


































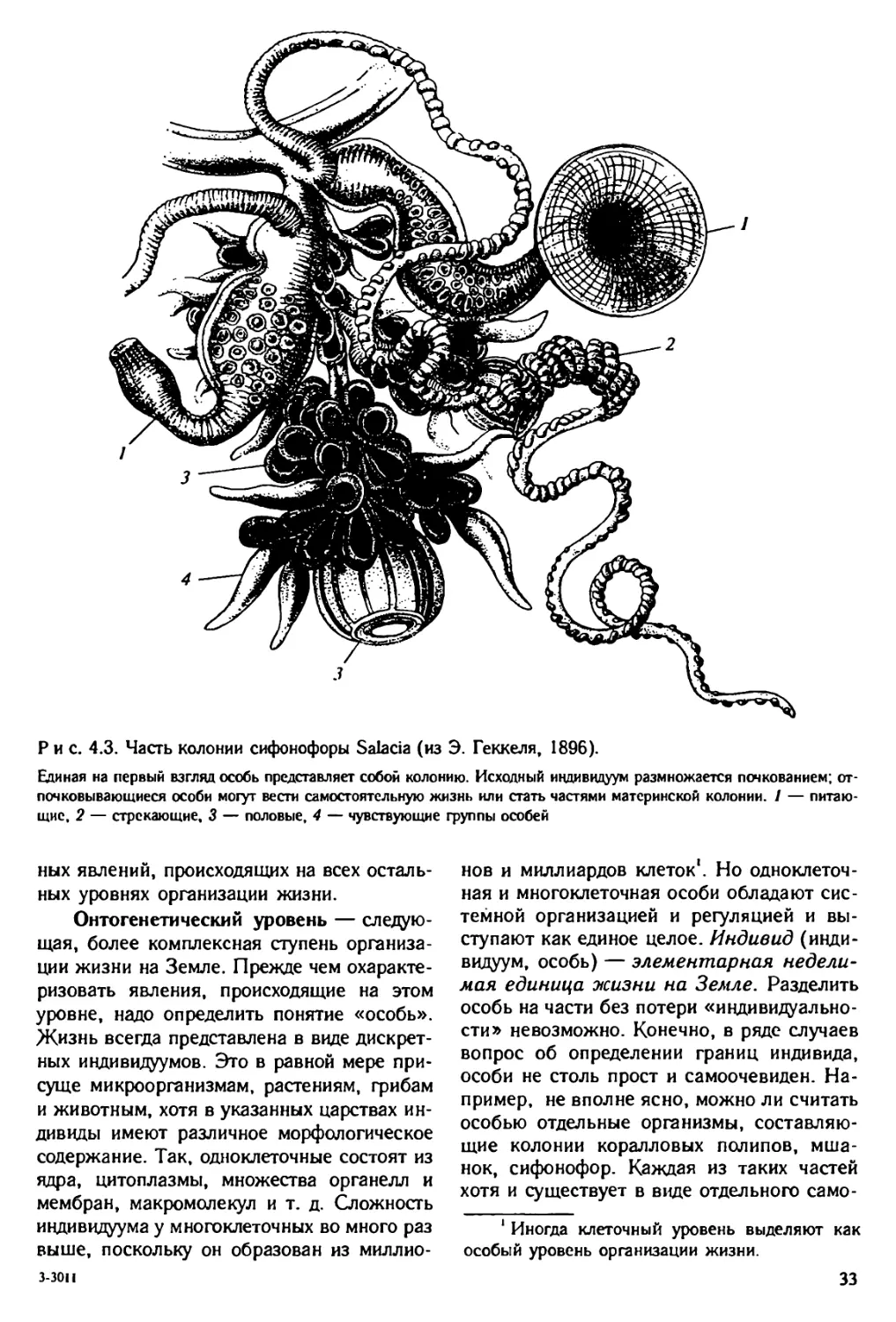









































































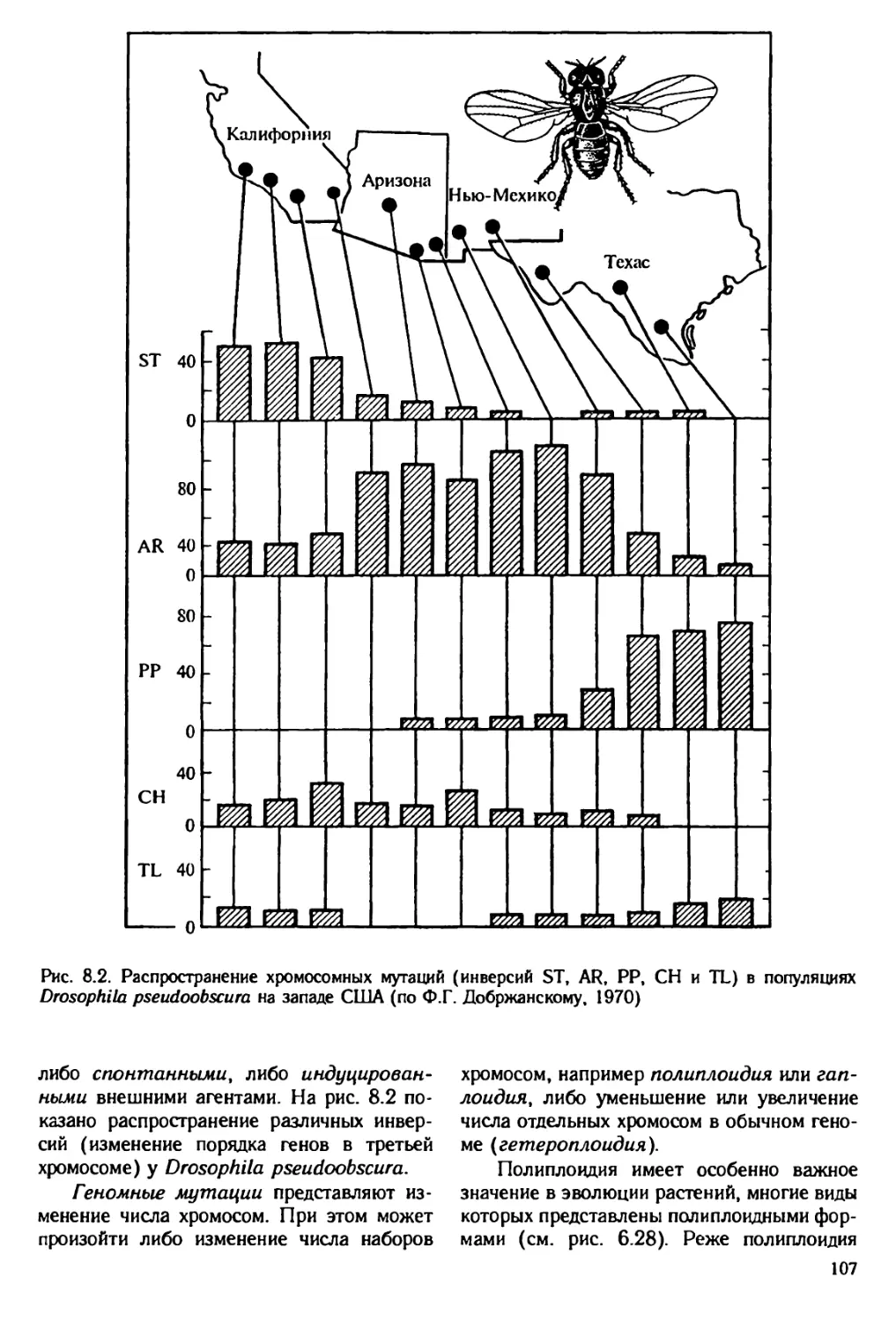












































































































































































































Author: Юсуфов А.Г. Яблоков А.В.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез экология и биогеография охрана живой природы учебное пособие теория эволюции
ISBN: 5-06-004584-6
Year: 2006
Author: Юсуфов А.Г. Яблоков А.В.
Tags: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез экология и биогеография охрана живой природы учебное пособие теория эволюции
ISBN: 5-06-004584-6
Year: 2006



