/







































































































































































Text
АКАДЕМИЯ НАУК СССР
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
ТОМ LXIV
Б. И. БОГОСЛОВСКИЙ
ДЕВОНСКИЕ АММОНОИДЕИ РУДНОГО АЛТАЯ
ИЗДАТЕЛЬСТВО АКАДЕМИИ ПАУК СССР
Москва 1958
АКАДЕМИЯ НАУК СССР
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА ТОМ LXIV
Б. И. БОГОСЛОВСКИЙ
ДЕВОНСКИЕ АММОНОИДЕИ РУДНОГО АЛТАЯ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва 1958
Работа Б. И. Богословского представляет описание девонских аммоноидей (вымерших головоногих моллюсков) Азиатской части СССР. Рассматриваются их систематика, происхождение, образ жизни и значение для определения геологического возраста слоев Земли. Рассчитана на палеонтологов и геологов.
ОТВЕТСТВЕННЫЙ РЕДАКТОР
В. Е. РУЖЕНЦЕВ
ПРЕДИСЛОВИЕ
Широко развернувшиеся в течение послевоенных пятилеток работы по геологическому исследованию Азиатской части СССР выдвинули в числе других проблем задачу всестороннего изучения важного в народнохозяйственном отношении и геологически сложного района Рудного Алтая. В связи с этим создалась необходимость разработки единой стратиграфической схемы палеозойских эффузивно-осадочных отложений данного района. Особенно большое практическое значение приобрел вопрос стратиграфии эффузивно-осадочного комплекса девонских отложений, для которых единой дробной стратиграфической схемы пока нет.
Предпринятое в последние годы Горно-металлургическим научно-исследовательским институтом и Геологическим институтом Академии наук Казахской ССР, Палеонтологическим институтом Академии наук СССР и другими учреждениями всестороннее изучение фауны эффузивно-осадочного комплекса Рудного Алтая с целью составления коллективной работы «Палеонтологическое обоснование стратиграфии палеозоя Рудного Алтая» несомненно должно содействовать успешному разрешению этой важнейшей задачи.
В связи с постановкой такой работы, согласно договору о социалистическом содружестве между Горно-металлургическим научно-исследовательским институтом Академии наук Казахской ССР и Палеонтологическим институтом Академии наук СССР, автору было поручено изучение девонских аммоноидей указанного района. Помимо решения чисто практических вопросов стратиграфического характера, необходимость изучения девонских аммонойдей диктовалась еще и тем, что до последнего времени в нашей палеонтологической литературе исследованию этой интересной группы древнейших цефалопод уделялось совершенно недостаточное внимание, в то время как всестороннее изучение древнейших представителей аммоноидей позволяет решить ряд вопросов, связанных с происхождением, эволюцией и угасанием всего отряда аммонитов в целом.
Материалом для настоящей работы послужили коллекции, собранные автором в течение летних периодов 1952 и 1953 гг. при участии практикантов Пермского государственного университета имени А. М. Горького — студентов А. К. Рогожникова и В. П. Телегина. Полевые работы были организованы Горно-металлургическим институтом Академии наук Казахской ССР и проводились в тесном контакте с Алтайской геологосъемочной экспедицией Треста аэрогеологии. Помимо собственных сборов, автор пользовался любезно переданными ему Д. В. Наливкиным небольшими коллекциями, собранными ранее Н. А. Ноздреевым, Д. М. Шилиным, Н. А. Елисеевым и другими исследователями. Некоторое количество образцов было получено также от старшего научного сотрудника Горно-металлургического научно-исследовательского института, доктора геологоминералогических наук Г. Н. Щербы и геологов Алтайской экспедиции
3
аэрогеологии Г. Ф. Яковлева, В. П. Курбатова и др. В результате в распоряжении автора оказалась коллекция, состоящая более чем из 1500 образцов, из которых около 1200 были пригодными для изучения и монографического описания.
Следует отметить, что большая часть материала была не очень хорошей сохранности и обработка его была сильно затруднена ввиду значительной твердости и плотности вмещающих пород, однако тщательная препаровка образцов позволила получить значительное количество удовлетворительной сохранности раковин и даже произвести в ряде случаев онтогенетические исследования.
При исследовании коллекций был применен метод изучения ископаемых головоногих, разработанный в Палеонтологическом институте Академии наук СССР, основанный на широком охвате изучаемой группы вымерших организмов, тесной взаимосвязи онто- и филогенеза и филогенетическом понимании систематики. Работа была проведена под руководством проф. В. Е. Руженцева.
При проведении полевых исследований помощь и содействие в работе были оказаны автору со стороны старшего научного сотрудника ГМНИИ доктора геолого-минералогических наук П. Ф. Иванкина, заведующего отделом геологии института Ж. А. Айталиева, геологов Алтайской экспедиции Треста аэрогеологии Г. Ф. Яковлева, В. А. Федоровского, Д. И. Горжевского и других, предоставивших необходимые материалы по геологии района. На протяжении всей работы всесторонняя помощь оказывалась автору старшим научным сотрудником ГМНИИ доктором геологоминералогических наук Н. Л. Бубличенко, проявлявшим большой интерес к проводимым исследованиям. Ряд ценных замечаний по работе был получен автором от доктора биологических наук В. Н. Шиманского.
Фотографии выполнены в фотолаборатории Палеонтологического института А. В. Скиндером.
Всем указанным лицам автор выражает свою глубокую признательность.
I. ИСТОРИЯ ИЗУЧЕНИЯ ДЕВОНСКИХ АММОНОИДЕИ СССР
Первые сведения о девонских аммоноидеях СССР относятся к 1844 г., когда А. Кейзерлингом (Keyserling, 1844) были описаны некоторые верхнедевонские аммоноидеи Печорского края из доманикового горизонта Южного Тимана: Goniatites аттоп Keyserling, G. cinctus Keyserling, G. strangulatus Keyserling, G. retrorsus Buch, G. bisulcatus Keyserling, G. uchtensis Keyserling, G. acutus Keyserling, G. wurmii Romer, относимые в настоящее время к родам Tornoceras, Manticoceras, Ponticeras и Tima-nites. Двумя годами позднее тот же автор среди представителей других групп организмов дает повторное описание указанных выше видов аммо-ноидей. Следует отметить, что Кейзерлинг рассматривает отложения доманикового горизонта как древнейшие в тиманском девоне, переходные от силура к девону. После сообщений Кейзерлинга о девонских цефалоподах Тимана никаких дальнейших исследований этих ископаемых не было произведено и опубликовано в течение полувека.
В течение 60—80-х годов прошлого столетия изучению девонских цефалопод уделил внимание А. П. Карпинский, которым в ряде мелких сообщений (1869, 1884, 1885) приводятся сведения о первых находках на Урале типичных верхнедевонских гониатитов и климений: Goniatites retrorsus Buch, G. аттоп Keyserling, G. globosus Munster, G. bronni Munster, G. falcifer (?) Munster, Goniatites sp. nov., Prolobites delphinus (Sand-berger), Clymenia cf. annulata Munster, Cl. striata Munster, относимых по современной систематике к родам Tornoceras, Manticoceras, S рог adorer as, Prolobites, Platyclymenia, Cymaclymenia.
В последние годы прошлого столетия ряд исследователей использовал результаты изучения аммоноидей для решения вопросов стратиграфии девона. Ф. Н. Чернышев (1887) в результате изучения фауны, собранной А. А. Краснопольским по рекам Пшикаю, Сикашты, Тереклы (Южный Урал), выделил среди верхнедевонских отложений два горизонта: нижний— D3, характеризующийся представителями Goniatites {Tornoceras) simplex (Buch), G. {Manticoceras) аттоп Keyserling, G. {Gephyroceras) bisulcatus Keyserling, G. {Manticoceras) intumescens^yxich, и верхний, содержащий Clymenia {Platyclymenia) annulata Munster, Cl. {Platyclymenia) fle-xuosa Minister, Cl. {Cyrtoclymenia) krasnopolskii Tschernyschew, Goniatites {Tornoceras) simplex Buch, G. {Cheiloceras) verneuili Minister. Чернышевым впервые на западном склоне Урала было обнаружено присутствие хейло-церового горизонта, который в следующем году был установлен П. И. Кротовым (1888) и на Северном Урале в бассейне р. Яйвы, где был найден Goniatites {Cheiloceras) Munster .В 1892 г. Ф. Ю. Левинсон-Лессинг
на восточном склоне Урала в Губерлинских горах обнаружил климение-выеслои с Clymenia undulata Munster, Cl. dunkeri Minister, Cl. angustisep-tata Minister, Cl. aff. flexuosavax. costulata Minister, Cl. laevigataWw&ter, Cl. cf. krasnopolskii Tschernyschew, Cl. paradoxa Minister, C/< cf. angulosa
5
Minister, Cl. cf. subarmata Munster, Cl. striata Miinster, Cl. speciosa Munster, Cl. cf. speciosa Miinster, Cl. barbarae Loewinson-Lessing, Cl. inostran-cevi Loewinson-Lessing, Cl. dubia Loewinson-Lessing, Goniatites cf. delphi-nus Sandberger, G. {Sporadoceras) miinsteri Buch, G. {Brancoceras) linearis Miinster, относимые в настоящее время к родам Prolobites, Sporadoceras, Platyclymenia, Cyrtoclymenia, Protoxyclymenia, Clymenia, Cymaclymenia и характеризующие пролобитовый горизонт фаменского яруса. Первое указание на присутствие аммоноидей {Anarcestes) в девонских отложениях Рудного Алтая и определение их возраста приводятся в опубликованной в следующем году работе Ф. Н. Чернышева (1893). Краткое описание отдельных находок аммоноидей из девонских отложений Мугоджар дается наряду с описанием иной фауны П. Венюковым в 1895 г.
В 1899 г. вышла работа Э. Гольцапфеля, посвященная изучению головоногих доманикового горизонта Южного Тимана, собранных ранее Кейзер-лингом, Цикендратом, Чернышевым и другими исследователями. В результате ревизии видов Кейзерлинга и обработки новых коллекций Гольцапфе-лем описаны: Tornoceras simplex (Buch), Т. cinctum (Keyserling), Mantico-ceras intumescens (Beyrich), M. ammon (Keyserling), M. retrorsum (Buch), ДГ. backlundi Holzapfel, Gephyroceras bisulcatum (Keyserling), G. tscherny-schewi Holzapfel, G. regale Holzapfel, G. uralicum Holzapfel, G. auritum Holzapfel, G. uchtense (Keyserling), G. lebedeffi Holzapfel, G. Keyser-lingi Holzapfel, G. domanicense Holzapfel, G. syrjanicum Holzapfel, Tima-nites acutus (Keyserling), T. stuckenbergi Holzapfel, Prolecanites timanicus Holzapfel, относимые в настоящее время к родам: Tornoceras, Ponticeras, Uchtites, Manticoceras, Timanites, Komioceras и Nordiceras.
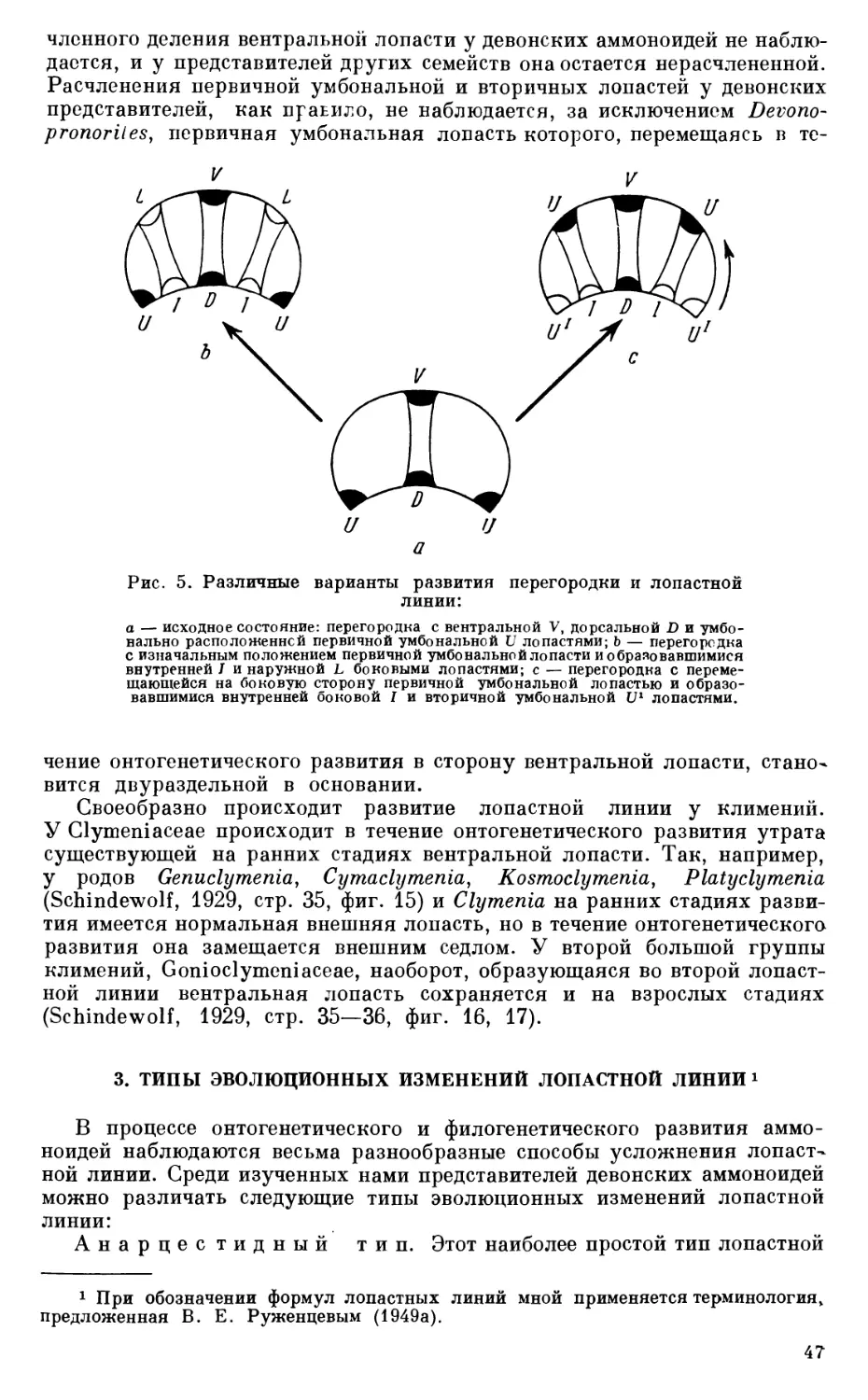
Работа Гольцапфеля выгодно отличается как от работ предыдущих исследователей, так и от многих (описательно-стратиграфических) работ более поздних авторов, так как в ней помимо описания, общей характеристики и стратиграфического значения изученной фауны затрагиваются вопросы фациальной приуроченности тиманских головоногих и впервые в отечественной литературе делаются попытки онтогенетического изучения девонских аммоноидей {Timanites acutus).
В первые годы текущего столетия (дореволюционный период) изучению девонских аммоноидей уделил внимание ряд исследователей, однако работы их преимущественно носят чисто описательный характер; представители аммоноидей описаны зачастую весьма кратко, попутно с описанием иной фауны. В 1901 г. Г. Петц описал единственного гониатита Beloceras multilobatum (Beyrich) из верхнедевонских отложений Кузнецкого угленосного бассейна. Двумя годами позднее Л. Токаренко (1903) были описаны из фаменских отложений района г. Верхнеуральска: Aganides sp., Prolobites delphinus (Sandberger), Sporadoceras miinsteri (Buch), Sp. miinsteri var. brachiloba Freeh var. varietatis uralensis Tokarenko, Manticoceras intumescens (Byerich) {=Sporadoceras inflexum^eAeWnA), Tornoceras planidor-satum (Miinster), Aphyllites frechi Tokarenko [=Cyrtoclymenia involuta var. jrechi (Tokarenko)], Clymenia krasnopolskii Tschernyschew, Cl. stuckenbergi Tokarenko, Cl. flexuosa Miinster, Cl. annulata Munster, Cl. annulata var. densicostata Freeh, ? Gephyroceras domanicense Holzapfel [=Cyrto-clymenia flexuosa var. orientalis (Perna)], Clymenia sp., относимые теперь к родам Prolobites, Tornoceras {Protornoceras), Sporadoceras, Cyrtoclymenia и Platyclymenia. Некоторые представители аммоноидей из фаменских Отложений Мугоджарских гор были кратко описаны в 1911 г. Г. М. Нахим-соном. В 1916 г. С. В. Обручевым сообщается о находке отдельных гониа-титов {Manticoceras intumescens) в Воронежском девоне.
Среди других исследований этого периода выделяется работа Э. Я. Пэр-на (1914), представляющая собой монографическое описание верхнедевонских гониатитов и климений восточного склона Южного Урала из окрест
6
ностей г. Верхнеуральска, пос. Спасского и дер. Кирсы. В результате исследования обширных собственных сборов и изучения монографических коллекций Левинсон-Лессинга и Токаренко, Парна было описано 48 видов и вариететов гониатитов и климений (из них 24 новых), относимых в настоящее время к родам Prolobites, Tornoceras (Protornoceras), Pseudocly-menia, Cheiloceras, Torleyoceras, Dyscheiloceras, Sporadoceras, Cyrtoclyme-nia, Platyclymenia, Rectoclymenia, Protoxy clymenia, Genuclymenia и характеризующих хейлоцеровый, пролобитовый и «аннулятовый» (плати-климениевый) горизонты фаменского яруса.
Работа Э. Я. Парна является существенным вкладом в дело познания девонской цефалоподовой фауны. Помимо описания, установления возраста и стратиграфического значения фауны, в ней освещаются вопросы фациальной приуроченности и образа жизни изучаемой группы, используется онтогенетический метод исследования для установления родственных связей между отдельными ее представителями.
За годы советской власти в результате широко развернувшихся комплексных геологических исследований в различных районах СССР было открыто значительное количество новых местонахождений девонских аммоноидей. Разнообразные гониатитовые фации различного возраста, пользующиеся значительным распространением, были найдены на Новой Земле, на Тимане, по всему Уралу, в Мугоджарах, в Центральном Казахстане, в Рудном Алтае, на севере Азиатской части СССР, в бассейне р. Колымы; отдельные находки аммоноидей были отмечены во многих пунктах на Русской платформе. Естественно поэтому, что изучением девонских аммоноидей, весьма важных для целей стратиграфии, занимался ряд авторов. В 1926 г. М. Э. Янишевский описывает Manticoceras intumescens var. latisellata Janischevsky, Gephyroceras aff. uchtense (Keyserling) (?), Gephyroceras sp., Tornoceras simplex (Buch) из верхнедевонских отложений Новой Земли. Несколько форм гониатитов было кратко описано Д. В. На-ливкиным (1930) среди фауны семилукских и воронежских слоев. В 1936 г. тем же автором при описании фауны верховьев рек Колымы и Хандыги кратко описываются Gephyroceras uchtense (Keyserling) [=Ponticeras uchtense (Keyserling)] и Pharciceras timanicum (Holzapfel) \= Nor dicer as timanicum (Holzapfel)], на основании которых устанавливается нижне-франский возраст вмещающих отложений. В 1936 г. А. К. Наливкина (1936а) впервые описывает весьма редкую для западного склона Урала хейлоцеровую фауну гониатитовых слоев р. Ай у Петропавловского завода, содержащую Tornoceras planidorsatum (Munster), Т. dorsoplanum Sobolew, T. escoti Freeh, T. haugi Freeh [ = 7\ simplex (Buch)], Cheiloceras subpartitum (Munster), Ch. subpartitum var. amblyloba (Sandberger), Ch. lagoviense Giirich, Ch. curvispinum (Sandberger), Ch. sacculum (Sandberger), Ch. circumflexum (Sandberger), Ch. praelentiformis Sobolew, Ch. umbili-catum (Sandberger), Ch. nehdense (Kayser), Ch. umbelifer Sobolew, Imitoce-ras lentiforme (Sandberger), Dimeroceras lentiforme Sobolew, Praeglyphio-ceras uralicum Nalivkina; кроме того, указываются Cheiloceras verneuili (Munster), Ch. acutum (Sandberger). В том же году А. К. Наливкиной (19366) описываются и некоторые верхнедевонские аммоноидей Новой Земли из мантикоцерового, хейлоцерового и пролобитового горизонтов: Manticoceras intumescens (Beyrich), М. ammon (Keyserling), M. carinatum (Sandberger), Gephyroceras sp., Tornoceras simplex (Buch), Cheiloceras acutum (Sandberger), Ch. verneuili (Munster), Dimeroceras sp., Cyrtoclymenia involuta Wedekind var. arctica Nalivkina.
В 1938 г. С. E. Колотухиной устанавливается наличие ортоклимение-вого и пролобитового горизонтов в фаменском ярусе Центрального Казахстана, откуда описываются: Praeglyphioceras pseudosphaericum Freeh var. itazuensis Kolotuchina, Varioclymenia varicata (Sobolew), V. callimorpha
7
Lange, V. cf. pompeckii (Wedekind), Cyrtoclymenia pinnata (Perna) var. kajraktensis Kolotuchina, Orthoclymenia cranoidejormis Kolotuchina, Pro-toxyclymenia cf. dunkeri (Munster), Sporadoceras sp. В следующем году H. Л. Бубличенко (1939) впервые отмечается верхнедевонский гониатито-вый горизонт с Manticoceras cf. carinatum (Sandberger), M. cf. schellwieni Wedekind, Gephyroceras sp., Beloceras multilobatum (Beyrich) [=B. sa-gittarium (Sandberger)] в Рудном Алтае.
Накопившиеся в результате предыдущих исследований сведения по девонским аммоноидеям дали возможность в 1947 г., в III томе «Атласа руководящих форм ископаемых фаун СССР» (А. К. Наливкина, 1947), привести краткие описания известных в СССР руководящих форм аммо-ноидей, относящихся к родам: Anarcestes, Agoniatites, Pinacites, Маепе-ceras (=Maenioceras), Manticoceras, Gephyroceras (=Ponticeras), Timani-tes, Beloceras, Pharciceras (= Nor dicer as), Tornoceras, Pseudoclymenia, Cheiloceras, Sporadoceras, Dimeroceras, Imitoceras, Prolobites, Prionoceras, Rectoclymenia, Cyrtoclymenia, Genuclymenia, Cymaclymenia, Platyclyme-nia, Laevigites (=Clymenia), Oxyclymenia (=Kosmoclymenia), Sellaclyme-nia.
В последующие годы наличие гониатитовых и климениевых фаций и списки находимых в них аммоноидей указываются А. И. Левенко (1951) для франских отложений Рудного Алтая, Н. Г. Чочиа и К. И. Андриановой (1952) для девона Колво-Вишерского края, В. Н. Тихим (1953) для девонских отложений Среднего Поволжья и Заволжья, С. М. Домрачевым (1952, 1953) для девона хребта Кара-Тау и прилегающих районов и для девона западного склона Среднего и Южного Урала, Е. Н. Ларионовой и Н. И. Петреневой (1953) для девона Пермского Прикамья и Удмуртской АССР, В. И. Троепольским (1953) для девона Татарской АССР и т. д.
Краткий обзор гониатитовых фаций СССР с выделением горизонтов и зон, для которых приводятся наиболее характерные формы, сделан в 1953 г. А. К. Наливкиной (1953а), дающей сводную таблицу гониатитовых горизонтов для отдельных регионов Европейской части СССР.
В том же году появилась работа А. К. Наливкиной (19536), представляющая специальную монографию по аммоноидеям фамепского яруса Мугоджар. Изученные коллекции характеризуют довольно полно фауну пролобитового горизонта, особенно богато представлена фауна верхней его части (зона III р). В работе описано 54 вида и разновидности гониатитов и климений (из них 12 новых), принадлежащих родам Tornoceras (с подродом Protornoceras), Lobotornoceras, Pseudoclymenia, Posttornoceras, Sporadoceras, Dimeroceras, Praeglyphioceras, Prolobites, Rectoclymenia, Cyrtoclymenia, Platy clymenia, Genuclymenia.
Заканчивая обзор истории изучения девонских аммоноидей СССР, следует, наконец, указать посвященные изучению фаменской фауны Казахстана и Кара-Тау работы Е. А. и 3. Г. Балашовых (1953, 1954) и А. И. Золкиной (1954), а также статьи автора (1954, 1955а, 19556, 1955в) о новых представителях аммоноидей из верхнего девона Рудного Алтая и Южного Урала.
Данный краткий обзор изучения девонских аммоноидей СССР исчерпывает все исследования, проводившиеся в этой области за более чем столетний период.
Несмотря на большое разнообразие гониатитовых фаун СССР, большой теоретический интерес и практическую важность изучения девонских аммоноидей, последние изучены еще далеко недостаточно. До сих пор главным образом описывались отдельные формы, найденные в различных горизонтах, но почти не давалось описания девонских аммонитовых фаун, почти не производилось монографического их изучения. Если фаменские аммоноидеи (далеко не все) более или менее изучены, то значительно мень
8
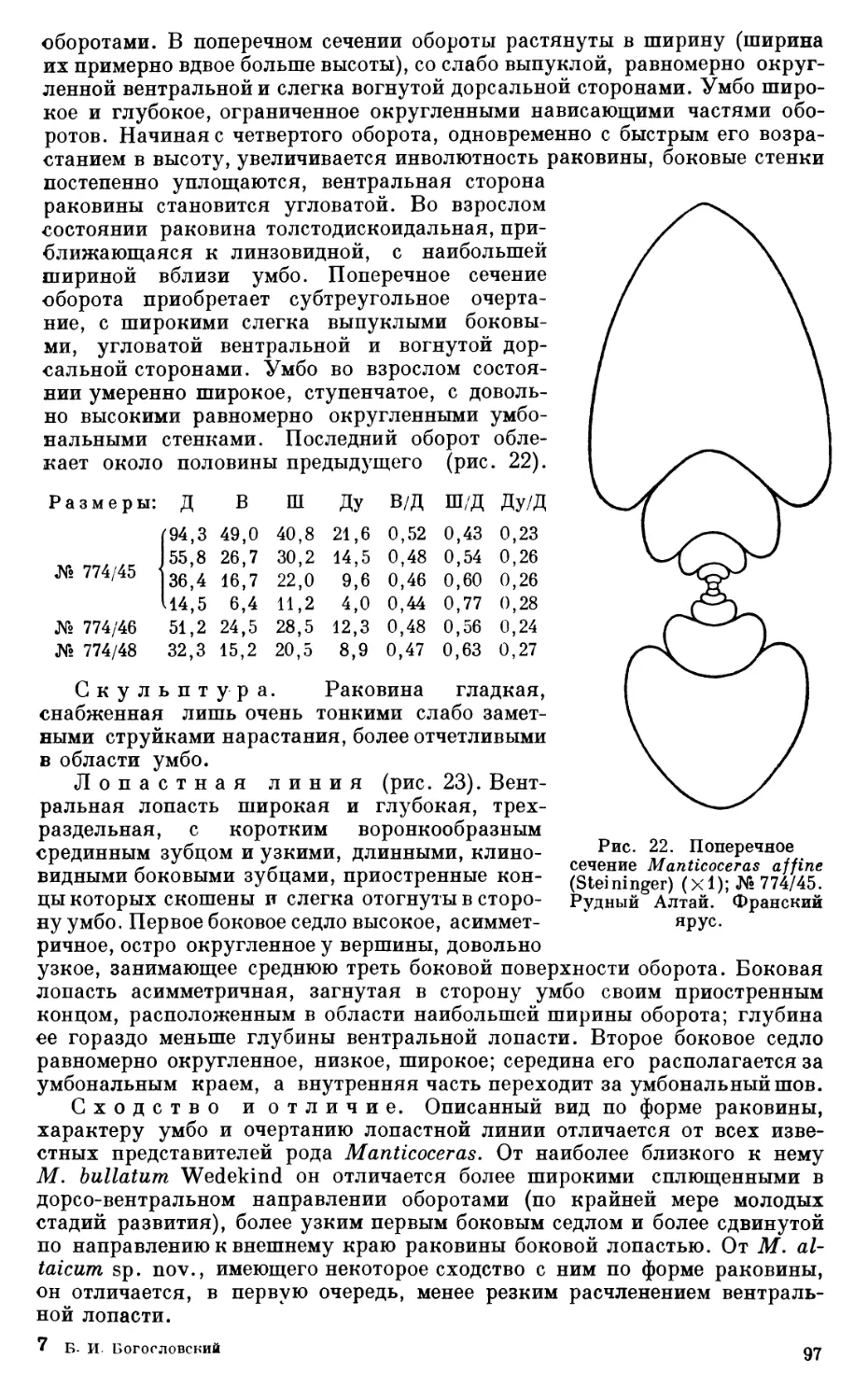
ше или почти не изучены аммоноидей франского яруса, не говоря уже о среднедевонских. Даже в тех немногочисленных случаях, когда изучаются аммонитовые фауны отдельных районов, работы носят чисто описательный (морфологический) характер, отличаясь односторонним стратиграфическим уклоном. В нашей литературе отсутствует широкое, разностороннее исследование девонских аммонитовых фаун, не затрагиваются и не освещаются важнейшие вопросы, связанные с происхождением, эволюцией, функциональными особенностями, фациальной приуроченностью, образом жизни этой весьма интересной и стратиграфически важной группы. При изучении девонских аммоноидей, к сожалению, совершенно не используется онто-филогенетический метод, с успехом применяемый при изучении вз хнепалеозойских аммоноидей. До сих пор в нашей палеонтологической литературе отсутствуют теоретически важные и практически необходимые сводки по девонским аммоноидеям СССР, какие, например, выполнены по девонским аммоноидеям Северной Америки.
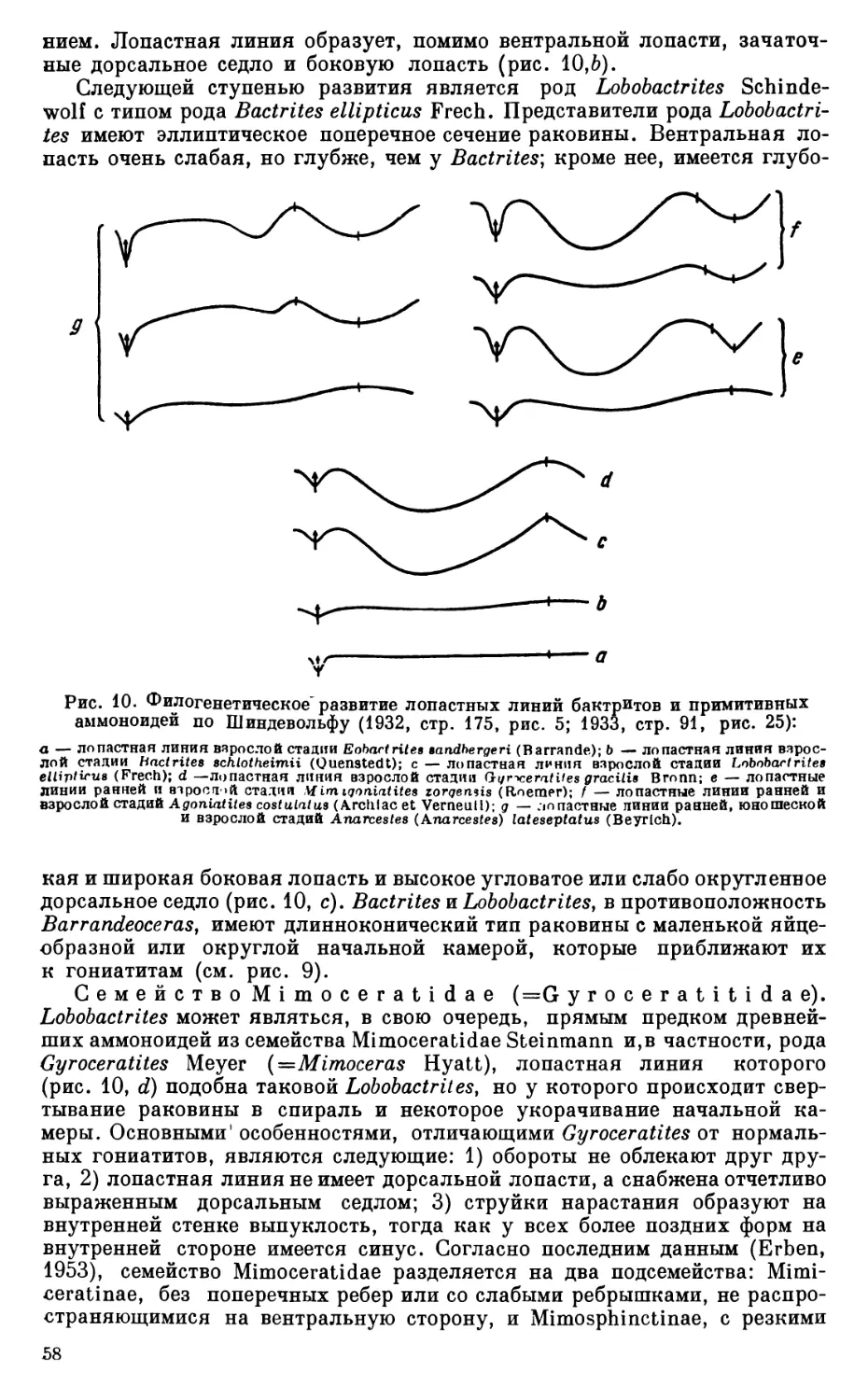
Тщательные поиски гониатитовых горизонтов и детальное, с применением онтогенетического метода, изучение как средне-, так и верхнедевонских фаун аммоноидей совершенно необходимы для разработки общей) стратиграфической схемы девонских отложений СССР, для построения филогенетической систематики девонских аммоноидей и всего отряда аммонитов в целом.
II. БИОСТРАТИГРАФИЧЕСКИИ ОЧЕРК
1. ОСНОВНАЯ БИОСТРАТИГРАФИЧЕСКАЯ ШКАЛА ЗАПАДНОЙ ЕВРОПЫ
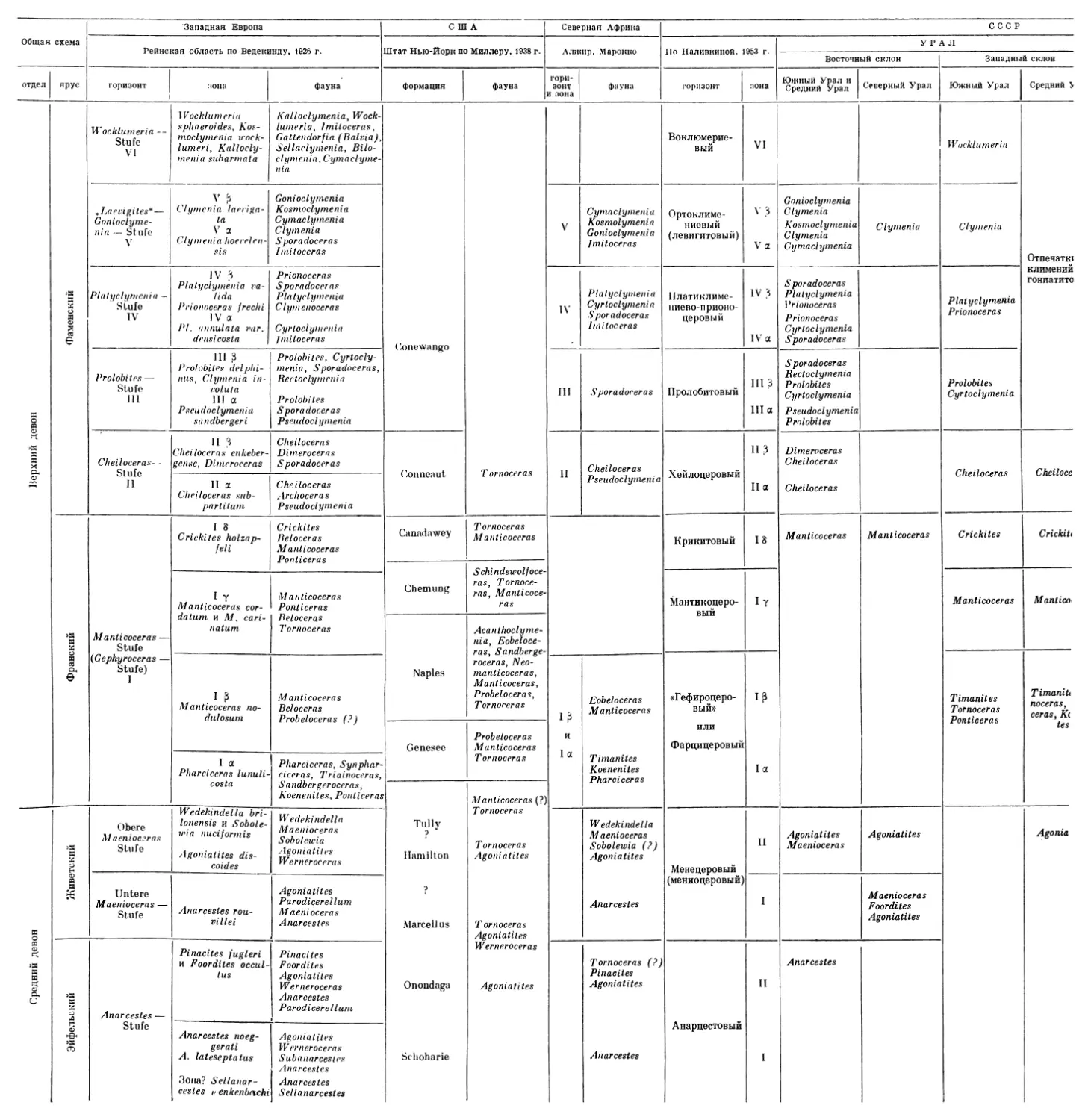
Для того чтобы правильно осветить стратиграфическое значение изученных коллекций аммоноидей, прежде чем перейти к вопросу о возрасте различных слоев с аммоноидеями в девонских отложениях Рудного Алтая, необходимо вкратце остановиться на биостратиграфических схемах девона по аммоноидеям, разработанных для других областей. Наиболее полно такая схема разработана для средне- и верхнедевонских отложений Рейнской провинции Западной Европы, для которых Ведекиндом (Wedekind, 1913, 1918, 1926) была предложена схема разделения на горизонты («Stufe») и зоны, несколько видоизмененная в 1926 г. Шиндеволь-фом (Schindewolf, 1926).
Среднедевонские отложения Западной Европы по аммоноидеям разделяются на три горизонта: горизонт с Anarcestes (Anarcestes-StvAe), соответствующий эйфельскому ярусу, и два горизонта с Maenioceras (Untere Maenioceras-SVafe и Obere Maenioceras-Stafe), отвечающие живетскому ярусу. Каждый из горизонтов разделяется на зоны. Внутри горизонта с Anarcestes выделяются зоны: Sellanarcestes wenkenbachi, Anarcestes late-septatus и A. noeggerati, Foordites occultus, Pinacites jugleri', нижний горизонт c Maenioceras характеризуется зоной Anarcestes rouvillei (низы); верхний горизонт с Maenioceras— зонами Agoniatitet discoides, A. fulgu-ralis и Wedekindella brilonensis.
Верхнедевонские отложения, содержащие богатую фауну аммоноидей, разделяются Ведекиндом на шесть горизонтов: Manticoceras-Sluie, <Cheiloceras-StVLfe, Prolobites-SVafe, Platy clymenia-Stafe, Laevigites-Stufe и Wocklumeria-SVaie. Эти шесть подразделений, первое из которых соответствует фрапскому ярусу, а остальные пять — фаменскому, часто именуются как Oberdevonstufe I, Oberdevonstufe II и т. д.; внутри этих горизонтов выделяются зоны: Oberdevonstufe la, Oberdevonstufe ip и т. д. Так, внутри горизонта с Manticoceras выделяются зоны: la (зона Pharciceras lunuli-costa), ip (зона Manticoceras nodulosum), ly (зона Manticoceras cordatum и M. carinatum), 18 (зона Crickites holzapfeliy, внутри горизонта c Cheiloceras — зоны Ila и lip и т. д.
Шиндевольф (1926), который продолжал работу Ведекинда, выделил еще верхнюю (седьмую) зону верхнего девона, или Gattendorfia-SVule, которая, однако, рассматривается Шмидтом и другими исследователями как нижнекаменноугольная. Позднее Шиндевольф (19346) объединил Prolobites-Stute и Platyclymenia-StvAe Ведекинда и различал шесть главных подразделений верхнего девона: Manticoceras-, Cheiloceras-, Prolobites-Platy clymenia-, Laevigites^Gonioclymenia-, Wocklumeria- и Gattendorfia-
1 Родовое название Laevigites, как установил Шиндевольф (1955), является синонимом Clymenia Munster.
10
Общая схема Западная Европа США Северная Африка СССР
Рейнская область по Ведекинду, 1926 г. Штат Нью-Йорк пс Миллеру, 1938 г. Алжир, Марокко По Наливкиной, 1953 г. УРАЛ
Восточный склон Западный склон
отдел ярус горизонт | зона фауна формация фауна горизонт и зона фауна горизонт зона Южный Урал и Средний Урал Северный Урал Южный Урал Средний У
Wocklumeria — Stufe VI Wocklumeria sphaeroides, Kos-moclymenia wock-lumeri, Kallocly-menia subarmala Kalloclymenia, Wocklumeria, Imitoceras, Gattendorfia (Balvia), Sellaclymenia, Bilo-clymenia, Cymaclymenia Воклюмерие-вый VI Wocklumeria
„Laevigites*— Gonioclymenia — Stufe V V Clymenia laeriga-ta \7 a Clymenia hoerelen-sis Gonioclymenia Kosnioclymen i a Cymaclymenia Clymenia S poradoceras Imitoceras V Cymaclymenia Kosmolymenia Gonioclymenia Imitoceras Ортоклиме-ниевый (левигитовый) V 3 V а Gonioclymenia Clymenia Kosmoclymenia Clymenia Cymaclymenia Clymenia Clymenia Отпечатю климений гониатито
Фаменский Platyclymenia - Stufe IV IV 3 Platyclymenia rail da Prionoceras frechi ]V a Pl. annulata rar. densicosta Prionoceras S poradoceras Platyclymenia Clymenoceras Cyrloclymenia Jmitoceras Conewango IV Platyclymenia Cyrloclymenia S poradoceras 1 mi toe eras Илатиклиме-пиево-прионо-церовый IV 3 IV а S poradoceras Platyclymenia Prionoceras Prionoceras Cyrloclymenia S poradoceras Platyclymenia Prionoceras
о л Prolobites — Stufe HI 111 (3 Prolobites delphi-nus, Clymenia itiro lut a III a Pseudoclytnenia sandbergeri Prolobites, Cyrtocly-menia, Sporadoceras, Recloclymeni a Prolobites Spora doceras Pseudoclytnenia III S poradoceras Пролобитовый 1П з III а S poradoceras Rectoclymenia Prolobites Cyrloclymenia Pseudoclytnenia Prolobites Prolobites Cyrloclymenia
эЯ S я и Рн Cheiloceras- -Stufe II 11 3 Cheiloceras enkeber-gense, Dimeroceras Cheiloceras Dimeroceras S poradoceras Conneaut T ornoceras II Cheiloceras Хейлоцеровый II 3 Dimeroceras Cheiloceras Cheiloceras Cheiloce
ф 11 a Cheiloceras subpar tituni Cheiloceras Archoceras Pseudoclytnenia Pseudoclytnenia Па Cheiloceras
1 8 Crickites holzap-feli Crickites Beloceras Manticoceras Ponticeras Canada wey Tornoceras M anticoceras Крикитовый 18 Manticoceras Manticoceras Crickites Crickiti
S chindew olfoce-ras, Tornoceras, Manticoceras
1 Y Manticoceras cor- M anticoceras ' Ponticeras Chemung Мантикоцеро-вый I Y Manticoceras Mantico
»я я я Manticoceras — Stufe datum и M. cari-natum Beloceras Tornoceras Acanthoclyme-nia, Eobeloce-ras, Sandbergeroceras, Neo-manticoceras, Manticoceras, Probeloceras, Tornoceras
о я со Рч е (Gephyroceras — Stufe) I I ? Manticoceras no-dulosum M anticoceras Beloceras Probeloceras (?) Naples и I а Eobeloceras Manticoceras «Гефироцеро-вый» I? Timanites Tornoceras Ponticeras TimaniU noceras, ceras, Kc
Genesee Probeloceras Manticoceras или Фарцицеровый tes
I a Pharciceras lunuli- Pharciceras, Synpharciceras, Triainoceras, Sandbergeroceras, Koenenites, Ponticeras Tornoceras Timanites Koenenites Pharciceras I а
costa Manticoceras (?)
>я я я Obere Л/aenioceras Stufe W edekindella bri-lonensis и Sobolewia nucifonnis Agoniatites dis-coides W edekindella Maenioceras Sobolewia Agoniatites Werneroceras Tully ? Hamilton Tornoceras Tornoceras Agoniatites W edekindella M aenioceras Sobolewia (?) Agoniatites Менецеровый (мениоцеровый) II Agoniatites Maenioceras Agoniatites Agonia
я о ф д я й Untere Maenioceras — Stufe Anarcestes rou-villei Agoniatites Parodicerellum M aenioceras Anarcestes ? Marcell us T ornoceras Agoniatites Anarcestes I Maenioceras Foordites Agoniatites
Средний дев »Я Я я л Anarcestes — Pinacites jugleri и Foordites occul-tus Pinacites Foordites Agoniatites Werneroceras Anarcestes Parodicerellum Onondaga Werneroceras Agoniatites Tornoceras (?) Pinacites Agoniatites Анарцестовый II Anarcestes
Эйфел Stufe Anarcestes noeg-gerati A. lateseptatus Зона? Sellanar-cestes a enkenbachi Agoniatites Werneroceras Subanarcestes Anarcestes Anarcestes Sellanarcestes Schoharie Anarcestes I
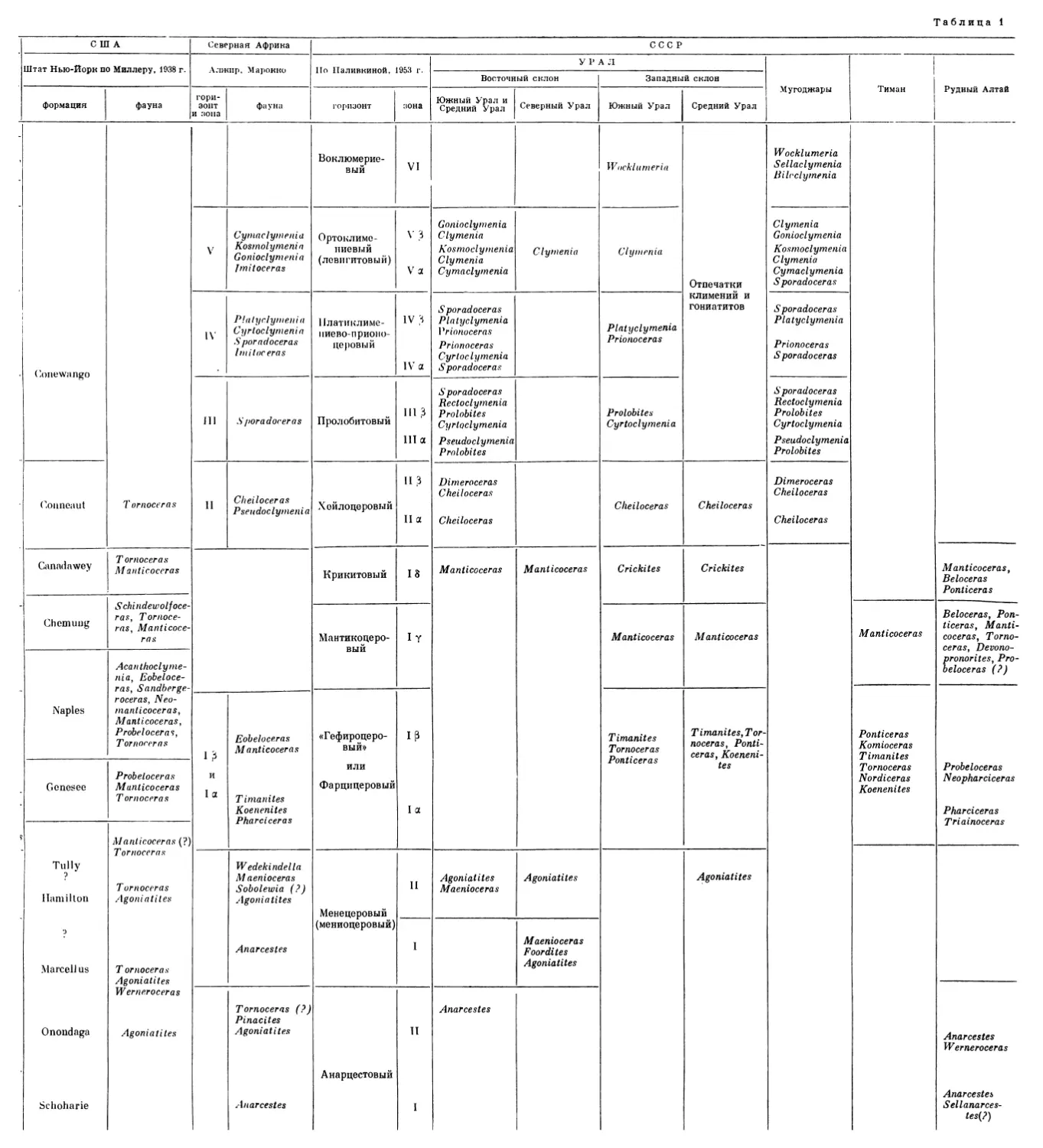
Таблица 1
США Северная Африка СССР
Штат Нью-Йорк по Миллеру, 1938 г. Алжир, Марокко По Наливкиной, 1953 г. УРАЛ Мугоджары Тиман Рудный Алтай
Восточный склон Западный склон
формация фауна горизонт и зона фауна горизонт зона Южный Урал и Средний Урал Северный Урал Южный Урал Средний Урал
I Conewango T ornoceras Воклюмерие-вый VI Wockl timer ia Отпечатки климений и гониатитов Wocklumeria Sellaclymenia Biloclymenia
V Cymaclymenia Kosmolynienia Gonioclymenia Jmitoceras Ортоклиме-пиевый (левигитовый) V 3 V а Gonioclymenia Clymenia Kosmoclymenia Clymenia Cymaclymenia Clymenia Clymenia Clymenia Gonioclymenia Kosmoclymenia Clymenia Cymaclymenia S poradoceras
IV Plalyclymenia Cyrtoclymenia S poradoceras I mi toe eras Нлатиклиме-пиево-прионо-церовый IV 3 IV а S poradoceras Plalyclymenia Prionoceras Prionoceras Cyrtoclymenia S poradoceras Plalyclymenia Prionoceras S poradoceras Plalyclymenia Prionoceras S poradoceras
III S poradoceras Пролобитовый III ,3 III а S poradoceras Rectoclymenia Prolobites Cyrtoclymenia Pseudoclymenia Prolobites Prolobites Cyrtoclymenia S poradoceras Rectoclymenia Prolobites Cyrtoclymenia Pseudoclymenia Prolobites
Conneaut II Cheiloceras Pseudoclymenia Хойлоцеровый II 3 Па Dimeroceras Cheiloceras Cheiloceras Cheiloceras Cheiloceras Dimeroceras Cheiloceras Cheiloceras
Manticoceras, Beloceras Ponticeras
Canada wey Tornoceras M anticoceras Крикитовый 18 Manticoceras Manticoceras Crickites Crickites
Chemung Schindewolfoce-ras, Tornoceras, Manticoceras
Мантикоцеро-вый I Y Manticoceras М anticoceras Manticoceras Beloceras, Ponticeras, Manticoceras, Tornoceras, Devono-pronorites, Probeloceras (?)
Naples Acanthoclyme-nia, Eobeloce-ras, Sandberge-roceras, Neo-manticoceras, Manticoceras, Probeloceras, Tornoceras
о и I а Eobeloceras Manticoceras Timanites Koenenites Pharciceras «Гефироцеро-вый» или Фарцицеровый I? I а T imanites Tornoceras Ponticeras Т imanites ,Т ornoceras, Ponticeras, Koenenites Ponticeras Komioceras Timanites Tornoceras Nor dicer as Koenenites Probeloceras Neo pharciceras Pharciceras Triainoceras
Genesee Probeloceras Manticoceras Tornoceras
? Tully ? Hamilton 9 Marcell us Onondaga Schoharie Manticoceras (?) Tornoceras Tornoceras Agoniatites T ornoceras Agoniatites Werneroceras Agoniatites
W edekindella M aenioceras Sobolewia (?) Agoniatites Anarcestes Менецеровый (мениоцеровый) II I Agoniatites Maenioceras Agoniatites Maenioceras Foordites Agoniatites Agoniatites
Tornoceras (?) Pinacites Agoniatites Anarcestes Анарцестовый II I Anarcestes Anarcestes Werneroceras Anarcestes Sellanarces-tes(?)
Stufe. Эти подразделения также именуются Oberdevonstufe I (или ОЬег-devon I), Oberdevonstufe II ит. д., причем Oberdevonstufe III Шиндевольфа эквивалентен как Oberdevonstufe III, так и Oberdevonstufe IV Ведекинда; Oberdevonstufe IV Шиндевольфа равняется Oberdevonstufe V Ведекинда и т. д. Принимая во внимание, что вопрос о возрасте слоев с Gatten-dorjia является до сих пор спорным в самой Германии и что схема Ведекинда деления этих слоев на горизонты используется значительно чаще, чем схема Шиндевольфа, я привожу в качестве стандартного деление верхнего девона Западной Европы по Ведекинду. Эта биостратиграфическая шкала с характерными для каждой зоны родами аммоноидей изображена на табл. 1. На этой же таблице дается сопоставление западноевропейской универсальной шкалы с биостратиграфическими схемами охарактеризованных фауной аммоноидей девонских отложений США (штат Нью-Йорк), Северной Африки и СССР, в том числе и девонских отложений Рудного Алтая.
2. ОПРЕДЕЛЕНИЕ ВОЗРАСТА ОПИСАННЫХ КОЛЛЕКЦИЙ
Среднедевонский отдел
Коллекции среднедевонских аммоноидей, находящиеся в нашем распоряжении, происходят из крюковской и сокольной свит среднедевонских отложений Лениногорского рудного поля и известны из четырех пунктов, которые указываются ниже в стратиграфическом порядке.
Эйфельский ярус Dj
Крюковская свита
1) Г. Лениногорск. Светло-серые известняки Терентьевской штольни (старые отвалы). Собрана фауна трилобитов, брахиопод, наутилоидей, кораллов и аммоноидей; из последних определены1:
Anarcestes praecursor Freeh......................2
W erneroceras ruppachense (Kayser)...............1
Sellanarcestes (?) wenkenbachi/or mis sp. nov....1
2) Г. Лениногорск. Выходы зеленовато-серых алевролитов крюковской свиты на ЮЗ склоне горы Сокольной, у ее подножья. Найдены аммо-ноидеи:
Anarcestes praecursor Freeh......................2
Два первых вида и близкий к третьему Sellanarcestes wenkenbachi (Kayser) известны из среднедевонских отложений Западной Европы. Согласно западноевропейским исследователям A. praecursor и S. wenkenbachi характерны для наиболее низких горизонтов среднего девона (низы Апаг-cestes-Stufe — зона Anarcestes wenkenbachi [=Sellanarcestes wenkenbachi]). Werneroceras ruppachense известен, правда, из несколько более высоких отложений AnarcestesStuie. Принимая во внимание сходство крюковской фауны с известняками Менани (Мнениан) Богемии [отмеченное еще Ф. Н. Чернышевым (1893)] и с Orthocerasschiefer Рупбаха и Грамберга, я считаю вполне возможным отнесение этих отложений (крюковской свиты) к низам эйфельского яруса (Anarcestes-SVoie I) — D2 (Сох Арденн).
Сокольная свита
3) Г. Лениногорск. Гора Сокольная. Выходы плотных толстослоистых зеленовато-серых и светло-серых алевролитов на СЗ склоне горы, в 400 м
1 Здесь и дальше цифры указывают количество экземпляров.
И
от ее вершины (у вентиляционной шахты). Собрана фауна брахиопод, трилобитов, аммоноидей и др. Среди последних определены:
Anarcestes (Latanarcestes) noeggerati (Buch)............34
Werneroceras ruppachense (Kayser).......................10
Werneroceras socolicum sp. nov...........................3
4) Кроме этих форм, имеется единственный экземпляр, найденный в алевролитах того же возраста на СЗ склоне горы Бахрушиной. Эта форма описана ниже как
Werneroceras altaicum sp. nov.................1
Из всех четырех указанных видов первые два известны из среднедевонских отложений Западной Европы, где они характерны для следующих, более высоких зон Anarcestes-SUife — зоны A. (Latanarcestes) noeggerati и зоны Pinacites jugleri и И7erneroceras ruppachense. Третий и четвертый виды являются местными, близкими, однако, к W. ruppachense.
Ведекинд (Wedekind, 1918, стр. 118), разбирая биостратиграфию среднего девона, отмечает, что в Западной Европе Anarcestes subnautilinus (Schlotheim), или — по современному — A. (Latanarcestes) noeggerati (Buch), занимает по сравнению с A. lateseptatus (Beyrich) более высокое» положение и характерен для верхов зоны A. subnautilinus, которая многими исследователями параллелизуется с нижней частью кувэнского (эйфельского) яруса Арденн (Coj). Werneroceras ruppachense (Kayser), синонимом которого Шиндевольф (Schindewolf, 1933, стр. 96) считает А~ (Werneroceras) subumbonale Wedekind, известен из верхней зоны Апагсе-stes-Stufe (зона Pinacites jugleri = зона Anarcestes ruppachensis) Западной Европы и Марокко, которая параллелизуется с верхней частью* кувэнского (эйфельского) яруса Арденн (Со2). Присутствие среди аммони-товой фауны нижней пачки алевролитов сокольной свиты A. (Latanarcestes) noeggerati и Werneroceras ruppachense позволяет, как мне кажется,, с полным основанием считать низы сокольной свиты соответствующими верхам Anarcestes-Stwfe I — низам AnarcestesStvSe II и относить их к средней части эйфельского (кувэнского) яруса (верхиСох— низы Со2 Арденн).
Верхнедевонский отдел
Местонахождения верхнедевонских аммоноидей известны в Рудном Алтае в большем количестве пунктов, нежели местонахождения среднедевонских аммоноидей, и собранные коллекции их значительно более обширны и разнообразны. Достаточно указать, что по сравнению со среднедевонскими аммоноидеями, представленными в коллекциях одним семейством и двумя-тремя родами, изученные представители верхнедевонских аммоноидей относятся к шести семействам и девяти родам. Перечень местонахождений верхнедевонских аммоноидей Рудного Алтая с указанием для каждого из пунктов фауны аммоноидей и обоснованием их возраста приводится ниже в стратиграфическом порядке.
Франский ярус D3
Фарцицеровый горизонт D1^ = зона 1а.
5) Шемонаихинский район. Правый берег р. Убы, в 2 км ниже ж.-д. моста. Выходы известково-глинистых алевролитов в 250 м севернее 25— 30-метровых обрывов, сложенных спилитами. В 6—8 м ниже контакта со спилитами найдена фауна аммоноидей плохой сохранности, среди которых определены:
12
Pharciceras kiliani (Freeh).............................30
Triainoceras sp.........................................56
6) Правобережье p. Убы. Выходы известняково-глинистых алевролитов в 1 км к СЗ от указанного выше обнажения. Фауна та же.
7) Золотушинский район. Правый берег рч. Грязнухи, в 1 км ниже с. Раздольного. Нижняя часть органогенных Неверовских известняков, -богатых разнообразной фауной. Из аммоноидей найдены:
Triainoceras gerassimovi sp. nov..........................3
Trlainoceras (?) sp.......................................1
Pharciceras kiliani Freeh, который рассматривается некоторыми исследователями как локальная южнофранцузская форма западногерманского Ph. lunulicosta (Sandberger), и близкий к Triainoceras gerassimovi Т. costatum {Archiac et Verneuil) описывались из самых низов верхнего девона — воны la (Prolecanitenschichten = Pharcicerasschichten), или зоны Ph. lunulicosta Германии и Франции; Ph. lunulicosta известен также из наиболее низких верхнедевонских отложений Северной Африки (Алжир, Марокко). В выше- и нижележащих отложениях представители указанных родов и видов не наблюдались, что, мне кажется, дает основание считать алевролиты правобережья р. Убы и нижнюю часть неверовских известняков, в которых найдены эти формы, наиболее низкими членами верхнедевон-•ского комплекса Рудного Алтая, соответствующими фарцицеровому горизонту (D*a) или зоне Pharciceras lunulicosta (зона la Рейнских провинций и, вероятно, Fi Арденн).
Зона 1р.
8) Золотушинский район. Правый берег рч. Золотухи. Известняковые линзы в 400 м восточнее дер. Горюновки. Известняки содержат фауну бра-хиопод, кораллов, наутилоидей и аммоноидей. Из последних определены:
Probeloceras (?) orientale sp. nov..................... 54
Neopharciceras kurbatovi Bogoslovsky....................15
Близкий к P. (?) orientale P.(?) forcipiferum (Sandberger) известен в Западной Европе из тех же слоев, что и упоминавшиеся выше Triainoceras costatum и Pharciceras lunulicosta (зона la = Prolecanitenschichten или Pharcicerasschichten), реже из несколько более молодых (зона ip— низы зоны Iy). С другой стороны нахождение такого высокоорганизованного представителя семейства Pharciceratidae, как Neopharciceras, указывает, по-видимому, на то, что отложения, содержащие данную фауну, должны считаться несколько более молодыми по сравнению со слоями, содержащими Pharciceras. Данные отложения должны, по всей вероятности, рассматриваться, как близкие по возрасту зоне 1а и соответствующие низам зоны ip (зона Manticoceras nodulosum) Рейнской провинции (Гга-с Арденн?).
Гониатитовый горизонт, или зона 1у.
9) Шемонаихинский район. Поселок Николаевского рудника. Известняковые линзы гониатитового горизонта на западном склоне гряды, отходящей от поселка в СВ направлении. Отсюда описаны:
Ponticeras altaicum sp. nov.............................15
» acutilobatum sp. nov...............................1
Probeloceras (?) forcipiierum (Sandberger).............. 2
Manticoceras affine (Steininger)........................15
» ammon (Keyserling)..................... 6
» carinatum (Sandberger)................ 26
13
Manticoceras bullatum Wedekind................... 6
» intumescens (Beyrich)................. 7
» sinuosum (Hall)......................22
» altaicum sp. nov..................... 1
» galeatum Wedekind.................... 3
Beloceras sagittarium (Sandberger).....................146
» stenumbilicatum sp. nov.............. 19
10) Шемонаихинский район. Известняковые линзы гониатитового горизонта в 3 км юго-восточнее предыдущего местонахождения по простиранию горизонта. В малиново-красных или бордово-красных известняках наряду с фауной криноидей, кораллов, мелких брахиопод, пелеципод и гастропод присутствуют аммоноидеи, из которых определены:
Tornoceras simplex (Buch).............................. 3
Manticoceras affine (Steininger)....................... 1
» intumescens (Beyrich)................ 1
» sinuosum (Hall)...................... 8
» artum sp. nov........................ 2
Beloceras sagittarium (Sandberger).....................30
Devonopronorites ruzhencevi Bogoslovsky...............193
Среди этих аммоноидей обращает на себя внимание присутствие в больших количествах Devonopronorites ruzhencevi Bogoslovsky, являющегося единственным представителем нового семейства Devonopronoritidae. Другие аммониты встречаются в значительно меньших количествах, и их видовой состав ничем существенно не отличается от такового первого местонахождения.
Анализ фауны аммоноидей гониатитового горизонта показывает, что наряду с наличием местных алтайских представителей она очень сходна с западноевропейской аммонитовой фауной, отчасти с североамериканской, уральской и тиманской. Большинство изученных видов известно из Man-ticoceras-Stuis Западной Европы, где они характеризуют собой третью (1у) зону Manticoceras cordatum и М. carinatum.
Присутствие среди изученной фауны Manticoceras intumescens, М. affine, М. galeatum, М. carinatum, характерных только для зоны 1у, позволяет рассматривать отложения гониатитового горизонта как соответствующие этой зоне или F2 (вероятно F2d — F2h) Арденн.
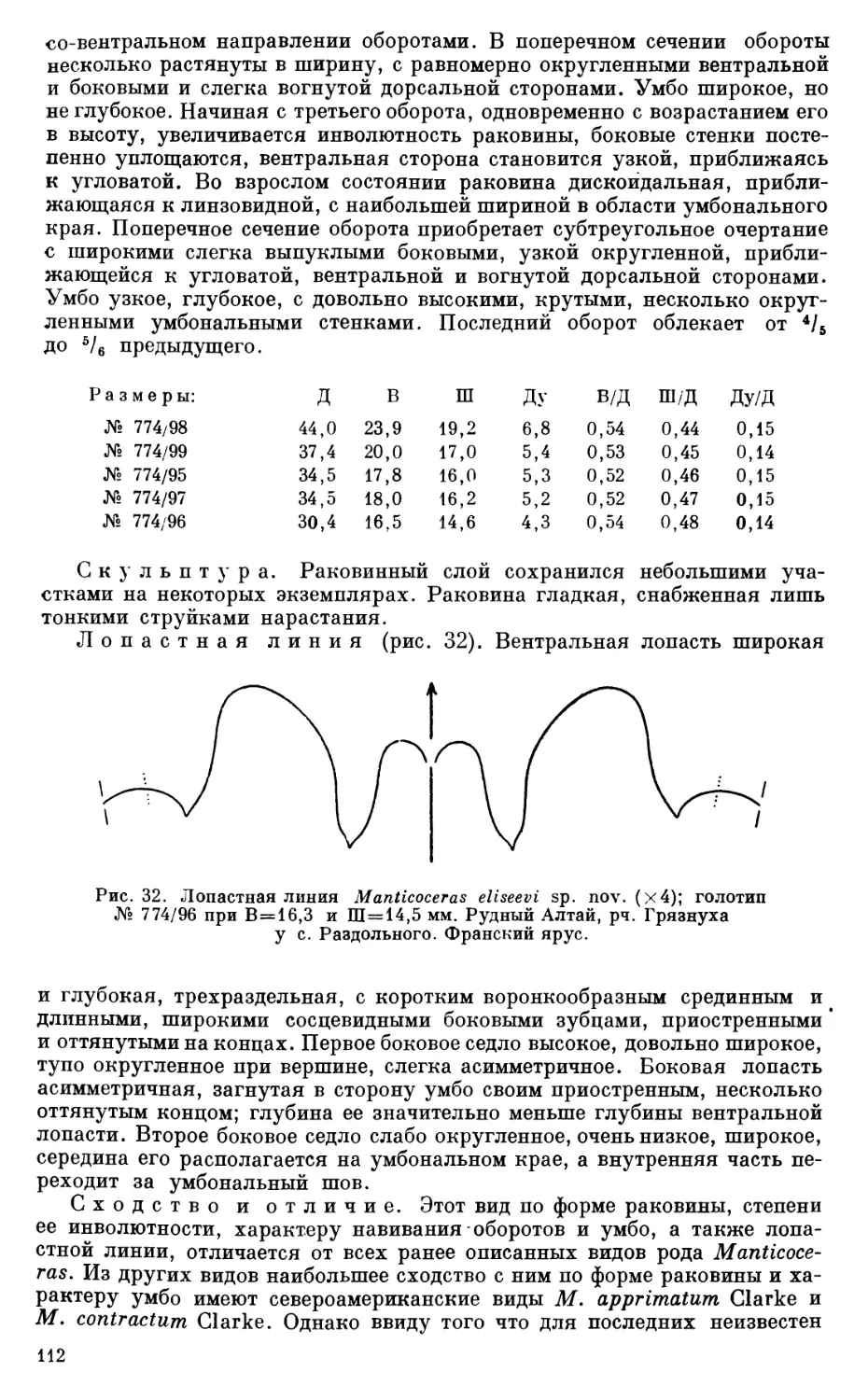
Зона 18.
11) Золотушинский район. Правый берег рч. Грязнухи, у нижней окраины с. Раздольного. Известняковые прослои песчано-сланцевого горизонта. Здесь собраны аммониты:
Ponticeras sp............................................ 9
Manticoceras bullatum Wedekind........................... 6
» neverovi sp. nov....................... 2
» sinuosum (Hall)........................14
» altaicum sp. nov....................... 1
» eliseevi sp. nov....................... 8
» drevermanni Wedekind................... 5
» carinatum (BeyriclP....................22
Beloceras sagittarium (Sandberger).......................78
Близость изученной фауны аммоноидей к фауне гониатитового горизонта (зона 1у) Николаевского месторождения, на что указывает наличиеобщих форм, и, с другой стороны, присутствие в ней Manticoceras drevermanni и Ponticeras sp., близкого к Ponticeras bickense Wedekind, тяготею
14
щих к верхней (13) зоне Manticoceras-Stuie Западной Европы, заставляет, полагать возраст этой фауны близким или несколько более молодым по сравнению с возрастом фауны гониатитового горизонта Николаевского месторождения и соответствующим примерно верхам зоны 1у — низам зоны 18 (слои с Manticoceras adorjense Wedekind Рейнской провинции или. F2i — F2j разреза Арденн).
Общий вывод
Подводя итог данным по распределению аммоноидей в девоне Рудного* Алтая и сопоставляя биостратиграфическую схему этих отложений по аммонитам с основной шкалой Западной Европы (табл. 1), можно установить несколько горизонтов и зон в среднем и верхнем девоне Рудного Алтая. В среднем девоне фауной аммоноидей охарактеризован эйфельский ярус (кувэнский), внутри которого можно различать две зоны. Нижняя, соответствующая примерно низам Anarcestes-Stufe Рейнской провинции, охарактеризована родами Anarcestes и Sellanarcestes (?); верхняя, соответствующая средней части Anarcestes-Stwiv — родами Anarcestes и Werneroceras. Более высокие зоны среднего девона, соответствующие верхам Апаг-cestesstufe и Maeniocerasstufe (Maenecerasstufe), фауны аммоноидей, по-видимому, не содержат (во всяком случае из этих отложений находок их пока не известно).
Наиболее охарактеризованы аммоноидеями франские отложения, внутри которых можно установить наличие элементов, свойственных в той или иной степени всем четырем зонам ManticocerasStvAe Рейнской провинции. Отложения, соответствующие зоне 1а, содержат роды Pharciceras и Triainoceras и представлены как известковистыми, так и песчано-глинистыми фациями неглубокого моря. Отложения, отвечающие, по всей вероятности, зоне ip, характеризуются присутствием родов Probeloceras и Neopharciceras, известных из органогенно-обломочных фаций мелкого моря. Наиболее богаты аммоноидеями отложения, соответствующие третьей (1у) зоне, представленные известняковыми, обычно с примесью довольно больших количеств глинистых частиц, фациями неглубокого моря и содержащие роды Ponticeras, Probeloceras, Manticoceras, Tornoceras, Beloceras и Devonopronorites. Отложений, полностью соответствующих четвертой (18) зоне Manticocerasstufe с типичной фауной аммоноидей {Crickites), на Рудном Алтае не известно, но, вероятно, к низам этой зоны следует отнести песчано-сланцевый горизонт района Гериховского месторождения (рч. Грязнуха у с. Раздольного), в известняковых прослоях которого встречены представители родов Ponticeras, Manticoceras и Beloceras.
Отложений с аммоноидеями, отвечающих более высоким горизонтам (фаменским) Рейнской провинции, в Рудном Алтае пока не известно, однако возможность нахождения их не исключается. В пользу этого говорит нахождение климений девонского облика в тарханской и бухтарминской свитах, которые относятся в настоящее время к карбону, что, может быть, не совсем правильно.
Анализ изученной фауны показывает, что, наряду с большим количеством местных форм, эта фауна обнаруживает большое сходство с девонской фауной Западной Европы, в меньшей степени с фауной Урала (возможно, ввиду недостаточной изученности последней) и Тимана, отчасти с фаунами Северной Америки и Северной Африки. Из 29 описанных видов общими для Алтая и Западной Европы являются 14; четыре алтайских вида близки к западноевропейским. Шесть видов известны из девона Урала, один — из Северной Америки; пять видов близки к североамериканским, пять — к североафриканским, два — к уральским, 14 видов —
15
Таблица 2
Стратиграфическое распределение изученных аммоноидей
Рудный Зап. Сев. Сев.
Вид Алтай Европа Урал Африка Америка
D2 D3 Dl А О 1 D^3 D2 D3 Рз D|
Anarcestes praecursor Freeh + - + - X —
A. (Latanarcestes) noeggerati (Buch) . + - + - — —
A. (Sellanarcestes? ) wenkenbachi]ormis
sp. nov + - X — — — — — — —
W erneroceras ruppachense (Kayser) . . + - + - — — — — — —
» socolicum sp. nov. . . . + - X — — — — — — —
» altaicum sp. nov. . . . + - X — — — — — — —
Tornoceras simplex (Buch) - + +? + - + — X — X
Ponticeras altaicum sp. nov. . . . - + — — — X --- — — —
» acutilobatum sp. nov. . - +
Probeloceras (?) forcipiferum (Sand-
berger) • - + - + — — — — — —
» orientale sp. nov. . . . - +
Manticoceras ajjine (Steininger) . . . - + - + — — — — — —
» ammon (Keyserling) . . - + — — - + — — — —
» carinatum (Sandberger) . - + - + - + — X — х
» bullatum (Wedekind) - + — + — — — — — —
Manticoceras intumescens (Beyricb) - + - + - + — — — X
„ sinuosum (Hall) .... — + - + — X — X - +
„ altaicum sp. nov. . . . - + — X
„ galeatum Wedekind . . - + — + - + — — — —
„ artum sp. nov. . . . - + — —
„ neverovi sp. nov. . . . - + — — — — — — — —
„ eliseevi sp. nov. . . . - + — X
„ drevermanni Wedekind . - + - + - + — — — —
Seloceras sagittarium (Sandberger) . . - + - + — — — — — —
» stenumbilicatum sp. nov. - + — —
Pharciceras kiliani (Freeh) — + - + — — — X — —
Neopharciceras kurbatovi Bogoslovsky - +
Triainoceras gerassimovi sp. nov. . . - + — X — — — — — —
Triainoceras sp - + — —
Devonopronorites ruzhencevi Bogoslov-
sky - +
новых, местных. Стратиграфическое распределение аммоноидей показано на табл. 2, на которой знаком + показано присутствие тех же видов в отложениях других регионов, а знаком X наличие в последних видов, близких к алтайским. Близость алтайской фауны к западноевропейской, наблюдающаяся и в других группах организмов (брахиоподы, трилобиты), указывает на имевшуюся в девоне связь между двумя бассейнами, которая осуществлялась, по всей вероятности, через зону Тетис. Наличие общих и близких к алтайским форм в девоне Урала и Северной Америки также указывает на связь бассейна Рудного Алтая с бассейнами этих двух регионов.
46
Обращает на себя внимание присутствие в верхнем девоне Рудного Алтая в больших количествах представителей весьма редкого в Западной Европе рода Triainoceras, а также таких высокоорганизованных и специализированных аммоноидей, как Neopharciceras, в массовых количествах представителей рода Beloceras и, наконец, рода Devonopronorites из не известного ранее в девоне семейства Devonopronoritidae, который встречается в довольно больших количествах. Алтайская фауна франских аммоноидей в целом по сравнению с аналогичными фаунами других областей выглядит более высоко организованной, содержащей в себе элементы, имеющие более молодой облик, что объясняется, по всей вероятности, тем, что она получила большие возможности для своего развития в обстановке неглубоких шельфовых зон свободно сообщающегося с геосинклинальным бассейном открытого моря, отдельные области которого с накопляющимися своеобразными кремнисто-глинисто-карбонатными осадками характеризовались благоприятными для жизни аммоноидей условиями.
3. НЕКОТОРЫЕ ЗАМЕЧАНИЯ ПО ПОВОДУ МЕСТНЫХ СТРАТИГРАФИЧЕСКИХ СХЕМ РУДНОГО АЛТАЯ
Вопрос стратиграфии эффузивно-осадочного комплекса девонских отложений Рудного Алтая, достигающих по некоторым данным мощности 4,5—5 км, имеет в настоящее время большое теоретическое и практическое значение. Несмотря на большие региональные исследования многих специалистов, доставившие значительное количество новых сведений по стратиграфии девона, разработка стратиграфической схемы еще далеко не завершена. Единой дробной стратиграфической схемы сейчас пока нет. Трудность разработки такой общей схемы объясняется метаморфизован-ностью и сложной тектоникой отложений, пестрым литологическим составом, спорадичностью находок и плохой сохранностью фауны и, отчасти, недостаточным до последнего времени вниманием к сборам и изучению фауны со стороны многих геологов-практиков.
Основные черты современной стратиграфии девонских отложений были намечены Н. Л. Бубличенко в результате работ Геологического комитета, проводившихся с 1919 по 1931 г. В этих работах принимали участие В. П. Нехорошев, Н. Н. Курек, Н. А. Елисеев и др.
В основе деления эффузивно-осадочного комплекса девона до последнего времени лежало три опорных горизонта: лосишинский, содержащий фауну брахиопод, мшанок и кораллов низов среднего девона, гониатито-вый и николаевский, содержащие фауну франского яруса. Граница средне- и верхнедевонских отложений устанавливалась в основании гониатитового горизонта и считалась «надежной» только там, где последний можно было установить по фаунистическим данным. Кроме горизонтов, В. Н. Даниловичем в 1938 г. (в результате работ 1935 г.) было предложено выделять на основании литологических признаков следующие свиты (снизу вверх):
Метаморфическая толща . •........................(?)
Березовская свита.................................D*
Лосишинский фаунистический горизонт...............D&
Таловская свита...................................D°
Каменевская свита.................................D2
Николаевский фаунистический горизонт .......... D2_3
Снегиревская свита .............................. D*
Боровиковская свита ............................. D£
Чудацкая свита.......................•............D£
2 б. и. Богословский
17
С некоторыми изменениями и добавлениями эта схема в дальнейшем обосновывается многими исследователями на основании преобладания в тех или иных свитах продуктов либо кислых, либо основных, лав. В основу подобного расчленения положена обычная для спилито-кератофировой формации закономерность: качественная периодичность излияний и выбросов, приводящая к напластованию резко отличных по составу серий лав и пирокластов, и количественная периодичность членения нормальных осадочных и туфогенно-осадочных продуктов морской седиментации. Периодам бурной вулканической деятельности отвечают мощные толщи эффузивно-пирокластического материала, периодам затухания — горизонты кремнисто-глинистых илов, органогенных осадков и продуктов подводного гильмерозиса (?) более древних туфов и лав.
Эта, в общем правильная, точка зрения (при рассмотрении отдельных регионов) не может, однако, в одинаковой степени использоваться при сопоставлении геологических разрезов смежных районов Рудного Алтая. Причина этому — разная степень проявления вулканической деятельности в различных районах, неравномерность накопления эффузивно-осадочных комплексов даже на соседних площадях, резкая смена и пестрота фациально-литологического состава выделяемых свит, подчас выпадение из разреза целых комплексов. Трудность корреляции местных разрезов объясняется отчасти и разной степенью изученности их.
В силу изложенного представляется более целесообразным, не вдаваясь в вопросы общей стратификации девонских отложений Рудного Алтая (хотя попытки установления обобщенной стратиграфической схемы имеются), рассмотреть более подробно стратиграфию отдельных районов, используя как новые данные других исследователей, так и собственные результаты, чтобы затем, сопоставляя разрезы этих районов со стандартными девонскими разрезами, установить более точно возраст рассматриваемых отложений и, по возможности, увязать последние между собой. Имея целью изучение девонских аммоноидей Рудного Алтая, мы, естественно, будем рассматривать стратиграфию районов, в пределах которых они найдены, а именно: Лениногорского рудного поля (средний девон) и Николаевского и Рериховского месторождений (верхний девон).
Лениногорское рудное поле
Первое упоминание о палеонтологических остатках из отложений Лениногорского рудного поля, а следовательно, и определение возраста относится к 1893 г., когда Ф. Н. Чернышевым была описана фауна из известняков Терентьевской штольни, среди которой был и один вид аммонитов — Goniatites (Anarcestes) lateseptatus Beyrich. Чернышев отнес указанные отложения к верхам нижнего девона, отметив сходство-данной фауны с фауной горизонта F2 Барранда и ряда слоев девона Гарца и Франции, считавшихся в то время нижнедевонскими. Однако он указывал и на сходство описанной им фауны с фауной некоторых заведомо среднедевонских отложений Западной Европы (стр. 234). Одним из доказательств нижнедевон-ского возраста крюковских известняков Ф. Н. Чернышев (стр. 233) считал сходство их фауны с фауной красного известняка в с. Покровском (восточный склон Урала), который он относил к нижнему девону.
В 1909 г. Д.Н. Соболев отнес крюковские известняки к низам среднего девона, присоединяясь к мнению ряда западноевропейских авторов, что известняки Грефенштейн Германии и Мнениан Богемии, с которыми Ф. Н. Чернышев параллелизовал крюковские известняки, должны быть отнесены к низам среднего девона (стр. 267).
Работы Н. Н. Курека в 1932 г., давшие схему стратиграфии девонских отложений Лениногорского рудного поля, не прибавили ничего существенного к фаунистической характеристике крюковских известняков.
18
Н. Н. Куреком приводится разрез небольшого участка девонских пород Риддерского (Лениногорского) рудного поля (снизу вверх):
1. Туфы преимущественно агломератового сложения, кислого состава с прослойками глинистых сланцев, в верхней части превращенные в мйкро-кварциты (т. наз. «роговики»). Фауна встречается спорадически. Мощность не менее 350 м.
2. Глинистые и известково-глинистые сланцы с редкими прослоями мергелистых известняков, мощностью в среднем от 10 до 50 м, в западной части — свыше 300 м (в этом горизонте встречена фауна, описанная Ф. Н. Чернышевым).
3. Красно-зеленые туфы основного состава мощностью 20—100 м.
4. Известково-глинистые и известковистые сланцы с редкими прослоями песчаников и туфов мощностью не менее 160 м.
В 1936 г. к выводу о среднедевонском возрасте крюковских известняков пришел Н. Л. Бубличенко, переизучивший коллекцию брахиопод Ф. Н. Чернышева.
Из современных исследователей кобленцский возраст крюковских известняков принят только Л. Л. Халфиным (1948).
В результате работ 1950—1951 гг. Г. Н. Щерба выделил в районе г. Лениногорска три свиты: крюковскую, ильинскую и сокольную, значительно дополнив разрез девона. В основании предположительно1 залегает известная по обломочным продуктам в агломератах риддерская туфогенноагломератовая толща, лежащая на метаморфических сланцах додевона. Мощность риддерской свиты неизвестна. Далее следуют:
1. Крюковская вулканогенно-осадочная свита. В основании — кислые фельзитовые кварцевые порфиры и альбит-порфиры, их туфобрекчии мощностью до 1С0 м. Выше — толща пестрых агломератовых и псеффито-агло-мератовых туфов, чаще кислого состава, мощностью 150—300 м, с прослоями туфосланцев. Перекрывается хлоритизированными глинистыми сланцами, аргиллитами, известковистыми сланцами и туфосланцами, мощностью от 20—50 до 350 м, с остатками фауны плохой сохранности. Сланцы содержат прослои мергелей и горизонты криноидных известняков и их брекчий, мощность которых в рифах достигает 220 м. Мощность сланцев в этом случае резко падает. Из известняковописана фаунаФ. Н. Чернышевым. Мсщность всей свиты 300—400 м.
2. Ильинская свита — эффузивно-пирокластическая с подчиненными прослоями осадочных пород (толща «красно-зеленых» туфов). Сложена чередующимися горизонтами агломератовых псефитовых и псаммитовых туфов с прсслоями туффитов преимущественно среднего и кислого состава. Мощность 100—300 м.
3. Сокольная свита — эффузивно-осадочная. Представлена известковистыми алевропелитами, мергелями, аргиллитами (известково-глинистые сланцы), мощностью 150—180 м с прослоями песчаников и диабазов (несколько маломощных горизонтов) и двумя-тремя горизонтами кварцевых альбит-порфиров и их лавобрекчий (кератофиров). Самый мощный имеет мощность 100—150 м, местами достигая 280 м. Известковистость обусловливает в большинстве случаев наличие фаунистических остатков. В алев-ропелитах на горе Сокольной собрана фауна, дополняющая список «крюковской фауны» Ф. Н. Чернышева.
Общая мощность всех последних трех свит достигает 1000 м.
В 1952 г. при составлении геологической карты Лениногорского рудного поля и прилегающих районов 3. В. Мусиенко и Г. Ф. Яковлев установили присутствие палеонтологически охарактеризованных отложений крюковской и сокольной свит и вне Лениногорского рудного поля. Данными этих исследователей существенно уточняется стратиграфия, и выработанную ими стратиграфическую схему можно применить для значитель
2*
19
ных площадей и сопоставления разрезов смежных районов. Вне Лениногорского рудного поля Мусиенко и Яковлевым выделяется эффузивнотуфовая успенская свита, нижняя часть которой рассматривается ими как фациальный аналог ильинской и сокольной свит, а верхняя часть считается более молодой по сравнению с сокольной свитой. Схематизированная стратиграфическая колонка среднедевонских отложений Лениногорского района на основании последних данных может быть представлена следующим образом (см. табл. 3).
На основании изучения трилобитов из отложений крюковской и сокольной свит, 3. А. Максимова приходит к выводу о близком возрасте обеих свит и считает, что отличия в составе их трилобитовых фаун вызваны, вероятно, несколько различными фациальными условиями накопления осадка. Однако данный автор указывает, что не исключено, что алевролиты сокольной свиты несколько моложе крюковских известняков. Сравнивая описанную фауну трилобитов с западноевропейской, Максимова отмечает, что из аналогичных форм в Западной Европе большинство встречается в верхах нижнего и низах среднего девона, однако, в среднем девоне они более распространены. Автор делает вывод, что комплекс •трилобитовой фауны несомненно тяготеет к среднему девону, так как среди описанных форм отсутствуют исключительно нижнедевонские •формы, большинство форм более распространены в среднем девоне, а часть форм является исключительно среднедевонскими. Ставя вопрос, «не является ли данная фауна переходной от D? к D2?», автор, отмечая неопределенность такого решения, считает, что имеется достаточно оснований относить крюковскую и сокольную свиты к низам среднего девона.
К подобному же заключению о возрасте и взаимоотношениях крюковской и сокольной свит приходит на основании изучения их брахио-подовых фаун и Н. Л. Бубличенко, который указывает на преемственность сокольной фауны от крюковской, поскольку в нее переходят трилобиты, по-видимому, аммоноидей; с другой стороны, в ней появляются новые формы. Изменение состава фауны, по Бубличенко, может быть связано с изменением фациальных условий обитания и несколько более молодым возрастом.
Для выяснения возраста крюковской и сокольной свит представляет интерес сравнение крюковской фауны с фауной лосишинского горизонта (слоев), выделенного в свое время Н. Л. Бубличенко (1928), который хотя и не наблюдается в Лениногорском районе, но широко распространен в других районах рудного Алтая. Отмечая в крюковской фауне наличие -близких к нижнедевонским форм и сравнивая ее с фауной лосишинского горизонта, распространенного в других районах, Н. Л. Бубличенко указывает на отличие крюковской фауны от лосишинской, которое, по его мнению, может быть объяснено различными фациальными условиями и более молодым возрастом лосишинской фауны по сравнению с крюковской. Относя лосишинскую фауну к верхам D2 (С02 Арденн), Н. Л. Бубличенко определяет возраст крюковской свиты как низы D*. К низам же среднего девона относится им и сокольная свита.
Наконец, нельзя не отметить результатов исследований Н. Я. Спасского, касающихся изучения кораллов из лосишинских слоев, с которыми, как указывалось выше, Н. Л. Бубличенко сравнивает отложения крюковской свиты. Изучение кораллов из лосишинских слоев, сравнение их с фаунами среднедевонских кораллов Кузбасса, Урала и Западной Европы также указывают на эйфельский (кувэнский) возраст лосишинских слоев.
Таким образом, в результате исследования ряда групп крюковскую и сокольную свиты нужно причислить к эйфельскому возрасту.
Что же дают для установления возраста этих свит аммоноидей?
20
Отдел | Ярус | Свита Литологический состав
Средний девон Живет-ский D2 Успенская (верхняя часть) Эффузивы и туфы кислого состава
Кувэнский (эйфельский) Сокольная Алевролиты с прослоями песчаников и туфов, в средней части пачка кератофиров
Ильинская Туфы среднего состава от псаммитовых до агломератовых
Крюковская Известняки, мергели, хлорито-серицитовые сланцы* (Известняки в основании и кровле)
Риддерская Туфы, гл. обр. агломератовые, кислого состава, эффузивы
Стратиграфическое несогласие. Базальный конгломерат
Додевон (Si?). Метаморфические сланцы
Таблица 3
Фауна
Брахиоподы
Leptostrophia perlana Conr., Nucleospira lens Schnur., Sowerbiella (Plec-todonta) minor Roemer, Dalmanella (Aucella) sokolica Bubl., Stropheodon-ta (?) solarissima Bubl., Chonetes (Plicochonetes) cf. davousti Oehlert, Uncinulus cf. nalivkini Bubl., Camarotoechia elliptica Schnur., Orbiculoidea sp., Trigeria sp.
Трилобиты
Phacops altaicus Tschern., Ph. sternbergi Corda, Ph. potieri ulbensis Maximova, Ph. oculeus Maximova, Phacopidella primitiva Maximova, Proetus altaicus Maximova, Proetus sp. Maximova
Аммониты
Werneroceras ruppachense (Kayser), W. socolicum sp. nov., W. altaicum sp. nov., Anarcestes (Latanarcestes) noeggerati (Buch)
Брахиоподы
Eospirifer orbitensis Tschern., Leptaena rhomboidalis Wibck , Dalmanella (Aucella) sokolica Bubl., Hypothyridina (Isopoma) brachyptycta Schnur., Merista (Dicamara) plebeja Sow., M. (?) altaica Tschern., Retzia promi-nula Roemer, Rhynchospirina (?) sp., Athyris undata Defr., A. (?) glassi Dav.
Три лобиты
Harpes reticulatus Corda var. altaica Tschern., Bronteus sibiricus Tschern., B. aff. scaber Goldf., Proetus oehlerti Bayle, Pr. cf. eremita Barr., Phacops altaicus Tschern.
Аммониты
Anarcestes praecursor Freeh., Sellanarcestes (?) wenkenbachiformis sp. nov. Werneroceras ruppachense (Kayser)
В нашем распоряжении имеются коллекции аммоноидей, собранные нами и другими исследователями из отложений крюковской и сокольной свит.
Небольшая коллекция крюковских аммоноидей собрана из светло-серых известняков Терентьевской штольни (старые отвалы) и зеленоватосерых алевролитов ЮЗ склона горы Сокольной. Среди изученных нами аммоноидей определены: Anarcestes praecursor Freeh, Sellanarcestes (?) wenkenbachiformis sp. nov. и Werneroceras ruppachense (Kayser). Как уже отмечалось выше, сравнение фауны крюковских аммоноидей со среднедевонской фауной Западной Европы и сходство ее с фауной известняков Менани (Мнениан) Богемии и Orthocerasschiefer Рупбаха и Грамберга дает возможность отнести крюковскую свиту к низам эйфельского яруса — D2 (Сох Арденн).
; Фауна аммоноидей из отложений сокольной свиты, большая по объему, но, к сожалению, плохой сохранности, собрана из низов свиты (нижняя пачка алевролитов) на СЗ склоне горы Сокольной и Бахрушиной. В верхней пачке алевролитов аммоноидей не обнаружено. Аммоноидей собраны из плотных, толстослоистых зеленовато-серых и светло-серых алевролитов, выходящих в коренном залегании по склону горы Сокольной, в 400 м от ее вершины. Несколько образцов было собрано из подстилающих алевро-пелиты зеленовато-светло-серых грубозернистых песчаников, выходящих в 50 м ниже по склону горы.
Среди изученной фауны определены: Anarcestes (Latanarcestes) noeg-gerati (Buch), Werneroceras ruppachense (Kayser), W. socolicum sp. nov. и altaicum sp. nov.
Отмеченное выше сходство аммонитовой фауны нижней пачки алевролитов сокольной свиты с фауной среднего девона Западной Европы и наличие в ее составе представителей A, (Latanarcestes) noeggerati (Buch) и W. ruppachense (Kayser), характерных для соответствующих зон Апаг-cestes-SJtufe Западной Европы, позволяют с полным основанием отнести низы сокольной свиты к верхам нижней — низам верхней части эйфельского (кувэнского) яруса (верхи Сох— низы Со2 Арденн). Сокольная свита .в целом должна, по-видимому, относиться к верхней половине эйфельского яруса.
Таким образом, результаты изучения аммоноидей из отложений крюковской и сокольной свит лишний раз подтверждают правильность отнесения этих отложений к нижней части (эйфельскому ярусу) среднего девона и необоснованность отнесения крюковских известняков к нижнему девону, как это делают некоторые исследователи (Л. Л. Халфин).
Более того, если по другим группам организмов возраст вышеуказанных свит определяется как соответствующий низам среднего девона, изучение аммоноидей позволяет более точно указать их стратиграфическое положение и относить крюковскую свиту к низам эйфельского яруса, а сокольную свиту — к верхней половине его.
Район Николаевского месторождения
В районе Николаевского месторождения (Шемонаихинский район), находящегося в центральной части Рудного Алтая, наблюдается наиболее детальный и полный разрез эффузивно-осадочного девонского комплекса. Этим обстоятельством, наряду с практической важностью, и объясняется неослабевающее внимание многочисленных исследователей к изучению геологии данного района. Именно здесь были заложены основы современной стратиграфии девонских отложений Рудного Алтая, выработана, в основном, схема стратификации последних.
Как уже отмечалось выше, основные черты современной стратиграфии девонских отложений Рудного Алтая были намечены Н. Л. Бубличенко,
22
В. П. Нехорошевым, Н. А. Елисеевым и др. в результате работ, проводившихся с 1919 по 1931 год. В качестве опорных горизонтов использовались два:
1) лосишинский горизонт известняков, мощностью 10—40 м, залегающий в основании среднего девона;
2) николаевский горизонт (кубоидные слои) в виде линз известняков среди глинистых сланцев, залегающих в основании верхнего девона.
Н. Л. Бубличенко приводилась следующая схема стратиграфии девонских отложений:
1. Метаморфическая свита — «додевонская метаморфическая свита». Залегает ниже среднего девона. Представлена хлоритовыми, эпидотовыми, кремнистыми и пр. (обычно зелеными) сланцами, нередко с хорошо выраженной слоистостью. Фауны не найдено. Возраст возможно —
2. Эффузивно-туфовая толща среднего девона — D2 Налегает несогласно на додевонскую метаморфическую толщу. Прослеживается полосой по правому берегу р. Убы от устья р. Кондаковки к с. Шемонаиха и мощной полосой по левую сторону р. Убы. Эта толща сложена эффузивами (преимущественно кислыми), туфами и песчаниками, имеются глинистые сланцы, редко известняки.
В самых низах наблюдается прослой известняка мощностью 10—20 м с обильной фауной (брахиоподы, кораллы, трилобиты, мшанки) лосишин-ского горизонта — низы кальцеоловых слоев Рейнского бассейна.
Следующий прослой известняка прослежен только по правую сторону р. Убы. Мощность приблизительно 15 м. Около пос. Березовки в нем наблюдается фауна: Spirifer cf. frequensBubl., Sp. cf . triplicatus, Fenestella, Poly рога, Alveolites, Pachypora (близка к лосишинской — низы D2). Мощность среднедевонских отложений достигает 2800 м (?). Средний девон без изменений переходит в верхний девон.
3. Верхний девон D3 прослеживается в виде полосы СВ-ЮЗ простирания от района пос. Усть-Таловского до района сопок Острухи-Убинской и Рулихи и уходит на ЮВ. Нижней границей верхнего девона приняты известняки Николаевского рудника, содержащие Uncinulus cuboides.
Кубоидный горизонт — известняки 10—20 м мощности, прослеженные в виде линз у Николаевского рудника. По имеющейся в известняках фауне: Pleurotomaria altaica Vern., Naticopsis primigena Eichw., Spirifer canali-jerus Valenc., Sp. undijerus F. A. Romer., Sp. curvatus Schloth., Sp. zick-zack Romer, Martinia glabra Mart., Atrypa aspera Schloth., A. arimaspus Eichw., Rhynchonella pugnus Mart., Rh. cuboides Schloth., Camarophoria microrhyncha C. F. Romer, Orthis striatula Schloth., Cyrtina heteroclyta Defr. H. Л. Бубличенко определяет возраст известняков — низы верхнего девона D3.
Над кубоидным горизонтом залегает мощная толща туфов, аркозовых песчаников и эффузивов.
Мощность отложений верхнего девона достигает 2300 м.
В результате работ 1935 г. В. Н. Данилович (1938) подразделил комплекс девонских отложений, наблюдающихся в районе, на основании литологии на ряд свит (см. выше стр. 17), которые были в дальнейшем положены в основу при составлении стратиграфических схем. В. Н. Данилович высказал предположение, что николаевский горизонт является переходным от средне- к верхнедевонским отложениям.
В 1938 г. при проведении геологической съемки района Николаевского месторождения Н. А. Ноздреевым вблизи пос. Николаевского рудника был впервые обнаружен маломощный гониатитовый горизонт, содержащий фауну аммоноидей, определенных А. К. Наливкиной как верхнедевонские (низы мантикоцерового горизонта франского яруса). Из коллекции
23
}{. А. Ноздреева А. К. Наливкиной были определены: Manticoceras intumescens Beyrich, М. cordatum Sandberger, M. inversum Wedekind, Beloceras sagittarium (Sandberger) и Altaites altaicum gen. et sp. nov.
Кроме указанных видов, ею же определены из сборов В. К. Катульского: М. cf. carinatum Beyrich, Gephyroceras, sp. nov. и M. cf. schellwini Wedekind.
А. И. Левенко в результате работ, производившихся в 1949 г. к югу от пос. Шемонаихи, пришел к выводу, что маломощный гониатитовый горизонт, обнаруженный Н. А. Ноздреевым к востоку от Николаевского рудника, «имеет значительное распространение по простиранию как к юго-востоку, так и, главным образом, к северо-западу, причем в последнем направлении мощность его резко возрастает». Была установлена значительная мощность гониатитового горизонта (самарского), являющегося составной частью каменевской свиты (по А. И. Левенко), достигающей мощности (по тому же автору) 850—900 м.
Согласно Левенко, каменевская свита сложена кислыми и основными эффузивами и их туфами, глинистыми сланцами, часто кремнистыми или известковистыми, алевролитами, глинисто-известковистыми алевролитами, песчаниками, в том числе аркозовыми, туфо-песчаниками и реже известняками.
Работавшим в этом районе в 1951 г. В. Ф. Масленниковым выделяются самостоятельные каменевская и гониатитовая (самарская) свиты. Более древняя гониатитовая свита сложена туфами диабазов и спилитов, основными эффузивами с многочисленными прослоями глинистых и кремнистых сланцев и известняков с фауной аммоноидей. Отмечаются линзообразные тела кислых эффузивов и их туфов. Мощность гониатитовой свиты, по Масленникову, достигает 400—500 м.
Содержащие почти исключительно фауну аммоноидей известняки прослеживаются в виде небольшой мощности линзовидных пропластков среди эффузивов на протяжении, по-видимому, нескольких километров.
В результате работ 1949 г. А. И. Левенко впервые обнаружил аммоноидей в плотных известково-глинистых алевролитах, которые наблюдаются южнее пос. Шемонаихи по правому берегу р. Убы, в 2 км ниже ж.-д. моста, и в виде довольно широкой полосы прослеживаются к северо-западу на правобережье р. Убы, севернее дер. Камышинки. Опираясь, по-видимому, на находки в алевролитах аммоноидей и сопутствие им, как и известнякам гониатитового горизонта района Николаевского рудника, основных эффузивов и их туфов, Левенко отнес известково-глинистые алевролиты правобережья р. Убы к гониатитовому (самарскому) горизонту (D |).
По правому берегу р. Убы, в 2 км ниже моста, 250 м севернее 25—30-метровых обрывов, сложенных темными, почти черными кварцевыми пор-
фиритами с белыми точечными включениями, наблюдается, по А. И. Левенко, следующий разрез сверху вниз:
1. Темно-зеленые кварцевые порфириты.....................
2. Буровато-зеленые кварцевые порфириты, ожелезненные по трещинам................................................2,3м
3. Темно-серые и синевато-серые неслоистые окремнелые глинистые сланцы (алевролиты)...........................15,0 м:
4. Светлые буровато-зеленые, преимущественно грубослоистые сланцы, ожелезненные по трещинам. Среди них встречаются и тонко-слоистые сланцы. Аз. пад. 210°/22° . . 16,4 м
В 9 м от контакта с кварцевыми порфиритами в сланцах найдена фауна гониатитов. Здесь же — ожелезненный прослой в 2 см мощностью из многочисленных стяжений
24
лимонита. В 15 м от контакта алевролиты становятся более темными, несколько окремнелыми, в алевролитах встречаются светлые мелкие многочисленные включения, вытянутые по слоистости. Здесь же встречена фауна . .
5. Светлая зеленоватая брекчия, в обломках которой — белые твердые зеленоватые сланцы, цементируется более мелкими обломками того же материала.....................78 см
В нижней части прослоя брекчии встречаются прерывистые глинистые прослойки мощностью около 2 см. К востоку брекчия сменяется светло-серыми кремнистыми сланцами. Мощность сланцев 80 см. . ................
6. Светло-зеленоватые неслоистые алевролиты............. 60 см
7. Светлые тонкослоистые кремнистые сланцы, образующие по простиранию микроскладки и микросбросы...............Q87 см
8. Алевролиты тонкослоистые со структурой течения. Аз. пад.
198°/ 30 . Алевролиты рассланцованы на правильные грубые плиты мощностью 25—30 см.....................90 см
9. Алевролиты светло-серые тонкозернистые и тонкополосчатые с микросбросами...................................50 см
10. Песчаники тонкозернистые, слоистые с точечной ожелез-ненной вкрапленностью, местами каолинизированные 50 см И. Темно-зеленые глинистые сланцы, сильно трещиноватые с неправильной овальной отдельностью, местами слоистые. В сланцах наблюдаются зеркала скольжения и более светлые песчаные прослои......................... 12—15 м
Склон покрыт осыпями и контакт между сланцами и известняками проследить не удалось ...................
12. Темные, почти черные известняки, местами слоистые, сильно трещиноватые; при выветривании дают тонкие длинные обломки, иногда караваи до 35 см в диаметре. В одном из караваев найдена фауна. В караваях — тонкие прожилки и вкрапления сульфидов. В нижней части аз. пад. 218°<^60°............................. 28—30 м
Известняки и сланцы срезаются интрузивом плагиогра-нит-порфиров........................................
В последнее время П. Ф. Иванкин (1952 г.), опираясь на данные бурения, подводя итоги собственным наблюдениям и исследованиям своих предшественников, считает ошибочным отнесение А. И. Левенко известково-глинистых алевролитов р. Убы к гониатитовому горизонту. Он выделяет их в нижнюю гониатитовую свиту (D i~r), отождествляя ее со среднедевонской надлосишинской свитой Н. А. Ноздреева, которая была установлена последним по левому берегу р. Убы.
П. Ф. Иванкиным дается следующая схема стратиграфии девонских отложений, развитых в районе (см. табл. 4 на стр. 26).
Верхняя граница девонской вулканогенной формации известна за пределами района, в бассейне р. Ульбы. По данным Н. Л. Бубличенко (1952) и В. А. Федоровского, она проводится в основании бухтарминской свиты известняков, отвечающей низам карбона.
Вдоль этого контакта устанавливаются базальные конгломераты и местами выпадение из разреза отдельных мощных его частей (тарханская терригенная свита).
Выделение П. Ф. Иванкиным среднедевонской нижнегониатитовой свиты не может, однако, не встретить серьезных возражений. Против такого выделения указанной свиты и определения ее среднедевонского возраста
25
Таблица 4
Возраст Свита Мощность, м Состав свиты Фауна
D3 Боровиковская 900 Основные, частью кислые лавы, их агломераты и туффиты —
Снегиревская 400— 500 Туфы кислых эффузивов, перемежающиеся с туфопесчаниками и кремнисто-глинистыми алевролитами —
Николаевский горизонт б— 250 Известняки, песчано-глинистые породы, частью туфы Имеется
Каменевская 300— 400 Туфолавы, их агломераты и среднеобломочные туфы кварцевых порфиров —
D>-r Гониатитовая „манти к о перовая" 100— 350 Пузыристые основные эффузивы. шаровые спилиты, их агломераты с известковистым цементом, кремнисто-глинистые породы, линзы известняков Имеется
Возможен размыв и незначительное несогласие
и о М IjS W W Таловская Нижнегониатито-вая («надлоси-шинская», по Н. А. Ноздрееву) 500 500— 600 Кварцевые порфиры и главным образом их агломераты Зеленовато-корйчневые алевролиты и туффиты с линзами известняков, перемежающихся с основными эффузивами и их плотными туфами Altaites sp. nov Tornoceras aff. simplex Buch.
о ьэ to в i Сугатовская (выделяется предположительно) 0— 800 Туфы, туфолавы и лавобрекчии кварцевых порфиров
Лосишинская 300— 350 Метаморфизованные известняки и песчано-глинистые алевролиты, частью кислые лавы и туфы Имеется
D3~6 Березовская 0— 900 Агломераты и лавобрекчии кислых эффузивов, частью песчаные породы, конгломераты —
Размыв и несогласие
М Метаморфическая толща Метаморфические сланцы —
говорят наши исследования наблюдающейся в алевролитах правобережья р. Убы фауны аммоноидей.
Наши наблюдения, произведенные во время летний полевых работ 1952 г. по правому берегу р. Убы, в 2 км ниже ж.-д. моста, дают в основном тот же разрез, что и приведенный выше по А. И. Левенко, с немногими лишь отличиями в мощности некоторых частей разреза. Так, верхняя пачка алевролитов (4-я по Левенко) составляет не 16,4 м, а около 12 м мощности, нижняя (11-я, по Левенко 12—15 м) — не более 8—10 м; преувеличена на наш взгляд и мощность подстилающих известняков. Сборы фауны аммо-26
ноидей произведены нами в слое 3-м, в интервале 6—8 м ниже контакта •со спилитами; в остальной части разреза аммоноидей не обнаружено.
Помимо этого, нами собрано относительно большое количество образцов из алевролитов зеленовато-серого и темно-серого цвета, выходы которых наблюдаются в 1 км к СЗ от рассмотренного выше обнажения по правому берегу р. Убы. Аммоноидей собраны из алевролитов мощностью до 60— 70см, залегающих почти горизонтально (аз. пад. 310°/А0°). Несколько форм было найдено в подобных алевролитах, наблюдающихся в 2,5 км севернее дер. Камышинки. Собранные из алевролитов образцы в большинстве своем деформированы и плохой сохранности, но ввиду отсутствия в алевролитах иной фауны приобретают немаловажное значение. Уже первое предварительное знакомство с наблюдающейся здесь фауной аммоноидей позволило отметить коренное отличие ее от фауны аммоноидей из известняков гониатитового горизонта Николаевского месторождения, говорящее о разновозрастности находящихся в районе Николаевского рудника и по правобережью р. Убы отложений.
На основании предварительного изучения собранных коллекций аммоноидей можно было заключить, что алевролиты правобережья р. Убы должны рассматриваться как относящиеся к наиболее низкому, фарцице-ровому горизонту (Вз~а) верхнего девона, т. е. имеющие несколько более древний возраст по сравнению с известняками гониатитового горизонта Николаевского рудника. Среди собранной фауны аммоноидей из алевролитов правобережья р. Убы определены: Pharciceras kiliani Freeh (? = Ph. lunulicosta (Sandberger)) и Triainoceras sp.
Pharciceras kiliani Freeh и близкий к Triainoceras sp. Triainoceras costatum (A. V.) описывались из самых низов верхнего девона — зоны 1а (Prolecanitenschichten = Pharcicerasschichten) Германии и Франции; Ph. lunulicosta (Sandberger) известен также из наиболее низких верхнедевонских отложенцй Северной Африки (Марокко, Алжир). В выше- и нижележащих отложениях представители указанных родов и видов не наблюдались, что, как мне кажется, позволяет считать алевролиты правобережья р. Убы, в которых найдены эти формы, наиболее низким членом верхнедевонского комплекса Рудного Алтая, соответствующим фарцице-ровому горизонту (Dga), (зона 1а Рейнских провинций и, вероятно, Fj Арденн). Использование же П. Ф. Иванкиным для палеонтологического обоснования среднедевонского возраста нижнегониатитовой свиты Altaites sp nov. и Tornoceras aff. simplex (Buch) (определения Г. П. Лященко) необоснованно.
Верхнедевонские аммоноидей собраны нами в результате полевых работ 1952 и 1953 гг. из известняковых линз, наблюдающихся в 3 км к юго-востоку от поселка Николаевского рудника по простиранию гониатитового горизонта (свиты). Здесь, среди кварцевых кератофиров, светло-серых, серых и зеленовато-серых туфов, на северном склоне гряды наблюдаются выходы небольших линз (мощностью 1,5—2,5 м, длиной до нескольких метров) органогенных или органогенно-обломочных известняков, крепких, перекристаллизованных малиновых и бордово-красных, с белыми пятнами окаменелостей. Известняки местами с обильной фауной энкринитов, кораллов, реже аммоноидей и мелких брахиопод, пелеципод, гастропод, трилобитов. Аммоноидей наблюдаются чаще небольшими гнездами в более рыхлых разностях известняков, встречаясь в таких случаях в довольно больших количествах. В более крепких разностях известняков, лишенных фаунистических остатков, наблюдаются мелкие пропластки и линзы песчаников и гальки и обломки эффузивных пород, местами тонкая слоистость. Известняки перемежаются с рыхлыми, выветрелыми известковистыми туфами и эффузицами. Элементы залегания известняков
27
неясные. Видимый азимут падения 220—230°/30—35°.Известняковые линзы прослеживаются на 100—120 м по простиранию при ширине полосы выхода известняков в 30—40 м. Среди собранных здесь аммоноидей определены: Manticoceras affine (Steininger), М. intumescens (Beyrich), M. sinuosum (Hall), M. artum sp. nov., Tornoceras simplex (Buch), Beloceras sagittarium (Sandberger) и Devonopronorites ruzhencevi Bogoslovsky.
Обращает на себя внимание присутствие в больших количествах представителей Devonopronorites ruzhencevi Bogoslovsky. Другие аммониты встречаются в значительно меньших количествах и их видовой состав ничем существенно не отличается от состава аммонитовой фауны известняковых линз гониатитоЕого горизонта, наблюдающихся непосредственно в районе самого Николаевского месторождения, а поэтому заключение о возрасте этих отложений будет дано нами после рассмотрения фауны аммоноидей гониатитового горизонта этого месторождения.
Основные сборы аммоноидей из района Николаевского месторождения, составляющие подавляющую часть коллекции из данного района, наиболее разнообразную по своему составу в видовом и родовом отношениях, были произведены из известняковых линз гониатитового горизонта, обнаруженного в свое время в районе месторождения еще Н. А. Ноздреевым.
Для вскрытия содержащих фауну аммоноидей линз известняков нами были пройдены на западном склоне гряды, отходящей от поселка в северо-восточном направлении, небольшие канавы со следующим разрезом снизу вверх (разрез сводный по простиранию 50—60 м):
1. Выветрелые светло-зеленые известковистые туфы с тонкими редкими прослоями глинистых сланцев темно-серого цвета...............................................до 2 м
2. Прослой плотного, неслоистого темного буровато-серого известняка с Manticoceras intumescens (Beyrich), М. sinuosum (Hall), Beloceras sagittarium (Sandberger) и др. . . .до 20 см
3. Чередующиеся тонкие (до 5 см мощности) прослои глинистых сланцев и известковистых туфов (по 2 прослойка)
4. Выветрелые туфы........................................1,5 м
5. Буровато-темно-серый, плотный неслоистый известняк (возможно глыбового залегания) с Manticoceras sinuosum (Hall), Beloceras sagittarium (Sandberger) и др................... 15 см
6. Выветрелые известковистые туфы........................до 4 м
7. Выветрелый прослой агломератовидного известняка с при-
месью туфового материала. Аз. пад. 220°/_60° . . . 10—15 см 8. Выветрелые известковистые туфы светло-зеленого цвета,
чередующиеся с прослоями плотных туфов................5,5 м
9. Буровато-серые перекристаллизованные известняки без видимой фауны....................................... 25—30 см
10. Туфы основных пород темно-зеленые........................ 1м
И. Известняки буровато-темно-серого цвета, трещиноватые, плохослоистые с Beloceras sagittarium (Sandberger), Manticoceras sp..................................................30 см
12. Туфы основных пород зеленого цвета....................до 1 м
13. Светло-серые сильно перекристаллизованные неслоистые
известняки без фауны............................ 50—60 см
14. Выветрелые известковистые зеленовато-белые туфы ... 1м 15. Темно-бурые метаморфизованные и перемятые известняки
с фауной деформированных гониатитов [Beloceras sagittarium (Sandberger)]...................................20 см
16. Известковистые туфы основных пород зеленого цвета . . до 1 м 17. Известняки буровато-темно-серые, плотные трещиноватые
28
c Ponticeras altaicum sp. nov., P. acutilobatum sp. nov., Manticoceras affine (Steininger), M. carinatum (Sandberger), M. ammon (Keyserling), M. sinuosum (Hall.), Manticoceras sp., Beloceras sagittarium (Sandberger), B. stenum-bilicatum sp. nov., реже с пелециподами, брахиоподами, трилобитами и др. Аз. пад. 220°z/60°.......................70 см
18. Туфы основных пород, плотные, с прослоями известкови-
стых туфов. Видимая мощность............................... 6 м
19. Плотные, темно-серые известняки, неслоистые трещино-
ватые с Probeloceras (?) forcipiferum (Sandberger), Ponticeras sp., Beloceras sagittarium (Sandberger), B. stenumbili-catum sp. nov. и др. Аз. пад. 160° / 40°...................40 см
20. Туфы основных пород (спилиты?) темно-зеленого цвета . . 40 см
Пройденные канавы дают представление о линзообразном залегании известняковых прослоев среди пород эффузивного происхождения. Многие из этих линзовидных прослоев содержат фауну, преимущественно гониа-титовую, причем в некоторых из них она довольно обильна. Подавляющую массу собранных здесь коллекций составляют роды Beloceras и Manticoceras, в значительно меньшей степени наблюдаются Ponticeras и Probeloceras. Отсюда определены: Ponticeras altaicum sp. nov., P. acutilobatum sp. nov., Probeloceras (?), forcipiferum (Sandberger), Manticoceras affine (Steininger), M. bullatum Wedekind, M. galeatum Wedekind, M. carinatum (Sandberger), M. intumescens (Beyrich), M. sinuosum (Hall), M. altaicum sp. nov., M. ammon (Keyserling), Beloceras sagittarium (Sandberger), B. stenumbilicatum sp. nov.
Анализ фауны аммоноидей гониатитового горизонта показывает, что наряду с наличием в ней местных алтайских форм она обнаруживает большое сходство с западноевропейской аммонитовой фауной, отчасти с североамериканской. В некоторой степени проявляется связь с Уралом и Тиманом. Большинство изученных видов известно из Manticoceras-SVafe Западной Европы, характеризуя третью (1у) зону Manticoceras cordatum и М. carinatum.
Присутствие среди изученной фауны Manticoceras intumescens (Beyrich), М. affine (Steininger), M. galeatum Wedekind, M. aff. carinatum (Sandberger), характерных только для зоны I у Западной Европы, позволяет считать отложения гониатитового горизонта соответствующими третьей зонеManticoceras-SVais (зона I?) или F2 (вероятно F2d — F2h Арденн). Более точная параллелизация гониатитового горизонта с разрезом Арденн затруднена в виду отсутствия в последнем достаточного количества характерных видов аммоноидей.
Подводя итог данным по стратиграфии девонских отложений района Николаевского месторождения, можно на основании изучения аммоноидей выделить два палеонтологически охарактеризованных горизонта: горизонт алевролитов правобережья р. Убы в основании верхнего девона, соответствующий фарцицеровому горизонту (1а), и гониатитовый горизонт известняковых линз Николаевского месторождения, соответствующий ман-тикоцеровым слоям (1у) франского яруса Западной Европы. Возможно, что оба эти «горизонта» следует рассматривать в пределах одной самарской (не употребляя наименования гониатитовая) свиты, относя один из них к основанию, а другой к верхам ее. Подобного взгляда, в частности, придерживаются в последнее время геологи Алтайской экспедиции треста Аэрогеологии (Яковлев, Зорова и др.), выделяющие гониатитовую свиту нижнефранского возраста, достигающую мощности до 500 м и заключающую в нижней и верхней своих частях охарактеризованные фауной аммоноидей горизонты.
29
Таким образом, результаты изучения аммоноидей позволяют уточнить стратиграфическое положение отдельных членов разреза и сделать следующие выводы:
1. Наиболее низким членом верхнедевонского эффузивно-осадочного^ комплекса, развитого в районе, являются алевролиты правобережья р. Убы, в основании которых должна проводиться граница между средним и верхним девоном.
2. Параллелизация указанных алевролитов с известняковыми линзами гониатитового горизонта и, тем более, отнесение их к среднему девону («нижнегониатитовой» или «надлосишинской» свите) необоснованно и ошибочно.
3. Гониатитовый горизонт, содержащий известняковые линзы с фауной аммоноидей, залегает не в основании франского яруса, а занимает более высокое положение, характеризуя собой мантикоцеровый горизонт (зону I?) франского яруса (среднефранский подъярус — D32 при делении яруса на три подъяруса, или, согласно другой точке зрения, верхнефран-ский подъярус D32 при делении яруса на два подъяруса).
4. Залегающий выше гониатитового горизонта николаевский фаунистический горизонт должен относиться не к нижней половине франского-яруса и, тем более, не к основанию его, а к верхней половине франского яруса (верхнефранский подъярус).
Район Гериховского месторождения
Несмотря на то, что район Гериховского и соседних с ним месторождений многократно посещался исследователями, начиная с XVIII столетия (Фалон, Ренованц, Герман, Шуровский, Богданов, Петц и др.), первые обстоятельные сведения о геологии района относятся к сравнительно позднему периоду и связаны с именем Н. А. Елисеева, которым в 1936 г. была проведена геологическая съемка и заложены основы понимания геологии данного района. Он выделил в районе метаморфические сланцы кембро-силура, эффузивно осадочные толщи среднего и верхнего девона и интрузивные породы змеиногорского комплекса. Важным результатом работ Елисеева явилось, на основании фаунистических находок вблизи с. Раздольного, определение возраста известняков, отнесенных им к среднему и верхнему девону.
В 1939 г. И. П. Коропец, проведя геологическую съемку участка между Гериховским и Титовским месторождениями, выделил здесь три свиты: известняковую (кубоидный горизонт D3), туфогенную (D2) и свиту красно-бурых туфов, лежащую на известняках, для которой им допускался возраст от D3 до Р включительно. Известняки, развитые южнее туфогеновой свиты, Коропец также относил к кубоидному горизонту и допускал существование антиклинали, к разным крыльям которой он приурочивал Ти-товское и Гериховское месторождения. В дальнейшем, однако, эти построения не подтвердились. В 1940 г. И. Ф. Трусова составила карту района на основании работ А. С. Егорова, однако она приводит стратиграфию и тектонику месторождения в очень упрощенном виде.
Л. Н. Краевская, определяя фауну, собранную А. С. Егоровым из нижнего горизонта известняков, установила франский возраст последних. Собранная Егоровым в верхах горизонта известняков фауна бухиол и гастропод была определена Л. Л. Халфиным как нижнефранская (кубоидный горизонт). У дер. Горюновки И. Ф. Трусова выделила два самостоятельных горизонта, отнеся их к среднему девону. Здесь, по А. С. Егорову, среди туфогенно-осадочного комплекса наблюдается следующий разрез, известняково-песчанистых отложений снизу вверх:
1. Туфы кварцевых порфиров зеленовато-серые, пятнистые (Z)2?)..................................1,5 м
30
2. Серые массивные известняки с криноидеями, трилобитами, брахиоподами . . . .................8 м
3. Задерновано (туфогенные породы)
4. Кварцевые порфиры плотные, серого цвета ... 16 м
5. Кремнисто-глинистые алевролиты плотные, темно-зеленого или зеленоватого цвета . . . . • . 40 м
6. Туфы кварцевых порфиров грубослоистые зеленоватые ............... ..................... . . 18 м
7. Ракушняковый сильно окремнелый известняк, розовый или светло-серый с обильной фауной криноидей, кораллов, мшанок, брахиопод . . . 4—Зм
8. Титовско-гериховская свита
Более поздними исследованиями было, однако, установлено, что приведенный выше разрез А. С. Егорова не наблюдается, а отложения, отнесенные И. Ф. Трусовой к среднему девону, должны, как показало исследование брахиопод и аммоноидей, относиться к франскому ярусу (см. ниже стр. 35).
В результате работ 1950—1951 гг. П. Ф. Иванкин разделил все палеозойские породы, наблюдающиеся в районе, на три комплекса:
1. Метаморфические сланцы кембро-силура и диоритовые гнейсы.
2. Эффузивные образования среднего (возможно и частично нижнего) девона.
3. Эффузивные, туфогенно-осадочные и органогенные образования (биогермы) верхнего девона.
Полностью выпадает из разреза S2 и, вероятно, Dj.
Весь комплекс девонских отложений подразделяется П. Ф. Иванкиным на четыре свиты сверху вниз:
1. Агломератовая свита............................D|
2. Туфогенно-обломочная свита...................D3
3. Титовско-гериховская свита, включающая Неверовский риф известняков •.D3
4. Свита кислых эффузивов с прослоями известковопесчаных отложений D1-|-D2
Агломератовая свита распространена севернее и восточнее Герихов-ского месторождения, отличается буро-красной окраской и лиловым оттенком. Представлена агломератовыми туфами и эффузивами основного состава (манделыптейны), преобладающими в нижней ее части, и кислыми эффузивами в верхней. В западной части площади распространения свиты известен прослой известняковых туфов. Мощность, вероятно, не превышает 100—150 м.
Туфогенно-обломочная свита лежит в основании агломератовой и, в свою очередь, налегает на титовско-гериховскую свиту и рифовые известняки франского яруса. Возраст — верхи франского яруса. Состав свиты отличается большой пестротой. К северу от Неверовского массива известняков в разрезе свиты преобладают зеленоватые, отчетливо слоистые туфопесчаники, перемежающиеся с плотными псаммитовыми туфами; восточнее, у с. Раздольного, господствуют агломератовые и псефитовые туфы и туфобрекчии с включениями крупных лапиллей и бомб кислых эффузивов. Общая мощность не превышает 80 м.
Титовско-гериховская свита. За верхнюю границу свиты П. Ф. Иванкиным принимается пласт известняков Рериховского месторождения, который прослеживается в широтном направлении, вплоть до рч. Грязнухи и переходит на восточный борт ее долины, приобретая G3 простирание. На юге нижняя граница менее определена и условно проводится по кон
31
такту основных туфов с кислыми туфами и порфирами коричневого цвета (D2). Это — наиболее сложная свита в районе. Вся свита разделяется на четыре толщи (А, Б, В, Г), отличающиеся фациально-литологическими особенностями и стратиграфическим положением. Каждая толща разделяется на пласты, которые состоят из нескольких слоев и линз. Свита в целом сложена эффузивно-осадочными образованиями с резким преобладанием вулканогенных образований (различного рода эффузивов и их туфов) над осадочными (алевролиты, туфо-песчаные известняки). В восточном направлении свита фациально замещается рифовыми известняками, которые считаются П. Ф. Иванкиным одновозрастными с титовско-гериховской свитой. Принимая большую мощность Неверовских известняков и отмечая периодичность накопления их в условиях циклической вулканический деятельности, П. Ф. Иванкин намечает 14—15 стадий роста рифа и отложений эффузивно-пирокластического материала. Риф рисуется им в виде веерообразно расходящихся линзовидныхобразований,которые сближаются и «срастаются» в восточном направлении в виде крупного массива известняка с редкими прослоями эффузивов и туфов. Риф, по мнению И. Ф. Иванкина, должен быть отнесен к барьерным (большая мощность) или к атоллам, вырастающим на склоне погружающегося вулканического острова. Прекращение роста рифа, вероятно, вызвано было поступлением туфогенно-обломочного материала и тонких илов (туфогенно-обломочная свита), которые покрывают риф.
Возраст рифа считается исследователем верхнедевонским, хотя он и оговаривается, что нижняя часть рифа, может быть, имеет и среднедевонский возраст. Истинная мощность свиты принимается равной 1200—1300 м.
Свита кислых эффузивов с прослоями известково-песчаных отложений. Выходы свиты наблюдаются южнее дер. Горюновки и с. Раздольного и представлены кварцевыми альбитофирами, содержащими прослои туфов и известково-песчаных отложений с обильной фауной. (Разрез известковопесчаных отложений у дер. Горюновки см. выше —разрез А. С. Егорова стр. 30). Мощность свиты принимается равной 900—1000 м.
Таким образом, в результате своих исследований П. Ф. Иванкин приходит к заключению: 1) о среднедевонеком возрасте известняковых линз (свита кислых эффузивов с прослоями известково-песчаных отложений), наблюдающихся у дер. Горюновки; 2) о столбообразном залегании мощных рифовых известняков вкрест простирания туфогенных пород, на основании выклинивания отдельных линз известняков.
Такая точка зрения оспаривается рядом исследователей. Так, Н. Л. Бубличенко, принимая несомненный рифовый характер некоторой части неверовских известняков (Tabulata), указывает, что известняки эти согласно пластуются с вмещающими породами. Их изгиб в виде сигмоида, при наличии местами сравнительно пологих залеганий, обусловливает в некоторых местах большие площади распространения и кажущиеся прихотливые очертания выходов известняков. Для известняковых линз, наблюдающихся у дер. Горюновки, Н. Л. Бубличенко на основании определения брахиоподовой фауны указывает верхнедевонский возраст. К аналогичным заключениям приходят в результате своих работ и другие исследователи (В. П. Курбатов, В. А. Комар и др.). Ими на основании геологической съемки Гериховского месторождения в 1951—1952 гг., по аналогии с районом Николаевского месторождения, принимается следующая схема стратиграфии девонских отложений:
Березовская свита.................•..............| t
Лосишинская » . . . •........................./ -
Таловская » .................................D2
32
Самарская (гониатитовая)......... .............1
Каменевская свита.............................j- D*
Снегиревская » ............................J
Боровиковская » ............................
Чудацкая » ............................[D3
Порфиритовая » ............................)
В основании девонских отложений района, по В. П. Курбатову, залегает мощная (до 2000 м), предположительно нижнепалеозойского (?) возраста метаморфическая свита, представленная метаморфическими сланцами, песчаниками, филлитовидными глинистыми сланцами с линзами мрамо-ризованных известняков, кварцево-хлорито-эпидотовыми и амфиболитовыми сланцами, метаморфизованными туфами и туфопесчаниками.
На метаморфической свите залегает с размывом березовская свита D23, представленная песчаниками, аргилитами, кремнистыми сланцами, туфами кислого состава с прослоями конгломератов и кислых эффузивов и линзами известняков мощностью 300—450 м. Лосишинская свита—D*6 представлена алевролитами, аргилитами с прослоями туфопесчаников, песчаников, известняков и линзами туфов кварцевых альбитофиров мощностью от 40 до 600 м. Таловская свита — D1^2 представлена псефитовыми, агломератовыми туфами кварцевых альбитофиров с прослоями лавобрекчий кислого состава и линзами аргилитов и туфопесчаников, мощностью 800—1000 м.
Общая мощность среднего девона по некоторым данным достигает приблизительно 2400 м.
Разрез верхнедевонских отложений начинается каменевской (+са-марекой) свитой. Последняя в виду плохой обнаженности в самостоятельную свиту в районе не выделяется.
Каменевская (+ самарская) свита—D13^-0. Пестрая толща туфогенноосадочных пород свиты представлена туфами кислого, среднего и основного состава с прослоями и линзами спилитов, аргилитов и кремнистых сланцев. Свита обнажается в районе восточнее и юго-восточнее дер. Горюновки в виде 3 выходов. В районе с. Раздольного верхняя граница свиты определяется залеганием на ней известняков франского яруса. Мощность свиты, по Курбатову, составляет 400—500 м.
Снегиревская свита —D1® представлена туфами кислого состава с линзами кварцевых альбитофиров, туфами среднего и основного состава с прослоями спилитов, тонкослоистыми аргилитами, песчаниками, кремнистыми сланцами,массивными рифовыми известняками николаевского горизонта. В районе с.Раздольного наблюдается следующий разрез снегиревской свиты:
1. В основании разреза — массивные рифовые (неверовские) известняки буро-красного, розового, серовато-розового и розовато-светло-серого цвета (максимальная мощность известняков 150 м) с линзовидными прослоями известковистых туфов, туффитов и кремнистых аргилитов. В известняках обильна фауна брахиопод (определены Н. Л. Бубличенко), мшанок, кораллов, гастропод и др. В прослоях кремнисто-глинистых аргилитов А. С. Егоровым собрана фауна пелеципод, определенная Л. Л. Халфиным (Buchiola retrostriata Buch, В. cf. conversa Clarke, В, altaica Chalf.) и указывающая на новый фациальный тип нижнефранских отложений. Горизонт известняков по простиранию быстро выклинивается и на левом борту рч. Грязнухи замещается туфогенной толщей с редкими линзами светлосерых известняков, содержащими франскую фауну (неправильно относимые А. С. Егоровым к D2 линзы известняков у дер. Горюновки).
2. На известняках, согласно с ними, залегает маломощный песчаносланцевый горизонт, состоящий из тонкого, частого чередования туфо-3 Б. II. Богословский on
генных песчаников, кремнисто-глинистых сланцев и аргилитов, туффитов, реже туфов кислого состава. Туфопесчаники и туффиты представлены тонко и мелкообломочными разностями желто-серого и зеленовато-серого цвета. Обломочный материал состоит из зерен плагиоклаза, кварца и обломков туфогенных пород. Встречаются обломки известняков. Цемент — известково-туфовый. Кремнисто-глинистые сланцы и аргилиты — толсто-и тонкоплитчатые породы, иногда листоватые, желтовато-серого и коричневатого цвета. К ним приурочены тонкие (2—5 см) прослои, реже линзы темно-серых и серых, часто кремнистых известняков с редкой, плохо сохранившейся фауной гониатитов. Мощность горизонта — 30—50 м.
3. Выше песчано-сланцевого горизонта залегает грубообломочный туфогенный горизонт туфов кислого состава с подчиненными прослоями эффузивов, кремнистых сланцев, туфопесчаников и туффитов. Между рч. Грязнухой и Золотухой наблюдается туфоконгломерат, в котором изредка встречаются глыбы и небольшие линзы серых известняков без фауны. Мощность горизонта — 100—150 м.
4. Горизонт туфогенно-эффузивных пород основного состава, представленный главным образом диабазовыми и андезитовыми порфиритами, спилитами, их туфами, изредка линзовидными прослоями известковистых туфов и кремнистых сланцев. Мощность горизонта — 150—200 м.
5. Разрез заканчивается горизонтом туфогенно-эффузивных пород кислого состава, главным образом гетеропластических туфов кварцевых альбитофиров, реже кварцевых альбитофиров и вулканических брекчий. У северной окраины с. Раздольного наблюдаются прослои пузыристых кислых лав. Для горизонта характерна буровато-лиловая окраска, реже коричневая, серая. Мощность горизонта достигает 600 м.
Первые указания на присутствие отложений гониатитового горизонта в районе Гериховского месторождения относятся к 1936 г., когда Н. А. Елисеевым были обнаружены аммоноидей в известняках рч. Грязнухи, ниже с. Раздольного. Позднее отдельные находки аммоноидей из района Гериховского месторождения указывались геологом В. П. Курбатовым, работавшим в этом районе в 1951—1952 гг. Результатом наших работ в 1952 и 1953 гг. в районе месторождения явились довольно обширные коллекции аммоноидей, собранные в ряде пунктов. Обследование массива неверовских рифовых известняков (толща 1-ая снегиревской свиты), богатых фауной брахиопод, кораллов, мшанок, гастропод и др., показало, что в этих известняках аммоноидей отсутствуют или находки их спорадичны. Несколько экземпляров, имеющихся в нашем распоряжении, принадлежат одному виду Triainoceras gerassimovi sp. nov., близкому к Т. costatum (А. V.), который известен в Западной Европе по единичным находкам из наиболее низких слоев (1а — фарцицеровый горизонт) франского яруса Рейнской провинции. Данная форма была найдена в органогенных известняках правого берега рч. Грязнухи, в 1 км ниже с. Раздольного.
Как уже отмечалось выше, нижняя часть неверовских известняков должна относиться к основанию франского яруса — зоне 1а.
Более обширная коллекция аммоноидей (правда, весьма однообразная по видовому составу) была собрана нами из известняковых линз у дер. Горюновки, которые рассматриваются в настоящее время как продолжение нижнего горизонта неверовских «рифовых» известняков по простиранию последних. Здесь, на правом берегу рч. Золотухи, в 400 м восточнее дер. Горюновки, на туфы и туфолавы кислого состава ложатся:
1. Кремнистые алевролиты.....................1,5 м
2. Туфы кислого состава, псефитовые с прослоями кремнистых известняков.......................... 20 м
3. Кремнистые известняки......................... 5 м
34
4. Алевролиты.................................. 20 м
5. Туфы кислого состава, псефитовые............ 10 м
6. Линза известняка с фауной D*. Среди сборов В. II. Курбатова Н. Л. Бубличенко определены: Gipl-dula globa Brown., Uncinulus ex gr cuboides Sow., Schizophoria striatula Schloth., Meristella sp., Retziasp.-, из аммоноидей нами определены: Probeloceras (?) orientate sp. nov., Neopharciceras kur-batori Bogoslovsky.............................1,5—2 м
7. Туфы кислого состава, псефитовые, анологичные слою 5......................................... 10 м
8. Грубые агломератовые туфы кислого состава . . 25 м
9. Слоистые, от мелко-до крупнозернистых, туфы кислого состава.
Как уже отмечалось выше, эти отложения должны, по всей вероятности, рассматриваться как близкие по возрасту зоне 1а и соответствующие низам зоны 1р (зона Manticoceras nodulosum) Рейнской провинции (Гга-е Арденн?). В районе Николаевского месторождения к ним близки по возрасту алевролиты правобережья р. Убы, а верхняя часть последних, возможно, соответствует данным отложениям.
В результате работ 1953 г. нами была собрана довольно большая коллекция аммоноидей из более высокого песчано-сланцевого горизонта отложений франского яруса, во 2-й толще снегиревской свиты. Выходы этого горизонта наблюдаются по правому берегу рч. Грязнухи, ниже с. Раздольного, где на массивных «рифовых» известняках согласно с ними залегает песчано-сланцевый горизонт чередующихся между собой туфогенных песчаников, кремнисто-глинистых сланцев, аргилитов, туффитов и туфов кислого состава. К кремнисто-глинистым сланцам приурочены тонкие (2—5 см) прослои, реже линзы темно-серых и серых, часто кремнистых известняков, с редкой, плохо сохранившейся фауной аммонитов. В 150 м вверх по течению речки, у окраины с. Раздольного, известняковые прослои достигают 10—15 см мощности. В верхней части песчано-сланцевого горизонта здесь наблюдаются линзообразно залегающие, чаще выветрелые бордово-красные и темно-бурые известняки, переслаивающиеся с тонкослоистыми глинистыми сланцами, песчаниками и известковистыми туфами. Известняковые прослои (5—10 см мощности) содержат фауну аммоноидей. (Здесь собрана большая часть коллекции). Общая мощность линзы достигает 2—2,5 м, протяженность по простиранию — до 20 метров. В 500 м по простиранию горизонта на восток в правом берегу рч. Грязнухи известняки выходят в виде глыб неясного залегания мощностью до 1 м бурого известняка, крепкого, иногда выветрелого, содержащего фауну аммонитов. Известняковые глыбы наблюдаются и в русле речки. Известняки перекрываются выветрелыми известковистыми туфами фиолетово-красного цвета, подстилаются кремнисто-глинистыми сланцами красно-бурого цвета. Среди выветрелых разностей известняков собрана фауна аммоноидей.
Подавляющую массу собранных аммоноидей составляют роды Manticoceras и Beloceras, в меньшей степени Ponticeras. Отсюда определены: Ponticeras sp., Manticoceras drevermanni Wedekind, M. altaicum sp. nov., M. sinuosum (Hall), M. neverovi sp. nov., M. eliseevi sp. nov., Beloceras sagittarium (Sandberger).
Близость этой фауны к фауне гониатитового горизонта Николаевского месторождения (Manticoceras-Stnio, зона 1у) и, с другой стороны, присутствие в ней таких форм, как Ponticeras sp., близкого к Ponticeras bickense Wedekind и Manticoceras drevermanni Wedekind (две последние формы тяготеют в Западной Европе к верхней 13 зоне Manticoceras-Stnio)*
3*
35
заставляют полагать возраст данной фауны близким или несколько более молодым по сравнению с возрастом фауны гониатитового горизонта Николаевского месторождения. Отложения, содержащие эту фауну, могут, на наш взгляд, параллелизоваться примерно с верхами зоны — низами зоны 13 (слои с Manticoceras adorjense) или соответственно с зоной F2i— F2j разреза Арденн. В районе Николаевского месторождения им, возможно, будут отвечать по возрасту известняковые линзы, наблюдающиеся среди отложений каменевской свиты, залегающей выше гониатитового горизонта (самарской свиты).
Подводя итог рассмотрению стратиграфии девонских отложений, развитых в районе Гериховского месторождения, можно на основании изучения аммоноидей уточнить стратиграфическое положение некоторых частей разреза и сделать следующие выводы:
1. Наиболее низким членом разреза верхнедевонских отложений, развитых в районе, должна считаться нижняя часть неверовских известняков, в основании которых следует проводить границу между средним и верхним девоном.
2) Отнесение неверовских известняков к низам снегиревской свиты и тем более параллелизация их с николаевским фаунистическим горизонтом района Николаевского месторождения вряд ли обоснованы.
3) Известняковые линзы, наблюдающиеся в туфогенной толще по правому берегу рч. Золотухи, восточнее дер. Горюновки, действительно являются верхнедевонскими, близкими по возрасту или несколько более молодыми по отношению к низам неверовских известняков, и могут рассматриваться как продолжение их по простиранию, соответствуя, вероятно, низам зоны 13 франского яруса.
4) Неверовские известняки в целом должны, по всей вероятности, относиться к нижней части франского яруса (нижнефранский подъярус Dy), соответствуя зонам 1а, 13 и, возможно, низам зоны I7.
, 5) Залегающий выше неверовских известняков песчано-сланцевый го-
ризонт с прослоями и линзами содержащих фауну аммоноидей известняков должен рассматриваться как близкий по возрасту (или несколько более молодой) к гониатитовому горизонту Николаевского месторождения и отвечающий, по всей вероятности, верхам зоны — низам зоны 1g франского яруса.
4. ФАЦИАЛЬНАЯ ПРИУРОЧЕННОСТЬ ИЗУЧЕННЫХ АММОНОИДЕЙ
В нижнем палеозое Алтай представлял собой часть геосинклиналь-ного бассейна, в котором были заложены глубинные структурные элементы, простиравшиеся параллельно юго-западному краю Сибирской платформы и в дальнейшем предопределившие направление каледонских и варисцийских складчатых сооружений. В результате каледонских тектонических движений отложения кембрия и нижнего силура были дислоцированы, и в верхнесилурийскую эпоху район испытал поднятие и долгое время представлял собой сушу, которая размывалась в течение верхнесилу-рпйской и нижнедевонской эпох.
В результате наступившей в среднем девоне трансгрессии эродированные каледонские сооружения образовали подвижный шельф, на котором шло в дальнейшем накопление продуктов вулканической деятельности, перемежающихся с осадочными образованиями.
В начале верхнедевонской эпохи, когда трансгрессия достигала мак симума, Алтай представлял собой область неглубокого открытого моря с серией островов и подводных складчатых сооружений, где накапливались вулканогенные и туфогенно-осадочные породы. Образовавшийся морской бассейн, имевший свободное сообщение с геосинклинальными средиземного
морской, уральской и тихоокеанской областями (на что указывает общность фауны), отличался довольно разнообразными фациальными условиями. В прогибающихся участках морского дна происходило быстрое накопление эффузивно-пирокластических масс в результате интенсивного проявления вулканизма. В условиях неглубокого моря и подвижной среды накапливались туфогенно-обломочные отложения, имеющие отчетливые признаки сортировки и перемыва, кремнисто-глинистые и известково-глинистые алевролиты, реже известняки. В условиях небольших глубин (35—40 м) постепенно погружающихся участков в чистых и теплых водах происходило формирование рифовых известняков барьерного типа. Внутри впадин и на участках, защищенных от прибоя и сильных течений подводными скалами и выступами рифов, осаждались морские илы, давшие неправильные по форме линзы кремнисто-глинистых алевролитов и глинистых известняков, залегающих среди грубого пирокластического и туфового материала. В более или менее удаленных от берега участках при наличии течений, приносивших достаточно большое количество терригенного материала, происходило накопление хорошо отсортированных слоистых известково-глинистых и глинистых осадков, давших довольно мощные и широко распространенные толщи алевролитов. В небольших впадинах при меньшем приносе терригенного материала происходило накопление кремнистых и глинистых известняков линзовидного залегания. При прекращении поступления терригенного материала глинистые отложения сменялись известняковыми, богатыми остатками нормальной бентосной фауны, преимущественно брахиоподовой. Различные области морского бассейна отличались разнообразными условиями и различными фаунами. В чистых теплых водах неглубокого моря, при наличии течений, существовала богатая бентосная фауна, представленная колониальными кораллами, строматопорами, криноидеями, мшанками, брахиоподами, гастроподами, трилобитами, рецептакулитами и т. д. В местах накопления терригенных и глинисто-известковистых осадков бентосная фауна значительно беднее и представлена мелкими брахиоподами, редкими пелециподами, трилобитами, одиночными кораллами, но зато богато представлены аммоноидей.
Аммоноидей наблюдаются как в среднем, так и в верхнем девоне, в терригенных песчано-глинистых отложениях и в мергелистых и глинистых известняках.
В среднедевонских отложениях они встречаются чаще в алевролитах, в той или иной степени известковистых, слоистых, хорошо отсортированных, накапливавшихся в условиях быстрой седиментации при наличии течений, на которые указывает правильная ориентировка окаменелостей вдоль плоскости напластования. Аммоноидей, встречающиеся в алевролитах, в большинстве своем деформированы и сплющены в плоскости напластования. Массовых скоплений их не наблюдается, хотя остатки раковин и приурочены к отдельным прослоям и гнездам. Помимо аммоноидей, в алевролитах присутствуют остатки наутилоидей, брахиопод, одиночных кораллов, пелеципод, трилобитов, что указывает на фацию неглубокого, открытого моря.
Находки аммоноидей в известковистых среднедевонских отложениях (крюковские известняки) значительно более редки. Отдельные экземпляры собраны из известняков наряду с кораллами, брахиоподами, гастроподами, криноидеями, трилобитами, наутилоидеями, что опять-таки указывает на фацию неглубокого, свободно сообщающегося с геосинклинальным бассейном моря. Условия захоронения фауны не совсем ясны, ввиду того что коллекции собраны из отвалов.
Местонахождения верхнедевонских аммонитов известны из терригенных и известняковых фаций. Терригенные фации представлены хорошо отсортированными, тонкослоистыми алевролитами (глинистыми сланцами)
37
низов франского яруса, накапливавшимися в условиях быстрой седиментации. Аммоноидей в алевролитах приурочены к отдельным прослоям и попадаются иногда в сравнительно больших количествах, никогда, однако, не переполняя породу. Остатки раковин деформированы,сплющены и ориентированы в плоскости напластования. Кроме аммоноидей, встречаются весьма редкие, плохо сохранившиеся остатки пелеципод, брахиопод, трилобитов, которые наряду со следами ползания червей и следами течения указывают на фацию неглубокого моря и зону течения.
В чистых слоистых и рифовых известняках остатки аммоноидей отсутствуют или весьма редки; в кремнисто-известковистых и глинисто-известковистых отложениях они более часты и скопления раковин нередко весьма многочисленны, иногда переполняют породу.
Скопления раковин аммоноидей приурочены к известковым линзам и прослоям среди песчано-глинистых и туфогенно-осадочных отложений. В отдельных линзах аммоноидей многочисленны и довольно разнообразны. Раковины ориентированы в плоскости напластования пород, преимущественно полные, сохранившие свою форму, реже деформированные. Кроме аммоноидей, наблюдаются редкие остатки мелких брахиопод, пелеципод, трилобитов, одиночных кораллов, криноидей, наутилоидей, указывающие на фацию неглубокого открытого моря. Сравнительно большое содержание в известняках глинистых частиц и сопутствие им глинисто-песчанистых и известково-глинистых отложений указывают на довольно большое количество материала, приносимого с суши.Наблюдающиеся в известняках следы течений и ориентировка раковин говорят о подвижности водной среды, наличии течений, не особенно, однако, сильных, так как захоронявшиеся остатки организмов недостаточно отсортированы и в одних и тех же местах наблюдаются как крупные, так и мелкие раковины.
Известняковые линзы и прослои, содержащие фауну, перемежаются с известково-глинистыми, кремнисто-глинистыми и глинисто-песчанистыми отложениями, эффузивами и их туфами; частая смена литологического состава пород, отображающая меняющиеся фациальные условия, указывает на колебание береговой линии, смену направления и силы течений, колебания морского дна. Указываемые для некоторых участков конгломераты нижнефранского возраста говорят за то, что в некоторые промежутки времени происходили временные поднятия отдельных участков над уровнем моря и размыв образовавшихся ранее пород. Сравнение мощностей сопоставимых по возрасту нижнефранских слоистых известняков и глинисто-песчанистых и известково-глинистых терригенных осадков указывает на значительно более быстрое накопление последних, обладающих большей мощностью по сравнению с известняками (вероятно, не менее 300 м против 150 м). Значительно более мощными являются и соответствующие песчано-глинистые отложения среднего девона Лениногорского рудного поля при сравнении их с известняковыми и песчанисто-известковистыми отложениями лосишинской свиты. Отложения, содержащие наибольшее количество остатков аммоноидей, характеризуются, таким образом, значительно большими мощностями по сравнению с отложениями, в которых аммоноидей отсутствуют или представляют редкость и которые характеризуются присутствием богатой бентосной фауны.
В результате рассмотрения фациальной приуроченности изученных аммоноидей можно сделать следующий вывод: девонские аммоноидей жили в областях неглубокого моря, в более или менее удаленной от берега зоне шельфа, в условиях сравнительно быстрой седиментации терригенных осадков.
III. ОНТОГЕНЕТИЧЕСКОЕ РАЗВИТИЕ РАКОВИНЫ ДЕВОНСКИХ АММОНОИДЕЙ
И ЕГО ЗНАЧЕНИЕ ДЛЯ ФИЛОГЕНИИ
1. ЭМБРИОНАЛЬНАЯ РАКОВИНА ДЕВОНСКИХ АММОНОИДЕЙ
Строение раковины аммоноидей на наиболее ранних стадиях развития изучалось многими специалистами и поэтому хорошо известно. Эмбриональная раковина представляла почти микроскопическое образование, состоящее из протоконха и жилой камеры, разделенных аселлатной, лати-селлатной или ангустиселлатной перегородкой.
а) Протокой х, или начальная камера. Аммоно-идеи произошли, по всей видимости, от бактритоидей. Начальные камеры бактритоидей описывались Баррандом (Barrande, 1877), Бранко (Branco, 1880, 1885), Кларком (Clarke, 1894, 1899), а в более позднее время Шин-девольфом (Schindewolf, 1932, 1933, 1934). Форма начальной камеры бактритоидей частью яйцеобразная, с высотой больше ширины, частью каплевидная или грушеобразная (рис. 1, d). Длина ее составляет 0,7—1,6 мм, ширина — 0,4—1,0 мм. Начальная камера часто отделена от следующих камер более или менее выраженным пережимом.
Следующая стадия развития протоконха наблюдается у древнейших аммоноидей — у родов Gyroceratites Meyer (=Mimoceras Hyatt) и Mimago-niatites Eichenberg. Начальная камера у первого была изучена Зандберге-рами (С. Sandberger, 1842, стр. 228; G. u F. Sandberger, 1851, стр. 304, табл. III, фиг. 26, и т. д.), у второго—Шиндевольфом (1933, стр. 29, табл. 3, фиг. 12—13). У указанных родов (рис. 1, е) протоконх подобен бактри-товому, т. е. яйцеобразный, но несколько округленный, со слегка укороченной длиной, более широкий по сравнению с протоконхами бактритов, слегка свернутый, так что его внешняя сторона плавно переходит в изгиб спирали первого оборота раковины, а внутренняя образует с внутренней стороной оборота раковины отчетливый пережим.
Следующей стадией в эволюции начальной камеры девонских аммоноидей является протоконх рода Anarcestes Mojsisovics. (рис. 1, /). У этого, еще низко организованного, раннего среднедевонского представителя начальная камера близка к шаровидной и более расширена по сравнению с начальными камерами Gyroceratites и Mimagoniatites. Она располагается ретортообразно по отношению к первому обороту, не соприкасаясь с первым его полуоборотом, так что образуется еще отчетливое умбональное отверстие. Этот тип начальной камеры является морфологическим переходом к начальным камерам родов Werneroceras Wedekind, Agoniatites Meek, и т. д., с плотно прилегающим первым оборотом раковины, без умбонального отверстия (рис. 1, g). В этих случаях внешняя сторона начальной камеры продолжается в начало первого оборота, который полностью облекает ее. Форма начальной камеры субсферическая, с высотой и шириной в среднем
39
равными 1,0 мм. Подобная форма начальной камеры свойственна почти всем без исключения девонским аммонитам. В ходе дальнейшей эволюции начальной камеры происходит сокращение размеров и некоторое относительное расширение ее, когда ширина протоконха становится больше его высоты. У позднепалеозойских и мезозойских аммоноидей начальная камера (рис. 1, Л) растянуто-эллиптическая, с диаметром в плоскости симметрии до 0,3 мм и шириной до 0,5 мм.
Таким образом, эволюция протоконха аммоноидей шла от бактритов через стадии, соответствующие протоконхам Gyroceratites, Mimagoniati-tes, Anarcestes и Werneroceras, к более поздним гониатитам, климениям и аммонитам.
Рис. 1. Филогенетическое развитие начальных камер Nautiloidea, Bactritoi-dea и Ammonoidea по Шиндевольфу (1932, стр. 169, рис. 2):
а — высоко коническая начальная камера палеозойских широко конических орто цератид; Ь — уменьшенная чашкообразная начальная камера мезозойских ширококонических на-утилид; с — яйцеобразная начальная камера длинно конических ортоцератид с центральным положением сифона; d — яйцеобразная начальная камера некоторых бактритов с краевым положением сифона-е — слегка свернутая начальная камера гироператитид; f — сильнее свернутая начальная камера некоторых гониатитов (Anarcestes) с небольшим умбональным отверстием; д — начальная камера некоторых инволютных гониатитов (Werneroceras) с округлым поперечным сечением протоконха; h — начальная камера мезозойских аммонитов с эллиптическим поперечным сечением протоконха.
В ходе эволюции девонских аммоноидей, одновременно с приобретением протоконхом субсферической формы происходило сокращение размеров его. Если у некоторых ранних среднедевонских представителей, напримерг Agoniatites vanuxemi (Hall), Werneroceras plebeiforme (Hall) ширина протоконха по наблюдениям Кларка (Clarke, 1899) превышает 1,5 мм (1,7 мм и 1,8 мм соответственно), а диаметр в плоскости симметрии доходит почти до 2,0 мм, у остальных среднедевонских аммонитов диаметр равняется в среднем 1,0—1,2 мм, то у большинства более поздних представителей Manticoceras и Tornoceras диаметр протоконха уменьшается до 0,7—0,8 мм, у Devonopronorites ruzhencevi Bogoslovsky — до 0,64—0,66 мм, у Imito-ceras—до 0,6 мм; у наиболее древней климении Acanthoclymenia пеаро-litana (Clarke) протоконх сравнительно большой (по данным Кларка, 1899, стр. 132, рис. 102—104), ширина его равна 1,1 мм, диаметр— 1,0 мм, в 2,5—3 раза превышающий размеры начальных камер более поздних кли-мений — Platyclymenia spinosa (Munster) и Clymenia laevigata Munster, изученных Шиндевольфом (1933, табл. 3, фиг. 15, 16).
В отличие от молодых представителей аммоноидей, имеющих гладкую начальную камеру и лишенный заметной скульптуры первый оборот раковины, у наиболее древних девонских аммонитов отмечается отчетливая радиальная струйчатость протоконха, переходящая и на первый оборот раковины. Подобная струйчатость указывалась Гольцапфелем (Holzapfel, 1895, стр. 77, табл. III, фиг. 19) для Cabrieroceras rouvillei (Коепеп)?
40
[ = Anarcestes karpinskyi Holzapfel (pars)], Кларком для Sandbergeroceras syngonum Clarke (1899, стр. 107, рис. 74—81), Werneroceras plebeiforme (Hall) (там же, стр. 165, фиг. 1—2) и Agoniatites expansus Vanuxem [ = A.va-nuxemi (Hall)] (там же, стр. 168, фиг. 6), Шиндевольфом (1933, стр. 45, табл. 4, фиг. 15 и фиг. 13) для Agoniatites costulatus (Archiac et Verneuil) и Anarcestes (Latanarcestes) noeggerati (Buch); радиальная струйчатость установлена и на протоконхе Tornoceras uniangulare (Conrad) Бичером (Beecher, 1890, стр. 74, табл. 1). В отличие от наутилоидей, у аммонитов струйчатость значительно менее выражена, и у более молодых представителей протоконх гладкий.
Внутреннее строение протоконха аммоноидей описано в русской литературе М. И. Шульга-Нестеренко (1926). Внутри протоконха помещается «caecum»— зачаточный сифон в виде пузырька, прилегающего изнутри к первой перегородке, и отходящий от него к внутренней стенке эмбриональной камеры (протоконха) «prosipho» («сифональный конус» по Хайэтту). «Caecum»—шаровидное полое образование, отверстие которого обращено к первой перегородке. Он похож на колбочку, суженная часть которой охвачена (не всегда) первой перегородкой, а длинная шейка является уже самим сифоном первой, второй и т. д. воздушных камер. По мнению большинства исследователей, образование «caecum» предшествует образованию первой перегородки.
б) Первая перегородка. Параллельно с развитием и усложнением начальной камеры у аммоноидей происходит усложнение отгораживающей начальную камеру первой перегородки и ее лопастной линии. Эта зависимость исследовалась Бранко (Branco, 1879, 1880) и позднее с некоторыми изменениями и дополнениями обосновывалась Шиндевольфом (1929). Бранко, уделивший много внимания изучению начальных стадий развития аммоноидей, разделил их всех на основе изучения начальной камеры и первых перегородок на Asellati, Latisellati и Angustisellati. К первой группе, у которой край перегородки проектируется в виде почти прямой линии, относится большинство девонских представителей аммоноидей. Ко второй группе, характеризующейся широким седлом первой перегородки, принадлежит большая часть палеозойских и часть триасовых аммонитов. Последнюю группу, у которой первая лопастная линия состоит снаружи из узкого вентрального седла, боковой лопасти, бокового седла и умбональной лопасти, образует большинство мезозойских аммонитов. В процессе эволюции аммоноидей происходит усложнение первой перегородки и ее лопастной линии и переход от Asellati к Latisellati и от последних к Angustisellati.
Девонские представители аммоноидей обладают аселлатной или лати-селлатной лопастной линией первой перегородки или «просутуры».Аселлат-ная просутура никогда, однако, не бывает полностью аселлатной, образующей совершенно прямую линию, так как она в большинстве случаев-образует хотя и очень слабо выраженные, сглаженные внутреннее (дорсальное) и внешнее (вентральное) углубления («лопасти»), так же как и слабые углубления, расположенные в районе умбонального шва. Подобный характер лопастной линии первой перегородки указывался Бранко для некоторых видов рода Manticoceras (1880, табл. VI, фиг. Ig, Ilg, Illg, IVa), Anarcestes lateseptatus (Beyrich) (?) (табл. VI, фиг. V), Agoniatites evexus (Buch) (табл. VII, фиг. I Id); такой же характер лопастной линии первой перегородки изображается Кларком для Manticoceras fasciculatum Clarke (Clarke, 1899, стр. 72, фиг. 46—48), вероятно, Sandbergeroceras syngonum Clarke (1899, стр. 107, фиг. 79—81) и обнаружен мной у Deco-nopronorites ruzhencevi Bogoslovsky (см. рис. 44, а).
В некоторых случаях, в противоположность такой форме первой лопастной линии, на внешней и внутренней сторонах образуются отчетливые,
4}
выступающие округленные седла, разделенные лопастью в районе умбо-нального шва. В этих случаях внешнее седло имеет на вершине слабое уплощение, которое, однако, никогда не переходит во внешнюю лопасть. Подобный тип первой лопастной линии указывался Шиндевольфом (1929) для Werneroceras ruppachense (Kayser) (стр. 18, рис. 1) и Tornoceras simplex (Buch) (стр. 18, рис. 2).
«Просутура» Tornoceras simplex (Buch) ( = «Goniatites retrorsus»)p&ccM&x-рпвалась Бранко (1880, стр. 25, 32) в качестве переходной формы к лати-селлатному типу, но, по мнению Шиндевольфа, она должна считаться еще как вполне нормальная и, возможно, даже наиболее широкая развивающаяся форма аселлатного типа, которая еще в значительной степени отличается от более поздних латиселлатных типов.
К латиселлатному типу, характеризующемуся широкими и довольно высокими внешним и внутренним седлами и более узкой умбональной лопастью первой перегородки, относятся Бранко (стр. 22, 31) представители родов Cheiloceras, Sporadoceras, Imitoceras, Prolobites, вероятно, Pharciceras и др. Латиселлатный тип первой перегородки указывался Бранко для Sporadoceras miinsteri (Buch) (стр. 22, 31), Imitoceras linearis (Munster) (стр. 22, 31, табл. V, фиг. VI), Prionoceras divisum (Munster) {? стр. 31). Латиселлатными являются и все климении, за исключением наиболее древней североамериканской Acanthoclymenia neapolitana (Clarke) (Clarke, 1899, стр. 133). Несмотря на то, что Бранко (1880, табл. VII, фиг. III, табл. VIII, фиг. /,в) указывался для Oxyclymenia cf. undu-lata (Munster) [ — Kosmoclymenia cf. undulata (Munster)] латисселатный тип первой перегородки, в более поздней литературе установилось ошибочное мнение, что Бранко доказывал у климений аселлатную начальную камеру. Исследование Шиндевольфом климений из верхнего девона Польши (1933, стр. 59) и, в частности, Cymaclymenia camerata Schindewolf (1923, стр. 25) показало наличие у них латиселлатной первой перегородки. Ла-тиселлатные первые перегородки обнаружены мной и у некоторых южноуральских климений (Platyclymenia, Genuclymenia, Kosmoclymenia), у которых были изучены начальные стадии развития. Вероятно, мнение Шиндевольфа (1933, стр. 58) о наличии у всех европейских климений латиселлатной начальной камеры правильно.
в) Длина жилой камеры. Вопрос о длине жилой камеры эмбриональной раковины аммоноидей до последнего времени решался на основании ее размера до слабого пережима, развитого на расстоянии приблизительно одного оборота от первой перегородки. В русской литературе этот вопрос был впервые рассмотрен М. И. Шульга-Нестеренко (1926), исследовавшей начальные стадии развития некоторых верхнепалеозойских аммоноидей. Первичный пережим наблюдается на первом обороте у всех аммонитов. Пережимы отображают задержки роста, так как под ними раковина утолщена в виде линзы. Задержка роста животного, связанная с выходом его из яйцевой капсулы и переходом к личиночному состоянию, характеризуется появлением первичного пережима раковины, который предположительно может указывать на местонахождение устья эмбриональной раковины. Первичный пережим по отношению к первой перегородке у различных групп аммоноидей имеет несколько различное положение. По данным Бёмерса (Bohmers, 1936), у древнейших девонских родов Anarcestes и Agoniatites он находится на расстоянии двух третей оборота от первой перегородки. По данным В. Е. Руженцева и В. Н. Шиманского (1954), первичный пережим у более молодого девонского рода Manticoceras расположен значительно дальше от перегородки, приблизительно на расстоянии 3/4 оборота от нее; это наблюдение согласуется с данными Бёмерса по роду Timanites. Исследования многих специалистов показали, что у еще более молодых аммоноидей, например у всех пермских предста-42
ъителей, первичный пережим находится еще дальше, на расстоянии одного оборота или более от первой перегородки. Наличие и положение первичного пережима на первых оборотах раковин различных представителей аммоноидей позволяли и раньше судить о длине трубки эмбриональной раковины, но все же более или менее предположительно. Недавно В. Е. Руженцевым и В. Н. Шиманским (1954) опубликованы интересные данные по строению эмбриональных раковин на основании непосредственного их наблюдения. В распоряжении указанных авторов были мельчайшие раковинки меловых и артинских аммонитов, имеющие единственную перегородку. Как у меловых, так и у артинских аммонитов раковины эти состоят из протоконха, одной перегородки и трубки, которая после целого оборота перекрывает перегородку в ее вентральной части. Эти раковины, по мнению авторов, безусловно принадлежали эмбрионам аммонитов, погибших сразу после выхода из яйцевой капсулы. Таким образом, если раньше можно было предполагать, что длина жилой камеры эмбриональных раковин аммоноидей достигала одного оборота, то после этих наблюдений доказано не только это положение, но и другое,—что эмбриональная раковина аммонитов имела только одну воздушную камеру (протоконх) и одну перегородку, отделявшую протоконх от жилой камеры.
Поперечное сечение эмбриональной раковины несколько изменялось в процессе эволюции аммоноидей. Исследования показывают, что если у большинства девонских аммонитов первый оборот раковины является совершенно эволютным (а у наиболее древних — даже не полностью охватывающим протоконх, так что имеется умбональное отверстие), то уже у некоторых девонских {Devonopronorites, например), каменноугольных и пермских — несколько объемлющий, у меловых — сильно объемлющий. Одновременно в процессе эволюции аммоноидей происходит сокращение абсолютных размеров эмбриональной раковины. Так, если у наиболее ранних среднедевонских представителей: Anarcestes {Anarcestes) simulans (Barrande), A. {Latanarcestes) noeggerati (Buch), Agoniatites co-stulatus (zirchiac et Verneuil), изображенных Шиндевольфом (1933, табл. 4, фиг. 11, 13, 16), диаметр эмбриональной раковины составляет, по-видимому, 1,8—2,0 мм, то у Sandbergeroceras singonum Clarke (Clarke, 1899, стр. 107, рис. 79—81) уменьшается до 1,6—1,8 мм (?), у различных видов рода Manticoceras составляет от 1,2 мм [М. sinuosum (Hall)] до 1,5 мм {М. rhyn-chostoma Clarke), у Tornoceras bicostatum (Hall) —1,4 мм (Clarke, 1899, стр. 119, фиг. 98), у Devonopronorites ruzhencevi Bogoslovsky — около 1,3 мм, у Platyclymenia spinosa (Munster) и Clymenia laevigata Munster (Schindewolf, 1933, табл. 3, фиг. 15 и фиг. 16) уменьшается до 1,2 мм, у Imitoceras sp. (Schindewolf, 1951, табл. I, фиг. 4)— почти до 1,0 мм. Еще меньшими диаметрами обладают эмбриональные раковины более молодых верхнепалеозойских и мезозойских аммонитов. У пермских аммоноидей размер эмбриональных раковин колеблется в пределах 1,3—0,9 мм (Руженцев и Шиманский, 1954), у меловых равен 0,6 мм. Приведенные данные говорят о том, что «эмбриональные раковины аммоноидей были исключительно малы (как зерна грубозернистого песка) и что в ходе эволюции их размер, как правило, в общем уменьшался» (Руженцев и Шиманский, 1954).
Подводя итог рассмотрению строения эмбриональной раковины девонских аммоноидей на основании исследований В. Е. Руженцева и В. Н. Шиманского, можно применительно к девонским представителям отметить следующие особенности:
1. Эмбриональные раковины девонских аммоноидей представляли почти микроскопические образования, состоящие из протоконха и жилой камеры, разделенных аселлатной или латиселлатной перегородкой.
2. Сифона у эмбрионов еще не было, однако существовал «caecum»,
43
т. е. шаровидное полое образование — основание будущего сифона и сифонного шнура.
3. В ходе эволюции: а) размер протоконха и всей раковины в общему хотя и очень слабо, уменьшался; б) форма протоконха изменялась от яйцевидной до округлой; в) строение первой перегородки (ее рельеф) усложнялось; г) длина жилой камеры и степень инволютности оборота постепенно возрастали.
2. РАЗВИТИЕ ПЕРЕГОРОДКИ И ЛОПАСТНОЙ ЛИНИИ
Лопастная линия второй перегородки («primasutur») у всех девонских как аселлатных, так и латиселлатных гониатитов и климений состоит из трех лопастей и разделяющих их седел (рассматривается обычно половина двусторонне-симметричной лопастной линии от внутренней до наружной лопастей). Две лопасти — наружная (вентральная) и внутренняя (дорсальная) — расположены в плоскости симметрии. Третья лопасть находится во второй лопастной линии и на наиболее ранних стадиях развития лопастной линии почти всегда в районе умбонального шва, иногда частично переходя на внутреннюю вогнутую сторону оборота раковины. Такое положение этой лопасти установлено Бранко для Anarcestes (Branco, 1880,. табл. VII, фиг. V), Tornoceras (табл. V, фиг. VII,Л), Manticoceras (табл. VI, фиг. I, ПЛ, I, ШЛ, IV\с) и Oxyclymenia (=Kosmoclymenia) (табл. VIII, фиг. 1с, d), Кларком — для Sandbergeroceras (Clarke, 1899, стр. 107, фиг. 79, 81), Manticoceras (1899, стр. 58, фиг. 28), Бичером для Tornoceras (Beecher, 1890, табл. I, фиг. 14, Ь, с), Фрехом для Pharciceras (Freeh, 1902, стр. 56, фиг. 17; с, стр. 64, фиг. 21, а), Гольцапфелем — для Timanites (1899, стр. 42, фиг. 12), Пэрна — для Prolobites (1914, стр. 23, фиг. 8У 6, с) и Sporadoceras (1914, стр. 33, фиг. 14, Ь, с), Шмидтом (Schmidt, 1921, стр. 541, средняя фиг. ) для W edekindella, Шиндевольфом — для Anarcestes (Schindewolf, 1927, стр. 184, фиг. 1,8), Tornoceras (1929, стр. 31, фиг. 13), Manticoceras (1929, стр. 30, фиг. 12, стр. 68, фиг. 34, а, стр. 74% фиг. 35, е), Cheiloceras (1929, стр. 32, фиг. 14) w Cymaclymenia (1923, стр. 25у фиг. 2, d)-, такое же положение этой лопасти наблюдается и у Devonop-ronorites (рис. 43, 5).
Таким образом, у девонских гониатитов и климений эта лопасть на ранних стадиях развития занимает умбональное положение. Исключение представляют nnnibMimoceratitidae и Agoniatitaceae, у которых эта лопасть, располагается, по-видимому, посредине боковой стороны раковины.
Вентральная, дорсальная и умбональная лопасти образуют первичные элементы лопастной линии. В то время как вентральная и дорсальная лопасти занимают свое постоянное положение (за исключением незначительного смещения вентральной лопасти из плоскости симметрии), умбональная лопасть может либо занимать умбональное положение на протяжении всего онтогенетического развития, либо смещаться в ходе онтогенеза на боковую сторону раковины. В некоторых случаях она сохраняет свое положение при умбональном шве, как у Anarcestes (Anarcestes), Subanarcestes (рис. 2), Archoceras, Ponticeras и др. В этом случае на взрослых стадиях небольшая умбональная лопасть лежит, как и в начале онтогенетического развития, в районе умбо. Снаружи к ней примыкает широкое седло, занимающее середину боковой поверхности раковины.
У других родов, как например Anarcestes (Latanarcestes), Werneroceras (рис. 3), Prolobites и др., она перемещается от умбо и, сильно расширяясь и углубляясь, располагается на середине боковой поверхности раковины, в то время как наружное седло сильно сужается. Первичная умбональная лопасть становится морфологической боковой лопастью в широком смысле слова, занимая соответствующее положение.
44
Помимо первичных вентральной, дорсальной и умбональной лопастей, на более поздних стадиях развития многих девонских аммоноидей в лопастной линии образуются новые лопасти. Такое увеличение количества элементов лопастной линии происходит за счет расчленения внутреннего бокового седла (Manticoceras, Costaclymenia) или за счет деления внешнего
Рис. 2. Лопастные линии Anarcestes (Anarcestes) lateseptatus(ReyAA}) и Subanarcestes macrocephalus Schindewolf по Шиндевольфу (1951, стр. 16, рис. 1):
a, b — две стадии онтогенетического развития лопастной линии Anarcestes (Anarcestes) lateseptatus (Beyrich); с — лопастная линия взрослой стадии развития Subanarcestes macrocephalus Schindewolf.
бокового седла (Tornoceras, Cheiloceras) или, наконец, за счет сочетания этих процессов, когда новые лопасти возникают в двух местах (Sporadoceras, Dimeroceras, Gonioclymenia, Kalloclymenia).
Рис. 3. Онтогенетическое развитие лопастной линии Werneroceras ruppachense (Kayser)no Шиндевольфу (1933, стр. 97, рис. 28).
Перемещение первично-умбональной лопасти наружу обычно происходит тогда, когда между ней и дорсальной лопастью образуются дополнительные умбональные лопасти в районе умбонального шва за счет расчленения внутреннего бокового или умбонального седел (Koenenites, Tima-nites, Pharciceras, Devonopronorites и т. д.).
У других гониатитов первичная умбональная лопасть занимает умбо-нальное положение и образуется одна или несколько боковых лопастей между умбональной и вентральной лопастями путем расчленения внешнего бокового седла. Подобным путем происходит усложнение лопастной линии у Tornoceras и Cheiloceras (рис. 4).
45
Аналогично Tornoceras и Cheiloceras развивается наружная часть лопастной линии Sporadoceras. Согласно Пэрна (1914, стр. 33, фиг. 14, Ь, с),. первичная умбональная лопасть сохраняет свое положение в районе умбо на протяжении всего хода онтогенетического развития, в то время как боковая лопасть является вторичной и образуется на внешнем седле, так же как у Cheiloceras, вблизи его вершины; рядом с ней аналогичным путем возникает вторая боковая лопасть, которая у некоторых видов остается плоской, у других становится глубокой. Совершенно иначе образуются боковые лопасти Prolobites и Wedekindella. Онтогенетическое
Рис. 4. Онтогенетическое развитие лопастной линии Cheiloceras sp. из верхнего девона Германии по Шиндевольфу (1929, стр. 32, рис. 14):
а — лопастная линия второй перегородки; Ь —лопастная линия десятой перегородки; с — лопастная линия взрослой стадии.
развитие лопастной линии для первого рода показано Пэрна (1914, стр. 33г фиг. 19), для второго — Шмидтом (Schmidt, 1921, стр.541, средняя фиг.). У этих родов первичная умбональная лопасть перемещается вторично-на середину бока. Аналогичное явление наблюдалось Шиндевольфом для Cymaclymenia (1923, стр. 25, фиг. 2,6), Бранко для Oxyclymenia ( = Kos-moclymenid) (Branco, 1880, табл. VIII, фиг. VII,й). Морфологическая боковая лопасть этих форм не гомологична боковой лопасти Tornoceras, Cheiloceras, Sporadoceras.
Таким образом, исходным пунктом в образовании лопастей является положение умбональной лопасти, которая или остается на месте и образуются наружные боковые лопасти, или смещается на боковую поверхность раковины и образуются внутренняя боковая и умбональные лопасти, возникающие при вершине внутреннего или умбонального седел (рис. 5).
Усложнение лопастной линии может происходить у девонских аммоноидей за счет расчленения не только седел, но и лопастей.
В некоторых редких случаях (Sellanarcestes, Wedekindella, Dyscheilo-ceras) происходит расчленение дорсальной лопасти срединным седлом. Значительно чаще наблюдается большее или меньшее расчленение вентральной лопасти, становящейся иногда очень широкой и состоящей из большого числа элементов, распространяющихся на значительную часть боковых сторон раковины. Трехчленное деление вентральной лопасти наблюдается у представителей девонских семейств Manticoceratidae, Sandbergeroceratidae, Pharciceratidae и рода Devonopronorites.TaKoe же деление ее наблюдается и у рода Praeglyphioceras. Особенно резкому расчленению подвергается вентральная лопасть белоцератид, которая у Beloceras, например, может иметь до 13—15 и даже более зубцов. Дву-46
членного деления вентральной лопасти у девонских аммоноидей не наблюдается, и у представителей других семейств она остается нерасчлененной. Расчленения первичной умбональной и вторичных лопастей у девонских представителей, как правило, не наблюдается, за исключением Devono-pronoriles, первичная умбональная лопасть которого, перемещаясь в те-
Рис. 5. Различные варианты развития перегородки и лопастной линии:
а — исходное состояние: перегородка с вентральной V, дорсальной D и умбо-нально расположенной первичной умбональной U лопастями; Ъ — перегородка с изначальным положением первичной умбональной лопасти и образовавшимися внутренней I и наружной L боковыми лопастями; с — перегородка с перемещающейся на боковую сторону первичной умбональной лопастью и образовавшимися внутренней боковой I и вторичной умбональной U1 лопастями.
чение онтогенетического развития в сторону вентральной лопасти, становится двураздельной в основании.
Своеобразно происходит развитие лопастной линии у климений. У Clymeniaceae происходит в течение онтогенетического развития утрата существующей на ранних стадиях вентральной лопасти. Так, например, у родов Genuclymenia, Cymaclymenia, Kosmoclymenia, Platyclymenia (Schindewolf, 1929, стр. 35, фиг. 15) и Clymenia на ранних стадиях развития имеется нормальная внешняя лопасть, но в течение онтогенетического развития она замещается внешним седлом. У второй большой группы климений, Gonioclymeniaceae, наоборот, образующаяся во второй лопастной линии вентральная лопасть сохраняется и на взрослых стадиях (Schindewolf, 1929, стр. 35—36, фиг. 16, 17).
3. ТИПЫ ЭВОЛЮЦИОННЫХ ИЗМЕНЕНИЙ ЛОПАСТНОЙ ЛИНИИ 1
В процессе онтогенетического и филогенетического развития аммоноидей наблюдаются весьма разнообразные способы усложнения лопастной линии. Среди изученных нами представителей девонских аммоноидей можно различать следующие типы эволюционных изменений лопастной линии:
Анарцестидный тип. Этот наиболее простой тип лопастной
1 При обозначении формул лопастных линий мной применяется терминология, предложенная В. Е. Руженцевым (1949а).
47
линии характерен для семейства Anarcestidae. У наиболее просто организованных представителей семейства, составляющих подсемейство Апаг-eestinae, вся лопастная линия состоит лишь из трех простых, нерасчле-ненных лопастей: одной вентральной, одной боковой (возникающей в районе умбонального шва) и одной дорсальной. Формула ее — VU : D. У несколько более организованных представителей, группирующихся в подсемейство Pinacitinae, лопастная линия испытывает некоторое, мало существенное усложнение путем образования в районе умбонального шва дополнительной, обычно небольшой округленной умбональной лопасти. В этом случае формула лопастной линии несколько изменяется — VUU1 : D. В некоторых редких случаях (у родов Sellanarcestes и Wede-kindelld) наблюдается расчленение дорсальной лопасти невысоким округленным срединным седлом. Аммониты с лопастной линией анарцестидного типа обычны в среднедевонских отложениях. В верхнем девоне известен только один редкий род с лопастной линией такого типа — Archoceras.
Торноцератидный тип характерен для семейств Тогпо-ceratidae и Cheiloceratidae. Усложнение лопастной линии происходит в этом случае за счет образования наружных и внутренних боковых лопастей в результате расчленения внешнего и внутреннего боковых 'седел. Формула лопастной линии у наиболее просто организованных представителей — VLU : D, у более организованных — VL1LU .* ID. В некоторых редких случаях (у родов Dyscheiloceras и Torleyoceras) наблюдается более или менее выраженное двучленное или трехчленное деление дорсальной лопасти (VLU : D^h VLU : D2D!D2 соответственно). Аммониты с такой лопастной линией известны из среднего девона, обычны в верхнедевонских и нижнекаменноугольных отложениях и редко встречаются даже в верхнем карбоне и в пермских отложениях.
Гефуроцератидный тип усложнения лопастной линии характерен для семейства Gephuroceratidae. Усложнение лопастной линии происходит в этом случае в результате трехчленного деления вентральной лопасти и образования дополнительных клиновидных лопастей в районе умбонального шва, которые в процессе эволюции смещаются на боковую сторону. Общее количество лопастей в процессе эволюции возрастает от четырех до десяти и более. В результате трехчленного расчленения вентральной лопасти, которое, правда, никогда не бывает полным, срединный зубец этой лопасти очень сильно укорачивается, а боковые разрастаются, становясь зачастую по величине значительно больше первого. Формула лопастной линии менее высокоорганизованных представителей, родов Probeloceras и Ponticeras'. (V2V1V2) U : D, у более прогрессивного Manticoceras'. (V2V!V2) U : ID. У следующих представителей эволюционного ряда усложнение лопастной линии происходит за счет образования новых лопастей в районе умбонального шва, которые смещаются на боковую стенку. Формулы лопастных линий родов Koenenites и Timanites соответственно:
(V2V1V2)UU1 : ID и (V^V^UU^^D;
Komioceras, вероятно:
(V2V1V2)UU1U3U5U7 : U6U4U21D.
К этому же типу усложнения лопастной линии следует отнести и лопастную линию рода Triainoceras, формула которой сходна (при одинаковом количестве боковых лопастей) или близка к формуле Timanites и может быть изображена:
(V^V^UUHJ2... : ID.
Аммониты с лопастной линией гефуроцератидного типа обычны во •фра неких отложениях верхнего девона.
Фарциц-ератидный тип характерен для семейства Phar-•ciceratidae, представители которого существовали в течение нижнефран-48
ского времени. Этот тип усложнения лопастной линии близок к манти-коцератидному, однако отличается от него большим количеством возникающих в районе умбонального шва ланцетовидных лопастей, которые в процессе развития смещаются как на наружную, так и на внутреннюю стенки. Формула лопастной линии наиболее просто организованного представителя семейства, рода Pharciceras’. (V^ViVgJUWU1 * 3 : U2ID. В ходе эволюции вентральная лопасть становится все более дифференцированной; срединный зубец ее укорачивается и расширяется, принимая форму широкой воронки, а расходящиеся на бока боковые зубцы становятся ланцетовидными, не отличаясь по форме от боковых лопастей; значительно увеличивается количество боковых наружных и внутренних лопастей, приобретающих в общем ланцетовидную форму; общее количество лопастей вокруг извилины доходит примерно до пятидесяти. Формула лопастной линии наиболее высокоорганизованного рода Neopharciceras становится довольно сложной:
V2V1V2UU1U2U4U6U8U10U12U14U16U18U20U22..
. U21U19U17U16U13U11U9U7U5U3ID.
Белоцератидный тип. Этот оригинальный тип усложнения лопастной линии характерен для семейства Beloceratidae, представители которого существовали в течение франского века верхнедевонской эпохи. Характерной особенностью этого типа является то, что усложнение лопастной линии в ходе эволюции происходит как за счет образования новых лопастей в районе умбонального шва и смещения их на внешнюю и внутреннюю стенки, так и, в особенности, за счет расчленения широкой вентральной лопасти различным количеством дополнительных лопастей, возникающих по бокам срединного зубца и постепенно смещающихся на боковые стенки. Формула лопастной линии наиболее примитивного рода Neomanticoceras может быть изображена следующим образом1:
V^V^V1) V2U : ID
или, возможно:
V2 V2U : ID.
У более прогрессивного члена филогенетического ряда—рода Eobeloceras, вентральная лопасть остается на той же стадии развития, но увеличивается количество боковых лопастей. Формула лопастной линии этого рода:
V2 (VWjV1) У2иишз: U2ID
или, возможно:
V2 (V^Vi.iV^) V2UU1U3: U2ID.
Резкое усложнение лопастной линии наблюдается у рода Beloceras. Количество элементов вентральной лопасти, которая становится очень широкой, доходит до 13; возрастает резко и количество боковых лопастей. Формула лопастной линии получает очень сложный характер:
1 Для данного типа усложнения лопастной линии приводятся два варианта фор-
мул, так как неясно, возникают ли дополнительные зубцы вентральной лопасти из вершины седел, прилегающих к срединному зубцу ее (первый вариант), или происходит периодическое трехчленное деление срединного зубца вентральной лопасти (вто-
рой вариант).
4 Б. И. Богословский 49
... (V3... Vi... V3) VWWgUUUJ3U2ID
или, возможно:
. • . (Vm.2. .. Vbbi.!. . . W VUA2WU3...:... U2ID.
Девонопроноритидный тип усложнения лопастной линии* характерен для семейства Devonopronoritidae, единственный представитель которого известен из франских отложений верхнего девона. Вентральная лопасть сохраняет свою нераздельность, хотя и оканчивается тремя зубцами. Первичная умбональная лопасть претерпевает симметричное деление на две части, которые, однако, не превращаются в самостоятельные лопасти. Новые лопасти появляются в области умбонального шва и постепенно смещаются на наружную и внутреннюю стенки. Полная формула единственного представителя семейства рода Devonop-ronorites выглядит следующим образом:
v (UiUi) U1U2U4UeU8U9: U7U6U3ID.
4. ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ ПЕРЕГОРОДОК АММОНИТОВ
Вопрос о функциональном значении перегородок аммонитов давно привлекал к себе внимание многих исследователей. В отечественной литературе этому вопросу впервые уделил внимание А. А. Чернов (1907; 1922), выявивший связь между спиралью, формой раковины и расчлененностью' лопастной линии. В дальнейшем В. Е. Руженцев (1946; 1949 б), используя эволюционно-онтогенетический метод, указал на структурную и функциональную зависимость между лопастной линией, перегородкой и всей раковиной аммоноидей. Последнее было подтверждено исследованием эволюции перегородки у представителей семейств Pronoritidae и Medlicot-tiidae.
Рельеф перегородки изменяется как в онтогенезе, так и в филогенезе* аммонитов, в зависимости от изменения поперечного сечения извилины.
Взаимоотношение лопастей и седел во всех случаях обеспечивает оптимальные условия прочности стенок раковины и самой перегородки.
Закономерности, выявленные при исследовании эволюции перегородок, верхнепалеозойских аммоноидей, полностью подтверждаются при изучении девонских аммонитов, особенно их более высоко организованных представителей, обладающих сложными перегородками и лопастными линиями.
Действительно, как можно видеть, например, на рис. 6 и рис. 17, в процессе эволюции семейства Gephuroceratidae происходит постепенное усложнение перегородки и лопастной линии. Перегородки, изображенные на рис. 6, отображают основные стадии эволюции семейства. В пределах основного филогенетического ряда Ponticeras —> Manticoceras —> Koenenites —> Honinghausia —> Timanites раковина изменялась от слабо инволют-ной дискоидальной до полностью инволютной линзовидной. Количество-элементов лопастной линии вокруг извилины увеличивалось с четырех до десяти, причем возникновение новых лопастей сопровождалось укреплением как инволютных, так и эволютных частей раковины. Происходило перестраивание всего рельефа перегородки. Наиболее простая перегород-50
ка наблюдается у слабо инволютного Ponticeras (рис. 6, а), который имеет лишь четыре лопасти, не связанные между собой. Вентральная лопасть еще не сильно расчленена, хотя и является трехзубчатой. Внутренние боковые лопасти отсутствуют. Следующую стадию представляет перегородка рода Manticoceras (рис. 6, Ь). Увеличение инволютности раковины и появление инволютных ее частей, которые требуют дополнительного крепления, сопровождаются развитием и усложнением перегородки и лопастной линии. Вентральная лопасть разрастается, связанные между собой боковые зубцы ее, расходясь на боковые стороны, укрепляют сводообразную вентральную часть раковины. На внутренней стенке возникают внутренние боковые лопасти (по одной с каждой стороны), которые, связываясь с наружными боковыми лопастями, укрепляют инволютные части извилины. В ходе дальнейшей эволюции при переходе к Koenenites, а затем к Honinghausia и Timanites в результате изменения формы раковины
Рис. 6. Перегородки ряда представителей семейства Gephuroceratidae: а — Ponticeras-, b—Manticoceras; с — Koenenites; d—Honinghausia; е — Timanites.
рельеф перегородки и лопастная линия претерпевают дальнейшие изменения. У Koenenites (рис. 6, с), в результате образования второй наружной боковой лопасти и смещения первой боковой лопасти в вентральном направлении, происходит изменение рельефа перегородки. Связь между внутренней боковой и первой наружной боковой лопастями утрачивается и последняя остается свободной. Внутренняя боковая лопасть в свою очередь связывается со второй наружной боковой. Перегородка рода Honing-hausia (рис. 6, d), обладающего линзовидной, уплощенной с боков раковиной, при общем сходстве в соотношении элементов с Koenenites является уже более адаптированной. В этом случае узкая килеватая вентральная часть извилины надежно укрепляется трехраздельной вентральной лопастью. Первые наружные боковые лопасти полностью переключаются на укрепление широких уплощенных боковых сторон. Возникшая у Koenenites связь между второй наружной и внутренней боковой лопастями еще более увеличивается и обеспечивает достаточно прочное укрепление узких инволютных частей. Наконец, у Timanites (рис. 6, е) с узкой, полностью инволютной линзовидной раковиной узкие и высокие инволютные части перегородки получают еще более прочное крепление за счет дополнительно возникающих умбональных лопастей. Все элементы перегородки в этом случае располагаются наиболее рационально, надежно укрепляя как эволютную, так и инволютные части перегородки и всей раковины.
Подобную же картину усложнения рельефа перегородки и увеличения количества элементов лопастной линии можно наблюдать и при эволюции семейства Beloceratidae (см. рис. 35, ниже), в несколько меньшей степени — при эволюции семейств Tornoceratidae и Gheiloceratidae.
4*
51
Особенно резкое усложнение перегородки и лопастной линии наблюдается у представителей семейства Pharciceratidae (см. рис. 41, ниже), в процессе эволюции которого при переходе от слабо инволютного Pharciceras к полностью инволютному Synpharciceras и от последнего — к Neopharciceras общее количество лопастей вокруг извилины увеличивается с 12 до, примерно, 50.
Усложнение рельефа перегородки и очертания лопастной линии, обусловленное изменением формы поперечного сечения извилины при возрастании ее относительной высоты и увеличении инволютности раковины, в свою очередь, как правило, сопровождается в ходе филогенетического
Рис. 7. Онтогенетическое развитие перегородки Devonopronorites ruzhencevi Bogoslovsky:
а—№ 774/111 при В -1,2 и 111=2,3 мм (Х8); Ь—№ 774/112 при В=5,9 и Ш=7,3 мм (ХЗ); с—№ 774/113 при В —11,7 и 111=10,0 мм (ХЗ); d—№ 774/109 при В-=16,1 и 111=13,1 мм (ХЗ). Рудный Алтай. Фран-ский ярус.
развития относительным утончением стенок раковины, что увеличивает плову честь последней.
Интересные изменения рельефа перегородки в связи с изменением формы извилины можно наблюдать в процессе онтогенеза рода Devonoprono-rites. На рис. 7, а — d показаны перегородки Devonopronorites ruzhencevi Bogoslovsky, зарисованные на разных стадиях развития раковины. При высоте оборота, равной 1,2 мм и ширине 2,3 мм, что соответствует примерно второму-третьему обороту (рис. 7, а), перегородка, вытянутая в латеральном направлении, имеет восемь лопастей. Дорсальная и внутренняя боковая лопасти соединены бороздками соответственно с вентральной и первой наружной боковой (первичной умбональной) лопастями, а вторая боковая (умбональная) лопасть остается свободной, укрепляя невысокую боковую стенку. Такое взаимоотношение лопастей наилучшим образом обеспечивает прочность вентрального свода на данной стадии онтогенеза. Обращает на себя внимание и тот факт, что уже на этой стадии развития расположенная на перегибе от вентральной стороны к боковой первая наружная боковая лопасть является двураздельной в основании, что способствует укреплению угловатого вентро-латерального края раковины.
По мере дальнейшего развития извилина возрастает больше в вентральном направлении, чем в латеральном. Разрастающиеся боковой стенки и появляющиеся инволютные части раковины требуют дополнительного крепления, которое достигается постепенным увеличением числа лопастей, возникающих в районе умбо и смещающихся на наружную и внутреннюю 52
стенки раковины. Этот процесс усложнения перегородки можно наблюдать на рис. 7, Ъ — d, на которых видно, как по мере разрастания боковых стенок в высоту эволютная часть извилины укрепляется постепенно возрастающим количеством лопастей. Быстро разрастающиеся в длину инволютные части извилины укрепляются связанными парами лопастей, в результате чего они приобретают поперечную гофрировку, которая обеспечивает прочность соответствующих частей боковых стенок раковины. В то же время по мере возрастания извилины в вентральном направлении первая внутренняя боковая лопасть теряет связь с каждой предыдущей лопастью, расположенной на боковой стороне и устанавливает ее с новой. Меняет свое взаимоотношение с наружными лопастями и вторая внутренняя боковая лопасть. Происходит сложная перестройка рельефа перегородки: первая внутренняя боковая лопасть в процессе онтогенетического развития приобретает связь с первой, второй и, наконец, третьей наружной боковыми лопастями; вторая внутренняя боковая лопасть — с третьей и четвертой наружными боковыми. Первая и вторая наружные боковые лопасти, постепенно теряя связь с внутренними лопастями, становятся свободными; каждая из них располагается симметрично одноименной лопасти другой стороны раковины. Этот интересный процесс онтогенеза перегородки, теснейшим образом связанный с изменением формы извилины, можно представить в виде следующего ряда формул1:
a) VQU1 Г I.D ,
b) VUU4^4Pj_U^lD , с) VUUfU2U4jeU7". UsU3IDl
3ID.
При рассмотрении приведенных выше рисунков можно видеть, что во всех случаях гофрировка перегородки расположена таким образом, чтобы внешняя поверхность раковины имела достаточную опору и могла противодействовать давлению воды. Взаимоотношение наружных и внутренних элементов перегородки находится, согласно В. Е. Руженцеву, в прямой зависимости от формы извилины, точнее, от высоты ее эволютной части. Количество элементов инволютной части перегородки, изменяясь в процессе онтогенетического развития, находится в прямой зависимости от ее высоты: чем выше инволютная часть извилины, тем больше лопастей нужно для придания ей прочности и тем больше их наблюдается в действительности. На любой стадии форма перегородки изменяется адаптивно к ее функции.
Сходство поперечного сечения извилины приводит подчас к формированию весьма близких по форме перегородок, что связано с выполнением одинаковых функций. Поразительный пример конвергентного развития перегородки наблюдается, например, у морфологически сходных (и даже относимых в свое время к одному семейству), но генетически далеких друг от друга представителей франского рода Beloceras и артинского рода Ме-dlicottia. Перегородки представителей указанных родов изображены на рис. 8. Как видно на рисунке, перегородки указанных родов действи-
1 В этих формулах для простоты U показана нерасчлененной. Двоеточие обозначает место расположения умбонального шва, т.е. линию схождения наружной и внутрей* ней поверхностей раковины. Скобки соединяют лопасти, связанные одна с другой;
53
а b
Рис. 8. Перегородки представитеЛей'родов Beloceras и Medlicottia:
a —Beloceras sagittarium (Sandberger) (XI »7). Рудный Алтай, район Николаевского месторождения, франский ярус; Ъ — Medlicottia orbignyana (Ver-neuil) (X2). Актюбинская обл. p. Жаксы-Каргала, гора Жиль-Tay. Артинский ярус (по Руженцеву, 1946, стр. 695, рис. 12, в).
тельно имеют много общего; различия между ними проявляются лишь в деталях, обусловленных несколько отличными формами поперечного сечения извилины. Как у Beloceras sagittarium, так и у Medlicottia orbignyana, обладающих линзовидной, уплощенной с боков, сильно инволют-ной раковиной, узкие и высокие эволютные и сильно объемлющие довольно тонкие инволютные части извилины требуют достаточно прочного крепления, которое достигается целой серией поперечных борозд, приводящих к интенсивной гофрировке перегородки. На данном примере мы видим, как сходная форма поперечного сечения извилины приводит к близкой форме перегородки, обусловленной выполнением одинаковой функции, однако, один и тот же эффект достигается у Beloceras sagittarium и Medlicottia orbignyana совершенно различными путями. Резко различные пути усложнения перегородки и лопастной линии указанных двух представителей четко проявляются при сравнении формул эволют-ных частей их перегородок, которые выглядят следующим образом:
1) Beloceras sagittarium: Vp.. V3V2V'V2U...
2) Medlicottia orbignyana: Vv1v2v3v4s2s1u5u4u3u2u1U1U1U2U3. ..
У Medlicottia orbignyana в эволютной части перегородки первые четыре наружные боковые лопасти (U,!}1, U2, U3) одной стороны расположены против соответствующих лопастей другой стороны. Вследствие сближения боковых стенок к вентральному краю лопасти первой пары связаны глубокой бороздой. Дальше взаимосвязь становится менее глубокой, и лопасти четвертой пары (U3) уже разобщены дорсальной лопастью. Первая внутренняя боковая лопасть (I) связана с пятой наружной боковой (U4). Взаимоотношение остальных лопастей можно видеть на рисунке.
У Beloceras sagittarium функцию укрепления эволютной части перегородки выполняет необычайно разрастающаяся вентральная лопасть, расходящиеся зубцы которой на одной стороне расположены против соответствующих им зубцов другой стороны и соединены с ними бороздами, глубина которых постепенно убывает по направлению к сифону. Первая пара боковых лопастей (U) разобщена дорсальной лопастью и отвечает по своему положению четвертой паре боковых лопастей Medlicottia orbignyana. Первая внутренняя боковая лопасть (I) связана со второй наружной боковой лопастью (U1). Вторая, третья и следующие внутренние боковые лопасти связаны соответственно с третьей, четвертой и т. д. наружными боковыми.
Приведенные примеры свидетельствуют о самой тесной структурной и функциональной зависимости между лопастной линией, перегородкой и всей раковиной. Рельеф перегородки изменяется как в онтогенезе, так 54
и в филогенезе аммонитов в зависимости от изменения поперечного сечения извилины. Поэтому топография перегородки и особенно взаимоотно-лпение ее различных элементов приобретают важное систематическое значение, а лопастные линии, отображающие основные морфологические особенности как перегородки, так и всей раковины, являются надежными своего рода онтограммами эволюции. Изучение онтогенетического развития перегородки и лопастной линии помогает разобраться в запутанных вопросах систематики и филогении аммоноидей.
5. ЗНАЧЕНИЕ ИЗУЧЕНИЯ ОНТОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ДЛЯ ФИЛОГЕНИИ
Для правильного понимания филогенетических связей между отдельными представителями аммоноидей, а также для решения вопроса о предках отдельных групп, огромное, если не первостепенное значение имеет проведение детальных онтогенетических исследований, начиная с самых ^ранних стадий. Такие исследования, основанные на возможности наблюдения в онтогенезе более высокоорганизованных представителей черт организации, в той или иной степени свойственных взрослым представителям их возможных предков, вскрывают возникновение и развитие тех или иных морфологических черт каждой отдельной группы и помогают разобраться в запутанных вопросах филогении и систематики. Детальные онтогенетические исследования делают возможным составление генетических формул, отображающих ход онтогенетического развития лопастной линии и подчеркивающих те или иные морфологические особенности в их историческом развитии. Изучение онтогенеза и составление генетических формул лопастной линии способствуют решению важнейшего вопроса о гомологии и аналогии различных сутурных элементов. Так, например, изучение начальных стадий развития лопастных линий агониатитид и некоторых представителей анарцестид, имеющих во взрослом состоянии боковые лопасти, расположенные латерально, показывает совершенно различные пути онтогенетического развития этих двух групп. У Agoniatitidae боковая лопасть появляется с самого начала на середине бока, в то время как у Anarcestidae эта лопасть является первично умбональной и уже в ходе онтогенеза перемещается у некоторых представителей на боковую стенку. Формулы лопастной линии поэтому будут различными: у Agoniatitidae— VO : D, у Anarcestidae — VU : D. Боковая лопасть агониатитид, таким образом, не гомологична боковой лопасти анарцестид.
Другой, еще более наглядный пример можно привести при сравнении развития морфологически сходных между собой родов Pharciceras и Рго-lecanites, которые многими исследователями даже объединялись под одним родовым названием. Однако их генетические формулы с полной очевидностью говорят о том, что у указанных родов происходит резко различное развитие лопастной линии, заключающееся в первую очередь в резко различном характере вентральной лопасти. Формулы этих двух родов должны быть изображены следующим образом:
1) Pharciceras — (V2V1V9) UUlU3 : U2ID, 2) Prolecanites — V UU4J2U3 : ID.
Таких примеров при желании можно привести много. По внешнему аналогичному сходству исследователи часто объединяют в роды и семейства генетически разные формы. Детальное же изучение онтогенеза и фиксация хода развития лопастных линий в виде генетических формул дает возможность правильно понять гомологию различных сутурных элементов, без чего невозможны ни филогенетические построения, ни естественная ^филогенетическая) систематика.
IV. ФИЛОГЕНИЯ ДЕВОНСКИХ АММОНОИДЕЙ
Происхождение аммоноидей. Общая филогения аммоноидей является довольно сложным, далеко еще неразрешенным вопросом. Немногие попытки выяснения вероятных предков и разработки филогении наиболее древних девонских аммоноидей еще не осветили всех вопросов, связанных с происхождением, эволюцией и угасанием этой некогда обширной группы. Различные исследователи в этой области приходят к разным выводам. Фрех (Freeh, 1902, стр. 111) думал, что гониатиты поли-филетичны, а в последнем издании «Основ палеонтологии» Циттеля и Бройли (Zittel und Broili, 1924) указано, что «до сих пор» неясно из какой формы, или из каких форм среди наутилоидей произошли аммониты. Тем не менее обычно полагают, что род Bactrites произошел от прямых наутилоидей и что. он дал начало слабо свернутому Gyroceratites (=Mimo-ceras), который, в свою очередь, дал начало инволютным гониатитам, из которых развились цератиты и аммониты. Эта точка зрения оспаривалась Спэтом (Spath, 1933), считавшим, что аммониты развились из некоторых силурийских свернутых наутилоидей типа Barrandeoceras, который он считает близким к Agoniatites. Однако начало первого оборота у Ваггап-deoceras чрезвычайно массивное, тупоконечное, с очень большим умбо-нальным отверстием. Начальная камера сравнительно большая, тупоконическая, быстро увеличивающаяся в ширину и непосредственно переходящая в оборот ширококонической раковины. Сифон лежит вблизи середины извилины и не перемещается никогда на внешнюю сторону. Лопастная линия лишена внешней и внутренней лопастей. У представителей рода Tarphyceras, который, по Спэту, также относится к этой группе наутилоидей, умбональное отверстие несколько уже, но начало оборота также массивное, и начальная камера крупная, конусообразная, как и у Barrandeoceras. В противоположность Barrandeoceras, у Agoniatites начало оборота сравнительно маленькое и тонкое. Начальная камера округленная, субсферической формы. Раковина с тонкими, очень медленно увеличивающимися в ширину длинноконическими оборотами. Умбональное отверстие отсутствует. Лопастная линия на всех стадиях онтогенетического развития с внутренней и внешней лопастями. Различия между Barrandeoceras и Agoniatites настолько разительны, что какая-либо родственная связь между ними вряд ли возможна (см. рис. 9, d, ё). Спэт считает, что Gyroceratites и Lobobactrites являются отклоняющимися от тесно свернутых гониатитов и полагает, что Bactrites s. s. представляет собой наутилоид с краевым положением сифона. Другие авторы рассматривают Bactrites, так же как и Gyroceratites, как вторично развернутую из инво-лютных гониатитов форму [Sandberger, 1841, 1843; Beyrich (в работе Branco, 1885, стр. 2)]. Вторичное происхождение Bactrites и Gyroceratites было поддержано позднее Гюрихом (Giirich, 1909), Соболевым (1914), Долло (Dollo, 1922) и другими авторами. Хайэтт (Hyatt, 1900), Смит
56
(Smith, 1903), Шиндевольф (Schindewolf, 1932, 1933, 1934a), А. К. Миллер (Miller, 1938) и некоторые другие авторы рассматривают Bactrites и Gyroceratites в качестве древних форм аммонитов, развившихся из длинноконических наутилоидей с яйцеобразной начальной камерой. Наконец, в 1954 г. В. Н. Шиманский выделил бактритов в самостоятельный отряд, отличающийся как от наутилоидей, так и от аммоноидей. Предками бактри-
е
Рис. 9. Схематизированные медианные срезы начальных стадий развития некоторых представителей бактритоидей, аммоноидей и наутилоидей по Шиндевольфу (1934, стр. 275, рис. 3—7):
а — Bactrites из низов нижнего девона с прямой раковиной и яйцеобразной начальной камерой (подобная картина наблюдается и у Lobobactrites (Х6); b —Gyroceratites из верхов нижнего девона со свернутой раковиной, слегка укороченной яйцеобразной начальной камерой и большим умбональным отверстием (Х6); с — Mimagonia-tites из низов среднего девона с более свернутой раковиной, сильнее укороченной начальной камерой и небольшим умбональным отверстием (Х6); d — Agoniatites из верхов среднего девона с объемлющими оборотами, субсферической начальной камерой и без умбонального отверстия (Х6); е — Barrandeoceras tyrannum (Barrande) из верхнего силура с массивной широко конической раковиной, конусовидной начальной камерой и в корне отличным положением сифона (XI ,25).
тоидей и, в частности рода Bactrites, возможно, явились некоторые орто-цератиды, например ордовичский род Bactroceras Holm, у которого сифон уже расположен очень близко к внешней стенке, хотя образования вентральной лопасти еще не произошло. Bactroceras и Bactrites в известной степени являются переходными формами от наутилоидей к аммоноидеям. Установлено, что наиболее древним представителем бактритоидей нужно признать Bactrites sandbergeri Barrande из ордовика Богемии, который теперь относится к роду Eobactrites Schindewolf. Этот род имеет округлое поперечное сечение и отчетливо выраженную вентральную лопасть (рис. 10, а). Следующую ступень развития представляет род Bactrites Sandberger с типом В. subconicus Sandberger со слегка эллиптическим поперечным сече
57
нием. Лопастная линия образует, помимо вентральной лопасти, зачаточные дорсальное седло и боковую лопасть (рис. 10,6).
Следующей ступенью развития является род Lobobactrites Schindewolf с типом рода Bactrites ellipticus Freeh. Представители рода Lobobactrites имеют эллиптическое поперечное сечение раковины. Вентральная лопасть очень слабая, но глубже, чем у Bactrites', кроме нее, имеется глубо-
Рис. 10. Филогенетическое развитие лопастных линий бактритов и примитивных аммоноидей по Шиндевольфу (1932, стр. 175, рис. 5; 1933, стр. 91, рис. 25):
а — лопастная линия взрослой стадии Eobadrites sandbergeri (Barrande); b — лопастная линия взрослой стадии Had rites schlotheirnii (Quenstedt); с — лопастная линия взрослой стадии Lobobad rites ellipticus (Freeh); d —лопастная линия взрослой стадии (хgr^ceratifes gracilis Bronn; e — лопастные линии ранней и взроет >й стадии М imigoniatites zorgensis (Roemer); / — лопастные линии ранней и взрослой стадий Agoniatites costulatus (Archlac et Verneuil); g — лопастные линии ранней, юношеской и взрослой стадий Anarcestes (Anarcestes) lateseptatus (Beyrich).
кая и широкая боковая лопасть и высокое угловатое или слабо округленное дорсальное седло (рис. 10, с). Bactrites и Lobobactrites, в противоположность Barrandeoceras, имеют длинноконический тип раковины с маленькой яйцеобразной или округлой начальной камерой, которые приближают их к гониатитам (см. рис. 9).
Семейство Mimoceratidae (=G yroceratitidae). Lobobactrites может являться, в свою очередь, прямым предком древнейших аммоноидей из семейства Mimoceratidae Steinmann и,в частности, рода Gyroceratites Meyer (=Mimoceras Hyatt), лопастная линия которого (рис. 10, d) подобна таковой Lobobactrites, но у которого происходит свертывание раковины в спираль и некоторое укорачивание начальной камеры. Основными1 особенностями, отличающими Gyroceratites от нормальных гониатитов, являются следующие: 1) обороты не облекают друг друга, 2) лопастная линия не имеет дорсальной лопасти, а снабжена отчетливо выраженным дорсальным седлом; 3) струйки нарастания образуют на внутренней стенке выпуклость, тогда как у всех более поздних форм на внутренней стороне имеется синус. Согласно последним данным (Erben, 1953), семейство Mimoceratidae разделяется на два подсемейства: Mimi-ceratinae, без поперечных ребер или со слабыми ребрышками, не распространяющимися на вентральную сторону, и Mimosphinctinae, с резкими
58
поперечными ребрами, пересекающими вентральную сторону раковины. К первому подсемейству относится род Gyrocer alites, ко второму — Mi-mosphinctes и, вероятно, Anetoceras и Palaeogoniatites. Представители семейства Mimoceratidae прослеживаются с низов нижнего до середины среднего девона.
Хронологически должно было бы казаться более логичным происхождение Gyroceratites от Eobactrites, так как Gyroceratites известен из нижней части нижнего девона, в то время как Bactrites и Lobobactrites появляются лишь первый в среднем девоне, второй — в верхах нижнего девона. Однако Eobactrites появляется очень рано в ордовике, и никаких гониатитов не известно из верхней части ордовика или из силура. Более того, при определении эволюционного развития аммонитов следует учитывать неполноту геологической летописи, в силу чего наши коллекции в большинстве своем являются далеко неполными. С другой стороны, для выяснения путей эволюции аммоноидей мы должны использовать все возможные критерии, включая строение и положение сифона, природу поверхностной скульптуры, форму раковины и начальной камеры, очертание лопастной линии. Как видно на рис. 10, лопастная линия Bactrites s. s. очень похожа на лопастную линию Eobactrites и по своей форме является более или менее промежуточной между лопастными линиями Eobactrites и Lobobactrites. В свою очередь, лопастная линия Lobobactrites очень сходна с лопастной линией слабо свернутого Gyroceratites, который почти наверняка близко родственен бактритам. Так как Lobobactrites не был найден ниже верхов нижнего девона, в то время как Gyroceratites известен в самых низах нижнего девона, может казаться, что Gyroceratites является предком Lobohac-trites, который следовало бы в таком случае считать вторично развернутой формой, но Шиндевольф и Миллер находят, что такое предположение неправильно, и убеждены, что будущие сборы покажут, что Lobobactrites прослеживается во всем нижнем девоне и, вероятно, в силуре.
Эволюция древнейших семейств аммоноидей. Помимо Gyroceratites и близких к нему родов, в нижнедевонских отложениях найдены представители различных родов древнейших тесно свернутых гониатитов: Mimagoniatites, Anarcestes, Sellanarcestes и, возможно, Agoniatites, Werneroceras и Foordites. Шиндевольф (1933, стр. 52, 82, 88, 93) показал, что лопастные линии ранних стадий развития Mimagoniatites и Anarcestes почти полностью подобны Gyroceratites, и это привело его к заключению, что эти два рода развились из Gyroceratites. Более того, Шиндевольф (1933, стр. 88) указывает, что Gyroceratites встречается в девонских отложениях на более низком уровне, чем тесно свернутые аммониты, и этот факт указывает на то, что он является предковым для них. Лопастные линии юношеских стадий Agoniatites весьма напоминают лопастные линии взрослых представителей Mimagoniatites и поэтому весьма возможно, что Agoniatites произошел от ствола, представленного родом Mimagoniatites, а не от свернутых наутилоидей, подобных Barrandeoceras, как считает Спэт. На основании положения сифона, очертания лопастной линии, формы и размеров начальной камеры можно заключить, что род Agoniatites служит окончанием эволюционного ряда Lobobactrites —> Gyroceratites —> Mimagoniatites. Род Anarcestes и семейство Anarcestidae в целом представляют собой, по-видимому, другую ветвь, параллельную ветви Mimagoniatites —> Agoniatites и развивавшуюся независимо от нее из Gyroceratites.
Таким образом, уже в нижнем девоне от гироцератитов произошли две параллельно развившиеся в течение среднего девона ветви: семейства Mi-magoniatitidae Agoniatitidae и семейство Anarcestidae. Судьба и место этих семейств в родословном древе аммонитов, однако, в корне различны.
Семейства Mimagoniatitidae->Agoniatiti-d а е, характеризующиеся постоянным в течение онтогенеза положе-
59
нием широкой боковой лопасти,не испытали сколько-нибудь существенных эволюционных изменений и представлены в среднем девоне сравнительно немногими родами и видами. В настоящее время в составе надсемейства выделяются два семейства: Mimagoniatitidae, представленное единственным родом Mimagoniatites, и Agoniatitidae с родами Agoniatites и Paraphyllites, Представители первого рода характеризуются наличием небольшого умбонального отверстия; у представителей родов Agoniatites и Paraphyllites умбональное отверстие отсутствует и первый оборот раковины полностью охватывает субсферическую начальную камеру. Эволюция внутри надсемейства Agoniatitaceae, как уже указано выше, шла от их предка Gyroceratites через Mimagoniatites к Agoniatites и Paraphyllites. С концом среднедевонской эпохи агониатиты вымирают, не давая начала ни одной из известных групп более молодых аммоноидей. Широкая боковая лопасть, занимавшая всю боковую поверхность раковины и не менявшая своего положения в течение онтогенетического развития, по-видимому, препятствовала дальнейшему расчленению и усложнению лопастной линии и связанного с этим укрепления перегородки и раковины и ограничивала тем самым возможность эволюционного развития семейств.
Семейства Anarcestidae-^Prolobitidae. Совершенно иную картину представляет собой семейство Anarcestidae, у представителей которого первичная умбональная лопасть или сохраняла свое положение вблизи умбо или перемещалась в процессе онтогенетического развития на боковую сторону раковины, оставаясь, однако, в большинстве случаев значительно меньше боковой лопасти агониатитид и создавая возможности для образования новых сутурных элементов либо в районе умбонального шва, либо на боках раковины, либо, наконец, за счет дополнительного расчленения расширяющейся вентральной лопасти.
Семейство Anarcestidae, возникнув в нижнедевонскую эпоху, достигает своего расцвета и наибольшего разнообразия в среднем девоне, частично переходит в верхний девон и в середине верхнедевонской эпохи, вероятно, заканчивает свое существование, давая в конечном счете начало всем известным в настоящее время многочисленным и разнообразным семействам аммоноидей.
Во второй половине среднедевонской эпохи (живетский век) появляются первые представители (род Sobolewia) широко распространенного в фаменский век семейства Prolobitidae, берущего, вероятно, начало от анарцестид с перемещающейся на боковую сторону первичной умбональ-ной лопастью. Во франских отложениях представителей семейства не найдено, но в фаменском веке оно достигает своего расцвета и представлено родами: Prolobites, Prionoceras, Clymenoceras, Paralytoceras, Cycloclymenia, По мнению Шиндевольфа (1922) и Миллера (1938), от пролобитид берет начало нижнекаменноугольное семейство Prolecanitidae, к которому Миллер относит также зандбергероцератид и, предположительно, род Pharciceras, но подобная филогенетическая связь, на наш взгляд, едва ли возможна. У эволютных пролобитид типа Clymenoceras, Cycloclymenia происходит резко отличное от пролеканитид развитие внутренней (дорсальной) части лопастной линии, исключающее, на наш взгляд, возможность филогенетической связи между семействами Prolobitidae и Prolecanitidae. Представители семейства Sandbergeroceratidae, как и входящий в самостоятельное семейство Pharciceratidae род Pharciceras, по-видимому, происходят от одного корня с семейством Gephuroceratidae, с которым их объединяет общий характер расчленения вентральной лопасти (см. ниже).
Семейства Anarcestidae и Prolobitidae характеризуют собой определенное направление в эволюции девонских аммоноидей, достаточно отличное от путей развития остальных групп. Характерными особенностями 60
представителей обоих семейств являются наличие простой, обычно небольших размеров, нерасчлененной вентральной лопасти и отсутствие лопастей, возникающих на боковых сторонах раковины.
Семейства Tornoceratidae^Cheiloceratidae —>А ganididae (= I m i t о с е г a t i d а е) характеризуют собой другое направление в эволюции девонских аммоноидей, отличительной особенностью которого является образование лопастей (L), возникших из вершины первого бокового седла на боковых сторонах раковины, при наличии простой нерасчлененной вентральной лопасти.
Уже в первой половине среднедевонской эпохи от анарцестид с умбо-нальным положением первичной умбональной лопасти берет начало семейство Tornoceratidae, которое прослеживается до середины верхнедевонской эпохи, не испытывая, однако, существенных эволюционных изменений. Семейство представлено в среднем девоне родом Tornoceras с подродами Paradoceras, Protornoceras и Tornoceras, из которых два последних переходят и в верхний девон, когда возникают еще подроды Aulatornoceras и Polonoceras.
Помимо Tornoceras, в верхнедевонскую эпоху появляются более высокоорганизованные роды Pseudoclymenia, Lobotornoceras и Posttornoceras, характеризующиеся образованием дополнительных элементов лопастной линии за счет возникновения внутренней и второй наружной боковых лопастей. Семейство заканчивает свое существование в конце фаменского века. Еще в среднем девоне от Tornoceratidae берет начало монотипное семейство Maenioceratidae, существовавшее в течение живетского века. В начале фаменского века от торноцератид, вероятно, ответвляется новое, широко распространенное в фаменский век семейство Cheiloceratidae, наиболее характерными представителями которого являются роды: Cheiloceras, Sporadoceras, Dimeroceras и др. Шиндевольф (1934 б) и Миллер (1938) склонны были считать предками семейства Cheiloceratidae анарцестид; эти филогенетические построения основывались на некотором сходстве лопастной линии наиболее просто организованного представителя семейства, рода Ray mondicer as, с лопастной линией среднедевонского представителя анарцестид рода Subanarcestes. Таким образом, эти исследователи считали, что корни фаменского семейства Cheiloceratidae уходят в средний девон. Мне кажется, что такая точка зрения не может не вызывать сомнения и что наиболее вероятными предками семейства Cheiloceratidae являются Во многих отношениях (форма раковины, очертание лопастной линии) близкие к представителям его торноцератиды, а не анар-цестиды. Подобного же мнения, в частности, придерживаются в последнее время Миллер и Фёрниш (Miller and Furnish, 1954, стр. 688, 689), объединяющие семейства Tornoceratidae и Cheiloceratidae в одно надсемейство Cheilocerataceae. Семейство Cheiloceratidae, заканчивая свое существование в конце фаменского века, дает, в свою очередь, начало семействам Praeglyphioceratidaen Aganididae ( = Imitoceratidae). Первое из них будучи монотипным, существует в первой половине фаменского яруса и характеризуется трехчленным делением расширяющейся вентральной лопасти. (Эта особенность резко отличает род Praeglyphioceras от других представителей семейства Cheiloceratidae, к которому он ранее относился, и заставляет выделить самостоятельное семейство Praeglyphioceratidae). Семейство Aganididae, представители которого появляются впервые еще в первой половине франского века, достигает расцвета уже в каменноугольном периоде, давая начало всем так называемым «гониатитам» каменноугольного и пермского периодов.
С началом верхнедевонской эпохи наступает резкий перелом в развитии фауны аммонитов. Полностью вымирают агониатитиды. Анарцестиды, хотя и продолжают еще свое существование, но роль их резко падает.
61
Появляются и быстро эволюционируют новые семейства Sandbergerocera-tidae, Gephuroceratidae, Pharciceratidae, характеризующиеся новым своеобразным типом усложнения перегородочной линии за счет появления дополнительных сутурных элементов в районе умбонального шва и,что особенно характерно для этой группы семейств, за счет трехчленного деления расширяющейся вентральной лопасти. Все три семейства имеют, вероятно, в конечном счете общее начало и уходят, возможно, своими корнями в средний девон. Предками их, по-видимому, были некоторые наиболее эволютные (типа Anarcestes и Archoceras) представители семейства Anarcestidae, обладавшие дискоидальной раковиной, простой лопастной линией, вентральная лопасть которой не была разделенной на части. В процессе эволюции такая простая лопасть испытала трехчленное деление, что-одновременно с образованием дополнительных лопастей в районе умбо и постепенным смещением их на боковые стенки привело к лопастным линиям гефуроцератид, зандбергероцератид и фарцицератид. Несмотря на некоторые общие черты строения представителей указанных семейств и вероятное происхождение их от общих предков, пути эволюции их значительно разнятся друг от друга.
Семейство Gephuroceratidae. Представители семейства Gephuroceratidae составляют наиболее многочисленную и одну иа самых характерных групп аммонитов франского яруса. Это семейство принадлежало к числу быстро эволюционировавших и пользовалось необычайно широким распространением в верхнедевонских (франских) морях всего земного шара. Возникнув в начале верхнедевонской эпохи, гефу-роцератиды испытали быструю эволюцию и в конце франского века закончили свое существование. Эволюция семейства (см. стр. 84) протекала в общем в направлении Ponticeras —> Manticoceras —> KoenenitesHonin-ghausia —> Timanites.
Семейство Beloceratidae. От гефуроцератид уже в первой половине франского века берет начало семейство Beloceratidae (см. стр. 116). В ходе эволюции, в пределах филогенетического ряда Neomanti-coceras —> Eobeloceras —> Beloceras одновременно, с возрастанием инво-лютности тонкой линзовидной раковины происходило увеличение количества элементов лопастной линии. Резкое возрастание расчлененности лопастной линии, достигающее предела у некоторых представителей рода Beloceras, способствовало укреплению утоньшающихся стенок раковины (что вызывало уменьшение веса ее), становящейся тонкой линзовидной, почти полностью инволютной, хорошо приспособленной для передвижения в водной среде. С другой стороны, резкое расчленение лопастной линии и приобретение раковиной линзовидной формы обусловили высокую одностороннюю специализацию белоцератид и быстро завело их в морфологический «тупик», т. е. такое состояние организации, при котором «формы, достигают крайней степени одностороннего развития, идти далее которого для организма невозможно» (В. Ковалевский, 1873). Просуществовав сравнительно короткий промежуток времени, белоцератиды полностью вымирают в конце франского века. Резкое вымирание белоцератид, вероятно, объясняется появлением новых, более приспособленных конкурентов — климений, проноритид и других групп аммоноидей. От белоцератид возможно, берет начало семейство Prodromitidae, представители которого известны из турнейских отложений Северной Америки.
Семейство Sandbergerocaretidae. Совершенно иное положение в составе верхнедевонской аммонитовой фауны по сравнению с Gephuroceratidae занимает одновременно с ним возникающее и генетически связанное семейство Sandbergeroceratidae.
Представители семейства Sandbergeroceratidae, одновременно с более или менее трехчленным делением вентральной лопасти и образов.нием
62
дополнительных, возникающих в районе умбо лопастей, пошли по пути’ук-репления раковины за счет образования резко выраженной (по крайней мере у молодых экземпляров) богатой скульптуры. Развитие подобной скульптуры и слабо-инволютная форма раковины, вероятно, поставили зандберге-роцератид в невыгодное положение по сравнению с более приспособленными к активному передвижению представителями других семейств с гладкими, более или менее инволютными раковинами и сильно ограничили возможности эволюционного развития семейства. В результате зандбер-героцератиды не получили широкого распространения, и, просуществовав сравнительно короткий промежуток времени, вымерли, не дав, вероятно, начала какой-либо группе более молодых представителей аммоноидей.
Семейство Pharciceratidae. На рубеже средне- и верхнедевонской эпох, помимо семейств Sandbergeroceratidae и Gephuro-ceratidae, возникает генетически связанное с ними семейство Pharciceratidae, уходящее, вероятно, как и первые два, своими корнями в средний девон. Эволюция внутри семейства (см. стр. 130) протекала как в направлении изменения формы раковины, так и в направлении усложнения лопастной линии за счет значительного увеличения количества сутурных элементов. Общее количество элементов лопастной линии возрастало в ходе эволюции от двенадцати (считая трехраздельную вентральную лопасть за один элемент) до, примерно, пятидесяти вокруг извилины. Возникшее и довольно широко распространенное в начале франского века семейство Pharciceratidae в результате быстрой эволюции вскоре достигает крайней степени своего эволюционного развития и, постепенно сокращая ареал своего распространения, уже к середине франского века, по-видимому, полностью вымирает, уступая место более приспособленным к широким условиям существования группам аммонитов.
Семейство Devonopronoritidae, От фарцицератид, по-видимому, еще в начале франского века берет начало семейство Devonopronoritidae (см. ниже, стр. 136), единственный представитель которого — род Devonopronorites известен из средней части франского яруса.
Семейство Prolecanitidae. Вопрос о происхождении нижнекаменноугольного1 семейства Prolecanitidae еще не вполне ясен и требует своего разрешения на основе тщательных онто-филогенетических исследований. Как уже указывалось выше, некоторые авторы склонны рассматривать в качестве предков данного семейства пролобитид, однако подобная филогенетическая связь вряд ли возможна. Возможными предками пролеканитид, на наш взгляд, могли явиться наиболее эволютные (типа Archoceras) представители семейства Anarcestidae, с дискоидальной раковиной и простой лопастной линией, вентральная лопасть которой была не разделенной на части. В процессе эволюции эта лопасть осталась простой, а образование дополнительных лопастей в районе умбо и постепенное смещение их на боковые стенки привели к формированию лопастных линий пролеканитид. Из пролеканитид впоследствии развиваются проноритиды, медликоттииды и др.
Надсемейства Gonioclymeniaceae—>Glymeni-а с е а е. Разбирая филогенетические связи девонских аммоноидей, следует, наконец, коснуться вопроса о систематическом положении довольно обширной группы климений. Климении являются типичными представителями аммоноидей и образуют среди них группу, которая отличается от гониатитов своим внутренним (дорсальным) положением сифона. Субсферическая начальная камера, фиксированное положение сифона на спин
1 Некоторые авторы (А. К. Миллер и др.) считают, что древнейший представитель семейства Prolecanitidae, родProtocanites, встречается и в наиболее верхних отложениях (если считать горизонт с Gattendorfia еще верхнедевонским) верхнего девона.
63
ной стороне раковины делают невозможным объединение климений с нау-тилоидеями, как это делали некоторые авторы: Анштед (Ansted, 1838), Гибель (Giebel, 1852), Квенштедт (Quenstedt, 1836), Рёмер (Roemer, 1856) и др. У климений не наблюдается переходных от наутилоидей форм, наподобие гониатитам; климений с умбональным отверстием и не полностью свернутым первым оборотом не известно. Барранд (Barrande, 1867), позднее Гюрих (Giirich, 1896) и другие исследователи полагали, что климении представляют собой эндогастрически свернутые формы, у которых внутреннее положение сифона — унаследованное. В большинстве случаев у климений развивается внешнее седло лопастной линии, отсутствующее у гониатитов. Gyroceratites, однако, имеют дорсальное седло, которое как бы гомологично вентральному седлу климений. Однако дальнейшие исследования (Schindewolf, 1923) показали, что внешнее (вентральное) седло климений не первичное, а образуется на месте начальной вентральной лопасти. С другой стороны, возможность перемещения сифона у климений на дорсальную сторону в процессе развития отмечалось еще Бранко (1880). Спэт (1919) утверждал, что климении развились из гониатитов, обладавших непостоянным положением сифона, однако это мнение, по Шиндевольфу, является ошибочным, так как гониатитов с непостоянным положением сифона в девоне не известно. Шиндевольф, уделивший внимание изучению начальных камер и положению сифона на ранних стадиях развития климений, установил, что в этот период у них сифон занимает внутреннее положение вблизи дорсальной стенки. Исключение составляют, вероятно, лишь древние климении. У единственной, известной до сих пор наиболее древней Acanthoclymenia neapolitana (Clarke) из нижней части верхнего девона Северной Америки сифон в первой перегородке начинается на вентральной стороне и на протяжении первого оборота перемещается на дорсальную сторону. Медианные срезы европейских климений, которые известны впервые из верхов верхнего девона, показывают всегда внутреннее положение сифона. Вентральное положение сифона на начальных стадиях онтогенетического развития древнейших климений говорит за происхождение их от типичных гониатитов.
Различные исследователи подходят различно к вопросу о возможном предковом роде для климений. Фрех, например, (Freeh, 1902, стр. 43). считал за их предка род Gyroceratites. Шиндевольф и некоторые другие исследователи считают такое толкование мало вероятным, так как древнейшие климении лишены умбонального отверстия и их боковая лопасть зарождается всегда вблизи умбо, а не на середине боковой стенки, как у Gyroceratites. Боковое положение ее на более поздних онтогенетических стадиях у климений вторично.
Соболев (1914) рассматривает климений как внутренне-сифонную мутацию различных ветвей нормальных гониатитов (Cyrtoclymenia пара ллелизуется, например, с Tornoceras), но эта идея пользуется малой поддержкой. В противоположность Соболеву, ряд других исследователей считают, что климении с внутренним положением сифона, представляющие собой большую группу аммонитов, произошли от одного общего корня — нормальных гониатитов с внешним положением сифона.
Хайэтт, Шиндевольф и другие исследователи считают, что климении произошли от полностью свернутых гониатитов группы Anarcestes. Наличие у Acanthoclymenia neapolitana (Clarke) сравнительно большой начальной камеры подтверждает это мнение и сближает этот вид с анар-цестидами. У наиболее древних климений, как уже указывалось выше, первая перегородочная линия аселлатная, у более молодых европейских климений — латисел латная.
Таким образом, в начале верхнедевонской эпохи или, возможно, еще в среднем девоне от анарцестид, в связи с утратой умбонального отверстия
64
Рис. 11. Филогенетическая схема^девонских аммоноидей:
1— семействоMimoceratidae(~Gyroceratitidae); 2 — сем. Mimagoniatitidae; 3 — сем. Agoniatitidae-4— сем. Anarcestidae; <5— сем. Prolobitidae; 6 — сем. Gephuroceratidae (^Manticoceratidae); 7— сем.’ Beloceratidae; 8 — сем. Prodromitidae; 9 — сем. Sandbergeroceratidae; Ю — сем. Pharciceratidap-11 — сем. Devonopronoritidae; 12 — сем. Prolecanitidae; 13 — сем. Tornoceratidae); 14 — сем’ Maenioceratidae; 15 —сем. Cheiloceratidae; 16 — сем. Aganididae ( = Imitoceratidae- 17 — сем
Praeglyphioceratidae; _18 — надсемейство Gonioclynieniaceae; 19 — надсемейство Clymeniaccae
и полным свертыванием раковины вокруг начальной камеры, вероятно, произошли монофилетически древние формы климений. Древнейшие кли-мении, уходящие, возможно, своими корнями в средний девон, относятся к надсемейству Gonioclymeniaceae, которое характеризуется наличием во взрослом состоянии вентральной лопасти. От Gonioclymeniaceae, вероятно, уже в первой половине верхнего девона берут начало представители надсемейства Glymeniaceae, характеризующиеся вторичной утратой вентральной лопасти и заменой ее вентральным седлом.
Шиндевольф (1923, стр. 366—367; 1931), отмечая, что уже на ранних стадиях онтогенетического развития настоящих аммонитов (наподобие Agathiceras из перми) сифон постепенно мигрирует из дорсального в вентральное положение, полагает, что эти аммониты вероятнее развились из климений скорее, чем из внешнесифонных гониатитов, как считалось до указанного исследователя. Более того, так как у юрских и меловых аммонитов, которых он расшлифовал, сифон вначале субцентральный, а затем с ростом раковины мигрирует на вентральную сторону, Шиндевольф заключает, что все послетриасовые аммониты произошли от климений (а не от Phylloceratidae, как считалось ранее). Спэт (1933, стр. 430—439), однако, нашел, что в течение ранних стадий развития у большинства юрских и меловых аммонитов положение сифона варьирует, и этот факт, вместе с общим сходством лопастных линий ранних внешнесифонных гониатитов с лопастными линиями Agathiceras и других форм с мигрирующим сифоном, позволяет, по мнению Миллера, считать, что Шиндевольф уделяет слишком много внимания положению сифона в течение юношеских стадий онтогенетического развития для указания филогенетического родства. Спэт, Миллер и другие исследователи считают, что климении заканчивают свое существование в конце девонского периода и что предками более поздних аммонитов являются внешнесифонные гониатиты, а не климении. Шмидт (1925, стр. 522) полагает, например, что все каменноугольные аммониты произошли от Aganides (=Imitoceras). Однако это мнение критикуется Смитом (1932, стр. 23) и не разделяется Миллером (1938, стр. 18) и другими исследователями. Действительно, как можно видеть из приведенной схемы филогенетических связей, границу девонского и каменноугольного периодов переходят не только представители семейства Agani-didae. От Beloceratidae отходит ветвь, дающая начало семейству Prodro-mitidea, от Anarcestidae — ветвь, дающая начало пролеканитидам, а следовательно, и проноритидам, медликоттиидам, дарэлитидам и др. На данной схеме можно видеть также, что все более поздние аммониты развились из анарцестид, а не из агониатитид.
ВЫВОДЫ
Подводя итог рассмотрению филогенетических связей и путей развития девонских аммоноидей, можно сделать следующие заключения:
1. Все аммоноидей развились монофилетически из бактритоидей.
2. Древнейшее среди девонских аммоноидей — семейство Mimocera-tidae.
3. От семейства Mimoceratidae берут начало еще в нижнем девоне два преимущественно среднедевонских семейства: Mimagoniatitidae и Anarcestidae; от первого из них происходит близко родственное семейство Agoni-atitidae, которое вымирает, не давая начала более молодым группам; семейство Anarcestidae является предковым для всех более молодых групп аммоноидей, эволюция которых идет различными путями.
4. Семейство Anarcestidae и близко родственное ему семейство Prolobitidae характеризуют собой определенное направление в эволюции де
66
вонских аммоноидей, достаточно отличающееся от путей развития остальных групп наличием простой нерасчлененной вентральной лопасти и отсутствием лопастей, возникающих на боковых сторонах раковины.
5. Семейства Tornoceratidae -> Cheiloceratidae -> Aganididae характеризуют собой другое направление в эволюции. Они обособляются в первой половине среднедевонской эпохи от семейства Anarcestidae; их характерной особенностью является образование лопастей (L), возникающих из вершины первого бокового седла на боковых сторонах раковины, при наличии простой нерасчлененной вентральной лопасти; представители этого направления эволюции при последующем двучленном делении вентральной лопасти дают начало подавляющему большинству верхнепалеозойских аммоноидей; в живетском веке от этого основного направления формируется монотипное семейство Maenioceratidae; в фаменском веке от него в результате трехчленного деления вентральной лопасти отделяется семейство Praeglyphioceratidae.
6. С началом верхнедевонской эпохи от анарцестид отделяется группа аммоноидей, характеризующаяся в той или иной степени трехчленным делением расширяющейся вентральной лопасти и образованием новых су-турных элементов в районе умбонального шва; представители этого особенно характерного для франского века направления эволюции относятся к семействам Gephuroceratidae, Sandbergeroceratidae, Pharcicerati-dae, Beloceratidae, Devonopronoritidae
7. В конце девонского периода от анарцестид берут начало пролека-нитиды, от которых в верхнем палеозое происходят семейства Pronori-tidae, Medlicottiidae, Daraelitidae, последнее из которых является, вероятно, предковым для почти всех постпалеозойских групп аммоноидей.
8. Климении представляют собой большую группу аммонитов с внутренним (дорсальным) положением сифона, происходящую монофилети-чески, вероятно, в начале верхнедевонской эпохи из нормальных полностью свернутых гониатитов группы Anarcestes и оканчивающую свое существование с концом девонского периода или, возможно, незначительно переходящую и в каменноугольный период.
V. ОПИСАТЕЛЬНАЯ ЧАСТЬ
СЕМЕЙСТВО ANARCESTIDAE STEINMANN, 1890
Диагноз. Раковина субсферическая или дискоидальная, иногда приближающаяся к линзовидной, преимущественно с сильно объемлющими оборотами, иногда с умбональным отверстием. На вентральной стороне раковины иногда наблюдаются отчетливые парные продольные борозды. Умбо у низших представителей широкое, у более организованных суженное. Струйки нарастания образуют узкие синусы на вентральной и широко-округленные на боковых сторонах раковины. Лопастная линия с V-образной вентральной, округленной боковой и округленной или угловатой дорсальной лопастями; у некоторых родов имеется в дополнение небольшая лопасть вблизи умбонального шва.
Филогения. Филогения семейства Anarcestidae и вопрос о возможных его предках, как и о предках аммоноидей вообще, еще не вполне ясны и требуют для своего разрешения дополнительных исследований как ранних, наиболее примитивных аммонитов, так и их возможных предков. Вопрос о возможных предках аммонитов более подробно освещен в главе о филогении аммонитов, и я поэтому не останавливаюсь на нем при описании данного семейства. Семейство было более или менее детально изучено Шиндевольфом (1933), который отметил его родство с семейством Agoniatitidae и происхождение их обоих от Gyroceratites. Однако несмотря на возможное происхождение Anarcestidae и Agoniatitidae от одних и тех же предков, они резко различаются по характеру развития лопастных линий и, на наш взгляд, не могут объединяться в одно надсемейство, как это делает А. К. Миллер (1938). У Agoniatitidae боковая лопасть появляется на ранней стадии вблизи середины боковых сторон раковины и занимает это положение в течение всего онтогенетического развития, являясь, таким образом, боковой по своему происхождению, в то время как у Апаг--cestidae боковая лопасть появляется в районе умбонального шва на спинно-боковой стороне раковины и затем у некоторых форм мигрирует вентрально в течение онтогенетического развития, так что на взрослой стадии она занимает положение вблизи середины боковой стороны, как у Agoniatitidae; у других форм боковая лопасть остается субумбональной по своему положению в течение всей жизни. Возможные филогенетические связи внутри семейства Anarcestidae указаны ниже при описании подсемейства Апаг-cestinae.
Ревизия семейства. На основании формы раковины, размеров умбо, степени развития лопастной линии, Шиндевольф (1933) разделяет формы, которые относятся к этому семейству, на два подсемейства: Anarcestinae и Pinacitinae. Характеристика и состав этих подсемейств приводятся ниже.
68
ПОДСЕМЕЙСТВО ANARCESTINAE STEINMANN, 1890
Диагноз. Раковина субсферическая или толстодискоидальная, реже тонкодискоидальная, низкоапертурная, в большинстве случаев с широким умбо. Лопастная линия образует вентральную, боковую и дорсальную лопасти. Формула лопастной линии: VU : D.
Общие замечания. Подсемейство включает следующие роды: Anarcestes, Subanarcestes, Cabrieroceras, Werneroceras, Sellanarcestesк Archo~ ceras. Роды, входящие в подсемейство, характеризуются различной степенью свернутости первого оборота вокруг начальной камеры. Крайними членами ряда являются Anarcestes и Werneroceras. У Anarcestes начальная камера яйцевидная до шаровидной, с не полностью спирально свернутым первым оборотом, так что протоконх не соприкасается с первым полуоборотом раковины и поэтому имеется умбональное отверстие. У Subanarcestes, Cabrieroceras, Werneroceras, Sellanarcestes и Archoceras протоконх субсферический и первый оборот раковины тесно свернут вокруг него, так что умбональное отверстие отсутствует. У Subanarcestes и Cabrieroceras боковые лопасти даже во взрослом состоянии маленькие и находятся вблизи умбонального шва, в то время как у W erneroceras и Sellanarcestes боковые лопасти во взрослом состоянии широкие и расположены вблизи середины боковых сторон раковины. Cabrieroceras легко отличим от Subanarcestes по форме эволютной бочковидной раковины. Sellanarcestes отличается от Werneroceras тем, что его дорсальная лопасть разделена во взрослом состоянии срединным седлом. Archoceras легко отличим от всех остальных родов по форме своей тонкодискоидальной, очень эволютной раковины. Шиндевольф (1933) и А. К. Миллер (1938) считают, что, по-видимому, как Anarcestes, так и Werneroceras произошли от одной из ветвей, отходящих от Gyroceratites. От Anarcestes затем произошел Subanarcestes и Sellanarcestes, а от Werneroceras— Parodicerellum из Pinacitinae. Краткая характеристика родов, входящих в подсемейство, такова:
1. Anarcestes Mojsisovics, 1882. Описание этого рода приводится ниже.
2. Subanarcestes Schindewolf, 1933. Раковина субсферическая, с узким умбо, без внешних борозд, с шаровидной начальной камерой, без умбонального отверстия. Боковая лопасть даже во взрослом состоянии маленькая, слабо выраженная, расположенная на умбональной стенке. Формула лопастной линии: VU : D. Тип рода — Subanarcestes тасгосе-phalus Schindewolf (1933). Низы среднего девона Германии. Других видов пока не известно.
3. Cabrieroceras gen. nov. Описание этого рода дано ниже.
4. Werneroceras Wedekind, 1918. Описание этого рода приводится ниже.
5. Sellanarcestes Schindewolf, 1933. Описание этого рода приводится ниже.
6. Archoceras Schindewolf, 1937. Раковина тонкодискоидальная, эво-лютная, с низкими, округленными, очень слабо объемлющими оборотами. Лопастная линия состоит из широкой и глубокой, слегка округленной в основании V-образной вентральной лопасти, широкой сглаженной боковой лопасти, расположенной в районе умбонального шва, и довольно глубокой и широкой, несколько округленной в основании дорсальной лопасти. Формула лопастной линии: VU : D. Тип рода — Archoceras paeckel-manni Schindewolf (1937) из верхнего девона (fiheiloceras-Stuiv) Германии. Кроме типичного вида, известного также из верхнего девона Северной Африки, к данному роду относится, возможно, Archoceras (?) varicosum (Drevermann), известный из франского яруса (мантикоцеровые слои, зона IS) Германии.
69
Род Anarcestes Mojsisovics, 1882
Anarcestes: Mojsisovics, 1882, стр. 181 (pars); Hyatt, 1884a, стр. 309 (pars); Holzapfel, 1895, стр. 69 (pars); Foord and Crick, 1897, стр. 38 (pars); Freeh, 1913, стр. 14 (pars); Schindewolf, 1933, стр. 90; Miller A. K., 1938, стр. 56; Наливкина,1947, стр. 159.
Anarcestes (Clarkloceras): Wedekind, 1918, стр. 108.
Тип рода — Ammonites lateseptatus Beyrich, 1837, стр. 25, табл. 1, фиг. 2—4. Низы среднего девона Западной Германии.
Диагноз. Раковина обычно толстодискоидальная, обороты довольно низкие, округленные с вентральной и боковых сторон, гладкие, без отчетливых внешних борозд. Умбо обычно довольно широкое, с умбо-нальным отверстием, так как первый оборот не полностью охватывает яйцевидный или субсферический протоконх. Струйки нарастания образуют широкие сглаженные синусы на боковых сторонах и узкие округленные синусы на вентральной стороне раковины. Лопастная линия во взрослом состоянии состоит из V-образной вентральной, округленной в основании боковой, располагающейся субумбонально или латерально, и широкой округленной дорсальной лопастей.
Формула лопастной линии: VU : D.
Общие замечания. В качестве типа рода Anarcestes Фурдом и Криком (Foord and Crick, 1897, стр. 38) был принят Goniatites plebeius Barrande (1865/1867). Но, как правильно отметил Флигель (Fliegel, 1896, стр. 417), эта форма является лишь вариететом ранее установленного Ammonites lateseptatus Beyrich (1937), который поэтому должен рассматриваться в качестве типа рода Anarcestes.
В 1918 г. Ведекинд (1918, стр. 106—108) на основании различного положения боковой лопасти выделил в составе рода Anarcestes три подрода: Anarcestes s. s. с боковым, Werneroceras—с субумбональным и Clarkeo-ceras — с умбональным положением боковых лопастей. Шиндевольф (1933), имевший возможность изучить образцы, которые послужили материалом для выделения Ведекиндом подродов, пришел к заключению, что A. (Clarkeoceras) umbonale Wedekind, установленный в качестве типа подрода Clarkeoceras, идентичен Anarcestes lateseptatus var. plebeia (Barrande) и подродовое название Clarkeoceras является поэтому синонимом Anarcestes s. s. В то же время установленный Ведекиндом A. (Werneroceras) subumbonale Wedekind в качестве типа подрода Werneroceras является, согласно исследований Шиндевольфа, лишь юным экземпляром ранее установленного Goniatites ruppachensis Kayser, который должен поэтому рассматриваться как тип подрода W erneroceras, который выделен Шиндевольфом в самостоятельный род.
В составе рода Anarcestes Шиндевольфом выделяются два подрода: Anarcestes. s. s. и Latanarcestes.
Подрод Anarcestes s. s. характеризуется наличием субумбонально расположенной (в области умбонального края раковины) маленькой и плохо выраженной боковой лопасти; боковые стороны заняты широким первым боковым седлом.
К данному подроду, помимо типичного вида A. (Anarcestes) lateseptatus (Beyrich), относятся A. (Anarcestes) praecursor Freeh A. (Anarcestes) simulans (Barrande).
Подрод Latanarcestes характеризуется широкими боковыми лопастями, расположенными на боковых сторонах раковины. Тип подрода — Ammonites noeggerati Buch (Buch, 1832) из низов среднего девона Германии. К этому же подроду относятся A. (L.) latisellatus Erben и, возможно, A. (L.)? applanatus Freeh.
70
Anarcestes (Anarcestes) praecursor Freeh
Табл. I, фиг. 1
Goniatites plebeius: Barrande, 1865—1870, стр. 37, табл. 5, фиг. 5; табл. 7, фиг. 3—9.
Goniatites (Anarcestes) lateseptatus: Чернышев, 1893, стр. 214, табл. Ill, фиг. 4—6.
Anarcestes praecursor: Fligel, 1896, стр. 414—420; Freeh, 1897, стр. 169, табл.30a, фиг. 9; 1913, стр. 14.
Anarcestes lateseptatus: Наливкина, 1947, стр. 161, табл. XLI, фиг.1, рис. в тексте 7.
Голотип происходит из низов среднего девона Богемии.
Форма. Раковина толстодискоидальная, у молодых экземпляров приближающаяся к субсферической, с равномерно округленными боко-
Рис. 12. Лопастная линия Anarcestes (Anarcestes) praecursor Freeh (x8); № 774/604 приШ=9 мм. Рудный Алтай, Лениногорский район. Низы среднего девона, крюковская свита.
выми и вентральной сторонами. Поперечное сечение молодых оборотов полулунное, с шириной, примерно вдвое превышающей высоту; с возрастом обороты возрастают в высоту и поперечное сечение их становится по-луэллиптическим. Умбо умеренно широкое и глубокое, ступенчатое, ограниченное крутыми, отчетливыми умбональными стенками. Умбональный край довольно резкий.
Размеры: Д В Ш Ду В/Д Ш/Д Ду/Д
№ 774/604 12,7 4,5 9,1 4,2 0,35 0,72 0,33
№ 773/4 18,0 6,4 11,4 6,0 0,36 0,64 0,33
Скульптура. На имеющихся в моем распоряжении экземплярах раковинный слой не сохранился. Ядро совершенно гладкое. На экземплярах, описанных Ф. Н. Чернышевым, скульптура представлена весьма тонкими струйками нарастания.
Лопастная линия (рис. 12). Наружная часть лопастной линии состоит из воронкообразной узкой вентральной лопасти, широкого, плоского, слабо-округленного наружного седла и маленькой округленной, расположенной в районе умбонального края боковой лопасти.
Сходство и отличие. Описанный вид, имея сходство с A. (A.) lateseptatus (Beyrich) по общей форме толстодискоидальной раковины и характеру лопастной линии, отличается, однако, от последнего значительно более инволютной раковиной, имеющей более узкое умбо.
Геологический возраст и распространение. Встречается в низах эйфельского яруса Богемии и Рудного Алтая.
Местонахождение. Два экземпляра найдены в алевролитах крюковской свиты Лениногорского рудного поля. Два экземпляра этого же вида, хранящиеся в Центральном научно-исследовательском геологоразведочном музее им. Ф. Н. Чернышева, описаны Ф. Н. Чернышевым из светло-серых известняков той же свиты.
71
Anarcestes (batanarcestes) noeggerati (Buch)
Табл. I, фиг. 2, 3
Ammonites subnautilinus: Beyrich,1837, стр. 24 (pars); Giebel, 1852, стр. 479 (pars). Ammonites noeggerati: Buch, 1832, стр. 166, табл. I, фиг. 6—8.
Goniatites noeggerati: Archiac et Verneuil, 1842, стр. 337, табл. XXV, фиг. 1, a.
Goniatites subnautilinus: Quenstedt, 1846—1849, стр. 63, табл. Ill, фиг. 4, a, G. Sandberger, 1851, стр. 304, табл. Ill, фиг. 27.
Goniatites subnautilinus var. typus: G. u. F. Sandberger, 1850—1856, стр. 114, табл. XI, фиг. 1, a — f] Beyrich, 1884, стр. 206.
Anarcestes subnautilinus: Hyatt, 1884a, стр. 309; Steinmann u. Doderlein, 1890, стр. 373, фиг. 432; стр. 392, фиг. 469, 470А; Freeh, 1913, стр. 15; Wedekind, 1918, стр. 109.
Anarcestes noeggerati: Foord and Crick, 1897, стр. 42.
Anarcestes (Latanarcestes) noeggerati: Schindewolf, 1933, стр. 93.
Голотип происходит из нижней части среднедевонских отложений Германии.
Форма. Раковина дискоидальная, среднеинволютная, с несколько уплощенными, слабо выпуклыми боковыми и округленной или слегка уплощенной вентральной сторонами, с наибольшей шириной вблизи умбо. Обороты во взрослом состоянии удлиненно-овальные в поперечном сечении, с высотой больше ширины; инволютная часть оборота составляет от х/4 до х/3 его высоты. Высота наружного оборота составляет от 2/5 до 4/9 диаметра раковины. Умбо умеренно широкое, глубокое, ступенчатое; ширина его варьирует в пределах от х/4 до 2/3 диаметра. Умбональные стенки отчетливые, несколько округленные, почти перпендикулярные к боковым сторонам. Умбональный край отчетливый.
Размеры. Ввиду плохой сохранности имеющегося в наших коллекциях материала произвести соответствующие измерения затруднительно. Единственный экземпляр, позволяющий произвести измерения его параметров, дает следующие результаты:
Размеры: Д В Ш Ду В/Д Ш/Д Ду/Д
№ 774/605 35,0 16,0 13,0 9,0 0,46 0,37 0,26
В коллекции имеется несколько экземпляров, обладающих более широким умбо, составляющим около х/3 диаметра раковины.
Скульптур а. У большинства экземпляров раковинный слой не сохранился. Отдельные участки его, наблюдающиеся на некоторых экземплярах, показывают, что раковина была гладкой, покрытой лишь очень тонкими струйками нарастания.
Лопастная линия образует узкую V-образную вентральную лопасть, округленное, довольно широкое, но не высокое первое боковое седло, широкую округленную в основании, занимающую собой всю боковую поверхность раковины боковую лопасть, расположенное в районе умбонального шва второе боковое седло и широкую, широко-округленную в основании дорсальную лопасть. В силу плохой сохранности наших образцов, точное графическое изображение даже наружной части лопастной линии дать не представляется возможным.
Сходство и отличие. От наиболее близкого к нему Л. (L.} latisellatus Erben описанный вид отличается более уплощенной с боков раковиной, имеющей более высокие в поперечном сечении обороты, менее широким и глубоким умбо и менее широким седлом в области умбонального шва. Различные авторы относят к данному виду экземпляры, несколько отличающиеся друг от друга шириной умбо. Среди наших коллекций имеются, как уже указывалось выше, как широкопупковые, так и более узкопупковые экземпляры. Возможно, что целесообразно было бы выделить 72
их в различные виды или, по крайней мере, подвиды, но не обладая достаточным количеством материала удовлетворительной сохранности, я воздерживаюсь в настоящее время от их разделения и отношу как узко-,, так и широкопупковые экземпляры к одному виду.
Геологический возраст и распространение. Часто в эйфельском ярусе Германии и Рудного Алтая.
Местонахождение. 34 экземпляра и большое количество фрагментарных обломков, относящихся, по-видимому, к этому виду, найдены в алевролитах на северо-западном склоне горы Сокольной, в нижней части сокольной свиты.
Род Cabrleroceras gen. nov.
Anarcestes: Freeh, 1913, стр. 14 (pars).
Anarcestes (Anarcestes): Wedekind, 1918, стр. 106 (pars).
Werneroceras: Schindewolf, 1923, стр. 96 (pars).
Тип рода — Goniatites rouvillei Koenen, 1886, стр. 166. Средний девон Южной Франции.
Диагноз. Раковина у небольших экземпляров бочковидная, эво-лютная, с очень широкими, низкими оборотами, у крупных экземпляров— толстодискоидальная. Вентральная и боковая стороны сливаются в одну слабо выпуклую поверхность. Умбональный край отчетливый. Умбональная стенка крутая. Умбо широкое и глубокое. Лопастная линия состоит из вентральной, боковой и дорсальной лопастей. Вентральная лопасть очень широкая, воронкообразная, с узким оттянутым концом. Слабо выраженная боковая лопасть находится на умбональной стенке.
Формула лопастной линии: VU : D.
Видовой состав и геологическое распространение. Кроме типичного вида, известного из среднего девона Южной Франции, Северной Африки и, вероятно, Западной Германии, к этому же роду, по-видимому, относится Goniatites crispijormis Kayser, из среднего девона Западной Германии.
Общие замечания. Новый род легко отличим от других представителей семейства по форме раковины и лопастной линии.
Род Werneroceras Wedekind* 1918
Anarcetes: Mojsisovics, 1882, стр. 181 (pars); Hyatt, 1884a, стр. 309 (pars); Holzapfel, 1895, стр. 69 (pars); Ford and Crick, 1897, стр. 38 (pars); Freeh, 1913, стр. 14 (pars).
Anarcestes (Anarcestes): Wedekind, 1918, стр. 106 (pars).
Anarcestes (Werneroceras}: Wedekind, 1918, стр. 108 (pars).
Werneroceras: Schindewolf, 1933, стр. 96; Miller, 1938, стр. 57.
Тип рода — Goniatites ruppachensis Kayser, 1879, стр. 304. Нижняя часть среднего девона Германии.
Диагноз. Раковина обычно толстодискоидальная, обороты довольно низкие, округленные с вентральной и боковых сторон. Умбо различных размеров, без умбонального отверстия, так как первый оборот раковины плотно свернут вокруг субсферпческого протоконха. Струйки нарастания образуют широкие сглаженные синусы на боковых сторонах и узкие округленные синусы на вентральной сторбне раковины. Во взрослом состоянии лопастная линия состоит из V-образной вентральной, широкой округленной боковой и широкой округленной дорсальной лопастей; лопасти разделены широкими округленными седлами, которые расположены в вентро-латеральной зоне раковины и вблизи умбонального шва. Боковые лопасти появляются на ранних стадиях развития в районе умбонального шва (на наружной его стороне), но перемещаются на бо-
7 3
новые стороны в течения онтогенетического развития и во взрослом состоянии располагаются вблизи середины боковых сторон раковины.
Формула лопастной линии: VU: D.
Видовой состав и геологическое распространение. Кроме Goniatites ruppachensis, известного из нижней части среднего девона Германии, Марокко и Рудного Алтая, в настоящее время к роду Werneroceras могут быть отнесены Anarcestes karpinskyi Holzapfel из среднего девона Германии, Goniatites plebeiformis Hall из среднего девона штата Нью-Йорк США, Goniatites wabashensis Kindle из верхнего девона штата Индиана США, возможно Goniatites crispus Barrande из верхней части нижнего девона Богемии и Ammonites subnautilinus Buch из среднего девона Германии. К этому же роду относятся Werneroceras socolicum sp. nov. и W. altaicum sp. nov. из среднедевонских отложений (сокольная свита) Лениногорского рудного поля, описанные ниже.
Werneroceras ruppachense (Kayser)
Goniatites subnautilinus: Roemer, 1850, стр. 19, табл. HI, фиг. 28, a, 6, c.
Goniatites subnautilinus чат. vittiger: G. u. F. Sandberger, 1850—1856, стр. 114, табл. XI, фиг. 3, 3, a — / (non vittiger Phillips); Kayser, 1878, стр. 57; Branco, 1880, табл. VII, фиг. 1.
Goniatites ruppachensis: Kayser, 1879, стр. 304.
Goniatites vittatus: Kayser, 1884, стр. 46, табл. V, фиг. 11—17.
Anarcestes ruppachensis: Food and Crick, 1897, стр. 45.
Anarcestes vittiger: Freeh, 1913, стр. 15; Wedekind, 1918, стр. 109, табл. XV, фиг. 3.
Anarcestes (Werneroceras) subumbonale: Wedekind, 1918, стр. 108, табл. XV, фиг. 6. Werneroceras ruppachense: Schindewolf, 1933, стр. 96.
Голотип происходит из нижней части среднедевонских отложений Германии.
Форма. Раковина во взрослом состоянии толстодискоидальная, с наибольшей шириной вблизи умбонального края, с широкими, выпуклыми, сходящимися наружу боковыми и умеренно широкой уплощенной, слегка выпуклой вентральной сторонами. Вентральный край отчетливый, несколько угловатый. Поперечное сечение оборота во взрослом состоянии субэллиптическое, с высотой больше ширины. Высота последнего оборота около 112 диаметра раковины. Умбо довольно узкое, глубокое. Последний оборот почти полностью облекает предыдущий. Ввиду плохой сохранности экземпляров произвести измерения параметров раковин не представляется возможным.
Скульптура. На имеющихся в нашем распоряжении экземплярах раковинный слой не сохранился. Ядро совершенно гладкое. Раковина была снабжена, вероятно, лишь струйками нарастания.
Лопастная линия. Наружная часть лопастной линии состоит из узкой V-образной вентральной лопасти, округленного умеренно широкого первого бокового седла, довольно широкой округленной в основании боковой лопасти, которая у небольших экземпляров располагается субум-бонально, а у взрослых перемещается на середину боковой стороны, и умеренно широкого и высокого второго бокового седла, округленная вершина которого располагается в районе умбонального шва.
Сходство и отличие. Этот вид отличается от Werneroceras (?) crispus(Barrande) из верхов нижнего(?) девона Богемии более узким умбо я большим возрастанием оборотов в высоту. От W. karpinsKyi (Holzapfel) и W. plebeiforme (Hall), описанных из среднедевонских отложений Германии и США, он легко отличим как по более инволютной раковине, так и по более выраженным боковым лопастям его лопастной линии.Отличия 74
его от описанных ниже ИЛ s ос о li cum sp. nov. и W. altaicum sp. nov. указаны при описании последних.
Геологический возраст и распространение. Встречается в низах среднего девона Западной Германии и Рудного Алтая.
Местонахождение. Десять неполной сохранности образцов найдены в известковистых алевролитах на северо-западном склоне горы •Сокольной, в низах сокольной свиты; один экземпляр найден в отвалах Терентьевской штольни, в светло-серых известняках крюковской свиты.
W erneroceras altaicum sp. nov.
Табл. I, фиг. 5
Голотип — ПИН № 773/2; Рудный Алтай, Лениногорское рудное поле, северо-западный склон горы Бахрушиной; низы сокольной свиты эйфельского яруса.
Форма. Раковина во взрослом состоянии толстодискоидальная, приплюснутая с боков, сильно инволютная, с наибольшей шириной вблизи умбо, с выпуклыми, несколько уплощенными в области умбо боковыми сторонами, широкой, слабо округленной вентральной стороной и сильно вогнутой дорсальной. Вентральный край отчетливый, несколько угловатый. Поперечное сечение оборота серповидное. Умбо относительно узкое и глубокое, ограниченное невысокими округленными умбональными стенками. Последний оборот полностью облекает предыдущий. Жилая камера составляет, по крайней мере, половину оборота.
Размеры: Д В Ш Ду в/д Ш/Д Ду/Д
№ 773/2 j 55,3 24,7 27,0 11,0 0,45 0,49 0,20
1 45,3 20,0 25,0 10,4 0,44 0,55 0,23
Скульптура. Поверхность раковины гладкая, несущая на себе лишь тонкие сглаженные струйки нарастания, образующие широкие синусы на боковых сторонах, узкие угловатые выступы на вентральном крае и более узкие синусы на вентральной стороне раковины. Струйки нарастания пересечены очень тонкими, перпендикулярными к ним штрихами.
Лопастная линия. На рис. 13 изображена наружная часть лопастной линии нового вида. Вентральная лопасть глубокая, очень узкая в нижней части и несколько расширяющаяся вверху; в целом дудкообразная. Первое боковое седло широкое и высокое, несколько асимметричное, трапециевидное, очень слабо округленное вверху, с очень крутой наружной и более пологой внутренней сторонами. Боковая лопасть широкая, довольно глубокая, округленная в основании, расположенная на середине боковой стороны раковины, слегка асимметричная. Второе боковое седло широкое, сравнительно невысокое, широко округленное; вершина его располагается на умбональном крае раковины, а внутренняя часть переходит за умбональный шов. Внутренняя часть лопастной линии неизвестна.
Сходство и отличие. Описанный вид отличается от типа рода, к которому он наиболее близок по форме раковины и характеру лопастной линии, узкой, глубокой вентральной лопастью, значительно большей шириной первых боковых седел и более узкими боковыми лопастями. Он имеет также нескблько более толстодискоидальную, несколько
75
приплюснутую с боков раковину. От остальных видов он легко отличим по характеру лопастной линии.
Геологический возраст и распространен е. Редко в эйфельском ярусе Рудного Алтая.
Рис. 13. Лопастные линии W erneroceras altaicum sp. nov.:
a—№ 773/2 при Ш=25,0 и В=20,0 мм (Х1,5); Ь — то же при 111=27,0 и В=24 мм (XI,5). Рудный Алтай. Сонольная свита низов среднего девона.
Местонахождение. Один экземпляр найден в известковистых алевролитах на северо-западном склоне горы Бахрушиной, в низах сокольной свиты.
Werneroceras socolicum sp. nov.
Табл. I, фиг. 4
Голотип —ПИН № 774/701; Рудный Алтай, Лениногорское рудное поле, северо-западный склон горы Сокольной; низы сокольной свиты эйфельского яруса.
Форма. Раковина толстодискоидальная, приближающаяся к эллипсоидальной, сильно инволютная, с наибольшей шириной вблизи умбонального края, со слабо выпуклыми, сходящимися наружу боковыми и умеренно широкой, округленной вентральной сторонами. Поперечное сечение во взрослом состоянии субтреугольное, несколько округленное. Вентральный край сглаженный на ранних оборотах и несколько более отчетливый во взрослом состоянии. Умбональный край отчетливый, но не резкий. Умбо довольно узкое, ступенчатое, ограниченное округленными умбо-нальными стенками. Последний оборот почти полностью облекает предыдущий.
Размеры: д В Ш Ду В/Д Ш/Д Ду/Д
v0 774/701 Г 37,4 18,2 20,3 6,6 0,49 0,54 0,18
tu / • т/ 9 vl 1 30,9 14,1 18,0 6,2 0,46 0,58 0,20
№ 774/702 16,0 6,3 9,2 (?) 3,6 0,40 0,58(?) 0,22
Скульптура. На имеющихся в нашей коллекции экземплярах раковинный слой не сохранился. Ядро совершенно гладкое.
76
Лопастная линия (рис. 14). Наружная часть лопастной линии состоит из узкой, воронкообразной вентральной лопасти; очень широкого широко округленного, асимметричного первого бокового седла с крутой наружной и пологой внутренней сторонами; широкой, умеренно глубокой округленной боковой лопасти и довольно высокого и широкого второго бокового седла, округленная вершина которого располагается на умбональном крае, а внутренняя часть переходит за умбональный шов.
Рис. 14. Лопастная линия Werneroceras socolicum sp.nov. (хЗ); голотип № 774/701 при Ш=1о,0 и В=13,6 мм. Рудный Алтай, Лениногорский район. Низы среднего девона, сокольная свита.
Сходство и отличие. Описанный вид отличается от типичного вида, с которым он имеет некоторое внешнее сходство, характером лопастной линии, имеющей очень широкое, сравнительно низкое первое боковое седло. Кроме того, наш вид имеет более низкие и менее быстро возрастающие в высоту обороты и, возможно, более узкое умбо. От других видов новый вид легко отличим по форме раковины и лопастной линии.
Геологический возраст и распространение. Редко в эйфельском ярусе Рудного Алтая.
Местонахождение. Три экземпляра найдены в известковистых алевролитах на северо-западном склоне горы Сокольной в низах сокольной свиты.
Род Sellanarcestes Schindewolf, 1933
Anarcestes: Freeh, 1913, стр. 14 (pars); Wedekind, 1918, стр. 106 (pars).
Sellanarcestes: Schindewolf, 1933, стр. 98; .Miller, 1938, стр. 56.
Тип рода — Goniatites wenkenbachi Kayser, 1884, стр. 42, табл. 4, фиг. 1—6. Нижняя часть среднего девона Германии.
Диагноз. Раковина толстодискоидальная, образованная низкими равномерно округленными с боков и с вентральной стороны оборотами; без внешних борозд. Умбо широкое. Боковая лопасть умеренно широкая, расположенная во взрослом состоянии латерально. Дорсальная лопасть разделена срединным седлом.
Формула лопастной линии: VU : DiDi.
Видовой состав и геологическое распространение. Кроме типичного вида, известного из верхов нижнего и низов среднего девона Германии, к этому же роду, возможно, относится S. (?) wenkenbachiformis sp. nov. из низов среднего девона Рудного Алтая, описанный ниже.
Sellanarcestes (?) wenkenbachifor mis sp. nov.
Табл. I, фиг. 6
Голотип — ПИН № 773/Г, Рудный Алтай, Лениногорское рудное поле, отвалы Терентьевской штольни; крюковская свита эйфельского яруса.
77
Форма. Раковина дискоидальная, среднеинволютная, с наибольшей шириной вблизи умбонального края, образованная довольно сильно объемлющими, нависающими над предыдущими оборотами. Боковые стороны умеренно выпуклые, сходящиеся наружу и постепенно переходящие в сравнительно неширокую округленную вентральную сторону и несколько подгибающиеся внутрь к умбональному краю. Умбональный край отчетливый, угловатый. Умбо довольно широкое и глубокое, ступенчатое^ ограниченное вогнутыми умбональными стенками, нависающими над предыдущими оборотами. Последний оборот облекает около 4/5 предыдущего.
Размеры: д В Ш Ду в/д Ш/Д Ду/Д
№ 773/1 1 72,0 34,0 31,0 18,0 0,47 0,43 0,25
1 56,2 22,4 27,0(?) 16,0 0,40 0,48(?) 0,28
Скульптура. Раковинный слой не сохранился. Ядро гладкое. Раковина была покрыта, по-видимому, лишь струйками нарастания.
Лопастная линия. Наружная часть лопастной линии изоб-
д
Рис. 15. Лопастная линия Sellanarcestes (?) wenKenbachiformis sp. nov. .(X2); голотип № 773/1 при В = 18,5мм. Рудный Алтай, Лениногорский район. Крюковская свита низов среднего девона.
ражена на рис. 15. Вентральная лопасть V-образная, довольно глубокая, но не широкая. Первое боковое седло широкое, асимметричное, слаба округленное в верхней части и более круто опускающееся к смежным лопастям; в целом трапециевидное. Боковая лопасть широкая, умеренно глубокая, округленная в основании и плавно переходящая в широкое и высокое равномерно округленное второе боковое седло, вершина которого располагается в области умбонального края, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Описанный вид имеет по форме раковины и очертанию внешней части лопастной линии некоторое сходство с S. wenkenbachi (Kayser), отличаясь от него, однако, более инволютной раковиной. Некоторое отличие наблюдается и в очертании лопастной линии, имеющей у нашего вида несколько менее глубокую боковую лопасть. В силу вышеизложенного я называю описанный вид Sellanarcestes (?) wenkenbachi formis sp. nov., ставя его родовое название под вопросом, так как внутренняя часть лопастной линии (имеющая у Sellanarcestes срединное седло дорсальной лопасти) у нашего экземпляра неизвестна.
Геологический возраст и распространение. Редко в нижней части эйфельского яруса Рудного Алтая.
Местонахождение. Единственный экземпляр найден в светло-серых известняках отвалов Терентьевской штольни Лениногорского рудного поля. Крюковская свита низов эйфельского яруса.
ПОДСЕМЕЙСТВО PINACITINAE SCNINDEWOLF, 1933
Диагноз. Раковина в большинстве случаев узкодискоидальная, реже — дискоидальная, обороты обычно довольно высокие, умбо узкое, и во взрослом состоянии лопастные линии образуют в дополнение к вент
78
ральной, дорсальной и первой боковой лопастям, еще одну лопасть, обычно неглубокую, в районе умбонального шва.
Формула лопастной линии: VUU1 : D.
Общие замечания. Подсемейство включает следующие роды: Parodicerellum, Wedekindella, Foordites и Pinacites. Отнесение к этому подсемейству Шиндевольфом и другими исследователями рода Maenioceras вряд ли обоснованно, так как у этого рода наблюдается резко отличное от других представителей семейства образование боковой лопасти при вершине первого бокового седла. Род Maenioceras должен, на наш взгляд, быть поэтому исключен как из состава подсемейства Pinacitinae, так и семейства Anarcestidae вообще и выделен в отдельное семейство, родственное Tornoceratidae. Краткая характеристика родов, входящих в подсемейство Pinacitinae, такова:
1. Parodicerellum Strand, 1929 (= Parodiceras Wedekind, 1913, non Hyatl, 1884; Holzapfeloceras Miller, 1932). Раковина более или менее толстодискоидальная, с низко- или главным образом высокоапертурными оборотами, с выпуклыми боками и округленной вентральной стороной, без отчетливых внешних борозд. Лопастная линия с формулой VUU1 : D, без внутреннего седла. Тип рода — Tornoceras convolutum Holzapfel (1895) из живетского яруса Германии. К этому же роду относятся известные из среднего девона Западной Европы: Parodicerellum circumflexiferum (Sandberger), Р. beushauseni (Wedekind), P. inversum (Wedekind), P. angulato-striatus (Kayser), P. djemeli (Petter) и P. numismalis (Petter) из среднего девона С-3 Африки.
2. Wedekindella Schindewolf, 1928. Этот род отличается от Parodicerellum наличием внутреннего срединного седла. Тип рода—Goniatites retrorsus var. brilonensis Kayser (1872) из верхов среднего девона Германии. К этому же роду относятся, вероятно, W. psittacinum (Whidborne) и W. clarkei (Holzapfel), известные из тех же отложений.
3. Foordites Wedekind, 1918. Раковина дискоидальная, сильно инво-лютная, с уплощенными боковыми и вентральной сторонами и отчетливыми внешними бороздами, ограничивающими последнюю. Лопастная линия, имеющая формулу VUU1 : D, без внутреннего срединного седла. Тип рода — Aphyllites occultus var. platypleura Freeh (1902). Нижняя часть среднего девона Германии. К этому же роду относятся F. annulatus (Maurer) и F. occultus (Barrande) из тех же отложений.
4. Pinacites Mojsisovics, 1882. Раковина линзовидная, уплощенная с боков, с широкими слабо выпуклыми боковыми и приостренной вентральной сторонами. Умбо более или менее узкое. Парные внешние борозды во взрослом состоянии отсутствуют. Лопастная линия, имеющая формулу VUU1 : D, без внутреннего срединного седла. Тип рода — Goniatites jugleri Roemer (Roemer, 1843). Низы среднего девона Западной Германии. Кроме типичного вида, известного также из средне девонских отложений Богемии и Северной Африки (Марокко и Алжир), к данному роду относится Р. irideum (Freeh) из среднего девона Германии. Р. emaliatus (Barrande, 1865—1867), который был указан Мойсисовичем (Mojsisovics, 1882, стр. 181) в качестве единственного представителя описанного им рода Pinacites и затем установлен Хайэттом как тип этого рода, по-видимому, является полностью идентичным установленному Ремером в 1843 г. Goniatites jugleri Roemer = Pinacites jugleri (Roemer).
СЕМЕЙСТВО TORNOCERATIDAE ARTHABER, 1911
Диагноз. Раковина дискоидальная от широко-до узкоумбональ-ной, часто полностью инволютная. Струйки нарастания образуют синусы на боковых и вентральной сторонах раковины. Лопастная линия образует
79
маленькую неразделенную вентральную лопасть, одну или две боковые лопасти, возникающие из вершины первичного бокового седла, первичную умбональную и дорсальную лопасти; у двух родов (Posttornoceras и Lobotornoceras), кроме того, между дорсальной и умбональной лопастями развивается одна внутренняя боковая лопасть.
Ревизия семейства. Представители семейства Тогпосега-tidae географически широко распространены п известны из девонских отложений Европы, Северной Америки, Азии, Африки и Австралии. Стратиграфически семейство ограничено средним и верхним девоном, отсутствуя лишь в самых низах среднего девона и наиболее верхних частях (верхи Oberdevonstufe VI) верхнего девона. Предками семейства, по всей вероятности, были наиболее инволютные представители ранних среднедевонских анарцестид,обладавшие простой лопастной линией с умбональ-но расположенной первичной умбональной лопастью.
В составе семейства Tornoceratidae в настоящее время известны роды Tornoceras, Pseudoclymenia, Lobotornoceras и Posttornoceras. В пределах первого рода, кроме того, выделяются подроды Tornoceras s. s., Paro-doceras, Protornoceras, Aulatornoceras и Polonoceras. Краткая характеристика указанных родов такова:
1. Tornoceras Hyatt, 1884. Описание этого рода дано ниже.
2. Pseudoclymenia Freeh, 1897. Раковина слабоинволютная, дискои-дальная, уплощенная с боков и округленная с вентральной стороны. Лопастная линия образует одну очень слабо выраженную маленькую и вторую приостренную топоровидную довольно большую боковые лопасти с каждой стороны вентральной лопасти. Тип рода — Clymenia pseudogoniatites Sandberger (1853) из верхнего девона Германии. К этому же роду относятся Pseudoclymenia fundifera (Perna), Р. applanata Schindewolf, P. australis Delepine, P. dillensis (Drevermann), P. orensis Na-livkina.
3. Lobotornoceras Schindewolf, 1936. Раковина дискоидальная, подобная Tornoceras. Отличается от Tornoceras тем, что внутренняя часть лопастной линии образует внутренние боковые лопасти (по одной с каждой стороны), отсутствующие у Tornoceras. Тип рода — Goniatites ausavensis Steininger (1853) из верхнего девона Германии. К этому же роду относится, кроме того, Tornoceras bilobatum Wedekind и, вероятно, Т. escoti Freeh.
4. Posttornoceras Wedekind, 1910. Раковина дискоидальная, полностью инволютная, с закрытым умбо, как у типичного Tornoceras, но как наружная, так и внутренняя части лопастной линии образуют на одну лопасть больше, чем внутренняя и наружная части лопастной линии Tornoceras. Тип рода — Posttornoceras balvei Wedekind (1910) из верхнего девона Германии. К этому же роду относится Р. contiguum Sobolew из верхнего девона Польши и Мугоджар.
Род Tornoceras Hyatt, 1884
Tornoceras: Hyatt, 1884а, стр. 320; Holzapfel, 1882, стр. 14 (pars); 1895, стр. 80; Foord and Crick, 1897, стр. 91 (pars); Clarke,1899, стр. 109; Freeh, 1902, стр. 45 (pars); 1913, стр. 15 (pars); Wedekind, 1908, стр. 577 (pars); Schindewolf, 1922, стр. 188; Miller A. K., 1938, стр. 140.
Parodiceras: Hyatt, 1884a, стр. 219 (pars).
Epitornoceras: Freeh, 1902, стр. 51—53; 1913, стр. 19.
Protornoceras: Dybczynski, 1913, стр. 512—513.
Polonoceras: Dybczynski, 1913, стр. 519—520.
Pernoceras: Schindewolf, 1922, стр. 188.
Aulatornoceras: Schindewolf, 1922, стр. 188.
Тип рода — Goniatites uniangularis Conrad, 1842, стр. 268, табл. 16, фиг. 4. Формация Гамильтон штата Нью-Йорк США.
80
Диагноз. Раковина дискоидальная или чечевицеобразная с уплощенными с боков, вогнутыми -со спинной стороны и округленными, угловатыми или уплощенными с вентральной стороны оборотами. Умбо закрытое или открытое, воронкообразное. Струйки нарастания образуют синусы на боковых и вентральной сторонах и выступы на вентро-латеральных и дорсо-латеральных частях раковины. Наружная часть лопастной линии состоит из маленькой неразделенной вентральной лопасти, вентро-латерального седла, значительно более широкой асимметричноокругленной боковой лопасти, широкого бокового седла и узко округленной или угловатой умбональной лопасти. Внутренняя часть лопастной линии состоит из выдающейся дорсальной лопасти и широко-округленного асимметричного седла, которое переходит в умбональную лопасть.
Формула лопастной линии: VLU: D.
Общие замечания. Род Tornoceras был установлен в 1884 г. Хайэттом (1884а, стр. 320—321), который отнес к нему несколько европейских и американских видов и установил в качестве типа рода Goniatites uniangularis Conrad из девонских отложений штата Нью-Йорк. Последующие определения Ведекиндом (1913, стр. 83, сноска) и Шиндевольфом (1922, стр. 188) Goniatites simplex Buch в качестве типа рода не имеют значения до тех пор, пока не будет доказано, что G. uniangularis является синонимом G. simplex,
К роду Tornoceras в разное время относились довольно разнообразные формы, многие из которых впоследствии были использованы для выделения различных родов и подродов.
В настоящее время род Tornoceras разделяется на пять подродов: Tornoceras s. s., Parodoceras, Protornoceras, Polonoceras и Aulatornoceras, которые могут быть различимы при помощи легко узнаваемых признаков. Краткая характеристика указанных подродов такова:
1. Tornoceras s. s. Hyatt, 1884. Умбо закрытое (или почти закрытое), вентро-латеральные зоны раковины округлены и не отмечены продольными желобками. Лопастная линия образует высокие выдающиеся седла на вентро-латеральных участках раковины. Тип подрода Goniatites uniangularis Conrad (1842), описанный из девонских отложений штата Нью-Йорк США. К этому же подроду могут быть отнесены известные из средне- и верхнедевонских отложений Западной Европы, отчасти Урала, Тимана и других областей: Tornoceras (Tornoceras) bertrandi Freeh, T. (T.) verae Freeh (средний девон), Т. (Т.) frechi Wedekind, Т. (Т.) pompecku Wedekind, Т.(Т.) holzapfeli Freeh, Т.(Т.) simplex (Buch), Т.(Т.) stachei Freeh, T.(T.) mithracoides Freeh, T.(T.) acutum Freeh, T.(T.) curvidorsatum Sobolew. Из североамериканских видов к данному подроду могут быть отнесены известные из верхнего девона: Т.(Т.) crebriseptum Raymond, Т.(Т.) edwin-halli Clarke, Т.(Т.) iowaenseMi\\er, Т.(Т.) mithrax (Hall), Т.(Т.) ре-racutum (Hall), Т.(Т.) rhysum Clarke и среднедевонские Т.(Т.) whitfieldi Miller, Т.(Т.) buttsi Miller.
2. Parodoceras Hyatt, 1884. Умбо закрытое, вентро-латеральные зоны раковины округлены и не отмечены продольными желобками; лопастная линия образует очень низкие, едва различимые седла на вентро-латеральных зонах раковины. Тип подрода — Goniatites discoideus Hall (Hall, 1860), описанный из среднедевонских отложений штата Нью-Йорк США. Других видов не известно.
3. ProtornocerasDybczynski, 1913 ( = PernocerasSchindewolf, 1922). Умбо открытое, воронкообразное, вентро-латеральные зоны раковины округлены и не отмечены продольными желобками; лопастная линия образует выдающиеся седла на вентро-латеральных зонах раковины. Тип подрода— Tornoceras (Protornoceras) polonicum Dybczynski (1913), описанный из верхнего девона Польши. К этому же подроду относятся Т.(Р.) latum 6 Б. И. Богословский см
Dybczynski, T.(P.) mirabile Dybczynski, T. (P.) kochi (Wedekind), T. (P.) г о tundatum Perna, T. (P.) dorsatum (Wedekind), T.(P.) evolutum Sobolew, T. (P.) sinuvaricatum Sobolew, T. (P.) planilobum Sobolew, T. (P.) sim-plijicatum Sobolew и др.
4. Aulatornoceras Schindewolf, 1922. Умбо различное, открытое или закрытое, вентро-латеральные зоны раковины несут продольные желобки; лопастная линия образует высокие выдающиеся седла на вентро-латеральных зонах раковины. Тип подрода — Goniatites auris Quenstedt (1846—1849), описанный из верхнего девона Германии. К этому же подроду могут быть отнесены верхнедевонские 7\ (A.) paucistriatum (Archiac et Verneuil), T. (A.) bicostatum (Hall), T. (A.) cinctum (Keyserling), T. (A.) constrictum (Steinmann), T. (A.) eifelensis (Steinmann), T. (A.) subundu-latum Freeh, T. (A.) loeschmanni Freeh, Z.(A.) undulatum (Sandberger) и среднедевонский T.{A.)westfalicum Holzapfel.
5. Polonoceras Dybczynski, 1913. Умбо закрытое; вентро-латеральные зоны раковины угловатые и не отмечены продольными желобками; лопастная линия образует высокие выдающиеся седла на вентро-латеральных зонах раковины. Тип подрода — Polonoceras planum Dybczynski (1913) из верхнего девона Польши. К этому же подроду относятся Т. (Р.) dor sop lanum Sobolew, T. (P.) planidorsatum (Munster), T. (P.) weissi Wedekind.
Возможно, что подроды Protornoceras, Aulatornoceras и Polonoceras^ достаточно отличные от Tornoceras s. s., должны рассматриваться в качестве самостоятельных родов, но, не имея в своем распоряжении достаточного количества материала, я в настоящее время воздерживаюсь от их выделения и оставляю в составе рода Tornoceras.
Tornoceras (Tornoceras) simplex (Buch)
Табл. II, фиг. 1, 2
Ammonites simplex'. Buch, 1832, стр. 42, табл. 2, фиг. 8.
Goniatites ovatus: Munster, 1832, стр. 16, табл. 4, фиг. 1.
Goniatites retrorsus: Keyserling, 1844, стр. 230, табл. А., фиг. 5; 1846, стр. 278, табл. 12, фиг. 5.
Goniatites retrorsus var. typus: Sandberger G., Sandberger F., 1850—1856, стр. 100, табл. 10, фиг. 14—16.
Goniatites simplex: Kayser, 1873, стр. 620, табл. 19, фиг. 6; Beyrich, 1884, стр. 212.
Goniatites (Tornoceras) simplex: Чернышев, 1887, стр. 23, табл. I, фиг. 22; табл. II, фиг. 9.
Goniatites hughesi: Whidborne, 1890, стр. 69, табл. 6, фиг. 1—3.
Tornoceras simplex: Holzapfel, 1895, стр. 95, табл. 6, фиг. И; табл. 7, фиг. 9; Голь-дапфель, 1899, стр. 14, табл. 8, фиг. 1—9; Freeh, 1902, стр. 16; Wedekind, 1918, стр. 135, табл. XVI, фиг. 12; Янишевский, 1926, стр. 83, табл. II, фиг. 14, 16, 17; Наливкина, 1947, стр. 167, табл. XLII, фиг. 6, 7, 8, рис. 21 в тексте.
Tornoceras simplex Buch mut. ovata: Freeh, 1902, стр. 47, табл. 2, фиг. 21, a, b. Tornoceras haugi: Freeh, 1902, стр. 47, табл. 2, фиг. 20, a, d; 1913, стр. 17.
Голотип происходит из девонских отложений Германии.
Форма. Раковина дискоидальная, полностью инволютная, с наибольшей шириной вблизи умбо, с широкими уплощенными, слабо выпуклыми, сходящимися наружу боковыми и сравнительно узкой, округленной вентральной сторонами.
Размеры: Д В Ш в/д ш/д
№ 774/257 18,0 10,8 80, 0,60 0,44
№ 774/258 10,0 6,0 50, 0,60 0,50
Скульптура. Поверхность раковины гладкая, несущая на себе лишь тонкие струйки нарастания.
Лопастная линия (рис. 16). Наружная часть лопастной ли
82
нии состоит из узкой V-образной вентральной лопасти, относительно широкого округленного при вершине внешнего седла, широкой и довольно глубокой, несколько асимметричной боковой лопасти и широкого и высокого асимметричного с крутой слегка вогнутой наружной и покатой выпуклой внутренней сторонами бокового седла, переходящего в умбо-нальную лопасть. Внутренняя часть лопастной линии для описываемых экземпляров неизвестна.
Сходство и отличие. Этот вид широко распространен в девонских отложениях Тимана и, особенно, Западной Европы, где в составе
Рис. 16. Лопастные линии Tornoceras (Tornoceras) simplex (Buch):
a — № 774/257 при B=7,5 и Ш=6,5 мм (Х6); Ъ — № 774/258 при В=5,5 и Ш=5,0 мм (Х6). Рудный Алтай, район Николаевского месторождения. Франский ярус, пониатитовый горизонт.
его различными исследователями выделяются различные «вариететы» и «мутации», отличающиеся друг от друга формой раковины и лопастной линией. Стратиграфически он прослеживается от верхней части среднего девона до хейлоцеровых слоев верхнего девона. Возможно, что отдельные уклоняющиеся «разновидности» его заслуживают выделения в отдельные виды или подвиды или, наоборот, некоторые из известных близких к нему видов должны быть объединены, но, не располагая достаточным материалом, я воздерживаюсь как от первого, так и от второго. Имеющиеся в моем распоряжении алтайские экземпляры по форме раковины и лопастной линии имеют наибольшее сходство с Т, simplex mut. ovata (Munster), изображенным Фрехом (1902, стр. 47, табл. III, фиг. 21, а, Ь), и весьма близки к тиманским представителям Т. simplex (Buch), если не аналогичны им, и поэтому, обладая ограниченным количеством материала, я рассматриваю их как относящиеся к Tornoceras (Tornoceras) simplex (Buch). От близкого по форме типа рода Tornoceras — Т. (Т.) uniangulare (Conrad) наши экземпляры отличаются характером лопастной линии, имеющей более высокое и широкое внешнее седло, более резко выраженную боковую лопасть и асимметричное боковое седло.
Геологический возраст и распространение. Известен из среднего и верхнего девона Западной Европы, Урала, Тимана, Рудного Алтая и других областей.
Местонахождение. Три экземпляра найдены в 3 км юго-восточнее поселка Николаевского рудника в известняковых линзах гониатитового горизонта франского яруса.
6*
83
СЕМЕЙСТВО GEPHUROCERATIDAE FRECH, 1902
Диагноз. Раковина изменяется от дискоидальной, слабо инво-лютной до линзовидной, полностью инволютной. Поверхность раковины гладкая, несущая лишь тонкие струйки нарастания, преимущественно двояковыпуклые. Вентральная лопасть широкая трехраздельная, с коротким воронкообразным срединным и обычно длинными приостренными боковыми зубцами. Дорсальная лопасть сравнительно небольшая, клиновидная. Общее количество боковых наружных и внутренних лопастей изменяется от одной до четырех (с каждой стороны) и более.
Филогения. Филогения семейства Gephuroceratidae, особенно вопрос о предковом роде его, в настоящее время далеки от ясности. Шиндевольф (1938) рассматривает в качестве такого рода Archoceras из семейства Anarcestidae, однако указанный род известен из более молодых фамен-ских («Cheiloceras-Stuie») отложений и поэтому не может рассматриваться в качестве предка развивавшегося в течение франского века семейства Gephuroceratidae. Это семейство, зародившееся в начале верхнедевонской эпохи, должно было иметь в качестве своего предка слабо инволютного, обладавшего простой лопастной линией аммонита. Таким возможным предком могли, действительно, явиться некоторые, наиболее эволютные представители семейства Anarcestidae, обладавшие дискоидальной раковиной, простой лопастной линией, вентральная лопасть которой была не разделенной на части. В процессе эволюции такая простая лопасть испытала трехчленное деление, что одновременно с образованием дополнительных умбональных лопастей привело к формированию лопастной линии Gephuroceratidae.
Вопрос о конкретных филогенетических связях внутри семейства еще не вполне ясен и требует для своего решения детального онто-филогене-тического изучения всех представителей гефуроцератид. Нет сомнения в том, что развитие этого обширного семейства (в составе которого, вероятно, будут установлены и выделены новые роды) шло различными путями. Эволюция семейства протекала в общем в направлении Ponticeras-^ Uchtites —> Manticoceras —> Koenenites —> Honinghausia —> Timanites. Помимо этого основного направления, как, вероятно, и внутри его, существовали боковые ветви. Такими боковыми ветвями были, вероятно, Ponticeras Probeloceras и Manticoceras —> Komioceras. От Gephuroceratidae уже в первой половине франского века происходит семейство Beloceratidae.
Ревизия семейства. Семейство Gephuroceratidae включает роды Ponticeras, Probeloceras, Uchtites, Manticoceras, Honinghausia, Koenenites, Timanites, Komioceras и, вероятно, Crickites. Систематика его может основываться на различиях в форме раковины, скульптуры и лопастной линии. В ходе эволюции раковина в различных филогенетических линиях изменялась весьма существенно от дискоидальной, слабо инволютной до линзовидной, полностью инволютной. Лопастная линия, сохраняя в общем форму лопастей, изменялась, однако, в отношении их количества. Достаточно отметить, что общее число лопастей вокруг извилины возрастало от 4 до 10 и более. Скульптура раковины, состоящая из тонких струек нарастания, испытывала гораздо меньшие изменения. Струйки нарастания Ponticeras, Probeloceras, Uchtites, Manticoceras, Honinghausia, Koenenites, Timanites и Komioceras образуют синусы на боковой и вентральной сторонах раковины; у Crickites же синус развит лишь на вентральной стороне. Различное сочетание признаков позволяет выделить указанные выше роды, краткая характеристика которых такова:
1) Ponticeras Matern, 1929. Описание этого рода дано ниже.
2) Probeloceras Clarke, 1899. Описание этого рода дано ниже.
£4
Рис. 17. Лопастные линии представителей семейства Gephuroceratidaet a—Probeloceras orientale Bogoslovsky, sp. ncv. (X6). Франский ярус. Рудный Алтай; b—Ponticeras tschernyschewi (Holzapfel) (X2). Франский ярус. Тиман (Гольц-апфель, 1899); с —Uchtites syrianicus (Holzapfel) (Х2). Франский ярус. Тиман (Гольцапфель, 1899); d — Manticoceras sinuosum (Hall) (XI). Франский ярус. Рудный Алтай; е —Koenenites lamellosus (Sandberger) (Х2). Франский ярус. Зап. Германия (Г. и Ф. Зандбергеры, 1850—1856); / — Honinghausia uchlensis
G. Ljaschenko(Xl )• Франский ярус. Тиман.; д — Timanites Keyserlingi Miller (xl). Франский ярус. Тиман (Гольцапфель, 1899); h —Komioceras stucnenbergi (Holzapfel) (ХЗ). Франский ярус. Тиман.
85
3) Uchtites. gen. nov. Описание этого рода дано ниже.
4) Manticoceras Hyatt, 1884. Описание этого рода дано ниже.
5) Koenenites Wedekind, 1913. Раковина дискоидальная, слабо инво-лютная, с уплощенными, слабо выпуклыми боковыми и округленной вентральной сторонами. Струйки нарастания образуют синусы на боковой и вентральной сторонах раковины. Вентральная лопасть широкая, трехраздельная, с довольно узкими, глубокими зубцами, разделенными высокими округленными при вершинах седлами. Наружных боковых лопастей две. Внутренняя боковая лопасть одна. Дорсальная лопасть клиновидная. Тип рода — Goniatites lamellosus Sandberger (1850—1856) из нижней части верхнего девона Германии. К этому роду относятся, кроме того, Koenenites sublamellosus (Sandberger) из тех же отложений Германии и Koenenites cooperi Miller из формации Антрим штата Мичиган США. В 1931 г. Матерн (Matern, 19316, стр. 75—76) отнес к этому роду Koenenites galeatus Matern из низов верхнего девона Германии, но этот вид, как полагает А. К. Миллер (1938, стр. 126), вероятно, должен быть причислен к Honinghausia.
6) Honinghausia Giirich, 1896. Этот род в общем сходен с типом рода Koenenites и, может быть, представляет собой его синоним, как считают, например, Ведекинд и Фрех. Однако вентральная сторона его раковины угловата и в настоящее время лучше оставить оба родовых наименования. Тип рода — Goniatites hoeninghausii Archiac et Verneuil (1842) из низов верхнего девона Германии.
7) Timanites Mojsisovics, 1882. Раковина линзовидная, полностью инволютная, со слабо выпуклыми, сходящимися кверху боковыми и угловатой или килеватой вентральной сторонами. Поверхность раковины гладкая, покрытая тонкими струйками нарастания, образующими синусы на боковых и вентральной сторонах раковины. Лопастная линия состоит в общем из десяти лопастей. Вентральная лопасть широкая, состоит из трех зубцов, разделенных широкими и высокими седлами. Дорсальная лопасть клиновидная, узкая и глубокая; боковые лопасти простые, приост-ренные на конце. Боковых наружных лопастей две, третья лопасть находится на умбональном шве. Внутренняя боковая лопасть одна. Тип рода — Timanites key serlingi Miller (1938) ( = Goniatites acutus^eyserXhig (1844) из дома-никового горизонта Южного Тимана. К этому роду относится, кроме того, Т. occidentalis Miller et Warren из формации Минневанка штата Альберта США.
8) Komioceras gen. nov. Описание этого рода дано ниже.
9) Crickites Wedekind, 1913. Этот род по форме раковины и очертанию лопастной линии сходен с Manticoceras и отличается от последнего лишь характером струек нарастания, образующих синусы лишь на вентральной стороне раковины при отсутствии таковых на боковых сторонах. А. К. Миллер указывает, что по этому признаку Crickites, возможно, может быть отнесен к Prolobitacea (?)или, возможно, должен быть выделен в самостоятельное подсемейство из семейства Gephuroceratidae, как считал Ведекинд (1913, стр. 70), но он соединяет его с Manticoceras, так как имеется прямое сходство его лопастной линии с этим родом. Тип рода — Crickites holzap-efli Wedekind (1913) из верхней части франского яруса Германии. К этому же роду относятся: Crickites acutus (Sandberger), С. expectatum Wedekind и C. koeneni (Clarke), известные из тех же отложений.
Помимо указанных родов, многие исследователи относят к семейству Gephuroceratidae род Pharciceras Hyatt, однако, как будет указано далее, последний должен рассматриваться совместно с другими родственными ему родами в составе самостоятельного семейства Pharciceratidae.
86
Род Probeloceras Clarke, 1899
Gephuroceras: Hyatt 1884a, стр. 316 (pars).
Gephyroceras: Wedekind, 1913, стр. 51 (pars); Freeh, 1913, стр. 21 (pars); Wedekind, 1918, стр. 122 (pars).
Probeloceras: Clarke, 1899, стр.90(pars); Freeh, 1902, стр. 60(pars); 1913, стр.25(pars); Miller A. K., 1938, стр. 63.
Тип рода — Goniatites lutheri Clarke, 18856, стр. 50, табл. II, фиг. 8. Формации Дженизи и Нейплс штата Нью-Йорк США. Франский ярус.
Диагноз. Раковина тонкодискоидальная, слабо инволютная, сдавленная с боков. На ранних стадиях вентральная и боковые стороны округленные, но во взрослом состоянии на вентральной стороне и вентральных частях боковых сторон раковины развиваются вогнутые зоны. Поверхность раковины покрыта струйками нарастания, образующими очень широкий, сглаженный синус на боковой и узкий и довольно глубокий синус на вентральной сторонах раковины. Лопастная линия во взрослом состоянии состоит из широкой трехраздельной вентральной лопасти, угловатого первого бокового седла, небольшой угловатой или округленной первой боковой лопасти, широкого округленного дорсо-латераль-ного седла, которое переходит через умбональный ’шов к более или менее клиновидной дорсальной лопасти. Внутренних боковых лопастей нет.
Формула лопастной линии: (V2ViV2)U : D.
Видовой состав и геологическое распространение. Представители рода Probeloceras считались до последнего времени неизвестными в восточном полушарии. В западном полушарии, кроме типичного вида Р. lutheri (Clarke), других представителей также неизвестно. Возможно, однако, что к этому роду относятся Goniatites forcipifer Sandberger и Goniatites planorbis Sandberger, форма раковины и лопастная линия которых не отличаются существенно от таковых рода Probeloceras. Probeloceras (?) forcipiferum (Sandberger) известен из фран-ских отложений Западной Европы и Рудного Алтая. К этому же роду, по-видимому, относится и описанный ниже Probeloceras (?) orientate sp. nov. из франского яруса Рудного Алтая, а также, возможно, к нему следует отнести Gephyroceras domanicense Holzapfel и Gephyroceras keyserlingi Holzapfel, описанные из доманика Южного Тимана.
Общие замечания. Кларк и Фрех относили к описываемому роду различные виды, отличающиеся, однако, от типа рода и выделенные впоследствии в самостоятельные роды Neomanticoceras и Eobeloceras. Кларк, вероятно, думал, что род Probeloceras является предковым по отношению к Beloceras, но Миллер (1938, стр. 63) с этим не согласен и относит Probeloceras к гефуроцератидам. Лопастная линия типа рода Ponticeras, Р. aequabile (Beyrich) сходна с лопастной линией Probeloceras lutheri Clarke и во взрослом состоянии образует также в общем лишь четыре лопасти и различие между Ponticeras и Probeloceras заключается лишь в наличии вогнутых зон на вентральной и боковых сторонах раковины последнего.
87
Probeloceras (?) forcipiferum (Sandberger)
Goniatites forcipijer: Sandberger G., Sandberger F., 1850—1856, стр. 43, табл. VI, фиг 13; Holzapfel, 1882, стр. 246.
Ammonites forcipifer: Giebel, 1852, стр. 467.
Gephuroceras forcipiferum'. Hyatt, 1884a, стр. 317.
Goniatites (Gephyroceras) forcipifer: Freeh, 1887, стр. 428.
Gephyroceras forcipiferum'. Wedekind, 1913, стр. 51; Freeh, 1913, стр. 21; Wedekind, 1918, стр. 122, рис. 28, f в тексте.
Голотип происходит из франских отложений Западной Германии.
Форма. Раковина тонкодискоидальная, сжатая с боков, полуинво-лютная, с наибольшей шириной вблизи умбонального края. В поперечном сечении обороты вытянуты в высоту, с широкими уплощенными, очень
Рис. 18. Лопастная линия Probeloceras (?) forcipiferum (Sandberger) (X 8);
№ 774/371 при В = 7,2 и Ш = 4,5 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
слабо выпуклыми боковыми, узкой уплощенной вентральной и вогнутой дорсальной сторонами. Вентральный край резкий, угловатый, окаймленный слегка вогнутыми зонами вентральной и боковых сторон раковины. Умбональный край отчетливый, несколько округленный. Умбо умеренно широкое, ступенчатое, ограниченное слабо округленными умбональными стенками. Последний оборот облекает около половины предыдущего.
Размеры: Д В Ш Ду В/Д Ш/Д Ду/Д
№ 774/371 18,0 7,8 5,0 4,3 0,43 0,28 0,24
Скульптура. Сохранившиеся участки раковинного слоя лишены скульптуры. Раковина, вероятно, гладкая, снабженная лишь струйками нарастания.
Лопастная линия (рис. 18). Вентральная лопасть очень широкая, трехраздельная. Срединный зубец ею сравнительно неглубокий воронкообразный; боковые зубцы — широкие и глубокие, клиновидные, приостренные на концах. Первое боковое седло высокое и широкое, асимметричное, остро округленное при вершине, с выпуклой наружной и несколько вогнутой внутренней сторонами. Боковая лопасть асимметричная, широкая, но не глубокая, угловатая, несколько округленная на конце; глубина ее гораздо меньше глубины вентральной лопасти. Второе боковое седло невысокое, широкое, равномерно округленное, вершина его находится на умбональной стенке, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Описанный вид отличается от Р. 1и-theri Clarke большей инволютностью раковины и несколько иным очертанием лопастной линии. От Р. orientale sp. nov., описанного ниже, он отличается более широким умбо и большей величиной срединных седел, разделяющих ветви вентральной лопасти. Лопастная линия алтайских образ
88
цов несколько отличается от таковой G. forcipifer Sandberger, изображенной Зандбергерами (1850—1856, табл. VI, фиг. 3, в), некоторой угловатостью ее элементов, но, не наблюдая других признаков, отличающих наши экземпляры от изображенного Зандбергерами, я склонен в настоящее время относить их к одному виду.
Геологический возраст и распространение. Встречается во франских отложениях Западной Германии и Рудного Алтая.
Местонахождение. Два экземпляра найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта.
Probeloceras (?) orientate sp. nov.
Табл. II, фиг. 3—5.
Голотип — ПИН № 774/325; Рудный Алтай, правый берег рч. Золотухи, известняковая линза в 400 м восточнее дер. Горюновки; нижняя часть франского яруса.
Форма. Раковина тонкодискоидальная, сжатая с боков, сильно инволютная, с наибольшей шириной вблизи умбонального края. В поперечном сечении обороты вытянуты в высоту, с широкими уплощенными, слегка выпуклыми, сходящимися наружу боковыми, уплощенной узкой вентральной и вогнутой дорсальной сторонами. Вентральная сторона на ранних стадиях округлена. На взрослых стадиях вентральный край становится резко угловатым, а на прилегающих к нему участках боковых сторон и на уплощающейся вентральной стороне образуются слегка вогнутые зоны. Умбональный край отчетливый, несколько округленный. Умбо во взрослом состоянии узкое, ступенчатое, ограниченное отчетливыми слегка округленными умбональными стенками, на ранних стадиях — сглаженное. Последний оборот облекает до 4/5 предыдущего.
Размеры: Д В Ш Ду В/Д ш/д Ду/Д
№ 774/327 16,6 8,9 5,0 2,4 0,54 0,3 0,14
№ 774/325 16,0 8,6 4,8 2,3 0,54 0,3 0,14
№ 774/334 14,0 7,6 4,5 2,0 0,54 0,32 0,14
Скульптура. Поверхность раковины покрыта тонкими струйками нарастания, образующими широкие, довольно глубокие синусы на боковых и узкие синусы на вентральной стороне раковины, разделенные узким выступом, тяготеющим к вентральному краю и окаймляющей его вогнутой зоне боковой стороны раковины. В области умбо струйки нарастания становятся более отчетливыми, переходя в тонкие морщинки раковинного слоя.
Лопастная линия (рис. 19). Вентральная лопасть очень широкая, трехраздельная, с укороченным воронкообразным срединным и глубокими, умеренно широкими приостренными на концах клиновидными боковыми зубцами. Первое боковое седло асимметричное, широкое, округленное при вершине, с выпуклой наружной и вогнутой внутренней сторонами. Боковая лопасть асимметричная, широкая, но не глубокая (глубина ее гораздо меньше глубины вентральной лопасти), тупо округленная в основании, располагающемся в районе умбонального края. Относительно высокое округленное второе боковое седло располагается своей вершиной в районе умбонального шва, переходя внутренней своей частью за умбональный шов.
89
Сходство и отличие. Описанный вид наиболее близок к Probeloceras (?) jorcipiferum (Sandberger), отличаясь, однако, от последнего более узким умбо и некоторой угловатостью и большей асимметричностью элементов лопастной линии. От типа рода Probeloceras — Р. 1и-theri Clarke, данный вид, как и указанный выше Р. (?) forcipiferum, отличается существенно лишь большей инволютностью раковины, и я поэтому
Рис. 19. Лопастная линия Probeloceras (?) orientale sp. nov. (x 10); голотип № 774/325 при В=4,6 мм. Рудный Алтай, рч. Золотуха у дер. Горюновки. Низы франского яруса.
отношу его к Probeloceras, приводя, правда, родовое название под вопросом.
Геологический возраст и распространение. Встречается в нижней части франского яруса Рудного Алтая.
Местонахождение. 54 большей частью неполных экземпляра найдены в известняковой линзе правого берега рч. Золотухи, в 400 м восточнее дер. Горюновки, в нижней части франского яруса.
Род Ponticeras Matern, 1929
Gephuroceras: Hyatt, 1884a, стр. 316 (pars).
Gephyroceras: Foord and Crick, 1897, стр. 59 (pars); Holzapfel, 1899, стр. 17 (pars); Freeh, 1902, стр. 56 (pars); 1913, стр. 21 (pars); Wedekind, 1913, стр. 46 (pars); Налив-кина, 1947, стр. 164.
Manticoceras: Hyatt, 1884a, стр. 317 (pars); Чернышев, 1887, стр. 23 (pars).
Ponticeras: Matern, 1929, стр. 151; Miller A. K., 1938, стр. 70.
Тип рода — Ammonites aequabilis Beyrich, 1837, стр. 34, табл. 2, фиг. 1. Нижняя часть франских отложений Западной Германии.
Диагноз. Раковина дискоидальная, слабо-инволютная, с округленной вентральной, округленными или слегка уплощенными боковыми и слабо вогнутой дорсальной сторонами. Умбо большое. Умбональные стенки округленные. Поверхность раковины покрыта струйками нарастания, образующими синусы на боковых и вентральной сторонах раковины. Лопастная линия состоит из вентральной, боковой и дорсальной лопастей. Внутренняя боковая лопасть отсутствует. Вентральная лопасть трехраздельная, с коротким срединным и более длинными приостренными боковыми зубцами. Боковая лопасть обычно округлена, реже является заостренной. Дорсальная лопасть в общем клиновидная. Седла, разделяющие лопасти, округлены.
Формула лопастной линии: (V2ViV2) U: D.
Видовой состав и геологическое распространение. Кроме типичного вида Ponticeras aequabile (Beyrich), описанного из низов верхнего девона Германии, в настоящее время к роду Ponticeras могут быть отнесены известные из верхнего девона Германии: Р. ge-
90
rolsteinense (Steininger), P. bickense (Wedekind), P. sandbergeri (Wedekind); описанные из доманика Южного Тимана: Р. bisulcatum (Keyserling), Р. tschernyschewi (Holzapfel), P. regale (Holzapfel), P. uralicum (Holzapfel), P. auritum (Holzapfel), P. lebedeffi (Holzapfel), P. uchtense (Keyserling). Из нижней части верхнего девона штата Айова («Independence shale») США известен Ponticeras stainbrooki Miller. Из франских отложений Рудного Алтая известны описанные ниже Р. altaicum sp. nov., Р. acutilobatum sp. nov.
Общие замечания. Род Ponticeras был установлен Матерном (1929, стр. 151) для части видов, которые относились ранее либо к Gephyroceras, либо к Manticoceras. В качестве типа рода был установлен A mmo nites aequabilis Beyrich из низов верхнего девона Германии. Полная лопастная линия A. aequabilis не была изображена, но Ведекинд (1918, стр. 121) указывает, что она образует в общем лишь четыре лопасти, в отличие от шести лопастей типов родов Manticoceras и Gephuroceras. Боковое седло внутренней части лопастной линии переходит через умбональный шов к первой боковой лопасти наружной части ее, а в области умбонального шва лопасти не образуется. Отсутствие внутренней боковой лопасти отличает Ponticeras от Manticoceras (синонимом которого является Gephuroceras), Honinghausia, Koenenites, Timanites и других. От Probeloceras, с которым он сходен! по лопастной линии, описываемый род отличается отсутствием вогнутых участков на вентральной и боковых сторонах раковины.
Ponticeras altaicum sp. nov.
Табл. II, фиг. 7
Голотип — ПИН № 774/305; Рудный Алтай, Николаевское месторождение, западный склон гряды, отходящей на северо-восток от поселка; линзы известняков гониатитового горизонта франского яруса.
Форма. Раковина дискоидальная, слабо-инволютная, уплощенная с боков, с наибольшей шириной вблизи умбонального края. В поперечном сечении обороты вытянуты в высоту, со слабо выпуклыми, сходящимися кверху боковыми, узкой, угловатой на ранних стадиях и округленной на более взрослых, вентральной, и вогнутой дорсальной сторонами. Умбональный край округленный, более отчетливый на последних оборотах. Умбо широкое, но не глубокое, сглаженное, особенно на первых оборотах. Последний оборот облекает около половины предыдущего.
Размеры: д В Ш Ду В/Д ш/д Ду/Д
№ 774/304 20,7 8,0 6,4 7,6 0,39 0,31 0,37
№ 774/305 18,7 7,0 5,5 7,0 0,37 0,30 0,37
№ 774/306 17,0 6,3 5,3 6,0 0,37 0,31 0,35
Скульптура. Поверхность раковины гладкая, несущая на себе лишь тонкие струйки нарастания.
Лопастная линия (рис. 20). Вентральная лопасть широкая, трехраздельная, с коротким сглаженным срединным и небольшими при-остренными в виде широких клиньев боковыми зубцами. Первое боковое седло широкое, занимающее всю боковую сторону оборота, асимметричное, широко округленное при вершине, со слегка выпуклой наружной и почти прямой, очень слабо вогнутой внутренней сторонами. Боковая лопасть широкая, неглубокая, широко округленная в основании, расположенном в области умбонального края. Второе боковое седло низкое,
91
широкое; вершина его располагается на умбональном шве, а внутренняя часть переходит на внутреннюю стенку.
Сходство и отличие. Описанный вид по форме раковины и очертанию лопастной линии наиболее близок к Р. tschernyschewi (Holzapfel), отличаясь от него лишь несколько иным очертанием элементов лопастной линии. Если у Р. tschernyschewi первое боковое седло имеет более крутую внешнюю сторону, то у описываемого вида более крутой является, напротив, внутренняя сторона. Кроме того, молодой экземпляр Р. tschernyschewi, изображенный Гольцапфелем (1899) на табл. 6, фиг. 8,
ДО* Лопастная лнння Ponticeras altaicum sp. nov. (X10); голотип № 774/305 при В—4,3 и Ш=3,6 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
соответствующий по размерам нашим образцам, имеет более угловатую вентральную сторону, окаймленную с боков несколько вогнутыми зонами. Различным у сравниваемых видов является, по-видимому, и характер струек нарастания, а следовательно, очертание апертуры раковины. Не исключена возможность, что тиманские и алтайские экземпляры должны быть отнесены к одному виду, но, не имея возможности детального их сравнения, я воздерживаюсь в настоящее время от их объединения.
1\ИЗпДРуГИХ видов наиболее близки к описанному Р. uralicum (Holzapfel) и Р. bisulcatum (Keyserling). От указанных видов новый вид, однако, отличается более широким умбо. По общей форме раковины он занимает промежуточное положение между указанными двумя видами. От Р. ига-hcum он отличается более уплощенной раковиной и узкой угловатой, несколько округленной вентральной стороной, от Р. bisulcatum напротив, несколько более широкой раковиной с менее вытянутыми в высоту оборотами и менее угловатой вентральной стороной последних оборотов.
1еологический возраст и распространение Встречается во франских отложениях Рудного Алтая.
Место на хождение. Пятнадцать различной сохранности экземпляров найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта франского яруса.
Ponticeras acutilobatum sp. nov.
Табл. II, фиг. 6
Г о л о т и п ПИН №774/322; Рудный Алтай, Николаевское месторождение, западный склон гряды, отходящей от поселка на северо-восток^ известняковые линзы гониатитового горизонта франского яруса.
о р м а. Раковина дискоидальная, полуинволютная, немного уплощенная с боков, с наибольшей шириной вблизи умбонального края. Поперечное сечение оборота субтрапецоидальное, вытянутое в высоту, 92
с уплощенными, слегка выпуклыми боковыми, узкой округленной вентральной и вогнутой дорсальной сторонами. Умбо широкое, ступенчатое, ограниченное отчетливыми, несколько округленными умбональными стенками. Последний оборот облекает до 3/5 предыдущего.
Размеры:
№ 774z322
д В Ш Ду в/д Ш/Д Ду/Д
68,5 25,4 20,0 23,5 0,37 0,29 0,34
51,4 20,1 15,5 18,5 0,39 0,30 0,36
Скульптура. Сохранившаяся часть раковинного слоя лишена скульптуры. Раковина, вероятно, гладкая, несущая лишь струйки нарастания.
Лопастная линия (рис. 21). Вентральная лопасть умеренно широкая, занимающая собой лишь вентральную сторону раковины, трехраздельная, с коротким срединным зубцом и узкими клиновидными, со слегка отогнутыми наружу концами, острыми боковыми зубцами. Первое
Рис. 21. Лопастная линия Ponticeras acutilobatum sp. nov. (x4); голотип № 774/322 при B=20,l и Ш = 15,5 мм. Рудный Алтай. Франский ярус.
боковое седло широкое, занимающее большую часть боковой поверхности оборота, асимметричное, субтрапецоидальное, округленное при вершине, с выпуклой, образующей перегиб в области вентрального края, наружной и прямой внутренней сторонами. Боковая лопасть несколько асимметричная, умеренно широкая, приостренная на конце, расположенном в области наибольшей ширины оборота.На более ранних стадиях боковая лопасть округленная. Глубина боковой лопасти значительно меньше глубины вентральной. Второе боковое седло умеренно широкое, невысокое, равномерно округленное; вершина его находится на умбональной стенке, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Описанный вид отличается от всех известных видов рода Ponticeras по очертанию лопастной линии, имеющей во взрослом состоянии приостренную на конце боковую лопасть. От представителей рода Manticoceras новый вид отличается слабо инволютной, уплощенной с боков раковиной.
Геологический возраст и распространение. Редко во франских отложениях Рудного Алтая.
Местонахождение. Единственный экземпляр найден на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковой линзе гониатитового горизонта.
Ponticeras sp.
Среди коллекций аммонитов, собранных нами из известняковых прослоев песчано-сланцевого горизонта в районе с. Раздольного, имеется некоторое количество образцов, несомненно принадлежащих к роду
93
Ponticeras, но плохая сохранность не позволяет определить их до вида. По форме своей тонкодискоидальной, очень слабо инволютной раковины^ имеющей невысокие округленные, несколько уплощенные с боков обороты и широкое сглаженное умбо, указанные образцы имеют сходство с Ponticeras bickense (Wedekind), описанным из верхней зоны мантикоцеровых слоев Германии (зона 18), но плохая сохранность, не дающая возможности наблюдать лопастную линию, заставляет меня в настоящее время определять их как Ponticeras sp.
Род Uchtites gen. nov.
Gephyroceras: Гольцапфель, 1899, стр. 27 (pars); Freeh, 1913, стр. 21 (pars).
Тип рода — Gephyroceras syrjanicum Holzapfel, 1899, стр. 40, табл. 6, фиг. 9. Доманиковый горизонт Южного Тимана.
Диагноз. Раковина линзовидная, уплощенная с боков, с узким умбо и резким вентральным килем, ограниченным у взрослых экземпляров слабыми бороздками. Лопастная линия состоит из трехраздельной вентральной, округленной внешней боковой, слабой сглаженной внутренней боковой и клиновидной дорсальной лопастей (рис. 17, с).
Формула лопастной линии: (V2VYV2) U : ID.
Видовой состав и геологическое распространение. Кроме типичного вида, известного из доманикового горизонта Южного Тимана, других видов не известно.
Общие замечания. Новый род отличается от родов Ponticeras и Manticoceras, занимая как бы промежуточное между ними положение. От Ponticeras его отличает форма раковины и характер лопастной линии с намечающимся образованием внутренней боковой лопасти. От Manticoceras его отличает отсутствие резко выраженной внутренней боковой лопасти. От других представителей семейства новый род также легко отличим по форме раковины и лопастной линии.
Ркод Manticoceras Hyatt, 1884
Gephuroceras: Hyatt, 1884a, стр. 316 (pars).
Gephyroceras: Foord and Crick, 1897, стр. 59 (pars).
Manticoceras: Hyatt, Д884а, стр. 317 (pars); Clarke, 1899, стр. 42; Holzapfel, 1899r стр. 19; Freeh, 1902, стр. 56; 1913, стр. 22; Wedekind, 1913, стр. 46; 1918, стр. 123;. Miller A. К., 1938, стр. 72; Наливкина, 1947, стр. 166.
Тип род а— Goniatites simulator Hall, 1874, стр. 2—3; 1876, табл. 69, фиг. 1, 2; табл. 74, фиг. 8. Слои с Liorhynchus формации Нейплс штата Нью-Йорк США.
Диагноз. Раковина дискоидальная, иногда приближающаяся к линзовидной, среднеинволютная, с округленными или уплощенными боками, округленной или угловатой вентральной и вогнутой дорсальной сторонами. Умбо умеренно широкое, реже узкое, ступенчатое, ограниченное отчетливыми несколько округленными умбональными стенками. Поверхность раковины гладкая, реже слабо скульптированная поперечными ребрами. Струйки нарастания образуют широкий, но не глубокий синус на боковой стороне и более глубокий узкий синус на вентральной сторона раковины. Лопастная линия состоит у взрослых экземпляров из широкой трехраздельной вентральной лопасти, более или менее клиновидной дорсальной лопасти и двух боковых лопастей, из которых одна является внутренней по своему положению. Боковые лопасти небольшие, простые, приостренные, реже округленные на конце. Седла, разделяющие эти лопасти, округлены.
94
Формула лопастной линии: (УгУ^г)!} : ID.
Видовой состав и геологическое распространение. Род Manticoceras широко распространен в Европе, Северной Америке, Азии и встречается также в Северной Африке. К этому роду принадлежат почти две пятых всех известных видов девонских аммонитов Америки. В Европе (и, возможно, в Азии и Африке) род ограничен стратиграфически нижней частью верхнего девона («Oberdevonstufe I») или франским ярусом.
Кроме типичного вида Manticoceras simulator (Hall), описанного из нижней части верхнего девона США, к роду Manticoceras могут быть отнесены известные из тех же отложений: М. accelerans Clarke, М. (?) amplexum (Hall), М. apprimatum Clarke, M. cataphractum (Clarke), M. clarkei Miller, M. contractum Clarke, M. cordiforme Miller, M. delphiense (Kindle), M. fasciculatum Clarke, M. genundewa (Clarke), M. holzapfeli (Clarke), M. kindlei Miller, M. nodifer (Clarke), M. oxy Clarke, M. perla-tum (Hall), M. regulare Fenton et Fenton, M. rhynchostoma Clarke, M. sep-tentrionale Miller, M. sinuosum (Hall), M. sororium Clarke, M. styliophi-lum Clarke, M. tardum Clarke, M. unduloconstrictum Miller, M. vagans Clarke. Из франского яруса Западной Европы (зоны ip—Io «Oberdevonstufe») известны: М. calculiforme (Beyrich), М. carinatum (Sandberger), M. intermedium (Sandberger), M. schellwieni Wedekind, M. cordatum (Sandberger), M. crassum Wedekind, M. galeatum Wedekind, M. inversum Wedekind, M. drevermanni Wedekind, M. intumescens (Beyrich), M. adorfense Wedekind, M. retrorsum (Buch), M. affine (Steininger), M. bullatum Wedekind; из доманикового горизонта южного Тимана описан: М. аттоп (Keyserling). Новыми алтайскими видами являются: М. artum sp.nov., М. altaicum sp. nov., M. neverovi sp. nov., M. eliseevi sp. nov. Некоторые из указанных видов, как, например, М. sinuosum, М, intumescens, М. cordatum и др., пользуются широким распространением. М. sinuosum известен, например, из франских отложений Северной Америки, Европы, Алтая, вероятно Урала; М. intumescens — из тех же отложений Западной Европы, Урала, Алтая и других областей; М. cordatum — из Западной Европы и Северной Африки; М. drevermanni, М. affine, М. galeatum — из Западной Европы, Алтая, Урала и т. д. Некоторые виды с другой стороны являются несомненно родственными между собой и, может быть, должны рассматриваться в составе одного вида (американские виды М. sinuosum, М. regulare, М. cordiforme, М. septentrionale, например, близкие к М. intumescens), но, не имея возможности детального сравнения их, я в настоящее время воздерживаюсь от их объединения.
Общие замечания. В литературе на протяжении продолжительного времени существовали различные мнения в отношении родства между родами Gephuroceras ( = Gephyroceras) и Manticoceras, Оба эти рода были установлены Хайэттом в 1884 г. в его работе «Genera of Fossil Cephalopods» (роды ископаемых цефалопод), первый на стр. 316, второй на стр. 317. Типом рода Gephuroceras является Goniatites sinuosus Hall из верхнего девона штата Нью-Йорк, типом рода Manticoceras—Goniatites simu-lator Hall из того же горизонта той же провинции. Раковины этих двух гониатитов не отличаются во взрослом состоянии по форме и имеют сходные умбо; различия наблюдаются лишь в очертаниях лопастных линий. Сравнение лопастных линий двух указанных представителей не обнаруживает существенной разницы между ними, но у наружной части лопастной линии G. simulator все лопасти округлены и второе боковое седло маленькое, в то время как у G. sinuosus все лопасти заострены, второе боковое седло относительно большое; большим и широким является и седло, разделяющее боковые ветви вентральной лопасти (ср. рис. 17,с? и рис.
95
35,а). Лопастная линия G. sinuosus действительно более развита, чем лопастная линия G. simulator, но различия между ними видовые, а не родовые, или даже подродовые. А. К. Миллер (1938, стр. 72), отмечая эти особенности, считает оба вида принадлежащими к одному роду. Кларк (1899, стр. 44) считал возможным рассматривать G. sinuosus Hall как особый способ сохранения G. intumescens Beyrich. Последний вид Кларк, как и многие другие исследователи, отождествляет с G. patersoni Hall, а Хайэтт относит его к Manticoceras. Таким образом, типом рода Gephuroceras является настоящий Manticoceras. Остальные формы, соединенные под названием Gephuroceras ( = Gephyroceras), представляют смесь видов родов Honinghausia, Timanites, Glyphioceras, Manticoceras, Ponticeras, Probeloceras и должны быть исключены из Gephuroceras.
Родовые названия Gephuroceras и Manticoceras, следовательно, являются синонимом один другого. Хотя название Gephuroceras ( = Gephyroceras) и имеет приоритет по сравнению с Manticoceras, А. К. Миллер (1938, стр. 72) предлагает ликвидировать его и считать синонимом Manticoceras, так как единственный образец, который был изображен в связи с первоначальным описанием типа рода Gephuroceras, очень неполный и на нем можно проследить характер лишь небольшой части одной из лопастных линий и поэтому идентичность образцов, относимых к этому виду, с голотипом его может вызывать сомнение. Более того, наименование Manticoceras теперь широко используется в стратиграфической терминологии и ликвидировать его будет трудно.
Manticoceras отличается от Ponticeras и Probeloceras тем, что каждая из его полных лопастных линий состоит из шести лопастей, тогда как у последних их лишь четыре. У Honinghausia и Koenenites, также тесно родственных с Manticoceras, полные лопастные линии образуют, в свою очередь, в общем восемь лопастей вокруг извилины. Timanites, который некоторыми авторами считается синонимом Manticoceras (Zittel, 1884, стр. 418; Foord and Crick, 1897, стр. 59), отличается от последнего угловатой вентральной стороной, полностью закрытым пупком и лопастной линией, образующей в общем десять лопастей вокруг извилины.
М anticoceras affine (Steininger)
Табл. II фиг. 8
Ammonites primordialis: Buch, 1832, стр. 168, табл. 1, фиг. 15—17 (pars) (non Schlotheim).
Goniatites affinis'. Steininger, 1853, стр. 42, табл. I, фиг. 4, 4a; Holzapfel, 1882, стр. 243, табл. XLVI (III), фиг. 3, За, b\ Beyrich, 1884, стр. 208.
Goniatites lamed var. latidorsalis'. Sandberger, 1850—1856, стр. 90, табл. VIII, фиг. 8; 1851, стр. 304, табл. Ill, фиг. 30.
Manticoceras latidorsatum'. Hyatt, 1884a, стр. 318.
Goniatites (Gephyroceras) affinis: Freeh, 1887, стр. 377, 428.
Gephyroceras affine: Foord and Crick, 1897, стр. 82.
Gephyroceras (Manticoceras) affine'. Freeh, 1902, стр. 59, фиг. 18, d.
Manticoceras affine'. Freeh, 1913, стр. 23; Wedekind, 1913, стр. 56, табл. IV, фиг. 10, 11, рис. в тексте 7, д]_2; 4918, стр. 126, табл. XXII, фиг. 12, рис. в тексте 36, а.
Голотип происходит из' франских отложений Западной Германии. Форма. Ввиду отсутствия более или менее удовлетворительного описания вида в работах предыдущих исследователей, считаю необходимым дать его характеристику возможно более подробно, тем более.что в изученной коллекции находятся экземпляры, дающие представление о более поздних стадиях его развития.
На ранних стадиях развития раковина эволютная, субсферического очертания, образованная уплощенными в дорсо-вентральном направлении
96
оборотами. В поперечном сечении обороты растянуты в ширину (ширина их примерно вдвое больше высоты), со слабо выпуклой, равномерно округленной вентральной и слегка вогнутой дорсальной сторонами. Умбо широкое и глубокое, ограниченное округленными нависающими частями оборотов. Начиная с четвертого оборота, одновременно с быстрым его возрастанием в высоту, увеличивается инволютность раковины, боковые стенки
постепенно уплощаются, вентральная сторона раковины становится угловатой. Во взрослом состоянии раковина толстодискоидальная, приближающаяся к линзовидной, с наибольшей шириной вблизи умбо. Поперечное сечение оборота приобретает субтреугольное очертание, с широкими слегка выпуклыми боковыми, угловатой вентральной и вогнутой дорсальной сторонами. Умбо во взрослом состоянии умеренно широкое, ступенчатое, с довольно высокими равномерно округленными умбо-нальными стенками. Последний оборот облекает около половины предыдущего (рис. 22).
Размеры: Д В Ш Ду В/Д Ш/Д Ду/Д
№ 774/45
№ 774/46
№ 774/48
(94,3 55,8 36,4 14,5 51,2 32,3
49,0
26,7
16,7
6,4
24,5
15,2
40,8
30,2
22,0
11,2
28,5
20,5
21,6
14,5
9,6
4,0
12,3
8,9
0,52 0,48 0,46 0,44 0,48 0,47
0,43 0,54 0,60 0,77 0,56
0,63
0,23 0,26 0,26
0,28 0,24 0,27
Скульптура. Раковина гладкая, снабженная лишь очень тонкими слабо заметными струйками нарастания, более отчетливыми в области умбо.
Лопастная линия (рис. 23). Вентральная лопасть широкая и глубокая, трехраздельная, с коротким воронкообразным срединным зубцом и узкими, длинными, клиновидными боковыми зубцами, приостренные концы которых скошены и слегка отогнуты в сторону умбо. Первое боковое седло высокое, асиммет-
Рис. 22. Поперечное сечение Manticoceras affine (Steininger) (X1); № 774/45. Рудный Алтай. Франский ярус.
ричное, остро округленное у вершины, довольно
узкое, занимающее среднюю треть боковой поверхности оборота. Боковая лопасть асимметричная, загнутая в сторону умбо своим приостренным концом, расположенным в области наибольшей ширины оборота; глубина ее гораздо меньше глубины вентральной лопасти. Второе боковое седло
равномерно округленное, низкое, широкое; середина его располагается за умбональным краем, а внутренняя часть переходит за умбональныйшов.
Сходство и отличие. Описанный вид по форме раковины, характеру умбо и очертанию лопастной линии отличается от всех известных представителей рода Manticoceras. От наиболее близкого к нему М. bullatum Wedekind он отличается более широкими сплющенными в дорсо-вентральном направлении оборотами (по крайней мере молодых стадий развития), более узким первым боковым седлом и более сдвинутой по направлению к внешнему краю раковины боковой лопастью. От М. altaicum sp. nov., имеющего некоторое сходство с ним по форме раковины, он отличается, в первую очередь, менее резким расчленением вентральной лопасти.
7 Б- И Богословский
97
Геологический возраст и распространение. Встречается во франских отложениях Западной Европы и Рудного Алтая.
Местонахождение. Пятнадцать экземпляров найдено на западном склоне гряды, отходящей на северо-восток от поселка Николаев-
Рис. 23. Лопастная линия Manticoceras ajjine (Steininger) (х2) 774/47 при В=17,3 и 111=22,0 мм. Рудный Алтай. Средняя часть франского яруса.
ского рудника, в известняковых линзах гониатитового горизонта; один экземпляр найден в трех км к юго-востоку от поселка в известняковой линзе того же горизонта.
Manticoceras bullatum Wedekind
Manticoceras bullatum: Wedekind, 1913, стр. 56, табл. IV, фиг. 12, 13, рис. в тексте 7 ai_2; 1918, стр. 126, табл. XXII, фиг. 13, рис. в тексте 36, а.
Голотип происходит из франских отложений Западной Германии.
Форма. Раковина толстодискоидальная, приближающаяся к линзовидной, с наибольшей шириной вблизи умбонального края. Поперечное сечение оборота субтреугольного очертания, с широкими слабо выпуклыми боковыми, узкой угловатой вентральной и вогнутой дорсальной сторонами. Ширина оборота несколько больше высоты. Умбо умеренно широкое, ступенчатое, с довольно высокими, равномерно округленными
умбональными стенками. Последний оборот облекает около 2/3 преды-
дущего.
Размеры: Д В Ш Ду в/д ш/д Ду/Д
№ ,774/44 30,0 4,2 16,2 7,8 0,47 0,54 0,26
№ 773/38 19,7 8,2 10,5 5,1 0,42 0,53 0,26
№ 774/9 19,0 8,0 10,0 5,0 0,42 0,53 0,26
Скульптура. Раковина гладкая, снабженная лишь тонкими
струнками нарастания.
Лопастная линия. При величине диаметра раковины в 19,7мм (рис. 24, а) вентральная лопасть широкая, но не особенно глубокая, трехраздельная, с коротким округленным срединным зубцом и сравнительно широкими и короткими, несколько асимметричными сосцевидными боковыми зубцами, концы которых слегка оттянуты. Первое боковое седло высокое, асимметричное, равномерно округленное у вершины, умеренно широкое, занимающее собой почти всю боковую сторону оборота, с очень слабо выпуклой наружной и более крутой незначительно вогнутой внешней сторонами. Боковая лопасть асимметричная, округленная на конце; глубина ее гораздо меньше глубины вентральной лопасти. Округленный
98
конец боковой лопасти лежит в области умбонального края. Второе боковое седло низкое, равномерно округленное, умеренно широкое; середина его располагается на умбональной стенке, а внутренняя часть переходит за умбональный шов.
При высоте оборота в 17 мм (рис. 24,6) боковые зубцы вентральной лопасти становятся узкими, более длинными, и приостренные, несколько оттянутые концы их слегка отгибаются в сторону умбонального края. Первое боковое седло приобретает более широко округленную форму.
Рис. 24. Лопастные линии Manticoceras bullatum Wedekind:
а — № 773/38 при В=8,2 и Ш=10.5 мм (ХЗ); Ь — № 774/39 при В=17,0 мм (Х2). Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
Боковая лопасть становится сосцевидной формы, с приостренным оттянутым концом, расположенным в области умбонального края.
Сходство и отличие. Описанные экземпляры по форме раковины имеют сходство с М. bullatum Wedekind. Отличие от указанного вида проявляется лишь в несколько ином характере лопастной линии, именно в характере боковой лопасти. На приведенном Ведекиндом рисунке (1913, стр. 55, фиг. 7,а2) боковая лопасть изображена менее приост-ренной, ее конец лежит ниже конца вентральной лопасти. Более детальное сравнение наших экземпляров, которые к тому же далеко не полной сохранности, с голотипом Ведекинда затруднено ввиду отсутствия у данного автора достаточно полного описания и удовлетворительного изображения выделенного им вида. Отмечая сходство описанного вида с голотипом Ведекинда, я отношу его к М. bullatum Wedekind, воздерживаясь от выделения нового вида ввиду недостаточной сохранности имеющихся у меня экземпляров. Отличие данного вида от М. affine (Steininger) и М. altaicum sp. nov., близких к нему, указано при описании последних.
Геологический возраст и распространение. Встречается во франских отложениях Западной Германии и Рудного Алтая.
Местонахождение. Шесть различной сохранности экземпляров найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта; шесть экземпляров найдены по правому берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта франского яруса.
7*
99
Manticoceras altaicum sp. nov.
Табл. IV, фиг. 1.
Г о л о т и п—ПИН№ 773/15; Рудный Алтай, Николаевское месторождение, западный склон гряды, отходящей на северо-восток от поселка; известняковые линзы гониатитового горизонта франского яруса.
Форма. Раковина во взрослом состоянии толстодискоидальная, приближающаяся к линзовидной, с наибольшей шириной вблизи умбонального края; инволютная, с широкими слегка выпуклыми боковыми, узкой несколько угловатой вентральной и вогнутой дорсальной сторонами. Умбо довольно широкое, ступенчатое, с равномерно округленными умбо-нальными стенками. Последний оборот облекает около половины предыдущего. Поперечное сечение оборота во взрослом состоянии субтреугольное с высотой, несколько превышающей ширину:
Размеры: д В Ш Ду в/д Ш/Д Ду/Д
179,5 37,0 33,0 20,5 0,47 0,42 0,26
№ 773, In [67,3 30,4 29,2 17,8 0,45 0,43 0,26
№ 774/40 61,0 27,4 28,0 16,6 0,45 0,46 0,27
Скульптура. Сохранившиеся на ядре небольшие участки раковинного слоя лишены заметных следов скульптуры. Раковина, по-видимому, гладкая.
Лопастная линия (рис. 25). Вентральная лопасть широкая и
Рис. 25. Лопастная линия Manticoceras altaicum sp. nov. (У 1); голотип № 773/15 при В=35,0мм. Рудный Алтай. Франский ярус, гониатитовый горизонт.
очень глубокая, трехраздельная, с укороченным воронкообразным сре динным и длинными боковыми зубцами. Боковые зубцы вентральной л о" пасти умеренно широкие, длинные, с несколько оттянутыми приострен” ными концами, в целом клиновидные. Первое боковое седло асимметрия" ное, высокое, умеренно широкое, остро округленное у вершины, занимаю" щее примерно среднюю треть боковой поверхности оборота, с выпуклой, ско” шейной у вершины наружной и почти прямой внутренней сторонами. Бо" ковая лопасть асимметричная, приостренная; глубина ее много меньше глубины вентральной лопасти. Второе боковое седло равномерно округлое, низкое, широкое, с вершиной, расположенной в области умбонального края.
Сходство и отличие. Этот вид по форме раковины, характеру умбо и, в особенности, лопастной линии отличается от описанных ранее представителей рода Manticoceras. По форме поперечного сечения оборотов он имеет сходство с М. bullatum Wedekind, отличаясь, однако, 100
от последнего характером и степенью развития лопастной линии и более широким умбо. От М. affine (Steininger) он отличается характером лопастной линии, в особенности вентральной лопасти, обладающей очень длинными боковыми зубцами. Он имеет по сравнению с М. affine и менее широкую раковину.
Геологический возраст и распространение. Встречается в средней части франского яруса Рудного Алтая.
Местонахождение. .Один экземпляр найден на западном склоне гряды, отходящей на сев’еро-восток от поселка Николаевского месторождения, в известняковой линзе гониатитового горизонта; один экземпляр, по-видимому, также принадлежащий этому виду, найден на правом берегу рч. Грязнухи у с. Раздольного, в известняковом прослое песчано-сланцевого горизонта франского яруса. Возможно, что к этому же виду относятся и несколько мелких экземпляров, найденных на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта.
Manticoceras neverovi sp. nov.
Табл. Ill, биг. 1
Г о л о т и п— ПИН № 774/62; Рудный Алтай, правый берег рч. Грязнухи у с. Раздольного; известняковые прослои песчано-сланцевого горизонта верхней половины франского яруса.
Форма. Раковина во взрослом состоянии толстодискоидальная, с наибольшей шириной вблизи умбонального края, инволютная, с уплощенными, слегка выпуклыми боковыми, узкой равномерно округленной вентральной и вогнутой дорсальной сторонами. Умбо умеренно широкое, с довольно высокими округленными умбональными стенками. Поперечное сечение оборота субтреугольное, с тупо-округленными углами и почти равными высотой и шириной.
Раз меры: д в ш Ду В/Д Ш/Д Ду/Д в/ш
№ 774/62 55,5 30,0 31,5 10,3 0,54 0,57 0,19 0,95
№ 774/62 а — 33,0 34,5 — — — — 0,96
Скульптура. Сохранившиеся на ядре небольшие участки раковинного слоя лишены заметных следов скульптуры. Раковина, по-видимому, совершенно гладкая.
Лопастная линия (рис. 26). Вентральная лопасть широкая и глубокая, трехраздельная, с очень коротким округленным срединным и длинными и широкими боковыми зубцами. Боковые зубцы ножеобразные, скошенные в основании с вентральной стороны, с приостренными, несколько оттянутыми утолщенными концами. Первое боковое седло высокое, почти вдвое превышающее по высоте седло, разделяющее боковые зубцы вентральной лопасти, широкое, равномерно округленное у вершины, со слегка выпуклой наружной и несколько вогнутой внешней сторонами. Боковая лопасть асимметричная, загнутая в сторону умбо своим приострен-ным концом, расположенным вблизи умбонального края; глубина ее меньше глубины вентральной лопасти. Второе боковое седло низкое, широкое, равномерно-округлое, середина его располагается за умбональным краем, а внутренняя часть немного переходит за умбональный шов. Внутренняя часть лопастной линии состоит из небольшой, несколько асимметричной, приостренной внутренней боковой лопасти, располагающейся непосредственно за умбональным швом, относительно высокого, узкого внутреннего бокового седла и узкой и глубокой клиновидной заостренной дорсальной лопасти.
101
Сходство и отличие. Этот вид по форме раковины и характеру лопастной линии, особенно боковых ветвей вентральной лопасти ее, отличается от описанных ранее, и я считаю возможным выделить его в качестве самостоятельного.
Геологический возраст и распространение. Встречается во франских отложениях Рудного Алтая.
Рис. 26. Лопастная линия М anticoceras* never ovi sp. nov. (X1); голотип № 774/62 при В=33,0 и 111=34,5 мм: Рудный Алтай, район Гериховского месторождения. Франский ярус.
Местонахождение. Один неполной сохранности экземпляр и один обломок взрослого оборота найдены на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта франского яруса.
Мanticoceras carma£umT(Beyrich)
Табл. VI, фиг. 1
Ammonites carinatus: Beyrich, 1837, стр. 35, табл. II, фиг. 2 а, Ь, с.
Goniatites carinatus: Sandberger G., Sandberger F., 1850—1856, стр. 88, табл. VI, фиг. £.
Gephyroceras (Manticoceras) complanatum: Freeh, 1902, стр. 59, рис. 18, Ь в тексте.
Manticoceras carinatum: Wedekind, 1913, стр. 63, табл. V, фиг. 3, 4, рис. 10, &]_2 в тексте; 1918, стр. 124—125, табл. XXI, фиг. 9, рис. 32, Ь в тексте; Freeh, 1913, стр. 22; Наливкина, 1947, стр. 167, табл. XLIII, фиг. 4, рис. 20 в тексте.
Голотип происходит из франских отложений Западной Германии.
Форма. У молодых экземпляров раковина эволютная, дискоидальная, образованная субтреугольными в поперечном сечении оборотами, с узко округленной вентральной, слабо выпуклыми боковыми и вогнутой дорсальной сторонами. Умбо широкое, сглаженное, в целом широко-коническое. Начиная с 3—4-го оборота, одновременно с возрастанием их в высоту, увеличивается инволютность раковины, вентральная сторона становится более широкой, боковые стенки постепенно уплощаются. Во взрослом состоянии раковина дискоидальная, с наибольшей шириной вблизи умбонального края. Поперечное сечение оборота приобретает субтрапе-цоидальное очертание, с широкими упрощенными, слегка выпуклыми боковыми, широко округленной вентральной и сильно вогнутой дорсальной сторонами. Умбо во взрослом состоянии умеренно широкое, ступенчатое, с довольно высокими, округленными умбональными стенками. Умбональ-ный край округленный. Последний оборот облекает около 2/3 предыдущего.
Размеры: д В Ш Ду В/Д Ш/Д Ду/Д
77Л/71 J 93,6 45,3 31(?) 20,0 0,48 0,33(?) 0,21
1/4//1 (75,5 36,6 28,7 13,8 0,48 0,38 0,18
(68,8 34,4 24,0 12,5 0,50 0,35 0,18
№ 774/72 (43,1 22,0 15,3 7,0 0,51 0,35 0,16
102
(66,3 №774/43 |61;3 № 774/42 55,5 32,0 29,6 28,0 24,0 21,8 20,0 12,5 0,48 11,5 0,48 9,3 0,50 0,36 0,36 0,36 0,19 0,19 0,17
Скульптура. Раковина гладкая, снабженная лишь «струйками нарастания, более отчетливыми в области умбо. ТОНКИМИ
Лопастная лини я (рис. 27). Вентральная лопасть широкая,
умеренно глубокая, трехраздельная, с очень коротким, округленным
Рис. 27. Лопастная линия Manticoceras carinatum (Beyrich) (Xl,5); № 774/43 при В=23,5 и Ш = 18,0 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
«срединным и более длинными, узкими, клиновидными, приостренными на концах боковыми зубцами. Первое боковое седло высокое, почти симметричное, широко округленное у вершины, очень широкое, занимающее всю боковую поверхность оборота. Боковая лопасть асимметричная, широкая, слегка загнутая в сторону умбо своим приостренным концом, расположенным вблизи умбонального края; глубина боковой лопасти меньше глубины вентральной лопасти. Второе боковое седло равномерно округленное, симметричное, очень низкое, умеренно широкое; середина его располагается за умбональным краем, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Наши экземпляры наиболее близки к М. carinatum (Beyrich), отличаясь от него лишь незначительными отклонениями формы поперечного сечения оборотов (слабо выпуклые, слегка сходящиеся кнаружи боковые стороны в отличие от уплощенных, параллельных друг другу боковых сторон М. carinatum) и, возможно, немного более тесным умбо. Помимо вышеуказанных особенностей, лопастная линия М. carinatum, изображенная Зандбергерами (1850—1856, табл. VI, фиг. 4,6), показывает округленную боковую лопасть. Возможно, что эта округленность объясняется степенью сохранности, тем более что Ведекинд подобного закругления для М. carinatum не указывает. Не имея возможности детального сравнения наших экземпляров с голотипом М. carinatum и не наблюдая на них резко отличных признаков, я воздерживаюсь от выделения их в новый вид и отношу к М. carinatum XBeyrich). Некоторое сходство наши экземпляры обнаруживают и с М. intumescens var. latisellati Janischevsky (Янишевский, 1926) из франского яруса Новой Земли, отличаясь, однако, от него меньшей глубиной боковых зубцов вентральной лопасти и более уплощенной с боков раковиной.
Геологический возраст и распространение. Встречается во франских отложениях Западной Европы, Урала, Рудного Алтая.
Местонахождение. Двадцать шесть экземпляров разной
103
сохранности найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта; 22 экземпляра и некоторое количество фрагментарных обломков обнаружены на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта) франского яруса.
Мanticoceras аттоп (Keyserling)
Табл. IV, фиг. 2.
Goniatites аттоп: Keyserling, 1844, стр. 226, табл. А, фиг. 1; 1846, стр. 283, табл. 12^ фиг. 1; табл. 13, фиг. 2.
Gephyroceras intumescens: Foord and Crick, 1897, стр. 287 (pars).
Manticoceras аттоп: Гольцапфель, 1899, стр. 23, табл. I, фиг. 4—8, 10, Налив-кина, 1947, стр. 166, табл. XLII, фиг. 10, рис. 18 в тексте.
Голотип происходит из доманикового горизонта южного Тимана.
Форма. На ранних стадиях развития раковина эволютная, дискоидальная, образованная равномерно округленными в поперечном сечении оборотами, со слегка вогнутой дорсальной стороной. Умбо широкое, но неглубокое. Начиная с третьего оборота, одновременно с возрастанием его в высоту, увеличивается инволютность раковины, боковые стенки постепенно уплощаются, вентральная сторона становится широко округленной. Во взрослом состоянии раковина дискоидальная, с наибольшей шириной вблизи умбонального края. Поперечное сечение оборота субтрапецоидаль-ного очертания, с широкими, слегка выпуклыми боковыми, широко округленной вентральной и вогнутой дорсальной сторонами. Умбо умеренно широкое, ступенчатое, с довольно высокими, несколько округленными умбональными стенками, отграниченными от боковых сторон довольно отчетливым перегибом в области умбонального края. Последний оборот облекает немногим более половины предыдущего.
Размеры: Д В Ш Ду В/Д И1/Д Ду/Д
№ 774/70 57,7 26,5 23,0 13,2 0,46 0,40 0,23
№ 774/20 39,0 18,6 16,3 8,5 0,48 0,42 0,22
№ 774/76 34,6 16,9 14,1 8,0 0,49 0,41 0,23
№ 774/77 33,8 16,7 14,3 7,2 0,49 0,42 0,21
№ 774/75 33,3 16,4 14,2 7,6 0,49 0,43 0,23
№ 774 35 30,7 14,6 13,0 7,0 0,48 0,42 0,23
Скульптура. Раковина гладкая, снабженная лишь тонкими струйками нарастания, более отчетливыми в области умбо, где они переходят в морщинки умбональной стенки.
Лопастная линия (рис. 28). Вентральная лопасть довольно широкая, глубокая, трехраздельная, с очень коротким, почти незаметным, округленным срединным и узкими, сравнительно длинными клиновидными боковыми зубцами, приостренные концы которых слегка отогнуты в сторону умбо. Первое боковое седло высокое, асимметричное, тупо округленное при вершине, широкое, занимающее всю боковую поверхность оборота, со слегка округленной внешней и прямой внутренней сторонами. Боковая лопасть асимметричная, загнутая в сторону умбо своим приострен-ным концом, расположенным в области наибольшей ширины раковины у умбонального края; глубина ее гораздо меньше глубины вентральной лопасти. Второе боковое седло равномерно округленное, умеренно широкое, низкое; середина его располагается на умбональной стенке непосредственно за умбональным краем, а внутренняя часть переходит за умбональный шов.
104
Сходство и отличие. Этот вид имеет наибольшее сходство с М. intumescens (Beyrich), но, однако, отличается от него менее резко выраженной расчлененностью и глубиной вентральной лопасти. Кроме того, у М. ammon струйки нарастания менее изогнуты и образуют менее глубокие синусы. При хорошей сохранности раковины хорошим отличительным признаком может служить морщинистый слой, который у М. ammon
Рис. 28. Лопастная 'линия Manticoceras ammon (Keyserling) (X 1,5); № 774/70 приВ=25,0и 111=23,0 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
(Keyserling) ограничивается спинной стороной, не распространяясь на боковые стороны. Помимо этих отличий, М. ammon имеет более толстую раковину. Имеющий некоторое сходство с описываемым видом М. wurmii (Romer) отличается от него закругленной боковой лопастью, более уплощен с боков и имеет более широкое умбо. Из североамериканских видов ближе других к описываемому стоит М. simulator (Hall), но приведенное Голлом изображение не позволяет произвести более точное сравнение, и лопастная линия при общем совпадении с лопастной линией М. ammon показывает сильно округленную боковую лопасть.
Геологический возраст и распространение. Встречается во франских отложениях (мантикоцеровые слои) Новой Земли, Тимана, Урала, Рудного Алтая.
Местонахождение. Шесть полных экземпляров и некоторое количество фрагментарных обломков найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта франского яруса.
Manticoceras drevermanni Wedekind
Табл. V, фиг. 3
Manticoceras drevermanni'. Wedekind, 1913, стр. 61, табл. V, фиг. 5, 6, рис. И &i-2 в тексте; 1918, стр. 125, табл. XXII, фиг. 11, рис. 34, 62 в тексте.
Голотип происходит из отложений зоны 1о мантикоцеровых слоев франского яруса Германии.
Форма. На ранних стадиях развития раковина эволютная. Поперечное сечение оборотов округлое с равномерно закругленными вентральной и боковыми и слегка вогнутой дорсальной сторонами. Умбо широкое. Начиная с третьего оборота, одновременно с его возрастанием в высоту, увеличивается инволютность раковины, боковые стенки уплощаются. Во взрослом состоянии раковина дискоидальная, с наибольшей шириной вблизи умбо. Поперечное сечение оборота приобретает субтреугольное очертание с широкими, слегка выпуклыми боковыми, округленной вентральной и вогнутой дорсальной сторонами. Умбо узкое, с доволь
105
но высокими крутыми округленными умбональными стенками. Последний оборот облекает около 3/4 предыдущего.
Размеры: Д В Ш Ду В/Д ш/д Ду/Д
/ 28,5 15,8 12,1 4,6 0,55 0,42 0,16
№ 774/26 | 17,0 8,6 8,3 3,8 0,51 0,49 0,22
№ 774/27 27,6 14,8 11,7 4,3 0,54 0,42 0,16
№ 774/28 25,7 13,2 10,6 4,1 0,55 0,41 0,16
Скульптура. Раковина гладкая, снабженная лишь тонкими
слабо заметными струйками нарастания, более отчетливыми в области умбо.
Лопастная линия (рис. 29). Вентральная лопасть широкая, трехраздельная, с очень коротким срединным и длинными боковыми зубцами. Боковые зубцы сосцевидные, довольно глубокие и широкие,
Рис. 29. Лопастная линия Manticoceras drevermanni Wedekind (Х13); № 774/27 при В=10,7 и Ш=8,8 мм. Рудный Алтай, рч. Грязнуха у с. Раздольного. Франский ярус.
с приостренными, слегка оттянутыми концами. Первое боковое седло высокое, асимметричное, остро округленное при вершине, со слегка выпуклой, скошенной вверху наружной и слабо вогнутой внутренней сторонами. Боковая лопасть асимметричная, слегка загнутая в сторону умбо своим приостренным, несколько оттянутым концом, расположенным в области наибольшей ширины раковины; основания боковой и вентральной лопастей лежат на одном уровне. Второе боковое седло равномерно округленное, низкое, широкое; середина его располагается на умбональ-ном крае, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Согласно Ведекинду (1913, стр. 62), М. drevermanni Wedekind является промежуточной формой между М. cordatum (Sandberger) и М. carinatum (Beyrich). М. cordatum отличается от описанного вида уплощенными, сходящимися под углом боковыми сторонами и большим возрастанием оборотов в высоту. Помимо этого, при равных размерах раковин представителей того и другого вида М. cordatum характеризуется меньшей степенью развития ветвей вентральной лопасти. М. carinatum отличается от М. drevermanni очень низким срединным седлом слабо развитой вентральной лопасти и широким, занимающим всю боковую сторону оборота округленным первым боковым седлом; наблюдаются различия и в форме поперечного сечения оборотов указанных видов.
Геологический возраст и распространение. Встречается в верхнефранских отложениях Западной Германии, Урала и Рудного Алтая.
Местонахожден ие. Пять экземпляров найдены на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта франского яруса.
106
Manticoceras intumescens (Beyrich)
Табл. V, фиг. 1,2
Ammonites intumescens: Beyrich, 1837, стр. 36, табл. II, фиг. 3.
Goniatites intumescens: Roemer F. A., 1843, стр. 33, табл. IX, фиг. 8, 9, 15; 1879, стр. 659; Branco, 1880, табл. VI, фиг. 4, а — d\ Holzapfel, 1882, стр. 242, табл. XLVI, фиг. 2; Чернышев, 1887, стр. 2, табл. I, фиг. 1.
Manticoceras intumescens: Hyatt, 1884а, стр. 318; Гольцапфель, 1899, стр. 21, табл. 1, фиг. 1—3, 9; табл. 2, фиг. 1—2, 5; Freeh, 1913, стр. 22; Wedekind, 1913, стр. 52, табл. IV, фиг. 9, рис. 6 в тексте; 1918, стр. 126, рис. 35, а, Ь, с в тексте; Наливкина, 1947, стр. 166, табл. ХЕШ, фиг. 1—3, рис. 17 в тексте.
Gephyroceras intumescens: Foord and Crick, 1897, стр. 62—63 (pars).
Gephyroceras (Manticoceras) intumescens: Freeh, 1902, стр. 59, рис. 18, с в тексте.
Голотип происходит из верхнедевонских отложений (зона I7 мантикоцеровых слоев) Германии.
Форма. У небольших экземпляров раковина дискоидальная, инво-лютная, довольно толстая. Поперечное сечение оборота субквадратное, с примерно равными высотой и шириной, с выпуклыми боковыми, широкой округленной вентральной и вогнутой дорсальной сторонами. Степень завивания равна 2/з—3/4« вследствие чего умбо узкое и глубокое, с довольно высокими, несколько округленными умбональными стенками. У крупных экземпляров в связи с большим возрастанием оборотов в высоту, нежели в ширину, раковина несколько уплощается с боков. Во взрослом состоянии раковина дискоидальная, инволютная, с наибольшей шириной вблизи умбо. Поперечное сечение оборота приобретает субтрапецоидаль-ное очертание, с широкими, слабо выпуклыми боковыми, равномерно округленной вентральной и вогнутой дорсальной сторонами. Умбо умеренно широкое, ступенчатое, с довольно высокими округленными умбональными стенками. Последний оборот облекает около 2/3 предыдущего.
Размер ы: Д В Ш Ду в/д Ш/Д Ду/Д
№ 774/1 /118,0 53,5 43,0 24,0 0,46 0,37 0,20
| 87,5 42,5 — 17,0 0,49 — 0,20
№ 774/82 I 47,0 22,5 — 10,5 0,49 — 0,22
I 37,6 18,0 17,9 8,9 0,48 0,48 0,24
Скульптура. На исследованных экземплярах, ввиду недостаточной сохранности раковинного слоя, скульптура не видна. Раковина гладкая. По наблюдениям других исследователей, гладкая поверхность раковины снабжена тонкими струйками нарастания, слабо заметными. На боковых сторонах и в области умбо при хорошей сохранности может наблюдаться морщинистый слой.
Лопастная линия (рис. 30). Лопастная линия характеризуется очень высокими, сравнительно широкими седлами и узкими глубокими лопастями. Особенно сильно развито вентральное седло. Лопастная линия мелких экземпляров характеризуется еще слабо развитыми округленными лопастями. При величине диаметра в 33 мм (рис. 30,а) вентральная лопасть широкая, трехраздельная, с коротким воронкообразным срединным зубцом и длинными, умеренно широкими, приостренными на концах боковыми зубцами. Первое боковое седло высокое, довольно широкое, округленное у вершины, асимметричное, с выпуклой, покатой наружной и крутой, слегка вогнутой внутренней сторонами. Боковая лопасть асимметричная, широкая, загнутая в сторону умбо своим приострен-ным оттянутым концом, расположенным в области умбонального края; глубина ее много меньше глубины вентральной лопасти. Второе боковое седло широкое, низкое, равномерно округленное, середина его располагается на умбональной стенке, а внутренняя часть переходит за умбональ-
107
ный шов. С ростом раковины происходит дальнейшее усложнение лопастной линии, проявляющееся главным образом в увеличении глубины и приостренности лопастей. Форма лопастной линии при диаметре в 80 мм изображена на рис. 30,6. На рисунке можно видеть, что вентральная лопасть широкая и глубокая, трехраздельная, с укороченным воронкообразным срединным и длинными, узкими, асимметричными боковыми зубцами. Первое боковое седло широкое и очень высокое, асимметричное, скошенное вверху с наружной стороны, округленное при вершине, с выпук-
Рис. 30. Лопастные линии Manticoceras intumescens (Beyrich): а— № 774/82 при В =14,0 и Ш=15,6мм (Х2); Ь—№ 774/1 при В=39,0 и Ш=30,5 мм (XI). Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
лой наружной и почти прямой, слегка вогнутой внутренней сторонами. Боковая лопасть асимметричная, широкая, свернутая в сторону умбо своим оттянутым острым концом, расположенным на боковой стороне вблизи умбонального края; глубина ее много меньше глубины вентральной лопасти. Второе боковое седло широкое, несколько асимметричное, умеренно высокое, вершина его располагается за умбональным краем, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Вид Manticoceras intumescens (Bey-rich) многими исследователями, начиная с Зандбергеров, рассматривается слишком широко. Некоторые исследователи относят к нему М. ammon (Keyserling) и М. wurmii (Romer) (Foord and Crick, 1897), другие (Ремер) причисляют к нему форму с острой режущей стороной. Однако экземпляр, принятый Бейрихом за голотип, имеет толстую раковину с выпуклыми боковыми сторонами и широкой округленной внешней стороной. Изображение его воспроизводится Гольцапфелем (1899, табл. I, фиг. 9). Кроме экземпляров, непосредственно примыкающих к типу, наблюдаются более или менее уклоняющиеся от него формы. Гольцапфель (1899, стр. 22), который причисляет к М. intumescens в качестве вариетета М. cordatum (Sandberger), различает внутри вида три разновидности: 1) нормальную форму с сильно и равномерно выпуклыми боковыми сторонами, 2) формы с плоско-выпуклыми параллельными и широкой вентральной сторонами [М. intermedium (Sandberger)], 3) формы с очень плоско-выпуклыми, кнаружи слабо сходящимися боками и поэтому с узкой, но равномерно округленной вентральной стороной — М. sinuosum (Hall). Две первые разновидности, по Гольцапфелю, могут быть более толстыми и более тон-108
кими, тогда как у третьей ширина оборотов, по-видимому, менее изменяется. Ведекинд (1913) рассматривает в качестве самостоятельных видов М. cordatum и М. intermedium, а Миллер (1938) считает обособленной, хотя и близкой к М. intumescens, целую группу сходных между собой североамериканских видов, группирующихся вокруг М. sinuosum.
М. intumescens (Beyrich) отличается от М. аттоп (Keyserling) более высокими вентральными седлами, более узкими и глубокими лопастями. Кроме того, у М. аттоп струйки нарастания менее изогнуты и образуют менее глубокие синусы.
При хорошей сохранности раковины отличительным признаком может служить морщинистый слой, который у М. аттоп ограничивается спин_ ной стороной, а у М. intumescens распространяется также и на боковые стороны. М. wurmii (Romer) отличается от описываемого вида закругленной боковой лопастью и, будучи более плоским, имеет широкое умбо. Североамериканская группа видов, тяготеющих к М. sinuosum, отличается от М. intumescens, по существу, лишь формой раковины, имеющей узкую округленную вентральную сторону и субтреугольное поперечное сечение оборотов.
М. intermedium (Sandberger), выделенный Ведекиндом (Wedekind, 1913, стр. 64), отличается от М. intumescens большей шириной раковины и формой поперечного сечения оборота с плоско-выпуклыми параллельными боковыми и широкой внешней сторонами. Иных отличий между указанными двумя видами не наблюдается и, возможно, что при наличии переходных форм, их следует объединять в один вид. В один же вид с М. intumescens, вероятно, следует объединять и М. buchi (Archiac et Verneuil).
Геологический возраст и распространение. Часто во франских отложениях Западной Европы, Урала, Тимана, Рудного Алтая и других областей.
Местонахождение. Один крупный экземпляр найден на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника; шесть более мелких экземпляров, частью неполной сохранности, найдены в 3 км на юго-восток от поселка в известняковых линзах гониатитового горизонта.
Manticoceras sinuosum (Hall)
Табл. Ill, фиг. 2—4
Goniatites sinuosus: Hall, 1843, стр. 244, 246, рис. в тексте 106 (6), 107(9); 1876, стр. 72, фиг. И; 1879, стр. 460—463, табл. 70, фиг. 13—15; табл. 72, фиг. И; табл. 74, фиг. И.
Goniatites patersoni: Hall, 1860, стр. 99, рис. 9, 10 в тексте; 1879, табл. 72, фиг. 1—5; табл. 74, фиг. 15.
Goniatites (Clymenia?) Nundaia: Hall, 1875, стр. 134; 1876, табл. 70, фиг. 13—15.
Gephuroceras sinuosum: Hyatt, 1884a, стр. 317.
Manticoceras patersoni: Hyatt, 1884a, стр. 318; Clarke, 1899, стр. 42, 45—62, 80, 101, рис. в тексте 1—28, 63, 64, табл. I, фиг. 1—12; табл. II, фиг. 1—4, 6; табл. IV, фиг. 14—18; Freeh, 1913, стр. 23—24.
Goniatites (Manticoceras) intumescens: Grabau, 1899, стр. 299, рис. в тексте 235, Manticoceras intumescens: Grabau and Shimer, 1910, стр. 136, фиг. 1389, a, 1389, b 1390, а в тексте.
Manticoceras sinuosum: Miller A. K., 1938, стр. 106, табл. 18, фиг. 4; табл. 19, фиг. 1—9; табл. 20, фиг. 6—10; табл. 33, фиг. 2, рис. 20, 21, 22 в тексте.
Голотип происходит из франских отложений (формация Нейплс) штата Нью-Йорк США.
Форма. На ранних стадиях развития раковина эволютная, образованная уплощенными в дорсо-вентральном направлении оборотами.
109
Поперечное сечение первых двух оборотов субтрапецоидальное, со слабо выпуклой вентральной, несколько вогнутой дорсальной и почти перпендикулярными к ним уплощенными боковыми сторонами. Углы, образованные двумя смежными сторонами, равномерно округлены. Умбо широкое, блюдцеобразное. Поперечное сечение третьего оборота, остающегося эволютным, становится уже более округленным. Начиная с четвертого оборота, одновременно с большим его возрастанием в высоту, увеличивается инволют-ность раковины, боковые стенки уплощаются, вентральная сторона раковины становится узко округленной, приближаясь к угловатой. Во взрослом состоянии раковина дискоидальная, с наибольшей шириной вблизи умбонального края. Поперечное сечение оборота приобретает субтреугольное очертание, с широкими слегка выпуклыми боковыми, узкой округленной, иногда угловатой вентральной и вогнутой дорсальной сторонами. Умбо узкое, ступенчатое, с относительно высокими равномерно округленными, умбональными стенками. Последний оборот облекает около 3/4 предыдущего. Форма раковины (соотношение высоты и ширины оборота), характер вентральной стороны и умбо в пределах вида изменчивы.Отношение В/Д колеблется у крупных экземпляров в пределах 0,50—0,55, ш/д-о ,40—0,50, Ду/Д — 0,13—0,19. Ввиду наличия в наших коллекциях переходных форм, связывающих между собой на первый взгляд разнящиеся друг от друга экземпляры, мне представляется возможным рассматривать эти колебания как индивидуальную изменчивость в пределах одного вида.
Раз меры: Д В Ш Ду В Д Ш/Д Ду/Д
95,5 49,0 27,4 16,0 0,51 0,30 0,17
№ 774/81 j 70,2 37,0 22,0 13,0 0,53 0,31 0,18
№ 773/12 95,0 49,0 29,0 16,5 0,52 0,31 0,17
№ 773/16 66,0 35,7 30,0 9,6 0,54 0,45 0,15
№ 774/15 54,8 28,7 22,3 9,0 0,52 0,41 0,16
№ 774/60 52,0 28,0 21,0 8,5 0,54 0,40 0,16
№ 774/29 49,5 25,8 22,4 8,9 0,52 0,45 0,18
(49,2 26,0 21,0 8,0 0,53 0,43 0,16
774/68 30,6 15,6 14,5 6,2 0,51 0,47 0,20
№ 18,5 8,9 10,0 5,2 0,48 0,54 0,28
.11,4 4,7 6.7 4,0 0,41 0,59 0,35
Скульптура. Раковина гладкая, снабженная лишь тонкими струйками нарастания, более отчетливыми в области умбо и на вентральной стороне раковины. На вентральной стороне раковины иногда наблюдаются очень тонкие продольные штрихи, образующие со струйками нарастания весьма нежную сеть. На некоторых раковинах сохранился слабо заметный невооруженным глазом морщинистый слой, покрытый густой сетью нежных дихотомически ветвящихся жилок.
Лопастная линия (рис. 31). Вентральная лопасть широкая и глубокая, трехраздельная, со сравнительно коротким, широким воронкообразным срединным и узкими, длинными клиновидными боковыми зубцами, приостренные концы которых несколько скошены и слегка отогнуты в сторону умбо. Первое бовокое седло высокое, асимметричное, умеренно широкое (ширина его может несколько варьировать), занимающее примерно половину боковой поверхности оборота. Наружная сторона седла, слегка скошенная у вершины, равномерно выпуклая, внутренняя — почти прямая, лишь слегка вогнутая. Боковая лопасть асимметричная, загнутая в сторону умбо своим приостренным концом, тяготеющим к области наибольшей ширины оборота; глубина боковой лопасти гораздо
ПО
меньше глубины вентральной лопасти. Второе боковое седло равномерно округленное, низкое, широкое, середина его располагается на умбональ-ном крае, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Этот вид весьма близок к М. intumescens (Beyrich), и, возможно, должен быть даже объединен с ним в один вид, как это делают многие исследователи. Однако голотип М. intumescens отличается от М. sinuosum эллиптическим поперечным сечением оборотов и несколько более открытым умбо. Лопастные линии обоих видов
Рис. 31. Лопастные линии Manticoceras sinuosum (Hall):
а — № 774/15 приВ=26,0 и 111=22,3 мм (XI); в —№ 773/2 при В =30,7 и Ш=24,0 мм (XI)- Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
также несколько разнятся. Не имея возможности детального сравнения голотипов обоих видов, а также не наблюдая в наших коллекциях переходных между этими видами форм, я воздерживаюсь в настоящее время от их объединения. Из североамериканских видов к М. sinuosum близки М. regulare Fenton et Fenton, M. cordiforme Miller, M. septentrionale Miller, которые, возможно, должны быть с ним объединены или даже отнесены вместе с ним к М. intumescens.
Геологический возраст и распространение. Встречается во франских отложениях Северной Америки, Северной Африки, Западной Европы, Тимана, Рудного Алтая и, вероятно, Урала.
Местонахождение. Двадцать два экземпляра найдены на западном склоне гряды, отходящей на северо-восток от Николаевского рудника, в известняковых линзах гониатитового горизонта; восемь экземпляров найдены в трех км к юго-востоку от поселка в известняковых линзах того же горизонта; 14 экземпляров и большое количество обломков найдены на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта франского яруса.
Manticoceras eliseevi sp. nov.1
Табл. IV, фиг. 3, 4
Голотип — ПИН № 774/96; Рудный Алтай, правый берег рч. Грязнухи у с. Раздольного; известняковые прослои песчано-сланцевога горизонта верхней половины франского яруса.
Форма. Раковина на ранних стадиях эволютная, дискоидальная, образованная равномерно округленными, несколько уплощенными в дор-
1 Вид назван в честь Н. А. Елисеева, впервые обнаружившего местонахождение гониатитов в районе Гериховского месторождения.
111
со-вентральном направлении оборотами. В поперечном сечении обороты несколько растянуты в ширину, с равномерно округленными вентральной и боковыми и слегка вогнутой дорсальной сторонами. Умбо широкое, но неглубокое. Начиная с третьего оборота, одновременно с возрастанием его в высоту, увеличивается инволютность раковины, боковые стенки постепенно уплощаются, вентральная сторона становится узкой, приближаясь к угловатой. Во взрослом состоянии раковина дискоидальная, приближающаяся к линзовидной, с наибольшей шириной в области умбонального края. Поперечное сечение оборота приобретает субтреугольное очертание с широкими слегка выпуклыми боковыми, узкой округленной, приближающейся к угловатой, вентральной и вогнутой дорсальной сторонами. Умбо узкое, глубокое, с довольно высокими, крутыми, несколько округленными умбональными стенками. Последний оборот облекает от 4/5 до 5/6 предыдущего.
Размеры: Д В Ш Ду в/д П1/Д Ду/Д
№ 774/98 44,0 23,9 19,2 6,8 0,54 0,44 0,15
№ 774/99 37,4 20,0 17,0 5,4 0,53 0,45 0,14
№ 774/95 34,5 17,8 16,0 5,3 0,52 0,46 0,15
№ 774/97 34,5 18,0 16,2 5,2 0,52 0,47 0,15
№ 774/96 30,4 16,5 14,6 4,3 0,54 0,48 0,14
Скульптура. Раковинный слой сохранился небольшими участками на некоторых экземплярах. Раковина гладкая, снабженная лишь тонкими струйками нарастания.
Лопастная линия (рис. 32). Вентральная лопасть широкая
Рис. 32. Лопастная линия Manticoceras eliseevi sp. nov. (x4); голотип № 774/96 при B=16,3 и Ш=14,5 мм. Рудный Алтай, рч. Грязнуха у с. Раздольного. Франский ярус.
и глубокая, трехраздельная, с коротким воронкообразным срединным и длинными, широкими сосцевидными боковыми зубцами, приостренными и оттянутыми на концах. Первое боковое седло высокое, довольно широкое, тупо округленное при вершине, слегка асимметричное. Боковая лопасть асимметричная, загнутая в сторону умбо своим приостренным, несколько оттянутым концом; глубина ее значительно меньше глубины вентральной лопасти. Второе боковое седло слабо округленное, очень низкое, широкое, середина его располагается на умбональном крае, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Этот вид по форме раковины, степени ее инволютности, характеру навивания оборотов и умбо, а также лопастной линии, отличается от всех ранее описанных видов рода Manticoceras. Из других видов наибольшее сходство с ним по форме раковины и характеру умбо имеют североамериканские виды М. apprimatum Clarke и М. contractum Clarke. Однако ввиду того что для последних неизвестен
112
характер лопастной линии, я воздерживаюсь от отождествления описанного вида с тем или другим из них и выделяю его в самостоятельный.
Геологический возраст и распространение. Встречается во франских отложениях Рудного Алтая.
Местонахождение. Восемь полных экземпляров и несколько фрагментарных обломков найдены на правом берегу рч. Грязнухи у с. Раздольного, в известняковых прослоях песчано-сланцевого горизонта франского яруса
Manticoceras galeatum Wedekind
Табл. Ill, фиг. 5
Manticoceras galeatum: Wedekind, 1913, стр. 60, табл. IV, фиг. 3, 4, рис. в тексте 8, а; 1918, стр. 126, табл. XXII, фиг. 8, рис. в тексте 33, а.
Голотип происходит из отложений верхней зоны мантикоцеровых слоев франского яруса Германии.
Форма. Ввиду отсутствия достаточно полного описания данного вида в работах Ведекинда считаю необходимым дать его характеристику подробнее. Раковина во взрослом состоянии инволютная, дискоидальная, приближающаяся к линзовидной с наибольшей шириной, тяготеющей к умбо. Поперечное сечение оборота субтреугольного очертания, с широкими, слегка выпуклыми боковыми, узкой угловатой вентральной и вогнутой дорсальной сторонами. Умбо узкое, ступенчатое, с довольно высокими, почти прямыми, слегка округленными умбональными стенками. Последний оборот облекает от 4/б до б/6 предыдущего.
Размеры: д В Ш Ду В/Д ш/д Ду/Д
№ 774/30 40,0 21,8 16,0 6,4 0,55 0,40 0,16
№ 774/14 35,5 19,5 15,0 6,0 0,55 0,42 0,17
№ 773/14 24,0 13,2 11,0 4,8 0,54 0,46 0,20
Скульптура. Раковина гладкая, снабженная лишь частыми тонкими струйками нарастания, более заметными в области умбо.
Лопастная линия (рис. 33). Вентральная лопасть широкая, довольно глубокая, трехраздельная. Срединный зубец ее очень короткий, узкий, воронкообразный, отделенный от боковых довольно высокими, но не широкими округленными при вершине седлами. Боковые зубцы вентральной лопасти сравнительно широкие, длинные, слегка асимметричные, с несколько оттянутыми приостренными концами, в целом сосцевидные. Первое боковое седло высокое, асимметричное, округленное у вершины, с выпуклой наружной и слегка вогнутой, почти прямой внутренней сторонами. Боковая лопасть асимметричная, широкая, тупоконечная; стороны ее образуют между собой близкий к прямому тупой угол, тяготеющий к области наибольшей ширины оборота. Концы боковой и вентральной лопастей располагаются на равном уровне. Второе боковое седло округленное, низкое, широкое, середина его располагается за умбональным краем, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Описанный вид отличается от близких к нему М. inversum Wedekind и М. cordatum (Sandberger) наличием во взрослом состоянии узкой угловатой вентральной стороны раковины, которая у М. inversum наблюдающаяся на молодых экземплярах, сменяется у взрослых округленной вентральной стороной, когда поперечное сечение оборота становится аналогичным сечению М. cordatum. От близкого по форме М. koeneni (Holzapfel) описанный вид отличается отсутст-в Б. И. Богословский цд
вием резко выраженной скульптуры раковины, а от Crickites acutus (Sandberger) наличием струек нарастания, образующих синусы на боковой и вентральной сторонах, не свойственных роду Crickites.
Рис. 33. Лопастная линия Manticoceras galeatum Wedekind (х4,5); № 774/30 при В=9,7 и Ш = 10,5 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
Геологический возраст и распространение. Встречается во франских отложениях (зона I7) Западной Германии, Урала, Рудного Алтая.
Местонахождение. Три экземпляра найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта.
Manticoceras artum sp. nov.
Табл. VI, фиг. 2
Голотип — ПИН № 774/80; Рудный Алтай, район Николаевского месторождения, в 3 км юго-восточнее поселка; известняковые линзы гониатитового горизонта франского яруса.
Форма. Раковина тонкодискоидальная, уплощенная с боков, инво-лютная, с наибольшей шириной вблизи умбо. Поперечное сечение оборота субтреугольного очертания, с широкими, уплощенными боковыми, узкой округленной, приближающейся к угловатой, вентральной и вогнутой дорсальной сторонами. Умбо довольно узкое, ступенчатое, с невысокими, равномерно округленными умбональными стенками. Последний оборот облекает около 2/з предыдущего.
Размеры: д В Ш Ду в/д П1/Д Ду/Д
JN® 774/80 (63,6 33,4 19,5 12,3 0,53 0,31 0,19
V 1— • < тх/ (53,0 27,0 16,2 9,5 0,51 0,31 0,18
Скульптура. Раковина гладкая, снабженная лишь чрезвычайно тонкими, еле заметными струйками нарастания.
Лопастная линия (рис. 34). Вентральная лопасть широкая, умеренно глубокая, трехраздельная, с коротким срединным и более длинными боковыми зубцами. Боковые зубцы вентральной лопасти умеренно глубокие и широкие, с приостренными, слегка оттянутыми концами, в целом сосцевидные. Первое боковое седло широкое, занимающее собой почти всю боковую поверхность оборота, но невысокое, асимметричное со скошенной вверху выпуклой наружной и слегка вогнутой, почти прямой внутренней сторонами, остро округленное при вершине. Боковая лопасть асимметричная, несколько загнутая в сторону умбо своим при-остренным концом, расположенным в области наибольшей ширины обо-114
рота; глубина ее гораздо меньше глубины вентральной лопасти. Второе боковое седло равномерно округленное, низкое, неширокое; середина его располагается за умбональным краем, а внутренняя часть переходит за умбональный шов.
Сходство и отличие. Описанный вид по форме раковины и характеру лопастной линии отличается от всех ранее описанных пред-
Рис. 34. Лопастная линия Manticoceras artum sp. nov. (xl,5); голотип № 774/80 при B=28,2 и 111=16,3 мм. Рудный Алтай, район Николаевского месторождения. Франский ярус, гониатитовый горизонт.
ставителей рода Manticoceras. Он наиболее близок к видам из группы М. cordatum, не имея, однако, ни с одним из них полного сходства. От М. cordatum его отличает более уплощенная с боков раковина, от М. galeatum— отсутствие характерной для последнего угловатой вентральной стороны; кроме того, описанный вид отличается от указанных выше характером лопастной линии, особенно наличием широкого, сравнительно невысокого первого бокового седла; имеется отличие и в форме вентральной лопасти. Сравнение нашего вида с М. sinuosum (Hall), с которым его несколько сближают некоторые относительные размеры раковины, показывает, что описанный вид отличается от М. sinuosum более уплощенной с боков тон-кодискоидальной раковиной и иным характером лопастной линии, в особенности первого бокового седла, более широкого и низкого у описанного вида.
Геологический возраст и распространение. Редко во франских отложениях Рудного Алтая.
Местонахождение. Два экземпляра найдены в 3 км к юго-востоку от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта.
Род Komioceras gen. nov.
Timanites'. Гольцапфель, 1899, стр. 41 (pars); Freeh, 1913, стр. 25 (pars); Wedekind 1918, стр. 127 (pars); Miller, 1938, стр. 130 (pars).
Тип рода — Timanites stuckenbergi Holzapfel, 1899, стр. 45, табл. 7, фиг. 8,9. Доманиковый горизонт Южного Тимана.
Диагноз. Раковина дискоидальная, уплощенная с боков, с широкими слегка выпуклыми боковыми и узкой полого-крышеобразной, ограниченной тупыми кантами, вентральной сторонами. Умбо узкое. Наружная часть лопастной линии состоит из трехраздельной вентральной и четырех-пяти округленных или угловатых боковых лопастей. Первая боковая лопасть обычно большая, остальные маленькие. На внутренней стенке развиты клиновидная дорсальная и четыре маленькие округленно-угловатые боковые лопасти (рис. 17,А).
8*
115
Формула лопастной линии, вероятно: (V^V^ULPLPIPU7 : U6U4U2ID.
Видовой состав и геологическое распространение. Кроме типичного вида, известного из доманикового горизонта Южного Тимана, другие виды не известны.
Общие замечания. Исследование имеющихся в распоряжении автора экземпляров Timanites stuckenbergi позволило установить резкие отличия его от типа рода Timanites. Эти отличия, проявляющиеся в первую очередь в характере перегородки и лопастной линии, заставляют выделить указанный вид в самостоятельный род. Новый род легко отличим от других представителей семейства по лопастной линии, имеющей большее количество боковых лопастей, в особенности внутренних.
CjE хМ Е Й С TjB О BELOCERATIDAE FRECH, 1902
Диагноз. Раковина линзовидная, сдавленная с боков, с широкими, уплощенными, слабо выпуклыми боковыми, очень узкой угловатой вентральной и сильно вогнутой дорсальной сторонами. Умбо различных размеров, от довольно большого до узкого, почти закрытого. Поверхность раковины гладкая, несущая лишь тонкие струйки нарастания, образующие широкие, относительно неглубокие синусы на боковых сторонах, угловатые вентро-латеральные выступы и глубокие синусы на вентральной стороне раковины. Лопастная линия у всех, кроме наиболее примитивных, представителей образует дополнительные, возникающие в районе умбонального шва боковые лопасти, и вентральная лопасть в различной степени расчленена адвентивными лопастями.
Филогения. Вопрос о предковом роде для семейства, как и филогения последнего, в настоящее время не вполне ясны. Кларк, устанавливая (1899, стр. 90) род Probeloceras, по-видимому, думал, что этот род является предком Beloceras. Однако такая точка зрения не поддерживается более поздними исследователями. А. К. Миллер (1938, стр. 134) указывает, что тип рода Probeloceras, Р. lutheri Clarke, близко родствен Ponticeras и оба эти рода являются примитивными представителями семейства Gephuroceratidae, а не Beloceratidae. Хотя Beloceratidae почти без сомнения произошли от Gephuroceratidae, изучение внутренних и наружных частей лопастных линий Probeloceras lutheri показало, по мнению Миллера, что существует одна или две отвечающие родовым категориям стадии между Probeloceras и большинством примитивных белоцератид и что родовое наименование Probeloceras поэтому до некоторой степени неправильно. Кроме того, указанный исследователь отмечает, что онтогенетическое развитие лопастной линии Eobeloceras iynx (Clarke) указывает, что Beloceratidae произошли скорее от рода Koenenites нежели Manticoceras и, возможно, Neomanticoceras naplesense не является прямой линией развития остальных белоцератид.
Проведенное мной изучение онтогенетического развития лопастной линии Beloceras sagittarium (Sandberger) показало (см.рис. 36), что на ранних стадиях онтогенетического развития (при В = 0,9 мм) лопастной линии его наблюдается стадия, соответствующая лопастной линии Manticoceras, что указывает на близкое родство двух семейств и возможность происхождения Beloceratidae от наиболее просто организованных Gephuroceratidae. Если у Gephuroceratidae дальнейшее усложнение лопастной линии в процессе эволюции происходило за счет образования дополнительных элементов лопастной линии в районе умбо, то у Beloceratidae наблюдается новый тип усложнения лопастной линии не только за счет возникновения новых элементов ее в районе умбо, но и за счет расчлене
116
ния вентральной лопасти рядом адвентивных лопастей, возникающих при вершине срединного седла (рис. 35). Роды Neomanticoceras, Eobe-loceras и Beloceras образуют, по-видимому, филогенетический ряд, который достаточно отличен от всех других групп девонских аммонитов, для того чтобы рассматриваться как отдельное семейство. Для выяснения конкретных филогенетических связей между семействами Gephuroceratidae и Beloceratidae, так же как и между отдельными родами последнего, нужны
Рис. 35. Филогенетическое развитие лопастной линии представителей семейства Beloceratidae:
а — Manficocerae simulator (Hall) (Х2.7) из формации Нейплс США. По Голлу и Миллеру (1938, стр. 133, фиг. 28,а); b—Neomanticoceras naplesense (Clarke) (увеличение не указано) из формации Нейплс США. По Кларку (1899, стр. 105, фиг. 78); с —Eobeloceras iynx (Clarke) (увеличение не указано) из формации Нейплс США. По Кларку (1899, стр. 104, фиг. 77); d — Beloceras sagittarium (Sandberger) (увеличение не указано) из франских отложений Зап. Германии. По Зандбергерам (1850—1856, табл. IV, фиг. 3, /).
дополнительные углубленные онтогенетические исследования представителей обоих семейств.
Ревизия семейства. В основу систематики рассматриваемого семейства положена главным образом степень расчлененности лопастной линии, в меньшей степени форма раковины и характер умбо. В ходе эволюции семейства Beloceratidae, существовавшего в течение франского века, раковина изменялась от слабо инволютной до почти полностью инволютной. Особенно большим изменениям подверглась лопастная линия, количество элементов которой изменялось в широких пределах.
Семейство Beloceratidae включает роды Neomanticoceras, Eobeloceras и Beloceras, краткая характеристика которых такова:
1К NeomanticocerasSchindewolf, 1936. Раковина дискоидальная, с уплощенными с боков и вдавленными с дорсальной стороны оборотами. Умбо довольно большое. Струйки нарастания образуют синусы на боковой и вентральной сторонах раковины. Наружная часть лопастной линии состоит из очень широкой вентральной лопасти (которая подразделена
117
на пять более меньших лопастей) и одной боковой лопасти. Тип рода —Manticoceras paradoxum Matern (19316) из верхнего девона Германии. К этому же роду относится рассматриваемый часто как синоним первого Neoman-ticoceras naplesense (Clarke) из формации Нейплс штата Нью-Йорк США.
2. Eobeloceras Schindewolf, 1936. Раковина дискоидальная. Обороты вдавлены с дорсальной стороны и, по-видимому, уплощены с боков. Умбо большое. Раковина гладкая, покрытая струйками нарастания, образующими широкий, но неглубокий синус на боковой и глубокий на вентральной сторонах. Наружная часть лопастной линии состоит из очень широкой вентральной лопасти, разделенной на пять более мелких лопастей, и трех боковых лопастей. Седла, разделяющие лопасти, при вершинах заострены. Тип рода — Ammonites multiseptatus Buch (1832), описанный из низов верхнего девона Германии. К этому же роду относится Eobeloceras iynx (Clarke) из формации Нейплс штата Нью-Йорк США.
3. Beloceras Hyatt, 1884. Описание этого рода дано ниже.
Род Beloceras Hyatt, 1884
Pinacoceras: Mojsisovics, 1873, стр. 43, 69 (pars).
Beloceras'. Hyatt, 1884a, стр. 333; Foord and Crick, 1897, стр. 273; Clarke, 1899, стр. 102 (pars); Freeh, 1902, стр. 61; 1913, стр. 26; Wedekind, 1913, стр. 47; 1918, стр. 129; Miller A. К., 1938, стр. 132; Наливкина, 1947, стр. 167.
Тип рода — Goniatites Sagittarius Sandberger, 1850—1856, стр. 77, табл. IV, фиг. 3,а—h (not 3,z—Z), стр. 56, фиг. 7. Нижняя часть (зоны ip—у) верхнего девона Западной Германии.
Д и а г н о з . Раковина линзовидная, сдавленная с боков, с широкими уплощенными слабо-выпуклыми, сходящимися кнаружи боковыми, очень узкой, часто килеватой вентральной и сильно вогнутой дорсальной сторонами. Умбо небольшое, иногда очень узкое, почти закрытое. Поверхность раковины гладкая, несущая лишь тонкие струйки нарастания, образующие широкий синус на боковой и узкий, глубокий на вентральной сторонах. Лопастная линия очень необычна для девонских аммонитов, так как она образует большое количество дополнительных лопастей на боковых частях раковины, тяготеющих к умбо, а вентральная лопасть исключительно широкая и подразделена на более мелкие лопасти, расположенные на вентро-латеральных частях раковины. Число элементов вентральной лопасти достигает 13—15 и более.
Формула лопастной линии:
(V3...V1...V3) VWW^IPU3... : ...U21D
или, возможно:
(V^.2... Уыдд ... V1,1.1.2)V1.1.2V1.2V2UU1U3... : ...U2ID
Видовой состав и геологическое распространение. Кроме типичного вида, известного из верхнего девона Европы, Северной Африки, Алтая, Урала, в настоящее время к роду Beloceras могут быть отнесены: Beloceras praecursor Freeh, описанный из низов верхнего девона Карнийских Альп (Австрия), В. kayseri (Holzapfel) из низов верхнего девона Германии; В. denckmanni Wedekind из тех же отложений. К этому же роду относится В. stenumbilicatum sp. nov. из франских отложений Рудного Алтая, описанный ниже.
Общие замечания. В 1884 г. Хайэтт, устанавливая род Beloceras, отнес к нему один вид Ammonites multilobatus Beyrich (1837) из низов верхнего девона Германии. Фурд и Крик (1897, стр. 274) и Фрех (1913, стр. 26) установили, что формы из верхнего девона Германии, ко
118
торые Г. и Ф. Зандбергеры описали как Goniatites Sagittarius (1850—1856, стр. 77, табл. IV, фиг. 3,а—h (not 3,i—Z), стр. 56, фиг. 7), идентичны образцам, описанным Бейрихом, и название Goniatites Sagittarius Sandberger — синоним Ammonites multilobatus Beyrich.
Однако название Ammonites multilobatus было уже ранее использовано для другой формы Бронном (1832, стр. 159—160). Таким образом, согласно правилам зоологической номенклатуры, видовое название Ammonites multilobatus Beyrich должно быть ликвидировано как гомоним Ammonites multilobatus Bronn, который имеет приоритет. Поэтому для типа рода Beloceras должно употребляться наименование Зандбергеров.
Beloceras sagittarium (Sandberger)
Табл. VI, фиг. 3, 4; табл. VII, фиг. 1
Ammonites multilobatus'. Beyrich, 1837, стр. 33, табл. I, фиг. 6.
Goniatites tenuistriatus: Archiacand Verneuil, 1842, стр. 343, табл. XXVI, фиг. 7, 7 а, Б', 8, 8, а.
Goniatites multilobatus'. Archiac and Verneuil, 1842, стр. 383; Branco, 1880, табл. VI, фиг. VI, a — d\ Holzapfel, 1882, стр. 13, табл. 45, фиг. 2—6.
Goniatites Sagittarius: G.u.F. Sandberger, 1850—1856, стр. 77, табл. IV, фиг. 3, a — h (not 3, i — l), стр. 56, фиг. 7.
Beloceras multilobatum: Hyatt, 1884a, стр. 333; Freeh, 1902, табл. 32, фиг. 9; 1913, стр. 26; Wedekind, 1913, стр. 48, рис. 5, а в тексте; 1918, стр. 129, табл. XXI. фиг. 13, рис. 38, а в тексте; Наливкина, 1947, стр. 167, табл. XLIII, фиг. 4, рис. 20 в тексте.
Beloceras sagittarium: Foord and Crick, 1897, стр. 274, рис. 133 в тексте; Miller А. К., 1938, стр. 132, рис. 28, d в тексте.
Голотип происходит из франских отложений Германии.
Форма. Раковина линзовидная, сильно уплощенная с боков, с широкими, слабо выпуклыми, сходящимися наружу боковыми сторонами, очень узкой, слегка вогнутой у молодых экземпляров и слабо округленной у взрослых форм вентральной и сильно вогнутой дорсальной сторонами. Умбо относительно узкое, сглаженное, ширина его и степень ин-волютности раковины в пределах вида изменчивы. Последний оборот облекает от 3/4 до 7/8 предыдущего.
Размеры: д Формы В с более Ш широк Ду им умбо В/Д Ш/Д Ду/Д
№ 774/730 107,3 55,4 19,3 17,2 0,52 0,18 0,16
№ 774/729 103,3 53,5 19,0 17,0 0,52 0,18 0,16
№ 774/728 91,5 46,7 16,0 16,0 0,51 0,17 0,17
№ 774/727 59,3 30,0 11,2 10,2 Формы с более узким умбо 0,51 0,19 0,17
№ 774/725 64,0 34,0 13,0 8,0 0,53 0,20 0,13
№ 774/726 60,0 33,0 11,5 6,0 0,55 0,19 0,10
Скульптура. Раковина гладкая, покрытая лишь тонкими струйками нарастания. На некоторых раковинах сохранился легко заметный невооруженным глазом морщинистый слой, покрытый густой сетью нежных, дихотомически ветвящихся жилок.
Лопастная линия. Некоторые стадии онтогенетического развития лопастной линии изображены на рис. 36. При высоте оборота раковины, равной 0,9 мм, лопастная линия (наружная ее часть) имеет «ман-тикоцеровый» характер, состоя из широкой трехраздельной вентральной и одной небольшой округленной боковой лопасти. Первое боковое седло широкое, тупо округленное. Следующая стадия развития лопастной линии характеризуется тем, что вентральная лопасть становится более
119
широкой, боковые ветви ее расходятся в стороны, а при вершине по краям срединного зубца происходит закладывание адвентивных лопастей. Одновременно в районе умбонального шва образуются дополнительные боковые лопасти, которые постепенно смещаются на боковую сторону; боковых лопастей две, основания их округлены. При высоте оборота
Рис. 36. Онтогенетическое развитие лопастной линии Beloceras sagittarium (Sandberger):
а — №774/738 при В =0,9 мм (Х22,5); Ъ—тоже при В=2,3 мм (хЮ); с— № 774/737 при В =2,7 мм (X10);d — №774/736 при В—5,Змм(Х8); I—№774/739 при В=7,5 мм (Х7,5);/ —№774/727 при В=22,8 мм(ХЗ); д — то же при В=28,7 мм (X3)- Рудный Алтай. Франский ярус.
раковины, равной 2,7 мм (рис. 36,с), широкая вентральная лопасть состоит из пяти неравных частей: срединный зубец ее воронкообразный, боковые — клиновидные, довольно глубокие. Адвентивные лопасти (по одной с каждой стороны) маленькие, клиновидные. Боковые лопасти простые, округленные в основании, число их до умбонального шва равно трем. Рисунки 36, d, е, /, g показывают следующие стадии развития и усложнения лопастной линии. Это усложнение происходит как за счет увеличения количества боковых лопастей, возникающих в районе умбонального шва и смещающихся на боковую сторону, так и за счет дальнейшего расчленения вентральной лопасти и образования ряда адвентивных лопастей, возникающих по бокам срединного зубца.
120
На рис. 36, g изображена наружная часть лопастной линии экземпляра, имеющего высоту оборота 28,7 мм. На этой стадии лопастная линия имеет уже все элементы, свойственные взрослому экземпляру. Вентральная лопасть состоит из 13 частей. Основания адвентивных лопастей и вершины седел, их разделяющих, приострены. Боковые лопасти простые, первые три из них в основании приострены, остальные — округлены. Общее число боковых лопастей до умбонального шва равно семи. Внутренняя часть лопастной линии не известна.
Перегородка (рис. 8,а). Эволютная часть перегородки занята разрастающейся вентральной лопастью, расходящиеся зубцы которой на одной стороне расположены против соответствующих им зубцов другой стороны п соединены с ними бороздами, глубина которых постепенно убывает к сифону. Первая пара боковых лопастей (U) разобщена дорсальной лопастью. Первая внутренняя боковая лопасть (I) связана со второй наружной боковой (U1). Вторая, третья и следующие внутренние боковые лопасти связаны соответственно с третьей, четвертой и т. д. наружными боковыми.
Сходство и отличие. Описанный вид по форме раковины и характеру лопастной линии имеет сходство с В. praecursor Freeh и В, stenumbilicatum sp. nov., являясь до некоторой степени промежуточным между ними. В. sagittarium отличается от В. praecursor (согласно Фреху) большим количеством элементов наружной части лопастной линии (13 вместо 10), более заостренными адвентивными лопастями и меньшими размерами боковых лопастей. От В. stenumbilicatum его отличает более широкое, сглаженное умбо, отсутствие резких умбональных стенок и меньшее количество элементов лопастной линии при одинаковых размерах раковин.
Геологический возраст и распространение. Вид обычен во франских отложениях Западной Германии, Рудного Алтая и Урала.
Местонахождение. 146 экземпляров различной сохранности найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта; 30 экземпляров найдены в 3 км к юго-востоку от поселка в аналогичных отложениях; 78 экземпляров найдены на правом берегу рч. Грязнухи в известняковых прослоях песчано-сланцевого горизонта франского яруса.
Seloceras stenumbilicatum sp. nov.
Табл. VI, фиг-. 5; табл. VIII, фиг. 1, 2
Голотип — ПИН № 774/703; Рудный Алтай, Николаевское месторождение, западный склон гряды, отходящий на северо-восток от поселка; известняковые линзы гониатитового горизонта франского яруса.
Форма. Раковина линзовидная, сжатая с боков, сильно инволют-ная, с широкими, уплощенными, слабо выпуклыми боковыми сторонами, которые несколько сближаются к умбо и сильно сходятся к вентральной стороне. Наибольшая ширина раковины тяготеет к средней части боковых сторон, несколько ближе к умбо. Вентральная сторона раковины очень узкая, уплощенная, отграниченная от боковых сторон отчетливыми угловатыми, слегка округленными, вентральными краями. Умбо узкое, ступенчатое, ограниченное перпендикулярными к боковым сторонам, резкими, но не высокими умбональными стенками. Последний оборот полностью облекает предыдущий.
121
Размер ы : д В Ш Ду в/д Ш/Д Ду/Д
№ 774/707 69,3 40,0 13,4 5,1 0,55 0,20 0,08
№ 774/703 49,3 27,7 10,3 4,8 0,56 0,21 0,10
№ 774/705 48,2 27,2 9,8 4,7 0,56 0,20 0,10
Ks 774/706 44,2 24,5 8,5 4,6 0,55 0,19 0,10
Скульптура. Поверхность раковины гладкая, покрытая лишь тонкими, слабо различимыми струйками нарастания. При достаточно хорошей сохранности можно наблюдать тонкий морщинистый слой, покрытый густой сетью нежных, дихотомически ветвящихся жилок, более отчетливых в области умбонального края, где они загибаются кзади.
Лопастная линия. На рис. 37 изображена наружная часть лопастной линии голотипа. При величине диаметра раковины 39,0 мм и
Рис. 37. Лопастная линия Beloceras stenumbilicatum sp. nov.; голотип № 774/703 при В=20,7 мм (х 4). Рудный Алтай, Николаевское месторождение.
Франский ярус, гониатитовый горизонт.
высоте оборота 20,7 мм вентральная лопасть очень 'широкая, состоящая из одиннадцати элементов. Адвентивных лопастей по четыре с каждой стороны; форма их ланцетовидная, концы приострены. Седла, разделяющие их, узкие, приостренные при вершинах. В районе широкого воронкообразного срединного зубца намечается образование пятой адвентивной лопасти. Боковые лопасти небольшие; первая из них ланцетовидная, при-остренная на конце, остальные конические, несколько округленные на концах. Всех боковых лопастей до умбонального края 10. Зачаточных умбональных лопастей две.
Сходство и отличие. Этот вид легко отличим от других видов Beloceras по своей сильно инволютной раковине, имеющей узкое умбо, окаймленное четкими умбональными стенками, не свойственными представителям других видов. Из других известных видов рода Beloceras к нему наиболее близок В. sagittarium (Sandberger), который отличается, однако, от описанного вида характером умбо и лопастной линии; при одинаковых размерах раковин общее количество элементов наружной части лопастной линии В. sagittarium (около 13) меньше, чем у В. stenumbilicatum (15, не считая умбональных).
Г еологическийвозраст и распространение. Встречается в средней части франских отложений Рудного Алтая.
Местонахождение. Девятнадцать различной сохранности экземпляров найдены на западном склоне гряды, отходящей на северо-восток от поселка Николаевского рудника, в известняковых линзах гониатитового горизонта.
122
СЕМЕЙСТВО SANDBERGEROCERATIDAE MILLER, 1938
Диагноз. Раковина дискоидальная, слабо инволютная, с широким открытым умбо, скульптированная на боках выдающимися поперечными ребрами и двумя продольными желобками на вентральной стороне. Вентральная лопасть в той или иной степени трехраздельная. Количество боковых лопастей изменяется от 1 до 5—6.
Ревизия семейства. Представители рассматриваемого семейства Sandbergeroceras(-\~ Triainoceras), Schindewolfoceras и Pseudarietites, обладающие богато скульптированными раковинами, были выделены в 1938 г. Миллером (стр. 178) в самостоятельное подсемейство Sandberge-roceratinae, которое рассматривается указанным автором совместно с подсемейством Prolecanitinae в составе семейства Prolecanitidae. Сравнение онтогенетического развития лопастных линий пролеканитид и представителей описываемого семейства показывает, однако, коренное их отличие, проявляющееся в разном развитии вентральной лопасти, простой у пролеканитид и трехчленной у зандбергероцератпд. Характер развития лопастной линии у последних указывает скорее на более тесное родство их с Pharciceratidae, нежели с Prolecanitidae. От фарцицератид, имеющих гладкие раковины, их, однако, отличает свойственная им богатая скульптура. В силу изложенного я считаю необходимым выделить подсемейство Sandbergeroceratinae из семейства Prolecanitidae и возвести его в ранг семейства Sandbergeroceratidae Miller как естественную, вполне самостоятельную группу, родственную семейству Pharciceratidae.
Филогения и систематика семейства Sandbergeroceratidae еще очень далека от ясности. Для того чтобы приблизиться к разрешению этого вопроса, необходимо тщательное исследование онтогенетического развития представителей семейства. Для решения вопроса о предковом роде семейства необходимы дополнительные данные по ранним верхнедевонским представителям аммонитов и углубленное исследование среднедевонских групп. В настоящее время можно лишь предполагать, что предком семейства был какой-то среднедевонский или ранний верхнедевонский представитель, обладавший дискоидальной, широкопупковой, развернутой, возможно скульптированной раковиной и вероятно простой лопастной линией. В настоящее время к семейству относятся верхнедевонские роды Sandbergeroceras, Triainocefas, Schindewolfoceras и Pseudari-etites. В основу их систематики положено очертание лопастной линии, степень расчлененности вентральной лопасти и количество боковых лопастей. Видовой состав каждого из указанных родов не вполне выяснен и некоторые виды относятся к тому или иному роду условно.
Краткая характеристика относимых к семейству родов такова:
1. Sandbergeroceras Hyatt, 1884. Раковина дискоидальная. Обороты сдавленные в дорсо-вентральном направлении, с округленными вентральной и боковыми и слегка вогнутой дорсальной сторонами. Умбо большое и открытое. Струйки нарастания образуют синусы на вентральной стороне раковины. Раковина скульптирована поперечными выдающимися боковыми ребрами или поперечными продолговатыми бугорками вблизи умбонального края. Лопастная линия образует более или менее трехчленную воронкообразную вентральную лопасть и четыре округленные простые боковые лопасти; пятая лопасть расположена в районе умбонального шва. Внутренняя часть лопастной линии состоит из округленной простой дорсальной и одной (?) внутренней боковой лопасти. Седла, разделяющие лопасти, округлены. Тип рода — Goniatites tuberculoso-costatus Sandberger (1850—1856) из верхнего девона Германии.
2. Triainoceras Hyatt, 1884. Описание этого рода дано ниже.
3. Schindewolfoceras Miller, 1938. Раковина субдискоидальная, с округ
123
ленными вентральной и боковыми и слегка вогнутой дорсальной сторонами. Умбо большое. Боковые стороны раковины несут выдающиеся поперечные ребра. Наружная часть лопастной линии состоит, согласно Миллеру, из небольшой воронкообразной вентральной лопасти и шести приостренных на концах простых боковых лопастей. Возможно, однако, что «первая боковая» лопасть (по Миллеру) является в действительности боковым зубцом широкой трехраздельной вентральной лопасти. В таком
Рис. 38. Лопастные линии S andbergeroceras tuberculoso-costatum (Sandberger) и Schindewolf oceras chemungense (Vanuxem):
a — Schindewolfoceras chemungense (Vanuxem) из формации Чемунг штата Нью-Йорк США. По Миллеру (1938, стр. 182, фиг. 38, Ь); (Ь — Sandbergeroceras tuberculosocostatum (Sandberger) из верхнего девона Германии. По Зандбергерам (1850—1856, табл. IV, фиг. 1, /).
случае количество боковых лопастей равно пяти. Тип рода — Goniatites chemungensis Vanuxem (1842) из формации Чемунг штата Нью-Йорк США.
4. Pseudarietites Freeh, 1902. Раковина дискоидальная, эволютная, богато скульптированная выдающимися поперечными ребрами. На вентральной стороне наблюдаются два продольных желобка, ограничивающих срединную килеватую зону. Лопастная линия состоит из глубокой вентральной и широкой, но не глубокой угловатой боковой лопасти. Тип рода — Pseudarietites silesiacus Freeh (1902) из верхнего девона (Clyme-nienkalk) Германии. К этому же роду относится Pseudarietites westfalicus Schmidt из тех же отложений.
Род Triainoceras Hyatt, 1884
Triainoceras («Triaenoceras»): Hyatt, 1884a, стр. 336; Freeh, 1902, стр. 63 (pars); Drevermann, 1903, стр. 85 (pars); Freeh, 1913, стр. 27 (pars); Wedekind, 1913, стр. 47 (pars); 1918, стр. 129 (pars).
Sandbergeroceras: Foord and Crick, 1897, стр. 242 (pars); Miller A. K., 1938, стр. 17S (pars).
Тип рода — Goniatites costatus Archiac et Verneuil, 1842, стр. 340, табл. XXXI, фиг. 1, l,a. Низы верхнего девона Западной Германии.
Диагноз. Раковина на ранних стадиях (первых пяти оборотах) дискоидальная, совершенно эволютная, образованная низкими и широкими, уплощенными в дорсо-вентральном направлении оборотами, ширина которых примерно вдвое больше высоты. На следующих двух-трех оборотах, одновременно с возрастанием в высоту, несколько увеличивается инволютность раковины, принимающей во взрослом состоянии, вероятно, линзовидную форму (рис. 39). Поверхность раковины покрыта
124
Рис. 39. Поперечное сечение Triainoceras costatum (Archiac et Verneuil (xl) из нижней части франского яруса Зап.Германии. По Ф. Древермапну (1903, табл. V, фиг. 4).
редкими выдающимися поперечными ребрами, во взрослом состоянии серповидно изогнутыми; на вентральной стороне раковины развиты две довольно глубокие, округленные продольные бороздки, постепенно сглаживающиеся и менее резко выраженные во взрослом состоянии. Лопастная линия во взрослом состоянии состоит из глубокой и широкой резко трехзубчатой вентральной лопасти и трех-пяти простых, приостренных или несколько округленных, резко убывающих по величине, боковых лопастей. На внутренней стенке развиты довольно глубокие, узкие, неколько прио-стренные дорсальная и одна (?) внутренняя боковая лопасти. Формула лопастной линии и характер перегородки неизвестны.
Формула лопастной линии, вероятно: (V2ViV2) UU1 : ID.
Видовой состав и геологическое распространение. Кроме типичного вида, описанного из низов верхнего девона (зона 1а) Рейнского бассейна Западной Германии, в настоящее время к роду Triainoceras может быть отнесен описанный ниже Т. gerassimovi sp. nov. из нижней части франских отложений Рудного Алтая. К этому же роду, по-видимому, должен быть отнесен и известный из тех же отложений Triainoceras <?) sp.
Общие замечания. В 1842 г. Аршиак и Вернейль описали несколько богато скульптиро-ванных форм из нижнефранских отложений Рейнской провинции: Goniatites costatus, G. costulatus, G. tuberculosus и G. incertus, из которых особенно первый часто упоминается в более поздней литературе. Так как лопастная линия была известна только у первого из этих видов, то вполне понятно, что при обсуждении большинство авторов пришли к различным результатам. Братья Зандбергер (1850— 1856) соединили G. tuberculosus и G. costatus и описали форму под названием G. tuberculosocostatus Archiac et Verneuil (1850—56, стр. 64, табл. IV, фиг. 1; табл. VIII, фиг. 2). Этого же мнения придерживались Фурд и Крик. Все остальные исследователи рассматривали G. costatus и G. tuberculosus как различные виды. Таким образом, существовали разные мнения в отношении систематической принадлежности G. costatus Archiac et Verneuil и G. tuberculosocostatus Sandberger.
В 1884 г. Хайэтт установил первый в качестве типа рода Triainoceras, второй — типа рода Sandbergeroceras. Таким образом, типом рода Sand-bergeroceras стал сборный вид «tuber culo so-costatum», представленный Зандбергерами на двух различных таблицах двумя разными формами. Эти две формы, по всей вероятности, не являются представителями одного вида. Кроме того, одна из них (табл. IV, фиг. 1), по которой Хайэтт установил род Sandbergeroceras, не аналогична изображенным типам G. costatus или G. tuberculosus, так как ее раковина более сильно сжата в дорсо-вентральном направлении. А. К. Миллер (1938, стр. 178) считает поэтому, что номенклатурное положение термина «tuberculoso-costatum» является отчасти неверным, но так как Зандбергеры не стремились пред-
125
дожить новое наименование (т. к. они указывали для своего нового названия авторами Аршиака и Вернейля), тип рода Sandbergeroceras не имеет действительного видового наименования и Миллер предлагает называть его S. sandbergerorum. Однако в 1952 г. тот же автор (Miller А. К., 1952, стр. 357—358) отказывается от своего первоначального заключения и считает, что братья Зандбергер установили для вполне самостоятельного вида (табл. IV, фиг. 1) новое наименование tuberculoso-costatum (или лучше tuberculosocostatum), авторами которого они и должны считаться. Название же S andbergeroceras sandbergerorum Miller должно рассматриваться в качестве синонима 5. tuberculosocostatum (Sandberger). Образец, изображенный Зандбергерами на их табл. VIII, фиг. 2, 2,а, не аналогичен образцу, представленному фигурами 1—1, а—/ на табл. IV [5. tuberculoso-costatum (Sandberger)], но, может быть, представляет S. (?) tu-berculosum (А. V.). Миллер, считает, что тип рода Triainoceras — G. costatus отличается от типа рода Sandbergeroceras лишь тем, что его раковина менее сильно сдавлена в дорсо-вентральном направлении и более инволютна и вентральная лопасть его лопастной линии трехзубчатая. Так как вентральная лопасть S. tuberculosocostatum показывает начинающееся трехчленное деление, Миллер считает, что различия между типами двух родов не являются, по-видимому, родовыми, или даже подродовыми, и рассматривает Sandbergeroceras и Triainoceras как синонимы, как это делали Гольцапфель, Фрех, Циттель, Фурд и Крик и позднее Хайэтт (1900). Так как род Sandbergeroceras имеет приоритет по сравнению с Triainoceras, Миллер предлагает восстановить его и ликвидировать последний. Такая точка зрения, однако, не может не вызвать серьезного возражения. К сожалению, лопастная линия S. tuberculosocostatum, зарисованная Зандбергерами (1850—56, табл. 4, фиг. 1, /)и воспроизведенная Миллером (1938, стр. 182, фиг. 38А), лишена указаний увеличения и параметров соответствующего ей оборота, что затрудняет сравнение ее с соответствующей ей лопастной линией Triainoceras, характер которой на ранних стадиях развития указывался Фрехом (1902, стр. 62, фиг. 20,с) и Древерманном (Drevermann, 1903, стр. 87). Указанные исследователи, наблюдая на ранних стадиях развития лопастной линии Triainoceras costatum Sandbergeroceras"овый характер вентральной лопасти, считали возможным рассматривать G. tuberculosocostatus и с ним род Sandbergeroceras как стадию развития Triainoceras costatum, к чему, однако, нет достаточных оснований в силу резко различного количества элементов лопастной линии (боковых лопастей) на одинаковых стадиях развития вентральной лопасти.
Исследование имеющихся в моем распоряжении алтайских экземпляров Triainoceras gerassimovi sp. nov. показывает, что в ходе онтогенетического развития лопастной линии вентральная лопасть действительно проходит стадию, во время которой она имеет воронкообразную форму с длинным концом и двумя округленными боковыми выступами, напоминая вентральную лопасть Sandbergeroceras tuberculosocostatum. (Подобную стадию в развитии вентральной лопасти проходят впрочем в той или иной степени на разных этапах все аммониты, вентральная лопасть которых испытывает трехчленное деление). Однако в отличие от лопастной линии S. tuberculosocostatum, имеющей четыре наружные боковые лопасти (см. рис. 38,а), лопастная линия Т. gerassimovi насчитывает лишь одну наружную боковую лопасть и намечающуюся в районе умбонального шва вторую, еще зачаточную лопасть. В дальнейшем, одновременно с увеличением количества боковых лопастей (в нашем случае лишь до трех) происходит и быстрое развитие вентральной лопасти, становящейся резко трехзубчатой, в корне отличной от таковой S. tuberculosocostatum. Различны по своей форме и боковые лопасти: у S. tuberculosocostatum они округленные, посте
126
пенно уменьшающиеся в направлении умбонального шва, у Т. gerassim-ovi—заостренные, резко убывающие по величине.
В силу изложенного, я считаю нужным разделить роды Sandbergeroceras и Triainoceras, как вполне самостоятельные.
Triainoceras gerassimovi sp. nov.1
Табл. VIII, фиг. 3
Голотип — ПИН № 773/65; Рудный Алтай, правый берег рч. Грязнухи в двух км ниже с. Раздольного; нижняя часть Неверовских известняков франского яруса.
Форма. На ранних стадиях своего развития раковина дискоидальная, полностью, эволютная, образованная уплощенными в дорсо-вентраль-ном направлении оборотами, ширина которых примерно вдвое больше высоты. Поперечное сечение оборотов субовальное, с уплощенной, очень слабо выпуклой вентральной, округленными боковыми и слегка вогнутой дорсальной сторонами. На вентральной стороне развиваются две продольные бороздки, еще не очень сильно выраженные. Умбо широкое, блюдцеобразное. Начиная с пятого-шестого оборота, одновременно с постепенным возрастанием оборотов в высоту, несколько возрастает инво-лютность раковины; вентральная сторона раковины становится более выпуклой; ее срединная зона, ограниченная двумя резкими продольными желобками, постепенно приобретает угловатую форму; резкие на молодых экземплярах продольные желобки с возрастом постепенно сглаживаются. При величине диаметра примерно в 50 мм раковина дискоидальная, слабо пнволютная, с округленно-угловатой вентральной, равномерно округленными боковыми и вогнутой дорсальной сторонами; Умбо широкое, но не глубокое, округленно-ступенчатое, блюдцеобразное. Последний оборот облекает около 1/3 предыдущего. На имеющихся в моем распоряжении экземплярах жилая камера не сохранилась и форма вполне взрослого экземпляра неизвестна, но более быстрое возрастание последнего сохранившегося оборота в высоту, сопровождающееся приобретением вентральной стороной угловатой формы и увеличением инволют-ности раковины, дает, по-видимому, основания предполагать наличие во взрослом состоянии дискоидальной, близкой к линзовидной, более инволютной раковины.
Размеры: д В Ш Ду В/Д Ш/Д Ду/Д
[52,5 14,0 15,5 28,7 0,27 0,30 0,55
№ 773/65 41,5 10,0 13,4 23,2 0,24 0,32 0,56
1зо,б 7,0 11,0 18,4 0,23 0,36 0,60
Скульптура. Поверхность раковины покрыта резкими, выступающими, поперечными, серпообразно изогнутыми ребрами, которые отражаются и на ядре. Ребра, начинаясь от умбо, проходят перпендикулярно на боковых сторонах, затем, приближаясь к вентральному краю, серпообразно загибаются кпереди и исчезают, не доходя до продольных борозд, окаймляющих срединную зону вентральной стороны. Указанные продольные борозды на молодых экземплярах представляют собой глубокие округленные желобки, постепенно сглаживающиеся с возрастом и менее отчетливо выраженные во взрослом состоянии. Струйки нарастания проходят параллельно ребрам, загибаясь на вентро-латеральных участках раковины вперед, но, переходя за продольные борозды, образуют на вентральной стороне довольно широкий и глубокий синус.
1 Вид назван в память покойного учителя автора профессора Н.П. Герасимова.
127
Лопастная линия. Некоторые стадии развития лопастной линии в онтогенезе показаны на рис. 40. При ширине оборота 4,3 мм и высоте 2 мм (рис. 40,а) наружная часть лопастной линии состоит из вентральной и одной боковой лопастей. Вентральная лопасть глубокая и широкая, воронкообразная, с длинным узким концом и крутыми боковыми
Рис. 40. Онтогенетическое развитие лопастной линии Triainoceras gerassimovi sp. nov.:
a — № 774/405 при B=2,0 и Ш = 4,3 мм (х5); Ъ—голотип № 773/65 при В =2,8 и 111=6,0 мм (Х4); с — то же при В=4,6 и 111=9,3 мм (Х4); d—то же при В = 10,0 и Ш=13,7 мм (ХЗ); е — то же при В=13,5 и Ш = 15,6 мм (ХЗ). Рудный Алтай. Низы франского яруса.
сторонами; боковая лопасть небольшая, округленная. Следующая стадия (рис. 40,6) характеризуется появлением в районе умбонального шва зачаточной второй боковой лопасти и дальнейшим развитием вентральной лопасти; на этой стадии намечается уже трехчленное ее деление путем образования округленных боковых выступов. Дальнейшее развитие лопастной линии происходит очень быстро. Уже на следующем обороте при ширине его 9,3 мм и высоте 4,6 мм (рис. 40,с) лопастная линия имеет все элементы, характеризующие взрослую стадию, и состоит из вентральной и трех боковых лопастей. Вентральная лопасть широкая и глубокая, резко трехзубчатая, с длинным срединным и значительно более короткими приостренными боковыми зубцами. Первая боковая лопасть языковидная, несколько приостренная на конце; глубина ее значительно меньше
128
глубины вентральной лопасти. Две другие боковые лопасти маленькиеу вторая приостренная, третья — округленная на конце. Седла, разделяющие лопасти, округленные. Рис. 40, d, е характеризуют лопастную линию вполне взрослого экземпляра. На рис. 40,е изображена полная лопастная линия голотипа при высоте оборота 13,5 мм и ширине 15,6 мм. Наружная часть лопастной линии состоит из вентральной и трех боковых лопастей. Вентральная лопасть широкая и глубокая, резко трехзубчатая, с длинным, сравнительно узким срединным и ножеобразными несколько более короткими боковыми зубцами, приостренные концы которых слегка отогнуты в сторону умбо. Первая боковая лопасть довольно глубокая, но не широкая, сосцевидной формы, с приостренным, несколько оттянутым концом. Вторая боковая лопасть значительно меньшая по величине, клиновидная, приостренная на конце; третья боковая лопасть маленькая, широкая, асимметричная, на конце заостренная; на внутренней ее стороне, примыкающей к умбональному шву, наблюдается выпуклость. Внутренняя часть лопастной линии состоит из узкой и глубокой, заостренной на конце клиновидной дорсальной и узкой ножевидной первой внутренней боковой лопастей. Седла, разделяющие лопасти, округленные.
Сходство и отличие. Описанный вид отличается от типа рода характером лопастной линии и формой раковины. В отличие от лопастной линии Т. costatum, имеющей четыре боковые лопасти, лопастная линия Т. gerassimovi имеет лишь три боковые лопасти, а зубцы вентральной лопасти более заострены, нежели у Т. costatum. Кроме того, раковина Т. gerassimovi sp. nov. обладает гораздо менее объемлющими оборотами, более широким умбо и богаче скульптирована (количество поперечных ребер на один оборот около 30 вместо 24) по сравнению с раковиной Т. costatum. В силу указанных особенностей я выделяю новый вид рода Triainoceras, называя его Triainoceras gerassimovi sp. nov.
Геологический возраст и распространение. Встречается в низах франского яруса Рудного Алтая.
Местонахождение. Два экземпляра и один обломок жилой камеры найдены на правом берегу рч. Грязнухи, в двух км ниже с. Раздольного, в нижней части неверовских известняков франского яруса.
Triainoceras sp.
Табл. VIII, фиг. 4—7
Среди коллекций аммонитов, собранных из алевролитов правобережья р. Убы в двух км ниже ж.-д. моста и севернее дер. Камышинки, имеется довольно большое количество образцов, принадлежащих, по всей вероятности, к роду Triainoceras, но плохая сохранность исключает возможность более точного их определения. Указанные образцы деформированы, что не позволяет произвести замеры параметров раковины. Представить себе былую форму раковины,однако,можно.Раковина дискоидальная, почти полностью эволютная, с округленными боковыми и вентральной и несколько вогнутой дорсальной сторонами. Умбо широкое, открытое. Поверхность раковины покрыта резкими выступающими поперечными ребрами, серповидно изогнутыми кпереди и сглаживающимися вблизи вентрального края раковины. На вентральной стороне развиты два продольных желобка, окаймляющих срединную, несколько выпуклую зону. Струйки нарастания проходят параллельно ребрам, загибаясь на вентро-латеральных участках раковины вперед, но, переходя за продольные желобки, образуют на вентральной стороне широкие и глубокие синусы. Лопастная 9 Б. И. БОГОСЛОВСКИЙ лил
линия не известна. Данные экземпляры имеют по форме раковины и скульптуры большое сходство с Triainoceras gerassimovi sp. nov., отличаясь от последнего, возможно, лишь более резкой скульптурой взрослых оборотов, однако, не зная лопастной линии описываемых форм, я воздерживаюсь от их объединения с Т. gerassimovi и определяю как Triainoceras sp.
Местонахождение. 56 экземпляров и большое количество фрагментарных обломков найдены в алевролитах правобережья р. Убы, в 2 км ниже ж.-д. моста и севернее дер. Камышинки, в низах франского яруса.
СЕМЕЙСТВО PHARCICERATIDAE HYATT, 1900
Диагноз. Раковина изменяется от дискоидальной, слабо инволютной до линзовидной, полностью инволютной. Поверхность раковины гладкая, несущая лишь тонкие струйки нарастания, образующие широкий сглаженный синус на боковой и резкий на вентральной стороне. Вентральная лопасть трехзубчатая, широкая; в процессе развития средний зубец ее укорачивается, становясь воронкообразным, а боковые расходятся на боковые стороны раковины. Количество боковых лопастей в процессе развития изменяется в широких пределах (от 3—4 до 13—15). Новые лопасти зарождаются в зоне умбо и затем постепенно смещаются на внутреннюю и наружную поверхности раковины.
Филогения. Вопрос о предковом роде семейства еще далеко не ясен. Ведекинд (1913) и Шиндевольф (1936) рассматривают Pharciceras как прямое продолжение филогенетического ряда гефуроцератид Ponticeras —» Manticoceras -» Koenenites -» Timanites, увязывая его непосредственно с Timanites. За основу этих связей принималась степень развития лопастной линии; остальные признаки, и в первую очередь форма раковины, остаются фактически без внимания. Не исключается возможность, что род Pharciceras действительно произошел от гефуроцератид или имеет с ними родственные связи, но непосредственная связь его с инволютным Timanites, к тому же наблюдающимся в несколько более молодых отложениях, не может не вызывать сомнения. Для решения этого вопроса необходимы дополнительные исследования филогении семейства Gephuroceratidae. Представители семейства Pharciceratidae известны из отложений нижней части франского яруса; в более молодых отложениях их пока не обнаружено. Несмотря на сравнительно непродолжительное время существования семейство претерпело значительные изменения, что говорит о быстрой его эволюции. Семейство Pharciceratidae развивалось в направлении Pharciceras -»Synpharciceras —> Neopharciceras, между которыми легко устанавливается непосредственная генетическая связь. Роды Sphaeropharciceras и Nordiceras представляют, вероятно, боковые ветви этого основного направления. Эволюция внутри семейства протекала как в направлении изменения формы раковины от широкопупковой, слабо инволютной к полностью инволютной, так и в направлении усложнения лопастной линии. Лопастные линии, показанные на рис. 41, отображают основные этапы эволюции перегородочной линии Pharciceratidae. Лопастная линия слабо инволютного Pharciceras tridens (Sandberger) (рис. 41, а) характеризуется наличием широкой трехзубчатой вентральной и трех (из них одна умбональная) боковых лопастей, округленных в основании. Срединный зубец вентральной лопасти узкий, глубокий, основание его лежит примерно на одном уровне с округленными концами боковых зубцов. В дальнейшем, в ходе эволюции лопастной линии, при переходе к инволютному Synpharciceras и от него к Neopharciceras (рис. 41, Ъ, с) лопастная линия испытывает значительные изменения: увеличивается количество боковых лопастей, при
130
обретающих в общем ланцетовидную форму; срединный зубец вентральной лопасти укорачивается и расширяется, принимая форму широкой воронки, а расходящиеся боковые зубцы становятся ланцетовидными, не отличаясь по форме от боковых лопастей.
Ревизия семейства. Представители описываемого семейства Pharciceras и Synpharciceras относятся одними авторами к Gephuroceratidae (Ведекинд, Шиндевольф), другими (А. К. Миллер), предположительно, к Prolecanitidae. Исследование родов Pharciceras, Sphaeropharciceras,
Рис. 41. Филогенетическое развитие лопастной линии представителей семейства Pharciceratidae:
а — Pharciceras tridens (Sandberger) (Х5) иа низов франского яруса Зап. Германии. По Зандбергерам (1850—1856, табл. IV, фиг. 2, е); Ь—Synpharciceras clavilobum (Sandberger) (Х7> из низов франского яруса Зап. Германии. По Зандбергерам (1850—1856, табл. VIII, фиг. 3, а;
с — N eopharciceras kurbatovi Bogoslovsky (Х4»5); голотип № 774/374 при Д -33,0 и Ш=14,0 мм. Рудный Алтай. Нижняя часть франского яруса.
Nordiceras, Synpharciceras и N eopharciceras показывает, что они представляют собой естественную, генетически связанную группу, существенно отличающуюся от мантикоцератид и заслуживающую выделения ее в самостоятельное семейство Pharciceratidae. Отнесение представителей описываемого семейства к Prolecanitidae, с представителями которого имеет некоторое сходство Pharciceras, на наш взгляд, не обосновано, так как сравнение онтогенетического развития лопастных линий фарцицератид и пролеканитид показывает коренное их отличие, которое проявляется в разном развитии вентральной лопасти. Вентральная лопасть пролеканитид сходна лишь со срединным зубцом вентральной лопасти фарцице-ратид, боковые зубцы которой соответствуют первой боковой лопасти пролеканитид. Для решения вопроса о возможных родственных связях между семействами Pharciceratidae и Prolecanitidae необходимы дополнительные онтогенетические исследования последнего.
Семейство Pharciceratidae состоит из родов: Pharciceras, Sphaerophar-ciceras, Nordiceras, Synpharciceras и TVeopharciceras, в основу выделения, которых положены форма раковины и степень развития лопастной линии. Краткая характеристика указанных родов такова:
1) Pharciceras Hyatt 1884. Описание этого рода дано ниже.
2) Sphaeropharciceras Bogoslovsky, 1955. Раковина субсферическая, среднеинволютная, образованная широкими низкими оборотами. Вентральная и боковые стороны сливаются в одну выпуклую, правильно округ
9*
131
ленную поверхность. Умбональный край округленный. Умбо средних размеров. Внутренняя поверхность извилины почти концентрична внешней. Лопастная линия состоит из трехраздельной вентральной, двух наружных боковых, умбональной,двух внутренних боковых и дорсальной лопастей. Вентральная лопасть умеренно широкая и сравнительно неглубокая. Тип рода—Sphaeropharciceras sandbergerorum Bogoslovsky (Богословский, 1955), из низов верхнего девона Западной Германии. Других видов не известно.
3. Nordiceras Bogoslovsky, 1955. Раковина тонкодискоидальная, с широкими уплощенными боками и очень узкой вентральной стороной. Вентральный край отчетливый, окаймленный плоской слабо выраженной бороздкой. Умбо широкое, плоское. Наружная часть лопастной линии состоит из неглубокой трехраздельной вентральной, двух боковых и умбональной лопастей; внутренняя часть не исследована. Тип рода — Рго-lecanites timanicus Holzapfel (Гольцапфель, 1899) из низов верхнего девона Южного Тимана. Других видов не известно.
4. Synpharciceras Schindewolf, 1940. Раковина полностью инволют-ная, дискоидальная у молодых и линзовидная у взрослых экземпляров, с широкими, умеренно выпуклыми боковыми стенками. Вентральная сторона — округленная у молодых и узкая, угловатая у взрослых форм. У молодых она несет на себе два неглубоких продольных желобка, ограничивающих слабо выпуклую вентральную зону. Поверхность раковины гладкая, покрытая лишь струйками нарастания. Вентральная лопасть широкая, трехраздельная, с глубоким воронкообразным срединным и ланцетовидными боковыми зубцами. Боковые лопасти простые, маленькие; число их не менее шести. На внутренней стенке развиты дорсальная и не менее трех внутренних боковых лопастей. Тип рода — Goniatites clavilobus Sandberger (1850—1856), из низов верхнего девона Западной Германии. Других видов не известно.
5. N eopharciceras Bogoslovsky, 1955. Описание этого рода приводится ниже.
Р о д Pharciceras Hyatt, 1884
Pharciceras-. Hyatt, 1884 а, стр. 333 (pars); Kayser, 1907, стр. 23 (pars); Wedekind, 1913, стр. 47 (pars); 1918, стр. 127 (pars); Наливкина, 1947, стр. 164.
Prolecanites-. Foord and Crick, 1897, стр. 244 (pars); Freeh, 1902, стр. 62 (pars); 1913, стр. 26 (pars).
Тип рода —Goniatites tridens Sandberger, 1850—56, стр. 66, табл. IV, фиг. 2. Низы верхнего девона (зона 1а) Западной Германии.
Диа г н о з. Раковина на ранних стадиях дискоидальная, слабо ин-волютная, образованная низкими, округленными или слегка угловатыми оборотами, с низкими, равномерно округленными или несколько уплощенными боковыми и широкой, округленной, иногда слегка уплощенной, вентральной сторонами. Умбо широкое, ступенчатое, ограниченное отчетливыми, несколько округленными умбональными стенками. Во взрослом состоянии раковина дискоидальная, приближающаяся к линзовидной, с широкими уплощенными боковыми и узкой, округленной или несколько угловатой вентральной сторонами. Последний оборот облекает до 4/5 предыдущего. Поверхность раковины гладкая, покрытая лишь струйками нарастания, образующими широкий сглаженный синус на боковой стороне и более глубокий на вентральной. Вентральная лопасть широкая, трехраздельная, с узким, глубоким воронкообразным срединным и более широкими ланцетовидными, чаще приостренными, боковыми зубцами. Наружные боковые лопасти простые, небольшие, приостренные «ли округленные на конце. Всех боковых лопастей три-четыре, из них 132
одна умбональная. На внутренней стенке развиты дорсальная и две внутренние боковые лопасти.
Формула лопастной линии:
(V2V1V2) и ию3 : U2ID.
Видовой состав и геологическое распространение. Кроме типичного вида, описанного из фарцицеровых слоев (зона 1а Ведекинда) верхнего девона Германии, Алжира и Марокко, в настоящее время к роду Pharciceras могут быть отнесены: известный: из тех же отложений Ph. lunulicosta (Sandberger) и его локальная южнофранцузская форма Ph.kiliani (Freeh), а также известные из низов франского яруса Германии (зона la) Ph. becheri (Buch), Ph. galeatum Wedekind, Ph. kayseri Wedekind, Ph. lateseptatum (Freeh), Ph. multiseptatum (Quenstedt).
Общие замечания. Хайэтт, устанавливая этот род, отнес к нему морфологически разные виды: эволютный Pharciceras tridens (Sandberger) и инволютный Ph. clavilobum (Sandberger). Более поздними исследователями видовой состав Pharciceras был дополнен рядом новых форм, при этом одни исследователи признавали самостоятельность данного рода, другие относили представителей его к роду Prolecanites (Freeh, 1902),, объединяя подчас далекие не только морфологически, но и генетически виды. Однако сравнение онтогенезов лопастных линий Pharciceras и Рго-lecanites показывает коренное отличие в развитии этих родов. У каменноугольных представителей пролеканитид, онтогенез лопастной линии для которых был прослежен еще А. П. Карпинским, вентральная лопасть в процессе онтогенеза не испытывает деления и остается простой, в то время как у Pharciceras в ходе онтогенеза эта лопасть рано испытывает трехчленное деление и становится затем настолько широкой, что ее боковые зубцы смещаются на боковые стороны, приобретая вид нормальных боковых лопастей, а укорачивающийся воронкообразный срединный зубец напоминает собой вентральную лопасть пролеканитид. Подобное трехчленное деление вентральной лопасти в процессе онтогенеза, прослеженное мной у Neopharciceras, несомненно имеет место и у Pharciceras] на это указывают изображенные Фрехом (1902, стр. 64, фиг. 21,а) некоторые стадии развития лопастной линии Pharciceras (Prolecanites) tridens. Ввиду изложенного я считаю вполне обоснованным выделение Pharciceras в качестве самостоятельного рода. Вопрос о том, какова степень родства между Pharciceras и пролеканитидами, для своего разрешения требует детального исследования их онтогенезов. Goniatites clavilobus Sandberger, резко отличающийся от остальных представителей Pharciceras своей полностью инволютной раковиной, я, присоединяясь к мнению Шиндеволь-фа, считаю необходимым выделить в самостоятельный род Synpharciceras. Род Pharciceras легко отличим от других представителей семейства Pharciceratidae по форме слабо инволютной раковины и характеру лопастной, линии.
Pharciceras kiliani (Freeh)
Prolecanites kiliani'. Freeh, 1902, стр. 64, табл. II, фиг. 14, a — f] 1913, стр. 26.
Г о л о т и п происходит из нижней части франских отложений Южной Франции.
Форма. Описываемые экземпляры имеют слабо инволютную раковину, форма и относительные размеры которой ввиду ее деформированно-сти не могут быть точно определены. Вентральная сторона раковины довольно узкая, округленная у молодых и уплощенная, ограниченная резко угловатыми вентральными краями у взрослых экземпляров. Боко
133-
вые стенки умеренно выпуклые. У крупных экземпляров вблизи вентрального края раковины наблюдается вогнутая зона. Умбональный край отчетливый, несколько округленный. Умбо довольно широкое, ступенчатое, ограниченное отчетливыми умбональными стенками.
Скульптура. Поверхность раковины покрыта тонкими струйками нарастания, более отчетливыми в области вентрального края, где некоторые из них, утолщаясь, образуют у крупных экземпляров низкие сглаженные ребрышки.
Лопастная лпния. Наружная часть лопастной линии, наблюдаемая на фрагментарных обломках, состоит из трехзубчатой широкой вентральной и трех-четырех боковых лопастей. Срединный зубец вентральной лопасти короткий, воронкообразный, боковые — длинные, ланцетовидные, приостренные на концах. Боковые лопасти простые, ланцетовидной формы.
Сходство и отличие. Имеющиеся в коллекции экземпляры по форме раковины и лопастной линии имеют наибольшее сходство с Pharciceras kiliani Freeh (Freeh, 1902, стр. 64, табл. II, фиг. 14, а—f), описанным из девонских отложений Южной Франции и рассматриваемым зачастую как локальная форма Ph. lunulicosta Sandberger (Sandberger G. nnd Sandberger F., 1850—1856, стр. 69, табл. Ill, фиг. 14), известного из нижнефранских отложений (зона 1а) Западной Германии, Алжира и Марокко. Более детальное сравнение наших экземпляров с указанными видами не представляется возможным сделать ввиду плохой сохранности алтайских форм. Возможно, что алтайские формы должны рассматриваться как принадлежащие новому виду, но, не обладая достаточным материалом, я воздерживаюсь от их выделения в самостоятельный вид и отношу к Ph. kiliani Freeh.
Геологический возраст и распространение. Встречается в низах франского яруса Западной Европы и Рудного Алтая.
Местонахождение. Более 30 деформированных плохой сохранности экземпляров и большое количество фрагментарных обломков найдено в алевролитах правобережья р. Убы, в двух км ниже ж.-д. моста; несколько экземпляров обнаружено в подобных алевролитах в 2,5 км севернее дер. Камышинки; низы франского яруса.
Род Ifeopharciceras Bogoslovsky, 1955
N eopharciceras: Богословский, 1955a, стр. 95; 19556, стр. 1104.
Тип рода — N eopharciceras kurbatovi Bogoslovsky, 1955a, стр. 95; 19556, стр. 1105, рис. 1, 2. Рудный Алтай; низы франского яруса.
Д и а г н о з. Раковина полностью инволютная, линзовидная, с широкими, умеренно выпуклыми боковыми стенками, которые сближаются в умбональном направлении и сильно сходятся к сифону, и очень узкой угловатой вентральной стороной. Умбо узкое у молодых экземпляров и полностью закрытое у взрослых. Поверхность раковины гладкая, покрытая лишь очень тонкими струйками нарастания, образующими неглубокий сглаженный синус на боковых и узкий, глубокий на вентральной сторонах раковины. Лопастная линия характеризуется большим количеством (до 13—15) наружных боковых лопастей, имеющих в общем ланцетовидную форму; первые пять-шесть из них приостренные, остальные в основании округлены. Вентральная лопасть трехраздельная, с коротким воронкообразным срединным и длинными ланцетовидными боковыми зубцами. Внутренняя часть лопастной линии состоит из глубокой клиновидной дорсальной и 12—14 небольших, в общем ланцетовидных боковых лопастей.
134
Формула лопастной линии:
V2V1V2UU1U2U4U6U8UiaU12U14U16U18U20U22..
U23U21U19U17U15U13UnU9U7U5U3ID.
Видовой состав и геологическое распространение. Кроме типичного вида, известного из нижней части франского яруса Рудного Алтая, других видов не известно.
Общие замечания. Род N eopharciceras легко отличим от других родов семейства по лопастной линии, имеющей большое количество боковых лопастей. Из других родов более близок к нему по форме раковины и характеру лопастной линии Synpharciceras, имеющий, однако, значительно меньшее количество лопастей.
N eopharciceras кигЪ atari Bogoslovsky 1
Табл. IX, фиг. 1—3
N eopharciceras kurbatovi] Богословский, 1955а, стр. 95; 19556, стр. 1105, рис. 1, 2.
Голотип — ПИН № 774/374; Рудный Алтай, правый берег рч. Золотухи, известняковая линза в 400 м восточнее дер. Горюновки; нижняя часть франского яруса.
Форма. На ранних стадиях развития раковина эллипсоидальная, сильно инволютная, образованная равномерно округленными, серповидными в поперечном сечении оборотами. Умбо узкое, глубокое, ограниченное округленными умбональными стенками. Начиная с четвертого оборота, одновременно с возрастанием его в высоту, инволютность раковины еще более увеличивается. Молодые экземпляры, имеющие раковину из четырех оборотов, приобретают толстодискоидальную, приближающуюся к линзовидной форму, с широкими, умеренно выпуклыми боками и узкой угловатой вентральной стороной. Умбо очень узкое. Во взрослом состоянии раковина полностью инволютная, линзовидная, с широкими, слабо выпуклыми боковыми стенками и очень узкой, округленно-угловатой вентральной стороной.
Размеры: Д В Ш в/д ш/д Ш/В
(74,0 45,0 20,0 0,60 0,27 0,44
№ 774/374 134,0 19,2 14,0 0,57 0,32 0,73
№ 774/373 61,0 35,0 17,5 0,57 0,29 0,50
№ 773/67 33,0 19,4 13,6 0,58 0,41 0,70
№ 774/375 31,4 17,8 13,4 0,57 0,43 0,75
№ 774/376 23,6 11,9 11,1 0,50 0,47 0,93
№ 774/380 (16,0 8,0 8,4 0,50 0,53 1,05
111,0 5,8 6,7 0,50 0,58 1,17
Ск ульптура. Раковина гладкая, несущая на своей поверхности
лишь тонкие струйки нарастания. На одной из раковин заметен тонкий морщинистый слой, покрытый слабо волнистыми, почти прямыми, изредка дихотомически ветвящимися морщинками.
Лопастная линия (рис. 42). Некоторые стадии онтогенетического развития лопастной линии изображены на рис. 42. При величине диаметра раковины 0,9 мм и ширине оборота 0,6 мм наружная часть лопастной линии (рис. 42,а) состоит из воронкообразной вентральной и округленной в основании боковой лопасти. При диаметре раковины 2,0 мм (рис. 42,в) боковых лопастей уже две; вентральная лопасть расширяется и образует с боков округленные выступы. На следующей стадии при ширине оборота в 2,8 мм (рис. 42,с) вентральная лопасть становится трехзубчатой, боковых лопастей уже четыре. На следующих стадиях (рис. 42,
1 Вид назван в честь геолога В. П. Курбатова, нашедшего первый экземпляр его.
135
d, ё) можно наблюдать дальнейшее развитие вентральной лопасти, когда одновременно с развитием боковых зубцов ее происходит укорачивание срединного. При диаметре раковины в 11,2 мм и ширине оборота 6,6 мм (рис. 42, /) вентральная лопасть резко трехраздельная, широкая; основание всех трех зубцов находится на одном уровне. Всех боковых лопастей восемь. При ширине оборота раковины 8 мм (рис. 42,g) количество боковых лопастей доходит до десяти; срединный зубец вентральной
Дл'гДЛ7 е
Рис. 42. Онтогенетическое развитие лопастной линии Neopharciceras kurbatovi Bogoslovsky:
а — № 774/380 при Д=0,9 и Ш=0,6 мм (><22,5); Ъ—то же при Д=2,0 мм (Х15); с —то же при Ш=2,8 мм (X9);d — то же при Д=6,0 и Ш=4,3мм (Х9);е — то же при 111=4,6 мм (х9): /—при Д=11,2 и Ш = 6,6 мм(х9); g— 774/377 при Д=18,7 иШ = 9,5 мм (Х9); h — голотип № 774/374 при Д -«33,0 и Ш = 14,0 мм (Х4,5); г — внутренний отрезок лопастной линии Кв 774/378 при Ш =12,2 мм (Х4,5)- Рудный Алтай. Нижняя часть франского яруса.
лопасти укорачивается, а боковые зубцы приобретают ланцетовидную форму и расходятся в стороны. Во взрослом состоянии при диаметре раковины в 33 мм и ширине оборота в 14 мм (рис. 42, /г) вентральная лопасть широкая, трехраздельная, с широким, коротким воронкообразным срединным зубцом и переходящими на боковые стороны ланцетовидными боковыми зубцами. Наружных боковых лопастей 13—15; форма их в общем ланцетовидная. Первые пять-шесть из них приостренные, остальные — в основании округленные. Внутренняя часть лопастной линии во взрослом состоянии состоит из очень глубокой клиновидной дорсальной лопасти и значительно более мелких ланцетовидных внутренних боковых лопастей, количество которых достигает 12—14.
136
Сходство и отличие. В составе рода Neopharciceras других видов пока не известно.
Геологический возраст и распространение . Встречается в нижней части франского яруса Рудного Алтая.
Местонахождение. Пятнадцать экземпляров разной сохранности найдены в известняковой линзе, в 400 м восточнее дер. Горюновки, на правом берегу рч. Золотухи. Низы франского яруса.
СЕМЕЙСТВО DEVONOPRONORITIDAE FAM. NOV.
Диагноз. Раковина дискоидальная, более или менее инволютная, гладкая, с плоскими или слегка выпуклыми боками и округленной или слегка уплощенной вентральной стороной. Вентральная лопасть широкая, трехзубчатая. Наружных боковых лопастей (до умбонального края) четыре-пять; основания их округлены. Первая боковая лопасть широкая, двураздельная, с округленными частями.
Общие замечания. В 1954 г., устанавливая род Devonopro-norites, я отнес его к семейству Pronoritidae. Такое заключение было основано на действительно большом сходстве нового рода (особенно молодых его экземпляров) с типичными представителями каменноугольных и пермских проноритид.
Однако несмотря на большое сходство между Devonopronorites и проноритидами, в их строении наблюдается и ряд существенных различий. Как уже указывалось мной ранее (1954, стр. 324), род Devonopronorites по сравнению с наиболее ранними каменноугольными представителями проноритид выглядит значительно более высокоорганизованным, •обладающим более инволютной и лучше обтекаемой раковиной, более развитыми перегородками и лопастными линиями. Округленная вентральная сторона раковины и широкая вентральная лопасть девонского рода также существенно отличают его от представителей семейства Pronoritidae. С другой стороны, большой перерыв во времени между Devonopronorites и простейшими Pronoritidae, отсутствие связующих форм в фаменских и т рнейских отложениях не могут не поставить под сомнение близкую родственную связь между ними. Род Devonopronorites, вероятно, не является предком типичных проноритид. Очевидно, мы имеем в данном случае дело с двумя разобщенными во времени, разнородными, поразительно сходными конвергентными группами, берущими начало от разных корней. Что же это за корни?
Со времен классических исследований А. П. Карпинского (1890, 1891, 1896) считается доказанным, что предками проноритид были представители семейства Prolecanitidae. Однако пролеканитиды известны из нижнекаменноугольных отложений и, естественно, поэтому не могут рассматриваться в качестве предков франского рода Devonopronorites. Возможными предками его, на наш взгляд, могли явиться некоторые, наиболее эво-лютные представители семейства Pharciceratidae, имевшие дискоидаль-ную, слабо инволютную раковину наподобие Pharciceras lunulicosta (Sandberger), трехзубчатую вентральную лопасть и простые, возникающие в районе умбо боковые лопасти. Изучение ранних стадий онтогенетического развития представителей Pharciceratidae и Devonopronorites (см. рис. 42 и 44) позволяет установить общий характер развития их лопастных линий. Начальные стадии развития лопастной линии как фарцицератид, так и Devonopronorites характеризуются наличием воронкообразной вентральной лопасти, которая испытывает затем трехчленное деление, и простых, округленных в основании, возникающих в районе умбо боковых лопастей. В дальнейшем, однако, пути эволюционного развития их заметно расходятся. У фарцицератид, одновременно с изменением формы рако
137
вины, становящейся полностью инволютной, значительно увеличивается количество элементов лопастной линии, доходящее примерно до пятидесяти вокруг извилины, вентральная лопасть становится очень широкой, срединный зубец ее укорачивается и расширяется, принимая форму широкой воронки, а расходящиеся боковые зубцы становятся ланцетовидными, не отличаясь по форме от боковых лопастей. У Devonopronorites же подобного резкого расчленения вентральной лопасти не наблюдается и она остается более или менее компактной, но, с другой стороны, укрепление раковины достигается путем расчленения первой боковой лопасти, расположенной на вентральном крае раковины. Начало двучленного расчленения первой боковой лопасти, имеющей формулу (Uj UJ, характеризует момент становления нового рода, а с ним и семейства. Род Devonopronorites, таким образом, имеет совершенно иное по сравнению с каменноугольно-пермскими проноритидами происхождение и заслуживает выделения в самостоятельное монотипное семейство Devonopronori-tidae fam. nov.
Род Devonopronorites Bogoslovsky, 1954
Devonopronorites'. Богословский, 1954, стр. 323.
Тип рода — Devonopronorites ruzhencevi Bogoslovsky, 1954y стр. 324; Рудный Алтай; гониатитовый горизонт средней части франского яруса.
Диагноз. Раковина во взрослом состоянии дискоидальная, сред-неинволютная, со слабо выпуклыми боками и равномерно округленной вентральной стороной. У крупных экземпляров, достигающих диаметра 45—50 мм, вентральная сторона в области жилой камеры уплощается, и вентральный край становится угловатым. Умбо довольно широкое, особенно у молодых особей. Поверхность раковины гладкая, несущая на себе тонкие струйки нарастания, слегка выпуклые на боковых сторонах и образующие широкий и глубокий синус на вентральной. Вентральная лопасть трехзубчатая. Первая боковая лопасть широкая, двураздельная, остальные — простые, нерасчлененные. Всех боковых лопастей четыре-пять. Умбональных лопастей одна-две. На внутренней стороне развиты длинная, узкая, приостренная на конце дорсальная лопасть и четыре (с каждой стороны) боковые лопасти.
й линии:
V (UiUi) U7U5U3ID.
Видовой состав и гео логическое распространение. Кроме типичного вида, других представителей не известно.
Общие замечания. Род Devonopronorites легко отличим от всех девонских аммоноидей по лопастной линии, имеющей двураздельную первую боковую лопасть. Этот признак сближает его с каменноугольными и пермскими представителями семейства Pronoritidae. Юные экземпляры
Ъ
Рис. 43. Поперечные разрезы представителей родов D evonopronorites и Sakmarites:
а — Sakmarites vulgaris (Karpinsky). Южный Урал. Артинский ярус. По Руженцеву (1949, стр 83, рис. 41, b); b — Devonopro-norites ruzhencevi Bogoslovsky; № 774/108. Рудный Алтай. Франский ярус. Увеличение в обоих случаях (Х4).
Т Н О
138
Devonopronorites по форме раковины и навиванию оборотов имеют большое сходство со взрослыми представителями самого простого пермского рода Sakmariles (рис. 43). Несмотря на столь большое конвергентное сходство девонского рода с проноритидами, он легко отличим от последних по форме раковины, очертанию лопастной линии (особенно, широкой вентральной лопасти) и характеру перегородки.
Devonopronorites ruzhencevi Bogoslovsky
Табл. IX, фиг. 4—6.
Devonopronorites ruzhencevi: Богословский, 1954, стр. 324, рис. 1—3.
Голотип — ПИН № 774/103; Рудный Алтай, район Николаевского месторождения, в трех км юго-восточнее поселка; известняковые линзы гониатитового горизонта франского яруса.
Форма. В личиночной стадии раковина эволютная, толстодискои-дальная, образованная уплощенными в дорсо-вентральном направлении оборотами (см. рис. 43). Начиная с четвертого оборота, одновременно с быстрым возрастанием его в высоту, увеличивается инволютность раковины. Молодые экземпляры, имеющие раковину из четырех оборотов, приобретают толстодискоидальную форму с широкими, быстро возрастающими оборотами, угловатое поперечное сечение которых приближается к трапециевидному. Во взрослом состоянии раковина полуинволютная, дискоидальная, с широкими уплощенными, слабо выпуклыми боками, округленной вентральной и вогнутой дорсальной сторонами. Умбональный край отчетливый, слегка округленный. У крупных экземпляров, достигающих диаметра 45—50 мм, вентральная сторона в области жилой камеры уплощается и вентральный край становится угловатым. Умбо во взрослом состоянии умеренно широкое, ступенчатое, ограниченное отчетливыми, слабо округленными умбональными стенками. Последний обо-
рот облекает от 3/4 до 4/5 предыдущего.
Размеры: Д В Ш Ду в/д ш/д Ду/Д
№ 774/101 49,4 22,8 17,2 И,2 0,46 0,35 0,23
№ 774/103 42,3 19,8 16,5 9,5 0,47 0,39 0,22
(39,4 19,0 14,2 8,5 0,48 0,36 0,22
№ 774/104 j (26,4 12,2 11,3 7,0 0,46 0,43 0,27
123,5 10,7 9,1 6,6 0,46 0,39 0,28
№ 774/108 16,0 6,5 7,3 5,5 0,41 0,46 0,34
11,1 4,1 5,8 4,7 0,37 0,52 0,42
Скульптура . Раковина несет на себе тонкие, отчетливые
струйки нарастания, очень слабо выпуклые на боковых сторонах и образующие широкий и глубокий синус на вентральной стороне.
Лопастная линия. Онтогенетическое развитие лопастной линии изображено на рис. 44. На рис. 44 а, b и с изображены лопастные линии 1-й, 2-й и 5-й перегородок. Лопастная линия первой перегородки— аселлатная, слабо волнистая, образующая сглаженные вогнутости («лопасти») на вентральной стороне и в области умбонального шва. Лопастная линия второй перегородки состоит из глубокой воронкообразной, с длинным, узким концом, вентральной лопасти и относительно неглубокой, широко округленной умбональной лопасти. Лопастная линия пятой перегородки (рис. 44, с) отличается от второй лишь несколько более широкой вентральной лопастью и тем, что первичная умбональная лопасть ее начинает смещаться в сторону вентральной лопасти, становясь впоследствии по своему положению первой боковой лопастью. На рис. 44, с?, е
139
показаны следующие стадии развития лопастной линии. Вентральная лопасть постепенно расширяется в своей нижней части, и на ее сторонах намечается образование округленных боковых выступов. Первичная ум-бональная лопасть, смещаясь на боковую сторону и становясь по своему положению первой боковой лопастью, испытывает двучленное деление. В районе умбонального шва обособляется вторая боковая лопасть (U1). При Д = 3,6 и Ш = 2,5 мм (рис. 44, /) вентральная лопасть глубокая и широкая, с узким концом и двумя округленными боковыми выступами. Первая боковая лопасть более короткая, широкая, двураздельная, с округленными частями. Вторая боковая лопасть простая, маленькая, округленная. В районе умбонального шва намечается третья боковая лопасть. На рис. 44, g—I показаны другие стадии развития лопастной линии. Вентральная лопасть постепенно расширяется и становится трехраздельной. Первая боковая лопасть, оставаясь двураздельной, разрастается в длину. Количество следующих лопастей (до умбонального шва) увеличивается до пяти. Полная лопастная линия взрослого экземпляра изображена на рис. 44, к. Вентральная лопасть широкая, трехзубчатая, с равными по длине зубцами, узким срединным и более широкими округленными боковыми. Наружных боковых лопастей пять, из них первая довольно широкая, двураздельная, с округленными частями, остальные простые, в основании тоже округленные. Умбональная лопасть одна, маленькая. На внутренней стороне развиты весьма длинная, узкая, приостренная на конце дорсальная лопасть и четыре боковые лопасти, имеющие форму более или менее приостренных, несколько скошенных наружу зубцов. Еще одна маленькая лопасть расположена на умбональном шве. Первая внутренняя боковая лопасть более чем вдвое короче дорсальной. На рис. 44, I изображена наружная часть лопастной линии голотипа. Вентральная лопасть очень широкая, трехзубчатая, с узким, коротким срединным и гораздо более глубокими и широкими, округленными боковыми частями. Наружных боковых лопастей четыре, пз них первая широкая и глубокая, двураздельная, с округленными частями, остальные простые, округленные в основании. Умбональных лопастей две.
Очертание лопастной линии в пределах вида изменчиво (рис. 45). Особенно большим колебаниям подвержено очертание вентральной лопасти, которая может быть то более, то менее широкой, с более или менее развитыми округленными, реже несколько приостренными боковыми частями; наблюдаются колебания и в длине срединного зубца, а также и в общем количестве боковых лопастей, которых может быть четыре или пять. Но, не обладая достаточным количеством удовлетворительной сохранности материала, и не отмечая на имеющихся в моем распоряжении экземплярах других резко отличных признаков, я склонен рассматривать в настоящее время указанные изменения в лопастной линии отдельных форм как индивидуальные.
Перегородка. Эволюция перегородки в ходе онтогенетического развития видна на рис. 7. При высоте оборота в 1,2 и ширине 2,3 мм (рис. 7, а) расположенные недалеко одна от другой вентральная и дорсальная лопасти связаны между собой. Первая внутренняя боковая лопасть связана с первой наружной боковой, расположенной на перегибе от вентральной стороны к боковой. Вторая наружная боковая лопасть свободна. На следующей стадии (рис. 7,в) при возрастании оборота в высоту, появлении инволютных его частей и связанного с ними увеличения количества лопастей как наружных, так и внутренних, соотношения между элементами перегородки меняются. Связь между вентральной и дорсальной лопастями утрачивается. Первая внутренняя боковая лопасть, теряя связь с отодвигающейся кверху первой наружной боковой лопастью, связыва-
140
Рис. 44. Онтогенетическое развитие лопастной линии Devonopronorites ruzhencevi Bogoslovsky*.
а/ Ь — № 774/114 — первая и вторая перегородки (Х25); с — то же — пятая перегородка (Х25); d — № 774/111 при Ш^-1,2 мм (Х15); е—то же при Ш—1,5 мм (XI5); /—то же при Д=3,6 и Ш-2,5 мм (X12); g — то же при Д=5,7 и Ш =3,4 мм (XI0);h — №774/110 при Д=9,0 и Ш=4,7 мм (Х8); i — №774/107 при В =5.4 и Ш-6,5 мм •(Х6); к—№ 774/109 при В-16,1 и Ш=13,1 мм (Х3);<—голотип № 774/103 при В-=17,5 и Ш=16,0 мм (ХЗ). Рудный Алтай, франский ярус.
10 Б. И. Богословский
ется со второй наружной боковой лопастью. Вторая внутренняя боковая лопасть связана с третьей наружной боковой. Четвертая наружная боковая и маленькая умбональная лопасти свободны. При высоте оборота 11,7 и ширине 10 мм (рис. 7, с) наблюдается следующая стадия. Отодвигающа-
Рис. 45. Лопастные линии D evonopronorites ruzhencevi Bogoslovsky:
a — № 774/105 при B=12,2 мм; b — № 774/106 при В=14,8 и Ш=12,7 мм; с — голотип № 774/103 при В=»17,5 и Ш=16,0 мм. Увеличение во всех случаях (><4). Рудный Алтай.
Франский ярус.
яся кверху вторая наружная боковая лопасть еще сохраняет связь с первой внутренней боковой, третья наружная боковая лопасть свободна, вторая внутренняя боковая, утратив с ней связь, соединяется с четвертой наружной боковой, а третья внутренняя боковая — соответственно с пятой наружной боковой лопастью. Умбональная лопасть свободна. Наконец, на последней стадии (рис. 7, с?), характеризующей перегородку взрослых индивидуумов, первая внутренняя боковая лопасть утрачивает связь со второй наружной боковой и связывается с третьей наружной боковой. Вторая, третья, четвертая внутренние боковые лопасти связаны соответственно с четвертой, пятой и шестой наружными боковыми лопастями.
Таким образом, в ходе эволюции перегородки Devonopronorites происходит перестраивание связей между внутренними и наружными ее эле-
142
ментами: первая внутренняя боковая лопасть последовательно соединяется с первой, второй и третьей наружными боковыми; вторая боковая лопасть— с третьей и четвертой наружными; третья внутренняя боковая лопасть своей связи не меняет.
Сходство и отличие. В составе рода Devonopronorites других видов пока не известно.
Геологический возраст и распространение. Встречается во франском ярусе Рудного Алтая.
Местонахождение. 193 экземпляра различной сохранности найдены в трех км к юго-востоку от поселка Николаевского рудника в известняковых линзах гониатитового горизонта франского яруса.
10*
выводы
1. Изучение девонских аммоноидей имеет первостепенное значение как для теоретического обоснования ряда вопросов, так и для решения практических задач. Однако до последнего времени исследованию этой интересной группы цефалопод уделялось недостаточное внимание, в частности, совершенно не изученными оставались девонские аммониты восточных районов нашей страны. Настоящая работа, посвященная изучению девонских аммоноидей Рудного Алтая, является фактически первой монографией по девонским аммоноидеям Азиатской части СССР.
2. Исследование алтайской фауны девонских аммоноидей показало значительное ее разнообразие. В изученной коллекции установлены 7 семейств (Anarcestidae, Tornoceratidae, Gephuroceratidae, Beloceratidae, Sandbergeroceratidae. Pharciceratidae и Devonopronoridae), 12 родов и 29 видов, из которых многие новые.
3. Описанные девонские аммоноидей весьма своеобразны, особенно в верхнем девоне (франском ярусе). Наибольшее своеобразие франскому комплексу придают сложно организованные роды Beloceras, Triainoceras, Neopharciceras и Devonopronorites, представленные большим количеством особей. Исключительно интересен род Devonopronorites — единственный представитель семейства Devonopronoritidae, ранее не известного в девоне. Франские аммоноидей Рудного Алтая в целом, по сравнению с аналогичными фаунами других областей, выглядят более высоко организованными, содержащими элементы, имеющие более молодой облик, что объясняется, вероятно, более благоприятными условиями существования.
4. Анализ изученной фауны показывает, что, наряду с наличием большого количества новых местных форм, аммоноидей алтайского девона обнаруживают большое сходство с фауной синхроничных отложений Западной Европы, в меньшей степени с фауной Урала (возможно, ввиду недостаточной изученности последней), отчасти с фауной Северной Америки. Близость изученной фауны к западноевропейской указывает на имевшуюся в девонском периоде тесную связь между двумя бассейнами, которая осуществлялась, по всей вероятности, через зону Тетис. Наличие общих и близких с алтайскими форм в отложениях Урала и Северной Америки указывает также на связь девонского бассейна Рудного Алтая с бассейнами этих двух регионов.
5. В результате рассмотрения фациальной приуроченности изученных аммонитов можно сделать следующий палеоэкологический вывод: девонские аммониты района Рудного Алтая жили в областях неглубокого моря, в более или менее удаленной от берега зоне шельфа, в условиях сравнительно быстрой седиментации терригенных осадков.
6. Проведение детальных онтогенетических исследований, начиная с самых ранних стадий, имеет огромное значение для правильного понимания особенностей индивидуального развития, и, в результате, для обо-144
снованного решения вопросов филогении и систематики. В данной работе во всех возможных случаях были проведены онтогенетические наблюдения, позволившие разработать генетические формулы и обосновать шесть типов эволюционных изменений лопастной линии у изученных аммоноидей (анарцестидный, торноцератидный, гефуроцератпдный, фарцицера-тидный, белоцератидный и девонопроноритидный).
7. На основе онтогенетического принципа исследования удалось по-новому разрешить некоторые вопросы филогенетических связей и таксономического положения отдельных родов и семейств девонских аммоноидей: установлено новое семейство Devonopronoritidae, восстановлено в новом понимании семейство Pharciceratidae; возведено в ранг семейства подсемейство Sandbergeroceratinae; установлено происхождение двух последних семейств от одного общего корня с семейством Gephuroceratidae; установлено происхождение семейства Beloceratidae от Gephuroceratidae, семейства Devonopronoritidae— от семейства Pharciceratidae и др. Выявление родственных связей внутри отдельных семейств, как и между последними, позволило дать существенно отличающуюся от имевшихся до этого схему филогении девонских аммоноидей.
8. Изучение всей литературы по девонским аммоноидеям, а также онто-филогенетические наблюдения и выводы по алтайскому материалу, позволили разработать общую схему филогении девонских аммоноидей, существенно отличную от опубликованных ранее схем. От древнейшего ствола Gyroceratitidae берут начало семейства Mimagoniatitidae-> Agonia-titidae и Anarcestidae. Первые два из них не дают потомков. От Anarcestidae происходят: ряд Tornoceratidae-> Cheiloceratidae -> Aganididae, семейство Prolobitidae, группа тесно связанных между собою семейств Gephuroceratidae, Beloceratidae, Sandbergeroceratidae, Pharciceratidae, Devonopronoritidae, семейство Prolecanitidae и, наконец, все климении.
9. Изучение девонских аммоноидей указывает на пышное развитие их, особенно в верхнем девоне, на часто весьма высокую организацию их представителей, по сравнению даже с более молодыми группами. Филогенетические исследования древних групп позволяют решить некоторые вопросы происхождения и эволюции верхнепалеозойских аммоноидей. На границе девона и карбона большая часть девонских типов аммоноидей вымирает. Три филогенетических ствола переходят из девона в карбон. Семейство Aganididae дает начало всем так называемым гониатитам карбона и перми. От Beloceratidae, вероятно, происходит монотипное нижнекаменноугольное семейство Prodromitidae. Семейство Prolecanitidae, происходя от Anarcestidae, дает начало Pronoritidae и Daraelitidae.
10. Изучение девонских аммонитов Рудного Алтая положено в основу детализации и уточнения стратиграфии вмещающих пород, особенно в тех случаях, когда в соответствующих отложениях иная фауна редка или отсутствует. Проведенные исследования позволили более точно определить возраст среднедевонских слоев (крюковской и сокольной свит), доказать присутствие в этом районе отложений, в той или иной мере соответствующих всем четырем зонам мантикоцеровых слоев франского яруса Западной Европы, более точно определить возраст нижней границы верхнедевонского эффузивно-осадочного комплекса. Все эти стратиграфические выводы имеют важное значение для понимания геологии района и уже используются местными геолого-разведочными организациями.
ЛИТЕРАТУРА
Балашова Е. А. и Балашов 3. Г. 1953. Новая находка верхнедевонской фауны в Казахстане. — Докл. АН СССР, т. XCII, № 2, стр. 413—416.
— 1954. Материалы к познанию верхнефаменской фауны Казахстана. — Вести. ЛГУ, сер. биологии, географии, геологии, № 1, стр. 179—202, табл. I—II.
Богословский Б. И. 1954. Новый девонский представитель семейства Pronoritidae Freeh. — Докл. АН СССР, т XCVII, № 2, стр. 323—326, фиг. 1—3.
— 1955а. Девонские аммоноидей Рудного Алтая. (Автореферат доклада,прочит. 5 ноября 1955 г.) — Бюлл. Моск, об-ва йен. прир., отд. геол., т. 30, вып. 1, стр. 94—95.
— 19556. О семействе Pharciceratidae Hyatt, 1900.— Докл. АН СССР, т. 103, № 6, стр. 1103—1106, фиг. 1—2.
— 1955в. О семействе Biloclymeniidae fam. nov.— Докл. АН СССР, т. 104, № 1, стр. 134—137, фиг. 1—2.
Бубличенко Н. Л. 1928. О некоторых среднедевонских брахиоподах Алтая.— Изв. Геол, ком., т XXI, № 10.
— 1936. Основные тектонические линии Рудного Алтая — Сб. «Большой Алтай», т. 2. Изд. АН СССР.
— 1939. Материалы по геологии Рудного Алтая.— Изд. ЦНИГРИ, стр. 67—89.
— 1952. Палеонтологическое обоснование стратиграфии палеозоя Рудного Алтая. Вып. 7. Брахиоподы среднего и верхнего девона Рудного Алтая.— Рук. Фонды ГМНИИ АН Каз. ССР.
Венюков П. 1895. Осадки девонской системы в Мугоджарских горах.— Тр. СПб. об-ва естеств., т. XXIII.
ГольцапфельЭ. 1899. Головоногие доманикового горизонта Южного Тимана.— Тр. Геол, ком., т. XII, № 3, стр. 1—56. табл. 1—10.
Данилович В.Н. 1938. К геологическому строению юго-западного крыла антиклинория Рудного Алтая.— Изв. АН СССР, сер. геол., № 2, стр. 179—198.
Домрачев С. М. 1952. Девон хребта Кара-Тау и прилегающих районов Южного Урала. Девон Западного Приуралья. (Сб. статей). — Тр. ВНИГРИ, нов. сер., вып. 61, стр. 5—121.
— 1953. Девон западного склона Среднего и Южного Урала. Девон Русской платформы. (Сб. статей).— ВНИГРИ, стр. 190—210.
ЗолкинаА. И. 1954. Cheiloceras aff. umbilicatum Sand, из девонских отложений хребта Кара-Тау.—Тр. МГРИ, т. XXVI, стр. 235—237.
Иванкин П. Ф. 1952. Структура рудных полей Прииртышской зоны смятия.— Рук. Фонды ГМНИИ АН Каз. ССР.
Карпинский А. П. 1869. Разведки ископаемого угля, произведенные в Уфимской и Оренбургской губерниях в 1868 г. Горный журнал, издав. Горным Ученым Ком., т. 4, № И, стр. 213—219.
— 1884. О нахождении климений на Урале.— Изв. Геол. ком. т III, стр. 157—159.
— 1885. Геологические исследования, произведенные в Южном Урале летом 1884 г.— Изв. Геол, ком., т. IV, стр. 1—323.
— 1890. Об аммонеях артинского яруса и о некоторых сходных с ними каменноугольных формах. СПб., стр. 1—192, табл. I—V.
— 1891. Об аммонеях артинского яруса и о некоторых сходных с ними каменноугольных формах.— Зап. Мин. об-ва, 2-я сер., ч. 27, стр. 15—195, табл. I—V.
— 1896. О нахождении в Азии Prolecaniles и о развитии этого рода.— Изв. имп. Ак. наук, 4, № 2, стр. 179—194.
— 1945. Собрание сочинений, т. 1, изд. АН СССР, стр. 1—521, табл. I—XVIII, рис. в тексте 1—234.
Ковалевский В. О. 1873. Остеология A nchitherium aurelianense Cuv. как формы, выясняющей генеалогию типа лошади (Equus)) стр. 1—86, табл. I—III, Киев.
146
Колотухина G. Е. 1938. К вопросу о климениевых фациях неодевона Центрального Казахстана. — Изв. АН СССР, сер. геол., № 5—6, стр. 671—688, фиг. 1—12 в тексте, 1 табл.
Кротов П. И. 1888. Геологические исследования на западном склоне Соликамского и Чердынского Урала.— Тр. Геол, ком., т VI, № 1, стр. 1—563, табл. I—II.
К у р е к Н. Н. 1932. Геологические исследования вдоль трассы проектируемой железной дороги Риддер-Рубцовка на Алтае.— Тр. Всес. геол.-разв. объед., вып. 176.
— 1939. Материалы по геологии Рудного Алтая.— Изд. ЦНИГРИ, стр. 34—66.
Ларионова Е. Н. и Петренева Н. И. 1953. Стратиграфия девона и до девонских отложений Молотовского Прикамья и Удмуртской АССР. Девон Русской платформы. (Сб. статей). ВНИГРИ, стр. 211—228.
Левенко А. И. 1951. К стратиграфии девонских отложений Рудного Алтая.— Бюлл. Моск, об-ва исп. прир., отд. геол., т. XXVI (1), стр. 67—83.
Наливкин Д. В. 1930. Семилукские и воронежские слои. — Изв. ГГРУ, т. XLIX, вып. 1, стр. 40, 3 табл.
— 1936. Среднепалеозойские фауны верховьев рек Колымы и Хандыги.— Мат. по изуч. Охотско-Колымского края. Серия 1, вып. 4, стр. 1—28, табл. I—II.
Наливкина А. К. 1936 а. Верхнедевонские гониатитовые слои р. Ай. Южный Урал.— Тр. ЦНИГРИ, вып. 79, стр. 1—24., табл. I—II.
— 19366. О верхнедевонских гониатитах Новой Земли.— Тр. Арктич. ин-та, т. 28, стр. 91—108, табл. I—II.
— 1947. Отряд Аммоноидей. Ammonoidea. Атлас руководящих форм ископ. фаун СССР, т. III. Девонская система, стр. 159—180, табл. XLI—XLV, рис. в тексте 7—52.
— 1953а. Значение гониатитовых фаций для стратиграфии девона. Девон Русской платформы. (Сб. статей).— ВНИГРИ, стр 264—273.
— 19536. Верхнедевонские гониатиты и климении Мугоджар.— Тр. ВНИГРИ, нов. сер., вып. 72, стр. 60—125, табл. I—V, рис. 1—37 в тексте.
Нахимсон Г. М. 1911. Материалы к изучению девонских отложений в Мугод-жарских горах.— Изв. Томск, техн, ин-та, т. XXI, стр. 32, 3 табл.
Нехорошев В. П. 1934. Краткий геологический очерк территории Большого Алтая.— Б. Алтай, т. 1 Изд. АН СССР.
Обручев С. В. 1916. Воронежский девон и группа Spirifer verneuili Murch.— Зап. Геол. отд. об-ва люб. естеств., антроп. и этногр., стр. 21—60, 1 табл.
П е т ц Г. 1901. Материалы к познанию фауны девонских отложений окраины Кузнецкого угленосного бассейна.— Тр. Геол, части, т. IV, стр. 1—393, табл. I—VI.
Парна Э. А. 1914. Аммонеи верхнего неодевона восточного склона Урала.— Тр. Геол, ком., нов. сер., вып. 99, стр. 1—114, табл. I—IV, рис. в тексте 1—86.
Руженцев В. Е. 1946. Эволюция и функциональное значение перегородок аммонитов.— Изв. АН СССР, сер. биол., № 6, стр. 675—706, рис. 1—12.
— 1949а. Основные типы эволюционных изменений лопастной линии верхнепалеозойских аммонитов. — Тр. ПИН, т. XX, стр. 183—198, рис. 1—12 в тексте.
— 19496. Систематика и филогения семейств Pronoritidae Freeh и Medlicottiidae Karpinsky.— Тр. ПИН, т. XIX, стр. 1—204, табл. I—XVII, рис. 1—88 в тексте.
— 1950. Верхнекаменноугольные аммониты Урала.— Тр. ПИН, т. XXIX, стр. 1—220, табл. I—XV, рис. 1—84 в тексте.
Руженцев В. Е. и Шиманский В. Н. 1954. Нижнепермские свернутые наутилоидеи Южного Урала.— Тр. ПИН, т. L. стр. 1—150, табл. I—XV, рис. 1 — 28 в тексте.
Соболев Д. Н. 1909. Средний девон Келецко-Сандомирского кряжа.— Матер, для геол. России, т. XXIV.
— 1914. Наброски по филогении гониатитов, стр. 1—192, табл. I—IX. Варшава.
Тихий В. Н. 1953. Стратиграфия отложений девона Среднего Поволжья и Заволжья. Девон Русской платформы. (Сб. статей).— ВНИГРИ, стр. 121—131.
Токаренко Л. 1903. Фауна верхнедевонских известняков окрестностей г. Верхнеуральска Оренбургской губ.— Тр. об-ваестеств. приКаз. ун-те, т. XXXVII, вып. 2, стр. 1—40, табл. I—III.
Троепольский В. И. 1953. Стратиграфическая схема девонских отложений Татарской АССР. Девон Западного Приуралья. (Сб. статей).— ВНИГРИ, стр. 166—178.
Халфин Л. Л. 1948. Фауна и стратиграфия девонских отложений Горного Алтая.— Изв. Томск, политехи, ин-та, т. LXV, вып. 1.
Чернов А. А. 1907. Артинский ярус. 1. Аммониты бассейнов Яйвы, Косьвы и Чусовой.— Бюлл. Моск, об-ва исп. прир., нов. сер., т. XX, 3, 4, стр. 270—401, табл. 1, рис. в тексте 1—14.
— 1922. Основные черты в эволюции аммонитов.— Бюлл. Моск, об-ва исп. прир., нов. сер., т. XXXI, стр. 149—181.
Чернышев Ф. Н. 1887. Фауна среднего и верхнего девона западного склона Урала.— Тр. Геол, ком., т. III, № 3, стр. 1 —208, табл. I—XIV.
147
Чернышев Ф. Н. 1893. Материалы к изучению Алтайской девонской фауны. — Изв. Геол, ком., т. XI, № 9—10. стр. 199—240, табл. I—IV.
Чочиа Н. Г. и Андрианова К. И. 1952. Девон К олво-В питерского края. Девон Западного Приуралья. (Сб. статей)— Тр. ВНИГРИ, нов. сер., вып. 61, стр. 122—199.
Шиманский В. Н. 1954. Прямые наутилоидеи и бактритоидеи сакмарского и артинского ярусов Южного Урала.—Тр. ПИН, т. XLIV, стр. 1—156, табл. I—XII, табл. 1—5 в тексте.
Шульга-Нестеренко М. И. 1926. Внутреннее строение раковин артинских аммонитов.— Бюлл. Моск, об-ва йен. прир., отд. геол., 4 (1—2), стр. 81—100, табл. II, рис. в тексте 1—9.
Янишевский М. Э. 1926. Материалы к познанию палеозойской фауны Новой Земли.— Тр. Геол, и Мин. музея АН СССР, т. V, вып. 4, стр. 73—116, табл. I— IV.
Ansted D. Т. 1838. On a new Genus of Fossil Multilocular Shells found in the Slate-Rocks of Cornwall.— Trans. Cambridge philos. Soc., vol. 6, pp. 415—422, Cambridge.
Archiac D. V. and Verneuil d. M. E. 1842. On the fossils of the older Deposits in the Rheinish provinces.— Trans. Geol. Soc. London, sec. ser., vol. VI, pp. 303—410, nls. XXV—XXXVIII.
Barrande J. 1865—1870. Systeme Silurien du la centre de la Boheme, partie I, vol. 2, Prague, p. 10—50, pls. 1—12, 17, 241, 245, 413 (pls. 1—12, 17 in 1865; pls. 241—244 in 1866; texte in 1867; pl. 245 in 1868; pl. 413 in 1870).
— 1877. Cephalopodes. Etudes Generales, Extraits du Systeme Silurien du centre de la Boheme, vol. II, p. V, XVI; 1—253, pls. 1—4.
Beecher Ch. 1890. On the development of the shell in the genus Tornoceras Hyatt.— Amer. Journ. Sci., 3d ser., vol. 40, pp. 71—75, pl. I.
Bey ri ch E. 1837. Beitrage zur Kenntniss der Versteinerungen des Reinischen Ubergangsgebirges, SS. 1—44, Taf. I, II, Berlin.
— 1884. Erlauterungen zu den Goniatiten L. v. Buch’s. Zeitschr. deutsch. geol. Gesellsch., Bd. 36, SS 203—219.
В 6 h m e r s I. G. A. 1936. Bau und Struktur von Schale und Sipho bei permischen Ammoncidea; SS. 1—125, Taf. 1, 2. Abb. 1—67, Apeldorn.
Branco W. 1879—1880. Beitrage zur Entwicklungsgeschichte der fossilen Cephalopo-den. Teil I: Die Ammoniten.— Palaeontographica, Bd. 26, SS. 15—51, Taf. IV—XIII.
— 1880—1881. Idem. Theil II: Die Goniatiten u. s. w.—Ibid, Bd. 27, SS. 13—81. Taf. IV—XI.
— 1885. Uber die Anfangskammer von Bactrites.— Zeitschr. deutsch. geol. Gesellsch., Bd. 37, SS. 1—9, Textfigs. 1—3a.
Buch v. L. 1832. Uber Goniatiten — Physikal. Abhandl. Akad. Wiss., SS. 159—187, Taf. I—V, Berlin.
Clarke John M. 1885a. Die Fauna des Iberger Kalkes.— Neues Jahrb. Min., Geol. und Palaont., Bd. Ill, SS. 316—411, Taf. IV—VI.
— 18856. On the higher Devonian faunas of Ontario County, New York.— U. S. Geol. Survey, Bull. 16, pp. 1—80, pls. 1—3.
— 1894. The early stages of Bactrites.— Am. Geologist, vol. 14, pp. 37—43, pl. 2.
— 1899. The Naples fauna (fauna with Manticoceras intumescens) in western New York.— New York State Geologist, Ann. Rep. 16, pp. 29—161. (The report was also issued as New York State Museum, Ann. Rep. 50, vol. 2).
Conrad F. A. 1842. Observations on the Silurian and Devonian sistems of the United States with descriptions of new organic remains.—Acad. Nat. Sci. Philadelphia Journ., vol. 8, pp. 228—280, pls. 12—18.
D о 1 1 о L. 1922. Les Cephalopodes deroules et I’irreversibilite de 1’evolution.— Bijdr. tot de Dierk, k. zool. Genootsch. Natura Artis Magistra Amsterdam, vol. 22, pp. 215—226, pl. 7, Leiden.
Drevermann Tr. 1903. Uber Triaenoceras costatum A. V. sp.— Zeitschr. deutsch geol. Gesellsch., Bd. 55, SS. 85—92, Taf. V.
Dybczynski Tabeusz. 1913. Amonity gornego Dewonu Kielec, Wiadomosc tymczasowa (Les ammonites du Devonien superieur de Kielce en Pologne, Note preliminaire).— Kosmos, Rocznik 38, pp. 5f0—525, pls. 1, 2.
Erben H. K. 1953. Goniatitacea (Ceph.) aus dem Unterdevon und dem Unteren. Mitteldevon.— Neues Jahrb. Geol. u. Palaont., Abh. 98, № 2, SS. 175—225, Taf. 17—19, 14 Abb u. 1 Tab. im Text.
F 1 i g e 1 G. 1896. Uber Goniatites evexus v. Buch und Goniatites lateseptatus Beyrich.— Zeitschr. deutsch. geol. Gesellsch., Bd. XLVIII, H. 2, SS. 414—420, Taf. IX, Textfig. 1-5.
Foord A. H. and С r i c k G. C. 1897. Catalogue of the fossil Cephalopoda in the British Museum (Natural History), part 3, containing the Bactritidae and part of the suborder Ammonoidea; pp. I—XXXIV, 1—303. London.
148
Freeh F. 1887. Uber das Devon der Ostalpen, nebst Bemerkungen fiber das Silur und einem palaontologischen Anhang.— Zeitschr. deutsch. geol. Gesellsch., Bd. 39, SS. 659—738.
— 1897. Lethaea geognostica, Theil I, Lethaea palaeozoica, Bd. 2, SS. I—788, Taf. I—35, Stuttgart.
— 1902. Uber devonische Ammoneen.— Beitrage zur Pa^ontologie und Geologie. Osterreich-Ungarns und des Orients, Bd. 14, SS. 27—111, Taf. 2—5.
— 1913. Ammoneae Devonice (Clymeniidae, Aphyllitidae, Gephyroceratidae, Ghei-
_ loceratidae).— Fossilium Catalogus. I: Animalia, pars I, pp. 1—42.
G i e b e 1 G. G. 1852. Fauna der Vorwelt mit steter Beriicksichtigung der lebenden Thiere, Bd. Ill, I. Cephalopoden, SS. 1—856.
G r a b a u A. W. 1899. The paleontology of Eighteen Mile Greek and the lake shore sections of Erie Country, New York.— Buffalo Soc. Nat. Sci., Bull., vol. 6, pp. 97—390.
G r a b a u A. W. and S c h i m e r H. W. 1910. North American index fossils, Invertebrates, vol. 2, pp. 16—229.
Giimbel G. W. 1862. Revision der Goniatiten des Fichtelgebirges.— Neues Jahrb. Min., Geog., Geol. und. Petrefaktenkunde, SS. 284—326, Taf. 5.
— 1863. Ober Glymenien in den Ubergangsgebilden des Fichtelgebirges. — Paleonog-raphica, Bd. II, SS. 85—165, Taf. XV—XXI.
G u r i c h G. 1896. Das Palaeozoikum im Polnischen Mittelgebirge.— Зап. ими. СПб. мин. об-ва, сер. 2, часть 32, стр. 1—539, табл. I—XV.
— 1909. Leitfossilien des Devons.— Leitfossi’ien, Lief. 2, SS. 97—199, Taf. 29—52.
Hall J. 1843. Geology of New York, part IV, comprising the survey of the fourth geological district Albany, pp. 1—683, pls. 1—19.
— 1860. Notes and observations upon the fossils of the Goniatite limestone in the Marcellus shale of the Hamilton group...—New York State Cabinet Nat. Hist. Ann Rep. 13, pp. 95—112.
— 1874. Descriptions of new species of Goniatitidae, with a list of previously described species, Albany, pp. 1—4 (Printed, May 1874, in advance of the 27 th Report of the New York State Museum of Nat. Hist.)
— 1876. Illustrations of Devonian fossils: Gastropoda, Pteropoda, Cephalopoda, Crustacea and Corals of Upper Helderberg, Hamilton and Chemung groups.— New York Geol. Survey, pp. 1—7, pls. 1—74.
— 1879. Descriptions of the Gastropoda, Pteropoda and Cephalopoda of the Upper Helderberg, Hamilton, Portage and Chemung groups.—New York Geol. Survey, Paleontology of New York, vol. 5, pt. 2, pp. I—XV, 1—492, pls. 1—113.
Haug E. 1898. Etudes sur les Goniatites.— Soc. geol. France, Mem. Paleont., t.7, № 18, pp. 1—114, pls. I—XX.
Holz a p f e 1 E. 1882. Die Goniatitenkalke von Adorf bei Waldeck.— Paleontographi-ca, Bd. XXVIII, SS. 227—261, Taf. 44—49.
— 1895. Das Obere Mitteldevon im Reinischen Gebirge.— Abhandl. d. k. geol. Lande-sanst. N. F., H. 16, SS. 1—459, Taf. I—XIX (in Atlas).
Hyatt, Alpheus 1884a. Genera of fossil cephalopods.— Boston Soc. Nat. History, Proceedings, vol. 22, pp. 253—338.
— 18846. The Evolution of Cephalopoda.—Science, vol. 3, pp. 122—127, 145—149., Cambridge, Mass.
— 1900. Cephalopoda, in Zittel-Eastman Text-book of paleontology, vol. ed. 1, pp. 536—592, London.
Kayser E. 1872. Studien aus dem Gebiete des Rheinischen Devon. III. Die Fauna des Rotheisensteins von Brilon in Westfalen.— Zeitschr. deutsch. geol. Gesellsch., Bd. 24, SS. 653—690, Taf. 25—26, 27, Fig. 1—6.
— 1873. Ober die Fauna des Nierenkalkes von Enkeberge und der schifer von Nehden — Zeitschr. deutsch. geol. Gesellsch., Bd. 25.
— 1878. Die Fauna der altesten Devon-Ablagerungen des Harzes.— Abhandl. geol. Specialkarte preussen und thiiringischen Staaten, Bd. 2, H. 4, SS. 1—295, Taf. 1—36 (in Altas).
— 1879. Uber einige neue Versteinerungen aus dem Kalk der Eifel.— Zeitschr. deutsch. geol. Gesellsch., Bd. 31, SS. 301—307, Taf. 5.
— 1884. Orthocerasschifer zwischen Balduinstein und Laurenburg an der Lahn.— Jahrb. preuss. geol. Landesanst. (1883); SS. 1—56, Taf. I—VI, 2 Abb.
— 1907. Erlauterungen zur geologischen Specialkarte von Preussen, Blatt Oberscheld.
Keyserling A. 1844. Beschreibung einiger Goniatiten aus dem Domanik-Schifer.— Verb. d. kais. russ. Miner. Ges., SS. 217—238, Taf. A.
— 1846. Wissenschaftliche Beobachtungen auf einer Reise in das Petschora-Land, SS. 1—465, Taf. I—XXII.
К о e n e n A. 1886. Ueber Clymenienkalk und Mitteldevon resp. Hercynkalk? bei Montpellier.— Neues Jahrb. f. Min., Geol. u. Pal., Jahrg. 1886, Bd. I, 6. 163—167.
Loewi nson- Lessing F. 1892. Les ammonees de la sone a S poradoceras Mun-
149
s teri dans les Monts Gouberlynskaya Gory.— Bull. d. 1. Soc. Beige de la Geol. t. VI, pp. 15—25, pl. 2.
M a t e r n H. 1929. Die Gliederung der Adorf-Stufe, zugleich ein Beitrag zur Nomenkla-tur von Gephuroceras Hyatt (Ceph.)— Senkenbergiana, Bd. XI, SS. 142—152.
— 1931 a. Die Goniatiten-Fauna der Schistes de Matagne in Belgien.— Bull. Mus. Royal d’ Hist. Naturelle de Belgique, t. 7, № 13, pp. 1—15.
— 19316. Das Oberdevon der Dill-Mulde.—Abhandl. preuss. geol. Landesanst., N F., Heft 134, SS. 1—139. Taf. 1—4.
Miller A. K. 1932. New names for Devonian cephalopod homonyms. — Amer. Journ. Sci., 5 th. ser., v. 24, pp. 330—331.
— 1938. Devonian ammonoids of America.— Geol. Soc. Amer., Spec. Papers, № 14, pp. 1—262, pls. 1—39, text-figs 1—41.
— 1952. Proposed acceptance of the emendation to «S andbergeoceras» of the generic name «Sanbergeroceras» Hyatt, 1884 (Class Cephalopoda, Order Ammonoidea); and the establishment of «Coniatites tuberculosocostatus» G. and F. Sandberger, 1850, as its type species.— Bull. zool. Nomenci., vol. 6, August, 1952.
Miller A. K. and Furnish W. M. 1954. The classification of the Paleozoic ammonoids.— Journ. Paleont., vol. 28, № 5, pp. 685—692.
Mojsisovics E. 1873. Die Cephalopoden der Hollstatter Kalke. I.— Abhandl. geol. Reichsanst., Bd. 6, SS. 1—356, Taf. 1—93.
— 1882. Die Cephalopoden der mediterranen Triasprovinz.— Abhandl. geolog. Reich-sanstalt, Wien, Bd. X, SS. 1—322, Taf. 1—94.
Miinster G. 1832. Uber die Planuliten und Goniatiten im Ubergangs-Kalk des Fichtelgebirges. SS 1—38, Taf. 1—6, Bayreuth.
— 1843. Ober die Clymenien und Goniatiten im Uberganskalk des Fichtelgebirges.— Beitrage zur Petrefaktenkunde, Bayreuth. Beitr. 1,2 Aufl., SS. 1—32, Taf. I—VI.
Quenstedt F. A. 1836. De Notis Nautilearum Primariis,Diss., SS. 1—34.
— 1846—1849. Petrefaktenkunde Deutschlands, Bd. I, Cephalopoden, Tiibingen, SS 1—580, Taf. 1—36 (in Atlas).
Roemer F. 1843. Die Versteinerungen des Harzgebirges. SS. I—XX u. 1—40, Taf. 1—12, Hannover.
— 1856. Kohlen-Periode—In: Bronn H. G. u. Roemer. F. Lethaea geognostica oder Abbildung und Beschreibung der fiir die Gebiergs-Formationen bezeichnendsten Versteinerungen, 1.2. Palaeo-Lethaea. 3. Aufl., SS. 1—788, Stuttgart (Scheizerbart).
Sandberger G. 1841. Mittheilung an Professor Bronn gerichtet.—Neues Jahrb. f. Min., Geog., Geol. u. Petrefaktenkunde, SS. 238—241.
— 1842. Kugeliges Anfangs.-Glied der Goniatiten.— Neues Jahrb. Min. u. s. w., SS. 228—229.
— 1843. Schilderung der paliiontologischen Verhaltnisse der alteren Formationen Nas-saus-Amtlicher Bericht fiber die Zwanzigste Versamml. der Gesellsch. deutsch. Naturf. und Aerzte zu Mainz. Sept. 1842, SS. 154—160.
— 1851. Beobachtungen uber mehrere schwierigere Punete der Organisation derGoniatiten.— Jahrb. Nassau Ver. Naturk. Bd. 7, SS. 292—304, Taf. 2—3.
Sandberger G. u. Sandberger F. 1850—1856. Die Versteinerungen des Reinischen Schichtensystems in Nassau. Wisbaden. SS. 1—564, Taf. I—XXXIX.
Schindewolf О. H. 1922. Einige Randbemerkungen zu E. Perna’s Abhandlung. «Die Ammoneen des oberen Neodevon von Ostabhang des Siidurals».— Senckenber-giana, Bd. 4, SS. 185—196.
— 1923. Entwurf einer naturlichen Systematik der Clymenoidea.—Centralbl. f. Min., Geol. u. Paliiont., SS. 23—30; 59—64. 5 Abb., Stuttgart.
— 1926. Zur Kenntnis der Devon-Karbon-Grenze in Deutschland.—Zeitschr. deutsch. geol. Gesellsch., Bd. 78, Abhandl., SS. 88—133.
— 1929a. Zur Systematik und Stammesgeschichte der altesten Cephalopoden.— Zeitschr. Geschriebeforsch., Bd. 5, SS. 169—178., 3 Abb., Berlin.
— 19296. Vergleichende Studien zur Phylogenie, Morphogenie und Terminologie der Ammoneenlobenlinie.— Abhandl. preuss. geol. Landesanst., N. F. 115, SS. 1—102, 40 Abb., 1 Taf., Berlin.
— 1931. Uber den Ammonitensipho.— Sitzungsb. d. Preuss.geol.Landesanst., H. 6, SS. 197—209, Taf. 5.
— 1933. Vergleichende Morphologie und Phylogenie der An fangskammern tetrabran-chiater Cephalopoden. Eine Studie uber Herkunft, Stammesentwicklung und System der niederen Ammoneen.— Abhandl. preuss. geol. Landesanst., N. F., H. 148, SS. 1—115, Taf. 1—4.
— 1934a. Zur Stammesgeschichte der Cephalopoden.— Jahrb. preuss. geol. Landesanst., Bd. 55, SS. 258—285, Taf. 19—22, Abb. 1—7.
— 19346. Uber eine oberdevonische Ammoneen-Fauna aus den Rocky Montains.—Neues Jahrb. Min., Geol. u. Paliiont., Bd. 72, Abt. B, SS. 331—350.
— 1936. Neue Gattungen tiefoberdevonischer Goniatiten.— Zeitschr. deutsch. geol. Geselsch., Bd. 88, SS. 689—691.
150
Schi ndewolfO. H. 1937. Zwei neue bemerkenswerte Goniatiten Gattungen des reinischen Oberdevons.—Jahrb preuss. geol. Landesanst., Bd. 58, SS. 242—252, Taf. 19.
— 1940. Konvergenzen bei Korallen und bei Ammoneen.— Fortschr. Geol. u. Palaont., t. 12, Heft 41, SS, VII u. 389—492, Abb. 33, Taf. 1.
— 1949a. Zur Nomenklatur der Glymenien (Cephalop., Ammon.).— Neues Jahrb. fiir Min., Geol. u. Palaont., Monatshefte, Jahrg. 1949, Abt. В, H. 1—3, SS, 64—76.
— 19496. Zur Phylogenie der Clymenien (Cephalop., Ammon.) — Neues Jahrb. fiir Min., Geol. u. Palaont., Monatshefte, Jahrg. 1949, Abt. В, H. 7, SS. 197—209.
— 1951. Zur Morphogenie und Terminologie der Ammoneen — Lobenlinie. — Palaont. Zeitschr., Bd. 25, № 1/2, SS. 11—34, Taf. I, Abb. 19.
Schmidt H. 1921. Ober Goniatiten, eine Revision ihrer Systematik mit Beifiigung neuer Beobachtungen.— Gentralbl. Min. u. s. w., H. 17, SS. 538—544.
— 1924. Zwei Gephalopodenfaunen an der Devon-Carbongrenze im Sauerland.—Jahrb. preuss. geol. Landesanst., Bd 44, SS. 98—171, Taf. 6—8.
— 1925. Die carbonischen Goniatiten Deutschlands.— Jahrb. preuss. geol. Landesanst., Bd. XLV, SS. 489—609, Taf. 19—26.
— 1926. Beobachtungen liber mitteldevonische Zonen-Goniatiten.— Senckenbergiana Bd. 8, SS. 291—295.
Smith J. P. 1903. The Carboniferous Ammonoids of America.—U. S. Geol. Surv., vol. XLII, pp. 1—211, pls. I—XXIX.
— 1932. Lower Triassic ammonoids of North America.— U. S. Geol. Surv. Prof. Paper 167, pp. 1—199, pls. 1—81.
Spath L. F. 1919. Notes on Ammonites.—Geol. Mag., new ser., decade 6, vol. 56, pp. 27—35, 65—71; 115—122, 170—177, 220—225.
— 1933. The evolution of the Cephalopoda.— Biol. Reviews, vol. 8, pp. 418—462.
Steininger J. 1853. Geognostische Beschreibung der Eifel, mit einer Karte und einer Profil-Tafel, nebst neun Tafeln mit Abbildungen von Versteinerungen. Trier, SS. 1—143. Taf. I—IX.
Steinmann G. u. Doderlein L. 1890. Elemente der Palaontologie, SS. I— XIX u. 1—848, Leipzig.
Strand E. 1929. Zoological und Palaeontological Nomenclatorical Notes.— Acta Univ. Latviensis, XX, pp. 1—29, Riga.
V a n i x e m L. 1842. Geology of New York, Part III, comprising the survey of the third geological district, Albany, pp. 1—306.
Wedekind R. 1908. Die Cephalopodenfauna des Oberdevon am Enkeberge.— Neues Jahrb. fiir Min., Geol u. Palaont., Bd. 26, SS. 565—639.
— 1910. Posttornoceras balvei n. g. et. n. sp. Ein neuer Fall von Konvergenz bei Goniatiten.— Centralbl. Min., Geol. u. Palaont., Jahrg. 1910, SS. 768—771, Textfigs 1,2.
— 1913. Die Goniatitenkalken des unteren Oberdevon von Martenberg bei Adorf.— Sitzungsb. d. Geselsch. naturforsch. Freunde, № 1, SS. 23—77, 14 Abb., Taf. IV — VII, Berlin.
— 1918. Die Genera der Palaeoammonoidea (Goniatiten).— Pal aeon tographica, Bd. LXII, SS. 85—184, 54 Abb., Taf., XIV—XXII.
— 1926. Die devonische Formation, in Salomon: Grundziige der Geologie, Bd. IIvErd-geschichte, SS. 194—226, Taf. 1—6, Stuttgart.
W h i d b о r n e G. F. 1890. A Monograph of the Devonian Fauna of the South of England. Vol. 1.
Z i t t e 1 K. 1884. Hahdbuch derPalaeontologie, Abt. 1, Palaeozoologie, Bd. 2, Miinchen and Leipzig, SS. 392—491.
Z i t t e 1 K. und В г о i 1 i F. 1924. Grundziige der Palaontologie (Palaozoologie). Abt. 1, Invertebrata, ed. 6, SS. 528—598, Miinchen u. Berlin.
ОБЪЯСНЕНИЕ ТАБЛИЦ
Таблица I
Фиг. 1. Anarcestes (Anarcestes) praecursor Freeh; № 774/604; Лениногорский район, северо-западный склон горы Сокольной; алевролиты крюковской свиты низов среднего девона (х2).
Фиг. 2, 3. Anarcestes (Latanarcestes) noeggerati (Buch): 2—№ 774/605; 3—№ 774/606; Лениногорский район, северо-западный склон горы Сокольной; алевролиты нижней части сокольной свиты эйфельского яруса (в обоих случаях х 1).
Фиг. 4. Werneroceras socolicum sp. nov.; голотип (X1) №774/701; Лениногорский район, северо-западный склон горы Сокольной; алевролиты нижней части сокольной свиты эйфельского яруса.
Фиг. 5. Werneroceras altaicum sp. nov.; голотип (xl) № 773/2; Лениногорский район, северо-западный склон горы Бахрушиной; алевролиты нижней части сокольной свиты эйфельского яруса.
Фиг* 6. Sellanarcestes (?) wenKenbachiformis sp. nov.; голотип (xl) № 773/1; Лениногорский район, отвалы терентьевской штольни; крюковская свита эйфельского яруса.
Таблица II
Фиг. 1,2. Tornoceras (Tornoceras) simplex (Buch): 1—№ 774/257 (x2); 2—№ 774/258 (ХЗ); район Николаевского месторождения, линзы известняка в 3 км к юго-востоку от поселка; гониатитовый горизонт франского яруса.
Фиг. 3—5. Рrobeloceras (?) orientale sp. nov.: 3 — паратип № 774/324; 4— паратип № 774/326; 5 — голотип № 774/325; район Гериховского месторождения, правый берег рч. Золотухи, известняковая линза в 400 м и северо-восточнее дер. Горюновки; нижняя часть франского яруса (во всех случаях х2).
Фиг. 6. Ponticeras acutilobatumsp. nov.; голотип (х 1) № 774/322; Николаевское месторождение, линзы известняка на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Фиг. 7. Ponticeras altaicum sp. nov.; голотип (х2) № 774/305; Николаевское месторождение, линзы известняка на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Фиг. 8. Manticoceras affine (Steininger); № 774/46 (xl); Николаевское месторождение, линзы известняка на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Таблица III
Фиг. 1. Manticoceras neverovi sp. nov.;голотип (xl)№ 774/62; район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчано-сланцевый горизонт франского яруса.
Фиг. 2—4. Мanticoceras sinuosum (Hall): 2—№ 773/16, 3—№ 774/60; район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчано-сланцевый горизонт франского яруса; 4 — № 774/15; район Николаевского месторождения, известняковые линзы на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса (во всех случаях xl).
Фиг. 5. Manticoceras galeatum Wedekind; № 774/30 (xl);район Николаевского месторождения, известняковые линзы на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
152
Табл и ц а I
Т а блица II
7а Tb 8Ь
Т а б л и ц a III
Г а б л и ц а IV
Т а б л и ц а V
Т а б л и ц а VI
Т а б лица VII
Т а б л и ц а VIII
Табл ицa IX
Таблица IV
Фиг. 1. Manticoceras altaicum sp. nov.; голотип(х!)№ 773/15; район Николаевского месторождения, известняковые линзы на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Фиг. 2. Manticoceras аттоп (Keyserling); № (х 1); район Николаевского месторождения, известняковые линзы на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Фиг. 3,4. Manticoceras eliseevi sp. nov.: 3—голотип № 774/96; 4 — паратип № 774/100; район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчано-сланцевый горизонт франского яруса (в обоих случаях xl).
Таблица V
Фиг. 1,2. Manticoceras intumescens (Beyricb): 1—№774/1; 2—№ 774/82; район Николаевского месторождения; известняковые линзы гониатитового горизонта франского яруса (в обоих случаях xl).
Фиг. 3. Manticocerasdrevermanni Wedekind; № 774/27(х 1); район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчано-сланцевый горизонт франского яруса.
Таблица VI
Фиг. 1. Manticoceras carinatum (Beyricb); № 774/43 ( х 1); Николаевское месторождение, известняковые линзы на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Фиг. 2. Manticoceras artumsp. nov; голотип(х1)№ 774/80; район Николаевского месторождения, известняковые линзы в 3 км юго-восточнее поселка; гониатитовый горизонт франского яруса.
Фиг. 3,4. Beloceras sagittarium (Sandberger): 3—№ 774/725; 4—№ 774/726; район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчаносланцевый горизонт франского яруса (в обоих случаях xl).
Фиг. 5. Beloceras stenumbilicatum sp. nov.;голотип(х 1) № 774/703; Николаевское месторождение, линзы известняка на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса.
Таблица VII
Фиг. 1. Beloceras sagittarium (Sandberger); № 774/735 (xl); район Гериховского месторождения, правый берег рч. Грязнухи ниже с. Раздольного; песчано-сланцевый горизонт, франского яруса.
Таблица VIII
Фиг. 1. 2. Beloceras stenumbilicatum sp. nov.: 1 — паратип № 774/705; 2 — паратип — № 774/704; Николаевское месторождение, линзы известняка на западном склоне гряды, отходящей от поселка на север; гониатитовый горизонт франского яруса (в обоих случаях xl).
Фиг. 3. Triainoceras gerassimovi sp. nov.; голотип (Xl) № 773/65; район Гериховского месторождения, правый берег рч. Грязнухи в 1 км ниже с. Раздольного; Неверовские рифовые известняки нижней части франского яруса.
Фиг. 4—7. Triainoceras (?) sp. : 4—№ 774/407; 5 — № 774/408; 6 — № 774/409; 7— № 774/410; район Николаевского месторождения, правобережье р. Убы, в 2 км ниже ж.-д. моста; алевролиты нижней части франского яруса (во всех случаях х 1).
Таблица IX
Фиг. 1—3. Neopharciceras kurbatovi Bogoslovsky: 1 — паратип № 774/373; 2 —голотип № 774/374; 3 — паратип № 774/376; район Гериховского месторождения, правый берег рч. Золотухи, известняковая линза в 400 м северо-западнее дер. Горюновки; нижняя часть франского яруса (во всех случаях х 1).
Фиг. 4—6. Devonopronorites ruzhencevi Bogoslovsky: 4 — голотип № 774/103; 5 — паратип № 774/101; 6— паратип № 774/120; район Николаевского месторождения, известняковые линзы в 3 км юго-восточнее поселка; гониатитовый горизонт франского яруса (во всех случаях xl).
СОДЕРЖАНИЕ
Предисловие .................................................... 3
I. История изучения девонских аммоноидей СССР 5
II. Биостратиграфический очерк....................... 10
1. Основная биостратиграфическая шкала Западной Европы...... 10
2. Определение возраста описанных коллекций ............... 11
3. Некоторые замечания по поводу местных стратиграфических схем Рудного Алтая.................................................. 17
Лениногорское рудное поле................................ 18
Район Николаевского месторождения........................ 22
Район Гериховского месторождения......................... 30
4. Фациальная приуроченность изученных аммоноидей.......... 36
III. Онтогенетическое развитие раковины девонских аммоноидей и его значение для филогении.. 39
1. Эмбриональная раковина девонских аммоноидей................... 39
а) Протоконх, или начальная камера ........................... 39
б) Первая перегородка......................................... 41
в) Длина жилой камеры......................................... 42
2. Развитие перегородки и лопастной линии........................ 44
3. Типы эволюционных изменений лопастной линии................... 47
Анарцестидный тип............................................. 47
Торноцератидный тип........................................... 48
Гефуроцератидный тип.......................................... 48
Фарцицератидныи тип........................................... 48
Белоцератидный тип............................................ 49
Девонопроноритидный тип....................................... 50
4. Функциональное значение перегородок аммонитов................. 50
5. Значение изучения онтогенетического развития для филогении ... 55
IV. Филогения девонских аммоноидей.......................... 56
Происхождение аммоноидей........................................... 56
Семейство Mimoceratidae (=Gyroceratitidae)......................... 58
Эволюция древнейших семейств аммоноидей........................... 59
Семейства Mimagoniatitidae -+ Agoniatitidae........................ 59
Семейства Anarcestidae -► Prolobitidae............................. 60
Семейства Tornoceratidae->Cbeiloceratidae->Agani di dae (=Imitoceratidae) 61
Семейство Gephuroceratidae......................................... 62
Семейство Beloceratidae............................................ 62
Семейство Sandbergeroceratidae..................................... 62
Семейство Pbarciceratidae.......................................... 63
Семейство Devonopronoritidae....................................... 63
Семейство Prolecanitidae .......................................... 63
Надсемейства Gonioclymeniaceae->Clymeniaceae....................... 63
Выводы............................................................. 66
V. Описательная часть..................................................... 68
Семейство Anarcestidae Steinmann...................................... 68
Подсемейство Anarcestihae Steinmann................................ 69
Род Anarcestes Mojsisovics................................... *70
Anarcestes (Anarcestes) praecursor Freeh.................. 71
154
Anarcestes (Latanarcestes) noeggerati (Buch)..................
Род Cabrieroceras gen. nov..........................................
Род W erneroceras Wedekind..........................................
Werneroceras ruppachense (Kayser).............................
W erneroceras altaicum sp. nov................................
Werneroceras socolicum sp. nov................................
Род Sellanarcestes Schindewolf .....................................
Sellanarcestes (?) wenkenbachiformis sp. nov..................
Подсемейство Pinacitinae Schindewolf..................................
Семейство Tornoceratidae Arthaber.....................................
Род Tornoceras Hyatt..............................................
Tornoceras (Tornoceras) simplex (Buch) . . •..................
Семейство Gephuroceratidae Freeh......................................
Род Probeloceras Clarke ..........................................
Probeloceras (?) forcipiferum (Sandberger)....................
Probeloceras (?) orientale sp. nov............................
Род Ponticeras Matern.............................................
Ponticeras altaicum sp. nov...................................
Ponticeras acutilobatum sp. nov. . . . . .....................
Ponticeras sp.................................................
Род Uchtites gen. nov.............................................
Род Manticoceras Hyatt............................................
Manticoceras affine (Steininger)..............................
Manticoceras bullatum Wedekind................................
Manticoceras altaicum sp. nov.................................
Manticoceras neverovi sp. nov.................................
Manticoceras carinatum (Beyricb)..............................
Manticoceras аттоп (Keyserling)...............................
Manticoceras drevermanni W’edekind............................
Manticoceras intumescens (Beyricb)............................
Manticoceras sinuosum (Hall)..................................
Manticoceras eliseevi sp. nov.................................
Manticoceras galeatum Wedekind................................
Manticoceras artum sp. nov....................................
Род Komioceras gen. nov...........................................
Семейство Beloceratidae Freeh.........................................
Род Beloceras Hyatt...............................................
Beloceras sagittarium (Sandberger)............................
Beloceras stenumbilicatum sp. nov.............................
Семейство Sandbergeroceratidae Miller.................................
Род Triainoceras Hyatt............................................
Triainoceras gerassimovi sp. nov..............................
Triainoceras sp...............................................
Семейство Pharciceratidae Hyatt.......................................
Род Pharciceras Hyatt.............................................
Pharciceras kiliani (Freeh)...................................
Род Neopharciceras Bogoslovsky....................................
Neopharciceras kurbatovi Bogoslovsky..........................
Семейство Devonopronoritidae fam. nov.................................
Род Devonopronorites Bogoslovsky..................................
Devonopronorites ruzhencevi Bogoslovsky........................
72
73
73
74
75
76
77
77
78
79
80
82
84
87
88
89
90
91
92
93
94
94
96
98
100
101
102
104
105
107
109
114
115
116
118
119
121
123
124
127
129
130
132
133
134
135
137
138
139
Выводы............................................................... 144
Литература........................................................... 146
Объяснение таблиц. Таблицы I—IX..................................... 152
Борис Иванович Богословский
Девонские аммоноидей Рудного Алтая
(Труды Палеонтологического института, том LXIV)
Утверждено в печать Палеонтологическим институтом Академии наук СССР
Редактор издательства К. Б. Корда
Технический редактор Ю. В. Рылина
РИСО АН СССР № 63—46В. Сдано в набор 26/VHI 1957 г.
Подп. в печать 7/II 1958 г. Формат бум. 70X1084*
Печ. л. 9,75(13,35)4-10 вклеек. Уч.-изд. лист. 13,9
Т-00264 Тираж 1500. Изд. № 1945. Тип. зак. 2082
Цена 3 р. 10 к.
Издательство Академии наук СССР.
Москва, Б-64, Подсосенский пер., д. 21
2-я типография Издательства АН СССР Москва, Г-99, Шубинский пер., д. 10
ОПЕЧАТКИ П ИСПРАВЛЕНИЯ
Страница Строна Напечатано Должно быть
24 5 св. schellwini schellwieni
26 Табл. 4, графа 1 и и сс ьэсо 1 1 о\ н н ю 1 1 Q Q
44 23 сн. 43 44
83 Подпись под рис. 16, 1 сн. пониатитовый гониатитовый
151 19 сн. V a n i х е m V а n и х е m
153 6 сн. северо-западнее северо-восточнее
Б. И. Богословский