

























































































































































































































































































































































Author: Дорофеев В.Ф. Филатенко А.А. Мигушова Э.Ф. Удачин Р.А. Якубцинер М.М.
Tags: хлебные злаки зерновые культуры география растений фитогеография флора географическое распространение растений зерновые и зернобобовые культуры биология биохимия физиология ботаника морфология иммунология издательство колос пшеница ботаническое разнообразие
Year: 1979




Г..........
h. i ?x j
/ s’ ,
I =
Под общим руководством
академика ВАСХНИЛ Д. Д. Брежнева
5 I И ? ? Т
HHU л
Редакторы тома чл.-кор. ВАСХНИЛ
В. Ф. Дорофеев,
канд. биол. наук О. Н. Коровина
ЛЕНИНГРАД • «КОЛОС»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ• 1 9 7 9
Published under the general editorship
of D. D. Brezhnev, Academician of VASK.HNIL
Editors of the volume
N. F. Dorofeev
Corresponding member of VASKHNIL,
O. N. Korovina
Candidate of biol. sci.
L E N I N G R A D • «К О L О S>
LENINGRAD BRANCH. 1979
ББК42.112
К90
УДК 633.11:581.9
Авторы: В. Ф. ДОРОФЕЕВ, А. А. ФИЛАТЕНКО,
Э. Ф. МИГУШОВА, Р. А. УДАЧИН, |М. М. ЯКУБЦИНЕР
К книге дано описание рода и всех видов лшеиипы, с указанием ботанического разнообразия,
географического распространения, морфологических, физиологических, биохимических, иммунологи-
ческих особенностей. Разновидности сгруппированы в таблицы, что облегчает их определение. Кни-
га предназначена для научных работников.
40401—323
К---------------.262—79. 3803030101
035(01)—79
ББК 42.112
633.1
© Издательство «Колос», 1979
ПРЕДИСЛОВИЕ
За последние годы ВИР провел эффективные сборы мировых рас-
тительных ресурсов. Экспедиции института вновь посетили практически
все страны с развитой селекцией, а также страны, входящие в первич-
ные центры происхождения культурных растений. Были обследованы
все земледельческие районы СССР. В результате мировая коллекция
ВИРа пополнилась образцами культурных голозерных и пленчатых ви-
дов пшеницы, а также их диких сородичей, многочисленными местными
и селекционными сортами — носителями генов высокой продуктивности,
короткостебельности, зимостойкости, засухоустойчивости, высоких тех-
нологических качеств, повышенной концентрации белка и лизина, устой-
чивости к грибным болезням и др. Только за годы 9-й пятилетки миро-
вая коллекция пшениц пополнилась 17 тыс. новых образцов. В результате
того, что все мировое богатство пшеницы широко и глубоко изучается
в тесном содружестве отдела пшеницы ВИРа с отделами генетики, фи-
зиологии, иммунитета, молекулярной биологии, биохимии и качества,
анатомии и цитологии, получен новый ценный материал к познанию
филогенетических связей в роде Triticum L. и селекционной ценности ви-
дового и сортового разнообразия пшеницы.
Настоящее издание «Пшеница» заметно отличается от издания 1935 г.
За прошедшее после 1935 г. время были открыты новые виды: Т. urartu,
Т. sinskajae, Т. militinae, Т. ispahatiicutn, Т. jakubzineri, Т. petropavlovs-
kyi, Т. zhukovskyi, Т. kiharae. Кроме того, ряд таксонов получили статус
вида '(Т- araraticum, Т. karamyschevii, Т. turanicum, Т. vavilovii). Наука
выдвинула идею о дифилетическом происхождении тетраплоидной пше-
ницы (Е. Н. Синская, 1955) и подтвердила ее (В. Г. Конарев и др., 1976).
Все это позволило нам, основываясь на законе гомологических рядов в
наследственной изменчивости Н. И. Вавилова (1920), разделить род Tri-
ticum на 2 подрода согласно геномному составу видов.
Описание видов пшеницы в каждом подроде начинается с диких ди-
плоидов. Для облегчения ориентировки в огромном внутривидовом гено-
фонде пшеницы нами предложены удобные для пользования таблицы,
где разновидности полиморфных видов определяются по табличному
ключу, составленному из рядов гомологичных признаков. Такой ключ
облегчает поиски нужной разновидности во множестве признаков.
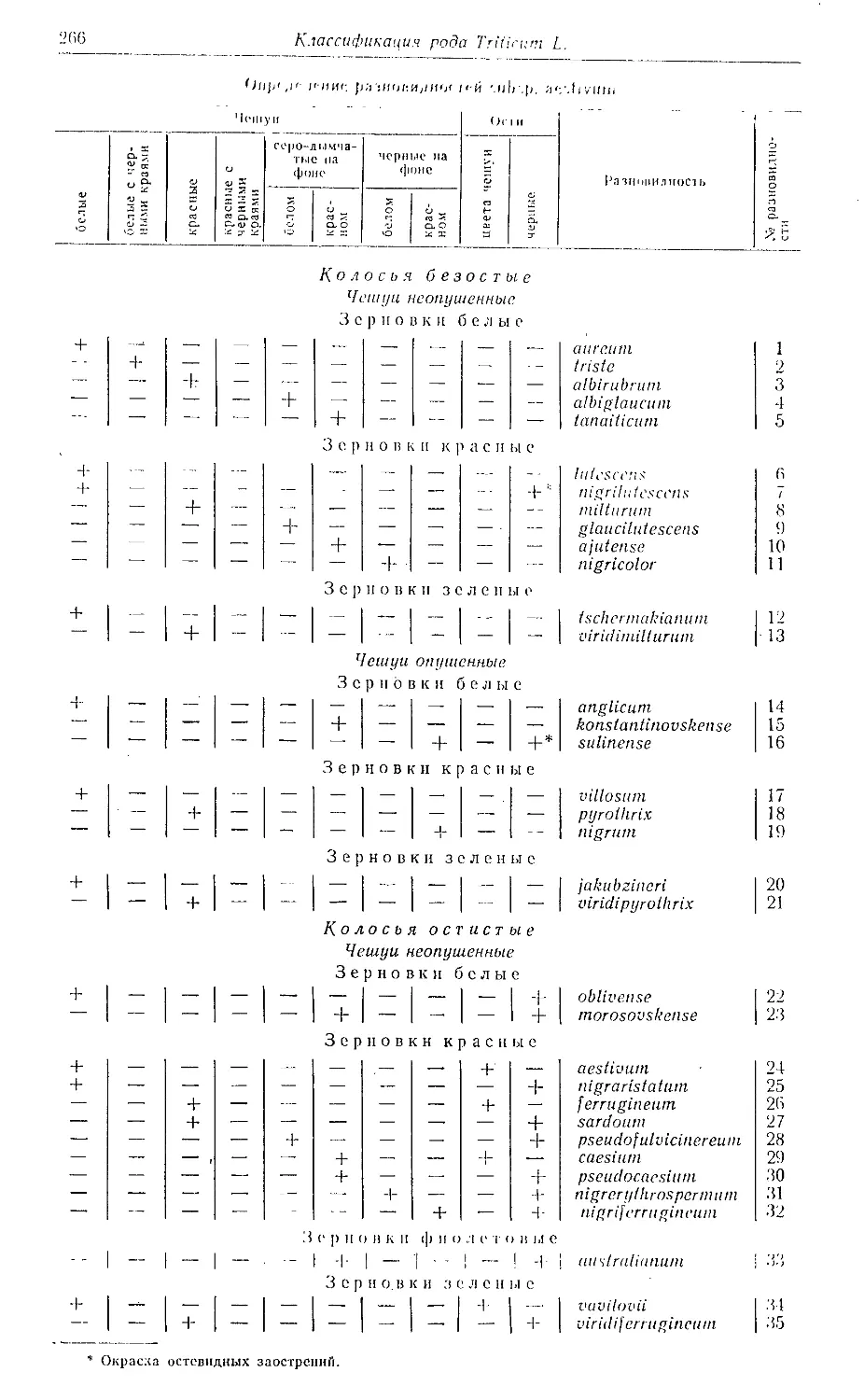
Для того чтобы установить название разновидности, нужно по табли-
це отыскать сумму признаков колоса (наличие признака обозначено
«+», отсутствие — знаком «—»). Разновидности приводятся в алфавит-
ном порядке, (^ведения об авторе каждой разновидности, литературном
источнике, енпонимных названиях и ареале завершаются заключенной
в скобки цифрой — порядковым номером разновидности, под которым
она находится в таблице. По этой цифре можно быстро отыскать в таб-
лице ее диагностические признаки.
Для определения подвидов, групп разновидностей, а также отдельных
разновидностей и форм, если они у вышестоящего таксона представле-
ны лишь двумя-тремя, использован дихотомический ключ. В тексте при-
няты следующие сокращения родовых названий: Т, —Triticum, Ле.—
Aegilops, А. —Agropyron.
6 Предисловие
В связи с ограниченным объемом книги в пей нс даны сведения по
эмбриологии пшеницы, которые сравнительно полно освещены в моногра-
фии Т. Б. Батыгиной «Эмбриология пшеницы» (1974). По той же причи-
не в указатель литературы помещены лишь основные источники. Кроме
того, исключено описание районированных сортов пшеницы, так как эти
сведения даются в «Руководстве по апробации сельскохозяйственных
культур», периодически издаваемом Государственной комиссией по сор-
тоиспытанию сельскохозяйственных культур с участием ВИРа.
Сведения об ареале тех разновидностей и экологических групп, кото-
рые послевоенными экспедициями ВИРа не выявлены на местах прежне-
го обитания, даются по времени их наибольшего распространения в пер-
вой половине XX в.
Чл.-кор. ВАСХГ1ИЛ В. Ф. Дорофеев и канд. биол. наук Л. А. Фила-
тенко разработали для настоящего издания систематику 23 видов пше-
ницы: Т. boeoticum, Т. urartu, Т. tnonococcum, Т. sinskajae, Т. dicoccoides,
Т. araraticum, Т. limopheevii, Т. militinae, Т. dicocctim, Т. ispahanictim,
Т. karamt/schevii, Т. durum, Т. turgidum, Т. ac.thiopictim. Т. poloiiicum,
Т. pcrsicutn, Т. zhukovski/i, Т. madia, Т. spelta, Т. vavilovii, Т. aesti-
vum, Т. compactum, Т. kiharae. Доктор с.-х. наук М. М. Якубцинер принял
участие в разработке систематики видов Т. boeoticum, Т. dicoccoides и
Т. aethiopicum. Систематика видов Т. turanicum, Т. iakubzineri, Т. sphae-
гососсшп, Т. pelropavlovskyi, сведения по их истории и географическому
распространению дапы доктором с.-х. паук Р. А. Удачиным.
Общая характеристика каждого вида, их географическое распростра-
нение, происхождение и история написаны В. Ф. Дорофеевым, Э. Ф. Ми-
гушовой, А. А. Филатенко и М. М. Якубципером; сведения об иммуните-
те — Э. Ф. Мигушовой. В разделе «Обзор рода Triticum L.» доктор с.-х.
паук М. М. Якубцинер изложил подразделы «Ботаническая характерис-
тика рода Triticum L.» и «История систематики рода Triticum L.»,
канд. биол. наук Э. Ф. Мигушова является автором подразделов «Совре-
менная система рода Triticum L.» и «Происхождение полиплоидной пше-
ницы».
За ценные замечания по рукописи, на основании которых внесены
соответствующие изменения, выражаем благодарность академику
ВАСХНИЛ В. Г. Копареву, докторам биол. паук В. С. Лехповичу,
Т. Я. Зарубайло, В. И. Кривченко, Г. В. Удовенко, Л. И. Орел, канд.
биол. паук Л. Л. Жестянниковой. Выражаем признательность за по-
мощь в работе над рукописью сотрудникам отдела пшениц А. Н. Брыко-
вой, О. А. Ляпуновой, Т. Н. Самовар и Н. Т. Шайдуко.
7
ОБЗОР РОДА TRITICUM L.
БОТАНИЧЕСКАЯ ХАРАКТЕРИСТИКА РОДА
TRITICUM L.
Triticum L. 1753, Sp. Pl. cd. 1: 85; id. 1754, Gen. Pl. ed. 5: 37; Dum. 1823,
Observ. Gram. Fl. Belg.: 94; Bcntli. ct Hook. f. 1883, Gen. PL 3: 1204, p.p. —
Aegicon Adans. 1763, Fam. pl. 2:36. — Bromus Scop. 1777, Introd.: 74,
non L. — Crithodium Link, 1834, Linnaea 9, 3: 132. — Gigachilon Seidl.
1836, Oecon. — techn. Fl. Bohm. 1: 425, nom. — Triticum a) Spelta Endl.
1836, Gen. PL: 103.— Triticum sect. Cerealia Koch, 1837, Syn. Fl. Germ. ecl.
1:823, p. p. — Triticum L. et Spelta (Bauh. et Cherl.) * Ser. et Nivieria
Ser. 1841, Descript, fig. Cor. Europ. 4: 66 (104), 76 (114). — Cryptopyruin
Heynh. 1846, Nomenci. 2: 174.— Triticum sect. Enlriticnm Godr. in Grcn. ct
Godr. 1856, Fl. France, 3: 598. — Triticum sect. Pyros et sect. Zea Doell,
1857, FL Grossh. Baden, 1: 123, 124. —Deina Alef. 1866, Landw. FL: 335.—
Triticum sect. Sitopyros Doell ct sect. Eremopyros Doell in Martins, 1880,
FL Bras. 2,3: 223, 227.— Triticum sect. Sitopyros Hackel in Engl, et Prantl,
1887, Nat. Pflanzenfam. 2,2: 80. —Frumenlum subgen. Triticum E. Krause,
1898, Bot. Zentralbl. 73, 10: 339.
Пшеница — растение однолетнее**. Это относится и к культивируе-
мым озимым формам и дикорастущим видам. 2п = 14, 28, 42. Геномный
состав Au, Ab, AUB, AbG, AUBD, AbGD.
Корневая система мочковатая, придаточные корни отходят от подзем-
ных стеблевых узлов пучками (мочками). Зародышевые корни сохраня-
ются. Главного корня нет.
Наземная часть растения травянистая, высотой 0,5—2 м. Стебель —
соломина, полая или выполненная рыхлой паренхимной тканью, с узлами
из плотной ткани. Твердая механическая ткань обеспечивает прочность
стебля, его устойчивость к полеганию. Этому способствует и выполнен-
ность соломины. Промежутки между стеблевыми узлами (междоузлия)
в молодом возрасте очень короткие; в дальнейшем они постепенно удли-
няются за счет вставочного роста. Стебель обычно имеет 4—6 узлов. При
корневом полегании пшеницы приземный узел, утолщаясь, содействует
выпрямлению растений. Подземные стеблевые узлы дают начало боко-
вым побегам, т. е. кущению стебля.
От стеблевых узлов отходят листья, состоящие из листового влагали-
ща и листовой пластинки. Листья у пшениц очередные, сидячие. Влагали-
ще почти до основания расщепленное, охватывающее междоузлие в виде
трубки, нижним своим основанием прикреплено к стеблевому узлу. Ме-
ханическая ткань придает прочность влагалищу, защищающему стебель
от сгибания и поломки над узлами и под ними. Листовая пластинка ли-
нейная, с параллельными рядами жилок. На границе между влагалищем
и пластинкой листа имеется язычок — лигула, представляющая собой
узкую тонкую ткань, лишенную проводящих пучков и плотно прилегаю-
щую к стеблю. Язычок защищает внутренние части влагалища от про-
никновения воды. По обеим сторонам язычка Соломину охватывают ли-
нейные ушки — отростки краев влагалища в месте отгиба листа. Ушки
* Указанные авторы предпочитали называть эту группу пшениц Zea, а не Spelta
(Bauhin, Cherler, 1G51), как это дано N. Seringe (1842).
** Среди пшенично-пырейных гибридов, созданных II. В. Цициным в Главном
ботаническом саду ЛИ СССР, и пшенично-ржаных, выведенных М. Садыховым в Ин-
ституте животноводства ЛИ ЛзССР, имеются и многолетние формы.
8
Обзор рода Triticum L.
Рис. 1. Колосок мягкой пше-
ницы:
а — общий вид; б— схема раз-
реза его в вертикальной плос-
кости; / — членик; 2 — колоско-
вая чешуя; 3 — наружная (ниж-
няя) цветковая чешуя; 4 —
внутренняя (верхняя) цветковая
чешуя; 5 — ость; £ —зерновка;
7 — рыльце пестика; 8 — тычин-
ка
нередко с ресничками. У некоторых пшениц Таджикистана, Афганистана,
Кипра язычок и ушки отсутствуют (безлигульные формы).
Верхняя часть стебля заканчивается соцветием, представляющим со-
бой колос. Стержень (ось) колоса у пшеницы состоит из члеников. На
верхней части каждого членика расположено по одному сидячему колос-
ку. Каждый колосок (рис. 1) с 2—5 (7) цветками, из которых 1—3 (4)
плодущие. Колоски образуют два ряда с боковой (двурядной) стороны.
С лицевой стороны колоски расположены черепицеобразно — своей верх-
ней частью колосок налегает на основание выше расположено™ колоска.
Колосок имеет 2 колосковые чешуи (у Т. jakubzineri — 4). Форма их —
продолговато-овальная, овальная, яйцевидная, лопатчатая, ланцетовид-
ная, консистенция — от грубой (деревянистой) до нежной (травянистой).
Колосковые чешуи защищают от повреждений заключенные между ними
части колоска. Колосковая чешуя имеет киль, килевой зубец, плечо п
зубец боковой жилки (рис. 2), иногда у Т. persicutn и некоторых форм
Т. aethiopicum и Т. aeslivutn вместо килевого зубца образуется ость. Меж-
ду колосковыми чешуями находятся цветки, которые являются обоепо-
лыми, неправильными (зигоморфными). Самый верхний цветок колоска
чаще выступает над колосковыми чешуями. Каждый цветок защи-
Рис. 2. Колосковая чешуя
мягкой пшеницы:
I — киль; 2 — килевой зубец;
3 — главная боковая жилка
щен и прикрыт двумя цветковыми чешуями
(рис. 1)—наружной (нижней) и внутрен-
ней (верхней). Наружные цветковые чешуи
продолговатые и продолговато-яйцевидные,
выпуклые, более или менее килеватые, со
сходящимися кверху жилками. Они несут
ость или (у безостых форм) более или ме-
нее короткий остевидпый придаток, который
изредка может отсутствовать. По краю ча-
сто расположены реснички. Внутренние цвет-
ковые чешуи — без ости и без остевидного
образования. Они имеют 2 киля и прикры-
вают зерновку с брюшной стороны. Между
наружной и внутренней цветковыми чешуя-
ми находятся главные части цветка — за-
вязь с перистым двухлопастным рыльцем и
3 тычинки. Завязь верхняя, одпогнездпая, с.
одной семяпочкой. Каждая тычинка состо-
ит из двух пыльцевых мешков и топкой,
очень короткой тычиночной нити, которая
при цветении резко удлиняется. Пыльники
качающиеся, легко разбрасывающие пыль-
цу. Они растрескиваются во время цветения
па всю длину или реже — только в верх-
ней части. У основания завязи за тычинками
История систематики рода Triticum Л. 9
находятся 2 бесцветные пленочки — лодикулы, которые во время цвете-
ния набухают, способствуя открыванию цветка.
Пшеница — самоопыляющееся растение. Однако она не является
облигатным самоопылителем: наблюдаются случаи перекрестного опы-
ления при помощи ветра. Пыльцевые зерна шаровидные, мелкие, глад-
кие. Часто самые верхние цветки в колоске недоразвиты и поэтому сте-
рильны. В неблагоприятных же условиях нередко наблюдается стериль-
ность других цветков, что вызывает череззерницу.
Плод — зерновка, не срастающаяся с цветковыми чешуями. По фор-
ме она может быть шаровидной, округлой (бочонковндной), яйцевидной,
удлиненной, очень удлиненной, серповидной и горбатой. У однозернянок
зерновка сплюснутая. На брюшной стороне зерновки имеется бороздка.
Зародыш расположен у основания зерновки на ее спинной стороне. На
противоположном конце зерновка покрыта волосками (хохолок). Основ-
ная часть зерновки — эндосперм. Зародыш прямой. Он соединяется с
эндоспермом при помощи щитка, всасывающие клетки которого при про-
растании транспортируют запасные питательные вещества для зародыша.
Важной составной частью последнего является зародышевая почка, со-
стоящая из первичного листа (колеоптиль), 2—4 зародышевых корешков
и точки роста. Колеоптиль имеет удлиненно-конусовидную форму и проч-
ную заостренную верхушку, помогающую защитить молодые зародыше-
вые листья при продвижении к поверхности почвы. После выхода колеоп-
тиля из повчы он отмирает, а верхушечная почка продолжает рост.
Лектотип: Т. aestivum L.
ИСТОРИЯ СИСТЕМАТИКИ РОДА
TRITICUM L.
Понятие Triticum — пшеница — имеет более чем двухтысячелетнюю
давность. Им пользовались еще в Древнем Риме. Его (наряду с термином
siligo) употребляет Катон (234—149 гг. до н. э.) в сочинении «De agricul-
tural. Название triticum для голозерной пшеницы (пленчатая именова-
лась far, odoreum) дают также Варрон (116—27 гг. до н. э.) и агроном
Колумелла (1 в.). Это наименование встречается позднее и у Плиния
Старшего (23—79 гг.) в «Naturalis historiae».
Предшественник К. Линнея—-французский ботаник Ж. Турнефор
(J. Р. Tournefort, 1700) дал характеристику рода Triticum с указанием
ряда видов пшеницы. В этот же род он включил и пырей (Elytrigia).
К. Линней описал род Triticum L. в 1737 г. в «Genera plantarum), и три-
тикологи многих поколений поныне рассматривают его как автора дан-
ного рода. Однако и К. Линней включал в состав Triticum формы пырея.
На протяжении более двух столетий в понятие Triticum L. разные ав-
торы прошлого и настоящего вкладывали различное содержание. У мно-
гих систематиков под этим наименованием были объединены наряду с
пшеницей также другие злаки, в частности житняк и пырей. Такого ши-
рокого толкования термина Triticum придерживались вслед за Ж. Турие-
фором и К. Лцннеем также другие исследователи (Р. Forskal, 1775;
J. С. D. Schreber, 1779, 1796; П. С. Паллас, 1781; D. Villar, 1785;
J. В. Р. Л. Lamarck, 1786; Р. Schrank, 1789; Л. W. Е. С. Wibel, 1790;
С. L. Willdenow, 1800; A. W. Roth, 1800; П. С. F. Schumacher, 1801;
Н. A. Schrader, 1803, 1806; F. Brotcro, 1804; R. L. Desfonlaines, 1801;
С. IL Persoon, 1805; J. В. P. A. Lamarck, Л. P. De Candolle, 1806; M. ISii-
берштейн, 1808; D. Viviani, 1808; Ф. Б. Фишер, 1815; 1821; J. Sadler, 1818:
J. H. Hornemann, 1819; J. H. F. Link, 1821; К. Ф. Ледебур, 1825; M. J. Bluff,
K. A. Fingerhuth, 1825; E. G. Steudel, C. F. Hochstetler, 1826; G. Gussone,
1827; C. S. Kunth, 1829; К- А. Мейер, 1831; W. D. J. Koch, 1837; H. С. Typ-
чапипов, 1838; W. J. Hooker, 1840; R. Visiani, 1842; Л. Г. Шренк, 1842;
Г). Don, L. Mitterpacher, 1848; E. G. Steudel, 1855; D. A. Godron, 1855;
J. C. Docll, 1857; R. Visiani et J. Pancic, 1862; A. Kerner, 1863; A. Gray,
1864; L. F. Celakovsky, 1867; J. Dnval-Jouvc, 1870; Д. И. Литвинов и
И. Ф. Шмальгаузен, 1880; J. Е. Т. Aitchison, W. В. Hemsley, 1886; Н. Loret,
A. Barandon, 1886; Е. Hackel, 1887; А. Н. Краснов, 1887; Р. F. A, Ascher-
soji, Р. Graebner, 1902; О. W. Paulsen, 1912; А. Г. Снятков и др., 1913).
Некоторые виды, ныне рассматриваемые по классификации Н. Н. Цве-
лева (1973, 1976) как представители рода Elymus, или, согласно С. Нев-
скому (1934), относимые к злаку Roegtieria С. Koch, также включались
различными авторами в состав Triticum L. (N. Т. Host, 1797, 1802, 1805;
Н. A. Schrader, 1808; J. Н. F. Link, 1829, 1843; К. A. Meyer, D. A. Bunge,
по К- Ф. Ледебуру, 1829; К,- А. Триниус, 1833, 1835; A. Bertoloni, 1841;
А. Г. Шренк, 1845; Ф. Б. Фишер и К. А. Мейер, 1845; А. Н. R. Grisebach,
1852; Н. Т. Турчанинов, 1854, 1856; L. М. Neuman, 1901; F. J. Hermann,
1936).
Ботаники прошлого века к роду Triticum относили is свое время виды
Elytrigia Desv. (N. Т. Host, 1805, 1809; II. A. Schrader, 1806; R. L. Desfon-
taines, 1815; К. A. Tpmniyc, 1829, 1835; X. 1'. Лессинг, 1834; Г. С. Карелин,
И. П. Кириллов, 1841; J. Н. F. Link, 1843; А. Н. R. Grisebach, 1852; Е. Bois-
sier, 1854; А. Н. Краснов, 1897), а также некоторые виды злака Егето-
pyrutn (Ledeb.) daub, et Spach (M. Биберштейн, 1808; A. W. Roth, 1802).
Некоторые авторы включили формы Dasypyrum (Coss, et Dur.) Borb.
(Haynaldia Schur) в состав Triticum (M. Биберштейн, 1809; J. H. F. Link,
1821; E. G. Steudel, C. F. Hochstetler, 1826; C. S. Kunth, 1829; M. J. Bluff
e. a., 1836; J. Sadler, 1840; R. Visiani, 1842; W. D. J. Koch, 1843; К. Ф. Ле-
дебур, 1853; R. Visiani, J. Pancic, 1862; A. Kerner, 1863; P. F. A. Ascher-
son, P. Graebner, 1902; O. W. Paulsen, 1912; A. Hayek, F. Markgraf, 1933)
как особую секцию или как подрод в'отличие от настоящей пшеницы,
именуемой ими Eutriticum (J. С. М. Grenier, D. A. Godron, 1856; A. Fiori,
G. Paoletti, 1896; A. Thellung, 1918).
Ряд ботаников (J. J. Roemer, J. A. Schultes, 1817; J. II. F. Link, 1853;
Р. F. A. Ascherson, 1864; Р. F. A. Ascherson, Р. Graebner, 1893, 1902)
включали в понятие Triticum формы ржи (Secale L.), которая еще в
1753 г. была выделена Линнеем в самостоятельный род. Кроме того, род
Triticum объединяли с белоусником — Nardurus (Bluff, Nees et Schauer)
Godr. (D. Villar, 1787; J. Banks, D. C. Solander, 1794; И. И. Геогри, 1800;
С. H. Persoon, 1805; D. Viviani, 1808 и др.) и с коротконожкой — Brachy-
podium Beauv. (A. W. E. C. Wibel, 1790; F. Brotero, 1804 и др.).
Особого внимания заслуживает вопрос, о систематическом родстве
между родами Triticum и Aegilops. R. L. Desfontaines (1700) рассматри-
вал Aegilops как отдельный род. К. Линией (1753) также выделил эги-
лопсы в самостоятельный род. Ряд последующих авторов объединяли его
с Triticum (Р. Forskal, 1775; F. V. Raspail, 1825; J. Н. F. Link, 1830;
И. Ф. Шмальгаузен, 1897; Р. F. A. Ascherson, Р. Graebner, 1901; К. W. De
Dalia Torre, Н. Harms, 1907; A. Thellung, 1907; F. Parlatore, 1908;
P. IO. Рожевиц, 1923 и др.), нередко обосабливая эгилопсы в особую сек-
цию (J. С. М. Grenier, D. A. Godron, 1856; Е. Hackel, 1887).
В конце 50-х 'годов текущего столетня под влиянием новейших успе-
хов цитогенетики, установившей исключительную роль некоторых видов
Aegilops как допоров геномов тетраплоидпых и гексанлопдпых видов
пшеницы, вновь появились тенденции к объединению под названием Tri-
ticum пшеницы и эгилопса. Такую концепцию выдвинул канадский бота-
ник W. М. Bowden (1959). Она получила затем поддержку в труде цито-
генетиков США (R. Morris, Е. R. Sears, 1967).
Некоторые авторы предлагают к роду Triticum отнести не все виды
эгилопса, а лишь входящие в состав секции Sitopsis, с которой пшеница
связана своим происхождением (М. S. Chennaveeraiah, 1960) или рас-
История систематики рода Triticum L.
11
сматривают Triticum л Aegilops как отдельные роды, генетически объе-
диненные в группу Triticum—Aegilops (D. Zohary, M. Feldman, 1962).
Близость указанных двух родов отмечает также видный генетик и мор-
фолог G. L. Stebbins (1956).
Противниками тезиса «пшеница и эгплопс— один род» являются вы-
дающиеся отечественные (II. И. Вавилов, Л. Л. Декапрслевпч, II. М. Жу-
ковский, К. А. Фляксбергер, В. Ф. Дорофеев, II. А. Гаидилян, Г. А. Ле-
витский, В. Л. Менабдс, И. Д. Мустафаев, С. А. Невский, В. Е. Писарев,
Е. Н. Сипская, II. II. Цвелев, II. В. Цицин, G. К. Черепанов, М. М. Якуб-
цинер) и зарубежные ангоры (М. Gokgol, F. Kornicke, A. Love и D. Love,
R. Mansfeld, E. Schiemann п др.). Этот принцип сохраняется и в настоя-
щем издании при рассмотрении границ рода Triticum.
До 50-х годов XX в. в специальной литературе первым, предложив-
шим термин Triticum для обозначения лишь культурных и дикорасту-
щих форм пшеницы (исключая Agropyron и Aegilops'), считался F. Kor-
nicke (1885). В действительности приоритет в ограничении рамок рода
Triticum (с исключением из его состава злаков Aegilops и Agropyron)
принадлежит ботанику В. С. Dumortier (1823). На этом основании ныне
в литературе иногда (J. Mac Key, 1966) употребляется термин Triticum
(L.) Dum. Кроме того, как правильно подчеркивал 11. М. Жуковский
(1971), вряд ли можно согласиться с присвоением авторства рода Triti-
cum не К. Линнею, a A. Thellung (1918), что допускается некоторыми
авторами (например, L. Briggle, 1976 и др.). За К. Линнеем как осново-
положником научной классификации и номенклатуры растений, произ-
ведшим переворот в ботанике, должно быть сохранено авторство рода
Triticum L.
Вопрос об объеме рода Triticum L. продолжает привлекать внимание
трнтикологов и после изъятия из его состава других злаков. К- Линней
первоначально (1737) описал 5 видов пшеницы: Т. aestivum L. (яровые
остистые формы), Т. hybernum L. (озимые безостые), Т. turgidum L.,
Т. spelta L. и Т. monococcum L. Впоследствии (1764) он приводит допол-
нительно вид Т. polonicum, которому дал такое название L. Plukonet
(1726).
F. Kornicke (1885) рассматривал в роде пшеницы 3 основных вида:
Т. polonicum L., Т. monococcum L., Т. vulgare Vill. s. 1. с делением послед-
него на виды в узком смысле [Г. vulgare Vill., Т. compactum Host, Т. tur-
gidum L., T. durum Desf., T. spelta L., T. dicoccum (Schrank) Schuebl.].
В первой четверти минувшего века (N. Seringe, 1819) положено нача-
ло устранению чрезмерного дробления вида. Дальнейшее развитие этого
принципа продолжалось на протяжении всего XIX в. (J. Metzger, 1824;
Н. Vilmorin, 1850; F. G. С. Alefeld, 1866; G. Henze, 1872).
В 1919 г. Н. И. Вавилов увеличил объем рода Triticum L., включив в
него новый вид тетраплоидной пшеницы — Т. persicum Vav. (Т. carthlicum
Nevski). Затем J. Percival (1921) впервые описал тетраплоидпые голо-
зерные пшеницы Т. orientate Perciv. (Т. turanicum Jakubz.), Т. pyramidale
Perciv. и гексаплондный вид Т. sphaerococcum Perciv.
В дальнейшем род Triticum L. пополнился видами: Т. timopheevii
(Zhuk.) Zhuk. (1928), Т. macha Dekapr. et Menabde (1932), T. vavilovii
Jakubz. (1933), T. abyssinicum Vav. (1939), T.aelhiopicuin Jakubz. (1947),
T. palaeocolchicum Menabde (1940), T. karamyschevii Nevski (1935),
T. araraticum Jakubz. (1947), T. ispahanicum Fleslot (1958), T. zhukovskyi
Menabde et Ericzjan (I960), T. militinae Zhuk, et Migusch. (1969), T. pet-
ropavlovskyi Udacz. et Migusch. (1970), T. sinskajae A. Filat, et Kurk.
(1975), T. jakubzineri Udacz. et Schachm. (1976), T. kiharae Dorof. et Mi-
gusch.
Морфологическая специфика однозернянок и их резкая обособлен-
ность от других пшениц привели к мысли исключить однозернянки из со-
става Triticum (J. Н. F. Link, 1834). Впервые в самостоятельный род
Ofi.nip ptnhr Tfih'ctttu
дикорастущие однозернянки были выделены иод названием Crilhodiutn
Link. Аналогично N. Scringe (1811- 1842) также рассматривал дикие и
культурные однозернянки, как особый род под названием Nivieria Scrin-
ge (1841). Через 89 лет к подобному заключению пришел также 10. Л. Фи-
липпенко (1930), предложивший для однозернянок новый термин—род
Мопососсит Philiptsch. Примечательно, что такую же позицию занял
J. Mac Key (1968), восстановивший для однозернянок старое наимено-
вание Crithodium Link. Он в составе рода Triticum сохранил тетраплоид-
пые и гексаплоидпые виды, представляющие собой аллополиплоиды—
результат межродовой гибридизации. В настоящей книге однозернянки
оставлены в пределах рода Triticum. В его состав ио принятой в данном
труде классификации входит также Т. polonicum L. (резко обособленный
по морфологии колоса от других видов), который ПО лет назад был вы-
делен в самостоятельный род Deina Alef. (1866).
Во флористической литературе на протяжении десятков лет пшеница
описывалась как часть Tribus Hordeaceae Benth. (Е. Н. Синская, 1955;
Р. 10. Рожевиц, 1935; С. А. Невский, 1934, 1937). Эту трибу впервые опи-
сал G. Bentham (1881). В ней С. А. Невский (1933) выделил subtribus
Aegilopitiae Nevski, куда отнес роды Triticum, Aegilops, Secale, llaytial-
dia, Eremopyrum.
До G. Bentham (1881) были известны трибы liordeaceae Kunth (1829)
и Hordeinae Nees (1843). Однако наименования Tribus Hordeaceae
Benth., как и Tribus Hordeaceae Kunth, на основании «Международного
кодекса ботанической номенклатуры» (1966) являются незаконными из-
за приоритета более раннего наименования Tribus Triticeae Dum., данно-
го В. С. Dumortier еще в 1823 г. и соответствующего правилам, уста-
новленным Международным кодексом. Род Triticum является родовым
типом этой трибы, включающей и другие злаки, что было забыто на про-
тяжении свыше 130 лет, пока к нему не привлек внимания R. Pilger
(1954).
В пределах трибы Triticeae Dum. Н. И. Цвелев (1973) выделил под-
трибу Triiicinae Trim ex Griseb., куда отнес Aegilops, Triticum, Secale n
другие роды. Между тем В. С. Dumortier еще в 1829 г. описал в трибе
Triticeae подразделение Frumentaceae, куда отнес роды Triticum L., Aegi-
lops L., Secale L., т. e. те, которые приводятся даже в новейшей литера-
туре под незаконным термином Subtribus Triticitiae Trim ex Griseb.
Странно, что наименование подтрибы Frumentaceae Dum. до настоящего
времени на протяжении почти 150 лет осталось незамеченным среди бо-
таников мира. Мы относим род Triticum L. к подтрибе Frumentaceae
Dum., входящей в состав трибы Triticeae Dum.
На протяжении свыше 160 лет внимание тритикологов привлекал во-
прос о внутриродовой группировке видов пшеницы. Впервые деление ро-
да Triticum L. па секции осуществлено в начале минувшего века G. Вау-
le-Barelle (1809), который отнес голозерные виды к Section I, а плен-
чатые к Section II. Деление рода Triticum на секции по характеру
вымолота зерна (голозерность— пленчатость) сделал также N. Seringe
(1818). Позднее в классификации F. Kornicke (1885) было отражено
различие между видами по признаку голозерность — пленчатость. Тот
же принцип лег в основу классификации, предложенной Е. Hackel (1887).
Первую внутриродовую классификацию пшеницы на филогенетичес-
кой основе дал A. Sthulz (1913). Он разделил пшеницу на 3 ряда: 1) Ein-
kornreihe, 2) Emmerreihe и 3) Dinkelreihe. Каждый из этих рядов он раз-
бил на 3 класса: а) дикий сородич — родоначальник рода; б) производ-
ные пленчатые, виды; в) производные культурные виды (эта схема полу-
чила название девятиклсточной).
К. А. Фляксбергер (1915) рассматривал ряды А. Шульца как сборные
виды. В монографии J. Percival (1921) эти ряды шпили свое выражение
в соответствующих обозначениях — Species I Perciv. и Species II Perciv.
История систематики рода Triticum L.
1.3
В. Kajanus (1927) нашел целесообразным вышеуказанные, ряды
Л. Шульца рассматривать как виды scnsu amplo: Т. monococcum (L.)
Kajanus (диплоиды), T. acuminatum Kajanus (теграллоиды), T. obtusa-
tum Kajanus (гсксаплоиды), которые он разделил на подвиды, оставив за
последними видовые наименования.
Значительным шагом вперед была последующая классификация
К- А. Фляксбергера (1928). В роде Triticum он установил секции: 1) Мо-
пососса Flaksb., 2) Dicoccoides Flaksb. и 3) Spelloidca Flaksb. Такая груп-
пировка получила признание многих тритикологов.
Однако еще в прошлом веке была обнародована секция Мопососсоп
Dum. (В. С. Dumortier, 1823), и этот термин имеет приоритет как секцион-
ное наименование. Впоследствии К- А. Фляксбергер (1935) отказался от
термина Sectio, заменив его не предусмотренным «Международным ко-
дексом ботанической литературы» термином Congregatio.
Классификация К,- А. Фляксбергера (1928) была позднее «модифици-
рована» С. А. Невским (1934), который дикие и культурные однозернян-
ки объединил как Sectio Crithodium (Link) Nevski, тстраплоидные ви-
ды— Sectio Orthatherum Nevski, а гексаплоидпые виды — Sectio Spelta
Nevski. Кроме того, им выделена дополнительно Sectio Gigachilon Nevs-
ki для вида T. polonicum L.
Необходимо указать на вклад других биологов в развитие морфоло-
го-филогенетической схемы классификации пшеницы. Н. И. Вавилов
(1919) обнаружил, что 3 ряда рода Triticum, которые установил A. Schulz
на основании морфологического анализа, четко различаются между собой
по отношению к грибным паразитам. A. Tschermak (1914) указал на от-
сутствие или наличие генетической совместимости между видами этих
рядов при межвидовой гибридизации, а Т. Sakamura (1918) установил
различие их по числу хромосом.
В ряде последующих работ исследователи придерживались в общих
чертах группировки классиков систематики пшеницы —А. Шульца и
К. А. Фляксбергера. Это относится, в частности, и к известной моногра-
фии Е. Schiemann (1948). Исключение составляют монографии В. Л. Ме-
набде, Е. Н. Синской и новейшие работы Н. Н. Цвелева. В. Л. Менабде
(1948) отказался от деления рода на секции и сгруппировал виды пше-
ницы на филогенетической основе в 4 ряда (группы) видов: 1) сегеталь-
ные и дикорастущие; 2) первичные; 3) вторичные; 4) географические
расы. Он в своей классификации игнорирует также критерий уровня пло-
идиости. Кроме того, гексаплоид Т. macha он относит к первичным ви-
дам. Е. Н. Синская (1955) разделила род Triticum на 3 секции. При этом
она отвергла критерий пленчатости—голозерности и уровни плоидности
вида. Кроме того, стремясь отразить историзм вида и его генетические
особенности, она ввела дополнительный термин Subsectio.
FI. Н. Цвелев (1976) разбил род Triticum на секции: 1) Мопососсоп
Dum.; 2) Triticum и 3) Pyrachne Dum. В этой классификации игнориро-
ван уровень плоидности и признак голозерности—пленчатости.
J. Mac Key (1975) на основе полученных им генетических данных раз-
делил род Triticum на секции: 1) Мопососса Flaksb.; 2) Dicoccoidea
Flaksb.; 3) Speltoidea Flaksb., сведя все многообразие пшеницы к 6
видам.
Современный уровень работ по цитогенетике и электрофорезу пшени-
цы потребовал внесения определенных корректив в классификацию рода
Triticum L. В частности, стало необходимым отразить особое положение
видов Т. araraticum и Т. timopheevii.
Еще Е. Н. Синская (1955) в пределах секции II выделила подсекцию
закавказских двузернянок, подчеркнув обособленность Т. araraticum от
Т. dicoccoides не только по комплексу морфологических признаков и аре-
алу, нон по генетической совместимости при гибридизации (стерильность
гибридов и др.).
14
Обзор рода Triticum I..
h\. М. Якубцпвер (1961) установил особую секцию Andropogon, объе-
диняющую t. araraticum и Т. timopheevii. Эта секция состоит из двух
серий (Моп/апае Jakubz. и Slrigosa Jakubz.), отражающих различия
между двумя генетически родственными видами — дикорастущим Т. ara-
raticum и культурным Т. timopheevii. Ошибочным является отнесение по-
следнего вида к числу диких (Е. Schiemann, 1948)'.
При классификации секции гексаплоидных видов, однако, часто не
принималось во внимание серьезное различие между обладателями гено-
мов AUBD и видом Т. zhukovskyi (геномы AbG). Это упущение было
устранено в последующей классификации В. Ф. Дорофеевым (1969), ко-
торый выделил самостоятельную серию Zatidurianum Dorof. в составе
Congregatio Quasihexaploidea Dorof. В этом аспекте Н. Н. Цвелев (1976)
выделил в пределах секции Triticum подразделение Т. aggregatio timo-
pheevii (Zhuk.) Zhuk., включающее T. araraticum, T. timopheevii, T. zhu-
kovskyi и T. militinae.
Следует отметить, что В. Ф. Дорофеев (1969) включил в состав Cong-
regatio Octoploidea 2 искусственно созданных межвидовых 56-хромосом-
ных амфидпплоида. Им, однако, не учтены октоплоидпые межвидовые
амфидиплоиды, созданные А. Р. Жебраком, Э. В. Тавриным и др., а так-
же не выделена конгрегация, включающая обнародованные (например,
А. Р. Жебраком, 1949) 70-хромосомные амфидиплоиды.
Н. Н. Цвелев (1976) приводит в своей классификации не только
Т. fungicidum Zhuk., но и другие межвидовые гибриды, обозначив их как
Т. X fungicidum Zhuk., Т. X edwardii Zhebrak, Т X soveticum Zhebrak,
T. X borisovii Zhebrak. Однако он не учел межвидовые амфидиплоиды,
созданные Э. В. Тавриным и другими генетиками. Кстати, Д. Мак-Кей
(1968) включил в состав рода Triticum в качестве самостоятельных ви-
дов межродовые амфидиплоиды, созданные при участии Secale (Тriticale)
и Agropyron, что не вполне оправдано. W. М. Bowden (1959) естественно
возникшие аллополиплоидные пшеницы рассматривает как гибриды.
Мягкую пшеницу он обозначает Triticum X aestivum. Такое обозначение
также не вполне оправдано.
Классификация пшеницы, принятая в настоящем издании, базируется
на концепции вида, сформулированной Н. И. Вавиловым (1931), и прин-
ципе К- А. Тимирязева об историческом методе в биологии.
Нам представляется недопустимым в свете принципа историзма вклю-
чение в состав единого вида пшеницы пленчатых и голозерных форм, как
дикорастущих, так и возделываемых. По этой причине мы не принимаем
классификации диплоидного ряда пшениц, предложенной Р. F. A. Ascher-
son и Р. Graebner (1898—1902), Н. С. Haussknecht (1899), A. Thellung
(1918), К- А. Фляксбергером (1925), A. Hayek и F. Markgraf (1933),
A. Love и D. Love (1961), J. Mac Key (1966). Они рассматривают дикую
и культурную однозернянки как один вид. Мы также не согласны с си-
стематиками, объединяющими культурную полбу с дикой (К. А. Флякс-
бергер, 1915; Р. F. A. Ascherson, Р. Graebner, 1901).Неприемлема для нас
и классификация тетраплоидных и гексаплоидных видов, данная A. Thel-
lung (1918), W. М. Bowden (1959, 1966), A. Love и D. Love (1961),
J. Mac Key (1966), R. Morris и Е. R. Sears (1967), нарушившими выше-
указанный принцип при описании ими границ Т. turgidum и Т. aestivum.
Дикорастущие виды более древние, чем культурные. По правилам но-
менклатуры пет препятствий к возведению культурного вида в ранг более
высокий, чем у дикорастущего. Однако в историческом плане вряд ли
оправдано представление дикорастущих видов Т. araraticum и Т. dicoccoi-
des в качестве разновидностей (W. М. Bowden, 1959) или подвидов
(A. Love, D. Love, 1961), подчиненных более молодому по происхожде-
нию культурному виду Т. turgidum L. С исторической точки зрения нело-
гично рассматривать эти дикорастущие виды в качестве подвидов про-
изошедших от них культурных видов.
переменна я
система рода Triticum Л.
Исходя из 110/1,01]устлмости обI,единения голозерных и пленчатых
форм, мы, вслед за 11. 11. Вавиловым и К. Л. Фляксбергером, отвергаем
классификацию J. Percival (1921), относившею голозерные пшеницы
Т. aethiopicum и Т. persicutn к пленчатому виду Т. dicoccum. W. М. Bow-
den (1959), со ссылкой на статью 28 «Восьмого Международного кодек-
са ботанической номенклатуры» (Utrecht-Netherlands, 1956), ошибочно
отрицает правомерность отнесения диких п культурных растений в пре-
делах рода Triticum к различным видам. Указанная статья гласит: «Рас-
тения, введенные из дикой флоры в культуру и ничем существенно не от-
личающиеся от своих родоначальников, имеют те же названия, что и
соответствующие виды и внутривидовые таксоны в природе». Между тем
культивируемые виды рода Triticum существенно отличаются от соот-
ветствующих диких сородичей.
Нам представляется неприемлемым чрезмерное укрупнение видов
пшеницы, которое сделано генетиками W. М. Bowden, J. Mac Key, Л. Lo-
ve, D. Love, R. Morris и E. R. Sears, которые группу полиморфных видов
рассматривают как разновидности одного вида. С другой стороны, так-
же следует остерегаться необоснованного дробления вида (например.
Т. dicoccum), допущенного С. Л. Невским.
Данные морфологии, генетики и филогении оправдывают возведе-
ние в ранг видов открытых в последние годы новых форм пшеницы
Т. sinskajae A. Filat, et Kurk., Т. militinae Zhuk, et Migusch., T. jakubzi-
neri Udacz. et Schachm., T. petropavlovskyi Udacz. et Migusch., T. kiha-
rae Dorof. et Migusch.
СОВРЕМЕННАЯ СИСТЕМА РОДА
TRITICUM L.
Пшеница — основной хлеб. В результате расселения ее из очага
происхождения в различные эколого-географические регионы и вслед-
ствие активной селекционной деятельности человека мировой гено-
фонд пшеницы оказался не только огромным, но и чрезвычайно много-
образным. Селекционер для удачного подбора исходных форм для
скрещивания должен свободно ориентироваться в системе рода.
Систематика пшеницы со времен Ж. Турпефора (1700) и К. Лин-
нея(1737) претерпела ряд модификаций. 14. И. Вавилов (1935) и
К. А. Фляксбергер (1935) в результате изучения мировой коллекции
пшеницы пришли к пониманию системы рода Triticum, как включаю-
щей все виды дикой и культурной пшеницы, но исключающей виды
пырея и эгилопса. Они понимали этот род как секции Eutriticum Gren.
et Godr. (J. С. M. Grenier, D. A. Godron, 1855—1856) или Sitopyros
Hackel (E. Hackel, 1887). Ныне такое толкование рода Triticum приня-
то Н. Kihara и его школой генетиков в Японии. В нашей стране ва-
виловского понимания системы рода придерживаются большинство
тритикологов, в том числе Л. Л. Декапрелевич, В. Л. Менабде,
И. Д. Мустафаев, П. А. Гандилян, а также авторы этой книги.
В 60-х годах в зарубежной литературе появились работы, в кото-
рых изложены результаты ревизии системы рода Triticum. W. М. Bow-
den (1959) поместил все виды эгилопса в этот род, а все многообразие
пшеницы объединил в 3 вида — Т. motiococcum L., Т. turgidum L..
Т. aestivum L. R. Morris п Е. R. Sears (1967), приняв в целом систему
W. М. Bowden, восстановили в ранге вида Т. timopheevii. М. S. Chen-
naveeraiah (1960) ввел в род Triticum не все эгплопсы, а лишь виды
секции Sitopsis. J. Mac Key (1954, 1968) выделил диплоидные виды в
16
Обзор рода Triticum L.
самостоятельный род Crithodium Link, а аллополиплоидныс пшеницы
объединил в 4 вида — Т. timopheevii Zhuk., Т. turgidum (L.) Thell.,
T. zhukovskyi Men. et Er., T. aestivum (L.) Thell. В род Triticum он ввел
также Triticale и пшенично-пырейные гибриды. Позднее (1975) в до-
кладе XII Международному ботаническому конгрессу он возвратился
к прежней своей классификации (1963, 1966), добавив к названным
видам еще 2-—Т. monococcum (L.) МК и Т. urartu Thum.
Многолетнее полевое изучение мировой коллекции пшеницы и эги-
лопса, сравнительно-морфологическое обследование вегетативных орга-
нов и колосьев привело нас к убеждению о нерациональности объеди-
нения их в один род, о неправильности сведения всего многообразия
пшеницы к 3—6 видам, об отсутствии оснований к выделению однозер-
нянок в отдельный род, о неудобстве для селекционеров систем
W. М. Bowden, R. Morris и Е. R. Sears, а также J. Alac Key.
Наиболее рациональной, как нам кажется, является та система ро-
да Triticum, основы которой разработаны рядом исследователей
прошлого, в том числе F. Kornicke (1885) и J. Percival (1921). Их сис-
темы были переработаны и дополнены II. И. Вавиловым (1935) и
К. А. Фляксбергером (1935). Воззрения последних являются основой
принятой нами системы. Существенное влияние на наши представле-
ния о ней оказали также исследования, проведенные в отделе молеку-
лярной биологии, биохимии и качества ВИРа под руководством акаде-
мика ВАСХНИЛ В. Г. Копарева. Предлагаемая нами система базиру-
ется на учете геномного состава видов и наличия или отсутствия ряда
главных генов в доминантном состоянии.
20-летний опыт межвидовой гибридизации убедил нас в том, что
каждый из тетраплоидных видов (геномы АиВ) при гибридизации с мяг-
кой пшеницей дает специфический для него спектр расщепления в по-
томстве. В качестве примера возьмем реципрокные скрещивания близ-
ких по морфологическим признакам колоса Т. turgidum и Т. durum, с
мягкой пшеницей. В комбинациях с участием первого вида в Fa наблю-
дается обильное выщепление типичной спельты, тогда как в комбина-
циях с участием второго вида ее не было (выщеплялись растения, ко-
лосья которых по внешнему облику были сходны со спельтой, но име-
ли легкий обмолот).
При обследовании отделом пшеницы ВИРа (В. Ф. Дорофеев и др.)
земледельческих районов Советского Закавказья, Средней Азии, Аф-
ганистана, Ирана, Турции, Ирака и Сирии было отмечено, что хлебо-
пашцы разделяют пшеницу именно по той системе, которая была
принята Н. И. Вавиловым. Например, они предпочитают сеять не
Т. turgidum, а Т. durum, хотя растения этих видов чрезвычайно сход-
ны. Учет их практического опыта показал, что у названных тетраплоид-
ных видов пшеницы признаки фенотипа сцеплены с рядом хозяйственных
признаков, которые учитываются при возделывании в производственных
условиях.
J. Мас 1<еу (1966, 1968) считает, что различие по одному гену не
может быть достаточным критерием для возведения формы пшеницы
в ранг вида. Рднако, если такой ген относится к числу главных, то его
действие резко меняет не только фенотип колоса, но и хозяйственную
характеристику пшеницы. Например, появление доминантной аллели
гена Q лишает Т. dicoccum и Т. spe.Ua трудного обмолота, способству-
ет упрочнению их колоса, меняет степень его плотности, т. е. превра-
щает эти виды соответственно в Т. persicum и Т. aestivum. Наличие
доминантной аллели гена Р ответственно за превращение Т. durum в
Т. polonicum, а Т. aestivum в Т. petropavlovskyi. К числу главных ге-
нов относят Q, Р, С, S и ген, определяющий ветвление колоса у Т. va-
vilovii и Т. jakubzineri (ген «вавилоидпости»). Переход этих генов из
доминантного в рецессивное состояние (или наоборот) кардинально
CoripcMeiuiau система рода Trilietiin I..
17
Система рода Tritit tun 1„
Подрод (Subgen.) Секция (Sect.) Группа видов Вил (sp.) 2il ГеномЫ Число разнока- чественных геномов
Triticum Urartu Do- rof et Л. Fi- lat. Однозер- нянка Т. urartu Thum, ex Gandil. 11 Au 1
Dicoccoides Flaksb. Полбы T. dicoccoides (Koern. ex Aschers. et Gra- ebn.) Schweinf. T. dicoccum (Schrank) Schuebl. T. karamyschevii Nevski T. ispahanicum Fleslot 28 28 28 28 I » » » 2 2 2 2
Голозерные тетрапло- иды T. turgidum L. T. jakubzineri Udacz. et Schachm. T. durum Desf. T. turanicum Jakubz. T. polonicum L. T. aethiopicum Jakubz. T. persicum Vav. (T. carthlicum Nevski) 28 28 28 28 28 28 28 2> » 2> 2> » О 2 2 2 2 2 2
T riticum Спельты T. madia Dekapr. et Menabde T. spelta L. Г. vavilovii (Thum.) Jakubz. 42 42 42 AUBD » 3 3 3
Голозерные гексапло- иды T. compactum Host T. aestivum L. T. sphaerococcum Per- civ. T. petropavlovskyi Udacz. et Migusch. 42 42 42 42 AAA A 3 3 3 3
Boeoiicuin Migusch. et Dorof. Monococcon Dum. Однозер- нянки T. boeoticum Boiss. T. monococcum L. 14 14 Ab » 1 1
Голозерный дпплоид T. sinskajae A. Filat, et Kurk. 14 1
T imopheevii A. Filat, et Dorof. Полбы T. araraticum Jakubz. T. timopheevii (Zhuk.) Zhuk. T. zhukovskyi Menab- de et Ericzjan 28 28 42 A"G » A”A''G 2 2 2
Голозер- ный тет- раплоид T. militinae Zhuk, et Migusch. 28 A"G 2
Kiharae Dorof. et Migusch, Спельта T. kiharae Dorof. et Migusch. 42 A»GD 3
2 762
18
Об:юр рода Triticum
меняет хозяйственную характеристику пшеницы, делая ее рентабель-
ной или, наоборот, невыгодной для возделывания.
Становление гипотезы о дифилетическом происхождении тетрапло-
идной пшеницы начиналось на результатах гибридологических и цитоло-
гических исследований (А. А. Ерпцян, 1928; В. В. Светозарова, 1929;
F. Lilienfeld, Н. Kihara, 1934). Дальнейшее развитие ее связано с воз-
зрениями М. Г. Туманяна (1936, 1939), Е. И. Сипской (1955) и
В. Л. Менабде (1971, 1972). Закон гомологических рядов в наследст-
венной изменчивости Н. И. Вавилова (1920, 1922, 1935), результаты
сравнительного изучения видов пшеницы и эгилопса методами биохи-
мии (В. Г. Конарев и др., 1970, 1976), иммунологии (В. И. Кривчен-
ко и др., 1976), физиологии (Э. А. Барашкова и др., 1977), проведен-
ного за последние годы в ВИРе, литературные данные (F. Lilienfeld,
Н. Kihara, 1934; М. Tanaka, S. Ichikawa, 1972; В. Яаска, 1974 и др.), а
также наши исследования (отдел пшениц ВИРа) привели нас к при-
знанию гипотезы о дифилетическом происхождении тетраплоидпой
пшеницы. В разное историческое время, в различных географических
районах Ближнего Востока, от разных материнских и отцовских ви-
дов в природе на тетранлоидпом уровне возникли 2 первичных амфи-
диплоида, различающихся по геномному составу и цитоплазме. Ими
были дикие полбы Т. dicoccoides (геномы АиВ) и Т. araraticum (гено-
мы AbG). В пашем понимании они были первыми полиплоидными ви-
дами двух подродов, основными геномами которых являются А11 и Аь.
Соединение этих геномов с тремя геномами (В, G, D) рода Aegilops
дало полиплоидные пшеницы.
В зависимости от геномного состава виды в подродах группируются
в 6 секций. Филогенетическое развитие рода шло как по пути появле-
ния видов-гомологов в двух гомологичных подродах, так и по пути
становления видов-аналогов внутри каждого из этих подродов. Виды-
гомологи имеют разный геномный состав, одинаковый уровень плоид-
ности и сходные хозяйственные особенности, контролируемые главны-
ми генами. Например, полбе Т. dicoccuni гомологична полба Т. timop-
heevii: они имеют разный геномный состав, 2п=28, трудный вымолот
зерна и ломкий стержень колоса. Виды-аналоги имеют одинаковый
геномный состав, один и тот же уровень плоидности и различные хо-
зяйственные свойства, контролируемые главными генами. Например,
Т. persicum является легкообмолачивающимся аналогом полбы 7’. di-
coccutn: они имеют одинаковый геномный состав, 2п=28, но в первом
случае вымолот зерна легкий, стержень колоса прочный, во втором —
вымолот зерна трудный, а стержень колоса ломкий.
Виды подродов Triticum и Boeoticum имеют 3 уровня плоидности
(2/7.= 14, 28, 42) являются одно-, дву- и трехгеномными. W. М. Bowden
(1959), как мы уже отмечали выше, разбил пшеницу па 3 группы, со-
ответствующие распределению по числу хромосом. Исторически такие
группы впервые выделил A. Schulz (1913). Деление рода Triticum по
J. Mac Key (1966, 1968, 1975) и R. Morris и Е. R. Sears (1967) факти-
чески является попыткой увязать систему рода с геномным составом
видов и дифилетическим происхождением тетраплоидов.
В хозяйственном отношении виды с геномами АиВ и AbG делятся
на полбы и голозерные тетраплоиды, виды с геномами A"BD - па
спельты и голозерные гексаплопды. В обоих подродах дикие виды име-
ются как средн однозернянок, так и среди полб. Диких видов нет у иле
гаплоидов.
Дикие виды секций Urartu (А") и Мопососсоп (А11) имеют плоский
колос (боковая стропа его шире лицевой), ломкий его стержень, труд-
ный вымолот зерна и одну зерновку (редко 2) н колоске. Одомашни-
вание коснулось лишь однозернянок с геномом А1'. При этом дикий вид
Т. boeoticum утерял спонтанную ломкость колоса и очень трудный вы-
Coii/ii'Mi’iunut система pi><)a Tritiruin I..
19
молот .зерна. Возникшая п.з нею культурная однозернянка Т. топо-
coccum более урожайная (в основном за снег некоторого укрупнения
зерновки и увеличения продуктивной кустистости). Кульминация фи-
логенетического развития секции Мопососсои -- появление естествен-
ного мутанта Т. sinskajae. On является легкообмолачиваемым анало-
гом Т. monococcum.
Виды секции Dicoccoides (AUB) прошли путь развития от дикой
полбы к культурной и затем к голозерным тетраплоидам. Все полбы
обычно имеют плоский колос с ломким стержнем, трудным вымолотом
зерна и двумя зерновками в колоске. Одомашнивание дикой полбы
Т. dicoccoides шло по липни утери колосом спонтанной ломкости и
очень сильного одревеснения колосковой чешуи. Все культурные пол-
бы имеют менее трудный, чем у дикой полбы, вымолот зерна, а коло-
сья их ломаются в основном лишь при надавливании. Дальнейший
процесс окультуривания тетраплоидной пшеницы шел по пути приоб-
ретения легкого вымолота зерна, прочного колосового стержня и уве-
личения числа зерновок в колоске. Последнее привело к появлению ко-
лосьев, у которых лицевая сторона равна или шире боковой. Потеря
сильного одревеснения колосковыми чешуямп, ответственного за труд-
ный обмолот, связана, по J. Mac Key (1968), с мутационным появле-
нием фактора Q (?'. persicutn) и со становлением полигенной системы
легкого обмолота (Т. durum и близкие к нему виды).
В секции Timopheevii (AbG) процесс одомашнивания шел путем,
сходным с гомологичной секцией Dicoccoides: от дикой полбы Т. агага-
ticum к культурной полбе Т. timopheevii. Филогенетический ход разви-
тия этой секции увенчался появлением в паши дни голозерного анало-
га Т. timopheevii, которому дано наименование Т. militinae. У послед-
него вида легкий вымолот зерна обеспечен мутационно возникшим
фактором Qm, действие которого сходно с действием фактора Q у
Т. persicutn. Полигонная система легкого обмолота у тетраплоидов —
носителей геномов AbG — не сформировалась. Это, видимо, связано с
их сравнительно недавним происхождением и с отсутствием массово-
го возделывания, что ограничило их селекционную обработку.
Виды секции Triticum (AUBD) возникли при спонтанном скрещи-
вании культурного тетраплоида с Ае. tauschii. Если исходным для них
видом пшеницы был Т. persicutn (IT. Kihara, F. Lilienfeld, 1949), то
первичная гексаплоидпая форма имела легкий обмолот и достаточно
прочный стержень колоса. Гексаплоидиые виды с трудным обмолотом,
возможно, появились в результате спонтанной гибридизации мягкой
пшеницы с тетраплоидными видами (Н. И. Вавилов, 1926; С. Flaksber-
ger, 1930; Е. Schiemann, 1932; Е. Н. Сииская, 1955; П. М. Жуковский,
1957), т. е. они являются вторичными по происхождению. Следует от-
метить, что пленчатые виды секций Мопососсоп, Timopheevii и Dico-
ccoides являются первичными. Пленчатые гексаплоидиые виды (спе.ть-
ты) имеют специфический тип ломкости: кроме типа ломкости, кото-
рый характерен для полб, стержень их колоса может ломаться, как
у Ае. tauschii. Последнее особенно характерно для европейских форм
Т. spelta, у которых членик стержня (после разлома колоса) прилега-
ет к брюшной, стороне колоска. Такой тип разлома связан с. наличием
генома D. В группе спелы- ломкость стержня колоса отсутствует у но-
сителя гена «вавилондпостп»— у Т. vavilovii. У гексаилоидпых видон
легкий обмолот обеспечивает фактор Q. В зависимости от селекцион-
ной проработки форм--носителей этого фактора- --лицевая сторона ко-
лоса может быть меньше боковой, равна ей или больше ечм Число зер-
новок в колоске в последнем случае достигает 5-6. Такое количество
зерновок могло разместиться в одном колоске после того, как была
обеспечена возможность раздвижения колосковых чешуй. Последнее
связано с потерей сильного их одревеснения из-за действия фактора Q.
2*
ЙО Об.чпр />111)11 ТгiIiniin I..
Секция Kiharae (гомологична секции Triticum') включает лишь
один вид, который синтезирован в лабораторных условиях. Этот вид —
Т. kiharae (AbGD) является гомологом Т, spelta. Он в своем геноти-
пе имеет все гены иммунитета, характерные для Т. timopheevii. Суще-
ствование Т. kiharae как высокофсртилыюго растительного организма
свидетельствует о возможности создания гомолога мягкой пшеницы.
Главный аргумент в пользу реальности идеи создания такого гомоло-
га— закон гомологических рядов в наследственной изменчивости
Н. И. Вавилова.
ПРОИСХОЖДЕНИЕ ПОЛИПЛОИДНОЙ ПШЕНИЦЫ
Мягкая пшеница (Т. aestivum) обеспечивает хлебом большую
часть человечества. Второе место после псе по объему возделывания
занимает твердая пшеница (Т. durum). Оба вида — полиплоидные. Ис-
следователей давно интересует вопрос установления источника гено-
мов полиплоидной пшеницы. Точная их идентификация позволит со-
вершить вторичный целенаправленный синтез мягкой пшеницы с
целью улучшения ее генетического содержания за счет проверенных
доноров геномов.
История науки знает 2 основные гипотезы о происхождении тетра-
плоидной пшеницы, понимающие ее возникновение как автополипло-
идное или как аллополнплоидпос. Ныне большинство исследователей
признало аллополиплонднос происхождение полиплоидной пшеницы, а
также пришло к убеждению, что не менее пяти диплоидных видов,
дающих пищевое зерно, участвовали в ее становлении. В связи с тем,
что род Triticum L. обладает лишь двумя собственными геномами, при-
надлежащими диким диплоидным пшеницам Т. boeoticum и Т. urartu,
остальные 3 генома были заимствованы им у рода Aegilops L. Послед-
ний значительно богаче, чем род Triticum, диплопдами — носителями
хлебных геномов. Существует еще не менее 6—9 (в зависимости от
классификации) диплоидных видов эгплопса, геномы которых еще не
привлечены к синтезу нищи для человечества.
В работах по филогении пшеницы применялись морфолого-ботани-
ческий, географический, цитологический, биохимический, иммунологи-
ческий и другие методы. Серьезные исследования были проведены в
Канаде, США и Японии. В нашей стране в начале 70-х годов сделан
значительный вклад в. решение проблемы происхождения полиплоид-
ной пшеницы. Основные работы по идентификации ее геномов осу-
ществлены отделом молекулярной биологии, биохимии и качества
ВИРа и в лаборатории геобпохимип Института зоологии и ботаники
Эстонской ССР.
Исследователи придерживались двух точек зрения па происхожде-
ние пшеницы: одни считали его монофилетическим (L. Sachs, 1953;
R. Riley и др., 1958; Е. R. Sears, 1956; Е. В. Wagenaar, 1961, 1966;
Д. Мак-Кей, 1968; М. Tanaka, И. Ichii, 1973 и др.), другие — полифи-
летическим (F. Lilienfeld, Н. Kihara, 1934; D. Zohary, М. Feldman, 1962;
В. Яаска, 1969;' В. Giorgi, A. Bozzini, 1969; G. Kimber, R. S. Athwal,
1972 и др.). Еще в 1935 г. И. И. Вавилов и К. А. Фляксбергер выска-
зались в пользу последней точки зрения. Исследования 70-х годов,
проведенные в нашей стране, в которых использовались биохимичес-
кие, морфолого-ботанические, генетические и иммунологические мето-
ды, позволили сделать заключение о дифилетическом происхождении
тетраплопдиой пшеницы (А. В. Коиарев и др., 1971, 1974; В. Л. Ме-
набде, 1971; Т. И. Пенева, 1974; В. Яаска, 1974; Э. Ф, Мигушова,
А. В. Коиарев, 1975; В. Г. Копарев и др., 1976; В. И. Кривченко и др.,
1976). За рубежом, на основании данных иммупохимического ана-
лиза белков, такая же точка зрения высказана A. Bozzini и др.
П/нин хим ih'ii не iio.'iiin лот hit >й niiii’i i iiifiii 21
(1973). В нс ri ;|i и'lei к ii различное время в i eoi рафпчески разобщенных
регионах от разных материнских и отцовских форм возникли виды сек-
ций Dicoccoides и Timopheevii, имеющие соответственно геномы Л1|В
и A!’G.
По мнению исследователей, первичный дикий амфиднплонд секции
Dicoccoides возник значительно раньше, чем первичный амфиднплонд
секции Timopheevii (К. Л. Фляксбергср, 1935; Л. Л. Ерицяп, 1941;
В. Giorgi, Л. Bozzini, 1969; В. Г. Конарев и др., 1976). Очаг происхож-
дения первых—Юго-Западная Сирия и прилегающие к ней районы
Израиля и Иордании (D. Zohary и др., 1969; В. Яаска, 1974), очаг про-
исхождения вторых ----- Северный Ирак и примыкающие к нему терри-
тории Ирана и Турции (М. Танака, S. Ichikawa, 1972; В. Яаска, 1974;
Э. Ф. Мигушова, Л. В. Конарев, 1975).
Рассмотрим происхождение отдельных геномов полиплоидной пше-
ницы.
Донор первого генома мягкой и твердой пшеницы. Первоначально
донором этого генома считали культурную однозернянку Т. monococ-
cum L. (II. Kihara, 1919, 1924; К. Sax, 1922). В последующих исследо-
ваниях (Ю. А. Фплпнчепко, 1930; A. Longlei, W. Sando, 1930; Д. Кос-
тов, 1940; Т. Б. Вакар, 1948; Е. Schiemann, G. Standi, 1958 и др.) выска-
зывались сомнения в правильности этого положения. R. Riley и
G. D. Bell (1958) при изучении гибридов между однозернянками и тет-
раплоидными пшеницами наблюдали развитие ослабленных растений в
комбинациях, в которых участвует Т. monococcum или Т. boeoticum
subsp. boeoticum. Подобного явления нс было у гибридов, полученных
с участием Т. boeoticum subsp. thaoudar. Был сделан вывод о возмож-
ном родстве между последним и геномом А тетраплоидпых видов сек-
ции Dicoccoides (R. Riley, G. D. H. Bell, 1958). С того времени утверди-
лось мнение, что геном А в твердую пшеницу привнесен дикой однозер-
нянкой Т. boeoticum.
Биохимические исследования G. Wolf, В. Lerch (1973) и В. Яаски
(1974) отвергли Т. boeoticum как донора генома А твердой пшеницы.
В 1974 г. была опубликована работа А. В. Копарсва и др., использовав-
ших для идентификации генома Л. твердой и мягкой пшеницы сравнитель-
ный иммунохимнческий анализ глиадипа. Оказалось, что полиплоидные
виды секций Dicoccoides и Triticum имеют антиген, специфичный только
для Т. urartu, что Т. boeoticum, Т. monococcum и тетраплоидпые виды сек-
ции Timopheevii не имеют этого антигена. На этом основании сделан вы-
вод, что донором генома А у твердой и мягкой пшеницы был предок сов-
ременного Т. urartu или близкая к нему форма (А. В. Конарев и др., 1974;
А. В. Конарев, 1974, 1975; В. Г. Конарев и др., 1976). Это открытие под-
тверждено следующими методами: 1) сравнительно-морфологическим —
Т. urartu имеет бархатистое на ощупь опушение листьев, сходное с опу-
шением у некоторых биотинов всех тетраплоидпых видов секции Dicoccoi-
des (А. В. Конарев и др., 1974); 2) иммунологическим — характер разви-
тия мицелия пыльной головни в тканях зародыша у зерновок Т. urartu
сходен с развитием гифов этого гриба в зародышах твердой и мягкой
пшеницы, по отличается от Т. boeoticum и Т. timopheevii (В. И. Кривчен-
ко и др., 1976),; 3) цитологическим — дителоцептрическнс и дважды дите-
лоцентрпческие (по всем хромосомам геномов А и В) линии мягкой пше-
ницы Chinese Spring были опылены пыльцой Т. zzrorZzz; обследование по-
ведения хромосом гибридов в мейозе показало, что хромосомы Т. urartu
близки к хромосомам генома А мягкой пшешщы (V. Chapman и др.,
1976). Подобный результат получен также при скрещивании Т. urartu с
линиями мягкой пшеницы, дптолосомпыми по хромосомам геномов А и В
(J. Dvorak, 1976); 4) биохимическим (Т. Nagayoshi, 1978).
Таким образом, донором первого генома твердой и мягкой пшеницы, а
также родственных им других полиплоидных видов следует признать
22 Обзор рода Triticum I,.
предки современного Т. urarlu, ядернып юном которого обозначается
символом Л". Геномный состав тетраплоида, произошедшего с его учас-
тием, обозначается символами Л'’В (геномная формула ЛПЛ’1ВВ).
Донор генома В. Вопрос о происхождении этого генома у твердой и
мягкой пшеницы вызвал оживленную дискуссию, которая не утихает и
поныне. Многократные попытки идентификации породили сомнения в
возможности его установления (D. Zohary, М. Feldman, 1962; G. Kimber,
R. S. Athwal, 1972; G. Kimber, 1974).
Источник генома В пытались искать в основном среди представителей
родов Triticum, Aegilops и Agropyron. Легкая обмолачивасмость колоса
/1. triticeum Gaerth. [Eremopyrum triticeum (Caerth.) Nevski], по мнению
E. S. McFadden и E. R. Sears (1944), свидетельствует о том, что этот злак
мог быть донором генома В пшеницы. Однако морфология его хромосом
не соответствует хромосомам генома В (II. П. Авдулов, 1931; Е. R. Sears,
1956; Р. Sarkar, G. L. Stebbins, 1956; R. Riley и др., 1958), как и содержа-
ние ДНК в ядре (И. Rees, 1963; Н. Rees, М. R. Walters, 1965).
В связи с изучением конъюгации хромосом в мейозе Б. А. Вакар
(1935), F. Н. Peto (1936), S. Matsumura (1951) пришли к предположе-
нию, что геном В произошел от A. elongalum (Host) Beauv. [Elytrigia.
elongata (Host) Nevski], Однако гомология хромосом в мейозе у гиб-
ридов от скрещивания его с тетраплоидными видами пшеницы секции Di-
coccoides оказалась незначительной (В. С. Jenkins, A. Mochizuki, 1957).
Предположение, что геном В произошел от диплоидной пшеницы, сде-
лано А. Camara (1935, 1944). В. L. Johnson (1972) поддержал эту гипоте-
зу. Позднее (В. L. Johnson, 1974, 1975; Н. S. Dhaliwal, В. L. Johnson,
1976) на основании изучения электрофоретических спектров
белков зерна и некоторых морфологических признаков растений
была высказана мысль, что донором генома В пшеницы секции Dicoccoi-
des был Т. urartu, а донором генома А— Т. boeolicum. А. В. Конарев и др.
(1976) методом электрофореза глиадинов зерна в полиакриламидном ге-
ле изучили популяции диких однозернянок Закавказья, Ирака, Ирана и
Турции, но не выявили среди них сходства с геномом В твердой и мягкой
пшеницы. Подобная работа по ферментам проростков проведена В. Яас-
кой (1974), также пришедшего к заключению об отличии генома В от ге-
номов диких однозернянок. Т. urartu как донор генома В отвергнут био-
химическими исследованиями К. Chen и др. (1975), L. Vittozzi и V. Sila-
по (1976).
Многие исследователи указывают на секцию Sitopsis (Jaub. et Spach.)
Zhuk, рода Aegilops L. как на возможный источник генома В. Еще
G. N. Pathak (1940) отметил сходство между спутничными хромосомами
Ае. speltoides и Т. turgidum. Затем Р. Sarkar и G. L. Stebbins (1956) на
основании изучения морфологических признаков колоска указали на Ае.
speltoides как па возможного донора генома В пшеницы. При наблюде-
ниях над прохождением мейоза у гибридов от скрещивания эгилопсов
этой секции с тетраплоидными пшеницами, имеющими геномный состав
АиВ, наиболее высокая степень конъюгации (до 8—10 бивалентов па 1
клетку) была установлена для комбинаций с участием Ае. speltoides
Tausch (J. A. Jenkins, 1929; F. Lilienfeld, H. Kihara, 1932; О. H. Сорокина,
1934; W. P. Thompson и др., 1943; E. S. McFadden, E. R. Sears, 1947;
R. Riley и др., 1958). На этом основании в течение многих лет донором
генома В считали Ае. speltoides.
R. Riley и V. Chapman (1958) установили, что Ае. speltoides обладает
способностью подавлять активность хромосомы 5В пшеницы, тормозящей
конъюгацию гомеологичных хромосом. В результате у гибридов, получен-
ных при скрещивании этого эгилопса с твердой и мягкой пшеницами, воз-
растает уровень бивалентизации, создающий иллюзию родства их гено-
мов. Впоследствии были открыты также биотипы Ае. speltoides, которые
не обладают способностью содействовать гомеологичпой конъюгации
Происхождение полиплоидной пшеницы
23
(J. Dvorak, 1972; G. Kimber, R. S. Athwal, 1972). У гибрида между такой
формой Ле. speltoides и мягкой пшеницей истинная степень конъюгации
хромосом в мейозс низкая: по данным G. Kimber и R. S. Athwal (1972),
она равна 2,9 бивалента на одну клетку. При изучении мейоза у гибридов
от скрещивания Ле. variabilis Eig с видами секции Sitopsis J. М. Ruben-
stein и Р. J. Salle (1973) получили косвенное подтверждение отсутствия
генома Ае. speltoides в генотипе мягкой пшеницы.
По содержанию ДПК в ядре геному В наиболее соответствует Ае.
speltoides (FI. Rees, 1963; Н. Rees, М. R. Walters, 1965). Электрофорети-
ческое обследование ферментов у проростков показало, что Ле. speltoides
и геном В тетраплопдных пшениц секции Dicoccoides обладают сходными
изоэстеразами и изофосфатазамп (В. Яаска, 1969, 1974, 1976). Электро-
форетические спектры изоферментов пероксидазы зерновок Т. durum, и
Т. aestivum оказались сходными также с Ае. speltoides (К. Kobrehel,
M.-F. Gautier, 1974). На этом основании исследователи сделали вывод,
что донором генома В был Ае. speltoides. Однако параллельно шли рабо-
ты по электрофорезу эстераз зерна (A. Bozzini и др., 1973), фосфодиэсте-
раз листьев (G. Wolf, В. Lerch, 1973), которые показали противополож-
ный результат. Экстраполированная зимограмма изоэпзима спирта
(ADH) не имела полного сходства с Ле. speltoides (R. К. Mitra, С. R. Bha-
tia, 1971).
Электрофоретическое обследование белков зерна, проведенное О. Hall
и В. L. Johnson (1966) отвергло Ае. speltoides как донора генома В, так
же как изучение гимза С-исчерчепности хромосом в опыте В. S. Gill,
G. Kimber (1974) и G. Hadlaczky, A. Belea (1975). В то же время харак-
тер исчерченности хромосом, по данным А. Т. Natarajan и N. Р. Sarma
(1974), свидетельствовал в пользу того, что Ае. speltoides был донором
генома В.
Иммунохимнческий анализ белка зерновок в опыте A. Aniol (1973)
показал, что белки, кодируемые геномом В у твердой пшеницы, антиген-
но близки к белкам Ае. speltoides. A. Bozzini и др. (1973) при иммунохи-
мическом анализе белков зерновок нашли, что реакция па специфические
белки отвергает Ае. speltoides как донора генома В. Иммунохимнческий
анализ глиадинов зерновок, проведенный Т. И. Пеневой (1974), подтвер-
дил последний вывод: Ае. speltoides имеет специфический антиген, отсут-
ствующий у видов секции Dicoccoides.
К. исследованиям по идентификации генома В была привлечена гибри-
дизация, при которой стремились синтезировать тетраплоидную пшени-
цу. Однако гибриды создавались с участием Т. boeoticum, а не Т. urartu.
Видимо, это одна из причин отсутствия у них сходства с видами секции
Dicoccoides.
Таким образом, большая часть современных исследований отвергла
Ае. speltoides как донора генома В. Тщательное электрофоретическое об-
следование ферментов, проведенное В. Яаской (1974) в ранней фазе рос-
та растений, свидетельствует о том, что донор генома В и современный
Ае. speltoides филогенетически родственны. Поэтому возможно, что мето-
ды исследований, примененные R. К. Mitra, С. R. Bhatia (1971) и Т. И. Пе-
невой (1974), улавливали наличие частичного сходства генома В с гено-
мом этого эгилопса.
М. Tanaka (1955) нашел, что по морфологическим признакам расте-
ний имеется сходство между Ле. bicornis (Forsk.) Jaub. et Spach и видами
секции Dicoccoides. Действительно, как нам кажется, средн эгилопсов
секции Sitopsis виды подсекции emarginata Eig, куда относятся Ле. bicor-
nis, Ле. lotigissima и Ле. sharonensis, имеют наибольшее сходство с твер-
дой пшеницей по общему облику растений.
Синтез тетраплоидов с участием Т. boeoticum показал, что амфидипло-
ид с ядерным геномом Ае. bicornis более похож на виды ряда твердой
пшеницы, чем амфидиплоид с геномом Ае. speltoides (Е. R. Sears, 1956,
2-1
Обаор рода Trilictun I..
1969; II. Kiliara, 1965). Методом тонкослойной хроматографии выявлено,
что Ле. bicornis сходен с. геномом В пшеницы (Н. С. Dass, 1972). Однако
оказалось, что у гибридов, полученных при скрещивании Ле. bicornis с
видами секции Dicoccoides, в мейозе наблюдается низкий уровень бпва-
лентпзацип (R. Riley и др., 1958; К- A. Siddiqui, J. К. Jones, 1968), сви-
детельствующий о наличии межгеномных различий. Кроме того, R. Riley
и др. (1958) показали, что геном В пшеницы и Ле. bicornis имеют разли-
чия по морфологии хромосом.
О. Hall и В. L. Johnson (196(1) нашли, что но электрофореграммам
белков зерна Ле. bicornis не может быть назван донором генома В. Экст-
раполированная зимограмма пзоэпзпма спирта (ADII) этого эгилопса со-
ответствовала геному В пшеницы, по по быстромигрирующей эстеразе
(EST-1) сходство отсутствовало (R. К. Mitra, С. R. Bhatia, 1971). Элек-
трофоретическое обследование ферментов также указало на отсутствие
подобия с геномом В пшеницы по изофосфатазам и и.зоэстеразам про-
ростков Ле. bicornis (В. Яаска, 1974). Иммунохимический анализ глиадп-
нов зерна показал, что маркера генома Ле. bicornis нет у видов секции
Dicoccoides (Т. И. Пснсва, 1974; В. I'. Копиров и др., 1976). G. Kimber
(1974) отрицает участие Ле. bicornis в становлении последних.
Таким образом, результаты большинства исследований отвергли Ле.
bicornis как донора генома В. Наличие морфологического сходства рас-
тений этого эгилопса и твердой пшеницы, как п эксперимент И. С. Dass
(1972), возможно, свидетельствуют о филогенетическом родстве донора
генома В пшеницы и Ле. bicornis.
По внешнему облику растений среди эгилопсов более всего сходен с
видами ряда твердой пшеницы Ле. sharonensis Eig. Однако при скрещи-
вании его с Т. turgidum L. наблюдались низкий уровень конъюгации хро-
мосом в мейозе, стерильность цветков и пыльны (М. Tanaka, 1955). По
результатам изучения мейотической бивалеитизации у гибридов G. Kim-
ber (1961, 1973, 1974), J. М. Rubenstein и Р. J. Salle (1973) сделали за-
ключение, что Ле. sharonensis не был донором генома В пшеницы.
По содержанию ДНК в ядре этот эгилопс также не соответствует ге-
ному В пшеницы (Н. Rees 1963; И. Rees, М. R. Walters, 1965). В. Яаска
(1974) пришел к заключению, что пет электрофоретического соответствия
между зимограммами изофосфатаз и изоэстераз Ле. sharonensis и гено-
мом В пшеницы.
Иммунохимический анализ глиадппов зерна показал, что все. специ-
фические для генома В компоненты имеются в спектрах Ле. sharonensis,
по у него присутствует еще один добавочный антиген, которого нет у тет-
раплоидной пшеницы секции Dicoccoides (Т. И. Пенева, 1974; В. Г. Ко-
нарев и др., 1976), т. е. Ле. sharonensis не соответствует донору генома В.
Установлено, что диплоид Ле. sharonensis возник при межвидовой ин-
трогрессивной гибридизации между Ае. bicornis и Ле. longissima
(J. G. Waines, В. L. Johnson, 1969, 1972; Т. И. Пснсва, 1973, 1974; В. Г. Ко-
нарев, Т. И. Пенева, 1975; В. Г. Конарсв и др., 1976). Ареал его неболь-
шой. Возможно, что он возник сравнительно недавно — позднее первич-
ной тетраплоидной пшеницы, имеющей геномы АиВ, Несомненно, он близ-
кородствен виду, давшему пшенице, геном В.
Ле. longissima Schweinf. et Muschl., как и все перечисленные виды сек-
ции Sitopsis, дает низкий уровень конъюгации хромосом в мейозе у гибри-
дов от скрещивания с твердой пшеницей (О. Н. Сорокина, 1934). По со-
держанию ДПК в ядре Ле. longissima не соответствует геному В пшени-
цы (II. Rees, 1963; II. Rees, М. R. Walters, 1965). По данным G. Martini
и A. Bnmori (1973) содержание ДНК У Ле. longissima ис соответствует
тому, которое должно быть у донора генома В полбы. В. Яаска (1974)
показал, что при электрофоретическом анализе ферментов у проростков
не выявлено соответствия между зимограммами изофосфатаз и изоэсте-
раз у Ле. longissima и у генома В пшеницы.
111>оисхождеиие полиплоидиии пиичищы
25
В. I,. Johnson (1972) отметил наличие сходства в электрофоретичес-
ких спектр ах белков зерновок Ле. longissima и видов секции Dicoccoides.
При иммунохимическом анализе глиадина зерновок все компоненты, спе-
цифичные для генома В пшеницы, обнаружены у Ле. longissima
(Т. И. Исаева, 1974; В. Г. Конарев и др., 197G). На спектрах преципитации
глиадина этот вид эгилопса наиболее соответствовал геному В пшеницы.
Учитывая, что иммунохнмическпй анализ глиадина относится к числу
наиболее чувствительных серологических методов идентификации гено-
мов (В. Г. Конарев, 1971, 1973; В. Г. Конарев и др., 1972), ио результа-
там описанного эксперимента сделан вывод, что донором генома В пше-
ницы был предок современного Ае. longissima пли близкая к нему форма.
При сравнительном иммунохимическом анализе белков спор пыльной
головни также установлено, что по наличию идентичных компонентов
наиболее близки между собой белки Ustilago tritici f. duri и пыльной го-
ловни, поражающей Ае. longissima. Причем характер поражения заро-
дышей зерновок, особенности локализации и морфология мицелия пыль-
ной головни в тканях зародыша зерновок у Ае. longissima и Т. durum
также имели сходство (А. М. Ямалссв, 1974; В. П. Кривченко и др., 1976).
Таким образом, пммунохимпчсским методом обнаружено, что среди
ресурсов диплоидных видон пшеницы и эгилопса наиболее близким к ге-
ному В пшеницы является Ае. longissima. Возможно, что при тщательном
обследовании ареала этого эгилопса будет обнаружен биотип, который
даст повышенный уровень конъюгации хромосом в мейозе у гибрида от
скрещивания с твердой пшеницей. L. Vitlozzi и V. Silano (1976) экспери-
ментально подтвердили, что Ае. longissima мог быть донором генома В
пшеницы.
Донор цитоплазмы твердой пшеницы. Изложенное выше свидетель-
ствует о том, что первичный тетраплоид секции Dicoccoides возник при
скрещивании диплоидных видов — дикой пшеницы и эгилопса — путем
соединения их ядерных геномов. Решение вопроса, какой из этих дипло-
идных видов был материнским растением при этом скрещивании, даст
ответ на вопрос, какому виду принадлежит цитоплазма твердой пшени-
цы. В ходе исследований по идентификации генома В было высказано
предположение, что при становлении первичного тетранлоида источником
его цитоплазмы был донор генома В (S. S. Маап, К- A. Lucken, 1967,
1968; Н. Suemoto, 1968; В. А. Крупнов, 1973). Цитоплазма эта определе-
на как эммероподобная (S. S. Маап, 1973), привнесенная эгплопсом сек-
ции Sitopsis (Н. Suemoto, 1973). Н. Kihara (1966) указал на Ае. speltoi-
des как на донора цитоплазмы твердой пшеницы. И. Suemoto (1973)
предположила, что цитоплазма этого эгилопса лишь родственна цито-
плазме полиплоидной пшеницы. Эксперименты по введению ядра твер-
дой пшеницы в цитоплазму Ае. speltoides показали, что при этом не про-
исходит гармонического совмещения ядерного генома и плазмона (цито-
плазмы): исследователи наблюдали полную стерилизацию пыльцы
(S. S. Маап, К. A. Lucken, 1969, 1971; S. S. Маап, 1973; К. Гоцов, 1-975;
И. Панайотов и др., 1976) или резкое снижение ее фертильности (до
33,8%, по Н. Suemoto, 1973). Рост и развитие стерильных растений с ци-
топлазмой Ае. speltoides и ядром твердой пшеницы были нормальными.
Совмещение ядра твердой пшеницы с цитоплазмой Ле. bicornis пол-
ностью или частично стерилизовало пыльцу и ослабляло мощность расте-
ний (S. S. Маап, К- A. Lucken, 1971, 1972; S. S. Маап, 1973). В подобном
опыте Н. Suemoto (1973) фертильность пыльцы у растений с ядром Т. tur-
gidum была высокой (88,4%), по у сеянцев наблюдалась пестролист -
посты
Таким образом, по степени стерилизации пыльцы можно предполо-
жить, что цитоплазма Ае. bicornis филогенетически ближе к цитоплазме
твердой пшеницы, чем цитоплазма Ле. speltoides. Однако пестролист-
ность сеянцев и ослабление мощности растительных организмов комби-
26
Обзор рода Triticum L.
нации «цитоплазма Ac. bicornis | ядро 7'. durum» свидетельствуют об от-
сутствии полного генетического соответствия между ними.
Опыты по переводу ядра твердой пшеницы в цитоплазму Ac. sharo-
nensis показали, что при этом вызывается полная или частичная стерили-
зация пыльцы в зависимости от условии выращивания (G. Bozzini и др.,
1976). Частичную фертильность растений комбинации «цитоплазма Ае.
sharonensis + ядро Т. durum», сопровождаемую пестролистностыо сеян-
цев, ослаблением растений и замедлением их роста, наблюдали S. S. Ма-
ли (1973) и II. Siiemoto (1973). По данным последней, фертильность
пыльцы у растений достигла лить 30,4%, т. е. как у твердой пшеницы,
имеющей цитоплазму Ле. speltoides.
Таким образом, цитоплазма Ае. sharonensis может быть квалифици-
рована лишь как имеющая неполное родство с цитоплазмой твердой
пшеницы.
Растения с цитоплазмой Ае. longissima и ядром Т. durum или Т. turgi-
dum могут быть стерильными или частично фертильными. S. S. Маап
(1973) определяет эту стерильность как полную, а фертильность как до-
статочно высокую, ио сопровождающуюся ослаблением растений. По
Н. Suemoto (1973), фертильность пыльцы достигает 77,4%, по наблюда-
ется пестролпстпость сеянцев и слабый рост растений. По изложенным
данным можно заключить, что есть какая-то степень родственности меж-
ду цитоплазмами Ае. longissima и твердой пшеницы. Однако следует от-
метить, что в названных экспериментах число беккроссов пыльцой тетра-
плоидной пшеницы было недостаточно большим.
Цитоплазма Т. boeoticum и ядро твердой пшеницы несовместимы:
у растений наблюдается стабильная стерилизация пыльцы в любых усло-
виях выращивания (S. S. Маап, К. A. Lucken, 1967, 1968, 1970; Т. Hori,
К. Tsunewaki, 1967; М. А. Федин, 1974). Кроме того, эта цитоплазма вы-
зывает ослабление роста, утончение соломины, узость и укороченпость
листьев, увеличение числа побегов. Цитоплазма Т. monococcum в присут-
ствии ядра Т. durum также дает мужскую стерильность и угнетает рост
растений (S. S. Маап, К. A. Lucken, 1967, 1968, 1970, 1971).
Цитоплазма Т. urartu не изучена. Пока не установлено, есть ли у нее
отличия от цитоплазмы Т. boeoticum или они сходны. Имеются сведения,
что при реципрокном скрещивании этих видов можно получить фертиль-
ные (J. Mac Key, 1969; А. А. Филатенко, личное сообщение) или стериль-
ные гибридные растения (В. L. Johnson, 1975). Однако такие данные не
проливают света на вопрос родственности цитоплазм Т. urartu и Т. durum.
Таким образом, на уровне современных знаний можно сделать пред-
положение, что наиболее близкородственными цитоплазме твердой пше-
ницы являются цитоплазмы Ае. bicornis и Ае. longissima. По имеющимся
данным трудно судить о степени сходства или отличия цитоплазмы этих
эгилопсов от цитоплазмы твердой пшеницы. Возможно, что цитоплазма
у названных видов однозначна. Отсутствие полного гармоничного разви-
тия у растений с цитоплазмой Ае. bicornis или Ае. longissima и ядром
твердой пшеницы может свидетельствовать о некотором изменении, про-
изошедшем у современных эгилопсов или твердой пшеницы по сравне-
нию с их формами, жившими несколько десятков тысяч лет назад.
Донор геноМа D. Переход пшеницы с тетраплоидного двухгепомпого
на гексаплоидный трехгеномпый уровень обеспечил ей место главного
хлеба человечества. Еще О. Stapf (1909) предполагал, что в становлении
мягкой пшеницы принял участие носитель генома D — Ае. ctjlindrica Host.
Такая же мысль отражена в монографии J. Percival (1921). К. и Н. Sax
(1924) обнаружили цитологическое сходство между 7 хромосомами Ае.
ctjlindrica и мягкой пшеницы. По результатам изучения мейоза у гибри-
дов Н. Kihara (1930) определил, что эти виды имеют один общий геном.
G. N. Pathak (1940) показал, что морфология хромосом свидетельствует
IIеоисхождение noAUiciomhioi'i ttiiieitti
i(i>i
27
о том, что донором трои.его геном;! мягкой пшеницы был диплоидный вид
Ле. tauschii Coss. (=Ле. squarrosa L.).
В 1944 г. сообщалось о синтезе гексаплоидной пшеницы нутом удвое-
ния хромосом у гибрида от скрещивания тетраплоидной пшеницы секции
Dicoccoides с Ле. tauschii (E.S. McFadden, Е. R. Sears, 1944; II. Kihara,
1944). Но результатам иммунохимпческого и электрофоретического ана-
лизов глиадииов зерна уточнено, что донором генома D гексаплоидной
пшеницы был Ле. tauschii subsp. strangulata (Eig) Tzvcl. (А. Г. Хакимо-
ва, И. И. Гаврилюк, 1973; В. Г. Коварен и др., 197(1). Зимограммы «-ами-
лазы также указали на этот подвид как на источник генома D мягкой
пшеницы (К. Nishikawa, 1973; К- Nishikawa и др., 1975). М. Tanaka и
Н. Kihara (1965) еще ранее показали, что у гибридных растений, полу-
ченных при скрещивании тетраплоидной пшеницы секции Dicoccoides с
Ае. tauschii subsp. strangulata, в F, озернепиость колосков выше, чем при
скрещивании с subsp. tauschii (=eusquarrosa Eig), т. с. реальность вы-
живания в природе у гибрида, возникшего с участием первого подвида
эгилопса, больше, чем во втором случае. Это косвенно свидетельствует в
пользу того, что именно Ле. tauschii subsp. strangulata мог в природе ско-
рее войти в контакт с тетраплоидной пшеницей, чем subsp. tauschii.
Донор компонента А"В гексаплоидной пшеницы. Источником тетра-
плоидного компонента для мягкой пшеницы сначала считали дикую пол-
бу— Т. dicoccoides (Koern. ex Achers. et Graebn.) Schweinf. (O. Cook,
1913; J. Percival, 1921 и др.), роль которой в становлении этой пшеницы
была отвергнута позже (М. Tanaka, 1961; Н. Kihara, 1965).
По результатам электрофоретического анализа белков и изоперокси-
даз зерновок В. L. Johnson (1972) и J. Brad и др. (1972) сделали заклю-
чение, что донором компонента А”В у гексаплоидной пшеницы была не
дикая, а культурная полба — Т. dicoccum (Schrank) Schuebl.
При гибридизации Т. dicoccoides, Т. dicoccum, Т. durum, Т. turgidum
и Т. polonicutn с носителями генома D — Ае. tauschii или Ае. cqlindrica —
Н. Bleier (1927, 1930), О. Н. Сорокина (1934), Н. Kihara (1949), Е. S. Mc-
Fadden и Е, R. Sears (1947) наблюдали низкий уровень конъюгации хро-
мосом в мейозе (0—4 бивалента на одну клетку). Это свидетельствовало
о значительной трудности соединения геномов названных тетраплоидных
пшениц и генома D.
По данным М. Tanaka (1959) и Н. Kihara (1965), при скрещивании
Т. dicoccoides, Т. dicoccum, Т. durum и Т. persicutn с Ае. tauschii subsp.
strangulata самофертильность цветков у гибридов в F] соответственно
равнялась 0; 2,4; 7,1 и 73,3%; полноценной пыльцы было 24,7; 48,1; 93,8 и
96,8%. Таким образом, наибольшая вероятность выживания в природных
условиях могла быть в случае, когда донором компонента АиВ при воз-
никновении первичного гексаплоида был Т. persicutn.
В. И. Кривченко и др. (1976) указывают па четкую специализацию
Ustilago tritici f. aestivi к поражению гексаплоидных видов с геномами
AUBD и авирулентность к ним возбудителя f. duri. Это, видимо, говорит
о том, что тетраплоидный вид, участвовавший в становлении первичного
гексаплоида, был устойчивым к f. duri. Именно такие формы найдены у
Т. dicoccum и Т. persicutn (В. И. Кривченко, 1973; А. М. Ямалеев, 1974).
Компонент ДиВ был выделен из мягкой пшеницы (Е. R. Kerber, 1964;
Р. J. Kaltsikes и др., 1968). Причем методом микрофотометрии было до-
казано, что Tetra-Canthatch (тетраплоидный компонент, полученный из
сорта Canthatch) не имеет существенных отличий по содержанию ДИК
в ядре от тетраплоидных видов секции Dicoccoides (Y. Furuta и др.,
1974).
В условиях Ташкентской области были изучены морфологические при-
знаки тетраплоидных производных от сортов Canthatch, Prelude, Rescue
и Thatcher (Э. Ф. Мпгушова, 1975). Оказалось, что все они отличались от
исходных сортов большей плотностью колоса,, более нежными колоско-
28
Об:top роди Triticum L.
ними чешуями, отсутствием плеча и поперечиой вдавленное!и у основа-
ния колосковой чешуи, выполненной (паренхимой) под колосом соломи-
ной, более топким и узким стержнем колоса, т. с. напоминали Т. persi-
сшп. Однако плотный колос не характерен для современного Т. persicum.
Возможно, что первичная форма 7'. persicum, возникшая мутациоппо у
Т. dicoccum, имела плотный колос. Косвенное подтверждение этому — на-
личие плотного колоса у тетранлоидного вида Т. militinae, который му-
тационио возник у Т. timopheevii (П. М. Жуковский, Э. Ф. Мигушова,
1969). Приняв 7’. militinae как модель первичного возникновения факто-
ра Q, можно предположить, что первичная форма Т. persicum также име-
ла плотный колос. Эта точка зрения была поддержана Р. А. Удачиным
(1975). Первичный Т. persicum скрестился с subsp. strangulata. В ареале
последнего возникла первая гексаплоидпая пшеница, которая дала нача-
ло пшенице свайных построек Т. antiquorum Heer. Первоначальная до-
гадка о подобном пути возникновения гексаплоида с геномами AUBD при-
надлежит И. Kihara и F. Lilienfeld (1949).
Присоединение генома D к пшенице обеспечило ей огромное внутриви-
довое разнообразие, повышенную приспособленность к условиям среды,
расширение ареала, приобретение озимого образа жизни, увеличение зи-
мо- и морозостойкости, повышение хлебопекарных свойств, а также сни-
жение устойчивости к грибным болезням, падение концентрации белка
в зерне, приобретение летальных генов Ch2 и Db
Донор цитоплазмы мягкой пшеницы. П. Kihara (1965) сообщил, что
скрещивание между тетраплоидным видом пшеницы (АиВ) и Ае. tauschii
(D) удается лишь в случае, когда эгилоис является опылителем. Н. Ki-
hara (1965, 1966) сделал заключение, что донором цитоплазмы у гекса-
плоидных видов был тетраплондный вид. К- Chen и др. (1975) определи-
ли полипептндный состав S-карбоксиметилировапиой фракции I белков,
контролируемых хлорапластным геномом, методом изоэлектрического
фокусирования. Оказалось, что хлоропластный геном мягкой пшеницы
был привнесен тетраплоидным видом пшеницы — донором компонента
АиВ. Последний при скрещивании с Ае. tauschii был материнской фор-
мой, т. е. цитоплазма у мягкой и твердой пшеницы одинаковая.
Огромная гибридизационная работа, выполненная селекционерами по
межвидовому реципрокному скрещиванию между гексанлоидными
(AUBD) и тетраплопднымн (АиВ) видами пшеницы, свидетельствует об
однозначности их цитоплазмы. В опытах по переводу ядра мягкой пше-
ницы в цитоплазму других видов показано, что цитоплазма твердой пше-
ницы похожа па цитоплазму мягкой пшеницы, что их цитоплазма одина-
кова или сходна с цитоплазмой Ае. tauschii (S. S. Маап, К. A. Liicken,
1971; S. S. Маап, 1973, 1975). Когда ядро тетраплоидпых видов пшени-
цы (АиВ) было переведено в цитоплазму Ае. tauschii, то обнаружилось
полное несоответствие между ядром и цитоплазмой, сопровождавшееся
полной стерилизацией пыльцы, частичной женской стерильностью, по-
зднеспелостью, ослаблением мощности растений (S. S. Маап, К.A. Luc-
ken, 1971, 1972; Н. Kihara, 1973). Иными словами, было доказано, что ци-
топлазма у видов секции Dicoccoides иная, чем у Ле. tauschii. После
большого числа беккроссов несоответствие цитоплазмы этого эгилопса
с ядром Т. dufuin выразилось в нежизнеспособности зерновок (S. S. Ма-
ап, 1973). Цитоплазма Ае. tauschii теряла стерилизующее действие лишь
в присутствии генома D, видимо, обладающего генами восстановления
фертильности для этой цитоплазмы (И. Kihara, 1973; S. S. Маап, 1973).
Таким образом, на основании перечисленных фактов следует за-
ключить, что донором цитоплазмы видов секции Triticum был исходный
для них тетраплондный вид, который в свою очередь эту цитоплазму
получил от донора генома В.
Донор первого генома видов секции Timopheevii. А. В. Конарев и др.
(1971, 1974) методом иммунохимического анализа глиадипов зерновок
IIpoucxmicdciiite полиплоидной пи/еницы
29
показали, что донором первого генома (Л1') у видов секции Timopheevii
была дикая однозернянка Т. boeoticum. То же нашел В. Яаска (1974) при
электрофоретическом обследовании изофосфатаз и изоэстераз у про-
ростков.
Донор генома G. В начале 30-х годов стало ясным, что Т. timopheevii
генетически отличается от видов ряда твердой пшеницы (Л. А. Ерицян,
1928; А. Г. Хипчук, 1929; Л. Л. Декапрслевич, В. Л. Мепабде, 1932).
F. Lilicnfeld и II. Kihara (1934), изучив ход мейоза у межвидовых гиб-
ридов, пришли к заключению, что второй геном Т. timopheevii иной, чем
у Т. durum. Они дали ему символ G.
Д. Костов (1937, 1940) по результатам наблюдений над поведением
хромосом в мейозе пришел к выводу о частичном сходстве гепомов В и
G, дав последнему символ р. Данные R. М. Love (1941), L. Sachs (1953)
и Е. В. Wagenaar (1961, 1963, 1966) также свидетельствовали о наличии
некоторого сходства между геномами В и G. На этом основании утвер-
дилась гипотеза о монофилетическом происхождении Т. timopheevii и
Т. durum, вызвавшая замену написания генома G символом В.
М. Г. Туманян (1937, 1938, 1939), основываясь па том, что Т. ti-
mopheevii является составной частью популяции зандури, высказал до-
гадку о его автополиплоидпом происхождении от Т. monococcum. В по-
следующем его точка зрения была поддержана Е. II. Списков (1955) и
грузинской школой тритикологов. Изучение морфологии хромосом при-
вело Л. К- Чхаидзе (1967, 1969) к заключению, что у Т. timopheevii и
Т. zhukovskyi кратно повторяется набор хромосом, характерный для
Т. monococcum. Эксперимент по ионизирующему облучению убедил
А. Д. Горгидзе (1968, 1973) в автополиплоидпом происхождении Т. ti-
mopheevii. На основании общности ареалов дикорастущей однозернянки
Т. boeoticum и полб Т. araraticum и Т. timopheevii, морфологического
сходства между ними, узкого размаха изменчивости в F2, появления при
скрещивании указанных полб в F2 растений типа Т. boeoticum А. А. Фи-
латенко (1968) подтверждает гипотезу о происхождении Т. araraticum
путем автополиплоидии Т. boeoticum. В. Л. Мепабде (1971, 1972) также
связывает происхождение Т. timopheevii с автополиплоидией.
Вопрос о генетической однозначности или разнокачественное™ гено-
мов В и G или А и G мог быть решен после точной идентификации доно-
ров этих гепомов.
В. Giorgi и A. Bozzini (1969) сообщили о значительном сходстве меж-
ду хромосомами Ае. speltoides и Т. timopheevii. Затем А. В. Коиарев и др.
(1971), Т. И. Пенева (1973), В. Г. Коиарев и др. (1976), применив имму-
нохимический и электрофоретический анализы глпадннов зерновок, на-
шли, что вторые геномы Т. durum и Т. timopheevii разнокачественны,
что донором генома G был Ае. speltoides, а не диплоидная пшеница.
Биохимические исследования В. Яаскп (1974), L. Vittozzi и V. Silano
(1976) также указали на Ае. speltoides как па источник генома G. Такое
же заключение можно сделать из эксперимента К- Chen и др. (1975).
S. S. Маап, К. A. Lucken (1971) и Н. Suemoto (1973), изучая ЦМС,
пришли к заключению, что Ае. speltoides был донором генома G. По
уровню конъюгации хромосом в мейозе у гибридов Т. timopheevii X
ХАе. speltoides (форма, у которой отсутствует способность вызывать
конъюгацию гомеологпчных хромосом) Н. L. Shands и G. Kimber (1973,
1974) пришли к такому же выводу.
Таким образом, па современном этапе развития филогенетических
знаний о пшенице следует признать аллотетраилоидиую природу видов
секции Timopheevii, произошедших при гибридизационном объединении
геномов Т. boeoticum и Ае. speltoides.
Донор цитоплазмы Т. timopheevii. Биохимические исследования
К. Chen и др. (1975) показали, что хлоропластный геном в Т. timophe-
evii был внесен вместе с ядерным геномом G, т. е. материнским расте-
30
ОДю/? рода Trilicum L.
пнем при гибридизации Ле. speltoides с Т. boeoticum был эгилопс.
Н. Suemoto (1968) указала, что в такой комбинации скрещивание уда-
стся лишь в случае, если в качестве материнской формы был взят эти-
лоне. В результате опытов по ядериому замещению II. Suemoto (1973)
пришла к заключению о родственности цитоплазмы Ае. speltoides и
Т. timopheevii. Эксперименты по переводу ядра Т. aestivum и Т. durum
в цитоплазму Ае. speltoides, Т. araraticum (из Армении и Ирака),
Т. timopheevii и 7'. zhukovskyi вызывали одинаковый результат: стерили-
зовали пыльцу, но не давали отрицательного фенотипического эффекта
у мужски стерильных растений твердой и мягкой пшеницы (И. Панайо-
тов, К. Гоцов, 1975 и др.). Эго косвенно свидетельствует о том, что
у Т. araraticum, Т. timopheevii и Т. zhukovskyi цитоплазма сходна с Ае.
speltoides. В дальнейшем (S. S. Маап, 1973) было выяснено, что между
системами «цитоплазма Ac. speltoides -ф ядро Т. timopheevii» и «цито-
плазма Т. araraticum (из Армении и Ирака)-)-ядро Т. timopheevii» есть
различия, так как в первом случае индуцируется полная мужская сте-
рильность, а во втором — фертильность.
Закон гомологических рядов в наследственной изменчивости Н. И. Ва-
вилова и возможный путь полиплоидизации пшеницы. Выше обсужден
вопрос о дифилетическом происхождении тетраплоидпой пшеницы и об
исторически меньшем возрасте видов с геномами AbG по сравнению
с видами, имеющими геномы АиВ. Последние в процессе эволюции пере-
шли на трехгеномиый гексаплоидный уровень (A"BD), дав начало про-
цессу возникновения основного хлеба человечества — мягкой пшеницы.
Среди видов с геномом G природа не создала еще трехгепомного гекса-
плоида: единственный естественно возникший гексаплоидный вид Т. zhu-
kovskyi имеет 2 разнокачественных генома. Трехгеномиый гсксаплоид-
ный уровень у пшеницы с геномом G достигнут искусственно: амфи-
диплоид с геномным составом AbGD был создан при скрещивании
Т. timopheevii X Ле. tauschii. По своему генетическому содержанию, по
морфолого-ботаническим и хозяйственным признакам он является го-
мологом Т. spelta L.: имеет трудный вымолот зерна и ломкий стержень
довольно рыхлого колоса. Ему дано название Т. kiharae (В. Ф. Доро-
феев, Э. Ф. Мигушова, 1977).
Присоединение к гепокомплексу тетраплоидпой пшеницы (АиВ) ге-
нома D снизило общий иммунологический тип гексаилопдпой пшеницы.
Селекционеры делали многочисленные попытки путем гибридизации пе-
ренести гены иммунитета к грибным болезням от Т. timopheevii в мягкую
пшеницу. Однако, несмотря на большие усилия, производство до сих пор
не. получило устойчивого к патогенам сорта и несет огромные потери
из-за болезней, поражающих растения пшеницы. Еще никто не сумел
привлечь на службу человеку всего комплекса генов иммунитета, кото-
рыми обладает Т. timopheevii — уникальный по устойчивости к грибным
болезням и эпдоврсдителям вид пшеницы.
Путь привлечения иммунитета Т. timopheevii па службу человеку,
очевидно, состоит в создании гомолога мягкой пшеницы. При этом мож-
но привлечь в скрещивания вид Т. militinae, являющийся естествен-
ным легкообмолачивающимся мутантом Т. timopheevii.
Скрещивание его с максимально устойчивой к грибным болезням
формой Ле. tauschii (с последующим удвоением числа хромосом) теоре-
тически должно дать трехгеномиый гексаплоид (AbGD) с легким обмо-
лотом. Закон гомологических рядов в наследственной изменчивости
позволяет сделать вывод о возможности создания гомолога мягкой пше-
ницы. Если удастся решить эту задачу, то к созданию хлеба для челове-
ка окажутся привлеченными 5 геномов (Аи, Аь, В, G, D) из родов Triti-
cum п Aegilops.
Ключ
дли определении
ипдои пшеницы
КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПШЕНИЦЫ *
1. В каждом колоске но 2 колосковые чешуи................2.
+ В каждом колоске по 4 нормально развитые колосковые чешуи;
колосья короткоостистые, ложповетвистыс (удлинены оси колосков);
зерновки стекловидные Т. jakubzineri Udacz. et Schachm.
2. Наружная (нижняя) цветковая чешуя первого (самого нижнего)
цветка в колоске длиннее внутренней (верхней) цветковой чешуи; зер-
новки стекловидные..........................................3.
-(-Наружная цветковая чешуя первого цветка в колоске равна или
почти равна по длине внутренней цветковой чешуе; зерновки стекловид-
ные или мучнистые...........................................5.
3. Колосья после созревания распадаются па колоски при надавлива-
нии; зерновки плотно заключены в чешуях (пшеница пленчатая); в
верхней части члеников стержня у основания колосков бугорки (мозоли-
стые утолщения) отсутствуют...............Т. ispahanicum Heslot.
+ Колосья после созревания не распадаются на колоски даже при
надавливании; зерновки неплотно заключены в цветковых чешуях (пше-
ница голозерная); в верхней части члеников стержня у основания ко-
лосков бугорки (мозолистые утолщения) четко выражены .... 4.
4. Колосковые чешуи травянистые, длинноланцетные; зерновки удли-
ненно-овальные .................................Т. polonicum L.
Колосковые чешуи грубые (жесткие), удлиненно-овальные; зернов-
ки овальные....................Т. petropavlovskyi Udacz. et Migusch.
5. Внутренняя (верхняя) цветковая чешуя после созревания зерно-
вок расщепляется на 2 продольные части; в колоске развивается 1, реже
2 зерновки..................................................6.
+ Внутренняя цветковая чешуя после созревания зерновок не рас-
щепляется па 2 продольные части; в колоске развиваются 2, реже боль-
ше зерновок.................................................9.
б. Стержень (ось) колоса без развитых сочленений, при надавливании
разламывается на членики, реже наблюдается спонтанная ломкость;
членики стержня колоса голые или лишь в верхней половине ребер по-
крыты волосками.............................................7.
+ Стержень (ось) колоса легко распадается па членики по сочлене-
ниям; членики стержня колоса по ребрам по всей длине густо покрыты
волосками...................................................8.
7. Зерновки неплотно заключены в чешуях (пшеница голозерная);
колоски 12—13 мм длины и 5—6 мм ширины; колосковые чешуи 10—
13 мм длины, нежные, па верхушке с остевидным отростком до 7 мм
длины................................Т. sinskajae A. Filat, et Kurk.
+ Зерновки плотно заключены в чешуях (пшеница пленчатая); ко-'
лоскп 10—11 мм длины и 4 мм ширины; колосковые чешуи 6—10 мм
длины, грубые, на верхушке с коротким и широким в основании киле-
вым зубцом 0,8—1,5 мм длины...................Т. inonococcuin I..
8. Листья все или только нижние бархатистые на ошупь из-за корот-
ких, почти невидимых невооруженным глазом волосков или голые; колос-
ковые чешуи 8—11 мм длины; килевой зубец 2—2,5 мм длины; зубец
главной боковой жилки значительно короче основного килевого зубца;
ость наружной (нижней) цветковой чешуи второго от основания цветка
в колоске длиннее, чем ость первого (самого нижнего) цветка и заклю-
чена между двумя ланцетными зубцами, равными 1,5 - 2,5 мм длины .
Т. urartu Tliuin. ex Gandil.
4-Лпстья рассеянно коротко щетинистые с. ирпмесыо более длинных
волосков; колосковые чешуи (> - 8 мм длины; килевой зубец 1-1,5 мм
длины; зубец главной боковой жилки почти равен или немного короче
Составлен каид. бпол. наук О. II. Коровиной.
<.6v,p p/j-)a Triticun [.
основного килевого зубка; ость наружной (нижней) цветковой чешуи
второго от основания цветка в колоске короче или почти короче, чем
ость первого (самого нижнего) цветка, и заключена между двумя ши-
роколапцетными зубцами, равными 1,5 мм длины . Т. boeoticum Boiss.
9. Стержень (ось) колоса после созревания распадается или разла-
мывается на членики спонтанно или при надавливании или сгибании;
зерновки плотно заключены в чешуях (пшеница пленчатая) реже (у
Т. militinae') пе плотно....................................10.
+ Стержень (ось) колоса после созревания не распадается и не раз-
ламывается на членики даже при надавливании или сгибании, а оста-
ется гибким; зерновки неплотно заключены в цветковых чешуях (пше-
ница голозерная), реже (у Т. vavilovii) зерновки плотно заключены в
чешуях и тогда колос ложповетвистып....................... 19.
10. Членики в средней части колоса (4) 5—8 мм длины, отчего ко-
лосья более рыхлые; лицевая сторона колоса равна или почти равна
боковой стороне, очень редко она уже (у Т. macha)...........11.
4-Членики в средней части колоса до 4 мм, реже 5 мм длины, от-
чего колосья более плотные; лицевая сторона колоса всегда значитель-
но уже боковой его стороны..................................13.
11. Листья голые, бархатистые или шероховатые; членики при раз-
ломе стержня колоса либо остаются прикрепленными верхним концом
к основанию колоска, либо прилегают и остаются прикрепленными
к брюшной стороне его.......................................12.
4-Листья щетпнпстоопушенпые; членики при разломе стержня ко-
лоса всегда остаются прикрепленными верхней своей частью к основа-
нию колоска...........................Т. kiharae Dorof. et Migusch.
12. Созревший колос легко отваливается от соломины; членики
средней части стержня (оси) колоса 4 реже 5 мм длины; основной зубец
на верхушке колосковой чешуи обычно более длинный, а иногда корот-
кий и тупой......................... Т. macha Dekapr. et Menabde.
4-Созревший колос не отделяется от соломины; членики средней
части стержня (оси) колоса 5—8 мм длины; основной зубец колосковой
чешуи короткий, почти пе возвышается над верхушкой самой чешуи .
......................................................Т. spelta L.
13. Членики стержня (оси) колоса голые или в верхней части по бо-
ковым ребрам опушенные, а иногда и по всей длине, но с полоской очень
коротких волосков, подобных шипикам . Т. dicoccum (Schrank) Schuebl.
4-Членики стержня (оси) колоса по всей длине боковых ребер по-
крыты короткими или длинными волосками, расположенными во много
рядов.......................................................14.
14. Стержень (ось) колоса легко спонтанно распадается на членики
(реже лишь при надавливании) по хорошо развитым сочленениям; по-
верхность колосковых чешуи плавно переходит в киль; пшеницы дико-
растущие ...................................................15.
4-Стержень (ось) колоса разламывается при надавливании или сги-
бании; сочленения неразвиты или развиты слабо; поверхность колоско-
вых чешуй резко переходит в киль; пшеппцы культивируемые . . .16.
15. Колосья 8—10 см длины, с длинными толстыми грубыми сильно
зазубренными остями; членики осн колосьев по всей длине боковых ре-
бер покрыты длинными волосками; листья бархатистые, реже шерохова-
тые или гладкие . Т. dicoccoides (Kocrn. ex Aschors. et Gracbn.) Schwcinf.
4-Колосья 5-8 (10) см длины, co сравнительно тонкими, не очень
грубыми и несильно зазубренными остями; членики стержня (оси) ко-
лосьев по всей длине боковых ребер покрыты короткими рассеянными
волосками; листья щетинисто опушены, довольно длинными рассеян-
ными волосками..............................Т. araraticum Jakubz.
Ключ для определения пидон пшеншрл 33
|Ь. Основной (к I i.ic lit >й) зубец колосковых чешуй па верхушке пере-
ходи! в короткую осн»; членики оержня (оси) колосьев до 1,5 (2) мм
длины; колосья (2) 3- 5 (7) см длины . . Т. militinae Zhuk, et Migusch.
+ Основной (килевой) .зубец колосковых чешуй короткий; членики
стержня (оси) колосьев 1,5 (2) мм длины; колосья (5) 6-9 см длины . 17.
17. Листья густо бархатистые; колосковые чешуи 7—9 мм длины, на
верхушке обычно с немного внутрь отклоненным основным (килевым)
зубцом, равным 0,5—1 мм длины, и почти незаметным зубцом главной
боковой жилки...........................Т. karamyschevii Nevski
4-Листья щетинистоопушенные; колосковые чешуи 8—10 см длины,
на верхушке с прямым или немного кнаружи отклоненным основным
(килевым) зубцом равным 1,3— 2,5 мм длины и с хорошо выраженным
зубцом главной боковой жилки ...............................18.
18. Членики стержня (оси) колоса по боковым ребрам с относитель-
но узкой 0,2—0,3 мм ширины полоской волосков, у основания ясно от-
шпурованпые от следующего более нижнего членика; колосья плотные
(D = 30—54) *..........................Т. timopheevii (Zhuk.) Zhuk.
+ Членики оси колоса по боковым ребрам с более широкой 0,4—
0,5 мм ширины полоской волосков, у основания постепенно переходящие
в более нижний членик (вследствие чего колосья мопсе ломкие); ко-
лосья менее плотные (D = 32—40). .Т. zhukovskyi Menabde et Ericzjan
19. Колосья с ложной ветвистостью вследствие удлинения оси у час-
ти колосков; зерновки плотно заключены в чешуях . Т. vavilovii Jakubz.
+ Колосья простые, реже ветвистые, но тогда колосковые чешуи
с сильно выступающим до самого основания килем; зерновки неплотно,
реже плотно заключены в чешуях..............................20.
20. Колосковые чешуи по всей длине с сильно выступающим ки-
лем 21.
+ Колосковые чешуи только в верхней половине с сильно выступаю-
щим килем, реже с заметным килем по всей длине чешуи .... 23.
21. Членики стержня (оси) колосьев 5—6,5 мм длины; колосковые че-
шуи 13—16 мм длины; колосья 11—14 см длины, простые, довольно рых-
лые (D = 16—18)..............................Т. turanicum Jakubz.
-(-Членики стержня (оси) колосьев 2,5—5 мм длины; колосковые че-
шуи 8—15 мм длины; колосья 4—13 см длины и более, нередко ветви-
стые, рыхлые или плотные (D = 16—50)....................... 22.
22. Колосковые чешуи 8—11 мм длины, примерно в 1,5 раза короче
прилегающих к ним цветковых чешуй; ость нижней цветковой чешуи
спонтанно обламывается у основания; зерновка мучнистая, 5,5—9 мм
длины.............................................Т. turgidum L.
+ Колосковые Чешуи 8—15 мм длины, обычно менее чем в 1,5 раза
короче прилегающих к ним цветковых чешуй; ость нижней цветковой че-
шуи спонтанно не обламывается у основания, очень редко обламывается;
зерновка стекловидная, 7—12 мм длины..............Т. durum Desf.
23. Колосковые чешуи на верхушке с 2 туповатыми зубчиками около
0,2 мм длины, между которыми выходит длинная ость, почти равная по
длине ости нижней цветковой чешуи, отчего весь колос имеет удвоенное
число остей; членики оси колоса 1,5 мм ширины; соломина под колосом
заполнена сердцевиной целиком или с небольшим просветом в цент-
ре .... '. .......................................Т. persicum Vav.
-(-Колосковые чешуи на верхушке обычно лишь с одним хорошо раз-
витым зубцом, иногда переходящим в короткую ость, более короткую
чем ость нижней цветковой чешуи, реже равную ей; членики стержня
(оси) колоса 2—3 мм ширины; соломина под колосом обычно полая,
* D — плотность колоса, определяется числом колосков, расположенных на отрез-
ке в 10 см длины стержня колоса.
3 762
34 Классификация рода Triticum
очень редко заполненная или с небольшим просветом в центре, по югда
колосковая чешуя с выраженным килем по всей длине чешуи . . . 24.
24. Соломина под колосом заполнена сердцевиной сплошь или с не-
большим просветом в центре; поперечная вдавленность в основании ко-
лосковой чешуи отсутствует....................Т. aethiopicum Jakubz.
+Соломина под колосом полая; поперечная вдавленность в основа-
нии колосковой чешуи четко выражена, очень редко отсутствует . . 25.
25. Колосья рыхлые (D — 10—22, реже 23—38), 5—17 см длины; дли-
на колоса превышает ширину его более чем в 4—5 раз; колоски в ко-
лосе расставленные, косо вверх направленные; зерновки 5—9 мм длины,
от удлиненно-овальной до яйцевидной формы, длина их обычно в 2 раза
превышает ширину, реже она равна ширине и тогда зерновки округ-
лые Т. aestivum L.
+ ,Колосья плотные (D = 33—54), 3—7 (8—10) см длины: длина ко-
лоса превышает ширину его в 3 раза и более; колоски в колосе соприка-
саются или прижаты друг к другу и направлены в сторону перпендику-
лярно стержню (оси) колоса; зерновки 4—7 см длины, овальные пли
почти шаровидные, длина их равна ширине или превышает ее, но менее
чем в 2 раза...................................................26.
26. Колосья более чем с 30 колосками; колоски с 3—4 плодущими
цветками; зерновки свальной формы; листья поникающие...............
...............................................Т. compactum Host
+ Колосья с 13—20 колосками, колоски с 4—5 плодущими цветка-
ми; зерновки почти шаровидные; листья под острым углом вверх направ-
ленные ....................................Т. sphaerococcum Perciv.
КЛАССИФИКАЦИЯ РОДА TRITICUM L.
ПОДРОД I. TRITICUM
Подрод I. Subgen. Triticum. Листовые пластинки молодых и взрос-
лых растений опушены очень короткими волосками (бархатистые на
ощупь) или голые (гладкие или шероховатые). 2п — 14, 28, 42. Геном-
ные формулы — AUAU, AUAUBB, AUAUBBDD.
Тип: Т. aestivum, L.
Геогр. распр. По всему ареалу рода Triticum L.
Видовой состав: Т. urartu Thum, ex Gandil., T. dicoccoides
(Koern. ex Aschers. et Graebn.) Schweinf., T. dicoccum (Schrank) Schucbl.,
T. karamyschevii Nevski, T. ispahanicum Heslot, T. turgidum L., T. jakub-
zineri Udacz. et Schachm., T. durum Desf., T. turanicum Jakubz., T. polo-
nicum L., T. aethiopicum Jakubz., T. persicum Vav. (-T. carthlicum
Nevski). T. spelta L., T. macha Dekapr. et Menabde, T. vavilovii Jakubz.,
T. aestivum L_, T. compactum Host, T. sphaerococcum Perciv., T. petro-
pavlovskyi Udacz. et Migusch.
СЕКЦИЯ 1. URARTU DOROF. ET A. FILAT.
Sect. 1. Urartu Dorof. et A. Filat, in Dorof. et Migusch., 1978, Tp.
прикл. бот. ген. сел. 62,2:151.— Triticum sect. Monococcon auct. non
Dum.: Цвелев, 1973, Новости сист. высш. раст. 10: 39, р.р.; Цвелев, 1976.
Злаки СССР: 163, р.р.
Листья с бархатистым па ощупь опушением из коротких волосков
или голые. Колосья двуостыс, неопушеппые, колоски с тремя цветками,
из которых плодоносит лишь 1 (редко 2). На колосковой чешуе зубец
главной боковой жилки значительно короче килевого зубца. Верхняя
цветковая чешуя имеет 2 киля, при созревании она расщепляется на
Подрод I. Triticum
35
2 продольные доли. Пылышки короткие (2,0 -2,7 мм). Зерновки крас-
ные. 2п — 14. Геном—А11.
Геогр. распр. Армения, Турция, Ирак, Иран, Ливан.
Тип: Т. urartu Thum, ex Gandil.
T. URARTU THUM. EX GANDIL. — ПШЕНИЦА УРАРТУ
T. urartu Thum, ex Gandil. 1972, Бот. жури. 57,2: 176; Туманян, 1938,
Tp. Арм. фил. АН СССР, сер. биол. 2: 211, descr. ross. — Т. armeniacum
Thum ex Flaksb. 1939, Опред. наст, хлебов: 27, nom. nud., non T. arme-
niacum. Nevski, 1933, non T. armeniacum Jakubz. 1938 et non T. armenia-
cum Makusch. ex Menabde, 1945. — T. monococcum subsp. urartu (Thum.)
A. et D. Love, 1961, Bot. Not. (Lund) 114, 1: 49, nom. illeg. — T. boeoticum
Boiss. subsp. urartu (Thum.) Vav. 1964, Миров, ресурсы хлебн. злаков:
21, nom. illeg. — T. boeoticum subsp. urartu (Thum.) V. Dorof. 1968,
Euphytica, 17: 453, nom. illeg. — T. monococcum subsp. michaelii An.
Theod. et Takht. ex Zhuk. 1968, Бот. журн. 53, 4: 442, nom. nud. — T. mi-
chaelii Zhuk. 1971, Культ, раст. и их сород.: 96, nom. nud.
Тип: АрмССР, юго-восточная окраина Еревана близ селений Вох-
чаберд и Гехадир, 30.VI. 1968, П. А. Гандилян.
Общая характеристика. Всходы фиолетовые или темно-зеленые. Куст
стелющийся. Соломина тонкая, упругая, высотой до 145 см. Стеблевые
узлы фиолетовые. Листья бархатистоопушенные или голые, линейно-
ланцетные (длина 15 см, ширина 0,7—1,0 см). Листовые влагалища бар-
хатистоопушенные или голые. Ушки белые, с ресничками. Колосья
удлиненные (рис 3,а), узкие (длина 7—9 см, ширина 0,6—0,7 см), имеют
спонтанную ломкость. Колоски двуостые, однозерные (реже двузерные).
Боковая (двурядная) Сторона колоса примерно вдвое шире лицевой.
Опушение члеников стержня колоса такое же, как у Т. boeoticum. Ко-
лосковые чешуи шероховато-бугорчатые, по консистенции более нежные,
чем у последнего вида. Килевой зубец длинный с широким основанием.
Зубец главной боковой жилки выражен значительно слабее, чем киле-
вой зубец: он короче последнего в 8—10 раз. Пыльники мелкие, длиной
2,0—2,7 мм (П. А. Гандилян, 1972).
Т. urartu имеет стебель значительно менее прочный, чем у Т. boeoti-
cum: у первого сопротивление на излом первого междоузлия достигает
456 г, у второго — 2000—3000 г (О. Д. Градчанинова, 1967).
Экологический фон вида в Армении — сухие склоны предгорий.
Все формы Т. urartu — озимые. Фаза колошения при выращивании на
поливном фоне в Ташкентской области очень растянута. Как в Арме-
нии, так и в местообитаниях вида на Ближнем и Среднем Востоке
Т. urartu встречается только в сообществе с Т. boeoticum.
У Т. urartu в отличие от Т. boeoticum наблюдается менее интенсив-
ное утреннее цветение. Имеет место и предвечернее цветение (В. И. Жу-
ков, 1969). По сведениям Э. Ф. Мпгушовой, в условиях Ташкента
у Т. urartu цветение начинается до полного выхода колоса из листового
влагалища. На своей родине в Армении, по наблюдениям А. Г. Араратян
и Г. А. Сурменян (1939), начало цветения Т. urartu наступает на не-
сколько дней позднее, чем у Т. boeoticum. По данным Н. S. Dhaliwal и
В. L. Johnson (1976), Т. urartu обособлен от других диплоидных видов
и по морфологии пыльников. У него пыльники необычайно короткие
(в среднем 2,2 мм, у Т. boeoticum 3,6). Раскрывание их происходит в ви-
де продольной щели, а после цветения они скручиваются. У Т. urartu
пыльники находятся на одном уровне с рыльцем, а цветковые чешуи
с трудом раздвигаются (А. Г. Араратян, Г. А. Сурменян, 1939).
Согласно данным В. Г. Коиарсва и др. (1971), Т. urartu, как и другие
дикорастущие виды пшеницы, выделяется высокой белковостью зерна
(23,7 и 25,0%). При этом содержание лизина в них относительно высо-
3*
36
Классификации potla Triticum J,.
Рис. 3. Колосья (co спонтанной ломкостью) и зерновки диких однозер-
нянок:
а — Т. urartu Ilium, ex Gandil.; б — T. boeoticum Bolss.
Подрод 1. Triticum
37
кос: 2,67 и 2,48% в белке; 0,632 и 0,6<Sl mi на 100 г зерня. По сведениям
М. М. Якубцпнера и 11, Ф. Покровской (1971), в среднем за 3 года со-
держание белка в зерне у этого вида (репродукция низменного Даге-
стана при орошении) составило 19,1—23,7%, крахмала 53,0—58,0%.
Иммунитет. Один из признаков, по которому тритикологн отличали
Т. urartu от Т. boeoticum, — это сильная его восприимчивость к желтой
ржавчине (В. О. Гулкапян, 1938; II. И. Вавилов, 1957, 1964; М. Д1. Якуб-
цппер, 1969). По наблюдениям Э. Ф. Мпгушовой (1976), в условиях Таш-
кентской области в 1968—1973 гг. поражение этим патогеном достигало
3 баллов (по 5-балльной шкале), а мучнистой росой — 4 баллов.
Э. X. Суханбердина (1977) также выявила сильное поражение мучни-
стой росой как в фазе проростков, так и у взрослых растений.
О. Г. Григорьева (1975) сообщила, что реакция па заражение популя-
цией рас бурой и стеблевой ржавчины в фазе проростков оказалась бо-
лее сильной (тин IV), чем у Т. boeoticum. При этом взрослые растения
в полевых условиях были практически устойчивыми. М. Якубцинер
(неопубликованные данные) отмечает, что в его совместном с Г. С. Ту-
ровым опыте в Южном Казахстане интенсивноегь поражения бурой
ржавчиной у взрослых растений не превышала 10%, хотя реакция па
заражение грибом характеризовалась типом IV.
По данным А. М. Ямалеева (1974), В. И. Кривченко и др. (1976),
искусственное заражение пыльной головней, относящейся к f. aestivi и
f. duri, вызвало поражение соответственно 10,0 и 11,5% колосьев. Ми-
целий гриба поразил все органы зародыша зерновки. Т. urartu, по за-
ключению этих исследователей, резко отличается от Т. boeoticum тем,
что не имеет ни полевой, ни эмбриональной расоспецифической устойчи-
вости к пыльной головне.
В целом Т. urartu неустойчив к грибным болезням.
Географическое распространение. По уточненным данным П. А. Гаи-
диляна (1976), ареал вида в Армении — юго-восточная окраина г. Ере-
вана, окрестности селений Шорбулак, Гехадир, Вохчаберд, Ацаван
в Абовянском районе.
В последние годы появились сведения о наличии форм Т. urartu
в Иране (В. Яаска, 1974), а также в Турции, Ираке и Ливане
(В. L.Johnson, 1975).
Происхождение. Ряд исследователей представляет ход филогенети-
ческого развития пшеницы в виде разветвленного дерева, исходящего
из единого родового типа. По К- А. Фляксбергеру (1935), последний дал
начало как диплоидным, так и полиплоидным видам пшеницы. С. А. Нев-
ский (1933) высказал мысль, что род Triticum произошел от предковой
трибы Protohordeae, точнее от такой ее формы, растения которой имели
соцветие метелку, несущую колоски, расположенные по 2—3 вместе.
Е. Н. Синская (1955) считала, что трибу Protohordeae составляли мно-
голетние биотипы с 2п = 14, которые эволюционировали и перешли
к однолетнему образу жизни, приобретая признаки ксерофильностн.
Н. Н. Цвелев (1976) пришел к заключению, что местом формирования
из фестукоидных злаков высокоспециализированных однолетников были
берега древнего Тетиса (древнее Средиземье). Здесь злаки-эфемеры
дали начало первой диплоидной пшенице. По нашему мнению, ею могла
быть древняя форма Г. urartu.
Систематика. Т. urartu был открыт М. Г. Туманяном в 1934 г., а пер-
вое описание на русском языке было обнародовано автором вида как
Т. urartu в 1938 г. В коллекцию ВИРа колосья его были доставлены
Н. И. Вавиловым с обозначением Т. armeniacum Thum., и это наимено-
вание появилось в печати в 1939 г. Указанный эпитет незаконен, ибо
armeniacum приводилось в 1933 г. для другого вида двузернянок, а в 1938
и 1945 гг. как более поздний омоним Т. araraticum. Предложенный
Ан. А. Федоровым и А. Л. Тахтаджяпом (1968) эпитет michaelii также
38
Классификация рода Triticum L.
является поздним омонимом. Этим самым в номенклатуру была внесена
путаница.
Диагноз для Т. urartu дал П. Л. Гандилян (1972). В ботанической
и генетической литературе ряд лет Т. urartu рассматривался как подвид
в пределах Т. boeoticum. (И. И. Вавилов, 1964) или в составе Т. mono-
coccum (A. Love, D. Love, 1961).
Определение разновидностей Т.urartu Thum, ex Gandil.
Чешуи Ости Разновидность № разновид- ности
бел ые белые с черными краями крас- ные красные с черными краями черные на фоне цвета чешуи черные
белом крас- ном
+ — — — — — + — spontaneoal- bum 1
— + — — — — 4- .— binartulutriru 9
— — + — — — + — spontanco- rubrum 3
— —. — + — — — + urartu 4
— —• — — + — — 4- albonigricans 5
— — — — — + — + nigrum 6
Var. albonigricans Thum. 1938, Tp. Арм. фил. АН СССР, сер. биол.
2:214, descr. ross. (5). А Т. urartu var. spontaneoalbum glumis aristi-
sque nigris differt.
Typus: Armenia. Reproductio stationis Daghestanskajae fulcracea, WIR,
k-33871, 3.VI. 1968, leg. Soskov (WIR).
Геогр. распр. Армения.
Var. binartulutriru Gandil. 1975, Tp. Арм. НИИЗ, сер. Пшеница (Эч-
миадзин), 3: 74, descr. ross. (2). Spica alba marginibus glumarum nigris.
Typus: Armenia, prope pagum Gegadir, 30.VI. 1968, leg. P. A. Gandi-
Ijan (WIR).
Геогр. распр. Армения.
Var. nigrum Thum. 1938, 1. c.: 214, descr. ross. (6). A T. urartu var.
urartu glumis nigris differt.
Typus: Armenia. Reproductio stationis Daghestanskajae fulcracea,
WIR, k-33870, 29.VI. 1968, leg. Soskov (WIR).
Геогр. распр. Армения.
Var. spontaneoalbum Thum. 1938, 1. c.: 214, descr. ross. (1). Spica
aristaque albae.
Typus: Armenia. 30.VI. 1968, leg. P. A. Gandiljan (WIR).
Геогр. распр. Армения.
Var. spontaneorubrum Thum. 1938, l.c.: 214, descr. ross. (3). Spica
aristaque rubrae.
Typus: Armenia. Reproductio stationis Daghestanskajae fulcracea,
WIR, k-33869, 3.VI. 1968, leg. Soskov (WIR).
Геогр. распр. Армения.
Var. urartu. — T. urartu var. binarturutriru Gandil. 1972, Бот. журн.
57,2: 177. (4).
Геогр. распр. Армения.
СЕКЦИЯ 2. DICOCCOIDES FLAKSB.
Sect. Dicoccoides Flaksb. 1928, Изв. Гос. ин-та опытн. агрономии,
6,2: 39. — Triticum sect. Frumentum Dum. 1823, Observ. Gram. Fl. Belg.:
94, quoad T. turgidum. — Triticum sect. Spelta Dum. 1823, l.c.: 94, quoad
T. dicoccum. — Triticum Etnmerreihe A. Schulz, 1913, Geschichte kult.
Подрод I. Ttilictim
39
Gctr.: 22, — Triticum consp. eudicoccoides I-’laksb. 1915, Опред, шлеи.: 8,
175. — Triticum turgidum L. emend. Thell. 1918, Naturw. Wochenschr. 17,
33: 470. — Triticum Series II (The Emmer series) Perciv. 1921, The Wheat
plant: 338. — Triticum acuminatum Kajanus, 1927, Bibliogr. genet. 3: 144,
145, species collect. — Triticum sect. Orthatherum Nevski, 1934, Фл. СССР,
2:681, excl. T. timopheevii. — Triticum congregatio tetraploidea Flaksb.
1935, Культ, фл. СССР, 1: 31, excl. T. timopheevii. — Triticum sect. Dicoc-
coidea subsect. Etidicoccoidea Sinsk. 1955, Пробл. бот. 2: 52, — Triticum
sect. Eutriticea subsect. Duriuscula Sinsk. 1955, l.c.: 52 et subsect. Pro-
totriticea Sinsk. 1955, l.c.: 52, nom. illeg. p.p. — Triticum sect. Triticum,
Цвелев, 1973, Новости сист. высш. раст. 10: 39, р.р.; он же, 1976. Злаки
СССР: 164, р.р.
Листья голые или опушенные короткими волосками (бархатистые
па ощупь). Колосья различной формы. Стержень колоса при созревании
ломается спонтанно (у дикорастущих форм) пли при надавливании
(у культурных пленчатых форм). Значительная часть видов имеет проч-
ный стержень колоса (у культурных голозерных форм). Колоски с 3—
5 (7) цветками, нз которых 2—4 (5—6) плодущие; верхняя цветковая
чешуя при созревании не расщепляется. 2п = 28. Геномы А"В.
Геогр. распр. СССР, Средний и Ближний Восток, Северная Аф-
рика, Европа, Южная и Юго-Восточная Азия, Северная и Южная Аме-
рика и Австралия.
Тип: Т. dicoccoides (Koern. ex Aschers. et Graebn.) Schweinf.
T. DICOCCOIDES (KOERN. EX ASCHERS. ET GRAEBN.)
SCHWEINF. — ПОЛБА ДИКАЯ
T. dicoccoides (Koern. ex. Aschers. et Graebn.) Schweinf. 1908 in Ber.
Deutsch. Bot. Ges. 26 a: 309. — T. vulgare var. dicoccoides Koern. 1889,
Verh. nat. Ver. Pr. Rheinl. 46: 21, nom. nud. — T. sativum subsp. dicoccum
var. dicoccoides (Koern.) Aschers. et Graebn. 1901, Synops. 2: 679.—
T. dicoccoides (Koern.) Aaronsohn, 1910, VerhandL Zool. —bot. Ges. Wien,
59: 491. — T. dicoccoides (Koern.) A. Schulz, 1913, Zeitschr. Naturw. 84,6:
414; idem, 1913, Bericht. Deutsch. Bot. Ges. Berlin, 31, 4: 226.-7’. her-
monis Cook, 1913, Bureau of Pl. Ind. (USA) Bull. 274: 13. — T. dicoccum
dicoccoides (Koern.) Flaksb. 1915, Опред. пшен.: 50. — T. turgidum subsp.
dicoccoides (Koern.) Thell. 1918, Naturw. Wochenschr. (Jena) 17,33:
470. — T. dicoccoides f. syriacae Flaksb. 1926, Tp. прикл. бот. сел. 16,3:
202. — T. dicoccoides subsp. palestinicum Jakubz. 1932, Tp. прикл. бот.
ген. сел. Прилож. 53: 160; он же, 1932, Тр. прикл. бот. ген. сел. сер. 5,1:
164. — Т. dicoccoides subsp. syrio-palestinicum Flaksb. 1935, Культ, фл.
СССР, 1: 325. — Т. turgidum var. dicoccoides (Koern.) Bowden, 1959,
Canad. Journ. Bot. 37: 671.
Общая характеристика. Всходы зеленые или фиолетовые. Колеоптиль
с двумя сосудистыми пучками. Куст стелющийся или прямостоячий. Вы-
сота растений от 50 до 100 см. Растения чаще без воскового налета.
Соломина полая или выполненная в верхней части под колосом. Узлы
зеленые или фиолетовые, крупные, голые или опушенные очень корот-
кими волосками. Влагалища нижних листьев опушены очень короткими
незаметными или довольно длинными (щетинистыми) волосками, иногда
они почти голые. Влагалища верхних листьев чаще голые. Листья ше-
роховатые или опушенные очень короткими волосками (бархатистые на
ощупь). Язычок и ушки зеленые пли фиолетовые. Колосья плоские
(двурядпая сторона шире лицевой, рис. 4), длиной 8—10 см, рыхлые
или среднерыхлые (D = 16—20). Членики стержня узкие (1—2 мм ши-
рины), но краям сильно опушены бесцветными или рыжими волосками,
которые у основания колосков образуют бородки. Слабо выраженное
опушение члеников встречается редко. Верх членика на месте соедине-
40
Классификации ро<)а Tiilicuiu
Рис. 4. Колосья, колоски и зерновки дикой полбы Т. dicoccoides (Koern. ex Aschers.
et Graebn.) Schweinf.:
a — convar. judaicuin Vav.; 6 — convar. incertum Jakubz.
ния его co следующим члеником имеет выемку (подковку), которой
в нижней части следующего членика соответствует бугорок (валик).
Колос ломается спонтанно или при надавливании. Колоски 11—20 мм
длины (длина превосходит ширину в 3 раза). Опп обычно имеют 3 цвет-
ка, из которых 2 плодоносят (3 зерновки в колосе развиваются редко).
В поперечном разрезе колосок имеет форму трапеции с рогами, обра-
зуемыми килем колосковых чешуи.
Колосковые чешуи одревесневшие, шероховатые, гладкие или опу-
шенные, 8—15 мм длины, 2,8—4,3 мм ширины, крыловидные. Плоскость
Подрод I. Triticum
II
чешуи постепенно переходит в резко выраженный киль и в его зубец.
Зубец колосковой чешуи широкий, треугольный, прямой или отогнутый
назад, острый. Главная боковая жилка ясно выражена, образует в вер-
ху чешуи небольшой зубец с широким основанием; между зубцами
остроугольная или закругленная выемка. Наружная цветковая чешуя
несет длинную (8—14 см и больше), грубую, зазубренную, иногда очень
толстую ость. Внутренняя цветковая чешуя двукилевая, при созревании
не расщепляется на 2 доли. Зерновки красные, длинные, узкие, отноше-
ние длины к ширине и толщине равно приблизительно 4 : 1 : 1, т. е. длина
превосходит ширину и толщину приблизительно в 4 раза. Пыльники
фиолетовые. Формы озимые, редко яровые, по времени созревания —
ранние или поздние.
Отличительные признаки («радикалы») колосьев Т. dicoccoides1.
1) чрезвычайная ломкость колосового стержня: колос при созревании
распадается на отдельные колоски; 2) волоски бородки на членике длин-
ные и густые; 3) наличие на колосковой чешуе сильно развитых киля и
главной боковой жилки; 4) наличие сильно развитой «подковки» на
членике стержня колоса (на месте сочленения). Первые два и последний
признаки в большей пли меньшей степени свойственны диким предста-
вителям других хлебных злаков.
Основной отличительный признак для растений вида — стелющаяся
форма куста. Типичные палестинские формы сохраняют «лежачее» поло-
жение почти до фазы колошения, поднимаясь лишь за несколько дней
до нес; после созревания вновь ложатся. Характерное для Т. dicoccoides
бархатистое опушение листовых влагалищ (особенно нижних) свойст-
венно также некоторым расам мягкой пшеницы Средней Азии. Оно чаше
встречается у яровых узкоколосых форм. На существование различных
степеней опушения и наличие голого листового влагалища указывал
J. Percival (1921). Опушение всходов дикой полбы Палестины он харак-
теризует как мягкое, бархатистое, сходное с опушением Т. dicoccum и
Т. turgidum. Все типичные формы Т. dicoccoides, по нашим данным, ха-
рактеризуются выполненной соломиной; форм с полой соломиной, упо-
минаемых J. Percival (1921), мы не встречали.
По данным В. Г. Александрова и О. Г. Александровой (1948), у
Т. dicoccoides перикарпий зерновки сильно редуцирован и состоит толь-
ко из двух слоев (поперечные клетки и эпикарпий). В процессе созрева-
ния мезокарпий в базальной части зерновки облитерируется. Согласно
исследованиям Л. Л. Жестянниковой (1954), у Т. dicoccoides наружная
ткань плодовой оболочки однослойная, иногда со следами исчезнувших
(облитерированных) рядов клеток. Пористость оболочек поперечных
клеток ясно обозначена в отличие от Т. boeoticum, Т. urartu и Т. mono-
coccum. У крупноколосых форм Т. dicoccoides группы judaicum из Из-
раиля структурные элементы более крупные. Мелкоколосые сирийские
формы группы dicoccoides по структуре перикарпия приближаются к
Т. boeoticum.
В анатомическом строении листа, как показали исследования
О. Д. Градчаниновой (1967), у Т. dicoccoides явно выражены черты ксе-
рофильности. Мезофилл, прилегающий к поверхности листа, столбчатый,
края листа острые, их структурные элементы сильно склерифицирова-
ны. Склерификация гидроцптной ткани наступает уже в фазе колошения.
На 1 мм2 листовой пластинки (верхняя сторона) приходится 59,0+0.72
устьиц (фаза колошения, Дербент, 1966), а длила устьиц 62,6+ 1,84 мкм.
Угол отклонения листа от стебля почти прямой (75—100°).
Дикие эммеры являются светолюбивыми растениями. По данным
D. Zohary (1970), па своей родине они произрастают как в низинных
(100 м ниже ур. м.), так и в горных районах, где поднимаются до вы-
соты 1200—1400 м над ур. м. Причем скороспелые формы обитают на
низменностях, а позднеспелые — па прохладных склонах горы Хермон.
•12 Классификации рода Triticum Л.
Л. Aaroiisolin (1908) находил иолбу дикую и расщелинах скал, 1де слой
почвы незначителен, в бесплодных местах, не защищенных от палящего
солнца. И. И. Вавилов (1957) собирал ее на каменистых местах, на
обочине полей, а также на межах. D. Zohary отмстил, что полба дикая
редко засоряет посевы.
Опытами ВИРа выявлены средн Т. dicoccoides среднепоздние фор-
мы, выколашивающиеся при осеннем посеве в Дербенте позднее стан-
дарта (пшеница твердая Дербентская чериоколосая) на 10 дней. Откры-
тое цветение у Т. dicoccoides и перекрестное опыление его с другими ви-
дами или родами обнаружено рядом авторов (A. Aaronsohn, 1910;
О. F. Cook, 1913; М. ЛА. Якубцинер, 1932; Л. И. Вигоров, 1960; В. М. Бер-
лянд-Кожевннков, 1965; А. А. Филатспко, 1969). J. Percival (1921) под-
черкивает, что этот вид легко скрещивается и с твердой и с мягкой
пшеницами.
Поступившие в коллекцию ВИРа от F. Kornicke некоторые нетипич-
ные константные формы являются продуктами естественных скрещива-
ний этого вида с другими формами в Западной Германии.
У Т. dicoccoides первыми открываются цветки на вершине колоса
(М. М. Якубцинер, 1932). Такой характер цветения подтвердился и трсх-
летними опытами В. И. Жукова (1969) па Дагестанской опытной стан-
ции ВИРа. Им уточнено, что цветение начинается немного ниже верши-
ны колоса и распространяется вниз. Такой порядок соответствует
порядку распадения колоса по мере созревания в нем зерновок. Т. dicoc-
coides цветет круглосуточно. Максимум цветения у него наблюдается
с 6 до 10 ч и с 14 до 18 ч. Этот вид заканчивает цветение в течение
3—5 дней. Т. dicoccoides превосходит другие виды пшеницы по содер-
жанию белка в зерне (М. М. Якубцинер, 1932). При испытании в лабо-
ратории биохимии ВИРа оригинального зерна выявлено содержание
белка в нем 23% (против 13—19% у твердой пшеницы и 9—1.1% у мяг-
кой), а при анализе репродукций осеннего посева в Кировабаде —
24,3%. Дальнейшие исследования ВИРа и других учреждений (а также
данные зарубежных авторов) подтвердили особую ценность этого вида
по химическому составу зерна. По анализам зерна, выращенного при
орошении в Дербенте (в среднем за 1958—1960 гг.), содержание белка
составило 24,4—27,7% (у других тетраплоидпых видов 11,1—24,0%),
крахмала 52,6—57,7%, золы 1,7—2,3%. В зерне репродукции Ташкент-
ской области (осенний посев, полив) содержится белка в среднем 20,8%,
крахмала 59,1 %, золы 2,1 % (М. М. Якубцинер, Н. Ф. Покровская, 1969).
В отдельные годы у Т. dicoccoides зафиксировано содержание в зер-
не белка до 37,0% (географические посевы ВИРа), а по зарубежным
источникам — от 26,0 до 32,0%. В опытах ВИРа не подмечено сущест-
венных различий между отдельными экологическими группами. Все же
у образца к-17157 хораиской группы содержится более высокий про-
цент белка и клейковины.
Результаты исследования последних лет в лаборатории белка и
нуклеиновых кислот ВИРа также показали, что дикорастущий вид Т. di-
coccoides по сравнению с другими тетраплоидами наиболее богат бел-
ком (24,3—26,5%). У отдельных представителей этого вида его количе-
ство достигает'28,6—31,6%. К. ним относятся образцы var. spontaneovil-
losum, var. kotschyi и var. aaronsohnii (к-17256, к-17259, к-23664).
На Среднем Урале, по данным опытов Л. И. Вигорова (1960, 1966)
в Уральской лесотехнической академии, белковость зерна Т. dicoccoides
достигает 24—27%, т. е. в 1,5—2 раза больше, чем у других (культур-
ных) видов. Кроме того, отмечено повышенное содержание витаминов —
рибофлавина (Вг) и никотиновой кислоты (РР). Этим же автором по-
лучены гибриды Т. dicoccoides с пшеницей твердой, отличающиеся вы-
сокой (18—20%) белковостью зерна и повышенным содержанием вита-
минов Вг и РР.
ГЬмТ/юд I. Trillium -13
Условия выращивания влияют па химический состав зерна Т. di-
coccoides. Так, у образца к-17157 в зависимости от места выращивания
содержание белка варьирует от 18,8 до 31,0%, сырой клейковины —
от 33,5 до 60,0%, сухой клейковины — от 14,0 до 23,0%- При выращи-
вании в условиях орошения обычно упругая клейковина становится де-
фектной.
У Т. dicoccoides наблюдаются более высокие, чем у культурных
тетраплоидных пшениц, показатели спирторастворимых белков. Содер-
жание же щелочерастворимых белков очень низкое. Несмотря па вы-
сокую белковость, количество триптофана в зерне полбы дикой (0,123 мг
с колебаниями 0,080—1,170 мг) значительно ниже, чем у других видов.
Содержание крахмала составляет 62,1% с колебанием 61,6—63,6%.
Отмечено наиболее низкое содержание амилазы в крахмале зерна
Т. dicoccoides (15,0% иа сухое вещество с колебанием 10,6—15,6%).
Иммунитет. На наличие у Т. dicoccoides поражаемых грибными бо-
лезнями форм указывали Н. И. Вавилов (1913, 1918), М. М. Якубцинер
(1932), К. А. Фляксбергер (1935). Отмечалось также присутствие
устойчивых к видам ржавчины форм. Последующие исследования под-
твердили эти данные.
В опыте Г. С. Турова и М. М. Якубципера (псоиубликовано) в усло-
виях Южного Казахстана восприимчивость Т. dicoccoides к бурой ржав-
чине (расы 20 и 77) в фазы кущения и колошения оценена как сильная
(тип реакции IV), хотя интенсивность поражения не превышала 20%>-
При изучении коллекции полбы дикой О. Г. Григорьева (1975) выявила
лишь один образец (к-17256), устойчивый к комплексу рас возбудителей
видов ржавчины. В полевых условиях на инфекционном фоне 45,4%
изученных ею образцов были восприимчивы к бурой и 36,4% к стебле-
вой ржавчине. А. М. Камелина (1973) определила, что Т. dicoccoides
имеет устойчивые и восприимчивые к стеблевой ржавчине формы.
В упомянутом эксперименте Г. С. Турова и М. М. Якубцинера полба ди-
кая значительно поражалась популяцией рас 21, 34, 40 и 77 стеблевой
ржавчины: интенсивность поражения превышала 60% при IV типе реак-
ции. К местной популяции желтой ржавчины около 50% образцов ока-
зались восприимчивыми (интенсивность поражения 15—80%, тип реак-
ции III и IV), остальные образцы были устойчивыми (тип 0 и II).
И. П. Шитова (1968) нашла, что в условиях Дагестана в 1954—1965 гг.
интенсивность поражения желтой ржавчиной в среднем достигала
1,8 балла (по 5-балльной шкале). Z. К. Gerechter-Amitai и др. (1970)
рекомендуют устойчивые к желтой ржавчине формы полбы дикой из по-
пуляций Израиля для использования в селекции.
В. И. Кривченко и др. (1979), изучив коллекцию полбы дикой, наш-
ли, что на провокационном фоне при естественном заражении в усло-
виях Ленинградской и Ташкентской областей 86,3% образцов были
иммунными, 3,4%—восприимчивыми к мучнистой росе. При ис-
кусственном заражении проростков сборным инокулюмом из смеси по-
пуляций гриба различных областей СССР Э. X. Суханбердина (1977)
определила иммунность у 40% и восприимчивость у 26% образцов.
Опыты В. И. Кривченко (1973) свидетельствуют о наличии у полбы
дикой устойчивых к пыльной головне форм. А. М. Ямалеев (1974) пока-
зал, что полба дикая восприимчива к специализированным расам f. duri
пыльной головни. При искусственном заражении мицелий достигает
зародышевой почки, поражая до 34,3% из общего их числа. В полевых
условиях при этом зарегистрировано до 18% заболевших колосьев. Пыль-
ная головня I. acstivi даст псспецифпческос поражение эмбриона зер-
новки: мицелий не достигает зародышевой почкй, полевое поражение
колосьев отсутствует. Искусственное заражение показало, что 81,8% об-
разцов полбы дикой были иммунными, остальные — высокоустойчивыми
к твердой головне (В. И. Кривченко, 1973).
Географическое распространение. Первый экземпляр Т. dicoccoides
собрал в Палестине Т. Кочи и 1855 г. В 1906-1908 гг. Л. Aaronsohn
обнаружил огромное количество этой пшеницы па терри горни совре-
менных Израиля, Иордании и Пивала, Bo.'ibinoe число форм (до 100 об-
разцов) было найдено здесь И. И. Вавиловым. Сборы полбы дикой
в пределах довоенной Палестины осуществили также местные флористы
(J. Е. Dinsmore, Л. Aaronsohn, A. Eig и др.). В. В. Маркович обнаружил
полбу дикую там в 1925 г. В послевоенные годы здесь сделаны большие
сборы (Н. Oppenheimer, 1963; D. Zohary, 1965; J. R. Harlan, D. Zoharv,
1966).
Ареал T. dicoccoides заходит в Юго-Восточную Турцию, где он най-
ден в популяциях совместно с Т. araraticum (Р. S. Rao, Е. L. Smith,
1968; D. Zohary. 1970; S. S. Маап, 1973; M. Tanaka,. Н. Ishii, 1973;
К. Rawal, J. R. Harlan, 1975).
Происхождение. Первоначально происхождение T. dicoccoides связы-
вали с Т. monococcum (Н. Kihara, 1919, 1924; R. Sax, 1922). Затем было
высказано сомнение в истинности этого положения (IO. А. Филиппенко,
1930; Л. Longlci, W. Sando, 1930; Д. Костов, 1940, и др.). К. А. Флякс-
бергер (1935) считал, что дикие полба и однозернянка произошли or
общего предка или же первая возникла от Т. boeoticum subsp. ihaoudar.
R. Riley и G. D. II. Bell (1958) по результатам межвидовых скрещиваний
сделали- заключение, что именно последний подвид был предком полбы
дикой. Биохимические исследования G. Wolf, В. Lerch (1973) и В. Яаска
(1974) отвергли дикую однозернянку как донора генома этой иолбы.
Л. В. Коиарев и др. (1974) по результатам иммупохимнческого анализа
глиадинов зерновок показали, что донором этого генома у видов секции
Dicoccoides, в том числе у полбы дикой, был Т. urartu. Сравнительно-
морфологический анализ подтвердил это открытие: многие биотины
Т. dicoccoides имеют сходный с Т. urartu тип опушения листовых пласти-
нок. В последующих исследованиях иммунологический и цитологический
методы подтвердили участие Т. urartu в становлении полбы’ дикой
(В. И. Кривченко и др., 1976; V. Chapman е. а., 1976).
Вопрос о доноре второго генома (В) тетраплоидпой пшеницы вызвал
наиболее острые дискуссии. Его пытались искать средн диплоидных
пшениц, эгилопсов и пыреев. В качестве источника генома В были на-
званы, например, Ае. bicornis (М. Tanaka, 1955; Н. С. Dass, 1972), Ае.
speltoides (Р. Sarkar, G. L. Stebbins, 1956; II. Rees, 1963; В. Яаска, 1969,
1974), Т. urartu (В. L. Johnson, 1974, 1975). Иммунохимнческий анализ
глиадинов зерновок показал, что наиболее близок к геному В пшеницы
вид секции Sitopsis Ае. longissima (Т. И. Пенева, 1974; В. Г. Коиарев
и др., 1976; L. Vittozzi, V. Silano, 1976). Иммунологический анализ пыль-
ной головни подтвердил это открытие (Л. М. Ямалеев, 1974; В. И. Крив-
ченко и др., 1976).
Т. urartu и Ае. longissima — дикие виды. При соединении их геномов
мог возникнуть тетраплоид лишь дикого типа, каким является Т. di-
coccoides. Ряд исследователей указывает, что при скрещивании между
донорами геномов тетраплоидпой пшеницы материнским компонентом
был носитель г.епома В (S. S. Маап, К- A- Lucken, 1967, 1968; И. Suemoto,
1968; В. А. Крупнов, 1973), который дал свою цитоплазму полбе дикой —
предку твердой и мягкой пшениц.
В опытах ряда исследователей, вводивших ядра тетраплоидпой пше-
ницы в цитоплазму Т. boeoticum, Т. monococcum и эгилопсов секции
Sitopsis, стабильная стерилизация пыльцы отсутствовала при цитоплазме
эгилопсов подсекции Dmarginata. Eig. По данным II. Suemoto (1973),
полученные ею гибриды с. цитоплазмой Ае. bicornis и Ае. longissima и
ядром Т. turgidum имели соответственно 88,4 и 77,4% фертильных пыль-
цевых зерен. Возможно, что эго свидетельствует о близости цитоплазм
эгилопсов и видов секции Dicoccoides, в том числе полбы дикой.
Под/мх) I. Triticum 45
Нет оснований отрицать, что в очаге происхождения полбы дикой —
в Юго-Западной Сирии и прилегающих районах Израиля и Иордании
(D. Zohary и др., 1969) па стыке ареалов Т. urartu и эгилопсов подсек-
ции Emarginata, куда относится Ле. longissima, — произошел синтез ди-
кой полбы с геномами ЛиВ.
И. И. Вавилов (1927) указывал, что полба дикая включает ряд ги-
бридных форм, возникших вследствие спонтанной гибридизации с куль-
турной пшеницей. М. М. Якубцинер (1932) даже выделил подобные
формы в convar. incerium. Это формы Т. dicoccoides вторичного про-
исхождения. К ним относятся также биотипы без бархатистого опушения
листовых пластинок. Иммунологически они могут отличаться от форм
первичного происхождения, например по восприимчивости к мучнистой
росе (В. И. Кривченко и др., 1979).
Систематика. Первоначально F. Kornicke (1889) описал дикую пше-
ницу, найденную в 1855 г. Т. Kotschy, под названием Т. vulgare var.
dicoccoides Koern. Впоследствии P. F. A. Ascherson и P. Graebner (1901)
эту пшеницу отнесли к Т. dicoccum dicoccoides (Koern.) Aschers. et
Graebn.
В литературе встречаются неправильные сведения о дате возведения
этой пшеницы в ранг вида, а также об авторе, чья заслуга в этом деле
должна быть отмечена. К- А. Фляксбсргер (1926) ошибочно считал, что
A. Schulz (1913) первый перенес двузернянку дикую из положения раз-
новидности в ранг вида и рекомендовал обозначить ее Т. dicoccoides
A. Schulz. Между тем С. А. Невский (1934, 1936, 1937) приписывал
присвоение видового статуса этой пшенице A. Aaronsohn, основываясь
па опубликованной им в 1910 г. статье, где приводится наименование
Т. dicoccoides, а не Т. dicoccum, как в предыдущей его работе 1906 г.
Такого же написания вида придерживался П. М. Жуковский (1964).
Однако в действительности же право приоритета в этом отношении
имеет не A. Schulz и не A. Aaronsohn, a G. A. Schweinfurth, который
в 1908 г. в своем докладе Германскому Ботаническому обществу назвал
двузернянку дикую Т. dicoccoides, что нашло отражение и в заголовке
статьи.
Объем вида Т. dicoccoides в нашем понимании иной, чем он пред-
ставлялся в прошлом.
П. М. Жуковский (1923) открытую им пшеницу Тимофеева первона-
чально относил к Т. dicoccum dicoccoides var. timopheevii Zhuk. Ныне
эта пшеница признана им и многими систематиками самостоятельным
видом Т. timopheevii (Zhuk.) Zhuk. Полбу дикую Закавказья сначала
также относили к Т. dicoccoides f. transcaucasicae Flaksb. (К. А. Флякс-
бергер, 1926) или subsp. armeniacutn Jakubz. (M. M. Якубцинер, 1932).
Позднее она была признана самостоятельным видом.
Многообразие форм Т. dicoccoides хорошо укладывается в 2 основ-
ные группы, выделенные Н. И. Вавиловым (М. М. Якубцинер, 1932):
grex horanum Vav. (мелкоколосые формы, ныне рассматриваемые нами
как типичная группа разновидностей — convar. dicoccoides) и grex
judaicum Vav. (крупноколосые формы, ныне рассматриваемые нами как
группа разновидностей — convar. judaicum Vav.). Впоследствии первая
группа была возведена Н. И. Вавиловым в ранг подвида — subsp. hora-
num а вторую он приводит под названием subsp. paleslinicum
Jakubz.
Группа incerium Jakubz., установленная в 1932 г., объединяет яровые
формы со слабовыражеппыми чертами дикой пшеницы и представляет
как бы связующее звено между дикой и культурной полбами. Ныне мы
эти 3 группы рассматриваем в ранге convarictas.
Изучение Т. dicoccoides ведется уже свыше 60 лет. У нас впервые
начал изучение этого вида Н. И. Вавилов в 1911 г. в Московском с.-х.
институте. Он выделил 4 формы — 3 с неопушенным белым, красным и
4G
Клас/ ификацин рода Trilicuni /..
черным колосом и 1 с опушенным колосом. A. Aaroiisohn (1909) указ;,л
на наличие у 7'. dicoccoides черноколосых форм с белыми остями. Не-
обходимо отмстить, что последние нами не обнаружены нс только среди
диких эммеров, по и в роде Triticum вообще. Как правило, ости в по-
добных случаях при проверке посевом выявляют свою черноту. Точно
так же отмеченные J. Percival (1921) формы с черной полоской по краю
чешуи в наших опытах оказались чсрноколосымп пли белоколосыми.
Установлены различия по размеру и форме чешуи, размеру колоса и
зубца колосковой чешуи (A. Aaronsohn, 1910; J. Percival, 1921;
К. А. Фляксбергер, 1915). Чаще встречаются формы с зубцом колос-
ковой чешуи 1,5—3 мм (denialшп), реже с зубцом меньше 1 MM(sabden-
tatum) и с зубцом больше 3 мм (macrodentatum), а также с зубцом,
отогнутым назад (recurvatum).
Формы со щетинистым опушением листовых влагалищ (ciliata) у
Т. dicoccoides довольно редки. Чаще встречаются формы, листовые вла-
галища которых опушены очень короткими волосками, дающими ощу-
щение бархатистости. Подавляющее число форм имеет неопушепные
листовые влагалища, на ощупь слабошершавые (scabra). Последние
встречаются чаще у convar. dicoccoides, чем у convar. judaictitn. У рас-
тений Т. dicoccoides восковой налет на листьях часто отсутствует (eprui-
nosa), реже он имеется (pruinosa). Причем у узкоколосой группы форма
epruinosa встречается реже, чем у ширококолосой. Подавляющее боль-
шинство форм Т. dicoccoides являются озимыми (hibernantes), яровые
(aestivates) —крайне редки.
Ключ для определения групп разновидностей Т. dicoccoides
1. Куст стелющийся; колосья ломкие, спонтанно распадающиеся на
колоски; членики стержня густо опушенные........................2.
+ Куст прямостоячий; колосья ломкие, не распадающиеся на ко-
лоски, боковая сторона 6—10 мм ширины; членики стержня слабоопу-
шенные, со слабовыраженными редкими бородками; колосковые чешуи
более округлые, чем у предыдущей группы (длиной 8—12 мм и шириной
3—5 мм); ости менее грубые, 8—14 см длины; зерновка среднего раз-
мера и крупная............................convar. incertum Jakubz.
2. Колосья широкие, боковая сторона до 8 мм; колоски иногда много-
цветковые; колосковые чешуи 8—15 мм длины, часто широкие (более
4 мм); зерновка длинная, крупная.............convar. judaicum Vav.
+ Колосья узкие, боковая сторона до 5 мм, колосковые чешуи 8—
12 мм длины, узкие (шириной около 3 мм), зерновка среднего раз-
мера . ..........................................convar. dicoccoides.
1. Convar. dicoccoides — хоранская группа разновидностей.
Convar. dicoccoides.-—Т. dicoccoides grex horanum Vav. in Jakubz.
1932, Tp. прикл. бот. ген. сел. Прилож. 53: 161. — Т. dicoccoides prol.
horanum Vav. in Фляксбергер, 1935, Культ, фл. СССР, 1: 326. — Т. di-
coccoides subsp. horanum Vav. 1964, Миров, ресурсы хлебн. злаков.
Пшеница: 24.
Геогр. распр. Преимущественно на юге Сирии, в Иордании, Из-
раиле, Турции( Киликийский Тавр).
Var. aaronsohnii (Flaksb.) Perciv. 1921, The Wheat plant: 183.—
T. dicoccum var. aaronsohnii Flaksb. 1914, Tp. Бюро прикл. бот. 7: 767.—
T. dicoccoides var. aaronsohnii (Flaksb.) Flaksb. 1935, Культ, фл. СССР,
1: 328. (3).
J. Percival (1921) относит к этой разновидности и формы по описа-
нию явно гибридного происхождения, соответствующие convar.
incertum.
Геогр. р ас и р. Израиль, Сирия.
Подрод I. Trilicum
‘17
Определение разпопидностеи convar. dicoccoides
Чешуи Ости Разновидность № разновид пости
белые крае- вые черные на фоне ссро-лымча^ые на фоне цвета чешуй черные
белом крас- ном белом крас- ном
Колосья неопушенные
kotschyi
pseudokot-
schyi
aaronsohnii
hermonicum
namuricum
dicoccoides
1
2
3
4
5
6
Колосья опушенные
palestinicu m
schweinf urthii
jordanictim
pseudojordanicum
spontaneovillosum
7
8
9
10
11
Var. dicoccoides. — T. dicoccum var. spontaneonigrum Flaksb. 1914.
l.c.: 768. — T. dicoccoides var. spontaneonigrum (Flaksb.) Perciv. 1921,
Lc.: 184, — T. dicoccoides var. spontaneonigrum (Flaksb.) Flaksb. 1935,
l.c.: 329. (6).
При просмотре подлинного экземпляра дикорастущей двузернянки,
собранного Т. Кочи в 1855 г. и хранящегося в Венском гербарии за
№ 1054 («crescit ad marginis aridas vinctorum pagi alt. 4000 ped. Die
Fuss Raschaja, Antiliban, 1855»), К. А. Фляксбергер (1929) отнес его
к разновидности spontaneonigrum Flaksb. На этикетке имеются собст-
венноручные определения F. Kornicke от 1873 и 1889 гг., как Т. vulgare
dicoccoides. Несомненно, этот гербарный лист можно считать типовым.
Геогр. распр. Сирия, Израиль, Иордания, реже в Турции (Ки-
ликийский Тавр).
Var. hermonicum Jakubz. 1932, Тр. прикл. бот. ген. сел. Прилож. 53:
163. (4).
Геогр. распр. Сирия, гора Хермон.
Var, jordanicum Vav. in Flaksb. 1922, Опред. наст, хлебов, 2 изд/.
23. (9).
На этикетке гербарного экземпляра, полученного К. А. Фляксбер-
гером (1922) от F. Kornicke— var. antilibani Koern. nom nud.
Геогр. распр. Сирия.
Var. kotschyi Jakubz. 1932, 1. c.: 162. (1).
Геогр. распр. Сирия, Израиль, Иордания.
Var. namuricum Jakubz. 1932, l. c.: 163. (5).
Геогр. распр. Сирия.
Var. palestinicum Vav. in Jakubz. 1932, l.c.: 164. (7).
Геогр. распр. Сирия, Израиль.
Var. pseudojordanicum Jakubz. 1932, 1. c.: 164. (10).
Геогр. распр. Сирия.
Var. pseudokotschyi Jakubz. 1932, 1. c.: 162. (2).
Геогр. распр. Сирия.
Var. schweinfurthii Jakubz. 1932,1. c.: 164. (8).
Геогр. распр. He выяснено. Разновидность выделена в сборах
A. Aaronsohn. Материал получен от F. Kornicke.
-18
Классификация рода Triticum L.
Var. spontaneovillosiim (I ’’la ksb.) Perciv. 1921, l.c.: 184. — T. dicoccutti
var. spontaneovillostim I-’laksb. 1914, 1. c.: 768. (11).
Г e о г p. расы p. Сирия.
2. Convar. judaicutn Vav. — эздрелопская группа разновидностей.
Convar. judaicum Vav. in Jakubz. 1932, Tp. прикл. бот. ген. сел. При-
,юж. 53: 161, 165. --- Т. dicoccoides prol. judaicum Vav. in Фляксбергер,
1935, Культ, фл. СССР, I: 330. ---7'. dicoccoides subsp. palesiinicum auct.
non Jakubz.: Вавилов, 1964, Миров, ресурсы хлебп. злаков. Пшеница:
18, 24.
Т и п: Т. dicoccoides var. arabicum Jakubz.
Геогр. распр. Израиль, Иордания, Сирия, Турция (Киликий-
ский Тавр).
Определение разновидностей convar. judaicum Vav.
Чешуи Ости Разнови шесть № ратшшнд ности
белое крас- ные черные на ф оне серо-лы мчатые на фоне цвета чешуи черные
белом крас* ном белом крас- ном
arabicum
pseudoarabicum
macraaronsohnii
koernickei
safedicum
vavilovii
1
2
3
4
6
Колосья неопушенные
Чешуи гладкие
Чешуи слабошероховатые
+ I — I — I — I — I — I + 1 — 1 ferearabictim
— | •— I — I + I — I — I — I + | ferevavilovii
Колосья опушенные
— I + I — I fulvovillosum
— I + I — I rufovillostim
9
10
Var. arabicum Jakubz. 1932, Tp. прикл. бот. ген. сел. Прилож. 53:
165. (1). Одна из наиболее распространенных разновидностей в преде-
лах группы.
Геогр. распр. Сирия, Израиль, Иордания.
Var. ferearabicum Jakubz. 1932,1. с.: 167. (7).
Геогр. распр. Израиль.
Var. ferevavilovii Jakubz. 1932,1. с.: 167. (8).
Геогр. распр. Израиль.
Var. fulvovillosum Perciv. 1921, The Wheat plant: 183. (9).
Геогр. распр. Сирия, Израиль.
Var. koernickei Jakubz. 1932, l.c.: 167. (4).
Геогр. распр. Сирия.
Var. macraaronsohnii Jakubz. 1932, 1. c.: 165. (3).
Геогр. рас и p. Сирия.
Var. pseudoarabicum Jakubz. 1932, l.c.: 165. (2).
Геогр. распр. Иордания.
Var. rufovillosum Jakubz. 1932, 1. c.: 169. (10).
Геогр. распр. Сирия.
Var. safedicum Jakubz. 1932, 1. c.: 167. (5).
Геогр. распр. Израиль.
Var. vavilovii Jakubz. 1932, l.c.: 167. (6).
Ih><)p<><) I. Triticum
49
Геогр. pa cup. Одна из наиболее распространенных разновидно-
стей дикой двузернянки в Сирии, Израиле, Иордании.
3. Convar. incertum Jakubz.--гибридная группа разновидностей.
Convar. .incertum Jakubz. 1932, Тр. прикл. бот. ген. сел. Прилож.
53: 170.
Установлена на образцах коллекции ВИРа и на материале F. Kor-
nicke, J. Percival, L. Trabut. Основные отличия этой группы от пере-
численных выше: 1) слабая по сравнению с типичными дикими эммера-
ми ломкость стержня колоса, который лишь частично распадается на
отдельные колоски (как у культурных полб или у Т. monococcum);
2) бородка выражена очень слабо; 3) куст прямостоячий; 4) соломина
полая; 5) по внешнему облику растения этой формы напоминают куль-
турные полбы, что свидетельствует о происхождении при спонтанном
скрещивании Т. dicoccoides с культивируемой пшеницей на его родине
или при репродуцировании в коллекционных питомниках.
О гибридном происхождении говорит также наличие в составе con-
var. incertum крайне редких, отсутствующих в природе форм: например,
короткоостистой (semiaristatum). Соответствующие, но по. тождествен-
ные формы (безостые) обнаружены в 1930 г. в потомстве от естествен-
ного скрещивания форм Т. dicoccoides.
Тип: Т. dicoccoides var. kotschyanum (Л. Schulz) Perciv.
Геогр. p а с п p. Сирия, Израиль, Иордания.
Определение разновидностей convar. incertum Jakubz.
белые ’lei красные ty и черные белом ia фоне красном О циста чешуи ти черные Разновидность К? разновнт- носги
Колосья неопушенные
1
2
3
kotschyanum
incertaaron-
sohnii
spontaneum
Колосья опушенные
- I + I
syriacuin
Var. incertaaronsohnii Jakubz. 1932, Tp. прикл. бот. геи. сел. При-
лож. 53: 171. — Т. dicoccoides f. incertaaronsohnii (Jakubz.) Flaksb. 1935,
Культ, фл. СССР, 1: 333. (2).
Обнаружена при пересевах сборов А. Ааронсоиа в Ботаническом са-
ду в Бонне (материал L. Traubut и F. Kornicke). По К- А. Фляксбергеру
(1935), формы гибридные или модификационные.
Var. kotschyanum (A. Schulz) Perciv. 1921, The Wheat plant: 183.—
T. dicoccoides f. kotschyana A. Schulz, 1913, Bericht Deutsch. Bot. Ge-
sellsch. Berlin, 31, 4: 229. — T. dicoccum var. kotschyanum (A. Schulz)
Flaksb. 1914, Tp'. Бюро прикл. бот. 7: 768. (1).
К. А. Фляксбергер (1914) и J. Percival (1921) предполагают гибрид-
ное происхождение дайной разновидности или обособление се в резуль-
тате влияния культуры.
Г е о г р. р а с и р. 11с выяснено.
Var. spontaneum Jakubz. 1932, l.c.: 172.— T. dicoccoides f. spontaneum
(Jakubz.) Flaksb. 1935, l.c.: 333. — 7’. dicoccoides var. spontaneonigrum
auct. non Flaksb.: Perciv. 1921, The Wheat plant: 184, cum tab. 122. (3).
В коллекцию ВИРа var. spontaneum поступила от F. Kornicke в
1921 г. и отнесена М. Г. Туманяном и П. М. Жуковским к var. sponta-
!>0 Кла< 1пф111<11ци.'1 Tiilicuiii
ueotiigrum. В посевах этой формы в Гандже (Кировабад) наблюдалось
расщепление с появлением типов, близких к 7'. durum.
Геогр. распр. Пе выяснено.
Var. syriacum Jakubz. 1932, l.c.: 170, - T. dicoccoides f. syriacum
(Jakubz.) Flaksb. 1935, l.c.: 333. — T. dicoccoides var. fulvovillosutn auct.
non Pcrciv.: Flaksb. 1926, Tp. прикл. бот. сел. 16,3: 213. (4).
Обнаружена в репродукции сбора Л. Aaronsohn в Сирии, полученной
от F. Kornicke в 1921 г.
Геогр. р а с п р. Сирия.
Выше была отмечена сильная склонность Т. dicoccoides к перекрест-
ному опылению. Многообразная группа гибридных форм условно была
объединена М. М. Якубцинером (1932) под названием grex hybridum.
Обзор разновидностей этой группы в данной работе не приведен из-за
малой константности большей части таких форм.
Т. DICOCCUM (SCHRANK) SCHUEBL. — ПОЛБА
(ДВУЗЕРНЯНКА, ЭММЕР)
Т. dicoccum (Schrank) Schijebl. 1818, Char, et Doser. Cereal. Ilort.
Tubing.: 29. — T. amyleum Ser. 1818, Melanges bot: 124. — T. zea Wagini,
1819, Anb. Getreid.: 33.—Spelta amylea Ser. 1841 —1842, Cer eur.: 76
(114). — T. vulgare var.-gr. dicoccum Alef. 1866,. Landw. Fl.: 331. — T. sa-
tivum dicoccum Hack, in Engler et Prantl, 1887, Nat. Pflanzenfam. 2: 81,
84; Aschers. et Graebn. 1901, Syn. Mitteleiirop. 2: 679. — T. turgidum
subsp. dicoccum (Schrank) Thell. 1918, Naturw. Wochenschr. (Jena) 17,
33: 470; Mac Key, 1966, Hercditas, Suppl. 2: 267.- 7’. turgidum gr. dicoc-
con Bowden, 1959, Canad. Journ. Bot. 37: 670. — T. turgidum subsp. di-
coccum (Schrank) A. et D. Love, 1961, Bot. Not. (Lund) 114, 1: 56.—
T. turgidum convar. dicoccon (Schrank) Morris et Sears, 1967, Wheat and
Wheat improv.: 36.
Общая характеристика. Всходы опушенные. Куст сомкнутый или
полуразвалистый. Кустистость различная. Растения от низкорослых
до высокорослых (длина соломины от 50 до 150 см). Соломина почти
выполненная в верхнем междоузлии или полая, большей частью неполе-
гающая, хотя имеются формы, в различной степени склонные к поле-
ганию. Влагалище опушенное или голое. Листья опушенные (бархати-
стые на ощупь) или шероховатые, иногда голые. Язычок и ушки ясно
выражены, но различной величины. Колосья 5—12,5 см длины, более
или менее плотные (D = 21—40, реже более), плоские, т. е. двурядная
сторона (0,8—1,2 см) значительно шире лицевой (0,5—0,8 см), остистые
(рис. 5), длина остей 3—17 см. Членики стержня голые, но по боко-
вым ребрам часто слабоопушепные, с более пли менее выражен-
ными бородками у основания колосков пли почти голые. Колоски раз-
личной формы, от удлиненных до овальных, в поперечном разрезе оваль-
ные, но со стороны, прилегающей к членику стержня, слабовыпуклые,
обычно развивающие только 2 зерна. При распадении колоса па ко-
лоски членик стержня своим верхним концом остается прикрепленным
к основанию колоска. Распадение колоса па колоски не такое легкое,
как у полб дйких, ввиду отсутствия валика и подковки, так что про-
исходит разрыв в месте соединения члеников, как у других пшениц (по
пе перелом членика). По этой причине колосья можно легко убирать
целыми. Колосковые чешуи различной формы (овальные, яйцевидные,
широколаицетные), с более или мопсе ясно выраженным килем (пе кры-
ловидные), переходящим вверху в зубец; плечо закругленное или широ-
кое. Главная боковая жилка обычно переходит вверху в тупой зубец.
Килевой зубец различной формы и величины, от короткого тупого до
крупного острого, прямого или клювовидно изогнутого. Наружная цвет-
ковая чешуя выпуклая, почти всегда несущая ость. Внутренняя цвегко-
//<»)/»)<) /. Tiiliitiiii
г>1
Рис. 5. Колосья, колоски и зерновки полбы Т. dicoccum (Schrank)
Schuebl.:
а — subsp. dicoccum-, б — subsp. asiaticuin Vav.
вая чешуя такой же длины, как наружная, двукилевая. Зерновка 6.5—
10 мм длины, с узкой продольной бороздкой с брюшной стороны, сжатая
с боков, обычно красная. Этот полиморфный вид характеризуется одно-
типной окраской колеоптиля: не содержит в своих тканях антоциана.
Е. Н. Столетова (1924) отмечала аптоциановую окраску колеоптиля
у индо-эфиопских и русских поволжских полб. Однако у этих полб ярко
окрашен антоцианом первый настоящий лист всходов, который виден
сквозь неокрашенный и прозрачный колеоптиль.
Аналогичное сочетание бесцветного колеоптиля и фиолетового пер-
вого настоящего .листа мы отметили у Т. dicoccoides. Прозрачный коле-
оптиль отмечен также у колхидской полбы. 11сокрашенпым и пот и та-
ким же прозрачным (беловатым) колеоптилем обладают всходы Т. рег-
sicum и образцы ветвистых форм Т. turgidum. Все эти виды пшеницы
сходны между собой также ио другим вегетативным признакам (барха-
тистое опушение листьев и др.).
4*
Перикарпии зерновки Т. dicoccum но своему просиню в общем in-
пичсн для пленчатых шнепиц. Эпи-мезокарпий представляет собой тон-
кую почти бесструктурную пленку, которая, однако, несколько толще,
чем у однозернянок и диких двузернянок. Стенки поперечных клеток
могут, хотя и в небольшой степени, одревесневать; поры в них всегда
четко выражены.
Из всех групп полб резко выделяется горносвропейская ветвистая
(к-20747, var. melanurum), отличающаяся от остальных анатомическим
строением перикарпия и морфологическими признаками. Структура ее
перикарпия сходна с таковой голозерных пшениц и в первую очередь
ветвистых форм Г. turgidum. Зерновка указанного образца в неветви-
стой части колоса по своей форме и анатомическому строению типична
для Т. dicoccum, а в ветвистой части колоса по тем же признакам зер-
новка сходна с ветвистыми образцами Т. turgidum (Л. Л. Жестянпико-
ва, 1954).
Таким образом, выявлено сходство строения перикарпия зерновки и
характера окраски колеоптиля у Т. dicoccum, Т. karamyschevii и ветви-
стых форм Т. turgidum.
Древность культуры полбы, широкий ареал возделывания в прошлом
обусловили большое разнообразие ее экологических типов. По
Е. Ф. Пальмовой (1935), экологическая дифференциация Т. dicoccum
параллельна мягкой пшенице. У полб находим крайние варианты среди
мирового разнообразия пшеницы по быстроте развития. К ним относят-
ся типичные полбы-эфемеры, отличающиеся крайней скороспелостью
(IT. И. Вавилов, 1964). Исключительной скороспелостью и зпоевыноелп-
востыо характеризуются полбы Индии. Позднеспелые ее формы из За-
падной Европы, приуроченные к горным районам, преимущественно
мезофиты или гигрофиты. Они развивают мощную надземную массу,
являются наиболее крупнозерными и продуктивными среди полб. Более
подробная эколого-морфологическая характеристика приводится при
описании групп разновидностей различных подвидов Т. dicoccum.
Полба представлена преимущественно яровыми формами. Лишь
в Западной Европе отмечено широкое распространение озимых, полу-
озимых и поздних яровых форм. По периоду вегетации полбы различа-
ются от очень ранних (приблизительно 80 дней) до очень поздних
(120 дней) форм.
У полбы выявлена большая склонность к открытому цветению. Прак-
тически все цветки в колосе (96—97%) открываются (В. И. Жуков,
1969; И. Л. Максимов, 1969). К открытоцветущим полбу относят также
Л. 3. Латыпов и В. И. Апель (1971), однако в их опыте имелись образцы
полбы с закрытым типом цветения.
Открытое цветение у полбы начинается в верхней части колоса, за-
тем распространяется вниз. В условиях Белоруссии максимум цветения
приходится на полуденные часы, а в Московской области И. Л. Макси-
мов наблюдал открытое цветение двузернянки и спельты на протяже-
нии всего дня.
Известны случаи спонтанной гибридизации полбы с рядом тетрапло-
пдных видов пшеницы — с Т. turgidum, Т. persicutn, Т. polonicum.
Поданным ВИРа, Т. dicoccum — один из высокобелковых видов пше-
ницы из числа возделываемых. М. М. Якубципером и II. Ф. Покровской
(1969) были изучены представители всех эколого-географических групп
полбы. Среднее содержание белка в зерне этого вида составляет 18,3%
при выращивании в условиях южной части плоскостного Дагестана
(г. Дербент) и 15,0% в Ташкентской области. Высокобелковые формы
встречаются среди групп ипдостапской (к-14928, 19475), иранской
(к-14322), гибридной (к-33226), марокканской (к-15837), западноевро-
пейской (к-7349, 37639), закавказской (к-31592, 28224, 38152), апеннин-
ской (к-21181) и волжской (к-2951). Эти формы содержат высокую
Подрод J. Triticum 53
концентрацию (33 Г>0%) клейковины. Такие же данные получены
С. Л. Тютеревым, 3. В. Числовой (1073) и др. Среди полб выявлены
формы с высоким содержанием (0,534 г) лизина па 100 г зерна
(И. И. Мойса, 1974).
Иммунитет. Один из ценнейших признаков 7’. dicoccum— наличие
устойчивых к болезням форм. Н, И. Вавилов (1931) указал на невоспри-
имчивость к ржавчине и мучнистой росе образцов subsp. dicoccum.
М. М. Якубцинер (1969) наблюдал устойчивость полбы обыкновенной
в условиях эпифитотии в Дагестане. Еще в 1916 г. G. М. Reed при ис-
кусственном заражении проростков выделил сорт Khapli и образец пол-
бы из России, как иммунные к мучнистой росе. Э. X. Сухапбердина
(1977) изучила 129 образцов полбы, инфицируя их проростки сборным
ниокулюмом из смеси популяций гриба Ленинградской, Куйбышевской,
Алма-Атинской, Новосибирской областей и Северной Осетии. 19,4% об-
разцов были иммунными, 42,6% — устойчивыми и 38% —восприимчивы-
ми к мучнистой росе. У взрослых растений на провокационном фоне при
естественном заражении эти показатели были соответственно равны
24,0; 42,7 и 33,3%. В. И. Кривченко и др. (1979) пришли к заклю-
чению, что устойчивость к мучнистой росе у полбы обусловлена
геномом В. Эти исследователи предполагают, что восприимчивые образ-
цы у полбы имеют вторичное происхождение (в результате спонтанной
гибридизации).
И. П. Шитова (1968) отнесла полбу к числу относительно устойчи-
вых к ржавчине видов пшеницы, указав на пиренейскую экологическую
группу сортов как на самую восприимчивую. Накопление восприимчи-
вых к ржавчине форм она связывает с озимым образом жизни. Макси-
мальный показатель поражения за 1954—1965 гг., по ее данным, в сред-
нем по бурой ржавчине равен 2,3 балла, по желтой ржавчине—1,6, но
стеблевой ржавчине— 1,3 балла (по 5-баллыюй шкале).
Устойчивыми к видам ржавчины оказались многие из закавказских
полб. В условиях сильной эпифитотии в Дагестане ’ некоторые формы
этой группы характеризовались слабой восприимчивостью к бурой,
желтой и стеблевой ржавчине и пыльной головне (В. Ф. Дорофеев,
1972).
О. Г. Григорьева (1975) сообщила, что в коллекции полбы полевая
устойчивость к бурой и стеблевой ржавчине варьирует широко. Из
123 изученных образцов иммунными и высоко устойчивыми к бурой
ржавчине были 67,5, к стеблевой — 74,8% образцов. При заражении про-
ростков агрессивными расами этих видов ржавчины О. Г. Григорьева
выявила значительную восприимчивость. А. М. Камелипа (1973) ука-
зала на наличие у полбы возрастной устойчивости к стеблевой ржавчи-
не. М. М. Якубцинер (1969) неоднократно обращал внимание селекцио-
неров па индийский сорт Khapli как на источник иммунитета к этой
ржавчине.
Д. В. Мягкова (1968) и В. И. Кривченко (1973) отнесли полбу обык-
новенную к числу видов, наиболее богатых формами, иммунными к ви-
рулентным и агрессивным расам пыльной головни. При искусственном
заражении расами f. duri и f. aestivi большая часть образцов полбы
в опыте В. И. Дривчепко имела высокий иммунологический тип.
А. М. Ямалеев (1973) выделил как наиболее устойчивые к пыльной го-
ловне subsp. asiaticuin Vav., в том числе convar. serbiettm (A. Schulz)
Flaksb., которые имели (соответственно) 95,4 и 78,9% иммунных образ-
цов. Наибольшее число восприимчивых к f. duri образцов выявлено у
subsp. abyssinicuin Vav. В целом при искусственном заражении из I ^об-
разцов иолбы is его опыте не имел пораженных колосьев 71 % образцов.
У части последних гифы гриба проникали в эмбрион, локализуясь в от-
дельпых частях зародыша, в том числе в зародышевой ночке. По элек-
трофоретическим спектрам запасных белков зерна, коллекция- полбы
51
Классификации роди Trilicum J..
обыкновенном были нм разделена на 5 i рули. Услойчивые к пыльной
головне образцы имели обедненные в ы- или у-зопе спектры. Ио элек-
трофоретическим спектрам можно было определить,, какой формой
(f. duri или f. aestivi) пыльной головни склонен поражаться исследуе-
мый образец полбы.
При искусственном заражении твердой головней из 131 образца пол-
бы обыкновенной В. И. Кривченко (1973) выявил 45,1% иммунных,
52,3% с различной степенью устойчивости и лишь 2,3% восприимчивых.
Устойчивые к грибным болезням образцы полбы используются как
исходный материал при выведении возделываемых сортов. С участием
полбы получены известные сорта мягкой пшеницы — Тулуп 197, Цезиум
94, Н-44, Hope, Ottawa, твердой пшеницы — Безснчукская 115, Леуку-
рум 19, Ракета, Харьковская 46, Харьковская 51, Jay, Lakota, Langdon,
Well, Yuma.
История. Полба является одной из первых хлебных культур на зем-
ле, имевших важнейшее хозяйственное, значение в древности. Ре зерна
обнаружены археологами в доисторических слоях во многих местах
Египта (G. Buschan, 1895; A. Schulz, 1916). Этим же временем (V ты-
сячелетие до н. э.) датируются находки смеси колосков однозернянки,
полбы и ячменя в Ираке (J. Harlan, 1955). Здесь же обнаружены про-
межуточные по морфологическим признакам формы между дикорасту-
щей и культурной полбами. При раскопках в горном Курдистане (Ирак)
в слоях, относящихся к началу VII тысячелетия до п.э., также обнару-
жили зерна типа Т. dicoccum в смеси с зернами диких однозернянок и
Т. dicoccoides (И. Helbaek, 1965).
Полбу выращивали в древней Вавилонии в IV тысячелетии до н. э.,
в Малой Азии, Сирии, Южной Аравии. Ныне опа сохранилась местами
лишь па юге Аравийского полуострова и в Турции.
Культура Т. dicoccum была известна в древности па территории
Закавказья. Остатки этого вида, датируемые III тысячелетием до и. э.,
обнаружены в Армении (близ г. Ереван) раскопками, проведенными
в 1936—1938 гг. под руководством К. А. Байбуртяна. Единичные зерна
полбы, относящиеся к концу IV — началу III тысячелетия до п.э., выяв-
лены в Грузии раскопками С. И. Надымашвили близ г. Гори.
Как показали исследования И. Д. Мустафаева (1955), полба возде-
лывалась в III—II тысячелетии до и. э. па территории городища № 1 в
Ханларском районе Азербайджана.
В Краснодарском крае, по сообщению В. Д. Блаватского (1948),
культура Т. dicoccum была известна еще в V в. до н. э. в древней Фана-
гории (Таманский полуостров).
Несомненно, Передняя Азия является первичным очагом развития
Т. dicoccum. Н. И. Вавилов (1964) предполагал, что в древности рас-
пространение пшеницы шло отсюда в Европу через Кавказ и Балканы.
Поэтому он искал следы реликтовых культур и действительно нашел
в Карпатах полбу — остаток шнрокораспрострапеппой в IV—11 тысяче-
летиях до и. э. в Поднепровье и на сопредельных территориях триполь-
ской культуры.
Описанные О. Heer (1871) полба и однозернянка из стоянок неолита
не были похожи на современные европейские полбу и однозернянки,
отличаясь большей плотностью колосьев. В бронзовом веке полба все
более вытесняется голозерной пшеницей. G развитием земледелия куль-
тура полбы принимала все более островной характер. В пашей стране
выявлена древность культуры Т. dicoccum в различных зонах. В южной
лесостепи (па современной территории Украины) земледелие, а с ним
и культура полбы появились у племен трипольской культуры еще в ка-
менном веке. Как показал проведенный М. М. Якубнинером (1956) ана-
лиз материала из раскопок в Хмельницкой области (селение Лука Вруб-
левецкая), проведенных под руководством С. Н. Бибикова, здесь еще
Подрод I. Trilii'tiin
н IV II । ысячеле.1 ни до и. >. наряду с 7'. durum и Т. aesHvum в культуре
была полба.
Па территории России наиболее древние следы культуры иолбы
обнаружены около г. Приморска (селение Пилконсп) Калининград-
ской области. Они относятся к III тысячелетию до п.э. (ДА. М. Якубци-
нер, 1956). До нашего века сохранились очаги возделывания иолбы
в волжско-камском районе, где начало ее. культуры датируется VI в.
до и. э. В XVIII в. полба возделывалась в ряде других районов европей-
ской части России, в XIX в. — в Литве, Белоруссии (ранние находки
здесь относятся к VIII в. и.э.). Полба широко распространилась на се-
вер. Ископаемая полба Старой Ладоги (VII в. п.э.) очень похожа на
волжскую.
Известны местные названия полбы в СССР. В Грузии ее называют
асли, в Армении и Азербайджане — атчар, париндж, гатча, в Чува-
шии— пыри; в Удмуртии — возь, в Башкирии и Татарии — борай,
в Крымской области встречалась под названием каплываддай, хаплы-
будай, в прикарпатских районах Украиныоркпш, лускппца.
Географическое распространение. Ныне иолба возделывается в очень
малых размерах в горных районах Болгарии, Югославии, Испании.
В Швеции эммер вместе со спельтой и однозернянкой встречается в по-
севе пшеницы мягкой как примесь, иногда довольно значительная
(Н. Hjelmquist, 1966). Полбы культивируется в Эфиопии, на юге Ара-
вийского полуострова, встречается в Марокко, Индии, Турции. По сооб-
щению Н. Kuckuck и Е. Schicmann (1957), очажки культуры эммера
в Иране тесно связаны с армянскими поселениями. В СССР возделыва-
ется в некоторых горных районах Азербайджана, Восточной Грузии и
Дагестана. В Армении встречается в качестве примеси и изредка—в
чистых посевах, как и в Татарии, Удмуртии, Чувашии.
Происхождение. Большинство исследователей связывают происхож-
дение Т. dicoccum с Т. dicoccoides (F. Kornicke, 1888; A. Aaronsohn, 1906;
G. Schweinfurth, 1908; A. Schulz, 1911, 1913; Д. К. Ларионов, 1914;
Г. И. Танфильев, 1923). К- А. Фляксбергер (1935, 1939 и др.) присое-
диняется к этой точке зрения, по делает оговорку, что Т. dicoccum и
Т. dicoccoides могли независимо друг от друга произойти от общего пред-
ка. Гипотеза о происхождении культурной полбы из иолбы дикой обос-
новывалась не только на сходстве их морфологических признаков и
правильности хода мейоза у гибридов между ними. Н. Rees и М. R. Wal-
ters (1965) показали, что у этих полб сходно среднее количество ядер-
ной ДНК- В. Яаска (1971) нашел сходство эпзимограмм кислой фос-
фатазы, полученных при электрофорезе вытяжки из проростков. В. Gior-
gi и A. Bozzini (1969) указали на сходство кариотипов: транслокаций
или других перестроек они не обнаружили. Исследователи часто наблю-
дали выщепление растений с признаками полбы дикой (например,спон-
танная ломкость колоса) среди гибридов, полученных при скрещивании
Т. dicoccum с другими видами культурной пшеницы.
Д. Мак-Кей (1968) считает, что полбу обыкновенную от дикой отде-
лил постепенный процесс одомашнивания, ответственный за полпгенные
изменения, произошедшие при этом. В. Ф. Дорофеев (1969, 1972) согла-
шается с таким толкованием, видя путь возникновения полбы обыкно-
венной из дикой в генетических перестройках наследственного материа-
ла. В процессе окультуривания полба дикая потеряла ряд признаков
(например, сильную одревеспелость колосковой! чешуи, спонтанную лом-
кость стержня колоса, развалистый тип куста) и приобрела другие.
Наличие различий между иолбами дикой п культурной дало повод
к сомнению о происхождении последней от первой (К. Yainashita, 1968).
Еще ранее И. II. Вавилов (1931, 1935) и Е. II. Снискан (1955) высказа-
ли мысль о том, что Г. dicoccoides не был родоначалыюи формой для
Т. dicoccum, эти виды возникли в параллельных рядах от общего предка.
5G Классификации рода Triticum
11. Kiickiick (196'1) связал происхождение полбы культурной с 7'. рег-
sicum. А. Д. Горгпдзе (1973) пришла к выводу, что эта полба появилась
вследствие мутации, произошедшей у колхидской полбы. В нашем пред-
ставлении Т. dicoccum является первой окультуренной полиплоидной
пшеницей, возникшей в далеком прошлом из первичной формы Т. dicoc-
coides. Современный генофонд культурной полбы включает, кроме
форм — потомков первичного Т. dicoccum, также формы вторичного
происхождения, возникшие вследствие межвидовой гибридизации.
В. И. Кривченко и др. (1979) связывают сильную восприимчивость к
мучнистой росе, которая обнаружена ими у 38% образцов коллекции
культурной полбы, ннокулировашюй патогеном в фазе проростков, с их
вторичным происхождением.
Систематика. Культурная полба включает 4 подвида в соответствии
с их ареалами и морфолого-экологическими особенностями: 1) subsp.
abyssinicum Vav. (эфиопская полба); 2) subsp. asiaiicum Vav. (восточ-
ная полба); 3) subsp. dicoccum (европейская полба); 4) subsp. marocca-
tiutn Flaksb. (марокканская полба).
Ключ для определения подвидов Т. dicoccum
1. Колеоптиль с тремя и более проводящими пучками; фиолетовая
окраска ясно проявляется на всходах и в дальнейшем на всем растении
(стеблях, узлах, ушках). Растения низкорослые (36—НО см). Куст
сомкнутый; соломина тонкостенная, под колосом выполненная, ниже —
с просветом. Листовая пластинка жесткая, шероховатая, с редкими ши-
пиками на верхней поверхности. Колосья мелкие (5—7 см), рыхлые
(D = 23—30). Колосковые .чешуи с коротким тупым килевым зубцом;
киль выражен слабо.........................subsp. abyssinicuin Vav.
+ Колеоптиль с двумя проводящими пучками; фиолетовая окраска
на всходах проявляется слабо....................................2.
2. Растения низкорослые и средней высоты (75—120 см). Куст сом-
кнутый. Соломина тонкостенная. Листовая пластинка короткая (17—
20 см длины), полужесткая, опушенная (па ощупь бархатистая). Ко-
лосья мелкие и средние (5—8 см), ости длиннее колоса; колосковые че-
шуи удлиненно-овальные; киль выражен слабо, килевой зубец короткий,
тупой или острый; главная боковая жилка хорошо или слабо обозна-
чена . ......................................subsp. asiaticuin Vav.
+ Растения средней высоты и высокорослые (до 150 см). Куст чаще
полуразвалистый. Соломина средне- и толстостенная. Листовая пластин-
ка длинная (20—30 см), мягкая, опушенная (бархатистая на ощупь)
или голая. Колосья крупные (7—13 см); ости от укороченных до длин-
ных; колосковые чешуи от ланцетных до широколапцетпых; киль ясно
выражен, килевой зубец острый или притупленный, прямой или клюво-
видно загнутый; главная боковая жилка ясно выражена.............3.
3. Колосья рыхлые (D=21—30), от плоско сжатых до овальных.
Колосковые чешуи ланцетные, плечо покатое или почти отсутствует;
киль ясно выражен; килевой зубец прямой или клювовидно загну-
тый subsp. dicoccum.
+ Колосья средней плотности (D=30—40) и очень плотные (D бо-
лее 41), плоские. Колосковые чешуи широколапцетныс, иногда с ши-
роким плечом; киль к основанию часто сходит на нет, образуя только
ребро чешуи, килевой зубец укороченный, слегка клювовидно загну-
тый .................................... subsp. maroccanum Flaksb.
I. Subsp. dicoccinii — подвид европейских нолб.
Subsp. dicoccum. — 7’. dicoccum subsp. curopaeum Vav. 1931, 'Гр.
прикл. бот. ген. сел. Прилож. 51: 224; он же, 1964, Миров, ресурсы хлеби.
злаков. Пшеница: 18, 30. — 7’. dicoccum gr. 2 European Emmers Perciv.
1921, Wheat pl.: 192, p.p.
Подрод I. Triticum
Всходы зеленые, опушенные. Колеоптиль с двумя проводящими пуч-
ками. Куст нолуразвалнстый или прямостоячий (реже). Кустистость
большая. Стебель зеленый (не фиолетовый), толстый (2,5—4 мм). Рас-
тение высокорослое (до 150 см). Листья крупные (длина 20—30 см.
ширина 1,0—1,6 см), опушенные (бархатистые на ощупь). Влагалище
слабоопушенное. Ушки бесцветные или зеленые (редко фиолетовые).
Колосья крупные, длинные (7,5—12,5 см), различной ширины (до 1,2 см
и больше), рыхлые (D =21—23). Ости различной длины, сравнительно
мягкие. Колосковые чешуи удлиненно-овальные или широколанцетпые.
с ясно выраженным килевым зубцом, прямым острым или клювовидно
загнутым внутрь. Плечо обычно ниспадающее или почти отсутствует,
зубец главной боковой жилки сближен с килевым. Зерновки крупные,
в среднем 9 мм длины. Растение мощное, хорошо облиственное. Формы
яровые и озимые, позднеспелые (вегетационный период 115—120 дней);
от посева до колошения приблизительно 65—75 дней; при посеве на
юге — поздние по колошению и созреванию; на севере сокращают пе-
риод от всходов до колошения. Влаголюбивые, страдающие от воздуш-
ной и почвенной засухи. Е. Ф. Пальмова (1935) относит их к типу влаж-
ного климата. Внутри этого подвида различается много разновидностей,
однако ныне возделывают только несколько из них.
Геогр. распр. Западная Европа. Редко в Закавказье.
В пределах subsp. dicoccum выделены 2 группы разновидностей, раз-
личающиеся по колосовым и вегетативным признакам, что связано с воз-
действием географической изоляции их в древности — convar. dicoccum
и convar. euscaldunense Flaksb.
Ключ для определения групп разновидностей
1. Куст прямостоячий, сомкнутый, фиолетовая окраска особенно
сильно проявляется на ушках; ушки с длинными ресничками. Соломи-
на толстая (до 6 мм), сравнительно прочная. Колосья средине и длин-
ные (до 11 см), в поперечном сечении овальные. Членики колосового
стержня почти голые; у основания колоска имеется бородка из немного-
численных волосков; колосковые чешуи ланцетные со скошенным пле-
чом, килевой зубец прямой, короткий, острый .......................
......................................convar. euscaldunense Flaksb.
+ Куст нолуразвалнстый. Фиолетовая окраска проявляется редко.
Соломина среднетонкая (2,5—4,0 мм), прочная или склонная к поле-
ганию. Колосья до 13 см длины, плоские. При основании колосков и по
сторонам члеников колосового стержня имеются волоски, по не по всей
длине; колосковые чешуи ланцетные пли более пли менее овальные, ки-
левой зубец острый прямой или клювовидно изогнутый.................
. . . ...........................................convar. dicoccum.
1. Convar. dicoccum — западноевропейская группа разновидностей.
Convar. dicoccum. — Т. dicoccum subsp. europaeum prol. iardoeuro-
paeum Flaksb. 1935, Культ, фл. СССР, 1: 305, descr. ross. — T. dicoccum
subsp. europaeum prol. meridionale Vav. et prol. bavaricum Vav. 1964.
Миров, ресурсы хлебп. злаков. Пшеница: 32, nom. invalid.
Растения большей частью высокие (150 см), силыюкустящисся, хо-
рошо облиственные. Листья длинные (20—30 см), широкие (10—16 мм).
Характерна высокая устойчивость к бурой и желтой ржавчинам, муч-
нистой росе (исключение составляют некоторые поздние сорта Бава-
рии, сильно поражающиеся бурой и желтой ржавчиной. Формы яровые
поздние и озимые. К этой группе относятся наиболее зимостойкие фор-
мы Т. dicoccum.
Геогр. распр. Рапсе возделывалась в Европе (ГДР, ФРГ.
Швейцарии, Франции, Швеции). Ввезена в Америку. Редко — в Закав-
казье.
58 Классификации /»><)</ 7’r iIicuш Г.
Определение разновидностей convar. dicoccum
’loin у и Осги Разновидность К» разновид- ности
белые красные черные поста чешу и черныс
Л' о л о с ь я простые
Колос остистый
Чешуи н е о п у ш е н и ы е
+ — — + — dicoccum 1
— — + iiguliforme 2
<— + — + — rufum 3
— + — — + pseudorujum 4
+ — — + — laschkentum * 5
Чешуи опушенные
+ ' 1 — 4- — fictesemicanum 6
+ — — — semicanum 7
— + — + — macratherum 8
— + — — + pscutlomacratherum 9
— + — atratum 10
+ — — + diploleucum * 11
Колос полцостистый
Чешу и neon у ш e n n ы e
+ — + — tricoccum 12
+ — — — 4- subliguliforme 13
— + — + — [uchsii 14
Чешуи опушенные
+ — — — + submajus 15
— + bauhinii 16
— + subatratutn 17
Колос безостый
Чешуи неопушенные
— + — novicium 18
Чешуи опушенные
+ 1 + 1 + Коло сь я с д с о й ны м muticum hybridum decussatum и колосками 19 20 21
Колос остистый
Чешуи неону ш сп пы е
- + — + — schuebleri 22
Чешуи опушенные
+ 1 - 1 “ | + | — | mazzucattii Колос полуостистый Чешуи неону ш е н и ы е | 23
— 1 + I — | +| — | dodonaei 1 24
Чешуи опушенные
_ 1 + ' - » | + | — 1 tragi Колос безостый Чешуи неопушенные \ 25
+ 1 — ! — | | | bispiculalum | 26
К о л о с ь и в с т и и с т ы с
Колос остистый
Чешуи II О И у HI (‘ II II ы с
+ + — albiramosum 27
-1- — + erythrurum 28
— + — — + pseudoerylhrurutn 29
Подрод Г. Trilicttm
59
П р одол же и и с
Чешуи Ости Разновидность № p:i3H0ii»i,T НО( ти
6С-1ЫС красные черные инета чешуи •U'pll Г.1С
Ч с ш у и о П у III С Н II ы с
— + — 4- — rubriraiiiosiim 30
— + — — . + pseiidorubriramosum 31
— —— -1- — + mclanurum 32
Колос полуостистый
Ч е in у и нсопуи е и и ы е
— + — -г — cladurum 33
Чешуи опушенные
- + — 4- kratisei 34
- + — 4- psetidokrausei 35
Колос безостый
Ч е ш у п И С О п У III 0 Н II ы с
4- - leticocladtitii 3(i
- + - subclad iirutn 37
Чешу и опушенные
+ tnetzgeri 38
4- tnelanocladuin 39
1 Разновидности зерновками белого цвета, у •(стальных разновидностей цвет .зерновок
красный.
Var. albiramosum Koern. 1908, Arch. Biontol.: 411; Perciv. 1921, Wheat
pl.: 198. (27).
Обнаружена Ф. Кёрпике в посеве Ботанического сада в Поппельсдор-
фе в 1901 г.; не возделывается.
Var. atratum (Host) Koern. in Koern. et Wern. 1885, Handb. Getreideb.
1: 89. —• T. atratum Host, 1809, Gram, austr. 4:5. (10).
Полба озимая, но незимостойкая. Устойчива к полеганию и пора-
жению видами ржавчины.
Аборигенное происхождение не выяснено. Высевалась в США, куда
попала из Франции.
К. А. Фляксбергер наблюдал появление двух колосков на одном чле-
нике стержня. Подобные формы были им выделены в var. modificatum
Flaksb. 1935, Культ, фл. СССР, 1: 319, descr. ross. Позднее оказалось,
что эти изменения пе наследственные и поэтому рассматриваются как
f. modificatum Flaksb. 1939, Опред. наст, хлебов: 63, nom illeg.
Var. bauhinii (Alef.) Koern. in Koern et Wern. 1885, l.c.: 87. — T. vul-
gare var.-gr. dicoccum bauhini Alef. 1866, Landw. FL: 332. (16).
Известна только из ботанических садов.
Var. bispiculatum Koern. 1908, Arch. Biontol. 2: 407, 410. (26).
Разновидность гибридного происхождения.
Известна тдлько из Ботанического сада в Поппельсдорфе.
Var. cladurum (Alef.) Koern. in Koern. et Wern. 1885, 1. c.: 90.— T. vul-
gare var.-gr. dicoccum cladura Alef. 1866, 1. c.: 333. (33).
Ф. Кёрпике отмечает, что ветвление выражено несильно и даже от-
сутствовало.
Известна только из ботанических садов.
Var. decussatuin Koern. 1908, l.c.: 409. (21). Гибридного происхож-
дения.
Не возделывается. Обнаружена Ф. Кёрпике в Ботаническом саду
в Поппельсдорфе.
GO
Классификация рода Triticum I..
Var. dicoccum. T. spelta dicoccum Schrank, 1789, Baier. 1'1. 1; 389
(’dicoccon’). — T. farrum Baylc-Barelle, 1809, Monogr. Ccr.: 50. —T. di-
coccum album Schuebl. 1818, Char, et Descr. Cereal. Hurt. Tubingen: 29.--
T. dicoccum var. farrum (Baylc-Barelle) Koern. in Koern. et Worn. 1885,
l.c.: 87; Persiv. 1921, l.c.: 197, excl. Slav, emmer (=var. volgcnse
Flaksb.). — T. dicoccum gr. farrum (Baylc-Barelle) Flaksb. 1935, Культ,
фл. СССР, 1: 306, р.р. (1).
Геогр. распр. Наиболее распространенная разновидность евро-
пейских полб в древности. В конце прошлого века еще возделывалась
в Средней Европе, главным образом в Южной Германии и Швейцарии,
а также Австрии, Франции. В коллекцию ВИРа от доктора Hjelmqvist
поступили образцы с о. Готланд (Швеция), где полба возделывалась
в смеси с Т. spelta и Т. aestivum 2 десятилетия назад.
Var. diploleucum Koern. 1908, l.c.: 409. (11). Гибридного происхож-
дения.
Не возделывается. Обнаружена Ф. Кёрнике в Ботаническом саду
в Поппельсдорфе.
Var. dodonaei Koern. 1908, l.c.: 410. (24).
Известна только по Ф. Кёрпике, обнаружившему разновидность в
посеве var. cladurum в Ботаническом саду в Поппельсдорфе.
Var. erythruruin Koern. in Koern. et Wern. 1885,. l.c.: 91. (28). Гиб-
ридного происхождения.
Г e о г р. р а с п р. Не выяснено.
Var. fictesemicanum Flaksb. 1929, Тр. прикл. бог. ген. сел. 21,5:
480. (6).
Известна по гербарию в Гейдельберге.
Var. fuchsii (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 87.--T. vul-
gare var.-gr. dicoccum fuchsii Alef. 1866, 1. c.: 332. (14).
Геогр. рас и p. He выяснено.
Var. hybridum Koern. 1908, l.c.: 409. (20).
Обнаружена Ф. Кёрнике в Ботаническом саду в Поппельсдорфе.
Var. krausei Koern. in Koern. et Wern. 1885, 1. c.: 91. (34).
Ф. Кёрнике указывает на гибридное происхождение.
Обнаружена в Ботаническом саду в Поппельсдорфе.
Var. leucocladum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 90,—
T. vulgare var.-gr. dicoccum cladura Alef. 1866, 1. c.: 333. (36).
Геогр. распр. He выяснено.
Var. liguliforme Koern. in Koern. et Wern. 1885, l.c.: 90. (2).
Для разновидности характерна большая плотность колоса к его вер-
шине (capitatum).
Известна из ботанических садов, не возделывается.
Var. macratherum Koern. in Koern. et Wern. 1885, 1. c.: 89. (8).
Известна из ботанических садов.
Var. mazzucattii Koern. 1908, l.c.: 410. (23).
Ф. Кёрнике предполагает гибридное происхождение, зерновки’ не-
плотно заключены в чешуях и легко обмолачиваются.
Обнаружена Ф. Кёрнике в Ботаническом саду в Поппельсдорфе.
Var. melanocladum Koern. 1908, l.c.: 410 (’melanocladon’). (39).
Обнаружена Ф. Кёрнике в Ботаническом саду в Поппельсдорфе.
Var. melanurum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 91.—
T. vulgare var.-gr. dicoccum melanura Alef. 1866, l.c.: 333. (32).
Колосья слабо ветвистые. Форм а поздняя.
Геогр. распр. Пе выяснено.
Var. melzgeri (Alef.) Koern. in Koern el Wern. 1885, l.c.: 91.-- T. vul-
gare var.-gr. dicoccum metzgeri Alef. 1866, 1. c.: 332. (38).
Геогр. p а с п p. He выяснено.
Var. nintictiin (Baylc-Barelle) Koern. in Koern. et Wern. 1885, l.c.:
86. — T. muticum Baylc-Barelle, 1809, I. c.: 52. (19).
Подрод I. Triticum С>1
Геогр. распр. 11с выяснено.
Var. noviciiun Koern. 1908, l.c.: 408. (18).
Обнаружена Ф. Кёрпике в Ботаническом саду в Поппельсдорфе.
Var. pseudoerythrurum Must. 1973, Опред, пшен. Лзсрб.; 30, descr.
ross. (29).
Обнаружена И. Д. Мустафаевым в 1970 г. По-видимому, спонтан-
ный гибрид полбы с Т. turgidum.
Геогр. распр. Азербайджан (Мардакертский район).
Var. pseudokrausei Flaksb. 1929, 1. с.: 480. (35).
Обнаружена К- Л. Фляксбергером и коллекции А. Шульца (Галле).
Var. pseiidoniacratheriun Flaksb. 1929, 1. с.: 480. (9).
Обнаружена К- А. Фляксбергером в коллекции А. Шульца (Галле)
и в гербарии в Гейдельберге.
Var. pseudorubriramosurn Flaksb. 1929, 1. с.: 480. (31).
Обнаружена К- А. Фляксбергером в коллекции А. Шульца (Галле)
и в гербарии в Гейдельберге.
Var. pseudorufum Flaksb. 1935, l.c.: 311, descr. ross. (4).
Обнаружена К- А. Фляксбергером в коллекции А. Шульца (Галле).
Var. nibriramosum Koern. 1908, l.c.: 411. (30).
Обнаружена Ф. Кёрнике в Ботаническом саду в Поппельсдорфе.
Var. rufutn Schuebl. 1818, l.c.: 29; Koern. in Koern. et Wern. 1885,
l.c.: 88.— T. vulgare var.-gr. dicoccum brunneum Alef. 1866, l.c.: 331. (3).
Г e о г p. p а с и p. Средняя и Западная Европа.
Var. sclniebleri Koern. 1908, 1. c.: 410. (22).
Геогр. распр. Пе выяснено. Обнаружена Ф. Кёрнике в Ботани-
ческом саду в Поппельсдорфе.
Var. seinicanum Koern. in Koern. et Wern. 1885, l.c.: 89. (7).
Геогр. распр. He выяснено. Ф. Кёрпике были получены семена
var. semicanum из Ботанического сада (Йена).
Var. subatratum Koern. 1908, 1. с.: 409. (17).
Геогр. распр. Не выяснено. Обнаружена Ф. Кёрнике в Ботани-
ческом саду в Поппельсдорфе.
Var. subcladurutn Koern. 1908, l.c.: 408, (37).
Геогр. распр. Не выяснено. Обнаружена Ф. Кёрнике в Ботани-
ческом саду в Поппельсдорфе и при посеве музейного образца из Лат-
вии па опорном пункте ВИРа в Гандже (Кировабад).
Var. subligtiliforme Flaksb. 1929, l.c.: 480. (13).
Геогр. распр. Не выяснено. Обнаружена К- А. Фляксбергером
в коллекции А. Шульца (Галле).
Var. submajus Koern. 1908, 1. с.: 409. (15).
Геогр. распр. Не выяснено. Обнаружена Ф. Кёрпике в Ботани-
ческом саду в Поппельсдорфе.
Var. taschkentum Udacz. 1976, Тр. прикл. бот. ген. сел. 56, 2: 149. (5).
Геогр. распр. На Среднеазиатской опытной станции ВИРа об-
наружена и выделена Р. А. Удачиным из гибридной комбинации Т. aes-
tivum X Т. turanicum.
Var. tragi Koern. in Koern. et Wern. 1885, l.c.: 90. (25).
Геогр. распр. He выяснено. Обнаружена Ф. Кёрнике в Ботани-
ческом саду в Поппельсдорфе.
Var. tricoccum (Schuebl.) Koern. in Koern. et Wern. 1885, 1. c.: 86.—
T. tricoccum Schuebl. 1818, Char, et descr. Cereal. Ilort. Tiibiiigcn: 33. - -
T. vulgare var.-gr. dicoccum subtricoccuni Alef. 1866, l.c.: 332. (12).
Геогр. p а с и p. He выяснено, по-видимому, Средняя Европа.
2. Convar. euscaldunense Flaksb. — пиренейская группа разновидно-
стей.
Convar. euscaldunense Flaksb. 1928, 'Гр. прикл. бот. ген. сел. 19, 1: 504.
Растения высокорослые, сильно кустящиеся, хорошо облиственные.
Характерен комплексный иммунитет к бурой и желтой ржавчине. Труп-
62
Классификация рода Triitcutn
па представлена преимущественно озимыми, реже полуозимымп и позд-
ними яровыми формами (самая позднеспелая полба).
Т и п: Т. dicoccum var. vasconicum (Stole).) Flaksb.
Геогр. распр. Испания (Наварра).
Пока известна только одна разновидность.
Var. vasconicum (Stole).) Flaksb. 1935, Культ, фл. СССР, 1: 305.—
Т. dicoccum var. jarrum f. vasconica Stole). 1925, Tp. прикл. бот. сел.
14,1: 91. Колосья белые, иеопушепные, ости белые, зерновки красные.
Геогр. распр. Испания (Наварра).
II. Subsp. maroccanum Flaksb.'—подвид марокканских полб.
Subsp. maroccanum (Flaksb.) Flaksb. 1935, Культ, фл. СССР 1:
302.— Т. dicoccum группа maroccanum Flaksb. 1928, Тр. прикл. бот.
ген. сел. 19,1: 504.
Тип: Т. dicoccum var miegei Flaksb.
Колеоптиль с двумя проводящими пучками. Листья опушенные
(бархатистые па ощупь). Соломина выполненная или полувыполненная
(70—80 см длины), жесткая, ломкая. Колосья 5—8 см длины, одно- и
двуостые (ости белые), неопушенныс, бледно-желтые, с двурядпой сто-
роны шире (0,8—1,2 см), чем с лицевой (0,5—0,8 см), несколько пира-
мидальной формы, средней плотности (D=30—40) и плотные (D бо-
лее 41). Стержень колоса ломкий, хотя и менее, чем у диких форм;
членики по боковым краям имеют средней длины волоски и бородки
вверху у основания колоска. Ости топкие, длинные (12—17 см), Ко-
лоски 4-цветковые, по зерновки развиваются лишь в первом и втором
цветках. Колосковые чешуи короче цветковых, с ниспадающим или при-
поднятым округлым и иногда широким плечом. Киль ясно выражен.
Килевой зубец короткий, прямой или слегка изогнутый, острый или
притупленный. Главная боковая жилка проходит но средней линии че-
шуи и .заканчивается вверху очень коротким зубчиком (бугорком).
Зерновки красные, симметричные, на спинке выпуклые, с боков сжатые
(длина 7—8 мм, ширина и высота 3—4 мм). Формы яровые, теплолю-
бивые, засухоустойчивые, раннеспелые (77—85 дней при яровом посе-
ве), силыювосприпмчнвые к желтой ржавчине.
Геогр. распр. Марокко.
Ключ для определения разновидностей
1. Колоски с двумя остями...................................2.
+ Колоски с одной остыо, колосья средней плотности (D = 30—40)
................................var. unimiegei A. Filat, et Migusch.
2. Колосья очень плотные (D более 41) . . var. compactomiegei Flaksb.
+ Колосья средней плотности (D = 30—40) . . . var. miegei Flaksb.
Var. compactomiegei Flaksb. 1935, Культ, фл. СССР, 1: 303. — T. di-
coccum var. liguliforme auct. non Koern.: Meige, 1925, Bull. Sok Nat.
Maroc. 5,3:108. — Растения с колосьями короткими (5—6 см), очень
плотными (D = 40 и более).
Г е о г р. р а с п р. Марокко.
Var. miegei Flaksb. 1935, l.c.: 303. — T. dicoccum var. farrum auct.
non. Bayle-Bajelle, nec Koern.: Miege, 1925, l.c.: 108. — Растения с ко-
лосьями длиной 5—11 см, средней плотности (D=30—-40).
Г с о г р. р а с п р. Марокко.
Var. unimiegei A. Filat, et Migusch. var. nov.--7'. dicoccum var.
unifarrttm Migusch. 19G7, Сб. тр. aciinp. мол. науч. corp. Bill’, 8, 12:
13, sine typo. — A T. dicocco var. miegei Flaksb. spicula arista una
differ!.
Typus: Alarocco, k-15836 (WIR).
II. И. Вавилов (1964) в марокканском подвиде выделяет 2 эколого-
географические группы: горную марокканскую (proles motilatium Vav.),
сильновосприимчивую к желтой ржавчине, по устойчивую к бурой ржав-
I. TtUictun (>:;
чипе и мучнистой росе и пред! орпую марокканскую (proles oasiculum
Vav.), силыювосприимчивую ко всем этим болезням.
III. Subsp. asiaticuin Vav. — подвид восточных полб.
Subs]>. asiaticuin Vav. 1931, Тр. прикл. бот, юн. сел. Прилож. 51:
224; Цвел. 1973, Новости сист. высш. раст. 10: 40; он же 1976, Злаки
СССР: 166. — Т. dicoccum subsp. euroum Flaksb. 1935, Культ, фл. СССР,
1: 296.
Всходы зеленые или слабо-фиолетовые, опушенные. Колеоптиль с
двумя проводящими пучками. Куст сомкнутый, кустистость небольшая.
Стебель перед созреванием приобретает фиолетовую окраску. Соломи-
на полая, под колосом выполненная, тонкостенная, диаметром большей
частью до 2,5 мм, обычно неполегающая, хотя иранские формы имеют
склонность к полеганию. Высота средняя (75—120 см). Листья корот-
кие (длина 17—20 см, ширина 0,7—1,3 см), па ощупь бархатистоопу-
шениые. Влагалище опушенное или голое. Ушки короткие, часто фиоле-
товые. Колосья мелкие пли средней величины, довольно рыхлые (D —
= 23—27), реже плотные (D = 40—50). Ости обычно длиннее колоса,
довольно грубые. Колосковые чешуи удлиненно-овальные, с коротким
(тупым или острым) килевым зубцом, с различно выраженным плечом
(слегка скошенным или ниспадающим), с ясно выраженной главной бо-
ковой жилкой, оканчивающейся зубцом. Зерновки средней величины.
Формы яровые (скороспелые и среднеспелые). Период до колошения
около 50 дней, весь период веютации до 95—100 дней. Мезофиты, неко-
торые засухоустойчивые. Не вполне устойчивые к трем видам ржавчины.
Т и п: Т. dicoccum var. haussknechtianum A. Schulz.
Геогр. распр. Ранее был широко распространен в районах Сред-
него Поволжья, Крыму, высокогорных районах Закавказья (СССР),
на Балканах, в Турции, гга северо-западе Ирана.
Значительный ареал подвида позволяет выделить 2 группы разно-
видностей, обусловленные эколого-географической и морфологической
дифференциацией: convar. transcaucasicum Flaksb. и convar. serbicum
(A. Schulz) Flaksb.
Ключ для определения групп разновидностей
1. Колосковая чешуя широкая, укороченная; килевой зубец тупой,
слабовыраженный, едва превышает короткий зубец главной боковой
жилки, между зубцами незначительная выемка..........................
................................convar. serbicum (A. Schulz) Flaksb.
+ Колосковая чешуя удлиненно-овальная, иногда обратнояйцевид-
пая; килевой зубец короткий острый или широкий тупой, или длинный
слегка крючковато изогнутый; зубец главной боковой жилки отсутству-
ет или хорошо выражен; в последнем случае выемка между зубцами
глубокая.............................convar. transcaucasicum Flaksb.
1. Convar. transcaucasicum Flaksb. — азиатская группа разновидно-
стей.
Convar. transcaucasicum Flaksb. 1928, Tp..прикл. бот. геи. сел. 19,1:
503.— Т. armeniaetttn (Stolet.) Nevski, 1926, Tp. Бот. ин-та АН СССР,
сер. 1, вып. 2: 67.!—Т. dicoccum prol. persicum Flaksb. 1935, Культ, фл.
СССР, 1: 300. — Т. dicoccum subsp. azerbajdzhanicum Dorof. el Lapt.
1967, Вести, с.-х. науки, 4: 30.— T. dicoccon subsp. subsponlaneum Tzvcl.
1973, Новости спет. высщ. раст. 10: 41.
Всходы опушенные, зеленые пли слабо окрашены ашоциаиом. Кус-
тистость слабая у полб из Ирана, несколько более сильная у армяно-
анатолийских и нагорно-карабахских. Растения низкорослые пли сред-
ней высоты, Соломина топкая, хотя сравнительно прочная. Листья бар-
хатпетоопушеппые, влагалище листа и стеблевые узлы опушенные,
особенно сильно у иранских полб. Для последних характерен сильный
>Л 1\лагсификацич /iot)<i Trilii'tim
восковой налет па растениях. Средн мелкоколосых азиатских полб бо-
лее крупными размерами колоса, колосков и зерна выделяются нагорно-
карабахские. Колосковая чешуя удлиненная, плоская, без плеча или
более или мопсе вздутая, или широкая, вздутая, с приподнятым плечом
('последнее чаще у закавказских полб). Килевой зубец короткий острый
или тупой широкий в основании, или длинный!, слегка крючковато
изогнутый; главная боковая жилка хорошо пли слабо выражена, се зу-
бец отсутствует или хорошо выражен.
Членики колосового стержня голые или опушенные по краям. Плен-
чатость сильная,средняя и слабая.
Т и п: Т. dicoccum var. haussknechtianum A. Schulz.
Азиатская группа разновидностей представлена в основном яровыми
формами. Скороспелые и среднеспелые (период вегетации до 100—
105 дней). Неустойчивы к поражению желтой и бурой ржавчиной и муч-
нистой росой; исключительно восприимчивы к этим заболеваниям и
твердой головне иранские формы. Среди азиатских полб наиболее устой-
чивы к бурой и желтой ржавчине карабахские.
Азиатскую полбу можно определить как сложную эколого-морфоло-
гическую группу.
Н. II. Вавилов (1964) среди азиатских полб выделяет 3 эколого-
географические группы: 1) армяно-анатолийскую (proles armeno-ana-
tolicum Vav.), для которой характерны скороспелость, тонкая соломи-
на, среднего размера колос, мягкие ости, мелкие зерновки; 2) нагорно-
карабахскую (proles carabachicum Vav.), отличающуюся от предыдущей
более крупными размерами колоса, зерновок, более прочной соло-
миной; 3) иранскую (proles iranicum Vav.) — низкорослую, исключи-
тельно восприимчивую к грибным болезням.
Морфологическая дифференциация азиатских полб затруднена из-за
значительного захождения рядов изменчивости признаков, что говорит
об их тесной взаимосвязи и близком происхождении.
По Е. Ф. Пальмовой (1935), азиатские полбы экологически относят-
ся к степному типу, относительно засухостойки и зноевыносливы, со
значительной реакцией на длину дня.
Геогр. распр. Азербайджан, Армения, Грузия, Дагестан, Тур-
ция, Иран.
Var. aeruginosum Flaksb. 1939, Оиред. паст, хлебов: 55. — Т. dicoc-
cum gr. aeruginosum var. aestivate (Stolet.) Flaksb. et var. autumnale
(Stolet.) Flaksb. 1935, Культ, фл. СССР, 1: 300. — T. dicoccum var. rtiftim
f. aestivalis Stolet. et f. autumnalis Stolet. 1925, Tp. прикл. бот. сел. 14,1:
91. — T. dicoccum var. rufum subvar. pschaviense Menabde, snbvar.
bolnissiense Menabde, subvar. dmanissicum Menabde et subvar. thiatie-
ticutn Menabde, 1948, Пшен. Груз. 40, 41, descr. ross.' (5).
E. H. Столетова (1925) делит красноколосые закавказские полбы по
продолжительности вегетационного периода. Более дробное деление на
подразновидности произведено В. Л. Менабде (1948) в зависимости от
длины и плотности колоса, формы и величины килевого зубца, опуше-
ния колосового стержня.
Геогр., распр. Грузия, Азербайджан, Армения.
Промежуточная форма (var. carabachicum Dorof. et Lapt.), возник-
шая при естественной гибридизации между Т. dicoccum и Т. durum, вы-
делена В. Ф. Дорофеевым и IO. П. Лаптевым (1967). Она имеет ряд
признаков первого вида, по легко обмолачивается. Подобная форма
отобрана также 11. А. Гандиляном (1972) -var. arnurual Gandil., ко-
торый считает се результатом спонтанной гибридизации между Т. du-
rum и Т. aeslivum.
Var. arptilunial Gandil. 1975, l.c.: 78, descr. ross. (10).
Выделена П. А. Гапдпляном на коллекционном посеве Ботанического
сада в Армении.
Vnr nrntirnnial Gnnrlil IQ?.^ 1 г • 7Q rlo^rr гп<;ч М
Ч е ш // it неону in с н н ы в
Колосок с одной остыо
3 с р п о в к п к р а с н ы с
“Г — — — — — 3- — iiniluteotin- 1
ctum
— _L — — — —. + — uniaertigino- 2
sutn
Колосок с двумя остями
3 о р н о в к и к р а с н ы с
-1- — - — — -1- — huiiKsbnech- 3
t ioiiu III
д_ — — — — — —- !- jii /ги b ~inoi i I
—. + •— — — — 4- _— ucriiginostim 5
— — + — — — + — chcusuriciim 6
— — — -f- — — — jlaksbergeri 7
Ч с ш у и о п у ш е н н ы с
Колосок с двумя остями
Зори о в к и к р а с п ы е
+ I — I — I — I — I — I + I — I gunbadi | 8
-- I + I — I — I — I — I — 1 + 1 rubrogiinbadi | 9
3 е р п о в к и б о л ы с
— I — I — I — 1 + 1 — 1 + 1 — I arpidiinial I 10
— I — I — I — I — I + I + I — I arpurunial | 11
Выделена И. Л. Гапдпляпом на коллекционном посеве Ботаническо-
го сада в Армении.
Var. chevsuricum Dekapr. 1954, Тр. Ип-та полевод. 8: 16, descr.
ross. (6).
Геогр. распр. Грузия.
Var. flaksbergeri Dekapr. in Flaksb. 1935, Культ, фл. СССР, 1: 300,
in lilt., 1928; Декапр. 1954, l.c.: 16. (7).
Геогр. распр. Грузия, Азербайджан.
Var. gunbadi Palm, in Flaksb. 1935, 1. c.: 301, in lilt. (8).
Геогр. распр. Иран (Курдистан).
Var. haussknechtianum A. Schulz, 1915, Berichte Deutsch. Bot. Ge-
sellsch. 33: 236, tab. 6,2.— T. dicoccum var. farrum f. iranica Stole!. 1925,
l.c.: 90, tab. 7. — T. dicoccum var. farrum f. caucasica Stolet. 1925, l.c.:
90. — T. dicoccum var. farrum f. armeniaca Stolet. 1925, l.c.: 91, tab. 4.—
T. dicoccum gr. luteo-iincium var. armetiiacum (Stolet.) Flaksb. 1935,
1. c.: 300.—T. dicoccum gr. luteo-tinctum var. caucasicum (Stolet.) Flaksb.
1935. l.c.: 299. — T. dicoccum var. luteo-tinctum Flaksb. 1939, l.c.: 54.—
T. dicoccum vaf. iranictim (Stolet.) Flaksb. 1935, l.c.: 301. — T. dicoccum
var. farrum osseticum Dekapr. 1954, Tp. Ип-та полевод. 8: 15, descr.
ross. — T. dicoccum var. farrum subvar. laeviglumarum Menabde, subvar.
thrialeticum Menabde, subvar. raruin Menabde, subvar. carthlicum Menab-
dc, 1948, l.c.: 40, descr. ross. (3).
Var. haussknechtianum очень изменчива по форме колосовых че-
шуй —от удлиненно-овальных, широких в верхней половине у f. cau-
casica Stolet. и f. iranica Stolet. (у первой килевой зубец притуп-
ленный, короткий, прямой, у второй — острый, короткий) до широких,
762
66
Классификация рода TrUicum L.
выпуклых колосковых чешуи с более длинным, слегка клювовидно за-
гнутым внутрь килевым зубцом (f. armetiiaca StoleL, I. osselicum
Dckapr.).
Среди белоколосых двузернянок В. Л. Мепабде (1948) выделил по
длине и плотности колоса, длине остей, форме киля колосковой чешуи
подразновидности, отображающие полиморфизм полб Грузии.
Геогр. распр. Грузия, Армения, Азербайджан, Иран.
Var. jakubzineri Udacz. 1976, Тр. прикл. бот. геи. сел. 56,2: 149. (4).
Геогр. распр. Выявлена и отобрана на коллекционном посеве
Среднеазиатской опытной станции ВИРа из местной полбы Ирана.
Var. rubrogunbadi Palm, in Flaksb. 1935, l.c.: 301, in litt. (9).
Геогр. распр. Иран (Курдистан).
Var. uniaeruginosum Dorof. 1968, Бюл. ВИРа, 13: 72. (2).
Геогр. распр. Азербайджан.
Var. uniluteotinctum Dorof. 1968, Бюл. ВИРа, 13: 72. (1).
Геогр. р а с и р. Азербайджан.
Среди азербайджанских полб И. Д. Мустафаев (1973) выделил под-
вид тургидоидных полб с рядом неэффективно обнародованных разно-
видностей (не приведены латинские диагнозы и типы). По-видимому, их
следует рассматривать как группу разновидностей гибридного проис-
хождения.
2. Convar. serbicum (A. Schulz) Flaksb. — волжско-балканская груп-
па разновидностей.
Convar. serbicum (A. Schulz) Flaksb. 1928, Тр. прикл. бот. ген. сел. 19,1:
503. — Т. dicoccum serbicum A. Schulz, 1915, Bcricht. Deutsch. Bot. Ge-
sellsch. 33: 240, 241, tab. 6,7. — T. dicoccum группа rosso-lartaricum
Flaksb. 1928, l.c.: 503.— T. dicoccum subsp. euroum prol. tartarobalcani-
cuin Flaksb. 1935, Культ, фл. СССР, 1: 297.—T. volgense (Flaksb.) Nev-
ski,1934, Фл. СССР, 2: 683. — T. turgidum subsp. volgense (Flaksb.)
A. et D. Love, 1961, Bot. Not. (Lund), 114, 1: 49.— T. dicoccum subsp.
volgense (Nevski) Vav. 1964, Миров, ресурсы хлебн. злаков. Пшеница:
32, comb, invalid. — Т. dicoccon subsp. volgense (Flaksb.) 1'zvel. 1973,
Новости спет. высш. раст. 10: 41.
Всходы силыюопушенные, часто окрашены антоцианом. Кустистость
слабая. Растения средней высоты (80-120 см). Листья бархатистоопу-
шеппые, влагалище голое или опушенное. Узлы на соломине опушенные
или почти голые.
Колосья небольшие, ости тонкие. Колосковая чешуя широкая, уко-
роченная, наиболее широкая в средней части. Килевой зубец короткий,
тупой, слабовыраженпыи, едва превышает размер зубца главной боко-
вой жилки, между зубцами незначительная выемка. Главная боковая
жилка выражена слабо. Членики стержня колоса в верхней части (у ос-
нования колосков) с бородками. Нленчатость невысокая. Формы скоро-
спелые (от посева до колошения 52—71 день) пли среднеспелые (95—
НО дней), довольно засухоустойчивые, яровые.
Т и п: Т. dicoccum var. serbicum A. Schulz.
Геогр. распр. Возделывается в Татарии, Чувашии и в прилегаю-
щих к ним районах (в остальных районах СССР встречается редко),
в Югославии, Болгарии и, вероятно, в Турции.
Ключ для определения разновидностей и форм
convar. serbicum (A. Schulz) Flaksb.
1. Колосья красные исоиушеиныс, ости красные, зерновки красные
var. serbicum A. Schulz
-|- Колосья белые, iieoiiyniennые, остн белые, зерновки красные . . .
...................................var. volgense (Flaksb.) Flaksb. 2.
Подрод I. Triticum
C>7
2. Влагалище .чиста и стеблевые узлы иеоиушеииые . . f. volgense
Flaksb.
+ Влагалище листа и стеблевые узлы опушенные f. tataricum Stolet.
Var. serbicum Л. Schulz, 1915, Bericht. Deutsch. Bot. Gesellsch. 33:
240, quoad f. rufum. — T. dicoccum rufum maturatum Flaksb. 1915, Опред.
пшен.: 60. — T. dicoccum var. maturatum Flaksb. 1935, Культ, фл. СССР,
1:298.
Встречается в смеси с белоколосой полбой и весьма сходна с пей.
Геогр. распр. По всему ареалу convar. serbicum.
Var. volgense (Flaksb.) Flaksb. 1935, l.c.: 297. — T. dicoccum farrum
volgense Flaksb. 1922, Опред. наст, хлебов: 24. — T. dicoccum serbicum
album A. Schulz, 1915, l.c.: 240. — T. dicoccum var. farrum 3. Slav emmer
Perciv., 1921. Wheat pl.: 198.
Преобладающая разновидность в татаро-балканской группе с не-
сколько большим диапазоном изменчивости признаков по сравнению
с var. serbicum. У var. volgense есть формы с почти голыми влагалищем
листа и стеблевыми узлами, которые выделила Е. Л. Столетова (1925),
отнеся их к f. volgense Flaksb. 1923; Опред. паст, хлебов: 24, p.p. (syn.
Т. dicoccum var. volgense f. [ere-ntidtim Flaksb. 1935, Культ, фл. СССР,
1: 298, descr. ross.) и формы с сильноопушеиными влагалищем листан
стеблевыми узлами — f. tataricum Stolet. 1925, Тр. прикл. бот. сел.
14,1: 90.
У var. serbicum по выявлено форм с неонушепными влагалищем лис-
та и стеблевыми узлами.
Геогр. распр. По всему ареалу convar. serbicum.
IV. Subsp. abyssinicum Vav. —• подвид эфиопских полб.
Subsp. abyssinicum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51:
223; он же 1964, Миров, ресурсы хлебп. злаков. Пшеница: 18, 29.—
Т. dicoccum gr. indo-abyssinian emmers Sect. A. Speltae Perciv. 1921,
Wheat pl.; 192.
Всходы слабоопушениые, густо окрашены в фиолетовый цвет. Ко-
леоптиль имеет 3 и более проводящих пучков. Куст сомкнутый, кусти-
стость слабая. Растения низкорослые (36—110 см). Стебли перед со-
зреванием фиолетовые. Листья светло-зеленые, с верхней поверхности
покрыты редкими шипиками, с нижней — голые; жесткие, блестящие,
небольшие (длина 18—20 см, ширина 0,9—1,3 см). Влагалище листа
голое. Соломина тонкостенная, под колосом почти выполненная (с про-
светом), обычно не полегающая; узлы большей частью фиолетовые.
Верхние листья подходят почти к самому колосу (образуя иногда как
бы кроющий лист). Колосья плоские (двурядпая сторона шире лицевой),
мелкие или средней длины (5—7 см), рыхлые (D — 23- -30). Колоски
ромбовидные (обычно средний цветок выступает над боковыми). Ко-
лосковые чешуи удлиненно-овальные, наиболее широкие в верхней части
чешуи. Килевой зубец слабовыражепиый, притупленный. Зубец главной
боковой жилки по размеру немного меньше. Плечо слабоскошеппос.
Зерновки мелкие (длина 9,5 мм, ширина и толщина 2—2,5,мм).
Формы только яровые, очень скороспелые (до колошения 40—45 дней
и от колошения до созревания 35—40 дней), с известной требователь-
ностью к влаге. Поражаются желтой, бурой и стеблевой ржавчиной.
Довольно устойчивы к мучнистой росе и пыльной головне.
Тип: Т. dicoccum var. arras (Ilochst.) 1\штп.
Геогр. распр. Индия, Эфиопия, юг Аравийского полуострова.
,В целом эфиопские полбы вместе с йеменскими и индийскими состав-
ляют определенный тип с рядом общих морфологических и биологиче-
ских свойств (Н. И. Вавилов, 1964). Однако в пределах подвида легко
выделяются 3 эколого-географические группы (прежде всего по их био-
логическим особенностям), почти не отличающиеся морфологически.
5*
(>8 l\.Ull'l 11<1>111«1ЦИ!1 )>Ot)(l Tritil'iun Д.
1. ( л)бс'| r.eiiHo )ф11<>iicкiie полбы | 7'. iliciicriiin pTcx abt/ssiiiica Slolc
lova in Vavilov, 1931, Tp. прикл, бог. ivii. сел. ] I рилож. 51: JG(>.
T. dicoccum prol. h/picum Jakubz.. in Flaksb. 1939, Опред. нас г. хлебов: 51,
descr. ross. -- T. dicoccum prol. proprioabi/ssinicum Vav. 1964, Мирон,
ресурсы хлеба, злаков. Пшеница: 30, nom invalid.] наиболее разнооб-
разны в ботаническом отношении. Средн них наряду с краспоколосымп
и белоколосымн разновидностями всгречаюк'я черноколосые и разно-
видности с черной окраской ио краю чешуй. По Е. Ф. Пальмовой
(1935), они относятся к высокогорному типу со средней реакцией на
длину дня, с малой потребностью в температуре, по с большей требо-
вательностью к влаге. Растения низкорослые, очень скороспелые. Боль-
шей частью устойчивы к. бурой и желтой ржавчине, пыльной головне и
мучнистой росе, относительно восприимчивы к твердой головне, а также
повреждаются шведской и гессенской мухами и зеленоглазкой.
Геогр. распр. В высокогорных районах Эфиопии (до высоты
2000—3000 м над ур. м.).
2. Пемепские полбы (Г. dicoccum var. arras f. jcmcni Flaksb. 1935.
Культ, фл. СССР, 1: 295. T. dicoccum prol. arabiciim Jakulv. in Flaksb.
1939, l.c.: 51, descr. ross. - T. dicoccum prol. jcmciiicum Vav. 1961, l.c.:
30, nom invalid.) более скороспелые и низкорослые, чем эфиопские. По
словам П. II. Вавилова (1964), они типичные эфемеры, крайний вариант
среди мирового разнообразия по быстроте развития. Е. Ф. Пальмова
(1935) относит их к полупустынному тину, отличающемуся значитель-
ной зиоевыпосливостыо. Восприимчивы к мучнистой росе.
Геогр. р а с и р. Юг Аравийского полуострова.
3. Индийские полбы (Т. dicoccum farrum indiciim Vav. in Flaksb. 1923,
l.c.: 25. — T. dicoccum prol. indosianicum Jakubz. in Flaksb. 1939, l.c.:
51, descr. ross.; Вавилов, 1964, цит. лит.: 30) более требовательны к теп-
лу. Характерна большая устойчивость к стеблевой и бурой ржавчине,
пыльной головне и мучнистой росе, восприимчивы к желтой ржавчине,
твердой головне, поражаются шведской и гессенской мухами, зелено-
глазкой.
Геогр. распр. Индия (очень небольшие островки культуры).
Определение разновидностей subsp. abyss!nicuin Vav.
белые Чен красные у и белые с черной каймой по краю черные (фон белый) Ос цвета чешуй ти черные Разновидность Л? ра.знови 1- НОС 1 и
Чешi/u иеопушениые
3 e p и о в к и к р а с н ы е
-к — — — + arms 1
-1- — — — — -к pseudoarras 2
— 4- _—. + — praecox 3
— + — — — pseudopraecox I
— — -1- — — + nigroajar 5
— — — + + tii pfru/fi 6
е р н о в х и ф И О л сто в ы с
-г — — — — violaeeoarras 7
Чешуи опушеи/юе (чернчнки красные)
-|- | — I — ' -|- — pubescent ' 8
Var. arras (Ilocbst.) Koern. in Koern. el Wern. 1835, Ilandb. Ge.treideb.
1: 88. — T. arras Ilocbst. 1848, Flora, 31: 450,- -T. dicoccum var. ajar
Perciv. 1921, Wheat pl.: 193. — T. dicoccum var. farrum indicum Vav. in
Подрод I. Triliciini 69
Flaksb. 1923, Онред. finer, хлебов: 25. /'. (licoccum var. ]arrum f. arras
(Ilochst.) Vav. et f. allins Slolel. in Vav. 193!, Tp. нрпкл. бог. юн. сел.
Прплож. 51: 16(5. (1).
Наиболее распростриценная разновидность иолбы. Обычное эфиоп-
ское название — «ajar» или «agga» В Индии встречается под названием
Khapli. Наряду с типичными скороспелыми (от всходов до колошения
43 дня в условиях весеннего посева в Крыму) f. arras выделены формы
более позднеспелые (от всходов до колошения 55 дней) и несколько
более высокорослые 1. altiiis Slolel. и f. jemeni I-’laksb. 1935, Культ,
фл. СССР, 1: 295, closer, ross.
Геогр. распр. Индия, Эфиопия, юг Аравийского полуострова.
Var. nigroajar Perciv. 1926, Journ. Bot.: 205. (5).
Геогр. распр. Эфиопия.
Var. nigrum Stolet. 1925, Tp. прикл. бот. сел. 14,1: 91. (6).
Геогр. распр. Эфиопия.
Var. praecox (Stolet.) Flaksb. 1935, l.c.: 295. — Т. dicoccum var. riijum
f. praecox Stolet. 1925, l.c.: 91. — T. dicoccum var. ru/um f. asiatica Vav.
in Stolet. 1925, I. c.: 91. (3).
Геогр. распр. Эфиопия.
К этой же разновидности К. А. Флякгберюр (1935 )огпосиг древне-
египетскую полбу, известную только ио раскопкам и выделенную
А. Шульцем как Т. dicoccum f. aegyptiaca rufa Schulz, 1915, Bericht.
Deutsch. Bot. Gesellscli. 33: 240. А. Шульц считает древнеегипетскую
полбу сходной с иранской (персидской), а К. Л. Фляксбсргер - с эфи-
опской полбой.
Var. pseudoarras Flaksb. 1935, Культ, фл. СССР, 1: 295, descr.
ross. (2).
Геогр. распр. Эфиопия.
Vav. psetidopraecox Palm, et Jakubz. in Flaksb. 1935, l.c.: 296, descr.
ross. (4). • .
Геогр. распр. Эфиопия.
Var. pubescens Jakubz. in Flaksb. 1939, l.c.: 52, descr. ross. (8).
Геогр. распр. Эфиопия.
Var. violaceoarras Jakubz. in Flaksb. 1939, l.c.: 52, descr. ross. (7).
Геогр. распр. Эфиопия.
T. KARAMYSCIIEVII NEVSKI — ПШЕНИЦА КАРАМЫШЕВА
(ПОЛБА КОЛХИДСКАЯ, ПШЕНИЦА ГРУЗИНСКАЯ)
Т. karainyschevii Nevski, 1935, Сов. бот. 6: 127. — 7’. dicoccum grex
georgicum Dekapr. et Menabde, 1932, Tp. прикл. бот. ген. сел. 5,1: 14,
37. — Т. dicoccum subsp. georgicum (Dckapr. et Menabde) Flaksb. 1935, t
Культ, фл. СССР, 1: 301. — T. madia subsp. georgicum (Dekapr. et Me-
nabde) Flaksb. 1939, Опред. паст, хлебов: 202. — T. pa'laebcolchicuni
Menabde, 1940, Сообщ. Груз. фил. АН СССР, 1, 9: 680, nom. illeg.—
Т. georgicum (Dckapr. et Menabde) Dekapr. 1941, Сообщ. АН ГССР,
2, 10: 920.— Т. turgidum gr. palaeocolchicum (Menabde) Bowden, 1959,
Canad. Journ. Bot. 37: 670, nom. illeg. — T. turgidum subsp. palaeocol-
chicum (Menabde) A. et D. Love, 1961, Bot. Not. (Lund), 114, 1: 50, nom
illeg.; Mac Key, 1966, lleredilas, Slippl. 2: 238.
Общая характеристика. Всходы co слабобархаi нет ым опушением, зе-
леные или темио-зелеиыс. Колеоптиль бесцветный. Об.тпст венное гь cii.uu-
пая. Листья широкие (до 2 3 см), барха i пот ые на ощупь, и период
колошения г восковым палпом. Листовое r.nai алите слабоопушениое.
Куст полусгелющпйся. Колосья средней длины (6—-S см), широкие
(в наиболее широкой части 1,4 — 1,5 см), суживающиеся к вершине и
основанию (рис. 6,и), плоские, плотные (D — 40 50). Соломина иод
колосом выполненная. Узлы бархатнетоопушенпые, ушки с короткими
70
Классификация рода Triticum L.
Рис. 6. Колосья, колоски и зерновки полб:
а — Т. karatnyschevii Nevski: б—Т. ispahariiciim Hcslot
ресничками. Членики колосового стержня опушенные, короткие (1,5—
2,0 мм) и узкие (1,4—1,6 мм). Из-за очень большой плотности колос
изогнут. В колоске 4—5 цветков. Колосковая чешуя короче цветковой
(па 2,2—3,5 мм), киль хорошо развит, доходит до основания чешуи.
Килевой зубец короткий (1,0—1,5 мм), острый, направлен в сторону
цветка. Ости негрубые, короткие (4,0—5,0 см). Растения до 130 см высо-
той. Колеоптиль имеет 2 проводящих пучка. Зерновки красные. По на-
блюдениям В. М. Супаташвили (1929) и Л. Л. Декапрелсвича (1956),
для этого вйда характерна склонность к образованию ветвистоколосых
форм. 2п=28. Геномы А"В. Т. karatnyschevii входил в качестве примеси
в состав ценоза Т. madia. Зона возделывания колхидской полбы, соглас-
но классификации Г. Т. Селянпнова (195(5), по термическом}' режиму
является умеренной и частью теплой, избыточно влажной. Экологически
эта полба — мезофит.
Формы Т. karaniyschevii в массе озимые и полуозпмые. По данным
В. Л. Мепабде и Л. Л. Декапрелсвича (1940), изредка встречаются яро-
вые формы. Па своей родине зимует удовлетворительно, в других зонах
зимостойкость слабая.
Подрод I. Triticum
71
JI. JI. Декапрелевпч (1956) характеризует эту пшеницу как пластич-
ную. Несмотря на то, что ее местообитание, связано с влажным клима-
том нагорья Западной Грузии, при переносе в восточную часть опа вы-
нослива к воздушной засухе.
Характер цветения по данным, полученным в 1966—1968 гг. В. И. Жу-
ковым (1969) в различных вертикальных зонах Дагестана, примерно
такой же, как у других пленчатых видов секции Dicoccoides. В первые
дни имеет место ускоренное отцветание верхушечных колосков. Цвете-
ние наступает прежде всего в первом (нижнем) цветке колоска, а затем
распространяется на вторые и последующие цветки. Как и у других
тетраплоидных видов, наблюдается круглосуточное цветение. Как пра-
вило, оно протекает быстро (за 3—5 дней). У этого вида, подобно дру-
гим тетраплоидный пшеницам, отмечено 93—100% открытоцветущих
цветков.
По сведениям М. М. Якубцинера, II. Ф. Покровской (1969), В. Г. Ко-
нарева с соавторами (1972), у Т. karamyschevii наблюдается сочетание
сравнительно высокой (15,9—18,8%) белковости зерна с повышенным
содержанием (2,70—2,91%) лизина в белке.
Иммунитет. Колхидская полба выделяется устойчивостью к видам
ржавчины и головни (М. М. Якубцинер, 1957). В опыте О. Г. Григорье-
вой (1975) се проростки при искусственном заражении были устойчивы-
ми (тип реакции 0—II) к расам 77, 20, 122 бурой ржавчины; в полевых
условиях па естественном инфекционном фоне поражение не превышало
2 балла (ио 5-баллыюй шкале). В условиях Дагестана максимальный
балл поражения за 1954—1965 гг., по данным И. П. Шитовой (1968),
в среднем был равен 2.
М. М. Якубцинер (1969) сообщал, что в отдельные годы в полевых
условиях полба колхидская устойчива к стеблевой ржавчине, но при
искусственном заражении популяцией рас 21, 40, 15 проростки ее сильно
восприимчивы. По данным О. Г. Григорьевой (1975), на естественном
инфекционном фоне полба колхидская устойчива к этой болезни
(балл 1).
Э. X. Суханбердина (1977) при искусственном заражении проростков
популяцией мучнистой росы из различных географических районов СССР
нашла, что образцы полбы колхидской устойчивы или восприимчивы
(балл 1—3), а взрослые растения иа провокационном фойе устойчивы
к патогену (балл 0—2).
По данным В. И. Кривченко (1973), полба колхидская более устой-
чива к пыльной головне f. aestivi (тип реакции 0—I), чем к f. duri (тип
реакции 0, II). А. М. Ямалеев (1974) подтвердил эти данные, указав,
что гифы последней формы (при искусственном заражении) проникают
во все части зародыша зерновки, в том числе в зародышевую почку.
При этом в полевых условиях у взрослых растений пыльная головня
поражает до 15% колосьев. Выявлена иммунность полбы колхидской
при искусственном заражении твердой головней (В. И. Кривченко, 1973).
История. Следы Т. karamyschevii, датируемые эиеолитическпм перио-
дом, были найдены В. Л. Менабде в 1940 г. среди растительных остатков
древней Колхиды. На этом основании В. Л. Менабде пришел к заклю-
чению, что эта, пшеница является сохранившимся реликтом культуры
доисторического прошлого.
По II. И. Вавилову (1964) Т. karamyschevii в прошлом имел более
широкое распространение.
Географическое распространение. Т. karamyschevii — эндем Запад-
ной Грузии.
Происхождение. В. Л. Менабде (1940) сообщил, что при раскопках
кургана Диди-Гудзуба в Мегрелии была обнаружена колхидская иолба
неолита. Он рассматривает ее как аборигенное растение древней Ко-ч-
хиды, возделывавшееся на территории, ныне занимаемой Грузией и
72
!\.шссификпци.ч ро<)п Triliciiin 1..
Турцией. I 1роисхождепие колхидской подои В. Л. Методе (1971) свя-
зывает с первичной полигонной популяцией Т. protomacha. Е. II. Син-
ская (1969) также принимала колхидскую полбу за рудимент интеграль-
ного вымершего вида Т. protomacha. Л. Л. Дскапрелевич (1941) склонен
видеть происхождение колхидской полбы и Т. dicoccum от одного пред-
ка. Он понимает в колхидской полбе тот вид, который замещает в За-
падной Грузии Т. dicoccum и ареал которого отсечен от основной части
Грузии Сурамскнм хребтом. ,
По мнению В. Ф. Дорофеева (1971), колхидская полба возникла на
заре примитивного земледелия из Т. dicoccoides путем значительных
генетических перестроек типа транслокаций, инверсий и пр. А. Д. Гор-
гпдзе (1973) пе отрицает возможности происхождения колхидской
полбы из полбы дикой, но не исключает также варианта, предполагаю-
щего происхождение ее с участием дикой однозернянки. Она считает,
что колхидская полба могла возникнуть раньше, чем Т. macha, пли
одновременно с этим видом. Подтверждение более раннего происхожде-
ния колхидской полбы, чем Т. dicoccum. она видит в результатах экспе-
римента по ионизирующему облучению семян, когда в М2 из первого
вида возник второй. G. Mandy (1970) предполагает, что колхидская
полба возникла от скрещивания между дикой однозернянкой Закавказья
и Ае. speltoides.
Сорок с лишним лет назад 1\. А. Фляксбергер (1935) высказал мне-
ние, что колхидская полба скорее гибридного происхождения, чем след-
ствие мутации. Естественно поэтому ее морфологическое сходство с
Т. macha, которое замечено рядом исследователей. Д. Мак-Кен (1963,
1969) даже полагает, что эти виды имеют один и тот же ген скверхсд-
пости.
Н. И. Вавилов (.1964) обратил внимание на то, что колхидская полба
имеет своеобразные признаки, свойственные Т. compactum. Подобные
признаки свойственны также Т. macha, в сообществе с которой открыта
колхидская полба (В. М. Супаташвили, 1929). Поэтому нельзя отрицать
возможность появления последней при спонтанном опылении цветков
Т. macha пыльцой какого-либо тстраплоидпого вида. Сходное выщеплс-
нпе растений с признаками колхидской полбы мы наблюдали в F2 у
подобного гибрида, полученного с участием Т. macha subsp. itnereticiun.
Систематика. Вид Т. karamyschevii сначала рассматривался
В. М. Супаташвили (1929) как разновидность (var. chwamlicum Supat.)
в составе Т. dicoccum. Позднее Л. Л. Дскапрелевич и В. Л. А1еиабде
(1932) выделили ее как особую группу grex georgicum в пределах того
же вида. К. А. Фляксбергер (1935) перевел эту группу в ранг подвида.
Первый, кто признал эту пшеницу самостоятельным видом, был
С. А. Невский (1935). Он па основании четких морфологических отличий
от Т. dicoccum и географической обособленности описал его как Т. kara-
myschevii Nevski, дав название в честь русского ботаника, ученика
К. Линнея А. Карамышева. До 1940 г. колхидская полба фигурировала в
литературе в ранге подвида Т. dicoccum subsp. georgicum (К. А. Флякс-
бергер, 1935; Н. И. Вавилов, 1964; В. Ф. Дорофеев, 1969), и даже как
особый подвид подтем же названием в составе Т. macha (К. Фляксбер-
гер, 1939). В1 1940 г. В. Л. Менабде возвел колхидскую полбу в ранг
вида Т. palaeocolchicum Menabde (синоним Т. karamyschevii).
Ключ для определения разновидностей Т. karamyschevii Nevski
1. Колосья пеоиушеппыс....................................2.
+ Колосья опушенные, черные (красный фон)........................
...........................................var. nigresccns Menabde
Подрод I. Triticum 73
2. Колосья белые (соломенно-желтые) .... var. karainyschcvii.
+ Колосья красные...............var. rubidum (Menabde) Dorof.
Var. karamyschevii. — T. dicoccum var. chwatnlicum Supat. 1929,
Вести. Ии-та экспер. агрономии Груз. 1: 92, tab. 2 и 3. — Т. palaeocolchi-
cum var. chwamlicum (Supat.) Menabde, 1910, Сообщен. Грез. фил. АН
СССР 1,9: 686.
Геогр. распр. Грузия (в прошлом встречалась на посевах
Т. madia).
Var. rubidum (Menabde) Dorof. comb.- nov. — T. palaeocolchicum
var. rubidum Menabde, 1940, 1. c.: 686.
Геогр. распр. Грузия (обнаружены единичные растения в по-
севах Т. madia).
В Институте ботаники АН ГССР (Тбилиси) в результате скрещива-
ния Т. karamyschevii XT. durum var. libycum получена константная
разновидность Т. palaeocolchicum var. nigrescens Menabde, 1948, Пшеп.
Груз. 43, descr. ross.
T. ISPAHANICUM HESLOT—ПШЕНИЦА ИСФАХАНСКАЯ
(ПОЛБА ИСФАХАНСКАЯ)
Т. ispahanicum lieslot, 1958, Compt. Rend. Acad. Sci. (Paris), 247:
2477—2479.— T. dicoccum subsp. ispahanicum (Heslot) Dorof. 1969,
Вести, с.-х. пауки, 3: 27.
Общая характеристика. Куст прямостоячий, редко полустелющиися.
Растения 90—100 см высоты. Соломина топкая, в верхнем междоузлии
под колосом выполненная. Узлы соломины густо, а листовое влагалище
редко опушены мягкими короткими волосками. Листья 24—30 см длины
и 0,7—1,2 см ширины, бархатпетоопушеппые. Язычок ясно выражен.
Колосья 7,6—10,6 см длины, неопушенные, одноостые (рис. 6,6).
Двурядная сторона колоса шире лицевой. Плотность колоса средняя
(D = 27—30). Стержень созревшего колоса распадается при надавли-
вании. Колоски удлиненные, двуцветковые, оба цветка плодущие. Чле-
ники колосового стержня опушены по бокам, у основания колоска име-
ются 3 бородки из мягких волосков. Колосковые чешуи удлиненные,
ланцетовидные, склернфицироваииые, со слаборазвитым килем, пере-
ходящим на верхушке в маленький тупой зубец; плечо слабо развито.
Наружная цветковая чешуя первого цветка имеет тонкую ость 3—4 см
длины. Внутренняя двухкилевая цветковая чешуя первого цветка при-'
близптелыю на 5 мм короче наружной. Наружная цветковая чешуя
второго цветка безостая, внутренняя — по длине почти равна наружной.
Для колосьев Т. ispahanicum характерна ориентация колосков в од-
ну сторону. При этом центральные оси колоса и колосков не совмеща-
ются в вертикальной плоскости, а двурядпые стороны колоса оказыва-
ются различными (стержень колоса на одной из его сторон обнажен,
а на другой — прикрыт колосками). Такая ориентация колосков наблю-
дается еще до выхода колоса из листового влагалища. Одиоостость ко-
лосков, характерная для 7'. ispahanicum, встречается также у некото-
рых форм Т. dicoccoides convar. dicoccoides, у 7'. dicoccum subsp. marocca-
num и subsp. asiaticum convar. transcaucasicum. Сходство с T. polonicum
по следующим признакам: удлиненные наружные цветковые и колоско-
вые чешуи; укороченная в сравнении с наружной внутренняя цветковая
чешуя; односторонняя ориентация колосков в колосе и удлиненная зер-
новка. Сходство с 7’. dicoccum — по морфологическим признакам вегета-
тивных органов, типу разлома колоса и трудности обмолота. У Т. ispa-
hanicum колеоптиль имеет 2 проводящих пучка.
П. М. Жуковский (1964) относит полбу исфаханскую к диуручкам,
так как па родине (Иран) она высевается весной или осенью. При вы-
ращивании ее в горах Дагестана без полива наблюдается слабое куще-
74 Классификация рода Triticum L.
лис, ;i прл орошении п осеннем посеве в Ташкентской области и в низ-
менном Дагестане — сильное (до 12—15 продуктивных стеблей). При
этом имеет место затенение нижней части растения, в результате —
склонность к сильному полеганию. В Ленинградской области полба
исфаханская имела 2—5 продуктивных стеблей на одно растение и прак-
тически не полегала.
При осеннем посеве на орошаемых участках в Ташкентской области
и Дагестане полба исфаханская колосится позднее местных стандартов
на 3—12 дней, а созревает одновременно с ними или па 1—5 дней рань-
ше за счет более укороченного прохождения межфазного периода ко-
лошение— восковая спелость. В Ленинградской области при весеннем
севе (без полива) колошение происходит одновременно со стандартом,
а вызревание — па 4—7 дней раньше, т. е. в этих условиях Т. ispahani-
cum является раннеспелой пшеницей.
У Т. ispahanicum отмечена склонность к прорастанию зерна па кор-
ню вследствие короткого периода покоя у семян. В условиях влажного
Северо-Запада России они прорастают в колосьях до уборки. В урожае,
выращенном в Ленинградской области в 1960, 1961 н 196'2 гг., соответ-
ственно наблюдалось 21,7; 84,6 и 68,1% проросших зерен.
По наблюдениям Л. 3. Латыпова и В. II. Апеля (1971), в Белорус-
сии свыше 50% цветков Т. ispahanicum цветут открыто. Суточный мак-
симум цветения — в полуденные часы.
Иммунитет. Полба исфаханская, по данным Э. Ф, Мигушовой,
П. М. Жуковского (1969) и Э. X. Сухапберднной (1977), сильно пора-
жается мучнистой росой. Первые исследователи также сообщили, что
в условиях Ташкентской области и плоскостного Дагестана в 1959—
1972 гг. полевое поражение ее бурой ржавчиной не превысило двух бал-
лов (по 5-балльной шкале). О. Г. Григорьева (1975) при искусственном
заражении расами 77, 20, 122 бурой ржавчины наблюдала сильную вос-
приимчивость проростков, хотя взрослые растения полбы исфаханской
при естественном заражении на провокационном фоне были устойчивы-
ми к патогену (балл 1). Опыт Г. С. Турова и М. М. Якубцинера (не-
опубликовапо), проведенный в условиях Южного Казахстана, также
указывает на устойчивость полбы исфаханской к бурой ржавчине (ра-
сы 72 и 20) в фазах кущения и молочной спелости, тогда как интенсив-
ность поражения популяцией рас 11, 21, 34, 40, 77 стеблевой ржавчины
была высокой (80%), как и тип реакции. В горном Дагестане также
наблюдалось сильное поражение стеблевой ржавчиной, а в условиях
Ташкентской области отмечена очень сильная восприимчивость полбы
исфаханской к желтой ржавчине (Э. Ф. Мпгушова, П. М. Жуковский,
1969).
В. И. Кривченко (1973) полевую устойчивость полбы исфаханской
к пыльной головне па расовом уровне оцепил как высокую, отметив
эмбриональную расоспецифическую восприимчивость к f. aestivi. Его
данные подтверждены экспериментом А. М. Ямалеева (1974), который
наблюдал, что в зародышевой почке у зерновки развитие мицелия
f. aestivi может быть сильным. Однако полевое поражение колосьев
взрослых растений отсутствовало. Эмбриональная устойчивость полбы
исфаханской к пыльной головне f. duri высокая. Полба исфаханская
иммупна к твердой головне (В. И. Кривченко, 1973).
Географическое распространение. 7'. is paha п ictim был обнаружен в
1957 г. французской экспедицией в горах близ города Исфахан (высота
2000—2500 м над ур. м.). По сведениям автора вида, эта пшеница ис-
стари возделывается бахтиарами в горных селениях Ирана при оро-
шении.
Происхождение. По результатам циклических межвидовых скрещи-
ваний К- Ай. Жпжилашвили (1974) пришел к заключению о генетиче-
ской близости Т. ispahanicum, Т. dicoccum н Т. karamyschevii. Еще ра-
llot)/>od I. Trituum
75
нее подобное же .заключение было сделано Э. Ф. Мнгушовой и
II. М. Жуковским (1969). Гибридизация, электрофоретический анализ
водорастворимых белков зерновок и изучение цитологического поведе-
ния гибридов привели A. Belea и др. (1975, 1976) к выводу о большей
близости полбы исфаханской к 7’. polonicum, чем к 7’. dicoccum. Еще
ранее Н. Kihara и др. (1956) по морфологическим признакам колоса
включили полбу исфаханскую в вид Т. polonicum. По данным M..G. Joshi
и Л. К. Kaul (1971), электрофоретические спектры белков .зерновок этих
видов близки, но характерный для Т. polonicum компонент отсутствовал
у полбы исфаханской.
К. М. Жижплашвилп (1974) свидетельствует, что в его эксперименте
в комбинации от скрещивания полбы исфаханской с твердой пшеницей
выщепились растения типа Т. polonicum. В подобных комбинациях на-
шего опыта такие особи отсутствовали. Па основании результатов цик-
лических межвидовых скрещиваний мы пришли к заключению, что пол-
ба исфаханская произошла от полбы обыкновенной, видимо, в связи
с мутационным возникновением фактора, ответственного за удлинен-
ность зерновки, колосковых и наружной цветковой чешуй (обозначаем
его символом Р1). Мутация, давшая сходный с полбой исфаханской
растительный организм, была получена Л. Д. Горгидзе (1973) после
ионизирующего облучения полбы колхидской.
Между эффектами воздействия факторов Р' и Р, носителем которого
является Т. polonicum, есть сходство (вызывают удлиненность чешуй и
зерновок) и различие (у полбы исфаханской отсутствует пергаментная
консистенция колосковой чешуи, характерная для Т. polonicum). Если
предположить возможность происхождения полбы исфаханской при
гибридизации Т. polonicum с Т. dicoccum., то как объяснить разрыв сцеп-
ления между признаками удлиненности чешуи и их псргаментпостыо?
Возможно ли, что такое качество чешуй является следствием взаимодей-
ствия гена Р с полигенноп системой легкого обмолота, которую Т. polo-
nicum унаследовал от твердой пшеницы? Тогда понятно, что действие
фактора Р1, наложенное на генотип полбы обыкновенной, даст расти-
тельный организм с трудным обмолотом, каковым обладает полба ис-
фаханская. Но вопрос об аналогичности или гомологичиостп факторов
Р и Р1 пока остается открытым из-за отсутствия их экспериментального
изучения.
Систематика. В. Ф. Дорофеев (1969) рассматривает Т. ispahanicum
в ранге подвида Т. dicoccum. К последнему виду исфаханскую двузер-
нянку относит и Н. Kuckuck (1959). Однако определенный комплекс
признаков обусловливает положение ее в роде Triticum в качестве са-
мостоятельного вида.
Этот вид включает всего 2 разновидности *.
Ключ для определения разновидностей Т. ispahanicum Heslot
1. Колосковые чешуи белые......................var. ispahanicum.
+ Колосковые чешуи красные .... var. ispahanorufum Udacz.
Var. ispahanicum. — T. ispahanicum var. uniaristaium Heslot, 1958,
Comp. Rend. Acad. Sci (Paris), 247: 2479.
Геогр. распр. Иран (Исфахан).
Var. ispahanorufum Udacz. 1976, Tp. прикл. бот. ген. сел. 56, 2: 150.
Геогр. распр. Выделена на Среднеазиатской опытной станции
ВИРа в популяции Т. ispahanicum из Ирана.
* Дпуостая разновидность var. biarishdtim Ileslol (1958) оказалась расщепляю-
щимся гибридом.
76 K.itici ификцци.ч /)<х)ч Triticum L.
T. HIRGIDUM I.. ПШГ.НИЦА ТУ1Ч ИДУМ
(ТУЧНАЯ, ВЗДУТАЯ, АНГЛИЙСКАЯ)
Т. turgidiini I,. 1753, Sp. 1’1.: 36. 'Г. sativum turgiduin Паек., 1837,
NaturL Pflanzenfam. 2, 2: <85. - - T. sativum b. sativum turgidum Richter,
1890, 1’1. Enr. 1: 130,- T. tenux I> 1 turgidum Ascliers. et Graebn. 1901,
Synops. Milteleiirop. I 'I. 2: 689. /'. turgidum subsp. simieniivum Rassc
turgidum Thell. 1918, Naturw isseiisch. Wochensclir. 17, 33: '170.
T. aestivum subsp. turgidum Dornin, 1935, Preslia, 13—15: 43.— 7’. durum
subsp. turgidum (L.) V. Dorof. 1968, Euphytica, 17: 456, comb, invalid.—
T. dicoccum convar. turgidum (L.) Mandv, 1970, Acta agronom. 19.3—
4:416.
Общая характеристика. Всходы сиренево-зеленые, темно-зеленые.
У молодых н взрослых растеши'! листовые пластинки опушены волоска-
ми длиной 120- 150 мкм (на ощупь бархатистые, как у Т. dicoccum) пли,
реже, голые (на ощупь шероховатые). Листовые пластинки взрослых
растений крупные (длиной 20- 31 см. шириной 1.0 -1,5 см). Ушкн боль-
шей частью с ресничками. Растения обычно мощные, с небольшой ку-
стистостью (3- 5 стеблей на 1 раст сине). Соломина длинная (120
180 см), толстая (диаметром до 5 мм), большей частило полая, реже
выполненная, но с небольшим просветом в центре. Число междоузлий
6—7. Узлы крупные.
Колосья простые или ветвистые (рис. 7 и 8, а), обычно крупные (дли-
ной до 15 см), в поперечном разрезе квадратные или плоские (ширина
боковой стороны колоса 1,0—1,5 см, лицевой — 1,5 см), поникающие,
тяжелые (иногда в колосе, до 70 и более зерновок), рыхлые (D =
= 20—40), среднеплотные (D = 30 —40) пли плотные (D свыше 40).
Число колосков в колосе 19 -33. В колоске 5-7 цветков, в которых раз-
вивается до 5 зерновок. Стержень колоса прочный. Изредка встречают-
ся формы с ломким, как у 7’. dicoccum, стержнем. Членики стержня по
ребрам обычно опушены длинными волосками. У основания колосков —
бородка из щетинок длиной до 1 мм (у верхушечного колоска щетинки
бородки длиннее). Колосковые чешуи сравнительно короткие (длиной
8—11 мм), вздутые, асимметричные, с 5—7 жилками (у верхушечного
колоска — обычно симметричные, округлые, овальные). Киль широкий,
четко выраженный. Килевой зубец небольшой (высотой 1 —1,5 мм),
заостренный. Главная боковая жилка у типичных форм ясно выражена.
Наружная цветковая чешуя округлая, выпуклая, вздутая (не лодочко-
образная, как у твердой пшеницы). Опа длиннее колосковой чешуи и
прикрывается ею обычно лишь па 2/з. Ости длиной 8—20 см, толстые,
треугольные в поперечном разрезе, шероховатые. Зерновка короткая
(длина 5,5—9,0 мм), широкая (шириной 3—4,1 мм), округлая n.ni
овальная, мучнистая (стекловндпость зерновок встречается редко).
Основные морфологические отличия Т. turgidum от Т. durum: ко-
лосковые чешуи выпуклые, они значительно короче цветковых чешуй;
главная боковая жилка резко обозначена; киль четко выражен; киле-
вой зубец короткий; у основания колоска на членике колосового стержня
густая бородку из щетинок; зерновка широкая, овальная, вздутая, муч-
нистая. По крупности колоса, числу колосков в колосе и по высоте со-
ломины (до 2 м) 7'. turgidum превосходит 7'. durum. У последнего вет-
вистоколосые формы встречаются реже, чем у первого вида.
У иеветвнстоколосых форм Г. turgidum строение плодовой оболочки
зерновки типичное, для тетраплоидных голозерных видов секции
Dicoccoides. Согласно характеристике Л. Л. /Кестяннпковой (1954), -ла
оболочка мало редуцирована и одревесневает- пезпачпIелыю. Структура
перикарпия зерновок у различных форм -лого вида довольно однооб-
разна. У вет впстоколосых форм перикарпий в общем развит слабее,
чем у форм с простым колосом. Причем у зерновок из ветвистой части
IhttlpDi) I, Trim inn 77
Рис. 7. Колосья, колоски и зерновки Т. turgidum I..
колоса перикарпии по строению почти пе отличается от перикарпия
7’. dicoccum (В. Г. Александров, Л. Л. Жсстяпиикова, 1952). По данным
О. Д. Градчаниновой (1957), стебель Т. turgidum отличается поздней
дифференциацией механических элементов. Деление клеток иернцпкли-
чсского происхождения и укрупнение проводящих пучков происходит
в фазе выхода в трубку.
Генофонд Т. turgidum включает озимые, иолуозпмые и яровые фор-
мы. Первые локализованы в Западной и К )i о-Восгочной Европе, вю-
рые- -в Дагестане и Закавказье, ipeii.n в Малой Азии и Южной
Европе (Западное Средиземноморье). 'Г. turgidum обладает большей
степенью озимостп, чем твердая пшеница.
Т. turgidum — пшеница приморскою климата (в отличие от 7'. du-
rum— пшеницы степного климата). Е. Ф. Пальмова (1935) отмечала,
78
Классификация рода Trilicufii 1.
что местообитание этого вида характеризуется малой годовой амплиту-
дой колебания температур (11 — 16°С), отсутствием сильных морозов
и зноя, значительной влажностью воздуха в продолжение всего периода
развития. Преобладающая масса полей этого вида тяготеет к морско-
му побережью, занимая низменные территории. Исключение — высоко-
горная группа из Тибета — бассейна р. Нуцзяп (М. М. Якубцинер,
1959).
У Т. turgidum преобладают формы, требовательные к теплу, но
влаголюбивые, хорошо отзывающиеся па орошение. Это самый влаго-
любивый в роде Triticum вид. Он очень чувствителен к почвенной и воз-
душной засухе. В засушливые годы его урожай резко падает. При де-
фиците тепла Т. turgidum задерживается в своем развитии, в резуль-
тате зерно не дозревает.
Подрод J. Triticum 79
По данным И. Д. Мустафаева (1975), 7’. turgidum в Азербайджане
встречается до высоты 1630 м над ур. м. Он установил также связь
вертикальной зональности с ботаническим составом этого вида. На по-
лях, расположенных на высоте 200—220 м над ур. м., локализованы раз-
новидности var. fumidum, var. joduruin, var. nigrogluinarum, ниже
(100—150 м надур. м.) —var. turgidum, выше (430—440 м над ур. м.) —
var. herrerae и var. megalopolitaniim, еще выше (780—800 м над
ур. M.)--var. rubroatrum. Исключительно широкое высотное распро-
странение— от крайне нижнего предела (50 м над ур. м.) до самого
высокого — характерно для var. nigrobarbaium. За этой разновидностью
следуют var. melanatherum (200—1530 м над ур. м.) и var. plinianutn
(200—1300 м над ур. м.).
Наряду с господствующим экотипом Т. turgidum в литературе опи-
саны экологические группы, которые различаются главным образом по
биологическим признакам. Е. Ф. Пальмова (1935) называет западно-
и южноевропейскую экологические группы; Н. И. Вавилов (1964) —
центральноевропейскую, восточпо- и западносредиземно морскую;
М. М. Якубцинер (1957) для вствистоколосых форм- южную. запад-
ную, китайскую, коупрадскую, кавказскую нолуозпмую и .закавказскую
позднюю. Л. Л. Дскапрелевич (1948) у Т. turgidum Грузии выделил
3 экотипа; 1) настоящие поливные формы — влаголюбивые, очень вы-
носливые к сухости воздуха и теплолюбивые; 2) формы приморского
увлажненного климата, требовательные к высокой влажности воздуха;
3) своеобразный месхетский экотип высокогорной ветвистой пшеницы,
обитающей на высоте 1200 м над ур. м.
По уровню открытого цветения, по данным В. И. Жукова (1969),
Т. turgidum в приморской низменности /Дагестана занимает промежу-
точное положение между другими видами секции Dicoccoides. В зави-
симости от условий выращивания доля хазмогамных цветков составля-
ет 56,6—96,7%. В менее увлажненных условиях количество от крытоцве-
тущих цветков увеличивается. Склонность к открытому цветению
Т. turgidum косвенно подтверждают случаи межвидовой спонтанной
его гибридизации, которые в условиях Дагестана наблюдал В. Ф. До-
рофеев (1968). Здесь же В. М. Берляид-Кожевников и др. (1975) выя-
вили у Т. turgidum образцы, являющиеся облигатными самоопылите-
лями, а также факультативными перекрестпиками и склонными к пере-
кресту.
На низкие хлебопекарные качества 7'. turgidum неоднократно указы-
вал Н. И. Вавилов. Опытами ВИРа выявлено, что у этого вида имеются
формы, интересные по биохимическому составу зерна. В среднем со-
держание белка в нем равно 12,5—17,0%, крахмала 58,0—66,2%, золы
1,6—2,2%. Озимые формы богаче белком, чем яровые. Лучшие показа-
тели по концентрации белка в зерне выявлены у образцов пиренейской
(к-5764) и западноевропейской (к-41930) групп (М. М. Якубцинер,
Н. Ф. Покровская, 1969).
Иммунитет. Н. И. Вавилов (1964) отнес Т. turgidum к числу видов,
относительно устойчивых к комплексу грибных болезней. М. М. Якуб-
цинер (1969) нашел, что этот вид полиморфен по отношению к бурой
ржавчине. К наиболее восприимчивым к ней он отнес образцы Афгани-
стана и Казахстана, а в составе южноевропейскпх сортов он отметил
преобладание устойчивых к большинству рас пли ко всему их комплек-
су. И. П. Шитова (1968) по наблюдениям над коллекцией в 1954—
1965 гг, (Дагестан) 83,2% сортообразцов отнесла к восприимчивым;
иммунные к бурой ржавчине отсутствовали. 0. Г. Григорьева (1975)
нашла, что расоспсцифическая устойчивость к патогену у 7'. turgidum
варьирует в широких пределах. В. М. Берлянд-Кожевников и др. (1973)
у отдельных образцов установили наличие групп генов Х|, Х2 и Х3,
80 ICun i ифимщич 1»><)ч Triliitiin /.
а также ichob I,г 3 n Lr 13, мнорые oi bci e н'.енны за ye i <>ii'iiino<-1 ь к ра-
сам 77, 151 и 202 бурой ржавчины.
J. Percival (1921) наблюдал поражение Т. turgidum желтой ржав-
чиной. По данным И. П. Шитовой (1968), в условиях Дагестана этот
вид пшеницы в целом сравнительно устойчив к названному заболева-
нию. М. М. Якубцинер (1969) не относит Т. turgidum к числу иммунных
к стеблевой ржавчине видов пшеницы. Он отметил, что восприимчивость
к пей характерна для образцов из Нагорного Карабаха, Ирана, Сирии,
Турции и Китая, а устойчивость — для образцов южносвропейского про-
исхождения.
G. М. Reed (1916) нашел, что Т. turgidum восприимчив к мучнистой
росе. В опыте Э. X. Сухаибсрдипой (1977) при искусственном зараже-
нии коллекции популяцией рас патогена из 5 географических районов
СССР показано, что в фазе проростков почти все образцы были вос-
приимчивыми, а на провокационном фоне в поле такое поражение уста-
новлено у 97,2% образцов. Иммунитет не обнаружен ею пи у одного
образца.
Т. turgidum, по М. М. Якубцпнсру (1969), устойчив к популяции рас
пыльной головни. В. II. Кривченко (1973) нашел, что большая часть
коллекции при искусственном заражении имеет высокий иммунологиче-
ский тип. Лишь у отдельных образцов отношение к иылыюй головне он
квалифицировал как практическую устойчивость или слабую восприим-
чивость. А. М. Ямалеев (1974), также при искусственном заражении,
выявил, что иммунитет (87,5% образцов) преобладает среди сортов
Западной Европы, а среди сортов Дагестана, Азербайджана и Ирана
их мало. Анализ данных по расоспецпфической устойчивости в его опы-
те показал, что Т. turgidum преимущественно поражается пыльной го-
ловней f. duri. При этом электрофоретические спектры глиадипа зерно-
вок у восприимчивых к патогену образцов имеют гетерогенную у-зоиу;
у устойчивых образцов эта зона обеднена.
При искусственном заражении коллекции выявлено, что 59,2% об-
разцов се иммунны к твердой головне; восприимчивость обнаружена
лишь у 3% образцов (В. И. Кривченко. 1973). П. Г. Чесноков (1936)
отнес Т. turgidum к числу видов, менее устойчивых к гессенской мухе,
чем твердая пшеница.
История. Древность культуры Т. turgidum подтверждается находка-
ми зерновок его в египетских пирамидах. Среди археологического ма-
териала неолитического периода из Швейцарии (III тысячелетие до
и. э.) обнаружены обугленные зерна, отнесенные к Т. turgidum (О. Heer,
1866). Сходство с этим же видом выявлено среди растительных остат-
ков пшеницы из Италии. По свидетельству В. Л. Мепабде (1964), па
территории Грузии в эпоху неолита сеяли по только древние примитив-
ные виды, по и Т. turgidum. На севере Испании, судя по доставленным
в Институт археологии АН СССР в I960 г. обугленным зернам пз раско-
пок, датируемых серединой! II тысячелетня до н. э., в тот период встре-
чался Т. turgidum.
Имеются недостаточно убедительные сведения А. Декандоля, что
в египетских саркофагах найдены зерновки ветвистоколосой 7’. turgidum
(J. Percival, 1921). Ветвистоколосые формы Т. turgidum были известны
еще около 2000 лет назад Плинию Старшему. Первый рисунок Т. turgi-
dum под названием «Welsch» приводит L. Buchs (1512), По данным
J. Percival, еще в XVI—XVIII вв. в Англии возделывали Т. turgidum
под названием Cone, Rivet. В Ферганской области Узбекистана (рас-
копки В. Ф. Гайдукевич в 1944 г.) Т. turgidum возделывался в X - XII вв.
(М. М. Якубцинер, 1956). Па основании археологического исследования
материала из раскопок А. Я. Якубовского (Пеиджикепт Ленинабадской
области) был сделай вывод, что Т. turgidum встречался в Таджикиста-
не еще в VI—VII вв.
11(><)р<к) /. Trilictiin 81
Ветвистоколосые формы Т. turgidum встречаются i.чанным образом
в Закавказье, где они являются аборигенными вопреки предположению
j\. П. Овсрипа (1874) об анатолийском происхождении грузинской се-
миколоски. В Среднюю Азию такие формы Т. turgidum проникли из
Закавказья, а также были занесены из Китая ('Г. Бурнашев, 1824).
В Западную Сибирь Т. turgidum занесен преимущественно из Ка-
захстана— из Семипалатинска и Кокчетава (по сведениям Н. Л. Скало-
зубова, 1899). Не исключено захождение его из Восточной Сибири.
В европейскую часть России Т. turgidum проник в XIX в. из За-
кавказья или из Средней Азии. В отдельных случаях эта пшеница зано-
силась сюда из Сибири. Так, в начале 30-х годов в Удмуртию семена
ветвистоколосой пшеницы завезены из Восточной Сибири, в Оренбург —
тогда же с Алтая.
Географическое распространение. Ареал Т. turgidum очень широкий.
Как подчеркивает Н. И. Вавилов (1964), он охватывает главным обра-
зом страны Балканского полуострова и Южной Европы, заходя в За-
падную Европу. В Англии в конце 40-х годов вид занимал до 5% всей
площади под пшеницей. В Ираке, Иране, Афганистане (в районе Ге-
рата), Малой Азии, Восточном Китае Т. turgidum возделывается мес-
тами. Ошибочны указания в литературе на отсутствие этого вида в Аф-
рике. В коллекции ВИРа имеется образец Т. turgidum из Алжира. Этот
вид пшеницы довольно часто встречается в Закавказье в основном
в виде примеси к мягкой или твердой пшенице. В недавнем прошлом
возделывался в Дагестане, Казахстане, Узбекистане и Таджикистане.
Завезенный из стран Старого Света 7'. turgidum. спорадически возделы-
вается в США (сорта Titanik, Alaska) и в Австралии.
Происхождение. Давно было замечено, что Т. turgidum и твердая
пшеница очень близки между собой (М. W. Beijcrink, 1884; F. Kornicke,
1908; A. Schulz, 1911; И. И. Вавилов, 1926, 1935 и др.). Специалисты,
с одной стороны, объединяют их в один вид (Е. Н. Снискан, 1969;
J. Mac Key, 1967 и др.), с другой стороны, усматривают между ними чет-
кие морфологические различия и дают Т. turgidum статус вида
(К- А. Фляксбергер, 1935; М. М. Якубцинер, 1957 и др.).
Несомненные различия имеются в полигонной системе обеспечения
легкого обмолота у Т. turgidum и пшеницы твердой. При их скрещива-
нии с Т. compactum в F2 настоящая спельта выщеилялась лишь в ком-
бинациях с участием первого вида (Э. Ф. Мигушова, пеопубликовано).
Возможно, что полпгенпая система легкого обмолота у пшеницы твер-
дой такова, что она подавляла трудный обмолот у растений F2, имев-
ших внешний облик спельты.
Многие исследователи пришли к заключению о наличии у Т. turgidum
и отсутствии у Т. durum ряда признаков, характерных для пшеницы мяг-
кой (К- А. Фляксбергер, 1911; Д. К- Ларионов, 1914; II. И. Вавилов,
О. В. Якушкина, 1925; А. А. Ерицян, 1928 и др.). Еще Ph. Vilmorin
(1906) и Ph. Н. Stoll (1907) отметили, что растения Fi, полученные при
скрещивании Т. turgidum с Т. aestivum, являются промежуточными, но
по морфологическим признакам больше похожи па пшеницу мягкую.
Б. А. Вакар (1932) нашел, что у растений Fj такой комбинации мейоз
в МКП проходит с меньшими нарушениями, чем при скрещивании
с другими гекс'аплоидпыми видами. Н. И. Вавилов (1964) подчеркивал,
что по мучнистости зерна и консистенции листа Т. turgidum занимает
промежуточное место между твердой и мягкой пшеницей. Известно, что
Т. turgidum в культуре сопутствует твердой пшенице, часто в виде при-
меси. Этот факт является веским аргументом в пользу гипотезы о про-
исхождении первого вида от второго. Возможно, что '/’. turgidum имеет
нолифилетическое происхождение.
Систематика. Некоторые генетики (W. М. Bowden, 1959; A. Love,
1). Love, 1961; J. Mac Key, 1966 и др.) включают в состав Т. turgidum L.
6 762
82
Классификация рода Triticum L.
ряд пленчатых и голозерных тетраплоидпых лидов пшеницы. Более
того, согласно их классификации древние дикие виды Т. dicoccoides и
Т. araraticum, а также культурные пленчатые - Т. dicoccum, Т. kara-
myschevii— рассматриваются как внутривидовые таксоны Т. turgi-
dum L. В контрасте с учением II. II. Вавилова (1931), изложенным в
труде «Линиеевский вид как система», где подчеркивается принцип
историзма в классификации растений, находится и классификация, пред-
ложенная A. Thellung (1918), R. Morris и Е. R. Sears (1967) и др.
Классификация G. Mandy (1970), в которой Т. turgidum в ранге conva-
rietas перенесен в состав пленчатого вида Т. dicoccum, неприемлема.
Н. И. Вавилов (1931) расширил границы Т. turgidum, включив в его
состав инфлятные формы пшеницы эфиопской. К настоящему времени
накопились факты, свидетельствующие о целесообразности выделения
всех тетраплоидпых пшениц Эфиопии в самостоятельный вид Т. aethi-
opicum. Кроме того, Н. И. Вавилов (1964) включил в состав Т. turgidum
новый подвид subsp. sinicuin Vav. Более, тщательный анализ показал,
что этот мономорфный уникальный подвид следует отнести к Т. durum..
Е. Н. Синская (1955) считала Т. turgidum нс самостоятельным видом,
а составной частью Т. durum. В. Ф. Дорофеев (1969) рассматривает
Т. turgidum в качестве подвида Т. durum, объясняя это их генетической
близостью и наличием промежуточных форм. Однако в целом оба вида
эколого-морфологически довольно хорошо дифференцированы.
Изменчивость признаков Т. turgidum гораздо уже, чем у пшеницы
твердой, но все же разнообразие его форм требует разделения па 2 груп-
пы разновидностей: convar. turgidum и convar. compositum (L. f.)
A. Filat.
Ключ для определения групп разновидностей Т. turgidum L.
1. Колосья простые, не ветвистые............convar. turgidum.
+ Колосья ветвистые или с двойными колосками, т. е. на члениках
стержня сидит не по одному, а по два колоска......................
................................convar. compositum (L. f.) A. Filat.
1. Convar. turgidum — группа разновидностей с простым (певетви-
стым) колосом.
Convar. turgidum — Т. vulgare var.-gr. II turgidum Alef. 1866,
Landwirtsch. Fl.: 325.— T. turgidum mediterraneum Flaksb. 1922, Опред.
паст, хлебов: 62, quoad I gr.
В целом для группы характерны мощное развитие растений, круп-
ный колос, толстая соломина, пе всегда достаточно прочная в связи
с высокой продуктивностью колоса. Колосковые чешуи широкие, взду-
тые. Ости длинные, параллельные. Зерновка округлая, мучнистая, редко
стекловидная. Разновидности этой группы представлены в большинстве
полуозимыми, озимыми и поздними яровыми формами.
Геогр. распр. Средиземноморье (Северная Африка, Малая Азия,
Балканы, страны Южной Европы) и Англия. Спорадически возделыва-
ется в Ираке, Сирии, Иране, Афганистане, а также в Китае. В СССР
эта пшеница распространена в Закавказье, отчасти в предгорном Да-
гестане в основном в виде примеси к твердой и мягкой пшенице. В Сред-
ней Азии встречается на поливных землях.
Convar. turgidum наряду с convar. compositum включает основную
массу разновидностей по Ф. Кёрпике, который разделил виды на разно-
видности по одному или нескольким морфологическим признакам, без
учета экологической характеристики. К. А. Фляксбергер (1935, 1939)
считал необходимым переход на генетико-экологическую систему, но
не смог его осуществить. Разновидности по Ф. Кёрпике чрезвычайно
удобны для определения и инвентаризации огромного внутривидового
разнообразия пшеницы. Однако деление экотипов или эколого-геогра-
фических групп на формальные разновидности увеличивает число по-
IIikI/hhI !. Tri til uni
83
слсдпих в многократном размере и делает сип ему 1ромоздкой и не-
удобной. Е. II. Спискам (1961) права, называя такие разновидности
«ненужным пережитком», по, к сожалению, внутривидовая система
у пшеницы еще недостаточно разработана, чтобы отказаться от разно-
видностей в понимании Ф. Кёрнике. В настоящем издании еще сохра-
няется тот ботанический «абсурд» (К- А. Фляксбергер, 1935, Культ, фл.
СССР, 1: 56), когда одна и та же разновидность отнесена к разным
эколого-географическим группам или экотипам. Таким образом, едини-
цы экологической классификации оказываются вспомогательными.
С точки зрения филогении и распределения фондов исходного материа-
ла для селекции более важно установление экогрупп или экотипов.
Широкий ареал возделывания Т. turgidum обусловил его значитель-
ное экологическое разнообразие, что нашло отражение в установлении
эколого-морфологических (К- А. Фляксбергер, 1935, 1939) или эколого-
географических (Н. И. Вавилов, 1964) групп, представители которых
по ряду морфологических признаков весьма сходны: различия между
этими группами можно выявить только в полевых условиях по ком-
плексу биологических свойств и с учетом районов их распространения.
Группы разновидностей convar. turgidum и convar. compositum (L.)
A. Filat, соответствуют следующему экологическому подразделению.
1. Средиземноморский экотип или южноевропейская экогруппа по
Н. И. Вавилову (Г. turgidum prol. meridionale Flaksb. 1935, Культ, фл.
СССР, 1:240, descr. ross.; Вавилов, 1964, Миров, ресурсы хлебп. злаков.
Пшеница: 53).
Растения мощные, высокорослые, богатооблиствепные. Колосья
крупные, обычно квадратные в поперечном сечении, более или менее
плоские. Большое разнообразие по плотности колоса. В. К. Кобелев
(1932) для Турции отмечает крайние варианты: плотпоколосые, с очень
коротким колосом (Т. turgidum turgidocompactum тип 4 marashii Kob.
1932, Хлебп. злаки Турции: 16. — Т. turgidum prol. hapolocompactum
Goekg. 1955, Ziraat vekal. 716: 104) и рыхлоколосые с длинным колосом
(T. turgidum mediterraneum тип 3 cristatum Kob. 1932, 1. c.: 16). Плотно-
колосые формы нередки также в Греции, Испании, Португалии.
Большая вариабельность наблюдается по высоте растений. Формы
из Италии отличаются высокорослостью. Наряду с озимыми имеются ти-
пичные яровые формы.
Ввиду значительной амплитуды количественных различий средизем-
номорских Т. turgidum II. И. Вавилов (1964) находит целесообразным
более дробное их деление на следующие подгруппы*: 1) западносреди-
земноморская группа — наиболее однообразная, представлена мощно-
развитыми растениями с толстой соломиной, хорошей кустистостью и
облпственностыо, в массе это формы озимые, а в Испании — яровые;
приурочена к странам Пиренейского полуострова, Италии, островам
Сардиния и Сицилия; 2) центрально-южноевропейская группа, к кото-
рой относится Т. turgidum из Югославии, характеризующаяся сравни-
тельной скороспелостью и некрупным колосом, озимым образом жизни
(за редким исключением); 3) восточносредиземпоморская группа — от-
личается особой пестротой состава и наибольшей амплитудой изменчи-
вости. К этой подгруппе относится Т. turgidum Малой Азии, Сирии,
Родоса, Греции, отчасти Югославии (Македония). Здесь особенно много
переходных форм от Т. turgidum к Т. durum.
2. Западноевропейский экотип (7'. turgidum prol. imglicum Flaksb.
1935, 1. с.: 242, descr. ross. - 7'. turgidum prol. ocridentute-europacum Vav.
1964, l.c.: 53, nom. illeg.).
В посмертной работе II. II. Вавилова (1IIG4) наблюдается некоторое necooiнет-
стние в подразделениях. Па с. 55, несомненно, имеется в виду не в челом средиземно-
морский подвид (subsp. mediterraneum Flaksb.), а только prol. meridionale Flaksb.
6*
84
Классификации рода Triticum L.
Растения мощные, хорошо облиственные, высокорослые. Колосья
крупные/уплотненные, обычно квадратные в поперечном сечении. Наи-
более позднеспелые в предела?; вида. Только озимые, приуроченные к
влажному прохладному климату. Многие формы с упругой соломиной.
Группа малополиморфная, представленная почти исключительно разно-
видностями jodurum и dinurtim.
Геогр. распр. Англия (типа сорта Rivet), ФРГ, Франция, Швей-
цария.
3. Закавказский экотип (Т. turgidum prol. transcaucasicum Flaksb.
1935, l.c.: 242, descr. ross.— T. turgidum prol. caspicum Vav. 19(54, l.c.:
53, nom. illeg.).
Растения характеризуются сравнительно широкой листовой пластин-
кой, высокой соломиной, крупным колосом, уступая в этом отношении
лишь Т. turgidum из Западной Европы. В общем экотип дагестанских
и азербайджанских Т. turgidum очень выдержанный — высокорослый,
среднеоблпственпый (листовые пластинки средней ширины — около
17 мм в верхних ярусах), соломина среднепрочная. Колос сравнительно
мало варьирует по плотности, занимая как бы промежуточное положе-
ние между рыхлым и плотным.
Разнообразие Т. turgidum в Азербайджане (включая Ленкорань) п
Дагестане исключительно велико. Здесь изредка встречаются ветвистые
формы. Особенность — сравнительно слабая зимостойкость (по сравне-
нию с западноевропейской группой). Средпеполиморфпая группа, при-
уроченная к поливным землям, расположенным не выше 600 м над ур. м.
Геогр. распр. Закавказье, Дагестан.
4. Восточный экотип (Г. turgidum subsp. mediterraneum prol. orien-
tate Flaksb. 1935, l.c.: 243, descr. ross. — T. turgidum subsp. mediterra-
neum prol. iranicutn Vav. 1964, I. c.: 54, nom. illeg.).
Растения крупные. Колосья крупные, квадратные в поперечном се-
чении. Чувствительны к недостатку почвенной влаги, но переносящие
воздушную сухость. Формы яровые и полуозимые, почти исключительно
представленные разновидностью turgidum.
Геогр. распр. Небольшие посевы в Прайс, в Афганистане
(в районе г. Герат), в Средней Азии (поливные земли), редко в Сибири
и Китае.
5. Тибетский экотип (Г. turgidum subsp. montanum Jakubz. 1957,
Бюл. ВИРа, 4: 54, nom. invalid.).
Растения 70—80 см высоты. Всходы зеленые, листовые пластинки
молодых растений покрыты нежными волосками. Стебли прямостоячие,
голые, узлы слабоопушенпые. Колос неветвиетый, более или менее ко-
роткий (6—8 см). Ости средней длины или почти отсутствуют. Колос-
ковые чешуи слабовздутые. Зерновки мелкие, мучнистые. Образ жи.зпп
яровой. За исключительно своеобразный ареал эта мономорфная пше-
ница была выделена М. М. Якубцппером (1957) в отдельный подвид.
В ВИРе изучалось несколько высокогорных эндемичных форм Ти-
бета. Одна из них получена из 'Чэнду (3100 м над ур.м.), другая — из
бассейна р. Нуцзян (Салуин).
Геогр. распр. Тибет. Крайний высотный предел Т. turgidum па
земном шаре 3700 м над ур. м. (Лхаса).
6. Крупнозерный экотип (Г. turgidum subsp. giganlosemineum Jakubz.
1901, Видов, сорт. раст. ресурсы, доклад-обобщен.: 33, пот.• invalid.).
Растения мощные, высокорослые (более 120 см длины); колосья
крупные, с 25—27 колосками, ости часто короткие; зерновка длинная
(8—9 мм и больше); исключительная крупность зерновок (масса
1000 зерен в благоприятных условиях достигает 70 г и выше) состав-
ляет главную специфику этой группы Т. turgidum. Экотип представлен
сортом Узбекистан.
Геогр. распр. Узбекистан.
Подрод !. Triticum
PJ34 овиднос ги
Ч с ш у п пс о п у in с н п ы е
1. Зерновки белые
+ — — — — — — — + — turgidum 1
— — — — — — — — ”Г inelanatheriini 9
— 4 — — — — — — -1- — nigroglumarum i 3
— 4 — — — — — — — + miscibile 1 1
— —. 4- — — — •—- — -1 — dreischinnum 1 5
— — 4 — — — — — — 4 sgeciosissimum ' l2
— — 4- — — — — — -- 4 + f erespeciosissimnm । /
— — ~— — 4 —• — — —- л lien ei ac 1 s
— — — — — + — — — t ubrokerrrrne . 9
2. Зерновки красные
4- — — — — — — — + — gentile 10
+ — — — — — — — — 4- nigrobarbuhim 11
— 4 — — — — — — — 4 striatum 12
— — 4 — — — — —. . + — speciosum 13
— — 4- — — — — — —. 4- martensii 14
— — — -h — — — — — + nigromartensii 15
— — — — — — — — 4- a I bof i; mid и in 1G
— — — — — + — — — 4- jumidum 17
Ч с ш у и о п у hi е н п ы с
1. Зерновки белые
L. 4- — 4- — — — — — 4- + 4 4 megalopolitaniim sedomonis pseudomirabile rubroalbum is 10 20 21
— — — + — — — — 4- — n igropseudo mirabile 22
— — —- — 4- — — — — 4- albojodurum 23
— — — — — — 4- — 4- —. griseomegalo- politanum 21
— —_ — — — — + — — 4- griseosedomonis 25
— — — — — — + 4- griscopseudomi rabile 26
— — — — — — — 4- — 4- griseorubroalbum 27
2. Зерновки красные
— — — — — — — + — buccale 2S
+ — —. .— — — — — 1 - pscudosiilomonis 20
— 4- __ — — — — — 4- — nigrobucctile 30
— -4- — — — — — — — 4- nigropseudosalo'noe.iy 31
— — — — — — —- + — dinurum
— — 4- — — — — 4- rubroatrum •
— — — 4 — — — — 4- — tristedinurum 31
— — — 4- — — —. — 4- frislerubroulrutn 35
— — — + — — — — 4- falsejodttrum 36
— — — — 4- — — — 4- jodurum 37
— — —. — — — 4- — — 4- abchasicum 3S
— — — — — — 4 4- .— i inereum 30
— — — — — — — 4 4- griseorubroatrum 10
86
Классификация рода Triticum L.
Колосья безостые
Чешуи опушенные (зерновки красные)
— I — I — I -|- I — I — I — I — I subjodurum I И
+ | — I ~• I — I — I — I — I — I muticospcciosum | 42
Колосья короткие, очень плотные (turgido-conipactiim)
Чешуи иеопушспиые (зерновки белые)
+
+
+
pseudomelan-
atherum
eompactoherrcrae
Ч е in у и о п у in е и н ы е (зерновки красные)
— | — ! + I — ' — — coinpactojodurum
43
44
15
* Остн слабошероховатые.
Var. abchasicuin Menabde, 1948, Пшеи. Груз.: 76, descr. ross. (38).
Геогр. распр. Грузия.
Var. albofumidum Ljapun. var. nov.— T. turgidum gr. nigriglumaceum
Flaksb. 1935, l.c.: 251. (16). descr. ross. — Л T. Ittrgidoo var. herrerae
Koern. caryopsi rubra differt.
Typus: Turcia, k-16066. Reproductio statione Daghestanica WIR, VI.
1972 (WIR).
Геогр. распр. Малая Азия, Югославия, Сардиния, редко в Азер-
байджане и Грузии.
Var. albojoduruin Flaksb. 1939, Опред. наст, хлебов; 126, descr. ross.
(23).
Геогр. распр. Азербайджан (Ленкоранский и Кахский районы).
Var. buccale (Alef.) Koern. in Koern. et Wern. 1885, Flandb. Gctrcideb,
1: 60. — T. vulgare buccale Alef. 1866, Landw. FL: 326. — T. turgidum gr.
buccale (Alef.) Flaksb. 1935, Культ, фл. СССР, 1: 253. (28).
Геогр. распр. Страны Средиземноморья, Англия, ГДР, ФРГ.
Var. cinereum Desv. 1833, Trav. Soc. d’Angers: 156; Фляксбергер,
1929, Tp. прикл. бот. ген. сел. 21,5: 472. (39).
Геогр. распр. Франция, Турция.
Var. compactoherrerae Flaksb. 1935, l.c.: 251, descr. ross. (44).
Геогр. распр. Сицилия и Родос; Турция.
Var. compactojodurum Jakubz. in Flaksb. 1935, l.c.: 259, descr. ross.
(45).
Г e о г p. p а с и p. Малая Азия.
Var. dinurum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 61.—
T. vulgare dinura Alef. 1866, l.c.; 326.— T. turgidum gr. diiiuruui (Alef.)
Flaksb. 1935, l.c.: 256. (32).
Геогр. распр. ГДР, ФРГ, Франция, Англия, Испания, Италия,
Швейцария, Болгария, Турция.
Var. dreischianuin Koern. in Koern. el Wern. 1885, l.c.: 60. — T. turgi-
dum gr. dreischianum (Koern.) Flaksb. 1935, 1. c.: 248. (5).
Геогр. распр. Средиземноморские страны, Малая Азия.
Подрод I. Triticum
87
Var. falsejoduruin Flaksb. 1929, I. c.: 480. (36).
Геогр. распр. Обнаружена К. Л. Фляксбергом в гербарии
в Бонне без указания места нахождения, а также М. Гекгелем в Турции.
Var. ferespeciosissimum Jakubz. 1932, Тр. прикл. бот. ген. сел. При-
лож. 53: 101. (7).
Геогр. распр. Сирия.
Var. fumidum Dekapr. 1924, Изв. Тифл. гос. политехи, ин-та. 1:4. (17).
Гео гр. р а с и р. Азербайджан, Грузия, Югославия, Италия (редко).
Var. gentile (Alef.) Koern. in Koern. el Wern. 1885, l.c.: 58. -T. vul-
gare gentile Alef. 1866, l.c.: 326. — T. turgidum gr. gentile (Alef.) Flaksb.
‘1935, l.c.: 246. (10).
Геогр. распр. ГДР, ФРГ, Испания, Португалия, Южная Фран-
ция, Италия, Сицилия, Греция, Турция, Азербайджан, Грузия.
Var. griseomegalopolitanum Gokg. 1955, Ziraat vekal. 716: 105, nom.
invalid. (24).
Геогр. распр. Турция.
Var. griseopseudomirabile Gokg. 1955, l.c.: 106, nom. invalid. (26).
Геогр. распр. Турция.
Var. griseorubroalbum Gokg. 1955, l.c.: 106, nom. invalid. (27).
Г e о г p. p а с п p. Турция.
Var. griseorubroatrum Gokg. 1955, l.c.: 106, nom. invalid. (40).
Геогр. распр. Турция.
Var. griseosalomonis Gokg. 1955, l.c.: 105, nom. invalid. (25).
Геогр. pa cup. Турция.
Var. herrerae Koern. in Koern. et Wern. 1885, l.c.: 60. — T. turgidum
var. izmiricum Ilaciz. 1932, Ziicht. 4,3: 76. nom. dub. — T. turgidum gr.
herrerae (Koern.) Flaksb. 1935, 1. c.: 251. (8).
Геогр. распр. Европейские средиземноморские страны, Малая
Азия, Закавказье (очень редко), Дагестан.
Var. jodurum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 61.—
T. vulgare jodura Alef. 1866, l.c.: 326. — T. turgidum gr. jodurum (Alef.)
Flaksb. 1935, l.c.: 258. (37).
Геогр. распр. Средиземноморские страны, Англия, ГДР, ФРГ,
Франция, США, Южная Африка, Грузия, Латвия, Эстония.
Var. martensii Koern. in Koern. et Wern. 1885, l.c.: 60. — T. turgidum
gr. martensii (Koern.) Flaksb. 1935, l.c.: 250. (14).
Г e о г p. p а с п p. Южная Европа, Турция, Закавказье.
Var. megalopolitanum Koern. in Koern. et Wern. 1885, l.c.: 60.—
T. turgidum gr. megalopolitanum (Koern.) Flaksb. 1935, l.c.: 252. (18).
Геогр. распр. Испания, Португалия, Сицилия, Крит, Греция,
Болгария, Турция, Египет, Чили.
Var. nielanatherum Koern. in Koern. et Wern. 1885, Handb. Gctreideb.
2: 396. — T. turgidum var. nemausense Wittm. ex Koern. et Wern. 1885,
1. c.: 59. (2).
Геогр. 'распр. Средиземноморские страны, Сицилия, Родос, Лес-
бос; Индия.
Var. miscibile Haciz. 1932, Ziicht. 4,3: 76. (4).
Геогр. распр. Малая Азия, Закавказье (редко).
Var. muticospeciosum Jakubz. in Flaksb. 1935, l.c.: 250, descr. ross.
(42).
Геогр. распр. Азербайджан (редко).
Var. nigrobarbatum Desv. 1833, Trav. Soc. d’Angers: 156. (11).
Геогр. распр. Средиземноморские страны, Азербайджан, Грузия
(редко).
Var. nigrobuccale Jakubz. in Flaksb. 1935, l.c.: 254, descr. ross. (30).
Геогр. распр. Испания (редко).
Var. nigrogluinarum Ilaciz. 1932, l.c.: 76. (3).
Г e о гр. p а с и p. Малая Азия, Азербайджан.
8В Классификация рода Triticum Л.
Var. nigromarleiisii Jakubz. in llaksb. 1935, l.c.: 251, descr. ross. (15).
Геогр. распр. Азербайджан.
Var. nigropseudomirabile Flaksb. 1935, l.c.: 255, descr. ross. (22).
Геогр. распр. Испания.
Var. nigropseudosalomonis Flaksb. 1935, l.c,: 254, descr. ross. (31).
Геогр. распр. Испания.
Var. pseudomelanatherum Schreib. 1934, Tp. прикл. бот. ген. сел. 5,2:
102. (43).
Геогр. распр. Сицилия.
Var. pseudomirabile Perciv. 1921, Wheat pl.: 25(5. (20).
Геогр. распр. Испания, Португалия, Югославия.
Var. pseudosaloinonis Papad. 1929, Formes Grecques de Ble.; 36.—
T. turgidum var. caryopsirubrum Ilaciz. 1932, l.c.: 76. — T. turgidum gr.
pseudosalomonis (Papad.) Flaksb. 1935, l.c.: 254. (29).
Геогр. p а с п p. Присредпземноморские европейские страны, Крит,
Сардиния, Малая Азия, Азербайджан, Грузия (редко).
Var. rubroalbum Flaksb. 1916, Тр. бюро прикл. бот. 9,2: 67. — Т. tur-
gidum var. rubroatrum auct. non Koern.: Papad. 1929, l.c.: 11. (21).
Геогр. распр. Испания, Италия, Греция, Родос, Югославия, Бол-
гария, Малая Азия, Азербайджан.
V ar. rubroatrum Koern. in Koern. et Wern. 1885, 1. c.: 61. — T. turgidum
pseudorubroairum Papad. 1929, l.c.: 41. — T. turgidum gr. rubroatrum
(Koern.) Flaksb. 1935, l.c.: 257. (33).
Геогр. распр. Испания, Португалия, Италия, Сицилия, Греция,
Крит, Малая Азия, Азербайджан, Грузия.
Var. rubroherrerae Jakubz. in Flaksb. 1935. 1. c.: 252, descr. ross. (9).
Геогр. распр. Азербайджан, Южная Африка.
V ar. salomonis Koern. in Koern. et Wern. 1885, l.c.: 61. — T. turgidum
gr. salomonis (Koern.) Flaksb. 1935, l.c.: 253. (19).
Геогр. распр. Присредпземноморские европейские страны, Гре-
ция, Турция, Сирия, Израиль, Пакистан, Иран, Азербайджан, Грузия,
Узбекистан.
V ar. speciosissimum Koern. in Koern. et Wern. 1885, l.c.: 60.—
T. turgidum gr. speciosissimum (Koern.) Flaksb. 1935, l.c.: 249. (6).
Геогр. распр. Средиземноморские страны, Закавказье, Дагестан.
V ar. speciosum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: (iO.—
T. vulgare speciosum Alef. 1866, l.c.: 326. — T. turgidum gr. speciosum
(Koern.) Flaksb. 1935,1. c.: 250. (13).
Геогр. распр. Присредпземноморские страны, Закавказье.
V ar. striatum Kob. 1932, Хлебн. злаки Турции: 17; Фляксб. 1935, l.c.:
248. (12).
Геогр. распр. Малая Азия, Азербайджан.
Var. subjodurum Koern. 1908, Arch. Biontol. 2: 401; Фляксб. 1935,
l.c.: 259. (41).
Геогр. распр. Получена путем искусственной гибридизации.
Var. tristedinurum Flaksb. 1935, 1. с.: 257, descr. ross. (34).
Геогр. распр. Редко в Испании.
Var. tristerubroatrurn Gokg. 1955, 1. с.: 106, nom. invalid. (35).
Геогр. распр. Турция.
Var. turgidum. — Т. turgidum var. lusitanicum Koern. in Koern. ct
Wern. 1885, Lc.; 59. — T. turgidum gr. lusitanicum (Koern.) Flaksb. 1935,
l.c.; 245. (1).
Геогр. распр. Средиземноморские страны, Пран, Афганистан,
Китай, Сибирь, Закавказье.
2. Convar. compositum — группа разновидностей с ветвистым ко-
лосом.
Convar. compositum (L. f.) A. Filat, comb. nov. — T. compositum L. f.
1774, Syst. Pl. ed. 13. 118. — T. linnaeanum Й spica ramosa Lagasca, 1816,
Надрод I. Triticum 89
Nov. gen. spec. pi.: 6 7’. turgidum |> compositum Doell, 1857, Fl.
Grossh. Baden, J: 124. — T. vulgare var-gr. 3 compositum Alef. 1866,
Landwirtsch. Fl.: 326. - T. turgidum mediterraneum Flaksb. 1922, Опред.
наст, хлебов: 62, quoad II gr.
Ветвистоколосая группа разновидностей сходна с предыдущей ио
многим признакам. Этой группе также присущи мощное развитие расте-
ний, хорошая облиственпость, толстая соломина. Колос крупный, ветви-
стый или с двойными колосками (т. с. на члениках стержня сидит не
по одному, а по два колоска). Зерновка мельче, чем у convar. turgidum.
Т и и: Т. turgidum var. compositum Doell.
Геогр. распр. Ареал convar. compositum совпадает с ареалом
основной группы разновидностей — convar. turgidum.
Экологически convar. compositum дифференцируется сходным обра-
зом с convar. turgidum.
Ветвистоколосая пшеница под названием ramosum и centigranium
упоминается еще в трудах Плиния (23—79 гг. и.э.).
Ветвистоколосость известна у ряда других видов пшеницы, однако
у Т. turgidum ее образование было особенно интенсивным, проявляю-
щимся в многообразии форм. В 1774 г. К. Линией (сын) выделил вет-
вистоколосую пшеницу в особый вид Т. compositum. В дальнейшем си-
стематики рассматривали ее как особую группу вида Т. turgidum или
как его ветвистоколоеые разновидности.
Против выделения ветвистых Т. turgidum в особый вид говорят сле-
дующие доводы: I) между ветвистыми и обычными Т. turgidum пет
резкой грани — при оптимальных условиях возделывания неветвистые
формы нередко приобретают ветвистость колоса и наоборот; 2) кроме
ветвистости, не существует других резких отличий этой группы от обыч-
ных форм Т. turgidum-, 3) ареалы обеих групп почти совпадают.
Экологическая характеристика convar. compositum в общем соответ-
ствует таковой convar. turgidum. Однако среди экотипов тибетского и
крупнозерного отсутствуют ветвистоколосые формы.
Var. alibekliense Thum. 1934, Тр. прикл. бот. ген. сел. 5,2: 269. (3).
Геогр. распр. Армения.
Var. caucasicum Jakubz. 1949, Тр. прикл. бот. ген. сел. 28,3: 49, descr.
ross. (26).
Геогр. распр. Азербайджан (редко).
Var. centigranium Koern. in Koern. et Wern. 1885, Handb. Gctrcideb.
1: 63. (20).
Ф. Кёрнике указывает на гибридное происхождение описанной им
разновидности.
Геогр. распр. Редко в ГДР, ФРГ, Испании, Азербайджане.
Var. cervinum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 63.—
T. vulgare cervinum Alef. 1866, Landwirtsch. FL: 327. (12).
Геогр. распр. Испания (редко).
Var. coeleste (Alef.) Koern. in Koern. et Wern. 1885, Lc.: 64.—
T. vulgare coeleste Alef. 1866, 1. c.: 327. (24).
Геогр. распр. He выяснено, предположительно страны Европы.
Известна по ботаническим описаниям (J. Metzger, 1824; N. С. Scringe,
1842). ' ,
Var. columbinum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 63.—
T. vulgare columbinum Alef. 1866, l.c.; 327. — T. turgidum var. diiplis
Koern. 1908, Archiv Bionl. Berlin, 2: 402. — T. turgidum gr. columbinum
(Alef.) Flaksb. 1935, Культ, фл. СССР, 1: 247. (6).
Геогр. распр. Предположительно в странах Европы, извеснш
ио ботаническим описаниям (J. Metzger, 1824; N. С. Scringe, 1842).
Var. compositum Doell, 1857, FL Grossh. Baden, 1: 124. — 7'. vulgare
linnaeanum Alef. 1866, l.c.: 327.--T. turgidum var. linnaeanum (Alef.)
Koern.. in Koern. et Wern. 1885, 1. c.: 63. — T. turgidum var. dubium Koern.
90
Классификация роди Trillium I..
Определение разновидное гей convar. compositum (I. ) Л. I'ilat.
Чешуи неопушенные
Зерновки белые
+ — — — — — — — 4- — ramosoltisilaniciim 1
-4- — — — — — — — nachitsclicvanicuin 2
— + —• —. — — — ——. — + alibekliense 3
— — + — — — — — ч- — pseiidocervinu m 4
— — + — — — — — — + plinianum 5
3 с р II о в к и к » а с п ы е
— — — — — — — — coltimbiiitim 6
— — — — 4- — — — — + ctibinum 7
+ — — — — — — — — + pavoninum 8
— + — — —- — — — 4- — ramosostriatum 9
+ — — — — — — — — + levipnvotiintim * 10
— — •— — •—• — — Ч- — leviramosostriatum * 11
— — -1- — — — — — + — cervinum 12
— — + — — — — + schemachinicum 13
Чешуи опушенные
Зерновки белые
+ + ramosomegalopo- liianum ramososalomouis 11 15
+ —. mirabile 16
— + falseniirabile 17
—. + lencoranicuin 18
— + gigant cum 19
Колосья
безостые (чешуи acoiiyincniibie, зерновки белые)
| 26
— 1 + 1 — i — I — ! — I — | — ! + ' caucasicum
* Ости гладкие.
in Koern. et Wern. 1885, l.c.: 62. — T. turgidum var. subdubium Koern.
1908, l.c.: 40?. (22).
Геогр. распр. Испания, Швейцария, Франция, Англия, США,
редко в Азербайджане.
Var. cubinutn Dorof. 1968, Бюл. ВИРа, 13: 74. (7).
Геогр. распр. Азербайджан.
Var. falseniirabile Flaksb. ex Jakubz. 1932, 'Гр. прикл. бог. ген. сел.
Ирилож. 53: 144. — Т. turgidum var. pseudomirabile auct. non Perciv.:
Flaksb. 1929, Tp. прикл. бот. ген. сел. 21,5: 460. — Т. turgidum var.
eandiense Flaksb. et Schreib. in Sclireib. 1934, Tp. прикл. бот. ген. сел.
5,2: 181. (17).
Подрод I. Triticum
91
Геогр. p а с п p. Страны Средиземноморья, Сирия, редко в Азер-
байджане.
Var. giganteum Jakubz. in Dorof. 1972, Tp. прикл. бот. геи. сел. 47,1:
73, descr. ross. (19).
Г с о г р. р а с п р. Азербайджан (редко).
Var. lencoranicum Schreib. in Flaksb. 1935, Культ, фл. СССР, 1:258,
descr. ross. (18).
Геогр. распр. Азербайджан (Ленкоранский район).
Var. levipavoninuin Dorof. 1970, Тр. прикл. бот. ген. сел. 43,1:
310. (10).
Обнаружена в посеве, представленном сложной популяцией из раз-
новидностей Т. durum, Т. turgidum, Т. aestivum, Т. compactum.
Геогр. р а с пр. Азербайджан (Кубинский район).
Var. leviramosostriatum Dorof. 1970, 1. с.: 309. (11).
Обнаружена в том же посеве, что и var. levipavoninuin.
Геогр. распр. Азербайджан (Кубинский район).
Var. mirabile Koern. in Koern. et Wern. 1885, l.c.: 03. — T. turgidum
var. subdubium Koern. 1908, l.c.: 402. (10).
Геогр. распр. Страны Средиземноморья и Западной Сиропы
(редко), в гХ.зербайджапс (редко).
Var. modigenituin Koern. 1908, l.c.: 403. (25).
Геогр. распр. Известна из Ботанического сада в Поипельсдорфе.
Var. nachitschevanicum Kulcsch. 1927, Тр. прикл. бот. ген. сел. 17,4:
163. — Т. turgidum caspicum Berg, 1929, Тр. Всесоюзи. съезда ген. сел.
сем. 3: 384. (2).
Геогр. распр. Малая Азия, Израиль, Дагестан, Азербайджан,
Армения, Грузия (редко).
Var. pavoninuni (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 63.—
T. vulgare pavoninum Alef. 1866, 1. c.: 327. (8).
Геогр. распр. Предположительно в странах Европы, известна
по ботаническим описаниям (J. Metzger, 1824; F. G. С. Alefcld, 1866),
а также очень редкой Азербайджане, Армении, Грузии.
Var. plinianuni Koern. in Koern. et Wern. 1885, 1. c.: 63. (5).
Ф. Кёрпике была известна лишь ио описанию N. С. Scringe (1842).
Геогр. распр. Греция, Сицилия, Малая Азия, Закавказье, Ка-
захстан.
Var. pseudocentigranium Flaksb. 1929, 1. с.: 480. (21).
Геогр. распр. Италия, Португалия, Азербайджан.
Var. pseudocervinum Koern. in Koern. et Wern. 1885, l.c.: 63. (4).
Геогр. распр. Редко в Испании, Португалии и других южно-
европейских странах, в Северной и Южной Африке, Малой Азии, также
редко в Азербайджане.
Var. pseudolinnaeanum Flaksb. 1929, 1. с.: 480. (23).
Геогр. распр. Испания, Греция, очень редко в Азербайджане.
Var. ramosolusitanicum Flaksb. 1935, Культ, фл. СССР, 1: 245, descr.
ross. (1).
Выделена по материалам заграничных гербариев (Австрия, Фран-
ция).
Геогр. распр. Казахстан, Азербайджан (редко).
Var. ramosomegalopolitanuin Perciv. 1926, Jotirn. BoE: 209. (14).
Геогр. распр. Описана по образцу Т. turgidum из Египта, хотя
автор не считает ее эндемичной для этой страны (J. Percival, 1926).
Var. ramososaloinonis Schreib. in Flaksb. 1935, Культ, фл. СССР, 1:
253, descr. ross. (15).
Геогр. pa сир. Крит.
Var. rainosostriatum Dorof. 1972, Tp. прикл. бот. геи. сел. 47,1: 72,
nom. invalid. (9).
Геогр. распр. Азербайджан.
92
Классификации /тда Triticum
Var. scheinachiniciim Dckapr. 1924, Нан. Гп<|>.1. юс. Политехи. hip ! ;i ,
1: 5. (13).
Геогр. p а с п p. Азербайджан (редко).
T. JAKUBZINERI UDACZ. ET SCHACHM.— ПШЕНИЦА ЯКУБЦИНЕРА
T. jakubzineri (Udacz. et Schachm.) Udacz. et Schachm. 1976, Tp.
прикл. бот. ген. сел. 56,2: 147; они же, 1977, Вести, с.-х. науки, 2: 43.—
Т. turgidum subsp. jakubzineri Udacz. et Schachm. 1972, Бюл. ВИРа,
23: 4.
В 1970 г. в процессе изучения мировой коллекции пшениц на Сред-
неазиатской опытной станции ВИРа (влизи г. Ташкента) Р. А. Удачи-
ным и И. Ш. Шахмедовым среди растений полуознмого образца Т. tur-
gidum var. turgidum (к-11597), привезенного еще в 1924 г. из Афгани-
стана И. И. Вавиловым, было найдено одно с ветвлением колоса по
тину Т. vavilovii Jakubz. Последующее изучение потомства этого рас-
тения показало константность этого типа ветвления колоса. По опреде-
лению Г. С. Ширяевой, 2п — 28.
Вавиловпопдпую форму Т. turgidum Р. А. Удачин и 11. 111. Шахме-
дов (1972) описали вначале как Г. turgidum subsp. jakubzineri Udacz.
et Schachm. Однако последующее изучение выявило присутствие у нее
совершенно уникального, неизвестного прежде в мировой ботанической
литературе для рода Triticum признака — наличие, в каждом колоске
четырех колосковых чешуй (по 2 справа и слева). Следует отмстить, что
подобный признак не установлен пока во всей трибе Triticeae Dum.
семейства Poaceae Barnh. (И. И. Цвелев, 1972, 1976) и только у ячменя
известна форма с тремя колосковыми чешуями, тоже из Афганистана
(И. И. Вавилов, Д. Д. Букивич, 1929; К. В. Иванова, 1937). Следует
также упомянуть данные Ch. Hochstetter (1848) о том, что он наблюдал
у одной певствистоколосой формы Т. turgidum из Аравии колос, у ко-
торого верхушечный колосок был окружен четырьмя колосковыми
чешуями.
В семействе злаков наличие четырех колосковых чешуй в каждом
колоске характерно для трибы Phleeae Dum., куда входят зубровка
(Hierocftloe R. Вг.), пахучеколосник (Anthoxanthum L.), канареечник
(Phalaris L.) и двукисточпнк (Phalaroides N. М. Wolf). Особенно резко
это выражено у зубровки альпийской [Н. alpina (Sw.) Roem. et Schult.].
Учитывая уникальность формы, описанной прежде как Т. turgidum
subsp. jakubzineri, Р. А. Удачин и И. III. Шахмедов (1977) выделили
се как самостоятельный вид Т. jakubzineri. Таким образом, в семействе
злаков выявлен новый пример действия закона гомологических рядов
в наследственной изменчивости И. И. Вавилова па уровне, триб. Т. jakub-
zineri имеет одинаковые с Т. turgidum (восточный экотип) признаки и
свойства, отличаясь лишь ветвлением колоса по типу Т. vavilovii и на-
личием в каждом колоске четырех колосковых чешуй. Образ жизни
полуозпмый.
Новый вид пшеницы голозерный, что опровергает вывод Е. Ausonius
и др. (1967) о том, что вавнловиондный тип ветвления колоса у пшени-
цы может появиться только в сочетании со спельтондпостыо. Эти авто-
ры утверждают, что гоп, контролирующий ветвление колоса по типу
Т. vavilovii, заключен в длинном плече хромосомы 5А, где также нахо-
дится и ген Q, подавляющий этот признак.
Ныне известна лишь одна разновидность, морфологически сходная
с 7'. turgidum var. turgidum.
Подрод I. Tiiticiini
93
T. DURUM DESF. - ПШЕНИЦА ТВЕРДАЯ
T. durum Desf. 1798, Fl. Atlant. 1: 111. — T. alatum Peterm. 1819,
Deutschl. Flora, 27,1: (547. — T. vulgar? var.-gr. durum Alef. 18(56, Landw.
Flora: 324. — T. sativum tenax 6 sativum durum Hack. 1887, Nat. Pflan-
zenfam. 2: 85. — T. sativum tenax В. II durum Aschers. et Graebn. 1901,
Synops. 2: 692. — T. turgidum subsp. semeutivum Rasse durum (Desf.)
Thell. 1918, Naturw. Wochenschr. (Jena) 17,33: 470. — T. aestivum subsp.
durum Dornin, 1935, Prcslia, 13—15: 43. — T. turgidum gr. durum (Desf.)
Bowden, 1959, Canacl. Journ. Bot. 37: 670. — T. turgidum subsp. durum
A. et D. Love, 1961. Bot. Not. (Lund) 114, 1: 49. — T. turgidum subsp.
turgidum convar. durum (Desf.) Mac Key, 1966, Flereditas, Suppl. 2:
268.
Общая характеристика. Всходы сине-зеленые, темно-зеленые, жел-
товато-зеленые, у некоторых форм фиолетовые, неопушенные, шерохо-
ватые или (редко) бархатистоопушенные. Куст прямостоячий. Стебель
темно-зеленый. Соломина выполненная пли полая высотой от 40 до
160 см. Кустистость большей частью 1.2—3 стебля па одно растение.
Листья сиповато-зеленые, темпо-зеленые, же.тговаro-зеленые, голые
(гладкие пли шероховатые) пли опушенные (бархатистые па ощупь)..
Длина листовой пластинки 16—35 см, ширина 0,7—1,1 см. Влагалище
обычно голое, шероховатое; ушки и язычок с ресничками или без них
(встречаются безлпгулыгые формы). Колосья простые или, реже, вет-
вистые, по форме от удлиненных тонких до коротких толстых (длина
колоса 5—13 см); боковая сторона колоса шире или равна лицевой
(колосья плоские или квадратные в поперечном сечении). Ширина бо-
ковой (двурядной) стороны колоса колеблется от 1 до 2 см. Колосья
от рыхлых до плотных (D = 16—50, большей частью 25—35). Стер-
жень колоса обычно прочный. Колоски по форме от ромбовидных до
округлых, 1,0—2 см ширины, многоцветковые, с 2—5 (реже до 6) зер-
новками. Число колосков в колосе 12—30. Членики колосового стержня
по краям опушенные пли почти голые со слабовыражснными бородками
у основания колосков. Колосковые чешуи более пли менее грубые, от
коротких и широких до длинных (8—15 мм длины при ширине до 4 мм),
голые или опушенные (степень опушенпостп может быть различной),
белые (соломенно-желтые), красные или черные па желтом или крас-
ном фоне, а также (редко) серо-дымчатые на белом или красном фоне.
У некоторых форм край чешуи черный. Киль большей частью хорошо
выражен, часто широкий, доходящий до основания чешуи, где морщи-
нистость и вдавленпость отсутствуют; в верхней части киль переходит
в широкий, четко выраженный обычно туповатый зубец. Наружная
(нижняя) цветковая чешуя менее выпуклая, чем у Т. turgidum, без ки-
ля, от короткой до удлиненной, переходит в ость постепенно. Ости от
коротких до очень длинных (7—23 см), шероховатые, слабошерохова-
тые или гладкие на ощупь, изредка спонтанно обламывающиеся.
Безостые формы встречаются редко. Их остевидпые заострения обыч-
но окрашены в цвет колосковой чешуи, изредка они черные. У основа-
ния ости с каждой ее стороны имеются мягкие слабовыражепные 2 зуб-
ца. Внутренняя (верхняя) цветковая чешуя двукилевая. При созревании
она не расщепляется вдоль на 2 части. Зерновка от короткой округлой
до длинной узкой (длина 6—12 мм, ширина 3-6 мм, толщина 3—4 мм),
большей частью стекловидная, в поперечном срезе у типичных форм
треугольная с округленными углами или округло-треугольная, со слабо-
выраженным хохолком (в отличие от мягкой пшеницы), с четкой про-
дольной бороздкой. Зерновки с брюшной стороны часто почти плоские,
здесь может быть поперечная вдавленпость, иногда придающая зернов-
ке серповидную форму. Зерновка заключена в чешуях неплотно (пше-
ница голозерная).
9-1 Классификация рода Tri/icuni I..
Веге га циоп и ы й период колебле i см <л 90 до 130 дней; бывают более
ранние формы. Обычно пшеницы твердые не осыпаются или слабо осы-
паются. В фазе колошения растения с восковым налетом па колосе,
стебле и листьях. Встречаются формы без воскового налета (epruinosa).
В отличие от Т. aelhiopicum пшеница твердая (М. С. Яковлев,
Е. И. Николаенко, 1936) представлена в основном (95,4% растений)
формами, характеризующимися двумя проводящими пучками в колеоп-
тиле. Лишь единичные растения этого вида (3,2%) отличаются тремя
пучками, еще меньше (1,3%)—четырьмя. По болеечем у0,1 % растений
пшеницы твердой выявлено 5 пучков. По данным В. Г. и О. Г. Алек-
сандровых (1948), для перикарпия зерновки пшеницы твердой харак-
терно одревеснение (различной интенсивности) поперечных и мешко-
видных трубчатых клеток. Отсутствие одревеснения иногда наблюда-
ется па ее спинной стороне вблизи зародыша.
По строению эпи-мезокарпия пшеница твердая отличается от плен-
чатых тетраплоидпых видов наличием трех (а пе двух) слоев остатков
клеточных.полостей мезокарпия п особенно толщиной всей пленки эпи-
мезокарпия.
Согласно результатам исследований Л. Л. Жссгянниковой (1964),
общими для различных эколого-морфологических групп пшеницы твер-
дой являются следующие признаки: присутствие нескольких слоев кле-
ток эпи-мезокарпия, налично одревеснения (но менее сильно выра-
жепого, чем у Т. aestivum) почти всех структурных элементов по-
крова. У кипрской и египетской экологических групп обнаружено не-
которое сходство в структуре перикарпия с Т. dicoccum.
Согласно данным В. Ф. Дорофеева и О. Д. Градчаииповон (1971),
поверхность листа взрослых растений пшеницы твердой снабжена ши-
ппками, причем на нижней стороне число их меньше, чем на верхней.
Иногда они отсутствуют.
Наиболее полная экологическая характеристика Т. durum, как и
многих других видов пшеницы, приводится в работе Е. Ф. Пальмовой
(1935). Пшеница твердая экологически является степным растением.
Ес культура приурочена в основном к каштановым почвам жарких су-
хих областей Средиземноморья. Осадки (500—800 мм) здесь выпадают
главным образом зимой и весной. Летом обычна малая облачность.
Пшеница твердая в общем влаголюбивее мягкой. Первые фазы ее
роста очень растянуты и приходятся па.относительно прохладное и
влажное осеннее время года, так как во многих областях возделывания
она выращивается при осеннем посеве. В период созревания пшеница
твердая требовательна к высокой температуре и сухости воздуха. Ти-
пичным мезофитом является пшеница твердая влажного климата гор-
ных районов юга Малой Азии. Опа здесь имеет короткий вегетационный
период п относительно низкую потребность в тепле.
Большая континентальность климата (меньшее количество осадков,
выпадающих зимой и весной, быстрое наступление летней засухи и жа-
ры, большая годовая амплитуда температур) в восточном Средизем-
номорье привела к формированию здесь полупустынного (сирийского,
по Е. Ф. Пальмовой) типа пшеницы твердой. Все фазы ее роста па
Ближнем Востоке и в прилегающих районах Турции протекают очень
быстро. Для пшеницы твердой этого типа характерна крайняя требо-
вательность к влажности весной и высокой температуре во время со-
зревания. Требовательна к таким же условиям развития пшеница твер-
дая Глинта, близкая к сирийской по экологическому типу.
На крайнем побережье Аравийского полуострова в субтропических
степях сформировался типичный эфемер - - феноменально скороспелая
твердая пшеница, отрицательно относящаяся к повышенной влажности,
но весьма требовательная к высоким температурам в период созре-
вания.
Подрод 1. Triticum 9."
Закавка.зскуio пшен ип.у твердую Е. Ф. Налимова (1935) счш ала
восточной ветвью североафриканской. Под влиянием особых климати-
ческих условий здесь у некоторой части форм пшеницы твердой вырабо-
тался озимый и полуозимый образ жизни. Это пшеница в основном по-
ливная, к воздушной засухе и зною выносливая.
На черноземных и каштановых почвах СССР возделывается пшени-
ца твердая, отнесенная Е. Ф. Пальмовой (1935) и II. И. Вавиловым
(1964) к степному типу. Она довольно требовательна к теплу, отзывчи-
ва на действие длинного дня, отличается повышенной засухоустойчи-
востью (И. А. Стефановский, 1950; М. И. Руденко, 1960). Ранее на
европейской части СССР возделывались исключительно яровые сорта,
В настоящее время здесь в связи с развитием селекции созданы сорта
озимой твердой пшеницы, которые пока что не получили широкого рас-
пространения.
Степной тип яровой твердой пшеницы сформировался в США и Ка-
наде. В селекции его большую роль сыграли русские сорта (Арнаутка,
Кубанка и др.). В общем здесь Т. durum повторяет экологические типы
пшениц Старого Света.
Крайне низкая повысотпая граница возделывания пшеницы твердой
отмечена в Иордании (230—300 м ниже ур. м.). К- А. Фляксбергер
(1935) указал на испанские образцы пшеницы твердой, собранные на
высоте 1800 м над ур. м. В массе 7’. durum не поднимается выше 1000 м
над ур. м. В Закавказье занимает низменности и предгорья в основном
до высоты 500—900 м над ур. м.
Несмотря на глубокую древность культуры пшеницы твердой, дол-
гие годы не было известно о существовании у этого вида зимующих
полуозимых форм и тем более о возможности создания настоящих ози-
мых его форм. А. М. Бажанов (1856) первый описал озимую твердую
пшеницу из Дагестана. Через 40 лет агроном А. Филиппенко (1895)
сообщил о нахождении им в Дагестане двух сортов озимой твердой пше-
ницы. Речь шла, очевидно, о зимующих в условиях мягких зим полу-
озимых сортах народной селекции.
О зимующих популяциях пшеницы твердой из Туркмении сообщил
Н. И. Вавилов (1926). Многие такие формы из горного Сумбарского рай-
она б. Закаспийской области (на границе с Ираном) были испытаны
в течении трех лет под Саратовом, где они ежегодно погибали при
перезимовке, как и две кавказские формы озимой твердой пшеницы.
Опыты ВИРа показали, что на фойе мирового разнообразия пшени-
цы твердой дагестано-азербайджанская группа выделяется максималь-
ной степенью озимости (к-35047, к-35052, к-35110, к-35115 и др.). В част-
ности, это относится к популяциям предгорной лесостепи Дагестана и
северо-восточного предгорья Большого Кавказа, а также к популяциям
Нахичевани и Нагорного Карабаха. Меньшей степенью озимости обла-
дают балканские и некоторые молдавские формы, например к-41134 из
Кагула. Ряд новых озимых форм пшеницы твердой выявлен в после-
военные годы в Азербайджане и Дагестане (В. Ф. Дорофеев, 1972).
Однако даже наиболее озимые формы Закавказья и Дагестана заметно
уступают по зимостойкости озимой мягкой пшенице. Созданные в СССР
за последние г од;.я (Ф. Г. Кириченко, И. П. Лукьяненко, Д. И. Малюта,
В. В. Костин, А. Ф. Шулындип, В. М. Пыльнов, II. И. Буюкли и др.)
формы озимой твердой пшеницы по зимостойкости также уступают ози-
мой мягкой пшенице, что сдерживает их продвижение в производство.
Согласно данным ВИРа, по отзывчивости па свет выделяются полу-
озимые формы пшеницы твердой балканской группы (к-22879 из Ма-
кедонии), азербайджанские популяции (к-16813, к-15473, к-16884) и
яровые формы средиземноморской группы (convar. jalcaium). В усло-
виях короткого дня они запаздывают с колошением па 21—30 дней.
Классификация рода Triliciini I..
Наименее отзывчивы па снег (запаздывание копии i i ей и я па 1 5 дней)
яровые твердые пшеницы с. о. Кипр.
Среди возделываемых в СССР твердых пшениц осеннего сева сор-
та Шарк, Лг-буыа 13, Ллтып-бугдай, Шавпха местная являются био-
логически яровыми.
В отличие от Е. В. Черняева (1874) Л. II. Стебут (1916), П. Н. Кон-
стантинов (1925), Б. А. Вакар (1927), Е. Ф. Пальмова (1935),
Л. С. Инякина (1947), А. II. Шехурдин (1946), В. II. Мамонтова (1946),
П. II. Богданов (1946) и многие другие считают пшеницу твердую ме-
нее засухоустойчивым видом, чем мягкая. Это подтвердилось также в опы-
тах ВИРа в Заволжье, па Алтае, в Северном и Западном Казахстане.
Как показывают многолетние данные (1961 —1975) Куйбышевского
НИИСХа и Госкомпссии по сортоиспытанию в Заволжье, пшеница твер-
дая способна давать более высокие урожаи, чем мягкая, в годы с влаго-
обеспеченным вегетационным периодом. Эта культура менее устойчива
к почвенной засухе, в большей степени повреждается вредителями и
более требовательна к агрофопу. В условиях же орошаемого земледелия
этой зоны пшеница твердая по жаростойкости и урожайности не усту-
пает лучшим сортам мягкой, а в годы сильного распространения бурой
ржавчины превосходит ее (М. И. Руденко, В. Д. Артамонов, 1978).
В Центрально-Черноземной зоне сравнительное испытание райони-
рованных сортов на госсортоучастках зоны (1969—1975) также показа-
ло, что в годы с повышенным количеством осадков наибольший урожай
дает пшеница твердая, в засушливые—мягкая, что свидетельствует о
превосходстве пшеницы мягкой по засухоустойчивости (II. Л. Кузьмин,
Ф. Л. Никуличева и др., 1978). На Кубани пшеница твердая также
уступает мягкой, в особенности в фазах выхода в трубку и колошения.
Согласно Н. И. Вавилову (1931), пшеница твердая в период созре-
вания отличается стойкостью к засухе, а в период начального роста
не обладает ею. Из сортов степной волжской группы к наиболее засухо-
устойчивым Н. И. Вавилов относил некогда популярнейший сорт Меля-
попус 69. Высокой устойчивостью к засухе характеризуются также сор-
та Мелянопус 26 и новые сорта Саратовская 40, Саратовская 41, Лсу-
курум 43, Краспокутка 6.
Исследования И. К- Иванова (1975) показали, что в развитии кор-
невой системы мягкой и твердой пшеницы имеются существенные раз-
личия: у пшеницы твердой корни растут несколько медленнее, а узловые
появляются па 3—6 дней позже, чем у мягкой; этим и объясняется осо-
бая чувствительность пшеницы твердой к запаздыванию с посевом. Уста-
новлено (А. С. Устименко, 1975), что в фазе цветения пшеница твердая
уступает мягкой по развитию надземной массы и содержанию корней
в слое почвы 0—50 см, что отрицательно сказывается па урожае.
Есть сведения о довольно широком распространении открытого цве-
тения у Т. durum. Опытами ВИРа в Омске (Л. П. Аладова, 1958) было
установлено, что у яровой твердой пшеницы в зависимости от сорта и
метеорологических условий раскрывается от 23,1 до 97,0% цветков. Во
влажный и прохладный год количество цветков с открытым цветением
увеличивается. Выявлена способность к цветению пшеницы твердой при
температурах воздуха +4 и 4-42°С.
В условиях Молдавии, согласно данным В. Р. Челака (1969), у пше-
ницы твердой преобладает открытое цветение над закрытым. М. А. Фе-
дин (1974) установил, что существуют различия в отношении степени
открытого цветения между сортами пшеницы твердой и условиях Цели-
ноградской и Московской областей.
В литературе приводится ряд свидетельств о спонтанной гибридиза-
ции пшеницы твердой (М. Ф. Терновский, 1929; II. Ц. Вавилов, 1935;
И. Д. Мустафаев и М, М. Якубцинер, 1952; В. Ф. Дорофеев, 1972 и др.).
Явление спонтанной гибридизации у пшеницы твердой шире распростра-
Подрод I. Triticum
97
нено. чем эго предполагалось раньше (В. М. Берлянд-Кожевников и
др., 1975).
Зерно пшеницы твердой характеризуется высокой стекловидностыо
эндосперма и клейковиной, обладающей высокой упругостью вследствие
повышенного содержания белка. В ее зерне, содержится белка больше,
чем в зерне пшеницы мягкой. Анализ большого количества образцов
пшеницы твердой, выращенных в различных зонах СССР, показал ши-
рокое варьирование содержания белка в пределах различных эколого-
географических групп. Так, в районах Среднего Поволжья, по многолет-
ним данным (1971 —1974), содержание белка в зерне коллекционных
образцов пшеницы твердой колебалось от 13,3 до 21,3% (В. Ф. Доро-
феев и др., 1977). Содержание белка в зерне репродукции Дагестанской
опытной станции ВИРа колебалось от 13 до 19,8%. При выращивании
коллекционных образцов Т. durum па Среднеазиатской опытной станции
ВИРа концентрация белка в зерне, по данным В. Г. Коиарева, 3. В. Чис-
ловой, Р. Л. Удачина и др. (1976), составляла 12,1—21,9% в зависимости
от экологической принадлежности сорта.
Испытания местных пшениц Закавказья (экспедиционные сборы
1961 —1964 гг.) в условиях орошения па Дагестанской опытной станции
ВИРа выявили значительные колебания содержания белка в зерне
(12,0—20,0%) и позволили выделить высокобелковые продуктивные
образцы (В. Ф. Дорофеев, 1972). При посеве по бобовому предшествен-
нику ла Кубани некоторые закавказские формы накапливают в зерне
до 21—23% белка (А. Ф. Мережко, 1970).
Иммунитет. 14. И. Вавилов (1964) относил пшеницу твердую к числу
видов, устойчивых к комплексу грибных болезней, выделяя по сравни-
тельной устойчивости к бурой и желтой ржавчине, мучнистой росе и
твердой головне средиземноморские ее формы. М. М. Якубцинер (1957)
писал, что у пшеницы твердой относительно устойчивых к бурой ржав-
чине форм больше, чем у мягкой.
В условиях плоскостного Дагестана, по наблюдениям 1954—1965 гг.
над 1038 образцами пшеницы твердой, И. П. Шитова (1968) определила,
что азербайджано-дагестанские образцы большей частью восприимчи-
вы к бурой ржавчине. В опыте О. Г. Григорьевой (1975) расоспецифиче-
ская устойчивость образцов коллекции пшеницы твердой варьировала
(обнаружены все типы устойчивости и восприимчивости). При изучении
888 образцов коллекции пшеницы твердой В. М. Берляид-Кожсвнпков
и др. (1973) выделили 142 образца, которые проявили устойчивость пе
менее чем к одной из тестируемых рас (151, 77, 202). Они установили,
что 15 образцов имели полигонную устойчивость к этим расам, что у
пшеницы твердой есть сорта — носители группы генов Хц Х2, Хз, а так-
же генов Lr 1, Lr 2, Lr 24, Lr 3, Lr 17.
Превосходство устойчивости к желтой ржавчине у пшеницы твердой
по сравнению с мягкой найдено при изучении мировой коллекции пше-
ницы (И. П. Шитова, 1968; О. Г. Григорьева, 1975). М. М. Якубцинер
(1969) указал на сорта Lakota и Yuma как на устойчивые к 7 расам
желтой и многим расам стеблевой ржавчины.
Еще G. М. Reed (1916) отметил варьирование устойчивости к муч-
нистой росе среду сортов пшеницы твердой. Э. X. Суханбердина (1977)
при искусственном заражении проростков популяцией рас этого пато-
гена (из 5 регионов СССР) нашла, что 89,5% образцов коллекции были
восприимчивыми, 1,4% — иммунными. В полевых условиях на провока-
ционном фоне эти показатели были соответственно равными 51,5 и 3,5%.
Она сделала заключение о том, что пшеница твердая в целом восприим-
чива к мучнистой росе.
В. И. Кривченко (1973) при искусственном заражении показал, что
патогенный эффект для ншсиицы твердой имеет пыльная головня f.
duri, а расы f. aestivi поражают ее значительно слабее. Степень поле-
7 762
98
Классификации рода Triticum I..
вой устойчивости к лим явно различат j ся: к I. duri бы,ян сильно вос-
приимчивы 26,5% и иммунпы 42,7% образцов, а к f. aestivi соответст-
венно 0 и 62,2%. Наиболее богаты высокоустойчивыми к пыльной го-
ловне f. duri были образцы с Ближнего Востока, из Средиземноморских
стран, Северной Африки, Северной и Южной Америки.
С. П. Зыбина (1939) при искусственном заражении популяцией рас
твердой головни из 17 районов СССР нашла, что пшеница твердая бо-
лее устойчива к этому патогену, чем мягкая. По данным В. И. Кривчен-
ко (1973), заражение твердой головней коллекции пшеницы твердой
выявило 14,7% иммунных образцов; сильная восприимчивость обнару-
жена у 20,8% образцов. Основная часть сортимента пшеницы твердой
была слабо или средне восприимчива к этой болезни.
Н. Harding (1972) нашел, что пшеница твердая более или менее
восприимчива к корневым гнилям. Н. И. Вавилов (1964) отметил не-
устойчивость ряда географических групп пшеницы твердой (в том числе
пшениц Египта, Эфиопии, ПАР, НДРЙ, Ирана, Индии и Китая) к гес-
сенской и шведской мухам.
История. Исследованиями Е. Schicmann (1948), В. Л. Мепабде
(1948), И. Д. Мустафаева (1955), II. Ilelbaek (1959, 1965) и др. уста-
новлено, что введение в культуру пшеницы твердой относится к IV—
III тысячелетию до п.э. Подробная сводка о распространении в прош-
лом культуры пшеницы твердой в нашей стране дана М. М. Якубцинс-
ром (1956).
Географическое распространение. Крупным производителем пшени-
цы твердой является СССР. Площадь посевов се is нашей стране около
5 млн. га, что составляет свыше 40% мировой площади под этой пше-
ницей. Основные площади Т. durum па территории нашей страны со-
средоточены в Поволжье, Казахстане, Западной Сибири, на Северном
Кавказе, в Закавказье (главным образом в Азербайджане) и Дагестане.
В Поволжье наибольшее распространение пшеница твердая получила
в Саратовской области. Большие массивы этого вида находятся в Куй-
бышевской, Волгоградской и Оренбургской областях, в степных и лесо-
степных зонах Сибири. Значительная часть площадей под пшеницей
твердой сосредоточена в Ростовской области. В европейской части на-
шей страны северная граница возделывания пшеницы твердой проходит
по южным районам Горьковской области.
За рубежом пшеница твердая широко распространена в Италии,
Алжире, Турции, Сирии, Ираке, США, Канаде, Мексике, Аргентине
и др.
Происхождение. К- А. Фляксбергер (1935) считал пшеницу твердую
древнейшим видом, ведущим свое происхождение от общего с Т. dicoc-
cum предка. Более ранняя (1911) его точка зрения —пшеница твердая
непосредственно возникла от Т. dicoccum. Последнего воззрения придер-
живались F. Kornicke (1908), A. Schulz (1911), Д. К. Ларионов (1914),
Е. Tschermak (1914) в прошлом, а в паши дни — Е. Р. Моррис и
Э. Р. Сирс (1970), В. Ф. Дорофеев (1972) и др. Б. А. Вакар (1932)
близость пшеницы твердой к Т. dicoccum усматривал в том, что их гиб-
рид имеет особенно мало неправильностей в мейозе. А. П. Щапова
(1969) нашка, что различия кариотипов Т. dicoccum и Т. durum связаны
с морфологией лишь двух хромосом.
П. М. Жуковский (1971) вс считал задачу происхождения пшеницы
твердой решенной, так как пи с каким из существующих диких пли
культурных видов пшеницы он не мог установить ее преемственности.
Наша точка зрения па происхождение пшеницы твердой сложилась
под влиянием высказываний Д. Мак-Кея (1968) о полигонном обеспече-
нии легкого обмолота у нее. Эта генетическая система, возможно, обра-
зовалась постепенно путем накопления микромутаций, приведших к пе-
реходу от трудного обмолота колоса, который имеет Т. dicoccum, к лег-
llo<)ptn) !. Tritiriini 99
кому обмолоту у пшеницы твердой. Переход этот ( эпровождалея не
только погерей сильною одревеснения колосковой чешуи, по и упрочне-
нием стержня колоса. По мнению В. И. Кривченко и др. (1979), одно-
временно также произошла потеря устойчивости к мучнистой росе. Пше-
ница твердая—результат отбора среди растений Т. dicoccum особей
с наименее трудным обмолотом, который произвела народная селекция
далекого прошлого. II. И. Вавилов (1964) происхождение пшеницы твер-
дой связывает с Восточным Средиземноморьем.
Систематика. Т. durum — второй по распространению па земном ша-
ре вид пшеницы. Огромный ареал вида с хорошо выраженными почвен-
но-климатическими различиями обусловил его широкую экологическую
дифференциацию. Однако внутривидовая классификация, которая пол-
ностью отвечала бы географо-морфологическому принципу, у Т. durum
весьма затруднена.
К. А. Фляксбергер (1935) отмечал необходимость использования
различных принципов при построении эколого-морфологической группи-
ровки Т. durum. Для удобства пользования в практической работе
К. А. Фляксбергер (1935, 1939) выделяет группы по отдельным морфо-
логическим признакам: плотности колоса (рыхлые при D до 30, плотные
при D — 30—40, очень плотные при D более 40), форме колоса (удли-
ненной, типично пирамидальной или с тенденцией к пирамидальности),
шероховатости остей (гладкие, слабошероховатые, шероховатые, силь-
нозазубренные), выполненности соломины (полая, выполненная парен-
химой), присутствию и отсутствию язычка и ушков листа и т. д. Отдель-
ные морфологические признаки проходят по многим экологическим
группам, некоторые носят узколокальный характер.
Точное внутривидовое разграничение отдельных морфологических
групп трудно дать из-за захождения признаков. Наиболее крупные под-
разделения Т. durum исследователи, начиная с N. Scringe (1842), не
случайно основывали прежде всего па признаке плотности колоса. Хотя
у Т. durum наблюдается значительная изменчивость плотности коло-
са— от крайне рыхлых (D = 16—20) до очень плотных (D =40 и бо-
лее) колосьев — со многими переходными формами, однако крайние
варианты по плотности колоса характеризуются свойственным нм опре-
деленным комплексом признаков. Одним из таких вариантов некоторые
исследователи считают Т. turanicum Jakubz. — рыхлоколосый вид, рас-
сматривая его в качестве подвида или более мелкой таксономической
единицы Т. durum. Однако специфичность многих признаков в какой-то
степени оправдывает видовой статус Т. turanicum. Вторым крайним ва-
риантом по плотности колоса является Т. durum subsp. horanicum Vav.—
плотноколосая пшеница с комплексом морфологических признаков, по
которым довольно отчетливо отличается от других групп пшеницы
твердой.
В то же время в упомянутых выше крайних группировках сущест-
вуют ряды переходных форм к типичному подвиду собственно твердых
пшениц (Т. durum subsp. durum}. Ареалы выделяемых нами подвидов
Т. durum в культуре обычно накладываются друг на друга. Следует
отметить, что, кроме морфологических различий subsp. durum и subsp.
horanicum, выявляется некоторая генетическая разнородность: при их
скрещивании могут наблюдаться аномалии и стерильность потомства.
В разнос время у Т. durum выделяли другие подвиды, при этом их
авторы руководствовались различными принципами выбора признаков
для характеристики одних и тех же таксонов, не обращая внимания на
ареал. Мы рассматриваем у Т. durum 2 подвида: subsp. durum и subsp.
horanicum. Собственно твердые пшеницы (subsp. durum) нами диффе-
ренцированы иа группы разновидностей.
Отдельные отличительные признаки или их комплексы у этих групп
носят локальный характер, т. е. имеют географическую приуроченность.
7*
100
Классификация рода Triticum
Ключ для определения подвидов Т. durum Desf.
1. Растения большей частью высокорослые (до 160 см высоты и боль-
ше), средперослые (80—100 см), реже низкорослые (до 40—70 см);
листья длинные, широкие, поникающие и скручивающиеся; колосья от
коротких (5—6 см) до очень длинных (13 см), рыхлые (D = 16—30),
средней плотности (D = 30—40) и реже очень плотные (D ='40—50),
но тогда растения средне- или высокорослые, ости параллельные, длин-
ные (до 10—23 см), реже сильноукорочепные (у китайской экологиче-
ской группы), колосковые чешуи от укороченных овальных до удлинен-
ных, с более или менее выраженным килем; зерновки большей частью
овальные, овально-удлиненные и длинные (длина превышает ширину
более чем в 1,5 раза)...............................subsp. durum.
+ Растения низкорослые (60—70 см высоты); листья короткие, сла-
бопопикающие, колосья короткие (5—6 см длины), очень плотные (D
около 40 и более); ости несколько расходящиеся, сравнительно укоро-
ченные; колосковые чешуи укороченные, широкие, с широким выдаю-
щимся килом; зерновки близкие к округлым (длина превышает ширину
приблизительно в 1,5 раза)...................subsp. horanicum Vav.
I. Subsp. durum — подвид собственно твердых пшениц.
Subsp? durum—T. durum subsp. expansum Vav. 1931, Tp. прикл. бот.
ген. сел. Прилож. 51: 216; он же, 1964, Миров, ресурсы хлебп. злаков.
Пшеница: 18.
Описание-собственно твердых пшениц совпадает с характеристикой
вида в целом. Для subsp. durum характерна большая вариация в от-
ношении плотности колоса, размеров и формы зерновки и ряда других
признаков. Некоторым ее формам свойственны безлпгулыюсть листа
или сильное опушение его и другие географически приуроченные при-
знаки, что позволяет у subsp. durum выделить группы разновидностей.
Геогр. распр. По всему ареалу Т. durum.
Ключ для определения групп разновидностей subsp. durum
1. Колос плотный (D около 40 и больше).....................2.
+ Колос рыхлый (D = 16—25) или средней плотности (D = 25—35,
иногда до 40).................................................3.
2. Колос 5—8 см длины, колосковые чешуи округлые, всегда значи-
тельно короче цветковых; ости короткие, на нижних колосках их нет
совсем............................convar. caucasicum (Dorof.) Dorof.
4- Колос около 6 см длины, колосковые чешуи овальные, приблизи-
тельно одинаковой длины с цветковыми или несколько короче их; ости
длинные по всему колосу..............convar. durocompactum Flaksb.
3. Растения высотой 40—160 см, богатооблиствепиые; колос средней
плотности (D = 25—40), длинный (9—12 см) или средней длины
(6—9 см) . !..................................................4.
+ Растения низкорослые (60—100 см), облнственность небольшая,
колос рыхлый’ (D около 25), 6—8 см длины......................5.
4. Колосковые чешуи различной формы, большей частью овальные,
6—-10 мм длины; зерновки 7—8 мм длины..............convar. durum.
’ + Колосковые чешуи удлиненные, 10 -12 мм длины; зерновки очень
длинные (до 10 мм) с характерным серповидным углублением с брюш-
ной стороны.............convar. falcatum (Jakubz.) Dorof. et A. Filat.
5. Всходы, листовая пластинка, влагалище листа густо опушены, опу-
шение сохраняется почти до конца периода вегетации; язычок и ушки
листа развиты ..... convar. villosum (Jakubz.) Dorof. et A. Filat.
+ Всходы с коротким редким опушением, иногда пеопушеиные, на
влагалище и пластинке листа редкое опушение исчезает по мере разви-
H(K)poi) /. Trilit uni 101
тия растений; язычок и ушки /шста очcyirinyioг или паходяюя в зама-
точном состоянии...............convar. aglossicon Dorof. et A. Filat.
1. Convar. durum -группа разновидностей собственно твердых
пшениц.
Convar. durum. — Т. durum commune Flaksb. 1922, Опред. наст, хле-
бов: 52. —Т. durum commune series prol. mediterranea (Vav.) Flaksb.
1935, Культ, фл. СССР, 1: 181. — T. durum commune series prol. europeae
(Vav.). Flaksb. 1935, 1. c_: 183, excl. subprol. eligulatum et prol. villosum. —
T. durum convar. thumanjanii Gandil. 1976, Бюл. ВИРа, 65: 74.
Распространен по всему ареалу пшеницы твердой. Обширный ареал
распространения собственно твердых пшениц обусловил чрезвычайно
широкую экологическую дифференциацию их па следующие экологиче-
ские группы.
1. Собственно средиземноморская экологическая группа — Т. durum
sect, mediterranea Vav., 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 216. —
Т. durum series prol. mediterranea (Vav.) Flaksb. 1935, Культ, фл. СССР,
1: 181. — T. durum prol. eumediterraneum Vav. 1964, Миров, ресурсы
хлебн. злаков. Пшеница: 38.
Растения большей частью высокорослые, соломина толстая, обычно
с выполненной сердцевиной, листья широкие.
Колосья крупные со средней плотностью (D =30—40), редко рых-
лые, ости грубые, длинные, обычно в 2 раза превышающие длину ко-
лоса, зерновки крупные (рис. 9, «), стекловидные.
Средиземноморские твердые пшеницы сравнительно устойчивы к бу-
рой и желтой ржавчине, к твердой и пыльной головне, отчасти к муч-
нистой росе; слабо поражаются шведской и гессенской мухами. Преиму-
щественно яровые, встречаются полуозимые и озимые (Дагестан, За-
кавказье), сравнительно позднеспелые.
Геогр. распр. Алжир, Тунис, Марокко, южная часть Испании и
Португалии, острова Сардиния, Сицилия, Корсика, Крит, прибрежная
полоса о. Кипр, Малой Азии, Сирии и Ливана.
Среди средиземноморских твердых пшениц выделены экологические
подгруппы с комплексом признаков, характерным в целом для этой
группы, и, кроме того, с рядом специфических особенностей, присущих
довольно узколокализованпым подразделениям.
а. Иорданская подгруппа [7. durum subgr. jordanica Jakubz. 1932.
Tp. прикл. бот. ген. сел. Прилож. 53: 52. — Т. durum gr. syriaca Jakubz.
1932, l.c.: 62. — T. durum mediterranea prol. jordanica (Jakubz.) Flaksb.
1935, l.c. 182.— T. durum mediterranea prol. syriaca (Jakubz.) Flaksb.
1935, l.c.: 182.— T. durum prol. palestinicum Vav., 1964, l.c.: 41, p.p.].
Растения низкорослые, соломина сравнительно тонкая, сравнительно
устойчивая к полеганию, облиственность средняя, листья с восковым на-
летом, реже без палета. Колосья средней плотности, редко рыхлые, ости
грубые, длинные, зерновка среднего размера, несколько мельче, чем
у средиземноморской группы, напоминает зерновки хорапскпх пшениц,
но более удлиненные, стекловидные. Устойчивы к грибным заболева-
ниям, отличаются значительной скороспелостью. М. М. Якубцинер (1932)
эти пшеницы подразделяет па иорданские и сирийские. Последние от-
личаются несколько большей позднеспелостью.
Г corp, распр. Сирия, Иордания, Израиль.
б. Ближневосточная низинная [7'. durum endemica Jakubz. 1932, Тр.
прикл. бот. ion. сел. Ирплож, 53: 52, nom. improp. 7'. durum prol.
endemicum (Jakubz.) Flaksb. 1935, Культ, фл. СССР, 1: 186. --T. durum
prol. melitense Flaksb. 1935, l.c.: 187, descr. ross. — T. durum prol. pa-
lestinicum Vav. 1964, Миров, ресурсы хлебн. злаков. Пшеница: 41, р.р.].
Растения низкорослые, соломина тонкая. Листья молодых и взрос-
лых растений и их влагалища неопушспные, листовые пластинки узкие,
102
Рис. 9. Колосья, колоски и зерновки Т. durum Desf.:
а — convar. durum; б — convar. cattcasicum (Dorof.) Dorof.
ярко-зеленые, без воскового палета. Восковой палет отсутствует па всех
частях растения. Колос топкий, рыхлый; ости параллельные, длинные,
топкие, более нежные, чем у остальных пшениц твердых. Зерновка длин-
ная, узкая. Скороспелые, относительно устойчивы к желтой и бурой
ржавчине.
От иорданской подгруппы отличается меньшей устойчивостью к по-
леганию, более рыхлым колосом и полным отсутствием воскового на-
лета. Формы без воскового палета обнаружены не только па Ближнем
Востоке, но и в Северной Африке, Малой Азии, однако ближневосточ-
ные отличаются от других подгрупп и морфологически и экологически.
Подрод /. Triticum
103
К ближневосгочпой подгруппе относятся также пшеницы твердые
о. Мальта, характеризующиеся отсутствием воскового палета, но с не-
сколько более крупным колосом и более длинной солом и пой, а также
слабым опушением молодых растений.
Геогр. распр. Преимущественно в низменных приморских и
долинных районах Ливана, Иордании, Сирии.
в. Сардинская подгруппа [Г. durum 1 сардинский тип и 2 сардинский
тип, Шрейбер, 1934, Тр. прикл. бот. геи. сел. 5, 2: G0.—Т. durum prol.
sardinicum (Schreib.) Flaksb. 1935, Культ, фл. СССР, 1: 182]. Растения
высокорослые. Соломина толстостенная, устойчивая к полеганию. Ко-
лосья крупные, 7—9 см длины, средней плотности пли рыхлые. Зерновка
крупная, слегка горбатая. Слабо поражаются грибными заболеваниями.
Относительно засухоустойчивы, при отсутствии осадков могут приоста-
навливаться в росте и затем быстро отрастать при последующих
дождях.
Геогр. распр. Сардиния.
г. Португальская подгруппа. Крайне позднеспелые пшеницы Порту-
галии, отличающиеся удлиненным колосом и сравни голыш топкой по-
легающей соломиной (П. П. Вавилов, 1961). Отстают ио созреванию
на 5— 6 дней от остальных средиземноморских твердых пшениц в усло-
виях предгорного Дагестана.
д. Горная атласская подгруппа (Г. durum prol. berbericutn Vav.
1964, 1. с.: 40, descr. ross.).
В горах Атласа возделывается пшеница твердая с наиболее легким
обмолотом, что нередко сочетается с легкостью осыпания зерна. Это
выделяет ее среди средиземноморских пшениц. Распространена па вы-
сотах до 2000 м над ур. м.
2. Европейская (степная) группа-— Т. durum sect, europeae Vav. 1931,
1. с.: 216. — T. durum prol. eustepposum Vav. 1964, 1. c.: 44.
Растения большей частью высокорослые, колосья крупные пли сред-
ней величины, среднерыхлые (D = 23—28, при колебаниях 21—30), ости
длинные, зерновки несколько укороченные.
Европейская группа, позднее названная Н. И. Вавиловым (1964)
степной, характеризуется комплексным иммунитетом к бурой и желтой
ржавчине. Между отдельными сортами имеются различия по устойчи-
вости к желтой ржавчине в разные фазы роста растений. Яровые, сред-
неспелые.
Геогр. распр. Балканы, Испания (частично), Малая Азия, Се-
верная Америка, Аргентина, Австралия, в СССР — степные и лесостеп-
ные районы европейской части, Закавказье, Сибирь, Казахстан.
К. А. Фляксбергер (1922, 1935) делит европейскую группу пшениц
твердых па 2 типа в зависимости от плотности колоса: 1) рыхлоколосый
[Т. durum taganrocense Ser. 1841, Сёг. Eur. 4.: 127 (165). — Т. durum
oblongum Orlov, 1923, Tp. прикл. бот. сел. 13: 373. — T. durum prol.
laxiusculum Flaksb. 1935, l.c.: 184]—с удлиненными, рыхлыми (D =
= 23—26) колосьями, удлиненными чешуями и зерновкой; 2) плотпоко-
лосый [Г. durum compactum Ser. 1841, l.c.: 126 (164).-—T. durum
densiusculum Flaksb. 1922, Опрсд. наст, хлебов: 52. — T. durum prol.
densiusculum (Flaksb.) Flaksb. 1935, l.c.: 183]—с небольшим колосом
средней плотности (D = 26—28), с чешуями и зерновкой несколько
укороченными. Однако колебание плотности колоса в обеих группах
одинаково и составляет 21—30 колосков на 10 см длины колоса. Первый
тип легче переносит недостаточное увлажнение и более скороспелый,
чем второй. В зависимости от влагообеспечеипости в посевах преобла-
дает тот или иной из них.
В степных и лесостепных районах СССР преобладают рыхлоколосыс
формы. Возделываемые здесь еще в 50-х годах пшеницы твердые отно-
сятся к следующим подгруппам.
104
Классификация рода Triticum L.
а. Стенная волжская (stepposum votgense Dorof.). Растения средней
высоты, средней кустистости и облиственности. Среднеспелые, довольно
засухоустойчивые. Стенной тип пшеницы явился основополагающим для
культуры пшеницы твердой в США и Канаде.
Гео г р. р а с и р. СССР (Поволжье.).
б. Степная южная (austrostepposum Dorof.). Близкая предыдущей,
но менее устойчивая к засухе.
Геогр. распр. Украина, Северный Кавказ.
в. Степная восточная (orientalistepposum Dorof.). Срсдиспоздняя,
влаголюбивая.
Геогр. распр. Зауралье, Сибирь.
г. Лесостепная волжская (silvistepposum volgense Dorof.). Близка
к степной южной.
Геогр. распр. До северного предела района возделывания пше-
ницы твердой (лесостепь Горьковской области и Удмуртия).
Несмотря на значительный ареал степных твердых пшениц, разио-
видностиый состав их беден (2—3 разновидности).
3. Эгейская группа — Т. durum тип aegettm Kob. 1932, Хлебн. злаки
Турции: 8. Растения высокорослые, об.тпсгвеипость средняя, соломина
слабоустойчивая к полеганию; колос длинный, плоский (сжатый с дву-
рядной стороны), большей частью рыхлый; колосковые чешуи удлинен-
ные, плоские, обычно почти лишенные плеча.
В целом эгейская группа иммунна к бурой и желтой ржавчине,
позднеспелая, представлена яровыми и полуозпмыми формами.
Геогр. распр. Прибрежные районы Турции, Греция, Югославия
(Македония).
4. Кипрская, группа — Т. durum cyprium Vav. 1931, l.c.: 218; Флякс-
бергер, 1935,1. с.: 185.
Растения низкорослые (50—90 см), кустистость слабая, облиствен-
ность небольшая, листья узкие. Колосья узкие, цилиндрически-веретено-
видные, 6—8 см длины, рыхлые (D = 24—28), иногда более плотные
(D =34); колосковые чешуи удлиненные, киль узкий, колосковый зу-
бец остевидный (до 3 см); зерновка длинная (10 мм).
Сильно поражаются ржавчиной, особенно желтой. Плотноколосые
формы обычно менее восприимчивы к этому патогену.
Геогр. распр. Кипр.
5. Азиатская группа — Т. durum f. asiatica Jakubz. 1932, l.c.: 67.—
T. durum тип asiaticum (Jakubz.) Kob. 1932, l.c.: 6. — T. durum prol.
aslaticum (Jakubz.) Flaksb. 1935, l.c.: 186.
Растения низкорослые, соломина топкая, листья узкие. Колосья ко-
роткие, 6—7 см длины, рыхлые, сжатые с двурядной стороны, реже квад-
ратные в поперечном сечении; колосковые чешуи узкие, зерна мелкие,
реже средней величины.
Яровые, скороспелые, устойчивы к фузарпозу.
Геогр. распр. Малая Азия, Сирия, Иордания, СССР (на юго-
востоке Русской равнины, редко в Средней Азин).
6. Каспийская группа-—Т. durum prol. caspicum Vav. 1964, l.c.: 43.
Растения высокорослые, 150—160 см длины, соломина толстая, вы-
полненная или с утолщенными стенками, листья широкие, облиствен-
ность хорошая. Колосья крупные, средней плотности, ости длинные,
грубые, в колоске обычно 3—4 крупные зерновки.
В каспийской экологической группе выделены 2 подгруппы (азер-
байджано-дагестанская и всрхпекарталпиская лесостепная).
а. Азербайджано-дагестанская подгруппа (7’. durum prol. azerbajdz-
hanicutn Groben. 1948, Изв. Азерб. с.-х. ин-та, 3,13: 5, descr. ross; Must,
et Kovd. 1970, Вести, с.-х. пауки, 15, 6: 17, descr. ross. — T. durum prol.
borcalinicum Dekapr. 1954, Tp. Ип-та полевод. 8: 28, descr. ross.). Пред-
ставлена озимыми, полуозпмыми и яровыми формами. Яровые высева-
Подрод I. Triticum
ются осенью. Все формы слабозпмостойкие, позднеспелые. ТреС>ов;иеди-
ные к почвенной влаге и теплу, они выносливы к воздушной засухе и
зною, возделываются в основном па поливе. Слабо или в средней сте-
пени устойчивы к бурой ржавчине, твердой головне, относительно устой-
чивы к желтой ржавчине и пыльной головне. Зерно отличного качества.
Высокий урожай дают только па богатом агрофоне.
Некоторые отличия (более рыхлый колос, более топкая соломина)
существуют между твердыми пшеницами, возделываемыми в Дагестие
и северо-восточных районах Азербайджана (7’. durum prol. daghestani-
cum Greben.. 1948, 1. c.: 6, descr. ross.) и собственно азербайджанскими.
Геогр. распр. Низменности и предгорья (до 500 м над у]), м.)
Азербайджана и сопредельных районов Грузин и Армении, плоскостные
районы Дагестана.
б. Верхнекарталипская лесостепная подгруппа (Г. durum pro!.
carthlicum Dekapr. 1954, l.c.: 28, descr. ross.). Представ,,тепа преимуще-
ственно var. caerulescens и var. libycum. Наименее требовательные к теп-
лу пшеницы твердые. В отличие от предыдущей подгруппы эти пшени-
цы яровые, высеваются с осени, но хорошо переносят местные зимы.
Геогр. распр. Грузия, Южная Осетия па высотах 500 -900 м.
над ур. м.
7. Индийская группа — Т. durum prol. indicum Vav. 1964, l.c.: 46.
descr. ross.
Растения низкорослые, редко высокорослые, соломина прочная,
листья широкие. Колосья короткие, рыхлые (у высокорослых форм бо-
лее длинные и плотные); ости незначительно превышают длину колоса;
зерновки мелкие. Близки к европейской степной группе, но имеют более
короткий вегетационный период.
Геогр. распр. Индия (сравнительно в небольшом количестве,
чаще встречаются в виде примеси).
8. Кашгарская группа — Т. durum prol. kaschgaricum Vav., 1964.
1. с.: 46, descr. ross.
Растения высокорослые, листья широкие. Колосья крупные, плотные.
Средне- и позднеспелые. Близки к европейской степной и средиземно-
морской группам.
Геогр. распр. Западный Китай (оазисное поливное земледелие
на высоте 1200—2000 м над ур. м.).
9. Китайская группа-—Т. durum subsp. siuicum Vav. 1964, l.c.: 49.
Растения высокорослые, соломина прочная. Колосья крупные, безостые,
колоски многоцветковые. Позднеспелая, по качеству зерна значительно
уступает другим группам.
Обнаруженную в образцах из Восточного Китая безостую пшеницу
твердую Н. И. Вавилов (1964) вначале рассматривал как спонтанный
гибрид между Т. durum и безостой формой Т. aestivum. Основываясь
на фактах формообразования и отбора в Китае в направлении безосто-
сти у других видов, он принял вероятным возникновение здесь на основе
занесенных извне пшениц твердых своеобразных безостых Т. durum.
Н. И. Вавилов выделяет эти пшеницы в подвид на том основании, что
отдельные гибридные семьи, полученные от скрещивания с разнообраз-
ными формами, Г. durum, в F3 имеют значительную стерильность. Однако
ввиду недостаточности экспериментального материала безостые китай-
ские пшеницы мы выделяем в особую эколого-географическую группу.
При описании разповидностпого состава Т. durum convar. durum
считаем возможным выделить в этой группе разновидности гибридного
происхождения с характерными признаками пшеницы твердой и с при-
знаками, привнесенными путем искусственной или спонтанной гибриди-
зации от других видов. В одном случае (subconvar. mulicutri) это будет
безостость, переданная от пшеницы мягкой, в другом — ветвистоколо-
сость, переданная пшенице твердой от 7’. turgidum (subconvar. durora-
1 ОС Классификация рода Triticum I..
mosum). Мы полагаем, что выделение подгрупп позволит избежать из-
лишней многоступенчатости в таблице по определению разновидное! oil
и что оно оправдано их происхождением.
Среди пшениц канадского происхождения выделены новые разновид-
ности пшеницы твердой, характеризующиеся опушением стебля под
колосом. Этот признак, не свойственный роду Triticum, передан пше-
нице твердой путем гибридизации ее с рожью. Указанные константные
разновидности в остальном совершенно соответствуют subconvar. durum
и потому они не выделяются в отдельную подгруппу.
Ключ для определения подгрупп разновидностей
1. Колос простой ...............................................2.
+ Колос ветвистый....................subconvar. duroraniosum Dorof.
2. Колос остистый.................................subconvar. durum.
+ Колос безостый . subconvar. muticuni (Orlov) Dorof. et A. Filat.
a. Subconvar. durum -- пшеница собственно твердая.
Subconvar. (lurtiin. - - T. durum u) aristatum Orlov. 11)23, 'Гр. прикл.
бог. сел. 13, 1: 372, excl. T. durum В. durocumpaclum Flaksb.
Обыкновенная остистая твердая пшеница, составляющая основную
массу в полевой культуре во всех частях света.
Var. aegyptiacum Koern. in Koern. et Wern. 1885, Ilandb. Gttreideb.
1: 73. — T. durum gr. aegyptiacum (Koern.) Flaksb. 1935, Культ, фл.
СССР, 1: 215. (31).
Геогр. распр. Средиземноморские страны, СССР'—Закавказье,
Дагестан, Северный Кавказ, Крым — редко как примесь.
Var. affine Koern. in Koern. et Wern. 1885, l.c.: 70.— T. durum var.
compylodon Koern. in Koern. et Wern. 1885, l.c.: 70-— T. durum gr. affine
(Koern.) Flaksb. 1935, l.c.: 199. (7).
Геогр. распр. Средиземноморские страны и острова, СССР —
Закавказье, Дагестан, Северный Кавказ и Украина'—редко как при-
месь.
У этой разновидности различаются 2 формы:
f. epruinosaffine Jakubz. et A. Filat, f. nov.—Tota planta epruinosa.
Typus: Jordania, 1926, leg. Markov, k-15057 (WIR),
Растения без воскового палета. Выделена из материала, полученного
от В. В. Марковича в 1926 г.
Геогр. распр. Иордания, Сирия, Малая Азия, Египет, о-в Мальта.
f. fereaffine Dorof. et A. Filat, f. nov. — Spica aristis scabriusculis vel
fere levibus.
Typus: Albania. Reproductio stationis Daghestanskajae fulcracea WIR,
1973, k-43179 (WIR).
Ости слабошероховатые. Описана no образцу из Албании.
Var. africanum Koern. in Koern. et Wern. 1885, l.c,: 73. — T. durum
gr. africanum (Koern.) Flaksb. 1935,1. c.: 212. (30) .
Геогр. распр. Средиземноморские страны, СССР (редко как при-
месь) .
К этой разновидности относится f. epruinosafricanum Jakubz. et
A. Filat, f. nov. — Tota planta epruinosa.
Typus: Marocco, prope pag. Tachdirt, Reproductio stationis Persijanov-
ka WIR, 1928, k-16467 (WIR).
Растения без воскового налета.
Геогр. распр. Северная Африка, Испания (Пиренеи).
Var. albobscurum Flaksb. var. nov. — T. durum var. albobscurum
Flaksb. 1935, l.c.: 207, descr. ross. — T. durum var. obscurum auct. non
Koern.: Vase. 1933, Bolet. Agric. I, 1—2, ser. 1: 78.—Л T. duro var.
reichenbachii Koern. spica nigra differt. (11).
Подрод I. Triticum
107
Определение ра шовидпос ieii snl>< onv.ir. durtini
Чешуи
6c.ll.iO крас- ные черные на фоне серо-ды мчатые на фоне цвета чешуи чер- ные
белом крас- ном белом крас- ном
Соломина под колосом н е о п у ш е н н а я
О Чешуи неопушенные с тп шероховаты с
1. Зерновки белые
4 — — — — — 4- — leucuriim 1
+ — — — — — — 4 leucomelan 2
— 4 — — — — 4- hordeiforme 3
— + —— — — — — 4 erythromelan 4
— — 4" — — — — 4- alboprovincialc
+ — 2. 3cpi овки к 4 :> ас in j е provinciate 6
4 — .— — — — 4- — uHine 7
4 — — — — — — 4 reichenbachii 8
— 4- — — — — 4- — murciense 9
— 4- — — — — — 4 pseudoalexandrinum 10
— — 4- — — — — 4 albobscurtim 11
— 4- 3 Зерно вки фи 4 алетов! obscuriim ic 12
+ - - 4 corvalanii 13
— 4- - Ост 1. 3ef и г л а )НОВКИ 4 д к и е белые alicnde 14
+ — — — — — — levileucurum 15
4- — — — —. —. — 4 levilcucomclan 16
— + — -— — — 4- — leuihordeiforme 17
4 — 2. 3epi 1ОВКИ к 4 расные levierythromelan 18
4- — - — — Д- levireichenbachii 19
4- - о — 1 — 1 4- 1 levialexandrinum Чешуи опушенные : т и шероховатые 1. Зерновки белые 20
+ — — — — — 4 — valenciac 21
— — — — — — 4 melanopus 22
— 4 —- —— — — 4- ' — italicum 23
— 4- — — — — — 4 apulicum 24
—— — 4- — —• — — 4 boeufii 25
— — —— 4- — — — 4- caeriilcscens 26
— — — —— 4- — 4 — cineraceum 27
— — — — + 2, 3epi овки к 4- расиыс rubromurinum 28
4- — — — — — . 4- — durum 29
4- — — — — — — 4“ africanum 30
— + — — — — 4 —• aegyptiacum 31
— 4- — — — — 4 nilolicuni 32
— —- 4 — — — — 4- inelaleuciitn 33
— — — + — . -1 liliycuni 34
О ст ii гладкие (зерновки белые)
4 — — — — — — 4 levimclanopiis 35
— 4- - — — — — — 4 leviapulicum 36
— — — 4 — — — 4 levicaerulesccns 37
108
Классификация рода Triticum L.
О к о и ч и и е
Чешуи Ос гн Разновидность раз- новидное! и
белые крас- ные черные на фоне серо-дымча- тые на фоне цвета чешуи чер- ные
белом крас- ном белом крас- ном
+ С о.ю л и н а под колосая опушенная Чешуи, неопушенные 1. Зерновки белые - - 1 + 1 — 1 jakubzineri 38
4- pilosohord ciforme 36
4' 2. Зерновки к - 1 + расньи pilosaffine I /10
+ + — pilosomurcicnsc 1 41
+ 1 — — 1ешуи опушенные (и грпонки белые) + pilosom eh i по ри s 1 и
Typus: Portugal. Reproductio stationis Daghestanskajae fulcracca,
WIR, k-42273 (WIR).
Геогр. распр. Португалия, Кипр, Египет, Азербайджан (редко).
Var. alboprovinciale Flaksb. var. nov.— T. durum var. alboprovinciale
Flaksb. 1935, l.c.: 206, descr. ross. — T. durum var. provinciate auct. non
Alef.: Vase. 1933, l.c.: 78. — A T. duro var. leucomelan (Alef.) Koern.
spica nigra differ!. (5).
Typus: Cyprus. Reproductio stationis Daghestanskajae fulcracea WIR,
k-47689 (WIR).
Геогр. распр. Португалия, Кипр, Азербайджан (редко).
Var. aliende Ruden. 1978, Бюл. ВИРа, 81: 41. (14).
Константная гибридная форма, выделенная нз образцов, полученных
из Чили.
Var. apulicum Koern. in Koern. et Wern. 1885, l.c.: 73. — T. durum gr.
apulicum (Koern.) Flaksb. 1935, I. c.: 214. (24).
Геогр. распр. Средиземноморские страны, СССР — Закавказье.
У этой разновидности различаются 3 формы:
f. epruinosapulicum Jakubz. et A. Filat, f. nov. — Tota planta eprui-
nosa.
Typus: Jordania, 1928, leg. Vavilov, k-18130 (WIR).
Растения без воскового налета. Привезена Н. И. Вавиловым из
Иордании.
Геогр. распр. Иордания, Египет.
f. fereapulicum Gokg. 1955, Ziraat Vck. 716: 92, descr. hire.
Ости слабошероховатые. Тип, вероятно, в Турции.
f. nigrapulicum Dekapr. f. nov. — T. durum var. nigroapulicutn
Dekapr. 1954, Tp. Ин-та полевод., 8: 30, descr. ross.— T. durum var.
apulicum f. nigroapulicuin Gokg. 1955, 1. c.: 92, descr. hire. Форма с чер-
ной окраской,края колосковой чешуи. Тип, вероятно, в Грузии.
Геогр. распр. Турция, Грузия.
Var. boeufii Flaksb. var. nov. — T. durum var. boeufii Flaksb. 1935,
1. c.: 217, descr. ross. — A T. duro var. melanopus (Alef.) Koern. spica
nigra differt. (25).
Typus: Morocco. Reproductio stationis Daghestanskajae fulcracca WIR,
k-44735 (WIR).
Г c o г p. p а с и p. Тунис, Марокко, Азербайджан.
Var cineraceuni Thum. 1934, Опред. хлебн. злаков (колос.): 207. —
Т. durum f. griseo-melanopus Gokg. 1955,1. c.: 92. (27).
Подрод I. Triticum
109
Гeo гр. распр. Армения, Турция.
Var. caerulesccns (Bayle-BarelIe) Koern. in Koern. et Wern. 1885,
I. c.: 73. — T. coerulescens Bayle-Barellc, 1809, Mon. agr. Cor.: 39. — T. du-
rum gr. coerulescens (Bayle-Barelle) Flaksb. 1935, 1. c.: 217. (26).
Геогр. распр. Средиземноморские страны. Закавказье, Даге-
стан, европейская часть СССР.
К этой разновидности относится:
f. ferecaerulescens Jakubz. et Anan. f. nov.— Spica aristis scabriusculis
vel fere levibus.
Typus: In stationc experimental! daghcstanica WIR e populatione culti-
var! Derbentskajae (k-32453) isolatum est (WIR). Ости слабошерохо-
ватые.
Геогр. распр. Выделена из посева сорта Дербентская черноко-
лосая на Дагестанской опытной станции ВИРа.
Var. corvalanii Rudcn. 1978, l.c.: 41. (13).
Геогр. распр. Константная гибридная форма пз коллекции пше-
ниц Чили.
Var. durum, — Т. durum var. faslttosum (Lag.) Koern. in Koern. et
Wern. 1885, l.c.: 72.— T. fastuosum Lag., 1816. Nov. gen. sp. pl.: 6.—
T. vulgare erion Alef. 1866, Landw. FL: 325. — T. durum gr. fastuosum
(Lag.) Flaksb. 1935, l.c.: 211. (29).
Геогр. распр. Средиземноморские страны, СССР — редко в Азер-
байджане и Дагестане.
Var. erythromelan Koern. in Koern. et Wern. 1885, 1. c.: 71. — T. durum
gr. erythromelan (Koern.) Flaksb. 1935, l.c.: 203. (4).
Геогр. распр. Средиземноморские страны, СССР — Закавказье,
Украина (редко).
К этой разновидности относится f. fereerythromelan (Nicol.) Dorof.
et A. Filat, comb. nov. — T. durum var. fereerythromelan Nicol, in Flaksb.
1929, Tp. прикл. бот. ген. сел. 22,2: 116. Форма со слабошероховатыми
остями.
Геогр. распр. Сирия.
Var. hordeiforme (Host) Koern. in Koern. et Wern. 1885, l.c.: 71.—
T. hordeiforme Host, 1809, Gram. Austr. 4: 3. — T. siculum Room, et Schult.
1817, Syst. veget.: 765. — T. durum gr. hordeiforme (Host) Flaksb. 1935,
1. c.: 201. (3).
Геогр. распр. Встречается и возделывается почти всюду, где
имеет место культура пшеницы твердой. Редко в Чехословакии (сорт
Adur).
К этой разновидности относится f. ferehordeiforme (Nicol.) Dorof. et
A. Filat, comb. nov. — T. durum var. ferehordeiforme Nicol, in Flaksb. 1929,
1. с.: 116. Форма co слабошероховатыми остями.
Геогр. распр. Малая Азия.
Var. italicum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 73. — T. vul-
gare italicum Alef. 1866, l.c.: 325. — T. durum gr. italicum (Alef.) Flaksb.
1935, l.c.: 213. (23).
Геогр. распр. Средиземноморские страны, СССР — Закавказье,
Дагестан.
Var. jakubzineri Kurk. 1976, Тр. прикл. бот. ген. сел., 58,2: 158. (38).
Геогр. рас'пр. Коллекция ВИРа. Отбор константных форм пше-
ницы твердой с опушенным под колосом стеблем из гибридного материа-
ла доктора Б. Ч. Дженкинса (Канада).
Var. leucomelan (Alef.) Koern. in Koern. et Wern. 1885, l.c,: 70.—
T. vulgare leucomelan Alef. 1866, l.c.: 324. — T. durum gr. leucomelan
(Alef.) Flaksb. 1935, 1. c.: 198. (2).
Геогр. распр. Средиземноморские страны, СССР — Закавказье.
К этой разновидности относятся следующие формы:
f. epruinosileucomelan Jakubz. et A. Filat, f. nov. — Tota planta eprui-
nosa.
110
Классификация рода Triticum L.
Typus: Jordania, 9. VI. 1926, leg. Vavilov, k-17230 (WIR).
Растения без воскового налета. Привезена Н. И. Вавиловым из
Иордании.
Геогр. распр. Иордания.
f. fereleucomelan (Palm.) Dorof. et A. Filat, comb. nov. — T. durum
var. fereleucomelan Palm, in Flaksb. 1929, l.c.: 117. Растения co слабо-
шероховатыми остями.
Г e о г р. р а с и р. Тунис, Марокко.
f. tristeleucomelan Gokg. 1955, 1. с.: 89, descr. tore.
Тип, вероятно, в Турции.
Форма с червой окраской края колосковой чешуи.
Геогр. распр. Турция, Азербайджан.
Var. leucurum (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 69.—
T. vulgare leucura Alef. 1866, l.c.: 324. — T. candidissinium Bayle-Barelle,
1809, l.c,: 42. — T. durum gr. leucurum (Alef.) Flaksb. 1935, l.c.: 196. (1).
Геогр. распр. Средиземноморские страны, Малая Азия, СССР —
Закавказье, Поволжье, Киргизия и как примесь в других районах.
К этой разновидности относятся формы:
f. epruinosileucurum Jakubz. et A. Filat, f. nov.—Tota planta
epruinosa.
Typus: Jordania, h-100 m, 1926, leg. Vavilov, к-17250 (WIR).
Растения без воскового налета. Привезена И. И. Вавиловым из
Иордании.
Геогр. распр. Иордания, Израиль, Малая Азия, Египет. Испа-
ния (Пиренеи).
f. fereleucurum Gokg. 1955, 1. с.: 89. descr. turc.
Ости слабошероховатые.
Тип, вероятно, в Турции.
Геогр. распр. Турция.
Var. levialexandrinum Jakubz. et Nicol, in Flaksb. 1929, l.c.: 116. (20).
Геогр. распр. Сирия.
Var. leviapulicum Dorof. 1968, Бюл. ВИРа, 13: 73. — T. durum f. le-
viapulicum Gokg. 1955, I. c.: 92, descr. turc. (36).
Геогр. распр. Турция, СССР — Азербайджан.
Var. levicaerulescens Jakubz. ct A. Filat, var. nov. —A T. duro var.
caerulescens (Bayle-Barelle) Koern. aristis levibus differ!. (37).
Typus: In stationc experimental! daghestanica WIR c populatione cul-
tivari Derbentskajae isolatum cst. 6. VII. 1973, к-47690 (WIR).
Формы с гладкими остями.
Геогр. распр. Выделена па посеве сорта Дербентская черноко-
лосая на Дагестанской опытной станции ВИРа.
Var. levierythrotnelan Dorof. var. nov. — T. durum var. erythromelan
f. levierythromelan Gokg. 1955, l.c.: 90, descr. turc. — A T. duro var.
erythromelan Koern. aristis levibus differt. (18).
Typus: Azerbajdzhan, regio Kubinski, 240 m, 1. VII. 1962, V. Dorofeev
(WIR).
Геогр. распр. Турция, Азербайджан.
Var. levihordeiforme Dorof. 1968, l.c.: 73. — T. durum f. levihordeiforme
Gokg. 1955, 1. c.,: 90, descr. turc. (17).
Геогр. распр. Турция, СССР — Азербайджан.
Var. levileucomelan Dorof. 1968. l.c.: 73. (16).
Геогр. распр. Азербайджан (Кубинский район).
Var. levileucurum Dorof. 1968, 1. с.: 72. (15).
Геогр. распр. Азербайджан (Дивичпискпй район).
Var. levimelanopus Dorof. var. nov. - 7’. durum var. levimelanopus
Dorof. 1972, 1. c.: 83, descr. ross. — Л T. duro var. melanopus Alef. aristis
levibus differt. (35).
Подрод I. Triticum 1 I 1
Typus: A zerl >;i j d/.l1 a 11, regio Scl i e 111 а с 11 i 11 s I; i, (>K() in, 4. VII. HM>1.
V. Dorofeev, (WIR).
Геогр. распр. Азербайджан (Шемахппский район).
Var. levireichenbachii Jakubz. 1932, l.c.: 38. (19).
Г e о г p. p а с н p. Единично в Сирии.
Var. libyctim Koern. in Koern. et Wern. 1885, l.c.: 73. — T. durum gr.
libycum (Koern.) Flaksb. 1935, l.c.: 218. (34).
Геогр. распр. Средиземноморские страны, СССР — Закавказье
(редкая примесь).
К этой разновидности относится форма f. epruinosilibyctun Jakubz.
et A. Filat, f. nov. —-Tota planta epruinosa.
Typus: Portugal. 1927, leg. Vavilov (WIR).
Растения без воскового налета.
Геогр. распр. Португалия.
Var. inelaleucum Dorof. var. nov. — T. durum var. melaleucum Dorof.
et Lapt. 1963, Вести, с.-х. науки, 11: 110, sync typo. — T. durum var.
nazilliense Gokg. 1955, l.c.: 94, descr. turc. — Л T. duro var. boeufii
Flaksb. caryopsis rubra differt. (33).
Typus: Azerbajdzhan, regio Schamchor, 650 m s. m., VII. 1961, leg.
Dorofeev et Laptev (WIR).
Геогр. распр. Малая Азия, Азербайджан.
Var. tnelanopus (Alef.) Koern. in Koern. et Wern. 1885, l.c.: 72.—
T. vulgare tnelanopus Alef. 1866, 1. c.: 325.— T. durum gr.tnelanopus (Alef.)
Flaksb. 1935, l.c.: 209. (22).
Геогр. распр. Средиземноморские страны, СССР.
К этой разновидности относятся формы:
f. epruinosimelanopus Jakubz. et A. Filat, f. nov. — Tota planta
epruinosa.
Typus: Jordania, 1926, к-17278, leg. Vavilov (WIR).
Геогр. распр. Иордания (редко).
f. feremelanopus (Jakubz.) Dorof. et A. Filat, comb. nov. — T. durum
var. feremelanopus Jakubz. 1932, 1. c.: 42.
Геогр. распр. Сирия.
V ar. murciense Koern. in Koern. et Wern. 1885, l.c.: 71.— T. durum
gr. murciense (Koern.) Flaksb. 1935, l.c.: 204. (9).
Геогр. распр. Средиземноморские страны (мало), СССР (редко
как примесь).
К этой разновидности относится f. feremurciense (Jakubz.) Dorof. et
A. Filat, comb. nov. — T. durum var. feremurciense Jakubz. 1932, l.c.: 101.
Геогр. распр. Турция, Югославия (Македония).
V ar. niloticum Koern. in Koern. et Wern. 1885, l.c.: 73. — T. durum
gr. niloticum (Koern.) Flaksb. 1935, l.c.: 216. (32).
Геогр. распр. Средиземноморские страны (редко как примесь).
К этой разновидности относится f. fereniloticum (Jakubz.) Dorof. et
A. Filat, comb. nov. — T. durum var. fereniloticum Jakubz. 1932, l.c.: 43,
Геогр. рас и p. Сирия.
V ar. obscuruin Koern. in Koern. et Wern. 1885, l.c.: 72; Jakubz. 1932,
l.c.: 42. — T. durum var. fulvoobscurum Vase. 1933, Bolet. Agric. I, 1—2,
ser. 1: 78. (12). ,
Геогр. распр. Северная Африка, Сирия, Израиль, Греция, Малая
Азия, о-в Сицилия, Южная Италия, Франция, Испания, Южная Африка
(ввезенная), СССР — редко как примесь.
Var. pilosaffine Kurk. 1970, Сб. тр. аеппр. ВИР, 17: 40. (40).
Геогр. распр. Коллекция ВИРа. Отбор константных форм
Т. durum с опушенном стебля под колосом из гибридного материала
доктора Б. Ч. Дженкинса (Канада).
Var. pilosohordeiforme Kurk. 1976, Тр. прикл. бот. ген. сел., 58,2:
158. (39).
И? Классификация рода Triticum I..
Геогр, р а с и р. Коллекция ВИРа, 1010 же происхождения, чю и
\ ar. pilosaffine.
Var. pilosoinelanopus Kurk. 1970, l.c.: 41. (42).
Геогр. распр. Коллекция ВИРа, того же происхождения, что
и var. pilosaffine.
Var. pilosomurciense Kurk. 1970, 1. с.: 41. (41).
Г e о г р. р а с и р. Коллекция ВИРа, того же происхождения, что
и var. pilosaffine.
Var. provinciale (Alef.) Koern. in Koern. et Wern. 1885, 1. c.: 71; Jakubz.
1932, l.c.: 41. — T. vulgare provinciale Alef. 1866, l.c.: 325. — T. durum
gr. provinciale (Alef.) Flaksb. 1935, l.c.: 206. — T. durum var. fulvopro-
vinciale Vase. 1933, 1. c.: 78. (6).
Геогр. распр. Средиземноморские страны, СССР — редко в За-
кавказье. К этой разновидности относится f. fereprovinciale Jakubz. f.
nov. — T. durum var. fereprovinciale Jakubz. in Flaksb. 1935, l.c.: 207,
descr. ross. — Л T. duro var. provinciale (Alef.) Koern. aristis fere levibus
differt.
Typus: Syria, 1928, к-17195 (WIR).
Г e о г p. p а с и p. Сирия.
Var. pseudoalexandrinum Flaksb. 1922, Опред. наст, хлебов: 55, non
Flaksb. in Schreib. 1934, nec Flaksb. et Schreib. in Flaksb. 1935.—
T. durum var. alexandrinum (Koern.) Perciv. 1921, Wheat pl.: 225, p.p.—
T. durum gr. alexandrinum (Koern.) Flaksb. 1935, l.c.: 205, p.p. (10).
Вместо названия pseudoalexandrinum нередко используется var.
alexandrinum Koern. (J. Papadakis, 1929; J. Vasconcelos, 1933 и др..),
хотя последнее наименование было дано Ф. Кёрнике для разновидности
с коротким плотным колосом из Египта (convar. durocompactum).
К var. pseudoalexandrinum относятся формы:
f. epruinosalexandrinum Dorof. et A. Filat, f. nov. — Tota planta
epruinosa.
Typus: Syria, к-17103, 1926, leg. Vavilov (WIR).
Растения без воскового налета.
Геогр. распр. Сирия, Иордания (редко).
f. ferealexandrinum (Jakubz. et Nicol.) Dorof. et A. Filat, comb. nov.—
T. durum var. ferealexandrinum Jakubz. et Nicol, in Flaksb. 1929, 1. c.:
116. — Ости слабошероховатые.
Геогр, распр. Сирия.
f. variegatum (Kob.) Dorof. et A. Filat, comb. nov. — T. durum var.
variegatum Kob. 1932, Хлебн. злаки Турции: 13. — Формы с черной ок-
раской края колосковой чешуи.
Геогр. распр. Турция.
Var. reichenbachii Koern. in Koern. et Wern. 1885, 1. c.: 71. — T. durum
gr. reichenbachii (Koern.) Flaksb. 1935, l.c.: 200. (8).
Геогр. распр. Средиземноморские страны, СССР — Северный
Кавказ, юг Украины, Крым, Закавказье.
К этой разновидности относятся формы:
f. epruinosireichenbachii Jakubz. et A. Filat, f. nov. •—Tota planta
epruinosa.
Typus: Syria, к-17101, 1926, leg. Vavilov (WIR).
Растения без воскового налета.
Геогр. рас п р. Сирия, Иордания, Египет.
f. ferereichenbachii (Jakubz. et Nicol.) Dorof. et A. Filat, comb. nov.—
T. durum var. ferereichenbachii Jakubz. et Nicol, in Flaksb. 1929, l.c.: 116.
Ости слабошероховатыс.
Геогр. распр. Сирия.
f. discolor (Kob.) Jakubz. et A. Filat, comb. nov. — T. durum var.
discolor Kob. 1932, l.c.: 12. Форма с черной окраской края колосковой
чешуи.
Подрод 1. Trilicum 113
Гео г р. р а с п |>. Турция.
Var. rubromurinum Menabde, 1948, Ишеп. Груз.: (i(i, descr. ross.—
T. durum f. griseoapulicum Gokg. 1955, 1. c.: 92, descr. tore. (28).
Геогр. распр. Турция, Грузия (Коди, Асурети).
Var. valenciae Koern. in Koern. et Wern. 1. c.: 72. — T. durum gr. va-
lentine (Koern.) Flaksb. 1935, 1. c.: 208. (21).
Геогр. pa сир. Как редкая примесь в ареале var. melanopus.
В Турции М. Гекгель выделил ряд форм с серо-дымчатой окраской
колоса (Г. durum f. griseoafricanum Gokg. 1955, 1. с.: 92, descr. turc.;
T. durum f. griseoitalicum Gokg. 1955, l.c.: 92, descr. turc.; T. durum
f. griseoleucomelan Gokg. 1955, l.c.: 89, descr. turc.; T. durum f. griseoni-
loticum Gokg. 1955, l.c.: 92, descr. turc.), которые следовало бы рассмат-
ривать разновидностями пшеницы твердой. Однако новые комбинации
невозможны из-за неэффективного обнародования этих таксонов и от-
сутствия в коллекции ВИРа экземпляров для выбора лектотипов.
б. Subconvar. muticum (Orlov) Dorof. et A. Filat. — пшеница твердая
безостая.
Subconvar. muticum (Orlov) Dorof. et A. Filat, subconvar. nov.—
T. durum p) muticum Orlov, 1923, Tp. прикл. бот. сел. 13,1: 372.
Безостые твердые пшеницы, полученные селекционерами от скрещи-
вания обыкновенной остистой твердой пшеницы с различными формами
других видов пшениц, а также в результате спонтанной межвидовой
гибридизации.
Т и и: Т. durum var. ausirale Perciv.
Геогр. распр. Селекционные учреждения СССР, Австралии, Ту-
ниса.
Определение разновидностей subconvar. muticum (Orlov) Dorof. et Л. Filat.
Чешуи
черные на
фоне
серо-лым»
Остистые
заострения
на (|юре
Разновидность
X
X
Чешуи неопушенные
Зерновки белые
4
candicatis
subaustrale
muticalbopro-
vincialc
australe
1
2
9
10
И
12
muticovalenciae
muticitalicum
miilicoboeulii
inuticocaenilesccns
Зерновки красные
— | — i — | muticonazilliense | 13
8 762
Var. anstrale Perciv. 1921, Wheal pl.: 229. T. durum gr. mulii о
provinciate Flaksb. 1935, Культ. ф.л. СССР, I: 222. (4). Описана Дж.
Персивалем по австралийскому’ сорту Huguenot, полученному путем
скрещивания пшеницы твердой с мягкой.
Геогр. распр. Незначительные хозяйственные посевы были от-
мечены в начале XX в. в Австралии и Тунисе,
Получена путем скрещивания Т. durum var. subausfrale Perciv.
с местными твердыми пшеницами в Азербайджане.
Var. candicans Meist. 1922, Жури, опыта, агрои. Юго-Востока 1.1:
55. — Т. durum gr. mu.ticoleucurum Flaksb. 1935, l.c.: 219. (1).
Получена па Саратовской обл. с.-х. опыт, станции (ныне HIIIICX
Юго-Востока) путем скрещивания Г. durum с Т. aestivum. В 1924 г.
обнаружена П. Гребенниковым is хозяйственных посевах Донской сел.
станции (ныне Донской НИИСХ). Формы этой разновидности выделены
в Тунисе из гибридного сорта Huguenot. В. Ф. Дорофеевым отмечена
как редкая примесь в сложных популяциях пшениц Закавказья.
Var. nniticalbobscurum Flaksb. var. nov. — T. durum gr. muticoal-
boobscurum Flaksb. 1935, l.c.: 222. descr. ross. — T. durum var. muti-
coalboobscurum Gokg. 1955, l.c.: 93. descr. lure. — A 7’. duro var. alboob-
scurum Flaksb. spica mutica differt. (7).
Typus: Tunis, 1919, к-15485 (WIR).
Выделена в Тунисе из сорта Huguenot как форма (NB) R-gM 14-1
(F. Boeuf, 1925).
Var. muticalboprovinciale Flaksb. 1935, l.c.: 222, descr. ross. (3).
Выделена в Tvnnce из сорта Huguenot как формы (NB) A-gM п
(BN) A-gM (F. Boeuf, 1925).
Var. inuticitalicuin Dorof. et A. Filat, var. nov. — T. durum gr. muti-
coitalicum Flaksb. 1935, l. c.: 223, descr. ross. — A T. duro var. italicum
(Alef.) Koern. spica mutica differt. (10).
Typus: Tunis. Reproductio stationis Daghestanskajae fulcracea WIR,
1978, к-16526 (WIR).
Выделена в Тунисе из сорта Huguenot как форма RA-PM (F. Boeuf,
1925). В Азербайджане акад. И. Д. Мустафаев выделил в гибридном
материале Т. durum var. subaustrale'X.T. durum var. apulicum.
Var. muticoboeufii Flaksb. 1935, 1. c.: 223, descr. ross. (II).
Выделена в Тунисе из сорта Huguenot как форма (NB) А-РМ
(F. Boeuf, 1925).
Var. muticocaerulescens Flaksb. 1935, l.c.: 223, descr. ross. (12).
Выделена в Тунисе из сорта Huguenot как формы (NB) А-РМ и
(RM) А-РМ (F. Boeuf, 1925). Очень редкая примесь в посевах пшеницы
твердой в Азербайджане.
Var. muticonazilliense Gokg. 1955, l.c.: 94, descr. hire. (13).
Тип, вероятно, в Турции.
Геогр. распр. Турция.
Var. muticobscurum Dorof. et A. Filat, var. nov. — T. durum gr.
muticoobscurum Flaksb. 1935, l.c.: 222, descr. ross. — A T. duro var.
obscurum Koern. spica mutica differt. (8).
Typus: Tunis. Reproductio stationis Daghestanskajae fulcracea WIR,
6. VI. 1968, к-1,6485, leg. Soskov (WIR).
Выделена в Тунисе из сорта Huguenot как форма (RN) R-GM
(F. Boeuf, 1925).
Var. iniiticovalenciае Dorof. et A. Filat, var. nov. — T. durum gr. muti-
covaleticiae Flaksb. 1935, 1. c.: 223, descr. ross. — i\ T. duro var. valenciae
Koern. spica mutica differt. (9).
Typus: Kazakhstan. Reproductio stationis Daghestanskajae fulcracea,
WIR, 6. VI. 1968, к-42335, leg. Soskov (WIR).
Iltulpoi) 1. Triliiiun J 15
Выделепи в Тунисе ч.ч сорта Huguenot как форма ЛЛ FM (I;. Boeuf,
1925), редко встречается в Казахстане.
V ar. schechurdinii Moist. 1922, l.c.: 55. - - T. durum gr. miiticoaljine
Flaksb. 1935, l.c.: 220. T. durum var. pscudoschechurdini Greb. 1929,
Опред. разной, шпон. Повочерк.: 26. (5).
Получена таким же образом, как и var. candicans Moist. (Саратов-
ская область). П. Гребенников (1929) указывает па редкое нахождение
в Ростовской области. В Тунисе выделена из сорта Huguenot как фор-
ма AR-GM (F. Boeuf, 1925).
V ar. slebutii Moist. 1922, l.c.: 55. — T. durum gr. muticomurdense
Flaksb. 1935, 1. c.: 221. (6).
Получена таким же образом, как и var. candicans Moist. (Саратов-
ская область). В Тунисе выделена из сорта Huguenot как форма RR-GM
(F. Boeuf, 1925).
V ar. subaustrale Perciv. 1921, l.c.: 229.— T. durum gr. muticohordei-
forme Flaksb. 1935, l.c.: 220. (2).
Происхождение и распространение таковы же, как у Т. durum var.
ausirale Perciv. Азербайджан (очень редкая примесь в посевах пшеницы
твердой).
Кроме указанных разновидностей, в литературе под латинскими на-
званиями приводится множество безостых гибридных форм пшеницы
твердой без указания их поколения и константности (П. Гребенников,
1926, 1929; М. Терповский, 1929).
в. Subconvar. duroramosuin Dorof.—пшеница твердая ветвистоко-
лосая.
Subconvar. duroramosuin Dorof. subconvar. nov. — T. durum convar.
duroramosuin Dorof. 1972, Tp. прикл. бот. ген. сел. 47,1: 76, nom. nud.—
A T. duro subconvar. durum spica ramosa differt.
Typus: T. durum var. dolgushinii Jakubz.
Отличается от T. durum subconvar. durum ветвнстоколосостыо.
Формы с крупными ветвистыми колосьями были обнаружены экспе-
дициями ВИРа в Закавказье. Происхождение их, возможно, связано со
спонтанной гибридизацией Т. durum с ветнпстоколосыми формами
Т. turgidum, произрастающими как примесь в посевах пшеницы твердой
(В. Ф. Дорофеев, 1970).
Геогр. распр. Предгорные районы Азербайджана и Грузии, Ка-
захстан (редко). Кроме того, выделены па коллекционных посевах Да-
гестанской и Павловской опытных станций ВИРа.
Var. dolgushinii Jakubz. 1950, Тр. прикл. бог. ген. сел. 28,3: 66.—
Т. durum var. ramosomelanopus Dorof. 1970, 1. с.: 306. (4).
Геогр. распр. Закавказье (редко).
Var. ramosafricanum Dorof. 1970, Тр. прикл. бот. ген. сел. 43,1:
307. (6).
Геогр. распр. Грузия (редко).
Var. ramosapulicum Jakubz. 1953, Докл. ВАСХНИЛ, 1: 46. (5).
Выявлена на посеве сорта Акмолинка 2 (Казахстан) и найдена
в Закавказье (редко).
Var. ramosohordeiforme Jakubz. 1953, l.c.: 46. (2).
Выявлена в Ленинградской области на посеве сорта Гордеиформе 10
и местного сорта из Гиндукуша, найдена в Азербайджане.
Var. ramosoleucomelan Jakubz. 1953, 1. с.: 46. (1).
Выявлена па посевах copra Mrari (Иордания) на Дагестанской опыт-
ной станции Bl IРа.
Var. rainosobsciirum Jakubz, 1953, I. с.: 46. (3).
Л'.-inci'iiфиuui{ii.4 рода Trilicum I..
Определение p;i шопидпостеи subconvar. clnroraiuosnin Dorof.
Чешуи Ости Ра нпжиАность № разно- видности
белые белые с черными краями крас- ные красные С черными краями черные на фоне инета чешуй чер- ные
белом крас- ном
4etui/u iieonytiieiimiie
Зерновки бел ы e
Зерновки к p а с н ы с
Г Г I - I +
Чешуи ош/шенные
3 е р и о в к и б е ,ч ы е
Зерновки красные
ramosoleu-
comelan
ramosohor-
deiforme
raniosobs-
ciirum
1
2
d ul у и shi nii
ramosapuli-
cum
1
5
ramosafri-
сипит
6
Выделена на посеве турецкого сорта Чехил-бугдай в Дагестане,
найдена в Закавказье (редко).
2. Convar. durocompactum Flaksb. — пшеница твердая плотноколосая.
Convar. durocompactum Flaksb. 1922, Опред. наст, хлебов: 52.—
Т. durum sect, africana Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51:
216, р.р. — Т. durum durocompactum Flaksb. 1935, Культ, фл. СССР, 1:
189, excl. prol. horanicum.
Растения низкорослые (меньше 90 см) и среднерослые (90—120 см).
Колосья очень плотные (D больше 40), 5—6 см длины. Длина колоса
лишь в 3—5 раз более его толщины. Колос иногда пирамидального по-
строения; ости длинные, зерновки овальные или удлиненно-овальные
(6—9 мм длины, 3—4 мм ширины).
Т и п: Т. durum var. alexandrinum Koern.
Геогр. распр. Алжир, Тунис, Марокко, Египет, Сирия, Иорда-
ния, ЛАалая Азия, Азербайджан.
Пшеница твердая плотноколосая дифференцирована на следующие
эколого-географические группы.
1. Африканская группа — Т. durum, prol. africanum Vav. 19S4, Миров,
ресурсы хлебн. злаков. Пшеница: 40. Характеризуется мощным разви-
тием растений с прочной соломиной и хорошей облпствепностью. Колос
нередко пирамидальной формы, короткий, плотный. Устойчива к бурой
и желтой ржавчине.
Среди плотноколосых твердых пшениц по ряду морфологических
признаков выделяется подгруппа с булавовидным колосом [Г. durum
тип cuspididimtatum Kob. 1932, Хлебн. злаки Турпин: К).- Т. durum
subprol. cuspididentatum (Kob.) Flaksb. 1935, l.c.: 191], т. с. с короткими
колосьями, плотность которых увеличивается к вершине; колосковые
чешуи широкие, с острым зубцом, вверху колоса переходящим в корот-
кую ость (остевидный зубец). Ости длинные.
ПоО/юО !. Triticum
117
Геогр. распр. Малая Азия, Сирия, Л\арокк<>, Азербайджан.
К африканекой группе относятся и переходные формы между ти-
пичными (convar. durum) и плотноколосыми (convar. ditrocompaclum)
пшеницами твердыми промежуточная подгруппа \Т. durum inn rurum
Kob. 1932, 1. с.: 11. — T. durum subprol. rarum (Kob.) Flaksb. 1935, l.c.:
190].
Промежуточная подгруппа отличается от средиземноморских твер-
дых пшениц (convar. durum) более коротким и плотным колосом.
В Турции встречается как очень редкая примесь. Судя по неполному
описанию, к промежуточной подгруппе относятся также переходные
формы между convar. durum и convar. durocompactum, выделенные как
Т. durum prol. marocccmum Flaksb. el Jakubz. in Flaksb. 1935, l.c.: 191,
closer, ross., распространенные по всему ареалу convar. durocompactum.
2. Египетская группа — T. sativum pyramidale Delile, 1813, Descrip,
egypt. 9: 181. — T. pyramidale Perciv. 1921, Wheat pl.: 262, quoad plantam
Egypt.— durum aegyptiaca Vav. 1931, Tp. прикл. бот. ген. сел. При-
лож. 51: 227. — Т. durum prol. aegyptiacum Vav. in Flaksb. 1935, Культ,
фл. СССР, 1: 193; Вавилов, 1964, l.c.: 40. Растения низкорослые, со-
ломина тонкая. Колосья короткие, плотные, большей частью пирами-
дальные. По плотности колоса имеются различия между иижнесгииег-
ской подгруппой (преимущественно плотноколосые) и верхиеегипетской
(более рыхлоколосые). Восприимчивы к желтой ржавчине, скороспелые.
К данной группе близки плотноколосые пшеницы Синайского полуост-
рова и Иордании, которые, по-видимому, имеют египетское происхож-
дение. Они были выделены М. М. Якубципером (1932) как сипайская
группа (Т. durum subprol. sinaicum Jakubz. in Flaksb. 1935, l.c.: 193)
и в отличие от типичных египетских пшениц являются более устойчи-
выми к бурой и желтой ржавчине. Меньшей восприимчивостью к чтим
видам ржавчины характеризуются плотноколосые твердые пшеницы
Кипра, по комплексу признаков близкие к египетским.
Геогр. распр. Египет, Кипр, редко в Иордании.
3. Степная группа—Т. durum prol. durocompactum-stepposum Must,
et Kovdysch. 1970, Вести, с.-х. науки, 6: 18. Растения 80—НО см высоты.
Колос 4—7 см длины, плотный, пирамидальный или почти пирамидаль-
ный; ости почти в 2 раза длиннее колоса, расходящиеся; колосковые
чешуи овальные или удлиненно-овальные, преимущественно с острым
прямым зубцом. Зерновки овальные или удлиненно-овальные.
Геогр. распр. Предгорные районы Азербайджана.
Var. alexandrinum Koern. in Koern. et Wern. 1885, Handb. Getreideb. 1:
71. — T. pyramidale var. pseudomudanianum Haciz. 1932, l.c.: 76.—
T. durum var. pseudoalexandrinum auct. non Flaksb.: Schreib. 1934, Tp.
прикл. бот. ген. сел. 5, 2: 154. — Т. durum gr. alexandrinum var. pseudo-
alexandrinum Schreib. et Flaksb. in Flaksb. 1935, Культ, фл. СССР, 1:
205. (10).
Геогр. распр. Средиземноморские страны (редко), Малая Азия.
К этой разновидности относится f. tristemudanianum Gokg. 1955, l.c.:
91, descr. hire. — форма с черпоокрашенными краями колосковых чешуй
описана М. Gokgol (1955) по образцам из Турции.
Var. aydinien$e (Haciz.) Gokg. 1955, Ziraat Vekal, 716: 89. — T. pyra-
midale var. aydiniense Haciz. 1932, Ziicht. 4,3: 76. — T. durum var. pseu-
doleucomelan Schreib. 1934, 1. c.: 148. (2).
Г co г p. p a c ii p. Киир и Сардиния, .Малая Азия.
К этой разновидности относится 1. tristeaydinien.se Gokg. 1955. l.c.:
89 —форма с черпоокрашенными краями колосковых чешуй. Ее выде-
лил М. Gokgol (1955) по образцам н.з Турции.
Var. flaksbergeri (Zhuk.) Jakubz. comb. nov. — T. durum gr. libycum
f. flaksbergeri Zhuk. 1924, Зап. науч,- прикл. отд. Тифл. Бот. сада 3:
15. (20).
118
Классификация рода Trifictiui К.
Определение разнопидностей convar. durocompactuni I'laksb.
Чешуи Ости Разновидность № разно- видности
белые крас- ные черные на фоне серо-дым- чатые на фоне цвета чешуи чер- ные
белом крас- ном белом крас- ном
К О Л о с. и я остист ы с
Чешуи неопушенные
Ости ще р о х о в а т ые
1. Зерновки белые
+ — — — — — + — serengei 1
+ — — — — — — -1- aydiniense 2
— + — — — — + — pseudohor deiforme 3
— + — — — — — + rubrinigruin 4
— — + — — — — -1- pscudoalboprovin dale 5
— — — + — — — -1 pseiidoprovinciale 6
2. Зерновки красные
.+ — — — — — -1- — pseudoaffine i
.+ — — — — — — 4- pseiidoreichen bachii 8
— + — — — — + — pseudoiniircien.se 9
— + —»- — — — — 4- alexandrinum 10
— — + — — — — 4- pscudoalbobsciiriim 11
—- — — — — — 4- pseudoobscurum 12
Чешуи опушенные
Ости ш е ро х о в а т ыо
1. Зерновки белые
+ — — — — — 4- — pseudoualenciue 13
4- — — — — — — -1- pseudomelanopus 14
— + — — — — 4- — pse.udoiialicum 15
— + — — — — — 4- pscudoapidicum 16
— -— — — — — 4- pseudocaerulescens 17
2. Зерновки красные
4- — — — — — -1- — pseudofasliiosum 18
4- — — — — — — 4- 1 virsaviacum 19
— — — — — — 4- 1 flaksbcrgcri 20
Колосья безостые
Чешуи неопушенные
О с т н in е р о х о в а т ы е (верновкп белые)
3' I — ; — i — ! — | — । — । yesilkoense 21
Ге о г р. р а с п р. Восточная Грузия.
Var. pseudoaffine (Haciz.) Schreib. 1934, l.c.: 147, non. Greb. 1829.—
T. pyramidale var. pseudoaffine Ilaciz. 1932, l.c.: 76. — T. durum var.
affine f. pseudoaffine (Haciz.) Gokg. 1955, 1. c.: 90. (7).
Геогр. распр. Кипр, Турция.
Var. pseudoalbobsctirum Flaksb. cl Schreib. in Flaksb. 1935, 1. c.:
207. — T. durum var. pseudoobscurum f. 2 Schreib. 1934, l.c.: 157. (11).
Геогр. рас и p. Кипр.
Var. psetidoalboprovinciale Flaksb. el Schreib. in Flaksb. 1935, l.c.:
206. — T. durum var. pseiidoprovinciale 1. 2. Schreib. 1931, l.c.: 156. (5).
Геогр. распр. Кипр, Азербайджан (Зака ra/incKini и К у би иски ii
районы).
Var. psetidoaptilicuin Flaksb. 1922, Опред. паст, хлебов: 59. - - Т. pyra-
midale var. pseudocopticum Perciv. 1921, Wheat PI.: 264, quoad pl.
aegypt. — T. pyramidale var. falsoapulictim Slot. 1929 -1930, Tp. прикл.
бот. геп. сел. 23,4: 126. (Ki).
Подрос) !. Triticum
119
Геогр. распр. I? liner, Испания, Кипр, Армения.
Var. pseiidocaeriilescens (Haciz.) Schreib. 1934, l.c.: 162 - - T. pyrami-
dale var. pseiidocoerulescens Haciz. 1932, l.c,: 76. -T. durum var. coeru-
lescens f. psetidocoertilescens (Haciz.) Gokg. 1955, l.c.: 93. (17).
Геогр. распр. Кипр, Турция.
Var. psetidofastiiosiiin Orlov, 1922 -1923, Tp. ирикл. бот. сел. 13,1:
375. — 7’. pyraniidale var. compression Perciv. 1921, l.c.: 264, quoad pl.
aegypt.- -7’. pyramidale var. arabietim Perciv. 1926, Jonrn. Bot.: 209. (18).
Гeо гp. p a сп p. Средиземноморские с траны.
Var. pseudohordeifornie Flaksb. 1922, l.c.: 59. — T. pyramidal? var.
rubrospicalnm Stol. 1929-1930, l.c.: 125. — T. pyraniidale var. diyarbeki-
ricuin Haciz. 1932, l.c.: 72. (3).
К этой разновидности относится f. ferepseudohordeiforine Gokg. 1955,
l.c.: 92, descr. turc. co слабошероховатыми остями, описана по образ-
цам из Турции.
Геогр. распр. Северная Африка, Турция.
Var. pseudoitalicuni Orlov, 1922 -1923, l.c.: 376. - -T. pyramidale var.
copt icn in Perciv. 1921, l.c.: 264, quoad, pl. aegypt. - 7'. pyraniidale var.
rubroi'eluliniim Stol. 1929 1930, l.c.: 126. (15).
Геогр. p a c ii p. Северная Африка, в СССР встречаются в юрпом
Крыму, Армении.
Var. serengei (Aschers. et Graebn.) Dorof. et A. Filat, comb. nov. —
7'. tenax B. 11 durum leiiciirum serenpei Aschers. et Graebn. 1901, Syn.
Mitteleur. 2: 693. -T. durum var. pseudo!euetirtiin Orlov, 1922 1923, l.c.:
375. — 7’. pyraniidale var. recognition Perciv. 1921, l.c.: 263, quoad pl.
aegypt. (1).
Геогр. распр. Египет, Сирия, Израиль, Северная Африка, Ма-
лая Азия. К этой разновидности относится f. niustafaevii Jakubz. f.
nov. — Spica compacta, clavata.
Typus: Azerbajdzhan, 30.VI. 1975, leg. Mustafaev (BAK). Форма
с булавовидным (скверхедным) колосом. Признак булавовидное!н ко-
лоса неустойчив и при пересеве, часто пе сохраняется.
Var. pseudomelanopus Flaksb. 1922, l.c.: 59. - - T. pyraniidale var.
pseudo-compressum Perciv. 1921, 1. c.: 261, quoad pl. aegypt. — 7’. durum
var. nielanopus f. pseudomelanopus Gokg. 1955, I. c.: 92. (14).
Г e о г p. p а с и p. Египет, Малая Азия.
Var. psetidomurciense Flaksb. 1922, l.c.: 59. — 7'. pyramidale var.
rtibroseniineuni Stol. 1929—1930, 1. c.: 125.--7’. pyraniidale var. niudania-
num Haciz, 1932, 1. c.: 76. (9).
Г e о г p. p а с п p. Египет, Турция, Армения.
Var. pseudoobscuruui Schreib. 1934, l.c.: 157. (12).
Г с о г p. p а с и p. Кипр.
Var. pseudoprovinciale Schreib. 1934, l.c.: 156. (6).
Геогр. распр. Кипр, Греция.
Var. pseudoreichenbachii (Haciz.) Schreib. 1934, l.c.: 151. — T. pyra-
midal? var. pseudoreichenbachii Haciz. 1932, I. c.: 76. (8).
Геогр. распр. Кипр, Турция.
Var. pseudovalenciae Orlov, 1922 — 1923, l.c.: 375. — T. pyramidale
var. compressuni, Perciv. 1921, 1. c.: 264, quoad pl. aegypt. (13).
Г e о г p. p а с п p. Северная Африка.
Var. riibrinigrtiin (Stol.) Dorof. et A. Filal. comb. nov. 7'. pyramidale
var. riibronigriim Slol. 1929 1930, l.c.: 125. /'. pyraniidale var. pseudo-
diyarhehiricum Haciz. 1932, I. <-.: 76. (I).
Геогр. рас up. Малая Азия, Кипр, Eiiinci; в СССР Армения.
Var. virsaviactitu Jakubz. 1932, Тр. прикл. бог. icn. сел. Прилож. 53:
64. -7'. durum var. pseudoaf ricanum Schreib. 1934, l.c.: 161. (19).
Геогр. p а с п p. Киир, Сирия, Иордания.
Var. yesilkoense Gokg. 1955, I. c.: 93, descr. turc. (21).
120
Кмгсификиi{ii:i рода Triticiini
Г с о г р. р а с. и р. Турция.
М. Гскгель обнаружил в Турции плотноколосую торцую пшеницу
с серо-дымчатой окраской колоса, которую выделил как Т. durum f. grt-
seocotnpresstim Gokg. 1955, I. с.: 91, descr. lure.
3. Convar. aglossicon (Flaksb.) Dorof. et A. Filat. — пшеница твердая
безлигульная.
Convar. aglossicon (Flaksb.) Dorof. et A. Filat, comb. nov. T. durum
eligulatum Flaksb. 1926, Tp. прикл. бот. ген. сел. 16,3: 135. — Т. durum
subprol. eligulatum Flaksb. 1935, Культ, фл. СССР, 1: 185.
Растения низкорослые (60—90 ем) с тонкой непрочной соломиной.
Облиствеипость небольшая. Листовое влагалище без перегиба перехо-
дит в листовую пластинку. Листовая пластинка узкая, яЛычок и ушки
отсутствуют или сильно редуцированы. Колосья веретеповидиыс, рых-
лые или средней плотности.
Тип: Т. durum var. aglossicon Flaksb.
Экологически соответствуют кипрским пшеницам Т. durum convar.
durum. Скороспелые, восприимчивы к поражению желтой ржавчиной.
Продуктивность невысокая как па Кипре, так и при испытании в южных
районах СССР.
Геогр. распр. Кипр.
Определение разновидностей convar. aglossicon (Flaksb.) Dorof. et A. Filat.
Чешуи Ости Разновидность № разно- видное ти
белые белые с черными краями крас- н ые красные с мерными краями черные на фоне цвета чешуи крас- ные
белом крас- ном
Чешуи неопушенные
Зерновки белые
1
quasilcucurum
quasileucome-
lun
quasiliordei-
forme
quasieryth.ro-
mclan
3
4
Зерновки красны
10
quasiaf rica-
mim
e
+
Var. aglossicon Flaksb. 1926, Tp. прикл. бот. сел. 16,3: 131. (6).
Геогр. распр. Кипр.
Var. quasiaffine Flaksb. ex Schreib. 1934, Tp. прикл. бот. юн. сел. сер.
5, 2: 143. (5).
Геогр. распр. Кипр.
Var. qtiasiafricantiin Flaksb. 1929, Тр. прикл. бот. ген. сел. 20: 97;
Schreib. 1934, l.c.: 145. (10).
Подрод I. Triliaim
121
Геогр. распр. Кипр.
Var. qnasialexandrintiin F’laksb. ex Schreib., 1934, I. c.: 145. (8).
Г e о г p. p а с и p. Кипр.
Var. quasierythromelan Flaksb. var. nov.- -T. durutn var. quasieryth.ro-
inelan Flaksb. 1935, l.c.: 204, descr. ross. — Л T. duro var. erythromelan
Koern. foliis eligulatis differt. (4).
Typus: Cyprus, к-17849, И.IV. 1926, leg. Vavilov (WIR).
Геогр. распр. Кипр.
Var. quasihordeiforme Flaksb. 1929, l.c.: 97; Schreib. 1934, l.c.:
145. (3).
Г e о г p. распр. Кипр.
Var. quasileucoinelan Flaksb. 1929, 1. c.: 97, Schreib. 1934, 1. c.: 143. (2).
Геогр. распр. Кипр.
Var. quasileucurum Flaksb. 1929, l.c.: 97; Schreib. 1934, l.c.: 143. (1).
Геогр. распр. Кипр.
Var. quasimelanopus Flaksb. 1929, l.c.: 97; Schreib. 1934, l.c.: 145. (9).
Геогр. p а с п p. Кипр.
Var. quasiintirciense Flaksb. 1929, l.c.: 97; Schreib. 1931, l.c.: 145. (7).
Г e о г p. p а с п p. Кипр.
4. Convar. villosum (Jakubz.) Dorof. et Л. Filat. — грубоколосая па-
лестинская группа разновидностей.
Convar. villosum (Jakubz.) Dorof. et A. Filat, comb. nov. — T. durum
villosa. Jakubz. 1932, Tp. прикл. бот. ген. сел Прилож. 53: 56. — Т. durum
prol. villosum (Jakubz.) Flaksb. 1935, Культ, фл. СССР, 1: 187. — Т. du-
rum prol. palestinicum Vav. 1964, Миров, ресурсы хлебн. злаков. Пшени-
ца: 41, р.р.
Всходы п листья опушенные, покрыты сильным восковым налетом.
Опушение листвы сохраняется па все время вегетации. Колосья рыхлые
(D=25), ости толстые, грубые. Колосковые чешуи опушены густыми
и длинными волосками. Восковой налет сохраняется па колосе и после
созревания. Зубец колосковой чешуи у многих растений на вершине
колоса остевидный. По грубости-колоса и остей эта группа разновидно-
стей аналогична ксероморфным грубоколосым мягким пшеницам из
Юго-Западной Азии, отличаясь от последних легкой вымолачивае-
мостыо зерна.
Грубоколосые палестинские твердые пшеницы экологически одно-
родны, биологически нередко отличаются от иорданской подгруппы
(convar. durum)-, скороспелые, восприимчивые к грибным заболева-
ниям.
Геогр. распр. Прибрежные равнины и предгорья Сирии, Иорда-
нии, Ливана, очень редко на Кипре.
Var. villosaffine Dorof. et A. Filat, var. nov.—A T. duro var. affine
Koern. pubescentia totae plantae densiore differt. (4).
Typus: Israel, k-17379, 1928, leg. Vavilov (WIR).
Геогр. распр. Сирия, Иордания, Ливан.
Var. villosafricannm Dorof. et A. Filat, var. nov. — A T. duro var.
africanum Koern. pubescentia totae plantae densiore differt. (8).
Typus: lordania, k-18127, 1928, leg. Vavilov (WIR).
Геогр. рас up. Сирия, Иордания, Ливан.
Var. villosapulicum Dorof. et A. Filat, var. nov. — A. T. duro var.
apulicum Koern. pubescentia totae plantae densiore differt. (7).
Typus: Israel, k-18130, 1928, leg. Vavilov (WIR).
Геогр. распр. Сирия, Иордания, Ливан.
Var. villosileucomelan Dorof. et A. Filat, var. nov. — A T. duro var.
leucomelan (Alef.) Koern. pubescentia totae plantae densiore differt. (2).
Typus: Israel, k-10055, cult. Kaf-el-Rachrnan (WIR).
Геогр. распр. Сирия, Иордания (очень редко).
122
Классификации рода Triticuin L.
Определение pa )iioini;iiiocicii convar. villosiini ( laktibz.) Dorof. et Л. I ilat.
Чешуи Ост Разновидное!ь № разно- видное।и
белые белые с черными краями крас- ные красные с черными краями черные на фоне цветя чешуи чер- ные
бел ом крас- ном
Чешуи пропущенные
3 с р П о в к и б е л ы г
-1- -L — — — -1- + + villosileticu- rum villosiletico- meSim villosiprovin- ciale 1 2 3
3 Ср ИО в к и к р а с и ы е
+ + - 7 ail j i e.e i’illosum 1
Чешуи опушенные 3 ерн о и к и бе л ы е
+ — 4- — — — — 1- + t'illosimela- nopus villosaptili- cum 6 7
Зерновки краев ы e
I — I — I — I + I villosafrica- | 8
Illi num
Var. villosileucuruni Dorof. et A. Filat, var. nov. — A T. duro var.
leucurum (Alef.) Koern. pubescentia totae plantae densiore differt. (1).
Typus: lordania, k-15059, 1926, leg. Markovich (WIR).
Ге о г p. p а с п p. Сирия, Иордания (очень редко).
Var. villosimelanoptis Dorof. et A. Filat, var. nov. • - Л T. duro var.
melanopus (Alef.) Koern. pubescentia totae plantae densiore differt. (6).
Typus: Syria, k-18126, 1926, leg. Vavilov (WIR).
Геогр. распр. Наиболее распространенная разновидность в Си-
рии, Иордании, Ливане, редко па о. Кипр.
Var. villosiprovinciale Dorof. et A. Filat, var. nov. — Л. T. duro var.
provinciate (Alef.) Koern. pubescentia totae plantae densiore differt. (3).
Typus: lordania, k-15063, 1926, leg. Markovich (WIR).
Геогр. распр. Сирия, Иордания (редко).
Var. villosum Dorof. et A. Filat, var. nov. — Л T. duro var. reichen-
bachii Koern. pubescentia totae plantae densiore differt. (5).
Typus: lordania, k-17311, leg. Vavilov (WIR).
Геогр. рас up. Сирия, Иордания, Ливан.
5. Convar. falcatum (Jakubz.) Dorof. et A. Filat. — серповидная (фаль-
катиая) группа разновидностей.
Convar. falcatum (Jakubz.) Dorof. et A. Filat, comb. nov.— T. durum
gr. falcata Jakubz. 1932, Tp. прикл. бот. ген. сел. Прнлож. 53: 65.—
Т. durum typus falcata (Jakubz.) Kob. 1932, Хлеби. злаки Турции: 8.—
Т. durum prol. falcatum (Jakubz.) Flaksb. 1935, Культ, фл. СССР, 1:
188.:—7'. luranicum subsp. falcatum (Jakubz.) Gokg. 1955, Ziraal Vckal.
716: 95. — T. durum subsp. falcatum (Jakubz.) Jakubz. 1959, Вот. жури.
44, 10: 1425; он же, 1961, Вид. сорт. раст. ресурсы пшеп.: 27.
Всходы и листья неопушенные, с восковым налетом. Растения вы-
сокорослые, соломина толстая. Колос длинный (до 12 см), средней плот-
ности, встречаются формы с уклоном колоса к иирамидалыюсти. Ко-
Подрод I. Trilli'iuu
Рне. 10. Колосья, колоски и зерпонки:
<7 — Т. durum cotivar. lalcatum (Jakubz.) Porof. ct Л. Filat.: 6 — T. lurumciiin Jakuliz..
досковые чешуи удлиненные с зубцом до 2—4 мм; ости длинные
(рис. 10, и), неопадающие. Зерновки длинные (до 10 мм). Брюшная
сторона их с характерным полулунным углублением (серповпдпость).
Группа включает яровые позднеспелые формы. На родине их носов
производится с осени. Слабо поражаются желтой и бурой ржавчиной.
Экологически эта группа соогветсi вует собственно средиземномор-
ским твердым пшеницам (convar. durum) и потому распределена почти
по всему ареалу последних, по отдельными небольшими локусами.
Геогр. р.аспр. Алжир, Тунис, Марокко, Сгниет, Греция, Сарди-
ния, Сицилия, /Мальта и Кипр, Турция, Праи, Афганистан, Китаи, Азер-
байджан, Киргизия, Таджикистан.
124
Классификация роди Triticum I..
IIccmoi|)я на сравнительно давнее (M. M. Якубцинер, 1932) выделе
пне серповидных пшениц в особую группу (grex falcala), они рассмат-
ривались как подразделения ряда разновидностей обыкновенных твер-
дых пшениц. Так, К. Л. Фляксбергер (1935) писал, что var. mclatiopiis
включает в себя pro parte формы, относящиеся к proles syriaca, jordani-
са, orientate, falcata и т. д.
М. Gokgol (1955) рассматривал группу фалькатпых твердых пшениц
как подвид Т. turanicuni Jakubz., дав неполное описание их разновидно-
стей. Поэтому трудно установить, какие из его новых разновидностей
относятся именно к фалькатпым формам. При анализе пшениц Ирана
М. Gokgol (1961) рассматривал почти все найденные там разновидно-
сти как принадлежащие к Т. luranictini subsp. falcatuin. Однако, судя
по названию, большинство их являются новыми комбинациями разно-
видностей, приводимыми М. Hacizade (1932) для Т. orientate Perciv.
(= Т. turanicuni Jakubz.) и, следовательно, должны быть отнесены
к Т. turanicuni subsp. orientate (Perciv.) Gokg.
Определение разновидностей convar. falcatuin (Jakubz.) Dorof. et A. I'ilat.
Чешуи Ости Разновидность № разно- видности
белые белые с черными краями крас- ные красные с черными. краями черные на фоне цвет чешуи чер- ные
белом крас- ном
Колосья неопушенные
Зерновки белые
Колосья опушенные
Зерновки белые
+ falcatimela-
noptis
+ falcafapuli-
cum
Зерновки красные
+
falcatifastuo-
sum
falcataf ri-
canum
Var. falcatafricanum Dorof. et A. Filat, var. nov. — T. luranictini var.
pseudobandirmaictun (Haciz.) Gokg. 1955, Ziraat Vekal. 716: 95, descr.
turc. quoad T. turanicuni subsp. falcatuin (Jakubz.) Gokg. — A T. duro
var. africanum Koern. caryopsis longis, falcatis differt. (8).
Typus: Cyprus, k-17777, 1927, leg. Vavilov (WIR).
Геогр. p a c it p. Праи, Турция, Израиль, Киир.
Var. falcatapulicuin Udacz. 1979, Вюл. ВНР, 84: 30. — T. durum var.
falcatoaputicuni Udacz. 1975, Ilineii. Средн. Азии: 18, nom. mid.—
A T. duro var. apulicuni Koern. caryopsis longis, falcatis differt. (6).
Typus: RSS Tadjik, k-51897, 1965, leg. Udaczin (WIR).
Геогр. рас и p. Таджикистан, Киир.
I. Triticum
12.5
Var. fa lea ti fast tiosuiii Dorof. of A. J'ilal. var. now turanicum var.
bundii maiciim (Ilaciz.) Gokg. 1955, l.c,; 97, descr. lure, quoad T. luiani
cum subsp. [alcatum (Jakubz.) Gokg. - - A T. duro var. durum caryopsis
longis, falcatis differt. (7).
Typus: Cyprus, k-17777, 1927, leg. Vavilov (WIR).
Геогр. распр. Иран, Кипр.
Var. falcatihordeiforme Dorof. et A. Filat, var. nov. - Л T. duro var.
hordeiforme (Host) Koern. caryopsis longis, falcatis differt. (3).
Typus: Ttircia, k-14386, Reproductio stationis Daghestanskajae fulcra-
cea WIR, 1976 (WIR).
Геогр. распр. Турция (редко), Азербайджан.
Var. falcatileucomelan Dorof. et A. Filat, var. nov. — T. turanicum
var. rarissimum Gokg. 1955, l.c.: 97, descr. turc. quoad 7’. turanicum subsp.
[alcatum (Jakubz.) Gokg. — Л T. duro var. leucoinelan (Alef.) Koern.
caryopsis longis, falcatis differt. (2).
Typus: Cyprus, k-17778, 1927, leg. Vavilov (WIR).
Г с о г p. рас п p. Киир, Турция, Иран.
Var. falcatileucurum Jakubz. et A. Filat, var. now- -7'. turanicum
var. gazi-musta[a-kemali (Haciz.) Gokg. 1955, l.c.: 97. descr. turc. quoad
7’. turanicum subsp. [alcatum (Jakubz.) Gokg. A 7'. duro var. leucu-
rum (Alef.) Koern. caryopsis longis, falcatis differt. (1).
Typus: Marocco, Reproductio stationis Daghestanskajae fulcracea WIR,
k-16361, 3.VI1. 1973 (WIR).
Геогр. распр. Турция, Праи, Афганистан, Сирия, Ливан, Египет,
Алжир, Тунис, Марокко, Кипр и Мальта, редко в Азербайджане.
Var. falcatiinelanopus Jakubz. et A. Filat, var. now — A 7’. duro var.
melanopus (Alef.) Koern. caryopsis longis, falcatis differt. (5).
Typus: Marocco, k-16399, 1926, leg. Vavilov (WIR).
Геогр. распр. Сирия, Израиль, Иордания, Египет, Кипр и Маль-
та, Турция, Афганистан, Китаи (Тибет), Азербайджан, Киргизия.
Var. falcatum Dorof. et A. Filat, var. nov. — A 7’. duro var. reicheti-
bachii. Koern. caryopsis longis. falcatis differt. (4).
Typus: Cyprus, k-17776, 1927, leg. Vavilov. (WIR),
Геогр. распр. Кипр, Турция.
6. Convar. caucasicum (Dorof.) Dorof. — кавказская группа разно-
видностей.
Convar. caucasicum (Dorof.) Dorof. comb. nov. — T. durum subsp.
caucasicum Dorof. 1968. С.-х. биол. 3,3: 347.
Растения высокорослые, соломина толстая, листья крупные, неопу-
шенные. Колосья 5—8 см длины, плотные (D = 28—30), хорошо озер-
пенные (30—50 зерновок в колосе); членики колосового стержня средпе-
опушены; колосковые чешуи округлые, всегда короче цветковых, без
плеча или со слабовыраженным плечом, зубец короткий, широкий, клю-
вовидно изогнутый (рис. 9,6); ости короткие, па нижних колосках их
пет совсем, па верхних — длина их наибольшая, но никогда не превос-
ходит длины колоса; зерновки округлые, стекловидные, крупные (масса
1000 зерен —44—58 г), бороздка их широкая, по неглубокая. Яровые,
возделываются при подзимнем севе.
Т и п: Т. durum var. cauliordei[orme Dorof.
Геогр. распр. Закавказье (предгорные районы и низменности —
100—600 м над ур. м.).
Кавказская группа разновидностей имеет гибридное происхожде-
ние: выявлена в сложных по составу популяциях, в которых всегда при-
сутствует большое число разновидностей 7'. durum и Т. turgidum. Опа
обнаруживает сходство с Т. turgidum (колосковые чешуи округлые,
всегда короче цветковых).
Тургидоподобпые пшеницы были замечены давно. В. К. Кобелев
(1932) обнаружил их в Турции. Однако тургидондность последних вы-
126
l\.ia< сификицип pixie Triticum I.
ражена значительно сл;iбее, чем у convar. caucasictim. Колосковые че-
шуи и.\ округлые, но в отличие ог /'. turgidum iiohiii равны цветковым
(В. К- Кобелев, 1932). Они являются формами разновидностей обычных
твердых пшениц, как собственно и рассматривал их В. К- Кобелев, на-
зывая подобные мелкие подразделения типами.
Определение разновидностей convar. caiicasiciim (Dorof.) Dorof.
Чешуи Ости -Разновидность № разно впдности
бел ы е белые с черными краями крас- ные красные с черными краями черные на фоле цвета чешуи чер- ные
бело м крас- ном
Чешуи неопушенные
3 е р н о в к и б е л ы е
Чешуи опушенные
Зерновки бел ы е
;+! — i — | — i — + сана pit I icu in
Var. catiaffine Dorof. 1968, С.-х. бнол. 3, 3: 348. (4).
Геогр. распр. Азербайджан.
Var. cauapulicum Dorof. 1968, l.c.: 348. (5).
Геогр. распр. Азербайджан (Сабирабадский п Шекппский рай-
оны).
Var. cauhordeiforme Dorof. 1968, I. c.: 348. (3).
Г e о г p. p а с и p. Азербайджан (Заката.тьский район).
Var. cauleucomelan Dorof. 1968, l.c.: 347. (2).
Геогр. распр. Азербайджан.
Var. cauleucurum Dorof. 1968, 1. c.: 347. (1).
Геогр. рас п p. Азербайджан (Закатальский район).
II. Subsp. horanicum (Vav.) Vav.— хорапский подвид твердых пшениц.
Subsp. horanicum (Vav.) Vav. 1964, Миров, ресурсы хлебп. злаков.
Пшеница: 47.— Т. durum grl horanica Vav. 1931, Tp. прикл. бот. ген. сел.
Прилож. 51: 216; Якубцинер, 1932, Тр. прикл. бот. ген. сел. Прплож. 53:
43. — Т. durum typus horanica (Vav.) Kob. 1932, Хлебп. злаки Турции:
10.— Т. durum prol. horanicum (Vav.) Flaksb. 1935, Культ, фл. СССР,
1: 191.
Растения низкорослые, с толстостенной, несклонной к полеганию
соломиной. Листья короткие (16—17 см), слабопоиикающие. гладкие
или с редкими шиппкамп; листья, стебель и колос покрыты восковым
налетом, нередко'восковой палет на этих частях растения отсутствует.
Колос короткий, очень плотный (D = 40 и более); ости слегка расходя-
щиеся, на 4—5 см короче, чем у обыкновенных твердых пшениц, легко-
опадающне после созревания. Колоски многоцветковые и миогозерные.
Колосковые чешуи укороченные, широкие, с. широким килем; главная
боковая жилка сильно развита, образует как бы второй киль. Форма
зерновки близка к округлой (длина лишь в полтора раза превышает
ширину), зерновки стекловидные, с неглубокой бороздкой.
В целом подвид характеризуется значительной скороспелостью,
сравнительно восприимчив к бурой и желтой ржавчине. У подвида вы-
llmlpm) 1. Triticum
127
явлены образцы, устойчивые к твердой солонце, ;Г шоке к шведской
мухе.
Т н и: Т. durum var. horanoleucurtim Vav. et Jakubz.
Экологически хоравский подвид однороден.
Геогр. распр. Сирия, Иордания, редко в Египте и па островах
Средиземного моря, в Малой Азии встречается в местностях с засушли-
вым летом и ранним наступлением высоких температур. Индивидуаль-
ный отбор из образцов хоравского подвида позволил ввести его в непо-
ливную культуру в предгорьях Азербайджана, где в дальнейшем путем
гибридизации с местными азербайджанскими твердыми пшеницами
созданы ценные сорта (Лк-бугда 13, Марк), относящиеся к subs]), hora-
nicum.
Определение разновидное гей subsp. bornnicnrn (Vav.) Vav.
Чешуи Ости Разновидность № p.i НЮ- НИ ТНОС 1 и
бел ыо красные черные белом на фоно красном пне га чешуи черные
Чешуи неопушенные
Зерновки белые
•г I — I — I — I + I — I horanolciiciirum I 1
— I + | — I — I + I — I horanohordeiforme | 2
Зерновки красные
+ | — — — I + ! — | horanatfine ; 3
4euiytt onушенные
3 e p и о в к и бе л ы е
4-1 — I — I — I — I + I horanomelanopus I I
— 1+1 — I — I — | +1 horanapulicum ] 5
Зерновки красные
+ | — I — I — I — I + 1 horanafricanum I 6
— 1+1 — I — I — I + | horanonilolicitin | 7
Var. horanaffine Vav. et Jakubz. in Jakubz. 1932, Tp. прикл. бот, ген.
сел. Прилож. 53: 44. (3).
Геогр. распр. Сирия, Израиль, Турция, Азербайджан.
Var. horanafricanum Vav. et Jakubz. in Jakubz. 1932, l.c.: 45. ((5).
Гео гр. p а с в p. Иордания, Азербайджан.
Var. horanapulicum Vav. et Jakubz. in Jakubz. 1932, l.c.: 45. (5).
Г e о г p. расп p. Иордания.
Var. horanohordeiforme Vav. et Jakubz. in Jakubz. 1932, l.c.: 44. (2).
Г e о г p. расп p. Сирия, Турция, Азербайджан.
Var. horanoleucurtim Vav. et Jakubz. in Jakubz. 1932, l.c.: 44. (1).
Геогр. pa сир. Сирия, Иордания, Турция, Азербайджан.
Var. horanomelanopus Vav. et Jakubz. in Jakubz. 1932, 1. c.: 45. (4).
Геогр. расп p. Сирия, Иордания, Азербайджан.
Var. horanoniloticum Vav. et Jakubz. in Jakubz. 1932, l.c.: 45. (7).
Геогр. p a<c п p. Сирия, Азербайджан.
T. TURANICUM JAKUBZ, — ПШЕНИЦА ТУРАНСКЛЯ
T. turanicuni Jakubz. 1947, Селеки, и семеновод. 5: 46, он же, 1943,
Тр. прикл. бот. ген. сел. 28, 1: 218.—-7’. orie.nlale Perciv.; 1921, Wheat
Plant: 204—205, non Biebcrstein, 1808, Elora Tatiro-Caucasica, 1: 8(5;
С. Кудряшов, 1957, во Фл,- ТаджССР, 1: 334. — T. durum commune orien-
tate (Perciv.) Flaksb., 1923, Опред. паст, хлебов: 14. — 'Г. durum subsp.
128
Классификация /юс)а Trilicuui
cxpaiisiim (ур. duroolilongum prol. orientate (Perciv.) b’laksb., 1935,
Культ. фл. СССР, 1: 137,- T. percivalii llubb. ex Sdiieniaiiu, 1948,
Ziichter 19: 323, nom. illeg. — T. percivalianum Parodi, 1959, Descr. Pl.
Cult., 1: 143, nom. illeg. — T. turgidum gr. turanicum (Jakubz.) Bowden,
1959, Canad. Journ. Bot. 37, 4: 670—671. — T. turgidum subsp. turanicum
(Jakubz.) A. et D. Love, 1961, Bot. Not. 114: 49. — T. durum subsp. orien-
tate (Perciv.) Vav., 1964, Миров, ресурсы хлебп. злаков. Пшеница: 36.—
Т. turgidum subsp. turgidum convar. turanicum (Jakubz.) Mac Key, 1966,
Ilereditas, suppl. vol. 2; 268. — T. georgicum convar. turanicum (Jakubz.)
Mandy, 1970, Acta Agronomica 19, 3—4: 415.
Общая характеристика. Всходы зеленые, неопушенные. или слабо-
шероховатые. Куст сомкнутый, при осеннем посеве на юге иногда про-
межуточный. Листья (к моменту выхода в трубку) опушены короткими
волосками; листовое влагалище голое. Листовая пластинка отходит от
стебля под углом 30—40° (25—35 см длины и 1,2—1,5 см ширины), лан-
цетной формы. Поверхность листовых пластинок главного стебля в фазе
колошения 90—120 см2. Всзлигульных форм не найдено. Ушки длинные,
с ресничками, бесцветные. Восковой палет па стебле и листьях слабой
или средней интенсивности. Растения в условиях весеннего сева в Евро-
пе достигают 106—НО см, а при осеннем севе, на поливе па юге СССР —
в среднем 115 —125 см (иногда 135 см) высоты. Неустойчивые к полега-
нию. Соломина 3,5—4 мм толщины, без антоциана, как правило, с 6 меж-
доузлиями; узлы утолщенные, светло-коричневые, опушены очень корот-
кими волосками. Междоузлия снизу вверх расположены на расстоянии
5—7, 11 —14, 13—15, 17—19, 19—21 и 35—45 см. Под колосом соломина,
как правило, выполненная, реже — полая, по толстостенная. Колосья
сильнопоппкающпе, удлиненные, 11 —14 см длины, в поперечном разрезе
почти квадратные (ширина двурядпой стороны 1,0—1,1 мм и лицевой
0,8—0,9), очень рыхлые (D = 16—18). В колосе по 16—20 колосков и
34—37 зерновок. Колосовой стержень гибкий; членики стержня по реб-
рам опушены белыми волосками, с «бородкой» у основания колоска
(с его фронтальной стороны), узкие, клиновидные, около 1,5 мм ширины
у основания и до 3 мм ширины в верхней части. Колоски вытянутые,
17—20 мм длины, 8—10 мм ширины при толщине около 4 мм, 3—4-цвет-
ковые. Колосковая чешуя лодочкообразпая, 13—16 мм длины и 3—4 мм
ширины, со слегка зазубренным килем, доходящим до основания, с ко-
ротким тупым зубцом (0,8—1,0 мм), одинаковым по всей длине колоса,
и ясно выступающей главной боковой жилкой; плечо узкое, слегка при-
поднятое из-за продления средней жилки. Цветковые чешуи немного
возвышаются над колосковой, довольно плотно охватывают зерновку.
Наружная цветковая чешуя вытянутая, с 7—8 жилками, 13—15 мм
длины, по краям опушенная. Ости 14—17 см длины, сильнозазубрен-
ные, грубые, более или менее легкоопадающпе, поэтому перед уборкой
иногда создается впечатление безостостп колоса. Внутренняя цветковая
чешуя первого цветка несколько короче наружной, окаймлена волос-
ками вдоль обоих килей. Третий цветок в колоске также несет ость, по
она нежная, 5—10 мм длины. Зерновки очень длинные. (И —12 мм),
стекловидные, сходные с зерновками Т. polonicum и с наиболее круп-
ными зерновками дикорастущей полбы Т. dicoccoides. Зародыш круп-
ный, с ясновыр^женной вдавленностыо. Спинка зерновки заостренная,
брюшная часть слегка вогнута, с ясповыраженной бороздкой.
Анатомические исследования вегетативных органов (О. Д. Градча-
нипова, 1967) показали, что строение листовой пластинки Т. turanicum
очень сходно с таковым у Т. durum. В фазу колошения Т. turanicum
отличается мощной но толщине листовой пластинкой (около 585 мкм) и
самым крупным среди видов рода Triticum радиальным сечением про-
водящих пучков по главной жилке (около 175 мкм). На 1 см2 нижней
поверхности листовой пластинки насчитывалось около 40 устьиц, длина
IIoilpill) I. Trilictllll
129
koi орых д<>ст11ra. i;i 63 mo. В отличие <ri опальных i страплоидных голо-
зерных видов у Т. turanicum на верхней и нижней стороне .чистовой
пластинки располагаются волоски различной длины (от 15 до 500 мкм).
Стебель, обладая высоким сопротивлением па излом (около 1700 г
в среднем но трем нижним междоузлиям), склонен к полета пню из-за
слабого развития механической ткани в трех нижних междоузлиях
(средняя толщина ее 85 мкм). Число проводящих пучков 50, причем
расположены они главным образом в паренхиме.
Согласно данным Л\. С. Яковлева и В. И. Николаенко (1936), в ко-
леоптиле имеются лишь 2 проводящих пучка.
Анатомическое исследование колосового стержня Т. turanicum вы-
явило большое сходство с пшеницей твердой (Е. Schroder, 1931). По
данным Л. Л. Жестянниковой (1954), в перикарпии спинной стороны
средней части зерновки бросается в глаза причудливое расположение
поперечных клеток, соединенных друг с другом своими длинными сторо-
нами. В местах сочленений ясно заметны поры, об'ьедпн.яющие отдель-
ные поперечные клетки в общую проводящую систему, направленную по
окружности поперечного сечения зерновки. Поры же, объединяющие
поперечные клетки в продольную систему, мелкие и расположены груп-
пами. Поперечные клетки боковой стороны зерновки более правильной
формы; поры в них короткие и широкие, окаймленного типа. Эпи-мезо-
карпий довольно сильно редуцирован и состоит из 2—3 слоев клеток, из
которых достаточно четко выражен лишь один наружный. Характер по-
перечных клеток на боковой стороне перикарпия (приподнятость их
концов, утолщение оболочек внутренней стороны) напоминает эту же
ткань в зерновке Т. aethiopicum. Сходство с последним видом также
в одинаковой степени редукции слоев эпи-мезокарпия, по в отличие от
Т. aethiopicum у Т. turanicum имеются здесь мощные отложения клет-
чатки.
Являясь резко выраженным экотипом оазисного земледелия районов
со знойным сухим климатом, вид Т. turanicum характеризуется высокой
устойчивостью к жаре и атмосферной засухе, но абсолютной неустойчи-
востью к почвенной засухе. В исследованиях ВИРа под Ташкентом уро-
жайность этой пшеницы при отсутствии почвенной засухи, по при жаре
и атмосферной засухе была лишь па 30% ниже, чем при оптимальных
условиях возделывания (осенний посев на поливном фойе). При соче-
тании же высоких температур воздуха с атмосферно-почвенной засухой
депрессия урожайности (главным образом, за счет резкого снижения
продуктивной кустистости и озернепности колоса) была почти в 8 раз
сильнее. По сообщению Ю. М. Иванова и Г. В. Удовенко (1970),
Т. turanicum отличается солеустойчивостыо. По данным Е. Ф. Пальмо-
вой (1935), эта пшеница характеризуется как вид с. малой приспособ-
ленностью ко всем жизненным факторам. В некоторые годы на отдель-
ных пунктах (Хибины в Мурманской области, Таловая в Воронежской
области) наблюдалась стерильность колосьев Т. turanicum. Эта пшени-
ца, согласно результатам исследования Ф. М. Куперман (1956), в Под-
московье часто чувствительна к повышенной влажности воздуха и из-
быточной облачности.
По оценке Е. Ф. Пальмовой (1933), для форм Т. turanicum из Тур-
ции характерно отрицательное отношение к длинному дню, дающему
позднее выколашивапие на северных пунктах. Характерна также требо-
вательность к высоким температурам в период созревания. М. М. Якуб-
цпнер (1938, 1958) отмечает, что в ряде случаев некоторые формы при
весеннем посеве не выколашиваются полностью. Отмечается особенно
сильная отзывчивость па свет у киргизских форм и меньшая — у узбек-
ских. Н. И. Вавилов j( 1964) сообщил об особой популяции из Ирана,
представленной озимыми и яровыми формами. Зафиксированы случаи
географической изменчивости отдельных образцов Т. turanicum по оира-
9 762
130 Классификации рода Triticum /..
зу жизни. Так, одна 13 турецких популяций (к-15912) в условиях Ле-
нинградской области вела себя как полуозимая, на других пунктах -
как яровая. У иранского образца к-14350 в условиях Московской и Ле-
нинградской областей образ жизни был яровым, а в Краснодарском
крае—озимым. В общем Г. turanicuni представлен is основном форма-
ми яровыми.
По характеристике J. Percival (1921), формы Т. turanicuni ранне-
спелые; по оценке И. К. Васильева (1899), — средцепоздние; по' много-
летним данным ВИРа, — среднеспелые.
При осеннем посеве в Ташкенте па поливном участке период от
всходов до созревания у Т. turanicuni равен 233—-238 дням, при посеве
весной— НО—113 дням. Период от всходов до колошения растянутый.
В европейской части СССР формы Т. turanicuni при осеннем посеве вы-
мерзают. В условиях юга хорошо перезимовывают даже в годы со
сравнительно суровой зимой.
Клейстогамный характер цветения Т. turanicuni отметил еще J. Per-
cival (1921). Уровень открытого цветения туринской пшеницы в Даге-
стане составлял около 60% (В. И. Жуков, 1969). По данным М. Л. Фе-
дина (1975). среди образцов Т. turanicuni открытое цветение отмечено
у 68—89% цветков колоса, причем «взрывной» тип цветения домини-
рует.
Согласно сведениям лаборатории белка и нуклеиновых кислот ВИРа
(В. Г. Конарев и др., 1971, 1972, 1976), содержание белка is зерне образ-
цов Т. turanicuni (репродукция осеннего посева па поливном участке
в Ташкенте) составляет 18,7—22,7%, но лизина в белке лини, 1,66—
1,84%.
По неопубликованным данным Г. 11. Ярины (ВНР), 'Г. turanicuni
характеризуется хорошими макаронными свойствами (прочность мака-
рон 882 г, цвет кремовый, разварпмость 3,2 раза).
По сведениям П. Е. Гребенникова (1948), в Азербайджане из зерна
Т. turanicuni получают прекрасную манную крупу. По данным
Г. И. Соляико (1948), лепешки из муки этой пшеницы характеризуются
высокой пористостью, отличным вкусом, долго пе черствеют и дают вы-
сокий припек (85% против 40—50% У пшеницы мягкой!). Согласно
сообщению Е. Г. Черняковскои (1930), в Пране колосья Т. turanicuni
срываются в фазу молочно-восковой спелости и употребляются затем
в пищу в печеном виде (как початки кукурузы).
Иммунитет. На сильную восприимчивость Т. turanicuni к твердой
головне, мучнистой росе, бурой, желтой п стеблевой ржавчине указы-
вали Е. Ф. Пальмова (1933) и М. М. Якубцинер (1938). По мнению
последнего (1957), по поражаемое™ грибными болезнями этот вид пше-
ницы более сходен с. пшеницей мягкой, чем с близкородственной ему
твердой. Р. А. Удачин (1973) отметил, что в условиях Ташкента Т. turci-
nicuni в годы эпифитогий грибных заболеваний резко выделяется среди
остальных тетраплоидпых голозерных видов резким падением урожай-
ности (масса 1000 зерен снижается почти вдвое).
По наблюдениям II. П. Шитовой (1968), в условиях Дагестана
Т. turanicuni очень сильно поражается бурой ржавчиной. При искусст-
венном заражении проростков популяцией рас бурой и стеблевой ржав-
чины О. Г. Григорьева (1975) отметила восприимчивость соответственно
у 65 и 85% образцов мировой коллекции этого вида пшеницы. 1< числу
особенно неустойчивых к последнему патогену ЛЕ М. Якубцинер (1969)
от.пес образцы Средней Азии, к числу менее восприимчивых образцы
1ipana п Турции.
На провокационном фоне в полевых условиях и при заражении про-
ростков популяцией рас из пяти географических регионов СССР
Э. X. Суханбердипа (1977) наблюдала восприимчивость к мучнистой ро-
се соответственно у 100 и 88,9% образцов..
Подрод I. Triticum
131
В. II. Кривченко (1973) при заражении пыльной головней установил
высокий иммунологический тип у большей части коллекционных образ-
цов. Причем патогенным эффектом обладала f. duri этого болезнетвор-
ного гриба, восприимчивость к которому обнаружена у 27,3% образцов,
тогда как восприимчивые к f. aestivi образцы отсутствовали. Л. М. Яма-
леев (1974) показал, что электрофоретические спектры глиадпна зер-
новок у восприимчивых к пыльной головне образцов гетерогепны
в и-зопс, у устойчивых эта зона обеднена. Т. turanicum, по данным
В. И. Кривченко (1974), имеет как устойчивые, так и восприимчивые
к твердой головне образцы.
Т. turanicum восприимчив к стеблевой головне (Е. Ф. Пальмова,
1933; Е. П. Муромцев, 1936), гсльминтоспориозу листьев и зерна
(Р. А. Удачин, 1973), сильно повреждается шведской мухой (П. Г. Чес-
ноков, 1956), хлебной полосатой блохой и пьявпцеп (Л. Г. Щелко, 1964).
По сообщению L. Defosse (1969), Т. turanicum отличается устойчи-
востью к церкоспориозу. Н. К- Васильев (1899) и Н. А. Стефановский
(1931) наблюдали лишь слабое поражение бурой ржавчиной у некото-
рых форм Т. turanicum. И. Д. Мустафаеву (11)64) в Азербайджане уда-
лось выявить, а также, создать при гибридизации формы, устойчивые
к видам ржавчины и головни.
История. До последнего времени считалось, что именно J. Percival
(1921) был первым ученым, обратившим внимание на систематические
отличия вида и описавшим его по образцам из Хорасана (Иран) как
Т. orientate Perciv. (К- А. Фляксбергер, 1935; И. И. Вавилов, 1964;
П. М. Жуковский, 1971). М. М. Якубцинер (1947), исходя из того, что
эпитет «orientale» уже был использован ранее (1808) в ботанической
литературе, дал виду новое название—Т. turanicum. Р. А. Удачин
(1973) установил, что честь первоначального выявления этой пшеницы
принадлежит русскому ботанику II. И. Васильеву (1899). Последний
в своей статье «Растения Азии и Кавказа в Уманском питомнике» дает
описание стародавнего среднеазиатского сорта пшеницы Ляйляк-бугдай.
Из описания, а также из помещенного в статье рисунка ясно видно, что
.речь идет именно о Т. turanicum. Кроме того, образцы этой пшеницы
под названием Ляйляк-бугдай, Туя-тыш, Китайская и Мпсри-бугдай из
Средней Азии и Дагестана имеются в мировой коллекции ВИРа.
Заслугой другого русского ученого Р. Р. Шредера (1913) является
оценка этого вида как самостоятельного ботанического таксона.
Географическое распространение. J. Percival (1921) считал обсуж-
даемый вид эндемичным для Ирана. К- А. Фляксбергер (1935, 1939) и
И. И. Вавилов (1964), обобщив имевшиеся в отечественной и .зарубеж-
ной литературе к концу 30-х годов данные, включили is ареал Т. turani-
cum также Среднюю Азию, Казахстан, Дагестан, Турцию, Афганистан,
Ирак, Индостан, Северную Африку, Судан, Сирию и Иорданию. Здесь
же И. И. Вавилов указывает, что характерной особенностью Т. turani-
cum является, несмотря на разбросанность его отдельными оазисами ио
широкому ареалу, большая выдержанность как в колосовых, так и в ве-
гетативных признаках, а также в биологических особенностях, в част-
ности, в отношении к паразитическим грибам. В сводке Р. А. Удачина
(1973) в ареал Т. turanicum включено Закавказье. П. М. Жуковский
(1971) ошибочно'включпл Северо-Западный Китай в ареал Т. turanicum
(там культивируется не этот вид пшениц, а Т. petropavlovskyi).
В настоящее время чистые посевы Т. turanicum встречаются редко.
Они размещены небольшими участками в Пране, Турции, Сирии, Афга-
нистане, а также в нашей стране---в Самаркандской, Бухарской и
Сурхандарьинской областях Узбекистана п в Чарджоуской области
Туркмении. Обычно же эта пшеница встречается только как незначи-
тельная примесь в посевах Т. durum.
9*
132
К.шссификпцип рода Tritictun I..
Происхождение. J. Percival (1921) считал, что Т. /iiranieum произо-
шел при । ибридизацип между пшеницей i верной и Т. polonicum.
II. II. Вавилов (1931) писал, что Т. turanicum действительно имеет сход-
ство с подобным гибридом. Н. Kuckuck (1970) поддержал точку зрения
первого, а П. М. Жуковский (1971) отверг ее. По мнению G. Mandy
(1970), становление Т. turanicum связано с мутацией у полбы колхид-
ской. Мысль о происхождении Т. turanicum непосредственно из дикой
полбы Т. dicoccoides при се окультуривании высказал Р. А. Удачин
(1973).
К. А. Фляксбергер (1935, 1938) понимал путь возникновения этого
вида, как обособление восточной группы Т. durum, отклонившейся от
средиземноморских твердых пшениц, по имеющей с последними много
общих признаков. Эта концепция была поддержана В. Ф. Дорофеевым
(1972).
Ряд экспериментальных данных свидетельствует о возможной фило-
генетической связи между 7'. turanicum и пшеницей твердой. Например,
при гибридизации последней с пшеницей мягкой в опыте Е. Д. Есыре-
вой (1970) наблюдалось выщеилеиие растений типа Т. turanicum. Гене-
тическая, экологическая, географическая, морфологическая, цитологи-
ческая и биохимическая близость Т. turanicum к пшенице твердой не-
сомненна. О. Д. Градчанинова (1967) показала их анатомическую
тождественность. Часто Т. turanicum в виде примеси сопутствует пше-
нице твердой, являясь ее спутником. Д. Мак-Кей (1968) считает, что
гены-модификаторы делают Т. turanicum более схожим с последней, чем
с Т. polonicum. Нам кажется, что наиболее соответствующей истине
является точка зрения К. А. Фляксбергера о происхождении 7’. turani-
cum из пшеницы твердой. В ходе становления первого вида микромута-
ции привели к появлению у последней тех признаков, которые отличают
эти виды друг от друга.
Систематика. Как отмечалось выше, эпитет «orientale» был исполь-
зован в 1808 г. М. Биберштейпом для обозначения одного из видов пы-
рея. На этом основании М. М. Якубцинер (1947) переименовал этот вид
в Т. turanicum. Годом позднее этот вид фигурировал на том же основа-
нии в литературе под названием Т. percivalii (II. Hubbard, Е. Schiemann,
1948). П. М. Жуковский (1971) ошибочно принял его за приоритетное
наименование.
В свое время Н. И. Вавилов (1923, 1935), следуя Д. Персивалю, рас-
сматривал эту пшеницу как самостоятельный вид, однако в дальнейших
его трудах она фигурирует уже как подвид твердой пшеницы. Т. turani-
cum включают в состав Т. durum также К- А. Фляксбергер (1923, 1935,
1939), Е. Н. Синская (1959), В. Ф. Дорофеев (1969); относят к Т. turgi-
dum— W. Bowden (1959), А. и D. Love (1961), J. Mac Key (1966, 1968,
1975). G. Alandy (1970) отнес его к Т. georgicum (?’. karamyschevii').
У Т, turanicum различаются две эколого-географические группы:
prol. anatolico-caspicum Jakubz. et Udacz. (Дагестан, Закавказье, Тур-
ция) и prol. irano-turkestanicum Jakubz. cl Udacz. (Иран, Ближний Во-
сток, Северная Африка, Средняя Азия). Первая отличается от второй
меньшей поражаемостыо грибными болезнями, более выраженной склон-
ностью к полурзимому образу жизни и меньшей крупностью вегетатив-
ных и генеративных органов. Эти группы включают 20 разновидностей.
Var. bandirmaicum (Haciz.) Gokg. 1955, Ziraat Vekal. Nes. Haberl.
Miidur., 716: 97. — T. orientale var. bandirmaicum Haciz. 1932, Zuchter,
4,3: 76. (16).
Геогр. распр. Турция, Иран.
Var. daralagesi (Thum.) Udacz. comb. nov. - T. orientale var. darala-
gesi Thum. 1935, Tp. Арм. СХИ, 1: 3. — T. turanicum var. griseoinsigne
Gokg. 1955, Ziraat Vekal. Nes. Haberl. Miidur., 716: 97. (14).
Геогр. распр. Армения, Турция.
Подрод I. Trilicum
) 33
Чешуи неопушенные
3 е р п о в к и белые
T —, gasimustafakemalii 1
— + rarissimum о
+ — generosum 3
— + pseudogenerosum '1
+ — wimshurstii 5
+ — turaniprovinciale 6
3 е р н о в к и к р а с и ы е
+ 1 1 i + + 111 + 1 -г- 4 liu anaffine tiiranimiii dense turanalcxaiidriiiiiin turanobsairnm 7 S <) 10
4- 4 3И euiyu II О в окупи к II 6 ННЫС С Л l.I 1 4- insigne 11
+ — — — — — — — — Р notabile 12
— — 4- — — —— — — — 4- ferghanicum 13
— — — — — — — — + daralagesi 14
— — — — — 4- — — — + hiraninigrum 15
-b 3 c p новь И К ] а с н 4- л с bandirmaicum 16
+ — — — — — — — — pseudobandirmaiciim 17
— —• 4- — — — — — 4- — jalovanianuin 18
— — 4- — — — — — — + pseudo jalovanianuni 19
— — — — + — — — — + odsisst'anuin 20
Var. ferghanicum (Kob.) Udacz. comb. nov. — T. orientate var. fergha-
nictun Kob. 1928, Пшен. Афганист.: 102.— T. turanicum var. turanoapu-
/icH/п Mustaf. 1964, Пшен. Азерб.: 24. (13).
Геогр. распр. Узбекистан, Азербайджан, Сирия.
Var. gasimustafakemalii (Haciz.) Gokg. 1955, Ziraat Vekal. Nes. Ha-
berl. Miidiir., 716: 97. —T. orientate var. gasimustafakemalii Haciz. 1932,
Ziichter, 4, 3: 76. — T. turanicum var. turanoleucurum Mustaf. 1964, Пшен.
Азерб.: 24. (1).
Геогр. p а с п p. Узбекистан, Азербайджан, Сирия.
Var. generosum (Haciz.) Gokg. 1955, Ziraat Vekal. Nes. Haberl.
Miidiir., 716: 97. — T. orientate var. generosum Haciz. 1932, Ziichter, 4, 3:
76. — T. turanicum var. turanohordeiforme Mustaf. 1964, Пшен. Азерб.:
24. (3).
Геогр. распр. Азербайджан, Туркмения, Турция, Пакистан.
Var. insigne (Perciv.) Gokg. 1955, Ziraat Vekal. Nes. Haberl. Miidiir.
716: 97. — T. orientate var. insigne Perciv. 1921, Wheat Pl.: 205. (11).
Геогр. распр. Средняя Азия, Казахстан. Азербайджан, Армения,
Дагестан, Пран, Турция, Ближний Восток, Прак, Афганистан, Га инет,
Ливия, Судан.
Var. jalovanianuin (Itaeiz.) (iokg. 1955, Ziraat Vekal. Nes. Haberl.
Miidiir., 716: 97,- T. orientate var. jalovaniantim Haciz. 1932, Ziichter,
1, 3: 76. (18).
Г e о г p. p а с п p. Турция.
134
Классификация рода Triticum L.
Var, notabile (Perciv.) Gokg. 1955, Ziraal Vckal, Nos. Haberl. Miidiir.,
716: 97. - T. orientate var. notabile Perciv. 1921, Wheat Pl.: 205. (12).
Геогр. распр. Средняя Азия, Казахстан, Азербайджан, Армения,
Дагестан, Иран, Турция, Ближний Восток, Ирак, Афганистан, Египет,
Ливия, Судан.
Var. odsissianuni (Zhuk.) Udacz. comb. nov. - T. durum var. libycum
f. odsissianuni Zhuk. 1923, Зан. Науч.-прикл. отд. Тифл. бот. сада, 3:
8. (20).
Геогр. р а сир. Грузия.
Var. pseudobandinnaicuni Gokg. 1955, l.c.: 97, descr. turc. (17).
Описана M. Гекгелем по образцу из Турции.
Геогр. распр. Турция, Пран.
Var. pseudogenerosuni Gokg. 1955, 1. с.: 97, descr. turc. (4).
Описана M. Гекгелем по образцу из Турции.
Г с о г р. р а с п р. Турция.
Var. psetidojalovanianuin Gokg. 1955, l.c.: 97, descr. turc. (19).
Описана M. Гекгелем по образцу из Турции.
Геогр. распр. Турция.
Var. rarissinium Gokg. 1.955, l.c.: 97, descr. turc. (2).
Описана M. Гекгелем по образцу из Турции.
Геогр. ра сир. Иран, Турция.
Var. turanaffine Mustaf. var. nov, — T. turanicum var. turanoajfihe
Mustaf. 1973, Опред. пшен. Азерб.; 59, descr. ross. — A T. turanicum var.
gasimustafakemalii (Ilaciz.) Gokg. caryopsis rubris differt. (7).
Typus: Azerbajdzhan, 1963, leg. Mustafaev (BAK).
Г e о г p. p а с и p. Азербайджан.
Var. turanalexandrinum Mustaf. var. nov. — T. turanicum var. tura-
noaffinc Mustaf. 1973, Опред. пшен. Азерб.: 60, descr. ross. — A T. turani-
cum var. gasimustafakemalii (Haciz.) Gokg. aristis nigris, caryopsis rubris
differt. (9).
Typus: Azerbajdzhan, 1963, leg. Mustafaev (BAK)-
Г e о г p. p а с п p. Азербайджан.
Var. turaninigrum Mustaf. var. nov. — T. turanicum var. turunonigrum
Mustaf. 1964, Пшеи. Азерб.: 24, descr. ross. — А T. turanicum var. notabile
(Perciv.) Gokg. spica nigra in colore sublito rubro differt. (15).
Typus: Azerbajdzhan, 1963, leg. Mustafaev (BAK)-
Геогр. рас п p. Азербайджан.
Var. turanobscurum Mustaf. var. nov. — T. turanicum var. turano-
obscurum Mustaf. 1973, Опред. пшен. Азерб.: 60, descr. ross. — A 7’. tura-
nicum var. generosuin Gokg. spica nigra in eolore sublito rubro, caryopsis
rubris differt. (10).
Typus: Azerbajdzhan, 1963, leg. Mustafaev (BAK).
Г e о г p. распр. Азербайджан.
Var. turanimurciense Adustaf. var. nov. — 7’. turanicum var, turano-
murciense Mustaf. 1973, Опред. пшен. Азерб.: 59, descr. ross. — A T. tura-
nicum var. generosuin Gokg. caryopsis rubris differt. (8).
Typus: Azerbajdzhan, 1963, leg. Mustafaev (BAK).
Геогр. распр. Азербайджан.
Var. turantprovinciale Mustaf. var. nov. — 7’. turanicum var.-turaiw-
provinciale Mustaf. 1973, Опред. пшен. Азерб.: 60, descr. ross. — A 7'. tu-
ranicum var. generosuin Gokg. spica nigra in colon1 sublito rubro dif-
fer!. (6).
Typus: Azerbajdzhan, 1963, leg, Mustafaev (BAK).
Геогр. p а с и p. Азербайджан.
Var. winishurstii Udacz. 1965, Вести, с.-х. науки, 10: 129. (5).
Г e о г p. p а с и p. Ирак.
Подрод I. Triticum
135
T. POLONICUM L. - ПШЕНИЦА ПОЛОНИКУМ
T. polonicum L. 1762, Sp. PI. ed. 2: 127. — T. maximum Vill. 1787,
Hist. Pl. Dauph. 2: 158.— T. glaucutn Moench, 1794, Method, plant.: 174.—
Deina polonica (L.) Alef. 1866. Landwirt. Fl.: 336. — T. turgidum gr.
polonicum Bowden, 1959, Canad. journ. Bot. 37: 670. — T. turgidum subsp.
polonicum (L.) Thell. 1918, Naturw. Wochenschr. (Jena) 17, 33: 470;
A. et D. Love, 1961, Bot. Not.: 50. — T, turgidum subsp. turgidum convar.
polonicum (L.) Mac Key, 1966, Heredit. Suppl. 2: 268.
Общая характеристика. Всходы сине-зеленые, желто-зеленые, у эфи-
опских образцов — фиолетовые, голые или бархатпстоопушениые. Куст
прямостоячий или (редко) полуразвалистый. Кустистость слабая (1—3).
Растения 100—160 см высоты. Соломина в верхнем междоузлии под
колосом выполнена паренхимой, а остальные междоузлия полые, с па-
ренхимой по стейкам. Листья у взрослых растений большей частью
сппе-зелспые, иногда желто-зеленые, голые пли опушенные (бархатис-
тые па ощупь), 15—33,5 см длины, 0,85—2,0 см ширины; язычок доволь-
но короткий, притупленный, зазубренный; ушки средней величины, с
ресничками. Листовое влагалище голое. Колос прямостоячий (рис. 11),
крупный (10—20 см длины, изредка до 4—8 см), различной формы —
квадратный, округлый, сжатый (двурядпая сторона шире боковой).
Стержень колоса по краям опушенный или почти голый, у основания
колоска с пучком волосков или без .пего. Под колосковыми чешуями
на членике стержня ясно выраженные бугорки, отсутствующие у других
видов пшеницы, кроме Т. petropavlovskyi. Колосья рыхлые, срсдисрых-
лые или плотные (D = 14—40). Колоски широкие (1,2—1,5 см), длин-
ные (2—3 см), многоцветковые. Верхний их цветок бесполый. Наружные
цветковые чешуи первого и второго цветков несут ости длиной 7—20 см,
нижняя цветковая чешуя третьего цветка имеет укороченную ость или
остевидное заострение. В колоске 3—4 зерновки. Колосковые чешуи
травянистые по консистенции, длинные или узкие (от 2,5 до 4 см длины
при ширине 0,3—0,5 см) ланцетные, опушенные (часто опушение слабое)
или голые. Киль слабовыраженпый, килевой зубец короткий или длин-
ный (до остевидного). Возле килевого зубца (с одной или с обеих его
сторон) расположен короткий зубец. Наружная цветковая чешуя, не-
сущая ость, ланцетная, лодочковидная. Внутренняя цветковая чешуя
двукилевая. У первого цветка она значительно короче нижней. Зерновка
стекловидная, обычно удлиненная (7—13 мм длины, дрп ширине и тол-
щине около 3—4 мм). Длина ее превосходит ширину приблизительно
в 3 раза (у форм, близких к Т. durum, — приблизительно в 2 раза).
Исследованиями М. Яковлева и Е. Николаенко (1931) в колеоптиле
Т. polonicum выявлено 2 проводящих пучка. Исключение — эфиопские
формы, у которых 4—6 таких пучков.
Т. polonicum является спутником пшениц твердых (И. И. Вавилов,
1964). По определению Е. Ф. Пальмовой (1935), этот вид песет экологи-
ческие черты соответствующих групп пшеницы твердой. Он тяготеет
к предгорьям. На Северо-Западе Китая на высоте 1350—1500 м над ур. м.
обнаружена эндемичная безостая форма Т. polonicum (М. М. Якубци-
нер, 1959). В Тибете A. Lein (1949) также констатировал наличие по-
добной формы Т. polonicum. Безостость колосьев характерца и для дру-
гих видов пшеницы Китая — Т. durum, Т. aestivum, Т. compactum
(И. И. Вавилов, 1964). Очевидно, в зоне муссонного климата, как под-
черкивала Е. II. Синская (1955), признак остистости является лишним
и устраняется естественным отбором.
У Т. polonicum, по данным ВИРа, известны только яровые формы.
Период вегетации составляет около 125—130 дней.
Содержание белка в зерне 7’. polonicum довольно значительно. Оно
в среднем составляет 16,9%, варьируя от 12,8 до 20,2% при выращива-
I 3f>
Классификация рода Triticum L.
Рис, 1!. Колосья, колоски и зерновки Т. polonicum L,
пни па орошении в плоскостной зоне Дагестана (М. М. Якубцинер,
Н. Ф. Покровская, 1969).
Иммунитет. Н. И. Вавилон (1961) отнес Т. polonicum к числу отно-
сительно устойчивых к комплексу грибных болезней. М. М. Якубцинер
(1969) нашел, что наиболее устойчивыми к бурой ржавчине являются
европейские формы, гетерогенными- -формы Эфиопии, а восприимчи-
выми- формы Ближнего Востока. Изучение коллекции Т. polonicum
позволило О, Г. Григорьевой (1975) сделать вывод о широком размахе
нолевой устойчивости к популяции бурой ржавчины, но с преобладанием
форм, восприимчивых к ее расам. В. М. Берлянд-Кожевников и др.
(1973) нашли у 7'. polonicum из Эфиопии группу генов Х3, обеспечиваю-
щих ус тойчивость к расе 77.
По/рюО /. Trilicum 137
11. II. Шигова (1968) oiHec.'ia T. polonitum к 4iic.iv видов, услойчи-
вых к желтой ржавчине: в условиях плоскостного Дагестана в среднем
за 1954—1965 гг. показатель Bi.icine.ro поражения патогеном был равен
баллу 2,3 (по 5-баллыюй шкале). II. И. Вавилов (1961), Е. Г. Рассадина
к др. (1963), А. М. Камелипа (1973) отнесли Т. polonicum к числу ви-
дов, восприимчивых к стеблевой ржавчине, проявляющих устойчивость
лишь к ее отдельным расам.
По данным М. At. Якубцннера (1961), Т. polonicum не относится
к числу видов, устойчивых к мучнистой росе. Еще в 1916 г. G. М. Reed
при искусственном заражении проростков наблюдал поражение Т. polo-
nicum этой болезнью. Заражение коллекции популяцией рас (из 5 гео-
графических районов СССР) в опыте Э. X. Суханбердппой (1977) вы-
явило восприимчивость к патогену у всех образцов (балл 3—4).
В. И. Кривченко (1973) и А. М. Ямалеев (1974) отнесли Т. polonicum
к слабовосприимчивым к пыльной головне видам пшеницы. Причем
устойчивость к f. aestivi патогена оказалась выше, чем к f. duri. Однако
исследователи отметили, что восприимчивые образцы у Т. polonicum
встречаются чаще, чем у Т. dicoccum, Т. turgidum, Т. turanicum, Т. persi-
cum. При искусственном заражении коллекции выявлены 41% иммун-
ных и 59% практически устойчивых к твердой головне образцов
(В. И. Кривченко, 1973). Н. Harding (1972) нашел, что Т. polonicum
более или менее восприимчив к корневым гнилям.
История. Как предполагал J. Percival (1921), первое описание об-
суждаемого вида («Triticum speciosum grano longo») было сделано по
материалу из Ботанического сада в Штутгарте (J. Bauhin et J. Cherler.,
1651). Эта своеобразная пшеница под названием Т. polonicum впервые
упоминается в каталогах и гербариях ботанических садов Европы во
второй половине XVII в. (Р. Hermann, 1687; J. Bobart, 1690; L. Plukenet,
1692; R. Morison, 1699). Это неудачное название (в Польше она не
возделывается) было принято и К- Линнеем. Основной территорией фор-
мообразования Т. polonicum являются Средиземноморские страны, где
он, как правило, сопутствует Т. durum, встречаясь в небольших количе-
ствах и спорадически.
На наличие небольших посевов в Англии указывали еще J. Bobart
(1690) и R. Morison (1699). К- А. Фляксбергер (1935) считал, что точ-
ных сведений о культуре Т. polonicum в Германии не имеется, хотя
A. Haller (1768) сообщал о незначительном возделывании этого вида
в Швейцарии и Тюрингии.
Нередко Т. polonicum принимали за рожь, о чем говорят также ее
местные названия: русское — «ассирийская рожь», немецкие—Riesen-
roggen, Riesenkorn и др.
Географическое распространение. Т. polonicum распространен по все-
му ареалу Т. durum. В мировой коллекции ВИРа есть образцы этого
вида из СССР (РСФСР, Украина, Узбекистан, Казахстан, Закавказье),
Эфиопии, где составляет примесь в посевах Т. aethiopicum (Н. И. Вави-
лов, 1931), Турции (В. К- Кобелев, 1933; М. Gokgol, 1955), Сирин, Ли-
вана, Иордании (М. М. Якубцинер, 1932), с островов Средиземного мо-
ря, из Марокко, Алжира, Италии, Испании (К. А. Фляксбергер, 1935;
И. И. Вавилов, 1964; Л. Шрейбер, 1933), а также из Южной Америки
(Аргентина, Бразилия). А. М. Горский собрал образцы Т. polonicum
в Китае. Им были обнаружены безостые формы, подобные описанным
ранее из Тибета (Л. Eein, 1949). В последние i оды получены образны
из .Монголии п Прака.
Происхождение. В филогенетической близости Т. polonicum и пше-
ницы твердой были убеждены Е. Tsclierinak (1907, 1913, 1914),
•F. Kornicke (1908) и др. И. И. Вавилов (1913) близость этих видов
усматривал не только в сходстве ряда морфологических признаков, ио
и в однозначной реакции па поражение грибными болезнями. Д. К. Ла-
138 Классификация рода TrUicum I,.
риоиов (1914) не связывал происхождение T. polonicum с твердой пше-
ницей. Однако близость этих двух видов столь значительна, что Л. Де-
кандоль, Г. Вильморен, Л. И. Стебут и др. (по R. Л. Фляксбсргеру, 1935)
даже объединили их >в один вид. Л. Schulz (1911, 1913) воспринимал
Т. polonicum как уродливую форму пшеницы твердой. Но Т. polonicum
нс урод, а полноценный растительный организм, у которого в МКП при
прохождении мейоза формируются 14 бивалентов, пыльники функцио-
нируют нормально, а оплодотворение после самоопыления проходит без
нарушений (Т. Б. Вакар, 1948).
J. Percival (1921), К- А. Фляксбергер (1935, 1938), И. Kuckuck (1970)
и др. происхождение Т. polonicum связывают с мутацией, произошедшей
у пшеницы твердой. Н. И. Вавилов (1931) предполагал, что Т. poloni-
cum— мутационные формы, возникшие у различных географических и
экологических групп и соответствующие по морфолого-экологическому
облику исходным типам пшеницы твердой. А. А. Ериияп (1941) по ре-
зультатам цитологического анализа сделала вывод о том, что Т. poloni-
cum— более молодой вид, чем пшеница твердая. В. Р. Челак (1976)
высказался за гибридное происхождение 7'. polonicum в результате спон-
танной гибридизации твердой или другой тетраплоидной пшеницы
с Т. ispahanicum.
М. Gokgol (1963) указывает па Переднюю Азию как на место возник-
новения Т. polonicum из пшеницы твердой. Н. И. Вавилов (1931),
К. А. Фляксбергер (1935, 1938), Е. Schiemann (1948), Е. Н. Синская
(1969) стояли на позициях признания полифилегического происхожде-
ния Т. polonicum из Т. durum. По нашему убеждению, это соответствует
истине. Мутация привела к появлению у пшеницы твердой фактора Р
(R. Н. Biffen, 1905; W. О. Backhouse, 1918; F. L. Englcdow, 1923;
|S. Matsumura, 1936; J. R. M. Almeida, 1937). Этот фактор ответствен
за удлиненность и травянистую консистенцию колосковых чешуи. Он
обеспечивает удлиненность зерновок и наружных цветковых чешуи ко-
лоска, не влияя на длину внутренней цветковой чешуи.
Закономерности наследования признака удлиненности колосковых
чешуи изучали A. S. С. Сарогп (1918), F. L. Engledow (1920), Е. Mali-
nowski (1927), Т. К. Лепин (1929). Последний (1932) пришел к заклю-
чению, что доминантный фактор Р оказывает множественное влияние,
которое проявляется также в уменьшении опушенностп чешуи и плот-
ности колоса, в укорочении остей и зубца колосковой чешуи. Еще рань-
ше Н. И. Вавилов и О. В. Якушкина (1925) высказали мнение о почти
абсолютном сцеплении белого цвета колоса е признаками Т. polonicum.
Удлиненная наружная цветковая чешуя характерна для Т. polonicum,
Т. ispahanicum и Т. pelropavlovskyi. Однако у второго вида нет сцепле-
ния между этим признаком и пергаментной консистенцией колосковой
чешуи. Возможно, что факторы, удлиняющие чешуи у Т. polonicum и
Т. ispahanicum, чем-то различаются. У Т. pelropavlovskyi действие фак-
тора Р немного подавлено, возможно, присутствием фактора Q и гено-
ма D. Наибольшая выраженность действия фактора Р характерна для
Т. polonicum. Генетически этот фактор мало изучен.
Систематика. По ряду признаков Т. polonicum соответствует видам,
которым сопутствует в посевах и повторяет их эколого-географическую
дифференциацию.
Т, polonicum делится иа 2 подвида: subsp. abyssinicum (Stem!.) Vav.
(примесь в посевах Т. aelhiopicum) и subsp. polonicum (встречается
в пределах ареала Т. durum).
Ключ для определения подвидов Т. polonicum
1. Колеоптиль с тремя и более проводящими пучками; фиолетовая
окраска ясно проявляется па всходах. Растения низкорослые (60—
Подрт) Л Trilicum
139
100 см), солом п и а гонкая, лисгья опушенные......................
...................................subsp. abyssinicum (Steud.) Vav.
+ Колеоптиль с двумя проводящими пучками, фиолетовая окраска
па всходах по проявляется. Растения средне- и высокорослые (100—
160 см), соломина толстая, листья неопушенные (у всходов опушен-
ность слабая).....................................subsp. polonicum.
I. Subsp. abyssinicum (Steud.) Vav. — эфиопский подвид пшеницы
польской.
Subsp. abyssinicum (Steud.) Vav. 1931, Tp. прикл. бот. ген. сел. При-
лож. 51: 159. — Т. abyssinicum Steud. 1855, Synops. plant, gram.: 342.
Растения низкорослые (60—100 см), кустистость слабая. Соломина
тонкая. Листья слабо опушены короткими шиниками. Колосья 8 — 14 см
длины, рыхлые (D=30). Стержень колоса гибкий, у сочленений опу-
шенный. Ости нормально развиты, средней длины (около 15 см). Ко-
лоски многоцветковые; колосковые чешуи около 2,5 см длины, с боль-
шим количеством жилок, с килевым зубцом — коротким или длинным
(1,8 мм), тупым или острым. Зерновка около 7 мм длины.
Для подвида характерна скороспелое гь за снег короткого межфаз-
ного периода колошение — созревание и восприимчивость к бурой ржав-
чине.
Геогр. распр. Эфиопия (встречается как примесь, не более 5%,
в провинциях Шоа, Тигре, Годжам, Харэр, Эритрея).
Var. abyssinicum (Steud.)
Koern. in Koern. et Wern.
1885, Handb. Getreideb. 1: 103. — Т. abyssinicum Steud. 1855, Synops. plant, gram.: 342. — T. polonicum var. kor- nickei Flaksb. 1935, Культ, фл. СССР, 1: 264. (1). Опре, а юление разновидностей subsp. byssinicum (Steud.) Vav.
Чешуи Ости Разновидность № разно- видности
белые чер- ные белые чер- ные
Геогр. распр. Эфио-
пия.
Var. deviatum Vav. 1931,
Tp. прикл. бот. ген. сел.
Прилож. 51: 160. (2).
Геогр. распр. Эфио-
пия (очень редкая примесь;
провинции Тигре и Эрит-
рея).
Var. pseudosporadicum
Vav. 1931, 1. с.: 160. (5).
Геогр. распр. Эфио-
пия (очень редкая примесь;
провинции Шоа и Эритрея).
Var. rarovillosum Vav.
1931, 1. с.: 160. (6).
Геогр. распр. Эфиопия
Чешуи неопушенные
Зернов к и б е лы е
1 Г I — I abyssinicum I
I — 1 + 1 deviatum I
Зерновки красные
I + I — I rubrosemi- I
I I I neum I
Чешуи опушенные
3 e p н о в к и б е л ы е
+ — sporadicum
— + pseudospo-
radicum
3 е р н о в к и к р а е н ы е
| + | — \rarovillosum\
жам — близ Дапглы, и Эритрея).
Var. rubrosemineum Vav. et Fortun. in Vav.
(очень редкая примесь; провинции
6
Год-
1931, 1. c.: 160. (3).
Ге or p. распр. Эфиопия (очень редкая примесь; провинция Шоа).
Var. sporadicum Vav. et Fortun. in Vav. 1931, l.c.: 160. (4).
Геогр. распр. Эфиопия (редкая примесь; около Харэра, Чэрчэ-
ра и Дапглы).
1
2
3
I
5
11. Subsp. polonicum — средиземноморский подвид и шеи и цы поль-
ской.
Subsp. polonicum. — Т. polonicum subsp. mediterraneum Vav. 1931,
Tp. прикл. бот. ген. сел. Прилож. 51: 222.
1'10 Kouciiiiliiii,iii(ini Tnlicitni I..
Растепия средперослые и высокорослые (100 160 см длины) со сла-
бой кустистостью. Соломина юлсгая. Листья неопушенные (опушение
бывает только на всходах), с восковым налетом и без пего. Колосья
длинные (10—20 см), рыхлые (D = 14 20) и среднеплотные (D —
— 20—30) пли укороченные (4- 7 см), плотные (1) — 30—40), редко -
ветвистые. Ости по длине равны, превосходят или значительно короче
колоса. Сеть безостые формы. Колосковые, чешуи от 25 до 40 мм длины
и 3- -5 мм ширины, с коротким или длинным (иногда ос1 евпдным) зуб-
ном; голые или опушенные, иногда опушение весьма слабое (волоски
стирающиеся, в этом случае опушение можно заметить только под лу-
пой). Зерновки 9—13 мм длины, 3—4 мм ширины, стекловидные. Фор-
мы яровые, позднеспелые, относительно устойчивые к бурой ржавчине.
Геогр. распр. Встречается спорадически (в виде единичных рас-
тений или небольших посевов) в ареале возделывания пшеницы твердой.
Как уже. отмечалось ранее, Т. polonicum subsp. polonicum является
спутником Т. durum и несет экологические черты его групп (В. Ф. Паль-
мой;!, 1935). В районах с наибольшей встречаемоегыо 7’. polonicum
II. II. Вавилов (19(14) выделяет 3 эколого-географические группы.
1. Собственно средиземноморская prol. eumediterraneum Vav. 1961,
Миров, ресурсы хлебп. злаков. Пшеница: 50.
Растения высокорослые (120—160 см), соломина толстая, но недо-
статочно устойчивая к полеганию, листовая пластинка широкая с воско-
вым налетом. Колосья плотные, с длинными остями. Формы сравни-
тельно позднеспелые, слабо поражаются желтой и бурой ржавчиной,
пыльной и твердой головней, восприимчивы к мучнистой росе.
Геогр. р а с п р. Страны Средиземноморья.
2. Палестинская группа — prol. paleslinicum Vav. 1964, I. с.; 51.
Растения НО—140 см 'высоты, соломина слабовыполпеппая, устойчи-
вая к полеганию. Листовая пластинка узкая, с восковым налетом и без
пего. Колосья рыхлые, 12—14 см длины. Ости равны по длине, колосу
или короче его. Формы сравнительно скороспелые.
Геогр. распр. Редко в Сирии, Иордании, Израиле.
3. Южная степная группа — prol. stepposum Vav. 1964, l.c,: 51.
Растения высокорослые (120 см и выше), соломина средней толщи-
ны, недостаточно устойчивая к полеганию. Облиствепиость слабая, ли-
стовая пластинка узкая, с восковым налетом. Колосья длинные, рых-
лые. Формы позднеспелые, до все же более ранние, чем у средиземно-
морской группы. Довольно устойчивые к бурой и желтой ржавчине.
Геогр. распр. Казахстан, Узбекистан.
У subsp. polonicum по плотности колоса выделены 2 группы разно-
видностей: 1) convar. polonicum и 2) convar. compactum Ser.
Деление по плотности колоса у Т. polonicum не вполне соответствует
таковому у других -видов пшеницы. У плотноколосой группы разновид-
ностей Т. polonicum convar. compactum редко бывает больше 40 колос-
ков па 10 см колосового стержня. С плотностью колоса связан ряд при-
знаков: нысокорослость, длинные колосья и ости у convar. polonicum-,
низкорослость, короткие колосья и ости у convar. compactum Ser.
У переходных между этими группами форм можно наблюдать захожде-
ние признаков; отнесение разновидностей к первой или второй группе
зависит от степени превалирования тех или иных из них.
Ключ для определения групп разновидностей subsp. polonicum
1. Колосья 9 13 см длины, рыхлые (1) — 14 20) или сроднен, ил иые
(I) = 20 30), двурядная сторона их почти равна лицевой.............
...............................................convar. polonicum.
Колосья 4- 10 см длины, плотные (12 = 30-40, редко больше),
двурядная сторона шире лицевой.................convar. compactum.
По()(хн) /. I’fi/iciiin
141
i. Convar. polonicum собственно среди и'миоморская ipvnna разно-
видное reii.
Convar. polonicum. T. polonicum oblongum Ser. 18-11, Descript, fipj.
Cereal, eiirop. 4: 147. T. polonicum ipuulralum Ser. 184 1, l.c.: 145. —
7’. polonicum [rr. oblongo-quadratum Flaksb. 1922, Опред. паст, хлебов:
(18. -T. polonicum simplex Flaksb. 1935, Культ, фл. СССР, 1: 259.
Растения высокорослые (120—1G0 см). Колосья длинные (рыхлые
или среднеплотные), двурядная сторона их почти равна лицевой. Ости
длинные, редко укороченные. Безостая форма var. gorskyi эндемична
для Северо-Западного Китая; имеются безостые разновидности, полу-
ченные искусственным скрещиванием (П. Е. Гребенников, 1929).
Г с о г р. р а с п р. По всем)' ареалу Т. polonicum.
Определение разновидностей convar. polonicum
Чешуи Ости Разновидность К? разно- видности
белые красные черные инета чешуи черные
К о .1 о с ь я и р о с т ы е
Колосья остистые
Чешу и н е о и у ш еи и ы о
1. Зерновки белые
+ + polonicum 1
+ + psetidolevissimum 2
2. Зерновки красные
+ — — + — cliri/sosperuium .3
+ — — — -1-. pseudoclirysospermum •1
— 4- — + — rufescens 5
Чешуи о п у ш е н ны е
1. Зерновки белые
+ .— — + — villosum 6
4- — — — + pseudovillostim 7
— + — — + czernjajevii 8
2. Зерновки красные
4- — — 4- — nouissimum 9
+ — — — 4- smirfiovii 10
— 4- — 4- — rubrouelutinum 11
— 4 —- — 4- vilmorinii 12
— — + — 4- nigrescens 13
Колосья безостые
Чешуи н е о п 1’. Зстяттл-. ушей н ы е
i ~ 1 - | eorskm I 14
— ^ + I — 1 - 1 - 2. Зерновки 1 prjunischnikovii | 15 красные
+ I - | - 1 = 1 = I skalosubovii 1 16
- | + 1 - t Ч е ш у и о п у 1. Зерновки | pissarevii | 17 Hl е и и ы е белые
+ 1- 2. Зерновки I tibeticum ' 18 красные
+ 1 — 1 — — — I miczurinii 1 19
— 1 + 1 — 1 — 1 — Колосья ветвистые (чешуи | talanovii | 20 неопушенные, зерновки белые)
+ I - I - 1 1 1 ramosum 1 21
+ 1 - - 1 — 14- [ kiritchenko | 22
142
Классификации рода Triticum I..
Var. clirysospermum Koern. in Koern. el Winn. I<485, Handl), Gelrenleb.
1: 101; Фляксбергер, 1935, Кулы. фл. СССР, 1: 270. (3).
Геогр. распр. Средиземноморские страны (редко), Польша,
СССР (Северный Кавказ).
Var. czernjajevii Grob. 1929, Опред. разной, union. Сев.-Кавк, края:
31.(8).
Геогр. распр. СССР (Северный Кавказ).
Var. gorskyi Jakubz. var. nov. — T. polonicum var. gorskyi Jakubz.
1959, Бот. журн. 44, 10: 1427, descr. ross.— А T. polonicum var. levissimum
(Haller) Koern. spica mutica differt. (14).
Typus: China (k-43319). Reproductio statiouis Dagheslanskaja fulcra-
cea WIR, leg. Л. Filatcnko (WIR).
Г corp, p a c ii p. Северо-Западный Китай.
Var. kiritchenko Jakubz. 1950, Tp. прикл. бот. геи. сел. 28, 3: 66. (22).
Выявлена в 1947 г. Ф. Г. Кириченко на посеве сорта .Кахетинская
ветвистая (Одесса), а также получена II. Д. Мустафаевым путем гиб-
ридизации 7'. polonicum с ветвистой формой 7’. turgidum, которой
М. М. Якубцинер (1950) дал название f. muslafaevii.
Var. miczurinii Greb. 1929, 1. c.: 31. (19).
Автор разновидности не дал сведений о ее происхождении и рас-
пространении. По сообщению К. Л. Фляксбергера (1935), она была по-
лучена путем искусственного скрещивания.
Var. nigrescens Koern. in Koern cl Wern. 1885, l.c.: 101, descr.
incompl.; Greb. 1929, l.c.: 31, corr. — 7’. polonicum var. violaceum Koern.
in Koern. et Wern. 1885, l.c.: 101. — T. polonicum var. anomalum Koern.
in Koern. et Wern. 1885, l.c.: 100. — T. polonicum gr. nigrescens (Koern.)
Flaksb. 1935, l.c.: 274. (13).
Геогр. распр. II. E. Гребенников (1929) указывает на очень ред-
кое нахождение этой краспозерпой разновидности па Северном Кавказе.
Более ранние неполные описания (F. Kornicke, Н. Werner, 1885) сделаны
по образцам из ботанических садов, среди которых выявлены также
короткоостистые формы (var. anomalum Koern.) неизвестного проис-
хождения.
Var. novissimum Koern. 1908, Archiv Biontol, 2: 404. (9). Описана no
образцам неизвестного происхождения. В коллекции ВИРа имеются се
образцы из Польши и Латвии.
Var. pissarevii Greb. 1929, l.c.: 32. (17).
Автор разновидности пе дал сведений о ее происхождении и распро-
странении. По сообщению К- Л. Фляксбергера (1935), опа получена пу-
тем искусственного скрещивания.
Var. polonicum. — 7’. levissimum Hall., 1768, Hist, stirp. indig. Helve-
tiae. 2: 209. — T. polonicum barbatum Desv. 1833, Mem. from. cult. France:
146. — Deina polonica alba Alef. 1866, Landwirt. Fl.: 336. — 7’. polonicum
var. levissimum (Hall.) Koern. in Koern. et Wern. 1885, l.c.: 100; Flaksb.
1935, l.c.: 269. — T. polonicum var. incerium Koern. in Koern. et Wern.
1885, 1. c.: 100, descr. incompl. (1).
Одна из наиболее распространенных разновидностей.
Геогр. распр. Средиземноморские страны, ГДР, ФРГ, Англия,
Китай, Монголия, США, СССР--Алтайский край и Азербайджан.
F. Kornicke (1885) указывает, что первые описания 7’. polonicum
различными авторами относились именно к этой разновидности.
Наряду с длинноостистыми типичными выделены формы с укорочен-
ными остями — f. submuiicum (Link) Flaksb. 1935, l.c.: 269. — 7’. poloni-
cum B. submuiicum Link, ' 1827, llarl. Berol. 1: 28. • Deina polonica
semiarislala Alef. 1866, l.c.: 336. • -7'. polonicum var. submuiicum (Link)
Koern. in Koern. et Wern. 1885, 1. c.: 100. Описывалась по материалам из
ботанических садов.
Геогр. рас и р. Пе выяснено.
1. Triticlllll
113
Var. prjanischnikovii Greb. 1929, l.c.: 32. (15).
Автор разновидности не дал сведений о ее происхождении и распро-
странении. Но сообщению К. Л. Фляксбергера (1935), она получена пу-
тем искусственного скрещивания.
Var. pseudochrysospermum Gokg. 1955, Ziraat Vekal. 71 (>: 1), descr.
lure. (4). Описана M. Гекгелем но образцам из Турции.
Г е о г р. р а с и р. Турция.
Var. pseudolevissimum Jakubz. var. nov. — T. polonicum van psendo-
levissinium Jakubz. in Flaksb. 1935, l.c.: 269, descr. ross. — A T. polonicum
var. polonicum aristis nigris differt. (2).
Typus: Syria k-43209. Reproductio stationis Daghesianskaja fulcracea
WIR, 1973, leg. A. Filatcnko (WIR).
Г corp, распр. Англия, Сирия, Китай, Казахстан, Азербайджан.
Var. pseiidovillosum Flaksb. var. nov. — T. polonicum gr. pseudovillo-
sum Flaksb. 1935, l.c.: 272, descr. ross. — T. polonicum var. pseiidovillo-
sum Flaksb. 1939, Опред. паст, хлебов: 136, descr. ross. —A T. polonicum
var. villosum Desv. aristis nigris differt. (7).
Typus: RSSUkr. k-23562. Reproductio stationis Daghestanskaja fulcra-
cea WIR, 1973, leg. A. Filatenko (WIR).
Геогр. p а с n p. Редко в Малой Азин, США, а также в Грузин. Ка-
захстане, на Украине.
Var. ramosum (Alef.) Dorof. et A. Filat, comb. nov. —Deina polonicum
ramosa Alef. 1866, I. c.: 336. — T. polonicum gr. ramosum (Metzg.) Flaksb.
1935, l.c.: 270.— T. polonicum B. Metzg. 1824, Europ. cereal.: 24, sine
nom.— T. polonicum dejormatum Ser. 1841, l.c.: 149, quoad spccim. ra-
niosuni. (21).
Геогр. распр. I le выяснено.
Var. rubrovelutinum Koern. in Koern. et Wern. 1885, l.c.: 100, descr.
incompl.; Гребенников, 1929, l.c.: 31, corr. — T. polonicum var. seringei
Koern. in Koern. et Wern. 1885, l. c.: 101. — T. polonicum gr. rubroveluti-
num (Koern.) Flaksb. 1935, l.c.: 273. (11).
Известна по образцам из ботанических садов, единичные колосья
обнаружены П. Е. Гребенниковым (1929) па Северном Кавказе.
Var. rufescens Koern. in Koern. et Wern. 1885, l.c.: 100; Flaksb.
1915, Опред. пшен.: 109; Greb. 1929, 1. c.: 31. (5).
Геогр. распр. Описана Ф. Кёрнике по образцу из Ботанического
сада в Поппельсдорфе. Единичные колосья обнаружены Н. Е. Гребен-
никовым (1929) на Северном Кавказе.
Var. skalostibovii Greb. 1929, l.c.: 31. (16).
По сообщению К. Л. Фляксбергера (1935). получена путем искусст-
венного скрещивания.
Var. smirnovii Vatz. 1929, Тр. Всесоюзи. съезда ген. сел. семей., январь,
1929 (Ленинград), 3: 122. — Т. polonicum var. meczetinskense Greb. 1929,
l.c.: 31.— T. polonicum var. pseudonovissimum Flaksb. 1935, l.c.: 273. (10).
Геогр. распр. Очень редко в СССР (Северный Кавказ. Украи-
на, Латвия).
Var; talanovii Greb. 1929, 1. с.: 32. (20).
По сообщению К. А. Фляксбергера (1935), получена путем искус-
ственного скрещивания.
Var. tibeticum Udacz. 1979, Бюл. ВИР, 84: 74. (18).
Геогр. распр. Китай.
Var. villosum Desv. 1833, Mem. from. cull. France: 146; Koern. in
Koern. et Wern. 1885, l.c.: 101. - Deina polonica velutina Alef. 1866, I. c;:
336. (6).
Геогр. распр. Наиболее распространенная разновидность I. po-
lonictim, встречающаяся по всему ареалу. В посевах этого вида является
преобладающей (var. polonicum преобладает реже).
Классификации рода Trilit ит 1..
К. Л. Фляксбергер (1929) обнаружил в гербарном материале (Herb.
Sprcngel) короткоостистые колосья этой разновидное!и, которые мы
рассматриваем как f. gracile (Flaksb.) Dorof. et A. Filat, comb. nov.—
T. polonicum var. gracile Flaksb. 1929, Tp. прикл. бот. ген. сел. 21, 5; 480;
он же, 1935. 1. с.: 273.
Г е о г р. расп р. Не выяснено.
Var. vilmorinii Koern. in Koern. et.Wern. 1885, l.c.: 101, descr. incompl.;
Greb. 1929, l.c. :31. — T. polonicum gr. vilmorinii (Koern.) Flaksb. 1935,
l.c.: 274. (12).
Геогр. распр. Известна ио образцам из ботанических садов.
П. Е. Гребенников (1929) указывает па нее как на очень редкую при-
месь на Северном Кавказе.
2. Convar. compactum Ser. — плотноколосая группа разновидностей.
Convar. compactum Ser. 1841, Descript, fig. Cereal, europ. 4: 147.—
T. polonicum gr. compactoides Flaksb. 1922, Опред. паст, хлебов; 69.
Растения средперослые (100—120 см). Колосья короткие (4—7 см)
или средней длины (до 10 см), плотные (D= 30—40, редко больше),
несколько сжатые с боковых сторон; ости большей частью укороченные,
реже длинные. Сюда же относятся переходные (по плотности и длине
колоса) формы.
Т и п: Т. polonicum var. compactum Link.
Геогр. распр. Средиземноморские страны (редко).
Var. aegeicum Gokg. 1955, Ziraat Vekal. 716: 112, descr. turc. (4). Опи-
сана M. Гекгелем по образцу из Турции с заходящими признаками
обеих групп разновидностей (false-compactoides).
Г е о г р. расп р. Турция.
Var. anatolicum Gokg. 1955, l.c.: 112, descr. turc. (3). Описана
AL Гекгелем по образцу из
Турции с заходящими при-
знаками обеих групп разно-
видностей.
Геогр. распр. Тур-
ция.
Var. caryopsirubrum
Gokg. 1955, l.c.: 112, descr.
turc. (5). Описана AY. Гек-
гелем по образцу из Тур-
ции.
Геогр. рас п р. Тур-
ция.
Var. compactum Link,
1827, Hort. Bertol. 1: 28;
Koern. in Koern. et Wern.
1885, Handb. Getreideb. 1:
102. — Deina polonica cla-
vata Alef. 1866, Landwirt.
Fl.: 337. T. polonicum var.
elongatum Koern. in Koern.
et Wern. 1885, l.c.: 102.--
T. polonicum var. attenua-
tum Koern. in Koern. et
Worn. 1885, 1. c.: 104, descr.
incompl.— T. polonicum var.
intermedium Koern. in Ko-
ern. et Wern. 1885, l.c.: 104,
descr. incompl.— T. poloni-
cum var. eucompactum
Aschers. et Graebn. 1901,
Synops. Mitteleurop. Fl. 2:
Определение разновидностей convar.
compactum Ser.
Чсип и Ости Разновид- ность № разно- видности
белые крас- ные чер- ные цвета чешуи чер- ные
Чешуи неопушенные
Зерновки белые
4- — — + — compactum 1
— — — pseudocom- 2
pactum
— + — + — anatolicum 3
— — — + aegeicum •1
Зерновки красные
+ — — + — caryopsi- rubrum 5
+ — — pseudocary- opsirub- rum 6
geuiyu опушенные
Зерновки белые
+ — — + — martinari 7
4- — — — + heidclbergi 8
— + — —• zaferanicum ‘)
— + — — 1 pseudozafe- ranicum 10
Зерновки краев ы е
halleri
homsicum
zonguldaki
cum
11
12
1.3
+
Подрод 1. Trillium
145
699.— Т. polonicum var. crassispicatum (Koern.) Flaksb. 1935, Культ, фл.
СССР, I: 275. T. polonicum tongiaristatum b. crassispicatum Koern.
1873. Syst. Uebers. Cereal.: 15, nom. mid. (1).
Г с о г ]>. p a e n p. Испания, Марокко, Иордания, Липан.
F. Kornicke (1885) и К- А. Фляксбергер (1915, 1935) перечисляют
ряд плотпоколосых длинпоостых разновидностей, ссылаясь на описание
N. Seringe (1841) и A. Desvaux (1833). Эти разновидности немного от-
личаются от var. compactum по тем или иным признакам, как-то: удли-
ненные полуостпстые колосья, изогнутые с двурядиой стороны (var.
elongatum Koern.); колосья, суживающиеся от основания к вершине
(var. attenuatum Koern. = Т. polonicum var. longiaristatuni b. crassispi-
catum Koern.); колосья булавовидные (var. intermedium Koern.). Ввиду
неполного описания и отсутствия указаний па места нахождений по-
добных форм мы оставляем их в качестве синонимов var. compactum.
У Т. polonicum Сирии и Иордании М. М, Якубцинер (1952) обнару-
жил плотноколосые формы без воскового налета на стебле, листьях и
колосе: var. compactum f. saronicum (Jakubz.) Dorof. et A. Filat, comb,
nov. — T. polonicum var. saronicum Jakubz. 1932, Tp. прикл. бот, ген. сел.
Прилож. 53: 140.
Var. halleri Koern. in Koern. el Wern. 1885, l.c.: 101. T. polonicum
var. rubroveslitum Flaksb. 1935, l.c.: 277, descr. ross. (11). Описана no
длинноостым образцам из ботанических садов.
Геогр. распр. Польша.
Var. homsicum Jakubz. 1932, Тр. прикл. бог. ген. сел. Прилож. 53:
142. (12).
Геогр. рас и р. Сирия.
Var. heidelbergi Flaksb. 1929, Тр. прикл. бот. ген. сел. 21, 5: 480,
emend. — Т. polonicum var. psetidomarlinari Flaksb. in Jakubz. 1932, l.c.:
142. — T. polonicum gr. psetidomarlinari Flaksb. 1935, l.c.: 276. (8).
Впервые короткоостистые формы этой разновидности были обнару-
жены К. А. Фляксбергсром в гербарии Гейдельбергского университета.
Геогр. распр. Сирия, Иордания, Кипр, Италия, Франция, Испа-
ния, Португалия, Марокко, Турция, Аргентина.
Var. martinari Koern. in Koern. et Wern. 1885, l.c.: 104, emend.—
T. polonicum var. vestitum Koern. in Koern. et Wern. 1885, l.c.: 103.—
T. polonicum gr. martinari (Koern.) Flaksb. 1935, l.c.: 276. (7).
Выявлены как длинноостистые, так и короткоостистые формы.
Геогр. распр. ГДР, ФРГ, Италия, Португалия, Испания, Марок-
ко, Иордания, Израиль, Турция, Аргентина.
Var. pseudocompactum Flaksb. 1935, l.c.: 276, descr. ross. —T. poloni-
cum var. nigrobarbatum Desv. 1833, l.c.: 149, descr. incornpl. — T. polo-
nicum var. aristinigrum Flaksb. 1915, Опред. пшен.: 116, descr. incompl.
(2). Выявлены как короткоостистые, так и длинноостистые формы.
Геогр. распр. Марокко.
Var. pseudocaryopsirubrum Gokg. 1955, descr. turc. (6).
Описан M. Гекгелем по образцу из Турции.
Var. pseudozaferanicum Gokg. 1955, l.c.: 113, descr. turc. (10).
Описан M. Гекгелем по образцу из Турции.
Var. zaferanicum Gokg. 1955, 1. c.; 113, descr. turc. (9).
Описан M. Гекгелем по образцу из Турции.
Var. zonguldakicum Gokg. 1955, 1. с.: 113, descr. turc. (13).
Описан M. Гекгелем no образцу из Турции.
T. AETHIOPICUM JAKUBZ — ПШЕНИЦА ЭФИОПСКАЯ
Т. aethiopicum Jakubz. 1947, Сел. и сем. 14, 5: 46. — 7'. abyssinicum
(Vav.) Vav. in Flaksb. 1939, Опред. наст, хлебов: 92, non Steud. 1855.—
T. durum abyssinicum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51:
26.-7'. turgidum abyssinicum Vav. 1931, l.c.: 61, — T. durum acutidenti-
10 762
1 16 К.шатфикиции роОи Trillium I,.
culatum Flaksb. 1929, 'Гр. прпкл. бот. теп. сел. 21, 5: 45(>. - - 7’. turgidum
цт. aethiopicum (Jakubz.) Bowden, 1959, Canad. journ. Bot. 37; 671.
Общая характеристика. Всходы фиолеювые, опушенные. Колеоптиль
имеет обычно больше двух проводящих пучков. Куегис1оеть слабая (2-
3), с небольшим числом подгонов. Растение невысокое (60—100 см, г. е.
в 1—2 раза ниже средиземноморских твердых пшениц). Соломина тон-
кая, гибкая, под колосом выполненная или с небольшим просветом.
Влагалище слабоопушеппое. Листовые пластинки взрослых растений
шероховатые. Ушки обычно фиолетовые, с очень короткими ресничками
или без них. Колосья от средних до укороченных (до 10 см) и от рыхлых
до плотных (D = 16 59). Колос па ощупь мягче и нежнее, чем у Т. du-
rum. Колосья некоторых форм имеют большое сходство с пшеницей мяг-
кой: часто их лицевая сторона (как и у мягких пшениц) бывает слегка
шире боковой стороны. Колосковые чешуи голые или опушенные, раз-
личной формы, от плоских до выпуклых (вздутых), среднежесткие или
мягкие (отличие от Т. durum). Киль колосковой чешуи выражен в разной
степени. У основания этой чешуи вдавленность и продольная морщини-
стость отсутствуют. Килевой зубец — от короткого острого до остевид-
ного. Наружная цветковая чешуя у остистых форм несет ость. Ости
сравнительно нежные, расходящиеся от оси колоса, как у пшеницы мяг-
кой, от длинных (длиннее колоса) до укороченных и коротких. У без-
остых форм — остевидные заострения. Внутренняя цветковая чешуя при
созревании не расщепляется вдоль на 2 части. Зерновки белые, крас-
ные и фиолетовые. Все формы яровые, скороспелые, с коротким перио-
дом от всходов до колошения (44—55 дней) и несколько растянутым —
от колошения до созревания (30—37 дней). Весь период вегетации ко-
леблется в зависимости от района высева от 73 до 91 дня.
Анатомическое строение плодовой оболочки зерновки типичное для
голозерных тетраплондпых пшениц (Л. Л. Жестяннпкова, 1954). Вместе,
с тем выявлено и некоторое сходство с. Т. dicoccum и Т. persicum.
В сравнении с последним видом структура плодовой оболочки у пшени-
цы эфиопской более упрощенная, редуцированная.
Для целей систематики рода Triticum J. Percival (1921) первый ис-
пользовал такой признак, как число проводящих пучков в колеоптиле.
Он выявил, что некоторые формы тетраплоидпой пшеницы из Эфиопии
(var. arraseita и var. schimperi) имеют от 3 до 6 таких пучков (Т. durum
имеет 2 пучка). Исследования М. С. Яковлева и Е. И. Николаенко (1931)
показали, что у подавляющего числа образцов пшеницы эфиопской
в колеоптиле больше двух проводящих пучков. Почти 30% образцов
у subsp. aethiopicum, более половины у subsp. vitvilovianum и subsp.
turgidoides имеют 4 пучка в колеоптиле. У некоторых форм число, пуч-
ков достигает 5 (иногда больше).
Исследования В. Ф. Дорофеева и О. Д. Градчаниповой (1971) по-
казали, что анатомическое строение листовой пластинки пшеницы эфи-
опской сходно с Т. persicum: опа у них тонкая, с почти одинаковым чис-
лом устьиц и количеством шипиков па 1 мм2, сходны размер и форма
моторных клеток. Однако в отличие от последнего вида у Т. aethiopicum
листья не бархатистые на ощупь: они снабжены шипикамп, отчего ше-
роховатые.
Эфиопская агро-экологнческая область, ио И. И. Вавилову (1964),
характеризуется обильными осадками (около 1000 мм в год), приуро-
ченными главным образом к первым периодам развития растений пше-
ницы. Посев обычно производи тся в июне июле, а уборка -в нояб-
ре--декабре. Пшеница в горных районах Эфиопии возделывается без
полива.
Вид Т. aethiopicum разделен II. И. Вавиловым (1964) на 2 эколого-
географические группы: 1) собственно эфиопская и 2) йеменская горно-
аравийская. Первая группа представлена яровыми формами, сравни-
П<>()/к») I. Ttiticum
1 17
TMbiio скороспелыми, с быстрым прохождением периода от колошения
до созревания, малой требоваге. 1i,11осi1,10 к теплу в (разе созревания,
влаголюбием, отсутствием засухоустойчивости и сравнительной устойчи-
востью к полеганию. По данным Е. Ф. Пальмовой (1935), она способна
переносить заморозки до —15°С.
Йеменская группа включает также биологически яровые формы, ио
менее влаголюбивые и более требовательные к высокой температуре.
В пределах собственно эфиопской группы II. И. Вавилов (1931) вы-
делил 3 экотипа, каждый из которых приурочен к определенной верти-
кальной зоне. На высоте 1800- -2000 м над ур. м. распространен эко-
тип I, отнесенный К. А. Фляксбергером (1935) к prol. expansoides. Этот
экотип менее разнообразен по ботаническому составу и представлен
преимущественно разновидностями с опушенными колосьями, белыми и
красными зерновками. Па высоте 2400—3000 м локализован основной
экотип II, отнесенный 1\. А. Фляксбергером к prol. tenerum Vav., разно-
видности которого имеют зерновки преимущественно фиолетовые, сте-
бель топкий, колосковые чешуи с очень длинным, иногда переходящим
в ость килевым зубцом. В наиболее высоко!орпом районе Гопдэр сосре-
доточены безостые и полуостпстые формы. На меньших высотах
(2000—2400 м над ур. м.) обитают формы, промежуточные между выше-
описанными тинами (экотип III).
Пшеница эфиопская (исключая йеменские формы), как растение вы-
сокогорного влажного климата, не приспособлена к засушливым усло-
виям. По данным И. А. Стсфановского (1950), опа очень чувствительна
к сухости почвы и зною. Ее посевы в засушливых условиях за период
вегетации сильно (до 40%) изреживаются, а рост растений задержива-
ется. При выращивании в Заволжье в засушливые годы высота соломы
не превышала 50—60 см, а продуктивная кустистость составляла лишь
1,1-—1,3 стебля па одно растение. При суховее урожай снижался па 94%
главным образом вследствие недостаточной выполненности зерна. Ко-
лосья при сильной засухе формировались частично или полностью сте-
рильными. Наименее устойчива пшеница эфиопская к ранней засухе.
К поздней засухе — сравнительно устойчива.
В отличие от собственно эфиопских пшениц йеменские формы лучше
приспособлены к засухе. В засушливых условиях Заволжья выпадение
растений не превышало 5%. Засухоустойчивость была средняя пли вы-
ше средней. Выносливость к суховеям в фазы колошения и налива зер-
на — средняя. Наиболее засухоустойчив местный сорт к-25856.
Как отмечал II. И. Вавилов (1964), пшеница эфиопская выделяется
повышенной склонностью к открытому цветению. Факты ее спонтанной
гибридизации на коллекционных питомниках ВИРа в различных гео-
графических условиях зарегистрированы Н. И. Вавиловым, П. М. Жу-
ковским, М. М. Якубципером. По данным В. М. Берлянд-Кожевникова
и др. (1975), у некоторых образцов (к-19289, к-19600) на долю облигат-
ных самоопылителей приходилось 30—40%, факультативных псрекрест-
ников —50,0%, склонных к перекрестному опылению—10—20% рас-
тений.
По результатам опытов М. М. Якубципера и П. Ф. Покровской (1969),
биохимический состав зерна пшеницы эфиопской выражен в следую-
щих цифрах: белковость—14,4—18,3%; содержание крахмала — 62,3—
66,8%; золы — 1,9—2,3 %.
Следует отметить, что зерно, репродуцированное непосредственно в
Эфиопии, отличается невысокой белковостью (9—11%). Близкие по-
казатели имеет зерно сухумской репродукции. При посеве в Ташкент-
ской области па поливе эти же образцы содержали 19—20% белка
в зерне.
Иммунитет. II. И. Вавилов (1931) охарактеризовал Т. cie.lhiopictim
как более восприимчивый к бурой ржавчине вид, чем пшеница твердая.
10*
1-18 Кмссифштци.ч рода Trilicum I..
Однако on отметил, что генофонд T. aethiopicum включает также формы,
иммунные к этому заболеванию. В опытах Л1. М. Якубцинсра и В. В. Шо-
пиной (1970) некоторые образцы оказались устойчивыми к 14 расам
бурой ржавчины. О. Г. Григорьева (1975) выявила в коллекции
Т. aethiopicum 4,3% иммунных и 26,2% восприимчивых к популяции
бурой ржавчины форм. В. М. Берлянд-Кожсвпиков и др. (1973) нашли,
что у этого вида пшеницы есть образцы — носители генов Lr 3 и Lr 17,
а также групп генов Х2 и Х3, обеспечивающих устойчивость к расам
77,151,202.
Т. aethiopicum, по данным II. II. Вавилова (1931), относительно
устойчив к желтой ржавчине. М. М. Якубцинер (1969) выявил как
восприимчивые, так и иммунные к стеблевой ржавчине формы, что под-
тверждено исследованием О. Г. Григорьевой (1975) при искусственном
заражении коллекции Т. aethiopicum.
Устойчивость к мучнистой росе у Т. aethiopicum отсутствует
(Н. И. Вавилов, 1931; М. М. Якубцинер, 1969). При инокуляции про-
ростков популяциями рас (из 5 географических регионов СССР) этого
патогена все образцы коллекции оказались сильно восприимчивыми; на
провокационном фоне в полевых условиях взрослые растения 95,7%
образцов имели такое же поражение (Э. X. Суханбердина, 1977).
По данным В. И. Кривченко (1973), Т. aethiopicum полиморфен по
отношению к пыльной головне, однако число устойчивых к патогену
образцов преобладает над числом восприимчивых. При искусственном
заражении в опыте Л. М. Ямалеева (1974) мицелий пыльной головни
f. dnri у восприимчивых образцов чаще поражал зародышевую почку
ипокулироваппых зерновок, чем мицелий f. aestivi. Причем электрофо-
ретические. спектры глиадинов зерновок у устойчивых образцов имели
обедненную зону медленных компонентов, а у восприимчивых образцов
эта зона спектра включала 5—6 компонентов. При искусственном зара-
жении большая часть образцов коллекции Т. aethiopicum была воспри-
имчива к твердой головне (В. И. Кривченко, 1973). Иммунность к кор-
невой гнили, вызываемой Cercosporella herpotrichoides Fron., отмечена
М. М. Якубципером (1969).
История. В 1931 г. Н. И. Вавилов писал о пшенице эфиопской как
о культуре очень древней. По его словам, Эфиопия является областью
максимального разнообразия тетраплоидпых пшениц и первичной миро-
вой базой генов этой группы. Однако в дальнейшем (1964) ему стало
ясно, что речь может идти лишь о вторичном гепцентре. Такого же взгля-
да придерживалась Е. Schiemann (1948). Первичным гепцентром тетра-
плоидных пшениц Н. И. Вавилов (1931) признавал Переднюю Азию,
включая Закавказье. Е. Н. Синская (1969) также рассматривала Эфио-
пию как область влияния африканской и древпссредпземпоморскоп
культурных флор, а эндемичную эфиопскую пшеницу считала результа-
том проникновения этой культуры в древние времена из Передней Азии.
Географическое распространение. Пшеница эфиопская возделывает-
ся в пределах горной Эфиопии и южной части Аравийского полуострова.
Происхождение. Первоначально Н. И. Вавилов (1926, 1931) считал,
что первичный центр становления тетраплоидиой пшеницы находится
в Эфиопии, предполагая, что пшеницы этой страны были исходными для
остальных тетраплоидпых видов. Позднее (в 1940 г.) он понял, что
тстраплоидная пшеница из Передней Азии расселилась ио берегам Сре-
диземного моря, обретя вторичный центр формообразования в Эфиопии.
К. А; Фляксбергер (1935, 1938) выводил происхождение Т. aelhiopi-
cum от пшеницы твердой, пришедшей в Африку с Востока. Е. И. Син-
ская (1969) связывала становление Т. aethiopicum с древней предпола-
гаемой популяцией «протомаха», возникшей в Закавказье. Однако,
учитывая эндемизм эфиопских голозерных тетраплоидов, с равным пра-
вом можно предположить их происхождение из полбы обыкновенной,
Ilodpoi) /. Triticum
149
пришедшей в Эфиопию и.з Передней Азии и издревле возделывавшейся
в Северной Африке. Безусловно, что здесь шел самостоятельный процесс
накопления генов, приведших к появлению тетраплоидных форм с поли-
гонной системой легкого обмолота. Становление этих форм—процесс
независимый от формирования 7'. durum. Нет ничего противоестествен-
ного в том, что в горах Эфиопии появились даже формы (subsp. uethiopi-
ctim), имеющие сходство с кавказским видом Т. persicum. По существу
подвид aethiopicum морфологически является формой, признаки которой
стоят между Т. durum и Т. turgidum, с одной стороны, и Т. persicum,
с другой стороны. Д. Мак-Кей (1968) считает, что признаки выделяющие
Т. aethiopicum из ряда других тетраплоидных видов, наложены па гено-
тип видов Т. turgidum и Т. durum. Нам кажется, что такого наложения
признаков не было, а эфиопские и европейско-азиатские голозерные
тетраплоиды произошли независимо друг от друга. Причем первые
возникли от 7’. dicoccum, который имел в колеоптиле 3 пли более про-
водящих пучков, а вторые—от Т. dicoccum с двумя такими пучками.
Признак этот, как уже отмечалось, до сих пор отделяет эфиопские тет-
раплоиды от европейско-азиатских.
Систематика. Эфиопские голозерные те граплопдпые пшеницы
Н. II. Вавилов в 1931 г. впервые выделил как самостоятельные подвиды
в пределах видов Т. durum и 7'. turgidum. /фо пего английский трнтиколог
J. Percival (1921) отнес пшеницу эфиопскую к виду Т. dicoccum па осно-
вании их сходства по числу проводящих пучков в колеоптиле. 11. И. Ва-
вилов подчеркивал своеобразно эфиопских тетраплоидных голозерных
пшениц (наличие фиолетовозервых форм, специфика по числу проводя-
щих пучков в колеоптиле). По характеру колоса, разнообразию колос-
ковых чешуй, наличию безостых, иолуостистых и пнфлятных форм,
шероховатости листовых пластинок пшеница эфиопская сходна с мяг-
кой. Некоторые плотноколосые формы ее напоминают Т. compactum.
Сходство с Т. persicum выражается в развитии остей па колосковых че-
шуях, тонкости колосового стержня (однако не достигающей той, ко-
торая характерна для 'Г. persicum), морщинистости спинки .зерновки.
Наличие форм с хорошо выраженным килем колосковой чешуи, с четко
выраженной двурядностыо боковой стороны колоса, выполненной соло-
миной сближает пшеницы эфиопские с 7’. durum. Основываясь на по-
следнем сходстве, а также па принадлежности пшениц эфиопских к
тетраплоидам, Н. И. Вавилов (1931) отнес их к Т. durum subsp. abyssi-
nicum Vav., подчеркивая при этом их резкое отличие от обычных типов
Т. durum и Т. aestivum. При описании эфиопских пшениц из европейских
гербариев К. А. Фляксбергер (1929) выделил среди них подгруппу vul-
garioides, а позднее (придавая особое значение сходству с пшеницами
мягкими)—subsp. vulgarioides Flaksb. (1939), включающий proles expan-
soides Flaksb. и proles tenero-expansoides Flaksb., которые более сходны
по ряду признаков с пшеницей твердой (ныне subsp. aethiopicum).
Тургидоидные эфиопские пшеницы, ранее относимые к Т. turgidum
subsp. abyssinicuin Vav., имеют сходство с Т. turgidum L. по признакам
колосковой чешуи (более короткая, чем цветковая; вздутая) и зерновки
(горбатая). Однако Н. II. Вавилов (1931) отмечал резкое отличие сре-
диземноморских Т. turgidum от тургцдопдпых эфиопских пшениц и ука-
зывал па большую общность всех групп эфиопских пшениц по экологи-
ческому и морфологическому облику. Наличие у Т. aethiopicum призна-
ков Т. aestivum, Т. durum, Т. turgidum II. И. Вавилов рассматривал как
черту примитивности культуры, хотя и очень древней.
К- А. Фляксбергер (1939) указывал па неоднократные высказывания
11. И. Вавилова о необходимости выделения пшеницы эфиопской в само-
стоятельный вид. Кроме того, он считал, что эта пшеница имеет комплекс
признаков, дающих право для возведения ее в ранг вида Т. abyssinicum.
М. М. Якубцинер в 1947 г. дал этому виду наименование Т. aethiopicum.
I .г>0 Классификации рода Triiictuu I,.
Для 7'. <t<'ictiнг харакчгреи исключи-!e.'H.iiblii по. шморфигм. В ря-
бого 11. 11. Вавилова (1931) приведена ciicieMa изменчивоеi и miieiiiiцы
эфиопской ио многим признакам с указанием места н степени распрост-
ранения ее форм. Наибольшая амплитуда изменчивости выявлена но
признакам колоса (форма и плотность колоса, форма и величина колос-
ковых чешуи, степень развития киля, длина и форма .зубца колосковой
чешуи с. крайними вариантами - - -от почти полного его отсутствия до
перехода в ость длиной до 7 см).
Наследственные различия по длине п плотности колоса были поло-
жены II. И. Вавиловым в основу подразделений вида па секции. Хотя
выраженность этих признаков зависит от места и времени посева и от
наличия переходных форм, тем не менее они являются ключевыми при
внутривидовой классификации 7’. aethiopicum. Этот вид памп подразде-
лен на 3 подвида: 1) subsp. acthiopicti/и (обнаруживает сходство
с Т. aestivum и Т. persicu.ni); 2) subsp. vavilovianum Jakubz. et A. Filat,
(имеет признаки 7'. durum); 3) subsp. turgidoides A. Filat, (по ряду при-
знаков сходен с Т. turgidum).
Ключ для определения подвидов
1. Колосковые чешуи короче цветковых, вздутые; зубец колосковой
чешуи всегда загнут внутрь; зерновка короткая, па спинной стороне
с ясно выраженной горбинкой................subsp. turgidoides Л. Filat.
+ Колосковые чешуи почти равны по длине цветковым, плоские или
выпуклые; зубец колосковой чешуи прямой или загнут внутрь, реже на-
ружу; зерновка удлиненная или округлая, реже горбатая ........... 2.
2. Колосья веретеповидпые, в поперечном сечении овальные, 8—
12 см длины, рыхлые (D около 17), колоски удлиненные, колосковые, че-
шуи со слабовыражсппым килем по всей длине колосковой чешуи; зер-
новка узкая, удлиненная, слегка морщинистая; бороздка зерновки очень
мелкая, широкая; волоски в верхней части зерновки (хохолок) длин-
ные .............................................subsp. aethiopicum.
+ Колосья цилиндрические или суживающиеся к вершине (пирами-
дальные), в поперечном сечении квадратные или прямоугольные, сред-
ней длины или короткие (5—9 см), плотные пли очень плотные (D =
= 30—59); колоски короткие; колосковая чешуя с хорошо выраженным
килем, зерновки короткие, округлые, иа спинной стороне с горбинкой;
бороздка зерновок глубокая, узкая; волоски хохолка зерновок корот-
кие ........................subsp. vavilovianum Jakubz. et A. Filat.
1. Subsp. aethiopicum—.подтип Синди.
Subsp. aethiopicum. — T. durum subsp. abi/ssinicum sect, elongata Vav.
1931, Tp. прикл. бот. гоп. сел. Прилож. 51: 27. — Т. durum subsp. abyssi-
nicum tenerum (экотип II) Vav. 1931, l.c.: 189. — T. abyssinicum subsp.
vulgarioides Flaksb. 1939, Опред. наст, хлебов: 94, quoad prol. tenerum,
nom. illeg.
Колосья 8—12 см длины, рыхлые (D около 17), большей частью ве-
ретеновидшуе (рис. 12, б), в поперечном сечении овальные, остистые или
безостые. Колосковые чешуи гладкие, (блестящие или со слабым воско-
вым налетом) пли опушенные, с остью (до 7 см длины) или с острым
килевым зубцом. Ости тонкие, длинные, слаборасходящпеся, почти па-
раллельные оси колоса. Зерновка узкая, удлиненная, слегка морщини-
стая, с брюшной стороны более пли менее плоская, чаще фиолетовая,
реже красная и еще реже белая. Фиолетовая окраска проявляется почти
на всех частях растения. Опушение всходов густое. Соломина топкая,
упругая. Очень скороспелые; восприимчивые к бурой ржавчине.
Г еогр. распр. Эфиопия (на высоте 2400—3000 м лад ур. м.).
Подрод I. Trilicum
Определение разпппидностсй subsp. aethiopicum
' 1ешуи О< иг Разновидность разновид- ности
белые белые с черными краями крас- ные красные с черными краями че тыс на фоне цвета чешуи черные
белом крас- ном
Колосья остистые
Колосья неопушенные
3 с р но в к и ф и о л с т о в ы е
4 — — — — — -1- — arraseita 1
-ф- — — — — —- — + schweinf urtliii 2
— -1- — — -— — — + idgriniargina- tII11) 3
— — 4 •— — — 4 — scliimpcri 4
— — 4 — -—. — — + sieudclianuni 5
— — — 4 — — — + pseudoschim- pcri 6
— — •— — + — — + niyrieiola- CCHIH 7
— — — — Зерно 4 в к и к расп + ы е fortuiiatiwac 8
4 — — — — -1- — uncinatum 9
— + — — — — — + nigrimenelikii 10
— — + — — — + — rufesccns 11
— — + — — — — + pseudodomi- nans 12
— — — — — — + psetidorufes- cens 13
— — — — 4 — — + negroide.s 14
— — — — Зори 4 0 в к и белы + е harruricum 15
+ — — — — — 4 + brownii albisemine- iiin 1G 17
— + — — — — — + pscudobrow- nii 18
— — + — —. — 4 — rubescens 19
— — + — —• — — 4 pscudoprae- hordeiforine 20
— . — — 4 — — — 4 pseudorubes- ccns 21
— — — — + — — 4 albinigrum 22
3 RoAOl С р Н О В К 4 'ЬЯ опу и ф и шенньи о л с т 4 ) в ы с rubrinigriiin 23
4 — — — — — 4 — albiviolaceum 21
— — — — — — 4 grabhami 25
— — + — — — + — aelhiopiciim 2G
4 Зерно в к н к Р .'1 С И 4 ы с psettdorubri- violaceum 27
4 — — — —. — + — ainharicutn 28
4 — '— — — — — 4 pseudoruriiiil 29
— — 4 — —. — -1- — rubriirisla- ititn 30
— — 4 — — — — 4 psciidorttbr- uristahun 31
— — — + 3 е р н овки белы 4 е rartssimum 32
+ — —• — — — + — praevalenciae 33
-1- — — — — — — -4 protomela.no- pus 34
152
Классификация рода Triticum Л.
Чешуи Остн Разновидность ,\j pj 1H1IBII 1- 1‘OCl H
белые белые с черными краями крас- ные красные с черными краями черные на фоне цвета чешуи черные
белом крас- ном
— + — — — — .— + cingulum 35
— + — — — <— + praeapulicum 36
— — — — + — — + kirae 37
— — — — — + — + prospicuum 38
К о л о с ья nо л у остистые Колосья неопушенные
3 с р и о в к и ф и о ,т е т о в ы с
— - + — 3 С Р II о В К И к + р 11 (' н ы с subechimperi 3'i
+ — + — — — + 1- — submenelibii goildai icuin 10 II
+ 3 е р п 9 В К И б е л ы с pseudogonda- ricum 12
+ — — — — — + — subbrownii 13
+ 3 Колос е р в о ЬЯ O1UJ в к и к + ценные р а с п ы с subpraehordei- forme 44
+ — — — — — — 4- subpseudora- rum 45
— — 4- — 3 е р п О в к и бель 4- с taborense 16
4- — — — — — + — subpraevalen- ciae 47
4“ ,— — — — — — 4- sttbprofome- lanopus 18
Колосья безостые
Колосья неопушенные
Зернов к и ф ноле т о в ые
+1 - I - I - I “ I - I..................... I 49
Зерновки к р а с н ы с
— I = 1 r 1 r | “ 1 figrfii.se 1 axumicuin | 50 1 51
Зерновки бе л ы е । rubrimuticum 1 52
Колосья опушенные (зерновки красные)
+ * galiasoruni | 53
* Окраска остсвндиых зубцов на цветковых чсшуях.
Var. aethiopicum. Т. dicoccuin var. elhiopicuin Perciv. 1926, Journ.
Bot.: 207. — 7’. durum subsp. abyssinicuin var. rubroviolaceum Vav. 1931.
Tp. прикл. бот. ген. сел. Прилож. 51: 31. — Т. abyssinicuin var. rubrnvio-
laceuni Vav. in Flaksb. 1939, Опред. паст, хлебов: 99. (2G).
Геогр. распр. Эфиопия (редкая примесь в Дэбрэ-Маркос).
Подрод 1. Triticum
153
Рис. 12. Колосья, колоски и зерновки Т. aethiopicum Jakubz.:
а—subsp. vavilovianum Jakubz. et Л. Filat.; 6 — subsp. aethiopicum
Var. albinigrum (Vav.) A. Filat, comb. nov.— T. durum var. albonig-
rum Vav. 1931, l.c.: 38.— T. abyssinicum var. albonigruni Vav. in Flaksb.
1939, l.c.: 104. (22).
Геогр. распр. Эфиопия (редко).
Var. albiseininetim (Vav.) A. Filat. comb. nov. T. durum var. adxi-
semineum Vav. 1931, l.c.: 37.- T. abyssinicum var. albosemineum Vav.
in Flaksb. 1939, l.c.: 103. (17).
Геогр. рас. п p. Эфиопия (редко в районах Харэра и Аддис-Абебы).
Var. albiviolaceum (Vav.) Л. Filat, comb. nov. — T. durum var. albn-
violaceum Vav. 1931, l.c.: 31. — T. abyssinicum var. alboviolaceum Vav.
in Flaksb. 1939, l.c.: 98. (24).
151
Классификация />о<)а Triticum !..
Г с oi р. распр. Эфиопия (редко и районе Аддис-Абебы и и горной
Эритрее).
Var. ainharicuni (Perciv.) Л. I'ilat. comb. nov. --- 'Г. dicoccum var.
amharicum Perciv. 1926, l.c.: 206.-- T. durum var. rarum Vav. 1931, l.c.:
35. — T. abyssinicum var. rarum Vav. in Flaksb. 1939, l.c.: 101. (28).
Геогр. распр. Эфиопия (редко).
Var. arraseita (Hochst. ex Koern.) A. Filat. comb. nov. — T. durum
var. arraseita Ilochst. ex Koern. 1885, in Koern. et Worn, llanclb. Gctrcidcb.
1: 70; Вавилов, 1931, l.c.: 27. - - T. durum var. hildebrandti Wiltm. 1879,
Bericht. Deutsch. Naturforsch. Baden-Baden: 211, nom. dnbiiirn.—
T. eloboni Caporn, 1918, Journ. Genet. 7: 259. — T. dicoccum var. arraseita
Perciv. 1921, Wheat pl.: 195. (1).
Var. arraseita, как и большинство других разновидностей вида, в за-
висимости от длины килевого зубца колосковой чешуи делится Н. И. Ва-
виловым (1931) на 3 подразиовидности: subvar. brevidentatum Vav. (зу-
бец короче 1 мм), subvar. longidentatum Vav. (зубец до 5 мм длины),
subvar. aristiforme Vav. (зубец более 5 мм, часто переходит в остевидные
заострения).
Геогр. распр. Наиболее распространенная разновидность, осо-
бенно в высокогорной зоне Эфиопии.
Var. axumicum (Vav.) A. Filat, comb. nov. — - T. durum var. axumicum
Vav. 1931, l.c.: 59.— T. abyssinicum var. axumicum Vav. in Flaksb. 1939,
l.c.: 106. (51).
Геогр. распр. Эфиопия (в районах Гопдэра и Аксума, редко
в горной Эритрее).
Var. brownii (Perciv.) A. Filat, comb. nov. — T. dicoccum sect, tenaces
var. brownii Perciv. 1926, Journ. Bot.: 205.— T. durum var. brownii
(Perciv.) Vav. 1931, l.c.: 36. — T. abyssinicum var. brownii (Perciv.) Vav.
in Flaksb. 1939, 1. c.: 103. (16).
Геогр. распр. Эфиопия (редко).
Var. fortunatovae A. Filat, nom. nov. — T. durum var. amharicum Vav.
1931, l.c.: 30, non Perciv., nom. praeocup. — T. abyssinicum var. atnhari-
cumNav. in Flaksb. 1939,1. c.: 98. (8).
Геогр. распр. Эфиопия (редкая примесь в районах Аддис-Абебы,
Харэра и Фиче).
Var. gallasorum (Vav.) A. Filat, comb. nov. — T. durum var. gallasorum
Vav. 1931, l.c.: 59. — T. abyssinicum var. gallasorum Vav. in Flaksb.
1939, l.c.: 106. (53).
Геогр. распр. Эфиопия (редкая примесь около Гондэра и Фиче).
Var. gondaricum (Vav.) A. Filat, comb. nov. — T. durum var. gondari-
cum Vav. 1931, 1. c.: 35. — T. abyssinicum var. gondaricum Vav. in Flaksb.
1939, l.c.: 100. (41).
Геогр. распр. Эфиопия (особенно часто встречается около Гон-
дэра, где нередко является преобладающей, а также около Фиче и
в горной Эритрее).
Var. grabhami (Perciv.) A. Filat, comb. nov. — T. dicoccum var. grab-
hami Perciv. 1926, l.c.: 206. — T. durum var. pseudoalboviolaceum Vav.
1931, l.c.: 31.— T. abyssinicum var. pseudoalboviolaceum Vav. in Flaksb.
1939, Lc.: 99. (25).
Геогр. распр. Эфиопия.
Var. harraricum (Vav.) A. Filat, comb. nov. — T. durum var. harrari-
cum Vav. 1931, l.c.: .35. - - T. abyssinicum var. harraricum Vav. in I’laksb.
1939, l.c.: 101. (15).
Геогр. распр. Эфиопия (преобладающая разновидность в рай-
оне Харэра, как примесь в районах Лнкобэра, Аддис-Абебы, Фиче).
Var. kirae (Vav.) Л. Filat, comb. nov. — T. durum var. kirae Vav. 1931,
l.c.: 39. — T. abyssinicum var. kirae Vav. in Flaksb. 1939, l.c.: 106. (37).
IIo<)po<) 1. Tttlicuin
155
Геогр. распр. Эфиопия (редко в районе Харэра).
Var. negroides (Vav.) j\. Filat. comb. nov. T. durum var. negroides
Vav. 1931, l.c.: 34. — T. abijssiiiictini var. negroides Vav. in Flaksb. 1939,
1. c : 101. (14).
Геогр. распр. Эфиопия (редкая примесь iiobccmcci ио) .
Var. nigrimarginatum (Chiov.) A. Filat, comb. nov. - T. durum var.
nigromarginaittm Chiov. 1912, Monogr. Rapp, colon. (Roma), 19: 15; Ва-
вилон, 1931, l.c.: 29,- -7’. dicoccum var. pseudoarraseita Perciv. 1926,
l.c.: 205. (3).
Геогр. распр. Эфиопия (широко распространена, главным обра-
зом в районах Аддис-Абебы, Апкобэра, Харэра, Фиче, Дэбрэ-Маркоса).
Var. nigrimenelikii (Vav.) A. Filat, comb. nov. — - T. durum var. nigro-
meneliki Vav. 1931, l.c.: 33. — T. dicoccum var. pseudouncinaltim Perciv.
1926, 1. c.: 205, quoad f. I. — T. abi/ssinicum var. nigromeneliki Vav. in
Flaksb. 1939, l.c.: 100. (10).
Геогр. распр. Эфиопия (повсеместно редкая примесь).
Var. nigriviolaceum (Orlov) A. Filat, comb. nov. — T. durum var.
nigroviolaceum Orlov, 1922—1923, Тр. прикл. бот. сел. 13: 374; Вавилов,
1931, l.c.: 30.— Т. abi/ssinicum var. nigroviolaceum (Orlov) Flaksb. 1939,
l.c.: 98. (7).
Геогр. распр. Эфиопия (широко распространена).
Var. praeapulicum (Vav.) A. Filat, comb. nov.— T. durum var. praeapu-
licum Vav. 1931, l.c.: 39. — T. abyssinicuin var. praeapulicum Vav. in
Flaksb. 1939, l.c.: 106. (36).
Геогр. распр. Эфиопия (редко).
Var. praevalenciae (Vav.) A. Filat, comb. nov. — 7’. durum var. prae-
valenciae Vav. 1931, l.c.: 38. — T. dicoccum var. lomentosum f. II Perciv.
1926, l.c.: 206.— T. abi/ssinicum var. praevalenciae Vav. in Flaksb. 1939,
l.c.: 105. (33).
Геогр. распр. Эфиопия (районы Харэра, Апкобэра, Аддис-Абебы,
Фиче, Гондэра, Дапглы).
Var. prospicuum (Vav.) A. Filat, comb, nov. — T. durum var. prospi-
ciiutn Vav. 1931, l.c.: 39. — T. abyssinicuin var. prospicuum Vav. in Flaksb.
1939, l.c.: 106. (38).
Геогр. распр. Эфиопия (редкая примесь в районе Харэра).
Var. protomelanopus (Vav.) A. Filat, comb. nov. — T. durum var. pro-
iomelanopus Vav. 1931, l.c.: 39. — T. abyssinicuin var. protomelanopus
Vav. in Flaksb. 1939, l.c.: 105. (34).
Геогр. распр. Эфиопия (редко).
Var. pseudobrownii (Perciv.) A. Filat, comb. nov. — T. dicoccum var.
pseudobrownii Perciv. 1926, l.c.: 205. — T. durum var. nigrobrownii Vav.
1931, l.c.: 37. •—T. abyssinicum var. nigrobrownii Vav. in Flaksb. 1939.
1. c.: 103. (18).
Г e о г p. p а с п p. Эфиопия (редко).
Var. pseudodominans (Vav.) A. Filat, comb. nov. — T. durum var.
pseudoclominans Vav. 1931, l.c.: 34. — T. abyssinicum var. pseudodomi-
nans Vav. in Flaksb. 1939, l.c.: 100. (12).
Геогр. распр. Эфиопия (районы Харэра, Аддис-Абебы, Апкобэ-
ра, Гондэра, Дапглы, оз. Таны).
Var. pseudogondaricum (Vav.) A. Filat, comb. nov. — T. durum var.
pseudogondaricum Vav. 1931, Re.: 55.-— T. abyssinicum var. pseudogon-
daricumVav. in Flaksb. 1939, I. c.: 100. (42).
Геогр. ]) а си p. Эфиопия (редко в районе Гондэра).
Var. pseudopraehordeiforme (Vav.) A. Filat, comb. nov. -- 7’. durum
var. pseudopraehordeiforme Vav. 1931, l.c.: 37—38. — T. abyssinicum var.
pseudopraehordeiforme Vav. in Flaksb. 1939, I. c.: 104. (20).
Геогр. распр. Эфиопия (районы Харэра, Аддис-Абебы и Лнко-
бэра).
1 56 K.uu i ификация роди Trilicuin Л.
Var. pseudorarum (Vav.) Л. Filat. comb. nov. T. durum var. pseudo -
rarum Vav. 1931, l.c.: 35.--7'. abyssinicum var. pseiidorariim, Vav. in
Flaksb. 1939, I. c.: 101. (29).
Геогр. распр. Эфиопия.
Var. pseudorubescens (Perciv.) A. Filat, comb, nov.-- T. dicoccum var.
pseudorubescens Perciv. 1926, l.c.: 203.— T. durum var. nigropraehordei-
forme Vav. 1931, l.c.: 38. — T. abyssinicum var. nigropraehordeiforme Vav.
in Flaksb. 1939, l.c.: 104. (21).
Геогр. распр. Эфиопия (районы Харэра, Ликобэра, Аддис-Абебы
и Эритрея).
Var. pseudorubraristatuni (Vav.) A. Filat, comb. nov. — T. durum
var. pseudorubroaristatum Vav. 1931, l.c.: 36. — T. abyssinicum var.
pseudorubroaristatum Vav. in Flaksb. 1939, I. c.: 102. (31).
Геогр. распр. Эфиопия (редкая примесь в районах Харэра, Ад-
дис-Абебы, Фиче и Гопдэра).
Var. psetidorubriviolaceum (Vav.) A. Filat, comb. nov. — T. durum var.
pseudorubroviolaceum Vav. 1931, l.c.: 31.— 7’. abyssinicum var. psetido-
rubroviolaceiim Viw. in Flaksb. 1939. l.c.: 99. (27).
Геогр. распр. Эфиопия (очень редкая примесь в районах Аттис-
Лбебы и Апкобэра).
Var. pseudorufescens (Perciv.) A. Filat, comb. nov. — T. dicoccum var.
pseudorufescens Perciv. 1926, l.c.: 207. — T. durum var. nigrodominans
Vav. 1931, l.c.: 34. — T. abyssinicum var. nigrodominans Vav. in Flaksb.
1939, l.c.: 101. (13).
Геогр. распр. Эфиопия (районы Харэра, Аддис-Абебы, Лпко-
бэра).
Var. pseudoschimperi (Perciv.) A. Filat, comb. nov. — T. dicoccum var.
pseudoschimperi Perciv. 1926, l.c.: 207. — T. durum var. nigrosteudelia-
num. Vav. 1931, l.c.: 30. — T. abyssinicum var. nigrosteudelianum Vav. in
Flaksb. 1939, l.c.: 98. (6).
Геогр. распр. Эфиопия.
Var. rarissimuni (Vav.) A. Filat, comb. nov. —T. durum var. rarissi-
mum Vav. 1931, l.c.: 36.— T. abyssinicum var. rarissimuni Vav. in Flaksb.
1939, 1. c.: 102. — T. dicoccum var. persicutn Perciv. 1921, l.c.: 196,
p.p. (32).
Геогр. распр. Эфиопия (редко в районе Харэра, Фиче, Аддис-
Абебы).
Var. rubescens (Perciv.) A. Filat, comb. nov.— T. dicoccum var. rubes-
cens Perciv. 1926, l.c.: 206, quoad f. I. — T. durum var. praehordeiforme
Vav. 1931, l.c.: 37. — T. abyssinicum var. praehordeiforine Vav. in Flaksb.
1939, 1. c.: 103. (19).
Геогр. распр. Эфиопия.
Var. rubraristatum (Vav.) A. Filat, comb. nov. — T. durum var. rubr.o-
aristatum Vav. 1931, l.c.: 35—36.— T. abyssinicum var. riibroaristatum
Vav. in Flaksb. 1939, 1. c.: 102. (30).
Геогр. распр. Эфиопия (редко в районах Харэра и Гопдэра).
Var. rubrimuticum (Vav.) A. Filat, comb. nov. — T. durum var. rubro-
muticum Vav. 1931, l.c.: 59.— T. abyssinicum var. rubromuticum Vav.
in Flaksb. 1939„ l.c.: 106. (52).
Геогр. распр. Эфиопия (очень редко в paiione Гопдэра).
Var. rtibrinigrum (Vav.) Л. Filat, comb. nov. — 7’. durum var. rtibro-
nigrum Vav. 1931, l.c.: 38. — T. abyssinicum var. rubronigrum Vav. in
Flaksb. 1939, 1. c.: 104. (23).
Геогр. распр. Эфиопия (районы Харэра, Аддис-Абебы, Анкобэ-
ра и в Лдуа).
Var. rufescens (Perciv.) Л. Filat, comb. nov. — T. dicoccum var. ru-
fescens Perciv. 1921, l.c.: 196. — T. durum var. dominans Vav. 1931, l.c.:
33. — T. abyssinicum var. dominans Vav. in Flaksb. 1939, l.c.: 100. (11).
Подрод I. Triticum
157
Ге or p. расп p. Эфиопия. Одна из наиболее распространенных раз-
новидностей.
Var. schiinperi (Koern.) Л. Filat, comb. nov. T. durum var. schimpcri
Koern. in Koern. et Wern. 1X85, l.c.: 71; Вавилов, 1931, l.c.: 29.---7’. di-
coccum var. schiinperi (Koern.) Perciv. 1921, l.c.: 196.— 7’. abyssinicum
var. schimperi (Koern.) Flaksb. 1939, l.c.: 98. (4).
Геогр. p a с H p. Эфиопия.
Var. schweinfurthii (Vav.) A. Filat, conib. nov. T. durum var. schwein-
fiirthii Vav. 1931, l.c.: 27.— 7'. abyssinicum var. schweinfurlhii Vav. in
Flaksb. 1939, l.c.: 97. (2).
Геогр. распр. Эфиопия (редко в районах Аддис-Абебы, Харэра
и Анкобэра).
Var. singulum (Vav.) A. Filat, comb. nov.-^7'. durum var. singulum
Vav. 1931, l.c.: 39. — T. abyssinicum var. singulum Vav. in Flaksb. 1939,
l.c.: 105. (35).
Геогр. распр. Эфиопия (редко в районе Гопдэра).
Var. steudelianuin (Vav.) Л. Filat, comb. nov.— T. durum var. steude-
lianuni Vav. 1931, l.c.: 30. — 7'. abyssinicum var. steudclianum Vav. in
Flaksb. 1939,1. c.: 98. (5).
Геогр. распр. Эфиопия (редко в районах Харэра, Анкобэра, Ад-
дис-Абебы, Гопдэра).
Var. subbrownii (Vav.) A. Filat, comb. nov. — T. durum var. subbrow-
nii Vav. 1931, l.c.: 55. — T. abyssinicum var. subbrownii Vav. in Flaksb.
1939, l.c.: 103. (43).
Геогр. рас. н p. Эфиопия (редко в районе Гопдэра).
Var. submenelikii (Vav.) A. Filat, comb. nov. — T. durum var. subme-
neliki Vav. 1931, l.c.: 55.— T. abyssinicum var. submenelikii Vav. in
Flaksb. 1939, l.c.: 99. (40).
Геогр. распр. Эфиопия (редко в районе Гопдэра па высоте
2600 м над ур. м.).
Var. subpraehordeiforme (Vav.) Л. Filat, comb. nov. — 7’. durum var.
subpraehordeifoime Vav. 1931, l.c.: 55.— T. abyssinicum var. subpraehor-
deifortneVav. in Flaksb. 1939, l.c.: 104. (44).
Геогр. распр. Эфиопия (в районе Гопдэра, редко в Фиче).
Var. subpraevalenciae (Vav.) A. Filat, comb. nov. — T. durum var.
subpraevalenciae Vav. 1931, l.c.; 51. — T. abyssinicum var. subpraevalen-
ciae Vav. in Flaksb. 1939, l.c.: 105. (47).
Геогр. распр. Эфиопия (горная Эритрея).
Var. subprotomelanopus (Vav.) A. Filat, comb. nov.— T. durum var.
subprotomelanopus Vav. 1931, l.c.; 57. — T. abyssinicum var. subproto-
melanopus Vav. in Flaksb. 1939, l.c.: 105. (48).
Геогр. распр. Эфиопия (редко в районе Гопдэра).
Var. subpseudorarum (Vav.) A. Filat, comb. nov. — T. durum var.
subpseudorarutn Vav. 1931, l.c.: 55. — T. abyssinicum var. subpseudorarum
Vav. in Flaksb. 1939, 1. c.: 101. (45).
Геогр. распр. Эфиопия (редко в районах Гопдэра и Фиче).
Var. subschimperi (Vav.) A. Filat, comb. nov. — T. durum var. stib-
schimperi Vav. 1931, l.c.: 55. — T. abyssinicum var. stibschimperi Vav. in
Flaksb. 1939, l.c.: 98. (39).
Геогр. pacrtp. Эфиопия (редко около Гопдэра).
Var. taborense (Vav.) A. Filat, comb. nov. — T. durum var. taborense
Vav. 1931, l.c.: 55. — 7'. abyssinicum var. taborense Vav. in Flaksb. 1939,
1. c.: 102. (46).
Геогр. распр. Эфиопия (районы Гопдэра и Дэбрэ-Табор).
Var. tigrense (Vav.) A. Filat, comb. nov. — T. durum var. tigrense Vav.
1931, l.c.: 57. — T. abyssinicum var. tigrense Vav. in Flaksb. 1939, l.c.:
106. (50).
Геогр. распр. Эфиопия (провинция Тигре, па высоте 2620 м над
УР- м.).
158 Классификация рода Trilicum I,.
Var. uncinatum (Perciv.) A. I’ilal. comb. nov. T. dicoccitm var. unci-
naltun Perciv. 1921, l.c.: 194, quoad form 1; idem, 1926, l.c.: 205, quoad
f. I, II spec, aethiop. /'. durum, var. menelikii Vav. 1931, l.c.: 32 33. -
T. abyssinicuin var. menelikii Vav. in Flaksb. 1939, l.c.: 99. (9).
Геогр. распр. Широко распространенная разновидность в Эфио-
пии (чаще в районах Аддис-Абебы и Харэра), иногда преобладает
в посеве.
Var. violaceomuticrim (Vav.) A. Filat, comb. nov. - - T. durum var.
violaceomuticum Vav. 1931, l.c.: 57. - - T. abyssinicuin var. violaceomuti-
cum Vav. in Flaksb, 1939, l.c.: 106. (49).
Геогр. распр. Эфиопия (редко около Гондэра).
II. Subsp. vavilovianum Jakubz. et A. Filat.- • подвид Вавилова.
Subsp. vaviloviannm Jakubz. et A. Filat, subsp. nov. — T. durum subsp.
abyssinicuin экотип I и III Vav. 1931, Tp. прикл. бот. ген. сел. Прилож.
51: 188, 189. — Т. durum subsp. abyssinicum prol. expansoides Flaksb.
1935, Культ, фл. СССР, 1: 156, nom invalid, el prol. teneroexpansoides
Flaksb. 1935, l.c.: 157, nom. invalid. — T. abyssinicum subsp. vulyarioides
Flaksb. 1939, Опред. наст, хлебов: 94, 96, excl. pro!, tenerum, nom. illeg. --
Spica cylindrica vol pyramidata, 5 -9 cm longae, sectione quadrangularis
vel rectangulata. D = 30—40 et ultra; spicula brevis; carina glumis dis-
tincta; caryopsis brevis, rotunda, dorso convexa.
Typus: T. aelliiopiciim var. densarraseita (Vav.) A. Filat.
Колосья от средних до укороченных (5-9 см длины), средней плот-
ности (D = 30—40) или плотные (D — 40—59), удлиненные, цилиндри-
ческие или иногда пирамидальные. Колосковые чешуи голые (блестящие
или с восковым налетом) или опушенные, разнообразные по форме и
размеру, нежные или грубоватые по консистенции; киль ясно выражен,
но сравнительно узкий, доходит до основания колосковой чешуи, где
отсутствуют поперечная вдавленность и продольная морщинистость. Ки-
левой зубец острый, от короткого до остевндпого (до 7 см длины). Зер-
новки короткие, округлые, нередко па спинной стороне с горбинкой, бе-
лые, красные пли фиолетовые. Кустистость слабая (2—3), с небольшим
числом подгонов. Растения невысокие. Соломина топкая, гибкая, вы-
полненная или с небольшим просветом. Влагалище слабоопушспное.
Листовые пластинки шероховатые. Ушки фиолетовые, с короткими рес-
ничками или без них. Растения яровые, раннеспелые (длина периода
вегетации 73—91 день), с небольшой продолжительностью периода от
всходов до колошения (44—55 дней) и еще более коротким от колоше-
ния до созревания (30—37 дней). Восприимчивы к бурой ржавчине,
менее восприимчивы к желтой ржавчине. Поражаются пыльной и мок-
рой головней.
Геогр. распр. Эфиопия (преобладает в районах Харэра и Чэрчэ-
ра) — от 1800 до 2400 м над ур. м.
Ключ для определения групп разновидностей
subsp. vavilovianum Jakubz. et A. Filat.
1. Колосья цилиндрические, 5—9 см длины, плотные (D = 30—40)
............'..........................convar. vavilovianum A. Filat.
+ Колосья суживающиеся к вершине, длиной до 5 см, плотные
(D более 40)......................convar. compactum (Vav.) A. Filat.
1. Convar. vavilovianum- группа плотноко.тосы.х разповидносюи.
Convar. vavilovianum A. Filat, nom. nov. — T. durum subsp. abyssini-
cum sect, densa Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 39, 57,
nom. pracocup.
Тип: Т. aethiopicum var. densarraseita (Vav.) A. Filat.
Геогр. pa сир. Эфиопия (па высоте 1800—2400 м над ур.м.).
Подрод I. Triticum
Определение разновидностей convar. vavilovianum Л. Filat.
Чешуи Ости PaiHOfinuiocib № разновид- ное 1 и
белые белые с черными краями крас- ные красные с черными краями черные на фоне цвета ЧСШ) к черные
белом крас- ном
Колосья о с 'г ист ы е
Колосья неопушенные
Зернов к и ф ио лето вые
+ — — — — — 4- — densarra- seita 1
— + — — — — — + nigridens- arruseila 2
— •— 4- — — — 4- —- dcnsischim- peri 3
—• __ — 4- — — — + nigridensi- schiiiiperi 4
3 ер и о в к и к расы + ы е d enxaiii hari- ctlin 5
+ — —- — — — + — densimeneli- kii 6
+ — — — — — • — + psettdodensi- menelikii 7
+ 4- — j — + pseudotincina- tiiin praescmiticum 8 9
— . — ч-- — — — — + pseudopraese- miticum 10
— — — ч- — — —. + nigripi aesemi- ticum 11
— — — — + — — + bicolor 12
— — —. — — + — + densiharrari- cuin 13
Зерновки белые
4- + + — — + 1 + 4- bialbum marginatum somalense 14 15 16
— — + — — — — 4- pseudosoma- Icnse 17
— — — 4~ — — — ч- nigrisomalen- se 18
— — — —- 4- — — -1- densalbinig- rum 19
3et Колос НОНК + ья on у и ф и ценные о л е т Д- в ы е densirubri- nigrum 20
4- — — — — — — + densiviola- ceuni 21
4- 3 e p и о В К II к р а с и -1- 4- 1.1 с abbaicuin Ithertihoiii uni 22 23
1 — — — — + (oiniltina 21
— 4- — — — + rubi ivillosum 25
— — — — — — + pscudorubri- ptibeciens 26
— i — 1 — । — -I- — 4- pilosini gr II III 27
н >0
Подрод I. Trilicum
О к о н ч а и и е
Чешуи Ости Разновидность № разновид- ности
белые белые с черными краями крас- н ы е красные с черными краями черные на (рои е цвета чешуи чер- ные
белом крас- ном
3 С Р И О 1! К и б с л ы е
+ — — 1 — — + — tomentosum 28
т — — — — — — + pseudotomen- 29
tosum
— — + - - - + — vulpinum 30
— — + - - - — + godjainicutn 31
Колосья полуостистые (колосья опушенные).
Зерновки красные
— 1+1 densisemiaris- 1 32
_ + | - |_|_ 1 1 tiihim |
Зернов к н б о л ы e
-1- — — — — — — : subnegustia- 33
num
— — + — — - — + antiquoaet- 34
hiops
Var. abbaicum (Vav.) A. Filat, comb. nov. — T. durum ,var. abbaicum
Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 40. — Т. abyssinicum var.
abbaicum Vav. in Flaksb. 1939, Опред. хлебн. злаков: 99. (22).
Геогр. распр. Эфиопия (редко в Годжаме, Дэбре-Маркосе и Ад-
дис-Абебе) .
Var. antiquoaethiops (Vav.) Л. Filat, comb. nov. — T. durum var.
antiquoaethiops Vav. 1931, l.c.: 57.— T. abyssinicum var. antiquoaethiops
Vav. in Flaksb. 1939, l.c.: 105. (34).
Геогр. распр. Эфиопия (редко в Гопдэре).
Var. bialbum (Vav.) Л. Filat, comb. nov. — T. durum var. bialbum Vav.
1931, Lc.: 46.— T. abyssinicum var. bialbum Vav. in Flaksb. 1939, l.c.:
103. (14).
Геогр. распр. Эфиопия (повсеместно как редкая примесь).
Var. bicolor (Chiov.) A. Filat, comb. nov. — T. durum var. bicolor
Chiov. 1912, Monogr. Rapp, colon. (Roma), 19: 15; Вавилов, 1931, l.c.:
42. — T. abyssinicum var. bicolor (Chiov.) Vav. in Flaksb. 1939, l.c.:
101. (12).
Геогр. распр. Эфиопия (северная и западная).
Var. comitans (Vav.) A. Filat, comb. nov. — T. durum var. comitans
Vav. 1931, 1. c.: 44. — T. abyssinicum var. comitans Vav. in Flaksb. 1939,
l.c.: 102. (24).
Геогр. распр. Эфиопия (повсеместно).
Var. densalbinigrtim (Vav.) A. Filat, comb. nov. — T. durum var.
densoalbonigrutn Vav. 1931, l.c.: 47. — T. abyssinicum var. densoalbonig-
rumVav. in Flaksb. 1939, l.c.: 104. (19).
Геогр. р'аспр. Эфиопия.
Var. densamharicum (Vav.) A. Filat, comb. nov. — T. durum var.
densoamharicum Vav. 1931, l.c.: 40.— T. abyssinicum var. densoatnhari-
cum Vav. in Flaksb. 1939,1. c.: 98. (5).
Геогр. распр. Эфиопия (редко в районеЧэрчэра).
Var. densarraseita (Vav.) A. Filat, comb. nov. — T. durum var. den-
soarraseita Vav. 1931, Lc.: 39. — T. abyssinicum var. densoarraseita Vav.
in Flaksb. 1939, Lc.: 97. (1).
Геогр. распр. Эфиопия (повсеместно как редкая примесь).
Подрод I. Triticum
161
Var. densiharraricum (Vav.) A. Filat, comb. nov. T. durum var.
densoharrariciim Vav. 1931, l.c..: 32. T. abyssinicutn var. densoharrari-
mtVav. in Flaksb. 1939, l.c.: 101. (13).
Геогр. распр. Эфиопия (районы Фиче и Чэрчэра).
Var. densinienelikii (Vav.) A. Filat, comb. nov. — 7’. durum var. den-
someneliki Vav. 1931, l.c..: 41. — T. abyssinicum var. densomeneliki Vav.
in Flaksb. 1939, l.c.: 99. (0).
Геогр. распр. Эфиопия (повсеместно).
Var. densirubrinigrum (Vav.) A. Filat, comb. nov. — T. durum var.
densorttbronigriim Vav. 1931, l.c.: 47. — 7'. abyssinicum var. densorubro-
nigrum Vav. in Flaksb. 1939, l.c.: 104. (20).
Геогр. p a cup. Эфиопия (редко в провинциях Харэр, Шоа и в гор-
ной Эритрее).
Var. densischimperi (Vav.) A. Filat, comb. nov. — T. durum var. den-
soschimperi Vav. 1931, I. c..: 40. — T. abyssinicum var. densoschimperi Vav.
in Flaksb. 1939, I. c.: 98. (3).
Геогр. распр. Эфиопия (Гондэр, Дэбрэ-Маркос, Шоа).
Var. densisemiaristatum (Vav.) Л. Filat, comb. nov. — T. durum var.
densosemiaristatum Vav. 1931, l.c.: 57. - - T. abyssinicum var. denso-
semiaristatum Vav. in Flaksb. 1939, l.c.: 102. (32).
Г e о г p. p а с п p. Эфиопия (редко в Гопдэрс).
Var. densiviolaceum (Vav.) A. Filat, comb. nov. — 7’. durum var. den-
soviolaceum Vav. 1931, l.c.: 40. — T. abyssinicum var. densoviolaceum
Vav. in Flaksb. 1939, l.c.: 99. (21).
Геогр. распр. Эфиопия (редко околоФиче,Харэра,Дэбрэ-Мар-
коса и в Эритрее).
Var. godjamicum (Vav.) A. Filat, comb. nov. — T. durum, var. godjami-
cum Vav. 1931, l.c,: 48. — T. abyssinicum var. godjamicum Vav. in Flaksb.
1939, l.c.: 106. (31).
Геогр. распр. Эфиопия (около Апкобэра, Аддис-Абебы, Адуа,
Харэра и в провинции Годжам).
Var. marginatum (Chiov. ex Vav.) A. Filat, comb. nov. — 7’. durum
var. marginatum Chiov. ex Vav. 1931, l.c.: 46. — T. abyssinicum var.
marginatum (Chiov.) Vav. in Flaksb. 1939, l.c.: 103. (15).
Геогр. распр. Эфиопия.
Var. nigridensarraseita (Vav.) A. Filat, comb. nov. — T. durum var.
nigrodensoarraseita Vav. 1931, l.c.: 40.— 7’. abyssinicum var. nigrodenso-
arraseita Vav. in Flaksb. 1939, l.c.: 97. (2).
Геогр. распр. Эфиопия (редко около Харэра и в Эритрее).
Var. nigridensischimperi (Vav.) A. Filat, comb. nov. — T. durum var.
nigrodensoschimperi Vav. 1931, l.c.: 40. — T. abyssinicum var. nigroden-
soschimperiVav. in Flaksb. 1939, 1. c.: 98. (4).
Геогр. распр. Эфиопия (редкая примесь около Аддис-Абебы).
Var. nigripraesemitictim (Vav.) A. Filat, comb. nov. — 7’. durum var.
nigropraesemiticum Vav. 1931, l.c.: 42. — 7’. abyssinicum var. nigroprae.se-
miticum Vav. in Flaksb. 1939, l.c.: 101. (11).
Геогр. распр. Эфиопия (повсеместно, по редко).
Var. nigrisomalense (Vav.) A. Filat, comb. nov. — T. durum var. nigro-
somalense Vav. 1931, l.c.: 46. — T. abyssinicum var. nigrosotnalense Vav.
in Flaksb. 1939, l.c.: 104. (18).
Геогр. распр. Эфиопия (повсеместно).
Var. pilosinigrum (Vav.) A. Filat, comb. nov. — T. durum var. piloso-
nigrurn Vav. 1931, l.c.: 44. - - 7’. abyssinicum var. pilosonigruin Vav. in
Flaksb. 1939, l.c.: 102. (27).
Геогр. распр. Эфиопия (редко в Харэре).
Var. praesemiticum (Vav.) A. Filat, comb. nov.— T. durum var. pra-
esemiticum Vav. 1931, l.c.: 41. — T. abyssinicum var. praesemiticum Vav.
in Flaksb. 1939, l.c.: 100. (9).
Геогр. распр. Эфиопия.
1 1 7S.9.
162
Классификация рода Triticum Т.
Var. psciidodensimcnclikii (Vav.) A. Filat. comb. nov. T. durum var.
pseudodensomenelikii Vav. 1931, l.c.: 41. 7'. abyssinicum var. pseudo-
dcnsomenelikii Vav. in Flaksb. 1939, l.c.: 99. (7).
Геогр. ]> а с и p. Эфиопия (редко в Шоа и Гопдэре).
Var. pseudopraesemiticum (Vav.) Л. Filat. comb. nov. - - 7’. durum var.
pseudopraesemiticum Vav. 1931, l.c.: 42. — T. abyssinicum var. pseudo-
praesemiticum Vav. in Flaksb. 1939, l.c.: 100. (10).
Гео гр. ]> а сир, Эфиопия (редко около Харэра, Чэрчэра и г. Дэбрэ-
Маркосс).
Var. pseudorubriptibescens (Vav.) A. Filat, comb. nov. — 7’. durum
var. pseudorubropubescens Vav. 1931, l.c.: 44. — T. abyssinicum var.
pseudorubropubescens Vav. in Flaksb. 1939, l.c.: 102. (26).
Геогр. распр. Эфиопия (повсеместно).
Var. pseudosomalense (Vav.) A. Filat, comb. nov. — 7’. durum var.
pseudosomalense Vav. 1931, l.c.: 46. — T. abyssinicum var. pseudosoma-
lense Vav. in Flaksb. 1939, I. c.: 104. (17).
Геогр. распр. Эфиопия (редко в районе Харэра и Чэрчэра).
Var. pseudotomentosum (Perciv.) Л. Filat, comb. nov. — 7'. dicoccum
var. pseudotomentosum Perciv. 1926, I. c.: 206. — 7’. durum var. negus-
tianum Vav. 1931, l.c.: 17. — T. abyssinicum var. negustianum Vav. in
Flaksb. 1939, l.c.: 105. (29).
Геогр. распр. Эфиопия (повсеместно; в восточной части стра-
ны — преобладающая разновидность).
Var. pseudouncinatum (Perciv.) Л. Filat, comb. nov. — 7'. dicoccum
var. pseudouncinatum Perciv. 1921, l.c.: 195; idem, 1926, l.c.: 205, quoad
form II. — T. durum var. nigrodensomeneliki Vav. 1931, 1. c.: 41. - 7’. abys-
sinicum var. nigrodensomeneliki Vav. in Flaksb. 1939, l.c.: 100. (8).
Геогр. распр. Эфиопия (редкая примесь около Харэра, Дэбрэ-
Маркоса, Гопдэра, Лдуа, Аддис-Абебы и в горной Эритрее).
Var. rubrivillosum (Perciv.) A. Filat, comb. nov. — T. dicoccum var.
rubrivillosum Perciv. 1926, l.c.: 207. — T. durum var. rubropubescens Vav.
1931, l.c.: 44. — T. abyssinicum var. rubropubescens Vav. in Flaksb. 1939,
l.c..: 102. (25).
Геогр. распр. Эфиопия.
Var. somalense (Vav.) A. Filat, comb. nov. — T. durum var. somalense
Vav. 1931, l.c.: 46.— T. abyssinicum var. somalense Vav. in Flaksb. 1939.
l.c.: 104. (16).
Геогр. распр. Эфиопия (редко около Харэра, Аддис-Абебы и
Дэбрэ-Маркоса).
Var. subnegustianum (Vav.) A. Filat, comb. nov. — T. durum var.
subnegustianum Vav. 1931, l.c.: 57. — 7’. abyssinicum var. subnegustianum
Vav. in Flaksb. 1939, l.c.: 105. (33).
Геогр. расп p. Эфиопия (единично в Годжаме).
Var. tchertchericum (Vav.) A. Filat, comb. nov. — T. durum var.
tchertchericum Vav. 1931, l.c.: 44.— 7’. abyssinicum var. tchertchericum
Vav. in Flaksb. 1939, l.c.: 101. (23). '
Геогр. распр. Эфиопия (редко в районе Чэрчэра).
Var. tomentosum (Perciv.) A. Filat, comb. nov. — T. dicoccum var.
tomentosum Perciv. 1921, l.c.: 195. — T. durum var. densopraevalenciae
Vav. 1931, l.c'.: 47. — 7’. abyssinicum var. densopraevalenciae Vav. in
Flaksb. 1939, l.c.: 105. (28).
Геогр. p аспр. Эфиопия (редко в районе Харэра и Чэрчэра).
Var. vulpinum (Perciv.) Л. F'ilat. comb. nov. — 7’. dicoccum var. vulpi-
num Perciv. 1926, l.c.: 207. — 7’. durum var. densorubrum Vav. 1931, l.c.:
48.-7’. abyssinicum var. densorubrum Vav. in Flaksb. 1939, l.c.:
105. (30).
Геогр. распр. Эфиопия (редко в районе Харэра и Чэрчэра и в
горной Эритрее).
Ilotlpoi) I. Triticum
2. Convar. compactum (Vav.) A. Filat. группа y.ni>i раплотпоколосых
разновидное i ей.
Convar. compactum (Vav.) A. Filat, comb. nov. — T. durum subsp.
abyssinicum sect, compacta Vav. 1931, Tp. прикл. бот. ген. сел. Прилож.
51:48.
Т ип: Т. aethiopicum var. typicum (Vav.) Л. Filat.
Геогр. распр. Эфиопия (главным образом в Харэре и севернее
Дэбрэ-Маркоса, в Эритрее).
Определение разновидностей convar. compactum (Vav.) Л. Filat.
Чешуи Ости Разновидность № разновнл ностн
белые белые с черными краями крас- ные красные с черными краями черные на фоне цвета чешуи черные
белом крас- ном
simiense
1
Колосья неопушенные
3 е р н о в к и ф и о л с т о в ы е
syrovatskyi
pseudoptolo-
mcieum
nigriptoloma-
eitm
jakovlevii
compacto-
negroides
Колосья опушенные
Зерновки красные
typicum
pseudotypicum
nigritypicum
carneum
pseudocariie-
um
takussicuin
asmaricum
pahnpvae
bojanovskajae
critraeum
Зерновки белы о
nubicum
stoleiovae
nikolucnkinis
2
3
4
5
6
7
8
9
10
It
12
13
14
15
16
17
18
19
Var. asmaricum (Vav.) A. Filat, comb. nov. — T. durum var. asmari-
cum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 52.— Т. abyssinicum
var. asmaricum Vav. in Flaksb. 1939, Опред. наст, хлебов: 104. (13).
Геогр. pactup. Эфиопия (редко в горной Эритрее).
Var. bojanovskajae A. Filat, nom. nov. — T. durum var. psetidoarabi-
cumNav. 1931, l.c.: 51, non Perciv., nom. praeoctip. — T. abyssinicum
var. pseiidoarabicum’Vax'. in Flaksb. 1939, l.c.: 102. (15).
Г corp, распр. Эфиопия (редко, около Харэра, Гопдэра и в Эрит-
рее).
Var. carneum (Vay.) A. Filat, comb. nov. — 7. durum var. carneum
Vav. 1931, l.c.: 52.— T. abyssinicum var. carneum Vav. in Flaksb. 1939,
l.c.: 104. (10).
11*
I (i'1 !\,iiu снфикици.'! TriHinni
Г e о г |>. p а с и p. Эфиопия.
Var. coinpactonegroides (Vav.) A. I'ilal. comb. nov. /'. durum var.
tompacionegroides \r;w. 1931, l.c.: 51. --T. abyssinicuin var. compacto-
negroides Vav. in Flaksb. 1939, l.c..: 101, ((>).
Геогр. распр. Эфиопия (редко в Гопдэрс и горной Эритрее).
Var. eritraeum (Vav.) Л. Filat, comb. nov.— T. durum var. eritraeum
Vav. 1931, l.c.: 51. — T. abyssinicuin var. eritraeum. Vav. in Flaksb. 1939,
I. c.: 102. (16).
Геогр. распр. Эфиопия (редко в lopiioii Эритрее, а также в
окрестностях Харэра и Дэбрэ-Маркоса).
Var. jakovlevii Л. Filat. nom. nov. --Г. durum var. thebaicuin Vav.
1931, Lc.: 50, non Perciv., nom. pracocup.— T. abyssinicuin var. ihebai-
cum Vav. in Flaksb. 1939, 1. c.: 100. (5).
Геогр. распр. Эфиопия (редко в Харэре, Гопдэрс, близ Голубо-
го Нила).
Var. nigriptolomaeum (Vav.) Л. Filat, comb. nov. — T. durum var.
nigroplolomaeutn Vav. 1931, Lc.: 50. — T. abyssinicum var. nigroptolo-
tnaeutn Vav. in Flaksb. 1939, 1. c.: 100. (4).
Геогр. p а с и p. Эфиопия (Гондэр).
Var. nigritypicum (Vav.) A. Filat, comb. nov. - T. durum var. nigro-
lypicum Vav. 1931, l.c..: 52. — T. abyssinicum var. nigrotypicum Vav. in
Flaksb. 1939, 1. c.: 103. (9).
Геогр. распр. Эфиопия (редко в Лксуме, Гопдэрс, Харэре, гор-
ной Эритрее).
Var. nikolaenkinis Л. Filat, nom. nov. — T. durum var. pseudocopticum
Vav. 1931, l.c.: 54, non Perciv., nom pracocup. — T. abyssinicum var.
pseudocopticum Vav. in Flaksb. 1939, 1. c.: 106. (19).
Геогр. распр. Эфиопия (повсеместно; в Харэре и около Чэрчэра
часто преобладает в посевах).
Var. nubicum (Vav.) A. Filat, comb. nov. — T. durum var. nubicum
Vav. 1931, 1. c.: 52.— T. abyssinicum var. nubicum Vav. in Flaksb. 1939,
l.c.: 105. (17).
Геогр. распр. Эфиопия (южнее Харэра, в Аксуме и Эритрее).
Var. palniovae Л. Filat, nom. nov. — T. durum var. compression Vav.
1931, Lc.: 51, non Koern., nom. praeocup. — T. abyssinicum var. conipres-
sum Vav. in Flaksb. 1939, l.c. : 101. (14).
Геогр. распр. Эфиопия.
Var. pseudocarneum (Vav.) A. Filat, comb. nov. — T. durum var. pseu-
docarneum Vav. 1931, l.c.: 52. — T. abi/ssinicum var. pseudocarneum Vav.
in Flaksb. 1939, l.c.: 104. (11).
Геогр. распр. Эфиопия (очень редко около Дэбрэ-Маркоса п
Харэра).
Var. pseudoptoloinaeuni (Vav.) Л. Filat, comb. nov. — T. durum var.
pseudoptolomaeum Vav. 1931, l.c.: 50. — 7’. abyssinicum var. pseudoptolo-
maeum Vav. in Flaksb. 1939, 1. c.: 99. (3).
Геогр. p а с п p. Эфиопия (очень редко в Гопдэре).
Var. pseudotypicum (Vav.) A. Filat, comb. nov. — T. durum var. psett-
dotypicum Vav. 1931, l.c.: 51. — T. abyssinicum var. pseudotypicum Vav.
in Flaksb. 1939,,l.c.: 103. (8).
Геогр. рас n p. Эфиопия (очень редко около Чэрчэра).
Var. simiense (Vav.) Л. Filat, comb. nov. — T. durum var. simiense
Vav. 1931, l.c.: 50,- T. abyssinicum var. simiense Vav. in Flaksb. 1939,
l.c.: 98. (1).
Геогр. распр. Эфиопия (редко к севе.ру от Гондэра).
Var. stoletovae Л. Filat, nom. nov. — T. durum var. coplicutn Vav. 1931,
1. c.: 54, non. Koern., nom. praeocup. — 7’. abyssinicum var. coplicum Vav.
in Flaksb. 1939, 1. c.: 105. (18).
Геогр. распр. Эфиопия.
П<к~)р1>() I. Triticum 1 (in
Var. syrovatskyi A. I'ilat. noin. nov. T. durum var. pttdomaeum Vav.
1931, l.c.: 50, non. Perciv., nom. praeocup.— T. abt/ssinicum var. ptolo-
maeuniNav. in Flaksb. 1939, Lc.: 99. (2).
Геогр. распр. Эфиопия (очень редко около Гопдэра).
Var. takussicum (Vav.) Л. Filat, comb. nov. — T. durum var. takussi-
cum Vav. 1931, Lc.: 52. — T. abyssinicum var. takussicum Vav. in Flaksb.
1939, Lc.: 104. (12).
Геогр. распр. Эфиопия (около Аддис-Абебы, Дэбрэ-Маркоса,
Аксума).
Var. typicuin (Vav.) Л. Filat, comb. nov. — T. durum var. typicum Vav.
1931, Lc.: 51. — T. abyssinicum var. typicum. Vav. in Flaksb. 1939, l.c.:
103. (7).
Геогр. распр. Эфиопия (редкая примесь в Гоидэре и Харэре).
III. Subsp. turgidoides Л. Filat. — подвид тургидоидиых пшениц.
Subsp. turgidoides A. Filat, subsp. nov. — T. turgidum subsp. abys-
sinicum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 61. — Т. abyssi-
nicum subsp. turgidoides Flaksb. 1939, Опред. паст, хлебов: 94, йот.
illeg. — Gluniae palcis breviores inflate; dentis gluniis inflexae; caryopsis
brevis, dorso convexa.
Тип: T. aethiopicum var. purpureum (Vav.) Dorof. et A. Filat.
Колосья удлиненные (6—9 см), короткие (5—7 см) или очень корот-
кие (до 5 см), большей частью сравнительно рыхлые (D около 25), плот-
ные (D до 40) или очень плотные (D больше. 40), многоцветковые,
с остями, которые значительно короче колоса. Ости тонкие, расходящие-
ся, часто изогнутые, искривленные, иногда недоразвитые (колосья без-
остые). У основания остей иногда образуются выросты (фурки). Колос-
ковые чешуи вздутые, с изогнутыми вовнутрь зубцами. Зерновки корот-
кие, с горбинкой, иолустекловидпыс или мучнистые, белые, красные пли
фиолетовые. Всходы фиолетовые. Куст прямостоячий. Соломина отно-
сительно короткая и тонкая. Листовые пластинки светло-зеленые, шеро-
ховатые на ощупь, довольно короткие и узкие. Восковой налет слабый.
Все формы яровые, скороспелые (как и subsp. vavilovianum), с быстрым
прохождением периода колошение — созревание. Экологически сходны
с subsp. aethiopicum и subsp. vavilovianum.
Геогр. распр. Эфиопия. Очень редко занимает господствующее
положение в посеве.
Ключ для определения групп разновидностей
subsp. turgidoides A. Filat.
1. Колосья удлиненные (6—9 см длины), сравнительно рыхлые (D
около 25).................convar. elongation (Vav.) Dorof. et A. Filat.
4- Колосья короткие (5—7 см длины)............................2.
2. Колосья 5—7 см длины, цилиндрические, плотные (D = 30 - 10) .
. .....................................convar. densum (Vav.) Dorof.
-(-Колосья очень короткие, (до 5 см длины), часто плоские, иногда
пирамидальной формы, очень плотные (D > 40) .........................
................. convar. recognituni (Steud.) A. Filat, et Dorof.
1. Convar. elongatum (Vav.) Dorof. et A. Filat. — группа рыхлоколо-
сых разновидностей.
Convar. elongatum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
subsp. abyssinicum sect, elongata Vav. 1931, Tp. прикл. бот. ген. сел.
Прилож. 51: 62.
Т и и: 7'. aethiopicum var. purpureum (Vav.) Dorof. et A. F'ilat.
Геогр. p а с и p. Эфиопия (главным образом в районе Аддис-Абе-
бы на высотах 2400—3000 м над ур. м.).
166
Классификация рода Trilicum Л.
Определение разновидностей convar. eloiigaliini (Vav.) Dorof. et Л. I ilat.
Чешуи Ос <и Разновидность К? разновид- ности
белые белые ' с черными краями крас* ные красные с черными краями черные на фоне цвета чешуи черные
белом крас- ном
Колосья остистые
3 с Колосья неопушенные и о в к и ф и о л е т о в ы е
+ + + + II 1 1+1 1 II 1 II +1 II 1 II 1 + + + | + + |Т + + purpureum pseiidopurpu- reum nigripurpure- iim sangiiincuiii nigi isangui- neum striatipurpu- reiun atratisangui- neutn 1 2 3 1 6 7
3 е р н о в к и к р а с и ы е
+ + Г 1 1 1 II 1 1 1. 1 1 + 1 1 + + II 1 1 1 + 1 1 II 1 II 1+1 + 1 1 1 1 1 1 1 1 1 1 1+1 1 + + + + + + + dispersion pseudodisper- sion nigridisper- sum coloration pseudocolora- tion nigricolora- tum melailOCCpha- lli III puliation 8 9 10 11 12 13 14 15
3 е р и О в к и бель е
++ 1 1 1 III 1 1 1 1 1+ II + + . 11+ III II II III 1 + 1 II III 11+ + 1 1 + 1 III + + + + + + decoloration pseudodeco- loration kusliianii ill rubidton pseudoriibi- dum nigrirtibidum atratbion atrirubrion 16 17 18 19 20 21 22 23
• 3 Колосья опушенные с р и о в к и фиолетовые
+ 1 ; 1 1 1 (iciilum | anlioberiHise 1 g
.3 С р JJ О в к л к р а (' и ы е
-1- -1- + — — 1 1- + idbii'il losuni pseudiiulbi- vil losil ill iiilvuin 26 27 28
Подрод I. Triticum
167
О к о и ч а и и е
Чешуи Ости Разновидность № разнонид-
белые белые крас- красные с черные на фоне ное ia
с черными КроЯМИ ные черными краями , крас- белом иом чешуи черные ности
Зерновки белые
+ — — — — — + — pilosum 29
+ — — — — — — + pseudopilosum .'10
— — -1- —• — — — + rubripilosum 31
Колосья безостые
Колосья неопушенные
Зор II О В К и ф и ) .Т 0 'Г ( в ы с
+ — — — — — — uiolacinf la- 3'2
tum
— — — — — — + addisabebae .33
— — + — — — + — rubrinf latum .34
— — — — — -— pscudorubr- ,35
iiiflatuin
— — — + —— — — + nigrirubrin- 36
flatum
— — —— — + — — + sabae 37
— — — — — — -и nigrinflatiim .38
3 С Р II О в к и к р а с и ы е
+ — — — — + — laxinflatum 39
— — — — — + duplirubrum 40
— — — ч- — — — + nigridupltriib- 41
rum
3 е р и О в к и бель о
+ 1 + 1 percivalii 42
— + 1 + 1 furcation 43
Колосья опушенные
3 ерно в к и краев ы с
— — — — — + ftdvinflatum 44
3 е р н О в к и белые
— — — — — + pilosinflahim 45
Окраска остеппдпых образований.
Var. acutum (Vav.) A. Filat, comb. nov. — T. turgidum. var. acutum
Vav. 1931, Tp. прукл. бот. геи. сел. Прилож. 51: 66. — Т. abyssinicum var.
acutum Vav. in Flaksb. 1939, Опред. наст, хлебов: 109. (24).
Геогр. распр. Эфиопия (редко около Харэра и в Эритрее).
Var. addisabebae (Vav.) A. Filat, cornb. nov. — T. turgidum var. addis-
abebae Vav. 1931, l.c.: 82.— T. abyssinicum var. addisabebae Vav. in
Flaksb. 1939, l.c.: 116. (33).
Геогр. распр. Эфиопия (в районе Аддис-Абебы и в Эритрее).
Var. albivMlostim (Vav.) Dorof. el A. Filat, comb. nov. — T. turgidum
var. alboviltosum Vav. 1931, l.c.: '68.— T. abyssinicum var. albovillosum
Vav. in Flaksb. 1939, l.c.: 111. (26).
168 Классификация роди Triliciuit L.
Геогр. p а с и p. Эфиопия (редко около Гопдэра, Фиче, Харэра и
в горной Эритрее).
Var. ankoberense (Vav.) Dorof. et A. Filat, eomb. nov. — T. turgidum
var. ankoberense Vav. 1931, l.c.: 66. — T. abyssinicum var. ankoberense
Vav. in Flaksb. 1939, l.c.: 109. (25).
Геогр. распр. Эфиопия (редко в районах Аддис-Абебы и Лпко-
бэра).
Var. atralbum (Vav.) Л. Filat, comb. nov. — T. turgidum var. atroal-
bum Vav. 1931, Lc.: 70. — T. abt/ssinicum var. atroalbum Vav. in Flaksb.
1939, l.c.: H3. (22).
Геогр. распр. Эфиопия (в провинциях Шоа, Харэр, близ Гопдэ-
ра, Лдуа).
Var. atratisanguineum (Vav.) Dorof. et A. Filat, comb. nov. — T. tur-
gidum var. atratosanguineum Vav. 1931, l.c.: 66. — T. abyssinicum var.
atralosanguineum Vav. in Flaksb. 1939, l.c.: 109. (7).
Геогр. распр. Эфиопия (в районах Аддис-Абебы и Фиче).
Var. atrirubnnn (Vav.) A. Filat, comb. nov.— 7’. turgidum var. alro-
rubrum Vav. 1931, l.c.: 70. —T. abt/ssinicum var. atrorubrum Vav. in
Flaksb. 1939, l.c.: 114. (23).
Геогр. распр. Эфиопия (очень редко около Харэра, Аддис-Абе-
бы, Анкобэра и в Годжаме).
Var. coloratum (Vav.) A. Filat, comb. nov.— T. turgidum var. coloratum
Vav. 1931, Lc.: 67. — T. abyssinicum var. coloratum Vav. in Flaksb. 1939,
1. c.: 110. — T. dicoccum var. rufescens Perciv. 1926, Journ. Bot. (London):
206, quoad f. II. (11).
Геогр. распр. Эфиопия (повсеместно).
Var. decoloratum (Vav.) A. Filat, comb. nov. — T. turgidum var. deco-
loration Vav. 1931, l.c.: 69.— T. abyssinicum var. decoloratum Vav. in
Flaksb. 1939, l.c.: 112. (16).
Геогр. распр. Эфиопия (редко около Харэра, Аддис-Абебы, Чэр-
чэра, в горной Эритрее).
Var. dispersum (Vav.) A. Filat, comb. nov.— T. turgidum var. disper-
sion Vav. 1931, Lc.: 66. — T. abyssinicum var. dispersum Vav. in Flaksb.
1939, l.c.: 110.-—T. dicoccum tenaces var. uncinatum Perciv. 1926, l.c.:
205, quoad f. III. (8).
Геогр. распр. Эфиопия (повсеместно).
Var. duplirubrum (Vav.) A. Filat, comb. nov. — T. turgidum var. dttplo-
rubrurn Vav. 1931, Lc.: 83. — T. abyssinicum var. duplorubrum Vav. in
Flaksb. 1939, 1. c.: 115. (40).
Геогр.. распр. Эфиопия (очень редко около Дэбрэ-Маркоса и
Фиче).
Var. fulvinflatum (Vav.) A. Filat, comb. nov. — T. turgidum var. ful-
voinflatum Vav. 1931, Lc.: 83.— T. abyssinicum var. fulvoinflatum Vav. in
Flaksb. 1939, Lc.: 115. (44).
Геогр. p а с п p. Эфиопия (около Гопдэра).
Var. fulvum (Vav.) A. Filat, comb. nov. — T. turgidum var. fulvum Vav.
1931, Lc.: 68. — T. abyssinicum var. fulvum Vav. in Flaksb. 1939, l.c.:
112. (28).
Геогр. р'аспр. Эфиопия (редко около Фиче и в горной Эритрее).
Var. furcatuni (Vav.) Л. Filat, comb. nov. — T. turgidum var. furcatuni
Vav. 1931, Lc.: 84. — T. abyssinicum var. furcatum Vav. in Flaksb. 1939,
Lc.: 116. (43).
Сильно выражена фуркатность (выросты ткани у основания остсвид-
ных придатков).
Геогр. распр. Эфиопия (очень редко около Харэра и Аддис-
Абебы).
IIixI/hh) I. Trilii tun
l(?i
Var. kiishiantiin (Va\.) A. Filat. comb, now T. turgidum var. kusltia-
nttm Vav. 1931, l.c.: 69. - T. abt/ssinicum var. kushianum Vav. in Flaksb.
1939, l.c.: 113. (18).
Геогр. p а с и p. Эфиопия (редко в районе /Анкобэра, Гопдэра, Ха-
рэра, Аксума, Дапглы и в горной Эритрее).
Var. laxinflatuni (Vav.) Л. Filat, comb. nov. — T. turgidum var. laxin-
flatum Vav. 1931, l.c.: 83.— T. abt/ssinicum var. laxinjlatum Vav. in
Flaksb. 1939, l.c.: 115. (39).
Геогр. распр. Эфиопия (редко около Аддис-Абебы и Фиче).
Var. melanocephalutn (Vav.) Л. Filat, comb. nov. — 7’. turgidum var.
melanocephalutn Vav. 1931, l.c.: 68. — T. abyssinicum var. melanoccpha-
lumVav. in Flaksb. 1939, l.c.: 111. (14).
Геогр. распр. Эфиопия (повсеместно).
Var. nigricoloratum (Vav.) A. Filat, comb. nov.— T. turgidum var.
nigrocoloratum Vav. 1931, Lc.: 67. — T. abyssinicum var. nigrocoloratum
Vav. in Flaksb. 1939, 1. c.: 111. (13).
Геогр. распр. Эфиопия (редко близ Аддис-Абебы, Анкобэра и
Харэра).
Var. nigridispersutn (Vav.) A. Filat, comb. nov. - - Г. turgidum var.
nigrodispersum Vav. 1931, l.c.: 67. T. ubyssinicttm var. nigrodispersiini
Vav. in Flaksb. 1939, 1. с.: 110. (10).
Геогр. расп p. Эфиопия (повсеместно).
Var. nigriduplirubrtini (Vav.) A. Filat, comb. nov. — T. turgidum var.
nigroduplorubrum Vav. 1931, 1. c.: 83.— T. abyssinicum var. nigroduplorub-
rumVav. in Flaksb. 1939, l.c.: 115. (41).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. nigrinflatum (Vav.) A. Filat, comb. nov. — T. turgidum var.
nigroinflatum Vav. 1931, 1. c.: 83.— T. abyssinicum var. nigroinf latum Vav.
in Flaksb. 1939, 1. c.: 115. (38).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. nigripurpureum (Vav.) A. Filat, comb, nov.-—T. turgidum var.
nigropurpureum Vav. 1931, Lc.: 63. — T. abyssinicum var. nigropurpureum
Vav. in Flaksb. 1939, 1. c.: 109. (3).
Геогр. распр. Эфиопия (центральная и восточная часть).
Var. nigrirubidum (Vav.) A. Filat, comb. nov. — T. turgidum var.
nigrorubidum Vav. 1931, 1. c.: 69.— T. abyssinicum var. nigrorubidum Vav.
in Flaksb. 1939, Lc.: 113. (21).
Г corp, распр. Эфиопия (редко близ Аддис-Абебы, Анкобэра, Ха-
рэра и в Годжаме).
Var. nigrirubrinflatum (Vav.) A. Filat, comb. nov. — T. turgidum
var. nigrorubroinflatutn Vav. 1931, Lc.: 83.— T. abyssinicum var. nigro-
rubroinflatumNav. in Flaksb. 1939, Lc.: 115. (36).
Геогр. расп p. Эфиопия (близ Аддис-Абебы).
Var. nigrisanguineum (Vav.) Dorof. et A. Filat, comb. nov. --- T. tur-
gidum var. nigrosanguineum Vav. 1931, 1. c.: 63. — T. abyssinicum var.
nigrosanguineum Vav. in Flaksb. 1939, Lc.: 109. (5).
Геогр. распр. Эфиопия (в районах Аддис-Абебы и Анкобэра).
Var. percivalii (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum var.
percivalii Vav. 1931, Lc.: 83. — T. abyssinicum var. percivalii Vav. in
Flaksb. 1939, 1. c.: 116. (42).
Геогр. распр. Эфиопия (очень редко около Дэбрэ-Маркоса).
Var. pilosinf 1 atuni (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. pilosoinjlatum Vav. 1931, l.c.: 81. T. iibi/ssinicum var. pilosoinflu-
Ittttt Vav. in Flaksb. 1939, I. e.: 115. (45).
Геогр. распр. Эфиопия (очень редко близ Фиче).
Var. pilosum (Vav.) Л. Filat, comb. nov.— T. turgidum var. pilosum
Vav. 1931, Lc.: 70. — T. abt/ssinicum var. pilosum Vav. in Flaksb. 1939,
l.c,: 114. (29).
170 Классификации /)ш)а Trilicmn К.
Геогр. распр. Эфиопия (редко в районе Харэра).
Var. pseudoalbiviIlosum (Vav). A. Filat, comb. nov. — T. turgidum var.
I)seiidoalbovillosum Vav. 1931, l.c.: 68.— T. abyssinicum var. pseudoalbo-
villosum May. in Flaksb. 1939, l.c.: Ill, (27).
Геогр. распр. Эфиопия (близ Гопдэра, Харэра, Аксума и в гор-
ной Эритрее).
Var. pseudocoloratum (Vav.) A. Filat, comb. nov.— T. turgidum. var.
pseudocoloratum Vav. 1931, l.c.: 67. — T. abyssinicum var. pseudocolora-
tum Vav. in Flaksb. 1939, l.c.: 110. (12).
Геогр. распр. Эфиопия (редко близ Аддис-Абебы и Гопдэра).
Var. pseudodecoloratum (Vav.) A. Filat, comb. nov. — T. turgidum var.
pseudodecoloratum Vav. 1931, l.c.: 69. — T. abyssinicum var. pseudodeco-
loratum Vav. in Flaksb. 1939, 1. с.: 112. (17).
Геогр. распр. Эфиопия (редко в Дапгле).
Var. pseudodispersum (Vav.) A. Filat, comb. nov.— T. turgidum var.
pseudodispersum Vav. 1931, l.c.: 66. — T. abyssinicum var. pseudodisper-
sum Vav. in Flaksb. 1939, 1. с.: 110. (9).
Геогр. распр. Эфиопия (повсеместно как редкая примесь в по-
севах) .
Var. pseudopilosum (Vav.) A. Filat, comb. nov. — T. turgidum var.
pseudopilosum Vav. 1931, l.c.: 70. — T. abyssinicum var. pseudopilosum
Vav. in Flaksb. 1939, 1. с.: 114. (30).
Геогр. распр. Эфиопия (редко в районе Харэра).
Var. pseudopurpureum (Vav.) Dorof. et A. Filat, comb. nov. — T. tur-
gidum var. pseudopurpureum Vav. 1931, l.c.: 63. — T. abyssinicum var.
pseudopurpureum Vav. in Flaksb. 1939, 1. c.: 109. (2).
Геогр. распр. Эфиопия (редко в районе Фиче).
Var. pseudorubidum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. pseudorubidum Vav. 1931, 1. c.: 69.— T. abyssinicum var. pseudo-
rubidum Vav. in Flaksb. 1939, l.c.: 113. (20).
Геогр. распр. Эфиопия (редко близ Харэра).
Var. pseudorubrinflatum (Vav.) Dorof. et A. Filat, comb. nov.— T. tur-
gidum var. pseudorubroinflatum Vav. 1931, 1. c.: 82. — T. abyssinicum var.
pseudorubroinjlatumNav. in Flaksb. 1939, l.c.: 115. (35).
Геогр. распр. Эфиопия (редко близ Аддис-Абебы).
Var. pullatum (Vav.) Dorof. et A. Filat, comb. nov.—7. turgidum var.
pullatum Vav. 1931, 1. c.: 68. — T. abyssinicum var. pullatum Vav. in Flaksb.
1939, 1. c.: 111. (15).
Геогр. распр. Эфиопия (редко близ Аддис-Абебы, Гондэра, Хар-
эра).
Var. purpureum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum var.
purpureum Vav. 1931, l.c.: 63. — T. abyssinicum var. purpureum Vav. in
Flaksb. 1939, 1. c.: 109. (1).
Геогр. распр. Эфиопия (близ Харэра, Аикобэра, Аддис-Абебы,
Фиче, Гопдэра и по долине р. Аббай).
Var. rubidum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum var.
rubidum Vav. 1931, 1. c.: 69.— T. abyssinicum var. rubidum Vav. in Flaksb.
1939, 1. c.: 113. (19).
Геогр. рарпр. Эфиопия (близ Гопдэра, Аддис-Абебы, Харэра,
Дэбрэ-Маркоса).
Var. rubrinflatum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. rubroinflatum Vav. 1931, l.c.: 82. — T. abyssinicum var. rubro-
inflatum Vav. in Flaksb. 1939, I. c.: 115. (34).
Геогр. распр. Эфиопия (в районе Аддис-Абебы).
Var. rubripifosum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. rubropilosum Vav. 1931, l.c.: 70—T. abyssinicum var. rubropilosum
Vav. in Flaksb. 1939, l.c.: 114. (31).
Гсогр.раси p. Эфиопия (редко в районе Аддис-Абебы и Аикобэра).
Подрод I. Triticum
171
Var. sabae (Vav.) Dmof. et A. Filat. comb. nov. T. turgidum var. so
bae Vav. 1931, l.c.: S3. — 7'. abyssinicum var. sabae Vav. in Flaksb. 1939,
1. c.: 115. (37).
Гe огр. p а с п p. Эфиопия (очень редко в районе Аддис-Абебы).
Var. sanguineuni (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. sanguineum Vav. 1931, l.c.: 63. — T. abyssinicum var. sanguineum
Vav. in Flaksb. 1939, l.c.: 109. (4).
Геогр. распр. Эфиопия (около Аддис-Абебы, Фиче, Дэбрэ-Мар-
коса и Гондэра).
Var. striatipurpureuni (Vav.) Dorof. et A. Filat, comb. nov.— T. turgi-
dum var. striatopurpureum Vav. 1931, 1. c.: 63. — 7'. abyssinicum var. stria-
topurpureum Vav. in Flaksb. 1939, 1. c.: 109. (6).
Геогр. распр. Эфиопия (повсеместно).
Var. violacinflatum (Vav.) Dorof. et A. Filat, comb. nov. — 7’. turgi-
dum var. violaceoinflatum Vav. 1931, 1. c.: 82. — T. abyssinicum var. viola-
ceoinflatum Vav. in Flaksb. 1939, l.c.: 114, (32).
Геогр. распр. Эфиопия (очень редко в районе Аддис-Абебы).
2. Convar. densuni (Vav.) Dorof. -- группа п.тотиоко.тосых разновид-
ностей.
Convar. densuni (Vav.) Dorof. comb. nov. — 7’. turgidum subsp. abyssi-
nicum sect, densa Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51 : 70.
Typus: Т. aethiopicum var. densipurpureum (Vav.) Dorof. et A. Filat.
Геогр. распр. Эфиопия (главным образом на высотах 1800—
2400 м над ур. м.).
Var. atrinfiatum (A7av.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. atroinj latum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51: 85.—
Т. abyssinicum var. atroinjlatum Vav. in Flaksb. 1939, Опред. наст, хле-
бов: 116. (35).
Геогр. р а с и р. Эфиопия (редко в районе Аддис-Абебы).
Var. atrimarginatum (Chiov.) Dorof. et A. Filat, comb. nov.— T. durum
var. atromarginatum Chiov. 1912, Monogr. Rapp, colon. (Roma), 19: 16. —
T. turgidum var. atromarginatum (Chiov.) Vav. 1931, 1. c.: 73. — T. abyssi-
nicum var. atromarginatum (Chiov.). Vav. in Flaksb. 1939, l.c.: 110. (8).
Геогр. распр. Эфиопия (в северных районах, а также близ Хар-
эра и Аддис-Абебы).
Var. dangalense (Vav.) Dorof. cl A. Filat, comb. nov.— T. turgidum var.
dangalense Vav. 1931, l.c.: 76. — T. abyssinicum var. dangalense Vav. in
Flaksb. 1939, 1. c.: 113. (18).
Геогр. распр. Эфиопия (около Харэра, Чэрчэра, в Эритрее).
Var. densatratipurpureum (Vav.) Dorof. et A. Filat, comb. nov.—
T. turgidum var. densoatratopurpureum Vav. 1931, l.c.: 72. — T. abyssini-
cum var. densoatratopurpureum Vav. in Flaksb. 1939, l.c.: 109. (5).
Геогр. распр. Эфиопия (в районе Аддис-Абебы).
Var. densatratisanguineum (Arav.) Dorof. et A. Filat, comb. nov.—
T. turgidum var. densoatratosanguineum Vav. 1931, 1. c.: 72. — T. abyssini-
cum var. densoatratosanguineum Vav. in Flaksb. 1939, l.c.: 109. (6).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. densatrinubrum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. densoatrorubrum Vav. 1931, I. c.: 76.— T. abyssinicum var. denso-
alrorubrumVav. in Flaksb. 1939, l.c.: 114. (19).
Геогр. распр. Эфиопия (около Аддис-Абебы, Гондэра, Харэра,
Апкобэра, в горной Эритрее).
Var. densidecoloratum (Vav.) Dorof. el A. Filat, comb, nov. - -7’. turgi-
dum var. densodecoloralum Vav. 1931, I. c.: 75. - - 7’. abyssinicum var. den-
sodecoloratum Vav. in Flaksb. 1939, l.c.: 112. (14).
Геогр. распр. Эфиопия (в районе Аддис-Абебы, в Эритрее).
Var. densifulvuin (Vav.) Dorof. et A. Filat, comb. nov. — 7’. turgidum
172
К.шссифштцип рода Triticum I..
Определение разновидное i ей convar. ilcnsiiin (Vav.) Dorof.
Чешуи О< 1И
белые белые крас- красные черные на фоне цвета Разновидность .V' разновид'
с черными . краями иыс с черными краями белом крас- ном чешуи черные ностп
1\ о л о с. Ь и о с т и с т w с
Колосья неопушенные
Зернов к и ф и о л с т о в ы е
4- — — — — — + — densipurpure- tim 1
— 4- — — — — — -о nigridensi- purpurcum 2
— — + — — — 4- — densisangui- neum 3
— — — 4- — — — + nigridensi- saitgiiineiim 4
— — — — 4- — — 4- deitsatruti- purpureuni
2 е р н о 4- В К И I расы + ы е densatrati- sanguineum 6
4- — — — — — -L — spectabile 7
—• 4- — — — — — + atrimargina- tum 8
— — + .— — — 4- — densiplanum 9
— — + — — — — + pseudodensi- planum 10
— — — 4- — — — + nigridensi- platium 11
— — — — 4- — — + heterochro- mtun 12
— — — — 3 е р и 4- 0 в к и белы + е densipullatum 13
4- — — — — — 4- — densidecolo- rattim 11
— —— — — — — + ricliardianunt 15
— — 4- — — — 4- — densiriibidum 10
— — — 4- — — — + diffusion 17
— — — — 4- — — + dangalense 18
3 ер Колос по вк 4- ья опу II ф II ценные о л о т 0 + в ы с densatririib- rum 19
— + — — — — — + marginali- villosum 20
— + — 3 е р н о в к и к р а с и + ы е rufivillosum 21
4- — — — — — 4- — incamun 22
4- — — — — — — 1- pseiidoinca- num 23
3 с р и > в к и 6 С Л 1 >1 1 с ileiisifuhmm 21
4- — — — — — 4- — densipilosum 25
+ — — — — — — + pubighimatum 20
— — 4- — — — — -1- solitariiiin 27
Ilodpoi) I. Triticum
173
Чешуи Ос I и О к ( Разновидное!ь н ч .ч if и е К.' разновид- ности
белые белые с черными краями крас- ные красные с черными краям и черные на фойе цвета чешуи черные
белом крас- ном
К о л о с ь .4 б с :< о с т ы с
Колосья неопушенные
Зернов к и ф и о л с т о в ы е
— --'!’ — flaksbcroeri 28
— — + sparsum 29
— + sanguinin- 30
j latum
* Окраска остевидиых образований.
var. densojulvum Vav. 1931, l.c.: 75.-7'. abyssinicum var. denso[ulvum
Vav. in Flaksb. 1939, 1. с.: 112. (24).
Геогр. распр. Эфиопия (редко в Эритрее и районе Харэра).
Var. densinflatum (Vav.) Dorof. et A. Filat, comb. nov. — 7'. turgidum
var. densoinflatum Vav. 1931, l.c.: 84. — T. abyssinicum var. densoinf la-
tumMav. in Flaksb. 1939, 1. c.: 116. (32).
Геогр. распр. Эфиопия (редко около Аддис-Абебы).
Var. densipilosum (Vav.) Dorof. et A. Filat, comb. nov. — 7'. turgidum
var. densopilosum Vav. 1931, 1. c.: 77. — T. abyssinicum var. densopilosum
Vav. in Flaksb. 1939, 1. s.: 114. (25).
Геогр. распр. Эфиопия (редко в районе Чэрчэра).
Var. densiplanum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. densoplanum Vav. 1931, l.c.: 73. — T. abyssinicum var. densoplanum
Vav. in Flaksb. 1939, l.c.: 110. (9).
Геогр. распр. Эфиопия (близ Аддис-Абебы, Харэра, в горной
Эритрее).
Var. densipullatum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. densopullatum Vav. 1931, l.c.: 74. — 7’. abyssinicum var. denso-
pullatum Vav. in Flaksb. 1939, l.c.: 111. (13).
Геогр. распр. Эфиопия (в районе Анкобэра, Харэра и в Год-
жаме).
Var. densipurpureum (Vav.) Dorof. et A. Filat, comb. nov. — 7’. turgi-
dum var. densopurpureum Vav. 1931, l.c.: 70. — T. abyssinicum var. den-
sopurpureumVav. in Flaksb. 1939, l.c.: 109. (1).
Геогр. растр. Эфиопия (в районе Аддис-Абебы).
Var. densirubidum (Vav.) Dorof. et A. Filat, comb. nov. -- T. turgidum
var. densorubidum Vav. 1931, I. c.: 76. — T. abyssinicum var. densorubidum
Vav. in Flaksb. 1939, 1. c.: 113. (16).
Геогр. распр. Эфиопия (в районе Харэра и Аддис-Абебы).
Var. densisanguineum (Vav.) Dorof. cl A. Filat, comb. nov. — 7'. turgi-
dum var. densosanguineum Vav. 1931, 1. c.: 72. — T. abyssinicum var. den-
sosanguineum Vav. in Flaksb. 1939, 1. c.: 109. (3).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
174
Кмссификацич рода Trilicum Л.
Var. difftistiin (Vav.) Dorof. et Л. F’ilal. comb. nov. 7'. turgidum var.
diffusum Vav. 1931, l.c.: 76. — T. ubbyssinicum var. dijfutiinti Vav. in
Flaksb. 1939, 1. c.: 113. (17).
Геогр. распр. Эфиопия (повсеместно).
Var. flaksbergeri (Vav.) Dorof. et A. Filat. comb. nov.— T. turgidum
var. flaksbergeri Vav. 1931, l.c.: 84. — T. abyssinicum var. flaksbergeri
Vav. in Flaksb. 1939, 1. c.: 115. (28).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. harlanii (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum var.
harlani Vav. 1931, l.c.: 84. — T. abyssinicum var. harlani Vav. in Flaksb.
1939, 1. c.: 116. (33).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. heterochroiniim (Chiov.) Dorof. et A. Filat, comb. nov. — T. durum
var. heterochromum Chiov. 1912. l.c.: 16. — T. turgidum var. heterochro-
mum (Chiov.) Vav. 1931, l.c.: 74. — T. abyssinicum var. heterochromum
(Chiov). Vav. in Flaksb. 1939, I. c.: 111. (12).
Геогр. распр. Эфиопия (повсеместно).
Var. incanum (Vav.) Dorof. et A. Filat, comb. nov. — 7’. turgidum var.
incanum Vav. 1931, I. c.: 75. — 7'. abyssinicum var. incanum Vav. in Flaksb.
1939, 1. c.: 111. (22).
Г e о г p. p а с п p. Эфиопия (редко в горной Эритрее).
Var. marginativillosuin (Vav.) Dorof. et A. Filat, comb. nov.— T. turgi-
dum var. marginatovillosum Vav. 1931, 1. c.: 72. — 7'. abyssinicum var. mar-
ginatovillosum Vav. in Flaksb. 1939, 1. c.: 109. (20).
Геогр. распр. Эфиопия (редко в восточной части страны и в Гонд-
оре), юг Аравийского полуострова (редко).
Var. nigridensiplanum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. nigrodensoplanum Vav. 1931, l.c.: 74. — T. abyssinicum var. nig-
rodensoplanum Vav. in Flaksb. 1939, 1. c.: 111. (11).
Геогр. распр. Эфиопия (близ Аддис-Абебы, Харэра, Апкобэра, в
Годжаме и Эритрее).
Var. nigridensipurpureum (Vav.) Dorof. et A. Filat, comb. nov.—
T. turgidum var. nigrodensopurpureum Vav. 1931, L c.: 72. — T. abyssini-
cum var. nigrodensopurpureum Vav. in Flaksb. 1939, l.c.: 109. (2).
Геогр. распр. Эфиопия (редко близ Аддис-Абебы, Лдуа и Анко-
бэра).
Var. nigridensisanguineum (Vav.) Dorof. et A. Filat, comb. nov.—
T. turgidum var. nigrodensosanguineum Vav. 1931, 1. c.: 72. — T. abyssini-
cum var. nigrodensosanguineum Vav. in Flaksb. 1939, l.c.: 109. (4).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. pseudodensiplanum (Vav.) Dorof. et A. Filat, comb. nov. — T. tur-
gidum var. pseudodensoplanum Vav. 1931, l.c.: 74. — 7'. abyssinicum var.
pseudodensoplanum Vav. in Flaksb. 1939, 1. c.: 110. (10).
Геогр. распр. Эфиопия (редко в районе Чэрчэра).
Var. pseudoincanuni (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. pseudoincanum Vav. 1931, l.c.: 75. — T. abyssinicum var. pseu-
doincanumN av.m Flaksb. 1939, l.c.: 112. (23).
Геогр. распр. Эфиопия (спорадически повсеместно, чаще в гор-
ной Эритрее).
Var. pubigltimatum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum
var. pubiglumatum Vav. 1931, l.c.: 77. — T. abyssinicum var. pubigluma-
tum Vav. in Flaksb. 1939, 1. с.: 114. (26).
Геогр. распр. Эфиопия (встречается в чистых посевах в районе
Харэра, составляет более или менее значительную долю в посевах близ
Лксума, Дапглы и в горной Эритрее).
Var. richardianuin (Vav.) Dorof. et A. Filat, comb. nov. — 7’. turgidum
var. richardianum Vav. 1931, 1. c.: 75. — 7’. abyssinicum var. richardianum
Vav. in Flaksb. 1939, 1. c.: 113. (15).
Ilodpoi) I. Trihi inn 17.",
Гео i’ P- p а с и p. Эфиопия (около Xap>p;i, Гопд эра, Аддис Абебы и
и горной Эрпipee).
Var. rubidinflatum (Vav.) Dorof. comb. nov. — T. turgidum var. rubi-
doinflalutn Vav. 1931, I. c.: 85. — 7'. abyssinicum var. rtibidoinllatum Vav
in Flaksb. 1939, l.c.: 116. (34).
Геогр. p а си p. Эфиопия (редко в ]?aiiojie Аддис-Абебы).
Var. rufivillosuin (Vav.) Dorof. et A. Filat, comb. nov. - T. turgidum
var. rufovillosum Vav. 1931, I. c.: 72. — T. abyssinicum var. rufovillostim
Vav. in Flaksb. 1939, I. c.: 109. (21).
Геогр. распр. Эфиопия (редко в районе Адшс \ Г>< а > i ,i).
Var. sanguininflatum (Vav.) Dorof. et A. Filat, couib. nov. luigi
dum var. sanguineoinf latum Vav. 1931, 1. c.: 84. — T. abyssinicum var. san-
guineoinflalum Vav. in Flaksb. 1939, 1. c.: 115. (30).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. solitarium (Vav.) Dorof. et A. Filat, comb. nov. — T. turgidum var.
solitarium Vav. 1931, I. c.: 77. — T. abyssinicum var. solitarium Vav. in
Flaksb. 1939, 1. c.: 114. (27).
Геогр. распр. Эфиопия (редко в Харэре и Эритрее).
Var. sparsum (Vav.) Dorof. et A. Filat comb. nov. — T. turgidum var.
sparsum Vav. 1931, 1. c.: 84.— T. abyssinicum var. sparsum Vav. in
Flaksb. 1939, 1. c.: 115. (29).
Геогр. распр. Эфиопия (редко в районе Аддис-Абебы).
Var. spectabile (Vav.) Dorof. et A. Filat, comb. nov. — 7'. turgidum var.
speciabile Vav. 1931, l.c.: 73. — T. abyssinicum spectabile Vav. in Flaksb.
1939, 1. c.: 110. (7).
Геогр. распр. Эфиопия (в районе Аддис-Абебы).
Var. unicum (Vav.) Dorof. et A. Filat, comb: nov. — 7'. turgidum var.
unicum Vav. 1931, 1. c.: 84. — T. abyssinicum var. unicum Vav. in Flaksb.
1939, 1. c.: 115. (31).
Геогр. распр. Эфиопия (очень редко в районе Аддис-Абебы).
3. Convar. recognitum (Steud.) Dorof. et A. Filat. — группа ультра-
плотноколосых разновидностей.
Convar. recognitum (Steud.) Dorof. et A. Filat, comb. nov. — T. recog-
nitum Steud. 1855, Sinops. Plant. Gram. 1: 342. — 7’. pyramidale Perciv.
1921, Wheat pl.: 262, quoad plantam Aethiopia. — T. turgidum pyraniidale
(Perciv.) Flaksb. 1922, Опред. паст, хлебов: 62, 66. — T. turgidum
gr. turgidocompactum Flaksb. 1928, Изв. Гос. ин-та. опытп. агрономии, 6,
2: 43, nom. mid.— Т. turgidum subsp. abyssinicum sect, compacta Vav.
1931, Tp. прикл. бот. ген. сел. Прилож. 51 : 77.
Геогр. распр. Эфиопия (в зоне 1800—2000 м над ур. .м.).
Var. atricompactum (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. atrocompactum Vav. 1931, Tp. прикл. бот. ген. сел. Прилож. 51:
81. — Т. abyssinicum var. atrocompactum Vav. in Flaksb. 1939, Опред.
паст, хлебов: 114. (11).
Геогр. рас п р. Эфиопия (редко в Гопдэре и Эритрее).
Var. atrirubricompactum (Vav.) Dorof. et A. Filat, comb. nov. -- 7'. tur-
gidum var. atrorubrocompacliim Vav. 1931, 1. c.: 82. — T. abyssinicum var.
atrorubrocompactumNax. in Flaksb. 1939, l.c.: 114. (12).
Геогр. p а с,п p. Эфиопия (редко в горной Эритрее).
Var. brevispicatum (Vav.) Dorof. et A. Filat, comb. nov.. — T. turgidum
var. brevispicatum Vav. 1931, l.c.: 77. — T. abyssinicum var. brevispica-
tum Vav. in Flaksb. 1939, l.c.: 110. (2).
Геогр. распр. Эфиопия (район Аддис-Абебы и горная Эритрея).
Var. compressum (Koern.) A. F'ilat. comb. nov.— 7’. compactum var.
compressum Koern. in Koern. et Wern. 1885, Ilandb. Gclreideb. 1: 55.—
T. pyramidale compressum auct. non Koern.: Perciv. 1921, Wheat
pl.: 264. — T. durum var. compressum auct non Koern.: Vav. 1931, l.c.:
51. — 7’. turgidum var. compressum (Koern.) Flaksb. 1935, Культ, фл.
176
К.1111'гиф111МЦ11'1 ро<)а Tiiticiim
Определение pa ’невидное i ей
cunvar. recognihiili (Slcixl.) Dorol.et Л. I'ilat.
белые белые с черным краем 4ei крас- ные ) у и красные с черным краем черные на фоне Ос нвет.ч чешуи и черные Разновидность №, разновил- МОСТИ
белом крас- ном
Колосья неопушенные
3 e ]> и о в к и ф и о л е т о в ы е
+ - + purpureocom- pactum
3 е р н о в к и к р а с и ы с
+ + — + + — 111 + + + + brevispicatum nigribrevi- spicatum planocom pac- tum notabile 2 3 I r>
3 е р и О в к и белые
+1 II 1 1 1 1 + 1 1 1 1 1 + + 1 1 + 1 1 II + + + + + + + + + recognitutn striaticompac- tum copticum pseudorubidi- compactum nigrirubidi- compactum atricompac- tum atriritbricom- pactiun 6 7 8 9 10 11 12
Колосья опушенные
Зерновки красны е
+ + 1 —' + — — — + + + compressum incanicompac- tum fulvicompac- tum 13 14 15
Зерновки белы е
+ — + — — — + + pilosicompac- tiim. ruficompac- tuni 16 17
СССР, 1:236. — Т. abyssinicum var. compressiun (Koern.) Flaksb. 1939,
1. c.: 111. (13).
Геогр. распр. Эфиопия.
Var. copticum (Koern.) A. Filat, comb. nov. — T. compactum var. copli-
cum Koern. in Koern. et Wern. 1885, 1. c.: 55. —T. pyraniidale copticum
Perciv. 1921, l.c.: 264. — T. durum var. copticum Vav. 1931, l.c.: 54.—
T. turgidum var. rubidocompaclum Vav. 1931, 1. c.: 81. — T. turgidum var.
copticum (Koern.) Flaksb. 1935, 1. c.: 239. — T. abyssinicum var. copticum
(Koern.) Flaksb. 1939, 1. c.: 113. (8).
Геогр. распр. Эфиопия (редко в районе Фиче и в горной Эрит-
рее).
Var. fiilvicoinpactiini (Vav.) Л. Filat. comb. nov.— 7. turgidum var. ful-
vocompactum Vav. 1931, 1. c: 78.—-7'. abyssinicum var. julvocompactum
Vav. in Flaksb. 1939, l.c.: 112. (15).
Геогр. распр. Эфиопия (редко в районе Гондэра и в Эритрее).
Htidpod I. Triticum
177
Var. incanicoinpacluni (Vav.) Dorof. et A. I'ilat. comb. nov. - - T. turgi-
dum var. iticanocompaclum Vav. 1931, l.c.: 78. — T. abyssinicum var.
incanocompactum Flaksb. 1939, l.c.: 112. — T. compactum var. compres-
sum auct. non Koern.: Chiov. 1912, Monogr. Rapp, colon. 24: 14. — T. du-
rum var. compression auct. non Koern.: Chiov. 1912, Monogr. Rapp, colon.
19: 17. (14).
Г e о г p. p а с и p. Эфиопия (в Гондэре и горной Эритрее).
Var. nigribrevispicatum (Vav.) Dorof. et A. Filat, comb. nov. — T. tur-
gidum var. nigrobrevispicatitm Vav. 1931, l.c.: 78. — T. abyssinicum var.
nigrobrevispicatum Vav. in Flaksb. 1939, 1. с.: 110. (3).
Геогр. распр. Эфиопия (редко в Гондэре и Эритрее).
Var. nigrirubidicompactum (Vav.) Dorof. et Л. Filat, comb. nov.—
T. turgidum var. nigrorubidocompactum Vav. 1931, l.c. 81. — T. abyssini-
cum var. nigrorubidocompactum Vav. in Flaksb. 1939, l.c.-. 113. (10).
Ге о г p. p а с и p. Эфиопия (очень редко в горной Эритрее и Харэре).
Var. notabile (Vav.) Л. Filat, comb. nov. — T. turgidum var. nota-
bile. Vav. 1931, 1. c.: 78. — T. abyssinicum var. notabile Vav. in Flaksb.
1939, 1. c.: 111. (5).
Ге. огр. распр. Эфиопия.
Var. pilosicoinpactuni (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. pilosocompactum Vav. 1931, 1. c.: 82.— T. abyssinicum var. pilo-
socompactum Vav. in Flaksb. 1939, l.c.: 114. (16).
Геогр. рас п p. Эфиопия (редко в Харэре и горной Эритрее).
Var. pianocompactum (Vav.) Dorof. et Л. Filat, comb. nov. — T. turgi-
dum var. pianocompactum Vav. 1931, 1. c.: 78. — T. abyssinicum var. piano-
compactum Vav. in Flaksb. 1939, 1. c.: 111. (4).
Геогр. распр. Эфиопия (редко в горной Эритрее и в Гондэре).
Var. pseudorubidicompactum (Vav.) Dorof. et Л. Filat, comb. nov.—
T. turgidum var. pseudorubidocompactum Vav. 1931, l.c.: 81. — T. abyssi-
nicum var. pseudorubidocompactum Vav. in Flaksb. 1939, 1. c.: 113. (9).
Геогр. распр. Эфиопия (очень редко в Гондэре).
Var. purpureocoinpactum (Vav.) Dorof. et A. Filat, comb. nov. — T. tur-
gidum var. purpureocoinpactum Vav. 1931, 1. c.: 77. — T. abyssinicum var.
purpureocoinpactum Vav. in Flaksb. 1939,1. c.: 109. (1).
Геогр. распр. Эфиопия (очень редко в районе Аддис-Абебы).
Var. recognitum (Steud.) Dorof. et A. Filat, comb. nov. — T. compactum
var. recognitum (Steud.) Koern. in Koern. et Werm 1885, 1. c.: 55. — T. tur-
gidum var. decoloratocompactum Vav. 1931, l.c.: 78. — T. turgidum var.
recognitum (Koern.) Flaksb. 1935, l.c.: 238. — T. abyssinicum var. decolo-
ratocompactum Vav. in Flaksb. 1939, l.c.: 112. — T. abyssinicum var.
recognitum (Koern.) Flaksb. 1939, l.c.: 112. (6).
Геогр. распр. Эфиопия (около Аддис-Абебы и в Эритрее).
Var. ruficompactum (Vav.) Dorof. et A. Filat, comb. nov.— T. turgidum
var. rufocompactum Vav. 1931, 1. c.: 82. — T. abyssinicum var. rufocompac-
tum Vav. in Flaksb. 1939, 1. c.: 114. (17).
Геогр. распр. Эфиопия (редко в Лксуме и горной Эритрее).
Var. striaticompactinn (Vav.) Dorof. et A. Filat, comb. nov. — T. turgi-
dum var. striatocompactum Vav. 1931, 1. c.: 80. — T. abyssinicum var.
striatocompactum Vav. in Flaksb. 1939, 1. c.: 113. (7).
Геогр. распр. Эфиопия (редко в Гондэре и Эритрее).
Т. PERSICUM VAV. — ПШЕНИЦА КЛРТЛД И НСКАЯ, ДИКА
Т. persiciun Vav. 1919, Изв. Пстровск. с.-х. акад. 1918, 1—4: 177—178,
nom. conserv. ргороп.; Жуковск. 1923, Тр. прикл. бот. сел. 13,1: 46, 47, поп
Aitch. et Hemsl. 1886. — t. dicoccum var. persiciun Perciv. 1921, Wheat pl.:
196, p.p. — T. carthlicum Nevski, 1934, Фл. СССР, 2: 685. — T. paradoxum
12 762
178
К.'Ш1<11<1>11К(1цич Triliitim I..
Parodi, 1910, Rev. Arpenl. Apron., 7, I: 19, nom. illep. T. ibericuin ,\\c-
nabde, I linen. Груз.: 50, нош. illep. - 7'. turgidum pr. curthlicum (Nevski)
Bowden, 1959, Canad. Jotirn. Bot. 37: 671. T. Iurgidtim subsp. carth/icиin
(Nevski) A. et D. Love, 1957, Bot. Not. (Lund) IM, 1: -'Ll; .Mac Key, 1966,
llcredit. Suppl. 2: 208. — T. turgidum convar. carlhlicum (Nevski) Morris
et Sears, 19(17, Wheat and wheat improv.: 3(1. T. cartldictim subsp. carth-
ticujn Dorof. 1968, Вести, с.-х. наука, 3: 33. - 7'. georgicum convar. carth-
licum (Nevski) Mandy, 1970, Acta Apron. Acad. Sci. I limp. 19. 3 4: 4 16.
Согласно правилам «Международного кодекса ботанической номен-
клатуры» (1974) действительным названием вида является Т. cartlilicum
Nevski. Однако в память о великом биологе II. II. Вавилове мы сохраня-
ем то наименование вида, которое он ему дал - Т. persicum.
Общая характеристика. Всходы зеленые, опушенные. Колеоптиль с
двумя проводящими пучками. Куст прямостоячий. Стебель и все расте-
ние с характерной зеленой окраской. Соломина почти выполненная, тон-
кая (диаметр 2—4 мм). Стеблевые узлы опушенные (иногда лишь в
верхней части) пли голые. Листовые влагалища опушены очень корот-
кими волосками. Листовые пластинки бархатпегоопушенньн'. Язычок
бахромчатый, ушки с ресничками. Колосья по внешнему облику похожи
на колосья пшеницы мягкой (рис. 13). Лицевая сторона колоса шире
боковой пли равна ей. Длина колоса 7,5—16 см, ширина лицевой сторо-
ны— 5—9 мм, боковой — 6—9 мм, D = 11—25. Окраска колосьев желтая,
светло-красная (иногда более интенсивная) или черная. Стержень колоса
отличается от такового других видов пшеницы. Он узкий и тонкий, что
обеспечивает гибкость колоса. Ширина членика приблизительно 1,5 мм
(у пшеницы мягкой — 2,5—3 мм). Членик стержня по краям голый или
очень слабо опушенный. На членике под колоском имеется бородка из во-
лосков, которая очень слабо выражена. Иногда она отсутствует. Колос-
ки ромбовидные, многоцветковые, чаще трехзерные. Имеются осыпаю-
щиеся и пеосыпающпеся формы. Колосковые чешуи по форме сходны с
таковыми пшеницы мягкой. У основания их отсутствуют продольная мор-
щинистость и поперечная вдавлеппость, которые характерны для пше-
ницы мягкой. Жилкование на поверхности колосковой чешуи не выраже-
но, сама она по консистенции нежная, а у пеопушенпых форм гладкая,
блестящая. Киль колосковой чешуи узкий, доходящий до основания че-
шуи. В верхней части он переходит в длинный остевидный отросток пли
в ость длиной до 12 см. Наличием длинных остей на колосковых чешуях
Т. persicum отличается от всех остальных видов пшеницы. Ости цветко-
вых чешуй нежные, зазубренные, как у пшеницы мягкой. На колосковых
и цветковых чешуях по обеим сторонам ости имеются 2 маленьких зуб-
чика. Между остыо и зубчиками — остроугольная выемка (синус). Зер-
новка обычно овальная, в поперечном разрезе округлая. Ее цвет — крас-
ный, редко белый. Длина зерновки 6—8 мм, консистенция чаще стекло-
видная. В последнем случае па спинке .зерновки поперечная морщини-
стость с амальгамным отблеском. Если этой морщинистости и такого от-
блеска нет, то зерновка мучнистая, схожая с зерновкой пшеницы мягкой.
Е. Ф. Пальмова (1935) отнесла Т. persicum к пшеницам высокогорно-
го умеренного пояса, где она возделывается на высоте 900—2000 м над
ур. м. Здесь она в чистом виде высевается очень редко: обычно возделы-
вается в популяциях с полбой (Армения), пшеницей мягкой (Грузия
и Дагестан) или с пшеницей твердой (Дагестан).
К. А. Фляксбергер (1935) характеризовал этот вид как единый эко-
лого-морфологический тип с ограниченным полиморфизмом. Л. Л. Де-
капрелевич (1954) выделил 2 экотипа: горно-лесной (caucasionis dika
Dekapr.) и горно-степной (dzhavachetis dika Dekapr.). Первый экотип
возделывается в нижней лесной полосе Главного Кавказского хребта
(900-1400 м над ур. м.). Почвы здесь лесные, климат умеренно холод-
ный с избытком осадков (800—1200 мм в год). Этот экотип представлен
Ilotlpcx) I. Tritiiiiin
Рис. 13. Колосья, колоски и зерновки Т. persicutn Vav.
( = Т. carthlicum Nevski)
в основном более влаголюбивыми формами (var. persiciun'}, которые со-
зревают на 5—9 дней позднее, чем горно-степной экотип. К горно-лесно-
му экотипу близки дагестанские формы Т. persiciun.
Формы горно-степного экотипа возделываются в Грузии (Джавахст-
ское плато и примыкающие к нему районы) па высоте 1400—2100 м над
ур. м. Климат здесь сравнительно континентальный, почвы— горно-чер-
ноземные, каштановые, чернозем невидные. Среднегодовое количество
осадков — до 500 мм (сравнительно засушливая зона). Вегетационный
период у растений этого экотипа короче, чем у форм горно-лесного эко-
типа, а высота растений ниже. В горно-степной зоне локализованы по-
пуляции из Т. persicutn (представлены красноколосыми формами —
var. rubiginosum) и пшеницы мягкой, формы которой значительно усту-
19*
180 Классификация рада Triticum I,.
итог осрiniMу компонспiу популяции ио \ с ioii'iiii’.ori и к i рибным болез-
ням. Близок к этому экотипу Г. persicum /Армении, а также Нахичевани
и Нагорного Карабаха. В двух последних регионах этот вид представ-
лен в качестве примеси.
Экотипы, описанные Л. Л. Декаирелевичем (1954), согласуются с дву-
мя группами Т. persicum, установленными II. II. Вавиловым (1961, 1964):
prol. daghestanicum Vav. и prol. armetiogeorgictim Vav. Первая отли-
чается наличием преимущественно черпоколосых разновидностей с опу-
шенными чешуями, вторая — белоколосыми и краспоколосыми с гладки-
ми чешуями.
У Т. persicum, несмотря па тщательные поиски па Кавказе, предпри-
нятые II. И. Вавиловым, Л. Л. Декаирелевичем, М. Г. Туманяном,
М. М. Якубцнпсром, И. Д. Мустафаевым и В. Ф. Дорофеевым, формы с
озимым образом жизни не обнаружены.
Поданным И. И. Вавилова (1935, 1964), Е. Ф. Пальмовой (1935),
М. М. Якубцпнера. (1938), И. А. Стефановского (1950) и Л. Л. Декаире-
левича (1967), Т. persicum имеет формы средпсспслые-средпспозднеепе-
лые (экотип горно-степной) и средпераиние-среднеспелые (экотип горно-
лесной). При весеннем посеве длительность периода от всходов до
колошения в Закавказье равна 80—120 дням (реже 77—127 дням), в За-
волжье — 78—83 дням. На своей родине хорошо удается и при осеннем по-
севе. При позднем сроке сева в условиях Поволжья не всегда вызревает.
Засухоустойчивость Т. persicum недостаточная, а регенерационная
способность высокая. Требовательность к теплу в период созревания низ-
кая. Отзывчивость на орошение и высокий агрофон хорошая. Выявлены
формы, устойчивые к весенним заморозкам. Период послеуборочного до-
зревания длительный. Поэтому Т. persicum устойчив к прорастанию
зерна на корню и в снопах.
По данным В. И. Жукова (1969), при осеннем посеве и орошении в
условиях низменного Дагестана у Т. persicum наблюдается высокий уро-
вень открытого цветения (до 100% хазмогамных цветков). Причем боль-
шая часть цветков открывается во второй половине дня. Открытое цве-
тение может стать причиной спонтанной гибридизации. М. М. Якубцинер
(1946) и В. Ф. Дорофеев (1972) выявили гибриды от естественного скре-
щивания Т. persicum с Т. durum.
В. М. Берляпд-Дожевников и др. (1975) в условиях низменного Даге-
стана установили, что формы горио-степпого экотипа более стабильны —
проявляют меньшую склонность к перекрестному опылению, чем формы
горно-лесного экотипа.
По данным В. Г. Допарева и др. (1972), у большей части образцов
Т. persicum выявлено высокое содержание белка в зерне (до 23%), осо-
бенно у образцов горно-лесного экотипа из Грузии.
Исследования Д. М. Чинго-Чипгаса (1925) показали, что мука из
зерна Т. persicum обладает низкой хлебопекарной способностью. Хлеб из
нее по мягкости корки и коричневому цвету сходен с хлебом из пшеницы
твердой.
Иммунитет. Характерная особенность Т. persicum — устойчивость к
мучнистой росе. Именно но этому признаку, как уже отмечалось,
И. И. Вавилов. (1918) выделил его из ряда сходных с ним образцов, пше-
ницы мягкой. В последующем (1935, 1964) он неоднократно обращал
внимание селекционеров па это свойство Т. persicum. По данным
М. М.. Якубцпнера (1969), некоторые образцы этой пшеницы могут по-
ражаться мучнистой росой. И. М. Жуковский (1971) указал, что в Гру-
зии, опа поражается специфической расой этого патогена. В. И. Кривчен-
ко и др. (1979) изучили коллекцию Т. persicum при искусственном зара-
жении проростков популяцией рас из 5 географических регионов СССР
и при естественном заражении -на провокационном фоне в поле. Боль-
шая часть образцов была иммунной к патогену. Около 30% их оказа-
Подрод I. Triticum 181
лись восприимчивыми. С,дела no пред положение, что иммунные к мучиис-
|oii росе образцы Т. pctsicum пмекн первичное происхождение (возникли
от иммунной формы Т. dicoccum), а восприимчивые- вторичное (в ре-
зультате спонтанной гибридизации между пшеницей мягкой и каким-
либо тетраплоидным ни,том секции Dicoccoides).
К бурой ржавчине большая часть образцов Г. persicum восприимчива
(М. М. Якубцинер, 1969). И. П. Шитова (1968) по результатам наблюде-
ний 1954—1965 гг. нашла, что в условиях Дагестана местные формы это-
го вида в основном устойчивы к бурой ржавчине, а образцы из Армении
большей частью восприимчивы к ней. Но данным О. Г. Григорьевой
(1975), в полевых условиях па провокационном фоле 46,6% образцов
коллекции были восприимчивы к этому заболеванию (балл 3—4), 10,3%
имели лишь следы поражения, иммунные образцы отсутствовали. В со-
ставе мировой коллекции Т. persicum В. М. Верлявд-Кожевников и др.
(1973) выявили образцы—носители генов Lr 1, Lr 2, Lr 2*, Lr 3, Lr 17,
а также образцы, имеющие группы генов X!, Х2, Х3. Перечисленные гопы
и группы их обеспечивают устойчивость к расам 77, 202 и 151. Выявлен
образец с полигонной устойчивостью к этим расам.
Т. persicum, но данным IL II. Шитовой (1968) и А\. Д\. Якубцннера
(1969), относится к числу видов устойчивых или слабо поражаемых жел-
той ржавчиной. Н. И. Вавилов (1935) отмечал устойчивость этой пшени-
цы также и к стеблевой ржавчине. М. М. Якубцинер (1961) сообщил о
сильном поражении этим патогеном в условиях Северного Кавказа и За-
кавказья. Л. М. Камслпна (1973) свидетельствует о наличии у Т. per-
sicum как устойчивых, так и восприимчивых к стеблевой ржавчине форм.
В полевых условиях па провокационном фоне 22,4% образцов коллек-
ции в опыте О. Г. Григорьевой (1975) были восприимчивы, 13,8% имели
следы поражения этой ржавчиной, иммунные образцы отсутствовали.
Этот исследователь пишет о большей устойчивости Т. persicum к стебле-
вой ржавчине, чем к бурой.
Н. И. Вавилов (1961) отнес Т. persicum к числу устойчивых к пыль-
ной головне видов, а М. М. Якубцинер (1969) отметил его восприимчи-
вость к этой болезни в условиях юга. Искусственное заражение коллек-
ции ипокулюмом пыльной головни в опытах В. И. Кривченко (1973) и
А. М. Ямалеева (1974) показало, что большинство образцов Т. persicum
имеет высокий иммунологический тип. К. пыльной головне L dtiri иммун-
ными были 79,2%, к f. aestivi — 73,1% изученных образцов, восприимчи-
выми (балл 3—4) оказались лишь (соответственно) 2,1 и 1,9% образцов.
Возможно, что последние образцы имеют вторичное происхождение.
Электрофоретический анализ белков зерна показал, что восприимчивые
образцы в со-зопе имеют. 6—8 компонентов, а устойчивые — 2—3- компо-
нента (А. М. Ямалеев, 1974).
Н. И. Вавилов (1961) и С. П. Зыбина (1939) отметили наличие у
Т. persicum неустойчивых к твердой головне образцов. При искусствен-
ном заражении В. И. Кривченко (1973) в коллекции этого вида выявил
42% иммунных и 12% восприимчивых образцов. Н. Harding (1972) отнес
изученные формы Т. persicum к числу более или менее восприимчивых к
корневым гнилям.
История. По некоторым данным (G. Mandy, 1970) возникновение
Т. persicum относится к 6500—6000 гг. до и. э. По результатам исследо-
ваний Л. Л. Декапрелевича (1938) Т. persicum — вид древнейшего про-
исхождения. В грузинской литературе V в. упоминается пшеница «дика».
Это народное название для 7’. persicum существует в Грузии исстари.
В отечественной литературе нового времени видовое название «,тика<>
впервые было приведено еще в 1798 г. И. Г. Георги в числе сортов, куль-
тивируемых в Имеретии. Это свидетельствует о возделывании Т. persi-
cum в тот период в Западной Грузии, т. е. за пределами eiо современного
ареала (М. М. Якубцинер, 1956).
182
К.'кк'сификици.ч po()<i
Trilicttm I..
I'eoi рафическое распространение. II. II. Вавилов (1918) iipe;nio.aai a.i,
'По вид T. persiiuiii Vac. pacnpoc i ранен лишь и I I ране. II. AV /Гуковский
(1923) первый обнаружил его в Грузии. В дальнейшем, благодаря науч-
ным поискам Л. Л. Декапрелевича (1925), М. Г. Туманяна (1934),
Е. А. Столетовой (1'930), М. М. Якубцппера (1938), II. Г.. Гребен-
никова (1948), И. Д. Мустафаева (1961), В. Ф. Дорофеева (1972),
уточнено, что в ареал вида входят также Армения, Нахичевань, Нагор-
ный Карабах, Северная Осетия и сопредельные Закавказью районы Тур-
кин. Этот вид возделывается в Праке при орошении (N. L. Вог, 1968).
Персидская пшеница обнаружена в Северо-Западном Иране (II. И. Ва-
вилов, 1961).
ГГ Д. Мустафаев (1961) в Азербайджане выявил б разновидностей
Т. persiciun в качестве примеси в посевах твердой и мягкой пшеницы.
По данным Н. ЕЕ Ксц.ховелп (1928), дика занимает в Грузин пояс от
650 до 2000 м над ур. м.; в Армении, по исследованиям Е. А. Столетовой
(1930). М. Г. Туманяна (1944), Г. X. /Агаджаняна (1931) н Б. П. Гарасе-
феряпа (1939), —от 1600 до 2500 м над ур. м.
Происхождение. 7'. persiciun в хромосоме 5А имеет фактор Q, кото-
рый ответствен за легкий обмолот колоса н прочность его стержня
(J. Mac. Key, 1954; AT Ahiramatsii, 1961, 1963; IE Kuckuck, 1961). Этот
фактор имел огромное эволюционное значение при переходе пшеницы с
двугеномного тетраплоидпого на трехгеномный гексаплоидный уровень,
обеспечив голозерность главного хлеба человечества — пшеницы мягкой.
К- А. Фляксбергер (1935, 1938) полагал, что Т. persiciun сформирова-
ли высокогорные условия и изменчивость. М. Г. Туманян (1944, 1948)
и В. О. Гулканян (1952), не отрицая возможности гибридного пути про-
исхождения, связывали возникновение этого вида из Т. dicoccum или
Т. durum с влиянием факторов внешней среды. Причем М. Г. Туманян
предполагал полифилетическое происхождение разновидностей Т. per-
sicum. В. Ф. Дорофеев (1969) причину возникновения Т. persicutn видит
в мутации, происшедшей у пшеницы твердой или полбы. Л. Л. Декапре-
левич (1971) местом возникновения такой мутации у полбы считает Вос-
точную Грузию. Экспериментально вызванная мутация с появлением Q
у Т. dicoccum (Н. Kuckuck, 1964) действительно дала Т. persiciun. Еще в
1922—1923 гг. П. М. Жуковский указывал на генетическую близость этих
видов. Видимо, J. Percival (1921) понимал это и потому квалифицировал
7'. persiciun как разновидность Т. dicoccum. В. Г. Конарев и др. (1970), а
также В. L. Johnson (1972) отметили сходство электрофоретических
спектров белков зерна у этих видов.
Существуют 2 точки зрения о происхождении фактора Q па тетра-
илондном уровне.
1. Фактор Q мутационпо возник у Т. dicoccum. В результате появился
Т. persicutn (первичное происхождение).
2. Фактор Q — мутация у Т. spelta. В результате появилась пшеница
мягкая. Последняя вступила в спонтанную гибридизацию с каким-то
тетраплоидный видом пшеницы. При расщеплении в гибридном потом-
стве этой комбинации выщегшлея Т. persiciun (вторичное происхожде-
ние).
С вопросом, какой из этих путей становления фактора Q у тетраплои-
да использовала природа, связано решение проблемы происхождения
Т. persicutn. Е. S. McFadden, Е. R. Sears (1944) и II. Kihara (1944) созда-
ли пленчатый гексаплоидный вид, минуя голозерный тстраплоид—
Т. persicutn (путем удвоения числа хромосом у гибридов от скрещива-
ния пленчатого тетраплоида с Ле. tauschii). Однако до сих пор паука не
представила факта, свидетельствующего о получении голозерного гекса-
плопдиого мутанта из созданного таким образом пленчатого гексанлоп-
да или из Т. spelta. Между тем такие авторитетные трптикологи, как
II. ЕЕ Вавилов ’(1918, 1926, 1935), К. А. Фляксбергер (1930) и Е. Schie-
Ihxlpot) I. Tri lit пт
in,-mu (I 932), были убеждены во в i орпчпом и । >< >1 к \ < >жд < 11 и и вида T. xpc/-
l;i (из пшеницы мягкой или lompaclitm). Фим eci ec i веиши о возпикно-
венпя голозериостп у пленчатого тетранлоида был зарегистрирован у пол-
бяной пшеницы — Т. timopheevii, которая является гомологом Т. dicoc-
cum. При этом появилась форма Т. militinae, колосковые чешуи которой
очень сходны с чешуями Т. persicum var. persicum (II. M. Жуковский,
Э. Ф. Мигушова, 19(19). Появление Т. militinae символизирует для пас
модель рождения Т. persicum из Т. dicoccum. Характер растепления в F2
при гибридизации между Т. persicum 11 Т. dicoccum в пашем опыте также
свидетельствует о том, что между этими видами не было посредников, что
генетически Т. persicum - это 'Г. dicoccum с фактором Q.
Автор вида Т. persicum—И. 11. Вавилов (1925, 1926, 1929, 1931) —
считал, что наличие у .этого вида признаков пшеницы мягкой свидетель-
ствует о происхождении его при гибридизации между Т. dicoccum и
Т. aestivum. В. Л. Мепабде (1948, 1971) предполагает место гибридного
происхождения 7’. persicum в горах Грузии. М. А. Арошпдзе (1953, 1956)
показала, что при скрещивании между пшеницей мягкой и 7'. dicoccum
в гибридном потомстве можно отобрать типичные формы 7'. persicum.
Е. Р. Aloppnc и Э. Р. Сирс (1970) также поддержали гипотезу о гибрид-
ном возникновении этого вида при скрещивании iексаилоида с тетра-
плоидом. Опп пришли к выводу о сравнительно недавнем происхождении
7'. persicum. П. А. Гандиляи (1972) описывает факты появления в усло-
виях Армении типичных форм этого вида в результате спонтанной гибри-
дизации между мягкой и твердой пшеницами.
Результаты изучения коллекции Т. dicoccum и Т. persicum па устойчи-
вость к мучнистой росе, пыльной и твердой головне (В. И. Кривченко,
1973; Э. X. Сухапберднпа, 1977), возможно, свидетельствуют о том, что
современный Т. persicum включает формы как первично!о (из 7’. di-
coccum), так и вторичного (с участием Т. aestivum) происхождения. Пер-
вые сохраняют иммупость к патогенам, унаследованную от Т. dicoccum,
вторые стали восприимчивыми, так как это качество они унаследовали
от пшеницы мягкой.
Систематика. Первоначально Т. persicum был определен К. А. Флякс-
бергером (1915) как разновидность пшеницы мягкой (7'. aestivum var.
fuligitlosuin Alef.) по строению колоса, форме цветковых чешуй и т. д.
Исследования по иммунитету, детальное морфологическое и анатоми-
ческое изучение позволили И. И. Вавилову (1919) выделить эту пшеницу,
полученную им от фирмы Гааге и Шмидт под названием «персидской», в
особый вид Т. persicum Vav.
Впоследствии И. А1. Жуковский (1923—1924) наряду с чериоколосы-
ми формами выявил в Грузин другие разновидности этого вида. Затем
ряд исследователей установил его разновидиостный состав в Армении,
Северной Осетии, а также в Турции и Иране.
Var. arnual (Gandil.) Gandil. comb. nov. — T. carthlicum var. arnual
Gandil. 1976, Бюл. ВИР, 65: 75. (6).
Получена И. А. Гандиляиом в результате гибридизации Т. durum с
Т. aestivum на Экспериментальной базе Института земледелия МСХ Ар-
мянской ССР (Эчмиадзип).
Var. arnurual (Gandil.) Gandil. comb. nov. — T. carthlicum var. arnu-
rual Gandil. 1976, 1. c.: 75. (7).
Получена И. А. Гандиляиом там же в результате спонтанной гибри-
дизации.
Var. arpual (Gandil.) Gandil. comb. nov. — T. carthlicum var. arpual
Gandil. 1976, 1. c.: 76. (13).
Получена 11. А. Гандиляиом там же и сходным образом.
Var. arpurual Gandil. 1975, Тр. Арм. ИИИЗ, сер. пшеница, 3:91, descr.
ross. (15).
Г с о г р. р а с и р. Армения.
184
Классификация роди Triticum I..
Колосья неопушенные.
3 с р и о в к и к р а с и ы с
4- — — — — — —. -1- — s/гн mi п ей in I
+ — — -— — — —- — ч- pscudostramineutn 2
— — + -— — — — -ь — rubiginosum 3
— —• + -— — — •— — ч- pseudorubiginosutil I
— — — + —- — — — + nigrorubiginosutn J
3 e p п о в к и бе л ы е.
+ + arnual
+ + arnttrtial
Колосья опушенные
Зерновки красные
+ —* — — — — -1- — rarissimum 8
— —- + — — — — — + zhukovskyi 9
— — + —. — — — .+ — ismeticum 10
— —- — —— + — — — + persicum 11
— — — — — + — — + darginicum 12
3 e p н о в к I бел ы е
+ — — — —. — — + — arpual 13
+ — — — —- — —— — + narpual 1 1
— — + — — — — + — arpurttal 15
— — + — — — — — + narpurual 16
— — — •— + — — — + osseticum 17
— — — — ~~ + — — + arpurunial 18
Var, arpurunial Gandil. 1975, 1. с.: 94, descr. ross. (18).
Геогр. распр. Армения.
Var. darginicum Berg et Muizhn. 1929, Tp. Всесоюзн. съезда ген. сел.
сем. 3:383. (12).
Геогр. распр. Дагестан, Грузия (редко).
Var. ismeticum Gokg. 1941, Zeitschr. Pflanzenziicht. A 23: 578. (10).
Геогр. распр. Турция (редко).
Var. narpual (Gandil.) Gandil. comb. nov. — T. carlhlicum var. narpual
Gandil. 1976, 1. c.: 76. (14).
Получена П. А. Гапдиляном в результате спонтанной гибридизации
на Экспериментальной базе Института земледелия МСХ Армянской
ССР (Эчмиадзин).
Var. narpurual (Gandil.) Gandil. comb. nov. — T. carlhlicum var. nar-
purual Gandil. 1976, 1. c.: 77. (16).
Г eo гр. распр. Получена П. А. Гапдиляном в результате гибриди-
зации Т. durum и Т. aestivum на Экспериментальной базе Института зем-
леделия МСХ Армянской ССР.
Var. nigrorubiginosutn Flaksb. 1935, l.c.: 284, descr. ross. (5).
Геогр. расп p. Армения, Грузия, Азербайджан.
Var. osseticuin Greb. 1930, Tp. прикл. бог. гоп. сел. 21, 2: 15. (17).
Г corp, ра си p. Северная Осетия, Армения, Да! естаи.
Var. persicum. - - Т. persicum var. juliginosum Zhuk. 1923, Tp. прикл.
бот. сел., 13, 1: 50. - 'Г. persicum var. cinereum Zhuk. 1923, l.c.: 49.—
T. ibericum var. fuliginosum f. albifuliginosum Menabde, 1948, Ilmen.
Груз.: 54.— T. ibericum var. fuliginosum f. laxiusculum Menabde et f. do-
Подрод 1. Triticum
minans Menabde, 1948, I. c.: 54. - T. carthlicum var. [uligitiostim f. capitu-
lum Dekapr. 1954, 'Гр. Пита полевод. 8:23, descr. ross. (11).
Довольно значительный полиморфизм var. fuliginosum позволь i
В. Л. Менабде (1948) выделить форму с очень длинными (13,0 см) и рых-
лыми (D = 14—17) колосьями — f. laxiusculum Menabde, а Л. Л. Дека-
ирелевичу (1954)—форму с очень плотными (D = 25—30) булавовид-
ными колосьями — Д capitatuni Dekapr. Кроме того, В. Л. Менабде выде-
лил форму с. совершенно неопушенными члениками колосового стерж-
ня — f. dominans Menabde.
Геогр. распр. Дагестан, Грузия; в Азербайджане и Армении
редко.
Var. pseudortibiginosum Zhuk. 1924, Изв. Тифл. гос. политехи, ин-та.
1: 140, nom. nud.; Flaksb. 1935, 1. с.: 284, descr. ross. — Т. ibericum var .ж-
biginosum f. nigriaristatum Menabde, 1948, 1. c.: 53, descr. ross. (4).
Г e о г p. p а с и p. Грузия, Азербайджан.
Var. pseudostraniineum Flaksb. 1935, l.c.: 283, descr. ross. — ?’. ibe’i-
cum var. slramineum f. nigriaristatum Menabde 1948, l.c.: 52, var. inva-
lid. (2).
Геогр. распр. Грузия и Азербайджан (редко).
Var. rarisshnuin Flaksb. 1935, l.c.: 284, descr. ross.--?’. ibericum var.
pilosostramineum Menabde, 1948, 1. c.: 53, var. invalid. (8).
Геогр. распр. Дагестан и Грузия (редко).
Var. rtibiginosum Zhuk. 1923, l.c.: 49. — T. persicutn var. rtibiginosum
f. capitatuni Flaksb. 1935, 1. c.: 283, descr. ross. — ?’. ibericum var. rubigino-
suni f. elongatum Menabde, f. pilosum Menabde, f. longiaristatum Menab-
de, f. uniaristatum Menabde, f. cauliplenum Menabde, f. duriusculum Me-
nabde, f. nigriaristatum Menabde, f. brevispicatum Menabde, 1948, 1. c.:
52—53, descr. ross. (3).
Одна из наиболее распространенных разновидностей Т. persicutn.
В пределах разновидности наблюдается большое разнообразие форм.
К- А. Фляксбергер (1935) отмечает форму с булавовидными плотными
(D около 30) колосьями. В. Л. Менабде (1948) выделил, ряд новых
форм: с.колосьями удлиненными (до 13,5 см), рыхлыми (D== 14—17); с
негрубыми колосьями и остями длиной 16—18 см; с колосьями плотными
(D=30—36), ости па колосковых чешуях которых укороченные (0,7—
1,8 см); с колосьями рыхлыми (D—16—18), с остями грубыми, у основа-
ния утолщенными; с колосьями рыхлыми (Г) — 17—20), длинными (8—
10 см длины), с остями длинными (15—17 см), нежными; с колосьями ко-
роткими (5—5,5 см), плотными (D = 34—36); с густоопушенными члени-
ками колосового стержня [подобная форма несколько рапсе (1946) опи-
сана М. А. Сихарулидзе]; с голыми члениками колосового стержня.
К. А. Фляксбергер (1935) у var. rtibiginosum (сборы Г. Ковалевского в
Армении па берегу озера Севан, 1960 м над ур. м) выделил форму
(Г. persicutn f. griseorubiginosum Flaksb. 1935, l.c.: 283, descr. ross.) co
слабовыраженной серо-дымчатой окраской колосьев. На нахождение по-
добных форм в Грузии указывает также Л. Л. Дскапрелевич (1954).
Геогр. распр. По всему Закавказью и в Дагестане.
Var. stramineum Zhuk. 1923, 1. с.: 49. — ?’. ibericum var. slramineum i.
nudum Menabde, f. pilosum Menabde, f. breviaristatum Menabde, f. duriu-
sculum Menabde, f. laxiusculum Menabde, f. densiusculum Menabde, 1948.
1. c.: 52, descr. ross. (1).
Геогр. распр. По всему Закавказью и в Дагестане.
Var. zhukovskyi Flaksb. 1935, 1. с.: 284, descr. ross. (9).
Выделена из гибридного материала комбинации Т. persicutn X Т ti-
mopheevii.
1 86
Клаа ификиция роди Triticum 1..
СЕКЦИЯ 3. TRITICUM
Sect. 3. Triticum; Ц целей, Новости сист. высш. раст. 10: 40, р.р. он же,
1976, Злаки СССР: 164, р.р. - Triticum sect. Frumentutn Dum. 1823, Ob-
serv. Gram. Fl. Belg.: 91, excl. T. turgidum cl T. compositum. — Triticum
sect. Spelta Dum. 1823, l.c.: 94, cxcl. T. dicoccum.— Triticum Dinkclrcihe
A. Schulz, 1913, Gcschichte knit. Getr.: 22. — Triticum consp. speltoides
Flaksb. 1915, Опред. union.: 10. — Triticum Scries III (The Bread Wheat
series) Perciv. 1921, Wheat pl.: 341. — T. obtusatum Kajanus, 1927, Bib-
liogr. Genet. 3: 146, species collect. — Trilicum sect. Speltoidea Flaksb.
1928, Изв. Гос. nn-та опыта, агрономии, 6, 2: 10. — Triticum sect. Spelta
Nevski, 1934, Фл. СССР, 2: 686, non Dum. 1823. — Trilicum congrcgatio
tiexaploiclea Flaksb. 1935, Культ, фл. СССР, 1: 30. — Triticum sect. Eutriti-
cca subsect. Vulgaria Sinsk. 1955, I IpoG.’i. бот. 2 : 52.
Листья голые (шероховатые) или опушенные (бархатистые па
ощупь). Соломина под колосом полая, редко выполненная. Колосья раз-
личной формы, простые или ветвистые, преимущественно двуоетые. Стер-
жень колоса в зрелом состоянии прочный или ломается при надавли-
вании. В последнем случае колоски остаются npii.iei ающпмп к брюшной
стороне колоска (прикреплены у своего основания к нижней части чле-
ника колосового стержня) или, реже, насаженными па верхний конец
членика (членик колосового стержня является как бы рукояткой колос-
ка). У пленчатых видов зерновка плотно заключена в чешуях, при обыч-
ной молотьбе вымолачивается с трудом. У голозерных видов зерновка
легко вымолачивается. Колоски 4—6(8)-цветковые, из которых 2—5(6)
плодущие. Верхняя цветковая чешуя у остистых форм несет ость. Ниж-
няя цветковая чешуя при созревании не расщепляется вдоль па 2 части.
Колосковая чешуя имеет килевой зубец (от короткого до длинного, из-
редка переходящего в укороченную ость). 2п = 42. Геномный состав
A"A"BBDD.
Геогр. распр. Границы ареала возделывания видов секции Trili-
сит в северном полушарии местами доходят до Полярного круга, в юж-
ном— до оконечностей материков Австралии, Африки и Южной Америки.
Т. MACHA DEKAPR. ET MENABDE — ПШЕНИЦА МАХА
Т. macha Dekapr. et Menabde, 1932, Tp. прикл. бот. ген. сел. Сер. 5,
1 : 14.— Т. aestivum gr. macha Bowden, 1959, Canad. Journ. Bot. 37 : 675.—
T. aestivum subsp. macha Mac Key, 1954, Sv. Bot. Tidskr. 48,2: 586.—
T. spelta subsp. macha (Dekapr. et Menabde) V. Dorof. 1966, Генетика,
3: 30, comb, invalid. — T. aestivum convar. spelta (L.) Morris et Sears
1967, Wheat and wheat improvement: 36, p.p., comb, invalid. — T. aestivum
convar. macha (Dekapr. et Menabde) Mandy, 1970, Acta Agron. 19, 3—4:
416, comb, invalid.
Общая характеристика. Первый лист молодых растении узкий, зеле-
ный, темпо-зеленый пли фиолетовый. В фазе кущения куст стелющийся
пли полустелющийся. Облнствениость растении хорошая. Листья голые
или опушенпйе (бархатистые на ощупь), в период колошения с воско-
вым налетом. У плотиоколосых форм лист широкий. Ушки зеленые пли
фиолетовые, с ресничками или без них. Влагалище голое или опушенное.
Узлы соломины опушенные. Соломина полая, около 100 см высотой!. Ко-
лосья от коротких до длинных, от плотных D = 30—52 до довольно рых-
лых (D = 18—30). У рыхлоколосых форм членики стержня длиной 4—
5,5 мм, у плотиоколосых — 2—3 мм. У первых в верхней части членика
развита полость, выполненная паренхимой, у вторых опа почти отсутст-
вует. Стержень колоса ломается, как у полбы (членик стержня верхним
Под/ю<) I. Trilicum
187
концом остле I (",i 111) 11 к ] >i'! i ле 11111,1 м к ос 11 о i >; i j 111 io колоска ) или, роже, как у
c 11 e. 11 > 11.1 (ч.кшик стержня в ннжисн части o< iacic;i прикрепленным к ос-
нованию колоска, прилегая к ею брюшной стропе). Колос обычно ло-
мается при надавливании, но у некоторых форм наблюдав гея спонтанная
ломкость верхних колосков. Созревший колос отламывается v основа-
ния. Иногда, например в Ташкентской области, такой колос может цели-
ком спонтанно отломиться и упасть па землю. Членики пержпя срсдне-
нлп cn.'jbiiooiiyiiieiiiibie. Ости обычно короче колоса (от 3 до >3 см длиной).
Встречаются также формы с деформированными остями. Колосковая че-
шуя по форме от лопатчатой, как у спелы ы, до ланцетной, как у пше-
ницы мягкой.
Имеется разнообразие по характеру' ее жилкования, по форме плеча
(иногда плечо отсутствует), выраженности киля, форме килевого зубца
(от тупого до острого; от прямого до загнутого). Зубец главной боковой!
жилки почти всегда имеется. Зерновки красные, с брюшной стороны
плоские или вдавленные, с широкой и глубокой бороздкой!. По рыхлости
колоса, по длине остей Т. macha уступает спелые. Кроме того, у нее от-
сутствуют черпоколосыеи яровые формы.
Число проводящих пучков в колеоптиле пе больше двух. Иногда
отделительная ткань, обусловливающая ломкость колосовою стержня,
бывает в редуцированном состоянии (G. Polilemll, 1958). У зародыша
сильно редуцирован эпибласт (М. С. Яковлев, 1950). Стенки соломины
тонкие, со слаборазвитой механической тканью (О. Д. Градчапинова,
1967). По данным Л. Л. Жестяпликовой (1954), .зерновка Т. macha по
структуре перикарпия не отличается существенно от других видов гекса-
плопдпой пшеницы. Для него характерны сильная редукция мезокарпия
и связанное с этим сохранение в эпи-мезокарпии лишь одного слоя эпи-
дермальных клеток, полное отсутствие отложения полуклетчатки. У con-
var. macha в эни-мезокарнни одни ряд эпидермальных клеток, тапгеп-
талыю вытянутых и сильно облитерированных. У convar. densiusculum
эти клетки короче, шире и на спинной стороне зерновки слабо облитери-
рованы, а сам перикарпий несколько мощнее, чем у первого подвида.
Л. Л. Жестянпикова пришла к выводу, что перикарпий Т. macha по строе-
нию близок к перикарпию пшеницы мягкой.
Внд Т. macha возделывался в горных условиях па высоте 900—1200 м
над ур. м. в зоне избыточного увлажнения, которая по термическому ре-
жиму является умеренной и частью теплой (Г. Т. Селяпппов, 1967). Об-
раз жизни его форм — озимый или полуозимый. Согласно трехлетним
данным А. М. Горского (1960), при весеннем посеве в условиях Адыгеи
и Ленинградской области эти формы не выколашиваются.
Преобладают позднеспелые формы, которые в Западной Грузии вы-
зревают на 5 дней раньше, чем Т. timopheevii. В целом Т. macha является
влаголюбивым и влаговыносливым видом. Обычно не страдает от поле-
гания. Чувствителен к воздушной засухе.
По данным А. 3. Латыпова и В. И. Апеля (1971), в Белоруссии у
Т. macha 57,6% цветков имели открытое цветение. Максимум цветения
наблюдался в период от 8 до 12 ч дня, минимум — от 21 до 5 ч утра. На-
блюдения М. А. Федина (1974) в Подмосковье показали, что Т. macha по
типу цветения имеет сходство с Т. aestivum (от 59 до 90% цветков в ко-
лосе цветет открыто). В приморской полосе в Дагестане В. И. Жуков
зарегистрировал у Т. macha 56,8—99,0% хазмогамных цветков. Здесь, по
данным В. М. Берляид-Кожсвпикова и др. (1978), в популяциях этого
вида преобладают растения- - облшатные самоопылители. На их долю у
convar. macha приходится 60%, а у convar. densiti sen la in — 80% расте-
ний. Для факультативных перекрест инков соответственно составляла 40
и 20%.
В .зерне 7’. macha содержание белка несколько ниже, чем у спельты
(М. М. Якубцинер, II. Ф. Покровская, 1971). Но исследованиям В. Г. Ко-
188
Классификации рода Triticum
нарова и др. (1971), ряд образцов Т. madia выделяв гея высоким сидер
жапием лизина в белке даже при выращивании в условиях орошения
в Дагестане. Сравнительно хорошее сочетание белковости зерна и высо-
кого содержания в нем лизина установлено у образцов к-28175 (соответ-
ственно 14,4 и 2,84%) и к-31688 (14,2 и 2,85%).
Иммунитет. При искусственном заражении проростков у Т. madia не
найдено устойчивых к видам ржавчины форм; взрослые растения ее в
восточных районах СССР сильно поражаются бурой и стеблевой ржав-
чиной (М. М. Якубцинер, 1957, 1969; О. Г. Григорьева, 1975). У Т. madia
обнаружена группа генов Х5, обеспечивающая устойчивость к расе 202
(В. М. Берляпд-Кожевпиков и др., 1973). Полевые наблюдения в усло-
виях Дагестана выявили относительную устойчивость Т. madia к мест-
ной популяции желтой ржавчины. Инокуляция проростков, смесью рас
мучнистой росы из различных географических районов СССР, как и на-
блюдения над взрослыми растениями на провокационном фоне показа-
ли, что образцы ее в слабой пли сильной степени восприимчивы к патоге-
ну (Э. X. Сухаибердипа, 1976). Искусственное заражение коллекции
также выявило, что образцы Т. madia устойчивы пли слабо восприимчи-
вы к пыльной головне (В. И. Кривченко, 1973; А. М. Ямалеев, 1971).
И. II. Вавилов (1961) отнес Т. madia к числу видов со средней и сильной
восприимчивостью к твердой головне. Он отметил также ее устойчивость
к зеленоглазке и слабую иоражаемость гессенской мухой.
История. Согласно Л. Л. Деканрелсвичу (1954), 7'. madia — древней-
ший вид гексаплоидной пленчатой пшеницы. II. И. Вавилов (1935)
назвал этот вид «прасиельтой» и подчеркнул ее своеобразие как полуди-
кой пшеницы. По данным В. Л. Менабде (1948), обугленные зерновки
пшеницы маха были найдены среди растительных остатков конца эпохи
неолита в холме. Диди-Гудзубп (Зугдидский район Грузии).
По предположению В. Л. Менабде к виду Т. madia относятся обуг-
ленные зерновки из раскопок государства Урарту (VII—VI вв. до и. э.),
а также Т. antiquorum из свайных построек в Швейцарии.
Е. Morris и Е. Sears (1967) считают 7'. macha видом более древним,
чем пшеница мягкая. В противоположность этому некоторые авторы, на-
пример Р. Mangelsdorf (1953), а за ним В. Ра! (1966), относят возник-
новение пшеницы махи к XX в. и. э.
В грузинской литературе наименование маха встречается в словаре
Сулхан-Саба-Орбелиани (1675—1685), а в кратком латино-русско-гру-
зинском ботаническом словаре (Р. Эрнстов, 1871) приводится под назва-
нием Т. atnileum.
Географическое распространение. Вид Т. madia эндемичен для За-
падной Грузии, его ареал узколокализован. Ко времени открытия этот
вид сохранился в культуре па изолированных горных участках возле се-
лений Рача и Лечхуми на площади около 50 га (Л. Л. Декапрслевич,
В. Л. Менабде, 1932; Л. Л. Декапрслевич, 1954). Ныне не возделыва-
ется.
Происхождение. Л. Л. Декапрслевич и В. Л. Менабде (1932),
В. Ф. Дорофеев (1969) и А. Д. Горгидзе (1973) считают Т. madia пер-
вичной гексаплоидной пшеницей—живым реликтом, дожившим до
наших дней со времен первоначальной земледельческой культуры.
И. И. Вавилов (1935) не исключал возможности занесения ее в Грузию
из Северо-Восточной Турции. В. Л. Менабде (1948) связывает происхож-
дение Т. madia с древним гипотетическим видом Т. protomadia, который
сосредотачивал в себе признаки многих видов. Е. II. Сипская (1969)
соглашалась с этой точкой .зрения. В ее понимании 'Г. madia в прошлом
из места своего возникновения расселилась по Средней и Северной Ев-
ропе.
Г. В. Канделаки (1967) па основании нахождения у 7'. madia углево-
да трпфруктозаиа пришла к выводу о возникновении convar. madia с уча-
Подрод I. Triticum
189
ci нем ржи. I Io («с мнению, convar. ileiisiusciiltim произошел e учасзием
полбы колхидской. С последней появление Т. macha связывает также
М. М. Якубцинер (1961). Л. Л. Декапрелевич (1954, 1961) и А. Д. Гор-
гидзе (1970) путь становления махи ведут от гибридизации полбы кол-
хидской с Ае tauschii, т. с. они считают Т. macha первичным гексаплоид-
ным видом. А. Д. Горгидзе (1973) синтезировала форму с признаками
Т. macha, удвоив число хромосом у гибрида Т. karamyschevii X Ае. tau-
schii. Т. С. Chin, С. S. Chwang (1944) по результатам изучения мейоза в
МК.П у гибрида Т. spelta X Т. macha и морфологических признаков рас-
тений высказали предположение о возникновении Т. macha при скрещи-
вании тетранлоидпого вида (геномы АиВ) с эгилопсом.
Японские ученые (К. Tsunewaki, 1970; К. Tsunewaki, Y. Nakai, 1973)
нашли, что ген гибридного хлороза Ch] у Т. macha является продуктом
интрогрессии от тетранлоидпого вида (геномы А" 13) вследствие его гиб-
ридизации с каким-то видом. Нам кажется, что последним не может быть
Ле. tauschii, который имеет ген Ch2. Гибридизация тетранлоидпого ви-
да— носителя гопа Cht — с этим эгилопсом должна иметь летальный
исход.
К. А. Фляксбергер (1935) связывал происхождение Т. macha с гибри-
дизацией, но, ио его мнению, этот гексаплоид является вторичным ви-
дом, возникшим при скрещивании пшеницы мягкой с полбой, П. А. Ган-
диляп (1972) неоднократно наблюдал появление форм с признаками
Т. macha при расщеплении спонтанных гибридов, возникших от гибри-
дизации пшеницы мягкой с эгилопсами и пшеницей твердой. Способность
пшеницы мягкой давать мунтанты с колосом типа Т. macha установлена
также М. S. Swaminathan (1965) при воздействии мутагенами.
Мы пришли к убеждению, что нельзя исключать точку зрения
К. А. Фляксбергера о вторичном происхождении 7’. macha. Оказалось,
что ряд комбинаций от скрещивания Т. compactum с тетраплондпыми
видами (геномы АиВ) уже в F2 обильно выщепляют растения с колосья-
ми типа convar. densiusculum п convar. macha, которые очень быстро
приобретают константность. У этих форм обычно отсутствует та легкость
обламывания у основания колоса, которая присуща Т. macha и безуслов-
но приобретена ею благодаря народной селекции, связанной с уборкой
колосьев в корзины с помощью двух палочек — шпаквп.
Систематика. Пшеница маха, несмотря па узкий ареал и экологичес-
кую однородность, отличается заметным полиморфизмом. Чрезвычайно
широка амплитуда изменчивости Т. macha по плотности колоса (D = 18—
60). По этому признаку В. Л. Мепабде и Л. Л. Декапрелевич (1932) раз-
делили вид на 2 группы. В большинстве случаев рыхлоколосая и плотно-
колосая пшеница маха возделывалась в отдельных популяциях под
разными названиями: гваца-маха (рыхлоколосая) и челта-маха (плотно-
колосая). Изредка встречались в смеси (Л. Л. Декапрелевич, 1954).
В отличие от авторов вида К. А. Фляксбергер (1939) рассматривал обе
группы в качестве подвидов, которые Л. Л. Декапрелевич (1954) возвы-
сил до статуса вида, от чего позднее отказался, оставив за ними ранг
подвидов. Мы считаем, что морфологическое разнообразие обусловлива-
ет деление этого экологически и биологически однородного вида на 2
группы разновидностей, не имеющих самостоятельных ареалов, и, следо-
вательно, основания для выделения их в качестве подвидов или видов.
Кроме того, между рыхлоколосой и плотноколосой группами разновид-
ностей не наблюдается гнатуса по плотности колоса, за условную грани-
цу между ними принята плотность I) —30.
190
К.111С( ификицич /Ю<)<1 Trilillllll
Ключ для определения iруин разновидностей
Г. macha Dekapr. et Menabde
1. Колосья рыхлые или средпеплотпые (D = 18— 30), длинные (7—
14 см), безостые, короткоостпетые п остисгые, колосовой стержень весь-
ма ломкий.............................................convar. macha.
+ Колосья плотные и очень плотные (D > 30), короткие (6—10 см)
короткоостпстыс, несколько менее ломкие, чем у convar. macha . . .
...........................convar. densiuscuhnn Dekapr. et Menabde
1. Convar. macha группа рыхлоколосых разновидностей.
Convar. macha.— T. macha группа normale Dekapr. et Menabde, 1932,
Tp. прикл. бот. ген. сел. 5,1 : 20. —- 7’- macha subsp. normale (Dekapr. et
Menabde) Flaksb. 1939, Опред. паст, .хлебов: 201, comb, invalid. — T. tu-
balicum Dekapr. 1949, в Гросс г. Опред. раст. Кавк.: 718, descr. ross; De-
kapr. 1954, Тр. Ин-та полеводства АН ГССР, 8: 55, descr. ross. — Т. macha
subsp. tubalicum (Dekapr.) Dekapr. 1961, Zeitschr. Pflanzcnziicht. 45, 1: 20,
Рис. 14. Колосья, колоски и зерновки
Т. macha Dekapr. ct Menabde:
a —convar. macha; 6 — convar. densiusculum Dekapr. ct Menabde
Подрт) !. Tt Hicum
comb. invalid. /’. s/ielia subsp. macha convar. Inbalicam (Dekapr.)
V. Dorof. 1966, Генетика, 3: 30, comb, invalid.
Колосья 7—14 см длины, плоские (боковая сторона их шире лице-
вой раза в полтора), рыхлые или средней плотности (D = 18—30). Ко-
лосовой стержень весьма ломкий
Геогр. распр. Грузия.
Определение разновидностей Т. macha
convar. madia
(рис. 14а). Зерновки красные.
Var. colchicum Dekapr. et Mt-
nabde, 1932, Tp. прикл. бот. ген.
сел. 5, 1:21. (9).
Г е о г р. р а с и р. Грузия
(очень редко).
В зависимости от размеров и
строения зубца колосковой че-
Колосья безостые
Колосья полуостистые
(ости 3—4 см длины)
+ —. + — + — sublet- shcliumi 4
cum
— + + — + — submeg- relicum 5
+ — — + + — stibcol- cliicum 6
Колосья остистые
(ости 7—8 см длины)
-1- + + : + + — macha megre- licum 7 8
+ — — + + — colchi- ctuu 9
— + — + + — ibericum 10
cea WIR, 1967, leg. V. Dorofeev (WIR).
туи В. Л. Мсиабде (1948) выде-
лил ряд форм (с зубцом до 1 мм,
с тупым зубцом, с острым зуб-
цом, с зубцом 5 8 мм, а также с
иифлятпыми остями).
Var. ericzjanae Menabde, var.
nov. - T. macha var. ericzjanae
Menabde, 1948, Пикш. Груз.: Ill,
descr. ross. — A T. macha var. col-
cliict/m spica mutica differt. (3).
Typus: Georgia, Reproductio
stationis Daghestanskajae fulcra-
cea WIR, 1967, leg. Dorofeev
(WIR).
Геогр. рас п p. Грузия,
(очень редко).
Var. georgicum Menabde, var.
nov. — T. macha var. georgicum
Menabde, 1948, l.c.: Ill, descr.
ross. — A T. macha var. megreli-
cuin spica mutica differt. (2).
Typus: Georgia, Reproductio
stationis Daghestanskajae fulcra-
Геогр. распр. Грузия (очень редко).
Var. ibericum Dekapr. et Menabde, 1932, 1. c.: 23. (10).
Геогр. распр. Грузия. Описан по одному колосу.
Var. macha. — Т. macha var. letshchumicum Dekapr. et Menabde, 1932.
1. c.: 20. (7),
Геогр. распр. Грузия. Преобладающая разновидность в посевах
вида.
По строению килевого зубца колосковой чешуи и его размерам
В. Л. гМенабде (1948) выделил формы с коротким зубцом, с ту-
пым, острым, с зубцом 5—8 мм длины, с остевпдным зубцом более 8 мм
длины, а также формы с инфлятно искривленными, укороченными в ниж-
ней части колоса,остями.
Var. mergelicum Menabde, var. nov. — T. macha var. megrelicnm Me-
nabde, 1948, descr. ross. — AT. macha var. macha spica rubra differt. (8).
Typus: Georgia RSS, Reproductio stationis Daghestanskajae fulcracea
WIR, 1971, leg. A. 1'ilalenko (WIR).
Геогр. распр. Грузия (редко).
Var scharaschidzei Menabde, var. nov. T. macha var. scharaschidzei
Menabde, 1948, l.c.: 110. descr. ross. — A 7'. macha var. macha spica mtrli-
ca differt. (I).
Классификация рода 1 riticum I..
Typus: Georgia, Reproductio stationis Daghestanskajae fulcracea WIR,
1971, leg. A. Filatenko (WIR).
Геогр. рас п p. Грузия (редко).
Var. subcolchicuni Dekapr. et Menabde, 1932, 1. c.: 21. (6).
Геогр. распр. Грузия (редко).
Var. subletshchumicum Dekapr. et Menabde, 1932, 1. c.: 21. (4).
Геогр. распр. Грузия (одна из часто встречаемых разновид-
ностей).
Var. submegrelicum Dekapr. et Menabde, 1932, 1. c.: 21. (5).
Геогр. распр. Грузия (редко).
2. Convar. densiusculum Dekapr. et Menabde — группа плотноколосых
разновидностей.
Convar. densiusculum Dckapr. et Menabde, 1932, Tp. прикл. бот. геи.
сел. 5, 1: 20. — Т. macha subsp. densiusculum (Dekapr. et Menabde) Flaksb.
1939, Опред. паст, хлебов: 202, comb, invalid.—T. imeretictim Dekapr. 1949,
в Гроссг. Опред. раст. Кавк.: 718, descr. ross.; Декапр. 1954, Тр. Ин-та
полевод. АП ГССР, 8: 57, descr. ross. — Т. macha subsp. imcreticum (De-
kapr.) Dekapr. 1961, Zeitschr. Pflanzenzucht. 45, 1: 20. comb, invalid.—
T. spelta subsp. macha convar. imeretictim (Dckapr.) V. Dorof. 1966, Гене-
тика, 3: 30, comb, invalid.
Колосья 6—9 см длины, плоские (двурядная сторона шире лицевой
более чем в 2 раза), весьма плотные (D=30—57), короткоостистые
(рис. 14, б) *. Колосовой стержень менее ломкий, чем у convar. macha.
Зерновки красные.
Геогр. распр. Грузия (по ареалу вида).
Var. palaeocolchictirn Dekapr.
et Menabde, 1932, 1. c. 5, 1: 23. (3).
Геогр. распр. Грузия
(очень редко).
Var. palaeoimereticum Dekapr.
et Menabde, 1932, Tp. прикл. бот.
ген. сел. 5, 1: 23. — Т. macha var.
planocotnpressum Menabde, 1948.
Пшен. Груз.: 113, descr. ross. (1).
Геогр. распр. Грузия.
Наиболее распространенная и
весьма полиморфная разновид-
ность. По строению килевого зуб-
ца колосковой чешуи и его раз-
мерам В. Л. Мепабде (1948) вы-
деляет формы с коротким зубцом.
Оределение разновидностей Т. macha
convar. densiusculum Dekapr. et Menabde
Чешуи Ости Разновид- кость № разно- видности
ф ф крас- ные hs О з Л X Э X ф X с = © 3 цвета чешуи о X сх ф
+ — + — + — palaeoimcrc- ticum 1
— 4- + — т — rubigi- nosum 2
+ — — + + — pataeo- colchi- cum 3
— -ф- + + — rubrive- lutinum •4
тупым и острым, с зубцом около 8 мм длины, а также формы с коротки-
ми изогнутыми остями. Редко встречающиеся растения с очень плотными
(D = 45—60) короткими (3—5 см) колосьями В. Л. Менабде выделяет
как var. planocopressum Menabde.
Var. rubiginosum Menabde, var. nov. — T. macha var. rubiginosum Me-
nabde, 1948, 1. c.: 113, descr. ross. — A T. macha var. palaeoimereticum
spica rubra dijfert. (2).
Tupus: Georgia. Reproductio stationis Daghestanskajae fulcracea WIR,
1967, leg. V. Dorofeev (WIR).
Геогр. распр. Грузия (очень редко).
Var. rubrivelutinuni Menabde, var. nov. — T. macha var. rubroveluti-
nuin Menabde, 1948, I. с.: 113, descr. ross. — A T. macha var. palaeocolchi-
cum Dekapr. et Menabde spica rubra differt. (4).
В. Л. Мепабде экспериментально получил безостую разновидность махи — var.
album Menabde, descr. ross. — путем скрещивания T. macha с безостой разновидностью
пшеницы мягкой. В культуре не обнаружена.
Iloilpod I. Tritisiun
193
Typus: Georgia. Reproductio stationis Daghestanskajae fulcracca WIR,
1967. leg. M. Ananjeva (WIR).
Геогр. распр. Грузия (очень редко).
I. SPELTA L. — ПШЕНИЦА СПЕЛЬТА
Общая характеристика. Всходы опушенные, в начальных фазах зе-
леные, темно-зеленые (спельта Испании) или фиолетовые (спельта ФРГ).
Куст прямостоячий, пол устелю щи и с я или стелющийся. Соломина полая
или по стенкам с паренхимной тканью, по всегда с просветом, длиной
60—155 см. Узлы голые или слабо опушенные. Влагалище листа голое
пли бархатистоопушеппое, иногда слегка фиолетовое. Листья зеленые
или темно-зеленые, шероховатые или опушенные, длиной 16—34 см, ши-
риной 0,8—-2,3 см. Язычок и ушки всегда имеются. Ушки с ресничками,
часто переходящими на края листа. Колосья грубые, жесткие, тонкие,
в поперечном сечении чаще квадратные. Длина колоса колеблется от 6 до
20 см при толщине 0,8-—0,9 см. Число колосков в колосе 12—24. Колосья
с разной степенью рыхлости (D = 11—25). Членики стержня широкие, по
краям опушенные, клиновидные (ширина верхней части членика пре-
восходит ширину нижней приблизительно в 2 раза). Толщина членика
в верхней части почти равна ширине основания колоска, в нижней — зна-
чительно уже (иногда в 4 раза). Характерна также вогнутость членика с
той его стороны, которая прилегает к колоску. Иногда членик в верхней
части полый. Колосок часто отламывается от верхней части членика, об-
нажая па месте сочленения плоский овал, заметный па месте разлома,
как на членике, так и па основании колоска. В этом случае колосок отва-
ливается вместе с прилегающим к его брюшной стороне члеником. Такой
тип ломкости является характерным для спсльты. Вследствие утончен-
ности членика в нижней части перелом может произойти около его осно-
вания. В этом случае разлом колосового стержня бывает сходным с
распадением колоса полбы и однозернянок. Изредка членики переламы-
ваются и в средней части. У азиатских и закавказских форм спсльты ко-
лосовой стержень распадается как но типу полбы, так и по типу спсльты.
Ломкость стержня от легкой до трудной. Колоски плотные, жесткие,
удлиненно-овальные (длина 0,8—1,3 см, ширина 0,6—0,9 см); часто дли-
на колоска превосходит ширину в 2 раза и больше. Число цветков в ко-
лоске до 5 (обычно меньше), число зерен 2—3 (редко больше). Колос-
ковые чешуи овально-лопатчатые, с почти необособленным килем, широ-
ким плечом, причем главная боковая жилка образует на плече бугорок.
У азиатских форм спсльты плечо может быть .косым, отчего лопатчатость
отсутствует. Наружная цветковая чешуя приблизительно такой же шири-
ны, как колосковая, но длиннее ее (иногда па 2 мм), у безостых форм —
с остевидиым заострением, у остистых — с шероховатой остью, длиной
4—13 см. Зерновки плотно охвачены чешуями, крупные, длиной 7—10 мм,
шириной 3—3,5 мм, стекловидные, реже мучнистые; их бороздка широ-
кая. При обычной молотьбе зерновки не вымолачиваются.
Анатомический анализ колосового стержня показал, что членик за-
кавказской.спельты имеет такое же строение, как п у европейской: про-
водящие пучки концентрируются по его периферии, а внутренняя часть
заполнена рыхлой паренхимой с полостью в центре (G. Pohlcnd, 1958;
В. Ф. Дорофеев, 1966).
По данным В. Ф. Дорофеева и О. Д. Градчапииовой (1971), в услови-
ях орошения на нолях низменного Дагестана в фазе колошения у /'. spelta
subsp. spelta толщина механической ткани двух нижних междоузлий со-
ставляет 103,8 и 87 мкм; в кольце механической ткани 18 и 14 проводящих
пучков, а сопротивление на излом составляет 850—670 г. Сделан вывод,
что анатомическое строение стебля свидетельствует об относительной
13 762
194
К.итификицин /юди
Tritii'tiiii
усгойчиносiи спелыы к полеганию. Установлена также несколько мень-
шая склерификация тканей по сравнению с Т. vavilovii.
Исследования Л. Л. Жсстяшшковоп (1956) показали, что наружная
часть плодовой оболочки зерновок спелым сильно редуцирована, в ней
лишь один ряд клеток. Перикарпий выделяется слабо вкрапленной по-
ристостью, как у диких пшениц. Толщина его больше, чем у Т. dicoccum.
По образу жизни четко различаются между собой западноевропейская
и астурийская эколого-морфологические группы. В составе первой груп-
пы преобладают озимые формы, второй — яровые. Примечательно, что
для этих групп специфичны и некоторые признаки колоса. Западноевро-
пейские формы представлены в основном безостыми разновидностями, а
астурийские—всегда остистые. Для азиатской спелым характерно на-
личие как озимых, так и яровых форм.
Одним из достоинств спельты является малая требовательность к
условиям произрастания, что связано с се формированием в горных рай-
онах. Согласно Е. Ф. Пальмовой (1935), спельта ФРГ экологически до-
вольно однообразна. Приуроченная к горным п предгорным районам,
эта пшеница отличается относительно длинным вегетационным периодом,
влаголюбием, свстолюбием, малой требовательностью к агрокультуре.
В Иране спельта высевалась в условиях высокогорья (2300—2500 м над
ур. м.) на границе возделывания хлебов. Климат здесь холодный, зима
затяжная, снежная, весной дуют продолжительные холодные ветры.
Естественно, в этих условиях выращивание других пшениц крайне за-
труднительно.
По оценке И. А. Стефановского (1950) характерная черта subsp.
spelta — влаголюбие. Устойчивость к атмосферной и почвенной засухе
очень слабая. Продуктивная кустистость и облпствеипость сильные. За-
падноевропейские формы отличаются замедленным темпом развития па
ранних его фазах.
Склонность спельты к спонтанной гибридизации отметил еще F. Kor-
nicke (1885). В условиях Московской области у нее 65—80% цветков цве-
тут открыто; наблюдается 2—3 максимума цветения в продолжение су-
ток (АЕ А. Федин, 1974). В Молдавии у европейской спельты отмечено
62,7—67,5% хазмогамных и 7,3- -8,3% клейкостогамных цветков
(В. Р. Челак, 1969). В Белоруссии в дополуденные и полуденные часы по-
вышается количество открытоцветущих цветков (А. 3. Латыпов.
В. И. Апель, 1971). В условиях низменного Дагестана закавказская фор-
ма спельты склонна к закрытому цветению (В. И. Жуков, 1969). Ио
данным В. М. Берлянд-Кожевникова и др. (1978), здесь в популяции
спельты ФРГ 20% растений были облигатными самоопылителями, 60% —
факультативными перекрестниками и 20%—склонными к перекрестно-
му опылению; у спельты из Астурии выявлено соответственно 60, 30 и
10% таких растений. Облигатные перекрестпики не обнаружены.
Концентрация белка в зерне спельты при репродуцировании на оро-
шаемых участках в условиях прибрежной полосы Дагестана достигала
15,6—18,8%, в Ташкентской области—14,8—18,3% (АЕ АЕ Якубцинер.
Н. Ф. Покровская, 1971). По данным В. Г. Копарева и др. (1972), в белке
зерна спельты -в этих условиях содержание лизина доходит до 2,68%.
Иммунитет,. И. И. Вавилов (1961, 1964) считал спельту Испании устой-
чивой, а спельту ФРГ — восприимчивой к бурой ржавчине. В целом при
искусственном заражении и в нолевых условиях Дагестана, Приморского
края и Ленинградской области образцы коллекции спельты оказались
средней сильно восприимчивыми к бурой ржавчине (И. П. Шитова, 1968;
А. АЕ Камелииа, 1973; О. Г. Григорьева, 1975). Данные 11. П. Вавилова
(1961, 1964) и АЕ М. Якубцииера (1957) об устойчивости спельты к жел-
той ржавчине подтверждены наблюдениями в условиях Дагестана, где за
1954—1965 гг. высшая оценка поражения в среднем оказалась равной
1,1 балла по 5-балльной шкале (И. П. Шитова, 1968). Спельта воспри-
UihI/xx) I. Trilirtitn l‘)5
имчива к стеблевой ржавчине (11. 11. Вавилов, 196 1; Л. N\. Камелнпа,
1973).
Еще G. М. Recd (191(1) отметил ее неустойчивость к мучнистой росе.
Искусственное заражение проростков сборным ипокулюмом из смеси по-
пуляций этого гриба, собранных в различных географических регионах
СССР, и полевые наблюдения на провокационном фоне показали, что
взрослые растения у 79,6% образцов коллекции восприимчивы к пато-
гену; иммунные образны етсугсгвошл-щ (?. X. (Л хш'.бср шчл. 1976).
Спе/.з:то.те >с‘. to;;'?.: а ныны:?;; f. ц чем к L d;n ;
(В. И. Кривченко, 1973). Указание Н. II. Вавилова (1961) об устойчивос-
ти спсльты к пыльной головне подтвердил опыт Л. М. Ямалеева (1974)
по искусственному заражению: в коллекции выявлено 78% иммунных
образцов. Причем электрофорез глиадииа зерновок показал, что спектры
этих образцов имеют 0—6 компонентов в (о-зоне, а спектры восприимчи-
вых образцов — до 8 компонентов. У спсльты пет восприимчивых к твер-
дой и стеблевой головне образцов (И. II. Вавилов, 1913, 1961; В. 11. Крив-
ченко, 1973). В опыте И. Harding (1972) спельта сильно поражалась кор-
невой гнилью.
История. До недавнего времени iосподсгвовала концепция, согласно
которой древность спсльты связана с эпохой бронзы (начало II тысяче-
летия до и. э.). Именно в слоях этой эпохи на территории Западной Евро-
пы— в Голландии, Дании — были найдены ее остатки (II. Helbaek, 1952).
К тому же времени относятся обугленные остатки из свайной постройки
Сумпф (К. Bertsch, F. Bertsch, 1949). Новейшие же данные вносят серь-
езные коррективы по этому вопросу. Так, в Польше (верховья Вислы)
М. Klichowska (1959), К. Wasylikowa (1956), W. Gizbcrt (1965) обнару-
жили следы культуры этого вида, относящиеся к раннему неолиту:
Неолитическим временем датируются, по сведениям 3. В. Янушевнч
(1976), отпечатки колосков спсльты из поселений в Молдавии.
В археологических материалах из Передней Азин спельты не обна-
ружено, однако И. Helbaek (1966) не отрицает возможности существова-
ния ее здесь в прошлом. Впервые предположение о передиеазиатском
происхождении спсльты высказал И. И. Вавилов (1926). Позднее экспе-
диционными обследованиями выявлены чистые посевы спельты в Иране
(И. Kuckuck, 1957, 1959) и Средней Азии (Р. А. Удачин, 1969), обнару-
жено большое разнообразие спельты в Закавказье (И. Д. Мустафаев,
1961, 1964; В. Ф. Дорофеев, 1966, 1970). В свете этих исследований под-
тверждается азиатское происхождение единичных колосьев спельты, в
прошлом найденных в Иране (A. Michaux, 1783, G. Olivier, 1867,
П. М. Жуковский, 1927), Турции (М. Hacizade, 1932), Средней Азии (F.
Baziner, 1848), Армении (А1. Г. Туманян, 1929, 1933). К. А. Фляксбер-
гер (1935) полагал, что спельта в эти регионы была занесена из других
областей земного шара или же явилась результатом неправильного бо-
танического определения. Одним из доказательств впеазиатского проис-
хождения спельты он считал отсутствие у древних народов Азии специ-
ального слова для обозначения этой пшеницы. Однако в Средней Азии
местное население долины р. Исфара (Таджикистан) издавна культиви-
рует спельту, называя ее «гапдуми обджувозп», т. е. пшеница, обмолачи-
ваемая па водяных крупорушках, в то время как для пшеницы мягкой
существует название «гапдуми галлоговн», т. е. пшеница, обмолачивае-
мая прогоном быков (Р. А. Удачин, 1971).
Географическое распространение. В начале XX в. посевы спельты бы-
ли довольно распространены в горных районах некоторых стран Цент-
ральной Европы (на юго-западе ФРГ, в Швейцарии, Австрии, реже во
Франции и Италии). Особая группа спельты сохранилась в северной
Испании (Астурия). В небольшом количестве спельта встречалась в
Югославии, Бельгии. На острове Готланд (Швеция) спельта возделы-
валась в смеси с другими пленчатыми видами пшеницы еще в 1965 г., а
196 J\ .тпифитщии poTla Trilicum /,.
до 1924 г. здесь is иекоюрых приходах наблюдались се чистые посевы
(II. Hjelmqvist, 1966).
Впервые о произрас i а п пи спельты в .Азии сообщил Л. Miehaiix (ио
,1. Lamarck, 1784), обнаруживший несколько ее растений в горах Хама-
дапа (Иран). К Ирану относятся и другие находки (G. Olivier, 1867), в
том числе типичные колосья спельты, обнаруженные II. М. Жуковским
(1927). В Иране обнаружены чистые посевы спельты в окрестностях
г. Шехре-Корд (2300- 2500 м над ур. м.), в сущности, па границе возде-
лывания хлебов (И. Kuckuck, 1957, 1959).
Несколько колосьев спельты обнаружены в пшеничных посевах в Тур-
ции (М. Hacizade, 1932).
Спельта встречается и в республиках Средней Азии. О нахождении
спельты в Хивинском оазисе (Узбекистан) сообщал F. Basiner (1848).
Однако Р. А. Удачин (1975) считает, что этот нсследовагель сделал лишь
предположение о возделывании здесь спельты, а сам лично не собрал
ее колосьев. Позднее спельта была найдена в посевах пшеницы мягкой
в Самаркандской области Узбекистана С. 11. Кудряшовым (1912) и в
Ташаузской области Туркмении В. II. гМоргенсепом (1916). Отдельные
колосья спельты были собраны Р. Л. Удачиным (1969, 1971, 1975) в Тад-
жикистане (Шаартузский район), Туркмении (Ташаузская область), Уз-
бекистане (Хорезмская и Бухарская области). Впервые на территории
СССР Р. Л. Удачин (1971) нашел чистые посевы спельты в Исфарипском
районе Ленинабадской области Таджикистана.
Большое разнообразие спельты в Азербайджане обнаружили
И. Д. Мустафаев (1961, 1964) и В. Ф. Дорофеев (1966, 1970). О нахожде-
нии спельты в Армении сообщил П. Л. Гапдилян (1967), в Грузии —
И. М. Сакварелидзе и К. II. Жижилашвпли (1974).
L. Ducellier (1923, 1929) указывает па возделывание спельты в оази-
сах северной Сахары. Он приводит описание разновидности спельты
var. saharae Due., особенностью которой является инфлятность колоса.
Следует отметить, что К. А. Фляксбергер (1935) считал эту разновид-
ность ипфлятпой пшеницей мягкой, хотя var. saharae имеет типичные
черты спельты.
Происхождение. Гипотеза первичного происхождения спельты (по
сравнению с пшеницей мягкой выдвинута Д. 1\. Ларионовым (К. А. Фляк-
сбергер, 1911), A. Schulz (1913) и Е. Tschcrmak (1914). Первые два ис-
следователя предполагали, что спельта произошла от неизвестного ди-
кого гексаплоидпого предка. J. Percival (1921) высказал догадку о ее
гибридном происхождении с участием тетраплоидпой пшеницы и Ае. су-
lindrica. В 1944 г. II. Kihara в Японии, Е. S. McFadden и Е. R. Sears в
США, введя Ле. tauschii в скрещивания с тетраплоидпой пшеницей, син-
тезировали спельту. Этот эксперимент, стал сильным аргументом в поль-
зу гипотезы о первичном происхождении спельты. После этого па пози-
цию признания первичности спельты—вторимпости пшеницы мягкой
встали L. Sachs (1953), Е. Schiemann (1958), Н. Kuckuck (1959, 1971),
Л. Л. Декапрелевич (1961), М. Muramatsu (1963), А. С. Andrews (1964),
J. Mac Key (1969) и др.
Следует упомянуть, что существует гипотеза происхождения спельты
из Т. macha. Ее в 1932 г. высказали Л. Л. Декапрелевич и В. Л. Мепабде.
Возможность такого пути становления спельты обсуждал К. А. Флякс-
бергер (1935), отрицали М. М. Якубцинер (1961) и В. Е. Писарев (1964),
признал М. Tanaka (1965).
F. Bertsch (1939) полагал, чю спельта альпийских гор произошла в
неолите при скрещивании Т. inonococcuin с Г. dicocciini. Е. Schiemann
и G. Standi (1958) синтезировали такой амфидиплопд, но он совершенно
не был похож па спельту. Наблюдения над появлением сиельтоидиых му-
тантов из пшеницы мягкой вызвали к жизни серию исследований, которые
стали основой гипотезы о вторичном происхождении спельты. Н. Nilsson-
llodpot) J. Trilicum
197
Ellie (1917) и M. I'Г\'м ;i 11я и (1932) наблio/iали появление снель-соидов в
чистых линиях пшеницы мшкой. Подобное явление, изучили A. Akerman
(1923), Е. Lindhard (1923), V. LathoHwcrs (1924), О. Winge (1924),
G. Nilsson-Leissner (1925), IO. Л. Филипченко (1927), Л. IL Делоне
(1934) и др.
E. Tscherinak (1914), J. Percival (1921) и многие другие исследовате-
ли зарегистрировали факты выщенлепия спельты при межвидовой гиб-
ридизации, которые привели II. И. Вавилова (1918, 1926) к убеждению
о вторичном гибридном происхождении спельты. Ее выщеплепис наблю-
дали при скрещивании пшеницы мягкой с Т. dicoccum (С. Е. Leighty,
S. Boshnakian, 1921; Е. Malinowsky, 1926; В. Kajanus, 1923 и др.), с Т. tur-
gidum (И. Vilmorin, 1880; С. Е. Leighty, S. Boshnakian. 1921; I. Райхати.
1954 и др.), с Т. durum (М. J. Vilmorin, 1883; А. Ф. Шулындпп, 1955 и
др.); при скрещивании Т. compactum с Т. dicoccum (Р. Mathis, 1925),
Т. sphaerococcum с 7’. dicoccoides, Т. dicoccum, Т. karamijschewii, Т. ispa-
hanicum (14. III. Шахмедов, 1972). О. Tscherinak (1914) отметил, что уже
со второго поколения некоторые вытопившиеся формы спельты были
константными.
Как упоминалось, при обследовании производственных посевов пше-
ницы мягкой примесь спельты была обнаружена II. Д. Мустафаевым,
В. Ф. Дорофеевым, Р. /А. Удачиным, II. А. Гапдиляном, 11. М. Сакваре-
лидзе и К. II. Жпжилашвпли. Все эти исследователи убеждены, что при-
месь спельты па полях Закавказья и Средней Азии является результатом
спонтанной гибридизации тетраплоидпой пшеницы секции Dicoccoides
с пшеницей мягкой, что это — спельта вторичного происхождения. При-
чем В. Ф. Дорофеев (1972) склонен признать, что в природе также су-
ществовала первичная спельта, которая произошла от Г. macha. Первич-
ная спельта, приобретя доминантность фактора Q, дала начало пшенице
мягкой, которая при спонтанной гибридизации с тетраплоидными видами
служит базисом возникновения вторичной спельты.
В ином понимании гипотеза вторичного происхождения спельты вос-
принималась A. De Candolle (1883), II. И. Вавиловым (1918, 1926),
Е. Hartmann (1923), Е. Schroder (1931), А. Р. Жебраком (1957),
Е. Н. Синской (1955, 1969), П. М. Жуковским (1971), которые считали,
что спельта исторически появилась позднее голозерной гексаплоидпой
пшеницы.
Гипотеза о вторичном происхождении западноевропейской спельты,
с указанием пути ее возникновения, выдвинута К. А. Фляксбергером
(1930) и Е. Schiemann (1932). Они полагали, что эта спельта возникла
в бронзовом веке при гибридизации древней голозерной гексаплоидпой
пшеницы Т. antiquorum Heer (сходна с современным видом Т. compac-
tum) с Т. dicoccum. Одним из доказательств правильности такого пред-
положения К. А. Фляксбергер (1935) считал наличие голозерной гекса-
плоидпой пшеницы и отсутствие спельты в археологических слоях, отно-
сящихся к каменному веку (неолит). О том же свидетельствует исследо-
вание Н. Helbaek (1964, 1965): оказалось, что нахождение спельты при
раскопках относится ко II тысячелетию до и. э., а находки пшеницы
мягкой па 3—4 тыс. лет старше.
Многочисленные работы по воздействию мутагенами на пшеницу
мягкую (М. V. Р. Rao, М. S. Swaminathan, 1963- К. Djclepov, G. Chav-
darov, 1967; J. V. Good, 1967; M. S. Swaminathan, 1965; P. B. Pal, 1965
и др.) также свидетельствуют о существовании у псе функциональной
способности, как выразился К. А. Фляксбергер (1935), «рождать» снель-
ту. М. V. Р. Rao п М. S. Swaminathan (1963) обратили внимание па то,
что в их обширном опыте облучение спельты не привело к появлению
пшеницы мягкой. А. В. Пухальский и II. Л. Максимов (1967) после гам-
ма-облучения спельты зарегистрировали выщеилепис пшеницы мягкой,
которое они связывают с продолжительным открытым цветением облу-
1 w
Классификации /юда Triticum I..
ченпой спельты, приведшим к нереопi>i.ichпк> с пшеницей мягкой, a lie
с переходом q в Q.
J. Mac Key (1951, 1968) считает, что доминантный фактор Q подавля-
ет признаки спельты, имеющей рецессивную аллель q. Снельтоидпыс му-
тации у пшеницы мягкой могут возникнуть при потере Q, расположенного
на длинном плече хромосомы 5А. М. Muramat.su (1963) показал, что 5 доз
гена q по эффекту действия равны 2Q. Потеря одной хромосомы 5А отра-
жается па фенотипе пшеницы мягкой: моиосомикп по этой хромосоме
становятся сиельтоидами (В. С. Josi, S. Kumar, 1967). II. Kuckuck (1964)
также считает, что степень эффективности сложного локуса Q зависит
от уровня дупликации q. С. К. Bhatia и М. S, Swamiuatlian (1962) связы-
вают разную степень спельтоидности с мутированием отдельных участ-
ков сложного гена (цистропа) Q. По мысли II. С. Мелпя (1972) появле-
ние. спельты, как примеси в песетах пшеницы мягкой, не мутация, а ре-
зультат ослабления силы действия Q пли <], так как эта пшеница несет
обе эти аллели.
По результатам 20-летней работы с межвидовыми гибридами пшени-
цы мы пришли к убеждению, что вся спельта Европы п Азии является
продуктом спонтанной гибридизации пшеницы мягкой или 7'. compactum
с тетраплоидными видами секции Dicoccoides. Спелый но происхожде-
нию— вид вторичный, имеющий полифилстпчсское происхождение.
Спельты Астурии, Алььпийских гор, Ирана, Закавказья, Таджикистана и
Туркмении возникли независимо друг от друга.
По данным А. Г. Хакимовой (1973), «-зона электрофоретических спек-
тров глиадинов испанской спельты идентична компонентам a-зоны испан-
ской полбы, что, возможно, свидетельствует о происхождении первой при
участии второй. О полифилетическом происхождении спельты свидетель-
ствует географическая локализация генов восстановления фертильности,
а также генов гибридных некроза и хлороза. Насыщенность ресурсов
спельты сортами — носителями генов Rf, восстанавливающими фертиль-
ность у стерильных аналогов с цитоплазмой пшеницы Тимофеева, в раз-
личных географических регионах различна: они установлены у 7,2%
сортов Испании, у 81,6—92,3% сортов Центральной Европы, у 1,7% об-
разцов Азербайджана; в Иране они пе обнаружены (А. И. Бороданепко,
1974; В. Ф. Дорофеев, А. И. Бороданепко, 1975). Спельты Испании име-
ют только ген Ch2, спельты Центральной Европы — только ген Neb спель-
ты Ирана и Азербайджана — гены Net и Ch2 (К. Tsunewaki, 1969, 1970;
Л. Л. Декапрелевич, Г. Г. Яшагашвили, 1970).
Систематика. До недавнего времени считалось, что полиморфизм
спельты невелик и ограничен двумя эколого-морфологическими группа-
ми: иберийской и баварской.
Находки спельты в Иране, Закавказье и Средней Азии значительно
обогатили ботанический состав вида Т. spelta. Азиатская спельта по ря-
ду признаков существенно отличается от европейской, что дало основа-
ние М. Gokgol (1961) выделить ее как subsp. ktickuckianum Gokg.
Ключ для определения подвидов Т. spelta L.
1. Колосья рыхлые (D=13—22), ломкие, спонтанно пли при легком
нажатии распадаются на колоски, при этом членик колосового стержня
остается прикрепленным к основанию колоска нижней своей частью
(ломкость типа спельты); колосковые чешуи овально-лопатчатые, с ши-
роким плечом, прямым пли приподнятым; в колоске 2, редко 3 зерновки,
плотно заключенные в чешуях, вымолот зерновок трудный...............
.......................................................subsp. spelta.
1 + Колосья рыхлые или более плотные (D= 11—25), менее ломкие,
чем у subsp. spelta, распадаются на колоски при нажатии, реже спонтан-
но, главным образом в верхней части колоса, при этом членик стержня
/Синсифпкиции pot)u 'ГгШаин Л. 199
<>c I .'id ся прикрои,iciiiii.im к основанию колоски uepxiiHi своей частью
(ломкосгь I ппа полбы), реже нижней (ломкость сне.илы); колос-
ковые чешуи с довольно узким косым плечом, характерным для пшеницы
мягкой, реже овально-лопатчатые с широким плечом; в колоске 2—3
зерновки; степень плотности заключения зерновок в чешуях различная;
вымолот зерновок от средне- до очень трудного .......................
.........................................subsp. kuckuckianuni Gokg.
I. Subsp. spelta — подвид европейской спельты.
Subsp. spelta. — 7'. spelta L. 1753, Sp. Pi.: 86. — T. zea Host, 1805,
Gram. Austr. 3: 20. — Spelta vulgaris Scringe, 1841, C6r. Europ.: 76
(114). — T. vulgare var.-gr. spelta Alef. 1866, Landw. Fl.: 334. — T. sati-
vum spelta Hackel, 1887, Nat. Pflanzenfam. 2: 81. — T. aestivum subsp.
spelta (L.) Thell. 1918, Naturw. Wochenschr. N. F. 17, 33; 471. — T. aesti-
vum L. group spelta Bowden, 1959, Canad. Journ. Bot. 37: 674.— T. spel-
ta subsp. europeum Dorof. 1966, Генетика, 3: 30, nom. illeg.
Растения 60—120 см высоты; лист с развитым язычком у основания
пластинки; колосья (рис. 15, а) длинные (10—20 см), очень рыхлые или
рыхлые (D—13—22), ломкие, спонтанно или при легком нажатии распа-
даются па колоски, при этом членик стержня остается прикрепленным к
основанию колоска нижней своей частью; колосковые чешуи овально-ло-
патчатые, с широким плечом, прямым или приподнятым; в колоске 2, ре-
же 3 зерновки, плотно заключенные в чешуях, вследствие чего вымолот
их трудный; зерновки в поперечном разрезе округло-треугольные, с ши-
рокой бороздкой, большей частью стекловидные, реже — мучнистые.
Геогр. распр. Горные районы Центральной Европы — ФРГ (Ба-
вария), Швейцария, Австрия, Франция, Испания, Италия, Швеция.
Описана по культивируемому экземпляру европейского проис-
хождения.
Европейская спельта подразделяется па 2 эколого-географические
группы: иберийскую и баварскую.
1. Иберийская спельта (Т. spelta prol. ibericum Flaksb. 1935, Культ,
фл. СССР; 1: 135; Т. spelta prol. asturicum Vav. 1964, Миров, ресурсы
хлебп. злаков. Пшеница: 60). Первый лист растения светло-зеленый, опу-
шенный. Растения высокорослые, хорошо облиственные, соломина проч-
ная, колос крупный. В целом группа характеризуется устойчивостью к
бурой и желтой ржавчине, известны исключительно яровые формы.
Геогр. распр. Испания (Астурия).
2. Баварская спельта (Т. spelta prol. allemanum Flaksb. 1935, l.c.:
135; T. spelta prol. bavaricum Vav. 1964, 1. c.: 60). Первый лист растения
темно-зеленый, часто с антоцианом, опушенный. Взрослые растения сред-
не- и высокорослые, с хорошей облиствеппостыо, колос их несколько
мельче, чем у иберийской спельты.
В целом группа характеризуется восприимчивостью к грибным забо-
леваниям, нетребовательностью к почвенным условиям, позднеспелостью
в сравнении с иберийской группой. Преобладают озимые формы.
Геогр. распр. Бавария (ФРГ), Швейцария, откуда спельта рас-
пространилась в другие страны Европы (Австрия, Бельгия, Швеция,
Франция, Италия), где возделывалась в незначительных размерах.
Var. albivelu(inum Koern. 1885, in Koern. et Wern. Handb. Getreideb.
1: 80. — T. tnacrodon Harz, 1885, Landwirt. Samenk.: 1214. (12).
Обнаружена F. Kornicke (1885) в Ботаническом саду в Поппельсдор-
фе па посеве спельты. Описана по экземпляру со слаболомким колосом,
как он предполагал, гибридного происхождения (от скрещивания сиель-
ты с. пшеницей мягкой). Редко как примесь встречается в Испании (Ас-
турия), ФРГ (Бавария), Швейцарии.
С. О. Harz (1885) описывает для южной Германии (ФРГ) под назва-
нием-7'. macrodon Harz форму с острым длинным зубцом (до 3 мм) на
колосковых чешуях — признаком, редким для спельты.
200
Классификации рода Trilicuui I..
Рис. 1,г>. Колосья, колоски И зерновки Т. spelta L.:
а-- subsp. spelta и i Ilcu.iiuiii; 6 subsp. kttcktickianiim G6kt{. m Азербайджана
Подрод I. Trilicum
201
Определение разновидностей subsp. spelta
Чешуи Ости РаДнони зное и. Л? ра »'<(- вилнос1 и
белые красные серо дымчатые или черные* пне i а чешуй черные
К о л о C b Я б С 3 о с т ы с
Чешуи неопушенные
3 с р п о в к и к р а с н ы с
+ + а- — — album d uhameliaiui in ainissum 1 2
+ — Зерновки зеле и ы е — I — ’ viridalbispiсаtши 1
+ Чешуи- энушенныс (зерновк it красные) recens
— + — — — neglectum (>
— — + — — alefeldii
+ \ О Л О С Ь 1 Чешуи н е р и о в к + о с т и с еопушенш и к р а с ы e te i i.t e ardtiini 8
— — + — vulpinum 9
— — + — schenkii 10
+ е р н о в к + и з с л о и ы е — | viridarduini 11
+ Чешуи с пушенные. (зерновк t красные) albivelutin um 12
— + — + — rubrivelutitium 13
— — + — i + caeruleum 14
* Черная окраска чешуй проявляется в зависимости от условий по-разному: от серо-дымчатых
(красный фон) или даже красных чешуй в одних условиях до черных в других, т. е. различия фе-
нотипические, и выделять отдельные разновидности по этому признаку, как считал К. Л. Флякс-
бергер (1935), не следует. Такой резкий переход серо-дымчатой окраски в черную у азиатского под-
вида спельты, как и у других видов пшеницы, наблюдается реже.
Var. album (Alef.) Koern. 1885, in Koern. et Wern. l.c.: 79; Perciv.
1921, Wheat pl. 333. — T. vulgare var.-gr. spelia album Alef. 1866, l.c.:
335. — T. vulgare var.-gr. spelta fringillarum Alef. 1866, 1. c.: 335 — T. al-
burneum Harz, 1885, l.c.: 1211. — T. ovatum Harz, 1885, l.c.: 1212.—
T. spelta var. albis plcat urn Flaksb. 1915, Опред. пшеп.: 120, nom. inval. (1).
Геогр. распр. Швейцария, южные районы ФРГ.
В начале XX в. в Баварском институте селекции растений (Weihens-
tephan) выведен ряд сортов с плотным колосом, которые были затем
завезены в Бельгию, Данию, США, Канаду.
В Испании var. album обнаружена как редкая примесь в посевах на
высоте 1100 м над ур. м. В коллекции ВИРа имеется образец этой разно-
видности из Франции под названием Epeautre ordinaire blanc Sans barlcs.
Var. alefeldii Koern. 1873, System. Uebersiclit Cer.: 13; idem. 1885, in
Koern. et Wern. l.c.: 80. — T. fuscocoeruleum Harz, 1885, l.c.: 1212.—
T. pseudoduhatneUanutn Harz, 1885, 1. c.: 1213. (7).
Известна по коллекциям ботанических садов и НИИ.
Var. ainissum Koern. 1885, in Koern. et Wern. 1. c.: 79.--7'. vulgare
var.-gr. spelia dtihameliaiiiim auct. non Mazz.: Alef. 1866, l.c.: 335. (3).
Известна только по описаниям. Ф. Кёрпике указывал на возделыва-
ние ее в Швейцарии до начала XIX в.
Var. arduini (Mazz.) Koern. 1885, in Koern. et Wern. 1. c.: 80. — T. ar-
?02
Классификации рода Tri/icuni
littini Mazz. 1807, Sopra ale. sp. friiin. Meiuor. botanicogeogr. 4: 50.--
I. vulgare var.-gr. urduini (Mazz.) Alef. 1866, 1. c.: 334. (8).
Геогр. рас. up. Возделывалась как редкая примесь к другим раз-
новидностям в ФРГ, Франции, I-Iiалии, Испании, Швейцарии. В коллек-
ции ВИРа имеются образцы из Польши и СПОР (Западная Украина,
Латвия).
Var. caeruleum (Alef.) Koern. 1885, in Koern. et Wern. 1. c.: 80. — T. vul-
ture var.-gr. spelta caeruleum Alef. 1866, 1. c.: 335. — 7’. caerulescens Harz,
1885, 1. c.: 1214. — T. caeruleum Harz, 1885, 1. c.: 1214. (14).
Ге or]>. распр. Испания, ФРГ, Франция.
Var. duhamelianiiin (Mazz.) Koern. 1885, in Koern. el Wern. l.c.: 79.—
T. duhamelianum Mazz. 1807, l.c.: 55. — T. vulgare var.-gr. spelta rufum
Alef. 1866, 1. c.: 335. — T. rufescens Harz, 1885, l.c.: 1213. (2).
Геогр. распр. ФРГ, Швейцария, Франция (горы Вогезы и Юра),
Австрия, Югославия, Бельгия.
Var. neglectuni Koern. 1885, in Koern. et Wern. l.c.: 80. — T. hirlum
Harz, 1885, 1. c.: 1212. (6).
Известна только по коллекциям ботанических садов и НИИ.
Var. recens Koern. 1885, in Koern. et Wern. 1. c.: 80. (5).
Известна no коллекциям ботанических садов. Описана Ф. Кёрнике по
растениям, как он полагал, гибридного происхождения.
Var. rubrivelutinum Koern. 1873, l.c.: 14; Koern. 1885, in Koern. et
Wern. l.c.: 80. — T. spelta var. erubescens Koern. 1873, l.c.: 14. (13).
Описана по растениям такого же происхождения, что и var. recens.
Геогр. распр. Н. И. Вавилов собрал в Астурии (Испания), где
она часто встречалась как примесь; редко — на юге ФРГ.
Var. schenkii Koern. 1885, in Koern. et Wern. l.c.; 80. — T. spelta nig-
rescens Schuebl. in Krause, 1836, Abbild. Beschr. Getreide, 4: 9. — T. glau-
uitn Harz, 1885, 1. c.: 1214. (10).
Геогр. распр. Известна по описаниям и в коллекциях. Обнаруже-
на как очень редкая примесь к другим разновидностям спельты в сборах
Н. И. Вавилова в Испании, произведенных в 1927 г.
К- А. Фляксбергер (1935, 1939) по описанию и рисунку J. Krause
(1935) выделил у этой разновидности ветвнстоколосую форму rainoso-
schenkii Flaksb., каковую обнаружить в посевах европейской спельты
больше не удавалось.
Var. viridalbispicatum Jakubz. 1971, Puchalski, Tp. прикл. бот. ген.
сел. 44, Г. 55. (4).
Выявлена в образце, присланном в ВИР из сельскохозяйственной
школы г. Брно (Чехословакия). Кроме того, экспериментально получе-
на А. В. Пухальским и И. Л. Максимовым под воздействием мутагена.
Var. viridarduini Jakubz. et Puchalski, 1971, in Puchalski I. c.: 55. (11).
Получена А, В. Пухальским и И. Л. Максимовым на Московском
отделении ВИРа экспериментально под воздействием мутагена.
Var. vulpinum (Alef.) Koern. 1885, in Koern. et Wern. l.c.: 80. — T. vul-
gare var.-gr. spelta vulpinum Alef. 1866, 1. c.: 335. (9).
Первые авторы разновидности описали ее по материалам из ботани-
ческих садов. Сборы Н. И. Вавилова указывают па широкое возделыва-
ние ее в Испании в 1927 и 1928 гг. В коллекции ВИРа имеется образец
var. vulpinum из Львовской области (Украина).
II. Subsp. kuckuckianum Gokg. — подвид азиатской спельты.
Subsp. kuckuckianum Gokg. subsp. nov. — T. spelta subsp. kuckuckia-
num Gokg. 1961, Zcilschr. Pflanzenzucht. 45, 3/4: 320, descr. germ, sine ty-
po.— Plantae 90—150 cm alt.; spicae 6—20 cm 1g., laxae vel densae (D =
— 11—25); spicarum rachis fragilis, arliculi rachidis basin spicularum
apicifixa (ad instar. T. dicoccum), rarius basifixa (ad instar T. spelta)-,
spiculae bi-triflora fertilis; caryopsis glumac arete vel arctiusculus inclusus.
Typus: T. spelta var. subbaktiaricum Dorof.
Ihxlji'x) J. Tnlitiini 2.03
Растения 90 — 155 см высоты; лист с лигулой (язычок у основания
HHICTUUKH развит) н.ш без лигулы |язы>ч>к чсысспеО; ко.’ \".я 1рис
15. б) ср-..-;.ИИ ЗЛИИЫ И л СИ'о. 2- ''). р - ;1 ;; у, ; ;( I) --
11 25), менее ломкие, чем у subsp. spelta, раснадаюн я на колоски при
нажатии главным образом в верхней части колоса; при этом членик
стержня остается прикрепленным к основанию колоска верхней своей
частью (ломкость по типу полбы), реже нижней (ломкость по тину спель-
ты); колосковые чешуи овально-лопатчатые, с прямым или косым пле-
чом (последнее характерно для пшеницы мягкой); колоски 4—5-цветко-
вые; в колоске 2—3 зерновки; степень плотности заключения зерновок is
чешуях различная — вымолот их от средпелегкого до очень трудного.
Зерновки от круглых до сильно удлиненных, с широкой бороздкой, стек-
ловидные.
Геогр. распр. Иран (в окрестностях г. Шехре-Корд па высоте
2300—2500 м над ур. м.). Как сообщал И. Kuckuck (1968), открывший
чистые посевы спельты в Иране, в настоящее время опа полностью
заменена сортами пшеницы мягкой.
В Закавказье спельта обнаружена как редкая примесь, иногда в ви-
де единичных растений, в популяциях шпеипны со сложным ботаничес-
ким составом (преобладает, как правило, Т. aestiviim) в предгорных рай-
онах Азербайджана, реже в Армении и Грузии.
Как примесь в посевах пшеницы мягкой спельта обнаружена в рес-
публиках Средней Азии. В долине р. Исфара на высоте 1000—1500 м
над ур. м. спельта высевалась в чистой культуре (Р. А. Удачин, 1971).
Впервые азиатскую спельту (Г. vulgare var. michauxi Alef. 1866,
Landwirt. Fl.: 334, descr. incompl.) F. Alefcld описал по растениям, соб-
ранным A. Michaux в горах Хамадана (Иран), и рассматривал ее как
единственную дикую форму, родопачальиую для всех культурных разно-
видностей. Подлинный экземпляр A. Michaux К- А. Фляксбергеру (1915)
разыскать не удалось.
В европейских гербариях среди обширных сборов пшеницы Ирана
спельта также не обнаружена (Н. Kuckuck, Е. Schiemann, 1957).
Безлигульпые формы пшеницы мягкой, овса, ржи и кукурузы были
обнаружены на Памире (Н. И. Вавилов, 1922/1923). Нахождение здесь
безлпгульной спельты — еще одно доказательство положений закона
гомологических рядов в наследственной изменчивости, выдвинутого
Н. И. Вавиловым. Безлигульная пшеница мягкая и Т. compactum с Па-
мира, а также безлигульная пшеница твердая с Кипра составляют у
этих видов особые географические группы. Параллелизм изменчивости
видов пшеницы дал основание для деления азиатского подвида спельты
на 2 группы разновидностей: convar. kuckuckianum Gokg. и convar. iara-
kanovii A. Filat, et Dorof.
Ключ для определения групп разновидностей
subsp. kuckuckianum Gokg.
1. Лист с лигулой (язычок и ушки у основания пластинки листа раз-
виты) ........................convar. kuckuckianum A. Filat, et Dorof.
+ Лист без лигулы (язычок и ушки у основания пластинки листа от-
сутствуют) .<•....................convar. tarakanovii A. Filat, et Dorof.
1. Convar. kuckuckianum A. Filat, et Dorof. — группа разновидностей
собственно азиатской спсльты.
Convar. kuckuckianum A. Filat, et Dorof. convar. nov. - - T. spelta con-
var. transcaucasicum Dorof. 1966, С.-х. бпол. 1,1: 137, nom. illeg. — T. spel-
ta convar. iranicum Mandy, 1970, Acta Agron, 19, 3/4: 416, nom. illeg.—
Folium ligulatum, patents.
Typus: T. spelta var. subbakliaricum Dorof.
Описание convar. kuckuckianum, куда относятся разновидности с раз-
витыми язычком и ушками, соответствует приводимому для подвида.
201
Классификация рода Trilicum I..
Определение разновидностей convar. kitckuckianuiii Л. Eilat, et Dorof.
Чешуи Ости Разновидное i ь № ра то- видное гн
белые крас- ные серо-дымчатые на фоне черные па фоне цвета чешуи черные
белом крас- ном белом крас- ном
К о л о с ь я и и ф л я т н ы е
Колосья безостые
Ч е ш у и опушены ы е.
1. Зерновки белые
+ [ — — • — | — 1 — . j vaoilovii 1
2. Зерновки красные
+ 1 — I — I — I — I — I — I — I ispharalbispicahun I 2
— I + I — I — I — | — I -— I — | asiduhatnelianum | 3
Коло сь si остистые
Чешуи н e о и у ш е и и ы е
(зерновки красные)
+ 1 — । — ! •— I — | — | + — | ispharobaktiaricum I I
Колосья обычного типа
Колосья безостые
Чешуи н е о и у ш е н н ы е
1. Зерновки белые
+ I — I —I — I — I — I — I — I ispharaecum I 5
— | + I ’— I — I — I — I — I — I albiduhamelianum | 6
2. Зерновки красные
+ — — — —- — — — asialbispicatum 7
— + . — — — — — — . dorofeevii 8
— — + — — — — jakubzineri 9
— — — + — — — — thumanianii 10
— — — — + — — — marinae 11
— — — — — + — — asiamissum 12
ч е ш у и О п у IJI о н и ы с
1. Зерновки белые
+ 1 — I — I — I —- I — I — I — I hiranalborecens I 13
— I + I — | — I — I — I — t — I mupurual | 14
2. Зерновки красные
+ — — — — — — — asirecens 15
— + — — — — — — asineglectum 16
— — + — — — — — griseoturanorecens 17
— —- — — — — — niupuluniru 18
— — — — — + — — turanalefeldii 19
Колосья остистые
Ч е in у и и е о и у ш е и н ы е
I. Зерновки белые
+ — — — — — 1 — subbuldojii 20
+ — — — — — — 1- pseudobuldojii 21
— -1- — — — — — muslaphaeuii 22
— — + — — — — -1- karabachicum 2.1
— — — — + — — T [ laksbergeri 21
IIo<)po<) I. Triticum
203
Чешуи Ости Ра июни 'шесть № разно- видности
белые крас- ные серо-дымчатые на фоне черные на фоне иве га чешуи черные
Гелом крас- ном белом крас- ном
2. Зерновки красные
4 — .— — — — — subbaktiai iciun 25
4- — — — — — — 4- pseudobaktiaricum 26
—— 4- — — — — + — subsharkordii 27
— 4- — — — — — 4- pseudosharkordii 28
— — 4- — — — — 4- samuricum 29
— — — — — — 4- shemachinictim 30
— — — — — + — 4- sinskajae - 31
ч е in у и О п у ш с н н ы с
I. Зерновки белые
Геогр. распр. Шемахипский район Азербайджана. Выявлена
В. Ф. Дорофеевым в виде единичных колосьев в популяции с богатым
разновидностным составом мягкой и твердой пшеницы на высоте 900 м
над ур. м. Эта разновидность получена им также экспериментально от
скрещивания Т. spelta var. subsharkordii с Т. aestivum. var. aureutn.
Var. arpurual Gandil. 1975, Tp. Арм. НИИЗ, сер. пшеница, 3: 82, nom.
invalid. (34).
Геогр. распр. Армения.
Var. asialbispicatum Dorof. 1970, 1. c.: 283. (7).
Геогр. pa сир. Единичные растения найдены В. Ф. Дорофеевым в
посевах пшеницы Масаллинского, Хачмасского, Кубинского районов
Азербайджана.
На редкое нахождение этой разновидности в Армении указывал
М. Г. Туманян (1929, 1933). Он приводит ее под названием var. albispica-
tuin Flaksb. Как очень редкая примесь обнаружена в Самаркандской об-
ласти Узбекистана и Исфарииском районе Таджикистана (С. Н. Кудря-
шев, 1942; Р. А. Удачин, 1975).
Var. asialbivelutinum Dorof. 1970, 1. с.: 294. (32).
Геогр. распр. Хачмасский и Шемахипский районы Азербайджа-
на (единичные растения в популяциях мягкой и твердой пшеницы).
Var. asiamissum Dorof. 1970, 1. с.: 288. (12).
Геогр. распр. Шемахипский район Азербайджана (единичные
растения в производственных посевах пшеницы).
Var. asicaeruleum Dorof. 1970, 1. с.: 297. (40).
Геогр. распр. Шемахипский район Азербайджана (II. Д. Муста-
фаев в 1964 г. обнаружил единичные растения в производственных посе-
вах пшеницы).
206
Класс ифшкщия /xxla Ti it ii и ill L.
Var. asididiainelianiini Dorof. 1970, l.c.: 285. (3).
Геогр. распр. 111 ем axi11ickini район Азербайджана, Исфаринскпй
район Таджикистана (единичные растения в посевах мягкой и твердой
пшеницы).
Var. asineglectum Dorof. 1970, 1. с.: 289. (1(5).
Геогр. распр. Хачмасский район Азербайджана (единичные рас-
тения в производственных посевах пшеницы).
Var. asirecens Dorof. 1970, 1. с.: 288. (15).
Гео гр. р а с п р. Хачмасский и Кубинский районы Азербайджана.
Обнаружена в посеве пшеницы твердой (сорт Лрандапы) со значи-
тельной примесью пшеницы мягкой. И. Д. Мустафаев (1973) нашел ее
также в Шекинском районе Азербайджана и приводит под названием
var. recens Koern.
Var. asirubrivelutinum Dorof. 1970, 1. c.: 295. (36).
Геогр. распр. Кубинский район Азербайджана.
Var. dekaprelevichii Dorof. 1970, l.c.: 296. (39). Константная форма от
спонтанного скрещивания Т. macha с Т. durum.
Геогр. распр. Выявлена на коллекционном посеве Дагес га некой
опытной станции BIIРа.
Var. dorofeevii Udacz. var. nov. — A T. spelta var. asiduhatneliantim
Dorof. spica non iflata differt. (8).
Typus: Tadzhikistan, reg. Isphara, pag. Czorku, 1100 m, 15.VII. 1969,
leg. R. Udaczin, N 069778 (WIR).
Геогр. распр. Таджикистан (Исфарннскнй район).
Var. flaksbergeri Dorof. var. nov. — T. spelta var. flaksbergeri Dorof.
1966, С.-х. биология, 1,1: 135, sine typo. — Plantae spicis rudibus, impu-
bibns griscis-nigeris, pruino, cerino, inanibus (D = 11-13); aristis nige-
ris, caryopsis alba. (21).
Typus: Azerbajdzhan, regio Lachinskyi in populatione culta T. durum et
T. aestivum, 780 m, 4.VII. 1961, N 168, leg. V. Dorofeev (WIR).
Геогр. распр. Азербайджан (Лачинский район).
Var. griseoturanorecens Udacz. 1979, 1. с.: 70. (17).
Геогр. распр. Таджикистан (Исфарннскнй район).
Var. ispharalbispicatum Udacz. var. nov. — A T. spelta var. asialbispi-
catuni Dorof. spica inflata differt. (2).
Typus: Tadzhikistan, reg. Isphara, pag. Nauglem, 1000 m, 13.VII. 1969,
leg. R. Udaczin, N 070110 (WIR).
Геогр. распр. Таджикистан (Исфарннскнй район).
Var. ispharaecum Udacz. var. nov. — A T. spelta var. asialbispicatum
Dorof. caryopsi albo differt. (5).
Typus: Tadzhikistan, reg. Isphara, pag. Nauglem, 1000 m, 13.VII. 1969,
leg. R. Udaczin, N 070106 (WIR).
Геогр. распр. Таджикистан (Исфарннскнй район).
Var. ispharobaktiaricum Udacz. var. nov.— A T. spelta var. bactiaricum
Gokg. spica inflata differt. (4).
Typus: Tadzhikistan, reg. Isphara, pag. Woruch, 1150 m, 13.VII. 1969,
leg. R. Udaczin, N 070299 (WIR).
Геогр. распр. Таджикистан (Исфарннскнй район).
Var. jakub^ineri Dorof. 1970, 1. с.: 286. (9).
Геогр. распр. Азербайджан (единичные растения собраны в
сложных ио составу популяциях пшеницы, сорно-нолевой ржи и эгилоп-
са); в Нахичевани в посеве пшеницы мягкой па высоте 1130 м
над ур. м.
Var. karabachictim Dorof. 1970, I. с.: 291. (23).
Г corp, распр. Азербайджан ([Путинский и Лачинский районы).
Var. marinae Dorof. 1970, 1. с.: 287. — Т. spelta var. alboamisstiin Dorof.
1966, С.-х. биол. 1,1: 134, sine typo. (11).
Под/юд 1. Tritisum
Г е о 1 р. распр. Масалл11пск11ii pinion A:<< pП;iи„чж;i па (0,1111111'1111,1
растения в сложных по составу посевах).
Var. menabdii Dorof. 1970, 1. с.: 296. (38). Константная форма от спон-
танного скрещивания Т. macha с Т. durum.
Выявлена на коллекционном посеве Дагестанской опытной станции
ВИРа.
Var. mupuluniru Gandil. 1975, 1. с.: 82, nom. illcg. (18).
Г e. о г р. р а с п р. Армения.
V а г. mupurual Gandil. 1975, 1. с.: 82, nom. illeg. (14).
Геогр. р а снр. Армения.
Var. mustaphaevii Dorof. 1970, 1. с.: 289. — Т. spelta var. laczinicum Do-
rof. 1. c:. 290. (22).
Геогр. распр. Азербайджан, как редкая примесь в посевах Шс-
махинского, Шушинского и Лачинского районов.
Var. pseudobaktiaricum Dorof. 1970, 1. с.: 291. (26).
Геогр. распр. Азербайджан (редко в Кусарском и Хачмасском
районах).
Var. pseudobuldojii Dorof. 1970, l.c.: 290. — T. spelta var. psetidoalb’ -
arduini Dorof. 1966, I. c.: 135, sine typo. (21).
Геогр. pa cup. Азербайджан (редко встречается в посевах Лачин-
ского района).
Var. pseudosharkordii Dorof. 1970, 1. с.: 291 — Т. spelta var. pseudovul-
pinum Dorof. 1966, 1. c.: 134, sine typo. (28).
Геогр. распр. Азербайджан (редко встречается в Исмапллинском
районе).
Var. samuricum Dorof. 1970, 1. с.: 292. (29).
Геогр. распр. Азербайджан (единичные растения обнаружены в
посевах Кусарского района).
Var. schaartusicum Udacz. 1969, Тр. прикл. бот. ген. сел. 40, 2: 44. (35).
Геогр. распр. Таджикистан (единичные растения обнаружены в
посевах пшеницы Шаартузского и Исфарипского районов), а также в
Азербайджане (Хачмасский и Шемахипский районы).
Var. shemachinicum Dorof. 1970, 1. с.: 292. — Т. spelta var. psetidoscheu-
kii Dorof. 1966, 1. c.: 134, sine typo. — T. spelia var. kusaricuni Dorof. 197<T
1. c.r 293. (30).
Геогр. распр. Азербайджан (редкая примесь и посевах Шема-
хинского, Шаумянонского и Кусарского районов).
Var. sinskajae Dorof. 1970, 1. с.: 293. (31).
Геогр. распр. Азербайджан (очень редкая примесь в посевах
пшеницы, представленных смесью разновидностей мягкой и твердой пше-
ницы в нескольких районах республики).
Var. subbaktiaricum Dorof. 1970, 1. с.: 289. — Т. spelta var. bakllaricum
Gokg. 1961, I. c.: 322, descr. germ, sine typo. (25).
Геогр. распр. Преобладающая разновидность спельты в Иране
(И. Kuckuck, 1959; М. Gokgol, 1961). Единичные растения найдены г
Азербайджане в Хачмасском и Кусарском районах (В. Ф. Дорофеев.
1972), а также в Ханларском районе на высоте 1120 м над ур. м.
(И. Д. Мустафаев, 1973); в Средней Азии в Шаартузском и Исфарин-
ском районах Таджикистана, Ташаузской области Туркмении, Бухарской
области Узбекистана (В. Н. Мортенсен, 1946; Р. А. Удачин, 1969, 1971.
1975).
Var. subbuldojii Udacz. 1979, Бюл. ВИРа, 81: 70. — Т. spelta var. bul-
dojli Gokg. 1961, Zeitsclir. Pflanzenziicht. 45, 3/4: 322, descr. germ, sine
typo. — T. spelta var. alboarduitii Must. 1971, Опред. iiuieii. Азерб.: 75.
nom. inval. (20).
Геогр. распр. Редко в Иране, Азербайджане, Таджикистане.
Var. subsharkordii Dorof. 1970, l.c.: 290. — T. spelta var. sharkordii
Gokg. 1961, 1. c: 322, descr. germ, sine typo. (27).
2(18 Классификации рода Tiiliciini L.
Г с <i г |>. распр. 1!рал (редко), Л.н рб а йдж ;i 11 (редкая примесь в
посевах мягкой и твердой пшеницы), Таджикистан (преобладающая раз-
новидность в чистых посевах спельты в Исфаринском районе).
Var. tashausicuin Udacz. 1971, Тр. прикл. бот. ген. сел. 45,2: 47. (33).
Геогр. распр. Туркмения (Ленинский район Ташаузской облас-
ти) и Узбекистан (Свердловский район Бухарской области и Хивинский
район Хорезмской области).
Var. thunianianii Dorof. 1970, I. с.: 287. (10).
Ге о rj). распр. Азербайджан (единичные растения обнаружены
в посевах Шемахивского района Азербайджана).
Var. turanalborecens Udacz. 1979, I. с.: 70. (13).
Геогр. распр. Таджикистан (Исфаринский район).
Var. turanalefeldii Udacz. 1979, 1. с.: 70. (19).
Геогр. распр. Таджикистан (Исфаринский район).
Var. vavilovii Dorof. 1970, 1. с.: 282. (1).
Геогр. распр. Азербайджан (в популяции мягкой и твердой
пшеницы па высоте 780 м над ур. м.) и Таджикистан (редко в Исфарин-
ском районе).
Var. zhukovskyi Dorof. 1970, l.c.: 295. (37). Константная форма or
спонтанного скрещивания Т. macha X 7’- durum.
Выделена па коллекционном посеве Дагестанской опытной станции
ВИРа.
2. Convar. tarakanovii A. Filat, et Dorof. — группа разновидностей
безлигулыюй спельты.
Convar. tarakanovii A. Filat, et Dorof. convar. nov. Folium eligulatum,
surstim spectantium.
Typus: T. spelta var. tarakanovii Udacz.
Растения с листьями без язычка и ушек. Листовая пластинка не отог-
нута от стебля, листья направлены вверх.
Геогр. распр. Памир (Рушанский район).
В последние годы открыто много новых разновидностей безлигулыюй
мягкой и карликовой пшеницы, эндемичных для советского Памира. Это
говорит об активном здесь процессе формообразования. Безлигульная
спельта представлена одной типовой разновидностью (var. tarakanovii
Udacz.). Однако, исходя из закономерностей изменчивости видов, вполне
вероятно ожидать па Памире существование других разновидностей без-
лигульпой спельты.
Var. tarakanovii Udacz. 1976, Тр. прикл. бот. ген. сел. 56, 2: 150. Колос
обычного типа, остистый, чешуи неопушенные, серо-дымчатые (фон
красный); ости черные; зерновки красные.
Г е о г р. р а с п р. Памир (Рушанский район).
П. А. Гаидиляп (1975) в азиатском подвиде выделил махопдную
группу разновидностей — convar. machoides Gandil. 1975, Тр. Арм.
НЙИЗ, сер. пшеница, 3: 80, descr. ross., sine typo. Для группы характер-
ны короткие (4—10 см), плотные (D = 30—40) колосья; боковая сторона
шире лицевой стороны колоса; колосковые чешуи почти вдвое короче
цветковых. Судя по приводимому автором рисунку, колосья convar.
machoides сходны с колосьями Т. macha.
Т. (VAVILOVII (THUM.) JAKUBZ. — ПШЕНИЦА ВАВИЛОВА
(ПШЕНИЦА ВАНСКАЯ)
Т. vavilovii (Thum.) Jakubz. 1933, в Жуков. Земледелья. Турция: 705;
Якубц. 1933, Социалист, растениевод., Сер. А, 7: 222, Якубц. 1933, При-
рода, 11: 73. — Т. vulgare var. vavilovii Thum. 1933, в Жуков. Земледелья.
Турция: 705.-— 7’. vulgare compositum Timm. 1933, Опред. хлебп. злаков
(колос).: 193, excl. var. kurduculense Thum.; он же, 1934, Tp. прикл. бот.
Подрод i. Tiitisum
209
!
Рис. 16. Колосья, колоски и зерновки:
а — Т. spelta subsp. kuckuckianum Gokg. из Таджикистана; б — Т. vauilovii (Thum.)
Jakubz.
ген. сел. Сер. 5, 2: 273, excl. var. kurduculense Thum. — T. vulgare prol.
thospitense Flaksb. 1935, Культ, фл. СССР, 1: 47, nom. illeg. — T. vavilovi-
anum Jakubz. 1935, у Вавилова, Теорет. основы селекции, 2: 10, nom.
illeg.; Jakubz. 1964, у Вавилова, Миров, ресурсы хлебп. злаков. Пшеница:
60, nom. illeg. Т. vulgare subsp. mulabile Thum. 1936, Tp. С.-х. ин-та
Армении, 1: 27, nom. illeg.; он же, 1939, Tp. Арм. фил. АН СССР, сер.
биол. 1; 71, nom. illeg. — Т. aestivum subsp. vavilovii (Thum.) Sears. 1959,
in Hanclb. Pflanzenziicht. cd. 2, 2: 164, comb, invalid. — T. aestivum group
vavilovii Bowden, 1959, Canad. Journ. Bot. 37: 675. — T. spelta subsp. va-
vilovii (Jakubz.) Dorof. 1966, Генетика, 3: 30, comb, invalid. — T. aesti-
vum convar. vavilovii (Thum.) Morris et Scars, 1967, Wheat and wheat
improvem.: 36, comb, invalid. — T. spelta subsp. vavilovii (Jakubz.) Must.
1973, Опред. пшои. Азерб.: 76, comb, invalid.
14 762
210 К-inсгифи/,7/ции роди Tritii tun I,.
Общая характеристика. 14< \<>,|ы зе. юные, с к<>р<>i кi>м и pr iiui\i опу-
шением. Куст и молодом возрасю прямостоячий, а и дальнейшем iio.iv-
развалнстый. Листовое влагалище в фазе кущения покрыто едва замет-
ным опушением; а в фазу выхода в трубку оно почти голое. Соломина
без антоциана, толстая и прочная, под колосом слабо выполнена парен-
химой. Высота стебля при орошении па юге СССР около 80 см. Стебле-
вые узлы короткие, выпуклые. Стебель, листья и узлы в фазу колошения
с восковым налетом. Кустистость средняя (около 5 стеблей на одно рас-
тение). Длина колоса варьирует от 10 до 15 см (чаще 13 см). Колос
ложповетвнстый (рис. 16, б). На значительной части колоса пли ночи,
по всей его длине колоски (за исключением 1—2 нижних и верхушечных)
обычно с удлиненными члениками оси и хорошим развитием каж-
дого цветка. Как показали многолетние исследования ВИРа, в различ-
ных географических условиях этот признак стабилен. Число колосков
(включая вторичные) достигает 19—20, плотность — около 16 колосков
па 10 см длины колоскового стержня, число зерновок в колосе — от 50
до 95, в колоске от 3 до 5. Длина ложноветвпето! о колоса от 23 до
25 мм. Стержень колоса неломкий. Подковка (сочленение) незаметна.
Опушение членика колосового стержня сильнее развито v основания ко-
лоска, несколько слабее — по бокам. Волоски бесцветные, иногда - с
желтоватым оттенком.
Колосковая чешуя лопатчатая или ромбовидная, опушенная или не-
опушепная, крупная, более одревеспелая, чем у спельты. Соотношение
длины чешуи к ее ширине (около 1,5) меньше, чем у большинства форм
пшеницы мягкой и спельты. Жилкование колосковых чешуй резко выра-
жено (4—5 слабовынуклых, грубо зазубренных, доходящих до основания
чешуи жилок). Киль колосковой чешуи сильно развит и зазубрен по всей
длине чешуи. Килевой зубец очень короткий--до 1 мм, прит\ пленный,
направленный в сторону цветка. Плечо прямое. Колосковые чешуи очень
плотно охватывают зерновки. Вид труднообмолачпвающийся. Зерновка
яйцевидно-овальной формы, в поперечном сечении менее округлая, чем
у большинства форм пшеницы мягкой, средних размеров: длина 7—8,
ширина 3,5, толщина 2,5 мм. Окраска желтовато-белая и красная. Бо-
роздка довольно широкая. Хохолок хорошо развит, со сравнительно
длинным, по редким опушением.
Колеоптиль у Т. vavilovii с двумя проводящими пучками. По данным
В. Ф. Дорофеева и О. Д. Градчанпповой (1971), у растений Т. vavilovii
в фазе колошения сильно развито кольцо механической ткани, в первом
нижнем междоузлии формируется 18, во втором—14 проводящих пуч-
ков. Е. Ф. Пальмова (1935) указывала на сильное одревеснение стебля
Т. vavilovii.
По характеристике Н. И. Вавилова (1935, 1964) и Е. Ф. Пальмовой
(1935), Т. vavilovii - крайний ксерофит. Облпствеппость его слабая, лист
жесткий, колос очень грубый. Этот вид выделяется зпоевыпосливостыо,
требовательностью к теплу. Образ жизни — озимый. Этот вид следует
отнести к средперапнпм. На юге при орошении Т. vavilovii созревает
примерно на 7 дней раньше спельты, па 11 дней раньше западноевропей-
ских и па 5—6 дней — степных и лесостепных украинских форм пшеницы
мягкой. Выделяется исключительной устойчивостью против осыпания
зерна.
По наблюдениям М. т\. Федина (1974), открытое цветение у этой
пшеницы — редкое явление. Лишь 1% цветков в колосе цветет открыто,
а раскрытие цветковых чешуй весьма кратковременное. В. М. Берляид-
Кожевников и др. (1978) указывают, что в условиях низменного Дагеста-
на при орошении завязываемость зерна у этого вида резко снижается
при строгой изоляции колосьев. Содержание белка в зерне у Т. vavilovii
достигает 17,2% (В. Г. Коиарев и др., 1972).
IIo<ll><>(3 I. Trihriuii 211
Иммунитет. Пшеница Baвилова очень r,oi-u[)нuмчнu.;i к грибным бо-
лезням. II. И. Вавилов (19(51, 19(51) назвал этот вид аккумулятором ин-
фекции, отметив его ус iойчивосгь лишь к (зеблевоб i оловне и .зелено-
глазке. М. М. Якубцинер (1957, 1969) подтвердил згу оценку. В условиях
Дагестана отмечено сильное поражение пшеннны Вавилова бурой, жел-
той и стеблевой ржавчиной (II. II. Шитова, 1968). Сходные результаты
показали также исследования по стеблевой и бурой ржавчине на расо-
вом уровне (О. Г. Григорьева, 1975). Полю синая устойчивость к расам
77, 151, 202 последнего патогена выявлена у одного образна (В. М. Бср-
ляпд-Кожсвипков и др., 1973). При искусственном заражении популяци-
ей рас из различных регионов СССР Э. X. Сухапбсрдппа (1976) подтвер-
дила данные II. И. Вавилова (1961) о сильной восприимчивости пшеницы
Вавилова к мучнистой росе. Н. И. Вавилов, указывал также на силь-
ное поражение этой пшеницы шведской и гессенской мухами, твердой и
пыльной головней. При пиокулпровапип популяцией рас В. И. Кривчен-
ко (1973) и Л. М. Ямалсев (1974) нашли, что f. duri пыльной головни
поражает ее слабее, чем f. aestivi.
История. Впервые единичные растения пшеницы Вавилова были об-
наружены в 1929—1930 гг. М. Г. Туманяном (1931) как примесь в старо-
местном армянском сорте, пшеницы мягкой под названием Дир.
Географическое распространение. Исходная форма —сорт Дир —
возделывалась преимущественно па Армянском нагорье, северо-восточ-
нее озера Ван (Турция, вилайет Ван, 1780 м над ур. м.). Ветвистую пше-
ницу, выделенную из этой популяции, М. Г. Туманян (1934) назвал
одним из замечательных эпдемов Армянского нагорья, характеризующе-
гося засушливо-континентальными условиями. Пшеница Вавилова была
также обнаружена в Азербайджане (И. Д. Мустафаев, 1973) и г. Арме-
нии (П. А. Гандилян, 1976).
Происхождение. При межвидовой гибридизации исследователи неод-
нократно наблюдали появление растений пшеницы с ветвящимся (по ти-
пу пшеницы Вавилова) колосом (R. Н. Biffcn, 1916; А. Кег/.ег, В. Boyack,
1918; G. Nilsson-Leissncr, 1925; S. Borosevic, 1959; В. C. Sharman, 1962;
М. V. Р. Rao, М. S. Swaminathan, 1963; J. Mac Key, 1966; А. А. Фплатеп-
ko, 1968; II. А. Гандилян, 1963, 1974, 1976). Появление таких «вавнлои-
дов» зарегистрировано также после воздействия мутагенами на пшеницу
мягкую (М. S. Swaminathan, 196.5; J. V. Goud, 1967; К. Nagi, 1968;
М. Р. Singh, 1969; R. S. Rana, 1970), пшеницу твердую (G. Т. Scarascia
и др., 1961), Т. persicutn. (Z. Barabas, 1959). К. А. Фляксбергер (1935)
допускал возможность естественного мутационного возникновения Т. va-
vilovii из грубоколосых спсльтоидных форм пшеницы мягкой.
Предполагается, что генетический контроль удлинения оси колоска
у Т. vavilovii осуществляется длинным плечом хромосомы 5А (М. S. Swa-
minathan, М. V. Р. Rao, 1961; J. Mac Key, 1966; Г. Р. Моррис, Э. Р. Сирс,
1970). Есть сведения (П. В. Singh и др., 1957), что не один геи, а два
рецессивных гена ответственны за удлинение осп колоска. Эта система
эффективна лишь при отсутствии фактора Q. Полигонная система легко-
го обмолота, которую, имеет пшеница твердая, не подавляет действия
генов (гена), ответственных за удлинение осп колоска. П. П. Лукья-
ненко и В. В. Koqthh (1970) выделили константный «вавилоид» с легким
обмолотом из комбинации, полученной при скрещивании твердой и мяг-
кой пшеницы, назвав его Т. durum subsp. uuicum. У. К. Куркиев (псопуб-
ликовапо) отобрал сходную форму среди растений фалькатной пшеницы
твердой. Р. А. Удачин и II. III. Шахмедов (1972, 1976, 1977) от крыли ва-
вплоидпую константную форму, спонтанно возникшую в образце /’. lurpi-
dum афганского происхождения. Они возвели се в ранг вида Т. jaktibzi-
neri на том основании, что она имеет не только вавилоидпость колоса при
легком обмолоте, но и обладает четырьмя колосковыми чешуями в каж-
дом колоске.
И*
212
Клшч ифпкнци.ч Tiitiiiim
Д. Мак-Кей (1969) считает, что апачиюльпаяпрочность колосовою
стержня у пшеницы Вавилова может быть связана с фактором сквер-
хедности, который исидептичен факторам Q и С, характерным для пше-
ницы мягкой и для 7’. compactum. Он предполагает, что если вавилоид-
ность колосков обеспечивается не одним, а большим числом генов, то
остальные гены по фенотипическому проявлению слабее, чем первый ген,
и, вероятно, кумулятивны по своему эффекту.
Исследователи связывают происхождение пшеницы Вавилова с мута-
цией у пшеницы мягкой (М. Г. Туманян, 1936, 1939; К. А. Фляксбергер,
1935 и др.). В. Ф. Дорофеев (1969) полагает, что такая мутация могла
произойти после спонтанной гибридизации тетраплоида (возможно,
Т. dicoccum) с пшеницей мягкой.
Систематика. Полиморфизм Т. vavilovii до недавнего времени был
ограничен тремя разновидностями, выделенными AI. Г. Туманяном
(1933). Позднее Р. Л. Удачин (1976) в одной из популяций Т. vavilovii
из Армении (к-29533), представленной var. vavilovii и var. mirabile.
выявил еще 2 новые разновидности. П. А. Гандилян (1976) относит к
Т. vavilovii разновидности, полученные в результате спонтанной гибри-
дизации Т. durum с Т. aestivum и дальнейшей их мутации. Он (1975) де-
лит Т. vavilovii па 2 группы разновидностей: convar. vavilovii и convar.
speltoramosum Gandil. Для последней характерно ветвление колоса с
образованием вторичных колосьев или сдвоенных колосков на одном ус-
тупе колосового стержня. Эту группу можно рассматривать как спельту
с ветвистыми колосьями, а не Т. vavilovii — вид с ложным ветвлением
колоса.
Определение разновидностей Т. vavilovii (Thum.) Jakubz.
Чешуи
белые
крас-
ные
серо-дымчатые
на фоне
белом
крас-
н ом
черные на
белом
фоне
цвета
чешуи
черные
Разновидность
Xi1 разно-
видности
Колосья безостые
Чешуи неопушенные (зерновки красные)
>— I f— I vavilovolutcscens
— intinuru
1
2
Чешуи опушенные
3 е р н о в к и б с л ы е
— I — I — I vavilovii
+ | — — mirabile
3
4
Зерновки крас н ы е
— I —1 I — I шириги
+ | — | — I rubromirabile
5
6
Колосья остистые
(чешуи неопушенные, зерновки белые)
+ I — | | | -- I + | — i mravianii [ 7
Var. mravianii (Thum.) Jakubz. 1933, Природа, 11: 73. — T. vulgare
var. mravianii Thum. 1933, Опред. хлебн. злаков (колос.): 194. (7).
Геогр. распр. Армения. Выделена М. Г. Туманяном па опытном
участке кафедры частного земледелия Гос. ун-та (ныне Армянский СХИ)
на посеве сорта мягкой пшеницы Дир.
Var. mirabile (Thum.) Jakubz. 1933, Социалист, растениевод. Сер. Л,
7: 222; он же, 1933, Природа, l.c.: 73. - Т. vulgare var. mirabile Thum.
1933, l.c.: 194; он же, 1934, Тр. прикл. бот. ген. сел. Сер. 5, 2: 273. (4).
Iloti/ttx) I. Tula inti 13
Геогр. распр. Армения. Найдена вчом же посеве, чта и var. тга-
vianii Thum. Единичные рапепия обнаружена И. Д. Мусiафаевым (1973)
в посевах Азербайджана.
Var. inuniirii Gandil. 1976, Bio.а. ВИРа, 65: 77. I. wioih>cii var. i’avi-
lovomilturum Udacz. 1979, Вюл. ВНР, 84: 73. (2).
Геогр. распр. Армения. Получена в результате спонтанной ги-
бридизации Т. durum с Т. aestivum и дальнейшей мутации в условиях
Экспериментальной базы Института земледелия МСХ Армянской ССР.
Var. mupiiru Gandil. 1976, I. с.: 78. (5).
Геогр. распр. Армения. Получена в результате спонтанной гиб-
ридизации на Экспериментальной базе Института земледелия МСХ Ар-
мянской ССР (Эчмиадзин).
Var. rubromirabile Udacz. 1976, Tp. прикл. бот. ген. сел. 56,2: 150. (6).
Выделена па коллекционном посеве Среднеазиатской опытной станции
ВИРа из популяции Т. vavilovii (к-29533).
Var. vavilovii. — T.vulgare var. vavilovii Thum. 1933, у Жуков. Земле-
делья. Турция: 705; idem. 1933, Определ. хлебн. злаков (колос.): 194,
idem. 1934, Тр. прикл. бот. ген. сел. Сер. 5, 2: 273. — Т. vavilovii var. va-
neum Jakubz. 1933, Социалист, растениевод. Сер. А, 7: 222; он же. 1933,
Природа, 11: 73. (3).
Геогр. распр. Армения. Выделена на опытном участке кафедры
частного земледелия Гос. ун-та па посеве сор та Дир.
И. Д. Мустафаев (1973) находил единичные растения var. vavilovii
в посевах Азербайджана.
Var. vavilovolutescens Udacz. 1976, 1. с.: 150. (1).
Геогр. распр. Узбекистан. Выделена из той же популяции Т. va-
vilovii (k-29533), что и var. rubroinirabile.
Т. COMPACTUM HOST — ПШЕНИЦА КОМПАКТНАЯ (ЕЖОВКА)
Т. compactum Host, 1809, Gram. Austr. 4: 4, tab. 7.— 7'. vulgare var.-gr.
compactum Alef. 1866, Landwirt. FL: 327. — T. sativum compactum Hack,
in Engl, et Prantl, 1887, Natilrl. Pflanzenfam. 2: 85. — T. tenax A. II com-
pactum Aschers. et Graebn. 1901, Synops.: 686. — T. aestivum subsp. aesti-
vum. (Host) Rasse compactum Thell. 1918. Naturwissensch. Wochenschr.
N. F. 17, 33: 471. — T. aestivum subsp. compactum (Host) Mac Key, 1954,
Sv. Bot. Tidskr. 48, 2: 586. — T. aestivum gr. compactum Bowden, 1959,
Canad. Journ. Bot. 37: 674.
Общая характеристика. Всходы зеленые пли синевато-зеленые. Лис-
товые пластинки покрыты шипиками (шероховатые па ощупь) или опу-
шены очень короткими волосками (бархатистые па ощупь), различной
длины (10—35 см) и ширины (0,7—2,4 см). Листовое влагалище голое
или опушенное. На месте его перехода в лист имеются язычок и ушки,
последние иногда с длинными ресничками. Имеются также формы без-
лигульные. Колосья плотные (D = 33—54, чаще 40—50), опушенные или
неопушенные, длиной 3—7 (реже до 8—10) см, шириной 1—2 см
(рис. 17). Часто боковая сторона колоса немного шире лицевой. Колосо-
вой стержень укороченный, его членики очень короткие, по их ребрам —
реснички из волосков. Колосковые чешуи с очень узким килем, пе всегда
доходящим до основания чешуи, где обычно имеется поперечная вдав-
ленность с продольной морщинистостью. Киль в верхней части перехо-
дит в килевой .зубец, который может быть по длине от короткого до осте-
видпого. Жилки па колосковой чешуе слабо nmcrvnaioi над ее поверх-
ностью. Главная боковая жилка выражена сильнее остальных. Она за-
капчивается бугорком пли коротким зубном. Плечо колосковой чешуи от
горизонтально-приподнятого до скошенного, иногда широкое, как у
спельты. Наружная цветковая чешуя несет ость, остсвидное заострение
или зубец (ее вершина изредка бывает притупленной, без зубца). Внут-
K.UI< < ll</llllull(ll!l 7 I Пиши /,.
Рис. 17. Колосья, колоски и зерновки Т. compactum Host
ренняя цветковая чешуя двукнлевая, при созревании не расщепляется
вдоль на 2 части. В колоске 3 -4 зерновки, длина зерновок до 7 мм
(обычно их длина превышает ширину менее чем в 2 раза).
По основным признакам анатомического строения листа, стебля^ ко-
лоса и зерновки Т. compactum сходна с пшеницей мягкой. В колеоптиле
2 проводящих пучка. Отличается от предыдущих гексаплоидных видов
некоторыми особенностями тканей колосового стержня. Анатомическое
изучение его (О. Д. Градчанппова, 1973) показало, что к моменту созре-
вания колоса у основания колосков формируется тонкостенная мелкокле-
точная паренхима. Она граничит с паренхимой сердцевины членика,
образуя с пей единую ткань, которая, не прерываясь, переходит из одного
членика в другой, внося свою долю в обеспечение прочности колосового
стержня. В месте перехода одного членика в другой прозенхпмные клеши
одревесневшего (внешнего) слоя, как и паренхимные клетки «разрыв-
ной» ткани, имеют не тангептальное, а продольное направление, что так-
же способствует упрочению колосового стержня.
Экологически Т. compactum является типичной горной пшеницей, ко-
торая имеет, по И. И. Вавилову (1935), формы влаголюбивые, с малой
требовательностью к теплу в период созревания, устойчивостью к весен-
нему понижению температуры и малой требовательностью к почвенным
и агротехническим условиям. Скороспелость и сравнительная выносли-
вость к низким температурам особенно присущи образцам из Якутии.
По образу жизни разделяется па озимые, полуозпмые и яровые (позд-
ние и ранние) формы. Опыт показал, что при промораживании (— 1(>°С)
образцов мировой коллекции морозостойкость Т. compactum невысокая.
По сравнению с сортом пшеницы мягкой Мироновская <308 она колеба-
лась в пределах 0 — 28% (Г. В. Удовенко и др., 1979). Е. Ф. Пальмова
(1935) указывала на слабую зимостойкость 'Г. compactum. Иамиро-ба-
дахшанские формы опа характеризовала как пустынные, поливные, ко-
торые при перенесении в иные эколо! ические условия вымирают.
Hix)i)i>i) I. Triliciuii ‘21 Г>
AV AV Як\бцинер (1970) указывавг iia 3;icyxoy("i ойчивос'1 ь иекторых
форм из Армении.
Т. compactum xapaKiepiiayt'icn пониженным уровнем открытое цве-
тения по еравн('ник) е ди- и тетраплоидными видами. В селениях Haie-
егаиа в опыте В. II. Жукова (1069) отмечено в зависимое! и oi сорта
1,6—71,7% хазмогамных цветков. Он оз метил, что значите, тьпые колеба-
ния в количестве -лих цветков связаны с местом происхождения сорта.
В условиях Белоруссии ширина щели, образующейся при расхождении
цветковых чешуи во время цветения, достигала лини, 2,1--2,4 мм, про-
должительность открытого состояния цветка — 12,6-23,5 мин (Л. 3. Ла-
тыпов, В. II. Апель, 1971).
Зерновки Т. compactum стекловидные, полустскловп.лпыс или муч-
нистые. Копнен грация белка в. них в условиях полнвнот \ чистка (плос-
костной Дагестан) определена в пределах 15,5—22%, содержание лизина
в белке— 1,94--2,44% (В. Г. Копарев и др., 1972).
Иммунитет. II. II. Вавилов (1913, 1961, 1964) указал па восприимчи-
вость Т. compactum к видам ржавчины, мучнистой росс, пыльной и твер-
дой головне, шведской и гессенской мухам, а также па устойчивость к
стеблевой головне. М. AV Якубцинер (1969) подтерли.! >ти данные но
бурой п стеблевой ржавчине, ио пыльной и твердой головне. Причем он
охарактеризовал Т. compactum как пшеницу, которая в целом восприим-
чивее мягкой. О. Г. Григорьева (1975) отметила, что образцы Т. com-
pactum средне пли сильно поражаются бурой и стеблевой ржавчиной па
всех фазах оптогенеза. При искусственном заражении популяциями рас
из различных районов СССР Э. X. Сухапбердина (1976) в коллекции
Т. compactum выявила 86,7% восприимчивых образцов (балл 3 и 4 по 5-
баллыюй шкале); иммунные, образцы отсутствовали. Инокуляция кол-
лекции хламидоспорами пыльной головни, проведенная В. И. Кривченко
(1973), выявила 66,7% образцов иммунных и устойчивых к f. duri этого
патогена, и 14,3%—к f. aestivi. Этот исследователь характеризует
Т. compactum как наиболее восприимчивый вид пшеницы по проявлению
заболевания в полевых условиях и по реакции органов зародыша на
внедрение мицелия. А. АС Ямалеев (1974) показал, что мицелий гриба
проникает во все органы зародыша зараженной зерновки. Причем
f. duri поражала 32%, a f. aestivi 66% зародышевых почек. В опыте
Н. Harding (1972) Т. compactum показал себя как очень восприимчивый
к корневым гнилям вид пшеницы.
История. Многие данные (J. Percival, 1921; !< Фляксбергер, 1923,
1935; G. Mandy. 1970 и др.) свидетельствуют о древности культуры
Т. compactum. Е. Schiemann (1948), F. Kornicke (1885) и G. Bnschan
(1895) считали, что в эпоху неолита в районе свайных построек
Швейцарии возделывался вид Т. compactum. О. Heer (1865) опи-
сал под названием Т. compactum-muticum форму, которая, по его словам,
найдена в неолите Швейцарии и отличается от Т. antiquorum. Е. II. Спн-
ская (1955) отнесла ее к Т. antiquorum, отметив ее кр\ пнозерность. По
данным F. Bertsch (1939) и J. Mac Key (1966), в эпоху бронзы па терри-
тории современной ФРГ возделывался Т. compactum.
В нашей стране обнаружены следы весьма древней культуры Т. com-
pactum. В частнрсти, судя по анализу археологических материалов из
раскопок 1954—1955 гг., обугленные зерна Т. compactum (III тысячеле-
тие до и. э.) найдены в Нахичевани в одном из погребений (М. АЕ Якуб-
ципср, 1956). Плетя культуры Т. compactum в Закавказье, относящиеся
к VII VI вв. до и. отражены археолш ическпми исследованиями в
Армении (близ г. Ереван), проведенными Б. Б. IIiioiровским в 1939 —
1941 гг. и продолженными им в послевоенный период. Она выявлена в
европейской части СССР в районе г. Костромы. Здесь в жилище, относи-
мом к концу I тысячелетия до и. э., найдены растительные остатки, кото-
рые представляют собой обугленные зерна пшеницы Т. compactum.
1 С> Кушссифинация роди Trilit uni I,.
К IX -X вв. п. относятся находки обугленных зерен Т. compactum
в районе Цимлянского водохранилища (на территории современной Рос-
товской области), где в тот период существовала хазарская крепость.
Позднее, после занятия этой крепости Святославом в 965 г., посевы
Т. compactum были вытеснены пшеницей мягкой как более ценной куль-
турой.
Географическое распространение. В недавнем прошлом в нашей стра-
не распространение Т. compactum было ограничено рядом районов
(Средняя Азия, Сибирь, Якутия, Ростовская область, Азербайджан,
включая Нахичевань). В 1970 г. посевы сохранились лишь в яровой
культуре па территории Армении, где районированы сорта Вардепнк 9 и
Норкопдпк. Вместе с тем посевы Т. compactum, по сообщению Р. А. Уда-
чипа (1974), обнаружены экспедицией ВИРа в Ташаузской области Турк-
мении, в Хорезмской и Кашкадарьинской областях Узбекистана, в Ош-
ской области Киргизии и Горно-Бадахшапской автономной области Тад-
жикистана. В Средней Азии зафиксировано 52 разновидности Т. compac-
tum. Общая площадь посева этого вида в Средней Азии исчислялась
лишь десятками гектаров.
Ареал вида в зарубежных странах, по данным Н. II. Вавилова (1935,
1964) и К. А. Фляксбергера (1935), охватывает Турцию, Афганистан, Ки-
тай, Монголию, Сирию, США, а по сведениям J. Percival (1921) — также
Южную Африку. Встречается и в Австралии. ЛА. L. Вог (1970) указыва-
ет на наличие Т. compactum в Ираке.
Происхождение. A. Schulz (1911), Д. К. Ларионов (1914) и др. не от-
вергали возможности происхождения Т. compactum из пшеницы мягкой.
В. Л. Мепабде (1948, 1972) предком голозерных гексаплоидных пшениц,
в том числе Т. compactum, считает Т. macha. К- А. Фляксбергер (1939)
предполагал, что Т. compactum возник при скрещивании эгилопса с пше-
ницей твердой. К- и F. Bertsch (1947) возникновение Т. compactum свя-
зывали с гибридизацией между Т. monococcum и Т. dicoccum.
Место происхождения Т. compactum Н. И. Вавилов (1926, 1929) сов-
мещал с районом его наибольшего разнообразия — с Афганистаном. Того
же мнения придерживается Н. Kihara (1958), который теоретически
допускает, что предшественник вида Т. compactum мог 'возникнуть при
гибридизации и иметь генотип вида Т. macha, генные мутации в хромосо-
мах которого дали начало Т. compactum и Т. spelta. Мнение сторонников
появления Т. compactum из последнего вида не подтвердилось палеобо-
таническими данными (A. Helbaek, 1964, 1965).
Не следует исключать возможности происхождения Т. compactum при
гибридизации Ле. tauschii с тетраплоидным носителем фактора Q—Т. per-
sicum. Н. Kihara и F. Lilienfelcl (1949) экспериментально доказали воз-
можность возникновения голозерного гексаплоида таким путем (К. Tsu-
newaki, 1966).
Многие исследователи неоднократно получали Т. compactum экспери-
ментально. А. Р. Жебрак (1949) зафиксировал выщепление этого вида
при гибридизации амфидиплоида Т. turgidum X Т, timopheevii с пше-
ницей мягкой. Б. Бочев и П. Павлов (1969) наблюдали его появление
при скрещивании мягкой и твердой пшениц. Опыты по воздействию му-
тагенами на пшеницу мягкую указывают на мутабилыюсть гена С, кото-
рый легко переходит в доминантное состояние из рецессивного, давая
Т. compactum (S. Н. К- Rao, Е. R. Sears, 1964; В. Р. Pal, 1965; С. II. Ма-
карова и др., 1966).
Известно, что Т. compactum отличается от пшеницы мягкой одним
главным геном С, который вызывает укорачивание члеников стержня
колоса (R. II. Biffen, 1905; Н. N i 1 sson - Eh I e, 1911; j\. А. Сапегнн и др.,
1916; E. F. Gaines, 1917; B. Kajanus, 1918, 1923; W. Arciszewski, 1924;
Ю. А. Филнпчсико, 1927, 1934 и др.). Этот геи локализован в хромосоме
2D (J. Unrau, 1950). Он не только увеличивает плотность колоса, но и
lloripoi) I. Triticum 217
обусловливает укорочение стебля и остей, влияет па массу зерен; д.|пн.о
1-го листа уменьшатся под его влиянием па (> 10% (Gul Azam,
R. Е. Allan, 1972).
J. Mac Key (1954) отметил, что ген С имеет способность укреплять
прочность колосового стержня наряду с фактором Q. Появление комнак-
тоидов он связывает также с дупликацией фактора Q. Компактоидные
формы могут возникнут!? даже при внутривидовых скрещиваниях пшени-
цы мягкой в результате дупликации Q и транслокации (J. В. Панг,
G. М. Wright, 1968).
С видом Т. compactum отождествляют древнейшую голозерную пше-
ницу Т. antiquorum Heer, открытую в виде целых колосьев при раскопках
свайных построек в Швейцарии. Е. И. Синская (1969) в современном ви-
де Т. compactum видит переходную форму между этой древней пшеницей
и Т. aestivum. В ее понятии Т. antiquorum из Закавказья проник в Евро-
пу, Египет, Месопотамию. В. Ф. Дорофеев (1972) также считает, что эта
пшеница прошлого имела обширный ареал в горных районах Закавказья.
Н. Kuckuck и R. Peters (1964) при воздействии мутагенами получили тет-
раплоидную пшеницу, сходную с пшеницей Т. antiquorum. Поэтому они
предположили, что последняя имела 28 хромосом.
Т. compactum мог возникнуть только при наличии фактора Q, так как
иначе легкий обмолот и прочность колосового стержня не были бы обес-
печены. Возможно, что фактор Q не мог быть продуктом интрогрессии от
диких диплоидных доноров, давших свои геномы пшенице. Если это так,
то он мог возникнуть мутационпо па тстранлоидном или гексаплоидпом
уровне. В первом случае возникновение Т. compactum могло быть связа-
но с гибридизацией тетраплоида, имеющего фактор Q, с Ае. tauschii. Во
втором случае Т. compactum мог возникнуть из первичной формы пшени-
цы мягкой после мутации рецессивной аллели в доминантную у гена С.
Предварительные данные опыта отдела пшеницы ВИРа по созданию го-
молога пшеницы мягкой пе отвергли возможности происхождения Т. com-
pactum последним путем. Если же становление этого вида шло при гиб-
ридизации Т. persicum X Ае. tauschii, то очаг возникновения Т. compac-
tum должен располагаться в ареале Ае. tauschii subsp. slrangulata (Eig)
Tzvel.
Систематика. Пшеница компактная широко культивировалась в
прошлом, о чем свидетельствуют многочисленные археологические на-
ходки. Ареал Т. compactum незначительно уступал ареалу Т. aestivum.
Оба вида формировались в одних и тех же условиях, и полиморфизм
одного из них в сущности повторяет полиморфизм другого. Однако у пше-
ницы компактной нет четкого обособления подвидов, что мы наблюдаем
у пшеницы мягкой. По предположению К- А. Фляксбергера (1935, 1939),
процесс дифференциации Т. compactum остановился, так как культура
пшеницы компактной начала вытесняться еще в далекие исторические
времена более продуктивной пшеницей мягкой. Возможно, сокращение
посевов пшеницы компактной и сосредоточение их в основном в горных
зонах привело к элиминации резко контрастных групп.
В пределах Т. compactum Host мы различаем 3 группы разновиднос-
тей: convar. compactum, convar. rigidicompactum, convar. inflation.
Ключ для определения групп разновидностей Т. compactum Host
1. Листья с язычками и ушками у основания пластинки пли без них;
колосья чаще остистые или с укороченными остями (иолуослистые), реже
безостые; колосковые чешуи жесткие или полужесткие; зерновки доволь-
но плотно заключены в чешуях...................................2.
+ Листья с язычками и ушками у основания пластинки; колосья ча-
ще безостые, реже остистые; колосковые чешуи тонкокожпстыс; зерновки
легко осыпаются.................................convar. compactum.
218
Классификации /юг)а Triliitim
2. К< >.'i < >< к < > I! 1.1 е чеш\и широкие, |',ып_\ к. ii.ii', с ,Ш1И\1Ыми bobhvi|h, ки-
левыми зубцами и осiеиндиыми заострениями, ости цветковых чешуи
обычно изогнутые, деформированные, часто у основания расширенные
......................................convar. inflation Vav. et Kob.
+ Колосковые чешуи более пли менее плоские, различной формы, с
прямым килевым зубном; ости цветковых чешуй прямые*.................
..................convar. rigidicompactum (Kudr.) Л. Filat. el Dorof.
I. Convar. rigidicompactum (Kudr.) A. Filat. et Dorof. — азиатская
группа разновидностей.
Convar. rigidicompactum (Kudr.) Dorof. et A. Filat, comb. nov.—
7'. compactum subsp. rigidicompactum Kudr. 1942, Питен. Узбекнст/. 48.—
T. compactum subsp. semiarislatocompactum Kudr. 1942, l.c.: 52, nom.
illeg. — T. compactum subsp. muticocompactum Kudr. 1942, l.c.: 53.—
T. compactum subsp. iranoasiaticum Flaksb. 1935, Культ, фл. СССР, 1:
101, quoad prol. rigidum Flaksb. et prol. subrigidum Flaksb., descr. ross. —
T. compactum subsp. armenoturkestanicum Vav. 1964, Миров, ресурсы
хлебн. злаков. Пшеница: 61, descr. ross.
Тип: Т. compactum var. splendidorigidum Kudr.
Колосья остистые, иолуосгпетые; реже безостые, колосковые чешуи
жесткие, чаще полужесткие; зерновки плотно заключены в чешуях, но
обычно легко обмолачиваются, реже вымолот затруднен.
Геогр. распр. Горные и предгорные зоны Средней и Юго-Запад-
ной Азии, Закавказье, Монголия.
По наличию или отсутствию язычка (лигулы) .у основания пластинки
листа convar. rigidicompactum делится па 2 подгруппы: subconvar. rigidi-
compactum и subconvar. eligulatum.
Ключ для определения подгрупп разновидностей convar.
rigidicompactum (Kudr.) A. Filat, et Dorof.
1. Листья с язычками и ушками у основания пластинки; листовая
пластинка отогнута от стебля .......................................
.......................subconvar. rigidicompactum A. Filat, et Dorof.
~Г Листья без язычков и ушек у основания пластинки; листовая плас-
тинка почти не отогнута от стебля . . . subconvar. eligulatum Flaksb.
a. Subconvar. rigidicompactum A. Filat, et Dorof. — азиатская под-
группа разновидностей.
Subconvar. rigidicompactum A. Filat, et Dorof. subconvar. nov.— Folia
ligulata, potens.
T и n: T. compactum var. splendidorigidum Kudr.
К subconvar. rigidicompactum относятся разновидности с развитыми
язычком и ушками у листьев. В остальном описание соответствует при-
водимому для convar. rigidicompactum.
Геогр. распр. По всему ареалу convar. rigidicompactum. В Сред-
ней и Западной Азии основные посевы пшеницы компактной, менее тре-
бовательной, чем пшеница мягкая, к условиям произрастания приуроче-
ны к высокогорным районам.
Азиатская пшеница компактная подразделяется на следующие эколо-
го-географические группы.
а) Кабульская группа (prol. cabulictim Vav. 1964, Миров, ресурсы
хлебп. злаков. Пшеница: 62, descr. ross.) отличается наиболее затруд-
ненным вымолотом зерна и резко выраженной компактностью колоса,
которая уменьшается при посеве в низменной зоне плоскостного Дагес-
тана. Зерно довольно крупное. Растения в основном среднерослые, лис-
тья короткие, широкие. Выявлена склонность к поражению желтой и бу-
рой ржавчиной. Озимые посевы этой группы сосредоточены в Юго-Вос-
точном Афганистане па высоте около 2000 м над ур. м. Зимостойкость
слабая, влаголюбивость большая.
/lorl/iod 1. Triticiuu
2 14
6) Хорезмская i руина (prol. khoresmiciun Vav. l.c.: 62, doser, ross.)
нредстав.чеиа низкорослыми растениями с коротким узким листом. Ис-
ключительно восприимчива к грибным заболеваниям. Относительно хо-
лодостойкая. Посевы малочисленны.
в) Палестинская группа (prol. palesiitiicum Vav. 1964, l.c.: 62, descr.
ross.) встречается как примесь среди посевов пшеницы твердой (Г. durum
subsp. horanicum Vav.). H. II. Вавилов (1961) рассматривал эгу группу
как типичный случай уподобления одного вида другому при засорении
первым последнего. Чистых посевов пшеницы компактной на Ближнем
Востоке не обнаружено.
г) Армяно-апатолпнская группа (prol. armeno-anatolicum Vav. 1964,
I. с.: 62) самая крупная по занимаемому ареалу и ботаническому разно-
образию. Ригидность у растений этой группы гораздо менее выражена,
чем у предыдущих. Характерные признаки: высокая кустистость и малая
облпсгвеппость растений, короткий колос, мелкое зерно, непрочная со-
ломина.
Отмечается восприимчивость к грибным заболеваниям.
Subconvar. rigidicompactuni отличается большим полиморфизмом н
продет а вл ей а значительным кол и чес гвом разновидностей.
Var. albiceps Koern. in Koern et Wern. 1885. llandb. Getrcidcb. 1: 51.—
T. aestivum var. albiceps (Koern.) Mansi. 1951, Dor Zuchter, 21, 1/2:
50. (32).
В Иордании выявлена форма без воскового налета — 1. epruinosa
Jakubz. 1932. 'Гр. прикл. бот. ген. сел. Прилож. 53: 147.
Геогр. распр. Средняя Азия, Закавказье, Афганистан, Малая
Азия, Иордания, Кипр.
Var. armeniacum Thum. 1928, Тр. прикл. бот. геи. сел. 19,1: 271.—
Т. aestivum var. armeniacum (Thum.) Mansf. 1951, l.c.: 51. (26).
Г e о г p. p а сп p. Армения.
Var. asiaticicterinum Kudr. 1942, Пшен. Узбскнст.: 49. (17).
Геогр. распр. Средняя Азия, Афганистан, Сирия, Иордания.
Var. atriceps Koern. in Koern. ct Wern. 1885, l.c.: 54, emend. Flaksb.
1935, l.c.: 115. — T. aestivum var. atriceps (Koern.) Mansf. 1951, l.c.:
52. (23).
Геогр. распр. Армения. По предположению Al. Г. Туманяна
(1928), завезена сюда пз Ирака.
Var. atrocyanum Thum. 1928, l.c.: 270. — T. aestivum var. atrocya-
niim (Thum) Mansf, 1951, 1, c.: 51. (30).
Геогр. распр. Закавказье (Армения), Малая Азия.
Var. cabristanicum Dekapr. 1924, 1-1зв. Тифл. политехи, ин-та, 1: 3.—
7". aestivum var. cabristanicum (Dekapr.) Mansf. 1951, l.c.: 52. (21).
Геогр. распр. Азербайджан, Средняя Азия, Казахстан.
Var. cyanorubrum Thum. 1933, Опред. хлсбп. злаков (колос.):
199. (20).
Геогр. распр. Армения.
Var. echinoideum Koern. in Koern. et Wern. 1885, l.c.: 54, nom. mnta-
tum — T. aestivum var. echinopsis Mansf. 1951, I. c.: 43. (34).
Геогр. распр. Афганистан, Средняя Азия, Закавказье, Малая
Азия, Китай.
Var. erinaceuin (Desv.) Koern. in Koern, et Wern. 1885, I. c.: 54 — T. sa-
tivum erinaceum Desv. 1833, From.: 161. — T. aestivum var. erinaceuiu
(Desv.) Mansf. 1951, l.c.: 52. (18).
Геогр. p а с в p. Испания, Сицилия, Греция, Средняя Азия, Вос-
1очиый Казахстан, Афганистан, Заканкаше, Малая Азия, Израиль, Кид
гай, Монголия, изредка в других pci ионах.
Var. erivanense Thum. 1928, l.c.: 272.-- T. aestivum var. erivanense
(Thum.) Mansf. 1951, l.c.: 49. (29).
Геогр. j) а спр. Армения.
К о л о сья безо с 7 ые
Чешуи неопушенные (зерновки белые)
+ I — ! — ! — ' — I — i — I — | — | — I karsiense ' 1
Чешуи опушенные (зерновки белые)
+ I — — I — I — ! — I — | — | — i — ; ntulicolinuza
Колосья по л у о сти стые
Чешуи неопушенные
3 е р и о в к и б е л ы е
+ I — I — | — I — I — I — i I I- I — I subsplendcns
— I — 1+ I — I — I — I — | — |+ I — I subfetissovii
Зерновки красные
+ — I — I — I — I — I — I — I + I — I stibiclerinum
— I — I + I — I — I — I — I — | + ! — I suberinaceum
4
Чешуи опушенные
Зерновки белые
+I--I-I-I-I-I- 1 + 1 — I subsericeum
-1-1 +1 - 1 - 1 - 1 - 1 - 1 + | — 1 sub rtibrice ps 1 8
Зерновки крас и ы е
1 subalbiceps 1 9
-l-l + l- l- l- l- l- l + l- 1 subechinoideum 1 10
Колосья остистые
Чешуи неопушенные
Зерновки бе л ы е
+ — — — — — — — — splendidorigidutn 11
— — + — — —— — — + — fetissovii 12
— — + — — — — — — -i- pseudo}etissovii 13
— — — — + — — — + — griseosplendens 14
— — — — — -1- — — + — griseofetissovii 15
— — + setosum 16
3 e p H 0 в к и < p а с Il Ы c
+ — 4- — asiaticicterinum 17
— — + — — — — + — erinaceum. 18
— — + — — — — — -1- pseiidoerinaceum 19
— — — — + — — — + — cyanorubrtun 20
— — — — — + — — — + cabristanicuin 21
— — — — — + — — 4- glabrocyanum 99
— — — — — — — + — + atriceps 23
Чешуи опушенные
3 е р п о в к и б е л ы с
+ — — — — — — — + — sericctini 21
+ — — — — — — — 4- siirchiiinuin 25
—. 4 — — — — -— — 4- —• armeniaciiin 26
— + — — — — — + — rubriccps 27
— — -1- — — — — — — ; pseudorubriceps 28
— — — + —. — — — 4 — erivanense 29
— — — — — — 4- — — + atrocyunum 30
— — — — — — — + — 4- flaksbergeri 31
Ilodpor) 1. TriHeum
221
Зернов к и к р а с н ы е
+ — — — — — — — -1- — albiceps 32
+ — — — — — — — + kanaschii 33
— — 4~ — — — — — 4- — echinoideum 34
— — — — .— — — — 4 kerkianum 35
— — — d- — — — — — orient ale 36
— — — — — — — 4- — + nigriveliitinum 37
Var. fetissovii Koern. in Koern. et Wern. 1885, l.c.: 54. — T. aestivum
var. fetissowii (Koern.) Mansf. 1951, 1. c.: 53. (12).
Геогр. распр. Средняя Азия, Афганистан, Малая Азия, Закав-
казье, Казахстан, Монголия, Китай.
Var. flaksbergeri Thum. 1928, 1. с.: 272. — 7'. aestivum var. flaksbergeri
(Thum.) Mansf. 1951, 1. c.: 49. (31).
Геогр. распр. Закавказье (Армения), Малая Азия.
Var. glabrocyanum Thum. 1933, 1. с.: 198. (22).
Геогр. распр. Армения. По предположению М. Г. Туманяна
(1933), завезена сюда из Ирака.
Var. griseofetissovii Flaksb. 1935, Культ, фл. СССР, 1: 114, descr.
ross. — Т. aestivum var. griseofetissovii (Flaksb.) Mansf. 1951, l.c.: 53,
comb, invalid. (15).
Геогр. распр. Средняя Азия.
Var. griseosplendens Flaksb. 1915, Опред. пшен.: 166. — T. aestivum
var. griseosplendens (Flaksb.) Mansf. 1951, 1. c.: 55. (14).
Геогр. распр. Средняя Азия.
Var. kanaschii Kob. 1928, Tp. прикл. бот. ген. сел. 19, 1: 92. — Т. com-
pactum var. anatolicum Haciz. 1932, Der Ziichter, 4,3: 76—T. aestivum
var. kanaschii (Kob.) Mansf. 1951, 1. c.: 50. (33).
Геогр. распр. Афганистан, Средняя Азия, Сирия, Малая Азия,
Закавказье.
Var. karsiense (Flaksb.) Л. Filat, comb. nov. — T. compactum var. huin-
bolclti karsiense Flaksb. 1922, Опред. наст, хлебов: 47. (1).
Геогр. распр. Средняя Азия, Западная Азия.
Var. kerkianuni (Flaksb.) Thum. 1928, l.c.: 270. — T. vulgare var. ker-
kianum Flaksb. 1922, Опред. наст, хлебов: 49. — T. aestivum var. kerkia-
num (Flaksb.) Mansf. 1951,1. c.: 43. (35).
Геогр. распр. Афганистан, Средняя Азия, Армения, Сирия.
Var. muticolinaza Kudr. 1942, 1. с.: 53. (2).
Геогр. распр. Средняя Азия, Малая Азия, Закавказье, Восточный
Казахстан, Монголия.
Var. nigrivelutinuin Thum. 1933, I. с.: 201. (37).
Геогр. распр. Закавказье (Армения), Малая Азия.
Var. orientate Thum. 1933, l.c.: 201. — T. aestivum var. orientate
(Thum.) Mansf. 1951, I. c.: 43. (36).
Геогр. распр. Армения.
Var. pseudoerinaceum Haciz. 1932, l.c.: 76. T. aestivum var. pseudoe-
rinaceum (Haciz.) Mansf. 1951, l.c.: 52. (19).
Геогр. p a c ii p. Малая Азия.
Var. pseudofetissovii Kob. 1932, Хлебп. злаки Турции: 33. — T. aesti-
vumvar. pseudofetissovii (Kob.) Mansf. 1951, I. c.: 53. (13).
Г e о г p. p а с п p. Малая Азия.
222 К/иитификиция /к>д(1 Ti itiruiii /..
V;ir. psendorubi iceps (I'I;iksb.) TI111111. 1924, I. c.: 269. T. milpui e \ ar.
pseudorubriceps Flaksb. 1922, l.c.: 49.-- /'. aestivum var. pscudortibnceps
(Flaksb.) Mansf. 1951, 1. c.: 49. (28).
Геогр. распр. Средняя Азия, Афганистан, Закавказье, Малая
Азия, Израиль, Иордания.
Var. rtibriceps Koern. in Koern. ct Worn. 1885, 1. c.: 54. — T. aestivum
var. rtibriceps (Koern.) Mansf. 1951, l.c.: 49. (27).
Геогр. распр. Средняя Азия, Восточный Казахстан, Афганистан,
Индия, Закавказье, Малая Азия, Кипр.
Var. sericeum (Alef.) Koern. in Koern. cl Wern. 1885, l.c.: 54.—
T. vulgare var. sericeum Alef. I860, Landwirtsch. Fl.: 328. — T. aestivum
var. sericeum (Alef.) Mansf. 1951, 1. c.: 51. (24).
Описана по материалам из (ботанических садов.
Геогр. распр. Закавказье (Армения), Средняя Азия. Турция, Аф-
ганистан, Индия.
Var. setosum Thum. 1933, 1. с.: 201. (16).
Геогр. распр. Армения.
Var. splendidorisridtini Kudr. 1912, l.c.: 19. (11).
Геогр. распр- Средняя Азия, Афганистан, Армения.
Var. subalbiceps Vav. et Kob. 1928, Tp. прикл. бот. ген. сел. 19. 1: 93.—
Т. aestivum var. subalbiceps (Vav. et Kob.) Mansf. 1951, l.c.: 51. (9).
Геогр. распр. Афганистан (1400—1900 м над ур. м.).
Var. subechinoideum Vav. el Kob. 1928, l.c.: 93, nom. mutatum.—
T. aestivum var. subechinodes (Vav. et Kob.) Mansf. 1951, l.c.: 49. (10).
Г еогр. распр. Афганистан (1400 -2100 м над ур. м.).
Var. suberinaceuni Vav. et Kob. 1928, 1. c.: 92. — T. aestivum var. suberi-
naceuni (Vav. et Kob.) Mansf. 1951, I. c.: 52. (6).
Геогр. распр. Афганистан (1400—2900 м над ур. м.), Восточный
Казахстан, Таджикистан.
Var. subfetissovii Vav. et Kob. 1928, 1. c.: 92. — T. aestivum var. subfe-
tisowii (Vav. et Kob.) Mansf. 1951, 1. c.: 53. (4)-
Геогр. распр. Афганистан, Средняя Азия, Восточный Казахстан.
V ar. subicterinuni Vav. el Kob. 1928, 1. c.: 92. — T. aestivum var. subic-
terinum (Vav. et Kob.) Mansf. 1951, I. c.: 54. (5).
Г corp, pa си]). Афганистан (1400—2000 м над ур. м.), Узбекистан.
V ar. subrubriceps Vav. et Kob. 1928, I. c.: 93. — T. aestivum var. subru-
briceps (Vav. et Kob.) Mansf, 1951, 1. c.: 50. (8).
Геогр. распр. Афганистан, Туркмения.
V ar. subsericeuin Vav. ct Kob. 1928, 1. c.: 93. — T. aestivum var. subseri-
ceum (Vav. et Kob.) Mansf. 1951, 1. c.: 52. (7)..
Геогр. распр. Афганистан, Таджикистан, Узбекистан.
V ar. subsplendens Vav. et Kob. 1928, l.c.: 92. — T. aestivum var. sub-
splendens (Vav. et Kob.) Mansf. 1951, 1. c.: 55. (3).
Геогр. распр. Афганистан (1400—1900 м над ур. м.). Казахстан.
Var. surchianum (Flaksb.) Thum. 1928, 1. с.: 262. — 'Г. vulgare var. sur-
chianum Flaksb. 1922, l.c.: 48. — T. aestivum var. surchianum (Flaksb.)
Mansf. 1951, 1. c.: 51. (25).
Г corp, распр. Таджикистан, Афганистан, Малая Азия, Армения.
б. Subconvar. eligulatum Flaksb. — азиатская безлигульная подгруп-
па разновидностей.
Subconvar. eligulatuin Flaksb. 1929, Тр. прикл. бот. ген. сел. 20: 98,
cxcl. plantae inflatae.
Растения с колосьями нолугрубого построения, с листьями без языч-
ков и ушек у основания пластппк I. Яровые.
Тип: 7’. compactum var. quasialbiceps Flaksb.
Геогр. распр. Таджикистан (редко па высоте 2000 м над ур. м.).
Var. bardarai Nipjni. 1973, Докл, АН ТаджССР, 16, 9: 72. (4).
Геогр. рас и р. Памир.
К.ии сифшащи:! i>i>i)u Tiilii iuii
4 сш у и и con I/ in ci 111 hi c
3 e p и о в к ii 6 c J ы e
— — -I- | — — — — — ; r ' -- I llliasijclissovii 1
3 e p ii о в к и крас п ы с
+ I — ' — [ — —- , — — 1 — | + — , sarezicitin 2
Чешуи опушенные (зерновки красные)
+ I — | -- I - I I — I -- I — I + I - I c/iiusiiilln'ceys I 3
— I — I + I - ~ I — I - I — I " | + I — I bnrdiirai I I
Var. quasialbiceps Flaksb. 1929, Tp. прикл. оот. ген. сел. 20: 101.—
T. aestivum var. quasialbiceps (Flaksb.) Mansf. 1951, l.c.: 51 (3).
Геогр. распр. Памир (2000 м падур. и.).
Var. quasifetissovii Flaksb. 1929, l.c.: 101. — T. aestivum var. quasi-
fetissovii (Flaksb.) Mansf. 1951, 1. c.: 53. (1).
Геогр. распр. Памир (2000—3000 м над ур. м.).
Var. sarezicuni Nigm. 1973, I. с.: 72. (2).
Г с о г р. р а с п р. Памир.
2. Convar. inflatiim Vav. et Kob.— инфлятпая группа разновидностей.
Convar. inflatiim Vav. et Kob. 1928, Tp. прикл. бот. ген. сел. 19, 1: 93,—
Т. compactum subsp. iranoasiaiictun Flaksb. 1935, Культ, фл. СССР, 1:
101, quoad prol. inflatiim. — T. afghanicum Kudr. 1941, Бот. мат. (Таш-
кент) 4: 19. — T. compactum prol. pamirobadakschanicum Vav. 1904, Ми-
ров. раст. ресурсы. Пшеница: (53, descr. ross.
Растения полугрубого построения, средиерослые или высокорослые,
с соломиной средней прочности. Листья короткие, широкие, у перехода
влагалища листа в пластинку имеются язычок и ушки пли они отсутст-
вуют. Колос плотный, широкий. Колосковые чешуи вздутые, укорочен-
ные и широкие; килевой зубец загнут внутрь. Нижние цветковые чешуи
с расширенными у основания остевидпыми заострениями пли остями,
обычно изогнутыми, деформированными. Зерновки плотно заключены
в чешуях, не осыпаются, по легко обмолачиваются.
Инфлятпая группа исключительно восприимчива к грибным заболе-
ваниям при посеве в лесных, лесостенных и стенных районах СССР.
Т и п. Т. compactum var. roshanum Korzh.
Геогр. распр. Высокогорные районы Афганистана и Таджикиста-
на. В Таджикистане инфлятпая группа ныне практически исчезла.
Ключ для определения подгрупп разновидностей
convar. inflatiim Vav. et Kob.
1. Листья с язычками и ушками у основания пластинки...................
..............' . . . subconvar. roshanum (Korzh.) A. Filat, el Dorof.
+ Листья без язычков и ушек у основания пластинки ....................
..............................subconvar. bartangiense A. Filat. el Dorof.
a. Subconvar. roshanum (Korzh.) A. Filat, et Dorof. инфляция ли-
гул иная подгруппа разновидностей.
Subconvar. roshanum (Korzh.) A. Filat, et Dorof. comb. nov. —
T. compactum prol. roshanum (Korzh.) Flaksb. 1939, Опред. паст., хле-
бов: 181.
224
Классификация рода Trilicum L.
Определение разновидностей subconvar. roslianum (Korzli.) Л. f'ilat. et Dorof.
К о л о с ь я бе я о с т ci е Чешуи неопушенные
Зерновки бел ы е
+ 1 “ 1 — 1 — 1 - 1 humboldtinflatum 1 1
- 1 - 1 + 3 е рновки крас и ы с rufulinf latum | 2
+ 1 — — - 1 werncrinflafum 3
+ 1 3 Чешуи опушенные е р н о в к и бел ы е roshanum 4
4- — — —- — — — — — — vaeilooianum 5
— — + — — — — — — — crassicepsinf latum 6
— 4- 3 е Р II О к и р а с н ы е griseovavilovianum 7
+ — — — — — — — — — wittmackinflatum 8
— — + — — —- — — — — rubrinf latum 9
Ко — 1 + лосья с 3 У к Jeiuyu е р н р о ч неоп Э В К II в Н Н 0 JIUCHH бел 1 м и ые ы с nepalicum ост я м II 10
+ - - - + — subsplendensinf latum и
- + - - Зе р и о в к и + 1 “ красные subfetissovinflatum 12
+ — - - 1 + - i subicl erinf lai um 13
+ - - 3 — 1 — 1 + 1 — Чешуи опушенные е р н о в к и бел ы е | creliciroshanum 14
+ — — — — — — — + — subsericinf latum 15
— — + — — —. — 1 •• — + montanum 16
— — + — — Зе Р II О в к и + к р а с п ы е subrubricepsinf latum 17
4- — — — — — — —- + —- subalbicepsinflalu m 18
— — + — — — — —. + — subechinoidinf latum 19
+ — — —-• — — — — —— + miguschovae 20
+ к О ло с Чешу1 е р и Ь Я О неоп о В К I стис утенн б с л + т ы е ые ы с bukiniezii 21
— — + - — - Зе - 1 ~ рновки + < р а с и ы е fetissovinflatum ; 22
+ — - 1 — 1 — 1 + 1 •"" 1 icierinflalum 23
+ - 1 -1 -1 + 1 - Чешуи опушенные 5 е р и о и к и б е л ы о | erinacinflatum 24
— 1 - 1 + 1 - j — 3 р Н 0 в к и 1 + крас 1 - н ы е | rubricepsinjlatum ! 25
4- . __ — — — + — albicepsinf latum 26
+ .— — + — echinoidinflatum 27
— — + — — — — - — — + kerkianinjlatum 28
I. T/itiriiiii 225
Растения с обычным для пнкчншы с троенном листа па месте пе-
рехода влагалища листа в пластинку ясно выражена перетяжка; листо-
вая пластинка отогнута от стебля; у основания листовой пластинки рас-
положены язычок и ушки.
'Г и и: 7\ compactum var. roshaiium Korzh.
Геогр. распр. По всему ареалу Т. compactum convar. inf tat um.
Var. albicepsinflatum Vav. et Kob. 1928, Tp. прикл. бот. ген. сел. 19,
1: 94. — Т. aestivum var. albicepsinflatum (Vav. et Kob.) Mansf. 1951,
Der Ziichter, 21, 1/2: 51. (26).
Г с о г p. p а с п p. Афганистан. Таджикистан.
Var. bukiniezii Vav. ct Kob. 1928, I. c.: 97. — T. aestivum var. bukiniezii
(Vav. et. Kob.) Mansf. 1951, 1. c.: 49. (21).
Геогр. распр. Афганистан.
Var. crassicepsinflatuin Vav. et Kob. 1928, l.c.: 95. — T, aestivum var.
crassicepsinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 56. (6).
Геогр. распр. Афганистан (1400—2400 м над ур. м.).
Var. creticiroshanum Kob. et Vav. 1928, l.c.: 96. — T. compactum var.
suberinaceuminf latum Palm, in Flaksb. 1935, Культ, фл. СССР. 1: 115,
descr. ross. (14).
Г e о г p. p а с п p. Афганистан, Таджикистан, Монголия.
V ar. echinoidinflatum Vav. et Kob. 1928, l.c.: 94. — T. aestivum var.
echinoidinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 49. (27).
Геогр. распр. Афганистан (1400—2400 м над ур. м.).
V ar. erinacinflatum Vav. et Kob. 1928, l.c.: 94. — T. aestivum var. eri-
naceuminflatum (Vav. ct Kob.) Mansf. 1951, 1. c.: 52. (24).
Г e о г p. распр. Афганистан, Узбекистан.
V ar. fetissovinflatum Vav. et Kob. 1928, 1. c.: 94. — T. aestivum var. [e-
tissovinflatuin (Vav. et Kob.) Mansf. 1951, 1. c.: 53. (22).
Геогр. распр. Афганистан (1400—2400 м над ур. и.).
V ar. griseovavilovianum Udacz. 1979, Бюл. ВИРа, 84: 71. (7).
Геогр. распр. Таджикистан.
V ar. humboldtinflatum Vav. et Kob. 1928, l.c.: 93. — T. aestivum var.
humboldtinflatum (Vav. et Kob.) Mansf. 1951, l.c.: 58. (1).
Геогр. распр. Афганистан (1400—1900 м над ур. м.).
V ar. icterinflatum Vav. et Kob. 1928, 1. c.: 94. — T. aestivum var. icteri-
numinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 54. (23).
Геогр. распр. Афганистан (1400—1900 м над ур. м.).
V ar. kerkianinflatum Vav. et Kob. 1928. 1. c.: 95. — T. aestivum var. ker-
kiatiuminf latum (Vav. et Kob.) Mansf. 1951, 1. c.: 49. (28).
Геогр. распр. Афганистан.
V ar. miguschovae Dorof. et A. Filat, nom. nov. — T. compactum, var.
album Vav. et Kob. 1928, 1. c.: 97, non Link, 1825. (20).
Геогр. p а с п p. Афганистан.
V ar. montanum Vav. et Kob. 1928, l.c.: 97. — T. aestivum var. monta-
num (Vav. et Kob.) Mansf. 1951, 1. c.: 50. (16).
Геогр. распр. Афганистан, Таджикистан.
Var. nepalicum Udacz. 1976, Tp. прикл. бот. ген. сел. 56,2: 149. (10).
Геогр. распр. Непал.
Var. roshanum Korzh. 1898, Очерк Рошана и Шугпапа, СПб.: 13,
quoad plantam exaristatam. — Т. compactum var. creticuminflatum Vav.
et Kob. 1928. l.c.: 94. — T. compactum var. korshinskianum Flaksb. 1939,
Опред. пшен.: 184. — T. aestivum var. roschanum (Korsh.) Mansf. 1951,
I. c.: 57. (4).
Г corp, рас ii]>. Афганистан, Таджикистан.
Var. rubricepsinflatum Vav. et Kob. 1928, l.c.: 94. — T. aestivum var.
rubricepsinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 50. (25).
Геогр. распр. Афганистан.
15 762
226 Клш сифшащи:) porta Tritiium
Var. riibrinfkituii) Vav. et Kob. 1928, l.c.: 94. — 7'. aestivum var. rnh-
ruminflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 55. (9).
Геогр. распр. /Афганистан. (MOO-- 1900 м над ур. м.).
Var. rufulinflatum Vav. et Kob. 1928, l.c.: 94. — T. aestivum var. rufti-
luminf latum (Vav. et Kob.) Mansf. 1951, 1. c. 57. (2).
Г e о г p. p а с н p. Афганистан.
Var. subalbicepsinflatum Vav. et Kob. 1928, 1. c.: 9G. — T. aestivum var.
subalbicepsinflalum (Vav. et Kob.) Mansf. 1951, 1. c.: 51. (18).
Геогр. распр. Афганистан.
Var. subechinoidinflatum Vav. et Kob. 1928, I. c.: 96. T. aestivum var.
subechinodesinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 49. (19).
Геогр. распр. Афганистан.
Var. subfetissovinflatum Vav. et Kob. 1928 1. c.: 96. — T. aestivum var.
subfetissoviinflatum (Vav. et Kob.) Mansf. 1951, 1. c.: 53. (12).
Геогр. рас и p. Афганистан, Таджикистан.
Var. subicterinfIatnni Vav. et Kob. 1928, I. c.: 95. — T. aestivum var. su-
bicterinuminf latum (Vav. et Kob.) Mansf. 1951, 1. c.: 54. (13).
Г e о г p. p а с п p. Афганистан.
Var. subrubricepsinflatum Vav. et Kob. 1928, 1. c.: 96. T. aestivum var.
subrubricepsinf latum (Vav. et Kob.) Mansf. 1951, l.c.: 50. (17).
Геогр. распр. Афганистан.
Var. subsericinflatum Vav. et Kob. 1928, l.c.: 96. — T. aestivum var.
subsericeuminflatum (Vav. et Kob.) Mansf. 1951, l.c.: 52. (15).
Геогр. распр. Афганистан.
Var. sabsplendensinflatum Vav. et Kob. 1928, l.c.: 95. — T. aestivum
var. sabsplendensinflatum (Vav. et Kob.) Mansf. 1951, l.c.: 55. (11).
Геогр. распр. Афганистан.
Var. vavilovianum Udacz. 1969, Tp. прикл. бот. ген. сел. 40, 2: 42. (5).
Геогр. распр. Таджикистан.
Var. wernerinflatum Vav. et Kob. 1928, l.c.: 93. — T. aestivum var.
wernerianuminflatum (Vav. et Kob.) Mansf. 1951, l.c.: 57.. (3).
Геогр. распр. Афганистан.
Var. wittmackinflatum Vav. et Kob. 1928, l.c.: 94. — T. aestivum var.
wittmackianuminf latum (Vav. et Kob.) Mansf. 1951, l.c.: 56. (8).
Геогр. распр. /Афганистан (1400—1900 м над ур. м.).
б. Subconvar. bartangiense Л. Filat, et Dorof. — инфлятпая безлигуль-
пая подгруппа разновидностей.
Subconvar. bartangiense A. Filat, et Dorof. subconvar. nov. — T. com-
pactum eligulatum Flaksb. 1929, Tp. прикл. бот. ген. сел. 20: 98, quoad
plantain inflatam. — T. compactum subprol. eligulatum Flaksb. 1935,
Культ, фл. СССР, 1: 102.—T. compactum subprol. eligulatoioschanum
Flaksb. 1939, Опред. паст, хлебов: 181, descr. ross. — Folia eligulata,
sursum spectantes.
Растения с ппфлятиыми колосьями, с листьями без язычков и ушек
у основания пластинки.
Тип: Т. compactum var. bartangiense Flaksb.
Геогр. распр. Высокогорные районы Памира (2000—2900 м над
ур. м.).
Var. bartangiense Flaksb. 1929, Тр. прикл. бот. ген. сел. 20: 100.—
Т. aestivum var. bartangiense (Flaksb.) Mansf. 1951, Der Ziichter, 21,
1/2:57. (1).
Геогр. распр. Таджикистан (2200 2900 м над ур. м.).
Var. gorbnnovii Flaksb. 1929, l.c.: 99. T. aestivum var. gorbuiiovii
(Flaksb.) Mansf. 1951, I. c.: 57. (2).
В долине p. Язгулем па высоте 2000 м над ур. м. обнаружена форма
с признаком инфлятиости — f. quasifalsefetissovii Flaksb. 1929, l.c.: 101.
Г e о г р . рас и р. Таджикистан.
11 (>()/!<>() I. Triticillll
227
Определение разновидностей subconvar. bartanpieiise Л. Filat, et Dorof.
- ,' + |- | — | — | — I — I — I — I bartangiense । 1
Зерновки красные
— I — I + I — I — I — I — I — I — I — I gorbunovii I 2
— ) — I — | — I — I + I — I — I — I — I karakurganiense | 3
Колосья остистые (чешуи неопушенные, зерновки белые)
— | — I + I — ; — f — I — | — ! + I — | jiisgulciuiciise j -I
Var. jasgulemiense Flaksb. 1929, l.c.: 101. — T. aestivum var. jasgule-
miense (Flaksb.) Mansf. 1951, 1. c.: 53. (4).
Геогр. распр. Таджикистан.
Var. karakurganiense Flaksb. 1929, l.c.: 100. — T. aestivum var. kara-
kurgani (Flaksb.) Mansf. 1951, 1. c.: 57. (3).
Геогр. распр. Таджикистан.
3. Convar. compactum — европейская группа разновидностей.
Convar. compactum. — T. compactum subsp. compactum-, Цвелев, 1973,
Новости спет. высш. pact. 10: 44. — Т. compactum subsp. indoeuropaettm
Flaksb. 1935, Культ, фл. СССР, 1: 102, descr. ross. — T. compactum subsp.
eurasiaticuni Vav. 1964, Миров, раст. ресурсы. Пшеница: 63, nom. illeg.
Растения и колосья нежного построения, колосковые чешуи обычно
тонкокожистые, у основания с поперечной вдавленпостыо. Зерновки не-
плотно заключены в чешуях, легко обмолачиваются и осыпаются.
Геогр. распр. Рассеянно по всему Старому Свету. Почти совер-
шенно выпала из культуры в Европе. В начале века редко возделывалась
в Сибири, на Дальнем Востоке, в Якутии, Монголии, Китае. В прошлом
веке была завезена в Южную и Северную Америку.
Пшеница компактная, с тонкокожистыми чешуями, нс связана с опре-
деленной территорией и представляет, по словам II. И. Вавилова (1964),
в значительной степени реликты. Однако Н. II. Вавилов выделял не-
сколько эколого-географических групп.
а. Западноевропейская группа (prol. occidentate-europaeum\Jav. 1964,
1. с.: 64, descr. ross.) характеризуется позднеспелостью н относительной
восприимчивостью к желтой и бурой ржавчине. Растения низкорослые,
хорошо облиственные, соломина прочная. В колосках нередко формиру-
ется по 4—5 зерен. Преимущественно озимые формы.
б. Якутская группа (prol. jacuticum Vav. 1964, l.c.: 63, descr. ross.)
довольно распространена в Сибири. Отличается скороспелостью, вынос-
ливостью к низким температурам. Соломина тонкая, но устойчивая к по-
леганию, средней длины. Образ жизни яровой.
в. Тяпыпапьская группа (prol. tiatiscliatiicum Vav. 1964, 1. с.: 63,
descr. ross.) позднеспелая, средпсвоспрпимчпна к желтой ржавчине. Со-
ломина прочная. Образ жизни преимущественно озимый. Приурочена
главным образом к Западному Тянь-Шаню.
г. Маньчжурская группа (prol. manchschurictim Vav. 1964, l.c.: 63,
descr. ross.) скороспелая, сравпптелыю устойчива к бурой и желтой
15*
228
I нфнн/щин poilll Till ill! Ill I..
J) жа B’u 11 le. Возделывается в небольшом количес гве в Северо-Восточном
Китае.
д. Ки । a lie кая i руина выделялась 11. II. Вавиловым как подвид ('I. сонь
paclu.nL subsp. siniciiin Vav. 1964, l.c.: 64, descr. ross.) ввиду исключи-
тельного разнообразия этой пшеницы по плотности колоса. У от oil груп-
пы, так же как у внутреппеапаюлпйской группы мягких пшениц, наблю-
дается большая амплитуда колебаний по плотности колоса, много форм,'
переходных от пшеницы мягкой к компактной. В целом группе свойст-
венны скороспелость, короткая, топкая соломина, колос, мелкий, часто
многоцветковый.
Велико разнообразие по форме колоса, длине остей. Зерновка мелкая.
Кроме того, Н. И. Вавилов выделяет американскую группу гибридно-
го происхождения, которую ввиду нечеткого выражения признаков труд-
но определенно отнести к той или иной группе разновидностей.
Колосья безостые.
Чешуи неопушенные
Зернов '< п б е л ы е
i|z|y|2|z| = |z| = | = |z|
1
2
3 е р и о и к и к р а с н и е
wern erianum
creticum
.3
4
linaza
(iraraticum
crcissicepx
pseudor rnssiceps
urarticum
lirasdanicnin
wagarschapali
witiniackianum
rubrum
pseudoriibrttm
clavatum
atrum
6
7
8
9
10
11
12
13
14
15
16
Колосья остистые
Чешуи неопушенные
Зерновки бе л ы е
— I — I"— I — I — I — I + I — i -f- i atrocrinaceuni
23
Окраска остспидпых заострении.
Jloilpoi) I. I'l'iUruni 329
У (-и ] > (> i le lie к <) li 1111 [ <' 1111111.1 ком и а к 11 loii 111 > с ч >бл; ।; i ;i к 11 6e <oc I ijo разно
видное i и. ,Y азиатских представителей згою вида пшеницы разновид-
ностпып состав богаче, чем у европейских.
Var. araraticum Thum. 1928, Тр. прикл. бог. геи. сел. 19, 1: 248.—
Т. aestivum var. araraticum (Thum.) Mansf. 1951, Dcr Ziichter, 21, 1/2:
56. (6).
Геогр. распр. Армения (редкая примесь в посевах выше 1000 м
над ур. м.).
V ar. atroerinaceuin Koern. 1908, Arch. Biontol. 2: 400. — T. aestivum
var. atroerinaceuin (Koern.) Mansf. 1951. 1. c.: 49. (23).
Описана по гибридному материалу из ботанического сада.
V ar. atrum Koern. 1908, 1. с.: 398; Фляксбергер, 1935, Культ, фл. СССР,
1: 109. — Т. aestivum vaf. atrum (Koern.) Mansf. 1951, l.c.: 55. (16).
Описана по гибридному материалу из ботанического сада.
По сообщению J. Percival (1921), изредка встречалась в США.
V ar. clavatum (Alef.) Flaksb. 1915, Опред. шлеи.: 165.— Т. vulgare
clavatum Alef. 1866, Landwirtsch. Fl.: 328.— T. aestivum var. clavatum
(Alef.) Mansf. 1955, 1. c.: 55. (15).
Геогр. распр. Встречается редко, точных указаний, где именно,
нс имеется.
Var. compactum. — Т. compactum var. aureuni Link, 1825, Hort. Berol.
1: 25. — T. compactum var. album Link, 1825,. l.c.: 25.— T. vulgare var.
splendens Alef. 1866, l.c.: 328.— T. compactum var. splendens (Alef.).
Koern. in Koern. et Wern. 1885, Handb. Getreidcb. 1: 53. — T. aestivum var.
splendens (Alef.) Mansf. 1951, 1. c.: 54. (17).
Геогр. расп p. Западная Европа, острова Средиземного моря, Ма-
лая Азия, Грузия, Северный Кавказ, Канада.
Var. crassiceps Koern. in Koern. et Wern. 1885, l.c.: 53. — T. aestivum
var. crassiceps (Koern.) Mansf. 1951,1. c.: 55. (7).
Геогр. распр. Редко в Монголии, Армении, Малой Азии, США.
Var. creticum (Mazz.) Koern. in Koern. et Wern. 1885, l.c.: 52.—
T. creticum Mazz. 1807, Sopra ale. sp. frum.: 2. — T. vulgare var. creticum
Alef. 1866, l.c.: 328. — T. aestivum var. creticum (Mazz.) Mansf. 1951,
I. c.: 57. (4).
Геогр. распр. По всему ареалу Т. compactum.
Var. griseoicterinum Flaksb. 1935, l.c.: 112, descr. ross.— T. aestivum
var. griseoicterinum (Flaksb.) Mansf. 1951, l.c.: 54, comb, invalid. (21).
Геогр. рас и p. Сибирь, Дальний Восток.
Var. hrasdanicum Thum. 1933. Опред. хлебн. злаков (колос.):
196. — Т. aestivum var. hrasdanicum (Thum.) Mansf. 1951, 1. c.: 56. (10).
Геогр. распр. Армения.
V ar. humboldtii Koern. 1873. Syst. Uebers. Ccr.: 12; idem, in Koern. ct
Wern. 1885, 1. c.: 52. — T. aestivum var. humboldtii (Koern.) Mansf. 1951,
l.c.: 58. (1).
Геогр. p а с и p. США, Чили, редко в Европе.
V ar. icterinum (Alef.) Koern. in Koern. ct Wern. 1885, l.c.: 53.—
T. vulgare var. icterinum Alef. 1866, 1. c.: 325. — T. aestivum var. icterinum
(Alef.) Mansf. 1951, 1. c.: 53. (19).
Геогр. рарпр. Западная Европа, Сибирь, Закавказье (па высоте
2000 м над ур. м.), Китай, Южная Африка.
V ar. linaza Koern. in Koern. et Wern. 1885, 1. c.: 53. — T. aestivum var.
linaza (Koern.) Mansf. 1951, I. c.: 56. (5).
Геогр. распр. Чили, Монголия.
V ar. percivalii Flaksb. 1935, l.c.: 115,- -7’. compactum var. atriceps
Koern. 1885, l.c.: 54, quoad plantain gliirnis nigris, colore siiblitus al-
bo. (22).
Выделена из французского copra пшеницы компактов ’Ilerisson bar-
bu’ (J. Percival, 1921).
230 Кмсгификаци.ч ро<)а Tritiriim I,.
Var. psetidocrassitcps Kob. I932, Хлебн. г шкп Турпин.: 33. T. aesli
vumvav. pseudocrassiceps (Kob.) Mansf. 1951, I. c.: 56. (3).
Г e о i- p. p а с и p. Малая Азия.
Var. pseudoicterinum Greb. 1929, Опред. разило, пшен.: 15; Haciz.
1932, Dor Ziichter, 4: 76. — T. aestivum var. pseudoicterinum (Greb.)
Mansf. 1951, 1. c.: 54. (20).
Геогр. распр. Северный Кавказ, Закавказье (Армения), Малая
Азия.
Var. pseudorubrum Kob. 1932, I. с.: 33. - - T. aestivum var. pseudorub-
rum (Kob.) Mansf. 1951, l.c.: 55. (14).
Геогр. распр. Малая Азия (редко).
Var. pseudosplendens Ilaciz. 1932, l.c.: 76. — T. aestivum var. pscu-
dosplendens (Haciz.) Mansf. 1951, l.c.: 55. (IS).-
Геогр. распр. Малая Азия.
Var. rubruin Koern. in Koern. et Wern. 1885, 1. c.: 53. -- Г. aestivum var.
rubrum (Koern.) Mansf. 1951, 1. c.: 55. (13).
Геогр. распр. CILIA, Китай, ЛАалая Азия, очень редко в Закав-
казье.
Var. rtifulum Koern. in Koern. et Worn. 1885, l.c.: 52. --- T. aestivum
var. rufulum (Koern.) Mansf. 1951, 1. c.: 57. (2).
Геогр. распр. США, ЛАопголия, Китай, как примесь в Малой
Азии, Закавказье, редко в Европе.
Var. urarticum Timin. 1928, l.c.: 249. — T. aestivum var. urarticum
(Thum.) Mansf. 1951, 1. c.: 56. (9).
Геогр. распр. Армения (редкая примесь в посевах выше 1000 м
над ур. м.).
Var. wagarschapati Thum. 1928, l.c.: 248. — T. aestivum var. wagar-
schapati (Thum.) Mansf. 1951, 1. c.: 56. (11).
Геогр. распр. Закавказье (Армения), Малая Азия (редко).
Var. wernerianum Koern. in Koern. et Wern. 1885, 1. c.: 52. — T. aesti-
vum var. wernerianum (Koern.) Mansf. 1951, 1. c.: 57. (3).
Геогр. распр. США, Чили, Китай, Малая Азия, редко в Европе.
Var. wittniackianum Koern. in Koern. et Wern. 1885, 1. c.: 53. — T. aes-
tivum var. wittmackianum (Koern.) Mansf. 1951, I. c.: 56. (12).
Геогр. распр. Восточный Казахстан, США (редко).
Т. AESTIVUM L. — ПШЕНИЦА МЯГКАЯ
(ХЛЕБНАЯ, ХЛЕБОПЕКАРНАЯ, ОБЫКНОВЕННАЯ)
Т. aestivum L. 1753, Sp. Pl.: 85, emend. Fiori et Paol. 1896. Fl. Anal.
Ital. 1: 107. — T. hybernum L. 1753. l.c.: 86. — T. sativum Lam. 1778, Fl.
Fr. 3: 625. — T. vulgare Vill. 1787, Hist. Pl. Dauph. 2: 153, nom. illeg.—
T. cereale Schrank, 1789, Baier. Fl. 1: 387, non Salisb. — T. muticum
Schuebl. et T. aristatum Schuebl. 1818. Char, et descr. Cereal.: 10. — T. vul-
gare var.-gr. muticum (Schuebl.) Alef. 1866, Landwirtsch. Flora: 328. —
T. vulgare var.-gr. aristatum Alef. 1866, l.c.: 330. — T. sativum vulga-
re Hack, in Engl, et Prantl, 1887, Naturl. Pfianzenfam. 2, 2: 85. — Frumen-
tum triticum E. Krause, 1898, Bot. Centralbl. 73, 10: 339. — T. tenax, A. I.
vulgare Aschefs. et Graebn. 1901, Synops. 2: 683. — T. aestivum subsp. aes-
tivum Rassen vulgare (Vill.) Thell. 1918, Nalurwissensch. Wochcnschr. N. F.
17, 33: 471. — T. amplissifolium Zhuk. 1949, Докл. AH СССР, 69, 2: 261. --
T. aestivum subsp. vulgare (Vill.) Mac Key, 1954, Sv. Bot. Tidskr. 48:
586. — 7’. aestivum gr. aestivum Bowden, 1959, Canad. Journ. Bot. 37: 674.
Общая характеристика. Всходы темпо-зеленые, .зеленые, сипевато-
илп желтовато-зеленые. Листовые пластинки длиной 8—35 см (иногда
больше), шириной 0,7—2,5 см, голые (гладкие), покрыты шпиикамп (ше-
роховатые) или опушены очень короткими волосками (бархатистые на
ощупь). Листовые влагалища голые пли опушенные. На месте их пере-
I. Triiidim
хода в липовую плас, ивку имением язычок и ушки. Последние имеют
или пе имеют длинные реснички, иногда переходящие на края листовой
пластинки. Есть безлпгульные формы. Соломина высотой 30—200 см,
полая по всей длине, редко выполнена паренхимой под колосом. Узлы
соломины голые или опушены короткими, сгпраклцимпси волосками.
Колосья опушенные или неопушенные, длиной 5—17 см, от рыхлых
(D = 10—22) до плотных (D=23—38), с. легким вымолотом зерновок.
Форма колоса веретеповидвая, цилиндрическая, пирамидальная или бу-
лавовидная. Окраска колоса белая, желтая (соломенная), красная, серо-
дымчатая, черпая (иногда с синеватым отливом). Есть формы с черной
каймой по краю чешуи. Число колосков в колосе 14—28 (редко до 32).
Колоски 3—8-цветковые. В них развиваются 2—3, реже 5—6 зерновок.
Колосковые чешуи по форме овальные, ромбические или ланцетовидные.
Киль узкий, часто не доходит до основания колосковой чешуи, где обыч-
но имеется поперечная вдавлепиость с продольной морщинистостью,
вдавленпость отсутствует у ригидных форм. В верхней части киль пере-
ходит в килевой зубец, который может быть ог короткого тупого до ост-
рого длинного, иногда остсвидного (длиной до 5 см). Жилки слегка вы-
ступают над поверхностью колосковой чешуи. Главная боковая жилка
выражена более четко, она вверху закапчивается бугорком пли корот-
ким зубцом. Плечо колосковой чешуи скошенное, приподнятое или пря-
мое, иногда широкое. Наружная цветковая чешуя выпуклая, песет ость
или острый зубец (изредка вершина его тупая, без зубца). Внутренняя
цветковая чешуя двукилевая, при созревании не расщепляется вдоль на
2 части.
В колеоптиле пшеницы мягкой 2 проводящих пучка. Исследование
О. Д. Градчаниповой (1967) показало, что первая листовая пластинка
молодого растения с верхней поверхности имеет гофрировапность, ко-
торая становится меньшей в фазе выхода в трубку. В фазе колошения
второй лист сверху имеет хорошо дифференцированные проводящие
пучки, и механические элементы. Нижняя сторона листа (за исключени-
ем центральной жилки) имеет выравненную поверхность. На верхней
стороне листа имеются выросты эпидермиса — шипики (у сорта Диа-
мант их количество в среднем равно. 130,5 на 1 мм2) или единичные во-
лоски, а также бугорки. Анатомическое строение колосового стержня
сходно с тем, которое выше приведено для Т. compactum. Анатомичес-
кое строение вегетативных органов в целом сходно с таковым других
гексаплоидиых видов секции Triticum.
Пшеница мягкая — экологически наиболее пластичный вид. Опа су-
мела приспособиться к разнообразным климатическим условиям и по-
тому возделывается па всех континентах земного шара. П. М. Жуков-
ский (1971) отмечал, что пшеница мягкая имеет огромный экологичес-
кий потенциал. Если посевы пшеницы твердой в нашей стране ограни-
чены территорией плодородных степей и лесостепью юга, то посевы
пшеницы мягкой внедрились в солонцеватые степи, пустыни, полупус-
тыни и освобожденные от древостоя регионы лесной полосы Нечерно-
земья. Озимая мягкая пшеница тяготеет к более южным теплым и уме-
ренно прохладным районам, где устойчивый снежный покров, яровая —
возделывается 6 местах с континентальным климатом.
Пшеница мягкая имеет озимый, полуозимый н яровой (поздние, и
ранние) образ жизни. Обширный эксперимент, предпринятый по про-
мораживанию при —16°С образцов мировой коллекции, показал, что
генофонд пшеницы мягкой включает сорта, морозостойкость которых
колеблется в пределах 0-170% <л морозос!ойкости copia Миронов-
ская 808, что селекция в роде только у пшеницы мягкой продвинула мо-
розостойкость до уровня отечественных сортов Алабасская, Альбидум 114
и Лютесценс 329 (Э. А. Барашкова и др., 1977; 1'. В. Удовенко и др., 1978).
232 Классификации рода Triticum К.
В условиях Дю остана, но дивным В. II. Жукова (1969), уровши,
о! крытого цветения у пшеницы мягкой в целом ниже, чем у ди- и тетра-
плоидных видов. В зависимости от сорта он колебался от 18,4 до 98,6%,
Количество зацветших цветков нарастает постепенно, достигая макси-
мума в дополуденные и полуденные часы. Цветение продолжается 7—
8 дней (до 10 дней при прохладной погоде). Наблюдения, проведенные
Л. 3. Латыповым и В. И. Лпслем (1971) в Белоруссии, зафиксировали
расхождение цветковых чешуй при открытии цветка лишь па 2.1 мм;
цветок остается открытым в среднем 18 мин. А1. А. Федин (1974) счи-
тает, что склонность пшеницы мягкой к открытому цветению относится
к сортовым наследственно обусловленным признакам.
. Зерно пшеницы мягкой мучнистое, полустск.товидное и стекловид-
ное. Изучение, мировой коллекции в орошаемых условиях Ташкентской
области (В. Г. Коиарев и др., 1975) показало, что концентрация белка
в зерне колебалась от 8,8 до 23,9% в зависимости от сорта и года ре-
продукции, лизина в белке высокобелковых сортов 2,05—3,04 %. Содер-
жание лизина зарегистрировано доходящим до 4,28 (сорт Украинка) и
4,31% (к-13274 из Австралии). Масса 1000 зерен в mix условиях до-
ходила до 60 г (сорт Хозанэ из Таджикистана) и 60,9 г (сорт Cologna
из Италии). В богарных условиях в Поволжье (Кинель) в зерне накап-
ливалось 17—21,3% белка, 1,62—2,49% лизина в белке, масса 1000 зе-
рен колебалась в пределах 22—46 г (В. Ф. Дорофеев, и др., 1977).
Иммунитет. Пшеница мягкая неустойчива к комплекс}' болезней и
вредителей, ежегодно уносящих значительную часть се урожая. Тита-
нические усилия селекции до сих пор не дали производству сорта, по-
добного пшенице Тимофеева по иммунитету. Причина этого феноме-
на— в генетическом содержании пшеницы мягкой, в се геномном
составе. Меньшая устойчивость к грибным болезням гексаплоидных ви-
дов секции Triticum по сравнению с тетраплоидными видами секции
Dicoccoides связана с геномом D.
По данным Н. И. Вавилова (1961, 1964), сортимент пшеницы мяг-
кой Средней Азии, Афганистана, Внутреннего Ирана и Кашгара выде-
ляется по восприимчивости к грибным заболеваниям, а также по пора-
жаемое™ шведской и гессенской мухами. Он указал на пшеницу
мягкую средних и южных увлажненных районов Китая как на устойчи-
вую к бурой и желтой ржавчине. Однако последующие исследования
М. М. Якубцинера (1961, 1967) выявили у них отсутствие иммунитета
к этим патогенам.
Целенаправленная селекция создала ряд сортов, которые устойчивы
к некоторым расам бурой ржавчины. Поражаемое™ ею шедевра совет-
ской селекции озимого сорта Безостая 1 оценивается как поражаемое™
ниже среднего уровня. Ряд сортов Аргентины, в том числе Magni! MG,
выделяется в условиях СССР устойчивостью к бурой ржавчине. При-
влечение их в скрещивания способствовало выведению сортов Осетин-
ская 3, Осетинская 4, Эритроспермум Г 720, имеющих полевую устой-
чивость к бурой ржавчине («Пшеницы мира», 1976).
В полевых условиях Дагестана, по данным И. П. Шитовой (1968),
в период 1954—1965 гг. из 872 коллекционных образцов пшеницы мяг-
кой не выделено ни одного иммунного пли слабо поражаемого бурой
ржавчиной; Й7,2% образцов оказались восприимчивыми (баллы 3 и 4
по 5-балльной шкале). Заражение вирулентными расами 77, 20, 21,
116, 52, 57, 143, 174, 185 этого патогена коллекции пшеницы мягкой
(1323 образца) различных стран мира выявило лишь 5% устойчивых
образцов (О. Г. Григорьева, 1975). Исследование В. М. Берляил-Кожев-
ипкова и др. (1973) показало, что из 1900 образцов коллекции устой-
чивостью к расам 151, 77, 202 обладают 387 образцов, относящихся в
основном к сортам современной сслскпии. Причем мопогсипая устой-
чивость была характерна для 16,7%, дигеипая — для 3,7% образцов.
J. Trili( um
II. II. Вавилов (1961, 1991) в иолом характеризовал шиешшу тон-
кую как вил, восприимчивый к желтой и сюблевой ржавчине, а также к
мучнистой росе. При иноку.тяпни 1317 образцов коллекции расами 21,
34, 40, 11, 16 и 17 стеблевой ржавчины остойчивость обнаружена лини, v
6,5% образцов (О. Г. Григорьева, 1975). Заражение пророст ков 2189
образцов коллекции сборным ипокулюмом из смеси популяций мучнис-
той росы Алма-Атинской, Куйбышевской, Ленинградской, Новосибир-
ской областей п Северо-Осетинской АССР выявило 90,3% восприимчи-
вых образцов (балл 3 и 4); иммунность отмечена лини, у 1,4% образ-
цов. Все. иммунные и высокоустойчивые образцы (9.7%) в этом опыте
оказались продуктом межвидовой гибридизации (Э. X. Ссханбердппа,
1976).
В. И. Кривченко (1973) в опыте по заражению пыльной головней
выявил, что 5,2% изученных образцов были восприимчивыми к I. duri
и 33,9% — к f. aestivi, иммунных образцов обнаружено соответственно
62,4 и 10,5%. Всего из 2693 образцов пшеницы мягкой устойчивыми (0 и
1 балл) к пыльной головне были 815 образцов. А. М. Ямалеев (1971)
показал, что у устойчивых к пыльной головне образцов в искусственно
зараженных зерновках мицелий гриба не достигает зародышевой почки.
Ряд исследователей (А. А. Ячевский, 1910, 1930; I\. Е. Мурашкпи-
ский. 1925, 1932; Э. Э. Гешеле, 1935, 1964; АЕ 3. Ампи.тогов, 195% 1965;
Д. В. Мягкова, 1968) указали на варьирование устойчивости к твердой
головне у пшеницы мягкой. Искусственное заражение 3419 образцов
коллекции выявило 72,8% восприимчивых (3 и 4 балла) и 0,7% иммун-
ных образцов (В. И. Кривченко, 1973).
История. Хотя пшеница мягкая считается более молодым видом в
сравнении с тетраплоидными, данные археологических исследований сви-
детельствуют о глубокой ее древности. Так, па территории Иранского
Курдистана, как сообщает Н. Helbaek (1966), обнаруженные при архео-
логических раскопках следы пшеницы мягкой датируются 5500—5000 гг.
до н. э., а на территории Ирака, согласно его исследованиям (1965),—
5800—5600 гг. до н. э. Там же найдены растительные остатки Т. monococ-
cum и Т. dicoccum.
К доисторическому периоду (5550 г. до и. э.) относятся и находки в
в Турции (Юго-Восточная Анатолия). Вместе с пшеницей мягкой здесь
также были найдены обугленные остатки Т. monococcum и Т. dicoccum
(Н. Helbaek, 1964).
На о. Крит, судя по археологическим данным (J. В. Evans, 1964).
пшеница мягкая и Т. dicoccum возделывались в 6100 г. до н. э.
Многие сведения свидетельствуют о том, что пшеница мягкая была
одомашнена в глубокой древности не в Европе, как считали многие ав-
торы, а на родине рода Triticum — в Передней Азии, на Ближнем и Сред-
нем Востоке.
По палеоботаническим материалам последних лет, миграция куль-
турных растений, в том числе и пшеницы, началась в крице VI — начале
V тысячелетия до и. э. (J. Renfrew, 1973).
Географическое распространение. Пшеница мягкая — комсомополнт:
опа возделывается на всех материках земного шара, за исключением Ан-
тарктиды. В северном полушарии местами доходит до Полярного Круга.
Происхождение. Фенотип пшеницы мягкой определяет наличие фак-
тора Q. Он относится к числу так называемых главных генов. При утере
или дупликации его пшеница мягкая «исчезает», вместо пес появляются
(соответственно) 7’. или Т / отти 1чт fl У> т '9'/ '/
,;ж.О,:-.:/- , г.', у г/, , О 3.1 ж > ф;; X Г О
ром Q. По вопросу о пути возникновения его у этой пшеницы нет единого
мнения.
Происхождение гексацлоидпой пшеницы связывают с присоединени-
ем генома I) (Ае. tauschii) к генетическому комплексу тетраплопдного
234 Классификации роди Triticum I..
вида ((}. N. Palhak, 1910; II. Kihara, 1911; I7.. S. Me I ;a <1 < lei i и E. I?. Scars,
1944, 1946). Система легкого обмолота, контролируемая фактором Q, не
могла быть привнесенной донором генома D, так как не обнаружена ни
одна его форма, которая имела бы в своем генотипе этот фактор. Он мог
возникнуть у пшеницы па гексап.топдпом или тетраплоидпом уровне.
Известен только одни тетраплопднын вид с фактором Q и геномами,
однозначными первому (А11) и второму (В) геномам пшеницы мягкой —
это Т. persicum. Н. Kihara и F. Lilienfeld (1949) скрепили этот тстрапло-
идпый вид с Ле. tauschii, у полученных гибридов Fi удвоили число хромо-
сом— получили амфпдпнлоид с легким обмолотом колоса (К. Tsunewaki,
1966), Этот эксперимент свидетельствует о том, что первичный гексапло-
ид с легким обмолотом мог произойти с непосредственным участием тст-
раплоида. Такой путь возникновения пшеницы мягкой возможен лишь в
случае, если фактор Q первоначально появился на тетраплоидпом уров-
не, например, у Т. dicoccum. Д. Мак-Кей (1968) подчеркивал, что фактор
Q маскирует за «фасадом» Т. persicum фенотип этой полбы. Появление
фактора Q у тетраплоида— реальная возможность. Моделью, иллюстри-
рующей это, является мутационное возникновение подобного фактора
у гомолога упомянутой полбы — у Т. timopheevii, давшее новый вид --
Т. militinae.
Возникновение пшеницы мягкой с участием тетранлоидпого вида ста-
вится под сомнение теми, кто считает, что Т. persicum произошел не мута-
циоино, а является продуктом расщепления гибридов, возникших при
межвидовой спонтанной гибридизации между тетраплоидом и гексаило-
идом ряда пшеницы мягкой. Е. S. McFadden и Е. R. Sears (1946),
Н. Kuckuck и R. Peters (1964) высказали предположение, что пшеница
свайных построек Т. antiquorum Heer была тетраплоидным видом с фак-
тором Q, который мог дать начало пшенице мягкой. Между тем рисунки
пшеницы времен свайных построек, приведенные в работах О. Heer
(1865) и К. А. Фляксбергера (1930), как нам кажется, убедительно сви-
детельствуют о ее гексаплоидной природе.
В случае, если пшеница мягкая возникла на гексаплоидном уровне,
то исходной формой для нее могла быть гексаплоидная пшеница с фак-
тором Q или без него. В последнем случае такой формой могли быть но-
сители рецессивной аллели q — Т. spelta или Т. macha. Грузинская шко-
ла тритикологов последний вид считает родоначальным для пшеницы
мягкой (Л. Л. Декапрелевич, 1938, 1962; В. Л. Мепабде, 1946, 1948, 1961,
1971; А. Д. Горгидзе, 1973). В. Л. Мепабде и А. Д. Горгидзе происхожде-
ние пшеницы мягкой связывают с гибридизацией между Т. macha и
Т. monococcum. Однако П. М. Жуковский (1971) считал такое заключе-
ние необоснованным. В. Ф. Дорофеев (1972) также не соглашается, что
из Т. macha могла произойти первичная спельта, давшая затем начало
пшенице мягкой.
A. Schulz (1913), Е. Tschermak (1914), М. Muramatsu (1963) считают
спельту исходной формой для голозерных гексаплоидов. Последний
предполагал, что при этом произошла мутация гена q в Q. Теория пер-
вичного происхождения спельты получила экспериментальную опору
после синтеза ее из Fj гибридов комбинации тетраплоид (AUB) * X Ле.
tauschii (D), проведенного Н. Kihara (1944), Е. S. McFadden и Е. R. Se-
ars (1944, 1946). Однако эта точка зрения не нашла подкрепления при
археологических изысканиях; гексаплоидная пшеница из наиболее древ-
них слоев является голозерной (Н. Helbaek. 1964, 1965).
Многочисленные эксперименты по облучению ншепнпы мягкой пока-
зали, что из нее легко возникает спельта (.1. Mac Key, 1954; М. V. Р. Rao,
М. S. Swaminathan, 1963; М. Р. Sinph, 1969; В. В. Хвостова, 1969 и др.).
Однако нам неизвестны литературные данные, свидетельствующие о по-
добном появлении пшеницы мягкой из спельты.
* Исключая Т. persicum.
235
Подрод 1. I rilicutn
Гипотеза вторичного происхождения спельi bi (t. e. первичною про-
исхождения ншеиппы мягкой) имеет мио,очисленных сторонников (Л. Do
Candolle, 1887; II. И. Вавилов, 1922, 1926; 1\. Л. Фляксбергер, 1930, 1932;
М. Г. Туманян, 1935, 193(1; Е. II. Синская, 1955 и др.). К. Л. Фляксбергер
(1930, 1932) и Е. Sdiieinaiin (1932) пришли к .заключению, что баварская
спельта моложе древней пшеницы Т. antiquorum. Примеры появления
спельты при расщеплении гибридов, полученных от скрещивания Т. aes-
tivum пли Т. compactum с тетраплопднымн видами, многочисленны пе
только в экспериментальных условиях, по н в природе (II. Д. Мустафа-
ев, 19G0; В. Ф. Дорофеев, 1961; Р. Л. Удачин, 1965; П. Л. Гапдилян, 1967).
В. Ф. Дорофеев (1972) указывает, что вытопившаяся гибридная спельта
может иметь различную степень ломкости колосового стержня: более
сильную, когда в скрещивания с пшеницей мягкой была вовлечена пол-
ба обыкновенная, меньшую, когда в гибридизации участвовавала пше-
ница твердая или Т. turgidum.
При вовлечении последнего вида в принудительное скрещивание с
пшеницей мягкой мы наблюдали, что уже в Е2 вытопилась в большом
количестве настоящая спельта, значительная часть которой оказалась
константной. Подобная спельта вытопилась также при гибридизации
Т. compactum, Т. aestivum и Т. spliaerococcum с. полбами (Т. dicoccoides,
Г. dicoccum, Т. ispahanicum, Т. karamyschevii).
Есть указания, что легкий обмолот у пшеницы мягкой может быть
обеспечен лишь при комплементарном взаимодействии фактора Q с ре-
цессивной аллелью tg (Е. R. Kerber, Р. L. Dyck, 1969; Р. L. Dyck,
Е. R. Kerber, 1970; Е. R. Kerber, G. G. Rowland, 1974). Доминантный ген
Tg расположен па хромосоме 2D у Ле. tauschii. Он обеспечивает одревес-
нение колосковой чешуи. Авторы гипотезы считают, что легкий обмолот у
гексаплоидпой пшеницы осуществляется лишь в случае, если произошла
мутация этого гена в рецессивную аллель 1g. Синтезированные ими ам-
фидиплоиды Т. persicum X Ае. tauschii имели трудный обмолот, так как
включали ген Tg, подавляющий действие фактора Q. В то же время они
отметили, что степень трудности вымолота зерна у амфпдиплоидов раз-
личных комбинаций варьировала. К- Tsunewaki (1966) писал, что подоб-
ные амфидиплоиды, созданные в Японии, имеют легкий обмолот. Изуче-
ние в условиях Ташкентской области показало, что пи одни из этих
амфидиплондов не имеет той легкости обмолота колоса, какая характер-
на для исходного вида Т. persicum. Степень обмолачиваемости их варьи-
рует от более или менее легкой до затрудненной (по нетрудной). Прак-
тик-селекционер Г. А. Айрапетов (Узбекская ССР), ознакомившись с
этими японскими амфидиплоидами, сделал заключение о том, что ко-
лосья их пе имеют трудного обмолота. По всей вероятности, таким был и
первичный гексаплоид Т. persicum X ^е- tauschii, спонтанно возникший
в далекие времена. Вторичный процесс—переход гена Tg в рецессивное
состояние — способствовал формированию той степени легкого вымоло-
та зерна, которая характерна для современной пшеницы мягкой.
Н. И. Вавилов (1964) указал на Переднюю Азию как на единый
центр происхождения пшениц всех трех уровней плоидности. Извест-
ные тритикологп нашей страны В. Л. Мепабде (1948), Л. Л. Декапрелс-
вич (1961), П. Д1. Жуковский (1964), И. Д. Мустафаев, (1964), Е. II. Спп-
ская (1969), В. Ф. Дорофеев (1972) и II. А. Гапдилян (1973) признава-
ли и признают правомерность этой теории. Той же точки зрения придер-
живаются зарубежные ученые М. Gokgol (1961), J. R. Harlan, D. Zohary
(1966), H. Kuckuck (1970) и др. II. Helbaek (1964, 1965) отметил, что
именно здесь при археологических раскопках найдена голозерная гекса-
плоидная пшеница, относящаяся к V VII тысячелетиям до и. э.
П. М. Жуковский (1971) колыбелью гексаплоидпой пшеницы назвал За-
кавказье, Иран, Афганистан. Здесь совпадают центры разнообразия пше-
ницы мягкой и Т. compactum. В работе В. Г. Конарева и др. (1976) пока-
236 K.uicitiil>iu;(ii{ii:i iun)ti Tritii uni I..
зано, ч го 11;i11 6<),nее вероятным донором i < 11ом;i 1) hb.ihcich .Ic. huisi lilt
subsp. strangulata. Ареал этого подвида сравните.tihio неболышш (Азер-
байджан, Армения, Юго-Западная Туркмения и примыкающая к ним
территория Ирана). Он включен внутрь ареала второго подвида — subsp.
tauschii. Вероятно, именно в пределах ареала subsp. strangulata произо-
шел спонтанный синтез гексаплоидной пшеницы.
По первоначальному предположению (Э. Ф. Мигушова, 1975, 1976) .
в этом географическом регионе произошло спонтанное скрещивание
эгилопса (D) с первичной плотпоколосон формой Т. persiciun (А" В).
Однако гибридизация плогноколосого гомолога последнего вида —
Т. militinae с этим эгилопсом, проведенная отделом пшениц ВИРа
(пеопубликовапо), показала, что гибрид Ip не имеет плотного коло-
са. Рассматривая этот опыт как иллюстрацию гомологичного про-
цесса создания природой в прошлом первичного гексаплоида с легким
обмолотом, мы пришли к предположению, что Т. aestivum мог возник-
нуть, минуя ступень плотноколосости. Иными словами пшеница мягкая
является первичным гексаплопдным видом, появившимся не из Т. com-
pactum. Судя по вышеупомянутым японским амфпдиплоидам, первичная
форма Т. aestivum имела признаки ригидности и обладала более трудным
обмолотом колоса, чем те, которые И. И. Вавилов (1923) объединял в
индо-европейском подвиде. Нежная колосковая чешуя последних сфор-
мировалась как следствие закрепленных отбором естественных мутаций
в сложном по своему строению факторе Q.
Систематика. Т. aestivum L. — один из крупнейших и древнейших ви-
дов культурных растений с ареалом, охватывающим все континенты, за
исключением Антарктиды. Не удивительно, насколько велико его разно-
образие.
Первые внутривидовые классификации мягкой пшеницы (J. Metzger,
1824; N. С. Seringe, F. Alefeld), из которых наиболее детально разрабо-
тана F. Kornicke (1885), строились па отдельных признаках (остистость,
цвет и опушение чешуй, цвет зерновок и т. п.). Эти системы, удобные и
легкие в обращении, не отражали филогенетические особенности отдель-
ных групп. Первые попытки естественной классификации пшеницы мяг-
кой сделаны в начале нашего века (К- А. Фляксбергер, 1915; J. Percival,
1921). д
Представления о внутривидовой системе Т. aestivum в корне измени-
лись со времени экспедиций И. И. Вавилова, по Средней Азии, Ирану,
Афганистану, Индии.
Поразительное разнообразие здесь пшениц заставило И. И. Вавило-
ва пересмотреть достоинство признаков для систематических целей. Он
обратил внимание на то, что несмотря на большое количество разновид-
ностей (в понимании Kornicke), аборигенная пшеница мягкая характе-
ризуется общим комплексом признаков, приуроченным к определенному
ареалу. К таким комплексным, или групповым, признакам у пшеницы
мягкой относится плотность заключения зерновок в чешуях, связанная
с трудностью обмолота и грубостью колоса. Этим признакам неизменно
сопутствует ряд других. Грубоколосые, грубоостые формы трудно обмо-
лачиваются, ксерофильпы, свойственны в первую очередь Западной Азии.
В противоположность им мягкоостые формы с колосом нежного построе-
ния легко обмолачиваются, свойственны Европе и северным умеренным
районам Азии.
Правильный выбор комплексных признаков позволил выявить 2 круп-
ные географические группировки, которые впоследствии К. А. Фляксбер-
гер и 11. И. Вавилов под разными названиями приводили как подвиды.
Широкое изучение изменчивости признаков в пространственном распре-
делении выявило более мелкую структуру, дало возможность охаракте-
ризовать. вид как целостную эколого-географическую систему. Первым
внутривидовым классификациям пшеницы мягкой Свойственна некоторая
//г><>!><><I I. I iilii urn 23/
двойственность, что объясняется действителып>1Ми iрудносгями при со.ч-
Д.11П1И естественной сисюмы.
Выделяя естественные группы пшеницы мяч кой, лирикiеризующиеся
комплексами признаков, связанными с определенным ареалом, более
мелкие подразделения — разновидности-- II. И. Вавилов (192(5) и
К. Л. Фляксбергер (1935, 1939) приводят в целом по виду, что нарушает
соподчиненпость групп. В настоящей книге мы старались по возможнос-
ти преодолеть указанное противоречие.
Следуя 11. 11. Вавилову и К. Л. Фляксбергеру, вид Т. aestivum L. раз-
деляем па азиатский и европейский подвиды. Подвиды имеют ряд хо-
рошо заметных морфологических отличий. Однако для выделения
отдельного вида (С. Н. Кудряшов, 1942, дал азиатскому подвиду ранг
вида Т. asiaticum) оснований мало, так как между азиатским и европей-
ским подвидами существует много переходных форм. Эти крупные ре-
гиональные экотипы могут рассматриваться в качестве подвидов, не-
смотря па то, что ареал европейского налегает па ареал азиатскою под-
вида. В этом значительную роль как факторы сыграли и играют человек
и большая пластичность более молодого европейского подвида. Азиат-
ский подвид отличается лучшей приспособленноетыо к засушливым усло-
виям. Эго в основном пшеница коро1коостисгая или остистая, реже ип-
флягная, т. е. со вздутыми чешуями и изогнутыми остевидпыми придат-
ками или короткими остями и безостая.
Азиатский подвид делится на 3 группы разновидностей с определен-
ными комплексами признаков, обособленных географически. У convar.
rigidttm сильно выражена жесткость колосковых чешуй и остей, как пра-
вило, укороченных. Эта группа хорошо приспособлена к засушливым
условиям, особенно стойка к воздушной засухе, часто встречается среди
поливных пшениц, редко заходит высоко в горы.
Ареал второй группы разновидностей (convar. semirigidum) нс огра-
ничен Западной Азией, где также поднимается высоко в горы, встреча-
ется в Китае, Индии, возделывался в присредпземноморскпх странах,
в СССР, кроме республик Средней Азии, — в южном Казахстане, Закав-
казье, на Нижней Волге. К. А. Фляксбергер (1935) рассматривал эту
пшеницу среди азиатского подвида как pro-1, stibrigidttm, для растений
которого характерно нолугрубое построение. Зерновки плотно заключе-
ны в чешуях, но легко обмолачиваются. Эта группа в значительной мере
является переходной между азиатским и европейским подвидами.
Проанализировав в целом разповидпостный состав Т. aestivum, мы
пришли к заключению, что большинство остистых разновидностей отно-
сятся к пшеницам именно полугрубого построения. Такой крупный зна-
ток экологии пшеницы, как Е. Ф. Пальмова (1935), отмечает определен-
ную закономерность в географическом распределении остистых и безос-
тых форм и считает этот признак как систематическим, так равным
образом и экологическим. Остистые пшеницы имеют 2 ареала: 1-й (боль-
шой)— области степей и полупустынь с высокой температурой и не-
достаточной влажностью, и 2-й — области тайги и лесотундры у пределов
земледелия. Для convar. semirigidum характерно преобладание остистых
форм, по именно в сочетании с достаточной грубостью построения всего
растения.
Если не брать во внимание специфический комплекс признаков, опре-
деляющих инфлятпость, в значительной степени эндемичный для Припа-
мпрья, вторая группа разновидностей азиатского подвида (convar. infla-
tum) близка полужеегко-чешуйиой группе разновидностей (convar. se-
mirigidum).
В 1916 г. II. И. Вавилов среди пшениц Ирипамирья обнаружил в
большом количестве безлигульпую пшеницу мягкую, нигде более на зем-
ном шаре не встречающуюся. У этой пшеницы отсутствуют или находят-
ся в зачаточном состоянии ушки и язычок у основания пластинки листа.
238
Классификации />о()а Triticum L.
Безлпгульиая пшеница выделена в качестве нот рун и разновидноетой
у convar. setnirigidum и convar. inf latum.
У каждого из указанных таксонов азиатского подвида имеются свои
разновидности. Эти систематические единицы строятся на основе парал-
лелизма признаков и их часто встречающихся комбинаций.
Таким образом, азиатский подвид отличается большим сист е.матичес-
ким полиморфизмом, а Западная Азия является центром интенсивного
формообразования пшеницы мягкой.
Для европейского подвида в отличие от азиатского с крупными мор-
фолого-географическими подразделениями и небольшим числом эколош-
ческих групп характерно исключительное разнообразие последних, резко
контрастных по длине вегетационного периода, зимостойкости, иммуни-
тету и ряду других биологических свойств.
Европейский подвид пластичен, с более высокой, чем у азиатского,
амплитудой изменчивости признаков.
Разнообразие стародавних местных сортов, по словам Н. И. Вавило-
ва (1964), выкристаллизовалось в определенные эколого-географические
группы, различающиеся по комплексам биологических и физиологичес-
ких свойств, тесно связанных с конкретными условиями среды.
В начале нашего века быстро развивалась селекция пшеницы мягкой.
Новые селекционные сорта значительно повысили урожайность культу-
ры. В настоящее время староместные популяции почти повсеместно вы-
теснены селекционными сортами. Однако исторически сложившиеся в
определенных условиях эколого-географические группы с совокупностью
свойств, определяющей тип и темпы протекания важнейших физиологи-
ческих процессов (Е. Н. Синская, 1961), играют большую роль при созда-
нии сортов для различных почвенно-климатических условий.
Путем отдаленных эколого-географических скрещиваний, значитель-
но расширяющих норму реакции гибридных популяций, созданы ценней-
шие сорта интенсивного типа. Площади под этими сортами нередко зна-
чительно превышают ареал экологической группы, к которой их можно
отнести по ряду биологических свойств. Большую роль в этом играет
современная сортовая агротехника.
Экологические группы пока не являются узаконенными таксонами,
но представляют ценнейший вспомогательный материал, позволяющий
четко представить структуру вида, его целостность. Кроме того, эколого-
географические группы хорошо характеризуют сортовой состав пшеницы.
Нами приводятся главнейшие подразделения пшеницы мягкой по их
типичным представителям, но между ними всегда имеются и промежу-
точные группы и формы—также продукты формообразовательного про-
цесса, дающие возможность проследить изменчивость признаков как в
широтном,так и в меридиональном направлениях.
Ключ для определения подвидов Т. aestivum L.
1. Колосковые чешуи жесткие, толстокожистые, у основания без по-
перечной вдавлегшости, зерновки трудно обмолачивающиеся . . . .
....................: . . . subsp. hadropyrum (Flaksb.) Tzvel.
+ Колорковые чешуи менее жесткие, обычно тонкокожистые, у осно-
вания с поперечной вдавлеппостыо; зерновки легко обмолачивающиеся
.................................................subsp. aestivum.
I. Subsp. hadropyrum (Flaksb.) Tzvel.--подвид азиатский.
Subsp. hadropyrum (Flaksb.) Tzvel. 1973, Новост сист. высш. paci.
К): 43; Цвел. 1976, Злаки СССР: 163. — 7'. vulgare subsp. hadropyrum
Flaksb. 1928, Изв. Г'ос. ин-та онытп. агрономии, 6,2: 47. - Т. vulgare
subsp. iranoasiaticum Flaksb. 1935, Культ, фл. СССР, 1:42, nom. illeg.--
7. asiaticum Kudr. 1941, Бот. мат. (Ташкент), 4: 16, рис. 1. — 7’. vulgare
Подрод I. Triticum
239
subsp. iraiioturliestanicum Vav. 196'1, Миров. ресурсы хлсбн. злаков.
Пшеница: 69, nom. illeg.- -7'. vulgare subsp. indicum Vav. 1964, l.c.: 73,
descr. ross. — 7’. vulgare subsp. abyssinicum Vav. 1964, I. c.: 100, descr. ross.
Растения и колосья грубого и полугрубого построения; колосья рых-
лые, редко плотные, остистые, реже безостые; колосковые чешуи различ-
ной формы, плоские и вздутые (ипфлятныс), у основания без поперечной
вдавленностп. Жилкование чешуи и киль хорошо развиты, жилки покры-
ты зубчиками. Ости грубые в разной степени. Зерновки плотно заключе-
ны в чешуях, обмолачиваются трудно или легко, часто стекловидные или
полустекловидные. Соломина нетолстая, полая, по встречаются формы с
соломиной, выполненной паренхимой по всей длине. Пшеница яровая,
озимая, часто полуозимая, обычно скороспелая или среднеспелая, чувст-
вительна к недостатку влаги в начале роста, в период от колошения до
созревания переносит сухость воздуха.
Тип.: Т. aestivum convar. rigidum (Vav.) Л. Filat.
Геогр. распр. Средняя Азия, засушливые районы Казахстана,
Закавказья, наблюдалось захождение в Нижнее Поволжье, Праи, Па-
кистан, Афганистан, Сирия, Индия, Китай, Средиземноморье.
Ключ для определения групп разновидностей
subsp. hadropyrum (Flaksb.) Tzvel.
1. Колосковые чешуи широкие, вздутые; килевой зубец загнут внутрь;
цветковые чешуи с расширенными у основания остевиднымн заострения-
ми или остями, сравнительно тонкими, обычно изогнутыми, деформиро-
ванными ..................convar, inflatum (Vav.). Л. Filat, et Dorof.
+ Колосковые чешуи более или менее плоские, различной формы;
килевой зубец прямой; ости прямые................................2.
2. Колосья полуостпстые, редко безостые, ости грубые, ломкие, к ос-
нованию утолщенные; колосковые чешуи грубые, с хорошо выраженным
жилкованием; килевой зубец всегда прямой, длинный (более 7 мм);
зерновки плотно заключены в чешуях, обмолот трудный.................
..............................convar. rigidum (Vav.) A. Filat, et Dorof.
+ Колосья остистые или безостые, полугрубого построения, ости бо-
лее тонкие, длинные; колосковые чешуи менее грубые, чем у convar. ri-
gidum, с менее выраженным жилкованием; килевой .зубец короткий (до
3 мм)...........................convar. semirigidum A. Filat, et Dorof.
1. Convar. rigidum (Vav.) A. Filat, et Dorof. — ригидная группа раз-
новидностей.
Convar. rigidum (Vav.) A. Filat, et Dorof. comb. nov. — T. vulgare
группа rigidum Vav. 1923. Tp. прикл. бот. сел. 13, 1:194. — T. vulgare
группа breviuristata Vav. 1923, l.c.: 168. — T. vulgare группа speltijorme
Vav. 1923, 1. c.: 195. — T. vulgare prol. rigidum. (Vav.) Flaksb. 1935.
Культ, фл. СССР, 1:43. — T. vulgare subprol. speltiformc (Vav.) Flaksb.
1935, 1, c.: 43. — T. vulgare subprol. breviarisiatum (Vav.) Flaksb.
1935, 1. c.: 43. — T. asiaticum subsp. semiaristatum Kudr. 1942, Пшен: Уз-
бекист.: 32, nom. illeg. — T. asiaticum subsp. speltiforme (Vav.) Kudr.
1942, l.c.: 31. — T. yutinanense King, 1957, Виды кит. пшен. их распро-
стри 15, descr. chin. — 7'. aestivum subsp. transcaucasicuni Dorof. ct Lapt.
1963. Вести, с.-х. пауки, 11: 111.
T и и: T. aestivum var. subgraecum. (Vav.) Mansf.
Растения грубого построения, колосья полуостпстые (рис. 18 щ 19 л);
ости грубые, ломкие, к основанию утолщенные; колосковые чешуи iру-
бые, с хорошо выраженными жилками. Зерновки плотно заключены в
чешуях, трудно обмолачиваются.
Н. И. Вавилов (1923) среди грубоколосых пшениц mhikiix выделяет
группу speltiforme, которая отличается от остальных ригидных разно-
2'10
Классификации porta Trilli нт I..
Рис. 18. Колосья (боковая сторона) и зерновки Т. aestivum L.:
a — subsp. hadropyrum (Flaksb.) Tzvel.; 6—subsp, aestiuum
видностей только лопатчатыми, усеченными сверху колосковыми чешуями,
отчего грубый колос очень напоминает таковой у европейской спель-
ты. С открытием спельты в Азии, характеризующейся большим разно-
образием колосковых чешуй — от сильно лопатчатых до полного отсут-
ствия лопатчатости, спсльтоидпой можно рассматривать всю convar.
rigidum. Нс только форма колосковых чешуи, но также грубоколосость,
плотное заключение зерновок в чешуях, удлиненный рыхлый колос н не-
которые другие признаки приближают- ее к Т. spelia. Ригидную группу
разновидностей можно рассматривать как клип- выражение после-
довательной изменчивости совокупности признаков Т. aestivum и Т. spel-
ta в засушливых условиях Юго-Западной Азии.
Геогр. распр. Средняя Азия, редко в Казахстане и Закавказье;
Иран, Афганистан, Пакистан, Китай.
I. ПИ и ит
Рис. 19. Колосья (лицевая сторона) и зерновки Т. aestivum L.:
а — subsp. hadropyrum (Flaksb.) Tzvel.; б — subsp. nesitvuni
Чистые поливные посевы этой пшеницы приурочены к высотам от
1000 м над ур. м. На больших высотах (2000—3200 м над ур. м.) в яро-
вых посевах встречаются в виде редкой примеси, как и на поливных по-
лях яровых и озимых посевов до 1000 м над ур. м.
Грубоколосая группа разновидностей приспособлена к засушливым
условиям и экологически довольно однородна. И. И. Вавилов (1964)
выявил среди грубоколосой азиатской пшеницы 3 эколого-географичес-
кие группы.
а. Богарная группа, или группа обеспеченной богары, характеризу-
ется сравнительной скороспелостью, устойчивостью к воздушной засу-
хе, исключительной восприимчивостью ко всем заболеваниям. Посевы
яровые и озимые. Приурочена в основном к склонам гор.
б. Поливная группа в период созревания очень требовательна к теп-
лу, устойчива к воздушной засухе. К этой группе относятся формы с
недостаточно прочной соломиной, восприимчивые к грибным заболе-
16 762
242
Классификации рода Triticum L.
нациям. Возделываются па пониженных высотах в Сроднен Азии, Л<]н а-
нистапе, Иране.
в. Группа районов необеспеченной богары сильно поражается ржав-
чиной. Соломина тонкая, средней прочности. Преимущественно яровые
формы горных и предгорных районов.
Определение разновидностей convar. rigidum (Vav.) A. Filat, et Dorof.
Чешуи
ссро-ды мча-
t ые на фойе
черные на
фоне
Разновидное।ь
’« разн( видности
Колосья безостые (чешуи неопушенные)
Зерновки белые
— — |-р[— | — | — | — | — [ navrozkyi 1
3 ерн о в к и крас п ы е
— I — l + i — I — I" i — ! — I — i — I schrcderi 2
Колосья полуостистые
Чешуи неопушенные
Чешуи опушенные
Зерновки белые
1 ++ + — — — — + + 1 +1 submeridionale subpseudomeridionale subhamadanicum 11 12 13
— — + — — —- — — — — subturcicum 14
— — + — — — — — — + subpseudoturcicum 15
— — + — — — — + — subkermanschachi- cum 16
— — - -» — + — — — — + caesiospeltiforme 17
— — — — — — 4- — — + submesopotamicum 18
— — — — — — — + — + subiranicum 19
Зерновки красные
4. — — — — — .— — + — subtiosfianum 20
+ — — — — — — — — + subpseudohostianum 21
- + — — — — — — subbarbarossa 22
— — — — — — — — -4- sub pseudobarbarossa 23
— — — — — — 4- — — + subfuliginosum 24
Var. caesiospeltiforme (Thum.) Mansf. 1951. Der Ziichler, 21, 1/2:
52. — T. vulgare var. caesiospeltiforme Thum. 1929, Tp. прикл. бот. ген.
сел. 22, 2:310. (17).
Геогр. распр. Армения, Турция (редко).
Var. kudrjaschevii Udacz. var. nov. — Differ! a var. cinerosiun Flaksb.
spicis speltiforrnibus et grants rubris. (9).
Typus: Uzbekskaja SSR, prov. Surchandariya, distr. Baysun, prope
pag. Tamczibulak, in populalionc culta T. aestivum L., 1800 m, 9.V1I.
1971, spicae, N 069820, R. Udaczin (WIR).
llodpoi) I. Triticum 243
Nomen var. in hoiioreni S. N. Kiidrjaschevii datur.
Отличается or var. cinerosum Flaksb. строением колосьев по типу
spelttforme и красными зерновками.
Т и и: Узбекская ССР, Сурхандарьинская область, Вайсунскпй рай-
он, близ селения Тамчнбулак, в посеве популяции Т. aestivum L., 1800 м
над ур. м., 9.V11. 1971, колосья, № 069820, Р. Удачин (ВИР). Название
разновидности дано в честь С. Н. Кудряшева.
Геогр. распр. Сурхандарья, Западный Памир.
Var. navrozkyi Udacz. 1976, Тр. прикл. бот. ген. сел. 56, 2: 147. (1).
Геогр. распр. Таджикистан.
Var. schrederi Udacz, 1976, 1. с.: 148. (2).
Геогр. распр. Таджикистан.
Var. subbarbarossa (Vav.) Mansf. 1951, Der Zuchter, 21, 1/2: 49.—
T. vulgare var. subbarbarossa Vav. 1923. Tp. прикл. бот. сел. 13,1:
173. (22).
Геогр. распр. Средняя Азия, Казахстан, Азербайджан, Иран,
Афганистан, Турция (редкая примесь).
Var. subcaesioides (Flaksb.) Mansf. 1951, l.c.: 53. — T. vulgare var.
subcaesioides Flaksb. 1929. Tp. прикл. бот. ген. сел. 20: 104. (5).
Гео гр. р а с и р. Таджикистан, Казахстан, Китай.
Var. subcaesittni (Flaksb.) Mansf. 1951, l.c.: 52. — T. vulgare var.
subcaesiuni Flaksb. 1929. l.c.: 106. (10).
Геогр. распр. Таджикистан и Туркмения.
Var. suberythroleucuni (Vav.) Mansf. 1951, l.c.: 53, «suberythroleu-
con». — T. vulgare var. suberythroleucon Vav. 1923, l.c.: 172. — T. aesti-
vum var. golbekii Udacz. 1976, 1. c.: 149. (4).
Геогр. распр. Средняя Азия, редко в Казахстане, Азербайджане,
Армении; Иран, Афганистан, Монголия, Китай.
Var. suberythrospermuni (Vav.) Mansf. 1951, l.c.: 54. — T. vulgare
var. suberythrospermuni Vav. 1923, l.c.: 172. — T. asiaticum var. formo-
sum Kudr. 1942, 1. c.: 32. (6).
Геогр. распр. Таджикистан, Узбекистан, Казахстан, Армения,
Азербайджан и Грузия (редко), Иран, Афганистан, Китай.
Var. subferrugineum (Vav.) Mansf. 1951, l.c.: 52. — T. vulgare var.
subferrugineum Vav. 1923, l.c.: 173. — T. asiaticum var. kvjatkovskyi
Kudr. 1942, 1. c.: 32. (7).
Геогр. распр. Средняя Азия, Казахстан, Закавказье (редко),
Иран, Афганистан, Китай.
Var. subfuliginosum (Vav.) A. Filat, comb. nov. — T. vulgare var. sub-
fuliginosum Vav. 1923, l.c.: 173. — T. vulgare var. subrenovalum Flaksb.
1935, Культ, фл. СССР, 1:98, nom. illeg. — T. aestivum var. subrenova-
tum (Flaksb.) Mansf. 1951,1. c.: 51. (24).
Геогр. распр. Иран, Индия.
Var. subgraecum (Vav.) Mansf. 1951, l.c.: 55.—• T. vulgare var. sub-
graecum Vav. 1923, l.c.: 172. — T. asiaticum var. lipskyi Kudr. 1942,
Пшен. Узбекист.: 32. (3).
Геогр. распр. Средняя Азия, редко в Казахстане, Азербайджане
и Армении; Иран, Афганистан, Китай.
Var. subhampdanicum (Thum.) Mansf. 1951, l.c.: 52. — T. vulgare
var. subhaniadanicum Thum. 1929, Tp. прикл. бот. ген. сел. 22,2:
311. (13).
Геогр. распр. Армения (редко).
Var. subhostianuni (Vav.) Mansf. 1951, l.c.: 51.- -T. vulgare var.
subhostianumVav. 1923,1. c.: 173. (20).
Геогр. распр. Средняя Азия, Закавказье .(редко), Афганистан,
Иран, Пакистан, Китай.
Var. subiranicum (Thum.) Mansf. 1951, 1. с.: 50. — T. vulgare var. sub-
iranicum Uhum. 1929, 1. c.: 311. (19).
1 t Классификации juxhi Tiiluum I..
Геогр. p ;| c II p. Армения, Чурция (i-.h.i;iйег Ban), Knraii.
Var. subkermanschachicinn (Thum.) Mansf. 1951, l.c.: 50,- T. vulga-
re var. subkernianschachii Thum. 1929, 1. c.: 311. (1G).
Геогр. распр. Армения и Турция (вилайет Ban).
Var.- submeridionale (Vav.) Mansf. 1951, l.c.: 52. — T. vulgare var.
submeridionale Vav. 1923, l.c.: 173. (11).
Геогр. распр. Узбекистан, Туркменистан, Армения и Азербай-
джан (редко), Пран, Пакистан, Афганистан.
Var. submesopotamicum (Thum.) Mansf. 1951, l.c.: 52. — T. vulgare
var. submesopotamicum Thum. 1929, l.c.: 310. — T. vulgare var. asiati-
cum Thum. 1933, Опред. хлебных злаков (колос.): 186. (18).
Геогр. распр. Азербайджан, Армения, Турция (вилайет Ban).
Var. subpseudobarbarossa (Vav. et Kob.) Mansf. 1951, l.c.: 49.--
T. vulgare var. subpseudobarbarossa Vav. et Kob. in Kob. 1928. 'Гр. прикл.
бот. ген. сел. 19, 1: 89. (23).
Геогр. распр. Узбекистан, Туркменистан, Казахстан, Афгани-
стан.
Var. siibpseudoliostianum (Vav. cl Kob.) Mansf. 1951, l.c.: 51.--
T. vulgare var. subpseudohosliaiium Vav. el Kob. in Kob. 1928. l.c.: 88.
(21).
Г e о г p. p а с н p. Туркменистан, Афганистан.
Var. subpseudomeridionale (Vav.) Mansf. 1951, l.c.: 52. — T. vulgare
var. subpseudomeridionale Vav. 1923, l.c.: 173. (12).
Геогр. распр. Туркменистан, Таджикистан, Афганистан, Пран,
Пакистан.
Var. subpseudoturcicuni (Vav. et Kob.) Mansf. 1951, l.c.: 50. — T. vul-
gare var. subpseudoturcicuni Vav. et Kob. in Kob. 1928, l.c.: 89. (15).
Геогр. распр. Туркменистан, Узбекистан, Казахстан, Афгани-
стан.
Var. stibsardoum (Vav. et Jakubz.) Mansf. 1951, l.c.: 52. — T. vulgare
var. subsardoum Vav. et Jakubz. 1931, l.c.: 184. (8).
Г e о г p. p а с п p. Эфиопия.
Var. subturcicum (Vav.) Mansf. 1951, l.c.: 50. — T. vulgare var. sub-
turcicum Vav. 1923, 1. c.: 173. (14).
Геогр. распр. Средняя Азия, Казахстан, Армения, Иран, Афга-
нистан.
2. Convar. inflatiim (Vav.) A. Filat, et Dorof. — инфлятпая группа
разновидностей. .
Convar. inflatiim (Vav.) A. Filat, et Dorof. comb. nov. — T. vulgare
группа inflatiim Vav. 1923, Tp. прикл. бот. сел. 13,1: 160. -T. vulgare
prol. inflatum. (Vav.) Flaksb. 1935. Культ, фл. СССР, 1: 46. — T. infla-
tion Kudr. 1941, Бот. мат. (Ташкент), 4: 18. — Т. aestivum subsp. infla-
tum (Kudr.) Tzvel. 1973, Новости спет. высш. раст. 10: 43.
Т и и: Т. aestivum var. khorassanicum (Vav.) Mansf.
Растения полугрубого построения. Листья голые или с редким опу-
шением, у перехода влагалища листа в пластинку располагаются язычок
и ушки или они отсутствуют. Колосья кверху суженные или почти
цилиндрические, безостые, реже остистые и полуостистые. Колосковые
чешуи вздутые, укороченные и широкие, килевой зубец загнут внутрь.
Цветковые чешуи с расширенными у основания остевпдными заостре-
ниями или остями, сравнительно топкими, обычно изогнутыми, дефор-
мированными. У основания остевидпых заострений или остей иногда
имеются выросты, напоминающие фуркп безостых ячменей. Зерновки
плотно заключены в чешуях, не осыпаются, по легко обмолачиваются.
Инфлятпая группа пшениц, выработавшаяся в условиях высокогор-
ных пустынь, сильно поражается видами ржавчины (II. И. Вавилов,
1964).
fiiuipot) 1. Triticum 15
Г <’ о । p. распр. Горные районы Средней и Юго Западной A.iini.
lie i речае i ся в Индии, Миш одни, Kiriae, редко в 3; i к а вк а з ье, Эфио-
пии и Алжире. В Афганистане обнаружены чистые посевы пнфлятной
пшеницы па высотах от 1000 до 2000 м над ур. м. (озимые поливные)
и выше--до 3200 м (яровые поливные). Неполивные яровые посевы
сосредоточены на высотах от 1500 до 3000 м. Ниже 1000 м над ур. м. ин-
флятная пшеница встречается как примесь.
Ключ для определения подгрупп разновидностей
convar. inflatuni (Vav.) A. Filat, et Dorof.
1. .Листья с язычками и ушками v основания пластинки . . . .
.....................subconvar. inflatuni (Vav.) A. Filat, et Dorof.
+ Листья без язычков и ушек у основания пластинки...................
................subconvar. eligulatinflatum (Flaksb.) A. Filat, ct Dorof.
a. Subconvar. inflatuni (Vav.) A. Filat, et Dorof. — иифлятпая .ти-
тульная подгруппа разновидностей.
Subconvar. inflatum (Vav.) j\. Filat, et Dorof. comb. nov. — T. vulgare
подгруппа inflatum Vav. 1923, Tp. прикл. бот. сел. 13,1: 173,- -T. vulga-
re prol. mulicorigidum Flaksb. 1935, Культ, фл. СССР, 1:45, descr.
ross. — T. vulgare subprol. oasicolum (Due.) Flaksb. 1935, l.c.: 47.
Растения с обычным для пшеницы строением листа — па месте пе-
рехода влагалища листа в пластинку ясно выражена перетяжка; листо-
вая пластинка отогнута от стебля. У основания листовой пластинки рас-
положены язычок и ушки.
Т и п. Т. aestivum var. khorassanicum (Vav.) Mansf.
Геогр. распр. По всему ареалу Т. aestivum convar. inflatum.
Var. albinflatum (Vav.) Mansf. 1951, Der Ziichter, 21, 1/2: 58.—
T. vulgare var. albinflatum Vav. 1923, Tp. прикл. бот. сел. 13,1: 174.—
T. vulgare var. albiduni subvar. inflatum Flaksb. 1910, Tp. Бюро прикл.
бот. 3: 67. — T. vulgare var. albidum f. inflatum Flaksb. et f. bucharicutn
Flaksb. 1915, Опред. пшен.: 129. (1).
К var. albinflatum относятся формы: f. albinflaticapitatum (Udacz.)
Udacz. comb. nov. — T. aestivum var. albinflaticapitatum Udacz. 1979,
Бюл. ВИРа, 84: 71. — с булавовидными колосьями и f. falsealbidum
(Udacz.) Udacz. comb. nov. — T. aestivum var.- falsealbidum Udacz. 1975,
Tp. прикл. бот. ген. сел. 55, 3: 48 — форма по построению колоса пере-
ходная от инфлятного к обычному типу (Фергана).
Геогр. распр. Средняя Азия, Афганистан, Иран, Монголия, Ки-
тай, Эфиопия, Алжир.
Var. albirubrinflatum (Vav.) Mansf. 1951, l.c.: 57. — T. vulgare var.
albortibrinf latum Vav. 1923. 1. c.: 174. — T. vulgare var. alborubrum orz'en-
tale Flaksb. 1915, 1. c.: 134. (3).
К var. albirubrinflatum относится f. falsealbirtibrum (Flaksb.) Dorof.
et A. Filat, comb. nov. — T. vulgare var. falsealborubrum Flaksb. 1929, Tp.
прикл. бот. ген. сел. 20: 106 — форма по построению колоса переходная
от инфлятного к обычному типу.
Геогр. распр. Средняя Азия, Афганистан, Эфиопия.
V ar. araxicum (Jakubz.) Jakubz. comb. nov. — T. vulgare var. araxi-
cum Jakubz. 4932, Tp. прикл. бот. ген. сел. Сер. 5,1: 173. (33).
Геогр. распр. Нахичевань.
V ar. bengalinflatum (Vav. et Jakubz.) Mansf. 1951, l.c.: 52. — 7'. vul-
gare var. bengalensinflalum Vav. et Jakubz. 1931, Tp. прикл. бот. ген.
сел. Прилож. 51: 186. (30).
Геогр. р а с и р. Эфиопия.
V ar. choresinum Jakubz. var. nov. - -7’. vulgare var. clioresmuiii
Jakubz. in Flaksb. 1935, Культ, фл. СССР, 1: 65, in adnot., descr. ross.
T. vulgare var. choresinum Jakubz. in Flaksb. 1939, Опред. паст, хлебов:
246
Классификации рода Triticum L.
Определение разновидностей subconvar. iiiflaltiiii (Vav.) Л. Ijlat. ct Dorof.
Разновидность
1> разновидности
Колосья бeзо стые
Чешуи неопушенные
3 с р н о в к и бел ы е
+ — — — — — — — — —. а 1 bin flat urn 1
--- + — — — — — — — simpani 2
— — + — — — — — — — albirubrinflatum 3
— — — — + —, — —. — — glaucilutinjlatum 4
— — — — — + — — — — choresmum o
Зерновки красные
4- — — — — — — — — — lutinflatum 6
— -h — — — — — — — +* nigrilutinflatiim 7
— + — — — — •— — — rufinflatum 8
— — — + — — — — —-II — nigrirtiKitflalum 9
— — — — — — + — — — nigricolorinjlatum 10
Чешуи опушенные
Зерновки белые
+ — — — — —. — — — — khorassanicum 11
+ +* pseudokhorassanicum 12
— + — — — — — — — — teheranicum 13
— — 4- — — — — — — — iranscaspicum 14
— — 4- — — — — — — +* pseudotranscaspicum 15
— — — — — + — — — — kobelevii 16
3e рновки крас п ы e
+ — — heraticum 17
+ +* pseudoheraticum 18
— — + — — — — — — —. turcomanicum 19
— — -r — — — — — — +* pseudoturcomanicum 20
— —— — — — — — + — — nigrinf latum 21
— — — — — + — —• — — cyanothrixlnf latum 22
Колосья полуостистые
Чешуи неопушенные
Зерновки белые
+ — — — — — — — + — sub graecinf latum 23
— — 4- — — — — — — suberythroleucinf latum 24
— — — — — 1- — — 4- —- subcaesioidinj latum 25
— — — — — — — + + — indicinf latum 26
Зерно в к и красные
+ — — — — — — — 4- — suberythrospermin- 27
f latum
— — + — — — — 4- — subferrugininf latum 28
— — — — — 4- — — 4- — subcaesi/ф latum 29
— — — — — — — + 4- — bengalinflatum 30.
t Чешу и ontj luennt яе
Зерновки бел ы е
+ — — — — — — — 4- — submeridionalinf latum 31
-+• + subpscuilomeridionalin- 32
f latum
—- + — — — — — — 4- — araxicuin 33
— + — — — — — + — subturcinf latum 34
— — + — — — — — 4- sub pseudot urdu] Iatan, 35
— — — — — + — — — + subsogdianinf latum 36
Окраска остеоидных заострении.
I. Tiitiiiim
Зерновки крас н ы e
subhostinf latum
sub pseuilohosiinf latum
subbarbarossinf latum
37
38
39
Колосья остистые
Чешуи неопушенные
Зерновки белые
graecinflatum
40
+ i —
+ I -
Зерновки крас н ы е
— I — | — I + I — | crythrosperminflatum 1 41
Чешуи опушенные
3 е р н о в к и б е л ы е *
— | — | — 1 | — [ meridionalinflatum 42
166, descr. ross. — Т. aestivum var. choresmum (Jakubz.) Mansf. 1951,
1. c.: 57, comb, invalid. — A T. aestivo var. cinerescens (Flaksb.) Mansf.
foliis ligulatis differt. (5).
Typus: Uzbekistan, reg. Fergana, pag. Rischtan, 600 m, 19. VII. 1969,
N 069906, leg. R. Udaczin (WIR).
Геогр. распр. Узбекистан (редко).
Var. cyanothrixinflatum Palm. var. nov. — T. vulgare var. cyanothri-
xinflatum Palm, ex Flaksb. 1935, Культ, фл. СССР, 1: 72, descr. ross.—
T. aestivum var. cyanothrixinj latum (Palm, ex Flaksb.) Mansf. 1951, l.c.:
55, comb, invalid. — A T. aestivo var. cyanothrix (Koern.) Mansf. spicis
inflatis differt. (22).
Typus: Mongolia, 1923, k-8073, leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. erythrosperminflatum Palm. var. nov. — T. vulgare var. erythros-
permuminflatum Palm, ex Flaksb. 1935, 1. c.: 80, descr. ross. — T. aestivum
var. erythrospermuminflatum (Palm, ex Flaksb.) Mansf. 1951, Lc.: 54,
comb, invalid. — AT. aestivo var. aestivum spicis inflatis differt. (41).
Typus: Mongolia, 1925, k-15613, leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. glaucilutinflatum Dorof. 1968. Бюл. ВИРа, 13: 74. — T. aestivum
var. popoviae Udacz. 1975,1. c.: 48. (4).
Геогр. распр. Азербайджан (редкая примесь в посевах пшеницы
мягкой на высоте 970 м над ур. м.), Узбекистан (в посеве пшеницы
мягкой на высоте 600 м надур. м.).
Var. graecinflatum Udacz. 1969, Тр. прикл. бот. ген. сел. 40,2:
41. (40).
Геогр. распр. Таджикистан.
Var. heraticuin (Vav. et. Kob.) Mansf. 1951, 1. c.: 56. — T. vulgare var.
heralicum Vav. et Kob. 1928. Tp. прикл. бот. ген. сел. 19,1: 89. - Т. vul-
gare var. lunangense King, 1957, Виды кит. пшен. их распростри 11,
descr. chin. (17).
Геогр. распр. Средняя Азия, Казахстан (очень редко), Афгани-
стан, Эфиопия, Китай, Монголия.
248
Классификация рода Trilictim
Var. indicinllatum (Vav. el Jakubz.) A'Vinsf. 1951, l.c.: 53. T. uiilg.it-
re var. indicinf lai tun Vav. et Jakubz. 1931, l.c.: 186.- 7'. vulgare var. la-
gendreanum Hosono, 1935, Mem. Coll. Agr. Kyoto Imp. Univ. 3-1, 4: 2.
(26).
Г e о г p. p а с и p. Эфиопия, Китай.
Var. khorassanicum (Vav.) Mansf. 1957, l.c.: 56. — T. vulgare var.
khorassanicum Vav. 1923, l.c.: 174. (11).
Г e о г p. p а с п p. Средняя Азия, Афганистан, Иран, Эфиопия.
Var. kobelevii Udacz. 1975, 'Гр. прикл. бот. ген. сел. 55, 3; 48. (16).
К var. kobelevii относится f. falsekonstaniinovsken.se (Udacz.) Udacz.
comb. nov. — T. aestivum var. falsekonstantinovskense Udacz. 1975, l.c.:
48 — форма по построению колоса переходная от ипфлятного к обычному
типу.
Геогр. распр. Узбекистан (Фергана, 600 м над ур. м.).
Var. lutinflatum (Flaksb.) Mansf. 1951, l.c.: 57. — T. vulgare var. lu-
tescens subvar. lutinflatum Flaksb. 1911, Tp. Бюро прикл. бот. 4: 3.—
T. vulgare var. lutinflatum (Flaksb.) Vav. 1923, l.c.: 174. (6).
В Китае обнаружена инфлятпая форма с полным отсутствием остей —
f. huangyangense (Hosono) Dorof. et A. Filat, comb. nov. — T. vulgare
var. huangyangense Hosono, 1935, Mem. Coll. Agr., Kyoto Imp. Univ.
34: 1. К этой же разновидности относится форма из' Таджикистана, по
построению колоса переходная от ипфлятного к обычному типу-—f. fal-
selulescens (Udacz.) Udacz. comb. nov. — T. aestivum var. falseluiescens
Udacz. 1976, Tp. прикл. бот. геи. сел. 56,2: 147.
Геогр. распр. Средняя Азия, Афганистан, Иран, Эфиопия, Мон-
голия, Китай.
Var. meridionalinflatum Nigm. 1977, Докл. АН ТаджССР, 20, 12:42.
(42).
Геогр. распр. Таджикистан.
Var. nigricolorinflatum Udacz. 1976, Тр. прикл. бот. геи. сел. 56, 2:
149. (10).
Среди var. nigricolorinflatum нз Китая имеются формы, по построе-
нию колоса переходные от ипфлятного к обычному типу-—f. falsenigri-
color (Udacz.), Udacz. comb. nov. — T. aestivum var. falsenigricolor
Udacz. 1976, l.c.: 149.
Teorp.’pacnp. Непал, Китай.
Var. nigrilutinflatum (Vav. et Jakubz.) Mansf. 1951, l.c.: 57. — T. vul-
gare var. nigrolutinflatum Vav. et Jakubz. 1931. Tp. прикл. бот. геи. сел.
Прилож. 51: 185. (7).
.Геогр. распр. Эфиопия.
Var. nigrinflatum (Vav.) Mansf. 1951, l.c.: 55. — T. vulgare var. nig-
rinf latum Vav. 1923, l.c.: 175. (21).
Геогр. распр. Иран.
Var. nigrirufinflatum (Vav. et Jakubz.) Mansf. 1951, l.c.: 57. — T. vul-
gare var. nigroruf inf latum Vav. et Jakubz. 1931, 1. c.: 186. (9).
Геогр. распр. Эфиопия.
Var. pseudoheraticum (Vav.) Mansf. 1951, Lc.: 56. — T. vulgare var.
pseudoheraticumVav. 1931, l.c.: 186. (18).
Геогр. распр. Эфиопия.
Var. psetidokhorassanicum Udacz. 1975, 1. c.: 48. (12).
Геогр. распр. Узбекистан (Фергана, 600 м над ур. м.).
Var. pseiidotranscaspicum Kob. ex Kudr. 1942, Пшен. Узбекпст.:
43. (15).
Геогр. распр. Узбекистан (Хорезм).
Var. pseiidoturcomanicum (Vav. et Kob.) Mansf. 1951, 1. c.: 55. T. vul-
gare var. pseiidoturcomanicum Vav. et Kob. 1928, l.c.: 89. (20).
Геогр. распр. Афганистан, Эфиопия.
Var. rufinflatum (Flaksb.) Mansf. 1951, l.c.: 57. — T. vulgare var. mil-
]lo<)pii() I. 1'iihttitn 2 I')
turum subvar. riifinfla/itm Flaksb. 1911, I. c.: 4. T. vulgare var. miUurum
I. ruf in) hit и/// (Flaksb.) Flaksb. 1915, l.c.: 135. -T. vulgare var. rufinfla-
tum (Flaksb.) Vav. 1923, 1. c.: 174. — T. vulgare var. menhsiengense King,
1957, I. c.: 11. (8).
Среди var. rufinf latum обнаружены формы, ио построению колоса пе-
реходные от инфлятного к обычному типу — f. jalserufinf lot uni (Flaksb.)
Dorof. et A. Filat, comb. nov. — T. vulgare var. falserufinf latum Flaksb.
1929, 1. c.: 106.
Геогр. p а с и p. Средняя Азия, Афганистан, Эфиопия.
Var. subbarbarossinflatum Palm. var. nov. — T. vulgare var. subbarba-
rossinflatum Palm, ex Flaksb. 1935, Культ, фл. СССР, 1: 97, descr. ross.—
T. aestivum var. subbarbarossinflatum (Palm, ex Flaksb.) Mansf. 1951,
l.c.: 49, comb, invalid. — A T. aestivo var. subbarbarossa (Vav.) Mansf.
spicis inflatis differt. (39).
Typus: Mongolia, 1923, k-8276. leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. subcaesioidinflatum Frenk. var. nov. — T. vulgare var. subcaesioid-
inflatum Frenk. ex Flaksb. 1935, l.c.: 83, descr. ross., in adnot. — T. aesti-
vum var. subcaesioidesinflatum (Frenk.) Mansf. 1951, l.c.: 53, comb, inva-
lid.— A T. aestivo var. subcaesioides (Flaksb.) Mansf. spicis inflatis
differt. (25).
Typus: Tuva, 1934, k-30966, leg. A. Frenkel et H. Necziporenko (WIR).
Геогр. распр. Тува.
Var. subcaesinflatum Frenk. var. nov. — T. vulgare var. subcaesiuinin-
flatum Frenk. ex Flaksb. 1935. 1. c.: 88, descr. ross., in adnot. — T. aestivum.
var. subcaesiuminflatum (Frenk. ex Flaksb.) Mansf. 1951, l.c.: 53,
comb, invalid. — Л. T. aestivo var. subcaesium (Flaksb.) Mansf. spicis
inflatis differt. (29).
Typus: Tuva, 1934, k-30944, leg. A. Frenkel et H. Necziporenko (WIR).
Геогр. распр. Тува.
Var. suberythroleucinflatum Frenk. var. nov.-—T. vulgare var. sube-
rythroleuconinf latum Frenk. ex Flaksb. 1935, l.c.: 82, descr. ross., in
adnot. — T. aestivum var. sober у throleuconinf latum (Frenk. ex Flaksb.)
Mansf. 1951, 1. c.: 53, comb, invalid. — A T. aestivo var. suberythroleucum
(Vav.) Mansf. spicis inflatis differt. (24).
Typus: Tuva, 1934, k-30947, leg. A. Frenkel et H. Hecziporenko (WIR.)
К var. suberythroleucinflatum относится форма, по строению колоса
являющаяся переходной от инфлятного к обычному типу — f. subfalse-
rythroleucon (Udacz.) Udacz. comb. nov. — T. aestivum var. subfal-
seerythroleucon Udacz. 1979, Бюл. ВИРа, 84: 71.
Геогр. распр. Тува, Таджикистан.
Var. suberythrosperminflatum Palm. var. nov. — T. vulgare var. sub-
erythrospermuminflalum Palm, ex Flaksb. 1935, 1. c.: 80, descr. ross.—
T. aestivum var. suberythrospermuminflatum (Palm, ex Flaksb.) Mansf.
1951, 1. c.: 54, comb, invalid. — A T. aestivo var. suberythrospermuni
(Vav.) Mansf. spicis inflatis differt. (27).
Typus: Mongolia, 1922, k-7988, leg, V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. subferrugininflatum Palm. var. nov. — T. vulgare var. subferrugi-
nettminflaium Palm, ex Flaksb. 1935, l.c.: 87, descr. ross. — T. aestivum
var. subferrugineuminflatum (Palm.) Mansf. 1951, l.c.: 53, comb, inva-
lid.—-A T. aestivo var. ferrugineum (Alef.) Mansf. spicis inflatis diff-
ert. (28).
Typus: Mongolia, 1923, k-8043. leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. subgraecinflaluin Palm. var. nov. — T. vulgare var. subgraecum-
inflatum Palm, ex Flaksb. 1935, l.c.: 76, descr. ross. — T. aestivum var.
subgraecuminflatum (Palm, ex Flaksb.) Mansf. 1951, l.c.: 55, comb, inva-
2.г>0 Классификация рода Triticum
lid. - - Л Т. aeslivo var. sttbgraeciim (Vav.) Mansf. spicis inflatis di-
ffer!. (23).
Typus: Mongolia, 1922, k-8276, leg. V. E. Pissarev (WIR).
К var. sub graecinf latum относятся формы, по сгросншо полоса яв-
ляющиеся переходными от ипфлятного к обычному типу - - /. subfalse-
graecum (Udacz.) Udacz. comb. nov. —- 7'. aestivum var. subfalsegraecum
Udacz. 1979, Бюл. ВИРа, 84: 71.
Геогр. распр. Монголия, Таджикистан.
Var. subhostinflatum Palm. var. nov. — T. vulgare var. subhoslianu-
minflatum Palm, ex Flaksb. 1935. 1. c.: 93, descr. ross. — T. aestivum var.
subhostianuminf latum (Palm, ex Flaksb.) Mansf. 1951, 1. c.: 51, comb,
invalid. — Л T. aestivo var. hostianuni (Clem.) Mansf. spicis inflatis dif-
fert. (37).
Typus: Mongolia, 1923, k-8043, leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. snbmeridionalinflatuni Palm. var. nov. — 7’. vulgare var. subme-
ridionalinflat um Palm, ex Flaksb. 1935, l.c.: 91, descr. ross. — T. aesti-
vum var. subnieridionalinf latum (Palm, ex Flaksb.) Mansf. 1951, l.c.: 52,
comb, invalid. — A T. aestivo submeridionale (Vav.) Mansf. spicis inflatis
differt. (31).
Typus: Mongolia, 1922, k-8277, leg. V. E. Pissarev (WIR).
Геогр. распр. Монголия.
Var. subpseudohostinflatum Udacz. 1975, l.c.: 49. (38).
Геогр. распр. Узбекистан (Фергана, 600 м над ур. м.).
Var. subpseudomeridionalinflatum Udacz. 1975, Lc.: 49. (32).
Геогр. распр. Узбекистан (Фергана, 600 м над ур. м.).
Var. subpseudoturcinflaturn Udacz. var. nov.— Differt a var. subpseudo-
turcicum Vav. et Kob. spicis inflatis. (35).
Typus: Uzbekskaya SSR, prov. Ferghana, dislr. Rischlan, propc pag.
Rischtan, in populatione culta T. aestivum L., 600 m, 19. VIE 1969, spicae,
N 070633, R. Udaczin (WIR).
Геогр. распр. Фергана.
Var. subsogdianinflatum Udacz. 1975, 1. c.: 49. (36).
Геогр. распр. Узбекистан (Фергана, 600 м над ур. м.).
Var. subturcinflatum Udacz. var. nov. — Differt a var. subturcicum Vav.
spicis inflatis. (34).
Typus: Tadzhikskaya SSR, distr. Pendzhikent, prope pag. Margidon,
in populatione culta T. aestivum L., 1250 m, 11. VIE 1969, spicae,
N 070309, R. Udaczin (WIR).
Геогр. распр. Таджикистан (долина Зеравшана).
Var. sunpani (Flaksb.) Mansf. 1951, 1. c.: 58. — T. vulgare var. sunpani
Flaksb. 1911, Tp. Бюро прикл. бот. 4:2. — T. vulgare var. triste f. sun-
pani Flaksb. 1915, Опред. пшен.; 138. — T. vulgare var. nigrohuanyangen-
se King, 1957,1. c.: 11, descr. chin. (2).
Геогр. распр. Китай, Эфиопия.
Var. teheranicum (Vav.) Mansf. 1951, l.c.: 56. — T. vulgare var. te-
heranicumNav. 1923, 1. c.: 174. (13).
Геогр. распр. Иран, Афганистан.
Var. transcaspicum (Vav.) Mansf. 1951, l.c.: 56. — 7’. vulgare var.
transcaspicum Vav. 1923, 1. c.: 174.. — T. vulgare var. oasicolum Due. 1930,
Espec. et var. Cereal, cult, en Algerie: 39. — T. vulgare var. chiugiengen-
se King, 1957, l.c.: 11. (14).
К var. transcaspicum относятся формы, ио построению переход-
ные от ипфлятного к обычному типу - f. Ialsedclfii (Udacz.) Udacz.
comb. nov. — T. aestivum var. falsedelfii Udacz. 1976, I. c.: 148.
Геогр. распр. Туркменистан, Афганистан, Эфиопия, Алжир.
Var. turcomanicum (Vav. et Kob.) Mansf. 1951, l.c.: 55. •—7’. vulgare
var. turcomanicum Vav. et Kob. 1928, 1. c.: 89. (19).
Геогр. распр. Средняя Азия, Казахстан (редко), Афганистан.
litxlpoil I. Tlitillllll
(>. Subconvar. eligulatinflatinn (Flaksb.) Л. Filat, et Dorof. — инфлят-
пая безлигульная подгруппа разновидностей.
Subconvar. eliqnlatinflatum (Flaksb.) A. Filat, el Dorof. comb. nov.
T. vulgare subprot. eligulato-injlatum Flaksb. 1935, Культ, фл. СССР, I:
46.— T. vulgare eligulaluni Vav. 1923, Тр. прикл. бот. сел. 13,1: 175,
quoad plantain inflatani.
Растения с инфлятными колосьями, с листья ми без язычков и ушек
у основания пластинки.
Т ин; Т. aestivum var. Iiorogense (Vav.) Mansf.
Геогр. рас и р. Высокогорные районы Памира.
Яровые поливные посевы в Горном Бадахшане не встречаются ниже
2100 м над ур. м., а в Афганистане — ниже 2400 м над ур. м., преобла-
дая на высотах 2900—3500 м над ур. м.
Определение разновидностей subconvar. eligulatinflatum (Flaksb.) A. Filat, et Dorof.
Колосья безостые
Чешуи неопушенные
Зерновки белые
pamiricum
Iiorogense
cinerescens
1
2
3
Зерновки красные
4-1 — I — | — I — I — I — I — 1 — I — I sc/iugnanicum
— I — I + I — I — I — I — I — I — I — I quasiruf inf latum
Чешуи опушенные (зерновки белые)
+ I — I — I — I — I — I — I — I — I — 1 oxianum
— I — I + I — | — I — I — I — I — I — I petropavlovskianum
Колосья остистые
Чешуи неопушенные
Зерновки белые
4-1 — I — I — I — I — I — I — I + | — | korshinskyi
— I — I + I — I — I — I — I — — I nicolai
8
9
Зерновки красные
+ I — I — I — I — I — | — I — | + I — I ruchczianum
— I — 1 + 1 — I — I — I — I — 1+1 — I suchobrusianum
Чешуи опушенные
Зерновки белые
+ 1 — 1 — I — I — | — .1 — I — I + I — I raikoviae
— | — 1 + 1 — I — I — I — I — 1 + 1 — I pjandshicum
10
11
12
13
' 3 е р н о в к и к р а с н ы е
— I — 1+ I — I — | — | — I — I +1 — 1 baranovii
1 I
I
Var. baranovii Udacz. 1969, Tp. прикл. бог. ген. сел. 40,2: 40. (14).
Геогр. распр. Таджикистан.
Var. cinerescens (Flaksb.) Mansf. 1951, Der Ziichtcr, 21, 1/2: 57.—
T. vulgare var. cinerescens Flaksb. 1929, Tp. прикл. бот. ген. сел. 20:
101. (3).
Г с о г р. р а с п р. Таджикистан.
252
Клиссификаци.ч podu Triticum I..
Var. horogense (Vav.) Mansf. 1951, l.c.: 57.-7'. vulgare var. horogii
Vav. 1923, Tp. прикл. бот. сел. 13,1; 175. (2).
Одна из наиболее распространенных разновидностей безлигульных
пшениц. Основная разновидность сорта Бабило.
Геогр. распр. Таджикистан.
Var. korshinskyi Udacz. 1976, Тр. прикл. бот. ген. сел. 56,2: 148. (8).
Ге-огр. распр. Таджикистан.
Var. nicolai Udacz. 1976, l.c.: 149. (9).
Геогр. распр. Таджикистан.
Var. oxianum (Vav.) Mansf. 1951, l.c..: 56. — T. vulgare var. oxianum
Vav. 1923, l.c.: 175. (6).
Геогр. распр. Таджикистан, Афганистан.
Var. pamiricum (Vav.) Mansf. 1951, l.c.: 58. — T. vulgare var. pamiri-
cum Vav., 1923, l.c.: 175. (1).
Геогр. распр. Таджикистан, Афганистан.
Var. petropavlovskianum Udacz. 1969, l.c.: 40. (7).
Геогр. распр. Таджикистан.
Var. pjandshicum Udacz. 1969, 1. c.: 41. (13).
Геогр. распр. Таджикистан.
Var. quasirufinflatum (Flaksb.) Mansf. 1951, l.c.: 57. — T. vulgare
var. quasirufinflatum Flaksb. 1929, 1. c.: 102. (5).
Геогр. распр. Таджикистан.
Var. raikoviae Udacz. 1969, l.c.: 40. — T. aestivum var. qtiasimeridio-
nale-inflatum Nigm. 1977, Докл. АН ТаджССР, 20, 12: 42. (12).
Геогр. распр. Таджикистан.
Var. ruchczianum Nigm. 1972, Докл. ТаджССР, 15, 12:47. (10).
Геогр. распр. Таджикистан.
Var. schugnanicum (Vav.) Mansf. 1951, l.c.: 57. •—T. vulgare var.
schugnanicumVav. 1923, l.c.: 175. (4).
Геогр. распр. Таджикистан, Афганистан.
У аг. suchobrusianum Udacz. 1969, 1. с.: 41. (11).
Теогр. распр. Таджикистан.
3. Convar. semirigidum A. Filat, et Dorof. — полуригидная группа
разновидностей. :
Convar. semirigidum A. Filat, convar. nov. — T. vulgare prol. subrigi-
dum Flaksb. 1935, Культ, фл. СССР, 1:45, descr. ross. — T. asiaticum
subsp. rigidum Kudr. 1942, Пшен. Узбекист.: 19. — T. asiaticum
subsp. muticorigidum Kudr. 1942, l.c.: 36. Spica semirigida, aristata vel
exaristata; glumis quarn convar, 'rigidum minus rigidis glumarum ner-
vatione minus differt.
Листья с язычком и ушками у основания пластинки пли без них.
Колосья остистые или безостые, полугрубого построения; колосковые
чешуи менее грубые, чем у convar. rigidum, с менее выраженным жил-
кованием, зерновки менее плотно заключены в чешуях, не осыпаются,
по легко обмолачиваются.
Т и и: Т. aestivum var. graecum (Koern.) Mansf.
Геогр. распр. Западная Азия, Индия, Китай, Средиземноморье,
Нижнее Поволжье, Закавказье, Центральная Америка.
Ключ для определения подгрупп разновидностей
convar. semirigidum A. Filat, et Dorof.
1. Листья с язычками и ушками у основания пластинки; листовая
пластинка отогнута от стебля ......................................
. . ....................subconvar. semirigidum A. Filat, ct Dorof.
+ Листья без язычков и ушек у основания пластинки; листовая плас-
тинка отогнута от стебля ..........................................
....................subconvar. eligulatimi (Vav.) A. Filat, et Dorof.
lh><)pot) 1. Tritictun
a. Subconvar. eligiilatiiin (Vav.) A. I 'ilat. cl Dorof. безлигульпая
подгруппа разновидностей.
Subconvar. eligulatum (Vav.) A. Filat, et Dorof. comb. nov. — 7'. vulga-
re группа eligulatum Vav. 1923, Tp. нрпкл. бог. сел. 13,1; J75, excl. plantar
inflatac. - T. vulgare subprol. eligulatum (Vav.) Flaksb. 1935, Культ. фл.
СССР, 1:45.
T и п: Т. aestivum var. tadjicorum (Vav.) Mansf.
Растения с колосьями полугрубого построения, с листьями без языч-
ков п ушек у основания пластинки.
Геогр. распр. Высокогорные районы Памира. Известны исклю-
чительно яровые поливные посевы не ниже 1900 м над ур. м.
Колосья безостые (чешуи неопушенные, зерновки красные)
+ — — — ; — , — I — ' — ; tnaksumovii 1
Колосья по л у о сти с гыс
Чешуи неопушенные
Зерновки белые
+ I — I — I — I — I — I — I — I + I — I subgunticum I 2
— I — I + I — I “ I — I — I — 1 + 1 — I subajghanicum | 3
Зерновки красные
+ । — ’ — : — I • — — I — } — I + ' — I subkabuliciim | 4
Чешуи опушенные
Зерновки белые
+ I — I — I — I — I — I — 1 — 1 + 1 - I flaksbergeri I 5
— I — 1 + | — I — I — | | — I + I ~ I quasisubtuicicuin | 6
Колосья остистые Чешуи неопушенные
3 е р н о в к и бел ы е.
— — 1 — — + — 1 gunlicum 7
— — — — — 4- — af ghanicutn 8
— — + — — + — buruschonicuni 9
3 с р и о в к и к р а с н ы о
— — — — — + kabulicum 10
— — — — — + — tadjicorum 11
— — i' — — + — murgabicum 12
ММ.;
Чешуи опушенные
Зерновки белые
Зернов к и крас и ы р
gorbunovii 13
quasiturcicutn 14
japschorvi 15
quasibarbarossa 1(>
badakschanicum 17
Var. afghanicum (Vav.) Mansf. 1951, Der Ziichter, 21, 1/2: 53.--
T. vulgare var. afghanicnm Vav. 1923, Tp. прикл. бот. сел. 13, 1: 176. (8).
Основная разновидность сорта Джальдак.
Геогр. распр. Средняя Азия, Афганистан.
251 Классификации рода Ttidtum L.
Var. badakscliaiiicinii (Vav. el Kob.) Mansf. 1951, I. c.: 49. T. vulgii
re var. badakschanicum Vav. et Kob. 1928, 1. c.: 89. (17).
Г e о г p. p а с и p. Таджикистан, Афганистан.
Var. banischonicum Udacz. 19(19, 'Гр. прикл. бог. ген. сел. 40.2: 41. (9).
Геогр. распр. Таджикистан.
Var. flaksbergeri Udacz. 1969, 1. с.: 40. (5).
Геогр. распр. Таджикистан.
Var. gorbunovii Udacz. 1969, 1. с.: 40, nom. praeocup. (13).
Геогр. рас и р. Таджикистан.
Var. gunticum (Vav.) Mansf. 1951, l.c.: 55. — T. vulgare var. gunti-
cum Vav. 1923, 1. c.: 176. (7).
Геогр. распр. Таджикистан, Афганистан.
Var. japschorvi Nigm. 1972, Докл. АН ТаджССР, 15, 12: 47. (15).
Геогр. распр. Таджикистан.
Var. kabulicum (Vav.) Mansf. 1951, l.c.: 54. — 7'. vulgare var. kabu-
licum Vav. 1923, 1. c.: 176. (10).
Основная разновидность ярового сорта Сафедрогз.
Геогр. распр. Таджикистан, Афганистан.
Var. maksumovii Udacz. 1969, 1. с.: 40. (1).
Гео г р. р а с и р. Таджикистан.
Var. inurgabicum (Flaksb.) Mansf. 1951, l.c.: 52.-7’. vulgare var.
murgabicum Flaksb. 1929, l.c.: 101. (12).
Геогр. распр. Таджикистан.
Var. quasibarbarossa Udacz. 1976, Tp. прикл. бот. ген. сел. 56,2:
149. (16).
Г е о г р. р а с и р. Таджикистан.
Var. qiiasisubturcicum (Flaksb.) Mansf. 1951, l.c.: 50. — T. vulgare
var. quasisubturcicum Flaksb. 1929, l.c.: 102. (6).
Геогр. распр. Таджикистан.
Var. quasiturcicum (Flaksb.) Mansf. 1951, l.c.: 49. — T. vulgare var.
quasiturcicum Flaksb. 1929, l.c.: 102. (14).
Геогр. распр. Таджикистан.
Var. subafghanicum (Flaksb.) Mansf. 1951, Lc.: 53. — T. vulgare var.
subafghanicum Flaksb. 1929, Tp. прикл. бот. ген. сел. 20: 104. (3).
Геогр. рас и р. Таджикистан.
Var. subgunticum (Vav.) Mansf. 1951, l.c.: 55. — T. vulgare var. sub-
gunticum. Vav. 1923, 1. c.: 176. (2).
Геогр. рас пp. Таджикистан.
Var. subkabulicum (Flaksb.) Mansf. 1951, l.c.: 54.— T. vulgare var.
subkabulicum Flaksb. 1929, Tp. прикл. бот. ген. сел. 20: 104. (4).
Геогр. распр. Таджикистан.
Var. tadjicorum (Vav.) Mansf. 1951, l.c.: 52. — T. vulgare var. tadji-
corum Vav. 1923, l.c.: 176. — T. vulgare var. bucharicum Vav. in Flaksb.
1923, Опред. наст, хлебов: 37. (И). Основная разновидность сорта
Руштак.
Геогр. распр. Таджикистан, Афганистан.
б. Subconvar. semirigidum A. Filat, et Dorof. — полуригидпая под-
группа разновидностей.
Subconvar. semirigidum A. Filat, et Dorof. subconvar. nov. — T. vul-
gare prol. subrigidum Flaksb. 1935, Культ, фл. СССР, 1: 45, descr. ross. —
T. vulgare subprol. indicum Flaksb. 1935, l.c.: 45, descr. ross. — Plantae
foliis ligiilatis, reclinalis.
T и и: T. aestivum var. graecum (Koern.) Mansf.
Описание subconvar. semirigidum, к которой относятся разновидно-
сти с развитыми язычком и ушками, по остальным признакам соответ-
ствует приводимому в целом для convar. semirigidum.
Г е о г р. р а с и р. По всему ареалу convar. semirigidum.
Широкий ареал возделывания обусловил значительное экологиче-
ское разнообразие subconvar. semirigidum.
Подрод I. Tritu inn
Подробное деление на эколого-географические группы и их описание
приведены в работах И. И. Вавилова (1957, 1964).
К полуригндпой а.пia iсков пшенице отпоен гея ряд групп, выделенных
II. И. Вавиловым в Юго-Западной Азии, Индии, Закавказье, а также
впутреппеапатолийская, пиренейская(внутренняя испанская но И.И. Ва-
вилову), аргентинская гибридная группы, рассматриваемые II. И. Вави-
ловым (1964) в европейском подвиде, и эфиопская группа, соответствую-
щая subsp. abyssinicum Vav.
Приводим краткое описание основных эколого-географических групп
subconvar. semirigidum.
а) Кашгарская группа отличается восприимчивостью к поражению
желтой и бурой ржавчинами и высокой холодостойкостью озимых форм
в степных малоснежных районах; сформировалась в оазисах Западного
Китая (Кашгарский, Яркендский, Учтурфанский, Кульджинскпй).
б) Белуджистанская группа развилась в горных пустынных и полу-
пустынных районах Пакистана и восточного Ирана, где земледелие
основано па поливе. Растения высокорослые, с прочной соломиной;
восприимчивые к ржавчинам, ио выявлены формы, отличающиеся им-
мунностью; продуктивные, е довольно крупным иеосынающпмгя зерном.
Ригидность растений более выражена, чем у индийской группы,
в) Индостанская труппа характеризуется исключительной скороспе-
лостью, прочностью соломины, устойчивостью к заболеваниям. Растения
низкорослые или среднерослые, зерно стекловидное, просыпаемое. Ин-
дийская группа сыграла важную роль в селекции современных продук-
тивных сортов пшеницы мягкой. Если subconvar. semirigidum является
переходным между азиатским и европейским подвидами пшеницы мяг-
кой, то индийская группа отражает связь азиатского подвида с восточ-
ной группой европейского. Из агроэкологических групп азиатского под-
вида у индийской признаки ксерофилыюсти выражены слабее.
г) Горпокавказская степная (армяпо-нахичевапская) группа харак-
теризуется устойчивостью к почвенной и воздушной засухе, ранне- и
среднеспелостыо, восприимчивостью к болезням.
д) Ашпероио-мугапская группа возделывалась на поливных землях,
исключительно стойка к воздушной засухе; скороспелая, формирует
низкий урожай, по устойчивый по годам. Для растений этой группы ха-
рактерны низкорослость, узколистность, мелкий колос н мелкое зерно.
Группа приурочена к засушливым районам Апшеронского полуост-
рова и Муганп.
е) Сирийская группа — низкорослая, скороспелая, сравнительно стой-
кая к желтой и бурой ржавчине пшеница, приурочена к нагорьям Сирии,
Израиля и Иордании.
ж) Впутреппеапатолийская ксерофнльная группа представлена глав-
ным образом яровыми формами, от низкорослых до сравнительно высо-
корослых, с соломиной среднеустойчивой или склонной к полеганию,
нередко выполненной паренхимой по всей длине, крупным грубым коло-
сом с трудным обмолотом и плотным заключением зерновок в чешуях.
Это наиболее устойчивая к поражению ржавчинами эколого-географиче-
ская группа азиатского подвида.
Географически находясь па стыке азиатского и европейского подви-
дов (Малая Азия), впутреппеапатолийская группа несет элементы евро-
пейского подвида: относительный иммунитет к бурой и желтой ржавчи-
не и довольно прочная соломина. Среди этой группы чаще появляются
безостые разновидности (II. II. Вавилов, 1964).
з) Пиренейская группа является переходной от азиатского к евро-
пейскому подвиду. Наряду с признаками, присущими азиатскому под-
виду— чешуи грубые с сильно выраженным жилкованием, ости обычно
довольно короткие, ригидные или полуригпдныс, грубость колоса хо-
рошо выражена у безостых форм; плотное заключение зерновок в чешу-
.’56 Классификации 1>оЛ) Tii/iiiun I..
ях, хотя обмолот средней грудное, и; засу\о\с i oii'iHiioci 1, хорошая, -
песет черты европейского подвида — растения мощно развитые, с широ-
кими поникающими листьями, сравнительно иммунные к желтой и бурой
ржавчине, средне- или позднеспелые, с длительным прохождением ста-
дии созревания, требовательные к влаге. Для пиренейской группы ха-
рактерна соломина с утолщенными стенками, нередко все междоузлия
выполнены паренхимой.
Приурочена к сухим нагорьям внутренней Испании, Португалии,
Алжира, Туниса и Центральной Америки, куда была завезена испан-
цами.
и) Эфиопская группа в общем близка индостанской, характеризуется
очень коротким вегетационным периодом, сравнительно мало поража-
ется бурой ржавчиной, сильнее — желтой. Соломина короткая (50—
80 см). В эфиопской группе довольно ярко отражается переходный ха-
рактер subconvar. semirigidum — сочетание признаков азиатского под-
вида (грубость общего построения колоса) с европейским (довольно
легкий обмолот).
В основном возделывается на высоте 2100—2400 м над ур.м., но
п здесь не является преобладающей.
к) Аргентинская гибридная группа близка к пиренейской как ио
происхождению (исходным материалом для этой группы послужили пше-
ницы Испании при участии итальянских, китайских и японских сортов),
гак и по морфологическим и биологическим особенностям (грубость ко-
лоса, скороспелость, высокая стекловидность зерна и др.).
К данной группе относятся гибридные сорта Аргентины, Уругвая и
других латиноамериканских стран.
Как уже указывалось ранее, в настоящее время эколого-географиче-
ские группы пшениц в большей части ареала своего исторического раз-
вития вытеснены продуктивными современными сортами интенсивного
типа. Но сохраненные, главным образом в коллекциях, местные сорта
различных экологических групп играют большую роль в селекции пше-
ницы.
Var. almaatense (Anik.) Mansf. 1951, Dor Zuchter, 21, 1/2: 55.—
T. almaatense Anik. 1929, Tp. Всесоюзп. съезда ген. сел. сем. январь,
1929 (Ленинград), 3: 48. — Т. vulgare var. alienuni Vav. et Jakubz. 1931,
Tp. прикл. бот. ген. сел. Прилож. 51: 182. — Т. vulgare var. setosomeso-
polamicum Thum. 1933, Опред. хлебн. злаков (колос.): 192. (28).
Геогр. распр, Эфиопия, Казахстан, Азербайджан и Армения
(редко).
Var. ardjeschicum (Thum.) Mansf. 1951, l.c.: 56. — T. vulgare var.
ardjeschicum Thum. 1929, Tp. прикл. бот. ген. сел. 22, 2: 310. — Т. vulga-
re var. nigroatrum Hasiz. 1932, Der Zuchter, 3/4: 76. (14).
Геогр. распр. Армения, Восточная Турция (редко).
Var. barbarossa (Alef.) Mansf. 1951, l.c.: 43. — T. vulgare var. bar-
barossa Alef. 1866, Landwirtsch. FL: 330; Koern. in Koern. et Wern. 1885,
Handb. Getreideb. 1: 48. — T. vulgare var. subvelutinum. Koern. 1873,
System. Ubers. Cer.: 12. (53).
Геогр. распр. Средняя Азия, Малая Азия, Афганистан, Пран,
Пакистан, Ицдия, Монголия, Китай, Эфиопия, Средиземноморье, Закав-
казье, Дагестан.
Var. bengalense (Howard) Mansf. 1951, l.c.: 52. — T. vulgare var.
bengalense Howard, in Howard et Khan, 1922, The wheat of Bihar and
Orissa (Calcutta): 19. (39).
Геогр. распр. Индия, Афганистан, Эфиопия, Казахе! аи, Узбеки-
стан, редко в Закавказье.
Var. caeruleovelutinum (Koern.) Dorof. et A. Filat, comb. nov.—
T. vulgare var. caeruleovelutinum Koern. 1873, l.c.: 12; Koern. in Koern.
et Wern. 1885, I. c.: 48, quoad plantain spiels griseis; Vav. 1923, l.c.:
11 оОрод 1. Triticum
'267
Определение разноиндноеиТ: subconvar. seniiripiduin Л. l ilaf. et Dorof.
Чешуи Ости Разновидность № разновидно- 1 сги 1 J
3 о 'О белые с черными краями красные красные с черными краям и ССрО- чаты ф<> 6 о 'О ды м- е на «с а§ Е черн! фо о •о е на нс a S а х цвета чешуи 1 черные |
Колосья безостые
Чешуи неопушенные
Зерновки белые
1 1 + + + — 3 е р II 0 В 1\ И К I а с и и с insigne turanicum nigralbirubrum 1 2 3
+ —. — — — — — — — — pulchrum 4
— — + —— — — — — — — uzbeliistanictim 5
— — + — — — — — — +i: psetidoalbinibrum 6
— — • — + — — — cinereum 7
ч 3 е 31UIJU р И 0 I 4- опуше к и иные е л ы с introitum s
+ — — — — — — — — — leucospermum <)
+ — — — —- — —. — - — +* pseudoleucospermum 10
— + —— —• — — — — — — vanense 11
— — + — — — — — — — delfii 12
— — 4- — — — — — — 4-* pseudodelfli 13
— — — — — — + — — — ardjeschicum 14
•— — — — 3 е р НОВ] + и к а с н ы е melanorubruin 15
+ — — — — — — — — +* pseudoveluiinutn 16
— — + — — — — — — — inagnificum 17
— — + — — — — — — -1 * pscudopyrothrix 18
4- К о ч Зс — го с ь ?tuyu р Н 0 I Я О С' ieonyi к и ГИСТ ленны о л ы ы в о е cyimothrix 19
+ — — — — — — — + — graccum 20
— + — — — — — — — + nigrigraecum 21
+ — + pseudograecum 22
— — 4- — — — — — 4- — ert/throleucum 23
— — + — — — —. — — + pseudoerythroleucum 24
— — — + — — — — — + chiovendae 25
— ,— — — — + — — — + caesioides 26
— — — — 4- — —- — — + cinerosum 27
— — — — — — + — — + almaatense 28
— — — — — 3 е р нов + < и к в а с н 4- ы е indicum 29
+ — — — — — — — + — pseudoerythros per- mum 30
+ — — — •— — — — — + melanopogon 31
— + — — — — — — — 4- ingrediens 32
— — + — — —— — + — var labile 33
— — + — —. — — — + hissaricutn 34
— —. — + — — — — 4- nigrisardoum 35
— — — — -I- — — — — 1- pgintsi harovii .36
— — —‘ —- I- — — 1- i acsiorigidum 37
- - — — — —. -1- — — 4- nigricans 38
— — — — — — — -1- — 4- bengalense 39
* Окраска остеоидных заострений.
17 шч
25S
Классификация por)a Triticum L.
Чешуи опушенные
Зс J 1! О В к п бе л ы е
+ — — — — — — — + — meridionale -10
+ — — — — — — + pseudomeridionede 41
— + —— —. —— — — — 4- hamadanicum 42
— -— + — — — — — + — turcicum 43
— — + — — — — — — + pseudoturcicum 4 1
— —- — 4- — — — — + — kermanschactucum 45
— — —— — + — — — — + griseum 46
— — — + — — — + sogdianiun 47
— — — — — — + — — + inesopotamicum 48
— — — — — — — + — + iraiiictini 49
Зерновки красные
+ — — — — — — — 4- .— hostianum 50
+ — — — — —- — — — + pseiidohostianam 51
— 4- — — — — — 4- — trurvinicum 52
— — 4- — — — — 4- — barbarossa <)• J
— — + - — — - — — — + pseiidobarbarossa 54
— — — + — — 4- — lurislriniciim 55
— — — — 4- — — + caeriileoL'eliitiniiin 56
,—. — — — — + — — — 4^ rubr'muriniim 57
— — —. — —- — + — — 4- renoeatum 58
— — — — — — + + [idiginosinn 59
170, descr. emend. — T. vulgare var. inurinuni Flaksb. 1935, l.c.: 94, descr.
ross. — T. asiaticum var. iranoturanicum Kudr. 1942, l.c.: 30. (56).
Геогр. распр. Иран, Индия, Пакистан, Монголия, очень редко
в Армении.
Var. caesioides (Flaksb.) Mansf. 1951, 1 с.: 53. — T. vulgare var.
caesioides Flaksb. 1929, Tp. прикл. бот. геи. сел. 20: 104. — Г. vulgare
var. Ichujense Anik. 1929, l.c.: 48. (26).
Геогр. распр. Памир, Казахстан, Туркмения, Таджикистан, Уз-
бекистан, Азербайджан.
Var. caesiorigiduni (Kudr.) Л. Filat, comb. nov. — T. asiaficum var.
caesiorigiduni Kudr. 1942, Пшен. Узбскпст.: 27. (37).
Геогр. распр. Средняя Азия, Индия, Иран, Афганистан.
Var. chiovendae (Vav. et Jakubz.) Mansf. 1951, Lc.: 53. — T. vulgare
var. chiovendae Vav. et Jakubz. 1931, Tp. прикл. бот. ген. сел. Прилож.
51: 182. (25).
Геогр. распр. Эфиопия.
Var. cinereum (Dekapr.) Mansf. 1951, l.c.: 57. — 7. vulgare var. cine-
reum Dekapr. 1924, Изв. Тифл. Политехи, ин-та, 1: 3. — 7’. vulgare var.
dshungaricum Orlov, 1935, Пшен. Казахст.: 14. (7).
Геогр. рас и р. Закавказье, Казахстан, Туркмения.
Var. cinerosiiin (Flaksb.) Mansf. 1951, l.c.: 55. — 7’. vulgare var. cine-
rosum Flaksb. 1929, 1. c.: 104. (27).
Геогр. распр. Таджикистан.
Var. cyanothrix (Koern.) AAansf. 1951, l.c.: 55. — T. vulgare var.
cyanothrix Koern. cl Wern. 1885, 1. c.: 46. (19).
Геогр. распр. Иран, Закавказье.
Var. delfii (Koern.) Mansf. 1951, Lc.: 55. 7'. vulgare var. ilcljii Koern.
in Koern. et Wern. 1885, Lc.: 46. 7’. asiaticum var. chivense Kiidr. 1942,
l.c.:37. (12).
Геогр. распр. .Средняя Азия, Закавказье, Афганистан. Индия,
Иран, Северная Африка, Эфиопия.
11 ОС/Пп) 1. Tlilil lint
259
Var. erythroleucum (Koern.) Mansf. 1951. l.c.: 53, ’eryihroleticon .—
T. vulgare var. erythrolcucon Koern. in Koern. et Worn. 1835, l.c.: 47. (23).
Геогр. распр. По всему ареалу азиатского подвида и небольших
размерах.
Var. fuliginosum (Alef.) Mansf. 1951, l.c.: 43. — T. vulgare var. fuligi-
nosum Alef. 1866, l.c.: 330; Koern. in Koern. et Wern. 1885, l.c.: 48;
Flaksb. 1935, l.c.: 98. — T. vulgare var. kurdistanicum Vav. 1923, Tp.
прикл. бот. сел. 13, 1: 171. — T. asiaticum var. komarovii Kudr. 1942, l.c.:
31. (59).
Геогр. распр. Иран, Афганистан, Средняя Азия, редко в Закав-
казье и Казахстане.
Var. gontscharovii (Kudr.) Л. Filat, comb. nov. — T. asiaticum var.
gontscharovii Kudr. 1942, 1. c.: 26. (36).
Геогр. распр. Узбекистан, Таджикистан.
Var. graecum (Koern.) Mansf. 1951, 1. c.: 54. — T. vulgare var. grae-
cuni Koern. 1893, l.c.: 11; Koern. in Koern. et Wern. 1885, l.c.: 46. (20).
Выделены формы с соломиной, выполненной паренхимой по всей
длине'—f. karbulakianum (Anik.) Dorof. et A. Filat, comb. nov. — T. vul-
gare var. karbulakianum Anik. 1929, I. c.: 48.
Геогр. распр. Одна из самых распространенных разновидностей
в ареале азиатского подвида — Юго-Западная Азия, Средняя Азия, Ка-
захстан, Китай, Монголия, Средиземноморье, Закавказье, Центральная
Америка.
Var. griseum (Vav.) Mansf. 1951, l.c.; 51. — T. vulgare var. griseum
Vav. 1923, l.c.: 170. — T. vulgare var. tnuriniim Vav. in Flaksb. 1922,
Опред. наст, хлебов: 43, descr. incompl., non Flaksb. 1935. (46).
Геогр. распр. Таджикистан, Узбекистан, Афганистан, Азербайд-
жан, Армения.
Var. hamadanicum (Vav.) Mansf. 1951, l.c.: 51.— T. vulgare var.
liamadaniciimVav. 1923, l.c.: 172. (42).
Геогр. распр. Иран, Азербайджан (Нахичевань).
Var. hissaricum (Kudr.) A. Filat, comb. nov. — T. asiaticum var.
hissaricum Kudr. 1942, l.c.: 26. — T. vulgare var. subsardoiun Vav. et
Jakubz. 1931, l.c.: 184. (34).
Среди тунисских образцов выявлена форма с соломиной, выполнен-
ной паренхимой по всей длине, описана как var. plenosardoum Flaksb.
1935, 1. с.: 86, descr. ross.
Геогр. распр. Средняя Азия, Иран, Малая Азия, Эфиопия, Сре-
диземноморье.
Var. hostianum (Clem.) Mansf. 1951, l.c.: 50. — T. hostianum Clemente
in Herrera, 1818. Agricult, gen.: 1. — T. vulgare var. hostianum Koern. in
Koern. et Wern. 1885, Handb. Getreideb. 1: 47. — T. vulgare var. velutinum
(Schuebl.) Alef. 1866, I. c.: 330, comb, invalid. (50).
Геогр. распр. Средняя Азия, Иран, Индия, Афганистан, Эфиопия,
Греция, Испания, Португалия, Закавказье (редко).
Var. indicum (Howard) Mansf. 1951, l.c.: 53. — T. vulgare var. indicum
Howard, in Howard et Khan, 1922, 1. c.: 18. (29).
Геогр. распр. Индия, Таджикистан, Узбекистан.
Var. ingrediens (Vav. et Jakubz.) Mansf. 1951, I. c.: 54. — T. vulgare
var. ingrediens Vav. et Jakubz. 1931, l.c.: 179,— T. vulgare var. pseudoin-
grediens Flaksb. 1935, 1. c.: 79, descr. ross. (32).
Геогр. p а сп p. Эфиопия, Ирак, Малая Азия, Туркмения.
Var. insigne (Kudr.) A. Filat, comb, now /'. asia/icinn var. insigne
Kudr. 1942, lliuen. Узбекист.: 36. (1).
1 corp, p а с и p. Средняя Азия, Монголия, .Афганистан.
Var. introitum (Vav. ct Jakubz.) Mansf. 1951, l.c.: 57.-7'. vulgare
\-armntroitumNa\.&Sa\<M'oz. 1931, l.c.: 181. (8).
Геогр. распр. Эфиопия, Узбекистан, Азербайджан.
17
2(>0 l\A<ii:cti(liiii;antin /xx)<i 77 itii um I,.
Var. iranicmn (Vav.) Mansf. 1951, l.c.: 49. T. vulpure var. iranicnm
Vav. 1923, l.c.: 171. (49).
Геогр. распр. Иран, Афганистан, Казахстан, Узбекистан, редко
в Заканказьс.
V ar. kazvinicum (Vav.) Mansf. 1951, l.c.: 50.— T. vulgare var. kazvi-
nicumVav. 1923, l.c.: 172. (52).
Г e о г p. p а с и p. Иран, о. Сардиния, Туркмения.
V ar. kermanschachicuni (Vav.) Mansf. 1951, l.c..: 49.— T. vulgare var.
kertnanschuchii Vav. 1923, Tp. прикл. бот. сел. 13, 1: 172. (45).
Геогр. распр. Иран, Афганистан.
V ar. leucospermum (Koern.) Mansf. 1951, l.c.: 43, quoad subsp. hadro-
pyrum.— T. vulgare var. leucospermum Koern. in Koern. et Worn. 1885,
1. c.: 45, p.p. (9).
Геогр. распр. Индия, Иран, Афганистан, Ирак, Эфиопия, Ма-
лая Азия, Испания.
V ar. luristanicuni (Vav.) Mansf. 1951, l.c,: 43. — T. vulgare var. luri-
sianicumMav. 1923, l.c.: 172. (55).
Гео г p. p а с п p. Иран, Азербайджан, Дагестан.
V ar. magnificuin (Kudr.) A. Filat, comb. nov.--Г. asiaticuin var.
magnijicum Kudr. 1942, l.c.: 37. (17).
Г еогр. распр. Средняя Азия, Афганистан, Монголия, Эфиопия;
V ar. melanopogon (Chiov.) Mansf. 1951, l.c.: 53. — T. vulgare var.
melanopogon Chiov. 1912, Monogr. Rapp. col. (Roma) N 19: 13. (31).
Геогр. распр. Средняя Азия, Закавказье, Сирия, Сардиния и
Сицилия.
V ar. melanorubruin (Thum.) Mansf. 1951, l.c.: 56.— T. vulgare var.
melanorubrum Thum. 1929, l.c.: 310. (15).
Геогр. распр. Армения, Восточная Турция, Узбекистан.
Var. meridionale (Koern.) Mansf. 1951, l.c.: 51. — T. vulgare var.
meridionale Koern. in Koern. et Wern. 1885, 1. c.: 47. (40).
Геогр. распр. Средняя Азия, Средиземноморье, Эфиопия, Закав-
казье (редко), Иран, Афганистан, Малая Азия.
Var. mesopotamicum (Vav.) Mansf. 1951, l.c.: 51.— T. vulgare var.
inesopolaniicuni'Vav. 1922—1923, l.c.: 171. (48).
Геогр. распр. Иран, Афганистан, Малая Азия, Туркмения, редко
в Закавказье.
Var. nigralbirubrtim (Vav. et Jakubz.) AJansf. 1951, l.c.: 57. — T. vul-
gare var. nigroalborubruni Vav. et Jakubz. 1931, l.c.: 180. T. vulgare
var. tarbagalanicum Orlov, 1935, Пшен. Казахст.: 14. (3).
Геогр. распр. Эфиопия, Казахстан.
Var. nigricans (Howard) Mansf. 1951, l.c.: 54.— 7'. vulgare var. nig-
ricans Howard. 1922, I. c.: 16. (38).
Геогр. распр. Индия, Афганистан, Пакистан, Средняя Азия,
Эфиопия, Северный Кавказ, редко в Закавказье.
Var. nigrigraeciim (Perciv.) Mansf. 1951, l.c.: 55. — T. vulgare var.
nigrograecum Perciv. 1926, Journ. Bot.: 210. — T. vulgare var. graecoimar-
ginatum Stol. 1922, Tp. прикл. бот. сел. 23, 4: 84. (21).
Геогр. распр. Иран, Армения, Эфиопия.
Var. nigrisardomn (Vav. et Jakubz.) Mansf. 1951, l.c.: 52. — T. vulgare
var. nigrosarclouniVav. et Jakubz. 1931, l.c.: 179. (35).
Геогр. p а с п p. Эфиопия.
Var. pseiidoalbirtibrum (Kob.) Alansf. 1951, l.c.: 57. — T. vulgare var.
pseiidoalboriibruni Kob. 1932, Хлебн. злаки Турции: 28. (6).
Г с о г р. р а с и р. Турция, Азербайджан.
Var. psendobarbarossa (Vav.) Mansf. 1951, l.c.: 43. — T. vulgare var.
pseudobarbarossa V av. 1923, l.c.: 170. (54).
Геогр. распр. В ареале возделывания var. barbarossa.
Ihjdpoi) 1. Tiiticiim 261
Var. psendodelfii (Kob.) M.insf. 1951, l.c.: 55. T. vulgare var. pseu-
dodelfii Kob. 1932, l.c.: 29. (13).
Г с о г p. p а с и p. Турция.
Var. psetidoeryihroleiicuin (Perciv.) Mansf. 1951, I. c.: 53 ' pwudoeryth-
roleucon'.— T. vulgare var. pseudoerylhroleucon Perciv. 1926, Journ. Bot.:
210. (24).
Ге or p. p a c ii p. Малая Азия, Афганистан, Ирак, Узбекистан, редко
в Закавказье.
Var. psetidoeryihrospernitini (Kudr.) Л. Filat, conib. nov. -- 7’. asiati-
cum var. pseudoerythrospermum Kudr. 1942, 1. c.: 23. (30).
Возделывается преимущественно на богарных землях, реже на по-
ливе.
Среди тунисских образцов выявлена форма с соломиной, выполнен-
ной паренхимой по всей длине, описана как var. plenoerythrospermum
Flaksb. 1935, l.c.: 79, descr. ross. В Сирии обнаружена форма без воско-
вого налета, описана как var. epruinosa Jakubz. in Flaksb. 1935, l.c.: 79,
descr. ross.
Геогр. распр. Средняя Азия, Западная Азия, Средиземноморье.
Var. psetidograecuin (Flaksb.) Mansf. 1951, l.c.: 54. -T. vulgare var.
pseudograecum Flaksb. 1929, 1. c.: 104. (22).
Геогр. распр. Таджикистан, Казахстан, Туркмения, Сирия, Иор-
дания, Югославия, Азербайджан, Армения.
Var. pseiidohostianuni (Flaksb.) Mansf. 1951, l.c.: 50. — T. vulgare
var. pseudohostianum Flaksb. 1915, Опред. пшен.: 157. (51).
Геогр. распр. Средняя Азия, Иран, Афганистан, Индия, Эфиопия,
о. Сардиния, редко в Закавказье.
Var. pseudoleucospermum (Kob.) Mansf. 1951, l.c.: 56. — T. vulgare
var. pseudoleucospermum Kob. 1932, 1. c.: 28. (10).
Геогр. распр. Турция.
Var. pseudomeridionale (Flaksb.) Mansf. 1951, l.c.: 51. — T. vulgare
vav.pseudomeridionale Flaksb. 1915, l.c. :155. (41).
Геогр. pa cup. Средняя Азия, Малая Азия, Афганистан, Иран,
Эфиопия, редко в Закавказье.
• Var. pseudopyrothrix (Kob.) Mansf. 1951, l.c.: 55. — T. vulgare var.
pseudopyrothrix Kob. 1932, l.c.: 29. (18).
Геогр. распр. Азербайджан, Турция.
Var. pseudoturcicum (Vav.) Mansf. 1951, l.c.: 49. — T. vulgare var.
pseudolurcicutnNav. 1923, l.c.: 170. (44).
В Испании и Турции обнаружена форма с выполненной соломиной.
Геогр. распр. В ареале var. turcicum.
Var. pseudovelutinum (Kob.) Mansf. 1951, l.c.: 56. — T. vulgare var,
pseudovelutinum Kob. 1932, 1. c.: 28. (16).
Геогр. распр. Турция.
Var. pulchrum (Kudr.) A. Filat, comb. nov. — T. asiaticum var. pulch-
rum Kudr. 1942, l.c.: 37. (4).
Геогр. распр. Средняя Азия, редко.
Var. renovatum (Flaksb.) Mansf. 1951, l.c.: 50.— 7’. vulgare var.
renovatum Flaksb. 1935, l.c.: 98. — T. vulgare var. fuliginosum non Alef.:
Vav. 1923, l.c.: 171. (58).
Геогр. рас п p. Иран, Индия, Пакистан, Малая Азия, редко в За-
кавказье.
Var. rubrimiirinimi Flaksb. var. nov. — - T. vulgare var. rubromurinum
Flaksb. 1935, Культ, фл. СССР, 1: 97, descr. ross. T. aestivum var. rtib-
rouiuritium (Flaksb.) Mansf. 1951, l.c.: 43, comb, invalid.- Л T. acsiivo
var. pseudobarbarossa spicis gri.seis differt. (57).
Typus: Tadzhikistan (k-52551), Reproductio stationis Daghestanskajae
fulcracea WIR, 1973, leg. A. Filatenko (WIR).
Геогр. распр. Средняя Азия, ^Монголия, редко в Закавказье.
'!(>2 К.пптпфпкпцип ТI iticutn I..
Var. sogdianum (Kudr.) A. Filat. comb. nov. T. asiaticum var. sog-
dianum Kudr. 19-12, l.c.: 30. (47).
Геогр. распр. Узбекистан.
Var. turanicum (Kudr.) A. l-'ilat. conib. tiov. - T. asiaticum var. tura-
nicum Kudr. 1942, l.c.: 37. (2).
Геогр. распр. Средняя Азия, Афганистан, Индия, Иран, Турция.
V ar. turcicum (Koern.) M.ansf. 1951, l.c.: 49. — T. vulgare var. turci-
cutn Koern. in Koern. et. Worn. 1885, 1. c.: 48. (43).
Геогр. распр. Средняя Азия, Иран, Афганистан, Малая Азия,
Эфиопия, Закавказье, Монголия.
V ar. uzbekistanicum (Kudr.) Л. Filat, comb. nov. — T. asiaticum var.
uzbekislanicum Kudr. 1942, l.c.: 37. (5).
Геогр. распр. Редко в Средней Азии, Афганистане, Иране, Ин-
дии.
V ar. vanense (Thum.) Mansf. 1951, l.c.: 56. — T. vulgare var. vanense
Thum. 1929,1. c.: 310. (11).
Геогр. распр. Армения, Турция (редко).
Var. variabile (Kudr.) A. Filat, comb. nov. — T. asiaticum var. variabi-
le Kudr. 1942, l.c.: 26. (33).
Среди тунисских образцов выявлена форма с соломиной, выполнен-
ной паренхимой по всей длине, описана как var. plenoferrugineum Flaksb.
1935, 1. с.: 86, descr. ross.
Геогр. распр. Средняя Азия, Западная Азия, Л-Тоиголпя, Китай,
Закавказье.
В Турции выявлена своеобразная плотиоколосая пшеница, которая
по признакам колоса и некоторым вегетативным является переходной от
Т. aestivum к Т. compactum. В. К- Кобелев выделил ее в особую группу
(тип по В. К- Кобелеву)— Т. vulgare compactoides Kob. 1932, Хлебн. зла-
ки Турции: 27. — Т. vulgare interjectum Flaksb. 1935, Культ, фл. СССР,
1: 55, nom. illeg. — с 13 разновидностями. М. Gokgol (1939, 1941) зна-
чительно увеличил число плотноколосых разновидностей, которые позд-
нее (М. Gokgol, 1955) рассматривал как редкие формы с соответствую-
щими гомологичными признаками распространенных в Турции разновид-
ностей с меньшей плотностью колоса. Подобные формы особенно часто
встречаются у var. variabile, var. turcicum и var. pseudoturcicum. To же
относится к большому количеству (113) разновидностей, которые
М. Gokgol выделил по мелким систематическим признакам. Такая дроб-
ная классификация независимо от ранга таксонов вскрывает широкий
полиморфизм 'пшеницы мягкой в Турции. Ввиду недостаточно четкого
систематического положения многих из этих неэффективнообнародовап-
ных таксонов, в настоящей работе они не приводятся. Обстоятельные
исследования пшениц позволили М. Gokgol (1939) установить в Турции
гораздо более частую встречаемость форм с соломиной, выполненной
паренхимой по всей длине.
II. Subsp. aestivum — подвид европейский.
Subsp. aestivum. — 7'. vulgare subsp. hapalopyrum Flaksb. 1928, Изв.
Гос. ин-та опытн. агрономии, 6, 2: 47. — Т. vulgare subsp. indoeuropaeum
(Vav.) Flaksb. 1935, Культ, фл. СССР, 1: 49, descr. ross. — T. vulgare
группа indoeuropaeum Vav. 1923, Tp. прикл. бот. сел. 13,1: 195.—
T. vulgare subsp. eurasiaticum Vav. 1964, ЛАиров. ресурсы хлебн. злаков.
Пшеница: 77, nom. illeg. — Т. vulgare subsp. sinicam Vav, 1964, l.c.: 76,
nom. illeg.
Растения и колосья нежного построения (рис. 18,6, 19, б), колоско-
вые чешуи обычно топкокожистыс, самой различной формы, у основания
с поперечной вдавлсппостыо. Зерновки неплотно заключены в чешуях,
легко обмолачиваются, много форм с осыпающимся зерном, плотность
заключения зерновок в чешуях обусловлена нс грубостью чешуи, как
П<к)ро<1 I, Tritiiitm
263
у subsp. hadropyrum, а зависит от степени выпуклости нижней половины
колосковой чешуи.
Геогр. распр. Но всему ареалу Т. aestivum. В XX в. сорта евро-
пейского подвила нередко возделываются в ареале азиатского.
Для subsp. aestivum характерно исключительное разнообразие эко-
логических групп, резко контрастных по длине вегетационного периода,
зимостойкости, засухоустойчивости, иммунитету и ряду других биоло-
гических свойств и морфологических признаков.
а) Кашмирская группа сформировалась на высотах около 2000 м
лад ур. м. западных Гималаев.
Растения с топкой соломиной, узкими листьями, колос рыхлый, топ-
кий, с нежными остями. Зерна мелкие, легко обмолачиваемые. Кашмир-
ские пшеницы сравнительно устойчивы к бурой и желтой ржавчине; за-
сухоустойчивость слабая. По биологичсскНхМ свойствам они близки яро-
вым сибирским, которые, по предположению Е. Ф. Пальмовой (1935),
родственны по происхождению.
б) Восточная группа сложилась в условиях влажного теплого кли-
мата. На основе некоторого сходства с индийскими пшеницами II. II. Ва-
вилов (1964) считает, что пшеница в Китай и далее в Японию занесена
из ее первичного центра развития через Индию. Древнее обособление
в своеобразных условиях произрастания привело к формированию пше-
ницы мягкой с рядом эндемичных признаков (исключительно нежные
колосковые чешуи; совершенная безостость— отсутствуют даже остс-
впдныс придатки; остн короткие по всей длине колоса; многоцветко-
вость; чрезвычайно слабое заключение зерновок в чешуях). Для пшениц
этой группы характерна очень широкая амплитуда изменчивости по
плотности колоса: от необычной для европейских пшениц рыхлоколосо-
сти (D — 13) до плотности (D = 33—40), характерной Т. compactum.
Среди группы чаще, чем среди других групп пшеницы мягкой, встре-
чаются ветвистоколосые формы. Две из них описаны М. М. Якубцинером
как разновидности Т. aestivum var. tarakanovii Jakubz. 1959, Бот. жури.
44, 10: 1430, descr. ross. и T. aestivum var. ramosoerythrospermum Jakubz.
1959, l.c.: 1430, descr. ross., а также формы с булавовидным колосом
(в Европе такие формы пшеницы мягкой получены путем гибридиза-
ции). Восточной агро-экологической группе присуща также низкорос-
лость. Стародавний японский сорт послужил основой для получения до-
нора генетически обусловленной короткостебельности— сорта Norin 10,
а в дальнейшем — серии знаменитых мексиканских короткостебельпых
сортов пшеницы. В своем большинстве пшеницы восточной группы вос-
приимчивы к грибным заболеваниям, но среди них выявлены очень
ценные по болезнеустойчивости. При посеве осенью отличаются скоро-
спелостью, крайне быстро проходят период налива зерна, при этом
в засушливых условиях формируется нормальное, с высокой стекло-
видиостью зерно. Большая часть китайских мягких пшениц полуозимая.
имеются озимые и яровые.
Несмотря на исключительный полиморфизм группы, опа проявляет
экологическое единство (Н. И. Вавилов, 1964).
Восточная группа приурочена в основном к Восточному, Центрально-
Южному и Юго-Западному Китаю, Японии и Корее.
На западе и севере от места первичного происхождения пшеницы
мягкой сформировались иные в экологическом и биологическом от ноше-
нии группы.
в) Кавказская горная озимая группа (prol. Iiicmale-immlaiiocaurasi-
ciim Vav., 1964, Миров, растит, ресурсы. Пшеница: 80, descr. ross.)
представлена почти исключительно озимыми формами, среднерослыми,
относительно скороспелыми, реже позднеспелыми. Стержень колоса
тонкий, ости несколько укороченные, нежные; колосковые чешуи с длин-
ными остевидными зубцами; зерно некрупное, его вымолот легкий.
2(>4 Классификицич р<ч<)а Tiituuui
Срсдлевосприимчивы к буроИ и желтой р/кавчине, сравнительно ycioii-
чивы к пыльной головне. Эта пшеница стенных районов горного Закав-
казья в Грузии известна под названием долис-иури; приурочена также
к высокогорной степной Армении и предгорным районам Азербайджана.
К кавказской горной озимой группе II. И. Вавилов относил праба-
натки предгорий Северного Кавказа и местные предгорные (300—
1000 м над ур. м.) озимые пшеницы Дагестана и Азербайджана— гюль-
гери. Прабаиатки в отличие от долис-иури имеют более широкий коло-
совой стержень, длинные, несколько более грубые ости, несколько за-
трудненный вымолот зерна, колос и зерно более крупные.
Гюльгери — местные популяции с более грубым колосом, пеосыпаю-
щимся зерном, вымолот зерна легкий; соломина средней длины, доволь-
но прочная.
г) Кавказская горная яровая группа (prol. aestivo-montanocaucasi-
cutn Vav. 1964, l.c.: 81, descr. ross.) сформировалась в условиях высоко-
горья (выше 1000 м над ур.м.) Дагестана, Грузии, Армении, Северной
Осетин и Чечено-Ингушетии. АГенее требовательна к теплу и более вла-
голюбива по сравнению с предыдущей. К горной яровой группе отно-
сятся персикоиды * (subprol. persicoidcs Vav.), по комплексу призна-
ков очень сходные с Т. persicum (топкий колосовой стержень, колос
рыхлый, чешуи топкокожистые, ости нежные, па колосковых чешуях
развиты ости, сравнительно устойчивы к грибным заболеваниям).
К кавказской горной яровой группе II. И. Вавилов относит также
прарусаки и праполтавки. Первые—аборигенные для горного Кавка-
за (в основном Кабардино-Балкария, Северная Осетия), близки по ря-
ду биологических и морфологических особенностей лесостепным и степ-
ным пшеницам, известным под названием русаков.
Яровые пшеницы Свапетии (безостые, преимущественно белоколосые
и краснозерные, с рыхлым колосом) за сходство с лесостепными пол-
тавками Украины названы ГГ И. Вавиловым праполтавками.
д) Степная озимая группа (prol. hiemale-stepposum Vav. 1964, 1. с.:
83, descr. ross.), известная еще под названием бапатки, отличается хо-
рошей зимостойкостью. Представлена преимущественно var. aestivum.
Экологически приурочена к лесостепным и степным районам СССР,
Средней Европы, Балкан, а также Северной Америки и отчасти Арген-
тины.
Широким ареалом возделывания объясняется большое многообразие
банаток, отраженное более дробным их подразделением Н. И. Вавило-
вым (1964). Пшеницы этой группы сыграли большую роль в создании
ряда продуктивных сортов современной селекции.
е) Степная яровая группа (prol. aeslivo-stepposum Vav. 1964, l.c.:
89) представлена русаками (остистые, преимущественно краспозерные
пшеницы, среднерослые, скороспелые, восприимчивые к ржавчинам) и
полтавками (безостые, краспозерные, со средней засухоустойчиво-
стью) — наиболее пластичными яровыми пшеницами. Большое значение
они имели в создании современных продуктивных яровых сортов По-
волжья, Украины, Сибири.
ж) Североевропейская лесная группа (prol. hiemale-boreale Vav.
1964, 1. с.: 90, descr. ross.) устойчива к низким температурам зимой и
избыточному увлажнению в осенний и весенний.периоды.
Растения выше средней высоты, хорошо облиственные, с довольно
крупным колосом и зерном, яровые и озимые. Сформировалась в лесной
полосе европейской части СССР, Полыни, Чехословакии, США. Ввиду
обширного ареала группа неоднородна в определенных частях его.
* II. А. Гапдилян выделил персикоиды и отдельную труппу разновидностей
(Г. aestivum convar. tetriiristatum Gandil. 1976, Гяол. ВИРа, 65: 78), ио ра.тновидност-
пый состав се не приведен.
Jlodpot) I. Тritii’um
26.5
з) Восточносибирская i руина (prol, orientale-sibiriiиm Vav. 196-1,
l.c.: 92, descr. ross.) - яровая скороспелая пшеница, связанная своим
происхождением с Монголией. Интересна зимостойкая озимая пшеница
згой группы, менее позднеспелая, с хорошей засухоустойчивостью.
и) Южноевропейская крупнозерная группа (prol. meridionale-euro-
paeum Vav. 1964, l.c.: 92, descr. ross.) стародавних сортов Южной
Европы отличается крупным колосом, крупным и очень крупным (мас-
са 1000 зерен до 60—70 г) зерном, прочной, толстой соломиной, широ-
кими листьями. Сорта преимущественно озимые, слабозимостойкие,
скороспелые, устойчивые к бурой ржавчине.
к) Западноевропейская группа (prol. occidentale-eiiropaeum Vav.
1964, l.c.: 94, descr. ross.) —гидрофильная, реже мезофильная, поздне-
спелая. Одна из первых групп, па местных сортах которой уже в XIX в.
шла интенсивная и успешная селекционная работа.
Аборигенные озимые и яровые сорта Западной Европы представле-
ны пшеницами высокорослыми, с прочной соломиной, с относительно
рыхлым колосом. Скандинавские местные озимые copra этой экологи-
ческой группы хорошо зимуют в условиях севера.
В настоящее время значительно улучшен культурный тип растения;
имеется богатый селекционный материал, полученный на основе сортов
западноевропейской группы.
Кроме экологических групп стародавних местных сортов пшеницы
мягкой, Н. И. Вавилов (1964) приводит ряд групп сложного гибридного
происхождения (канадская, австралийская, итальянская, французская,
южноамериканская и др.), сыгравших большую роль в современной
селекции пшеницы.
Разповидпостный состав европейского подвида гораздо беднее, чем
у азиатского.
Var. aestivum. — Т. vulgare var. erylhrospermtim Koern. 1873, System.
Ubers. Cer.: 11; Koern. in Koern. et Wern. 1885, Handb. Getreideb. 1:
46. (24).
Геогр. распр. Главным образом в Восточной Европе, реже
в Средней и Северо-Западной, в Китае, Америке, Австралии.
Var. ajutense (Nosatov.) Л. Filat, comb. nov. — T. vulgare var. ajuten-
se Nosatov. in Greb. 1926, Нов. формы яр. пшеп, на Дону: 4. (10).
Геогр. распр. Северный Кавказ.
Var. albiglaucum (Vatz.) Mansf. 1951, Der Zflchter, 21, 1/2: 58.—
T. vulgare var. alboglaucum Vatz. 1929, Tp. Всесоюзи. съезда ген. сел.
сем. Январь, 1929 (Ленинград), 3: 122. (4).
Геогр. распр. Украина.
Var. albirtibruni (Koern.) Mansf. 1951, l. c.: 57.— T. vulgare var.
alborubrum Koern. in Koern. et Wern. 1885, l.c.: 45. (3).
В Закавказье выявлена форма с булавовидным колосом — f. dekap-
relevitschii Menabde.
Геогр. распр. По всему ареалу европейского подвида.
Var. anglicum (Aschers. ct Graebn.) A. Filat, comb. nov. — T. ienax
A. I. vulgare var. anglicum Ashers, ct Graebn. 1901, Syn. Mitteleur.
Fl. 2.: 684; Flaksb. 1915, Опред. пшен.: 139. — T. anglicum iAl\7.z. 1807,
Sopra als. sp. 'frum.: 39. — T. vulgare var. leucospermum Koern. in Koern.
et Wern. 1885, 1. c.: 45, p.p. (14).
Геогр. распр. Западная Европа, редко Восточная, Китай, Юж-
ная Африка, Северная Америка, Австралия.
Var. aiireinn (Link) Mansf. 1951, l.c. ;58. 7’. vulgare var. aureum
Link, 1829, JIandb. Erk. Gewachse, 1: 10.—-7’. vulgare var. albiclum Alef.
1866, I. c.: 329; Koern. in Koern. cl Wern. 1885, 1. c.: 45. (1).
В Армении выявлены формы с двойными колосками па одном
уступе колосового стержня — f. kurductilense (Thum.) Dorof. ct A. Filat.
266
Классификация рода Tritirum L.
Колосья безостые
Чешуи неопушенные
Зерновки бе лы е
+ 1 i 1 ; 4- 11 + 11 — 1 1 1 + 1 + 3 е р IIOB1 1 ! 1 11 н|и i Ы С аигеиш triste albirubrum albiglaucum tanaiticum 1 2 3 4 5
ч- + — — - — — —. — — 4-:: lutcsccns nigrilutesccns 6 1
— — + — — — — — — milturum <3
— — — — 4- — — — — • — glaucilutescens 9
— — — — — + — — — — ajutense 10
— — — — — — + — — nigricolor 11
+ 3 е р НОНКИ 3 С Л С II 1.1 0 - 1 - 1 -- 1 fschermakianu m 12
+ — — - viridimilturum •13
4- — L Зе leiuyu р Н О Ofiyill к и енные е л ы с anglicum 14
— — —— —• — + — — — — konstantinovskense 15
—- — — — — —. — + — +* sulinense 16
4- — — — 3 е р новь И К | а с и ы е villosum 17
— — + — — — — — — — pyrolhrix 18
— — — — — — — _1_ — nigrum 19
+ 3 е р н о в к и зелен ы с jakubzineri 20
+ — — — viridipyrothrix 21
4- Колосья остист Чешуи неопушенньи Зерновки белы ы е е + oblivense 22
— — + + morosovskense 23
4- 3 е р Н О в <нк ) ас н ы е aestivum 24
4- — — — — — — — — + nigraristatum 25
— — 4- — — — — — 4- — ferrugineum 26
— — + — — — — — — + sardoum 27
— — — — 4- —. — — — + pseudo fulvicinereum 28
— — J — —. 4- — — _д_ — caesium 29
— — — — — 4- — — — + pseudocacsium 30
— — — — — —• 4- — — 4- n ig rerythrosperm и m 31
— — — •— — 4- — Ч- nigrijerrugineum 32
-- - - - 3 (‘ р и -I- > в к и 1 - ф л о 1 л е т о в ы е 1 - । 4- auslralianum . J
+ 3 е р и о. в < И 3 с л с н — 1 -I- >1 е vavilovii 34
4- + viridiferrtigincuin 35
* Окрасха остевидных заострений.
HoOpoi) I. 'Гi itii iim
Чешуи опушенные
Зерновки красные
pseiidocaeruleovelu- I 36
tinum
3 eрновки зеле н ы е
+ I — | -- I — | — I — I — I — I + 1 — I viridihostianum I 37
— I — I + | — I — | — I — I — | +| — I viridibarbarossa [ 38
comb. nov.— 7’. vulgare var. kurd и cu I ensc Thum. 19.33, Опред. хлебн.
злаков (колос.): 194.
В Туве обнаружена пшеница европейского подвида с признаками
инфлятпости— f. araticum Frenk. ex Flaksb. 1935, Культ, фл. СССР,
1: 59.
Геогр. распр. Западная Европа, редко Восточная, Закавказье,
Иран, Китай, Южная Африка, Австралия.
Var. australianum Udacz. et Schachm. 1979, Бюл. ВИРа, 84: 71. (33).
Фиолетовозерная разновидность выделена из австралийского сорти-
мента пшениц. По-видимому, получена гибридизационным путем при
участии фполстовозерпых разновидностей Т. aethiopicum.
Var. caesium (Alef.) Mansf. 1951, l.c.: 52. — T. vulgare var. caesium
Alef. 1866, 1. c.: 330; Koern. in Koern. et Wern. 1885, 1. c.: 47. (29).
Описана по материалам из ботанических садов.
Геогр. распр. Европа, Сибирь, Закавказье, Северный Кавказ,
Дагестан, Аргентина.
Var. ferrugineiim (Alef.) Mansf. 1951, l.c.: 52. — T. vulgare var. fer-
rugineum Alef. 1886, l.c.: 330. — T. vulgare var. vulpinum Alef. 1866, l.c.:
330. (26).
Геогр. распр. Широко распространена в Северной Европе и
Восточной Сибири; в остальных районах встречается реже.
Var. glaucilutescens (Vatz.) Mansf. 1951, l.c.: 57. — T. vulgare var.
glaucolutescens Vatz. 1929, 1. c.: 122. (9).
Геогр. распр. Украина.
Var. jakubzineri Puch. et Maksimov, 1971, Tp. прикл. бот. ген. сел.
44,1:55.(20).
Выявлена в образцах, поступивших в ВИР из Чехословакии.
Var. konstantinovskense (Greb.) Mansf. 1951, l.c.: 56.— T. vulgare
var. konstantinovskense Greb. 1929, Опред. разнов. пшен. Сев. Кавказ,
края: 13. (15).
Геогр. распр. Северный Кавказ.
Var. lutescens (Alef.) Adansf. 1951, 1. с.: 57. — T. vulgare var. lutescens
Alef. 1866, 1. c.: 329. — T. vulgare var. xanthura Alef. 1866, l.c.: 329.—
7’. vulgare var. rufescens Wittm. 1876, Landwirtsch. Jahrb. 5: 617, non
Link. (6).
Геогр. распр. Одна из распространенных разновидностей евро-
пейского подвида, главным образом в Европе, Китае, реже в остальных
частях Азин, Австралии, Африке.
Var. milturum (Alef.) Mansf. 1951, l.c.': 56.--7’. vulgare var. miltura
Alef. 1866, l.c.: 329.— T. vulgare var. milturum (Alef.) Koern. in Koern.
et Wern. 1885, Lc.: 45. — T. aegyptiacum Mazz. 1807, Lc.: 39. (8).
268
Классификации рода Triticum L.
Один и.) паибо. ice pfieiipoci раненных разновидное ей, европейскою
подпила.
Var. morosovskense (Greb.) A. Filat, coinb. now- T. vulgare var.
morosovkense Greb. 1929, l.c.: 11. (23).
Геогр. рас п p. Северный Кавказ.
Var. nigraristatum (Flaksb.) A. Filat, comb. nov. — T. vulgare var.
nigraristatum Flaksb. 1915. Опред. пшен.: 149. — T. vulgare var. sub-
nigriaristatumN^ii. 1929, 1. c.: 120. (25).
Г еогр. p а с и p. Северный Кавказ, Закавказье, Украина.
Var. nigrerythrospernium (Jakuschk.) Mansf. 1951, l.c.: 54. — T. vul-
gare var. nigroerythrospermum Jakuschk. 1923, Tp. прикл. бот. сел. 13, 1:
96. (31).
Геогр. распр. Украина (редко).
Var. nigricolor (Flaksb.) Mansf. 1951, l.c.: 57, comb, invalid. --T. vul-
gare var. nigricolor Flaksb. 1935, 1. c.: 67, descr. ross. (11).
Геогр. pa снр. Франция (редко).
Var. nigriferrugineum (Jakuschk.) Mansf. 1951, l.c.: 52.-- T. vulgare
var. nigroferrugineum Jakuschk. 1923, Tp. прикл. бот. сел. 13, 1: 95. (32).
Геогр. распр. Украина (редко).
Var. nigrilutescens (Greb.) Mansf. 1951, l.c.: 57. — T. vulgare var.
nigrolutescens Greb. 1929, 1. c.: 12. (7).
Геогр. распр. Северный Кавказ (очень редко).
Var. nigrum (Koern.) A. Filat, comb. nov. — T. vulgare var. nigrum
Koern. in Koern. et Wern. 1885, l.c.: 46; Vav. 1923, Tp. прикл. бот. сел.
13, 1: 169, descr. ampl. (19).
Геогр. распр. Редко в Западной Европе, на Северном Кавказе.
Var. oblivense (Greb.) Л. Filat, comb. nov. — T. vulgare var. oblivense
Greb. 1929, l.c.: 10. (22).
Геогр. распр. Северный Кавказ (редкая примесь).
Var. pseudocaeruleovelutinum (Greb.) Л. Filat, comb. nov. — T. vul-
gare var. pseudocaeruleovelutinum Greb. 1929, 1. c.: 11. (36).
Геогр. распр. Северный Кавказ.
Var. pseudocaesium (Greb.) Mansf. 1951, l.c.: 52.— T. vulgare var.
pseudocaesium Greb. 1929, 1. c.: 11. (30).
Геогр. распр. Северный Кавказ (очень редко).
Var. pseudofulvicinereum (Vatz.) Mansf. 1951, l.c.: 54. — T. vulgare
var. pseudo}ulvocinereum Vatz. 1929, l.c.: 122. — T. vulgare [ulvocinereum
Flaksb. 1935, l.c.: 81, descr. ross. (28).
Геогр. распр. Украина.
Var. pyrothrix (Alef.) Mansf. 1951, l.c.: 55. — T. vulgare var. pyrothrix
Alef. 1866, 1. c.: 329. (18).
Геогр. распр. Западная Европа, редко на Северном Кавказе, Ки-
тай, Северная Америка.
Var. sardoum (Koern.) Mansf. 1951, l.c.: 52. — T. vulgare var. sar-
doum Koern. in Koern. ct Wern. 1885, 1. c.: 47. (27).
Геогр. распр. Редко в районах возделывания var. ferrugineum.
Var. sulinense (Greb.) A. Filat, comb. nov. — T. vulgare var. sulinense
Greb. 1929, l.c.: 13. (16).
Геогр. p a c n p. Северный Кавказ (очень редко).
Var. tanaiticum (Greb.) Mansf. 1951, l.c.: 57. — T. vulgare var. lanaiii-
cum Greb. 1926, 1. c.: 7. (5).
Геогр. распр. Северный Кавказ, Сибирь (очень редко).
Var. triste (Flaksb.) Mansf. 1951, l.c.: 58. — 7’. vulgare var. triste
Flaksb. 1915, Опред. пшен.: 137. (2).
Г e о г p. p а с и p. Северный Кавказ, Украина (редко).
Var. tscliermakianum Mansf. 1949, Ind. sein. Inst. Gatersleb.: 42- idem,
1951, l.c.: 58. (12).
Выделена из гибридного материала Е. Tschcrmak.
Подрод I. Triticum
269
Var. vavilovii Jakubz. v I lyxa.iii.CKoi o, 1971, 'I']). npiiK.i. Гют. icn. сел.
1 I. 1: 56. (34).
Выявлена в образцах из Баварии (ФРГ).
Var. villosum (Alef.) Mansf. 1951, l.c.: 56. -- T. vulgare var. villosum
Alef. 1866, l.c.: 329. — 7’. velutinum Schuebl. 1818, Char, et descr. cer.:
7. — 7'. vulgare var. velutinum Koern. in Koern. et Wern. 1885, l.c.: 45,
non Alef. (*1866). (17).
Геогр. распр. Средняя и Западная Европа, Америка (редко).
Var. viridibarbarossa Puch. et Max. 1975, Tp. прикл. бот. ген. сел.
55,3:122.(38).
Отобрана из гибридного материала, полученного па основе скрещи-
вания пшеницы мягкой со спельтой (Московское отделение ВИРа).
Var. viridiferrugineum Max. et Puch. 1975, 1. c.: 122. (35).
Получена таким же путем, как и var. viridibarbarossa Puch. et Max.
Var. viridihostianum Puch. et Max. 1975, l.c.: 122: (37).
Получена таким же путем, как и var. viridibarbarossa Puch. et Max.
Var. viridimilturum Max. et Puch. 1975, 1. c.: 122. (13).
Получена таким же путем, как и var. viridibarbarossa Puch. et Max.
Var. viridipyrothrix Max. et Puch. 1975, l.c.: 122. (21).
Получена таким же путем, что и var. viridibarbarossa Puch. et Max.
T. SPHAEROCOCCUM PERCIV. ПШЕНИЦА ШАРОЗЕРНАЯ
T. sphaerococcum Perciv. 1921, Wheat Plant: 321. — 7'. aestivum subsp.
sphaerococcum (Perciv.) Mac Key, 1954, Bot. Tidskr. 48: 586. — 7'.. aesti-
vum group sphaerococcum (Perciv.) Bowden, 1959, Canad. Journ. Bot. 4:
674. — 7'. aestivum convar. sphaerococcum (Perciv.) Morris et Sears, 1967,
Wheat and wheat improv.: 36. — T. compactum auct. non. Host.: A. et
G. Howard, 1910, Wheat in India: 166.
Общая характеристика. Всходы светло-зеленые, слабо опушенные,
без антоциана (реже густоопушениые, окрашенные антоцианом). Куст
прямостоячий, Листовые влагалища со слабым восковым налетом, без
опушения. Листовые пластинки взрослых растении также со слабым
восковым налетом, шероховатые па ощупь, отходят от стебля иод углом
15—16°, сравнительно короткие (длина 15—20 см, ширина 1,3—1,6 см),
резко суживающиеся к вершине. Листовая поверхность главного стебля
в фазу колошения 65—70 см2. В массе — формы лигульныс (имеют язы-
чок и ушки). Ф. Г. Нигматуллин (1972) создал безлигульиую (cligula-
tum) форму. Ушки длинные, узкие, бесцветные, с несколькими реснич-
ками. Растения невысокие, в среднем 70—85 см. Соломина по всей длине
полая, исключительно прочная (проволочная), трудно сгибаемая, пря-
мая, толщиной 2,5—2,7 мм, под колосом иногда волнообразно изогнутая.
Узлы снизу вверх по стеблю расположены па расстоянии 4—5, 6—9,
13—14, 16—20, 30—32 см. Содержание сухого вещества в 1 пог. см стеб-
ля 38—40%.
Колосья короткие (рис. 20), 4—7 см длины, почти цилиндрической
формы, с 13—19 колосками. Боковая сторона колоса обычно шире лице-
вой. Членики колоскового стержня по бокам и у основания колосков опу-
шены белыми волосками. Колос плотный (D = 30—34, в особо благо-
приятных условиях до 38—42).. Колоски 3—4-цветковые (иногда 6—
7-цветковые). Колос содержит 35—40 зерновок. Колоски короткие
(длина и ширина около 10 мм, толщина около 5 мм). Колосковая чешуя
широкая (длина 7—8 мм, ширина 5 -6 мм), часто почти полушаровид-
иая, с килем, доходящим до ее основания. Килевой зубец изогнут внутрь
(клювовидный). По обеим сторонам киля расположены 6 7 жилок,
которые слегка выступают. Наружная цветковая чешуя вздутая, округ-
лая, с 2—9 жилками.
270
ly.itii i ифииацич рода Triticum L.
Рис. 20. Колосья, колоски и зерновки Т. sphaerococcum Perciv.
Цветковая чешуя у безостых разновидностей заканчивается наверху
жестким остевидпым заострением длиной 3—4 мм, а у остистых форм —
жесткими, сильно зазубренным!!, короткими (15—20 мм) остями. Внут-
ренняя цветковая чешуя окаймлена волосками вдоль обоих килей; она
несколько короче наружной. Но форме колосковой и цветковой чешуи,
загнутостп килевого зубца и внешнему облику колосков Т. sphaerococ-
cum имеет сходство с ипфлятпой мягкой пшеницей. Поэтому, видимо,
не случайно последняя выщспилась при скрещивании шарозерпой пше-
ницы с Т. compactum в опыте Р. А. Удачина (1959). Зерновки стекловид-
ные, почти шаровидной формы: у типичных форм длина их 4—5 мм.
ширина и толщина 3,4—3,5 мм. Хохолок короткий, бороздка ншлубо-
кая (изредка — глубокая). Масса 1000 зерен 30—32 г.
Анатомические исследования листа и стебля (О. Д. Градчанинова,
1967) показали, что клетки эпидермы, is том числе моторные, крупные.
Число проводящих пучков на поперечном разрезе листа равно 36—37.
Размер пучка главной жилки 142—144 мкм. На нижней поверхности
листа насчитывается 42—43 устьица на 1 см2. Оболочка склеренхимных
волокон листового влагалища сильно утолщены, дают реакцию на лиг-
нин. Стебель' характеризуется хорошим развитием механической ткани
в трех нижних междоузлиях (толщина ее 71 —123 мкм), с числом про-
водящих пучков 11 —16 в механической ткани и 24—26 в паренхиме. При
сравнительно топком нижнем междоузлии (диаметр около 3,5 мм) со-
противление его па излом равно почти 1 кг. 11. Sdirotler (1931) писал,
что анатомически колосовой стержень /’. sphaerococcum имеет признаки
полбы и спельты.
Л. Л. Жестяииикова (1964) показала, что эии-мс ижарпий зерновки
7’. sphaerococcum имеет 2 слоя к.теюк (изредка оенноюя участи клеток
Подрод I. Triticum 271
третьего слоя). Отложения иолуклел чат ки настолько мощные, что за-
полняют почти нее клетки эии-мезокариня. Поперечные клетки правиль-
ной формы, причем внутренняя их стенка толще наружной. В местах
сочленений поперечных клеток видны поры. Трубчатые клетки встреча-
ются лишь па спинной стороне зерновки. Одревеснению подвергаются
оболочки лишь этих клеток. Таким образом, плодовая оболочка зернов-
ки Т. sphaerococcum сходна с пшеницей мягкой.
К. Miczynski (1930), проведя реципрокные скрещивания Т. sphaero-
coccum с Т. aestivum, пришел к выводу, что весь комплекс признаков,
характерных для первого вида, контролируется одним геном s, для ко-
торого S. Matsumura (1954) предложил символ sp. Е. Sears (1947),
М. Rao (1971, 1977) определили локализацию гена s в хромосоме 3D и
подтвердили его аллелытоегь гену s пшеницы мягкой. J. Schmidt и др.
(1963) отрицают это. J. Mac Key (1954)- доказал, что голозсрпость
Т. sphaerococcum определяется тем же фактором Q, что и у остальных
голозерных гексаплоидных видов пшеницы.
II. Singh и др. (1957) показали, что Т. sphaerococcum имеет главные
гены Qcs (у Т. aestivum — QcS, у Т. compactum — QCS). Изучение фено-
типического эффекта обработки семян шарозерпой пшеницы рентгенов-
скими и Й-лучами провели М. Rao и М. Swaminathan (1963). В М2 3,65%.
мутаций оказались спельтоидами с топкой соломиной и меньшей жест-
костью листьев. Типичная штыкообразность последних исчезла. Расте-
ния стали более высокорослыми, имели рыхлые колосья ригидного типа
и колосковую чешую менее округлую, чем у исходной формы. Зерно
было мелким, сферическим, сходным с зерном пшеницы шарозерной.
Небольшой процент спельтоидных мутаций у этой пшеницы показывает,
что мутационное изменение фактора Q у пес бывает сравнительно ред-
ким, в отличие от пшеницы компактум и мягкой. Кроме спельтоидов,
в М2 было выделено 2 мутантных компактоида, имевших в 1,5 раза бо-
лее плотные колосья, чем пшеница шарозерпая. Подобные супсркомпак-
тоиды наблюдались и в исследованиях Р. А. Удачина (1959) при скре-
щивании Т. compactum с Т. sphaerococcum.
Как подчеркивал II. И. Вавилов (1964), пшеница шарозерная пред-
ставляет собой вполне выдержанную экологическую группу. По оценке
Е. Ф. Пальмовой (1935) этот вид имеет ряд признаков, свойственных
пшенице мягкой Индии. Это относится, в частности, к элементам ксеро-
фитиости, что находит свое выражение в жесткости и вертикальном
положении листа, слабой облиствсиностп растеши'!. Это пшеница оро-
шаемого земледелия.
Т. sphaerococcum имеет яровой образ жизни. Недавно Р. А. Удачин
(1975) обнаружил озимую форму этого вида — var. kibraicum Udacz.
А. Ценов и К, Джслспов (1976) также сообщили о выделении озимой
формы пшеницы шарозерной. Р. В. Карпшпсв (1958), М. П. Аладова
(1960), М. М. Якубцинер (1961), Н. И. Вавилов (1964), Р. А. Удачин и
Э. Ф. Мигушова (1965) в комбинациях от скрещивания между яровыми
формами пшеницы шарозерпой и мягкой наблюдали случаи выщепления
озимых растений.
В литературе (J. Percival, 1921; К. А. Фляксбергер, 1935; И. И. Ва-
вилов, 1964; II. М. Жуковский, 1971) пшеница шарозерпая описывается
как скороспелая. По данным М. М. Якубцинера (1955, 1961), при осен-
нем посеве на юге она является одной из наиболее скороспелых форм,
в более прохладных условиях — позднеспелой. Являясь биологически
яровым видом, пшеница шарозерпая в ряде опытов не выколашивалась
па юге при позднем весеннем посеве, что указывает на потребность
в пониженных температурах и чувствительность к длине дня.
При осеннем посеве под Ташкентом на поливном участке период от
всходов до созревания равен 195—225 дням, а при посеве весной —
80—85 дням. Па юге пашей страны в обычные зимы хорошо псрезпмо-
1\м1т1фик(щи>1 рода Triticum 1
вываст (па (SO ‘)0 (Х>), а в суровые вымерзав г. В Волгарии, eoi.aaciio
с । в >01 це 1111 ю 11. Станкова п И. CiaiiKor.a (1975), jioi вид морозосшпкий.
Являясь резко выраженным экотипом поливной пшеницы жаркого
южного климата, Т. sphaerococcum характеризуется требовательностью
к теплу при созревании, высокой устойчивостью к жаре и атмосферной
засухе, по педостаточной устойчивостью к почвенной засухе. Р. Л. Уда-
чин и И. 111. Шахмедов (1974) показали, что урожайность этой пшеницы
при отсутствии почвенной засухи и при повышенных температурах воз-
духа (-R35°C) была па 30% выше, чем при почвенной засухе. Падение
урожайности в основном происходило за счет резкого снижения про-
дуктивной кустистости и озерпепности колоса.
Е. И. Преображенская (1971) указывает, что Т. sphaerococcum наи-
более устойчив к у-облучепню по сравнению с другими видами рода
Triticum. М. S. Swaminathan и М. Р. Rao (1969) установили, что этот
вид пшеницы по мутабилыюсти близок Т. aestivum п Т. compactum.
Согласно исследованиям В. Д. Карпенко (1970), в условиях При-
аралья количество первичных корней в фазу кущения у пшеницы шаро-
зерной достигало в среднем 3,5, а число вторичных при колошении —
27,8. Максимальный суточный прирост первичных корней в периоды
всходов составил 3,2 см, кущения --5 ем. Наибольшая глубина проник-
новения корней в фазы всходов — 40 см, кущения — 66 см, начала нали-
ва зерна— 160 см, причем последний показатель отражал наибольшую
глубину проникновения корней в почву по сравнению с другими видами
пшеницы. Л. А. Вельсовская (1972) указывает, что в Павлодарской об-
ласти Казахстана суточный прирост первичных корней был равен
2,01 см, а вторичных — 1,85 см.
По наблюдениям В. И. Жукова (1969), пшеница шарозерпая склон-
на к клейстогампому цветению: цветков с открытым цветением 19,8—
50%, т. е. меньше, чем у других гексаплоидных видов. По данным
А. 3. Латыпова (1971), у нее при цветении наблюдалось наименьшее
(1 мм) по сравнению с другими видами расхождение цветковых чешуй,
цветки в открытом состоянии находились всего около 1 ч. Видимо, по-
этому гибриды от свободного перекрестного переопыления у Т. sphaero-
coccum практически пе встречаются.
Результаты анализов, проведенных В. Г. Копаревым и др. (1972),
выявили, что содержание белка в зерне Т. sphaerococcum в условиях
орошения составляет 15.4—21,1% при сравнительно богатом содержании
лизина (от 0,230 до 0,470 г на 100 г зерна).
В работе Л. В. Милова новой и Р. А. Удачина (1960) были показаны
высокие хлебопекарные качества пшеницы шарозерной (объем хлеба
из 100 г муки, полученной из зерна, выращенного па орошаемом участ-
ке, достигал 520 мл). Изучение качества зерна образцов коллекции
этого вида (Л. В. Семенова, И. Ш. Шахмедов, 1975) также выявило, что
показатель набухаемости муки в уксусной кислоте у пшеницы шарозер-
ной ниже среднего или средний (до 48 мл). Стекловидпость зерна, вы-
ращенного при орошении, была очень высокой (95—100%); оценка тес-
та на микрофарипографе показала, что для пшеницы шарозерной харак-
терна невысокая устойчивость теста к замесу (до 3,5 мин), а испытание
«силы» па альвеографе с микроприставкой выявило довольно широкую
амплитуду '(до 313-10-4 Дж). Хлеб отличался хорошей пористостью
(до 70 баллов), отличным вкусом и довольно долго не черствел; припек
до 52%. Исследование Г. И. Ярины и И. Ш. Шахмедова (1977) показа-
ло, что среди образцов пшеницы шарозерпой есть такие, которые не усту-
пают пшенице твердой по прочности макарон, их разварнмости и про-
центу сухого остатка. Способность шарозерпой пшеницы к передаче по
наследству высокого содержания белка в зерне при межвидовой гибри-
дизации показана в работах Н. Ф. Покровской и Р. А. Удачина (1970).
П. П. Попова и др. (1973).
1!o<)/iui) I. Tiitii uni 273
Иммунитет. Сц.’пшая ш > p а ж ;i о м< >c т i> T. sphtmrococcum бурой, желтой
и стеблевой ржавчиной, пыльной и 'твердой головней, мучнистой росой,
а также повреждаемость зеленоглазкой, шведской п leecencKoii мухами
отмечена II. И. Вавиловым (1961. 1961). 11о ею данным, эта пшеница
устойчива к стеблевой колонне. Восприимчивость к бурой и стеблевой
ржавчине подтверждена опытами М, М. Якубцинера (1969), А. М. Ка-
мелиной (1973) и О. Г. Григорьевой (1975). Иммунные образцы не обна-
ружены как при искусственном заражении (в ([тазе проростков) кол-
лекции 7'. sphaerococcum популяциями рас мучнистой росы из различных
географических районов СССР, так и па инфекционном фоне в полевых
условиях (Э. X. Сухаибердина, 1976).
В опыте В. И. Кривченко (1973) ипокулироваиие .хламидоспорами
пыльной головни коллекции Т. sphaerococcum выявило устойчивость к
f. aestivi у 4,3%, к f. duri — у 20,9% образцов. Большая часть образцов
оказалась восприимчивой к патогену. А. М, Ямалсев (1974) показал,
что мицелий пыльной головни обильно распространяется во всех органах
зародыша, поражая до 37,2% зародышевых почек у зараженных зерно-
вок. С. П. Зыбина (1939) нашла, что популяции твердой головни из
17 регионов СССР сильно поражали Т. sphaerococcum, в том числе
твердая головня, собранная с Т. timopheevii в Грузии, которая поразила .
до 25% колосьев. Изучение коллекции показало, что 87,3% ее образцов
восприимчивы к твердой головне, что иммунных к licit образцов нет
(В. И. Кривченко, 1973). Т. sphaerococcum восприимчив к корневым
гнилям (И. Harding, 1972), к хлебной полосатой блохе, пьявице и впут-
ристебельпым вредителям (Л. Г. Щелко, 1964).
История. Впервые пшеница с шаровидными зерновками была описа-
на О. Heer (1865) как Т. antiquorum. Он исследовал найденные при рас-
копках свайных поселений Швейцарии (эпоха неолита, III тысячелетие
до и. э.) целые колосья, отдельные колоски и много зерна этой пшеницы. '
У нее был плотный, мелкий, мпогозерпый, безостый колос длиной около
4,5 см, толщиной 1 см. В колоске развивалось по 3—4 зерновки. Колос-
ковая чешуя округлая, с хорошо выраженным килем, доходящим до ее
основания, с изогнутым внутрь острым зубцом; рядом с. килем с обеих
сторон проходят более или менее ясно выраженные продольные жилки.
Наружная цветковая чешуя возвышается над колосковой и оканчивает-
ся коротким, изогнутым внутрь заострением. Зерновки округлые, с глу-
бокой бороздкой, длина их в среднем 5 мм, толщина и ширина 3,5 мм.
О. Heer писал, что зерно и колоски этой пшеницы, относящиеся к ран-
нему периоду свайных поселений, найдены в единичных экземплярах.
Количество их резко возрастает в поздний период этих построек.
L. Wittmack (1881), F. Kornicke (1885) и G. Buschan (1895) описали
последующие археологические находки Т. antiquorum. Последний автор
наряду с Т. antiquorum выделил среди ископаемых неолитических пше-
ниц формы с наиболее крупным шаровидным зерном, описав их как
Т. globiforme Buschan. Однако К. А. Фляксбергер (1930), изучив вопрос
о Т. antiquorum и Т. globiforme, пришел к выводу, что они относятся
к одному виду.
G. Saraun (1900), J. Hoops (1905), G. Schweinfurth (1908), H. Messi-
kommer (1913), E. Neuweiler (1924), H. Peake (1927), E. Schiemann
(1931), J. Percival (1936), E. Werth (1939), B. Pal (1948, 1966),
В. Л. Менабде (1948), E. H. Сипская (1955, 1969) и M. M. Якубцинер
(1956, 1961) дали подробный обзор археологических находок этой шаро-
видной пшеницы. В период с V по II тысячелетие до и. э. ее ареал про-
стирался от Индостана и Средней Азии до Швеции и Испании. В эпохи
неолита и ранней бронзы она была одним из основных хлебных злаков
па этой обширной территории. Затем, вследствие похолодания климата,
она постепенно была вытеснена из культуры в Европе, Закавказье, Пе-
18
274 i ифиьации liitiium
редней и Юго-Западной 7\.ши более сухолюбивыми и .холодостойкими
урожайными пшеницами 'Г. compactum и Т. aesliviini.
Была ли эта неолитическая пшеница идентичной современному
Т. sphaerococcum? .1. Percival (1921) указывает, чго Г. sphaerococcum
очень близок к Т. antiquorum, а В. Pal (1948, 1966) прямо отождест-
вляет находки 7'. antiquorum в Индостане (Мохенджо-Даро, III тыся-
челетие до п.э.) с Т. sphaerococcum. Е. II. Синская (1955) писала, чго
по форме зерна эта типичная ископаемая пшеница свайных построек
больше всего напоминает современную индийскую круглозерную пше-
ницу Т. sphaerococcum. М. М. Якубцинер (1956) указывал, что все
археологические находки круглозерных пшениц следует отнести к древ-
ним формам Т. compactum и Т. aestivum. Е. McFadden и Е. Sears (1946)
отнесли, однако, Т. antiquorum к числу тетраплоидных пшениц. Этот
вывод поддержал Н. Kuckuck (1959).
Р. Л. Удачин (1975) пришел к заключению, что 7'. antiquorum —
гексаплоидпая пшеница, произошедшая от скрещивания Т. prolopersi-
cum Udacz., имевшего главные гены QCs, с Ле. tauschii. В irroie в ге-
нотип Т. antiquorum оказались включенными аллели комплекса глав-
ных генов, пе имеющиеся ни у одного современного вида пшеницы.
Затем в древности при мутациях последовательно возникли из Т. anti-
quorum-^T. compactum (QCS)->7'. aestivum (QcS)->7’_ sphaerococ-
cum (Qcs). Очевидно, только зерновки из археологических находок на
полуострове Индостан, па котором располагался ареал Т. sphaerococ-
cum, действительно можно отождествлять с этим видом.
Географическое распространение. Т. sphaerococcum — эпдем полу-
острова Индостан. Он возделывался па территории Пакистана. Были
указания о нахождении его в Иране. (J. Percival, 1921), Ираке (S. ЕПег-
ton, 1939) и Северной Африке (G. Morris, 1933; R. Porleres, 1958), что
можно объяснить завозом с полуострова Индостан. Еще в 40—50-х го-
дах были сообщения о наличии площадей посева этой ншенины (В. Pal,
1966). Затем ее вытеснила более урожайная мягкая пшеница. Экспе-
диции ВИРа пе обнаружили в ареале прежнего возделывания
Т. sphaerococcum пи одного его колоса (Д. В. Тер-Аванесян, 1961;
В. Ф. Дорофеев, 1971).
Происхождение. При гибридизации пшеницы шарозерпой с видами
секций Dicoccoides и Triticum расщепление гибридного потомства со-
провождается появлением растеши'! типа Т. aestivum (И. Станков, 1972;
И. 141. Шахмедов, 1972). Это свидетельствует о филогенетической бли-
зости шарозерпой и мягкой пшеницы. О том же говорит высокая само-
фертильность гибридов F], полученных при скрещивании этих видов
друг с другом (Н. Kihara, 1937; Р. Л. Удачин, 1959). Близкую родст-
венность их отмечают Б. А. Вакар (1932), П. Попов и Д. Бояджиева
(1965), Е. Н. Синская (1969) и др.
Главный ген S у пшеницы мягкой находится в доминантном состоя-
нии, у шарозерной — в рецессивном. II. П. Зоз (1973) высказала пред-
положение, что локус. S представляет собой блок генов. М. V. Р. Rao и
М. S. Swaminathan (1963) определили, что действие рецессивной
аллели s ослабляется, если шарозерная пшеница под воздействием
мутагенов теряет фактор Q. При этом опа приобретает трудный обмо-
лот, соломина ее становится тоньше и исчезает жесткость листьев.
Воздействие на пшеницу мягкую мутагенами приводит к возникнове-
нию признаков пшеницы шарозерной, что вызвано, видимо, утерей до-
минантности геном S (С. И. Макарова, II. II. Зоз, 1965; М. S. Swarni-
nathan, 1966; J. V. Goad, 1967; К. П. Джелепов, 1’. Л. Чавдаров, 1969;
10. И. Абрамович, 1976). Ф. Г. Нигматуллин (1972) при мутагенной
обработке западнопамирского сорта безлигулыюй пшеницы мягкой
Бабило получил безлигульный тип пшеницы шарозерной. Подобная
обработка пшеницы шарозерпой в свою очередь ведет к появлению
Ill nil I. I I till Ilin
пшеницы мягкой (M. V. P. Kao, M. S. Swaminathan, 1963; В P, Pal,
1965).
Перечисленные эксперименты свпдетельсi вуют о возможное!и пе-
рехода гепа S из доминантною в рецессивное состояние и обратно.
Причем пшеница шарозерпая с большей частотой мутировала в мяг-
кую и с меньшей частотой — в Т. compact tun; облучение последнего ви-
да не привело к появлению пшеницы шарозерной (М. V. Р. Rao,
М. S. Swaminathan, 1963). Поэтому предположение J. Percival (1921)
и К. Л. Фляксбергера (1935) о возникновении последней из 7'. compac-
tum не получило экспериментального подтверждения. Значительная
часть исследователей склонна согласиться с другой догадкой
К. А. Фляксбергера — о происхождении пшеницы шарозерной из мяг-
кой. S. Kumar (1970) отметил, что гомозиготное рецессивное состояние
локуса s дает фенотип пшеницы шарозерной, а гетерозиготное пли го-
мозиготное доминантное состояние обеспечивает фенотип пшеницы
мягкой.
Ген S отсутствует у диплоидных доноров гепомов пшеницы
(Е. Р. Моррис, Э. Р. Сирс, 1970). Его пет у тетраплоидпой пшеницы.
Он, видимо, возник на гсксаплоидном уровне. По мысли Н. 11. Вави-
лова (1926). при продвижении вида из центра происхождения ряд ге-
нов теряет доминантность, переходя в рецессивное состояние. Если это
так, то пшеница шарозерная, имеющая рецессивную аллель названного
гена, должна была произойти позднее носителей его доминантной
аллели—Т. aestivum и Т. compactum. Как известно, последний вид
имеет ген С в доминантном состоянии, а мягкая и шарозерпая пшени-
ца— его рецессивную аллель. Поэтому более вероятно, что вид
Т. sphaerococcum возник из Т. aestivum, а не из Т. compactum.
Существует еще несколько гипотез происхождения пшеницы шаро-
зерпой. Н. Bleier (1928) считал, что опа возникла при гибридизации
между тстраплоидпым и гексаплоидным видами. G. Mandy (1970) свя-
зывает ее появление со скрещиванием между Т. turanicum и Ае. tau-
schii. В. Л. Мепабде (1971) полагает, что становление пшеницы шаро-
зерной происходило на базе древней гипотетической популяции Т. pro-
to mac ha.
Систематика. А. и G. Howard (1910) первыми описали Т. sphae-
rococcum как разновидности Т. compactum, a J. Percival (1921) их вы-
делил в указанный самостоятельный вид. Статус Т. sphaerococcum
как вида признают виднейшие тритпкологи (К- А. Фляксбергер, 1935,
1939; Н. И. Вавилов, 1964; П. М. Жуковский, 1971 и др.). Противопо-
ложного мнения, однако, придерживаются J. Mac Key (1954), W. Bow-
den (1959), Е. Morris и Е. Sears (1967).
Var. echinatum Perciv. 1921, Wheat Plant.: 323. (8).
Геогр. рас и p. Индия.
Var. globosum Perciv. 1921. Wheat Plant.: 324. (4).
Геогр. распр. Пакистан.
Var. ischkaschimicum Nigm. 1972, Докл. АН ТаджССР, 10: 58. (17).
Получена Ф. Г. Нигматуллиным в Памирском биологическом ин-
ституте АН ТаджССР при воздействии химическим мутагеном па старо-
давний памирский сорт безлигулыюй мягкой пшеницы Вабило.
Var. jakubzineri Stank. 1977, Докл. Болгар. АН, 30. 1: 111, sine
typo. (7).
Выделена И. Станковым на с.-х. опытной станции им. Малкова
(Болгария) из межвидового гибрида, полученного при скрещивании
Т. sphaerococcum с Т. durum.
Var. kibraicum Udacz. 1976, Tp. прикл. бот. юн. сел. 56, 2: 149. (13).
Выделена Р. А. Удачиным из коллекционного образца Т. sphaerococ-
cum (Индия) па Среднеазиатской опытной станции ВИРа (Ташкент-
ская область).
18”
/\ ан । ti<lnii.<iци 4 po<)u Ti ilи iiiii
К о л о с ь я и с о ii у ш с „ н ы е
1. Зерновки белые
+ I — , — । — | — : — ! — । — tumidiiin
2. Зерновки краевые
+ I — 1—1 — | *" | — I — I — I — I — I rotiiiidutuni
-| -|+|— — | — malko[[ii
К о л о е ъ я о и у in е и и ы с
I. Зерновки белые
+ I ~ I I ~ I ~ I Ч Ч ~ I Ч " I dlobosuI"
— | — | + |— )— |— |-Ч~1~1“Ч rubralbuin
2. Зерновки красные
- I - I + I -- I — I “ I - I -- I — I - I juliubTineri
Колосья с короткими остями
К о л о с ь я п е о п у ш е п и ы е
1. Зерновки белые
+ I ~ I - - I — I -- I — I — I — I + I — I cchinafuin
— | — I +| — | — I -- I — | — I +| — | rubriglabruin
2. Зерновки красные
4- I — I — I — I — | — I — | — I 4- I — | spiccituin
— I — | -f- I — | — I — I — I — I f I — I rtibitriuosuin
К о л о с ь я о и у ш е и н ы е
1
о
3
6
7
8
9
1. Зерновки белые
2. Зерновки красные
randhawaii
Idbraiciiiu
pakislanicum
II?
12
13
1-1
popovii I 15
rubraristaium | 1(>
P а с т с н и я б e з л и г у л ь н ы е
(колосья безостые, неопушенные, зерновки белые)
+
ischkaschimictim | 17
Var. kostoffii Stank, et Tsik. 1973, Докл. Болгар. АН, 26, 6: 810. (6).
Выделена И. Станковым и Д. Циковым в Институте генетики и се-
лекции Болгарской АН из межвидового гибрида комбинации Т. sphae-
rococcum. X Т. durum.
Var. malkoffii Stank. 1977, Докл. Болгар. AH, 30, 1:111, sine
typo. (3).
Выделена И. Станковым на с.-х. опытной станции им. Малкова
(Болгария) из межвидового гибрида от скрещивания Т. spliaerococ-
cum X Т. durum.
Var. pakistanicum Udacz. 1976, Tp. прикл. бот. ген. сел. 56, 2:
149. (14).
Геогр. распр. Пакистан.
11 ex)/>г)д I. Triticuin 277
Var. popovii Stank. cl J’sik. 1973, Доклады Болар. AH, 26, (i. 309.
(15).
Выделена И. Станковым и Д. Циковым в Институте генетики и се-
лекции Болгарской АН из межвидового гибрида комбинации Т. sphaero-
coccum X Т. durum.
Var. randhawaii R. Singh 1959, Indian J. Genet., 19: 200. (12).
Геогр. распр. Пакистан.
Var. rotundatuin Perciv. 1921, Wheat Plant: 324. (2).
Геогр. распр. Пакистан.
Var. rubiginosum Perciv. 1921, Wheat Plant: 323. (11).
Геогр. распр. Пакистан.
V ar. rubralbum Stank. 1977, Докл. Болгар. АН, 30, 1: 111, sine
typo. (5).
Выделена И. Станковым па с.-х. опытной станции им. Малкова
(Болгария) из межвидового гибрида комбинации Т. sphaerococcum^
X Т. durum.
V ar. rubraristatum Stank. 1977, Докл. Болгар. All, 30, 1: 111, sine
typo. (16).
Выделена П. Станковым па с.-х. опытной станции им. Малкова
(Болгария) из гибридов, полученных при скрещивании Т. sphaerococ-
cum X Т. durum.
V ar. rubriglabrum R. Singh, 1959, Indian J. Genet., 19: 200. — T. sphae-
rococcum var. alborubiginosum Jakubz. 1939, у К. А. Фляксбергера,
Опред. наст, хлебов: 194, descr. ross. (9).
Геогр. распр. Пакистан.
Var. spicatum Perciv. 1921, Wheat Plant: 323. (10).
Г e о г p. p а с и p. Индия.
Var. tuniiduni Perciv. 1921, Wheat Plant: 324. (1).
Геогр. p а с п p. Индия, Пакистан.
T, PETROPAVLOVSKYI UDACZ. ET MIGUSCH.—
ПШЕНИЦА ПЕТРОПАВЛОВСКОГО
T. petropavlovskyi Udacz. et Migusch. 1970, Вести, с.-х. пауки, 9:
20. — T. turanicum convar. montanostepposum f. arislijorme Jakubz. 1959,
Бот. жури. 10: 1428, nom. illeg.
Общая характеристика. Всходы зеленые, бархатистоонушеиные. Про-
дуктивная кустистость 3—6 стеблей на 1. растение. Соломина высо-
кая— при осеннем посеве на орошаемом участке в Ташкентской об-
ласти достигает 145—150 см. Стеблевые узлы опушены короткими
нежными стирающимися волосками. Под колосом соломина очень сла-
бо выполнена паренхимой или полая. Листовая пластинка длинная
(до 35—38 см), средней ширины (1,4—1,6 см). Листья взрослого рас-
тения шероховатые. Язычок (лигула) четко выражен. Восковой налет
на стебле и листьях слабый. Колос остистый, грубый, в фазах колоше-
ния и цветения покрыт восковым налетом; в поперечном сечении квад-
ратный; рыхлый (19 = 11—12). Колоски удлиненные, трехцветковые,
членики колосового стержня опушены короткими, редкими волосками
(рис. 21,я). Колосковые чешуи удлиненно-овальные (как у Т. turani-
cuiii). Киль колосковой чешуи доходит до ее. основания. Плечо колоско-
вой чешуи очень слабо выражено. Килевой зубец длинный, переходящий
в верхней части колоса в короткую ость длиной до 2,5 см. У основания
колоска заметны 2 бугорка, как у 7'. polonicum. Наружная цветковая
чешуя длиннее внутренней на 4 5 мм. с двумя килями, при созревании
не расщепляется. Ости длинные (8 И) см), грубые. Зерновка оваль-
ная, средней длины (9 К) мм), стекловидная.
В колеоптиле пшеницы Петропавловского 2 проводящих пучка.
Образ жизни яровой. Вегетационный период продолжительный, как
278
Классификация рода Triticum
Рис. 21. Колосья, колоски и зерновки:
а~ Т. petropavlavskyi Udacz. cl Migusch.; б — T. kiharae Dorof. cl Migusch.
у позднеспелых форм пшеницы мягкой. Засухоустойчивость ниже сред-
ней, жаростойкость высокая. Является ярко выраженным экотипом
оазисного поливного земледелия районов со знойным сухим климатом.
Иммунитет. М. М. Якубцинер (1969) отметил, что пшеница Петро-
павловского сильно поражается бурой и стеблевой ржавчиной. Эти
данные подтверждены экспериментом О. Г. Григорьевой (1975) при
искусственном заражении проростков и наблюдениями над взрослыми
растениями в полевых условиях па инфекционном фоне. По наблюде-
ниям авторов вида (неопубликовано), в Ташкентской и Ленинградской
областях они очень сильно поражаются бурой и желтой ржавчиной,
а также мучнистой росой. При ииокулировапии популяцией рас из раз-
личных географических регионов СССР Э. X. Су.ханбердипа (1976)
также выявила сильную восприимчивость к мучнистой росе. В. И. Крив-
ченко (1973) указал на восприимчивость к твердой и пыльной головне.
Подрод I. Tiiticum
279
A. /М. Ямалсен (197-1) при пекут венном зарижснии выявил, что
к пыльной головне Г. aeslivi этот вид пшеницы средне восприимчив,
а к f. diiri слабо восприимчив как в эмбриональном, так и во взрослом
состоянии.
История. При изучении образцов пшеницы, собранных в 1937 г.
А. М. Горским в Китае (Синьцзян), Р. Л. Удачин обратил внимание на
сходство зерновок нескольких форм, описанных AV. М. Якубципером
как Т. turanicum, с мягкой пшеницей. Электрофоретический анализ
глиадипов зерновок показал, что в генотипе этой пшеницы есть геном D
(А. Г. Хакимова, 1972). При подсчете числа хромосом, проведенном
Л. II. Оре.ч и О. Д. Градчапиновой, было установлено, что 2п = 42.
Все это дало основание рассматривать указанные формы как самосто-
ятельный гексаплоидный вид Т. petropavlovskyi Udacz. et Migtiseh.
Происхождение. Пшеница Петропавловского имеет гены Р и Q, от-
носящиеся к числу главных генов. Ген Р маскирует за фенотипом
этого вида генотип пшеницы мягкой. Действие гена Р выражается
в первую очередь is удлинении наружной цветковой чешуи но сравне-
нию с внутренней, но в меньшей мере, чем у Г. polonii'iim. У шиеницы
Петропавловского есп> признаки Т. persiciun (удлинение килевого зуб-
ца до размера укороченной ости, отсутствие опушения в средней
части поверхности колосковой чешуи). Именно поэтому П. А. Ганди-
ляп (1972) назвал ее «неренкоидом» азиатской спельты. По мнению
авторов вида, пшеница Петропавловского могла возникнуть прп му-
тационном появлении гена Р у пшеницы мягкой пли же в результате
спонтанной гибридизации между Т. aestivum и Т. polonicum. Произо-
шло это, наиболее вероятно, именно в Западном Китае. Это район
древних земледельческих оазисов, изолированных высокогорными хреб-
тами Тянь-Шаня, Памира, Каракорума. Куньлуня, а также обшир-
нейшими пустынями Такла-Макан и Гоби. В подобных изоляторах, как
показал Н. И. Вавилов (1927), возможно появление специфических
эндемичных форм пшеницы, одной из которых и является пшеница,
Петропавловского.
Систематика. Вид узко локализован и, возможно, произошел сравни-
тельно недавно. Поэтому разнообразие его форм очень ограниченное.
Определение разновидностей Т. petropavlovskyi
Колосья неопушенные
Зерновки белые
— , — ! Д-| — I — ! — | — | — ; Д-| — I pctroerythroleucam I
3 е р п о в к н к р а с и ы е
— | — 1 Д- ! — — ; — I — I - ; -I- i — I petrofcrriif>inenm , 2
Колосья опушенные
3 е р п о и к и б е л ы с
— I — I -|~ ! — ' '— । — I — I — I — I I petropavlovskyi | 3
3 срн о в к н крас н ы е
— | — | -(- | — I I I I - I | I pclropscudobarba- I 4
2Ж)
Классификация рода Tcilicum
Var. pclroeryIhioleiiciim Udacz. id Mij'ir.cli, 1976, Ip npiii..i. ddi hii.
сел. 56, 2: 150. (1).
Г e о г p. p а с и p. Западный Китай.
Var. petroferrugineuni Udacz. ct Migusch. 1976, l.c.: 150. (2).
Геогр. распр. Западный Китай.
Var. petropavlovskyi. (3).
Геогр. распр. Западный Китай.
Var. petropseudoharbarossa Udacz. et Aligiiseli. 1979 Бюл. ВИРа, 8-1:
74. (4).
Геогр. распр. Западный Китай.
ПОДРОД II. BOEOTICUM MIGUSCH. ET DOROF.
Подрод II. Subgen. Boeoticum Migusch. et Dorof. 1978, Tp. прикл.
бот. ген. сел. 62, 2: 151. — Листовые пластинки взрослых растений опу-
шены щетинистыми волосками. 2п = 14, 28, 42. Геномные формулы —
A1’Ab, AbAbGG, A”AbAbA”GG, AbA”GGDD.
Т и и: Т. boeoticum Boiss.
Геогр. распр. Балканский полуостров, Закавказье, Израиль,
Иордания, Иран, Сирия и Турция.
Видовой состав: Т. boeoticum Boiss., Т. monococcum L., Т. sin-
skajae A.. Filat, et Kurk., T. araraticum Jakubz., T. timopheevii (Zhuk.)
Zhuk., T. militinae Zhuk, et Atigusch., T. zhukovskyi Menabde et Ericzjan,
T. kiharae Dorof. et Migusch.
СЕКЦИЯ 4. MONOCOCCON DUM.
Sect. 4. Monococcon Dum. 1823, Observ. Gram. Fl. Belg.: 94; Нев-
ский, 1936, Tp. Бот. ин-та АИ СССР, сер. I, 2: 66; Цвелев, 1973, Новости
сист. высш. раст. 10: 39, р.р.; idem. 1976, Злаки СССР: 163, р.р.—
Criihodium Link, 1834, Linnaea, 9: 132. — Nivieria Ser. 1841, Descr. et
Fig. Cer. Europ. 4: 66. (104). — Triticum sect. Eremopyros Doell, 1880
in Martius, Fl. Bras. 2, 3: 227. — Triticum Einkornreihe A. Schulz, 1913,
Geschichte knit. Getr.: 21. — T. consp. monococcum Flaksb. 1915, Опред.
пшен.: 8. — Triticum series I Perciv. 1921, The wheat Plant: 338.—
T. monococcum (L.) Kajanus, 1927, Bibliogr. genet. 3: 144, s. a.— Triti-
cum sect. Monococca Flaksb. 1928, Изв. Гос. ин-та опытп. агрономии,
6, 2: 39. — Monococcum Philiptsch. 1930, Zeitschr. induct. Abst. 54, 1/2:
314. — Triticum sect. Crithodium (Link) Nevski, 1934, Фл. СССР, 2:
677.— Triticum congr. diploidea Flaksb. 1935, Ку-тьт. фл. СССР, 1: 131.—
Triticum sect. Monococca Schiem. у Списков, 1955. Пробл. бот. 2: 52.
Листовые пластинки взрослых растений опушены щетинистыми во-
лосками. Колосья одпоостые или двуостые, колоски с 2—3 цветками,
из которых 1—2 плодущие. На колосковой чешуе имеются 2 хорошо
выраженных зубца. Внутренняя двукнлевая цветковая чешуя расщеп-
ляется на 2 продольные доли при созревании. Пыльники длинные
(6 мм). 2п = 14. Геномная формула — А1’А1’.
Геогр. распр. Балканский полуостров, Закавказье, Израиль,
Иордания, Иран, Сирия и Турция.
Т и п: Г. monococcum L.
Т. BOEOTICUM BOISS. — ОДНОЗЕРНЯНКА ДИКАЯ
Т. boeoticum Boiss. 1853, Diagi). PI. Or., ser. 1, 13: 69; Fpocci. 1939,
Фл. Кавк. изд. 2,1: 349; Schiemann, 1943, Ergebn. Biol. 19: 453, 515.
Crithodium aegilopoides Link, 1834, Linnaea, 9, 3: 132. - Aeyilops critho-
dium Steud. 1855, Syn. Gram.: 355.- 7'. (letfilopoidcs (Link) Bal. ex
//. НогиНгшп Mi.i’iisch rt 1)пк>1 ,'S|
Koern. 1885, in Koern. et Wern. Ilandb. Getrcideb. 1: 109, non Forsk.
1775. — T. monococcum lasiorrachis Boiss. in Nym. 1882. Consp. Fl.
Furop.: 840; Boiss. 1884, Fl. Or. 5: 673.- 7'. aegilopoides (Link) llausskn.
1899, Atilt. Thiiring. Bol. Ver., N. F. 13—14: (>(>.- -7'. monococcum A.
aegilopioides Ascliers. et Graebn. 1901, Synops. 2: 701. — T. monococcum
subsp. aegllopoides (Link) Tliell. 1918, Nalurw. Wochensc.hr. (Jena)
17, 33: 470. —T. monococcum boeoticum (Boiss.) Hayek et Markg., 1932,
Rep. sp. nov. regni veget. 30, 3: 228.— T. spontaneum Flaksb. 1935,
Культ, фл. СССР, 1: 341, nom. illeg.— T. monococcum subsp. boeoticum
(Boiss.) A. et D. Love, 1961, Bot. Not. (Lund), 114, 1: 49. — T. monococ-
cum subsp. boeoticum (Boiss.) Mac Key, I960, Hereditas, Suppl. 2: 267.
Описан по образцу из Греции.
Общая характеристика. Всходы темно-зеленые с антоцианом. Кусти-
стость сильная. Куст молодых растений стелющийся, позднее стебли
коленчато приподнимаются. Соломина тонкая, упругая, прочная, не по-
легающая. Узлы соломины щетниистоопушеипые, часто фиолетовые.
Высота растений 40—100 см (в благоприятных условиях больше). Ли-
стовое влагалище большей частью щетпппстоопушешюе, ио внешнему
краю с длинными волосками. Листья узкие (7—13 мм), средней длины
(14—27 см), темно-зеленые, щетинистоопушенные. Ушки фиолетовые,
с ресничками. Колосья 5—12 см длины, плоские, (ширина боковой
стороны 4,5—9 мм, лицевой 2—2,5 мм). D = 25—50. Колос при созре-
вании спонтанно распадается на колоски (см. рис. 3,6). Последние
в верхней части колоса созревают раньше, чем в нижней. Членики ко-
лоскового стержня по ребрам сильно опушенные, причем на верхнем
конце их волоски. (длиной 1,5—3 мм) образуют густую бородку. Ко-
лоски с одной или двумя остями, в поперечном разрезе имеют форму
трапеции с рогами, образуемыми килями колосковых чешуи. Эти че-
шуи узкие, удлиненные (длина 6—8 мм, ширина 2—2,5 мм), короче
цветковых, крыловидные, поверхность их постепенно переходит в киль.
Килевой зубец (длиной в 1,2—2 мм) в виде треугольника с острой вер-
шиной. Главная боковая жилка хорошо развита, зазубренная, вверху
переходит в острый зубец, который почти равен длине килевого зубца
или несколько короче его. Между зубцами остроугольная выемка. По-
верхность чешуи шероховатая или почти гладкая. Внутренние цветко-
вые чешуи выпуклые, немного длиннее наружных. У одноостых форм
наружная цветковая чешуя нижнего плодоносящего цветка песет ость
длиной в 6—11 см, у второго цветка—остевидпос заострение. У дву-
остых форм наружные цветковые чешуи обоих цветков несут ости оди-
наковой длины пли одна из них короче другой. Ости довольно жесткие,
зазубренные. Внутренняя цветковая чешуя двукилевая, в зрелом со-
стоянии расщепляющаяся на 2 продольные части: расщепление начина-
ется от основания чешуи. В колоске 1 (2) плодущий цветок; обычно
развивается 1 зерновка, редко — 2. Зерновки длиной 5,6—7,9, шириной
1 —1,5 мм и толщиной 2—2,6 мм, плоские (ложатся на боковую сторону,
а не на брюшную). Бороздка у них узкая, хохолок ясно обозначен.
Масса 1000 зерен 16—27 г, стекловидпость высокая, цвет красный пли
изредка зеленый. Белозерные формы не обнаружены.
По данным В. Г. Александрова и О. Г. Александровой (1948), зер-
новки Т. boeoticum отличаются сильно редуцированным перикарпием.
Плодовая оболочка (у зрелых зерновок) очень топкая. Исследования
Л. Л. Жсстяпниковой (1954, 1976) показали, что эии-мезокарипй око-
лоплодника представлен всего одним слоем клеток. Поперечные клетки
короткие, без видимых нор. Трубчатые клетки расположены более или
менее равномерно но всей поверхности зерновки. В пределах вида раз-
личий по анатомической структуре перикарпия зерновок не подмечено.
2Й2 Классификация рода Tritiruia L.
(J. Д. Градчапипова (1967) установила, что у Т. boeoticum в отличие
от других видов наблюдаются ранняя дифференциация анатомической
структуры листа и стебля, сильная склерификация механических эле-
ментов, мелкоклеточность эпидермы и мезофилла, минимальная среди
всех видов пшениц длина устьиц (48 мкм) и максимальное их число
(82) па 1 мм2 верхней стороны листовой пластинки.
Т. boeoticum -- горный ксероморфпый вид, наиболее характерный
для пологих склонов и плоскогорий; он спускается в долины, по ые вы-
ходит на равнины. В Армении и Азербайджане (Нахичевань) поднима-
ется is горы до высоты 1600 м над ур. м. В южных частях ареала про-
израстает до высоты 1700 м над ур. м. В Болгарии опускается до 200—
300 м над у]). м.
Все формы Т. boeoticum поздно- пли среднеспелые (скороспелые
формы пе выявлены). Па территории пашей страны (Закавказье) оби-
тают только озимые формы, южнее государственной Гранины наряду
с озимыми есть формы яровые.
Зимостойкость Т. boeoticum пе достигает степени .зимостойкости
пшеницы мягкой. В суровую зиму 1971/72 г. в Полтавской области не
было полной гибели всех растений пи у одного его образца (3). А. Ба-
рашкова и др., 1979). Морозостойкость п засухоустойчивость невысо-
кие (Г. В. Удовенко и др., 1978).
Для Т. boeoticum в условиях низменности Дагестана характерен
один четко выраженный максимум цветения (В. И. Жуков, 1969).
Основное количество цветков (70—90%) отцветает в течение 15—20 ч.
Цветение продолжается ночью и заканчивается к 7 ч утра. Последую-
щая пауза в цветении продолжается с 7 до 14 —15 ч.
Изучение коллекции Т. boeoticum в орошаемых условиях (Даге-
стан) показало, что концентрация белка в его зерне достигает 30,6%,
лизина в белке — 2,7% (В. Г. Конарев и др., 1972). В отдельных слу-
чаях (Ташкентская область, богарные условия) зарегистрировано со-
держание белка в зерне, достигающее 37% (Н. Ф. Покровская, 1971).
Иммунитет. Т. boeoticum известен как вид, устойчивый к грибным бо-
лезням, по с исключением для некоторых биотипов (J. Percival, 1921;
М. Г. Туманян, 1936: В. О. Гулканян, 1938; II. И. Вавилов, 1940;
И. П. Шитова, 1968; М. М, Якубцинер, 1969; П. М. Жуковский, 1971).
Заражение проростков агрессивными расами 77, 20, 122 бурой ржав-
чины выявило их высокую расоспецифическую устойчивость (тип 0—2);
при выращивании в полевых условиях на провокационном фоне обна-
ружены лишь следы поражения популяцией бурой ржавчины
(О. Г. Григорьева, 1975). Об устойчивости к желтой ржавчине свиде-
тельствует исследование И. П. Шитовой (1968). Иммунитет к стебле-
вой ржавчине подтвержден О. Г. Григорьевой (1975): при изучении
коллекции Т. boeoticum ею обнаружен лишь один образец со следами
поражения патогеном.
На наличие у Т. boeoticum высокоустойчивых к мучнистой росс
форм указали С. И. Ригина-Трайпипа и И. Г. Одинцова (1974). Искус-
ственное заражение проростков 35 образцов сборным ипокулюмом из
смеси популяций гриба Ленинградской, Куйбышевской, Новосибирской,
Алма-Атинской областей и Северной Осетии показало, что восприимчи-
вых к патогену образцов среди них пе было; па провокационном фоне
в полевых условиях взрослые растения лишь одного образца оказались
восприимчивыми (3. X. Сухапбердина, 1977).
В. 11. Кривченко (1973) и А. М. Ямалеев (1974) при искусственном
заражении расами 2 и 38 (I. aeslivi и 1. dnri) выявили высокую поле-
вую расоспсцифическую устойчивость к пыльной головне. Причем пос-
ле заражения мицелий был обнаружен во всех частях зародыша зер-
новки некоторых образцов; он достигал даже зародышевой почки, по
растения продуцировали свободные от инфекции зерновки. Устойчи-
П/Н^Мм) //. U Ilin AIh'UuIi cl /Jt/l/l)
bocib к 11i,i,!iмн>ii головие у 7'. boeoticum ii;i:ih;iiiiimmii исследователями
квалифицирована как устопчив<>сiь типа 7’. /imo/iheeoii. Иммунность
7’. boeoticum к твердой головне выявлена также в опыте В. II. Кривчен-
ко (1973).
В целом Г. boeoticum следует отнести к числу видов, устойчивых
к грибным болезням.
История. Наиболее древние, находки растительных остатков Т. boeoti-
сшп отмечены в раскопках, проведенных па севере Сирии близ р. Ев-
фрат, а также в археологическом материале (II. Ilelbaek, 1959, 1966) из
Ирака. Первая из них датирована 8050—-7542 гг. до н. э., вторая —
7500—6750 гг. до и. э. К 7000 г. до и. э. относится находка в Анатолии
(Турция).
О нахождении однозернянки дикой па Восточном Кавказе имеется
нечеткое указание с обозначением ее под названием Т. monococcum
(ЛЕ Bicberstein, 1809). Впоследствии здесь в Дпвпчпнском и Шсмахпн-
ско.м районах Азербайджана Т. boeoticum был обнаружен экспедицией
II. Д. Мустафаева (1964) и в последующем экспедициями ВИРа.
В Крыму дикие однозернянки были отмечены ЛЕ Bicberstein (1S09),
Д. К. Ларионовым (1913), Е. II. Барулиной (1922), И. А. Дроздовым
(1922), А. Е. Коварскпм (1929). А. И. Мордвинкина (иеопубликовапо)
наблюдала здесь произрастание их одноостых форм в 1967 г. Поиски
последних лет не дали положительных результатов в обнаружении
однозернянки дикой в Крыму.
В Армении они были открыты ЛЕ Г. Туманяном (1929). Сбор их
в 30-х годах произвели II. И. Вавилов, Б. Гарасеферяп и М. ЛЕ Якуб-
цинер, а в послевоенные годы — II. Д. Мустафаев, В. Ф. Дорофеев и
II. Л. Гандилян. В Азербайджане однозернянка дикая выявлена
ЛЕ ЛЕ Якубципером (1932), И. Д. Мустафаевым (1961, 1967) и
В. Ф. Дорофеевым (1972). Места обитания ее отмечены в Турции
(В. Balansa, 1857; Е. Bourgeau, I860; J. Bornmuller, 1889; П. ЛЕ Жу-
ковский, 1926), Сирии (С. Haussknecht, 1865; Е. Bourgeau, I860;
Т. Kotschy, 1843; A. Aaronsohn, 1906), Греции (II. F. Link, 1833;
Н. И. Вавилов, 1935), Югославии (И. Страпски, 1929), Болгарии
(Н. Стоянов, Б. Стефанов, 1921; Т. Лйптков, 1939), Иране (И. Strauss,
1910), Ираке (И. HandeKMazzelti, 1910; С. Haussknecht, 1865; F. Na-
bclek, 1935) и Израиле (Л. Aaronsohn, 1906).
Географическое распространение. Одпоостые формы сосредоточены
на Балканском полуострове, в Крыму (после 1967 г. нет сведений об
их нахождении), Армении (Абовянскни, Азизбсковскнй, Араратский и
Ехегпадзорский районы), Азербайджане (Дивпчинскпй, Шемахппскнй,
Джебраильский, Нахичеванский, Запгслапский, Гадрутский районы),
Югославии, Греции, Болгарии, Иране, Ираке, Сирии, Иордании. Сведе-
ния о нахождении их в Грузии и Испании не подтверждены, а сборы
единичных растений носят случайный характер и, видимо, являются
результатом заноса (И. Кецховели, 1928; К. А. Фляксбергер, 1935).
Двуостые формы приурочены большей частью к Армении, Азер-
байджану, Ираку, Ирану, Сирии, изредка встречаются в Болгарин и
Югославии. И. И. Вавилов (1964), К. А. Фляксбергер (1925),
ЛЕ М. Якубцинер (1961) и И. ЛЕ Жуковский (1964) отмечали, что фор-
мы subsp. boeoticum приурочены к Балканскому полуострову и частич-
но к западу Малой Азии, к Закавказью и Ближнему Востоку. По
II. ЛЕ Жуковскому, почти вся ЛЕалая Азия, за исключением лесной зоны
Северной Анатолии (Турция), является единой областью массового
произрастания диких двуостых однозернянок subsp. thuoudar.
Происхождение. G. D. II. Bell (1965) возникновение Т. boeoticum
связывает с диплоидным прототипом ишеппцы. В. Ф. Дорофеев (19.69)
становление этого вида объясняет мутациями, произошедшими у
2SI l\.lill I inlllll.l>m)u 7 util HUI I..
вымерим о родовою uina пшеницы t'roloti iticiim. Пи нашим наблюде-
ниям, в условиях Ленинградской области молодые растения Т. boeoli-
cum неотличимы or Т. ururtw. .аистовые плаегннкп обоих имеют барха-
тистое на ощупь опушение. С началом фазы выхода г. гребку па .thciо-
вой пластинке и влагалище листа Т. boeoticum появляется новый при-
знак— щетинистое опушение. У Т. urartu с возрастом тип опушения не
меняется. Подобная общность в опушении листовой пластинки у моло-
дых растений может свидетельствовать о происхождении Т. boeoticum
от древней формы Т. urartu. Причина дивергенции этих двух однозерня-
нок— интрогрессивная гибридизация или мутация, приведшие к появ-
лению на эпидерме листовой пластинки и влагалища v Т. boeoticum
щетинистого опушения и усиления такого опушения на стеблевом
узле.
Систематика. Систематическое положение однозернянок вызывало
немало споров среди тритикологов. J. Н. F. Link (1834) выделил дикие
однозернянки в самостоятельный род Crithodium Link. N. Scringe
(1841) отдельным родом Nivieria Ser. считал культурные однозернянки.
Генетик 10. А. Филиппенко (1930) предложил все дикие и культур-
ные однозернянки объединим, в одном роде Monococcum Philiptsch.
Е. Boissier (1884), A. Fiori, G. Paolelti (1896), P. F. A. Ascherson cl
P. Graebncr (1901), T. Sakamura (1918), A. Thclking (1818), A. Ilavek
et F. Markgraf (1932), A. el D. Love (1961), J. Mac Key (1966),
R. Morris et IL R. Sears (1967), как и К.. Л. Фляксбергер (1925), объ-
единили все однозернянки, как дикие, так и культурные, под видовым
названием 7’. monococcum L. Это не вполне правильно, ибо 1\. Линией
так называл лишь культурные формы, о чем свидетельствует и лекто-
тип (каталог № 1044 Линнсевского гербария в Лондоне). Название
Т. aegilopoides (Link) Bal. введено в литературу F. Kornicke (1885),
и оно приводилось в разное время многими авторами для обозначения
диких однозернянок. Такого термина придерживались J. Percival
(1921), II. И. Вавилов (1935), Е. Ф. Пальмова (1935), М. Л\. Якубци-
нер (1947, 1957), П. М. Жуковский (1950), Е. II. Сипская (1955),
рассматривавшие Т. aegilopoides (Link) Bal. как дикорастущий вид,
давший начало Т. monococcum L. Другое, видовое наименование
Т. aegilopoides (Link) Hausskn., предложенное в 1899 г., должно быть
отвергнуто независимо от границ его па основании статьи №64 «Меж-
дународного кодекса ботанической номенклатуры» как более поздний
омоним. В литературе известно название, ранее действительно обнаро-
дованное,— Т. aegilopoides Forsk., ио в современном понимании оно
обозначает растение Rotboellia hirsuta. Незаконным является и другое
название диких однозернянок, предложенное К. А. Фляксбергером
(1935) г., — Т. spontaneum Flaksb., ибо оно противоречит статье №63
«Кодекса», т. е. является более поздним омонимом, поскольку известно
более раннее название таксона — Т. boeoticum Boiss. (1853).
Мы рассматриваем все дикие однозернянки с геномом А1’ как осо-
бый вид, отличный от культурных форм, т. е. как вид Т. boeoticum
Boiss., по в более широком понимании, чем принимал его сам Е. Bois-
sier. Мы включаем в него в качестве подвидов одиоостые и двуостые
формы (хот'я граница между ними не всегда четкая). Некоторые си-
стематики под наименованием Т. monococcum boeoticum (Boiss.) Hayek
et Markg. подразумевают как одиоостые, так и двуостые формы (A. Ha-
yek el F. Markgraf, 1932). Примерно так понимает 'Г. boeoticum ряд
современных тритикологов. Е. Scliieinanii (19'18) даже обозначила груп-
пу пшениц как 'Г. boeoticum Boiss. eiiiciid. Scliieni. В действи i е. ibiioci и
задолк) до псе наш соотечественник А. Гроссгей.м (1939) для рода
Triticum приводит вид Т. boeoticum Boiss. именно в таком понимании.
Ряд систематиков не включали дикие однозернянки в состав вида
Т. monococcum L., а рассматривали их как единый вид с выделением
По()р<к) JI. I>of<)tii uni Ali/ч/ .< h fl DoK/j.
одноосных и двуос|ых форм в качестве отдельных рас (II. ('. Ilaiis-
sknechl, 1899; Л. 'I'll е 111111 g, 1918), подвидов (A. A. J'pocci cii m, 1939) ила
разновидностей (К. Л. Фляксбергер, 1913; J. Percival, 1921). C;i,mo-
стоятельные виды- T. boeofictun Boiss. (одноосные формы) и Т. tliaou-
dar Pout, (двуослые) приводит II. М. Жуковский (1971).
К- Л. Фляксбергер (1925) в составе 7’. monococcum Л. aegilopioides
Aschers. et Graebn. различал группы uniaristala Flaksb. (одноостые) и
biniaristata Flaksb. (двуостыс) дикие однозернянки. Затем он (1935)
у однозернянки дикой Т. spontancum выделяет subsp. иещ/<>poide.s
(Bal.) Flaksb. n subsp. Ihaomlar (Kent.) J’lnksb. Такие наименования
подвидов закрепились затем во многих литературных источниках. Как
подразделения вида Т. boeoticum. они впервые приводились Л. А. Гросс-
геймом (1939), Н. II. Вавиловым (19(31) и др. Е. Schiemaim (1948)
также приводит subsp. aegilopoides (Eink) Schiem. и subsp. lliaoudar
(Rent.) Schiem. в пределах вида T. boeoticum.
Следует отметить, что в понятии Т. boeoticum Boiss. подразумева-
лась его автором именно одпоостая однозернянка дикая. Двуостые
дикие однозернянки он относил к другому виду. Действительно, в гер-
барии Ботанического института АН СССР хранится лектотип двуосгой
однозернянки дикой с собственноручной надписью Е. Boissier кTriticum
thaoudar n. sp. Boiss.».
Нами принимается деление Т. boeoticum Boiss. на subsp. boeoticum
и subsp. thaoudar (Rent, ex Hausskn.) Grossh.
В разработке внутривидовой классификации Т. boeoticum большую
роль сыграли советские тритикологи, особенно К- А. Фляксбергер и
ЛЕ Г. Туманян. Ряд новых разновидностей п форм установлены
Б. Гарасеферяном, М. ЛЕ Якубцинером, П. Гапдиляном, А. Е. Ковар-
скнм и П. М. Жуковским.
Дикие однозернянки со слабошероховатымп, а также с бугорчаты-
ми чешуямп иногда (ЛЕ Г. Туманян, Б. Гарасеферян и др.) рассматри-
вали как самостоятельные разновидности. Они в соответствии с прин-
ципом К. Фляксбергера (1935) нами не выделяются в особые разновид-
ности, а рассматриваются как формы соответствующих разновидностей
с неопушепными чешуямп колоса.
Ключ для определения подвидов
1. Колосья длиной 5—6 см (реже больше), колоски несут по одной
ости (обычно вместо второй ости—остсвидное заострение 0,3—2,5 см);
колосковые чешуи 6—6,5 мм длины. В колоске пе более I плодущего
цветка. Пыльники 3 мм длины...................'. . subsp. boeoticum.
+ Колосья длиной 5,5—12 см (изредка больше), колоски несут по
2 ости, одна из них до И см длины, вторая немного короче. Колосковые
чешуи 7,5—8 мм длины. В каждом колоске по 1—2 плодущих цветка.
Пыльники 3,9 мм длины................................................
.......................subsp. thaoudar (Rent, ex Hausskn.) Grossh.
1. Subsp. boeoticum — подвид беотийский.
Subsp. boeoticum.--?’. aegilopoides (Link) Hausskn. Rasse b)
T. boeoticum (Boiss.) Hausskn. 1899. Mitt. Thiiring. Bot. Ver. 13: 67.—
T. monococcum aegilopoides grex uniaristala Flaksb. 1925, Tp. прикл.
бот. сел. 15, 1: 208. -7’. thaoudar auct. non Rent, ex Hausskn.; Nevski,
1934, Фл. GGGP, 2; 677, p.p.- -7’. spontaneum subsp. aegilopoides (Bai.)
Flaksb. 1935, Культ, фл. GGGP, 1: 349,- T. boeoticum subsp. aegilopoi-
des (Bal.) Grossh. 1939, Фл. Кавк. изд. 2,1: 350.
Геогр. распр. Турция, Ирак, Армения, Азербайджан (включая
Нахичевань), Балканский полуостров.
286
1\ЛШЧ ш/И1М1ЦШ1 /><>()(! I'lHlCUIII 1..
Определение разновидностей subsp. boeoticum
Чешуи Огги Р.ГШН1П1ЛН0СТ1. 1 № разновидности j
белые белые с чер- ными краями красные красные с черными краями черные на <|>оне серо- лымча тыс н; крас- ном фон? цвета чешуи черные
белом крас- ном
+ Коло Чеш 3 е р и С b Я 0 уи neot 0 в к и С Т и с ушеин к р а с + т ы е я в н ы с boeoticum 1
+ — — — •— — — — + pseudoboeoticum 2
+ — — — — — + — albi nigrum 3
— — + — — — — + — sumbolonense 4
— — + — — — — 4- lariotiovii 5
— + + — — + + theydjerabaki pantchitchii 6 7
—— — — — — + — — + baydariaim 8
— — — — — — -1 — + avdalaricum 9
+ 3 с р н ) В К 11 3 с л о + [ ы е viridiboeoticum 10
+ — — — — — — — + aznabtirticum 11
— + — — — — + — viridisymboloner.se 12
— — + — — — — — + buluchevskajae 13
— — — + — — — — + larisae 14
— — — — + — — — + nigrichlorococcum 15
— — — — — + — + — gandil janii 10
+ 4et Зори lytl Ofll, э в к и шенны к р а с 4’ с I ы с ziiccarinii 17
+ — — — — — — — + pseudozu ccarinii 18
—- 4“ — — 1 1 — — + — garnicnse 19
— — + — — — — + — mayssurianii 20
— — + — —- — — — + pseiidomayssurianii 21
— — — —- + — —— —. + piibescentinigrtim 22
— — — — — + — — + helenae 23
— — — — — — + — + cinereuni 24
4“ 3 е р п ) в к и зеле 4- I ы е kovarskii 25
— + — — — — + — chochlovae 26
— — + — — — — — + garasseferianii 27
— — + — — — — + daralagezictim 28
— — — — + — — — + kurbagalense 29
— — — — — + — —. + mikoiani 30
— — — — — — + — + ainazuri 31
+ К С лось Чеш 3 с р и я поя /и iicoi о в к п У ОСТ ушеша к р а с + исты яе и ы е e subalbum 32
+ — — — — — — + subpseudoalbum 33
— — — + — — - - + sub pantchitchii 34
— — — — + — — + subbaydarictim 35
+ 1 — — -- - Зерновки зелены е + 1 - niicrospermum 36
Чешуи. опушенные (зерновки красные)
I I I I I subpubescentinig-
I - - - I- гит
Ih.ti/ioi) IL 1 )'<?<•<>iic iim Mii>u\i li. et Driroj. 287
Var. ainazuri Garass. var. nova. T. aegilopoides var. uinaztiri
Garass. 1939, 'Гр. Лрм. фил. ЛИ СССР, сер. биол. 1: <88, descr. ross.--
Т. boeoticum var. unnarpurucivir Gandil. 1975, Tp. Лрм. НПНЗ, cep.
пшеница, 3: 72, descr. ross. A T. boeotico var. cinereum (Thum.) Jakubz.
caryopsibus viridibus differt. (31).
Typus: Armenia, 1935, к-25808 (WIR).
Геогр. pa сп]). Армения.
Var. aibinigrum (Thum.) Л. Filat, et Dorof. comb. nova. — T. aegilo-
poides var. albotiigrttm Thum. 19<31, Tp. прикл. бог. ген. сел. 5,2: 257. -
Т. sponlaneutn var. boeoticum f. albonigrtun (Thum.) Flaksb. 1935,
Культ, фл. 1: 350. (3).
Геогр. распр. Армения.
Var. avdalaricum (Thum.) Л. Filat, el Dorof. comb. nova. — T. aegi-
lopoides var. avdalaricum Thum. 1934, l.c.: 257. — T. aegilopoides var.
tuberculatum Thum. 1934, l.c.: 257.— T. spontaneum var. symbolonense
f. avdalaricum (Thum.) Flaksb. et f. tuberculatum (Thum.) Flaksb. 1935,
I. c.: 352. (9).
Геогр. распр. Армения.
Var. aznabtirticum (Jakubz.) Л. Filat, et Dorof. comb. nova. /’. aep'-
lopoides var. aznabtirticum Jakubz. 19,32, Tp. прикл. бог. ген. сел. 5,2;
171. — -Т. aegilopoides var. chlorococcum Thum. 1934, l.c.: 257.— T. spon-
taneum var. pseudoboeoticum f. aznabtirticum (Jakubz.) Flaksb. et
f. chlorococcum (Thum.) Flaksb. 1935, l.c.: 351.— T. aegilopoides var.
pseudogelapi Garass. 1939, l.c.: 88, descr. ross. (11).
Геогр. распр. Азербайджан (включая Нахичевань).
Var. baydaricum (Flaksb.) A. Filat, et Dorof. comb. nova. — T. mono-
coccum A. aegilopioides var. baydaricum Flaksb. 1925, Tp. прикл. бот.
сел. 25, 1: 224. — T. spontaneum var. baydaricum Flaksb. 1935, l.c.:
353. (8).
Геогр. распр. Азербайджан (Нахичевань), Армения, Крым.
Var. boeolictiin. — Т. monococcum var. boeoticum (Boiss.) Flaksb.
1915, Опред. пшен.: 42. — T. aegilopoides var. boeoticum Perciv. 1921,
Wheat pl.: 166. — T. aegilopoides var. album. Thum. 1934, l.c.: 255.—
T. spontaneum var. boeoticum (Boiss.) Flaksb. 1935, l.c.: 349. — T. boeo-
ticum var. euboeotictun Grossh. 1939, Фл. Кавк.: 346. (1).
Г e о г р. рас п р. По всему ареалу диких однозернянок.
У var. boeoticum К- А. Фляксбергер в сборах С. Haussknecht из Гре-
ции обнаружил колосья типа карликовых булавовидных пшениц и вы-
делил их как Т. aegilopoides var. boeoticum f. breviata Flaksb. 1929. Tp.
прикл. бот. ген. сел. 21,5: 469.
Var. buluchevskajae A. Filat, et Dorof. var. nov. — T. boeoticum var.
unnarnuruvir Gandil. 1967, 1. c.: 96, sine typo; Gandil. 1975, 1. c.: 72, descr.
ross. Л T. boeotico var. viridisymbolonense (Jakubz.) A. Filat, et Dorof.
aristis nigris differt. (13).
Typus: Armenia, VI. 1967, leg. P. Gandiljan (WIR).
Геогр. рас п p. Армения.
Var. chochlovae A. Filat, et Dorof. var. nov.— T. boeoticum var.
unarpurivir Gandil. 1967, l.c.: 96, sine typo; Gandil. 1975. l.c.: 72, descr.
ross. A. T. boeotico var. mayssurianii (Zhuk.) A. Filat, et Dorof. caryopsi-
bus viridibus dif,fert. (26). Typus: Armenia, VI. 1967, leg. P. Gandiljan
(WIR).
Г e о г p. p а с и p. Армения.
Var. cinereum (Thum.) Л. Filat, et Dorof. comb. nova. T. aegilopoi-
des var. cinereum Thum. 1934, l.c.: 257,- T. sponlaneum var. inayssuri-
unii f. cinereum (Thum.) Flaksb. 1935, l.c.: 354. (24).
Геогр. распр. Армения.
Var. daralagezicum A. Filat, et Dorof. var. nov. — T. boeoticum var.
unnnrpurutrivir Gandil. 1967, l.c.; 96, sine typo; Gandil. 1975, l.c.: 72,
28<S /\.кк < т1/ик,щп:1 /ялМ ToHium
descr. ross. Л T. boeotico var. larisae A. Filat, et Dorof. glumis pubcs-
ceiilibus differ!. (28). Typus: Armenia, VI. 196/, leg. I’, Gandiljan (WIR).
Гео i' p. p а с и p. Армения.
Var. gandiljanii A. Filat, var. nov. T. boeoticum var. иnarunrnnivir
Gandil. 1975. l.c.: 72, descr. ross. — Л T. boeotico var. baydaricum caryop-
sibus viridibus differt. (1G). Typus: Armenia, VI. 1967, leg. P. Gandiljan
(WIR).
Г e о г p. p а с п p. Армения.
Var. garasseferianii A. Filat, et Dorof. var. nov.- T. boeoticum var.
unnarpurttvir Gandil. 1967, l.c.: 96, sine typo. Л T. boeotico var. pseudo-
mayssurianii Dorof. caryopsibus viridibus differt. (27).
Typus: Armenia, VI. 1967, leg. P. Gandiljan (WIR).
Геогр. распр. Армения.
Var. garniense (Thum.) A. Filat, et Dorof. comb. nova. — T. aegilopoi-
des var. garniense Thum. 1934, l.c.: 257. — T. spontaneum var. zuccarinii
f. garniense (Thum.) Flaksb. 1935, l.c.: 354. (19).
Геог p. p а с п p. Армения.
Var. helenae (Flaksb.) A. Filat, et Dorof. comb. nova. — T. monococ-
cum A. aegilopioides var. helenae Flaksb. 1925, l.c.: 221.— T. sponianeum
var. helenae Flaksb. 1935, l.c.: 355. — T. spontaneum var. villosonigrum
Flaksb. 1935, l.c.: 355. (23).
Геогр. распр. Азербайджан, Армения, Крым, Югославия.
Var. kovarskii Dorof. et A. Filat, var. nov. — J', spontaneum var,
psetidozuccarinii f. kovarskii Flaksb. 1935, l.c.: 354, descr. ross. Л T. boe-
otico var. psetidozuccarinii (Kovarsk.) A. Filat, ct Dorof. caryopsibus viri-
dibus differt. (25). Typus: Armenia, VI. 1962, leg. Dorofeev (WIR).
Геогр. распр. Армения, Украина (Крым).
Var. kurbagalense (Thum.) A. Filat, el Dorof. comb. nova. — T. aegi-
lopoides var. kurbagalense Thum. 1934, l.c.: 257. — T. spontaneum var.
pubesceniinigrum f. kurbagalense (Thum.) Flaksb. 1935, l.c.: 355. (29).
Геогр. распр. Армения.
Var. larionovii (Flaksb.) A. Filat, et Dorof. comb. nova. — T. mono-
coccum var. larionowii Flaksb. 1913, Tp. Бюро прикл. бот. 6: 682.—
T. aegilopoides var. larionowii Perciv. 1921, Wheat pl.: 167. — T. aegilo-
poides var. pseudostjmbolonense Thum. 1929—1930, Tp. прикл. бот. ген.
сел. 24, 2: 3. — Т. spontaneum var. larionovii Flaksb. 1935, l.c.: 352. (5).
Геогр. распр. Закавказье, Крым, Балканы, Малая Азия.
Var. larisae A. Filat, et Dorof. var. nov. — T. boeoticum var. unnarnu-
rutrivir Gandil. 1967, 1. c.: 96, sine typo. — T. boeoticum var. unnarrutrivir
Gandil. et var. unnarlurutriuir Gandil. 1975, 1. c.: 72, descr. ross. Л. T. boe-
otico var. theydjerabakii (Thum.) A. Filat, et Dorof. caryopsibus viridibus
et aristis nigris differt. (14). Typus: Armenia, IV. 1967, leg. P. Gandiljan
(WIR).
Геогр. p а с и p. Армения.
Var. mayssurianii (Zhuk.) A. Filat, ct Dorof. comb. nova.— T. mono-
coccum aegilopoides var. mayssurianii Zhuk. 1924, Зап. науч.-прикл. отд.
Тифл. бот. сада, 3: 5. — Т. spontaneum var. mayssuriani (Zhuk.) Flaksb.
1935, Lc.: 354. (20).
Геогр. распр. Закавказье, Крым, Балканы, Малая Азия.
Var. iniclospenninn (Thum.) A. Filat, et Dorof. comb. nova. — T. aegi-
lopoides var. microspermum Thum. 1934, l.c.: 258. — T. spontaneum
var. boeoticum f. microspermum (Thum.) Flaksb. 1935, l.c.: 350. (36).
Г с о г p. p а с и p. Армения.
Var. inikoiani Garass. var. nova. — 'T. aegilopoides var. mikoiani Ga-
rass. 1939, l.c.: 88, descr. ross.— T. boeoticum var. unarpuruni (vir)
Gandil. 1967, l.c.: 96, sine typo. Л T. boeotico var. helenae (Flaksb.)
A. Filat, et Dorof. caryopsibus viridibus differt. (30).
Typus: Armenia, VI. 1967, leg. P. Gandiljan (WIR).
Л. lioi'ofiсиin Mirniiih. et Doroj.
Гео г p. p ;i с и p. Армения.
Var. nigrichlorococciim (Tlnini.) A. Filal. et Dorof. comb. nova.
7'. aegilopoides var. nigrochlorococcum Tlnini. 1934, l.c.: 257. — T. spon-
taneum var. pancicii Flaksb. f. nigrochlorococcum (Thum.) Flaksb. 1935,
1. c.: 353. (15).
Геогр. распр. Армения.
Var. pantchitchii (Flaksb.) A. Filat, et Dorof. comb, nova. —Г. mono-
coccum var. pancici Flaksb. 1913, Tp. Бюро прикл. бот. 6, 10: 682.—
T. aegilopoides var. pancicii Perciv. 1921,1. c.: 167. — T. nigrescens Pane,
in Koern. et Wern. 1885, Handb. Getreid, 1: 109. — T. aegilopoides var.
luteonigrum Thum. 1929—1930, Tp. прикл. бот. ген. сел. 24, 2: 3.—
7'. sponlaneutn var. pancicii f. litteonigrum (Thum.) Flaksb. 1935, 1. c.:
353. — T. aegilopoides var. nigrescens Thum. 1934, 1. c.: 257. (7).
Геогр. распр. Закавказье (Армения), Крымский и Балканский
полуострова, Малая Азия.
Var. pseudoboeoticum (Flaksb.) Л. Filat, et Dorof. comb. nova.—
T. monococcum A. aegilopioides var. pseudoboeoticum Flaksb. 1925, l.c.:
207. — T. aegilopoides var. pseudoalbum Thum. 1931, l.c.: 257. - 7'. spon-
taneum var. pseudoboeoticum Flaksb. 1935, l.c.: 351. (2).
Геогр. распр. Азербайджан, Армения, Крым, Югославия.
Var. pseudomayssurianii Dorof. 1968, Бюл. ВИРа, 13: 72. — Т. aegi-
lopoides var. Ihumanianii Garass. 1939, l.c.: 88, descr. ross. (21).
Геогр. распр. Армения.
Var. pseudozuccarinii (Kovarsk.) A. Filat, et Dorof. comb. nova.—
T. aegilopioides var. pseudozuccarinii Kovarsk. 1929, Tp. прикл. бот. ген.
сел. 22, 2: 72. — Т. spontaneum var. pseudozuccarinii (Kovarsk.) Flaksb.
1935, 1. c.: 354. (18).
Геогр. pa с. п p. Армения, Украина (Крым).
Var. pubescentinigrum (Flaksb.) A. Filat, et Dorof. comb. nova.—
T. monococcum A. aegilopioides var. pubescentinigrum Flaksb. 1925, l.c.:
209. — T. aegilopoides var. stramineonigrum Thum. 1929, Tp. прикл. бот.
ген. сел. 24, 2: 3; Якубцинер, 1932, Тр. прикл. бот. ген. сел. 5, 2: 171.—
Т. spontaneum var. pubescentinigrum Flaksb. 1935, l.c.: 355. (22).
Геогр. распр. Армения, Украина (Крым), Югославия.
Var. subalbum (Thum.) Л. Filat, et Dorof. comb. nova.— T. aegilopoi-
des var. subalbum Thum. 1933, Опред. хлебн. злаков (колос.): 220.—
Т. spontaneum var. boeoticum f. stibalbum (Thum.) Flaksb. 1935, l.c,:
350. (32).
Геогр. p а с п p. Армения/
Var. subbaydaricum (Thum.) A. Filat, et Dorof. comb. nova.—
T.. aegilopoides var. subbaydaricum Thum. 1934, l.c.: 259. — T. sponta-
neum var. baydaricum f. subbaydaricum (Thum.) Flaksb. 1935, 1. c.:
353. (35).
Геогр. pa с и p. Армения.
Var. subpantchitchii (Thum.) A. Filat, et Dorof. comb. nova. — T. aegi-
lopoides var. subpancicii Thum. 1934, l.c.: 259. — T. spontaneum var.
pancicii f. subpancicii (Thum.) Flaksb. 1935, 1. c.: 353. (34).
Геогр. распр. Армения.
Var. subpseudoalbum (Thum.) A. Filat, et Dorof. comb. nova.—
T. aegilopoides 'var. subpseudoalbum Thum. 1934, l.c.: 258. — T. sponta-
neum var. pseudoboeoticum f. subpseudoalbum (Thum.) Flaksb. 1935,
l.c.: 351. (33),
Геогр. распр. Армения.
Var. subpubescentimgrum (Thum.) A. Filal. et Dorof. comb. nova. --
T. aegilopoides var. subpubesccniinigrum Thum. 1934, l.c.: 259. -T. spon-
taneum var. pubescentinigrum f. subpubesc.entinigrum (Thum.) Flaksb.
1935, l.c.: 355. (37).
19 762
290
К.чассификчци'! /ю<)а Tnliiitin /,.
Г е о । р. р .ч с 11 р Армения.
Var. syinbolonciise (Flaksb.) Л. Filat. et Dorof. comb. nova. T. ino-
nococctim aegilopioides var. symbolonense Flaksb. 1925, l.c.: 207.—
T. spontaneum var. symbolonense Flaksb. 1935, 1. c.: 351. (4).
Геогр. распр. Закавказье (Азербайджан — в Нахичевани, Ар-
мения), Крымский и Балканский полуострова, Малая Азия.
Среди образцов Малой Азии К. Л. Фляксбергер обнаружил булаво-
видные формы этой разновидности — Т. spontaneum var. symbolonense
f. capitata Flaksb. 1929, Tp. прикл. бот. ген. сел. 21, 5: 4(>3.
Var. theydjerabaki (Thum.) A. Filat, ct Dorof. comb. nova. — T. aegi-
lopoides var. theydjerabaki Thum. 1934, l.c.: 257. — T. spontaneum var.
symbolonense f. theydjerabaki (Thum.) Flaksb. 1935, l.c.: 352. (6).
Геогр. распр. Армения.
Var. viridiboeoticum (Jakubz.) A. Filat, et Dorof. comb. nova. —
T. aegilopoides var. viridoboeoticum Jakubz. 1932, l.c.: 171. — T. aegilo-
poides var. albochlorococctim Thum. 1934, l.c.: 258. — T. spontaneum
var. boeoticum f. viridoboeoticum (Jakubz.) Flaksb. 1935, l.c.: 351.—
T. aegilopoides var. getapi Garass. 1939, l.c.: 88, descr. ross. (10).
Геогр. p а с и p. Азербайджан (11ахпчеваиь), Армения.
Var. viridisymbolonense (Jakubz.) A. Filat, et Dorof. comb. nova. -
T. aegilopoides var. viridosymbolonense Jakubz. 1932, l.c.: 171.— T. spon-
tane'um var. symbolonense f. viridosymbolonense (Jakubz.) Flaksb. 1935,
l.c.: 352. — T. boeoticum var. tinarnuru (vir) Gandil. 1967, l.c.: 96, sine
typo. (12).
Геогр. рас ир. Армения, Азербайджан (Нахичевань).
Var. zuccarinii (Flaksb.) A. Filat, ct Dorof. comb. nova.— T. aegilo-
poides var. zuccarinii Flaksb. 1929, l.c.: 463. — T. monococcum var. zuc-
cariorii Flaksb. 1913, l.c.: 683. — T. sponianeum var. zuccarinii (Flaksb.)
Flaksb. 1935, 1. c.: 353. (17).
Геогр. распр. Испания, Сирия, Ирак, Армения, Крымский и Бал-
канский полуострова, Малая Азия.
Среди образцов Малой Азин К- А. Фляксбергер выделил форму
с булавовидными колосьями (колоски в верхней части колоса распола-
гаются настолько плотно, что принимают положение почти под прямым
углом к оси колоса) —Т. spontaneum var. zuccarinii f. bourgeani Flaksb.
1935, l.c.: 354.
II. Subsp. thaoudar (Reul, ex Hausskn.) Grossh.--подвид таудар.
Subsp. thaoudar (Rent, ex Hausskn.) Grossh. 1939, Фл. Кавк. изд.
2, 1: 350. — T. thaoudar Rent, ex Hausskn. 1899, Mitt. Thiiring. Bot. Ver.
N. F. 13—14: 66. — T. thaoudar Rent, ex Bourgeau, 1860, Pl. exs.: n° 281
(nomen); Koern. 1885, in Koern. et Wern. Handb. Getreideb. 1: 109;
Невский, 1934, Фл. СССР, 2: 677, р.р. — Т. monococcum aegilopioides
grex biniaristafa Flaksb. 1925, Tp. прикл. бот. сел. 15. 1: 208. — T. spon-
taneum subsp. thaoudar (Rent.) Flaksb. 1935, Культ, фл. СССР, 1: 344.
Геогр. распр. Турция, Ирак, Сирия, Армения, Азербайджан,
очень редко на Крымском и Балканском полуостровах.
Var. abovjanii A. Filat, et Dorof. var. nov. — T. boeoticum var. biarnu-
alvir Gandil. 1967, Биол. жури. Арм. 20, 4; 96, sine typo. A T. boeotico
var. thaoudar caryopsibus viridibus differt. (10).
Typus: Armenia, VI. 1967, leg. Gandiljan (WIR).
Геогр. распр. Армения.
Var. albinigrescens (Flaksb.) A. Filat, et Dorof. comb. nova. -
T. thaoudar var. albinigrescens Flaksb. 1929, Tp. прикл. бот. геи. сел. 21,
5: 465. — 7’. thaoudar var. atratum Thum. 1934, Tp. прикл. бот. геи. сел.
5, 2: 259. — Т. spontaneum var. albinigrescens Flaksb. 1935, Культ, фл.
СССР, I: 346. (7).
Геогр. pa сп]). Малая Азия, Северный Ирак, Сирия, /Армения.
Подрод II. lioeoticum Mifiusdi. et Doroj.
291
Определение pa шоппдиостсй subsp. thaoud.ir (Rent, ex llaiisskn.) Grossli.
Чешуи
Ости Рязпонл 1Н0С/ь овидности
цвета мер-
чешуи ные X ге
%
Чешуи неопушенные
Зерновки крас I ы с
+ — — — — — — + thaoiidar 1
+ — — — — — — — pseudoreuteri 2
—— + — —• — — — + — nigrireuteri 3
— — + — — —. — + — balansae 1
— — + — — — — balaclavicum s
— — — + — — — 4 — bicolor 6
— — — — + — — 4 albinigrescens 7
— — — — 4- — — 4 fusctirn 8
— — — — — — + — 4- ftimidum 9
3 С [4 II ) В К ] I з о л II ы с
4- — — — — — — — abovjanii 10
+ — — — — — — — 4 azerbajdjanicum 11
— — 4- — — •— —— + — gegadiricum 12
— — -1- •• — — — — — + harnicutn 13
— -— — 4- —. — — — + banasuricum 14
— — — — — + — + — viridifiiscum 15
Чешуи опушенные
Зерновки красные
+ — — — — — + — borinnuclleri 16
— + — — — — -1- — hausskne chili 17
— -— + — — — — — + mazzettii 18
— — — — + — — + — luteinigruni 19
— — — — — + — + — rufinigrutn 20
Зерновки зелены о
+ — — — — — — — + viridalb urn 21
— — — 4- — — — — — + tchivi i'll ill 22
—. — — — + —• — + — uiridinigrum 23
— — — — — + — + — biarpurunivir 24
Var. azerbajdjanicuin (Jakubz.) A. Filat. et Dorof. comb. nov.—
T. thaoiidar var. azerbajdjanicum Jakubz. 1932, Tp. прикл. бот. ren. сел.
5. 2: 172. — Т. thaoiidar var. virescens Thum. 1934, Tp. прикл. бот. ген.
сел. 5, 2: 259. — Т. spontaneum var. pseudoreuterii f. azerbajdjanicutn (Ja-
kubz.) Flaksb. 1935, Культ, фл. СССР, 1: 345. - 7'. thaoiidar var.
rindi Garass. 1939. Tp. Лрм. фил. All СССР, сер. биол. 2: 93, descr.
ross. (11).
Геогр. распр. Азербайджан (в том числе Нахичевань), Армения.
Var. balaclavicum (Kovarsk.) Filat, ct Dorof. comb. nova. — T. aegi-
lopoides var. balaclavicum Kovarsk. 1929, Tp. прикл. бот. ген. сел. 22, 2:
72. — Т. thaoiidar var. psetidobalansae Thum. 1934, l.c.: 259. --T. spon-
taneum var. balaclavicum (Kovarsk.) Flaksb. 1935, l.c.: 346. (5).
Геогр. распр. Армения, Украина (Крым).
Var. balansae (Flaksb.) A. Filat, et Dorof. comb. nova.— T. thaoiidar
var. balansae Flaksb. 1929, l.c.: 463. — T. thaoiidar var. rosetun Thum.
1934, l.c.: 259. — T. spontaneum var. balansae Flaksb. 1935, l.c.: 346. (4).
Геогр. pa си]). Малая Азия, Иран, Северный Прак, Армения.
Var. baiiasiiricuiti Л. Filat, el Dorof. var. nov. T. boeoticum var.
binarniirutrivir Gandil. 1967, l.c.: 96, sine tipo. Л T. boeotico var. bicolor
(Thum.) A. Filat, et Dorof. caryopsibus viridibus differt. (14).
Typus: Armenia, VI. 1968, leg. P. Gandiljan (WIR).
Геогр. распр. Армения.
19*
292
Классификация рода Triticum
V а г. bi a ip u run i vi г Gandil. 1967, l.c.: 97, sine I уро. (21).
Г e о г р. р а с п р. Армения.
Var. bicolor (Thum.) A. Filat, ct Dorof. comb, nova.-- T. Ihaoudur
var. bicolor Thum. 1934, l.c.: 260. — T. sponluneum var. balansae f. bico-
lor (Thum.) Flaksb. 1935, I. c.: 346. (6).
Геогр. распр. Армения.
Var. bormnuelleri (Flaksb.) A. Filat, et Dorof. comb. nova. — 7'. thaou-
dar var. bornmitlleri Flaksb. 1929, l.c.: 464. — T. spontaneum var. born-
miilleri Flaksb. 1935, l.c.: 347. (16).
Геогр. распр. Малая Азия, Ирак, Пран, Сирия, Армения.
Var. fumidum (Thum.) Л. Filat, et Dorof. comb. nov. — T. thaotidar
var. fumidum Thum. 1934, Tp. прикл. бот. геи. сел. 5, 2: 260. — 7’. thaou-
dar var. torosofumidiim Thum. 1934, l.c.: 260.- 7’. spontaneum var.
fuscum f. fumidum (Thum.) Flaksb. et f. torosofumidum (Thum.) Flaksb.
1935, l.c.: 346, 347. (9).
Геогр. p а с п p. Малая Азия, Армения.
Var. fuscum (Zhuk.) A. Filat, ct Dorof. comb. nova. T. aegilopoides
var. fuscum Zhuk. 1927, Tp. прикл. бог. геи. сел. 17, 4: 1 13. Т. thaotidar
var. nigritubcrculatum Thum. 1931, l.c.: 260. 7’. ihaoudur var. rufinig-
rum Flaksb. 1932, in Jakubz. l.c.: 172. — 7’. spontanetim var. fuscum
(Zhuk.) Flaksb. 1935, 1. c.: 346, p. p. (8).
Геогр. распр. Закавказье (Азербайджан — в Нахичевани, Ар-
мения), Крымский полуостров, Малая Азия.
Var. gegadiricum Л. Filat, var. nov. — T. boeoticum var. biarnurtivir
Gandil. 1967. l.c.: 96, sine typo. Л T. boeotico var. balansae (Flaksb.)
A. Filat, ct Dorof. caryopsibus viridibus differt. (12).
Typus: Armenia, VI. 1967, leg. P. Gandiljan (WIR).
Геогр. распр. Армения.
Var. harnicum A. Filat, et Dorof. var. nov.— T. boeoticum var. binar-
nuruvir Gandil. 1967, l.c.: 96, sine typo; Gandil. 1975, l.c.: 73, descr.
ross.— T. spontaneum var. balaclavicum f. kovarskii Flaksb. 1935, l.c.:
346. A. T. boeotico var. balaclavicum (Kovarsk.) A. Filat, et Dorof. caryop-
sibus viridibus differt. (13).
Typus: Armenia, VI. 1966, leg. P. Gandiljan (WIR).
Геогр. распр. Армения, Украина (Крым).
Var. haussknechtii (Flaksb.) A. Filat, ct Dorof. comb. nova. — T. thao-
ndar var. haussknechtii Flaksb. 1929, l.c.: 463. — T. spontaneum var.
haussknechtii Flaksb. 1935, I. c.: 348. (17).
Г e о r p. p a c n p. Малая Азия, Сирия, Армения.
Var. luteinigrum (Kovarsk.) A. Filat, ct Dorof.. comb, nova.- - T. aegi-
lopoides var. luteinigrum Kovarsk. 1929, Tp. прикл. бог. ген. сел. 22, 2:
72.— T. aegilopioides var. biilgaricum. Stransk. 1929, За Димена
в Българ.: 202. — 7’. Ihaoudur var. schorbtilachicttm Thum. 1934, l.c.:
259.- 7’. spontaneum var. luteinigrum (Kovarsk.) Flaksb. 1935, l.c.:
348.' (19).
Геогр. p a с n p. Крым, Армения, Болгария.
Var. mazzettii (Flaksb.) A. Filat, ct Dorof. comb. nova. — T. thaotidar
var. mazzettii Flaksb. 1929, l.c.: 465. — T. spontaneum var. mazzettii
Flaksb. 1929, I. ,c.: 348. (18).
Геогр. распр. Северный Ирак, Украина (Крым), Армения.
Vdr. nigrireuteri (Jakubz.) A. Filat, et Dorof. comb. nova. — T. thao-
udar var. nigroreuteri Jakubz. 1932, Tp. прикл. бот. ген. сел. Прилож.
53*: 180. — 7’. thaotidar var. albiatralum Thum. 1934, l.c.: 260.— 7’. spon-
lanetim var. reuleri f. nigroreuteri (Jakubz.) Flaksb. et f. albiatratum
(Thum.) Flaksb. 1935, I. c.: 345. (3).
* В цитируемой работе ее автором сделана опечатка — вместо nigroreuteri напе-
чатано nigroliaussknecliti.
II. Iloeoticnm ЛЬЛыЛ <’l Dtunj /ИЗ
Геогр. p а с и p. ЛI ;i .1 а я Аиш, Армения.
Var. pseudoreuteri (Thum.) A. F'ilat. cl Dorof. comb. nova. — T. thao-
udar var. pseudoreuteri Thum. 1934, l.c.: 259. - T. spontaneum var.
pseudoreuteri (Thum). Flaksb. 1935, 1. c.: 345. (2).
Г co г p. p асп p. Армения, Азербайджан.
Var. rufinigrum (Thum.) A. Filat, et Dorof. comb. nova. — T. thaoudar
var. rufinigriini Thum. 1934, l.c.: 2G0.— T. spontaneum var. rufinigrum
(Thum.) Flaksb. 1935, l.c.: 349. (20).
Геогр. распр. Армения.
Var. tchivictim A. Filat, et Dorof. var. nov. — T. boeoticum var. binar-
purutrivir Gandil. 1967, l.c.: 97, sine typo; Gandil. 1975, l.c,: 73, descr.
ross. A T. boeotico var. banastiricum A. Filat, et Dorof. glumis pubescen-
tibus differt. (22).
Typus: Armenia, VI. I960, leg. P. Gandiljan (WIR).
Геогр. распр. Армения.
Var. thaoudar. — T. thaoudar var. reuteri Flaksb. 1929, l.c.: 4G4.—
T. sponianeum var. reulerii Flaksb. 1935, I. c.: 345. (1).
Геогр. распр. Малая /Азия, Прак, Иран, Сирия, Армения, Азер-
байджан. Украина (Крым).
Var. viridalbum A. Filat, var. nov. — T. boeoticum var. binarpitalvir
Gandil. 1975, l.c.: 73, descr. ross. Л T. boeotico var. bornmuelleri (Flaksb.)
A. Filat, ct Dorof. caryopsibus viridibus et aristis nigris differt. (21).
Typus: Armenia, VI. 1966, leg. P. Gandiljan (WIR).
Г e о г p. p а с и p. Армения.
Var. viridifuscum A. Filat, var. nov. — T. spontaneum var. fuscum f. zhtt-
kovskyi Flaksb. 1935, 1. c.: 347, nom. illeg. A T. boeotico var. fuscum (Zhuk.)
A. Filat, et Dorof. caryopsibus viridibus differt. (15).
Typus: Armenia, VI. 1966, leg. P. Gandiljan (WIR).
Геогр. распр. Закавказье (Армения), Малая Азия.
Var. viridinigrum Flaksb. var. nov. — T. spontaneum var. luteinigrum
f. viridonigrum Flaksb. 1935, l.c.: 349, nom. illeg. A T. boeotico var.
luteinigrum (Kovarsk.) A. Filat, et Dorof. caryopsibus viridibus differt. (23).
Typus: Armenia, VI. 1966, leg. P. Gandiljan (WIR).
Геогр. распр. /Армения.
T. MONOCOCCUM L. — ОДНОЗЕРНЯНКА КУЛЬТУРНАЯ
Т. monococcum L. 1753, Sp. PI. cd. 1: 86. — T. pubescens Bicb. 1800,
Beschr. Land. Casp. Meerc: 81.--7’. hornemannii Clementi. 1818, in Her-
rera, Agric. gen. I: 3. — Nivieria monococcum Ser. 1841, Descr. et fig.
Cer. Eur. 4.: 66 (104). — T. vulgare monococcum Alef. 1866, Landw. Fl.:
333. — T. monococcum B. cereale Aschers. et Graebn. 1901, Synops, 2:
702. — T. monococcum subsp. cereale (Aschers. et Graebn.) Thell. 1918,
Naturw. Wochenschr. N. F. 17, 33: 470. — Monococcum Philipchcnko,
1930, Zcilschr., indukt. Abst. 54, 1, 2: 314. — T. monococcum eumonococ-
cum Hayek et Markgr. 1932, Rep. spec. nov. regni veget. 30, 3: 228.—
T. monococcum subsp. monococcum A. ct D. Love, 1961, Bot. Not. (Lund),
114,1:49.
Тип: описан по культивируемым экземплярам из Европы.
Общая характеристика. Всходы темпо-зеленые, с антоцианом, слабо
опушенные. Кустистость высокая, кует наше, прямостоячий, полураеки-
дистый, реже — развалистый, причем стебли поднимаются от перво! о
узла соломины вертикально вверх. Первое нижнее междоузлие корот-
кое (4 6 см). Растение 60 130 см высоты. Соломина тонкостенная,
полая, гибкая, нс полегает. Узлы соломины опушенные, под ними часто
фиолетовый пояс. Листья короткоопушенвыс, шероховатые, узкие (дли-
на 18—35 см, ширина 0,6-0,8 см). Колосья (рис. 22,а) длиной 4,5
9 см (короче, чем у дикой однозернянки, во более широкие и плоские),
294
К.ИЦгифиКИЦИ.Ч 'j'l ilicitlll
боковая сторона их значительно шире (0,5—1,0 см) лицевой. Колосья
плотные (I) = 40—55). Членики стержня от слабоопушспных до пеону-
шеппых. Стержень колоса при надавливании ломается. Ишида наблю-
дается спонтанная ломкость верхних колосков. Ости длинные или уко-
роченные, шероховатые. Колоски удлиненно-овальные, контуры попе-
речного разреза их напоминают сегмент окружности (а нс трапецию,
как у диких однозернянок), хорда которого обращена к стержню коло-
са; рога сегмента выражены слабо. Колосковые чешуи вытяиуто-обрат-
нояйцевндиые, слегка выпуклые, короче цветковых. Поверхность этой
чешуи постепенно переходит в киль, который закапчивается треуголь-
ным зубцом. Зубец главной боковой жилки по размеру меньше послед-
него. Между рубцами более или менее остроугольная выемка. Поверх-
ность чешуи бугорчатая, шероховатая, на вид точечная пли гладкая,
блестящая. Все культурные однозернянки остистые (безостых форм
не выявлено), причем ость песет только наружная цветковая чешуя
первого цветка, у второго цветка опа имеет лишь остевпдное заострение
(редко-• укороченную осп,). Как и у однозернянки дикой, внутренняя
цветковая чешуя при созревании расщепляется (начиная от основания)
па 2 продольные части. Зерновки узкие (отношение длины к ширине и
толщине равно приблизительно 16:3:4), более крупные, чем у одно-
зернянки дикой, стекловидные, красные (белозерные формы не обна-
II /йл <i;:i А! D-.-r
РУ А ЗШ< ) . . > сулеек,; , /',’.4:10 р .’к ВЦ Юс 1 к Ч lo.ll.KO В Первом Цветке В С.1\
чае развития зерновки ио втором цвете опа бывав! оолее мелкой, чем
зерновка первого цветка.
Для Т. monococcum характерна редуцированная структура перикар-
пия, свойственная пленчатым пшеницам. Однако от однозернянки ди-
кой этот вид отличается более сильным развитием эпи-мезокарпия
(более толстые стенки клеток с заметной слоистостью), более крупны-
ми поперечными клетками, наличием одревеснения клеточных оболочек
отдельных участков перикарпия спинной стороны зерновки, а также
некоторым внутривидовым разнообразием строения этой ткани, соот-
ветствующим морфобиологическому разнообразию (Л. Л. Жестяппи-
кова, 1976).
Для образцов средиземноморской группы (Италия, Испания) ха-
рактерны короткие и широкие поперечные клетки, для армяно-анато-
лийской группы, напротив, типичны удлиненные поперечные клетки.
Т. monococcum рассматривается Е. Ф. Пальмовой (1935) как экотип
сухой степи. Однако однозернянка культурная западпогрузинская из
ценоза запдури, как и горноевропсйскпс формы, представляет со-
бой, несомненно, мезофитпый тин. Однозернянку заиадногрузппекую
В. Л. Менабде (1918) выделил в особый экогцц. Все формы однозер-
нянки - нагорные, по не высокогорные. Как пояснял 11. М. Жуковский
(1950, 1964), в орошаемых древних высокоземледельческих районах
Месопотамии опа не могла конкурировать с полбой и ячменем и пото-
му локализировалась в более холодных горных районах, тде нет дефи-
цита естественных осадков.
Однозернянка культурная в Грузии поднималась до высоты 900—
1600 м, в Чечено-Ингушетии—до 1500, в Испании—до 750 м над ур.м.
По данным Г. Г. Давидяна и др. (1971), в Югославии ее посевы спуска-
ются до 80 м над ур. м. Однозернянка культурная нетребовательна
к условиям произрастания. Она может возделываться на бедных не-
плодородных горных почвах. Однако на высоком агрофопе дает отлич-
ные результаты.
Т. monococcum встречается ныне в качестве примеси к Т. dicoccum.
И. Д. Мустафаев (1961) собрал ее образцы на полях Нагорного Кара-
баха и Нахичевани. В Западной Грузии однозернянка входила в состав
популяции зандури, которая включала также Т. timopheevii и Т. zhu-
kovskyi.
В Югославии чистые посевы однозернянки культурной были при-
урочены к районам с неблагоприятными природными условиями
(Е. Schiemann, 1955; S. Borojcvic, 1956; Л. Павечсвич, 1963, 1973). Та-
кие посевы ее встречались также в Болгарии и Албании (М. М. Якуб-
цинер, 1961). Иа о. Готланд в Швеции в недавнем прошлом сохраня-
лась чистая культура Т. monococcum. Ныне эта пшеница встречается
здесь лишь как примесь на нолях пшеницы мягкой (И. Hjeltnquist,
1963).
В Крыму в прошлом чистые посевы этой однозернянки встречались
редко (Е. Барулина, 1924). Н. А. Дроздов (1922—1923) и А. Е. Ковар-
ский (1929) собрали здесь растения этого вида, произраставшие в ка-
честве примеси па поле пшеницы мягкой. По К.. А. Фляксбергеру (1935),
однозернянка культурная дифференцирована на 3 эколого-морфологи-
ческие группы: восточную, среднеевропейскую и испанскую. 11ервая
сформировалась в Малой Азии, Закавказье и Крыму, вторая в юж-
ной горной части ФРГ, северной Швейцарии, третья в Псшаипп, Юж
ной Франции и Марокко. Наиболее крупноколосые, позднеспелые
(включая озимые) формы относятся к среднеевропейской группе; самые
раннеспелые — к испанской.
Подавляющее большинство культурных однозернянок в отличие от
дикорастущих имеет яровой образ жизни. Нередко в Западной Европе
29(i Ктитифитщис /nidti Triticum I..
и 3.'Iк;1 вказье, где климат мш кий, яровые однозернянки высева,шен
с осени, что дало основание. J. Percival (1921) писать о возделывании
в Европе озимых форм. В коллекции ВИРа имеются единичные образ-
цы однозернянки культурной из ФРГ (к-20746, к-20399) с озимым
образом жизни. Но степень озимостн у них ниже, чем у сорта пшени-
цы мягкой Украинка. Об озимой ее форме сообщал cine II. Еашп
(1933). Среди культурных однозернянок обнаружены и полуознмые
формы из Югославии, Албании, ГДР и ФРГ.
В массе культурные однозернянки позднеспелые. При осеннем посе-
ве в Дагестане и Узбекистане период от всходов до колошения варьиру-
ет от 193 до 214 дней, до созревания — от 227 до 233 дней. Прп весеннем
посеве в Поволжье эти периоды составляют соответственно 60—70 и
90—100 дней. От фазы кущения до колошения наблюдается замедлен-
ный темп роста и развития. Среднеспелыми являются формы испанские,
срсднепозднимп — дагестанские, крымские, армяно-анатолнйскпс.
По утверждению И. А. Стефаиовского (1950), культурные однозер-
нянки экологически слабо приспособлены к засушливым условиям За-
волжья. Они неустойчивы к суховею в межфазный период от колоше-
ния до налива зерна, а также к почвенно-атмосферной засухе. Ослаб-
ление и гибель Т. monococcum могут быть вызваны также чрезмерной
кустистостью и обилием подгонов, быстро иссушающих почву.
В условиях плоскостного Дагестана цветение у Т. monococcum про-
должается с 3 ч ночи до 7 ч утра. В первый час отцветает 71—88%
цветков от числа отцветших в течение суток. Отмечено определенное
сходство в темпах цветения у культурной и дикой однозернянки. У обо-
их видов (в отличие от большинства других видов) цветение протекает
наиболее быстро — в течение 3—4 дней; имеется один четко выражен-
ный максимум цветения: у Т. monococcum — утренний, а у Т. boeoii-
сиш— предвечерний.
В Швеции на о. Готланд (Н. Hjemqvist, 1966), местами в Югославии
(Л. Павечевич, 1963) однозернянка культурная ценилась за качество
зерна. Противоположная оценка ей дана в Болгарии за обилие отру-
бей при помоле зерна, в Японии — за низкое качество клейковины
(К. Jamashita, 1953).
В Испании, по данным П. ГЕ Вавилова (1964), однозернянка куль-
турная возделывалась преимущественно на корм для свиней, а в Ба-
варии использовалась на крупу. По сведениям Л. Павечевича (1963),
в Югославии зерно Т. monococcum используется па корм птицам и как
фуражная культура на корм лошадям. С последним связано п одно из
местных названий этой культуры — овне.
По данным В. Г. Копарева и др. (1972), в зерне однозернянки куль-
турной в условиях поливного земледелия в Дагестане и Узбекистане
накапливается 15,6—23,4% белка, лизина в нем — 2--2,59°/о. II. Ф. По-
кровская (1971, 1974) указывает, что концентрация триптофана в зерне
в этих условиях колебалась в пределах 0,73—1,05%. Она отмечает так-
же низкое качество клейковины.
Иммунитет. Высокая устойчивость Т. monococcum к грибным болез-
ням отмечена еще у R. Н. Biffen (1907), который советовал использо-
вать этот вид при создании устойчивых к патогенам сортов продоволь-
ственной пшёницы. Н. И. Вавилов (1961) назвал однозернянку куль-
турную «аккумулятором комплексного иммунитета», хотя и отмечал,
что некоторые формы ее могут поражаться грибными болезнями.
Т. monococcum является составной частью грузинской популяции
заидури, которая выделяется иммунностью к грибным болезням. Устой-
чивость однозернянки культурной к поражению ими выше, чем у одно-
зернянки дикой.
Т. monococcum устойчив к ряду рас бурой, желтой и стеблевой ржав-
чины и пыльной головни, отмечается также его невосприимчивость
Подрод II. lioi'ot i с и т Mipiis,i/i. of Doioj.
247
к мучнистой росе (M. M. Якубцинер, 19,59, 19(51, 1961). Однако в неко-
торые юды ржавчина и мучнистая роса moivt поражать представите-
лей ряда экологических групп этого вида. I. Nover и Cli. Lehmann (1967)
к числу рас бурой ржавчины, поражающих однозернянку культурную,
отнесли 15 и 20/31.
И. П. Шитова (1966) изучила коллекцию Т. monococcum г, различ-
ных высотных зонах Дагестана. Независимо от места изучения была
выявлена высокая устойчивость к бурой, желтой и стеблевой ржавчине.
О. Г. Григорьева (1975) обследовала 55 образцов в полевых \словия<
на инфекционном фоне и выявила иммунность и высокую устойчивое in
к бурой ржавчине у 49 образцов. Кроме того, проведенное ею зараже-
ние расами 21, 34, 17 стеблевой ржавчины показало восприимчивость
большей части образцов в фазе проростков, хотя взрослые растения
в полевых условиях не поражались. Подобные результаты по стеблевой
ржавчине получены также Л. М. Камслипой (1973).
Устойчивость отдельных образцов Т. monococcum. к твердой голов-
не отмечена еще Н. М. Wollman к N. Humphrey (1924). О том же сви-
детельствует опыт Т. В. Ярошенко, II. Я. Зубко (1966) и др. Воспри-
имчивость к спецна.ш шрованпым формам гвердой головни orMein.ur
Д. II. Тетеревнпкова-Бабаяп (1911) и В. Тодорова (1969). Исследование
В. II. Кривченко (1973) выявило у Т. monococcum доминирование им-
мунности к грибу: из 51 образца лишь 3 имели слабую или среднюю
восприимчивость к твердой головне.
Д. В. Мягкова (1963), В. И. Кривченко (1973) и А. М. Ямалсев
(1974) обследовали мировую коллекцию Т. monococcum па устойчи-
вость к пыльной головне (Ustilago tritici) при искусственном заражении
f. duri и f. aestivi. Выявлена высокая расоспецифическая устойчивость
к патогену. Зародыши зараженных зерновок у большей части образцов
были иммунными, реже — слабочувствптельными. Мицелий гриба не
достигал зародышевой почки не только у Т. monococcum, но п у амфи-
диплоидов, полученных от скрещивания его с Т. durum и Т. persicum.
Зараженные зерновки продуцировали свободные от инфекции колосья.
Исследователи квалифицировали устойчивость Т. monococcum как
устойчивость типа Т. timopheevii.
Наличие значительного числа устойчивых к мучнистой росе форм
у Т. monococcum отмечено Н. И. Вавиловым (1964), П. М. Жуковским
(1957), М. М. Якубцинером (1969), В. Ф. Дорофеевым (1972). С. И. Ри-
гпна-Трайнипа (1972) сообщила о наличии у Т. monococcum внутри-
видовой дифференциации по устойчивости к мучнистой росе. Э. X. Су-
ханбердииа (1977) изучила 51 образец этого вида при искусственном
заражении проростков популяцией пыльной головни Ленинградской,
Куйбышевской, Новосибирской и Алма-Атинской областей, а 1,зкже
Северной Осетии. Кроме того, были обследованы взрослые растения
тех же образцов на провокационном фоне в ноле. Выявлено, что подав-
ляющая часть образцов была иммунной и высокоустойчивой к грибу
в обеих возрастных фазах.
Корневой гнилью растения этого вида поражаются меньше, чем ви-
ды секций Dicoccoides ц Triticum (И. Harding, 1972).
Таким образом, Т. monococcum в целом характеризуется как вы-
сокоустойчивый ' к грибным болезням вид пшеницы. II. II. Вавилов
(1964) отметил также устойчивость этого вида к зеленоглазке, гессен-
ской и шведской мухам.
История. Древнейшие находки обугленных .зерен однозернянки куль-
турной, поданным П. llelbaek (1966), отмечены в Иране (Курдистан).
Опп отнесены к 7500- 6700 гг. до и. э. В долине р. Иордан они дати-
руются VII—VIII тысячелетием до н. э. Археологические раскопки
выявили этот вид в Грециивозраст культурного слоя, к которому
они отнесены, —6000 -5000 гг. до и. э., Турции (Анатолия)—5800—
248 К.шссифитщи.ч /imhi Triticum !..
5000 11.,io !i. I*. 5000 5000 ii. v> п.э,, Bo. n арии 50’00 ii.
до и. j. (J. Л1. Renfrew, 1968). Очень дрсг.иш- неходки зафик< пропаны,
согласно сводке Z. Teinpir (I960), в Чехословакии (остаiкп неолити-
ческого периода). Эти данные свидетельсгнуют о значительных мас-
штабах возделывания однозернянки культурной в древности. Ныне она
почти целиком вытеснена из производства более урожайными видами
полиплоидной пшеницы.
Географическое распространение. Ареал однозернянки культурной
в прошлом был более широким, чем у дикой. В пределах ареала по
следпеп она возделывалась в Турции, Болгарии, IOiославии, Хлерблс
джапе (Нахичевань, Нагорный Карабах) и па Украине (Кры ) 6 у
того, она выращивалась на о.. Крит, в Италии, Албанш!. о- .
(куда предположительно попала из Испании), в АХ,чр • i 1
была сюда завезена из Франции), Австрии (Тироль), Швейцарии,
Швеции (о. Готланд), Дании, Чехословакии, Грузни, Дагестане и Че-
чено-Ингушетии.
Происхождение. В 1884 г. /М. W. Bcijerinck высказал предположение,
что Т. monococcum произошел от Т. boeoticum. A. Schulz (1913),
К. А. Фляксбергер (1935) и др. придерживались также этой точки зре-
ния. В этом же была убеждена Е. И. Синская (19-18, 1955), считавшая
Т. monococcum вторичной культурой, возделыванию которой предшест-
вовало сорняковое состояние. А. Д. Горгпдзе (1974) возникновение
Т. monococcum связывает с вымершей популяцией протомаха (2п =
= 14. 28, 42), из которой человеком была отобрана первоначальная
форма однозернянки культурной.
Н. Kuckuck (1959, 1970) предположил, что образование однозер-
нянки культурной из дикой сопровождалось генными мутациями во
многих локусах. На этих позициях стоит также М. Tanaka (1965).
В. Ф. Дорофеев (1969) считает, что с началом земледельческой куль-
туры создались благоприятные условия для мутаций, приведших к
микроэволюциопиым сдвигам и явившихся причиной возникновения
культурных однозернянок из диких.
L. Smith (1936) пришел к заключению, что ген, контролирующий
ломкость колоса у однозернянки культурной, был привнесен от Т. boe-
oticum. О близости между Т. monococcum и однозернянкой дикой
свидетельствует наличие у них сходного содержания ядериого ДИК
(Н. Rees, М. R. Walters, 1965), двух нар спутппчных хромосом
(G. N. Pathak, 1940), высокоплодовитого потомства от их скрещивания
друг с другом (А. А. Филатепко, 1968), нормального прохождения мей-
оза в МК.П (Н. Kihara е. а., 1929; J. Percival, 1932) и порциалыюго
взрывного характера открытия цветков при зацветании (В. И. Жуков,
1969). Кроме того, иммунохимическнй анализ растворимых белков об-
наружил в спектре однозернянки культурной почти все белковые ком-
поненты однозернянки дикой (В. Г. Конарев и др., 1970). Растительные
организмы с цитоплазмой 75 monococcum и ядром Т. boeoticum фер-
тильны и имеют нормальный рост (S. S. Маап, К- A. Lucken, 1967).
М. А. Федин (1974) считает эти виды изоплазматпчпыми.
Все перечисленное даст основание согласиться с мнением В. Часки
(1974), Н. Н,. Цвелева (1976) и ряда вышеназванных исследователей
о происхождении Т. monococcum от Т. boeoticum.
Дивергенция этих видов шла в первую очередь по линии потери
сильного одревеснения колосковой чешуи и элиминации еиоитаииоп
ломкости колосового стержня. В. Г. Конарев и др. (1970) свидетель-
ствуют, что в процессе эволюции усложнилась та часть наследственного
комплекса, которая контролирует синтез глиадина. И. Ясуо и др. (1969)
наблюдали наличие подобного усложнения по зимограммам эстераз.
Методом гибридологического анализа ДИК также показано, что у
Т. monococcum число повторов ДПК больше, чем у Т. boeoticum
Ilo<)i>oi) JI. IJoi-oliriun Miци sch I't Dorof. 299
(P. Ф. Ma x плева, C. JI. Iio'i ерев, 1973). Дпг.ер| eiiinia лих видов отра-
зилась также и изменении морфоло! нл двух хромосом (А. Н. Щапова,
1969), в приуроченноегв цветения диких однозернянок к предвечерне-
му времени, а ку.пътурпьгх - - к раппеутронному периоду суток
(В. И. Жуков, 1969).
Генетическое содержание геномов ликон и культурной однозерня-
нок, видимо, изменилось в такой степени, которая нренятс!вуег обиль-
ному образованию нолнвалеитов в мейозе. В результат о i оксаилоидпый
вид Т. zhttkovskyi, несущий 4 генома А1’, является высокофертильным
организмом. Этот признак характерен также для амфидинлоидов, по-
лученных Д. Костовым (1936) и Э. В. Тавриным (1963) от скрещива-
ния Т. monococcum с Т. timopheevii (один геном последнего, по совре-
менным воззрениям, произошел от Т. boeoticum).
Попытка воздействием мутагена (рентгеновские лучи) получить из
Т. boeoticum однозернянку культурную не увенчалась успехом: полу-
ченные мстанты не имели признаков пшеницы культурной (К. Yama-
shita, 1956).
Очаг возникновения Т. monococcum и место его первоначального
вхождения в культуру ,1. R. Harlan и IX Zoliary (1966) связыв.-iio.i
с Юго-Восточной Турцией.
Систематика. По Линнею Т. monococcum L. объединяет различные
формы однозернянки культурной. Его видовой лектотип, по свидетель-
ству S. Savage (1945), хранится в Линпеевском гербарии в Лондоне
нод номером 1044. Однозернянка культурная рассматривается как вид
F. Kornicke (1885), К. А. Фляксбергером (1935), II, II. Вавиловым
(1935, 1964), П. М. Жуковским (.1950, 1964), М. М. Якубципсром (1961),
В. Ф. Дорофеевым (1969) и др. в отличие от N. Seringc (1841) и
IO. А. Филипченко (1930), которые возвели диплоидную группу (куль-
турную н дикую однозернянки) в ранг рода. Мы также нс согласны
с W. М. Bowden (1959), который объединяет дикие п культурные дип-
лоидные виды пшеницы под названием Т. monococcum L. К диплоид-
ным видам пшениц он относит ряд диплоидных видов рода Aegilops L.
К. А. Фляксбергер (1935) делит культурные однозернянки на 3 груп-
пы, а Н. И. Вавилов (1961, 1964) —на 6 экологических групп.
Приводим описание трех групп* культурных однозернянок по
К. А. Фляксбергеру.
1. Heothinum Flaksb. 1935, Культ, фл. СССР, I: 359. Восточная груп-
па (или тип). Колосья сравнительно мелкие, колосковые чешуи но килю
и по главной боковой жилке с зубчиками; членики стержня большей
частью слабо опушенные, со слабо выраженными бородками у основа-
ния колосков. Растения некрупные, довольно позднеспелые.
Геогр. распр. Малая Азия, Закавказье, Крым.
2. Alemanum Flaksb. 1935, Культ, фл. СССР, 1: 359. Среднеевропей-
ская группа. Колосья сравнительно крупные; колосковые чешуи слабо
зазубренные; членики стержня почти голые. Растения сравнительна
крупные, позднеспелые (вегетационный период до 135 дней), яровые
формы (высеваемые с осени как озимые).
Геогр. распр. Южные горные районы ФРГ, северная Швей-
цария.
3. Ibericum Flaksb. 1935, Культ, фл. СССР, I: 359. Иберийская, или
испанская, группа. Колосья мелкие, растения некрупные, ранние (со-
зревают на 14 дней раньше группы alemanum).
Г е о г р. рас и р. Испания, Южная Франция, Марокко.
В группировке К- А. Фляксбергера не нашли отражения некоторые
формы однозернянок, возделывавшиеся в отдельных районах СССР
* Название proles, которое употребляют К. А. Фляксбергер и II. И. Вавилов, нс
фигурирует ныне среди принятых «Международным кодексом» таксонов.
300
Классификация рода Triticum L.
(например, па CeoepuoM Канюке) и на рубежных cipaii (Италия, Ал-
бапня). Кроме тою, некоторые i руины К. А. Фляксбергера елишком
укрупнены. 'Гак в ipyiniy heolhitiiiiu вошли наряду с закавказскими
также крымские формы. 11. II. Вавилов (19(H) приводит бо.чее дробное
деление иа экологические группы.
Согласно II. II. Вавилову, горная западноевропейская группа (топ-
tano-occidenlale-europaeinn Vav. 1964, ЛГиров. ресурсы хлебп. злаков.
Пшеница: 23) объединяет однозернянки Швейцарии (группа nletnu-
пит), а также Чехословакии п Югославии. Пам представляется целе-
сообразным отнести сюда формы Албании, Австрии (Тироль), Швеции.
II. И. Вавилов различает в составе этой группы ряд подгрупп во про-
должительности вегетационного периода, длине соломины и, что осо-
бенно важно, по образу жизни—яровые, полуозимые и озимые.
Средиземноморская группа (accident aletnedilerrcmeiim Vav. 1964.
l.c.: 23) включает наряду с однозернянками Испании п сопредельных
зон также однозернянки Италии. Характерные отличия группы — ши-
рокий короткий колос, прочная соломина с заметно утолщенными стен-
ками, средняя высота растений, наличие форм с. двузерными колосками,
устойчивость к видам ржавчины и головни, а также к повреждениям
гессенской и шведской мухами и зеленоглазкой, слабая восприимчи-
вость к мучнистой росе. В опытах ВИРа испанские формы были склон-
ны поражаться желтой ржавчиной в слабой, а иногда и в средней сте-
пени. В пределах этой группы Н. И. Вавилов рекомендовал выделить
подгруппы по крупности колоса и скороспелости.
Выделена группа крымская (lauricum Vav. 1964, l.c.: 23), формы
которой входили раньше в группу heolhititun Flaksb. Характерные от-
личия ее, по Н. И. Вавилову: низкорослость, скороспелость, мелкий
колос, топкая, упругая соломина, иммунитет практически ко всем за-
болеваниям.
Однозернянки Северного Кавказа составляют, по II. И. Вавилову,
2 особые группы — предгорнодагестанскую (daghestanicum Vav. 1964,
1. с.: 22) и северокавказскую (ciscaucasictim Vav. 1964, l.c.: 22). Пер-
вая отличается широкими листьями, высокой соломиной, удлиненным
колосом, сравнительной скороспелостью при посеве осенью и слабой
зимостойкостью. Вторая включает формы Чечено-Ингушетии — более
позднеспелые, сильно кустистые, с более длинным тонким колосом.
Восточную группу — heothinum Flaksb. —- II. И. Вавилов расчленил
на группы: нагорно-карабахскую (carabachicum Vav. 1964, I. с.: 22)
и армяно-анатолийскую (armenoanalolicuin Vav. 1964, l.c.: 22). Первая
встречалась в Нагорном Карабахе, где из-за своеобразных климатиче-
ских условий и сравнительно высокой культуры земледелия формирова-
лись растения с продуктивным колосом (нередко с двузерпыми колос-
ками), среднеспелые, болезнеустойчивые, по восприимчивые к твердой
головне.
В отличие от этих однозернянок армяно-анатолийская группа из
Горной Армении и Внутренней Анатолии (Турция) характеризуется
средним или мелким колосом, более мелкими листьями, средпеспело-
стыо.
В общем, же. резких морфологических различий между упомянуты-
ми группами этого древнего вида не обнаружено.
Классификация II. II. Вавилова (1964) более полно характеризует
видовое разнообразие и обособленность отдельных эколого-географи-
ческих групп по комплексу признаков. Современное разнообразие, поч-
ти исчезнувшего из производственных посевов вида Т. monacacciitn не-
велико. Вид представлен всего 14 разновидностями.
llcxlpoil ll. Htieoliciiin Migiisch. et Dnrnj.
301
Определение p.TiiioniiyiHoc i ей T. iiioiiocoeciiin I..
бел ые красные ешун черные белом на фоне красном Ос цвета чешуи г и черные Разнонидностъ № рззновид ности
Чешуи неопушенные
Чешуи шероховат ы е, бугорчатые, матовые
+ + — — — + + monococcum tauricum 1 9
— + — — -г — flavescens 4
— + — — — + pseudoflaves- cens 4
— — + 4 е hi у и I л а д к и с б л е с т sofianum 1 1Ц и c 5
+ — — — + — macedonicum G
-1- + — — 4- + pseudomacedo- nicutn vul puce 7 8
— -1- — — — -Г atriaristalum 9
— — + — — Д- symphacropoli- taniim 10
— — — ешуи onyt + ценные nigriciiltum 11
+ — — + — hohensteinii 12
—. + — — + — hornemannii 13
— 4- — — — + pseudohorne- mannii 1 1
Var. atriaristalum Flaksb. 1925, Тр. прикл. бот. сел. 15, 1: 224.—
Т. monococcum var. nigroatrum Flaksb. in Кобелев, 1932, Хлебн. злаки
Турции: 39. — Т. monococcum gr. atriaristalum Flaksb. 1935, l.c.:
365. (9).
Геогр. расп p. Франция, Болгария, Малая Азия.
Var. flavescens Koern. in Koern. et Worn. 1885, Ilandb. Getreideb. 1:
112. — T. monococcum gr. flavescens (Koern.) Flaksb. 1935, l.c.: 363. (3).
Геогр. распр. Малая Азия, Балканы, Швейцария, ФРГ, Фран-
ция. Испания, Марокко.
К. А. Фляксбергер выделил у gr. flavescens форму bigranum Flaksb.
(1935, Lc.: 363), известную во Франции под названием engrain double,
а в Испании — escana minor, esprilla, espelta comuna и характеризую-
щуюся развитием двух зерновок и второй укороченной ости в колосках
средней части колоса. Созревает приблизительно на 14 дней раньше
западноевропейских форм.
Var. hohensteinii Flaksb. 1929, l.c.: 477. — T. monococcum var. lata-
ricum Kovarsk. 1929, Tp. прикл. бот. ген. сел. 22, 2: 68. — Т. monococcum
var. ratschinicum Dekapr. et Menabde. 1929, Зап. Науч.-прикл. отд. Тифл.
бот. сада, 6: 236. — Т. monococcum gr. albohornemannii f. hohenslenii
Flaksb. et f. ratschinicum (Dekapr. et Menabde) Flaksb. 1935. 1. c.: 366,
367. (12).
Геогр. распр. ФРГ, Грузия, Украина (Крым).
Var. hornemannii (Clem.) Koern. in Koern. et Worn, 1885, Ilandb.
Getreideb. 1: 112. — T. hornemannii Clem, in Herrera, 1818, Agric. gen.
I: 3,-—T. pubcscens Bicb. 1800, Beschr. Land. Casp. Месте: 81. — T. mo-
nococctim var. pubcscens (Bicb.) Koern. 1877, Nath, mid Thiel, Landw.
Jahrb. 6: 1044; Koern. in Koern. et Wern. 1885, Ilandb. Getreideb. 1:
112. — T. monococcum var. sangesuri Thum. 1933, Опред. хлебн. злаков
(колос.): 217. — Т. monococcum gr. hornemannii (Clem.) Flaksb. 1935,
I. c.: 367. (13).
302
Классификация рода Trilicum 1..
Геогр. распр. Испания, ФРГ, Швейцария, Австрия. Балкан-
ский полуостров, Кавказ, Крым.
У разновидности hornemannii К. Л. Фляксбергер установил 2 фор-
мы: 1) f. capiiata Flaksb. 1935, l.c.: 368, descr. ross. — форма с булаво-
видными колосьями, выделенная в Weihenslcphan (Бавария) под на-
званием WW38; 2) f. sangesuri (Thum.) Flaksb. 1935, l.c.: 368. — Г. mo-
nococcum var. sangesuri Thum. 1934, l.c.: 217 -форма с черными но
краям цветковыми чешуями.
Var. macedonicum Papag. 1919, Descr. hies Greqncs: 124. T. mo-
nococcum var. eredviantim Zhuk. 1924, Зан. Науч.-прикл. отд. Тифл. бот.
сада, 3: 6, р.р. — Т. monococcum var. eredviantim f. punctata Stransk.
1929, l.c.: 210. — T. monococcum gr. macedonicum f. eredviantim (Zhuk.)
Flaksb. 1935, l.c.: 362 et f. punctatum (Stransk.) Flaksb. 1935, l.c.:
362. (6).
Геогр. распр. Балканский полуостров, Малая Азия, Грузия, Ук-
раина (Крым).
Var. monococcum. — Т. monococcum var. laetissimum Koern. in Koern.
et Wern. 1885, Handb. Getreideb. 1: 113. (1).
Геогр. распр. Малая Азия. Болгария, Югославия, Испания.
Франция, Марокко.
Var. nigricultum Flaksb. 1916, Тр. Бюро прикл. бот. 9. 2: 68.—
Т. monococcum gr. nigricultum Flaksb. 1935, l.c.: 366. (11).
Геогр. распр. Малая Азия, Балканский полуостров. Италия,
ФРГ, Крым.
Var. pseudoflavescens Flaksb. var. nov. — T. monococcum gr. pseudo-
flavescens Flaksb. 1935, l.c.: 364, descr. ross. A T. monococca var. fla-
vescens aristis nigris differt. (4).
Typus: Hispania, 1926, k-20498, leg. N. I. Vavilov (WIR).
Геогр. распр. ФРГ, Испания, Югославия, Малая Азия.
Var. pseudohornemannii Dekapr. et Menabde, 1932, Tp. прикл. бот.
ген. сел. 5, 1: 11. (14).
Геогр. распр. Грузия, Украина (Крым), ФРГ.
Var. pseudomacedonicum Flaksb. var. nov. — T. monococcum var.
pseudomacedonicum Flaksb. 1935, l.c.: 362, descr. ross. — T. monococcum
var. psetidoeredvianum Zhuk, in Flaksb. 1935, l.c.: 362, nom. mid.—
T. monococcum var. macedonicum f. psetidoeredvianum (Zhuk.) Flaksb.
1939, Опред. паст, хлебов.: 33, descr. ross.—Л T. monococca var. mace-
donicum aristis nigris differt. (7).
Typus: Asia Media, 1927, k-27171, leg. P. Zhukovsky (WIR).
Геогр. распр. Турция, Болгария, Украина (Крым).
Var. sofianum Stransk. 1929, За Лимеца в България: 208. — Т. mo-
nococcum gr. sofianum (Stransk.) Flaksb. 1935, l.c.: 366. (5).
По И. Странски (1929) членики стержня по ребрам густо опушены
короткими волосками. У var. sofianum он различает 2 формы: 1) f. den-
siuscula Stransk. 1929, l.c.: 209 — колосья узкие, короткие, ости парал-
лельны колосу, шероховатость на чешуях слабая; 2) f. laxitiscula
Stransk. 1929, l.c.: 209 — колосья широкие, рыхлые, остп несколько
расходящиеся.
Геогр., распр. Болгария.
Var. symphaeropolitanum Drosd. 1922—1923, Тр. прикл. бот. сел. 13,
1: 520. (10).
И. Странски отмечает для Болгарии 2 формы этой разновидности:
1) f. parallelica Stransk. 1929, Lc.: 218—с параллельными колосу остя-
ми и 2) f. patula Stransk. 1929, l.c.: 218 — с расходящимися остями.
Геогр. распр. Малая Азия, Балканский и Крымский полуост-
рова.
Var. tauricum Drosd. 1922—1923, Тр. прикл. бот. сел. 13, 1: 520.—
Т. monococcum gr. tauricum (Drosd.) Flaksb. 1935, l.c.: 360. (2).
Подрод II. Boeoticum Migusch. et Dorof.
303
Геогр. распр. .Малая Азия, Крымский и 1 >а.1 к;i 1 ick 11 й iio.'iytic 1 -
рона, Италия, 1 киаиля, Марокко.
Var. vulgare Koern. in Koern. et Wern. 1885, Handb. Getrcideb. 1:
112. — T. monococcum. var. pseudovulgare Flaksb. 1922, Опред. паст,
хлебов, изд. 2: 20. — Т. monococcum gr. vulgare (Koern.) Flaksb. 1935,
1. c.: 364. (8).
Геогр. распр. Малая Азия, Крымский и Балканский полуост-
рова, Австрия, ФРГ, Франция, Испания, Закавказье.
Т. SINSKAJAE A. FILAT. ET KURK. -ПШЕНИЦА СИНСКОЙ
Т. sinskajae A. Filat, et Kurk. 1975, Тр. прикл. бот. геи. сел. 54,
1:239.
Всходы темпо-зеленые или фиолетовые. Кустистость высокая. Кус1'
прямостоячий пли иолустелющийся. Соломина при осеннем посеве и
орошении довольно высокая (120 см), тонкостенная, полая, под коло-
сом выполненная, без воскового налета. Узлы соломины густо опушен-
ные, под ними — бурый пояс. Листья взрослых растений гладкие или
слабошероховатые, узкие, без воскового налета. Листовое влагалище
опушенное, голое или шероховатое. Язычок (лигула) короткий, каем-
чатый. Колосья безостые (см. рис. 22,6), длиной 5—6 см, плоские (бо-
ковая сторона их значительно шире лицевой). Колос плотный (D —
= 55). Ломкость колосового стержня (при надавливании на пего)
выражена слабее, чем у Т. monococcum. Колоски однозерные, удлинен-
но-овальные. Колосковые чешуи длинные (10—13 мм), почти равные
по длине цветковым чешуям. Киль колосковой чешуи узкий, резко вы-
раженный до основания чешуи и покрытый короткими щетинками. Ки-
левой зубец в верхней части колоса длиной до 7 мм. Внутренняя цвет-
ковая чешуя при созревании расщепляется вдоль на 2 части. Пыльники
желтые. Зерновки красные, сплюснутые по бокам, длиной 7—9 мм,
имеют легкий вымолот. Важнейшие морфологические отличия от
Т. monococcum — голозерность и безостость колоса, меньшая его дли-
на, более нежная консистенция колосковых чешуи, большая плотность
колоса, наличие длинного остевидпого килевого зубца.
Строение перикарпия зерновки сходно с таковым у Т. boeoticum и
Т. monococcum. Отличительная особенность — наличие в стенках попе-
речных клеток спиипой стороны зерновки мелких пор (Л. Л. Жестян-
никова,1976).
Биологические признаки пшеницы Сииской и культурной однозер-
нянки сходные. По данным авторов вида, при естественном заражения
в условиях прибрежной полосы Дагестана он был иммунен к бурой,
желтой и стеблевой ржавчине, мучнистой росе, пыльной и твердой
головне. Э. X. Сухапбердипа (1977) подтвердила эти данные в отноше-
нии мучнистой росы как при полевой оценке на провокационном фоне,
так и при заражении проростков сборным ииокулюмом из смеси попу-
ляций гриба Ленинградской, Куйбышевской, Новосибирской, Алма-
Атинской областей и Северной Осетии.
Пшеница Сииской возникла в результате спонтанной мутации у рас-
тений Т. monococcum — образец к-20970, интродуцированный из Тур-
ции. Зерновки -этого вида были обнаружены в репродукции опытных
станций ВИРа — Среднеазиатской (урожаи 1963—1967 гг.) и Дагестан-
ской (урожаи 1963—1970 гг.). Возможно, что и на родине исходного
образца в северных причерноморских районах Турции обитают подоб-
ные мутанты.
304
Классификация рода Triticum L.
СЕКЦИЯ 5. TIMOPHEEVII Л. EILAT. ET DOROF.
Sect. 5. Timopheevii A. Filat, ct Dorof. 1978, Tp. прикл. бот. геи. сел.
62, 2: 151. — Triticum sect. Dicoccoidea subscct. Transcaucasica Sinsk.
1955, Пробл. бот. 2: 55, nom. illeg. — Triticum sect. Transcaucasicum
(Sinsk.) Dorof. 1969, в кп.: Вавилов и с.-х. паука: 282, nom. illeg. —
Triticum sect. Andropogon Jakubz. 1962, Symp. gen. and wheat breed.
Martonvasar: 407, 409, nom. illeg. — Triticum sect. Orthalherum Nevski,
1934, Фл. СССР, 2: 681. quoad T. timopheevii. — Triticum congregatio
Tetraploidea Flaksb. 1935, Культ, фл. СССР, 1: 31, quoad T. timophee-
vii.— Triticum sect. Triticum-. Цвелев, 1973, Новости спет. высш. расг.
10: 40, quoad Т. aggr. timopheevii (Zhuk.) Zhuk., sensu Tzvel.; он же, 1976,
Злаки СССР: 164, quoad Т. aggr. timopheevii (Zhuk.) Zhuk., sensu Tzvel.
Листовая пластинка взрослых растений всегда щетинистоопушен-
иая. Колосья пирамидальные, плоские, в зрелом состоянии спонтанно
распадаются па колоски (дикорастущие формы) пли ломаются лишь
при надавливании (культурные); колоски с 3—4 цветками, из которых
2 плодущие. Наружная цветковая чешуя на 1-1,5 мм короче внутрен-
ней. Последняя при созревании не расщепляется вдоль на 2 части.
2п — 28. Геномные формулы AbAbGG, AbAbAbAbGG.
Т и п: Т. araraticum Jakubz.
Геогр. распр. Закавказье, Ирак, Иран, Турция.
Т. ARARATICUM JAKUBZ. — ПОЛБА ДИКАЯ АРАРАТСКАЯ
Общая характеристика. Всходы слабо-фиолетовые пли зеленые, опу-
шенные. Растение с восковым налетом, сизо-зеленое. Стебель сравни-
тельно топкий, 60—120 см высоты, без фиолетовой окраски; слабая
фиолетовая окраска наблюдается на узлах соломины и на ушках листь-
ев. Узлы слабо или густо опушенные. Листовая пластинка узкая, ли-
нейная или линейно-ланцетная, щетиннстоопушепная (с нижней поверх-
ности сильнее). Листовое влагалище щетиннстоопушенпое. Колосья
остистые, длиной 5—10 см, плоские (боковая сторона 0,7—1,5 см, ли-
цевая 0,3—0,5 см). Ости 12—15 см длины. Колосовой стержень очень,
ломкий (рис. 23). Членики стержня негусто опушенные, с короткими
белыми бородками. Зерновки красные, 8—11 мм длины, с коротким
хохолком.
В колеоптиле Т. araraticum имеется 2 проводящих мучка. Устыща
длиной 39—51 мкм (короче, чем у Т. dicoccoides). Характерна сильная
склернфикация листа в области жилок и по краям. На эпидермисе
верхней поверхности листа'расположены (только в области жилок)
длинные (до 400 мкм) волоски, па нижней — по всей поверхности. Ли-
стовая пластинка весьма топкая— до 236,6 мкм (О. Д. Градчаннпова,
1967). Среди длинных клеток эпидермиса нижней стороны листа до-
вольно часто встречаются короткие клетки изодпаметрической формы,
над жилками обычно подобные клетки-выросты не образуются
(Ю. И. Берхин, 1963). Ткани перикарпия зерновки резко редуцирова-
ны. Эпи-мезокарпий состоит из одного слоя тангенталыю удлиненных
клеток с тонкими стенками. В отличие от однозернянок в стенках по-
перечных клеток полбы араратской хорошо выражены поры.
Т. araraticum — пшеница горных степей. Произрастает па сухих
горных склонах на высоте 1200—1600 м в Армении, спускается значи-
тельно ниже (300—700 м над ур. м.) в Азербайджане.
Полба дикая араратская Закавказья имеет формы средне- и поздне-
спелые, со средней требовательностью к теплу н невысокой зимостой-
костью. Морозостойкость ее выше, чем у полбы дикой Сирии и Израи-
ля (Г. В. Удовенко и др., 1978). Засухоустойчивость высокая, зноевы-
носливость средняя.
//)()po<) U. util Mipuscll. et /)<uo[. ?,U”)
Рис. 23. Колосья (co спонтанной ломкостью) и зерновки дикой полбы Г. arara-
ticum Jakubz.
я —sttb'-p. <iraraticum\ 6—subsp. kuriPstaiutiim Doiof. ct Mii’nsch.
В низменном Дагестане у Т. и rural icn tn цветение начинается в ко-
лосках верхней части колоса и зачем распространяется ниже. В таком
же порядке происходит спонтанное распадение колоса при его созре-
вании. У этого вида наблюдается один резко выраженный максимум
>/420 па
.’!()(> Клисгпфикиццч роди Tt Uii uni I..
цветения в 6 7 ч утра при температуре воздуха I'.) 2ГС, За ) ч oi-
ивсгает 43 --83% цветков (В. И.Жуков, 1969).
Концентрация белка в зерне образцов коллекции в условиях оро-
шаемого .земледелия установлена в пределах 21,8---30,6% (Ташкент-
ская область) и 20.9—26,9% (Дагестан), лизина в белке соответствен-
но 1,54—1,82 и 1,76—2,17%; в зерне образцов из Ирака 23,4—31,6%
белка и 1,62—2,05% лизина в нем (В. Г. Коиарев и др., 1972).
Полба дикая араратская имеет формы, которые не скрещиваются
(тип I) или скрещиваются (тип II) с пшеницей Тимофеева, давая в 1%
стерильное или фертильное потомство. При гибридизации типов I и
II между собой растения F] фертильные. В природе растения этих ти-
пов обитают обычно в единой популяции (М. Tanaka, S. Ichikawa, 1968,
1972; Э. Ф. Мигушова, А. В. Коиарев, 1975).
Иммунитет. М. М. Якубцинер (1932, 1957, 1969) характеризует
Т. araraticum как вид, низко- и среднеустойчивый к грибным болезням
и поражаемый всеми видами ржавчины. Последнее отметил также
П. А. Гапдилян (1973). По данным И. П. Шитовой (1968), в условиях
Дагестана в 1954—1965 гг. максимальное поражение бурой ржавчиной
в среднем было равно 4 баллам (но 5-бал,тьной шкале) у форм из Пра-
ка и 1,7 балла у форм из Закавказья. В условиях Ташкентской и Ле-
нинградской областей отмечено сильное поражение дикой араратской
полбы бурой и желтой ржавчиной («Пшеницы мира», 1976). Макси-
мальное поражение стеблевой ржавчиной за 12 лет наблюдений в усло-
виях Дагестана в среднем было равно 2,5 балла (И. II. Шитова, 1968).
При искусственном заражении проростков сборным ипокулюмом из
смеси популяций мучнистой росы из разных географических районов
СССР Э. X. Сухапбердипа (1977) нашла, что 78,9% образцов коллек-
ции Т. araraticum иммунны к патогену, 21,1% —устойчивы (1—2 балла).
У взрослых растений при естественном заражении па провокационном
фоне в поле эти показатели соответственно были равны 83,9 и 16,1%.
По данным А. М. Ямалеева (1974), при искусственном заражении у
Т. araraticum типа I из Закавказья и образцов иракского происхожде-
ния выявлен иммунитет к специализированным расам пыльной головни
на всех фазах роста растений. Т. araraticum типа II из Закавказья си.ть-
повоспрннмчпв к патогену в эмбриональной стадии. Причем мицелий
гриба обнаружен во всех частях зародышей, даже в их почках. Мице-
лий пыльной головни f. duri поражал до 77%, a f. aestivi до 88% заро-
дышевых почек. Несмотря на это, взрослые растения не имели поражен-
ных колосьев. А. М. Ямалеев пришел к заключению, что устойчивость
Т. araraticum к пыльной головне сходна с устойчивостью пшеницы Ти-
мофеева. В. И. Кривченко и др. (1976) отнесли Т. araraticum к числу ви-
дов, обладающих высокой возрастной расоспецифической устойчивос-
тью к этому патогену, а характер развития гифов гриба в эмбрионе
отнесен к неспецифическому поражению.
Географическое распространение. Полба дикая араратская была от-
крыта М. Г. Туманяном в 1925—1930 гг. близ г. Ереван па высоте 1200 -
1500 м над ур. м. В 1931 г. М. М. Якубцинер обнаружил ее на террито-
рии Азербайджана (Нахичевань—между селениями Азнабюрт и Папе)
на высоте 1300—1400 м над ур. м. В 1949 г. он открыл ее обитание
в Ахсуинском районе Азербайджана, а позднее И. Д. Мустафаев (1973)
нашел заросли этого вида в Шемахииском районе на высоте 600—700 м
над ур. м. и в Нагорном Карабахе на высоте 410 м. В Армении Б. Гара-
сефсряи (1939) обнаружил крупные массивы Т. araraticum в Ехегиадзор-
ском (на высоте 1200 -1300 м), а также в Араратском районах.
Ареал полбы дикой араратской нс ограничивав!ся горной Армени-
ей, Нахичеванью и Ахсуппско-Шемахппским нагорьем па северо-вос-
токе Азербайджана. Она произрастает также в Ираке, Иране и Турции.
Сходство дикой полбы, найденной Т. Strauss в 1910 г. (по J. Percival,
11. Hocohi tun <‘t Dotoj 307
I 921 ), c .'{JiK.'iHiGi.'tcKiiMii oTM(i4ciH) AV AV Я к \ бп и пером (1432) и
К. Л. < 1>л я к<< >г J) । ером ( 1935). Внос.je,i.c । нии было показано, но среди
диких полб Ирики, Ирана и Турции есть формы с геномным сосаном
AhG, г. е. относящиеся к виду Т. araraticum (L. Sachs, 1953; L. В. Wa-
цепааг, 1961; J. R. Harlan, 1). Zohari, 196(5; В. L. Johnson, 1967; Л. В. Ко-
нарев и др., 1971, 1972; В. Яаска, 1971; S. S. Маап, К. A. Lucken, 1973;
М. Tanaka, Н. Ishii, 1973; И. Д. Мустафаев, 1975).
Происхождение. Т. ага rat iciini в Закавказье почти всегда обитает в
сообществе с Т. boeoticum, с которым сходен но ряду признаков, в част-
ности щетинистому опушению листьев. .Эти факты вызвали к жизни тео-
рию автополиплопдпого происхождения полбы дикой араратской из
Т. boeoticum, которую ныне поддерживают Л. А. Филатеико (1968),
В. Л. Мепабде (1971) и А. Д. Горгидзе (1973). В. Ф. Дорофеев (1969),
пе отрицая возможности авгоиолиплоидпого пути ее возникнове-
ния, отдает предпочтение гибридизационному пути ее появления. По
его мнению, при этом скрестились диплоидная пшеница и Ле. speltoides.
А. В. Конарев и др. (1971, 1974), Т. II. Ненова (1973, 1971) и В. Г. Ко-
варен и др. (1976) методом иммупохимическо!о анализа глиадпнов зер-
новки установили, что донором первого генома полбы дикой араратской
был Т. boeoticum. К такому же выводу пришел В. Яаска (1974) при
электрофоретическом обследовании изофосфатаз и изоэстераз про-
ростков.
Исследования А. В. Конарева и др. (1971) также показали, что ан-
тиглпадиновая сыворотка Ае. speltoides интенсивно проявляет в глиади-
пе полбы дикой араратской почти все компоненты, характерные для
Ае. speltoides, в том числе главный компонент, специфичный только для
этого вида эгилопса и отсутствующий у однозернянок. Т. И. Пенена
(1973, 1974), истощив сыворотку полбы дикой араратской глиадинами
Т. boeoticum, получила в реакции преципитации с глиадшюм Т. arara-
ticum 3 слабых компонента, специфичных для Ае. speltoides, в том числе
главный компонент, имеющийся только у этого вида эгилопса и отсут-
ствующий у диплоидных видов пшеницы. Перечисленное свидетельству-
ет о том, что Т. araraticum имеет 2 разнокачественных генома, донором
одного пз которых был Ac. speltoides. В. Яаска (1974) также свидетель-
ствует о том, что некоторые образцы Ае. speltoides по электрофорети-
ческой подвижности изофосфатаз и изоэстераз проростков соответству-
ют одному из геномов полбы дикой араратской.
В запасных белках зерновок Т. araraticum пммупохи.мический ана-
лиз не выявил специфических компонентов Т. urartu. Поэтому гипотеза
Н. S. Dhaliwal и В. L. Johnson (1976) о происхождении полбы дикой
араратской с участием Т. urartu нам представляется необоснованной,
как и предположение L. Sachs (1953) о возникновении полбы дикой
Ирака, относящейся к Т. araraticum, при скрещивании между полбой
дикой с геномом В и Т. timopheevii.
Дикие полбы Закавказья и Ирака, имеющие геномный состав AbG,
генетически родственны. В Северном Ираке и прилегающих районах
Турции и Ирана эти полбы имеют ряд доминантных признаков (черный
цвет и опушенность чешуй колоска, наличие биотипов с яровым образом
жизни), отсутствующих у полбы Закавказья. М. Tanaka и S. Ichikawa
(1972) полагают, что наиболее вероятным очагом возникновения полб с
геномом G является территория, охватывающая Восточную Турцию,
Северный Ирак и Закавказье. По нашему мнению, последний географи-
ческий район пе может быть местом происхождения Т. araraticum из-за
отсутствия в нем Ле. speltoides. Сюда эта полба попала при миграции
из центра своего происхождения.
Систематика. ДА. Г. Туманян (1930) первым обратил внимание па
морфологические отличия диких полб Армении от сирийско-палестин-
ских, выделив первые в особую секцию — армянскую, а М. М. Якубцп-
'/< + 20 Ж2
308 Классификация /юра Triticum К.
пер (1932) (>бособ11/I ее в подвид Т. dicoccoides snbsp. iirmeuiaciim
Jakubz.., куда отнес и иранские формы, собранные 'Г. Strauss.
В свете учения II. И. Вавилова (1931) неприемлемы позиции пекою-
рых генетиков, относящих полбу дикую араратскую в качестве подвида
к Т. turgidum (W. М. Bowden, 1959; Л. et L). Love, 1961) пли к Т. timo-
pheevii (J. Mac Key, 1966; R. Morris et Е. R. Sears, 1967). Такой подход
означает в сущности обратный ход эволюции. Пшешша Тимофеева име-
ет ряд морфологических черт, общих с Т. araraticum, и близка с ним ге-
нетически. Т. araraticum рассматривается ныне как родоначальник
7'. timopheevii.
Подробный анализ оригинального материала из Ирака и выявление
генетической близости этих форм к закавказским образцам дали осно-
вание для включения их в состав вида Т. araraticum в ранге подвида.
Т. araraticum в свете новых данных представлен двумя подвидами:
subsp. araraticum и subsp. kurdistanicum Dorof. et Migusch.
Ключ для определения подвидов
Т. araraticum Jakubz.
1. Растения 60-120 см высотой; колосья 6-10 см длиной, в колосе
12—18 развитых колосков, ости сравнительно тонкие; колоски 14—
20 мм длиной и 4—5 мм шириной; колосковые чешуи неопушенные. Ки-
левой зубец 1 —1,5 мм длиной, притупленный (редко-—заостренный),
между ним и зубцом главной боковой жилки остроугольная, выемка час-
то отсутствует. Плечо колосковой чешуи прямое . . subsp. araraticum.
+ Растения 60—100 см высотой (при орошении — до 130 см); коло-
сья 5—8 см длиной, в колосе 11 —13 развитых колосков, ости грубые;
колоски 16—25 мм длиной и 6—7 мм шириной; колосковые чешуи опу-
шенные или неопушенные. Килевой зубец 1—4 мм длиной, заостренный
или притупленный, между ним и зубцом главной боковой жилки — остро-
угольная выемка. Плечо колосковой чешуи приподнятое.................
.............................subsp. kurdistanicum Dorof. et Migusch.
I. Subsp. araraticum — подвид араратский.
Subsp. araraticum. — T. araraticum Jakubz. 1947, Сел. и сем. 5: 46;
Якубц. 1948, Тр. прикл. бот. ген. сел. 28, 1 : 218; Якубц. 1950, Бот. жури.
СССР, 35,2: 191. — Т. dicoccoides subsp. armeniacum Jakubz. 1932. Гр.
прикл. бот. ген. сел. сер. 5, 1 : 164; idem, 1932, Тр. прикл. бот. ген. сел.
прилож. 53:160. — Т. armeniacum (Jakubz.) Makush. 1938, Докл. АН
СССР, Нов. сер. 21,7: 350, non Nevski, 1934, пес Thum, in Flaksb. 1939. —
Т. montanum Makush. 1948, Учен. зап. Лешшгр. под. ин-та, 66, Бот. 8: 138,
nom. illeg. — Т. chaldicum Menabde, 1948, Пшен. Груз.: 196. — Т. turgi-
dum var. tumanianii (Jakubz.) Bowden, 1959, Canad. Journ. Bot. 37:
670. — 7'. turgidum subsp. armeniacum (Jakubz.) A. et D. Love, 1961, Bot.
Not. (Lund) 114, 1:50. — T. timopheevii subsp. araraticum (Jakubz.) Mac
Key, 1966, Hereditas, Suppl. 2: 267, nom illeg. — T. nikolai An. Fed. et
Takht. ex Zhuk. 1968, Бот. жури. 53, 4: 442, nom. nud.
Геогр. распр. Армения, Азербайджан, Ирак, Иран, Турция.
Var. araraticum.— Т. dicoccoides f. straussiana Schulz, 1913. Bericht.
Deutsch. Botan. Gesellsch. 31, 4:229. — 7’. dicoccoides var. straussianum
(Schulz) Thum. 1929—1930, Tp. прикл. бот. ген. сел. 24,2: 8; Туманян,
1934, Тр. прикл. бот. ген. сел. 5, 2: 255. (6).
Геогр. распр. Армения, Азербайджан (включая Нахичевань),
Ирак, Иран, Турция.
Var. araxicum (Thum.) Jakubz. comb. nov. — T. dicoccoides var. araxi-
num Thum. 1934, l.c.: 255. — T. dicoccoides var. nachitschevanicum f.
araxicum (Thum.) Flaksb. 1935, Культ, фл. СССР, 1: 335. (3).
Геогр. распр. Армения, Азербайджан (включая Нахичевань).
Ilnilpocl II. Iloeoticnm Л!йЦ|ч< h. el Dnroj.
Определение p.Tiiioinwiocicii T. ar;ir;itiviiui subsp. araraticum
1 ешун Остн I'a'HfwBii топь ра.нонн нос г и
болы о кр;ц-ные черные ОС Ю.М r;t фоне красном ине i a чешуи McpHi ie
Чешуи / i (4 ш jч in e 11 aide d.uu): tie
-I- + — — — j .j. llminnniimii pscndoai тема- 1 2
— .— — — cum araxicum 3
— -1- — — — j- psciiilonruxicum 4
— — J. — + nigricans 5
+ — Чешуи z tepoxoeciThic, + бугор1/ 1ТЫС arnrniicnm C>
— — — — -I- pseudoslraiissi- i
— + — — + - nnnm nacliitschecani- 8
— 4- — — — 4- cum psetiilouiiron- 9
— — — + — + sohnii nigrum 10
Var. nachitschevanicum (Jakubz.) Jakubz. comb. nov. — T. dicoccoides
var. tiachiischevanicum Jakubz. 1932, Tp. прикл. бог. ген. сел. 5, 1: 163. --
Т. dicoccoides var. aaronsohnii auct. non Flaksb.: Туманян, 1929—1930,
1. c.: 8; on же, 1934, 1. c.: 255. (8).
Геогр. распр. Армения, Азербайджан (включая Нахичевань).
Var. nigricans (Thum.) Jakubz. comb. nov. — T. dicoccoides var. nigri-
cans Thum. 1929—1930, 1. c.: 8; Туманян, 1934, l.c.: 255. — T. dicoccoides
var. nigrum f. nigricans (Thum.) Flaksb. 1935, 1. c.: 335. (5).
Геогр. распр. Армения (экспедиции ВИРа в 1961 —1976 гг. та-
кую форму не обнаружили).
Var. nigrum (Thum.) Jakubz. comb. nov. — T. dicoccoides var. nigrum
Thum. 1934. l.c.: 255.— T. dicoccoides var. spontaneonigrum auct. non
Flaksb.: Туманян, 1929—1930, l.c.: 8. (10).
Геогр. распр. Армения (экспедиции ВИРа в 1961 —1976 гг. та-
кую форму не обнаружили).
Var. pseudoaaronsohnii (Thum.) Jakubz. comb. nov. — T. dicoccoides
var. pseudoaaronsohnii Thum. 1929^-1930, l.c.: 8. Туманян, 1934, l.c.:
255. — T. dicoccoides var.- pseudonachitschevanicum Flaksb. 1935. l.c.:
335, descr. ross. (9).
Геогр. распр. Армения.
Var. pseudoaraxicum (Thum.) Jakubz. comb. nov. — T. dicoccoides var.
pseudoaraxicum Thum. 1934, l.c.: 255. — T. dicoccoides var. pseudonachi-
tschevanicum f. pseudoaraxicum. (Thum). Flaksb. 1935, l.c.: 335. (4).
Геогр. распр. Армения.
Var. pseudoarmeniacum (Thum.) Jakubz. comb. nov. — T. dicoccoides
var. pseudoarmeniacum Thum. 1934, l.c.: 255. — T. dicoccoides var. pseu-
dothumanianii f. pseudoarmeniacum (Thum.) Flaksb. 1935, l.c.: 335. (2).
Геогр. распр. Армения.
Var. pseudostraussianum (Thum.) Jakubz. comb. nov. — T. dicoccoi-
des var. pseudostraussianum Thum. 1929—1930, 1. c.: 8; Туманян, 1934,
1. c.: 255. — T. dicoccoides var. pseudo! Iiumanianii Flaksb. 1935, l.c.: 334,
descr. ross. p. p. (7).
Геогр. распр. Армения, Азербайджан (включая Нахичевань).
Var. ihumanianii (Jakubz.) Jakubz. comb. nov. — T. dicoccoides var.
thumanianii Jakubz. 1932, l.c.: 169. — T. dicoccoides var. artneniacuni
Thum. 1934, l.c.: 255. — T. turgidum var. Ihumanianii (Jakubz.) Bowden,
1959, Canad. Journ. Bot. 37: 672. (1).
310 К лас спфи л а ци ч Ti i/i< нт I..
Ген i p. распр. Армения. Л’.грбзй.'Ы,ап ( включая I la11 ч < • ii i 11.),
lip ак.
II. Subsp. kurdistaniciiin Dorof. el Migusch. - под.г.и i \\ рдпс'гашчшii.
Subsp. kurdistanicum Dorof. el Migiis< 11. 1977, Б10.1. ВИ Pa, 72: 76.
Растения (>()-—100 см высотой, длина колотьен 5 -IS см, колосковые
чешуи опушенные пли неопушенные. Килевой зубец острый или тупой,
прямой пли несколько отогнут назад, короткий пли средней длины (1---
4 мм). Зубец главной боковой жилки слабо развит (не выше 1 мм),
между этими зубцами—остроугольная выемка.
Геогр. р а с н р. Прак, прилегающие районы Ирана и Турции.
Определение разновидностей subsp. kurdistanicum Dorof. et Migusch.
’ I сш ум Oci н Разновидности У- ра (повил- И’ ги
белые красные черные на фоне цвета чешуи черные
белом красном
+ . Чешуи неопушенные - + kurdistanicum 1
Чешуи опушенные
+ 1 - 1 - 1 - 1 + 1 - I nabelicum
- 1 - 1 + 1 - 1 - 1 + 1 hirtiglumis 1 ~
Var. hirtiglumis (Nabel.) Migusch. el Dorof. 1977, Бюл. ВИРа, 72:
76. — T. dicoccoides f. hirtiglumis Nabel. 1929, Publ. Fac. Masaryk Univ.
Brno, 111: 27. (3).
Геогр. распр. Ирак.
Var. kurdistanicum. — T. dicoccoides f. nudiglumis Nabel. 1929, l.c.:
27. — T. timopheevii auct. non Zhuk.: Morris et Sears, 1967, Wheat and
wheat improvem.: 34, quoad T. dicoccoides var. nudiglumis. (1).
Геогр, распр. Ирак, Иран, Турция.
Var. nabelicum Migusch. et Dorof. 1977, Бюл. ВИРа, 72: 76. (2).
Геогр. распр. Ирак.
Т. TIMOPHEEVII (ZHUK.) ZHUK — ПШЕНИЦА ТИМОФЕЕВА
Т. timopheevii (Zhuk.) Zhuk. 1928, Тр. прикл. бот. ген. сел. 19,2: 64; Нев-
ский, 1934, Фл. СССР, 2: 682. — Т. dicoccum dicoccoides var. timopheevii
Zhuk. 1923, Зап. Науч.-прикл. отд. Тифл. бот. сада, 3: 1; Столетова, 1925,
Тр. прикл. бот. сел. 14,1:42. — Т. turgidum var. timopheevii (Zhuk.) Bow-
den, 1959, Canad, Journ. Bot. 37: 67L— T. turgidum subsp. timopheevii
(Zhuk.) A. et D. Love, 1961, Bot. Not. (Lund), 114, 1: 50.
Описал из Грузии (Карталпппя, близ села Азарми).
Общая характеристика. Всходы опушенные. Листовые пластинка и
влагалище разреженно опушены щетинистыми волосками длиной в
1,5—2 мм. Длина листовой пластинки приблизительно 20 см, ширина —
1 см. Язычок,и ушки бесцветные; ушки с длинными ресничками. Соло-
мина длиной до 140 см, полая, тонкостенная, ее узлы щетииистоопушеп-
пые. Колосья остистые (рис. 24, о), плотные (D = 30—54), длиной 4—
7 см, плоские (ширина их боковой! стороны до 2 см). Стержень колоса
при надавливании ломается. Тип его ломкости как у полб. Созрев-,
шпй колос отделяется от соломины, обламываясь у своего основания.
Членики стержня слабо опушенные, в верхней части (у основания ко-
лоска) с бородками. Ости мягкие, тонкие, немного длиннее колоса, ше-
роховатые. Колоски удлиненные, обычно двузерпые, в поперечном раз-
резе овальные, с выступающими килями колосковых чешуи (рога).
11(>Г)ро(1 П. Itoi'ol iiu т Mieut-c/i. el Dot <»/.
Вымолот зерновок трудный. По спинной стороне зерновок проходит
очень узкое, неглубокое (в виде линии) углубление, с брюшной сторо-
ны— узкая бороздка. Хохолок небольшой. Зародыш узкий, удлинен-
ный. Пыльники короткие (2,8 мм).
По типу опушения листовой пластинки, по консистенции и длине во-
лосков, а также по ряду анатомических признаков пшеница Тимо-
феева сходна с полбой дикой араратской. Некоторые их различия
отражены в том, что у первого вида клетки эпидермы и мезофилла лис-
та крупнее, рдзмер устьиц больше, а число их па единицу листовой по-
верхности меньше, чем у последнего. На 1 мм2 поверхности листа у
пшеницы Тимофеева насчитывается около 31 волоска, у полбы дикой
араратской — около 9(5. У первого вида стенки соломины тоньше, про-
водящих пучков в механической ткани меньше (О. Д. Градчанинова,
1967; В. Ф. Дорофеев, О. Д. Градчанинова, 1971). Близость этих видов
отражена также is строении плодовой оболочки зерновки. У них отмече-
на тангептальиая удлиненность клеток эпидермиса, их топкостенность,
а также наличие четко выраженных пор в стенках поперечных клеток
(Л. Л. Жестяппикова, 1954).
312 K.ini сифш.ици.ч рода Tiitiriim I..
I 111 ic 11111 i,;i T и м <)(|)e1' i i ;i имеет яровой ofipa.i жизни. ()ii;i хар;ш i ери iv-
ется позднеспелоеti.io и златолюбием. Места ее общ аипя -зона влаж-
ного прохладного климата со среднегодовым количеством осадков
800—1200 мм. В Грузии обычно высевалась с осени, но давала урожаи
также и при весеннем посеве.
Для Т. iimopheevii в условиях низменною Даюстапа характерно
ускоренное ответчице цветков в верхней части колоса. Цветение коло-
са протекает быстро (3 5 дней). Отмечен резко выраженный утренний
максимум цветения, наступающий при температуре воздуха 19- 21°С
(В. II. Жуков, 19(59). Известны случаи спонтанной iибрндпзации пше-
ницы Тимофеева. В. Л. Мепабде (1948) в Грузии выявил спонтанные
гибриды от скрещивания Г. timopheeuii с Т. iiioiiococcinn и Г. aesiiviim.
В зерне пшеницы Тимофеева в условиях орошаемого земледелия по
данным В. Г. Конарева и др. (1972), концентрация белка колебалась
в пределах 19,7—22%, лизина в белке - 1,56—1,88%.
Иммунитет. Еще в 1930 г. II. II. Вавилов обратил внимание па устой-
чивость пшеницы Тимофеева к ржавчине (М. М. Якубцинер, 1931). Гео-
графические опыты 1923—1931 гг. подтвердили то наблюден и'
(Н. II. Вавилов, 1935). Вид оказался уникальным но иммунитету к муч-
нистой росе, бурой и стеблевой ржавчине, пыльной и твердой головне,
по устойчивости к фузариозу, гессенской и шведской мухам, клопу-че-
репашке, зеленоглазке и пьявпце (М. М. Якубцинер, 1932, 1934, 1969;
Н. И. Вавилов, 1935; II. М. Жуковский, 1914, 1950, 1964; В. Л. Мепаб-
де, 1948; Л. Л. Декапрелевич, 1954 и др.). Существующие расы бурой
ржавчины, очевидно, приспособлены к паразитированию на видах пше-
ницы с геномным составом А11В. Они генетически не запрограммирова-
ны для жизни за счет таких растительных организмов, каким является
пшеница Тимофеева. При искусственном заражении пли в условиях
естественных эпифитотии у нее лишь изредка обнаруживали единичные
мелкие пустулы бурой ржавчины (Н. И. Вавилов, 1961; М. М. Якубци-
пер, 1934, 1969; Э. Ф. Мигушова, Л. А. Смельцова, 1970 и др.). В усло-
виях Дагестана И. П. Шитова (1968) оцепила поражаемость пшеницы
Тимофеева этим патогеном как равную в среднем 0,04 балла (по 5-бал-
льной шкале) при реакции типа I. О. Г. Григорьева (1975) при искус-
ственном заражении наблюдала слабую восприимчивость проростков
этой пшеницы к расам 77, 20, 122. В. М. Бсрляпд-Кожевипков и др.
(1973) установили, что подавляющее число образцов коллекции пшени-
цы Тимофеева имеет полигонную устойчивость к расам 77, 151, 202.
Наблюдения, начатые ВИРом в 1923 г. над коллекцией, высевав-
шейся в широком географическом диапазоне (от Ленинградской облас-
ти до Дальнего Востока), показали, что пшеница Тимофеева иммуииа
к желтой ржавчине (Н. И. Вавилов, 1935; М. М. Якубцинер, 1969;
А. М. Камелипа, 1973; О. Г. Григорьева, 1975). Следы этого патогена на
ее листьях обнаруживались очень редко (М. М. Якубцинер, 1934;
Н. И. Вавилов, 1961). Несмотря на эпифитотии, в условиях плоскостно-
го Дагестана в 1954—1967 гг. поражение желтой ржавчиной отсутство-
вало (И. П. Шитова, 1968; Э. Ф. Мигушова, Л. А. Смельцова, 1970).
Пшеница Тимофеева слабо поражается стеблевой ржавчиной — до
балла 2 (М. М. Якубцинер, 1934, 1969; Н. И. Вавилов, 1961; И. П. Ши-
това, 1968; Э. Ф. Мигушова, Л. А. Смельцова, 1970). При искусствен-
ном заражении проростков ипокулюмом этого патогена А. М. Камелипа
(1973) наблюдала реакцию типа IV. Восприимчивость проростков к ра-
сам 34 и 17 (тип реакции I—II) отметила О. Г. Григорьева (1975).
На иммунность пшеницы Тимофеева к мучнистой росе указывали
М. М. Якубцинер (1957), II. М. Жуковский (1969) и др. Очень редкое
появление единичных пустул этого патогена отметили II. II. Вавилов
(1961) и II. А. Скурыгппа (1963, 1970). Высокая устойчивость к боль-
шинству физиологических рас возбудителя мучнистой росы пшеницы в
llo<)i>o<) 11. Hiieoticnnt Л1!»iisell. el Doioj. 313
СССР и Европе определена и а бл io.t.ci и i я м 11 Н. Sniiljakovie (1962),
11. Рнгпноп Tpaiii.. и II. Г. Одинцовой (1966), Р. A. McIntosh,
Е. Р. Baker (1966), J. Vallega (1967). При искусственном заражении
проростков сборным ииокулюмом гриба Алма-Атинской, Куйбышевской,
Ленинградской, Новосибирской областей и Северной Осетин Э. X. Су-
хапбердипа (1977) наблюдала иммунность у 71,1 % и высокую устойчи-
вость у остальных образцов коллекции пшеницы Тимофеева. Взрослые
растения на провокационном фоне в Ленинградской области также
были иммунными.
Пшеница Тимофеева иммунпа к пыльной головне (А\. А\. Якубцинер,
1957, 1969; В. И. Кривченко, 1972). Прп изучении коллекции установ-
лено, что практически все образцы выдерживают самые жесткие испы-
тания на естественном и искусственном инфекционных фонах .зараже-
ния пыльной головней. При этом устойчивость пе зависела от специали-
зации и рас. патогена (В. И. Кривченко, 1973). А. М. Ямалеев (1974)
при искусственном заражении наблюдал мицелий f. aeslivi в зародыше-
вых почках у 10% обследованных зерновок, f. dnri — у 1,7%. Несмотря
па это, выросшие из заразившихся зерновок растения были свободны от
поражения пыльной головней. Освобождение от мицелия патогена у
пшеницы Тимофеева происходило вследствие лизиса гифов гриба. При
электрофоретическом изучении запасных белков зерна А. М. Ямалеев
показал, что те образцы пшеницы Тимофеева, у которых w-зона спектра
имеет 5—6 компонентов, являются эмбрионально восприимчивыми к
пыльной головне. Если эта зона спектра редуцирована до грех компо-
нентов, то образцы характеризовались устойчивостью к грибу.
Высокий иммунологический тип характеризует отношение пшеницы
Тимофеева к твердой головне (М. М. Якубцинер, 1934, 1969). Внедряю-
щийся внутрь клеток мицелий патогена подвергается угнетению и начи-
нает дегенерировать: цитоплазма этих клеток является неблагоприят-
ной средой, ведущей к лизису мицелия гриба (Т. Д. Страхов, II. В. Греч-
ко, 1960). При искусственном заражении популяциями рас твердой го-
ловни из 17 географических районов СССР пшеница Тимофеева пора-
жалась лишь формой патогена, собранного в Грузии на колосьях этой
пшеницы (С. П. Зыбина, 1939). Подобные сведения сообщает также
В. Л. Мепабде (1948). В опыте В. И. Кривченко (1973) искусственное
заражение спорами твердой головни коллекции пшеницы Тимофеева
(27 образцов) выявило иммунность 96,3% образцов.
В восточных районах СССР пшеница Тимофеева сильно поражается
корневой гнилью, а в Грузии — септориозом и гсльмиптоспорпозом
(М. М. Якубцинер, 1961, 1969). Н. Harding (1972) пришел к выводу,
что она все же более устойчива к корневой гнили, чем тетраплоидные
виды секции Dicoccoides. В условиях провокационного фона пшеница
Тимофеева средне поражается вирусом полосатой мозаики (М. М. Якуб-
цинер, Ю. В. Велянчпкова, 1970).
Иммунитет пшеницы Тимофеева к ряду грибных болезней до сих
пор мало использован в селекции. Огромные усилия селекционеров,
пытавшихся соединить комплекс генов иммунитета-пшеницы Тимофеева
с генотипом пшеницы мягкой, не увенчались созданием иммунного про-
изводственного сорта Т. aestivum. Причина в первую очередь в генети-
ческой отчужденности, существующей между видами с геномами В и G,
которая, очевидно, связана с их дифилетическим происхождением. Со-
гласно положениям закона гомологических рядов в наследственной
изменчивости II. И. Вавилова (1920), возможно создание гомолога
пшеницы мягкой, который будет иметь геномы АЪ(Л). Только таким пу-
тем, вероятно, можно успешнее использовать комплексный иммунитет
пшеницы Тимофеева к грибным болезням для защиты урожая.
История. В VIII в. популяция зандури представляла собой сообще-
ство двух видов - Т. timopheevii и 7’. monococcum (В. Л. /Мепабде,
•Я ! /' Л(и < ш/шкици:! pr»)n Tri/ictiiii
1918). 11ослед||ПЙ вит з :i 11 и м ал в ней । оено.ю i вующее положение, Bno-
следе i ни и < >e 11 < > hi i у io naeii, 1 i.ei i o. i ;i eiii.i coe , ;i i;.i-,i 11, rei ранлои ijii.iij вид.
Компоненты популяции зандури имели народные пап минования - чел-
та зандури (пшеница Тимофеева) и гваца зандури (культурная одно-
зернянка).
Географическое распространение. Пшеница Тимофеева -узколока-
лизовапный вид пшеницы. II. М. Жуковский (1923) обнаружил ее в
1922 г. в Западной Грузни. Г. Л. Столетова (1925) уточнила места воз-
делывания этой пшеницы. Грузинские трптпкологи jl. Л. Декаирелевич
п В. Л. .Менабде в ряде своих работ указывали, что северная граница
возделывания пшеницы Тимофеева лежала на липни селений Нагори —
Орбелп—-Лайлапш--Опп, южная граница проходила по селениям Мек-
вена и Дгпориса. Иными словами, ареал ее располагался южнее Леч-
хумского хребта Главного Кавказа па территории современных Амбро-
лаурского, Опекой), Цагерского районов. Ныне пшеница Тимофеева
там не возделывается.
Экологически пшеница Тимофеева приурочена к предгорной части
Западной Грузии (400—800 м над ур. м.), где климат влажный и про-
хладный.
Происхождение. Еще в 1932 г. М. М. Якубцинер подметил наличие
сходных морфологических признаков у ишеппцы Тимофеева и дикой
араратской полбы. Позднее В. В. Светозарова (1939) пришла к заклю-
чению о сходстве мейотического поведения хромосом у гибридов этих
видов, что нашло подтверждение в работах Е. II. Макушипой-Горощеп-
ко (1948), L. Sachs (1953), Е. В. Wagenaar (1961, 1966), J. Mac Key
(1966). В близкой родственности пшеницы Тимофеева и полбы дикой
араратской убеждены В. Л. Менабде и Л. Л. Ерицяп (1941). Е. II. Сни-
скан (1955), Л. Л. Фплатеико (1968), В. Ф. Дорофеев (1969), В. Г. Ке-
наров и др. (1970). В. Е. Johnson (1967), М. G. Joshi и Л. К- Kaul (1971)
подтвердили генетическую близость этих видов при электрофоретиче-
ском обследовании белков. Качественно одинаковыми оказались также
эпзимограммы их кислых фосфатаз в опыте В. Яаска (1970, 1971,
1974). Введение ядер твердой и мягкой пшеницы в цитоплазму пше-
ницы Тимофеева и полбы дикой араратской сходным образом стерили-
зовало пыльцу гибридов (И. Д. Мустафаев и др., 1968; Н. А. Скуры-
гина, 1968; S. S. Маап, К- A. Lucken, 1972). A. Bozzini и В. Giorgi
(1969) нашли, что кариотипы этих видов также сходны.
Наряду с перечисленными фактами найдены некоторые генетичес-
кие различия между пшеницей Тимофеева и полбой дикой араратской.
В. Яаска (1971, 1974) указал на различия в зимограммах изоэстераз
у проростков этих видов. Отсутствует полное сходство также в зимо-
граммах пзоэнзимов а-амилазы (К. Nishikawa, М. Nobuhara, 1971).
А. В. Конарев и др. (1971) нашли, что в иммупохимпческих реакциях
аптиглиадиновая сыворотка Т. boeoticum у полбы дикой араратской вы-
являет глиадипы значительно сильнее, чем у пшеницы Тимофеева.
Наличие подобных различий — свидетельство дивергенции, которая
привела к становлению пшеницы Тимофеева из полбы дикой арарат-
ской.
М. Tanaka и S. Ichikawa (1968) показали, что полба дикая арарат-
ская типа 1Г из Закавказья наиболее близка к пшенице Тимофеева.
К. Nishikawa и Y. Sawai (1969) нашли, что эти пшеницы имеют сходное
количество ядерпого ДНК. Сходство их продемонстрировано также па
электрофоретических спектрах белка зерновок (J. G. Wains, 1970;
А. В. Копиров, 1974).
Кариотип пшеницы Тимофеева оказался сходным с Т. araraticum
из Ирака (R. Giorgi и др., 1971). При гибридизации последнего с пше-
ницей Тимофеева Е. Sachs (1953), Е. R. Wagenaar (1966), Р. S. Rao и
Е. Е. Smith (1968) обнаружили стерильность их гибридов при правили-
Подрод II. Boeoticum Migusch. et Dorof.
315
ном ходе мейоза в 1д. Дальнейшие последования показали, что Т. ara-
raticum Ирака, подобно закавказской форме, имеет пе менее двух гене-
тически разнокачественных форм: I — нескрещивающуюся с видом
Т. timopheevii и II — скрещивающуюся с ним (Э. Ф. Мнгушова, Л. В, Ко-
варов, 1975).
Логично предположить, что исходной формой для пшеницы Тимофе-
ева была полба дикая Ирака типа II с опушенным колосом (var. nabe-
licum). Среди подобных форм этой полбы имеются морфологически
сходные с пшеницей Тимофеева, по обладающие спонтанным распаде-
нием стержня колоса на отдельные колоски. Возможно, что они пред-
ставляют собой переходный биотип между истинно дикой полбой и
Т. iimopheevii.
Пшеница Тимофеева в Западной Грузии, возможно, является занос-
ной из Северо-Восточной Турции (II. И. Вавилов, 1935). В. Л. Менабде
и Л. Л. Ерицяп (1942) связывали происхождение Т. timopheevii с террито-
рией древнего государства Урарту, откуда предки картвельских племен
при переселении завезли этот вид в Западную Грузию. Безусловно,
нельзя категорически отвергать возможность заноса пшеницы Тимофе-
ева в Грузию с юга. Нахождение се. в Западной Сербии в долине реки
Лима в селении Бело-баба (Г. Г. Давидяп и др., 1971) можно рассмат-
ривать как свидетельство миграции этого вида не только в Грузию.
Наука пока не обладает достаточным количеством информации для
окончательного решения вопроса о месте происхождения Т. timopheevii.
Говоря об исключительном иммунитете этого вида к грибным болез-
ням, Н. И. Вавилов (1964) подчеркивал, что он мог выработаться и в
условиях теплого, влажного климата Западной Грузии, Именно здесь
мыслят место становления пшеницы Тимофеева Г. И. Абесадзе (1929)
и В. Л. Менабде (1948).
Мысль о возможности интрогрессивной гибридизации полбы дикой
араратской с Т. monococcum была высказана В. Л. Менабде (1948).
Вопрос об участии этой однозернянки в становлении пшеницы Тимофе-
ева обсуждался рядом исследователей (Г. И. Абесадзе, 1929;
К. А. Фляксбергер, 1935; Е. Н. Синская, 1955; А. А. Филатенко, 1968;
А. Д. Горгидзе, 1973, 1974).
G. Kimber (1973) высказал мысль о возникновении пшеницы Тимо-
феева при скрещивании между Т. monococcum и Ае. speltoides. Такой
путь возникновения Т. timopheevii пока тоже нельзя категорически от-
вергнуть, ибо при этом мог соединиться генетический материал двух
названных иммунных диплоидных видов, дав начало тетраплоидному
виду с рекордной устойчивостью к грибным болезням, какую имеет
Т. timopheevii. В. Яаска (1974) пе отрицает возможность интрогрессив-
ной гибридизации с Т. monococcum при возникновении пшеницы Тимо-
феева. Спонтанная гибридизация между 7’. araraticum и Т. monococcum
var. hornemannii обеспечила интрогрессию генов, ответственных у одно-
зернянки за щетинистое опушение узлов соломины, иммунность и, мо-
жет быть, яровой образ жизни (Э. Ф. Мнгушова, А. В. Конарев, 1975).
Существующая гипотеза об автополиплоидном происхождении пше-
ницы Тимофеева от Т. monococcum в свете ряда современных исследо-
ваний методами биохимии, ядерпо-цитоплазматпческого замещения, ци-
тогенетики представляется нам необоснованной. II. И. Вавилов (1935,
1962) стоя,/! на позициях признания аллополиплоидпого происхождения
Т. timopheevii.
Систематика. П. М. Жуковский (1923) сначала рассматривал пшени-
цу Тимофеева как дикорастущую разновидность Т. dicoccoides (Koern.)
Schweinf. Е. А. Столетова (1925) установила, что эта пшеница является
культурной. Она отнесла ее к виду Т. dicoccum. К. А. Фляксбергер
(1926) указал, что пшеница Тимофеева имеет ряд промежуточных
(между Т. dicoccum п 7’. dicoccoides) признаков. П. М. Жуковский пере-
.416
Классификации рода Trilicum I..
Hi'.'i ее в ринг вида в 1928 г. из-за наличия у нее резких морфоло! нисских
отличий от других возделывавшихся тетраплоидных видов. Правиль-
ность этого была затем подтверждена исследованиями по генетике и ци-
тогенетике (Л. Г. Хиннен, 1929; F. Lilienfeld, II. Kihara, 1934; Д. Костов,
1936). ’
Вызывает недоумение отрицание некоторыми зарубежными исследо-
вателями правомочности выделения этой пленчатой пшеницы.в само-
стоятельный вид и определение ее как подвида голозерной пшеницы
Т. turgidum (A. et I). Love, 1961) или даже как ее разновидности
(W. М. Bowden, 1959).
Определение разновидностей Т. timopheevii (Zhuk.) Zhuk.
Чешуи Ости Разновидность № разновид- ности
белые красные черные иа фоне цвета чешуи черные
белом красном
Чешуи неопушенные
rubi gi ncisuni
Чешуи опушенные
+
+
tinioplieevii
viticulosum
nigrum
Var. nigrum Ericzjan, 1939, Tp. Тбил. бот. нп-та, 7: 172. (4).
Получена А. А. Ерицяп в Институте ботаники АН ГССР при гибри-
дизации пшеницы Тимофеева с Т. durum и Т. persicum.
Var. rubiginosum Ericzjan, 1939, I. c.: 172. (1).
Получена А. А. Ернцян в Институте ботаники АН ГССР при скре-
щивании пшеницы Тимофеева с Т. persicum.
Var. timopheevii. — Т. timopheevii var. typicutn Zhuk. 1928, Tp. прикл.
бот. ген. сел. 19, 2: 64. (2).
Геогр. распр. Грузия.
Описана из Грузии (селение Азармн).
Var. viticulosum Zhuk. 1928, 1. с.: 64. (3).
Геогр. р а с п р. Грузия.
Кроме того, А. А. Ерицян (1939) и В. Л. Мепабде (1948) гибридо-
генно получили еще ряд константных разновидностей (Т. timopheevii
gr. planosupercompactum Menabde. 1948, 1. c.; 47, descr. ross.) с плотны-
ми колосьями (D = 50—70), которые нехарактерны для пшеницы Ти-
мофеева в культуре.
Т. ZHUKOVSKYI MENABDE ET ERICZJAN —
ПШЕНИЦА ЖУКОВСКОГО
Т. zhukovskyi Menabde et Ericzjan, 1960, Сообщ. AH ГССР, 25,6:
732. — T. timopheevii var. zhukovskyi (Menabde et Ericzjan) Morris e.t
Sears, 1967, Wheat and wheat improvem.: 36, comb, invalid.
Общая характеристика. T. zhukovskyi морфологически весьма схо-
ден с Т. timopheevii. Опушение листовых пластинок и влагалища щети-
нистое. Листья широкие, 16—20 см длины. Соломина 100—125 см дли-
ны. Стеблевые узлы густо опушены щетинистыми волосками. Or Т. ti-
mopheevii отличается более удлиненным и более крупным, но менее
плотным колосом (D=32—40). Колосья остистые (рис. 25, а), ост
негрубые, 5—7 см длины, с меньшей ломкостью стержня, чем у пшени-
цы Тимофеева. Колосковые чешуи более удлиненные, почти равные по
длине нижним цветковым чешуям или незначительно короче последних.
Dodpor) I/. Hoeoticum Minnich, et Dorof.
317
Рис. 25. Колосья, колоски и зерновки:
а— Т. zliuko-.'skyi Menabde ct Eric/jan; б — T. fungicidum Zhuk.
Зубец главной боковой жилки на колосковой чешуе хотя и значительно
короче килевого зубца, но хорошо развит. Между зубцами — остро-
угольная выемка. Членики колосового стержня опушены менее густо,
чем у пшеницы' Тимофеева. Зерновки красные, вымолот их трудный.
2п = 42. Геномная формула A1’A1,A1’AbGG. Образ жизни — яровой.
По экологическому облику пшеница Жуковского сходна <• пшеницей
Тимофеева, в сообществе которой она была открыта. Оба вида пше-
ницы влажного, прохладного климата предгорий. Первая из них более
позднеспелая, чем вторая: разница во времени созревания в условиях
Ташкентской области и Дагестана составляет 9—10 дней. Пшеница Жу-
ковского весьма влаголюбива и плохо переносит воздушную засуху.
Кроме того, она характеризуется сильной склонностью к открытому цве-
21 762
318
Классификация /юда Tiilicum I..
Tvnitto: у нес отмечено почти 100% огкрын) цветущих ннешш (М. Л. Фе-
дин, 1974).
Концентрация белка в зерне пшеницы Жуковскою достигает при
возделывании па орошаемых участках в условиях Ташкентской области
23,6%, лизина в белке — 2,9% (В. Г. Коиарев и др., 1972). Она получила
хорошую оценку по хлебопекарным качествам. Объем хлеба, выпечен-
ного из урожая, выращенного в условиях Ленинградской области, до-
стигал 500 см3 (Э. В. Таврин, 1963).
Иммунитет. По отношению к ряду грибных болезней пшеница Жу-
ковского характеризуется высоким иммунологическим т ипом. ЛА. М. Якуб-
ципер (1969) и А. М. Камелипа (1973) сообщили о невосприим-
чивости ее к ряду рас бурой ржавчины. Высокую расоспецифпчес-
кую устойчивость к этому виду ржавчины на сильнейшем инфекцион-
ном фоне выявила О. Г. Григорьева (1975) в различных возрастных
фазах растений. Заражение проростков сборным ипокулюмом мучнис-
той росы из 5 географических пунктов нашей страны показало иммуни-
тет к этому грибу ие только у пшеницы Жуковского, по и у ее анало-
гов— Т. timococcum Kost, п амфидиплопда Э. В. Таврина, имеющих
одинаковый геномный состав (Э. X. Сухапбердппа, 1977).
При искусственном заражении пыльной головней 1. duri мицелий
гриба достигал зародышевых почек у 1,7% зерновок, а при заражении
f. aestivi — 46,5% зерновок, однако при этом колосья пшеницы Жуков-
ского были свободны от поражения патогеном (В. П. Кривченко, 1973;
А. М. Ямалеев, 1974). Иммунность к твердой головне выявлена в опы-
тах Т. В. Ярошенко, И. Я. Зубко (1966) и В. И. Кривченко (1973).
Иммунность к вышеназванным грибным болезням, а также к жел-
той ржавчине наблюдалась при естественном заражении пшеницы Жу-
ковского в условиях Дагестана, Краснодарского края, Ленинградской и
Ташкентской областей в 1958—1978 гг. В экстремальных условиях
(поздний весенний посев в приморской полосе Дагестана) зарегистри-
рована восприимчивость к стеблевой ржавчине, доходившая до 3 бал-
лов (по 5-балльпой шкале), при иммунности к бурой и желтой ржавчи-
не (Э. Ф. Мигушова, пеопубликовано). А. М. Камелипа (1973) также
отметила поражение этим патогеном в условиях эпифитотии па Даль-
нем Востоке.
Происхождение. Пшеница Жуковского обнаружена в Грузин в попу-
ляции запдурп (В. Л. Мепабде, А. А. Ерицян, 1958, I960). А. Д. Гор-
гидзе (1968), Л. К- Чхаидзе (1969), В. Л. Мепабде (1971, 1972) счита-
ют ее результатом автополиплопдизацви Т. monococcum в популяции
зандури. В их понимании в соматических клетках пшеницы Жуковско-
го присутствуют хромосомы 6 гепомов однозернянки культурной. Экс-
перименты Э. В. Таврина (1963), М. 1). Upadhya и М. S. Swaminathan
(1965) показали, что пшеница Жуковского является аллогексаилоидом,
имеющим геномы Т. monococcum и Т. limoplieevii. Еще в 1959 г.
W. М. Bowden представил геномную формулу пшеницы Жуковского
как сложение геномов этих видов. Подтверждением такого пути проис-
хождения пшеницы Жуковского является сходство морфологических
признаков ее колоса с колосьями амфидиплопдов п гибридов Fi, полу-
ченных при спонтанной или принудительной гибридизации Т. timophee-
vii с Т. monococcum Д. Костовым (1936), Г. В. Канделаки (1946),
М. D. Upadhya и Al. S. Swaminathan (1963), а также Э. В. Тавриным
(1963). Последний скрестил созданный им амфидиплопд с пшеницей
Жуковского и получил фертильное потомство. П. S. Dhaliwal и
В. L. Johnson (1976) наблюдали нормальную конъюгацию хромосом с
образованием бивалентов в мейозе и нормальное плодоношение у гиб-
ридов от скрещивания одинаковых ио геномному составу амфпдиплои-
дов — Т. araraticum X Т. boeoticum и Т. zhиkovskyi.
Подрод II. Hoeot i< и in Mi p и sell. el Doroj.
319
Иммунохимический и Элек J рофоре i лчсск ini а н;iл 11.41,1 i./iiiayuiiiou зер-
новки указыh.’iiot на участие T. motioiocciim и Т. titnopheevii в станов-
лении пшеницы Жуковского (j\. В. Конарев и др., J971; A. Aniol, 1973;
В. Г. Конарев и др., 1976). Таким образом, пшеница Жуковского — ре-
зультат спонтанной гибридизации между диплоидным и тетраплоид-
ным компонентами ценоза зандури, приведшей к возникновению алло-
гексаплоидпой пшеницы.
Г. MILITINAE ZHUK. ET MIGUSCH. — ПШЕНИЦА МИЛИТИНЫ
Т. militinae Zhuk, et Migusch. 1969, Вести, с.-х. науки, 2 : 16.
Общая характеристика. Всходы фиолетовые. Листовые пластинки и
влагалища опушены щетинистыми волосками. Язычок и ушки бесцвет-
ные; ушки с длинными ресничками. Соломина 100—120 см длины. Сте-
бель и узлы покрыты восковым налетом. Стеблевые узлы опушены ще-
тинистыми волосками. Колосья остистые (рис. 24, б), длиной 3,5—
4,5 см, широкие и плоские, очень плотные (D = 50—70). Стержень коло-
са почти неломкий. Его членики короткие, иногда ломающиеся не по
сочленению. Ости пегрубые, длиной 6- 8 см. Килевой зубец остевпд-
пый. Колоски с 3 цветками, из которых третий неразвит. Наружная
цветковая чешуя короче внутренней на 1 —1,5 мм. Внутренняя цветко-
вая чешуя при созревании не расщепляется на 2 части. Зерновки крас-
ные, 8—9 мм длины, сплющенные, иногда деформированные, стекловид-
ные, с хохолком. Вымолот зерновок легкий. По морфологии вегетатив-
ных органов пшеница Мплитппы сходна с исходным видом Т. timo-
pheevii. В условиях орошаемого земледелия (плоскостной Дагестан и
Ташкентская область) имела низкую продуктивность. Масса 1000 зерен
достигала лишь 27 г, число колосков в колосе — 23, а число зерновок
в колосе — 26.
Экологический облик пшеницы Мплптипы такой же, как у пшеницы
Тимофеева. Это—тип влажного климата предгорий. Пшеница Милити-
ны — биологически яровая, по плодоносящая и при осеннем посеве в
условиях мягких зим (Закавказье, Дагестан, Средняя Азия). Для пес
характерна позднеспелость. В низменном Дагестане при осеннем посе-
ве и орошении период от всходов до колошения в среднем равен 237
дням. Исходная форма — Т. timopheevii—созревала почти на 9 дней
раньше (Э. Ф. Мигушова, Л. Л. Смельцова, 1970).
Иммунитет. Пшеница Мплитппы — иммунный к ряду грибных болез-
ней вид. Ее невосприимчивость к ним оценена как превосходящая устой-
чивость пшеницы Тимофеева (П. М. Жуковский, Э. Ф. Мигушова, 1969).
При искусственном заражении проростков агрессивными расами бурой
ржавчины и при изучении взрослых растении па провокационном фоне
в полевых условиях О. Г. Григорьева (1975) подтвердила иммунитет
к этому патогену. На иммунитет к мучнистой росс указывает исследо-
вание Э. X. Сухапбердппой (1977), которая инфицировала проростки
пшеницы Мплитппы популяцией рас Алма-Атинской, Куйбышевской,
Ленинградской, Новосибирской областей и Северной Осетии, а также
оцепила взрослые растения па провокационном фоне в полевых усло-
виях Ленинградской области.
В. И. Кривченко (1973) и А. М. Ямалеев (1974) показали, что при
заражении пыльной головней мицелий 1. duri и f. aestivi достигает за-
родышевых почек у 2,2 и. 5,0% зерновок (cooiнетс.твепио). Однако при
этом взрослые растения не. имеют пораженных колосьев: освобождение
от гпфов гриба у них происходит вследствие лизиса. Высокий иммуно-
логический тип установлен также при искусственном заражении твер-
дой головней (В. И. Кривченко,1973).
Иммунитет пшеницы Мплитппы к перечисленным болезням, а также
к желтой ржавчине подтверждают наблюдения в условиях Дагестана,
21 762
320 Каш t ификици'! />о<)и Tiiliium I..
Ленинградской n Ташкен некой облает ей, проведенные па eeicc i венном
инфекционном фоне is 1963—1978 гг. (Э. Ф. Мнгушова, пеопубликова-
ио). Отмечены лишь следы стеблевой ржавчины, поражение которой у
пшеницы Тимофеева в тех же условиях достигало балла 2 (по 5-балль-
ной шкале).
Происхождение. Пшеница Милитииы — естественный мутант, возник-
ший у пшеницы Тимофеева (II. М. Жуковский, Э. Ф. Мнгушова, 1969).
Подобный мутант, появившийся независимо от пшеницы Милитииы и
имеющий белый цвет колоса, был отобран среди гибридов, полученных
от скрещивания между пшеницей Тимофеева и Т. polonicum
(В. И. Аиель, А. 3. Латыпов, 1974). Форма, подобная пшенице Милити-
ны, была получена также в Венгрии (D. Szalay, 1977).
Происхождение пшеницы Милитииы связано со спонтанным появле-
нием фактора, который обеспечивает легкий обмолот и действует как
доминантный в системе генов, которыми обладают геномы AhG. Он воз-
ник независимо от сходно действующих факторов Т. sinskajae и Т. per-
sicum. В связи с этим фактор легкого обмолота пшеницы Милитииы
обозначен символом Qm.
Систематика. Как упоминалось, появление форм, сходных по призна-
кам колоса с пшеницей Милитииы, отмечалось неоднократно в условиях
Армении, Белоруссии, Грузии, Венгрии, а также Болгарии. Так, в Инсти-
туте пшеницы и подсолнечника им. генерала Тошева (София) С. М. Се-
вов получил в 1975 г. на посеве пшеницы Тимофеева в М, спонтанный
ультраплотноколосый мутант с черными опушенными и белыми, чешуя-
ми, подобный Т. militinae. Такие же формы созданы в ВИРе при гиб-
ридизации пшеницы Милитины с пшеницей Тимофеева.
Ключ для определения разновидностей Т. militinae Zhuk, et Migusch.
1. Колосковые чешуи черные па красном фоне . . . var. militinae.
+ Колосковые чешуи белые ......................................
..............................var. albimilitinae A. Filat, et Migusch.
Var. albimilitinae A. Filat, el Migusch. var. nova - A T. militinae var.
militinae spica alba differt.
Typus: In statione experimental! dagestanica WIR e populatione
T. militinae Zhuk, et Migusch. X T. timopheevii Zhuk. 2.VII. 1970. leg.
E. F. Miguschova (WIR).
Геогр. распр. В коллекционных питомниках.
Var. militinae. — Т. militinae Zhuk, et Migusch. 1969, Вести, c.-x.
науки, 2: 16.
Геогр. распр. В коллекционных питомниках.
СЕКЦИЯ 6. KIHARAE DOROF. ET MIGUSCH.
Sect. Kiharae Dorof. et Migusch. 1978, Tp. прикл. бот. ген. сел.
62,2 : 151.
Листовые пластинки и влагалище опушены щетинистыми волосками.
Колосковая'чешуя имеет киль. На ее поверхности выраженная главная
боковая жилка. Внутренняя цветковая чешуя па 1 —1,5 мм длиннее на-
ружной, при созревании не расщепляется вдоль па 2 части. 2п=42, ге-
номная формула AbAhGGDD.
Геогр. распр. В коллекциях научно-исследовательских институ-
тов и других учреждений СССР, Японии, США, Канады и друтх стран.
Тип: Т. kiharae Dorof. et Migusch.
Секция 6 включает пока один синтезированный наукой вид—пше-
ницу Кихары.
T. KIHARAE DOROF. ET MIGUSCH. — ПШЕНИЦА КИХАРЫ
T. kiharae. Dorof. et Migusch. 1977, Bioji. ВИРа, 83: 71. Листовые
пластинки и влагалища взрослых растений опушены, как у Т. timo-
pheevii, щетинистыми волосками. Высота растении 123--125 см (ниже
Т. timopheevii па 5—6 см). Кустистость — 4—6 стеблей па 1 растение.
Соломина под колосом полая. Узлы соломины опушены очень тонкими
короткими волосками, которые стираются, поэтому узлы становятся го-
лыми. Колос желтовато-белый, жесткий, остистый (см. рис. 21, б), опу-
шенный, со средней рыхлостью (D = 17—18), легко обламывается у ос-
нования. Ости зазубренные, черноватые или белые (в зависимости от
условий созревания). Стержень колоса ломается при сгибании. При
этом членики стержня верхней своей частью остаются прикрепленными
к основанию колоска. Колосковая чешуя плавно переходит в выражен-
ный, но немощный киль. Килевой зубец острый, длиной 2—2,5 мм. Глав-
ная боковая и соседняя с ней жилки выражены четче, чем остальные.
У верхнего края чешуи эти 2 жилки сливаются, образуя бугорок. Плечо
колосковой чешуи приподнятое, довольно широкое у нижних и неболь-
шое у верхних колосков. Наружная цветковая чешуя короче внутренней
на 1,5—2 мм. Последняя при созревании не расщепляется вдоль на 2
части. Фертильность колосьев нормальная. Зерновка продолговатая,
красная, с выраженным хохолком и трудным вымолотом.
Пшеница Кихары имеет высокобелковое зерно. В условиях поливно-
го земледелия Ташкентской области и Дагестана концентрация белка в
ее зерне достигала соответственно 23,3 и 22,4%. Содержание лизина в
белке в последнем случае было равным 1,7%, масса 1000 зерен 39 г.
Пшеница Кихары иммуппа к желтой ржавчине, практически не по-
ражается бурой ржавчиной, во восприимчива к мучнистой росе. Послед-
нее унаследовано по линии генома D. Этот вид пшеницы создан при
скрещивании Т. timopheevii X Tie. tauschii. В коллекцию ВИРа посту-
пил из Японии. С гексаплондпыми видами секции Triticum генетически
несовместим: имеет иные, чем пшеница мягкая, ядро и цитоплазму.
Поддерживается в живом состоянии в коллекциях научно-исследова-
тельских институтов и других учреждений СССР, Японии, США, Кана-
ды и других стран.
УКАЗАТЕЛЬ ЛИТЕРАТУРЫ1
Абрамова 3. В. Цветение, оплодотворение п формирование зерновки пшеницы
в зависимости от сорта и условий произрастания. Автореф. дне. на сопск. учен, сте-
пени д-ра с-.х. наук. Л.. 1969.
Батыгина Т. Б. Эмбриология пшеницы. Л., 1974.
Брежнев Д. Д. Мировые растительные ресурсы и советская селекция. — В кН.:
Мир идей Вавилова, М., 1968, с. 41—48.
Вавилов Н. И. Лпннеевскпй вид как система. — Тр. по прикладной ботанике, ге-
нетике и селекции, 1931, т. 26, вып. 3, с. 109—134.
Вавилов Н. И. Ботанико-географические основы селекции. — В кп.: Теоретические
основы селекции растений, т. 1, М.; Л„ 1935, с. 17—75.
Вавилов II. II. Научные основы селекции пшеницы. — В кп.: Теоретические основы
селекции, т. 2, М.; Л.. 1935, с. 3—244.
Вавилов 11. II. Учение о происхождении ку.тыурных р,-iciciiнii после Дарвина. •
Советская паука, 1940, №2, с. 55—75.
Вавилов Н. И. Мировые ресурсы сортов хлебных злаков, зерновых бобовых, льна
и их использование в селекции. М.; Л., 1957.
Вавилов 11. И. Азия — источник видов. — Растительные ресурсы, 1966, т. 2, вып. 4:
577—80.
Ганди л ян II. А. Разнообразие диких пшениц Армянской ССР, вопросы нх систе-
матики и использования. — Учен. зап. Азерб. СХИ, 1966, т. 2. с. 61—64.
Г ан ди л ян II. А. О дикорастущих видах Triticum Армянской ССР. — Бот. жури,
1972, т. 57, №2, с. 173—181.
Г р о с с г е й м А. Флора Кавказа. Т. 1. Баку, 1939.
Декапрелевич Л. Л. Ботанический состав, экология и хозяйственные свойства
пшениц Грузии, Азербайджана и Армении. — В кн.: Пшеница в СССР. М/ Л.,
1959, с. 107—122.
Декапрелевич Л. Л. Виды, разновидности и сорта пшениц Грузии. — Тр. Ин-та
полеводства АН ГССР, 1954, т. 8, с. 3—58.
Дорофеев В. Ф. Проблемы современной филогении и систематики пшеницы.—
Вести, с.-х. науки, 1969, № 3, с. 25—35.
Дорофеев В. Ф. Пшеницы Закавказья. — Тр. но прикладной ботанике, генетике и
селекции, 1972, т. 47, вып. 1, с. 3—206.
Мак-Кей Дж. Генетические основы систематики пшениц.— С.-х. биол., 1968, т. 3, № 1.
с. 112—23.
Мустафаев И. Д. Материалы по изучению пшениц, ржи, ячменя и эгилопсов
Азербайджана. Баку, 1961.
Невский С. Triticum L. — Пшеница. Л.. 1934 (Флора СССР; т. 2), с. 675—688.
Н о с а то в с к и й А. И. Пшеница (биология). М., 1965.
Пшеницы мира/ В. Ф. Дорофеев, М. М. Якубцинер, М. И. Руденко и др. Л., 1976.
Роже виц Р. 10. Система злаков в связи с их эволюцией. — Сб. науч, работ 1941 —
1943 гг. Л., 1946.
СикскаяЕ И. Происхождение пшеницы. — В кн.: Проблемы ботаники. М.; Л., 1955.
т. 2, с. 5—73.
С инс кая Г:. Н. Исторический обзор работ ВИР по систематике. — Тр. по приклад-
ной ботанике, генетике и селекции, 1968, т. 39, вып. 2, с. 3—38.
Списка я Е. И. Историческая география культурной флоры. Л., 1969.
Туманян М. Г. Ботанический состав диких пшениц Армении и условия их про-
израстания в природе. — Тр. по прикладной ботанике, генетике и селекции, 1934.
сер. 5, №2, с. 241—268.
Удачин Р. А., Мигушова Э. Ф. Новое в познании рода Triticum L. Вести, с.-х.
науки, 1970, № 9, с. 20—24.
Фил а теп к о А. А., Кур к не и У. К. Пшеница Сипской — новый вид. — Тр. но при-
кладной ботанике, генетике и селекции, 1975, т. 54, вып. 1, с. 239—241. * I. 2
* Библиография мировой литературы по филогении, генетике, селекции и отдель-
ным вопросам биологии шиеницы периодически и щастся в специальных указателях:
I. Библиография мировой литературы ио генетике и селекции пшениц. Тр. по при-
кладной ботанике, генетике и селекции, 1935, сер. 15А, № 1, с. 1—135.
2. Селекция и генетика пшеницы. Бнблиогр. указат. отеч. лит. за 1935—1970 гг.
Л., 1977, ч. 1; 1978, ч. 2.
У/иикпель ли Г1'1><гг1Ц1ы .’,23
Ф л я к с б е |> г е р К. A. 1111 к' 1111111.(. Л\.; 1935
Фляксбергер К. A. I lnieiiiiu.i. AV; JI., 1335. (Кулц| урная ф.’гора СССР, i. I).
Ц велев II. 11. Злаки СССР./I.. 1976.
Цицин II. В. Новый вид и новые разновидноеiи пшеницы. — Бюл. Гл. Бот. сада,
I960, вын. 38. с. 38—41.
Якубцинер АЕ АЕ К познанию пшениц Китая, —Бот. жури., 1959, т. 44, № 10,
с. 1425—1436.
Bor N. L. Flora of Iraq. Baghdad, 1968 (Gramineae, vol. 9), p. 194—208.
Bowden \V. AL The taxonomy and nomenclature of the wheats, barleys and ryes and
their relatives. — Canad. Journ. Bot., 1959, N 37, p. 657—684.
Gokgol AV Tiirkiye bugdaylari. Istanbul. 1939.
G о k g <> 1 AV Bugdaylarin tasnif Anahtari. Istanbul, 1955.
Harlan J. R.. Zoharv D. Distribution of wild wheats and barley. — Science. 1966,
N 153, p. 1074—1080.
Helbaek H. Comnientarv on the phylogenesis of Triticum and Hordeuni. — Econ. Bot.,
1966. N 20, p. 350—360.
Hjelmqvist H. Some notes of the old wheat species of Gotland. — Hereditas, 1966,
Bd. 56, p. 2—3.
Kihara H. Interspecific relationship in Triticum and Aegilops. — Seiken Ziho, 1963,
N 15, p. 1 — 12.
Kihara II., Ya ma shit a K., Tanaka M. Alorphological, physiological, genetical
and cytologieal studies in Aegilops and Triticum collected from Pakistan. Afghanistan
and Iran. Results Kyoto Univ. Sci. Exp., Karakorum and Ilindukush, 1965, Kvoto, vol.
l.p. 1 — 118.
Kimber G.. Riley R. The 1 elalionship ol (he diploid progenilois oi hexapioid
wheat. — Canad. Journ. Genet. Cytol., 196.3, N 5, p. 83—88.
Kuckuck IL, S c h i e m a n n E. Uber das Vorkommen von Spelz. und Emrner (Triticum
spelta L. und T. dicoccum Schuebl.) in Iran.—Zeitschr. fiir Pflanzenziicht. Berlin,
1957, Bd. 38, H. 4, S. 383—396.
Al а с К e у J. The Taxonomy of hexapioid wheat. — Swensk Botanisk Tidskrift, 1954,
Bd. 48, II. 2, p. 579—587.
Al a ns f eld R. Das morphologische System des Saatweizens, Triticum aestivum L. s. I.
Der Zuchter, 1951, Bd. 21, II. 1/2, p. 41—60.
Renfrew J. M. Palaeoethnobotany the prehistoric food plants of the Near East and
Europe. London, 1973.
Vavilov N. I. La probleme de L’origine des plantes cultivees. — Ann. Inst. Nat. Agron.,
1934, N 26, p. 239—246.
Vavilov H. I. The new systematics of cultivated plants. — The New Systematics.
Oxford, 1940, p. 549—566.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
РАСТЕНИЙ * *
А
kegicon ** 7
Aegilops 5, 10, 12, 18, 20, 22, 30, 299
— Sitopsis sect, 10, 15, 22—25, 44
— Emarginata subsect. 23, 44, 45
— bicornis 23—26, 44
— crithodium 280
— cylindrica 26, 27, 196
— longissima 23, 24, 26, 44, 45
— sharonensis 23, 24, 26
— speltoides 22, 23, 25, 26, 29, 30, 44,
72, 307, 315
— squarrosa 27
— — eusquarrosa 27
— tauschii 19, 27, 28, 30, 182, 189, 196,
217, 234—236, 274, 275
---strangulate subsp. 27, 28, 217,
236
----tauschii subsp. 27, 216, 321
— variabilis 23
Agropyron 5, 9, 11, 22
— elongatum 22
— triticeum 22
Anthoxanthum 92
В
Brachypodiuni 10
Broinus 7
-----ramosa 143
— — semiaristata 142
— — velutina 143
E
Elymus 10
Elytrigia 9
— elongata 22
Eremopyrum 10, 12
— triticeum 22
F
Frumentum 7, 230
— Triticum subgen. 7, 230
G
Gigachilon 7
H
Haynaldia 10, 12
Hierochloe 92
— alpina 92
C
Crithodium 7, 12, 16, 280, 284
— aegilopoides 280
Cryptopyrum 7
D
Dasypyrum 10
Deina 7, 12
- polonica 135
- • • alba 142
-- — clavata 144
M
Monococcum 12, 280, 284, 293
N
Nardurus 10
Nivieria 7, 12, 280, 284
— monococcum 293
P
Phalaiis 92
Phalaroides 92
* Указатель составлен А. А. Филатеико
* Курсивом отмечены синонимы.
Указатель латинских незнаний
325
I ’o.'icc.'i < 92
Hordern ear lril>. I 2
- llordeinae trib. 12
— Phleeae trib. 92
— Protohordeae trib. 37
— Triticeae trib. 12, 92
— Aegilopinae subtrib. 12
— Frumentaceae subtrib. 12
— Triticinac subtrib. 12
P
Prototriticum 284
R
Roegneria 10
Rotboellia hirsuta 284
Secale 10, 12, 14
Spelta 7
— amylea 50
— vulgaris 199
T
Triticale 16
Triticum 5—7, 9—18, 20, 22, 30, 34, 37,
75, 78, 92, 106, 128, 146
— Boeoticum subgen. 17, 18, 280—321
— Eutriticum subgen. 10
— Triticum subgen. 17, 34—280
— Andropogon sect. 14, 304
— Cerealia sect. 7
— Crithodium sect. 13, 280
— Dicoccoidea sect. 13, 39, 304
— Dicoccoides sect. 13, 17, 19, 21—24,
27, 28, 38—185, 197, 198, 232, 274,
297, 313
— Eremopyros sect. 7, 280
— Eutriticea sect. 39, 186
— Eutriticum sect. 7, 10, 15
— Frumentum sect. 38, 186
— Gigachilon sect. 13
— Kiharae sect. 17, 20, 320—321
— Monococca sect. 13, 280
— Мопососсоп sect. 13, 17—19, 34
280—303
— Orthatherum sect. 13, 39, 304
— Pyrachne sect. 13
— Pyros sect. 7
— Silopyros sect. 7, 15
— Spelta sect. 13, 38, 186
— Speltoidea sect. 13, 186
— Timopheevii sect. 17, 19, 21, 28, 29,
304—320
— Transcaucasicum sect. 304
— Triticum sect. 13, 14, 17, 19 21, 28
39, ,186—280, 297, 304
- - Zea sect. 7
— Urartu sect. 17, 18, 3-1—38
- Dtiriuscula subsect. 39
— Eudicoccoidea subsect. 39
— Prototriticea subsect. 39
— Transcaucasira subsect. 304
— Vulgaria subsect. 186
Montunue mt. 11
S/rigosa s<-r. 1'1
Zandurianum ser. 14
— diploidca congr. 280
— hexaploidea congr. 186
— octoploidca congr. 14
— quasihexaploidca congr. 14
— tetraploidea congr. 39, 304
— eudicoccoides consp. 39
— monococcum consp. 280
— speltoides consp. 186
— timopheevii aggr. 14, 304
— Spelta a) 7
— abyssinicum 11, 139, 145, 149
----turgidoides subsp. 165
----vulgarioides subsp. 149, 150, 158
----expansoides prol. 147, 149
— — tenero-expansoides prol. 149
— — tenerum prol. 147, 158
----abbaicum var. 160
----addisabebae var. 167
— -- acutum var. 167
-- albonigrum var. 153
albosemineum var. 153
albovillosum var. 167
— — alboviolaceum var. 153
----amharicum var. 154
----ankoberense var. 168
----antiquoaethiops var. 160
----asmaricum var. 163
----atraiosanguineum var. 168
----atroalbum var. 168
----atrocompactum var. 175
----atroinflatum var. 171
----atromarginatum var. 171
----atrorubrocompactum var. 175
----atrorubrum var. 168
----axumicum var. 154
----bialbum var. 160
----bicolor var. 160
------- brevispicatum var. 175
----brownii var. 154
— — earneum var. 163
----compactonegroides var. 164
----coloratum var. 168
----comitans var. 160
----compressum var. 164, 176
— — copticum var. 164, 176
----dangalense var. 171
----decoloratocompactum var. 177
- - — decoloratum var. 168
— — densoalbonigrum var. 160
----densoamharicum var. 160
— — densoarraseita var. 161
— — densoatratopurpureuni var. 171
----densoatratosanguineum 171
— -- densoatrorubrutn var. 171
— — densodecoloraturn var. 171
----densofulvum var. 173
— — densoharraricum var. 161
----densoinjlatum var. 173
----densomenelikii var. 160
----densopilosum var. 173
----densoplanum var. 173
----densopraevalenciae var. 162
— - densopullatum var. 173
- - • densopurpureum var. 173
----densorubidum var. 173
----densorubronigrum var. 161
— — densorubrum var. 162
- densosanguineum var. 173
-- — densoschimperi var. 161
Указатель латинских иазттий
<1 t‘ll lilt iiJuliiin \ a i I li I
ill'll I, ovinliu cum va I 11 > I
- - di]Iusum var. 174
- dispersum var. 168
- - - dominans var. 156
----duplorubrum var. 168
- — erithre.um var. 164
— — flaksbergeri var. 174
— — fulvocompaclum var. 176
- - - - lulvoinflatiim var. 168
fulvuin var. 168
- - I urea him var. 168
— — gallasoruni var. 154
godjamicum var. 161
- -- gondaricum var. 154
----harlanii var. 174
— — harraricum var. 154
- - — lieterochromum var. 174
----incanocompactiim var. 177
- - — incanum var. 174
- kirae var. 154
----ktishianum var. 169
— — laxinflatum var. 169
- - -- marginaioeillosum var. 174
— — marginatum var. 161
— — melanoceplialum var. 169
— — menelikii var. 158
----negroides var. 155
— -- negustianum var. 162
----nigrobrcvispicatum var. 177
----nigrobrownii var. 155
----nigrocoloratum var. 169
— — nigrodensoarraseita var. 161
- - — nigrodensomenelikii var. 162
— - - nigrodensoplanum var. 174
— — nigrodensopurpureum var. 174
----nigrodensosanguineum var. 174
— — nigrodensosclumperi var. 161
----nigrodispersum var. 169
----nigrodominans var. 156
----nigroduplorubrum var. 169
----nigroinflatum var. 169
------- nigromenelikii var. 155
— — nigropraehordeiforine var. 156
— — nigropraesemiticum var. 161
----nigroptolomaeum var. 164
— — nigropurpureum var. 169
------- nigrorubidocompactum var. 177
----nigrorubidum var. 169
----nig rorubroinf latum var. 169
----nigrosanguineum var. 169
— — nigrosomalense var. 161
----nigrosteudelianum var. 156
----nigrotypicum var. 164
----nigroviolaceum var. 155
----notabile var. 177
— — nubicum var. 164
----percivalii var. 169
— — pilosocompactum var. 177
------- pilosoinflatum var. 169
— — pilosonigrum var. 161
----pilosum var'. 169
----pianocompactum var. 177
—• — praeapulicum var. 155
— — praehordeijorme var. 156
— — praesemiticum var. 161
- • - praevalenciae var. 155
prospicuum var. 155
----protomelanopus var. 155
----pseudoalbovillosum var. 170
----pseudoalboviolaceum var. 154
• — -- pseudoarabicum var. 163
pseudocopticum var. 164
- - — pseudodecolor alum var. I/O
— — pseudodensomenelikii var. 162
— — pseudodensoplanum var. 174
- - — pseudodispersum var. 170
- - - - pseudodominans var. 155
- - - /meudogoiidariciini var. 155'
— — pseudoincanum var. 174
psi'tidopraehordei]oi me var. 155
— -- pseudopraesemiticum var. 162
— — pseudopilosum var. 170
— — pseiidoptolomaeum var. 164
— — pseudopurpureum var. 170
— - pseudorarum var. 156
- - - • pseudorubidocompactum var. 177
-- — pseudorubidum var. 170
— — pseudorubroaristatum var. 156
— — pseudorubroinflatum var. 170
- - - pscudorubropubescens var. 162
- - -- pseiidorubroeiolaceiim var. 156
- - - pscudosomalense var. 162
- pseudotypicum var. 161
— - - pfolomaeum var. 165
— — pubiglumatum var. 174
----pullatum var. 170
- purpureocompactum var. 177
----purpurcum var. 170
----rarissimum var. 156
— — rarttm var. 154
----recognition var. 177
— — richardianum var. 174
—- — rubidoinjlatum var. 175
---- rubidum var. 170
— -- rubroaristatum var. 156
----rubroinjlatum var. 170
— — rubromuiicum var. 156
----rubronigrum var. 156
----rubropilosum var. 170
— — rubropubescens var. 162
— — rubroviolaceum var. 152
— — rufocompactum var. 177
— rufovillosum var. 175
— — sabae var. 171
---- sanguineoinflatum var. 175
----sanguineum var. 171
- - — schimperi var. 157
-- — schweinfurthii var. 157
— — simiense var. 164
----singulum var. 157
----somalense var. 162
— — solitarium var. 175
- - sparsum var. 175
----spectabile var. 175
— — steudelianum var. 157
— — striatocompactum var. 177
----striatopurpureum var. 171
----subbrownii var. 157
— submenelikii var. 157
— — subnegustianum var. 162
— — subpraehordeiforme var. 157
— — subpraevalenciae var. 157
— — subprotomelanopus var. 157
----subpseudorarum var. 157
- - - subschimperi var. 157
- — taborense var. 157
....takussicum var. 165
- - - - tchertchericum var. 162
— — (hebaicum var. 164
- - - - - tigrense var. 157
• - - lypicum var. 165
Указатель латинских imaimmii'i
327
null ши var. I 75
violai eomulii um vai. 158, ]7I
acuminatum 13, 39
— aegilopoides 281, 284, 285
----ainazuri var, 287
- - — albochlorococcum var. 290
- - - albonigrum var. 287
• album var. 287
- - - avdalaricum var. 287
— - - aznaburticum var. 287
— - -- balaclavicum var. 291
- - -- boeoticum var. 287
— breviuta f. 287
- - --- Imlgaricum var. 292
- - - - chlorococcum var. 287
----cinereum var. 287
- - — fusciim var. 292
— — garniense var. 288
----getapi var. 290
----kurbagalense var. 288
----larionowii var. 288
----luteinigrum var. 292
— — luteonigrum var. 289
- — microspermum var. 288
----mikoiani var. 288
----nigrescens var. 289
----nigrochlorococcum var. 289
----pancicii var. 289
— — pseudoalbum var. 289
----pseudogetapi var. 287
----pseudosymbolonense var. 288
------pseudozuccarinii var. 289
------stramineonigrum var. 289
----subalbum var. 289
-- — subbaydaricum var. 289
— — subpancicii var. 289
----subpseudoalbum var. 289
— — subpubescentinigrum var. 289
----theydjerabaki var. 290
— — thumanianii var. 289
----tuberculatum var. 287
------viridoboeoticum var. 290
----viridosymbolonen.se var. 290
----zuccarinii var. 290
— aegyptiacum 267
— aestivum 6, 8, 9, 11, 14—17 20 23.
30, 34. 55. GO, 64, 81, 91, 94, 105 114
135, 149, 150, 183, 184, 187, 203, 205,
212, 213, 217, 230—269, 271, 272, 274,
275, 279
----aestivum subsp. 230, 238, 240,
241, 262—269, 312
----compactum subsp. 213
----durum subsp. 93
----hadropyrum subsp. 238—262, 263
----inflatum subsp. 244
----macha subsp. 186
----spelta subsp. 199
— sphaerococcum subsp. 269
----transcaucasicum subsp. 239
------turgidum subsp. 76
----vavilovii subsp. 209
----vulgare subsp. 230
----inflatum convar. 237, 238, 239,
244—252
— — macha convar. 186
----rigidum convar. 237, 239—244,
252
— - - semirigidum convar. 237, 238.
239, 252—262
— — spelta convar. 186
sphaerococcum convar. 269
- loti ai istutum convai. 2(11
mimlocii convai. 209
- el i gu la 1 i nl 1 a turn subconvar. 245,
251 -252
— - eligulatum subconvar. 252—254
-- — inflatum subconvar. 215—250
semirigidum subconvar. 252,
251 262
— -- aestivum gloup 230
- compactum group 213
- - - macha group 186
— — spelta group 199
sphaerococcum group 269
- - vavilovii group 209
- - compactum Rasse 213
----- vulgare Rasse 230
— - aeslivo-montano-caucasiciim prol.
264
-• — aeslivo-stepposum prol. 264
-----hiemale-boreale prol. 264
-----hiemale-montano-caucasicum prol.
263
- - liiemale-stcpposum prol. 261
- meridioiialc-i'uiopaeum prol. 265
— - - occidetitale-curopaeum prol. 265
----- orientale-sibiricum prol. 265
-----aestivum var. 247, 264. 265. 266
- — afghanicum var. 253
----- ajutense var. 265, 266
-----albiceps var. 219
— — albicepsinf latum var. 225
-----albiglaucum var. 265, 266
----- albinflatocapilalum var. 245
— - albinflatum var. 245, 246
— -- albinflaticaptatum f. 245
-----falsealbidum f. 245
-----albirubrinflatum var. 245, 246
-----falsealbirubrum 1. 245
-----albirubrum var. 265, 266
-----dekaprelevitschji f. 265
-----almaatense var. 256, 257
-----anglicum var. 265, 266
-----araraticum var. 229
- - — araxicum var. 245, 246
-----ardjeschicum var. 256, 257
— — armeniacum var. 219
— - atriceps var. 219
— atrocyanum var. 219
— — atroerinaceum var. 229
-----atrum var. 229
aureuin var. 206, 265, 266
• — - araticuni f. 267
— — kurduculense f. 265
-----auslralianum var. 266, 267
— — badakschanicum var. 253, 254
- - - baranovii var. 251
• - - - barbarossa var. 256, 258, 260
- - - - barlangiensc var. 226
- - - - baruschonicum var. 253, 254
----- bcngalense var. 256, 257
— — bengalinflatum var. 245, 246
— — bukiniezii var. 225
— — cabristanicum var. 219
— — caeruleovelutinuin var. 256, 258
- - cacsioidcs var. 257, 258
- - - - caesiorigidum var. 257, 258
caesiospeltifornie var. 242
- - caesium var. 266, 267
- - chiovendae var. 257, 258
clioresmum var. 245, 246, 247
cinerescens var. 247, 251
cinereum var. 257, 258
328
Указатель латинских названии
ciiierosmii var. 212, 243, 257, 258
chiatilum v;ir. 229
- - crassiceps var. 229
— — crassicepsinf latum var. 225
— — creticum var. 229
----- cyanolhrix var. 247, 257, 258
— - cvanothri.xinflatuni var. 246, 247
-----(ielfii var. 257,258
-----echinoidinf latum var. 225
— - ecliinopsis var. 219
erinaceum var. 219
-----erinaceuminflatum var. 225
— — erivanense var. 219
— — erythroleucum var. 257, 259
-----erythrosperminflatum var. 247
-----erythrospcnnuni var. 265
— — jalsealbidum var. 245
-----jalsedelfii var. 250
— — falsekcmstantinovskense var. 248
— — falselutescens var. 248
-----falsenigricolor var. 248
-----ferrugiiieum var. 249. 266, 267
— — fetissovii var. 221
— — fetissovinf latum var. 225
-----flahsbergeri var. 221, 253, 254
-----fuliginosum var. 258, 259
-----glaucilutescens var. 266, 267
— — glaucilutinflatum var. 246, 247
— — golbckii var. 243
— — gontscharovii var. 257, 259
-----gorbunovii var. 226, 253, 254
-----graecinflatum var. 247
-----graecum var. 252, 254, 257, 259
-----karbulakianum f. 259
— — griseofetissovii var. 221
— — griseoictcrinum var. 229
-----griseosplendens var. 221
-----griseum var. 258, 259
-----gunticum var. 253, 254
— — hamadanicum var. 258, 259
-----heraticum var. 246, 247
-----hissaricum var. 257, 259
— — horogiense var. 251, 252
— — hostianum var. 250, 258, 259
-----hrasdanicum var. 229
— — humboldtii var. 229
— — humboldtinflatum var. 225
-----icterinum var. 229
-----icterinuminflatum var. 225 .
-----indicinflatum var. 246, 248
-----indicum var. 257, 259
— — ingrediens var. 257, 259
-----insigne var. 257, 259
-----introitum var. 257, 259
-----iranicum var. 258, 260
-----jakubzineri var. 266, 267
-----japschorvi var. 253, 254
-----jasgtilemi var. 227
-----kabulicum var. 253, 254
-----kanascliii var. 221
— — karakurgani var. 227
-----kazvinictim Var. 258, 260
— — kerkianum var. 221
— — kerkianuminflatum var. 225
— — kermanschachicum var. 258, 260
— — khorassanicum var. 244, 246, 248
-----kobelevii var. 246, 248
— — falsckonstantinovskense f. 248
-----konstantinovskense var. 266, 267
— — korshinskyi var. 251, 252
— — kudrjaschcvii var. 242
-----leucospcrimnn var. 257, 260
- - - linac a var. 229
1111asI a11icinn var, 258, 260
-- lutescens var. 266, 267
— -- 1 ii tin 11 a turn var. 246.248
— -- falselutescens f. 248
- - - hmmgyangen'se f. 248
magiiificiiin var. 257, 260
— maksumovii var. 253, 254
- - iiielanopogon var. 257, 260
- melanoriibniiii var. 257,260
-- - meridionale var. 258, 260
----- nieridionalmflalum var. 217, 248
— — mesopotamicum var. 258, 260
— — milturum var. 266, 267
— — montanum var. 225
-- — morosovskensc var. 266, 268
— — murgabiciun var. 253, 254
- - • - navrozkyi var. 242, 243
— nicolai var. 251, 252
— - nigralbirubrutn var. 257, 260
-----nigraristatum var. 266. 268
— — nigrerythrospernium var. 266, 268
—- — nigricans var. 257. 260
-- • nigricolor var. 260. 268
-----nigricolorinflaluin var. 246, 248
— — falsenigricolor f. 248
— — nigriferrugineum var. 266. 268
— — nigrigraecum var. 257, 260
— — nigrilutescens var. 266, 268
-----nigrilutinflatinn var. 246, 218
-----nigrinflatum var. 246, 248
— nigrirufinflatum var. 246. 248
-----nigrisardoum var. 257, 260
-----nigrum var. 266, 268
— — oblivense var. 266, 268
orientale var. 221
— — oxiantnn var. 251, 252
----- painiricuin var. 251, 252
— — petropavlovskianum var. 251, 252
-----pjandshicum var. 251, 252
-----popoviae var. 247
— — pseudoalbirubrmn var. 257, 260
— — pseudobarbarossa var. 258, 260.
261
— — pseudocaeruleovehitinum var. 267,
268
— — pseudocaesium var. 266. 268
— — pseudocrassiceps var. 230
-----pseudodelfii var. 257, 261
— —- pseudoerinaceum var. 221
— -- pscudoerythroleucuni var. 257.
261
----- pscudoerytlirosperinum var. 257,
261
— — pseudofetissovii var. 221
— — pseudofulvicinereum var. 266, 268
-----pseudograecum var. 257. 261
— — pseudoheraticum var. 246, 248
-----pscudohostianum var. 258, 261
-- — pseudoicterinum var. 230
— - pseudokhorassanicum var. 246,
248
— -- pseudoleucospermum var. 257,
261
— — pscudomeridionalc var. 258. 261
— — pseudopyrothrix var. 257, 261
— pseudorubriceps var. 222
- - - pseudorub rum var. 230
— — pseudosplendens var. 2,30
-- -- pseudotranscaspicum var. 246,
248
pseudoturcicuin var. 258, 261, 262
Указатель латинских iiauuumu'i
329
I>•,!ч।<I<>11lie>iи,-i11 н 11111 var. 21G, 213
pseu<1 ove 1 иti1111111 var. 257, 261
- - - - pukhrum var. 257, 261
- — pyrothrix var. 266, 268
quasialbiceps var. 222
quasibarbarossa var. 253, 254
-- - qmisimeridionalinf latum var. 252
- - - quasifelisowii var. 223
- cpiasiriifinflatum var. 251, 252
qiiasisubturcicum var. 253, 254
- quasiturcicuin var. 253,25.4
• - raikoviae yar. 251,252
-----ramosoerythrospertnum var. 263
- renovatum var. 258, 261
- roschanum var. 225
- - rtibriceps var. 222
— .— rtibricepsinflahim var. 225
-----rubrimurinum var. 258, 261
-----rtibrtim var. 230
-----rubruminflatum var. 226
-- -- ruchczianiim var. 251, 252
- - riifinflatum.var. 246, 248
- - - falserufintlatum f. 249
- rujtilitijlatиm var. 226
nijtiliim var. 230
— - - sardotim var. 266. 268
— - schrcdcri var. 242, 243
— -- sclnignanicum var. 251, 252
-----sericeum var. 222
--------------------------sogdianttm var. 258, 262
--------------------------spletulens var. 229
— — subafghanicum var. 253, 254
-- —- subalbiceps var. 222
-----subalbicepsinflalum var. 226
- — subbarbarossa var. 242, 243, 249
— — subbarbarossinflatum var. 247,
249
-----subcacsinllatuni var. 246, 249
-----subcaesioides var. 24.2, 243, 249
------subcaesioidinflatum var. 246, 249
------subcaesium var. 242, 243, 249
------subechinod.es var. 222
------subechinodesinflatum var. 226
------ suberinaceum var. 222
-----suberythroleucinflatum var. 246,
249
- -- subfalseerythroleucum f. 249
— — suberythrolcucuni var. 242, 243.
249
-----subcrythrospenninflatum var. 246
249
-----subervthrospermum var. 242, 243,
249
— — subjalseeryihroleucon var. 249
------ subfalsegraecum var. 250
-----subferrugineum var. 242, 243
----- subferrugininflatum var. 246, 249
— -- subfetissovii var. 222
— — subfetissowiinflatum var. 226
-----subfuliginosum var. 242, 243
-----subgraecinflatuin var. 246 249
250
— — subfalsegraecum f. 250
-----subgraecum var. 239, 242 243,
250
-----siibgiinticum var. 253, 254
- subhamadanicimi var. 242, 243
- - • sublioslianum var. 242, 243
-----subhostinflatum var. 247, 250
-----subicterinum var. 222
-----subicterinuminflatum var. 226
----- subiranicum var. 242, 243
• 111 > к :i I >i 111 c 11111 var. 253, 254
subkermaiischacliicum var. 242
214
submeridionale var. 242, 244, 250
siibmeridionalinflatum var. 246,
259
stibiiiesopolamicuin var. 242, 244
- • - suhpseudobarbarossa var. 242, 244
.subpseudohostiamini var. 242; 24 1
- --siibpseudoho.stinflatnm var. 247,
250
stibpseudomeridionale var. 242,
244
- subpseudomeridionalinflatum var.
246, 250
— - subpseudoturcicum var. 242, 244.
250
- — subpseudoturcinflatum var. 246.
250
- —• subrenovutuin var. 243
- -- - siibrubriceps var. 222
- subrtibricepsinflatum var. 226
- siibsardoum var. 242. 24 1
subsericeum var. 222
subsericeuminjlalum var. 226
- subsogdianiiiflatinn var. 246, 250
- sttbsplendens var. 222
-----siibsplendcnsiiiflaliim var. 226
----- subturcicum var. 242, 244, 250
-----subturcinflatum var. 246, 250
— --- suchobrusianum var. 251, 252
- - - sulinense var. 266, 268
-----sunpani var. 246, 250
— — siircliiaiium var. 222
— — tadjicorum var. 253, 254
— — tanaiticuin var. 266, 268
-----tarakanovii var. 263
— — tehcranicum var. 246, 250
— — transcaspicum var. 246, 250
-----falsedelfii f. 250
-----triste var. 266, 268
- - — tschermakianum var. 266, 268
-----turanicum var. 257, 262
-----turcicum var. 258, 261, 262
-----turcomanicum var. 246, 250
— — urarticum var. 230
-- — uzbekistanic.um var. 257, 262
vanense var. 257,262
variabile var. 257, 262
— vavilovii var. 266, 269
villosum var. 266, 269
— — viridibarbarossa var. 267, 269
— — viridiferrugineum var. 266, 269
----- viridihostianum var. 267. 269
— viridimilturum var. 266, 269
-- — viridipyrothrix var. 266, 269
-----wagarschapati var. 230
-- — wernerianum var. 230
— — wernerianuininflalum var. 226
-----wittmackianum var. 230
— •- wittmackianuminflatum var. 226
— aethiopicum 6, 8, 11, 15, 17, 34 82,
94, 138, 145—177, 267
----- aethiopicum subsp. 146, 149,
150 -158, 165
turgidoides subsp. 146, 150,
165 177
- -• vavilovianum subsp. 146, 150,
153, 158-165
•— — compactum convar. 158, 163—165
dcnsum convar. 165, 171 —175
-----clongatum convar. 165, 166—171
3:50
Указатель латинских iirui/iainii'i
11 '< < 111I11111111 convar. I(|(>. 173, 17/
v ;i v i I < > v i :i 11 u 111 convar. 158, 15!)
163
-----abbaicum var. 159, 160
-----acutun) var. 166, 167
- • — addisabebae var. 167
----- aethiopicum var. 151, 152
-----aIbinigruin var. 151, 153
-• albiseminemn var. 151. 153
— — albivillosum var. 166. 167
- — albiviolaceum var. 151, 153
-----ainharicimi var. 151, 151
— — ankoberense var. 166, 168
-----antiquoaethiops var. 160
-----arraseita var. 146, 151, 154
— — aristiforme subvar. 154
-----breviclcntatum subvar. 154
-----longidentatum subvar. 154
-- — asmaricuni var. 163
-----atralbum var. 166, 168
-----artralisanguineum var. 166, 168
— — atrinflatuin var. 171, 173
-----atriconipactum var. 175, 176
— — atrimarginatum var. 171, 172
-----atriiiibrieompactiini var. 175, 176
— -- atririibriini var. 166, 168
---axiiiniciini var. 152, 154
-- — bialbum var. 159, 160
— .... bicolor var. 159, 160
-----bojanovskajae var. 163, 164
-----brevispicatum var. 175, 176
— — brownii var. 151, 154
— - carnemn var. 163
- coloratmn var. 166, 168
-----coniitans var. 159, 160
— — compactonegroides var. 163, 164
- — coiiiprcssuin var. 175, 176
- — copticum var. 176
.....dangalense var. 171, 172
decoloraium var. 166, 168
- - • - densamliaricuin var. 159, 160
densalbinigrum var. 159, 160
- densarraseita var. 158, 159, 160,
161
-----densatratipurpureum var. 171, 1/2
- densatratisanguinemn var. 171, 172
• - — densatririibruni var. 171. 172
densidecoloratum var. 171, 172
-- densifulvum var. 171, 172
- - - - densiharraricum var. 159, 161
— densiuflatiun var. 173
- — dens.inienclikii var. 159. 160
- — dcnsipilosum var. 172. 173
- densiplanum var. 172. 173
-- — deusipu I latum var. 172. 173
• - - - densipurptiretim var. 172, 173
-----densirubidum var. 172, 173
-----densirubrinigrum var. 159, 161
— — densisanguineum var. 172. 173
-----densischimperi var. 159, 161
— •— densisemiarist,atum var. 160, 161
— — dcnsiviolaceum var. 159, 161
-----diffusion var. 172, 174
-----dispersum var. 166, 168
-----duplirubrmn var. 167, 168
-----eritraeum. var. 163, 164
-----flaksbergeri var. 173, 174
- - — lortunalovae var. 151, 154
- - fiilvinflatum var. 167, 168
- - — fulvicompactum var. 176
- - - fnlviun var. 166, 168
-- — furcatum var. 167. 168
I’..i II.I'.OI uni r .11 1.52. I '.11
- g' id j a 111 ic 11111 vai. 160, I 61
— - gondaricum var. 152, 154
grabhami-var. 151, 154
— * - harlanii var. 173, 174
liarraricmn var. 151, 154
—- - heterochromum var. 172. 174
- - iiicanicompactuiu var. 176. 177
- — incanum var. 172, 174
----- jakovlevii var. 163, 164
kirae var. 152, 154
- - - kushianum var. 166, 169
---laxinflatum 168, 169
- - - niarginativillosum 172, 174
----- marginatum var. 159, 161
-----iiielanoc.ephalum var. 166. 169
- - - negroides var. 151, 155
— iiigrinflatimi var. 167, 168. 169
— nigrinflatum var. 167, 168. 169
- nigricoloratuni var. 166, 16!)
iiigridensarraseita var. 159. 161
— nigridcnsiplanmn var; 1 / 2 174
- 11 igridensipurptiretuiI var. 17: ’. 174
- - nigridensisangiiinemn 174 var. 172,
nigridensischimperi var. 159 . 161
nigridispersum var. 166, 16!)
nigridiipliriibrum var. 167. 168,
169
----- nigrimarginatum var. 151. 155
- - nigrimenelikii var. 151, 155
— - nigripraesemiticum var. 159, 161
— - - nigriptolomaemn var. 163. 161
----- nigripurpureum var. 167, 169
----- nigrirubidiconipactum var. 176,
177
.... ._ nigrirubidum var. 166. 169
-----nigrirubrinflatum var. 167. 168,
169
— -- nigrisangiiineuin var. 166, 169
. .. - nigrisomalense var. 159. 161
- nigritypieum var. 163, 164
nigriviolaceum var. 151, 155
nikolaenkinis var. 163, 164
notabile var. 176, 177
nubicum var. 163. 164
pahnovae var. 163, 164
percivalii var. 167, 169
pilosiuf latum rar. 167, 169
pilosicoinpactmii var. 176, 177
|>iIosi11ign11n var. 159. 161
— -- pilosum var. 167, 169
- -- planocompactum var. 176, 177
- - --- praeapulicum var. 152. 155
- - praesemiticum var. 159, 161
----- praevalenciae var. 151, 155
— — prospicuum var. 152, 155
-----protomelanopus var. 151, 155
— — psetidoalbivillosuin var. 166, 170
-----pseudobrownii var. 151. 155
-----pseudocarneum var. 163, 164
-----pseudocoloratmn var. 166, 170 .
-----pseudodecoloratum var, 166. 170
— — pse.udodciisimeiiclikii var. 159, 162
-----pseudodensiplanuin var. 172, 174
— — pseudodispersum var. 166, 170
— — pseudodoininans var. 151, 155
— — pseudogondaricum var. 152, 155
-----pseudoincanuni var. 172, 174
— — psetidopraehordeiforme var. 151,
155
— — psetidopraeseiniticimi var. 159, 591
Указатель латинских названии
331
psi'iiih>рi|<(Sinn vrir. I67, 170
| >sei I < I < >р to h >i I) aei 1111 var. 163, 164
---pscudopurpureum var. 166, 170
— — pseudorarum var. 151, 156
— — pseudorubescens var. 151, 156
— — pseudorubidicompactum var. 176,
177
— - -- pseudorubidum var. 166, 170
- - — pseudorubraristatmn var. 151, 156
pseiidonibriiiflatuni var. 167, 170
- - - pseudonibripubescens var. 159
162
— — pscudorubriviolaceum var. 151,
156
— — pseudorufescens var. 151, 156
— — pseudoschimperi var. 151, 156
-- — pseudosomalense var. 159, 162
-----pseudotomentosum var. 160, 162
— — pseudotypicum var. 163, 164
- - — pseudouncinatum var. 159, 162
— pubighimatinn var. 172, 174
- pullatum var. 166, 170
- purpureocompactum var. 176, 177
-----purpureum var. 165, 166. 167, 170
- -- rarissimum var. 151, 156
— — recognitum var. 176, 177
-----richardianum var. 172, 174
— — rubescens var. 151, 156
- - rubid inf la turn var. 173, 175
-- rubidum var. 166, 170
-----rubraristatum var. 151, 156
- -- rubriinuticum var. 152, 156
- — rubriiiflatuin var. 167. 168, 170
• - - • rubrinigrum var. 152, 156
- • - - rubripiiosum var. 167, 170
- - - rubrivillosum var. 159, 162
- - rufcscens var. 151, 156
-----ruficompactum var. 176, 177
- - — rulivillosuui var. 172, 175
-----sabae var. 167, 171
-----sanguineuin var. 166, 171, 172
-----sanguininflatum var. 173, 175
- schiinperi var. 146, 151, 157
schweinfurthii var. 151, 157
- - - simiense var. 163. 164
singulum var. 152, 157
solitarium var. 172, 175
-----somalense var. 159, 162
-----sparsum var. 173, 175
spectabile var. 172, 175
steudelianum var. 151,157
- — stoletovae var. 163, 164
- • -• striaticompactum var. 176, 177
- — striatipurpureum var. 166, 171
— • - subbrownii var. 152, 157
submenelikii var. 152, 157
-----subnegustianum var. 160, 162
-----subpraehordciforme var. 152, 157
-----subpraevalenciae var. 152, 157
- — subprotomelanopus var. 152, 157
-----subpseudorarum var. 152, 157
-----subschimperi var. 152. 157
-----syrovatskyi var. 163. 165.
— -- taborense var. 152, 157
-----takussicum var. 163, 165
— — tchertchericum var. 159, 162
tigrense var. 152, 157
-- tomentosum var. 160, 162
-----typicum var. 163, 165
-----tmeinatum var. 151, 158
- - - - unicum var. 173, 175
- - — vioiaceomuticum var. 15.2, 158
\ i < 11 a c i 11 f I a 111111 var. 167, I 71
v 11111 i I u 1111 var. 160, 162
— a[ghanicum 223
— albtirneum 201
— almaatense 256
- - alatiim 93
— amplissifolium 230
— amyleum 50, 188
aiigliaim 265
— antiquarum 28. 188. 197, 215 217.
234, 235, 273, 274
— araraticum 5. 6. 11, 13. 14, 17—19.
29. 3(1, 32. 44, 82, 280, 304 -310 314,
315, 318.
-- — araraticum subsp. 305, 308—310
----kurdistanicum subsp. 305 308.
310
----araraticum var. 308, 309
— ara.xicum var. 308, 309
----hirtiglumis var. 310
kurdistanicum var. 310
- - nabelicum var. 310
-- - nachitschevanicum var. 309
....nigricans var. 309
----nigrum var. 309
— — pseudoaaronsolmii var. 309
----pseudoaraxicum var. 309
- - -- pseudoarmeniacum var. 309
— - pseudostraussianum var. 309
----thumanianii var. 309
— ariluini 201
— aristalum 230
- - armeuiacum 35, 37, 63, 308
- arras 68
- - asiaticum 238, 252
— — imiticorigiduin subsp. 252
- -- rigidum subsp. 252
— — semiaristaliim subsp. 239
----speltiforme subsp. 239
----caesiorigidum var. 258
— — chivense var. 258
----[ormosum var. 243
— — gontscharovii var. 259
----hissaricum var. 259
----insigne var. 259
— - iranoturanicum var. 258
— - komarovii var. 259
----kujatkovskyi var. 243
----lipskyi var. 243
— — niagiiijiciim var. 260
— — pseudoerythrospermum var. 261
----pidchrum var. 261
----sogdianum var. 262
----turanicum var. 262
— -- uzbelcistanicum var. 262
---- variabile var. 262
— atratum 59
— boeoticum 6, 17, 18, 20—23, 26. 29.
30, 32, 35, 36, 37, 38, 41, 44, 280 293,
298, 299, 303, 307, 314, 318
- - — aegilopoides subsp. 285
— — boeoticum subsp. 21. 285—290
----thaoudar subsp. 21, 4 1, 283, 285.
290—293
-- — urartu subsp. 35
----abovjanii var. 290, 291
— — ainazuri var. 286, 287
----albinigrescens var. 290,291
----albinigrum var. 286, 287
----avdalaricum var. 286, 287
----azerbajdjanictnn var. 291
----aznaburticum var. 286, 287,
332
Указатель латинских назпании
li.i hie 1 ;i v ii'ii 111 v:ir. 29| , 292
I>;11 ;i 11 s;u- v;i г. 291 , 292
I>a 11a sur icuи I var. 291, 293
— — baydaricum var. 286, 287, 288
— — biarnualvir var. 290
— — biarnuruvir var. 292
- -- biarpurunivir var. 291. 292
binarnurutrivir var. 291
binarnuruvir var. 292
- binarpiialuir var. 293
-- — binarpurutrivir var. 293
- - — bicolor var. 291, 292
-----boeoticum var. 286, 287
-----bornmuelleri var. 291, 292, 293
— — buluchevskajae var. 286, 287
-- — chochlovac var. 286,287
— -• cinereum var. 286, 287
----- daralagezicum var. 286, 287
— — euboeoticum var. 287
-----fumidum var. 291, 292
-----fuscuin var. 291, 292, 293
— — gandiljanii var. 286, 288
— - - garasseferianii var. 286, 288
— — garniense var. 286, 288
— — gegadiricum var. 291, 292
-----harnicum var. 291, 292
— — haussknechtii var. 291, 292
-----helenae var. 286, 288
— — kovarskii var. 286, 288
— — kurbagalense var. 286, 288
-----larionovii var. 286, 288
— — larisae var. 286, 288
— — luteinigrum var. 291. 292, 293
-----maz.zettii var. 291, 292
— — maysstirianii var. 286, 287, 288
— — microspermum var. 286, 288
-- — mikoianii var'. 286, 288
- -- nigrichlorococcum var. 286, 288
-----nigrireuteri var. 291, 292
-----pantchitchii var. 286, 289
-----pseudoboeoticum var. 286, 289
-----pseudomayssurinaii var. 286, 288,
289
-----pseudoreuteri var. 291, 293
— — pseudozuccarinii var. 286, 288.
289
-----pubescentinigrum var. 286, 289
-----rufinigrum var. 291, 293
----- subalbum var. 286, 289
— — subbaydaricum var. 286, 289
-----subpantchitchii var. 286, 289
-----subpseudoalbum var. 286, 289
— — subpubescentinigrum var. 286,
289
-- symbolonense var. 286, 290
— — thaoiidar var. 290, 291, 293
-----tchivicum var. 291, 293
-----theydjerabakii var. 286, 288, 290
-----unarntirutiivir var. 288
-----unarntiruvir var. 290
-----unarpurivir, var. 287
— — unarpurunivir var. 288
- - — unnarnurutrivir 288
— — unnarnuruvir var. 287
-----unnarpurucivir var. 287
- - — unnarpurutrivir var. 287
— — untiarpuruvir час. 288
-----iinnarrutrivir var. 288
— — unnarturutrivir var. 288
-----viridalbum var. 291,293
-----viridibocticum var. 286, 290
-----viridifuscum var. 291, 293
v ii 111i111(>i urn var. 291, 29,3
vii idi'OiiiboloiieiL'.c var. 286, - 287,
290
— • - zuccarinii var. 286, 290
— x borisovii 14
• — caerulescens 109, 202
— caeruleum 202
— candidissimum 110
-- carthlicum 11, 17, 34, 177, 179
- - carllilictim subsp. 178
arnual var. 183
~ - arnurual var. 183
----arpual var. 183
— — fuliginosum var. 185
— -- capitatuni f. 185
— -- narpual var. 184
narpurual var. 184
— cercale 230
- - chaldicum 308
— creticum 229
- compactum 6, 11, 17, 34, 72. 81, 91,
135, 149. 183, 189, 197, 198. 203, 212,
213—2.30, 231. 233, 235, 236, 269. 270,
27J. 272, 274, 275
— — armcnoturkeslanicum subsp. 218
— — compactum subsp. 227
----eurasiaticum subsp. 227
— — indoeuropaeum subsp. 227
----iranoasiaticum subsp. 218, 223
----muticocompactuin subsp. 218
----rigidicompactum subsp. 218
----semiaristatocompactum subsp. 218
----sinicum subsp. 228
----compactum convar. 217, 227—230
----inflatum convar. 217, 218, 223—
227
— — rigidicompactum convar. 217,
218—223
----bartangiense subconvar. 223,
226—227
— — eligulatum subconvar. 218, 222—
223
— — rigidicompactum subconvar. 218—
222
— — roshanum subconvar. 223 -226
— — armeno-anatolicum prol. 219
----cabulicum prol. 218
----eligulatoroschanum subprol. 226
— — eligulatum subprol. 226
----inflatum prol. 223
— — jacuticum prol. 227
— — khoresmicum prol. 219
----mancliscliuricum prol. 227
— — vccidentaleeuropaeum prol. 227
— .... palestinicum prol. 219
— - - pamirobadakschanicuin prol. 223
.... ._ rigidum prol. 218
----roshanum prol. 223
----subrigidum prol. 218
— — tianschanicum prol. 227
----albiceps var. 219, 221
----epruinosa f. 219
----albicepsinflatum var. 224, 225
----album var. 225, 229
----anatolicum var. 221
----araraticum var. 228, 229
----armeniacum var. 219, 220
---- asiaticicterinum var. 219, 220
— — atriceps var. 219, 220, 229
— atrocyanum var. 219, 220
— — atrocrinaceuin var. 228, 229
----atrum var. 228, 229
Указатель латинских названий
333
uuiciim v.ii. 229
- bardarai var. 222
— — bartangiense var. 226, 227
— — bukiniczii var. 224, 225
— — cabristanicum var. 219, 220
— - clavatuni var. 228, 229
— — compactum var. 228, 229
-----compressum var. 175, 177
-----copticum var. 176
-----crassiceps var. 228, 229
— — crassicepsinflatum var. 224, 225
— — creticiroshanum var. 224, 225
-----creticum var. 228, 229
— — creticuminflatum var. 225
— — cyanorubrum var. 219, 220
— — echinoideum var. 219, 221
— — echinoid inf latum var. 224, 225
— — erinaceum var. 219, 220
— — erinacinflatuin var. 224, 225
-----erivanense var. 219. 220
-----fctissovii var. 220, 221
-----fetissovinflatum var. 224, 225
-----flaksbergeri var. 220. 221
— — glabrocyanum var. 220, 221
— — gorbunovii var. 226, 227
-----quasifalsefetissovii f. 226
— — griseofetissovii var. 220, 221
-----griseoicterinum var. 228, 229
-----griseosplendens var. 220, 221
-----griseovavilovianum var. 224, 225
-----hrasdanicurn var. 228, 229
— — humboldtii var. 221, 228, 229
-----humboldtinflatum var. 224, 225
-----icterinflatum var. 224, 225
-----icterinum var. 228, 229
-----jasgulemiense var. 227
— — kanaschii var. 221
-----karakurganiense var. 227
— -- karsiense var. 220,221
- — kerkianinflatum var. 224, 225
— — kerkianum var. 221
-----korschinskianutn var. 225
- - — linaza var. 228, 229
-----miguschovae var. 224, 225
-----montanum var. 224, 225
— miiticolinaza var. 220, 221
----- nepalicum var. 224, 225
-----nigrivehitinum var. 221
- — orientale var. 221
- - pcrcivalii var. 228, 229
— — pseudocrassiceps var. 228, 230
— — pseudoerinaceum var. 220, 221
— — pseudofetissovii var. 220, 221
-----pseudoicterinum var. 228, 230
-----pseudorubriceps var. 220, 222
-----pseudorubruin var. 228, 230
-----pseudosplendens var. 228, 230
-----quasialbiceps var. 222
-----qnasifetissovii var. 223
-----recognitum var. 177
-----roshanum var,. 223, 224, 225
-----rubriceps var. 220, 222
-- — rubricepsinflatum var. 224, 225
-----rubrinflatum var. 224, 226
-----rubruin var. 228, 230
- - rnfulinflatuni var. 224, 226
rufilium var. 228, 230
- - sarcziciiui var. 223
sericeiiin var. 220, 222
- - • setosuni var. 220, 222
------splendcns var. 229
'.pleudidorigidum var. 218 220,
222
— - subalbiceps var. 220,222
— — subalbicepsinflatum var. 224, 226
- -- subechinoidcmii var. 220, 222
— — subechinoidinflatum var. 224. 226
— — suberinaceum var. 220, 222
-- — suberinaceuminflatum var. 225
— -- subfetissovii var. 220, 222
---- subfetissovinflatum var. 224, 226
- - - - subicterinflatum var. 224, 226
- • ••- subicterinum var. 220, 222
----snbrubriceps var. 220, 222
—- -- subrubricepsinflatum var. 224, 226
— - subsericeum var. 220, 222
---- subscricinflaluin var. 224, 226
- - - - subsplendens var. 220, 222
— -- subsplcndensinflatum var. 224, 226
- - — surchianurn var. 220, 222
----urarticum var. 228, 230
— — vavilovianum var. 224, 226
wagarschapali var. 228, 230
wernerianuni var. 228,230
— — wernerinflatum var. 224, 226
— — witlmaekianum var. 228, 230
— — wittmackinflatum var. 224, 226
— compositum 88, 89, 186
— creticum 229
— dicoccoides 6, 13, 14, 17—19, 27, 32.
34, 39—50. 51, 54, 55, 56, 72, 82, 128,
132, 197, 235, 304, 315
----armeniacutn subsp. 45, 308
— — horanum subsp. 45, 46
----palestinicum subsp. 39, 45, 48
----syrio-palestinicum subsp. 39
----dicoccoides convar. 41, 45, 46—
48, 73
----incertum convar. 40, 45, 46. 49
----judaicum convar. 40, 41, 45, 46.
48
---- horanum prol. 46
— -- judaicum prol. 48
— — horanum grex 45, 46, 50
:— — judaicum grex 45
— — aaronsohnii var. 42, 46, 47, 309
----antilibani var. 47
---- arabicum var. 48
— — armeniacutn var. 309
— -- araxicum var. 308
— — dicoccoides var. 47
— — ferearabicum var. 48
— — ferevavilovii var. 48
— — fulvovillosum var. 48, 50
hermonicum var. 47
incertaaronsohnii var. 49.
incertaaronsohnii f. 49
— — jordanicum var. 47
----koernickei var. 48
----kotschyanum var. 49
----kotschyana f. 49
----kotschyi var. 42, 47
— — macraaronsohnii var. 48
— — nachitschevanicum var. 308, 309
— - araxicum f. 308
- namuricum var. 47
nigricans var. 309
nigrum var. 309
- - nigricans f. 309
— nudiglumis var. 310
— - - palestinicum var. 47
- - — pseudoaaronsohnii var. 309
----pseudoarabicum var. 48
on
331
Указатель латинских назпиниИ
pseudoaiaxicuin var. .309
— - pseudoarmeniacum var. 309
- - — pseudojordanicum var. 47
— — pseudokotsciiyi var. 47
- - — pseudonachilschevanicum var. 309
— — pseudoaraxicum f. 309
----pseudastraussianum var. 309
— — pseudothumanianii var. 309
pseudoarmeniacum f. 309
rufovillosum var. 48
----safedicum var. 48
----schweinfurthii var. 47
- — spontaneonigrum var. 47, 49, 309
— — spontaneovillosum var. 42, 47, 48
— — spontaneum var. 49
----spontaneum f. 49
— — straussianum var. 308
— — syriacum var. 49, 50
- —.. syriacum f. 50
• - - - tumanianii var. 309
— — vavilovii var. 48
— — hirtiglumis f. 310
— — nudiglumis f. 310
— — siriacae f. 39
— — straussiana f. 308
— — transcaucasicae f. 45
— dicoccum 6, 11, 15-18, 27, 28, 32,
34, 38, 41, 45, 50-69, 72. 73, 74, 75,
76, 77, 82, 94, 98, 99, 137, 146, 149,
181, 182, 183, 186. 194, 196, 197, 202,
212, 216, 233, 234, 315
— — abyssinicum subsp. 53, 56, 67—69
----asiaticum subsp. 51, 53, 56, 63—
67
— — azerbajdzhanicum subsp. 63
----dicoccum subsp. 51, 56, 57—62
----europaeum subsp. 56, 57
— — euroum subsp. 63, 66
— — georgicum subsp. 69, 72
— — ispahanicum subsp. 73
----maroccanum subsp. 56, 62 -63, 73
— — subsponianeum. subsp. 63
— — volgense subsp. 66
- - — tenaces sect. 154, 168
— — dicoccum convar. 57
— -- e.uscaldtinense convar. 57, 61—62
— — serbicum convar. 53, 63, 66—67
-- —- transcaucasicuni convar. 63—66,
73
----turgidum convar. 76
----arabicum prol. 68
— — armeno-anatolicum prol. 64
----bavaricum prol. 57
----carabachicum prol. 64
— — indostanicum prol. 68
------- iranicum prol. 64
— — jemenicum prol. 68
— —• meridionale prol. 57
— — oasiculum prol. 63
---- persicum prol. 63
— — proprioabyssinicum prol. 68
----tardo-europaeum prol. 57
— — tartarobalcanicum pro!. 66
— — typicum prol. 68
---- dicoccoides группа 39, 45, 310
— --- abyssinica gr. 68
- - — acruginosum gr. 64
----jarrum gr. 60
----georgicum gr. 69, 72
----luteatinctum gr. 65
----maroccanum gr. 62
----rossotartaricum группа 66
wiliti um 66, 67
- наrinisi>hnii var. 46
aerugiuosuiii var. 64, 65
— aesiivale var. 64
----ajar var. 68
— — albiraniosuni var. 58, 59
— — album var. 60
— — amharicum var. 154
- - armeniacum var. 65
- - - arniirual var. 61
- arpulunial var. 64, 65
— — arpurunial var. 64, 65
— - - arras var. 67, 68
— — arraseita var. 154
— — atratum var. 58, 59
— — autumnale var. 64
-- — bauhinii var. 58, 59
-----bispiculatum var. 58, 59
brownii var. 154
- carabacfiicum var. 61
— — caucasicum var. 65
- - -- chcvsuricinn var 65
— - - clraiamlicum var. 72. 73
.....claduruni var. 59, 60
— — compactomiegei var. 62
-----decussatum var. 58, 59
— - dicoccum var. 58, 60
•— - - diploleucum var. 58, 60
— — dodonaei var. 58, 60
— — erythrurum var. 58, 60
-----ethiopicum var. 152
-----farrum var. 60, 62. 65, 67, 68. 69
-----fictescmicanum var. 58, 60
— — flaksbergcri var. 65
-----fuchsii var. 58, 60
-----grabhami var. 154
-----gunbadi var. 65
-----haussknechtianum var. 63, 64, 65
— — hybridum var. 58, 60
-----iranicum var. 65
----- jakubzineri var. 65, 66
— — kotschyanum var. 49
-----krausei var. 59, 60
— — leucocladum var. 59, 60
-----liguliforme var. 58. 60, 62
— — luteo-tinctum var. 65
-----macratherum var. 58, 60
— — maturatum var. 67
-----mazzucattii var. 58, 60
- - — melanocladum var. 59, 60
-----melanurum var. 52, 59, 60
— — metzgeri var. 59, 60
-----miegei var. 62
— — modificatum var. 59
-----muticum var. 58, 60
-----nigroajar var. 68, 69
-----nigrum var. 68, 69
-----novicium var. 58, 61
-----persicum var. 156, 177
— — praecox var. 68, 69
-----pseudoarras var.. 68, 69
-----pseudoarraseita var. 155
— — pseudobrownii var. 155
-----pseiidocrylhrurum var. 58, 61
— - - pseudokrausei var. 59, 61
----- pseudomacratheruin var. 58, 61
— - - pseudopraecox var. 68, 69
— — pseudorubescens var. 156
— — pseudorubriramosum var. 59, 61
- - - pseudorufescens var. 156
pseudorufum var. 58, 61
— — pseudoschimperi var. 156
Указатель латинских названий
335
pscudotomentosum v;ir, 162
- pseudouncinatnm var. 155, |62
— — pubeseens var. 68, 69, 156
-----rubrigunbadi var. 65, 66
— •— rubriramosum var. 59, 61
-----rubrivillosum var. 162
-----rufescens var. 156, 168
-----rufum var. 58, 61, 64, 67, 69
-----schiinperi var. 157
- - - scliuebleri var. 58, 61
— - - semicanuin var. 58, 61
-----serbicum var. 66, 67
-----spontaneonigrum var. 47
-----spontaneovillosum var. 48
-----subatratum var. 58, 61
-----subcladurum var. 59, 61
-----subliguliformc var. 58, 61
-----submajus var. 58, 61
-----taschkentum var. 58, 61
-----timopheevii var. 45, 310
-----tomentosum var. 155, 162
-----tragi var. 58, 61
— — tricoccum var. 58, 61
— — uniaerugiuosutn var. 65, 66
unci mil и m var. 158, 168
— — unifarruni var. 62
-----uniluteotinctum var. 65, 66
-----unimiegei var. 62
— — vasconicum var. 62
-----violaceoarras var. 68, 69
-----volgense var. 60, 66, 67
— — vulpinum var. 162
— — bolnissietise subvar. 64
— — carthlicum subvar. 65
-----dmanissimum subvar. 64
-----laevi'gltimartim subvar. 65
-----pscllaviense subvar. 64
— — rarum subvar. 65
-----thianeiicum subvar. 64, 65
— — thrialeticum subvar. 65
-----aegyptiaca f. 69
-----aestivalis f. 64
------- altius f. 68
-----album f. 67
-----armeniaca f. 65, 66
-----arras f. 68, 69
-----asiatica f. 69
-----aulumnalis f. 64
-----caucasica f. 65
-----fere-nudum f. 67
-----irartica f. 65
-----jemeni. f. 68, 69
-----modificatum f. 59
-----osseticum f. 65, 66
-----rufum f. 67
----- tataricum f. 67
-----vasconica f. 62
-----volgense f. 67
-----indicum 68
— duhamelianum 202
— durum 6, 11, 16, 17, 19, 20, 25—30,
34, 55, 64, 76, 77, 81, 83, 91, 92—127,
131, 132, 135, 137, 138, 140, 146, 149,
150, 180, 182, 184, 197, 206, 207, 208,
212, 213, 275, 276, 277, 297, 316
-----abi/ssinicum subsp. 145, 149, 150
158, 160
-----caucasicum subsp. 125
-----durum subsp. 99, 100—126
-----expansum subsp. 100,128
-----falcatum subsp. 122, 124
hoi aiiicinn Mibsp. 99, 100, 126
12/, 219
orientate subsp. 128
- - sinicum subsp. 105
— — turgidum subsp. 76
- — africana sect. 116
- - — compacta sect. 163
— — densa sect. 160
- - elongate sect. 150
europeac sect. 103
- - mediterranca sect. 101
-----aglossicum convar. 101. 120—121
— -- caucasicum convar. 100, 102,
125—126
— — durocompactum convar. 100, 112,
116—120
— — duroramosuin convar. 115—116
-----durum convar. 100, 123, 101 —
116, 117, 120, 121
-----falcatum convar. 95, 100, 122 —
125
- - - - thumanjanii convar. 101
- villosum convar. 100, 12b 122
dtirorainosuni subconvar. 105,
106, 115—116
-----durum subconvar. 106—113
-----muticum subconvar. 105, 106,
113—115
— — europeac ser. prol. 101
-----mediterranca ser. prol. 101
-----commune группа 101, 127
-----duro-oblongum группа 128
-----aegyptiacum prol. 117
-----africanum prol. 116
— — asiaticum prol. 104
-----azerbajdzhanicum prol. 104
-----berbericum prol. , 103
— — borcalinicum prol. 104
— — carthlicum prol. 105
-----caspicum prol. 104
— — daghestanicum prol. 105
-----densiusculum prol. 103
-----durocompactum-stepposum prol.
117
•----cndcmicum prol. 101
-----eumediterraneum prol. 101
-----eustepposum prol. 103
-----expansoides prol. 158
-----falcatum prol. 122
-----horanicum prol. 116, 126
-----indicum prol. 105
-----jordanica prol. 101, 124
-----Haschgaricutn prol. 105
-----laxiusculum prol. 103
-----maroccanum prol. 117
— — melitense prol. 101
-----orientate prol. 124, 128
-----palestinicum prol. 101, 121
-----sardinicum prol. 103
-----syriaca prol. 101,124
-----teneroexpansoides prol. 158
-----tenerum prol. 150
-----villosum prol. 101, 121
-----cuspididentatum subprol. 116
• • - - eligulatum subprol. 101, 120
— rarum subprol. 117
-----sinaicuin subprol. 117
-----aegeum тип 104
-----asiaticum тип 104
-----cuspididentatum гни 116
-----falcata тип 122
-----horanica тип 126
22*
3,36
Указатель латинских названии
rurum । iiп 1 17
uep.yptiiicum gr. 106
afline gr. 106
africanum gr. 106
— alexandrinum gr. 112, 117
— — apulicum gr. 108
-----caerulescens gr. 109
— — erythromelan gr. 109
- — falcata gr. 122
- - — fastuosum gr. 109
— -- horanica gr. 126
— — hordeiforme gr. 109
-----italicum gr. 109
- — leucomelan gr. 109
— — leucurum gr. 110
— — libycum gr. Lil, 117
-----tnelanopus gr. Ill
— — murciense gr. Ill
-----muticoaffine gr. 115
-----muticocilbobscurum gr. 114
— — muficoliordeiforme gr. 115
-----muticoleiicurum gr. 114
-• —- muticomurcicnse gr. 115
— -- muticoprovincalc gr. 114
-----muficoitalicum gr. 114
— — muticovalenciae gr. 114
— — mulicoobscurum gr. 114
-----niloticum gr. Ill
— — provinciate gr. 112
-----reichenbachii gr. 112
— — valenciae gr. 113
-----syriaca gr. 101
-----jordanica subgr. 101
- - — abbaicum var. 160
— - aegyptiacum var. 106, 107
--- affine var. 106,107,121
-----epruinosaffine f. 106
— pseudoaf fine f. 118
-----fereaffine f. 106
africanum var. 106, 107, 121, 124
— — epruinosafricanum f. 106
-----griseoafricanum f. 113
-----aglossicon var. 120
-• — albiprovinciale var. 107, 108
----- albobscurum var. 106, 107, 114
-----albonigrum var. 153
— — albosemineum var. 153
-----alboviolaceum var. 153.
-----alexandrinum var. 112, 116, 117,
118
— — tristemudanianum f. 117
— — aliende var. 107, 108
— — amharicum var. 154
— — anliquoaethiops var. 160
-----apulicum var. 107, 108, 114. 121,
124
-----epruinosapulicum f. 108
-----fereapulicum f. 108
-----griseoapulicum f. 113
-----leviapulicum f. 110
-----nigrapulicum f. 108
— — arraseita var. ' 154
-----asmaricum var. 163
--- — atromarginatum var. 171
- -- australe var. 113, 114, 115
axumicum var. 154
-----aydiniensc var. 117, 118
-----tristeaydin'iense f. 117
-----bialbum var. 160
-----bi col or var. 160
boeufii var. 107, 108, 111
— brownii var. 154
larrnlevee 11var. 105, 107, 109
j <o г-i-.-i<-i 11Jem i-i).s f. 109
pseiiilocaerulesci-ns f. 11!)
- candicans var. 113, 114, 115
- - carneum var. 163
-----cauaff inc var. 126
-----cauapulicuin var. 126
-----cauhordeiforme var. 125, 126
-----caulcucomelan var. 126
-- — cauleucuruni var. 126
— — cineraceum var. 107, 108
-- - - comilans var. 160
— —• compactonegroides var. 164
- - compressum var. 164. 175, 177
— compylodon var. 106
.. — copticum var. 164, 176
— —- corvalanii var. 107, 109
-- — densoharraricum var. 161
-----densoalbonigrum var. 160
-----densoamharicutn var. 160
— - densoarraseita var. 161
densomenelildi var. 160
densopraevalenciae var. 162
-- - - densorubronigrum var. 161
- - densorubrum var. 162
— — densoschimperi var. 161
— — densosemiaristatum var. 161
— — densoviolaceum var. 161
— — discolor var. 112
-----dolgushinii var. 115, 116
— — domitians var. 156
-----durum var. 107, 109, 125
— — eritraeum var. 164
-----erythromelan var. 107, 109, 110,
121
-----fereerythromelan f. 109
— — levierythromelan f. 110
— — falcatafricanum var. 124
— — falcatapulicum var. 124
— — falcatifastuosuni var. 124, 125
— — falcatihordeiforme var. 124, 125
— — falcatileucomelan var. 124, 125
-----falcatileucurum var. 124, 125
— — falcatimelanopus var. 124, 125
— -- falcatum var. 124, 125
-- — fastuosum var. 109
-----ferealexandritium var. 112
-----fereerythromelan var. 109
-----ferehordeiforme var. 109
— — feremelanopus var. 111
feremurciense var. Ill
--- — fereniloticum var. Ill
— — fereprovinciale var. 112
— - ferereichcnbaclu var. 112
— — flaksbergeri var. 117, 118
— -- julvobscurum var. Ill
— — fulvoprovinciale var. 112
— — gallasorum var. 154
-----godjamicum var. 161
— — gondaricum var. 154
-----harraricum var. 154
— — heterochromum var. 174
— — hildebrandti var. 154
- - — horanaffine var. 127
horanafricanum var. 127
horanapulicum var. 127
.....horanohordciforme var. 107, 127
- - - • horanoleucuruni var. 127
lioranoine.lanopus var. 127
lioranoniloticum var. 127
hordeiforme var. 109, 125
ferehordeiforme f. 109
Указатель латинских незнании
337
leuiln>rdci]orme I 110
italiciini v.ir. 107, 109, 1 I I
- griseoilalicum f. I 13
- jakiibz.ineri var. 108, 109
—• kirtie var. 154
leucomclau var. 107, 108 109
121, 125
— epruinosileucomelan f. 109
— fercleucomelan f. 110
- griscolciicomclaii f. 113
- - tristeleucomelan f. 110
--- leuctirum var. 107, 110, 122, 125
- - epniinosileuctiruni f. 110
— fereleucurum f. 110
- serengei 119
-- levialexandrinuin var. 107, 110
— leviapiilicuni var. 107, 110
-- levicaciulesceiis var. 107. 110
— Jevierytliromelan var. 107, 110
— levihordeifonne var. 107, 110
— levileuconielan var. 107. 110
— levilciiciiruin var. 107. 110
levimelanopns var. 107, 110
levireichenbacliii var. 107, 1 11
— libycum var. 73, 105, 107 111
134
— epruinosilibycuni f. Ill
— flaksbergcri f. 117
— odsissianttm f. 134
— marginatum var. 161
— melaleucum var. 107, 111
— melanopus var. 107, 108 110 111
113, 122, 124, 125
— eprninoshnelanopiis 1. Ill
— feremelanopus f. Ill
— griseomelanopus f. 108
— pseudomelanopus f. 119
—- menelikii var. 158
-- murciense. var. 107, 111
- feremurciense f. Ill
-- multicalbobscurum var. 113, 114
— miticalboprovinciale var. 113, 111
- multicitaliciini var. 11,3, 114
- muticoboeufii var. 113, 114
— muticobscuruni var. 113, 114
— niuticocaeriilescens var. 113. 114
— inuticonazilliense var. 113, 114
— muticovalenciae var. 113, 114
— nazilliense var. 111
— negroides var. 155
— negusiiantim var. 162
-- nigrobrownii var. 155
— nigrodensoarraseita var. 161
— nigrodensomenelikii var. 162
— nigrodensoschimperi var. 161
— nigrodominans var. 156
— nigroinarginatum var. 155
— nigromenelikii var. 155
— nigropraehordeiforme var. 156
— nigropraesemiticum var. 161
— nigroptolomaeum var. 164
— nigrosomdlensc var. 161
— nigrosteudelianum var. 156
-- nigrotypicum var. 164
-- nigroviolaceum var. 155
— - niloiiciini var. 107. 1 11
fereniloticum f. Ill
- - griseoniloticum 1. 113
— nubt'eum var. 164
- obscurmn var. 106, 107, 111, 114
- pilosa 1 line var. 108, 111, 112
pilosohordeilorine var. 108, 111
j) i I < >*-r > 11 и-1 ;i i >< > | > I is va i 108, 112
pi I < >s() 11111 re 11111 st' var 108, 1 1 2
pdosonigrtim var. 161
- praeapulicum var. 155
- - - praehordeiforme var. 156
-- praesemilicum var. 161
- - praevalcnciae var. 155
prospicuum var. 155
protomelanupus var. 155
jirovinciale var. 107, 108, 1 12, 122
- - fereprovinciale f. 112
pseudoailiue var. 1 18
— -- psetidoafricanum var. 119
------- pseudoalbobscurum var. 118
- pseudoalboprovinciale var. 118
-- -- pseudoalbovirdaceum var. 15-1
pseudoalexandrinuni var. 107, 112,
117
-- — epruinosalexandrimim i. 112
— — ferealexandriniun f. 112
-- varicfjatum 1. 112
psei id oa p 111 ic i и 11 v.ir. 1 18
psen (I i ia r a Hi c 11 in var. 163
psetidocaeriilescens vai. 118. 119
— — pseudocarneum var. 164
— — pseudocopiicum var. 164
— pseudodensomenelikii var. 162
-- pseiidodomiiuins var. 155
-----pseudocaertilescens var, 118. 119
— — pseudogondaricum var. 155
— pseudohordeifonne var. 118. 119
-- — ferepseudohordeifonne f. 119
— -- pseudoitalicum var. 118, 119
- •— pseudoleucomelan var. 117
pscudoleucui urn var. 1 19
pseudomelanopus var. 118, 119
pseiidoinurcien.se var. 118, 119
pseudoobscurmn 118, 119
pseudopraehordeijorme var. 155
-- --- pseudopraesemiticum var. 162
pseudoprovinciale var. 118, 119
- - pseudoptolomaeum var. 164
- pseudorarum var. 156
----- pseudoreichenbachii var. 118. 119
— - pseudorubroaristaium var. 156
— — pseudorubropubescens var. 162
— --- pseudorubroviolaceum var. 156
— - pseudoschechurdinii var. 115
— pseudosomiden.se var. 162
— - pseudo! у picum var. 16'1
pseudovalenciae var. 118, 119
-- — ptolomaeum var. 165
— — quasiafiine var. 120
— -- qtiasiafricanum var. 120
-• — quasialcxandrinuni var. 120. 121
-----quasicrytliroinelan var. 120, 121
— — quasihordeiforme var. 120, 121
— — quasileucomelan var. 120, 121
— — quasileucuruiu var. 120, 121
-----quasiinelanopus var. 120, 121
- — quasimurcieuse var. 120, 121
— — ramosafricanum var. 115, 116
• - - • rainosapulicuin var. 115, 116
- ramosohordeifornie var. 115, 116
raniosoleucomelan var. 115, 116
ramosomelanopns var. 115
raniosobsciiriiiii var. 115, 116
rurissimum var. 156
rarum var. 154
reieheiibacli i i var. 106, 107, 112,
122, 125
discolor 1. 112
338
Указатель латинских нациииnil
epi uiiiosn I'ichei11>acliii 1 112
lei ('reichenbachii f. 112
— — rulirinifji’iin> var. 118, 11!)
— — rubroaristatum var. 156
— — rubromurinum var. 107, 113
— — rtibromulicum. var. 156
----- rubronigrum var. 156
-----rubropubescens var. 162
- -- rubroviolaceum var. 152
- schechurdini var. 113, 114
-----sehimperi var. 157
-----scliweinfurtliii var. 157
-----serengei var. 118, 119
— — mustafaevii f. 119
- — simiense var. 164
— — singulum var. 157
— — somalense var. 162
- — stebutii var. 113,115
-----steudeliaiuim var. 157
-----subauslrale var. 113, 114, 115
-----subbrownii var. 157
-- — submenelildi var. 157
— - - subiieguslianum var. 162
— subpraehordeifoi me var. 157
-----subpraevalenciae var. 157
-----subprolomelanopus 157
— — subpscudorarum var. 157
-----subschimperi var. 157
-----taborense var. 157
-----takussicum var. 165
-----fchertchericum var. 162
-----thebalcum var. 164
-----tigrense var. 157
-----typicum var. 165
-----valenciae var. 107, 113, 114
-----variegatum. 112
— — villosaffine var. 121, 122
-----villosafricanum var. 121, 122
— — villosapulicum var. 121, 122
-----villosileucomelan var. 121, 122
-----villosileucurum var. 122
-----villosimelanopus var. 122
-----villosiprovinciale var. 122
— — villosum var. 122
— — violaceomuticum var. 158
-----virsaviacum var. 118, 119
-----yesilkoense var. 118, 119
-----griseocompressum f. 120
-----acutidenticulatum 145
-----aegyptiaca 117
-----aristatum a 106
-----asiatica f. 104
-----austrostepposum 104
-----compactum 103
-----cyprium 104
-----densiusculum 103
-----durocompactum 106, 116
-----eligulatum 120
-----endemica 101
-----muticum p 113
-----oblongum 103
-----orlentalistepposum 104
-----silvistepposum volgense 104
— '— stepposum volgense 104
----- taganrocense 103
— — viltosa 121
-- X edwardii 14
— eloboni 154
— [arrum 60
-- fastuosum 109
— X fuagicidum. 14, 317
— fuscocoeruleum 201
!;eoi oil uni 6!), 1.32. I 78
< in lliliciim convar. I 78
turanicum coiiv.ar. 1 28
glaticum 135, 202
- globijorme 273
— hermonis 39
— hirtum 202
— hordeiforme 109
- hornemannii 293, 301
- - hostianum 259
-- liybernum I I, 230
ibericum 178, 184
— --- fuliginosum var. 184
albijuliginosum f. 184
- - - dominans f. 181, 185
- laxiusculum f. 184, 185
— -- pilosostramineam var. 185
— — rtibiginosum var. 185
- - — brevispicatum f. 185
- •• — cauliplenum f. 185
— — diiriusculum f. 185
- elongation f. 185
pi iscoi ubiginosion f. 185
longiaristaiiim I. 185
--- nigriaristatum f. 185
— — pilosum f. 185
— — uniaristatum f. 185
— ••• slramineum var. 185
- breviaristatum 1. 185
— - densiusculum f. 185
---- diiriusculum f. 185
---- laxiusculum f. 185
— — nigriaristatum f. 185
----nudum f. 185
----pilosum f. 185
— imercticum 192
- - inflatum 244
— ispahanicum 5, 6. 11, 17, 31, 34. 70,
73—75, 138. 197, 235
— — biaristatum var. 75
----ispahanicum var. 75
----ispahanorufum var. 75
— -- uniaristatum var. 75
— jakubzineri 5, 6, 8, 34. 78, 92, 211 11, 15—17, 31
- - karamyschevii 5, 6, 11. 17, 34. 52. 69—73, 74, 82, 132, 189, 197, 235 — — karamyschevii var. 73 — — nigrescens var. 72 rubidum var. 73 — kiharae 5, 6, 11, 15, 17, 20, 30, 32, 278, 280, 320, 321 — levissimum 141 — linnaeanum 88
— macha 6, 11, 13, 17, 32, 34, 70, 72.
73, 186—193, 196, 206, 207, 208, 216.
234
— — densiusculum subsp. 192
-----georgicum subsp. 69
— — imereticum subsp. 72, 192
— — normale subsp. 190
-----tubalicum subsp. 190
densiusculum । convar. 187, 189,
190, 192—193 macha convar. 187, 189, 190-192
normale группа 190
album var. 192 colchicum var. 191
ericz.janae var. 191
georgicum var. 191
ibericum var. 191
- — letshchumicum var. 191
Укапитель латинских mmuimuii
macha var. 191
megrelicmil var. 191
pa 1 a e< >co I ch icm 11 var. 192
- - palaeoimereticum var. 192
- -- pianocoin pressuni var. 192
- - rubiginosum var. 192
— rubri\eliitinuin var. 192
- - scharaschidzei var. 191
-----subcolchicum var. 191, 192
- - - - subletshchumicum var. 191, 192
submegrelicum var. 191, 192
- macrodon 199
-- maximum 135
— michaelii 35, 37
— militinae 5, 6, 11, 14, 15. 17, 19 28,
30, 32, 183, 234, 236, 280, 304, 311.
319—320
-----albimilitinae var. 320
-----militinae var. 320
— monococcum 6, 11, 13, 15—17, 19,
21, 26, 29, 31, 38, 41, 44. 48, 196, 216.
233, 234, 280, 281, 283, 284, 293—303.
312, 315, 318, 319
-----aegilopoides subsp. 281, 287, 288,
289, 290
—• — boeoticum subsp. 281
-----cercale subsp. 293
-----micliaelii subsp. 35
-----monococcum subsp. 293
— — urartu subsp. 35
-----alemanum группа 299, 300
—• —• armenoanatolicum группа 300
-----carabachicum группа 300
-----ciscaucasicum группа 300
-----daghestanicum группа 300
— — heothinum группа 299, 300
-----ibericum группа 299
-----montanooccidentale-europaeum
группа 300
— — occidentalemediterraneum
группа 300
-----tauricum группа 300
-----albohornemannii gr. 301
-----hohensteinii f. 301
-----ratschinicum f. 301
-----atriaristatum gr. 301
-----biniarisiata gr. 285, 290
— —- flavescens gr. 301
-----bigranum f. 301
-----hornemantiii gr. 301
-----macedonicum gr. 302
— — eredviantim f. 302
-----punctatum f. 302
-----nigricultum gr. 302
-----pseudoflavescens gr. 302
-----sofianum gr'. 302
-----tauricum gr. 302
-----uniaristata gr. 285
-----vulgare gr. 302
-----atriaristatum var. 301
-----baydaricum var. 287
-----boeoticum var. 287
— eredvianum var. 302
-----punctata f. 302
----- flavescens var. 301, 302
- -- - helenae var. 288
— — hohensteinii var. 301
-• — hornemannii var. 301, 302, 315
— — capitata f. 302
-----sangesuri f. 302
— — laetissimum var. 302
-----larionowii var. 288
macedonicum var. 301,302
psi'mlocredi'iimum. 1. 302
mat/ssurianii var. 288
monococcum var. 301, 302
nigricultum var. 301, 302
- nigroalrum var. 301
- pancici var. 289
- - pseudoboeoticum var. 289
psetidoeredvianum var. 302
pseudoflavescens var. 301. 302
pseudohoriiemannii var. 301, 302
pseudomacedonicimi var. 301, 302
pseudouulgare var. 303
pubescens var. 301
- --- pubescetilinigrtim var. 289
ratschinicum var. 301
- - - sangesuri var. 301, 302
— - sofianum var. 301. .302
— densiuscula f. 302
----laxiuscula f. 302
— — st/mbolonensc var. 289
symphaeropolitainim var. 301. 302
- - -- parallelica f. 302
----patula f. 302
----tatariciim var. 301
----tauricum var. 301, 302
----vulgare var. 301, 303
----zuccariorii var. 290
— — aegilopioides 281, 285
----boeoticum 281, 284
— - - cereale 293
---- ettmonococcum 293
----lasiorrachis 281
— montanum 308
— mutieum 230
— nigrescens 289
— nikolai 308
— obtusattim 13, 186
— orientate II, 124, 127, 131
— — bandirmaicum var. 132
----daralagesi var. 132
— — ferghanicum var. 13.3
— — gasimustafakemalii var. 133
----generosum var. 133
— — insigne var. 133
— — jalovanianuni var. 133, 134
----notabile var. 134
— ovatum 201
— palaeocolchicum II, 69, 72
— — chwamlicum var. 73
— — nigrescens var. 73
— •— rubidum var. 73
— paradoxum 177
— percivalianum 128
— perci valii 128, 132
— persicum 6, 8, 11, 15—19, 27, 28, 51
56, 137, 146, 149, 150, 177—185, 211,
216, 217, 234—236, 264, 279, 297, 316.
320
— — caucasionis dika группа 178
----dzliavachetis dika группа 178
— — armenogeorgicuin prol. 180
---- daghestanicum prol. 180
- — arnual var. 183, 184
- arnurual var. 183, 184
- arpual var. 183, 184
arpurual var. 18.3, 181
- • - - arpurunial var. 184
— — cinereuin var. 184
— — darginicum var. 184
— — fuliginosum var. 183, 184
310
Указатель латинских mciminiii'i
- — ismeticum var. 184
narpti.-il var. 184
narpuru.il var. 181
-- nigrirubiginosum var. 184
osseticum var. 184
persiciun var. 183, 184
-- --- pseudortibiginosum var. 184, 185
— pseudostraniineuin var. 184, 185
- - rarissimuni var. 184, 185
- - — rubiginosum var. 179, 184, 185
capitatuni f. 185
-----stramiiieuin var. 184, 185
— — zhukovskyi var. 184, 185
- pctropavlovskvi 5, 6, 11, 15—17, 31
34, 131, 135, 138, 277—280
-----petroerythroleuc.on var. 279, 280
-----petroferrugineum var. 279, 280
-----petropavlovskyi var. 279, 280
— petropseudobarbarossa var. 279,
280
— polonicum 6, 11 — 13, 16, 17, 27, 34,
52, 73, 75, 128, 132, 135—145, 150,
277, 279, 320
-- — abyssinicum subsp. 138, 139
-----mediierranetim subsp, 139
-----polonicum subsp. 138, 139—145
-----compactum convar. 140, 144—145
-----polonicum convar. 140, 141 —144
— — eumediterraneum prol. 140
— — palestinicum prol. 140
-----stcpposum prol. 140
— — compactoides gr. 144
— — martinari gr. 145
-----nigrescens gr. 142
— — oblongoquadratum gr. 141
-----pseudomartinari gr. 145
-----pseudovillosum gr. 143
-----ramosum gr. 143
-----rubrovelutinum gr. 143
-----vilmorinii gr. 144
-----abyssinicum var. 139
— — aegeicum var. 144
-----anatolicum var. 144
— — anomalum var. 142
-----aristinigrum var. 145
— — attenuation var. 141, 145
— — caryopsirubrum var. 144
— — chrysospernuini var. 141, 142
-----compactum var. 144, 145
-----saronicum f. 145
— — crassispicatum var. 145
— czernjaevii var. 141, 142
- — devialuni var. 139
-----elongatum var. 144, 145
— — eucompacium var. 144
-----gorskyi var. 141, 142
----- gracile var. 144
— — halleri var. 144, 145
-----heidelbergi var. 144, 145
-----homsicuin var. 144, 145
-----incertum var. 142
-----intermedium var. 144, 145
-----kiritchenko var. 141, 142
-----mustafaevii f. 141
— — koernickei 139
-- — levissiinum var. 141, 14 2
- - • - martinari var. 144, 145
— meczelinsbense var. 143
-- micz.ininii var. 141
- — nigrescens var. 141, 142
-----nigrobarbalum var. 145
— — novissinium var. 141, 142
— - - pissarevii var. 141, 142
- - - poionicum v.'ir. 14 1,14 2, 1 13
---- siibmiilicttm f. 142
prjanischnikovii var. Ill, 113
- pseiidocarvopsirubrum var. 1 14
145
— - - psetidoclirvsospermum var. 1 12
143
----pseudocoinpactum var. 144, 145
pseudolevissmiiiii) var. 142, 143
pseudomiirtimiri var. 145
- - - pseuilonoeissimiim var. 143
— — pseudosporadicum var. 139
----pseudovillosum var. 141, 143
— - pscudozaferanicum var. 144, 145
— — ramosum var. 141, 143
----rarovillosuin var. 139
--- —- rubrosemineinn var. 139
— — rubrovelutinum var. 141, 143
----rubrovestituni var. 145
— — rufescens var. 141, 143
----saronicum var. 145
— — seringei var. 143
— — skaiosubovii var. 141, 143
- - - siilirnovii var III, 14,3
— - sporadicum var. 139
— -- submuticum var. 142
---- talonovii var. 141, 143
— — tibeticum var. 141, 14,3
— - - vestitum var. 145
— — villosum var. 141, 143
----gracile f. 144
----vilmorinii var. 142, 144
----violaccum var. 142
— — zaleranicum var. 144, 145
----z.onguldakicum var. 144, 145
----bar bat uni 142
----deformation 143
— — longiaristatum 145
----crassispicatum b. 145
----oblongum 141
— — quadratum 141
- - — simplex 141
— protomacha 72. 188, 275
— protopersicum 274
— pseudodidiamelianum 201
— pubescens 293, 301
— - rufescens 202
— pyramidale 117, 175
— — arabicum var. 119
— — aydiniense var. 117
— — compression var. 119, 175
- copticum var. 119, 176
— — diyarbekiricum var. 119
— — [alsoapulicum var. 118
----mudanianum. var. 119
— — pseudoaffine var. 118
— — pseudocoerulescens var. 119
— — pseudocompresstim var. 119
— — pseudocopticum var. 118
— — pseudodiyarbekiricum var. 119
— — pseudomudanianum var. 117
----pseudoreiclienbachii var. 119
-- -- recognitum var. 119
rubi onigriim \ar. 119
• - riibrosemineiim var. I 19
i ubi ospicti (и in var. 119
rubi oveliilinum var. I 1 9
recognitum 175
— sativum 39, 230
— - dicoccum subsp. 39
dicoccoides var. 39
Указатель латинских lumuaiun'i
:м
s/tella a 19!)
dicoi i um Ii 5(1
tenax c. 7(>, 93
- compactum 213
iluriiin 93
erinacettm 219
piinimidale 117
- - spelta 199
turgidum 76
vulgare 230
siculum 109
siuskajac 5. 6. 11, 15, 17, 19, 31 230,
294, 303, 320
— X soveticum 14
— spelta L. 6, 11, 16, 17, 19, 20, 30, 32,
34, 60, 182, 183, 189, 193—208. 216,
233, 234, 240
-- --- europetim subsp. 199
-----kuckuckianum subsp. 198 199,
200, 202 -208. 209
macha subsp. 186, 191, 192
-----spelta subsp. 193, 194, 198, 199 —
202, 203
- vavilovii subsp. 209
—- imeretictim convar. 192
— — iranictim convar. 203, 205
- — kuckuckianum convar. 203—208,
209
— — machoides convar. 208
-----tarakanovii convar. 203, 208
- — transcaucasicum convar. 203
— — tubalicum convar. 191
-----alletnanutn prol. 199
-----asturicum prol. 199
— — bavaricum prol. 199
— — ibericum prol. 199
- - — albamissum var. 206
-----albarduini var. 207
-----albiduhamelianum var. 204, 205
-- — albispicatum var. 201, 205
-----albivelutinum var. 199, 201
-----album var. 201
-----alefeldii var. 201
-----amissum var. 201
----- arduini var. 201
-----arpurual var. 205
- - — asialbispicatum var. 204, 205, 206
— — asialbivelutinum var. 205
-----asiamissum var. 204, 205
- -- asicaeruleum var. 205
-----asiduhamelianum var. 204, 206
-----asineglectuin var. 204, 206
-- - - asirecens var. 204, 206
— — asirubrivelutinum var. 205, 206
— — baktiaricum var. 206, 207
— -- biildojii var. 207
— — caeruleum var. 201, 202
-----dekaprelevichii var. 205, 206
— — dicoccon var. 60
-----dorofeevii var. 204, 206
-----duhamelianum var. 201, 202
— — erubescens var. , 202
— — flaksbergeri var. 204, 206
-----griseoturanorecens var. 204, 206
— — isfaralbispicatum var. 204, 206
—- - - ispharaeciim var. 204, 206
-- — ispharobakliaricutn var. 204, 206
- jakubzineri var. 204, 206
-- karabachicum var. 204, 206
- - — kusaricum var. 207
-----laczinicum var. 207
marinae var. 204, 206
111 < и a I к I i i var. 205. 207
иiи|mlu11ii u var. 201. 207
111111>11r11a I var. 20 I, 207
iiiiisIaphacvii var. 201. 207
in mice I и m var. 20I, 202
uigiesceiis var. 202
- pseudoalboarduini var. 207
- - - pseudobaktiaricum var. 205. 207
- - -- pscudobuldojii var. 204, 207
---- - pseiidoschenkii var. 207
- psi-tidosharkordii var. 205, 207
- - pseudovulpinum var. 207
---- - recens var. 201, 202
-----rubrivelulinum var. 201, 202
— — rufum var. 202
-----saharae var. 196
-----samuricum var. 205, 207
-----schaartusicum var. 205. 207
— — sheinachinicum var. 205, 207
----- sclienkii var. 201, 202
— - - ramososctie.nki f. 202
— - sharkordii var. 207
— - sitiskajac var. 205, 207
siibbaktiaricum var. 202 203 205
207
-----subbuldojii var. 204, 207
---subsliarkordii var. 205, 207
-----tarakanovii var. 208
— — tashausicum var. 205, 208
-----Ihumanianii var. 204, 208
-----turanalborecens var. 204, 208
-----turanalefeldii var. 204, 208
-----vavilovii var. 204, 208
-----viridalbispicatum var. 201, 202
-----viridarduini var. 201, 202
-----vulpinum var. 201,202
-----zhukovskyi var. 205, 208
— sphaerococcum 6, 11, 17, 34 197,
235, 269 -277
-----alborubiginosum var. 277
-----echinatuin var. 275, 276
-----globosum var. 275, 276
-----ischkaschimicum var. 275, 276.
-----jakubzineri var. 275, 276
-----kibraicum var. 271, 275, 276
-- — kostoffii var. 276
----- malkoffii var. 276
pakislaniciiin var. 276
- - — popovii var. 276, 277
• - — randhawaii var. 276, 277
— — rotundatum var. 276, 277
-----rubiginosum var. 276, 277
— -- rubralbum var. 276, 277
----- riibraristatum var. 276, 277
—- - - riibriglabnnn var. 276, 277
spicatum var. 276, 277
-----tumiduin var. 276, 277
— spontaneum 281, 284
— — aegilopoides subsp. 285
-----thaoudar subsp. 285, 290
-----albinigrescens var. 290
-----balaclavicum var. 291, 292
— — kovarskii f. 292
-----balansae var. 291,292
- - - bicolor f. 292
baydaricum var. 287, 289
- - subbaydaricum f. 289
- boeoticum var. 287, 288, 289. 290
-- — albonigrum f. 287
-----microspermum f. 288
-----subalbum f. 289
- - viridohoeolictim 1. 290
Укаиатем латинских напиапий
bonimuelleri var. 292
- - Insulin var. 292
- jumidum f. 292
torosofumidum f. 292
zhukotciskyi f. 293
haussknechtii var. 292
• - - - helenae var. 288
----- larionovii var. 288
— — luteinigrum var. 292, 293
- - — viridonigrum f. 293
-----mayssuriani var. 287, 288
- cinereum f. 287
— — mazzettii var. 292
— pancicii var. 289
— — luteonigrum f. 289
-----nigrochlorococcum f. 289
-- — subpancicii f. 289
—• — pseudoboeoticum var. 287, 289
— — aznaburticum f. 287
-----chlorococcuin f. 287
— subpseiidoalbum f. 289
- - —- pseudoreuteri var. 291, 293
-----azerbajdjanicum f. 291
-----pseudozuccarinii var. 288, 289
- — kovarskii f. 288
— — pubescentinigrum var. 288, 289
-----kurbagalense f. 288
-----subpubescentinigrum f. 289
— — reuteri var. 292, 293
— — albiatratum f. 292
-----nigroreuteri f. 292
-----rufinigrum var. 293
:----symbolonense var. 287, 289, 290
-----avdalaricum f. 287
-----capitata f. 290
— — theydjerabaki f. 290
— — tuberculatum f. 287
-----viridosymbolonense f. 290
-----villosonigrutn var. 288
-----zuccarinii var. 288, 290
-----bourgeani f. 290
— — garniense f. 288
— tenax 213
-----compactum A II 213
-----durum В II 119
-----turgidum В I 76
-----vulgare Л I 230, 265
— — anglicum var. 265
— thaoudar 285, 290
-----albiatratum var. 292
-----albinigrescens var. 290
-----atratum var. 290
-----azerbajdjanicum var. 291
-----balansae var. 291
-----bicolor var. 292
-----bornmuelleri var. 292
— — jumidum var. 292
-----haussknechtii var. 292
-----mazzettii var. 292
-----nigrituberculatum var. 292
-----nigroreuteri var. 292
-----pseudobalansae var. 291
-----pseudoreuteri var. 293
-----reuteri var. 293
-----rindi var. 291
- - — roseum var. 291
-- — rufinigrum var. 292, 293
- schorbulachicum var. 292
-----toroxofumidum var. 292
— — virescens var. 291
— X timococcum 318
— timopheevii 6, 11, 13 21, 28 -30,39,
•15. 18.';. 185. 1.87, 216, 23-1. 273, 280.
283, 297, 299, 301, 307, 308, 310 316,
318, 319. 320. 321
aiariiticiun subsp, 308
- phinosupei louipaihim ^r. 316
- - 11ipГЧ1 n var. 31 6
-- nibiginosturi var. 316
— -- timopheevii var. 316
- - — typicum var. 316
— vi 1 iculosum var. 316
- -- zhukovskyi var. 316
- - tricoccum 61
- - tuhaliciim 190
— turanicum 5. 6, 11. 17, 34. 99. 123,
124, 127 134, 137, 275, 277, 279
------falcatuin subsp. 122, 124, 125
------orientale subsp. 124
— — montanostepposum convar. 277
------anatolico-caspicum prol. 132
— irano-turkestanicum prol. 132
- — aristiforme I. 277
— — bandinnaicuin var. 125, 132, 133
- - — daralagesi var. 132. 133
- - - ferghanictim var. 133
- - - pa z i in ii sta fa kem a I i i var. 125,
133. 1.34
------generosum var. 133, 134
— — griseoinsigne var. 132
— — insigne var. 133
------- jalovanianum var. 133
------nolabile var. 133, 134
------odsissianum var. 133, 134
------pseudobandirmaicurri var. 124,
133, 134
------pseudogenerosuin var. 133, 134
------ pseudojalovanianum var. 133, 134
------rarissimum var. 125, 133, 134
------turanaffine var. 133, 134
------turanalexandrinum var. 133, 134
— •— iuranoapulicum var. 133
— — turanohordeijorme var. 133
— — turanoleucurum var. 133
— — turanimurciense var. 133, 134
— Itiraninigruin var. 133, 134
- — turanobscurum var. 133, 134
------turaniprovinciale var. 133, 134 ’
------wimsnurstii var. 133, 134
— turgidum 6, 1 1 — 17, 22, 24- 27. 34,
38. 39, 41. 44, 51, 52, 61. 76 -92, 93,
106, 115, 125, 126, 132, 137, 141, 149,
150, 186, 197, 211, 216, 235, 308, 316
— — abyssinicum subsp. 145, 165, 166
17'1, 175
— — armeniacum subsp. 308
------carlhlicum subsp. 178
------dicoccoides subsp. 39
------dicoccum subsp. 50
------durum subsp. 93
------gigantosemineum subsp. 84
— — jakubzineri subsp. 92
------mediterraneum subsp. 83, 84
------montanum subsp. 84
— — palaeocolchicum subsp. 69
------polonicum subsp. 135
------sementivum subsp. 76, 93
------sinicum subsp. 82
- -- limopheevii subsp. 310
- — turanicum subsp. 128
— — volgense subsp. 66
------compacta sect. 175
— — densa sect. 171
- - elongata sect. 165
Указатель латинских mi.immin'i
( arthlicum <(>iiv;iг. I 78
compositum convar. 78, 82, 83,
88—92
- - dicoccon convar. 50
- durum convar. 93
- - polonicum convar. 135
-- turanicum convar. 128
— turgidum convar. 82—88, 89
— anglicum prol. 83
• - caspicum prol. 84
- - hapalocompactum prol. 83
iranicum prol. 84
— meridionale prol. 83
— occidentale-europaeum prol. 83
- -- orientate prol. 84
— transcaucasicum prol. 84
— cristafum тип 83
- - aethiopicum gr. 146
— buccale gr. 86
- - carthlicum gr. 178
— columbinum gr. 89
— dicoccon gr. 50
— dinurum gr. 86
— dreischianum gr. 86
— durum gr. 93
— gentile gr. 87
— herrerae gr. 87
— jodurum gr. 87
— lusitanicum gr. 88
— martensii gr. 87
— megalopolitanum gr. 87
— nigriglumaceum gr. 86
— palaeocolchicum gr. 69
— polonicum gr. 135
— pseudosalomonis gr. 88
— rubroatrum gr. 88
— salomonis gr. 88
— speciosissimum gr. 88
— speciosum gr. 88
— turanicum gr. 128 '
— turgidocompactum gr. 175
— abchasicum var. 85, 86
— acutum var. 167
— addisabebae var. 167
— albofumidum var. • 85, 86
— albojodurum var. 85, 86
— albovillosum var. 167
— alibekliense var. 89, 90
— ankoberense var. 168
— atratosanguineum var. 168
— atroalbum var. 168
— otrocompactum var. 175
— atroinflatum var. 171
— atromarginatum var. 171
— atrorubrocompactum var. 175
— atrorubrum var. 168
— brevispicatum var. 175
— buccale var. 85, 86
— candiense var. 90
— caryopsirubrum var. 88
— caspicum var. 91
— caucasicum var. ' 89, 90
— centigranium var. 89,90
— cervinum var. 89, 90
— cinereum var. 85, 86
coeleste var. 89, 90
coloratum var. 168
columbinum var. 89, 90
— compactoberrerae var. 86
compactojodurum var. 86
compositum var. 89, 90
— compressum var. 175
i ophi um var. 1 76
ct11>11111111 var. 90
- - - - dangalense var. 1 7 I
dccoloratocompactum var. 177
- - dccolorattun \ ar. 168
densoatrorubrum var. 171
----densoatratopurpureum var. 171
----densoatratosaguineum var. 171
— — densodecoloratum var. 171
-- — densofulvuni var. 173
---- densoinflatum var. 173
- densopilosum var. 173
-- -- densoplanum var. 173
----densopullatum var. 173
----densopurpureum var. 173
----densorubidum var. 173
— — densosanguineum var. 173
— — dicoccoides var. 39
----diflusum var. 174
----dinurum var. 84, 85, 86
- - — dispersum var. 168
— - dreiscliianum var. 85, 86
----dubium var. 89
— — d и pl is var. 89
— — duplorubrum var. 168
— — falsejodurum var. 85, 87
- — falscmirabile var. 90
- - — ferespeciosissimum var. 85, 87
- flaksbergeri var. 174
— - fulvocompactum var. 176
- • - julvoinflatum var. 168
- - — fulvum var. 168
-- •• fumidum var. 79, 85, 87
— — furcatum var. 168
----gentile var. 85, 87
----giganteum var. 90,91
----griseomegalopolitanum var. 85,
87
----griscopseudomirabile var. 85, 87
----griseorubroalbum var. 85, 87
— — griseorubroatrum var. 85, 87
----griseosalomonis var. 85, 87
-- — harlanii var. 174
— — herrerae var. 79, 85, 86, 87
— — heterochromum var. 174
— — incanocompactum var. 177
- incanum var. 174
— '— izmiricum var. 87
----jodurum var. 79, 84, 85, 87
— — laxinflatum var. 169
— — lencoranicum var. 90, 91
— — levipavoninum var. 90, 91
----leviramosostriatum var. 90, 91
----linnaeanum var. 89
— — lusitanicum var. 88
— — kushianum var. 169
— — marginatovillosum var. 174
— martensii var. 85, 87
----megalopolitanum var. 79, 85, 87
----melanatherum var. 79, 85, 87
— —- melanocephalum var. 169
- - — mirabile var. 90, 91
— -- miscibile var. 85, 87
- - - inidigenitum var. 90. 91
- - innticospeciosum var. 86, 87
nachitschevanicuin var. 90, 91
nemaiisense var. 87
nigrobarbatum var. 79, 85, 87
- - nigrobrevispicatum var. 177
- — nigrobuccale var. 85, 87
-- nigrocoloratum var. 169
nigrodcnsoplanum var. 174
Указатель латинских iimiiiaimii
иigi oth’iisopurptiiейin var. 17 1
- - nigtodeiisosangtiineiim var. 17 1
- - - nigrodispersum var. 169
nigroduplorubrum var. 169
n i g r< >g h 11 и an i in var. 79, 85, 87
ni groi n j I a I и in var. 169
- -- - - 11igroinarlensii var. 85, 88
— — nigropseudomirabile var. 85, 88
— — nigropsetidosalouioiiis var. 85, 88
— -— nigropurpureum var. 169
-- — nigroriibidocompactum var. 177
— - - nigrorubidum var. 169
— nigrorubroinf latum var. 169
— -- nigrosanguineum var. 169
— - - notabile var. 177
— - pavoninuni var. 90, 91
- - — persivalii var. 169
- - — pilosocompactum var. 177
— — pilosoinflatum var. 169
— — pilosum var. 169
- - -• pianocompactum var. 177
plitiianiim var. 90. 91
-- — pseudoalbovillosum var. 170
--- - pseudoccntigraniuin var. 90, 91
—- - - pseudocervinuin var. 90, 91
-----pseudocoloratum var. 170
-----pseiidodecoloratum var. 170
— — pseudodensoplanum var. 171
— -- pseudodispersum var. 170
— — pseudoincanum var. 174
— -- pseudolinnaeanum var. 90, 91
-----pseuclomelanathcrum var. 86, 88
— — pseudomirabile var. 85, 88, 90
-----pseudopilosum var. 170
-----psettdoptirpureum var. 170
— -- pseudorubidocompactum var. 177
— — pseudorubidum var. 170
-----pseudorubroatriim var. 88
— -- pseudortibroinflaliun var. 170
— — pseudosalomonis var. 85, 88
— — pubiglumatum var. 174
-----pullatum var. 170
— — purpureocom pact uni var. 177
-----purpureum var. 170
-----ramosolusitanicum var. 90, 91
-----rainosomegalopolitanum var. 90,
91
-----ramososalonionis var. 90, 91
— — ramosostriatum var. 90, 91
— — recognitum var. 177
— — richardianum var. 174
— — rubidocompaclum var. 176
-----rubidoinflatum var. 175
-----rubidum var. 170
-----rubroalbum var. 85, 88
-----rubroatrum var. 79, 85, 88
- - — nibroherrerae var. 85.88
-----ritbroinflalum var. 170
— — rubropilosiim var. 170
— — rufocompactum var. 177
-----rufovillosum var. 175
-----sabae var. 171
-----salomonis var. 85, 88
— — sanguineoinflatum var. 175
- - - - sanguineuni var. 171
— scheinachinicuin var. 90. 92
solitarium var. 175
sparsum var. 175
— - speciosissiiiuiiii var. 85, 88
— — speciosum var. 85, 88
spectabile var. 175
--- - strialocompactum var. 177
-di iutopur pin гит var. 171
.striatum var. 85, 88
subdtibiiim var. 90. 91
siibj(>iIiiru111 \ ar. 86, 88
timopheeeii var. 308. 310
I ri a I ei I ii 111 ri 1111 v.ii. 85, 88
IrislerulimatruIn var. 85, 88
— — tumanianii var. 309
— -- turgidum var. 81, 85, 88. 92
- unicum var. 175
• - --- violacinf latum var. 171
------mediterranean! 82, 89
— - - pt/ramidale 175
- - tiirgidocompactum 83
— — maraschii тип 83
- urartu 5, 6, 16, 17. 20 23. 26.
31. 34, 35. 36, 37, 38, 41, 14. 45.
284, 307
- - albonigricans var. 38
— - - binartulutriru var. 38
binarturutriru .38
- - nigrum var. 38
spuntaneoaIbuin var. 38
- - -- spontancoriibnim var. 38
- - - urartu var. .38
- vavilovianum 209
— vavilovii 5, 6, 11, 16. 17, 19. 32, .34,
92. 194. 208—213
- — vavilovii convar. 212
— — spelloramosum convar. 212
— — mirabile var. 212
— — mravianii var. 212, 213
------inunuru var. 212, 213
- — inupuru var. 212. 21.3
-- - rubromirabile var. 212, 21.3
— — vaneuni var. 213
— — vavilovolutcscens var. 212, 213
— -- vavilovomilturum var. 213
------vavilovii var. 212, 213
-- velutinum 269
-- volgense 66
---vulgare 11.202.230
— abt/ssinicum subsp. 239. 255
— - - curasiaticum subsp. 262
— — hadropyrum subsp. 238
hapalopyriim subsp. 262
- - - - indicum subsp. 239
------indoeuropueuin subsp. 262
- iranoasiaticum subsp. 238
- - —- iranoturkestanicum subsp. 239
— - mutabile subsp. 209
— — sinicum subs]). 262
- — — breviarisfaia группа 239
— --- compactoidcs группа 262
— — eligulatum группа 251, 253
— indoeuropaeum группа 262
— — inflatum группа 244
— — inflatum подгруппа 215
— — interjectum группа 262
•— rigidum группа 239
— — speltiforme группа 239
------inflatum prol. 244
— — muticorigidum prol. 245
------rigidum prol. 239
--- — siibrigidum prol. 237, 252. 251
- Ihospilense prol. 209
breviaristatuni subprol. 2.39
i-ligulatoinllatum subpro! 251
- eligulatum subprol. 253
- indicum subprol. 254
- - persicoides subprol. 264
Указатель латинских uasuautii'i
31
oii'.ii i >1 и nt si 11 ip । ol. ;* I,'I
s/hY/ijorsniipKil. 239
- compositum 208
arduini var.-gr. 202
— aristatum var.-gr. 230
coeruleum var.-gr. 202
coinpiicitim var.-gr. 213
compositum var.gr. 89
dicoccum var.-gr. 50
- durum var.-gr. 93
monococcum var.-gr. 293
miticum var.-gr. 230
spelta var.-gr. 199
- turgidum var.-gr. 82
ufghanicum var. 253
— ajatense var. 265
— albidum var 245, 265
— inflation subvar. 245
— bucharicum f. 245
- inflatum 1. 245
- - albinflatum var. 245
- alboglaucum var. 265
— alborubrinflatum var. 245
— alborubrum var. 245, 265
— orient ale 245
— album var. 201
— alienum var. 256
— araxicum var. 245
— ardjeschicum var. 256
— arduini var. 202
— asiaticum var. 244
— aureum var. 265
— badaksclianicum var. 254
— barbarossa var. 256
— bauhini var. 59
- - bengalense var. 256
— bengalinflaturn var. 245
— brunneum var. 61
— buccale var. 86
- - bucharicum var. 254
— caeruleovelutinum var. 256
— caesioides var. 258
— caesiospelliforme var. 242
- - caesium var. 267
— cervinum var. 89
— chiovendae var. 258
chiugiengense var. 250
— choresmum var. 245
— cinerescens var. 251
- - cinereum var. 258
— cinerosum var. 258
— cladura var. 59, 60
— clavatum var. 229
- coeleste var. 89
— coeruleum var. 202
— columbinum var. 89
— creticum var. 229
— cyanothrix var. 247, 258
— cyanothrixinf latum var. 247
— delfii var. 258
— dicoccoides var. 39, 45, 47
- - diiiura var. 86 <
— dshungaricum var. 258
— duhamelianum var. 201
— epruinosum var. 261
— erion var. 109
— erythroleucon var. 259
- erylhrosperminf latum var. 247
- - erythrospermum var. 265
- - falsealborubrum var. 245
— falserufinflatum var. 249
Iei i iip im-um var. 267
111 и/о II urn m 201
- - - jin hsii var. 60
f uliginosiim var. 259, 261
-- - ful-eocincreum var. 268
gentile var. 87
glaiicilutesccns var. 267
- - graceoimarginaluni var. 260
graecum var. 259
grist um var. 259
gunticiim var. 254
hamadanicum. var. 259
hcraticum var. 247
horogii var. 252
- — hostianum var. 259
— - huangyangense var. 248
-----icterinum var. 229
— indicinflatum var. 248
-- —- indicum var. 259
- ingrediens var. 259
introitum var. 259
-- -- iranicum var. 260
- italicum var. 109
— -- fodura var. 87
- - - - kabulicum var. 254
-----karbulakianum var. 259
-----kuzvinicum var. 260
— — kerkianutn var. 221
— — kermanschachii var. 260
-----khorassanicum var. 248
-----konstantinovskense var. 267
-----kurdistanicum var. 259
-----kurduculense var. 208, 209, 267
-----lagendreanum var. 248
----- leucomelan var. 109
-----leucospermum var. 260, 265
-----leucura var. 110
-----linnaeanum var. 89
-----lunangense var. 247
-----luristanicum var. 260
-----lutescens var. 248, 267
-----lutinflatum subvar. 248
-----lutinflatum var. 248
-----melanopogon var. 260
-----melanorubrum var. 260
-----melanopus var. 111
- - — melanura var. 60
-----menhsiengense var. 249
-----meridionale var. 260
-----mesopotamicum var. 260
— — metzgeri var. 60
— — michauxi var. 203
— — milturum var. 248, 267
-----rufinflatum subvar. 249
— — rufinflatum f. 249
— — mirabile var. 212
— — morosovskense var. 268
-----mravianii var. 212
— — murinum var. 258, 259
-----murgabicum var. 254
— — nigraristatum var. 268
-----nigricans var. 260
----- nigricolor var. 268
-----nigrinflaium var. 248
nigroalborubrum var. 260
— -- nigroatrum var. 256
- - - - nigroerythrospermum var. 268
nigroferrugineum var. 268
- - -• nigrograe'.uni var. 260
- - nigrohuanyangense var. 250
- nigrolutescens var. 268
346
Указатель латинских названий
ni groin I inf lain in var. 248
nigroi ujiiil/aliim var. 248
— — nigrosardoum var. 260
— — nigrum var. 268
— — oasicolum var. 250
- - -- obtivenae var. 268
---- oxianuni var. 252
----pamiricum var. 252
----pavoninum var. 91
-- — plenoerythrospermum var. 261
----plenoferrugineuni var. 262
— — plenosardoum var. 259
----provincials var. 112
— — pseudoalborubrum var. 260
----pseudobarbarossa var. 260
— —- pseudocaeruleovelitinum var. 268
— — pseudocaesium var. 268
----pseudodelfii var. 261
----pseudoerythroleucon var. 261
----pseudofulvocineretim var. 268
----pseudograecum var. 261
— — pseudoheraticum var. 248
— - - pscudohostiamun var. 261
— - - pseudoitigrediens var. 259
— — pseudoleucospermum var. 261
— — pseudomcridionale var. 261
----pseudopyrothrix var. 261
----pseudorubrieeps var. 222
— — pseudoturcicum var. 261
----pseudoturcomanicum var. 248
— — pseudovelutinum var. 261
— — pyrothrix var. 268
— — quasirufinflatum var. 252
----quasisubturcicum var. 254
----quasiturcicum var. 254
----renovatum var. 261
----rubromurinum var. 261
— — rufescens var. 267
----rufinflatum var. 248
----rufum var. 202
----sardoum var. 268
-- — schugnanicum var. 252
— — sericeum var. 222
----setosomesopotamicum var. 256
----speciosum var. 88
----splendens var. 229
----subafghanicum var. 254
— — subbarbarossa var. 243, 249
— — subbarbarossinf latum var. 249
----subcaesioides var. 243
----subcaesioidinjlatum var. 249
----subcaesium var. 243
-- — subcaesiuminflatum var. 249
— — suberythroleucon var. 243
siiberi/tliroleiiconinjlutiim var.
249
— - - suberythrospermum var. 243
- — subcrythrospermuminflatum var.
249
- subferrugitteumitijliditm var. 249
subferruginetim var. 243
- - -- subjuliginosum var. 243
— -- subgraecum var. 243
— — subgraecuminflatum var. 249
— — subgunticum var. 254
- - — subhamadaniciim var. 243
- - —- subliostianuni var. 243
- — subhostianuminjlatum var. 250
— — subira-nicum var. 243
— — subkabulicum var. 254
----- subkerinanscliacliii var. 244
— — submeridionale var. 244
— — submeridionalinflatum var. 250
— — submesopotamicum var. 244
— — subnigriaristatum var. 268
— — subpseudobarbarossa var. 244
--- subpscudohostianum var. 244
- subpseiidomeridionale var. 244
---- subpseudoturcicuni var. 244
— — subrenovatum var. 243
----subsardoum var. 244, 259
— — subtricoccum var. 61
— — subturcicum var. 24 1. 250
— — subvelutinum var. 256
— — sulinetise var. 268
----sunpani var. 250
— — surcfiianum var. 222
— -- tadjicorum var. 254
----tanaiticum var. 268
----larbagatanicum var. 260
----tchujense var. 258
----teheranicum var. 250
----transcaspicum var. 250
— — triste var. 250, 268
— — sunpani f. 250
— - - turcicum var. 262
turcomanicum var. 250
.. vanense var. '262
— - - vavilovii var. 208. 213
---- velutiniim var. 259, 269
- - villosum var. 269
----vulpinum var. 202
----xanthura var. 267
— quiinanense 239
- zea 50,199
— zhukovskyi 5, 6, II, 14, 16. 17, 29,
30. 280, 299, 316—319
СОДЕРЖАНИЕ
Предисловие................................................................
Обзор рода Triticum L......................................................
Ботаническая характеристика рода Triticum L..............................
История систематики рода Triticum L......................................
Современная система рода Triticum L......................................
Происхождение полиплоидной пшеницы.......................................
Ключ для определения видов пшеницы.......................................
Классификация рода Triticum L..............................................
Подрод I. Triticum.....................................................
Секция 1. Urartu Dorof. ct Л. Filat..................................
Т. urartu Thum, ex Gandil. — шпеница Урарту.......................
Секция 2. Dicoccoides Flaksb.........................................
T. dicoccoides (Koern. ex Aschers. ct Graebn.) Sclrwcinf. — полба дикая
T. dicoccum (Schrank) Schuebl. — полба (двузернянка, эммер)
T. karamyschevii Nevski — пшеница Карамышева (полба колхидская
пшеница грузинская)...............................................
Т. ispahanicum Heslot — пшеница исфаханская (полба исфаханская)
Т. turgidum L. — пшеница тургидум (тучная, вздутая, английская).
Т. jakubzineri Udacz. et Schachm. — пшеница Якубцнпера
Т. durum Desf. — пшеница твердая..................................
Т. turanicum Jakubz. — пшеница туранская..........................
Т. polonicum L. — пшеница полоникум . ..................
Т. aethiopicum Jakubz. — пшеница эфиопская........................
Т. persiemn Vav. — пшеница карталпнекая, дика.....................
Секция 3. Triticum...................................................
Т. macha Dekapr. et Menabde — пшеница маха........................
Т. spelta L. — пшеница спельта....................................
Т. vavilovii (Thum.) Jakubz. — шпеница Вавилова (пшеница ванская)
Т. compactum Ilost — пшеница компактная (ежовка) ....
Т. aestivum L. — пшеница мягкая (хлебная, хлебопекарная, обыкновен
пая)..............................................................
Т. sphaerococcum Perciv. — пшеница шарозерпая.....................
Т. pclropavlovskyi Udacz. et Migusch. - пшеница Петропавловского
Подрод II. Boeoticum Akigusch. et Dorof.................................
Секция 4. Monococcon Dum.............................................
T. boeoticum Boiss. — однозернянка дикая..........................
T. monococcum L. — однозернянка культурная........................
T. sinskajae A. Filat, et Kurk.— пшеница Сииской..................
Секция 5. Timopheevii A. Filat, et Dorof.............................
T. araraticum Jakubz. — полба дикая араратская ......
T. timopheevii (Zhuk.) Zhuk. — пшеница Тимофеева ....
T. zhukovskyi Menabde et Ericzjan <—пшеница Жуковского
T. militinae Zhuk, et Migusch. — пшеница Мплитппы ....
Секция 6. Kiharae Dorof. et Migusch..................................
T. kiharae Dorof. et Migusch. — пшеница Кнхары....................
Указатель литературы....................................................
Указатель латинских названий ..................................................
5
7
7
9
15
20
31
34
34
34
35
'38
39
50
G9
73
76
92
93
127
135
145
177
186
186
193
208
213
230
269
277
2,80
280
280
29.3
303
304
304
310
316
324
КУЛЬТУРНАЯ ФЛОРА СССР, ТОМ I
Владимир Филимонович Дорофеев, Анна Александровна Филатенко, Эмилия Филиппов-
на Мнгушова, Роальд Арсеньевич Удачин, Моисей Маркович Якубцинер
ПШЕНИЦА
Под общим руководством академика ВАСХНИЛ Д. Д. Брежнева
Редактор Г? А. Пенькова. Художественный редактор О. П. Андреев. Технические редакторы
Р. Н. Егоров а и Л. Б. Рез ннков а. Корректор Л. В. Вешнякова.
ИБ № 2034
Сдано в набор 18.04.79. Подписано к печати 15.11.79. Формат 70X108‘/ig. Бумага тин. № I. Гарни-
тура литературная. Печать высокая. Уел. поч. л. 30,45. Уч.-изд. л. 33,41. Изд. № 92. Тираж
2000 экз. Заказ № 762. Цепа 2 р. 60 к.
Отделение ордена Трудового Красного Знамени издательства «Колос», 191186, Ленинград, Д-186.
Невский пр., 28.
Белоцерковская книжная фабрика республиканского производственного объединения «Полиграфкнн-
га» Государственного комитета Украинской ССР по делам издательств, полиграфии и книжной тор-
говли, 256400, г. Белая Церковь, ул. Карла Маркса, 4.