром, с помощью животных).
Онтогенез (греч. ontos — сущее, genesis — происхожде-
ние) — индивидуальное развитие от рождения до смерти. Есте-
ственным завершением жизни многоклеточного животного явля-
ется смерть. Такая форма окончания жизни возникла в эволюции
животного мира вместе с дифференцировкой тела на соматиче-
скую и воспроизводящую части. Для протозоа при их одноклеточ-
ном теле жизнь особи длится до разделения материнского орга-
низма на дочерние особи, представляющие собой новое поколение,
которое растет и развивается до готовности к размножению.
63
У протозоа, имеющих в размножении стадию полового процесса,
жизненный цикл совершается с чередованием поколений. Вслед
за несколькими генерациями в результате бесполого размноже-
ния наступает появление половозрелого поколения. Так, инфузо-
рии парамеции производят до 60 поколений путем деления, прежде
чем появится поколение конъюгантов. Бесполое размножение
влечет возрастание числа особей! и тем самым способствует со-
хранению вида. Развитие с чередованием поколений встречается и
у метазоа, например у кишечнополостных.
ТИП САРКОДОВЫЕ И ЖГУТИКОВЫЕ
(SARCOMASTIGOPHORA)
КЛАСС САРКОДОВЫЕ (SARCODINA)
Характерная особенность саркодовых — способность тела, по-
крытого тонкой мембраной, образовывать временные выросты —
псевдоподии. Они служат для передвижения животных и для
захвата пищи. Принятие пищи и удаление ее непереваренных
остатков происходят в любом месте поверхности тела.
Из саркодовых наиболее известны различные виды голых амеб
(подкл. Корненожки Rhizopoda, отр. Amoebina). Это микроскопи-
чески малые животные, содержащие одно или реже несколько
ядер. Цитоплазма подразделяется на более плотную, прозрачную
эктоплазму и более жидкую, содержащую различные включения
эндоплазму. Размножаются амебы делением.
Амебы обычно встречаются на дне пресных водоемов и в почве,
где питаются микроорганизмами, гниющими остатками животных
и растений. Некоторые виды ведут паразитический образ жизни.
Такова дизентерийная амеба (Entamoeba histolytica), обитающая
в кишечнике человека. Она может быть возбудителем тяжелой
формы кишечного колита — амебиаза. Распространение этих
амеб осуществляется четырехъядерными цистами, которые в
огромном количестве выделяются с калом больного. В нового
хозяина цисты попадают обычно с загрязненной водой. В кишеч-
нике хозяина из цисты выходит паразит, который делится на четы-
ре особи соответственно числу ядер. Многие виды амеб парази-
тируют в кишках различных сельскохозяйственных животных.
Интересны пресноводные раковинные амебы (отр. Testacea),
тело которых заключено в раковинку (рис. 40). У обитающей в во-
доемах арцеллы (Arcella) раковинка напоминает шапочку гриба:
на ее нижней поверхности есть отверстие, через которое животное
выдвигает наружу псевдоподию. Раковина арцеллы образована
кремнийорганическим веществом, выделяемым телом амебы.
У другой раковинной амебы — диффлугии (Diffhipia) раковинка,
имеющая вид опрокинутого кувшинчика, построена из песчинок,
сцементированных органическим веществом. Раковинные амебы
64
Рис. 40. Раковншпяе амебы:
новинка арцеллы
В.
живут обычно па дне водоемов,
стеблях и листьях водных расте-
ний. По образу жизни близки
к голым амебам.
Многие голые и раковинные
амебы населяют почву, участ-
вуя в почвообразовании. По
видовому составу близки к прес-
новодным, но отличаются от них
мелкими размерами, что связа-
но с обстоятельствами жизни
в почве,— существованием
топком слое воды, окружающем твердые конкреции почвы. Во
влажной почве они живут в активном состоянии, в высыхающей —
ипцистируются. Питаются перегноем и почвенными'бактериями.
Поедая последних, в том числе и таких, которые обогащают
почву азотистыми соединениями, амебы могут несколько снизить
их численность. Но с другой стороны, они полезны тем, что,
питаясь остатками различных организмов, способствуют их разло-
жению. В благоприятных условиях почвенные амебы быстро
размножаются и их численность нередко составляет многие ты-
сячи особей в каждом грамме почвы.
В морях, находясь во взвешенном состоянии в толще воды или
у дна, обитают своеобразные саркодовые — фораминиферь! (отр\
1'oraminiIera, рис. 41). Их тело заключено в тонкостенную рако-
вину различной формы, стенки которой пронизаны большим
числом пор. Раковина образована органическим веществом,
обычно пропитанным углекислой известью. Через поры раковинки
наружу выходят тонкие и длиннпыс псевдоподии, образующие
вокруг животного протоплазматическую сеть, которая служит для
улавливания и переваривания пищи. Размножаются форамини-
феры как бесполым (деление), так и половым путем. В последнем
случае образуют многочисленные подвижные гаметы со жгути-
ками. Сливаясь попарно, они образуют зиготу.
После смерти фораминифер раковинки, оседая на дне моря,
образуют вместе с кокколитофоридами (из жгутиковых) фора-
мипиферно-кокколитовые мощные меловые породы. Виды, свой-
ственные отдельным геологическим эрам, служат руководящими
формами в определении возраста отложений и их принадлежности
к той или иной геологической эпохе. Более молодые нуммули-
товые известняки свидетели эоценового периода кайнозойской
эры, а фузулиповые — далекого каменноугольного периода палео-
зойской эры и т. д. Палеонтологический материал фораминифер
3 В. А. Кузнецов и др. 65
Рис. 41. Фораминифсра с сетью тончайших псевдоподий
позволяет проследить эволюцию отдельных групп, их расцвет и
угасание.
Фораминиферы вызывают не только теоретический, но и прак-
тический интерес, в частности при разведке полезных ископаемых.
Для работников сельского хозяйства полезны знания биологи-
ческих источников накопления извести в земле, причин и мест
залегания.
Отряд Пироплазмиды (Piroplastnida). Кровяные паразиты
домашних и диких млекопитающих. Положение пироплазмид
в системе протозоа остается неопределенным. Амебоидная форма
пироплазмид и ряд других черт позволяют включить их в класс
Саркодовые.
Жизнь этих опасных паразитов протекает последовательно
в теле двух хозяев: млекопитающего и иксодового клеща (паст-
бищного). В клещах пироплазмиды живут в разных органах, а
у млекопитающих — в крови. Паразиты, попав в кровь, прони-
кают в эритроциты, где растут и размножаются делением. У одних
видов в результате бесполого размножения образуются две до-
черние особи, у других — четыре. Они грушевидной формы
(рис. 42).
Сами клещи инвазируются в разном возрасте при сосании
крови больного животного. Заболевание передают, как правило,
взрослые клещи, но у собачьего клеща и ряда других паразитов
66
Рис. 42. Пироплазмы:
а -- Babesia canis (в крови собаки); б — Nuttalia (в кро-
ви лошади)
передают и их личинки, даже не сосавшие крови больного живот-
ного. Находясь в самке клеща, паразит может проникать в ес
яйца и сохраняться при развитии из таких яиц личинки.
ПОДКЛАСС РАДИОЛЯРИИ (RADIOLARIA)
Морские саркодовые со сложным внутренним скелетом из крем-
незема или сернокислого стронция. Наружные слои эктоплазмы
содержат крупные вакуоли, облегчающие парение лучевиков в
воде. Тонкие прямые псевдоподии расходятся лучеобразно (от-
сюда название). Живут радиолярии в поверхностных слоях морей.
Их скелеты образуют на дне отложения радиоляриевого ила, из
которого со временем слагаются кремнеземные горные породы
(рис. 43).
Радиоляриевая «горная мука» трепел идет для шлифовки
деталей и на изготовление особых сортов наждачной бумаги.
КЛАСС ЖГУТИКОВЫЕ (MASTIGOPHORA)
Жгутиковых (рис. 44) отличает от других одноклеточных способ
передвижения — при помощи одного или нескольких жгутиков.
Их тело одето оболочкой (пелликулой), имеет более или менее
постоянную форму, характерную для отдельных видов. При бес-
полом размножении тело жгутиковых делится продольно.
Известно более 6 тыс. видов жгутиковых. Большинство их
живут в морях, пресных водоемах и почве, но около 700 видов
ведут паразитический образ жизни. Некоторые обитают в рубце
жвачных млекопитающих или в слепой кишке других травоядных
(например, лошадей).
Рис. 43. Радиолярии
3*
67
Рис. 44. Жгутиковые. Эвглена зеленая (/); ворот-
ничковые жгутиковые (//):
1 -- ядро; 2 — зерна парамила; 3— хромагофоры, 4 -- гла-
зок; 5 — сократительная вакуоль; 6 — резервуар; 7 жгу-
тик, 8 — воротничок
Пресноводные окрашенные жгутиковые, имеющие хлорофилл
и другие пигменты (например, эвглена), при массовом размноже-
нии могут вызвать «цветение воды» в прудах, изменяя физико-
химические свойства воды (хотя чаще это вызывают микроскопи-
ческие водоросли).
Из паразитических жгутиковых особенно опасны различные
виды трипанозом (Trypanosoma, отр. Kinetoplastida), живущие
в плазме крови человека и различных животных (рис. 45). Они
имеют вытянутое веретеновидное тело. Длинный жгутик отходит от
базального тельца и тянется вдоль тела, обрамляя ундулирующую
мембрану. Близ него расположен кинетопласт. Волнообразные
движения мембраны способствуют движению в вязкой плазме
крови. Африканская трипанозома (Т. gambiense) вызывает сон-
ную болезнь человека. Этот паразит попадает в кровь человека
при укусе мухой цеце, в кишечнике и слюнных железах которой
трипанозомы проходят часть цикла своего развития. Сонная
болезнь — длительное и тяжелое заболевание, заканчивающееся
нередко смертельным исходом. Другой африканский вид трипа-
нозом (Т. brucei) вызывает повальное заболевание скота. Этот
паразит также разносится мухой цеце, но другого вида. Мухи
заражаются трипанозомами от антилоп, которые являются их
68
постоянными носителями, но сами не болеют. Следовательно,
антилопы служат «природным резервуаром» данного паразита.Для
скота, не имеющего иммунитета, это заболевание обычно смер-
тельно.
В СССР встречается трипанозома (Т. equiperdum), паразити-
рующая в крови лошадей. Она вызывает так называемую случную
болезнь, распространяющуюся при случке больной и здоровой
лошадей. Известна трипанозома (Т. euansi) — возбудитель сур-
ры болезни лошадей, ослов, верблюдов и крупного рогатого
скота. Передается болезнь обычно слепнями.
К кинетопластидам относятся также различные виды лейш-
маний (Leishmania). Один из них — тропическая лейшмания
(L. tropica) вызывает тяжелое заболевание человека - кожный
лейшманиоз, или пендинскую язву. Паразит живет в лейкоцитах
крови и клетках лимфатических желез, разрушая их. На коже
появляются большие, плохо заживающие раны, оставляющие
затем глубокие рубцы. Болезнь распространяется москитами.
Находясь в белых кровяных клетках, паразит лишен жгутика,
и форма тела его округлая. Но попав в жидкую среду содер-
жимого кишечника москита, он образует жгутиконосную веретено-
видную форму. Другой вид лейшмании (L. donowatii) вызывает
у человека болезнь внутренний лейшманиоз. Паразит поселяется
в клетках костного мозга, в пе-
чени и других внутренних органах,
вызывая их глубокие поврежде-
ния. Переносчиками являются
москиты и постельные клопы.
Отряд Фитомонады (Phytomo-
nadina). В отряде фитомонад
встречаются одиночные и коло-
ниальные формы. Среди колони-
альных форм наблюдается боль-
шое разнообразие по строению и
биологии размножения. Клетки,
как это характерно для фитомо-
над, с двумя жгутиками, глазком
и хроматофором. В одних коло-
ниях клеток мало (4—8, 16—32)
и они сходны по строению. У го-
ниум каждая из 16 клеток спо-
собна к воспроизводству КОЛО-
Рис. 46. Трихомонас (схема строения):
/—свободные жгутики; '2— ядро; Г/--
ядрышко; 4--опорная фибрилла; 5-- аксо-
стиль; 6 — рулевой жгутик, идущий ио краю
у 11 д у л и р у ю щ е й мембраны
69
нии. Колониальность — не случайность, а наследственный при-
знак. Сложнее эудорины. Они раздельнополы. В женской особи
при половом размножении каждая клетка развивается в крупную
без жгутиков макрогамету, а в мужской — из клеток созревают
двухжгутиковые микрогаметы. У вольвоксов число клеток около
20 тыс., в большинстве соматические, и лишь несколько десятков
клеток генеративные. Половые продукты: макрогаметы и микро-
гаметы наподобие яиц и сперматозоидов метазоа. Есть виды
раздельнополые, есть и гермафродиты. Колония вращается вокруг
оси, сохраняя передний полюс направленным вперед. Здесь четыре
клетки физиологически отличаются от других. Неоднородность
клеток подмечена и по оси тела. Характерная для многоклеточных
при развитии организма дифференциация клеток в разных направ-
лениях имеет место и у вольвоксов.
Отряд Многожгутиковые (Polymastigina) объединяет бес-
цветные формы с усложненным двигательным аппаратом, в боль-
шинстве паразитов. Их отличают двусторонняя симметрия тела,
наличие двух ядер, четырех пар жгутиков. Вогнутая брюшная
сторона служит присоской. Опасны трихомонасы — паразиты
кишечника и половых путей человека (рис. 46). Кишечные лямб-
лии выделены в особый отряд Diplomonadida.
ТИП СПОРОВИКИ (SPOROZOA)
Этот тип объединяет паразитических одноклеточных, носящих
явные черты упрощения организации в связи с образом жизни.
Форма тела разнообразна, часто амебовидная. Органоиды дви-
жения отсутствуют, лишь у половых клеток — гамет иногда
имеются жгутики. Обычно нет пищеварительных вакуолей, по-
скольку питательные вещества поступают в тело осмотически
через всю его поверхность. Споровикам свойственны сложные
циклы развития, нередко со сменой хозяев. Размножаются как
бесполым, так и половым путем. Для них характерно образование
в конце жизненного цикла массы мелких зародышей — спор, по-
крытых плотной защитной оболочкой. Образование их способ-
ствует распространению паразитов.
Кокцидии (подкл. Coccidiomorpha, отр. Coccidiida). Паразиты
эпителия кишечника и протоков печени кроликов, рогатого скота,
многих домашних животных и диких птиц, насекомых и других
животных. Особенно часто кокцидиоз поражает кроликов, вы-
зывая их массовую гибель. Кокцидиозом болеют и люди.
Циклы развития кокцидий весьма сложны. Примером может
служить цикл развития паразита кроликов эймерии (Eimeria
stiedae). Эймерия (рис. 47) паразитирует в клетках эпителия
протоков печени кроликов. Бесполое размножение этого споро-
вика протекает по типу шизогонии. Паразит, находясь в клетке
эпителия печени хозяина, растет, пока не достигнет зрелости,
70
Рис. 47. Жизненный цикл кокцидии:
/. 2 — рост молодых кокцидии в клетках эпителия кишечника хозяина; <У—<5 -- процесс
шизогонии и образования мерозоитов; 6, 10 — проникновение мерозоитов в клетки эпи-
телия хозяина; 7—9 — образование макрогамет из мерозоитов; 11, 12 — образование
макрогамет из мерозоитов; /Л — слияние макро- и микрогамет; 14 — ооциста (зигота,
окруженная оболочкой); 15—17 --образование из ооцисты четырех споробластов с дву-
мя спорозоитами в каждом; 18 — выход спорозоитов в кишечнике нового хозяина; 19
внедрение их в клетки эпителия нового хозяина
после чего его ядро рядом последовательных делений дает значи-
тельное число мелких ядер. Затем вокруг каждого из них обо-
собляется участок цитоплазмы, что ведет к образованию новых
мелких особей — мерозоитов. Последние покидают разрушенную
клетку эпителия печени хозяина и внедряются в соседние, еще
здоровые клетки. В результате возникает глубокое поражение
печени хозяина, что приводит его к гибели. После циклов бес-
полого размножения кокцидии путем шизогонии, в течение кото-
рых в теле хозяина происходит быстрое нарастание численности
паразитов, наступает подготовка, а затем и сам половой процесс.
При этом одни из мерозоитов развиваются в макрогаметоциты,
71
а другие — в микрогаметоциты. Первые, созрев, становятся
крупными округлыми макрогаметами, а вторые, делясь, превра-
щаются в мелкие вытянутые подвижные микрогаметы со жгути-
ками. После копуляции макрогамет с микрогаметами образуются
зиготы, которые инцистируются, превращаясь в ооцисту. Током
желчи ооцисты выносятся из печени в кишечник, а из него с калом
выбрасываются наружу. Во внешней среде содержимое ооцисты
делится на два споробласта, которые окружаются собственными
оболочками. Споробласты, в свою очередь, распадаются на четыре
спорозоита. Когда ооциста попадет с загрязненным кормом в
кишечник нового хозяина, спорозоиты освобождаются от оболо-
чек и внедряются в клетки эпителия протоков печени. Весь процесс
образования спорозоитов у вида, паразитирующего в печени кро-
ликов, занимает около трех суток.
Другой вид кокцидий (Е. perforans) паразитирует в клетках
эпителия кишечника кроликов. Каждому сельскохозяйственному
животному свойствен свой вид кокцидий. Так, у крупного рога-
того скота паразитирует кокцидия (Eimeria bovis), вызывающая
кровавый понос.
Паразиты красных кровяных клеток различных животных и
человека—гемоспоридии (Haemosporidia) составляют подотряд
в отряде Coccidiida. К ним относится, например, малярийный
плазмодий (Plasmodium) —возбудитель малярии человека
(рис. 48). Часть своего жизненного цикла они проводят в тка-
невых клетках (печени), затем в эритроцитах человека, а заклю-
чительную — в теле малярийных комаров. В эритроцитах чело-
века, больного малярией, можно обнаружить амебовидных пара-
зитов, которые растут здесь и размножаются бесполым путем
по типу шизогонии. Образующиеся мелкие мерозоиты выходят
из разрушенных эритроцитов в плазму крови и вскоре внедряются
в новые красные кровяные клетки того же хозяина.
Быстро размножаясь путем шизогонии, плазмодии накапли-
ваются в крови человека в короткий срок (100 тыс. и более в 1 мм3
крови). После ряда циклов бесполого размножения наступает
подготовка к половому процессу с образованием гаметоцитов.
Если в это время больной малярией человек будет укушен маля-
рийным комаром, то вместе с кровью в желудок насекомого
попадают микро- и макрогаметоциты плазмодия. Из одних
гаметоцитов образуются округлые крупные женские макрогаметы.
Из других путем ряда делений формируются мелкие подвижные
микрогаметы. После копуляции макрогамет с микрогаметами
образуются зиготы, которые за способность к движению называют
оокинетами. Они внедряются в стенку кишечника, инцистируются
и путем многократного деления дают по нескольку тысяч мелких
подвижных спорозоитов, которые проникают в слюнные железы
комара и накапливаются в них в столь большом количестве, что
при укусе насекомого в организм человека со слюной комара про-
никает множество паразитов. Попав в кровь человека, спорозоиты
72
19
Рис. 48. Жизненный цикл
малярийного плазмодия:
I - спорозоиты, выходящие из слюнных желез комара и попадающие в тело человека;
2, 3 - развитие спорозоита в клетке иечепи человека; 4 — распадение паразита путем
шизогонии на массу мерозоитон; .5--// - -цикл развития и размножения паразита путем
шизогонии в красных кровяных клетках люден; 12 -- макрогаметоцит; 13 —- зрелый макро-
гаметоцит; 14 — микрогаметоцит; 15 -- зрелый микрогаметоцит; 16 - образование макро-
гамет из микрогаметоцита; 17 - образование макрогаметы и ее слияние с макрогаметой;
18 внедрение образовавшейся зиготы - оокинеты в стенку кишки комара; 19..... образо-
вание внутри ложной цисты множества спорозоитов; 20 - выход из цисты; 21 - скопление
спорозоитов в слюнных железах комара; 22—26 -- цикл развития плазмодия в клетках
тканей человека. Стадии развития паразита от 2 до 15 и от 22 до 26 проходят в теле челове-
ка, а стадии от 16 до 21 и теле комара
обычно сначала внедряются в клетки стенок кровеносных сосудов,
затем опять выходят в плазму крови и потом входят в эритроциты.
Приступы малярии у людей начинаются в результате инток-
сикации организма при массовом выходе мсрозоитов из разру-
шенных эритроцитов в плазму крови. Промежуток времени между
приступами соответствует длительности периода развития и раз-
множения паразитов в эритроцитах. У Pl. viuax — возбудителя
73
трехдневной лихорадки он занимает около 48 ч. При заболевании
тропической малярией обычно один приступ накладывается на
другой из-за большой токсичности паразита.
Малярия до сих пор широко распространена в тропических
странах и уносит много человеческих жизней. Но в СССР на-
пряженная. и систематическая борьба с малярией дала хорошие
результаты. Очаги этой болезни ликвидированы.
В борьбе с малярией главное внимание было обращено на
уничтожение переносчиков этого заболевания — малярийных ко-
маров и их личинок.
Обнаружены плазмодии, паразитирующие в эритроцитах
домашних птиц. Много видов плазмодиев выявлено у птиц отряда
воробьиных. Распространяют их комары, но других видов.
Сравнение циклов развития различных споровиков убеждает
в том, что последние обусловливаются образом жизни паразита.
В частности, отчетливо выступают приспособительные черты
чередования бесполого и полового процессов размножения.
У больного малярией человека насыщенная паразитами —
малярийными плазмодиями — кровь обеспечивает заражение
комара-кровососа, а массовое накопление спорозоитов плазмодиев
в слюнных железах комара — передачу паразитов человеку.
Оба перехода совершаются, минуя внешнюю среду, и оокинеты не
имеют защитных оболочек. У кокцидий ооцисты выводятся из
организма хозяина во внешнюю среду, отсюда — наличие плот-
ных защитных оболочек; споробласты, развивающиеся в ооцистах,
также имеют оболочки.
Среди споровиков имеются паразиты, живущие не в клетках
хозяина, а в полостях его внутренних органов (рис. 49). Тако-
Рис. 49. Цикл развития грегарины:
1 - спора; 2 •- спорозоиты, вышедшие из раскрывшейся споры;
,7 внедрение спорозоита в клетку эпителия кишки; 4, 5 — разви-
тие спорозоита в грегарину; 6 — передний отдел тела; 7 — средний
отдел; <S' - задний отдел
74
вы различные представители подкласса Грегарины (Gretfari-
nida).
Паразиты встречаются в кишечнике и в полостях других ор-
ганов, главным образом у насекомых и кольчатых червей. Удли-
ненное тело грегарин подразделено на три участка: передний,
у некоторых видов несущий крючочки, которыми паразит за-
крепляется в стенке кишечника хозяина, средний и задний,
содержащий ядро. Наиболее крупные грегарины достигают в дли-
ну несколько миллиметров. У большинства видов размножение
происходит половым путем, и лишь у немногих — со сменой по-
лового и бесполого поколений. После копуляции гамет образуется
зигота, которая инцистируется, превращаясь в спору. Последняя
выводится из тела хозяина, и содержимое ее разделяется на
большое число спорозоитов. Споры заглатываются новым хозя-
ином с пищей.
Попав в кишечник, спорозоиты выходят из оболочек споры
и закрепляются в стенке кишки. Степень патогенности грегарин
для хозяев еще не установлена.
ТИП КНИДОСПОРИДИИ (CNIDOSPORIDIA)
Еще недавно книдоспоридий относили к классу Споровиков в
качестве особого подкласса. Но ряд своеобразных черт строения
этих животных определил выделение их в тип.
Так, споры у книдоспоридий образуются в течение всей жизни,
и нет правильного чередования бесполого и полового размно-
жения. Половой процесс проходит по типу аутогамии, самоопло-
дотворения, при этом копулируют дочерние ядра. Споры отли-
чаются сложным строением. Они многоядерные с двуядерным
зародышем внутри (рис. 50).
Споры чаще двустворчатые, реже покрыты несколькими створ-
ками. На обоих или только на одном полюсе находятся полярные
капсулы, содержащие спирально свернутую нить. Попадая в
кишечник хозяина, под воздействием ки-
шечных соков споры выстреливают эти
нити, которыми прикрепляются к стенке
кишки. В дальнейшем паразит проникает
внутрь тела хозяина. Книдоспори-
дии — эндопаразиты. Локализуются в раз-
ных органах — в мочевом пузыре, в жаб-
рах и т. д.
Рис. 50. Схема строения споры Myxobolus:
I — полярные капсулы; 2 — вакуоль; 3 — амебоидный за-
родыш; -/ --его ядро; 5 — ядра клеток — образователь-
пиц полярных капсул; 6 —стрекательная нить
75
Рис. 51. Усач, пораженный Mijxobolus pfeijferi
КЛАСС СЛИЗИСТЫЕ СПОРОВИКИ (MYXOSPORIDIA)
В него входят несколько сот видов, в большинстве паразитов рыб.
В хозяине локализуются в разных органах.
Тяжелые заболевания вызывают поселяющиеся в мышечной
ткани паразиты, вызывая ее разрушение с образованием крупных
опухолей. Болезнь заканчивается гибелью рыбы. На рис. 51 изо-
бражен усач, зараженный Myxobolus pfeifferi — возбудителем
шишечной болезни.
ТИП МИКРОСПОРИДИИ (MICROSPORIDIA)
Мелкие внутриклеточные паразиты насекомых и рыб. Споры
одеты сплошной (не створчатой) оболочкой и имеют одну поляр-
ную капсулу со стрекательной нитью. К этому отряду относятся
ноземы (Nosema), опасные ’паразиты пчел и шелкопрядов.
Пчелиная нозема (Nosema apis) вызывает «черный понос» взрос-
лых пчел, нередко влекущий их массовую гибель. Этот паразит
живет в клетках эпителия кишечника пчел, разрушая их. Нозема
тутового шелкопряда (N. bombycis) (рис. 52) нередко служит
причиной опасного заболевания гусениц этой бабочки. При силь-
ном размножении паразиты заполняют клетки стенок кишечника,
Рис. 52. Схема цикла развития нозе-
мы в эпителии кишечника тутового
шелкопряда:
слева — клетка эпителия кишечника гусе-
ницы шелкопряда, в которой происходят
бесполое размножение и образование спор
ноземы; справа — выход спор из клетки
хозяина и образование амебоидного заро-
дыша, покидающего спору и внедряюще-
гося в новые клетки эпителия
76
прядильных желез и других внутренних органов. Проникновение
их в яйца бабочки влечет передачу паразитов потомству в случае
выживания гусеницы.
Французский ученый Л. Пастер предложил в конце прошлого
столетия простой способ распознавания яиц шелкопряда (грены),
зараженных ноземой. После яйцекладки вскрывают самку и
самца шелкопряда, растирают их внутренности в ступке с водой
и рассматривают каплю этой кашицы под микроскопом. При
обнаружении в ней спор ноземы грена уничтожается. Если же
спор нет, то грена считается здоровой. В СССР разработан тер-
мический способ обработки грены шелкопряда для уничтожения
позем.
ТИП РЕСНИЧНЫЕ (CILIOPHORA)
КЛАСС ИНФУЗОРИИ (INFUSORIA)
Инфузории отличаются наиболее сложной организацией среди
одноклеточных животных (рис. 53). Тело их покрыто пелликулой
и поэтому имеет относительно постоянную форму, характерную
для разных видов. Органоидами движения служат реснички,
которые участвуют также в захвате пищи. Одна из характерных
черт инфузорий — ядерный комплекс, состоящий, как правило,
из макро- и микронуклеуса. У некоторых инфузорий имеется
несколько малых ядер. Бесполое размножение инфузорий про-
исходит поперечным делением и изредка почкованием. Половой
процесс протекает по типу конъюгации.
Описано около 6 тыс. видов инфузорий. Они населяют прес-
ные воды и моря, встречаются в почве, обитают в теле различных
животных в качестве паразитов или симбионтов. Среди свободно-
живущих видов большая часть плавает в толще воды, держась
близ дна. Некоторые инфузории, например стилонихии (Stylo-
tiichia, отр. Равноресничные Holotriclia), передвигаются по суб-
страту на утолщенных ресничках, как на ножках. Ряд видов
инфузорий ведет сидячий образ жизни. Так живут своеобразные
сувойки (Vorticella, отр. Спиральноресничные Spirotricha), бока-
лообразное тело которых прикрепляется к субстрату длинным сте-
бельком. При каком-либо раздражении последний закручивается
спиралью, подтягивая животное. По верхнему краю тела сувоек,
вокруг ротовой воронки, расположен ряд ресничек, подгоняющих
пищу.
Нередко сувойки живут целыми колониями. Распростра-
нение сувоек в пространстве осуществляется с помощью под-
вижных особей, отпочковывающихся от материнского орга-
низма.
Большинство инфузорий питаются различными бактериями,
водорослями, разлагающимися остатками разных организмов.
77
Рис. 53. Панцирная инфузория из рубца желудка
жвачных
Рис. 54. Паразитические инфузории:
а — балантидиум; б — ихтиофтириус
Среди инфузорий имеются и хищные виды, нападающие на других
микроскопических животных. Интересны сосущие инфузории,
снабженные особыми трубочками. Поймав с помощью последних
живую добычу (обычно инфузорию другого вида), эти животные
сначала убивают ее ядовитыми выделениями, а затем высасывают.
Питание многих свободнодвижущихся инфузорий бактериями
позволяет считать их санитарами водоемов. Одна инфузория
туфелька за 1 ч может поглотить тысячи бактерий.
Ряд инфузорий ведут паразитический образ жизни (рис. 54).
Так, инфузория балантидиум {Balantidium') поселяется в толстых
кишках человека. Инфузории ихтиофтприусы (Ichtyophthirius),
поселяясь на жабрах и в коже карпов, форелей и других
рыб, вызывают воспаление тканей и могут привести к гибели
рыб.
Особый интерес представляет большая группа инфузорий —
панцирные инфузории из семейства Ophryoscolecidae (отр. Спи-
ральноресничные), живущие в содержимом рубца желудка
жвачных животных. Некоторые из них покрыты плотной оболоч-
кой (отсюда название — панцирные инфузории). Реснички
обычно сохраняются только вокруг ротовой воронки и пояском
вокруг тела. У здоровых коров в 1 см3 содержимого рубца их
бывает до 2 млн. По-видимому, инфузории способствуют пере-
вариванию клетчатки пищи жвачных. Инфузории, попадая
из рубца с пищей в сычуг и кишечник, становятся белковой
пищей скота.
78
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ (METAZOA)
К подцарству относятся животные, тело которых состоит из мно-
гих клеток, разнообразных по строению и функции. Клетки, за
исключением половых, входят в состав тканей, системы тканей
образуют органы.
Онтогенез метазоа характеризуется сложными процессами
роста и развития. Взаимосвязь всех структур в многоклеточном
организме отражает единство, целостность организма.
ТИП ГУБКИ (SPONGIA)
Губки — неподвижные морские и пресноводные животные, оди-
ночные и колониальные (рис. 55). Форма их тела разнообразна.
Многие имеют форму более или менее определенную — чаше-
образную, бокаловидную и др. Однако особи ряда видов при вы-
растании приобретают неопределенную форму, нередко отражаю-
щую форму того субстрата (камня, ветви), на котором растут.
Необычайно простое для многоклеточных организмов строение их
тела и своеобразие эмбрионального развития послужили основа-
нием для выделения губок в особый тип животных.
Тело губок состоит из двух слоев клеток: эктодермы и эндо-
дермы (двухслойные животные).
Поверхность тела губок пронизана множеством пор, через
которые вода поступает в сложную систему каналов и камер.
Вода увлекается внутрь животного движением жгутиков особых
воротничковых клеток, выстилающих полость камер. Свое назва-
ние эти клетки получили за наличие на наружном конце цито-
плазматической кольцевой складки (воротничка), окружающей
длинный жгутик. Из каналов и камер вода попадает в центральную
полость, а оттуда выводится наружу через общее отверстие —
устье (у колониальных губок имеется несколько центральных
полостей и устьев, рис. 56).
Движение воды по каналам тела губок обеспечивает им луч-
шее дыхание и способствует выносу из тела остаточных продуктов
обмена веществ. С водой в тело губок попадает и пища — мель-
чайшие водные животные и растения, а также кусочки гниющих
организмов. Попавшие в губку пищевые частицы захватываются
псевдоподиями клеток жгутиковых камер и перевариваются в их
цитоплазме, а чаще транспортируются в мезоглею, где поступают
в амебоидные клетки. Следовательно, губкам свойствен внутри-
клеточный способ пищеварения. Большое значение в питании гу-
бок имеет также осмотическое поглощение органических веществ,
растворенных в воде.
Между покровами и выстилкой внутренних каналов и камер на-
ходится студенистое вещество — мезоглея, в которой содержатся
разнообразные по форме и функции клетки и скелетные эле-
менты (рис. 57). Среди клеток, находящихся в мезоглее, особое
79
Рис. 56. Строение губок с различной сложностью системы каналов и расположе-
нием жгутиковых камер:
/ - поры; 2 - жгутиковые камеры; J — устье. Стрелки указывают направление тока
воды
80
I
Рис. 57. Строение тела губок:
I разрез стенки тела губки; 11 различные формы спикул скелета;
1 - клетки, покрывающие поверхность тела губки и выстилающие ее каналы;
2 - жгутиковые воротннчковые клетки; ,4 — яйцеклетка в мезоглее; 4 — склеро-
бласт со спикулой в мезоглее; 5 - каналец; 6 — звездчатая клетка
значение имеют так называемые амебоциты (полибласты). Они
выделяют вещество мезоглеи, из них образуются половые клетки,
они же участвуют в бесполом размножении губок и дают начало
клеткам, образующим скелет животного. Подобная широкая по-
тенциальная возможность этих неспециализированных клеток
наряду с отсутствием обычных для многоклеточных животных
тканей характеризует губок как животных, уклоняющихся по своей
организации от других многоклеточных.
81
Скелет губок бывает известковым, кремневым и роговым.
Известковый и кремневый скелеты образованы многочисленными
скелетными элементами — спикулами, имеющими разнообразную
форму — иголочек, звездочек и др. Роговой скелет представлен
тонкими упругими волокнами спонгина — вещества, близкого к
роговому. У пресноводных губок бадяг скелет образован крем-
невыми иглами, связанными волокнами из спонгина. В теле губок
встречаются также клетки с сократительными волокнами —
миоциты. Они определяют способность губок отвечать на внеш-
ние раздражения очень слабыми и медленными движениями.
Нервных клеток у губок нет.
Размножаются губки как бесполым, так и половым путем.
Среди них есть и раздельнополые, и гермафродиты. Бесполое
размножение губок осуществляется либо почкованием, либо пу-
тем образования геммул. Геммулы образуются в мезоглее. Это
комочки клеток, окруженные оболочками, в цитоплазме которых
имеется запас питательных веществ. У наших пресноводных
губок бадяг геммулы образуются обычно в конце лета. Осенью
губка погибнет, а геммулы перезимовывают и весной разрастаются
в новую губку.
Половое размножение губок происходит путем образования
в теле из амебоцитов яйцеклеток и сперматозоидов. У герма-
фродитных видов те и другие формируются в мезоглее одной особи
и оплодотворение яиц происходит внутри тела. У разнополых
видов оплодотворение яиц происходит в женской особи, куда
сперматозоиды проникают с током воды. Из зиготы в материнском
организме развивается личинка. Личинка выходит наружу, не-
которое время активно плавает, а затем прикрепляется к суб-
страту и превращается в губку.
Известно более 5 тыс. видов губок, из которых подавляющее
большинство обитают в море. Часто наблюдается сожительство
губок с другими животными — крабами, раками-отшельниками
и др. Так, рачки Spongicola попарно, самец и самка, еще в личи-
ночном состоянии проникают в полость стеклянной губки Euple-
ctella, внутри которой и проводят всю жизнь. У краба Dromia
две задние пары грудных ног видоизменены и несут функцию
поддержания губки, которую сам краб помещает себе на спину.
Губка маскирует краба, а краб обеспечивает передвижение
губки. Это пример симбиоза двух различных организмов, т. е. отно-
шений взаимовыгодных для обоих сожительствующих животных.
У пресноводных губок бадяг наблюдается сожительство с водо-
рослями — хлореллами, которые, выделяя в процессе фотосинтеза
кислород, облегчают дыхание губок.
Практическое значение губок невелико. В Средиземном, Кра-
сном и некоторых других морях добывают туалетных губок. Вы-
ловленных животных подвергают гниению, после чего остается
их мягкий, упругий, пористый волокнистый скелет, используемый
в быту. Сверлящие губки клионы, выделяя особый секрет, содер-
82
жащий кислоту, разрушают известковые скалы. Некоторые из них
сверлят раковины устриц, принося вред устричным хозяйствам.
Филогенетически губки связаны с колониальными воротнич-
ковыми жгутиковыми из подцарства Протозоа.
ТИП КИШЕЧНОПОЛОСТНЫЕ (COELENTERATA)
Общая характеристика. Тип объединяет примитивных многокле-
точных животных, стенки тела которых образованы двумя слоями
клеток: наружным — эктодермой и внутренним — энтодермой.
Находящаяся между ними первичная полость тела заполнена
промежуточным веществом — мезоглеей, которая у некоторых
видов (например, у гидры) имеет вид тонкой бесструктурной
пластинки, а у других (медузы) представлена большой массой
студенистого вещества с включением отдельных клеток и волокон.
Как правило, тело кишечнополостных имеет радиальную сим-
метрию. Ротовое отверстие обрамлено щупальцами. Эктодерма
содержит особые стрекательные клетки, служащие для добывания
пищи и защиты. Кишечная полость заканчивается слепо, аналь-
ное отверстие отсутствует.
Размножаются кишечнополостные как бесполым (образова-
нием почек), так и половым путем. У многих форм наблюдается
чередование поколений, когда бесполая генерация полипов сме-
няется половым поколением медуз.
Кишечнополостные известны с древнейших времен палеозой-
ской эры.
Ныне живущих кишечнополостных описано около 9 тыс. видов.
Подавляющее большинство их обитают в море, и лишь гидры и
некоторые близкие к ним гидроидные обитают в пресных водо-
емах.
Кишечнополостные интересны как примитивные двухслойные
многоклеточные животные для уяснения путей эволюции более
высокоорганизованных животных.
Кишечнополостные имеют также и практическое значение.
Тип Кишечнополостные делится на три класса: Гидроидные
(Hydrozoa), Сцифоидные медузы (Scyphozoa), Коралловые по-
липы (Л п thozoa).
Строение и жизненные отправления. Для многих кишечно-
полостных характерны две жизненные формы: полип и медуза
(рис. 58). Полипы ведут неподвижный или малоподвижный образ
жизни, укрепившись на каком-либо субстрате. Тело их цилиндри-
ческой формы. Его нижняя часть обычно расширена в так назы-
ваемую подошву, которой животное удерживается на субстрате.
На верхнем конце находится ротовое отверстие, окруженное
щупальцами. Между экто- и эндодермой залегает тонкий слой
мезоглеи. Нервная система полипов обычно состоит из отдельных
клеток (диффузного характера). Форма полипа более распростра-
нена среди кишечнополостных. Медузы — свободноплавающие
83
Рис. 58. Схема строения полипа (I) и медузы (II):
1 — рот; 2 — щупальца; 3 — кишечная полость; 4 — ме-
зоглея; 5 - радиальный канал кишечной полости; 6 --
парус
организмы, передвигающиеся в воде (как
активно, так и пассивно) течениями и вол-
нами. Тело медуз имеет вид прозрачного
студенистого купола (зонтика). Рот рас-
положен посредине его нижней стороны:
он окружен предродовыми лопастями. От
кишечной полости отходят радиальные
каналы.
Между экто- и эндодермой расположен
мощный слой студенистой, насыщенной
водой, прозрачной мезоглеи.
Различия полипов и медуз отдельных групп кишечнополост-
ных указаны в систематическом обзоре. В жизненном цикле
некоторых кишечнополостных наблюдается чередование форм
полипа и медузы, когда ряд поколений полипов, размножающихся
почкованием, сменяется поколением половых особей медуз. Но у
многих кишечнополостных жизненная форма медузы отсутст-
вует, и полипы размножаются как половым, так и бесполым
путем.
Размеры кишечнополостных варьируют от нескольких милли-
метров (гидры) до нескольких метров (некоторые океанические
медузы). Разнообразна и окраска этих животных. У плавающих
в воде медуз тело обычно прозрачное, голубоватое, под цвет воды.
Покровы кишечнополостных образованы однослойным
эпителием эктодермы. Среди ее клеток расположены различные
специализированные клеточные элементы. Таковы эпителиально-
мышечные клетки полипов, которые у основания имеют вытянутые
вдоль тела отростки, содержащие миофибриллы. Их сокращение
вызывает укорочение тела. На щупальцах и вокруг рта, а в
меньшем количестве и по всей поверхности тела разбросаны чув-
ствующие клетки, служащие рецепторами, воспринимающими
различные раздражения внешней среды. Но особенно характерны
для покровов кишечнополостных стрекательные клетки, располо-
женные преимущественно на щупальцах, окружающих рот. Эти
клетки имеют на наружной поверхности осязательный волосок.
Внутри клетки находится капсула, оттесняющая цитоплазму и
ядро к периферии клетки. Капсула содержит спирально завитую
полую нить. Прикосновение к осязательному волоску клетки вы-
зывает выбрасывание стрекательной нити, которая при этом
выворачивается. У основания нити расположены крупные и
мелкие шипики. До выброса нити большие шипы сложены вместе
84
и направлены наружу. Они вонзаются в тело жертвы, затем рас-
правляются в стороны, расширяя ранку и удерживая добычу.
В рану вводится ядовитая нить, которая парализует или убивает
жертву или вызывает ожоги на теле врага. У других стрекатель-
ных клеток нити служат для опутывания и удерживания добычи.
Каждая стрекательная клетка «стреляет» один раз.
В покровах кишечнополостных имеются недифференцирован-
ные клетки, способные превращаться в половые, стрекательные,
чувствующие и др.
Нервная система полипов имеет вид нервного сплетения,
лежащего под слоем эпителия покровов. Ее образуют звездчатые
нервные клетки, соединяющиеся своими отростками. Медузы име-
ют более сложное строение нервной системы, что связано с их
подвижным образом жизни. Она усложняется наличием нерв-
ного кольца по краю купола со скоплениями нервных клеток около
глазков и статоцистов.
Органы чувств кишечнополостных примитивны и лучше
развиты у медуз, у которых по краю купола расположены органы
равновесия — статоцисты и глазки. Имеются чувствующие клетки,
которых особенно много на щупальцах и около рта.
Мускулатура имеет своеобразное строение. У полипов
изменения формы тела происходят вследствие движения располо-
женных в мезоглее мускульных отростков эпителиально-мышечных
клеток эктодермы и энтодермы. Более сложные движения медуз
обусловлены наличием особых мышечных волокон, залегающих
в мезоглее, главным образом на краю купола тела. У коралловых
полипов имеются также продольные и поперечные мышечные
волокна в перегородках кишечной полости,
Органы пищеварения у разных представителей типа
устроены различно, но всегда кишечная полость заканчивается
слепо, так как анальное отверстие отсутствует. У гидр и близких к
ним форм ротовое отверстие ведет прямо в цилиндрическую ки-
шечную полость. Но у большинства видов за ртом следует экто-
дермальная глотка, которая ведет в кишечник. У коралловых
полипов в кишечную полость вдаются радиально расположенные
продольные перегородки, что увеличивает поверхность кишеч-
ника. У медуз от кишечной полости внутрь купола расходятся
радиальные каналы, впадающие в кольцевой канал. Кишечная
полость продолжается в полости щупалец. Она выстлана одно-
слойным эпителием энтодермы. Высокие клетки его несут жгутики,
служащие для передвижения пищи. Часть клеток энтодермы
способна образовывать на своей свободной поверхности псевдо-
подии, которыми они захватывают кусочки пищи. Но одновременно
с подобным внутриклеточным пищеварением у кишечнополостных
наблюдается также и внеклеточное переваривание пищи в кишеч-
ной полости под действием соков, выделяемых железистыми клет-
ками ее стенок. У полипов большая часть клеток энтодермы имеет
мышечные отростки с миофибриллами, направленными поперек
«5
тела. Сокращение этих отростков клеток энтодермы вызывает
утончение и удлинение тела животного.
Органы дыхания у кишечнополостных отсутствуют, и
газообмен происходит через покровы.
Выделительная система. Выделительную функцию
несет эпителий как эктодермы, так и энтодермы.
Размножение у кишечнополостных бесполое и половое.
Большинство представителей типа — раздельнополые животные,
но среди них имеются и гермафродиты. У гидроидных половые
продукты образуются в эктодерме, а у остальных — в эндодерме.
Развитие обычно происходит со стадией личинки-планулы. Сна-
ружи планула покрыта ресничками, при помощи которых она
плавает.
КЛАСС ГИДРОИДНЫЕ (HYDROZOA)
Гидроидные представлены разнообразными одиночными и коло-
ниальными формами, обитающими преимущественно в морях, хотя
имеются и пресноводные виды. Для них характерно отсутствие
глотки, ротовое отверстие ведет прямо в обширную кишечную
полость. Эктодерма и эндодерма сходятся по краю рта. Кишечная
полость без продольных перегородок. Яйца и сперматозоиды
образуются в эктодерме.
Характерными представителями класса являются различные
виды пресноводных гидр (рис. 59). Это одиночные полипы, длина
тела которых обычно менее 3 см.
В теле гидры можно различить четыре отдела: головной с
ротовым конусом, окруженным щупальцами, туловище — рас-
ширенную верхнюю часть тела, стебель — утонченную гибкую его
часть и подошву — расширенное основание, которым животное
удерживается на субстрате. Внутри гидры находится обширная
кишечная полость без продольных перегородок. Мезоглея в виде
тонкой опорной пластинки. В ней расположены нервные клетки
и мышечные отростки эпителиально-мышечных клеток. Окраска
гидр обычно буроватая, но у зеленой гидры — зеленая, что обус-
ловлено обитанием в ее эндодермальных клетках многочислен-
ных одноклеточных водорослей — хлорелл. Летом обитают в прес-
ных водоемах со стоячей или малоподвижной водой. Передви-
гаются гидры медленным скольжением подошвы или «кувыр-
канием» через головной конец путем присасывания к субстрату то
подошвой, то ртом. Щупальца животного находятся в движении
для улавливания добычи. Пищей служат мелкие рачки, инфу-
зории, коловратки и другие планктонные животные. Зеленые
гидры способны переносить длительное голодание.
Размножаются как почкованием, так и половым путем. Фор-
мирование почки происходит путем выпячивания обоих слоев его
стенки. Полость материнского организма продолжается в почку.
Дочерние организмы, возникшие путем почкования, отделяются
86
1
Рис. 59. Гидра:
/ внешний вид животного; // - продольный разрез; /// - поперечный разрез тела;
/Г -участок разреза стенки тела; Г -эпителиально-мускульная клетка;
1 — эктодерма; 2 — эндодерма; 3 — опорная пластинка мезоглеи; 4 — кишечная полость;
.5 - эктодермальные эпителиально-мускульные клетки; б - интерстициальные клетки;
7 — стрекательные клетки; 8 — нервные
клетки; .9 — эндодермальные эпителиаль-
но-мускульные клетки; 10 — железистые
клетки; 11 — ротовое отверстие; 12 — ро-
товой конус; 13 — почка; /‘/—подошва;
15 яйцеклетка; 16 — мужские гонады;
17 — щупальца
Рис. 60. Колониальный гидроидный
иолип:
/ — отдельный полип; 2—4 — разные ста-
дии образования медуз; 5 — отделившая-
ся медуза (видны каналы, соединяющие
кишечные полости всех особей колонии)
от материнского, поэтому почкование у гидр не ведет к образова-
нию колоний. Почкование длится все лето, осенью гидры перехо-
дят к половому размножению. На поверхности тела образуются
выпуклости семенники и яичники. Семенников у одной гидры
бывает несколько — они располагаются в верхней части тела.
Яичники — один или два — находятся ниже семенников: в каждом
из них бывает лишь одно большое яйцо. Сперматозоиды выходят
в воду и оплодотворяют яйца. Часть гидр раздельнополы, часть —
гермафродиты. После оплодотворения развитие яйца сначала про-
исходит в яичнике, а затем зародыш выпадает и зимует на дне
водоема. Весной из него вырастает гидра.
Гидры обладают удивительной способностью регенерации —
даже из небольшого кусочка тела быстро восстанавливается
весь организм.
Среди морских представителей класса Гидроидные преоб-
ладают колониальные формы (рис. 60) со сложным жизненным
циклом, включающим чередование бесполого поколения прикреп-
ленных полипов и полового — обычно свободноплавающйх ме-
дузою Колонии их образуются путем многократного почкования
животных, причем дочерние особи не отрываются от материнского
организма, а остаются соединенными с ним. Периодически на
веточках колонии гидроидных полипов образуются особые почки,
из которых развиваются не новые полипы, а половые особи —
мелкие гидроидные медузки. Они отрываются от колонии и сво-
бодно плавают. Из развивающихся в них яиц выходят личинки —
планулы, плавающие благодаря движению покрывающих их
ресничек. Через некоторое время планулы опускаются на дно,
прикрепляются к субстрату и превращаются в неподвижные
полипы, дающие начало новым колониям. Плавающие медузы
и планулы способствуют расселению вида.
Особой сложностью отличаются колонии морских гидроидов —
сифонофор. Плавающие колонии этих животных обычно образо-
ваны полипами различного строения и выполняющими различные
жизненные функции. Одни из них, имеющие вид наполненных
газом пузырей, играют роль поплавков, другие, особенно бога-
тые стрекательными клетками, обеспечивают защиту колонии и
добывание пищи, третьи ее переваривают, в четвертых разви-
ваются половые продукты.
КЛАСС СЦИФОИДНЫЕ МЕДУЗЫ (SCYPHOZOA)
Сцифоидные медузы отличаются от гидроидных более сложной
организацией (рис. 61). У них имеются нервная и мышечная
ткани. В мезоглее купола имеются мышечные волокна, сокра-
щение которых обусловливает сжатие зонтика. Рот ведет в глотку,
которая открывается в кишечную полость. В нее с боков вдаются
четыре радиальные складки. От кишечной полости отходят ра-
диальные каналы, впадающие в кольцевой канал, идущий по
88
Рис. (И. Медузы:
/ корцерог, // схема строения аурелии;
/ рог, 2 ропалий; 3 ротовые лепестки; 4 -..........-кольцевой
канал; 5 радиальные каналы; 6 • щупальца; 7 — половые
железы
Рис. С>2. Развитие сцифоидной медузы аурелии:
/ - яйцо; 2 личинка (планула); .'/--полип (сцифиетома);
/ почкующаяся сцифиетома; 5 - деление ецнфистомы на
диски; 4 молодая медуза (эфира); 7—взрослая медуза
краю купола. Вокруг рта расположены большие лопасти с
многочисленными стрекательными и осязательными клетками.
Как правило, сцифоидные медузы раздельнополые. Половые про-
дукты у них развиваются в эндодерме. Размеры колеблются от
нескольких миллиметров до 2 м в диаметре.
Развитие проходит с превращениями (рис. 62). Из оплодот-
воренных яиц развиваются микроскопически малые личинки —
планулы. Сначала они плавают, передвигаясь при помощи ресни-
чек, а затем опускаются на дно и прикрепляются к субстрату, где
превращаются в мелкие одиночные полипы бокалообразной
формы — сцифистомы. Когда такой полип подрастет, на его теле
появляются поперечные перетяжки, делящие его на ряд дисков,
каждый из которых, отделившись от других, превращается в пла-
вающую медузу.
Сцифоидные медузы — обитатели морей и океанов. Большин-
ство их свободно плавают в поверхностных слоях водной толщи,
но есть и глубоководные формы. Могут приносить вред истреб-
лением рыбы, а некоторые идут в пищу человеку.
КЛАСС КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
Коралловые полипы — одна из древнейших групп животных.
Это одиночные и колониальные полипы, не образующие в своем
жизненном цикле медуз. От гидроидных полипов отличаются более
сложным строением. Их радиальная симметрия несколько нару-
шена двусторонней симметрией пищеварительной полости. Щеле-
видный или овальный рот ведет в сплющенную эктодермальную
глотку, которая открывается в обширную кишечную полость,
выстланную эпителием эндодермы. В кишечную полость с боков
вдаются продольно расположенные перегородки (их бывает восемь
или число, кратное шести) (рис. 63). Скелет коралловых полипов
бывает роговой и известковый.
Коралловые полипы размножаются почкованием или половым
путем. Половые продукты образуются в энтодерме. Из оплодо-
творенных яиц выходит планула, которая прикрепляется к
субстрату и превращается в полип.
У колониальных форм полип путем почкования дает начало
образованию колонии.
Примером одиночных коралловых полипов могут служить раз-
личные актинии. Они имеют мешковидное тело со ртом, окружен-
ным многочисленными щупальцами. Поражает их яркая разно-
образная окраска. Актинии вооружены сильными стрекательными
клетками.
Интересен симбиоз актиний и раков-отшельников. Эти раки
живут в пустых раковинах моллюсков, а актинии часто прикреп-
ляются к этим раковинам (рис. 64). Рак передвигает актинию по
дну моря, кроме того, актиния улавливает пищевые частицы,
90
Рис. 64, Актиния на раковине,
занятой р а ко м - Отт ел ы I и ко м
Рис, 63, Схематическое изображение
строения кораллового полипа:
/ -- щупальца; 2 - роговое отверстие;
3 -.глотка; 4 - перегородки кишечной по-
лости; 5— мезентериальные нити; 3 —
яйца
Рис. 65. Веточка колонии красного коралла:
1 — полни; 2 — кора веточки; 3 — осевой скелет
91
Рис. 66. Каралловый риф во время
отлива
когда рак разрывает клешнями
свою добычу. В то же время
она защищает рака своими
стрекательными клетками.
Особенно интересны коло-
ниальные коралловые полипы
(рис. 65), образующие слож-
ный известковый или роговой
скелет.
В тропических морях ске-
летами коралловых полипов об-
разованы огромные прибреж-
ные рифы (рис. 66) и целые ко-
ралловые острова. Ископаемые
остатки коралловых рифов име-
ют вид мощных отложений известняков, используемых в строи-
тельстве, в сельском хозяйстве и др.
Скелеты некоторых коралловых полипов используют для
изготовления художественных и ювелирных изделий.
ТИП ГРЕБНЕВИКИ (CTENOPHORA)
Своеобразная группа животных, которых ранее рассматривали
как класс Кишечнополостные.
Гребневики — морские одиночные животные. Большинство их
живут в толще воды, но некоторые ведут придонный образ жизни.
Гребневики обладают нежным полупрозрачным телом, вдоль
которого тянутся восемь рядов гребных пластинок, образованных
сросшимися ресничками клеток покровов (рис. 67). Расположен-
ный внизу рот ведет в глотку, а последняя при помощи короткого
пищевода — в сплющенную кишечную полость. От нее отходят
каналы, которые затем дважды ветвятся: от концов этих каналов
отходят вверх и вниз слепые отростки, тянущиеся под рядами
гребных пластинок. Снаружи тело покрыто эктодермой, а кишеч-
ная полость и ее каналы выстланы эпителием энтодермы. Между
экто- и эндодермой лежит студенистая мезоглея. Наличие в мезо-
глее многочисленных клеточных элементов позволяет рассматри-
вать ее как зачаточную, примитивную мезодерму. Этим и отсут-
ствием стрекательных клеток гребневики резко отличаются от
типичных кишечнополостных.
Теоретический интерес представляют донные ползающие греб-
невики, впервые описанные А. О. Ковалевским. Эти животные
92
сочетают признаки строения кишечнополостных, гребневиков и
ресничных плоских червей, что позволяет предположить о возмож-
ной филогенетической связи животных указанных групп.
ТИП ПЛОСКИЕ ЧЕРВИ (PLATHELMINTHES)
Общая характеристика. Плоские черви — двустороннесимметрич-
иыс животные с уплощенным в спинно-брюшном направлении
телом. Покровы и лежащие под ними слои мышечных волокон
образуют кожно-мускульный мешок. Первичная полость тела за-
полнена рыхлой массой клеток — паренхимой, в которой распо-
ложены различные внутренние органы (паренхиматозные черви).
Специальные органы дыхания отсутствуют. У форм, имеющих
пищеварительную систему, кишечник (обычно разветвленный)
закапчивается слепо: анальное отверстие отсутствует. У одной из
паразитических групп плоских червей — ленточных червей —
органов пищеварения нет и пища всасывается через покровы
осмотически. Органы выделения представлены протонефридиями.
Плоские черви, за редким исключением, гермафродиты. Развитие
происходит обычно с метаморфозом, реже — без него.
Описаны около 12 тыс. видов плоских червей. Часть из них
живут в морях, пресных водоемах и почве, но большинство яв-
ляются наружными или внутренними паразитами различных
животных и человека. Многие причиняют значительный вред
животноводству, вызывая заболевание, а иногда и гибель скота.
Некоторые плоские черви служат причиной серьезных заболеваний
л юдей.
Изучение плоских червей важно для разработки мер борьбы
с паразитами, для решения ряда вопросов эволюции животного
93
мира, поскольку они представляют низшую группу трехслойных
многоклеточных животных, имеющих ряд примитивных черт
организации. Плоские черви интересны также для познания
приспособлений животных к паразитическому образу жизни.
В тип Плоские черви входят четыре класса: Ресничные черви
(Turbellaria), Дигенетические сосальщики (Tremalodd), Моно-
генетические сосальщики (Monogenea), Ленточные черви (Се-
stoda).
Строение и жизненные отправления. Форма тела плоских
червей весьма разнообразна. Большинство их имеют листообраз-
ную или лентовидную форму. У лентовидных червей тело обычно
разделено на ряд члеников. Длина тела плоских червей колеб-
лется от нескольких миллиметров до 18 м.
Покровы плоских червей представлены однослойным эпи-
телием. У свободноживущих ресничных червей клетки покровов
несут на поверхности реснички, при помощи которых они плавают
в воде. У паразитических червей тело покрыто тегументом —
сложным цитоплазматическим образованием.
Мускулатура кожно-мускульного мешка слагается из
двух слоев мышечных волокон: наружного кольцевого и внутрен-
него продольного (рис. 68). Кроме того, отдельные пучки мы-
шечных волокон идут наклонно через паренхиму от спинной
стороны тела к брюшной. Такое строение мускулатуры позволяет
червю совершать различные движения, а также вытягивать
и сокращать тело в длину.
Полость тела заполнена рыхлой массой соединительно-
тканных клеток — паренхимой, в промежутках между которыми
циркулирует межтканевая жидкость. В паренхиме заложены
пищеварительные, выделительные и половые органы.
Нервная система у наиболее примитивных плоских чер-
вей имеет диффузный характер, представляя подкожное спле-
тение, образованное нервными клетками, контактирующими окон-
чаниями своих отростков. Но чаще она представлена скопле-
нием нервных клеток в головной части животного, от которых
вдоль тела тянутся нервные тяжи. У свободноживущих плоских
червей и эктопаразитов имеются органы зрения.
Пищеварительная система свойственна не всем
плоским червям. У большой группы их — ленточных червей —
в связи с паразитированием в кишечнике хозяина пищеваритель-
ные органы отсутствуют и пища всасывается покровами тела.
У других плоских червей пищеварительная система имеется и
кишечник обычно разветвлен на две или несколько ветвей,
которые заканчиваются слепо. Анальное отверстие отсутствует.
Органы выделения представлены протонефридиями
(рис. 69). Среди клеток паренхимы расположены крупные клетки
с просветом внутри. В последний вдается пучок длинных ресни-
чек, колеблющихся, как пламя свечи. В просвет клетки поступают
в растворенном виде продукты обмена веществ, а реснички «пла-
94
7.
1
Рис, 68. Разрез кожно-мускульного
мешка планарии:
/ — кожный эпителий; 2 кольцевые мы-
шечные волокна; 3 — косые мышечные
волокна; 4 — продольные мышечные во-
локна; 5—клетки паренхимы полости
тела; 6 — клетки, образующие рабдиты;
7 — рабдиты; 8 — кожная железа
Рис. 69. Клетка с «мерца-
тельным пламенем» прото-
неф риди альной выделитель-
ной системы плоских червей:
/ — ядро; 2 — «мерцательное
пламя»; 3 — канал клетки
мени» гонят их в тончайшие, находящиеся в выделительных
клетках, канальцы. Последние, соединяясь, образуют все более
крупные протоки, которые сливаются в непарный или парные
главные каналы, открывающиеся наружу 1—2 отверстиями. У не-
которых плоских червей выделительных клеток нет и моча вы-
деляется стенками выделительных канальцев. У ряда морских
форм экскреция происходит через покровы.
Органы дыхания отсутствуют. Свободноживущие виды
и эктопаразиты дышат через покровы, а у эндопаразитов, живущих
в среде, бедной кислородом, имеет место анаэробное дыхание.
Кровеносная система, как у всех первичнополостных
животных, отсутствует.
Половая система гермафродитная и очень сложна.
Обычно в эту систему, помимо семенников и яичников, входят
различные придаточные образования, обеспечивающие процесс
оплодотворения яйцеклеток сперматозоидами, снабжение яйца
необходимыми для" развития эмбриона питательными веществами
и создание вокруг яйца защитных оболочек.
Развитие плоских червей проходит в большинстве случаев
с метаморфозом, когда животное, прежде чем достигнуть полово-
зрелости, проходит ряд личиночных стадий. Значительно реже их
развитие идет без подобных сложных превращений.
КЛАСС РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA)
Общая характеристика. В большинстве случаев свободножи-
вущие в водной среде или почве плоские черви с нерасчле-
95
ненным телом, покрытым мерцательным эпителием. На спинной
стороне переднего конца тела имеется обычно несколько прими-
тивных глазков. Рот расположен у большинства видов посредине
брюшной поверхности тела.
Известно около 3 тыс. видов ресничных червей, из которых
несколько десятков видов ведут паразитический образ жизни. Оби-
тают в морях, озерах и реках, но иногда также на суше. Практи-
ческого значения почти не имеют, но интересны при изучении
процессов регенерации, сильно развитой у них, и при решении
вопроса о филогенетических взаимоотношениях классов тина
плоских червей.
Строение и жизненные отправления. Форма тела реснич-
ных червей различна (рис. 70). У некоторых по бокам тела тянут-
ся складки в виде оторочки. Длина тела колеблется от 0,2 мм до
35 см и более (у наземных).
Покровы образованы ресничным (мерцательным) одно-
слойным эпителием. Клетки его несут на своей поверхности
мелкие реснички, движение которых способствует плаванию червя.
Внутри клеток эпителия включены мельчайшие блестящие
тельца — рабдиты, выполняющие защитную функцию, иногда
служат для добывания пищи. Они периодически выбрасываются
наружу и, растворяясь в воде, одевают животное рыхлой клейкой
оболочкой. В покровах ресничных червей заложено много желе-
зистых клеток. Некоторые из них выделяют слизь, другие —
особые ядовитые белковые вещества. Так, у молочных планарий
ядовитых железок много на брюшной стороне тела. Найдя какое-
либо мелкое животное, червь накрывает его своим плоским телом
и убивает ядом.
Нервная система ресничных червей различна по степени
сложности. У некоторых примитивных форм она диффузная.
У других вдоль тела идет несколько нервных тяжей, дающих
ветви к различным органам. У третьих имеются (обычно в го-
ловной части тела) скопления нервных клеток—ганглии, от
которых отходят продольные нервные тяжи (рис. 71). Но, как
показали исследования, интегрирующая роль этих головных ганг-
лиев у ресничных червей невелика.
Органы чувств представлены примитивными глазками,
органами равновесия — статоцитами — осязательными клетками,
разбросанными в покровах. Число глазков и их строение варь-
ируют.
Органы пищеварения имеют весьма разнообразное
строение. У большинства видов рот расположен в средней части
брюшной стороны тела. Он ведет в объемистую глотку, способ-
ную у многих видов выпячиваться наружу, присасываться к до-
быче и высасывать ее. От глотки отходит кишечник, иногда
.прямой, но чаще ветвящийся на две, три или много ветвей. Аналь-
ное отверстие отсутствует, и остатки пищи выбрасываются через
рот. Некоторые ресничные черви лишены кишечника, и пища,
96
Рис. 70. Виды ресничных чер-
вей:
/-молочная нланарпя; 2 -- много-
глаз к а
Рис. 71. ^Пищеварительная и нервная
системы молочной плапарии:
1 — рот; 2 -- глотка; 3 -- передняя ветвь
кишечника; 4, 5 ..задние ветки кишеч-
ника; б — головной нервный узел; 7 -•
боковой нервный тяж; 8 — глазок
поступающая через рот, попадает в рыхлую массу клеток па-
ренхимы, которое ее поглощают и переваривают. У форм, имею-
щих кишечник, пища переваривается как в его просвете, так и
клетками стенок, захватывающими кусочки корма. Следователь-
но, ресничным червям свойственно как внеклеточное, так и вну-
т р и кл еточ ное п и ще в а ре ние.
Внутриклеточное пищеварение у ресничных червей впервые
было описано выдающимся русским ученым И. И. Мечниковым
в 1865 г. Позднее он установил, что внутриклеточное перевари-
вание пищи широко распространено среди многоклеточных живот-
ных. Он обнаружил, что у самых различных животных имеются
особые клетки — фагоциты, способные захватывать и перевари-
вать проникшие в их тело различные микроорганизмы (в том
числе и болезнетворные). На основе этих данных И. И. Меч-
4 Б. А. Кузнецов и др.
97
никовым была создана фагоцитарная теория борьбы организмов
с возбудителями болезней, имеющая большое значение для меди-
цины и ветеринарии.
Органы дыхания отсутствуют. Газообмен происходит
через покровы всей поверхности тела.
Органы выделения у большинства видов ресничных
червей представлены протонефридиями. У многих морских видов
особых органов выделения нет или они слабо развиты и остаточные
продукты обмена веществ удаляются из тела через покровы и
стенки кишечника.
Органы размножения ресничных червей имеют разное
строение. Большая часть этих животных — гермафродиты, но
самооплодотворение устраняется разновременным созреванием
яиц и сперматозоидов. Оплодотворение внутреннее.
Развитие ресничных червей происходит у морских форм
обычно с превращением, а у пресноводных — без него.
Ресничным червям присуща исключительная способность реге-
нерации. Недаром их назвали бессмертными под ножом хирурга.
У некоторых видов этих животных при их разделении на 1500 ча-
стей удавалось получить из каждой части новую особь.
После открытия А. О. Ковалевским в Красном море свое-
образных ползающих гребневиков (Coeloplana), имеющих упло-
щенное тело, ресничные покровы и ряд других признаков,
свойственных ресничным червям, филогенетическая близость
последних к гребневикам, а через них к кишечнополостным
представляется вероятной.
КЛАСС ДИГЕНЕТИЧЕСКИЕ СОСАЛЬЩИКИ
(TREMATODA)
Общая характеристика. Класс Сосальщиков состоит из парази-
тических плоских червей с уплощенным нерасчлененным телом
обычно листовидной формы. Как правило, имеются присоски для
присасывания к телу хозяина. Кишечник образует две ветви,
которые заканчиваются слепо. За редкими исключениями герма-
фродиты. Развитие происходит со сменой хозяев.
Известно, более 6 тыс. видов сосальщиков. Многие из них —
опасные и широко распространенные паразиты сельскохозяй-
ственных животных и человека.
Строение и жизненные отправления. Строение сосальщиков
во многом сходно со строением ресничных червей, от которых они
произошли. Форма тела разнообразная, нередко листовидная.
Характерно наличие присосок, служащих органом прикрепления
к телу хозяина. Обычно имеются две присоски, из которых одна
окружает рот, а другая расположена на брюшной стороне тела.
У некоторых видов встречается третья присоска около полового
отверстия. Иногда брюшная присоска отсутствует, а у некоторых
форм, паразйтирующих’в полости тела птицы, они редуцированы.
98
П о к р о в ы - тсгумент. Гиподерма представлена сетью удли-
ненных, вдающихся в паренхиму клеток. Снаружи тело у неко-
торых усеяно в передней части шипиками, облегчающими фик-
сацию паразита внутри тела хозяина. В покровах заложены
железки различного назначения. Личинки сосальщиков в началь-
ном периоде развития имеют покров из мерцательного эпителия
подобно покровам ресничных червей.
Мускулатура представлена слоями мышечных волокон,
лежащими под покровами и образующими с ними кожно-мускуль-
ный мешок. Наружный, более тонкий,слой образован волокнами,
тянущимися поперек тела, а более мощный внутренний — волок-
нами, идущими вдоль тела. Кроме того, имеются диагональные
пучки мышечных волокон и поперечные, проходящие через парен-
химу от спинной до брюшной стенки кожно-мускульного мешка.
Нервная система в типичном виде слагается из парного
головного ганглия и отходящих от него парных нервных тяжей,
тянущихся к заднему концу тела. От ганглия и тяжей отходят
ответвления к разным органам.
Органы чувств. Органы зрения у взрослых отсутствуют.
Имеются осязательные и другие нервные окончания в покровах.
Органы пищеварения развиты хорошо. Ротовое отвер-
стие ведет в глотку, выполняющую функцию сосания. Затем идет
пищевод, ветвящийся на левую и правую ветви кишечника. По-
следние иногда дают многочисленные боковые отростки. У неко-
торых форм концы обоих ветвей кишечника соединяются. Аналь-
ного отверстия нет. Переваривание пищи происходит в основном
в полости кишечника (т. е. внеклеточно). У немногих встречается
внекишечное переваривание с помощью ферментов, выделяемых
особым органом, расположенным на переднем конце тела, с после-
дующим всасыванием пищи отдельными участками покровов.
Наряду с обычным приемом пищи отмечена способность всасы-
вания через покровы ряда растворенных органических ве-
ществ. Остатки пищи выбрасываются через рот. Всосанные стенка-
ми кишечника питательные вещества разносятся по всему телу
межтканевой жидкостью и клетками паренхимы; движению ее
способствует сокращение мускулатуры тела. По данным электрон-
ной микроскопии, клетки паренхимы имеют выросты, контакти-
рующие с клетками кишечника и с тегументом.
Органы дыхания отсутствуют. У эндопаразитических
форм, обитающих в среде, очень бедной кислородом, энергети-
ческие ресурсы черпаются в процессе брожения пищи.
Органами выделения обычно служат протонефридии.
Их многочисленные начальные клетки разбросаны в паренхиме.
От них отходят канальцы, сливающиеся в более крупные каналы.
Вся сложная система выводных каналов открывается наружу на
заднем конце тела.
Органы размножения имеют очень сложное строение
(рис. 72).
4*
99
2 __ Рис. 72. Схема строения органов размножения
~^7жь дигенетнческих сосальщиков:
J /з 1 — семенники; 2— семяпроводы; 3 - - еемяизнергатель-
5- у (и J 1Н.1Й канал; 4 — копулятивный орган; 5 — его сумка;
77 ~ железы сумки; 7 — яичник; 8— тельце Мелиса;
Л] <) — Лауреров канал; /0 — желтом ники; И — оотип;
2^-2/ / 12 — семяприемник; 13 — матка
£ 1 in
(\[ S^-J 11 / в Почти все сосальщики — гермафро-
( у диты, лишь паразитирующие в крови
yL W 1 2г млекопитающих и птиц кровяные сосаль-
щики раздельнополы. Мужские половые
\ / Ж органы состоят из двух семенников и от-
* ходящих от них семявыносящих прото-
ков, продолжающихся в семяизверга-
тельный канал, который заканчивается
копулятивным органом. Женские половые органы представлены
непарным яичником. От него начинается яйцевод, в который
открываются придаточные образования женской половой системы:
протоки желточников и выводные отверстия особой железки —
тельца Мелиса. Место слияния яйцевода с протоками желточ-
ников и тельца Мелиса называется оотипом. От него наружу
тянется лауреров канал, назначение которого не вполне ясно.
От оотипа начинается длинная трубчатая извитая матка, конец
которой открывается близ мужского полового отверстия.
Сосальщикам свойственны как самооплодотворение, так и
перекрестное оплодотворение. Сперма, образующаяся в семен-
никах, выводится через семявыносящие пути и при помощи копу-
лятивного органа вводится в свою матку или матку другой особи.
По матке сперматозоиды проникают в расширение ее нижнего
отдела — семяприемник, где и накапливаются. Из него сперма
постепенно поступает в оотип, принимающий также яйца из яйце-
вода и продукцию придаточных желез. Здесь яйца оплодотворя-
ются и покрываются сначала желточными клетками, а затем проч-
ной скорлуповой оболочкой. Желточные клетки богаты желтком,
который служит питательным веществом для зародыша при его
развитии в яйце. Секрет тельца Мелиса способствует процессу
оплодотворения и формирования яиц. Оплодотворенные и покры-
тые скорлупой яйца поступают в матку, а затем выводятся на-
ружу. Яйца сосальщиков обычно имеют крышечку, которая от-
крывается при выходе из него личинки.
Развитие многих сосальщиков сопровождается рядом пре-
вращений и сменой хозяев. Нет сомнения в том, что сосальщики
произошли от свободноживущих ресничных червей, приобретя ряд
признаков приспособления к паразитическому образу жизни. Об
этом свидетельствует наличие многих общих черт строения этих
двух групп плоских червей.
Наибольшее значение как опасные паразиты сельскохозяй-
ственных животных и людей имеют следующие виды.
100
Печеночный сосальщик (Hasciola /юра! ica) имеет .листовидное
тело длиной до 5 см и шириной до 15 мм (см. рис. 73). На перед-
нем конце червя выдается небольшой выступ с ротовой присоской.
На брюшной стороне тела расположена брюшная присоска.
Поверхность передней части тела покрыта небольшими шипиками,
задняя — гладкая. Каждая из двух ветвей кишечника образует
многочисленные ответвления, которые, в свою очередь, дают
отростки. Выделительные органы сильно разветвленные. В средней
части тела лежат два ветвистых семенника, а перед ними распо-
ложен непарный яичник. По бокам тела находятся желточники.
Печеночный сосальщик живет в желчных протоках печени
многих растительноядных и всеядных животных, а также недове-
рие. 73. Строение половой (/) и пищеварительной (//) систем пече-
ночного сосальщика фаециолы:
1 — ротовая присоска; 2— брюшная присоска; .7--разветвленный кшпечнпк
(левая и правая ветви); 4 - копулятивный орган; .5 - яичник; 6 желточ-
ники; 7 — желточные протоки; Л' -- семенники; 9 - семяпровод; 10. матка
101
ка. Молодые мигрирующие особи встречаются в различных ор-
ганах хозяина. Более всего от поражения этим паразитом —
фасциолезом — страдают молодые овцы, но нередко им бывает
заражен и крупный рогатый скот. Печеночные сосальщики вы-
зывают расстройство деятельности пищеварительных органов,
отечность и истощение хозяина, и нередко болезнь закан-
чивается смертью. Основная пища — желчь печени хозяина.
Печеночные сосальщики обладают огромной плодовитостью.
Они способны к самооплодотворению, но более часто наблюда-
ется перекрестное оплодотворение. За неделю фасциола может от-
ложить более миллиона яиц, которые вместе с калом хозяина
выводятся наружу. Если яйцо попадет в воду, то спустя 3—6 не-
дель крышечка его открывается и наружу выходит микроскопи-
чески малая личинка — мирацидий (рис. 74). Она покрыта мер-
цательным эпителием, реснички которого служат для плавания
личинки. На переднем конце мирацидия находятся глазки и рот,
ведущий в зачаточный кишечник. В паренхиме заложены выде-
лительные органы и зародышевые клетки. Мирацидии могут
существовать в воде не более 2 сут. Дальнейшее развитие личинки
связано с проникновением в тело промежуточного хозяина, кото-
рым обычно является небольшой пресноводный моллюск — ма-
лый прудовик (Litnnaea truncatula). Внедрению паразита помо-
гает секрет особых железок, открывающихся около рта. Про-
никнув в моллюска, личинка сосальщика претерпевает ряд
упрощений в строении и превращается в следующую стадию раз-
вития — спороцисту. Она мешкообразной формы и лишена
ресничек, питается осмотически соками хозяина через покровы.
Внутри спороцисты сохраняются элементы нервной системы,
выделительные органы и зародышевые клетки, из которых путем
дробления и дифференцировки развивается следующее поколение
личинок — редии. Развитие в спороцисте длится 3—4 недели.
Редии имеют вытянутое тело с ротовым отверстием на переднем
конце, которое ведет в глотку, а она — в пищевод и зачаточный
кишечник. Питаются они, заглатывая ткани хозяина ртом. В теле
редий заложены зародышевые клетки, из которых развивается
второе поколение редий, а затем новая форма личинок — цер-
карии. Они имеют овальное тельце с двумя присосками и длинным
хвостом. Кишечник имеет две ветви. Есть выделительные и зача-
точные половые органы. Развитие церкарий продолжается 4—6 не-
дель. Выходя из тела своего промежуточного хозяина — прудо-
вика, они некоторое время активно плавают в воде при помощи
хвоста, прикрепляются затем к водным или прибрежным расте-
ниям, отбрасывают хвост и инцистируются, покрываясь плотной
оболочкой. Эта стадия развития сосальщика носит название
адолескарии. Они сохраняют жизнеспособность в воде и влажной
среде в течение многих месяцев.
Окончательные (дефинитивные) хозяева заражаются фасцио-
лезом, проглатывая адолескарий с водой или поедая зараженную
102
Рис. 74. Цикл развития
печеночного сосальщика фасцполы
ими прибрежную траву и сено с заливных лугов. В кишечнике
хозяина оболочка адолескарий растворяется и освободившийся
зародыш червя проникает в желчные протоки печени, где и
развивается в половозрелую форму. Весь цикл развития паразита
занимает 2—3 мес. Фасциолез более распространен в районах
со многими водоемами. Борьба с фасциолезом состоит из ряда
профилактических и лечебных мер: дегельминтизации больных жи-
вотных, предотвращения заражения яйцами печеночного сосаль-
щика пастбищ и водоемов, предупреждения возможностей ис-
103
пользования скотом воды из зараженных прудов, смены выпаса
скота (Скрябин, Шульц). Основанием служат расчеты: 1. Раз-
витие личинок в промежуточном хозяине (прудовике) длится от
30 до 80 дней в зависимости от температуры. 2. Созревание мариты
в дефинитивном хозяине происходит за 2—3 мес. Чтобы не до-
пустить заглатывания адолескарий, необходимо менять пастбище
3 раза за выпасной сезон, проводить мелиорацию заболоченных
земель и др.
Ланцетовидный сосальщик, (Dicrocoelium lanceolatum) —
небольшой червь длиной до 1 см, с удлиненным плоским телом
ланцетовидной формы (рис. 75). Имеет ротовую и брюшную при-
соски. Ветви кишечника без боковых ответвлений. Как яичник,
так и семенники компактные, неветвящиеся. Паразитируют в
печени мелкого и крупного рогатого скота и других травоядных
млекопитающих. Более распространен в засушливых районах.
11ервые стадии развития проводит в теле некоторых наземных
моллюсков, а конечные •— в теле муравьев, где инцистируется.
Скот заражается, проглатывая с травой
муравьев, содержащих инцистированных
личинок. Основная мера борьбы с дикроце-
лиозом — изгнание паразитов из больных
животных различными препаратами.
П ростогонимусы (Prosthogonimus) —
мелкие, длиной 3—6 мм сосальщики, пара-
зитирующие в яйцеводах домашних и диких
птиц. Жизнедеятельность их вызывает сна-
чала образование бссскорлупых яиц (птицы
«льют яйца»), а затем прекращение яйце-
кладки. Первым промежуточным хозяином
личинок простогонимусов служат пресновод-
ные моллюски, а дополнительным —личинки
стрекоз.
Кошачий сосальщик (Opistorchis feli-
neus) паразитирует в печени собак, кошек
и диких хищных млекопитающих, иногда
встречается и у человека. Длина червя до
13 мм. Характерны два больших компактных
семенника, расположенных в заднем отделе
тела. Личинки сначала развиваются в прес-
новодном моллюске битинии, а затем — в ры-
бах. Хищные звери заражаются, поедая рыб.
Рис. 75. Ланцетовидный сосальщик:
I — ротовая присоска; 2 — брюшная присоска; ,4 - глотка,
4 - ветви кишечника; 5— яичник; 6 — матка; 7 — семя-
приемник; 8 — желточники; 9—семенники; 10 — семяизвер-
гательный канал
104
Рис. 76. Моиогепетичсские сосаль-
щики:
/ - сиайинк; // - многоустка; ///
дакшлогнрус;
1 рот; У — прнеасывательпый диск
КЛАС С М. О Н О Г Е Н Е Т И -
Ч ЕСКИ Е СОС АЛ Ы ЦИ КИ
(MONOGENEA)
Плоские черви по организа-
ции близки к трематодам.
За единичными исключения-
ми, это эктопаразиты раз-
личных позвоночных (рыб,
амфибий) и беспозвоночных
(моллюсков). Они обладают
мощными органами прикреп-
ления к телу хозяина. Кроме
ротовой присоски, на заднем
конце тела имеется присасы-
вательный диск, вооружен-
ный присосками с крючья-
ми или только крючьями.
Обычно имеются глаза. Вы-
делительные органы откры-
ваются на переднем конце
тела. Гермафродиты. Разви-
тие происходит без смены хо-
зяина. Некоторые виды мо-
ногенез приносят существен-
ный вред рыбному хозяйству, вызывая истощение рыб. Из
опасных паразитов рыб надо отметить дактилогируса (Daciilo-
gyrus), поселяющегося на покровах и жабрах разных пресно-
водных рыб и питающегося их кровью. Своеобразен другой пара-
зит рыб — спайник (Diplozoon paradoxus), селящийся преимуще-
ственно на жабрах (рис. 76). У этого паразита две особи соеди-
няются средними частями крестообразно на всю жизнь так, что
возможно только перекрестное оплодотворение их половых про-
дуктов.
КЛАСС ЛЕНТОЧНЫЕ ЧЕРВИ (CESTODA)
Общая характеристика. Ленточные черви — эндопаразиты раз-
личных животных (преимущественно позвоночных) и людей. Взро-
слые черви обитают в кишечнике дефинитивного, а их личинки —
в разных органах промежуточного хозяина. Тело плоское, как
правило, лентовидное. У подавляющего большинства представи-
105
liWSSjsfl
Рис. 77. Цепень невооруженный:
I — внешний вид; II — гермафродит-
ный членик; Ill — зрелый членик;
1 — головка; 2 - шейка; 3 ~~ стробила;
4 . двураздельный яичник; 5 — яйце-
вод; 6 — тельце Мелиса; 7 -- желточ-
ник; 8 — семяприемник; 9 - влагали-
ще; 10 — половая клоака; 11 — устье
матки; 12 — матка; 13 — копулятивный
орган; 14 — семяпровод; /5 — семен-
ники; 16 — канал выделительной систе-
мы; 17 — нервный тяж
гелей этого класса оно подразделяется на головку (сколекс),
продолжающуюся в шейку, и тело (стробилу), (/гробила обра-
зована рядом члеников — проглоттид, число которых колеблется у
разных видов (у эхинококков до 3—4 члеников, у широких
лентецов — до нескольких тысяч). Эти членики постепенно отшну-
ровываются от шейки паразита. От заднего конца червя отде-
ляются зрелые членики, наполненные яйцами. У взрослых особей
имеются на головке различные приспособления для фиксации
паразита в кишечнике хозяина (рис. 77). Пищеварительные ор-
ганы отсутствуют. Все ленточные черви — гермафродиты.
Строение и жизненные отправления. Тело плоское, ленто-
видное. Его размеры от нескольких миллиметров до 15 м и более.
На переднем конце головка, несущая органы прикрепления:
у цепней округлые присоски, у лентецов щелевидпые ямки —
ботрии, своими краями ущемляющие стенку кишечника хозяина.
Иногда есть дополнительные придатки (хоботок с крючьями и др.)
(рис. 78). Головка продолжается в шейку, зону роста червя.
Здесь происходит отпочковывание члеников — проглоттид, из
которых состоит стробила — членистая лента. По-иному происхо-
дит рост ремнецов, не имеющих наружной членистости тела.
Молодые членики, недавно отделившиеся от шейки, еще не имеют
половых органов. Раньше в них формируется мужская половая
система, а затем и женская, и членики становятся гермафро-
дитными.
П о кров ы представляют собой видоизмененный цитоплазма-
тический слой погруженных клеток подобно тегумепту трематод
Рис. 78. Головки вооруженного
(/) и невооруженного (//)
цепней:
1 — головка; 2 — шейка; 3 - при-
соски; 4 — хоботок с крючьями
Рис. 79. Схема строения тегумента цестод:
/ — микротрихи; 2.-наружный цитоплазматический слой; 3- базальная мем-
брана; 4 — цитоплазматические отростки погруженных клеток тегумента; 5 -
кольцевые и продольные мускульные волокна; 6 -- погруженные клетки тегумента
107
(рис. 79). Покровы выполняют разнообразные функции: защи-
щают от действия ферментов кишечника хозяина благодаря
содержанию веществ, противодействующих ферментам, выделе-
нию слизистых веществ и т. п., всасывают пищу, выделяют на-
ружу свои ферменты, воздействующие на окружающую червей
пищу. В покровах цестод выявлены волоски и ворсинки, увели-
чивающие площадь всасывания пищи. В кутикуле встречаются
шипики.
Мускулатура состоит из наружного кольцевого слоя и
внутреннего продольного. Иногда есть еще мышцы диагональные
и в паренхиме. Полость члеников содержит паренхиму. К спе-
цифическим особенностям ее относится наличие в ее клетках
известковых телец.
Нервная система состоит из скопления нервных клеток
в головке червя и парных продольных тяжей, тянущихся до конца
тела.
Органы чувств у ленточных червей выражены слабо.
Органы пищеварения и дыхания отсутствуют.
Органы выделения протонефридиального типа. В парен-
химе заложены крупные клетки с мерцательным «пламенем».
От них отходят выносящие канальцы, которые у большинства
видов впадают в два крупных канала, тянущихся по бокам стро-
билы. В каждом членике эти каналы соединяются поперечным
протоком. Боковые каналы открываются на заднем крае по-
следнего членика.
Органы размножения ленточных червей во многом
сходны с таковыми сосальщиков (рис. 80). У перасчлепенных
ленточных червей (Cestodaria) в полости тела только один комп-
лект мужских и женских половых органов. У расчлененных (Ces-
toda) этот комплект повторяется в каждом гермафродитном
членике (иногда в одном членике имеются два набора половых
органов). Женская половая система образована яичником (обыч-
но двудольчатым), яйцеводом, желточниками, тельцем Мелиса и
маткой. Для проведения спермы служит влагалище, расширяю-
щееся у впадения в яйцевод (семяприемник). Мужская половая
система состоит из многочисленных мелких округлых семенников
и семявыносящих канальцев, которые, сливаясь, открываются в
семяпровод, заканчивающийся семяизвергательным каналом и ко-
пулятивным органом. Семяизвергательный канал и влагалище
открываются в углубление поверхности тела червя — половую
клоаку. У ленточных червей наблюдается как самооплодотворение,
так и перекрестное оплодотворение. Самооплодотворение про-
ходит двумя путями: когда копулятивный орган вводит сперму
во влагалище того же членика, на котором он расположен, или
когда сперма вводится им во влагалище другого членика. Опло-
дотворенное яйцо окружается желточными клетками и скорлупой,
чему способствует секрет тельца Мелиса. Сформированные
яйца выводятся в матку. У лентецов на конце матки имеется
108
Рис. 80. Схема строения полового аппарата гермафродитного членика пения
(/) и лептеца (//):
/ - семенники; 2 семяпровод; 14 - копулятивный орган; 4 -- его сумка; 5 яични-
ки; 6 - яйцевод; 7 - желточннк; Я оотип; 9 - тельце Мелиса; 11) семяприемник,
// --- влагалище; 12 . матка
отверстие, через которое яйца по мере созревания выводятся
наружу. У цепней матка замкнутая и яйца, попадающие в нее,
постепенно там накапливаются. По мере их накопления матка
увеличивается в размере и образует боковые ответвления,
которые также заполнены яйцами. В матке одного членика может
скопиться несколько тысяч яиц. У наполненных яйцами члеников
заднего конца стробилы, называемых зрелыми, происходит по-
степенное рассасывание половых органов, кроме матки с яйцами.
Они теряют связь со стробилой и выводятся с калом из тела
хозяина. В наружной среде членики разлагаются, освобождая
яйца.
Развитие ленточных червей сопровождается сложными
метаморфозами. Еще в теле хозяина в яйце формируется округ-
лая личинка с шестью острыми хитиновыми крючками (онко-
сфера). Если яйцо попадает в кишечник промежуточного хозяина,
онкосфера выходит из его скорлупы, проходит через стенку
кишки и проникает в кровеносные сосуды; лишь у немногих
форм она продолжает свое развитие в стенках кишечника. Попав
в кровяное русло, онкосферы разносятся током крови по телу
промежуточного хозяина. Оседая в различных органах последнего,
они преобразуются в новую форму личинки — финну. Строение
финн у разных цепней неодинаково. Финна-цистицерк имеет вид
109
небольшого овального пузырька с одной впяченной внутрь его
головкой будущего червя. Финна-ценур значительно больше —
с крупный орех или даже куриное яйцо; в его пузырь впячено
много головок. Финна-эхинококк отличается крупными размерами
и сложным строением; оболочка пузыря отпочковывает внутрь
его дочерние пузыри, а те, почкуясь, дают внучатые пузыри со
многими головками. Своеобразные личинки лентецов описаны
ниже.
Дефинитивный хозяин заражается ленточными червями, по-
едая промежуточного хозяина, в котором находятся финны. В ки-
шечнике под воздействием желчи, выделяемой печенью, головки
выворачиваются, прикрепляются к стенке кишки и начинается
рост стробилы.
Вопрос о происхождении ленточных червей является дис-
куссионным. Более вероятным является взгляд, согласно кото-
рому за исходную группу в эволюции ленточных червей прини-
мают турбеллярий. Ленточные черви в современной система-
тике делятся на два подкласса: Цестодообразных (Cestodaria)
и Цестод (Cestoda).
ПОДКЛАСС ЦЕСТОДООБРАЗНЫЕ (CESTODARIA)
К подклассу Цестодообразные относится небольшое число лен-
точных видов червей, у которых тело не подразделено на головку
и стробилу и нет членистости. В теле листовидной формы один
набор половых органов. Клдсс включает небольшое число
видов — паразитов осетровых рыб (например, амфилину).
ПОДКЛАСС ЦЕСТОДЫ (CESTODA)
К подклассу Цестод принадлежит подавляющее большинство
ленточных червей. Их лентовидное тело ясно разделено на го-
ловку, шейку и стробилу, которая, как правило, состоит из ря-
да члеников. Половой аппарат повторяется в каждом членике.
Подкласс разделяется на ряд отрядов, из которых мы опишем толь-
ко отряды Цепней и Лентецов, имеющих наибольшее значение для
животноводства.
Отряд Цепни (Cyclophyllidea) —ленточные черви, у которых
на головке расположены четыре большие округлые присоски.
Матки члеников не имеют наружных отверстий. Половая клоака
расположена, как правило, сбоку членика. Личинки онкосферы
и финны различного типа. Из многочисленных представителей
отряда особенно опасными паразитами сельскохозяйственных жи-
вотных являются бычий и свиной цепни, овечий мозговик, эхи-
нококк и мониезия.
Невооруженный (бычий) цепень (Taeniartiynchus sagitia-
tus) — крупный червь, длина тела которого иногда достигает
12 м. На голове расположёны четыре присоски, но хоботка с крю-
110
чьими нет. Стробила состоит из нескольких сотен и даже тысяч
члеников. Матка зрелого членика имеет до 35 ответвлении с
каждой стороны. Число яиц в ней достигает 100 тыс. Вышедшие
поодиночке из тела хозяина зрелые членики могут пере-
двигаться. Дефинитивным хозяином бычьего цепня является чело-
век, а промежуточным — крупный рогатый скот, у которого финны
обычно концентрируются в мышцах. Заражение человека происхо-
дит при употреблении плохо проваренного или прожаренного го-
вяжьего мяса. Заболевание вызывает истощение человека, про-
исходят интоксикация продуктами выделений червя,'расстройство
деятельности кишечника. Меры предупреждения заболевания
сводятся к ветеринарному надзору за заболеваемым скотом и
выбраковке финнозного мяса.
Вооруженный (свиной) цепень (Taenia solium) отличается от
бычьего тем, что на головке, помимо четырех присосок, имеет
хоботок с венчиком острых крючков. Длина тела 2—8 м. В стро-
биле до 1 тыс. члеников. Матка зрелого членика имеет 5—12 ответ-
влений с каждой стороны. Число яиц в ней обычно по превышает
50 тыс. Зрелые членики выходят из тела человека обрывками
стробилы: самостоятельно двигаться они не могут. Взрослый
червь живет в, кишечнике человека, а его развитие протекает в
теле свиней, кабанов, верблюдов, собак, кошек и кроликов.
Финны его чаще всего находятся в различных мускулах про-
межуточных хозяев. Длина их достигает 15 мм, а ширина -
до 8 мм. Развитие финны из онкосферы длится 3—4 мес. В органах
промежуточного хозяина они могут оставаться жизнеспособными
ряд лет. Люди заражаются обычно, употребляя в пищу мало
проваренное или прожаренное свиное мясо. Но человек может быть
для свиного цепня и промежуточным хозяином — это происходит
в тех случаях, когда в кишечник попадают яйца этого паразита
с уже развившимися в них онкосферами. Последние покидают
оболочки яиц и внедряются в кровеносное русло человека, а затем
превращаются в его мышцах в финны. Своим присутствием финны
могут вызывать тяжелые заболевания (рис. 81).
Овечий мозговик (Milticeps milticeps) достигает длины 80 см.
Головка имеет четыре присоски и хоботок с двумя рядами крючьев.
В стробиле около 200 члеников. Дефинитивными хозяевами его
являются собаки, которые рассеивают с фекалиями зрелые членики
червя с яйцами. При попадании в кишечник промежуточного
хозяина — овцы — из них выходят онкосферы, которые внедряют-
ся в стенки кишки и током крови заносятся в мозг животного, где
и превращаются в финну-ценур. Это пузырчатая финна мозговп-
ка достигает величины куриного яйца и содержит несколько
головок. При поражении одного из полушарий мозга овца начи-
нает двигаться по кругу («вертячка»), У больных овец извлекают
паразита путем трепанации черепа. Для профилактики заболе-
вания необходимо регулярно проводить дегельминтизацию собак
(особенно пастушьих).
111
Рис. 81. Цикл развития вооруженного цепня
Эхинококк (Echinococcus granulosus) — половозрелый червь
имеет длину всего лишь 5—6 мм (рис. 82). На его головке
имеются четыре присоски и хоботок с двумя рядами крючьев.
В стробиле всего лишь три членика: незрелый, гермафродитный
и зрелый, длина которого больше длины двух первых члеников.
Зрелый членик содержит до 800 яиц. Созрев, он открывается
от материнского тела и выносится вместе с фекалиями хозяина
наружу. Его место занимает гермафродитный членик, который
после оплодотворения яиц становится зрелым. Зрелые членики
эхинококка способны некоторое время ползать. Во взрослом со-
стоянии эхинококки живут в кишечнике собак, волков, лисиц и дру-
112
Рис. 82. Эхинококк:
/ - - половозрелая особь; II — схема строе-
ния эхинококкового пузыря;
1 — головка с присосками и хоботком
с крючьями; 2 — незрелый членик; 3 —
гермафродитный членик; 4 -- зрелый чле-
ник; 5 кутикула пузыря; 6 — произво-
дящая оболочка; 7—11 — внутренние до-
черние пузыри па разных стадиях раз-
вития; 12—15 — наружные дочерние пу-
зыри па разных стадиях развития
Рис. 83. Схема строения различных
типов финн:
I — цистицерк; // — ценур; III - эхино-
кокк; IV — цистицеркоид; V' -- плероцер-
коид;
1 — стенка пузыря; 2 — вывернутые го-
ловки; 3 — внутренние дочерние пузыри
гих животных. Промежуточными хозяевами червя служат крупный
и мелкий рогатый скот, свиньи, лошади, верблюды, кролики и мно-
гие грызуны. Личиночной стадией эхинококка поражается и чело-
век, заболевая тяжелым (иногда смертельным) эхинококкозом. Из
яйца, попавшего в кишечник промежуточного хозяина, выходит он-
косфера, проникающая через стенку кишки в кровеносные сосуды
хозяина, по которым мигрирует, оседая в разных органах.
Из них развиваются огромные пузырчатые финны, наиболее
часто встречающиеся в печени хозяина, реже — в легких,
трубчатых костях, мышцах и мозге. У крупного рогатого скота
масса пузырей эхинококка может достигать 60 кг. Внутри нахо-
дится большое число дочерних пузырей, одни из них содержат
несколько головок, а другие совсем не имеют. Финны эхинокок-
ка растут весьма медленно (рис. 83).
Главными распространителями эхинококкоза животных и че-
ловека являются собаки. Зрелые членики паразита, выползающие
из анального отверстия собаки, вызывают почесывание зудящего
места и переноса их на шерсть. Человек заражается при неосто-
рожном обращении с собаками. Больные эхинококкозом собаки,
охраняющие отары овец, служат источником заражения яйцами
паразита пастбищ. Профилактика заболевания — дегельминти-
зация собак, а для людей — личная гигиена.
Мониезия (Motiiezia expanse,) достигает в длину 5 м (рис. 84).
Ее головка несет четыре присоски, но хоботка с крючьями нет.
В каждом гермафродитном членике имеется двойной комплект
половых органов. Половые отверстия расположены на обеих сторо-
нах членика. Дефинитивными хозяевами червя служат крупный
рогатый скот, овцы и козы. Молодняк скота переносит мониезиоз
весьма тяжело, взрослые животные — легче. Промежуточными
хозяевами мониезии являются микроскопические малые панцир-
ные клещи, живущие в почве и на ее поверхности. Наиболее
действенная мера борьбы с мониезеией — дегельминтизация боль-
ных животных.
Отряд Лентецы (Pseudophyllidea). Имеют головку без присо-
сок, но с двумя щелевидными ямками — ботриями, которыми
они зажимают складку стенки кишки. Матка имеет наружное
отверстие, через которое выходят яйца. Половое отверстие нахо-
дится посредине брюшной стороны членика.
Отряд включает большое количество видов, из которых наи-
большее значение как опасный паразит человека имеет широкий
лентец.
Лентец широкий (Diphyllobothrium latum) в половозрелом
состоянии живет в кишечнике человека и хищных животных
(рис. 85). Промежуточных хозяев два: первый — веслоногие рачки
(циклопы и диаптомусы), второй — рыбы, главным образом щуки.
Длина взрослого червя до 15 м. Стробила широкая, из 3—-4 тыс.
члеников. Яйца с крышечкой. Если яйцо попадет в воду, из него
через некоторое время выходит личинка — корацидия (рис. 86),
114
Рис, 84. Монисзия:
/ -- сколекс; // - гермафро-
дитный членик; III — личин-
ки моннезии в теле панцир-
ного клеща
покрытая ресничным эпителием и снабженная шестью крючками.
Эта личинка может быть заглочена рачком, в теле которого она
теряет реснички и в течение двух недель развивается в следующую
стадию личинок — процеркоид длиной около 0,5 мм с крючками
на конце тела. Если рачка проглотит рыба, в ее теле формируется
новая личинка с двумя ботриями — плероцеркоид длиной 1—2 см.
Плероцеркоиды находятся в различных органах рыбы, но преиму-
щественно в мышцах. Дефинитивный хозяин заражается, поедая
рыбу. В нем плероцеркоиды превращаются в зрелых червей. У
людей дифилоботриоз встречается в районах, где рыбу потреб-
ляют в большом количестве, особенно строганину из мороженой
рыбы. Отсюда вытекают и меры профилактики этого заболевания.
115
Рис. 85. Лентец широкий:
/— головка с ботриями; // —-гермафродитный членик;
/ — копулятивный орган; 2 — семянровод; //--семенники; 4 — яичник; 5 —
оотип; 6 — желточник; 7—желточный проток; 8 — матка; 9 — влагалище;
10 — отверстие матки
Рис. 86. Цикл развития лентеца широкого
Важно также предупредить возможность попадания фекалий
человека в водоемы.
К числу лентецов относится также червь с нерасчлепепным
телом — ремнец (сем. Ligulidae). Длина его тела достигает 1 м и
имеет вид ровной ленты. Но комплекты половых органов повто-
ряются в нем многократно. Он паразитирует в кишечнике
водных и болотных птиц. Из яиц ремнеца, выделяемых этими
птицами, выходят плавающие личинки — корацидии, которые за-
глатываются мелкими рачками — циклопами и диаптомусами. В
них корацидии превращаются в процеркоид. Если зараженного
рачка съест какая-нибудь рыба, то в полости тела ее процеркоид
становится плероцеркоидом, имеющим лентовидное иерасчленеп-
ное тело длиной до 80 см. Поедая рыбу, водные и болотные
птицы становятся хозяевами половозрелой стадии ремнеца.
Плероцеркоиды ремнеца иногда ’ вызывают массовую гибель
различных пресноводных рыб.
ТИП КРУГЛЫЕ ЧЕРВИ (NEMATHELMINTHES)
Круглые черви имеют несегментированное тело. Первичная по-
лость тела обычно заполнена полостной жидкостью. Кишечный
канал (если имеется) неветвистый и заканчивается анальным
отверстием.
Нервная система представлена окологлоточным кольцом,
образованным скоплением нервных клеток и рядом отходящих
от него продольных нервных тяжей. Органы дыхания отсутствуют.
Выделение продуктов диссимиляции происходит при помощи про-
тонефридиев или внутриклеточными канальцами, но без мерца-
тельных клеток; у некоторых видов выделительных органов нет.
Как правило, круглые черви раздельнополы. У многих форм ясно
выражен половой диморфизм.
Описано свыше 12 тыс. видов круглых червей. Часть их ве-
дет свободное существование в водоемах и почве, другие— па-
разиты растений, животных и человека. Практическое значение
круглых червей очень велико. Почвенные виды играют существен-
ную роль в почвообразовательных процессах, паразитические -
наносят огромный вред сельскому хозяйству, резко снижая про-
дуктивность сельскохозяйственных растений и животных и нередко
вызывая их гибель.
Ряд паразитических круглых червей является причиной
серьезных заболеваний людей.
Тип Круглые черви включает классы: Собственно круглые чер-
ви, или Нематоды (Netnaioda); Волосатики (Nernatomorpha);
Брюхоресничные (Gastrotricha); Коловратки (Rotatoria).
Для сельского хозяйства наибольшее значение имеет класс
Нематод.
117
КЛАСС СОБСТВЕННО КРУГЛЫЕ ЧЕРВИ,
ИЛИ НЕМАТОДЫ (NEMATODA)
Общая характеристика. Тело в сечении округлое, сохраняет
постоянную ширину, покрыто плотной кутикулой. Передвигаются
изгибаясь, как тугая струна («струнцы»). Полное отсутствие кле-
ток с ресничками. К классу Нематод принадлежит большинство
видов типа. Свободноживущие обитают в морях, пресных водах и
в почве. При многочисленности видов и особей их участие в
круговороте веществ в природе значительно. Многие нематоды из-
вестны как часто встречающиеся паразиты человека, животных и
растений. Так, паразитов растений описано свыше тысячи видов.
Положительную роль играют почвенные нематоды, как сапрофиты,
так и хищные. Из паразитов полезны виды, имеющие своими
хозяевами вредных насекомых, а также паразитирующие на кор-
нях и тканях растений-сорняков.
Строение и жизненные отправления. Форма тела у боль-
шинства веретеновидная, нитевидная, реже — колбасовидная, но
встречается и округлая. На переднем конце рот, органы осязания
в виде папилл и щетинок и органы химического чувства — амфиды
(в виде углублений в кутикуле). Светочувствительные органы
лишь у немногих свободноживущих. Туловище заканчивается хво-
стом, так называют участок тела за анальным отверстием.
У самок ряда видов, ведущих неподвижный образ жизни, хвост
отсутствует. Тело самцов многих видов оканчивается бурсой, слу-
жащей органом прикрепления к телу самки при спаривании.
Хорошо выражен почти у всех видов половой диморфизм.
Длина тела от долей миллиметра до нескольких сантиметров
и, как исключение, до 1,5 м.
Покровы нематод образованы гиподермой и покрывающей
ее кутикулой. Толщина кутикулы может достигать 20 % диаметра
тела. В обширном классе нематод строение и химический состав
кутикулы неодинаковы. У кишечных паразитов — аскарид—она
из 8—10 слоев, у других количество слоев меньше. Поверхность
ее у одних гладкая, у других кольчатая благодаря кутикулярным
утолщениям.
Кутикула нематод сложная и по химическому составу. У па-
разитирующих в кишечном тракте в ней выявлены: белки (альбу-
мин, эластин, коллаген, кератин и др.), нуклеопротеиды, липо-
протеиды. Все эти вещества находятся в биологически активном
состоянии, что объясняет устойчивость кишечных паразитов к дей-
ствию пищеварительных ферментов. Кутикулярный покров мерт-
вых гельминтов переваривается в кишечнике хозяина.
Кутикула органически связана с выделяющей ее гиподермой.
У свободноживущих нематод и молодых паразитических она в виде
однослойного эпителия, у взрослых особей многих пара-
зитических представлена синцитием — протоплазматической мас-
сой с ядрами. На спинной и брюшной сторонах и по бокам гипо-
118
Рис. 87. Изолированные мышечные
клетки аскариды (продольный разрез):
1 — сократимая часть клетки; 2 — пла-
зматическая часть клетки е ядром; 3 -
плазматический отросток; 4 -- ядро
заиы одним слоем веретено-
вдоль тела червя и содержат
дерма образует утолщения в
форме валиков, простирающих-
ся вдоль тела.
Кожно-мускульный меток
заключает хорошо представлен-
ную первичную полость тела,
заполненную полостной жид-
костью. В полости тела распо-
ложены внутренние органы.
Мускулатура, как пра-
вило, образована четырьмя
полосами продольных мышеч-
ных волокон (рис. 87), разде-
ленных по бокам тела валика-
ми гиподермы, а на спине и брю-
хе —- нервными стволами.
Эти мышечные полосы обра
видных клеток, которые вытянут
большое число сократительных волокон — миофибрилл. От
мышечных клеток отходят ветвистые протоплазматические отрост-
ки, которые углубляются в полость тела. Одни из них, принадлежа-
щие клеткам спинных мышечных полос, направлены к спинному,
а отростки клеток брюшных мышц — к брюшному нервным тя-
жам. Подобное строение мускулатуры круглых червей, не имеющей
кольцевых мышц, определяет однообразие движений этих живот-
ных, выражающихся только в изгибах тела.
Нервная система круглых червей состоит из нервного
кольца, окружающего пищевод, и нескольких продольных нервных
тяжей. Она образована небольшим числом нервных клеток,
что свидетельствует о ее примитивных чертах. Наиболее развиты
спинной и брюшной стволы, соединенные комиссурами и дающие
ответвления к различным органам.
Органы чувств развиты Слабо. У некоторых свободножи-
вущих форм имеются глазки примитивного строения. Около рта
расположены чувствительные щетинки и осязательные бугорки.
Осязательные придатки имеются также обычно у самцов на зад-
нем конце тела, которым они прикрепляются к телу самок или
охватывают его при спаривании.
Пищеварительная система. Пищеварительный тракт
состоит из трех отделов: переднего, среднего и заднего (рис. 88).
Передний отдел обычно разделяется на ротовую полость, глотку
и пищевод: они выстланы эпителием эктодермального происхож-
119
.Рис. 88. Поперечный разрез тела самки аскариды:
/_ кутикула; 2— гиподерма; 3 — мышечные клетки; 4 - - сип........ нернпып
тяж; 5 — брюшной нервный тяж; 6
боковые валики гинодермы е каналом
выделительной системы; 7 - первичная
полость тела; 8 -- кишечник; .9 - яич-
ник; 10— яйцевод; II — матка
Рис. 89. Выделительная система
различных нематод
дения. Ротовое отверстие обычно прикрыто губами, в нем у многих
хищных видов имеются хитиновые зубчики, а у растительноядных
форм расположено острие — стилет, которым они прокалывают
ткани растений. За ротовой полостью следует пищевод, имеющий
одно или несколько расширений — бульбусов с сильно развитой
мускулатурой стенок. Пищеварение происходит в средней части
кишки, имеющей вид прямой трубки и выложенной однослойным
эпителием эндодермального происхождения. Задняя кишка экто-
дермальная, выстлана кутикулой. Кишечник заканчивается
а нал ь и ы м от вс р сти е м.
Орга н ы д ы х а и и я . У свободпоживущих нематод и парази-
тов растений газообмен происходит через покровы тела, а у боль-
шинства видов — паразитов животных — дыхание анаэробное.
Органы выделения (рис. 89). В головном отделе к
гиподерме прилегает шейная железа. Ее считают органом выделе-
ния, ио она прежде всего функционирует как орган осморегуляции.
Железу образуют 1—2 клетки с отростками и с каналом внутри,
открывающимся наружу. У многих нематод — лошадиной, свиной
и др.— шейная железа в виде гигантской клетки с двумя отрост-
ками, тянущимися вдоль тела в боковых валиках гиподермы.
Каналы, находящиеся внутри отростков, соединяются в теле клет-
ки. Наружу непарный канал открывается норой на брюшной
стороне в передней части тела. Обычно к отросткам прилегают
крупные фагоцитарные клетки (у аскарид—по две па каждом
отростке), в которых накапливаются некоторые остаточные про-
дукты обмена веществ, по передача их в каналы отростков не уста-
новлена.
Нематоды раздельнополы и большей частью обладают ясно
выраженным половым диморфизмом.
Органы р а з м н о ж е н и я. Половые органы самцов и самок
имеют трубчатое строение. Мужские половые органы имеют
форму непарной трубки, тонкий конец которой является семен-
ником, средняя часть — семяпроводом, а основной, наиболее
толстый отдел — семяизвергательным каналом, открывающим-
ся в конечный отрезок кишечника — клоаку. У самок половые
органы, как правило, парные. Нитевидные яичники продолжаются
в яйцеводы, которые, расширяясь, переходят в две матки,
открывающиеся в короткое непарное влагалище. Влагалище слу-
жит для приема спермы самца и вывода'наружу яиц. У некоторых
паразитических круглых червей в цикле развития имеется герма-
фродитное поколение. Оплодотворение у круглых червей внутрен-
нее. Большинство видов откладывают яйца, но есть и живородя-
щие формы.
Развитиеу одних видов прямое, у других — с метаморфо-
зом и сменой хозяев.
Среди форм нематод, имеющих большое значение для сель-
ского хозяйства и здравоохранения, выделяют следующие:
121
1. Свободноживущие формы, населяющие водоемы и почву и
питающиеся органическими остатками или мелкими почвенными
животными;
2. Паразиты растений, обитающие в течение всей жизни или
чаще только на определенной стадии развития в тканях различных
растений. Вне времени паразитирования они живут в почве;
3. Паразиты животных и человека.
СВОБОДНОЖИВУЩИЕ КРУГЛЫЕ ЧЕРВИ
Значительное число мелких круглых червей обитают в морях,
пресных водоемах и почве. Для сельского хозяйства большое
значение имеют различные почвенные нематоды, встречающиеся
преимущественно в почвах значительного увлажнения и богатых
органическими веществами. Нередко в верхних горизонтах почвы
на площади 1 м2 обитают десятки миллионов этих червей.
Большая часть их обитает в почве постоянно, проходя все стадии
развития, но среди них немало и таких форм, которые проводят
в почве только часть жизненного цикла, внедряясь затем в те
или иные растения и становясь их паразитами.
Почвенные нематоды питаются чаще всего гниющими органи-
ческими веществами. Но среди них есть хищные формы, поедаю-
щие других мелких почвенных животных. Роль нематод в почвооб-
разовательных процессах и, в частности, в гумификации почв
весьма значительна.
КРУГЛЫЕ ЧЕРВИ - ПАРАЗИТЫ РАСТЕНИЙ
Подавляющее большинство круглых червей, паразитирующих
в растениях, принадлежит к отряду , Шишкоиглых нематод
(Tylenchida). В ротовой полости их находится колющий орган —
стилет, которым разрушаются ткани растения. В стилете имеется
канал, через который в ранку растения поступает секрет железок
пищевода, растворяющий оболочки клеток. В средней ча-
сти пищевода расположено мускулистое расширение — бульбус,
с помощью которого происходит всасывание паразитом соков ра-
стения. Отряд включает ряд важных вредителей различных куль-
турных растений.
Семейство Разнокожие нематоды (Heteroderidae) характе-
ризуется тем, что самки имеют плотные покровы шаровид-
ного тела, а самцы — тонкое вытянутое тело с прозрачными
покровами.
Картофельная нематода (Heterodera rostochiensis) паразити-
рует на корнях и клубнях картофеля. Самец имеет веретеновид-
ное бесцветное тело с тонкими покровами длиной до 1,2 мм. Взрос-
лые самки шаровидной формы, с выдающимся головным концом,
диаметр их до 1 мм. Их плотные покровы окрашены в коричневый
цвет. Самцы живут внутри корней и клубней картофеля, а самки —
122
на их поверхности. Развитие происходит внутри тканей корня.
За лето, по-видимому, дает 1 — 2 поколения. Длительность раз-
вития каждого поколения до 50 дней. Плодовитость до 1 тыс.
яиц. Пораженное растение прекращает рост и часто погибает.
Главные способы борьбы: высокий уровень агротехники и исполь-
зование в севооборотах устойчивых растений.
Свекловичная нематода (Heterodera schachli) — паразит кор-
ней свеклы, приносящий нередко большие повреждения свеклови-
чным плантациям (рис. 90). Самцы имеют вытянутое прозрачное
тело длиной до 1,6 мм. Самки отличаются лимонообразной
формой тела, окрашенного в желтый цвет, длина его от 0,4 до
1,1 мм. Снаружи самки одеты беловатой слизистой оболочкой.
Самцы живут внутри корней свеклы, самки сидят на поверхности
корешков, внедрившись головным концом. Выросшие самцы вы-
ходят из корней в почву, разыскивают самок и оплодотворяют
их. Оплодотворенные самки откладывают 200—600 яиц в слизи-
стые яйцевые мешочки. Яйца сохраняют жизнеспособность в тече-
ние нескольких лет (Кирьянова). Личинки живут сначала в почве,
а затем внедряются в корешки свеклы. В почве мигрируют в
пределах 30—40 см. Развитие одного поколения длится 4—5 не-
дель. На юге за лето эти нематоды могут дать до пяти поколений.
Растения замедляют рост, легко привядают на солнце, рано от-
мирают. Урожайность и сахаристость корней резко падает. Меры
борьбы те же, что и с картофельной нематодой.
Рис. 90. Свекловичная нематода:
/ — самка; // — самки на корешке свеклы
123
1
Рис. 91. Галловая нематода:
/ - самка; II - самец; /// - галлы на корнях огурца;
1 -стилет; 2- бульбус; ,3 - пищеводная железа; -1 - нервное кольцо; .5 выдел тельное
отверстие; 6 кишка; 7 яичник; Л’ - яйцевод; 9- матка с яйцами, 10 половое
отверстие; 11 — анус; 12 - семенник; 1,3 — семяпровод; 14 — семяизвергательный канал;
/5 - спикулы
Галловая нематода (Meloidogyne marioni) — паразит корне-
вой системы широкого круга растений. Первостепенный вреди-
тель огородных культур (рис. 91). Взрослые самки шаровидной
формы, с выдающимся головным концом, тело белое, дли-
ной 0,4—1,9 мм. Самцы с длинным прозрачным телом длиной до
1,2 мм, шириной 0,03—0,04 мм. Паразитирует в паренхиме кореш-
ков растений, вызывая разрастание клеток корней, что влечет
образование бесформенных наростов — галлов величиной до 3 см
в диаметре. Самки откладывают яйца в слизистый мешочек,
выступающий над поверхностью корня. Личинки мигрируют в поч-
ве, а затем внедряются в корешки растений. Борьба с этой не^
матодой заключается в проведении севооборотов с исключением
бахчевых и овощных культур на данном поле в течение 2—3 лет,
уничтожении ботвы и корневых систем растений после сбора уро-
жая, химической обработке полей.
Семейство Настоящие шишкоиглые нематоды (Tylenchidae).
Тело самцов и самок вытянутое, тонкое. Пшеничная нематода
(Anguina iritici) живет в тканях надземных частей пшеницы
(рис. 92). Самец и самка имеют веретеновидное прозрачное тело.
Длина самок до 5 мм, самцов до 2,5 мм. Пораженный этими
Рис. 92. Пшеничная нематода:
/ — самка; II -молодое растение пшеницы, пораженное нематодой; ///—галлы;
IV — здоровый колос пшеницы; К — колос, пораженный нематодой
125
паразитами колос пшеницы содежит вместо зерен овальные гаЛлы.
Сначала они имеют гладкую поверхность и зеленую окраску и
содержат внутри рыхлую полужидкую массу с большим числом
мелких молодых' личинок нематод. Позднее галлы становятся
твердыми и коричневыми.
Часть зрелых галлов выпадает из колосьев еще до уборки
хлебов, но большинство их отделяются от колосьев лишь при
обмолоте пшеницы, засоряя зерно. При высеве зараженного
зерна или при самостоятельном выпадении галлов из колосьев они
попадают в почву. Оболочки их во влажной земле размягчаются,
и подросшие личинки нематод выходят наружу. Когда появляют-
ся всходы пшеницы, личинки внедряются в них, концентрируясь
Рис. 93. Стеблевая картофельная нематода
126
в точках роста стеблей и в пазухах листьев. При формировании
колосьев они проникают в цветки, поражение которых влечет
образование вместо зерен галлов. В них личинки быстро превра-
щаются во взрослых, интенсивно размножающихся нематод —
одна самка может дать 2 тыс. яиц и более. После откладки
яиц взрослые черви погибают, а из яиц выходят личинки, достигаю-
щие к моменту выпадения галлов из колосьев примерно 1 мм. Один
галл содержит несколько тысяч личинок паразита. Они способны
впадать в состояние анабиоза (покоя), в котором могут пре-
бывать до 28 лет, пока галл не попадет во влажную почву. В
одном колосе может быть до 50 галлов. Пораженные нематодами
растения пшеницы отстают в росте, листья приобретают «гофриро-
ванную» форму, стебель искривляется, в колосьях вместо зерен
формируются галлы. Главная мера борьбы от пшеничной немато-
ды — предпосевная очистка зерна и смена полей под посевы.
В последние годы степень зараженности полей пшеничной немато-
дой в СССР резко сократилась.
Стеблевая картофельная нематода (Dityletichus destructor)
повреждает клубни картофеля (рис. 93). Самец и самка имеют
бесцветное тонкое тело длиной до 1,3 мм. В начале лета парази-
тируют в стеблях и листьях картофеля, задерживая рост.
Позднее мигрируют в клубни, вызывая их разрушение. Клубни
картофеля поражаются также червями, проникающими из почвы.
В стеблях и клубнях картофеля происходит интенсивное размно-
жение паразитов, которые за лето успевают дать несколько
поколений. Осенью часть нематод остается в клубнях, остальные
уходят в почву. У пораженных кустов картофеля стебли
укорочены и утолщены, листья мелкие, бледноокрашенные, клуб-
ни с глубокими впадинами серого цвета. Методы борьбы со сте-
блевой нематодой сводятся к использованию только здорового
семенного картофеля, сменой полей под картофель.
Луковая нематода (Ditylenchus аШ) — паразит лука и чеснока.
Черви с тонким телом длиной до 1,5 мм. Мигрируют в почве и
внедряются в подземные части лука и чеснока. Размножаются
в луковицах и листьях. Пораженные растения отстают в росте и
часто гибнут.
КРУГЛЫЕ ЧЕРВИ - ПАРАЗИТЫ ЖИВОТНЫХ И ЧЕЛОВЕКА
Значительное число круглых червей паразитируют в различних
сельскохозяйственных животных. Они вызывают серьезные забо-
левания и снижают их продуктивность,причиняя значительный
убыток животноводству нашей страны. Некоторые круглые черви
являются паразитами человека, нанося вред его здоровью.
Аскариды (различные виды сем. Ascaridae) (рис. 94) парази-
та pyiof в кишечнике лошадей, свиней и многих других сельскохо-
зяйственных и диких млекопитающих, а также человека. В каждом
из указанных хозяев живет свой вид аскарид. Аскариды имеют
127
Рис. 94. Внешний вид лошадиной аскариды:
/ — самка; II — самец
веретеновидное тело, у самцов хвостовой отдел за-
W гнут крючком (рис. 95). Длина лошадиной аскари-
ды (Paraskaris equorum) до 37 см, свиной (Ascaris
11 suum) — до 30 см, человеческой (Ascaris Lumbrc-
li| coides) — до 20 см при диаметре до 3—5 мм.
1д Самка значительно крупнее самцов. Аскариды
1Я очень плодовиты — самка лошадиной аскариды
// откладывает за сутки до 200 тыс. яиц. Оплодотво-
3 рение внутреннее. Оплодотворенное яйцо аскари-
f ды одето четырьмя оболочками, которые надежно
// защищают его от различных неблагоприятных
воздействий среды (рис. 96). Яйца аскарид оста-
д ются живыми в течение 1,5 мес при погружении в
3 %-ный раствор формалина и в продолжение ме-
сяца — в 2—4 %-ный раствор едкого натра или
едкого калия. Но 5—10 %-ный раствор лизола
быстро их убивает, что позволяет использовать этот
препарат для обеззараживания помещения от яиц
аскарид. С той же целью может быть использо-
ван кипяток, так как при температуре выше
60 °C яйца аскарид погибают.
Дробление яиц аскарид начинается еще в теле матери, но
в основном развитие личинки происходит во внешней среде в те-
чение 10—15 дней в зависимости от температуры среды. Когда в
яйце разовьется личинка, оно становится инвазионным. Прогло-
тив такое яйцо, животное или человек может заболеть аскаридо-
зом. В .кишечнике хозяина из яиц выходят микроскопические
малые личинки, которые внедряются в стенку кишки, попадают в
кровеносные сосуды и с током крови мигрируют по телу хозяина.
Попав в лёгкие, они выходят в просвет легочных пузырьков,
а оттуда через бронхи и дыхательное горло достигают ротовой
полости. Из нее вместе со слюной и пищей уже подросшие личин-
ки вторично попадают в кишечник того же хозяина, где и за-
канчивают свое развитие. Весь цикл развития личинок аскарид в
теле хозяина длится около 3 мес.
Аскариды причиняют вред людям и животным. В случае мас-
сового скопления этих паразитов в кишечнике хозяина они мо-
гут вызвать его закупорку и даже разрыв, что влечет смерть
хозяина. Продукты выделений аскарид ядовиты, поэтому у боль-
ных аскаридозом часто наблюдаются температура, нарушения рит-
ма сердца и другие симптомы отравления. Личинки аскарид, ми-
грируя из кишечника, открывают путь бактериям к внутренним
органам. Иногда наблюдается проникновение личинок аскарид в
128
Рис. 95. Внутреннее строение аскариды:
- рот; 2 ~~ пищевод; 3 — кишка; 4 — окологлоточное нервное кольцо; 5 - брюшной
нервный тяж; 6 — боковой валик гиподермы с каналом выделительной системы; 7-
фагоцитарные клетки; 3 - яичники; 9 - яйцеводы; 10 матка; И влагалище; 12 -
анальное отверстие; 13- семенник; 14 — семяпровод; /5 — семяизвергательный капал
Рис. 96. Яйца различных круглых червей — паразитов животных:
1 —острицы; 2— власоглава; 3, 4— аскариды; 5 — анкилостомы (свайника)
5 Б. А. Кузнецов и др.
желчные протоки печени и поджелудочную железу, что влечет пов-
реждение этих органов. У людей бывают случаи кровохаркания,
вызываемого повреждением стенок легких мигрирующими личин-
ками аскарид. Аскаридозы людей, а также сельскохозяйственных
животных (особенно лошадей и свиней) имеют широкое рас-
пространение. В борьбе с ними наряду с дегельминтизацией
больных при помощи различных лекарственных препаратов важ-
но содержать помещение в чистоте, а людям — соблюдать пра-
вила личной гигиены.
Аскаридия (Ascaridia galli) — широко распространенный
паразит, живущий в кишечнике кур. По внешнему виду похожа на
аскариду. Длина самок до 11 см, самцов до 7 см. Яйца аскаридий
становятся инвазионными через 7—20 дней после выхода из тела
хозяина. Заражение кур и цыплят происходит при поедании с
загрязненным кормом инвазионных яиц аскаридий. Освобожда-
ющиеся в желудке хозяина из оболочек яиц личинки в течение
1—2 ч остаются в кишечнике, а затем внедряются в его стенки.
Спустя 17—19 дней они вновь выходят в просвет кишки, где и до-
стигают зрелости. Весь цикл развития длится 35—58 дней. Следо-
вательно, миграции личинок по телу хозяина, столь характерные
для личинок аскарид, не происходят.
Аскаридиоз кур особенно распространен среди цыплят, вы-
зывая их массовую гибель.
Острицы (различные виды сем. Oxyurldae) — мелкие паразиты
кишечника позвоночных животных и человека (рис. 97). Остриц
узнают по тонкому заостренному хвостовому концу и наличию
луковицеобразного вздутия пищевода — бульбуса. Длина челове-
ческой острицы (Enterobius vertnicularis) менее 2 см, лошадиной
(Oxyura equi) —до 18 см за счет очень длинного хвоста.
Живут они в толстой кишке хозяина. Оплодотворенные сам-
ки спускаются к анальному отверстию хозяина и откладывают
здесь яйца, после чего выходят наружу и погибают. Яйца во
внешней среде дозревают и вскоре становятся инвазионными:
у человеческой — через 5—6 ч, у лошадиной — недели через 3.
Попав в кишечник хозяина, инвазионные яйца раскрываются, и
из их'оболочек выходят микроскопически малые личинки, которые
скоро достигают зрелости.
Свайники (Strongylidae) — довольно крупные черви длиной до
10 см, с нитевидным телом белого цвета. Самцы имеют на хвосто-
вом конце особое колоколообразноё образование — бурсу, способ-
ствующую спариванию (рис. 98). Один из представителей рода
Dictyocaulus viviparus вызывает опасное заболевание дыхатель-
ных путей скота. Длина самки до 57 мм, а самца до 44 мм. Ро-
товая капсула простая, невооруженная. Личинки развиваются в
теле матери, и выходящие наружу яйца содержат развитую
личинку.
Особенность развития этих свайников в том, что из яиц,
отхаркиваемых хозяином, выходят личинки, которые затем начи-
130
Рис. 98. Свайник (Ancylostonui)
двенадцатиперстной кишки:
1 - ротовая полость с зубами; 2 —- пи-
щевод; <3—кишка; 4 — бурса; 5 —
спикулы (щетинки)
Рис. 97. Острица человеческая:
1 — рот; 2 -г пищевод; 3 — бульбус; 4 —
кишечник; 5—яичник; 6—матка; 7—
хвостовой конец; 8 — анальное отверстие;
9 — женское половое отверстие; 10—го-
ловное вздутие
нают вести самостоятельную жизнь в почве, питаясь гниющими
веществами. Почвенная стадия развития личинок свайника длит-
ся около 1 недели'с. двумя линьками, после чего они выползают
из почвы на траву, вместе с которой и поедаются скотом. По-
пав в кишечник нового хозяина, они сначала проникают через стен-
ки кишки в лимфатические узлы, а оттуда по сосудам мигрируют
в легкие, где и заканчивают свое формирование. Для предупреж-
дения заболевания скота диктиокаулезом важно следить за чи-
стотой скотных дворов и овчарен, периодически дезинфициро-
вать стойла и инвентарь, не допускать загрязнения корма и
воды. Больной скот необходимо подвергать дегельминтизации.
К этому отряду принадлежат также широко распространенные
Strongylus eqainus — паразит преимущественно слепой кишки
лошадей и Syngamus trachea — паразит трахей птиц: кур, фаза-
нов, гусей и др. (рис. 99).
5* 131
Рис. 99. Стронгилиды в толстых кишках лошади
Рис. 100. Власоглав:
/ — самка; // — самец (передняя часть
его погружена в стенку кишки хозяина)
Рис. 101. Трихинелла:
/ — самка и самец; II —- личинки,
инкапсулированные в мышцах свиньи
Власоглавы (Tric/iocephalus, сем. Trichocepfialidae) живут в
кишечнике мелкого и крупного рогатого скота, свиней, кроликов и
многих диких зверей (рис. 100), иногда встречаются в кишках лю-
дей. Каждому виду хозяина свойственна своя форма паразита.
Длина червя до 5 см. Форма тела своеобразна: передняя часть
его тонкая, нитевидная, а задняя — утолщенная, мешкообразная.
Вдоль передней части тянется узкий пищевод, а в задней рас-
положены кишечник и половые органы. Тонким передним концом
паразит глубоко внедряется в стенку кишки хозяина, а задняя,
расширенная часть его тела свободно свисает в просвет кишечни-
ка. Яйца, в огромном количестве откладываемые самкой, вы-
водятся вместе с калом хозяина, где остаются живыми в течение
нескольких месяцев. Животные и люди заглатывают их с водой или
пищей. В кишечнике хозяина из яиц выходят личинки, которые
закрепляются в стенке кишки и примерно через месяц превра-
щаются во взрослого червя. Заражение власоглавами вызывает
воспаление кишечника и глубокие нарушения деятельности пище-
варительной системы. Наблюдается отравление хозяина выделе-
ниями паразита.
Трихинелла {Тrichinella spiralis, сем. Trichiucllidae) —- мелкие
черви с веретеновидным телом, заостренным на переднем конце
и закругленным сзади. Длина самок до 4 мм, самцов до 1,5 мм
(рис. 101). Половозрелые черви живут в кишечнике, а личинки —
в мышцах свиней, медведей, крыс и других животных, а также
человека. Все стадии жизни этого паразита проходят в теле хозяи-
на без выхода в наружную среду. Заражение человека трихинел-
лезом происходит при употреблении в пищу недостаточно про-
жаренного или проваренного мяса свиньи. Свиньи заражаются,
поедая дохлых крыс или же отбросы с боен, а крысы — пожирая
павших от трихинеллеза собратьев. В желудке личинки освобо-
ждаются и укрепляются на стенках кишок нового хозяина.
Они быстро растут и через 2—3 дня становятся половозрелыми.
После спаривания самцы погибают, а самки углубляются в стенку
кишечника и там спустя 9—10 дней рождают до 1500 подвижных
личинок, которые проникают в кровеносные сосуды, где живут,
вырастая с 10 до 120 мкм. В дальнейшем подросшие личинки
проникают в мышцы, и вокруг каждой из них постепенно
образуется обизвествленная овальная капсула. Внутри капсулы
личинка лежит, свернувшись спиралью, и питается тканевой
жидкостью хозяина. В мышцах людей они могут жить до 14 лет.
Число капсул с личинками трихинелл может достигать нескольких
десятков в поле зрения малого увеличения микроскопа. При за-
ражении трихинеллами люди испытывают большие страдания.
Свиньи, больные трехинеллезом, плохо растут и откармливаются.
С целью предупреждения заражения людей мясо забиваемых на
бойнях свиней исследуется трихиноскопом. Зараженные туши сви-
ней уничтожают.
133
Ришта (Dracunculus meilinetisis) имеет тонкое нитевидное тело
длиной до 100 см. Взрослые черви живут в подкожной клетчатке
рук и ног человека и вызывают язвы, через которые высовывается
наружу передний конец червя. Самка рождает большое количество
личинок. Промывая язвы у водоема, человек смывает личинок в
воду, где они внедряются в мелких рачков циклопов. Здесь и
происходит ряд стадий развития. Человек заражается, заглатывая
зараженных циклопов с водой. Следовательно, развитие ришты
происходит со сменой хозяина.
КЛАСС ВОЛОСАТИКИ (NEMATOMORPHA)
Первичнополостные черви с волосовидным телом, почти черного
цвета. Длина тела до 1,5 м при ширине 1 —1,5 мм. Полость тела
заполнена паренхимой. Раздельнополы. Развитие со сменой хозяев
(моллюски, насекомые). Вбуравливаться в тело хозяина не могут.
КЛАСС КОЛОВРАТКИ (ROTATORIA)
Черви микроскопических размеров, длиной не более 2 мм
(рис. 102). Покровы у одних мягкие и тело может сжиматься на-
подобие телескопической трубы, у других заключено в панцирь
с отверстиями для головного отдела и ноги. Нога оканчивается
обычно двумя пальцами. На голове ресничный орган. Он служит
для плавания и одновременно для добычи микроорганизмов, ко-
торые осаждаются на дне воронки, образуемой коловращением
ресничек. Глазков два. Имеются группы осязательных щетинок.
Нервная система — нервные ганглии и нервные волокна. Муску-
латура в основном состоит из поперечнополосатых волокон.
Органы пищеварения — рот, мускулистая глотка, ее расшире-
ние — зоб с хитиновыми челюстями; за глоткой — пищевод, объ-
емистый желудок, кишка, заканчивающаяся клоакой. Анус у осно-
вания ноги на спинной стороне. Органы выделения — протонефри-
дии. Они открываются в клоаку.
Раздельнополы. Самки имеют яичник, желточник и яйцевод,
который открывается в клоаку; самцы семенник, семяпровод,
предстательные железы и копулятивный орган с острым хитино-
вым стилетом, которым прокалываются покровы самки. Сперма
вносится в полость тела и уже из нее проникает в яичник. Опло-
дотворенные яйца заключены в скорлупу. Самцы обнаружены не у
всех. Если они имеются, то размножение происходит с чередо-
ванием партеногенетических поколений и полового. Партеногене-
тические поколения развиваются из яиц с диплоидным набором
хромосом и представлены только самками. После ряда поколений
самки дают яйца с гаплоидным набором хромосом. Из таких
яиц развиваются самки и самцы. Оплодотворенные самки от-
кладывают покоящиеся яйца. Состояние покоя длится от несколь-
ких дней и более года. Причины этого не выяснены.
134
1
Рис. 102. Коловратки:
А — сидячая коловратка Slephatioiwros fimbrial us: 1 головное щупальце; 2 — глотка
с жевательным аппаратом; 3 — желудок; 4 — задняя кишка; 5 — мочевой пузырь;
6' — студенистый домик;
Б—Brachionus (самка): / -—щетинки коловращательпого аппарата; 2 — чувствитель-
ные щетинки; 3 - реснички коловращатсльпого аппарата; 4 — спинное щупальце;
5 — мозговой ганглий; 6 — ретракторы переднего конца тела; 7 — глотка с жевательным
аппаратом; 8 — пищеварительные железы; 9 — панцирь; 10 — капал протонефридия;
// — концевая клетка протонефридия с «мерцательным пламенем»; 12 — боковой нерв
в чувствительном щупальце; 13 — яичник; 14 -- кишка; 15 - анус; 16 - ножные железы;
17 — их протоки; 18 — пальцы ноги; 19 — мочевой пузырь; 20 — яйцо; 21 --желудок;
22 — пищевод; 23 -- глазок
У сидячих форм из яиц сначала выходит личинка свобод-
ноплавающая. Подобное наблюдается и у сидячих животных
других типов. Такие «бродяжки» способствуют распространению
вида в пространстве.
В наших внутренних водах коловратки многочисленны. Они
играют большую роль в биоценозах водоемов. Коловратки —
корм для планктоноядных рыб и мальков всех видов рыб.
Потребление коловратками бактерий указывает на значение их
как санитаров.
135
Рис. 103. Брюхоресничный червь
Способность переходить в состояние
анабиоза характерна у живущих в почве,
во мху и у обитателей придонной зоны
водоемов. Коловратки переносят дли-
тельное высушивание и промораживание
в высушенном виде. Они сохраняют жиз-
неспособность при нагреве свыше 100 °C
и охлаждение до крайне низких темпера-
тур. Характерно, что планктонные виды не
выносят высушивание даже нескольких
часов. У них стойкостью обладают покоя-
щиеся яйца.
ПРОИСХОЖДЕНИЕ КРУГЛЫХ ЧЕРВЕЙ
Вопрос о происхождении круглых червей
неясен. Наибольшего внимания заслужи-
вает теория об их происхождении от мор-
ских ресничных плоских червей. Подтвер-
ждения этому можно найти в строении
примитивных круглых червей, относящих-
ся к классу Брюхоресничных (Gastrol-
riclia, рис. 103), которые обитают па дне
водоемов. Они совмещают признаки
круглых и ресничных червей. Их округ-
лое в сечении тело покрыто на брюшной
поверхности ресничками. Кожно-мус-
кульный мешок имеет только продольные
мышечные волокна. Первичная полость
тела, как у всех круглых червей, выражена хорошо. Органами вы-
деления служат протонефридии, как и у ресничных червей. Воз-
можно, что брюхоресничные черви произошли от ресничных. Воз-
можно и то, что гастротрихи и нематоды имели общих предков.
ТИП СКРЕБНИ (ACANTHOCEPHALES)
КЛАСС СКРЕБНИ (ACANTHOCEPHALA)
Паразитические черви с цилиндрическим телом, на переднем
конце которого имеется втяжной хоботок, усаженный рядами ост-
рых направленных крючочков (рис. 104). Этим хоботком скре-
бни внедряются в стенки кишечника хозяина, удерживаясь в его
просвете. В кожно-мускульном мешке скребней мышечные волок-
на располагаются двумя слоями: в наружном они кольцевые, а во
136
Рис. 104. Скребни:
/ - великан; II - четковпдпый
внутреннем — продольные.
Кишечника нет, и пища по-
ступает в организм червя ос-
мотически через покровы.
Скребни раздельнополы. Раз-
витие личинок происходит
обычно со сменою хозяев.
Живут в кишечнике различ-
ных животных. Класс вклю-
чает большое число видов,
из которых многие являются
и а разитами сел ьс ко х оз я й-
ственпых животных, вызы-
вая их тяжелые заболевания. Так, например, скребень-великан
(Масгасаnthorliyiichns liiriidiiiaccns), достигающий в длину 70 см,
наносит большой урон свиноводству, вызывая заболевание и
падеж свиней. Последние заражаются, роясь и поедая личинок
майских жуков, в которых живут личинки скребней.
ТИП КОЛЬЧАТЫЕ ЧЕРВИ (ANNELIDA)
Общая характеристика. Кольчатые черви — двусторонпесимме-
тричпыс животные метамерного строения со вторичной полостью
тела — целомом. Метамерия кольчатых червей выражается в том,
что их тело снаружи расчленено на ряд сходных сегментов и из
сегмента в сегмент наблюдается повторность многих органов.
Кольчатые черви обладают кровеносной системой, состоящей из
сосудов, пронизывающих тело. Нервная система слагается из го-
ловных (надглоточного и подглоточного) нервных узлов и цепи
парных нервных ганглиев, соединенных продольными тяжами; от
нервных узлов отходят нервы ко всем органам. Эта нервная це-
почка расположена на брюшной стороне животного. Органами вы-
деления служат метанефридии.
Известно около 8 тыс. видов кольчатых червей, которые оби-
тают в морях, пресных водоемах и почве. Многие из них имеют
большое практическое значение: дождевые черви и энхитреиды
активно участвуют в почвообразовательных процессах, многие ви-
ды служат кормом промысловых рыб, некоторые пиявки использу-
ются в медицине.
Кольчатые черви интересны в теоретическом отношении, пос-
кольку у них некоторые органы, свойственные и высшим вторично-
полостным животным, имеют примитивные черты строения, что по-
137
могает выяснить путь эволюции многих групп более высокоор-
ганизованных животных.
Тип Кольчатые черви включает три класса: Многощетинковые
черви (Polychaeta), Малощетинковые черви (Oligochaeta),
Пиявки (Hirudinea').
Строение и жизненные отправления. Форма тела кольчатых
червей обычно вытянутая, слегка уплощенная. Тело разделено
на ряд более или менее сходных сегментов. Они имеют вид ко-
лец, соединенных тонкой кожей, что обеспечивает большую гиб-
кость тела. Сегментация тела у многощетинковых морских чер-
вей подчеркивается наличием на каждом сегменте особых парных
выростов-параподий, играющих роль органов передвижения при
ползании и плавании. Иногда у них сегментарно расположены
также жабры и другие придатки. У многих многощетинковых чер^
вей хорошо диффренцирована голова с щупальцами и глазами.
Размеры кольчатых червей колеблются от нескольких миллиметров
до 1 м и более в длину. Кожно-мускульный мешок образован кож-
ным эпителием и двумя лежащими под ним слоями мышечных
волокон.
Полость тела. Вторичная полость тела — целом — обра-
зуется в процессе эмбрионального развития червя как полость
внутри закладок третьего зародышевого листка — мезодермы.
Стенки целома выстланы эндотелием мезодермального происхо-
ждения. Он наполнен целомической жидкостью. В типичном
случае целом у кольчатых червей разделен посегментно попе-
речными перегородками на камеры. Но у многих посегментное
деление целома на камеры нарушено. Образование вторичной
полости тела у кольчатых червей имеет свои особенности.
В качестве примера ее развития можно проследить образование
и формирование целома у трохофор личинок некоторых’ морских
многощетинковых червей. Такое название им дано за наличие двух
поясков ресничек, опоясывающих тело. На определенной стадии
развития трохофоры ее тело образовано двумя зародышевыми
листками — эктодермой и эндодермой, между которыми залегает
первичная полость тела. Затем близ заднего конца тела от эндо-
дермы отделяются две большие клетки — телобласты. В дальней-
шем путем многократного деления из них образуются две полоски
клеток мезодермы, которые располагаются вдоль кишечника меж-
ду экто- и эндодермой. Затем клетки мезодермальных полосок де-
лятся и образуют ряд отдельных групп клеток, попарно лежащих
по бокам кишки. Внутри каждой из них путем расхождения клеток
возникают просветы — зачатки вторичной полости тела. На каж-
дый сегмент тела приходится одна пара целомических мешков.
Увеличиваясь в размерах, они вытесняют первичную полость те-
ла. Соответственно увеличивается и полость целомических меш-
ков — вторичная полость тела. Разрастание целомических мешков
приводит к смыканию их друг с другом. Из их стенок образуются
перегородки, разделяющие целом на камеры. Смыкаясь над кищеч-
138
пиком и под ним, целомические мешки левой и правой сторон
образуют спинную и брюшную продольные брыжейки, фиксирую-
щие положение кишечника. При этом между левым и правым
листками брыжеек остаются продольные просветы, превращаю-
щиеся в спинной и брюшной кровеносные сосуды, которые
следует рассматривать как остатки первичной полости тела,
вытесненной целомом.
Покровы кольчатых червей образованы однослойным эпите-
лием с тонкой кутикулой на поверхности. Кожа богата железами,
выделяющими слизь. Выделения кожных желез используются не-
которыми морскими червями при постройке домиков.
Мускулатура кольчатых червей входит в состав кожно-
мускульного мешка. В более тонком наружном слое мышечные
волокна кольцевые, а в более мощном внутреннем — продоль-
ные. Благодаря такому строению мускулатуры черви могут из-
гибаться в различных направлениях, а также удлинять или уко-
рачивать свое тело. У некоторых форм между слоями попереч-
ных и продольных мышечных волокон залегает слой волокон
диагонального направления. Мускулы входят также в структуру
некоторых внутренних органов.
Нервная система. В голове от пары крупных надгло-
точных ганглиев спускаются по обе стороны глотки два нервных тя-
жа, идущих к менее крупным парным подглоточным узлам.
Надглоточные ганглии, боковые тяжи и подглоточные нервные уз-
лы образуют окологлоточное нервное кольцо. За подглоточными
узлами следует брюшная нервная цепочка с посегментно рас-
положенными парами нервных узлов, соединенных поперечными
и продольными комиссурами. От ганглиев отходят нервы к раз-
личным органам. У многих кольчатых червей пары ганглиев в
каждом сегменте сближены в один нервный узел.
Органы чувств развиты у кольчатых червей в различной
степени. У почвенных дождевых червей глаза и щупальца от-
сутствуют, но у них в коже заложены многочисленные чувствую-
щие клетки и нервные окончания. У морских многощетинковых
червей, живущих на дне или плавающих в толще воды, хорошо
развиты глаза и щупальца.
Пищеварительная система начинается ротовым и за-
капчивается анальным отверстием. Кишечник состоит из трех от-
делов: переднего и заднего эктодермального происхождения и
среднего, образованного эндодермой. Строение отделов имеет
отличия у разных’представителей типа.
Органы’ дыхания. Многие кольчатые черви дышат через
покровы тела. Но у морских и некоторых пресноводных видов есть
жабры — тонкостенные выросты покровов различной формы, рас-
положенные метамерно по бокам сегментов тела на параподиях,
реже — пучком на голове или лишь на задних сегментах тела.
Кровеносная система у подавляющего большинства
кольчатых червей замкнутая, т. е. кровь у них движется по
139
Рис. 105. Строение метанефридия дождевого
червя:
I — воронка, открывающаяся в целом; 2 — меж-
сегментная перегородка; -- петлистый канал; 4 -
ресничный канал; 5—расширение канала; 6- же-
лезистый пузырек; 7 — мочевой пузырь; 8 - наруж-
ное моченое отверстие
сосудам, не выливаясь в полость тела.
Движение крови обусловливается пуль-
сацией сосудов, главным образом опоясы-
вающих пищевод. Кровяной поток обеспе-
чивает снабжение всех частей тела пита-
тельными веществами, всосанными стен-
ками кишечника, и кислородом, поступаю-
щим из внешней среды. Основные сосуды
системы — спинной, идущий над кишеч-
ником, и брюшной, тянущийся под ним.
По спинному сосуду кровь движется
1 к переднему, а по брюшному — в об-
ратном направлении. Оба сосуда посегментно соединены кольце-
выми сосудами, охватывающими кишечник. Из них выделяются
своей толщиной пульсирующие сосуды, окружающие пищевод,
называемые сердцами.
Органы выделения представлены расположенными по-
сегментно метанефридиями. Они имеют вид извитых трубочек
(рис. 105), начинающихся в полости тела воронкой с ресничками.
Внешний конец метанефридиев открывается либо непосредственно
наружу, либо в продольный общий выделительный канал. Ко-
нечный отдел метанефридия, или общего мочеточника, имеет рас-
ширение — мочевой пузырь.
Органы размножения. Одни кольчатые черви раздель-
нополы, другие — гермафродиты. Размножение, как правило, по-
ловое, но иногда образование новых особей происходит бесполым
путем — путем почкования или деления.
Развитие кольчатых червей прямое или с метаморфозом.
КЛАСС МНОГОЩЕТИНКОВЫЕ ЧЕРВИ (POLYCHAETA)
Общая характеристика. Многощетинковые черви — наиболее
древняя группа типа кольчатых червей, давшая начало другим
классам этого типа. Поводом к наименованию класса послужило
то, что у этих червей по бокам сегментов тела образовались
своеобразные органы — параподии, снабженные многочисленны-
ми щетинками.
Тело многощетинковых червей состоит из ряда внешне сход-
ных сегментов. Передние сегменты, сливаясь, образуют хорошо
выраженный головной отдел, на котором расположены рот и раз-
личные придатки, а также некоторые органы чувств. Животные
140
Рис. 106. Вилы многощетинковых кольчатых червей:
/ -пескожил (/Irezi/co/ti); // перепс (ЛДтеЫ); /// — филлодоце
(Phi/lodoce); IV .- амфитрпге (Д тpftitriteY, I'- сабелларпя (Sabellaria);
VI ~ еерпула (Ser/ш/п), VII морская мышь (Aphrodite)-,
I - щупальца; 2 — жабры; .'I .. параподпи
раздельнополые. Развиваются с метаморфозом (рис. 106).
Описано несколько тысяч видов. Имеют практическое значение
как корм рыб.
Строение и жизненные отправления. Длина тела многощетин-
ковых червей колеблется от нескольких миллиметров до 1 м
и более.
Органами движения служат параподии — парные бо-
ковые выросты, состоящие обычно из непарной основной части и
двух лопастей: спинной и брюшной (рис. 107). Каждая лопасть
141
содержит пучок упругих щетинок, а также обычно осязательный
усик. При недоразвитии спинной лопасти параподии становятся
одноветвистыми. Параподии используются червями при ползании
по дну водоема, а при плавании животного играют роль плавников.
У червей, роющихся в грунте или живущих в трубчатых
домиках, параподии полностью или частично редуцированы.
Покровы многощетинковых червей, ведущих активную
жизнь на дне водоема, отличаются хорошо развитой кутикулой.
Напротив, у червей, плавающих в толще воды, зарывающихся
в грунт или строящих трубчатые домики, кутикула очень тонкая.
Выделения покровов служат строительным и цементирующим ма-
териалом при постройке трубок, в которых живут некоторые много-
щетинковые черви.
Органы чувств у большинства могощетинковых червей
хорошо развиты. На голове (рис. 108) обычно имеются 1—2 пары
глаз, осязательные усики, щупальца и обонятельные ямки.
Органы дыхания —жабры. У некоторых они отсутст-
вуют, и дыхание происходит всей поверхностью тела.
Органы размножения. Многощетинковые черви обычно
раздельнополы, наружных отличий между полами нет. У отдель-
ных видов наблюдается партеногенез. Большинство откладывают
яйца, но встречаются и живородящие формы. Некоторые раз-
множаются почкованием, в результате которого могут образо-
вываться временные разветвленные колонии.
Развитие многощетинковых червей происходит с метамор-
фозом или без него. У большинства форм из яиц выходят микро-
Рис. 107. Параподий многощетинко-
вого кольчатого червя нереис:
/спинной усик; 2—спинная лопасть;
3 — брюшная лопасть; 4 — брюшной усик;
5 — щетинки; 6 — опорные щетинки
Рис. 108. Голова многощетинкового кольчатого червя нереис:
1 — щупальца; 2 — глаза; 3 — параподии
142
скопически малые плавающие4 личинки — трохофоры, имеющие
округлое несегментированное тело с поясками ресничек. Сначала
они имеют первичную полость тела, которая в процессе раз-
вития животного сменяется вторичной.
Многощетинковые черви населяют моря, обитая от мелководья
до больших глубин. Некоторые представители встречаются в пре-
сноводных бассейнах, например в оз. Байкал. Большинство видов
живут на дне, но некоторые обитают в толще воды. Одни
весьма подвижны и способны ползать по дну и плавать, другие
зарываются в грунт, третьи постоянно живут в трубкообразном
домике, построенном из извести или органических веществ, вы-
деляемых покровами червя.
У каждой из экологических групп многощетинковых червей
свои особенности организации, способы питания, защитные при-
способления. Обитающие в домиках обычно имеют недоразвитые
параподии, их головные щупальца, сливаясь, могут образовывать
крышечку, закрывающую вход в домик. Щупальца некоторых
сидячих видов приобретают перисторазветвленную форму; они
выполняют роль жабр и участвуют в добывании пищи.
Поверхность их покрыта ресничным эпителием, реснички которого
гонят воду со взвешенными в ней пищевыми частицами ко рту.
Для расселения таких прикрепленных червей служит свободно-
плавающая личинка.
У некоторых роющихся в грунте много щетинковых есть хо-
боток с зубчиками, помогающий им закапываться. Параподии
у них часто атрофированы, и короткие щетинки торчат прямо из те-
ла пучками.
Паразитические виды среди многощетинковых червей немно-
гочисленны. Встречается их симбиоз с другими животными:
раками, губками и пр.
Многие виды многощетинковых служат основной пищей для
промысловых морских рыб. Поэтому распространение и числен-
ность их учитывается при оценке биологической продуктивности
водоемов и разведке запасов промысловых рыб. Из кольчатых
червей, которые являются кормом для рыб, важны живущие на
мелководье нереиды (Nereis). По предложению проф. Л. А. Зен-
кевича в 1939—1941 гг. они были переселены из Азовского
моря в Каспийское, где ранее не водились. Нереиды хорошо при-
жились в новых условиях и ныне стали в Каспийском море цен-
ным кормом для осетровых рыб.
Своеобразный многощетинковый червь пескожил (Aretiicola
marina) в огромном количестве населяет песчаные отмели. Он
живет в заиленном песке, пропуская его через кишечник и
переваривая содержащиеся в нем органические вещества.
У тихоокеанского червя палоло (Eunice viridis) в период раз-
множения сегменты задней части тела, наполненные половыми
продуктами, отрываются и всплывают на поверхность моря.
Путем разрыва стенок сегментов содержащиеся в них яйца или
143
сперматозоиды выходят в воду, где и происходит их соеди-
нение. Из зигот развиваются плавающие личинки, а из них —
взрослые черви, которые опускаются на дно. Подобное размноже-
ние способствует расселению малоподвижных червей. В период
массового всплытия местное население добывает их в большом ко-
личестве и использует как продукт питания.
КЛАСС МАЛОЩЕТИНКОВЫЕ ЧЕРВИ (OLIGOCHAETA)
Общая характеристика. К этому классу принадлежат дожде-
вые черви и многие другие почвенные и водные формы. Для них
характерны отсутствие параподий и малое число щетинок, кото-
рые обычно сидят пучками по бокам сегментов (кроме передних и
задних). Головной отдел тела не обособлен. Щупалец у большин-
ства форм нет. Гермафродиты. Развитие происходит без метамор-
фоза.
Известно около 3 тыс. видов этого класса. Обитают преи-
мущественно в почве и на дне пресных водоемов. Почвенные
дождевые черви играют большую роль в почвообразовании
(рис. 109).
Строение и жизненные отправления. Тело малощетинковых
червей сильно вытянуто, цилиндрическое и состоит из разного
числа внешне сходных сегментов. На переднем конце
тела расположен рот, на заднем — анальное отвер-
zgk стие-
Размеры тела колеблются от нескольких мил-
диметров до 3 м (у некоторых тропических форм).
2—Покровы содержат большое количество кожных
;|Ц желез, выделяющих слизь. Каждый сегмент снабжен
|Ц[ щетинками, собранными в пучки.
У почвенных форм они играют большую роль при
передвижении червей.
ЕЕд Полость тела вторичная, хорошо развитая, запол-
ненная целомической жидкостью с клеточными элемен-
Лр тами (лимфоциты и др.). У большинства видов
она разделена посегментно на отдельные камеры
(рис. 110, 111).
Нервная система представлена надглоточ-
|||| ным нервным узлом, окологлоточным кольцом, подгло-
точным ганглием и нервной брюшной цепочкой.
Wa Органы чувств развиты слабо. Глаза и щу-
уЦа пальца у большинства форм отсутствуют. Имеются чув-
ствующие щетинки, обонятельные ямки и статоциты.
Рис. 109. Дождевой червь:
/ — женское половое отверстие; 2 — мужское половое отверстие; ,? -
поясок
144
Рис. 110. Анатомия дождевого червя:
/ ротовая лопасть; 2 надглоточный нервный узел;
3 - глотка; 4 - пищевод; 5. /.? - кольцевые сосуды;
6 спинной сосуд; 7 семенные мешки; Л' -семенники;
9 - семенные воронки; 10 семяпровод; II перего-
родки между сегментами; 12 - метанефридип; 14 — киш-
ка; 15 - желудок; Hi - зоб; 17 яйцевод; 18 - яйце-
вые воронки; 19 яичник; 20 семяприемники. Рим-
скими цифрами обозначены сегменты тела
П п 1.ц с в а р и т е л ь и ы е о р г а и ы
обычно велики и приспособлены к про-
хождению больших масс почвы и донного
грунта, которыми большинство червей
питаются. За ротовой полостью следуют
глотка, пищевод, мускульный желудок
и кишечник.
Все эти органы лежат вдоль тела,
не образуя изгибов.
К р о в с и о с п а я система. Глав-
ные сосуды спинной и брюшной. В покро-
вах густая сеть капилляров, из которых
окисленная кровь собирается в субнев-
ральный сосуд, лежащий под брюшной
нервной цепочкой.
Органы д ы х а и и я, за редкими
ис кл юче н и я м и, отсутству ют.
Кожа малощетинковых червей прони-
зана сетью капилляров, облегчающих га-
зообмен через покровы (рис. 112).
Половая с и с т е м а. Все малоще
фродиты, но им свойственно перекрестное оплодотворение. Строе-
ние гонад отличается у представителей разных групп.
Из большого числа малощетинковых червей для сельского
хозяйства особый интерес представляют различные виды дожде-
вых червей. Тело их слегка уплощенное и состоит из 50—248 внеш-
не сходных сегментов. На границе передней и средней третей тела-, ,
ясно выделяется поясок из нескольких утолщенных сегментов. ‘
Дождевые черви предпочитают жить в умеренно влажной поч-”.
ве, богатой гумусом. Кислых и засоленных почв они избегают.
Зиму проводят обычно в грунте на глубине 2—3 м (в условиях
умеренного пояса). Питаются органическими веществами, содер-
жащимися в почве, а также отмершими частями растений. -
Размножаются путем перекрестного оплодотворения. Гермаф-
родитные половые органы дождевых червей отличаются слож-
ностью строения. Яичников одна пара. Около них воронкой,
отороченной ресничками, начинаются короткие яйцеводы, другим
концом открывающиеся наружу в следующем сегменте. Семсн-
‘типковые черви герма-
145
3
Рис. 111. Поперечный разрез тела дождевого червя:
1 — метанефридий; 2 — воронка метанефридия; 3 — ганглий
брюшной нервной цепочки; 4 — кожный эпителий; 5 — попе-
речные мышцы; 6 — продольные мышцы; 7 — щетинки; 8 —
спинная складка кишки; 9, 10 — спинной и брюшной кровенос-
ные сосуды; 11 — субневральный сосуд
ников две' пары. Они находятся внутри больших семенных
мешков, в которых происходит накопление семенной жидкости.
К семенникам прилегают воронки семяпроводов, открывающихся
наружу позади отверстий яйцеводов. Обычно в 9—10-м сегментах
находятся две пары небольших мешочков — семяприемников, ко-
торые открываются наружу короткими протоками. При спаривании
два дождевых червя прикладываются брюшными сторонами.
При этом их головы обращены в противоположные стороны, а
отверстия спермопроводов одной особи прилегают к спермоприем-
нику другой. После этого происходит взаимный обмен семенной
жидкостью. Затем спаривавшиеся черви расходятся.
Когда созревают яйца, на пояске червя выделяется слизистое
кольцо. Кольцо червь сбрасывает через передний отдел тела; когда
оно проходит мимо отверстий яйцеводов и семяприемников, в
Рис. 112. Поперечный разрез стенки кожно-
мускульного мешка дождевого червя:
1 — кожный эпителий; 2 ~~ слой кольцевых мышц;
3 — слой продольных мышц; 4 — кровеносные
сосуды и капилляры
4
146
Рис. 113. Почвенные малощетинко-
вые кольчатые черви (энхитреиды)
в момент спаривания
Рис. 114. Пресноводный малоще-
тинковый кольчатый червь трубоч-
ник (Tubtfex)
него поступают яйца и спер-
матозоиды и происходит оп-
лодотворение яиц. Сброшен-
ное кольцо с заключенными
в нем яйцами затвердевает,
превращаясь в кокон. В нем
и происходит первоначаль-
ное развитие молодых чер-
вей, которое идет без мета-
морфоза.
Дождевые черви играют
огромную роль в почвообра-
зовательных процессах.
Ходы червей способству-
ют проникновению в почву
воды и воздуха, чем дости-
гаются важное для успеш-
ного роста растений равно-
мерное увлажнение и венти-
ляция почвы. Черви разрых-
ляют почву и удобряют ее,
затаскивая в норки расти-
тельные остатки, способству-
ющие образованию гумуса.
Численность дождевых чер
вей в почве иногда бывает огро-
мной, достигая 5 млн особей на 1 га, что по массе составляет
около 1 тыс. кг. Если в почве на площади 1 м2 обитают 50—100 до-
ждевых червей, то они выбрасывают на поверхность 1 га от 10
до 30 т земли, прошедшей за год через их кишечник. Отсюда оче-
видно огромное значение дождевых червей для создания пло-
дородия почвы.
В почвообразовательных процессах деятельное участие прини-
мают также мелкие малощетинковые черви из семейства Enchy-
treidae (рис. 113). Длина их обычно не превышает 1 см, а
толщина — 1 мм. Нередко в почвенном слое на площади 1 м2 мож-
но обнаружить десятки тысяч различных представителей этого
семейства. Особенно много их бывает около гниющих остатков
растений и животных.. Многие энхитреиды живут на дне пресных
и солоноватых водоемов. Их жизнь здесь проходит в иле, в почти
147
бескислородной среде. Во время заморов они впадают в анабиоз.
Обычные в озерах и прудах мелкие малощетинковые черви
трубочники (сем. TibificUlae) (рис. 114). Черви, зарывшись задним
концом в ил, совершают своим телом колебательные движения.
Замечено, что чем меньше в воде кислорода, тем сильнее они
вытягивают тело и чаще совершают колебательные движения,
усиливая газообмен через покровы.
У ряда водных малощетинковых червей наблюдается бесполое
размножение почкованием. Иногда образуются целые цепи поч-
кующихся червей. Водные малощетинковые черви служат важ-
ным кормом рыб. Они способствуют ускорению круговорота
веществ в грунте водоемов.
КЛАСС ПИЯВКИ (HIRUDINEA)
Общая характеристика. Пиявки — своеобразные кольчатые черви,
большинство которых являются эктопаразитами различных жи-
вотных (рис. 115). Тело обычно уплощенное, без ясно выражен-
ного головного отдела. Наружная кольчатость очень мелкая и не
соответствует более крупной внутренней сегментации тела. Пара-
подий нет. Шетинки обычно отсутствуют. У большинства видов
имеются присоски. Поло£ть тела частично редуцирована. Герма-
фродиты.
Всего известно около 250 видов пиявок, подавляющее боль-
шинство которых живут в пресных водоемах.
Строение и жизненные отправления. В организации пиявок
много черт приспособления к полупаразитическому образу жизни,
который ведет большая часть представителей этого класса
(рис. 116).
Покровы. Тело одето снаружи кутикулой. Лежащий под
ней эпителий богат железистыми слизистыми клетками, а у осио-
ш
и
148
Рис. 116. Анатомия медицинской пиявки:
/ - надглоточный нервный узел; 2 глотка; ,3 --
пищевод; 4 - желудок; 5 — задний вырост желуд-
ка; tf — средняя кишка; 7- задняя кишка; Л' —
анальное отверстие; 9 задняя присоска; К) --
ганглии брюшной нервной цепочки; 11 - мета-
нефридни; 12 — мочевой пузырь; 13 -- семенные
мешки; 14 - семяпроводы; /,5 — воронки мета-
нефридиев; 16 - влагалище; 17 яичники; 18
придаток семенника; /9 - копулятивный орган;
20 - предстательная железа; 21 боковая лакуна
вания эпителиальных клеток рассея-
ны многочисленные пигментные
клетки, обусловливающие окраску
пиявок.
Мускулатура развита очень
сильно. В кожно-мускульном мешке
три слоя мышечных волокон, кото-
рые тянутся в поперечном, диаго-
нальном и продольном направлениях
к оси тела.
Полость тела сильно реду-
цирована и имеет вид системы лакун.
Нервная система. Есть
брюшная нервная цепочка.
Органы чувств. Глаза, если
имеются, отличаются примитивным
строением.
В покровах чувствующие клет-
ки и нервные окончания.
Пищеварительная сис-
тема песет в себе черты приспособ-
ления к паразитическому образу
жизни.
Рот ведет в ротовую полость, в
которой у одних видов (например, у
медицинской пиявки) расположены
три челюсти, снабженные множеством зубчиков (челюстные
пиявки), у других имеется хоботок, которым они внедряются в по-
кровы жертвы (хоботные пиявки).
Ротовая полость ведет в глотку, играющую роль сосательного
аппарата. В глотку открываются одноклеточные слюнные железы.
У медицинских пиявок слюнные железы выделяют особое ве-
щество — гирудин, обладающий свойством препятствовать свер-
тыванию крови.
Передний отдел кишечного тракта имеет несколько пар кар-
манообразных боковых выростов, увеличивающих их объем, что
149
позволяет делать большой запас крови, которого хватает меди-
цинской пиявке на 2—3 мес. Благодаря примеси гирудина
кровь пиявок не свертывается и долго сохраняется в свежем ви-
де. Переваривание происходит в эндодермальном отделе кишеч-
ника.
Дыхание у большинства видов происходит через покровы
тела, но у некоторых видов есть жабры.
Органами выделения служат метанефридии.
Половая система. Пиявки — гермафродиты. Спаривание
медицинских пиявок происходит весной близ водоема в сырой зем-
ле выше уровня воды. Крупные коконы их напоминают желуди.
Они образуются к концу июня. Развитие пиявок в коконе длится
около 5 недель. Половой зрелости пиявки достигают к 5 годам. Жи-
вут до 20 лет.
Некоторые пиявки питаются кровью определенных животных.
Так, виды рода Protoclepsis паразитируют на водоплавающих пти-
цах, представители родов Acatilhobdella и Ichthyobdella на-
падают на рыб и т. д.
Практический интерес представляет медицинская пиявка,
используемая для лечения больных людей.
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
Общая характеристика. Наиболее богатый видами и разнооб-
разный тип животных. К нему относятся более 80 % известных
науке представителей животного мира: различные насекомые, пау-
ки, скорпионы, многоножки, раки.
Членистоногие — двустороннесимметричные животные с сег-
ментированным телом и членистыми конечностями. Их тело покры-
то прочной хитиновой кутикулой, служащей для защиты организма
и являющейся наружным скелетом. Полость тела смешанная —
она образуется слиянием первичной и вторичной полостей тела в
процессе эмбрионального развития. Центральная нервная система
в основном сходна с таковой кольчатых червей. Органами
дыхания у разных представителей служат жабры, легкие или
трахеи. Кровеносная система незамкнутая. Органы выделения —
либо видоизмененные метанефридии, либо специфические образо-
вания — мальпигиевые трубочки.
Членистоногие — группа высокоорганизованных животных,
играющая важную роль в жизни природы. Они встречаются на су-
ше и в морях, населяя самые различные места. Большинство чле-
нистоногих — свободноживущие организмы, однако среди них
встречаются экто- и эндопаразиты растений и животных. Членисто-
ногие имеют большое значение как для хозяйства человека, так и
здравоохранения.
Тип Членистоногие объединяют три подтипа: Жабернодыша-
щие (Branchiata), Хелицероносные (Chelicerata), Трахейноды-
шащие (Тгас heat а).
150
Рис. 117. Различная степень слияния сегментов тела у членистоногих:
/ - многоножка; II насекомое; /// клещ
Строение и жизненные отправления. Форма тела члени-
стоногих разнообразна. Тело слагается из ряда следующих друг
за другом сегментов (рис. 117). У некоторых членистоногих
(например, многоножек) членики сходны, но у большинства
сегменты различны, что позволяет выделять несколько отделов
тела — голову, грудь и брюшко. Передние членики, сливаясь,
образуют голову, у многих наблюдается
слияние сегментов других отделов тела.
Так, у пауков членики сливаются в пе-
расчленеппые головогрудь и брюшко, а
у многих клещей сегментация тела совсем
не выражена.
У членистоногих в типичном случае
каждый членик тела песет по одной паре
членистых конечностей (рис. 118). Но у
большинства представителей конечности
па отдельных сегментах пли отделах тела
атрофированы или превращены в различ-
ные специфические органы (ротовые ор-
га н ы, я й i io к л а д ы и др.).
Рис. 118. Конечности членистоногих:
а двухветвистая ножка ракообразного; б - однопет-
вистая ножка насекомого
151
Покровы членистоногих представлены гиподермой, образо-
ванной однослойным эпителием. Клетки гиподермы выделяют на
наружной поверхности кутикулу сложного химического состава,
содержащую стойкое вещество хитин. У ряда членистоногих
(у высших раков) покровы содержат углекислую известь. На
сегментах тела и члениках конечностей хитиновая кутикула
заметно утолщена, а в местах их соединения остается тонкой и гиб-
кой, что обусловливает подвижность частей тела. Рост членистоно-
гих сопровождается линькой. Хитиновый покров Препятствует
увеличению размеров тела, поэтому рост животного происходит
в тот период, пока новая кутикула мягка. Твердый и прочный по-
кров не только защищает членистоногих от неблагоприятных воз-
действий внешней среды, но и служит наружным скелетом, к ко-
торому прикрепляются мускулы.
Мускулатура членистоногих представлена отдельными
мышечными пучками — мышцами, прикрепляющимися к наруж-
ному покрову. Кожно-мускульного мешка, столь характерного
для червей, у членистоногих (за исключением некоторых прими-
тивных форм) нет. Мышцы, приводящие в движение части тела,
имеют поперечнополосатую структуру, а мышцы внутренних орга-
нов слагаются из гладких волокон.
Полость тела членистоногих смешанная. В процессе раз-
вития зародыша она претерпевает сложное развитие. У эмбрионов
сначала имеется первичная полость тела, затем по бокам пер-
вичного кишечника образуются парные выросты мезодермы с по-
Рис. 119. Нервная система таракана:
/ -надглоточный 1апглий; 2 подглоточный ганглий, обра-
зующий е надглоточным окологлоточное нервное кольцо;
;i—5 - грудные нервные ганглии; 6—11 — брюшные нервные
ганглии
Рис. 120. Сложный глаз на-
секомого:
/ — роговина; 2 — хрустальный
конус; -- светочувствительные
элементы глаза; 4 — пигменти-
рованные прослойки между ом-
матидиями; 5 —- зрительный
нерв
152
лостями внутри. Впоследствии стоики выростов частично нару-
шаются и находящиеся в них зачатки вторичной полости сли-
ваются с первичной полостью тела.
Н е р в н а я с и с т е м а членистоногих во многом сходна с
таковой кольчатых червей (рис. 119). В голове, над глоткой, нахо-
дится крупный парный надглоточный нервный узел. От него
отходят вниз два нервных тяжа, охватывающих с боков глотку
и соединяющихся под ней с парным подглоточным нервным
узлом, образуя окологлоточное кольцо. От подглоточного ганглия
начинается брюшная нервная цепочка, образованная расположен-
ными в каждом членике парой ганглиев, связанных между со-
бою продольными комиссурами с другими ганглиями цепочки.
От ганглиев отходят нервы к различным органам. У членистоногих,
сегменты которых слились, обычно бывают слиты и нервные узлы.
О р г а н ы ч у в с т в членистоногих нередко отличаются слож-
ностью строения и функций. У большинства имеются глаза, прос-
тые или сложные либо те и другие. Простые глаза имеют вид
бокалов, устье которых закрыто хрусталиком, направляющим луч
света в глубь глаза, а оно выстлано светочувствительными
Клетками, к которым подходит нерв, ("ложные глаза состоят из
множества отдельных глазков — омматидиев, которые имеют ци-
линдрическую 'или коническую форму (рис. 120). Наружную
часть омматидиев составляет прозрачная роговица, под которой
лежит хрустальный- конус, направляющий луч света в глубь глаз-
ка. С боков омматидии одеты пигментными клетками, а в глубине
их расположены светочувствительные клеточные образования.
Сложные глаза позволяют различать не только форму пред-
метов, но и цвет. У членистоногих также хорошо развиты органы
осязания и обоняния.
Органы пищеварения начинаются расположенным на
голове ртом, у отверстия которого находятся видоизмененные
конечности ~ ротовые органы, помогающие добывать, раз-
мельчать и заглатывать пищу. Форма и строение ротовых ор-
ганов разнообразны в зависимости от способа питания и характера
нищи животного. Пищеварительный канал, заканчивающийся зад-
непроходным отверстием, распадается на передний, средний и
задний отделы. Передний и задний отделы образуются впячива-
нием эктодермы и покрыты изнутри хитином. Переваривание
и усвоение пищи происходят в основном в среднем отделе
кишечника, имеющем эндодермальное происхождение. Он обра-
зует различные выросты (печеночные, пилорические и др.), кото-
рые имеют секреторную функцию и увеличивают всасывающую
поверхность пищеварительного тракта.
Органы д ы х а п и я большинства водных членистоногих —
жабры, у сухопутных и некоторых водных членистоногих органами
дыхания служат легкие и трахеи. Легкие имеют вид тонкостенных
мешков, открывающихся наружу, внутри которых расположены
многочисленные тонкие листочки; через покровы их происходит
153
Рис. 121. Трахеи насекомых:
/ — участок трахеи; // . микроскопическое строение трахеи
газообмен. Трахеи — ветвящиеся трубочки, открываются наружу
рядом отверстий — дыхалец (рйс. 121). Внутри они имеют спи-
ральную хитиновую нить, не позволяющую им спадаться. Конеч-
ные разветвления трахеи лише,ны.хитиновой спирали, они проника-
ют во все органы животного и приносят к тканям кислород воздуха.
У некоторых мелких членистоногих дыхание происходит осмоти-
чески через кожу.
Кровеносная система членистоногих незамкнутая, что
связано со смешанной'полостью тела. От пульсирующего сердца
(или ряда следующих друг за другом его камер), расположенного
на спинной стороне животного, к различным частям тела отходят
кровеносные сосуды, ио обратный ток крови к сердцу проходит
не по сосудам, а по лакунам — промежуткам между органами.
Кровь в сердце засасывается из полости тела через отверстия в
его стенках. У некоторых членистоногих кровеносная система от-
сутствует или представлена лишь сердцем. Кровь окрашенная или
бесцветная. У трахейнодышащих видов функции ее ограничены в
основном доставкой всем органам питательных веществ от ки-
шечника.
Органы выделения у ракообразных представлены ви-
доизмененными метанефридиями, расположенными в головном от-
деле животного. У насекомых и многоножек органами выделения
154
служат специфические образования — мальпигиевы трубочки —
тонкие слепые отростки кишечника. Через их стенки происходят
фильтрация излишков воды и удаление продуктов распада
тканей организма. У паукообразных имеются мальпигиевы тру-
бочки и измененные метанефридии. В обмене веществ многих чле-
нистоногих значительную роль играет жировое тело, лежащее в
полости тела. Ойо образовано скоплением крупных клеток, в плаз-
ме которых при обильном питании образуются капельки жира, слу-
жащего запасным энергетическим ресурсом. В клетках жирового
тела в течение жизни животного накапливаются отложения мо-
чевой кислоты, что указывает на выполнение ими выделительной
функции.
Органы размножения. Подавляющее большинство
членистоногих раздельнополы, лишь некоторые паразитические и
сидячие формы — гермафродиты. У отдельных видов наблюдается
чередование поколений: ряд партеногенетических генераций сме-
няется половой. При партеногенетическом размножении живот-
ные развиваются из неоплодотворенных яиц. У многих наблю-
дается резко выраженный половой диморфизм. Половые органы у
самок представлены парными (иногда сливающимися) яичниками,
яйцеводами, непарным влагалищем и спермоприемником, куда
поступает сперма при спаривании. Половые органы самца обычно
слагаются из парных семенников, двух спермопроводов, пред-
стательной железы, которая дает жидкий секрет, разбавляющий
и активизирующий сперму, и разного рода приспособлений для
передачи спермы самкам. Оплодотворение происходит внутри тела
самки.
Развитие членистоногих протекает прямым путем или путем
сложных превращений (с метаморфозом). При прямом развитии
из яйца выходит молодое животное, сходное со взрослой особью.
При развитии с метаморфозом животное проходит личиночную
стадию. Личинки обычно отличаются по морфологии и образу
жизни от взрослых особей.
ПОДТИП ЖАБЕРНОДЫШАЩИЕ (BRANCHIATA)
К этому подтипу из ныне живущих членистоногих принадле-
жит только один класс — Ракообразные. Поэтому приведенная
ниже характеристика этого класса является в то же время харак-
теристикой подтипа Жабернодышащие.
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
Общая характеристика. К этому классу принадлежат раки,
крабы, мокрицы, разнообразные мелкие рачки пресных и морских
вод. Подавляющее большинство ракообразных обитают в различ-
ных водоемах. Лишь немногие виды (например, мокрицы) живут
на суше. Тело ракообразных подразделяется на голову, грудь и
155
Рис. 122. Речной рак:
А — со спинной стороны; Б — с брюшной стороны;
/ —.антеннула; // — антенна; IX -- XIII — ходильные ноги; XIV—XIX — членики
брюшка;
1 — рострум; 2—головогрудной щит; 3 • кран щита, прикрывающий жабры; 4 —
головогрудь; .5 — брюшко; 6 — хвостовой плавник; 7 — половое отверстие самца;
8 — половое отверстие самки
брюшко; нередко голова и грудь сливаются, образуя головогрудь.
На голове расположены две пары антенн, играющие роль органов
осязания и обоняния. Конечности, как правило, двухветвистые.
Водные формы дышат жабрами. Органами выделения служат ви-
доизмененные метанефридии. Развитие в большинстве случаев
протекает с метаморфозом. Число известных видов превышает
20 тыс. Ракообразные имеют большое хозяйственное значение,
представляя важный пищевой продукт людей и корм рыб. Пара-
зитические виды наносят вред.
Строение и жизненные отправления. Размеры и форма тела
ракообразных разнообразны. Среди них имеются едва различимые
глазом рачки-циклопы и огромные крабы и омары массой до 8 кг.
Голова ракообразных образуется в процессе развития путем объе-
динения нескольких передних члеников. У большинства она несет
две пары антенн и три пары челюстей. Те и другие пред-
ставляют собой видоизмененные конечности животного. Членики
груди у некоторых свободные, но чаще слиты друг с другом или с
головой, образуя головогрудь (рис. 122). У высших раков (речные,
раки, омары, крабы) хитиновый щит, одевающий сверху голову и
грудь, спускается по бокам складками, прикрывая нежные жабры.
Каждый сегмент груди несет по паре членистых конечностей раз-
личной формы и строения в зависимости от выполняемых функций.
Сегменты брюшка обычно не сливаются друг с другом. У высших
раков каждый из них несет пару хорошо развитых ножек, у низ-
ших они редуцированы на брюшке. Конечности ракообразных
отличаются разнообразием формы и строения и выполняют раз-
личные функции (рис. 123). Некоторые из них служат опорой при
хождении, другие помогают животному при плавании, третьи
участвуют в добывании пищи и ее размельчении. Обычно ножки
животных имеют основную непарную часть — протоподит — и две
ветви: наружную — экзоподит и внутреннюю — эндоподит. Двух-
ветвистая форма конечностей ракообразных близка к двухло-
пастной форме параподий многощетинковых кольчатых червей. Но
если параподии этих червей не расчленены, то ножки ракообразных
состоят из ряда члеников. Деление конечностей на членики
обеспечивает их подвижность.
Покровы. Тело раков одето хитиновой кутикулой, образую-
щей наружный скелет. У высших раков скелет нередко утолщается
и укрепляется отложением углекислой извести, превращающей
кутикулу в твердый панцирь. У низших скелет тонкий. У многих
видов от хитинового покрова внутрь тела отходят отростки, к
которым прикрепляются мышцы.
Нервная система раков состоит из парного надглоточ-
ного узла, окологлоточного нервного кольца, парного подглоточно-
го ганглия и брюшной нервной цепочки с отходящими от всех
нервных узлов периферическими нервами (рис. 124). Если у ра-
ков произошло слияние отдельных сегментов, то происходит и
соединение их нервных узлов.
157
Органы чувств развиты хорошо и представлены простыми
и сложными глазами, органами осязания (главным образом на
усиках) и обоняния.
Пищеварительная система. Кишечный канал состоит
из передней, средней и задней кишок. Передняя кишка образована
коротким пищеводом, расширяющимся в желудок, выстланный
хитином. У ряда высших раков (например, у речных) желудок
разделен на два отдела: жевательный и пилорический. В жева-
тельном находятся три больших
выступа — «зуба», участвую-
щих в размельчении пищи.
Стенки пилорического отдела
образуют систему складок с ще-
тинками по краям, которые
создают фильтр, пропускаю-
щий в кишечник только раз-
дробленную пищу. Перевари-
вание и всасывание веществ пи-
щи происходят в короткой сред-
Рис. 123. Конечности речного рака:
/ — антеннула; 2 — антенна; 3 — верхние
челюсти; 4,5 — первая и вторая пары
нижних челюстей; 6—8 — ногочелюсти;
9—13— ходильные ноги; 14—19 — брюш-
ные ножки; 20 — нити; 21 — жабры
Рис. 124. Нервная система речного
рака:
1 — надглоточный ганглий; 2 — окологло-
точное нервное кольцо; 3— нервы первой
пары ходильных ног; 4 — подглоточный
ганглий; 5,6 — ганглии брюшной нервной
цепочки; 7 — отходящие от них нервы
158
Рис. 125. Анатомия речного рака:
/ некрытый рак; // - вскрытый же нательный желудок рака;
/ росгрум; 2 жевательный желудок; печень; 4 - кишка; 5 - сердце; 6’, 7 -
артерии; 8 - яичник; .9 - брюшная нервная цепочка; 10 - ~ жабры
ней кишке, в которую открываются протоки пищеварительной
железы — печени. Задняя кишка заканчивается анальным отвер-
стием (рис. 125).
О р г а п а м и д ы х а н и я у подавляющего большинства ра-
кообразных служат жабры, имеющие вид тонких нежных выростов
у основания ножек. Многие мелкие низшие рачки дышат всей по-
верхностью тела. Живущие на суше мокрицы дышат при помощи
тонких воздухоносных трубочек — трахей, находящихся в брюш-
ных ножках.
Кровеносная система имеет различное строение. У выс-
ших раков на спине лежит мешковидное или вытянутое вдоль тела
сердце с отверстиями, через которые кровь засасывается из полос-
ти тела. От сердца отходят артерии, из которых кровь выливается
в лакуны.. К сердцу кровь возвращается по лакунам и заса-
сывается через его отверстия. У некоторых низших раков крове-
носная система представлена трубчатым или мешковидным серд-
цем и сосудами или только сердцем или кровь перемещается
благодаря движению мышц.
Органы выделения представлены парными железами,
лежащими в головном отделе животного. Это видоизмененные
метанефридии, выводные протоки которых открываются наружу
в базальном членике антенн (антеннальные железы) или в основа-
нии нижних челюстей (максиллярные железы).
159
Половая система. Большинство раков раздельнополы.
Иногда имеется половой диморфизм. Половые органы у самок
представлены парными яичниками и яйцеводами, а у самцов —
парными семенниками и семяпроводами.
Лишь у немногих паразитических и сидячих форм наблюдается
гермафродитизм.
Развитие 'у ракообразных сопровождается метаморфозом
различной сложности, реже — без стадии личинок.
ПОДКЛАСС ЖАБРОНОГИЕ (BRANCHIOPODA)
К этому подклассу относятся мелкие ракообразные, обитающие
в морях и водоемах суши. Отличаются отсутствием конечностей
на брюшке. Грудные ножки двухветвистые, листовидные. Прото-
ки выделительных органов открываются у основания нижних
челюстей.
Органы дыхания — жабры в виде лепестков в основании
ног. Иногда функцию дыхания выполняют’ участки покро-
вов.
Отряд Ветвистоусые раки (Cladocera) — имеют мешковидное
тельце, заключенное в двустворчатую тонкостенную хитиновую
раковину. Голова несет две пары антенн, из которых первая мала,
а вторая достигает больших размеров и имеет двухветвистое строе-
ние; они участвуют в плавании. Имеется пара сближенных слож-
ных глаз. Грудной отдел тела состоит из 4—6 члеников, несущих
короткие листовидные ножки с рядами щетинок по краям и жабер-
ными лепестками у основания. Вода, увлекаемая движением
этих ножек, движется внутри раковинки, омывая жабры, а взве-
шенные в ней микроорганизмы и пищевые частицы задерживаются
щетинками ног и направляются
в рот. Брюшко сильно укорочено и
подогнуто к груди. У некоторых
хищных ветвистоусых раков ра-
ковина и жабры атрофированы.
Ноги изменены в хватательные ко-
нечности. Дыхание у них осущест-
вляется через покровы.
Наиболее яркими представите-
лями являются различные виды
Рис. 126. Дафния:
1 — антеннулы; 2 — двухветвистые антенны;
3—простой глаз; 4 — сложный глаз; 5 —
выделительная железа; 6' — сердце; 7 — груд-
ные ножки; 8— брюшко; 9 — выводковая
камера
160
дафний (рис. 126), в массе населяющие озера и пруды. Летом они
размножаются партеногенетически: из неоплодотворенных яиц
развиваются только самки, которые, в свою очередь, откладывают
нсоплодотворенные яйца. Партеногенетическое размножение обес-
печивает более быстрое воспроизводство популяции. Осенью появ-
ляются самцы, которые оплодотворяют яйца самок. Оплодотво-
ренные яйца перезимовывают, и весной из них выводится новое
поколение рачков.
ПОДКЛАСС МАКСИЛЛОПОДЫ (MAXILLOPODА)
Отряд Веслоногие раки (Copepoda) — мелкие рачки с удли-
ненным стройным телом.
На голове сильно развита первая пара одноветвистых ан-
тенн. На груди расположены двухветвистые, уплощенные и снаб-
женные щетинками ножки, движение которых толкает тело вперед.
Брюшко заканчивается вилкой. Дышат они всей поверхностью
тела.
Сердце и сосуды у большинства отсутствуют. Размножение
только половое. Самки носят оплодотворенные яйца в особых яйце-
вых мешочках. Характерные представители отряда циклопы
(Cyclops) и диаптомусы (Diaptonius), населяющие в огромном
количестве разные водоемы (рис. 127). Они служат кормом маль-
кам рыб и планктоноядным рыбам. Среди веслоногих раков
встречаются эктопаразиты рыб, вызывающие заболевания и ги-
бель последних. Некоторые из паразитических форм настолько из-
меняются, что совершенно утрачивают вид ракообразного. Только
развитие со стадией иауплиуса и наличие у самок парных яйцевых
мешков указывают на принадлежность их к веслоногим рачкам
(рис. 128).
Копеподы известны как промежуточные хозяева червей —
паразитов сельскохозяйственных животных и человека.
Отряд Усоногие раки (Cirripedia) — морские ракообразные,
ведущие во взрослом состоянии сидячий или паразитический
образ жизни, что вызывает упрощение и изменение их организации
(рис. 129). Наиболее известны морские желуди и морские
уточки, нередко в огромном количестве обрастающие подводные
камни и скалы. Сначала их личинки подвижны, по позднее они
прикрепляются головным отделом к субстрату и испытывают слож-
ные превращения. Их тело окружается складкой кожи — мантией,
которая выделяет на наружной поверхности несколько известко-
вых пластинок, образующих в совокупности домик-раковину с
подвижными створками. Грудные ножки удлинены. Движение
последних загоняет в раковину воду со взвешенными в ней пи-
щевыми частицами.
Усоногие — гермафродиты. Могут причинять убытки судо-
ходству: ими обрастают днища судов.
6 Б. А. Кузнецов н др.
161
Рис. 127. Свободпоживущие веслоногие рачки:
I — циклоп; // — диаптомус; III — каптокамптус;
/ --глаз; 2 — антеплулы; 8 - головогрудь; 4 свободные сегменты груди; 5
брюшко; 6 — яйцевые мешки; 7 — вилочка брюшка; 8 — кишка
Рис. 128. Веслоногие рачки — паразиты рыб:
/ — Laniftruiilenir, II — Ler налоге г tr, HI — Achteres' IV — Traelwliuster
ПОДКЛАСС ВЫСШИЕ РАКИ (MALACOSTRACA)
В большинстве случаев раки средней или большой величины.
Тело состоит из 20—21 сегмента: 5—6 головных, 8 грудных и 7
брюшных. У многих наблюдается тенденция к слиянию члеников с
образованием головогруди. Конечности расположены не только на
голове и груди, но и на брюшке. Протоки выделительных
органов обычно открываются у основания антенн. Развитие про-
исходит либо без превращения, либо со стадией своеобразной
личинки — зоеа, имеющей в отличие от науплиуса низших раков
расчлененное тело (рис. 130). Высшие раки населяют моря, озера,
пруды и реки, но некоторые из них (мокрицы) приспособились к
жизни на суше.
6*
163
Рис. 130. Личинки ракообразных:
/ - паунлиус—личинка низших раков,
// — зоеа - личинка высших раков
Рис. 131. Равноногие раки:
1 мокрица; 11 — водяной ослик
I Я
Из отрядов данного подкласса рассмотрим равноногих и деся-
тиногих раков, имеющих наибольшее значение для промыслового
и сельского хозяйства нашей страны.
Отряд Равноногие раки (Isopoda) — небольшие рачки с упло-
щенным телом (рис. 131). Голова слита с передним сегментом
груди, остальные членики грудного отдела свободны. Каждый сег-
мент груди и брюшка несет пару относительно коротких ножек.
К отряду относятся различные виды мокриц, обитающих на суше
(обычно во влажных местах), водяные ослики, живущие в боль-
шом числе в озерах и прудах, и многие морские формы.
Мокрицы местами приносят значительный вред культурным
растениям, особенно в теплицах и парниках.
Отряд Десятиногие раки (Decapoda) — наиболее высокоорга-
низованные ракообразные среднего или крупного размера
(рис. 132). Голова и все членики груди слиты в головогрудь,
прикрытую сверху и с боков головогрудным щитом, хитин которого
обычно пропитан известью. Грудных конечностей восемь пар,
из которых три передние — ногочелюсти — участвуют в добыва-
нии пищи, а остальные пять пар служат для передвижения
животного (ходильные ноги). У многих видов передняя пара
ходильных ног несет мощные клешни. Брюшко имеет шесть пар
ног. Десятиногие раки делятся на три группы: длиннохвостые,
мягкохвостые и короткохвостые.
Длиннохвостые раки — речные раки и морские раки — омары и
лангусты, креветки и др. У них хорошо развито брюшко, которое
несет на конце хвостовой плавник, образованный последним чле-
164
Рис. 132. Десятиногие раки:
1 - креветка; // - рак-отшельпик; II! краб
ником тела и широкими лопастями ножек предпоследнего
сегмента.
Речные раки селятся в реках с тихим течением, в прудах
и озерах. Днем они скрываются в норках или под корягами, а
ночью выходят на поиски пиши. Питаются мелкими донными жи-
вотными и падалью. Икру самки вынашивают, прикрепляя ее к
ножкам брюшка. Многие длиннохвостые раки имеют большое про-
мысловое значение.
165
Мягкохвостые раки включают раков-отшельников, обитающих
в морях. Брюшко у них средней величины, изогнутое, мягкое, без
конечного плавника. Такое строение обусловлено тем, что жи-
вут раки-отшельники в спирально изогнутых раковинах брюхоно-
гих моллюсков, в которые прячут свое брюшко. Хозяйственного
значения не имеют.
Короткохвостые раки (крабы) имеют широкую уплощенную
головогрудь, под которую подвернуто брюшко. Многие виды упо-
требляются как пищевой продукт. '
ПОДТИП ХЕЛИЦЕРОНОСНЫЕ (CHELICERATA)
Общая характеристика. Наземные членистоногие, ведут ро-
дословную от палеозойских водных предков, близких к предкам
ракообразных. В расчленении тела нет единообразия. Частый слу-
чай, когда сегменты головы и груди, сливаясь, образуют голо-
вогрудь. В наибольшей мере слияние сегментов тела представлено
у клещей. Антенны отсутствуют, глаза только простые в количест-
ве от одной до восьми пар. Членистых придатков шесть пар.
Из них первые две пары: хелицеры и педипальпы —
выполняют функцию антенн и челюстей. Остальные четыре пары —
ноги. Брюшко обычно лишено ног, но у пауков они видоизменены
в паутинные бородавки. У скорпионов ноги на брюшке заклады-
ваются в эмбриональном периоде, у взрослых они отсутствуют.
В подтипе два класса: Меристомовые— Мечехвосты (Xip/iosu-
га) и Паукообразные (Arachnida). В данном курсе рассматри-
ваются только паукообразные, широко представленные в фауне
СССР.
Строение и жизненные отправления. Форма тела разно-
образна, как и расчлененность.
Покровы хитиновые, нередко с многочисленными чувствую-
щими волосками. У мелких клещей тонкие покровы служат и для
дыхания.
Нервная система, как и у других членистоногих: брюш-
ная нервная цепочка с ганглиями. При слиянии члеников про-
изошло и объединение ганглиев. У пауков и клещей узлы груди
и брюшка слиты в единый нервный узел. Пауков отличает слож-
ность нервно-психических проявлений, рефлексов, инстинктов
(рис. 133).
Органы зрения. Имеется 1—6 пар простых глаз. Два
центральных глаза пауков способны различать форму и цвет пред-
метов.
Органы пищеварения имеют различное строение в
зависисмости от характера питания. У видов, питающихся жид-
кой пищей (кровью животных, соками растений), передний отдел
пищеварительного тракта слагается из мускулистой глотки и пи-
щевода, которые служат для всасывания пищи. Основные про-
цессы переваривания и всасывания пищи протекают в средней
166
кишке, выстланной железистым эпителием и имеющей слепые
выросты, что увеличивает вместимость и всасывающую поверх-
ность. У большинства паукообразных в брюшной отдел средней
кишки открываются протоки парной железы — печени, выделяю-
щей пищеварительные ферменты и всасывающей переваренные пи-
тательные вещества.
Органы дыха н и я разнообразны. У одних — это легочные
мешки, у других -- трахеи, у третьих — и те и другие. Легочные
мешки расположены в передней части брюшка и сообщаются с
внешней средой дыхальцами. Внутри легких имеются параллельно
расположенные тонкие листки, в лакунах которых течет кровь.
Газообмен происходит через тонкие покровы листочков. Трахеи
начинаются отверстиями в покровах брюшка, от которых уходят в
глубь тела в виде ветвящихся трубочек.
Кровеносная система слагается из лежащего па спин-
ной стороне мускулистого пульсирующего сердца и отходящих от
него сосудов, направляющихся к различным органам. Обратный
ток крови к сердцу проходит по лакунам. У некоторых видов
сердце имеет вид мускулистой трубки с многими отверстиями, у
других оно мешкообразно и имеет 1— 2 пары входных отверстий.
У клещей кровеносные сосуды частично редуцированы, а иногда
нет и сердца.
167
Орга и ы в ы д е л е н и я у паукообразных двоякого порядка.
Большая часть из них сохранила 1—2 пары метанефридиальных
органов, открывающихся у основания ног. Кроме того, в брюшке
имеется 1—2 пары тонких слепых трубочек— мальпигиевы сосу-
ды, открывающиеся в кишечник. Первые выделяют продукты рас-
пада из грудного, вторые — из брюшного отделов тела.
Половая система. Паукообразные раздельнополы. Яич-
ники самок расположены в брюшке, а яйцеводы сливаются в еди-
ный проток, открывающийся в передней части брюшного отдела.
Семенники самцов также лежат в брюшке. Отходящие от них спер-
мопроводы сливаются и одним отверстием открываются на нижней
стороне брюшка.
Развитие. Оплодотворение происходит в половых путях
самки. Большинство наукообразных откладывает яйца, у некото-
рых наблюдается живорождение. Развитие происходит без мета-
морфоза или со стадией личинки (клещи). У клещей иногда наб-
людается партеногенетическое размножение. Самцы паукообраз-
ных значительно мельче самок.
КЛАСС ПАУКООБРАЗНЫЕ (ARACHNIDA)
Класс Паукообразные делится на отряды, из которых наибольшее
значение имеют следующие: Скорпионы (Scorpiones), Фаланги
(Solpugida), Пауки (Aratiei), Клещи (Acari).
Отряд Скорпионы (Scorpiones) — внешне напоминают речных
раков (рис. 134). Длина наиболее крупных тропических видов
достигает 18 см. Сегменты головы и груди слились в нерасчлеиен-
ную головогрудь. Брюшко образовано рядом сегментов. Передняя
часть брюшка широкая, а задняя — тонкая, вытянутая. Послед-
ний членик брюшка заканчивается острым жалом, на конце кото-
рого открывается проток ядовитой железы, лежащей в теле члени-
ка. Ударом брюшка скорпион ранит жалом врага или жертву и
впускает в ранку яд. Уколы крупных скорпионов могут быть
смертельными даже для человека. Около рта расположены неболь-
шие клешневидные хелицеры и огромные, оканчивающиеся боль-
шими клешнями педипальпы. Эти конечности служат для улавли-
вания и удержания добычи.
Скорпионы обитают в тропических и субтропических странах,
в СССР встречаются в Средней Азии, Крыму и на Кавказе. Днем
скрываются под камнями, в норках и щелях, а ночью охотятся за
различными мелкими животными.
Отряд Фаланги (Solpugida)—довольно крупные животные,
внешне сходные с пауками (рис. 135). Тело состоит из голово-
груди и расчлененного брюшка. Последние два сегмента груди
свободны. На головном отделе расположены мощные клешневид-
ные хелицеры и длинные, сходные с ногами педипальпы. Ды-
шат трахеями. Паутинные бородавки отсутствуют. Обитают фа-
ланги в жарких и сухих местностях. В СССР встречаются в
168
Рис. 134. Скорпион:
А -- вид со спппной стороны; /> — вид с брюшной стороны;
/ - головогрудь; 2 - хелицеры; 3 - недпиалыня; 4 -- ноги; .5 конечный членик брюш-
ка; 6’--жало; 7 - передний отдел брюшка; Н - задний отдел брюшка; .9- анальное
отверстие; Щ-- легочная щель; II - гребенчатые органы; 12 — половые крышечки
{VIII. XIX -сегменты брюшка)
Средней Азии и Казахстане. Ночные хищники, поедают саранчу и
много других вредных насекомых. Укус неядовит, но остатки
пищи на хелицерах могут вызвать воспаление и оказаться
ядовитыми.
Отряд Пауки (Aranei). Тело большинства пауков состоит из
перасчлененных головогруди и брюшка. Хелицеры состоят из двух
члеников, из которых конечный имеет вид изогнутого острого
169
Рис. 135. Фаланга:
I — хелицеры; 2 — педипальпы; 3 — ноги;
4—задние свободные членики груди;
5 — брюшко; 6 — головогрудь; 7 — глаза
Рис. 136. Паук-крестовик:
1 — головогрудь; 2 — хелицеры; 3 —
педипальпы; 4 — ноги; 5 — брюшко
коготка. В основании хелицер залегает ядовитая железа, проток
которой открывается на острие коготка. Им пауки прокалывают
покровы жертв и вводят в ранку яд. Педипальпы пауков имеют вид
членистых щупалец. От основного членика педипальп в сторону
рта отходят челюстные отростки с щетинками по краям, которые
препятствуют попаданию твердых частиц при высасывании пау-
ком жертвы. У самцов конечный членик педипальп имеет
небольшой резервуар с копулятивным аппаратом. В период раз-
множения самец наполняет эти резервуары педипальп спермой,
вытекающей из семенного протока на брюшке. При спаривании
он вводит копулятивный аппарат в семяприемник самки и остав-
ляет в нем сперму. Все пауки имеют четыре пары ходильных ног.
На конце брюшка расположены паутинные бородавки с отвер-
стиями протоков паутинных железок, которые выделяют особые ве-
щества, быстро застывающие на воздухе, образуя паутинную
нить. У пауков имеется несколько типов желез разного назначения.
Их протоки на бородавках открываются трубочками и конуса-
ми. У крестовика на бородавках насчитывается до 560 трубочек
и до 20 конусов. Паутинные нити служат для различных целей:
170
изготовления ловчей сети, яйцевого кокона, жилого домика. На па-
утинке они спускаются с деревьев, а молодые паучки, распустив
паутинную нить по ветру, совершают далекие перелеты. Все пау-
ки — хищники. Они в значительных количествах уничтожают на-
секомых-вредителей, что определяет их полезное значение.
Некоторые из пауков ядовиты.
Домовые пауки (Tegenaria) селятся обычно в жилых поме-
щениях, где охотятся за мухами, тараканами и другими насекомы-
ми —- спутниками человека.
Науки-крестовики (Aratieus) легко узнаются по крестообраз-
ному рисунку на верхней стороне брюшка (рис. 136). Они
селятся в лесах и на кустарниках. На ветвях они делают красивые
радиальные сети, нити которых несут капельки клейкого вещест-
ва, приклеивающего насекомых.
Тарантулы (Lycosa) — самые крупные пауки фауны СССР
(рис. 137). Их длина достигает 3 см. Живут они в степных
районах в отвесной норке. Укус тарантула бывает болезненным,
но неопасен.
Каракурты (Lathrodeetus) пауки среднего размера, барха-
тно-черной окраски, с красными точками на брюшке (рис. 138).
Встречаются в степях Казахстана и Средней Азии. Укусы кара-
курта нередко смертельны для человека, крупного рогатого скота
и лошадей. Самки откладывают яйца группами в шаровидные
коконы из паутины, расположенными у земли.
Паук-серебрянка (Argyroneta) живет в водоемах. На подвод-
ных растениях он строит из паутины колоколообразный домик,
который наполняет воздухом. Охотится в воде за мелкими водными
животными.
Рис. 137. Тарантул
Рис. 138. Каракурт (сам-
ка в гнезде у яйцевых
коконов, /) и паук-пти-
цеед (//)
171
Отряд Клещи (Acari) — мелкие наукообразные, длиной от до-
лей миллиметра до 2 см и более. Для клещей характерно
слияние сегментов тела, лишь у немногих форм сохраняются
слабо заметное разделение тела на головогрудь и брюшко и
границы отдельных сегментов брюшка, У клещей, питающихся
жидкой пищей (кровью животных и соком растений), хелицеры и
педипальпы вытянуты и образуют колюще-сосущий хоботок.
Хелицеры других клещей зазубрены и приспособлены к раздроб-
лению твердой пищи. Клещи дышат трахеями или через кожу.
Развитие клещей происходит с метаморфозами. Из яиц выходят
личинки, похожие на взрослых особей, но с тремя парами ног.
Они превращаются в нимфу, имеющую уже четыре пары ног, но с
еще недоразвитыми половыми органами. За стадией нимфы сле-
дует стадия взрослого животного.
Известно более 10 тыс. видов клещей. Часть из них являются
паразитами людей и животных. Многие паразитируют на расте-
ниях (в том числе и на сельскохозяйственных). Ряд видов
ведет хищнический образ жизни, нападая на мелких насекомых и
других клещей. Многочисленны почвенные клещи, питающиеся
гниющими веществами почв.
Подотряд Сенокосцы ( Notostigmata). Малочисленная, наибо-
лее примитивная группа. Тело сегментировано. Практического
значения не имеют.
Подотряд Саркоптоидные (Sarcoptiformes). Дыхальца отсут-
ствуют и трахейная система развита не у всех (если трахеи
имеются, то открываются на поровых полях). Хелицеры клешне-
видные (грызущий тип). Тазики ног очень тесно слиты с хитино-
вым покровом. Это обширный подотряд со многими семействами.
Большое значение имеют панцирные клещи (Oribatei) как
почвообразователи, амбарные — как вредители. Есть и хищные, и
паразиты насекомых. Практическое значение большое.
Подотряд Краснотелковые (Trombidiformes) — наиболее круп-
ный из подотрядов. В нем свободноживущие и паразиты,
сухопутные и водные. Дыхалец одна пара, расположена в передней
части тела. Хелицеры колющие. Имеют большое практическое
значение.
Из растительноядных клещей большой вред культурным расте-
ниям причиняют паутинные клещики (Tetranychus telarius)
(рис. 139). Это название они получили за способность выделять
паутинные нити, которыми покрывают нижнюю поверхность листь-
ев. Тело самки яйцевидное, нерасчлененное, длиной до 0,4 мм, а
самца — округлой формы, длиной до 0,25 см. Из яиц выходят
личинки, превращающиеся в нимфы I и II стадий, которые затем
становятся взрослыми особями. Весь цикл развития клеща длится
обычно 12—28 дней. За лето может отродиться 8—10 поколений
клещей. Клещики сосут сок из листьев растений. Особенно
опасны они для огородных и бахчевых культур: огурцов, тыквы,
арбузов, дынь и др. На юге поражают хлопчатник, сою, фасоль и
172
щик
Рис. 139. Паутинный кле-
пр. На листьях появляются мелкие белые пятна, цветки, завязи
плодов опадают. Вред, причиняемый паутинными клещиками
растениеводству (особенно в условиях закрытого грунта), очень
велик. К мерам борьбы относятся: опрыскивание и опыление
растений различными химикатами, уничтожение сорняков, уборка
растительных остатков после сбора урожая.
Семейство Собственно краснотелки (более 300 видов) вклю-
чает паразитов животных и человека (кровососов), приносящих
вред, и полезных, паразитирующих на вредных насекомых, вклю-
чая саранчовых.
Железничные клещи (Demodicidae) паразитируют в волосяных
сумках, вызывая заболевание животных и человека — железни-
цу (рис. 140). Отличаются вытянутой, червеобразной формой
тела и короткими ножками. У мериносовых овец их находили
до 25 тыс. па 1 см2 кожи. На пораженных местах шерсть выпадает,
а в дерме образуются пузырьки — пустулы.
Подотряд Паразитоидные (Parasitiformes). Дыхалец одна па-
ра, открывающаяся позади 2, 3 и 4-й пары ног. Ротовой аппарат
у большинства колюще-сосущий. Пальцы 5-члениковые, без ко-
готков па лапках. В подотряде встречаются как хищные (свободпо-
живущие), так и паразитические виды. Практическое значение
большое. Некоторые виды клещей питаются продовольственными
продуктами.
Амбарные клещики (различные виды сем. Tyroglyphida)
относятся к числу опасных вредителей зерна и муки (рис. 141).
Это очень мелкие клещики овальной формы, длина тела до 0,5 мм.
На теле заметны отдельные щетинки. Размножаются в благо-
приятных условиях круглый год. Цикл развития длится 25 - (>()
дней. Клещики, как и их личинки и нимфы, питаются зерном,
173
мукой, крупой. Испражнениями загрязняют эти продукты. Для
борьбы с ними применяют дезинфекцию складских помещений,
огневую сушку зерна и муки, удаляют зерновую просыпь и др.
Из клещей того же подотряда — паразитов сельскохозяйственных
животных и человека — особенно опасны следующие.
Зудневые клещи (Sarcoptes) живут в коже диких млекопитаю-
щих и домашних животных (особенно лошадей и свиней), а также
человека (рис. 142). Это очень мелкие животные с овальным
тельцем и четырьмя парами коротких ножек, длина тела до 0,4 мм.
Самка откладывает в толще кожи хозяина до 50 яиц. Раз-
витие продолжается около 15 дней. Зудень вызывает заболева-
ние — чесотку, при которой кожа воспаляется и резко утолщается.
На ней развиваются струпья и роговые корки, скапливающие
гной. Волосы на пораженных участках кожи выпадают. Иногда
болезнь приводит к гибели животного.
Накожные клещи (Psoroptes) живут на поверхности кожи овец
и других домашних животных. Они крупнее часоточных, длина
их тела до 1 мм. Ножки развиты лучше. Впиваясь в кожу хозяина
хоботком, они сосут кровь, вызывая воспаление кожного покрова
и выпадение шерсти. При массовом размножении могут стать при-
чиной истощения и даже гибели овец. С целью уничтожения
клещей овец после стрижки купают в креолиновом растворе.
Иксодовые клещи (подотр. Паразитоидных, надсем. Пастбищ-
ных клещей Ixodea) — эктопаразиты диких и сельскохозяйствен-
ных животных, а также человека (рис. 143). На одной корове
одновременно может паразитировать до 1 тыс. клещей, которые
высасывают до 3—5 л крови. Болезненное состояние животно-
го вызывает при этом снижение молочной продуктивности на
40—75 %. По биологическому признаку различают 1-, 2- и 3-хозя-
инных клещей. Первые все ста-
дии своего развития проходят
на одном хозяине. Вторые, на-
сосавшись крови, покидают хо-
зяина и, претерпев линьку, на-
падают вновь. К 3-хозяинным
относится широко распрост-
раненный собачий клещ (Ixo-
des ricitius). Его личинка, насо-
савшись крови, сходит на зем-
лю, где совершает линьку, пре-
вращаясь в нимфу. Нимфа кор-
мится уже на новом хозяине.
Она тоже покидает хозяина и
после очередной линьки стано-
Рис. 142. Чесоточный зудень (самка)
174
вится зрелой, т. е. переходит в стадию имаго. Половозрелый клещ
должен опять найти хозяина (млекопитающего) и насосаться его
крови. Последняя кормежка обеспечивает созревание половых
продуктов.
После спаривания следует откладка яиц. Хозяевами иксодовых
клещей, помимо домашних животных, служат ежи, полевки,
зайцы и другие млекопитающие, а из пресмыкающихся — яще-
рицы.
Отдельные виды иксодовых клещей могут голодать на стадии
личинки и нимфы до 1,5—2 лет, а взрослые — до года, что создает
трудности в борьбе с клещами.
Иксодовыс клещи -- переносчики возбудителей опасных забо-
леваний сельскохозяйственных животных: пироплазмоза млекопи-
тающих, спирохетоза кур, гусей и уток. Они переносят возбуди-
телей и таких тяжелых болезней человека, как таежный энцефалит,
клещевой тиф, туляремия и др.
В подотряд Parasitiformes, семейство Plujloseiidae входят хищ-
ные клещи, многие из них питаются вредными клещами —
паутинным клещиком и др.
Один из видов Phytoseiulus persimilis ввезен для акклимати-
зации из Канады (Бегляров, 1963) и искусственно размножается
в ряде хозяйств.
175
ПОДТИП ТРАХЕЙ НОДЫШАШИ Е (TRACHEATA)
Подтип объединяет членистоногих, дышащих при помощи тра-
хей. У всех трахейнодышащих головной отдел ясно обособлен от
грудного. Голова несет одну пару антенн. В подавляющем боль-
шинстве они являются обитателями суши, лишь немногие вторично
приспособились к жизни в воде.
Подтип включает два класса: Многоножки (Myriapoda), На-
секомые (Insecta).
КЛАСС МНОГОНОЖКИ (MYRIAPODA)
Удлиненное тело многоножек слагается из головы и сегмен-
тов туловища (рис. 144). Все сегменты схожи, грудной и брюш-
ной отделы тела неразличимы. У одной из групп — кивсяков —
сегменты тела слиты попарно. На голове расположены пара антенн
и 2—3 пары челюстей. У большинства многоножек каждый членик
тела несет одну пару членистых ножек, а у кивсяков вследствие
попарного слияния сегментов — две пары. По бокам сегментов
тела расположены дыхальца, ведущие в пучки разветвляющих-
ся трахей. Органами выделения служат мальпигиевы трубочки,
открывающиеся в кишечник, и особые нижнегубные выделитель-
ные железы метаиефридиального происхождения. Развитие жи-
вотных из яиц проходит через стадию личинки или минуя ее.
Известно около 9 тыс. видов многоножек, разнообразных по
внешнему виду и образу жизни. Большинство их — ночные живот-
176
Рис. 145. Строение многоножки-костянки:
/ — голова; 2 - антенны; 3, 7 межсегментные
покровы; •/, 3, !) - ножки; 5 - анус; (> дыхальца;
/О — иогочелюсти; / Л/Л сегменты тела
ные, днем прячущиеся под камнями,
в лесной подстилке и других убежи-
щах. Имеются хищные и раститель-
ноядные формы.
Сколопендра — крупная, длиной до
15 см,, хищная многоножка, обитаю-
щая па юге СССР.
Первая пара ног превратилась в
иогочелюсти, служащие для захвата
и умерщвления добычи. На заострен-
ном последнем членике ее открывают-
ся протоки ядовитых желез, располо-
женных в основании сегмента. Питает-
ся насекомыми, червями и другими мел-
кими животными.
Яд сколопендр опасен даже для
человека.
Костянка, или сороконожка, сход-
на со сколопендрой, но значительно мень-
ше— длина до 5 см (рис. 145).
Широко распространена в СССР.
Питается мелкими беспозвоночными.
Кивсяк, или тысяченожка, отличается цилиндрическим телом
и большим числом тонких ножек, которые сидят по две пары на
каждом членике тела. Растительноядные формы, распространен-
ные по всей Европе. Часто повреждают различные огородные
культуры, особенно в теплицах.
КЛАСС НАСЕКОМЫЕ (1NSECTA)
Общая характеристика. В классе насекомых объединены трахей-
нодышащие членистоногие с характерным расчленением тела па
голову, грудь и брюшко (рис. 146). На голове находятся одна
пара антенн, глаз и рот е тремя парами челюстных придатков.
Грудь, как правило, состоит из трех сегментов, на каждом из ко-
торых по паре конечностей (отсюда другое название насеко-
мых — шестиногие). У большинства насекомых на втором и тре-
тьем члениках груди по паре крыльев. Брюшко, состоящее из
6—12 сегментов, лишено конечностей, и только у некоторых они
сохранились в измененном виде и выполняют специальные функ-
ции (яйцеклада и др.).
177
Рис. 146. Расчлененное насекомое (жук-олень l.itcanus seruus):
1 — нижняя губа; 2-- нижняя челюсть; Л--мандибула; 4 — верхняя губа; 5 - голова;
6 — антенны; 7 — переднегрудь; А, //, 14 -- грудные конечности; .9 - передняя пара
крыльев (надкрылья); 10- среднегрудь; 12 заднегрудь; 13 — задние крылья; 15
брюшко
К классу Насекомые принадлежит более половины всех оби-
тающих на Земле видов животных. Нет такого уголка суши, где
бы они не встречались. Роль насекомых в природе и хозяйстве
человека огромна.
Класс Насекомые включает подклассы: Первичнобескрылых
(Apterygota) и Крылатых (Pterygota).
Строение и жизненные отправления. Внешний вид насекомых
весьма разнообразен. Это касается как размера тела, окраски,
так и строения членистых придатков головы, груди и брюшка.
Так, антенны на голове бывают короткие и длинные, в виде ще-
тинок и перисторазветвленные, тонкие и пластинчатые и т. д.
Ротовые органы в количестве трех пар представлены верхними че-
люстями (мандибулами), нижними челюстями (максиллами) и
178
нижней губой, образованной срастанием второй пары нижних че-
люстей. Все они развились в процессе эволюции из членистых при-
датков головных сегментов.
Строение ротовых органов у насекомых различных система-
тических групп варьирует в широких пределах, отражающих
разнообразие в способах питания. Различают четыре основных
типа ротовых органов: грызущий, грызуще-сосущий, колюще-сосу-
щий и сосущий (рис. 147).
Грызущие ротовые органы свойственны насекомым, питающим-
ся преимущественно твердой пищей — жуками, тараканами, пря-
мокрылыми и др. У них верхние челюсти — жвалы — имеют вид
массивных хитиновых пластинок с зазубренными острыми вну-
тренними краями. Первая пара нижних челюстей (макеиллы)
состоит из нескольких члеников и несет нижнечелюстные щупики, а
на концах — парные лопасти, внутренняя из которых усажена хи-
тиновыми щетинками. Как жвалы, так и нижние челюсти служат
насекомому для отделения и размельчения пищи. Вторая пара
нижних челюстей срастается основными частями в единую рас-
члененную пластинку, образуя нижнюю губу, по боком которой
расположены нижнегубные щупики. На ротовых щупиках имеются
осязательные волоски и ямки органов осязания и вкуса.
Нижняя губа прикрывает .рот и его органы снизу; сверху же он
ограничен хитиновой пластинкой (складкой) — верхней губой.
Грызуще-сосущие ротовые органы имеются у насекомых, ко-
торые питаются как жидкой (нектар цветков), так и твердой пи-
щей. Таковы, например, ротовые органы пчел и шмелей. Их верх-
ние челюсти имеют вид пары зазубренных по внутреннему краю
лопаточек. Нижние челюсти вытянуты в ланцетовидные пластинки
с зачаточными щупиками. Нижняя губа продолжается вперед
тонким язычком, образованным сложенными вместе внутренними
Рис. 147. Ротовые органы насекомых:
/ — грызущие (таракана); // — грызуще-сосущие (пчелы); /// — сосущие (бабочки);
/V' — колюще-сосущие (самки комара);
/ — верхняя губа; 2 — верхние челюсти; 3— нижние челюсти; 4 — нижняя губа; 5 -
подглоточник; 6 — нижнечелюстные щупики; 7 — нижнегубные щупики
179
лопастями. Когда к язычку прикладываются нижние челюсти,
образуется трубочка - хоботок, при помощи которого насекомое
сосет нектар.
Колюще-сосущие ротовые органы свойственны комарам, кло-
пам, тлям и некоторым другим насекомым. Ротовые органы их
сильно удлинены и, складываясь, образуют хоботок, служащий
для высасывания крови или сока растений. Так, например, у самок
комаров нижняя губа имеет вид желоба, открытого сверху, за-
канчивающегося двумя лепестками. В углублении желоба лежат
остальные ротовые части.
Верхняя губа с загнутыми вниз краями, почти смыкающи-
мися, образует канал, по которому всасывается пища. Щель на
его нижней стороне закрыта длинным, па конце заостренным
подглоточником. Он имеет внутри слюнопроводящий канал. Рядом
залегают мандибулы и макспллы. Они в виде упругих колющих
щетинок. В нанесении укола участвуют верхняя губа, подглоточ-
ник и челюсти, а нижняя губа по мере погружения в рану колюще-
го аппарата изгибается.
Сосущие ротовые органы характерны для' бабочек (чешуе-
крылых) и устроены в виде сосательного хоботка. Верхняя и ниж-
няя губы, верхние челюсти сильно редуцированы. Нижние же че-
люсти велики и имеют вид гибких длинных желобков. Склады-
ваясь, желобки образуют длинный, полый внутри хоботок. В по-
коящемся состоянии он свернут спиралью. В расплавленном виде
хоботок погружается бабочкой в венчик цветка.
Строение ног у насекомых весьма различно в зависимости
от образа жизни и способа передвижения (рис. 148). Они состоят
из следующих члеников: основной — тазик, за ним расположен
небольшой членик — вертлуг, облегчающий движение конечно-
сти. Затем идут два длинных членика — бедро и голень, в которых
заложены сильные мышцы. Кончается ножка лапкой из несколь-
ких мелких члеников, последний из которых обычно имеет 1—2 ко-
готка.
У большинства насекомых второй и третий сегменты груди
несут по паре крыльев. Мухи, комары и близкие к ним формы
сохранили только переднюю пару крыльев, тогда как задняя ви-
доизменилась в своеобразные органы — жужжальца. Некоторые
насекомые не имеют крыльев: у примитивных форм отсутствие
крыльев является первичным, у более высокоразвитых — вторич-
ным, возникшим в результате редукции их в процессе эволюции.
Редукция крыльев характерна для паразитических видов — вшей,
пухоедов, блох, постельных клопов и др.
Крылья образуются как тонкие и плоские складки покровов
второго и третьего сегментов груди (рис. 149). Они представ-
ляют собой пластинки различной формы, образованные двумя сло-
ями кутикулы с подстилающей ее гиподермой. По крылу тянется
сеть жилок — трубчатых утолщений хитиновых покровов крыла,
придающих ему прочность. По жилкам в крыло заходят трахеи и
180
Рис. 148. Типы ножек насекомых:
Рис. 149. (/троение крыла насекомого:
/ жилкование крыла; // — разрез крыла;
/ . копалъиая жилка; 2-субкоегаль-
иая; ,3 радиальная; 4 медиальная;
5 . кубитальная; 6 - анальные; 7
югальные; Л1 трахея
/ бега гель пая (жужелицы); //
прыгательная (сверчка); III хиата-
Н'льная (богомила); /Г роющая
(медведки);
1 тазик; 2 вертлуг; ,'} ~ бедро;
4 голень; 5 лапка
нервы. Характер жилкования крыльев служит одним из важных
систематических признаков насекомых. У жуков передние кры-
лья - надкрылья - очень толстые и жесткие, их называют элн-
трами. Они служат как для полета, так и для защиты тонких
перепончатых крыльев.
Полет насекомых разнообразен: он может быть машущим,
парящим, порхающим и пр. Движение крыльев может быть очень
частым. Так, летящая пчела делает до 440 взмахов в секунду.
Брюшко насекомых состоит из 6- -12 сегментов. У взрослых
насекомых ножек на брюшке пет, но у некоторых личинок па
члениках брюшка расположено несколько пар печленистых лож-
ноножек. Самки многих насекомых имеют на конце брюшка яйце-
клад, при помощи которого откладывают яйца в почву или щели,
а паразитические наездники — в тело будущего хозяина личинки.
Некоторые насекомые имеют па конце брюшка жало, на котором
открывается проток ядовитой железы.
Покровы насекомых, как и других членистоногих, образо-
ваны однослойным эпителием — гиподермой и выделяемой ею хи-
тиновой кутикулой.
Н с р в и а я с и с т е м а состоит из сложного надглоточного
парного ганглия, соединяющегося с помощью окологлоточного
нервного кольца с меньшим по размеру подглоточным ганглием,
от которого вдоль тела по его брюшной стороне тянется цепочка
парных нервных узлов (по паре па каждом членике), соединен-
ных продольными нервными тяжами (рис. 150). Нередко наблю-
дается слияние нервных узлов соседних члеников. От нервных уз-
лов отходят к различным органам нервы.
О р г а н ы ч у в с т в у большинства насекомых отличаются
сложным строением. На голове расположены сложные и простые
181
1
Рис. 150. Нервная система пчелы:
/ — надглоточный ганглий; 2 -- ганглий I сегмен-
та груди; 3 — объединенный ганглий II, III сег-
ментов груди и I, II сегментов брюшка; 4 ганг-
лий III сегмента брюшка
Рис. 151. Внутреннее строение насекомого
(таракана):
/ — вид сверху; // — вид сбоку;
/ — глотка; 2 — пищевод; 3 — зоб; 4 — мус-
кульный желудок; 5 — слепые отростки кишеч-
ника; 6 — средняя кишка; 7 — задняя кишка;
8 — мальпигиевы трубочки; 9 — спинной сосуд
с рядом сердец; 10 — надглоточный ганглий;
//—брюшная нервная цепочка; 12 — семен-
ник; 13, 14 — придаточные железы полового
аппарата; 15 — трахеи; 16 — слюнная железа;
17 — ее резервуар; 18 — дыхальца; 19 — сим-
патическая нервная система
глаза. Сложные глаза состоят из многих, иногда до 20 и более
тысяч отдельных глазков (омматидиев) и могут воспринимать
форму и цвет предметов. Органы обоняния расположены на ан-
теннах, которые часто служат одновременно и органами осязания.
Органы слуха имеются далеко не у всех насекомых.
П и щ е в а р и т е л ь н а я с и с т е м а . Пищевой канал подраз-
деляется на передний, средний и задний отделы. В передний
отдел входят ротовая полость, в которую открываются слюнные
железы, глотка и пищевод. Задний отдел пищевода нередко
расширяется в зоб, служащий для накопления пищи (рис. 151).
У многих насекомых передний отдел кишечника заканчивается
мускульным желудком, в котором идет перетирание пищи. В сред-
ней кишке происходят переваривание и усвоение пищи. Часто в нее
впадает несколько слепых выростов кишечника, служащих для
увеличения его всасывающей поверхности.
Орга н а м и в ы д е л е и и я насекомых являются мальпигие-
вы сосуды — тонкие неветвящиеся трубочки, открывающиеся в
кишечник.
Органы дыхания. Дыхание у подавляющего большин-
ства насекомых осуществляется при помощи трахей. Воздух в них
поступает через отверстия но бокам тела —- дыхальца. Изнутри
трахеи выстланы тонкой хитиновой пленкой со спиральным утол-
щением (только мельчайшие трахеи лишены ее), придающим тра-
хее эластичность и препятствующим их сплющиванию. Трахеи
связаны рядом продольных стволов. Поступление и удаление воз-
духа из них происходит путем изменения объема брюшка сокраще-
нием мышц. Личинки некоторых насекомых (стрекоз, поденок
и др.), живущие в воде, дышат трахейными жабрами — тонко-
стенными (нередко ветвистыми) выростами брюшка, в которых
расположена сеть трахей. Иногда у личинок, живущих в воде,
встречаются жабры, лишенные трахей, газообмен происходит че-
рез их тонкие покровы.
Кровеносная система насекомых незамкнутая. Вдоль
спины в полости брюшка тянется длинная трубка, состоящая из
ряда пульсирующих камер — сердец. В грудном отделе сердце
продолжается крупным сосудом — аортой. Кровь попадает в серд-
це из полости тела через парные боковые отверстия с клапанами.
Из сердца кровь движется по аорте и через ее конечное отвер-
стие выливается в полость тела, омывая все органы.
Органы размножения. Насекомые раздельнополы.
У самцов в брюшке расположены парные семенники, от которых
отходят семяпроводы, соединяющиеся в непарный семяизвер-
гательный канал. Яичники самок имеют вид пучка трубочек,
постепенно расширяющихся книзу. Они открываются в парные
яйцеводы, которые ниже соединяются в единое влагалище, откры-
вающееся наружу. При спаривании семя самца вводится в сово-
купительную сумку самки, затем у одних насекомых по особому
каналу поступает во влагалище, где и происходит оплодотворение
183
Рис. 152. Развитие насекомых с непол-
ным превращением (саранча):
/ -- яйцо; 2 — личинки разных возрастных
стадий; 3 — взрослое насекомое
яиц, у других сперма из сово-
купительной сумки попадает в
семяприемник, где может сох-
раняться длительное время. От-
сюда семя отдельными порция-
ми поступает во влагалище и
оплодотворяет яйца.
У этих форм самка может в
течение долгого срока после
спаривания откладывать оплодотворенные яйца. Так, матка пчелы
после спаривания с трутнем в течение жизни (4—5 лет) дает ты-
сячи оплодотворенных яиц без повторного оплодотворения.
Развитие насекомых протекает либо без превращений, либо
с неполным или полным метаморфозом. При прямом развитии
(без превращений), что характерно для низших насекомых,
из яиц выходят особи, отличающиеся от взрослых преимуществен-
но малыми размерами и недоразвитыми половыми органами.
У насекомых с неполным превращением (рис. 152) из яиц
появляются личинки с чертами имаго, но отличающиеся от них
меньшей величиной, обычно зачаточными крыльями и еще слабо
развитыми половыми органами. Эти личинки претерпевают не-
сколько линек и со временем превращаются во взрослых насе-
комых, не проходя стадию куколки.
У насекомых с полным превращением (рис. 153) из яиц вы-
ходят червеобразные личинки, совершенно не похожие на имаго.
Достигнув определенного возраста и величины после ряда линек
покровов, они перестают двигаться и питаться и вскоре превра-
щаются в куколку. Куколка обычно неподвижна или может совер-
шать самые простые движения. Внутри ее тела происходит глубо-
кая перестройка организма с формированием тканей и органов
взрослого насекомого. Когда этот сложный процесс заканчивается,
покровы куколки лопаются и из них выходит имаго.
У многих насекомых наблюдается резко выраженный половой
диморфизм. Так, мелкие самцы зимней пяденицы имеют хорошо
развитые крылья и тонкое брюшко. Самки этой бабочки крупнее,
их крылья редуцированы, а брюшко раздуто.
У насекомых, живущих колониями, нередко имеет место по-
лиморфизм, при котором особи одного и того же вида имеют раз-
личное строение соответственно той роли, которую они играют в
жизни колонии. Так, в колониях многих термитов родоначальни-
ца семьи — матка отличается огромным брюшком, которое не
позволяет ей передвигаться, и ее кормят термиты-рабочие
(рис. 154). Ежедневно матка откладывает несколько тысяч яиц.
184
Рис. 153. Развитие насекомых с полным превращением (тутовый
шелкопряд):
1 - бабочка; 2 гусеница; <4 ...кокои; 4... куколка, изнлечеш1ая из ко-
кона
Рис. 154. Полиморфизм у термитов:
/-- рабочий; //солдат; III— крылатый самец; IV •- молодая самка,
сбросившая крылья после оплодотворения; У' — взрослая самка
Самцы термитов живут недолго. Они оплодотворяют матку. Ос-
новную массу членов колонии составляют термиты-рабочие, отли-
чающиеся мелкими размерами и небольшой головой: они строят
термитник, добывают пищу, выкармливают личинок. В термит-
никах обитают также термиты-солдаты, которых легко узнать по
огромной голове с мощными челюстями; они бесстрашно защи-
щают колонию от нападения врагов.
Экология насекомых. Подавляющее большинство насекомых —
обитатели суши. На суше насекомые населяют самые различные
места обитания. Многие из них являются настоящими подземными
животными, которые питаются подземными частями растений, гни-
ющими веществами или почвенными животными. Большое число
видов насекомых обитает в опавшей листве (лесной подстилке).
Многочисленны они и в травяном покрове, где находят убежище
и обилие пищи. Многие насекомые держатся на деревьях и кустах,
поедая листья, побеги, семена и плоды, высасывая соки или
вгрызаясь в древесину стволов. Наконец, некоторые насекомые
много времени проводят в полете, нередко поднимаясь с воздуш-
ными потоками на сотни, а то и на тысячи метров над поверхностью
земли. Водные насекомые обитают преимущественно в стоячих
водоемах и медленно текущих реках. В морях их почти нет.
По времени наибольшей активности насекомых делят на днев-
ных, сумеречных и ночных.
Замечено, что фауна насекомых тропических стран значи-
тельно богаче видами, чем энтомофауна умеренных и тем более
холодных зон. Это объясняется прежде всего тем, что насекомые
относятся к числу животных с переменной температурой тела. По-
этому в зонах умеренного и холодного климата почти все насе-
комые впадают в длительную зимнюю спячку, к которой смогли
приспособиться лишь относительно немногие виды. Но и в'теплое
время года климатические и погодные условия оказывают огром-
ное влияние на различные процессы жизнедеятельности насеко-
мых. Например, гусеница кукурузного мотылька при 20 °C пот-
ребляет 266 мм3/ч, при 12 °C — 95, а при 0 °C — лишь 22 мм3/ч
кислорода на 1 г массы. Бабочка совка-гамма на севере евро-
пейской части нашей страны за год дает только одну генерацию,
в средней полосе — две, а на юге — три генерации.
Погодные условия относятся к числу основных причин ко-
лебаний численности насекомых в разные годы. Это важно
учитывать при организации борьбы с насекомыми — вредителями
сельского хозяйства. Бороться с ними надо различными методами.
Колебания численности насекомых нередко обусловливаются так-
же обилием или недостатком кормов и миграциями животных.
В зависимости от численности насекомого область распро-
странения его может быть разделена на несколько зон:
зона постоянного обилия данного вида (а для вредителей —
также постоянной и активной вредной деятельности);
зона постоянного обитания, но лишь периодического обилия
186
в годы с благоприятными условиями жизни и размножения.
Вредоносная деятельность по годам выражена в разной степени;
зона, где насекомое редко встречается и не вредит расте-
ниеводству, но где в некоторые годы наблюдаются вспышки чис-
ленности и вредоносной деятельности;
зона, где данное насекомое обычно не встречается, но изредка
появляется, мигрируя из других зон в годы массового раз-
множения.
По образу жизни насекомые могут быть разбиты на следующие
группы:
свободноживущие, питающиеся растениями, животными, гнию-
щими веществами, трупами, навозом и т. и.;
паразиты растений: а) растительные эктопаразиты, живущие
на поверхности растения и сосущие их соки (тли, червецы и др.);
б) растительные эндопаразиты, живущие в растении и питающиеся
его тканями (личинки короедов и дровосеков);
паразиты животных: а) эктопаразиты — вши, блохи, пухоеды и
многие другие; б) эндопаразиты — личинки желудочных и кожных
оводов, наездников, яйцеедов и др.
По характеру питания среди насекомых различают:
фитофаги — растительноядные формы, которые, в свою оче-
редь, делятся на питающихся соками растений, листьями и побе-
гами, плодами, семенами, корнями, корнеплодами и т. д.;
зоофаги — насекомые, питающиеся животными;
копрофаги — насекомые, поедающие навоз и экскременты жи-
вотных;
иекрофаги -- насекомые, нищей которым служат трупы;
сапрофаги — насекомые, питающиеся гниющими раститель-
ными веществами;
паптофаги — всеядные насекомые.
Насекомые отличаются сложной нервной деятельностью.
В их поведении особое' значение имеют инстинкты — совокупность
подчас очень совершенных безусловных рефлексов, т. е. таких
реакций организма на раздражения внешней среды, которые не
приобретаются опытом данной особи, а сложились исторически в
течение длительного времени и стали наследственными, врожден-
ными. Иногда инстинкты отличаются большой сложностью и опре-
деляют весьма целесообразное поведение насекомого. Так, напри-
мер, песчаная оса аммофила выкармливает своих личинок
гусеницами бабочек. Найдя гусеницу, она вонзает жало в те места
тела, где расположены нервные узлы брюшной цепочки. Пораже-
ние нервных узлов не убивает гусеницу, но парализует ее движе-
ния. Затем оса затаскивает неподвижную гусеницу в вырытую за-
ранее норку и откладывает на нее свое яичко. Вышедшая личинка
долгое время питается живой парализованной гусеницей.
Особой сложности инстинкты достигают у общественных на-
секомых — муравьев, пчел, термитов и др. В их колониях (пред-
ставляющих потомство одной матки) обычно наблюдается диф-
187
Рис. 155. Полезные насекомые:
/ — муха-журчалка и ее хищная личинка; //--златоглазка и ее хищная личинка;
III, /Г — жук-красотел и его личинка; V — божьи коровки и их личинки; 17 — жуже-
лица; 17/ — етафилинуе
фереициация особей на несколько форм: маток, трутней, рабочих,
солдат и др. Каждая из этих групп выполняет в колонии свою
роль, имеет свои обязанности. В колониях некоторых обществен-
ных насекомых создаются иногда удивительно сложные и своеоб-
разные взимоотношения между их членами. Так, например, круп-
ные муравьи-амазонки нападают на колонии других муравьев и за-
хватывают их куколок. Выведшиеся из них муравьи превращаются
в «рабов» амазонок, обеспечивающих постройку муравейника, до-
бычу пищи, уход за молодняком амазонок и другие работы.
Некоторые тропические виды муравьев и термитов разводят в
своих гнездах «грибные сады», выращивая мицелий грибов на мас-
се мелко пережеванных листьев и древесины. Многие муравьи
культивируют корневых тлей, принося в свое гнездо и высаживая
на корни растений, они «доят» их, заставляя щекотанием усиков
выделять особые сахаристые вещества.
Хозяйственное значение насекомых. Значение насекомых для
хозяйства человека огромно и многообразно. Многие из них при-
носят большой вред сельскому хозяйству, уничтожая культурные
растения или снижая их урожайность. Насекомые-паразиты умень-
шают продуктивность сельскохозяйственных животных, вызывают
их тяжелые заболевания. Кровососущие насекомые — комары,
блохи, вши, клоны и др. — переносят возбудителей многих опас-
ных болезней человека. Есть среди насекомых и промежуточные
хозяева паразитических червей.
Но среди насекомых много видов, приносящих человеку боль-
шую пользу. Прежде всего надо помнить, что опыление многих
культурных растений производится пчелами, шмелями, цветочны-
ми мухами и другими насекомыми. Среди насекомых немало хищ-
ников, уничтожающих массу различных вредителей сельскохозяй-
ственных растений (рис. 155). Наездники, яйцееды и некоторые
другие перепончатокрылые насекомые откладывают яйца в тело
или яички вредных насекомых, что влечет их гибель. Пчелы
разводятся человеком для получения меда и воска, для опыле-
ния сельскохозяйственных растений. Гусеницы тутового шелкопря-
да дают шелковое волокно, служащее сырьем для выработки ес-
тественного шелка. Многие насекомые питаются трупами живот-
ных, их экскрементами, разными отбросами и гниющими вещества-
ми, очищая поля и леса. В тропических странах некоторых насеко-
мых (саранчу) употребляют в пищу.
ПОДКЛАСС ПЕРВИЧНОБЕСКРЫЛЫЕ (APTERYGOTA)
Первичнобескрылые — примитивные насекомые, у которых от-
сутствие крыльев первично, т. е. крыльев не было и у их пред-
ков (рис. 156). Челюсти погружены в капсулу (скрыточелюст-
ные). Органами зрения служат простые глаза, у ряда видов гла-
за отсутствуют. Развитие без превращений. Обитают в почве,
сырых местах, под камнями, во мхе, в погребах. В почве особен-
189
Рис, 156. Первичнобескрылые насекомые:
энтогнатные: 1 — отряд Бессяжковые; 2 — отряд
Ногохвостки; 3 — отряд Двухвостки; эктогнат-
ные: 4 — отряд Щетиикохвостки (чешуйница)
но много мелких (длиной до 1 мм) бесцветных прыгающих ного-
хвосток, которые играют некоторую роль в почвообразователь-
ных процессах.
ПОДКЛАСС КРЫЛАТЫЕ (PTERYGOTA)
Крылатые — более высокоорганизованная группа насекомых.
Челюсти расположены свободно (открыточелюстные). Подавляю-
щее большинство их имеют крылья. У остальных отсутствие крыль-
ев — явление вторичное; они были утрачены в процессе эво-
люции, чаще всего как приспособление к паразитическому образу
жизни. Глаза как простые, так и сложные. Развитие происходит
с полным или неполным превращением. К этому подклассу при-
надлежат большинство видов насекомых. Подкласс разделяется на
ряд отрядов, из которых ниже описаны преимущественно те,
представители которых имеют значение для сельского хозяйства
нашей страны. Отряды, входящие в данный подкласс, могут быть
объединены по особенностям развития представителей в два отде-
ла: насекомых с неполным и полным превращением.
ОТДЕЛ НАСЕКОМЫЕ С НЕПОЛНЫМ ПРЕВРАЩЕНИЕМ
(HETEROMETABOLA)
Отряд прямокрылые (Orthoptera). К ним относятся саранча,
кузнечики, сверчки, медведки и др. Передние крылья у них прямые,
кожистые, относительно узкие, складывающиеся вдоль тела. Они
закрывают широкие перепончатые крылья второй пары. Ротовые
органы грызущие. Задние ноги прыгательного типа: они отличаются
от ног двух передних пар большой длиной, зависящей от сильно уд-
линенных бедер и голеней. Благодаря такому устройству ног пря-
мокрылые способны делать большие прыжки. Большинство прямо-
крылых могут стрекотать — издавать характерные звуки путем
190
трения друг о друга некоторых частей тела; кузнечики и сверчки
при этом трут крыло о крыло, саранча и кобылки — бедро задних
пог о край крыла.
Многие прямокрылые сильно вредят сельскохозяйственным
растениям, к их числу относятся саранча, кобылки, кузнечики,
медведки.
Рис. 157.' Саранча перелетная:
1 -взрослые насекомые; // личинка (пешая саранча); /// кубышки
с яйцами
191
Рис. 158. Медведка
Рис. 159. Тараканы:
/— рыжий таракан-прусак; II -- черный таракан
Рис. 160. Богомол
Саранча - довольно крупные насекомые (до 10 см длиной),
похожие па кузнечиков, по отличающиеся от них короткими (до
2 см) усиками (рис. 157). Из многих видов саранчи, обитающих
в СССР, большое значение как вредитель сельского хозяйства
имеет саранча перелетная (Locusla niigraloria). Она размно-
жается в плавнях южных рек. Самка коротким яйцекладом,
находящимся на конце брюшка, делает в почве норку, куда
откладывает яйца вместе с пенистыми выделениями особых желез.
Эти выделения затвердевают, склеивая яйца в колбасовидную
кубышку. Вышедшие из яиц личинки похожи на взрослых на-
секомых, но крылья у них еще зачаточны. Стаи (кулиги) этих личи-
нок, именуемых пешей саранчой, могут совершать большие кочев-
ки, уничтожая ио пути посевы. Взрослая крылатая саранча хо-
рошо летает, и ее стаи делают далекие перелеты. Она может
наносить нолям еще большие повреждения, чем «пешая». В
СССР ведется упорная борьба с саранчой главным образом
опылением с самолетов различными ядами плавней. Благодаря
планомерной ликвидации очагов размножения численность саран-
чи в нашей стране резко сократилась.
Кобылки близки к саранче, но отличаются более мелкими
размерами и обычно яркой окраской задних крыльев. Многие ви-
ды кобылок наносят ощутимый вред сельскохозяйственным куль-
турам, лугам и пастбищам.
Кузнечики, которые представлены в нашей фауне многими
видами, легко узнаются по очень длинным (длиннее тела) тон-
ким усикам. Вредоносная деятельность их не столь велика. Не-
которые из них иногда повреждают огородные культуры и травы
па лугах, но многие полезны как хищники, поедающие вред-
ных насекомых.
Медведка (Grillotalpa) живет в норках, которые роет своими
мощными, укороченными, широкими передними ногами (рис. 158).
Приносит вред огородным культурам, повреждая корни и корне-
плоды растений.
Отряд Тараканы (Blattoidea). Имеют уплощенное тело
(рис. 159). Передние крылья кожистые, задние — тонкие,
перепончатые. У самок крылья нередко укорочены или полностью
редуцированы. Ротовые органы грызущие. Задние ноги лишь нем-
ного длиннее передних и средних. В домах живут крупный — чер-
ный таракан и мелкий — рыжий таракан-прусак. Они не только
портят продукты, но опасны распространением различных забо-
леваний человека.
Отряд Богомолы (Mantoidea). Хищные насекомые, ловят добы-
чу при помощи хватательных передних ног (рис. 160). Удлинен-
ная голень входит своим зазубренным краем в желобок длинных
бедер, что позволяет зажимать жертву. Крылья листовидные. Ро-
товые органы грызущие. Встречаются в южных областях СССР.
Полезные истреблением вредных насекомых.
7 Б. А. Кузнецов и др.
193
Рис. 161. Стрекоза-коромысло:
1—взрослое насекомое; 2—личинка; 3—ее голова с вытянутой маской
Отряд Стрекозы (Odonata). Голова стрекоз несет огромные
сложные глаза (рис. 161). Брюшко вытянутое, тонкое. Крыльев две
пары сходного строения, с густой сетью жилок. На передних
краях их обычно имеется утолщенный хитиновый прямо-
угольный участок. У большинства стрекоз крылья постоянно рас-
простерты в стороны, у некоторых же могут складываться
вертикально над спинкой. Ротовые органы грызущие. Стрекозы.-
дневные хищники, питаются насекомыми, которых ловят на
лету. Истребляют много комаров. Держатся преимущественно
близ водоемов. Личинки живут в воде. Их нижняя губа
превращена в снабженную щипцами маску, служащую для схва-
тывания добычи: мелких водных животных, в том числе мальков
рыб.
Отряд Вши (Anoplura). Накожные паразиты многих млеко-
питающих. Мелкие бескрылые насекомые с плоским телом и цепки-
ми лапками, несущими загнутые коготки (рис. 162, //).
Ротовые органы колюще-сосущие, своеобразного строения.
В спокойном состоянии колющий аппарат втянут в рот. Прокалы-
вая кожу хозяина, вши сосут его кровь. Распространяют среди
людей сыпной тиф.
Отряд Пухоеды и Власоеды (Mallophaga). Мелкие насекомые,
обитающие в перьевом покрове птиц (пухоеды) и волосяном по-
крове млекопитающих (власоеды). Похожи на вшей, по отли-
194
чаются от них грызущими ротовыми органами, а также очень боль-
шой головой, ширина которой превышает ширину груди (см.
рис. 162, /). Различные виды паразитируют па млекопитающих
и птицах. Встречаются почти на всех домашних животных. Пи-
таясь пухом, шерстью п роговым слоем эпидермиса кожи хозяев,,
они вызывают кожные заболевания, разрушают перьевой п воло-
сяной покров.
Отряд Равнокрылые (Homoptera). К этому отряду относятся
тли, червецы, цикады и др. Отличаются они двумя парами сходных
по форме и строению топких крыльев с небольшим числом жилок.
Ротовые органы колюще-сосущие.
Тли (разные виды сем. Aphidae) (рис. 163, ///)- -мелкие
насекомые с мягким раздутым телом. У многих тлей наблюдаются
сложные циклы развития, при которых бескрылые партеногене-
тические поколения сменяются крылатыми половыми. Тли сосут
соки растений, нередко вызывая их гибель. Особенно вредна
виноградная тля филлоксера (Viteus vi lift >lii), которая паразити-
рует па корнях виноградных лоз, приводя их к гибели. На капусте
живет капустная тля (Brevicoryne brassicae), па яблонях --
яблонная тля (Ap/iis pond) и кровяная тля (Eriosoma laniperutn),
устраивающая гнездо из красной пены.
Червецы (сем. Coccidae) — эктопаразиты южных растений,
особенно цитрусовых (см. рис. 163, /, //). Бескрылые самки их по-
крыты сверху щитком из особого воскообразного вещества. Очень
мелкие самцы у некоторых видов имеют тонкие крылья. Самки
прокалывают хоботком покровы растений и сосут их соки,
сильно истощая деревья.
Цикады (сем. Cicadidae и др.) значительно крупнее тлей
(иногда до 5 см). Обычно крылья имеют и самки, и самцы.
Многие цикады - вредители сельскохозяйственных растений.
Кроме того, они переносят некоторые вирусные болезни
Рис. 162. Пухоед собачий (/), вошь платяная (//), вошь свиная
7*
195
Рис. 163. Представители отряда Равнокрылые:
/—самец красного гигантского червеца; II — самка мучнистого
червеца; /// —капустная тля;
I —бескрылая самка; 2— крылатая самка; 3 -• нимфа; 5 —
личинки
Рис. 164. Полужесткокрылые:
/ — гладыш; II — постельный клоп; III — клоп-черепашка
Рис. 165. Переднее крыло полужесткокры- — )
лых: i /\————/
1 ~ утолщенная часть; 2 — перепончатая часть —
этих растений. Личинки живут в почве, повреждая корни де-
ревьев и кустов. Самцы многих цикад издают своеобразное
стрекотание.
Отряд Полужесткокрылые, или Клопы (Heteroptera). Обшир-
ная группа насекомых, включающая паразитов теплокровных жи-
вотных и большое число вредителей полезных растений (рис. 164).
Передние крылья в основной части жесткие, а к свободным
концам — тонкие, прозрачные. Задние крылья также тонкие, пере-
пончатые. В покое как задние, так и конечные половины пе-
редних крыльев складываются под жесткими основными частями
последних (рис. 165). У постельного клопа крылья редуцированы.
Ротовые органы колюще-сосущие.
Клоп-черепашка вредная (Eurygater integriceps)--вреди-
тель сельскохозяйственных растений. Длина насекомого до 15 мм.
Плоское овальное тело покрыто коричневым щитком передне-
спинки и основаниями передних крыльев (см. рис. 164,///).
Живут на хлебных злаках: взрослые сосут соки из молодых по-
бегов, личинки — из наливающегося зерна. Другие виды клопов
приносят вред различным полевым, садовым и огородным расте-
ниям. Постельный клоп (Cimex leciularius) паразитирует на чело-
веке и различных теплокровных животных.
ОТДЕЛ НАСЕКОМЫЕ С ПОЛНЫМ ПРЕВРАЩЕНИЕМ
(HOLOMETABOLA)
Отряд Жесткокрылые, или Жуки (Coleoptera). Самый богатый
видами отряд насекомых. Передние крылья жуков превратились
в надкрылья — твердые, жесткие, выпуклые хитиновые пластинки.
В покое они прикрывают сложенные тонкие, нежные, перепон-
чатые задние крылья. Ротовые органы грызущего типа. Личинки
червеобразные, обычно с тремя парами членистых ножек на трех
первых сегментах. Многие жуки относятся к числу опасных вреди-
телей лесных, садовых, огородных и полевых растений. Из вреди-
телей леса и сада большое значение имеют короеды, усачи и май-
ские жуки.
Короеды (сем. Ipidae) — мелкие жучки длиной менее 1 см
(рис. 166)'. Самки короедов вгрызаются в кору деревьев и делают
под ней длинный прямой маточный ход, к стенкам которого
прикрепляют свои яички. Выведшиеся из них личинки делают под
корой личиночные ходы, отходящие в обе стороны от маточного.
В нашей стране встречается много видов короедов, повреждающих
древесные породы, причиняя большой убыток лесному и садовому
хозяйствам.
197
Рис. 166. Короед (жук-типограф):
/ — взрослый жук; // — .ходы короедов
под корой ели
Рис. 167. Жук-усач сосновый
Усачи (сем. Ceratnbycidae) — жуки разной величины с очень
длинными, постепенно утончающимися к концу усиками (рис. 167).
Личинки многих видов живут в древесине различных деревьев,
проделывая в ней ходы, что вредит деревьям и снижает качество
лесных материалов.
Майские жуки (Meloluntha) во взрослом состоянии питаются
листьями деревьев и кустов, а их личинки, живущие в течение
трех лет в почве, повреждают корни (рис. 168).
Многочисленны жуки — вредители огородных и полевых куль-
тур. Из них наибольший вред приносят колорадский жук, свекло-
вичный долгоносик, жуки-щелкуны и хлебные жуки.
Колорадский жук (Leptinofarsa decemlineata) имеет тело ок-
руглой формы, длиной до 10 мм. Легко узнается по ярко-желтой
окраске с пятью черными продольными полосами на каждом
надкрылье (рис. 169). Родина его Северная Америка, откуда он
Рис. 168. Майский жук (/) и его
личинка (//)
Рис. 169. Колорадский жук (/) и
его личинка (//)
198
Рис. 170. Долгоносик све-
кловичный
Рис. 171. Щелкун посевной (/) и его ли-
чинка (//)
был завезен в ряд стран Европы. Опасный вредитель картофеля,
листьями которого питаются жуки и личинки.
Свекловичный долгоносик (Boihytioderes рипсiiventris) имеет
длину тела около 15 мм (рис. 170). Как у всех видов
долгоносиков, передняя часть головы вытянута в хоботок, на конце
которого расположены рот и усики. Окраска серая с буроватой
полосой поперек надкрыльев. Жуки повреждают всходы свеклы, а
безногие личинки, живущие в почве,— ее корни.
Жуки-щелкуны (сем. Elateridae) имеют вытянутое уплощенное
тело длиной до 17 см (рис. 171). Если щелкуна положить на
спину, то он, изгибаясь, с характерным щелканьем подпрыгивает
и перевертывается на ноги. Личинок щелкунов, имеющих очень
твердые хитиновые покровы желто-бурого цвета, называют про-
волочными червями. Они живут в почве, или под корой гниющих
пней. Некоторые (например, личинки посевного щелкуна) сильно
вредят злаковым культурам, подгрызая их корни.
Хлебные жуки (Anisoplia, сем. Scarabaeidae) имеют блестящую
зеленовато-черного цвета переднюю спинку и буроватые над-
крылья, обычно с темными пятнами. Все они — серьезные вре-
дители хлебов. Взросые особи поедают еще мягкие зерна хлебных
злаков, а личинки грызут корни.
Среди жуков немало видов, приносящих значительную пользу
сельскому хозяйству. Таковы, например, хищные жуки жужелицы
(сем. Carabidae), которые уничтожают большое количество
вредных насекомых; божьи коровки и их личинки, питающиеся
вредными тлями. Многие жуки (навозники, скарабеи, мертвоеды,
могильщики и др.) очищают поля и леса от трупов и
навоза.
199
Рис. 172. Блоха собачья:
/--личинка; // — куколка; III - взрослое
насекомое
Отряд Блохи (Aphaniptera).
Мелкие насекомые с уплощен-
ным с боков телом, без крыль-
ев, с удлиненными прыгательны-
ми задними ногами (рис. 172).
Ротовые части колюще-сосущие.
Эктопаразиты млекопитающих
и птиц. Различным видам зве-
рей и птиц свойственны опреде-
ленные виды блох. Взрослые
блохи питаются кровью хозяев.
Червеобразные личинки блох
живут в трещинах пола, мусо-
ре, гнездах хозяев, питаясь раз-
лагаю щи м и с я орган и ч е с к и м и
веществами. Блохи служат пе-
реносчиками различных забо-
леваний человека и животных.
Отряд Чешуекрылые, или Бабочки (Lepidoptera). Две пары
больших крыльев бабочек покрыты мелкими хитиновыми чешуй-
ками, черепицеобразно налегающими друга на друга (рис. 173).
У дневных бабочек в спокойном состоянии крылья сложены верти-
кально над спиной, а у ночных обычно лежат вдоль тела крыше-
образно. Ротовые органы сосущие, преобразованные в гибкий
хоботок, сворачивающийся в спираль. Им бабочки сосут нектар
цветков. Личинки бабочек — гусеницы имеют червеобразную
форму. Три первых членика их тела несут настоящие членистые
ножки, а остальные (или часть остальных) — нерасчленениые
ложные ножки. Ротовые органы гусениц в отличие от имаго грызу-
щего типа.
Тутового шелкопряда (Bombyx niori) разводят с целью полу-
чения натурального шелка. Гусеницы этих бабочек имеют особые
железы, выделяющие белковое вещество фиброин, которое па воз-
духе затвердевает, превращаясь в шелковую нить. Когда гусеница
достигает полного роста, она делает из нити кокои, в котором
и окукливается. На шелкомотальных фабриках из нити коконов
свивают шелковую пряжу. Разводят также дубового шелкопряда
(Antherea pernyi), из коконов которого получают более грубую
пряжу, идущую на изготовление ткани чесучи.
Среди чешуекрылых много видов, гусеницы которых являются
вредителями лесов и садов. Так, гусеницы непарного шелкопряда
(Porlhetria dispar), питаясь листьями различных деревьев, в годы
массового размножения могут уничтожить целые массивы лесов
и садов (рис. 174).
Кольчатый шелкопряд (Malacosonia neustria) откладывает
яйца колечком вокруг веточек деревьев (отсюда его название).
Гусеницы в годы большой численности причиняют огромный вред
лиственным деревьям, объедая листву.
200
Рис. 173. Чешуйки па крыле бабочки:
/ расположение чешуек Па крыле лисиной бабочки; //, 111
пых нидон бабочек
форма чешуек ра.ч-
Рпс. 174. Шелкопряд непарный:
/ самеп.; // самка,
IV кокон е куколкой
о1кла дынаюпшя яйпа, III
I усенни ы разного возраст,
Сосновый шелкопряд (Dendrolimus pini) — один из основных
вредителей сосны, нередко уничтожающий сосновые боры на боль-
шой площади.
Златогузка (Euproctis chrysorrhoea) — небольшая белая ноч-
ная бабочка, конец брюшка которой покрыт золотистыми волос-
ками.
Гусеницы сильно повреждают плодовые деревья, объедая
листья. Зимуют они в больших гнездах, построенных из листьев,
соединенных шелковыми нитями.
Боярышница (Aporia craiaegi) — крупная дневная бабочка бе-
лого цвета с черноватыми жилками крыльев. Гусеница держится
на плодовых деревьях. Вредитель фруктовых садов.
Яблоневая моль (Hypononieuta tnalinella) — мелкая белая, с
черными пятнышками бабочка, по величине и форме сходная
с обычной комнатной молью. Гусеницы живут группами на листьях
яблони под тонким слоем паутины. Серьезный вредитель ябло-
невых садов.
Яблоневая плодожорка (Laspeyresia- pomonella) — мелкая
ночная бабочка, гусеница которой живет в мякоти плодов яблони.
Яблоки с «червоточинкой» рано опадают, и ценность их резко
снижается.
Из бабочек, гусеницы которых наносят вред огородным куль-
турам, надо прежде всего указать широко распространенную
капустную белянку (Pieris brassicae), получившую название за
чисто-белую окраску с несколькими черными пятнышками на кры-
льях (рис. 175).
Гусеница сильно повреждает капусту и некоторые другие
огородные растения. Гусеницы более мелкой репной белян-
ки (Pieris гарае) вредят репе, брюкве, редису.
Рис. 175. Белянка капустная:
/ — бабочка; II — яйца; III — гусеница; IV — куколка
202
Рис. 176. Совка озимая:
/ - бабочка; // гусеница; Ш куколка
Гусеницы ряда бабочек на-
носят вред также и зерновым
культурам.
Так, гусеницы ночной ба-
бочки озимой совки (Aprotis
sepetum) (рис. 176) питаются
в основном всходами хлебных
злаков.
Наконец, гусеницы некото-
рых бабочек - молей, огневок
и др.— портят продукты пита-
ния и изделия из животного
сы рья.
Отряд Двукрылые (Diptera).
К ним относятся мухи, слепни,
оводы, мошки, москиты, комары. У них сохранилась лишь первая
пара крыльев, а вторая превратилась в жужжальца булаво-
видные придатки, расположенные позади основания крыльев.
Внутри жужжалец находятся органы равновесия, очень важные
для полета насекомых. Ротовые органы различного устройства.
Личинки безногие, обычно без ясно дифференцированной головы.
Комары (сем. Ctilicidae) имеют длинные усики и колюще-
сосущие ротовые органы. Самцы комаров питаются нектаром или
соком растений, а самки многих видов - кровью человека и жи-
вотных. Личинки и куколки живут в стоячих водоемах (рис. 177).
Малярийные комары (Anopheles) распространяют малярию.
Москиты (Phlebothomus) — мелкие двукрылые насекомые,
длина тела которых обычно не превышает 3 мм (рис. 178).
Тело покрыто волосками. Самцы сосут соки растений. Самки
питаются кровью человека и теплокровных животных. Очень мно-
гочисленны в тропических странах. В СССР встречаются в
Крыму, Средней Азии, на Кавказе. Укусы москитов весьма болез-
ненны и вызывают зуд кожи. Распространяют возбудителей
ряда заболеваний человека: лейшманиозы, летний грипп (заболе-
вание типа временной лихорадки).
Мошки (сем. Siniullidae) хорошо известны жителям тайги
(рис. 179). Они составляют основную массу гнуса — огромных
скоплений мелких кровососущих насекомых. Мошки, длина кото-
рых не превышает 5 мм, отличаются коротким телом с приподнятой
горбом передней грудкой. Усики их короче, чем у комаров, но
длиннее, чем у мух. Кровью теплокровных животных и человека
питаются только самки. Мошки переносят возбудителей ряда забо-
леваний человека и сельскохозяйственных животных.
203
Рис. 177. Комар обыкновенный:
/ — яйца; II — личинка; III — куколка; IV — взрослая самка
204
Галлицы (сем. /thouididae) вклю- \
чают большое число видов мелких
комариков с удлиненным телом, длин-
ными ногами и тонкими крыльями с
немногими продольными жилками
без поперечных соединений. Личинки /ff
галлиц, поселяясь в тканях расте-
ний, нередко вызывают образование рис. 179. Мошка
наростов -- галлов. Некоторые виды
галлиц причиняют значительный
вред сельскохозяйственным рас-
тениям. Такова, например, гессенская мушка (вернее, комарик)
(Mayetiola destructor), личинка которой живет в стеблях хлебных
злаков (рис. 180).
Мухи (сем. Muscidae) отличаются широким уплощенным те-
лом, полушаровидной головой с короткими усиками. Обыкновен-
ная комнатная муха опасна тем, что переносит на лапках и хоботке
яйца паразитических червей и возбудителей различных заболе-
ваний. Столь же опасны как распространители заболеваний круп-
ные зеленые и синие падальные мухи.
Слепни (сем. Tabaiiidae) — крупные или среднего размера кро-
вососущие мухи с огромными радужными глазами. Укусы слепней
беспокоят скот. Являются переносчиками сибирской язвы.
Оводы (сем. Oestridae) относятся к числу важных паразитов
сельскохозяйственных животных (рис. 181). Отличаются от слеп-
ней коротким, покрытым волосками телом и небольшими глазами.
У взрослых оводов недоразвиты ротовые органы, и они за недол-
гую жизнь ничего не едят. Личинки бычьего овода (liypoderma
bouts) и овода крупного рогатого скота (Hypodernia liueala) пара-
зитируют в теле коров и быков, в последних стадиях своего разви-
тия скапливаясь под кожей. Личинки овечьего овода (Oestrus)
живут в носовой полости и лобных пазухах овец, вызывая ложную
Рис. 180. Гессенский комарик:
/ — самец; II — самка; III - личинка
205
Рис. 181. Желудочный овод:
/-яйцо на волоске; II—IV—личинки
разных стадий развития; V — самец;
VI — самка; VII - личинки на стенке
желудка лошади
«вертячку». Желудочные оводы (сем. Gastrophilidae) похожи на
кожных оводов. Их личинки паразитируют в кишечнике и двенад-
цатиперстной кишке лошадей и ослов, нередко вызывая тяжелые
воспаления слизистой оболочки этих органов. Взрослые оводы от-
кладывают яйца на шерсть лошадей, откуда они слизываются
хозяином.
Отряд Перепончатокрылые ( Hymenoptera). Отряд объединяет
свыше 70 тыс. видов пчел, шмелей, ос, наездников, муравьев и не-
206
которых других групп насекомых (рис. 182). За исключением
бескрылых форм, перепончатокрылые имеют две пары перепон-
чатых, сравнительно небольших крыльев, из которых задние за-
метно меньше передних. На крыльях имеется небольшое число
жилок, образующих крупные ячейки. У большинства видов
крылья имеют своеобразный радужный переливчатый оттенок.
Часто грудь соединяется с брюшком тонким стебельком. На конце
брюшка самок обычно имеется яйцеклад или жало, являющееся
видоизмененным яйцекладом. Ротовые органы грызущие или гры-
зуще-сосущие.
Многие перепончатокрылые живут обществами, которые явля-
ются потомством одной матки. У многих общественных перепон-
чатокрылых наблюдается полиморфизм. Их семьи состоят из одной
или нескольких самок (маток), некоторого количества самцов и
многих рабочих особей (бесплодных самок). Этим насекомым
свойственна сложная нервная деятельность, в которой преобла-
дают инстинктивные действия.
Личинки многих перепончатокрылых паразитируют в яйцах
или личинках других насекомых, в том числе различных вредите-
лей растениеводства, чем они приносят большую пользу сельско-
му хозяйству. Ряд их принимает деятельное участие в опылении
Рис. 182. Перепончатокрылые насекомые:
/ - муравьи; II - крыжовниковый пилильщик; III - наездник парализует гусеницу
бабочки перед откладкой на нее яиц; IV --яйцеед-трихограмма;
/ - рабочий; 2 -- крылатый самец; .7 солдат; 4. взрослая самка; 5 — личинка
207
сельскохозяйственных растений. Пчелы дают мед, воск, пчели-
ный яд и др.
Из многих видов перепончатокрылых наибольшее значение
имеют следующие группы.
Пчелы. Наибольший интерес представляет медоносная пчела
(Apis mellifera), которая встречается в диком состоянии и раз-
водится в пчеловодческих хозяйствах. Дикие пчелы живут в дуп-
лах деревьев, трещинах скал и других убежищах. Домашних
пчел содержат в ульях различной конструкции. Пчелиная семья со-
стоит из матки, самцов — трутней и многих бесплодных самок --
рабочих пчел (рис. 183). Зимой в улье остаются только матка и
некоторое число рабочих пчел, народившихся в конце лета. Весной
рабочие пчелы начинают строить соты с шестигранными ячейка-
ми; материалом для их постройки служит воск, выделяемый особы-
ми железами, расположенными снизу брюшка^Часть ячеек рабо-
чие пчелы заполняют пищевыми продуктами — медом и цветочной
пыльцой, часть же служит камерами, в которые матка откладывает
яйца. Большинство ячеек мелкие и тонкостенные, в них матка кла-
Рис. 183. Медоносная пчела:
/ — мачка; II — ее голова; /// — рабочая пчела; IV - ее голова; И тру-
тень; I'/ — его голова
208
дет оплодотворенные яйца, из которых при особом режиме кормле-
ния развиваются рабочие пчелы. Некоторые ячейки, более крупные
и толстостенные, предназначены для выведения трутней из нео-
плодотворепных яиц. Наконец, в сотах устраивается несколько
особо крупных, сначала чашеобразных, а позднее желудепо-
добных ячеек — маточников, в которых выводятся матки. В такую
ячейку старая матка откладывает оплодотворенное яйцо, из него
выходит личинка. Рабочие пчелы выкармливают се особой пи-
тательной смесью — «пчелиным молочком», а затем медом и пыль-
цой, что и обусловливает превращение личинки (после стадии ку-
колки) в молодую матку. За день матка откладывает до 1 тыс. яиц
и более. Летом рабочие пчелы собирают нектар и пыльцу с цветков
различных растений. Нектар перерабатывается в расширенном
отделе пищевода — медовом желудочке — в мед. Когда из маточ-
ника появляется первая молодая матка, старая вместе с частью
рабочих пчел обычно покидает улей и образовавшийся рой ищет
нового убежища. Молодая же матка и выведшиеся к этому
времени трутни вскоре совершают брачный полет, во время кото-
рого происходит спаривание. Сперма, введенная в спермоирнемиик
матки, сохраняется весьма долго и служит для оплодотворения
многих тысяч яиц. После брачного полета матка возвращается в
улей, а трутни вскоре погибают.
Пчеловодство — важная отрасль сельского хозяйства СССР.
При хорошей организации дела от одной семьи пчел можно полу-
чить до 300 кг меда и много воска. Пчелы приносят также огром-
ную пользу, совершая опыление сельскохозяйственных растений.
Шмели (сем. Bonibidae) отличаются от пчел более крупными
размерами и относительно толстым телом, густо покрытым волос-
ками. Зиму переживают только матки шмелей, которые весной
строят гнездо в земле, под мхом пли в другом укромном
месте, делают там небольшие соты и выводят первых рабочих
особей. В дальнейшем же работы в гнезде --- постройку сотов, вы-
кармливание молодняка и др. — выполняют только рабочие шмели,
а матка занята откладкой яиц. К осени в гнезде появляются
трутни и молодые самки. После спаривания трутни погибают, а
матки с приближением холодов забиваются в укрытые места на зи-
мовку. Шмели относятся к числу важнейших опылителей растений,
особенно клевера. Поэтому часто по краям клеверищ ставят не-
большие ящички для привлечения шмелей, которые устраивают в
них гнезда.
Наездники (сем. / cliiieumotiidae) — стройные насекомые с
брюшком, сидящим обычно на длинном стебельке (см. рис.
182, ///). У самок имеется длинный яйцеклад, которым
они вводят яйца в тело жертвы (чаще всего гусениц бабочки).
Личинки паразитируют в теле насекомых, вызывая их гибель. По-
скольку многие наездники откладывают яйца в тело вредителей ра-
стениеводства, они приносят большую пользу сельскому хозяй-
ству.
209
Яйцееды (сем. Scelionidae и др.) близки к наездникам
(см. рис. 182, IV). Это мелкие перепончатокрылые, откладываю-
щие яички в яйца других насекомых. Выведшиеся личинки яйце-
едов поедают личинок хозяев. Яйцеед трихограмма откладывает
яички в яйца различных бабочек, а теленомус — в яйца клопа-
вредной черепашки.
В нашей стране работает ряд биофабрик, выводящих еже-
годно миллиарды этих яйцеедов, которые затем выпускаются
на поля для истребления вредителей.
Муравьи (сем. Fornu'cidae) живут в сложных постройках —
муравейниках. У некоторых видов муравьев (см. рис. 182, /) это
сложные лабиринты подземных ходов, у других муравейник имеет
еще и надземную часть, обычно в виде конуса из хвои и иных
материалов. В больших муравейниках обитает много тысяч му-
равьев. В отдельных камерах их находятся яйца, личинки и ку-
колки в коконах (неправильно именуемые муравьиными яйцами).
Подавляющее большинство обитателей муравейников составляют
неспособные к размножению бескрылые самки — рабочие му-
равьи.
Они строят муравейник, добывают пищу себе, матке и
личинкам, защищают гнездо от врагов. В середине лета на
муравейниках появляются крылатые особи — стройные самцы и
более массивные самки. После брачного полета самки освобож-
даются от крыльев и начинают строить новое гнездо, а затем
приступают к откладке яиц, из которых развиваются рабочие
особи. Возникает новый муравейник. Крупные лесные муравьи,
строящие конусовидные муравейники, весьма полезны истребле-
нием многих вредителей леса.
Среди перепончатокрылых имеется ряд видов, вредящих сель-
скохозяйственным культурам. Таковы различные пилильщики
(сем. Ceph.id.ae). В отличие от большинства других перепонча-
токрылых их брюшко имеет широкое основание без сужения на
месте соединения с грудью. Особенно вреден хлебный пилильщик
(Cephus pygniaeus). На конце брюшка самки этого насекомого
имеется пиловидный яйцеклад, которым она пропиливает в стеблях
хлебных злаков отверстия и откладывает туда яйца. Вышедшие
личинки питаются тканями стебля, передвигаясь вниз. Спустив-
шись к основанию стебля, они подпиливают соломку и остаются
зимовать в оставшейся части. Поэтому основным способом борьбы
с хлебным пилильщиком является лущение стерни.
ПРОИСХОЖДЕНИЕ ЧЛЕНИСТОНОГИХ
Не подлежит сомнению, что членистоногие произошли от кольча-
тых червей. Это доказывается наличием многих общих для
этих типов животных черт строения: сегментации тела, строения
нервной системы в виде брюшной нервной цепочки, сходства
кровеносной системы с главным сосудом, лежащим па спинной
210
стороне тела, присутствием у некоторых членистоногих видоизме-
ненных метанефридиев, сходством некоторых черт строения па-
раподиев многощетинковых кольчатых червей и конечностей рако-
образных. ;
ТИП МОЛЛЮСКИ (MOLLUSCA)
Общая характеристика. Моллюски (мягкотелые) — вторичнопо-
лостные животные с несегмснтированным телом, обычно заключен-
ным в раковину. Тело состоит из трех отделов: головы, туловища
и ноги. Для моллюсков характерна мантия — складка кожи с
многочисленными и разнообразными железами, выделяющими
секреты, идущие на построение раковины. Между мантией и телом
находится мантийная полость, в которой расположены органы
дыхания — жабры у водных и легкие у наземных видов. Покровы
моллюсков тонкие, мягкие, богатые слизистыми железами. Крове-
носная система незамкнутая. Центральная нервная система обра-
зована обычно нервными узлами, соединенными комиссурами.
Моллюски, как и кольчатые черви, членистоногие, иглокожие
и хордовые, относятся к группе высших вторичнополостных жи-
вотных. Это доказывается наличием у них целома, кровеносной
системы, метаиефридиальных органов выделения, ганглиозной
нервной системы. От других вторичнополостных животных они
отличаются отсутствием сегментации тела. Сходство начальных
стадий дробления яиц, наличие ряда общих черт строения личинок
и отмеченных выше особенностей взрослых животных сближают
моллюсков с кольчатыми червями. Возможно, что эти две группы
животных произошли от общих предков.
Известно более 100 тыс. видов моллюсков. Они встречаются
в морях, пресных водоемах и на суше.
Практическое значение моллюсков велико и разнообразно.
Многие идут в пищу человека и корм скоту. Из жемчужниц добы-
вают жемчуг. Раковины используются для выработки перламутро-
вых изделий. Ряд видов наносят значительный вред полевым
и огородным культурам. Некоторые служат промежуточными
хозяевами опасных паразитов животных и человека. В теоретиче-
ском отношении интересны ископаемые моллюски. Их раковины
в большом количестве сохраняются в древних осадочных породах.
Изучение их позволяет проследить эволюцию ряда видов.
Тип Моллюски включает несколько классов, из которых наибо-
лее распространены и представляют большой интерес для народ-
ного хозяйства следующие: Двустворчатые моллюски (Bivalvia),
Брюхоногие моллюски (Gastropoda), Головоногие моллюски
(Cephalopoda).
Строение и жизненные отправления. Форма и размер мол-
люсков крайне разнообразны в связи с особенностями мест оби-
тания и жизни. У одних тело имеет двустороннюю симметрию, но
у многих, имеющих спиральную раковину, оно асимметрично.
211
Тело состоит из туловища, в котором расположено большинство
внутренних органов, ноги, служащей для передвижения живот-
ного, и головы, которая несет органы чувств и содержит крупные
головные нервные узлы. У многих моллюсков голова не обособ-
лена от туловища. Размеры моллюсков от микроскопических до
гигантских (кальмары, тридакны).
Раковины моллюсков разнообразны. У одних она образована
двумя одинаковыми или различными по величине и форме створ-
ками, соединенными зубчатым замком или связкой, у других не
разделена на створки и обычно спирально закручена, у третьих
представлена рядом следующих друг за другом щитков. Как
правило, стенка раковины состоит из трех слоев: наружного,
образованного органическим веществом конхиолином; среднего,
сложенного из перпендикулярных к поверхности раковины извест-
ковых призмочек, и внутреннего, образованного из тонких листоч-
ков перламутра. У ряда моллюсков раковина может подвергаться
частичной редукции.
Полость тела смешанная, образована остатками первичной
полости и сильно редуцированным целомом. Полость заполнена
рыхлой массой клеток — паренхимой, в которой лежат внутренние
органы. Вторичная полость тела сведена к околосердечной сумке
и полости половых органов.
Покровы моллюсков мягкие, слизистые. Они образованы
однослойным, местами ресничным эпителием. В нем заложены
многочисленные железы, секрет которых увлажняет кожу сухо-
путных видов и облегчает скольжение ноги по субстрату, ('кладки
кожи образуют мантию.
Мускулатура большинства моллюсков слагается из глад-
ких мышечных волокон, и это сказывается в медленных движениях
тела. Но у головоногих моллюсков -- осьминогов, кальмаров и
других имеются поперечнополосатые мышцы, что связано с их
подвижным образом жизни. Особенно сильно развиты они в
ноге животного.
Нервная система у большинства моллюсков представ-
лена несколькими парами нервных узлов, которые расположены
в различных частях тела и связаны нервными тяжами; от них
отходят нервы к органам. Лишь у примитивных панцирных мол-
люсков нервная система образована окологлоточным нервным
кольцом, от которого вдоль тела тянутся нервные тяжи, содержа-
щие нервные клетки. Строение нервной системы этих моллюсков
напоминает таковую некоторых плоских червей.
Органы чувств развиты в разной степени. Большин-
ство имеют глаза. У головоногих моллюсков они сложного стро-
ения, имеются также органы осязания, химического чувства и
равновесия.
Органы пищеварения начинаются ротовым отверстием,
ведущим в глотку, которая переходит в пищевод, за ним сле-
212
дует желудок. Далее начинается кишечник, заканчивающийся
анальным отверстием. В кишку впадает проток печени.
Органами дыхания большинства водных моллюсков
служат жабры, расположенные в мантийной полости. Жабры
видоизмененные участки мантии. Они имеют вид лепестков, при-
легающих друг к другу и сидящих с двух сторон от осп жанры,
или иной формы. Вторичноводные и все сухопутные моллюски ды-
шат легкими, также представляющими участки стенки мантийной
полости, богато снабженные кровеносными сосудами.
К р о в е н о с н а я с и с т е м а незамкнутая и состоит из сердца
и кровеносных сосудов. В околосердечной сумке находится сердце,
которое состоит из одного желудочка и одного или нескольких
предсердий. От желудочка отходят артерии, которые ветвятся н
расходятся по всему телу. Из них кровь поступает в систему
лакун —- промежутки между тканями и органами, а затем засасы-
вается в венозные сосуды, которые несут ее к жабрам или легким.
Окисленная кровь по сосудам возвращается в сердце.
Органы выделения моллюсков — почки, представля-
ющие видоизмененные метанефридии. Каналец каждой! почки
начинается воронкой в околосердечной сумке (целоме), а другим
концом открывается в мантийную полость.
Органы размножения развиваются в целоме, протоки
открываются либо в мантийную полость, либо в протоки почек.
Большинство моллюсков раздельнополы. Оплодотворение яиц
происходит внутри материнского организма или в мантийной
полости.
Развитие моллюсков протекает с метаморфозом или без
него. Водные виды откладывают яйца в воду, а наземные в
почву или на ее поверхность. Некоторые моллюски живородящие.
КЛАСС ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ (BIVALVIA)
Общая характеристика. Двустворчатые моллюски с двуствор-
чатой раковиной (рис. 184). Голова не обособлена. Нога обычно
клиновидной формы, часто в той или иной степени редуцирована.
Жабры имеют вид пластин, расположенных в мантийной полости
с двух сторон ноги (другое название класса — Пластипчатожа
берные).
Описано более 15 тыс. видов двустворчатых моллюсков. Боль-
шинство их обитают в морях, меньшая часть — в пресных водо
емах. Обычно это малоподвижные или неподвижные животные,
обитающие на дне водоемов. Питаются пассивно, путем удержива-
ния пищевых частиц, поступающих вместе с водой в мантийную
полость. Развитие прямое (часть пресноводных форм) пли е
превращением.
Строение и жизненные отправления. Тело двустворчатых
моллюсков двустороннесимметричное. Оно состоит из туловища
и ноги, голова редуцирована. Нога обычно имеет клиновидную
5.
Рис. 184. Двустворчатый мол-
люск беззубка:
1 — нога; 2 — жаберный сифон;
5 — клоакальный сифон; 4 - вер-
шина раковины; 5 . связка створок
форму. У подвижных видов опа может высовываться из раковины и
зарываться в грунт водоема, подтягивая затем за собой все жи-
вотное. У прикрепленных или лежащих на грунте моллюсков нога
обычно в той или иной степени редуцирована.
Покровы двустворчатых моллюсков богаты различными
железами, выделяющими слизь и другие вещества. Так у моллю-
сков камнеточцев (Pholas) железы покровов ноги выделяют кис-
лоту, разрушающую известь, что позволяет животному углуб-
ляться в известковые скалы. У дрейссен и мидий кожные железы
выделяют особое вещество — биссус, затвердевающее в воде в
виде тонких нитей, которыми моллюски прочно прикрепляются к
субстрату.
Мантия свешивается с боков в виде двух складок. Они ог-
раничивают мантийную полость, в которой расположены нога и
жабры. В мантийную полость открываются задняя кишка, протоки
почек и половые пути. Вода в нее поступает через жаберный
сифон, а удаляется через клоакальный сифон, образованный
задним краем мантии. Ток воды создается движением ресничек
эпителия, покрывающего мантию, жабры и сифоны.
Раковина образована двумя створками, соединенными на вер-
шине эластичной связкой пли замком — зубами, расположенными
по верхнему краю створок (рис. 185, 186). У большинства видов
обе створки раковины одинаковы, но у ряда лежащих па дне мол-
люсков они отличаются размером и формой. Раскрытие створок
происходит автоматически под действием эластичных свойств
соединяющей связки. Сближение створок вызывается сокращени-
ем мощных мускулов — запирателей, связывающих обе створки.
Нарастание раковины происходит по наружному краю створок за
счет выделений из желез подстилающей мантии.'Зимой рост мол-
люсков почти прекращается, отчего на створках образуются годич-
ные полосы, по числу которых можно определить возраст живот-
ного.^
Нервная система образована тремя парами нервных
узлов, один из которых расположен над глоткой, второй — в ноге
и третий — в задней части тела. Узлы связаны нервными тяжа-
ми — комиссурами. От узлов отходят нервы к разным органам
(рис. 187).
214
Рис. 185. Раковина перловицы (с внут-
ренней стороны):
1 . кардинальные .чубы замка; 2 боко-
вой зуб в виде пластинки; 3 - отиечагки
мышц
Рис. 186. Разрез раковины и мантии
беззубки:
1 — конхиолиновый слой раковины; 2
фарфоровидный ее слой; ,? перламутро-
вый слой; 4 наружный эпителий мантии;
5 -• средний соединительнотканный слой
мантии; 6' - внутренний эпителий мантии
Рис. 187. Внутреннее строение беззубки:
1 — мантия; 2 нога; 3 — жаберный сифон; 4 -- клоакальный сифон; 5 » замыкательные
мышцы; С> — ротовые лопасти; 7 — ротовое отверстие; 8 желудок; .9 печень, 1»
кишка; /1 — анус; 12 —- сердце; 13 — околосердечная сумка; 11 почка; /5 Кеберов
орган; 16 - головной ганглий; 17 — ножной ганглий; 18 висцеральный ганглий;
19 — жабры; 20 -- мантийная полость
Органы чувств у двустворчатых моллюсков развиты
слабо. В покровах разных частей тела разбросаны чувствующие
клетки — органы кожного чувства. На жабрах имеются особые
органы химического чувства — осфрадии. В ноге расположены
органы равновесия — статоцисты. У ряда видов имеются много-
численные глазки по краю мантии.
Пищеварительная система. Рот расположен на пе-
реднем конце тела над основанием ноги. По бокам его располо-
жены две лопасти, покрытые мерцательным эпителием, реснички
которого подгоняют пищевые частицы к ротовому отверстию. Ко-
роткий пищевод проводит пищу в небольшой желудок, в который
открываются протоки печени. Отходящая от желудка кишка
образует ряд петель, поднявшись к спинной стороне тела, проходит
назад через околосердечную сумку и желудочек сердца и откры-
вается в клоакальный сифон.
Органами дыхания являются жабры, лежащие в ман-
тийной полости по обеим сторонам ноги.
Кровеносная система состоит из сердца и сосудов.
Сердце имеет один желудочек и несколько предсердий, число
которых соответствует количеству жабр, поскольку каждое из них
представляет расширение сосудов, идущих от органов дыхания.
Сердце лежит на спинной стороне тела.
Органы выделения представлены двумя почками, ле-
жащими под сердцем. Каждая из них начинается в околосердеч-
ной сумке воронкой, выстланной мерцательным эпителием. Моче-
точники открываются в мантийную полость. Эпителий околосер-
дечной сумки содержит особые перикардиальные железы (Кеберов
орган), также несущие выделительные функции.
Органы размножения. Большинство двустворчатых
моллюсков раздельнополы. Семенники и яичники парные. Протоки
открываются обычно в мантийную полость. Оплодотворение яиц
наружное. У пресноводных форм оно происходит в мантийной
полости самки, куда сперматозоиды проникают вместе с водой
через жаберный сифон.
Развитие у большинства видов протекает с метаморфозом.
Личинка морских видов парусник имеет сходство с личинками
кольчатых червей. Тело ее округлое, прозрачное, окруженное
поясками ресничек. Личинки пресноводных моллюсков беззубок
и перловиц, носящие название глохидиев, имеют двустворчатую
раковину с зазубренными шипами на краях (рис. 188). Глохидии
образуются осенью и всю зиму проводят в мантийной полости
материнского организма. Весной они покидают тело матери и
укрепляются на жабрах рыб. Ткани хозяина обрастают глохидиев,
и они становятся эндопаразитами, питаясь осмотически соками
хозяина. Через 2—3 мес моллюски покидают жабры рыб, опуска-
ются на дно и переходят к свободному существованию.
Значение двустворчатых моллюсков для хозяйства человс-
216
Рие. 188. Глохидия беззубки:
/ епюрки рикопнпы; 2 ее «убцы; 3
мускул замыкатель тюрок; 7 щетинки
органов чувств
ка велико. Многие из них упот-
ребляются в пищу. Таковы устри-
цы, гребешки, мидии и др.
Устрицы (Ost геи) - непод-
вижные морские моллюски с
а с и м метр и ч i юй р а ко в и и о й.
Большой выпуклой створкой 2
они соприкасаются с субстратом
(рис. 189, /). На мелких местах устрицы нередко образуют огром-
ные скопления — устричные банки, где их и вылавливают. Во мно-
гих странах устрицы высоко ценятся как деликатес. Мясо их
по калорийности не уступает мясу рыб и богато витаминами.
Во многих странах их разводят в специальных хозяйствах.
Гребешки (Pecten) имеют красивые округлые раковины с
радиальными ребрами (рис. 190). Они пе прирастают к субстра-
ту, а лежат на дне. При опасности гребешок быстро захлопывает
Рис. 189. Промысловые съедобные моллюски:
/ — устрицы; // — мидия
217
Рис. 190. Гребешок
Рис. 192. Древоточец и
его ходы в куске де-
рева
Рис. 191. Жемчужница:
/ — внешний вид; // - распил жемчуга
(видны концентрические слои вокруг ино-
родного тела)
218
раковину, выбрасываемая при этом из нее вода отбрасывает жи-
вотное, которое делает плавный скачок. Гребешки в большом коли-
честве добываются в морях Дальнего Востока СССР. Используют-
ся для приготовления консервов.
Мидии (Mytilus) имеют темную раковину из двух створок,
одна сторона которых почти прямая, а другая — выпуклая (см.
рис. 189, II). Моллюск прикрепляется к субстрату пучком нитей
биссуса. В Черном море имеются огромные запасы мидий, которые
широко эксплуатируются приморскими животноводческими хо-
зяйствами.
Для кормления сельскохозяйственных животных пригодны
также наши пресноводные двустворчатые моллюски — беззубки
и перловицы (Anodonta, Unio). Они широко распространены
по рекам и озерам. Раковины их удлиненно-овальные, крупные,
зслеповато-бурого цвета, состоят из двух одинаковых створок.
Моллюск передвигается, погрузив в песок клинообразную ногу.
У беззубки раковина тонкостенная и створки ее соединяются
эластичной связкой без замка. У перловицы раковина более вытя-
нутая и утолщенная, створки ее соединены замком из зубчиков.
Раковины перловиц служат сырьем для перламутрового произ-
водства.
Ряд видов двустворчатых моллюсков дает жемчуг. Он образу-
ется в результате отложения слоев перламутра вокруг какого-либо
инородного тельца (песчинки), попавшего между мантией и
раковиной животного. Наиболее крупные и ценные жемчужины
дает жемчужница (Pteria), обитающая в тропических и субтро-
пических морях (рис. 191). В Японии разводят жемчужниц в осо-
бых клетках, погруженных в море. Молодым жемчужницам в
мантию вводят инородное тельце, становящееся центром роста
жемчужины. Жемчуг добывается также из речной жемчужницы
(Margarilana), которая встречается в реках и озерах северных
областей СССР.
Вредным представителем двустворчатых моллюсков является
дрейссена (Dreissena), живущая в пресной и соленой воде. Опп
нередко скапливаются на шлюзах и защитных решетках гидро-
технических сооружений, нарушая нормальную работу. Корабель-
ное точило (Teredo) имеет длинное червеобразное тело и малень-
кую раковинку на переднем конце (рис. 192). Она выполняет
роль сверлящего аппарата. Глубоко внедряясь в древесину
подводных сооружений и днищ деревянных судов, они наносят
им сильные повреждения.
КЛАСС БРЮХОНОГИЕ МОЛЛЮСКИ (GASTROPODA)
Общая характеристика. Брюхоногие — моллюски, тело которых
разделено на голову, туловище и ногу с широкой ползательной
подошвой. Раковина, если она имеется, целая, спирально завитая.
Тело асимметричное. На голове имеются 1—2 пары щупалец.
219
У большинства хорошо развиты глаза. Дышат жабрами или
легкими.
Известно свыше 50 тыс. видов брюхоногих моллюсков. Они
обитают не только в морях и пресных водоемах, но и на суше. Пи-
таются животными и растениями, а также гниющими веществами.
Развитие проходит с метаморфозом или без него. Многие брюхо-
ногие имеют большое практическое значение как вредители рас-
тениеводства и промежуточные хозяева некоторых опасных пара-
зитов из числа плоских червей.
Строение и жизненные отправления. Форма тела брюхо-
ногих разнообразна, обычно асимметричная в силу закрученности
туловища спиралью. На голове имеются 1—2 пары способных
втягиваться щупалец и хорошо развитые глаза, расположенные
у некоторых видов на вершинах щупалец. Нога обычно широкая,
с плоской подошвой. Большинство брюхоногих передвигаются
скольжением по субстрату благодаря волнообразным изгибам
подошвы ноги.
Раковина нередко имеет причудливую форму и яркую окраску.
У брюхоногих, плавающих в толще воды морей, раковина в той или
иной степени редуцирована. Она отсутствует также у наземных
слизней, прячущихся на день в норки. В спокойном состоянии
животного внутри раковины помещается только его туловище, но
в случае опасности в нее втягивается все тело. Как правило, ра-
ковина брюхоногих изогнута спиралью, но у моллюска морского
блюдца она конической формы.
Мантийная полость находится в нижних витках раковины.
В нее открываются заднепроходное отверстие, мочеточники, ино-
гда проток половых органов. У водных в ней размещаются органы
дыхания — жабры. У воздушно-дышащих мантийная полость ста-
новится легким, открываясь наружу дыхательным отверстием.
В стенках полости находится густое сплетение кровеносных
сосудов.
Покровы брюхоногих богаты различными железами, в том
числе слизистыми, которые обильны на подошве ноги.
Нервная система этих моллюсков состоит из несколь-
ких пар ганглиев, связанных комиссурами.
Органы чувств. У брюхоногих имеются глаза, органы
равновесия — статоцисты, находящиеся в ноге, органы осязания
(щупальца) и химического чувства.
Органы пищеварения начинаются ротовым отверстием,
расположенным на нижней стороне головы, которое ведет в
глотку. В глотке находятся одна или две челюсти и терка (радула),
имеющая вид пластинки с множеством расположенных попе-
речными рядами мелких зубчиков. Благодаря ей моллюск может
отделять кусочки пищи и соскабливать обрастания (микрона-
селение с подводных растений и предметов). В глотку впадают
протоки слюнных желез. Глотка переходит в пищевод, который
открывается в желудок, принимающий протоки большой печени.
220
Из желудка пища попадает в среднюю, а затем в заднюю
киш ку.
О р г а и а м и д ы х а п и я служат жабры пли легкие. К жабер-
ным относятся все морские и часть пресноводных брюхоногих
моллюсков. К числу легочных брюхоногих моллюсков относятся
все наземные и многие пресноводные виды (прудовики, катушки
и др.). Последние вынуждены периодически подниматься к по-
верхности водоема, чтобы набрать воздух в мантийную полость.
Кров е и о с н а я с и с т е м а представлена сердцем, сосудами
и лакунами. Сердце лежит в околосердечной сумке. От не-
го отходят артериальные сосуды, которые изливают кровь в ла-
куны.
Органами в ы д с л е н и я являются почки, воронки которых
открываются в околосердечную сумку. Мочеточники заканчива-
ются в мантийной полости.
Органы размножения у брюхоногих имеют различное
строение. Морские формы обычно раздельнополые, а наземные
и многие пресноводные — гермафродиты. Оплодотворение яиц
происходит внутри материнского организма.
Развитие происходит без превращений или с наличием
стадии личинки. Есть виды живородящие.
Практическое значение брюхоногих моллюсков довольно ве-
лико. Они играют значительную роль в круговороте веществ в
водоемах. Обитая на дне и потребляя различные органические
отложения, они ускоряют их разложение. Многие служат кормом
промысловых рыб, китов и ластоногих. Морские трубачи — источ-
ник ценного черного и розового жемчуга, пурпурные улитки
имеют особые железы, из секрета которых получают пурпурную
краску. Велико значение брюхоногих как вредителей растение-
водства.
Виноградная улитка (Helix) — крупный моллюск с полосатой
раковиной (рис. 193). При массовом размножении вредит вино-
градной лозе. В ряде стран их разводят в пищевых целях.
Слизни (рис. 194) частично или полностью утратили рако-
вину. Тело вытянуто. Нога хорошо развита. На голове две пары
щупалец. Большинство — ночные животные. Днем прячутся в
Рис. 193. Виноградная улитка:
/ — раковина; 2 — голова со щупальцами;
3 — нога; 4 — дыхательное отверстие; 5 -
половое отверстие; 6 -- глаза
Рис. 194. Слизень оранжево-жел-
тый
221
почве. Питаются различными растениями, вредят огородным
культурам. Гермафродиты. Дают за лето несколько кладок от 9 до
50 яиц в каждой. Молодь вылупляется через 15 - 20 дней. Пере-
зимовывают на стадии яйца, а иногда и во взрослом состоянии.
Живут от 1 до 3 лет. Слизни семейства Arionidae отличаются
крупными размерами, на спине известковой пластинки нет. К мно-
гоядным относится Arion empiricorum, который часто встречается
в садах и лесах. Озимым больше других арионов вредит арион
окаймленный (Arion circumscriptus) —слизень оранжевого цвета
со светлой полоской на спине, длиной до 5 см. Слизни семейства
Limacidae меньших размеров. Под кожей на спине у них есть
небольшая известковая пластинка — рудимент раковины. Мио-
гоядны. Вредят различным сельскохозяйственным растениям.
К лимацидам относятся полевой, черный, сетчатый, большой
слизни. Они прожорливы и подвижны, быстро размножаются.
Наиболее вреден полевой слизень (Agriolimax agreslis). Он вредит
всходам озимых. Размножается все лето, откладывая за год до
500 яиц. Молодь выходит через 2—3 недели, а через 1,5 мес дости-
гает полного развития.
Слизни — промежуточные хозяева ланцетовидного сосаль-
щика, некоторых ленточных червей — паразитов кур и ряда не-
матод, паразитирующих в овцах и козах. Для борьбы с ними
применяют опыление табачной пылью в смеси с известью и раз-
личные ядохимикаты. Замечено, что при внесении в почву супер-
фосфата, селитры и калийной соли их число сокращается. Много
слизней уничтожают жабы.
Рис. 195. Раковины различных пресноводных брюхоногих мол-
люсков:
/ прудовик обыкновенный; 2— катушка; 3 — прудовик малый; /
лужайка живородящая; 5 — битиния
222
Прудовики и бигинии - промежуточные хозяева сосаль-
щиков — паразитов домашних животных (рис. 195). Наибольшее
значение имеет в этом отношении легочный моллюск малый
прудовик (Limnaea iruncalula). Раковина у него яйевидпо-
коничсская с 5—6 сильно выпуклыми оборотами. Устье раковины
довольно узкое, яйцевидное, высота 5—10 мм, ширина 3—5 мм.
Живет в болотах, прибрежной зоне прудов среди растений, во вре-
менных водоемчиках на заливных лугах. Известен как промежу-
точный хозяин печеночной фасциолы. Моллюск встречается
местами в количестве до 900 и более экземпляров на 1 м". По окра-
инам болот насчитывали до 3 млн особей па 1 га. Яйца откла-
дывают, начиная с весны, во влажную землю, редко в воду. В тече-
ние лета наблюдается несколько кладок. Молодь выходит дней
через 10—20 в зависимости от температуры. Половозрелости
достигает к 6—7 мес. Зимовать уходят в глубь водоема, где зары-
ваются в грунт. Летом переносят временное высыхание водоема.
КЛАСС ГОЛОВОНОГИЕ МОЛЛЮСКИ (CEPHALOPODA)
К этому классу относятся осьминоги, кальмары, каракатицы и др.
(рис. 196, 197). Головоногие — двустороннесимметричиые живот-
ные, тело которых разделено на голову и туловище. Голова несет
8—10 щупалец с присосками, они окружают ротовое отверстие,
22.3
Рис. 197. Наутилус и ископаемые головоногие моллюски:
/ — наутилус; II — белемнит и часть его скелета (чертов па-
лец); И! — реконструкция аммонита; IV аммонит (окаме-
нелость)
вооруженное обычно роговым клювом. На голове расположена
пара больших глаз. Туловище яйцевидной или цилиндрической
формы, нередко с хвостовым плавником на конце. Раковина
у некоторых видов хорошо развита, у большинства же рудимен-
тарна и скрыта под кожей спины. У осьминогов раковина пол-
ностью атрофирована. Мантийная полость открывается щелевид-
ным отверстием, способным замыкаться. Перед ним находится
воронкообразный орган — видоизмененная часть ноги животного.
Набрав воду в мантийную полость и закрыв ее отверстие, моллюск
выпускает воду сильной струей через эту воронку, что вызывает
движение тела в обратную сторону, задним концом вперед. В ман-
тийной полости расположены 1—2 пары жабр. У многих форм в
нее открывается также проток особой железы — чернильного меш-
ка, выделяющей черную жидкость. При опасности моллюск вы-
брасывает ее в воду и скрывается за образовавшейся завесой.
Живут эти моллюски у дна моря или в толще его вод. Все
они — хищники. Мясо моллюсков употребляется в пищу и идет на
изготовление консервов. В СССР промысел кальмаров и осьми-
ногов развит на Дальнем Востоке.
224
ТИП ИГЛОКОЖИЕ (ECHINODERMATA)
Общая характеристика. Иглокожие — вторичнополостные живот-
ные, во взрослом состоянии имеющие радиальную симметрию тела.
У большинства видов органы расположены по пяти радиусам,
но у некоторых число лучей иное. Если у кишечнополостных
радиальная симметрия тела является первичной, то у иглокожих
она будет вторичной, поскольку их предки имели двустороннюю
симметрию тела. Свободноплавающие личинки иглокожих двусто-
роннесимметричны. Для иглокожих характерно наличие амбуляк-
ральной системы, служащей для передвижения и участвующей
в процессах дыхания и выделения. Вторичная полость тела хорошо
выражена и наполнена полостной жидкостью. Иглокожие — жи-
тели моря. Это преимущественно донные животные, способные к
медленному передвижению по субстрату, реже прикрепленные к
нему. Некоторые иглокожие служат объектом промысла.
Иглокожие, как впервые показали исследования И. И. Мечни-
кова, интересны для выявления филогенетических связей беспоз-
воночных животных с представителями типа Хордовые. Несмотря
на радиальную симметрию тела взрослых особей, в организации
и развитии иглокожих много общих черт с хордовыми животными.
Вторичная полость тела у них, как и у хордовых, образуется
путем отделения мезодермальных мешков от кишечника. Как и
хордовые, они являются вторичнополостными животными, у кото-
рых в процессе развития гастропор зарастает или превращается
в анальное отверстие, а рот личинки образуется заново. У пред-
ставителей обоих типов имеются двухслойная кожа и скелетные
элементы мезодермальной природы. Эти черты сходства позво-
ляют предположить, что низшие хордовые филогенетически свя-
заны через общих предков с иглокожими. Остатки иглокожих
найдены в отложениях палеозойской эры.
Тип Иглокожие делится на пять классов: Морские звезды
(Asteroidea), Офиуры (Ophiuroided), Морские лилии (Crinoidea),
Морские ежи (Echitioidea), Голотурии (Holotfiuroidea).
Строение и жизненные отправления. Покровы иглокожих
состоят из двух слоев: наружного, имеющего характер однослой-
ного эпителия, и внутреннего, образованного волокнистой соедини-
тельной тканью. Во внутреннем слое развиваются различные эле-
менты известкового кожного скелета. У морских звезд они имеют
вид известковых пластинок, расположенных продольными (вдоль
лучей) рядами и обычно несущих выдающиеся наружу шипики.
У морских ежей тело заключено в известковый панцирь из рядов
плотно соединенных пластинок с сидящими на них длинными
иглами. У голотурии мелкие известковые тельца разной формы
разбросаны в коже.
Мускулатура развита в различной степени в зависи-
мости от подвижности и характера кожного скелета. Она слагает-
ся из отдельных мышц и мускульных полос.
8 Б. А. Кузнецов и др.
225
Амбулякральная система начинается пористой
мадрепоровой пластинкой, расположенной на спинной стороне
тела. От нее в глубь тела тянется каменистый канал, который
открывается в кольцевой канал, окружающий пищевод. Кольцевой
канал дает радиальные каналы в каждый луч тела. От радиальных
каналов ответвляются в обе стороны короткие канальцы, от кото-
рых внутрь полости отходят сократимые пузырьки — ампулы, а
наружу — сократимые трубчатые амбулякральные ножки с при-
сосками на концах. Амбулякральная система заполнена водой,
поступающей через мадрепоровую пластинку. При сокращении
ампул вода из них переходит в полость ножек, отчего они удли-
няются, вытягиваются. Находящиеся на концах ножек присоски
присасываются к субстрату, после чего длина ножек сокращается,
так как вода из их полости отводится обратно в ампулу. Общими
усилиями многих одновременно сокращающихся ножек тело игло-
кожего подтягивается, и животное медленно движется по дну.
Благодаря присоскам амбулякральных ножек иглокожие могут
ползти даже по вертикальной поверхности скалы.
Нервная система иглокожих имеет радиальное строе-
ние. От окологлоточного нервного кольца отходят радиальные
нервные тяжи по числу лучей тела.
Органы.чувств развиты слабо. Примитивные глазки рас-
положены у морских звезд на концах лучей, а у морских ежей —
на верхней части тела. Имеются также органы осязания и др.
Пищеварительная система. Ротовое отверстие рас-
положено у большинства посредине на нижей поверхности тела.
Рот ведет в короткий пищевод, за которым следуют средняя кишка
и короткая задняя. Анальное отверстие у некоторых отсутствует.
Органами дыхания у морских звезд и ежей служат кож-
ные жабры — тонкостенные выросты на верхней стороне тела.
По-видимому, в дыхательном процессе принимает участие и
амбулякральная система. У ряда иглокожих дыхание происходит
через покровы тела.
Кровеносная система обычно состоит из двух кольце-
вых сосудов, один из которых окружает рот, а другой —- анальное
отверстие, и радиальных сосудов, количество которых совпадает
с числом лучей тела. Оба кольцевых сосуда соединены крове-
творным осевым органом, пронизанным сетью кровеносных сосу-
дов.
Выделительные органы. Специальных органов выде-
ления у иглокожих нет. Выделение продуктов диссимиляции
происходит через стенки каналов амбулякральной системы и при
помощи особых амебовидных клеток крови, мигрирующих внутри
организма.
Половые органы имеют разное строение. Большинство
иглокожих раздельнополы, но имеются и гермафродитные формы.
Развитие происходит с рядом сложных превращений. Дву-
стороннесимметричные личинки иглокожих плавают в толще воды.
226
Многие иглокожие обладают удивительной способностью к ре-
генерации частей тела. Например, один луч морской звезды может
восстанавливать целое животное.
Наиболее распространены классы: Морские звезды, Морские
ежи и Голотурии.
КЛАСС МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA)
Тело имеет вид плоской звезды с пятью или большим числом
лучей (рис. 198, /). По нижней стороне лучей тянутся продольные
бороздки, в которых расположены многочисленные амбулякраль-
ные ножки. Передвижение осуществляется как с помощью ножек,
так и лучей. Рот расположен в центре нижней поверхности тела.
Короткий пищевод ведет в объемистый желудок (рис. 199, 2),
способный выворачиваться наружу. Найдя крупную добычу (на-
пример, моллюска), морская звезда накрывает ее телом, вывора-
чивает желудок и прижимает его к добыче. Соки, выделяемые
стенкой желудка, переваривают пищу. Короткая кишка ведет к
анальному отверстию, лежащему на верхней поверхности тела.
Морские звезды живут на дне морей, спускаясь на большую
глубину. Питаются донными животными, органическими остат-
ками. Приносят вред устричным хозяйствам.
КЛАСС МОРСКИЕ ЕЖИ (ECHINOIDEA)
Тело шаровидное или уплощенное (дискообразное или сердцевид-
ное) (см. рис. 198, IV). Оно заключено в сплошной панцирь из
плотно соединенных известковых пластинок, которые несут тон-
кие острые иглы у одних форм и толстые тупые — у других. На
нижней стороне тела расположен рот, вооруженный особым
челюстным аппаратом — аристотелевым фонарем — с пятью зу-
бами, выступающими изо рта. Кишечник заканчивается анальным
отверстием, расположенным на вершине тела. От рта к верхнему
полюсу тела тянутся пять рядов пластинок панциря с отверсти-
ями, через которые выступают амбулякральные ножки. Пользуясь
ими, морские ежи медленно передвигаются по дну и скалам.
Морские ежи — донные малоподвижные животные, обитающие
в море на различной глубине. Питаются животной и растительной
пищей. Их икра используется в пищу людям. Полостная жидкость
их обладает тонизирующими свойствами.
КЛАСС ГОЛОТУРИИ (HOLOTHUROIDEA)
Голотурии отличаются мешковидным или червеобразным телом с
венчиком ветвистых щупалец вокруг рта (см. рис. 198, III). Вдоль
тела тянутся пять полос амбулякральных ножек. В коже располо-
жены мелкие известковые тельца. Донные, реже плавающие мор-
8*
227
ш
Рис. 198. Различные иглокожие:
I — морская звезда; // — морская лилия; /// — голотурия; IV — морской
еж (с одной половины его тела удалены покровы и виден панцирь из изве-
стковых пластинок)
Рис. 199. Внутреннее стро-
ение морской звезды:
1 — рот; 2 — желудок; 3 -
анус; 4 — печень; 5 — мад-
репоровая пластинка; 6 —
осевой орган; 7 — камени-
стый канал; 8—кольцевой
канал амбулякральной си-
стемы; 9 — ее радиальный
капал; 10—амбулякраль-
ные ножки; // —- их ампулы;
12 — кольцевой сосуд кро-
веносной системы; 13 — ее
радиальный сосуд; 14 —
окологлоточное кольцо нерв-
ной системы; 15 — ее ра-
диальный ствол; 16 — поло-
вая железа
скиё животные. Обычно ползают по дну, разыскивая пищу — мел-
ких донных животных, водоросли, гниющие остатки. В ряде стран,
в том числе и в СССР, ведется промысел голотурий.
ТИП ХОРДОВЫЕ (CHORDATA)
К хордовым принадлежат вторичнополостные, двусторонне-
симметричные, вторичноротые животные метамерного строения.
Метамерия более полно выражена на ранних стадиях эмбриональ-
ного развития, чем у взрослых особей. У последних она в большей
мере сохраняется у первичноводных, чем у наземных видов. Как и
другие вторичнополостные животные, хордовые имеют кровенос-
ную систему, органы выделения метанефридиального типа, их по-
ловые железы развиваются в целоме.
Для хордовых характерны следующие, присущие только им
признаки:
надиже. хорды — упругого гибкого стержня, тянущегося вдоль
тела животного, ближе к спинной стороне. Хорда играет роль осе-
вого скелета. У примитивных хордовых она обычно сохраняется
в течение всей жизни, а у более высокоорганизованных форм
замещается в процессе развития позвоночником, позвонки кото-
рого формируются из скелетообразующей соединительной ткани,
окружающей хорду и расположенную над ней нервную трубку;
центральная „нервная система имеет вид нервной трубки с
узким’каналом (невроцелём) внутри. Она расположена на спин-
ной стороне тела животного над осевым скелетом (хордой или
телами позвонков). Из переднего отдела нервной трубки путем
сложных изменений формируется головной мозг, тогда как осталь-
ная ее часть образует спинной мозг;
образование в стенках глотки жаберных щелей, которые у
первичноводных животных сохраняУотся 'в течение всей жизни, а
у наземных и вторично перешедших к жизни в водной среде ви-
дов обнаруживаются только на определенных стадиях развития.
К хордовым относятся бесчерепные, оболочники, круглоротые
(миноги и миксины), рыбы, земноводные, пресмыкающиеся, птицы
229
и млекопитающие (звери). Известно около 50 тыс. видов этих
животных, обитающих как в водной среде, так и на суше. Неко-
торым присуща способность к полету. Большинство их ведут
свободный, подвижный образ жизни.
Тип Хордовые делится на подтипы: Бесчерепные (Acraiiia),
Личиночнохордовые {Urochordata), Позвоночные (Vertebrata).
Хордовые имеют огромное значение для хозяйства человека.
К этому типу принадлежат все сельскохозяйственные животные,
кроме пчел и тутового шелкопряда. Многие хордовые (рыбы,
птицы, звери) служат объектом промысла, давая большое коли-
чество мяса, жира, кожи, меха и других продуктов и промышлен-
ного сырья. Некоторые хищные виды полезны истреблением вре-
дителей сельского хозяйства. В то же время среди хордовых
немало видов, причиняющих значительный вред растениеводству
и животноводству. Некоторые из них участвуют в распростране-
нии опасных заболеваний человека.
ПОДТИП БЕСЧЕРЕПНЫЕ (ACRAN1A)
Общая характеристика. Бесчерепные — морские, преимуществен-
но донные животные, сохраняющие основные признаки типа в
течение всей жизни. Их организация представляет как бы схему
строения хордового животного. Бесчерепные представляют боль-
шой интерес для решения вопроса о происхождении животных.
Познанию бесчерепных наука, обязана прежде всего исследова-
ниям А. О. Ковалевского.
Строение и жизненные отправления. Из сравнительно неболь-
шого числа представителей бесчерепных (около 20 видов) наибо-
лее обычным и хорошо изученным является ланцетник (Amphio-
xus lanceolatus) (рис. 200). Это небольшое животное (длиной
до 8 см) обитает на мелководье морей, зарывшись в песок и выста-
вив наружу передний отдел тела. Питается мелкими пищевыми
частицами, опускающимися на дно.
Форма тела ланцетника вытянутая, сжатая с боков, заост-
ренная спереди и сзади. По спине тянется невысокая продольная
складка кожи — спинной плавник. На заднем конце тела имеется
копьевидный хвостовой плавничок. Парных конечностей нет.
Кожный покров образован однослойным слизистым эпи-
дермисом и соединительнотканной дермой.
Скелет представлен хордой, тянущейся вдоль тела, утон-
чаясь к концам. Хорда и лежащая над ней нервная трубка окру-
жены соединительнотканной оболочкой.
Мускулатура тянется лентами по обе стороны тела. Эти
мускульные ленты метамерно разделены тонкими соединитель-
нотканными перегородками (миосептами) на ряд миомеров.
Центральная нервная система примитивного стро-
ения (рис. 201). Она имеет вид трубки, невроцель которой в пе-
редней части образует полость, представляющую зачаток желу-
230
Рис. 200. Ланцетник (схема):
/ — предротовая воронка, окруженная щупальцами; 2 — хвостовой плавник; 3 — спин-
ной плавник; 4 — подхвостовой плавник; 5 — отверстие околожаберпой полости; 6 -
половые железы; 7 — мпомер; 8 -- миосепта
Рис. 201. Внутреннее строение ланцетника:
/ — продольный разрез тела; II — поперечный разрез тела (в области глотки и в области
кишечника);
/ — хорда; 2 — спинной мозг; 3 — спинной плавник; 4 — миомер; 5 — целом; 6 - глотка;
7 — жаберная щель; 8 — межжаберная перегородка; 9 — эндостиль; 10 — околожаберная
полость; // — отверстие околожаберпой полости; 12— печень; 13 — кишка; 14 — нефри-
дий; 15 — анус; 16 — спинная аорта; 17 — подкишечная вена; 18 — половые железы
дочка головного мозга. От центральной нервной системы попарно
отходят спинные — двигательно-чувствующие и брюшные - - дви-
гательные нервы, которые не соединяются в общие смешанные
нервы, как у позвоночных животных.
Органы чувств примитивные. Вдоль центральной нервной
системы заложены светочувствительные клетки — глазки Гессе.
На переднем конце тела находится обонятельная ямка. Около-
ротовые щупальца выполняют осязательную функцию.
Пищеварительная система начинается предротовой
воронкой, окруженной щупальцами. На дне ее расположен рот,
ведущий в обширную глотку. Сверху и снизу глотки тянутся
бороздки, выстланные ресничным эпителием. Движением ресничек
нижней борозды — эндостиля — частицы пищи, попавшие в глот-
ку, сначала передвигаются вперед, а затем по спинной борозде
глотки — к кишечнику. Эндостиль у позвоночных животных
видоизменяется в щитовидную железу внутренней секреции. Ки-
шечник тянется без изгибов и заметных расширений от глотки
до анального отверстия. От передней части кишечника отходит
печеночный вырост, который гомологичен печени позвоночных.
Органами дыхания служат перегородки между много-
численными жаберными щелями, прорезающими стенки глотки.
У некоторых глубоководных форм бесчерепных жаберные щели
открываются наружу. У обыкновенного ланцетника, живущего
в песке мелководья, они ведут в обширную околожаберную по-
лость. Последняя образуется у эмбриона путем срастания по
средней линии брюха двух боковых складок кожи. Вода, посту-
пающая через жаберные щели из глотки в околожаберную по-
лость, выводится из нее через непарное отверстие (антриопор) на
брюшной стороне тела.
Кровеносная система замкнутая (рис. 202). Имеется
один круг кровообращения. Сердце отсутствует, и кровь движется
благодаря пульсации некоторых крупных сосудов. Под глоткой
тянется брюшная аорта, от которой в обе стороны отходят прино-
сящие жаберные артерии, несущие венозную кровь к межжабер-
ным перегородкам. Через тонкие покровы последних происходит
поглощение кровью растворенного в воде кислорода. Окисленная
артериальная кровь через выносящие жаберные артерии посту-
пает в парные наджаберные сосуды — корни спинной аорты, ко-
торые позади глотки сливаются в спинную аорту. Спинная аорта
тянется назад над хордой, давая ответвления ч< различным орга-
нам задней половины тела. Наджаберные сосуды продолжаются
вперед сонными артериями, снабжающими кровью головной отдел
животного.
Венозная кровь оттекает от кишечника по подкишечной вене
к печеночному выросту и распадается в стенках на капилляры,
образуя воротную систему печени. Из печени кровь выходит
по печеночной вене, впадающей в венозный синус, лежащий
у корня брюшной аорты. В синус впадают также слева и справа
232
Рис. 202. Схема кровообращения ланцетника:
/ брюшная аорта; 2 расширения оснований приносящих жаберных артерий; 3 -
жаберные артерии; 4 - корни спинной аорты; 5 — сонные артерии; 6 — спинная аорта;
7 хвостовая вена; 8 подкишечная вена; !) - воротная система печени; 10 — около-
жаберная вена; 11 --передняя кардинальная вена; 12 - задняя кардинальная вена;
13 — кювьеров проток
крупные кювьеровы протоки. Они образуются слиянием передних
и задних парных кардинальных вен, несущих кровь из передней
и задней частей тела. Из венозного синуса кровь поступает в брюш-
ную аорту. Так замыкается круг кровообращения.
Органы выделения представлены видоизмененными
метанефридиями, расположенными метамерно в области глотки.
Наружными концами они открываются в околожаберную полость.
Органы размножения имеют вид двух рядов парных по-
ловых желез. Яичники самок и семенники самцов образуют ряды
бугорков на стенках полости тела в области жаберных щелей.
Половые продукты выводятся в околожаберную полость.
‘ПОДТИП ЛИЧИНОЧНОХОРДОВЫЕ (UROCHORDATA)
Группа примитивных хордовых животных, которые в личиноч-
ной стадии развития имеют все характерные для типа Chordata
черты строения, но при переходе во взрослое состояние теряют
хорду и испытывают глубокое преобразование центральной нерв-
ной системы, которая превращается из нервной трубки в компакт-
ный нервный узел (лишь аппендикулярии сохраняют хорду и
нервную трубку всю жизнь). Упрощение организма с возрастом
животных связано с переходом от подвижного существования ли-
чинки к неподвижному взрослых особей. Для личиночнохордовых
характерно также и то, что их тело заключено в оболочку, обра-
зованную веществами, близкими к клетчатке. Личиночнохордо-
вые — гермафродиты. Некоторые из них способны размножаться
и бесполым путем, образуя почки.
Известно около 1,5 тыс. видов. Все они — морские животные.
Большая часть их во взрослом состоянии ведет сидячий образ
жизни, прочно прикрепляясь к субстрату.
Наиболее многочисленная группа личиночнохордовых — асци-
дии. Большинство асцидий обитают на дне морей, прирастая
к камням и другим подводным предметам (рис. 203). Некоторые
асцидии образуют колонии, пассивно плавающие в водах океана.
233
Рис. 203. Асцидия:
1 — ротовой сифон; 2 - клоакальный
сифон; 3 — туника; 4, 5 -- мантия;
6 — глотка; 7 — полость глотки; 8 —
жаберные отверстия; 9—эндостиль;
10, 11—околожаберная полость; 12 - -
ее стенка; 13 — желудок; 14 — пече-
ночный вырост; 15 — анус; 16 — се-
менник; 17— яичник; 18 — протоки
половых желез; 19 — околосердечная
сумка; 20— сердце; 21 — нервный ган-
глий
Из яиц асцидии выходят
микроскопически малые под-
вижные личинки (рис. 204).
Их округлое тельце закан-
чивается длинным, сжатым
с боков хвостом.
Внутри хвоста тянется
хорошо развитая хорда ти-
пичного строения.
Над хордой лежит нерв-
ная трубка, чуть расширяю-
щаяся в передней части.
Глотка личинки пронизана жаберными щелями. Следовательно,
личинкам асцидий присущи все основные признаки хордовых
животных. Подвижная личинка вскоре прикрепляется передним
концом к подводному камню и претерпевает регрессивные пре-
вращения. Хвост с находящейся в нем хордой и большей частью
нервной трубки атрофируется. Передняя часть нервной трубки
Рис. 204. Развитие асцидии:
/— прикрепившаяся личинка; II — превращение личинки во взрослую асцидию; 1 --
хвост; 2—хорда; 3—гаиглий; 4 —- рот; 5 — эндостиль; 6—задняя кишка; 7—-жа-
берные щели; 8—околожаберная полость; 9 — сердце
234
видоизменяется в нервный узел взрослой асцидии. Складками
покровов образуется околожаберная полость. Снаружи тело по-
крывается туникой. Так подвижная личинка, несущая характер-
ные для хордовых животных признаки, постепенно превращается в
сидячую взрослую асцидию, утратившую большинство особен-
ностей представителей типа Хордовые. Именно поэтому опи-
сываемые животные получили название личиночнохордовых.
ПОДТИП ПОЗВОНОЧНЫЕ (VERTEBRATA)
Общая характеристика. К этому подтипу принадлежит подавляю-
щее большинство хордовых животных: круглоротые, рыбы, земно-
водные, пресмыкающиеся, птицы и звери. Позвоночные отличаются
более высокой организацией, чем описанные бесчерепные и
личиночнохордовые (рис. 205). .Для позвоночных особенно
характерны прогрессивное развитие центральной нервной си-
стемы, усложнение кровеносной системы и других систем органов,
образование хрящевого или костного внутреннего скелета. Услож-
нение организации позвоночных связано с тем, что эти животные
ведут подвижный образ жизни, активно добывая пищу. Совер-
шенствование центральной нервной системы выражается прежде
всего в дифференцировке спинной нервной трубки на головной и
спинной мозг и разделение головного мозга на ряд отделов с раз-
личными функциями. Усложнение кровеносной системы прояв-
ляется, в частности, в образовании сердца, а выделительной —
почек.
Позвоночные животные широко распространены на земном шаре.
Современная систематика предусматривает деление позвоноч-
ных животных на следующие систематические группы:
Раздел Бесчелюстные (Agnatha)
Класс Круглоротые (Cyclostomata)
Рис. 205. Схема строения позвоночного животного:
1— хорда; 2 — спинной мозг; 3—головной мозг; 4 — жаберные щели; 5 — сердце;
б — легкое; 7 — головная почка; 8 — туловищная почка; 9 — тазовая почка; 10 —
половая железа; //—желудок; 12 — кишечник; 13—поджелудочная железа; 14 —
печень; 15 — селезенка; /6‘ — мочевой пузырь; 17 — клоака
235
Раздел Челюстноротые (Gnathostomata)
Надкласс Рыбы (Pisces)
Класс Хрящевые рыбы (Chondrlchthyes)
Класс Костные рыбы (Osteichthyes)
Надкласс Наземные позвоночные (Tetrapoda)
Класс Земноводные (Amphibia)
Класс Пресмыкающиеся (Reptilia)
Класс Птицы (Л<у«)
Класс Млекопитающие (Mammalia)
Строение и жизненные отправления. Форма тела позвоноч-
ных разнообразна. В теле различают голову, шею, туловище,
хвост и конечности. Но у первичноводных видов шейный отдел
отсутствует. У некоторых нет парных конечностей — отсутствие
их бывает либо первичным, либо объясняется редукцией. Размеры
позвоночных колеблются от нескольких миллиметров (некоторые
рыбы) до 33 м (отдельные виды китов).
Покровы позвоночных образованы кожей, состоящей из
двух слоев: наружного — эпидермиса и внутреннего — дермы
(рис. 206). Эпидермис представлен многослойным эпителием.
У круглоротых, рыб и личинок земноводных, жизнь которых
тесно связана с водной средой, в .эпидермисе залегает много
железистых клеток, выделяющих слизь, облегчающую движение
животного в воде. У наземных позвоночных — пресмыкающихся,
птиц и млекопитающих — наружные слои клеток эпидермиса
ороговевают. На их коже обычно образуются различные роговые
придатки — чешуя, щитки, перья, волосы и др. Дерма кожи состо-
ит из плотной волокнистой соединительной ткани. В ней образу-
ются костная чешуя (у рыб), костные щитки и кожные (наклад-
ные) кости. В коже позвоночных залегают железы различного
строения и функций (слизистые, потовые, сальные и др.).
Скелет. У позвоночных первичным опорным образованием
служит хорда. Но у взрослых особей хорда в той или иной степени
замещается позвоночником (рис. 207). У миног (круглоротые)
хорда полностью сохраняется в течение всей жизни животного, но
в соединительнотканной оболочке ее и лежащего над ней спинного
мозга развиваются метамерно расположенные парные хрящи,
защищающие спинной мозг. Хорошо сохраняется хорда в течение
всей жизни у осетровых рыб, но у них вокруг хорды образуются
верхние и нижние хрящевые дуги позвонков. Хорда костистых
рыб в значительной степени вытесняется в процессе онтогенеза
позвоночником, образованным метамерно расположенными дво-
яковогнутыми позвонками, между которыми сохраняются четко-
образные или линзообразные остатки хорды. У взрослых наземных
позвоночных двояковогнутые позвонки встречаются редко и хорда
сохраняется во взрослом состоянии лишь в виде незначительных
остатков между позвонками. Отдельный позвонок обычно имеет
тело, верхнюю дугу (через канал которой проходит спинной мозг)
и нижнюю дугу. Если тело позвонка вогнуто с обеих сторон, то
236
4
Рис. 206. Схема строения кожи
/—роговой слой эпидермиса; 2 —
эпидермиса; .3 --дерма; 4 — волос;
ее протока; 7 — сальная железа
позвоночного (млекопитающего):
мальпигиев слой из живых клеток
5 — потовая железа; 6’ — отверстие
Рис. 207. Образование позвонка;
/ — стадия образования верхних и нижних дуг позвонка; // — стадия смыкания
концов верхних дуг и образования спинномозгового капала; III - стадия форми-
рования тела позвонка;
/ — нижняя дуга; 2— верхняя дуга; 3— боковые отростки позвонка; 4 — тело по-
звонка; 5 — хорда
позвонок именуется амфицельным, если он вогнут спереди, выпук-
лый сзади — процельным, если наоборот — опистоцельным,
а если спереди и сзади плоский — платицельным. К позвонкам
причленяются ребра.
Череп позвоночных животных бывает хрящевым, костно-хря-
щевым или костным. При развитии костно-хрящевого черепа
на хрящевую основу накладываются плоские кости, образующися
за счет окостенения окружающей соединительной ткани. При
формировании костного черепа часть костей возникает путем
окостенения хрящевого черепа эмбриона (первичные кости), а
другие накладываются на него снаружи (накладные кости). В че-
репе позвоночного различают мозговую коробку и висцеральный
скелет (рис. 208). Мозговая коробка защищает от механических
воздействий головной мозг и органы чувств головы.
Висцеральный скелет черепа образован у круглоротых хря-
щевой решетчатой коробкой, а у прочих позвоночных в основе
его лежит ряд парных жаберных дуг. В процессе эволюции одна
из передних пар жаберных дуг видоизменилась в челюсти, а сле-
дующая за ней — в подъязычную дугу. У костных рыб и наземных
позвоночных первичные хрящевые челюсти заменены вторичными,
образованными накладными костями. Остальные жаберные дуги
служат у рыб скелетом жаберного аппарата, а у наземных позво-
ночных редуцируются, частично входя в подъязычный аппарат.
У всех позвоночных, кроме миног и миксин, имеется скелет
парных конечностей и их поясов (иногда он бывает редуцирован
при атрофии конечностей). У рыб парные конечности имеют вид
плавников, а у наземных позвоночных они построены по типу
пятипалых ног (рис. 209).
В состав скелета передней пятипалой конечности (при ее
типичном строении) входят плечевая кость, две кости предплечья
(локтевая и лучевая), ряд костей запястья, пять костей пясти и
фаланги пальцев. Скелет типичной пятипалой задней конечности
образован бедром, двумя костями голени (большой и малой бер-
цовой), несколькими косточками предплюсны, пятью костями
плюсны и фалангами пальцев.
Плечевой пояс служит опорой для скелета передних конеч-
ностей. У наземных позвоночных он образован лежащими на спин-
ной стороне тела лопатками и расположенными на брюшной сто-
роне ключицами и коракоидами. Задние конечности сочленяются
с тазовым поясом, который у земноводных, пресмыкающихся,
птиц и млекопитающих слагается из трех пар костей: подвздош-
ных, лобковых и седалищных. У рыб пояса конечностей имеют
иное строение.
Мускулатура. У рыб и круглоротых с каждой стороны
тела тянутся мощные продольные мышцы, разделенные тонкими
соединительнотканными прослойками (миосептами) на ряд мио-
меров. У наземных позвоночных метамерность мускулатуры во
многом нарушена образованием многочисленных отдельных мышц.
238
Рис. 208. Череп акулы:
/ — слуховая капсула; 2 — глазница;
3 — обонятельная капсула; 4 — жабер-
ные дуги; 5 — верхний отдел подъязыч-
ной дуги; 6 — подъязычная дуга; 7 —
первичная верхняя челюсть; 8 — пер-
вичная нижняя челюсть; 9 — губные
хрящи
Рис. 209. Схема строения пятипа-
лой конечности наземных позво-
ночных:
1—передняя конечность; II—задняя
конечность;
1 — лопатка; 2 — подвздошная кость;
3 - прокоракоид; 4 — лобковая кость;
5 — коракоид; 6 — седалищная кость;
7 — плечевая кость; 8 — бедренная
кость; .9 — лучевая кость; 10 — боль-
шая берцовая кость; 11—локтевая
кость; 12- -малая берцовая кость;
13 — запястье; 14 — предплюсна; 15 —
пясть; 16 - плюсна; 17 — фаланги
пальцев; 18 — ключица
1
Рис. 210. Схема развития головного мозга позвоночных:
/—/// — стадии развития мозга; IV — продольный разрез мозга 111 стадии; V'— сфор-
мированный мозг с полушариями;
/ — зачаток переднего мозга; 2— зачаток среднего мозга; 3 — зачаток заднего мозга;
4 — спинной мозг; 5 — промежуточный мозг; 6— мозжечок; 7 — продолговатый мозг;
8 — обонятельные доли; 9 — гипофиз; 10— теменной орган; // — эпифиз; 12 — желу-
дочек переднего мозга; 13 — зрительные бугры; 14 — третий желудочек; /5 — желу-
дочек среднего мозга; 16 — желудочек мозжечка; 17 — четвертый желудочек; 18 — по-
лушария переднего мозга; 19 — невроцель
Нервная система позвоночных разделяется на централь-
ную, периферическую, симпатическую, парасимпатическую и веге-
тативную.
Центральная нервная система состоит из головного и спинного
мозга. У эмбрионов позвоночных передний конец нервной трубки
расширяется, образуя три первичных мозговых пузыря — за-
чатки головного мозга (рис. 210). Из переднего пузыря в даль-
нейшем образуется передний и промежуточный мозг, из средне-
го — средний мозг, а из заднего — продолговатый мозг, на крыше
которого путем выпячивания возникает мозжечок. Невроцель в
головном мозге представлен рядом полостей — желудочков мозга.
От головного мозга отходят 10—12 пар головных чувствующих,
двигательных и смешанных нервов:
обонятельные —• чувствующие нервы, передающие химические
раздражения с чувствующих клеток эпителия слизистой оболочки
обонятельной полости клеткам обонятельных луковиц переднего
мозга; зрительные — чувствующие нервы, проводящие световые
раздражения с сетчатки глаза в мозг животного; глазодвигатель-
ные нервы отходят от среднего мозга, иннервируя мышцы глаза;
блоковые — двигательные нервы, идущие от среднего мозга к не-
которым мышцам глазного яблока; тройничные — смешанные
нервы, отходящие (как и все последующие) от продолговатого
мозга, иннервируют многие мышцы, железы и участки кожи голо-
вы, а также слизистую оболочку носовой и ротовой полостей; отво-
дящие— двигательные нервы, иннервирующие некоторые мышцы
глаза; лицевые — двигательные нервы, идущие к некоторым
наружным мышцам головы; слуховые — чувствующие нервы, пе-
редающие раздражения, воспринятые органами слуха и равнове-
сия, в продолговатый мозг; языкоглоточные — смешанные нервы,
которые иннервируют часть слизистой оболочки ротовой и глоточ-
ной полостей и органа слуха, а также мышцы глотки и языка;
блуждающие — смешанные нервы, которые связаны анастомо-
зами с рядом головных нервов и участвуют в образовании нервных
сплетений сердца и некоторых других органов. У низших позво-
ночных иннервирует жаберные дуги (кроме первой) и органы
боковой линии, а у высших — кожу задней части головы, часть
глотки, щитовидную железу, дыхательные пути и легкие, печень,
переднюю часть кишечника, почки и некоторые другие органы;
добавочные — двигательные нервы, свойственные только высшим
позвоночным, иннервируют преимущественно мышцы шеи; подъ-
язычные — в основном двигательные нервы, но содержат также
и чувствительные волокна, иннервируют мышцы языка и подъ-
язычного аппарата.
Спинной мозг позвоночных животных в течение всех жизни
сохраняет трубчатое строение. Он состоит из расположенного внут-
ри серого и снаружи белого мозгового вещества. В сером веществе
находится большое количество нервных клеток и нервных воло-
кон, не одетых миэлиновыми оболочками. Белое вещество обра-
240
зовапо проходящими вдоль отростками нервных клеток — нейри-
тами, покрытыми изолирующими их миэлиновыми оболочками.
Периферическая нервная система позвоночных представ-
ляет сложную систему нервов, отходящих от головного и спинного
мозга к различным органам тела. Нервы слагаются из многих
нервных волокон, являющихся отростками нервных клеток мозга
и нервных узлов — ганглиев. Нервные волокна обычно одеты
миэлиновыми оболочками. Различают три категории нервов:
1) чувствующие нервы проводят раздражения, получаемые из
внешней среды или от внутренних органов, к нервным клеткам
мозга. По ним нервные импульсы идут по отношению к мозгу
центростремительно; 2) двигательные нервы передают нервные
импульсы центробежно от мозга к мускулам и другим органам,
вызывая реакцию соответственно их функциям: сокращение
мышц, секрецию желез и др.; 3) смешанные (чувствующе-двига-
тельные) нервы слагаются из чувствующих и двигательных нерв-
ных волокон. Эта группа нервов наиболее многочисленна.
От спинного мозга нервы отходят метамерно, попарно. Они на-
чинаются двумя корешками: спинным — чувствующим и брюш-
ным — двигательным. Эти корешки вскоре сливаются в единый
спинномозговой нерв, который вновь разветвляется, исключение
составляют миноги, у которых корешки спинномозговых нервов
не сливаются.
Вегетативная нервная система осуществляет регуляцию работы
внутренних органов животного - биение сердца, перистальтику
кишечника, секрецию желез и др. Деятельность вегетативной
нервной системы имеет некоторую степень автономности, но
все же контролируется центральной нервной системой. Вегетатив-
ная нервная система состоит из следующих элементов (рис. 211):
скоплений нервных клеток в сером веществе головного и спинно-
го мозга, образующих в нем особые центры; нервных волокон,
идущих к нервным узлам, расположенным по бокам позвоночника,
в полости тела и во внутренних органах; нервных волокон, идущих
от нервных узлов к иннервируемому
органу.
Таким образом, нервный импульс, по-
лучаемый органом через вегетативную
систему, обусловливается деятельностью
и центральной нервной системы.
Рис. 211. Схема вегетативной нервной системы
в связи со спинным мозгом:
1 -- серое вещество спинного мозга; 2 — его белое ве-
щество; 3 - двигательные волокна нерва; 4 -- брюшной
корешок ’спинномозгового нерва; 5 — прсганглионарпое
волокно нейрона; 6' — пограничный симпатический ствол;
7 — узел попншичного ствола; 8 - ганглий в стенке
кишки; 9... боковой столб серого мозгового вещества;
И) -- чувствительные волокна; 11—спинной корешок
спинномозгового нерва; 12 — смешанный спинномозго-
вой нерв; 13 - - ганглий; 14 - блуждающий нерв
241
Органы чувств в связи с активной жизнью позвоночных
обычно отличаются сложностью строения и функции. Глаза позво-
ночных имеют форму бокала, внутренняя полость которого за-
полнена студенистым стекловидным телом (рис. 212). Снаружи
глаз прикрыт прозрачной роговицей. Между краями бокала рас-
положен круглый или линзообразный хрусталик. Его форма у на-
земных позвоночных может меняться, это изменяет фокусное
расстояние линзы хрусталика и обусловливает аккомодацию гла-
за. У рыб аккомодация достигается путем перемещения хруста-
лика. Стенки глазного бокала состоят из трех слоев (оболо-
чек): 1) наружной сосудистой, богатой кровеносными сосудами;
2) средней пигментной, служащей для световой изоляции глаза;
3) внутренней — сетчатки, клетки которой (палочки и колбоч-
ки) способны воспринимать световые и цветовые раздражения.
Спереди сосудистая оболочка образует радужину, ограничиваю-
щую отверстие зрачка. Снаружи глазной бокал одет прочной за-
щитной капсулой — склерой.
Слуховые органы позвоночных животных и связанные с ними
органы равновесия (рис. 213) имеют различное строение, опи-
санное при обзоре отдельных групп этих животных. В передней
части головы расположена непарная (у круглоротых) или пар-
ная (у всех остальных позвоночных) обонятельная полость,
открывающаяся наружу ноздрями. У позвоночных, дышащих лег-
кими, обонятельные полости соединяются с ротовой полостью
внутренними ноздрями — хоанами, что позволяет им дышать, не
раскрывая рта. В коже позвоночных имеются рецепторы осяза-
ния, восприятия температуры, давления и иных раздражений
внешней среды. Имеются также рецепторы, воспринимающие раз-
дражения внутренней среды.
Органы пищеварения имеют различное строение.
Обычно пищеварительный тракт дифференцирован на ротовую
полость, глотку, пищевод, желудок и кишечник. Имеются пищева-
рительные железы — печень, поджелудочная железа и др.
Органы дыхания. Круглоротые и рыбы, а также личинки
земноводных дышат жабрами, взрослые же земноводные, пресмы-
кающиеся, птицы и млекопитающие — легкими. В газообмене
низших позвоночных большое значение имеет также кожное дыха-
ние. Жабры водных позвоночных — пластинчатые выросты стенок
жаберных щелей, в виде тонких жаберных лепестков, с богатой
сетью кровеносных сосудов. Легкие позвоночных возникают в
онтогенезе как парные выросты брюшной стороны глотки. У зем-
новодных они имеют вид тонкостенных мешков со складчатыми
стенками, у пресмыкающихся также мешкообразны, но внутри
разделены многочисленными перегородками и складками, у птиц
имеют губчатое строение. У млекопитающих конечные развет-
вления бронхов оканчиваются в легких мельчайшими пузырька-
ми — альвеолами. Стенки легких всех видов пронизаны сетью
кровеносных капилляров. Воздух поступает к легким через
242
Рис. 212. Схема развития и строения
глаза позвоночного животного:
/- IV — последовательные стадии разви-
тия глаза;
/ — промежуточный мозг; 2 — глазной бо-
кал; 3 — ножка глазного бокала; 4 —•
пигментная оболочка; 5—сетчатка; 6—
хрусталик; 7 — радужина; 8 — склера;
9 — роговица
6
Рис. 213. Схема строения слухо-
вого аппарата позвоночных:
1—овальный мешочек; 2— круглый
мешочек; 3 — передний полукружный
канал; 4 — горизонтальный и о — зад-
ний; 6 — эндолимфатический проток
Рис. 214. Схема кровообращения у различных позвоночных животных:
/— рыба; //-—земноводное; /// — млекопитающее (черное — венозная кровь, серое —
смешанная, белое — артериальная);
1 — предсердие; 2 — желудочек сердца; 3 — брюшная аорта; 4 — спинная аорта; 5 —
правое предсердие; 6 — левое предсердие; 7 — общий желудочек; 8 — правый желудочек;
9 левый желудочек; 10 — легочная артерия; //—легочная вена
ноздри в глотку, где начинается дыхательное горло — трахея,
которая в грудной полости делится на два бронха, идущих к
легким.
Кровеносная система замкнутая (рис. 214). Она сла-
гается из сердца, артериальных сосудов, несущих кровь от сердца
к различным органам, венозных сосудов, по которым кровь воз-
вращается в сердце, и капилляров — мельчайших сосудов, соеди-
няющих окончания разветвлений артерий и вен. Кровь позво-
ночных представляет собой бесцветную вязкую жидкость — плаз-
му, в которой взвешены красные и белые кровяные клетки. У круг-
лоротых, рыб и личинок земноводных, дышащих жабрами, один
круг кровообращения: от сердца венозная кровь идет к жабрам,
где окисляется и освобождается от углекислого газа, а затем раз-
носится артериальными сосудами по всему телу, возвращаясь
в сердце по системе венозных сосудов (рис. 214,/). Сердце у
этих животных имеет две камеры: предсердие и желудочек.
У взрослых земноводных, пресмыкающихся, птиц и млекопитаю-
щих в связи с переходом к легочному дыханию два круга крово-
обращения (рис. 214, II, III). Соответственно в сердце имеются
два предсердия. Малый круг образован легочными артериями,
несущими венозную кровь от сердца к легким, и легочными ве-
нами, возвращающими окисленную кровь в левое предсердие.
По артериям большого круга кровообращения обогащенная
кислородом кровь разносится от сердца по всему телу, где отдает
кислород и растворенные в ней питательные вещества тканям
и органам и поглощает углекислый газ и продукты распада орга-
низма. В сердце (в правое предсердие) кровь возвращается по ве-
нозным сосудам. У всех позвоночных вены, несущие кровь от
кишечника, входят в печень, где разветвляются в сложную
систему капилляров, образуя воротную систему печени. Воротная
система почек хорошо развита у рыб и слабо — у наземных поз-
воночных.
Сердце земноводных и большей части пресмыкающихся
трехкамерное: оно имеет два предсердия и один желудочек. В же-
лудочке происходит частичное смешение венозной крови, поступа-
ющей из правого предсердия с артериальной кровью, втекающей
из левого предсердия.
У птиц и зверей сердце состоит из четырех камер: двух пред-
сердий и двух желудочков. Этим устраняется смешение венозной
и артериальной крови.
Органы выделения — почки. Строение почек и проте-
кающие в них процессы отличаются у представителей разных групп
животных и изменяются в процессе онтогенеза (рис. 215). У заро-
дышей рыб и земноводных сначала закладываются головные поч-
ки, имеющие характер метанефридиев. Они образованы каналь-
цами, открывающимися одним концом, несущим воронку с мер-
цательным эпителием, в полость тела, а другим — в общий вы-
водной проток. Близ воронки стенки канальца имеют утолщение,
244
Рис. 215. Схема строения мочеполовых органон позвоночных:
1 — зародыш низшего позвоночного; // — самка низшего позвоночного; /// самец
низшего позвоночного; IV - зародыш высшего позвоночного; V' самка высшего позво-
ночного; VI — самец высшего позвоночного;
1— головная почка; 2 — туловищная почка; 3— тазовая почка; ‘/ — проток эмбрио-
нальной почки; 5 — мюллеров капал, служащий у самок яйцеводом; 6 - вольфов канал,
являющийся у низших позвоночных обоего пола мочеточником, а у самцов - - также семя-
проводом, у самцов высших -только семяпроводом; 7 — матка; 8 - мочеточник тазо-
вых почек; .9 — мочевой пузырь; 10 --клоака; 11 --мочеполовой синус; 12 задняя
кишка; 13 — половая железа; 14 -- кишка
в котором кровеносные капилляры образуют сосудистый клубочек.
Через воронки канальцев головных почек из полости тела удаляют-
ся излишки полостной жидкости с растворенными в ней продук-
тами азотистого распада. Часть же влаги и удаляемых из организ-
ма веществ фильтруется через стенки канальцев из крови сосу-
дистого клубочка.
По мере развития зародышей рыб и земноводных головные
почки сменяются туловищными почками. Последние располага-
ются позади головных, которые постепенно атрофируются. Они
имеют более сложное строение, чем головные почки. В них как
излишняя для организма вода, так и продукты распада выделяют-
ся из крови в особых боуменовых капсулах, имеющих вид округлых
полых телец, в которые глубоко впячивается часть стенки. В этом
впячиваиии находятся клубочки тонких сосудов, которые выделя-
ют влагу и продукты распада в полость капсулы, откуда они
стекают по коротким протокам в выводной канал почки.
С возрастом зародыша рыб и земноводных меняются также
выводные протоки почек. Эмбриональные выводные протоки
головных почек расщепляются вдоль на два канала: мюллеров
и вольфов. Вольфов канал становится мочеточником первичной
почки, тогда как мюллеров проток у самцов редуцируется, а у са-
мок выполняет функцию яйцевода. У самцов вольфов канал
выполняет также функцию семяпровода.
245
У пресмыкающихся, птиц и млекопитающих при эмбриональ-
ном развитии туловищные почки заменяются более сложного
строения тазовыми почками, лежащими в полости таза. В них
мочевые канальцы начинаются мальпигиевыми тельцами — боу-
меновыми капсулами с лежащими в их впячиваниях сосудистыми
клубочками. Мочеточники тазовых почек представляют собой
новообразование.
Органы размножения. Почти все позвоночные жи-
вотные раздельнополы. Лишь среди круглоротых и рыб имеется
небольшое число гермафродитных видов. Половые железы —
семенники у самцов и яичники у самок — обычно парные. У кругло-
ротых половые клетки из половых желез попадают в полость тела,
откуда выводятся через особые поры в стенках тела у мочевого
отверстия. У других позвоночных развиты специальные протоки,
выводящие половые продукты наружу. Нередко они дифференци-
рованы на ряд отделов.
Различия в особенностях строения, характере размножения и
развития эмбрионов, а также путей эволюции челюстноро-
тых позвоночных позволяют разделить их на две группы: Atiainnia
и A-mniota.
К Anamnia принадлежат рыбы и земноводные, т. е. животные,
проводящие всю жизнь или личиночную стадию развития в воде.
У подавляющего большинства видов этой группы наблюдается
наружное оплодотворение яиц в водной среде. Самки мечут яйца в
воду, а самцы поливают их семенной жидкостью. Развитие эмбри-
она проходит в воде без образования специальных зародышевых
оболочек — амниона и аллантоиса. Животные, относящиеся к этой
группе, имеют слизистую кожу и дышат всю жизнь или на стадии
личинок жабрами. Органами выделения у взрослых особей слу-
жат туловищные почки.
Группа Afntiiota включает наземных позвоночных живот-
ных — пресмыкающихся, птиц и млекопитающих. У них оплодо-
творение яиц происходит в половых органах самки (внутреннее).
При развитии зародыш образует зародышевую оболочку — ам-
нион, окружающую эмбрион (отсюда название группы) (рис. 216).
\ _ 4 jj/ Между амнионом и зародышем
находится околоплодная жидкость.
Из заднего отдела кишечника эм-
бриона развивается вторая заро-
дышевая оболочка — аллантоис,
в котором накапливаются моче-
вые выделения зародыша. На-
ружная стенка аллантоиса, бо-
Рис. 216. Зародышевые оболочки амниот:
/—/|/ — стадии развития зародыша;
/ —эктодерма; 2 — эндодерма; 3 — мезодер-
ма; 4 — полость кишечника; ,5 — впезароды-
шевая полость; 6—амнион; 7 — амниоти-
ческая полость; 8 — сероза; 9 — аллантоис;
10 - желточный мешок
246
гатая кровеносными сосудами, выполняет дыхательную функцию.
У животных этой группы кожа не выделяет слизи и наружные
слои се эпидермиса подвергаются ороговению. Дыхание на* всех
стадиях развития легочное. Ночки тазовые.
РАЗДЕЛ БЕСЧЕЛЮСТНЫЕ (AGNATHA)
Небольшая группа примитивных позвоночных животных с со-
сущим ротовым аппаратом без подвижных челюстей. Жаберных
дуг нет. Парные конечности отсутствуют. Имеется непарная
ноздря, ведущая в непарный обонятельный мешок.
КЛАСС КРУГЛОРОТЫЕ (CYCLOSTOMATA)
Общая характеристика. Примитивная группа позвоночных жи-
вотных, включающая миног и миксин. Тело круглоротых вытяну-
тое, цилиндрическое. Кожа голая, слизистая. Парные конеч-
ности отсутствую'!'. Рот без челюстей, он расположен в глубине
ротовой присоски. Ноздря непарная. Хорда полностью сохраняет-
ся в течение всей жизни животного. По бокам спинного мозга
метамерно расположены попарно небольшие хрящики — зачатки
верхних дуг позвонков. Череп слагается из нескольких хрящей.
Миксины и большая часть миног - обитатели морей, но часть
миног заходят в реки для икрометания, а некоторые из них посто-
янно живут в пресных водах.
Строение и жизненные отправления. Тело круглоротых вы-
тянутое, цилиндрическое в передней и средней частях и уплощен-
ное с боков, в задней! без резких границ разделенное на голову,
туловище и хвост (рис. 217). Парные плавники отсутствуют —
их нс было и у предков круглоротых. Хвост оторочен узким хвосто-
вым плавником. У мпно!' па спине возвышается непарный спин-
ной плавник, разделенный выемкой на передний и задний.
Покровы круглоротых тонкие, с большим количеством
сл и з и ст ы х ж ел ез о к.
Скелет представлен хорошо развитой хордой. По бокам
спинного мозга в толще соединительнотканной оболочки, окружа-
ющей его и хорду, расположены два ряда небольших хрящиков,
являющихся зачатками верхних дуг позвонков. Черен состоит из
нескольких отдельных хрящей, соединенных тонкой перепонкой.
Основанием черепа служит хрящевая пластинка, по бокам которой
лежат слуховые капсулы, а спереди — обонятельная капсула.
Скелет глоточной области имеет вид хрящевой решетки. Жабер-
ные дуги и челюсти отсутствуют.
Мускула т у р а ясно разделена миосептами на ряд мио-
. мерой. Продольной миосеиты пет.
Нервная система весьма примитивна. Головной мозг
мал. В крыше переднего мозга нет нервных клеток. Мозжечок
имеет вид валика на передней стенке продолговатого мозга.
247
Рис. 217. Минога:
/ — внешний вид; // — внутреннее строение;
1 — ротовая присоска; 2 — ноздря; 3 — глаз; 4 — жаберные щели; .5, 6 — спинные плав-
ники; 7 — хвостовой плавник; 8,9 — мочеполовой сосочек; 10 — хорда; 11, 12 — спинной
мозг; 13—его оболочка; 14— мышцы; 15— головной мозг; 16—18 — орган обоняния;
19—23 — хрящи черепа; 24— сосочки ротовой воронки; 25 —- роговые зубы; 26 —
подъязычный хрящ; 27 — ротовая полость; 28 — пищевод; 29— кишка; 30 — дыха-
тельный отдел глотки; 31 — внутренние отверстия жаберных мешков; 32 — вскрытый
жаберный мешок; 33 — его наружное отверстие; 34, 35 — сердце; 36 — печень; 37 -
семенник; 38 — задняя кишка; 39 — ночка; 40 ~ мочеточник
Органы чувств развиты слабо. Глаза малы, а у миксин
сильно редуцированы. Обонятельная полость непарная, она откры-
вается наружу одной ноздрей (но обонятельные нервы парные).
Обонятельный мешок продолжается в так называемый питуитар-
ный вырост. Ухо внутреннее; орган равновесия имеет только два
полукружных канала. Имеются кожные рецепторы различного
назначения.
Пищеварительная система начинается околоротовой
присоской, усаженной роговыми зубами. В глубине ее располо-
жен рот, который ведет в обширную глотку. Глотка делится го-
ризонтальной перегородкой на дыхательную трубку и лежащий
над ней пищевод, который переходит в кишку, кончающуюся
анальным отверстием. Желудок слабо выражен. Имеется большая
печень, желчного пузыря нет.
Органами дыхания служат жабры. Их строение отли-
чается от строения жабр рыб. В боковых стенках дыхательной
трубки глотки имеются парные жаберные отверстия, ведущие в
жаберные мешки, стенки которых несут многочисленные тонкие
лепестки. В них расположена сеть кровеносных сосудов. У миног
каждый жаберный мешок открывается наружу отдельным жабер-
ным отверстием. У миксин же наружные жаберные отверстия
ведут в продольный канал, открывающийся одной парой на боках
тела. Развиваются жабры у круглоротых из эндодермы, а не из
эктодермы, как у рыб.
248
Кровеносная система круглоротых близка к таковой
ланцетника. Имеется один круг кровообращения. Сердце состоит
из предсердия и желудочка. Селезенка отсутствует.
Органами выделения у взрослых миног являются туло-
вищные почки, а у некоторых миксин всю жизнь функционируют
головные почки.
Органы р а з м н о ж е н и я. Половые железы непарные. По-
ловые продукты выводятся через разрыв стенок гонад в полость
тела, а оттуда — через мочеполовой синус наружу.
Р а з в и т и е у миксин прямое, а у миног — со стадией личинки.
В класс Круглоротые входят два отряда: Миноги (Petrorny-
zoncs), Миксины (Myxiiioideci).
Отряд Миноги (Petromyzones) — свободноживущие живот-
ные. Имеют спинной плавник. Семь пар жаберных отверстий.
Обитают в морях, за исключением ручьевых миног, живущих в
ручьях и речках. Морские формы заходят в реки для икромета-
ния. Питаются мелкими донными животными и падалью. Икру
мечут один раз в жизни. Из нее выходят мелкие личинки — песко-
ройки, позднее уходящие в море. Мясо употребляется в пищу.
Промышляются во время хода на нерест.
Отряд Миксины (Myxinoidea) — полупаразиты. Спинной
плавник отсутствует. Глаза скрыты под кожей. Жаберные мешки
открываются в общий продольный капал, выходное отверстие
которого находится позади головы.
Живут в морях. Миксины вгрызаются в тело рыб и поедают
их внутренности.
РАЗДЕЛ ЧЕЛЮСТНОРОТЫЕ
(GNATHOSTOMATA)
Более высокоорганизованные позвоночные животные. Рот с
челюстями. Две пары конечностей имеют вид плавников или ног.
Ноздри и обонятельные полости парные. К данному разделу
относятся рыбы, земноводные, пресмыкающиеся, птицы и млеко-
питающие.
НАДКЛАСС РЫБЫ (PISCES)
Общая характеристика. Рыбы — водные позвоночные, органами
движения которых служат парные и непарные плавники. У боль-
шинства кожа покрыта чешуей различного строения. Рот ограни-
чен подвижными челюстями. Органами дыхания служат жабры.
Обонятельные отверстия парные. У всех рыб, кроме двоякодыша-
щих, имеется один круг кровообращения. Сердце имеет две ка-
меры: предсердие и желудочек. Температура тела переменная.
Гидростатическим органом служит плавательный пузырь.
У большинства рыб в коже заложены особые органы боковой
249
линии. Рыбы, как правило, раздельнополы, но имеются и герма-
фродиты. Размножаются они обычно икрометанием, но есть также
и живородящие.
В современной фауне насчитывается около 20 тыс. видов
рыб, большинство из которых живут в морях. Рыбы имеют огром-
ное значение как продуценты ценных пищевых продуктов и тех-
нического сырья.
Современных рыб обычно подразделяют на два класса:
Хрящевые и Костные. Каждый из них, в свою очередь, делится
на подклассы.
Класс Хрящевые рыбы (Chondrichthyes)
Подкласс Пластинчатожаберные (Elasmobran-
chia)
Класс Костные рыбы (Osieichthyes)
Подкласс Лучеперые (Actinopterygii)
Надотряд Костно-хрящевые (Chondrostei)
Надотряд Костистые (Teleostei)
Подкласс Кистеперые (Crossopterygii)
Подкласс Двоякодышащие (Dipnoi)
Строение и жизненные отправления. Размеры рыб колеблют-
ся в очень широких пределах: от 1 см (один из видов бычков)
до 16—20 м (китовая акула). Масса некоторых рыб достигает
1,5 т и более.
Форма тела рыб крайне разнообразна (рис. 218), что свя-
зано с многообразием мест обитания и образа жизни. У большин-
ства рыб тело обтекаемой формы, что облегчает их перемещение
в воде. У таких быстро плавающих рыб, как акулы, лососи, суда-
ки и др., тело удлиненное, торпедообразное. Рыбы, не соверша-
ющие больших перемещений (лещ, карась, карп), часто имеют
высокое, сжатое с боков тело. У донных рыб брюшная сторона
тела чаще уплощенная, что обеспечивает больший контакт с дном,
где находится пища.
Органы движения. Рыбы передвигаются в воде путем
изгибов тела и с помощью парных и непарных плавников (рис. 219).
Основанием плавников служит внутренний опорный скелет. На-
ружные лопасти поддерживаются костными или хрящевыми плав-
никовыми лучами. Последние бывают жесткими нерасчлененными
или мягкими членистыми, мягкие лучи бывают ветвистыми.
Парных плавников две пары: грудные и брюшные. Первые
расположены за жабрами, а вторые — в разных местах брюшной
поверхности. У тресковых они находятся на горле впереди грудных.
Отсутствие парных плавников обусловлено утратой их в процессе
эволюции.
Парные плавники поддерживают тело рыбы в горизонталь-
ном положении и служат рулями поворота и глубины.
Брюшные плавники бычков сращены и образуют присоску, при
помощи которой рыбы удерживаются на дне в местах с быстрым
течением или сильным прибоем. Очень длинные грудные плавники
250
Рис. 218. Форма тела рыб:
/ сарган; 11 скумбрия; /// лет; IV -- луна-рыба; V' -- камбала; Г/ -
угорь; VII сельдяной король; VIII -иглобрюх; /Л--скат
летучих рыб образуют несущие плоскости, на которых они плани-
руют, отделившись от воды.
К непарным плавникам относятся хвостовой, один или несколь-
ко спинных и один, реже больше анальных. В поступательном дви-
жении рыб основную роль играет хвостовой плавник: он же служит
рулем при поворотах и погружениях животного. Спинные и аналь-
ные плавники — это прежде всего стабилизаторы направления
движения рыбы, но они участвуют также в поворотах тела. Так,
лещ движением длинного анального плавника может наклонять
передний конец тела вниз, что облегчает нахождение пищи. У щук
спинной и анальный плавники отодвинуты к хвосту, что увеличи-
вает мощность удара хвостом и, следовательно, стремительный
251
Рис. 219. Внешний вид леща:
1 — спинной плавник; 2 — хвостовой плавник; 3— анальный плавник;
•4 — брюшные плавники; 5 — грудные плавники; 6 — рот; 7 — ноздри;
8 — глаза; 9 — жаберная крышка; 10 — боковая линия; 11—аналь-
ное отверстие
бросок на добычу из засады. У живущей на дне морей рыбы удиль-
щика удлиненный передний луч спинного плавника расположен
над верхней губой: его колебанием хищник привлекает добычу.
У рыбы прилипало спинной плавник изменен в присоску. Иногда
плавники представлены колючками.
Покровы образованы кожей из эпидермиса и дермы. На-
ружные эпидермиса не ороговевают. В нем заложены многочислен-
ные одноклеточные железки, выделяющие слизь, которая умень-
шает трение тела рыб о воду. Дерма имеет волокнистое строение
(рис. 220). У большинства рыб тело покрыто чешуей того или иного
строения. У акул и скатов чешуя плакоидного типа (рис. 221, /).
Она состоит из погруженной в кожу дентиновой основы и выдаю-
щегося наружу, покрытого эмалью шипа. На челюстях этих рыб
плакоидные чешуи видоизменены в зубы. У осетровых рыб по
хребту, бокам и границе боков и брюха тянутся пять рядов костных
пластинок — жучек, а хвост покрыт ромбической ганоидной чешу-
ей, характерной для древних рыб. Костистые рыбы обычно покры-
ты костной чешуей, имеющей ‘
вид тонких, налегающих друг на
друга пластинок различной фор-
мы (рис. 221, //). Они сидят
наклонно в коже животного,
образуя в большинстве случаев
правильные ряды. Неравномер-
ность роста чешуи в разные се-
Рис. 220. Строение эпидермиса кости-
стой рыбы:
1 —слизеотделительная одноклеточная
железа; 2 — колбовидная одноклеточная
железа; 3 — лимфатический сосуд
252
1
Рис. 221. Типы чешуи рыб и соотношение между скоростью роста рыбы и разме-
ром ее чешуи по годам (///):
/ — разрез кожи акулы с плакоидной чешуей; // — костная чешуя костистой рыбы
(видны годовые кольца);
Н
зоны года ведет к образованию на ней широких летних и узких зим-
них колец, по их числу можно судить о возрасте рыбы.
Поскольку есть зависимость между скоростью роста чешуи и
тела рыбы, то по ширине годовых колец на чешуе можно судить о
темпах роста рыб за прошлые годы. Иногда чешуя рыб видо-
изменена в иглы, шипы, костные щитки и другие кожные обра-
зования. Окраска рыб определяется находящимися в коже пиг-
ментными клетками. Серебристая окраска и металлический блеск
чешуи обусловливается наличием иглистых кристаллов особого
вещества — гуанина, который отражает падающий свет.
Скелет. Строение скелета связано с высотой организации и
особенностями биологии рыбы. У осетровых, двоякодышащих и
некоторых других рыб в течение жизни сохраняется хорошо
развитая хорда, а позвонки представлены только хрящевыми ду-
гами. У костистых рыб во взрослом состоянии остатки хорды
наблюдаются только между позвонками. Акулы и скаты имеют
скелет, состоящий из хрящевых элементов без костных частей.
У осетровых рыб череп, позвоночник и пояса плавников в основе
хрящевые, но хрящевой череп их покрыт наружным панцирем
из плоских накладных костей. Кости имеются и в поясе грудных
плавников. У костистых рыб скелет образован в основном кост-
ными элементами (рис. 222). Скелет рыб слагается из черепа,
позвоночника, связанных с ним ребер, костей и хрящей плавников
и их оснований. Череп состоит из мозговой коробки и висцераль-
ного скелета, который образован костными или хрящевыми челю-
стями, подъязычной дугой и жаберными дужками. У большинства
рыб жабры прикрыты костными жаберными крышками. Позвонки
хрящевых и костных рыб имеют двояковогнутое тело. Их верх-
ние дуги образуют спинномозговой канал, а нижние дуги хвосто-
вых позвонков — канал, по которому тянутся крупные кровенос-
Рис. 222. Скелет костистой рыбы (окуня):
/ — остистые отростки позвонков; 2 — нижние дуги позвонков; 3 — нижние остистые
отростки позвонков; 4 — ребра; 5 — мускульные косточки; б—основные косточки
(радиалии) лучей плавников; 7—лучи плавников; 8—кости плечевого пояса;
9 — кости тазового пояса; 10 — череп
254
Рис. 223. Мускулатура рыбы (окуня):
/ — миомеры; 2 — миосепты
ные сосуды. С позвонками туловища связаны ребра, свободно
оканчивающиеся другим концом в мускулатуре стенок тела. Пар-
ные плавники имеют пояса, образованные хрящами или костями,
лежащими в туловище рыбы.
Мускулатура туловища и хвоста рыб имеет метамерное
строение (рис. 223). Вдоль тела, слева и справа, тянутся широкие
полосы продольных мышц, разделенные соединительными
прослойками — миосептами на ряд мышечных сегментов — мио-
меров. Продольная миосепта разделяет их на спинные и брюш-
ные отделы. В голове, плавниках и их поясах расположены отдель-
ные мышцы.
Нервная система рыб более совершенна, чем у кругло-
ротых, но все же несет много примитивных черт. Головной мозг
рыб невелик: так, например, у крупных щук он составляет лишь
1/1300, а у акул— 1/3700 массы тела (рис. 224). Он состоит
из переднего, промежуточного, среднего и про-
долговатого мозга, а также мозжечка. Передний
мозг не образует полушарий (кроме двоякоды-
шащих рыб) и имеет только один желудочек, при-
крытый сверху тонким покровом без нервных кле-
ток. Передний конец его продолжается в обоня-
тельные доли с отходящими от них обонятельны-
ми нервами. Передний мозг рыб играет роль обо-
нятельного центра и участвует в координации
движений. Его удаление влечет потерю рефлекса
на восприятие запахов. Промежуточный мозг
Рис. 224. Головной мозг рыбы (окуня):
/—обонятельные, капсулы; 2 — обонятельные доли; 3 — передний
мозг; 4 — средний мозг; 5—мозжечок; 6 — продолговатый мозг;
7 — спинной мозг; 8—10 — головные нервы
255
рыб мал. На его крыше расположен вырост — эпифиз, который
выполняет роль железы внутренней секреции. Нижняя поверхность
промежуточного мозга рыб образует воронку, к вершине которой
прилегает другая железа внутренней секреции — гипофиз. От дна
этого отдела мозга отходят зрительные нервы. Средний мозг
развит относительно хорошо. На его верхней поверхности выдают-
ся два зрительных бугра, с которыми связаны волокна зрительных
нервов. Мозжечок, являющийся у рыб, как и у других позвоноч-
ных, центром регуляции движений, развит по-разному в зависи-
мости от степени подвижности животного. Продолговатый мозг
рыб прикрыт сверху эпителиальной пленкой. Боковые стенки его
утолщены. От головного мозга рыб отходят десять пар головных
нервов. Спинной мозг тонок, он тянется до конца позвоночника.
Органы чувств приспособлены к восприятию раздраже-
ний в условиях водной среды. Обонятельные органы рыб имеют
вид пары полостей, открывающихся наружу двумя ноздрями. Они
не соединены с ротовой полостью или глоткой хоанами, как у на-
земных позвоночных. Внутри они выстланы чувствующим эпите-
лием. Обоняние играет большую роль в отыскивании пищи. Так,
ослепленный налим находит червя по запаху на расстоянии до
30 см, но не может его обнаружить на расстоянии 1 см, если ему
закрыть ноздри.
Осязательную функцию у рыб несут покровы, усики на голове,
иногда удлиненные лучи парных плавников. Органами вкуса слу-
жат вкусовые сосочки, расположенные на губах и в ротовой
полости.
Глаза рыб приспособлены к видению на близком расстоянии,
что обусловлено относительно малой прозрачностью воды. Они
отличаются плоской роговицей и шаровидным хрусталиком. Боль-
шинство рыб хорошо видят на расстоянии до 1 м, но иногда акко-
модацией достигается дальность видения до 10—12 м. У глубоко-
водных и пещерных рыб глаза нередко редуцируются. Рыбы
различают как форму, так и цвет окружающих предметов. У видов,
наиболее жизнедеятельных днем и обитающих в освещенных
слоях водоема, в сетчатке глаз частично преобладают колбочки,
воспринимающие цвета. Сумеречные и глубоководные рыбы, на-
против, имеют в сетчатке глаз больше палочек, воспринимающих
свет различной интенсивности. У многих глубоководных рыб кол-
бочек в сетчатке глаз нет совсем. У рыб, живущих в поверхност-
ных слоях воды, обычно наблюдается положительный, а у боль-
шинства донных рыб — отрицательный фототаксис. У мальков
придонных рыб, живущих на первых стадиях развития в верхних,
хорошо освещенных слоях водоема, при переходе к жизни у дна,
где света меньше, положительный фототаксис сменяется отрица-
тельным. Световая реакция некоторых промысловых рыб позволя-
ет ловить их, привлекая ярким светом.
Органы слуха представлены только внутренним ухом, заклю-
ченным в костную капсулу черепа. Перепончатый лабиринт внут-
256
Рис. 225. Продольный разрез через капал боковой линии костистой
рыбы:
I .-боковой нерв; 2- чувствительные клетки органа; 3~ канал органа-
4 - наружное отверстие
репного уха рыб состоит из верхнего овального мешочка с тре-
мя взаимно перпендикулярными полукружными каналами и ниж-
него круглого мешочка, заполненных эндолимфой. Собственно
органом слуха служит лишь нижний мешочек; улитка уха, свойст-
венная наземным позвоночным, отсутствует. Сравнительно простое
строение органа слуха рыб связано с большой звукопроводностью
воды. Полукружные каналы выполняют функцию органов равнове-
сия. В их эндолимфе взвешены мелкие камешки — отолиты.
У костных рыб отолиты прикреплены к вершинам ресничек чувст-
вительного эпителия, выстилающего внутреннее ухо. При изме-
нении положения тела животного давление и натяжение ресничек
отолитами меняются, что и воспринимается клетками эпителия.
Органы слуха имеют большое значение при установлении
контактов между рыбами. Доказано, что многие рыбы издают
различные звуки зубами, трением плавников, при помощи плава-
тельного пузыря и другими способами. Расшифровка этих звуков
позволяет использовать их для обнаружения косяков рыбы.
Своеобразным органом чувств рыб является боковая линия
(рис. 225). В простейшем случае, как, например, у плащеносных
акул, это открытая борозда, тянущаяся вдоль боков тела. Но у
большинства рыб органы боковой линии имеют вид канала в коже,
сообщающегося с внешней средой рядом пор, прободающих
чешую. Ряд таких перфорированных чешуек тянется по бокам
рыбы, образуя пунктирную линию. В стенках канала расположены
окончания нервных волокон ветви блуждающего нерва. Органы
боковой линии воспринимают движение и давление воды. Они
чувствительны к низким звукам (ниже 100—150 колеб/с). У ряда
рыб органов боковой линии нет, но имеются особые чувствующие
канальцы. У сельдей они находятся на голове.
Органы пищеварения. Пищеварительный тракт рыб
делится на три отдела: передний, включающий ротовую полость,
глотку и пищевод; средний, состоящий из желудка, тонкой кишки
и пищеварительных желез — печени и поджелудочной железы, и
задний, представленный задней кишкой (рис. 226).
9 Б. Л. Кузнецов и др.
257
Рис. 226. Внутреннее строение рыбы (окуня):
/ — жабры; 2~ сердце; 3—печень; '/ — плавательный пузырь; 5 - селезенка;
6 — яичник; 7 — желудок; 8 — слепые отростки кишки; У— кишка; 10 - мочевой
пузырь; // — анус; 12 — мочеполовое отверстие
Рот рыб бывает различных типов: хватательный, когда челюсти
вооружены острыми зубами для схватывания добычи (хищные ры-
бы) ; всасывающий, способный вытягиваться в трубку для всасыва-
ния пищи (лещ и др.); дробящий — с тупыми большими зубами,
способными дробить твердую пищу (зубатка и др.). По положению
различают: рот нижний, расположенный на нижней стороне
головы (характерен для рыб, добывающих пищу на дне водоема);
конечный — на конце головы и верхний, когда нижняя челюсть
выдается за конец верхней (свойствен рыбам, которые питаются
падающими в воду насекомыми и мелкими животными, обитающи-
ми в верхних слоях воды). У большинства рыб на челюстях и на
небе расположены зубы. У рыб семейства Карповые зубов нет,
но на последней жаберной дуге у них находятся костные выросты,
вдающиеся в полость глотки — глоточные зубы. Они обращены
вершинами к находящемуся на верхней стороне глотки жерновку,
трением о который размельчается пища.
Важную роль в добывании пищи у рыб играют жаберные
тычинки — выросты вогнутой стороны жаберных дуг. У планктоно-
ядных сельдей, сигов, толстолобиков и др. длинные и многочислен-
ные жаберные тычинки образуют своеобразный цедильный аппа-
рат, задерживающий проходящие с водой через жабры мелкие
организмы. Короткий пищевод ведет в желудок. У карповых
желудок отсутствует. Длина кишечника зависит от характера
258
пищи: у хищных рыб он значительно короче, чем у растительно-
ядных. У ряда рыб он образует то или иное число пилорических
отростков, наличие которых увеличивает всасывающую поверх-
ность кишок. Обычно хорошо развиты печень и поджелудоч-
ная железа, которые вместе с железами стенок желудка и кишки
вырабатывают ферменты, необходимые для переваривания пищи.
Слюнных желез у рыб нет.
Плавательный пузырь, присущий большинству рыб, представ-
ляет собой тонкостенный полый орган различной формы и строе-
ния, наполненный газом. Он лежит в спинной части полости тела
и образуется у эмбриона как вырост глотки. У некоторых рыб пла-
вательный пузырь остается связанным каналом с глоткой в течение
всей жизни (открытопузырные). У других эта связь нарушается в
процессе развития малька (закрытопузырные). Плавательный пу-
зырь служит гидростатическим органом. Изменение его объема
влечет изменение удельного веса рыбы, что облегчает ее пребыва-
ние в определенном слое водной толщи. Объем газа в пузыре регу-
лируется выделением его из крови с помощью особого сплетения
капилляров кровеносных сосудов — так называемого красного
тельца и сети капилляров в стенках пузыря. Плавательный пузырь
отсутствует у многих донных рыб и рыб, совершающих быстрые
вертикальные перемещения в воде.
Органы д ы х а н и я рыб — жабры. У акул и скатов жабер-
ные щели разделены межжаберными перегородками и открывают-
ся наружу отдельными отверстиями. На передней и задней стенках
жаберных перегородок сидят рядами топкие жаберные лепестки,
имеющие густую сеть кровеносных сосудов. У костистых рыб
межжаберные перегородки редуцированы и жаберные лепестки
сидят попарно на жаберных дужках. Их жаберные полости не
разделены перегородками и прикрыты снаружи костными жабер-
ными крышками. Важную роль в газообмене рыб играет кожное
дыхание. Так, у годовалых карпов кожное дыхание обеспечивает
до '/а, а у карася и угря — даже '/з потребности организма в
кислороде. Некоторые рыбы способны также использовать для
дыхания кислород воздуха. Вьюны, живущие на дне водоемов, ре-
гулярно поднимаются к поверхности и заглатывают воздух, про-
пуская его через кишечник, в стенках которого находится сеть
кровеносных капилляров. У аквариумных рыбок макроподов в
задней части головы расположен особый лабиринтовый аппарат,
сообщающийся с глоткой, куда животное заглатывает воздух и
где усваивается кислород. У двоякодышащих рыб, обитающих в
пересыхающих водоемах тропиков, помимо жабр, есть особые
мешковидные легкие, при помощи которых они дышат, когда
впадают в анабиотическое состояние, зарывшись в ил высохшего
водоема.
Кровеносная система рыб в основных частях сходна
с таковой ланцетника (рис. 227). У всех рыб, кроме двоякодыша-
щих, один круг кровообращения. Сердце двухкамерное: оно со-
9*
259
6
4 5 Ю
7 9
Рис. 227. Кровеносная система хряще-
вых (4) и костистых (Б) рыб:
/ — схема кровеносной системы;//— строение
сердца;
1— сердце; 2 — брюшная аорта; 3 - прино-
сящие жаберные артерии; 4 — выносящие жа-
берные артерии; <5 — корни аорты; 6 — сон-
ная артерия; 7 — епиппая аорта; 8 -- хвосто-
вая вена; 9 — задняя кардинальная вена;
10 — передняя кардинальная вена; // — кю-
вьеров проток; 12 — воротная вена печени;
13 — печеночная вена; 14 — боковая вена;
/5 — предсердие; 16 — желудочек сердца;
17 — артериальный конус С клапанами; 18 -
луковица аорты
стоит из предсердия и желудочка. Перед, предсердием лежит
тонкостенный венозный синус, в который оттекает венозная кровь
из разных частей тела. У хрящевых и осетровых рыб желудочек
сердца продолжается вперед мускулистым артериальным конусом
е клапанами внутри. Впереди он переходит в брюшную аорту,
несущую венозную кровь к жабрам. У костистых рыб артериаль-
ный конус редуцирован, и у них брюшная аорта отходит непо-
средственно от желудочка сердца, образуя у основания утол-
щение-луковицу аорты.
Окисленная в жабрах кровь поступает в парные наджабер-
ные артерии, которые впереди продолжаются сонными артериями,
несущими кровь к голове, а сзади переходят в парные корни
аорты. Последние, сливаясь, образуют спинную аорту, которая
тянется под позвоночником и дает ответвления к различным ча-
стям тела.
Кровь, отдавшая тканям тела кислород и поглотившая угле-
кислоту, течет обратно к сердцу по венам. От головы венозная
кровь оттекает по парным кардинальным венам. Из хвостового
отдела она собирается в непарную хвостовую вену, которая, войдя
в полость тела, разделяется на две ветви; обе или одна из них
проходят через почки, образуя воротную систему почек. По вы-
ходе из почек венозная кровь течет к сердцу по двум задним
кардинальным венам. Сосуды, несущие кровь от кишечника с
растворенными в ней питательными веществами, соединяются в
подкишечную вену, которая идет к печени. Войдя в последнюю,
она образует воротную систему печени. Капилляры последней со-
260
бираются в крупную печеночную вену. Передние и задние кар-
динальные вены, сливаясь, образуют два кювьерова протока,
которые открываются в общий венозный синус, туда же впадает
печеночная вена. Из венозного синуса венозная кровь переходит
в предсердие и далее в желудочек.
Количество крови у рыб в сравнении с наземными позвоноч-
ными невелико (например, у карпа около 2 % массы тела). Эри-
троциты крови рыб имеют овальную форму и содержат ядра.
У рыб, живущих в водоемах, бедных кислородом или с резкими
колебаниями содержания его в воде (карп, карась, линь и др.),
гемоглобин крови способен поглощать кислород даже при малом
его количестве, тогда как у рыб, обитающих в воде, постоянно
богатой кислородом (форели, сиги и пр.), он может связывать
кислород только при значительном его количестве.
Сердце рыб сокращается сравнительно редко — 20—30 раз в
минуту. С повышением температуры среды сердцебиения уча-
щаются. У рыб, находящихся в зимнем оцепенении, число их па-
дает до 1—2 в минуту.
Кроветворными органами служат преимущественно селезенка
и соединительная ткань почек.
Лимфатическая система костистых рыб представлена парными
сосудами, идущими по бокам спинной аорты, и сетью мелких
сосудов, принимающих лимфу из тканей тела и проводящих
ее в основные спинные сосуды. Лимфатических желез у рыб нет.
Органы выделения эмбриона представлены сначала
головными почками, которые в процессе развития сменяются
туловищными ночками — двумя длинными и плоскими краснова-
тыми полосками, расположенными в средней части полости тела
по бокам от позвоночника. Моча, выделяемая туловищными поч-
ками, выводится мочеточниками, которыми служат вольфовы ка-
налы. Мочеточники в конце сливаются в единый проток, который
открывается у акул и скатов в клоаку, а у костистых рыб — в моче-
вой пузырь или наружу.
Органы размножения. Рыбы, как правило, раздельно-
полы, хотя среди них имеются и гермафродиты (морской окунь).
Яичники самок содержат многочисленные фолликулы, в каждом
из которых развивается яйцо. У акул, скатов, осетровых рыб
созревшие яйца выпадают из яичников в полость тела и увлекают-
ся движением ресничек в воронки яйцеводов, которыми являются
мюллеровы каналы. Оплодотворение яиц у акул и скатов проис-
ходит в половых путях самки, куда сперма вводится при помощи
особого копулятивного аппарата.
У костистых рыб в процессе онтогенеза мюллеровы каналы
с воронками обычно атрофируются и яйца выводятся вторично
образующимися яйцеводами. Последние одним концом срастаются
с яичником, а другим открываются особым отверстием наружу.
У некоторых лососевых рыб яйца выпадают из яичников в полость
тела, а оттуда выходят наружу через особые половые поры на
261
брюхе. Оплодотворение икринок костистых рыб, как правило, про-
исходит в воде. Размер яиц рыб колеблется от долей миллиметра
до 20 см (у некоторых акул).
Развитие у большинства рыб протекает с превращением..
В этом случае из икринок выходят личинки, отличающиеся от
взрослых особей рядом признаков. Сначала они питаются остаточ-
ным желтком, сохранившимся в желточном мешке, а позднее
переходят на активное питание. У некоторых рыб личинки имеют
наружные жабры, позднее исчезающие.
Экология рыб. Местом обитания рыб служат различные водо-
емы: моря, реки, озера, пруды и др.. Вода как физическое тело
имеет особые свойства, обусловливающие неоднородность водной
среды в разных зонах водоема. Вода относительно малопрозрач-
на, и в водоеме близ поверхности днем много света, но чем глубже,
тем его становится все меньше, а на больших глубинах морей царит
полный мрак. Вода обладает большой теплоемкостью, но мало-
теплопроводна. Верхние слои ее поглощают основную массу
тепловых лучей, поступающих из атмосферы. Это определяет
слоистое распределение температуры в водоемах летом с пониже-
нием ее от поверхности ко дну. Зимой в малых водоемах создается
обратная слоистость — от 0 °C у ледового покрова до 4°С в глу-
бине. В литре воды растворено кислорода примерно в 30 раз мень-
ше, чем содержится его в том же объеме воздуха. Диффузия
кислорода, поступающего главным образом из атмосферы, проис-
ходит в воде крайне медленно, что определяет понижение его
концентрации от поверхности к глубине (как летом, так и зимой).
Вода хорошо растворяет многие вещества. В ней (особенно в
морской) растворены многочисленные элементы, входящие в со-
став разных солей. Основное количество их находится в воде в
диссоциированном состоянии в виде ионов, что обусловливает
электропроводность воды. При отмеченных свойствах воды исклю-
чительное значение для жизни водных животных имеет ее дви-
жение: течения и перемешивание, вызываемые разными причина-
ми. Большое влияние на условия жизни в воде оказывают геогра-
фическое'положение водоема, водосборная площадь, с которой
поступают различные вещества, хозяйственная деятельность че-
ловека и другие природные и хозяйственные факторы. Все они
создают конкретные условия жизни в воде живых существ, в том
числе и рыб.
При всем разнообразии условий жизни в водной среде все же
можно наметить характерные экологические зоны водоемов:
пелагиаль, бенталь, литораль.
Пелагиаль — толща воды в водоеме. Здесь обитают многие
виды рыб, именуемые пелагическими. Одни из них питаются планк-
тоном (мелкими животными и растениями, взвешенными в воде),
другие поедают рыб, третьи ловят насекомых, упавших в воду.
Большинство пелагических рыб (среди морских, например, акулы,
тунцы, макрели, сельди; среди пресноводных судак, жерех, лососи,
262
сиги и др.) — хорошие пловцы, обычно обладающие стройным
удлиненным телом и сильным хвостом. Окраска их сверху, как
правило, темная, а снизу — светлая, серебристая.
Шенталь — придонная зона водоема. Придонные — бентальныс
рыбы (в морях скаты, камбалы, бычки, в пресных водах — сом,
стерлядь и др.) обычно менее подвижны. У лежащих на дне тело
обычно уплощено, верхняя сторона его окрашена под цвет окру-
жающего грунта. Другие бентальные рыбы плавают в придонном
слое воды, питаясь преимущественно донными животными.
Литораль — прибрежная зона водоемов. В морях обитают
различные бычки, морские собачки и многие другие рыбы. Не-
которые из них имеют особые приспособления, чтобы удерживать-
ся на грунте во время прибоя. В литорали пресных вод обитают
многие из семейства Карповые и др.
В зависимости от места обитания рыб делят на три группы:
1) морские, которые постоянно живут в море; 2) пресноводные,
всю жизнь проводящие в озерах, прудах и реках; 3) проходные,
обитающие в море, но для размножения поднимающиеся в реки
(многие лососевые, осетровые и др.) или, наоборот, живущие в
пресных водоемах, но для нереста уходящие в море (угорь).
Половозрелости рыбы достигают в различном возрасте. Хам-
са мечет икру впервые в годовалом возрасте, большинство карпо-
вых и окуневых — в возрасте 3—4 лет, осетры и севрюги — на 10—
15-м, а белуги -- иа 14—17-м и даже 20-м году жизни. Большая
часть рыб наших пресных вод мечут икру в теплое время года, но
щуки нерестятся ранней весной, а налимы — зимой. К наступле-
нию срока икрометания многие рыбы приобретают бра.чный на-
ряд — окраска их меняется и изменяется форма некоторых частей
тела. Так, у горбуши перед входом в реки для нереста окраска
меняется с серебристой на коричневую с темными пятнами, на
спине самцов вырастает горб, а челюсти загибаются крючком.
Многие рыбы перед нерестом совершают более или менее далекие
миграции: из озер в реки, из глубин на мелководье, из низовьев
в верховья рек и т. п. Проходные рыбы мигрируют из морей
в реки или реже из рек в моря. Весною, а иногда еще с осени рыбы
собираются в предустьевые пространства рек и с наступлением
определенных условий устремляются вверх по течению. Морские
рыбы совершают далекие путешествия с мест кормежки к местам
икрометания. Некоторые рыбы (дальневосточные лососи, сельдь-
залом и др.) мечут икру только один раз в жизни и вскоре после
икрометания погибают. Во время хода к местам нереста проходные
рыбы, как правило, не питаются.
Икра осетровых, многих карповых, окуневых и ряда других
рыб тяжелее воды; при откладывании она падает на дно или под-
водные предметы, к которым прикрепляется выделяемой икрой
клейкой слизью. У ряда рыб икра легче воды и плавает в верх-
них слоях водоемов (камбала, треска и др.). Некоторые рыбы
откладывают икру в особое гнездо (колюшка), другие закапывают
263
ее в песок или гравий дна (кета), третьи вынашивают ее в особых
сумках на теле (самцы морской иглы), что способствует большей
выживаемости икры и мальков.
Плодовитость рыб связана с условиями развития икры и
молоди. Живородящие рыбы родят лишь единицы или десятки
мальков. Всего несколько тысяч икринок мечут лососи, развитие
икры которых происходит в гнезде под слоем песка или гравия.
Карповые рыбы, откладывающие икру на дно водоема или водяные
растения, в теплое время года продуцируют за один нерест по не-
скольку сот тысяч икринок. Морские рыбы, дающие плавающую
икру, выметывают за нерест миллионы икринок (крупная самка
трески — до 9 млн, а луна-рыба — до 300 млн). Высокая плодо-
витость объясняется массовой гибелью икры, поедаемой другими
животными.
Развитие икры рыб нашей фауны, нерестящихся весной и ле-
том, длится обычно 3—8 дней (чем теплее вода, тем скорее),
а у рыб, мечущих ее осенью и зимой (лососи, налимы и др.), затяги-
вается до весны.
Способы добывания пищи у различных рыб весьма разнообраз-
ны. Некоторые (щука) подкарауливают добычу, укрывшись в
засаде, другие (судак, жерех) преследуют ее, третьи находят
пищу на дне водоема, четвертые ловят насекомых, упавших на
поверхность воды, многие улавливают планктон, пропуская воду
через жаберные щели, где он удерживается жаберными тычин-
ками, и т. д. Молодь рыбы питается в основном планктоном, а за-
тем постепенно переходит на пищу, свойственную взрослым осо-
бям. В теплое время года рыбы питаются интенсивнее, чем зимой.
Многие рыбы нашей страны зимой совсем не питаются, впадая
в зимнее оцепенение. Такая сезонная голодовка теплолюбивых
рыб является приспособлением к перенесению холодного периода.
Для ряда видов рыб установлена способность поглощать раство-
ренные в воде питательные вещества через жабры и покровы, а
личинки угрей в период миграции в океане питаются только этим
путем. У многих рыб наблюдаются миграции в поисках пищи.
Хозяйственное значение рыб. Практическое значение рыб
огромно. Их добывают прежде всего для получения ценных про-
дуктов питания. Из отходов рыбного промысла вырабатывают
рыбную муку и другие корма для сельскохозяйственных животных.
Из печени некоторых видов (тресковых, акул) получают лечебный
и технический жир. Из плавников акул, плавательного пузыря
осетровых и других рыб вырабатывают ценный клей. Из чешуи
ряда видов рыб- изготовляют искусственный жемчуг.
Советский Союз принадлежит к числу стран с высокоразви-
тым рыбным промыслом. Основная масса рыбы, добываемая на-
шими рыболовными судами, вылавливается в морях и океанах
всех частей земного шара. Но немало ее добывается и во внутрен-
них водоемах СССР.
В нашей стране ведется большая работа по обогащению фауны
264
рыб и по разработке путей рационального использования запа-
сов рыбы. Выполнение этой важной народнохозяйственной задачи
требует проведения ряда биотехнических мероприятий, из кото-
рых особое значение имеют следующие:
1. Охрана нерестилищ и производителей;
2. Мелиорация водоемов, удаление избытка ила и водной
растительности, расчистка водоемов от хлама. Все эти меро-
приятия способствуют повышению естественной продукции
биомассы населения водоема, которое служит пищей рыб и улуч-
шает условия их жизни;
3. Предохранение рек, озер и прудов от вредных сточных вод,
спускаемых промышленными предприятиями, от нефти и пр.;
4. Акклиматизация ценных промысловых пород в крупных водо-
емах, пригодных для жизни рыб, с целью повышения эффектив-
ности использования кормовых запасов в водоеме и получения
более ценного продукта и в большем количестве. В СССР успешно
проведена акклиматизация азово-черноморской кефали в Каспии,
сазана в оз. Балхаш, сигов в ряде уральских озер и т. д.;
5. Расширение сети рыбоводных заводов, задачей которых
является искусственное оплодотворение икры, инкубация ее в
специальных рыбоводных аппаратах и выращивание молоди, кото-
рая затем выпускается в естественные водоемы для дальнейшего
развития. В СССР имеются рыбоводные заводы для наиболее цен-
ных промысловых рыб: лососей, белорыбицы, сигов, осетровых
и ряда карповых. Работа этих заводов приобрела особенное
значение в связи с возведением гидроэлектростанций на реках,
по которым совершают миграции на нерест проходные рыбы.
Огромное значение для улучшения снабжения населения на-
шей страны имеют дальнейшее расширение и совершенствование
рыбоводства в специальных рыбоводных хозяйствах, совхозах и
колхозах. В рыбоводных хозяйствах широко применяют различные
зоотехнические приемы по разведению, кормлению и содержанию
разводимой рыбы. Так, в полносистемном прудовом карповом хо-
зяйстве имеется система прудов разного назначения: маточники
для содержания взрослых карпов-прозводителей; нерестовые
пруды, в которые сажают производителей для икрометания, пруды
для выращивания молоди (мальковые, выростные); зимовальные
и нагульные водоемы. Обычно пруды устраивают искусственно,
располагая их так, чтобы водоснабжение каждого из них было
независимым и пруд мог быть осушен или заполнен водой по мере
надобности. В прудах создается нужная степень проточности и
глубины. В выростные и нагульные пруды после зимовки сажают
определенное количество молоди, исходя из площади водоема, его
естественной кормиости и размера дополнительно вносимого в
водоем корма. Лучшие прудовые карповые хозяйства дают до
30—40 ц рыбы с 1 га в год, а в хозяйствах с использованием тер-
мальных вод (теплой воды с теплоцентралей и др.) продуктивность
измеряется центнерами с 1 м2 площади садков. Кроме карпа, в на-
265
Рис. 228. Рыбы — объекты прудового рыбоводства:
/ — карп; // — карась; III — толстолобик; IV — форель
шей стране разводят толстолобика, белого амура и другие виды те-
плолюбивых рыб, а из холодноводных — форель (рис. 228, IV).
Комбинированные карпо-утиные хозяйства дают возможность
получать повышенный выход не только мяса рыб, но и птиц и, кро-
ме того, увеличить яйценоскость птиц.
КЛАСС ХРЯЩЕВЫЕ РЫБЫ (CHONDRICHTHYES)
Древняя группа рыб с хрящевым (без костных элементов) ске-
летом. Кожа покрыта плакоидной чешуей. Группа включает два
подкласса: Пластинчатожаберные и Химеры, Поскольку химе-
ры — редкие морские рыбы, не имеющие практического значения,
описание их опускается.
' ПОДКЛАСС ПЛАСТИНЧАТОЖАБЕРНЫЕ (ELASMOBRANCH! А)
В современной фауне этот подкласс представлен акулами и ска-
тами (рис. 229). Верхняя лопасть хвоста значительно больше ниж-
ней, в нее заходит конец позвоночника. Рот расположен на нижней
стороне головы. Жаберные щели в числе 5—7 пар открываются
Рис. 229. Хрящевые пластинчатожаберные рыбы:
I — скат-хвостокол; // — гигантская акула; /// — яйцо ската в оболочке; IV — зародыш
одной из живородящих акул;
/ — брызгальце; 2 — жаберные щели; 3 — грудные плавники; '/—брюшные плавники;
5 — спинной плавник; 6 — анальный плавник; 7 — хвостовой плавник
267
наружу отдельными отверстиями. Желудочек сердца продолжает-
ся вперед артериальным конусом. В кишечнике имеется спираль-
ный клапан. У всех пластинчатожаберных оплодотворение яиц
внутреннее. Некоторые из них живородящие, другие окладывают
крупные яйца. У отдельных видов живородящих акул эмбри-
оны связаны со стенками половых путей самки, получая от мате-
ринского организма кислород и питательные вещества.
Акулы и скаты — морские животные. Акулы — превосходные
пловцы. Тело их имеет торпедообразную форму, а хвост отли-
чается сильным развитием. Они, за немногим исключением,— хищ-
ники. Некоторые опасны для купающихся в море людей. У скатов,
обитающих на дне, тело плоское, с большими грудными плавника-
ми. Питаются они различными донными животными. Мясо акул и
скатов вполне съедобно. Акулы во многих странах служат объ-
ектом промысла. Кожа крупных видов ценится как кожевенное
сырье. Из печени акул получают технический рыбий жир.
КЛАСС КОСТНЫЕ РЫБЫ (OSTEICHTHYES)
К этой группе принадлежит подавляющее большинство сов-
ременных видов рыб. Скелет их полностью или частично состоит из
костных элементов. Чешуя различного строения (ганоидная, кост-
ная), но не плакоидная. Жаберные щели прикрыты костными жа-
берными крышками. Оплодотворение обычно наружное.
ПОДКЛАСС ЛУЧЕПЕРЫЕ (ACTINOPTERYGII)
Скелет парных плавников образован веерообразно расположен-
ными твердыми и мягкими лучами. К подклассу относится около
90 % известных рыб, объединенных в несколько надотрядов.
Надотряд Костно-хрящевые (Chondrostei). Рыбы с удли-
ненным, уплощенным снизу телом, вдоль которого по хребту,
бокам и границе боков и брюха тянутся пять рядов костных
ромбических пластинок — жучек (рис. 230). Голова с удлинен-
ным рылом. Рот расположен на нижней стороне головы. Хво-
стовой плавник неравнолопастный, парные горизонтальные.
Скелет в основном хрящевой, но хрящевой череп покрыт
накладными плоскими костями. Кости входят также в состав поя-
са грудных плавников. Осевым скелетом служит сохраняющаяся
в течение всей жизни хорда. Позвонки представлены хрящевыми
верхними и нижними дугами: тела их не развиты. Сердце с арте-
риальным конусом. В кишечнике имеется спиральный клапан.
В современной фауне этот надотряд представлен семейством
Осетровые (Acipetiseridae), к которому принадлежат осетры,
белуга, севрюга, стерлядь, лопатоносы и др. Большинство видов
осетровых обитает в морях, особенно много их в Каспийском море.
Но некоторые виды, такие, как стерлядь, лопатоносы, постоянно
живут в реках и озерах. Все виды осетровых нерестятся в реках
268
весной и летом. Большинство питается различными донными жи-
вотными. Мясо и особенно икра этих рыб высоко ценятся.
Надотряд Костистые (Teleostei). К данному надотряду
относятся большинство лучеперых рыб. Форма тела разно-
образна. Рыбы обычно покрыты костной чешуей, имеющей
вид тонких, налегающих друг на друга пластинок. Верх-
няя и нижняя лопасти хвоста примерно одинаковой величины и
формы (равнолопастный хвостовой плавник). Грудные плавники
обычно вертикальные. Скелет костный. Хорда у взрослых особей
в той или иной степени редуцирована. Артериального конуса в
сердце и спирального клапана в кишечнике нет.
Многие костистые рыбы имеют большое промысловое значение.
Особенно велико оно у рыб, относящихся к семействам Сельде-
вые, Лососевые, Карповые, Сомовые, Щуковые, Угревые, Окуне-
вые, Камбаловые и Тресковые (рис. 231).
Семейство Сельдевые (Clupeidae). Тело вытянутое, стройное,
уплощенное с боков, невысокое. Брюшные плавники расположены
269
Рис. 231. Костистые рыбы:
/ —сельдь (сем. Сельдевые); II — лосось (сем. Лососевые); III — кари (сем. Кар-
повые); IV— сом (сем. Сомовые); V'— щука (сем. Щуковые); VI— угорь (сем. Угре-
вые); (/// — судак (сем. Окуневые); VIII - бычок речной (сем. Бычковые); IX
камбала (сем. Камбаловые)
посредине брюха, над ними находится один спинной плавник.
Боковой линии нет. На брюшной стороне имеется острый киль из
чешуек. Стайные рыбы. Питаются планктонными животными,
пользуясь цедильным аппаратом из жаберных тычинок. Одни из
них, как, например, океаническая сельдь, кильки, шпроты, сардины
и др., всю жизнь проводят в море. Другие (большинство сельдей
Каспийского, Азовского и Черного морей) на нерест поднимаются
в реки. Сельдевые имеют большое промысловое значение.
Семейство Лососевые (Sahnotiidae). По внешнему виду
сходны с сельдями. Отличаются жировым плавничком на хвосто-
вом стебле. Брюшные плавники расположены посредине брюшной
стороны. У собственно лососей рот большой, с зубами, чешуя
мелкая; у сигов рот маленький, беззубый, чешуя крупная. Морские,
проходные и пресноводные рыбы. Лососевые — очень ценные про-
мысловые рыбы с высокими вкусовыми качествами мяса и икры.
В реках Сибири и Дальнего Востока являются основным объектом
рыбного промысла. Дальневосточные лососи — кета, горбуша,
нерка и др/— ежегодно в большом количестве заходят в реки для
нереста. Европейский лосось — семга, а также кумжа нерестятся
в реках Европейского Севера. От кумжи произошли пресноводные
форели — обитатели горных рек и озер. Форелей разводят в спе-
циальных рыбоводных хозяйствах. Известны различные
виды сигов — байкальский омуль, чудский сиг, ряпушка и др.
Промысловое значение имеют морская корюшка и ее пресновод-
ные разновидности — снетки в наших северо-западных озерах.
Семейство Карповые (Cyprinidae). Отличаются от сельдевых
наличием боковой линии, а от лососевых — отсутствием жирового
плавника. Рот у карповых лишен зубов, и пища дробится особы-
ми глоточными зубами — костными выростами последней жабер-
ной дуги, вдающимися в полость глотки. Зубы обращены вершина-
ми к находящемуся на верхней поверхности глотки жерновку,
трением о который размельчается пища. Семейство Карповые,
самое обширное среди рыб, объединяет несколько десятков родов
и сотни видов пресноводных и морских рыб. Большинство карпо-
вых— теплолюбивые рыбы. Осенью они постепенно прекращают
питаться и на зиму впадают в оцепенение. Размножаются в конце
весны и летом. Мальки питаются обычно планктоном. Взрослые
особи поедают преимущественно различных донных беспозвоноч-
ных животных и водные растения, хотя среди карповых имеются
планктоноядные и хищные виды (жерех).
Карповые рыбы служат важнейшим объектом рыбного промыс-
ла наших внутренних водоемов. Наибольшее промысловое значе-
ние имеют из проходных форм рыбец, шемая, кутум, вобла, та-
рань, а из пресноводных — сазан, карась, лещ, язь, жерех, го-
лавль, линь, южные усачи, среднеазиатские маринки и др. Основ-
ным видом рыб, разводимых в рыбоводных прудовых хозяйствах
СССР, является карп. В прудах разводят также карасей, линей,
толстолобиков, белых амуров и другие виды карповых рыб.
271
Семейство Сомовые (Siluridae). Отличается голым телом,
большой головой с огромной пастью, очень длинным анальным
плавником. В СССР широко распространен обыкновенный сом —
крупная рыба, обитающая в южных реках страны. Сомы держатся
в омутах. Хищники. Зимой лежат в ямах. Растут быстро — к 5-лет-
нему возрасту достигают нередко 1 м. Живут до 100 и даже более
лет, достигая 300—400 кг.
Семейство Щуковые (Esocidae). Характерный представи-
тель — обыкновенная щука — крупная рыба с вытянутым, валько-
ватым телом, уплощенной сверху головой с пастью, усаженной
крупными острыми зубами. Спинной и анальный плавники сдвину-
ты далеко назад.
Щуки широко распространены по рекам и озерам нашей стра-
ны. Это прожорливые, сильные хищники, питающиеся в основ-
ном другими рыбами. Отличаются долголетием. Местами добы-
ваются в большом количестве.
Семейство Угревые (Anguillidae) представлено в нашей фауне
только речным угрем. Эта рыба отличается длинным змеевидным
телом без брюшных плавников. Чешуя очень мелкая. Спинной и
анальный плавники слиты с хвостовым. В СССР обитает преиму-
щественно в водоемах бассейна Балтийского моря и очень редко
в реках, впадающих в Черное море. Самки живут на дне заилен-
ных речек и пойменных озер. Самцы держатся в море. Взрослые
самки спускаются по рекам к устьям, где встречаются с самцами.
Затем они совместно мигрируют к местам нереста в Саргассово
море (Атлантический океан). Здесь на большой глубине самки
мечут икру. После нереста самки и самцы погибают. Молодь
переносится течением Гольфстрим через океан к берегам Европы.
Угри достигают 1,5 м длины. Их мясо высоко ценится.
Семейство Окуневые (Percidae) объединяет большую группу
морских и пресноводных рыб, у которых в парных и особенно в
непарных плавниках имеются колючие твердые лучи. Спинных
плавников два или реже один, но двураздельный; первый из
них поддерживается только колючими лучами. В анальном плав-
нике два колючих луча. Рот большой, с зубами. Кости жаберной
крышки зазубренные. К семейству относятся окуни, ерши, судаки,
морские судаки и другие виды. Многие из них являются объек-
том промысла.
Семейство Бычковые (Gobiidae) отличается от других тем, что
у его представителей брюшные плавники срастаются, образуя при-
соску, которой удерживаются на дне. Морские и реже речные ры-
бы. Многие виды промышляются.
Семейство Камбаловые (Pleuronectidae) — рыбы с уплощен-
ным с боков телом, лежащие на дне на одном боку. Глаза у взрос-
лых рыб смещены на одну сторону головы. Мальки камбалы, жи-
вущие сначала в толще воды, имеют симметричное тело и глаза по
обе стороны головы. Вырастая, переходят к донному образу жиз-
ни — тело их уплощается, а один глаз перемещается на другую
272
сторону головы. Различные виды камбал промышляются во многих
морях СССР.
Семейство Тресковые (Gadidae). Тресковые (кроме налимов)
имеют три спинных плавника и два анальных. Брюшные плав-
ники расположены на горле впереди грудных. На подбородке
есть непарный усик. Лучи всех плавников мягкие. К семейству
относится небольшое число видов северных рыб — треска, пикша,
сайда, навага, морской налим. Все живут преимущественно в мо-
рях Севера и Дальнего Востока. Лишь речной налим обитает
в пресных водах. Налим — единственная из наших пресноводных
рыб, нерестующая зимой. Треска — самая крупная (до 30 кг) и в
уловах наиболее многочисленная из тресковых. Распространена
в морях Европейского Севера и Дальнего Востока. Один из ее
отличительных признаков — изогнутая белая боковая линия.
Пикша промышляется в Баренцевом море. Боковая линия у нее
черная, прямая. Навага живет на Европейском Севере. Это не-
большая рыба длиной около 30 см. Значительно крупнее ее ро-
дич — дальневосточная вахня. У тресковых ценятся мясо и жирная
печень, из которой приготовляют консервы и медицинский рыбий
жир.
ПОДКЛАСС КИСТЕПЕРЫЕ (CROSSOPTERYGII)
Палеонтологи давно обнаружили в древних слоях осадочных по-
род, относящихся к палеозойской эре развития Земли, остат-
ки своеобразных рыб с особым строением парных плавников и
рядом других, как правило, примитивных признаков. Их считали
Рис. 232. Кистеперые и двоякодышащие рыбы:
/ — латимерпя; //— неоцератод
273
вымершими миллионы лет назад. И велико было удивление зооло-
гов, когда в 30-х годах XX в. в Индийском океане была выловле-
на рыба, относящаяся к данному подклассу. Позднее здесь было
добыто еще несколько особей этой замечательной рыбы, получив-
шей название латимерия (Latitnerid) (рис. 232,/).
У кистеперых рыб основанием каждого из парных плавни-
ков служит мясистая, вытянутая, покрытая крупной чешуей ло-
пасть, на конце которой располагаются лучи плавника. Располо-
жение скелетных элементов внутри основной лопасти плавника
несколько напоминает расположение костей пятипалых конеч-
ностей наземных позвоночных. К тому же ископаемые — палеозой-
ские представители подкласса — обладали, помимо жаберного,
также легочным дыханием. Все это позволяет предполагать бли-
зость древних кистеперых рыб к предкам четвероногих назем-
ных позвоночных.
ПОДКЛАСС ДВОЯКОДЫШАЩИЕ (DIPNOI)
Включает несколько видов крайне своеобразных пресноводных
рыб, обитающих в мелких, пересыхающих водоемах: в Африке —
протоптерус (Protopterus), в Южной Америке — лепидозирен
(Lepidosiren) и в Австралии — неоцератод (Neoceratodus)
(рис. 232,//). Наиболее характерный признак этих рыб — нали-
чие одного или двух тонкостенных, сообщающихся каналом с
глоткой, мешкообразных легких. Когда водоемы пересыхают,
двоякодышащие рыбы зарываются в ил и впадают в состояние
оцепенения, дыша воздухом при помощи легких. В связи с воздуш-
ным дыханием у них имеется два круга кровообращения, предсер-
дие разделено неполной перегородкой. Скелет двоякодышащих
костно-хрящевой. Хорда сохраняется всю жизнь.
НАДКЛАСС НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ (TETRAPODA)
КЛАСС ЗЕМНОВОДНЫЕ (AMPHIBIA)
Общая характеристика. Земноводные — четвероногие позвоноч-
ные из группы Anamnia. Температура их тела переменная, завися-
щая от температуры внешней среды. Кожа голая, с большим
количеством слизистых железок. Передний мозг имеет два полу-
шария. Носовая полость сообщается с ротовой внутренними
ноздрями — хоанами. Имеется среднее ухо, в котором расположе-
на одна слуховая косточка. Череп сочленяется с единственным
шейным позвонком двумя мыщелками. Крестец образован одним
позвонком. Органы дыхания личинок — жабры, а взрослых —
легкие. Большую роль в дыхании играет кожный покров. Имеется
два круга кровообращения. Сердце трехкамерное и состоит из
двух предсердий и одного желудочка с артериальным конусом.
Почки туловищные. Размножаются икрометанием. Развитие зем-
274
новодных проходит с метаморфозом. Икра и личинки развиваются
в воде, имеют жабры, у них один круг кровообращения. Взрос-
лые земноводные после метаморфоза становятся наземными
легочнодышащими животными с двумя кругами кровообращения.
Только немногие земноводные всю жизнь проводят в воде, со-
храняя жабры и некоторые другие признаки личинок.
Известно более 2 тыс. видов земноводных. Они широко распро-
странены по материкам и островам земного шара, но более много-
численны в странах с теплым, влажным климатом.
Амфибии служат ценными объектами физиологических опытов.
При их изучении было сделано много выдающихся открытий.
Так, И. М. Сеченов в опытах над лягушками открыл рефлексы
головного мозга. Земноводные интересны как животные, филогене-
тически связанные, с одной стороны, с древними рыбами, а с
другой - с примитивными пресмыкающимися.
Строение и жизненные отправления. Внешний вид земноводных
разнообразен (см. рис. 238). У хвостатых амфибий тело удли-
ненное, ноги короткие, примерно одинаковой длины, всю жизнь
сохраняется длинный хвост. У бесхвостых земноводных тело ко-
роткое и широкое, задние йоги прыгательныс, значительно длин-
нее передних, хвост у взрослых особей отсутствует. Червяги (без-
ногие) имеют длинное червеобразное тело без ног. У всех амфибий
шея не выражена или выражена слабо. В отличие от рыб голова у
них сочленяется с позвоночником подвижно.
Покровы. Кожа земноводных тонкая, голая, обычно по-
крытая слизью, выделяемой многочисленными кожными железами.
У личинок слизистые железы одноклеточные, у взрослых — много-
клеточные. Выделяемая слизь препятствует подсыханию кожи, что
необходимо для кожного дыхания. У некоторых земноводных
кожные железы выделяют ядовитый или жгучий секрет, защища-
ющий их от хищников. Степень ороговения эпидермиса у разных
видов земноводных далеко не одинакова. У личинок и тех взрос-
лых особей, которые ведут в основном водный образ жизни, орого-
вение поверхностных слоев кожи развито слабо, но у жаб на спи-
не роговой слой составляет (>() % всей толщи эпидермиса.
Кожа - важный орган дыхания земноводных, о чем свиде-
тельствуют цифры отношения длины капилляров кожи к длине
этих сосудов в легких; у тритона оно равно 4:1, а у жаб, имеющих
более сухую кожу, - 1:3.
Окраска земноводных часто носит покровительственный ха-
рактер. Некоторые, как, например, древесная квакша, способны
изменять ее.
Скелет земноводных состоит из позвоночника, черепа, костей
конечностей и их поясов (рис. 233). Позвоночник разделяется
на отделы: шейный, состоящий из одного позвонка, туловищный —
из ряда позвонков, крестцовый — из одного позвонка и хвостовой.
У бесхвостых амфибий рудименты хвостовых позвонков срастают-
ся в длинную косточку -- уростиль. У некоторых хвостатых земио-
275
Рис. 233. Скелет лягушки:
/ — общий вид; // — позвонок сверху; 111 — позвонок спереди;
1— шейный гюзгинок; 2 — крестцовый позвонок; 3 — уростиль; 4, 5 грудина; 6 -
предгрудина; 7 - коракоид; 8 — прокоракоид; .9, 10 - лопатка; 11 - подвздошная
кость; 12— седа •"чцная кость; 13 — лобковая кость; /-/• - плечевая кость; 15 - кости
предплечья; 16 — запястье; 17 — пясть; 18—20 — фаланги пальцев; 21 — бедро; 22 — ко-
сти голени; 23 — предплюсна; 24 — плюсна; 25 — шпора; 26 — фаланги пальцев задней
ноги; 27 — тело позвонка; 28 — спинномозговой канал; 29—сочленовная площадка;
30 — остистый отросток; 31 — поперечный отросток
водных позвонки двояковогнутые: между ними сохраняются остат-
ки хорды. У большинства же амфибий они либо выпуклые спереди
и вогнутые сзади, либо, наоборот, вогнутые спереди и выпуклые
сзади. Грудная клетка отсутствует.
Череп в основном хрящевой, с небольшим числом накладных
(вторичных) и основных (первичных) костей. С переходом от жа-
берного дыхания водных предков амфибий к легочному висцераль-
ный скелет изменился. Скелет жаберной области частично видо-
изменился в подъязычную кость. Верхняя часть подъязычной
дуги — подвесок, к которому у низших рыб прикрепляются челюс-
ти, у амфибий в связи со срастанием первичной верхней челюсти
с черепом превратилась в маленькую слуховую косточку — стре-
мечко, расположенное в среднем ухе.
Скелет конечностей и их поясов складывается из элементов,
характерных для пятипалых конечностей наземных позвоночных
животных. Число пальцев на ногах неодинаково у разных видов.
276
Мускула т у р а земноводных в связи с более разнообраз-
ными движениями и развитием конечностей, приспособленных к
движению по суше, в значительной степени теряет метамерное
строение и приобретает большую дифференцировку. Скелетная
мускулатура представлена множеством отдельных мышц, число
которых у лягушки превышает 350.
Нервная система претерпела значительные усложне-
ния по сравнению с таковой рыб. Головной мозг относительно
крупнее (рис. 234). Прогрессивными чертами его строения следует
считать образование полушарий переднего мозга и наличие нерв-
ных клеток не только в боковых стенках, но и в крыше полушарий.
В связи с тем что земноводные малоподвижны, мозжечок у них
развит слабо. Промежуточный мозг Сверху имеет придаток — эпи-
физ, а от дна его отходит воронка, с которой связан гипофиз.
Средний мозг развит слабо. От головного и спинного мозга отходят
нервы ко всем органам тела. Головных нервов десять пар. Спин-
номозговые нервы образуют плечевое и пояснично-крестцовое
сцепления, иннервирующие передние и задние конечности.
Органы чувств у амфибий получили в процессе эволюции
прогрессивное развитие. В связи с тем что воздушная среда менее
звукопроводна, в органах слуха земноводных усложнилось строе-
ние внутреннего уха и образовалось среднее ухо (барабанная
полость) со слуховой косточкой. Среднее ухо снаружи ограни-
чено барабанной перепонкой. Оно сообщается с глоткой каналом
(евстахиевой трубой), что позволяет уравновешивать давление
воздуха в нем с давлением
внешней среды. В связи с
особенностями видения в воз-
душной среде у земноводных
произошли изменения в стро-
ении глаз. Роговица глаза
выпуклая, хрусталик линзо-
образной формы, есть веки,
защищающие глаза. Органы
обоняния имеют наружные
и внутренние ноздри. У ли-
чинок и постоянно живу-
Рис. 234. Мозг лягушки:
/ - сверху; // — снизу; /// - сбоку;
/—полушария переднего мозга; 2 -
обонятельные доли; 3 — обонятельный
пера; 4 - промежуточный мозг; 5 —
зрительные нервы; 6 — воронка про-
межуточного мозга; 7—гипофиз; 8 —
средний мозг; 9 - ~ мозжечок; 10 —
продолговатый мозг; // — спинной
мозг ° 1
277
щих в воде земноводных сохранились характерные для рыб органы
боковой линии.
Органы пищеварения (рис. 235). Широкий рот ведет
в обширную ротовую полость: у многих земноводных на челюстях,
а также на нёбе расположены мелкие зубы, помогающие удер-
живать добычу. У амфибий имеется язык различной формы; у
лягушек он прикреплен к передней части нижней челюсти и может
выбрасываться изо рта, животные пользуются этим для ловли
насекомых. В ротовую полость открываются внутренние ноздри --
Рис. 235. Внутреннее строение лягушки:
1 — сердце; 2 —легкое; 3, 4 — печень; 5 — желчный пузырь; 6 -- желудок; 7 — подже-
лудочная железа; 8, 9 — топкая кишка; 10 — толстая кишка; 11 -- селезенка; 12 клоа-
ка; 13 — мочевой пузырь; 14—его отверстие в клоаке; 15 — почка; 16 - мочеточник;
17 — его отверстие в клоаке; 18 — яичник; 19 — жировое тело; 20, 21 — яйцеводы; 22 —
маточный отдел яйцеводов; 23 — отверстие яйцеводов в клоаке; 24 -- спинная аорта;
25—задняя полая вена; 26 — сонная артерия; 27 - душ аорты; 28- легочная артерия
278
хоаны, а в глотку - евстахиевы трубы. Интересно, что у ля-
гушки в проглатывании пищи принимают участие глаза; захватив
ртом добычу, лягушка сокращением мышц втягивает глаза в глубь
ротовой полости, проталкивая корм в пищевод. Через пищевод
пища попадает в мешкообразный желудок, а оттуда — в сравни-
тельно короткий кишечник, который разделен на тонкий и толстый
. отделы. В начало топкой кишки но особым протокам поступают
< желчь, вырабатываемая печенью, и секрет поджелудочной железы.
В конечную часть толстой кишки — клоаку — открываются
мочеточники, проток мочевого пузыря и половые протоки.
Органы дыхания изменяются с возрастом животного.
Личинки земноводных дышат наружными или внутренними жаб-
рами. У взрослых амфибий развиваются легкие, хотя у некоторых
хвостатых амфибий жабры сохраняются пожизненно. Легкие
имеют вид тонкостенных эластичных мешков со складками на
внутренней поверхности. Поскольку земноводные не имеют груд-
ной клетки, воздух в легкие поступает путем заглатывания: при
опускании дна ротовой полости воздух входит в нее через ноздри,
затем ноздри закрываются, а дно ротовой полости поднимается,
проталкивая воздух в легкие. Как было указано, в дыхании земно-
водных большую роль играет газообмен через кожу.
Кровеносная с и с т е м а. Амфибии в связи с воздушным
дыханием имеют два круга кровообращения (рис. 236). Сердце
амфибий трехкамерное, оно состоит из двух предсердий и желу-
дочка. Левое предсердие принимает кровь из легких, а в правое
поступает венозная кровь со всего тела с примесью артериаль-
ной крови, идущей от кожи. Кровь из обоих предсердий вливается
в желудочек через общее отверстие с клапанами. Желудочек
продолжается в крупный артериальный конус, за которым следует
короткая брюшная аорта. У бесхвостых амфибий аорта делится
на три пары симметрично отходящих сосудов, являющихся видо-
измененными приносящими жаберными артериями рыбообразных
предков. Передняя пара — сонные артерии, несут артериальную
кровь к голове. Вторая пара — дуги аорты, изогнувшись на спин-
ную сторону, сливаются в спинную аорту, от которой отходят ар-
терии, несущие кровь к разным органам и участкам тела. Третья
пара — легочные артерии, по ним венозная кровь течет в легкие.
По пути к легким от них ответвляются большие кожные артерии,
направляющиеся в кожу, где разветвляются на множество сосуди-
ков, обусловливая кожное дыхание, имеющее у амфибий большое
значение. Из легких артериальная кровь по легочным венам
движется в левое предсердие.
Венозная кровь от задней части тела частично проходит в
почки, где почечные вены распадаются на капилляры, образуя
воротную систему почек. Вены, выходящие из почек, образуют не-
парную заднюю (нижнюю) полую вену. Другая часть крови от
заднего отдела тела течет по двум сосудам, которые, сливаясь,
образуют брюшную вену. Она направляется, минуя почки, в пе-
279
Рис. 236. Кровеносная система лягушки:
1 — артериальная; II — венозная;
1—желудочек сердца; 2—правое предсердие; 3—левое предсердие; 4 - артериаль-
ный конус; 5—7 — ветви сонных артерий; 8— дуги аорты; 9 — подключичная артерия;
10 — легочная артерия; И — большая кожная артерия; 12 — спинная аорта; 13 - печень;
14— желудочная артерия; 15— кишечная артерия; 16 — почечная артерия; 17 - почка;
18—семенник; 19—подвздошная артерия; 20 — бедренная вена; 21 - седалищная
вена; 22—подвздошная вена; 23 — брюшная вена; 24 — воротная вена печени; 25 —
печеночная вена; 26 — яичник; 27 — почки; 28 — задняя полая вена; 29 — большая кож-
ная вена; 30—подключичная вена; 31, 32 — яремные вены; 33—передняя полая вена;
34 — легкое; 35 — легочные вены
чень и участвует вместе с воротной веной печени, несущей кровь
от кишечника, в образовании воротной системы печени. По выхо-
де из печени печеночные вены впадают в заднюю полую вену, а
последняя — в венозный синус (венозную пазуху) сердца, пред-
ставляющий расширение вен. Венозный синус принимает кровь от
головы, передних конечностей и кожи. Из венозного синуса кровь
изливается в правое предсердие. У хвостатых амфибий сохрани-
лись кардинальные вены от водных предков.
Органы выделения у взрослых амфибий представлены
туловищными почками (см. рис. 235). От почек отходит пара моче-
точников. Выводимая ими моча сначала попадает в клоаку, от-
туда — в мочевой пузырь. При сокращении последнего моча вновь
оказывается в клоаке, а из нее выделяется наружу. У зародышей
амфибий функционируют головные почки.
Органы размножения. Все земноводные раздельнопо-
лы. У самцов имеются два семенника, расположенных в полости те-
ла близ почек. Семявыносящие канальцы, пройдя через почку, впа-
280
Рис. 237. Развитие лягушки:
/—икринки в слизистой оболочке; II—VI!— стадии развития голо-
вастика; VIII, IX -- превращение головастика в лягушку; IVa — -го-
лова головастика с наружными жабрами; IV6 — жабры
дают в мочеточник, представленный вольфовым каналом, который
служит для выведения мочи и спермы. У самок большие парные
яичники лежат в полости тела. Созревшие яйца выходят в полость
тела, откуда попадают в воронкообразные начальные отделы
яйцеводов. Проходя по яйцеводам, икринки покрываются прозрач-
ной толстой слизистой оболочкой. Яйцеводы открываются в
клоаку.
Развитие у земноводых проходит со сложным метамор-
фозом (рис. 237). Из икринок выходят личинки, отличающиеся как
по строению, так и по образу жизни от взрослых особей. Личинки
земноводных — настоящие водные животные. Обитая в водной
среде, они дышат жабрами. Жабры у личинок хвостатых амфи-
бий наружные, ветвистые; у личинок бесхвостых земноводных
жабры сначала наружные, но вскоре становятся внутренними
вследствие обрастания их складками кожи. Кровеносная система
личинок земноводных сходна с таковой рыб и имеет только один
круг кровообращения. У них имеются органы боковой линии, как
и у большинства рыб. Передвигаются в основном за счет движения
уплощенного хвоста, отороченного плавником.
При превращении личинки во взрослое земноводное проис-
ходят глубокие изменения большинства органов. Появляются пар-
ные пятипалые конечности, у бесхвостых амфибий редуцируется
хвост. Жаберное дыхание заменяется легочным, жабры обычно
исчезают. Вместо одного круга кровообращения развиваются два:
большой и малый (легочный). При этом первая пара приносящих
жаберных артерий превращается в сонные артерии, вторая ста-
новится дугами аорты, третья в той или иной степени редуци-
руется, а четвертая преобразуется в легочные артерии. У мекси-
канской амфибии амблистомы наблюдается неотения -- способ-
ность размножаться на стадии личИнки, т. е. достигать половой
зрелости при сохранении личиночных черт строения.
Экология и хозяйственное значение земноводных. Места обита-
ная земноводных разнообразны, но большинство видов придержи-
вается влажных мест, а некоторые проводят в воде всю жизнь,
не выходя на сушу. Тропические земноводные — червяги — ведут
подземный образ жизни. Своеобразная амфибия — балканский
протей обитает в водоемах пещер; глаза у него редуцированы, а ко-
жа лишена пигмента. Земноводные относятся к группе холодно-
кровных животных, т. е. температура их тела непостоянна и зави-
сит от температуры окружающей среды. Уже при 10 °C их движе-
ния становятся вялыми, а при 5—7 °C они обычно впадают в оце-
пенение. Зимой в условиях умеренного и холодного климата
жизнедеятельность амфибий почти замирает. Лягушки зимуют
обычно на дне водоемов, а тритоны — в норках, во мху, под
камнями.
Размножаются земноводные в большинстве случаев весной.
Самки лягушек, жаб и многих других бесхвостых земновод-
ных выметывают икру в воду, где ее оплодотворяют самцы, ноли-
282
вая спермой. У хвостатых амфибий наблюдается своеобразное
внутреннее оплодотворение. Так, самец тритона откладывает на
водные растения комочки спермы в слизистых мешочках-спермато-
форах. Самка, найдя сперматофор, захватывает его краями
кл о а кал ьного от ве pc ti i я.
Плодовитость земноводных колеблется в широких пределах.
Обычная травяная лягушка выметывает весной 1—4 тыс., а зеле-
ная лягушка — 5—10 тыс. икринок. Развитие головастиков тра-
вяной лягушки в икринке длится в зависимости от температуры
воды от 8 до 28 дней. Превращение головастика в лягушонка
происходит обычно в конце лета.
Большинство земноводных, отложив икру в воду и оплодот-
ворив ее, не проявляют о ней заботы. Но некоторые виды забо-
тятся о своем потомстве. Так, например самец жабы повитухи,
широко распространенной в пашей стране, наматывает шнуры
оплодотворенной икры па задние ноги и плавает с ней, пока из
яиц не выведутся головастики. У самки южноамериканской (сури-
намской) жабы пипы во время икрометания кожа на спине сильно
утолщается и размягчается, клоака вытягивается и становится
яйцекладом. После выметывания и оплодотворения икры самец
накладывает ее на спину самки и брюшком вдавливает их в раз-
бухшую кожу, где и происходит развитие молодняка.
, Питаются земноводные мелкими беспозвоночными животными,
в первую очередь насекомыми. Они поедают много вредителей
культурных растений. Поэтому большинство земноводных очень
полезны для растениеводства. Подсчитано, что одна травяная
лягушка за лето может съесть около 1,2 тыс. вредных для сельско-
хозяйственных растений насекомых. Еще более полезны жабы,
поскольку они охотятся ночью и поедают массу ночных насекомых
и слизней, малодоступных для птиц. В Западной Европе жаб часто
выпускают в оранжереи и парники для истребления вредителей.
Тритоны полезны тем, что поедают личинок комаров. Вместе
с тем нельзя не отметить вред, который приносят крупные лягушки
истреблением молоди рыб. В природе лягушками питаются многие
животные, в том числе и промысловые.
Класс Земноводные делится на три отряда: Хвостатые амфи-
бии (Urodela), Бесхвостые амфибии (Anura), Безногие амфибии
(Apoda).
Отряд Хвостатые амфибии (Urodela). Наиболее древняя груп-
па земноводных, представленная в современной фауне примерно
130 видами (рис. 238). Тело удлиненное, вальковатое. Хвост
сохраняется всю жизнь. Передние и задние конечности примерно
одинаковой длины. Поэтому хвостатые амфибии передвигаются
ползанием или хождением. Оплодотворение внутреннее. Некото-
рые формы сохраняют жабры всю жизнь.
В нашей стране из хвостатых амфибий широко распростра-
нены тритоны. (Trilurus). Наиболее часто встречаются крупный
гребенчатый тритон (у них самцы черные с оранжевым брюхом)
283
Рис. 238. Различные представители класса Земноводные:
/ — огненная саламандра; // — протей; /// — травяная лягушка; IV — червяга
и более мелкий обыкновенный тритон (обычно самцы имеют
светлую пятнистую расцветку). Летом тритоны живут в воде, где
и размножаются, а зиму проводят на суше в состоянии оцепенения.
В Карпатах можно встретить довольно крупную огненную сала-
мандру (Salatnandra), которую легко узнать по черной окраске
с оранжевыми или желтыми пятнами. Гигантская японская сала-
мандра достигает 1,5 м длины. К семейству Протеев (Proteidae)
относится балканский протей, живущий в водоемах пещер и сохра-
няющий жабры всю жизнь. Его кожа не имеет пигмента, а глаза
рудиментарны, так как животное живет в темноте. В лабора-
ториях для проведения физиологических опытов разводят личинок
американских амблистом, именуемых аксолотлями. Эти животные,
как и все хвостатые амфибии, обладают замечательной особен-
ностью восстанавливать утраченные части тела.
Отряд Бесхвостые амфибии (Anura) — лягушки, жабы, квак-
ши. Для них характерно короткое, широкое тело. Хвост у взрос-
лых особей отсутствует. Задние ноги значительно длиннее перед-
них, что определяет движение скачками. Оплодотворение наруж-
ное.
У лягушек (Ranidae) кожа гладкая, .слизистая? Во рту есть
зубы. Преимущественно дневные и сумеречные животные. У жаб
(Bufonidae) кожа сухая, бугристая, во рту зубов нет,- задние
ноги относительно короткие. Квакши (Hylidae) отличаются не-
большими размерами, тонким стройным телом и ляпами с при-
сосками на концах пальцев. Присоски облегчают передвижение по
деревьям, где квакши охотятся за насекомыми. Окраска квакш
обычно ярко-зеленая, может меняться в зависимости от расцветки
окружающего фона среды.
Отряд Безногие амфибии (Apoda)—тропические земновод-
ные, ведущие подземный образ жизни. Имеют длинное, валысо-
ватое тело с коротким хвостом. В связи с жизнью в норках под
284
землей ноги и глаза у них подверглись редукции. Оплодо-
творение внутреннее. Питаются почвенными беспозвоночными
животными.
ПРОИСХОЖДЕНИЕ ЗЕМНОВОДНЫХ
Изучение земноводных представляет интерес для выяснения эво-
люции позвоночных животных, перехода их из водной среды на
сушу. Переход, хотя и частичный, древних земноводных к назем-
ному существованию сопровождался при сохранении ряда призна-
ков рыбообразных предков появлением новых приспособлений:
из парных плавников образовались пятипалые конечности, жа-
берное дыхание у взрослых заменилось легочным, произошли и
другие глубочайшие изменения организма.
Земноводные произошли от рыбообразных предков, вероятно,
близких к кистеперым рыбам. Древнейшие амфибии стегоце-
фалы (панцирноголовые) -- появились уже в девонское время.
Они были тогда в основном водными животными, по-видимому,
лишь ненадолго покидавшими водоемы. Об этом свидетельствует,
например, то, что у них были органы боковой линии. От современ-
ных земноводных они отличались, в частности, шитом из наклад-
ных костей на черепе.
КЛАСС ПРЕСМЫКАЮЩИЕСЯ (REPTILIA)
Общая характеристика» Пресмыкающиеся — наземные позво-
ночные из группы Amtiiota с. переменной температурой тела. Ряд
черт характеризует пресмыкающихся как типичных наземных
животных. Шея хорошо выражена. Конечности пятипалые, но у
некоторых они частично или полностью редуцировались. Кожа
сухая, с сильным ороговением эпидермиса. Ее поверхность покры-
та обычно роговыми чешуйками или щитками. Череп сочленяется
с позвоночником одним мыщелком. Позвонки туловища несут
ребра, которые составляют грудную клетку. Полушария перед-
него мозга хорошо развиты, в их крыше имеется серое мозговое
вещество, образующее кору мозга. В течение всей жизни дышат
легкими, вдох и выдох производятся путем, расширения и сжа-
тия грудной клетки. Сердце у большинства видов трехкамерное,
с неполной перегородкой в желудочке. У крокодилов она полно-
стью разделяет желудочек на две камеры. Артериального конуса
нет; две дуги аорты и легочная артерия отходят непосредственно
от желудочка сердца. Органами выделения у взрослых служат
тазовые почки. Оплодотворение внутреннее. У эмбрионов раз-
виваются оболочки — амнион и аллантоис.
Число видов современных пресмыкающихся превышает 6 тыс.
Большая часть их обитает в тропических и субтропических стра-
нах. В СССР пресмыкающиеся особенно многочисленны и разно-
образны в зоне пустынь.
285
Рис. 239, Различные представители класса Пресмыкающиеся:
I — гаттерия; II—желтопузик; III — гадюка; IV — хамелеон;
Г — степная черепаха; VI — нильский крокодил
Многие пресмыкающиеся полезны истреблением вредных на-
секомых и моллюсков. Некоторые из них (крокодилы, крупные
змеи) дают ценные шкуры, а черепахи — съедобное мясо. Яд змей
используется в медицине. Некоторые ядовитые змеи опасны для
человека.
Современные пресмыкающиеся относятся к одному из следую-
щих четырех подклассов: Первоящеры (Prosauria), Чешуйчатые
(Squamata), Черепахи (Chelonia), Крокодилы (Crocodllia}.
Строение и жизненные отправления. Форма тела пресмы-
кающихся в связи с обитанием в различных условиях среды весь-
ма разнообразна (рис. 239). Размеры колеблются от нескольких
сантиметров до 11 м (некоторые удавы). Окраска тела варьирует,
часто с рисунком из пятен и полос. Нередко она носит явно покро-
вительственный характер.
Покровы пресмыкающихся имеют черты приспособления
к жизни в воздушной среде (рис. 240). Наружные слои эпидер-
миса кожи подвергаются сильному ороговению, что обеспечивает
животному защиту от высыхания. У большинства пресмыкаю-
щихся кожа покрыта роговыми чешуйками, бугорками или щит-
ками. Поверхность ее сухая, так как кожные железы отсутствуют
(щюме некоторых желез особого назначения). В дерме у ряда
ш чов развиваются костные пластинки.
?86
Рис. 240. Продольный разрез кожи ящерицы:
1 эпидермис; 2 дерма; ,'У роговой слой эпидермиса; 4 -- пигмент-
ные клетки в мальпигиевом слое эпидермиса; .5--костные образования
в дерме
Скелет пресмыкающихся почти полностью образован ко-
стными элементами (рис. 241). Череп состоит как из окостенений
хрящей черепа эмбриона, так и из покровных костей. Позвоночник
подразделяется на шейный, грудной, поясничный, крестцовый щ
хвостовой отделы. Первый шейный позвонок — атлант, имеет вид
кольца, передняя поверхность которого снизу несет суставную
ямку для сочленения с единственным мыщелком черепа. Второй
позвонок — эпистрофей — имеет направленный вперед отросток,
который служит осью вращения головы. Такое строение обеспе-
чивает подвижность головы. Грудные позвонки пресмыкающихся
несут по паре хорошо развитых ребер, большая часть которых
нижними хрящевыми концами прикрепляется к грудине, образуя
грудную клетку. Ребра поясничных и задних грудных позвонков
не соединяются с грудиной. Крестцовых позвонков два, к ним
прикрепляются подвздошные кости таза.
Скелет передних и задних конечностей состоит из тех же ко-
стей, что и у других наземных позвоночных. У змей и безногих
ящериц конечности утрачены.
Рис. 241. Скелет ящерицы:
/ —- ключица; 2 —- лопатка; 3 плечевая кость; 4 — лучевая кость; 5 — локте-
вая кость; 6' — запястье; 7 — пясть; <9 — фаланги пальцев; 9 — ребра; 10 — таз;
11 — бедренная кость; 12 — большая берцовая кость; 13 — малая берцовая кость;
14 г- предплюсна; 15 — плюсна; 16 — фаланги пальцев; /7 — позвоночник; 18-
черен
287
Рис. 242. Головной мозг ящерицы:
1 — сверху; // — снизу; III, IV — сбоку;
1 — полушария переднего мозга; 2 - обо-
нятельные доли; 3 — полосатое тело; 4
промежуточный мозг; 5 — теменной орган;
6— гипофиз; 7—средний мозг; 8 — зри-
тельные нервы; 9—мозжечок; 10 -- чет-
вертый желудочек; 11 — продолговатый
мозг; 12—спинной мозг; 13 — третий
желудочек
Нервная система пре-
смыкающихся отличается от тако-
вой земноводных рядом прогрес-
сивных черт. Головной мозг их
значительно больше, полушария
переднего отдела хорошо разви-
ты и имеют на поверхности серое
мозговое вещество — кору
(рис. 242). Мозжечок довольно
большой, что соответствует слож-
ности движений большинства
рептилий.
Органы чувств. Глаза
пресмыкающихся имеют веки, но
у змей и некоторых ящериц они
срастаются, образуя прозрачную
пленку, защищающую глаз. У мно-
гих рептилий на темени распо-
лагается своеобразный теменной орган (теменной глаз), связан-
ный с промежуточным мозгом. Он может воспринимать световые
раздражения.
Орган слуха состоит из внутреннего и среднего уха, в котором
находится одна слуховая косточка — стремечко. Наружного уха
нет. Имеются также органы осязания, обоняния и вкуса.
Органы пищеварения. Пищеварительный тракт пре-
смыкающихся (рис. 243) начинается ротовой полостью, в которой
обычно находятся язык и зубы. Зубы либо прирастают основа-
нием к челюстям, либо (у крокодилов) сидят в ямках (альвеолах)
челюстных костей. Крокодилы имеют вторичное небо, образован-
ное внутренними отростками верхних челюстей, небными и кры-
ловидными костями. Благодаря образованию вторичного неба
хоаны у крокодилов отодвинуты назад и открываются в глотку.
Воздух, вдыхаемый через ноздри, поступает в глотку и далее
по трахее в легкие, минуя ротовую полость. Это позволяет живот-
ному дышать при заглатывании пищи. Пищевод ведет в хорошо
развитый желудок. Кишечник ясно подразделяется на более длин-
ную тонкую и относительно короткую толстую кишку. У места
288
соединения кишка образует слепой вырост. Протоки печени и
поджелудочной железы открываются в начальный отдел тонкой
кишки — двенадцатиперстную кишку. Закапчивается кишечник
расширением — клоакой, в которую впадают мочеточники и по-
ловые протоки.
О р г а н а м и д ы х а и и я пресмыкающихся в течение всей
жизни являются легкие. От глотки начинается трахея (дыхатель-
ное горло), которая делится на два бронха, ведущих в мешковид-
Рис. 243. Внутреннее строение яще-
рицы:
1 — бедренные норы; 2 ~ подъязычная
кость; ,3 - щитовидная железа; 4 — зобная
железа; 5 — трахея; 6 . легкое; 7,8-
сердце; .9- сонная артерия; 10 левая
дуга аорты; И, 12 — епннпая аорта; 13—
печеночная пена; 14- яремная вена;
15 - пищевод; 16 тонкая кишка; 17 —
желудок; 18 — прямая кишка; 19 — не-
чет,; 20 — желчный пузырь; 21 — желч-
ный проток; 22 — поджелудочная железа;
23 -- селезенка; 24, 25 — семенники; 26 —
надпочечники; 27 — семяпроводы; 28—
ночки; 29 - мочеполовое отверстие; 30 -
клоака; 31 — мочевой пузырь; 32— хвост;
33 — жировое тело
Рис. 244. Кровеносная система
ящерицы:
1 - левая дуга аорты; 2 - правая
дуга аорты; 3 - сонные артерии; 4 -
спинная аорта; 5 - внутренностная
артерия; 6 - кишечная артерия; 7
межпозвоночная артерия; 8- -артерии
задних конечностей; 9 — левая брюш-
ная артерия; 10 - мезентериальная
артерия; 11 — легочная артерия; 12-
легкое; 13, 14 - яремные вены; 15 —
вены задней конечности; /6 - - тазовые
вены; 17 -- брюшная вена; 18 - почки;
19 - задняя полая вена; 20 — кишка;
21 — воротная вена печени; 22 - пе-
чень
10 Б. Л. Кузнецов и др.
289
ные легкие. Стенки легких внутри имеют многочисленные складки,
увеличивающие дыхательную поверхность.
Кровеносная система пресмыкающихся имеет ряд
прогрессивных черт по сравнению с земноводными (рис. 244).
Хотя сердце у тех и других трехкамерное, но сложная струк-
тура желудочка (перегородки, гребни) в сердце рептилий способ-
ствует разделению в нем артериальной и венозной крови. У кроко-
дилов желудочек полностью перегорожен па правую и левую по-
ловины. Артериальный конус отсутствует, и три артерии (легоч-
ная и две дуги аорты) отходят непосредственно от желудочка. От
левой его половины, принимающей артериальную кровь, посту-
пающую из легких, отходит правая дуга (загибающаяся напра-
во). С ней связаны сонные артерии, питающие артериальной
кровью головной мозг. Ближе к правой стороне берет начало ме-
нее развитая левая дуга (загибающаяся налево). Обе дуги аорты
соединяются на спинной стороне в единую спинную аорту. Много-
численные сосуды, отходящие от нее, несут кровь к разным орга-
нам тела. Легочные артерии начинаются общим стволом от пра-
вой половины желудочка. По ним венозная кровь поступает в
легкие.
Строение венозной системы в основном сходно с таковой у бес-
хвостых амфибий, но венозный синус развит у пресмыкающихся
слабее.
Органы выделения у взрослых пресмыкающихся пред-
ставлены тазовыми почками. От почек отходят мочеточники, впа-
дающие в мочевой пузырь, который открывается в клоаку. У .змеи
и крокодилов мочевого пузыря нет.
Органы размножения. Семенники у самцов и яичники
у самок лежат в полости тела по бокам позвоночника. Оплодотво-
рение яиц происходит внутри тела самки.
Развитие пресмыкающихся происходит в яйце. Эмбрион ок-
ружен амнионом, наполненным амниотической жидкостью, что об-
условливает развитие зародыша в жидкой среде. Вырост заднего
конца кишечника эмбриона — аллантоис — является зародыше-
вым мочевым пузырем и в то же время служит органом дыхания.
Его наружная стенка, прилегающая к оболочке яйца, содержит
сеть капилляров, через которые и происходит газообмен.
Экология пресмыкающихся. Пресмыкающиеся широко рас-
пространены по земному шару, но большинство обитают в стра-
нах с жарким и теплым климатом. Это обусловливается зави-
симостью их жизнедеятельности как животных с переменной тем-
пературой тела от температуры окружающей среды.
Пресмыкающиеся, как правило,— наземные животные. Од-
нако ряд видов этих животных ведет водный образ жизни. Так,
в морях обитают морские змеи и черепахи, в реках и озерах --
крокодилы, некоторые змеи и черепахи. Все водпые пресмыкаю-
щиеся дышат воздухом и размножаются па суше (исключение
составляют живородящие морские змеи).
290
В умеренных и северных широтах пресмыкающиеся прово-
дят зиму в глубоком оцепенении, укрывшись в различные убе-
жища.
Большинство размножаются путем откладывания яиц. Яйца
пресмыкающихся имеют сходное строение с яйцами птиц. Они
богаты желтком и белком, снаружи покрыты прочной кожистой
оболочкой или известковой скорлупой. Рептилии откладывают
яйца обычно в почву или в трещины скал, под кору деревьев, в
навоз. Срок инкубации зависит от температуры внешней среды.
Некоторые пресмыкающиеся, например крокодилы, охраняют
кладку яиц. У ряда рептилий наблюдается яйцеживорождение:
в этом случае оплодотворенные яйца остаются в теле матери и в
них развиваются молодые животные, которые выходят из яйца
тотчас же после откладки. Имеются и живородящие пресмыкаю-
щиеся.
Мелкие пресмыкающиеся питаются преимущественно беспоз-
воночными, главным образом насекомыми. Более крупные напа-
дают на позвоночных животных. Ядовитые змеи убивают жертву,
вонзая в нее ядовитые зубы, по каналам или бороздкам которых
в ранку стекает яд, вырабатываемый специальными железами.
Имеются также и растительноядные пресмыкающиеся, поедаю-
щие траву и плоды (например, сухопутные черепахи).
В СССР и других странах созданы специальные питомники
ядовитых змей, от которых получают .яд, используемый для изго-
товления ряда лекарств.
ПОДКЛАСС ПЕРВОЯЩЕРЫ (PROSAURIA)
Древняя группа пресмыкающихся, представленная в современ-
ной фауне только одним видом — гаттерией. Это животное оби-
тает в Новой Зеландии. По внешнему виду гаттерия напоминает
крупную (до 75 см длиной) ящерицу (см. рис. 239, /), но отлича-
ется рядом примитивных черт строения. Так, ее позвоночник об-
разован двояковогнутыми позвонками, между телами которых
сохраняются всю жизнь остатки хорды. На брюхе, под кожей,
расположены брюшные ребра, которые, по-видимому, являются
рудиментами брюшного костного щита палеозойских амфибий —
стегоцефалов. Хорошо развит теменной глаз. Гаттерия тщательно
охраняется законом.
ПОДКЛАСС ЧЕШУЙЧАТЫЕ (SQUAMATA)
Подкласс объединяет около 6 тыс. различных видов ящериц, змей
и хамелеонов. Тело их покрыто роговыми чешуйками, бугорками
или щитками. Костные образования в коже встречаются редко.
Отверстие клоаки в виде поперечной щели. У многих представи-
телей отряда наблюдаются частичная редукция или даже полное
исчезновение конечностей.
ю*
291
Подкласс делится на три отряда: Ящерицы (Lacertilia), Змеи
(Ophidia), Хамелеоны (Chamaeleontes).
Отрад Ящерицы (Lacertilia)— животные с продолговатым
телом и обычно длинным хвостом. Ноги, как правило, хорошо
развиты, но у некоторых форм они редуцированы или совсем от-
сутствуют (веретеница, желтопузик); в последнем случае в теле
сохраняются рудименты поясов конечностей (см. рис. 239, //). Гла-
за у большинства видов снабжены подвижными веками, но у гек-
конов, гологлазов и некоторых других ящериц они срастаются
и превращаются в прозрачные пленки на глазах. Имеются бара-
банные перепонки. Многие ящерицы способны при преследовании
самопроизвольно отделять часть хвоста. В дальнейшем на месте
отпавшего вырастает новый хвост несколько измененной формы.
Большинство ящериц размножаются, откладывая яйца, но
некоторые виды яйцеживородящи (веретеница, живородящая
ящерица). Питаются мелкими животными, в том числе различ-
ными вредными насекомыми и моллюсками, чем приносят пользу
сельскому и лесному хозяйствам. Ядовитых видов среди ящериц
нашей фауны нет.
В Советском Союзе встречается большое число видов ящериц,
из которых большинство обитают на юге страны. Но некоторые
из них, как, например, живородящая и прыткая ящерицы (La-
certa vivipara, L. agilis), распространены далеко на север. В пу-
стынях Средней Азии обычны круглоголовки (Rlirynocephalus),
отличающиеся округлой головкой на подвижной шее, тело их по-
крыто мелкими роговыми бугорками. В домах и среди скал южных
районов СССР по ночам можно встретить своеобразных гекконов
(Geckonidae), ловко бегающих по стенам, а то и по потолку. В пу-
стынях Средней Азии обитает крупная ящерица — серый варан
(Varanus grlseus), длина которой достигает 1,5 м. Вараны, оби-
тающие на о-ве Комодо (Индонезия), достигают 3 м.
Отряд Змеи (Ophidia) имеют длинное цилиндрическое тело
со слабо выраженным делением на голову, шею, туловище и хвост
(см. рис. 239, ///). Конечности отсутствуют, лишь у удавов руди-
менты задних ног сохраняются в виде небольших крючочков по
бокам анального отверстия. Глаза змей покрыты прозрачной
пленкой сросшихся век. Барабанной перепонки нет. Кости обеих
челюстей и неба соединены связками, которые позволяют пасти
открываться настолько широко, что змеи могут заглатывать круп-
ную добычу целиком. В связи с вытянутой формой тела у змей
имеется ряд особенностей анатомического строения: одно легкое,
лентовидные почки и др.
Ползают змеи, изгибая тело. Щитки, покрывающие брюхо,
налегают друг на друга задними краями и препятствуют сколь-
жению тела назад.
Некоторые змеи (ужи) заглатывают живую добычу, другие
(удавы) душат ее кольцами тела, третьи (ядовитые змеи) — уби-
вают ядом. У ядовитых змей два зуба верхних челюстей резко
292
Рис. 245. Череп гремучей змеи с за-
крытой и открытой пастью (я - - ядо-
витые зубы)
выделяются размерами. Они
сращены с подвижными челюст-
ными костями. Ядовитые зу-
бы сидят на верхней челюсти в
вертикальном положении при
открытой пасти, а при закры-
той отогнуты назад (рис. 245).
•У одних ядовитых змей (га-
дюки, гремучие змеи) яд стекает по каналу внутри зуба, у других
(кобра, аспиды) — но борозде на его поверхности. Яд выраба-
тывается особыми верхнечелюстными железами. Для человека
особенно опасны укусы кобры, гюрзы, эфы, обитающих в южных
районах пашей страны. Укус широко распространенной у нас
гадюки болезнен, но не смертелен. Обычный во многих районах
страны уж ядовитых зубов не имеет.
Размножаются змеи либо откладывая яйца, либо живорож-
дением. На зиму змеи нашей фауны впадают в спячку.
Отряд Хамелеоны (Chamaeleontes) — своеобразные пресмы-
кающиеся, по форме тела напоминающие ящериц (см. рис. 239,
IV). Для них характерны черты приспособления к древесному
образу жизни. Пальцы лап срастаются но 2—-3 вместе и проти-
востоят друг другу. Такими клещеобразными лапами хамелеоны
крепко охватывают ветку дерева, по которой передвигаются.
Хвост длинный, тонкий, способный обвиваться вокруг сучков.
Огромные глаза подвижны и могут одновременно смотреть в раз-
ные стороны. Окраска хамелеонов быстро меняется в зависимости
от фона окружающей среды. Язык очень длинный — выбрасы-
вая его, они ловят насекомых. Около 90 видов хамелеонов насе-
ляют тропические и субтропические страны.
ПОДКЛАСС ЧЕРЕПАХИ (CHELONIA)
Уплощенное широкое тело черепах (см. рис. 239, (/) покрыто
прочным панцирем, образованным двумя щитами: более выпук-
лым спинным и уплощенным брюшным. По бокам тела они сра-
щены или соединены связками. Спереди и сзади между щитами
остаются широкие щели; из передней выступают голова и перед-
ние ноги, а из задней — хвост и задние конечности. При испуге
или опасности черепахи втягивают их под край панциря. Щиты
панциря состоят из костной основы, которая образуется как за
счет расширения и слияния остистых отростков позвонков, ребер,
грудины и ключиц, так и путем костных образований в дерме
кожи.
293
Костная основа панциря покрыта у кожистых черепах мяг-
кой кожей, а у других — роговыми щитками.
Для черепах характерно отсутствие зубов — их челюсти обыч-
но одеты роговым чехлом с режущим краем («клювом»). У мор-
ских черепах ноги превратились в ласты.
В настоящее время в мире насчитывается около 200 видов
черепах, большинство из которых встречаются в тропических
странах. На островах Тихого и Индийского океанов водятся ги-
гантские слоновые черепахи, масса которых достигает 200 кг.
Объектом промысла служит морская зеленая черепаха (Chelo-
nia mydas) длиной до 2 м и массой до 450 кг. Все морские чере-
пахи откладывают яйца в песок отмелей побережья.
В степях Средней Азии и Казахстана обитает степная чере-
паха (Testudo), медленно передвигающаяся на толстых ногах с
четырьмя когтями на передних конечностях. Мясо и яйца ее
съедобны. В болотах и озерах южных районов нашей страны во-
дится сильно уплощенная болотная черепаха (Emis orbicularis),
питающаяся водными животными.
ПОДКЛАСС КРОКОДИЛЫ (CROCODILIA)
Крупные пресмыкающиеся, некоторые виды достигают 10 м
(нильский крокодил). Отличаются вытянутой головой, короткими
лапами и мощным длинным хвостом, сплюснутым с боков (см.
рис. 239, К/). Тело покрыто крупными роговыми щитками, под
которыми в дерме кожи залегают костные пластинки. Пальцы
задних ног соединены плавательными перепонками. Огромная
пасть усажена многочисленными крупными зубами, сидящими в
альвеолах. Сердце четырехкамерное. Объемистые легкие имеют
губчатое строение. Вторичное небо отделяет носоглоточный ход
от ротовой полости. Поперечный валик в глубине рта при откры-
той пасти смыкается с небом, закрывая глотку, что позволяет
крокодилам хватать добычу под водой.
Крокодилы населяют реки, озера и болота тропических стран.
Большую часть времени проводят в воде, хорошо плавая и ныряя.
Но нередко они вылезают на берег погреться на солнце. Яйца
зарывают в прибрежных песках, где и проходит их развитие. Не-
редко самка охраняет кладку яиц.
Питаются крокодилы как водными животными, так и зверями,
приблизившимися к водоему. Крупные крокодилы опасны для
людей.
В ряде стран крокодилы промышляются ради ценной шкуры
и мяса. На Кубе крокодилов разводят с этой целью на специаль-
ных фермах.
ПРОИСХОЖДЕНИЕ ПРЕСМЫКАЮЩИХСЯ
Несколько десятков миллионов лет назад, в мезозойскую эру, на
Земле господствовали многие виды пресмыкающихся. Они были
294
Рис. 246. Ископаемые пресмыкающиеся мезозоя:
/--котилозавр; // птерозавр; ///--цератозавр; /Г - - неспдозухия; Г ихтио
завр; VI трицератопс
представлены большим разнообразием форм и на суше, и в воде.
По берегам водоемов бродили гигантские бронтозавры. В водо-
емах, близ берегов жили огромные диплодоки, длина тела кото-
рых достигала 25 м. На суше паслись носороговидные трицера-
топсы. Верхушками деревьев лакомились ходившие на задних но-
295
гах колоссальные игуанодоны. В море носились косяки похожих
на дельфинов ихтиозавров, плавали змеевидные мезозавры и
огромные плезиозавры. В воздухе парили на кожистых крыльях
летающие драконы — птеродактили. Уже тогда были крокодилы,
черепахи, ящерицы. Это время было подлинным веком рептилий
(рис. 246).
Но позднее пресмыкающиеся уступили господство более со-
вершенным организмам —- млекопитающим и птицам.
Данные сравнительной анатомии, эмбриологии и палеонтоло-
гии свидетельствуют о происхождении рептилий от древней па-
леозойской группы котилозавров, которая, в свою очередь, свя-
зана по происхождению со стегоцефалами — панцириоголовыми
земноводными. Подобно последним котилозавры тоже были цель-
ночерепными, имели массивное тело и пятипалые конечности.
Формирование котилозавров относится к концу каменноугольного
периода палеозойской эры и связано с изменением климатиче-
ских условий на больших пространствах суши: сменой теплого и
влажного климата на сухой, с суровыми сезонами. Котилозавры
вымерли в начале мезозойской эры, уступив место различным
ветвям рептилий, занявшим в мезозое доминирующее положе-
ние на суше.
КЛАСС ПТИЦЫ (AVES)'
Общая характеристика. Птицы — теплокровные позвоночные
животные из группы Amniofa, приспособленные к полету. Перед-
ние конечности видоизменены в крылья. Тело покрыто перьями,
которые образуют также несущую плоскость крыльев и хвоста.
Часть костей плюсны и предплюсны, слившись, образовали еди-
ную кость — цевку. Череп сочленяется с позвоночником одним
мыщелком. В полушариях мозга имеется кора, но поверхность
их. гладкая. Мозжечок хорошо развит. Легкие губчатые, соеди-
нены с системой воздушных мешков. Сердце четырехкамерное.
Имеется только правая дуга аорты, левая атрофируется еще при
эмбриональном развитии. Органами выделения являются тазовые
почки. Оплодотворение внутреннее. Размножаются, отклады-
вая яйца.
В настоящее время на Земле обитает около 9 тыс. видов птиц,
населяющих все материки и острова. В СССР обитает примерно
750 видов птиц.
Современных птиц подразделяют на три обособленных над-
отряда: Килегрудые птицы (Carinatae), Бескилевые птицы (Ra-
titae), Пингвины (Inipennes).
Строение и жизненные отправления. Внешний вид птиц отра-
жает их приспособленность к полету (рис. 247). Туловище об-
текаемой яйцеобразной формы, отличается компактностью. Шея
большинства птиц тонкая, гибкая. На голове вперед выдается
клюв, состоящий из надклювья и подклювья. Для полета служат
296
измененные передние конечности —
крылья. Большую часть их несущей
плоскости образуют большие упру-
гие маховые перья. Ноги птиц при-
нимают всю тяжесть тела-при пере-
движении по земле, лазании по де-
ревьям, взлете и посадке. Ноги имеют
четыре отдела: бедро, голень, цевку
и пальцы. Обычно ноги птицы четы-
рехпалые, но иногда число их сокра-
щается до трех и даже двух (афри-
канский страус). Из четырех паль-
цев в большинстве случаев три
Рис. 247. Внешний вид птицы
(полевой лунь)
направлены вперед, а одни — назад.
Покровы. Кожа птиц тонкая, сухая. Кожные железы от-
сутствуют. Лишь над основанием хвоста у большинства птиц
расположена особая копчиковая железа, секрет которой исполь-
зуется для смазки перьев, что препятствует намоканию. Для птиц
характерен перьевой покров. Перья присущи всем видам птиц и
не встречаются у других животных. Перья птиц развились из ро-
говых чешуй пресмыкающихся.
Перо — производное эпидермиса кожи (рис. 248). Оно об-
разовано роговым веществом — кератином. Отдельное перо со-
стоит из очина (часть, погруженная в кожу), стержня и опахала.
Рис. 248. Строение пера птицы:
1 — стержень; 2 - наружное опахало; 3 внутреннее опа-
хало; 4 — ствол; .5 — очин; 6 -- отверстие очина; 7 дужка
Рис. 249. Строение крыла птицы:
1 — плечевая кость; 2 — локтевая кость; 3 . лучевая кость;
4 — пяетно-заняегпая кость; 5 ... часть запястья; 6’, 7
фаланги пальцев; Л — крылышко; .9 крыловая перепонка;
10 — основания маховых перьев; 11 - первостепенные ма-
ховые перья; 12 -- второстепенные маховые перья
297
Стержень представляет собой плотную роговую трубку с рыхлой
роговой сердцевиной. Опахало образовано отходящими от стерж-
ня в обе стороны бородками первого порядка, от которых, в свою
очередь, отходят короткие бородки второго порядка. Бородки
второго порядка несут мелкие крючочки, сцепляющие бородки
друг с другом, отчего образуется упругая легкая пластинка опа-
хала пера. У нежных пуховых перьев стержень укорочен и несет
тонкие нежные, не сцепленные крючочками бородки. У пуха стер-
жень не развит и бородки отходят пучком от общего основания.
Крупные упругие перья, образующие основную часть несу-
щей плоскости крыла, называются маховыми. Их опахало асим-
метрично — передняя сторона узкая, а задняя — широкая. Такое
строение допускает при поднятии крыла прохождение воздуха
между перьями, а при опускании крыла под давлением воздуха
вызывает плотное соединение перьев. Более крупные маховые
перья, опирающиеся на кости кисти крыла, называются перво-
степенными маховыми, а меньшие по размеру и не столь упругие
перья, соединенные с костями предплечья,— второстепенными
маховыми (рис. 249). Рулевые перья, слагающие хвост и направ-
ляющие полет птиц, отличаются большими размерами, упруго-
стью и асимметрией опахал. Более мелкие перья, покрывающие
тело птиц, называются контурными, они придают телу обтекаемую
форму. Участки, на которых они находятся, называются птери-
лиями, а участки кожи, лишенные их,— аптериями (рис. 250).
Аптерии расположены по средней линии груди, в подмышечной
области, вдоль лопаток, т. е. в тех местах тела, где кожа над мыш-
цами напрягается при полете. Аптерии прикрыты соседними кон-
турными перьями. У многих птиц, особенно водных, между кон-
турными перьями расположены пуховые перья и пух, согреваю-
щие организм.
Роль перьевого покрова в жизни птиц велика и разнообразна.
Маховые и рулевые перья образуют большую часть несущей по-
верхности крыльев и хвоста, следовательно, они необходимы для
полета. Перьевой покров придаст телу птиц обтекаемую форму,
что облегчает их полет. Благодаря высоким теплозащитным свой-
ствам перьев и воздушных прослоек между ними перьевой покров
способствует сохранению тепла тела птиц и, следовательно, уча-
ствует в терморегуляции организма. Он также защищает птицу
от различных механических воздействий. Разнообразные пиг-
менты перьев придают птицам ту или иную окраску, которая часто
носит покровительственный характер.
Периодически, обычно один или два раза в год, перьевой по-
кров птиц полностью или частично обновляется путем линьки;
при этом старые перья выпадают, а на их месте развиваются но-
вые (иногда другой окраски). У большинства птиц линька опере-
ния протекает медленно и постепенно, благодаря чему они сохра-
няют способность полета, но у водоплавающих птиц она протекает
столь быстро, что они временно не могут летать.
298
Рис. 250. Птерилии и аптерии
птицы (голубя)
Рис. 251. Скелет птицы (голубя):
/ - шейные позвонки; 2 — грудные по-
звонки; 3 — хвостовые позвонки; 4 —
копчиковая кость; 5, 6 — ребра; 7 —
грудина; 8 — киль; 9 лопатка; 10 —
коракоид; //—ключица (вилочка);
12 -- плечевая кость; 13 - - лучевая
кость; /^/—локтевая кость; 15 —
пясть; 16..18 — фаланги пальцев;
19—21 — тазовые кости; 22— бедрен-
ная кость; 23 -- кость голени; 24 —
цевка; 25, 26 — фаланги пальцев
Скелет птиц легок и в то же время прочен, что важно для
полета (рис. 251). Легкость его достигается тонкостью состав-
ляющих его костей и наличием полостей в трубчатых костях
передних конечностей. Прочность скелета в значительной степени
обусловлена срастанием многих костей.
Череп птиц отличается большой тонкостенной мозговой ко-
робкой, огромными глазницами, беззубыми челюстями. У взрос-
лых птиц кости черепа полностью срастаются, что обеспечивает
его прочность. Череп сочленяется с первым шейным позвонком
одним мыщелком.
Шейные позвонки, число которых у разных птиц варьирует,
сочленяются друг с другом седлообразными суставными поверх-
ностями, что придает шее большую гибкость. Грудные позвонки
у взрослых птиц сращены между собой. Ребра нижними концами
прикрепляются к большой грудной кости; на заднем крае они
имеют крючковидные отростки, которые налегают концами на
ребра следующей пары; это придает грудной клетке прочность.
Грудина птиц, за исключением тех, которые утратили способ-
ность к полету, несет на передней поверхности высокий костный
киль, к которому с обеих сторон прикрепляются мощные грудные
и подключичные мышцы, приводящие в движение крыло.
Задние грудные, поясничные, крестцовые Ц передние хвосто-
вые позвонки у взрослых птиц срастаются между собой и с тон-
299
кими подвздошными костями таза в единый крестец, который
служит прочной основой для ног. Задние хвостовые позвонки
сливаются, образуя копчиковую кость, имеющую вид вертикаль-
ной пластинки. Она служит опорой для рулевых перьев.
Плечевой пояс состоит из трех пар костей: саблевидных ло-
паток, лежащих вдоль позвоночника; тонких ключиц, которые
срастаются нижними концами в вилочку, распирающую основа-
ния крыльев; коракоидов — массивных костей, соединяющихся
одним концом с лопатками и основаниями плечевых костей, а
другим — с грудиной.
Скелет крыла состоит из крупной, полой внутри кости плеча,
двух костей (локтевой и лучевой) предплечья, ряда сросшихся
косточек запястья и пясти и сильно редуцированных и измененных
фаланг II, III и IV пальцев, I и V пальцы атрофированы, II имеет
только одну фалангу, служащую опорой для обособленного пучка
перьев на внешнем крае крыла, так называемого крылышка.
Тазовый пояс скелета образован тонкими подвздошными, лоб-
ковыми и седалищными костями, срастающимися у взрослых птиц
в единую кость. Задние концы лобковых и седалищных костей у
большинства птиц (кроме некоторых страусов) не сходятся, по-
этому таз остается снизу открытым.
Скелет каждой из задних конечностей состоит из крупной
кости бедра, двух костей голени (большой и малой берцовой),
цевки и фаланг пальцев. Малая берцовая кость сильно редуциро-
вана и приращена к большой берцовой кости. В процессе онтоге-
неза к нижнему концу голени прирастают косточки основного
ряда предплюсны. Остальные косточки предплюсны и три ко-
сточки плюсны сливаются в единую вытянутую кость — цевку.
К нижнему концу цевки причленяются фаланги пальцев.
Мускулатура. Особенно развиты грудные и подключич-
ные мышцы, приводящие в движение крылья. Мощны также
мышцы ног, выполняющие большую работу при хождении птицы
и передвижении по веткам деревьев, при взлете и посадке.
Нервная система, особенно центральный отдел, у птиц
имеет более сложное строение, чем у рептилий, что соответствует
более высокому уровню жизнедеятельности. Головной мозг птиц
отличается большими размерами полушарий переднего мозга,
сильным развитием зрительных бугров среднего мозга и огром-
ным складчатым мозжечком (рис. 252). Крыша полушарий имеет
гладкую поверхность, и серое мозговое вещество в ней выражено
слабо. Сильное развитие зрительных бугров среднего мозга, не-
сущих зрительную функцию, обусловлено значением зрения в
жизни птиц. Мозжечок велик и имеет сложное строение. Его сред-
няя часть — червячок — передним краем почти соприкасается с
полушариями, а задним концом прикрывает продолговатый мозг.
Червячок покрыт характерными поперечными бороздами. Разви-
тие мозжечка связано с полетом, требующим точно координиро-
ванных движений. Головных нервов у птиц 12 пар.
зоо
Рис. 252. Головной мозг ппщы (го-
лубя) :
/ -- енерху; II снизу; III ..- сбоку;
/ -- полушария передне!о мозга; 2 -
обоншельные доли; .‘I - iлазные яб-
локи; 4 эпифиз; .5 зрительные до-
ли; 6 мозжечок; 7 продолговатый
мозг; 8 цнпельные нервы
О р г а и ы ч у в с г в раз-
виты у птиц в различной сте-
пени. Наибольшее значение
в их жизни имеет зрение, по-
этому глаза велики и сложно
устроены. У большинства ви-
дов также хорошо раз-
вит слух. Органы слуха состоят из внутреннего и среднего уха,
барабанная перепонка несколько углублена. Слуховая косточка
одна. Органы обоняния развиты слабо.
Орга и ы и и те в а р е и и я начинаются ротовой полостью.
Зубы у современных птиц отсутствуют —- их частично заменяют
острые края рогового чехла клюва, которым птица захватывает,
удерживает и иногда размельчает пишу (рис. 253). Длинный пи-
щевод у многих птиц расширяется в зоб; здесь пища, подверга-
ясь обработке слюной, набухает и размягчается. Из пищевода
корм попадает в железистый желудок, где смешивается с пище-
варительными соками. Из железистого желудка пища переходит
в мускулистый желудок. Стенки его сложены мощными мышцами,
а в полости, выстланной твердой оболочкой, обычно находятся
мелкие камешки, заглоченные птицей. Эти камешки и складки
стенок желудка при сокращении мышц стенок перетирают
пищу.
Кишечник птиц относительно короткий. В нем различают более
длинный тонкий и менее протяженный толстый отделы. На гра-
нице этих отделов от кишечника отходят два слепых выроста.
Прямая кишка не развита, поэтому фекалии не накапливаются
в кишечнике, что облегчает массу птицы. Заканчивается кишеч-
ник расширением — клоакой, в которую открываются мочеточ-
ники и протоки половых желез. Секреты большой двухлопастной
печени и поджелудочной железы, поступающие в двенадцати-
перстную кишку, способствуют перевариванию пищи.
Затрата птицами во время полета огромного количества энер-
гии и высокий уровень обмена веществ обусловливают необхо-
димость поглощения больших масс пищи. Так, мелкая птичка
наших лесов королек за день потребляет количество пищи, превы-
шающее '/4 массы тела. Процессы пищеварения протекают у
птиц очень быстро: у свиристели ягоды рябины проходят весь
кишечник за 8—10 мин, а у утки, вскрытой через 30 мин после
301
Рис. 253. Внутреннее строение птицы (голубя):
[ — вскрытый голубь; // — разрез желудка голубя;
/ — трахея; 2 — пищевод; 3 — зоб; 4 — легкое; 5 — воздушные мешки;
6 — сердце; 7 — железистый желудок; 8 — мускульный желудок
того, как она проглотила карася длиной 6 см, уже нельзя было
обнаружить в кишечнике его остатков.
Органы дыхания птиц также несут признаки приспособ-
ления к полету, во время которого организм нуждается в усилен-
ном газообмене (рис. 254). От глотки птицы отходит длинная
трахея, которая в грудной полости делится на два бронха. На
месте разделения трахеи на бронхи имеется расширение — ниж-
няя гортань, в которой расположены голосовые связки; стенки
ее имеют костные кольца. Нижняя гортань играет роль голосового
аппарата и особенно сильно развита у птиц, поющих или издаю-
щих громкие звуки.
Легкие птиц имеют губчатое строение. Бронхи, входя в легкие,
распадаются на все более мелкие ветви. Последние заканчива-
ются тончайшими слепыми канальцами — бронхиолами, в стенках
которых проходят капилляры кровеносных сосудов.
Часть разветвлений бронхов выходит за пределы легких, про-
должаясь в тонкостенные воздушные мешки, расположенные
302
между мышцами, средн внутренних органов и в полостях трубча-
тых костей крыльев. Эти мешки играют большую роль в дыхании
птицы во время полета. У сидящей птицы дыхание осуществля-
ется путем расширения и сжатия грудной клетки. В полете же,
когда движущимся крыльям нужна твердая опора, грудная клет-
ка остается почти неподвижной и прохождение воздуха через
легкие обусловливается в основном расширением и сжатием воз-
душных мешков. Этот процесс получил название двойного дыха-
ния, поскольку отдача кислорода в кровь происходит как при
душные мешки
Рис. 255. Кровеносная система птицы (голубя):
/ правое предсердие; 2 - правый желудочек сердца; 3— левая легочная артерия;
4 правая легочная артерия; 5 -левое предсердие; 6—левый желудочек сердца;
7 - правая дуга аорты; W, 9 - безымянные артерии; 10—12 --- сонные артерии;
13 — подключичная артерия; 14 -- левая грудная артерия; /5 — аорта; 16 - правая
бедренная артерия; 17 почечная артерия; 18 - седалищная артерия; 19 -- под-
вздошная артерия; 20 задняя брыжеечная артерия; 21 — хвостовая артерия;
22 хвостовая вена; 23- воротная вена почек; 24 - бедренная вена; 25 - под-
вздошная вена; 26 задняя полая вена; 27 — кишечная вена; 28 - иадкишечная
вена; 29 почечная вена; 30 — яремная вена; 31 — подключичная вена; 32— передняя
полая вена
303
вдохе, так и при выдохе. Чем быстрее машущий полет, тем интен-
сивнее дыхание. При подъеме крыльев они растягиваются и воз-
дух засасывается в легкие и далее в мешки. При опускании крыль-
ев происходит выдох, причем через легкие проходит воздух из
мешков, что способствует окислению крови в легких.
Кровеносная система птиц имеет два круга кровооб-
ращения (рис. 255). Большое по объему сердце полностью раз-
делено на правую и левую половины и имеет левое и правое пред-
сердия и левый и правый желудочки. Этим достигнуто полное
разделение потоков артериальной и венозной крови. Артериаль-
ная кровь, идущая от легких по легочной вене, попадает в левое
предсердие, а оттуда — в левый желудочек, из которого уходит в
аорту. Венозная кровь со всего тела поступает в правое предсер-
дие, а из него — в правый желудочек, с тем чтобы потом напра-
виться по легочной артерии к легким.
У эмбрионов птиц, как и пресмыкающихся, закладываются
как левая, так и правая дуги аорты, но в процессе эмбриональ-
ного развития животного левая атрофируется. Начинаясь от
левого желудочка сердца, правая дуга аорты изгибается направо
(почему и называется правой), поворачивает назад и продолжа-
ется стволом аорты, тянущимся под позвоночником. От дуги
аорты отходят большие парные безымянные артерии, которые
вскоре делятсяиа сонные артерии, несущие кровь к голове, и мощ-
ные грудные и подключичные артерии, идущие к грудным мыш-
цам и крыльям. От спинной аорты ответвляются артерии к раз-
личным частям туловища птиц и к ногам. Венозная система птиц
в основном сходна с таковой пресмыкающихся.
Высокая активность процесса обмена веществ у птиц делает
необходимым быструю и обильную доставку питательных веществ
и кислорода во все части организма. Поэтому кровообращение
у них совершается очень быстро, что обеспечивается энергичной
работой сердца. Так, у многих мелких птиц сердце сокращается
более 1 тыс. раз в минуту (у человека 60—80 раз).
Органы выделения птиц также приспособлены к интен-
сивному обмену веществ в организме, вследствие чего увеличи-
вается объем продуктов распада, подлежащих удалению. Почки
у птиц отличаются большими размерами и лежат в углублениях
тазовых костей. От них отходят мочеточники, открывающиеся
в клоаку. Густая моча поступает в клоаку, откуда выводится на-
ружу вместе с калом.
Органы размножения. Два семенника, лежащих в
брюшной полости, имеют бобовидную форму. От них отходят се-
мяпроводы, открывающиеся в клоаку. У некоторых птиц (гуси)
самцы имеют совокупительный орган. У самок обычно имеется
только один, левый, яичник, лежащий близ почки. Вышедшее из
яичника яйцо попадает в непарный яйцевод, в верхней части ко-
торого происходит оплодотворение. Пройдя по яйцеводу, яйцо
приобретает белковую оболочку, а попав в более широкую матку,
304
Рис. 256. Строение яйца птицы:
/ . скорлупа; 2 -.подскорлуновая оболочка; 3 -
воздушная камера; 4 белок; 5 желточная
оболочка; 6 желток; 7 - зародышевый диск;
8- белый желток; 9 - желтый желток; 10 -
халазы
6
покрывается известковой скорлупой. Через конечный отдел поло-
вых путей самки влагалище-- яйцо попадает в клоаку, а от-
туда выводится наружу.
Яйцо птиц имеет (относительно к величине животного) очень
крупные размеры, так как содержит много питательных веществ
в виде желтка и белка (рис. 256). Зародыш развивается из не-
большого зародышевого диска, находящегося на поверхности
желтка.
На тупом конце яйца под скорлупой и подскорлуповой обо-
лочкой находится полость, наполненная воздухом; она помогает
дыханию зародыша. Развитие птенца в яйце показано па рис. 257.
Рис. 257. Развитие зародыша птицы:
/ - IV - последовательные стадии развития эмбриона;
/ - зародыш; 2 — желток; 3— белок; 4 -- амниотическая складка; 5 -- внезароды-
шевая полость; 6 — воздушная камера; /--скорлупа; 8 - сероза; .9—амнион;
10 -- полость амниона; // -- аллантоис; 12 - желточный мешок
305
Экология птиц. Основная форма передвижения большинства
птиц — полет. Приспособление к полету вызвало ряд описанных
изменений в строении организма этих животных, а также наложи-
ло отпечаток на все виды их жизнедеятельности. Благодаря спо-
собности к полету птицы обладают огромными возможностями
для далеких миграций и расселения: именно полет позволил им
заселить все океанические острова, нередко лежащие в сотнях
километров от материка. Полет помогает птицам избегать врагов.
Многие птицы во время полета добывают пищу или высматри-
вают ее па земле.
Характер полета разных видов птиц далеко не одинаков -
он всегда находится в соответствии с их образом жизни. Разли-
чают два основных вида полета птиц: парение и гребной полет.
Парением называется полет птиц на более или менее неподвиж-
ных, распростертых крыльях. Этот полет может осуществляться
при постепенном снижении птицы в воздухе. Но часто птица путем
парения может сохранять набранную высоту над землею или
даже подниматься вверх (достигается благодаря использованию
птицей восходящих потоков воздуха). Гребной полет осуществля-
ется взмахами крыльев. У многих птиц такая активная форма
полета чередуется с парением в воздухе. Ворона при спокойном
гребном полете делает в среднем 2,9, а чайка —• 2,2 взмаха крыла
в секунду. Максимально возможная скорость полета ласточки
28 м, глухаря— 16 и лебедя — 14 м в секунду. Некоторые птицы
могут летать без остановки для отдыха более 3 тыс. км.
Способность к активному полету, теплокровность и высокий
уровень развития центральной нервной системы обеспечили пти-
цам возможность широкого распространения на Земле. С приспо-
соблением птиц в ходе эволюции к жизни в различных условиях
(леса, открытые пространства, водоемы) связано образование
разных экологических групп, отличающихся внешним обликом
и специфическими чертами строения.
Древесные птицы — обитатели разнообразных лесов и кустар-
никовых зарослей. К этой группе относятся дятлы, попугаи, по-
ползни, пищухи, кукушки, скворцы, дрозды, голуби, глухари, ряб-
чики и др. Добывают корм и гнездятся обычно на деревьях, реже
на земле. У наиболее специализированных, приспособленных
к лазанию по деревьям птиц (попугаи, дятлы, поползни) лапы
сильные, вооруженные загнутыми когтями. У дятлов два пальца
направлены вперед, два — назад, что позволяет им ловко лазать
по стволам деревьев, опираясь при этом на жесткие и упругие
перья хвоста. Попугаи используют при передвижении по ветвям
деревьев не только задние конечности, но и клюв.
Наземные птицы — обитатели открытых пространств — лу-
гов, степей и пустынь. В эту группу входят страусы, дрофы, стре-
петы, некоторые кулики. Кормятся и гнездятся на земле. В поис-
ках корма передвигаются преимущественно путем хождения и
бега, а не полета. Это крупные и средних размеров птицы с мас-
306
сивным и широким туловищем и длинной шеей. Ноги длинные
и сильные, с короткими и толстыми пальцами, число которых мо-
жет сокращаться до трех, а у африканского страуса — до двух.
Болотные птицы населяют заболоченные луга, болота, заросли
побережий водоемов. Типичные представители: цапли, аисты, жу-
равли, многие кулики. Корм обычно собирают на земле. Гнезда
устраивают на земле или на деревьях. Это крупные или средней
величины птицы. Большинство имеют длинные тонкие ноги с уд-
линенными пальцами, при помощи которых они легко передвига-
ются по вязкому грунту или мелководью. Голова небольшая, с
длинным твердым клювом. Крылья хорошо развиты. Хвост корот-
кий. Оперение рыхлое, со слабо развитым пухом.
Водоплавающие птицы значительную часть жизни проводят
на водоемах. К данной группе относятся гагары, поганки, кайры,
чистики, пингвины, бакланы, пеликаны, утки, гуси, лебеди. Они
хорошо плавают, а многие и ныряют, но ходят по суше и летают
обычно плохо, а некоторые вообще не летают (пингвины). Мно-
гие птицы добывают корм (рыба, моллюски, ракообразные) в
воде, другие кормятся на суше вегетативными частями растений
и семенами. Гнездятся по берегам водоемов, на земле, па деревь-
ях, в зарослях тростника, на скалах и в их расщелинах, в норах.
Это крупные и средней величины птицы с несколько уплощенным
с брюшной стороны туловищем и коротким хвостом. Ноги отне-
сены далеко назад, что обеспечивает почти вертикальное положе-
ние тела при хождении. Имеют плотное оперение с хорошо раз-
витым пухом, плавательные перепонки на лапах, большинство —
развитую копчиковую железу.
Воздушно-водные птицы в отличие от предыдущей группы
в меньшей степени связаны с водоемами. В группу входят чайки,
крачки, буревестники. Они обычно хорошо летают, плавают, но
плохо ныряют. Полет парящий с использованием завихрений
воздуха над волнами или разной скорости воздушных потоков.
Питаются в основном рыбой, которую высматривают во время
полета, затем стремительно бросаются на нее и вытаскивают из
воды своим крепким и длинным клювом, загнутым на конце. Гнез-
дятся часто па берегах рек, озер, морей, на каменистых уступах
морских берегов. Это крупные и средних размеров птицы с вытя-
нутым туловищем, длинными, острыми крыльями и короткими
лапами, на которых три передних пальца соединены плаватель-
ной перепонкой. Оперение густое, с большим количеством пуха.
Воздушно-наземные птицы значительную часть светового
времени суток проводят в воздухе, где ловят насекомых своим
коротким, по широко раскрывающимся клювом. Типичные пред-
ставители: стрижи, ласточки, козодои. Это отличные летуны, об-
ладающие быстрым и маневренным полетом. Гнездятся обычно в
постройках, в норах по берегам рек, на земле. Их туловитце вытя-
нутой формы, шея короткая, крылья длинные и узкие. Ноги ко-
роткие, что затрудняет хождение по земле.
307
Питание птиц. Большинство птиц животиоядиы, другие рас-
тительноядны или всеядны. Имеются виды, питающиеся преиму-
щественно вегетативными частями растений (гуси), ягодами
(дрозды, свиристели), семенами (воробьи, клесты), нектаром
(колибри), насекомыми (кукушки, дятлы, многие воробьиные),
рыбой (чайки, бакланы, пеликаны), лягушками (утки, аисты,
цапли), ящерицами и змеями (аисты, некоторые дневные хищ-
ники), птицами (ястреба), грызунами (совы, многие дневные
хищники). Некоторые хищники предпочитают поедать падаль
(грифы, стервятники, сипы). Характер пищи может изменяться
в зависимости от возраста: большинство зерноядных птиц вы-
кармливают своих птенцов насекомыми. Состав пищи варьирует
и по сезонам года. Например, тетерев летом кормится зелеными
частями растений, ягодами и насекомыми, а зимой — в основном
хвоей сосны, почками, побегами и сережками березы и ольхи.
Годовая периодичность в жизни птиц. У птиц, как и у других
животных, годовая периодичность жизнедеятельности тесно свя-
зана с сезонными изменениями условий существования и имеет
большое приспособительное значение. Она позволяет приурочить
наиболее ответственный момент в жизни каждого вида — раз-
множение — к определенному сезону, когда условия для выкарм-
ливания птенцов будут наиболее благоприятны. Можно выде-
лить следующие этапы годового цикла птиц: подготовка к раз-
множению, размножение, линька, подготовка к зиме, зимовка.
Подготовка к размножению выражается в образовании пар.
Объединение в брачное время в пары (м о н о г а м и я) свойствен-
но большинству видов птиц. Однако продолжительность суще-
ствования пар у разных птиц существенно различна. На несколько
лет или возможно даже пожизненно образуют пары лебеди,
аисты, орлы. Другие птицы образуют пары на сезон размножения,
а многие утки держатся парами только до начала яйцекладки.
У меньшего числа видов птиц пары не образуются и самец за пе-
риод размножения оплодотворяет несколько самок, которые несут
всю заботу о потомстве. Такое явление получило название и о л и-
гамии (многоженство). Оно свойственно тетеревам, фазанам,
глухарям, домашним курам. У этих птиц особенно хорошо выра-
жен половой диморфизм.
Разбивка на пары у птиц сопровождается токованием: птицы
принимают различные позы, необычно держат оперение, издают
особые звуки, у некоторых полигамных видов происходят драки
между самцами. Брачное поведение птиц облегчает встречу осо-
бей разного пола и образование пар, стимулирует синхронное
созревание половых продуктов обоих партнеров.
Период размножения начинается с гнездостроения. Устрой-
ство гнезд — характерная черта размножения птиц, повышаю-
щая сохранность потомства. Гнезда птиц очень разнообразны.
Лишь немногие виды откладывают яйца прямо на уступ скалы
(кайры) или на землю (стрепеты), предварительно не изменяя
308
естественный субстрат. Большинство птиц строят гнезда. У одних
видов (курообразные, кулики) гнездо представляет ямку в земле,
которую эти птицы обычно выстилают скудной подстилкой из
сухой травы или веток. У других (воробьиные) гнезда более слож-
ные, имеющие вид чаши из сухой травы, мха, перьев и другого
материала и размещающиеся обычно в развилке ветвей или па
земле. Многие птицы (дятлы, синицы, мухоловки, скворцы, совы
и др.) гнездятся в дуплах деревьев. Ласточки часто сооружают
лепные гнезда из комочков грязи, которые они скрепляют слюной.
Зимородки, щурки, береговые ласточки и ряд других птиц роют
в обрывах рек и оврагах глубокие норы, в которые и откладывают
яйца. Нс строят собственных гнезд только птицы, которым свой-
ствен гнездовой паразитизм (кукушки). Они откладывают яйца
в гнезда других птиц, которые их насиживают и выкармливают
птенцов.
Плодовитость птиц значительно меньше, чем рептилий, что
связано с наличием у птиц разнообразных форм заботы о потом-
стве (гпездостроение, насиживание и выкармливание птенцов).
Количество яиц в кладке колеблется от 1 (пингвины, кайры) до
22 (серая куропатка). Большинство птиц насиживают свою клад-
ку. У полигамных видов насиживание осуществляется только
самкой (курообразные, гусеобразные), у моногамных — попере-
менно самцом и самкой (голуби, чайки, многие воробьиные)
или только самкой, а самец кормит ее и охраняет гнездовой уча-
сток (совы, дневные хищники, некоторые воробьиные).
Длительность инкубации у разных птиц различна и зависит
от размеров яйца и птицы, типа гнезда и интенсивности насижи-
вания. Мелкие воробьиные насиживают в течение 11 —12 сут, во-
рона — 17, лебеди — 35—40. Продолжительность инкубации у
домашних птиц: у курицы 21 сут, у утки — 28, у гусыни .. 30,
у индейки — 28, 29 сут.
В зависимости от степени развития птенцов, только что вы-
лупившихся из яиц, птиц делят на выводковых, нолувыводковых
и птенцовых (рис. 258). Птенцы выводковых птиц опушенные,
зрячие, способны через короткое время самостоятельно питаться
(курообразные, гусеобразные, страусы). Птенцы иолувыводко-
вых птиц вылупляются зрячими и опушенными, но выкармлива-
ются родителями до приобретения способности к полету (чайки,
чистики, буревестники). У птенцовых птиц птенцы голые, слепые,
длительное время остаются в гнезде (воробьиные, дятлы, голуби),
где их интенсивно кормят родители. Так, пара мухоловок, синиц
или пеночек приносит корм своим птенцам до 450- 500 раз в
сутки.
После завершения выкармливания птенцов семья обычно рас-
падается и птицы объединяются в стаи. Самая высокая смерт-
ность наблюдается в первый год жизни птиц. Иногда опа может
достигать более 50 % числа особей, вылетевших из гнезда. Поло-
вой зрелости птицы достигают в разном возрасте. Большинство
зон
Рис. 258. Птенцы различных птиц в одном возрасте:
/ — птенцовых (конек); 11 — иолувыводковых (орел); ///—выводковых (куропатка)
птиц мелких и средних размеров (многие воробьиные) начинают
размножаться уже на следующий год жизни, более крупные пти-
цы (серая ворона, утки, мелкие хищники и чайки) — на 2-м году,
а гагары, орлы, буревестники — на 3—4-м, страусы — на 4—
5-м году.
Средняя продолжительность жизни мелких воробьиных птиц
составляет 1 — 1,5 года, а предельная — 8—10 лет. Более крупные
виды птиц могут жить 40 и более лет.
Линька различно протекает у разных птиц. У одних видов
(воробьиные) она постепенная, у других (курообразные, гусе-
образные) — бурная. Линные особи гусеобразных на 2—5 недель
теряют способность к полету. Линька обычно начинается сразу
после размножения. У самцов многих видов птиц, которые не уча-
ствуют в выводе потомства, линька наступает раньше, чем у са-
мок. Линяющие самцы глухарей и тетеревов держатся поодиночке
в глухих участках леса, а селезни уток на период линьки скапли-
ваются в больших количествах в труднодоступных заболочен-
ных местах.
Подготовка к зиме. В этот период птицы начинают кочевать
в поисках корма. Интенсивное питание обеспечивает накопле-
ние жира. Некоторым птицам свойственно запасание кормов,
что облегчает их зимовку. Сойки собирают и закапывают в почву
или под лесную подстилку желуди, а кедровки — орешки. Зимой
птицы используют эти запасы лишь частично. Другая часть се-
мян поедается мышевидными грызунами и насекомыми или, сох-
ранившись до весны, прорастает. Поползни и синицы прячут се-
мена различных деревьев в трещинах коры, обеспечивая себя кор-
мами на 50—60 %. Мелкие совы (воробьиный и мохноногий
сычи) заготавливают на зиму тушки мышевидных грызунов и
складывают их в дупла деревьев. Свои кладовые птицы находят,
по-видимому, благодаря памяти и обонянию.
310
Зимовка. В зимний период птицы сталкиваются с большими
трудностями при добывании нужного количества корма. В поис-
ках мест обитания, которые наиболее полно могут обеспечить
тот или иной вид кормом и защитными условиями, многие птицы
начинают совершать направленные перемещения (кочевки и миг-
рации). Лишь оседлые птицы остаются в тех местах, где прохо-
дило их размножение, а если и меняют места обитания, то улетают
не далее чем за несколько десятков километров (глухари, ряб-
чики, дятлы, воробьи, синицы). Кочующие птицы могут улетать
за сотни километров, по обычно держатся в пределах одной при-
родной зоны (свиристели, чечетки, снегири). Самые дальние миг-
рации совершают перелетные птицы, зимующие в других при-
родных зонах, находящихся за тысячи километров от мест гнез-
дования.
Деление птиц на оседлых, кочующих и перелетных осложнено
тем, что один и тот же вид в разных частях своего ареала может
вести себя различно. Так, серая ворона па юге европейской ча-
сти СССР - оседлый вид, а па севере — перелетный. Изменение
погодных и кормовых условий но годам также сказывается па
характере подвижности птиц. В теплые зимы при достаточной
обеспеченности кормом некоторые перелетные для данного района
виды остаются зимовать в местах своего размножения (утки,
грачи, дрозды). Эго свидетельствует о том, что основная причина
перелетов птиц заключается в сезонных изменениях условий су-
ществования. В районах, где эти изменения ио сезонам выражены
резче, число перелетных видов больше. 'Гак, в СССР из 750 видов
птиц 600 - перелетные, зимующие в основном на Британских
островах, в Южной Европе, в Средиземноморье, в Африке и Азии.
Миграционные пути птиц огромны. Пролетный путь наших
пеночек и ласточек, зимующих в Африке, составляет 9—10 тыс. км,
а полярной крачки от побережий Варенцова моря до побережий
Африки 16 18 тыс. км. Пролетные пути водоплавающих и
болотных птиц приурочены к речным долинам и морским побе-
режьям, где имеются условия, подходящие для их отдыха и кор-
межки, Многие птицы летят широким фронтом. Мелкие воробьи-
ные покрывают за сутки расстояние 50—100 км, утки 100—
500, аисты - 250, вальдшнепы 500 км. В сутки па полет птицы
обычно тратят 1—2 ч, используя остальное время па остановки
для отдыха и кормежки. Пересекая водные пространства, они без
отдыха пролетают расстояние в тысячи километров. Веспой оста-
новки птиц более редки и кратковремеипы, чем осенью, поэтому
весенние миграции обычно происходят с большей скоростью, чем
осенние.
Перелеты птиц - один из интереснейших и слабоизученных
вопросов биологии птиц. До сих пор не изучен до конца механизм,
определяющий ориентацию птиц во время перелетов. На основе
наблюдений в природе и постановки экспериментов удалось вы-
явить, что мигрирующие птицы могут ориентироваться по поло-
зы
жению солнца, луны, звезд, по особенностям ландшафта. Важную
роль в миграционном поведении птиц и выборе общего направле-
ния во время полета играет врожденный миграционный ин-
стинкт. Однако он проявляется при наличии определенной суммы
факторов внешней среды. Под влиянием условий среды возможно
изменение этого врожденного инстинкта.
Перелеты птиц сложились в течение тысячелетий. Бесспорно
влияние ледникового периода на формирование миграционных
путей птиц в Северном полушарии. Современные пролетные пути
некоторых птиц повторяют исторический путь их расселения в
послеледниковое время.
Большое значение для изучения перелетов птиц имеет метод
кольцевания, когда птенцам перед вылетом из гнезда или взрос-
лым птицам одевают на лапу металлическое кольцо с номером
и обозначением учреждения, проводящего мечение. В нашей стра-
не все сведения о кольцевании и добыче окольцованных птиц на-
правляются в Центр кольцевания Академии наук СССР (Мо-
сква). Ежегодно в мире кольцуют около 1 млн птиц, из них в
СССР — более 100 тыс. Кольцевание позволяет проследить миг-
рационные пути, скорость полета, продолжительность жизни и
другие важные вопросы экологии птиц.
Хозяйственное значение птиц. Роль птиц в хозяйственной
деятельности человека велика и многообразна. Птицы, одомаш-
ненные человеком (куры, гуси, утки, индейки, цесарки, голуби),
издавна используются для получения от них мяса, яиц, пуха,
пера и других ценных продуктов и промышленного сырья. В нашей
стране птицеводство — важнейшая и быстро развивающаяся
отрасль животноводства. Многие виды диких птиц (курообраз-
ные, гусеобразные, некоторые кулики) служат объектами спор-
тивной и промысловой охоты, что позволяет дополнительно во-
влекать в хозяйственный оборот значительное количество вкус-
ного мяса.
Велика роль птиц в истреблении насекомых и мышевидных
грызунов — вредителей сельского хозяйства. Значение синиц,
мухоловок, поползней, скворцов, дроздов и многих других птиц
как регуляторов численности вредных насекомых особенно воз-
растает в период выкармливания птенцов. Так, семья обыкновен-
ного скворца за гнездовой период уничтожает 8—10 тыс. майских
жуков и их личинок или свыше 15 тыс. гусениц зимней пяденицы.
Многие хищные птицы, совы, чайки, аисты и ряд других истреб-
ляют мышей, полевок, сусликов, крыс, хомяков и других вредных
грызунов. Полезность птиц связана с их способностью быстро
находить и концентрироваться в очагах массового размножения
вредителей, а для многих видов птиц —- переходить на многочис-
ленный, хотя часто не свойственный им корм. Так, в годы массо-
вого размножения мышевидных грызунов ими начинают питаться
грачи, чайки и др.
Некоторые птицы выступают в роли распространителей рас-
312
Рис. 259. Различные типы домиков
для гнездования полезных насекомояд-
ных птиц
тений. Так, в тайге Восточной
Сибири на гарях восстановле-
ние кедра часто связано с дея-
тельностью кедровки. Сойки
участвуют в расселении дуба.
Свиристели, дрозды, рябчики и
многие другие разносят семена
рябины, черемухи, терна, бузи-
ны, калины, бересклета, черни-
ки, малины, брусники и т. д.
Для повышения численнос-
ти и привлечения полезных
птиц создают благоприятные
условия для их гнездования,
развешивают искусственные
гнездовья: скворечники, синич-
пики, дуплянки (рис. 259),
проводят зимнюю подкормку нт. д. При развешивании искусствен-
ных гнездовий численность птиц-дунлогнезднпков (мухоловки,
синицы, скворцы) резко возрастает.
В некоторых случаях птицы могут причинять определенный
ущерб. Грачи, полезные уничтожением почвенных насекомых,
местами вредят посевам сельскохозяйственных культур (особенно
кукурузе), выклевывая семена и выдергивая проростки. Кочую-
щие скворцы расклевывают зрелые плоды вишни и винограда.
В южных районах нашей страны местами серьезный вред урожаю
зерновых причиняют воробьи. Золотистая щурка, истребляющая
пчел, может приносить вред пчеловодству. Охотничьему хозяй-
ству местами вредят камышовый лунь и серая ворона. При столк-
новении в воздухе со скоростными самолетами птицы иногда ста-
новятся причиной серьезных аварий, что обусловливает создание
системы отпугивания птиц с территории аэродромов. Необходимо
учитывать также роль птиц в распространении некоторых опас-
ных для человека и сельскохозяйственных животных заболеваний
(орнитозы, грипп, энцефалит и др.).
НАДОТРЯД КИЛЕГРУДЫЕ ПТИЦЫ (CARINATAE)
К этому надотряду относятся большинство современных птиц,
в том числе все птицы фауны Советского Союза. Почти все киле-
.грудые птицы способны летать, и поэтому у них хорошо выражены
различные приспособления к полету. Они имеют развитые крылья.
На передней поверхности грудной кости выдается киль, к кото-
313
рому прикрепляются мощные грудные мышцы. Плечевые кости
полые. Контурные перья имеют хороню выраженную пластинку
опахала, бородки которого сцеплены друг с другом.
Килегрудые птицы распространены по всем материкам и ост-
ровам земного шара. Надотряд включает много отрядов. Ниже
дается описание только тех из них, которые распространены в
СССР и имеют значение для сельского и охотничьего хозяйства
нашей страны.
Отряд Курообразные (Galliformes). Птицы средней величины
(рис. 260,/). Они отличаются плотным телом, небольшой голо-
вой с коротким клювом, довольно короткими, широкими крыль-
ями и сильными, средней длины четырехпалыми ногами с боль-
шими тупыми когтями. Оперение плотное, но почти без пуха. Мясо
высокого качества. Курообразные встречаются во всех зонах
нашей страны. Полет этих птиц сильный, но тяжелый, с частыми
взмахами крыльев, без парения. Большинство ведет оседлый об-
раз жизни, но некоторые (белые куропатки) совершают далекие
сезонные кочевки, а перепела улетают на зиму далеко на юг. Все
курообразные — выводковые птицы. У большинства видов самцы
резко отличаются от самок размерами, окраской и характером
оперения. Гнезда устраивают на земле в виде неглубокой ямки.
Кладки обычно содержат много яиц. Питаются преимущественно
растительной пищей, но в корме птенцов преобладают насекомые.
В СССР обитает 20 видов куриных птиц, которые относятся
к двум семействам: Фазановые и Тетеревиные.
Фазановые (Phasianidae) отличаются тем, что цевка у них не
оперена и по бокам пальцев нет роговой бахромы. У самцов мно-
гих видов на ногах имеются шпоры. Из диких птиц к этому семей-
ству принадлежат фазаны, серые и каменные куропатки, пере-
пела, а из разводимых в СССР домашних птиц — куры, индейки,
павлины, цесарки. Домашние куры произошли от обитающих в
Индии банкивских диких кур (Gallus gallus), а домашние ин-
дейки — от живущих в Северной Америке диких индеек (Meleag-
ris gallopavo) (рис. 261). Куры являются основным объектом
птицеводства нашей страны: они дают большое количество цеп-
ного мяса и яиц. Дикоживущие фазановые служат предметом
охоты (рис. 262).
Тетеревиные (Tetraonidae) характеризуются оперенной цевкой
и наличием по бокам пальцев рядов роговых зубчиков, обеспе-
чивающих более прочный захват ветви. Шпор у самцов пет. К
этому семейству относятся глухари, тетерева, белые куропатки,
рябчики. Все эти птицы являются важными объектами спортив-
ной и промысловой охоты.
Отряд Голубеобразные (Columbiformes) — средней величины
и небольшие птицы. Голова с коротким сильным клювом; основа-
ние надклювья покрыто мягкой утолщенной кожей (восковицей),
которую прорезают ноздри (см. рис. 260,//). Крылья длинные,
острые. Ноги короткие, с четырьмя пальцами, один из которых
314
Рис. 260. Представители надотряда килегрудых птиц:
/..глухарь (отр. Курообразные); II — скалистый голубь (отр. Голубеобразные);
/// кроншнеп (nixlorp. Кулики); IV— чайка (подотр. Чайки); V' - кайра
(поботр. Чистиковые); VI - серый гусь (отр. Гусеобразные); VII - баклан
(отр. Веслоногие); VIII -- аист (отр. Листообразные); IX -- дрофа (отр. Журавле-
образные).
Рис. 260. (Продолжение)
А'—орлан (отр. Дневные хищные птицы)', XI — неясыть (отр. Совообразные);
XII — пестрый дятел (отр. Дятлообразные); XIII грач (отр. Воробьинообразные)
Рис. 261. Предки различных сель-
скохозяйственных птиц:
I - - банкнвский петух (Индия); И —
дикая индейка (Северная Америка);
HI - утка-кряква
направлен назад. Когти туповатые. Хвост средней длины. Опере-
ние плотное, по пух развит слабо. Держатся па деревьях, скалах,
постройках человека. Живут обычно колониями. Хорошо летают.
Некоторые виды ведут оседлый образ жизни, другие на зиму от-
летают па юг. Птенцовые птицы. В кладке только два яйца. Пи-
таются в основном растительной пищей (семенами, ягодами и
т. и.). Местами вредят посевам. Мясо высокого качества.
В городах и селах широко распространены полудомашние
сизари. В лесах и полях обитают близкие виды — клинтух, вяхирь
и др. В садах и лесах южных частей страны обычны мелкие го-
лубки - горлицы.
Отряд Ржанкообразные (Charadriiformes) включает три под-
отряда.
Подотряд Кулики (Charadrii) — мелкие и средней величины
птицы, большинство которых имеют удлиненную шею, острые,
но не очень длинные крылья, короткий хвост, длинные ноги с голой
цевкой, а то и голенью (см. рис. 260, III). Клюв обычно длинный,
тонкий. Плавательных перепонок на ногах, как правило, пет, или
они слабо развиты. Оперение рыхлое, почти без пуха.
Обитатели болот и лугов, сырых лесов и перелесков, берегов
рек, озер и морей; реже они встречаются в степи и пустыне. На
зиму улетают па юг. Гнездятся обычно на земле. Самка отклады-
вает в небольшую ямку четыре яйца. Выводковые птицы. Пита-
ются преимущественно мелкими беспозвоночными. Вальдшнепы,
бекасы, дупеля и многие другие кулики служат объектом спортив-
ной охоты.
317
Рис. 262. Породы кур. Пример образования новых форм путем се-
лекции:
/ — дикий банкивский петух; II — петух и курина породы леггорн; III .. бойцо-
вый петух; IV—петух длиннохвостой породы Иокогама; V'— петух и курица
орловской породы
Подотряд Чайки (Lari). К этому подотряду принадлежат раз-
личные виды чаек и крачек. Птицы различной величины, с вытя-
нутым телом, короткими ногами, длинными острыми крыльями и
хвостом средней длины (см. рис. 260, IV). Ноги четырехпалые,
три передних пальца связаны плавательной перепонкой. Клюв
большой, прямой, часто с крючком на конце. Оперение плотное,
с развитым пухом.
Живут по берегам морей, на озерах и реках. Большую часть
дня летают над водой, выхватывая из нес корм. Крупные чайки
нападают па мелких зверьков и птиц. В стенных районах прино-
сят пользу истреблением сусликов и полевок. Хорошо плавают,
но не пыряют. Некоторые оседлы, но большинство видов кочу-
ющие или перелетные птицы. Гнездятся на земле или на скалах
близ водоемов. В кладке обычно три яйца. 11олувыводковые
птицы. Мясо низкого качества.
Подотряд Частиковые (Alcae). Сюда относятся кайры, га-
гарки, чистики, тупики и др.
Морские птицы среднего размера с вальковатым вытянутым
телом, короткими, по острыми крыльями, небольшим хвостом
(см. рис. 260, V). Ноги отодвинуты далеко назад, поэтому сидя-
щая птица держится почти вертикально. На ногах только три
пальца, соединенных плавательной перепонкой. Спина темная,
брюшко обычное белое. Оперение очень плотное.
Населяют берега морен Северного Ледовитого п Тихого оке-
анов. Живут большими колониями, составляя основную часть
птиц птичьих «базаров». Гнездятся чаще всего на карнизах скал,
откладывая 1-3 яйца прямо в камень; некоторые живут в порах.
Питаются главным образом рыбой, которую ловят, пыряя в воду.
Мясо низкого качества.
Отряд Гусеобразные (Anseriformes). Объединяет различные
виды уток, гусей и лебедей.
Крупные и средней величины водоплавающие птицы с плот-
ным, несколько уплощенным туловищем, короткими, отнесенными
назад ногами, небольшим хвостом, крыльями среднего размера
(см. рис. 260,1/7). Передние пальцы соединены плавательной
перепонкой. Клюв обычно уплощенный, с рядами поперечных
роговых пластинок или острых роговых зубчиков по краям. Опе-
рение плотное, с густым пухом.
Обитают на водоемах различных типов. Хорошо плавают,
много пыряют. Гнездятся на земле, но берегам водоемов, в порах
и дуплах. В кладке бывает до 20 яиц. Выводковые птицы. Пита-
ются мелкими беспозвоночными, водными и береговыми расте-
ниями, травой на лугах.
Дикие гуси - серый и сухопос и утка-кряква (см. рис. 261) —
считаются исходными формами, от которых были выведены раз-
нообразные породы домашних гусей и уток. Разведение этих птиц,
дающих ценное мясо, яйца и пух, имеет большое народнохозяй-
ственное значение. Дикие гусиные служат важным объектом спор-
319
тивной и промысловой охоты. Высоко ценится нежный и теплый
пух, выбираемый из гнезд морских нырковых уток — гаг.
Отряд Веслоногие (Pelecaniformes). Сюда относятся пели-
каны, бакланы и некоторые другие водоплавающие птицы. Для
них характерно, что все четыре пальца коротких ног соединены
плавательной перепонкой (см. рис. 260, VII).
Обитают по берегам морей, на озерах и реках. Питаются в
основном рыбой. Бакланы ловят ее, ныряя в глубь водоема, а пе-
ликаны — выхватывая на мелководье. Местами бакланы прино-
сят вред рыболовству. Мясо бакланов и пеликанов низкого ка-
чества.
Отряд Листообразные (Ciconiiformes). Характерные предста-
вители— аисты и цапли. Большинство из них — довольно круп-
ные птицы на длинных тонких йогах (см. рис. 260, VIII). Цевка
и нижняя часть голени лишены оперения. Длинная тонкая гибкая
шея несет небольшую голову у большинства с прямым удлинен-
ным острым клювом. Ноги с четырьмя длинными пальцами: пе-
редние соединены узкой перепонкой. Оперение рыхлое, почти
без пуха.
Большинство видов отряда держится по берегам водоемов,
на болотах, пойменных лугах. Гнездятся на деревьях или па
земле, белые аисты часто делают гнезда на крышах домов. В клад-
ке 2—3 яйца. Птенцовые птицы. Питаются мелкими животными.
На зиму обычно улетают на юг. Мясо аистов и цапель низкого
качества.
Отряд Журавлеобразные (Gruiformes) объединяет журавлей,
дроф, стрепетов и др.
Журавли — крупные птицы с длинными шеей, ногами и клю-
вам. Гнездятся в труднодоступных районах, на больших болотах
и лугах, реже в степях. Гнездо устраивают на земле. В кладке
1—2 яйца. Выводковые птицы. Питаются преимущественно ра-
стительной пищей. Все виды перелетные. Охота на журавлей за-
прещена.
Дрофы и стрепеты (см. рис. 260, IX) — птицы с массивным
телом, довольно длинной шеей, большой головой, несущей корот-
кий конический клюв. Крылья короткие, округлые. Ноги крепкие,
трехпалые; цевка покрыта шестигранными роговыми щитками.
Обитатели степей и полупустынь Восточного полушария. Дер-
жатся преимущественно на участках целинной степи. Гнездо на
земле. Выводковые птицы. В связи с быстрым сокращением чис-
ленности включены в Красную книгу СССР.
Отряд Дневные хищные птицы (Falconiformes). К ним отно-
сятся орлы, ястребы, луни, коршуны, соколы, грифы. Для всех
их характерны острые изогнутые когти и сильный, хотя и не длин-
ный, загнутый на конце клюв (см. рис. 260, X). Основание над-
клювья покрыто тонкой голой кожей — восковицей. Оперение
упругое, плотное, но почти без пуха.
Места обитания хищных птиц разнообразны. Они живут обыч-
320
но парами. Гнезда устраивают па скалах, деревьях, иногда на
земле. Птенцовые птицы. Пищей им служат преимущественно
различные позвоночные животные, хотя мелкие соколиные' (коб-
чики, пустельги) поедают также' много насекомых. Грифы, сипы
и стервятники пожирают падаль. Многие' виды полезны истреб-
лением вредных грызунов и поэтому подлежат охране. Некото-
рые хищные птицы вредны истреблением дичи и полезных насе-
комоядных птиц.
Отряд Совообразные (Strigiformes). Как и дневные хищные
птицы, совы имеют острые' загнутые когти и клюв. Но они резко
отличаются от дневных хищников рядом признаков, связанных
с ночным образом жизни (см. рис. 260, XI). Огромные глаза сов
направлены вперед. Перья вокруг глаз расходятся радиально,
образуя плоский лицевой диск. Ушные отверстия велики. Шея
столь гибкая, что позволяет птице поворачивать голову почти
на 300°. Наружный палец пог может обращаться как вперед, так
и назад. Оперение рыхлое, очень мягкое.
Совы -.ночные хищники, днем они скрываются в дуплах, пе-
щерах, развалинах, на чердаках. Ночью, а некоторые виды также
на рассвете и закате охотятся на различных мелких позвоночных.
Почти все совы приносят большую пользу, истребляя огромное
количество вредных грызунов. Одна сова неясыть за год уничто-
жает около тысячи мышей и полевок, сохраняя сельскому хо-
зяйству до 0,5 т зерна. Самая крупная сова — филин — может
вредить истреблением дичи.
Отряд Дятлообразные (Piciformes). Своеобразные лесные
птицы, приспособившиеся к лазанию по стволам деревьев (см.
рис. 260, XII). Ноги у них короткие, с острыми когтями. Обычно
два пальца ноги направлены вперед, а два — назад. Перья хвоста
очень жесткие, заостренные на концах; находясь на дереве, птица
упирается хвостом в ствол. Клюв прямой, острый, долотообраз-
ный, приспособленный к долблению древесины. Язык очень длин-
ный и тонкий, что позволяет извлекать насекомых из щелей и
отверстий в деревьях.
В СССР наиболее обычны пестрые дятлы — большой, сред-
ний и малый, несколько реже встречаются черный, зеленый, трех-
палый и др.
Отряд Воробьинообразные (Passeriformes) птицы малой и
средней величины (см. рис. 260, XIII). Туловище стройное. Ноги
тонкие, с четырьмя пальцами, расположенными на одном уровне,
из которых три направлены вперед, а один — назад. Когти острые.
Плавательной перепонки нет. Клюв разной формы.
К отряду принадлежит более половины всех птиц. Сюда от-
носятся вороны, галки, грачи, сороки, скворцы, иволги, дрозды,
славки, пеночки, мухоловки, трясогузки, ласточки, жаворонки,
синицы, поползни, щеглы, чижи, зяблики, вьюрки, клесты, ов-
сянки и многие другие.
Образ жизни воробьиных разнообразен. Большинство из
1 1 Б. Л. Кузнецов и др.
321
них — дневные птицы, реже — сумеречные. Птенцовые птицы.
Многие воробьиные питаются насекомыми. Почти все зерноядные
виды выкармливают птенцов насекомыми. Поэтому большинство
воробьиных птиц очень полезны для сельского и лесного хозяй-
ства. Так, семья розовых скворцов за лето может уничтожить
свыше 100 кг саранчи. А самая маленькая птичка наших лесов —
королек истребляет за год до 4 млн лесных насекомых и их яиц.
На полях массу вредных насекомых поедают жаворонки, коньки,
скворцы и др. С огородных растений вредных насекомых склевы-
вают трясогузки, горихвостки, чеканы и пр. Для лесов и садов
особенно полезны различные синицы, корольки, пищухи, славки,
пеночки, дрозды, мухоловки, иволги.
Но некоторые воробьиные (например, воробьи) местами при-
носят вред, поедая семена культурных растений.
НАДОТРЯД БЕСКИЛЕВЫЕ ПТИЦЫ (RATITAE)
К этой группе птиц относятся страусы и близкие к ним формы,
утратившие способность к полету, что вызвало редукцию и изме-
нения ряда органов (рис. 263). Крылья короткие, недоразвитые,
у киви они почти исчезли. Киль на грудине не выражен (отсюда
наименование отряда). Грудные мышцы развиты относительно
слабо. Перья рыхлые, без сцепления бородок опахала в пла-
стинку. Ноги, ставшие единственным органом передвижения,
мощно развиты. Число пальцев ног в связи со способностью быст-
рого бега у большинства видов (кроме бескрыла) сократилось
до трех, а у африканского страуса — до двух.
Крупные африканские страусы, достигающие высоты 3 м, на-
селяют степи и пустыни Африки. Способны к очень быстрому бегу.
Самка откладывает в яму до 15 яиц, массой до 1,5 кг каждое.
Высиживают птенцов оба родителя. В Южной Африке страусов
разводят ради мяса, яиц и ценных перьев.
Более мелкие трехпалые страусы нанду живут в Южной Аме-
рике. Самец насиживает 20—35 яиц, отложенных 5—7 самками
его «гарема». Австралийский страус эму отличается сильной ре-
дукцией крыльев, тяжелым телосложением и относительно ко-
роткими ногами. Высота его до 1,8 м. Ведет моногамный образ
жизни. Высиживает яйца и выводит птенцов самец. В лесах Се-
верной Австралии и Новой Гвинеи обитают казуары, по форме
тела напоминающие эму, но пестрой окраски и с роговым шлемом
на голове. В Новой Зеландии водится самая малая из бескилевых
птиц — бескрыл, или киви, величиной с курицу.
НАДОТРЯД ПИНГВИНЫ (IMPENNES)
Небольшая группа своеобразных птиц, утративших способность
к полету и приспособленных к жизни в водах океана (рис. 263,
///). Тело вальковатое, вытянутое. Крылья превратились в ласты,
322
Рис. 263. Бескилевые птицы и пингвины;
/ - африканский страус; II бескрыл; III - пингвин
помогающие птице двигаться в воде. Развиты киль и грудные
мышцы.
В костях нет воздушной полости. Оперение плотное, стерж-
ни перьев уплощенные. На ластах перья имеют вид чешуек.
Ноги сдвинуты далеко назад, поэтому посадка птицы на земле
вертикальная. Пальцы ног соединены перепонкой.
Пингвины обитают в Антарктиде, у южных берегов Южной
Америки, Австралии и Африки. Живут колониями. Большую часть
времени проводят в море, по гнездятся на суше, устраивая гнезда
в виде ямки или поры. В кладке 1—2 яйца. Питаются рыбой и
другими морскими животными. Промыслового значения не имеют.
Известно около 20 видов пингвинов. Наиболее крупный
императорский пингвин — достигает в высоту 1 м.
11*
323
ПРОИСХОЖДЕНИЕ ПТИЦ
Несомненно, что птицы произошли от одной из древних групп
пресмыкающихся. Об этом свидетельствует наличие у них ряда
общих черт строения с рептилиями. Первые представители класса
Птиц появились в мезозойской эре. В юрских отложениях обна-
ружены остатки первоптицы археоптерикса (рис. 264), совме-
щающего признаки пресмыкающихся и птиц. Внешний вид его
напоминал птицу, но имевшийся длинный хвост состоял из многих
позвонков, по бокам располагались попарно крупные перья. Че-
люсти несли ряд мелких острых зубов. На крыльях сохранились
три длинных, свободно двигающихся пальца с когтями. На гру-
дине’ киля не было. Вероятно, археоптериксы жили па деревьях,
перепархивая с ветки на ветку. Пальцы передних конечностей
помогали при лазании по сучкам. Археоптерикс — боковая ветвь
в эволюции птиц.
В конце мезозойской эры появились настоящие птицы. Однако
у них еще сохранялись мелкие зубы, головной мозг был очень
мал. Беззубые птицы с роговым клювом возникли только в палео-
гене кайнозойской эры.
Рис. 264. Археоптерикс:
/ — отпечаток на сланце; // — реконструкция
324
КЛАСС МЛЕКОПИТАЮЩИЕ (MAMMALIA)
Общая характеристика. Млекопитающие теплокровные позво-
ночные животные из группы Amniota. Тело покрыто волосами
(у некоторых видов волосяной покров редуцирован). В коже есть
сальные и потовые железы. Головной мозг отличается сильным
развитием коры полушарий. Череп сочленяется с позвоночником
двумя мыщелками. В среднем ухе расположены три слуховые
косточки. Зубы различаются по форме и назначению, они сидят
в ячейках челюстей. Легкие альвеолярного строения. Сердце че-
тырехкамерное. Единственная дуга аорты загибается влево. Груд-
ная полость отделена от брюшной мускульной перегородкой
диафрагмой. Почки у взрослых тазовые. Как правило, живоро-
дящие животные, но имеются также яйцекладущие формы. Самки
выкармливают детенышей молоком, выделяемым молочными
железами.
Млекопитающие -- наиболее высокоорганизованная группа
позвоночных животных. Прогрессивные черты их организации
выражены во многих органах, но особенно наглядно: 1) в высоком
развитии центральной нервной системы и органов чувств; 2) в
сложной системе терморегуляции, определяющей относительное
постоянство температуры тела (у большинства млекопитаю-
щих 37--39 °С); 3) в приспособлении к живорождению и вы-
кармливанию детенышей матерью своим молоком, что создает
благоприятные условия для выживания потомства.
Высокая организация млекопитающих и сложная высшая
нервная деятельность обеспечили широкое их распространение.
Ныне на земном шаре насчитывается более 4 тыс. видов млеко-
питающих, из которых около 300 обитает на территории СССР.
Класс Млекопитающие делится на три подкласса: Яйцекла-
дущие (Protolheria), Сумчатые (Metatheria), Плацентарные
(Eutheria}.
Строение и жизненные отправления. Размеры и форма тела
млекопитающих разнообразны. Самый мелкий зверек белозубка-
крошка (отряд Насекомоядные) имеет длину всего лишь около
2 см, масса его примерно 2,5 г, тогда как синий кит достигает
33 м длины и массы 150 т. В теле млекопитающих можно разли-
чить голову, шею, туловище, хвост (иногда отсутствует), передние
и задние конечности (рис. 265).
Покровы. Кожа имеет особые трубчатые потовые и альвео-
лярные сальные железы (рис. 266). Первые несут выделительные
функции, и их секрет — пот — способствует охлаждению тела при
перегреве. Сальные железы выделяют жировую смазку, покры-
вающую волосы и поверхность кожи, предохраняя их от хими-
ческого воздействия среды и намокания. Но особенно характерны
для млекопитающих молочные железы, выделяющие молоко, ко-
торое служит пищей детенышам. Число сосков молочных желез
у различных млекопитающих колеблется от одной до десяти и
325
Рис. 265. Внешний вид млекопитающего (тигра)
Рис. 266. Строение кожи млеко-
питающего:
1 — роговой слой эпидермиса; 2 —
мальпигиев слой; 3 — дерма; 4 - по-
товая железа; 5 — ее устье; 6 — волос;
7—сальная железа; 8—кровеносные
сосуды; .9 — нервы; 10 — подкожные
жировые отложения
Рис. 267. Строение волоса (продольный разрез корня волоса):
1 — роговой слой эпидермиса; 2 -- его мальпигиев слой; 3 - дерма; 4 -- потовая железа;
,5 -- ее устье; 6 — волос; 7 - сальная железа; 8 — жировые отложения; 9 — сердцевина
волоса; 10 — корковое вещество волоса; II — стекловидная оболочка между влагали-
щем волоса и волосяным мешочком; 12— волосяной мешочек; 13— влагалище волоса;
14 — сосочек волоса; 15 — мышка волосяного мешочка
даже более пар. Обычно они расположены па груди и брюхе,
реже — на боках тела. Протоки молочных желез открываются на
вершине соска или в его углублении.
Тело большинства млекопитающих одето волосяным покро-
вом: лить у немногих он в той или иной степени подвергается
редукции. Волосяной покров играет большую роль в терморегу-
ляции млекопитающих, уменьшает испарение влаги с поверхности
тела, смягчает механические воздействия, обусловливает окраску
животного. Особенно сильно развит он у зверей зон умеренного
и полярного климата. Волосяной покров млекопитающих состоит
из огромного количества отдельных волос (например, у песца па
1 см~ кожи спины растет до 20 тыс. волос). Волос - нитевидное
роговое образование, которое формируется из клеток глубоких
слоев эпидермиса (рис. 267). Он состоит из стержня, выдающе-
гося из кожи, и корпя, погруженного в нее. Корень закапчивается
луковицей, размножение клеток которой обусловливает рост
волоса. Корень волоса находится в так называемом мешочке
волоса.
Мех большинства млекопитающих слагается из волос двух
категорий: более длинных, толстых и грубых - - остевых и нежных,
топких пуховых. Густой слой пуховых волос определяет в ос-
новном теплозащитные свойства волосяного покрова, а остевые
защищают пуховые от вытирания и свойлачивают. На голове,
лапах и других местах расположены особо длинные упругие во-
лосы - вибриссы, корпи которых окружены нервными оконча-
ниями. Они играют роль органов осязания.
Для млекопитающих зон умеренного и холодного климата
характерны сезонные смены волосяного покрова — линьки.
Осенью редкий и низкий, летний волосяной покров сменяется гу-
стым и высоким зимним мехом, а весной происходит смена зим-
него покрова на летний. Нередко зимний и летний волосяные по-
кровы отличаются и окраской: так, например, зимний мех зайца-
беляка белый, а летний — буровато-серый. У тропических мле-
копитающих волосы сменяются постепенно, без выраженной се-
зонности.
Конечные фаланги пальцев почти у всех млекопитающих не-
сут роговые образования - когти, ногти, или копыта, которые
являются производными эпидермиса (рис. 268). По строению
эти роговые образования сходны, но отличаются степенью раз-
вития отдельных частей. Все они состоят из прочной роговой пла-
стинки, подошвенной пластинки и подушечки пальца. Когти име-
ют толстую выпуклую и изогнутую роговую пластинку, которая
выдается вперед острым выступом; подошвенная пластинка хо-
рошо развита, подушечки пальцев большие и мягкие. У ногтей
роговая пластинка высокая, относительно тонкая, подошвенная
пластинка недоразвита, подушечка пальцев большая, мягкая.
У копыт толстая роговая пластинка охватывает конец фаланги
пальца; подошвенная пластинка утолщенная, твердая.
327
Рис. 268. Продольный разрез конечной фаланги пальцев:
1 — обезьяны; 11 — собаки; 111 — лошади;
/1 — ноготь; Б коготь; Б .- копыто;
1 - подушечка пальца; 2 подошвенная пластинка; 3— ногтевая пластинка;
4 — ногтевой валик; 5 -- конечная фаланга пальца
У многих млекопитающих развиваются рога, в образовании
которых участвуют покровы.
Скелет взрослых млекопитающих образован костными эле-
ментами (рис. 269). Хорда хорошо развита только на ранних ста-
диях эмбрионального развития. Позвоночник состоит из позвон-
ков, обычно с плоскими сочленовными поверхностями. Между
ними лежат хрящевые прослойки с остатками хорды. Шейных
позвонков, как правило, семь. Первый из них — атлант — имеет
вид кольца с двумя сочленовными поверхностями для сочленения
с мыщелками черепа. Второй — эпистрофей — имеет направлен-
ный вперед отросток, который входит в кольцо атланта. Подобное
Рис. 269. Скелет кролика:
/ — шейные позвонки; 2— грудные позвонки; 3— поясничные позвонки; 4 --крестец;
5 — хвостовые позвонки; 6 — ребра; 7 — грудина; 8— лопатка; .9 - ее акромиальный
отросток; 10 - коракоидиый отросток; 11—13 — таз; /-/ — отверстие таза; 15 пле-
чевая кость; 16 — локтевая кость; 17—лучевая кость; 18— запястье; 19 — пясть; 20 -
бедренная кость; 21 — коленная чашечка; 22— большая берцовая кость; 2?---малая
берцовая кость; 24 — пяточная кость; 25 — таранная кость; 26 - плюсна
328
соединение позвонков обеспечивает подвижность головы. Груд-
ных позвонков от 12 до 15. К ним причленяются хорошо развитые
ребра, большая часть которых нижними хрящевыми концами
срастается с грудиной, образуя грудную клетку. Поясничных
позвонков от 2 до 9. Они несут рудиментарные ребра; крестцо-
вых 3 4, они срастаются в единую крестцовую кость, что
обеспечивает прочную основу для прикрепления костей таза.
Число хвостовых позвонков различно.
Череп млекопитающих отличается большой мозговой короб-
кой. С позвоночником он сочленяется двумя мыщелками. Нижняя
челюсть образована одной зубной костью, которая причленяется
непосредственно к височной области черепа.
У предков млекопитающих - пресмыкающихся нижняя че-
люсть соединяется с черепом квадратной костью; у млекопитаю-
щих эта кость превращается в одну из слуховых косточек сред-
него уха - наковальню. Задний угол нижней челюсти рептилий
образован отдельной сочленовной костью, которая у млекопитаю-
щих становится другой слуховой косточкой — молоточком.
Третья слуховая косточка млекопитающих — стремечко пред-
ставляет собой измененную верхнюю часть подъязычной дуги
низших позвоночных.
Плечевой пояс скелета млекопитающих, как правило, состоит
из двух нар костей: лопаток и ключиц. Коракоидные кости почти
у всех млекопитающих рудиментарны и еще в процессе эмбрио-
нального развития прирастают к лопаткам, образуя коракоидные
отростки. Лишь у утконосов п ехидн коракоидные кости сохра-
няют самостоятельность. Тазовый пояс млекопитающих образо-
ван тремя парами костей: седалищными, лобковыми и под-
вздошными.
Млекопитающие имеют пятипалые конечности обычного для
наземных позвоночных типа. Но у быстро бегающих форм число
пальцев сокращается (у лОшади до одного на каждой ноге). Раз-
личают млекопитающих стопоходящих — опирающихся при хож-
дении па всю стопу (медведи), полустопоходящих, у которых с
землей соприкасается только передняя часть стопы (многие гры-
зуны и хищники), и пальцеходящих, опирающихся только на кон-
цы пальцев (копытные).
Мускул а т у р а млекопитающих достигает большой слож-
ности и образована многочисленными отдельными мышцами.
Скелетная мышца образована поперечнополосатой мышечной
тканью. Во внутренних органах заложены гладкие мускульные
волокна.
И с р в и а я с и с т е м а отличается высокой степенью слож-
ности. Головной мозг большого размера: особенно велики полу-
шария переднего мозга и мозжечок, что соответствует сложности
высшей нервной деятельности и разнообразию движений млеко-
питающих (рис. 270). Кора полушарий мозга имеет сложное
строение. У многих кора полушарий образует сложную систему
329
Рис. 270. Мозг млекопитающих:
/ - кролика; II шимпанзе;
/ — полушария переднего мозга; 2 --
обонятельные доли; 3 - эпифиз; 4.сред-
ний мозг; 5 мозжечок; 6......... продолгова-
тый мозг; 7 - извилины полушарий
борозд и извилин, что значи-
тельно увеличивает ее поверх-
ность. Головных нервов 12 нар.
Органы чувств млекопитающих развиты хорошо.
Органы обоняния играют в жизни млекопитающих большую
роль; у большинства хорошее чутье позволяет находить пищу,
опознавать особей своего вида, избегать врагов. Поэтому у зве-
рей, как правило, имеются большие обонятельные полости, по-
верхность которых значительно увеличена наличием складок —
обонятельных раковин.
Органы слуха обычно также хорошо развиты, в их состав вхо-
дят наружное, среднее и внутреннее ухо (рис. 271). Наружное
ухо образовано ушной раковиной и наружным ушным проходом.
Барабанная перепонка, лежащая в глубине этого прохода, отде-
ляет наружное ухо от среднего. В полости среднего уха распо-
ложены три слуховые косточки. С барабанной перепонкой связана
первая из них — молоточек, с которым подвижно соединена на-
ковальня, сочлененная с третьей косточкой — стремечком, упи-
Рис. 271. Схема строения органа слуха и равновесия млекопитающих:
1 — овальный мешочек; 2 — полукружный капал; 3 ~~ эндолимфатический проток;
4 - круглый мешочек; 5 — улитка; 6— перилимфатическая полость; 7 круглое окно;
8 — полость среднего уха; 9 -- евстахиева труба; 10 — стремечко; 11 - наковальня;
12—молоточек; 13— барабанная перепонка; 14 —наружный слуховой проход; 15 —
слуховое отверстие; 16 — ушная раковина; //--каменистая кость; 18 -чешуйчатая
кость
330
Рис. 272. Схема строения глаза млекопитаю-
щего:
1 - зрительная ось; 2 - роговина; .7 радужи-
на; -/ ресничный мускул; 5 - хрусталик; 6
стекловидное тело; 7 сенника; Л' волнистая
складка, .9 ши меитная оболочка; 10 соеу-
дипая оболочка; 11 склера; 12 желтое пят-
но; 111 зршельпый нерв
рающимся в перепонку овального ок-
на внутреннего уха. Эти косточки слу-
жат для передачи звукового раздра-
жения от наружного уха к внутрен-
нему. Звуковое раздражение воспри-
нимается кортиевым органом внутреннего уха, образованным
тысячами тончайших волокон, натянутых в канале улитки уха
и начинающих вибрировать при определенной длине звуковой
волны.
Глаза млекопитающих имеют в основном типичное для орга-
нов зрения позвоночных строение (рис. 272). Для них характерны
выпуклая роговица, линзообразный хрусталик, наличие век. Зре-
ние у зверей развито обычно слабее, чем у птиц.
Полость тела млекопитающих отличается тем, что она
разделена эластичной мускульной перегородкой — диафраг-
мой - на два отдела: грудной и брюшной. В грудной полости
находятся сердце и легкие, а в брюшной —- желудок, кишечник,
печень, почки и ряд других органов (рис. 273).
О р г а п ы и и щ е в а р с п и я. Пищеварительный тракт млеко-
питающих начинается ротовой полостью. На верхней и нижней
челюстях подавляющего большинства зверей расположены зубы,
сидящие в альвеолах. Характерна дифференцировка зубов на
резцы, клыки, передпекоренные и собственно коренные (рис. 274).
Резцы чаще имеют долотообразную, а клыки — коническую
форму..
Коренные зубы у хищников обычно уплощены с боков и несут
ряды острых бугорков, а у растительноядных форм они имеют
уплощенную поверхность со складками эмали или тупыми бугор-
ками, что облегчает перетирание жесткой пищи.
Число зубов у млекопитающих обозначают зубной формулой.
Обычно записывают их количество в верхней и нижней челюстях.
Резцы обозначают буквой I (iticisivi), клыки ---с (catuni), перед-
некоренные — pm (praeinolares) и заднекоренные т (molares).
Наибольшее число зубов у плацентарных млекопитающих, кроме
зубатых китов,— 44 (у свиней, кротов и немногих других). Зуб-
ная формула этих животных такова:
i с Ь pm т — -Ц-• 2 = 44.
3 1'4 ,3 11
331
Знание полной зубной формулы помогает разобраться в осо-
бенностях зубного состава у других животных. Так, например,
у коровы зубная формула такова:
Из этой формулы видно, что у коровы произошли редукция
резцов и клыков верхней челюсти и сокращение числа передне-
коренных зубов.
В молодом возрасте звери обычно имеют молочные зубы, ко-
торые в дальнейшем сменяются постоянными. В число молочных
Рис. 273. Внутреннее строение кролика:
/ — пищевод; 2 — желудок; 3 . печень; 4 — поджелудочная железа; 5 — тонкая кишка;
6 — слепая кишка; 7— червеобразный отросток; 8 — толстая кишка; 9 — прямая кишка;
10 — анус; 11 — селезенка; 12 — трахея; 13 — легкие; 14 — сердце; 15 -- аорта; 16—18 -
подключичная артерия; 19 — наружная яремная вена, 20— задняя полая вена; 21-
диафрагма; 22— почки; 23 — мочевой пузырь; 24 яичники; 25 -- яйцеводы; 26 -мат-
ка; 27 — влагалище; 28— мочеполовой синус; 29 -- мочевое отверстие; 30—желчный
пузырь
332
Рис. 274. Зубная система различных млеко
питающих:
/ насекомоядного (землеройки); // хищного
(собаки); /// - непарнокопытного (лошади);
/V зайцеобразного (кролика)
1 резцы; 2 - клыки; .7 - предки репные и ко-
ренные зубы
Рис. 275. Строение желудка различных
млекопитающих:
/ собаки; // - крысы; III - мыши; IV - нерб-
люда; I' короны; / III - простой желудок;
/Г, Г сложный желудок;
/ рубец; 2 сетка; 3 книжка; •/ сычуг
зубов входят резцы, клыки и переднекоренные; задпекореппые
развиваются позднее и смене не подлежат. Некоторые млекопи-
тающие в силу специфики питания утратили зубы, как, например,
питающиеся мелкими насекомыми муравьеды или фильтрующие
планктон из воды беззубые киты.
В ротовую полость млекопитающих открываются протоки
слюнных желез, секрет которых способствует перевариванию уг-
леводов пищи, превращению крахмала в сахар.
Ротовая полость переходит в глотку, за которой следует пи-
щевод, открывающийся в желудок. У большинства млекопитаю-
щих желудок однокамерный (рис. 275). В стенках его заложены
железы, которые выделяют желудочный сок, содержащий фер-
мент пепсин и соляную кислоту, воздействующие прежде всего па
белки пищи. Сложное строение имеет желудок жвачных (коров,
овец, коз и др.). Он разделен на четыре отдела: рубец, сетку, книж-
ку и сычуг. Три первых отдела желудка жвачных являются видо-
измененными частями пищевода, а собственно желудком следует
считать только сычуг. Столь сложное строение желудка жвачных
связано с особенностями их пищеварения — наличием вторичного
пережевывания пищи — жвачки.
ззз
Из желудка пища продвигается в кишечник. У хищных и на-
секомоядных млекопитающих он относительно короткий (у лету-
чих мышей он лишь в 2—3 раза длиннее тела), у травоядных он
может достигать большой длины (у коровы он примерно в 20 раз
превышает длину тела). Кишечник делится на длинный тонкий
и короткий толстый отделы. У многих форм от места их соедине-
ния отходит слепая кишка, размер которой изменчив.
У большинства млекопитающих анальное отверстие отделено
от мочеполового промежностью. Лишь у утконосов и ехидн име-
ется клоака.
В пищеварении зверей большую роль играют желчь, образую-
щаяся в большой печени, а также секреты поджелудочной железы
и железок стенок кишечника.
Органы дыхания млекопитающих состоят из дыхатель-
ного горла—трахеи, бронхов и легких. Трахея начинается гор-
танью, открывающейся в глотку. В ее стенках заложен ряд хря-
щей, движение которых препятствует попаданию пищи в трахею
при заглатывании. Внутренняя оболочка гортани имеет голосовые
связки, участвующие в образовании звуков, издаваемых живот-
ным. В стенке трахей заложены неполные хрящевые кольца, не
позволяющие ей спадаться. В грудной полости трахея разделя-
ется на два бронха, идущих к легким.
Рис. 276. Строение легочных альвеол
(справа вскрытая альвеола е сетью капил-
ляров)
Рис. 277. Схема кровеносной системы мле-
копитающих:
/ — внешняя сонная артерия; 2 — внутренняя сон-
ная артерия; 3 —- подключичная артерия; 4
дуга аорты; 5 — легочная артерия; 6'— левое
предсердие; 7—правое предсердие; S — левый
желудочек сердца; 9 — правый желудочек серд-
ца; 10 — спинная аорта; 11 — внутренностная
артерия; 12—почечная артерия; 13 — подвздош-
ная артерия; 14 — яремная вена; 15 — подключичная вена; 16 —- левая непарная вена;
17 — правая непарная вена; 13 — задняя полая вена; 19 — печеночная вена; 20 — ворот-
ная вена печени; 21 -- печень; 22 — почки; 23 — повздошная вена
334
Легкие млекопитающих имеют альвеолярное строение. Бронхи
в легких распадаются на множество все более утончающихся
разветвлений, которые переходят в тончайшие трубочки — брон-
хиолы, заканчивающиеся микроскопически малыми пузырь-
ками — альвеолами (рис. 276). Число альвеол в легких исчисля-
ется миллионами, что обусловливает их большую дыхательную
поверхность. Вдох и выдох осуществляются вследствие измене-
ния объема грудной клетки, вызываемого как поднятием и опуска-
нием ребер, так и движением диафрагмы.
Кровеносная система млекопитающих характеризу-
ется следующими основными признаками (рис. 277). Сердце че-
тырехкамерпос: оно состоит из двух предсердий и двух желудоч-
ков, поэтому смешения артериальной и венозной крови не проис-
ходит. Имеются большой и малый круги кровообращения. От
левого желудочка сердца, как и у птиц, отходит только одна дуга
аорты, по она загибается не на правую, а па левую сторону (ле-
вая дуга аорты). Дуга аорты продолжается в спинную аорту, тя-
нущуюся назад вдоль позвоночника. От дуги аорты отходят ар-
терии, несущие артериальную кровь к различным органам тела.
Венозная кровь, оттекающая от заднего и среднего отделов
тела животного, попадает по венам в мощную заднюю полую
вену, тянущуюся вперед вдоль позвоночника. Кровь из передней
части тела и головы у некоторых млекопитающих собирается в
левую и правую передние полые вены, которые, проходя к сердцу,
сливаются друг с другом. Ио большинство зверей имеют непар-
ную короткую переднюю полую вену, в которую вливаются все
вены, несущие кровь от передней части тела. Вены, несущие кровь
от кишечника, образуют воротную вену печени, которая, войдя
в печень, распадается па сеть капилляров воротной системы пе-
чени. От печени кровь отводится печеночной веной, впадающей
в заднюю полую вену.
Малый круг кровообращения состоит из легочных артерий,
несущих венозную кровь от правого желудочка сердца к легким,
из сети капилляров, обволакивающих альвеолы легких, и из ле-
гочных вен, по которым уже окисленная кровь возвращается в
сердце (в левое предсердие).
Органами выделения у млекопитающих служат тазо-
вые почки, лежащие в поясничной области. Обычно они имеют
бобовидную форму. Каждая из почек состоит из поверхностного
коркового и внутреннего мозгового слоев. Корковый слой состоит
из массы тончайших трубочек, начинающихся округлыми полыми
расширениями -- боуменовыми капсулами. В них вдаются клу-
бочки капилляров, через стенки которых происходит выделение
из крови излишков воды и продуктов распада. Мозговой слой
состоит из огромного количества топких собирательных каналь-
цев, в которые впадают трубочки коркового слоя. Эти канальцы
открываются в полость почки — почечную лоханку, куда и посту-
пает выводимая моча. Из почечных лоханок моча стекает по мо-
335
источникам в мочевой пузырь, а из него периодически выводится
наружу.
Органы в н у т р е н н ей секреции играют важную роль
в жизни млекопитающих, регулируя важнейшие физиологические
процессы. Особое положение среди желез внутренней секреции
занимает гипофиз, связанный с нижней поверхностью промежу-
точного мозга. Эпифиз расположен на верхней поверхности про-
межуточного мозга.
Щитовидная железа имеет обычно вид двухлопастного тела,
лежащего перед дыхательным горлом. Выделяемые ею гормоны
(в первую очередь тироксин) участвуют в регулировании про-
цессов обмена веществ, роста и дифференцировки тканей. Над-
почечники— овальные или бобовидные тельца, лежащие близ
почек, состоят из двух слоев: наружного —• коркового и внутрен-
него — мозгового. Гормоны коркового слоя регулируют водный,
минеральный и, главное, углеводный обмен животного, а также
влияют на функцию половых органов.
В теле млекопитающего имеются и другие железы внутрен-
ней секреции. Гормоны выделяются также половыми железами,
поджелудочной железой и некоторыми другими органами.
Органы размножения представлены у самцов парными
семенниками, расположенными либо в полости тела, либо в особой
складке кожи — мошонке. Сперма выводится из семенников на-
ружу по семяпроводам через копулятивный орган. Семенные пу-
зырьки и предстательная железа выделяют секрет, который обра-
зует жидкую часть спермы и активизирует сперматозоиды. Ор-
Рис. 278. Схема строения яйцеводов и матки различных млекопитающих:
/ --- клоачные; //, III - сумчатые; IV — плацентарные с двойной магкой; К — с двурогой;
VI — с простой маткой;
1 — яйцеводы; 2 — матка; 3 — влагалище; 4 — мочеполовое отверстие; 5 -- мочевой пу-
зырь; 6 — клоака; 7 — прямая кишка
336
гапы размножения самок состоят из парных яичников, лежащих
в полости тела, яйцеводов с фаллопиевыми трубами, матки и
влагалища. Строение матки у разных млекопитающих неодина-
ковое. У одних имеются две матки, открывающиеся во влагалище
отдельными отверстиями — двойная матка (кролики). У других
тоже имеются две матки, но они открываются во влагалище
одним общим отверстием — такая матка называется двураздель-
ной (свиньи, многие грызуны). У ряда копытных парные матки
слиты нижними отделами, а верхние части остаются свободными
(двурогая матка). Наконец, у многих млекопитающих произошло
полное слияние обеих маток в единый полый орган — простую
матку (обезьяны, летучие мыши, человек, рис. 278).
Оплодотворение яиц происходит в верхних отделах половых
путей самки. Все млекопитающие (за исключением утконоса и
ехидны, откладывающих яйца) живородящие. У сумчатых дете-
ныши рождаются слаборазвитыми и дальнейшее развитие их про-
исходит в сумке на животе матери, У остальных млекопитающих
эмбрионы связаны со стенками матки плацентой, через которую
получают от материнского организма кислород и питательные
вещества.
Экология млекопитающих. Общий прогресс организации мле-
копитающих позволил им заселить практически всю Землю и в
высшей степени приспособиться к разнообразной жизненной об-
становке. В ходе эволюции млекопитающие приобрели большое
разнообразие форм, которые по характеру связи со средой могут
быть объединены в несколько экологических групп.
Наземные млекопитающие — наиболее многочисленная груп-
па, объединяющая лесных зверей и обитателей открытых про-
странств (ежи, зайцы, волки, лисицы, тигры, антилопы, олени,
бараны, жирафы и др.). Они имеют стройное тело, хорошо раз-
витую шею, часто длинные ноги, сокращенное число пальцев.
Многих характеризует быстрота передвижения, необходимая при
добывании пищи и спасении их от врагов. У видов, не способных
к быстрому бегу, свои приспособления. Так, у ежей произошло
частичное видоизменение волосяного покрова в иглы, служащие
защитой.
Древесные млекопитающие обнаруживают тесную связь с
древесной растительностью. К этой группе относятся белки-
летяги, сони, ленивцы, большинство обезьян. Они приспособлены
к лазанию по деревьям с помощью длинных цепких конечностей
и хвоста. Пальцы лап вооружены удлиненными, иногда изогну-
тыми когтями. Многие древесные млекопитающие способны со-
вершать прыжки с дерева на дерево, а некоторые — планировать
в воздухе благодаря наличию кожных складок по бокам тела
между передними и задними конечностями (летяги).
Летающие млекопитающие представлены летучими мышами.
Они приспособлены к полету — основному способу их передви-
жения. Во время полета большинство видов добывает себе пи-
12 Б. Л. Кузнецов и др.
щу — разнообразных насекомых. Их крылья представляют собой
кожистые летательные перепонки, натянутые между сильно вытя-
нутыми пальцами передних конечностей, передними и задними
конечностями и хвостом.
Норные млекопитающие используют почву для рытья нор.
Норы служат им жилищем, где они размножаются и спасаются
от врагов. Кормятся они на поверхности земли. Таковы кролики,
суслики, сурки, хомяки, барсуки. Они имеют мешковидное тело,
укороченные конечности с развитыми тупыми когтями.
Подземные млекопитающие хорошо приспособлены к подзем-
ному роющему образу жизни. В эту группу входят кроты, сле-
пыши, цокоры, слепушонки. Вся жизнь этих млекопитающих свя-
зана с почвой, где они размножаются, прячутся от врагов и до-
бывают себе пищу. На поверхности появляются редко. Туловище
у них вальковатой формы, шея выражена слабо, ноги короткие,
хвост сильно редуцирован. Волосяной покров низкий, без ворса.
Ушных раковин нет. Глаза маленькие, у слепышей они скрыты
под кожей. Рытье ходов осуществляется с помощью передних
конечностей (кроты, цокоры) или выступающих изо рта крупных
резцов (слепыши, слепушонки).
Полу водные млекопитающие значительную часть жизни про-
водят в воде, но при этом сохраняют тесную связь с сушей (выдра,
норка, выхухоль, ондатра, бобр и др.). Хорошо плавают и ныряют.
Имеют короткие конечности, пальцы часто соединены плаватель-
ной перепонкой. Хвост обычно уплощен в горизонтальной пло-
скости или сжат с боков. Ушные раковины маленькие.
Водные млекопитающие имеют еще большую, чем предыдущая
группа, связь с водной средой. Одни из них значительную часть
жизни проводят в воде, но для размножения и линьки выходят
на сушу или на лед (ластоногие). Их конечности превращены
в ласты, причем у большинства видов задние ласты не принимают
участия в движении по твердому субстрату, а служат локомотор-
ным органом при плавании и нырянии. Ушные раковины у многих
видов отсутствуют. Волосяной покров в той или иной степени
редуцирован. Имеется слой подкожного жира. Другие предста-
вители данной группы полностью утратили способность выходить
на сушу (китообразные). Тело у них рыбообразной формы, за-
канчивается хвостовым плавником. Шея короткая и голова сли-
вается с туловищем. Передние конечности превратились в ласты,
а задние полностью редуцировались. Ушных раковин нет. Воло-
сяной покров редуцирован. Термоизоляционную роль выполняет
толстый слой подкожного жира.
Все названные экологические группы млекопитающих связаны
между собой рядами переходных форм.
Питание. Состав пищи млекопитающих крайне разнообразен.
Многие из них используют многочисленные растительные корма
(грызуны, зайцеобразные, копытные, обезьяны). Среди зверей
немало и плотоядных видов, которые приспособились либо к пое-
338
данию в основном беспозвоночных животных (насекомоядные,
рукокрылые, беззубые киты), либо к добыванию различных поз-
воночных животных (хищные, ластоногие, зубатые киты). Однако
большинство млекопитающих используют как растительные, так
и животные корма, долевое значение которых часто меняется в
зависимости от места обитания, от сезона года и т. д.
Годовой цикл жизни млекопитающих состоит из нескольких
этанов, отражающих сезонные изменения во взаимоотношениях
организма и условий его существования и отличающихся своеоб-
разием поведения зверей и их приуроченностью к определенным
местам обитания,
Подготовка к размножению сопровождается у моногамных
видов разбивкой на пары, а у полигамных (тюлени, котики, оле-
ни) — формированием гаремов и косяков, драками между сам-
цами. Многие звери меняют места обитания.
В период размножения млекопитающие обычно держатся в
малодоступных местах и ведут оседлый образ жизни. Спаривание
у разных зверей происходит в различные сроки: зимой (собачьи),
весной (грызуны, насекомоядные), летом (куньи), осенью (копыт-
ные, рукокрылые). Деторождение чаще приурочено к концу вес-
ны — началу лета, т, е. к наиболее благоприятному для выращи-
вания потомства времени года. Продолжительность беременности
варьирует от 2—4 недель (мыши, полевки) до'2 лет (слоны). Дли-
тельность беременности у домашних животных, дней: у осли-
цы— 380 (360—390); верблюдицы — 365 (335—395); кобылы —
340 (307—412); буйволицы — 307 (300—315); коровы — 285
(240—311); козы, овцы— 150 (146—160); свиньи — 114 (110—
140); собаки —62 (59—65); кошки — 58 (55—60); крольчихи —
30 (28—33). Многие грызуны при благоприятных условиях могут
размножаться в течение всего года, давая за год 3—4 помета.
У крупных зверей (белый медведь, тигр, слон) детеныш рожда-
ется 1 раз в 3 года. В конце этого периода у многих млекопитаю-
щих происходят распадение семей и расселение молодняка (на-
секомоядные, многие грызуны). Животные переселяются в места,
хорошо обеспеченные кормом.
В период подготовки к зиме млекопитающие интенсивно пи-
таются, накапливают жировые запасы, линяют, меняя короткий
и редкий летний мех на длинный и густой зимний. Млекопитаю-
щие запасают на зиму корм, особенно грызуны. Так, белки заго-
тавливают орехи, желуди, шишки, сушат шляпки грибов, засовы-
вая их в развилки между ветвями. Зимние запасы желтогорлой
мыши, состоящие в основном из желудей и орехов лещины, рас-
полагаются в дуплах и могут достигать 10 кг. Бобры делают
запасы веточного корма, складывая его в воду вблизи жилища.
В этот период некоторые млекопитающие (мыши, полевки, многие
копытные, волки) объединяются в большие группы, что облегчает
им зимнее существование, миграции и добывание пищи. Многие
звери (китообразные, ластоногие, летучие мыши, копытные) со-
12*
339
вершают далекие миграции, которые имеют сезонный характер,
ежегодно повторяются и проходят по определенным маршрутам.
Однако в целом миграции у млекопитающих не получили такого
большого распространения, как у рыб или птиц.
Зимовка. В этот период происходит ухудшение условий добы-
вания млекопитающими необходимого количества корма. Одним
из приспособлений для переживания этого неблагоприятного
времени года, получившим широкое распространение у млекопи-
тающих, является спячка. Спячка характерна для многих зверей,
но степень ее глубины у разных видов неодинакова. Так, у медве-
дей, енотовидных собак, барсуков она принимает вид зимнего
сна, при котором у неподвижного животного уровень обмена
веществ, температура тела и частота дыхания меняются мало.
Такого зверя очень легко разбудить, после чего он может больше
не уснуть. У медведей в период зимнего сна происходит рождение
детенышей. В настоящую зимнюю спячку с сильным оцепенением,
резким снижением температуры тела и частоты дыхания впадают
суслики, сурки, сони, ежи и многие летучие мыши.
Численность многих видов млекопитающих колеблется по
годам, что связано с изменением кормовых условий, заболева-
ниями, количеством врагов, погодными явлениями и другими
факторами. Так, численность белки в кормные и голодные годы
резко меняется. Периодически повторяющиеся неурожаи семян
хвойных деревьев, от которых зависит кормовое благополучие
белки, обусловливают снижение ее численности, что вызывается
гибелью зверьков от голода и уменьшением числа детенышей.
Хорошо известно резкое увеличение численности мышей и полевок
в результате их массового размножения в так называемые «мыши-
ные годы».
Прогнозирование массового появления вредных грызу-
нов имеет большое значение для сельского хозяйства, позволяет
своевременно и эффективно вести с ними борьбу.
Хозяйственное значение млекопитающих. Велико и многооб-
разно значение млекопитающих в народном хозяйстве нашей
страны. К этому классу принадлежат более 60 % домашних жи-
вотных (крупный и мелкий рогатый скот, свиньи, верблюды, ло-
шади, кролики, собаки, кошки и др.), а также все объекты пуш-
ного звероводства (серебристо-черные лисицы, голубые песцы,
норки, соболи, нутрии и др.). Некоторые виды млекопитающих
разводят для лабораторных целей (мыши, крысы, морские свинки
и др.).
Многие млекопитающие служат объектами промысла, обеспе-
чивая совместно с клеточным разведением пушных зверей пот-
ребности страны в пушнине. По добыче пушнины СССР занимает
первое место. Основные промысловые виды нашей фауны: белка,
соболь, ондатра, лисица, песец и др. При промысле диких копыт-
ных (северные олени, лоси, сайгаки, кабаны) используются мясо
и шкура. Продуктами морского зверобойного промысла служат
340
шкуры животных, представляющие собой хорошее кожевенное п
меховое сырье, жир и другие ценные продукты.
Большой вред народному хозяйству Советского Союза при-
носят многие виды грызунов, уничтожающие и повреждающие
различные культурные, полезные дикорастущие растения и раз-
ные виды сельскохозяйственной продукции. Различные виды сус-
ликов, мышей и полевок вредят полеводству, поедая высеянные
в почву семена, подгрызая побеги, уничтожая большое количе-
ство зерна. Плодовым насаждениям иногда немалый вред причи-
няют сони, водяные и обыкновенные полевки, зайцы, а овощным
культурам — крысы, полевки, мыши и хомяки. Крысы и мыши
уничтожают и портят большое количество пищевых продуктов
на складах и в домах. Волки местами наносят ущерб животно-
водству, нападая на сельскохозяйственных животных.
Оценивая отрицательную роль млекопитающих, нельзя забы-
вать о том, что многие звери, особенно грызуны, являются пере-
носчиками и хозяевами возбудителей опасных болезней человека
и домашних животных.
ПОДКЛАСС ЯЙЦЕКЛАДУЩИЕ (PROTOTHERIA)
Древняя группа примитивных млекопитающих, представленная
в современной фауне только несколькими видами, населяющими
Австралию, Новую Гвинею и Тасманию (рис. 279).
В отличие от других млекопитающих размножаются, откла-
дывая яйца, которые либо высиживают (утконосы), либо вынаши-
вают в сумке, образованной складкой кожи брюха (ехидны).
Рис. 279. Яйцекладущие млекопитающие:
1 — ехидна; 11 утконос; 11! - проехпдпа
341
Выкармливают детенышей молокообразной жидкостью, которая
выделяется многочисленными примитивными молочными желе-
зами, открывающимися на железистых полях покровов брюшка.
Сосков нет, и детеныши слизывают капельки секрета желез, сте-
кающие по волосам. Трубчатое строение молочных желез яйце-
кладущих указывает на их происхождение от потовых желез
кожи.
У взрослых яйцекладущих зубы отсутствуют и морда одета
роговым чехлом—клювом. Наличие клоаки — признак, сохра-
нившийся от рептильных предков этих зверей. Кора полушарий
головного мозга развита слабо. Коракоидная кость плечевого
пояса сохраняет самостоятельность. Терморегуляция недоста-
точно совершенна, поэтому температура тела колеблется от 25
до 37 °C.
Известны три рода современных яйцекладущих млекопитаю-
щих: утконосы, ехидны и проехидны. Тело утконосов покрыто
густым мягким мехом. Характерен широкий, похожий на утиный
клюв. Пальцы ног соединены плавательной перепонкой. Утконосы
живут в норах по берегам рек, хорошо плавая и ныряя. Питаются
мелкими водными животными. Ехидны иглистым покровом спины
и боков тела несколько напоминают ежей. Клюв короткий, кони-
ческий. Самка откладывает лишь одно яйцо, которое вынаши-
вает в складке кожи брюха. Питаются насекомыми. Проехидна
отличается от ехидны длинным тонким клювом.
Яйцекладущие — боковая ветвь, рано отделившаяся от основ-
ного ствола родословного древа млекопитающих.
ПОДКЛАСС СУМЧАТЫЕ (METATHERIA)
Сумчатые — древняя группа млекопитающих (рис. 280). Оби-
тают в Австралии и на прилегающих островах; небольшое число
видов встречается в Северной, Центральной и Южной Америке.
Плацента отсутствует или слабо развита, в связи с чем детеныши
рождаются после короткого срока эмбриогенеза слабо развитыми.
У подавляющего большинства сумчатых дальнейшее развитие
детенышей протекает в сумке, представляющей собой складку
кожи брюха самки (рис. 281). Молоко впрыскивается в рот дете-
нышам сокращением особых мышц молочных желез. Зубы обычно
слабо дифференцированы. Задняя кишка отделена от мочеполо-
вых протоков.
Сумчатые обитают в различных угодьях: лесах, степи, горах.
Среди них встречаются лазающие, бегающие, норные и подзем-
ноживущие виды, есть хищные, насекомоядные, растительнояд-
ные и всеядные формы. У сумчатых Австралии и плацентарных
млекопитающих других континентов, обитающих в сходных усло-
виях и ведущих сходный образ жизни, нередко наблюдается кон-
вергенция — внешнее сходство по ряду признаков различных по
происхождению и систематическому положению животных.
342
Рис. 280. Сумчатые млекопитающие:
/ — опоссум; II -- сумчатая белка; /// - - гигант-
ский кенгуру с детенышем в сумке; /Г - - сумчатый
медведь коала; |/- бандикут; 17 сумчатый
крот; VII - сумчатый волк
Рис. 281. Вскрытая сумка кенгуру с дете-
нышем
Из сумчатых широко известны различные виды кенгуру, кото-
рые передвигаются прыжками на сильно развитых задних ногах;
укороченные передние конечности служат для захвата пищи.
Хищный сумчатый волк, ныне почти истребленный, внешне напо-
минает крупную собаку. На эвкалиптовых деревьях живет медли-
тельный сумчатый медведь коала, питающийся листьями. Сум-
чатые куницы, сумчатые белки и сумчатые летяги ведут древесный
образ жизни. В почве роют сложные и глубокие норы слепые сум-
чатые кроты. В Америке встречаются своеобразные сумчатые —
опоссумы.
Многие сумчатые дают ценную пушнину. Мясо кенгуру хоро-
шего качества.
В палеогене сумчатые были широко распространены по мате-
рикам земного шара, но позднее почти везде (кроме Австралии
и Америки) были вытеснены более высокоорганизованными пла-
центарными.
ПОДКЛАСС ПЛАЦЕНТАРНЫЕ (EUTHERIA)
К данному подклассу относятся большинство современных мле-
копитающих. У плацентарных питательные вещества и кислород
поступают в организм зародыша из тела матери через особый
временный орган — плаценту, которая образуется путем соеди-
нения хориона со стенкой матки. Хорион — губчатое тело
(рис. 282), возникает в результате срастания внешней стенки
аллантоиса с наружной оболочкой плода — серозой. От хориона
в глубь утолщенной стенки матки врастают многочисленные тон-
кие выросты — ворсинки, богатые кровеносными сосудами и ка-
пиллярами. Сложная сеть последних приходит в тесное соседство
с капиллярами и кровеносными лакунами утолщенной стенки мат-
ки матери, что позволяет питательным веществам и кислороду ос-
мотически поступать из крови матери в кровь эмбриона. Из пла-
центы они переносятся по кровеносным сосудам пупочного кана-
тика в тело зародыша. Другие сосуды канатика, несущие кровь
от зародыша к плаценте, выносят продукты диссимиляции эм-
бриона.
Различают несколько типов плаценты: диффузная, когда вор-
синки распределяются по всей поверхности хориона равномерно
(китообразные, многие копытные); дольчатая (котиледонная),
когда ворсинки собраны на отдельных участках хориона в виде
пятен (большинство жвачных); зональная (кольцевая, когда
сосочки располагаются широким поясом (некоторые хищные,
слоны); дискоидальная, когда ворсинки собраны на одном резко
ограниченном участке хориона, имеющем форму диска (грызуны,
обезьяны, человек). Плацента может быть отпадающей и неот-
падающей. В первом случае, в связи с тем что ворсинки хориона
прочно соединяются со слизистой оболочкой матки, отторжение
плаценты при родах сопровождается отпадением части стенки
344
Рис. 282. Развитие зародышевых
оболочек и плаценты у плацен-
тарных млекопитающих (схема):
/ --/// - последовательные стадии раз-
вития;
1 впезародышевая полость; 2 - ал-
лантоис; 3 - амнион; •/ желточный
пузырь; 5 зародыш; б — плацента
матки и кровотечением (хищ-
ные, грызуны, обезьяны). У
свиней, китообразных, вер-
блюдов, лошадей, многих
жвачных — неотпадающая
плацента, так как при родах
ворсинки хориона выходят
из углублений слизистой
оболочки матки, не повреж-
дая ее, без кровотечения.
Наличие связи через плаценту с организмом матери позволяет
эмбриону относительно долго оставаться в матке самки и дости-
гать в ней значительно большей стадии развития, чем зародышам
сумчатых. Новорожденные плацентарные способны самостоя-
тельно сосать молоко из молочных желез матери, имеющих хо-
рошо развитые соски.
Головной мозг плацентарных имеет более сложное строение,
чем мозг сумчатых. Зубы, как правило, хороню дифференциро-
ваны на резцы, клыки и коренные. Клоаки пет. Коракоидная кость
стала отростком лопатки.
Подкласс Плацентарные включает ряд отрядов.
Отряд Насекомоядные (Insectivora) — небольшие зверьки с
относительно примитивными признаками. Их остробугорчатые
зубы слабо дифференцированы, или деление их па группы имеет
другой характер, чем у прочих плацентарных. Головной мозг мал,
полушария имеют гладкую поверхность. Внешний вид насекомо-
ядных разнообразен, но для всех характерен вытянутый подвиж-
ной хоботок на конце морды (рис. 283).
В нашей стране отряд Насекомоядные представлен различ-
ными видами ежей, землероек, кротов, а также выхухолью. Ежи
и землеройки приносят пользу, истребляя вредных насекомых.
Кроты уничтожают почвенных животных (личинок майских жу-
ков), но в то же время они питаются полезными дождевыми чер-
вями. Кроме того, их роющая деятельность приносит вред лесным
посадкам, огородам и лугам. Шкурки выхухолей и кротов исполь-
зуются как пушное сырье. Выхухоль охраняется законом.
Отряд Рукокрылые, или Летучие мыши (Chiroptera). Эти
шерьки приобрели способность активного полета. Передние ко-
нечности у них видоизменились в крылья, несущей поверхностью
345
Рис. 283. Представители различных отрядов плацентарных млекопитающих:
о с ряс) Насекомоядные: I -- выхухоль; // - крот; отряс) Летучие мыши: II/ крылан;
отряд Хищные: IV -- соболь; Г - рысь; 17 — бурый медведь; осрчд Лсиюиоеие:
17/— тюлень; отряс) Китообразные: VIII -- дельфин; IX - скелет гренландского
кита
которых служит тонкая кожистая летательная перепонка, натя-
нутая между очень длинными пальцами передних ног и между
передними и задними ногами; у многих летучих мышей эта пере-
понка соединяет также задние ноги и хвост. Грудные мышцы,
двигающие крылья, прикрепляются к грудине, на которой имеется
небольшой киль. Первый палец передних конечностей и кисть
задней остаются свободными, что помогает зверьку при лазании
и цеплянии (рис. 283,///).
346
Летучие мыши — ночные животные. День они проводят па
чердаках. На зиму некоторые виды улетают на юг, другие впадают
в спячку.
Полету в темноте летучим мышам помогает своеобразная зву-
ковая локация. В полете они издают прерывистые звуки высокой
частоты, которые, распространяясь, отражаются от находящихся
на пути зверька предметов и воспринимаются его слуховым ап-
паратом. Поэтому летучие мыши даже в полной темноте избе-
гают во время полета столкновения со встречными предметами.
Летучие мыши относятся к числу полезнейших животных —
они истребляют огромное количество вредных ночных насекомых.
Поэтому рекомендуется устанавливать в садах специальные до-
мики для их привлечения.
Отряд Грызуны (Rodentia) — наиболее богатый видами отряд
млекопитающих. Это мелкие или средней величины наземные или
реже полуводные растительноядные млекопитающие различного
внешнего вида (рис. 284). Общим признаком всех грызунов яв-
ляется своеобразное строение зубной системы, приспособленной
Рис. 284. Представители отряда грызунов:
/ — белка; // — тушканчик; /// — нутрия; /V'— серая полевка; К—
лесная мышь; VI — заяц-русак {отр. Зайцеобразные)
347
к разгрызанию и разжевыванию твердой растительной пищи.
В верхней и нижней челюстях имеется по одной паре больших
долотообразных, лишенных корней и постоянно растущих, резцов.
Стачиваясь на концах, они постоянно отрастают. Твердая эмаль
покрывает только переднюю поверхность резцов, поэтому, стачи-
ваясь, они всегда остаются острыми. Клыки у грызунов отсут-
ствуют. Резцы отделены от коренных зубов широким промежут-
ком— диастемой. Коренные зубы приспособлены к перетиранию
жесткой растительной нищи. Они имеют широкую жевательную
поверхность, обычно плоскую или несущую тупые бугорки или
гребни. Как и резцы эти зубы часто не имеют корней и растут
в течение всей жизни зверька. Сочленовные поверхности нижней
челюсти устроены так, что последняя может несколько сдвигаться
вперед и назад. Когда зверек грызет пищу, он выдвигает нижнюю
челюсть немного вперед, так что концы резцов верхней и нижней
челюстей сходятся; но коренные зубы при этом не соприкаса-
ются. Когда же грызун приступает к перетиранию отделенного
куска пищи, нижняя челюсть сдвигается несколько назад, что вле-
чет соприкосновение жевательных поверхностей коренных зубов
и расхождение концов резцов. Большинство грызунов имеют силь-
но развитую слепую кишку, в которой проходят процессы сбра-
живания пищи. Полушария головного мозга обычно гладкие,
без извилин. Пальцы несут когти. Грызунам свойственны раннее
половое созревание и высокая плодовитость.
Грызуны имеют большое хозяйственное значение. Многие из
них являются серьезными вредителями сельского и лесного хо-
зяйств. Ряд грызунов служат переносчиками и вирусоносителями
некоторых опасных заболеваний человека (чумы, туляремии). Но
многие дают ценную пушнину и потому служат важным объектом
охотничьего промысла (белки, бобры, ондатры, нутрии и др.).
Из грызунов — вредителей сельского и лесного хозяйств наи-
большее значение имеют следующие.
Суслики (род Citellus, сем. Беличьи—Sciuridae) — зверьки
величиной с белку, с вальковатым телом, короткими ногами и
обычно небольшим хвостом. Населяют преимущественно лесо-
степные, степные и пустынные районы. Только своеобразный
длиннохвостый суслик обитает в Восточной Сибири на севере
вплоть до Арктики, селясь на открытых местах среди тайги. Сус-
лики живут в норах, часто колониями. На зиму впадают в спячку.
Начинают размножаться в годовом возрасте, весной. Самки при-
носят 4—10 детенышей. Питаются различными растениями, нано-
сят огромный вред посевам. Являются носителями ряда заболе-
ваний, опасных для человека (в первую очередь чумы).
Сони (сем. Myoxidae} внешне напоминают белок, но значи-
тельно меньше их. Живут в дуплах деревьев и гнездах, которые
сооружают на деревьях и кустах. Преимущественно ночные жи-
вотные. На зиму залегают в длительную спячку (откуда назва-
ние). Крупные виды сонь — соня-полчок и садовая соня — причи-
348
няют местами вред садоводству, уничтожая плоды и ягоды.
В СССР наибольший убыток от этих грызунов терпят сады Кав-
каза.
Тушканчики (сем. Dipodidae) —своеобразные грызуны, пере-
двигающиеся прыжками на длинных задних ногах; передние ко-
нечности у них сильно укорочены и служат преимущественно
для держания пищи. Ряд видов населяет степную и пустынную
зоны СССР. Некоторые из них вредят бахчевым культурам.
Хомяки (род Cricetus, сем. Хомяковые— Cricelidae) —зверь-
ки среднего размера (величиной с большую крысу) с мешкова-
тым телом, короткими лапами и небольшим хвостом. Окраска
верха ржаво-серая, низа — черноватая, по бокам тела располо-
жены большие белые пятна. Хомяки широко распространены на
юге нашей страны, населяя поля, огороды, заросли бурьяна, ов-
раги, перелески, поймы рек. Живут в норах, обычно поодиночке.
Самки приносят за год 1—2 помета из 3—18 детенышей. Местами
сильно вредят посевам зерновых и огородам. На зиму делают
большие запасы зерна и корнеплодов.
Хомячки (род Cricetulus, сем. Хомяковые — Cricelidae) по
форме тела похожи на хомяков, по значительно мельче их (обычно
с крупную мышь). Окраска различная - серая, бурая, рыжева-
тая, песочная, часто с черной полоской по спине. Встречаются
в степных, лесостепных и пустынных зонах СССР, где часто се-
лятся на полях, бахчах, плантациях, в огородах, нанося большой
вред посевам и культурам. По образу жизни похожи на хомяков.
Песчанки (роды Rhombomys и Meriones, сем. Хомяковые —
Cricelidae). Большая группа грызунов, внешне похожих на-крыс
или крупных мышей, но с хвостом, густо покрытым волосами,
образующими на конце кисточку (у^крыс одет кольцами роговых
чешуек и редкими отдельными волосками). Обитатели пустынь.
Живут как среди песков, так и на участках с твердым грунтом,
охотно селятся на культурных землях оазисов, где причиняют
большой вред посевам. Опасные передатчики чумы.
Полевки (подсем. Microtiuae, сем. Хомяковые — Cricelidae)
похожи на мышей, но отличаются более коротким хвостом, длина
которого, как правило, короче ’/г длины тела, и широкой, тупой
мордочкой. Зубы с плоской жевательной поверхностью и глубо-
кими боковыми складками эмали. Живут в неглубоких порках.
За год самки приносят несколько пометов из 3 —10 детенышей.
На зиму в спячку не впадают. Наиболее распространена обыкно-
венная полевка (Microtus arvalis), которая обитает преимуще-
ственно на полях, лугах и в огородах. Она питается зелеными
частями растений, зерном и корнеплодами, местами принося боль-
шой вред растениеводству.
Рыжие полевки (род. С let brio ио ту s, сем. Хомяковые -- С/7-
celidae) похожи на обыкновенную полевку, но отличаются рыже-
ватой окраской спины. Обитатели леса. Вредят уничтожением
семян и коры древесных пород, повреждением лесных посадок.
349
Мыши (сем. Мышиные — Миridae). В нашей стране встре-
чается много видов мышей. Наиболее обычны домовая мышь
(Mus tnusculus) грязно-серого цвета, с относительно коротким
хвостом; лесная мышь (Apodemus sylvaticiis) коричневатая и
чисто-белая снизу, с длинным тонким хвостом; полевая мышь
(Apodemus agrarius) с коричневато-рыжей спинкой, вдоль кото-
рой тянется черная полоска, и мышь-малютка (Mlcromus minu-
tus), отличающаяся малыми размерами. Домовая мышь в север-
ных районах живет преимущественно в домах, питаясь продук-
тами. Но на юге этот вид часто живет и на полях, принося большой
вред посевам. Полевая и лесная мыши держатся в природных
угодьях и местами также вредят сельскохозяйственным куль-
турам.
Крысы (род Radius, сем. Мышиные — Muridae) отличаются
от мышей более крупными размерами. В СССР обитает несколько
видов, из которых наиболее распространена серая крыса пасюк.
Крысы наносят большой вред, уничтожая и повреждая продо-
вольственные продукты. Все они являются носителями и распро-
странителями возбудителей некоторых опасных инфекционных
болезней для людей.
В Советском Союзе проводятся широкие работы по истреб-
лению вредных грызунов с использованием авиации, сложных
химикатов и аппаратуры. Наибольшее распространение имеют
химические способы истребления этих вредителей. Но нужно пом-
нить, что химические меры борьбы с грызунами требуют значи-
тельных затрат и часто, особенно при нарушениях инструкции их
использования, влекут гибель многих полезных животных. По-
этому они по возможности должны заменяться механическими,
биологическим и бактериальным методами истребления грызунов-
вредителей.
В настоящее время широкое применение из химических мер
борьбы с этими животными приобрел приманочный метод. Он
заключается в раскладке в местах обитания грызунов съедобных
приманок, заранее отравленных каким-либо ядохимикатом. В ка-
честве приманок используют зерна злаков, куски корнеплодов,
хлеба, кусочки дынь и тыкв и другие привлекательные для гры-
зунов продукты.
В борьбе с сусликами, хомяками, водяными полевками и не-
которыми другими грызунами огромную роль играет отлов их
капканами для получения шкурок и технического жира. Промысел
этих зверьков дает ежегодно несколько миллионов шкурок, ис-
пользуемых меховой промышленностью для изготовления раз-
личных меховых изделий, и в то же время в значительной сте-
пени способствует истреблению важных вредителей растение-
водства.
В борьбе с мышами и крысами, а также против некоторых
других грызунов иногда используют приманки, зараженные бак-
териями мышиного и крысиного тифа. Этот бактериальный метод
350
особенно рационален в условиях человеческих поселений, где
применение многих химических препаратов недопустимо.
Отряд Зайцеобразные (Lagomorpha). К этому отряду отно-
сятся зайцы, кролики и небольшие зверьки — пищухи. Зубная
система этих животных напоминает таковую грызунов, по в верх-
ней челюсти за передней парой больших долотообразных рез-
цов находятся еще маленькие столбиковидные резцы (вторая
пара). Дикие кролики встречаются на юге Украины. Домашних
кроликов разводят ради мяса, шкурки и пуха. Зайцы служат пред-
метом охоты. Местами зайцы наносят вред плодовым деревьям,
объедая зимой кору.
Отряд Хищные (Carnivora) объединяет наземных и полувод-
пых млекопитающих, питающихся в основном различными поз-
воночными животными; некоторые из них всеядны (медведь).
Зубы четко дифференцированы па резцы, клыки и коренные. Рез-
цы довольно мелкие. Клыки, напротив, большие, конические,
острые. Коренные зубы обычно остробугорчатые. Почти у всех
хищных последний переднекоренной зуб верхней челюсти и пер-
вый заднекорешюй пижней челюсти выделяются большими раз-
мерами --- эти зубы называются хищными. Число коренных зу-
бов обычно сокращено.
Из ряда семейств отряда наибольшее значение имеют семей-
ства: Псовые, Медвежьи, Куньи, Кошачьи.
Семейство Псовые (Сипidae) включает домашних собак, вол-
ков, шакалов, лисиц, песцов п некоторые другие виды. Эти звери
среднего размера, с уплощенным с боков телом, высокими ногами,
удлиненной мордой и пышно опушенным хвостом. Пальцеходя-
щие животные. На передних ногах по пять, а па задних — по че-
тыре пальца. Когти невтяжные, тупые. Хищники. Добывают пищу
преимущественно преследованием. Ведут (кроме времени раз-
множения) бродячий образ жизни. Размножаются 1 раз в году —
весной.
Семейство Медвежьи (Ursidae) объединяет крупных стопохо-
дящих зверей с массивным туловищем на мощных, но относи-
тельно коротких лапах, с очень коротким, спрятанным в мехе
хвостом. Когти невтяжные. Медведи — всеядные животные. На зи-
му (кроме самцов белых медведей) впадают в спячку. В СССР, в
Арктике, обитает белый медведь, в лесной зоне — бурый медведь
и на Дальнем Востоке -- черный медведь.
Семейство Куньи (Mustelidae) включает большое число жи-
вотных: соболей, куниц, горностаев, ласок, хорьков, норок, выдр,
барсуков и др. Это звери средней или малой величины различного
телосложения. Стопоходящие или полустопоходящис. Когти пе-
втяжные, но острые. Большинство -- настоящие хищники, но есть
и всеядные виды (барсук). Размножаются весной. В спячку впа-
дает только барсук.
Семейство Кошачьи (Felidae), кроме домашней кошки, вклю-
чает львов, тигров, леопардов, рысей, различные виды диких ко-
351
тек. Кошачьи обладают гибким вытянутым телом с округлой
головой. Пальцеходящие звери. Лапы (кроме гепарда) с очень
острыми втяжными когтями. Типичные хищники, добывающие
своих жертв путем подкарауливания.
Многие хищные млекопитающие нашей фауны (соболь, ку-
ница, норка, горностай, лисица, песец и др.) принадлежат к числу
ценных путных зверей, промысел которых дает высококачествен-
ную пушнину. Некоторые из них (серебристо-черная лисица, го-
лубой песец, соболь, норка) разводятся в звероводческих хозяй-
ствах. Ряд видов хищных (степной хорек, ласка, горностай) по-
лезны истреблением вредных грызунов. Так, семья степных хорь-
ков за лето уничтожает до 300 сусликов, а одна ласка — до 2 тыс.
мышей и полевок. Некоторые хищники, напротив, вредны для
сельского хозяйства. Например, волки там, где их численность
еще высока, режут много скота и диких копытных. К числу вред-
ных хищников относятся также шакалы, истребляющие мелкий
скот, домашнюю и пернатую дичь. Некоторые хищники являются
носителями вируса бешенства, опасного для человека и домашних
животных.
Отряд Ластоногие (Pinnipedia). Отряд объединяет тюленей,
моржей, сивучей, морских котиков и некоторых других живот-
ных. Жизнь этих животных тесно связана с водной средой, что
накладывает отпечаток на их организацию. Тело ластоногих вы-
тянутое, торпедообразное. Конечности имеют вид ластов. У тю-
леней задние ласты направлены всегда назад и не участвуют в
передвижении животных по суше или льду. У моржей, сивучей и
морских котиков они могут подгибаться вперед и помогают жи-
вотным двигаться по твердому субстрату. Волосяной покров мор-
ских котиков густой, с плотным пухом. У тюленей он редкий и
жесткий, состоящий у взрослых только из грубых остевых волос.
У моржей на коже имеются только редко расположенные оди-
ночные грубые волосы. Ластоногие имеют толстый слой подкож-
ного жира, который сохраняет внутреннее тепло тела.
Промысел тюленей дает значительное количество технического
жира, кожевенных и меховых шкур. Моржей добывают ради мяса,
жира и огромных клыков (моржовая кость), идущих на косте-
резные изделия. Шкуры молодых морских котиков ценятся как
высококачественное меховое сырье. Морские котики в пределах
СССР размножаются на островах Дальнего Востока, где и произ-
водятся их охрана и забой.
Отряд Китообразные (Cetacea). Отряд включает китов и дель-
финов. Организация этих животных носит явные признаки при-
способления к жизни в водной среде. Тело торпедообразной фор-
мы, шея не выражена, передние конечности имеют вид ластов,
задние атрофированы, на конце туловища имеется горизонталь-
ный плавник, служащий основным органом движения животного.
Кожа голая, под ней лежит толстый жировой слой, играющий
роль теплоизолирующей прослойки. Легкие отличаются большой
352
вместимостью. Так, синий кит, ныряя, может захватить в легкие
до 14 тыс. л воздуха. Большой объем легких позволяет китооб-
разным долго находиться под водой и нырять на большую глу-
бину. Например, кашалоты могут пребывать под водой, не обнов-
ляя воздуха в легких, более часа, погружаясь в глубь океана
до 1 тыс. м.
Китообразные делятся на два подотряда: Зубатые и Беззубые
киты. У зубатых китов (Odontoceti) имеются конусовидные зубы,
помогающие добывать рыбу и головоногих моллюсков, которыми
они питаются. У беззубых китов (Mystacoceti) зубов нет, но во рту
имеется своеобразный цедильный аппарат из роговых пластин
(китовый ус), несущих по нижнему краю бахрому из волокон.
Пластины свисают с наружных краев неба. Забрав в пасть воду,
киты процеживают ее обратно через ряды пластин китового уса,
и находящиеся в воде мелкие животные задерживаются бахро-
мой пластин и заглатываются. К зубатым китам относятся огром-
ные кашалоты, крупные белухи и касатки и небольшие дельфины.
К беззубым китам принадлежат гренландский кит и различные
виды китов-полосатиков. Синий кит, также относящийся к подот-
ряду Беззубых китов, достигает в длину 33 м — это самое круп-
ное животное мира.
Добыча всех видов китов в настоящее время запрещена.
Отряд Парнопалые (Artiodactyla) — растительноядные или
всеядные животные. Сюда относятся свиньи, бегемоты, дикие
быки, горные бараны и козлы, антилопы, олени, жирафы и многие
другие (рис. 285). Для всех представителей характерно то, что
конечные фаланги пальцев одеты роговыми чехлами -- копытами.
Как на передних, так и па задних ногах наибольшее развитие
получили III и IV пальцы, между которыми проходит ось тела.
I палец атрофирован, а II и V короче двух средних или совсем
недоразвиты. Конечности двигаются в основном в плоскости, па-
раллельной оси тела. В связи с этим ключицы отсутствуют. Ве-
дут наземный образ жизни, лишь бегемоты тесно связаны с водо-
емами. Многие способны к быстрому бегу.
Парнопалые делятся на два подотряда: Нежвачные и Жвачные.
Подотряд Нежвачные (Noriruniitiatiiia) включает свиней и бе-
гемотов. Это животные с массивным грузным телом, короткой
толстой шеей, невысокими ногами и коротким хвостом. На ногах
по четыре пальца, из которых два средних развиты сильнее бо-
ковых. Лишь у южноамериканских пекари на задних ногах только
три пальца. Кожа толстая, с хорошо развитым слоем подкожного
жира. Зубная формула полная:
рш^ щ | = 2 = 44.
Коренные зубы многобугорчатые, клыки сильно развиты. Же-
лудок однокамерный. У самок на брюхе два ряда сосков. Пла-
цента диффузная.
353
К семейству Свиньи относятся кабаны, бородавочники, кис-
теухие свиньи и др. Представители этого семейства отличаются
массивным, сжатым с боков телом, короткой шеей, удлиненной
конической головой с хрящеватым «пятачком» на конце, корот-
кими сильными ногами. Это всеядные животные. Пища переже-
вывается во рту и лишь после этого поступает по пищеводу в желу-
док (жвачка не образуется).
Домашние свиньи произошли от дикого кабана (Sus scrofa),
широко распространенного в Европе и Азии. От дикого предка они
отличаются редукцией щетинного покрова, большой массой, высо-
кой мясной и сальной продуктивностью. Свиньи очень плодовиты.
Беременность маток длится около 4 мес.
К семейству Бегемоты принадлежат только два вида: беге-
мот обыкновенный и бегемот карликовый. Это крупные живот-
354
ные, массой до 3 т, с грузным цилиндрическим телом, короткими
толстыми ногами и огромной уплощенной головой. Кожа голая.
Живут в водоемах Африки. Питаются водной и прибрежной
растительностью.
Подотряд Жвачные (Ruminantia) включает дикие и одомаш-
ненные формы животных. Из представителей подотряда надо
отметить домашний крупный и мелкий рогатый скот, а из диких
зверей — зубров, бизонов, буйволов, яков, горных баранов и коз-
лов, антилоп, оленей, жирафов.
Жвачные характеризуются прежде всего своеобразным про-
цессом пищеварения — наличием жвачки. Грубо пережеванная
пища сначала попадает в первый отдел сложного желудка — ру-
бец, где под влиянием слюны и деятельности микроорганизмов
подвергается брожению. Из рубца пища перемещается во второй
отдел желудка — сетку с ячеистым строением стенок. Отсюда она
отрыгивается обратно в ротовую полость, где подвергается
размельчению зубами и обильно смачивается слюной. Образо-
вавшаяся полужидкая масса вновь заглатывается и попадает в
третий отдел желудка — книжку, стенки которой образуют парал-
лельные складки — листочки. Здесь пища несколько обезвожива-
ется и переходит в последний отдел желудка — сычуг, где подвер-
гается воздействию желудочного сока.
Для жвачных характерно отсутствие резцов в верхней че-
люсти; их функционально заменяет твердый поперечный валик.
Зубная формула у большинства видов такова:
i". с-2-1; рт~; //Ц==±Г1.2=32—34.
На коренных зубах имеются складки эмали лунчатой формы.
Кишечник жвачных очень длинный. Молочные железы образуют
вымя, расположенное в паху самки, с 2—4 сосками. У большин-
ства видов на лобных костях черепа самцов (а иногда и самок)
сидят рога различной формы и строения. Обычно это стройные
животные, способные к быстрому бегу. II и V пальцы у них зача-
точные или полностью редуцировались. Пястные кости III и IV
пальцев на передних конечностях и плюсневые на задних сращены
в массивные кости, что вместе с частичной редукцией одной
из костей предплечья и голени придает конечностям стерж-
невидное строение — признак, выработавшийся как приспособ-
ление к бегу (так же как и сокращение числа пальцев).
В фауне СССР подотряд Жвачные представлен двумя семей-
ствами: Бычьи и Оленьи. У представителей семейства Бычьи (Во-
uidae) самцы, а иногда и самки имеют рога, образованные ко-
ническими (прямыми или изогнутыми) костными выростами лоб-
ных костей черепа, одетыми роговыми чехлами. Почти у всех
видов (кроме американского вилорога) они не подвержены еже-
годной смене. Клыков в верхней челюсти нет.
Из диких животных фауны нашей страны к этому семейству
355
относятся зубры, горные козлы и бараны, сайгаки, джейраны,
дзерены, серны и горалы. Могучие дикие быки — зубры прежде
были широко распространены в лесах Европы, но позднее были
почти полностью истреблены. В настоящее время удалось их
вновь размножить, и ныне стада зубров пасутся в ряде заповед-
ников.
Несколько видов диких горных козлов обитают в пределах
СССР на Кавказе, в горах Средней Азии и на Алтае. Они насе-
ляют высокогорную зону, держась по скалам и на альпийских
лугах. Пасутся обычно небольшими табунами. В Советском Союзе
обитают два вида диких баранов: один из них — горный баран
(Ovis аттоп) встречается в горах и предгорьях Южной Сибири,
Средней Азии и Закавказья, акклиматизирован в Крыму. Он на-
селяет высокогорные степи (сырты), увалы предгорьев, горные
останцы среди степи; другой — толсторог (Ovis canadensis), от-
личающийся от горного барана толстыми рогами, обитает в горах
северных районов Дальнего Востока, Якутии и Таймыра. Оба
вида — ценные охотничьи животные. В степях Нижнего По-
волжья и Казахстана ныне бродят огромные стада сайгаков
(Saiga tatarica), которые еще 50 лет назад были здесь весьма
редкими животными. Теперь они служат объектом интенсивного
промысла. В пустынях Средней Азии обитает стройная газель —
джейран (Gazella gutturosa). В связи с резким сокращением чис-
ленности внесен в Красную книгу СССР.
Разводимый человеком крупный рогатый скот произошел от
широко распространенного в Европе и Азии тура (Bos taurus),
истребленного уже в историческое время. В Закавказье разводят
также буйволов, которые отличаются от крупного рогатого скота
почти голой кожей и огромными полулунными рогами. Эти жи-
вотные — одомашненная форма дикого индийского буйвола
(Bubalus агпее). В горах Памира и Алтая можно встретить стада
одомашенных быков — яков (Bos mutus). Наши домашние овцы
ведут свою родословную от диких горных баранов (Ovis аттоп),
а козы — от своеобразного дикого безоарового козла (Capra
aegagrus), и сейчас встречающегося в горах Закавказья и Пе-
редней Азии (рис. 286).
Виды семейства Оленьи (Cervidae) характеризуются тем,
что их самцы, а у северного оленя и самки носят на голове ветви-
стые костные рога, сменяемые ежегодно. Из диких представите-
лей этого семейства в СССР водятся лоси, северные, благородные
и пятнистые олени и косули. В северных районах страны и на юге
Сибири разводят одомашненных северных оленей, которых ис-
пользуют как транспортных животных, от них получают мясо,
молоко, меховые и кожевенные шкуры. На юге Дальнего Востока
и Алтае разводят пятнистых оленей и маралов (разновидность
благородного оленя) для получения пантов — молодых рогов,
отрастающих после ежегодной смены и еще не успевших окосте-
неть. Из пантов изготовляют ценное лекарство — пантокрин.
356
Рис. 286. Предки сельскохозяйственных млекопитающих:
I тур вымерший предок рогатого скота (реконструкция); II м\ф.иои; ///
архар; IV сгонной баран предок домашних овец; И - Пезоаровый козел пре-
док ко I
Отряд Мозоленогие (Tylopoda). Этот отряд объединяет вер-
блюдов (Camelus) и лам (Lama). Для представителей отряда
характерно отсутствие I, II и V пальцев, на длинных стройных
ногах лишь ио два пальца. Животное опирается не па концы фа-
ланг пальцев, как нарноналые, а на все суставы пальцев. Снизу
ног имеются большие эластичные мозолистые подушки, что обес-
печивает передвижение верблюдов в песках. На концах пальцев
выдаются небольшие тупые роговые выросты. Зубов 32-34. Зуб-
ная формула следующая:
/•Ь щ^12±.2«32 34.
В верхней челюсти имеется с каждой стороны но одному клы-
коподобному резцу. Предкоренные и коренные с лунчатыми склад-
ками эмали. Желудок трехкамерный, распадается па рубец, сетку
и сычуг. Вымя с двумя сосками.
Верблюды отличаются крупными размерами и наличием на
спине одного или двух горбов. В Африке, на Ближнем Востоке п
357
в Туркмении разводят одногорбых верблюдов — дромадеров, а в
Центральной, Средней и Южной Азии — двугорбых верблюдов.
Дикие двугорбые верблюды в небольшом числе встречаются еще
в настоящее время. Верблюдов используют как транспортных
животных, приспособленных к жизни в пустыне, от них получают
шерсть, молоко, мясо.
Ламы отличаются более мелкими размерами и отсутствием
горбов. Домашние ламы разводятся в Южной Америке как вьюч-
ные животные и продуценты шерсти и молока.
Отряд Непарнопалые (Perissodactyla). Это крупные расти-'
тельноядные животные. Конечные фаланги пальцев покрыты ко-
пытами. Ось конечностей проходит по среднему пальцу, который
Рис. 287. Непарнопалые млекопитающие:
/ — чепрачный тапир; // — индийский носорог; III — зебра
358
развит сильнее остальных. Желудок однокамерный. Ключицы от-
сутствуют.
Отряд делится на три семейства: Лошадиные, Тапировыс и
Носороговые (рис. 287).
Семейство Лошадиные (Equidae) объединяет, помимо домаш-
них лошадей и ослов, дикую лошадь Пржевальского, сохранив-
шуюся в Монголии, африканских зебр и ряд видов диких ослов.
У представителей этого семействе! на каждой ноге хорошо развит
и одет копытом только средний (III) палец, тогда как II и IV реду-
цированы и превратились в тонкие, лежащие под кожей грифель-
ные косточки; I и V пальцы полностью атрофированы. Зубная
формула такова:
Зубы, постоянно растущие с плоскими жующими поверхно-
стями, имеют сложнораскладчатую коронку. Желудок однокамер-
ный. Слепая кишка велика.
Рис. 288. Происхождение
дошили:
А 1 армии - предок совре-
менных лошадей; Ь’ изме-
нение величины п строения
черепа в ряду предков ло-
шади (/ -югипнус; II
протогиппус; /// - мерпгнп-
пус); В- изменение перед-
них конечностей в эволю-
ционном ряду предков ло-
шади (/--5 -- нумерация
пальцев)
Б
359
Длина кишечника в 12 раз больше длины тела. В паху распо-
ложены два соска.
Домашние лошади произошли от вымершей дикой лошади —
тарпана (Equus caballus), прежде широко распространенного
в Евразии (рис. 288). В СССР сохранились (в Туркмении и Ка-
захстане) куланы, совмещающие признаки лошадей и ослов.
Ослы отличаются от лошадей узкими копытами, длинными ушами
и короткой шерстью на хвосте, кроме концевой кисти удлинен-
ных волос. Домашний осел произошел от дикого африканского
осла.
В сельском хозяйстве ряда стран используют в качестве уп-
ряжных и верховых животных помесей осла и кобылы — му-
лов и помесей жеребца и ослицы — лошаков. И те и другие бес-
плодны.
Тапиры (сем. Tapiridae) отличаются тем, что у них на передних
ногах по четыре, а на задних —- по три пальца с небольшими ко-
пытцами. Морда с коротким подвижным хоботком. Ряд ви-
дов тапиров населяет леса Южной Америки и Юго-Восточной
Азии.
Носороги (сем. Rhinoceratidae) —- гигантские трехпалые звери
с толстой, почти голой кожей. На морде расположены один или
два рога из слитых роговых волокон. Длина этих рогов иногда
достигает 1,5 м. Обитают в Африке и Южной Азии.
Отряд Хоботные (Proboscidea) включает африканских и ази-
атских слонов. Для этих гигантских животных характерны длин-
ный подвижной хобот, образованный удлиненным носом и верх-
ней губой, огромные резцы верхней челюсти, выступающие изо
рта в виде пары бивней, клыков нет. Ноги пятипалые, каждый
палец одет копытцем. Кожа почти голая. Коренные зубы функ-
ционируют посменно. Азиатских слонов приручают и используют
в сельскохозяйственных и транспортных работах. На севере Си-
бири, в вечной мерзлоте, иногда находят туши вымерших сло-
нов — мамонтов, живших в Евразии в ледниковое время.
Отряд Обезьяны (Primates). Для обезьян характерно мощ-
ное развитие полушарий головного мозга, кора которых образует
сложную систему извилин и борозд (рис. 289). Это связано с вы-
соким уровнем их высшей нервной деятельности. Конечности
хватательного типа с противопоставлением большого пальца
остальным, что облегчает охват ветвей деревьев при лазании.
Пальцы конечностей несут плоские ногти. Одна пара молочных
желез расположена на груди. Зубная формула обезьян' Старого
света такова:
4 4 рт1' 4=г2==32-
Обезьяны обитают в тропических и субтропических странах
Старого и Нового света.
360
Рпс. 289. Отряд полуобезьян: I — долгопят; отряд обезьян:
11 - черный ревун; III - павиан-бабуин; IV шимпанзе
Особенно интересны обитающие на юге Азии и в Африке че-
ловекообразные обезьяны •— гориллы, шимпанзе, орангутаны,
гиббоны.
ПРОИСХОЖДЕНИЕ МЛЕКОПИТАЮЩИХ
Предполагают, что млекопитающие произошли от какой-то при-
митивной малоспециализированной группы палеозойских пре-
смыкающихся. Среди рептилий того времени было несколько
групп, носивших отдельные признаки, характерные для млеко-
питающих (рис. 290).
Первые млекопитающие появились, по-видимому, в триасо-
вом периоде. В отложениях юрского периода найдены остатки
своеобразных животных, получивших наименование трехбугорча-
тых (Trituberculata). Это были небольшие зверьки, по ряду приз-
наков близкие к насекомоядным млекопитающим. По-видимому,
Ани дали начало сумчатым и плацентарным млекопитающим.
Происхождение яйцекладущих млекопитающих, сохранивших по-
361
Человекообразные
Узконосые
Человек
I Спбакопбрпзные
I Кишкообразные
ХЛаапенпгив
'Зубатые киты
Хищные
Обезьяны
fНепарноко-
пытные у
Насекомоядные \ Первичные
< \ хищные
Первичные
копытные
Рукокрылые
[рызуны -
Первичные плацентарные
Сумчатые /
Клоачные /
Первичные млекопитающие
Рис. 290. Родословное древо млекопитающих
(схема)
ныне ряд признаков пресмыкающихся, до сих пор не совсем ясно.
Предполагают, что они ведут родословную от древних многобу-
горчатых (Multitube г culata).
РОДОСЛОВНОЕ ДРЕВО ЖИВОТНОГО МИРА
Живая природа находится в непрерывном движении, она по-
стоянно изменяется и развивается. Изучение многочисленных
данных зоологической науки подводит к материалистическому
и диалектическому пониманию путей развития живой природы.
Всестороннее исследование убеждает в происхождении жи-
вотных от общего корня, в монофилетическом развитии всего жи-
вотного мира. Поэтому филогенетические отношения животных
разных систематических групп могут быть изображены в виде раз-
ветвленного древа, в основании которого находятся примитив-
ные одноклеточные животные (рис. 291). Выше в разделе о фило-
гении было отмечено, что установлению филогении животных,
выявлению их родственных взаимоотношений способствуют три
пути исследования: 1) сравнительное изучение морфологии со-
временных животных (сравнительная морфология); 2) изучение
их эмбриологии также в сравнительном освещении (сравнитель-
ная эмбриология); 3) изучение по ископаемым остаткам живот-
ных прошлых геологических эпох (сравнительная палеозоология).
Сравнительно-морфологический метод изучения животных
362
Костистые Амфибии Рептилии Птицы Млекопитающие
рыбы
Киствперые
Первичные
Хрящекостные рыбы
Хрящевые рыбы
НасЕкимые
Диплоподо
Первично-
кордовые
Иглокожие
Круглоротые
Первичные черепные
Оболочники—...
Первичные бвсчерепуые
Первичные костные
рыбы
бесчерепные
Паукообразные
Симоррлы
Первичнотраквйные
'Цетинконогие
Втпричнопплоапные
втаричнаротые \ Вторичнополастные Пиявки
\ пврвич/шрптые
Г •' —-Ресничные
- Трематоды
Цестоды
Пврвичнопа'лостные
двустороннесииметричные
$>, Коралловые полипы
Инфузории
Протозоа
Рис. 291. Родословное древо животного мира (схема)
Жгутиковые
Споровики
Корненожкр
позволяет вскрыть разную степень сложности их строения и обна-
ружить те или иные признаки сходства и различия, что помогает
составить представление о возможных переходных формах и гипо-
тетических предках той или иной группы животного мира. Напри-
мер, выяснено, что плоские гребневики — животные, близкие
363
к кишечнополостным, в то же время имеют ряд черт сходства
с ресничными червями из тина плоских червей, сочетая в строении
признаки двуслойных животных с радиальной симметрией тела
с признаками трехслойных животных с двусторонней симметрией.
Подобное сходство позволяет предположить о филогенетической
связи плоских червей с кишечнополостными.
Использование данных сравнительной эмбриологии основы-
вается на биогенетическом законе (правиле), согласно которому
прошлое не проходит в эволюции животных бесследно и живот-
ные в индивидуальном развитии повторяют в той или иной сте-
пени этапы своего исторического развития —- онтогения отражает
филогению. Основанием для формулировки этого закона изве-
стным немецким дарвинистом Э. Геккелем послужили много-
численные исследования корифеев нашей отечественной науки
А. О. Ковалевского и И. И. Мечникова — по эмбриологии беспоз-
воночных и низших хордовых и К. М. Бэра — по эмбриональному
развитию позвоночных. Сходство в эмбриональном развитии жи-
вотных может служить дополнительным аргументом при решении
вопроса о филогенетических связях.
Палеонтологический путь исследования филогении животных
также дает веские доказательства для установления родословной
тех животных, останки которых сохранились в слоях земли.
Одноклеточным организмам со всеми характерными для них
чертами сложного строения — наличия цитоплазмы, ядра и раз-
личных органоидов, выполняющих жизненные отправления —
предшествовали живые существа значительно более простой
организации, неклеточного строения. Первые организмы облада-
ли способностью воссоздавать живую материю тела путем асси-
миляции из окружающей среды неорганических и органических
веществ небиологического происхождения. От них произошли
одноклеточные животные, возможно, амебоидного строения.
Среди жгутиковых появились организмы, способные к построе-
нию тела из неорганических соединений с использованием солнеч-
ной энергии в процессе фотосинтеза. Зеленые жгутиковые, соеди-
няющие в строении и биологии черты животных и растений,
свидетельствуют о происхождении двух царств органической
природы от общих предков.
Эволюция одноклеточных протекала в разных направлениях,
но одноклеточная организация тела у большинства сохранялась.
Древние амебоидные организмы дали начало современным сар-
кодовым и паразитическим книдоспоридиям. Одна из ветвей
родословного древа от жгутиковых идет к инфузориям — живот-
ным наиболее сложного строения, другая — к паразитическим
споровикам, хотя о происхождении споровиков и книдоспоридий
нет единого мнения.
Развитие одноклеточного животного мира привело через ко-
лониальных жгутиковых к многоклеточным животным. Жгути-
ковые отличаются среди Protozoa наибольшим разнообразием,
364
сложностью приспособлений к питанию и жизни в разных усло-
виях. Есть среди них простые и сложные организмы колониальной
формы, давшие начало двум ветвям современных многоклеточных
животных: губкам и остальным многоклеточным — через первич-
ных двухслойных предков.
Существует несколько гипотез о пути происхождения много-
клеточных, и все они объединены общим признанием происхож-
дения многоклеточных от колониальных жгутиковых. От ворот-
ничковых жгутиковых произошли губки, неподвижный образ
жизни которых не способствовал их прогрессивному развитию,
поэтому они сохранили много примитивных черт строения. От
других колониальных жгутиковых произошли двухслойные предки
кишечнополостных животных. Большинство их перешло к непод-
вижному или малоподвижному существованию, как это наблю-
дается у современных кишечнополостных.
Сравнительное изучение двухслойных кишечнополостных при-
водит к представлению о возможных путях происхождения и
эволюции тканей, дифференцировки их функций и специализации.
Кишечнополостные имеют в мезоглее клеточные структуры раз-
личной сложности и назначения, которые помогают представить
этапы развития третьего зародышевого листка — мезодермы.
Зачатки мезодермы, имеющиеся у целентерат, еще более отчет-
ливо выражены у гребневиков, животных по происхождению
и организации, близких к кишечнополостным. Вопрос о происхож-
дении плоских червей продолжает оставаться дискуссионным.
Гипотеза Ланга о филогенетической близости их с плоскими пол-
зающими гребневиками оспаривается, но ряд новых гипотез так-
же не решает вопроса. Ланг основывает свою гипотезу на мор-
фологическом сходстве гребневиков и ресничных червей. Послед-
ние подобно некоторым плоским гребневикам имеют уплощенное
тело, покрытое ресничным эпителием, нервную систему в виде
сети нервных клеток или собранных в нервных тяжах; кишечный
канал тех и других лишен анального отверстия. У некоторых тур-
беллярий наряду с двусторонней симметрией тела сохраняются
черты радиальной симметрии в расположении отдельных орга-
нов: центральное положение рта на брюшной стороне, радиальное
расхождение от глотки ветвей кишечника, расположение нервного
узла почти над глоткой с радиально расходящимися от него нерв-
ными тяжами.
В родстве с плоскими червями состоят круглые черви и скреб-
ни. Тип Круглые черви включает класс Брюхоресничные, предста-
вители которых имеют как во взрослом состоянии, так и в период
эмбрионального развития ряд сходных черт строения с реснич-
ными плоскими червями (наличие ресничного эпителия на брюш-
ной стороне, типичных протонефридиев и др.). Это дает основа-
ние сближать круглых червей с плоскими.
От предков современных первичнополостных отделились вто-
ричнополостные животные (целомата), которых с полным ос-
365
нованием называют высшими многоклеточными животными.
Основанием этому служат такие черты их организации, как кон-
центрация нервных клеток в ганглиях — прогрессивное развитие
нервной системы, образование кровеносной сосудистой системы,
выделительных органов метанефридиального типа, сегментар-
ность тела.
Одну из ветвей вторичнополостных животных составляют
организмы, у которых во взрослом состоянии сохраняется пер-
вичный рот (первичноротые). Раньше других от первичных вто-
ричнополостных животных отделилась ветвь, приведшая к воз-
никновению типа моллюсков. Несмотря на то что моллюски имеют
на всех этапах онтогенеза несегментированное тело, родство их
с другими вторичнополостными животными не вызывает сомне-
ния. Как у тех, так и других имеются нервные ганглии, кровенос-
ная система, метанефридиальные выделительные органы. Такое
глубокое сходство организации не может быть случайным. Кроме
того, типичная для морских моллюсков личинка парусник сходна
с личинкой многощетинковых кольчатых червей трохофорой.
Среди сегментированных вторичнополостных животных исход-
ной группой следует считать древних первичных многощетинко-
вых червей, близких к типу Кольчатые черви. Они дали начало
малощетинковым червям, а от последних произошли пиявки.
Сравнение кольчатых червей с членистоногими убеждает
в большом сходстве их строения: у тех и других имеются вторич-
ная полость тела, метамерность, нервная система в виде надгло-
точных ганглиев, окологлоточного кольца и брюшной нервной
цепочки, кровеносная система с главным сосудом на спинной
стороне и другие общие признаки; посегментное расположение
конечностей, которые у многощетинковых кольчатых червей
имеют вид нерасчлененных параподиев, а у членистоногих вместе
с приобретением твердого покрова преобразовались и стали чле-
нистыми ножками. Общие черты строения кольчатых червей и
членистоногих указывают на близкое родство тех и других. Чле-
нистоногие — прогрессивная ветвь, берущая начало от общего
предка с современными многощетинковыми кольчатыми червями.
Среди членистоногих ракообразные, продолжающие жить
в воде, где жили и их предки, сохранили многие примитивные
признаки строения, сближающие их с кольчатыми червями.
Паукообразные во многом сходны с ракообразными, ведут
происхождение от древних жабернодышащих членистоногих.
В родстве с предками кольчатых червей находятся и трахейно-
дышащие членистоногие, среди которых наиболее прогрессивной
группой являются насекомые.
Другая ветвь родословного древа развития животного мира
ведет от древних вторичнополостных червеобразных животных
к вторичноротым животным — иглокожим и хордовым.
Иглокожие — древняя группа животных, которые вторично
приобрели радиальную симметрию тела. Она была у палеозой-
366
ских представителей этого типа в связи с переходом к прикреплен-
ному образу жизни. Личинки иглокожих имеют двусторонпесим-
метричное тело.
Низших хордовых с иглокожими сближают вторичное образо-
вание рта в процессе онтогенеза, сходство процесса образования
вторичной полости тела, двухслойное строение покровов и ряд
других признаков.
Хордовые животные - одна из наиболее высокоорганизован-
ных групп животного мира. Происхождение хордовых остается
в деталях еще неясным. Возможно, они произошли от общего
корня с иглокожими.
Первичные хордовые, как показали исследования А. Н. Се-
верцева, представляли собой существа, довольно близкие к со-
временному ланцетнику. Это были вторичнополостныс, вторично-
ротые животные с признаками внутренней сегментации, имевшие
хорду, нервную трубку вдоль спины и жаберные щели в стенках
глотки. От этих примитивных первичных хордовых произошли,
с одной стороны, современные бесчерепные (ланцетники и близкие
к ним формы), а с другой — первичные черепные (позвоночные),
у которых передняя часть спипиого мозга преобразовалась в го-
ловной мозг и защищать его стал сформировавшийся черен.
Боковую ветвь низших хордовых образуют личиночпохордовые,
в организации взрослых особей которых произошел регресс в
связи с переходом к сидячему образу жизни при пассивном пи-
тании.
В эволюции позвоночных одна из ветвей привела к появлению
бесчелюстных (миноги и миксины), а другая дала начало челю-
стноротым, к которым относятся все остальные классы Vertebral а.
У животных этой ветви в связи с активным добыванием пищи
и общим нарастанием активности поведения развились парные
конечности, челюстной аппарат, сложный скелет и ряд других
усложнений в строении. Все это обеспечивало им развитие и ши-
рокое распространение. Первыми представителями челюстноро-
тых позвоночных были рыбы, в первую очередь хрящевые (акулы
и скаты). По уже в конце палеозоя широкое распространение
получают различные рыбы с кос.тным или костно-хрящевым ске-
летом. От древних палеозойских рыб ведут родословную двояко-
дышащие и кистеперые рыбы. Последних надо считать родона-
чальниками наземных позвоночных, древнейшими из которых
были панцирпоголовые земноводные — стегоцефалы. От древних
примитивных амфибий произошли как земноводные, так и первые
палеозойские пресмыкающиеся, достигшие в мезозойскую эру
большого разнообразия форм.
В мезозое от пресмыкающихся отделились два высших класса
позвоночных: Млекопитающие и Птицы. Как уже было сказано,
в юрских отложениях найдены остатки своеобразных существ ----
археоптериксов, которые совмещали признаки рептилий и птиц.
Это боковая ветвь в эволюции птиц. Настоящие птицы появились
3(57
в меловом периоде и достигли большого разнообразия в палеогене
и неогене кайнозойской эры.
Млекопитающие произошли от древних рептилий в самом на-
чале мезозойской эры, т. е. раньше птиц. Но развитие, приведшее
к современному богатству форм этого класса позвоночных, отно-
сится, как и у птиц, к кайнозойской эре (см. рис. 290).
ЖИВОТНЫЙ МИР СОВЕТСКОГО СОЮЗА
Территория Советского Союза расположена в пределах Палеарк-
тической области. Поэтому фауна нашей страны носит все харак-
терные черты животного мира этой области. Но различные зоны
СССР населяют далеко не одни и те же животные. Зональность
в их распределении связана с природными зонами Евразийского
материка. Каждой ландшафтной зоне его свойственна характер-
ная для нее фауна.
Зона тундр занимает Крайний Север страны. Ее безлесые про-
странства расстелились по северной части Кольского полуост-
рова, равнинам низовьев рек Мезени и Печоры, Ямальскому,
Гыданскому и Таймырскому полуостровам, северным районам
Якутии и Чукотки и по островам Северного Ледовитого океана.
Для зоны тундр характерен суровый арктический климат
с длительной морозной и ветреной, но относительно малоснеж-
ной зимой и коротким прохладным летом. Болотистые, песчаные
или каменистые почвы тундры покоятся на мощном слое вечной
мерзлоты. В низменных районах преобладают болотистые тундры,
в растительном покрове которых господствуют различные влаго-
любивые травы, мхи и лишайники. На более возвышенных местах
расстилаются песчанистые и каменистые тундры со скудной
растительностью. Местами тундру прорезают горные хребты.
Животный мир тундр своеобразен (рис. 292). Здесь обитает
сравнительно немного видов животных, приспособившихся к
местным суровым условиям. Преобладают наземные, водные и
норные животные. Многие из них на зиму приобретают белую
окраску под цвет снега.
Из млекопитающих характерны северный олень, тундровый
волк, песец, белый медведь (по побережью и на островах), своеоб-
разные грызуны—лемминги. Заходят лисица, росомаха, бурый
медведь.
Из птиц для этой зоны наиболее типичны белая и тундровая
куропатки, полярная белая сова, ряд гусей-казарок, мелкие птич-
ки — пуночки. На скалах морских побережий расположены
птичьи «базары», где гнездятся кайры, чистики, чайки, поморники
и другие птицы. Пресмыкающиеся и земноводные в тундре отсут-
ствуют.
Между зонами тундр и лесов лежит полоса лесотундры, фауна
которой образована представителями животного мира обеих зон.
368
Рис. 292. Характерные представители Голарктической области (зоны тундры):
/--тундровая куропатка; // -- белая куропатка; Ш - мохноногий канюк; IV - лапланд-
ский подорожник; V - пупочка; VI — белая сова; VII — белолобая казарка; VIII чер-
пая казарка; IX -- черпошаночпый сурок; Л' — краепозобая казарка; XI .копытный
лемминг; XII - обский лемминг; XIII — снежный баран; XIV-северный олень; Л’Г—
песец
13 Б. Л. Кузнецов и др.
Зона лесов тянется широкой полосой от западной границы
СССР до побережья Тихого океана. К югу она спускается при-
мерно до линии Львов — Киев — Брянск — Тула—Рязань —
Горький — Казань — Уфа — Оренбург — Свердловск — Тю-
мень — Новосибирск — Алтай и степи, языками заходит на
Алтай, в Туву и Южное Забайкалье.
Климат зоны лесов не столь суровый, более умеренный, чем
зоны тундр. Зимы здесь короче, мягче и многоснежнее. Лето более
продолжительное и теплое. Вечная мерзлота характерна только
для северных частей зоны. Господствуют подзолистые и болотные
почвы. Прежде большая часть зоны была покрыта могучими ле-
сами различного типа. Но в настоящее время значительные пло-
щади, ранее занятые лесами и болотами, преобразованы в поля
и пастбища, в населенные пункты.
Животный мир зоны лесов более богат видами, чем фауна
зоны тундр. Здесь много древесных животных. Из млекопитающих
характерны лось, благородный олень, косуля, кабарга, бурый
медведь, соболь, лесная куница, росомаха, рысь, белка, летяга,
бурундук, сони, бобр, лесные мыши. Типичными для зоны лесов
птицами являются глухари, тетерева, рябчики, различные дятлы,
лесные совы — неясыти, кукушки, кедровки, сойки, клесты, сви-
ристели, дрозды и многие другие птицы.
Пресмыкающиеся здесь немногочисленны (гадюка, обыкно-
венный уж, некоторые ящерицы). Из земноводных отметим раз-
личных тритонов и лягушек.
Обширная лесная зона естественно делится на подзону хвой-
ных лесов (тайги) и подзону смешанных и широколиственных
лесов. Первая занимает всю азиатскую часть зоны лесов и север
Европы на юг до линии Петрозаводск — Вологда — Киров —
Казань. Вторая простирается на лесные районы Европы на север
до указанной линии. Для таежной подзоны характерны лось,
соболь, росомаха, бурундук, летяга, кедровка, кукша и другие
звери и птицы. Для зоны смешанных и широколиственных лесов
типичны благородный олень, косуля, кабан, лесная куница, лес-
ной хорек, разные сони, иволги, зеленый и седой дятел и др.
(рис. 293, 294).
В последнее столетие в зоне лесов вслед за вырубкой лесных
массивов расселилось много животных, характерных для полей
и пастбищ: заяц-русак, еж, черный хорь, серая куропатка и пр.
К югу от зоны лесов, отделяя ее от зоны степей, тянется поло-
са лесостепи, в лесных островах которой обитают различные
лесные, а на степных участках —- степные и полевые животные.
Зона степей занимает южные части Украины, Дон, большую
часть Северного Кавказа, Среднее Поволжье, Южный Урал,
Зауралье, Северный Казахстан, южные районы Забайкалья.
Климат здесь еще более теплый, но менее влажный, чем в зоне
лесов. Большая часть зоны покрыта черноземными почвами.
Для степной зоны характерно отсутствие лесов, если не считать
370
Рис. 293. Характерные представители Голарктической области (зоны широко-
лиственных и смешанных лесов Европы):
/ — лесная куница; II - черный хорь; /// — садовая сопя; IV — полчок; V - иволга;
VI -- квакша; VII орешниковая соня; VIII — зеленый дятел; IX — дубонос; А' — евро-
пейская неясыть; А7 белка; XII - зубр; XIII - европейская косуля
13*
Рис. 294. Характерные представители Голарктической области (зоны тайги):
/ — трехпалый дятел; // — желна; III - глухарь; /Г — кедровка; I* — свиристель; 17
бородатая неясыть; VII— белокрылый клест; VIII — мохноногий сыч; IX — соболь,
V — колонок; XI — белка; XII — лось; XIII — летяга; А'/V — бу рундук; XV — кабарга;
Х17 — росомаха
лесные искусственные посадки. Прежде почти вся зона была по-
крыта целинными ковыльными или ковыльно-полынными степями.
Но в настоящее время они почти полностью распаханы под раз-
личные сельскохозяйственные культуры. Поэтому ныне для зоны
характерен так называемый культурный ландшафт, в котором
бескрайние поля чередуются с пастбищами, лесными посадками,
садами и поселениями людей.
Прежде на целинных степях паслись стада сайгаков, диких
лошадей — тарпанов. Сейчас тарпаны истреблены, а сайгаки
оттеснены в зону полупустынь и пустынь. Особенно многочислен-
ны в степной зоне различные грызуны: сурки, суслики, тушкан-
чики, слепцы, хомяки, различные полевки и мыши. Многие из них
являются серьезными вредителями растениеводства. Из зайце-
образных в степях водятся зайцы-русаки и степные виды пищух,
из хищных зверей — волки, лисы, корсаки, степные хорьки
(рис. 295). Из типично степных птиц — дрофы, стрепеты, журав-
ли-красавки, степные орлы, степные луни, степные жаворонки.
В степной зоне значительно больше, чем в лесной, различных
пресмыкающихся: полозов, гадюк, ужей, разных ящериц и др.
Зона пустынь и полупустынь занимает огромные пространства
в Восточном Азербайджане, Калмыкии, Нижнем Поволжье, Ка-
захстане и на равнинах Средней Азии. Эта зона отличается за-
сушливым климатом. Почвы скудные — сероземы, пески, солон-
чаки и солонцы. Растительность не образует сплошного покрова.
Местами имеются заросли пустынных кустарников: саксаулов,
гребенчуков, пустынных акаций. Северная часть зоны занята
глинистой полынной или злаково-полынной полупустыней, слу-
жащей переходной полосой между степной и пустынной зонами.
В Средней Азии огромные площади занимают песчаные пустыни,
в которых можно различить все переходы от голых барханных
песков до закрепленных растительностью ровных песчанистых
участков. В предгорьях широкое развитие получают каменистые
пустыни с щебнистыми почвами и скудной растительностью.
Животный мир пустыни крайне оригинален (рис. 296). В спе-
цифических условиях этой зоны могут существовать только хо-
рошо приспособленные виды и формы, хорошо уживающиеся
в самых различных условиях среды. Из копытных характерны
куланы (дикие ослы), джейраны и сайгаки (последние преиму-
щественно держатся в полосе полупустыни). Из хищных млеко-
питающих обитают барханные и степные кошки, пустынные ры-
си — каракалы, степные лисицы -- корсаки, волки, лисы и другие
виды. Очень многочисленны грызуны: различные формы тушкан-
чиков, песчанок, тонкопалые и песчаные суслики и многие другие.
Из пустынных птиц наиболее характерны рябки, саджи, дрофы-
красотки, саксаульные воробьи и сойки, пустынные вьюрки и
славки. Очень многочисленны различные пресмыкающиеся:
огромные вараны, агамы, круглоголовки, различные змеи, степ-
ные черепахи и прочие виды. Земноводных здесь мало.
373
Рис. 295. Характерные представители Голарктической области (зоны степей):
/ — обыкновенный хомяк; II — крапчатый суслик; /// — степная мышевка; IV — слепыш,
I — степная пеструшка; 1-7 — заяц-русак; VII — дрофа; VIII — большой тушканчик;
/X— байбак; X— степная пищуха; А7 — степной жаворонок; X//— черный жаворонок;
XIII — краснощекий суслик; XIV — стрепет; XV' —сайгак; XVI — полоз желтобрюхий
Рис. 296. Характерные представители Голарктической области (зоны пустынь
и полупустынь):
/ — тонкопалый суслик; II -саксаульная сойка; III гребенчатый тушканчик; JV ....
варан; V--ушастая круглоголовка; VI - белобрюхий рябок; VII ---каракал; I///
барханный кот; IX ушастый еж; X кулан; XI — джейран
Почти все звери пустынь — наземные и норные животные.
Многие из них имеют защитную песчаную окраску.
По долинам больших рек в зоне пустынь местами развиты
тугайные леса, в которых находят себе убежище тугайный олень,
шакалы, камышовые коты, кабаны, прежде здесь жили также
тигры.
Особый зоогеографический район нашей страны представ-
ляет южная часть Приморского края. Здесь по горам Сихотэ-
Алиня далеко к югу спускаются таежные леса с их характерными
обитателями: лосями, бурыми медведями, рысями, росомахами,
соболями, белками, летягами, бурундуками и другими таежными
животными. А с юга по долинам рек в этот район распростра-
няются широколиственные и смешанные леса маньчжурского типа,
для которых типичны такие южные животные, как тигр, леопард,
гималайский медведь, енотовидная собака, куница — харза,
маньчжурский заяц, фазан, белая цапля, колпица и другие
формы. Смешение южных и северных по происхождению видов
животных образует крайне своеобразные сообщества.
Зоогеографически выделяются также области высокогорья
Кавказа, гор Средней Азии и Южной Сибири. На высокогорных
альпийских лугах Кавказа пасутся стада горных козлов — туров
и серн, держатся табунки горных индеек и каменных куропаток,
летают стайки альпийских галок и клущиц, поют горные вьюрки,
завирушки и другие мелкие птички.
На высокогорных лугах, среди скал и ледников гор Средней
Азии и Южной Сибири (Алтая и Саян) можно встретить горного
козла — сибирского козерога, горных баранов — архаров и арга-
ли, снежных барсов, горных красных волков, горных сурков,
.пищух, горных индеек — уларов, каменных куропаток — кекли-
ков, клущиц, горных голубей и прочих представителей высоко-
горной фауны.
В настоящее время, когда природа нашей страны быстро
преображается хозяйственной деятельностью человека, меняется
также и характер фауны различных ее областей. Вслед за про-
движением на север земледелия расселяются в том же направле-
нии и многие животные, жизнь которых связана с полями и луга-
ми. По мере распашки целинных степей исчезают типично степ-
ные виды и пашню заселяют различные грызуны, в том числе
многие вредители растениеводства. В городах и селах поселяются
крысы, мыши, голуби, воробьи и др.
ОХРАНА И РАЦИОНАЛЬНОЕ ИСПОЛЬЗОВАНИЕ
ЖИВОТНОГО МИРА В СССР
Животный мир СССР богат и разнообразен. Многочисленные
виды промысловых животных обеспечивают народное хозяйство
страны пищевыми продуктами, техническим и лекарственным
.376
сырьем. Промысловые животные — огромный резерв для разве-
дения. Генофонд диких видов может быть использован для улуч-
шения пород домашних животных.
Установление в нашей стране социалистического строя с его
общественной собственностью на средства производства и пла-
новым ведением хозяйства создало благоприятные условия для
рационального и бережного отношения к природным богатствам.
Охрана природы в СССР стала делом большой государственной
важности.
В современных условиях под влиянием быстро развивающихся
производительных сил и роста народонаселения усилилось воз-
действие хозяйственной деятельности человека на природную
среду, что сделало проблему охраны природы, в том числе живот-
ного мира, особо актуальной. Сегодня на численности диких жи-
вотных сказывается не только перепромысел. Гораздо чаще со-
кращение численности животных происходит под воздействием
таких косвенных факторов, как разру шейпе местообитания,
интродукция (вселение) новых видов, сокращение и ухудшение
кормовой базы, применение пестицидов и гербицидов, загрязне-
ние среды и т. д.
Формы охраны природы в СССР различны. Велика роль в деле
охраны природы нашей страны государственных заповедников,
призванных сохранить особо ценные природные комплексы. Пер-
вые заповедники (Астраханский — 1919 г., Ильменский - -
1920 г.) были созданы по инициативе В. И. Ленина. В настоящее
время в нашей стране действуют 143 заповедника общей пло-
щадью 13,7 млн га. В них исключены всякая хозяйственная дея-
тельность (кошение травы, рубка леса, выпас скота, охота) и
другие вмешательства человека, не связанные с целями охраны
природы. Заповедники служат хорошей базой для выполнения
многолетних комплексных научных исследований и работ, связан-
ных с поддержанием численности исчезающих видов животных и
растений. Во многом благодаря заповедникам были восстанов-
лены до промыслового уровня запасы таких ценных животных,
как соболь и бобр, а зубр, кулан и тигр спасены от угрозы исчез-
новения.
Важное значение для сохранения многих животных и рас-
тений имеют охраняемые леса, выполняющие водоохранную,
защитную, санитарно-гигиеническую и оздоровительную функ-
ции.
Другой формой охраны естественных местообитаний в СССР
являются заказники разнообразного назначения, в которых до-
пускаются определенные формы хозяйственной деятельности. Они
способствуют восстановлению и увеличению запасов многих охот-
ничьих зверей и птиц, а также ценных рыб. В СССР существует
более 1 тыс. заказников общей площадью около 25 млн га. К кате-
гории охраняемых естественных местообитаний относятся и созда-
ваемые в нашей стране национальные парки, которые наряду
377
с охраной живой природы служат местом отдыха и массового
туризма.
За годы Советской власти в нашей стране созданы плановое
социалистическое охотничье и рыбное хозяйства, в которых ве-
дется рациональная эксплуатация ресурсов животных. Она обес-
печивает сочетание добычи животных, производимой на строго
научной основе, с широкими мероприятиями ио их охране и обо-
гащению. Охрана промысловых животных осуществляется путем
соблюдения сроков и способов охоты и рыболовства. Советским
законодательством запрещено применение хищнических способов
охоты (с автомашин, самолетов, разорение нор, гнезд, логовищ,
сбор яиц и т. д.) и рыболовства (с помощью взрывчатых веществ,
остроги и т. д.), приводящих к массовому истреблению животных.
В положениях об охоте и охотничьем хозяйстве (по каждой
союзной республике) указаны виды зверей и птиц, охота на кото-
рых полностью запрещена, и виды, добыча которых разрешена
только по лицензиям — особым разрешениям на отстрел или
отлов строго определенного числа особей. Рыболовство в СССР
регламентируется положением об охране рыбных запасов и о ре-
гулировании рыболовства в водоемах СССР и республиканскими
правилами рыболовства. Важными мерами по охране рыбных
запасов являются: охрана нерестилищ ценных видов рыб, уста-
новление минимального размера промышляемых рыб того или
иного вида с целью сохранения молоди, борьба с загрязнением
рек и озер отходами заводов, фабрик и других промышленных
предприятий и сточными водами. Нарушение существующих за-
конов и правил охоты и рыболовства наказывается в администра-
тивном и уголовном порядке. Ведется борьба с браконьерством.
В комплекс проблем по охране животного мира СССР входит
задача сохранения и восстановления редких и исчезающих видов
животных. Еще в первые, годы Советской власти (1919—1922)
были изданы декреты об охоте, охране таких редких уже в то вре-
мя животных, как зубр, пятнистый олень, сайгак, выхухоль, бобр,
белая цапля, фламинго. Несколько позже к этому списку были
добавлены: кулан, джейран, бухарский олень, горал, винторогий
козел, тигр, гепард, белый медведь, гага, турач, лебеди, некото-
рые гуси и др. Полный запрет на отстрел и отлов редких животных
в сочетании с мероприятиями по их расселению в местах прежнего
обитания во многих случаях дали хорошие результаты. Имеется
немало примеров возвращения ставших редкими видов в число
ведущих промысловых животных фауны Советского Союза. Так,
в период гражданской войны резко сократилась численность лося,
особенно в центральных районах европейской части СССР. В ре-
зультате введения полного запрета на отстрел численность его
постепенно возрастала, так что через 25 лет была разрешена
охота на лося по лицензиям. В настоящее время в нашей стране
ежегодно добывается более 75 тыс. лосей. Значительные успехи
были достигнуты в увеличении численности сайгака, превратив-
378
шегося из исчезающего животного в важный объект промысла.
Выросло поголовье зубра, также находившегося на грани исчез-
новения. К началу XX в. в большинстве районов тайги был хищ-
нически истреблен соболь. Введение полного запрета на отлов
соболя в сочетании с широким расселением его в местах прежнего
обитания позволило не только восстановить, но и довести чис-
ленность вида до промыслового уровня. Ежегодно в нашей стране
заготавливается шкурок соболя в 15 раз больше, чем в дореволю-
ционной России. Удалось также увеличить поголовье речного
бобра, калана, куницы, выдры и некоторых других ценных пушных
зверей.
Постоянное внимание к проблемам охраны природы нашло
свое отражение в Основных направлениях экономического и со-
циального развития СССР на 1986—1990 годы и на период до
2000 года, принятых XXVII съездом КПСС и в Законе СССР об
охране и использовании животного мира (1980 г.). В Законе
большое внимание уделено редким и находящимся под угрозой
исчезновения видам животных. С целью инвентаризации и учета
этих животных в нашей стране в 1974 г. была учреждена Красная
книга СССР, первое издание которой вышло в свет в 1978 г.,
второе — в 1984 г. В зависимости от степени угрожаемой опас-
ности все виды и подвиды животных, внесенных в Красную книгу
СССР, подразделены на пять категорий: 1 — исчезающие; 2 —
сокращающиеся; 3 — редкие; 4 — неопределенные малоизве-
стные, очевидно, находящиеся под угрозой; 5 -- восстановленные.
Всего в Красную книгу СССР внесено 94 вида и подвида млеко-
питающих, 80 видов птиц, 37 видов пресмыкающихся, 9 видов
земноводных, 9 видов рыб, 204 вида членистоногих, 19 видов
моллюсков и 11 видов кольчатых червей. Красная книга СССР —
одновременно и научно обоснованная программа действий по
спасению редких видов животных и растений, напоминающая о
бережном отношении к живой природе. Она нацеливает на реше-
ние одной из основных задач, разработанных во Всемирной стра-
тегии охраны природы: сохранить все генетическое многообразие
биосферы, ибо сегодня нет возможности определить значимость
того или иного вида для будущего.
Перспективным является создание специальных центров по
разведению редких видов животных в неволе. В нашей стране
уже имеется опыт такой работы. Численность некоторых ценных
осетровых рыб бассейна Азовского и Каспийского морей (белуга,
севрюга, русский осетр) и лососевых (белорыбица) сохраняется
и увеличивается исключительно благодаря искусственному вос-
производству их на рыборазводных заводах. Это же относится
к разведению дальневосточных лососевых рыб. Разведение фаза-
нов в специальных фазанариях имеет большое значение для
обеспечения охотничьих хозяйств страны этими ценными птицами.
Большая роль в охране редких видов животных отводится зоо-
паркам.
379
В нашей стране проводится большая работа но обогащению
фауны, преследующая цель — повысить продуктивность угодий.
Она включает мероприятия по искусственному расселению жи-
вотных с целью акклиматизации ряда ценных представителей
отечественной и зарубежной фауны и реакклиматизации ранее
истребленных видов в местах их прежнего обитания.
Работы по акклиматизации многих промысловых живот-
ных были начаты в нашей стране в 20-х годах. Ряд видов пуш-
ных зверей был завезен в СССР из-за границы. Успешно про-
шла акклиматизация североамериканского грызуна ондатры,
начатая в 1928 г. В настоящее время ондатра распространилась
на значительной части территории Советского Сюза и занимает
одно из ведущих мест в пушном промысле. Хорошие результаты
получены по расселению южноамериканского грызуна нутрии
в Закавказье и других местах.
Было также проведено искусственное расселение в целях
акклиматизации ряда промысловых зверей и птиц отечественной
фауны, обеспечившее значительное расширение их ареалов. Так,
заяц-русак, ранее обитавший только в европейской части СССР
и в Зауралье, был выпущен во многих местах Южной Сибири,
в некоторых из них стал объектом охоты. Белка была расселена
в лесах Кавказа, Крыма и Восточного Тянь-Шаня, где успешно
акклиматизировалась. Пятнистые олени, обитавшие в Примор-
ском крае, были акклиматизированы в ряде заповедников и охот-
ничьих хозяйств европейской части СССР. В некоторых из них
также прижились завезенные с Алтая олени маралы. В охотничьих
хозяйствах страны были расселены фазаны и другие ценные
охотничьи птицы.
Для обогащения фауны промысловых рыб морей и пресных
водоемов СССР проведена акклиматизация многих видов рыб
в тех водных бассейнах, где они раньше не встречались. Так,
в Каспийском море была успешно акклиматизирована черномор-
ская кефаль, в оз. Иссык-Куль — севанская форель, в оз. Бал-
хаш — аральский усач, судак, лещ, в озерах Урала, Зауралья,
Армении и Красноярского края — сиги, что позволило повысить
рыбопродуктивность этих водоемов. Акклиматизация завезенных
из рек бассейна Амура в реки, водохранилища и каналы европей-
ской части СССР и Средней Азии растительноядных рыб — тол-
столобика и белого амура — не только обеспечила повышение
рыбопродуктивности, но и позволила решить проблему зараста-
ния этих водоемов, в частности Каракумского канала.
В СССР проводится также реакклиматизация ряда промысло-
вых зверей, направленная на восстановление их прежних ареалов.
Так, речной бобр в начале настоящего столетия был почти пол-
ностью истреблен и сохранился лишь в наиболее глухих районах
Белорусии, Украины, в Воронежской, Тюменской областях и Ту-
винской АССР. С 1930 г. началось искусственное расселение
бобра. В настоящее время бобр распространен от Белоруссии и
380
Карелии до Амура. Увеличение его численности позволило с
1963 г. начать промысел этого ценного зверя но лицензиям. Реак-
климатизация соболя, выпущенного более чем в 100 районах, где
он ранее обитал, ио был истреблен, позволила почти полностью
восстановить ареал этого ценного пушного зверя. Проводится
также реакклиматизация кабанов, благородных оленей, косуль
в охотничьих хозяйствах ряда центральных и западных обла-
стей СССР.
Экономический эффект, полученный от акклиматизации и ре-
акклиматизации в СССР многих ценных видов животных, оче-
виден. В то же время опыт предыдущих лет показал, что вселение
новых видов в естественные биогеоценозы может быть целесооб-
разным лишь при уверенности, что «вселенцы» не станут причи-
ной гибели местных видов.
Большое значение' для обогащения фауны Советского Союза
имеют охрана и воспроизводство промысловых зверей, птиц, рыб
и других полезных животных. С этой целью в заповедниках и
охотничьих хозяйствах СССР реализуется широкая программа
работ, направленных па улучшение условий обитания животных.
Известно, что численность последних часто лимитирована числом
мест, пригодных для постройки гнезд, нор, убежищ, для отдыха,
а также запасами кормов, наличием солонцов, водопоев и т. д.
В этой связи важное значение приобретает организуемая охот-
ничьими хозяйствами подкормка диких копытных, пушных зве-
рей и охотничьих птиц, особенно в наиболее трудное для животных
время года. Зимняя подкормка не только спасает полезных жи-
вотных от голода, но и позволяет им избежать контакта с пести-
цидами и сохранить хорошую упитанность к периоду размноже-
ния. В ряде случаев зимняя подкормка птиц может послужить
причиной появления оседлых популяций.
Численность полезных птиц-дуплогнездников удается увели-
чить путем развески около домов, в садах, парках, лесных полосах
искусственных гнездовий (дуплянок, скворечников). Создавая
искусственные укрытия и гнездовья по берегам водоемов, можно
увеличить число гнездящихся уток-крякв.
Среди различных биотехнических работ, проводимых в нашей
стране, многие направлены на воспроизводство и увеличение
запасов промысловых рыб. Они включают следующие мероприя-
тия: улучшение мест нереста, спуск молоди по каналам из отде-
лившихся во время половодья стариц, подкормку рыб в прудах
и озерах, очистку водоемов от сорной растительности и т. д. В ры-
боразводных заводах СССР производятся искусственное оплодо-
творение и инкубация икры рыб с дальнейшим выпуском выра-
щенных мальков в естественные водоемы. На реках, течение ко-
торых перекрыто плотинами, для проходных рыб сооружаются
рыбопропускники и рыбоподъемники.
В силу неделимости биосферы Земли успех в решении многих
проблем охраны природы, в том числе животного мира, может
381
быть достигнут только благодаря объединению усилий всех го-
сударств планеты. Заинтересованность многих государств в деле
охраны природы нашла свое отражение в принятии системы ши-
роких международных соглашений по охране и использованию
животного мира под эгидой ООН и развития двусторонних согла-
шений между странами, связанными общими интересами ио рас-
сматриваемой проблеме.
Советский Союз с первых лет своего существования постоянно
выступает за развитие и укрепление международного сотрудни-
чества по охране природы и рациональному использованию ее ре-
сурсов.
Государство активно участвует в заключении международ-
ных договоров, соглашений и конвенций, имеющих природоохра-
нительное значение. Так, СССР заключил специальные соглаше-
на о регулировании рыболовства с Финляндией, Ираном, Турцией
и Японией. Приняты и согласованы национальные законода-
тельства СССР, Болгарии и Румынии по охране морских млеко-
питающих Черного моря, подписана конвенция по охране пере-
летных и редких птиц между СССР и Японией, СССР и США,
а также соглашения СССР с Данией, Норвегией, Канадой и США
о сохранении белых медведей и др.
Активные действия СССР, направленные на разностороннее
международное сотрудничество, уже принесли немалую пользу
делу охраны и рационального использования животного мира.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ И ТЕРМИНОВ
Адолескарии 102, 104
Анеты 307, 308, 320
Аксолотль 284
Актинии 90, 91
Акулы И, 254, 257, 259, 262, 267, 268
Аллантоис 246, 285, 290
Амблистома 282, 284
Амбулакральная ножка 226, 227
— система 225, 226
Амеба 45, 58, 59, 60, 64
голая 64, 65
-- дизентерийная 45, 60, 64
раковинная 53, 64, 65
Аммонит 48, 224
Аммофила 187
Амнион 246, 285, 290
Амфибии 48, 274, 275
безногие 284, 285
- бесхвостые 48, 275
-- хвостатые 48, 275, 283, 284
Амфилнпа 110
Анабиоз 60, 127, 136, 148
Анизогамия 61
Антилопа 355
Аппарат Гольджи 25, 55
Аптерии 298
Ареал 15, 20, 311
Архар 354
Археоптерикс 324, 367
Арцелла 53, 64
Аскариды 45, 118, 127, 128
Аскаридия 130
Аскаридоз 128
Аспиды 293
Асцидия 233, 234
Атлант 287, 328
Аутогамия 62, 75
Бабочки 200, 202
Байбак 374
Баклан 307, 320
Бандикут 343
Баран 356
Барсук 351
Бегемоты 354, 355
Беззубка 219
Бекас 317
Белемнит 48
Белка 343, 347, 371, 372
Белозубка-крошка 325
Белуга 269
Бента ль 15, 263
Бентос 15
Беспозвоночные 4
Бесчелюстные 247
Бесчерепные 230
Бизон 355
Биотоп 15
Биоценоз 15
Биссус 214
Битинии 104, 223
Бластомер 39
Бластопор 40
Бластоцель 39, 40
Блефаропласт 57
Блохи 200
Бобр 338
Богомолы 193
Бородавки паутинные 166, 170
Ботрии 114
Бронтозавр 295
Брыжейка 139
Буйволы 355, 356
Бульбуе 120, 122, 130
Буревестник 307, 309
Бурундук 372
Бычковые 270, 272
Бычьи 355, 356
Варан 292, 375
Вальдшнеп 311, 317
Верблюды 354, 357
Веретеница 292
Веслоногие 320
Вибриссы 327
Власоглав 133
Власоеды 194, 195
Волк 344
383
Волоса тки 134
Вольвокс 50
Воробьипообразпые 321, 322
Ворона 321
Восковица 320
Вторичнополостные 42, 211, 225, 229,
365, 366
Вторпчнорогые 41, 229, 366
Вши 194
Вьюрок 321
Выдра 338, 351
Выхухоль 338, 345, 346
Вяхирь 317
Гага 320
Гагара 307
Гадюка 286, 293
Галка 321
Галлицы 205
Гальванотаксис 55
Ганглий 96, 220, 241, 366
Гастропор 225
Гастроцель 41
Гаструла 40
Гаструляция 40, 41
Гаттерия 286, 291
Геккон 292
Гемоспоридип 72
Геммула 82
Гиббон 361
Гепард 352
Гидра 86, 87, 88
Гидроидные 83, 86
Гиподерма 99, 118, 152
Гирудин 149, 150
Глазки Гессе 232
Глохидий 216
Глухарь 306, 372
Головастики 281
Голотурии 227, 228, 229
Голубеобразные 314, 317
Горилла 361
Горихвостка 322
Горлицы 317
Горностай 351
Грач 311, 321
Гребешки 217, 218
Гребневики 92, 93
Грегарины 52, 75
Грена 77
Гриф 320, 321
Грызуны 347, 348
Губки 48, 79, 80
- - бадяги 82
- клионы 82
- стеклянные 82
туалетные 82
Гусеница 187, 202, 203
Гусеобразные 319, 320
Гюрза 293
Да кти логи рус 105
Дафния 161
Двоякодышащие 274
Двукрылые 203
Двусторонпесимметричные 213, 223,
226, 229
Дельфин 11, 346
Джейран 356
Дзсреп 356
Диаптомус 161
Дикроцелиоз 104
Диктиокаулез 131
Диморфизм половой 19, 117, 118, 155,
184, 308
Диплодок 295
Диффлугия 52, 64
Долгопят 361
Дрейссена 214, 219
Дробление оплодотворенного яйца 39,
40
Дрозд 306, 308
Дрофа 306, 320, 374
Дубонос 371
Дупель 317
Дятел 306
Дятлообразные 321
Еж 345
— морской 227
Ехидна 341
Жабернодышашие 155
Жаброногие 160
Жабры 139, 142, 211, 213, 226, 242,
248, 259
Жабы 283, 284
Жаворонок 321, 374
Жвалы 179
Жвачка 333
Жгутиковые 55, 56, 64, 67
Желна 372
Желтопузик 286, 292
Жемчужница 218, 219
Жерех 264
Жесткокрылые 197
Жизненный цикл грегарины 74
— — кокцидии 71
— - лептеца широкого 116
— — ноземы 76
— — плазмодия малярийного 72, 73
— — фасциолы 103
- — цепня -вооруженного 1 12
Жирафа 354
Жужелицы 199
Жужжальца 180
Жуки 197
Журавлеобразные 320
Жучки 252 •
Зайцеобразные 351
384
3 а к p ы то пу з ы р н ые 2 59
Заказники 377
Заповедники 377
Заяц 347
Звезда морская 227
Зебра 359
Землеройка 345
Земноводные 274, 275
Зигота 39, 72
Змеи 292, 293
Зоны ландшафтные 15, 16
Зоогеография 15
Зоология 4, 6, 15
Зоофаги 187
Зубр 354, 355, 356, 371
Зяблик 321
Иволга 321, 322, 371
Иглобрюх 251
Иглокожие 225, 228, 366, 367
Игуанодон 296
Изогамия 61
Имаго 175, 184
Индейки 317
Инфузория 55, 56, 77
— балантидиум 78
— ихтиофтириус 78
— панцирная 78
— стилонихия 57, 77
— туфелька 54, 59, 78
Ихтиозавр 11, 296
Кабан 354
Кабарга 372
Казарка 369
Казуар 322
Кайра 308, 315
Кальмар 223
Камбала 251, 270
Камбаловые 272
Камнеточец 214
Канюк мохноногий 369
Каракал 375
Каракурт 171
Карась 266
Кариоплазма 26
Карп 266, 270
Карповые 271
Категория систематическая 18, 52
Квакша 284, 371
Кедровка 310, 372
Кенгуру 343, 344
Киви 322
Кивсяк 176, 177
Киль 299, 313
Кинетосома 57
Кит 353
Китообразные 352, 353
Кишечнополостные 83
Классификация 18
Клест 308, 372
Клетка 23, 24, 25, 27
— амебовидная 79, 226
— веретеновидная 119
— выделительная 95
— железистая 26, 96, 149, 236
— желточная 100
— звездчатая 29, 85
— мышечная 53
- нервная 23, 37, 85, 108, 119, 212
— осязательная 89
Клетка пигментная 30, 149, 254
— стрекательная 54, 55, 83, 84, 85, 89
— фагоцитарная 121
— эпителиальная 27, 84
Клещи 172, 173, 174, 175
Клещики 172, 173
Клинтух 317
Клопы 197
Книдоспоридии 75
Коала 343, 344
Кобра 293
Кобылки 191, 193
Когти 327, 348
Козел безоаровый 356
Козерог сибирский 354
Козодой 307
Кокцидии 45, 70
Коловратки 134, 135
Колонок 372
Комары 203, 204
Комиссура 211, 214, 220
Комменсализм 44
Конвергенция 11
Коиек 310
Конъюгация 62, 63, 77
Копрофаги 187
Копуляция 63, 72
Копыта 327
Коракоид 238
Корацидия 114, 117
Короеды 197
Королек 301, 322
Коршун 320
Костянка 177
Косуля 356, 371
Котик морской 352
Котилозавр 296
Кошачьи 351, 352
Крабы 82, 155, 166
Краспотелковые 172
Крачка 307
Креветки 164
Крокодилы 294
Кролик 337
Кроншнеп 315
Кроты 338, 345, 346
Круглоголовка 292, 375
Круглоротые 247
Крылан 346
385
Крылатые 190
Крысы 350
Кузнечики 191, 193
Куколка 184
Кукушка 306, 309
Кулан 360, 375
Кулики 306, 317
Куница 351, 371
Куньи 351
Курообразные 309, 314
Куропатка 369
Кутикула 53, 118, 139
Лакуны 154, 213, 221, 344
Лама 357, 358
Лангусты 164
Ланцетник 230, 231
Ласка 351
Ластоногие 352
Ласточка 307, 321
Латимерия 273, 274
Лебедь 307
Лев 351
Легкие 153, 211, 242
Лейкоциты 33, 34
Лейшмании 69
Лейшманиоз 69
Лемминги 368
Лентецы 114, 115
Леопард 351
Лепидозирен 274
Летяга 337
Лещ 251
Лимфа 34, 35, 261
Линь 271
Линька 152, 174, 298, 310, 327
Литораль 15, 263
Личинка 82, 234, 249, 275, 282, 367
Личиночнохордовые 233, 234
Лопатонос 269
Лососевые 271
Лосось 270
Лось 354, 356, 372
Лошадиные 359
Лошадь Пржевальского 359
Лошак 360
Лунь 297, 320
Лягушки 282, 283, 284
Макрогаметоциты 71
Макрогаметы 69
Максиллоподы 161
Максиллы 178, 179
Мандибулы 178
Мантия 161, 211, 212, 214
Марал 356
Медведка 191, 192, 193
Медвежьи 351
Медуза гидроидная 88
— сцифоидная 83, 88, 89
Мезенхима 29, 33
Мезоглея 79, 83, 84, 92
Мезодерма 41, 43, 92, 138, 365
Мезозавры 295
Мембрана 23, 25
Мерозоит 39, 71
Метаморфоз 140, 142, 184, 213, 216,
275
Мета нефридий 137, 140, 150, 213, 233
Мешок кожно-мускульный 93, 95, 99,
119, 138, 149
— целомический 138
— чернильный 224
Мидии 214, 219
Микрогаметоциты 72
Микрогаметы 69, 72
Микроспоридии 45, 76
Миксины 247, 249
Миноги 247, 249
Миомеры 230, 240, 247, 255
Мионемы 53
Миосепты 230, 240, 247, 255
Миофибриллы 35, 53, 84, 119
Мирацидий 102
Митохондрии 25
Млекопитающие 20, 48, 325, 337, 361,
362
— водные 338
— древесные 337, 344
— летающие 337
— наземные 337
— норные 338
— подземные 338
— полуводные 338
Многобугорчатые 362
Многожгутиковые 70
Многоклеточные 79, 364, 365
Многоножки 176, 177
Мозг головной 37, 235, 239, 240, 245,
277, 288, 300
— костный 33
— спинной 37, 235, 240, 256, 277
Мозговик овечий 111
Мозоленогие 357
Мокрицы 155, 164
Моллюски 211
— брюхоногие 219, 220
— головоногие 212, 223, 224
— двустворчатые 213
Мониезиоз 114
Мониезия 114, 115
Моногамия 308
Морж 352
Морфология 10, 11, 362
Москиты 203
Мошки 203
Мул 360
Муравьи 210
Мутуализм 44
Мухи 205
386
Мухоловка 30$), 321
Мышевка степная 374
Мыши 350
Наездники 209
Насекомые 177, 178
Н а секо м оя д и ые 345
Наутилус 224
Невроцель 229, 230
Нейрофибриллы 30
Нскрофаги 187
Нематоды 118, 122, 123, 125, 126
Неоцератод 274
Непарнопалые 358, 359
Нервная система позвоночных 235
- - — вегетативная 240, 241
- - - — парасимпатическая 240
- периферическая 240, 241
— - симпатическая 240
- - - центральная 240
Нервы 240, 241, 256, 257
Нереиды 143
Нереис 142
Неясыть 316, 321, 371
Нимфа 172, 173
Ноземы 45, 76
Норка 351
Носороги 360
Номенклатура бинарная 18
Нутрия 347
Обезьяны 360
Области зоогеографические 16
Оводы 205
Овсянка 321
Одноклеточные 5, 49, 362
Окуневые 272
Оленьи 355, 356
Омары 157, 164
Омматидии 153, 183
Ондатра 338
Онкосфера 109, 110
Онтогенез 12, 14, 63, 79, 236, 367
Оокинеты 72
Оотип 100
Ооциста 72
Опахало 297
Опоссум 343, 344
Орангутан 361
Организмы автотрофные 22
- гетеротрофные 22
Органы аналогичные 10
- боковой линии 240, 257, 278, 282,
285
- гомологичные 11
Орел 308, 320
Орлан 316
Осел 359
Осетровые 268, 269
Ослик водяной 164
Острицы 45, 130
Осфрадии 216
Осьминог 223
От к р ыто 11 узы р п ые 2 59
Отолиты 257
Очпи 297
Павиан-бабуин 361
Палеозоология 362
Палеонтология 15, 16, 17, 47, 48, 296
Налео-жология 17
Палоло 143
Паразитизм 45
Паразитоидпые 173
Паразитология 51
Параподии 138, 140
Паренхима 93, 94, 99, 212
Парнопалые 353
- жвачные 355
- нежвачные 353, 354
Партеногенез 142
Пауки 169, 170, 171
Паукообразные 168
Педипальпы 166, 168
Педогамия 62
Пелагпаль 15, 262
Пеликан 307, 320
Пелликула 53, 67
Пеночка 309, 321
Первичпобеекрылые 189, 190
Первичпополостные 11
Первичноротые 41, 366
Первоящеры 291
Перепончатокрылые 45, 206, 207
Перловица 219
Перья птиц 297, 298
-- - - контурные 298
— -- маховые 298
- -- рулевые 298
Песец 369
Пескоройки 249
Пеструшка стенная 374
Песчанки 349
Пилильщики 210
Пингвины 307, 322, 323
Пироплазмиды 66
Пищуха 306, 322, 374
Пиявки 148, 149
Планктон 15, 262
Плантофаги 187
Планула 86, 88, 90
Пластинчатожаберные 213, 267
Плацента 337, 344
— дпекоидальная 344
- диффузная 344
— дольчатая 344
— зональная 344
Плацентарные 344
Плезиозавры 296
Плероцеркоид 115, 117
387
Поганка 307
Подорожник лапландский 369
Позвоночные 235, 236, 249, 274
Полевки 347, 349
Полигамия 308
Полиморфизм 19, 20, 184, 185
Полип 83, 84
— гидроидный 87, 88
— коралловый 83, 85, 90
Полоз желтобрюхий 374
Полость тела вторичная 225
— — первичная 212
Полужесткокрылые 197
Полчек 371
Поползень 306, 321
Попугай 306
Пресмыкающиеся 285, 286, 294, 295
Проглоттиды 107
Проехидна 341, 342
Простейшие 49
Простогонимус 104
Протей балканский 282, 284
Протозоология 51
Протонефридий 94, 98, 99, 134
Протоподит 157
Протоптерус 274
Процеркоид 115, 117
Прудовики 102, 223
Прямокрылые 190, 192, 193
Псевдоподия 56, 58, 64
Псевдозухия 295
Псовые 351
Птерилии 298
Птеродактиль 296
Птицы 48, 296, 297, 298, 299, 324
— бескилевые 322
— болотные 307
— водоплавающие 307, 320
— воздушно-водные 307
— воздушно-наземные 306, 307
—- выводковые 309, 314, 317, 319, 320
— древесные 306
— килегрудые 313, 314
— наземные 306, 307
— полувыводковые 309, 319
— птенцовые 309, 317, 320, 321
— хищные 320, 321
Пузырь плавательный рыб 249, 259
Пуночка 369
Пухоед 194, 195
Пчелы 35, 208, 209
Рабдиты 96
Равнокрылые 195
Радиолярии 48, 67
Радула 220
Раки 155, 160, 161, 164, 165, 166
—- высшие 157, 161, 163
— низшие 157
Ракообразные 155, 157
Редия 102
Ревун черный 361
Реликты 16
Ремнец 116
Рептилии 288, 291
Ресничные 77
Ржанкообразные 317
Рибосомы 26
Ришта 134
Росомаха 372
Ротовые органы насекомых 179
- — — грызущие 179, 190
- — — грызуще-сосущие 179
— — _ колюще-сосущие 180, 197
— — — сосущие 180
Рукокрылые 345, 346
Рыбы 249, 250
— двоякодышащие 274
— кистеперые 273, 274
— костистые 48, 269
— костно-хрящевые 48, 268
— костные 268
— лучеперые 268
— морские 263, 264
—- пресноводные 263, 271, 274
— проходные 263, 271
— хрящевые 267
Рысь 346, 351
Рябок белобрюхий 375
Рябчик 306, 311
Сайгак 354, 356, 374
Саламандра 284
Сапрофаги 187
Саранча 189, 191, 193
Сарган 251
Саркодовые 64
Сарколемма 35
Саркоптоидные 172
Свайник 130, 131
Сверчки 191
Свиньи 353, 354
Свиристель 301, 308, 372
Севрюга 269
Сельдевые 269, 270
Сенокосцы 172
Серна 356
Сивучи 352
Симбиоз 44, 45, 90, 143
Симметрия 43, 90
— двусторонняя 44, 211
— радиальная 44, 83, 225
Синица 309, 321
Синцитий 29
Сипы 321
Систематика 18, 48, 235
Сифон 214
— жаберный 214, 216
— клаокальный 214, 216
— ротовой 214, 216
388
Сифонофора 88
Скат 254, 259, 267, 268
Скворец 306, 321
Сколекс 107
Сколопендра 177
Скорпион 168, 169
Скребни 136, 137
Скумбрия 251
Славка 321
Слеппи 205
Слепыш 338, 374
Слепушонки 338
Слизни 221, 222
Слоны 360
Соболь 346, 351, 372
Сова 321, 369
Совообразные 321
Совка озимая 203
Сойка 310, 375
Сокол 320
Сом 263, 270
Сомовые 272
Сопи 348, 349
Сороки 321
Сосальщик кошачий 104
— ланцетовидный 104
- - печеночный 101
Сосалыцики дигенетичеекие 94, 98
- - монотеистические 94, 105
Сосуды мальпигиевы 168, 183
Спайник 105
Спикула 82
Спонгина 82
Спора 70, 75, 76
Споробласт 72
Споровики 45, 70
Спорозоит 72
Спороциста 102
Спячка 293, 340
Статоцисты 56, 85, 96, 216, 220
Стегоцефал 48, 285, 291, 367
Стервятник 321
Стержень 297
Стерлядь 263, 269
Стигма 55
Страусы 306, 322
Стриж 307
Стрекозы 194
Стрепет 308, 320, 374
Стробила 107
Судак 264, 270
Сумчатые 342, 343
Сурки 369
Суслик 348, 374, 375
Сцифистомы 90
Сыч мохноногий 372
Таксис 55
- отрицательный 55
• - положительный 55
Тапир 360
Тараканы 193
Тарантул 171
Тарпан 360
Тегумент 94, 99
Телобласты 138
Тельце Мелиса 100, 108
Термотаксис 55
Тетеревиные 314
Тигмотаксис 55
Тигр 326, 351
Ткань жировая 29
— костная 29, 31
— мышечная 27, 35
— нервная 27, 36
—- ретикулярная 29, 30
— соединительная 27, 29, 30, 236
— хрящевая 29, 31
— эпителиальная 27, 43
Тли 195
Толстолобик 266
Толсторог 356
Тонофибриллы 27
Трахейнодышащие 176
Трахея 153, 167, 242
Тресковые 273
Трипанозомы 68, 69
Тритоны 283, 284
Трихинелла 45, 46, 133
Трихинеллез 133
Трихоцисты 54
Трицератопс 295
Тромбоциты 33, 34
Трохофора 143
Трубочки мальпигиевы 155
Трубочник 148
Трясогузка 321
Туника асцидий 235
Тупик 319
Тушканчик 347, 349, 374, 375
Тюлень 346, 352
Угорь 251, 270
Угревые 272
Удавы 292
Уж обыкновенный 292
Улитки 221
Уростиль 275
Усачи 198
Устрицы 217
Утки 302
Утконос 341
Фагоциты 34, 97
Фазановые 314
Факторы среды 12, 14
— — абиотические 14
— _ антропогенные 15
— — биотические 15
— — географические 14
389
Факторы среды климатические 14
— — эдафические 14
Фаланга 168, 169
Фасциола 46, 47, 101
Фасциолез 102, 103
Фибриллы 53
Физиология 12
Филин 321
Филогенез 12, 14
Филогенетика 17
Финна 109, НО, 111, 113
— ценур 110, 111
— цистицерк 110
— эхинококк 110, 114
Фитомонады 69
Фитофаги 187
Фораминифера 51, 65, 66
Форель 266, 267
Фототаксис 55
— отрицательный 256
— положительный 59, 256
Хамелеоны 286, 293
Хелицероносные 166
Хелицеры 166, 168
Хемотаксис 55
Хищные 351, 352
Хоаны 242
Хоботные 360
Хоботок 180, 345
Хомяки 349, 374
Хомячки 349
Хордовые 229, 367
Хорион 344
Хорек 351
Хорь черный 371
Хромосомы 26
Хрящ 31, 32, 33, 236
Цапли 307, 320
Цевка 296, 300
Целом 42, 137, 211
Цепни 110, Ill
Цератозавр 295
Церкарии 102
Цестодообразные ПО
Цестоды НО
Ценур 110, 111
Цикады 195
Цистицерк 110, 111
Цитокариоплазма 26
Циклоп 161
Цитоплазма 23, 25
Цокор 337
Чайки 307, 309, 319
Чекан 322
Челюстноротые 249
Червецы 195
Черви брюхоресничные 136
— дождевые 144, 145
— кольчатые 137, 138
— круглые 117, 118, 122, 123, 127, 136
— ленточные 94, 105, 107
— малощетинковые 144
— многощетинковые 140, 141
— плоские 93, 94
— ресничные 94, 95, 96
Червяга 282
Черепахи 293, 294
Чесотка 174
Чешуекрылые 200
Чешуйчатые 291, 292
Чешуя рыб 252
— — ганоидная 252, 268
— — костная 252, 268, 269
— — плакоидная 252, 267
Чиж 321
Чистиковые 309, 319
Членистоногие 150, 210, 211
Шелкопряд 200, 201
Шизогония 39, 70, 72
Шимпанзе 361
Шмели 209
Щегол 321
Щука 251, 270
Щуковые 272
Эвглена зеленая 22
Эволюция 4, 22, 48, 63, 238, 364
Эймерия 70
Экзоподит 157
Экология 4, 14, 15
— земноводных 282, 283
— млекопитающих 337—340
— насекомых 186—189
— пресмыкающихся 290, 291
— птиц 306—312
— рыб 262, 263, 265, 267
Эктодерма 41, 43, 79, 83
Эктоплазма 26, 53
Элитры 181
Эмбриология 12, 296, 362, 364
Эмбрион 12, 95, 238, 285, 290, 344
Эндемики 16
Эндодерма 41, 43, 79, 83
Эндокринология 12
Эндоплазма 26, 53
Эндоподит 157
Эндостиль 232
Энхитреиды 147
Эпидермис 236
Эпистрофей 287, 328
Эпителий 43
— железистый 29, 167
— мерцательный 29, 96, 99, 102, 216
— многослойный 27, 28, 29, 236
— однослойный 27, 28, 96, 118, 139, 181,
390
212, 225
- ресничный 115, 212, 232
- цилиндрический 27, 28
- чувствующий 256
Эра архейская 48
- - кайнозойская 48, 65, 324, 368
- мезозойская 48, 294, 324, 367
палеозойская 48, 65, 83, 225, 273
- протерозойская 48
Эритроциты 33
Этология 14
Эхинококк 112, 113
Эхинококкоз 114
Эфы 293
Ядро 23, 26, 27, 49, 61, 62
Ядрышко 26
Яйца гомолецитальные 40
— телолецитальные 40
— центролецитальные 40
Яйцееды 210
Яйцеклад 151, 177, 209
Яйцекладущие 325, 341, 342
Як 356
Ястреб 308, 320
Ящерицы 292
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Acanthobdella 150
Acanthocephala 136
Acanthocephales 136
Acari 172
Acipenseridae 268
Acrania 230
Actionopterygii 268
Agnatha 247
Agriolimax agrestis 222
Alcae 319
Amniota 246, 296
Amoebina 64
Amphibia 274
Amphioxus laceolalus 230
Anamnia 246, 247
Anguillidae 272
Anguina tritici 125
Anisoplia 199
Annelida 137
Anodonta 219
Anopheles 203
Anoplura 194
Anseriformes 319
Antherea pernyi 200
Anthozoa 83, 90
Anura 284
Aphidae 195
Aphis pomi 195
Apis mellifera 208
Apoda 284
Aporia crataegi 202
Apterygota 189
Arachnida 168
Aranei 168, 169
Araneus 171
Arcella 64
Arenicola marina 143
Argyroneta 171
Arion circumscriptus 222
— empiricorum 222
Arionidae 222
Artiodactyla 353
Ascaridae 127
Ascaridia galli 130
392
Ascaris lumbrieoides 128
— suum 128
Asteroidea 227
Aves 296
Balantidium 78
Bivalvia 213
Blattoidea 193
Bombidac 209
Bombyx niori 200
Bos niutus 356
— taurus 356
Bothynoderes puiietiventris 19!)
Bovidae 355
Branchiala 155
Branchiopoda 160
Brevicoryne brassieae 195
Bubalus arnee 356
Camelus 357
Canidae 351
Capepoda 161
Capra aegagrus 356
Carinatae 313
Carnivora 351
Cephalopoda 223
Cephidae 210
Cephus pygmaeus 210
Cerambycidae 198
Cervidae 356
Cestoda 105, 108, 110
Cestodaria 108
Cetacea 352
Chamaeleontes 293
Charadrii 317
Charadriiformes 317
Cheliccrata 166
Chelonia 293
— mydas 294
Chiroptera 345
Chondrichthyes 267
Chondrostei 268
Chordata 229
Cicadidae 195
Cicouiilormes 320
Ciliophora 77
Cimex lectularius 197
Cirripedia 161
Ci tel iris 348
Cladocera 160
Clethrionoinys 34!)
Clupeidae 26!)
Cnidosporidia 75
Ciccidae 70, 105
Coelenterata 85
Coeloplana 98
Coleoplera 197
Colmnbiformcs 314
Cricelidae 349
Crieetnlus eversmanni 349
Cricetns 349
Crocodilia 294
Crossoptergygii 273
Crustacea 155
Clenophora 92
Cyclophyllidea 110
Cyclops 161
Cyclostomata 247
Cyprinidae 271
Culicidae 203
Daetilogyrus 105
Decapoda 164
Deiidrolinms piui 202
Dia p loinus 161
Dicrocoelium laneeolalinn 104
Dictyocaulus viviparns 130
Diffugia 64
Diphyllobothriimi latum 1 14
Diplozoon paradoxus 105
Dipnoi 274
Dipodidae 349
Ditylenchus alii 127
destructor 127
Dracuuculus niedinensis 134
Dreissena 219
Droniia 82
Echinococcus granulosus 112
Ecluiioidea 227
Eiiueria perforate 72
- sliedae 70
Elasniobranchia 267
Elateridae 199
Ends orbicularis 294
Enchy treidae 149
Enterobius vermicularis 130
Equidae 359
Equus caballus 360
Esocidae 272
Eunice viridis 143
Eupleclella 82
Euproctis ehysorrlioea 202
Eurygaster integriceps 197
Eutheria 344
Ealeoniforrnes 320
Easciola hepatica 101
Eelidae 351
Eoraminifera 65
Gadidae 273
Gallifornies 314
Gallus gallus 314
Gastrophilidae 206
Gastropoda 219
Gastrotricha 136
Gazella guttorosa 356
Geckonidae 292
Gnathostomata 249
Gobi id ae 272
Gregarinida 75
Grillotalpa 193
Gruiforines 320
Haemosporidia 72
Helix 221
Heterodera rostoehieusis 122
schaehti 123
Heteroderidae 122
1 leteroinetabola 190
Heteroptera 197
Hirudinea 148
I lolonietabola 197
Holothurioidea 227
Holotricha 77
Hydrozoa 83, 86
Hylidae 284
Hynienoptera 206
Hypoderma bovis 205
- 1 ineata 205
Hyponomeuta nialinella 202
Ichthyobdella 150
lehtyophthirius 78
Impennes 322
Infusoria 77
Insecta 177
Insectivora 345
Ixodes 174
Lacertilia 292
Lacerta agilis 292
~~ vivipara 292
Lagomorpha 351
Lama 357
Lari 319
Laspeyrcsia pomonella 202
Lathrodectus 171
Latimeria 274
Leishmania 69
— donowani 69
— tropica 69
Lepidoptera 200
393
Leptinotarsa decemlineata 198
Limacidae 222
Limnaea truncatula 102, 223
Locusta migratoria 193
Lycosa 171
Malacosoma neustria 200
Malacostraca 163
Mallophaga 194
Mammalia 325
Margaritana 219
Mastigophora 67
Mayetiola destructor 205
Maxillopoda 161
Meleagris gallopava 314
Meloidogyne marioni 125
Melolontha 198
Meriones 349
Metatheria 342
Metazoa 79
Microsporidia 76
Microtus arvalis 349
Mollusca 211
Moniezia expansa 114
Monogenea 150
Multiceps multiceps 111
Multituberculata 362
Muridae 350.
Muscidae 205
Mustelidae 351
Myoxidae 348
Myriapoda 176
Mystacoceti 353
Mytilus 219
Myxinoidea 249
Myxosporidia 76
Namathelminthes 117
Nematoda 118
Nematomorpha 134
Nonruninantia 353
Nereis 143
Nosema 76
— apis 76
— bombycis 76
Notostigmata 172
Odonata 194
Odontoceti 353
Oestridae 205
Oestrus 205
Oligochaeta 144
Ophidia 292
Ophryoscolecidae 78
Opistorchis felineus 104
Orthoptera 190
Osteichthyes 268
Ovis ammori 356
— canadensis 356
Oxyuridae 130
Parasitiformes 173, 175
Passeriformes 321
Pecten 217
Pelecanifornies 320
Percidae 272
Perissodactyla 358
Petromyzones 249
Pholas 214
Phlebothomus 203
Picifonnes 321
Pieris brassicae 202
— rapae 202
Pinnipedia 352
Piroplasmida 66
Pisces 249
Plasmodium 72
Plathelminthes 93
Pleuronectidae 272
Polychaeta 140
Polymastigina 70
Primates 360
Proboscidea 360
Prosauria 291
Proteidae 284
Prosthogonimus 104
Protoclepsis 150
Protothcria 341
Protozoa 49, 364
Pseudophyllidea 114
Psoroptes 174
Pteria 219
Pterygota 190
Radiolaria 67
Ranidae 284
Ratitae 322
Rattus 350
Reptilia 285
Rhinoceratidae 360
Rhombomys 349
Rodentia 347
Rotatoria 134
Ruminantia 355
Saiga tatarica 356
Salamandra 284
Salmonidae 271
Sarcodina 64
Sarcoptiformes 172
Scelionidae 210
Sciuridae 348
Scorpiones 168
Scyphozoa 83, 88
Siluridae 272
Simulidae 203
Solpugida 168
Spongia 79
Spongicola 82
Sporozoa 70
394
Squamata 291
Strigiformes 321
Strongylidae 130
Stylonichia 77
Sus scrota 354
Tabanidae 205
Taeniarhynchus saginatus 110
Taenia solium 111
Tapiridae 360
Tegenaria 171
Teleostei 269
Teredo 219
Testudo 294
Tetranidae 314
Tetrapoda 164, 274
Tracheata 176
Trematoda 98
Trichinella spiralis 133
Trichocephalus 133
Trituberculata 361
Triturus 283
Trombidiformes 172
Trypanosoma 68
— brucei 68
— equiperdum 69
— gambiensis 68
— evansi 69
Tubificidae 148
Turbellaria 95
Tylenchidae 122, 125
Tyroglyphida 173
Unio 219
Urochordata 233
Urodela 283
Ursidae 351
Varanus griseus 292
Vertebrata 235
Viteus vitifolii 195
Vorticella 77
СОДЕРЖАНИЕ
Предисловие к четвертому изданию......................................... 3
Введение ................................................................ 4
ОБШАЯ ЧАСТЬ............................................................. 22
Основные отличительные черты животного организма........................ 22
Клетка.................................................................. 23
Ткани................................................................... 27
Органы.................................................................. 37
Размножение животных.................................................... 37
Начальные этапы развития многоклеточных животных........................ 39
Симметрия тела животных................................................. 43
Симбиоз в животном мире................................................. 44
Развитие животного мира................................................. 47
СИСТЕМАТИЧЕСКИЙ ОБЗОР ЖИВОТНЫХ.......................................... 49
Подцарство Одноклеточные (Protozoa)..................................... 49
Тип Саркодовые и Жгутиковые (Sarcomastigophora)..................... 64
Класс Саркодовые (Sarcodina).................................... 64
Подкласс Радиолярии (Radiolaria)............................. 67
Класс Жгутиковые (Mastigophora)................................. 67
Тип Споровики (Sporozoa)............................................ 70
Тип Книдоспоридии (Cnidosporidia)................................... 75
Класс Слизистые споровики (Myxosporidia)........................ 76
Тип Микроспоридии (Microsporidia)................................... 76
Тип Ресничные (Ciliophora).......................................... 77
Класс Инфузории (Infusoria)..................................... 77
Подцарство Многоклеточные (Metazoa)..................................... 79
Тип Губки (Spongia)................................................. 79
Тип Кишечнополостные (Coelenterata).............................. 83
Класс Гидроидные (Hydrozoa).................................. 86
Класс Сцифоидные медузы (Scyphozoa).......................... 88
Класс Коралловые полипы (Anlhozoa)........................... 90
Тип Гребневики (Ctenophora)......................................... 92
Тип Плоские черви (РI a t hel m inthes)............................. 93
Класс Ресничные черви (Turbellaria)............................ 95
Класс Дигенетические сосальщики (Trematoda)..................... 98
396
Класс Мопогенетические сосальщики (Monogenea)................ 105
Класс Ленточные черви (Cestoda).............................. 105
Подкласс Цестодообразные (Cestodaria)...................... 110
Подкласс Цестоды (Cestoda)................................. 110
Тип Круглые черви (Nemathelminthes)............................... 117
Класс Собственно круглые черви, или Нематоды (Nematoda). . . 118
Свободноживущие круглые черви............................. 122
Круглые черви — паразиты растений......................... 122
Круглые черви — паразиты животных и человека.............. 127
Класс Волосатики (Neniatomorpha)............................. 134
Класс Коловратки (Rotatoria)................................. 134
Тип Скребни (Acanthoeephales).................................... 130
Класс Скребни (Aearithocephala).............................. 136
Тип Кольчатые черви (Annelida)................................... 137
Класс Многощетинковые черви (Polychaeta)..................... 140
Класс Малощетинковые черви (Oligochaeta)..................... 144
Класс Пиявки (Hirudinea)..................................... 148
Тин Членистоногие (Arthropoda)................................... 150
Подтип Жабернодышащие (Brancliiata)............................ 155
Класс Ракообразные (Crustacea)............................... 155
Подкласс Жаброногие (Branchiopoda)......................... 160
Подкласс Максиллоподы (Maxillopoda)........................ 161
Подкласс Высшие раки (Malacostraca)........................ 163
Подтип Хелицероноспые (Chelicerata)............................ 166
Класс Паукообразные (Arachnida).............................. 168
Подтип Трахейподышащие (Tracheata)............................. 176
Класс Многоножки (Myriapoda)................................. 176
Класс Насекомые (Insecta).................................... 177
Подкласс Первичнобескрылые (Apterygota).................... 189
Подкласс Крылатые (Pterygota).............................. 190
Отдел Насекомые с неполным превращением (Heteroine-
tabola).................................................. 190
Отдел Насекомые с полным превращением (Holometabola) . . 197
Тип Моллюски (Mollusca)...............................................211
Класс Двустворчатые моллюски (Bivalvia)...........................213
Класс Брюхоногие моллюски (Gastropoda)............................219
Класс Головоногие моллюски (Cephalopoda)..........................223
Тип Иглокожие (Echinodermata).........................................225
Класс Морские звезды (Asteroidea).................................227
Класе Морские ежи (Echinoidea)....................................227
Класс Голотурии (Holotliuroidea)..................................227
Тип Хордовые (Chordata)...............................................229
Подтип Бесчерепные (Acrania)........................................230
Подтип Личиночиохордовыс (Urochordata)..............................233
Подтип Позвоночные (Vertebrata).....................................235
Раздел Бесчелюстные (Agnatha)....................................... 247
Класс Круглоротые (Cyclostomata)................................ 247
Раздел Челюстноротые (Gnathostomata)................................ 249
Надкласс Рыбы (Pisces)............................................ 249
Класс Хрящевые рыбы (Chondrichthyes).............................267
Подкласс Пластинчатожаберные (Elasmobranchia)..................267
Класс Костные рыбы (Osteichthyes)................................268
Подкласс Лучеперые (Actinopterygii)............................268
Подкласс Кистеперые (Crossopterygii)...........................273
Подкласс Двоякодышащие (Dipnoi)............................... 274
397
Надкласс Наземные позвоночные (Tetrapoda)........................274
Класс Земноводные (Amphibia)..................................274
Класс Пресмыкающиеся (Reptilia)...............................285
Подкласс Первоящеры (Prosauria)...............................291
Подкласс Чешуйчатые (Squamata)................................291
Подкласс Черепахи (Chelonia)..................................293
Подкласс Крокодилы (Crocodilia)...............................294
Класс Птицы (Aves)...........................................29(i
Надотряд Килегрудые птицы (Carinatae).......................313
Надотряд Бескилевые птицы (Ratitae).........................322
Надотряд Пингвины (Iinpennes)...............................322
Класс Млекопитающие (Mammalia)................................325
Подкласс Яйцекладущие (Prototlieria)........................34 1
Подкласс Сумчатые (Metalheria)..............................342
Подкласс Плацентарные (Eutheria)............................344
Родословное древо животного мира......................................362
Животный мир Советского Союза............................................368
Охрана и рациональное использование животного мира в СССР................376
Указатель русских названий и терминов....................................383
Указатель латинских названий ........................................... 392
398
Кузнецов Борис Александрович,
Чернов Анатолий Зиновьевич,
Катонова Людмила Николаевна
КУРС ЗООЛОГИИ
Зав. редакцией В. И. Орлов
Художественный редактор В. Г. Прибегина
Технический редактор Т. Б. Платонова
Корректор В. //. Маркина
ИБ № 4972
Сдано в набор 30.11.87. Подписано к печати 23.08.88. Формат 60Х90‘/и>- Бумага
тин. № 1 . Гарнитура Литературная. Печать офсетная. Уел. печ. л. 25,0. Уел. кр.-отт.
25,0. Уч.-изд. л. 26,40. Изд. № 007. Тираж 31 500 экз. Заказ № 862. Цена 1 р. 20 к.
Зак. '62'3.
Ордена Трудового Красного Знамени ВО «Агропромйздат».
107807, ГСП-6, Москва, Б-78, ул. Садовая-Спасская, 18.
Диапозитивы изготовлены в Ленинградской типографии № 2 головном предприя-
тии ордена Трудового Красного Знамени Ленинградского объединения «Техни-
ческая книга» им. Евгении Соколовой Союзполиграфпрома при Государственном
комитете СССР по делам издательств, полиграфии и книжной торговли. 198052,
г. Ленинград, Л-52, Измайловский проспект, 29.
Отпечатано с диапозитивов в Ленинградской типографии № 6 ордена Трудового
Красного Знамени Ленинградского объединения «Техническая книга» им. Евгении
Соколовой Союзполиграфпрома при Государственном комитете СССР по делам
издательств, полиграфии и книжной торговли, 193144, г. Ленинград, ул. Моисе-
енко, 10.
В 1989 году
ВО «Агропромиздат»
выпустит книги:
Лукин Е. И. Зоология: Учебник для вузов.—
3-е изд., перераб. и доп.— М.: Агропромиздат, 1989
(I кв.) — 25 л.— ISBN 5—10-000722 2,—
1 р. 30 к.
Зоология в учебнике дана в применении к сель-
скохозяйственному производству.
В третье издание (второе — в 1981 г.) в перера-
ботанном виде включены «Введение», «Системати-
ческий обзор групп животных». Изложены наибо-
лее важные общебиологические и общезоологиче-
ские положения, новым материалом дополнены все
разделы учебника.
Для студентов по специальностям «Зоотехния»
и «Ветеринария».
Георгиевский В.И. Физиология сельско-
хозяйственных животных: Учебник для вузов.— М.:
Агропромиздат, 1989 (I кв.).— 45 л.— ISBN
5—10—000676—5. — 2 р. 10 к.
Рассмотрены функции и деятельность систем
организма: крови, крово- и лимфообращения, ды-
хания, пищеварения, а также обмен веществ и энер-
гии, выделительные процессы и внутренняя секре-
ция. Представлена физиология размножения, лак-
тации, работы мышц и нервов, центральной нервной
системы. В отдельном разделе даны сведения об
этологии животных.
Для студентов по специальности «Зоотехния».