/
























































































































































































































































































































































































































































Text
БКМОПЯ
НАВЧАЛЬНИЙ ПОСІБНИК
За редакцією 3. Д. Воробця
УДК 57(075.8)
ББК 28.0я73
Б63
Рекомендовано Міністерством освіти і науки України
(лист № 1.4/18-Г-1409 від 13 серпня 2007р.)
Автори — викладачі Львівського національного медичного
університету ім. Данила Галицького: З.Д. Воробець, доктор біол.
наук, проф., зав. каф. медичної біології, паразитології та генетики
ЛНМУ (ред., розд. 1, 2, 6); Н. М. Воробець, доктор біол. наук, проф.
(розд. 5, 7); О. С. Корчинська, канд. біол. наук, доц. (розд. 4, 7);
Я. В. Матвієнко, канд. біол. наук, доц. (розд. 6); О. І. Периіин,
канд. біол. наук, асистент (розд. 4); Л. М.Сергієнко, канд. біол.
наук, доц. (розд. 2, 3); О. Р. Ткаченко, канд. біол. наук, ст. викл.
(розд. 3); О. Я. Чупашко, канд. біол. наук, доц. (розд. 1, 3, 4, 6);
М. І. Калинський, доктор біол. наук, проф. Кентського університе
ту, США (розд. 1)
Р ец ен зен ти :
Л. І. Остапченко — доктор біологічних наук, професор Київ
ського національного університету імені Тараса Шевченка;
Б. П. Романюк — доктор біологічних наук, професор Луганського
державного медичного університету;
О. Є. Пахомов — доктор біологічних наук, професор Дніпропет
ровського національного університету
Біологія: Навч. посіб. / 3. Д. Воробець, О. Я. Чупашко,
Б63 Л. М. Сергієнко та ін.; За ред. 3. Д. Воробця. — К.: Знан
ня, 2010. — 436 с.
I8ВN 978-966-346-721-4
Навчальний посібник підготовлено відповідно до чинної на
вчальної програми з медичної біології. Видання містить найваж
ливіші відомості про основи життєдіяльності людини. Особливу
увагу приділено медичній генетиці та медичній паразитології,
зокрема спадковій патології та паразитарним хворобам людини.
Для студентів вищих медичних навчальних закладів IV рівня
акредитації, лікарів, магістрів, студентів біологічних факультетів.
УДК 57(075.8)
ББК 28.0я73
І8ВК 978-966-346-721-4
© 3. Д. Воробець, О. Я. Чупашко,
Л. М. Сергієнко та ін., 2010
© Видавництво“Знання” , 2010
ЗМІСТ
ПЕРЕДМОВА ..................................................................................
7
ВСТУП .............................................................................................. 9
Розділ 1. БІОЛОГІЧНІ ОСОБЛИВОСТІ ЖИТТЄ
ДІЯЛЬНОСТІ ЛЮДИНИ. МОЛЕКУЛЯРНОКЛІТИННИЙ РІВЕНЬ ОРГАНІЗАЦІЇ
ЖИТТЯ ....................................................................... 15
1.1. Структурні рівні організації життя, їх зна
чення для медицини ............................................... 15
1.2. Неклітинні форми органічного світу. Віруси.
Білки пріони .............................................................. 18
1.3. Клітина— найменша структурно-функціо
нальна одиниця живого .........................................25
1.3.1. Прокаріотичні клітини .......................................... 25
1.3.2. Еукаріотичні клітини. Цитоплазма та клі
тинні органели .......................................................... 27
1.3.3. Ядро клітини. Хроматин: рівні організації
спадкового апарату. Будова хромосоми .............36
1.4. Обмін речовин і енергії ........................................... 38
1.5. Молекулярний рівень організації спадкової
інформації у клітині. Нуклеїнові кислоти ....... 40
1.6. Механізми реалізації спадкової інформації
в клітині ......................................................................43
4
Зміст
1.6.1. Регуляція експресії генів у прокаріотів.
Теорія оперона ......................................................... 48
1.6.2. Екзонно-інтронна організація генома
еукаріотів .................................................................. 50
1.7. Розмноження — універсальна властивість
живої матерії ............................................................ 52
1.7.1. Життєвий цикл клітини. Мітоз ............................53
1.7.2. Статеве розмноження. Гаметогенез. Мейоз.
Статеві клітини .........................................................56
1.8. Клонування. Життя клітин поза організ
мом ............................................................................... 66
Розділ 2. ОРГАНІЗМОВИЙ РІВЕНЬ ОРГАНІЗАЦІЇ
ЖИТТЯ. ОСНОВИ ГЕНЕТИКИ ЛЮДИНИ ...... 73
2.1. Предмет і завдання генетики та медичної
генетики. Основні поняття ................................... 73
2.2. Закономірності успадкування ознак .................. 81
2.2.1. Перший і другий закони Менделя. Менделюючі ознаки людини ............................................ 82
2.2.2. Третій закон Менделя..............................................83
2.3. Взаємодія генів у детермінуванні та розвитку
ознак (захворювань) організму .............................88
2.3.1. Форми взаємодії алельних генів. Множин
ний алелізм. Успадкування груп крові ............. 89
2.3.2. Взаємодія неалельних генів ..................................96
2.4. Генетика статі. Зчеплене успадкування
генів .......................................................................... 103
2.5. Закономірності успадкування зчеплених
генів. Кросинговер. Хромосомна теорія
спадковості ............................................................. 112
2.6. Особливості людини як об’ єкта генетичних
досліджень .............................................................. 118
2.6.1. Методи вивчення генетики людини. Спад
кові хвороби ............................................................119
2.6.2. Принципи діагностики спадкової патології .... 143
2.6.3. Медико-генетичне консультування ...................150
2.7. Нехромосомна спадковість .................................. 154
Зміст
5
2.8.
2.9.
Генна інженерія та біотехнологія ....................155
Поняття про імуногенетику та фармакоге
нетику ........................................................................ 161
2.10. Мінливість у людини. Форми мінливості.
Мутагенні фактори ................................................ 168
2.11. Генетична небезпека забруднення середо
вища. Поняття про антимутагени і комутагени ........................................................................ 179
Розділ 3. ІНДИВІДУАЛЬНИЙ РОЗВИТОК ..................... 180
3.1. Основні закономірності ембріонального
розвитку .................................................................... 181
3.2. Основні закономірності постембріонального
розвитку ....................................................................191
3.3. Регенерація і трансплантація ............................. 199
3.4. Механізми гомеостазу ...........................................215
3.5. Біологічні ритми ....................................................225
Розділ 4. ПОПУЛЯЦІЙНО-ВИДОВИЙ РІВЕНЬ
ОРГАНІЗАЦІЇ ЖИТТЯ. БІОЛОГІЧНА
ЕВОЛЮЦІЯ ............................................................. 230
4.1. Вид, його критерії та структура ........................ 231
4.2. Основні напрями еволюційного процесу ......... 246
4.3. Закономірності розвитку органічного світу.
Теорії виникнення життя на Землі ...................249
4.4. Філогенез і антропогенез ..................................... 258
Розділ 5. ЕКОЛОГІЯ ЛЮДИНИ ТА БІОСФЕРА ............ 288
5.1. Життєве середовище людини ............................. 293
5.2. Стан середовища існування та здоров’ я
людини ..................................................................... 303
5.3. Отруйні організми. Лікарські рослини ............ 312
5.4. Основні екологічні закони ...................................319
5.5. Поняття про екосистеми .......................................326
Розділ 6. МЕДИЧНА ПАРАЗИТОЛОГІЯ ......................... 335
Є
_____ ____________________________ ______________ _______ Зміст
6.1.
6.2.
6.3.
6.4.
Підцарство Найпростіші— Ргоіогоа ...............344
Тип Плоскі черви — Ріаікеїтіпікез ................. 362
Тип Круглі черви — Nетаікеїтіпікез ............. 386
Тип Членистоногі — Агікгоройа ....................... 406
ЛІТЕРАТУРА .............................................................................. ...
ПЕРЕДМОВА
Біологія — наука про життя. Вона як фундаментальна при
роднича наука розкриває закономірності розвитку життя, є
теоретичною базою медицини й основою підготовки майбутніх
лікарів і провізорів. Біології належить провідна роль у навчан
ні студентів-медиків, у формуванні їхніх природничо-наукових
і світоглядних позицій.
Вивчення біології студентами медичних і фармацевтичних
факультетів має дві основні мети:
1. Забезпечити високий рівень загальнобіологічної підготов
ки, оскільки медицина і фармація органічно входить до систе
ми медико-біологічних наук.
2. Досягти чіткої медико-біологічної орієнтації студентів
першого курсу, заклавши фундамент їх навчання на профіль
них теоретичних кафедрах, а в подальшому — на кафедрах
клінічного та фармацевтичного профілю.
Одним із основних завдань медичної біології для лікарів і про
візорів є аналіз впливу на здоров’ я людини молекулярно-гене
тичних, клітинних, онтогенетичних і екологічних факторів.
Важливим розділом біології, якому приділяється значна
увага в медичних університетах, є генетика. Предмет вивчення
генетики — явища спадковості та мінливості, матеріальні носії
спадковості. Спадковість лежить в основі усіх життєвих проявів.
Без явищ спадковості та мінливості неможлива була б еволюція
життя на Землі. Оскільки людина є “ продуктом” довготривалої
8________________________________________________________ Передмова
еволюції живої природи, то їй властиві всі загальнобіологічні
закономірності життя. Генетика також вивчає роль спадковості
у патології людини, закономірності передавання із покоління
в покоління спадкових хвороб тощо, а це має суттєве значення
в підготовці лікаря чи провізора. Важливим аспектом у підго
товці лікарів і провізорів є також вивчення механізмів мута
генної дії різних хімічних речовин і медикаментів.
Вивчення питань медичної паразитології важливе тому, що
велика кількість паразитарних захворювань досить розповсю
джена серед населення і невпинно зростає. У навчальному
посібнику тваринний світ показано як компонент екологічно
го середовища людини. Приділено значну увагу різним формам
взаємовідносин між паразитами й організмом людини, похо
дженню та еволюції паразитизму, життєвому циклові паразитів,
методам діагностики та профілактики паразитарних хвороб.
Вивчення медичної біології студентами медичних, стомато
логічних і фармацевтичних факультетів ВМНЗО в Україні є
складовою частиною державного стандарту освіти.
Навчальний посібник підготовлено відповідно до програми
з медичної біології для студентів медичних університетів IV рів
ня акредитації.
ВСТУП
Біологія (Ьіоз — життя, Іо§оз — вчення) — наука, що вивчає
живу природу. Предметом вивчення біології є всі живі організ
ми, особливості їх будови, властивості та функції, а також
розповсюдження, розвиток, взаємовідносини.
Життя — це унікальне природне явище, для якого харак
терні деякі специфічні прояви, котрі відрізняють живі істоти
від неживої природи. До сьогодні суть життя залишається не
до кінця зрозумілою тому, що форми і прояви життя дуже
різноманітні, немає точного і повноцінного визначення життя.
З погляду природничих наук, життя — це якісно особлива
форма існування матерії, відмінна від хімічної та фізичної, але
включає їх у себе.
Життя широко розповсюджене по всій планеті, незважаючи
на дуже різноманітні фізичні та хімічні умови зовнішнього
середовища. Живі організми, будучи результатом пристосуван
ня й еволюції, є надзвичайно різноманітними за розмірами,
складністю будови, особливостями метаболізму та життєдіяль
ності. Це дає їм змогу займати будь-які екологічні ніші на
Землі — жити “ на” і “ під” землею, “ на” і “ під” водою, у повітрі,
на каменях, в інших організмах, у льодах, гарячих гейзерах,
у глибинах океану при великому тиску та при температурі
близько 100 °С. Субстанціями життя є молекули нуклеїнових
кислот і білків, властивості та функції яких (у водному сере
довищі) забезпечують усі форми життя. Тільки катастрофи
космічного масштабу зможуть призвести до зникнення води,
10
Вступ
а значить і життя, за умови підвищення температури на Землі
понад 100 °С. Навіть ядерна війна і спричинена нею “ ядерна
зима” зможуть знищити тільки “ розумне життя” і сотні чи ти
сячі різноманітних видів, які живуть на Землі. Але практично
без проблем переживуть цю катастрофу віруси, мікроорганізми
та складніші істоти, які населяють товщу землі й океану.
Отже, життя як якісно особлива форма руху матерії зако
номірно буде існувати стільки, скільки на Землі зберігатимуть
ся необхідні умови для “ водних розчинів нуклеїнових кислот
і білків” .
Життя на Землі зумовлене специфічним розміщенням пла
нети у Сонячній системі. Саме на цій відстані від Сонця існують
температурні умови для існування води у рідкому стані, а це
забезпечує існування “білкових тіл” .
Поняттю “життя” не можна дати точного визначення, бо
воно є досить абстрактне. Однак можна глибше зрозуміти зміст
цього явища, якщо вивчити його властивості й характеристи
ки. Відомо, що жива матерія добре організована, здатна три
валий час підтримувати складні хімічні реакції метаболізму.
Вона росте, відповідає на подразнення, здатна до розмноження,
адаптації до певних умов оточуючого середовища.
Сьогодні підхід до розуміння життя базується на відомій
інформації про молекули, їх взаємодію, а також про енерге
тичні й інформаційні процеси, які відбуваються у живих ор
ганізмах.
Життя може існувати в межах живого тіла, організму. Ж и
вий організм — це вже не абстрактне, а конкретне поняття.
Він є одиницею біологічної активності, “ носієм життя” . Кожен
організм має свої особисті, особливі характеристики, тобто
індивідуальність, тому його називають індивідуумом. Це склад
на організаційна одиниця виду, якій притаманні усі властиво
сті живого.
Організми дуже різноманітні за своєю природою, формами
і розмірами. Тіло може бути одноклітинним, багатоклітинним
і неклітинним (віруси). Розміри коливаються від 20 нм (вірус
СНІДу) до ЗО м (синій кит). Тіла можуть мати різноманітні
форми. Наприклад, одноклітинні мають сферичні, довгасті, віб
ріони, спірохети, паличкоподібні та ін.; можуть мати ворсинки,
Вступ
11
джгутики, ундулюючі мембрани, різні вирости, зовнішній
скелет тощо. Багатоклітинні бувають грибами, рослинами,
тваринами різноманітних форм і розмірів. Це є результатом
тривалої еволюції та пристосувань до конкретних умов існу
вання. Однак для всіх живих тіл характерні загальні власти
вості: вони здатні до розмноження, підтримки впорядкованості
молекул і клітин, а також до специфіки своєї організації про
тягом мільйонів років і поколінь.
Найпростішим організмом можна вважати вірус, який має
матеріальне тіло, котре складається з однієї молекули ДНК чи
РНК та невеликої групи білків. Його розміри становлять лише
кілька нанометрів. Потрапляючи у клітину, він набуває бага
тьох ознак живого. У бактерій значно більші розміри тіла — де
кілька мікрометрів. Вони мають цитоплазматичну мембрану,
значне число генів, багато різноманітних білків, складний
метаболізм. Одноклітинні організми у десятки разів більші,
ніж бактерії, — їх розміри сягають сотні мікрометрів. Вони
мають кілька хромосом, значне число генів, сотні різноманіт
них білків, оформлене ядро і органели. Тіла багатоклітинних
організмів представлені великою різноманітністю видів тварин,
рослин і грибів. Ці організми мають різні клітини, тканини,
органи та системи. Усі органи і тканини характеризуються
“доцільністю” структури, розмірів, місця розташування, а також
функціями, скерованими на виживання та розмноження.
Таким чином, усі властивості та прояви життя пов’ язані
з існуванням певний час високоорганізованої системи молекул
і клітин — живого тіла. Порушення у структурі та функціях
тіла і подальше його руйнування ведуть до припинення явища
життя.
Незважаючи на істотні відмінності живої та неживої при
роди, необхідно зауважити, що живі організми живуть і функ
ціонують за загальними принципами хімії та фізики. Живий
організм володіє рядом фізико-хімічних характеристик: диск
ретністю, ієрархічністю будови, взаємодією і взаємозалежністю
частин (цілісністю), має молекулярну основу будови і перетво
рення енергії. Властивості різних молекул, з яких складається
організм, не відрізняються від таких самих молекул у неживій
природі. Всі біохімічні реакції відбуваються в організмі на
12
Вступ
основі законів хімії. Для їх перебігу необхідні ферменти, які
функціонують за принципами хімічного каталізу. Такі факто
ри, як світло, температура, тиск мають такий самий вплив на
біохімічні реакції, як і на хімічні реакції поза біологічними
системами.
Рух молекул у живих системах базується на таких фізичних
процесах, як дифузія та осмос, в основі котрих лежить тепло
вий рух молекул. Багато молекул і надмолекулярних структур
клітини мають фізичні властивості полярності, гідрофобності,
мембрани клітин мають електричні потенціали, відростки не
рвових клітин проводять електричний струм. Суглоби та кіс
тки скелета, м’ язи діють на основі принципів механіки. Пере
творення енергії у живих організмах підпорядковуються зако
нам термодинаміки.
Отже, живі організми є частиною природи, мають молеку
лярну основу і за своїм хімічним складом та фізичними прин
ципами функціонування не відрізняються від неживих тіл.
Однак значна частина впорядкованості та високий рівень ор
ганізації молекул і їх комплексів є причиною появи нової
форми існування матерії — життя. А живий організм функціо
нує за складнішими законами, ніж його окремі “ неживі” ком
поненти. Живі тіла проявляють властивості, які характерні
тільки для них, — наприклад, розмноження, живлення, ди
хання, рух тощо
Живі організми складаються з безлічі частин, чітко впоряд
кованих у просторі та часі. Взаємодія цих частин зумовлює
цілісність і всі життєві процеси.
Дискретність означає перервність організації матеріальних
тіл. Дискретність будови організму означає те, що він як си
стема складається з багатьох окремих, складних, взаємозв’ яза
них і взаємодіючих частин. Дискретність є важливою характери
стикою будь-якої системи, оскільки забезпечує внутрішній рух
і взаємодію її частин. А це означає прояв різних якостей, вла
стивостей і функцій.
Впорядкованість — це закономірне розміщення матеріаль
них тіл у певному просторі. Важливість цієї характеристики
полягає в тому, що у конкретному просторі в певний час, всупереч
Вступ____________________________________________________________ 13
другому законові термодинаміки, збираються певні елементи
системи, закономірна взаємодія яких зумовлює появу якісно
нових характеристик і властивостей цієї системи (наприклад,
нової форми існування матерії — життя).
Взаємозалежність і взаємодія елементів упорядкованої си
стеми зумовлює її відносну незалежність від зовнішнього се
редовища. Така система виглядає як цілісний окремий організм.
Таким чином, упорядкованість і взаємодія дискретних елемен
тів системи веде до якісно нового результату — появи цілісності
системи. Стосовно живих систем — це прояв відособленості
живих тіл. Вони як єдине ціле реагують на різного роду под
разники, рухаються, мають стандартний набір біохімічних
реакцій і тотожність різноманітних функцій.
Основою організації живого і неживого є атом. З атомів скла
даються молекули та їх комплекси. У неживій природі вони
можуть утворювати розчини, кристали та суміші, у яких від
сутній рівень організації.
У живому світі молекули та макромолекули об’єднуються
в різні комбінації й утворюють кілька рівнів організації: мемб
рани, клітинні органели, цитоплазматичний матрикс, клітини,
тканини тощо. Живі організми можуть бути одноклітинними
та багатоклітинними. У багатоклітинних організмах клітини
організовані у тканини, тканини — в органи, а органи — у систе
ми органів.
У природі більшість речовин розсіяні хаотично, приблизно
у рівних співвідношеннях. Однак у живих організмах концен
трації деяких речовин у тисячі разів перевищують їх вміст
у навколишньому середовищі. Зокрема, у мікроскопічному
об’ємі клітини сконцентровані різноманітні білки, нуклеїнові
кислоти, амінокислоти, специфічні ліпіди, вуглеводи, яких
практично немає у зовнішньому середовищі. Молекули цих
речовин строго організовані у клітині й утворюють складні
спеціальні структури. Наприклад, біологічні мембрани скла
даються зі специфічних молекул фосфоліпідів, які чітко впо
рядковані один відносно одного, утворюючи у водному середо
вищі тонкий суцільний шар. Він повністю вкриває поверхню
клітини чи органели. У рідкий шар фосфоліпідів мембран
у певній послідовності занурені різноманітні (специфічні для
14____________ _ _ _ _ _ _ ______________________________________ Вступ
різних органел) структурні та функціональні білки. Завдяки
такій вибірковості та впорядкованості молекул утворюються
різноманітні органели, які можуть забезпечити більшість жит
тєвих процесів у клітині.
Дискретність і впорядкованість — є основою кількох струк
турно-функціональних рівнів організації цілих живих організ
мів. Кожен рівень має більший ступінь ускладнення і йому
притаманні нові властивості та функції.
Розділ 1
БІОЛОГІЧНІ ОСОБЛИВОСТІ
ЖИТТЄДІЯЛЬНОСТІ ЛЮДИНИ.
МОЛЕКУЛЯРНО-КЛІТИННИЙ РІВЕНЬ
ОРГАНІЗАЦІЇ ЖИТТЯ
1.1. Структурні рівні організації життя,
їх значення для медицини
Молекулярно-генетичний рівень. Елементарною одиницею
цього рівня є ген — фрагмент молекули ДНК, у якому записа
но визначений в якісному та кількісному співвідношенні об’ єм
біологічної інформації про синтез одного білка. Елементарне
явище полягає у процесі редуплікації ДНК. Біологічна інфор
мація, що міститься в молекулі ДНК, не бере безпосередньої
участі у процесах життєдіяльності. Вона переходить у діючу
форму завдяки механізмові матричного синтезу й утворення
мРНК, яка контролює синтез білків. На молекулярно-генетич
ному рівні дискретні одиниці одноманітні. Життєвий субстрат
для всіх живих організмів — приблизно 20 амінокислот і 5 азо
тистих основ, які входять до складу нуклеїнових кислот. Біо
логічна енергія запасається в макроергічних зв’ язках молекул
АТФ.
16_________________________________________________________ Розділ 1
Цей рівень вивчають у курсі молекулярної біології та моле
кулярної генетики.
Субклітинний рівень. Завдяки електронній мікроскопії та
вдосконаленню диференційного центрифугування і біохімічних
методів дослідження стало можливим пізнання субклітинної
організації: виявлено й ідентифіковано деякі субклітинні
структури (мітохондрії, ендоплазматичний ретикулум, комп
лекс Гольджі, лізосоми, рибосоми та ін.). За допомогою біохіміч
них методів з’ясовано їх функції.
Клітинний рівень. Клітина — елементарна одиниця цього
рівня, найменша структурно-функціональна одиниця живого.
Елементарне явище представлене реакціями клітинного мета
болізму, що становлять основу потоків речовин, енергії, інформа
ції. Речовини, що надходять ззовні, завдяки діяльності кліти
ни перетворюються в субстрати й енергію, які використовують
ся у процесі біосинтезу білків та інших сполук, потрібних
організмові.
Клітинний рівень вивчають такі науки, як цитологія, біо
хімія, клітинна біологія, молекулярна генетика.
Тканинний рівень. Сукупність клітин, подібних за будовою,
походженням і функціями, разом із тканинною речовиною
утворюють тканину. Тканинний рівень виникає з появою ба
гатоклітинних організмів.
Вивчення цього рівня проводиться здебільшого гістологіч
ними, біохімічними, імунологічними методами.
Рівень організму (онт огенет ичний). Елементарна одини
ця цього рівня — особина в її розвитку від моменту зароджен
ня до припинення існування як живої системи. Закономірні
зміни організму в індивідуальному розвитку становлять еле
ментарне явище цього рівня. Ці зміни забезпечують ріст ор-ганізму, диференціацію його частин і одночасно інтеграцію
розвитку в єдине ціле. У процесі онтогенезу в певних умовах
зовнішнього середовища відбувається втілення спадкової ін
формації в біологічні структури і процеси, на основі генотипу
формується фенотип організмів певного виду.
Основні науки, що вивчають цей рівень, — анатомія та
фізіологія.
Біологічні особливості життєдіяльності людини
17
Популяційно-видовий рівень. Популяція — елементарна
одиниця цього рівня. Популяція — це сукупність особин одно
го виду, які заселяють один ареал і відносно відокремлені від
інших популяцій цього ж виду. Об’єднання особин у популяцію
відбувається завдяки спільності генофонду, що використовуєть
ся у процесі статевого розмноження для створення генотипів
особин наступних поколінь. Популяція внаслідок можливості
міжпопуляційних схрещувань є відкритою генетичною систе
мою. Дія на генофонд популяції елементарних еволюційних
факторів (мутаційний процес, природний добір, коливання
чисельності особин) приводить до еволюційно значущих змін
генофонду.
Екосистемний (біогеоценотичний) рівень. Елементарна
одиниця цього рівня — екосистема (сукупність популяцій
різних видів, які заселяють територію з певними абіотичними
показниками і пов’язані між собою та навколишнім середови
щем обміном речовин, енергії й інформації). Екосистеми
об’ єднуються на планеті у єдиний комплекс — біосферу.
Біосферний рівень. Сукупність екосистем утворює біосферу.
Біосфера — це сукупність усього живого на Землі — всіх ор
ганізмів, які існують у тісному зв’язку з неживою природою
в атмосфері, гідросфері та землі. Біосферу складають мільйони
найрізноманітніших видів тварин, рослин, грибів, найпрості
ших і вірусів. Біосфера — це вищий рівень організації живої
природи. Єдина біосфера — це єдність усіх живих організмів,
свідоцтво єдиного джерела виникнення життя, еволюції та
тісного взаємозв’ язку всього живого.
Рівні організації живих організмів взаємозв’ язані за прин
ципом ієрархії: нижчі рівні організації обов’ язково входять
складовою частиною у вищі рівні.
Уявлення про організацію живих організмів на різних рів
нях сприяє кращому розумінню явища життя, причин виник
нення патологій, успішнішій розробці методів профілактики
захворювань і лікування хворих організмів.
Основними принципами організації всіх рівнів є об’єднання
і взаємодія. Наприклад, агрегація на молекулярному рівні веде
до об’єднання груп окремих одиниць і утворення макромолекул
певної структури, біомембран, ферментативних комплексів.
18
Розділ 1
Об’ єднання різних органел і їхнє узгоджене функціонування
зумовлює клітинний рівень організації живого. У свою чергу,
різні за структурою та функціями клітини об’ єднуються, взає
модіють між собою й утворюють функціональні одиниці тканин
і органів. Взаємодія комплексів веде до стабільності цієї си
стеми, а також до закономірних змін її структури і функції.
Висока впорядкованість молекул — одна з основних харак
теристик живих систем. Основною умовою життя клітини є
підтримка високої впорядкованості протягом усього життя. Це
можливо завдяки постійному використанню клітиною енергії
ззовні та координації біохімічних реакцій.
1.2. Неклітинні форми органічного світу.
Віруси. Білки пріони
До неклітинних форм належать віруси. Вони утворюють
групу Віра (Уіга). Віруси є примітивними і нескладними ор
ганізмами, які проявляють свою життєздатність тільки у стадії
внутрішньоклітинного “ паразитизму” .
Розміри вірусів варіюють у межах 10— 250 нм, вони у серед
ньому в 50 разів менші від бактерій, і їх неможливо розгляну
ти за допомогою світлового мікроскопа. Віруси перебувають на
межі між живим і неживим, і це свідчить про те, що існує
неперервний спектр всезростаючої складності, який починаєть
ся простими молекулами і закінчується складними замкнути
ми системами клітин.
Віруси побудовані досить примітивно — вони складаються
з ДНК або РНК, які містяться всередині вірусів і оточені за
хисною білковою оболонкою (капсидом). Залежно від типу
нуклеїнової кислоти розрізняють дезоксивіруси (ДНК двониткова або однониткова) та рибовіруси (РНК однониткова або
двониткова). Оболонка вірусів часто побудована з ідентичних
одиниць-капсомерів, які повторюються. Із капсомерів утворю
ються симетричні структури, здатні до кристалізації.
Віруси можуть утворювати собі подібних тільки всередині
живої бактеріальної або еукаріотичної клітини. Потрапивши
Біологічні особливості життєдіяльності людини
19
всередину клітини хазяїна, вони “ включають” свою нуклеїно
ву кислоту в ДНК клітини, і клітина, використовуючи ДНК чи
РНК віруса, синтезує нові копії цього вірусу.
Життєві цикли більшості вірусів подібні, але в клітину во
ни потрапляють по-різному:
— повітряно-крапельним шляхом — віруси грипу, кору,
віспи;
— аліментарним (з їжею) — віруси ентериту, ящуру;
— фекально-оральним (через брудні руки) — вірус гепатиту А;
— перкутанним (через шкіру) — віруси сказу, віспи, гер
песу;
— статевим — віруси герпесу, папіломи, ВІЛу;
— трансмісивним (через кровососних членистоногих) —
вірус енцефаліту;
— хірургічним — вірус ВІЛу, гепатиту В.
У процесі розмноження вірусів розрізняють такі стадії:
— адсорбція вірусів на поверхні клітини-хазяїна;
— проникнення у клітину;
— втрата вірусом білкової оболонки;
— латентний період (у клітині відбувається реплікація,
транскрипція, трансляція);
— формування вірусів, що супроводжується їх виходом із
клітини.
Віруси, які паразитують у бактеріях, називаються бактеріо
фагами, або фагами. їх відкрив д’ Херел 1917 року у Франції.
Фаги мають специфічних хазяїв, різноманітну форму, розміри
і структуру. Найбільш вивченим є бактеріофаг Т-4, який інфі
кує бацили, Е. Соїі. Він містить головку, коротку шийку та
комірець і сильний хвіст, у якому міститься гексагональна
базальна пластинка з хвостовими нитками. Хромосома фага є
лінійною молекулою ДНК, яка містить більш ніж 75 генів,
щільно розміщених усередині білкового капсиду полігедральної головки.
Усі віруси є “ паразитами” і тому можуть викликати у своїх
хазяїв хвороби, зокрема грип, віспу, свинку, краснуху, поліоміє
літ, енцефаліт, гепатит, жовту лихоманку, кір, ящур, СНІД, сказ
тощо. У людини й інших живих організмів у процесі еволюції
20
Розділ 1
виробилися захисні механізми проти вірусних інфекцій. Існу
ють гуморальний і клітинний імунітети. Гуморальний здійс
нюється завдяки виробленню в організмі антитіл (імуноглобулінів) і захисних білків (інтерферонів), які пригнічують розмно
ження вірусів — збудників багатьох захворювань. Клітинний
імунітет ґрунтується на здатності певних видів лейкоцитів
розпізнавати інфіковані вірусами клітини та знищувати їх.
Походження вірусів не з ’ясовано. Існує три основні гіпоте
зи щодо цього. Одні вважають віруси первинно примітивними
організмами, інші — що віруси походять від організмів, які
спростились у зв’язку з паразитичним способом життя, треті —
що віруси являють собою групу генів або фрагментів клітинних
структур, які набули автономії.
Окрім вірусів, відомі віроїди. Віроїди — це рослинні вну
трішньоклітинні паразити, які представлені у вигляді замкне
ної дволанцюгової РНК з каталітичним центром і не містять
у своєму складі білків (були відкриті 1971 року Теодором Діенером). За своєю структурою — це коротколанцюгові РНК,
найменший віроїд має лише 220 нуклеотидів, а в середньому
350— 500 нуклеотидів. Для порівняння: найменший РНК-вмісний вірус налічує дещо більше 2 тис. нуклеотидів. Послідовність
нуклеотидів у віроїдів не кодує жодного білка, але дуже ак
тивно конкурує з клітинними структурами за білковий фермент
РНК-полімеразу II, перешкоджаючи синтезу мРНК, а відповід
но й білків. Механізм інфікування рослин віроїдами ще не
розкрито, а симптоми віроїдних захворювань проявляються
у гальмуванні росту і спотворенні органів. Походження віроїдів
залишається великою загадкою для науковців; найбільш по
ширеною є гіпотеза “ інтрона-втікача” — структурної частини
інформаційної РНК, яка видаляється ферментами при її доз
ріванні та формуванні мРНК.
Білки пріони. В останні десятиріччя мікробіологи та ві
русологи відкрили низку нових інфекційних агентів — бак
терій, віроїдів, вірусів і віріно. Пріони серед виявлених збуд
ників захворювань посідають особливе місце. Пріони — це
збудники захворювань, які складаються лише з білка; вони не
містять нуклеїнової кислоти і продукуються самим організмом
(рис. 1.1).
Клітинна
будова
( наявність
оформленого
ядра)
Клітинна
будова
( доядерні)
Нуклеїнові
кислоти
Білок
Пріони
Віроїди
Віруси
Прокаріоти
І,.,: 'Iі :
'•.1•
ш ІЙ^ИІвяЯІ(№
ЯЙіІЖ *
«нчмгч
н
н
ш
и
св
X
Я
Я
=Я
■&
Я
V
•в*
Н
і
Я
есов
X
X
и
а
о
я
н
о
3
04
Еукаріоти
22
Розділ 1
РгР — пріоновий білок; РгРс — неінфекційний пріоновий
білок, який міститься в клітинах організму; РгР8с — інфекцій
ний пріоновий білок, який виявляється при пріоновій патоло
гії; РІШР — ген, який кодує синтез клітинного РгР°.
Перші повідомлення про пріонну хворобу під назвою зсгаріе
(“ вітрянка овець” ) датовані початком XVIII ст. Це захворюван
ня супроводжувалося порушенням координації, паралічем,
свербежем, агресивністю чи сонливістю, Захворювання постій
но прогресувало і призводило до летального кінця. Пізніше це
захворювання виявили у кіз і перенесли його на мишей. Так
була доведена інфекційна природа хвороби.
У 1920— 1921 рр. описано захворювання, яке пізніше отри
мало назву хвороби Кройцфельда — Якоба. Хвороба супрово
джувалася запамороченням, погіршенням рівноваги, зору,
слуху, порушенням мозкових функцій. У 30-х роках X X ст.
описана хвороба, яка тепер має назву синдрому Герстмана —
Страуслера —- Шайнкера. Цей синдром характеризується мозочковою атаксією, прогресуючим слабоумством, відсутністю
рефлексів на нижніх кінцівках і суцільними патологічно-амілоїдними бляшками у центральній нервовій системі. Хвороба
триває 2— 10 років.
У 1957 р. описана прогресуюча, летальна спонгіоформна
енцефалопатія під назвою “куру” . Хвороба супроводжується
нестійкістю пози та ходи, погіршенням мови, тремором, роз
ладом координації в нижніх кінцівках. Із розвитком хвороби
пацієнт не може вставати без підтримки, спостерігається більш
виражений тремор, посмикування м’язів, емоційна лабільність,
вибухи сміху, депресія, уповільнене мислення. На терміналь
ній стадії хвороби пацієнт не здатний сидіти без сторонньої
підтримки, відбувається повна втрата координації рухів, тре
мор, нетримання сечі та калу. В основі патогенезу лежить
порушення функції мозочку. За підтвердження інфекційної
природи куру, зокрема за “ відкриття нових патогенетичних
механізмів інфекційних захворювань” , у 1976 р. Д. Гайдужек
отримав Нобелівську премію.
У Великій Британії (1985) було вперше зареєстровано хво
робу під назвою “сказ корів” (тепер — спонгіоформна енцефа
лопатія великої рогатої худоби).
Біологічні особливості життєдіяльності людини
23
У 1886 р. описана пріонова хвороба під назвою фатальне
сімейне безсоння. Це генетично детермінована дегенеративна
хвороба, обмежена певними теламічними ядрами. Характери
зується прогресуючим безсонням, гарячкою, дисфункцією
сфінктерів. Згодом розвиваються сонний стан, тремор, а через
дев’ять місяців — коматозний стан і смерть.
Виявлена обширна група вікових міопатій (1993). Захворю
вання характеризується міозитом, повільно прогресуючою
слабкістю. Гістологічно виявляли некротичну міопатію з наяв
ністю амілоїдних включень, які ідентифікували як суміш
пріону, амілоїду АЬ та аполіпопротеїну Е.
У 2001 р. було ідентифіковано хворобу під назвою Хантінгтон-подібний нейродегенеративний розлад. Ця хвороба харак
теризується розладами переважно маніакального характеру.
Гістологічно було виявлено атрофію мозочкового молекуляр
ного шару, який містив пріонові бляшки.
Етіологія усіх вказаних вище хвороб, які нині об’єднують
під назвою “ трансмісивні спонгіоформні енцефалопатії” , три
валий час залишалася нез’ ясованою. У 1982 р. С. Прусінер
довів, що інфекційними агентами є білок. Цей білок отримав
назву “ пріон” . За відкриття пріонів С. Прусінер у 1997 р. от
римав Нобелівську премію. На сьогодні вивчено амінокислот
ний склад цього білка, клоновано ДНК.
Інфекційні пріони являють собою змінені білкові молекули
хазяїна. Вони не містять нуклеїнових кислот і цим кардиналь
но відрізняються від бактерій, вірусів і віроїдів. Сучасне уяв
лення про рівні організації інфекційних агентів дає змогу
розподілити їх на п’ ять основних груп: пріони, віроїди, віріно,
віруси, прокаріоти. Окремі види еукаріотів, які викликають
паразитарні захворювання, відносять до інвазійних агентів.
Пріоновий білок (РгР) — це сіалоглікопротеїн із молекуляр
ною масою 33—35 кД, який складається у людини з 254 амі
нокислот, включаючи А-термінальний сигнальний пептид із
22 амінокислот, котрий відсутній в активному зрілому білку.
На С-кінці розміщений глікозилфосфатидилінозитол, який
використовується для фіксації на плазматичній мембрані клі
тини. Пріон кодується єдиним геном, розташованим у людини
в короткому плечі 20-ї хромосоми, який позначається як РІШР.
Таким чином, пріон має дві ізоформи — нормальну клітинну
24
Розділ 1
(Ргрс) та інфекційну (РгР8С). Інші молекулярні форми можуть
бути протеолітичними фрагментами, які, також проникаючи
у клітини, спрямовують синтез собі подібних інфекційно-ак
тивних пріонів. Клітинна ізоформа РгРс у нормі входить до
складу зовнішніх клітинних мембран. Вона необхідна для
процесів ендоцитозу й катаболізму клітин. Найвищу концен
трацію РгРс виявлено у нервових клітинах, де вони, ймовірно,
беруть участь у нормальній синаптичній функції. Делеція гена
РІШР не призводить до смерті організму, й особини, позбавлені
цього гена не схильні до пріонових захворювань. Підвищення
експресії гена РІШР, навпаки, є сприяючим чинником для
виникнення пріонового захворювання.
Усі відомі на сьогодні випадки пріонових захворювань мож
на класифікувати на спорадичні, спадкові та власне інфекцій
ні. Спорадичні випадки стосуються здебільшого хвороби Крайцфельдта — Якоба, а також Герстмана — Страуслера — Шайнкера та фатального сімейного безсоння, і становлять основну
масу пріонових захворювань (9 0 % ). Ці випадки виникають,
коли відбувається спонтанна конверсія нормальної клітинної
ізоформи білка РгРс у патологічну РгР8С, або коли соматична
мутація гена РІШР призводить до утворення мутантного РгР,
який трансформується в РгР8С. Відомо близько 20 мутацій гена
РІШР, достовірно пов’ язаних із вродженими пріоновими за
хворюваннями.
Куру та ятрогенна (інфекційна) форма хвороби Крайцфельдта — Якоба являють собою приклади інфекційних форм пріо
нових захворювань. Відомі випадки інфікування внаслідок
використання контамінованих хірургічних і стоматологічних
інструментів чи електродів, мозкових трансплантатів, переса
док трупних тканин, уведення фармакологічних препаратів
мозкового походження тощо. Варто зазначити, що пріоновий
білок РгР8С контагіозний незалежно від причини його виник
нення, тому поділ на спорадичні, спадкові та інфекційні фор
ми є умовним і не відображає різниці у патологічній анатомії
та клініці захворювань.
Репродукція пріонів має свої особливості. Патологічна ізо
форма РгР8Сутворюється шляхом трансформації нормальної
клітинної ізоформи в ході посттрансляційного процесингу.
Вважається, що при цьому одна молекула РгР8С з’єднується
Біологічні особливості життєдіяльності людини
25
з однією молекулою РгРс з утворенням гетеродимерного про
міжного продукту, який розщеплюється на дві молекули РгРзс.
У наступному циклі кожна молекула РгР8Ссполучається з новою
молекулою РгРс, трансформуючи її, в результаті чого утво
рюється чотири молекули інфекційної форми і т . д . , тобто
відбувається експоненціальне зростання кількості РгР8С.
Незважаючи на значні успіхи у вивченні пріонових захво
рювань, питання їх патогенезу остаточно не з’ ясовано. Це є
основною причиною того, що ефективне лікування пацієнтів
із пріоновими захворюваннями досі не запропоноване. Значні
труднощі при розробці методів лікування становить той факт,
що збудники — пріони — являють собою модифіковані власні
білки організму людини.
1.3. Клітина — найменша
структурно-функціональна одиниця
живого
1.3.1. Прокаріотичні клітини
До прокаріотів відносять бактерії та синьо-зелені водорості.
Вони об’ єднані в підцарство Дроб’янки. Характеризуються тим,
що не мають типового ядра, оточеного ядерною мембраною.
Генетичний матеріал представлений кільцевою молекулою ДНК,
яка не утворює комплекси з білками гістонами.
Бактерії — це дрібні організми, які мають клітинну будову,
їх розміри коливаються в межах 0,1— 10 мкм, і тому їх можна
розглядати тільки під мікроскопом. Бактерії займають найріз
номанітніші екологічні ніші: живуть у ґрунті, воді, повітрі,
всередині тваринних організмів і на їхніх зовнішніх покривах.
Так, в 1 г ґрунту може міститися до 100 млн бактерій. Бактерії
викликають різноманітні захворювання. До найважчих захво
рювань людини і тварин належать харчові отруєння, викликані
сальмонелами.
Бактерії є також збудниками таких важких захворювань,
як дифтерія, туберкульоз, коклюш, сифіліс, холера, черевний
26
Розділ 1
тиф, бактеріальна дизентерія. Бактерії використовуються для
отримання необхідних продуктів, антибіотиків, розщеплюють
органічні речовини і беруть участь у кругообігу речовин у при
роді.
Клітинна стінка бактерій надає їм відповідної форми і твер
дості. Вона запобігає осмотичному набуханню та розриву клі
тин. Через неї легко проходить вода, відповідні іони. Полісаха
риди і білки надають клітинній стінці антигенних властивостей.
Багато бактерій рухливі. їх рухливість обумовлена наявністю
в них джгутиків.
Протоплазма бактерій оточена плазматичною мембраною,
яка не відрізняється від мембран еукаріотичних клітин. У де
яких бактерій плазматична мембрана має впнуті всередину
складчасті структури мезосоми, які беруть участь у процесі
дихання. Форма бактерій може бути різноманітна і є система
тичною ознакою. У цьому плані розрізняють чотири основних
типи клітин: коки (сферичні), бацили (циліндричні), спірили
(спіралеподібні) і вібріони (короткі палички у вигляді коми).
У свою чергу, коки поділяються на мікрококи (одиничні клі
тини), диплококи (дві клітини в одній капсулі), стрептококи
(ланцюжки клітин) і стафілококи (група клітин у вигляді ви
ноградного грона).
Генетичний матеріал бактерій, який представлений однією
кільцевою ниткою молекули ДНК, містить приблизно 5— 10 пар
нуклеотидів. Ця нитка не має складної будови, характерної
для хромосом, вона містить усього кілька тисяч генів, що при
близно у 500 разів менше, ніж у клітині людини.
У клітинах прокаріот відсутні ендоплазматичний ретикулум,
апарат Гольджі, мітохондрії, лізосоми, центріолі, пластиди,
однак вони містять 708 рибосоми, які у процесі біосинтезу
білка утворюють полірибосоми. Деякі фотосинтезуючі бактерії
мають хроматофори, в яких міститься бактеріохлорофіл.
Бактерії дуже швидко ростуть, їх ріст і розмноження зале
жить від температури, рН, поживних речовин, концентрації
іонів. Облігатним аеробам потрібен ще кисень, аоблігатним
анаеробам, навпаки, — повна його відсутність.
Діляться бактерії шляхом поділу. Перед поділом клітини
відбувається реплікація ДНК. Перехід до поділу залежить
від співвідношення об’ єму генетичного матеріалу і о б ’ єму
Біологічні особливості життєдіяльності людини
27
цитоплазми. Завдяки швидкому поділові бактеріальні клітини
легко адаптуються до змін оточуючого середовища.
Наприклад, у лабораторії популяція бактерій за кілька тиж
нів завдяки спонтанним мутаціям і природному доборові набу
ває здатності використовувати як джерело вуглеводів нові
типи цукрів.
Крім безстатевого розмноження, бактеріям притаманне
і статеве розмноження. Воно відрізняється від статевого роз
множення еукаріот тим, що у бактерій не утворюються гамети
і не відбувається злиття клітин.
Під час генетичної рекомбінації бактерій частина ДНК клітини-донора переноситься в клітину-реципієнта, ДНК якої
генетично відрізняється від ДНК донора. У процесі заміщення
ДНК беруть участь ферменти, які розщ еплю ю ть і знову
з’ єднують ланцюги ДНК. При цьому утворюється ДНК із вмі
стом генів обох батьківських клітин. Таку ДНК називають
рекомбінантною. У рекомбінантів спостерігається різноманіт
ність ознак, що дуже важливо для еволюції.
Відомі три шляхи отримання рекомбінантів: трансформація,
кон’ югація і трансдукція.
1.3.2. Еукаріотичні клітини. Цитоплазма та клітинні
органели
Клітинні структури лежать в основі будови всіх тваринних
і рослинних організмів, незалежно від їх різноманітності.
У прокаріот і найпростіших поняття “ клітина” й “ організм”
збігаються, їх називають одноклітинними. Однак більшість
тварин і рослин, які є еукаріотами, складаються із багатьох
клітин, і їх називають багатоклітинними. У багатоклітинних
організмів клітини складають тканини, які входять до складу
органів. Життєдіяльність клітин у багатоклітинних організмів
підпорядкована координаційному впливові цілого організму.
Ця координація здійснюється нервовою та гормональною си
стемами.
Уважають, що всі організми і всі клітини, з яких вони скла
даються, утворилися еволюційним шляхом від спільної пред
кової клітини. Переживши своїх конкурентів, ця клітина
28
Розділ 1
поклала початок процесові клітинного поділу та еволюції, який
врешті-решт створив зелений покрив землі, змінив склад ат
мосфери і створив розумне життя. Тільки так можна пояснити
феноменальну подібність між усіма організмами. Завдяки клі
тинній будові, організм є дискретним і зберігає цілісність.
Розчленування цілого організму на дрібні морфологічні одини
ці — клітини, які мають великі поверхні, дуже доцільне для
обміну речовин. Клітинна структура, не порушуючи ж иттє
діяльності цілого організму, сприяє поступовій заміні відмира
ючих або патологічно змінених клітин чи частин тіла новими.
Значення клітинної структури у всьому органічному світі
обумовлене тим, що тільки вона забезпечує найкраще збері
гання, репродукцію і передачу спадкової інформації. Крім
цього, поділ функцій між клітинами в багатоклітинному ор
ганізмі забезпечує широкі можливості пристосування організ
мів до навколишнього середовища.
До кінця XIX ст. накопичився обширний матеріал про бу
дову та життєдіяльність клітини, і до середини X X ст. цей
описовий морфологічний підхід домінував при вивченні клітин,
хоча використовувалися більш досконалі мікроскопи для ви
явлення нових клітинних структур. Цим структурам часто
давали назви, не маючи уявлення про їх призначення чи функ
цію. Тому багато вчених, уважаючи такий підхід незадовільним,
перейшли від морфологічного вивчення клітин до аналізу хі
мічних реакцій, які лежать в основі їх життєдіяльності. Швид
ко було з’ ясовано, що для побудови нових біологічних молекул
клітини використовують енергію молекул поживних речовин.
Таким чином, було показано, що клітини можуть рости і діли
тись, не порушуючи законів термодинаміки, згідно з якими
спонтанні реакції повинні супроводжуватися виділенням теп
ла і призводити до зростання невпорядкованості.
Однак незрозумілим залишилося місце генів у цій хімічній
картині, а саме їх роль є вирішальною у правильному зв’ я
зуванні між собою сотень амінокислот при утворенні молекул
білка. Потім за короткий період між 1953 р., коли Уотсон і Крік
відкрили структуру ДНК, і 1966 р. була розкрита природа
і шляхи передачі генетичної інформації. Коли була встановле
на домінуюча роль ДНК у передачі спадкової інформації, ви
никло відчуття, що вивчивши структуру і функцію нуклеїнових
Біологічні особливості життєдіяльності людини
29
кислот, людство зрозуміло суть живого і що найбільша загад
ка природи розгадана. Однак сьогодні вчені знову відчувають,
що це не так, що вони ще далекі від розуміння суті живого.
Складне переплетення метаболічних шляхів від АТФ до ДНК,
яким би дивним воно не було, ще не є живою клітиною. Навіть
найпростіші клітини є значно складнішими від будь-якого зі
сконструйованих до цього часу комп’ ютерів.
Тому кожен лікар повинен усвідомлювати складність будо
ви та функціонування живого і розуміти, наскільки ми ще
мало знаємо про саму клітину, її будову і процеси, які в ній
відбуваються. А всі хвороби якраз і полягають у певних тонких
структурних і функціонованих змінах тих чи інших клітин.
Усі відомі одноклітинні та багатоклітинні організми поді
ляються на дві великі групи — прокаріоти й еукаріоти. До
прокаріотів належать бактерії, синьо-зелені водорості, а до
еукаріотів — зелені рослини, гриби, тварини.
Структурними компонентами еукаріотичної клітини є:
• Клітинна оболонка (плазмалема), в основі будови якої —
цитоплазматична мембрана — вибірково проникний бар’єр,
що регулює обмін між клітиною і середовищем. Усі біологічні
мембрани складаються з подвійного шару ліпідних молекул,
у якому занурені різні мембранні білки. У плазматичній мем
брані є три основні класи ліпідних молекул — фосфоліпіди,
холестерин і гліколіпіди, однак ліпідний склад зовнішнього
і внутрішнього моношарів різняться між собою. Різний ліпід
ний склад характерний і для різних мембран однієї еукаріо
тичної клітини. Ліпідний бішар визначає основні структурні
особливості біологічних мембран, тоді як білки є відповідаль
ними за більшість мембранних функцій: вони є специфічними
рецепторами і ферментами, а також транспортують крізь мем
брану різноманітних речовин. Велика кількість мембранних
білків мають гідрофобні поверхні, тому вони нерозчинні у вод
них середовищах. Білки асоціюють із ліпідним бішаром різни
ми способами: деякі з них пронизують бішар наскрізь у вигляді
єдиної а-спіралі, інші містяться у більш згорнутій конформації
і, таким чином, перетинають бішар кілька разів. Також до
складу клітинної оболонки входить глікокалікс — надмембранна зона, утворена вуглеводневими радикалами глікопротеїнів
(рис. 1.2).
зо
Розділ 1
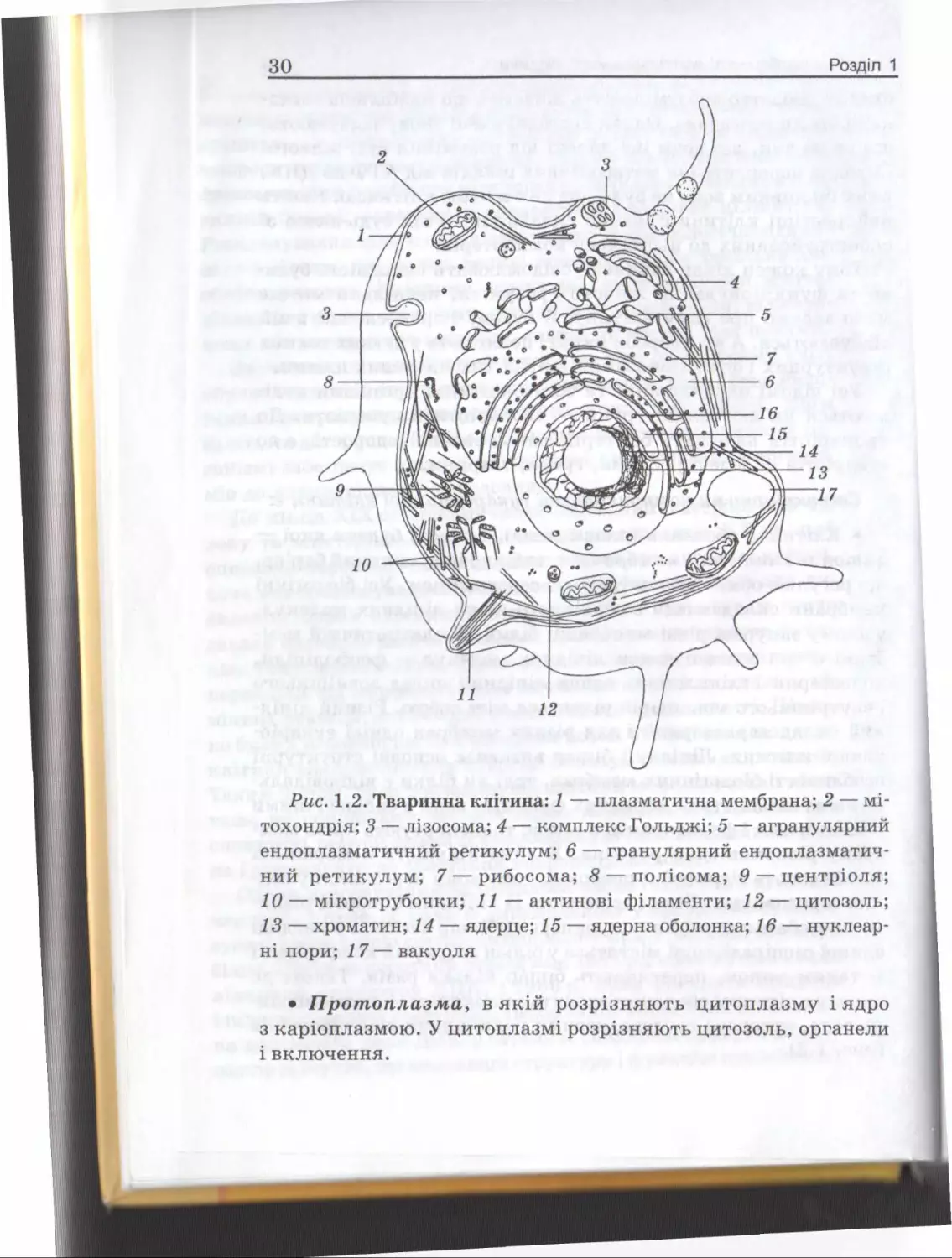
Рис. 1.2. Тваринна клітина: 1 — плазматична мембрана; 2 — мітохондрія; 3 — лізосома; 4 — комплекс Гольджі; 5 — агранулярний
ендоплазматичний ретикулум; 6 — гранулярний ендоплазматич
ний ретикулум; 7 — рибосома; 8 — полісома; 9 — центріоля;
10 — мікротрубочки; 1 1 — актинові філаменти; 12 — цитозоль;
13 — хроматин; 14 — ядерце; 15 — ядерна оболонка; 16 — нуклеарні пори; 17 — вакуоля
• П рот оплазма, в якій розрізняють цитоплазму і ядро
з каріоплазмою. У цитоплазмі розрізняють цитозоль, органели
і включення.
Ьюлогічні особливості життєдіяльності людини
31
Цитозоль утворений переважно білками, забезпечує колоїд
ні властивості цитоплазми, її в’ язкість, еластичність, скорот
нії вість, внутрішній рух. Функціонально є внутрішнім середо
вищем клітини, одним із місць здійснення внутрішньоклітин
ного метаболізму.
Ендоплазматичний ретикулум утворений системою ущіль
нених мембранних мішечків (цистерн) у вигляді трубочок
і пластинок (рис. 1.3).
Рис. 1.3. Гранулярний ендоплазматичний ретикулум: 1 — ве
лика субчастка рибосоми; 2 — мала субчастка рибосоми; 3 — ри
босома; 4 — агранулярний ендоплазматичний ретикулум
Зернистий (гранулярний) ендоплазматичний ретикулум
утворює комплекс із рибосомами і виконує функції синтезу
білків. Крім цього, у ділянках гранулярної частини ретикулуму відбувається синтез білків і ліпідів цитоплазматичних мем
бран та їх збирання. Агранулярний (гладенький) ендоплазма
тичний ретикулум функціонально пов’язаний з обміном вугле
водів, ліпідів та інших речовин небілкової природи, наприклад,
32
Розділ 1
стероїдних гормонів (у статевих залозах, кірковому шарі над
ниркових залоз). У гепатоцитах на гладенькому ендоплазма
тичному ретикулумі руйнуються і знешкоджуються токсичні
речовини, деякі ліки (барбітурати). У пухирцях і канальцях
гладенького ендоплазматичного ретикулуму посмугованих
м ’язів депонуються іони кальцію, і він бере участь у підтримці
гомеостазу кальцію в клітині.
Мітохондрй — це органели двомембранної будови у вигляді
паличок, гранул, ниток розміром 0,5— 7 мкм. Внутрішня мем
брана мітохондрій утворює кристи, простір, обмежений кристами, складає матрикс мітохондрій (рис. 1.4). Під час аероб
ного дихання на кристах відбувається окиснювальне фосфорилювання і перенос електронів, а у матриксі працюють ферменти
циклу Кребса. Основна функція мітохондрій — синтез АТФ.
У матриксі є також власні білоксинтезуючі системи, які за
основними властивостями подібні до апарату синтезу білка
прокаріотів (кільцева молекула ДНК, рибосоми, РНК), і цим
підтверджується симбіотична гіпотеза походження мітохондрій.
Кільцеву молекулу ДНК мітохондрій часто називають 25 хро
мосомою людини. ДНК мітохондрій становить близько 5 % від
загальної кількості ДНК в організмі. Серед інших функцій
мітохондрій — участь у синтезі стероїдних гормонів і деяких
амінокислот (глутамінової).
Рис. 1.4. Мітохондрія: 1 — зовнішня мембрана мітохондрії;
2 — внутрішня мембрана мітохондрії; 3 — кристи; 4 — матрикс
Біологічні особливості життєдіяльності людини
33
Комплекс Гольджі — це система ущільнених мембранних
цистерн, від країв яких відшаровуються пухирці (везикули).
Комплекс розташований переважно в навколоядерній зоні,
контактує з плазматичною мембраною. Він бере участь у кон
центрації, зневодненні й ущільненні продуктів внутрішньоклі
тинної секреції та речовин, які надходять ззовні та призначені
для виведення з клітини. З ним пов’ язаний також синтез поліса
харидів, ліпідів, формування лізосом, глікозилювання білків.
Лізосоми — органели одномембранної будови, мають кулясту
форму, діаметр 0,2— 1 мкм. Розрізняють первинні лізосоми,
які утворюються внаслідок відбрунькування мембранного мі
шечка від комплексу Гольджі та містять набір ферментів кис
лих фосфатаз, які каталізують при низьких значеннях рН
гідролітичне розщеплення нуклеїнових кислот, білків, жирів,
полісахаридів; прелізосоми (фагоцитуючі вакуолі) та вторинні
лізосоми, які утворюються у процесі ендоцитозу внаслідок
злиття первинних лізосом із прелізосомами. Вторинні лізосоми
поділяються на гетерофагосоми (перетравлюють субстрат, який
надійшов до клітини ззовні) і автофагосоми (руйнують власні
структури клітини, які завершили свою функцію). Продукти
розпаду поглинаються і засвоюються цитоплазмою, а неперетравлені рештки залишаються в постлізосомі.
Клітинний центр (центросома) утворений двома центріолями, оточеними ущільненою цитоплазмою — центросферою.
Кожна центріоля — це циліндричне тільце завдовжки 0,3—
0,5 мкм, діаметром 0,15 мкм і утворене дев’ ятьма триплетами
мікротрубочок (побудованих із полімеризованого білка тубуліну). Функція — утворення ахроматинового веретена під час
поділу клітини.
Рибосоми — це дрібні органели немембранної будови розмі
ром 15— 35 нм, утворені малою і великою субчастками, об’ єд
нання яких відбувається за наявності мРНК. Це є рибонуклеопротеїнові структури, що виконують функцію активного
синтезу білка. Кожна рибосома має два функціональні актив
ні центри: аміноацильний і пептидильний. Здебільшого одна
молекула мРНК об’ єднує кілька рибосом і утворює полісому.
Полісоми вільно розташовані в цитоплазмі або прикріплені до
34
Розділ 1
мембран гранулярної ендоплазматичної сітки. Доведено, що
на полісомах цитоплазматичного матриксу утворюються білки
для власних потреб клітини, а на полісомах гранулярної ендо
плазматичної сітки синтезуються білки, які виводяться з клі
тини і використовуються для потреб організму (наприклад,
травні ферменти, білки грудного молока).
Мікротільця — група одномембранних пухирців з дрібно
зернистим матриксом. До цієї групи належать пероксисоми.
Вони утворюються в комплексі Гольджі. їх діаметр стано
вить — 0,3— 1,0 мкм. Пероксисоми містять переважно фермен
ти для знешкодження пероксиду водню, який утворюється
в результаті окиснення деяких органічних речовин і є дуже
токсичним для клітини. Переважно пероксид водню розщеп
люється під дією ферменте каталази. Ці реакції включені до
різних метаболічних циклів, зокрема в обмін сечовини у клі
тинах печінки та нирок. Пероксисоми беруть участь також
у процесі окиснення жирних кислот. У гепатоциті кількість
пероксисом досягає 70— 100.
Включення — відносно непостійні компоненти цитоплазми,
які виконують функцію запасання поживних речовин (жир,
глікоген), продуктів, які підлягають виведенню з клітини (гра
нули секрету), баластних речовин (деякі пігменти).
Для рослинної клітини характерними є такі особливості
будови (рис. 1.5):
• Клітинна стінка, у складі будови якої — целюлоза.
• Вакуолі з клітинним соком, оточені мембраною (тонопластом).
• Пластиди (хлоропласти, лейкопласти, хромопласти); усі
види пластид мають двомембранну структуру (рис. 1.6). Усі
пластиди, як і мітохондрії, мають кільцеву молекулу ДНК
і систему біосинтезу білка;
Хлоропласти — це зелені пластиди, зовнішня мембрана
яких гладка, а внутрішня утворює тилакоїди гран у вигляді
стовпчика монет, з’ єднаних між собою ламелами. Вміст хло
ропласта — це строма, багата ферментами. На тилакоїдах
синтезується пігмент хлорофіл і здійснюється світлова фаза
Біологічні особливості життєдіяльності людини
35
Рис. 1.5. Рослинна клітина: 1 — плазматична мембрана; 2 — клі
тинна стінка; 3 —- серединна пластинка; 4 — мітохондрія; 5 — хло
ропласт; 6 — тилакоїди гран; 7 — центральна вакуоля; 8 — комп
лекс Гольджі; 9 — рибосома; 10 — полісома; 11 — ядерна оболон
ка; 12 — хроматин; 13 — ядерце; 14 — нуклеарні пори
Рис. 1.6. Хлоропласт: 1 — зовнішня мембрана хлоропласта;
2 — внутрішня мембрана хлоропласта; 3 — строма; 4 — грани;
5 — тилакоїди
36
Розділ 1
фотосинтезу, а у стромі хлоропласта відбувається темнова фаза
фотосинтезу.
Хромопласти — це кольорові пластиди, в яких синтезують
ся пігменти каротиноїди, ксантофіли, фікоціани, фікоеритрини.
Лейкопласти — це безбарвні пластиди, в яких відкладають
ся про запас поживні речовини рослинної клітини.
• Відсутність клітинного центру у клітинах вищих рослин.
• Плазмодесми — цитоплазматичні містки, які з’ єднують
клітини рослинного організму між собою.
1.3.3. Ядро клітини. Хроматин: рівні організації
спадкового апарату. Будова хромосоми
Ядро еукаріотичної клітини відмежоване від цитоплазми
двомембранною оболонкою. Зовнішня та внутрішня мембрани
ядерної оболонки містять пори. Зокрема, пори зовнішньої
мембрани з’ єднані з мембранами ендоплазматичної сітки і за
безпечують взаємозв’язок ядра та цитоплазми. Зовнішня мем
брана може бути вкрита рибосомами. Вміст ядра— це каріоплазма. У каріоплазмі є велика кількість ферментів, а також
спадковий апарат клітини — хроматин (деконденсовані хромо
соми). У ядрі також розрізняють ядерця — непостійні струк
тури. Вони складаються з білків і РНК та формуються вторин
ними перетяжками хромосом.
Ділянки хромосом, які інтенсивно фарбуються барвниками,
називаються гетерохроматином. Більш світлі, слабо забарвлені
ділянки хромосом, називаються еухроматином. Припускають,
що в еухроматині містяться гени, які експресуються, а гетерохроматин містить неактивні гени. Хромосоми можуть перебу
вати у двох структурно-функціональних станах: конденсованому
(спіралізованому) і неконденсованому (деспіралізованому). Найдрібнішими структурними компонентами хромосоми є нуклеопротеїнові фібрили, які складаються з ДНК і білків, переваж
но гістонів. Молекули гістонів комплексують з ДНК не по всій
довжині рівномірно, а окремими ділянками. Комплекс молекул
гістонів із ДНК має назву нуклеосоми. Припускають, що хро
Біологічні особливості життєдіяльності людини
37
мосома утворена однією величезною фібрилою нуклеопротеїну.
За іншими уявленнями, фібрили нуклеопротеїну попарно скру
чуються, утворюючи хромонеми, які входять до складу напівхроматиди, дві напівхроматиди утворюють хроматиду, а дві
хроматиди — хромосому. Кожна хромосома має первинну пе
ретяжку (центромеру), яка розмежовує її на два плеча. Залеж
но від місця розташування розрізняють три типи хромосом:
метацентричні (розташування центромери медіанне), субметацентричні (розташування центромери субмедіанне) і акроцентричні (розташування центромери термінальне). Кінці плечей
хромосоми називають теломерами. Це спеціалізовані ділянки,
які заважають сполученню хромосом між собою або з іншими
фрагментами. На плечах хромосом видно товстіші й інтенсив
ніше забарвлені ділянки — хромомери, які чергуються з міжхромомерними нитками.
Диплоїдний набір хромосом клітини — каріотип. Нормаль
ний каріотип людини включає 46 хромосом, або 23 пари; з них
22 пари аутосом і одна пара статевих хромосом. Для вивчення
складного комплексу хромосом каріотипу їх розташовують
у вигляді ідіограми. В ідіограмі хромосоми розташовуються
попарно у порядку зменшення розмірів, за винятком статевих
хромосом. Тільце Барра (Х-статевий хроматин) — утворення,
яке спостерігається у світловий мікроскоп в інтерфазному ядрі.
Тільце Барра може мати різноманітну форму (трикутну, оваль
ну, півмісяцеву) і щільно прилягає до ядерної оболонки. Вва
жають, що воно утворене однією Х-хромосомою, яка перебуває
у спіралізованому стані (гетерохроматизована). У жінок дві
статеві Х-хромосоми, одна з яких є активною, а друга утворює
грудочку статевого хроматину і є неактивною. У чоловіків є
статеві X і У-хромосоми.
Термінальні ділянки хромосом, названі теломерами, не під
даються реплікації і втрачаються під час кожного клітинного
поділу. У 1980-х роках було експериментально встановлено,
що теломери хромосом старих клітин справді коротші за теломери молодих клітин.
Тепер відомо, що теломерна ділянка нитки ДНК хромосом
складається більш як з 1000 нуклеотидних повторів — ТТАООО.
Оскільки під час реплікації ДНК фермент ДНК-полімераза не
може забезпечити реплікацію кінцевих нуклеотидів у нитці
38
Розділ 1
ДНК, то з кожним наступним поділом клітини довжина хро
мосоми вкорочується приблизно на 10— 20 теломерних фраг
ментів. При досягненні критичної довжини теломера відбува
ються такі порушення структури ДНК, які не сумісні з нор
мальним функціонуванням клітини.
Теломераза — це клітинний фермент рибонуклеопротеїнової
природи, який забезпечує відновлення довжини теломерної
ділянки хромосомної ДНК. Однак у більшості нормальних
клітин людини чи тварин теломераза відсутня. Тому за зви
чайних умов ці клітини піддаються апоптозу після 50— 100 мі
тотичних поділів. Якщо 1000 теломер, які містяться на одному
кінці хромосоми, поділити на 1—20 теломер, які втрачаються
під час одного мітотичного циклу, то це означає, що клітина
може пережити 50—100 мітотичних поділів.
Встановлено, що ген теломерази активний у багатьох клі
тинах злоякісних пухлин. Це пояснює те, що тривалість ж ит
тя злоякісних клітин практично не обмежена кількістю клітин
них мітозів.
1.4. Обмін речовин і енергії
Життєдіяльність клітини як одиниці біологічної активності,
найменшої структурно-функціональної одиниці живого забез
печується сукупністю взаємопов’язаних, упорядкованих у про
сторі та часі метаболічних процесів. Ці процеси утворюють
потоки речовин, енергії та інформації.
У живому організмі постійно відбуваються складні процеси
перетворення та переносу речовин, енергії, синтез, розпад, рух
тощо. Процеси синтезу називають анаболізмом (асиміляцією),
і являють собою пластичний обмін, а процеси розпаду — ката
болізмом — і являють собою енергетичний обмін. Сукупність
пластичного й енергетичного обмінів називається обміном ре
човин, або метаболізмом. Складні органічні речовини, які
надходять в організм, розщеплюються ферментами. Продукти
розщеплення білків — амінокислоти, жирів — гліцерин і жир
ні кислоти, вуглеводів— гл ю к оза— всмоктуються у кров
і використовуються організмом для подальших перетворень.
Біологічні особливості життєдіяльності людини
39
Спряжені реакції дихання й окисного фосфорилювання
в мітохондріях клітин не тільки постачають енергію, але й за
безпечують клітину речовинами, необхідними для синтезу
різноманітних молекул. Ними є різні продукти розщеплення
продуктів харчування. Особлива роль у цьому процесі належить
циклові Кребса, ферменти якого локалізовані на внутрішній
мембрані мітохондрій. Через цей цикл проходить шлях вугле
водневих атомів більшості речовин, які є проміжними продук
тами синтезу хімічних компонентів клітини. У циклі Кребса
відбувається вибір шляху перетворення тієї чи іншої сполуки,
а також переключення обміну клітини з одного шляху на ін
ший, наприклад з вуглеводного на жировий. Таким чином,
обмін у циклі Кребса — основна ділянка потоку речовин, що
об’ єднує метаболічні шляхи розщеплення й утворення вугле
водів, білків, жирів, нуклеїнових кислот.
Потік енергії у представників різних груп організмів забез
печується механізмами енергозабезпечення — диханням, фо
тосинтезом або хемосинтезом, бродінням. Основна роль у біо
енергетиці тваринних клітин належить диханню й окисному
фосфорилюванню. Ці спряжені процеси включають розщеп
лення низькокалорійного органічного “ палива” у вигляді глю
кози, жирних кислот, амінокислот, а також використання
енергії, що виділяється, і спрямовані на утворення висококало
рійного “палива” у вигляді аденозинтрифосфату (АТФ). Енер
гія АТФ — безпосередньо або через інші макроергічні сполуки
(наприклад, креатинфосфат) — у різноманітних внутрішньоклі
тинних процесах перетворюється в той чи інший вид роботи:
хімічну (синтез), механічну, осмотичну, електричну, регуля
торну.
Окрім мітохондрій, які відіграють основну роль у синтезі
АТФ, завдяки спряженим реакціям дихання й окисного фос
форилювання, важливе місце в синтезі АТФ належить також
анаеробному гліколізові, ферменти якого локалізовані з вну
трішнього боку плазматичної мембрани. Із двох механізмів,
які забезпечують життєдіяльність клітини енергією, анаеробний
гліколіз менш ефективний. Через відсутність оксигену відбу
вається неповне окиснення глюкози, тому в процесі гліколізу
для потреби клітини використовується не більше 10 % енергії.
Недоокиснені продукти гліколізу (піруват) далі надходять
40
Розділ 1
у мітохондрії, де в умовах повного окиснення, спряженого
з фосфорилюванням АДФ до АТФ, віддають для потреб кліти
ни залишену в їх хімічних зв’ язках енергію. Із перетворювачів
енергії хімічних зв’язків АТФ у роботу найкраще вивчена механохімічна система поперечносмугастих м ’ язів. Вона склада
ється зі скоротливих білків (актоміозиновий комплекс) і фер
менту аденозинтрифосфатази, який розщеплює АТФ зі звіль
ненням енергії.
Особливість потоку енергії в рослинних клітинах полягає
у наявності фотосинтезу — механізму перетворення енергії
сонячного світла в енергію хімічних зв’язків органічних речо
вин. Різні механізми енергозабезпечення клітини відрізняють
ся своєю ефективністю. Коефіцієнти корисної дії хлоропласта
і мітохондрії досягають 25 і 45—60 % відповідно.
Потоки речовин, енергії та інформації циркулюють безпе
рервно і становлять необхідну умову збереження клітини як
живої системи. Окрім структур і процесів, прямо включених
в обмін речовин та енергії, у клітині також функціонують інші
механізми, які є життєво необхідними. Так, лізосоми забезпе
чують гідролітичне розщеплення макромолекул, які надійшли
у клітину, до низькомолекулярних сполук. Вони також руй
нують внутрішньоклітинні структури, які втратили своє фун
кціональне значення. Утворені при цьому хімічні сполуки
включаються в потоки речовин, енергії, інформації. Пероксисоми розщеплюють токсичні для клітини пероксиди, які по
стійно утворюються в результаті біохімічних процесів. Органі
зація внутрішньоклітинних транспортних потоків забезпечуєть
ся також активністю мікротрубочок, мікрофібрил.
1.5. Молекулярний рівень організації
спадкової інформації у клітині.
Нуклеїнові кислоти
Завдяки наявності потоку інформації, клітина у процесі
еволюції стала структурою, яка відповідає всім критеріям живо
го, підтримує своє існування в часі, передає свої властивості
наступним поколінням. У потоці інформації беруть участь ядро
Біологічні особливості життєдіяльності людини
41
(конкретно ДНК хромосом), мРНК, тРНК, цитоплазматичні
апарати транскрипції та трансляції тощо. На завершальному
етапі цього потоку поліпептиди, які синтезовані на полісомах,
набувають третинної та четвертинної структури і використо
вуються як ферменти-каталізатори чи структурні білки. В еукаріотичних клітинах, окрім основного за обсягом уміщеної ін
формації ядерного геному, функціонують також геноми мітохондрій, а в зелених рослинах — і хлоропластів.
Дезоксирибонуклеїнова кислота (ДНК) — матеріальний
субстрат спадковості та мінливості. ДНК — це макромолекули,
полімери, що утворені нуклеотидами — аденіловим, гуаніловим, цитидиловим і тимідиловим. До складу кожного нуклеотиду входить пентоза (дезоксирибоза), ортофосфорна кислота
й азотиста основа (пуринова чи піримідинова). Види азотистих
основ, які входять до складу нуклеотидів ДНК: аденін (А),
гуанін (Г), цитозин (Ц), тимін (Т). Сполучення нуклеотидів
у макромолекулу ДНК відбувається через взаємодію гідрокси
лу одного нуклеотиду з фосфатом наступного з утворенням
фосфодіефірного зв’ язку. Збирання полінуклеотидного ланцю
га здійснюється за участю фермента ДНК-полімерази.
Особливістю структурної організації ДНК є те, що її молеку
ли утворені двома полінуклеотидними ланцюгами. Відповідно
до трьохвимірної моделі ДНК, запропонованої в 1953 р. амери
канським біофізиком Дж. Уотсоном і англійським біофізиком
та генетиком Ф. Кріком, ці ланцюги з’ єднуються один з одним
гідрогеновими зв’язками між їх азотистими основами за прин
ципом комплементарності: А з’ єднується зТ двома гідрогеноііими зв’ язками, а між Г і Ц різних ланцюгів утворюються три
гідрогенові зв’ язки. Таке сполучення азотистих основ забезпе
чує тривкий зв’ язок двох ланцюгів і збереження однакової
відстані між ними на всьому проміжку.
Інша важлива особливість об’єднання двох полінуклеотидних
ланцюгів у молекулу ДНК — антипаралельність: 5-кінець од
ного ланцюга сполучається зЗ-кінцем іншого і навпаки.
Дані рентгеноструктурного аналізу засвідчили, що двохланірогова молекула ДНК утворює спіраль, закручену навколо
власної осі; діаметр спіралі — 2 нм, довжина кроку — 3 ,4 нм,
до кожного закруту входить 10 пар нуклеотидів.
У живій клітині подвійна спіраль, яка становить вторин
ну структуру ДНК, не має вигляду розгорнутої молекули,
42
Розділ 1
а додатково згорнута у просторі, утворюючи третинні структу
ри, — суперспіралі. У суперспіралізованому стані молекула
ДНК в комплексі з певними клітинними білками входить до
складу нуклеоїду прокаріотів та ядерного хроматину еукаріотів.
Завдяки суперспіралізації довгі молекули ДНК формують ком
пактні утворення, зокрема хромосоми ядра.
Таким чином, у структурній організації молекули ДНК
розрізняють первинну будову — полінуклеотидний ланцюг,
вторинну — два комплементарні один до одного і антипаралельні полінуклеотидні ланцюги, з’єднані гідрогеновими зв’язками,
і третинну — трьохвимірну спіраль із певними просторовими
характеристиками.
Основними біологічними функціями ДНК є:
1. Збереження спадкової інформації. Кількість ДНК у сома
тичних і статевих клітинах організму людини є сталою вели
чиною, яку ці клітини отримують у процесі запліднення бать
ківських гамет і подальшого поділу зиготи.
2. Передавання спадкової інформації нащадкам через под
воєння молекул ДНК під час реплікації (рис. 1.7). Процес
реплікації напівконсервативний, у розплітанні подвійної спіралі
ДНК бере участь фермент геліказа. Утворені при цьому ділян
ки зв’ язуються спеціальними дестабілізуючими білками. В міс
цях розходження полінуклеотидних ланцюгів (у зонах репліка
ції) утворюються реплікаційні вилки. У кожній з цих ділянок за
наявності ферменту ДНК-полімерази синтезується ДНК двох но
вих дочірніх молекул. У процесі синтезу реплікаційна вилка
рухається вздовж материнської спіралі та захоплює нові зони.
Особливість ДНК-полімерази — її нездатність почати синтез
нового полінуклеотидного ланцюга через звичайне зв’язування
двох нуклеозидтрьохфосфатів: необхідний 3-ОН-кінець якогось
полінуклеотидного ланцюга, що спарений з матричним ланцю
гом ДНК, до якого ДНК-полімераза може лише додавати нові
нуклеотиди. Такий полінуклеотидний ланцюг називається
праймером (“ затравка” ). Функцію праймера для синтезу полі
нуклеотидних ланцюгів ДНК у процесі реплікації виконують
короткі послідовності РНК, що утворюються за наявності фер
менту РНК-праймази. Вказана особливість ДНК-полімерази є
свідченням того, що матрицею при реплікації може бути
Біологічні особливості життєдіяльності людини
43
З'
Рис. 1.7. Ділянка реплікаційної вилки в молекулі ДНК
лише той ланцюг ДНК, котрий несе спарений з ним праймер,
який має вільний 3-ОН-кінець. Механізм реплікації відзна
чається надзвичайно високою точністю відтворення структури
ДНК, і в її підтриманні важлива роль належить, перш за все,
ДНК-полімеразі.
1.6. Механізми реалізації спадкової
інформації в клітині
Ця біологічна функція здійснюється за рахунок передачі
закодованої в ДНК інформації молекулам інформаційних (мат
ричних) РНК (транскрипції) та подальшої розшифровки цієї
інформації при синтезі білків (трансляції). Сукупність зазна
чених біологічних функцій ДНК та механізмів їх реалізації
отримала назву центральна догма молекулярної біології.
Рибонуклеїнові кислоти (РНК) — посередники в перекодуван
ні та реалізації біологічної інформації. РНК представлені одним
иолінуклеотидним ланцюгом, який складається з чотирьох
44
Розділ 1
різновидів нуклеотидів, які містять пентозу (рибозу), ортофос
форну кислоту і одну з таких азотистих основ: А , Г, Ц,
У (урацил). РНК синтезується на молекулі ДНК за допомогою
ферментів РНК-полімераз із дотриманням принципу комплементарності й антипаралельності, зокрема, аденін ДНК комп
лементарний урацилу РНК.
Матричні (інформаційні) РНК (м РН К або ІРНК). Це
клас РНК, які становлять 2— 5 % від загальної кількості клі
тинної РНК. Матричні РНК (мРНК) виконують функцію пере
носників генетичної інформації від геному (ядерної ДНК) до
білоксинтезуючої системи клітини. Вони є інформаційними
матрицями, що визначають амінокислотні послідовності в моле
кулах поліпептидів, що синтезуються у рибосомах.
Синтез мРНК відбувається на відповідних ділянках ДНК.
Процес синтезу мРНК — транскрипція — починається з вияв
лення РНК-полімеразою особливої ділянки в молекулі ДНК —
про мотора. Після приєднання до промотори РНК-полімераза
розплітає закрут спіралі ДНК, який розташований поруч. Два
ланцюги ДНК у цьому місці розкручуються, і на одному з них
фермент здійснює синтез мРНК. Збирання рибонуклеотидів
у ланцюг відбувається з дотриманням їх комплементарності
нуклеотидам ДНК, а також антипаралельно щодо матричного
ланцюга ДНК. РНК-полімераза здатна збирати полінуклеотид
лише від 5-кінця, і тому матрицею для транскрипції може
бути лише один із двох ланцюгів ДНК, а саме той, котрий
обернений до фермента своїм 3-кінцем. Такий ланцюг нази
вається кодогенним. Просуваючись уздовж кодогенного лан
цюга ДНК, РНК-полімераза здійснює поступове точне перепи
сування інформації доти, поки не натрапить на специфічну
нуклеотидну послідовність — термінатора транскрипції.
У цій ділянці РНК-полімераза від’єднується і від матриці ДНК,
і від синтезованої мРНК. Таким чином, мРНК містить точну
копію інформації, записаної у відповідній ділянці ДНК. Фраг
мент молекули ДНК, який включає промотор, структурні гени
(цистрони) і термінатор, утворює одиницю транскрипції —
транскриптон.
Триплети нуклеотидів мРНК, розташовані поряд, — це кодони. Послідовність кодонів мРНК шифрує послідовність амі
нокислот у поліпептидному ланцюзі.
Біологічні особливості життєдіяльності людини
45
Транспортні РН К (т РН К ). На тРНК припадає 10— 20 %
клітинної РНК. їх молекули — де полірибонуклеотидні лан
цюги, довжина яких становить 70—90 нуклеотидів. Транспор
тна тРНК постачає необхідні амінокислоти до місця збирання
поліпептидних ланцюгів і, таким чином, виконує функцію
трансляційного посередника. Первинна структура тРНК, яка
визначається послідовністю нуклеотидів, формує вторинну
структуру тРНК. Це тривимірна третинна структура, для якої
характерне утворення двох перпендикулярно розташованих
подвійних ланцюгів. Один із них утворений акцепторною
зоною, другий — антикодоновою. До акцепторної зони приєд
нується амінокислота, а на антикодоновій розміщений антикодон (триплет нуклеотидів, який комплементарний кодонові
мРНК). Ці ділянки максимально віддалені одна від одної. Ста
більність третинної структури тРНК підтримується завдяки
виникненню додаткових гідрогенових зв’ язків між основами
полінуклеотидного ланцюга, що локалізуються в різних її ді
лянках, але просторово наближені у третинній структурі.
Одна з особливостей тРНК — наявність нетипових основ, що
виникають внаслідок хімічної модифікації вже після включен
ня нормальної основи в полінуклеотидний ланцюг. Ці змінені
основи зумовлюють структурну різноманітність тРНК при
спільному плані їх будови. На особливу увагу заслуговують
модифікації основ, які формують антикодон. Вони впливають
на специфічність взаємодії антикодона з кодоном. Наприклад,
нетипова основа інозин, яка інколи стоїть у першому положен
ні антикодона тРНК, здатна помилково сполучатися з трьома
різними третіми азотистими основами мРНК — У, Ц, А. Оскіль
ки одна із властивостей генетичного коду є його виродженість,
то більшість амінокислот шифрується кількома кодонами, які
відрізняються переважно третьою основою. Завдяки неспеци
фічному зв’язуванню модифікованої основи антикодоном, одна
тРНК впізнає кілька кодонів-синонімів. Встановлено також
існування кількох видів тРНК* які здатні сполучатися з одним
і тим самим кодоном. У результаті в цитоплазмі трапляється
близько 40 різних молекул тРНК, і цієї кількості достатньо,
щоб транспортувати 20 різних амінокислот до місця синтезу
білка.
Водночас із функцією точного впізнавання визначеного
кодона в мРНК, молекула тРНК постачає до місця синтезу
46
Розділ 1
поліпептидного ланцюга амінокислоту, зашифровану за допо
могою цього кодона.
Рибосомні РНК (рРНК). Це клас клітинних РНК, які вхо
дять до складу рибосом прокаріотичних і еукаріотичних клітин.
На рРНК припадає до 90 % загальної кількості клітинних
РНК.
Рибосомні рРНК і специфічні білки становлять основу струк
тури та функції рибосом, у яких відбувається процес трансля
ції — біосинтез поліпептидних ланцюгів на основі коду, що
доставляється мРНК.
У рибосомі є дві борозенки: одна утримує поліпептидний
ланцюг, який нарощується в процесі трансляції, а інша —
мРНК. Окрім цього, в рибосомі розрізняють дві ділянки, які
зв’ язують тРНК. В аміноацильній розташована аміноацилтРНК, в пептидильній — тРНК, яка навантажена ланцюжком
амінокислот. Рибосома постійно екранує сегмент мРНК за
вдовжки близько ЗО нуклеотидів (рис. 1.8).
Під час трансляції виділяють три фази:
1. Фаза ініціації (початок синтезу поліпептиду) — полягає
в об’ єднанні двох субчасток рибосом на визначеній ділянці
мРНК. Процес ініціації трансляції каталізується особливими
білками — факторами ініціації (еІГ-1, еІГ-2, еІГ-3) та ГТФ, які
лабільно зв’язані з малою субчасткою рибосоми. Після завершен
ня фази ініціації та утворення комплексу рибосома — мРНК з кодоном ініціації АУГ — аміноацил-тРНК ці фактори від’ єд
нуються від рибосоми.
2. Фаза елонгації (продовження пептиду) — включає всі
реакції від моменту утворення першого пептидного зв’ язку до
приєднання останньої амінокислоти.
3. Фаза термінації (завершення синтезу поліпептиду) —
пов’язана з упізнаванням специфічним рибосомним білком
одного з термінуючих кодонів (УАА, УАГ, УГА). При цьому
до останньої амінокислоти в поліпептидному ланцюзі приєд
нується Н20 , і її карбоксильний кінець від’ єднується від тРНК.
У результаті завершений поліпептидний ланцюг втрачає зв’язок
із рибосомою і вона розпадається на дві субчастки.
Он
о
з
ії біосинтезу білків у прокаріотів
48
Розділ 1
1.6.1. Регуляція експресії генів у прокаріотів.
Теорія оперона
Загальну схему роботи генів прокаріотів запропонували
французькі генетики Ф. Жакоб і Ж. Моно у 1961 р. (рис. 1.9).
Рис. 1.9. Схема функціонування лактозного оперона в Е. соїі:
а — включення Ьас-оперона за допомогою індуктора; б — репресія
£ас-оперона
Біологічні особливості життєдіяльності людини
49
Ця схема запропонована на основі вивчення механізмів ре
гуляції процесів транскрипції у Е. соїі (кишкової палички).
Ф. Жакоб і Ж. Моно вивчали молекулярно-генетичні механіз
ми контролю у клітинах Е. соїі біосинтезу р-галактозидази, що
забезпечує розщеплення лактози на глюкозу та галактозу. Щоб
пояснити механізми контролю функції генів в Е. соїі, які від
повідають за метаболізм у клітинах енергетичного субстрату
дисахариду лактози, Ф. Жакоб і Ж . Моно висунули теорію
оперона. Оперон — це комплекс генетичних елементів, які
відповідають за координований синтез групи функціонально
зв’ язаних ферментних білків. Зокрема, у разі І,ас-оперона Е. соїі
це є ферменти метаболізму лактози: р-галактозидаза, р-галактозидпермеаза та р-галактозидтрансацетилаза.
До складу оперона входять:
а) структурні гени, які містять інформацію відносно пер
винної структури поліпептидів, що транскрибуються з даного
оперона;
б) контрольні сайти, до яких належать:
— промотор — ділянка ДНК, яка первинно взаємодіє
з РНК-полімеразою;
— ген-оператор — ділянка ДНК, з якою може спе
цифічно зв’ язуватися білок-репресор; ген-оператор безпо
середньо прилягає до структурних генів, і його зв’язування
з репресором протидіє зчитуванню РНК-полімеразою ін
формації зі структурних генів;
— ген-регулятор — розташований на певній відстані
від оперона і кодує синтез білка-репресора.
За відсутності в середовищі, на якому вирощуються бактерії,
цукру лактози активний білок-репресор, що синтезується геном-регулятором (І), взаємодіє з оператором (О), чим переш
коджає з’ єднанню РНК-полімерази з промотором (Р) і транс
крипції генів 2, У, А. Поява в середовищі лактози інактивує
репресор, він не з ’ єднується з оператором, РНК-полімераза
взаємодіє з промотором і відбувається транскрипція поліцистронної мРНК. Остання забезпечує синтез одночасно всіх фер
ментів, що беруть участь у метаболізмі лактози ф-галактозидази,
50
Розділ 1
Р-галактозидпермеази і (3-галактозидтрансацетилази, що коду
ються генами 2, У, А). Зменшення вмісту лактози у результаті
її ферментативного розщеплення призводить до відновлення
здатності репресора з’ єднуватися з оператором і припинення
транскрипції генів 2, У, А.
1.6.2. Екзонно-інтронна організація генома
еукаріотів
В еукаріотів не встановлено оперонної організації генів
(рис. 1.10). Гени, що визначають синтез ферментів одного лан
цюга біохімічних реакцій, можуть бути розсіяні в геномі й, оче
видно, не мають, як прокаріоти, єдиної регулюючої системи
(ген-регулятор, оператор, промотор).
Транскриптон (одиниця транскрипції) еукаріотів утворений
акцепторною зоною (включає ген-промотор — Р і акцепторні
гени — А) й інформативною зоною, що включає структурні
гени — 8,, 82, 83. У процесі синтезу мРНК, який відбувається
в еукаріотичній клітині, утворюється первинний транскрипт
(про-мРНК), здатний до перетворення у функціонально повно
цінну молекулу мРНК у результаті процесингу (дозрівання).
Це пов’ язано з тим, що геноми еукаріотів і гени, що транслю
ються у відповідні білки, — екзони, — мають значну кількість
нуклеотидних послідовностей, які не несуть генетичної інфор
мації, — інтронів.
Процесинг первинного транскрипта включає:
а) приєднання до 5'-кінця молекули специфічної нуклеотидної структури — “ кепу” ;
б) приєднання до З'-кінця первинного транскрипта поліаденілатного “ хвоста” ;
в) вирізання інтронів із молекули про-мРНК за участю фер
ментів рестриктаз і зшивання внутрішніх кінців молекул екзонів — сплайсинг за участю ферментів лігаз.
Модифікування кінцевих І
° 1 / \!х‘уі\//////27//Ж і Ш//////л2 т Щ ш ш а
Первинний транскрипт
3
52
Розділ 1
1.7. Розмноження — універсальна властивість
живої матерії
Біологічна роль розмноження полягає в тому, що воно за
безпечує зміну поколінь. Залежно від характеру клітинного
матеріалу, що використовується з метою розмноження, розріз
няють різні форми та способи розмноження.
Нестатеве розмноження в одноклітинних організмів:
— поділ клітини навпіл: цьому процесові передує редуплі
кація ДНК (прокаріоти, одноклітинні, найпростіші), в основі
поділу лежить мітоз;
— шизогонія, або множинний поділ ядра: після утворення
певної кількості ядер цитоплазма розпадається на окремі фраг
менти, які зосереджені навколо утворених ядер (малярійний
плазмодій);
— ендогонія, або внутрішнє брунькування (трапляється
у токсоплазмі);
— брунькування: у певному місці на клітині утворюється
горбик, який містить дочірнє ядро або нуклеоїд, цей горбик
росте і відокремлюється від материнської клітини (бактерії,
дріжджі);
— спороутворення (зустрічається в одноклітинних тварин
і бактерій): спора як одна зі стадій життєвого циклу є клітиною,
яка оточена міцною оболонкою; спори у рослин — це один зі
способів безстатевого розмноження.
Нестатеве розмноження у багатоклітинних організмів:
— брунькування (характерне для кишковопорожнинних
і кільчастих червів);
— поліембріонія: на певному етапі розвитку зародок поді
ляється на дві або більше частин, і кожна з них розвивається
у самостійний організм (утворення монозиготних близнюків
у людини).
Біологічні особливості життєдіяльності людини
53
1.7.1. Життєвий цикл клітини. Мітоз
Закономірні зміни структурно-функціональних характери
стик клітини у часі становлять зміст життєвого циклу кліти
ни (клітинного циклу).
Клітинний цикл — це період існування клітини від момен
ту її утворення через поділ материнської клітини до власно
го поділу або смерті.
У клітинному циклі розрізняють:
1. Період спокою (О0) — клітина починає підготовку до
мітозу, або починається спеціалізація клітини у певному функ
ціональному напрямі.
2 . Інтерфазу— період виконання клітиною багатоклітин
ного організму специфічних функцій. Включає: С^-період
(постмітотичний, або пресинтетичний) — відновлюються риси
інтерфазної клітини, завершується формування ядерець, від
бувається біосинтез білка та РНК; 5-період (синтетичний) — від
бувається редуплікація ДНК, продовжується біосинтез білка
та РНК; Суперіод (постсинтетичний, або премітотичний) — від
бувається синтез РНК, білка та накопичення молекул АТФ.
3. Мітоз — непрямий поділ соматичних клітин, у результаті
якого з материнської клітини утворюються дві дочірні клітини
з таким самим набором хромосом, як і в материнській клітині
(рис. 1.11).
Мітоз включає в себе 4 фази:
— Профаза: хромосоми спіралізуються, ядерця руйнуються,
ядерна оболонка розпадається, центріолі розходяться до полюсів
і утворюють веретено поділу.
— Метафаза: утворення веретена поділу завершується, хро
мосоми розташ овуються в площ ині екватора й утворюють
метафазну пластинку, кожна хромосома розщеплюється на дві
хроматиди, що з ’ єднані в ділянці центромери.
— Анафаза: зв’ язок між хроматидами (дочірніми хромосо
мами) руйнується, і вони як самостійні хромосоми рухаються
до полюсів клітини; після завершення руху на полюсах збира
ються два рівноцінні набори хромосом.
П роф аза
Біологічні особливості життєдіяльності людини
55
— Телофаза: реконструюються інтерфазні ядра дочірніх
клітин, хромосоми деспіралізуються, поділ генетичного ма
теріалу завершується; руйнується веретено поділу, материнська
клітина ділиться на дві дочірні (цитокінез).
Основну роль у почерговій зміні періодів клітинного циклу
і, зокрема, фаз мітозу відіграють спеціальні протеїнкінази — так
звані циклінзалежні кінази. Кожна із них фосфорилює від
повідні білки, задіяні у певні фази циклу, і таким чином акти
вує чи інгібує їх. Для активації циклінзалежної кінази необ
хідне зв’язування з нею спеціального білка — цикліну. Відомо
кілька різних циклінів. Можуть існувати різні комплекси
конкретних циклінів і конкретних кіназ, причому кожен такий
комплекс характерний для строго відповідної фази мітозу чи
періоду клітинного циклу.
Втрата контролю над генетичною стабільністю та порушен
ня клітинного циклу відіграють ключову роль у розвитку
злоякісних новоутворень. Нині доведено, що продукти генів
р53, КЬ, Е2Г і ЦГ-кВ є ключовими компонентами у ланцюзі
регуляторів біохімічних подій, які регулюють цілісність гено
ма. Встановлено, наприклад, що у відповідь на пошкодження
структури ДНК різними генотоксичними чинниками продукція
білка р53 і його біологічна активність швидко зростають. Це
або супроводжується зупинкою клітинного циклу (якщо клі
тина може усунути незначне пошкодження ДНК шляхом ре
парації), або призводить до апоптозу (якщо пошкодження ДНК
несумісне з продовженням ж иттєдіяльності клітини). Про
важливу роль білка р53 у регуляції клітинного циклу свідчить
також той факт, що більше половини всіх злоякісних пухлин
людини мають мутаційно змінений ген р53.
Однак за нормального чи патологічного стану клітини не
можуть ділитися безмежно. Вони мають обмежений проліферативний потенціал. Так, для молодих фібробластів шкіри він
становить приблизно 50 поділів. Клітини можуть відмирати
різними шляхами. Найголовніші з них — апоптоз, автофагія
та некроз. Апоптоз (запрограмована клітинна смерть) супро
воджується конденсацією клітинних структур і активацією
нелізосомних протеолітичних ферментів каспаз або фрагмен
тацією клітини без конденсації її структур і без залучення
56
Розділ 1
лізосомальних систем до цитодеструкції. Найбільш характер
ними цитоморфологічними змінами під час апоптозу є агрега
ція хроматину, конденсація цитоплазми та ядра, фрагментація
ядра і цитоплазми на покриті плазматичною мембраною вези
кули (апоптичні тільця), які містять конденсований ядерний
матеріал, мітохондрії, рибосоми та інші. Відмирання клітин
шляхом апоптозу відбувається як за фізіологічних умов, так
і при патологічних процесах. Клітина при цьому активно спри
яє своїй смерті. Апоптоз добре виражений під час ембріогене
зу, формування та функціонування імунної системи, розвитку
нервової системи, тканинної атрофії, підтримання тканинного
гомеостазу тощо. Варто зазначити, що апоптоз не супрово
джується запальним процесом, який характерний для некро
тичного відмирання клітин. Автофагія (автофагоцитоз) поля
гає у руйнуванні внутрішньоклітинних біомолекул, зокрема
білків, за участю лізосом. Некроз, або патологічна смерть,
у більшості випадків супроводжується пошкодженням плазма
тичної мембрани, що призводить до порушення іонного гомео
стазу і руйнування клітини шляхом лізису. Некротична смерть
клітини іп оіио супроводжується пошкодженням тканини, що
сприяє розвиткові запального процесу. Здебільшого некроз
відбувається тоді, коли клітини піддаються дії екстремальних
чинників, наприклад гіпоксії, дії токсинів, гіпертермії, вірусів
тощо.
1.7.2. Статеве розмноження. Гаметогенез. Мейоз.
Статеві клітини
В основі статевого розмноження — статевий процес, суть
якого полягає в об’ єднанні у спадковому матеріалі для розвит
ку нащадка генетичної інформації від обидвох батьків. Уяв
лення про статевий процес дає кон’ югація у інфузорій.
Також у найпростіших трапляється копуляція — злиття
двох особин в одну, об’ єднання та рекомбінація спадкового
матеріалу.
Подальше ускладнення процесу розмноження пов’ язане
з диференціюванням гамет на великі та дрібні клітини, тобто
Біологічні особливості життєдіяльності людини
57
з появою анізогамії. У табл. 1.1 наведено відмінності між ста
тевим і безстатевим розмноженням.
Таблиця 1.1. Порівняльна характеристика нестатевого і ста
тевого розмноження
№
з/п
Показник
Нестатеве
розмноження
Багатоклітинні: одна
або кілька соматич
них клітин батька;
одноклітинні: клітина-організм як ціле
Одна особина
Генетично точна копія
батька
2
Клітинні дже
рела спадкової
інформації для
розвитку нащад
ків
Батьки
3
Нащадки
4
Основний клі
Мітоз
тинний механізм
1
5
Еволюційне зна
чення
Статеве розмноження
Батьки утворюють
статеві клітини,
спеціалізовані для
виконання функції
розмноження
Звичайно дві особини
Генетично відмінні від
обидвох батьків
Мейоз
Унаслідок генетичної
різноманітності ство
Сприяє підтриманню
рює передумови для
найбільшої присто
освоєння різноманіт
сованості в умовах,
них умов існування,
що практично не
дає еволюційні й
змінюються, підсилює
екологічні перспекти
роль стабілізуючого
ви, сприяє здійсненню
природного добору
творчої ролі природ
ного добору
На певному етапі еволюції у багатоклітинних організмів
статевий процес як спосіб обміну генетичною інформацією між
особинами в межах виду виявився пов’ язаний із розмноженням.
Для здійснення статевого процесу батьківські особини утворю
ють гамети — спеціалізовані клітини, які забезпечують генера
тивну функцію (рис. 1.12). Злиття материнської та батьківської
гамет приводить до утворення зиготи — клітини, яка є дочір
ньою особиною на найбільш ранній стадії розвитку. Утворення
зиготи в деяких організмів відбувається внаслідок злиття гамет,
що морфологічно не відрізняються між собою, тобто відбуваєть
ся ізогамія. У багатоклітинних має місце лише овогамія, тобто
злиття нерухомої яйцеклітини з рухомим сперматозоїдом.
58
Розділ 1
Розвиток гамет відбувається у статевих залозах — гонадах.
Жіночі статеві клітини (яйцеклітини) розвиваються у яйниках,
чоловічі статеві клітини (сперматозоїди) — у сім ’ яниках.
Статеві клітини високодиференційовані та спеціалізовані.
Мейоз, характеристика І і I I мейотичних поділів
Мейоз — біологічний процес, який спостерігається у період
дозрівання статевих клітин.
Його суть полягає в тому, що на певних стадіях розвитку
попередники статевих клітин із диплоїдним набором хромосом
діляться непрямим шляхом, у результаті чого утворюються
гамети з гаплоїдним набором хромосом. Мейоз включає І і II мейо
тичні поділи.
Біологічні особливості
59
ЖИТТЄДІЯЛЬНОСТІ
людини------------------- ------------------- -—
Перший мейотичний поділ (риє. 1.13).
Н а й в а ж л и в іш и м
і н ай тр и ва л іш и м п ер іод о м в п р о ф а з а п е р ш о г о п о д і л у .
Рис. 1.13. Мейоз І
60
Розділ 1
Тут розрізняють кілька стадій:
Лептонема — збільшення ядра і виявлення у ньому слабо
спіралізованих хромосом.
Зигонема — попарне об’ єднання гомологічних хромосом,
під час якого центромери і плечі точно наближаються один до
одного (явище кон’ югації).
Пахінема — спіралізація хромосом і їх о б ’ єднання у па
ри — біваленти. У хромосомах ідентифікуються хроматиди,
в результаті чого утворюються тетради. При цьому відбуваєть
ся обмін ділянками хромосом — кросинговер.
Диплонема — початок відштовхування гомологічних хро
мосом, однак у місцях кросинговеру зв’язок зберігається.
Діакінез — подальше розходження хромосом, які ще зали
шаються з’ єднаними в бівалентах своїми кінцевими ділянками
гомологічних хромосом. У результаті виникають характерні
кільцеві фігури. Ядерна мембрана розчиняється.
Особливістю мейозу І в овогенезі є наявність спеціальної
стадії — диктіотени, яка відсутня в сперматогенезі. На цій
стадії, яка спостерігається ще в ембріогенезі, хромосоми набу
вають особливої морфологічної форми “лампових щіток” і при
пиняють будь-які подальші структурні зміни на багато років.
При досягненні жіночим організмом репродуктивного віку під
впливом лютеїнізуючого гормону гіпофіза, як правило, один
овоцит І порядку відновлює мейоз.
У метафазі першого поділу хромосоми, з’ єднані в бівален
тах, розташовуються по екватору. В анафазі відбувається
розходження до діаметральних полюсів клітини, не хроматид,
а гомологічних хромосом із кожної пари. У цьому принципова
відмінність від аналогічної стадії мітозу.
Телофаза — формування двох клітин із гаплоїдним набором
хромосом, проте кількість ДНК ще відповідає диплоїдному
наборові (якщо позначити гаплоїдний набір хромосом п, а кіль
кість ДНК в гаплоїдному наборі с, то в телофазі утворюються
клітини з набором хромосом і ДНК п, 2с).
Зміст
61
Другий мейотичний поділ (рис. 1.14). Йому передує корот
ка інтерфаза, у якій синтез ДНК не відбувається. В анафазі
розходяться не гомологічні хромосоми, а лише їхні хроматиди.
Тому утворені дочірні клітини залишаються гаплоїдними і ма
ють відповідну до цього набору кількість ДНК, тобто вдвічі
меншу, ніж у соматичних клітинах (п, с).
Р ис.
1.14. Мейоз II
62
Розділ 1
Сперматогенез. Сім’ яник утворений великою кількістю канальців і на поперечному перерізі його видно кілька шарів
клітин. Це і є послідовні стадії розвитку сперматозоонів. Зов
нішній шар (зона розмноження) утворений сперматогоніями,
які інтенсивно діляться шляхом мітозу в період ембріонально
го розвитку, а в постембріональному періоді — до настання
статевої зрілості. Мітотичний поділ сперматогоній триває і далі,
однак деякі з них переміщуються у наступну зону — зону р о
сту, яка розташована ближче до просвіту канальця. Тут клі
тини ростуть за рахунок збільшення маси цитоплазми, їх на
зивають первинними сперматоцитами (2п, 4с).
Третій період розвитку чоловічих гамет — це період дозрі
вання. У результаті І мейотичного поділу з кожного первинно
го сперматоцита утворюються два вторинні сперматоцити (п,
2с), унаслідок II мейотичного поділу з кожного вторинного
сперматоцита утворюються дві сперматиди (п, с) овальної фор
ми і менших розмірів. Тоді сперматиди вступають у четвертий,
останній період розвитку — зону формування (ближче до про
світу канальця сім ’яника), і в результаті утворюються сперматозоони (п, с).
Овогенез. Періоди овогенезу подібні до періодів сперматоге
незу. У період ембріонального розвитку в зоні розмноження
утворюються овогонії (2п, 2с).
У зоні рост у утворюються первинні овоцити, і під час ста
тевої зрілості окремі овоцити вступають у період росту, тобто
клітини збільшуються, в них накопичується жовток, жир,
пігменти. У цитоплазмі, в її органелах і мембрані відбуваються
складні морфологічні та біохімічні перетворення. Кожен овоцит
оточений фолікулярними клітинами, які його живлять.
У період дозрівання відбуваються два послідовні мейози
з нерівномірним розподілом цитоплазми між дочірніми кліти
нами. У І мейотичному поділі з первинного овоцита утворюєть
ся вторинний овоцит (п, 2с). Під час II мейотичного поділу
утворюється яйцеклітина (п, с) і вторинний полоцит (п, с),
також з первинного полоцита утворюються два вторинні полоцити (п, с). У ссавців і людини періоди розмноження та росту
статевих клітин відбуваються у фолікулах, а період дозрівання
Біологічні особливості життєдіяльності людини
63
яйцеклітини — у маткових трубах (під час овуляції), де також
здійснюється і процес запліднення.
Будова статевих клітин:
• Яйцеклітина — це високодиференційована спеціалізована
жіноча статева клітина (гамета), яка містить гаплоїдний
набір хромосом, зокрема одну Х-хромосому. Яйцеклітина лю
дини кулястої форми діаметром 130— 200 мкм, з великою
кількістю цитоплазми і всіма органелами загального призна
чення, за винятком центросоми. Ядро яйцеклітини велике,
містить ядерце і хромосоми типу “ лампових щіток” . Зазвичай
ядро розташоване ексцентрично, тому яйцеклітина поляр
на — вона має анімальний і вегетативний полюси. Ззовні яйце
клітина людини оточена кількома оболонками: цитоплаз
матична мембрана, жовткова вітелінова мембрана, утворена
яйцеклітиною, блискуча оболонка (зона), утворена мукополісахаридами. Між жовтковою і блискучою оболонками може
утворюватися перивітеліновий простір, у якому часто розташо
вується полярне тільце. Існує також промениста оболонка —
зовнішня оболонка яйцеклітини, утворена фолікулярними
клітинами і міжклітинним матриксом.
• Сперматозоон — це високоспеціалізована чоловіча ста
тева клітина (гамета), яка містить гаплоїдний набір хро
мосом, зокрема X- чи У-хромосому. Морфологічно складається
з головки, шийки, середньої частини і хвостика. У головці
розміщене велике ядро, а на кінчику головки є акросома (ви
дозмінений комплекс Гольджі, який виділяє фермент гіалуронідазу, що руйнує оболонку яйцеклітини). Шийка утворена
двома центріолями, розміщеними перпендикулярно одна до
одної. Центріолі, потрапивши в яйцеклітину, утворюють ахроматинове веретено для першого мейотичного поділу зиготи.
Середня частина розміщена за шийкою і утворена з мітохондріальної спіралі. Хвостик — це рухома і найдовша частина
сперматозоона. Складається з аксонеми (осьової нитки, яка
утворена мікротрубочками) і цитоплазматичної мембрани.
Хвостик здійснює коливальні рухи, забезпечуючи просування
сперматозоона вперед. Довжина сперматозоона людини коли
вається в межах 52— 70 мкм.
64
Розділ 1
Осіменіння — це низка процесів, які забезпечують зустріч
яйцеклітини і сперматозоона.
Запліднення — це процес злиття ядер батьківської та ма
теринської гамет. У момент контакту з яйцеклітиною на вер
хівці головки сперматозоона плазматична мембрана і частина
мембрани акросоми розчиняється, вивільняється фермент гіалуронідаза, який розчиняє у певному місці оболонки яйця й утво
рює канал для проникнення ядра сперматозоона (рис. 1.15).
Рис. 1.15. Процес запліднення: 1 — акросома; 2 — ядро спер
матозоона; 3 — головка сперматозоона; 4 — шийка сперматозоона;
5 — хвостик сперматозоона; 6 — секреція акросомою фермента
гіалуронідази; 7 — плазматична мембрана; 8 — вітеліновий про
стір; 9 — жовткова мембрана; 10 — рецептори; 11 — жіночий
пронуклеус; 12 — чоловічий пронуклеус; 13 — синкаріон
Проникнення сперматозоона в яйцеклітину спричинює її ак
тивацію: змінюється обмін речовин, підвищується проникність
мембрани, інтенсифікується надходження з міжклітинного
Біологічні особливості життєдіяльності людини
65
середовища фосфору і калію, виводиться кальцій, активується
біосинтез білка, обмін вуглеводів. При контакті сперматозоона
з яйцеклітиною кортикальні гранули зливаються з мембраною
і вивільняють свій вміст назовні. Жовткова мембрана відша
ровується від поверхні цитоплазматичної мембрани, і виникає
перивітеліновий простір, що є непроникним для сперматозоонів.
Ці процеси необхідні, щоб запобігти поліспермії. Таким чином,
у яйцеклітину проникає лише ядро і середня частина сперма
тозоона. Ядро сперматозоона просувається до місця зустрічі
з жіночим ядром, перетворюється на чоловічий пронуклеус.
Його хроматин розпушується, ядерна оболонка розчиняються.
Одночасно чоловічий пронуклеус повертається на 180° і цен
тросомою вперед рухається у бік жіночого пронуклеуса; остан
ній також переміщується йому назустріч. Після зустрічі ядра
зливаються. Процес злиття ядер сперматозоона і яйцеклітини
має назву синкаріогамія. У результаті утворюється запліднена
яйцеклітина— зигота, з диплоїдним набором хромосом, яка
починає дробитись і дає початок новому організмові.
Сперматозоони чоловіка у статевих шляхах жінки мають
обмежену тривалість життя і ще меншу тривалість здатності
до запліднення. Так, звільнена з яйника яйцеклітина зберігає
спроможність до запліднення протягом 24 год. Сперматозоони
чоловіка у статевих шляхах жінки залишаються рухомими
понад 4 доби, але запліднювальну здатність втрачають уже
через 1— 2 доби.
У секулоті чоловіків міститься приблизно 350 млн сперма
тозоонів. Якщо їх кількість менша 150 млн, тобто менша 60 млн
в 1 мл, то ймовірність запліднення знижується. Швидкість
руху сперматозоонів у нормі становить 1,5—3,0 м м/хв. Відхи
лення від такого поступального руху викликає втрату здатності
до запліднення. До цього призводить також зміна рН середо
вища піхви, загальні явища, патологічні зміни морфології
сперматозоонів тощо.
66
Розділ 1
1.8. Клонування. Ж иття клітин поза
організмом
Клонування, незважаючи на те, що більшість населення
розвинених країн засуджує його, широко розповсюджене у при
роді. Прикладом клонування може бути популяція бактерій,
усі клітини якої утворилися внаслідок повторних поділів з од
нієї батьківської клітини. Всі бактерії такого клону успадкову
ють гени батьківської клітини. Клонування — це також, на
приклад, розмноження рослин вегетативним шляхом: отри
мання рослини із вусика полуниці, розділеного кореня ревеня,
вкоріненої гілки винограду чи шматочка листка бегонії. Не
рідко такий тип розмноження трапляється і у тварин. Для
губок, поліпів, планарій — це звичайний шлях розмноження,
який чергується зі статевим. Дощові черв’ яки так не розмно
жуються, але якщо черв’яка розрізати на кілька частин, то
кожен фрагмент “доросте” до цілого організму. Однак чим
складніший і досконаліший живий організм, тим менше він
здатний до вегетативного розмноження.
Клонами називають групу генетично ідентичних індивіду
умів, отриманих шляхом безстатевого розмноження або один
від одного, або від спільного предка.
Усі складні організми зароджуються у вигляді однієї кліти
ни, з якої поступово розвивається цілий організм. Започаткував
клонування організмів, яким не властиве вегетативне розмно
ження, німецький ембріолог Ганс Дриш (1892). Він узяв за
пліднену яйцеклітину морського їжака і після першого її
поділу раз поділилася, роз’єднав утворені бластомери. Із кож
ної половини розвинулася нормальна личинка, а потім і їжак.
Подібні експерименти широко почали проводитись у 50—60-х
роках X X ст. У 1952 р. американці Р. Брігс і Т. Кінг пересадили
в яйцеклітини ядра з клітин зародка на одній із ранніх стадій
розвитку і виростили здорових жаб. На початку 70-х років
X X ст. у Великій Британії вдалось отримати дорослих жаб із
яйцеклітин, у які були пересаджені ядра клітин кишок пуголов
ка. І навпаки, якщо донорське ядро бралося із дорослої жаби,
зародок вдавалося виростити тільки до стадії пуголовка.
Біологічні особливості життєдіяльності людини
67
Причини подібних невдач з’ясовувалися в міру того, як вчені
одержували інформацію про функціонування генів у процесі
розвитку зародка. Кожна клітина організму (за винятком ерит
роцитів людини, у яких відсутнє ядро) зберігає всі гени, отри
мані у спадок, але мРНК транскрибуються тільки з деяких, чиї
білки потрібні в цей момент. Інші гени “ мовчать” . Виявилося,
що на найбільш ранніх стадіях розвитку зародка “ мовчать” усі
його гени. У цей час вони подвоюються, щільно упаковуються
в хромосомах, пізніше розходяться у клітини, що діляться,
і знову починають подвоюватись. Між поділами клітини прак
тично не збільшуються в розмірах, але деякі білки у них всетаки синтезуються. Цей процес забезпечують мРНК, синтезовані
клітиною ще до запліднення. Досі не з’ ясовано, який фактор
у відповідний момент включає синтез мРНК на генах зародка
і який сигнал свідчить, що цей момент настав. Рано чи пізно
власні гени зародка переривають свою бездіяльність, транс
крибують РНК і скеровують їх у цитоплазму. У більшості тва
рин цей процес відбувається на стадії гаструли. У ссавців
включення власних генів відбувається раніше. Так, у зародка
миші деякі гени включаються вже після першого поділу зиго
ти. “ Мовчання” переривають тільки деякі строго відповідні
гени, потім до них додаються другі — строго відповідні, за
ним и—•треті, четверті...
Як у такому разі виглядає пересадка ядра зрілої клітини
у яйцеклітину? У такій клітині з пересадженим ядром відбу
ваються грубі порушення, не сумісні з життям зародка. Щоб
клонувати тварин, треба було навчитись узгоджувати, син
хронізувати стан цитоплазми зиготи та пересадженого ядра.
Такого узгодження було певною мірою досягнуто у 80-х
роках X X ст. Якщо культуру дорослих клітин помістити в се
редовище, де концентрація поживних речовин уп’ ятеро менша
від оптимальної, то клітини втрачають ознаки спеціалізації,
а їх ядра стають майже неактивними. Це дуже подібне на стан
зиготи. Таке ядро краще синхронізує свою діяльність із цито
плазмою.
Ґрунтуючись на цих даних, вчені для дослідів узяли кліти
ни молочної залози дорослої вівці. Після витримки їх у сере
довищі з низькою концентрацією поживних речовин вчені
об’ єднали ці клітини з яйцеклітинами овець іншої породи, із
68
Розділ 1
яких попередньо видалили власні ядра. “ Голодні” клітини
молочної залози і без’ ядерні яйцеклітини з’єднували і пропус
кали через них електричний струм. Мембрани клітин злива
лись, а утворена зигота починала дробитись. Певний час такий
зародок розвивався у пробірці, а на відповідній стадії розвитку
його пересаджували в матку сурогатної мами.
Навіть при такій складній процедурі ефективність клонування була надто малою. Використавши 277 яйцеклітин і ство
ривши 257 реконструйованих зародків, група І. Уїлмута зумі
ла виростити одну-єдину тварину-клона — овечку Доллі (1997).
Доллі була точною генетичною копією вівці, від якої взяли
клітини молочної залози. Інших генів, зокрема генів донора
яйцеклітини чи сурогатної мами, у неї не виявили. Принци
пова можливість клонування дорослих ссавців була доведена.
Вслід за клонуванням вівці Доллі в інших лабораторіях були
успішно клоновані макаки-резуси, миші, корови, свині, коти,
собаки. Список тварин, яких вдалося клонувати, продовжує
зростати. Водночас збільшуються знання про клонованих тва
рин і про можливості цього методу (рис. 1.16).
За останні роки ефективність клонування вдалося збільши
ти, але вона і далі залишається вкрай низькою. У найкращих
лабораторіях до народження доживає близько 3 % клонованих
ембріонів (табл. 1.2). Причини— непогодженість функціону
вання ядра зрілої клітини з цитоплазмою яйцеклітини, трав
матичні процедури видалення і пересаджування ядер та інших
маніпуляцій з клітинами. Самі тварини, народжені в результаті
клонування, мають більше вад розвитку, частіше хворіють.
Наприклад, Доллі прожила менше 7 років (близько половини
середнього віку для її породи), довго хворіла артритом і була
приспана ветеринарами, коли у неї розвинулося захворювання
легень.
Генетична подібність клонів із вихідним організмом і один
з одним є абсолютною: розрізнити їхні тканини не можуть
навіть їхні імунні системи. Водночас інколи клон відрізняєть
ся від оригінала навіть зовнішньо. Це пов’ язано з тим, що
деякі ознаки формуються під час індивідуального розвитку під
впливом випадкових факторів, у батьківського організму і клона можуть функціонувати різні алелі одного гена. Також,
наприклад, багато нейробіологів вважає, що тонка структура
Біологічні особливості життєдіяльності людини
69
Донор незаплідненої Донор соматичних клітин Донор заплідненої
( тварина, яку клонують)
яйцеклітини
яйцеклітини
Яйце
Донорські
клітина
}<£' клітини
<*>
ПЕРШЕ ПЕРЕСАДЖЕННЯ
_
(перший перенос ядра) Запліднена яйцеклітина
^
^
з двома ядрами —
>власним та одержаним
від сперматозоїда
Видалення ядра
Електричний розряд спричиняє
злиття соматичної клітини
з “пустою" яйцеклітиною
Активація генів у ядрі
соматичної клітини
Видалення ДНК
яйцеклітини та
сперматозоїда
Видалення ядра
Пуста" яйцеклітина
ДРУГЕ ПЕРЕСАДЖЕННЯ
(другий перенос ядра)
Клоновані
тварини
Р ис.
Активоване ядро соматичної клітини
зливається з “пустою” яйцеклітиною
Сурогатна ___
— мат襳?::=^ ь ==::*їьРдноклітинний зародок
підсаджується
сурогатній матері
1.16. Основні етапи клонування тварин
70
Розділ 1
Т аблиця
1.2. Ефективність клонування
Показник
Миша,
у%
Кіт,
у%
Вівця,
у%
Корова,
у%
Людина, у %
Пересадка ядра
Власне ядро
яйцеклітини
замінюють ядром
клітини організ
му, який клонують. Отриману
клітину спонука
ють до ділення
76
43
94
63
75
Дроблення клі
тин
Клітина, яка
ділиться, стає
бластоцистою,
попередником
зародка
49
28
23
34
4
Пересадка за
родка
Зародок, який
розвивається, пе
реносять у матку
сурогатної матері
та стежать за ви
никненням у неї
ознак вагітності
38
13
41
41
Успішне народ
ження
Якщо вагітність
проходить нор
мально, на світ
з’являється жит
тєздатний клон
2
1
7
19
Ефективність
клонування
Частина вдалих
спроб: від отри
мання яйцекліти
ни до народження
нащадків
1
0,5
2
1
Клони людини
ніколи не були
доведені далі
стадії бласто
цисти з етич
них міркувань
Біологічні особливості життєдіяльності людини
71
мозку і спеціалізація нейронів не строго генетично детерміно
вана, а формується в процесі індивідуального розвитку. Це
означає, що характер та індивідуальні особливості поведінки
клона можуть відрізнятись від оригіналу.
Успіхи клонування тісно пов’язані з медичними проблемами.
Так, 2005 р. в Японії народилося 12 клонованих телят, із гено
ма яких видалені гени пріонів — білків, що є субстратом для
губчастої енцефалопатії. Тим самим покладено початок породі,
несприйнятливій до цього важкого захворювання. Ще один
напрям, де клонування могло би бути корисним, — це збере
ження рідкісних видів тварин.
Отримавши інформацію про народження вівці Доллі, люд
ство зацікавилося можливістю клонування людини. Почали
з’ являтися прогнози та заяви стосовно отримання клонованих
дітей, однак підтвердження цього немає. Уряди і парламенти
практично всіх розвинутих країн прийняли закони, які забо
роняють клонування людини. Окрім цього, на заваді стоїть
низька ефективність клонування. Сподіватися на народження
хоча б однієї дитини можна лише тоді, якщо створити й імплан
тувати сурогатним матерям сотні реконструйованих ембріонів.
Знайти таку кількість жінок для явно незаконного експери
менту практично неможливо. Інша проблема, про яку вже
згадувалося, — те, що клоновані організми частіше хворіють
і менш життєздатні. Клонування людини сьогодні призвело б
до народження хворих дітей.
Паралельно з клонуванням організмів успішно проводяться
роботи зі створення культур клітин людини без маніпуляції
з ядрами. Нині медицина значно просунулась у цьому напрямі.
Зі стовбурових клітин вдається отримати не лише окремі тка
нини, але й такі складні багатоклітинні структури, як, напри
клад, ділянки судин, частини сечового міхура. Якщо переса
дити ядро зрілої клітини в яйцеклітину і дати їй кілька днів
розвиватися в пробірці, можна отримати бластоцисту із кількох
сотень клітин, значна частина яких — це ембріональні стовбу
рові клітини. Вирощування з цих клітин ізольованих органів
і заміна ними за необхідністю хворих органів вирішили би
одразу дві складні проблеми сучасної трансплантології: пробле
му імунної сумісності, оскільки всі клітини, вирощені “у про
бірці” , будуть нести гени їхнього майбутнього отримувача, та
72
Розділ 1
проблему нестачі органів. Для того щоб створити такий склад
ний орган, як серце, потрібні якраз ембріональні стовбурові
клітини, здатні перетворюватись у будь-яку тканину організму.
Перспективи такого терапевтичного клонування настільки
захоплюючі, що, незважаючи на прийняту 2005 р. декларацію
ООН, яка забороняє будь-яке клонування людини, та протести
релігійних організацій, розвинуті країни одна за одною при
єдналися до цих перегонів. Терапевтичне клонування дедалі
більше стає однією з доступних людству технологій.
Розділ 2
ОРГАНІЗМОВИЙ РІВЕНЬ ОРГАНІЗАЦІЇ
ЖИТТЯ. ОСНОВИ ГЕНЕТИКИ ЛЮДИНИ
2.1. Предмет і завдання генетики та медичної
генетики. Основні поняття
Генетика — наука, яка вивчає закономірності успадкуван
ня ознак, матеріальні носи спадковості та мінливість. Як
наука генетика розвинулась у зв’ язку з практичними потреба
ми сільського господарства, ветеринарії, необхідністю виведен
ня нових сортів рослин, порід тварин. Порівнюючи гібриди
з вихідними формами, практики давно помітили деякі особли
вості успадкування ознак.
Спадковість і мінливість забезпечують:
1. Дискретне успадкування і зміну конкретних ознак.
2. Відтворення в особин кожного покоління всього комплек
су морфофункціональних характеристик організмів конкрет
ного біологічного виду.
3. Перерозподіл у видів зі статевим розмноженням відтво
рення спадкових задатків, у результаті чого в нащадків спосте
рігається поєднання ознак, відмінне від їх поєднання у батьків.
74
Розділ 2
Етапи розвитку генетики. Основні закономірності успад
кування властивостей і ознак у поколіннях були відкриті
Г. Менделем. На засіданні товариства природодослідників
у м. Брно 1865 р. Г. Мендель зробив доповідь “ Досліди над
рослинними гібридами” , яку опублікував пізніше у журналі
“ Праці Брюнського товариства природодослідників” (1866).
Важливим у дослідженні було твердження про те, що спадкові
задатки не змішуються, а передаються нащадкам у вигляді
дискретних одиниць. Проте робота не була належно оцінена
науковою громадськістю.
Закономірності успадкування ознак, відкриті Г. Менделем,
підтвердили 1900 р. незалежно один від одного Г. де Фріз (Гол
ландія), К. Корренс (Німеччина), Е. Чермак (Австрія). Тому
1900 р. можна вважати роком народження генетики.
У XIX — на початку X X ст. передачу спадкових властиво
стей не пов’ язували з певними структурами.
У 80-х роках X IX ст. Ван-Бенден, Т. Бовері описали склад
ні процеси, які відбуваються при утворенні гамет, і встановили,
що у зиготі спостерігається відновлення диплоїдного набору
хромосом.
У 1902 р. Т. Бовері, Е. Вільсон, Д. Сеттон виявили, що при
передачі спадкових ознак існує паралелізм у поведінці хромо
сом при формуванні статевих клітин і заплідненні. Вони ви
словили припущення про зв’ язок спадкових факторів із хро
мосомами.
У 1909 р. датський вчений В. Йогансен дискретні одиниці
спадковості назвав генами.
Упродовж 1911— 1912 рр. Т. Морган сформулював хромо
сомну теорію спадковості. На плодовій мушці дрозофілі було
з’ ясовано основні закономірності успадкування зчеплених генів
і доведено, що дискретні одиниці (гени) містяться у хромосомах
і розташовані лінійно.
У 1928 р. М. К. Кольцов висловив гіпотезу про зв’ язок генів
із певною хімічною речовиною. На думку Кольцова, хромосо
ма — це велика білкова молекула, окремі радикали якої вико
нують функцію генів. Хоча теорія Кольцова була помилковою,
але в ній уперше зроблено спробу розглянути закономірності
спадковості на молекулярному рівні.
Організмовий рівень організації життя. Основи генетики людини_______ 75
У 1940 р. Д. Бідл і Е. Тейтем запропонували теорію “один
ген один фермент”: гени зумовлюють утворення ферментів,
які, регулюючи клітинний метаболізм, впливають на розвиток
структур і фізіологічних властивостей організмів.
У 1944 р. О. Ейвері, К. Мак-Леод і К. Мак-Карті вперше
встановили, що передача спадкової інформації пов’ язана з нук
леїновою кислотою.
Дослідженнями А . М. Білозерського було доведено, що
ДНК — обов’язковий компонент хромосом рослинних і тварин
них клітин, вивчено нуклеотидний склад ДНК багатьох видів.
На початку 40-х років X X ст. були запропоновані нові мето
ди дослідження: електронна мікроскопія, метод мічених атомів,
рентгеноструктурний аналіз. Виникла і успішно розвивається
молекулярна біологія.
У 1949 р. Е. Чаргафф відкрив правило комплементарності
пуринових і піримідинових азотистих основ. На початку 50-х
років X X ст. М. Уїлкінс за допомогою рентгенівських променів
одержав рентгенограми нитки ДНК.
У 1953 р. Д. Уотсон і Ф. Крік запропонували модель ДНК,
яка має вигляд подвійної спіралі. Почалося поглиблене ви
вчення спадковості на молекулярному рівні.
Таким чином, в історії генетики можна виділити три етапи:
1. Вивчення явищ спадковості на рівні організму.
2. Вивчення явищ спадковості на клітинному рівні.
3. Вивчення явищ спадковості на молекулярному рівні.
Генетика тісно пов’ язана з медициною. Відомо понад дві
тисячі спадкових хвороб і аномалій розвитку. їх вивчають на
молекулярному, клітинному рівнях, на рівні організму, попу
ляції. Генетичними дослідженнями одержані важливі дані про
характер успадкування спадкових захворювань, їх пенетрантність; розроблені методи ранньої діагностики спадкових захво
рювань, дефектів розвитку, злоякісних новоутворень.
Основні поняття генетики:
Спадковість — це властивість організмів передавати із
покоління в покоління подібні ознаки і забезпечувати специфічний
характер індивідуального розвитку в певних умовах середовища.
76_________________________________________________________ Розділ 2
Спадковість — це властивість живої матерїі, що полягає у її
матеріальності, дискретності, цілісності.
Успадкування — це спосіб передачі спадкової інформації,
який може змінюватися залежно від форм розмноження. При
безстатевому розмноженні — через вегетативні клітини, спори;
при статевому розмноженні — через статеві клітини.
Мінливість — це властивість організмів змінювати спад
кові задатки при взаємодії з навколишнім середовищем.
Елементарними дискретними одиницями спадковості є гени.
Ген — це ділянка молекули ДНК, яка містить інформацію
про послідовність амінокислот у білку (поліпептиді) або по
слідовність нуклеотидів у рРНК і тРНК.
Алельні гени — це парні гени, які розміщені в однакових
локусах гомологічних хромосом і відповідають за розвиток
альтернативної пари ознак.
Гомозиготний організм — це організм, у якого в однакових
локусах гомологічних хромосом розміщені однакові алелі гена.
Генотип організму: АА; аа.
Гетерозиготний організм — це організм, у якого в однако
вих локусах гомологічних хромосом розміщені різні алелі гена.
Генотип організму — Аа.
Генотип — це сукупність генів у соматичних клітинах
організму; в диплоїдів — сукупність генів у диплоїдному наборі
хромосом.
Геном — це сукупність генів гаплоїдного набору хромо
сом.
Генофонд — це сукупність усіх генів і алелей виду або по
пуляції.
Фенотип — це сукупність усіх зовнішніх і внутрішніх
ознак організму. Фенотип — результат взаємодії зовнішнього
середовища і генотипу.
Домінантна ознака — це ознака, яка виявляється у гібридів
першого покоління.
І
Організмовий рівень організації життя. Основи генетики людини_______ 77
Рецесивна ознака — це ознака, яка пригнічується і не ви
являється у гібридів першого покоління.
Класифікація генів та їх властивості. Ген — одиниця
функціонування спадкового матеріалу. За хімічною природою
ген, як відомо, — ділянка молекули ДНК, що містить інфор
мацію про послідовність амінокислот у молекулі білка або
нуклеотидів у молекулі рРНК, тРНК.
Термін ген запропонував В. Йогансен у 1909 р. для позна
чення відкритих Г. Менделем спадкових факторів.
Функції генів:
1.
2.
3.
4.
5.
6.
Збереження генетичної інформації.
Редуплікація, подвоєння ДНК в періоді інтерфази.
Репарація ДНК.
Забезпечення спадкової мінливості клітин, організмів.
Рекомбінація.
Контроль за розвитком і старінням клітин, організмів.
Класифікація генів:
1. Структурні гени контролюють розвиток конкретних оз
нак. Містять інформацію про амінокислотні або нуклеотидні
послідовності макромолекул. Утворені фракцією середніх пов
торів ДНК. Включають групи генів:
а) гени, що кодують амінокислотні послідовності струк
турних і ферментативних білків;
б) гени, що кодують амінокислотні послідовності білків,
які функціонують у всіх клітинах (рибосомні білки, гістони);
в) гени, що кодують послідовності нуклеотидів у моле
кулах рРНК, тРНК.
2. Гени-модулятори змінюють розвиток ознаки чи будь-яко
го генетичного явища. Включають групи генів:
а) інгібітори або супресори;
б) інтенсифікатори або гени-мутатори, які підвищують
частоту мутацій;
в) модифікатори або комплементарні гени.
78
Розділ 2
З.
Гени-регулятори координують активність структурних
генів, контролюють час включення різних локусів у процесі
індивідуального розвитку залежно від типу клітин багатоклі
тинного організму, а також від стану середовища.
Властивості гена:
1. Дискретність дії: розвиток різних ознак контролюється
різними генами, локалізація яких у хромосомі не збігається.
2. Стабільність (постійність) дії: за відсутності мутації гени
передаються у ряді поколінь без змін.
3. Специфічність дії: кожний ген зумовлює розвиток певної
ознаки або груп ознак.
4. Плейотропність дії: ген визначає розвиток кількох ознак
організму.
5. Експресивність і пенетрантність дії гена.
Плейотропність дії гена. Уперше явище описане Г. Менделем:
рослини з пурпуровими квітками мають листки з червоними
черешками і білу шкірку насінини.
У дрозофіли ген білого кольору впливає на колір тіла, дов
жину крил, будову статевого апарату, знижує плодючість,
зменшує тривалість життя.
У людини відомий синдром Марфана (арахнодактилії, доліхостеномелії). Ген локалізований у хромосомі 15ц21, відповідає
за синтез білка сполучнотканинних волокон фібриліну і фор
мування довгих, тонких пальців, викликає аномалії в будові
грудної клітки, порушення розвитку сполучної тканини, звих
кришталика ока, аномалії серцево-судинної системи, ектазію
(розширення) твердої оболонки мозку. Захворювання сполучної
тканини успадковується за аутосомно-домінантним типом.
Плейотропна дія гена може бути:
1.
Первинна. Розвиток відбувається за схемою зображеною
на рис. 2.1.
Хвороба Хартнепа. Мутація гена призводить до порушення
всмоктування триптофану в тонкому кишківнику і його реабсорб
ції в ниркових канальцях. Одночасно вражаються мембраниепітеліальних клітин кишківника і ниркових канальців, спостері
гаються розлади травної та видільної систем.
Організмовий рівень організації життя. Основи генетики людини
Р ис.
79
2.1. Схема розвитку хвороби Хартнепа
2.
Вторинна: спостерігається первинний фенотиповий про
яв гена, за яким розвивається ступінчастий процес вторинних
ознак, що призводять до множинних ефектів. Розвиток відбу
вається за схемою (рис. 2.2):
Ген
І
Білок-фермент
І
О знака
І
О знака
і
Ознака
І
Ознака
Р ис.
1
2
З
4
2.2. Схема розвитку серпоподібноклітинної анемії
Серпоподібноклітинна анемія. Утворення аномального НЬ
унаслідок заміни в 6-положенні р-ланцюга Сій —» Уаі зу
мовлює його кристалізацію, веде до зміни форми еритроцитів,
80
Розділ 2
їх злипання і руйнування, викликає зміну фізіологічних функ
цій еритроцитів, сприяє розвиткові анемії, дефектів у нирках,
серці, мозку.
Хвороба Тей-Сакса. Порушення обміну гангліозидів, нако
пичення у нейронах, сітківці ока, печінці ліпоїдоподібних
речовин спричинюють зниження гостроти зору аж до повної
сліпоти, затримку психічного розвитку, прогресуючий параліч.
Успадковується за аутосомно-рецесивним типом.
Експресивність і пенетрантність дії гена. У 1927 р. М. Тимофеєв-Ресовський ввів у наукову мову терміни експресив
ність і пенетрантність.
Експресивність — це ступінь прояву ознаки ( наприклад,
хвороби) при реалізації генотипу в різних умовах середовища.
Тобто експресивність зумовлюється сумарною дією генетичних
і середовищних чинників.
Пенетрантність — це кількісний показник фенотипового
прояву гена. Характеризується співвідношенням особин, у яких
певний ген виявляється у фенотипі, до загальної кількості
особин, у яких ген міг би виявити свою дію. Якщо у всіх осіб
цього генотипу з ’ являється очікуваний фенотип, то це вказує
на повну пенетрантність. Неповна пенетрантність з ’являється
у випадку багатьох домінантних алелів і у випадку гомозигот
ності за рецесивними алелями. Функція пенетрантності вира
жає ймовірність хвороби для кожного генотипу і залежить від
таких чинників, як стать, вік, вплив навколишнього середо
вища й ефект дії інших генів.
Наприклад, ризик захворіти на рак молочної залози у віці
40, 55 і 80 років становить 37, 66 і 85 % відповідно.
Пенетрантність деяких захворювань і ознак організму лю
дини, у % :
1. Вроджений вивих стегна— 20.
2. Ретинобластома — 60.
3. Цукровий діабет — 65.
4. Епілепсія — 67.
5. Групи крові за системою АВО — 100.
Організмовий рівень організації життя. Основи генетики людини_______ 81
Пенетрантність гена завжди варто враховувати при медикогенетичному консультуванні, щоб з’ ясувати ризик народження
дітей із певною ознакою (захворюванням).
Отже, передачею генів у поколіннях досягається матеріаль
на, структурно-функціональна спадкоємність, успадкування
нащадками ознак батьків.
2.2. Закономірності успадкування ознак
Основні закономірності успадкування ознак відкриті Г. Менделем завдяки розробленому ним методу гібридологічного
аналізу. Суть методу полягає в тому, що про генотип батьків
судять за фенотипом нащадків, одержаних у результаті схре
щування. При цьому:
1. Аналізується успадкування за окремими альтернативни
ми проявами ознак.
2. Проводиться точний кількісний підрахунок успадкування
кожного альтернативного вияву ознаки в наступних поколін
нях.
Схрещування, при якому батьківські особини аналізуються
за однією парою альтернативних виявів ознаки, називають
моногібридним-, двох ознак — дигібридним-, багатьох ознак —
полігібридним.
Класичним об’ єктом генетичних досліджень Г. Мендель
вибрав горох — рослина самозапильна, у якої чітко виявля
ються альтернативні ознаки, а саме жовте і зелене забарвлення,
гладка і зморшкувата поверхня насіння. На основі аналізу ха
рактеру успадкування ознак Г. Мендель відкрив три закони:
одноманітності гібридів першого покоління; розщеплення;
незалежного успадкування і комбінування ознак.
82
Розділ 2
2.2.1. Перший і другий закони Менделя. Менделюючі
ознаки людини
Перший закон М енделя — закон одноманітності гібридів
першого покоління: при схрещуванні двох гомозиготних осо
бин, які відрізняються за однією парою альтернативних виявів
ознаки, всі нащадки в першому поколінні одноманітні як за
фенотипом, так і за генотипом.
А
а
— алель гена кароокості;
— алель гена блакитноокості.
Рі: $ АА
С:
®
Г,:
Аа
х
в аа
©
Перше покоління:
— за генотипом (г/т) 100 % гетерозиготи;
— за фенотипом (ф/т) 100 % кароокі.
Другий закон М енделя — закон розщеплення: при схре
щуванні гібридів першого покоління, тобто двох гетерозигот
них особин, у нащадків у другому поколінні спостерігається
розщеплення за фенотипом у співвідношенні 3 : 1 ; за генотипом
1 : 2 : 1 за умови повного домінування ознаки.
С:
Р^
$ Аа
х $ Аа
@ ®
® @
АА; 2Аа; аа
Розщеплення за:
— г/т 1 : 2 : 1 (1 гомозигота за домінантними алелями гена :
: 2 гетерозиготи: 1 гомозигота за рецесивними алелями гена);
— ф/т 3 : 1 (3 кароокі : 1 блакитноокі).
Гіпотеза “чистоти гамет”: у гетерозиготного організму при
гаметогенезі гамети залишаються “чистими”, тобто вони несуть
лише один алель, що визначає один із альтернативних виявів
ознаки.
Організмовий рівень організації життя. Основи генетики людини_______83
Гіпотеза отримала цитологічне обґрунтування. Якщо особи
на гетерозиготна, то в одній із гомологічних хромосом розта
шований домінантний алель, у другій — рецесивний. При ут
воренні статевих клітин відбувається мейоз і в кожну з гамет
потрапляє лише одна із гомологічних хромосом. Тобто у гаметі
може бути лише один алель гена.
2.2.2. Третій закон Менделя
Третій закон М енделя — закон незалежного успадкуван
ня і комбінування ознак: при схрещуванні гомозиготних
особин, які відрізняються за двома (або більше) парами аль
тернативних виявів ознак, у другому поколінні спостерігаєть
ся незалежне успадкування і довільне комбінування ознак за
умови розміщення генів, які визначають розвиток цих ознак,
у різних парах гомологічних хромосом.
А
а
В
Ь
—
—
—
—
алель
алель
алель
алель
гена
гена
гена
гена
$ ААВВ
О:
кароокості;
блакитноокості;
праворукості;
ліворукості.
х
@
в ааЬЬ
@
АаВЬ — кароокі, праворукі
Р2: $ АаВЬ
х
$ АаВЬ
С:
9 X .
АВ
АЬ
аВ
аЬ
АВ
ААВВ
ААВЬ
АаВВ
АаВЬ
АЬ
ААВЬ
АаЬЬ
АаВЬ
АаЬЬ
аВ
АаВВ
АаВЬ
ааВВ
ааВЬ
аЬ
АаВЬ
АаЬЬ
ааВЬ
ааЬЬ
84
Розділ 2
генотипові групи
1/16
ААВВ
2/16
ААВЬ
2/16
АаВВ
4/16
АаВЬ
1/16
ААЬЬ
2/16
АаЬЬ
1/16
ааВВ
2/16
ааВЬ
1/16
ааЬЬ
фенотипові групи
кароокі, праворукі
кароокі, праворукі
кароокі, праворукі
кароокі, праворукі
кароокі, ліворукі
кароокі, ліворукі
блакитноокі, праворукі
блакитноокі, праворукі
блакитноокі, ліворукі
Проаналізуємо розщеплення за кожною парою альтернатив
них ознак:
1. За кольором очей:
12 карооких : 4 блакитноокі,
тобто 3 : 1 .
2. За ознакою кращого володіння рукою:
12 праворуких : 4 ліворукі,
тобто 3 : 1 .
Отже, ознаки успадковуються незалежно одна від одної. Це
можливе, оскільки під час мейозу розподіл хромосом у статевих
клітинах при дозріванні гамет відбувається незалежно, що
може привести до появи нащадків, які несуть ознаки у поєднан
нях, не властивих батьківським і прабатьківським особинам.
Т а б л и ц я 2.1. Кількісні закономірності утворення гамет гібрида
ми і розщеплення їх у нащадків при різних типах схрещування
Параметри
1
Кількість типів гамет,
утворених гібридом
Кількість комбінацій
гамет при утворенні Р2
Розщеплення за
генотипом у Г2
Типи схрещувань
Моногібридне
Дигібридне Полігібридне
2
3
4
2
2"
2Л
4
42
4"
1:2:1
( 1 : 2 : І)2
( 1 : 2 : 1)"
Організмовий рівень організації життя. Основи генетики людини_______85
3
4
3:1
(3 : І)2
(3 : 1)"
II
Кількість різних класів
за фенотипом
2
З2= 9
3"
2і = 2
22 = 4
2"
СО
1
Розщеплення за
фенотипом у Р?
Кількість різних класів
за генотипом
СО
Закінчення табл. 2.1
Ознаки, успадкування яких відповідає закономірностям,
встановленим Г. Менделем, називаються менделюючими. На
сьогодні їх відомо більше 2 500. Усі менделюючі ознаки диск
ретні, характер їх успадкування моногенний.
Таблиця 2.2. Менделюючі ознаки людини
№
з/п
1
Ознака
2
Нормальні
Домінантна (АА)
Рецесивна (аа)
3
4
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
Колір очей
Колір волосся
Розріз очей
Форма носа
Пігментація волосся
Ластовиння
Мочка вуха
Губи
Ямочки на щоках
Краще володіння
Карі
Темне
Монголоїдний
3 горбинкою
Біле пасмо
Є
Вільна
Повні
Є
Правою рукою
11.
Кров
Резус-позитивна
Блакитні
Світле
Європеоїдний
Прямий
Рівномірна
Відсутнє
Приросла
Тонкі
Відсутні
Лівою рукою
Резуснегативна
Патологічні
12.
13.
Будова емалі зубів
Посивіння волосся
14.
Гострота зору
15.
Гострота слуху
Гіпоплазія
Раннє (у 25 років)
Короткозорість
Далекозорість
Нормальна
Нормальна
Пізнє
Нормальна
Знижена
86
Розділ 2
Закінчення табл. 2.2
1
16.
17.
18.
19.
2
Пігментація шкіри
Наявність потових
залоз
Артеріальний тиск
Внутрішньоочний
тиск
3
Нормальна
4
Альбінізм
Є
Немає
Гіпертензія
Нормальний
Глаукома
Нормальний
20.
Розвиток скелета
21.
Будова кінцівки
22.
Будова кінцівки
23.
24.
25.
26.
27.
28.
29.
Зсідання крові
Товстий кишківник
Сприйняття кольорів
Шизофренія
Мігрень
Ретинобластома
Синдром Марфана
Карликова
хондродистрофія
Полідактилія
(шестипалість)
Брахідактилія
(короткопалість)
Нормальне
Поліпоз
Нормальне
Є
Є
Є
Є
ЗО.
Вуглеводний обмін
Нормальний
31.
32.
33.
34.
35.
36.
37.
Схильність до
ожиріння
Наявність пігментів
Засвоєння
фенілаланіну
Засвоєння лактози
Засвоєння фруктози
Здатність
відчувати смак
фенілтіокарбаміду
Пігментна
ксеродерма
Нормальний
Нормальна
Нормальна
Гемофілія
Немає
Дальтонізм
Немає
Немає
Немає
Немає
Цукровий
діабет
Є
Немає
Є
Нормальне
Нормальне
Альбінізм
Фенілкетонурія
Галактоземія
Фруктозурія
Є
Немає
Немає
Є
Нормальне
Організмовий рівень організації життя. Основи генетики людини
87
Типи успадкування ознак (рис. 2.3). На знаннях законів
генетики ґрунтується розуміння механізмів передачі спадкових
ознак від батьків до нащадків. Тому успішна діяльність сучас
ного лікаря неможлива без чіткого знання основних законо
мірностей спадковості. Це необхідно для правильного розумін
ня виникнення спадкових аномалій і захворювань. Студентимедики зобов’ язані засвоїти основи загальної генетики, що
допоможе їм глибоко та свідомо сприймати матеріал багатьох
клінічних дисциплін, а пізніше — використовувати ці знання
у практичній діяльності.
УСПАДКУВАННЯ
Моногенне
(визначається однією
парою алельних генів)
Полігенне
(визначається двома
парами алельних генів)
аутосомно-домінантне
аутосомно-рецесивне
зчеплене із статтю
зчеплене Х-хромосомою
домінантне
Р ис.
рецесивне
2.3. Типи успадкування ознак
зчеплене У-хромосомою
88
Розділ 2
2.3. Взаємодія генів у детермінуванні
та розвитку ознак (захворювань)
організму
Генотип людини визначається як сукупність усіх генів
(алелів) організму. Генотип — це система генів, які взаємодіють
між собою. Генні взаємодії можуть відбуватися на кількох
рівнях: безпосередньо у генетичному матеріалі клітин, між
іРНК і поліпептидами, що утворюються у процесі біосинтезу
білка, між білками-ферментами одного метаболічного циклу.
На сьогодні для більшості ознак неможливо точно вказати
рівень взаємодії генів, які контролюють їх розвиток. Лікар
повинен розуміти, що становлення у людини морфологічних,
фізіологічних, біохімічних, імунологічних, а також патологіч
них ознак (симптомів захворювання) має генетичну природу
і визначається взаємодією первинних і вторинних продуктів
.генів у певних умовах середовища.
Враховуючи інтерес лікарів-практиків до закономірностей
успадкування ознак людини, можна виділити такі форми взає
модії генів:
I. На рівні однієї алельної пари:
1.
2.
3.
4.
Повне домінування.
Неповне домінування.
Наддомінування.
Кодомінування.
II. На рівні різних алельних пар:
1. Комплементарність.
2. Епістаз.
3. Полімерія.
Організмовий рівень організації життя. Основи генетики людини_______8 9
2.3.1. Форми взаємодії алельних генів. Множинний
алелізм. Успадкування груп крові
Повне домінування — один алель гена повністю приховує
наявність іншого алеля.
Неповне домінування — домінантний ген не повністю при
гнічує рецесивний ген. Гетерозиготи мають власний фенотип.
Н аддомінування — домінантний ген у гетерозиготному
стані має сильніший вияв, ніж у гомозиготному.
Кодомінування — прояв у гетерозиготному стані ознак,
що детермінуються двома домінантними алельними генами,
кожен із яких окремо має самостійний прояв.
Неповне домінування характерне деяким генам, які викли
кають спадкові аномалії та хвороби людини. Так успадковуєть
ся серпоподібноклітинна анемія, атаксія Фрідрейха, цистинурія, таласемія.
Серпоподібноклітинна анемія — спадкова гемолітична ане
мія, зумовлена наявністю в еритроцитах патологічного гемо
глобіну 8; за умов гіпоксії еритроцити набувають серпоподібної
форми, легко руйнуються. Захворювання успадковується за
аутосомно-рецесивним типом. У гомозигот перебіг захворюван
ня тяжкий, закінчується смертю; гетерозиготи за звичайних
умов здорові та рідко хворіють на малярію.
8 — алель гена дископодібної форми еритроцитів;
8 — алель гена серпоподібної форми еритроцитів;
рг: $ Зз
X
в
38
О:
2 5 % 88 — здорові діти з дископодібною формою ерит
роцитів, можуть хворіти малярією;
50 % 8з — легка форма гемолітичної анемії, частина
еритроцитів має серпоподібну форму, стійкі до малярії;
25 % 88 — гемолітична анемія, еритроцити мають сер
поподібну форму, смерть у ранньому віці.
90______________________________________________________ Розділ 2
Множинний алелізм., причини виникнення
Значення явища. Ознаки, успадкування яких вивчав Г. Мендель, визначаються парою алельних генів. Проте більшість
ознак у популяції визначається не двома, а більшою кількістю
алелів. Це явище називають множинним алелізмом, а гени
утворюють серію множинних алелів.
Множинні алелі — це різні, відносно стійкі стани одного
і того ж гена, який займає певний локус у хромосомі та пред
ставлений у вигляді нормального алеля або мутації.
Множинні алелі виникають у результаті багаторазової му
тації одного і того ж локуса в хромосомі:
С > ссЬ > сь > с,
де С — алель гена чорного забарвлення шерсті у кролика; ссЬ—
алель гена шиншилового забарвлення шерсті; сь — алель гена
гімалайського забарвлення шерсті; с — алель гена білого забарв
лення шерсті.
При цьому виникає серія генотипів і фенотипів;
г/т
СС
СссЬ
ссЬссЬ
ссЬсн
сьсн
сс
ф/т
чорний;
темно-сірий;
сірий;
світло-сірий;
гімалайський;
білий (альбінос).
„
.
п(п + 1)
При /і-алелів м ож лива:---------------кількість генотипів;
2
п іп -1)
п — кількість гом ози гот;-------------- кількість гетерозигот.
Організмовий рівень організації життя. Основи генетики людини_______91_
Г. Меллер класифікував мутації, які призводять до виник
нення множинних алелів.
1. Гіпоморфи — мутації, які призводять до появи алелей із
послабленими властивостями. Різний колір шерсті у кроликів,
морських свинок — це результат утворення серії множинних
алелей, які контролюють утворення різної кількості меланіну.
2. Аморфи — мутації, які призводять до появи алелей,
пов’язаних із інактивацією гена (існування у природі тваринальбіносів).
3. Гіперморфи— мутації, що призводять до появи більш
сильних за дією алелей (успадкування у людини м ’язової ди
строфії, вроджених аномалій форми кісток, надколінка).
4. Неоморфи — мутації, які призводять до появи алелей
з новими функціями, властивостями (при опроміненні рентге
нівськими променями у дрозофіли виникає новий домінантний
алель, який визначає розвиток глибокої вирізки на крилах).
5. Антиморфи — мутації, які пригнічують ефект дії нормаль
них генів (ген білого опірення у курей виявляється в результаті
пригнічення гена, що контролює утворення пігменту).
Вивчення мутацій серії множинних алелів показало:
1. Будь-який алель зі серії може виникнути через мутацію
безпосередньо алеля дикого типу, або будь-якого іншого алеля.
2. Будь-який алель зі серії може мутувати в інший як у пря
мому, так і у зворотному напрямку.
3. Кожен алельний ген має свою частоту мутації.
4. Серії множинних алелів у різних локусах мають різну
кількість членів.
Успадкування множинних алелей відбувається відповідно
до законів Г. Менделя, але необхідно пам’ ятати:
1. У кожного диплоїдного організму серія множинних алелей
може бути представлена одночасно лише двома алельними
генами.
2. Кожен ген із серії може повністю або не повністю домі
нувати над іншими генами.
92__________________________________________________________Розділ 2
3. Гени однієї серії діють на одну і ту ж ознаку, одночасно
можуть мати множинний ефект дії.
4. Гетерозиготи, які мають різні алелі зі серії множинних
генів, називаються компаундами.
Значення явища: множинний алелізм має пристосувальне
значення, збільшує резерв мутаційної мінливості в еволюції.
Групи крові людини. Успадкування груп крові за системою
АВО та резус-фактором
Система груп крові АВО була відкрита 1900 р. К. Ландштейнером.
У 1911 р. Е. Дунгерн і Л. Гіршфельд підтвердили, що групи
крові успадковуються організмом і не змінюються протягом
усього життя людини. Берштейн 1924 р. встановив, що система
груп крові АВО контролюється серією множинних алелей од
ного локуса, який розміщений у довгому плечі 9-ї хромосоми.
Гени Iа, ів контролюють синтез білків-аглютиногенів, розміще
них на поверхні еритроцитів, відповідно до яких у сироватці
утворюються білки-аглютиніни (табл. 2.3).
і — алель гена І групи крові;
Iа — алель гена II групи крові;
Ів — алель гена III групи крові;
Iа > і ; Ів > і;
Iа = Ів.
Таблиця
2.3. Генотипи груп крові
Група крові
Аглютиногени
(антигени)
еритроцитів
Аглютиніни
(антитіла)
сироватки
Ймовірні
генотипи
1(0)
—
а, Р
іі
II (А)
А
ІАІА, ІАІ
III (В)
В
Р
а
IV (АВ)
А, В
—
ІАІВ
ІВР, Рі
IV група крові успадковується за типом кодомінування.
Організмовий рівень організації життя. Основи генетики людини
93
Серед європейців групи крові за системою АВО трапляють
ся з такою частотою, у % :
— 1(0) - 45;
- I I (А) - 41;
— III (В) — 10;
— IV (АВ) — 4.
Кодомінування — це форма взаємодії алельних генів, при
якій у гетерозиготному стані виявляються ознаки, що детер
мінуються двома домінантними генами.
Окрім системи груп крові АВО, відомі такі системи:
1. Система МІМ: Iм, Ін.
Серед європейців: ІМІМ = 36 % ;
ІМР _ 48 о/о.
імрі = 1 6 % .
2.
3.
4.
5.
Система
Система
Система
Система
Кісісі: Іка, Ікь.
Біе£о: Б і \ Біь.
Ьиіегап: Ілі\ Ьиь.
Биііу: Гуа, Гуь, Гус.
Ж и т т є в о важливо враховувати групи крові при підборі доно
рів для переливання крові, щоб запобігти агрегації еритроцитів
донора при потраплянні їх у кров реципієнта. Крім того, прин
цип успадкування груп крові за системою АВО використовується
у судовій практиці з метою експертизи батьківства.
Успадкування резус-факторної системи. У 1940 р. із ерит
роцитів мавп макака-резус був виділений антиген, який отри
мав назву резус-фактор. Білок знайдений і у крові людини.
Близько 85 % європейців є резус-позитивними, 15 % — резуснегативними.
Синтез білка Юі детермінується трьома домінантними гена
ми С, Б, Е, які абсолютно зчеплені між собою і локалізовані
в короткому плечі І хромосоми. Тому успадкування резус-фак
тора імітує моногенне успадкування.
У нормі в людей із резус-негативною кров’ ю не виробляють
ся антитіла до резус-фактора, але вони неодмінно почнуть
з’являтись у результаті переливання їм резус-позитивної крові
Розділ 2
94
як захисна реакція організму проти чужорідного антигену.
Тому варто пам’ ятати:
1. При шлюбі жінки з резус-негативною кров’ ю і чоловіка
з резус-позитивною кров’ ю за умови гомозиготності батька всі
діти будуть резус-позитивними.
2. При такому ж шлюбі за умови гетерозиготності батька
буде спостерігатися розщеплення в співвідношенні 1 : 1 .
Р : $
С:
33
@
х
6
оо
®
100 % БЗ
Р .:$
а:
Б\
(ІЗ
®
х
50 % Б 3 ,
6
03
50 % 3 3
Якщо у жінки із резус-негативною кров’ ю дитина успадкує
резус-фактор батька, то перша вагітність, як правило, завер
шиться цілком нормально. Але при цьому у кров’яному руслі
матері утворяться антитіла до резус-фактора. При наступній
вагітності антитіла через плаценту проникнуть у кров плода
і викличуть руйнування еритроцитів, які мають антиген Ші+.
З кожною наступною вагітністю, несумісною за антигенами,
кількість антитіл до Ші+-фактора у крові матері зростатиме,
що може спричинити викидні та мертвонародження. Внаслідок
проникнення у кров’ яне русло дитини антитіл у неї розвиваєть
ся гемолітична хвороба, що призводить до руйнування ерит
роцитів. Урятувати новонародженого може тільки термінове
переливання крові з повного її заміною.
Знання про успадкування резус-факторної системи крові
майбутні лікарі повинні використовувати у практичній діяль
ності при переливаннях крові, при медико-генетичному кон
сультуванні резус-негативних жінок.
Задача 1. У пологовому будинку переплутали двох хлопчи
ків. Батьки одного з них мають І і II групи крові, батьки ін
шого — II і IV. У дітей І і IV групи крові.
Визначте, хто чий син.
Організмовий рівень організації життя. Основи генетики людини_______95
Розв’язок:
у
40
і — алель гена І групи крові;
Iа — алель гена II групи крові;
Ів — алель гена III групи крові;
х
в Iа і
® 0
0:
©
Р^ Ілі; іі
Р2:
р2 :
$ Iа і х в Iа і в
© 0
0 ©
ІЛІА; Ілі; ІЛІВ; Іві
Відповідь: хлопчик із І групою крові належить першій бать
ківській парі; хлопчик з IV групою крові — другій батьківській
парі.
Задача 2. У карооких резус-позитивних батьків народилася
донька кароока, резус-позитивна і син блакитноокий резуснегативний.
Визначте генотипи батьків і дітей.
Розв’язок:
А — алель гена кароокості;
а — алель гена блакитноокості;
Б — алель гена резус-позитивності крові (Ші+);
сі — алель гена резус-негативності крові (гЬ ).
Р:
С:
$
АВ
Ай
аБ
асі
$АаБсіх
б'АаБб
АБ
Ай
аБ
асі
ААБ Б
ААВсі
АаББ
АаВсі
ААВсі
ААсИ
АаВсі
Аасісі
АаББ
АаВсі
ааББ
ааВсі
АаВсі
Аасісі
ааВй
аасісі
Відповідь: генотипи батьків — АаБб; АаБб; генотипи ді
тей — А-Б-; А-бсі; ааБ-; аадсі.
Розділ 2
96
2.3.2. Взаємодія неалельних генів
Комплементарна дія — це вид взаємодії неалельних генів,
при якому для формування ознаки необхідна наявність двох
або більше неалельних, як правило, домінантних генів, кожен
із яких окремо має самостійний прояв.
Нова ознака в організмі з ’являється за наявності домінант
них генів із різних алельних пар. Розщеплення за фенотипом
при дигібридному схрещуванні є видозміною менделівської
формули 9 : 3 : 3 : 1 і можливе у співвідношенні: 9 : 7 ; 9 : 3 : 4 ;
9 : 6 : 1; 9 : 4 : 3 .
Такий тип успадкування дуже поширений у природі.
1. У рослин:
а) успадкування кольору віночка в запашного горош ку:
А-Е---- пурпурові; А-ее, ааЕ-, ааее — білі;
б) успадкування форми плоду в гарбуза: А-В---- дископодіб
на; А-вв, ааВ-----сферична; аавв — видовжена;
в) успадкування забарвлення цибулини: А-В- — червоне;
А-вв — жовте; ааВ-, аавв — біле.
2. У тварин:
а) забарвлення кольору шерсті у мишей: А-В-----сірі; А-вв —
чорні; ааВ-, аавв — білі;
б) успадкування забарвлення кольору очей у дрозофіли:
А-В---- червоні; ааВ- — яскраво-червоні; А-вв — коричневі;
аавв — білі.
3. У людини:
а) синтез білка інтерферону: утворення білка пов’ язане
з комплементарною взаємодією двох неалельних генів, які
локалізовані у 2 і 5 хромосомах;
б) синтез гемоглобіну: кожен поліпептидний ланцюг гемо
глобіну кодується окремим незалежним геном, отже, для син
тезу молекули гемоглобіну необхідна наявність чотирьох ком
плементарних генів;
Організмовий рівень організації життя. Основи генетики людини_______9 7
в) синтез важких і легких поліпептидних ланцюгів імуноглобулінів: синтез контролюють три різні групи неалельних
генів;
г) розвиток нормального слуху у людини визначається дво
ма домінантними генами Б і Е, ген Б відповідає за розвиток
завитки вуха, а ген Е — за функціонування слухового нерва;
ґ) спадкування різних форм спадкової короткозорості.
Задача 1. У сім’ї глухих батьків народилася дівчинка з нор
мальним слухом. Яка ймовірність народження глухих онуків,
якщо дівчинка одружиться з юнаком із таким генотипом, як
у неї?
Розв’язок:
Б — алель гена нормального розвитку завитки вуха;
сі — алель гена аномалії розвитку завитки вуха;
Е — алель гена нормального функціонування слухового
нерва;
е — алель гена аномалії функціонування слухового нерва.
Рг : $
С:
ББее
©
х
$ сісіее
©
Р^ 100 % БсІЕе — нормальний слух
Р2 : $ БсІЕе
О:
\ с ?
х
в БсІЕе
БЕ
Бе
4Е
СІЄ
БЕ
ББЕЕ
БВЕе
БсІЕЕ
БсІЕе
Бе
ВБЕе
Б Б ее
БсІЕе
Всіее
ав
БсІЕЕ
Б сІЕ е
аавв
йсіЕе
СІЄ
БсІЕе
В йее
йсіЕе
сісіее
Відповідь: ймовірність народження глухих онуків становить
4 3 ,7 5 % .
У другому поколінні спостерігається розщеплення:
98
Розділ 2
9/16
Б-Е-
—
нормальний слух;
3/16
сІсІЕ-
—
глухі в результаті аномалії завитки вуха;
3/16
В-ее — глухі в результаті аномалії функціонуван
ня слухового нерва;
1/16
сісіее — глухі в результаті недорозвитку завитки
вуха і аномалії функціонування слухового нерва.
Епістаз — вид взаємодії неалельних генів, при якій ген із од
нієї алельної пари пригнічує дію домінантного гена іншої алельної пари, але кожен із них окремо має самостійний прояв.
Ген, який пригнічує дію іншого гена, називається епістатичним супресором. Його позначають І (і), або 8 (з).
Ген, дія якого пригнічується, називається гіпостатичним.
Розрізняють:
а) домінантний епістаз;
б) рецесивний епістаз.
Домінантний епістаз — це пригнічення геном-супресором
у домінантному стані дії гіпостатичного домінантного гена.
Прикладами домінантного епістазу є:
1) успадкування білого оперення у курей при схрещуванні
білих леггорнів з білими плімутроками (А-І-, ааі-, ааіі — біле
оперення; А-іі — забарвлене оперення);
2) сірої масті у коней (С-І-, ссі---- сіра масть; С-іі — корич
нева масть; ссіі — руда масть);
3) ферментопатій у людини (фенілкетонурії, альбінізму,
галактоземії, лактоземії, тирозинозу).
Фенілкетонурія — спадкова хвороба, зумовлена порушенням
обміну фенілаланіну, проявляється у відставанні фізичного та
розумового розвитку, розладах руху, м ’ язового тонусу. Успад
ковується за аутосомно-рецесивним типом.
С — алель гена синтезу ферменту фенілаланінгідроксилази;
Організмовий рівень організації життя. Основи генетики людини_______9 9
с — алель гена відсутності синтезу фенілаланінгідроксилази;
І — алель гена-супресора;
і — алель відсутності гена-супресора.
Р, : $ ССіі
О:
®
Р
х
в ссІІ
©
100 % СсІі — хворі на фенілкетонурію
Р2 : $ СсІі х
С:
в СсІі
СІ
Сі
сі
сі
сси
С С іі
СсІІ
Ссіі
Сі
с с іі
ССіі
С с іі
Ссіі
СІ
С с іі
Ссіі
с с іі
ССІІ
СІ
С с іі
Ссіі
ССІІ
ССІІ
СІ
У другому поколінні спостерігається:
13/16
С-І-, ссі-, с с і і — хворі на фенілкетонурію;
3/16
С-іі — здорові.
Рецесивний епістаз — це пригнічення геном-супресором
у рецесивному стані дії гіпостатичного домінантного гена.
Прикладом рецесивного епістазу є “бомбейський феномен”.
У 1952 р. в одній індійській сім ’ї, яка мешкала на околицях
Бомбея, був виявлений і описаний незвичний родовід за успад
куванням груп крові. “ Бомбейський феномен” трапляється
з частотою 1 : 13 000.
У сім’ ї, в якій мати мала III групу крові, а батько І групу,
народилася донька з І групою крові. Вона вийшла заміж за
юнака з II групою крові та народила дві дівчинки: першу — з IV
групою крові, другу — з і групою. Поява у третьому поколінні
дівчинки з IV групою крові від матері з І групою і батька з II
групою крові викликала сумнів. У літературі описано кілька
подібних випадків. За повідомленням Мак-К’ юсіка, деякі гене
тики пояснюють це явище дією рецесивного епістатичного гена,
здатного пригнічувати дію генів, які визначають II і III групи
100
Розділ 2
крові. Зважаючи на цю гіпотезу, установіть імовірні генотипи
всіх трьох поколінь, описаних у “бомбейському феномені” .
і
Iа
Ів
8
з
—
—
—
—
—
алель
алель
алель
алель
алель
Р,: $ Іві 8з
О:
гена І групи крові;
гена II групи крові;
гена III групи крові;
відсутності гена-супресора;
гена-супресора.
х
9
Ів8
ІВ8
і8
ІЗ
Р2 : $ ІВІ88
х
в И 8з
І8
ІЗ
Іві88
Іві8в
іі88
іі8з
Іві8з
ІВІ88
іі8з
ІІ88
в ІАі88
Причиною появи ферментопатій (захворювань, пов’язаних
із відсутністю певних ферментів) часто є епістатична взаємодія
неалельних генів, коли наявність або відсутність продуктів
реалізації будь-якого гена перешкоджає утворенню життєво
важливих ферментів, які кодує інший ген.
Полімерія — вид взаємодії неалельних генів, за якої декіль
ка домінантних неалельних генів впливають на прояв однієї
фенотипової ознаки, підсилюючи її прояв. При нагромадженні
домінантних полімерних генів їх дія сумується. Ці гени мають
кумулятивний ефект.
Полімерні гени прийнято позначати однією літерою латин
ського алфавіту з цифровим індексом:
А 2, А 3 ... а^ а2, а3.
Організмовий рівень організації життя. Основи генетики людини_____ 101
Уперше явище полімерії описав шведський генетик Г. Нільсон-Еле в 1908 р. при вивченні успадкування кольору зернівки
пшениці. Було встановлено, що забарвлення насіння у пшени
ці визначається двома неалельними полімерними генами.
Біологічне значення полімерії полягає втом у, що воно за
безпечує існування безперервних рядів кількісних змін. Озна
ки, які кодуються полімерними генами, стабільніші, ніж ті,
що кодуються одним геном. Організм без полімерних генів був
би дуже вразливим: будь-яка мутація або рекомбінація призво
дила б до зміни ознак і зумовлювала би мінливість організму.
У тварин і рослин чимало полігенних ознак, серед них є
цінні для господарства: інтенсивність росту, скороспілість,
яйценосність, кількість молока та його жирність, кількість
білка в ендоспермі зерна, вміст цукристих речовин і вітамінів
у рослин.
Багато морфологічних, фізіологічних і патологічних особ
ливостей людини визначається полімерними генами: зріст,
маса тіла, конституція, величина артеріального тиску, колір
шкіри, розумові здібності. Розвиток цих ознак підпорядко
вується загальним закономірностям полігенного успадкування
і дуже залежить від умов середовища.
Зокрема, пігментація шкіри у людини визначається п’ ятьма
або шістьма полімерними генами. У аборигенів Африки (не
гроїдна раса) переважають домінантні алелі, у представників
європеоїдної раси — рецесивні. Тому мулати мають проміжну
пігментацію, але при шлюбі мулатів у них можлива поява як
більш, так і менш інтенсивно пігментованих дітей.
А ]( А , А 3, А 4, А 5 — алелі генів темної пігментації шкіри
а,, а2, а3, а4, а5 — алелі генів світлої пігментації шкіри
?1 •
О:
9 9 1 а і 3 2 а 2 а3 а3 й4 а 4 а 5 а 5
Х
^ А 1^ 1А 2 А 2 А 3 А 3 А 4А 4 А 5 А 5
С&Га2 аз а4
А іаіА 2а2А зазА 4 аА
А 2 А З А 4 А Ц9->
а5
— мулати
Полігенне успадкування має такі характеристики:
1.
Мінливість кількісних ознак залежить від варіювання
кількості домінантних алелей полімерних генів у окремих осіб.
Мірою мінливості ознак є амплітуда варіювання.
102
Розділ 2
2. Межі коливань міри вираженості кількісної ознаки ге
нетично контролюються.
3. Амплітуда коливань тим більша, чим більше локусів
займає поліген.
Для полігенних (мультифакторіальних) спадкових захворю
вань характерне існування перехідних форм від прихованих
до клінічно виражених. Полігенні захворювання людини:
1) юнацька прогерія;
2) полікистоз нирок;
3) гастрит (2 гени);
4) діабет дорослих (5 генів);
5) подагра;
6) косоокість;
7) глухота (60 генів);
8) псоріаз.
Генетична схильність до деяких захворювань у людини успад
ковується полігенно. За сприятливих умов середовища дані
захворювання можуть не виявитись або виявитись незначною
мірою. Це такі захворювання:
1) шизофренія;
2) тромбофлебіт;
3) атеросклероз;
4) гіпертонія;
5) емфізема легенів;
6) бронхіальна астма;
7) туберкульоз легенів;
8) вроджений звих стегна.
Отже, формування фенотипу завжди є результатом взаємодії
генотипу і факторів довкілля. Процес становлення у людини
морфологічних, фізіологічних, біохімічних, імунологічних,
а також патологічних ознак (симптомів захворювання) має
генетичну основу і визначається взаємодією первинних і вторин
них продуктів експресії генів у певних умовах середовища.
Організмовий рівень організації життя. Основи генетики людини
103
2.4. Генетика статі. Зчеплене успадкування
генів
Хромосоми відіграють важливу роль у передачі спадкової
інформації та властивостей організму. Це було доведено від
криттям генетичного визначення статі, встановленням груп
зчеплення генів, які відповідають кількості хромосом, побудові
генетичних і цитологічних карт хромосом. Хромосомна теорія
спадковості вивчена й обґрунтована в роботах Т. Моргана, К. Бріджеса і А. Стертеванта.
Успадкування статі у різних організмів. Розрізняють чоти
ри рівні статевого диференціювання:
1.
2.
3.
4.
Хромосомне визначення статі.
Визначення статі на рівні гонад.
Фенотипове визначення статі.
Психологічне визначення статі.
Хромосомне визначення статі. Стать успадковується як менделююча ознака. У людини в ядрі соматичної клітини 46 хро
мосом, з них 44 аутосоми і 2 статеві хромосоми. Поєднання ста
тевих хромосом у зиготі визначає стать майбутнього організму:
а) у людини:
Р: $
44А + X X
х
Р:
44А + X X — жіноча стать
44А + ХУ — чоловіча стать
З
б) у коників-стрибунців:
Р:
$
XX
х
ХО
в
О:
Г:
X X — самки
ХО — самці
в) у метеликів, птахів:
Р:
?
21У
х
$
22
44А + ХУ
104
Розділ 2
С:
@ ®
Г:
2\У — самки
22 — самці
®
Гомогаметна стать — це стать, у якої при гаметогенезі
утворюються гамети одного типу.
Гетерогаметна стать — це стать, у якої при гаметогенезі
утворюються гамети двох типів.
Визначення статі на рівні гонад. Первинні статеві клітини
(гоноцити) з’являються в ектодермі жовткового мішка на тре
тьому тижні ембріонального розвитку. Статеві залози представ
лені генітальними валиками, у яких ще немає статевих клітин.
Гоноцити амебоподібними рухами мігрують у генітальні вали
ки: якщо міграція відбувається в кіркову периферійну частину
генітального валика, — формується яєчник; при міграції в моз
кову (центральну) частину — сім ’ яник. Міграція гоноцитів
зумовлена НУ-геном, локус якого розміщений у Х-хромосомі,
а в У-хромосомі локалізований ген, який регулює активність
НУ-гена.
Подальше формування статевого апарату відбувається під
впливом статевих гормонів, які виділяються статевими зало
зами. На ранніх стадіях розвитку ембріона генітальний тракт
є індиферентним (біпотентним) і представлений мюлеровими
і вольфовими каналами (утворюються з первинної нирки), які
впадають у сечостатевий синус. Із вольфового і мюлерового
каналів формуються внутрішні статеві органи. Сечостатевий
синус утворює статеві складки, статевий горбик і статевий
валик, із яких утворюються зовнішні статеві органи.
При формуванні жіночої статевої системи вольфові канали
редукуються, а із мюлерових утворюються фалопієві труби,
матка, піхва. Статевий горбик перетворюється на клітор, ста
теві складки — на малі, а статевий валик — на великі сороміт
ні губи.
При формуванні чоловічої статевої системи мюлерові кана
ли редукуються, а вольфові диференціюються на сім’ япровід,
сім ’ яні міхурці та придаток яєчка. Зі сечостатевого синуса
утворюється передміхурова залоза, зі статевого горбика — зов
нішній статевий орган, зі статевого валика — калитка.
Організмовий рівень організації життя. Основи генетики людини
105
Для формування жіночого фенотипу статеві гормони не
потрібні. Для формування чоловічого фенотипу потрібна наяв
ність двох факторів, які утворюються в сім ’ яниках:
1. Синтез горм ону— мюлерового інгібуючого фактора —
в клітинах Сертолі, який пригнічує розвиток мюлерових ка
налів.
2. Синтез гормонів тестостерону і 5-дигідротестостерону
в клітинах Лейдіга сім’ яників під впливом хоріонічного гонадотропіну матері. Тестостерон зумовлює диференціювання
вольфових каналів; 5-дигідротестостерон сприяє розвиткові
зовнішніх статевих органів.
Фенотипове визначення статі. Полягає у формуванні під
впливом жіночих (естрону, естріолу, естрадіолу) статевих гормо
нів і чоловічих (тестостерону, андрогенів) статевих гормонів
зовнішніх статевих ознак; жіночого або чоловічого фенотипу.
Психологічне визначення статі. Полягає у відповідності
паспортної статі психологічній. Стать початково визначається
генами, які розміщені у статевих хромосомах. Гени зумовлюють
розвиток гонад, гонади продукують відповідні гормони, які
впливають на розвиток соматичних ознак статі. На основі со
матичних ознак визначається громадянська, а потім психічна
та соціальна стать. За даними І. В. Голубаєвої, послідовність
визначення статі така: генетична (хромосомна) стать — гонадна стать — гормональна стать — соматична стать — громадянсь
ка стать — психічна стать — соціальна стать. Саме тому фор
мування психічної та соціальної статі при порушеннях стате
вого диференціювання повинно проводитися з урахуванням
хромосомної і гонадної статі.
Успадкування ознак, зчеплених зі статтю. Ознаки, гени
яких локалізовані у статевих хромосомах, успадковуються
зчеплено зі статтю. Гени можна розділити на три групи:
І.Гени, які локалізовані в гомологічних ділянках X- і Ухромосом — частково зчеплені гени:
Розділ 2
106
а)
б)
в)
г)
ґ)
д)
е)
ген
ген
ген
ген
ген
ген
ген
загальної кольорової сліпоти;
пігментного ретиніту;
хвороби Огуті;
епідермального бульозу;
геморагічного діатезу;
спазматичної параплегії;
судомних розладів.
2. Х-зчеплені гени, які розміщені в тій ділянці Х-хромосоми,
що не має гомологічної ділянки в У-хромосомі:
а)
б)
в)
г)
ґ)
д)
е)
ген
ген
ген
ген
ген
ген
ген
гемофілії;
дальтонізму;
гіпоплазії емалі зубів;
відсутності других різців;
фавізму;
м ’язової дистрофії Дюшена;
нецукрового діабету.
3. У-зчеплені гени (голандричні гени), які локалізовані
у непарній ділянці У-хромосоми:
а)
б)
в)
г)
ґ)
д)
ген іхтіозу;
ген гіпертрихозу;
ген гістосумісності;
ген чутливості до андрогенів;
ген, який зумовлює низький тембр голосу;
ген наявності шкірної перетинки між пальцями.
Успадкування захворювань, зчеплених із X- і У-хромосо
мами
Х-зчеплене успадкування: гемофілія — спадкове захворю
вання, зумовлене нестачею VIII або IX факторів зсідання
крові, яке проявляється симптомами підвищеної кровоточи
вості. Успадковується як рецесивна, зчеплена з Х-хромосомою
ознака.
Задача. У сім’ї здорових батьків народився син, хворий на
гемофілію. Яка ймовірність народження здорового сина?
Організмовий рівень організації життя. Основи генетики людини_____ 107
Розв’язок:
Х н — алель гена нормального зсідання крові;
X і1— алель гена гемофілії.
Рі: $ Х НХ Ь
С:
@ @
Г :
25%
х
в Х н¥
® ©
Х НХ Н— донька здорова;
25 % Х НХ Ь— донька фенотипово здорова, але носій
гена гемофілії;
25 % Х н¥ — син здоровий;
25 % Х ь¥ — син-гемофілік.
¥-зчеплене успадкування: гіперт рихоз — захворювання,
при якому спостерігається надмірний розвиток волосяного
покриву на вушних мушлях.
Задача. Батько страждає гіпертрихозом. Яка ймовірність
прояву даної ознаки у дітей?
Розв’язок:
¥ь — алель гена гіпертрихозу
Р,: $ X X
Б:
(§ )
50 %
50 %
х
в
Х¥ь
@ ©
X X — донька здорова
Х ¥ ь — син хворий, з ознаками гіпертрихозу.
Голандричні ознаки — це ознаки, що успадковуються через
У-хромосому.
Проблема статі:
І.
Мозаїцизм. Існування у природі гінандроморфів. Мозаїцизм — це явище, при якому хромосомна або генна мутації
розвиваються після першого мітотичного поділу зиготи
і пошкоджень зазнає тільки частина соматичних клітин.
Індивід при цьому буде мозаїчним за даним геном.
В одних клітинах буде функціонувати нормальний алель,
в інших — мутантний, або патологічний. При цьому, на відміну
108
Розділ 2
від летальності (при ураженні всіх клітин), у мозаїчному стані
не обов’ язково порушується життєдіяльність організму. Вини
кає функціональний мозаїцизм. Якщо мутація домінантна,
вона проявиться у відповідних клітинах і призведе до тяжкого
захворювання. Якщо мутація у клітинах на ранніх стадіях роз
витку зародка рецесивна, то її ефект проявиться тільки у го
мозигот. Частота виникнення мозаїчних форм невисока. Сома
тичний мозаїцизм доведено при ЗО генних хворобах (синдром
Олпарта, синдром Марфана, туберозний склероз тощо). Сома
тичний мозаїцизм виявляють при злоякісних новоутвореннях.
Явище мозаїцизму може спостерігатися при хромосомній анеуплоїдії — 47, Х Х (+13)/46, X X або 47, Х У (+13)/ 46, ХУ, тоб
то частина клітин у особини жіночої або чоловічої статі містить
нормальний набір хромосом (46, X X або 46, ХУ), а в части
ни — явище трисомії (47, ХХ(+13) або 47, ХУ(+13)).
Іноді під час другого мейотичного поділу направляючі тіль
ця не зникають, а зберігаються. Так виникає двоклітинне
яйце. Воно може бути запліднене X- і У-сперміями, і виникає
рідкісний тип гермафродитизму — мозаїцизм гонад. У суб’ єкта
міститься оваріальна тканина поряд із тестикулярною, з мозаїцизмом X X - і ХУ-клітин. Описані випадки мозаїцизму у па
цієнтів із хворобою Дауна — 46, Х У / 47, ХУ,+21 або 47, Х Х + 2 1 /
46, X X .
Патологія може виникнути із трисомної зиготи внаслідок
втрати однієї з 21 хромосоми в мітотичному поділі або з нормаль
ної зиготи в результаті мітотичного нерозходження. Наявність
у мозаїків клітин із нормальним каріотипом зумовлює кращий
психічний розвиток, ніж у хворих при чистій трисомії.
Сучасні молекулярно-генетичні методи дають змогу діагно
стувати мозаїцизм на клітинному і тканинному рівнях.
Гінандроморфи — це організми, у яких одна частина тіла
і статеві залози жіночого типу, а друга частина — чоловічого
типу.
Гінандроморфи життєздатні. Характеризуються різними
конституційними відхиленнями. Фенотиповий прояв мозаїч
ного хромосомного набору різноманітний: від мінімальних
відхилень (крипторхізм, гіпоспадія) до сильно вираженого
синдрому Шерешевського — Тернера. Всі гінандроморфні осо
би з 45, ХО/46, ХУ каріотипом розподіляють на три групи:
Організмовий рівень організації життя. Основи генетики людини_____ 109
1. З жіночими зовнішніми статевими органами. Такі жінки
фенотипово нагадують синдром Ш ерешевського — Тернера.
Соматичні аномалії відсутні. Піхва та похідні мюлерових ка
налів розвинені слабо. Через нестачу статевих гормонів молоч
ні залози недорозвинені, оволосіння на лобку, під пахвами
незначне або відсутнє. Якщо ж молочні залози розвиваються,
то переважно під впливом гонадобластоми або дисгерміноми,
що продукують естрогени. Пухлини трапляються у 15— 20 %
випадків мозаїцизму 45, ХО/46, ХУ і розвиваються до 20 років.
У зв’язку з цим існують прямі покази для видалення статевих
залоз із метою профілактики пухлин. Визначення Х-хроматину недостатнє, оскільки при каріотипі 45, Х0 і 46, ХУ Х-хромосома не конденсується.
2. Бісексуальна будова статевих органів. Асиметрична або
змішана дисгенезія статевих залоз спостерігається при наяв
ності одного недиференційованого зачаткового тяжа і одного
дисгенетичного сім’ яника. При мозаїчному каріотипі присутні
похідні мюлерових каналів (матка).
3. Майже нормальна будова чоловічих статевих органів. При
цьому матка відсутня, розвиток пухлин не передбачається.
II. Існування гермафродитизму (інтерсексуальності). При
цьому спостерігається порушення розвитку вольфових або
мюлерових каналів. У клініці розрізняють:
а) істинний гермафродитизм;
б) псевдогермафродитизм.
Істинний гермафродитизм виникає у результаті порушення
раннього процесу гонадогенезу. При цьому у людини одночас
но присутні і гонадні, і соматичні ознаки чоловічої та жіночої
статі.
Форми істинного гермафродитизму:
1.
Передзиготна— виникає в результаті кон’ югації при
сперматогенезі короткого плеча У-хромосоми, в якій локалізо
ваний НУ-антиген, з довгим плечем Х-хромосоми. У результаті
утворюється транслокаційна ХУ-хромосома, при заплідненні
якою яйцеклітини з Х-хромосомою утворюється зигота Х ХУ.
110________________________________________________________ Розділ 2
У зиготи спостерігається одночасний розвиток яєчників і сі
м’ яників.
2. Зиготна — виникає в результаті подвійного запліднення
яйцеклітини двома сперматозоонами з X- і У-хромосомою.
Каріотип зиготи 46, Х Х /4 6 , ХУ. Виявляється мозаїчний гер
мафродитизм.
3. Постзиготна — запліднена яйцеклітина починає діли
тись, і на стадії двох-трьох перших поділів хромосоми не роз
ходяться. У результаті нерозходження хромосом виникають
різнорідні мозаїки, в яких частина клітин має 47 хромосом
і комплекс статевих хромосом ХХУ, а частина — 46 хромосом
і комплекс X X . Мозаїків такого типу позначають Х Х У /X X .
Псевдогермафродитизм виникає в тому випадку, коли у лю
дини гонади однієї статі (чоловічі або жіночі), а зовнішні ста
теві органи — обидвох статей із різним ступенем вираженості
у різних індивідів. Причина — нечутливість тканин геніталь
ного тракту до гормонів або аномальне утворення статевих
гормонів.
III. Вплив генних взаємодій на процес формування фенотипу.
У 1953 р. описаний синдром Моріса (синдром тестикулярної
фемінізації). Народжуються дівчатка з чоловічим каріотипом
(46, ХУ) і сім ’ яниками. При настанні статевої зрілості спосте
рігається:
1. Повна або часткова відсутність оволосіння на лобку, під
пахвами.
2. Первинна аменорея.
3. Матка, фаллопієві труби недорозвинені, піхва закінчуєть
ся сліпо.
4. Сім’ яники локалізовані у великих соромітних губах або
в черевній порожнині.
5. Сперматогенез відсутній.
6. Утворюються чоловічі статеві гормони (андрогени, тесто
стерон). Але через гомозиготність за рецесивним геном і і т
у клітинах зародка не утворюється білок-рецептор, який робить
клітини тканин і органів, що розвиваються, чутливими до
андрогенів. У результаті відсутності взаємодії гормон — рецеп
тор розвиток за чоловічим типом припиняється.
Організмовий рівень організації життя. Основи генетики людини_____ 111
Синдром Моріса має аутосомно-рецесивний тип успадкуван
ня. У жінок проявляються чоловічі риси характеру: рішучість,
витривалість до фізичних навантажень, підприємливість, енер
гійність.
IV. Вплив факторів зовнішнього середовища на розвиток
статі. Феномен описаний на прикладі морського черв’ яка бонелії. Із яєць бонелії розвиваються личинки, які можуть стати
самцем або самкою. Якщо личинка прикріпиться до хоботка
самки, то під впливом її гормонів перетворюється на самця.
Якщо личинка веде самостійний спосіб життя, то перетворюєть
ся на самку.
V. Перевизначення статі. У 1953 р. вчений Ямомото провів
досліди на акваріумних рибках медаках (Огугіаз Іаіірез). До
мінантний ген червоного кольору тіла К локалізований в У-хромосомі, а рецесивний ген білого кольору г в Х-хромосомі. У ри
бок із покоління в покоління відтворюються білі самки (Х ГХ Г)
і червоні самці (Х г¥а). Якщо малькам упродовж восьми місяців
згодовувати жіночий статевий гормон — естроген, — все одер
жане покоління буде самками, половина з них червоні.
Х г — алель гена білого забарвлення тіла;
¥к — алель гена червоного забарвлення тіла.
ДОСЛІД
КОНТРОЛЬ
Р:
О:
Г:
$
Х ГХ Г х
б ?)
в
Х г¥к
Р: $
О:
Х ГХ Г х
<1 Х гуК
50 % Х ГХ Г— самки білі;
Г: 50 % Х ГХ Г— самки білі;
50 % Х г¥ к — самці червоні;
50 % Х г¥ к— самки чер
воні.
Отже, під впливом статевих гормонів можна змінити стать
майбутнього організму.
112
Розділ 2
2.5. Закономірності успадкування зчеплених
генів. Кросинговер. Хромосомна теорія
спадковості
Гени, локалізовані в одній хромосомі, утворюють групу
зчеплення генів. У кожного виду організмів кількість груп
зчеплення дорівнює кількості пар хромосом:
•
•
•
•
•
•
У дрозофіли $ 4; <$ 5.
У гороху 7.
У кукурудзи 10.
У томатів 12.
У шимпанзе $ 24; $ 25.
У людини
$ 23; <$ 24.
Закономірності успадкування зчеплених генів були вивчені
Т. Морганом і співробітниками. Протягом 1910— 1916 рр. на
плодовій мушці дрозофілі БгозорНИа теїаподазіег. У дрозофі
ли гени довжини крил (V — нормальні, V — зачаткові) і кольору
тіла (В — сіре, Ь — чорне) локалізовані в одній хромосомі й ут
ворюють групу зчеплення генів.
Можливі два види зчеплення генів:
1. Абсолют не (повне) зчеплення: кросинговер не відбу
вається, і у дигетерозиготи при гаметогенезі варто очікувати
утворення двох типів гамет.
У людини резус-фактор кодують три тісно зчеплені гени: С,
Б ІБ . Домінантні гени визначають резус-позитивні властивості
крові, а їх рецесивні алелі — резус-негативні.
Задача. Резус-позитивна дівчина (гомозигота) одружується
з резус-негативним юнаком. Яких дітей і внуків за резус-факторною системою варто очікувати?
Розв’язок:
С, Б, Е — алелі генів резус-позитивності;
с, сі, е — алелі генів резус-негативності.
СБЕ
ссіе
вСБЕ
ссіе
Організмовий рівень організації життя. Основи генетики людини_____ 113
Сх:
<ґГл5бГ)
("Ж >
СБЕ
Б\: 100 % -----ссіе
Р2: ?
С:
СБЕ
----- ссіе
х
— резус-позитивні;
в
ссіе
—
ссіе
50 % ССПІГ)
50 % рВсіе^
СБЕ
-----ссіе
Г„: 50 %
— резус-позитивні;
ссіе
— резус-негативні.
50%
ссіе
2 . Відносне (неповне) зчеплення: під час мейозу відбуваєть
ся кон’ югація хромосом і обмін ідентичними ділянками гомо
логічних хромосом. Кросинговер може відбуватись у будь-якій
ділянці хромосоми, навіть у кількох місцях однієї хромосоми.
Чим більша відстань між окремими локусами, тим частіше
відбувається перехрест і обмін алельними генами. У результаті
в дигетерозиготи утворюється чотири типи гамет: гамети з перех
ресними хромосомами (кросоверні гамети) і з неперехресними
хромосомами (некросоверні гамети). Некросоверних гамет зав
жди більше, ніж кросоверних.
Процес кросинговеру можна відтворити (рис. 2.4):
Б
Е
Рис.
2.4. Процес кросинговеру
Відстань між генами Б і Е дорівнює 3 морганіди.
114
Розділ 2
Задача. У людини ген еліптоцитозу і резус-фактора локалі
зовані у першій хромосомі на відстані 3 морганід. Мати резуспозитивна з еліптичними еритроцитами (гомозигота), чоловік —
резус-негативний з нормальними еритроцитами. Еліптоцитоз
і позитивний резус-фактор — домінантні ознаки, нормальна фор
ма еритроцитів і негативний резус-фактор — рецесивні ознаки.
Які генотипи і фенотипи дітей, онуків?
Розв’язок:
Е — алель гена еліптоцитозу;
е — алель гена дископодібної (нормальної) форми еритро
цитів;
Б — алель гена резус-позитивності;
сі — алель гена резус-негативності.
БЕ
Рл $
сіє
X
в
БЕ
СІЄ
С:
Гд
100%
БЕ
=
сіє
— резус-позитивні з еліптичними
еритроцитами
БЕ
Р2 $
бе
X
бе
в
бе
1,5 %
1 ,5 %
Е2: 48,5 % БЕбе — резус-позитивні, еліптоцитоз;
48.5 % йесіе — резус-негативні, дископодібна форма
еритроцитів;
1.5 %
Вебе — резус-позитивні, дископодібна форма
еритроцитів;
1.5 %
сіЕсіе — резус-негативні, еліптоцитоз.
Організми, які виникають унаслідок злиття кросоверних
гамет, називають кросоверами, або рекомбінантами.
Організмовий рівень організації життя. Основи генетики людини_____ 115
Організми, які виникають від поєднання некросоверних
гамет, називають некросоверами, або нерекомбінантами.
Хромосомна теорія спадковості. Закономірності, які вста
новлені школою Т. Моргана, а потім підтверджені й поглиблені
на численних об’ єктах, відомі під загальною назвою хромосом
ної теорії спадковості.
Основні положення хромосомної теорії спадковості:
1. Гени локалізовані у хромосомах. Кожна хромосома — це
група зчеплення генів. Кількість груп зчеплення у кожного
виду дорівнює гаплоїдному наборові хромосом.
2. Гени у хромосомах розміщені лінійно. Кожен ген у хромо
сомі займає певний локус.
3. Між гомологічними хромосомами може відбуватися кон’ ю
гація і кросинговер (обмін алельними генами).
4. Відстань між генами у хромосомі пропорційна процентові
кросинговеру між ними і виражається в морганідах:
1 морганіда = 1 % кросинговеру.
Генетичні та цитологічні карти хромосом. Роботи Т. Мор
гана і співробітників показали, що частота кросинговеру між
одними і тими ж генами — величина постійна і прямо пропор
ційна відстані між генами у хромосомах.
У 1911— 1914 рр. Т. Морган запропонував принцип побудо
ви генетичних карт хромосом. За одиницю відстані між генами
у хромосомі прийнята 1 морганіда, яка дорівнює 1 % кросин
говеру.
Генетична карта хр ом осом — це графічне зображення
хромосоми у вигляді лінії, на якій вказано взаємне розташу
вання генів.
Щоб побудувати генетичну карту хромосоми, необхідно:
1. Провести
падкування не
2. З’ ясувати
3. З’ ясувати
рекомбінантний аналіз (вивчити характер ус
менше трьох генів).
відстань між генами.
порядок розташування генів.
116
Розділ 2
Картовані хромосоми:
1. Мухи дрозофіли, курей, мишей.
2. Кукурудзи, томатів.
3. В Х-хромосомі людини картовано більше сотні генів;
в одній аутосомі — кілька десятків генів.
У 1919 р. Стертевант сформулював закон адитивності: гени
розташовані у хромосомі в лінійному порядку, і відстань між
ними обчислюється додаванням або відніманням проценту
кросоверних особин.
Задача. В Х-хромосомі людини розміщені гени:
а — алель гена очного альбінізму;
б т — алель гена нецукрового діабету;
ас — алель гена ангіокератоми.
У результаті проведеного рекомбінантного аналізу одержали
такі результати:
а — бт
а — ас
б т — ас
7 % кросоверних організмів;
10 % кросоверних організмів;
17 % кросоверних організмів.
Побудуйте генетичну карту ділянки Х-хромосоми людини.
Розв’язок:
$ А Б М абт
х
абтабш
$
О:
46,5 % САШГ)
С Ж у
IV
4 6 ,5 % АБ М абт;
46.5 % а б т а б т ;
3.5 %
А бтабт;
3.5 %
аБМ абт.
Отже, відстань між генами а і б т дорівнює 7 морганід.
Р2:
$ БМ АСбтас
О:
4 1 ,5 %
х
$ бтасбтас
ГбМ А їу Г^Пнас)
Організмовий рівень організації життя. Основи генетики людини
41,5%
8,5 %
8,5 %
117
СПліА^Р
41,5%
41,5%
8.5 %
8.5 %
БМАСсІтас;
сітассітас;
Б тассітас;
йтАСсІтас.
Отже, відстань між генами йш і ас дорівнює 17 морганід.
Р3: $ ААСаас
С:
Г3:
х
3
аасаас
45 %(2ШГ>
5 %<ГаЛС^)
45 % ААСаас;
45 % аасаас;
5 % Аасаас;
5 % аАСаас.
Отже, відстань між генами а і ас дорівнює 10 морганід.
На основі рекомбінантного аналізу будуємо генетичну карту
ділянки Х-хромосоми людини:
йт
а
ас
<--------------- 7--------------------- ► ◄------------------ 1 0 ------------------------ ■>
17 морганід
Цитологічні карти хромосом. Для їх побудови використо
вують політенні хромосоми, які утворюються в результаті
збільшення кількості ДНК у хромосомах під час ендомітозу
при збереженні диплоїдного числа хромосом. Кожна хромосо
ма характеризується строго визначеним чергуванням еу- і гетерохроматинових ділянок. Еухроматинові ділянки — це деспіралізовані ділянки ДНК, які слабо фарбуються барвниками
і містять структурні гени. Гетерохроматинові ділянки — це
спіралізовані ділянки ДНК, які добре фарбуються барвниками
і не містять структурних генів.
Відомі хромосомні аберації, при яких окремі ділянки хро
мосом подвоюються (дуплікації) або втрачаються (делеції). На
цитологічних препаратах політенних хромосом із делеціями
або дуплікаціями кінцеві точки таких аберацій можна визна
чити досить точно. Втрати генетичної інформації, пов’ язані
з делеціями, фенотипово проявляються у гетерозигот при ре
цесивних мутаціях. Зіставлення даних цитологічного і рекомбінантного аналізів свідчить про те, що генетичні карти хромосом
повністю відповідають реальній фізичній структурі хромосом.
Отже, поруч з ознаками, які успадковуються незалежно, є
ознаки, що успадковуються зчеплено. Експериментальне до
слідження зчепленого успадкування, проведеного Т. Морганом
і співробітниками, підтвердило хромосомну локалізацію генів
і покладене в основу хромосомної теорії спадковості. Кон’югація
гомологічних хромосом і обмін алельними генами в результаті
кросинговеру має велике значення для еволюції, оскільки
збільшує можливості комбінативної мінливості. Внаслідок
перехресту добір у процесі еволюції відбувається не за цілими
групами зчеплення, а за групами генів і навіть за окремими
генами. Адже в одній групі зчеплення можуть бути розміщені
гени, які кодують як адаптивні (пристосувальні), так і неадаптивні стани ознак. У результаті кросинговеру корисні для ор
ганізму алелі можуть бути відокремлені від шкідливих і, як
наслідок, виникнуть більш корисні для існування виду генні
комбінації.
2.6. Особливості людини як об’єкта
генетичних досліджень
Генетика людини вивчає явища спадковості та мінливості
у популяціях людей, особливості успадкування нормальних
і патологічних ознак, залежність захворювання від генетичної
схильності й факторів середовища. Завданням медичної гене
тики є виявлення і профілактика спадкових захворювань.
Академік І. П. Павлов, визначаючи важливе значення генети
Організмовий рівень організації життя. Основи генетики людини_____ 119
ки для фізіології та медицини, писав: “Наші лікарі повинні
як азбуку знати закони спадковості... Втілення в життя науко
вої істини про закони спадковості допоможе позбавити людство
від багатьох скорбот і горя” .
Засновником медичної генетики є видатний невропатолог
С. М. Давиденков. Він уперше застосував ідеї генетики у клініці,
проаналізував низку спадкових захворювань. Важливою заслу
гою вченого є розробка і практичне використання методів медико-генетичного консультування.
Людина є специфічним об’ єктом генетичних досліджень,
який має свої переваги та недоліки.
Переваги:
1. Високий ступінь вивченості фенотипу людини (анатоміч
ні, фізіологічні, біохімічні, імунологічні ознаки, поведінка).
2. Висока фенотипова мінливість людей, яка зумовлена ге
нетичною різноманітністю та значною варіабельністю еколо
гічних і соціальних умов.
Недоліки:
1. Неможливий цілеспрямований підбір шлюбних пар і екс
периментальні шлюби.
2. Невелика тривалість життя (генетик може спостерігати
не більше 1— 2 поколінь).
3. Невелика кількість дітей (2— 3), що недостатньо для ко
ректного статистичного аналізу розподілу ознак у нащадків.
4. Велика кількість груп зчеплення (23—24 групи). Сумар
не число генів у геномі людини 104— 10б.
Проте, незважаючи на всі ці труднощі, генетика людини
успішно розвивається.
2.6.1. Методи вивчення генетики людини.
Спадкові хвороби
При вивченні генетики людини використовуються такі ме
тоди, якіклініко-генеалогічнийі близнюковий; цитогенетичний,
біохімічний, популяційно-статистичний, метод гібридизації
соматичних клітин, моделювання і (дерматогліфіки )
120
Розділ 2
Клініко-генеалогічний метод. Цей метод ґрунтується на
простеженні будь-якої ознаки (захворювання) у ряді поколінь,
з вказанням типу родинного зв’ язку між членами родоводу.
Метод запропонований у кінці X IX ст. Ф. Гальтоном, умовні
позначення — Г. Юстом і М. Бочковим у 1931 р.
Метод включає два етапи:
1. Збір даних про сім’ ю (анамнез захворювання).
2. Генеалогічний аналіз.
Пробанд — особа, родовід якої складають і аналізують.
Сибси — рідні брати і сестри пробанда.
Легенда завжди додається до родоводу; лаконічно інформує
про хворобу того чи іншого члена сім’ї.
Генетична символіка
О
□
оо
0 -0
оо
□ ©
■ •
жінка;
чоловік;
шлюб;
неофіційний шлюб;
кровнородинний шлюб;
гетерозиготні носії;
мають досліджувану ознаку;
помер до 1 року;
ш
помер у п років;
121
монозиготні
близнята;
дизиготні близнята;
О
ґ 'Ь
&
6
Організмовий рівень організації життя. Основи генетики людини
медичний аборт;
бездітний шлюб;
викидень.
Клініко-генеалогічний метод вивчення спадковості людини
дає змогу:
1. З’ясувати тип успадкування ознаки (захворювання).
2. Визначити гетерозиготність організму.
3. Вирахувати ризик народження хворої дитини.
Типи успадкування ознак:
1. Аутосомно-домінантний тип (рис. 2.5):
а)
мутантний ген (ознака) виявляється як у гомозиготному,
так і в гетерозиготному стані;
122
Розділ 2
б) ознака (захворювання) виявляється однаковою мірою
у представників обидвох статей;
в) наявність ознаки (захворювання) у всіх поколіннях (по
вертикалі) та при відносно великій кількості сибсів (по гори
зонталі);
г) у гетерозиготних батьків ймовірність прояву ознаки (за
хворювання) у дітей становить 75 % .
II
III
IV
Рис. 2.5. Родовід сім’ї з аутосомно-домінантним типом успад
кування вродженої катаракти
2. Аутосомно-рецесивний тип (рис. 2.6):
а) мутантний ген (ознака) виявляється тільки в гомозигот
ному стані (за рецесивом);
б) відносно невелика кількість хворих у родоводі;
в) наявність хворих по горизонталі (хворіють сибси);
г) при гетерозиготності батьків ймовірність народження
хворої дитини становить 25 % ;
ґ) при появі рецесивних захворювань у нащадків часто трап
ляється кровна спорідненість батьків.
3. Захворювання, зчеплені зі статтю:
— Х-зчеплене успадкування:
1. При домінантному успадкуванні (рис. 2.7):
а)
захворювання виявляється як у чоловіків, так і у жінок,
але жінки хворіють удвічі частіше;
Організмовий рівень організації життя. Основи генетики людини_____ 125
б) сини успадковують ген від матері;
в) дочки можуть успадкувати ген як від матері, так і від
батька.
2. При рецесивному успадкуванні (рис. 2.8):
а) хворі з’ являються у родоводі через покоління;
б) хворіють, як правило, чоловіки;
в) жінки фенотипово здорові, але є носіями патологічного
гена.
— У-зчеплене успадкування (рис. 2.9):
а) хворіють лише особи чоловічої статі;
б) захворювання передається від батька синові.
Близнюковий метод. Запропонований у 1876 р. Гальтоном.
Близнята — це нащадки однієї матері, які спільно розви
вались і народились у процесі одних пологів.
Монозиготні (однояйцеві) близнята (МБ) розвиваються із
однієї зиготи завдяки поділу її на 2 бластомери. Близнята
мають однаковий генотип, завжди однієї статі. Якщо роз’ єд
нання бластомерів відбувається на стадії дроблення аж до стадії
морули, то кожен ембріон матиме повний набір зародкових
оболонок: 2 амніони, 2 плаценти, 2 хоріони. Якщо ембріон
ділиться після утворення трофобласта, але перед появою ам
ніона, то близнята матимуть спільні хоріон і плаценту, але
2 різних амніони. Якщо ембріон ділиться після появи амніона,
то близнята матимуть спільні плаценту, хоріон і амніон.
Дизиготні (різнояйцеві) близнята (ДБ) утворюються внаслі
док одночасного запліднення двох чи більшого числа яйцеклі
тин. Близнята матимуть приблизно 50 % однакових генів;
можуть бути однакової або різної статі. Кожен близнюк має
повний набір зародкових оболонок: амніон, плаценту, х о
ріон.
Нині на кожні 100 пологів народжується 1 пара близнят.
На дві-три пари дизиготних близнят народжується одна пара
монозиготних.
Рис. 2.8. Родовід сім’ї з Х-зчепленим рецесивним типом успадкування гемофілії
О1
II
0-1
1
III
11 в
0-1
2
1
3
2
1
5
--(о
4
/
О
і
IV
О
4
Рис. 2.9. Родовід сім’ї зУ-зчепленим типом успадкування гіпертрихозу
128________________________________________________________ Розділ 2
У 1811 р. в Таїланді народилися двоє близнят— хлопчики
Енг і Чанг — сіамські близнята, які зрослися грудьми.
У 1934 р. в Канаді народилися п’ятеро дівчаток.
У 1974 р. в Польщі (Гданськ) народилися три хлопчики і дві
дівчинки.
У 1980 р. Віталії народилися чотири хлопчики і дві дів
чинки.
У 1994 р. в Україні (Київ) народилися три хлопчики і дві
дівчинки.
У 2001 р. в Україні (Львів) народилися три хлопчики і одна
дівчинка.
Близнюковий метод дає змогу вивчити співвідносну роль
генотипу і зовнішнього середовища в реалізації ознак (захво
рювань) людини.
Конкордантні ознаки — це ознаки, які фенотипово вияв
ляються в обидвох близнят пари.
Дискордантні ознаки — це різні ознаки або відсутність
ознаки в одного із близнят.
Для оцінки ролі спадковості у розвитку тієї чи іншої ознаки
визначають коефіцієнт спадковості за формулою К. Хольцингера:
% подібності МБ - % подібності ДБ
100 % —% подібності ДБ
де Н — коефіцієнт спадковості; МБ — монозиготні близнята;
ДБ —дизиготні близнята.
Якщо Н = 1, то ознака (захворювання) повністю визначаєть
ся генотипом, якщо Н = 0,5, то на вияв ознаки (захворювання)
однаково впливають спадковість і фактори довкілля, а якщо
Н = 0, то ознака (захворювання) повністю визначається факто
рами довкілля.
Вплив факторів довкілля (С) на розвиток ознаки (захворю
вання) визначають за формулою:
С = 100 % - Я .
Організмовий рівень організації життя. Основи генетики людини_____ 129
За допомогою близнюкового методу проведені дослідження
схильності людей до серцево-судинних захворювань. У табл. 2.4
і 2.5 наведені деякі узагальнені дані з вивчення генетичної
компоненти в походженні захворювань.
Таблиця 2.4. Результати дослідження близнюковим методом
схильності до серцево-судинних захворювань
Конкордантність близнят, %
Монозиготних
Дизиготних
26,2
10,0
19,6
15,5
22,4
10,8
26,0
10,5
Захворювання
Гіпертонічна хвороба
Інфаркт міокарда
Інсульт
Ревматизм
Таблиця 2.5. Генетичний аналіз спадкової схильності до деяких
мультифакторіальних захворювань
Показник
ІХС
Р
ЦД
ВХ
Ш
Частота в загальній
популяції, %
19
2
0,6
0,6
1
Частота повторних
випадків серед
родичів 1-го ступеня
спорідненості, %
30—60
10
10
8
14
67
43
37
7
42
12
50
14
67
18
Конкордантність
близнят:
монозиготних, %
дизиготних, %
Примітка. ІХС — ішемічна хвороба серця; Р — ревматизм; ЦД — цук
ровий діабет; ВХ — виразкова хвороба; ПІ — шизофренія.
Дерматогліфіка. Це метод вивчення рельєфу шкіри на
подушечках пальців, долонях і підошвах стоп. Термін “дерма
тогліфіка” був запропонований Г. Каммінсоном і Ц. Мідло,
введений на 42 сесії А м ер и кан ськ ої асоц іац ії анатомів
(1926).
130
Розділ 2
Ще у 1892 р. Ф. Гальтон установив, що візерунки не зміню
ються упродовж життя людини.
Дерматогліфіка включає три розділи:
1 . Дактилоскопія — вивчення візерунків на подушечках
пальців.
2. Пальмоскопія— вивчення візерунків на долонях.
3. Плантоскопія — вивчення візерунків на підошвах стоп.
1.
Дактилоскопія. Шкірні візерунки формуються з ектодер
ми і мезенхіми в період між 6 і 19-тим тижнями внутрішньоутробного розвитку і залишаються незмінними протягом усьо
го життя. Візерунки на підошвах стоп формуються на 2— З
тижні пізніше. Індивідуальні особливості візерунків зумовлені
генотипом батьків або мутаціями в каріотипі при дії деяких
факторів довкілля в перші 4— 5 місяців вагітності. Гребені на
шкірі пальців рук відповідають сосочкам дерми, тому їх нази
вають папілярними лініями. Міжсосочкові заглибини утворю
ють борозенки. На поверхні гребенів відкриваються протоки
потових залоз, а у товщі сполучнотканинного сосочка розміщені
чутливі нервові закінчення.
Папілярні лінії формують візерунки (рис. 2.10):
1. Дуги:
А ( 6 %).
2. Петлі:
Ь (6 0 % ); Ьк ІІД.
3. Завитки:
(34 % ).
Рис. 2.10. Папілярні шкірні візерунки людини: А — дуги;
Ь — петлі; \У — завиток
Організмовий рівень організації життя. Основи генетики людини_____ 131
Гребеневий рахунок — це кількість папілярних ліній між
трирадіусом і центром візерунка. У чоловіків гребеневий раху
нок становить 144 ± 51; у жінок — 127 ± 52.
Трирадіус — це місце сходження папілярних ліній трьох
різних напрямків.
2. Пальмоскопія. На долоні розрізняють:
1. Долонні подушечки — 11: тенар, гіпотенар, 4 міжпальцеві
подушечки; 5 подушечок на нігтевих фалангах.
2. Долонні лінії— 5: браслетна, лінія великого пальця,
проксимальна, дистальна, фалангова.
3. Долонні поля— 14.
4. Кут аід:
а)
б)
в)
г)
д)
у нормі, не перевищує 57°;
синдром Клайнфельтера 42°;
синдром Шерешевського — Тернера 66°;
синдром Дауна 81°;
синдром Патау 108°.
Дерматогліфіка — науковий метод. Має важливе значення
як допоміжний тест при діагностиці хромосомних і генних
захворювань людини. Метод застосовують для діагностики вро
джених вад і для клінічно невиразних, стертих форм при різних
хромосомних синдромах.
Цитогенетичний метод. Цей метод дає можливість вив
чити будову хромосом і каріотип людини. Метод забезпечує:
1. Вивчення каріотипу людини.
2. Уточнення числа хромосомних наборів, кількість і мор
фологію хромосом.
3. Вивчення геномних і хромосомних мутацій.
4. Складання генетичних карт хромосом.
Цитогенетичний метод включає каріатицування, та визна
чення статевого хроматину.
Каріотипування — це вивчення каріотипу соматичної клі
тини. Каріотип досліджують у лімфоцитах периферійної крові
132________________________________________________________ Розділ 2
людини. Для цього венозну кров центрифугують, виділяють
лейкоцити. Лейкоцити переносять у поживне середовище з фітогемаглютиніном, який стимулює поділ одного з видів лейко
цитів — лімфоцитів. Одержану суспензію поміщають у термо
стат на 72 год. За 3 год до закінчення інкубації додають кол
хіцин (затримує поділ лімфоцитів на стадії метафази). До
культури лімфоцитів додають гіпотонічний розчин КСІ, клі
тини збільшуються в об’ ємі, руйнується ядерна оболонка,
хромосоми розміщуються в екваторіальній площині клітини.
Препарат фарбують ядерними барвниками і вивчають під мік
роскопом, використовуючи імерсійний об’ єктив.
Метафазна пластинка — це сукупність хромосом одного
лімфоцита на стадії метафази. Метафазну пластинку можна
сфотографувати й вивчити кількість і форму хромосом на мікро
фотографії.
Визначення статевого хроматину дає змогу побічно судити
про кількість статевих хромосом у соматичних клітинах лю
дини. Розрізняють X- і У-статевий хроматин.
Х-статевий хроматин — це конденсована Х-хромосома
в інтерфазному ядрі соматичної клітини. Відповідно до теорії
Лайон, у ядрі соматичної клітини жінки активна лише одна
Х-хромосома, друга інактивується ще під час ембріогенезу.
Інактивована Х-хромосома виявляється:
1. У вигляді грудки (тільце Барра) в ядрах клітин епітелію
слизової оболонки ротової порожнини.
2. У вигляді барабанної палички в нейтрофільних лейкоци
тах.
Кількість грудок Х-хроматину можна визначити за форму
лою:
а
=
п - 1,
де а — кількість грудок Х-хроматину; п — кількість Х-хромосом.
Організмовий рівень організації життя. Основи генетики людини_____ 133
У жінок:
1. У нормі а = 1.
2. При моносомії а = 0 (синдром Шерешевського — Тернера).
3. При трисомії а = 2 (синдром трипло-Х).
У чоловіків:
1. У нормі а = 0 (Х-статевий хроматин відсутній).
2. При трисомії а = 1 (синдром Клайнфельтера).
У-статевий хроматин — це У-хромосома, яка люмінесціює
в ядрі соматичної клітини чоловіків. Досліджують за допомогою
люмінесцентного мікроскопа, при цьому У-хромосома світить
ся “ діамантовим сяйвом” . У ядрі здорового чоловіка видно
одну ділянку флюоресценції.
Механізм виникнення моносомій і трисомій. Моносомії і три
сомії виникають внаслідок нерозходження однієї пари хромо
сом при першому або другому мейотичному поділах у процесі
гаметогенезу в одного з батьків, або на ранніх мітотичних поді
лах зиготи.
У нормі
Р: $
XX
О:
®
Г:
При патології
х
<3
ХУ
X X — здорова дівчинка;
ХУ — здоровий хлопчик;
Р: $
XX
х
О:
@ )©
З
ХУ
Ф®
Г: X X X — Х-трисомія;
Х Х У — синдром Клайнфель
тера;
ХО —
синдром Шерешевсько
го — Тернера;
УО —
зигота нежиттєздат
на.
’
/
134_____________________________________________________Розділ 2
Аномалії аутосом:
1.
Синдром К. Патау. Описаний К. Патау 1960 р. у дівчинки
з ціанозом.
Каріотип 47, 13+, трисомія за Б-групою хромосом. Частота
синдрому 1 : 7600. Можливі два варіанти синдрому:
а) трисомія за Б-групою хромосом (47, 13+);
б) транслокація і злиття двох хромосом Б-групи (46, 13).
Фенотипово синдром характеризується такими ознаками:
1. Нормальний зріст і маса тіла.
2. Незаростання верхньої губи і піднебіння.
3. Полідактилія (наявність 6 -го пальця на нозі).
4. Глухота (у 80—85 % хворих).
5. Різка розумова відсталість (мікроцефалія, аненцефалія,
гіпоплазія мозочка).
6 . Вади серця (дефекти міжшлункових і міжпередсердних
перетинок, розширені камери серця).
7. Вади статевих органів (крипторхізм, подвоєння матки,
піхви).
8 . Подвоєння сечоводів.
9. Аномалії очного яблука (мікрофтальм, анофтальм).
10. Тривалість життя — три місяці; методи лікування від
сутні.
2.
Синдром Едвардса. Описаний 1960 р. лікарем Дж. Едвардсом у дівчинки з вузьким фронтальним і широким потиличнотім’яним діаметром черепа.
Каріотип 47, 18+, трисомія за Е-групою хромосом. Частота
синдрому 1 : 8100; у дівчаток трапляється у 3 рази частіше,
ніж у хлопчиків.
У 90 % дітей смерть настає на першому році життя.
Синдром фенотипово характеризується такими ознаками:
1. Мікростомія, мікрогнатія.
2. Асиметрія вушних мушель.
3. Наявність шкірної перетинки між пальцями.
І
Організмовий рівень організації життя. Основи генетики людини_____ 135
4. Виражена рухливість плечових суглобів.
5. Гіпоплазія, або аплазія м ’ язів тенара.
6 . Вади серця, підковоподібна нирка, подвоєння сечоводів.
3. Синдром Дауна. Описаний 1866 р. лікарем Л. Дауном.
Каріотип 47, 21+, трисомія за О-групою хромосом. Частота
синдрому 0,15 % ; впливає вік матері.
Можливі два варіанти синдрому:
1. Трисомія за О-групою, нерозходження 21-ї пари хромосом
під час І мейотичного поділу в період гаметогенезу (47, 2 1 +).
2. Транслокація 21-ї хромосоми на 15-ту хромосому ( 2 Н —»
15). При цьому каріотип урівноважений 46, 211 (21+15).
Фенотипово синдром характеризується такими ознаками:
1. Маленька голова, плоске обличчя, ніс з широким плоским
переніссям, рот напіввідкритий, язик географічний.
2. Монголоїдний розріз очей.
3. Порушена координація рухів.
4. Різко виражена розумова відсталість.
5. Вади серця, шлунково-кишкового каналу, нирок, сечових
шляхів.
6 . Однаково часто хворіють як хлопчики, так і дівчатка.
7. Неплідні, хоча відомі випадки, коли жінки з синдромом
Дауна народжували дітей-Даунів.
8 . Дерматогліфічна діагностична ознака: поперечна чотирипальцева лінія на долоні — мавпяча складка. Долонний трирадіус зміщений дистально; кут аій = 81°.
Аномалії статевих хром осом :
1. Синдром Шерешевського— Тернера. Описаний 1925 р.
лікарями М. Шерешевським і Г. Тернером.
Каріотип 45, ХО; тільце Барра відсутнє. Частота синдрому
0,02 %.
Фенотипово синдром характеризується такими ознаками:
1. Низький зріст (139— 148 см).
2. Крилоподібна складка на шиї — шия сфінкса.
136
Розділ 2
3. Низький заріст волосся на потилиці.
4. Повільний статевий розвиток, дисгенезія гонад.
5. Первинна аменорея, неплідність.
6 . Аномалії видільної системи (підковоподібна нирка, под
воєння нирок, сечоводів).
7. Інтелект нормальний, інколи спостерігається психічний
інфантилізм, емоційна лабільність.
8. Дерматогліфічні дослідження показують, що на пальцевих
узорах зменшується кількість радіальних петель і дуг, а збіль
шується частота ульнарних петель і завитків. Долонний трирадіус зміщений дистально і ульнарно; кут аій = 66°.
9. Лікування гормональне, спрямоване на корекцію вторин
них статевих ознак.
2. Синдром Клайнфельтера. Описаний 1942 р. лікарем
Г. Клайнфельтером.
Каріотип 47, ХХУ; 48, Х Х Х У ; 49, Х Х ХХ У . 1 тільце — 2 тіль
ця — 3 тільця Барра. Частота синдрому 0,1 % .
Фенотипово синдром характеризується такими ознаками:
1. Астенічний, євнухоподібний тип будови тіла.
2. Високий зріст. Диспропорція в розмірах кінцівок і тулу
ба, вузький плечовий пояс, широкий тазовий пояс.
3. Слаборозвинена мускулатура.
4. Розміщення жирової клітковини й оволосіння за жіночим
типом, на щоках заріст відсутній.
5. Недорозвинені сім’ яники, сперматогенез відсутній. Чо
ловіки неплідні.
6 . Невисокий інтелект, розумова відсталість, дебільність.
7. Зайва Х-хромосома зумовлює порушення психіки: апа
тичні, параноїдні стани, психози.
8 . Дерматогліфічні дослідження вказують на збільшення
кількості дуг, зменшення гребенів, проксимальне зміщення
долонного трирадіуса;
9. Кут аій = 42°.
Організмовий рівень організації життя. Основи генетики людини_____ 137
3. Синдром трипло-Х. Описаний 1959 р. Р. Джекобсоном.
Каріотип 47, X X X ; 2 тільця Барра. Частота синдрому 0,1 % .
Аномалія дуже мінлива у фенотиповому прояві: від явно ви
раженої до клінічно стертої.
Фенотипово синдром характеризується такими ознаками:
1. Супержіночий тип.
2. Незначна дисплазія різних систем органів.
3. Спостерігається затримка росту, епікант, високе тверде
піднебіння, клинодактилія мізинців.
4. У 75 % жінок виявлена розумова відсталість, високий
ризик захворювання психозом, надто шизофренією.
5. Первинна, частіше вторинна аменорея.
6 .
Недостатня функція яєчників з відсутністю фолікулів
зумовлює неплідність і прискорює клімакс.
7. Зі збільшенням кількості Х-хромосом спостерігається
різке зниження інтелекту.
8 . Трапляються жінки без жодних конституційних, фізіо
логічних і психічних розладів, однак вони фертильні з поси
леним лібідо.
9. Дерматогліфічні дослідження вказують на підвищення
кількості завитків і дуг на пальцях, зменшення гребеневого
рахунку.
4. Синдром дубль-У. Описаний 1962 р.
Каріотип 47, ХУУ; 2 зони “діамантового сяйва” У-хроматину. Частота синдрому 0,45 % .
Фенотипово синдром характеризується такими ознаками:
1. Чоловіки високого зросту (вище 186 см).
2. Асиметрія в будові тіла, обличчя, кінцівок.
3. Гіпоплазія сім ’ яників, неплідність.
4. Підвищений рівень андрогенів та лютеїнізуючого гор
мону.
5. Інтелект нормальний, проте розумовий розвиток низь
кий.
6 . Психопатичні риси характеру, агресивність, неправильна
поведінка, схильність до асоціальних вчинків.
138
Розділ 2
7. Зайва У-хромосома більше впливає на поведінку, ніж на
інтелект: хворі самовпевнені, імпульсивні, афективно нестійкі,
гіперсексуальні.
8 . Дерматогліфічні дослідження не дають підстави для
діагностики синдрому.
Популяційно-статистичний метод. У загальнобіологічному значенні під “ популяцією розуміють сукупність особин
одного виду, яка протягом тривалого часу (великої кількості
поколінь) займає певний ареал, всередині якого практично
здійснюється той чи інший ступінь панміксй і немає поміт
них ізоляційних бар’єрів, яка віддалена від сусідніх популяцій
певного виду тим чи іншим ступенем тиску різних форм ізо
л я ц ії(Тимофеєв — Ресовський).
Популяційна структура людства:
1. Ізоляти — людські популяції з чисельністю не більше
1500 осіб, споріднені шлюби становлять понад 90 % .
2. Деми — людські популяції з чисельністю від 1500 до
4000 осіб, частота споріднених шлюбів 80— 90 % .
3. Популяції — людські популяції села, міста, області.
Гени, які розповсюджені в популяції:
• Універсальні гени трапляються в будь-якій популяції
в середньому з однаковою частотою. Зокрема, ген фенілкетонурії
у гетерозиготному стані трапляється в 1 % населення Європи;
ген дальтонізму в гомозиготному стані зустрічається у 7 % чо
ловіків і 0,5 % жінок, в гетерозиготному стані — у 13 % жінок.
• Локальні гени трапляються у певних районах. Ген серпо
подібно-клітинної анемії виявляється у мешканців Африки,
Середземномор’ я; ген вродженого звиху стегна — у місцевого
населення північно-східної частини Євразії.
Популяційно-статистичний метод дає змогу визначити:
1. Генетичну структуру популяції.
2. Частоти розповсюдження домінантного і рецесивного
алелей гена в популяції.
3. Тенденції до зміни генетичного складу популяції.
Організмовий рівень організації життя. Основи генетики людини_____ 139
Д. Харді і В. Вайнберг відкрили закон (1908), який є тео
ретичним обґрунтуванням популяційного-статистичного ме
тоду.
В ідеальній популяції співвідношення генотипів домінант
них гомозигот (АА), гетерозигот (Аа ) і рецесивних гомозигот
(аа) залишається сталим.
(р + д)2 - р2+ 2 рц + д2 = 1 (100 % );
Р + 9 = 1»
де р — частота домінантного алеля гена; <7 — частота рецесивного
алеля гена; р2= АА — частота домінантних гомозиготних особин;
2рд = Аа — частота гетерозиготних особин; (?2 = аа — частота реце
сивних особин.
Ідеальна популяція характеризується такими ознаками:
• Велика кількість особин.
• Вільне схрещування (панміксія).
• Відсутність відтоку генів за рахунок добору чи міграції
особин за межі певної популяції.
• Відсутність притоку генів за рахунок мутації чи міграції
особин у певну популяцію З З О В Н І.
• Однакова плодючість гомозигот і гетерозигот.
• Кодування ознаки в популяції парою алельних генів або
серією множинних алелей.
У медичній генетиці закон Харді — Вайнберга використо
вується для аналізу розповсюдження в популяції патологічних
генів, які знижують життєздатність і плодючість людини. Це
можливо тому, що в панміксичних популяціях, які існують
тривалий час, відтік патологічних алелей, викликаний при
родним добором (загибеллю особин зі зниженою життєздатні
стю), врівноважується притоком тих самих алелей, що відбу
вається внаслідок мутаційного процесу.
Задача. Альбінізм успадковується як рецесивна ознака
і трапляється з частотою 1 : 1 0 0 .
1. Визначити генетичну структуру популяції.
2. Визначити частоти домінантного і рецесивного генів.
140
Розділ 2
Розв’язок:
А — алель гена нормальної пігментації шкіри;
а — алель гена альбінізму;
аа = д1
2 = 1 : 100 = 0, 01 (1 % );
а = д = VI : 100 = ОД;
А - р = 1 — д;
А = 1 — 0,1 = 0,9;
АА = р 2 = 0,92 = 0,81 (81 % );
Аа = 2рд = 2 •0,9 •0,1 = 0,18 (18 %).
В ід п о від ь :
1. Генетична структура популяції:
А А = 81 %;
Аа = 18 % ;
аа = 1 % .
2. Частоти алелей генів: домінантного
рецесивного
А = 0,9;
а = 0,1.
Біохімічні методи. Використовуються для діагностики
хвороб обміну речовин, причиною яких є зміни активності
ферментів. За допомогою біохімічних методів відкрито близь
ко 5000 молекулярних хвороб, які є наслідком прояву мутан
тних генів. Методи дуже трудомісткі, вимагають спеціального
обладнання і тому не можуть бути використані для масових
популяційних досліджень із метою раннього виявлення хворих
зі спадковою патологією.
Біохімічна діагностика порушень обміну речовин проводять
у два етапи:
1. Скринінг-діагностика (просіювальна): за допомогою експрес-методів, які ґрунтуються на простих якісних реакціях
виявлення продуктів обміну в сечі та крові, серед великої
кількості обстежуваних виділяють імовірно хворих, що мають
певне спадкове відхилення від норми.
2. За допомогою точних хроматографічних методів визна
чення ферментів, амінокислот, мікробіологічних тестів прово
диться уточнення діагнозу або відхилення при хибно-позитив
ній реакції на першому етапі.
Організмовий рівень організації життя. Основи генетики людини_____ 141
Науковою групою ВООЗ розроблена та рекомендована до
практичного використання така класифікація спадкових за
хворювань обміну речовин:
1. Спадкові порушення обміну амінокислот (фенілкетонурія).
2. Спадкові порушення обміну вуглеводів (глікогенова хво
роба, галактоземія).
3. Спадкові порушення обміну ліпідів (хвороба Німанна — Пі
ка, хвороба Гоше).
4. Спадкові порушення обміну стероїдів (адреногенітальний
синдром).
5. Спадкові порушення обміну пуринів і піримідинів (синд
ром Леша — Наяна).
6. Спадкові порушення обміну сполучної тканини (мукополісахаридози, синдром Марфана).
7. Спадкові порушення гема-порфірину (гемоглобінопатії).
8. Спадкові порушення обміну в еритроцитах (анемія Мінковського — Шоффара).
9. Спадкові аномалії обміну металів (хвороба Коновалова — Вільсона).
10. Спадкові порушення обміну білірубіну (синдром Кріглера — Наджара).
11. Спадкові порушення всмоктування в травному каналі
(муковісцидоз, целіакія, лактоземія).
М етоди генетики соматичних клітин. Вивчають спад
ковість і мінливість соматичних клітин, які містять повний
об’ єм генетичної інформації. Соматичні клітини людини отри
мують із різних органів (шкіра, кістковий мозок, клітини крові
та сполучної тканини, тканини ембріонів) через біопсію або
аутопсію. Найчастіше використовують фібробласти (клітини
сполучної тканини) або лімфоцити крові.
Нині використовують такі методи генетики соматичних
клітин людини:
• Культивування клітин поза організмом.
• Клонування.
142
Розділ 2
• Селекція.
• Гібридизація.
Культивування —• розмноження клітин на поживних сере
довищах з метою одержання їх у достатній кількості для ви
вчення цитогенетичними, біохімічними, імунологічними ме
тодами. Таке дослідження може бути більш точним, ніж на
рівні організму, бо метаболічні процеси вдається виділити зі
складного ланцюга взаємопов’язаних реакцій, які відбувають
ся в організмі.
Клонування — одержання нащадків однієї клітини (клону).
Всі клітини будуть з однаковим генотипом. Наприклад, шляхом
злиття нормального лімфоцита і пухлинної клітини одержують
гібридоми.
Селекція — відбір клітин із наперед заданими властивостя
ми при культивуванні їх на селективних живильних середо
вищах.
Гібридизація — злиття клітин двох різних видів (типів).
При цьому утворюється гетерокаріон — гібридна клітина зі
спільною цитоплазмою і ядрами двох батьківських клітин.
Після мітозу і подальшого поділу цитоплазми із двоядерного
гетерокаріону утворюються дві одноядерні клітини, кожна
з яких є синкаріоном (справжня гібридна клітина, яка має
хромосоми обох батьківських клітин). Залежно від мети аналі
зу дослідження проводять на гетерокаріонних або синкаріонних
клітинах. Синкаріони звичайно вдається отримати при гібри
дизації у межах класу. Це справжні гібридні клітини, бо в них
відбулося поєднання двох геномів. Наприклад, гібридні кліти
ни людини і миші мають 43 пари хромосом: 23 — від людини
і 20 — від миші. При розмноженні цих клітин доля вихідних
геномів різна: відбувається поступова елімінація хромосом
того організму, клітини якого мають повільніший темп роз
множення. Через ЗО поколінь можна знайти клітини, які мі
стять 1—2 пари хромосом людини. Якщо у гібридній клітині
відсутня хромосома і не відбувається синтез білків, то можна
припустити, що гени, які визначають їх синтез, локалізовані
в цій хромосомі.
Організмовий рівень організації життя. Основи генетики людини_____ 143
Метод гібридизації дає можливість вивчати:
1. Механізми взаємодії генів і шляхи регуляції генної ак
тивності.
2. Мутагенну дію факторів довкілля, генні мутації.
3. Локалізацію генів у хромосомі, аналізувати групи зчеп
лення генів.
4. Первинні продукти активності окремих генів.
Упровадження методів генетики соматичних клітин дає змо
гу точно діагностувати спадкові захворювання ще у пренаталь
ному періоді.
2.6.2. Принципи діагностики спадкової патології
Спадкові хвороби становлять дедалі більшу частку у загаль
ній патології людини. Тому лікарі будь-якої спеціалізації
постійно стикаються зі спадковими хворобами, більшість із
яких мають хронічний перебіг. Особливо багато хворих зі спад
ковими формами захворювань потрапляють у спеціалізовані
клінічні та діагностичні відділення. Проте спадкові форми діа
гностуються у клінічних умовах не завжди, оскільки діагности
ка спадкової патології — це складний і трудомісткий процес.
Ускладнення діагностики зумовлені, перш за все, тим, що
нозологічні форми спадкових хвороб дуже різноманітні (близь
ко 2000) і кожна з них має широкий діапазон клінічної карти
ни. Так, у групі нервових хвороб відомо понад 200 спадкових
форм, а в дерматології й офтальмології їх понад 250. Деякі
форми вкрай рідкісні, і лікар у своїй практиці може не стика
тися з ними. Тому він повинен знати основні принципи, які
допоможуть запідозрити спадковий генез хвороби, а після додат
кових консультацій і обстежень встановити точний діагноз.
Діагностика спадкових хвороб базується на даних клінічно
го, параклінічного та спеціального генетичного обстеження.
Щоб не пропустити спадкову хворобу, лікар має пам’ ятати
про те, що здебільшого така патологія є супутником основного,
неспадкового захворювання, з приводу якого пацієнт звернув
ся до лікаря.
144
Розділ 2
Для того, щоб не пропустити спадкову хворобу, діагноз
треба ставити у два етапи:
— перший — загальне клінічне обстеження хворого відповід
но до сучасних вимог;
— другий — при підозрі певної спадкової хвороби необхідно
провести спеціалізоване діагностичне обстеження.
Лікар будь-якої спеціалізації при обстеженні пацієнта має
пам’ятати загальні принципи діагностики спадкових хвороб:
1. При обстеженні хворого необхідно застосовувати клінікогенеалогічний метод. Завдяки цьому методу лікар констатує
наявність або відсутність схожих захворювань у сім ’ ї, а також
виявляє інші “ сімейні” захворювання. Одержання відомостей
про родичів — завдання непросте, тому що не всі пацієнти
мають повну інформацію про своїх родичів. Часто приховують
сімейні випадки з етичних міркувань, або, навпаки, виявляю
чи їх у сім’ї партнера починають звинувачувати його у виник
ненні захворювання.
Тому лікар має особисто обстежити родичів хворого. За
наявності сімейних випадків захворювання або при підозрі на
спадковий характер хвороби необхідно провести другий етап
обстеження хворого, спрямований на диференціальну діагно
стику конкретного захворювання.
2. Рецидивуючі, хронічні захворювання, що тривалий час
погано лікуються, особливо в дитячому віці, часто належать
до спадкових форм. Так, хронічна пневмонія у дітей із легене
вою формою муковісцидозу, тривалі розлади кишківника ви
никають при целіакії, кишковій формі муковісцидозу, при
дисахаридозній нестачі та інше. Хронічний процес розвиваєть
ся в міру постійної дії мутантного гена, що сприяє утворенню
патологічних метаболітів. Багато форм спадкових хвороб були
відкриті під час обстеження груп хворих із хронічними захво
рюваннями.
3. Наявність у хворого специфічних симптомів, які рідко
трапляються, або їх поєднання є підставою думати про при
роджений чи спадковий характер захворювання. Так, вивих
або підвивих кришталика ока характерний для трьох спадко
вих синдромів: Марфана, Вейля — Марчезані та гомоцистинурії.
Організмовий рівень організації життя. Основи генетики людини_____ 145
Блакитні склери характерні для недосконалого остеогенезу
і деяких інших хвороб сполучної тканини. Кожен лікар пови
нен знати маркери, характерні для спадкових хвороб.
4. Патологічні зміни, які охоплюють багато органів і систем,
наводять на думку про спадкову (або вроджену) патологію.
Більшість мутантних генів, які викликають спадкові хвороби,
мають плейотропний ефект, унаслідок чого патологічні проце
си охоплюють багато органів. Особливо розповсюджений харак
тер уражень спостерігається при хромосомних хворобах. Для
них характерні не тільки численні мікроаномалії, але й тяжкі
вади розвитку. Наявність кількох мікроаномалій або ознак
дисплазії розвитку зумовлюють необхідність диференціальнодіагностичного обстеження хворого.
5. Спадковим захворювання можна вважати тоді, коли хво
роба має вроджений характер. Як зазначено вище, вроджений
характер патологічних ознак не завжди свідчить про спадкову
природу захворювання, але багато спадкових хвороб є резуль
татом хромосомних аномалій або генних мутацій і формуються
вже внутрішньоутробно. Коли дитина народжується з комп
лексом патологічних ознак, то вважають, що хвороба є вро
дженою. Прикладом уроджених спадкових хвороб є хромосом
ні синдроми, ахондроплазія, іхтіоз, Х-зчеплена гідроцефалія,
аутосомно-рецесивна справжня гідроцефалія та ін. Прикладом
уроджених, але неспадкових хвороб є талідомідний, сифілі
тичний, алкогольний і деякі інші синдроми, етіологію яких
встановлюють при цілеспрямованому зборі анамнезу. Якщо
при дотриманні вищезгаданих принципів у процесі обстеження
хворого точний діагноз усе-таки не встановлено, а підозра на
спадкове захворювання залишається, необхідно продовжити
дослідження з використанням спеціальних методів. Варто за
значити, що для багатьох хвороб інструментальні й лаборатор
ні параклінічні методи є вирішальними у диференціальній
діагностиці.
Пренатальна діагностика — напрям медичної генетики,
який виник на межі клінічної медицини і фундаментальних
наук: генетики, молекулярної біології, біохімії, цитогенетики.
Основне завдання — розробка комплексу методів, які дозволяють
146
Розділ 2
ефективно оцінити стан плоду людини вже на ранніх стадіях
розвитку й організувати раціональну профілактику народжен
ня дітей з тяжкими некорегованими вадами спадкової або
екзогенної природи.
Вона використовує ультразвукову діагностику (УЗД), опе
ративну техніку (хоріонбіопсію, амніо-і кордоцентез, біопсію
м ’ язів і шкіри плоду), лабораторні методи (цитогенетичні,
біохімічні, молекулярно-генетичні).
Пренатальна діагностика має винятково важливе значення
при медико-генетичному консультуванні, оскільки вона дає
змогу перейти від імовірного до однозначного прогнозування
здоров’я дитини у родинах із генетичним обтяженням. Нині
пренатальна діагностика здійснюється в І і II триместрах ва
гітності, тобто в періоди, коли у разі виявлення патології ще
можна перервати вагітність. На сьогодні можлива діагностика
практично всіх хромосомних синдромів і близько 100 спадкових
хвороб, біохімічний дефект при яких встановлений імовірно.
Рішення про проведення пренатального переривання вагіт
ності необхідно приймати тільки після оцінки таких кри
теріїв:
1. Хвороба повинна бути досить тяжкою, щоб було виправ
дане переривання вагітності.
2. Лікування хвороби плоду неможливе і незадовільне.
3. Згода на переривання вагітності родини, яка консуль
тується.
4. Наявність точного тесту для постановки пренатального
діагнозу.
5. Досить високий генетичний ризик несприятливого резуль
тату вагітності.
При організації та розвитку системи пренатальної діагно
стики повинні виконуватися такі умови:
1. Діагностичні процедури мають бути безпечні для здоров’ я
матері та плоду.
2. Частота ускладнень вагітності після пренатальної діагно
стики не має помітно підвищуватися зі спонтанним рівнем,
тобто процедура не має підвищувати ймовірність втрати плоду
відразу чи після її проведення у віддалений період.
Організмовий рівень організації життя. Основи генетики людини_____ 147
3. Лікарі, що володіють технікою пренатальної діагностики,
повинні знати ймовірність постановки псевдопозитивних чи
псевдонегативних діагнозів, — іншими словами, повинні добре
знати обмеження методу.
4. Пренатальна діагностика має включати два етапи:
— перший етап — виявлення ж інок (точніше, родин) із
підвищеним ризиком несприятливого в генетичному плані
результату вагітності при медико-генетичному консультуванні
чи первинному обстеженні всіх вагітних, у тому числі з вико
ристанням методів просіваючої діагностики;
— другий етап — власне пренатальна діагностика: аналізи
проводяться тільки жінкам, які мають фактори ризику.
5. Група фахівців із пренатальної діагностики (акушер-гінеколог, лікар-генетик, лікар-лаборант-генетик) повинні знати
діагностичні обмеження методу не взагалі, а в їхній лабора
торії.
6. Група фахівців повинна суворо дотримуватися стандартів
для процедур і лабораторних аналізів, здійснювати поточний
контроль якості роботи, а також мати статистику завершення
вагітностей і розбіжностей діагнозів (контроль після абортів чи
після народження).
Показання для пренатальної діагностики:
1. Вік матері понад 35 років.
2. Наявність у родині попередньої дитини з хромосомною
патологією, у тому числі зі синдромом Дауна (попередній анеусомік).
3. Перебудови батьківських хромосом.
4. Наявність у родині захворювань, успадкованих зчепле
но зі статтю.
5. Синдром фрагільної Х-хромосоми.
6. Гемоглобінопатії.
7. Природжені порушення метаболізму.
8. Різні спадкові захворювання, що діагностуються мето
дом зчеплення з ДНК-маркерами.
9. Дефекти нервової трубки.
10. Інші показання для цитогенетичної пренатальної діагно
стики.
148
Розділ 2
Методи пренатальної діагностики:
• Скринінг-методи.
• Неінвазивні.
• Інвазивні з наступною лабораторною діагностикою.
Скринінг-методи дають змогу виділити тих жінок, які ма
ють підвищений ризик народити дитину зі спадковою або
вродженою аномалією. До них належать:
1. Визначення концентрації а-фетопротеїну (АФП) у сиро
ватці крові вагітних. АФП — білок, який продукує печінка
під час внутрішньоутробного розвитку. Його концентрація
підвищена при вроджених дефектах нервової трубки і черевної
стінки; знижена при хворобі Дауна у плоду.
2. Визначення рівня хоріонічного гонадотропіну. В нормі
його концентрація знижується після І триместру вагітності та
залишається підвищеною при хромосомній патології.
3. Визначення рівня незв’ язаного естріолу. Рівень цього
гормону значно нижчий при виношуванні плоду із хворобою
Дауна.
Неінвазивним методом пренатальної діагностики є уль
тразвукове сканування (УЗС), яке дозволяє виявити до 80 %
всіх вроджених аномалій розвитку (табл. 2.6). УЗС викорис
товують як метод скринінг-діагностики, так і як уточнений
діагностичний метод. Покази для використання УЗС:
1. Підвищений рівень АФП у сироватці крові на 16— 18-й
тиждень вагітності.
2. Несприятливий перебіг вагітності, затримка розвитку
плоду.
3. Народження попередньої дитини з вродженими вадами
розвитку.
4. Наявність вроджених вад розвитку в подружжя.
Інвазивними методами отримують клітини і тканини ем
бріона, плоду в І і II триместрах вагітності. Хоріонбіопсію про
водять після 8-го тижня вагітності, а плацентобіопсію — після
11-го. Процедура здійснюється трансабдомінально або трансцервікально під контролем УЗС. Ворсинки хоріона використовують
для лабораторної (цитогенетичної, молекулярно-генетичної,
біохімічної) діагностики спадкових захворювань.
Організмовий рівень організації життя. Основи генетики людини_____ 149
2.6. Вроджені вади розвитку, які діагностуються за
допомогою УЗС
Таблиця
Система або орган
ЦНС
Кінцівки
Серце
Нирки
Кишково-шлунковий
тракт
Вада
Аненцефалія, спинномозкова грижа,
енцефалоцеле
Редукційні вади, тяжкі форми
карликовості із укороченням кінцівок,
остеогенез
Тяжка вада серця
Агенезія нирок, полікистоз нирок
Атрезія 12-палої кишки, дефекти
передньої черевної стінки
Амніоцентез використовують з метою одержання навколоплодової рідини, у якій містяться злущені клітини амніона
і плоду. Процедура здійснюється на 15— 18-му тижні вагітності.
Злущені клітини культивують 2— 4 тижні та використовують
для цитогенетичних і біохімічних досліджень.
Кордоцентез — взяття крові із пуповини під контролем УЗС.
Процедуру проводять у період 18— 22-го тижня вагітності.
Зразки крові використовують для цитогенетичних (культивують
лімфоцити), біохімічних і молекулярно-генетичних досліджень.
Кордоцентез використовують для діагностики хромосомних
захворювань, гематологічних спадкових захворювань (гемо
глобінопатії), імунодефіцитів, внутрішньоутробних інфекцій.
Біопсію тканин плоду здійснюють у II триместрі вагітності
під контролем УЗС. Для діагностики м ’ язової дистрофії Дюшена розроблений імунофлуоресцентний метод біопсії м ’ язів
плоду.
Фетоскопія — введення зонда в амніотичну порожнину й ог
ляд плоду. Проводять на 18— 23-й тиждень вагітності. Можна
діагностувати майже всі вади розвитку.
1 5 0 ________________________________________________________ Розділ 2
2.6.3. Медико-генетичне консультування
Медико-генетичне консультування — це спеціалізована
медична допомога для запобігання поширенню спадкових хво
роб. Генетичне консультування передбачає інформування лю
дини про ризик розвитку спадкового захворювання, передачі
його нащадкам, про діагностичні та терапевтичні дії. Досвід
роботи медико-генетичних консультацій показує, що велика
кількість звернень пов’язана з питанням прогнозу нащадків,
з оцінкою генетичного ризику. Генетичний ризик, який не пе
ревищує 5 % , розцінюється як низький, до 20 % — як підвище
ний і понад 20 % — як високий. Консультування з приводу
прогнозу нащадків можна розділити на дві великі групи: проспективне і ретроспективне.
Проспективне консультування — це найбільш ефективний
вид профілактики спадкових хвороб, коли ризик народження
хворої дитини визначається ще до настання вагітності чи на
ранніх її етапах. У цьому випадку подружжя, направлене на
консультацію, не має хворої дитини, але існує певний ризик
її народження, який ґрунтується на даних генеалогічного до
слідження, анамнезу чи перебігу вагітності.
Ретроспективне консультування — це консультування що
до здоров’ я майбутніх дітей після народження у родині хворої
дитини.
Завдання медико-генетичного консультування:
1. Встановлення точного діагнозу природженого чи спадко
вого захворювання.
2. Визначення типу успадкування захворювання в певній
родині.
3. Розрахунок величини ризику повторення захворювання
в родині.
4. Пояснення змісту медико-генетичного прогнозу тим лю
дям, які звернулися за консультацією.
5. Диспансерне спостереження і виявлення групи підвище
ного ризику серед родичів індивіда зі спадковою хворобою.
6. Пропаганда медико-генетичних знань серед лікарів і на
селення.
Організмовий рівень організації життя. Основи генетики людини_____ 151
Покази для медико-генетичного консультування:
1. Народження дитини з вродженими вадами розвитку.
2. Встановлена або підозрювана спадкова хвороба у родині.
3. Затримка фізичного розвитку або розумова відсталість
у дитини.
4. Повторні спонтанні аборти, викидні, мертвонародження.
5. Близькородинні шлюби.
6. Вік матері старше 35 років.
7. Несприятливі впливи факторів зовнішнього середовища
у ранній термін вагітності (інфекційні захворювання, особливо
вірусної етіології; посилена лікарська терапія; рентгенодіагно
стичні процедури; шкідливі умови праці).
8. Несприятливий перебіг вагітності.
Етапи медико-генетичного консультування. Консультування
повинно складатися з кількох етапів для того, щоб лікар-генетик міг дати обґрунтовану рекомендацію і підготувати людей до
правильного сприйняття поради. При цьому перед лікарем ви
никають не тільки генетичні, але й морально-етичні питання.
Медико-генетична консультація складається з чотирьох
етапів: діагноз, прогноз, висновок, порада. При цьому необхід
не відверте і доброзичливе спілкування лікаря-генетика з ро
диною хворого.
Перший етап консультування починається з уточнення
діагнозу хвороби. Це вимагає близького контакту між генети
ком і лікарем-фахівцем у галузі тієї патології, що є предметом
консультування (акушер, педіатр, невропатолог та інші). По
чатковим моментом діагностики є клінічний діагноз. У медикогенетичних консультаціях діагноз уточнюють за допомогою
генетичного аналізу (що і відрізняє лікаря-генетика від інших
фахівців), широко використовують генеалогічний і цитогенетичний методи, а також специфічні методи біохімічної генети
ки, які спеціально розроблені для діагностики спадкових
хвороб і не часто застосовуються у клінічній практиці.
На другому етапі консультування завдання лікаря-генетика
полягає у визначенні ризику народження хворої дитини. Почат
ковим моментом є родовід обстежуваної родини. Генетичний ри
зик виражає імовірність появи певної аномалії в обстежуваного
1 52 ________________________________________________________ Розділ 2
чи його нащадків. Він визначається двома способами: або шля
хом теоретичних розрахунків, які ґрунтуються на генетичних
закономірностях, або за допомогою емпіричних даних.
На третьому етапі консультування лікар-генетик повинен
зробити висновок про ризик виникнення хвороби у дітей об
стежуваних і дати їм відповідні рекомендації. Складаючи ви
сновок, лікар враховує тяжкість сімейної патології, величину
ризику народження хворої дитини і морально-етичний бік пи
тання.
Заключний етап консультування — порада лікаря-генетика — вимагає найбільш уважного ставлення. Як зазначають
деякі дослідники, багато обстежуваних не готові до сприйнят
тя генетичної інформації. Всі особи, котрі звертаються в кон
сультацію, хочуть мати дитину і чекають від консультантів
позитивної відповіді. Нерідко їх запити нереальні, оскільки
вони не знають про можливості консультанта-генетика й очі
кують від нього практичної допомоги. Консультант-генетик
завжди повинен враховувати мотиви, якими можуть керува
тися люди (емоційні, соціально-економічні тощо), оцінювати
інтелектуальний і освітній рівень подружжя (пробанда), пси
хологічний клімат у родині. Тільки за цієї умови він може
справді допомогти подружжю в аналізі ситуації. Родина само
стійно приймає остаточне рішення.
У кожному конкретному випадку за наявності більше одно
го фактора ризику всі бали додаються. Ґрунтуючись на бальній
оцінці обтяжуючих факторів, є реальна можливість вирахува
ти ступінь ризику народження дитини з вадами розвитку.
Правильне своєчасне виділення групи ризику з подальшим її
консультуванням дозволяє зменшити кількість народжених
дітей з вадами розвитку.
Нині дуже мало родин (не більше 1 0 %) , яким потрібна
порада лікаря-генетика, звертається за такою спеціалізованою
допомогою. Така ситуація пов’ язана з двома обставинами:
1) недостатній рівень медико-генетичних знань у суспільстві;
2) недостатнє розуміння організаторами охорони здоров’я
значення медико-генетичного консультування як методу про
філактики спадкових хвороб.
Організмовий рівень організації життя. Основи генетики людини
153
Медико-генетичне консультування повинно стати складовою
комплексу заходів, спрямованих на охорону здоров’ я матері
та дитини, на зниження пренатальної смертності, тому особ
ливого значення варто надавати пропаганді знань у галузі
клінічної генетики для лікарів-практиків — акушерів-гінекологів, неонатологів, невропатологів, терапевтів, онкологів.
Важливим профілактичним заходом уникнення різних ано
малій у нащадків є широка санітарно-просвітницька пропаганда
як серед дорослого населення, так і серед підлітків, які стануть
родоначальниками нових, фізично та розумово здорових по
колінь.
Фенокопії. При проведенні медико-генетичного консульту
вання варто пам’ ятати про можливість виникнення фено
копії — тобто фенотипу, схожого за характерними проявами
мутантного гена, зумовленого чинниками зовнішнього середо
вища.
Фенокопії —
ц е і н д у к о в а н і з м ін и о з н а к о р г а н і з м у , я к і з б е р і
г а ю т ь с я т іл ь к и п р о т я г о м й о го ж и т т я , н а щ а д к и т а к о г о о р
га н із м у при с т а т е в о м у р о з м н о ж ен н і в т р а ч а ю т ь ці о з н а к и .
Фенотипно однорідні аномалії можуть бути наслідком спад
кових або середовищних факторів, які діють на ембріон. Так,
уроджене помутніння кришталика може бути спадковим реце
сивним захворюванням, результатом інфекції (краснуха) або
впливом іонізуючого випромінювання на ранньому терміні ва
гітності.
Вживання вагітною синтетичних гормонів, яким властивий
маскулінізуючий ефект, може спричинити явище вірилізації
плоду жіночої статі (надмірне оволосіння), яке нагадує адреногенітальний синдром.
Деякі ліки (талідомід) хімічної природи, які призначалися
хворим вагітним, схильні до провокування вад розвитку (ат
резія слухового ходу, аплазія дванадцятипалої кишки, вади
серця та ін.), які нагадують фенокопії домінантного синдрому
Холта — Орама і рецесивної панмієлопатії Фанконі.
Хімічні речовини (етанол) порушують генетичні механізми
формування систем органів, кінцівок, голови і мозку на пізніх
154
Розділ 2
стадіях ембріонального розвитку, спричиняють появу фенокопій,
схожих на патологію спадкового генезу.
Усе це докази взаємодії генетичних і негенетичних чинників
у виникненні виродливостей та вад розвитку, зокрема фено
копій.
Генокопії. За умов виникнення генних мутацій у різних
локусах клінічна картина генного спадкового захворювання
формується внаслідок порушення різних патогенетичних ланок.
При цьому фенотипні ефекти мутацій різних генів об’єднуються
в одну групу, тобто до однієї групи входять різні генетичні
захворювання (мутації у різних локусах). Такі випадки нази
вають генокопіями.
2.7. Нехромосомна спадковість
Нехромосомна (цитоплазматична) спадковість вперше від
крита 1909 року К. Корренсом і Е. Бауером у квіткових рослин.
Пізніше було доведено присутність стабільних і постійних ге
нетичних детермінат, які перебувають поза ядром. Успадку
вання ознак і властивостей організму, зумовлених цитоплаз
матичними структурами, доведено для багатьох рослин, тварин
і людини.
Носіями спадковості можуть бути тільки органели клітини,
які здатні до самовідтворення. Серед них одні регулюють клі
тинний метаболізм (мітохондрії та пластиди), а інші — проду
кують фібрилярні утворення (клітинний центр, або центросоми,
і базальні гранули, або кінетосоми). Доведено, що в цих цито
плазматичних структурах наявна ДНК, яка використовується
для передавання в клітині багатократно (близько 100 пластид,
50—5000 мітохондрій, 100 000 рибосом). Завдяки такій кіль
кості цих органел, цитоплазма порівняно з ядром менш мінли
ва, а невеликі кількісні їх зміни не супроводжуються істотни
ми порушеннями ознак в організмі.
Під впливом різних зовнішніх чинників в організмі відбу
ваються специфічні зміни компонентів нехромосомної спадко
вості. Вони можуть істотно відрізнятись між собою за стійкістю
фенотипного прояву. Найменш стійкі зміни — фенокопй.
Організмовий рівень організації життя. Основи генетики людини______155
Отримано достатню кількість доказів того, що мітохондрії
людини мають власну спадковість. Ідентифіковано всі гени
мітохондріального геному, які успадковуються лише від матері
через цитоплазму яйцеклітини. У ньому налічується 15 690
нуклеотидних пар. Охарактеризовано унікальні властивості
організації мітохондріального геному, що відрізняються від
ядерного, мутації якого зумовлюють ряд хвороб людини. Виділе
но окремий розділ генетики — мітохондріальна генетика.
Доведено, що мітохондріальна ДНК передається тільки від
матері всім її нащадкам і від дочок всім її поколінням (мате
ринське успадкування). Оскільки мітохондріальна ДНК нале
жить тільки одному з батьків, то рекомбінаційні зміни, властиві
ядерній ДНК в мейозі, відсутні. Отже, відсутня і комбінативна
мінливість. Нуклеотидна послідовність зазнає змін від поколін
ня до покоління тільки за рахунок мутацій, які відбуваються
в 10 разів інтенсивніше, ніж у ядерній ДНК. Наприкінці X X ст.
виник новий розділ медицини — мітохондріальна патологія.
Класичними патолого-анатомічними проявами мітохондріальних
хвороб вважаються міопатії та енцефалопатії, лактат-ацитоз,
синдром Кернса — Сера, міоклонус-епілепсія та ін. Для хвороб
такого генезу властивий значний клінічний поліморфізм.
Останнім часом мітохондріальна ДНК використовується
в популяційних дослідженнях, вивченні поширення Н о т о
з а р і е п з на планеті, походження людських рас, походження та
генетичної структури окремих етносів і популяцій.
2.8. Генна інженерія та біотехнологія
Окрім терапевтичного клонування, для лікування деяких
хвороб, зокрема спадкових, широко використовується генна
терапія, що базується на досягненнях генної інженерії та біотехнології.
Г е н н а і н ж е н е р і я — галузь молекулярної біології та генети
ки, яка займається створенням організмів із новою генетичною
програмою. Основні методи генної інженерії було розроблено
у 70-х роках X X ст. Створення трансгенних організмів включає
три основні етапи (рис. 2.10):
Виділення
вбудованоїДНК
з очищених плазмід
Виділення білкового
продукту гену
мРНК 5',
ТТ
І
І
\
Зворотна
___
\
г и.н/іл
транскриптЬра Розділення
_
(ревертаза) ' МНК
Гібрид
мРНК-
Отримання гена
\
або білка, що/
Сумарна ним коди
мРНК*\ еться
кднк с ; ■. 1 ■ 1 15
*
' Одноланцюгова
кДНК
ІЛпилькоподібна
кДНК
НАРОЩ УВАН НЯ
Б ІО М А С И
а
Дволанцюгова 5'-,
кДНК З '- 1
©
Лужний
І гідроліз
і і і і і
Виділення
Бактерії
клону
Днк- 5' ДНК-копія к Д Н /КК Л/І''\
ТИНА
- полімераза
пр'окаріотична
1
і— г— г З ’
Ампліфікація
Дволанцюгова60
в клітині
Нуклеаза кд н К
реципієнта
ОТРИМ АННЯ
/
1— 1— 1-3"
Р Е К О М Б ІН А Н Т Н О Ї
ДНК
5'- та З'
__ модифікації
-------
Рис. 2.10. Генна інженерія: схема клонування генів
Вбудовування"
у вектор
Скринінг
по "+” бак
теріях і клонування
БактерїА
Ріст
у культурі
158
Розділ 2
1. Отримання генетичного матеріалу з біологічних об’єктів
або хімічний синтез генів.
2. Вбудовування генів у векторну молекулу (плазміду або фаг)
і створення таким чином рекомбінантної молекули ДНК.
3. Трансформація рекомбінантної молекули ДНК у клітинуреципієнт.
Для отримання генетичного матеріалу із однотипових клітин
організму виділяють ІРНК і знімають з неї за допомогою зво
ротної транскриптази (ревертази) ДНК-копії, які потім вводять
у реципієнтні клітини. ІРНК виділяють із клітини або тканин,
у яких експресується пошуковий ген. Так, для клонування
проінсулінового гена необхідно використовувати р-клітини
підшлункової залози, оскільки для них характерний найбіль
ший вміст проінсулінової ІРНК. Сумарну клітинну ІРНК за
допомогою центрифугування в градієнті густини сахарози роз
діляють на фракції. При такому грубому очищенні кількість
сторонніх молекул значно зменшується. Потім із молекул ІРНК,
які попали в необхідний нам діапазон молекулярних мас (для
проінсуліну 25 000 нукл), знімають ДНК-копії.
Синтез ДНК-копій здійснюється ферментом РНК-залежною
ДНК-полімеразою (зворотна транскриптаза, ревертаза). Після
завершення копіювання вихідний ланцюг ІРНК деполімеризують шляхом лужного гідролізу. Ланцюги ДНК стійкі до луж
ного гідролізу, а РНК повністю деполімеризуються. Отримана
в результаті такої процедури ДНК є одноланцюговою, лише на
кінці молекули утворюється шпилька з невеликою петлею.
Така шпилька утворюється в результаті того, що на 5'-кінці
більшості молекул ІРНК наявна послідовність нуклеотидів,
одна половина якої комплементарна іншій та яка в результаті
копіювання залишається і в ДНК. Дволанцюгову ДНК отриму
ють шляхом добудови олДНК до длДНК, що здійснюється
ферментом ДНК-полімеразою І. Одноланцюгова петля, яка
залишається в длДНК, видаляється за допомогою ферменту
нуклеази 81. Після такої обробки ДНК можна вбудовувати
у вектор.
Варто відзначити, що окрім отримання необхідних генів із
біологічних об’ єктів, можливий і хімічний синтез генів. Однак
для цього потрібно мати повністю розшифровану послідовність
нуклеотидів гена. Вперше ген був синтезований 1970 р. Г. Ко-
Організмовий рівень організації життя. Основи генетики людини
159
раною в Канаді. Це був ген аланінової тРНК дріжджів, який
складався зі 77 нуклеотидних пар і не мав регуляторних діля
нок. Потім, 1976 р. було синтезовано ген тирозинової тРНК,
який складався зі структурної ділянки (126 нуклеотидних пар),
промотора, термінатора. Цей ген було введено у бактеріальну
клітину, у якій він функціонував. Синтезовано ген, який відпо
відає за синтез лактатдегідрогенази, що розщеплює лактат.
Його було вбудовано в плазміду, а потім у Е. соїі, яка почала
засвоювати лактат. Хімічним шляхом вдається синтезувати
гени невеликих розмірів.
Вектор — це молекулярна структура, здатна переносити
чужорідну ДНК всередину бактеріальної чи еукаріотичної
клітини таким чином, щоб вона могла там реплікуватися. Іс
нують два основні типи векторів: бактеріальні плазміди та
бактеріофаги.
Плазміди — позахромосомні елементи бактеріальних клі
тин, які є замкнутими кільцевими молекулами ДНК. Вони
здатні реплікуватись незалежно від геномної ДНК бактерій.
Часто плазміди містять гени, білкові продукти яких забезпе
чують нечутливість до тих чи інших антибіотиків. Ці власти
вості використовуються для відокремлення бактерій, що містять
плазміди, від бактерій без плазмід. Щ оби включити ДНК
у плазміду, треба розірвати її замкнуте кільце. Для цього плаз
міди піддають дії рестриктаз. Ці ферменти розрізають длДНК
у відповідних нуклеотидних послідовностях, які називають
ділянками рестрикції. У плазмідах, які трапляються в природі,
часто буває багато таких ділянок для кожної рестриктази. Ос
кільки здебільшого потрібна одна-єдина ділянка розрізу, про
водять направлений відбір плазмід із такими властивостями.
Зшивання — процедура, у результаті якої чужорідна ДНК
вбудовується між двома кінцями плазмідної ДНК і зшивається
з ними за допомогою ферменту ДНК-лігази. Щоб ця операція
пройшла успішно, кінці ДНК і плазміди мають бути “липкими” .
Для цього на їх кінцях пришивають послідовності нуклеотидів,
які за рахунок комплементарності забезпечують з’єднання двох
пар кінців. Так утворюються плазміди, які називаються рекомбінантними.
1 60
Розділ 2
Трансформація відбувається після того, як рекомбінантні
плазміди додають до бактерій-реципієнтів. Плазміда проникає
всередину бактеріальної клітини і включається в її життєвий
цикл. Оскільки не всі бактеріальні клітини в реакційній суміші
будуть трансформовані, бажано провести їх відбір, щоби роз
множуватись могли тільки рекомбінантні бактерії. Відбір
ґрунтується на тому, що плазміди забезпечують резистентність
до тих чи інших антибіотиків. Тобто тільки ті бактерії, у яких
є плазміди, будуть розмножуватись за наявності відповідного
антибіотика.
Скринінг — процедура, яка полягає в пошуку необхідних
бактерій. Використання її обумовлено тим, що у вихідному
препараті представлено багато різних ІРНК, і лише частина
плазмід несе необхідний ген. Як тільки необхідну бактерію
вдалося виділити, її не важко клонувати і розмножити в куль
турі.
Ампліфікація— здійснюється завдяки тому, що водній
бактеріальній клітині може синтезуватися багато копій необ
хідної нам плазміди, а також за рахунок отримання великої
кількості клітин із такими плазмідами. Після виділення й очи
щення плазміди обробляють відповідною рестриктазою, яка
вирізає вбудовані в них копії необхідного гена. Ампліфікований
таким чином ген можна використовувати для подальших експе
риментів у галузі генної інженерії. Окрім цього, бактерії мож
на використовувати для отримання білкового продукту гена,
вбудованого у плазміду.
Отримання біологічної продукції з генно-інженерних об’єктів
почалося 1982 р., коли компанія Елі Ділі вперше випустила
у продаж комерційний препарат інсуліну людини, одержаного
методом генної інженерії з трансформованої Е. соїі. Пізніше
у США вивели свиней з трансформованим гормоном росту лю
дини (1986), а у Великій Британії (Единбурзі) — ген людини,
який відповідає за фактор IX зсідання крові, був уведений
у геном молочної залози овець (1987). Вівці набули здатності
давати молоко, яке містить фактор IX. Цей фактор легко очи
стити з молока і продавати як препарат для лікування гемофілії.
На сьогодні отримано тисячі генетичних конструкцій.
Організмовий рівень організації життя. Основи генетики людини_____ 161
Генна інженерія нині стала основою генної терапії. Генну
терапію в сучасному її розумінні можна визначити як сукуп
ність біомедичних технологій, що ґрунтуються на введенні
хворому генетичного матеріалу. Це перенесення генів ех оіио,
що проводиться зазвичай на клітинах крові; іп зііи, наприклад,
введення в клітини трахеї та бронхів генетичного матеріалу
при муковісцидозі; іп оіоо — введення генетичних конструкцій
безпосередньо в ембріон. Лікувальний ефект генно-терапевтич
них процедур досягається у результаті корекції чи заміни де
фектного гена, екстрахромосомної експресії введеного терапев
тичного гена або пригнічення функції дефектного гена. Першим
прикладом успішної соматичної генної терапії є лікування
у 1990 р. важкого імунодефіциту, обумовленого генетичною
хворобою — дефіцитом аденозиндеамінази. Вірусним вектором,
який містив аденозиндеаміназу, трансфікували Т-лімфоцити,
виділені з крові хворих, а потім вводили лімфоцити назад,
у результаті чого досягався терапевтичний ефект.
2.9. Поняття про імуногенетику
та фармакогенетику
Імуногенетика — наука, яка поєднує імунологію та гене
тику і, зокрема, вивчає спадкову зумовленість груп крові,
типи гемоглобіну, ферментів, білків сироватки крові, молока
тощо. Імуногенетика використовує методи імунології для ви
рішення генетичних завдань.
Так, кожен орган, тканина, клітина і біологічна рідина мі
стять тільки їм властиві антигенні речовини. Антигени успадко
вуються від батьків. Синтез антигенів визначається від батьків
окремими генами, які успадковуються незалежно один від
одного або зчеплено. Впродовж життя антигени залишаються
сталими; вони не змінюються з віком, не залежать від факторів
зовнішнього середовища. У різних організмів одного виду, за
винятком монозиготних близнюків, набір антигенів різний.
В організмі людини у відповідь на введений антиген виробля
ється специфічний захисний компонент — антитіло, яке зв’я
зується з антигеном і нейтралізує його. Антитіла — це білки,
162
Розділ 2
синтез яких контролюється генами. Тому всі імунологічні про
цеси, які відбуваються в організмі, зумовлені спадковістю.
Організм ніколи не виробляє антитіл проти тих антигенів,
які у нього є, а тільки проти чужорідних. Антитіла завжди
специфічні — вони взаємодіють тільки з тими антигенами,
проти яких вони утворилися в організмі. Для виявлення анти
генів використовують спеціальні сироватки, які містять певні
антитіла (моноспецифічні сироватки). Антиген і антитіло взає
модіють між собою, що супроводжується гемолізом, реакцією
осадження (преципітації), відторгнення трансплантату тощо.
Фармакогенетика — наука, яка вивчає генетичні механіз
ми індивідуальної чутливості організмів до лікарських препа
ратів. Вона виникла з розвитку генетики та фармакології і нині
інтенсивно розвивається. Незвичні реакції на лікарські препа
рати часто викликають побічні ефекти і можуть бути навіть
причиною смерті. Встановлено, що в багатьох випадках індиві
дуальні реакції на лікарські препарати обумовлені генетични
ми механізмами, які визначають особливості всмоктування,
розподілу, метаболізму та виведення ліків. Хоча більшість
ліків, які вживають хворі, є безпечні, однак через різноманіт
ність індивідуальної реакції на лікарську терапію побічні
ефекти трапляються нерідко. Індивідуальні особливості можуть
проявлятись як у відсутності терапевтичного ефекту, так і у ці
лому спектрі різноманітних побічних реакцій. Так, в останні
роки у Великій Британії виявлено близько 20 тис. серйозних
несприятливих реакцій на лікарські засоби. Протягом 1998 р.
в СІЛА було госпіталізовано 221,6 тис. осіб через несприятливі
реакції на лікарські засоби. Через такі реакції 1994 р. в СІЛА
померло понад 100 тис. осіб.
Можливість передбачити й уникнути несприятливих уск
ладнень дала би змогу призначати лікарські препарати в опти
мальних дозах, уникати використання інших препаратів.
Вивчення генетичних механізмів, які визначають індивіду
альну чутливість до ліків, і вдосконалення ДНК-технологій
у генетичних дослідженнях дозволяють проводити скринінг
хворих за чутливістю до того чи іншого препарату. Віднедавна
часу ці технології стали звичними у фармацевтичній індустрії
при плануванні й аналізі клінічних випробувань лікарських
засобів.
Організмовий рівень організації життя. Основи генетики людини______163
Рутинне використання генетичних методів для виявлення
індивідуальної чутливості до лікарської терапії стає реаль
ністю.
Фармакогенетична мінливість лікарської відповіді визнача
ється генами, які можна поділити на дві групи. Першу групу
становлять гени, що кодують білки-медіатори, які є частиною
внутрішньоклітинної системи. Ці білки — первинна мішень
лікарських засобів. Друга група генів бере участь у специфіч
них процесах надходження, метаболізму і накопичення. Саме
цю групу генів найбільш інтенсивно вивчає фармакогенетика.
Як уже зазначалось, ця група генів формувалась і розвивалась,
захищаючи багатоклітинні організми в процесі їх еволюції від
шкідливої дії природних токсинів, які надходять з їжею, та
була одним із важливих чинників еволюційного процесу. Також
ці гени беруть участь у надходженні, метаболізмі, накопиченні
лікарських препаратів, які потрапляють в організм ззовні.
Варто зазначити, що існує багато чинників, які не належать
до генетично детермінованої індивідуальної чутливості до фарма
котерапії. Зокрема, мінливість терапевтичного ефекту на ан
тибіотики, протипаразитарну і противірусну терапію пов’ язана
з генетичними змінами в організмі-мішені. Соматичні мутації,
що виникають в організмі, можуть бути причиною лікарської
стійкості при хіміотерапії злоякісних новоутворень. Проте
дослідження в цьому напрямі показали, що фармакогенетичний
поліморфізм може бути важливим чинником, який визначає
результат протимікробної та протипухлинної терапії.
Фармакологічна ефективність терапії залежить від необхід
ної концентрації препарату в заданій ділянці мішені протягом
достатнього для отримання необхідного ефекту часу без виник
нення побічних дій. Механізми взаємодії ліків з мішенню на
поверхні та всередині клітин також є критичними детермінан
тами ефективності ліків.
У людини є гени, що відповідають за захист організму від
зовнішніх хімічних чинників. Вони і визначають чутливість
або резистентність до фармакотерапії. Деякі вчені вважають,
що ці гени не мають значення для виживання виду, будучи,
мабуть, рудиментом, який зникає у процесі еволюції виду.
Інші дотримуються думки про те, що поліморфізм — це
відображення гетерогенності чинників зовнішнього середовища
164
Розділ 2
та наслідки тиску на окремі гени в процесі еволюції, щоб орга
нізми могли подолати певні провокуючі чинники середовища.
Різноманітні хімічні речовини, що надходять до організму
з їжею, повітрям і водою, стимулювали розвиток ферментних
систем детоксикації в процесі еволюції. Біологічна необхідність
їх розвитку полягала у вдосконаленні резистентності й адапта
ції до коливань чинників зовнішнього середовища, що сприяло
збільшенню тривалості життя, захисту від тератогенних, канце
рогенних впливів і хвороб, які індукуються ксенобіотиками.
Оптимальність життєдіяльності кожного організму, як ві
домо, визначається поєднанням його генотипу і довкілля. Вони
і обумовлюють певні рівні активності ферментних систем деток
сикації. Значне погіршення екологічної ситуації веде до пору
шення збалансованої системи “ генотип — середовище” і спри
яє розвитку хвороб навіть у гетерозиготних носіїв мутантних
алелів, особливо при мультифакторних спадкових хворобах.
Такі ж закономірності впливають і на результативність фар
макотерапії — варіантів взаємодії організму і лікарських ксенобіотиків.
На підставі численних спостережень А. Мотульські вперше
з’ ясував (1957), що спадкові дефекти метаболізму можуть по
яснити багато індивідуальних відмінностей в ефективності ліків
і побічних реакцій, які потрібно відрізняти від алергічних
реакцій.
Спочатку дослідження в галузі фармакогенетики не мали
якоїсь системності. Це були самостійні незалежні роботи. Тим
не менш вони мали велике значення в поясненні біохімічної
та генетичної основи багатьох реакцій організмів, які не пере
носять лікарської терапії.
Так, було доведено, що рівень активності сироваткової холінестерази — це ознака, яка генетично детермінована. Спостере
ження показали, що певна кількість пацієнтів пережили про
лонговану анестезію після призначення мієлорелаксанта суксаметоніуму (сукцинілдихоліну).
Суксаметоніум, або сукцинілдихолін широко використову
ють як релаксант м ’язів при хірургічних операціях. Псевдохолінестераза каталізує гідроліз препарату, тому в нормі його
дія нетривала. У інших хворих цей фермент має дуже низьку
спорідненість з препаратом, що призводить до тривалої затрим
Організмовий рівень організації життя. Основи генетики людини______165
ки дихання внаслідок пригнічення діяльності дихальних м’язів.
У таких випадках пацієнтові необхідно вводити очищений
фермент або плазму, що утримує псевдохолінестеразу, а доти
переводити його на штучне дихання.
Причиною пригнічення дихальних м ’ язів є різні мутації як
у гетерозиготному, так і в компаунд-гетерозиготному стані, які
змінюють активний центр псевдохолінестерази, що втрачає
здатність ефективно гідролізувати субстрат. Донині не охарак
теризовано деякі алельні варіанти цього гена і певні молеку
лярні механізми його поліморфізму. Основна атипова форма
ферменту виникає внаслідок заміни аспарагінової кислоти на
гліцерин у ділянці білка, який приєднує субстрат. У цьому разі
не відбувається електростатична взаємодія, і субстрат не при
єднується.
Інший приклад. Під час війни в Кореї на початку 50-х років
X X ст. усі американські солдати проходили профілактичний
курс лікування протималярійним препаратом примахіном.
У 10 % чорношкірих і 1— 2 % білошкірих солдатів із тисячі
у відповідь на прийом примахіну розвивалася судинна гемолі
тична реакція. Раніше схож і реакції спостерігалися при ліку
ванні чорношкірих хворих сульфаніламідами, а також у жи
телів Сардинії після вживання у їжу кінських бобів. Спочатку
це намагалися пояснити патологічними імунними реакціями,
але подальші дослідження показали, що побічна дія (гемоліз)
примахіну пов’язана з недостатністю глюкозо-6-фосфатдегідрогенази.
Незабаром була виявлена ще одна особливість — гемоліз
частіше трапляється у чоловіків, ніж у жінок. Подальше ви
вчення рівня глутатіону в еритроцитах показало, що в афро
американців спостерігається яскраво виражений бімодальний
характер кривої розподілу, причому в значній частині популя
ції рівень вмісту глутатіону був дуже низьким. У групі афроамериканських жінок крива зміщена вліво, а частка хворих із
низьким вмістом глутатіону набагато менша, ніж у групі чоло
віків. Виходячи з цього, було зроблено висновок, що ця ознака
зчеплена з Х-хромосомою. Подальші роботи стосувалися аналі
зу родоводів — генеалогічні дані також підтвердили Х-зчеплений характер успадкування реакції на ці препарати, причому
в цих роботах було використано і прямі тести дослідження
166
Розділ 2
активності ферменту Г-6-ФДГ. Надалі в популяціях людини
було виявлено декілька типів Г-6-ФДГ, які відрізняються ак
тивністю ферменту, електрофоретичною рухливістю в різних
буферних системах, залежністю активності ферменту від рН,
термостабільністю, субстратною специфічністю та іншими па
раметрами. Нині розрізняють близько 400 варіантів Г-6-ФДГ.
Генетичний аналіз ДНК дозволив виявити велику кількість
мутацій Г-6-ФДГ. Найменш активна форма ферменту виникає
в результаті однієї заміни (аспарагіну на аспарагінову кислоту).
Вона успадковується як Х-зчеплена ознака. Більше ЗО алелей
гена має змінену послідовність нуклеотидів. Мутації утворюють
неперервний ряд фенотипів від варіантів із практично незміненими біологічними функціями до тих, які виявляються
в несприятливих умовах, аж до варіантів, що зумовлюють
розвиток захворювання навіть за відсутності несприятливих
чинників. Окрім протималярійних препаратів, Г-6-ФДГ метаболізує широкий спектр лікарських препаратів, у тому числі
хлорамфенікол, сульфаніламіди.
Наступним кроком уперед у вивченні індивідуальної чутли
вості до лікарських препаратів було виявлення генетичної
варіабельності через окиснення лікарських препаратів, опосе
редкованих системою цитохрому Р-450. Після призначення
антипірину і біс-гідроксикумарину з ’явилися перші повідом
лення про індивідуальну мінливість у кліренсі препаратів.
Доказом того, що це генетично обумовлено, було вивчення
підвищеної чутливості до низки гіпотензивних і антиаритмічних засобів, до яких належать дебрисохін і спартеїн.
Індивіди, які здатні швидко окиснювати ці препарати, є
нормальними гомозиготами і гетерозиготами, тоді як повільне
окиснення властиве лише мутантним гомозиготам. Виявити
окиснювальний поліморфізм іп иііго, на жаль, неможливо. Для
визначення окиснювального статусу людини необхідний прийом
контрольного препарату і аналіз його метаболітів у сечі (ви
значення співвідношення 4-оксидебрисохіну до дебрисохіну).
Ці лікарські засоби метаболізуються за участю поліморфного
ферменту СУР2Б6 родини (гідроксилаза дебрисохина) Р-450.
Підвищена чутливість була пов’ язана з відсутністю функції
ферменту внаслідок мутації у відповідному гені. Основні типи
мутацій, які обумовлюють інактивацію ферментів, ідентифі
Організмовий рівень організації життя. Основи генетики людини______1 67
ковані недавно. На підставі отриманих результатів розроблені
ДНК -тести, для виявлення індивідуумів з порушенням мета
болізму СУР2Б6.
У вищенаведених прикладах відображено концептуальні
положення екогенетики загалом і фармакогенетики зокрема.
Основне в них таке:
— поліморфізм є генетичним субстратом, на якому форму
ються сприйнятливість або стійкість до захворювань;
— ефективність або безуспішність лікарської терапії;
— існування достатньо великої кількості мутацій генома
людини в окремих локусах, які обумовлюють патологічну ре
акцію на специфічний чинник середовища; лише деяка части
на мутацій призводить до розвитку хвороби або небажаної
реакції.
Моногенний характер патологічних реакцій існує разом із
полігенними системами, контролюючими екогенетичні від
повіді. У таких випадках докази участі багатьох генів у реакції
на конкретний екологічний чинник будуються на близнюкових
зіставленнях, внутрішньосімейному кореляційному аналізі
досліджуваних фенотипів і на порівнянні характеру реакції
в різних етнічних групах населення.
Нині основний блок наукових досліджень в галузі фармако
генетики складається із праць, які присвячені вивченню гене
тичного поліморфізму ферментів, відповідальних за метаболізм
лікарських засобів.
Таким чином, можливість ідентифікації молекулярних основ
фармакогенетичного поліморфізму і, як наслідок, сканування
індивідуумів на наявність відповідного алеля, відкриває пер
спективи передбачати результати лікарської терапії і дає змо
гу індивідуалізувати лікарську терапію на базі генетичної
конституції. Це найбільш перспективний шлях підвищення
ефективності лікарських засобів і скорочення кількості побіч
них ефектів.
1 68
Розділ 2
2.10. Мінливість у людини. Форми
мінливості. Мутагенні фактори
Мінливістю називають властивість організмів набувати
нових ознак під впливом умов зовнішнього середовища або
внаслідок змін спадкового апарату. Мінливість, як і спад
ковість, властива всій живій природі. Розрізняють неспадкову
і спадкову мінливість.
Неспадкова мінливість — здатність організмів реагувати
на умови зовнішнього середовища, змінювати свій фенотип
у межах норми реакції, заданої генотипом.
Спадкова мінливість — здатність до змін самого генетич
ного матеріалу, які можуть проходити на різних рівнях його
організації. Спадкова мінливість, поділяється на комбінативну і мутаційну.
Мінливість
Неспадкова
(фенотипна, модифікація)
Спадкова
(генотипна)
Комбінативна
Мутаційна
Рис. 2.11. Форми мінливості
Неспадкова мінливість — це фенотипна модифікаційна мінли
вість, яка виникає у процесі індивідуального розвитку організ
мів і не передається нащадкам, не викликає змін генотипу.
Мінливість забезпечує різноманітність за будовою і фізіоло
гічними функціями організмів. Вона є результатом різних
процесів. Деякі з них відбуваються у спадковому матеріалі
(генотипі). Інші обмежуються фенотипом.
Кожний організм у процесі розвитку і життя зберігає при
таманні виду певні властивості, які контролюються спадкові
стю. Вона ніби закріплює рівень розвитку, досягнутий видом
під дією природного добору. Кожна ознака формується у процесі
онтогенезу за дії не одного, а багатьох генів, і є результатом
численних і складних процесів. Так, колір волосся людини або
Організмовий рівень організації життя. Основи генетики людини
1 69
шерсті тварини залежить від складу пігменту, кількості його
у волоссі чи шерсті, характеру розподілу по довжині волосся
чи шерсті тощо.
У вищих тварин і людини на розвиток тієї чи іншої ознаки
ішливає багато генів. За участю значних ділянок молекули
ДІІК синтезуються відповідні ІРНК, а за участю останніх — від
повідні ферменти. Отже, у вищих форм процес розвитку озна
ки характеризується цілим рядом специфічних взаємодій генів,
які входять до генотипу організму. Тому залежно від генотипу
при одному і тому ж гені ознака може зазнавати значних змін.
І(«приклад, масть гомозиготного за геном чорного забарвлення
шерсті корови може бути суцільно чорною або різних відтінків,
сірою, тигровою, кавовою або плямистою у різних ділянках
тіла. Це залежить від інших генів і їх взаємодії між собою.
Проте генотип визначає тільки спрямування розвитку ознаки.
Залежно від умов середовища характер розвитку ознаки може
зазнавати змін. У вищих організмів і людини ембріональний
розвиток відбувається за відносно стабільних умов, тому озна
ки, які формуються до народження, не зазнають істотного
впливу зовнішнього середовища і розвиваються в основному
під впливом генотипу і після народження майже не змінюють
ся. До таких ознак варто віднести форму вух і носа, групи
крові, типи гемоглобіну тощо.
Проте відомі випадки, коли, здавалося б, сталі ознаки за
знають змін під впливом зовнішніх факторів. Так, у горностає
вих кроликів забарвлення тулуба біле, а вуха, кінчик мордоч
ки, хвіст і кінці лапок пігментовані. Таке забарвлення змінить
ся, якщо на різних ділянках тіла шерсть виголити й утримувати
тварин в умовах різної температури.
Висока температура може гальмувати прояв певних ознак
у личинок дрозофіли, викликати захворювання на екзему і за
палення шкіри голови в ягнят тощо.
Усі ознаки, які формуються в постембріональному періоді,
зазнають істотного впливу середовища. Це можна спостерігати
в монозиготних близнюків, які перебувають у різних умовах.
Незважаючи на ідентичність їх генотипу, навіть маса тіла за
знає змін.
Середовище може впливати і на прояв домінантної ознаки. Це
залежить як від спадковості, так і від впливу умов середовища.
170
Розділ 2
Проте існують і такі ознаки, ступінь домінування яких майже
не залежить від умов середовища.
Тому виняткового значення набуває знання критичних пе
ріодів розвитку ембріона та періодів, коли зигота чутлива до
дії різних чинників і легко пошкоджується, що призводить до
розвитку вад.
Кожен організм розвивається за участі генотипу і під, впли
вом зовнішнього середовища. Спадкові ознаки та властивості
виявляються по-різному і залежать від умов, у яких відбуваєть
ся розвиток. Спадковий матеріал при модифікаційній мінли
вості змін не зазнає.
Зміни ознак і властивостей організму в межах норми реакції,
що виникають внаслідок різних умов існування, називаються
модифікаціями.
Модифікації мають адаптивний (пристосувальний) характер
і є важливими для виживання організмів, виконуючи певну
роль у збереженні виду. Крім корисних (пристосувальних),
зміни бувають індиферентні.
Модифікаційна мінливість — явище масове. Вона стосуєть
ся не одного, а сукупності об’ єктів, і тому її ще називають
груповою, або визначеною.
На розвиток ознаки впливають регуляторні системи організ
му, насамперед, ендокринна система. Такі ознаки у півня, як
забарвлення пір’я, величина гребеня та сережок, характер співу
і тембр голосу, зумовлені дією статевого гормону тестостерону.
Введення півням жіночих статевих гормонів гальмує ці ознаки.
Отже, внутрішнє середовище організму має значний вплив на
прояв генів у формі ознак.
Не всі ознаки однаково зазнають модифікаційної мінливості.
Під впливом середовища змінюються маса і розміри тіла, ро
зумові здібності, артеріальний тиск. Проте, наприклад, мор
фологічні ознаки є більш стійкими і менше залежать від екзо
генних чинників. Модифікаційна мінливість характеризується
такими властивостями: 1) неуспадкування; 2) груповий харак
тер змін; 3) відповідність змін щодо дії певного чинника сере
довища; 4) обумовлені генотипом межі змін (хоча напрям змін
однаковий, ступінь їх прояву в різних організмів неоднако
вий).
і ірганізмовий рівень організації життя. Основи генетики людини_____ 171
Модифікаційна мінливість — це закономірне біологічне
чий іце, яке постійно супроводжує розмноження організмів.
Процес розвитку кожної ознаки або властивостей організму
здійснюється на основі генотипу за різних умов середовища.
Тому успадкування ознак або властивостей завжди проявляєтьгн у формі різних його модифікацій.
Модифікаційну мінливість вивчають за допомогою методів
математичної статистики. Вона характеризується варіаційним
рядом. Середня величина варіаційного ряду нагадує точку
ііого рівноваги. Діапазон модифікаційної мінливості обумов
лений нормою реакції.
Генотипна мінливість — основа різноманітності живих
організмів і головна умова їх здатності до еволюційного розвит
ку. Механізми генотипної мінливості різноманітні. Як зазна
чалося, розрізняють дві її основні форми: комбінативну і му
таційну.
Комбінативною називають мінливість, яка залежить від
иерекомбінування алелів у генотипах нащадків порівняно з ге
нотипами батьків, що призводить до появи нових ознак у дітей.
ІІа відміну від перерозподілу спадкового матеріалу нерегуляр
ного типу, при хромосомних абераціях така рекомбінація
н організмів, які розмножуються статевим шляхом, відбуваєть
ся закономірно.
Комбінативна мінливість — найважливіше джерело безмеж
но великого спадкового розмаїття, яке спостерігається у живих
організмів. В основі комбінативної мінливості лежить статеве
розмноження. Генотип нащадків являє собою поєднання генів,
які були властиві батькам. Кількість генів у кожного організ
му дуже велика. При статевому розмноженні комбінації генів
приводять до формування нового унікального генотипу і фено
типу. Комбінативна мінливість широко розповсюджена у при
роді. Навіть у мікроорганізмів, які розмножуються нестатевим
шляхом, виникли своєрідні механізми (трансформація і транс
дукція), що приводять до появи комбінативної мінливості. Все
це вказує на велике значення комбінативної мінливості для
еволюції та видоутворення.
Існує три механізми комбінативної мінливості. Два із них
пов’ язані з мейозом. Завдяки обмінові ідентичними ділянками
1 72
Розділ 2
між гомологічними хромосомами при кросинговері у профазі
мейозу алельний склад хромосом статевих клітин нащадків
зазнає змін порівняно з гаметами батьків. Хромосоми, успад
ковані від обох батьків, у метафазі мейозу комбінуються в різ
них поєднаннях.
Третій механізм комбінативної мінливості пов’ язаний із
заплідненням. Унаслідок випадкового підбору гамет, які беруть
участь у заплідненні, створюються різноманітні поєднання хро
мосом із різним набором генів, а відповідно, й алельного скла
ду зигот. Явище комбінативної мінливості пояснює причину
поєднання у дітей і онуків ознак родичів по батьківській або
материнській лінії. З урахуванням міграційних процесів серед
людей зростає кількість шлюбів між особами, які походять із
віддалених регіонів. Чим більша відстань, тим більша ймовір
ність того, що гамети будуть відрізнятися за набором генів.
Комбінативна мінливість сприяє більшій пристосованості до
мінливих умов середовища, підвищує виживання виду.
Мутаційна мінливість. Термін “мутація” вперше запровадив
Гуго де Фріз 1901 р.
Мутаційна мінливість зумовлена як впливом на організм
чинників зовнішнього середовища, так і його фізіологічним
станом. Частота виникнення мутацій залежить від таких фак
торів:
—
—
—
—
—
—
генотип організму;
фаза онтогенезу;
стать;
стадія гаметогенезу;
мітотичний і мейотичний цикли хромосом;
хімічна будова окремих ділянок хромосом тощо.
Властивості мутацій:
— мутації виникають раптово, стрибкоподібно;
— мутації успадковуються, тобто передаються від покоління
до покоління;
— мутації ненаправлені — зазнавати мутації може будь-який
локус, викликаючи зміни як незначних, так і життєво важли
вих ознак;
Організмовий рівень організації життя. Основи генетики людини
173
— одні і ті ж мутації можуть виникати повторно;
— за проявом мутації можуть бути корисними і шкідливими,
домінантними і рецесивними.
Класифікація мутацій:
1. Мутації за причинами виникнення. Розрізняють спон
танний та індукований мутаційні процеси. Індукований му
таційний процес — це виникнення спадкових змін під впливом
направленої дії факторів зовнішнього і внутрішнього середо
вища. Виникнення мутацій без встановлених причин прийня
то називати спонтанним мутаційним процесом.
2. Мутації за характером прояву. Більшість із мутацій ре
цесивні, тому не проявляються у гетерозигот. Як правило,
мутації шкідливі, бо порушують чітко збалансовану систему
біохімічних перетворень. Домінантні мутації проявляються
відразу у гомо- і гетерозиготного організму: здебільшого такі
особини, будучи нежиттєздатними, гинуть на ранніх стадіях
онтогенезу. Мутації часто знижують життєстійкість або пло
дючість. Мутації, які різко впливають на життєздатність і ча
стково або повністю зупиняють розвиток, називаються напівлетальними, а несумісні з життям — летальними. У людини
до таких мутацій належить рецесивний ген гемофілії.
3. Мутації за місцем виникнення. Мутації, що виникають
у соматичних тканинах, отримали назву соматичних мутацій.
Це зміни спадкового характеру в соматичних клітинах, які
виникають на різних етапах розвитку особини. Вони не пере
даються по спадковості, а залишаються, поки живе організм,
який зазнав мутаційного впливу.
Соматичні мутації обумовлюють генотипну різноманітність
тканин, часто не передаються по спадковості й обмежені тим ін
дивідуумом, у якому вони виникли. Соматичні мутації виникають
у диплоїдних соматичних клітинах, тому проявляються тільки
за домінантними генами або за рецесивними, але в гомозигот
ному стані. Що раніше в ембріогенезі людини виникла мутація,
то більша ділянка соматичних клітин відхиляється від норми,
і навпаки. Наприклад, забарвлення райдужної оболонки ока —
1 74
Розділ 2
білий або карий сегменти на блакитній райдужці — зумовлені
соматичною мутацією. Вважають, що наслідком соматичних
мутацій є ракове переродження. Злоякісний ріст викликаєть
ся канцерогенами, серед яких найбільш негативними є прони
каюча радіація й активні хімічні речовини.
І хоча соматичні мутації не успадковуються, вони знижують
репродуктивні можливості організму, в якому виникли.
Мутації, що виникають у гаметах або клітинах, з яких вони
утворюються, отримали назву генеративних, або гермінальних
мутацій. Що раніше у статевих клітинах виникає мутація, то
більшою буде частка статевих клітин, які нестимуть нову му
тацію. Верхня межа частки клітин, які міститимуть індукова
ну або спонтанну мутацію, становить 50 % .
Існує думка, що найбільша кількість мутацій у статевих
клітинах виникає в овоцитах. Оскільки сперматогонії зазнають
постійного поділу, то серед них може відбуватися відбір проти
мутацій, які зумовлюють шкідливий ефект, і частота мутацій
знижується до часу статевої зрілості. Жінка, навпаки, наро
джується майже з усіма мутантними змінами, в лінії статевих
клітин немає паралельного мітотичного відбору. Овоцити не
тільки не зазнають мітозу, вони залишаються малоактивними
впродовж десятиліть, доки не стануть яйцеклітинами. За цей
період овоцити старіють, стають непропорційно чутливими до
спонтанної мутації.
На статеві клітини найбільше впливають цезій-137, стронцій-90 івуглець-14.
Генеративні мутації при статевому розмноженні передають
ся наступним поколінням. Домінантні мутації з’являються вже
у п е р т о м у поколінні, а рецесивні— тільки у другому та на
ступних поколіннях, з переходом у гомозиготний стан.
4.
Мутації за характером зміни спадкового матеріалу. Ген
ні, або точкові, мутації — зміни, зумовлені заміною, вставкою
або випаданням одного чи кількох нуклеотидів у межах одно
го гена. Вони обумовлюють зміни як у будові білків, так і у функ
ціональній активності білкової молекули (див. розд. 1— 3, 7).
Хромосомні мутації (аберації) — зміни структури хромосом.
Такі мутації можуть виникнути в результаті втрати частини
Організмовий рівень організації життя. Основи генетики людини
1 75
хромосоми (делеція), подвоєння частини хромосоми (дупліка
ція), відриву і повороту частини хромосоми на 180° (інверсія).
Якщо зміна зачіпає життєво важливі ділянки гена, то така
мутація призведе до смерті. Так, втрата невеличкої ділянки
21-ї хром осоми у людини викликає тяж ке захворювання
крові — гострий лейкоз. В окремих випадках відірвана ділян
ка хромосоми може приєднатися до негомологічної хромосоми
(транслокація), що призведе до нової комбінації генів і зміни
їх взаємодії.
Геномні мутації — зміни каріотипу, кратні або некратні
гаплоїдній кількості хромосом. Унаслідок порушення розхо
дження пари гомологічних хромосом під час мейозу в одній із
утворених гамет міститься на одну хромосому менше, а в ін
шій — на одну хромосому більше, ніж за нормального гаплоїд
ного набору. Злиття такої аномальної гамети з нормальною
гаплоїдною гаметою при заплідненні призводить до утворення
зиготи з меншою або більшою кількістю хромосом порівняно
з диплоїдним набором, характерним для цього виду.
Геномні, хромосомні та генні мутації у соматичних клітинах
є наслідком дії мутагенних чинників. У людини це етіологічні
фактори спадкових хвороб. Захворювання, зумовлені геномними (зміна кількості хромосом) і хромосомними (зміна структу
ри хромосом) мутаціями, називаються хромосомними хвороба
ми. Коли зміна кількості хромосом визначається подвоєнням
або зменшенням усього набору хромосом, це призводить до
поліплоїдії або гаплоїдії (відповідно). Поява зайвої або вида
лення однієї чи кількох хромосом призводять до гетероплоїдії
або анеуплоїдії. Зміна структури хромосом — це перебудови
або аберації. При цьому порушується збалансованість набору
генів і нормальний розвиток організму. Як наслідок хромосом
ного дисбалансу відбувається внутрішньоутробна загибель
ембріона або плоду, виникають вроджені вади розвитку. Чим
більша кількість хромосомного матеріалу зазнала мутаційного
ефекту, тим раніше захворювання з’ явиться в онтогенезі і тим
суттєвішими будуть порушення фізичного та психічного роз
витку особини. Характерна риса хромосомного дисбалансу —
множинність вад розвитку різних органів і систем. Хромосом
ні хвороби становлять близько 0,5— 1 % усіх спадкових хвороб
людини.
176
Розділ 2
Генні, або точкові, мутації— це результат молекулярних
змін на рівні ДНК. У людини описані такі види генних мутацій,
які призводять до розвитку спадкових генних хвороб: нісенс,
нонсенс, зміщення рамки зчитування, делеції, вставки (інсерції), порушення сплайсингу, збільшення кількості (експансії)
тринуклеотидних повторів. Мутації ділянок, що транскрибу
ються (які визначають амінокислотну послідовність у молекулі
білка, що синтезується), призводять до синтезу аномального
продукту. Мутації ділянок, що транскрибуються, можуть при
звести до зниження швидкості синтезу білка.
Фенотипно генні мутації виявляються на молекулярному,
клітинному, тканинному і органному рівнях. Генних хвороб
на даний час виявлено близько 3500— 4500.
Генні мутації поділяють на односайтові і багатосайтові. Од
носайтові — це такі, які стосуються змін одного сайту, бага
тосайтові — охоплюють кілька сайтів генного локусу.
Розрізняють генні мутації прямі й обернені. Прямі мута
ції — це мутації, які інактивують гени дикого типу і зумовлю
ють появу мутантного типу. Обернені мутації, або зворот
ні, — це зміни до висхідної форми від мутантної.
Більшість генів стійкі до мутацій, але окремі гени зазнають
мутацій досить часто.
Соматичні мутації зумовлюють генотипну різноманітність
тканин однієї особини і здебільшого не передаються по спадко
вості при статевому розмноженні, якщо організм розвивається
з однієї клітини або групи клітин, у яких виникла мутація, —
такі зміни можуть передаватися нащадкам. Соматичні мутації
використовують в організмів, які розмножуються вегетативно.
Соматичні мутації становлять основу селекції культурних рос
лин.
Мутагенні фактори. Мутагенні фактори поділяються (кла
сифікуються) на фізичні, хімічні біологічні. Кожну із мутацій
викликає певна причина. Здебільшого ці причини невідомі.
Людина зазнавала і зазнає тиску мутацій, які виникають під
впливом тих чи інших чинників.
Чинники, які здатні проникати в соматичні зародкові
клітини живих організмів і викликати мутації, отримали
назву мутагенів.
Організмовий рівень організації життя. Основи генетики людини_____ 1 7 7
Для того, щоб агент вважався мутагеном, він повинен мати
певні властивості: легко проникати в клітини організму, зберіга
ти їх життєстійкість, досягати ядра клітини та впливати на хі
мічну структуру хромосом і процеси, які відбуваються в них.
Вони можуть мати хімічну, фізичну та біологічну природу.
Близько 90 % мутагенів є канцерогенами (здатні викликати
пухлинний ріст).
До фізичних мутагенів належать іонізуюча радіація, рент
генівське випромінення й ультрафіолетові промені. Вони вва
жаються найбільш активними мутагенами.
Усі види випромінення мають високу енергію і у тканинах
організму обумовлюють радіоліз води, утворення гідропероксидів органічних речовин, порушення структури ДНК.
Ультрафіолетові промені належать до електромагнітних
коливань, але іонізації вони не викликають. їхня дія на ор
ганізм пов’ язана з утворенням у тканинах збуджених молекул
і атомів. Залежно від довжини хвилі, УФ-промені мають різну
мутагенну активність. Найбільша вона при довжині хвилі
260 нм, оскільки ДНК поглинає саме цю частину спектру УФ.
Коротші та довші за 260 нм промені менш активні, проникна
активність їх низька. Для отримання мутацій у наукових цілях
широкого застосування набули гамма-промені, промені Рент
гена і нейтрони. Ці види іонізуючого випромінювання дуже
високу проникність.
Дія радіації на живу істоту визначається дозою випромінен
ня і дозою енергії, поглинутої тканинами організму. Доза
випромінення вимірюється у рентгенах, а доза поглинутої енер
гії — в радах.
Іонізуюче випромінення викликає переважно хромосомні
перебудови, які супроводжуються різкими змінами структури
і функції організму. Більшість їх шкідлива.
Хімічних мутагенів надзвичайно багато. Вони можуть впли
вати на спадковість різних експериментальних об’ єктів: фагів,
мікробів, рослин, комах, ссавців. Мутагенну активність про
являють чотири умовні групи хімічних факторів зовнішнього
середовища: пестициди, промислові отрути, харчові добавки,
ліки.
Серед хімічних мутагенів до найсильніших відносять: хло
ровані вуглеводні, етиленамін, діетилсульфат, диметилсульфат,
178
Розділ 2
нітрозоетилсечовину, нітрозометилсечовину, пероксид гідроге
ну, іприт тощо. їх ще називають супермутагенами, оскільки
вони викликають один і той же тип мутацій. Другу групу ста
новлять речовини, які близькі за будовою з азотистими основа
ми нуклеїнових кислот і діють на них: 5-бромурацил, 5-фтордіоксіуредин, 5-бромоксіурацил та ін. До третьої групи нале
жать акридини та їх похідні (акридиновий жовтий, етидій
броміду, профлавін акридинового смарагдового тощо). Вони
здатні зміщувати окремі основи ДНК, або викликати мутації
типу делецій (випадання ділянки ДНК). У четверту групу вхо
дять азотиста кислота, формальдегід, гідроксиламін тощо.
Кожен із хімічних мутагенів може викликати як хромосом
ні розриви, так і генні мутації. Але, на відміну від фізичних
мутагенів, хімічні переважно індукують генні мутації, їх дія три
валіша, мутації проявляються не відразу, а через певний час.
Характер спадкових змін, викликаних дією хімічних чин
ників, залежить від дози мутагену. Невеликі концентрації хі
мічної речовини-мутагену викликають здебільшого дрібні му
тації, які стосуються переважно кількісних ознак; зі збільшенням
дози мутагену зростає ефект мутацій, зумовлений перебудовою
хромосом.
Хімічні мутагени здатні впливати на спадкову основу як
прямо, так і опосередковано.
До хімічних мутагенів можна віднести і деякі лікарські
речовини. Лікарські препарати як хімічні речовини різної
структури є чужорідними для організму людини. Звідси вини
кає проблема їх мутагенності. Як нові, так і давно відомі лі
карські речовини, здатні викликати мутації генів у рослин,
мікроорганізмів, комах, а також у культурі клітин ссавців і лю
дини. Деякі з лікарських препаратів за своїм мутагенним по
тенціалом не поступаються іонізуючій радіації. Так, усі цитостатичні й антимітотичні препарати, які використовуються для
лікування злоякісних пухлин, здатні індукувати генні мутації,
хромосомні аномалії й інші біологічні ефекти, характерні для
іонізуючого випромінювання. Лікарські препарати можуть
викликати широкий спектр мутацій — від точкових до пошко
дження всього хромосомного набору.
Біологічні мутагени — це віруси, бактерії, найпростіші,
токсини гельмінтів, гриби злаків. Вони спричиняють зміни
такого ж характеру, як і фізичні та хімічні мутагени.
Організмовий рівень організації життя. Основи генетики людини
1 79
2.11. Генетична небезпека забруднення
середовища. Поняття про антимутагени
і комутагени
Забруднення середовища мутагенами хімічної чи біологічної
природи може призводити здебільшого до виникнення генних
мутацій. Близько 90 % мутагенів є одночасно канцерогенами,
здатними викликати злоякісні пухлини.
Водночас, відомо понад кілька сотень хімічних речовин, які
мають антимутагенний вплив, тобто здатні захищати геном від
дії мутагенів. Такі речовини — антимутагени — підвищують
стійкість клітин до негативного впливу мутагенів, знижують
кількість пошкоджених мутаціями клітин. Так, р-каротин,
вітамін Е, вітамін С у відповідних концентраціях і співвідно
шеннях знижують мутагенне пошкодження ДНК у лімфоцитах
периферичної крові. Поліфенольні антиоксиданти, які містять
ся у зеленому чаї, знижують частоту хромосомних аберацій.
Прийом вітамінів А і С у відповідних дозах протягом місяця
знижує аномально високий рівень спонтанного мутагенезу
у людей, які пов’ язані з виробництвом молібдену. Встановлено,
що захисний антимутагенний ефект тих чи інших речовин
залежить від часу їх введення, стадії клітинного циклу, кон
центрації тощо.
На відміну від мутагенів, комутагени — це речовини, які
посилюють дію мутагенів, хоча самі по собі не спричиняють
мутацій. До них можна віднести кофеїн, який активує індук
цію метотрексатом сестринських хроматидних обмінів; верапаміл, який посилює кластогенну дію блеоміцину; аскорбінову
кислоту — збільшує пошкодження ДНК циклофосамідом. На
явність у середовищі комутагенів може підвищувати негативні
ефекти фізичних, хімічних чи біологічних мутагенів.
Розділ З
ІНДИВІДУАЛЬНИЙ РОЗВИТОК
Онтогенез — індивідуальний розвиток живого організму
з моменту запліднення яйцеклітини (утворення зиготи) і до
смерті. При статевому розмноженні розрізняють передзародковий період — формування статевих клітин і їх запліднення
(або розвиток без запліднення — партеногенез), ембріональний
і постембріональний розвиток.
Існують непрямий (личинковий) і прямий (неличинковий,
внутрішньоутробний) типи онтогенезу.
Личинковий тип розвитку трапляється у тих видів організ
мів, яйця яких бідні на жовток (більшість безхребетних, деякі
хребетні). Личинка веде вільний або паразитичний спосіб жит
тя і перетворюється на дорослу форму.
Неличинковий тип розвитку характерний для риб, плазунів,
птахів, а також безхребетних, яйця яких мають багато жовтка,
що дає можливість завершити зародковий розвиток.
Внутрішньоутробний тип розвитку характерний для вищих
ссавців і людини. Яйцеклітини при цьому типі розвитку май
же не мають поживних речовин. Усі життєві функції зародка
здійснюються через материнський організм, який найкращим
чином забезпечує не тільки виживання зародка, але й необхід
ність вигодовування новонароджених молоком.
Індивідуальний розвиток
181
Онтогенезові передує гаметогенез — процес утворення чо
ловічих і жіночих статевих клітин-гамет, які необхідні для
виникнення нового організму. У разі злиття чоловічої та ж і
ночої статевих клітин утворюється одноклітинний зародок —
зигота. З цього моменту починається ембріональний розвиток,
або розвиток організму до народження, який у людини в нормі
триває близько 280 діб (40 тижнів). У ембріональному розвит
ку розрізняють початковий період, який охоплює 1— 7 діб.
У цей період багатоклітинний організм — бластоциста в кінці
початкового періоду вростає у стінку матки (здійснюється про
цес імплантації).
З другого до восьмого тижня триває зародковий період роз
витку, під час якого у складі зародка з ’ являються зачатки
тканин, органів і систем. Плодовий період пренатального он
тогенезу триває з третього до дев’ятого місяця і завершується
народженням дитини. У цей період здійснюються процеси
подальшого структурного і функціонального становлення тка
нин і органів плода, а також диференціації клітин, які їх ут
ворюють.
Постембріональний онтогенез розпочинається з моменту
народження дитини і закінчується смертю організму.
Онтогенез зумовлений тривалим процесом філогенетичного
розвитку. У кінці XIX ст. було сформульовано основний біоге
нетичний закон (закон Геккеля — Мюллера), згідно з яким
пренатальний онтогенез є коротким повторенням філогенезу,
тобто індивідуальний розвиток зародка у скороченому вигляді
повторює історичний розвиток свого виду.
3.1. Основні закономірності ембріонального
розвитку
Ембріональний період ділиться на стадії зиготи, дроблення,
бластули, утворення зародкових листків, гісто- і органогенезу.
Зародки ссавців і людини до утворення зачаткових органів
прийнято називати ембріоном, а потім — плодом.
Запліднення у людини здійснюється в матковій трубі. У резуль
таті злиття чоловічої (сперматозоїда) та жіночої (яйцеклітини)
1 82
Розділ З
статевих клітин виникає одноклітинний зародок. У цитоплазмі
заплідненого овоцита здійснюється низка послідовних змін.
Зокрема, завершується другий поділ дозрівання, у результаті
якого з овоцита II утворюється зріла яйцеклітина і одне поляр
не тільце. Ядро сперматозоїда після проникнення в овоцит
рухається вперед і перетворюється у чоловічий пронуклеус. Із
ядра яйцеклітини формується жіночий пронуклеус. Кожен із
пронуклеусів реплікує свою ДНК, після чого вони втрачають
ядерні оболонки й утворюють загальну метафазну пластинку
зиготи з веретеном поділу. Таким чином забезпечується дип
лоїдність клітин майбутнього зародка. Взаємодією пронуклеусів
завершується власне запліднення. У складі шийки спермато
зоїда в яйцеклітину потрапляє центріоль, необхідна для поділу
зиготи. Окрім батьківських хромосом, сперматозоїд вносить
у новий організм свій мітохондріальний геном разом із мітохондріями, а також сигнальний білок дроблення.
Дроблення — наступний після запліднення період життя
зародка, котрий включає низку послідовних мітотичних
поділів, у результаті яких зигота перетворюється в багатоклі
тинний організм.
Залежно від особливостей будови яйцеклітини дроблення
буває повним (охоплює всю зиготу) і неповним (борозна дроб
лення зміщується до одного з полюсів клітини). Неповне дроб
лення може бути поверхневим або дискоїдальним. Для яйцек
літини людини характерне повне нерівномірне асинхронне
дроблення.
Дроблення зиготи людини охоплює проміжок з першої до
кінця шостої доби. Характерною його особливістю є дуже ко
роткий період інтерфази між двома послідовними мітозами,
внаслідок чого розміри новоутворених клітин зародка у період
дроблення мають назву бластомерів. Дроблення завершується,
коли розмір бластомерів наближається до розміру соматичних
клітин організму.
Бластомери після третього поділу, приблизно через три до
би після запліднення, утворюють клітинну компактну масу.
На цій стадії зародок має назву морули. Периферійні клітини
утворюють бар’єр, який обмежує внутрішнє середовище мору
ли. Вони формують зовнішню клітинну масу (трофобласт). На
Індивідуальний розвиток__________________________________________ 1 83
четверту добу після запліднення зародок потрапляє у матку.
В цей час у ньому виникає порожнина — бластоцель (первинна
порожнина тіла), заповнена рідиною, яка транспортується
всередину зародка з його мікрооточення клітинами трофобласта. Ембріон тепер називається бластоциста. Він має вигляд
пухирця, стінку якого утворюють видовжені клітини трофобласта. Всередині міститься бластоцель, на одному з полюсів на
внутрішній поверхні трофобласта локалізована внутрішня
клітинна маса, з якої розвивається власне зародок і деякі позазародкові органи (часткове або повне розділення внутрішньої
клітинної маси приводить до утворення близнюків). До кінця
четвертої доби розвитку ензими розчиняють оболонку заплід
нення, створюючи цим передумови для наступної імплантації.
Імплантація — процес вростання зародка у слизову оболонку
матки, який здійснюється на шосту—сьому добу розвитку.
Гаструляція — період ембріогенезу, коли виникають за
родкові листки — ектодерма, ендодерма, мезодерма — і зародок
набуває тришарової будови.
Спосіб гаструляції у різних тварин залежить від структури
яйцеклітини (кількості та розміщення в ній жовтка) й умов
розвитку зародка. В одних випадках ендодерма утворюється
шляхом впинання (інвагінації) певної ділянки стінки бластули
всередину її порожнини або “ вселення” (імміграції) сюди окре
мих клітин із поверхні зародка. Якщо ж клітини майбутньої
ендодерми перевантажені жовтком і не здатні до пересування,
то відбувається обростання (епіболія) їх дрібнішими клітинами
протилежної півкулі бластули. У деяких тварин ектодерма
й ендодерма виникають шляхом повного та часткового розщеп
лення (делямінації) стінки бластули на два шари.
У людини гаструляція відбувається шляхом імміграції та
делямінації. Гаструляція у людини охоплює період з 14-ї по
17-ту добу пренатального онтогенезу.
У процесі гаструляції виділяють два етапи: а) утворення
екто- і ендодерми (двошаровий зародок); б) утворення мезодер
ми (тришаровий зародок). Вегетативний полюс бластули впи
нається всередину, протилежні полюси бластодерми майже
змикаються, бластоцель або зникає зовсім, або залишається
у вигляді незначної порожнини, а з кулі виникає двошаровий
184
Розділ З
зародок: ектодерма й ендодерма. Порожнина називається гастроцелем, або первинною кишкою, а вхід у кишку — бласто
пором, або первинним ротом. Його краї зближуються, утворю
ючи верхню і нижню губи. Гаструляція відбувається й іншими
способами: делямінацією (розшаруванням), епіболією (обро
станням) та імміграцією (проникненням усередину).
Третій, або середній, зародковий листок називається мезо
дермою. У хордових із двох боків від первинної кишки утво
рюються випини — кишені (целомічні мішки). Всередині ки
шень міститься порожнина. Целомічні мішки відшнуровуються від первинної кишки і розростаються між екто- і ендодермою.
Клітинний матеріал цих ділянок дає початок мезодермі. Дор
зальний відділ мезодерми, який лежить по боках нервової
трубки і хорди, розчленований на сегменти — соміти. Вен
тральний її відділ утворює суцільну бокову пластинку, яка
розташована з боків кишкової трубки.
Гістогенез і органогенез здійснюється в результаті розмно
ження, міграції, диференціації клітин, що його складають,
встановлення міжклітинних контактів і загибелі частини клі
тин. З 17-ї по 20-ту добу триває пресомітний період, із 20-ї
доби починається сомітний період розвитку, коли одночасно
дорсальні ділянки мезодерми зародка діляться на окремі сег
менти, розташовані з обох боків х ор д и — соміти. На 21-шу
добу в організмі зародка є дві—три пари сомітів, а у кінці
п’ятого тижня кількість сомітів в ембріоні досягає 43— 44 пар.
На основі підрахунку числа сомітів можна приблизно визна
чити термін розвитку (сомітний вік) ембріона.
Зі зовнішньої частини кожного соміта виникає дерматом, із
внутрішньої — склеротом, зі середньої — міотом. Дерматом
стає джерелом розвитку дерми шкіри, склеротом — хрящової
та кісткової тканин, міотом — скелетних м’ язів спинної частини
зародка. Вентральні ділянки мезодерми — спланхнотом — не
сегментуються, а поділяються'на вісцеральний і парієтальний
листки, із яких розвиваються серозні оболонки внутрішніх
органів, м ’ язова тканина серця та кора надниркових залоз.
З мезенхіми спланхнотома утворюються кровоносні судини,
клітини крові, сполучна та гладка м ’ язова тканини зародка.
Ділянка мезодерми, яка зв’ язує соміти зі спланхнотомом,
служить джерелом розвитку нирок і статевих залоз, а також
Індивідуальний розвиток__________________________________________ 1 8 5
парамезонефральних проток. З останніх утворюється епітелій
матки і яйцеводів.
У процесі диференціації зародкової ектодерми утворюється
нервова трубка, нервові гребені, плакоди, шкірна ектодерма
та прехордальна пластинка. Процес формування нервової труб
ки має назву нейруляції. Він полягає в утворенні щілиноподіб
ної заглибини на поверхні ектодерми; потовщені краї цієї за
глибини зростаються з утворенням нервової трубки. З краніаль
ної частини нервової трубки формуються мозкові пухирці, які
є зачатком головного мозку. Клітини нервових гребенів, мігру
ючи у напрямку дерматоми, дають початок пігментним кліти
нам — меланоцитам; мігруючи у напрямку черевної порожни
ни, дають початок симпатичним і парасимпатичним нервовим
вузлам, мозковій речовині надниркових залоз. Із клітин не
рвових гребенів, які не мігрували, утворюються гангліозні
пластинки, з яких розвиваються спинномозкові та периферій
ні вегетативні нервові ганглії. Із плакод формуються ганглії
голови та нервові клітини органів слуху та рівноваги. Прехор
дальна пластинка є джерелом розвитку епітелію трахеї, брон
хів, легень, а також ротової порожнини і стравоходу.
Зі шкірної ектодерми формуються епідерміс і його похідні,
емаль і кутикула зубів, епітелій пристінка ротової порожнини,
анального відділу прямої кишки та піхви. Ендодерма зародка
є джерелом утворення епітелію середньої частини кишкової
трубки, печінки та підшлункової залози.
Маса зародка до кінця восьмого тижня досягає 5 г, до цьо
го часу в ембріона вже сформовані зачатки всіх органів.
З початком третього місяця розвитку завершується зарод
ковий і розпочинається третій, плодовий, період пренатально
го онтогенезу.
Зв’ язок зародка зі середовищем забезпечується спеціальни
ми позазародковими органами, які функціонують тимчасово
мають назву провізорними. До них належать плацента, амніон,
пуповина, жовтковий мішок і алантоїс. Вони формують транс
портну систему, яка забезпечує постачання плода поживними
речовинами і киснем, видалення продуктів обміну речовин,
продукують гормони, виконують функцію імунного захисту.
Жовтковий мішок закладається на ранніх етапах ембріоге
незу, але не розвивається, а поступово редукується. Алантоїс
186
Розділ З
також не розвинений. Зачаток його входить до складу нового
пупкового канатика. Функцію зовнішньої зародкової оболонки
виконує хоріон, або ворсиста оболонка. Численні вирости,
ворсинки хоріона, вростають у слизову оболонку матки — спеці
ального органа материнського організму, який є тільки у ссав
ців. Місце найбільшого галуження ворсинок хоріона і найбільш
тісного контакту їх зі слизовою оболонкою матки має назву
дитячого місця, або плаценти.
Зв’язок тіла зародка з плацентою здійснюється через пупо
вину або пупковий канатик, який містить кровоносні судини.
Кровоносні капіляри тіла зародка галузяться у ворсинках
хоріона. Так встановлюється плацентарний кровообіг. Через
плаценту плід постачається поживними речовинами, киснем
і звільняється від продуктів життєдіяльності. Епітелій хоріона
і стінок судин утворює специфічний клітинний бар’єр; мікро
організми і ряд речовин із кровотоку матері у нормі не потрапля
ють у кровоток плоду. Плацента є бар’ єром для деяких лікар
ських речовин, у тому числі наркотиків, виробничих і харчових
отрут, чужорідних білків і антитіл.
Навколо плоду розташована суцільна оболонка, яка нази
вається амніоном і бере участь у виробленні навколоплодових
вод. Ці води містять антитіла, які беруть участь у захисті за
родка від дії хвороботворних чинників.
Хребетні, які мають амніон (плазуни, птахи і ссавці), об’єд
нуються у групу вищих хребетних, або амніот. Нижчі хребет
ні, які не мають амніону (круглороті, риби і земноводні), на
лежать до групи анамній.
Механізми розвитку організму. При поділі зиготи утворюєть
ся два бластомери, кожен із яких може розвинутися в повно
цінний організм. На цій стадії вони тотипотентні, тобто рівноспадкові. Було встановлено, що така тотипотентність у трито
на зберігається до стадії 16 бластомерів, у кроля — до стадії
4 бластомерів. Про існування подібної тотипотентності у бла
стомерів людини свідчать випадки народження двох, чотирьох
і навіть семи близнят. Надалі спостерігається диференціація,
тобто формування різноманітних структур і частин тіла. При
цьому клітини зберігають генетичну інформацію і кожна со
матична клітина, яку б спеціалізовану функцію вона не вико
Індивідуальний розвиток__________________________________________1 87
нувала, несе генотип цілого організму. На хід диференціюван
ня впливають три фактори — ядро, цитоплазма і зовнішні
фактори середовища.
Незважаючи на велику різноманітність клітин тіла, усі во
ни мають однакову кількість хромосом, тому що гени при
мітозі рівномірно розподіляються між утвореними клітинами.
В одній клітині — зиготі — закладена інформація розвитку
особини. Які механізми її реалізації?
Ще американський біолог Т. Морган (1866— 1945) припустив,
що в кожній клітині тіла функціонують не всі гени, а тільки
певна їх частина, решта генів неактивні. Ця теорія сьогодні
загальноприйнята. Її підтверджують генні мутації, які визна
чаються одним геном або групою генів. Різні летальні мутації
у дрозофіл призводять до смерті у різні періоди розвитку. Так,
одні з них викликають смерть у яйці, інші — на личинковій
стадії, треті — на стадії імаго. Це свідчить про активність різ
них генів на різних етапах індивідуального розвитку. Одні
гени “ включаються” , інші — “ виключаються” . Періодичну
активність окремих ділянок хромосом можна спостерігати під
мікроскопом на “ гігантських” хромосомах слинних залоз ли
чинок дрозофіл та інших двокрилих у вигляді здуттів (пуфів),
які виникають у результаті деспіралізації хромосомних ниток
в певних ділянках хромосом. Розташування цих здуттів змі
нюється на різних стадіях розвитку. Вони різні також і в клі
тинах різних тканин.
Отже, у різні періоди розвитку організму, а також у різних
частинах його тіла функціонують різні гени. Діяльність генів
регулюється білками-гістонами, які входять до складу хромо
сом. Ці білки вкривають значну частину молекул ДНК. Синтез
іРНК відбувається тільки у тих ділянках ДНК, які не закриті
гістонами. Речовини, які надходять із цитоплазми у ядро,
вивільняють певні ділянки ДНК від гістонів. Встановлено, що
на хромосомний апарат клітини мають вплив гормони.
Отже, гени регулюються білками-гістонами і гормонами.
У результаті діяльності генів утворюються білки-ферменти.
Процес онтогенезу — ланцюг реакцій, які регулюються за
принципом зворотного зв’ язку. В цьому ланцюзі нагромаджен
ня певних речовин, які утворюються в результаті діяльності
генів, може або гальмувати, або стимулювати функцію генів.
1 88
Розділ З
Гени діють через кодовані ними ферменти. Це можна сформу
лювати так: ген (ДНК) — іРНК — білок (фермент) — ознака.
На молекулярному рівні реалізація ознаки проходить такі
етапи: транскрипція — іРНК — процесинг — тРНК — транс
ляція — утворення білків і їх участь у формуванні ознаки. На
кожному з цих етапів можливий вплив інших генів. Саме цим
пояснюється існування генів-модифікаторів, епістазу, гено
копій.
У 1924 р. проведеними ембріологічними експериментами,
які стали класичними, Г. Шпеман і X . Мангольд показали, що
диференціювання значною мірою контролюється впливом цито
плазми клітин одного типу на клітини другого типу. В одному
зі своїх експериментів автори брали гаструлу амфібії, вирізали
шматочок тканини із дорсальної губи, які при нормальному
розвитку утворюють хорду та мезодермальні соміти (міотоми).
У цьому досліді в гаструли-реципієнта з тканин транспланта
(донора) розвивалася друга хорда і міотоми. Над ними із екто
дерми реципієнта виникала нова, додаткова нервова трубка,
що приводило до утворення органів другого пуголовка. Струк
тури, які утворилися в експерименті на незвичному для них
місці, називають атопічними.
На основі цих даних автори висунули гіпотезу про механізм
диференціювання, який отримав назву ембріональної індукції.
Згідно з цією гіпотезою, певні клітини діють як організатори
на інші клітини, які для цього підходять. Організат' о спонукає
такі клітини до розвитку у напрямі, відмінному від того, в яко
му вони розвивались би за відсутності організаторів.
У процесі ембріонального розвитку організму певна ділянка,
яка називається організаційним центром, детермінує подаль
ший розвиток. У амфібій — це дорсальна губа бластопора,
у птахів — ділянка, яка називається первинною смужкою.
Згодом були виявлені організаційні центри для розвитку інших
органів. Реалізація індукції можлива тільки за умови, що
клітини реагуючої системи здатні сприймати вплив, тобто є
компетентними. Компетенція виникає на певних стадіях роз
витку і зберігається протягом обмеженого часу.
Експерименти з пересадкою частин зародків, які перебували
на різних стадіях розвитку, показали важливу роль взаємо
впливу його частин. Виявилося, що частини зародка, із яких
Індивідуальний розвиток
189
у нормі формуються одні органи, при перенесенні на нове міс
це дають початок іншим органам, тобто тим, які мають утво
рюватися на цьому місці. Такий розвиток називається залеж
ною диференціацією. Згодом, коли диференціація уже почала
ся, пересадка зачатків органа на іншу ділянку зародка не
змінює його розвитку, бо характер його уже визначений. У та
ких випадках кажуть про незалежну, стабільну диференціацію.
У розвитку органів звичайно спостерігається перехід від залеж
ної диференціації до незалежної.
На формування частин зародка впливають фізичні та хіміч
ні фактори середовища, а також синтез гормонів і певних біл
ків, які характерні для морфологічних структур. Саме вони,
мабуть, на певному етапі розвитку визначають напрям морфогенезу.
Це переконливо показано в експериментах Г. В. Лопашова
(1968), присвячених розвиткові структури ока амфібій. Вия
вилося, що сітківка ока і пігментований епітелій тривалий час
можуть перетворюватися одне в одного. Напрям їхнього роз
витку залежить від певних зовнішніх умов. За нормального
розвитку очного міхура у зовнішній стінці, яка щільно приля
гає до ектодерми покривів тіла, відбувається нагромадження
продуктів життєдіяльності та ускладнюється постачання кис
ню. Цим визначається напрям розвитку очного міхура у сіт
ківку. Інші умови створюються у стінці міхура, оберненій до
мозку. Тут тонкий шар клітин омивається зовнішнім середо
вищем, продукти життєдіяльності видаляються, кисень має
вільний доступ. Це сприяє утворенню пігментної оболонки.
Також було доведено, що диференціюванню тканин і утво
ренню органів передує синтез гормонів і певних білків. Саме
вони, мабуть, на даному етапі розвитку визначають напрям
морфогенезу. Це можна проілюструвати на прикладі формуван
ня статевих клітин. У хребетних під час ембріогенезу спочатку
закладаються індиферентні зачатки гонад, але згодом відбу
вається диференціація або в одному, або в іншому напрямі. При
цьому у риб і амфібій жіночими і чоловічими статевими гор
монами можна не тільки прискорити розвиток відповідно
яєчників або сім ’яників, але також і гормонами протилежної
статі вдається перевизначити генетичну стать. Такий результат
був досягнутий у дослідах на акваріумних рибках гупі.
1 90
Розділ З
Критичні періоди розвитку. У процесі онтогенезу існують
періоди підвищеної чутливості організму до ушкоджувальної
дії чинників зовнішнього середовища. Ці періоди отримали
назву критичних періодів розвитку. Вперше поняття критичних
періодів розвитку було сформульоване австрійським лікарем
Н. Грегом у 1944 р. Значний внесок у розробку положень теорії
критичних періодів зробив ембріолог П. Г. Свєтлов.
Підґрунтям для виникнення критичних періодів є перехід
організму зародка від одного морфофункціонального етапу до
наступного, якісно відмінного від попереднього. Якісна пере
будова організму при цьому супроводжується проліферацією,
детермінацією та диференціацією клітин, що є його складни
ками. Такими періодами підвищеної чутливості у гаметогенезі
є мейоз (стадія дозрівання статевих клітин), а також процес
запліднення. У пренатальному онтогенезі до критичних періодів
відносять імплантацію (6— 7-ма доба), плацентацію і розвиток
осьових зачатків органів (3—8-й тиждень), період посиленого
розвитку головного мозку (15—20-й тиждень), період форму
вання основних функціональних систем організму (20— 24-й
тиждень), а також процес пологів. У постнатальному онтогенезі
до критичних періодів розвитку належить період новонародженості (перший рік життя дитини), період статевого дозрі
вання (11— 16 років).
Порушення природного ходу ембріогенезу призводить до
формування каліцтв або вад розвитку. Вадами називаються
стійкі відхилення в будові органа або цілого організму, які
призводять до функціональних порушень. Вони полягають
у відсутності органу або його частин, недорозвитку органів,
зменшенні маси тіла, непропорційному збільшенні маси орга
ну, збільшенні довжини тіла. Причини, механізми і шляхи
профілактики їх вивчає наука тератологія.
За даними ВООЗ, вади розвитку характерні для в 1— 2 %
людей.
Залежно від причини, яка призводить до неправильного
розвитку, розрізняють вади генетичної, екзогенної (зовнішньої)
і комбінованої (мультифакторіальної) природи.
В основі вад генетичної природи лежать зміни структури
і кількості хромосом, мутаційні зміни спадкового матеріалу.
Екзогенні вади виникають через з дією на зародок пошкоджу
ючих зовнішніх факторів, які поділяють на фізичні (іонізуюче
Індивідуальний розвиток__________________________________________ 191
випромінювання, вібрація, високі температури, механічні
впливи, гіпоксія), хімічні (лікарські препарати, гормони, ал
коголь, наркотики, гербіциди) і біологічні (неповноцінне хар
чування, хвороби матері, інфекційні захворювання: грип,
краснуха, токсоплазмоз; токсини гельмінтів, гриби злаків).
Причиною мультифакторіальних вад є поєднання дії на процес
розвитку генетичних і екзогенних факторів. Впливи, які спри
чинюють вади розвитку ембріона, називають тератогенними.
3.2. Основні закономірності
постембріонального розвитку
Після народження або виходу організму з яйцевих чи зарод
кових оболонок починається постембріональний, або постнатальний, етап онтогенезу, протягом якого відбувається подаль
ший його розвиток. У різних видів тварин постнатальний етап
життя може тривати від кількох днів до десятків років. Три
валість індивідуального життя — видова ознака, яка не зале
жить від рівня організації.
Постембріональний онтогенез людини можна розділити на
такі періоди: ювенільний (до статевого дозрівання); зрілий
(дорослий, статевозрілий стан); період старості, який закін
чується природною смертю.
Ювенільний період — залежно від типу онтогенезу харак
теризується прямим або непрямим розвитком.
При прямому розвитку організми виходять з яйцевих обо
лонок або є новонародженими, відрізняються від дорослих форм
переважно розмірами, а також недостатнім розвитком органів
і пропорцій тіла.
При непрямому розвитку личинка зазнає перетворень, які
називаються метаморфозом. Личинка, яка різко відрізняється
від дорослої форми, проходить стадії повного метаморфозу,
а та, яка схожа на статевозрілу форму та зазнає линяння, розви
вається з неповним метаморфозом. Метаморфоз досить пошире
ний у представників різних типів тварин. Трапляється не тіль
ки у безхребетних тварин (кишковопорожнинних, плоских і круг
лих червів, членистоногих), але і у хордових (земноводних).
192
Розділ З
Однією з найхарактерніших рис онтогенезу є збільшення
розмірів організму, який розвивається, тобто його ріст. Ріст
пов’ язаний зі збільшенням кількості клітин і нагромадженням
маси позаклітинних утворів. За характером росту всіх тварин
можна поділити на дві групи — з визначеним і невизначеним
ростом.
Невизначений ріст спостерігається у молюсків, ракоподіб
них, риб, земноводних, плазунів та інших тварин, які ростуть
протягом усього життя. Визначений ріст властивий організмам,
які при досягненні певного віку рости перестають, наприклад,
комахи, птахи, ссавці.
Ріст характеризується рядом закономірностей, які вивчали
зоологи К. Майнот і І. Шмальгаузен.
Досліджено, що інтенсивність росту більша на початку он
тогенезу, а потім поступово знижується, у різні періоди індиві
дуального розвитку вона неоднакова. В онтогенезі спостері
гається чергування періодів росту і диференціації. Коли в од
ному органі переважає ріст, то в іншому — процеси диференціації,
і навпаки. Диференціація спричинює певні якісні зміни клітин,
які зумовлюють зниження або повну втрату їх здатності до
розмноження (нервові клітини не здатні розмножуватись).
Онтогенез людини відрізняється від розвитку інших видів
і приматів тривалішим періодом дитинства, бо в цей час відбу
вається не тільки фізичний розвиток організму, але й станов
лення особистості.
Багато фізичних ознак людини формується у процесі роз
витку специфічних для нього функцій. Хребет новонародже
ного за формою відрізняється від хребта дорослої людини.
Типова форма хребта, яка має два вигини вперед (лордози) і два
вигини назад (кіфози), розвивається у процесі переходу до
прямоходіння. Першим виникає шийний л ор д оз— у віці
2,5— 3,0 місяці, коли дитина починає тримати голівку. На
ш остому місяці, коли дитина починає сидіти, формується
грудний кіфоз. При освоєнні стояння і ходіння починає утво
рюватися поперековий лордоз. У 1,5—2,0 роки повністю сфор
мовані всі вигини хребта. Дитина відрізняється від дорослої
людини відносно короткими ногами, довгим тул ’бом, великою
головою.
Ріст супроводжується збільшенням довжини і маси тіла.
Найінтенсивніший ріст спостерігається на першому році жит
Індивідуальний розвиток
193
тя, коли довжина тіла дитини збільшується в середньому на
23—25 см. У подальшому темп росту сповільнюється: 10— 11 см
протягом другого року і 8 см протягом третього року життя.
Подібна закономірність простежується і в нарощуванні маси
тіла.
Ріст — кількісна ознака, що має полігенний характер успад
кування. Такі ознаки здебільшого залежать від умов середо
вища. Для нормального росту необхідне повноцінне харчуван
ня, яке б містило повний набір амінокислот, вітамінів, зокре
ма ретинолу (віт. А), кальциферолу (віт. Б), вітамінів групи
В, мінеральні солі та мікроелементи, а також такі фактори
середовища, як кисень, температура і світло. На ріст організму
мають вплив і гормони, а саме гормон росту — соматотропін,
який виділяється передньою часткою гіпофізу. У людини може
спостерігатись як підвищення функції передньої частки гіпо
фізу, що призводить до гігантизму (у чоловіків більше 200 см),
так і зниження функції — розвивається гіпофізарна карли
ковість (нанізм). При нанізмі ріст уповільнюється, але частини
тіла зберігають нормальні пропорції.
На ріст і статеве дозрівання мають вплив також гормони
епіфізу. При його гіпофункції ріст затримується, відбувається
раннє статеве дозрівання з передчасним розвитком вторинних
статевих ознак і раннім розумовим розвитком. Гормони щито
подібної залози підвищують загальну інтенсивність обміну
речовин, впливають на ріст і розвиток хребетних тварин і лю
дей. При гіпофункції або повній атрофії щитоподібної залози
у дитячому віці спостерігається кретинізм, який проявляється
у затримці росту, недорозвитку статевих органів і вторинних
статевих ознак, недоумкуватості. Гіпофункція щитоподібної
залози викликає мікседему — захворювання, при якому спосте
рігаються знижений обмін речовин, порушений водно-сольовий
обмін. Гіперфункція зумовлює розвиток базедової хвороби, при
якій підсилюється обмін речовин, що призводить до різкого
схуднення, підвищення збудливості нервової системи.
Протягом останніх 100— 150 років спостерігається приско
рення соматичного розвитку та фізіологічного дозрівання дітей
і підлітків. Це явище називають акселерацією. Вже на стадії
внутрішньоутробного розвитку довжина тіла новонароджених
збільшується на 0,5— 1,0 см, а м аса— на 50— 100 г. Значне
194
Розділ З
прискорення розвитку спостерігається і в немовлят: подвоєння
маси відбувається швидше, раніше прорізуються зуби. У більш
ранньому віці збільшується маса м’ язів, закінчується ріст. На
сьогодні у більш ості дівчат ріст припиняється у віці 16—
17 років, у ю наків— 18— 19 років. Існують різні пояснення
явища акселерації: зміни магнітного поля Землі, підсилення
дії іонізуючої, сонячної радіації, поліпшення харчування, ус
піхи профілактики та гігієни тощо.
Зрілий вік настає у чоловіків у 22 роки, у жінок — у 21 рік.
Перший період зрілого віку — до 35 років, другий період — від
36 до 60 років у чоловіків і до 55 років у жінок.
У віці ЗО—35 років, коли завершується перший період зріло
го віку, виявляються деякі зміни фізіологічних реакцій, зміни
обміну речовин. Упродовж п’ятого десятиріччя відбуваються
зміни, які визначають процес старіння. Водночас із цими змі
нами вмикаються і механізми, які забезпечують перебудову
організму та його адаптацію. Похилий вік починається з 61 року
у чоловіків і з 56 років у жінок. Багато людей зберігають у цей
період достатню професійну працездатність.
Старість — загальнобіологічна закономірність, притаманна
всім живим організмам. Старість — заключний етап онтогене
зу, віковий період, який настає за зрілістю і характеризується
суттєвими структурними, функціональними і біохімічними
змінами в організмі, що обмежують його пристосувальні мож
ливості.
На рівні організму зміни при старінні проявляються перш за
все у зовнішніх ознаках: змінюються постава, форма тіла, змен
шуються його розміри, з’являється сивина, шкіра втрачає ела
стичність, що призводить до утворення зморшок. Спостерігається
ослаблення зору і слуху, погіршення пам’яті. Зменшуються
компактна і губчаста речовини кісткової тканини, зокрема, це
проявляється у зміні лицевого відділу черепа.
На клітинному рівні спостерігається зменшення вмісту води
у цитоплазмі, зміна активного транспорту іонів, зростання
процесу гліколізу, зменшення активності окислюваного фосфорилювання й активності деяких ферментів.
Функціональні зміни при старінні не є однотипними, старін
ня не можна розглядати як ослаблення всіх функцій. За су-
Індивідуальний розвиток
195
масними уявленнями, вікові зміни можуть бути різнобічними.
Одні функції прогресивно знижуються з віком, інші — суттєво
не змінюються, а деякі показники зростають. До першого типу
належать скоротливість серця, гормональна активність залоз
внутрішньої секреції (щитоподібна, статеві), гострота зору
і слуху, зниження активності багатьох ферментів; до другого
типу — рівень цукру у крові, кількість еритроцитів, лейкоцитів,
гемоглобіну; до третього — синтез гіпофізарних гормонів, чут
ливість клітин до гормональних і хімічних факторів, рівень
холестерину у крові.
Дві людини одного й того ж календарного віку можуть
відрізнятися за рівнем вікових змін. Для визначення біологіч
ного віку використовують систему різноманітних тестів: ар
теріальний тиск, холестерин крові, акомодацію ока, життєву
ємність легенів, м’ язову силу, показник остеопорозу п’ясткових
кісток (на рентгенограмі) тощо.
Теорії старіння. У геронтології існує понад 100 теорій та гі
потез причин та механізмів старіння. їх можна розділити на
дві великі групи: перша група теорій виходить з того, що
старіння — це генетично запрограмований процес; згідно з дру
гою групою уявлень, старіння є результатом випадкових про
цесів, які руйнують організм. Л. 3. Тель (1997) серед усіх тео
рій виділяє молекулярні (генетичні, метаболічні, конформаційні), клітинні, організмові та надорганізмові.
Одна із найпопулярніших груп теорій обґрунтовує уявлення
про генетичну основу старіння. Послідовники цих теорій до
водять думку про існування ювенільних генів, завдяки яким
розгортається програма раннього онтогенезу, а також генів
старіння, експресія яких і викликає старіння. Інші автори
вважають, що існує плейотропний механізм дії генів: одні й ті
к гени на ранніх етапах онтогенезу функціонують сприятливо
для розвитку, тоді як на пізніх етапах їх функція трансфор
мується у несприятливу. Недавно генетиками було доведено,
що при старінні частина молекули ДНК, вільна від генів, по
ступово зменшується; ймовірно, що така редукція молекули
ДІІК має певний вплив на старіння організму і його загибель.
Інші прихильники генної теорії, відкидаючи уявлення про
спеціалізовані гени, стверджують, що старіння є наслідком
196
Розділ З
появи у геномі порушень. На їхню думку, у процесі онтогенезу
в геномному апараті клітин під впливом найрізноманітніших
чинників виникають дефекти, частина яких не відновлюється
і призводить до порушення синтезу білкових молекул (теорія
соматичної мутації, генетична мутаційна теорія, теорія пошко
дження генетичного апарату вільними радикалами, теорія
нагромадження помилок). Найбільш визнаною генетичною
теорією старіння є генорегуляторна теорія В. В. Фролькіса
(1988), яка велику увагу приділяє дефектам на етапах транс
крипції й трансляції синтезу білка, а також процесові сплайсингу, завдяки якому фрагменти РНК, які залишаються в ядрі,
здійснюють регулюючий вплив на транскрипцію. Показано, що
речовини, які гальмують процес транскрипції, збільшують три
валість життя. Багато авторів основну причину старіння вбача
ють у тому, що з віком порушується механізм репарації ДНК.
Серед метаболічних теорій сьогодні в центрі уваги дослід
ників такі, як вуглеводна гіпотеза, за якою глюкоза визна
чається як “медіатор старіння” ; теорія накопичення пігментних
тіл, перш за все ліпофусцину; кальцієва теорія Ганса Сельє,
яка пояснює старіння зниженням роботи кальцієвих насосів;
теорія вільних радикалів.
Конформаційні теорії вважають старіння результатом конформаційних змін на рівні вторинної, третинної та четвертин
ної структур. У рамках цього напряму запропоновані мембран
ні теорії і теорія поперечних “зшивок” макромолекул. Згідно
з мембранними теоріями, при старінні змінюється структура
мембран, що суттєво впливає на їхні властивості, зокрема по
рушує транспорт речовин, а тим самим і функції клітин. При
пускають, що в основі вікових змін клітинних мембран лежать
вільнорадикальні процеси, а також “ зшивки” макромолекул.
Згідно з теорією поперечних “зшивок” макромолекул, вперше
запропонованою Ф. Верцар у 40-х роках X X ст., при старінні
за рахунок утворення дисульфідних містків посилюються між
молекулярні зв’ язки, що знижує функціональні можливості
макромолекул. Це доведено стосовно колагену й еластину, що
становлять основу сполучної тканини.
Клітинні теорії старіння вважають первинними власне клі
тинні механізми геронтогенезу. Основи цих теорій заклали
К. Бернар, А. Вейсман, І. Мечніков, О. Богомолець.
Індивідуальний розвиток__________________________________________ 1 97
І. Мечніков пов’язував старіння з підсиленням в організмі
процесів інтоксикації у результаті нагромадження продуктів
азотистого обміну і продуктів гниття у товстій кишці. Підкрес
лював, що “ людина має вік своєї сполучної тканини” . Але
сьогодні доведено, що основою клітинного старіння є молеку
лярні зміни, про які йшлося вище. У рамках клітинних теорій
була висунута імунологічна теорія, що пояснювала старіння
як наслідок імунодефіцитного стану організму, зниження
імунного нагляду і як результат активації продукції аутоантитіл
(аутоімунна теорія).
Організмові теорії розглядають старіння як функцію ціліс
ного організму. У рамках цього напряму запропоновані теорії
“ біологічного годинника” , теорії еволюційного походження
старіння, теорії, що розглядають старіння як результат роз
витку, диференціювання і спеціалізації тканин, а також адап
таційно-регуляторні теорії. Остання група теорій, на думку
Л. 3. Теля (1997), є найбільш перспективною, оскільки вона
інтегрує існуючі уявлення про геронтогенез і пов’ язує процеси,
що відбуваються на молекулярному і клітинному рівнях, з ме
ханізмами адаптації організму до мінливих умов зовнішнього
середовища. Старіння розглядається як процес інтеграції мікроиошкоджень, що виникають при кожному окремому акті адап
тації в системах негайної відповіді та в системах забезпечення.
Чим вища інтенсивність відповіді організму на адаптаційні або
стресові фактори, тим вища швидкість настання старості. Ві
домо, що довгожителям, як і короткожителям, властива певна
ендемічність. Так, довгожителі трапляються переважно у сіль
ській місцевості з помірним кліматом і сприятливим життєвим
укладом. Водночас у дуже жарких або в дуже холодних країнах
(у районах Півночі), де існують підвищені вимоги до адапта
ційних можливостей організму, спостерігаються більш ранні
старечі зміни. На думку Л. 3. Теля, інтенсивні фізичні наван
таження, характерні для спорту, як і інтенсивна розумова
діяльність, — це потужні фактори старіння організму, тоді як
помірне фізичне навантаження і помірне загартовування підви
щують адаптаційні можливості організму та протистоять роз
виткові патології. Таким чином, старіння, згідно зЛ . З.Телем, — це довгий і необхідний для організму процес адаптації,
198
Розділ З
який відбувається з постійним зниженням функціональних
здібностей, але саме за рахунок цього — зі збереженням рів
новаги й узгодженості у функціонуванні клітин, органів і си
стем, і в кінцевому рахунку — зі збереженням життя.
Надорганізмові теорії старіння стверджують, що основною
причиною старіння є несприятливі впливи навколишнього
середовища, в тому числі радіації.
Смерть — це припинення усіх життєвих функцій організму.
Розрізняють смерть природну, або фізіологічну, яка настає
у результаті природного старіння організму, і смерть передчас
ну, яка настає внаслідок різних захворювань, дії несприятли
вих умов зовнішнього середовища (переохолодження, перена
грівання, недостатність кисню та інші), внаслідок отруєння,
нещасного випадку або вбивства.
Фізіологічна смерть — це закономірне явище, що завершує
індивідуальний розвиток організму. Причина природної смер
ті багатоклітинних організмів — старечі зміни в органах і тка
нинах, а також порушення злагодженості в роботі усіх систем
організму.
Вмирання — це складний процес, який здійснюється по
ступово і завершується смертю. Розрізняють два типи смерті:
клінічну і біологічну.
Окрім того, виділяють два термінальні стани, що передують
клінічній смерті, — передагонія і агонія. Передагонія може
мати різну тривалість (години, доби). Свідомість у цьому стані
затьмарена, артеріальний тиск знижений, пульс прискорений,
ниткоподібний, дихання утруднене, ш кірні покриви бліді.
Передагонія поступово переходить в агонію. Агонія триває
2—4 хв, іноді довше. Це стан між життям і смертю. Мобілізу
ються всі захисні механізми. Дихання прискорюється, з’яв
ляються судоми. Настає клінічна смерть.
При клінічній смерті всі видимі ознаки життя зникають:
зупиняється робота серця, припиняються дихальні рухи, зни
кають рефлекси, свідомість відсутня. Але більшість клітин,
тканин і органів ще зберігають життєздатність. Обмін речовин,
хоча на низькому рівні, але ще триває. Клінічна смерть — про
цес зворотний. У цей період можливе оживлення організму.
Індивідуальний розвиток__________________________________________ 1 99
У людини клінічна смерть при звичайних умовах триває
4—6 хв.
За клінічною смертю настає смерть біологічна, при якій
відбуваються необоротні зміни у клітинах, тканинах і органах,
порушується обмін речовин, починається руйнування клітин.
3.3. Регенерація і трансплантація
Регенерація (від лат. ге§епегаііо — відродження, відновлен
ня) — це процес оновлення структурних елементів організму
у процесі життєдіяльності, а також тих структур, які були
втрачені внаслідок патологічних процесів.
У біологічному сенсі регенерація — це пристосувальний
процес, вироблений в ході еволюції та властивий усьому ж и
вому. Ступінь і форми вираженості регенераційної здатності
неоднакові у різних тварин. Найпростіші, Кишковопорожнин
ні, Плоскі та Кільчасті черви, Голкошкірі володіють здатністю
відновлювати з окремого фрагмента або частинки тіла цілий
організм. Інші організми відновлюють лише окремі втрачені
органи або їх частини: регенерація ампутованих кінцівок, ву
сиків, очей — у ракоподібних; частин ноги, мантії, голови,
очей, щупалець, раковин — у молюсків; кінцівок, хвоста, очей,
щелеп — у хвостатих амфібій; відновлення внутрішніх органів
печінки, м ’ язів, кісток, шкіри, окремих кл ітин— у людини
і ссавців. При регенерації відбувається самовідновлення живої
матерії, і це “ самовідновлення живого відбиває принцип ауторегуляції й автоматизації життєвих відправлень” (І. В. Давидовський).
Регенераторне відновлення структури може відбуватися на
різних рівнях — молекулярному, субклітинному, клітинному
й органному, однак завжди йдеться про компенсацію структу
ри, яка здатна виконувати спеціалізовану функцію. Регенера
ція — це відновлення як структури, так і функції, значення
якої полягає в матеріальному забезпеченні гомеостазу. Віднов
лення структури і функції може відбуватися за допомогою
клітинних або внутрішньоклітинних гіперпластичних процесів.
Розділ З
2 00
На цій основі розрізняють клітинну та внутрішньоклітинну
форми регенерації. Для клітинної форми регенерації характер
не розмноження клітин мітотичним шляхом; для внутріш
ньоклітинної — збільшення кількості (гіперплазія) і розмірів
(гіпертрофія) ультраструктур (ядра, ядерець, мітохондрій, ри
босом, комплексу Гольджі) та їх компонентів. Внутрішньоклі
тинна форма регенерації є універсальною, тому що властива
всім тканинам і органам. Однак структурно-функціональна
спеціалізація органів і тканин у філо- і онтогенезі відібрала для
одних переважно клітинну форму, для інших — переважно або
виключно внутрішньоклітинну, для ще інших — однаковою
мірою обидві форми регенерації (табл. 3.1).
Т а б л и ц я 3.1. Форми регенерації в органах і тканинах ссавців
(за Д. С. Саркісовим, 1998)
Клітинна регенерація
Кістки;
епідерміс;
слизова оболонка
шлунково-кишкового
тракту, дихальних і се
човивідних шляхів;
пухка сполучна тка
нина;
ендотелій;
кровотворна система;
лімфоїдна тканина;
мезотелій
Клітиннаі внутрішньо
клітинна регенерація
Печінка;
нирки;
підшлункова залоза;
ендокринні залози;
легені;
гладкі м’язи;
вегетативна нервова
система
Внутрішньоклітинна
регенерація
Міокард;
скелетні м’язи;
гангліозні клітини
цнс
При регенерації відбуваються такі процеси, як детермінація,
диференціювання, ріст, інтеграція. Морфогенез регенератор
ного процесу складається з двох фаз — проліферації та дифе
ренціації. У фазі проліферації розмножуються молоді, недиференційовані клітини. Ці клітини називають камбіальними,
стовбуровими клітинами, клітинами-попередниками (напр.,
стовбурова клітина кровотворної системи, лімфоїдної тканини).
У фазі диференціації молоді клітини визрівають, відбувається
їх структурно-функціональна спеціалізація.
Індивідуальний розвиток____________________
201
Джерелом регенерації можуть бути:
— резервні клітини — камбіальні або незрілі клітинні еле
менти (стовбурові клітини, клітини-попередники), які інтен
сивно розмножуються і диференціюються, відновлюючи втрати
високодиференційованих клітин органу і забезпечують його
специфічну функцію;
— високодиференційовані клітини органа, які в умовах па
тологічного процесу можуть перебудовуватися, втрачати части
ну специфічних органел і одночасно набувати здатність до мі
тотичного поділу з подальшою проліферацією і диференціацією.
Розрізняють три види регенерації: фізіологічну, репаративну та патологічну.
Фізіологічна регенерація відбувається протягом усього жит
тя та характеризується постійним і неперервним відновленням
структур на клітинному (зміна клітин крові, епідермісу шкіри,
рогівки ока, епітелію слизової кишківника) і внутрішньоклі
тинному (оновлення клітинних органел) рівнях, чим забезпе
чується функціонування органів і тканин. Разом із відновлен
ням клітин і субклітинних структур постійно відбувається
біохімічна регенерація, тобто відновлення молекулярного скла
ду всіх компонентів тіла.
Інтенсивність проліферації визначають за кількістю мітозів,
які відбуваються в 1000 підрахованих клітин. Якщо в соматич
ній клітині мітоз, у середньому, триває 1 год, а весь мітотичний
цикл 22—24 год, то для визначення інтенсивності оновлення
клітинного складу тканин необхідно підрахувати кількість мі
тозів упродовж однієї або кількох діб. Таким чином було
з’ясовано, що інтенсивність мітозу в різні години доби неодна
кова, і було відкрито добовий ритм клітинних поділів.
Репаративна регенерація спостерігають при різних патоло
гічних процесах, які призводять до пошкодження клітин
і тканин. До ушкоджуючих факторів належать: механічні трав
ми (оперативне втручання), опіки, обмороження, дія отруйних
речовин, радіації, хвороботворні агенти. Два види регенерації
по є відособленими і незалежними. Так, репаративна регене
рація відбувається на базі фізіологічної, тобто на основі тих
самих механізмів, і відрізняється лише більшою інтенсивністю
202
Розділ З
проявів. Тому репаративну регенерацію необхідно розглядати
як нормальну реакцію організму на пошкодження, яка харак
теризується значним підсиленням фізіологічних механізмів
відтворення специфічних тканинних елементів того чи іншого
органу. Однак у зв’ язку з тим, що репаративна регенерація
збуджується патологічними процесами, вона має якісні мор
фологічні відмінності та може бути повною або неповною.
Повна регенерація (реституція) характеризується запов
ненням дефекту тканиною, ідентичною втраченій. Відбуваєть
ся у тканинах, де переважає клітинна регенерація (сполучна
тканина, кістки, шкіра, слизові оболонки).
Неповна регенерація (субституція) характеризується за
повненням дефекту сполучною тканиною, рубцем. Характерна
для органів і тканин, у яких переважає внутрішньоклітинна
форма регенерації. Біологічний сенс субституції полягає в ком
пенсаторній гіперплазії елементів залишеної спеціалізованої
тканини, маса якої збільшується, тобто відбувається гіпертро
фія тканини.
Регенераційна гіпертрофія може здійснюватися двома шля
хами:
— за допомогою гіперплазії клітин печінки, нирок, підшлун
кової та надниркових залоз, легень і селезінки відбувається
відновлення початкової маси органа та його функцій;
— за допомогою гіперплазії клітинних ультраструктур міо
карда, головного мозку.
Відновлення втрачених органів здійснюється шляхом епіморфозу, морфолаксису, ендоморфозу та замісної регенерації.
Епіморфоз — це відновлення органа шляхом надбудови,
відростання від поверхні рани, яка при цьому не змінюється
і зберігає старі структури. Регенераційний процес виявляється
тільки в ушкодженому органі, процес розпочинається з лізису
тканин, які прилягають до рани, інтенсивного розмноження
клітин, із яких утворюється регенераційний зачаток — бла
стема. Розмноження клітин приводить до збільшення зачатка,
а диференціювання клітин — до формування органа. При цьо
му форма і розмір інтактних частин організму зовнішньо прак
Індивідуальний розвиток
2 03
тично не змінюються. Найбільш типовими прикладами реге
нерації шляхом епіморфозу є: відновлення кінцівок у хвостатих
амфібій (тритон, аксолотль) і у личинок безхвостих амфібій
після ампутації. Аналогічно регенерує передня частина тіла
у планарій і хвіст у ящірок.
Морфалаксис — це регенерація за рахунок перебудови,
перегрупування частини органа, що залишилася, з подальшим
значним розростанням і утворенням цілого органа, у процесі
якої він поступово набуває вихідну форму та розміри. У пере
будову залучений весь ушкоджений організм або орган. Цей
спосіб регенерації широко розповсюджений у безхребетних,
а саме у кишковопорожнинних (гідроїдні поліпи) і червів (пла
нарій).
Ендоморфоз — це регенерація, що відбувається всередині
органа за рахунок росту частини, що залишилася, без віднов
лення попередньої форми. Регенерація починається із загою
вання рани, потім відбувається збільшення маси та розмірів
ушкодженого органа (гіпертрофія). Через регенераційну гіпер
трофію відновлюються всі паренхіматозні органи (печінка,
нирки, легені, селезінка, підшлункова, щитоподібна, молочна,
слинні, статеві залози, наднирники) хребетних тварин. Реге
нераційна гіпертрофія має виражений адаптивний характер,
оскільки за будь-яких патологічних станів, незалежно від то
го, виникли вони в результаті захворювання, впливу екстре
мальних факторів довкілля чи оперативного втручання, для
організму важливою є нормалізація функції ушкодженого
органа, відновлення гомеостазу.
Замісна регенерація — це відновлення ушкоджених органів
за участю клітинних елементів, які не належать цим органам.
Зокрема, відновлення лімфоїдних органів (тимуса, лімфатичних
вузлів, селезінки) при введенні кортизону, після рентгенівсь
кого опромінення і резекції відбувається не тільки шляхом
репопуляції стовбурових клітин, але й за рахунок міграції
у регенеруючий орган лімфоцитів, які циркулюють у крові. До
замісної регенерації можна віднести трансплантаційну регене
рацію, коли пересаджені тканини (наприклад, фрагменти
посмугованих м ’ язів) беруть участь у відновленні новоутворе
ного органа, зокрема ушкодженого м’яза реципієнта.
2 04
Розділ З
Незалежно від рівня, на якому відбуваються процеси від
новлення, водночас із основними шляхами регенерації існують
різновидності способів регенерації (табл. 3.2).
Таблиця
3.2. Основні способи репаративної регенерації
Рівні регенерації
Організмовий
Способи регенерації
Різновидності способів
регенерації
Епіморфоз
Морфолаксис
Епіморфоз
Регенераційна і компен
саторна гіпертрофія
Ендоморфоз
Позарановий вставний
ріст
Регенерація шляхом
заповнення дефекту
Органний
Загоєння рубцем
Замісна регенерація
Трансплантаційна
регенерація
За участю клітин, які
мігрують
Загоєння рубцем
Замісна регенерація
Тканинний
За участю клітин, які
мігрують
Ендоморфоз
Клітинний
Субклітинний
Трансплантаційна
регенерація
Репаративний
ендоморфоз клітини
Гіпертрофічний
ендоморфоз клітини
Репаративний
мембраногенез
органел
Гіпертрофований
мембраногенез
органел
Регенерація шляхом
заповнення дефекту
Індивідуальний розвиток__________________________________________2 0 5
Патологічна регенерація спостерігається в тих випадках,
коли внаслідок тих чи інших причин відбувається спотворен
ня регенераторного процесу, порушення зміни фаз проліфера
ції та диференціації (табл. 3.2). Як правило, патологічна реге
нерація розвивається при порушеннях загальних і місцевих
умов регенерації: іннервації, білковому та вітамінному голоду
ванні, хронічному запаленні. Проявом патологічної регенерації
є надмірне або недостатнє утворення регенераторної тканини
(гіпер- або гіпорегенерація), а також перетворення у процесі
регенерації одного виду тканини в інший (метаплазія). При
кладами можуть бути гіперпродукція сполучної тканини з утво
ренням келоїду, надмірна регенерація периферичних нервів
і надмірне утворення кісткового мозоля при зростанні перело
му, повільне загоювання ран і метаплазія епітелію в осередку
хронічного запалення.
Наслідками процесу регенерації є:
— повна регенерація (реституція, гомоморфоз): завершуєть
ся формуванням органа, ідентичного втраченому за формою
і побудованого з такої ж тканини;
— гетероморфоз: утворення зовсім іншого органа (наприклад,
у ракоподібних утворення кінцівки замість вусика, у ящ ір
ки — хвостоподібного органа замість ампутованої кінцівки);
— гіпотипія: неповний розвиток органа, що регенерує (напри
клад, поява меншої кількості пальців на кінцівках у тритона);
— гіперморфоз (суперрегенерація): надлишкове розростання
тканин (наприклад, формування більшої кількості кінцівок
при розсіканні місця ампутації органа на кілька частин; над
лишкове новоутворення кісткової тканини у місці перелому);
— неповна регенерація (реституція): утворення в зоні по
шкодження тканини, не притаманної для цього органа, при
цьому сполучна тканина в подальшому рубцюється.
Регенераційна здатність у людини характеризується значним
різноманіттям проявів. Так, у кістковому мозку, покривному
епітелії, слизових оболонках, кістках фізіологічна регенерація
виражається в неперервному оновленні клітинного складу,
а репаративна — у повному відновленні дефекту тканин і ре
конструкції її вихідної форми шляхом інтенсивного мітотич
ного поділу клітин. У печінці, нирках, підшлунковій залозі,
206
Розділ З
легенях, органах ендокринної системи відновлення клітинного
складу відбувається значно повільніше, а ліквідація пош ко
джень і нормалізація ушкоджених функцій відбувається на
основі двох процесів — розмноження клітин і нарощування
маси органел у збережених клітинах. Унаслідок цього відбу
вається їх гіпертрофія, що збільшує функціональну активність.
При цьому вихідна форма органів після ушкодження не від
новлюється, а на місці травми формується рубець. Відновлення
втраченої частини відбувається за рахунок неушкоджених
відділів, тобто відновлювальний процес відбувається за типом
регенераційної гіпертрофії. Внутрішні органи людини мають
високу потенційну здатність до регенераційної гіпертрофії. Так,
структура печінки відновлюється протягом 3—4 тижнів після
резекції 70 % її паренхіми через доброякісну пухлину внаслі
док ехінококозу, відновлюється вихідна маса і в повному обсязі
функціональна активність. У ц. н. с. і в міокарді, клітини яких
не здатні до мітотичного поділу, структурне та функціональне
відновлення після пошкодження досягається за рахунок збіль
шення маси органел у збережених клітинах, їх гіпертрофії при
внутрішньоклітинній регенерації.
Сучасний етап вивчення проблеми відновлення органів
у тварин і людини характеризується зростанням інтересу до
слідників до питань регуляції репаративних процесів. Вста
новлено, що у тканині, яка регенерує, підвищується вміст
води, інтенсифікуються процеси редуплікації ДНК, синтезу
РНК і білків, нагромаджується АТФ, зростає виділення азо
тистих речовин. Процеси, що відбуваються у тканині під час
регенерації, регулюються нервовою і гуморальною системами,
функціональним станом органів. При денервації органа спосте
рігається сповільнення темпів регенерації, що, на думку деяких
дослідників (Л. Ліознер, М. Воронцової), пов’ язане з відсутніс
тю трофічного впливу нервової системи на початкові стадії
репаративної регенерації органа.
На регенераційний процес впливають залози внутрішньої
секреції. Соматотропний гормон гіпофіза зменшує тривалість
синтезу ДНК, РНК, таким чином прискорює процеси регене
рації. Гормони надниркових залоз, зокрема кортикостероїди,
підвищують проліферацію клітинних елементів сполучної тка
нини й епітелію, регенерацію кісток, прискорюють утворення
Індивідуальний розвиток
207
хрящового мозоля; глюкокортикоїди стимулюють синтез РНК
в печінці. Тиреоїдні гормони (кальцитонін) регулюють усі ета
пи остеогенезу, підвищують рівень білкового обміну, сприяють
осифікації кісткового мозоля. Статеві гормони (тестостерон,
естрогени) змінюють функціональну активність клітин через
активацію синтезу РНК. На процеси регенерації впливає фун
кціональний стан органа, який, виконуючи специфічну функ
цію (функції), виробляє і виділяє в гуморальне середовище
речовину або, навпаки, використовує у процесі роботи і вилучає
зі середовища певні речовини. Ці речовини є постійними ком
понентами гуморального середовища організму, а функціональ
на активність органа визначає стан гомеостазу.
При порушенні тканинної рівноваги, функції органа, іму
нологічної реактивності, нейроендокринної регуляції (а це
можливе при будь-якому пошкодженні) концентрація вказаних
речовин, гормонів, біологічно активних речовин у крові змі
нюється. Порушення гомеостазу, зумовлене однією із вказаних
причин, виявляється у вигляді функціонального запиту, пере
дача якого клітинам уш кодж еного органа за допомогою
біохімічних посередників через гуморальне середовище є уні
версальним механізмом регуляції регенерації. У відповідь на
функціональний запит клітини ушкодженого органа починають
активно функціонувати. Активація функції супроводжується
підвищенням рівня метаболічних процесів у клітині, гіпер
плазією і гіпертрофією внутрішньоклітинних структур, інтен
сифікацією їх оновлення. Функціональний запит і пов’ язані
з ним зміни в гуморальному середовищі можуть бути однією зі
складових частин регенераційного стимулу до проліферації
клітин, що виникає внаслідок одночасної дії кількох регуля
торних механізмів: внутрішньотканинного, нервового і гумо
рального.
Отже, формування функціонального запиту в умовах уш
кодження органа розвивається за типом множинної ланцюгової
реакції, дослідження механізмів якої є перспективним напря
мом сучасної регенерації.
Значення регенерації для організму визначається тим, що
на основі клітинного та внутрішньоклітинного оновлення орга
нів і структур забезпечується широкий діапазон пристосуваль
них коливань функціональної активності в умовах довкілля,
2 08
Розділ З
що постійно змінюється, а також відновлення і компенсація
ушкоджених функцій при дії різних патологічних факторів.
Регенерація забезпечує стійку роботу органів, гомеостаз як
у нормальних умовах, так і при різних патологічних процесах.
Трансплантація (від лат. ігапзріапіаге— пересаджува
ти) — це заміщення ушкоджених або відсутніх органів, тканин
власними або тканинами і органами, взятими від іншого орга
нізму. Проблеми трансплантації вивчає клінічна трансплантологія, яка розробляє методи консервування органів і тканин.
Уперше трансплантація нирки у клініці була виконана
1933 р. професором Ю. Вороним. Ця дата вважається початком
ери клінічної трансплантології. Першу трансплантацію серця
людині виконав 1964 р. американський хірург Харді, який
пересадив хворому серце шимпанзе.
Хірург К. Барнард у м . Кейптауні 1967 р. вперше в історії
пересадив хворій людині серце 25-річної дівчини, яка загину
ла в автомобільній катастрофі. Операція Барнарда започатку
вала цілу серію трансплантацій серця у всьому світі. У другій
половині X X ст. відбувся бурхливий розвиток трансплантацій
ної хірургії. Поліпшення результатів трансплантації нирки,
печінки, серця, підшлункової залози було пов’ язане не тільки
з удосконаленням хірургічної техніки і кращою підготовкою
хворого до операції, але й нагромадженням знань про механіз
ми імунного відторгнення, відкриттям методів оцінки гістосумісності та використанням нових імуносупресивних препаратів
селективної дії. Ці наукові відкриття значно збільшили терміни
функціонування пересаджених органів.
Особу, в якої за життя або після її смерті взято анатомічні
матеріали для трансплантації чи для виготовлення біоімплантатів, називають донором; особу, для лікування якої застосовують
трансплантацію, — реципієнтом.
Трансплант ант — тканини, орган, анатомічний утвір,
фетальний матеріал, який використовують для трансплантації.
Трансплантація як метод лікування застосовується суто за
наявності медичних показів і згоди об’єктивно інформованого
дієздатного реципієнта лише тоді, коли усунення небезпеки
для життя або відновлення здоров’я реципієнта іншими мето
дами лікування неможливе.
Індивідуальний розвиток_________________________________________ 209
Види трансплантації:
Аутотрансплантація — пересадка органів і тканин у ме
жах організму.
/ зотрансплантація — пересадка органів і тканин між ге
нетично ідентичними організмами (монозиготні близнюки) або
між тваринами чистої (інбредної) лінії.
Алот рансплант ація — пересадка органів і тканин між
нлогенними індивідами (між тваринами одного і того ж виду,
але різних генотипів).
Ксенотрансплантація — пересадка органів або тканин між
організмами, що належать до різних видів.
Залежно від джерела, звідки взято трансплантат, розріз
няють:
— аутогенну трансплантацію: трансплантат беруть з однієї
ділянки тіла пацієнта і пересаджують на іншу ділянку (пере
садка шкіри, кістки, хряща, частки легенів, пальців);
— ізогенну трансплантацію: трансплантат беруть від ізогенного стосовно реципієнта донора (від одного з однояйцевих
близнят);
— сингенну трансплантацію: трансплантат беруть від роди
чів першого покоління;
— алогенну трансплантацію: пересадка тканин і органів
у межах одного виду (від людини — людині);
— ксеногенну трансплантацію: пересадка тканин і органів
від одного виду іншому (від мавпи — людині);
— алопластичну трансплантацію: заміна частин тіла синте
тичними матеріалами (метал, пластмаса) — судинні протези,
клапани серця, штучні суглоби.
При трансплантації тканина або орган можуть бути роз
міщені в організмі реципієнта:
— у тій же анатомічній ділянці, що і відсутній або видалений
орган (ортотипічна трансплантація); так імплантують донорське
серце, при цьому залишають своєрідний футляр із правої та
лівої передсердних стінок реципієнта, до яких анастомозують
пересаджене серце;
2 10
Розділ З
— в іншій, не властивій йому анатомічній області (гетеротипічна трансплантація): печінку можна трансплантувати в че
ревну порожнину реципієнта на місце видаленої селезінки, при
цьому власну, але ушкоджену патологічним процесом печінку
реципієнта зберігають; пересадку нирки, як правило, здійсню
ють гетеротипічно в ліву або праву клубові ямки.
Залежно від виду пластики розрізняють:
— трансплантацію тканин і органів з однієї ділянки тіла на
іншу або від донора реципієнтові;
— реплантацію, коли уражені органи і тканини пересаджу
ють на своє місце (відірвана кінцівка, скальпована шкіра);
— імплантацію, коли клітини або тканини спеціально пе
ресаджують в іншу ділянку (імплантація ізольованих острівців
підшлункової залози в печінку або селезінку).
Трансплантаційний імунітет. На початку X X ст. були
розроблені практично всі технічні аспекти трансплантації ор
ганів, проте застосування методів у клініці не давало бажаних
результатів, оскільки пересаджений орган швидко втрачав свою
функцію і гинув. Пояснення цьому явищу дав І. Мечніков,
заклавши основи нової науки — трансплантаційної імуно
логії. П. Медавар установив зв’ язок між процесами відторгнен
ня органів при трансплантації та станом імунної системи. Пе
ресаджений орган втрачає свою функцію і дегенерує в організ
мі реципієнта внаслідок клітинної чи гуморальної реакції
відторгнення — імунної відповіді, спрямованої проти транс
плантаційних антигенів, що розташовані на мембранах клітин
донорського органа.
Трансплантаційний імунітет пояснюється тим, що транс
плантат генетично відрізняється від тканин і органів реципієн
та, тобто тканина, що вводиться, генетично чужорідна. Імуно
логічна несумісність зумовлюється клітинними і гуморальними
реакціями організму. Основу для підбору пар донор — реципієнт
становлять еритроцитарні антигени системи АБО, резус-фак
тора і НЬА-системи. Водночас, якщо система АБО потребує
повної сумісності донора і реципієнта, то НЬА-система робить
можливою ідентифікацію лише за найбільш вираженими ан
тигенами.
Індивідуальний розвиток__________________________________________211
Синтез трансплантаційних антигенів детермінується гене
тичними структурами, які дістали назву локусів гістосумісності — НЬА (Нитап Ьеисосуіе Апіідепз). НЬА-система є го
ловною системою тканинної сумісності людини. Головний
локус гістосумісності людини розміщений у 6-й парі хромосом.
У ньому є два тісно зчеплених і серологічно визначених локуси (НЬА-сублокуси). До офіційної номенклатури ВООЗ вклю
чено 22 основних локуси гістосумісності людини (НЬА 1 ...
ИЬА 22), а також 73 додаткових локуси (УУ 1 ... УУ 73). Най
більше значення мають чотири локуси: НЬА-А, НЬА-В, НЬАС, НЬА-Б. Антигени гістосумісності, що кодуються генами
локусів НЬА-А, НЬА-В, НЬА-С, визначають серологічно, а ан
тигени, які контролюються генами локусу НЬА-Б, виявляють
за допомогою лімфоцитів.
Трансплантаційний імунітет зумовлюється в основному
клітинною реакцією за типом сповільненої гіперчутливості.
Сенсибілізовані лімфоцити справляють цитопатогенну дію на
клітини, які різняться одним або кількома генами. Навколо
трансплантата й у стінках судин скупчується багато лімфоцитів
і лімфоцитоподібних клітин, макрофагів і плазматичних клітин,
потім відбувається інфільтрація лімфоцитами (Т-кілерами,
клітинами з Т-популяції, що атакують трансплантат), закупор
ка судин трансплантата і загибель його через ішемію.
Успіх трансплантацій залежить від біохімічної та імуноло
гічної сумісності тканин або органів донора і реципієнта, які
в генетичному аспекті повинні бути ідентичними. Повна суміс
ність тканин можлива тільки між однояйцевими близнюками
і між тваринами чистих ліній. Так, при аналізі випадків транс
плантації серця спеціалісти дійшли висновку, що тканини
реципієнта і донора не повинні різнитися більш ніж двома
групами генів, при більшій несумісності ймовірність відторг
нення дуже висока.
Запобігти розвиткові трансплантаційного імунітету можна
пригніченням активності імунної системи організму, викори
стовуючи специфічні та неспецифічні методи.
До специфічних методів відносять:
— підбір донора і реципієнта за тканинною сумісністю і су
місністю груп крові;
212
Розділ З
— пригнічення трансплантаційного імунітету;
— виховання толерантності у реципієнта до антигенів донора.
Неспецифічні методи діють на імунну систему організму.
Вони пригнічують не тільки трансплантаційний, але й інфек
ційний імунітет. До них належать:
— Фізичні фактори: рентгенівське опромінення, яке має
супресорний вплив на процеси імуногенезу. Дозоване опромі
нення лімфоїдної тканини з екрануванням кісткового мозку,
легенів, інших життєво важливих органів застосовується вже
більше 25 років для лікування хвороби Ходжкіна. Проте вар
то пам’ ятати, що великі дози радіації небезпечні для життя
реципієнта, тому для імунодепресії використовують місцеве,
а не загальне опромінення.
— Хімічні імунодепресори широко використовують у клініч
ній практиці. До них належать циклофосфамід, аміноптерин,
метотрексат, імуран, 6 меркаптопурин, гормональні препарати
(кортизон, гідрокортизон, преднізолон, адренокортикотропний
гормон). Дані препарати пригнічують розвиток лімфоїдної
тканини, блокують реакції обміну, пригнічують клітинний
і гуморальний імунітет, гальмують синтез антитіл, цитокінінів,
зв’язують комплемент.
— Біологічні імунодепресори: ефективним є використан
ня антилімфоцитарних цитотоксичних сироваток (АЛС), антилімфоцитарного у-глобуліну, циклоспорину А. АЛС має
тканинну специфічність, пригнічує, в основному, трансплан
таційний імунітет, не впливаючи на процеси кровотворення.
Циклоспорин А блокує імунну відповідь Т-лімфоцитів за раху
нок зменшення продукції інтерлейкіну (ІЬ-2), пригнічує генну
транскрипцію (ІЬ-2) і, на сьогодні, є методом вибору для при
гнічення імунітету при пересадці. Біологічними імунодепресорами є гепарин і хондріотинсульфат, ядерні білки — гістони.
Гепарин поліпшує кровопостачання у капілярах, запобігає
зсіданню крові в них, пригнічує процес утворення антитіл і не
впливає на імунний комплекс організму.
Отже, основними вимогами при трансплантації органів
і тканин є повна відповідність груп крові донора та реципієнта,
максимальна відповідність локусів гістосумісності, старанне
Індивідуальний розвиток
213
запобігання розвиткові інфекційних захворювань і різних ус
кладнень, пов’ язаних із застосуванням лікарських засобів, які
мають імунодепресивну дію.
Одна з важливих проблем трансплантації — одержання не
обхідного трансплантата. У більшості випадків для трансплан
тації використовують власні тканини організму. При транс
плантації органів донарами є кровні родичі хворого або трупи
людей, смерть яких настала внаслідок несумісних із життям
ушкоджень. На сьогодні найбільш поширена алогенна транс
плантація, тобто пересадка трансплантата від іншої людини.
Лише деякі тканини й органи можна взяти від живої людини,
не завдаючи їй шкоди (шкіра, нирки). Як же вийти зі стано
вища? Потрібний матеріал для пересадки беруть від трупа,
частіше від людини, що загинула в катастрофі або внаслідок
іншого нещасного випадку. У зв’ язку з цим виникають органі
заційні, правові, етичні проблеми, які потребують вирішення.
Найбільш важливими з них є такі:
— визнання суспільством стану “ смерть мозку” як соціаль
ного еквівалента “ біологічної смерті” ;
— утвердження законодавчої процедури ухвалення рішення
про вилучення донорських органів після констатації смерті
мозку, а також у живих родинних донорів;
— розподіл донорських органів серед реципієнтів, які очі
кують на трансплантацію, відповідно до принципів соціальної
справедливості та медичної обґрунтованості.
Оскільки трансплантація багатьох важливих органів є опе
рацією, що рятує життя, рівень розвитку цієї галузі медицини
свідчить про рівень морального розвитку суспільства. В Україні
1999 р. прийнято “ Закон про трансплантацію органів, тканин
і клітин” , у якому регламентуються морально-етичні, правові
та медичні аспекти цієї проблеми. В Україні напрацьовано
відповідні ін стр у к ц ії, створено спеціальні бригади, які
об’єднують фахівців різного профілю: хірургів-трансплантологів, реаніматологів, морфологів, біохіміків, імунологів — пра
цівників медичних установ одного міста або центрів трансплан
тації з різних міст.
Багато країн входять у міждержавні об’єднання: Євротрансплантат (Бельгія, Нідерланди, Австрія, Швейцарія, Німеччина),
Скандинавтрансплантат (Данія, Швеція, Норвегія, Фінляндія).
214
Розділ З
Консервування трансплантатів. Консервування тканин
проводять у лабораторіях, обладнаних необхідним устаткуван
ням і апаратурою. Для взяття матеріалу використовують осіб,
які загинули від пошкоджень або миттєвої смерті (інфаркт
міокарда, розрив аневризми). Не підлягають консервуванню
тканини осіб, які хворіли і померли від інфекційних захворю
вань або пухлин. Тканини й органи від трупа забирають в умо
вах суворої асептики і антисептики у перші 6 год після кон
статації факту загибелі мозку. Консервують їх різними спосо
бами: швидким заморожуванням за температури -1 9 6 °С
з подальшим зберіганням за температури від -2 5 °С до -ЗО °С;
у розчинах, які містять антибіотики або антисептики, з подаль
шим зберіганням в охолоджених розчинах, плазмі чи крові
реципієнта; у парафіні, розчині формальдегіду, шляхом “ х о
лодної перфузії” (перфузія судин органа колоїдним розчином
за температури 8 °С); ліофілізацією кісток, хрящів. Варто
пам’ ятати: чим складніший орган, тим менший термін він
може зберігатися без кровопостачання. Під час зберігання
і транспортування органи й тканини піддаються консерваційному ушкодженню. Допустимі терміни холодової ішемії органів
при консервації такі: для нирок не більше 36 год, для печін
к и — 15 год, для підшлункової залози— 12 год, для серця —
5 год, для легенів — 4 год, для кишківника — 2 год. Протягом
цього терміну органи повинні бути доставлені до центру транс
плантації, де здійснюється вибір реципієнта (селекція пари
донор — реципієнт) і проводиться операція.
Сьогодні багато реципієнтів живуть з трансплантованим
органом 10 і навіть 20 років, досягнувши стану активно набутої
імунної та фізіологічної толерантності. Якість життя реципієн
тів значно поліпшилася. Протягом першого року після транс
плантації від 25 до 40 % реципієнтів повертаються до праці,
в середньому ЗО—50 % мають обмежену працездатність, 10—
15 % займаються домашнім господарством. Одним із перекон
ливих доказів високої фізичної та соціальної реабілітації після
трансплантації є успішний виступ реципієнтів на Олімпійських
іграх у 2002 р. для людей, які перенесли трансплантацію ор
ганів.
Трансплантація перейшла зі стадії клінічного експерименту
і стала загальноприйнятим методом лікування багатьох хроніч
Індивідуальний розвиток__________________________________________2 1 5
них і деяких гострих захворювань, при яких порушується
функція життєво важливих органів. Розробка й ефективне
впровадження в клінічну трансплантологію неспецифічних
методів подолання бар’ єру біологічної несумісності дозволить
їй стати важливим засобом лікувальної та профілактичної
медицини, поліпшити якість і тривалість життя людини.
3.4. Механізми гомеостазу
Індивідуальний розвиток неможливий без нормального функ
ціонування різних гомеостатичних механізмів.
Організм можна визначити як фізико-хімічну систему, що
існує в навколишньому середовищі у стаціонарному стані. Са
ме ця здатність живих систем зберігати стаціонарний стан
в умовах безупинно мінливого середовища й обумовлює їх ви
живання. Для забезпечення стаціонарного стану всіх організ
мів — від морфологічно найпростіших до найбільш складних —
виробилися різноманітні анатомічні, фізіологічні та поведінкові
пристосування, які слугують одній меті — збереженню сталості
внутрішнього середовища.
Уперше думка про те, що сталість внутрішнього середовища
забезпечує нормальні умови для життя і розмноження організ
мів, була висловлена у 1857 р. французьким фізіологом К. Бернаром. “ Сталість внутрішнього середовища — умова вільного
життя” , — так сформулював це положення К. Бернар. Здатність
зберігати сталість внутрішнього середовища назвали гомеоста
зом, і цей термін запропонував В. Б. Кеннон для опису “ різно
манітних фізіологічних процесів, підпорядкованих відновлен
ню нормального стану, як тільки його було порушено” . В основі
гомеостазу лежать не статичні, а динамічні процеси, оскільки
сталість внутрішнього середовища безперервно порушується
і так само безперервно відновлюється. Організм — це ультрастабільна система, яка сама здійснює пошук найбільш стійкого
й оптимального стану, утримуючи різні параметри функцій
у межах фізіологічних коливань. Коливання складу внутрішньо
го середовища та параметрів функцій у межах фізіологічних норм
необхідні для досягнення оптимального рівня життєдіяльності
216
Розділ З
організму. Комплекс процесів, спрямованих на підтримання
сталості внутрішнього середовища, називають гомеокінезом.
Згідно зі загальноприйнятою теорією еволюції, зародження
та виникнення життя відбулось у водному середовищі. З появою
багатоклітинних організмів більшість клітин втратила безпо
середній контакт із зовнішнім середовищем. Вони існують,
оточені внутрішнім середовищем — позаклітинною речовиною.
Завдяки наявності систем крово- та лімфообігу, а також дії
органів і систем, які забезпечують надходження різноманітних
речовин ззовні (органи дихання і кровообігу), та органів, які
забезпечують виведення назовні продуктів обміну, у багатоклі
тинних організмів виникла можливість сталості внутрішнього
середовища організму. Внаслідок цього клітини організму іс
нують і виконують власні функції у відносно сталих (стабільних)
умовах. Завдяки діяльності низки регуляторних механізмів,
організм здатний зберігати сталість внутрішнього середовища
за різких змін різноманітних характеристик зовнішнього се
редовища — перепадах температури, тиску, вологості, освітлен
ня тощо. За класифікацією, запропонованою на початку XIX ст.
відомим французьким анатомом і фізіологом М. Біша, гомео
статичні функції організму відносять до вегетативних функцій.
Мається на увазі, що характер усіх цих процесів — обмін ре
човин, ріст, розмноження, забезпечення умов для збереження
структури і здійснення процесів життєдіяльності організму —
однаковий для тваринних і рослинних організмів.
Роль транспорту іонів у підтриманні гомеостазу. Діяль
ність клітин потребує адекватної функції постачання їх киснем
та ефективного вимивання з них вуглекислого газу й інших
метаболітів чи відпрацьованих речовин. Для відновлення зруй
нованих білкових структур і вилучення енергії клітини повин
ні отримувати пластичний та енергетичний матеріал, що над
ходить до організму з їж ею. Усе це клітини отримують із
мікросередовища, який їх оточує, через позаклітинну речови
ну. У тварин із замкненою судинною системою позаклітинна
речовина умовно розмежовується на два компоненти: інтерстиціальна рідина та циркулююча плазма крові. Концентрація
електролітів і колоїдних речовин суттєво відрізняються у плаз
мі, інтерстиціальній і внутрішньоклітинних рідинах. Клітинні
Індивідуальний розвиток__________________________________________ 217
мембрани відмежовують внутрішньоклітинне середовище від
позаклітинної рідини, а стінки капілярів — інтерстиціальну
рідину від плазми крові. Транспортування речовин через ці
бар’ єри може відбуватися пасивно (дифузія, осмос, фільтрація),
а також шляхом активного транспорту. Активний транспорт
здійснюється транспортними аденозинтрифосфатазами (АТФазами) і відбувається за рахунок енергії гідролізу АТФ (первин
но-активний транспорт) чи енергії, що обумовлена градієнтом
концентрації деяких іонів, найчастіше Натрію. Первинно-ак
тивний транспорт відбувається за участю АТФаз, що отримали
назву іонних помп. До них належать Иа+,К+-АТФаза (Иа+,К+помпа), Н+,К+-АТФаза (локалізується здебільшого у клітинних
мембранах слизової оболонки шлунка, ниркових канальцях),
а також Са+, М£+-АТФаза, що локалізується у плазматичних
мембранах і мембранах сарко(ендо)плазматичного ретикулуму.
Для вторинно-активного транспорту у клітин тваринних ор
ганізмів використовується енергія градієнта концентрації іонів
Натрію, що створюється роботою Ка+, К+-помпи. Наприклад,
у кардіоміоцитах і гладеньком’ язових клітинах із функціону
ванням Иа+, К +-АТФази пов’язаний транспорт Са2+через плаз
матичну мембрану, завдяки наявності у мембрані білка, що
здійснює протитранспорт (антипорт) Иа+ іСа+. Іони Кальцію
транспортуються через клітинну мембрану в обмін на іони
Натрію і за рахунок енергії концентраційного градієнта іонів
Натрію. У головному мозку робота Иа+,К+-помпи спряжена із
реабсорбцією медіаторів. Таким чином, сталість складу крові
та стан бар’єрів є умовами гомеостазу мікросередовища клітин.
Вибіркова проникність цих бар’ єрів забезпечує певну специфі
ку складу мікросередовища клітин, необхідну для їхніх фун
кцій. З іншого боку, позаклітинна речовина бере участь в ут
воренні лімфи, обмінюється з лімфатичними капілярами, а це
дозволяє ефективно вилучати з клітинного мікросередовища
макромолекули, що не здатні дифундувати через гістогематичні бар’ єри у кров. Таким чином, в організмі між рідинами
внутрішнього середовища відбувається безперервний обмін, що
є необхідною умовою гомеостазу.
Механізми захисту клітинного гомеостазу. У процесі
життєдіяльності організму у внутрішнє середовище можуть
218
Розділ З
потрапляти ззовні молекули та мікроорганізми, що здатні по
рушувати його сталість і ушкоджувати клітинні структури. Ці
речовини та мікроорганізми отримали назву сторонніх, оскіль
ки вони не характерні для конкретного організму, не могли
бути синтезовані у ньому, тобто несуть ознаки сторонньої ге
нетичної інформації. Надійшовши в організм, вони загрожують
його генетичній індивідуальності, тобто фундаментальним оз
накам, які відрізняють один організм від іншого.
Водночас зі сторонніми агентами в організмі постійно від
бувається утворення внутрішніх сторонніх речовин і клітин,
пов’язане з процесом мутації соматичних клітин. Структурні
гени, що зазнали змін, кодують синтез білків із атиповою для
організму амінокислотною послідовністю. Звісно, в організмі
повинна існувати система розпізнавання та вилучення клітин
і речовин, які стали сторонніми в результаті мутації. Існує ціл
ком аргументована теорія, що низька ефективність механізмів
вилучення змінених мутацією соматичних клітин може бути
однією із причин виникнення пухлин.
Таким чином, внутрішнє середовище має забезпечувати
реалізацію механізмів захисту, по-перше, від мікроорганізмів
та екзогенних сторонніх речовин і, по-друге, від сторонніх
речовин і клітин ендогенного походження. Ці механізми за
хисту умовно поділяють на неспецифічні та специфічні. Неспе
цифічними називають механізми захисту, що не мають пріо
ритету в протидії сторонньому началу, тобто ефективні проти
будь-яких сторонніх речовин. Специфічні механізми захисту
спрямовані проти конкретних, визначених сторонніх агентів,
забезпечують пріоритетну протидію цьому сторонньому началу.
Специфічні механізми захисту здійснюються імунною системою
за рахунок гуморального і клітинного імунітету. Розмежуван
ня механізмів захисту на специфічні та неспецифічні є умовним,
оскільки реалізація неспецифічних захисних механізмів вима
гає насамперед розпізнавання стороннього начала, а це є одним
із завдань імунологічного нагляду, що здійснюється імунною
системою. Також відомо, що ефективність неспецифічних чин
ників різко посилюється за рахунок імунних механізмів.
Індивідуальний розвиток__________________________________________2 19
Неспецифічні механізми захисту клітинного гомеостазу:
1. Шкіра та епітелій слизових оболонок. Бар’ єрна функція
шкіри та епітеліальних структур (ротової порожнини, носог
лотки, шлунково-кишкового каналу, легенів, ока, сечовивідних
шляхів) забезпечується не лише механічним шляхом, але й за
вдяки хімічним речовинам, які виділяються клітинами бар’єрів.
Зокрема, шкіра має бактерицидні властивості за рахунок ре
човин, що містяться у секретах потових і сальних залоз, нап
риклад, молочної та жирних кислот, утворення пероксиду
гідрогену. Соляна кислота і ферменти шлункового соку руйну
ють мікроорганізми, тому у здорових людей шлунковий сік
практично стерильний. Бар’ єрна функція підтримується і лі
зоцимом, який має потужну бактеріолізуючу дію. Лізоцим
міститься у слині, слізній рідині, слизі дихальних шляхів,
а також у крові, материнському молоці, синовіальній, перитонеальній і плевральній рідинах.
2. Гуморальні чинники внутрішнього середовища, які за
безпечують механізми неспецифічного захисту, переважно
представлені білковими речовинами плазми крові. Це, насам
перед, білкова система комплементу, яка здійснює лізис сто
ронніх клітин.
3. Клітинні механізми неспецифічного захисту характерні
запальною реакцією тканин і фагоцитозом. Запальна реакція
тканин є механізмом захисту внутрішнього середовища від
проникнення сторонніх макромолекул, що виробився у процесі
еволюції. Наприклад, мікроорганізми, що проникли у тканину,
фіксуються в місці проникнення, руйнуються і навіть вилуча
ються з тканини у зовнішнє середовище з рідким середовищем
вогнища запалення — ексудатом. У вогнищі запалення особ
ливо ефективно відбувається процес фагоцитозу. Фагоцитоз,
будучи механізмом неспецифічного захисту, також сприяє
імунологічним механізмам захисту.
Специфічні механізми захисту клітинного гомеостазу.
Специфічні механізми захисту клітинного гомеостазу станов
лять основу імунітету. Імунітет — це спосіб захисту організ
му від живих тіл і речовин, які несуть у собі ознаки генетично
220
Розділ З
сторонньої Інформації. До них належать мікроорганізми і віру
си (інфекційний імунітет), найпростіші, гриби і черви (пара
зитарний імунітет), а також клітини (зокрема ракові), тканини
(зокрема органи, які використовують для пересадки), білки та
їх сполуки з ліпідами і полісахаридами (неінфекційний імуні
тет). Розрізняють природний і штучний імунітети. Природний
імунітет означає, що в організмі від народження існують
механізми захисту від сторонніх тіл. Штучний імунітет
означає, що у процесі життєдіяльності організм, розпізнаючи
стороннє тіло, набуває механізмів протидії та захисту, що от
римали назву імунних.
Система організму, яка виконує цю функцію, називається
імунною системою. Вона представлена всіма видами лейкоци
тів: лімфоцитами, моноцитами, макрофагами, нейтрофілами,
базофілами, еозинофілами, а також органами, в яких вони утво
рюються (лімфоїдні органи). Лімфоїдні органи і тканини пред
ставлені в організмі вилочковою залозою (тимусом), лімфатични
ми вузлами, селезінкою, лімфатичною тканиною кишківника
(апендиксом і пейєровими бляшками), носоглотки (мигдалики),
кісткового мозку. Оскільки ці органи і тканини можуть забез
печувати імунітет, їх називають імунокомпетентними. Імунокомпетентними клітинами є лімфоцити і макрофаги.
Примітивний клітинний імунітет виявляється у целомічних
безхребетних (кільчасті черви, голкошкірі). Він забезпечуєть
ся целомоцитами — клітинами вторинної порожнини тіла, що
знищують сторонній матеріал. На цьому етапі імунні реакції
набувають властивості специфічності, з’ являється імунологіч
на пам’ ять.
На наступному етапі сформувалася система інтегрованого
клітинного і гуморального імунітету, для якої типовими є спе
цифічні гуморальні та клітинні реакції на стороннє тіло, на
явність спеціальних лімфоїдних органів імунітету, утворення
антитіл. Жоден із перелічених показників у достатньо розви
неному стані не трапляється в безхребетних, — цей етап ево
люції імунітету характеризує лише хребетних тварин.
На думку одних дослідників, імунітет формувався у процесі
філогенезу як механізм захисту від мікроорганізмів. Згідно
з іншою теорією (концепція імунологічного нагляду), розвиток
системи імунітету стимулювався необхідністю захисту організ
Індивідуальний розвиток__________________________________________221
му від внутрішньої небезпеки у вигляді власних соматичних
клітин, які стали генетично чужими внаслідок мутації. Пору
шення процесів життєдіяльності у зв’ язку зі соматичними му
таціями (мутаційний ризик) зростає зі збільшенням кількості
клітин в організмі. Зокрема, тіло людини утворене приблизно
із 1015 клітин, і не менше, ніж 1013 з них проліферують. За
таких умов при наявній частоті мутації генів у кожному по
колінні клітин, які діляться, має накопичуватися щодоби 10е
соматичних мутацій, тобто мутаційний ризик значно зростає.
У класах хребетних тварин спостерігається посилення імунної
відповіді в міру ускладнення структурно-функціональної ор
ганізації. Наприклад, круглороті здатні до відторгнення ало
трансплантату і синтезу антитіл, однак питання наявності у них
органа імунітету — вилочкової залози — залишається відкри
тим. Розвинений тимус з’ являється лише в акул та інших
нижчих риб. В акул уперше серед хребетних спостерігаються
типові плазматичні клітини, що виробляють антитіла.
Еволюційні попередники лімфоїдних органів ссавців і лю
дини (тимус, селезінка, скупчення лімфоїдної тканини) з’ яв
ляються у безхвостих амфібій. У нижчих хребетних (риби,
амфібії), на відміну від птахів і ссавців, тимус активно виділяє
антитіла.
Особливість імунної системи птахів полягає у наявності
фабрицієвої сумки — особливого лімфоїдного органа. В ембріо
генезі цьому органові належить провідна роль у розвитку клі
тинної лінії В-лімфоцитів, тобто здатності організму утворю
вати антитіла.
Ссавці відрізняються складністю й різноманітністю будови
і розташування в організмі лімфоїдних органів. Вважається,
що гомологом фабрицієвої сумки у ссавців є апендикс. Згідно
з даними порівняльно-морфологічних досліджень, лімфоїдні
структури хордових у процесі еволюції утворилися з елементів
видільної системи.
Таким чином, виходячи з вищесказаного, формування імун
ної системи ссавців і людини — це тривалий еволюційний
процес.
2 22
Розділ З
Специфічний і неспецифічний імунітет:
1.
Неспецифічний імунітет спрямований проти будь-якої
сторонньої речовини (антигена). Він виявляється у вигляді
гуморального, за рахунок продукції бактерицидних речовин,
і клітинного, у результаті якого здійснюється фагоцитоз і ци
тотоксичний ефект. Вилучення будь-яких сторонніх у генетич
ному сенсі тіл, частинок здійснюється гуморальними і клітин
ними механізмами. Гуморальні механізми представлені таки
ми факторами, як фібронектин, лізоцим, інтерферон, система
комплементу тощо. Фібронектин — це білок, який має здатність
приєднуватися до сторонніх частинок, клітин, мікроорганізмів,
при цьому сприяє полегшенню наступного етапу інактивації
сторонніх тіл — фагоцитозу. Цей білок продукується макро
фагами, ендотелієм, гладеньком’ язовими клітинами, гепатоцитами тощо. Лізоцим — фермент, який продукується нейтро
філами і макрофагами. Його функція полягає у руйнуванні
мембран бактерій і їх лізисі. Інтерферони — білки, що забез
печують гальмування синтезу білка у клітинах, уражених
вірусами, і внаслідок цього вони блокують розмноження вірусів.
Система комплементу налічує близько 20 білків, які належать
до глобулінів плазми крові. Усі компоненти комплементу про
дукуються макрофагами, і їх функція полягає у знищенні
сторонніх агентів клітинного типу (цитоліз), участі у розвитку
реакції запалення та модифікації імунних комплексів із подаль
шим їх виведенням з організму.
Неспецифічний клітинний імунітет за своїм походженням
є вродженим і здійснюється переважно нейтрофілами, основ
ними функціями яких є: 1) фагоцитоз; 2) внутрішньоклітинне
перетравлення; 3) цитотоксична дія; 4) дегрануляція з виділен
ням лізосомальних ферментів.
Фагоцитоз і внутрішньоклітинне перетравлення сторонніх
тіл відкриті у 1892 р. лауреатом Нобелівської премії І. Мечниковим. Фагоцитоз відбувається у три етапи: адгезія, поглинан
ня, перетравлення за участю лізосомальних ферментів. Важ
ливу роль у процесі фагоцитозу відіграє здатність нейтрофілів
утворювати активні форми кисню (супероксидний радикал,
пероксид водню тощо). Активні форми кисню ушкоджують
молекули бактеріальних клітин, а це і є основою фагоцитозу.
Індивідуальний розвиток__________________________________________2 23
Варто мати на увазі, що ушкоджуюча дія активних форм кис
ню поширюється і на нейтрофіли, тому вони також можуть
загинути, як і сусідні з ними клітини.
Цитотоксичний ефект відкритий у 1968 р. Він полягає в то
му, що у присутності імуноглобулінів І§С і за наявності комп
лементу нейтрофіли підходять до клітини-мішені, але не фагоцитують її, а ушкоджують на відстані. Це здійснюється за
рахунок виділення нейтрофілами активних форм кисню — пероксиду водню, гіпохлорної кислоти тощо.
За останніми даними, нейтрофіли також посилюють про
дукцію антитіл В-лімфоцитами, у малих концентраціях вияв
ляють інгібуючу дію на функції Т-супресорів, а у великих — сти
мулюючу дію на Т-супресори.
2.
Специфічний імунітет спрямований проти певної сто
ронньої речовини. Специфічний імунітет реалізується удвох
формах — гуморальний (продукція антитіл В-лімфоцитами
і плазматичними клітинами) та клітинний (за участю Т-лімфоцитів).
Т-лімфоцити отримали назву від тимуса, їх називають також
тимус-залежними. Ще у періоді ембріонального розвитку в зача
ток тимуса, який називають центральним органом імунітету,
проникають із крові попередники Т-клітин. Упродовж усього
життя кістковий мозок постачає у кров, а потім у тимус невели
ку кількість попередників Т-клітин. У тимусі Т-клітини набу
вають імунокомпетентних властивостей, тоді виходять у кров
і заселяють периферійні лімфоїдні органи. Тут ці ще незрілі
клітини можуть реагувати на антигени, до яких вони вже ма
ють поверхневі рецептори, проліферацією з наступною дифе
ренціацією в ефекторні Т-лімфоцити. Таким чином, у тимусі
незрілі Т-клітини проходять своєрідний цикл “ навчання” на
здатність розпізнавати стороннє начало, а в периферійних лімфоїдних тканинах першими впізнають антигени. Т-лімфоцити
безперервно мігрують із лімфи у кров і навпаки. Приблизно
60— 80 % лімфоцитів, які циркулюють у крові, становлять Тлімфоцити. З усіх Т-лімфоцитів близько 10 % без поділу цир
кулює в організмі до 10 років, вони зберігають інформацію про
всі антигенні впливи, і тому їх називають клітинами імунної
пам’ яті. При надходженні в організм антигену він зв’ язується
2 24 ________________________________________________________ Розділ З
з рецептором відповідних Т-клітин, які активуються, утворю
ють ефекторні клітини, що знищують антиген.
Серед ефекторних Т-клітин розрізняють такі класи: Т-хелпери, або клітини-помічники, які забезпечують стимуляцію
диференціації В-лімфоцитів; Т-кілери — забезпечують цито
токсичний ефект і руйнують сторонні клітини; Т-ампліфайєри — посилюють і розширюють проліферацію Т-кілерів; Т-супресори — пригнічують імунну відповідь на певний антиген.
Імунні реакції, що реалізуються за допомогою ефекторних
Т-лімфоцитів, отримали назву клітинного імунітету. Одним
із різновидів клітинного імунітету є трансплантаційний, що ви
являється реакцією відторгнення трансплантованого органа.
Другим типом лімфоцитів є В-клітини. Свою назву вони от
римали від фабрицієвої сумки птахів (Ьигза — сумка), де були
виявлені. У людини немає фабрицієвої сумки, аналогом цього
органа вважають лімфоїдну тканину кишківника (апендикс),
пейєрові бляшки чи кістковий мозок. В-лімфоцити також утво
рюються у кістковому мозку зі стовбурових лімфоїдних клітинпопередників. Після набуття антигенної специфічності ці ще
незрілі клітини розселяються переважно в лімфатичні вузли,
а також у селезінку та пейєрові бляшки. Тут при антигенній
стимуляції В-лімфоцити проліферують і диференціюються
в В-лімфоцити пам’ яті та плазматичні клітини, які секретують антитіла. Зв’ язуючись з антигенами, антитіла сприяють
руйнуванню сторонніх клітин, нейтралізації продуктів їх жит
тєдіяльності. Оскільки антитіла чи імуноглобуліни є білками
лімфи і крові, тобто транспортуються рідким середовищем, є
підстави вважати, що В-лімфоцити забезпечують гуморальний
імунітет.
Імунна система розпізнає чужорідні агенти, що отримали
назву антигенів. Антигени — це речовини, котрі при введенні
в організм здатні викликати утворення антитіл або іншу фор
му імунного реагування: це є високомолекулярні речовини,
структура чи просторова конфігурація яких (наприклад, амі
нокислотна послідовність) не типові для певного індивідууму.
Антигенами найчастіше є білки, але можуть бути і полісаха
риди, ліпіди, полімеризована нуклеїнова кислота. Звичайно
антигени локалізуються на мембранах клітин, причому анти
генними властивостями володіє не цілісна молекула антигену,
Індивідуальний розвиток
225
а лише специфічна структура зовнішньої частини, яка нази
вається детермінантною групою. Кількість детермінантних груп
на поверхні клітини може бути дуже великого і досягати кіль
кох сотень чи навіть тисяч. Найчастіше антигенами є глікопротеїни, що локалізуються у мембранах клітин. За своїм походжен
ням антигени можуть бути інфекційними і неінфекційними.
Суть теорії імуногенезу:
1. В ембріональному періоді закладається стільки лімфоцитів
(чи навіть більше), скільки є антигенів у середовищі. Кожен
лімфоцит містить антитіла проти передбачуваних антигенів.
Ці антитіла продукуються лімфоцитом у невеликих кількостях
і локалізуються на поверхні лімфоцита, виконуючи роль рецеп
тора антигену.
2. Коли в організмі з’ являється антиген, то він взаємодіє
лише з одним видом лімфоцитів, який відповідає йому за рецепторами-антитілами. У результаті починається проліферація
цього виду лімфоцитів, клонування окремих видів лімфоцитів,
напрацювання ними відповідних кількостей антитіл (відшнуровування рецепторів) і наступна елімінація антигену або
шляхом його зв’ язування, або за рахунок пошкодження клітини-антигену.
3. Лімфоцити, які мають рецептори до власних антигенів і які
були у контакті з ними в ембріональному періоді, не здатні до
проліферації, оскільки це заборонено їм відповідними Т-супресорами. Не виключено, що ця заборона здійснюється за
рахунок вироблення Т-супресорними клітинами антитіл до
власних антигенів, які й блокують рецептори на звичайних
лімфоцитах.
3.5. Біологічні ритми
Живий організм з усіми його фізіологічними системами, як
і будь-який вид матерії, має просторово-часову організацію.
Фізіологічні системи і складові елементи цих систем пов’ язані
один з одним різноманітними просторовими взаємовідносинами.
Життєдіяльність будь-якого організму можлива лише за умов
226
Розділ З
нормальної його пристосованості до періодичних змін умов
зовнішнього середовища, що мають геліогеофізичну природу.
Наприклад, ритм “сон — бадьорий стан” синхронізується з до
бовим обертанням Землі, ритм линяння у тварин — зі зміною
пір року. Здатність організмів орієнтуватись у часі, яка дає
змогу узгоджувати швидкість і спрямованість головних фізіо
логічних процесів із закономірними та, насамперед, цикліч
ними змінами умов існування, має назву “ біологічного годин
ника” . Зовнішнім виявом функціонування такого годинника є
ритмічні коливання функцій організму— біологічні ритми.
Галузь біології, що займається вивченням закономірностей
часової організації живих систем, має назву хронобіології. Своїм
корінням хронобіологія сягає стародавньої медицини — вчені
стародавнього світу не могли не відзначати змін, які відбува
ються в організмі протягом доби, протягом пір року. Недарем
но ще представники натурфілософії вважали, що “ макрокосм”
(світ) і “ мікрокосм” (людина) — єдині. Для того, щоб науково
обґрунтувати це поняття, потрібні були століття роботи фізиків,
астрономів, медиків і біологів. Дана проблема була предметом
наукових досліджень В. М. Бехтєрєва, І. П. Павлова, І. М. Сєченова, В. В. Паріна та багатьох інших вчених різних галузей
науки. На сьогодні відомо, що біологічні ритми властиві усім
рівням організації живої матерії — від внутрішньоклітинного
до популяційного. У клітинах і тканинах відбувається безперерв
ний перебіг процесів асиміляції та дисиміляції, що складаються
з дискретних хімічних реакцій. Усі фізіологічні системи функ
ціонують також дискретно: у вигляді замкнених циклів (на
приклад, дихання) або у вигляді послідовних етапів перебігу
(наприклад, травлення). При цьому як цикли, так і етапи про
цесів мають власні часові параметри. Для серцево-судинної
системи характерною часовою мірою є серцевий цикл (у серед
ньому 0,8 с). Процес дихання складається із циклічної зміни
вдиху і видиху та, загалом, дає власний ритм — 12 дихальних
рухів за 1 хв. Травна система також працює із власними часо
вими показниками, що характеризують швидкість переробки
їжі у кожному відділі та її переміщення до наступного. Для
кожної залози внутрішньої секреції характерне виділення
певної кількості гормону за одиницю часу. Тканини поглинають
близько 300 мл кисню за хвилину. Певними часовими харак
Індивідуальний розвиток
2 27
теристиками володіють і показники внутрішнього середовища
організму. Будь-який показник крові, як, наприклад, в’язкість,
осмотичний тиск, рН, вміст того чи іншого аніона є кінцевим
вираженням складних процесів, пов’язаних між собою. Напри
клад, вміст Натрію у крові залежить від надходження його
з їжею, від виділення зі сечею і потом. У свою чергу, вміст
ІІатрію пов’язаний з інтенсивністю роботи гіпофіза та наднирко
вих залоз, що секретують гормони, котрі регулюють концент
рацію іонів Натрію у крові. Численними є такі приклади у рос
ини: піднімання й опускання листя або рух пелюсток залежно
від часу доби, вихід спор зі спорангіїв у грибів і водоростей.
Біологічні ритми розвиваються у тісному взаємозв’ язку
;і навколишнім середовищем і є результатом пристосування до
тих його чинників, котрі змінюються з чіткою періодичністю
(обертання Землі навколо Сонця і власної осі, коливання освіт
лення, температури, вологості тощо). Формування біологічно
го ритму, на думку багатьох учених, забезпечується двома
основними механізмами — генетичним і метаболічним. Поши
реною на сьогодні є гіпотеза хронона, згідно з якою припус
кається генетична обумовленість відліку часу організмом. Ця
гіпотеза розглядає механізм білкового синтезу як клітинний
годинник (процес транскрипції триває близько 24 годин).
( Стосовно метаболічного механізму, то його обґрунтування ба
зується на основі математичного моделювання синтезу полі
ферментних комплексів, які беруть участь в обміні вуглеводів.
Виявилося, що деякі замкнені ферментні цикли, в яких відбу
вається розсіювання енергії макроергічних сполук, постійно
перебувають в автоколивному режимі. З усіх природних фак
торів у підтриманні стабільності й узгодженості біоритмів
найважливіша роль належить фотоперіодизмові та змінам тем
ператури навколишнього середовища. При класифікації ритміч
них процесів залежно від їх частоти біологічні ритми об’єднують
у декілька груп: високочастотні, коливання середньої частоти
і біологічні ритми низької частоти. Періоди коливань високо
частотних біоритмів перебувають у межах від частки секунди
до півгодини. Навкологодинні ритми характеризують часову
організацію деяких внутрішньоклітинних метаболічних про
цесів, наприклад, синтез і виділення білкового секрету клітина
ми деяких залоз, а також коливання біоелектричної активності
228
Розділ З
головного мозку, серця, м ’язів, ритмічність зовнішнього ди
хання. їх вивчення почалося відносно недавно. Біоритми з ко
ливаннями середньої частоти відбуваються з тривалістю від
півгодини до 28 годин, їх ще називають циркадними (навколодобовими). Серед них виділяють біоритми з періодом від
півгодини до кількох годин (ультрадіанні). Для найважливіших
із них характерний період близько 90 хвилин. Вони простежу
ються вже у немовлят, у яких приблизно кож ні 90 хвилин
активність змінюється відносним спокоєм. У дорослих із такою
періодичністю відбувається чергування різних стадій сну, а та
кож періодів підвищеної працездатності та відносного розслаб
лення. Встановлені циркадні коливання понад 300 фізіологіч
них функцій організму людини. У результаті цих досліджень
була розроблена і складена “ циркадна система людини” , згідно
з якою частота серцевих скорочень є максимальною у проміж
ку між 13 і 18-ю годинами, рівень систолічного артеріального
тиску — між 15 і 18-ю годинами, кількість еритроцитів у кро
в і — між 11І12-Ю годинами, лейкоцитів— між 21 і 23-ю го
динами, рівень гормонів у плазмі крові — між 8 і 12-ю годи
нами. Виражені циркадні ритми мають місце також з боку
факторів імунітету, зокрема фагоцитозу, вмісту у крові Т- і Влімфоцитів.
Добові ритми багатьох фізіологічних процесів є ендогенни
ми, тобто визначаються механізмами, які діють у самому ор
ганізмі. На користь цього свідчить, наприклад, збереження
ритму, яке залежить від фотоперіодичності, навіть після того,
коли організм помістити в умови постійного освітлення. Зокре
ма, миші, які існували протягом кількох поколінь при постійно
му освітленні, повернувшись в умови чергування світла і тем
ряви, відтворюють нормальну добову періодичність рухової
активності. Біоритми із низькою частотою — це навколотижневі, навколомісячні, сезонні, навколорічні, багаторічні тощо.
Зокрема, навколотижневому біоритмові відповідає рівень виді
лення зі сечею деяких фізіологічно активних речовин; навко
ломісячному — менструальний цикл у жінок; сезонним — змі
ни тривалості сну, м’язової сили; навколорічним і багаторіч
ним — темпи росту і фізичного розвитку дітей, показники
імунітету. Залежність стану фізіологічних систем організму
від пори року пояснюється сезонними змінами кліматичних
Індивідуальний розвиток
2 29
умов, складом продуктів харчування (наприклад, дефіцит ві
тамінів у зимовий період). Сезонну динаміку репродуктивної
функції організму пов’ язують із фотоперіодизмом. Провідна
роль у координації всіх циклічних процесів належить циркадним ритмам активності механізмів нервової та ендокринної
регуляції. Практично всі її ланки (вищі відділи ЦНС, вегета
тивної нервової системи, гіпоталамічна секреція, секреція
тропних гормонів гіпофіза, метаболізм і виведення гормонів)
мають власні біоритми і визначають добові коливання концен
трації гормонів, тим самим запускають біоритми інших фізіо
логічних показників. Підкоряючись біоритмам, кожний фізіо
логічний показник протягом доби може суттєво змінюватись,
і тому діагностика та лікування багатьох захворювань людини
повинні проводитися з урахуванням біоритмів.
Розділ 4
ПОПУЛЯЦІЙНО-ВИДОВИЙ РІВЕНЬ
ОРГАНІЗАЦІЇ ЖИТТЯ.
БІОЛОГІЧНА ЕВОЛЮЦІЯ
Еволюція органічного світу — це процес історичного роз
витку живої природи.
Одним із основних положень еволюційної теорії є положен
ня про те, що складні організми утворилися від простих форм,
які існували раніше, шляхом поступових змін пристосуваль
ного характеру і нагромадження в ряду поколінь. Цей процес
супроводжувався зміною генетичного складу популяцій, ви
робленням адаптацій, виникненням одних і вимиранням інших
видів, перетвореннями біогеоценозів і біосфери. На певному
етапі біологічної еволюції створилися передумови для появи
біосоціальної істоти — людини. Еволюція загалом — процес
поступовий, прогресивний, пристосувальний і незворотний.
Результати еволюції — величезна різноманітність видів, при
стосованість їх до середовища існування і складність будови
багатьох із них.
Еволюція відбувалася з часу виникнення життя на Землі
понад 3,5 млрд років. Тому до наших днів живі організми
перебувають у тісній єдності з неживою природою.
Доказами еволюції, що була в минулому, є викопні рештки
організмів. Еволюція має місце і в наш час, а темпи її не тіль
Популяційно-видовий рівень організації життя
231
ки не сповільнилися, а навіть прискорилися в результаті збіль
шення антропогенного тиску на природні системи. Свідченням
цього є зникнення багатьох видів рослин і тварин, наявність
зникаючих і рідкісних видів, занесених до Червоної книги,
виникнення штамів мікроорганізмів, резистентних до антибіо
тиків, популяцій комах-переносників збудників захворювань
і комах-шкідників, на яких не діють інсектициди, що були
раніше для них смертельними.
Необхідною умовою для опанування еволюційного процесу
є розуміння природи виду. Поняття виду є одним із основних
у біології.
4.1. Вид, його критерії та структура
Уперше термін “ вид” запровадив англійський ботанік Джон
Рей (1693).
Карл Лінней (1707— 1778) сформував уявлення про вид як
про універсальну, дискретну й об’ єктивно існуючу у природі
реальну категорію.
Згідно зі сучасними уявленнями, вид — це сукупність осо
бин, які подібні між собою за будовою, функціями, каріотипом,
екологічними потребами, мають спільне походження, населя
ють певну територію (ареал), у природних умовах схрещу
ються виключно між собою і дають плодюче потомство.
Особливе положення виду серед інших систематичних оди
ниць (таксонів) зумовлене тим, що в цьому угрупованні окремі
особини існують реально. На відміну від виду, таксони надвидового рівня (рід, родина, ряд, клас, тип) не є одиницями ре
ального життя організмів. Розподіл організмів за надвидовими
таксонами вказує на ступінь їх філогенетичної спорідненості.
Згідно з біологічною концепцією, види реальні характери
зуються певними біологічними властивостями, мають єдину
генетичну програму і складаються із популяцій. В и д — диск
ретна одиниця живої природи, репродуктивно ізольована від
інших видів, здатна до самовідтворення й еволюції. В ієрар
хічній системі рівнів організації органічного світу він утворює
популяційно-видовий рівень.
232
Розділ 4
Види виникли у результаті еволюції від видів, що існували
в минулі геологічні епохи. Одночасно вид — це якісний етап
еволюції, який реально існує, допоки не зміняться умови. У разі
їх зміни вид або дає початок новим видам і надвидовим таксо
нам, або вимирає. Як результат еволюції вид — дискретний,
репродуктивно ізольований від інших видів, стабільний, ціліс
ний, пристосований до певних умов середовища. Як етап ево
люції — вид динамічний, лабільний, має розпливчасті межі,
здатний змінюватися, пристосовуватися до мінливих умов
середовища, еволюціонувати.
Варто зазначити, що універсального визначення виду не
існує. Це пояснюється величезною різноманітністю видів (опи
сано 1,5 млн видів тварин і близько 500 тис. видів рослин)
і широкою внутрішньовидовою мінливістю. Ця характеристи
ка певною мірою властива тільки для організмів зі статевим
розмноженням і неприйнятна для агамних (безстатевого роз
множення), самозаплідних і облігатно партеногенетичних форм.
Не існує також універсального видового критерію. Види і на
лежність особини до певного виду визначають за сукупністю
критеріїв, які взаємно доповнюють один одного.
Критерії виду:
— морфологічний критерій полягає в тому, що подібність
особин, які належать до одного виду, не мають відрізнятися
одна від одної більше, ніж відрізняються діти одних батьків,
а також мають відновлювати свої ознаки у нащадків;
— фізіологічний критерій полягає у подібності процесів
життєдіяльності, насамперед процесу розмноження;
— генетичний критерій: вид є генетично замкненою репро
дуктивною ізольованою системою, яка складається з генетично
відкритих систем-популяцій;
— біохімічний критерій: подібність біохімічного складу
і єдність біохімічних процесів;
— географічний критерій полягає в адаптації специфічного
комплексу умов середовища: оскільки вид займає власну еко
логічну нішу, що забезпечує можливість існування його на
Популяційно-видовий рівень організації життя______________________ 2 33
певній території, то всі популяції займають одну видову еко
логічну нішу і е взаємозамінними;
— критерій несхрещуваності полягає в тому, що особини,
які дають плодюче потомство, належать до одного виду, а ті,
які не дають — до різних видів (Ж. Бюффон);
— еволюційний критерій полягає в існуванні виду як єдино
го цілого до того часу, поки всі популяції еволюціонують вза<мозв’ язано, тобто обмінюються генами і взаємно обумовлюють
існування одна одної.
Коли єдина система популяцій розпадається у процесі ди
вергенції, то утворюється кілька таких систем популяцій, які
можна вважати різними видами.
Генофонд. Біологічний вид, який складається з численних
особин, які мають генетичні родинні зв’ язки, але різняться
за комбінаціями спадкових ознак, становить цілісну біологіч
ну макросистему.
Завдяки дії природного добору генофонд виду збалансований
і має оптимальні якості щодо умов існування виду.
Генофонд утворюють особини одного виду тварин або рослин,
у генних локусах яких міститься багато алелів.
Унаслідок вільного схрещування виникають високогетерозиготні диплоїдні особини. Багато алелів кожного із локусів
уже зазнали в попередніх поколіннях добору на комбінативну
здатність у диплоїдних гетерозиготних генотипах. Стабільність
розвитку, або морфогенетичний гомеостаз, — результат гете
розиготності.
Новий вид виникає з популяції або групи популяцій бать
ківського виду. І тому генофонд популяції є складовою гено
фонду виду, який істотно різноманітніший, ніж генетична
інформація окремої популяції або навіть групи популяцій. Не
кожна популяція (група популяцій) перетворюється на новий
вид, оскільки не завжди складаються відповідні умови, які
були оптимальними для цієї популяції, створюються необхід
ні ізоляційні бар’єри, які захищають перетворення популяції
на вид тощо.
Генофондові виду належить виняткова роль в еволюції.
Мид — якісний етап еволюції, її вузловий момент. Проте вид
2 34
Розділ 4
еволюціонує, опосередковано через популяції, які здатні існу
вати необмежено в часі.
Популяції — головні складові одиниці виду. Вид має склад
ну внутрішню популяційну структуру, яка забезпечує присто
сування до оптимального використання різноманітних умов
у межах видового ареалу. Високий ступінь пристосування
і пристосованості виду забезпечується мутаційною, комбінативною і модифікаційною мінливістю. На підґрунті комбінативної мінливості формуються популяційна структура і полі
морфізм.
Усі види складаються з популяцій.
Популяція (від франц. рориіаііоп — населення) — група
особин одного виду, яка здатна до самовідновлення, населяє
певну територію (ареал). У популяції здійснюється велика
кількість випадкових схрещувань (панміксії) та вона відокрем
лена від інших певною формою ізоляції.
Популяція характеризується:
— величиною ареалу (залежить від радіуса індивідуальної
активності);
— чисельністю особин, яка коливається залежно від змін
екологічної ситуації;
— віковим складом, який залежить від тривалості життя,
інтенсивності розмноження, віку досягнення статевої зрілості;
— статевим складом, який зумовлений еволюційно закріп
леними механізмами формування первинного (у момент заплід
нення), вторинного (у момент народження) і третинного (у до
рослому житті) співвідношення статей.
Популяція характеризується генофондом, тобто генотипами
організмів цієї популяції. Генетична популяція — різноманіт
на, бо в генофонді одночасно присутні різні алелі окремих генів.
Мутації, як правило, рецесивні, вони зберігаються і накопичують
ся, створюючи резерв спадкової мінливості. Завдяки комбінативній мінливості, цей резерв використовується для створення
нових комбінацій алелів.
Популяції — генетично відкриті у межах виду угруповання
організмів.
Популяційно-видовий рівень організації життя______________________ 2 35
Комплекс генів однієї особини називається генотипом. За
гальний запас генів особин, які входять до складу популяції,
утворює генофонд певної популяції.
Видоутворення починається зі зміни генетичного складу
(генофонду) популяції шляхом появи генних, хромосомних
і геномних мутацій. Вони відбуваються постійно, і частота їх
збільшується при дії мутагенів різної природи.
Сукупність алелів, які виникли у результаті мутації, утворю
ють елементарний еволюційний матеріал для рекомбінацій
і добору. Природний добір усуває невдалі комбінації генів і збе
рігає генотипи, які не порушують пристосувальне формоутво
рення.
У популяціях виникають випадкові коливання частот алелів,
які не зумовлені дією природного добору, — це дрейф генів.
Інтенсивність дрейфу генів залежить від чисельності популяції.
1Іим менша кількість особин, тим більша інтенсивність дрейфу.
Це явище зумовлює втрату або закріплення алелів у гомо
зиготному стані в усіх членів популяції поза зв’ язком з при
стосувальною цінністю.
Міжпопуляційні міграції запобігають поглибленню відмін
ностей і об’ єднують популяції в єдину систему виду. Якщо
ізоляція популяції тривала, то це веде до репродуктивної ізо
ляції та, як наслідок, до утворення нового виду. Отже, попу
ляція— елементарна еволюційна одиниця, а ви д — якісний
етап еволюції, який закріплює її суттєвий результат.
Генетична характеристика популяцій. Генетично кожна
популяція характеризується певним генофондом (сукупність
усіх генів популяції). Носіями генів є особини популяції.
Гени існують у різних формах (алелях). У генотипі особини
представлені тільки два алелі певного гена і в рівних співвід
ношеннях (0,5А :0,5 а), а у генофонді популяції можуть бути
псі алелі певного гена, і ці алелі трапляються у різних співвід
ношеннях, із різною частотою (наприклад, 0,ЗА :0,7а). Гене
тична структура популяції характеризується якісним складом
генів і певними частотами алелів і генотипів. Частотою алелів
генотипу називається відносна кількість особин у популяції
з певним геном, генотипом. Частоти виражають у відсотках
або, що найчастіше, десятковими дробами.
236
Розділ 4
Генофонди природних популяцій характеризуються гене
тичною гетерогенністю, генетичною єдністю і динамічною
рівновагою частот алелів і генотипів. Генетична гетерогенність
(спадкова різноманітність, генетичний поліморфізм) зумовлена
наявністю в генофонді популяції одночасно різних алелів ок
ремих генів. Вона створюється мутаційним процесом (мутацій
на мінливість), процесом рекомбінацій (комбінативна мін
ливість), а також вноситься в популяцію міграціями з інших
популяцій, але і в цьому разі зумовлена вказаними вище про
цесами. Первинним є мутаційний процес, оскільки лише він
створює нові алелі. Але основна частина генетичної мінливості
представлена комбінативною мінливістю (рекомбінаціями), яка
створюється кросинговером і незалежним розходженням негомологічних хромосом у мейозі, а також випадковим поєднанням
гамет при заплідненні. Комбінативна мінливість — невичерп
не джерело спадкових змін. Розрахунки показують, що при
схрещуванні організмів, які відрізняються у загальному за 1000
локусів, кожен із яких представлений 10 алелями, кількість
можливих генотипів у нащадків становитиме 101000, що пере
вищує число електронів у Всесвіті. Мутаційний процес і гене
тична комбінаторика створюють таку різноманітність, що
в популяції, яка розмножується статевим шляхом, не існує
двох абсолютно ідентичних особин. Еволюційне значення генетичності полягає у тому, які мутації та рекомбінації є ма
теріалом для природного добору. У разі їх відсутності, як це
можливо у генетично однорідних популяціях (чисті лінії),
природний добір неефективний. Незважаючи на гетерогенність
особин (генотипів), популяція — це єдина генетична система,
цілісність якої забезпечується достатнім рівнем внутрішньопопуляційної панкмісії.
Генофонд (алелофонд) популяції. Природним популяціям
властива генетична гетерогенність. Завдяки цьому популяції
набувають необхідної пластичності щодо гетерогенного, мінли
вого середовища існування. Генетична гетерогенність популяції
дозволяє видові використовувати як нові спадкові зміни, що
виникли за життя одного покоління, так і ознаки, набуті у по
передніх поколіннях.
Популяційно-видовий рівень організації життя
2 37
Генофонд популяції — це різноманітність елементарних
спадкових ознак у межах сукупності особин, які становлять
популяцію. Генофонд — це сукупність алелів, які містить пев
на вибірка індивідів. Тому термін генофонд тотожний термінові
алелофонд. Генофонд локальної популяції містить, крім мономорфних, різні поліморфні гени. Популяція може набувати
нового алеля внаслідок мутації, яка виникла в окремої особи
ни, або в результаті міграції носія цього алеля з іншої популя
ції. Такий процес отримав назву потік генів. Він здійснюється
шляхом розселення, переміщення у просторі окремих особин,
їхніх спор, пилку (гамет) або окремих органів (насіння тощо).
Мутаційний процес і потік генів створюють у популяції
мінливість за окремими генами.
Кожна популяція характеризується специфічним генофон
дом із властивим тільки їй співвідношенням частот різних
алелів і з різним характером мінливості.
Завдяки дії природного добору популяції набувають опти
мальної пристосованості до умов існування та стійкості до
зміни цих умов. Це забезпечується широтою норми реакції
генотипів і генетичною гетерогенністю популяцій.
Облік генофонду популяції завжди залишається неповним,
іцо зумовлено недосконалістю методик, обмеженістю можли
востей вивчення всіх особин популяції тощо.
На підґрунті комбінативної мінливості виникає адаптивний
поліморфізм природних популяцій. Концепція генофонду шир
ша, ніж концепція поліморфізму. Генофонд популяції скла
дається зі всіх генів, що входять до її складу. Тому популяцію
можна описати кількісно, використовуючи типи генів, які мі
стяться в її генофонді, та їх частоти.
Способи видоутворення. Видоутворення полягає у поділі
первинного єдиного виду на два і більше нових.
Дивергентне видоутворення пов’ язане з виникненням міжпопуляційних ізоляційних бар’єрів і поглибленням різниці між
генофондами популяцій під дією природного добору до гене
тичної ізоляції.
Філетичне видоутворення полягає у поступовому перетво
ренні одного виду в інш ий протягом деякого часу. При
238
Розділ 4
алопатричному (географічному) видоутворенні перешкоди для
схрещування зумовленні просторовим розділом популяцій.
Генетична ізоляція розвивається вторинно.
Симпатричне (від гр. суш — разом) видоутворення веде до
появи нового виду в межах вихідного ареалу. Ізоляція є гене
тичною і проходить швидко. Цей тип видоутворення на основі
поліплоїдії характерний для рослин (різні види пшениці мають
14, 28, 42 хромосоми).
Основні типи еволюційного розвитку:
1. Дивергенція — виникнення відмінностей на основі однієї
і тієї ж організації. Причиною дивергенції є розходження по
різних екологічних нішах і міжгрупова конкуренція, у резуль
таті яких перевагу одержують в ході добору особини, які мак
симально відрізняються від особин іншої групи. Проміжні
форми не витримують конкуренції та елімінуються. Завдяки
дивергенції виникають види у межах родів.
2. Паралелізм — це виникнення подібних ознак у таксонів,
які походять від одної предкової форми на загальній організа
ційній основі, що відбувається в подібних умовах. При даній
формі еволюції ступінь подібності не свідчить про ступінь ро
динних зв’ язків. Наприклад, розвиток легенів у кистеперих
і двоякодихаючих риб еволюціонував незалежно і паралельно.
Наслідком був розвиток трикамерного серця і малого кола
кровообігу.
3. Конвергенція — виникнення подібних рис організації на
початково різній основі, тобто шлях розвитку аналогічних
пристосувань. В однакових умовах середовища добір виробляє
подібні пристосування навіть у різних філогенетичних груп
організмів. Наприклад, крило пінгвіна (птахи) і плавець акули
(риби), літальна перетинка кажанів (рукокрилі) і білки-летяги
(гризуни).
Абіотичне середовище подібно впливає на всі організми,
тому конвергенції, на відміну від паралелелізмів, завжди більш
поверхневі та стосуються лише небагатьох рис організації груп,
які конвергують.
Популяційно-видовий рівень організації життя______________________ 239
Мінливість. Ч. Дарвінові було важливо показати, яким чи
ном забезпечується різноманітність видів. Тому він насамперед
обґрунтував положення про мінливість живих організмів. Уче
ний виділив визначену і невизначену мінливість.
Визначена (гр уп ов а ) м ін л и віст ь— подібна зміна всіх
особин потомства в одному напрямі внаслідок впливу певних
умов (зміна росту при зміні кількості та якості їжі, товщи
на шкіри і густота покриву шерсті залежно від зміни кліма
ту і т. д.).
Невизначена (індивідуальна) мінливість — це поява різ
номанітних незначних відмінностей у особин одного виду,
якими в існуючих умовах одна особина відрізняється від інших
(нащадки однієї пари тварин не бувають цілком подібними,
хоча розвиваються в подібних умовах). Значна різноманітність
особин унаслідок індивідуальної мінливості є важливим мате
ріалом для еволюційного процесу. Водночас із незначними
змінами, Ч. Дарвін не виключав можливості появи різких
відхилень.
Корелятивна (співвідносна) мінливість. Організм — цілісна
система, окремі частини якої тісно пов’ язані між собою. Тому
зміна структури або функції однієї частини зумовлює зміну
іншої чи ряду інших (довжина шиї і ніг болотних птахів, дов
жина дзьоба і язика, розвиток м ’язів і розмір кісток, до яких
вони прикріплені).
Компенсаційна мінливість полягає в розвитку одних органів
чи функцій і одночасному пригніченні інших, тобто спостері
гається зворотна кореляція. Мінливість різних ознак організ
мів здійснюється в різних напрямах. У домашніх тварин і рос
лин розвиваються корисні в господарському розумінні ознаки,
а в диких видів добре розвинені адаптації, які забезпечують
краще виживання організмів.
Ч.
Дарвін розробив також вчення про штучний і природній
добір як ведучі та спрямовуючі фактори еволюції домашніх
форм і диких видів.
Штучний добір — це система заходів, спрямована на вдоско
налення існуючих і створення нових порід тварин і сортів
240
Розділ 4
рослин із корисними в господарському розумінні спадковими
ознаками. Творча функція штучного добору базується на су
купній взаємодії в ряді поколінь мінливості, спадковості, під
бору батьківських пар, направленого вирощування, переваж
ного розмноження особин із корисними ознаками й елімінація
небажаних індивідів. Завдяки цьому з покоління в покоління
підсилюється розвиток корисних ознак, а внаслідок кореля
тивної мінливості відбувається перебудова всього організму.
Штучний добір веде до дивергенції— розходження ознак
у порід і сортів, утворення великої їх різноманітності. Але де
коли подібні вимоги ведуть до появи однакових ознак у різних
за походженням організмів, що отримало назву конвергенції.
Наприклад, у м ’ ясних порід великої рогатої худоби і всіх порід
свиней зменшена довжина ніг, шиї, голови, збільшена довжи
на тулуба і загальна маса. Ч. Дарвін виділив дві форми штуч
ного добору — несвідомий і методичний.
Несвідомий добір є первинною формою добору, якою ко
ристувалися ще найдавніші люди. Водночас людина не має за
мету створення нових порід чи сортів, а лише переважно роз
множує кращі особини. У результаті диференційного підходу
в ряді поколінь поступово підсилюються певні ознаки, що зре
штою, хоч і повільно, приводить до утвореня нових порід чи
сортів. Ця форма добору має творчий характер. Сьогодні у гос
подарстві людини ще існує несвідомий добір. При цьому від
бувається елімінація (вибракування) особин із гіршими озна
ками.
Методичний (свідомий) добір полягає у цілеспрямованому
виведенні породи чи сорту. Використовуючи мінливість, важ
ливо провести підбір пар для спарювання, забезпечити макси
мальний розвиток і закріплення бажаних ознак у кожному
поколінні.
Фактори еволюції:
1. Спадкова мінливість — здатність змінюватись і передава
ти зміни потомству. Має випадковий характер. Зміни (мутації)
можуть бути корисні, ш кідливі або нейтральні для орга
нізму.
Популяційно-видовий рівень організації життя______________________ 2 41
2.
Природний добір— результат взаємодії з навколишнім
середовищем, при якому зберігаються форми з найбільш ко
рисними (відповідними до нього) спадковими змінами. Прояв
ляється у формі боротьби за існування: внутрішньовидової,
міжвидової та зі силами неживої природи. Розрізняють три
основні форми природного добору: рушійний, стабілізуючий
і дизруптивний (дивергентний) (за Ч. Дарвіном).
Згідно з теорією Ч. Дарвіна, дія добору веде до створення
нових адаптивних ознак; знищує ознаки, які втратили своє
пристосувальне значення; зберігає пристосування, корисні для
виду в певних умовах середовища.
І.
І. Шмальгаузен у монографії “ Фактори еволюції. Теорія
стабілізуючого добору” (1946) запропонував розрізняти дві
форми природного добору: рушійний (ведучий, або дарвінівський), який зумовлює адаптивні перетворення організмів у ході
еволюції, і стабілізуючий добір, який визначає стабільність
сформованих ознак, що зберігають своє адаптивне значення.
Стабілізуючий добір — це переважне виживання організмів,
яким притаманні ознаки, що не мають помітних відхилень від
норми, властивої певній популяції. Він проходить через елімі
націю будь-яких відхилень від норми. Мінливість ознаки в по
пуляції описується кривою нормального розподілу. Виділяють
дві зони мінливості: безумовної та умовної елімінації. У зону
безумовної елімінації потрапляють всі особини, які сильно
відхилені за певною ознакою і за певних умов середовища ста
ють нежиттєздатними. У зону умовної елімінації потрапляють
менш відхилені від норми особини, які здатні вижити в більш
сприятливих умовах середовища і гинуть при погіршені цих
умов. У результаті дії стабілізуючого добору на кривій ліміти
зближуються, а ексцес збільшується. Прикладом можуть бути
результати М. Бампаса, американського орнітолога. Він виз
начив, що домові горобці на вулицях Манхеттена взимку за
мерзали. При цьому виживали тільки ті птахи, ознаки яких
наближалися до середньої норми (довжина крила, цівки, висо
та дзьоба, маса і довжина тіла), тоді як птахи з відхиленнями
в бік зменшення чи збільшення розмірів і маси гинули. У 1952 р.
М. Карн і Л. Пенроуз встановили зв’ язок між масою новона
роджених дітей і їх смертністю: чим сильніше відхилення
2 4 2 ________________________________________________________ Розділ 4
в будь-який бік від середньої норми (3,6 кг), тим рідше діти
виживають.
Дією стабілізуючого добору пояснюються всі випадки збе
реження ознак на будь-яких рівнях організації. Прикладами
такого тривалого збереження адаптацій на морфологічному
рівні є кількість очей у хребетних (2), що не змінювалася про
тягом усієї історії; кількість променів парних кінцівок (5).
У тварин і у людини відомі мутації, які збільшують або змен
шують кількість пальців, а в ході еволюції хребетних неодно
разово відбувалася редукція кількості пальців (птахи, копитні).
Тому можна стверджувати, що збереження п’ ятипалості — ре
зультат дії стабілізуючого добору. На клітинному рівні органі
зації — це єдність будови клітинних органел у всіх еукаріотів,
на молекулярному — незмінна будова гена гемоглобіну всіх
хребетних. Стабілізуючий добір веде до накопичення рецесив
них мутацій, до підвищення гетерозиготності популяції. При
зміні умов середовища ця мінливість є матеріалом для пере
творення популяції під дією рушійного добору.
Альтернатива стабілізуючої форми добору — рушійний добір,
який веде до утворення еволюційно нових адаптивних ознак.
При направленій зміні середовища частіше виживають особи
ни, які мають індивідуальні особливості, що відповідають цій
зміні; особини з відхиленнями в протилежний бік, неадекватні
до зміни зовнішніх умов, частіше гинуть. У напрямі зміни се
редовища зона безумовної елімінації стискається, а зона умов
ної — зміщується. У протилежній частині кривої картина зво
ротна. У результаті — розподіл ознаки набуває асиметричної
форми, і відбувається направлене перетворення популяції.
Таким чином, рушійна форма добору веде до утворення но
вих пристосувань через направлену перебудову генофонду по
пуляції, а це супроводжується перебудовою генотипу особин.
У природі рушійна та стабілізуюча форми добору постійно
співіснують, і можна говорити лише про переважання тієї чи
іншої форми в певний період часу за певною ознакою.
Однак еволюціоністи виділили третю форму добору — дизруптивний добір. Він веде до розпадання (дивергенції) попе
редньої форми і формування двох і більше норм реакції. Ме
ханізм його протилежний до механізму стабілізуючого добору,
бо зберігаються крайні варіанти й елімінують проміжні. При
Популяційно-видовий рівень організації життя
2 43
кладом дизруптивного добору є виникнення рас сучасного
виду Ното заріепз.
У результаті сформувалася концепція трьох форм добору,
які плавно і поступово переходять одна в одну. Всі форми до
бору належать до індивідуального добору. Водночас існують
індивідуальна, родинна, групова форми елімінації, пов’ язані
з індивідуальною, міжродинною, міжпопуляційною і міжвидо
вою боротьбою за існування.
3. Хвилі життя (популяційні хвилі) — коливання чисельності
особин популяції, яке залежить від сезонних явищ або непе
ріодичних факторів (за С. С. Четвериковим). При цьому часто
та алелів різних генів у популяціях змінюється випадково,
незалежно від добору. Популяція, що різко зменшилася кіль
кісно, з часом відновлюється і досягає попереднього рівня за
рахунок особин, які вижили випадково. Внаслідок того, що
окремі особини не можуть бути носіями всього генофонду по
пуляції, відновлена у кількісному аспекті популяція має інший
генофонд. Коливання чисельності особин може приводити до
тимчасового розширення ареалу виду. Організми виявляються
у нетипових для них умовах, що може викликати підсилення
мутаційного процесу. Ріст чисельності популяцій приводить
до їх злиття й обміну генофондом. У невеликих популяціях
(при зниженні їхньої чисельності та на межах ареалу) велику
роль відіграє дрейф генів.
Випадкове підвищення концентрації деяких мутацій дає
новий матеріал для добору. Таким чином, популяційні хвилі
поряд із мутаційним процесом є джерелом еволюційного ма
теріалу.
4. Ізоляція — неможливість обміну генами (схрещувань) між
особинами різних популяцій. Розрізняють географічну, біоло
гічну, екологічну, генетичну репродуктивну ізоляцію. Ізоляція
не створює нові форми, але сприяє дивергенції.
Географічна ізоляція виникає у результаті фрагментації ареалу
материнського виду. Вона може бути наслідком розмежування
фізичними бар’ єрами (гірськими хребтами, водними простора
ми тощо).
Біологічна ізоляція забезпечується двома механізмами: докопуляційними і посткопуляційними. Докопуляційні механізми
244
Розділ 4
роблять неможливим схрещування. Наприклад, різна темпе
ратура води у період розмноження в жаби озерної та жаби
ставкової; зміщений період цвітіння розоцвітих запобігає пе
рехресному запиленню.
Екологічна ізоляція передбачає відмінності поведінки
у близьких форм (спів птахів, обмін звуковими і хімічними
подразниками). Посткопуляційні механізми передбачають ізо
ляцію при схрещуванні та після запліднення.
Генетична ізоляція (зміна кількості чи структури хромосом)
і неможливість розвитку зиготи веде до загибелі ембріона.
Навіть при утворенні життєздатних гібридів вони будуть не
плідні. Оскільки гібридний організм має у генотипі негомологічні хромосоми, які не вступають у мейоз, то гамети не будуть
утворені. Наприклад, гібрид між самкою коня і ослом — мул —
є неплідним.
Якщо гібриди дають потомство, то воно є малоплодюче і ос
лаблене. Конкуренція з іншими особинами призводить до заги
белі таких гібридів. У цьому полягає репродуктивна ізоляція.
Морфофізіологічні відмінності в органах розмноження також
є ізолюючим механізмом.
Боротьба за існування. Ч. Дарвін звернув увагу на складні
форми взаємовідносин між організмом і навколишнім середо
вищем, на різні форми залежності між собою, на пристосуван
ня до несприятливих умов. Такі форми залежності Ч. Дарвін
називав боротьбою за існування, або боротьбою за життя.
Складні взаємовідносини у природі можна згрупувати у три
форми:
1. Взаємовідносини з умовами середовища, абіотичними
факторами (клімат, температура, вологість, ґрунт тощо).
2. Міжвидові взаємовідносини — з особинами інших видів.
3. Внутрішньовидові взаємовідносини— між особинами
і групами індивідів одного виду.
Перша форма виникає залежно від кліматичних і ґрунтових
умов, інших абіотичних факторів, які впливають на життєдіяль
ність організмів. У процесі еволюції в організмів різних видів
виробляється ряд пристосувань до несприятливих умов. Однак
Популяційно-видовий рівень організації життя______________________ 2 45
відносність цих пристосувань і постійні зміни довкілля потре
бують удосконалення адаптації. З іншого боку, живі організми
також змінюють неживу природу.
Міжвидова боротьба за існування, за Ч. Дарвіном, веде до
витіснення одного виду іншим. Вона включає не тільки кон
куренцію, але і пряму боротьбу за життя, — відносини в лан
цюгах живлення. У результаті йде інтеграція виду, вироблен
ня морфофізіологічних кореляцій між особинами даного виду.
О. М. Сєвєрцов назвав такі кореляції конгруенціями. Тобто
загибель одних організмів у результаті прямої боротьби за іс
нування веде до відбору і збереження будь-яких ознак, які
знижують смертність, у тому числі ознак, які зменшують ін
тенсивність внутрішньовидової конкуренції. Іншим прикладом
є поява ознак, корисних для виду в цілому і шкідливих для
окремої особини. Тривалий внутрішньоутробний розвиток
шкідливий для самки-матері, але знижує вікову загибель. Так
само небезпечний ризик для самки, яка імітує поранення і від
водить переслідування від місця, де ховається маля.
Наслідками міжвидової боротьби є витіснення одних видів
іншими, з їх остаточною акліматизацією. Виникають захисні
забарвлення, явище мімікрії (подібність до якого-небудь пред
мета чи інших тварин за формою і поведінкою), з’ являється
імунітет до захворювань, здатність виробляти певні речовини
(фітонциди й антибіотики).
Міжвидова боротьба є причиною коеволюції (спряженої
еволюції). При цьому вдосконалення однієї групи організмів
обумовлює вдосконалення іншої, залежної групи: хижак — жер
тва, паразит — хазяїн, комахи — комахозапильні рослини.
Згідно з Ч. Дарвіном, найважливіша для еволюції внутріш
ньовидова боротьба за існування. Вона є особливо гострою
і визначає виживання або загибель організмів. При цьому від
значається, що чим більше подібні організми, тим гостріша
конкуренція між ними. Внутрішньовидова боротьба загалом
проходить через відбір особин, які ухиляються від виникнення
різновидів і видоутворення. Якщо особини, які більше подібні,
сильно конкурують між собою, то виживати (підлягати доборо
ві) будуть ухилені, менш подібні особини. У результаті почина
ється розходження ознак — дивергенція. Ті організми, які ухи
лилися, поступово пристосовуються до нових умов середовища,
246
Розділ 4
а проміжні форми (гірше пристосовані, ніж крайні варіанти)
продовжують конкуренцію між собою, вимирають більш ін
тенсивно. Зрештою різновиди, які ухиляються, відособлюють
ся одні від одних. Отже, внутрішньовидова (міжпопуляційна)
боротьба за існування веде до диференціації виду: виникнення
підвидів і видоутворення. У ряді випадків між особинами однієї
популяції відбувається конкуренція за територію, самців за
самку, що веде до статевого добору. Виробляється статевий
диморфізм за вторинними статевими ознаками, складні комп
лекси шлюбної поведінки і захисту території. Ці ознаки не
мають для самців захисного значення. Вони не мали б закріп
люватися добором, бо демаскують самця і знижують виживан
ня. Але добір за своєю суттю направлений не на виживання
окремої особини, а на повноцінне і чисельне потомство, яке
вона залишить. Таке потомство дають фізично розвиненіші
самці з добре вираженими вторинними статевими ознаками.
Вони перші вступають у розмноження, обмежуючи участь
у ньому інших самців. Дивергенція за розпізнавальними озна
ками самців і самок спочатку іде під дією звичайного природ
ного добору. Далі ці ознаки стають об’ єктом статевого добору,
кінцевий результат якого збігається з результатом природного
добору.
Внутрішньовидові взаємозв’язки є складними (між особи
нами різної статі, між батьківськими і дочірніми особинами,
між особинами одного покоління у процесі індивідуального
розвитку, відносини у стаді, зграї тощо). При значному збіль
шенні чисельності особин одного виду та обмеженні умов для
їх існування між окремими індивідами виникає гостра взає
модія, яка веде до загибелі частини або всіх особин чи до усу
нення їх від розмноження. Крайньою формою таких відносин
є канібалізм — поїдання особин свого виду.
4.2. Основні напрями еволюційного процесу
Теорія головних напрямів еволюційного процесу відповідає
на питання: якими шляхами може підвищуватися пристосо
ваність організмів у ході філогенезу? Згідно з О. М. Сєвєрцовим
найбільш очевидний напрям еволюції — біологічний прогрес,
Популяційно-видовий рівень організації життя______________________ 2 4 7
або збільшення пристосованості нащадків порівняно з предка
ми. Критеріями біологічного прогресу є:
1) збільшення чисельності;
2) розширення ареалу (екологічної ніші);
3) прогресивна диференціація — збільшення кількості си
стематичних груп, які утворюють даний таксон: видів у роді,
видів і родів у родині.
Виникнення нової адаптації знижує елімінацію, у результаті
середній рівень чисельності виду зростає. Це веде до розширен
ня ареалу й освоєння нових екологічних умов, викликає по
дальшу диференціацію виду, підсилює дивергенцію. Протилеж
ністю біологічного прогресу є біологічний регрес, який приз
водить до вимирання таксонів. Критерії біологічного регресу
діаметрально протилежні критеріям біологічного прогресу.
Основі напрями еволюційного процесу (за О. М. Сєвєрцовим
та 1.1. Шмальгаузеном):
1. Ароморфоз, або морфофізіологічний прогрес — стрибко
подібні зміни, які підвищують рівень організації організму.
Це веде до освоєння нових середовищ існування, джерел жив
лення. Ароморфози приводять до формування класів, типів.
Наприклад, виникнення хребта у хребетних, квітки у покри
тонасінних, парних кінцівок земноводних із парних плавців
риб, виникнення серця і його подальша еволюція. Поява за
родкових оболонок яйця забезпечила хребетним розвиток за
родків на суші, у результаті сформувалися наземні хребетні,
які в період розмноження не були пов’язані з водою.
2. Ідіоадаптація — вироблення часткових пристосувань до
конкретних умов, що не змінює рівня організації. Характерна
для нижчих систематичних груп: видів, родів, родин, рядів.
Прикладами можуть бути типи пристосувального забарвлен
ня, розвиток мімікрії, різна форма дзьоба птахів, ротових
органів комах, пристосування до запилення квіток чи до роз
повсюдження насіння покритонасінних.
3. Загальна дегенерація, або морфофізіологічний регрес —
вторинне спрощення організації. Спостерігається при переході
248
Розділ 4
організму до осілого або до паразитичного способу життя.
У паразитів стьожкових червів зникає травна система, але
добре розвинена статева. Двостулкові молюски ведуть прикріп
лений спосіб життя, тому у них з’ являються сифони для за
хоплення харчового матеріалу. У паразитичних рослин (омела)
замість кореня — присоски.
Мікроеволюція — це процес адаптивного перетворення по
пуляцій: від виникнення спадкових змін особин через форму
вання на їх основі нових адаптацій за дії природного добору
до виникнення нового виду.
Процеси, які приводять до формування надвидових систе
матичних категорій (родів, родин, рядів), називають макроево
люцією. При мікро- і макроеволюції діють однакові закономір
ності. Утворення видів пов’ язано з перетворенням генофонду
і дивергенцією у популяціях, боротьбою за існування і природ
ним добором. Макроеволюційні зміни — наслідок процесів на
мікроеволюційному рівні.
Дослідження вчених X X ст. (С. С. Четверикова, Р. Фішера,
1.1. Шмальгаузена, М. В. Тимофєєва-Ризовського та ін.) роз
ширили знання про закономірності макроеволюції. Встановле
но, що популяція — це елементарна еволюційна структура;
мутації — елементарний еволюційний матеріал, а мутаційний
процес, популяційні хвилі, ізоляції та природній добір — еле
ментарні еволюційні фактори.
Особини одного виду розміщені в межах ареалу у вигляді
популяцій, між якими є простори з низькою кількістю особин
одного виду. Особини в одній популяції вільно схрещуються
(панміксія), тому створюється особливий генофонд, який відріз
няється від інших популяцій. Особливості генетичних процесів
у популяціях див. у розд. 2.
Популяційно-видовий рівень організації життя
249
4.3. Закономірності розвитку органічного
світу. Теорії виникнення життя на Землі
Упродовж тисячоліть людям видавалось очевидним, що
жива природа була створена такою, якою ми її знаємо тепер,
і завжди залишалася незмінною. Водночас, ще у глибоку дав
нину були висловлені здогади про поступові зміни, розвиток
(еволюцію) живої природи.
Давньогрецький філософ Геракліт (VI—V ст. до н. е.) сфор
мулював положення про зміни в природі, які відбуваються
постійно: “ Все тече, все змінюється” . Його можна вважати
попередником еволюційних ідей. Інший давньогрецький мис
литель Емпедокл (V ст. до н. е.) висунув найдавнішу теорію
еволюції. Він уважав, що спочатку на світ з ’ явилися різні
частини організмів (голови, тулуби, ноги). Вони з’ єднувалися
в різних варіантах. Таким чином, на думку Емпедокла, з’ я
вилися кентаври (міфічні напівлюди-напівконі). Пізніше на
чебто всі нежиттєздатні комбінації загинули. Великий учений
Арістотель (384—322 рр. до н. е.) вибудував усі відомі йому
організми в ряд за ступенем їх ускладнення. Він проаналізував
загальний план будови вищих тварин, гомології та кореляції
органів. Фундаментальні твори Арістотеля “ Про частини тва
рин” , “Історія тварин” , “Про виникнення тварин” мали великий
вплив на розвиток біології. Арістотель є засновником теорії
“активного начала”, тобто самовільного спонтанного зароджен
ня, яка була поширена у Стародавньому Китаї, Вавилоні,
Єгипті. Дослідники вважали, що певні частини речовини мі
стять “ активне начало” , яке за сприятливих умов може ство
рити живий організм. Це “ активне начало” міститься у заплідне
ному яйці, в сонячному світлі, тіні, у гниючому м’ ясі.
Так, для зародження червів потрібне гниле м’ ясо. Однак
було зауважено, що черви у м ’ ясі з’являються після того, як
на м’ ясо сіли мухи. Прихильником теорії “ активного начала”
був Ж . Б. Ламарк (1744— 1829). Він припускав, що глисти
і кишковопорожнинні можуть самозароджуватись, а одноклі
тинні з ’ явились у результаті взаємного впливу тепла, вологи
й електрики.
250
Розділ 4
Лікар Франческо Реді 1688 р. піддав сумніву цю теорію
і припустив, що життя може виникати тільки з попередньо
існуючого життя (концепція біогенезу).
Ладзаро Спаланцані 1765 р. провів дослід, у якому м ’ясні
й овочеві відвари піддавали кип’ятінню протягом кількох го
дин. Подальші дослідження рідин через кілька днів не вияви
ли жодних ознак життя.
У 1860 р. Луї Пастер довів справедливість теорії біогенезу
і остаточно заперечив теорію спонтанного зародження. Він
довів, що раптове виникнення (“ спонтанне зародження”) мік
роорганізмів у різних видах гниючих екстрактів не є виник
ненням життя. Гниття і бродіння — результат життєдіяльності
мікроорганізмів, занесених ззовні.
У період середньовіччя природознавство контролювалося
церквою. Було заборонено висловлювати ідеї розвитку приро
ди, досліджувати давні наукові книги. Саме в цей час виникає
теорія креаціонізму (від лат. сгеаііоп — створення), яка ствер
джує, що органічний і неорганічний світ виник унаслідок акту
“ божественного творіння” . Ця теорія лежить в основі всіх
релігій. Вона заперечує зміну видів та їх історичний розвиток
і тісно пов’язана з теологією, ідеалізмом, катастрофізмом.
У 1865 р. X. Ріхтер розвинув ідею космічного посіву (панс
пермії). Згідно з нею, зародки життя були занесені на Землю
метеоритами або космічним пилом. Ідею розвинув шведський
фізикохімік Сванте Авенаріус у 1884 р., який припустив, що
життя на Землі походить від спор рослин або мікроорганізмів,
принесених з інших планет під дією світлового тиску або з ме
теоритами. Однак П. Беккерель і деякі інші вчені довели не
можливість переносу в життєздатному стані зародкового жит
тя. На нього згубно діють космічні промені, особливо корот
кохвильове ультрафіолетове випромінювання, яким пронизаний
Всесвіт.
Хоча до сьогодні ще не встановлений контакт з іншими
(позаземними) цивілізаціями, це не може бути доказом, що
життя існує тільки на Землі. Водночас, ймовірність існування
життя на інших планетах зовсім не означає, що “ зародки жит
тя” з таких позаземних “ плантацій” можуть безперешкодно
переноситися з одного космічного тіла на інше.
Популяційно-видовий рівень організації життя______________________ 251
Теорія біохімічної еволюції. У 1924 р. О. Опарін на зборах
Російського ботанічного товариства зробив доповідь “ Про ви
никнення ж иття” . Через 5 років незалежно від 0. Опаріна
подібні ідеї були розвинуті англійським вченим Дж. Холдейном.
Спільним у поглядах двох вчених є спроба пояснити походжен
ня життя як результату хімічної еволюції на первісній землі.
Як доповнення до цього Дж. Холдейн уперше висловив ідею
про відсутність кисню у первинній атмосфері Землі, або про
його мізерну кількість. Сучасна теорія походження життя
заснована на ідеї про те, що біологічні молекули могли виникну
ти в далекому геологічному минулому неорганічним шляхом.
Складну хімічну еволюцію можна зобразити узагальненою
схемою:
Атоми —> прості сполуки —» прості біоорганічні сполуки —>
-» макромолекули —» організовані біологічні системи.
Початком еволюції був нуклеосинтез у Сонячній системі,
коли утворювалися всі основні елементи (абіогенні та біогенні).
Перша умова хімічних перетворень — їх загальнокосмічний
характер, тобто, єдина хімічна основа цілого Всесвіту. Друга
(основна) умова — планетарна причина, яка визначається ма
сою планети (не більше 1/ 20 маси Сонця). Із планет Сонячної
системи масу, наближену до Землі, мають Венера і Марс, але
там існують інші умови.
Важливими умовами для виникнення і розвитку життя є
відносно постійна й оптимальна радіація та наявність води.
У Сонячній системі тільки Земля має гідросферу, а на Марсі є
лише незначна кількість води. Специфічні термічні особливості
води зумовлюють колообіг води у природі.
В океані у результаті хімічних реакцій виникли і нагрома
дились органічні речовини, які утворили “ первинний бульйон” ,
де могло зародитися життя.
Життя — особлива форма існування матерії, яка характе
ризується двома особливостями — самовідновленням та обміном
речовин із навколишнім середовищем. На цій основі визнані
коацерватна і генетична гіпотези.
О. Опарін, опираючись на експерименти Бунденберга де
Йонга, розвинув положення коацерватної гіпотези походження
252
Розділ 4
життя. Основою гіпотези є ствердження, що початкові етапи
біогенезу були пов’ язані з формуванням білкових структур.
Перші білкові структури (за О. Опаріним, протобіонти) з’я
вились у період, коли білкові молекули відмежовувались від
навколишнього середовища мембраною. Відбувався процес
коацервації — самовільне розділення водного розчину полімерів
на фази з різною концентрацією, що супроводжувалось утво
ренням мікроскопічних крапель із високою концентрацією
полімерів. Частина цих крапель поглинала зі середовища низь
комолекулярні сполуки: амінокислоти, глюкозу, каталізатори,
їх взаємодія означала виникнення найпростішого метаболізму
всередині протобіонтів.
Поступово об’ єми коацерватів збільшувались, і вони розпа
далися на дрібніші краплі, які знову продовжували рости. На
межі між коацерватами і зовнішнім середовищем вибудовува
лися молекули ліпідів, що привело до появи примітивної клі
тинної мембрани. Так виникла примітивна клітина. Усклад
нення протобіонтів здійснювалося добором таких коацерватних
крапель, які краще використовували речовини й енергію сере
довища.
Отже, центральним положенням у гіпотезі Опаріна є добір
як причина вдосконалення коацерватів.
Генетична гіпотеза. У 1929 р. Г. Мюллер висунув гіпотезу
про первинне виникнення нуклеїнових кислот як матричної
основи синтезу білків.
Основні етапи біогенезу:
— І етап — виникнення органічних речовин. Стенлі Міллер
(1953) змоделював умови, які теоретично існували на первісній
Землі. У створеній ним установці вдалося синтезувати такі
амінокислоти, як аспарагін, гліцин, глютамін. Разом із нук
леїновими основами були експериментально синтезовані пури
нові та піримідинові основи, цукор.
— II етап — утворення складних полімерів, які подібні до
нуклеїнових кислот і білків.
— III етап — поява перших живих організмів, які мали вну
трішню організацію протобіонтів і були пристосовані до умов
Популяційно-видовий рівень організації життя
253
навколишнього середовища завдяки стійкому обмінові речовин
та енергії, й успадкуванню цієї організації на основі реплікації
генетичного коду (матричного коду).
Гіпотези походження еукаріот:
1. Аутогенна гіпотеза (або гіпотеза інвагінації) — припускає,
що еукаріотична клітина виникла шляхом диференціації вихід
ної прокаріотичної клітини. Спочатку розвинулася зовнішня
клітинна мембрана з вп’ ячування всередину клітини, з якої
сформувалися окремі структури. Вони дали початок клітинним
органам.
2. Симбіотична гіпотеза (Л. Маргуліс) — еукаріотична клі
тина виникла внаслідок кількох актів симбіогенезу. Спочатку
об’ єдналася велика амебоподібна клітина з дрібними аеробни
ми бактеріями, які перетворилися в мітохондрії. Потім ця
симбіотична прокаріотична клітина включила в себе спірохетоподібні бактерії, з яких сформувалися кінетосоми, центросоми
і джгутики. Після відокремлення ядра в цитоплазмі (ознака
еукаріот) клітина з цим набором органел стала вихідною для
утворення Царств Гриби і Тварини. Об’ єднання прокаріотичної
клітини з ціанеями привело до утворення пластидної клітини,
що зробило можливим формування Царства Рослини.
Після виникнення одноклітинних другим ступенем еволюції
було утворення і прогресивний розвиток багатоклітинних
організмів. Цей ступінь включає такі стадії:
• Колоніальна одноклітинна. Приклад: колоніальні джгу
тикові Сопіит, Уоіоох, Рапдогіпа. Це об’ єднання клітин без
жодної спеціалізації їх функцій.
• Первинно-диференційована стадія — спеціалізація клітин
і їх поділ на соматичні (функція живлення і руху), генератив
ні (гонідії для розмноження). Спеціалізація відбувається на
тканинному, органному і системно-органному рівні. У киш ко
вопорожнинних сформована проста нервова система, яка ко
ординує діяльність інших типів клітин.
• Централізовано-диференційна стадія — ускладнення мор
фологічної структури через посилення тканинної спеціалізації.
254
Розділ 4
Виникає добре розвинута нервова система. Одночасно вдоско
налюються способи статевого розмноження — від зовнішнього
запліднення до внутрішнього, від вільної інкубації яєць поза
материнським організмом до живонародження.
Фіналом в еволюції багатоклітинної організації тварин була
поява організмів із поведінкою “розумного типу” . Тварини, які
мають високорозвинену умовно-рефлекторну діяльність, здат
ні передавати інформацію нащадкам не тільки через спад
ковість, але і надгаметним шляхом (передача молодим особинам
досвіду через навчання).
Порівняльно-анатомічні докази еволюції. Після виходу
у світ праць Ч. Дарвіна порівняльна анатомія продовжила ак
тивно розвиватись і суттєво доповнила еволюційне вчення.
Першорядне значення при з’ясуванні філогенетичної спорід
неності між організмами мало встановлене явище гомології
органів.
Гомологічними органами (від грец. цотопоз — подібний) —
називаються органи або структури, що мають спільне походжен
ня, але відрізняються деталями будови і функціями. Напри
клад: рука людини, крило птаха, передня кінцівка крота
гомологічні між собою. Також молочні залози гомологічні по
товим; щелепи ракоподібних — їх кінцівкам; жало бджоли —
яйцекладові інших комах; волосся ссавц ів— пір’ ю птахів
і лусці плазунів; зуби ссавців — лусці акули; слухові кісточки
середнього вуха ссавців (стремінце, ковадло і молоточок) — кіст
кам вісцерального черепа риб, амфібій; частини квітки (маточ
ка, тичинки, пелюстки) — листкам.
Доказами еволюції також є атавістичні та рудиментарні
органи.
Атавізм (від лат. аіаоиз — предок) — прояв у фенотипі ок
ремих особин виду рис далеких предків. Наприклад, надмірна
волохатість тіла, багатососковість або наявність хвоста у лю
дини, поява бічних пальців у коней, смугастість у домашніх
свиней тощо.
Рудимент (від лат. гидітепіит — зачаток) — це орган, який
втратив своє значення і зазнав часткової редукції. Він однако
Популяційно-видовий рівень організації життя
255
во розвинений у всіх особин певного виду. Наприклад, залишок
третьої повіки та червоподібний відросток сліпої кишки лю
дини, залишки листків (лусочки) на кореневищах рослин.
У людини рудиментарними органами є куприк (залишок хвос
тових хребців), зачаткові вушні м’ язи, які свідчать про те, що
предки людини мали рухливу вушну мушлю.
Порівняльно-анатомічні дослідження сучасних прогресивних
і примітивних форм дають змогу виявити “ перехідні форми” .
Це допомагає відтворювати шляхи еволюції. Наприклад, лан
цетник має ряд ознак, які наближають його з одного боку до
голкошкірих і напівхордових (баланоглос), а з другого боку до
хребетних, разом із якими він належить до одного типу хор
дових.
Серед сучасних ссавців існують однопрохідні (мають клоаку
і при розмноженні відкладають яйця, як плазуни), сумчасті та
плацентарні. Порівняння їх вказує на те, що ссавці споріднені
з плазунами і що еволюція ссавців йшла від тварин, які відкла
дали яйця, до живородних форм із ще недорозвиненою плацен
тою і, нарешті, до тварин, які народжують уже добре сформо
ваних малят.
Ембріологічні докази еволюції. Карл Бер перетворив емб
ріологію з описової науки в порівняльну. Він простежив усі
етапи розвитку пташенят курей від 1-го по 21-й день, від за
родкового диска в яйці, яке знесла курка, до прориву дзьобом
тонкої плівки, першого вдиху і першого писку. К. Бер спосте
рігав ту ранню стадію розвитку, коли на зародковій пластинці
утворюються два паралельних валики, які надалі змикаються
й утворюють мозкову трубку. Він вважав, що кожне нове утво
рення виникає збільш простої попередньо існуючої основи.
Таким чином, у зародку з’ являються спочатку загальні основи,
а з них відокремлюються більш спеціальні частини. Цей процес
поступового руху від загального до спеціального відомий як
диференціація.
К. Бер відкрив 1826 р. яйцеклітину ссавців (на прикладі
собаки), а 1834 р. — спинну струну, основу внутрішнього ске
лета ссавців.
Ембріологічні дослідження продовжили вчені О. О. Ковалевський та 1.1. Мечніков. Вони довели, що у процесі ембріонального
256
Розділ 4
розвитку в безхребетних, як і у хребетних, закладаються три
зародкові листки. Це підтверджує спільність походження всьо
го тваринного світу.
Саме О. О. Ковалевському належать відкриття стадії бла
стули у кишковопорожнинних, червів, голкошкірих, оболонників і ланцетника.
1.1. Мечніков описав інвагінаційну гаструлу. Порівняння
розвитку зародків усіх класів хребетних вказує на подібність
їх внутрішньої та зовнішньої будови на ранніх стадіях розвитку.
У ході їх подальшого розвитку подібність зменшується, з’являю
ться ознаки, характерні для певного класу, ряду, роду, виду.
Через десятки років було сформульовано біогенетичний
закон Ф. Мюллера і Е. Геккеля (незалежно один від одного) —
ряд форм, які проходить індивідуальний організм під час
свого розвитку від яйцеклітини до розвиненого стану, є корот
ким і стислим повторенням довгого ряду форм, який пройшли
предки того ж організму з давніх часів органічного творіння
до сьогодні. У стислій формі закон формулюється так: онтоге
нез є коротким повторенням філогенезу.
Пізніше 0. М. Сєвєрцов вивів теорію філембріогенезів. Філембріогенезами вчений називав зміни, які відбуваються у процесі
ембріонального розвитку, зберігаються в дорослому стані й
успадковуються нащадками. Він показав, що співвідношення
онтогенезу і філогенезу значно складніше, ніж вважав Е. Геккель. Насправді не філогенез творить індивідуальний розвиток
(нові еволюційні здобутки продовжують онтогенез, додають
нові стадії), як припускав Е. Геккель, а навпаки, спадкові змі
ни ходу онтогенезу ведуть до еволюційних перебудов (філоге
нез — це еволюція онтогенезу). Лише у випадку анаболії (над
ставки), коли еволюційна перебудова якого-небудь органу
проходить шляхом зміни пізніх стадій його індивідуального
розвитку, тобто нові ознаки формуються наприкінці онтогене
зу, дійсно спостерігається біогенетичний закон. Наприклад,
вторинна нирка хребетних пройшла стадію переднирки і пер
винної нирки.
При девіації (відхиленні) виникають новоутворення в серед
ніх стадіях розвитку. Наприклад, різні типи лусок плазунів
і риб.
При архалаксисі розвиток змінюється на початкових етапах
розвитку. Наприклад, розвиток волосяного покриву у ссавців,
Популяційно-видовий рівень організації життя______________________ 2 5 7
гомологічного до ембріональних закладок луски риб і пла
зунів.
Шляхом філембріогенезу досягається не тільки підвищення
організації, але і спрощення, як у паразитів (наприклад, сакуліна із ракоподібних) і сидячих форм (наприклад, оболонники).
Палеонтологічні докази еволюції. У результаті навколосвіт
ньої подорожі Ч. Дарвіна на кораблі “ Бігль” (1831— 1836) було
зібрано колекцію палеонтологічних знахідок. їх аналіз пока
зував, що вимерлі та сучасні тварини мають спільне походжен
ня, але останні суттєво змінені. Причиною могли бути зміни
земної поверхні, що могло призвести до вимирання видів, за
лишки яких лежать у пластах землі. Для остаточних висновків
даних Ч. Дарвіна було недостатньо.
О.
О. Ковалевському вдалося побудувати філогенетичні ряди
непарнокопитних, хоботних, хижаків, молюсків.
Доведено, що чим молодші пласти землі, тим рештки тварин
і рослин ближчі до сучасних форм. За розміщенням осадових
порід можна визначити їх відносний вік (суть методу закладе
на в XVIII—X IX ст. У. Смітом). Серед викопних рештків завжди
виділяють декілька найбільш звичайних, численних і широкорозповсюджених видів, які називають керівними копалина
ми. Абсолютний вік вивержених вулканічних порід можна
визначити за вмістом у них радіоактивних елементів і продук
тів їх розпаду. Виявлено викопні “ перехідні форми” (першоптах
археоптерикс, звірозубі ящури, насіннєві папороті). “Перехід
ні форми” поєднують ознаки організмів різних систематичних
груп. Наприклад, першоптах зберіг ряд ознак плазунів. Озна
ки плазунів: відсутність рогового дзьоба; наявність зубів; від
сутність кіля грудини; довгий хвостовий відділ хребта, що
складається з багатьох хребців. Ознаки птахів: наявність пір’я,
передні кінцівки перетворені на крила; ключиці зрослись
у “виделку” ; наявність цівки.
На дереві родоводу (графічному зображенні філогенезу) па
леонтологічні дані та методи недостатні. Тут допомагає метод
потрійного паралелізму, який ввів у науку німецький вчений
Е. Геккель. Він заснований на порівнянні палеонтологічних,
порівняльно-анатомічних та ембріологічних даних. Вивчення
індивідуального розвитку сучасних організмів дозволяє певного
258
Розділ 4
мірою міркувати про хід еволюційних змін їх далеких предків,
у тому числі тих, які не збереглися в палеонтологічному літо
писі.
Сучасні знання про еволюцію та її фактори склалися у так
звану синтетичну теорію еволюції, яка є результатом розвитку
дарвінізму, генетики, екології та інших біологічних наук.
4.4. Філогенез і антропогенез
Еволюція кровоносної системи хордових тварин. Наяв
ність рідкого динамічного внутрішнього середовища є необхід
ною умовою існування багатоклітинних організмів, оскільки
воно забезпечує інтеграцію організму в цілісну систему, вико
нуючи транспортні функції. Ці функції є основними для кро
воносної системи. Кровоносна система всіх хордових замкнена
і утворена двома основними артеріальними судинами: черевною
та спинною аортами. У ланцетника кровоносна система най
простіша. Коло кровообігу одне. Черевною аортою венозна
(деоксигенована) кров надходить до приносних зябрових ар
терій, насичується киснем, по виносних зябрових артеріях
проходить до коренів спинної аорти, котрі симетрично роз
міщені з обидвох боків тіла. Передні гілки цих судин — це
сонні артерії, а задні гілки на рівні заднього кінця глотки утво
рюють спинну аорту, котра розгалужується до численних ар
терій і, таким чином, постачає всі органи і тканини. Після
тканинного газообміну деоксигенована кров надходить до пар
них передніх і задніх кардинальних вен, які з кожного боку
впадають у кюверову протоку. Обидві кюверові протоки впа
дають до черевної вени. Від стінок травної системи венозна
кров надходить ворітньою веною печінки до печінкового виро
сту і тут формується система капілярів. Тоді капіляри знову
збираються й утворюють венозну судину — печінкову вену,
котра впадає в черевну аорту.
У риб більш активний спосіб життя передбачає інтенсифі
кацію метаболічних процесів. У зв’язку з цим на тлі олігомеризації їх артеріальних зябрових дуг до чотирьох пар в них
відмічається високий ступінь диференціації: зяброві судини
Популяційно-видовий рівень організації життя
259
розгалужуються до капілярів, які пронизують зяброві пелю
стки. У процесі інтенсифікації скоротливої функції черевної
порти частина її перетворюється на двокамерне серце, утворе
не передсердям і шлуночком. До передсердя прилягає венозний
синус, а до шлуночка — цибулина аорти. В іншому кровоносна
система риб відповідає її будові у ланцетника.
В амфібій, внаслідок їх виходу на сушу та формування ле
геневого дихання, виникає два кола кровообігу. Відповідно,
н будові серця й артерій з’являються пристосування, спрямовані
на розмежування артеріальної та венозної крові. Серце амфібій
трикамерне і складається з двох передсердь (правого та лівого)
і шлуночка. У шлуночку є три порції крові: венозна, змішана
й артеріальна. Від правої частини шлуночка бере початок єди
на судина — артеріальний конус, що розгалужується на три
мари судин: шкірно-легеневі артерії, дуги аорти, сонні артерії.
Як і у всіх більш високоорганізованих класів, до правого пе
редсердя впадають вени великого кола, котрі несуть деоксигеновану кров, до лівого— вени малого кола з оксигенованою
кров’ ю. Під час скорочення передсердь до шлуночка, у якого
внутрішня стінка має велику кількість м ’ язових перетинок,
одночасово надходять обидві порції крові. Повного змішуван
ня немає у зв’ язку зі своєрідною будовою стінки шлуночка,
тому при його скороченні перша порція венозної крові надхо
дить до артеріального конуса і за допомогою спірального кла
пана, що є в його стінці, проходить у шкірно-легеневі артерії.
Змішана кров із середини шлуночка надходить таким самим
чином до дуг аорти, а невелика кількість артеріальної крові,
яка потрапляє останньою до артеріального конуса, надходить
у сонні артерії. Дуги аорти огинають серце і стравохід дорзаль
но, утворюють спинну аорту, котра постачає всі органи тіла,
за винятком голови. Задні кардинальні вени редукуються, і їх
функціонально замінює непарна задня порожниста вена. Пе
редні кардинальні вени називають уже яремними венами,
п кюверові протоки, в які вони впадають разом із підключич
ними венами, — передніми порожнистими венами.
У рептилій виникають такі прогресивні зміни в будові кро
воносної системи: у шлуночку серця є неповна перетинка, котра
запобігає повному змішуванню крові, що надходить з правого
260
Розділ 4
та лівого передсердь, від серця відходять три судини, що ут
ворилися внаслідок поділу артеріального стовбура. З лівої
половини шлуночка бере початок права дуга аорти з оксигенованою кров’ю, з правої — легенева артерія, що несе деоксигеновану кров. Зі середини шлуночка, у ділянці неповної пере
тинки, бере початок ліва дуга аорти зі змішаною кров’ ю.
Обидві дуги аорти зростаються й утворюють спинну аорту,
у якій кров змішана, проте більш насичена киснем, ніж у ам
фібій. Окрім цього, сонні та підключичні артерії беруть поча
ток від правої дуги аорти, а, отже, оксигенованою кров’ ю по
стачається не лише голова, а також і передні кінцівки. Венозна
система рептилій не має принципових відмінностей від систе
ми вен амфібій.
У птахів і ссавців прогресивні зміни кровоносної системи
полягають в повному розмежуванні венозного й артеріального
плину крові. Це досягається, насамперед, завершеною чотирикамерністю серця, а, по-друге, редукцією лівої (у птахів) чи
правої (у ссавців) дуги аорти. В результаті цього всі органи
птахів і ссавців (крім печінки) постачаються артеріальною кро
в’ю. У венах великого кола кровообігу також спостерігаються
прогресивні зміни: виникла безіменна вена, яка об’ єднала ліві
яремну та підключичну вени з правими, тому залишилась од
на передня порожниста вена. Ліва кюверова протока у вигляді
рудиментарної судини зіпиз согопагіз збирає венозну кров ли
ше від міокарду, а непарна і напівнепарна вени — рудименти
задніх кардинальних вен, — мають суттєве значення переважно
у випадку формування обхідних шляхів венозного відтоку через
кава-кавальні анастомози, що формуються ними (рис. 4.1).
Ф ормування арт еріальних судин у ссавців і людини —
результат філогенезу артеріальних зябрових дуг хребетних
тварин. У ембріогенезі абсолютної більшості хребетних закла
дається шість пар артеріальних зябрових дуг, що відповідають
шести парам вісцеральних дуг черепа. У зв’язку з тим, що пер
ші дві пари вісцеральних дуг входять до складу лицьового че
репа, дві перші артеріальні дуги швидко редукуються. Решта
чотири пари функціонують як зяброві артерії. У наземних
хребетних третя пара зябрових артерій втрачає зв’ язок із ко
ренями спинної аорти і несе кров до голови (сонні артерії).
Популяційно-видовий рівень організації життя
261
Рис. 4.1. Будова серця хордових: а — риби: 1 — артеріальний
конус; 2 — передсердя; 3 — венозний синус; 4 — шлуночок; б —
земноводні: 1 — сонна артерія; 2 — легенева артерія; 3 — легене
ва вена; 4 — артеріальний конус; 5 — передсердя; 6 — шлуночок;
и — рептилії: 1 — права дуга аорти; 2 — легенева артерія; 3 — лі
ва дуга аорти; 4 — легенева вена; 5 — передсердя; 6 — шлуночок;
7 — міжшлуночкова перетинка; — ссавці: 1 — ліва дуга аорти;
2 — легенева артерія; 3 — легенева вена; 4 — передсердя; 5 — шлу
ночок; 6 — міжшлуночкова перетинка
г
2 62
Розділ 4
Судини четвертої пари розвиваються найбільше і разом з ді
лянкою кореня спинної аорти у постембріональному періоді
формуються в дуги аорти — основні судини великого кола
кровообігу.
У земноводних і плазунів обидві судини розвинені та беруть
участь у кровообігу. У ссавців і людини також закладаються
обидві судини 4-ї пари, а пізніше права дуга аорти редукуєть
ся таким чином, що від неї залишається невеликий руди
мент — плечоголовий стовбур. П ’ ята пара артеріальних дуг
тому, що вона функціонально дублює четверту, редукується
у всіх наземних хребетних, за винятком хвостатих амфібій.
Шоста пара, котра постачає кров’ ю, крім зябер, ще і плаваль
ний міхур, у кистеперих риб формується у легеневу артерію
(рис. 4.2).
Рис. 4.2. Еволюція артеріальних зябрових дуг хребетних:
а — закладка в ембріогенез: І—VI — зяброві дуги; б — зяброві ду
ги риби; в — зяброві дуги амфібій; г — зяброві дуги рептилій; д — зяб
рові дуги ссавців: І — сонні артерії; 2 — боталова протока; 3 — ар
теріальні дуги великого кола кровообігу; 4 — легеневі артерії
Популяційно-видовий рівень організації життя______________________ 2 6 3
Вроджені вади розвитку серця і судин у людини — наслі
док порушень ембріонального розвитку. Персистенція обидвох
дуг аорти четвертої пари — одна з найбільш поширених атаві
стичних вад розвитку судин, трапляється 1 випадок на 200 роз
тинів дітей, котрі померли від вроджених вад серця. При ць
ому обидві дуги зростаються позаду стравоходу і трахеї, утво
рюючи низхідну частину спинної аорти (подібно до земноводних
і плазунів). Вада проявляється порушенням ковтання (дисфа
гія). Дещо частіше (2,8 випадка на 200 розтинів) трапляється
порушення редукції правої дуги аорти з редукцією лівої. Ця
аномалія часто клінічно не проявляється. Одна з поширених
вад (0,5— 1,2 випадка на 1000 новонароджених) — персистенція
артеріальної (боталової) протоки, яка є частиною кореня спин
ної аорти між четвертою і шостою парами артерій зліва. Про
являється регургітацією артеріальної крові з великого кола
кровообігу в мале. Дуже важка вада — персистенція первин
ного артеріального стовбура, у результаті якої з серця виходить
лише одна судина, яка локалізується переважно над дефектом
у міжшлуночковій перетинці. Ця вада у більшості випадків
закінчується смертю. Порушення диференціації первинного
стовбура може призвести до транспозиції судин — виходу аор
ти з правого шлуночка, а легеневого стовбура — з лівого (1 ви
падок на 2500 новонароджених). Ця вада несумісна з життям.
Серед вад розвитку венозного русла можливою є персистенція
обидвох верхніх порожнистих вен. Якщо вони обидві впадають
до правого передсердя, то аномалія — без клінічного прояву.
За умови впадання лівої порожнистої вени до лівого передсер
дя відбувається викид венозної крові у велике коло кровообігу.
Така вада несумісна з життям і трапляється з частотою 1 % від
усіх вроджених вад серцево-судинної системи.
Рідкісна вроджена аномалія — редукція нижньої порожни
стої вени. Відтік крові від нижньої частини тулуба і нижніх
кінцівок здійснюється у цьому випадку через колатералі не
марної та напівнепарної вен, що є рудиментами задніх карди
нальних вен (рис. 4.3).
264
Розділ 4
І
II
в
Ш
Рис. 4.3. Вроджені вади розвитку серця і судин у людини:
а — персистенція боталової протоки у людини: 1 — легеневий
стовбур; 2 — ліва дуга аорти; 3 — боталова протока; б — онтофілогенетичні основи формування вад розвитку серця і судин у лю
дини: І — серце зародка людини у віці 5 тижнів; I I — те ж саме,
у розрізі; I I I — розмежування артеріального конуса зародка лю
дини у віці 6 тижнів; I V — серце риби; V — те ж саме, у розрізі;
VI — персистенція не розмежованого артеріального конуса люди
ни: 1 — артеріальні зяброві дуги; 2 — артеріальний конус; 3 — пе
редсердя; 4 — кардинальні вени; 5 — шлуночок; 6 — загальна
легенева артерія; 7 — ліва дуга аорти; в — аномалії міжпередсердної (І, II ) і міжшлуночкової (III) перетинок
Популяційно-видовий рівень організації життя
265
Еволюція нирки:
— Еволюція нирки у хордових тварин. Органом виділення
у хребетних є нирки — парні компактні органи, структурна
одиниця яких представлена нефроном. У найбільш примітив
ному вигляді — це лійка, яка відкривається в целом і з’ єднана
з видільним канальцем, який впадає у загальний вивідний ка
нал — сечову протоку. У філогенезі хребетних нирка пройшла
три етапи еволюції:
— переднирка, або головна нирка (пронефрос);
— первинна, або тулубова нирка (мезонефрос);
— вторинна, або тазова нирка (метанефрос).
Переднирка повністю розвивається і функціонує як само
стійний орган у личинок риб і земноводних. Вона міститься на
передньому кінці тіла, складається з 6— 12 нефронів, лійки
яких відкриті в целом, а вивідні канальця впадають у пронефричний канал, який з’ єднаний із клоакою. Переднирка має
сегментну будову. Продукти дисиміляції фільтруються у целом
із кровоносних судин, які формують клубочки і розташовані
біля нефронів.
У дорослих риб і земноводних позаду переднирок, у тулубо
вих сегментах тіла, формуються первинні нирки, у яких є кіль
ка сотень нефронів. Під час онтогенезу кількість нефронів
збільшується за рахунок їх брунькування один від одного з по
дальшою диференціацією. Вони вступають у зв’ язок із крово
носною системою, формуючи капсули ниркових клубочків.
Капсули мають вигляд двобічних чаш, у яких розміщуються
судинні клубочки, завдяки чому продукти дисиміляції можуть
надходити безпосередньо з крові у нефрон. Деякі нефрони пер
винної нирки зберігають зв’ язок із целомом через лійки, а ін
ші втрачають його.
Видільні канали видовжуються, і в них відбувається реаб
сорбція — зворотне всмоктування у кров води, глюкози й інших
речовин, тому концентрація продуктів дисиміляції у сечі збіль
шується. Тварини з такою ниркою можуть жити лише у вод
ному або вологому середовищі, оскільки води зі сечею втра
чається дуже багато. Первинна нирка зберігає ознаки метамерної будови.
266
Розділ 4
У плазунів і ссавців виникають вторинні нирки. Вони за
кладаються в тазовому відділі тіла і містять сотні тисяч нефронів ускладненої будови. У новонародженої дитини в нирці
налічується близько 1 млн нефронів. Вони утворюються за
рахунок багаторазового галуження нефронів, які розвивають
ся. Нефрони не мають лійки і тому втрачають зв’язок із целомом. Канадець нефрона видовжується і завдяки цьому тісніше
контактує з кровоносною системою, а у ссавців диференціює на
проксимальну та дистальну частини, між ними розміщується
петля Генле.
Така будова нефрона забезпечує не лише повноцінну філь
трацію плазми крові у капсулі, але, що дуже важливо, зворот
не всмоктування у кров води, глюкози, гормонів, солей та інших
необхідних для організму речовин. Тому концентрація продук
тів дисиміляції у сечі, яка виділяється вторинними нирками,
є високою, а сама кількість сечі — малою. У людини за добу
в капсулі нефронів двох нирок фільтрується близько 150 л
плазми крові, а сечі виділяється близько 2 л. Ці особливості
дозволяють тваринам, які мають вторинні нирки, не залежати
від водного середовища. У плазунів вторинні нирки протягом
життя зберігаються на місці їх початкового закладання — у та
зовому відділі. У них простежуються риси первинної метамерної будови (рис. 4.4).
Нирки ссавців розміщені у поперековому відділі, у більшості
сегментація не виражена. В онтогенезі людини спостерігається
рекапітуляція у розвитку нирки: закладання спочатку про-,
потім мезо-, а пізніше метанефроса. Останній розвивається
у тазовому відділі, а потім за рахунок різної швидкості росту
хребта, тазу й органів черевної порожнини переміщується
у поперековий відділ. На початкових етапах розвитку нирки
людини сегментовані. Пізніше метамерія зберігається лише
у вигляді ниркових пірамід.
Складність формування нирок сприяє появі відхилень від
нормального процесу. Крім того, у процесі росту зародка від
бувається переміщення вторинної нирки із тазової частини
у поперекову. Аномалії розвитку нирок бувають різними: одна
із нирок може не піднятись і залишитись у ділянці таза. При
низькому розташуванні обидвох нирок і зростанні їх нижніми
полюсами утворюється підковоподібна нирка. Обидві нирки
Популяційно-видовий рівень організації життя
267
Рис. 4.4. Еволюція нефрону: а — переднирка; б — первинна
нирка; в, г — вторинна нирка: 1 — збірна трубочка; 2 — видільний
канадець; 3 — нефростом; 4 — целом; 5 — капілярний клубочок;
в — капсула; 7,8 — звивистий канадець; 9 — петля нефрону
можуть розміщуватись по один бік від середньої лінії і зроста
тись у суцільну ниркову масу. Кількість нирок може бути
більшою або меншою від нормальної, дуже рідко зустрічається
третя нирка (яка лежить на хребті між двома нирками або
нижче однієї з них).
268
Розділ 4
— Еволюція сечостатевих проток у хордових тварин. В ем
бріогенезі всіх хребетних при розвитку переднирки вздовж
тіла, від головного відділу до клоаки, закладається канал, по
якому продукти дисиміляції з нефронів надходять у зовнішнє
середовище. Це пронефричний канал. Під час розвитку пер
винної нирки цей канал розщеплюється на два канали, які
розміщені паралельно, або другий канал утворюється у поз
довжньому потовщенні стінки першого. Один із них — вольфів
(мезонефричний) — зв’язується з нефронами первинної нирки.
Другий — мюллерів (парамезонефричний) — зростається пе
реднім кінцем з одним із нефронів переднирки й утворює яйцевід, що одним кінцем відкривається лійкою у целомі, а іншим
кінцем упадає в клоаку.
Незалежно від статі у всіх хребетних формуються обв’ язково
два канали: вольфів і мюллерів. Але подальший їх розвиток
у різних статей і представників різних класів відбувається не
однаково. У самок риб і земноводних (анамнії) вольфів канал
завжди виконує функцію сечової протоки, а мюллерів — яйцеводу. У самців мюллерів канал редукується, тому статеву і виділь
ну функцію виконує вольфів канал. Сім’ яні канали впадають
у нирку, а сперматозоїди при заплідненні виводяться зі сечею
У воду.
У плазунів і ссавців більша частина вольфового каналу не
бере участі у виведенні сечі, тільки найбільш каудальна частина
в ділянці впадання у клоаку утворює випин, який стає сечопроводом вторинної нирки. Сам вольфів канал у самців виконує
функцію сім’явипорскувального каналу. Мюллерів канал у них
зазнає редукції. У самок вольфів канал редукується, за винят
ком каудальної частини, з неї формується сечопровід, а мюл
лерів стає яйцеводом. У плацентарних ссавців (амніот) мюллерів
канал диференціюється на власне яйцевід, матку і піхву. Ос
кільки мюллерів канал є парним утвором, то він зберігає пар
ність у будові яйцекладних і частково у сумчастих ссавців,
тому у них є дві піхви, дві матки та два яйцеводи. Подальша
еволюція веде до зростання мюллерових каналів, і утворюють
ся одна піхва та одна матка. Матка може бути подвійною (у гри
зунів), двороздільною (у хижих), дворогою (у китоподібних) або
простою (у приматів і людини).
Популяційно-видовий рівень організації життя______________________ 2 6 9
В ембріогенезі людини закладаються парні вольфові та мюллерові канали. Пізніше залежно від статі здійснюється їх ре
дукція. Рудимент мюллерового каналу у чоловіків розміщуєть
ся в передміхуровій залозі (простаті). Канальці передньої ча
стини первинної нирки вступають у зв’ язок зі сім’ яниками та
перетворюються на придатки сім ’ яника — епідидиміс.
У плодів жіночої статі можливе порушення редукції вольфових каналів, які розміщуються по боках від піхви. Ця ано
малія тісно пов’ язана з можливістю утворення кістозів і злоя
кісних перероджень. Поширеними аномаліями розвитку є
різні форми подвоєння матки (частота аномалії 1 на 1000).
Вони розвиваються як результат порушення зростання мюл
лерового каналу. Порушення зростання парних зачатків ста
тевого члена в ембріогенезі людини може призвести до форму
вання такої аномалії розвитку, як його подвоєння (рис. 4.5).
Еволюція статевої системи. Статеві залози у хребетних
розвиваються у вигляді парних складок частини нефрогонотома в ділянці ніжки.
Диференціація клітин на статеві й соматичні у процесі фі
логенезу була найбільш ранньою.
У процесі еволюції репродуктивна система у хордових змі
нилася від подібних за будовою до спеціалізованих жіночих
і чоловічих залоз та до виникнення зв’ язку з різними для кож
ної статі частинами видільної системи, які стали статевими
протоками; від зовнішнього до внутрішнього процесу запліднен
ня і до пов’ язаних із ним пристосувань у особин обох статей.
У ланцетника (самця і самки) статеві залози метамерно роз
ташовані у стінках навколозябрової порожнини. Статевих
протоків немає. Дозрілі статеві клітини через розриви стінок
потрапляють у навколишнє середовище через атріопор.
У риб статеві органи досить різноманітні. Яєчник має фоліку
лярний тип будови, а у сім’яниках (частіше парних) з’ являються
сім’ явиносні канальці. У риб вивідні протоки статевих залоз
уже тісно пов’ язані з видільними каналами нирок. У багатьох
риб статеві залози сильно розростаються у зв’ язку з пристосу
ванням до відкладання величезної кількості ікринок. У біль
шості риб запліднення зовнішнє, у небагатьох живородних
риб — внутрішнє.
Рис. 4.5. Еволюція нирки та сечостатевих каналів: а — нейтральний зародковий стан; б — анам
нії; в — амніоти: І — самці; II — самки: 1 —- переднирка; 2 — вторинна нирка; 3 — канал переднир
ки; 4 — статева залоза; 5 — мюллерів канал; 6 — вольфів канал; 7 — сечовий міхур; 8 — клоака;
9 — вторинна нирка; 10 — матка; 11 — сечостатевий синус; 12 — задня кишка; 13 — статевий член;
14 — сечоводи вторинної нирки; 15 — чоловіча “маточка”
Популяційно-видовий рівень організації життя______________________ 2 71
У амфібій від сім’яників відходять виносні протоки, які
проникають у передній відділ нирки. Тут вони сполучаються
зі статевими канальцями і відкриваються у сечопровід (вольфів
канал), який функціонує і як сім ’ япровід, що впадає у клоаку.
У самок яйця із яєчників випадають у порожнину тіла, звідти
через звивисті яйцеводи (мюллерові канали) виводяться назов
ні через клоаку. У хвостатих амфібій запліднення внутрішнє,
а у безхвостих — зовнішнє.
У всіх вищих хребетних (амніоти) запліднення внутрішнє.
У рептилій і птахів статеві залози закладаються як парні ор
гани, але потім розвиваються нерівномірно. У багатьох рептилій
лівий яєчник розвинений слабше від правого. У птахів, навпа
ки, розвивається тільки лівий яєчник із дуже великим яйцеводом.
В однопрохідних ссавців статева система нагадує систему
рептилій (наявність клоаки, відкладання яєць). У інших пред
ставників ссавців відбувається подальше ускладнення будови
статевої системи, яке пов’ язане перш за все із розвитком при
стосування до розвитку зародків у матці й живлення їх за ра
хунок організму матері через плаценту. В результаті розши
рення задньої частини яйцеводів і зростання їх в один непарний
орган утворюється матка. Протилежні кінці яйцеводів відкри
ваються лійками у черевну порожнину поряд із яєчниками.
У деяких видів ссавців сім’ яники містяться у черевній порож
нині, але у більшості видів вони виходять назовні та лежать
у мошонці. З’ являються також складна система додаткових
залоз (сім ’ яні міхурці, передміхурова залоза) і копулятивні
органи.
Статеві залози ссавців розвиваються, я к ів інших хребетних,
спереду від закладання нирки, але потім переміщуються у та
зову ділянку. Яєчники залишаються у задній частині черевної
порожнини, а сім ’ яники опускаються з черевної порожнини
через паховий канал у зовнішній мішкоподібний виступ — мо
шонку, разом зі складкою черевини, яка їх огортає. Іноді
відбувається відхилення від звичайного шляху розвитку, один
із сім’ яників може не опускатись у мошонку, а залишатись
у черевній порожнині (крипторхізм). Це призводить до атрофії
сперматогенного епітелію і порушення функції сім ’ яника. При
виявленні у дитини крипторхізму застосовують хірургічну
корекцію.
2 72 ________________________________________________________ Розділ 4
Аномалії розвитку статевої системи людини. При пору
шенні ембріонального розвитку у людини дуже рідко трапля
ються випадки розвитку залоз обох статей — справжній гер
мафродитизм. Описані випадки, коли з одного боку були яєчник
і добре розвинена мюллерова протока, а з іншого — яєчко
і вольфова протока. При несправжньому гермафродитизмі ста
теві залози належать до якої-небудь однієї статі, хоч вони мо
жуть бути недорозвинені. Зовнішні статеві органи при цьому
слабо диференційовані, визначення статі за їх будовою немож
ливе. Іноді вони виявляються характерними для статі, проти
лежної тій, яку можна встановити при вивченні внутрішніх
статевих органів.
Можливі причини розвитку таких відхилень стають зро
зумілими на підставі вивчення ранніх стадій ембріонального
розвитку. Закладка статевої системи на початкових стадіях
у обох статей відбувається однаково, процес утворення гонад
і протоків носить індиферентний характер. Закладка статевих
залоз відбувається на стадії 3—4 тижнів у вигляді статевих
валиків — потовщень целомічного епітелію на поверхні первин
них нирок. Від цих валиків у первинну нирку вростають тя
жі — статеві шнури із зачатковими клітинами — гонобластами.
Диференціація індиферентної статевої залози за чоловічим
чи жіночим типом починається на 6—8 тижні. При розвитку
чоловічого ембріона статеві шнури перетворюються у сім ’ яні
канальці, а гонобласти утворюють сперматогенний епітелій.
При розвитку за жіночим типом відбувається ріст статевих
шнурів, які утворюють кіркову частину яєчника, а мозкова
його частина утворюється мезенхімними елементами — залиш
ками тіла первинної нирки (мезонефроса).
В ембріональному періоді маткові труби можуть недорозвиватися, що призводить до неплідності. У результаті порушення
зростання мюллерових каналів на ранніх стадіях розвитку
жіночого ембріона можуть виникати аномалії будови матки
і піхви: роздвоєння дна матки, а іноді і шийки. У деяких видів
ссавців це нормальне явище (дворога матка хижаків, парно
копитних, подвійна матка у багатьох видів гризунів). У при
матів, у тому числі в людини, у нормі зростання мюллерових
каналів відбувається на всьому проміжку нижче яйцеводів,
які залишаються парними. У результаті утворюється поодино
ка проста матка.
Популяційно-видовий рівень організації життя______________________ 2 73
Еволюція нервової системи у хордових тварин. Розвиток
нервової системи у хордових тварин в ембріогенезі відбуваєть
ся з ектодерми. На стадії гаструли формується нервова пла
стинка, яка у ході нейрули прогинається й утворює нервову
трубку. Покривна ектодерма змикається над нервовою трубкою,
яка заглиблюється у покрив (невроцель). Розширений кінець
нервової трубки, що розміщений у ділянці головного відділу
тіла, поділяється на три первинних мозкових міхурці (кефалізація). Порожнини шлуночків головного мозку і каналу спин
ного мозку є залишками просвіту ембріональної нервової
трубки.
Центральна нервова система виконує функції координації,
інтеграції та регуляції майже всіх видів нервової активності.
При цьому центральна нервова система працює у тісному кон
такті з периферичною нервовою системою.
У ланцетника — примітивного представника хордових —
центральна нервова система складається із нервової трубки.
Серед клітин нервової трубки є окремі світлочутливі утво
ри — вічка Гесса. Нервова трубка ланцетника виглядає як
жолоб, у якого передній кінець розширений. Від нервової труб
ки відходять спинномозкові нерви, які виконують функції
руху і сенсорних систем.
Кожен із відділів головного мозку залежно від класу орга
нізмів обумовлений рівнем життя та рівнем його організації.
Деякі відділи головного мозку збільшуються у розмірах, що
вказує на їх важливість для цього представника, а інші, навпа
ки, зменшуються. В ембріогенезі від міхура, який відповідає
проміжному мозкові, в обидва боки формуються очні міхурі,
а у середній його частині закладається задня частка гіпофіза.
Довгастий мозок переходить у спинний. Із підвищенням рівня
організації хребетних тварин відбувається диференціація голов
ного мозку на 5 відділів. Мозок дорослої хребетної тварини
складається з переднього, проміжного, середнього, заднього і дов
гастого. Тканини мозку утворені сірою речовиною (скупчення
нервових клітин) та білою (відростки нервових клітин). Про
відні шляхи зв’ язують між собою різні ядра, а також головний
мозок зі спинним.
Риби у пошуках їжі сильно залежать від нюху, тому у них
великі нюхові частки, які обробляють сенсорні сигнали. Великий
2 74 _________________________________________________________Розділ 4
мозочок здійснює у них координацію рухів, а досить розвинені
зорові частки мозку хоч беруть участь у зорових реакціях, але
в основному слугують головним координуючим центром мозку.
В проміжному мозку, з яким пов’ язані епіфіз і гіпофіз, розмі
щений гіпоталамус, який є центральним органом ендокринної
системи. Довгастий мозок забезпечує зв’ язок вищих відділів
головного мозку зі спинним і містить центри дихання та кро
вообігу. Головний мозок такого типу, де головним інтеграцій
ним центром є середній мозок, називають іхтіопсидним.
У земноводних головний мозок іхтіопсидний, але передній
мозок розділений на півкулі. Такі самі функції, як у риб, ви
конує середній мозок. Мозочок мало редукований у зв’ язку
з примітивними рухами. Спинний мозок у нижчих хордових
рівномірно тягнеться вздовж усього тіла. У них добре виражені
низхідні спинномозкові шляхи. Спинний мозок земноводних
починає вкорочуватися.
Наземне існування плазунів зумовлює складну організацію
мозку. Найкраще розвивається передній мозок, до нього пере
ходять функції інтегративного центру. Середній мозок втрачає
значення, і тому його розміри зменшуються. Мозочок добре
розвинений завдяки складній поведінці плазунів. Головний
мозок такого типу, де ведучим відділом є передній мозок, на
зивається зауроксидним.
У птахів півкулі переднього мозку досягають великих роз
мірів. Нюхові частки дуже малі. Мозочок дуже великий, що
пов’язано з координацією складних рухів при польоті.
Ссавці мають мамалійний тип мозку. Для нього характерний
розвиток переднього мозку за рахунок кори. Кора стає інте
груючим центром мозку. У ній розміщуються центри вищої
нервової діяльності, а також центри зорового, слухового, нюхо
вого та рухового аналізаторів. Кора має дуже складну будову,
у ній розміщуються тіла нейронів і асоціативні волокна, які
з’єднують різні її частини. Між півкулями виникають комісури, які з’ єднують їх в одне ціле. Проміжний мозок включає
гіпоталамус, гіпофіз, епіфіз. У середньому мозку розміщують
ся чотиригорбикові тіла, які відповідають за зір і слух. Дуже
добре розвинений мозочок.
Популяційно-видовий рівень організації життя______________________ 2 7 5
Спинний м о з о к ссавців на задньому кінці тіла має рудимент
у вигляді кінцевої нитки — / ііит іегтіпаїе. Нерви, які ідуть
до заднього кінця тіла, проходять по хребцевому каналу само
стійно, утворюючи так званий кінський хвіст — саисіа едиіпа.
Основні етапи еволюції центральної нервової системи відоб
ражаються в онтогенезі людини. На стадії нейруляції закла
дається нервова пластинка, яка перетворюється у жолоб, а потім
у трубку. Передній кінець трубки утворює спочатку три моз
кових міхурі: передній, середній, задній. Водночас, передній
мозок ділиться на двоє, і утворюються передній і проміжний
мозок. Середній мозковий міхур розвивається у середній мозок,
а задній — у задній і довгастий мозок.
Задній кінець спинного мозку редукується і перетворюється
на термінальну нитку. Швидкість росту спинного мозку і хреб
та є різною, тому в момент народження кінець спинного мозку
міститься на рівні третього хребця, а у дорослої людини вже
на рівні першого поперекового хребця.
Головний мозок швидко починає розвиватися за схемою,
характерною для ссавців і людини. Спочатку майже пряма
нервова трубка різко згинається в ділянці майбутнього довгас
того і середнього мозку. На цьому фоні великі півкулі перед
нього мозку ростуть дуже швидко. Тому головний мозок роз
ташовується над лицьовим відділом черепа. Диференціювання
кори приводить до розвитку закруток, борозен і до формуван
ня вищих сенсорних та рухових центрів.
Центральна нервова система є важливою ланкою всього
людського організму. Виходячи з цього, вроджені аномалії
розвитку нервової системи несумісні з життям. Серед вад спин
ного мозку, онтогенетичні механізми яких вивчені, варто на
звати рахісхізис — відсутність змикання нервової трубки. Ця
аномалія пов’ язана з порушенням клітинних переміщень і ад
гезією в зоні формування нервової трубки у процесі нейруляції.
Аномалія переднього мозку — прозенцефалія — виражена
в порушенні морфогенезу мозку, при якому півкулі залиша
ються з’єднаними, а кора не розвивається. Ця вада проявляєть
ся на четвертий тиждень ембріогенезу, в момент закладки
переднього мозку. Ці вади несумісні з життям, часто трапля
ються у мертвонароджених при різних хромосомних і генних
синдромах.
276
Розділ 4
Порушення диференціювання кори — агірія (відсутність
звивин) та олігосірія з пахігірією (мала кількість потовщених
звивин) — супроводжується спрощенням гістологічної будови
кори. У дітей із такими вадами виявляються олігофренія і по
рушення багатьох рефлексів. Такі діти переважно помирають
у перші роки життя.
Філогенез ендокринної системи. Ендокринна система разом
із нервовою забезпечують інтеграцію багатоклітинного організ
му, зокрема, гуморальну регуляцію функцій органів. Ця регу
ляція здійснюється гормонами — біологічно активними речо
винами різної хімічної природи, що продукуються залозами
внутрішньої секреції. Дія гормонів строго специфічна: різні
гормони діють на різні органи, спричиняючи певні зміни в їх
функціонуванні. Залози внутрішньої секреції не мають вивід
них проток і секретують гормони безпосередньо у кров, що
спрощує їх транспорт до органів-мішеней. Клітини цих органів
мають на своїх мембранах специфічні рецептори, з якими
зв’ язуються активні центри гормонів, при цьому відбуваються
зміни клітинного метаболізму. Гуморальна регуляція виникла
значно раніше, ніж нервова, оскільки вона є більш простою
і не потребує розвитку таких складних структур, як нервова
система.
Припускають, що вже стародавні одноклітинні організми
використовували біологічно активні речовини для міжклітин
них комунікацій. Деякі речовини такого роду, що володіли
регуляторними функціями, могли діяти як на окремі клітини
найпростіших, так пізніше і на клітини багатоклітинних ор
ганізмів. Потім вони стали виконувати функції гормонів.
В ембріогенезі багатоклітинних організмів деякі гормони
виявляють свою дію вже у перші години і дні розвитку. У про
цесі дроблення вони регулюють перебіг клітинного циклу.
Пізніше — переміщення клітин і утворення міжклітинних кон
тактів. Гормони набувають властивостей дистантних регулято
рів у філогенезі лише тришарових тварин, а в онтогенезі бага
токлітинних — відповідно на стадії первинного органогенезу.
При незмінності хімічної структури функції гормонів можуть
часто змінюватися. Наприклад, гормон пролактин, який секретується у ссавців і людини гіпофізом і регулює виділення
Популяційно-видовий рівень організації життя______________________ 2 7 7
молока молочними залозами, виявлений також у риб, земно
водних і птахів. У перших він регулює секрецію шкірними
залозами слизу, яким живляться мальки, у других — утворен
ня оболонок ікринок в яйцеводах, у третіх — деякі елементи
шлюбної поведінки, а також виділення “ пташиного молочка” .
Цей приклад є демонстрацією того, що еволюція дії гормонів
проявляється виконанням однієї і тієї ж функції у найширшому діапазоні. Справді, всі перелічені функції пролактину у тва
рин, що перебувають на різних рівнях організації, спрямовані
на створення умов для успішного розмноження.
Функції інших гормонів, наприклад адреналіну, можуть
у філогенетичному ряді хребетних практично не змінюватися,
забезпечуючи у всіх регуляцію енергетичного обміну.
Залози внутрішньої секреції, так само як гормони, мають різ
не походження, що дуже важливо для виявлення їх еволюції.
Деякі ендокринні залози пов’ язані за походженням з епі
телієм, що вистеляє глотку. До них належить щитоподібна
і паращитоподібна залози. Епіфіз розвивається як виріст моз
ку; гіпофіз, наднирники і підшлункова залоза мають складне
походження.
Серед хордових лише у безчерепних ендокринна система
існує у вигляді окремих клітинних комплексів, які розміщені
у різних відділах тіла й об’ єднані один з одним за рахунок гу
моральної взаємодії. У хребетних в основі проміжного мозку
розвивається гіпоталамус — нейросекреторне утворення, що
здійснює зв’ язок між двома системами інтеграції організму
в єдине ціле: нервовою та ендокринною. Разом з гіпофізом гі
поталамус утворює єдину гіпоталамо-гіпофізарну систему.
Еволюційним попередником гіпоталамуса є так званий інфундибулярний виріст ланцетника, що складається з нейросекреторних клітин і переднього кінця нервової трубки.
Починаючи від риб, гіпоталамус диференціюється на чис
ленні ядра, клітини яких за допомогою відростків контактують
як із нейронами мозку, так і з клітинами гіпофізу. Нейросекреторні клітини гіпоталамусу виділяють дві основні групи
гормонів: пептидні та моноамінові.
Перші — гормони, що впливають на функції внутрішніх
органів, — вазопресин (регулює артеріальний тиск) і окситоцин (впливає на мускулатуру матки). Друга група гормонів —
2 78
Розділ 4
дофамін, норадреналін, серотонін — регулює роботу передньої
частки гіпофіза. За дії цих гормонів стимулюється чи пригні
чується секреція гормонів відповідними гіпофізарними кліти
нами.
Розглянемо еволюцію гіпофіза. Ця залоза утворена трьома
частками — передньою (аденогіпофіз), середньою (проміжною)
і задньою (нейрогіпофіз). Гіпофіз з’єднується з гіпоталамусом
його виростом — лійкою, через яку проходять відростки ней
ронів гіпоталамуса і кровоносні судини, що забезпечують їх
гуморальну взаємодію.
Частини гіпофіза мають різне походження. Передня части
на розвивається з вип’ ячування ектодермального епітелію даху
ротової порожнини. Задня частина розвивається із задньої
частини лійки. Клітини, що входять до їхнього складу, за
походженням є гліальними. Проміжна частка — похідна від
передньої.
У хрящових риб в дорослому стані зберігається первісний
зв’язок з епітелієм ротової порожнини. За рахунок її задньої
частини формується також середня частка. Обидві частини
продукують гонадотропні гормони. У кісткових риб і личинок
земноводних є вже передня та середня частки, а у дорослих
амфібій, що переходять до наземного існування, з’являється
також і задня, що регулює водний обмін. Середня частка у них
припиняє продукувати гонадотропний гормон, однак виробляє
пролактин.
У зв’ язку з наземним способом життя рептилій і ссавців
у них найбільш прогресивно розвивається задня частка гіпо
фіза, і це пов’ язано з інтенсифікацією обміну води.
У людини в ембріогенезі розвиток гіпофіза відповідає ос
новним етапам її еволюції. Досить часто, у ЗО— 40 % нормаль
них людей під слизовою оболонкою даху глотки, в основі кли
ноподібної кістки, виявляється група клітин завдовжки 5—
6 мм і шириною 0,5— 1,0 мм, що за структурою та функціями
відповідає передній частці гіпофіза. Це є результатом пору
шення переміщення клітин при закладанні гіпофіза в ектодермі
ротової порожнини в ділянці турецького сідла. Ця аномалія
має назву ектопія аденогіпофіза, вона не супроводжується
патологічними виявами. Більш небезпечним є збереження по
рожнини в ділянці кишені Ратке — кіста кишені Ратке. Вона
Популяційно-видовий рівень організації життя
279
утворюється між передньою і середньою частками, містить слиз
і у деяких випадках має тенденцію до росту і навіть до перехо
ду в злоякісне новоутворення. Ростучі кісти стискають гіпофіз
і спричиняють його гіпофункцію. При цьому необхідне опера
тивне втручання.
Щитоподібна залоза, гормон якої — тироксин — регулює
основний обмін, серед хордових як компактний орган з’ яв
ляється вперше у риб. Однак вже у ланцетника окремі тироксиносинтезуючі клітини виявляють у жолобку на вентрально
му боці глотки. Щитоподібна залоза риб закладається також
у вигляді жолобка на вентральному боці глотки між першою
і другою зябровими щілинами в ділянці зачатка основи язика.
Пізніше цей клітинний матеріал занурюється під слизову обо
лонку і формує фолікули, характерні для щитоподібної залози.
У людини в ембріогенезі щитоподібної залози відбувається
рекапітуляція. Гетеротопія її здійснюється шляхом міграції
клітин у вигляді тяжу, порожнистого всередині, що має назву
щитоязикової протоки — сапаїіз ікугеоціоззиз. При нормаль
ному розвитку ця вада цілком і повністю редукується, а руди
ментом її є сліпий отвір (їогатеп саесит) на корені язика.
Персистування ділянок під’ язикової протоки в постнатальному періоді супроводжується накопиченням у них рідини
й утворенні серединних кіст шиї, які можуть локалізуватись
у будь-якому місці від кореня язика до верхньої межі щито
подібного хряща.
Інколи кісти нагноюються і проривають. Не менш відома
така вада розвитку, як ектопія щитоподібної залози, що може
розвиватись як наслідок порушення клітинної міграції.
Надниркові залози хребетних мають подвійне походження.
У риб і амфібій тканини, які відповідають мозковій і кірковій
речовині цих залоз, розміщені окремо. Зачатки мозкової речо
вини пов’ язані за походженням зі симпатичними нервовими
гангліями і розміщені метамерно. Зачаткй кіркової речовини
розвиваються з потовщень епітелію очеревини. У наземних
хребетних мозкова та кіркова речовини об’ єднуються в ком
пактні. У ссавців вони прилягають до переднього кінця нирок.
Мозкова речовина секретує переважно адреналін — регулятор
кровообігу й енергетичного обміну, а кіркова речовина — сте
роїдні гормони, що мають вплив на мінеральний, вуглеводний
обміни і функції нирок.
2 80
Розділ 4
Антропогенез. Одним із важливих об’ єктів біології є лю
дина. Людина як біологічний вид належить до типу хордових,
підтипу хребетних, класу ссавців, ряду приматів.
У X IX ст. деякі вчені, зокрема Бюффон, Ламарк, звернули
увагу на подібність людиноподібних мавп і людини, але вони
підкреслювали духовні здібності людини, які, на їхню думку,
могли мати тільки божественне походження. І лише Ч. Дарвін
у книзі “ Походження людини і статевий добір” (1871) спробу
вав довести походження людини від мавпоподібних предків
шляхом послідовних змін у результаті добору, однак учений
врахував основні біологічні фактори, які визначали еволюцію
людини. Соціальні фактори згодом розкрив Ф. Енгельс у праці
“ Роль праці в процесі перетворення мавпи у людину” (1876).
Ч.
Дарвін навів багато фактів, що підтверджували пох
дження людини від тварин, які пізніше були доповнені пра
цями багатьох учених. Людина має велику анатомо-фізіологічну подібність до вищих людиноподібних мавп: відносно
великі розміри тіла, редукцію хвостового відділу хребта, бу
дову кисті, папілярні візерунки на пальцях, долонях і стопах,
наявність нігтів, у людини така ж формула зубів (32 зуби).
Мавпи мають такі ж групи крові, як і людина. Вищі антропо
морфні мавпи мають добре розвинений мозок, маса якого до
сягає 420 г, спостерігається подібність і за будовою кори півкуль
великого мозку. У мавп трапляються ті ж паразити, що і в лю
дини, а також мавпи заражаються хворобами, які властиві
тільки людині (черевний тиф, віспа, сифіліс, туберкульоз,
грип). Каріотип шимпанзе має 24 пари хромосом, з яких 13 пар
хромосом практично однакових з людиною.
Зрозуміло, що ні горили, ні шимпанзе не є прямими пред
ками людини, насамперед тому, що вони її сучасники. Наука
доводить, що люди і сучасні людиноподібні мавпи виникли від
спільних предків, які жили у далекому минулому.
Істотною морфофізіологічною відміною людини від людино
подібних мавп є об’єм головного мозку. У людини він становить
у середньому 1400— 1600 см3, а у м авп— до 600 см3. Площа
поверхні півкуль переднього мозку у людини становить у се
редньому 1250 см2, а в людиноподібних м авп— у 3,5—4 рази
менше (табл. 4.1). Найважливішою якісною особливістю лю
дини як біологічного виду є її здатність до праці.
Таблиця 4.1. Основні етапи антропогенезу
282
Розділ 4
Сучасна людина сформувалася під впливом не тільки біоло
гічних, а й соціальних факторів. На ранніх етапах антропоге
незу в еволюції давніх людей провідну роль відігравали біоло
гічні фактори — мінливість і природний добір. Проте вже на
цих етапах добір відбувався не тільки за фізичними показни
ками (сила, витривалість), а й за рівнем розумового розвитку
(вміння виготовляти знаряддя праці, використовувати їх, здат
ність до спілкування з іншими членами племені, навички до
колективних дій тощо). Отже, вже у цю епоху добір сприяв
виживанню особин із вираженими соціальними рисами. У мі
ру того, як роль біологічних факторів зменшувалася, зростало
значення соціальних. Здатність до трудової діяльності, абстракт
ного мислення та членороздільної мови з ’ явилась і вдоскона
лювалась як специфічна якість людини розумної у результаті
впливу колективного способу життя на багато поколінь наших
предків (тобто сформувалась у філогенезі). Проте ця якість не
закріпилась у генотипі, і в кожної сучасної людини вона форму
ється у процесі її індивідуального розвитку (онтогенезу) тільки
під впливом виховання та навчання.
Об’ єктом антропологічних досліджень є людське тіло. Це
дослідження поєднує антропологію та медицину. В антрополо
гії проводиться опис ряду якісних ознак (антропоскопія) і кіль
кісне визначення особливостей будови тіла людини (антропо
метрія).
Дані антропології використовуються у судовій медицині для
ідентифікації особи, коли за черепом або за іншими навіть
незначними залишками необхідно встановити вік, стать, расо
ву приналежність та індивідуальні особливості суб’єкта. Осно
ву наукової антропометрії становить уніфікована методика. Ця
методика запропонована на основі робіт К. Верба, П. Брока,
Р. Мартіна, В. Кунака. Антропометричними точками називають
точки, що лежать в основному на виступах кісток, відростках,
які добре відчутні через м’ які тканини.
Основні антропометричні виміри голови зображено на
рис. 4.6.
Верхівкова — иегіех (о) — найвища точка тімені при розташу
ванні тіла, яке відповідає військовій виправці — голова фіксу
ється так, щоб край очниці й обидві козликові точки (над верх
нім краєм вуха) розташовувались в одній горизонтальній пло-
Популяційно-видовий рівень організації життя
283
Рис. 4.6. Краніометричні точки: а — спереду; б — збоку
щині (очно-вушна горизонталь). На верхівкову точку кладеться
планка ростоміра при визначенні зросту (довжини тіла).
Глабела — §ІаЬеІІа (§) — точка між бровами, яка найбільше
виступає вперед, лежить на носовому відростку лобної кістки,
де утворює помітну випуклість.
Метопіон — теіоріоп (т) — точка, яка лежить на перетині
серединної площини з лінією, що сполучає точки лобних горбів,
які виступають найбільше. Іноді через цю точку проходить так
званий метопічний шов, який поділяє луску лобної кістки на
дві (праву і ліву) частини. Це є варіантом норми.
Тім’яна — еигуоп (еи) — точка на латеральній поверхні
голови, яка найбільше виступає назовні, анатомічно не фіксо
вана, визначається шляхом вимірювання спеціальним цирку
лем найбільшої ширини голови. Її місце розташування залежить
від форми черепа.
Потилична— орізікокгапіоп (о р )— точка потилиці, яка
найбільше виступає назад (найбільш віддалена від глабел).
Звичайно лежить на зовнішньому потиличному підвищенні,
284
Розділ 4
визначається шляхом вимірювання найбільшого поздовжнього
діаметра голови.
Верхньоносова — пазіоп (п) — точка, яка лежить у медіаль
но-сагітальній площині на рівні носолобного шва.
Підборідна — дпаіНіоп (§п) — найнижча точка підборіддя
у медіально-сагітальній площині.
Вилична — гу§іоп (гу) — точка виличної кістки, яка най
більше виступає назовні, анатомічно не фіксована. Вона може
лежати як на відростку виличної кістки, так і на виличному
відростку скроневої кістки.
Нижньощелепна— §опіоп ( § о ) — точка на куті нижньої
щелепи (кут між тілом нижньої щелепи і заднім краєм гілки),
яка найбільше виступає назовні. Деякі точки можна визначи
ти тільки на черепі.
Брегма — Ьгедта (Ь) — точка на місці перетину сагітально
го і вінцевого швів.
Ламбда — ІатЬда (І) — точка на перетині ламбдоподібного
і сагітального швів.
Череп — найважливіший об’ єкт антропологічного дослід
ження як для вивчення сучасної людини, так і для характе
ристики викопних форм. Вивчення черепа, варіацій його
розмірів і форм становить спеціальний розділ антропології —
краніологію (лат. сгапіит — череп). Як відомо, у черепі розріз
няють два відділи: мозковий і лицьовий. Мозковий череп ут
ворений кістками, що входять до складу черепної коробки,
у якій міститься мозок.
Уявлення про відносний розвиток лицьового і мозкового
відділів черепа дає поперечний краніофаціальний (черепнолицьовий) показник (КФП). Це виражене у відсотках відношен
ня найбільшої ширини обличчя (відстань між точками гу) до
поперечного діаметра мозкового відділу черепа (відстань між
точками еи). У сучасної л ю д и н и в і н становить 87— 105 % .
За черепом можна визначити стать: чоловічий череп має
більш грубий рельєф (місце прикріплення м ’ язів), більший
розвиток надбрівних дуг і перенісся, масивнішу нижню щеле
пу; луска лобної кістки розташовується менш вертикально,
Популяційно-видовий рівень організації життя______________________ 2 85
ніж у жінок. Більший середній об’ єм черепа чоловіка корелює
з більшим середнім зростком. За кістками черепа можна виз
начити і вік індивідуума. Старечий вік характеризується за
ростанням (облітерацією) швів (сагітального, вінцевого, ламбдоподібного). Атрофуються альвеолярні краї щелеп. Для ха
рактеристики розмірів і форми мозкового черепа здійснюють
вимірювання його об’ єму, окружності та трьох основних діа
метрів — поздовжнього, поперечного і висотного.
Поздовжній діаметр — найбільша довжина черепа — ви
мірюється від глабели до опістокраніона по сагітальній пло
щині. Ця величина коливається у межах 167— 193 мм. Попе
речний діаметр вимірюється між точками евріон у фронтальній
площині, він становить 123— 153 мм.
Загальне уявлення про форму черепа дає поперечно-поздовж
ній індекс (черепний показник, ЧП):
тім’яна — тім’яна
ЧП = ---------------------------------- 100% .
глабела — потилична
Це виражене у відсотках відношення поперечного діаметра
до поздовжнього. Залежно від величини черепного показника
розрізняють череп видовженої форми (доліхокранія), кругля
стий, сферичний (брахікранія) і проміжні варіанти (мезокранія).
Найбільш поширену рубрикацію подано у табл. 4.2.
Таблиця 4.2. Рубрикація черепа
Ч ер е п н и й п о к а з н и к
Д о л іх о к р а н ія
На черепі
Д о 7 4 ,9
Н а гол ові
Д о 7 5 ,9
М е з о к р а н ія
7 5 , 0 — 7 9 ,9
7 6 ,0 — 8 0 ,9
Б р а х ік р а н ія
8 0 ,0 і б іл ь ш е
8 1 ,0 і б іл ь ш е
Краніофаціальний показник (КФП) більший, ніж черепний
(ЧП), за рахунок товщини м’ яких тканин.
Різні форми черепа у сучасних людей — це нейтральні оз
наки, варіанти норми. У межах однієї раси можуть трапляти
ся різні співвідношення цих показників. Спроби пов’ язати ці
морфологічні критерії з історичними долями народів і їх со
ціальним прогресом позбавлені будь-якої наукової основи.
286
Розділ 4
Раси людей. Великі історично сформовані ареальні групи
людей, які пов’язані єдністю походження, що виявляється
у спільних спадкових морфологічних і фізіологічних ознаках,
називають расами людей. Дані антропології свідчать, що
раси людей за основними біологічними особливостями і розу
мовими здібностями цілком рівноцінні та належать до єдино
го виду людина розумна (Ното варіепз). За останніми дослід
женнями, все людство складається з чотирьох великих рас,
формування ознак яких на перших етапах розвитку мало ви
разну кліматогеографічну адаптацію. Формування рас — яви
ще вторинне, оскільки всі основні ознаки людини як біологіч
ного виду з’ явились до поділу людей на окремі раси. Історія
формування рас, яка розпочалася 25— 40 тис. років тому,
пов’язана з міграцією людей у різні зони та континенти Землі.
Сьогодні внаслідок змішаних шлюбів триває процес стирання
відмінностей між окремими расовими групами людей.
Для європеоїдної раси властиві такі ознаки: світла пігмен
тація шкіри різних відтінків, м ’ яке волосся (пряме або хви
лясте), рясний третинний волосяний покрив, зокрема борода
і вуса, порівняно тонкі губи. Пігментація волосся й очей, варіює
від дуже світлої до дуже темної. Основний ареал поширення
цієї раси — Європа, частина Азії (Східна і Середня), Північна
Африка, крім того, вона поширилась на материках Америки
й Австралії.
Негроїдна раса характеризується темним кучерявим волос
сям, темним кольором шкіри і очей, повними губами, широким
носом. Розвиток покриву слабкий або середній. Лицьова ча
стина черепа дещо виступає по вертикальній площині (прогматизм). Основний ареал поширення цієї раси — Африка, Пів
нічна Америка (наслідок торгівлі рабами).
Монголоїди мають пряме жорстке темне волосся, темні очі,
шкіру жовтавого кольору, обличчя сплющене, вилиці дуже
видаються, плескате перенісся. Характерні особлива будова
передніх зубів (“ совкоподібні” різці) і наявність нависаючої
шкірної складки верхньої повіки — епікантуса. До монголоїд
ної раси близькі американські індіанці, хоча деякі риси, що
властиві цій расі, у них відсутні або трапляються рідко (епі
кантус).
Популяційно-видовий рівень організації життя______________________ 2 8 7
Проте не всі популяції сучасної людини можна віднести до
однієї з великих трьох рас. Аборигени Австралії відрізняють
ся від негроїдної раси деякими ознаками (хвилясте волосся,
густий третинний волосяний покрив на обличчі й тілі, менш
темна пігментація шкіри). Деякі антропологи розглядають їх
як четверту основну расову групу — австралоїдну. За іншою
класифікацією, їх поєднують із негроїдами в одну велику еква
торіальну расу (австралонегроїдну). За класифікацією, запро
понованою Я. Я. Рогінським, сучасне людство поділене на три
великі раси: екваторіальну (австралонегроїдну), євразійську
(європеоїдну), азіатсько-американську (монголоїдну) (табл. 4.3).
Таблиця 4.3. Схема класифікації рас (за Я. Я. Рогінським,
М. Г. Левін)
Є в р а зій сь к а р а са
А з іа т с ь к о -а м е р и к а н с ь
ка р аса
Е к в а т о р іа л ь н а р а са
А т л а н т о - б а л т ій с ь к а
П ів н іч н о а з іа т с ь к а
А в ст р а л ій сь к а
С ер ед н ьоєвр оп ей ськ а
А ркти чн а
В е д о їд н а
Ін д о се р е д н ь о м о р сь к а
Д а л е к о с х ід н а
М е л а н е з ій с ь к а
Б іл о м о р с ь к о - б а л т ій с ь к а
П ів д е н н о а з іа т с ь к а
Н егр ськ а
Б а л к а н о -к а в к а з ь к а
А м ер и к а н ськ а
Н егри л ьська
Б уш м ен ськ а
У межах великих рас є кілька малих. Так, європеоїдна ра
са розпадається на п’ ять расових груп. Азіатсько-американська
включає монголоїдів Азії, Далекого Сходу, ескімосів Арктики,
американських індіанців. До екваторіальної раси належать
чорні люди Африки й Америки і низькорослі племена Цен
тральної та Південної Африки, яким властиві низький зріст,
більш виступаючий ніс і тонші губи (пігмеї та бушмени), або
ригени Австралії.
Народи різних рас перебувають на різних рівнях культур
ного розвитку, проте ці відмінності визначаються суспільноекономічними факторами, а не біологічними особливостями
людей.
Розділ 5
ЕКОЛОГІЯ ЛЮДИНИ ТА БІОСФЕРА
Дослідження живого світу завжди було однією з важливих
сфер діяльності людини. Спочатку від цього залежало її ж ит
тя — необхідно було знати, які рослини, тварини, гриби мож
на вживати у їжу, для облаштування житла чи для лікування,
а які з них небезпечні та отруйні. І лише згодом людина змог
ла зайнятися наукою з метою пізнання. Термін екологія був
запроваджений у 1869 р. Ернстом Геккелем для позначення
науки, що вивчає взаємовідносини організмів з навколишнім
середовищем. Однак судячи з наскельних малюнків давньої
людини, можна дійти висновку, що екологія — одна із найдав
ніших наук.
Предмет екології відомий усім, однак наукова екологія не
проста, оскільки це фундаментальна наука, яка має свої тон
кощі та складність. Екологія — це фундаментальна біологічна
наука, яка має свою мету, об’ єкти і методи пізнання, свої
об’єктивні закони, які виконуються незалежно від знання чи
незнання їх людиною. Особливістю екології є те, що вона ще
на початку свого становлення набула певної самостійності,
коли пропонувалися перші екологічні концепції та вироблялись
основні екологічні підходи. Саме з цього часу, на думку деяких
вчених, в екології співіснують популяційний і екосистемний
підходи, які практично вичерпують у своїй сукупності основний
її зміст.
Екологія людини та біосфера
289
Існує багато визначень екології. На нашу думку, найбільш
повним і точним є таке: екологія — це фундаментальна біоло
гічна наука, яка вивчає взаємовідносини організмів між собою
та з навколишнім природним середовищем на надорганізменому рівні. Щоб розширити це визначення, необхідно зазначити,
що екологія займається трьома рівнями організації: окремими
особинами (індивідуумами) як членами популяцій, популяція
ми (які складаються з особин одного виду) та угрупованнями
(які складаються з певного набору популяцій), а також екоси
стемами.
Займаючись особинами, екологія виявляє, як на них впливає
абіотичне та біотичне середовище і як вони самі впливають на
середовище. Вивчаючи популяції, вона вирішує питання про
наявність або відсутність окремих видів, про їх стійкість, зміни
і коливання чисельності популяцій. При дослідженні на попу
ляційному рівні можливі два методологічні підходи. Перший
аналізує основні властивості окремих особин і лише після цього
намагається виявити форми співвідношення цих властивостей,
які визначають особливості популяції загалом. Другий звер
тається до властивостей популяції безпосередньо, намагаючись
пов’ язати ці властивості з параметрами середовища.
Екологія угруповань розглядає їх склад, структуру, а також
проходження через угруповання енергії, біогенних елементів
та інших речовин.
Екологія займає центральне місце серед інших біологічних
дисциплін, а тому з більшістю із них перекривається. Екологів
цікавлять не лише природні угруповання та популяції, але
й ті, що створені самою людиною, або під її впливом (сади
і парки, поля і зерносховища, заповідники і ставки). Крім того,
екологи часто вивчають експериментальні системи і матема
тичні моделі.
Визначний вчений X X ст. Ю. Одум писав, що будь-яка біосистема, яка включає всі організми, які функціонують разом
(біотичне угруповання) на певній ділянці та взаємодіє з фізич
ним середовищем таким чином, що потік енергії створює чітко
визначені біотичні структури та колообіг речовин між живою
і неживою частинами, являє собою екологічну систему, або
екосистему. Термін “ екосистема” вперше був запропонований
у 1935 р. А. Тенслі. На сьогодні більшість екологів уважають,
2 90 ________________________________________________________ Розділ 5
що екосистема — основна функціональна одиниця в екології,
оскільки до її складу входять і організми, і нежива приро
да — компоненти, які взаємно впливають на властивості один
одного і необхідні для підтримання життя в тій її формі, яка
існує на Землі.
А. І. Опарін висловив припущення, що 3,5 млрд років тому
в первісному океані Землі під впливом різних типів випромі
нювання й електричних розрядів почалось утворення перших
органічних сполук, які утворили “ органічний бульйон” . Зі
зростанням його концентрації деякі органічні молекули,
об’єднуючись, почали утворювати коацерватні краплі, ізольо
вані від навколишнього середовища, котрі використовували
елементи цього розчину для збільшення свого розміру. Так
виникли молекули, які здатні самовідтворюватись — виникло
життя, з’ явилися живі організми. Подальший еволюційний
розвиток живих організмів привів до їх видового різноманіття.
Внаслідок живлення одного організму іншим і внаслідок при
родного добору виник колообіг речовин і енергії, які характер
ні для екосистем нашої планети, з’явилася біосфера. Отже,
тварини, рослини та мікроорганізми своєю життєдіяльністю
протягом мільярдів років створювали і вдосконалювали умови,
сприятливі для існування, тобто біосферу, перш ніж з’явилася
людина. На основі праць В. І. Вернадського та його учнів і пос
лідовників використовується таке визначення біосфери:
Біосфера — це загальнопланетна оболонка, до складу якої
належать нижні шари атмосфери, вся гідросфера та верхні
шари літосфери; її склад, структура і властивості зумовлені
сучасною та минулою життєдіяльністю всієї сукупності
живих організмів. Вона є наслідком взаємодії живих і неживих
компонентів, акумуляції та перерозподілу в ній величезної
кількості енергії. Це термодинамічно відкрита, самоорганізова
на, саморегульована, динамічно зрівноважена, стійка, мозаїч
на (досиметрична), глобальна система.
Володимир Іванович Вернадський — великий український
натураліст і філософ X X ст. створив сучасне вчення про біосфе
ру та ноосферу. В. І. Вернадський одним із перших усвідомив
величезний вплив живих організмів на біосферу, тісну взаємо
дію і взаємозалежність усіх форм життя. Особливо В. Вернад-
Екологія людини та біосфера
291
ський виділив перетворювальний вплив на землю однієї з форм
життя — людини — через її розумну діяльність і передбачив
швидке зростання глибини та масштабів цього впливу. У роботі
“Декілька слів про ноосферу” (1944) В. І. Вернадський ствер
джує, що під впливом розвитку науки і пізнання біосфера має
стати ноосферою, тобто цариною розуму, де панують закони
мудрості й гармонії.
Серед величезного різноманіття видів живих організмів
у біосфері примати мають найвищу здатність до адаптації.
Найбільш характерні адаптивні ознаки приматів пов’ язані
з дуже високим розвитком деяких відділів нервової системи,
а особливо тих відділів головного мозку, від яких залежить
розумна поведінка і здатність м ’ язів до тонких дій. Людина є
особливим видом у біосфері — їй притаманне мислення і все,
що з ним пов’ язане. Людство збагнуло надзвичайну “ геологіч
ну могутність” , за В. І. Вернадським, свого інтелекту і “ керо
ваної ним праці” , які пронизують не лише біосферу та підпо
рядковані їй живі підсистеми, але і глибокі шари атмосфери,
інші геосфери й космічне довкілля Землі. З моменту свого
виникнення протягом одного-півтора мільйона років людина
перетворювала біосферу, однак лише у X X ст. стала “ геологіч
ною силою” . Знищення рослинного покриву і видів диких
тварин, розорювання земель на великих площах, потужне
використання горючих і негорючих корисних копалин, синте
зу великої кількості чужорідних для біосфери сполук (ксенобіотиків), забруднення усіх трьох сфер біосфери призвели до
суттєвих змін у ній. Тобто сили людства були спрямовані не
па формування сфери розуму (ноосфери), а внаслідок нераціо
нального природокористування та господарської діяльності
призвели до вичерпання і пошкодження природних ресурсів,
деформації процесів природного колообігу речовин і енергії,
що склалися протягом багатовікової еволюції, руйнування
механізмів саморегуляції біосфери.
Нині людство переживає вирішальний момент своєї історії.
Світ зіткнувся з проблемами посилення злиднів, голоду, хвороб,
неграмотності та тривалої деградації екологічних систем, від
шейх залежить наш добробут. Взаємовідносини людини з нав
колишнім середовищем— одна з глобальних проблем сучас
ності. Її вирішення стає необхідною передумовою розвитку
292
Розділ 5
економіки й самого існування людства. Для цього насамперед
необхідно знати основні екологічні закони. Адже закони еко
логії об’ єктивні. Порушення цих законів може призвести до
необоротних процесів у біосфері, поставити під питання саме
існування людини як виду.
Екологія людини — це розділ екології, який вивчає взаємодію
людини як біосоціальної істоти зі складним багатокомпонен
тним навколишнім середовищем, що постійно ускладнюється.
У 80-х роках X X ст. було сформульовано визначення екології
людини як науки, що вивчає “ закономірності взаємодії люди
ни з навколишнім космопланетарним середовищем, питання
розвитку народонаселення, збереження і розвитку здоров’ я,
удосконалення психофізичних можливостей людини” .
В екології людини можна виділити ряд напрямів:
— природне навколишнє середовище і здоров’я людей;
— негативний вплив на людину антропогенних порушень
біосфери;
— урбанізація та її негативні наслідки;
— тривалість життя як показник, що характеризує функ
ціональні резерви організму та якість життя, обумовлені при
родними та соцільними факторами тощо.
Екологія людини повинна допомогти у пізнанні механізмів
адаптації організму людини до нового для неї середовища та
розробці заходів, які полегшать її пристосування до навколиш
ніх умов. Отже, в центрі екології людина як біологічний об’єкт
зі специфічною морфо-фізіологічною організацією та соціальною
суттю. Екологію людини можна вважати однією з найважли
віших наук майбутнього.
Виділення екології людини як міждисциплінарної комплек
сної науки, що розкриває взаємовідносини між людиною і при
родою, обумовлене саме причинами, зазначеними вище, — тим,
що людина постала перед необхідністю вирішення головних
питань: яке місце вона займає як вид у глобальній екосисте
мі — біосфері — та які закони біосфери вона не має права по
рушувати, щоб вижити і процвітати на планеті Земля.
Упродовж біологічної еволюції з’ явилася висока пристосо
ваність організмів до природних умов. У той же час, на відмі-
Екологія людини та біосфера
2 93
ну від решти видів, людина, значною мірою за відсутності
факторів природного добору, увійшла в соціальну еволюцію
генетично недостатньо пристосованою до тривалих несприят
ливих нових факторів навколишнього середовища. Тому зов
нішні дії на людський організм лише частково компенсуються
певними фізичними і біологічними механізмами. Якщо ці ме
ханізми вичерпуються, то у найменш сильних особин популя
ції з’ являються відповідно гострі, а пізніше хронічні захворю
вання. Передусім це пов’язано з тим, що властивості організму
людини, які сформувалися протягом тривалого періоду еволю
ції, не можуть змінюватися з такою ж ш видкістю, з якою
змінюються умови оточуючого його природного середовища.
5.1. Життєве середовище ЛЮДИНИ
Природне середовище, яке оточує людину, — це всі фактори
природно-антропогенного походження. Поміж середовищем,
яке являє собою сукупність умов існування організмів, і цими
організмами встановлюється тісний взаємозв’ язок. Організми
та їх сукупності, з одного боку, у процесі своєї життєдіяльності
змінюють навколишнє середовище, пристосовують його до своїх
потреб, а з іншого — самі пристосовуються до середовища, за
знають змін. Ж иття будь-якого організму — тваринного чи
рослинного, в тому числі і людини, — залежить від навколишнього середовища. Між живими організмами і середовищем
існують тісні взаємозв’ язки, взаємозалежності та взаємовпли
ви. Біологічні системи на будь-якому ієрархічному рівні є
відкритими системами — вони отримують для свого існування
з навколишнього середовища речовини і хімічні елементи,
енергію (сонячну та хімічну) й інформацію і віддають у навко
лишнє середовище трансформовані речовини, енергію та ін
формацію, таким чином активно впливаючи на нього, зміню
ючи його. Хімічний склад навколишнього середовища є ре
зультатом складних і тривалих динамічних процесів, які
її донині відбуваються на нашій планеті. Саме тому параметри
кожного чинника навколишнього середовища постійно зміню
ються в часі, що потребує перманентної адаптації організмів.
294________________ _______________________________________ Розділ 5
У свою чергу, середовище обумовлює пристосувальні ознаки
організмів.
За ступенем впливу на організми та за походженням виді
ляють такі фактори середовища: абіотичні, біотичні й ан
тропогенні.
Абіотичні чинники поділяються на елементарні (температу
ра, вода, атмосферний тиск, повітря, електромагнітне поле, гра
вітація) та комплексні (хімічний склад субстрату, розчину, газу;
агрегатний стан субстрату, розчину, газу; сонячне світло).
Найбільш значущими кліматичними факторами, які впли
вають на організм людини, є освітлення, температура та воло
гість. Іноді виділяють окремий розділ екології — біокліматологію людини, яка вивчає реакції організму на вплив різно
манітних умов середовища, а також вимоги організму до цих
умов для збереження нормальної фізіологічної діяльності.
У цьому контексті особливо ретельно вивчають адаптивні мож
ливості людини до життя в екстремальних і у швидкозмінюваних умовах середовища. Іншими важливими кліматичними
екологічними чинниками для людини є тиск, вітер, кислотність
ґрунту і його сольовий склад, склад повітря. Часто хімічні
компоненти атмосфери, гідросфери та літосфери, якщо надхо
дять у надмірній кількості, також стають екологічними фак
торами. Крім основних кліматичних факторів і хімічних речо
вин, які можуть надходити у довкілля внаслідок антропогенної
діяльності, людина має адаптуватися до фізичних забрудників:
виробничих шумів, вібрації, світлових і теплових упливів,
іонізуючого випромінювання тощо.
Світло є найважливішим абіотичним чинником на планеті.
Лише сонячне випромінювання забезпечує трофічні потреби
всіх живих істот. Разом із гравітацією та радіаційним опромі
ненням цей чинник є тим, до якого жива природа адаптовується впродовж усього свого існування. Сонце випромінює в ши
рокому діапазоні хвилі всіх довжин (зокрема, від ультрафіолето
вих до інфрачервоних). Хвилі довжиною менше 290 нм є тією
чи іншою мірою згубними для більшості живих організмів; знач
на кількість цих хвиль відбивається озоновим шаром Землі.
Довші хвилі проходять крізь цей шар. Споживачами проме
нистої енергії Сонця є передусім автотрофні організми — рос
лини та ціанобактерії, які використовують її для фотосинтезу.
Екологія людини та біосфера
2 95
Тварини теж потребують сонячного світла. Зоровий аналізатор,
притаманний більшості тварин і людині, пристосований до
функціонування в діапазоні видимого світла. Орієнтація у про
сторі, одержання необхідної інформації для більшості тварин
неможливі без цього апарату, а для людини дуже утруднені.
Поверхня шкіри також може містити фотосенсибілізатори, які
здатні поглинати світлові промені видимої частини й утворю
вати біологічно активні речовини, які можуть призвести до
дерматитів, екзем і пухлин.
Інтенсивність ультрафіолетових променів коливається за
лежно від сезону, часу доби, висоти над рівнем моря, погодних
умов тощо. Наприклад, якщо промені Сонця спрямовуються
під кутом 30°, то інтенсивність ультрафіолетового опромінення
стає вдвічі меншою, ніж удень. Засмагу, яка виникає у людей
ЗІ світлим кольором шкіри після тривалого опромінення ультра
фіолетовими променями, можна розглядати як фенокопію
генетично визначеного темного кольору шкіри. Водночас це
явище належить до адаптивних реакцій організму. Роль такої
адаптації полягає у підвищенні порогової еритемної дози опро
мінення (еритемафізіологічне почервоніння шкіри). При цьому
потовщується роговий шар опромінених ділянок шкіри, нако
пичується меланін унаслідок посилення його вироблення і міг
рації з базальних клітин до поверхні. Крім того, людині воно
необхідне для синтезу вітаміну Б — кальциферолу (промені
в діапазоні 250— 300 нм), необхідного для нормальної жит
тєдіяльності організму, кальцієвого та фосфорного обміну, які
забезпечують розвиток і репарацію кісткової тканини.
Загальностимулююча дія УФ полягає в утворенні в організ
мі під дією БАР гістаміну, ацетилхоліну тощо. Під дією цих
сполук стимулюються робота нервової системи, синтез гемогло
біну та фагоцитоз, збільшується кількість антитіл і посилюєть
ся стійкість організму до шкідливих агентів.
Однак УФ і негативно впливає на стан здоров’ я людини.
Існує пряма кореляція між тривалістю й інтенсивністю ультра
фіолетового опромінення шкіри людини і частотою захворювань
па рак шкіри. Цей показник залежить від географічної широ
ти, на якій мешкають люди, тому що вона впливає на дозу
ультрафіолетового опромінення. В умовах гіперінсоляції мо
жуть загострюватися хронічні захворювання.
296
Розділ 5
У лікувальних закладах широко використовують бактери
цидну дію короткохвильового (180—280 нм) випромінювання,
яке згубно діє практично на всі види мікроорганізмів, на гри
би та віруси.
Інфрачервоні промені, залежно від довжини хвилі, можуть
проникати крізь шкіру на глибину від 0,1 до ЗО—40 мм. Унас
лідок цього розширюються судини і поліпшується кровообіг,
стимулюються метаболічні процеси у шкірі. Тому інфрачервоні
промені застосовуються у фізіотерапії і травматології.
Іонізуюче випромінювання. У природі існує багато речовин,
які містять радіоактивні елементи, або радіоактивні ізотопи
Карбону, Гідрогену, Калію та інших хімічних елементів. Ево
люція органічного світу і людини відбувалася на фоні певних
доз опромінення, яке отримує кожна жива істота. Ці дози
становлять природний радіоактивний фон. До їхнього впливу
організми тією чи іншою мірою адаптовані. Однак штучно
створені радіоактивні речовини, ядерні реактори, устаткуван
ня сконцентрували у собі набагато більші від природного обся
ги іонізуючого випромінювання, до чого природа виявилася
непристосованою.
Радіоактивні ізотопи можуть потрапляти у травну систему
людини разом із їжею, частина їх — через органи дихання,
шкіру. Окремі ізотопи (стронцій-90) швидко всмоктуються
й осідають у сполучній тканині. Радіоактивне випромінювання
є найсильнішим мутагенним чинником. У людини певна частка
мутацій виникає під впливом природних джерел радіації. Особ
ливо небезпечним є вплив радіоактивного випромінювання на
статеві клітини. Значна частина радіоактивних ізотопів, які
через трофічні ланцюги потрапили в живі організми, продов
жуватимуть циркулювати в екосистемах ще тривалий час.
Аварія на Чорнобильській АЕС у квітні 1986 р. спричинила
проникнення в атмосферу (згодом у воду та ґрунт) значної
кількості радіоактивних елементів. Метаболічні процеси в ж и
вих організмах, спроможність атмосфери та гідросфери розбавля
ти концентрацію ізотопів не можуть нейтралізувати негативні
наслідки цього лиха. Людина передусім стає об’єктом накопи
чення радіогенних мутацій, які є причиною багатьох захворю
вань.
Екологія людини та біосфера
2 97
Електромагнітне забруднення. Головними джерелами елек
тромагнітного випромінювання є радіо-, телевізійні та радіо
локаційні станції, засоби радіозв’ язку різного призначення.
Забруднення природного середовища електромагнітними ви
промінюваннями (полями) у таких районах може змінюватися
від 50— 100 Гц до 100 Гц, часто перевищує допустимі гігієніч
ні норми, що шкодить здоров’ ю. Передусім страждає нервова
система. Напруженість поля 1000 В /м спричиняє головний
біль, сильну втому, а це веде до безсоння, неврозів.
Температура є суттєвим чинником, який визначає мож
ливість перебігу біохімічних процесів, адже білки-ферменти,
що контролюють перебіг цих процесів, функціонують лише
у певних температурних межах. Механізми регулювання тем
ператури у рослин, грибів, дроб’ янок і тварин принципово від
мінні. У рослин при високих температурах процеси дихання
починають перебігати значно швидше, ніж реакція фотосин
тезу, і вони гинуть. У тварин високі температури викликають
“тепловий шок” , із якого вони можуть вийти при зниженні
температури.
Людина не має механізмів, які б допомогли їй пристосува
тися до життя в умовах постійно високих або низьких темпе
ратур без застосування допоміжних засобів і заходів. Наша
шкіра має незначний шар підшкірної жирової клітковини,
волосяний покрив майже відсутній. Тому людина більш чут
лива до температури, ніж інші ссавці. Однак, на відміну від
ч'варин, людина здатна захистити свій організм за допомогою
одягу та завдяки перебуванню у приміщеннях із комфортними
температурними умовами.
Температура тіла людини у стані фізіологічного комфорту,
або оптимуму, найбільш рівномірна. Чим більше відхилення
л’емператури в той чи інший бік від оптимальних значень, тим
більша частка енергії в усіх ендотермних організмів витра
чається на підтримання постійної температури тіла. Основне
тепло в організмі людини утворюють глибинні структури тіла,
звідки це тепло розповсюджується на периферію. Регуляція
температури тіла здійснюється передусім термостатичним мехамізмом мозку (гіпоталамусом). Нормальна температура тіла лю
дини 37 °С, летальні — 32—33 і 43 °С. Добові коливання темпе
ратури тіла у людей не перевищують 1,7 °С. Якщо створюється
298
Розділ 5
загроза рівновазі, вступають уд ію компенсаторні механізми.
Негайна реакція організму на перегрів — збільшення віддачі
тепла через систему кровообігу та шляхом потовиділення.
При незначному охолодженні серцева діяльність змінюєть
ся — збільшуються сила та кількість скорочень. При істотному
охолодженні порушується збудливість і провідність кардіоміоцитів. Якщо у зовнішньому середовищі холодно, значна части
на організму піддається переохолодженню, а температура
кінцівок може знижуватися на 10 °С порівняно з температурою
внутрішніх органів, однак температура мозку, органів грудної
клітки та черевної порожнини зберігається на постійному рів
ні. Пристосування до холоду досягається у людини різними
шляхами. Європейці реагують на зниження температури сере
довища за межу критичної підвищенням рівня метаболізму.
Водночас аборигени Австралії здатні провести холодну ніч, не
прикриваючи свого тіла, — при нічному охолодженні у них
спостерігається ізоляційна гіпотермія, яка супроводжується
зниженням температури поверхні тіла без інтенсифікації об
міну речовин (як у європейців).
У підвищенні стійкості організму людини до дії сублетальних високих і низьких температур суттєве значення має під
готовчий період (теплова або холодова акліматизація), для якої
потрібен певний період (місяць і більше).
Підтримання водного балансу організму людини зумовлене
передусім необхідністю водного середовища для перебігу всіх
біохімічних процесів. Водний баланс формується за рахунок
надходження води з їжею та питтям, за рахунок утворення
метаболічної води і втрати її через видільну, травну й дихаль
ну системи, а також із потом. Наявність близько 64 % води від
маси тіла забезпечує умови для фізико-хімічних процесів у клі
тинах організму. Вся система водно-сольового обміну повинна
забезпечувати постійний об’ єм, іонний склад, рН і осмотичний
тиск усередині організму.
Атмосферний тиск, тиск води, вітер, вміст іонів і елементів
у питній воді та повітрі також належать до екологічних фак
торів.
Атмосферний тиск у нормі на рівні моря дорівнює 1 атмо
сфері (це тиск стовпчика ртуті 760 мм з діаметром 1см 2). На
територіях нижче рівня моря тиск вищий, а вгору від рівня
Екологія людини та біосфера
299
моря тиск знижується. Зміни атмосферного тиску по вертикалі
та горизонталі тісно пов’ язані зі змінами температури повітря
та переміщенням повітряних мас. Атмосферний тиск впливає
на хід життєвих процесів, самопочуття і працездатність люди
ни. Так, на значних висотах, де знижений вміст кисню у повітрі,
з’ являється серцева недостатність, знижується працездатність.
Людина може відчувати дискомфорт, головний біль, безсоння,
запаморочення, порушення діяльності центральної нервової
системи. Можуть виникнути спазми легенів, порушення кро
вообігу, набряки. Важка гіпоксія, яка розвивається, як пра
вило, внаслідок різкого підняття на висоту, може спричинити
порушення психіки і призвести людину до загибелі.
Якщо людина піднімається в гори поступово, відбувається
адаптація до перебування у високогір’ ї, перебудовується її
газотранспортна система. Процеси в організмі стають подібни
ми до тих, які спостерігаються у постійних мешканців висо
когір’ я.
При зануренні людини у воду можуть виникнути значні
фізіологічні проблеми. Гідростатичний тиск із зануренням на
глибину 10 м збільшується на 1 атм. Збільшення тиску з занурен
ням у воду призводить до збільшення розчиненності газів у кро
ві. При підйомі людини з глибини на поверхню тиск зменшу
ється і може настати газова емболія — виділення міхурців газів,
серед яких найнебезпечнішим є азот. Міхурці азоту блокують
судини, що призводить до швидкої загибелі людини. Це необхід
но враховувати при роботі водолазів — створювати суміш для
дихання із відповідним співвідношенням газів і їх тиском.
Вплив вітру на стан організму. Повітряні маси поділяють
за температурою, вологістю, хімічним складом, напрямком
руху тощо. Залежно від географічних особливостей місцевості
формуються повітряні потоки певного регіону, які потім пере
міщуються. Зона контакту двох повітряних мас називається
атмосферним фронтом. Відомо такі типи вітрів, як мусони,
бризи, гірські та долинні вітри, фени, бори (містралі), шквали,
торнадо (смерчі). Напрямок і швидкість вітрів у надґрунтових
шарах повітря залежить від конфігурації території та структу
ри рослинності.
На організм людини вітер впливає безпосередньо або опосе
редковано через уплив на стан навколишнього середовища,
300
Розділ 5
а, відповідно, на умови життя людини. Існують науково обґрун
товані гігієнічні вимоги щодо складу повітря у житловій і ро
бочій зонах.
Великий вплив на здоров’ я людей мають і інші екологічні
чинники.
Так, коливання кислотності та сольового складу ґрунту
може позначатися і на стані здоров’ я людей, що мешкають
у відповідній місцевості. Рослинна продукція, вирощена на
лужних ґрунтах, містить менше заліза, марганцю і фосфору,
тому що ці елементи перебувають у таких ґрунтах у зв’ язаній
формі, недоступній кореням рослин. Через трофічні ланцюги
певні хімічні елементи, якщо вони є у ґрунті або питній воді
у надлишкових кількостях, потрапляють в організм людини,
накопичуються в ньому у значній кількості, і це може стати
причиною розвитку захворювань. Особливо небезпечними є
іони важких металів. Кадмій, зокрема, потрапляючи в організм
людини, накопичується в нирках і спричиняє підвищення нир
кового тиску та розвиток деяких хвороб. Арсен потрапляє
у навколишнє середовище в результаті спалювання кам’яного
вугілля, а також із пестицидами. Свинець із вихлопних газів
досить швидко потрапляє у поверхневі ґрунтові води, а звід
ти — в рослини та гриби. Залізо і марганець забруднюють се
редовище поблизу місць їхнього видобутку.
Повітря для людини не є лімітуючим фактором, якщо воно
чисте. Однак атмосфера містить певну кількість домішок, котрі
зумовлюються природними й антропогенними джерелами і ви
ступають екологічними факторами. Нині налічується понад
500 шкідливих речовин, котрі забруднюють атмосферу, і їхня
кількість зростає. Найбільш поширеними токсикантами є:
оксид карбону СО, діоксид сульфуру 802, оксид азоту N0, вугле
водні СтНп, сажа та пил. Усі ці сполуки виявляють пряму й опо
середковану дію на організм людини. Небезпека підвищення
концентрації СО полягає в тому, що його не можна визначити
органами чуттів. Головним чином він потрапляє у повітря
з вихлопних газів автомобілів. Високий вміст СО спричиняєть
ся до асфіксії (задухи), а малі концентрації — до поступового
збільшення у крові вмісту карбоксигемоглобіну. Ця речовина
надзвичайно стійка, внаслідок чого з гемоглобіном не може
зв’язуватися кисень або вуглекислий газ. У людей із підвище
Екологія людини та біосфера_____________________________________ ЗО Ї
ним вмістом карбоксигемоглобіну знижується рівень сприй
няття зовнішніх подразнень, порушуються процеси мислення,
координація рухів і адекватність прийняття рішень. Постійно
високий рівень карбоксигемоглобіну спостерігається у людей,
які багато курять. Вони перебувають у стані підвищеного ри
зику раптового порушення серцевої діяльності.
За чотири дні Лондонського смогу у 1952 р. загинуло понад
чотири тисячі людей. Сполуки азоту і сірки, потрапляючи
у хмари, випадають на землю у вигляді кислотних дощів, що
призводить до закислення водойм і ґрунтів. Поліхлоровані та
полібромовані біфеніли руйнують озоновий шар, що призводить
до посилення ультрафіолетового опромінення, зростання захво
рювань на рак шкіри.
Підвищення С 02 у повітрі посилює парниковий ефект, по
тепління в Арктиці й Антарктиці, а це з часом призведе до
зміни погодних умов на планеті в цілому.
Шумове забруднення виникає, якщо рівень шуму перевищує
60 дБ. Наслідок дії шумових забруднень — захворювання орга
нів слуху (до повної глухоти), ендокринної, нервової, серцевосудинної систем, у т. ч. гіпертонія, нервово-психічні захворю
вання. 90— 100 дБ — больовий поріг шуму. В той же час шум
листя, дзюрчання джерела, тиха музика (10— 15 дБ) мають
позитивний вплив на нервову систему людини, її здоров’я за
галом. Надмірний шум тривалої дії може спричинити затрим
ку розумового розвитку дітей, прискорити старіння організму.
Людський організм не здатний адаптуватися до шуму, тому
єдиним і обов’ язковим заходом є обмеження шуму.
Часто шумові виробничі забруднення супроводжуються віб
рацією. Звукові хвилі з частотою нижче 16 Гц людина сприй
має не як звук, а як вібрацію. Вібрація супроводжується стру
сами окремих частин тіла або тремтінням цілого тіла. Тривалі
вібрації можуть викликати порушення діяльності нервової та
серцевої системи, кровообігу, струс головного мозку.
Фотографії поверхні планети, зроблені в інфрачервоних
променях, свідчать про наявність так званих теплових ост
ровів на місцях розташування міст. Причиною цього, зокрема,
є покриття поверхні асфальтом та іншими матеріалами, які
нагріваються краще, ніж ґрунт. Потужні викиди тепла від
3 02
Розділ 5
працюючих агрегатів ще більше посилюють цей ефект. Зміни
газового складу атмосфери над промисловими містами призво
дять до формування смогу, що негативно позначається на
здоров’ї людей.
Сигнальний вплив антропогенних чинників виражається
у зміні місць проживання окремих видів або їх популяцій.
У зв’язку з тим, що умови природного навколишнього сере
довища змінюються, людський організм змушений постійно
адаптуватись. Для людини, яка не адаптована до коливань
параметрів абіотичних чинників, типовою є низька працездат
ність.
Виділяють кілька генетично зумовлених конститутивних
типів людей за їхніми адаптивними можливостями: “спринтер” ,
“ стайєр” , “ мікст” . “ Спринтер” може витримувати короткочас
ні потужні навантаження. Однак для відновлення працездат
ності потребує тривалого часу. “ Стайєр” має загалом невисокі
резервні можливості, однак витримує рівномірні навантажен
ня протягом тривалого часу і стійко відновлює працездатність.
“ Мікст” має проміжні адаптивні можливості й оптимальноадекватний спосіб реагування на зміни навколишнього середо
вища.
У процесі еволюції в кожному географічному районі скла
лися певні умови для життя людини, зв’ язки між життєвими
функціями організму людини і періодичними змінами зовніш
ніх природних умов середовища — біологічні ритми. Розріз
няють біологічні ритми як реакцію на зміну умов середовища
(іекзогенні) і такі, що генеруються самим організмом (ендоген
ні). Механізми, які дозволяють організмам циклічно змінюва
ти свій стан, орієнтуючись у часі, становлять так званий біоло
гічний годинник. У їх основі лежить періодичність фізикохімічних процесів, що відбуваються в організмах. Це ендогенні
біологічні ритми.
Коли порушуються ці адаптивні зв’язки, неминуче виника
ють розлади життєдіяльності й функціонування людського
організму, які потребують адаптації. Фізіологічна стійкість
організму змінюється навіть упродовж однієї доби. Наприклад,
хворі на ревматоїдний артрит уранці почувають себе значно
гірше, ніж удень. Це обумовлено тим, що уночі, коли немає
світлового подразнення, яке опосередковано, через нервову
систему, стимулює кортикостероїдну функцію кори наднирни-
Екологія людини та біосфера
3 03
ків, у хворих вона різко зменшується, і виникає гіпокортицизм.
Зниження рівня оксикортикостероїдів у крові призводить до
зниження рівня артеріального тиску, виникають ранкова ско
ваність і погане самопочуття як прояв добових ритмів організ
му. Крім того, варто зазначити, що вплив на таких хворих
у ранковий час електромагнітними хвилями сприяє посиленню
секреції стероїдних гормонів, зменшенню рівня транскортину
і послаблленню виявів клінічних ознак хвороби, поліпшує
самопочуття. Встановлення важливості біоритмів організму
людини дало змогу підвищити ефективність лікування різних
хвороб дією природних лікувальних грязей, мінеральних вод,
фармакотерапевтичними препаратами. Наприклад, встановле
но, що оптимальне значення артеріального тиску у гіпертоніків
дають хлоридні натрієві ванни, призначені з 14 до 19 год. У цей
час під їх впливом поліпшується геодинаміка, зменшуються
периферійний опір, систолічний і діастолічний тиск. Під впли
вом йодно-бромних ванн у хворих на ішемічну хворобу серця
в ці години зростає в 1,5 раза толерантність серця до фізичних
навантажень порівняно з хворими, котрі приймають ті ж самі
ванни вранці.
Дія лікарських препаратів і природних фізичних факторів
на організм людини у різні години доби по-різному стимулює
певні адаптивні системи організму. Тому успіх лікування знач
ною мірою залежить від урахування біоритмів людини. Завдан
ня лікаря в тому, щоб вибрати найефективніший час для ліку
вання тієї чи іншої системи організму. При плануванні оздо
ровчих заходів із позиції координації біологічних ритмів
людини та зовнішніх умов довкілля необхідно враховувати
досягнення сучасної хронобіології.
5.2. Стан середовища існування та здоров’я
людини
На думку експертів ВООЗ, 80 % хвороб людини пов’ язані
з екологічним станом середовища існування. Цілющ а дія
клімату, різних типів мінеральних вод, повітря хвойних лісів,
лікувальні грязі та мінерали ш ироко застосовуються для
304
Розділ 5
профілактики і лікування різних недуг. У терапії використо
вують реформовані фізичні фактори — гальванічний струм,
електричні та магнітні поля, імпульсні струми різної частоти,
ультразвук, лазерні промені. Більшість дослідників уважають,
що механізми дії фізичних факторів мають рефлекторну при
роду і остаточно проявляються в діяльності гуморальної й ен
докринної систем організму людини. Одночасно мають значен
ня специфічні та хімічні властивості природного фізичного
фактора, які проявляються при проходженні захисно-відновлювальних процесів в організмі, визначаючи терапевтичний
ефект. Нині відомо, що деякі фізичні фактори мають сигналь
ну дію. Тому доза впливу окремого фактори визначається за
лежно від його біохімічних та біофізичних властивостей, ха
рактеру захворювання, стану організму хворого. Загальний
неспецифічний вплив мінеральних ванн, лікувальних грязей
і кліматерапевтичних процедур полягає в тому, що вони обу
мовлюють різні геодинамічні, терморегуляційні, нейроцидинні реакції, змінюють тонус і реактивність скелетних м’язів,
кровонаповнення шкірних судин і периферійний опір капілярів
та прекапілярної сітки, внаслідок чого відбувається пере
розподіл крові, змінюється частота дихання, інтенсивність
сечовиділення. Необхідно враховувати, що при впливі кліма
тичних факторів важливу роль відіграє стан таких природних
параметрів, як рівень атмосферного тиску, напруга електрич
ного і магнітного полів, радіоактивність, швидкість вітру.
Людина є компонентом біосфери і виступає як особливий
біотичний чинник. Водночас є багато інших біотичних еколо
гічних чинників, які взаємодіють між собою. Біотичні чинни
ки — це дія живих організмів один на одного. Біотичні чин
ники є найскладнішою категорією екологічних чинників, що
зумовлено складністю самих проявів життєдіяльності.
Серед сукупності зв’язків між живими організмами можна
виділити кілька основних типів відносин, які мають багато
спільного серед організмів різних систематичних груп. В основі
такого поділу є необхідність використовувати спільні природ
ні ресурси. Відносини типу “хижак — ж е р т в а “паразит —
господар” — це прямі зв’язки, які для одного із партнерів мають
позитивне, а для іншого — негативне значення. У природі немає
видів, для яких людина була б основним джерелом харчуван
Екологія людини та біосфера
305
ня. Проте видів, які можуть бути хижаками щодо людини, є
чимало. Це великі хижаки з родини котячих (лев, тигр, леопард
тощо); крокодили, акули, баракуди тощо. У місцях їх поши
рення людині необхідно остерігатися прямого нападу. Пара
зитів, для яких людина може бути хазяїном, значно більше.
Серед них багато найпростіших, плоских і круглих червів.
Інвазивні хвороби, зумовлені патогенними найпростішими,
гельмінтами, вивчають відповідні розділи медичної біології.
Розділ екології, який вивчає біологію розвитку, шляхи поши
рення паразитів у природі та умови їх існування іноді виділя
ють в окремий розділ — екологічна паразитологія.
Основне місце у дослідженні екології людини займають
людські спільноти. Людина має декілька суттєвих відмінностей
від усіх інших видів, які існують у біосфері. Суспільна сутність
людини з її трудовими навичками та соціальними потреба
ми — властивість, не притаманна іншим живим істотам.
Кожне нове покоління формується під впливом різних вза
ємовідносин між людьми у великих і малих соціальних групах,
які включені в різні соціально-історичні процеси. Загалом по
пуляція як система виживання, саморозвитку з часом може
втрачати свої адаптивні властивості, можуть суттєво скорочу
ватися резерви здоров’ я і зростати загроза депопуляції. Дослі
дження прямих і зворотних зв’ язків, які відбуваються у процесі
формування народонаселення в умовах конкретного навколиш
нього середовища, а також причин, які змінюють поведінку
населення у різних соціально-економічних ситуаціях, вивчає
розділ екологія народонаселення.
Динаміка кількості населення на Землі визначається співвід
ношенням показників народжуваності та смертності. У деяких
країнах світу на зміну кількості населення суттєво впливають
міграційні процеси. Після великих географічних відкриттів
зросли еміграційні процеси, особливо в Австралію й Америку.
Для виявлення тенденцій популяційних змін необхідно знати
демографічну ситуацію і напрями її розвитку. Протягом остан
ніх 150 років населення Землі зростає дуже швидкими темпа
ми. Близько 1830 р. населення планети становило 1 млрд осіб,
протягом X X ст. зросло більш як на 3 млрд і наприкінці 1999 р.
сягнуло 6 млрд. Одночасно природні ресурси, якими мож
на забезпечити життя населення, неухильно зменшуються.
306
Розділ 5
Розміщується населення Землі вкрай нерівномірно. У шести
країнах із населенням понад 100 млн осіб кожна (Китай — по
над 1 млрд осіб; Індія, США, Індонезія, Бразилія, Японія)
живе понад половину населення планети. У 57 великих держа
вах світу мешкає понад 95 % населення Землі, а в 38 неве
ликих — усього понад 1 млн осіб. На початку X X ст. найбіль
ший приріст населення спостерігався у Європі (висока народжу
ваність при відносно низькій смертності), а також у Латинській
Америці. Однак згодом народжуваність у Європі зменшилась.
У країнах Азії й Африки у зв’ язку з високою смертністю (не
зважаючи на високу народжуваність) приріст населення був
незначний. Після закінчення Першої світової війни починаєть
ся “демографічний вибух” у країнах третього світу — Африки
та Азії. Друга світова війна завдала усім воюючим країнам
величезних людських втрат, у тому числі призвела до змен
шення народжуваності. Щонайменше на 175 млн людей стало
менше на планеті у результаті війни. Дещо скоротився приріст
і населення Азії та Африки. Лише в країнах Америки й А в
стралії, яких ця війна торкнулася мало, кількість населення
почала зростати.
Глобальний “ демографічний вибух” , який настав після Дру
гої світової війни, відрізнявся від усіх попередніх. Насамперед
це пов’ язано зі значним зменшенням смертності у багатьох
країнах (особливо у країнах третього світу) завдяки досягненням
медицини. Народжуваність у цей період змінилася не дуже
суттєво, зросла середня тривалість життя, а смертність із препродуктивної групи перемістилась до постпродуктивної та пе
рестала виступати як обмежуючий фактор народжуваності. За
рахунок цього зріс демографічний потенціал — відсоток моло
дих людей у країнах третього світу. Однак від 60-х років X X ст.
спостерігається стійке зниження темпів приросту населення.
Вважають, що 15 народжених на 1000 осіб населення є критич
ним показником, при якому відбувається просте відтворення
населення. У Японії, США, Канаді, Австралії та Новій Зелан
дії народжуваність упала до 13— 16 народжених на 1000 осіб.
Унаслідок нерівномірного приросту населення у різних ре
гіонах Землі змінилася їхня частка в загальній кількості на
селення. Так, населення Латинської Америки більш ніж под
воїлось, а Центральної та Західної Європи зменшилось більш
Екологія людини та біосфера
307
ніж на 40 % . За даними ООН, населення Землі до 2025 р. сяг
не 8 млрд осіб, до 2095 р. — 10,2 млрд, і на цьому практично
припиниться зростання населення планети.
В Україні згідно з останніми даними Держкомстату, на
1 вересня 2005 р. проживало 47,03 млн людей. В Україні дуже
низька щодо інших європейських країн очікувана тривалість
життя — 68,2 року. У чоловіків — 62,2, у жінок -— 74,1 року.
У світі розрив тривалості життя між статями становить 4—
5 років. Вікова структура європейських країн уміщує чотири
покоління, а в У країні— щонайбільше три. Неймовірно висо
кий в Україні ризик смертності для чоловіків у працездатному
віці — близько 38 % . З 1991 р. серед населення України смер
тність перевищує народжуваність, тобто відбувається депопу
ляція. За прогнозами Інституту демографії та соціальних до
сліджень Національної академії наук України, до 2050 р. чи
сельність населення України може зменшитися до 35 млн
осіб.
Негативні наслідки неправильного господарювання призвели
до забруднення навколишнього середовища, посилення нер
вово-емоційного напруження, до погіршення здоров’ я людей.
Отже, незважаючи на те, що розвиток медицини дозволив
суттєво знизити рівень дитячої смертності та скоротити захво
рюваність і смертність від інфекційних недуг, посилився нега
тивний вплив на здоров’ я людей забруднення природного
навколишнього середовища і його наслідків. Крім того, значно
зросли стресові навантаження, пов’ язані з прискоренням тем
пів життя. Сукупність цих факторів призвела до збільшення
кількості серцево-судинних захворювань, злоякісних новоут
ворень, травматизму. Хвороби вірусної природи розповсюджу
ються все швидше та набувають загрозливих форм.
В особливостях життєдіяльності кількох поколінь проявля
ються нові специфічні як індивідуальні, так і популяційні
закономірності. На рівні популяції сучасний процес біосоціального напруження і еволюції характеризується появою нових
механізмів адаптації організму людини, а також наростанням
конститутивної гетерогенності популяцій. З’являються механіз
ми, які регулюють генофенотипову гетерогенність популяції:
підтримання специфічних, рідкісних форм генофенотипових
здібностей (обдарувань), оскільки без таких людських феноменів
308
Розділ 5
популяція не може мати повноцінної суспільно-трудової ак
тивності. Вважають, що так, очевидно, формувались, відбира
лись, закріплювались вказані специфічні для людини нові
конститутивні варіанти, генофенотипові адаптивні програми,
можливості їх підтримання і заміни, потенційне видове актив
не довгожительство. Поява нових токсикантів, вірусів і бак
терій, а також фізичних агентів діє на найбільш еволюційно
давні механізми пристосування (наприклад, макрофаги). Зараз
в Україні, як і в усіх інших країнах світу, є необхідність роз
ширювати і поглиблювати наукові дослідження взаємодії фак
торів навколишнього середовища. Деградоване штучне міське
середовище шкідливо впливає на здоров’я населення внаслідок
забруднення атмосферного повітря, дефіциту сонячного про
міння, води, а також стресових факторів, зумовлених напру
женим ритмом життя, скупченістю населення. Мешканці ве
ликих міст п’ ють воду гіршої якості, ніж у селах. В Україні
в більшості великих індустріальних міст якість питної води не
відповідає санітарним нормам. У великих містах люди частіше
хворіють бронхітами, астмою, алергічними хворобами. Ступінь
поширення багатьох хвороб упродовж X X ст. був набагато
більшим у великих містах, ніж у малих містах або селах.
Однак останніми роками така закономірність починає змі
нюватись. У світі близько 25 тис. сільськогосподарських робіт
ників щороку отруюються пестицидами. Усі пестициди не
безпечні, адже вони створюються з метою знищення усіх форм
життя. Пестициди вражають печінку, викликають захворю
вання серцево-судинної системи, центральної нервової та ди
хальної систем, негативно впливають на репродуктивну фун
кцію людини.
У природі існує багато речовин, які містять радіоактивні
елементи або радіоактивні ізотопи Карбону, Гідрогену, Калію
та інших хімічних елементів. Еволюція органічного світу і лю
дини відбувалася на фоні певних доз опромінення, яке отримує
кожна жива істота. Ці дози становлять природний радіоактив
ний фон. До їхнього впливу організми тією чи іншою мірою
адаптовані.
Унаслідок Чорнобильської катастрофи територія України
сильно забруднена радіонуклідами. Вплив радіонуклідів на
організм людини дуже різний. Гостра біологічна дія радіації
Екологія людини та біосфера
3 09
проявляється у вигляді променевої хвороби, локальних уражень
шкіри, кришталика ока, кісткового мозку. Багато радіонуклідів
потрапляють в організм людини з повітрям через органи ди
хання, травлення, ш кіру. Основними радіонуклідами, що
визначають медичні наслідки аварії на ЧАЕС, були радіоак
тивні ізотопи Йоду. їх викид у біосферу призвів до зростання
частоти тиреоїдної патології, зокрема злоякісних новоутворень
щитоподібної залози. Радіоактивний Цезій відкладається пе
реважно у м’ язах. Радіоактивний Стронцій має здатність за
міщувати Кальцій і його сполуки в організмі. Тому він нагро
маджується в кістковій тканині і сприяє захворюваності на рак
або лейкемію.
Розрізняють пряму та непряму дію радіонуклідів і хімічних
молекул. Під прямою дією розуміють такі зміни, що виникають
унаслідок поглинання енергії випромінювання клітинами, а під
непрямою — зміни, які викликають продукти радіоактивного
розпаду речовин у живій клітині. Крім того, вплив радіонуклі
дів на організм людини може бути соматичним або генетичним.
Соматичний проявляється протягом усього життя людини.
Особливо небезпечним є вплив радіоактивного випромінювання
на статеві клітини. Вплив на генетичний апарат проявляється
у нащадків унаслідок порушень у генах і хромосомах. Винайден
ня рентгенівського випромінювання, а потім ядерної зброї, за
провадження атомної енергетики призвели до різкого підвищен
ня радіоактивного опромінення окремих категорій людей.
Наслідки аварії на ЧАЕС нині ще повністю на проявилися.
Однак, на думку багатьох учених, в Україні вже загинуло по
над 500 тис. осіб від її прямої чи опосередкованої дії.
Дуже гострим залишається питання про можливість про
філактики та лікування наслідків негативної дії радіоактив
ного забруднення організму людини, виведення з нього токсикантів. Розроблені та використовуються радіопротектори, ан
тиоксиданти, а також речовини, які адсорбують в організмі та
виводять з нього окремі радіоізотопи (адсобар, фероцин, пентацин) або підвищують неспецифічну резистентність організму,
посилюючи активність імунної системи (нероболіл, ретаболіл,
вітамінні препарати). Ефективними радіопротекторами є віта
мін А (оптимізує роботу з експресії генів, сприяє збільшенню
Т-кілерів), р-каротин та деякі інші каротиноїди (інгібують
310
Розділ 5
передракову лейкоплакію). Особливо позитивні результати їх
спільного застосування з аскорбіновою кислотою. Застосування
спеціальної дієти для попередження пострадіаційного канце
рогенезу передбачає зменшення калорійності їж і, введення
в неї у певних співвідношеннях вітамінів, антиоксидантів і ад
сорбентів.
Пестициди і радіонукліди, особливо з великим періодом
піврозпаду, можуть зберігатись у навколишньому середовищі
десятки років, продовжуючи свою негативну дію. Ефект спіль
ної дії пестицидів і радіонуклідів — поява найрізноманітніших
захворювань, включаючи онкологічні. Однак доведено, що
пестициди та радіонукліди внаслідок колообігу хімічних еле
ментів і речовин у природі поширюються на значні території,
які на тисячі кілометрів можуть бути віддалені від джерела
забруднення. У довкіллі зростає рівень ксенобіотиків — хіміч
них речовин, які у природі не утворюються, а тому в живих
організмів немає систем захисту від них. Отже, наслідки дії
найрізноманітніших хімічних елементів і радіонуклідів прояв
ляються в усіх регіонах нашої планети. Розробка методів ви
далення радіонуклідів із води, ґрунтів, повітря залишається
однією з найважливіших.
Серйозну небезпеку для здоров’я людини становлять важкі
метали: Кадмій, Свинець, Ртуть, Олово, Вісмут, Сурма, Ко
бальт, Мідь, Хром, Цинк, Селен та інші. Залежно від хімічної
форми і концентрації вони можуть спричиняти гострі або
хронічні захворювання.
Поверхнево-активні речовини, синтетичні мийні засоби,
синтетичні покриття можуть мати загальну токсичну дію,
а також алергенну, імунодепресивну, ембріотоксичну, мута
генну та канцерогенну дії.
Відомо, що одними з основних зв’ язків у екосистемах є хар
чові зв’язки, які об’єднують між собою організми за принципом
“ їжа — споживач” . В екології співвідношення біомас організмів
або зв’язаної в біомасі енергії зображують у вигляді екологічних
пірамід. Як правило, екологічні піраміди звужуються від ос
нови до верхівки. У трофічних ланцюгах усі види речовин
послідовно переходять від одного організму до іншого. Інша
закономірність притаманна тим речовинам і ксенобіотикам,
Екологія людини та біосфера
311
які не використовуються і не виводяться живими ланками
трофічного ланцюга, — при переході на кожен новий трофіч
ний рівень їх концентрація у розрахунку на одиницю біомаси
стає більшою. Цей ефект називається законом концентрування
речовин у трофічних ланцюгах. Людина стоїть у кінці трофіч
них ланцюгів. Тому забруднення, яке вважається незначним
при оцінці кількості забруднювача в навколишньому середо
вищі, стає катастрофічно небезпечним при оцінці його для
самої людини.
З огляду на вищесказане, у 1972 р. започаткована міжна
родна програма моніторингу навколишнього природного сере
довища для контролю за станом об’ єктів екосистеми, контролю
за джерелами поширення екологічної рівноваги, моделювання
та прогнозу екологічного стану екосистеми, керування еколо
гічними процесами. Важлива роль у питаннях контролю за
станом довкілля належить лікарям і Міністерству охорони
здоров’ я.
Певну небезпеку для здоров’я людини можуть становити
недостатньо апробовані лікарські препарати, харчові добавки,
харчові продукти, забруднені патогенними мікроорганізмами.
Сьогодні весь світ занепокоєний епізоотією пташиного гри
пу. Ситуація загрозлива. Внаслідок глобального потепління
змінилися звичайні маршрути міграції перелітних птахів.
Дикі птахи найчастіше лише переносять вірус (самі вони
хворіють менше). Однак від диких птахів збудник може пере
датися свійським птахам. У диких птахів за тривалий період
співіснування з вірусом виробився надійний імунітет, у домаш
ніх птахів його немає. Вірус також еволюціонує, виникають
нові його штами, високопатогенні. Українські вірусологи ви
явили велику кількість видів високопатогенних вірусів рослин.
Це призводить до зниження врожаю, цінних компонентів у на
сінні, яке споживають люди. Виявлено кілька варіантів вірусу
тютюнової мозаїки, який може вражати близько 400 видів
рослин, а також викликати зміни у клітинах ссавців. Висока
мінливість штамів в умовах радіації у Чорнобильській зоні
призводить до інфікування все ширшого спектра рослин. Сьо
годні збудники таких захворювань, як сказ у тварин, краснуха
у коропа, вазикулярний стоматит у ссавців знаходять у 60 видів
312
Розділ 5
рослин (пшениця, цукровий буряк, хміль, малина, чорна смо
родина тощо). Вони дуже подібні. А. Бойко вважає: “ Єдине,
що рятує людей зараз, — геном рослинних вірусів влаштований
інакше. Але немає гарантії, що в результаті дії якихось стре
сових факторів такі віруси від рослин не перейдуть на тварин
і людину” . Тобто нині всі найбільш вразливі ніші займають
віруси. Сьогодні необхідно якнайретельніше здійснювати кон
троль за сільськогосподарською продукцією і рослинного,
і тваринного походження.
Вивчення основних закономірностей існування та функціо
нування надорганізмових систем дало можливість виділити
понад сімдесят основних законів і правил. Ми розглянемо ли
ше ті з них, які найбільшою мірою необхідно враховувати
в екології людини.
5.3. Отруйні організми. Лікарські рослини
Отруйні тварини. Отруйними називають організми, у тілі
яких міститься отрута, здатна викликати постійні або тимча
сові розлади функцій у тварин і людини, а інколи — смерть.
Продукування отруйних речовин є специфічною видовою оз
накою.
Отруйними тваринами називають організми, що містять
речовини при різних формах контакту з якими в інших тварин
або людини виникають порушення життєвих функцій і навіть
смерть.
Розрізняють первинно- і вторинноотруйні тварини.
У первинноотруйних отруйність є біологічною властивістю,
тобто притаманна всім особинам певного виду. їх поділяють
на дві групи:
Активноотруйні мають особливі залози, що виділяють
отруйний секрет. Вони можуть бути озброєними і неозброєними.
Озброєні, окрім отруйної залози, мають жалкі клітини, жало,
ротовий апарат колючого або ріжучого типу, плавці, кісткові
шипи, отруйні зуби. Неозброєні отруйні організми мають
тільки отруйні залози — шкірні (у земноводних) та анальні
(у попелиці).
Екологія людини та біосфера_____________________________________ 3 1 3
Пасивноотруйні не мають отруйних органів, а їхня отруй
ність визначається токсичними властивостями тканин і ор
ганів.
Вторинноотруйні — це види тварин, серед яких тільки
окремі особини мають отруйні властивості, причому ці вла
стивості проявляються за певних умов: живлення, бактеріаль
ної інфекції, впливу зовнішнього середовища тощо. Отруєння
такими тваринами відбувається тільки при їх поїданні.
Універсальних отруйних тварин, які вражали б однаково
всі організми, немає. Отрута певної тварини неоднаково діє на
тварин різних видів і на людину. Отруйність залежить від
кількості введеної отрути, від віку тварин, їх фізіологічного
стану, пори року, від віку жертви, місця введення отрути то
що.
Отруйні тварини різноманітні за систематикою. Так, серед
кишковопорожнинних відомі отруйні медузи. Найнебезпечніша з них — хіронекс, або морська оса, діаметром 45 мм. При
контакті з її ногощупальцями у людини виникають корчі, па
раліч і смерть. Кишковопорожнинні озброєні кропив’ яними,
або жалкими, клітинами, які мають капсулу зі спірально за
крученою ниткою. При контакті з іншим організмом нитка
викидається назовні та проникає у покриви жертви, паралізу
ючи її. Деякі види медуз здатні завдавати опіків. Так, медуза
хрестовичок-гоніонема має 80 щупалець, її отрута викликає
пекучий біль, невеликі крововиливи, задишку.
Отруйні форми є і серед молюсків. У представника черево
ногих — великого конуса — отрута викликає параліч скелетних
м’ язів людини.
Із хребетних тварин отруйні види є серед риб, земноводних,
плазунів,
До отруйних риб належить хвостокол, або морський кіт,
який мешкає у Чорному морі. Його хвіст озброєний зазубреним
шипом, у який відкривається протока отруйної залози. На
людей, як правило, не нападає, але при контакті може пора
нити і спричинити запалення.
У Тихому океані трапляється скорпена (Зсогрепа риііаіа) —
риба із гострими променистими плавцями. Проколюючи ними
шкіру людини, впорскує у жертву отруту, яка викликає ура
ження і смерть.
314
Розділ 5
Серед риб є пасивноотруйні форми. Наприклад, фуга (во
диться у Тихому океані) має отруйну ікру, печінку та кров.
Серед фуг найбільш небезпечним видом є куля-риба, отрута
якої викликає параліч дихальних шляхів і, як наслідок, смерть.
Середньоазіатська риба маринка має смачне м’ ясо, але отруй
ні очеревину та статеві органи.
Серед земноводних отруйними є зелена жаба зі шкірними
бородавчастими залозами, секрет якої діє на роботу серця;
тропічна південноамериканська колумбійська жаба-дереволаз
плямиста, смертельну отруту якої використовували індіанці
для наконечників стріл.
Та найбільша кількість отруйних форм серед плазунів.
Змії — це активноотруйні тварини, загальна кількість їх сягає
2000 видів. Серед них 270 є отруйними.
У Європі трапляється 56 видів змій, 14 із них отруйні. Най
характерніша ознака отруйних змій — наявність двох зубів
і отруйних залоз. Найнебезпечнішими є кобри. Вони у збудже
ному стані можуть піднімати верхню частину тулуба і розши
рювати шию завдяки рухливим ребрам і шийним хребцям.
Отрута кобр містить токсин, який уражає дихальний центр
нервової системи, зумовлюючи параліч. До отруйних змій на
лежать види: гадюка звичайна (Уірега Ьегиз), г. степова, г. ро
гата тощо. Гадюки мають темну смугу вздовж спини. Звичай
на гадюка поширена у південній частині Європи, Сибіру.
До гадюкових належить американська гримуча змія (Сгоіаіиз
Ноггдиз). Свою назву вона отримала через наявність на хвості
рухливо сполучених рогових дисків (“ тріскачки” ). Збуджена
змія, розхитуючись, видає тріскотливі звуки. Укус її досить
небезпечний.
У Середній А зії живе щитомордник (Апсізігодоп Наїиз),
близький до групи гримучих змій. Видовою ознакою його є
правильно розташовані рогові щитки на голові. Отрута щито
мордника змінює склад крові та порушує кровообіг.
До родини гадюкових належать також гюрза (Уірега ІеЬесііпа)
і ефа пісщана (Ескіз сагіпаіиз), які водяться у Середній Азії,
Південному Сибіру і Закавказзі.
Залежно від дії, зміїну отруту називають “кров’яною отрутою”
(отрута гадюкових) і нервово-паралітичною отрутою (отрута
кобр). Отрути гадюк викликають як місцеву (сильний біль,
Екологія людини та біосфера
315
набряк, крововиливи), так і загальну токсичну дію на організм
(ослаблення серцевої діяльності, зниження артеріального тис
ку, гемоліз еритроцитів). У важких випадках смерть може
настати навіть через кілька днів після укусу. Отрута кобр діє
на нервові центри, насамперед на дихальний. Смерть настає
від паралічу дихальних м ’ язів. Сила вражаючої дії отрути
залежить від кількості, місця укусу та фізіологічного стану
тварин і людини.
Різні тварини неоднаково чутливі до отрути. Наприклад,
одна і та ж кількість отрути гримучої змії вбиває 24 собаки,
60 коней, 600 кроликів, 800 щурів, 2000 морських свинок. Є
види тварин, які мало чутливі до деяких отрут, тобто мають
природний імунітет. Наприклад, свині нечутливі до укусів
гримучої змії; їжак — до отрути гадюки. Вчені дослідили, що
сироватка крові їжака може нейтралізувати цю отруту. Гри
зуни пустель несприйнятливі до отрути скорпіонів. Деякі пта
хи (лелеки, ворони, кондори) можуть поїдати отруйних змій.
Встановлено, що людину і тварину можна зробити менш
чутливими до деяких тваринних отрут. Це досягається послі
довним введенням в організм отрути у зростаючій кількості (від
дуже малих до більших доз). При цьому у крові з’ являються
антитоксини, які нейтралізують її. Імунізований кінь може
витримати одноразове введення отрути кобри, яка у десятки
разів перевищує смертельну дозу. Лабораторно виготовляють
лікувальні протиотруйні специфічні сироватки. Для їх вико
ристання необхідно знати, отрутою якого виду змій була ура
жена людина.
Отруйні речовини тваринного походження в малих дозах ви
користовуються з лікувальною метою. Бджолину отруту засто
совують при лікуванні ревматизму, при захворюваннях сугло
бів, подагрі, невралгіях. Отруту гримучих змій використовують
при лікуванні епілепсії, змій (ВоіНгорз }агагаса) — при гемо
філії. Кобротоксин зменшує або знімає приступи бронхіальної
астми. Вчені Пастерівського інституту (Париж) виявили пози
тивний ефект отрути кобри при лікуванні деяких пухлин. Для
одержання отрути змій утримують у спеціальних розплідни
ках — серпентаріях, де умови є близькими до природних.
Змії є ланкою складного екологічного ланцюга. Вони при
носять велику користь, поїдаючи величезну кількість щурів,
316
Розділ 5
мишей, сарани, багатьох шкідливих комах. Відомо, що зни
щення змій у деяких місцевостях мало негативні наслідки — ма
сове розмноження мишей завдало великої шкоди сільському
господарству. Отже, людині необхідно раціонально ставитися
до природи. Тварини, які населяють нашу планету, повинні
бути збережені, у тому числі й отруйні види.
Отруйні рослини. Отруйними називають рослини, які міс
тять отруйні для людей і тварин речовини, що при потраплян
ні в організм викликають у ньому хворобливий стан і можуть
спричинити його загибель.
Розрізняють власне отруйні рослини (отруйність є видовою
специфічною ознакою) та випадкові отруйні (отруйними є
окремі екземпляри).
У кожного виду рослин отруйність виявляється у певній фазі
їх розвитку. В одних найбільша кількість отруйних речовин
нагромаджується до цвітіння, в інших — під час цвітіння, у ще
інших — у фазі дозрівання плодів. Нагромадження токсичних
речовин у рослині залежить від ґрунтово-кліматичних умов,
температури, освітлення, вологи й інших факторів.
Отруйні речовини нерівномірно розподіляються у різних
органах рослин. Наприклад, в аконіту і цикути найбільше
отруйних речовин нагромаджується у кореневищах і коренях,
у наперстянки — в листках і стеблі, у жабрію та кукілю — в на
сінні, у вовчих ягодах — в корі та плодах. Найбільї е отруйних
рослин серед покритонасінних: родини метеликових, макових,
лілійних, пасльонових, молочайних. Проте зустрічаються окремі
екземпляри і серед голонасінних, хвощів, папоротей.
Отруєння тварин або людей залежить не тільки від отруй
ності рослин, а й від сприйнятливості організму до дії отрути.
Відомо, що дорослий міцний організм менш чутливий до дії
отруйних рослин, ніж виснажений або хворий. Особливо чут
ливими є діти. Ще первісні люди збирали рослини та різні
плоди для харчування. Одні з них виявлялися непридатними
для їжі через погані смакові якості, інші викликали захворю
вання, а іноді і смерть людей.
У Східній Європі трапляється 400 видів отруйних рослин.
Основні діючі компоненти: алкалоїди, глікозиди, смоли, ефір
ні олії, органічні кислоти.
Екологія людини та біосфера
317
Серед рослин із цілющими властивостями трапляються от
руйні, які необхідно застосовувати обережно. Так, аїр звичай
ний є небезпечним при підвищеній кислотності, оман — при
захворюваннях нирок, черемша — при гастритах, виразковій
хворобі шлунка та дванадцятипалої кишки, гірчиця — при
туберкульозі, запаленні нирок, цибуля ріпчаста — при захво
рюванні серця, морська капуста — при фурункульозі та кро
вотечах, чебрець — при захворюваннях щитоподібної залози,
атеросклерозі, материнка, м’ята польова, оман — при вагіт
ності, а валеріана — при тривалому застосуванні й у великих
кількостях може викликати головний біль, нудоту, пригнічен
ня діяльності шлунково-кишкового тракту, роботи серця.
Отруйні гриби. Особливу групу отруйних організмів ста
новлять гриби. Білки грибів досить швидко розщеплюються
в організмі людини з утворенням отруйних азотистих основ,
тому отруєння може бути викликане навіть несвіжими гриба
ми. Серед шапкових грибів отруйними є бліда поганка, мухомор
червоний, пантерний, гадючий, несправжні опеньки та лисич
ки, сатанинський гриб. У 50 % людей, отруєних грибами,
настає смерть.
Отруєння можуть викликати ріжки (“ лихі корчі” ) або про
дукти життєдіяльності різних мікроскопічних грибів, зокрема
види фузаріїв (“ п’ яний хліб” ) — зерно, насичене отруйними
речовинами, що є продуктами життєдіяльності грибів. У разі
отруєння грибами необхідно терміново викликати швидку
медичну допомогу. Медпрацівники інтенсивно промивають
шлунок із використанням активованого вугілля, призначають
проносні препарати.
Гриби також можуть паразитувати на рослинах і тваринах,
викликаючи їх захворювання та відмирання, можуть розмно
жуватись і поширюватися на продуктах харчування і промисло
вих матеріалах.
У людини гриби можуть викликати такі захворювання, як
мікози (мукоро-мікози легенів, головного мозку й інших ор
ганів), кандидози, дерматомікози тощо.
Лікарські рослини. До категорії лікарських відносять рос
лини, які містять біологічно активні речовини, впливають на
318
Розділ 5
процеси обміну в організмі людини. Статус лікарської сирови
ни рослина отримує тоді, коли її властивості доведено в нау
кових лабораторіях, протестовано на піддослідних тваринах.
Такі види заносять у спеціальний реєстр — Державну фарма
копею.
Значення лікарських рослин у житті людини: задоволен
ня потреб у поживних речовинах; пригнічення хвороботворної
мікрофлори; мобілізація захисних функцій організму; антиалергічна дія; посилення припливу крові до окремих органів;
стимуляція поділу клітин; збудлива, тонізуюча, заспокійлива,
снодійна дія на нервову систему й організм загалом.
Лікарські рослини доцільно використовувати для профілак
тики та лікування більшості захворювань, а також для імунопідтримуючої терапії. Під час їх застосування в організм лю
дини надходить цілий комплекс біологічно активних речовин:
алкалоїди, глікозиди, сапоніни, флавони, ефірні масла, мікро
елементи, фітонциди, мінеральні солі, ферменти, органічні
кислоти, барвники, смоли, гіркоти тощо. Так, алкалоїди — це
органічні азотовмісні сполуки, що утворюються у процесі метаболізації білків. У рослинному організмі алкалоїди взає
модіють з органічними або мінеральними кислотами, утворю
ючи солі лимонної, винної, яблучної, оксалатної та інших
кислот. Варто пам’ятати, що алкалоїди є сильнодіючими, навіть
токсичними речовинами. Це атропін, хінін, морфін, лобелін,
кофеїн, папаверин, стрихнін, ефедрин, нікотин, нівалін, ерго
тамін та інші, які широко застосовують у практичній медицині.
Найбільше алкалоїдів у рослинах родин бобові, макові, пасль
онові та інших.
На кількісний склад активнодіючих речовин у рослині впли
ває ряд факторів: тип рослини (дикоросла чи культивована),
умови проростання (клімат, водний режим, склад ґрунту), час
заготівлі, умови зберігання. Тому лікарську рослинну сирови
ну необхідно чітко стандартизувати.
Екологія людини та біосфера
319
5.4. Основні екологічні закони
(Основні екологічні закони подано у викладі В. С. Джигирея,
2002).
Історично першим є закон Лібіха, який встановлює залеж
ність живих систем від факторів, які обмежують їхній розви
ток (так званих лімітуючих факторів). Одне з тлумачень цього
закону: якщо кількість і якість екологічних факторів близькі
до мінімуму, необхідного для організму, то він виживає, а якщо
менші за цей мінімум, то організм гине, екосистема руй
нується.
Закон толерантності ( Закон Ш елфорда) узагальнює за
кон мінімуму: відсутність або неможливість розвитку еко
системи визначається не лише нестачею, але й надлишком
будь-якого з факторів ( тепло, світло, вода тощо). Тобто лімі
туючим фактором процвітання організму може бути як мінімум,
так і максимум екологічного впливу, діапазон між якими ви
значає ступінь витривалості (толерантності) організму до цьо
го фактора.
Обидва згадані закони стосуються і людини. Адже вона мо
же жити лише за наявності ресурсів для існування (повітря,
води, світла, їжі, території тощо) і у певному діапазоні цих та
інших факторів (зокрема, кліматичних).
Закон конкурентного виключення формулюється так: два
види, що займають одну екологічну нішу, не можуть спів
існувати в одному місці нескінченно довго. Те, який із видів
перемагає, залежить від зовнішніх умов. Нездатність виду до
біотичної конкуренції призводить до його витіснення та не
обхідності пристосування до більш складних умов і факторів.
І цей закон працює в людському суспільстві. У біосфері не
має місця для розселення та немає надлишку ресурсів. Можна
говорити про екологічне суперництво між країнами і навіть
війни, спричинені екологічними причинами. Гітлер виправдо
вував агресивну політику нацистської Німеччини боротьбою
за життєвий простір. Ресурси нафти, вугілля тощо і тоді були
важливими. Надзвичайної ваги вони набувають на початку
X X I ст. Адже додалася потреба чистої питної води та територій
для захоронення радіоактивних відходів.
3 2 0 ________________________________________________________ Розділ 5
Закон біогенної міграції атомів (закон В ернадського):
міграція хімічних елементів на земній поверхні та в біосфері
загалом здійснюється під домінуючим впливом живої речовини,
організмів. Розуміння тих хімічних процесів, які відбувають
ся в біосфері, неможливе без урахування дії біогенних факторів.
Люди змінюють фізичний і хімічний склад біосфери, умови
збалансованої міграції елементів. Якщо ці зміни виходять за
рамки саморегуляції біосфери, то вони стають глобальними
і некерованими (спустелювання, деградація ґрунтів, катастро
фічні паводки і цунамі).
Закон внутрішньої динамічної рівноваги: речовина, енергія,
інформація та динамічні якості окремих природних систем
і їх ієрархи дуже тісно пов'язані між собою, тому зміна одно
го з показників неминуче призводить до функціонально-струк
турних змін інших, але при цьому зберігаються загальні якості
системи — речовинно-енергетичні, інформаційні та динамічні.
Наслідки дії цього закону виявляються в тому, що після
будь-яких змін елементів природного середовища (речовинно
го складу, енергії, інформації, швидкості природних процесів
тощо) обов’язково розвиваються ланцюгові реакції, які нама
гаються нейтралізувати ці зміни. Навіть незначна зміна одно
го показника може спричинити великі відхилення в інших
і в усій екосистемі. Зміни у великих екосистемах можуть бути
непоправними, а будь-які локальні перетворення природи
викликають у біосфері реакції-відповіді. Закон свідчить, що
при незначних втручаннях у природне середовище, його еко
системи здатні саморегулюватися та відновлюватися, а коли
ці втручання перевищують певні межі та вже не можуть
згаснути в ланцюзі ієрархії екосистем, вони призводять до
значних порушень енерго- і біобалансу на значних територіях
та в усій біосфері.
Закон генетичної різноманітності: все живе генетично
різне і має тенденцію до збільшення біологічної різнорідності.
Необхідність і значення закону особливо зросли у наш час,
коли біотехнологія розвивається швидкими темпами. Не зав
жди можна передбачити результат перенесення генів від одно
го виду до іншого, особливо якщ о ці гени відповідають за
стійкість тих чи інших сполук, які віддавна циркулюють в еко
системах.
Екологія людини та біосфера_____________________________________ 3 21
Закон константності (сформульований В. І. Вернадським):
кількість живої речовини біосфери, утвореної за певний час, є
величиною постійною.
За цим законом будь-яка зміна кількості живої речовини
в одному з регіонів біосфери неминуче призводить до такої ж
за обсягом зміни речовини в іншому регіоні, тільки зі зворот
ним знаком. Наслідком цього є правило обв’ язкового запов
нення екологічних ніш.
Закон максимуму біогенної енергії ( закон Вернадського — Б ауера): будь-яка біологічна та біонедосконала система,
що перебуває у стані стійкої нерівноваги (динамічно рухливої
рівноваги з довкіллям), збільшує, розвиваючись, свій вплив на
середовище.
У процесі еволюції видів виживають ті, котрі збільшують
біогенну геохімічну енергію. Живі системи ніколи не перебу
вають у стані рівноваги та виконують за рахунок своєї вільної
енергії корисну роботу проти рівноваги, якої потребують зако
ни фізики й хімії за існуючих зовнішніх умов. Цей закон разом
з іншими є основою розробки стратегії природокористування.
Закон оптимальності: жодна система не може звужува
тись або розширюватись до нескінченності.
Жоден цілісний організм не може перевищити певних кри
тичних розмірів, які забезпечують підтримку його енергетики.
Ці розміри залежать від умов живлення та факторів існування.
Закон оптимальності допомагає знайти оптимальні розміри для
ділянок полів, вирощуваних тварин і рослин. Ігнорування за
кону — створення великих площ монокультур, масові забудо
ви ландшафту — призводять до порушень у функціонуванні
екосистем, до екологічної кризи.
Закон піраміди енергій (сформульований Р. Ліндеманом):
з одного трофічного рівня екологічної піраміди на інший перехо
дить у середньому не більше 10% енергії. Зворотний потік із
більш високих на більш низькі рівні набагато менший — не
більше 0 ,5 — 0 ,2 5 % , і тому говорити про колообіг енергії в біоце
нозі не доводиться.
Цей закон дає можливість виконувати розрахунки земельних
площ, лісових угідь з метою забезпечення населення продо
вольством та іншими ресурсами.
322
Розділ 5
Закон рівнозначності умов життя: всі природні умови
середовища, необхідні для життя, відіграють рівнозначні ролі.
Звідси випливає закон сукупної дії екологічних факторів.
Аксіома адаптованості Ч. Дарвіна: кожен вид адаптований
до певної, специфічної для нього, сукупності умов існування.
Аксіома розповсюджується і на популяцію людей. Зусилля
американських учених щодо створення штучної екосистеми
(“ Біосфера-1” та “ Біосфера-2” ), яка була б повністю ізольована
від природного навколишнього середовища біосфери, зазнали
невдачі.
Закон розвитку довкілля: будь-яка природна система роз
вивається лише за рахунок використання матеріально-енер
гетичних та інформаційних можливостей навколишнього
середовища.
Абсолютно ізольований саморозвиток неможливий — це
висновок із законів термодинаміки. З цього закону випливають
такі висновки:
— абсолютно безвідходне виробництво неможливе;
— будь-яка більш високоорганізована біотична система
у своєму розвитку є потенційною загрозою для менш організо
ваних систем, тому в біосфері Землі неможливе повторне заро
дження життя — воно буде знищене існуючими організмами;
— біосфера Землі як система розвивається за рахунок внут
рішніх і космічних ресурсів.
Закон фізико-хімічної єдності живої речовини (сформу
льований за В. І. Вернадським): уся жива речовина Землі має
єдину фізико-хімічну природу.
З цього випливає, що шкідливе для однієї частини живої
речовини шкодить і іншій її частині, тільки різною мірою.
Через наявність у будь-якій популяції стійких до фізико-хімічного впливу видів, швидкість добору за витривалістю популя
цій до шкідливого агента прямо пропорційна швидкості роз
множення організмів і чергування поколінь. Внаслідок цього
тривале використання пестицидів є екологічно недопустимим,
бо шкідники, які розмножуються значно швидше, пристосо
вуються та виживають, а обсяги хімічних забруднень доводить
ся дедалі збільшувати.
Екологія людини та біосфера_____________________________________ 3 2 3
Це ж стосується і тривалого застосування одного і того ж
препарати з метою лікування пацієнта.
Закон історичної необоротності: розвиток біосфери й люд
ства як цілого не може відбуватися від пізніших фаз до по
чаткових, загальний процес розвитку однонаправлений.
Відомі також чотири закони екології американського вче
ного Б. Коммонера:
— усе пов’язане з усім;
— усе мусить кудись подітися;
— природа знає краще;
— ніщо не дається даремно.
Перший закон Б. Коммонера, на думку М. Реймерса, близь
кий за змістом до закону внутрішньої динамічної рівноваги,
другий — до цього ж закону та закону розвитку природної
системи за рахунок довкілля, третій — застерігає людство від
самовпевненості. Згідно з четвертим — ми маємо повертати
природі те, що беремо від неї, інакше катастрофа неминуча.
У 1991— 1993 рр. відомий американський еколог Д. Чірас
;і|)0 бив висновок, що природа існує вічно (з погляду людини)
і чинить опір деградації завдяки дії чотирьої екологічних за
конів:
— рециклізацй, або повторного багаторазового використан
ня найважливіших речовин;
— постійного відновлення ресурсів;
— консервативного споживання (коли живі істоти спожива
ють лише те і у тій кількості, що їм необхідно, не більше і не
менше );
- популяційного контролю ( природа не допускає “вибухо
вого" росту популяцій, регулюючи кількісний склад того чи
іншого виду шляхом створення відповідних умов для його існуаиння та розмноження) .
ІІайважливішим завданням екології Д. Чірас вважає вив....... структури та функції екосистем, їх урівноваженості або
мсурівноваженості, тобто причини стабільності й розбалансу111 1 IIмя екосистем.
324
Розділ 5
Внутрішнє середовище організму людини, цілісність і го
меостаз якого забезпечуються нервовими і гуморальними ме
ханізмами регуляції, тісно пов’язане з природним навколишнім
середовищем. Нормальний ріст, розвиток і розмноження лю
дини може відбуватися лише за певного комплексу умов сере
довища. Комплексним показником, що характеризує стан
будь-якої людської популяції, є рівень здоров’ я її представни
ків. За сучасними уявленнями, здоров’я — це природний стан
організму, що характеризується його певною рівновагою
з біосферою і відсутністю будь-яких хворобливих змін. Все
світня організація охорони здоров’ я визначає “ здоров’ я як стан
повного фізичного, духовного і соціального добробуту, а не
лише відсутність захворювання або фізичних дефектів” . Тобто
здоров’ я — це об’ єктивний стан і суб’єктивне відчуття благо
получчя в усіх його аспектах — генетикоанатомо-морфологічному, психолого-психічному, соціально-економічному та ін
формаційному. Якість життя складається із взаємодії людини
(людей) і середовища її (їх) існування. Забезпечення оптималь
ного стану фізичного середовища існування є однією з найваж
ливіших умов збереження здоров’ я людини. Адже від вирішен
ня екологічних проблем залежить здоров’ я і благополуччя
людини. З другої половини X X ст. в економічно розвинених
країнах змінилося співвідношення між гострими та хронічни
ми захворюваннями, зміщуючись у бік хронічних процесів. До
Другої світової війни інфекційні та паразитарні захворювання
були одними з головних причин смертності населення. їх міс
це зайняли з кінця X X ст. серцево-судинні розлади та злоякіс
ні новоутворення, які дають нині приблизно 70 % смертності.
Крім того, значно збільшилася кількість нервово-психічних
розладів, хвороб обміну, алергічних станів, вад розпитку і па
тологічних станів генетичної природи, зріс транспортний трав
матизм.
Певною мірою ці зміни у структурі захворювання пов’ язані
причинним зв’ язком зі змінами в демографічних процесах
(збільшення абсолютної та відносної кількості осіб старшого
віку, зниження народжуваності, зниження дитячої смертності).
Однак основну роль у цих змінах відіграють, очевидно, зміню
вані умови життя. Лікарі та фармацевти повинні чітко усві
домлювати про те, що причинами захворювань людини можуть
Екологія людини та біосфера
325
бути не тільки збудники інфекційних чи інвазійних хвороб,
а й забруднення навколишнього середовища, накопичення
в живих організмах деяких хімічних речовин, біотоксикантів,
дія на організм чинників фізичної природи, стресів тощо.
Порушення екологічної рівноваги у природі може спричи
нити екологічну небезпеку чи екологічну катастрофу. Воно
зумовлює появу невідомих раніше хвороб (генетичних, токси
кологічних, алергічних тощо). Усі ці питання є предметом
вивчення екології, зокрема екології людини. Знання екології
людини є обов’ язковим для кожного лікаря і фармацевта.
На завершення підрозділу хочемо нагадати, що усі біологіч
ні дисципліни розвиваються нині дуже швидкими темпами,
а одночасно з ними — й інші природничі науки: фізика, мате
матика, хімія. Це дає можливість моделювання. Згідно з ма
тематичною моделлю багаторічної динаміки Карбону в атмосфе
рі, ймовірність необоротних змін клімату і руйнування біосфери
цілком реальна. Не виключений і катастрофічний варіант,
коли доза або інтенсивність змін перевищить поріг стійкості
системи “ клімат — біосфера” . Тоді глобальні зміни можуть
стати лавиноподібно наростаючими і необоротними. Згідно
з моделлю, біосферу врятує повне припинення антропогенної
емісії Карбону в атмосферу в 2059 р. Якщо зробити це хоча б
на п’ ять років пізніше, то необоротні зміни почнуться з сере
дини X X IV ст. Якщо ж людство не припинить спалювати па
ливо до 2090 р., то не зможе дожити до 2200 р. Це лише прогноз
за одним, хоча і могутнім, екологічним фактором. Прогнози
не вельми оптимістичні. Що ж робити?
Уявлення людини про світ і про себе не змінювались упро
довж тисячоліть. Нещодавно наука вступила в фазу “ вибухо
подібного росту” . Визнаний у всьому світі український нейро
фізіолог О. Кришталь вважає, що вихід — в особливостях
людського мозку і мислення. “ Світовий розум знайде відповіді
на вічні запитання” . Ми, очевидно, повинні вивчити базові
елементи ноосфери і навчитися їх використовувати.
Продовжувати розвивати науку! Продовжувати вивчати
екологічні закони і, найголовніше, не порушувати їх. Упродовж
існування органічного світу на планеті продукти життєдіяль
ності та механічний вплив організмів на неживі об’єкти призве
ли до суттєвих змін у їхньому хімічному складі та фізичному
326
Розділ 5
стані. Вказані явища сучасної природи є результатом сукупної
дії абіотичних і біотичних чинників. Так, на формування типів
клімату й окремих погодніх явищ неабияк впливає рослинний
покрив.
Організми впливають один на одного і на навколишнє сере
довище.
Конкуренція — одна з найпоширеніших форм взаємодії
організмів, яка проявляється у взаємному обмеженні вико
ристання ресурсів.
Парціальна конкуренція передбачає взаємообмеження од
ного ресурсу, а інтегральна — сукупності ресурсів.
Внутрішньовидова конкуренція буває лише інтегральною,
оскільки представники одного виду займають одну й ту ж
еконішу.
Міжвидова конкуренція може бути або парціальною, або
інтегральною. Конкурентна боротьба спричиняє коливання
чисельності виду (популяційні хвилі), міграції й виступає як
другорядний еволюційний чинник.
Формально закономірності конкуренції висвітлює закон
конкурентного взаємовиключення: у випадку обмежених
ресурсів різні види або популяції з даною щільністю населення
не можуть співіснувати на певній території; у результаті
спостерігається витіснення конкурентів чи вимирання одно
го з них або частини перенаселеної популяції (якщо конкурен
ція внутрішньовидова ).
5.5. Поняття про екосистеми
Залежно від багатьох факторів існує відповідна класифіка
ція екосистем.
Природні екосистеми — це системи, у яких процеси відбу
ваються без прямої участі людини. Виділяють три групи при
родних екосистем: наземні, прісноводні та морські.
Екосистема — це сукупність популяцій різних видів,
пов’язаних між собою трофічними зв’язками, які взаємодіють
Екологія людини та біосфера
327
із фізичним середовищем існування. У фізичному середовищі
виділяють біотичну й абіотичну частини.
До складу абіотичної частини входять неорганічні й орга
нічні сполуки, кліматичний режим (температура, вологість,
рельєф місцевості тощо), які визначають умови існування орга
нізмів.
Біотичну частину біогеоценозу становлять різні екологічні
групи популяцій організмів, поєднані між собою трофічними
та просторовими зв’ язками:
— продуценти — популяції автотрофних організмів, здатних
синтезувати органічні сполуки з неорганічних;
— консументи — популяції гетеротрофних організмів, які
споживають інші організми або мертву органічну речовину
(хижаки, паразити);
— редуценти — популяції організмів, які розщеплюють (пе
рероблюють) органічну речовину (бактерії, гриби).
Унаслідок цих взаємозв’ язків виникають потоки енергії та
кругообіг речовин, які зв’ язують складові біогеоценозу в єдину
систему.
Екологічний гомеостаз — це здатність системи організмів
підтримувати динамічну стабільну рівновагу в мінливих умовах
навколишнього середовища.
Натурценози — екосистеми, які зазнали відносно незнач
ного антропогенного впливу. В них міститься досить багато
диких видів рослин і тварин, з якими людина вступає у певні
зв’язки, однак, негативні наслідки цих зв’ язків є незначними.
З натурценозами пов’язані природно-осередкові захворювання.
Проникнення людини в такі екосистеми зумовлює винесення
інфекції за межі природних осередків і навіть їх поширення.
Агроценози — це штучні біогеоценози, які виникають у ре
зультаті сільськогосподарської діяльності людини (в них збе
реглося небагато диких тварин і рослин, а культурних рослин,
навпаки багато).
До створюваних людиною штучних біогеоценозів належать
парки, плодові сади і ягідники, лісові насадження, рекульти
вовані землі. Істотного антропогенного впливу зазнають ті
328
Розділ 5
штучні екосистеми, де скупченість людей є найбільшою. Гос
подарська діяльність людей тут сприяє поширенню багатьох
захворювань. Забруднення середовища може бути причиною
професійних і алергічних хвороб, а надзвичайно висока скуп
ченість, шум тощо викликають нервові та серцево-судинні
розлади. Здатність екосистеми до самовідтворення залежить
від саморегулювальної взаємодії компонентів, які входять до
її складу. Завдяки цій взаємодії популяції організмів відтво
рюють свою чисельність і умови середовища існування. Стій
кість екосистем проявляється у протистоянні несприятливим
зовнішнім впливам. Порушення взаємозв’ язків організмів
у екосистемах унаслідок діяльності людини можуть призвести
до їх змін.
З усіх видів порушення природної рівноваги через госпо
дарську діяльність найбільшим за своїми наслідками є забруд
нення навколишнього середовища.
Відомо, що коли на поверхню ґрунту потрапляє 5 г палива,
знищуються всі організми на площі до 50 м2, а коли в 1 м3 во
ди міститься 10 г палива, то там гине усе живе.
Унаслідок потрапляння в атмосферу відпрацьованих газів
змінюється атмосферне повітря, тоді як найбільше значення
для всіх живих організмів має його відносно сталий склад.
Відпрацьовані гази — це суміш близько 200 речовин, серед
яких багато шкідливих. До них належать оксиди Карбону,
оксид сірки, альдегіди (в т. ч. й сильнодіючий формальдегід),
токсичні сполуки свинцю. Більшість цих газів важчі за повітря,
тому накопичуються біля поверхні Землі.
Щ ороку в біосферу викидається приблизно 5 млрд т твердих
відходів, 250 млн т дрібнодисперсних аерозолів, 200 млн т ок
сиду Карбону, 151 млн т оксиду сірки, 385 млн т оксидів азоту.
У Світовий океан щороку скидається понад 10 млн т нафти,
300 млн т заліза, 6,5 млн т фосфору, 7000 т ртуті. Щ ороку на
сільськогосподарських землях розпилюється понад 500 млн т
мінеральних добрив і 5 млн т пестицидів, третина яких зми
вається у водойми або залишається в атмосфері. Безпосередні
ми об’ єктами забруднення є атмосфера, вода, ґрунт, а жертва
ми цього забруднення стають людина, рослини, тварини.
Сірка потрапляє в атмосферу під час спалювання вугілля.
При спалюванні 1 млн т вугілля у повітрі додатково з’ являється
Екологія людини та біосфера
329
близько 25 тис. т сірки у вигляді її диоксиду. Сира нафта мі
стить у середньому 1 % сірки, значна кількість якої після пе
регонки переходить у мазут.
Середньодобовий вміст диоксиду сірки в повітрі має бути не
більшим, ніж 365 м кг/м 3. За незначного перевищення такої
кількості збільшується кількість випадків різноманітних за
хворювань, підвищується смертність, особливо серед літніх
людей. Унаслідок хімічної взаємодії сірковмісних речовин
у повітрі утворюються також сульфатні частки, які входять до
складу кислотних дощів і негативно впливають на стан здоров’ я
людей.
Ртуть трапляється у природі в кількох формах: металічній,
у складі неорганічних солей і алкільних сполук. Саме останні
є найнебезпечнішими. Металічна ртуть і її солі виводяться
з організму людини порівняно швидко, зате алкіли — дуже
повільно. Підвищення концентрації ртуті в організмі призво
дить до безсоння, підвищеної збудливості, страхів, а згодом і до
втрати зору, координації рухів. Ртуть, яка потрапляє у навко
лишнє середовище в нетоксичній формі, завдяки мікроорганіз
мам перетворюється в алкільну форму. По трофічних ланцюгах
вона переходить, наприклад, у тканини риб, при цьому її кон
центрація підвищується до небезпечної.
До комбінованих чинників варто віднести, наприклад, ерозію
і зсуви ґрунту, спричиненні сільськогоподарським виробниц
твом чи будівництвом, загальне потепління, викликане підви
щенням концентрації вуглекислого газу, та інші явища. У деградованих ґрунтах зменшується вміст гумусу, погіршуються
аерація та мікробіологічні показники.
Ланцюги живлення. Перенесення енергії від її джерела
(рослин) через низку організмів називають ланцюгом живлен
ня. Всі ланцюги беруть початок від зелених фотосинтезуючих
рослин, переходять до травоїдних тварин, а далі до хижаків
(організми, які поїдають тварин). Ланцюги живлення не ізо
льовані один від одного, а тісно переплетені.
Зелені рослини займають перший трофічний рівень (проду
центи). Другий трофічний рівень займають травоїдні тварини
(первинні консументи). Третій трофічний рівень належить
вторинним консументам (м ’ясоїдні та всеїдні тварини). Більше
трьох-чотирьох трофічних рівнів у екосистемах не існує, адже
3 3 0 ____________ ____________________________________________Розділ 5
чим вищий трофічний рівень, тим менше органічної речовини
(енергії) міститься в ньому.
Вивчення ланцюгів живлення актуальне і для медицини.
Так, усе більшого значення набуває застосування у тваринниц
тві лікарських речовин з метою стимуляції продуктивності
тварин.
Надмірне призначення сільськогосподарським тваринам
антибіотиків і гормональних стимуляторів дає негативні наслід
ки для здоров’я людини, яка вживає м ’ ясні вироби, молоко,
яйця та інші продукти тваринного походження.
Антибіотики, стимулятори росту тварин та інші лікарські
речовини по мірі просування у ланцюзі живлення не розсію
ються, а, навпаки, накопичуються.
Принцип біологічного накопичення необхідно враховувати
при надходженні забруднень у середовище. Загальновідомі
негативні наслідки накопичення радіонуклідних відходів або
осадів у ланках живлення, адсорбція ДДТ і пестицидів у тка
нинах риб і водоплавних птахів.
Хлоровмісні вуглеводні, що застосовуються як інсектициди,
можуть адсорбуватися на органічних речовинах. Така влас
тивість поліпшує їх перенесення і накопичення при передачі
в ланцюгах живлення — від рослин до всеїдних, а від них — до
хижаків. Повідомляється про негативний вплив хлоровмісних
вуглеводів на ендокринну систему людини, про можливість
виникнення злоякісних пухлин і шкідливих мутацій.
Цими проблемами займається наука екогенетика.
Загальне завдання екогенетики — вивчення спадкових від
мінностей у реакціях людей на певні фактори середовища.
У ширшому розумінні в центрі екогенетичної проблеми пере
бувають взаємозв’ язки генотипу й середовища.
Основні напрями екогенетики людини виходять як із харак
теру дій середовища, так і з тривалості впливу. Є багато фак
торів середовища, які потребують спеціального екогенетичного аналізу. До найважливіших із них належать природні
абіотичні фактори, природні біотичні фактори, штучні факто
ри, які дають високе екогенетичне навантаження.
ДІЮ природних абіотичних факторів можна ілюструвати
деякими ситуаціями, пов’ язаними з міграційною активністю
Екологія людини та біосфера
3 31
людей. Організоване переміщення людей за трудовими або
іншими угодами призводить до формування нових популяцій
у кліматичних параметрах, не властивих для них раніше.
Доведено, що в умовах Крайньої Півночі більш пристосова
ними і відповідно закріплюваними виявляються переселенці
з певними варіантами кислої еритроцитарної та лужної сиро
ваткової фосфатаз, генетипово адекватні за даними показни
ками аборигенам.
Про вплив природних біотичних факторів на генетичну
структуру свідчить зміна типів живлення на рівні акліматизо
ваних популяцій. Негативні тенденції виявляються при різкій
зміні традиційного харчування. За останні десятиріччя в ескі
мосів Західної півкулі значно зросла частота певних форм
діабету з тяжким клінічним перебігом. Дослідження показали,
що ріст цієї патології прямо залежить від зміни статусу жив
лення, а саме — від інтенсифікації вживання нетрадиційних
харчових продуктів з підвищеним вмістом глюкози, карбогідратів та інших легкозасвоюваних цукрів. Це і сприяло прояву
патологічних генів, які були раніше в прихованому стані та
віповідають за розвиток діабету.
До штучних факторів, які дають посилене екологічне на
вантаження, можна віднести підвищений радіаційний фон,
хімічне забруднення середовища та деякі інші прояви техні
зації, наслідки яких для організму й генетичного апарату
добре відомі. У суто медичному аспекті тут необхідно врахову
вати великий матеріал з фармакогенетики.
Адапт ація людей до екстремальних умов. Всесвітня ор
ганізація охорони здоров’ я зазначає, що здоров’я — це стан
повного фізичного, духовного і соціального благополуччя, а не
лише відсутність хвороб чи фізичних дефектів.
Якраз завдяки біосоціальній природі людини основою її
адаптації є біологічна інформація, отримана від батьків, (вона
забезпечує пристосувальні та захисні механізми), а також со
ціально-економічні механізми. Останні пов’ язані з освоєнням
людиною нових середовищ, екстремальних умов, удосконален
ням засобів і систем життєзабезпечення. Тобто людина повин
на постійно пристосовуватись не тільки до мінливих природних
умов, але і до антропогенних впливів.
332
Розділ 5
Так, вихідці зі зони помірного клімату, які потрапляють на
роботу в Арктику або Антарктиду, опиняються в умовах суво
рого клімату, незвичайних для середніх широт атмосферних
явищ, різкого зниження кількості мікроорганізмів у ґрунті та
повітрі, життя у відносно нечисленних, згрупованих колекти
вах. Зазвичай такі люди після прибуття на Заполяр’я тривалий
час перебувають у хворобливому стані, який посилюється на
приклад, під час зміни полярних дня та ночі. Хворобливий
стан виявляється підвищенням артеріального тиску та приско
ренням пульсу, що змінюються згодом зниженням тиску і час
тоти пульсу. Спостерігаються масові простудні та кишкові
захворювання. З’ являються ознаки втоми і навіть виснаження
нервової системи — погіршується оперативна пам’ ять, зни
жується надійність роботи людини, збільшується тривалість
періоду рухових реакцій. Ці явища трактуються деякими до
слідниками як метеоневроз.
Зазначена ситуація відображається в рекомендаціях гігі
єністів, які обмежують тривалість роботи для людей, щойно
прибулих у Заполяр’ я. З часом у частини людей функціональ
ні показники повертаються до нормального рівня. У інших
вони залишаються зміненими порівняно з вихідними значен
нями, але відновлюється працездатність і самопочуття. У таких
випадках говорять про акліматизацію людей до нових умов
існування. Для рослин і тварин, перенесених у незвичайне
середовище існування, критерієм їх акліматизації є виживан
ня та здатність до відтворення, для людей — відновлення ви
сокого рівня працездатності.
Сьогодні людині доводиться адаптуватись до зовсім незвич
них умов існування, наприклад, до умов невагомості.
Вивчаючи біологічні ефекти факторів космічного польоту,
насамперед мікрогравітації та важкого компонента космічної
галактичної радіації, з якими живі системи не стикалися на
Землі, космічна біологія одержує принципово нову наукову
інформацію. Ця інформація надзвичайно важлива для з’ ясу
вання фундаментальних проблем сучасної біології, — вона є
базовою для розробки космічних клітинних біотехнологій
і контрольованих екологічних систем життєзабезпечення лю
дини в космічних літальних апаратах, значення яких тепер
різко зросло в зв’язку з планами пілотованих польотів у далекий
космос.
Екологія людини та біосфера
333
В Україні до зон особливого контролю із різним ступенем за
брудненості віднесено 78 районів у 12 областях (більше 2000 насе
лених пунктів). Особливо небезпечне погіршення стану здоров’я
дітей. Падає опір їх організму до різних інфекційних хвороб,
зростає питома вага дітей із поганим фізичним розвитком,
прогресують хвороби, зумовлені генетичними порушеннями.
У населення забруднених районів спостерігається зростання
захворюваності за такими нозологіями, у % :
— захворювання печінки і жовчовивідних ш ляхів— 54,2;
— легенева патологія — 26,5;
— захворювання нервової системи — 20,5;
— астено-невротичний стан — 13,3;
— вегетативні дисфункції — 6,0.
У 42,9 % обстежених спостерігали симетричні й асиметрич
ні хромосомні аберації.
Поняття про стрес. Сучасне урбанізоване суспільство
випробовує здоров’я людини ще і стресовими ситуаціями. Кон
цепцію явища “зігезз” (з англ. — напруга, тиск, натиск) упер
ше виклав Г. Сельє (1936). За Г. Сельє, стрес — це неспецифіч
на відповідь організму на будь-яку вимогу. Стрес розвивається
у кілька стадій. Спершу виникає стадія неспокою, відбуваєть
ся мобілізація захисних сил організму. Для неї характерне
переважання процесів катаболізму. Далі спостерігається від
новлення порушеної рівноваги і перехід у стадію резистентності,
якій властива стійкість до дії подразника та стабілізація фун
кцій. Процес відображає прагнення організму підтримати
сталість внутрішнього середовища. Переважають тут анаболічні процеси. У тому разі, коли організм не може перебороти дію
патогенного подразника, настає стадія виснаження. Клінічним
виявленням стресу є загальний адаптаційний синдром. Реакції-відповіді людини, спрямовані на подолання стресу, за своєю
природою поділяють на фізіологічні і психологічні. У фізіоло
гічних реакціях провідну роль відіграє система гіпоталамус —
передня частка гіпофіза — кора наднирників. Стресорна реак
ція розвивається у певній послідовності. Під дією стресора
у кров викидається “ аварійний гормон” адреналін, який про
дукується мозковим шаром наднирників. Крізь відповідні
334
Розділ 5
ділянки гематоенцефалічного бар’ єру він проникає в гіпотала
мус і спричинює в чутливих до адреналіну клітинах стан збуд
ження. Цей стан передається нервовими шляхами в гіпоталамічні клітини, які виробляють кортикотропний рилізингфактор — кортилібеїн. Він контролює синтез у гіпофізі адренокортиконтропного гормону (АКТГ). Під дією АКТГ у корі
наднирників синтезуються глюкокортикоїдні гормони (кортикостерон, кортизол). Ці гормони впливають на обмін вуглеводів,
білків і ліпідів, підсилюють процеси фосфорилювання та спри
яють утворенню речовин, які акумулюють і вивільнюють енер
гію в клітинах. У результаті підвищується опірність організму
до дії шкідливих факторів, а також до екстремальних станів,
підсилюються захисні й компенсаторні реакції. Описана гор
мональна перебудова здійснюється протягом дуже коротких
інтервалів часу. Вивченню психологічних реакцій на стрес
присвячено багато досліджень. Основна проблема для людини
в тому, аби впоратися зі стресовою ситуацією. Важливим спо
собом психологічного захисту для людей є здатність адекватно
реагувати на екстремальну ситуацію, оцінюючи її з боку ра
ціонального виходу. Люди, які мають внутрішній контроль
над своєю діяльністю, “ інтернали” (які вірять у себе і надіють
ся лише на себе), краще вийдуть зі стресу, ніж “ екстернали”
(які потребують допомоги і покладаються на випадок). Г. Сельє
справедливо зазначав, що захисні реакції організму не завжди
оптимальні. Тому в багатьох випадках у результаті стресу роз
виваються хвороби дизадаптації. Основна причина їх або
в неправильних співвідношеннях гормонів, або у зміні реак
тивності організму, що зумовлена попередніми хворобами,
а також психоемоційними порушеннями, невротизацією особи
тощо.
Наведені приклади ще раз підтверджують вплив факторів
довкілля на функціонування організму. Порушення стабільності
екологічних систем спричинює зростання рівня захворювано
сті.
Розділ 6
МЕДИЧНА ПАРАЗИТОЛОГІЯ
Паразитизм як форма біотичних зв’язків і його основні
ознаки. Співжиття організмів, які належать до різних біоло
гічних видів, називається симбіозом.
Розрізняють такі форми симбіозу:
1. Мутуалізм— взаємовигідне співжиття організмів. Нап
риклад, у кишках людини живе кишкова паличка, яка спожи
ває їх вміст, зате вона діє як антагоніст на розвиток хворобот
ворних бактерій (черевного тифу, дизентерії).
2. Коменсалізм — співжиття, коли один організм споживає
залишки або відходи їж і іншого, не завдаючи йому шкоди.
Наприклад, існування ротової амеби в ротовій порожнині лю
дини.
3. Синойкія — співжиття, при якому один партнер викори
стовує іншого як житло. Наприклад, мальки деяких риб три
маються біля щупалець медуз, а у разі небезпеки ховаються
під їх тілом.
4. Паразитизм — співжиття, при якому один організм ви
користовує іншого як місце перебування і джерело харчування
та завдає шкоди (зумовлює захворювання).
1
336
Розділ 6
Паразитологія — розділ біології, який вивчає взаємозв’язок
у системі паразит-живитель і хвороби, які викликають парази
ти, а також заходи боротьби з ними. Значний вклад у розвиток
паразитології внесли вчені К. А. Рудольф, Р. Лейкарт, М. О. Холодковський, Є. Н. Павловський, К. І. Скрябін, В. А. Догель,
О. П. Маркевич.
Паразитологію поділяють на медичну (вивчає паразитів
людини), ветеринарну (вивчає паразитів свійських і промисло
вих тварин), агрономічну та фітопаразитологію (вивчає пара
зитів рослин).
Залежно від рівня організації паразита існують розділи:
— медична протозоологія — наука про паразитичних най
простіших;
— медична гельмінтологія — наука про паразитичних чер
вів — гельмінтів;
— медична арахноентомологія — наука про членистоногих
(кліщів і комах), які завдають шкоди здоров’ю людини і при
цьому передають збудників хвороб.
Назви паразитних хвороб утворюються від назви роду пара
зита і суфіксів “ -аз” , “ -оз” , “ -ез” (амебіаз, теніоз, скабіез).
Хвороби, викликані бактеріями і вірусами, називаються
інфекціями.
Хвороби, збудниками яких є паразитичні тваринні організ
ми, називаються інвазіями.
Паразитів поділяють на дві групи: тимчасові та стаціонарні.
Тимчасові паразити — це такі, що живуть і розмножуються
у довкіллі, а людину (або тварину) використовують тільки для
живлення (комарі, іксодові кліщі, людська блоха, блощиця).
Стаціонарні паразити живуть і розмножуються всередині
або на тілі людини тривалий час. Цих паразитів поділяють на
дві групи: постійні та періодичні.
Постійні паразити від яйця до смерті живуть усередині або
ззовні організму. Всі стадії життєвого циклу відбуваються у тілі
людини (трихінела, воші, коростяні кліщі).
Періодичні паразити заражають людину на певний час і пе
ребувають в організмі на стадії або личинки, або статевої зрі
лості (біогельмінти, муха вольфартова, аскарида).
Медична паразитологія
337
Залежно від локалізації паразитів поділяють на ектопара
зитів, які поселяються переважно на поверхні тіла людини
(воші), та ендопаразитів, що паразитують у внутрішніх органах
і тканинах людини (гострик, трихінела).
Несправжні паразити ті, які випадково опинились усередині
живителя, там паразитують, але, вийшовши з нього, ведуть
вільноживучий спосіб життя.
Дія паразита не є патогенною тоді, коли він не викликає
захворювання. Якщо ж паразит призводить до захворювання,
то його дія патогенна, вона може мати кілька проявів:
— механічний вплив — пошкодження тканин кишки гач
ками свинячого ціп’ яка або закупорювання просвіту кишки
клубком аскарид;
— токсичний вплив — гельмінти викликають зниження
апетиту, безсоння, нудоту, блювання, головний біль, знижен
ня працездатності;
— зміна хімічного складу крові або міжклітинної ріди
ни — живлення паразитів перетравленою їжею збіднює кров
людини на вміст органічних речовин і макроелементів;
— міграція личинок призводить до руйнування тканин,
появи запальних процесів, сприяє проникненню збудників
інфекції;
— трансмісивна передача збудника через укус членистоногих
кровососних викликає одночасно механічну, хімічну, токсич
ну дію.
Усі ці прояви дії паразита не викликають загибелі живите
ля, адже паразит не зацікавлений в його смерті. У процесі
еволюції виробилася нестійка рівновага між двома організма
ми — паразитом і живителем.
Часто розвиток паразитарних червів відбувається зі зміною
хазяїна.
Кінцевим, або дефінітивним, живителем називається ор
ганізм, у якому розвивається статевозріла стадія паразита
і відбувається його статеве розмноження. Проміжним живите
лем називається той, у якому розвивається личинка і відбу
вається безстатеве розмноження. При наявності двох проміж
них живителів другий з них називається додатковим. Черви,
розвиток яких проходить з обов’ язковою зміною живителя,
3 3 8 ________________________________________________________ Розділ 6
називаються біогельмінтами (сисуни, стьожаки). Коли ж для
їх розвитку необхідне перебування у ґрунті, черви називають
ся геогельмінтами (круглі черви). Звільнення від паразита
називають дегельмінтизацією, знищення паразита як виду —
це девастація.
Живителі поділяються на облігатних, факультативних і тран
зитних.
Облігатний живитель необхідний для розвитку паразита,
без нього розвиток не відбувається.
Факультативний живитель не є обов’язковим для розвит
ку, паразит потрапляє туди випадково. Наприклад, цикл роз
витку п’явки проходить без попадання в організм людини, але
коли п’явка потрапила у глотку, то деякий час живе там як па
разит.
Транзитний живитель нетиповий для паразита, в ньому
цикл розвитку не завершується. Наприклад, яйця котячої та
собачої аскариди можуть заражати людину, личинки здійсню
ють міграцію, а після повторного проковтування викидаються
з фекаліями у довкілля. В цьому випадку людина стосовно
собачої та котячої аскариди є транзитним живителем.
Найчастіше людина заражується паразитами такими шля
хами:
— аліментарно — занесення перорально цисти, яйця або
личинки з продуктами харчування при недотриманні особистої
гігієни;
— перкутанно — проникнення личинок в організм через
шкірний покрив (вугриця);
— конгеніально — внутріутробно (токсоплазма);
— контамінаційно — втирання через пошкоджену ділянку
шкіри збудників захворювання, які є в порожнині та гемолім
фі переносника.
Багаторазове зараження живителя паразитом називається
суперінвазією. Розвиток яєць паразита у дорослу форму без
виходу в зовнішнє середовище називається автоінвазією (гемінолепідоз, цистоцеркоз).
Трансмісивні хвороби передаються переносниками, вони
можуть бути викликані бактеріями та вірусами (інфекції) або
Медична паразитологія___________________________________________ 339
збудниками тваринного походження (інвазії). Серед трансмі
сивних хвороб розрізняють:
— облігатно-трансмісивні, що передаються від одного ж и
вителя до іншого лише через переносника;
— факультативно-трансмісивні — можуть передаватись через
переносника і безпосередньо від хворої людини, через забруд
нену фекаліями їжу чи воду (чума, туляремія), або при зніман
ні шкіри з тварини.
Трансмісивні хвороби поділяють на групи:
— зоонози — хвороби, притаманні лише тваринам (малярія
птахів);
— антропозоонози — збудники вражають людей і тварин
(чума);
— антропонози — хвороби, притаманні лише людині (амебіаз,
трихомоноз).
Є. Н. Павловський у 1939 р. виділив природно-осередкову
категорію хвороб, яка має такі особливості:
— збудники циркулюють у природі незалежно від діяльності
чи захворюваності людини;
— резервуаром збудників є дикі тварини;
— переносники і збудники утворюють біоценотичний ком
плекс;
— збудники поширені на обмеженій території з певним гео
графічним ландшафтом, де наявні різні компоненти біоценозу
(ліс чи степ, поширення кліщів, гризунів, москітів).
Коли людина потрапляє в такий осередок, вона заражається.
Прикладом природно-осередкової хвороби може бути лейшма
ніоз: збудник — Ь. ігоріса, природний резервуар — гризуни, облігатні переносники — москіти (комахи з ряду двокрилих).
Особливості імунітету при паразитозах. Різноманітні
чинники генетичної і негенетичної природи обумовлюють різну
чутливість організму живителя до паразита.
Серед негенетичних чинників виділяють вік, харчування,
гормональний статус, супутні захворювання й особливості
їх лікування. До прикладу, у дітей із порушеним білковим
3 4 0 ________________________________________________________ Розділ 5
метаболізмом важчий перебіг амебіазу, стронгілоїдозу, тоді як
перебіг малярії більш легкий.
Люди, які мають важкі форми злоякісних пухлин, зазвичай
не заражаються вісцеральним лейшманіозом. Залізодефіцитні
анемії практично забезпечують захист людини від малярії. При
злоякісних новоутвореннях товстої кишки і жіночої статевої
системи спостерігається більш важкий перебіг амебіазу і три
хомонозу урогенітального. Велике значення у сприйнятті лю
дини до паразитарних хвороб має її генетична конституція.
Зокрема, в осіб з II (А) групою крові виявлена висока чутливість
до лямбліозу. У Західній Африці у людських популяціях від
сутні особи з групою крові Баіїу, причому в цьому регіоні
дуже рідко трапляються збудники триденної малярії.
Захисна дія хазяїна проти паразитарної хвороби забезпе
чується, головним чином, імунними механізмами. Зважаючи
на сучасне розуміння ролі та значення імунної системи в ор
ганізмі людини, зрозуміло, що природжена стійкість до певних
інфекцій та інвазій забезпечується тісною взаємодією всіх ре
гуляторних і ефекторних систем організму, які беруть участь
у підтриманні імунного гомеостазу. Останній здійснюється
завдяки тісній взаємодії комплексу неспецифічних чинників
імунної резистентності та специфічних клітинних і гумораль
них реакцій імунітету.
Імунні реалізації хазяїна виникають у відповідь на дію ан
тигенів двох різних типів: тих, які входять до складу організ
му паразита, і тих, що виділяються паразитом у довкілля.
Антигени першого типу, за винятком антигенів, які входять
до складу покривів, вивільняються лише після загибелі пара
зита. Вони різноманітні, але часто у споріднених форм можуть
бути схожими. Антигени покривів різноманітні та специфічні.
Часто у них глікопротеїнова природа, і на різних етапах жит
тєвого циклу паразита вони можуть змінюватись, тому імуні
тет до них виробляється важко. Антигени другого типу спе
цифічні. Це компоненти слини кровососного паразита, що
секретуються різними залозами гельмінта.
Імунна відповідь організму хазяїна на протозойних паразитів
полягає в тому, що одноклітинні паразити, середовищем існу
вання яких є позаклітинний простір, вкриваються антитілами
і, таким чином, втрачають рухливість. При цьому вони стають
Екологія людини та біосфера
3 41
доступними для захоплення їх макрофагами. У деяких випад
ках антитіла забезпечують аглютинацію паразитів, які після
цього гинуть. Внутрішньоклітинні паразити, що живуть у мак
рофагах (лейшманії, токсоплазма), можуть перетравлюватися
на місці перебування у випадку активації макрофагів анти
тілами.
Такі механізми імунного захисту проти гельмінтних інвазій
не діють. Імунна відповідь при гельмінтозах має особливості,
обумовлені такими чинниками:
1. Гельмінти можуть не розмножуватися в організмі хазяїна,
але, проходячи складний цикл розвитку від яйця до дорослої
особини, значно змінюють свій антигенний склад.
2. Гельмінти, на відміну від інших паразитів людини, мають
великі розміри, а нематоди, крім того, мають щільну кутику
лу. Для їхнього ураження ефективною стає клітинна форма
боротьби з боку хазяїна.
3. Гельмінти — конгломерат антигенів, у відповідь на який
виділяється велика кількість антитіл.
4. При гельмінтозах імунна відповідь проходить за ТЬ2 (гу
моральним) шляхом, тому розвиваються алергічні реакції
негайного типу, частіше за реагіновим або цитотоксичним
типом. У поєднанні з медикаментозною або харчовою алергією
можливий імунокомплексний варіант (за Сеіі і СоотЬз).
5. На початкових етапах гельмінтної інвазії спрацьовують
неспецифічні механізми захисту: вмикання альтернативного
шляху активації комплементу, що захищає організм хазяїна,
але може сприяти проникненню личинок у тканини хазяїна.
Тому імунітет при гельмінтозах частковий і спрямований
в основному проти личинок паразитів.
Інвазійні личинки містять дві групи антигенів: а) антиген,
який належить до секреторно-екскреторних речовин; б) антиген,
який експресується на кутикулі тегумента. Першу групу ан
тигенів становлять цинковмісні цистеїнпротеїнази (СР). Кож
на з них відіграє важливу роль у взаємовідносинах системи
“ паразит-хазяїн” . Так, СР ТгісНіпеІІа зрігаїіз пошкоджує сли
зові оболонки. СР Апсуіозіопга сапіпит сприяє проникненню
у слизову оболонку шлунка, стимулюючи субепітеліальну по
пуляцію ТЙ2 і розвиток еозинофільного ентериту, вона володіє
також антиплазмовою активністю. СР Разсіоіа кераііса попе
реджує АТ-залежне прикріплення еозинофілів до ексцистованих личинок паразита.
Другу групу антигенів становлять молекули з детермінан
тами 33 кБ до рецептора Сіц-К системи комплементу на лімфо
цитах. Вони експресуються інвазійними личинками Тгіскіпеїіа
зрігаїіз, зв’язують С іц на епітеліальних клітинах тонкої киш
ки, сприяючи проникненню личинок у стінку кишки. До цієї
групи належить також глютатіон 8-трансфераза, яку секретують личинки Опскосегса аоіоиіиз та ін.
У процесі гельмінтозної інвазії в організмі хазяїна утворю
ються антитіла. Найбільш виражений синтез антитіл спостері
гається при паразитуванні тканинних і кишкових мігруючих
гельмінтів, особливо в період міграції та линяння личинок.
Установлено, що антитіла продукуються організмом хазяїна
при аскаридозі, цестодозах, філяріатозах, шистосомозах та ін.
Синтезовані антитіла належать, переважно, до І&М і І£С
класів.
При інвазії Разсіоіа кераііса зростає рівень сироваткових
І&С, І£М, і £Е. У половини хворих визначаються специфічні
І0Е-антитіла. Встановлено позитивну кореляцію між рівнем а
І^Е-антитіл і масивністю інвазії, клінікою, ступенем еозино
філи.
При інвазіях, що викликаються Орізі когокіз їеііпеиз і Сіопогскіз зіпепзіз, є характерним початкове збільшення І£М із по
дальшим зростанням рівня І£А і І^О. Рівень І&Е підвищується
у кілька разів. У інвазованих відзначається кореляція між рів
нем сироваткового І&Е і специфічних І^Е-Ат проти С. зіпепзіз.
Відомо, що і £Е фіксує еозинофіли на паразитах. Еозинофілія
характерна для всіх гельмінтозів. Вона може бути дуже висо
кою, що патогномонічно тільки для еозинофільних лейкозів.
Виражена еозинофілія при гельмінтозах зазвичай супровод
жується ознаками алергії: кропивницею, набряком Квінке,
лихоманкою, артралгіями, міалгіями, еозинофільними інфіль
тратами в легенях та ін. Еозинофілія при гельмінтозних алер
гіях — найбільш простий і показовий тест сенсибілізації ор
ганізму.
Еозинофіли — основні клітини-ефектори, що мають протигельмінтну спрямованість, оскільки система фагоцитозу не
спрацьовує проти паразитів великих розмірів.
Медична паразитологія
343
Еозинофіли містять велику кількість крупних гранул, до
складу яких входять різноманітні біологічно активні речовини.
Так, у ядрі гранул міститься головний основний білок — МВР
(та]ог Ьазіз ргоіеіп). У матриксі гранул локалізовані катіонні
білки, пероксидаза. Гранули також містять фосфоліпазу Д,
гістаміназу, кислу фосфатазу, р-глюкуронідазу, рибонуклеазу,
катепсин. При активації еозинофілів відбувається активація
дихання з продукцією активних метаболітів кисню.
Передбачається, що активовані еозинофіли виробляють гра
нули в навколишнє середовище, вміст яких має токсичну дію
щодо паразитів. Найбільшою активністю відрізняються голо
вний основний білок і, особливо, катіонні білки. Щодо актив
них форм кисню дані суперечливі.
Встановлено, що самі гельмінти мають імуносупресивну дію
на організм хазяїна. Це може істотно вплинути на перебіг ін
фекційних захворювань, особливо кишкових інвазій. Лімфоцитотоксичним ефектом володіють трихінели, ехінокок, шистосоми, печінковий сисун. Сироватка крові хворих на шистосомоз, опісторхоз пригнічує активність макрофагів іп оііго.
Очевидно, імуносупресивний ефект при гельмінтозах забезпе
чується багатокомпонентними механізмами, що запускаються
не тільки антигенами, але й іншими речовинами, які вироб
ляються паразитами. Велика кількість паразитів у процесі
еволюції виробила механізми, що дозволяють послаблювати
несприятливий вплив імунітету хазяїна. Тому тривалі та пов
торні інвазії можливі лише у разі зниження імунітету хазяї
на.
До прикладу, деякі одноклітинні — збудники протозойних
захворювань — паразитують всередині клітини, і це робить їх
малодоступними для антитіл хазяїна. Окрім цього, такі пара
зити розмножуються і розселяються по організму хазяїна, не
виходячи навіть за межі клітин. Переваги деяким паразитам
дає локалізація у тканинній рідині, де концентрація антитіл
звичайно у 5 разів нижча, ніж у плазмі крові.
На паразитів, які живуть у порожнині кишківника, не діють
ні антитіла плазми крові, ні механізм клітинного імунітету.
Деякі дорослі гельмінти вкриті кутикулою, яка у непошкодженому стані майже не викликає захисних реакцій з боку
хазяїна.
344
Розділ 6
Деякі паразити у процесі еволюції виробили своєрідний
засіб захисту — антигенне маскування. Вони синтезують по
верхневі антигени, що за своїм складом близькі до білків ха
зяїна, і, відповідно, хазяїн не ідентифікує їх як сторонні.
Імунний захист проти багатоклітинних паразитів усклад
нюється багатоступінчастим циклом їх розвитку, у якому кож
на стадія може виробляти свій антигенний комплекс.
Щ одо паразитичних одноклітинних організмів, то вони
здатні змінювати антигенну структуру своєї оболонки, причо
му різними шляхами. Це може бути або природний добір, який
формує виникнення популяцій паразитів з новими антигенни
ми властивостями, або почергова активація різних, але функ
ціонально подібних за походженням генів, які постійно змі
нюють антигенну конституцію організму.
6.1. Підцарство Найпростіші — Р г о іо г о а
Загальні риси будови:
— у морфологічному відношенні — одна клітина, у фізіоло
гічному — цілісний організм; тіло має мікроскопічні розміри
й утворене клітинною оболонкою, цитоплазмою з ядром і органелами;
—- найважливіші органели: травна вакуоля; видільна, або
скоротлива вакуоля; в інфузорій — цитостом, цитофаринкс,
порошиця;
— органоїди руху: псевдоподії, джгутики, війки;
— подразливість у формі таксисів (позитивні й негативні
фото-, хемо-, термотаксиси тощо);
— за несприятливих умов утворюють цисти;
— розмноження безстатеве (поділ клітини навпіл, цьому
процесові передує редуплікація ДНК; множинний поділ ядра)
та статеве (кон’югація, копуляція).
Медична паразитологія___________________________________________ 3 4 5
Тип Саркомастігофора — З а гсо т а зіі^ о р к о га
Клас Лобозеа — ЬоЬогеа
Амеба дизентерійна (ЕпіатоеЬа Нівіоіуііса) (рис. 6.1) —
збудник амебіазу
Місце паразитування: товста кишка, інколи внутрішні
органи (печінка).
Рис. 6.1. Амеба дизентерійна: а — велика вегетативна форма;
б — чотириядерна циста: 1 — ектоплазма; 2 — ядро; 3 — ендо
плазма; 4 —- травна вакуоля; 5 — цитоплазматична мембрана
Морфофізіологічні особливості та цикл розвитку (рис. 6.2):
в кишках людини трапляється у трьох формах:
— мала вегетативна, або просвітна, форма (іогта тіпиіа),
утворюється із форми цисти. Розміри 15—20 мкм. Трапляєть
ся у паразитоносіїв, а також у людей в період одужання. При
ослабленні організму переходить у велику вегетативну форму.
В нижньому відділі товстої кишки перетворюється на цисту;
— велика вегетативна, або тканинна форма ( /огта тацпа),
утворюється із просвітної форми. Розміри 20— 50 мкм. Цито
плазма розмежована на екто- і ендоплазму. В ендоплазмі трап
ляються фагоцитовані еритроцити. Хроматин ядра утворює
радіальну структуру, в центрі якої міститься велика грудка
346
Розділ 6
хроматину — каріосома. Виявляється в період загострення
хвороби, а при покращенні стану хворого перетворюється на
просвітну форму, цист не утворює;
— циста, або постійна форма (/огта сівііса), утворюється із
просвітної форми у нижніх відділах кишківника. Розміри
9— 13 мкм, нерухома, безбарвна, псевдоніжки видовжені. Зрілі
цисти чотириядерні, містять хроматоїдні тіла і глікогенові
вакуолі, при ексцистуванні з них у кишківнику здорової лю
дини виходить чотири просвітні форми.
Рис. 6.2. Цикл розвитку дизентерійної амеби: 1 — чотириядерна циста; 2 — ексцистування цисти у метацистні форми; 3 — роз
множення; 4 — мала вегетативна форма амеби; 5 — вихід із фе
каліями чотириядерної цисти; 6 ,7 — велика вегетативна форма
амеби; 8 — вихід великої вегетативної форми амеби у кровотік
Шляхи зараження: пероральний (заковтування цист із пит
ною водою або продуктами харчування).
Симптоми амебіазу: частий рідкий стілець (до 10— 20 разів
на добу) з домішками слизу та крові, абсцеси печінки й інших
органів, куди проникає амеба. При відсутності лікування смер
тність сягає 40 % .
Лабораторна діагностика: мікроскопічне дослідження фе
калій на наявність вегетативних форм і цист. У гострому періоді
в мазку містяться великі вегетативні форми з фагоцитованими
347
Медична паразитологія
еритроцитами, цисти переважно відсутні. При хронічній фор
мі у фекаліях виявляють чотириядерні цисти.
Профілактика: особиста гігієна, миття рук, овочів, фруктів,
санітарний нагляд за водопостачанням, знищення переносників,
лікування хворих, саносвітня робота серед населення.
Географічне поширення: всі області України, а також країни
з гарячим кліматом.
Амеба кишкова (ЕпіатоеЬа соїі) (рис. 6 .3 ) — непатогенна
форма
а
б
Рис. 6.3. Амеба кишкова: а — вегетативна форма; б — циста:
1 — ядро; 2 — травна вакуоля
Морфофізіологічні особливості: амеба кишкова існує у ве
гетативній формі та в формі цисти. Вегетативна форма має
розміри 20— 60 мкм. У цитоплазмі є вакуолі, бактерії, грибки,
але немає еритроцитів. Псевдоподії широкі й тупі. Ядро добре
помітне. Локалізується у верхньому відділі товстої кишки,
а в нижньому утворює цисти.
Циста округлої та овальної форми з товстою оболонкою
(1— 8 ядер). Хроматоїдні тіла і глікогенова вакуоля помітні рід
ко. Ексцентрично розміщена каріосома неправильної форми.
Лабораторна діагностика: мікроскопічне дослідження фе
калій на наявність вегетативних форм і восьмиядерних цист.
3 48
Розділ 6
Амеба ротова (ЕпіатоеЬа §іп£іоаІІ8) — патогенна дія сум
нівна
Морфофізіологічні особливості: розміри амеби ротової
10—ЗО мкм, у цитоплазмі багато вакуолей, лейкоцитів. Рух
повільний. Цист не утворює. Локалізується у білому налипан
ні на зубах та в карієсних порожнинах. Вважають, що амеба
ротова може бути однією з причин розвитку карієсу.
Тип Ціліофора — СіїіорКога
Клас Цітостоматеа — С ііозіот а їеа
Балантидій (Ваіапіісііит соїі) (рис. 6.4) — збудник балан
тидіазу
Місце паразитування: стінка товстої кишки.
Рис. 6.4. Балантидій: а — вегетативна форма; б — циста: 1 — цитостом; 2 — цитофаринкс; 3 — травна вакуоля; 4 — скоротлива
вакуоля; 5 —- макронуклеус; 6 — мікронуклеус; 7 — порошиця
Морфофізіологічні особливості: розміри ЗО— 200 х 20—
70 мкм. У життєвому циклі виділяють вегетативну форму і ци
сту. Тіло овальне, вкрите пелікулою, що несе війки. На перед
ньому кінці тіла є перистом, цитостом і цитофаринкс. Добре
виражені мікро- (ядро кулястої форми) та макронуклеус (ве
лике ядро, бобоподібної форми). У цитоплазмі є травні та ско
Медична паразитологія
349
ротливі вакуолі. Розмножується статевим шляхом (кон’югація)
і поперечним поділом. У нижніх відділах кишківника здатний
утворювати цисти. Цисти одноядерні, мають овальну або ку
лясту форму, розміри 50— 60 мкм.
Шляхи зараження: пероральний (проковтування цист), алі
ментарний.
Симптоми балантидіазу: балантидії можуть проникати
у слизову оболонку товстої кишки, утворювати там вогнища
запалення і виразки, спричиняти діарею, болі у животі, блю
вання, головний біль. У фекаліях кров і слиз. Можливий роз
виток перитоніту.
Лабораторна діагностика: мікроскопічне дослідження фе
калій на наявність вегетативних форм і цист.
Профілактика: дотримання правил особистої гігієни має
основне значення; боротьба із забрудненням середовища фе
каліями свиней, своєчасне виявлення та лікування хворих.
Географічне поширення: в усіх областях України, особливо
там, де розвинене свинарство.
Тип Саркомастігофора — З а гсот а зіі^ор Н ога
Клас Зоомастігофора — Х оот а зіі^орігога
Трихомонада кишкова (ТгісНотопаз Нотіпіз) (рис. 6.5) —
збудник кишкового трихомонозу
Місце паразитування: товста кишка людини.
Рис. 6.5. Трихомонада кишкова: 1 — джгутик; 2 — базальне
зернятко; 3 — ядро; 4 — ундулююча мембрана; 5 — аксостиль
350
Розділ 6
Морфофізіологічні особливості: форма тіла овальна, довжи
на 8—20 мкм. На передньому кінці 5 джгутиків. З одного боку
вздовж тіла розміщена ундулююча мембрана. Посередині про
ходить опірний стержень, кінець якого виступає на задньому
кінці тіла. Недалеко від ядра розміщений клітинний рот. У ци
топлазмі помітні травні вакуолі. Цист не утворює. Розмно
жується поздовжнім поділом.
Шляхи зараження: пероральний (проковтування вегетатив
них форм).
Симптоми трихомонозу кишкового: діарея, часто безсимптомно.
Лабораторна діагностика: мікроскопічне дослідження фе
калій на наявність вегетативних форм.
Профілактика: особиста гігієна, миття рук, овочів, фруктів,
санітарний нагляд за водопостачанням, знищення механічних
переносників, лікування хворих, саносвітня робота серед на
селення.
Географічне поширення: всі області України.
Трихомонадаурогеттальн&(Тгіскотопа8оа£іпаІІ8)(рис. 6.6) —
збудник урогенітального трихомонозу
Місце паразитування: сечостатеві шляхи чоловіків і жінок.
Рис. 6.6. Трихомонада урогенітальна: 1 — джгутик; 2 — базаль
не зернятко; 3 — ядро; 4 — ундулююча мембрана; 5 — аксостиль
Медична паразитологія
351
Морфофізіологічні особливості-, форма тіла грушоподібна,
завдовжки 14—ЗО мкм, на передньому кінці є 4 джгутики і ундулююча мембрана до середини тіла. На задньому кінці — дов
га шпичка, в цитоплазмі - вакуолі.
Шляхи зараження: через статеві контакти, дуже рідко по
бутовим шляхом.
Симптоми трихомонозу урогенітального: свербіж, пекучий
біль, серозно-гнійні виділення з піхви. Чоловіки можуть трива
лий час бути безсимптомними носіями. Згодом виникає запален
ня уретри, до інвазії приєднуються інфекції та грибкові захво
рювання, з’ являються затяжні запальні процеси, розвивається
простатит.
Лабораторна діагностика: мікроскопічне дослідження ви
ділень сечостатевих шляхів на наявність вегетативних форм.
Профілактика: особиста гігієна, уникнення контактів із
хворими, стерилізація гінекологічних інструментів, санітарноосвітня робота серед населення, виявлення і лікування хво
рих.
Географічне поширення: всі області України.
Трипаносома ( Т г і р а п о с о т а Ь г и с е і § а т Ь і е п з е ) — збудник
трипаносомозу — африканської сонної хвороби
Місце паразитування: плазма крові, лімфа, лімфатичні
вузли, спинномозкова рідина, тканини спинного і головного
мозку людини та хребетних тварин.
Морфоанатомічні особливості. Тіло видовжене, звужене на
кінцях, завдовжки 15— 40 мкм. Посередині тіла є ядро, на
задньому кінці блефаропласт і базальне зерно, від якого бере
початок джгутик. Між тілом і джгутиком утворюється ундулююча мембрана, джгутик вільно закінчується в передній частині
тіла. Живлення осмотичне. Розмноження шляхом поздовжньо
го поділу.
Природний резервуар — людина, рогата худоба, свині, со
баки.
Шляхи зараження: трансмісивний (укус мухи цеце (Сіоззіпа
раїраііз).
352
Розділ 6
Симптоми: потрапивши в організм людини при укусі мухи
цеце, трипаносоми накопичуються в лімфатичних вузлах
і судинах, виходять у кров, поширюються по організму, вра
жаючи головний мозок. Унаслідок загальних і аутоімунних
ушкоджень в тканинах внутрішніх органів розвиваються різ
ного характеру зміни: страждає клубочковий апарат нирок,
у серці — некрози та крововиливи, в печінці, селезінці — ди
строфічні зміни. У хворих спостерігається лихоманка, збіль
шення лімфатичних вузлів, болісні набряки, головний біль,
стомленість, апатія, сонливість удень, безсоння вночі, судоми,
які призводять до коми і смерті хворого.
Лабораторна діагностика: виявлення трипаносом у мазках
периферичної крові, у спинномозковій рідині, кістковом у
мозку.
Заходи боротьби та профілактика: знищення мухи цеце
у місцях її розмноження, використання інсектицидів, прий
мання лікарських препаратів, які запобігають зараженню при
укусі мухи цеце.
Географічне поширення: Західна і Центральна Африка.
Інші види трипаносом
Тгірапозота Ьгисеі гНодезіепзе морфологічно не відрізняєть
ся від Т. Ьгисеі §атЬіепзе. Для неї основним резервуаром є
антилопа, рогата худоба і людина.
Переносник — муха цеце (Сіоззіпа тогзііапз). Хвороба поши
рена у східних районах Африки.
Захворювання трипаносомозом переважно закінчується
смертю.
Тгірапозота сгигі — збудник хвороби Чагаса. Живе в орга
нізмі людини у двох формах: трипаносомній (у крові) та лейшманіоподібній (інтенсивно розмножується у клітинах м ’ язів,
серця, центральній нервовій системі).
Основним резервуаром є броненосці, опосуми, мурахоїди.
Переносник — поцілунковий клоп.
Гостра форма хвороби Чагаса уражає переважно дітей. Роз
виваються мцнінгоенцефаліт або корчі, стійкі розумові та фі-
Медична паразитологія________________________________________ 353
зичні вади. Значна кількість випадків захворювання закін
чується смертю. Захворювання поширене в країнах Північної
та Центральної Америки.
Лямблія кишкова (ЬатЬІіа іпіезііпаїіз) (рис. 6.7) — збудник
лямбліозу
Місце паразитування: 12-пала кишка, жовчні протоки
печінки.
1
2
3
4
5
6
Рис. 6.7. Лямблія кишкова: а — вегетативна форма; б — циста:
1 — клітинна оболонка; 2 — ядра; 3 — присмоктувальний диск;
4 — парабазальні ядра; 5 — аксостиль; 6 — джгутик
Морфоанатомічні особливості: розміри 9— 18 мкм, грушо
подібна форма, двобічна симетрія тіла. На черевному ввігну
тому боці розміщений присмоктувальний диск. У середній
частині тіла є дві опірні палички — аксостилі. По краях від
них розміщені 2 ядра і 2 парабазальні тіла, джгутиків чотири
пари. Лямблія має здатність у нижньому відділі товстої кишки
утворювати цисти, які з випорожненнями виділяються в зов
нішнє середовище. Цисти овальні, двох- або чотирьохядерні,
з подвійною прозорою оболонкою. При потраплянні на продук
ти харчування можуть заражати людину.
Шляхи зараження: пероральний (проковтування цист).
Симптоми лямбліозу: нудота і блювання, метеоризм, болі
у животі, розлади стільця.
354
Розділ 6
Лабораторна діагностика: мікроскопічне дослідження
вмісту 12-палої кишки на наявність вегетативних форм і фе
калій на наявність цист.
Профілактика: особиста гігієна, миття рук, овочів, фруктів,
санітарний нагляд за водопостачанням, знищення переносників,
лікування хворих, саносвітня робота серед населення.
Географічне поширення: всі області України.
Лейшманії — збудники лейшманіозів
Лейшманії дерматотропні (ЬеізНтапіа ігоріса та]ог, Ь. ігоріса тіпог, Ь. ігоріса техісапа) (рис. 6.8)— збудники дерматотропного лейшманіозу
Місце паразитування: шкіра людини.
Рис. 6.8. Лейшманія: а — лептомонадна форма; б —лейшманіальна форма: 1 — джгутик; 2 — базальне зернятко; 3 — ядро;
4 — клітинна оболонка
Морфофізіологічні особливості і цикл розвитку: розрізняють
безджгутикову (лейшманіальну) і джгутикову (лептомонадну)
форми лейшманій. Безджгутикові форми мають овально-видовжену форму тіла, розміри 2—6 мкм, округле ядро, блефаропласт, джгутик редукований, нерухомі, локалізуються в орга
нізмі людини й інших хребетних.
Медична паразитологія
355
Джгутикова форма має веретеноподібне тіло завдовжки
10—20 мкм, велике ядро, блефаропласт, джгутик до 15—20 мкм,
рухлива, локалізується в організмі комах-переносників (мос
кітів).
У життєвому циклі розвитку лейшманій існує джерело ін
вазії (гризуни) і переносник — москіт (РНІеЬоіотиз рарраіазіі),
який заражає людей при укусах.
Шляхи зараження: трансмісивний (укуси москіта).
Симптоми лейшманіозу дерматотропного: на шкірі роже
вого кольору папула, потім утворюється гнійна виразка.
Лабораторна діагностика: мікроскопічне дослідження
виділень із виразок на шкірі на виявлення лейшманіальних
форм.
Профілактика: боротьба з москітами, знищення природних
резервуарів (гризунів), профілактичні щеплення.
Географічне поширення: Південна Америка, Середня Азія,
південь Казахстану, Закавказзя. В Україні можуть бути лише
зввезені форми.
Лейшманії вісцеротропні ( Ь . д о п о о а п і ,
ники вісцеротропного лейшманіозу
Ь . іп ^ а п іи т )
— збуд
Місце паразитування: клітини внутрішніх органів (печінки,
селезінки, лімфатичних вузлів).
Морфофізіологічні особливості: розміри безджгутикової
стадії 2—4 мкм, джгутикової — 10—20 мкм. Природний резер
вуар — собаки, лисиці, шакали.
Шляхи зараження: трансмісивний (укус москіта).
Симптоми лейшманіозу вісцеротропного: інкубаційний пе
ріод від 15 днів до кількох місяців. Підвищується температура
до 40 °С, адинамія, кволість, знесилення, втрата апетиту, ане
мія, збільшується печінка і селезінка, шкіра темного, майже
чорного кольору.
Лабораторна діагностика: мікроскопія мазків пунктату
грудини, лімфатичних вузлів і крові на виявлення лейшманізмі.них форм, серологічні реакції (зв’ язування компліменту,
іштскс-аглютинація з культури лейшманій).
Розділ 6
356
Профілактика: захист тіла від укусів москітів, знищення
бродячих собак, раннє виявлення та лікування хворих людей.
Географічне поширення: відомі дві форми захворювання — ін
дійський кала-азар і середземноморський. В Україні не зареє
стровано.
Тип Апікомплекса — А р ісо т р іе х а
Клас Споровики — З р о го го а
Збудник
Збудник
Збудник
Збудник
3-денної малярії — Ріазтосііит оіоах
4-денної малярії — Ріазтосііит таїагіае
тропічної малярії — Ріазтосііит {аісірагит
за типом 3-денної малярії — Ріазтосііит ооаіе
Місце паразитування: еритроцити, клітини печінки, селе
зінки.
Морфофізіологічні особливості та цикл розвитку: розміри
4— 5 мкм, форма мінлива, є ядро, цитоплазма, вакуоля.
Людина для малярійного плазмодія — проміжний господар,
комар — кінцевий.
Ш изогонія в організмі людини складається із двох фаз:
екзоеритроцитарної, або тканинної (в клітинах печінки), і еритроцитарної людини.
Тканинна шизогонія. При укусі людини зараженим комаром
разом із його слиною у кров надходять спорозоїти (дрібні, вере
теноподібні клітини, довжиною 14—15 мкм) і заносяться у гепатоцити. Там вони ростуть і стають шизонтами, в яких ділиться
спочатку ядро, а потім цитоплазма, утворюючи тканинні ме
розоїти (1000—5000 особин). Тканинний цикл триває 8— 12 діб.
Мерозоїти руйнують гепатоцит і виходять із нього у кров.
Еритроцитарна шизогонія. Мерозоїти, які проникли в ерит
роцити, перетворюються на трофозоїти (ростучі форми), а остан
ні — на шизонти (форми, що діляться). Стадія розвитку, на
якій шизонт ділиться, називається морулою. При розпаді морули мерозоїти розривають оболонку еритроцита і разом з про
дуктами інтоксикації виходять у кров. У цей момент у хворого
спостерігають напади малярії, які залежно від виду збудника
повторюються через 48 або 72 год. У період еритроцитарної
Медична паразитологія___________________________________________ 357
шизогонії малярійні плазмодії проходять ряд стадій, які мають
важливе діагностичне значення.
Гаметогонія. Окрім шизонтів, у деяких еритроцитах розви
ваються чоловічі та жіночі незрілі статеві клітини (гамонти),
або жіночі — макрогаметоцити (великі, ядро компактне, роз
міщене скраю клітини, в цитоплазмі є пігмент) і чоловічі —
мікрогаметоцити (з великим пухким ядром у центрі).
Спорогонія. Кусаючи хворого на малярію, комар разом із
кров’ ю проковтує гамонти. У шлунку комара макрогаметоцити
перетворюються на макрогамети, а мікрогаметоцити діляться,
утворюючи 5— 6 мікрогамет. Після злиття макрогамети з мікрогаметою утворюється рухлива зигота (оокінета), яка в стінці
шлунка комара перетворюється на ооцисту. В результаті спо
рогонії всередині ооцисти утворюється до 1000 спорозоїтів, які
виходять із ооцисти в гемолімфу тіла і потім проникають
у слинні залози комара. Такий комар може заразити людину.
Шляхи зараження: трансмісивний (укус малярійного кома
ра роду АпорНеІег), а також при гемотерапії.
Симптоми малярії: лихоманка, спочатку з ’ являється слаб
кість, швидко підвищується температура, хворого морозить,
протягом двох годин температура сягає 40 °С. Хворий неспокій
ний, з’являється сильний головний біль. Стан важкий. Через
кілька годин температура знижується до 35— 36 °С. У хворого
спостерігається інтенсивне потовиділення, втома, знесилення.
Напади повторюються через кожні 48 або 72 години, їх кіль
кість 10— 15, потім вони припиняються. Без лікування напади
повторюються за кілька тижнів або місяців. При малярії спостері гається збільшення печінки і селезінки, малокрів’ я та вис
наження. Триденна, чотириденна та овалє-малярія не призво
дять до смерті.
Лабораторна діагностика: мікроскопія великої краплі кро
ві, взятої при клінічних проявах і у стані спокою. Плазмодій
має блакитну цитоплазму і червоне ядро (забарвлення за Ромаповським). Для диференційної діагностики кожного виду плаз
модія можна використати таблицю, де вказані відмінності
в картині крові при малярії, зумовленій паразитуванням між
різними видами малярійних плазмодіїв.
Медична паразитологія___________________________________________ 3 5 9
Профілактика: захист тіла від укусів комарів, виявлення
та лікування хворих на малярію і паразитоносіїв.
Географічне поширення: сьогодні малярія широко розповсю
джена хвороба. Кожного року у світі захворює 100 млн осіб.
В Україну малярію найчастіше завозять.
Токсоплазма (Тохоріазта ропсііі) (рис. 6.10)— збудник ток
соплазмозу
Місце паразитування: клітини головного мозку, печінки,
селезінки, лімфатичних вузлів.
Рис. 6.10. Токсоплазма: 1 — коноїд; 2 — роптрії; 3 — комплекс
Гольджі; 4 — ядро; 5 — мітохондрія; 6 — ендоплазматична сітка
Морфофізіологічні особливості та цикл розвитку: токсо
плазма має півмісяцеву форму, один кінець заокруглений,
другий — загострений, довжина 4— 7 мкм, ширина 3—4 мкм.
Зовні тіло вкрите оболонкою, під якою містяться мікротрубочки.
Па передньому кінці розташований орган прикріплення —
коноїд, від якого відходять трубчасті мішкоподібні органої
ди — роптрії. Ядро вкрите двоконтурною оболонкою з числен
ними порами.
Життєвий цикл розвитку токсоплазми відбувається за на
пій юсті кінцевого (родина котячих) і проміжного (птахи, ссав
ці, людина) господарів. Проникнувши в клітини кишки котя
чих, спорозоїти перетворюються на шизонти, які діляться
н утворюють мерозоїти. Через декілька шизогоній безстатеве
3 60
Розділ 6
розмноження замінюється статевим. Мерозоїти, які проникли
у клітину, утворюють макрогамети і мікрогамети. Внаслідок
злиття макро- і мікрогамет утворюється зигота, що перетво
рюється на ооцисту. Ооциста виділяється в зовнішнє середови
ще і є інвазійною стадією і для проміжного, і для кінцевого
господаря. З ооцисти, яка потрапила в організм проміжного
господаря, спорозоїти звільняються, проникають у клітини
(печінка, селезінка, мозок) і там розмножуються шляхом поді
лу. В результаті багаторазового поділу в клітинах господаря
утворюється багато особин, оточених однією щільною оболон
кою (справжня циста). Скупчення токсоплазм, оточених обо
лонкою ураженої клітини, називається псевдоцистою. При
руйнуванні оболонки спорозоїти звільняються і проникають
у нові клітини.
Шляхи зараження: токсоплазми потрапляють в організм із
погано просмаженим м ’ ясом ссавців і птахів (аліментарно) або
з некип’ яченим молоком, сирими яйцями. Можуть заноситися
брудними руками в рот при догляді за тваринами (перорально),
через слизову оболонку при зніманні шкір і розрізанні тварин
них туш (перкутанно), при заковтуванні ооцист, а також внутрішньоутробно від матері до дитини через плаценту.
Симптоми токсоплазмозу: при вродженому токсоплазмозі
дитина народжується із множинними вадами, не виключені
викидні та неплідність. Часто, новонароджені фізично або
конституційно здорові, однак, відстають у розумовому розвит
ку, мають розлади зору. При токсоплазмозі клінічна картина
дуже різноманітна і залежить від того, який орган уражений.
У хворого підвищується температура, збільшуються лімфатич
ні вузли, на шкірі з’являється висипання, збільшуються миг
далики. Часто спостерігаються рецидиви, інколи зараження
проходить безсимптомно.
Лабораторна діагностика: мікроскопічне дослідження пунктату лімфатичних вузлів на виявлення токсоплазм, вагінальних
мазків, навколоплідних оболонок, плаценти, трупного матеріа
лу. Забарвлення мазків за Романовським. Серологічні реакції.
Внутрішньошкірна проба з токсоплазміном дає можливість
виявити хворобу через чотири тижні після зараження. Вну-
Медична паразитологія___________________________________________ 361
трішньошкірна проба часто позитивна і у здорових людей, які
колись хворіли токсоплазмозом.
Профілактика: обмеження контакту з котами, дотримання
правил особистої гігієни, не вживання в їжу сирого та недо
статньо термічно обробленого м ’ яса птиці та ссавців.
Географічне поширення: в усіх областях України.
Ряд СоссіЛііЛа — кокцидії
Ізоспора — Ізозрога Ьеііі або І. Нотіпіз — збудник кокцидіозу
Місце паразитування: різні органи тварин, а також кишківник людини.
Морфофізіологічні особливості. Ооциста видовжено-овальної
форми, один кінець заокруглений, другий витягнутий, 25—
33 мм в довжину і 12— 16 мм в ширину. Оболонка подвійна
прозора. Протоплазма сконцентрована всередині. Ядро у вигляді
великої світлої плями.
Цикл розвитку кокцидій відбувається у кілька стадій без
зміни господаря:
безстатеве розмноження, або шизогонія;
— статеве — гаметогонія;
спорогонія.
Зі зрілої ооцисти, занесеної в організм господаря, виходять
<іюрозоїти і проникають в епітелій клітини, де ростуть, набуишочи круглої форми. На цій стадії вони називаються трофоюїтами. Ядро трофозоїтів ділиться, утворюються шизонти,
.1 шейх унаслідок множинного поділу формуються мерозоїти.
Киітеліальні клітини гинуть, мерозоїти виходять із них і прониііпють у нові клітини. Шизогонія повторюється багаторазово.
Ііезстатеве розмноження змінюється гаметогонією. Із деяких
мерозоїтів утворюються мікро- і макрогаметоцити. Із мікрогамстоцитів формуються мікрогамети, з макрогаметоцитів —
мшфогамети. Внаслідок копуляції виникає зигота, яка вкрииін ться щільною оболонкою, — ооциста. Вона виходить зепі!<■ Ш И Л Ь Н И Х клітин у просвіт кишки і з фекаліями виноситься
\ зовнішнє середовище. Спорогонія відбувається за межами
Розділ 6
362
господаря. За сприятливих умов у зовнішньому середовищі
всередині ооцисти утворюються спори (4), а всередині спор спорозоїти (2). На цій стадії ооциста є інвазійною для людини.
Шляхи зараження: аліментарний (занесення ооцист в орга
нізм).
Симптоми кокцидіозу: частий рідкий стілець, слабкість,
біль у животі.
Лабораторна діагностика: дослідження свіжих фекалій на
наявність ооцист.
Профілактика: охорона зовнішнього середовища від забруд
нень, особиста гігієна, виявлення і лікування хворих людей,
санітарно-просвітня робота серед населення.
Географічне поширення: в усіх областях України.
6.2. Тип Плоскі черви — РІаіНеїт іпіНез
Загальні ознаки:
— тришаровість (розвиток у ембріонів екто-, ендо-, мезодер
ми);
— білатеральна симетрія тіла; тіло сплюснуте у спинно-че
ревному напрямку;
— первинна порожнина тіла, заповнена паренхімою;
— покриви представлені шкірно-м’ язовим мішком;
— розвинені такі системи органів: нервова, травна, виділь
на, статева;
— складні цикли розвитку зі зміною господарів.
Клас Сисуни —
Т ге т а їо д а
Загальні риси будови: статевозріла гермафродитна стадія
паразита має назву марита.
Покриви тіла: ш кірно-м ’ язовий мішок, зовнішній шар
якого — це тегумент, утворений без’ ядерною цитоплазмою
з великою кількістю мітохондрій (верхній шар) і ядрами (ниж
ній шар). Під тегументом розміщена базальна мембрана, а під
Медична паразитологія
363
мембраною лежать гладенькі м ’ язи (кільцеві, діагональні,
поздовжні).
Травна система утворена переднім (ектодермального похо
дження) і середнім (ентодермального походження) відділами
кишківника і починається ротовим отвором, який переходить
у глотку, стравохід, кишку і закінчується сліпо.
Нервова система представлена навкологлотковим нервовим
кільцем і трьома парами нервових стовбурів уздовж тіла.
Видільна система протонефридіального типу починається
н паренхімі клітинами зіркоподібної будови (термінальні, кін
цеві); від клітин відходять видільні канальці, що зливаються
у бічні канали, які закінчуються видільними порами (відкри
ваються назовні).
Статева система: здебільшого представники класу Сису
ни — гермафродити. Чоловіча статева система утворена парою
сім ’ яників, сім ’ япроводів, які зливаються у с ім ’ явипорскувальний канал, і копулятивним органом (цирусом). Жіноча
статева система складається з яєчника, жовтівників, сім’яприймача, які відкриваються в оотип. Тут відбувається запліднення
та формування запліднених яєць. До оотипу надходить пожив
шій матеріал із жовтівників і виділення спеціальних залоз —
гілець Меліса. Запліднені яйця з оотипу надходять до матки
і виводяться назовні через статевий отвір. Розвиток сисунів
відбувається стадійно, зі зміною хазяїнів і кількома поколін
нями личинкових стадій: марита —> яйце —> мірацидій —> спо
роциста-» редія —> церкарія- » метацеркарія (адолескарія).
Сисун печінковий (Р а з с і о і а
фасціольозу
к е р а ііс а )
(рис. 6 .1 1 ) — збудник
Місце паразитування: жовчні протоки печінки, жовчний
м і х ур, підшлункова залоза переважно великої та малої рогатої
худоби, свиней, коней, зрідка людини.
Морфоанатомічні особливості: сірого кольору з коричневим
відтінком завдовжки 3—5 см, тіло листкоподібне, сплющене
у черевному напрямку. На передньому кінці добре помітний
і івіусоподібний виступ, на вершині якого розташований ротовий
присосок. Черевний присосок лежить на невеликій відстані
364
Розділ 6
Рис. 6.11. Сисун печінковий: 1 — ротовий присосок; 2 — кишківник; 3 — цирус; 4 — черевний присосок; 5 — матка; 6 — сім’я
проводи; 7 — оотип; 8 — сім’яники; 9 — жовтівники; 1 0 — яйник
від ротового, по середній лінії тіла біля основи конусоподібно
го виступу.
Особливості внутрішньої будови: короткий розгалужений
кишківник, закінчується сліпо; за черевним присоском роз
міщена багатолопатева матка і яєчник; численні жовтівни
ки — по краях тіла; середню частину тіла займають сім’ яники.
Органи чуттів, крім дотику, відсутні.
Цикл розвитку: проміжний хазяїн — ставковик малий СаІЬа
ігипсаіиіа, кінцевий хазяїн — ссавці, зрідка людина. Яйце
фасціоли потрапляє у воду, з нього виходить личинка — мірацидій (має нервовий ганглій, світлочутливе вічко, органи
Медична паразитологія
365
виділення, а в задній частині зародкові клітини; передній кінець
тіла містить залозу, що синтезує фермент, який здатний роз
чинити живі тканини при проникненні в проміжного хазяїна).
Мірацидій вкритий війками, завдяки яким вільно плаває у воді,
активно проникає в тіло молюска, далі в його печінку і пере
творюється в наступну личинкову стадію — спороцисту. Зі
спороцист шляхом партеногенезу утворюються нові личинкові
стадії — редії. У редій формуються рот, глотка, травна трубка
і отвір для виходу особин нового покоління. Всередині редій
також зі зародкових клітин партеногенетично утворюється
наступне покоління личинок — церкарії. У тілі церкарії роз
винені системи органів, які характерні для марити: присоски,
кишка, нервова та видільна системи, але, на відміну від мари
ти, церкарія озброєна довгим мускульним хвостом, який за
безпечує поступальний рух. Церкарія залишає молюска й ак
тивно пересувається уводі. Прикріпившись до стебла водної
рослини і вкрившись оболонкою, церкарія перетворюється на
адолескарію. При заковтуванні тваринами адолескарій із водою
вони потрапляють у кишку хазяїна: оболонка розчиняється,
паразит проникає у печінку і досягає там статевої зрілості.
Шляхи зараження: пероральний (проковтування адолескарій, які можуть бути у стоячих водоймах або на стеблах чи
листках рослин вологих лугів).
Симптоми фасціольозу: фасціола з кишківника проникає
у печінку. Розлади функцій печінки, які можуть при інтенсив
ній інвазії призвести до смерті. Частіше сисун локалізується
у жовчних протоках. Виникають запальні процеси, болі в ділян
ці епігастрію та правого підребер’ я, нудота, блювота. Печінка
збільшена, болюча, щільна. Спостерігається жовтяниця, збіль
шення селезінки, у крові лейкоцитоз, еозинофілія. У печінці
після запальних процесів розростається сполучна тканина
(цироз).
Лабораторна діагностика: овоскопія фекалій із попереднім
виключенням печінки з раціону. Зрілі яйця жовтаво-брунатні,
опальні, на одному з полюсів добре помітна кришечка, а на
протилежному — плоский горбочок, їх розміри близько 135 х
- КО мкм.
366________________________________________________________ Розділ 6
Профілактика: не споживати сиру воду, ретельно мити
овочі; велике значення має санітарно-освітня робота.
Географічне поширення: всі області України (здебільшого
Полісся).
Сисун ланцетоподібний (Рісгосоеііитіапсеаіит) (рис. 6.12) —
збудник дикроцеліозу
Місце паразитування: печінка великої та малої рогатої
худоби й інших тварин, зрідка людини.
Морфоанатомічні особливості: довжина паразита близько
10 мм, тіло звужене на кінцях, форма ланцетоподібна. Є рото
вий і черевний присоски. Травна система складається із рото
вого отвору, розташованого на дні ротового присоска, короткої
глотки, що переходить у стравохід, двох нерозгалужених гілок
кишок, які закінчуються сліпо, не доходячи до заднього кінця
тіла.
Спереду від черевного присоска є два округлі сім’ яники,
кулястий яєчник розміщений за сім’ яниками, розгалужена
матка займає задню частину тіла. У матці можна побачити
велику кількість дрібних яєць. Яйця можуть мати різне забар
влення — від світло-жовтого до темно-коричневого. Різне за
барвлення зумовлене різним ступенем зрілості. У середній
частині тіла по боках містяться жовтівники.
Цикл розвитку: проміжні господарі — наземні молюски
роду ЯеЬгіпа, Неіісеїа (яйце з мірацидієм —» мірацидій - » спо
роциста 1-го порядку-» спороциста 2-го порядку-» церкарія),
у вигляді збірних цист виходять із тіла молюска та потрапляють
до мурашки роду Рогтіса (метацеркарія), кінцевий господар —
травоїдні ссавці.
Шляхи зараження: пероральний (випадкове проковтування
мурашок, інвазованих метацеркаріями).
Симптоми дикроцеліозу: перебіг дикроцеліозу подібний до
перебігу фасціольозу.
Лабораторна діагностика: овоскопія фекалій. Яйця за за
барвленням варіюють від жовтуватого до темно-коричневого
кольору, мають овальну форму, на одному з боків — маленьку
кришечку, розміри яйця 35— 45 мкм.
367
Медична паразитологія
Рис. 6.12. Сисун ланцетоподібний: 1 — ротовий присосок;
:! - черевний присосок; 3 — сім’яники; 4 — яйник; 5 — оотип;
(1 — жовтівники; 7 — кишківник; 8 — матка
Профілактика: знищення молюсків і дегельмінтизація ху
доби.
Географічне поширення: в Україні скрізь, де є пасовища.
Китайський сисун
хозу
(С Іо п о г с К із з іп е п з із )
— збудник клонор-
Місце паразитування: печінка, жовчний міхур, підшлун
кова залоза.
Морфоанатомічні особливості: плоске видовжене тіло черво
нуватого кольору завдовжки 10— 20 мм. Має два присоски —
368
Розділ 6
ротовий і черевний. Кутикула без шпичок. В останній третині
тіла містяться галузисті сім ’ яники. Перед сім ’ яником лежить
грушоподібний сім’ яприймач. Жовтівники по боках тіла. Яєч
ник і матка спереду сім’ яників.
Цикл розвитку, яйця з мірацидіями від хворої людини
потрапляють у воду і захоплюються молюском Рагаїоззгиіиз
тапсйоигісиз, з яєць в и х о д я т ь мірацидії. У кишківнику мо
люска з мірацидій партеногенетично розвиваються спороциста,
редії, церкарії. Церкарії покидають тіло молюска і проникають
в організм другого проміжного господаря прісноводних риб
(зокрема коропових), де розвивається метацеркарія, яка є ін
вазійною стадією для людини.
Шляхи зараження: споживання погано посоленої або недо
статньо термічно обробленої риби з метацеркаріями (алімен
тарний).
Симптоми клонорхозу: алергічні реакції, жовтяниця, за
пальні процеси у печінці можуть викликати цироз, а згодом
рак печінки
Лабораторна діагностика: виявлення яєць у калі. Яйця
грушоподібної форми, світло-золотистого кольору. На одному
полюсі є кришечка, на протилежному— горбик. Розмір 27—
35 мкм.
Профілактика: не вживати в їжу термічно не обробленої
риби. Охорона водойм від забруднення фекаліями.
Географічне поширення: Далекий Схід Росії. Часто виявля
ються паразити в емігрантів з далекосхідних країн.
Сисун котячий (ОрізіНогсНіз / еііпеиз) (рис. 6.13) — збудник
опісторхозу
Місце паразитування: печінка, жовчний міхур, підшлун
кова залоза.
Морфоанатомічні особливості: тіло завдовжки 4— 13 мм,
звужене на передньому кінці та заокруглене на задньому. На
передньому кінці помітний слабо розвинений ротовий присосок,
черевний присосок малопомітний. Паразит блідо-жовтого ко
льору.
Медична паразитологія
369
Рис. 6.13. Сисун котячий: 1 — ротовий присосок; 2 — черевний
присосок; 3 — матка; 4 — жовтівники; 5 — кишківник; 6 — сім’яприймач; 7 — яйник; 8 — сім’яник; 9 —головний видільний канал
Канали кишок доходять до заднього кінця тіла і закінчу
ються сліпо.
У сисуна котячого статева система такої самої будови, як
і у ланцетоподібного, але розташування статевих органів інак
ше. Трубчаста матка розташована між каналами кишок у се
редній частині тіла. Два лопатеві сім’ яники лежать у задній
частині тіла, і між ними проходить звивистий видільний канал.
Яєчник і сім’ яприймач розміщені перед сім’ яниками. У серед
ні й третині тіла сисуна розміщені жовтівники. Статеві отвори
відкриваються на передньому краї черевного присоска.
Цикл розвитку: проміжні господарі — молюск роду Віікупіа
(яйце з мірацидієм —> мірацидій —> спороциста —» редія -* цері арії) -» риби (метацеркарія) - » кінцевий господар — людина,
м’ ясоїдні ссавці.
Шляхи зараження: аліментарний (споживання погано терміч..... Зроблених м ’ яса та риби, заражених метацеркаріями).
Симптоми опісторхозу: болі у верхній половині живота
і іррадіацією вліво до селезінки, болючість при пальпації під
ім лункової залози, збільшення жовчного міхура, застій жовчі,
іце призводить до цирозу печінки, а при тривалій інвазії — до
раку печінки. Перебіг хвороби з помітними симптомами холе
циститу і гепатиту.
3 70 ____________ ____________________________________________Розділ 6
Лабораторна діагностика: овоскопія фекалій і дуоденталь
ного вмісту. Жовтуваті овальні яйця звужені на передньому
полюсі, де міститься кришечка. Розміри їх 26— ЗО мкм.
Профілактика: споживання добре термічно обробленої ри
би, охорона водойм від забруднення фекаліями, виявлення та
лікування хворих людей, санітарно-освітня робота серед насе
лення.
Географічне поширення: природно-осередкове захворювання,
на Україні зареєстровані окремі випадки в басейнах Дніпра
і Південного Бугу.
Фасціолопсис (Газсіоіорзіз Ьизгі) — збудник фасціолопсидозу
Місце паразитування: верхній відділ тонкої кишки і шлу
нок людини, свині та собаки.
Морфоанатомічні особливості: листкоподібне, сплющене
тіло червоно-оранжевого кольору, довжиною до 5 см. На зву
женому передньому кінці тіла лежать ротовий і черевний при
соски.
За черевним присоском розміщені матка і яєчник: середньозадню частину тіла займає розгалужений сім ’ яник, а по боках
тіла — жовтівники.
Цикл розвитку: яйця паразита потрапляють у воду. Із яєць
виходить мірацидій, який проникає в молюска (РІапогЬіа
зегтепііпа) і проходить кілька стадій розвитку (спороциста,
редія, церкарія). Церкарії виходять із тіла молюска і перетво
рюються на адолескарії. Адолескарії осідають на листях і пло
дах водяного горіха.
Проміжний живитель: молюск (РІапогЬіа зегтепііпа).
Кінцевий господар: людина, свині та собаки.
Шляхи зараження: люди заражаються при обдиранні рука
ми свіжих водяних горіхів, на яких можуть бути адолескарії,
а свині — поїдаючи водяні горіхи.
Симптоми фасціолопсидозу: запалення слизової оболонки
шлунка і тонкої кишки, що призводить до розладу їх функцій.
Медична паразитологія
371
Лабораторна діагностика: дослідження фекалій на наяв
ність яєць. Яйця овальної форми зі звуженими полюсами, на
одному з полюсів є перемичка, на другому — горбик сіруватожовтуватого кольору. Розміри 130— 140 х 80—95 мкм.
Профілактика: використання в їж у водяних горіхів тільки
після термічної обробки або після витримування їх у 5— 20 %
розчині МаСІ протягом 2,5—3,0 годин.
Географічне поширення: центральні та північні райони Ки
таю, Гонконгу, Гуанди, Філіппіни.
Метагонімуз (Меіа£опітиз уого§ашаі) — збудник метагонімозу
Місце паразитування: верхній і середній відділ тонкої
кишки.
Морфоанатомічні особливості. Плоске видовжене тіло,
спереду звужене, ззаду загострене, довжиною 1,0— 2,5 мм.
Тіло вкрите дрібними шпичками. Ротовий присосок менших
розмірів, ніж черевний. У кінці тіла розташовані овальні сім’я
ники. Жовтівники розташовані по боках тіла. На передньому
краї черевного присоска відкривається матка зі сім ’ явипорскуішльним каналом.
Цикл розвитку: яйце разом із фекаліями потрапляє у воду,
де з нього виходить личинка мірацидій. Мірацидій проникає
н тіло проміжного господаря — молюска роду Меіапіа, в ор
ганізмі якого проходять стадії розвитку спороцисти, редії та
церкарії. Церкарії виходять із тіла молюска і проникають у ті
ло риби (коропових і лососевих), у м ’ ясі та на плавниках риби
формуються метацеркарії.
Проміжний живитель: молюск Меіапіа ІіЬегііпа, М. еЬепіпа.
Додатковий живитель: риби (коропові та лососеві)
Кінцевий господар: домашні та дикі ссавці, людина.
Шляхи зараження: аліментарний (споживання недостатньо
пропареної, прожареної або просоленої риби).
372
Розділ 6
Симптоми метагонімозу: алергія, гарячка, еозинофілія
крові, нудота, болі в животі, нестійкі випорожнення, діарея до
5—6 разів на добу.
Лабораторна діагностика: дослідження фекалій на наяв
ність яєць. Яйця лимоноподібної форми, світло-коричневого
кольору з кришечкою. Розміри 26—28 х 10— 15 мкм.
Профілактика: не споживати сиру, малосолену, в’ ялену
і недостатньо термічно оброблену рибу. Охорона зовнішнього
середовища від зараження, санітарно-освітня робота серед на
селення.
Географічне поширення: Японія, Китай, Корея, Далекий
Схід. В У країні— басейни річок Дніпра, Дунаю, Дністра.
Сисун легеневий (Р а г а £ о п і т и з
мозу
гіп д е гі)
— збудник парагоні-
Місце паразитування: легені людини, кота, собаки, свині.
Морфоанатомічні особливості: тіло завдовжки 4— 13 мм,
форма яйцеподібна. На тілі є шипи. Паразит коричневого ко
льору.
Цикл розвитку: яйця з харкотинням або калом потрапляють
у навколишнє середовище (у воду). З яйця виходить мірацидій,
який активно проникає в молюска роду Меіапіа, там проходить
стадії спороцисти, редії і церкарії. Церкарії виходять із молюс
ка і проковтуються прісноводними раками чи крабами, де
перетворюються в метацеркарії.
Шляхи зараження: споживання термічно необроблених раків
і крабів з метацеркаріями.
Симптоми: за характером перебігу нагадує туберкульоз —
хронічний кашель, харкотиння з домішками крові.
Лабораторна діагностика: виявлення яєць у харкотинні,
фекаліях. Форма яєць: овальні, золотисто-брунатного кольору,
розмір — 80— 100 мкм.
Профілактика: не вживати сирих і погано термічно оброб
лених раків, крабів.
Медична паразитологія
3 73
Географічне поширення: Китай, Індонезія, Філіппіни, Пів
денна Америка. В Україні не розповсюджені. Можуть бути
лише завезені випадки захворювання.
Сисун кров’яний (ЗсНізІозота НаетаіоЬіит) (рис. 6.14) —
збудник урогенітального шистосомозу
Місце паразитування: великі вени черевної порожнини
й органів сечостатевої системи.
Рис. 6.14. Шистосома: 1 — самець; 2 — гінекофорний канал;
З — самка
Морфофізіологічні особливості: роздільностатеві, довжина
самки 20 мм, самця — 10— 15 мм. У самця широке тіло містить
жолоб, у якому лежить тонка і довга самка. У самки є один
яєчник, коротка матка, жовтівник, який займає задню части
ну тіла.
Цикл розвитку: проміжний господар — прісноводні молюс
ки Виїепиз ігипсаіез, РІапогЬіз теіійепзіз, кінцевий госпо
дар — людина, мавпа. Розвиток відбувається таким чином:
марита -» яйце - » мірацидій -» молюск (спороциста І порядку,
спороциста II порядку, церкарія).
3 74 _______________________________ __ ______________________ Розділ 6
Шляхи зараження: перкутанний (проникнення церкарій
через шкірні покриви).
Симптоми урогенітального шистосомозу: від проникнення
церкарій через шкіру розвиваються дерматити, свербіж. На
тлі загальної слабкості, зниження апетиту, схуднення, болів
голови з’являється алергія. У сечовому міхурі розвиваються
запальні процеси, руйнування тканин, кровотечі, біль при
сечовиділенні від незначного ниючого до сильного за типом
ниркової коліки. У сечі багато еритроцитів. У жінок часто ви
являються запальні процеси піхви та матки.
Лабораторна діагностика: овоскопія сечі. Яйця безбарвні,
овальної форми, містять на одному із полюсів великий шип.
У яйці є мірацидій із зачатком кишки. У самця на рівні ста
тевого отвору міститься три — шість сім’ яників, мішок цирусу відсутній.
Профілактика: не споживати сиру воду, не купатись у за
боронених водоймах.
Географічне поширення: Єгипет, Південна Африка, А в
стралія, Іран.
Клас Стьожкові черви — Сезіоійеа
Ціп’як озброєний (свинячий) ( Таепіа зоїіит) (рис. 6.15) —
збудник теніозу
Місце паразитування: доросла форма паразитує лише в тон
кому кишківнику людини. Ф ін и — в мозку, нирці, очному
яблуці.
Морфоанатомічні особливості: На передньому кінці тіла
ціп’ яка озброєного міститься головка — сколекс, далі коротка
шийка (5— 10 мм) без члеників і потім — стробіла, що скла
дається із члеників — проглотид, у якій є незрілі, гермафро
дитні та зрілі проглотиди. Стробіла містить до 1000 проглотид.
Розміри тіла ціп’ яка озброєного, або свинячого, 1,5—2,0 м.
Головка свинячого ціп’ яка має чотири присоски і подвійний
віночок гачечків.
По боках гермафродитних проглотид гельмінта добре видно
канали протонефридій видільної системи, які тягнуться
Медична паразитологія
375
Рис. 6.15. Ціп’як озброєний (свинячий): а — сколекс; б — гер
мафродитна проглотида; в — зріла проглотида: 1 — гачечки;
2 — присоски; 3 — шийка; 4 — сім’ яники; 5 — матка; 6 — яйник;
7 — тільце Меліса; 8 — оотип; 9 — жовтівники; 10 — сім’явиносний
канал; 11 — піхва; 12 — розгалуження матки, заповнені заплід
неними яйцями
вздовж тіла і з’ єднуються поперечними анастомозами в кож
ному членику. У центральній частині розташована гермафро
дитна статева система. Чоловіча представлена численними
сім ’ яниками. Від них відходять сім ’ явивідні канали, які зби
раються у сім ’ явипорскувальний канал, який заходить у ста
теву клоаку. У задній частині членика розташований яєчник.
376
Розділ 6
У ціп’яка озброєного він має дві великі частки округлої форми
і одну маленьку, яка називається додатковою. Від яєчника
відходять яйцепроводи, що відкриваються в оотип, оточений
тільцями Меліса. Від нього починається трубчаста матка. Ни
жче від яєчника розташовані жовтівники.
Довжина члеників зрілих проглотид гельмінта є більшою
від ширини. Матка у ціп’ яка озброєного має від 7 до 12 бокових
відгалужень, сім ’ явипорскувальний канал і піхву.
Цикл розвитку: свиняче м ’ ясо з цистицерками потрапляє
в кишківник людини, під дією травних соків озброєний сколекс
вивертається назовні та гачками прикріплюється до стінки
кишки. Починається ріст стробіли. Приблизно через два міся
ці після зараження хворий виділяє з калом велику кількість
яєць і проглотид свинячого ціп’яка. Подальший розвиток мож
ливий тільки тоді, коли яйце потрапляє у травний канал свині.
У шлунку оболонка яйця руйнується і виходить зародок — онкосфера, що має 6 ембріональних гачків. За їх допомогою
проходить крізь стінку кишківника у кровоносні судини і стру
менем крові розноситься в м ’ язи; там протягом 60— 70 днів
перетворюється в цистицерк — міхурець, заповнений рідиною,
із ввігнутою головкою. Розміри цистицерка залежать від ло
калізації (10— 20 х 7—10 мм). У свині ларвоциста вражає м’язи.
У людини, крім скелетних м ’ язів, може локалізуватися в ли
цевому м ’язі, очному яблуці, центральній нервовій системі.
Шляхи зараження: аліментарний (вживання у їжу термічно
необробленої свинини, інвазованої цистицерками).
Симптоми теніозу: симптоматика різко варіює, інвазія
інколи спричиняє різкі розлади травлення, діарею, болі, слаб
кість, інколи нема жодних проявів захворювання. Основні
ознаки: нудота, блювання, діарея, втрата ваги й апетиту.
Симптоми цистицеркозу значно тяжчі й небезпечніші. При
локалізації ларвоцист в оці людина втрачає зір, при локаліза
ції в мозку цистерки розростаються і можуть досягати розміру
голубиного яйця, що призводить до смерті, при їх локалізації
у м’язах особливих скарг немає.
Лабораторна діагностика: виявлення в калі зрілих члени
ків ціп’яка. Для диференційної діагностики членик затискають
Медична паразитологія
3 77
між двома предметними скельцями і під мікроскопом досліджу
ють кількість відгалужень матки. Діагностика цистицеркозу
важка: імунологічні методи, комп’ ютерна томографія, енцефа
лографія.
Профілактика: не вживати в їж у термічно необробленої
свинини, здійснювати ветеринарний огляд м ’ яса, що реалі
зується. Дегельмінтизація хворих. Охорона ґрунту і водойм від
забруднення фекаліями. Забій свиней під ветеринарним нагля
дом. Санітарно-освітня робота серед населення.
Географічне поширення: трапляється рідко у всіх областях
України, найчастіше у людей, які вживають напівсире м’ ясо.
Ц іп ’ як н еозброєн и й (ТаепіагкупсНиз за^іпаіив) (рис. 6.16) —
збудн и к тен іа ри н хозу
Місце паразитування: тонка кишка людини.
Морфоанатомічні особливості:
На передньому кінці тіла ціп’ яка бичачого розташована
головка (сколекс), далі шийка і потім — стробіла, що скла
дається із члеників — проглотид. Розміри тіла 4— 10 м. Головка
ціп’ яка бичачого має 4 присоски. По боках гермафродитних
проглотид гельмінта добре видно канали протонефридій виділь
ної системи, які тягнуться вздовж тіла і з’ єднуються попереч
ними анастомозами в кожному членику. У центральній частині
розташована гермафродитна статева система. Чоловіча пред
ставлена численними сім’ яниками. Від них відходять сім’ янивідні канали, які збираються у сім ’явипорскувальний канал,
який заходить у статеву клоаку. У задній частині членика роз
ташований яєчник, який має лише дві округлі частки. Між
частками яєчника видно оотип в оточенні тілець Меліса. Від
нього починається трубчаста матка. Нижче від яєчника розта
шовані жовтівники.
Довжина члеників зрілих гельмінтів є більшою від ширини.
Із органів статевої системи видно лише матку (яка у ціп’яка
неозброєного має від 17 до 35 відгалужень з кожного боку),
сім ’ явипорскувальний канал і піхву.
Цикл розвитку: з калом людини у навколишнє середовище
виходять членики бичачого ц іп ’ яка. Подальший розвиток
378
Розділ 6
Рис. 6.16. Ц іп’як неозброєний: а — сколекс; б — гермафродит
на проглотида; в — зріла проглотида: 1 — присоски; 2 — шийка;
З — сім’яники; 4 — матка; 5 — яйник; 6 — тільце Меліса; 7 — оотип; 8 — жовтівники; 9 — піхва; 1 0 — сім’ явипорскувальний
канал; 11 — бокові відгалуження матки
відбувається тоді, коли їх проковтує велика рогата худоба.
У шлунку яйця звільняються від оболонки, утворюються онкосфери, які проникають крізь стінки кишківника в кровонос
не русло і заносяться у м’ язи. Кожна онкосфера перетворюєть
ся у фіну. При споживанні фінозного м’ яса в організмі людини
розвивається статевозріла форма ціп’ яка.
Медична паразитологія___________________________________________ 3 7 9
Шляхи зараження: аліментарно (вживання у їж у термічно
необробленої фінозної яловичини).
Симптоми теніаринхозу: симптоматика різко варіює, ін
вазія інколи викликає різкі розлади травлення, діарею, болі,
слабкість, інколи нема жодних проявів захворювання. Основ
ні ознаки: нудота, блювання, діарея, втрата ваги й апетиту.
Ускладнення: заповзання члеників у червоподібний відросток,
жовчні шляхи, непрохідність кишківника, перфорація 12-палої кишки, проникнення члеників у підшлункову залозу.
Лабораторна діагностика: наявність зрілих проглотид
ціп’ яка у калі людини.
Профілактика: не вживати в їж у термічно необробленої
яловичини. Дегельмінтизація хворих. Охорона ґрунту і водойм
від забруднення фекаліями. Санітарно-освітня робота серед
населення.
Географічне поширення: у всіх областях України, де розво
дять велику рогату худобу.
Ц іп ’ як к арл и к ови й (Н у т е п о іе р із п а п а ) — збудн ик гім ен ол еп ідозу
Місце паразитування: у ворсинках кишківника, в просвіті
тонкої кишки.
Морфоанатомічні особливості: ціп’ як карликовий має дов
жину від 1 до 5 см. У стробілі 200 і більше члеників. На гру
шоподібному сколексі розташовані 4 присоски і хоботок із
віночком гачків.
Цикл розвитку: яйця, які потрапили в зовнішнє середовище,
прилипають до дитячих іграшок, до ручок дверей, до дитячих
горщиків, забруднюють руки, переносяться мухами і тарганами
на їжу та потрапляють у кишківник людини. З яєць виходить
омкосфера, яка у ворсинці кишки розвивається у цистицеркоїд,
у якого на передньому кінці є здуття з головкою, а на задньому
кін ці— хвостоподібний придаток. За 6—8 днів ворсинка руй
нується, цистицеркоїди потрапляють у просвіт кишки, при
кріплюються до її стінки і виростають у статевозрілу форму.
380_______________________________________________
Розділ 6
Шляхи зараження: пероральний (проковтування яєць ціп’ я
ка карликового).
Симптоми гіменолепідозу: біль у животі, нудота, блювання,
діарея, біль голови, зниження пам’яті, втрата ваги й апетиту.
Лабораторна діагностика: виявлення яєць у калі людини.
Яйця кулясті, покриті двоконтурною прозорою оболонкою.
Всередині яйця міститься личинкоподібна онкосфера. Розмір
яйця 40 х 50 мкм.
Профілактика: дотримання правил особистої гігієни.
Географічне поширення: у всіх областях України
Ц іп ’як щ урячи й (Н у т е п о іе р із й іт іп и іа ) — збудник щ у р я
ч ого гем ін ол епід озу
Місце паразитування: кишківник гризунів.
Морфоанатомічні особливості. Колір білий, довжина 20—
40 см. На передньому кінці є головка з 4 присосками, гачки
відсутні, хоботок рудиментарний. У члениках від шийки посту
пово закладаються органи статевої системи. Зрілі членики
короткі й широкі, довжиною 5— 6 мм. Мішковидна матка ви
тісняє інші частини статевої системи.
Цикл розвитку: виділені яйця щурячого ціп’яка поїдають
борошняні хрущаки, в тілі яких розвиваються личинки.
Шляхи зараження: споживання недостатньо термічно об
роблених виробів із борошна (людина заносить у свій організм
борошняних хрущаків і стає кінцевим господарем паразита).
Симптоми: характерні для гемінолепідозу.
Лабораторна діагностика: дослідження фекалій на наяв
ність яєць.
Профілактика: не вживати в їжу погано термічно обробле
них борошняних виробів, боротьба з гризунами, що забрудню
ють зерно мучними хрущаками.
Географічне поширення: в усіх областях України.
Г а р б у зо в и й ц іп ’ як (О ір у ї ій іи т с а п іп и т ) — збуд н и к ди піл ідіум озу
Медична паразитологія___________________________________________ 381
Місце паразитування: тонкий кишківник котів, собак,
лисиць, вовків, а також людини.
Морфоанатомічні особливості. Ц іп’ як гарбузовий білува
того кольору, довжиною 15—50 см. Сколекс довжиною 0,3—
0,4 мм, має 4 присоски і хоботок з 3— 4 рядами гачків. Зрілі
членики за формою нагадують насіння огірків.
Цикл розвитку: яйця з’ єднуються в овальні кокони і виді
ляються у зовнішнє середовище. Іноді у фекаліях кокони мо
жуть руйнуватись, і тоді яйця трапляються поодинці. Личин
ка розвивається в організмі собачої блохи.
Шлях зараження: пероральний (проковтування личинок
собачої блохи, іноді яєць ціп’ яка).
Симптоми: як при гіменолепідозі.
Лабораторна діагностика: виявлення у фекаліях яєць і ко
конів із яйцями. Часто в калі макроскопічно виявляють рухливі
зрілі членики, подібні на насіння гарбуза.
Профілактика: дотримання особистої гігієни.
Географічне поширення: усі регіони України.
С тьож а к ш и р ок и й (БіркуІІоЬоІНгіит Іаіит) (рис. 6.17) —
збудн ик д и ф іл оботр іозу
Місце паразитування: тонкий кишківник людини.
Морфоанатомічні особливості: сколекс стьожака широкого
має видовжену форму і дві поздовжні щілини (ботрії) замість
присосок. Гермафродитний членик гельмінта має ширину біль
шу від довжини. Чоловіча і жіноча статеві системи відкрива
ються у статеву клоаку, яка розміщена на черевному боці пе
редньої частини членика. Особливістю будови зрілого членика
є наявність петлеподібної матки, що лежить у вигляді розетки
я середній частині членика.
Цикл розвитку: яйця стьожака виходять із калом хворої
пюдини або тварини; їх розвиток закінчується уводі. Корацидій — перша личинкова стадія — округла, вкрита війками,
проковтується першим живителем— циклопом. Корацидій
втрачає війки і перетворюється в онкосферу, а потім у другу
382
Розділ 6
Рис. 6.17. Стьоясак широкий: а — сколекс; б — гермафродитна
проглотида; в — зріла проглотида: 1 — ботрії; 2 — шийка; 3 — цирус; 4 — піхва; 5 — матка; 6 — жовтівники; 7 — яйник; 8 — оотип;
9 — тільця Меліса; 10 — сім’яники; 11 — розеткоцодібна матка
личинкову стадію — процеркоїда (0,5 мм). Хиж і риби проков
тують циклопів із процеркоїдами, і в їхніх м ’ язах, внутрішніх
органах, в ікрі протягом 3—4 тижнів формується остання ли
чинкова стадія — плероцеркоїд. Він має ботрії, але тіло не
почленоване. При поїданні заражених риб у кишці людини або
рибоїдних ссавців розвивається статевозрілий стьожак.
Шляхи зараження: аліментарний (при вживанні у їжу річ
кової риби, яка не пройшла термічної обробки і містить у м’язах
та ікрі плероцеркоїди).
Симптоми дифілоботріозу: слабкість, запаморочення, ну
дота, розлади стільця, болі в животі, анемія, іноді епілептичні
судоми, кишкова непрохідність, зменшення кількості еритро
цитів і гемоглобіну.
Лабораторна діагностика: мікроскопія калу на виявлення
яєць і проглотид, у яких матка має вигляд розеток у центрі
Медична паразитологія
3 83
членика. Яйця овальної форми, сірого або бурого кольору,
розміром 68—71 мкм.
Профілактика: не вживати в їжу термічно не оброблених
хижих річкових риб (щук, окунів, йоржиків); періодичні об
стеження рибалок, працівників річкового транспорту, санітар
ний благоустрій кораблів, населених пунктів, боротьба із за
брудненням водойм фекаліями, боротьба з бездомними котами
і собаками, які можуть розповсюджувати яйця і членики ши
рокого стьожака.
Географічне поширення: у басейнах рік Бугу, Дністра (через
забруднення рік вилов риби не набуває промислового масшта
бу, тому захворювання осередкове).
Е хінокок (ЕсНіпососсиз §гапиІозиз) (рис. 6.18) — збудник е х і
н ок ок озу
Місце паразитування: легені, печінка, мозок.
Морфоанатомічні особливості: ехінокок — дрібний гельмінт
довжиною до 5 мм, білого кольору. На сколексі містить 4 при
соски і 36—40 гачків. Стробіла складається із 3—4 проглотид,
із яких остання — зріла рухлива і містить до 800 яєць.
Фіна типу ехінококового міхура може мати розміри від
курячого яйця до голівки немовляти. Стінка міхура утворена
зовнішньою капсулою і внутрішньою паренхіматозною оболон
кою. На останній утворюються дочірні міхурці. Міхур запов
нений безбарвною рідиною, яка містить продукти життєдіяль
ності паразита. При розриві міхура сколекси та дочірні міхур
ці можуть призвести до рецидиву захворювання.
Цикл розвитку: в кишківнику собаки розвивається стате
возріла форма ехінокока. З випорожненнями яйця потрапляють
на шерсть, у ґрунт, траву, на всі предмети, до яких торкається
собака. Членики ехінокока забруднюють пасовиська. Потра
пивши у шлунок людини, з яєць вивільняється онкосфера, яка
потрапляє у кровоносні судини і заноситься в печінку, легені
чи мозок, де розвивається фіна, що має форму міхура. Міхур
має здатність рости протягом усього життя. При споживанні
заражених органів м ’ ясоїдними тваринами (вовк, ведмідь,
лисиця, собака, кіт) у їх кишківнику зі сколексів розвивають
ся статевозрілі форми ехінокока. Личинки (міхурі) гинуть
лише тоді, коли помирає живитель.
384
Розділ 6
Рис. 6.18. Ехінокок: 1 — гачечки; 2 — присоски; 3 — сім’яники;
4 — матка; 5 — яйник; 6 — матка, заповнена заплідненими яй
цями
Шляхи зараження: пероральний (проковтування яєць, які
можуть найчастіше потрапити на руки з шерсті собак).
Симптоми ехінококозу: біль, загальна слабкість і відчуття
тягаря, задуха, при легеневій формі — кашель, у харкотинні
кров. Нагноєння місць ураження може призвести до смерті.
При розриванні міхура заражаються сусідні тканини, тоді
починається множинний ріст міхурів. Лікування лише хірур
гічне.
Лабораторна діагностика: складна і неспецифічна. Діагноз
ставиться на основі рентгенологічного обстеження, реакції
латекс-аглютинації. Пункція міхура з подальшою мікроскопією
центрифугату дає можливість побачити уламки сколексів,
гачків, оболонок міхура.
Профілактика: при контакті зі собаками дотримуватися
правил особистої гігієни. Не торкатися руками собак при вжи
Медична паразитологія___________________________________________ 385
ванні їжі. Після торкання до собак мити руки. Дегельмінти
зація собак, ветеринарні огляди органів тварин на бійні, не
гайне спалювання заражених органів (щоб їх не поїдали гри
зуни і м ’ ясоїдні тварини); санітарно-освітня робота серед
населення, боротьба з бездомними собаками.
Географічне поширення: в усіх областях України.
А л ь в е о к о к (А іо е о с о с с и з т и ііііоси їа гіз) — збудник ал ьвеокок озу
Місце паразитування: печінка, мозок, легені.
Морфоанатомічні особливості. Будовою подібний до ехіно
кока. На сколексі має 4 присоски і подвійний віночок гачків.
Матка кулястої форми без бічних виростів, містить 130—
400 яєць. Статевий отвір у передній частині збоку членика.
Цикл розвитку: статевозрілі форми паразитують у тонкій
кишці собаки, песця, лисиці, іноді вовка і кота (кінцеві госпо
дарі). З випорожненнями у зовнішнє середовище виносяться
яйця та членики. Проміжний господар: людина, полівки, он
датра, інші гризуни. Із яєць в організмі проміжного господаря
розвивається ларвоциста паразита у вигляді дрібногорбкуватого міхура, який складається з великої кількості міхурців (у них
є сколекси і колоїдні маси). Об’ єднані міхурці фіброзною тка
ниною. Міхурі не містять рідини і брунькуються тільки назов
ні, даючи метастази в інші органи. Покриті міхурі зовнішньою
(кутикулярною) і внутрішньою (зародковою) оболонками.
Шляхи зараження: пероральний (занесення в організм яєць
із забрудненими продуктами), від контакту зі собаками, під
час знімання шкури з убитих лисиць і песців. Собаки заража
ються, поїдаючи інвазованих мишоподібних гризунів.
Симптоми: найчастіше уражається печінка, у неї “ кам’я
ниста” щільність. З’ являються відчуття тяжкості та здавлю
вання в правому підребер’ї, якщо вузол міститься в ділянці
воріт печінки, виникає жовтяниця. Величина вузлів сягає
в діаметрі 25 см, проростання вузлів в середні органи, метаста
зи в лімфатичні вузли, легені, мозок ускладнюють перебіг
захворювання. Лікування хірургічне. Хіміотерапія альвеококозу не розроблена.
386
Розділ 6
Лабораторна діагностика: вивчення гемограми на виявлен
ня еозинофілії, реакція аглютинації з латексом, реакція Кацоні,
застосовують рентгенографію, ультразвукову діагностику,
комп’ ютерну томографію для виявлення вогнища ураження.
Заходи боротьби та профілактика: така ж, як при ехіно
кокозі.
Географічне поширення: в усіх областях України.
6.3. Тип Круглі черви — N ет аїК еїт іпіК ез
Клас Власне круглі черви — N ет а їой а
А ск а р и д а л ю дська (Азсагіз ІитЬгісоідев) (рис. 6.1 9 ) — збу д
ник а ск ар и д озу
Місце паразитування: тонка кишка.
Морфоанатомічні особливості: тіло аскариди людської має
циліндричну форму, загострене на передньому та задньому
кінцях, вкрите кутикулою, гіподермою і поздовжніми м ’ я
зовими волокнами. Довжина самки ЗО—40 см, самця— 15—
25 см. У самця задній кінець тіла закручений.
На головному кінці тіла аскариди містяться три “губи” . По
боках уздовж всього тіла в гіподермальних валиках просвічу
ються видільні канали у вигляді двох тонких темних смужок.
Починаються канали однією — двома одноклітинними шкір
ними залозами у задній частині тіла, а в передній частині
сполучаються в один канал, що відкривається назовні порою
позаду губ.
Травна система починається ротом, який оточений трьома
“губами” . Травна трубка ділиться на три відділи — передній,
середній і задній, який закінчується анальним отвором. За
коротким стравоходом міститься кишка, обплетена трубочками
різного діаметру. Це різні відділи статевої системи. Статеві
органи трубчастої будови. У самки вони парні, а в самця — не
парні. Найтонші трубочки — це яєчники. За яєчниками тяг
нуться трубкоподібні яйцепроводи, які переходять у матковий
Рис. 6.19. А ск а р и д а лю дська : а — внутрішня будова.*1 навкологлоткове нервове кільце, 2
нер
вові стовбури; З — яйце; 4 — піхва; 5 — кишка; 6 — матка; 7 яйник; 8 — анальний отвір; б — цикл
розвитку: 1 —дорослі особини в організмі хазяїна; 2 — запліднене яйце; 3 — інвазійне яйце; 4 — фак
тор передачі
3 88
Розділ 6
відділ. Матки мають найбільший діаметр, у передній частині
тіла обидві матки з’єднуються у непарну коротку піхву, яка
відкривається зовнішнім статевим отвором.
У самця початковим відділом статевої системи є сім’ яник,
який має вигляд тонкої нитки, за яким лежить сім’япровід.
Сім’ япровід переходить у сім ’явипорскувальний канал, який
впадає у задній відділ кишки з утворенням клоаки.
Нервова система складається з навкологлоткового нервового
кільця, від якого відходять нервові стовбури — спинний, черев
ний і два бічні. Стовбури сполучені між собою комісурами.
Органи чуттів розвинені слабо і представлені органами дотику
та хімічного чуття.
Цикл розвитку, запліднення внутрішнє. Самка відкладає
близько 200 тис. яєць, які з фекаліями потрапляють у зовніш
нє середовище і за наявності вологи при температурі 24—25 °С
за три тижні стають інвазійними, тобто містять сформовану
личинку. При проковтуванні інвазійних яєць у шлунку личин
ка звільняється від оболонки і всвердлюється в капіляри, кров’ ю
заноситься в печінкову вену, через порожнисту вену потрапляє
у праве передсердя, правий шлуночок. Легеневою артерією ра
зом із кров’ ю заноситься в легені. У капілярах альвеол личин
ка два рази линяє, просвердлює стінки альвеол, потрапляє
у просвіт альвеоли і піднімається дихальними шляхами догори
(через бронхіоли, бронхи, трахею, ротову порожнину) і повтор
но проковтується. У кишківнику розвиваються статевозрілі
самки та самці. Тривалість їх життя 12 місяців.
Шляхи зараження: пероральний (проковтування інвазійних
яєць).
Симптоми аскаридозу: при легеневій стадії — кашель, біль
у грудях, підвищення температури. При кишковій стадії —
втрата апетиту, нудота, болі у животі, біль голови, неспокійний
сон, зниження уваги, працездатності. Аскариди можуть бути
причиною непрохідності киш ківника, інколи заповзають
у жовчний міхур, стравохід, трахею, що може викликати
смерть. При наявності молодих аскарид або самців яєць у калі
не буде.
Лабораторна діагностика: овоскопія фекалій. Одна самка
виділяє протягом доби 200 тис. яєць. Личинки виявляються
Медична паразитологія
3 89
в харкотинні. Форма яєць овальна. Заплідненні яйця вкриті
білковою оболонкою, яка має неправильні хвилясті контури
жовтого кольору, вона товста і непрозора. Під нею знаходять
ся безбарвна блискуча та волокниста внутрішні оболонки.
Інколи яйця можуть бути без білкової оболонки, всередині яких
округла зародкова клітина. Незапліднені яйця овальні, заповнені
жовтковими клітинами.
Профілактика: дотримування правил гігієни, не вживати
в їж у немитої городини, овочів, фруктів. Перед їдою і після
відвідування громадських місць (транспорт, туалети) мити
руки. Боротьба з мухами, тарганами, щурами.
Географічне поширення: в усіх областях України.
Т о к со к а р а соб а к , чи к отів (Т о х о с а г а са п ів ) — збудн и к токсок а р о зу
Місце паразитування: в організмі людини личинки уража
ють центральну нервову систему, очі, печінку, легені, серце.
Статевозрілі форми живуть у кишківнику собак і котів.
Морфоанатомічні особливості: за будовою токсокара нага
дує аскариду людини.
Цикл розвитку: людина є неспецифічним господарем, роз
виток паразита обмежується стадією мігруючих личинок. Ін
вазійні яйця потрапляють укиш ківник. З яєць виходять ли
чинки, які проникають у кровоносні судини. Кров’ ю личинки
заносяться в центральну нервову систему, очі, печінку, легені,
серце. Личинки не змінюючись, залишаються життєздатними
протягом місяців і років.
Шляхи зараження: джерело інвазії — коти та собаки. Мож
ливе внутрішньоутробне зараження плоду, перорально (про
ковтування інвазійних яєць, які подібні до яєць аскариди).
Личинки проникають у тканину, зумовлюючи місцеву гранульоматозну реакцію. В центрі гранульом розвивається некроз,
иерфокальна запальна реакція і алергічний набряк. Спостері
гається лихоманка.
Симптоми токсокарозу: лихоманка, збільшення печінки
та селезінки, лейкоцитоз, еозинофілія, кашель і свербіж шкі
ри. Можливе ураження очей із втратою зору.
3 9 0 ________________________________________________________ Розділ 6
Лабораторна діагностика (різні імунологічні реакції: імунодифузії, ензим мічених антитіл).
Профілактика: дегельмінтизація собак, знищення бродячих
тварин, упорядкування вигулювання собак, особиста гігієна.
Охорона дитячих майданчиків, пісочниць, дворів, парків від
забруднення собаками. Призначення домашнім собакам протиг
лисних засобів.
Географічне поширення: в усіх областях України.
Волосоголовець ( Т г і с К о с е р Н а ї и в
хоцефальозу (рис. 6.20)
іг іс Н іи ги з )
— збудник три
Місце паразитування: личинки у ворсинках тонкої кишки,
статево-зрілі форми у товстій кишці.
Морфоанатомічні особливості: передній кінець волосого
ловця дуже тонкий, нагадує волосину і становить 2/ 3 довжини
тіла. У завуженій частині тіла міститься стравохід. Усі органи
статевої системи й основні відділи травного каналу розташовані
в задньому, розширеному відділі. Статева система має вигляд
спірально закрученої трубки. Стать паразита визначають за
наявністю у самця на задньому кінці тіла спікули (копулятивного органу).
Цикл розвитку: яйця волосоголовця стають інвазійними
протягом 25— ЗО днів у ґрунті при 15—37 °С і за наявності
кисню та вологості. Зберігають інвазійність протягом двох ро
ків. При проковтуванні яєць у тонкій кишці личинки без мігра
ції перетворюються на статевозрілі форми. Личинки проника
ють у ворсинки кишки і там розвиваються протягом трьох — де
сяти діб. Ворсинки руйнуються, личинки знову потрапляють
у просвіт кишки, сповзають до товстої кишки, там передній
волоскоподібний відділ тіла всвердлюється в оболонку, а заднійкінець виступає у просвіт. За місяць статевозрілі самки
запліднюються і виділяють яйця. Тривалість життя — кілька
років.
Шляхи зараження: пероральний (проковтування інвазійних
яєць).
Симптоми трихоцефальозу: розлади травлення та дефекації,
нудота, болі в епігастральній ділянці, що симулюють виразкову
Рис. 6.20. Волосоголовець: а — самка, б — самець: 1 — стравохід; 2 — кишка; 3 — анальний от
вір; 4 — яєчник; 5 — піхва; 6 — матка; 7 — статевий отвір; 8 — сім’яник; 9 — сім’ япровід; в — цикл
розвитку: 1 — статевозрілі форми в організмі хазяїна; 2 — запліднене яйце; 3 — інвазійне яйце із
зрілою личинкою; 4 — фактор передачі
3 92
Розділ 6
хворобу, головний біль, зниження кислотності шлунку, запа
морочення.
Лабораторна діагностика: овоскопія фекалій. Яйце воло
соголовця жовтувато-коричневого кольору, за формою нагадує
бочечку, на полюсах якої є втулкоподібні світліші утворення.
Оболонка гладка, товста, багатошарова.
Профілактика: дотримання правил гігієни, не вживати
в їжу немитої городини, овочів, фруктів. Перед їдою і після
відвідування громадських місць (транспорт, туалети) мити
руки. Боротьба з мухами, тарганами, щурами. Санітарно-ос
вітня робота серед населення. Боротьба із забрудненням ґрун
ту фекаліями людини, компостування вигрібних ям.
Географічне поширення: в усіх областях України.
Гострик (ЕпіегоЬіиз оегтісиїагіз) (рис. 6.21) — збудник ен
теробіозу
Місце паразитування: тонка і товста кишки.
Морфоанатомічні особливості: гострик має циліндричне
тіло, довжина самок 10— 12 мм, сам ців— 2— 5 мм. У самця
задній кінець закручений. У передній частині тіла є кутикулярне здуття — везикула. В передній частині гострика добре
видно стравохід і бульбус, які утворюють передню кишку. За
передньою кишкою лежить середня, яка продовжується у за
дню. Статеві органи у самки трубчастої будови і представлені
парними яєчниками, яйцепроводами, непарною маткою та
піхвою. Матка заповнена яйцями.
Цикл розвитку: інвазійні яйця потрапляють у кишківник.
Личинки без міграції розвиваються у статевозрілих самок
і самців. Самки ротовими отворами прикріплюються до стінки
кишки. Самці запліднюють самок і гинуть, а самки спускають
ся вниз до сигмоподібної та прямої кишки. Частіше вночі,
коли підвищується вологість тіла, виповзають з анального от
вору і на шкірі перианальних складок відкладають близько
12 000 яєць. При відкладанні яєць тіло самки зигзагоподібно
рухається, спричиняючи свербіж. Людина (часто дитина) роз
чухує сверблячі місця, при цьому розносить яйця на руки,
тіло, постіль. Яйця потрапляють під нігті, на підлогу, на дитячі
3 3
з*
>3
о*
Є
0 3
1 .2
’З,
02 33
3 3
3й 1
с (М
3
.*
з
3
>з 00 4
(•2
а
з
З
н
§с ®
о з- і
а, -з .3
з
зм
&
§ .& з
з ® з
& £ *•
Є о &
■&
3
3
4
•г-і
&
о
зз
3і
З
<з3
&
з
>в 1
ю
9
в о
н Зі
хо
з
3і
й л
8 'о
Л 1
н
о
о
Рн • .
гН
счі
со
о
3
йі
а
4
з
0
32
V
»
Г-1
5
3
з
з
О
3
3
3'
1
1
Н ю
о •„
з
3
3
§< в*
5 3
з о
1
і >3
3
со 3
§■
е
3
сз
1
1
. „
3
З4
>3
3
3
>3
•>-»
со
3
3
3-і
•1
1
1
со
іграшки, посуд. Протягом 6—8 год. у яйці розвивається ли
чинка, і воно стає інвазійним. Тривалість життя гострика
в організмі людини 4 тижні, але внаслідок повторного самоза
раження (автореінвазії) можуть паразитувати в організмі кіль
ка місяців. У всіх випадках потрібне лікування.
Шляхи зараження: пероральний (проковтування інвазійних
яєць).
Симптоми ентеробіозу: механічна і токсично-алергічна дія.
Свербіж у промежині, печія, легеньке поколювання в ділянці
ануса, втрата апетиту, головний біль, неспокійний сон. Інколи
гострики заповзають у жіночі статеві органи, викликають їх
запалення. Можуть бути причиною онанізму в хлопчиків.
Лабораторна діагностика: мікроскопія зіскрібків з перианальних складок. Зіскрібок проводять вранці після сну і до
сліджують на наявність гостриків чи яєць. Форма яйця аси
метрична, одна сторона плоска, друга більш випукла. Всередині
яйця видно личинки на різних стадіях розвитку. Яйця гостри
ка безбарвні та прозорі, з тонкою гладенькою оболонкою.
Профілактика: дотримання правил особистої гігієни, зміна
білизни, миття рук, коротке обстригання нігтів. Два рази на
рік проводити обстеження дитини на виявлення яєць гостриків.
Санітарно-освітня робота серед населення (батьків, особливо
дітей). Профілактичні аналізи на виявлення яєць гостриків
у дитячих колективах. Якщо яйця виявлені у дитини, треба
перевірити всіх членів сім’ї. Вологе прибирання приміщень
дитячих садочків, ясел, шкіл-інтернатів.
Географічне поширення: всі області України.
Кривоголовка (Апсуіозіота йиойепаїе) (рис. 6.22) — збудник
анкілостомозу
Місце паразитування: личинки у легенях, статевозрілі фор
ми у 12-палій кишці.
Морфоанатомічні особливості: кривоголовка має довжину
тіла 10—20 мм, рожево-жовтуватого кольору. На передній
частині тіла є характерний згин. Головний кінець має ротову
капсулу, що містить чотири кутикулярні зубці, за допомогою
1 О, 3
з * 3
•• 3 3
2
« 3
4
л 2
з» 3
5 а 3
ч
а 8 3
£ 1 3
хо
З ''4 <3
'В ==> о
2 * 3
з
§ £
&
<^> <0 ч
ч
в О •|»4
-Сі.
о
о
о
З
ч
а
а
зіІ
хо
* «
І
І
.„
4
з
Ԥ
<М о
СО
о
З
а,
з
з
з
з
з
3
4
'о
з
3 96
Розділ 6
яких паразит прикріплюється до слизової оболонки кишки.
Самці відрізняються наявністю на хвостовому кінці особливо
го розширення кутикули — статевої бурси.
Цикл розвитку: яйця паразита з екскрементами виводять
ся назовні. За сприятливих умов через один-два дні з яйця
виходить рабдитна личинка з подвійним здуттям стравоходу.
Після кількох линянь вона стає філярієподібною (інвазійна
стадія), яка може бути занесена в організм через рот або про
никнути через шкіру. По кровоносних судинах доходить до
серця і через легеневу вену заноситься до легень. Далі через
бронхіоли, бронхи, трахею потрапляє у глотку, кишківник
і там перетворюється на статевозрілу форму. Тривалість життя
паразита в організмі людини 5— 15 років.
Шляхи зараження: пероральний (проковтування личинок)
і перкутанний (всвердлювання личинок через шкіру).
Симптоми анкілостомозу: у хронічній стадії загальна слаб
кість, головний біль, біль у животі, втрата апетиту, розлади
дефекації. Крім хронічної форми, є ще шкірна та легенева.
Лабораторна діагностика: овоскопія фекалій. Яйця прозорі,
безбарвні, з закрученими краями, довжиною 50—60 мкм. Фор
ма яйця овальна з вираженою тонкою оболонкою, всередині
4—8 кулястих бластомерів.
Профілактика: у місцевостях, де розташовуються шахти
або проводять земляні роботи, не ходити без взуття, не лягати
роздягнутим на ґрунт, який може бути заражений личинками.
Не вживати в їжу немитої городини. Дотримуватись особистої
гігієни. Обстеження населення, виявлення хворих, дегельмін
тизація, боротьба із забрудненням ґрунту людськими фекалія
ми, санітарний благоустрій шахт.
Географічне поширення: в Україні там, де теплий і вологий
клімат, де є шахти або ведуться глибокі підземні роботи. Спа
лахи анкілостомозу спостерігались у районі шахт Червонограда. Планомірне проведення обстеження, дегельмінтизації та
проведення профілактичних заходів дало змогу швидко лікві
дувати спалах гельмінтозу серед шахтарів.
Медична паразитологія___________________________________________ 3 9 7
Некатор американський —
П е с а іо г а т е гіс а п и з
Місце паразитування: личинки у легенях, статевозрілі
у 12-палій кишці.
Морфоанатомічні особливості: колір тіла червонуватий.
Самки довжиною 8,0— 13,5 мм, самці — 5,2— 10,0 мм. Ротова
капсула менша, ніж у кривоголовки. У ротовій порожнині за
мість зубів є дві гострі пластинки. Бурса (копулятивна сумка)
на кінці тіла самця вузька, але довша, ніж у кривоголовки.
Спікули бурси з’єднані та закінчуються гачком.
Цикл розвитку: як у кривоголовки.
Шляхи зараження: як у кривоголовки.
Симптоми: можливі запальні процеси на шкірі, живлення
паразита кров’ ю призводить до анемії, болі в області 12-палої
киш ки, розлади травлення, головний біль, апатія, у дітей
уповільнений розвиток. При відсутності лікування можлива
смерть.
Лабораторна діагностика: овоскопія фекалій. Яйця довжи
ною 54— 70 мкм нагадують яйця анкілостоми. Відрізняються
менш заокругленими полюсами та більшою довжиною при
однаковому поперечному діаметрі.
Профілактика: як у кривоголовки.
Географічне поширення: тропічні та субтропічні пояси, пе
реважно Азія та Південна Америка.
Трихінела
(Т гіс К іп е ІІа з р іга їіз )
— збудник трихінельозу
Місце паразитування: статевозрілі форми в тонкому кишківнику, личинки у м’ язах (жувальних, міжреберних, литкових).
Морфоанатомічні особливості: трихінела має циліндричне,
звужене на передньому кінці й потовщене у задній частині
тіло довжиною 2—4 мм (самка) і 1,4— 1,6 мм (самець). Довгий
тонкий стравохід утворює розширення і переходить у кишкову
трубку, яка відкривається на задньому кінці тіла анальним
отвором. Статеві органи самки непарні та представлені яєчни
ком, коротким яйцепроводом і широкою маткою, яка перехо
дить у піхву. У самця на задньому кінці тіла є два конусоподіб
ні шкірні вирости.
398
Розділ 6
Цикл розвитку: в кишківнику з личинок розвиваються
статевозрілі форми. Після копуляції самці гинуть, а самки
проникають у лімфатичні судини, в яких народжують до 1500
личинок. Личинки з кров’ю та лімфою розносяться по всьому
організму і проникають у посмуговані м’ язи (діафрагму, міжре
берні, шийні, литкові), навколо личинки утворюється капсула.
Така личинка життєздатна протягом багатьох років і гине зі
смертю господаря. Личинка перетворюється на статевозрілу
форму лише тоді, коли м’ ясо з личинками з’їсть інший госпо
дар. Людина для трихінели є біологічним кінцем, але паразит
спричиняє захворювання трихінельоз.
Шляхи зараження: аліментарний (споживання м ’ яса сви
нини з капсульованими личинками трихінели).
Симптоми трихінельозу: набряки повік, обличчя, болі
у м ’ язах, висока температура, еозинофілія. Своїм перебігом
захворювання нагадує черевний тиф і нерідко закінчується
смертю.
Лабораторна діагностика: біопсія посмугованих м ’ язів,
серологічна реакція (кільцепреципітація, зв’язування компле
менту). Серед м ’ язових волокон рожево-червоного кольору
розташовані капсули. В порожнині капсули можна побачити
спірально закручену личинку.
Профілактика: не вживати м ’ яса, яке не пройшло ветери
нарно-санітарної експертизи, чистота у свинарниках, боротьба
з пацюками, які можуть бути носіями личинок трихінел і дже
релом зараження свиней.
Географічне поширення: по всій території України, частіше
в північно-західних областях.
Вугриця кишкова (8 і г о п § у І о і д е з
стронгілоїдозу
зіе гс о га їіз )
— збудник
Місце паразитування: у тонкому кишківнику, жовчних
і панкреатичних протоках людини і деяких тварин.
Морфоанатомічні особливості: вугриця кишкова — нема
тода довжиною 1— 3 мм.
Цикл розвитку: самка за добу виділяє 50 яєць. Із них ще
у кишківнику виходять рабдитні личинки, у яких стравохід
Медична паразитологія
399
має два характерних здуття. Коли з калом рабдитні личинки
потрапляють у ґрунт, то перетворюються у філярієподібні,
з циліндричним стравоходом. Інвазійні личинки — філяріє
подібні, проникають у капіляри людини і розносяться кров’ ю:
печінкова і порожниста вени, праве предсердя і правий шлу
ночок, легенева артерія, легені та дихальні шляхи, глотка.
Личинки пронизують капіляри альвеол, проникають у порож
нину і там перетворюються на статевозрілі форми. Якщо про
тягом 24 год личинки, що вийшли з яєць, залишаються в кишківнику, то вони перетворюються у філярієподібну стадію,
проникають у кров’ яне русло і мігрують в легені, з легень
у глотку, ротову порожнину, повторно заковтуються і, потрап
ляючи в кишківник, утворюють статевозрілі форми. Весь цикл
триває близько місяця. У зовнішньому середовищі може спо
стерігатися чергування поколінь — вільноживучого і парази
тичного. Рабдитні личинки у теплому вологому ґрунті розви
ваються у вільноживучі статевозрілі форми, які за несприят
ливих умов дають паразитичне покоління.
Шляхи зараження: пероральний (проковтування філярієподібних личинок) і перкутанний (проникнення личинок через
шкіру).
Симптоми стронгіло'ідозу: на шкірній фазі — свербіж ш кі
ри, кропивниця, підвищення температури. На легеневій фа
з і — еозинофільні інфільтрати у легенях. При кишковій фа
зі — слабкість, біль голови, діарея, болі в животі, у печінці,
в ділянці жовчного міхура, втрата ваги.
Лабораторна діагностика: виявлення личинок при мікро
скопії калу і дуоденального вмісту. Висока еозинофілія вказує
на наявність гельмінтозу. Личинки розміром 0,3—0,5 мм.
Профілактика: в місцевостях, де розташовуються шахти
або проводяться земляні роботи, не ходити без взуття, не ля
гати роздягнутим на ґрунт, який може бути заражений личин
ками. Не вживати в їж у немитої городини. Дотримуватись
особистої гігієни. Профілактичне обстеження шахтарів і людей,
що працюють на земляних роботах, виявлення хворих, дегель
мінтизація, боротьба із забрудненням ґрунту людськими фе
каліями, санітарний благоустрій шахт. Санітарно-освітня ро
бота серед шахтарів.
400
Розділ 6
Географічне поширення: в Україні, регіони з теплим і воло
гим кліматом, де є шахти, або глибокі підземні роботи.
Трихостронгілідози — ТНсН08ІГ0П§ІШ08Є8 (ТгІско8ГОП§уІи8
соїитЬгЦогтіз, Т. охеі, Т. огіепіаіів)
Місце паразитування: верхній відділ тонкого кишківника,
переважно 12-пала кишка.
Морфоанатомічні особливості: форма тіла ниткоподібна,
довжиною до 5 мм, з тонким головним кінцем. Рот оточений
трьома губами. У самців добре розвинута статева бурса, розділе
на на дві гілки, які розщеплюються ще на дві, містять дві не
однакові спікули. У самок статевий апарат парний. У них добре
розвинута статева бурса, оточена латеральними лопатями.
Цикл розвитку: яйця паразита з фекаліями виділяються
у зовнішнє середовище, через 1—3 доби з них виходять рабдитні личинки, які двічі линяють і стають філярієподібними — ін
вазійними для людини. Тривалість розвитку личинок становить
4— 14 діб. Після попадання в організм людини личинки не
мігрують. Тривалість життя понад вісім років.
Шляхи зараження: пероральний (проковтування філярієподібних личинок). Через шкіру філярієподібні личинки не
проникають.
Симптоми: зниження апетиту, болі в животі, відригування,
загальна слабість, нестійкі випорожнення, головний біль,
дратівливість, блідість шкірних покривів.
Лабораторна діагностика: виявлення яєць паразита у ви
порожненнях. Яйця овальні, трохи витягнутої форми, один із
полюсів заокруглений, другий загострений. У яйці міститься
зародок із 8—ЗО бластомерів. Проводять також дуоденальне
зондування для виявлення яєць у дуоденальному вмісті.
Профілактика: дегельмінтизація носіїв інвазії — людей
і тварин. Охорона зовнішнього середовища від забруднення
фекаліями, особиста гігієна.
Географічне поширення: Австралія, Північна Америка, краї
ни Азії, Африка. В Україну можливе завезенення випадків.
Медична паразитологія
Ришта
____________________________________401
(О г а с и п с и їи в т е с ііп е п з із )
— збудник дракункульозу
Місце паразитування: підшкірна клітковина рук, ніг.
Морфоанатомічні особливості’, тоненькі білі паразити, роз
мір самця до 20 см, самки — до 120 см.
Цикл розвитку: при проковтуванні циклопа з личинкою під
дією травних соків мікрофілярія звільняється від тканин цик
лопа, перетворюється на статевозрілу форму, лімфатичною
системою розноситься у шкірну клітковину рук або ніг, там
згортається в клубочок, і на шкірі виникає висипання. При
контакті з водою шкіра на папулі тріскає, а в щілину потрапляє
частина нематоди, в якій міститься матка, уводу виходять
численні личинки. Вони потрапляють у тіло циклопа. Коли
циклоп із філярією заноситься в організм людини, починаєть
ся захворювання.
Шлях зараження: пероральний — проковтування циклопа
з мікрофілярією.
Симптоми: на пізній стадії розвивається слоновість.
Лабораторна діагностика: серологічні реакції.
Профілактика: не пити некип’ яченої води в заражених
районах, санітарно-просвітня робота серед населення. Осушен
ня водойм, де розповсюджені циклопи.
Географічне поширення: тропічні країни. В Україні можуть
виявлятися завезені випадки захворювання.
Томінкс
(Т к о т іп х а его р Н И и з)
— збудник томінксозу
Місце паразитування: слизова оболонка носової порожнини,
трахеї, бронхи собак, лисиць, котів, іноді людини.
Морфоанатомічні особливості: ниткоподібна нематода світ
ло-сірого кольору зі звуженим переднім кінцем. Самець дов
жиною 15— 18 мм, самка — 18—20 мм.
Цикл розвитку: яйця виділяються з харкотинням. У зов
нішньому середовищі стають інвазійними.
Шляхи зараження: пероральний (проковтування яєць).
Симптоми: кашель, біль у грудях, підвищення температури.
402_____________________________________________________ Розділ 6
Лабораторна діагностика: дослідження харкотиння і фе
калій на наявність яєць.
Профілактика: типова для боротьби з гельмінтозами.
Географічне поширення: усі регіони України.
Пальовик гігант (О і о с і о р К у т а
мозу
геп а їе)
— збудник диоктофі-
Місце паразитування: ниркові миски собак, деяких копит
них ссавців, іноді людина.
Морфоанатомічні особливості: колір тіла криваво-червоний.
Довжина самця 14— 20 см, самки — 20—ЗО см.
Цикл розвитку: яйця паразита виділяються незрілими зі
сечею господаря. У зовнішньому середовищі при оптимальних
умовах стають інвазійними.
Шлях зараження: пероральний (проковтування яєць).
Симптоми: ниючий біль у поперековій ділянці.
Лабораторна діагностика: виявлення яєць у сечі.
Профілактика: дотримання особистої гігієни, охорона зов
нішнього середовища від забруднення.
Географічне поширення: всі регіони України.
Родина філярії— (Рііа гіій а е ) — збудники
філяріозів
Паразити мають ниткоподібну форму тіла, яке звужується
на кінцях. Живородні. Біогельмінти. Передаються трансмісив
ним шляхом через кровосисних комах.
Вухереріоз (\ ¥ и с к е г е г і а
Ь а п с го Ц і)
— збудник вухереріозу
Місце паразитування: дорослі черви паразитують у лімфа
тичній системі, а личинки — у кровоносній.
Морфоанатомічні особливості: ниткоподібна форма тіла,
колір білий. Самка досягає у довжину 5— 10 см, в ширину
0,2— 0,3 мм, самець — 2—4 см в довжину і 0,1м м в ширину.
Самці та самки переплітаються між собою, утворюючи клубок.
Медична паразитологія
403
Живуть паразити до 17 років. Самка народжує личинок (мікрофілярії), які оточені чохликом, передній кінець тіла має стилет.
Цикл розвитку: із лімфатичної системи личинки мігрують
у кровоносну і живуть до 70 днів. Проміжний господар — комарі
родів Сиіех, АпорНеІез, Мапзопіа. При смоктанні крові людини
комаром личинки потрапляють в організм комара, залежно від
температури цикл розвитку триває 8—ЗО днів. Личинки мігру
ють у хоботок.
Шляхи зараження: укус комара, який вносить в організм
людини мікрофілярії.
Симптоми: дорослі гельмінти на ранніх стадіях спричиня
ють алергічний синдром, подальше запалення лімфовузлів,
закупорюють просвіт лімфатичних судин, утруднюють відтік
лімфи, що призводить до збільшення об’ єму органів (нижніх
кінцівок, статевих органів, у жінок — молочних залоз). Захво
рювання відоме під назвою “ слонова хвороба” .
Лабораторна діагностика: дослідження мазків крові на
наявність мікрофілярій. Кров беруть уночі. Застосовують також
імунологічні реакції.
Профілактика: захист від укусів комарів, осушення боліт,
лікування хворих людей.
Географічне поширення: тропічні країни. В Україні трапля
ються завезені випадки захворювання.
Бругія (В г и § і а
т а їа іа )
— збудник бругіозу
Місце паразитування: дорослі черви паразитують у лімфа
тичній системі, мікрофілярії у крові.
Морфоанатомічні особливості: самці в довжину 23 мм,
у ширину 0,09 мм, самки відповідно 55 мм і 0,16 мм. Живород
ні. Мікрофілярії на передньому заокругленому кінці мають
подвійний стилет.
Цикл розвитку: не відрізняється від \У. ЬапсгоЦі. Кінцевий
господар — людина, а також коти, собаки, мавпи. Проміжний
господар — усі види комарів, але найчастіше комарі роду
Мапсопіа.
4 0 4 ________________________________________________________ Розділ 6
Симптоми: подібні до вухеріозу, трапляються переважно
серед сільського населення.
Лабораторна діагностика: дослідження мазків крові на
наявність мікрофілярій.
Профілактика: захист від укусів комарів, осушення боліт,
лікування хворих людей.
Географічне поширення: Південно-Східна Азія. В Україні
можливі завезені випадки захворювання.
Лоа (Ьоа Іоа) — збудник лоалозу
Місце паразитування: підшкірна клітковина, під серозни
ми оболонками, іноді під кон’юнктивою ока людини.
Морфоанатомічні особливості: самець у довжину 34 мм,
у ширину 0,3 мм, самка — відповідно 60 мм і 0,5 мм — живо
родні. Мікрофілярії покриті малопомітним чохликом. Передній
кінець тіла широкий.
Цикл розвитку: мікрофілярії проникають у кров. Проміж
ний господар— ґедзі родини СНгузорз. В організмі ґедзів
мікрофілярії досягають інвазійної стадії через 7— 10 днів.
Шляхи зараження: укус ґедзів.
Симптоми: на початкових стадіях захворювання спостері
гають свербіж шкіри, пропасницю, пізніше вражається під
шкірна клітковина, з’ являються “ пухлини” , внутрішньоочна
міграція дорослих паразитів супроводжується набряками повік
і кон’ юктиви, сильними болями в очах.
Лабораторна діагностика: кров у хворого беруть удень
і досліджують мазки на наявність мікрофілярій. Імунологічні
реакції.
Профілактика: боротьба з переносниками, лікування хворих
людей.
Географічне поширення: Західна і Центральна Африка.
В Україні можливі завезені випадки захворювання.
Медична паразитологія
405
Акантохейлонема ( А с а п і к о с К е і І о п е т а
акантохейлонематозу
р егз^а п в)
— збудник
Місце паразитування: брижа, печінка, перикард людини,
порожнини серця, легені, селезінка.
Морфоанатомічні особливості: ниткоподібне тіло, білого
кольору, самець у довжину 45 мм, у ш ирину— 0,07 мм, сам
ка — відповідно 75 мм, і 0,13 мм.
Цикл розвитку: самка плодить личинки, які локалізують
ся у крові. Мокреці — проміжний господар.
Шляхи зараження: укус мокреця.
Симптоми: лихоманка, біль у животі, кінцівках, запаморо
чення, мікроабсцеси в печінці.
Лабораторна діагностика: виявлення мікрофілярій у мазках
крові.
Профілактика: захист від укусу мокреців, осушення боліт,
лікування хворих людей.
Географічне поширення: Африка, Південна Америка.
Онхоцерк
(О п с і їо с е г с а и о іи и іи з )
— збудник онхоцеркозу
Місце паразитування: біля колінного і стегнового суглобів,
на ребрах, біля хребта, у пахових впадинах людини.
Морфоанатомічні особливості: самки довжиною до 5 см,
шириною 0,4 мм, самці довжиною 3—4 см і шириною 0 ,1 —
0,2 мм.
Цикл розвитку: самка народжує личинки, які проникають
у кровоносні судини.
Проміжний господар — мошки роду Зітиііит. В організмі
проміжного господаря мікрофілярії досягають інвазійної стадії
та переміщуються у хоботок мошки.
Шляхи зараження: укус мошки — личинки потрапляють
на шкіру і занурюються в неї.
Симптоми: підшкірні пошкодження призводять до розро
стання навколо мертвих і живих гельмінтів, утворення вузлів,
406
Розділ 6
ураження очей личинками призводить до сліпоти, ураження
нервової системи.
Лабораторна діагностика: гістологічне дослідження зрізів
епідермісу, вузлів.
Профілактика: знищення переносників, використання ін
сектицидів у місцях розмноження мошок, лікування хворих
людей.
Географічне поширення: країни Африки. В Україні можливі
завезені випадки онхоцеркозу.
6.4. Тип Членистоногі — А гіК го р о А а
Характерні ознаки:
— наявність членистих кінцівок;
— зовнішній хітиновий скелет;
— тіло розмежоване на головогруди і черевце (у павукоподіб
них) або на голову, груди і черевце (у комах);
— травна система: диференційована передня кишка, серед
ня і задня, що закінчується анальним отвором;
— видільна система: зелені залози, або мальпігієві трубочки;
— дихальна система: у водних — зябра, у наземних — леге
неві мішки або трахеї;
— кровоносна система: незамкнена, серце на спинному боці;
— ріст членистоногих у процесі їх розвитку відбувається
через линяння.
Клас павукоподібні — АгасК поіЛ еа
Клас включає близько 35 000 видів. Для павукоподібних
характерне тією чи іншою мірою злиття сегментів у головог
руди і черевце, між якими є перетяжка. Не мають вусиків
і крил. Головогруди несуть 6 пар кінцівок: перша пара хеліцери (верхні щелепи) і друга пара педипальпи (ногощупальця),
чотири пари ходильних ніг. Тіло покрите хітином, під яким є
гіподерма.
Медична паразитологія___________________________________________ 4 0 7
Отруйні павуки
Скорпіон —
Е и з с о гр іи з іа и г іс и з
Великий павук, жовтого, або темно-коричневого кольору.
На кінці сегментованого черевця є твердий чорний гачок — жа
ло, на вершині якого відкривається протока отруйної залози.
Швидко рухається, може заповзти у житло людини, в одяг,
взуття. Після уколу у людини з ’ являється біль, який віддає по
ходу нервів, гіперемія, набряки, затруднене дихання, ковтан
ня, судоми. Усі ці явища можуть тривати кілька днів. В Україні
скорпіони поширені в Криму.
Каракурт —
2 а Іго Л е с іи з іг е й е с іт £ и Ііа Іи з
Самка довжиною 2 см. Черевце велике кругле чорного кольо
ру, на спинній стороні видно червоні плями у два ряди. Кін
цівки покриті дрібними чорними волосинками. Протоки от
руйних залоз зв’ язані зі щелепами.
Самці менших розмірів, ніж самки. У них подовгасте черев
це коричневого кольору, довгі, тонкі кінцівки.
У кінці літа самка будує гніздо, причому нитки павутиння
обплітають кущі, каміння, суху траву. У гніздо самка відкла
дає 100— 200 яєць, навесні з них вилуплюються маленькі па
вучки. Укус самки отруйний, з’ являється біль, підвищення
температури, оніміння рук і ніг, болі в ділянці живота, рук,
поясниці. Виникає відчуття страху, можлива смерть.
Поширені у степовій зоні, в передгір’ ях і пустелях, у Серед
ній Азії, на півдні України.
Тарантул —
Т госН оза з іп д о гіе п з із
Верхня поверхня тіла коричневого кольору, нижня — чор
ного. Кінцівки з темно-коричневими і жовтуватими попереч
ними кільцями. Самка восени відкладає 100— 400 яєць. Навес
ні з них вилуплюються павучки, яких самка певний період
носить на собі. У цей період вона найбільш агресивна.
При укусі в рану потрапляє отрута, яка викликає хворобливі
місцеві явища та симптоми загальної інтоксикації. Прогноз
сприятливий.
Поширені в сухих місцевостях Середньої Азії. В Україні
немає.
408
Розділ 6
Фаланги, або сольпуги — 8оІри§ае
Тіло розчленоване. Хеліцери у вигляді клешень, а педипальпи подібні до ніг. Отруйні залози відсутні, але укуси болючі,
можуть викликати запальні процеси, бо ранки забруднюються
залишками їжі із хеліцер. Поширені у Середній Азії, на Кав
казі, в Криму, Молдові.
Клас Павукоподібні — АгасК поіЛ еа
Ряд Кліщі — А с а гіп а
Кліщі паразитоформні— РагазіЩогтез (рис. 6.23)
Паразитоформних кліщів поділяють на родини: іксодові,
аргасові, гамазові.
Рис. 6.23. Кліщі: а — сита самка собачого кліща; б — тайговий
кліщ; в — степовий кліщ
Родина Іксодові кліщі — ІхоЛШ ае
Кліщ собачий — ІхоЛез гісіпиз (рис. 6.23, а)
Місце перебування: відкрита місцевість лісової та лісосте
пової зон.
Морфоанатомічні особливості: тіло несегментоване, оваль
ної форми, на спині є щиток. У самців щиток повністю вкриває
спину, а у самок, личинок і німф він є лише у передній частині
спини, решта частин тіла мають м’які покриви, що забезпечує
можливість розтягування і збільшення об’ єму тіла. Очі відсут
ні, хітиновий покрив пронизаний численними порами з волос
ками, які виконують функцію органів чуття. Колір брунатний,
Медична паразитологія___________________________________________ 4 0 9
довжина тіла 2,5—4,0 мм, а ситої самки — до 11 мм. Хеліцери
та педипальпи видозмінені у хоботок — колючо-сисний ротовий
апарат, розміщений на звуженому головному кінці. Основа
хоботка прямокутна. До країв тіла нерухомими коксами при
єднані чотири пари кінцівок, що мають перетинчастий прида
ток. Кожна кінцівка складається з основного членика (кокса),
вертлуга, стегна, гомілки, передньолапки, лапки з двома кіг
тиками, між якими є присосок у вигляді подушечки. Позаду
кокса четвертої пари ніжок з кожного боку міститься ущіль
нений хітиновий щиток — перитрема, який оточує дихальну
стигму з черевного боку, а на рівні 3-ї пари кінцівок — витяг
нутий в поперечну щілину статевий отвір (у самки дугоподіб
ний). Ближче до заднього кінця тіла міститься анальний отвір,
що має вигляд короткої поздовжньої щілини. Дугоподібна
анальна борозна огинає анальний отвір спереду і своїми кінця
ми доходить до заднього краю тіла.
Процес кусання кліща безболісний, оскільки він виділяє
особливі анестезуючі речовини, завдяки чому присмоктування
кліща залишається непоміченим живителем. Збудник захво
рювання потрапляє зі слиною кліща або переноситься ротови
ми органами. Дорослі кліщі нападають на гризунів, їжаків,
собак, лосів, овець, а також на людину.
Цикл розвитку: непрямий із повним метаморфозом: яйце —»
личинка (три пари кінцівок) —> німфа (чотири пари кінцівок) —>
імаго (після 1— 3 линянь). Термін розвитку 3— 7 років.
Епідеміологічне значення: переносник збудників туляремії,
енцефаліту західних районів, шотландського енцефаліту.
Ш «а.
'ШСЦ> „тгем-р&нз-
Заходи боротьби та профілактики: знищення кліщів у при
роді на всіх стадіях розвитку, своєчасне виявлення і зняття
кліщів із тіла людини, застосування репелентів.
Кліщ тайговий — Іходез регзиісаіиз (рис. 6.23, б)
Місце перебування: відкрита місцевість лісової та лісосте
пової зон.
Морфоанатомічні особливості: подібний за формою і роз
мірами до кліща собачого. Личинки та німфи нападають на
малих ссавців, статевозрілі форми — на людину або великих
Розділ 6
410
ссавців. Але у самця на коксах першої пари ніг відсутній пе
ретинчастий придаток, внутрішній зубець кокса короткий,
а кокси другої і третьої пари кінцівок однакові. У самок на
рівні другої пари ніжок на черевній стороні розміщена пряма
або хвиляста статева щілина, прикрита хітиновим щитком.
Розповсюджений за межами України, в Сибіру, на Далекому
Сході, виявлений у Європі.
Цикл розвитку: непрямий з повним метаморфозом: яйце —>
—> личинка —> німфа -» імаго.
Епідеміологічне значення: переносник збудника тайгового
енцефаліту.
Заходи боротьби та профілактики: знищення кліщів у при
роді на всіх стадіях розвитку, своєчасне виявлення і зняття
кліщів із тіла людини, застосування репелентів.
Родина Аргасові
Кліщ селищний —
О г п ііН о д о г и з р а р іИ ір еа
Місце перебування: печери, покинуті кам’ яні споруди, хлі
ви, нірки гризунів.
Морфоанатомічні особливості: продовгувато-овальна форма
тіла, розмір 5,8—8,2 мм. Задня частина тіла заокруглена, перед
ня загострена. Тіло вкрите суцільним дрібногорбкуватим шаром
хітину сірого кольору. Очі відсутні. Ротовий апарат розміщений
у заглибині, на черевному боці. Чотири пари кінцівок за довжи
ною не однакові, друга пара найкоротша, а четверта — найдов
ша. Позаду від першої пари кокс міститься статевий отвір,
а трохи далі до кінця тіла — анальний отвір. В Україні не ви
явлений. Найбільш поширений у Середній Азії, Афганістані,
на Уралі.
Цикл розвитку: непрямий із повним метаморфозом: яйце —>
—> личинка
німфа 1 -» німфа 2 -> німфа 3 -» німфа 4,5 ->
- » імаго. Живляться на гризунах, кажанах, жайворонках, со
баках, конях, котах, великій рогатій худобі, а також на людині.
Личинки можуть голодувати до 2 років, німфи — 3— 4 роки,
дорослі кліщі — до 13 років.
Медична паразитологія____________________________________ 411
Епідеміологічне значення: переносник кліщового поворот
ного тифу.
Заходи боротьби та профілактики: знищення кліщів у при
роді на всіх стадіях розвитку, своєчасне виявлення і зняття
кліщів з тіла людини, застосування репелентів.
Степовий к л іщ —
О е г т а с е п іо г т а г^ іп а іи з
(рис. 6.23, в)
Місце перебування: рівнини, гірські, степові та лісостепові
ділянки.
Морфоанатомічні особливості: тіло овальне, звужене до
переду, довжиною 6—8 мм. На світлому спинному щитку є ве
ликі темні плями, розміщені симетрично. По краю тіла наяві
фестони. Очі плоскі. Хоботок короткий, з прямокутною основою,
ширина якого у два рази перевищує довжину. Добре розвинуті
кокси четвертої пари кінцівок. На черевній стороні статевий
отвір оточений спереду статевою борозною. На рівні четвертої
пари кокс по боках тіла розміщені овальні перитреми.
Цикл розвитку: непрямий із повним метаморфозом (яйце —»
—» личинка -> німфа —» імаго). Личинки і німфи живуть на дріб
них ссавцях, переважно на гризунах, а імаго — на великих ссав
цях (верблюди, вовки, собаки, можуть нападати на людину).
Епідеміологічне значення: переносник кровопаразитів до
машніх тварин. Переносить вірус енцефаломієліту коней,
збудника кліщової висипнотифозної лихоманки. Довго збері
гають у своєму тілі збудників туляремії, чуми, бруцельозу.
Заходи боротьби та профілактика: знищення кліщів на
всіх стадіях розвитку, особистий захист, застосування репе
лентів.
Родина Гамазові — Сат азоіЛеа
Курячий кліщ — О е г т а п у з з і и з £ а 1 1 іп а е
Щурячий кліщ — О г п і і к о п у з з и з Ь а с о іі
Мишачий кліщ — А І І о д е г т а п у з и з з а п £ и і п е и з
Місце паразитування: гнізда, печери, нори гризунів,
ґрунт.
412
Розділ 6
Морфоанатомічні особливості: дрібні кліщі (0,2— 2,5 мм).
Забарвлення тіла від білуватого до коричневого. Тіло вкрите
спинними та черевними щитками і щетинками. Хеліцери у ви
гляді клешні або голки. Це факультативні кровососи, які на
короткий час заповзають на тіло живителя, щоб насмоктатися
крові.
Цикл розвитку: весь життєвий цикл проходить у гнізді.
Самка після смоктання крові протягом 1— 2 діб відкладає яй
ця. Цикл розвитку ділиться на такі стадії: яйце -» личинка -»
—> німфа (протонімфа) - » німфа II (дейтонімфа) —> імаго. Ці
кліщі витривалі до холоду і морозів. Після замерзання відтаюють і оживають. Можуть голодувати 6— 7 місяців.
Епідеміологічне значення: гамазові кліщі можуть нападати
на людину, викликати подразнення шкіри, висипання та інші
явища дерматиту. Також беруть участь у циркуляції збудників
серед тварин у природних осередках енцефаліту, геморагічної
лихоманки, туляремії, Ку-лихоманки, рикетсіозів, грипу тощо.
Заходи боротьби та профілактика: знищення кліщів на
всіх стадіях розвитку з використанням хімічних препаратів,
індивідуальний захист (захисний одяг, репеленти, аерозольні
балони), боротьба з гризунами, які є живителями для кліщів.
Рикетсіозний кліщ —
А І Іо д е г т а п у з з и з з а п §и іп е и з
Місце перебування: лісова підстилка, печери, під корою де
рев, на рослинах, комахах, у норах гризунів, гніздах птахів.
Морфоанатомічні особливості: будовою подібний до куря
чого кліща. Тіло овальне, довжиною 0,2— 0,5 мм, на покривах
є дві ущільнені ділянки — щитки: передній — вузький видов
жений, задній — маленький, округлий. На поверхні тіла, яка
не покрита щитками, є численні щетинки. Перитреми доходять
до середини кокс II пари ніг. Не має очей.
Цикл розвитку: непрямий з повним метаморфозом і скла
дається із таких фаз: яйце —>личинка —> німфа і (споронімфа) ->
-» німфа II (дейтонімфа) —» дорослі кліщі.
Епідеміологічне значення: кліщі паразитують на гризунах,
але часто нападають на людей, укуси їх спричиняють дерма-
Медична паразитологія________________________________________ 413
тити. Вони переносять рикетсіозні захворювання — визекулярний рикетсіоз і рикетсіозну віспу.
Заходи боротьби та профілактика: боротьба з гризунами,
знищення кліщів на всіх стадіях розвитку, використання ре
пелентів.
Саркомастігоформні кліщі
Свербун коростяний —
(рис. 6.24, а)
А са ги з
зіго
з ., З а г с о р і е з
зс а Ь іе і
Місце перебування: роговий шар епідермісу.
Рис. 6.24. Кліщі: а — коростяний свербун; б — залозиця вуг
рова; в — селищний кліщ
Морфоанатомічні особливості: дуже дрібні розміри: сам
ка — близько 0,4 мм; самець — близько 0,3 мм завдовжки. Має
округлу форму тіла, на поверхні якого помітні численні лусоч
ки, тупі шпички і спрямовані назад щетинки. Помітна попе
речна борозна, яка розмежовує тіло на головогруди і черевце.
Очі відсутні. У передній частині тіла міститься ротовий апарат
гризучого типу, по боках від нього, з кожного боку по дві пари
коротких ніжок. Задні дві пари кінцівок видно лише з черев
ного боку. Всі кінцівки короткі та мають щетинки, що пов’язано
з пристосуванням до життя у ходах всередині шкіри. На черев
ному боці помітні хітинові утворення — епімери, які тягнуть
ся від основи кінцівок до середини тіла. Дихання у кліща
здійснюється через усю поверхню тіла.
414
Розділ 6
Довжина ходу, який самка робить за день, сягає 2— 3 мм
(самці ходів не роблять). Живляться кліщі клітинами епідермі
су хазяїна. Тривалість життя дорослих кліщів 40—45 днів.
Захворювання проявляється сильним свербежем. Поширене
в усіх областях України. Діагноз ставиться при виявленні
кліщів у ходах, які вони прокладають.
Цикл розвитку: у ходах самки відкладають яйця (20 і біль
ше за життя), де відбувається метаморфоза, яка триває 1—
2 тижні: яйце -» личинка -> німфи І, II порядку -> імаго.
Епідеміологічне значення: збудник корости або скабієсу.
Заходи боротьби та профілактики: виявлення хворих,
періодичні перевірки у місцях скупчення людей, дезинфекція
постільної білизни та одягу.
Тирогліфоподібні кліщі, або акаридієві кліщі. Вони невели
ких розмірів, уражають продукти харчування і можуть із їжею
потрапити у шлунково-кишковий тракт людини, викликаючи
запальні процеси, а при вдиханні — катар верхніх дихальних
шляхів і астматичні напади. Часто живі кліщі виявляють у ви
порожненнях, у дуоденальному вмісті, слині, харкотинні.
Мікроскопічний кліщ Т у с а р к а £ и з п о х і и з живе у сіні, соло
мі, житлі людини (в кімнатному пилі), де поїдає бакалійні та
кондитерські вироби.
Борошняні кліщі — Т у г о § І у р Н у з ї а г і п а е , Т . п о х і и з , Т . Р е г п і Форма тіла яйцеподібна, молочно-білого кольору зі
скляним блиском, довжиною 0,35—0,67 мм. Потрапивши на
вологий ґрунт, виїдають зародки зерна, знижують його схо
жість. Екскременти кліщів сприяють склеюванню насіння та
його зараженню бактеріями.
с іо з и з .
Сирний кліщ ( Т у г о £ І у р к у з ) живе у сирі, а С а г р о § І у р К у з —
у кислому молоці, гниючих овочах, сухофруктах.
Винний кліщ (Іп8ііо£азіЄ8 ЬассУіиз) розмножується на по
верхні вина, викликаючи його псування в дубових бочках,
у природі живе на деревах, із яких витікає сік.
Медична паразитологія___________________________________________ 4 1 5
Цибульковий кліщ —
К к і г ^ і у р к у з е с к іп о р и з
Пошкоджує корені рослин, цибулю, бульби у місцях їх
зберігання. Значної шкоди здоров’ ю людини завдають так звані
порохові, або алергенні кліщі родини Ругодіуркісіае.
У поросі, зібраному в житлових приміщеннях із килимів,
доріжок, підлоги тощо виявлено близько 100 видів кліщів
розміром 0,2— 0,6 мм. Із них найбільш поширені “ліжковий”
кліщ И е г т а і о р к а ^ о і с і е з р і е г о п у з з і п и з . Продукти життєдіяль
ності та фрагменти мертвих кліщів і личинок містяться у по
росі.
Залозниця вугрова —
О е т о д е х іо іііс и їо г и т
Місце локалізації: шкіра, сальні залози та волосяні цибу
лини обличчя, шиї, тулуба і зовнішній слуховий прохід.
Морфоанатомічні особливості: дрібні розміри (0,2—0,5 мм),
овальна форма тіла, яке нагадує черв’ ячка, короткі кінцівки.
Ротові органи пристосовані до смоктання. Відкладають яйця.
Личинки, німфи, імагінальна стадія подібні. Діагностують
шляхом мікроскопічного дослідження виділень із гнійних
волосяних фолікул.
Цикл розвитку: непрямий із повним метаморфозом: яйце - »
—» личинка - » німфа -» імаго.
Епідеміологічне значення: збудник демодекозу у людини
і залозистої корости у тварин.
Заходи боротьби та профілактики: дотримання правил
особистої гігієни (не витискати вугрів брудними руками), ви
явлення та лікування хворих.
Кліщі — червонотільці належать до родини ТготЬісиІідае.
Вони мають такі стадії розвитку: яйце —> передличинка —» ли
чинка —> протонімфа —» дейтонімфа —> тритонімфа -» імаго.
Активні личинки, дейтонімфа й імаго. Розмір личинок 0,15—
0,4 мм, а дорослих кліщів — 2— 4 мм у довжину. Личинки
яйцеподібної форми оранжево-червоного кольору — густо вкриті
волосинками. Личинки живляться протягом кількох діб лімфою
та зруйнованими клітинами. Дорослі кліщі живуть у ґрунті.
Тіло овальне з перетяжкою на рівні третьої — четвертої пари
кінцівок, червонуватого кольору.
Розділ 6
4 16
Личинки при живленні на тілі людини викликають дерма
тити. При кровосмоктанні передають рикетсії лихоманки цуцу гаму ші.
Родина Хижі кліщі
Представник — пузатий кліщ — Р е & і с и ї о і й е з о е п і г і с о в и з
Кліщ жовтого кольору довжиною 0,2 мм. Самка живородна.
Цей кліщ забруднює зерно і борошно, при потраплянні на лю
дину викликає дерматит, свербіж, гіперемію. Захворювання
розвивається у суху погоду і пов’язане з контактом людини із
зерном або соломою. Назва хвороби — зернова короста.
Клас Комахи — Іп зесіа
Ряд Двокрилі — Б ір іе га
Родина Мухи — М изсіЛае
Осіння жигалка —
З і о т о х у з с а іс іїга п з
(рис. 6.25,
а)
Рис. 6.25. Осіння жигалка (а); муха Вольфартова (б)
Місце перебування: епідерма шкіри людини.
Морфоанатомічні особливості: тіло світло-сіре, складаєть
ся із 3 відділів: голова, груди і черевце. На голові у мухи є
складні очі, вусики, щупики і твердий довгий хоботок, який
служить для проколювання шкіри людини. На черевці є темні
округлі плями, на грудях чотири темні смужки, також у ділян
Медична паразитологія
4 17
ці грудей три пари кінцівок, дві пари крил і одна пара дзижчалець. На кінці має хітинові пластинки. Тертям хоботка по
шкірі муха зіскрібає епідерміс і живиться кров’ ю, одночасно
впускає отруйну речовину і викликає сильне подразнення.
Розмір мухи 6— 7 мм.
Цикл розвитку: яйце (відкладає у рослини, що гниють, або
у гній) -» личинка -> лялечка (утворюється в ґрунті) —» імаго.
Епідеміологічне значення: розповсюджувач збудників сибір
ки, поліомієліту, туляремії, гнійних інфекцій.
Профілактика: захист шкіри від укусів мух, використання
інсектицидів (хімічних речовин для знищення личинок і до
рослих мух).
Географічне поширення: Карпатський регіон, узбережжя
річок України.
Муха Вольфартова — \У^оНЦагііа та£пЦіса (рис. 6.25, б)
Місце перебування: м’ язи людини і тварин.
Морфоанатомічні особливості: великих розмірів (10— 13 мм
завдовжки), покрив тіла світло-сірий, три темні поздовжні
смужки на грудях. Черевце яйцеподібної форми, з округлими
і темними плямами по краях і чорною смужкою посередині.
Доросла муха живиться соком квітів і після запліднення на
роджує понад 100 живих личинок.
Цикл розвитку: самки живородні, живих личинок відкла
дають у ранки або на слизову оболонку носа, ока тварин чи
людини. Личинки мають жовтувато-коричневі шипики, про
никають у м ’ язи, прогризають ходи аж до кісток, спричиняють
відмирання тканин. Після кількох линянь через 3— 5 днів
личинка покидає тіло людини чи тварин і заляльковується
у ґрунті.
Епідеміологічне значення: збудник міазу.
Профілактика: захист тіла від нападу Вольфартової мухи.
Географічне поширення: середня і південна частина Європи.
Розділ 6
4 18
Хатня муха —
М и з с а й о т е з ііс а
Місце паразитування: покриви тіла людини.
Морфоанатомічні особливості: три відділи тіла: голова,
груди, черевце. Колір тіла темний. Голова напівкругла, з боків
великі фасеткові очі, спереду ротовий апарат лижучого типу.
Нижня губа перетворилася на хоботок із двома смоктальними
дольками, між ними є ротовий отвір. Верхня губа і язик розмі
щені на передній стінці хоботка. Верхня щелепа і перша пара
нижніх щелеп атрофовані. Черевце складається із п’ яти сегмен
тів. Тіло і кінцівки вкриті волосинками. По боках тіла є стигми.
На лапках кігтики та клейкі лопасті, які допомагають перемі
щуватись по плоскій поверхні. Сегменти грудей злиті. Є одна
пара крил і одна пара дзижчалець. Від грудей відходить три
пари кінцівок. Лапка має п’ять сегментів, останній із них за
кінчується двома кігтиками та двома подушечками.
Цикл розвитку: за один раз муха відкладає 100— 150 яєць.
Через добу з них вилуплюються личинки, ще через два тижні
утворюється лялечка. Розвиток від яєць до імаго при 25 °С
триває 16 днів.
Профілактика: використання інсектицидів, механічні за
соби (липучки, мухоловки), захист зовнішнього середовища
від забруднення і недопущення виплоду мух.
Епідеміологічне значення: механічний переносник різних
видів бактерій, яєць гельмінтів, цист найпростіших.
Географічне поширення: по всій Україні.
Сіра м’ясна муха —
З а гс о р Н о га с а гп а гіа
Місце паразитування личинок: виразки людини, очі, по
рожнини носа, вуха, кишківник.
Морфоанатомічні особливості: розмір до 14— 16 мм. Колір
тіла сірий, голова — жовтувато-біла, з чорною смужкою посе
редині, на грудях також є темні поздовжні смужки, кінець
черевця чорний.
Цикл розвитку: живородна. Самка відкладає личинки на
свіже м ’ ясо, а також на трупи людей і тварин у кількості 20 тис.
У ґрунті личинка перетворюється в лялечку -» імаго.
419
Медична паразитологія
Епідеміологічне значення: личинки викликають захворю
вання — міаз, а імаго є механічними переносниками збудників
інфекційних захворювань.
Заходи боротьби та профілактика: такі ж, як і з іншими
видами мух.
Географічне поширення: по всій Україні.
Сирна муха —
Р іо р Н й а са зеі
Місце паразитування личинок: кишківник людини.
Морфоанатомічні особливості: типова будова мух.
Цикл розвитку: самка відкладає свої яйця на сир, копчену
ковбасу, копчену і солену рибу, які споживає людина.
Епідеміологічне значення: кишковий міаз.
Заходи боротьби та профілактика: такі ж самі, як і з інши
ми видами мух.
Географічне поширення: по всій Україні.
Синя м’ясна муха —
С а ІІір Н о га ь іс іп а
Місце паразитування личинок: виразки і рани людини.
Морфоанатомічні особливості: велика муха, розміром до
14 мм. Колір тіла темно-синій із металевим відливом, нижня
частина голови червонувата, груди чорні, черевце з білуватим
нальотом.
Цикл розвитку: з повним перетворенням. Самка відкладає
450—600 яєць на свіже м’ясо, а також на трупи людей і тварин.
Личинка живиться тим субстратом, на якому були відкладені
яйця. Заляльковуються в землі, де лялечки і зимують.
Епідеміологічне значення: личинки викликають захворю
вання — міаз, а імаго є механічними переносниками збудників
інфекційних захворювань, зокрема сибірки.
Заходи боротьби і профілактика: санітарно-профілактичні
заходи спрямовані на недопущення виплоду мух у населених
пунктах та на використання інсектицидів.
Географічне поширення: по всій Україні.
4 20
Розділ 6
Домова муха — Мизсіпа зіаЬиІапз
Місце паразитування: переважно сільська місцевість.
Морфоанатомічні особливості. Тіло бурого кольору, а щ у
пики і кінцівки — жовтого. Копрофаг. Живиться фекаліями
та їжею людини.
Цикл розвитку: розмножується найчастіше у гної, туалетах,
гниючих відходах, деколи відкладає яйця на їжу людини.
Розвиток із повним перетворенням. Залітає у житло людини,
особливо поблизу неканалізаційних туалетів і хлівів.
Епідеміологічне значення: переносник збудників кишкових
інфекцій та інвазій. Відкладання яєць на продукти харчуван
ня людини призводить до розвитку кишкового міазу.
Заходи боротьби та профілактика: проведення санітарних
заходів, які забезпечують збір відходів і їх своєчасне видален
ня, закриття неканалізаційних туалетів, недопускання забруд
нення ґрунту фекаліями людини. Для знищення личинок
у місцях виплоду мух використання інсектицидів.
Географічне поширення: по всій Україні.
Муха цеце — Сіоззіпа раїраііз
Місце паразитування личинок: покрив тіла.
Морфоанатомічні особливості: розмір 10— 13 мм. Забарв
лення тіла темно-коричневе. На спинній стороні черевця є
кілька вузьких поперечних смужок жовтого кольору, а посе
редині — одна продовгувата. Між поперечними смужками
розташовані дві великі темні петлі. Спереду виступає хітинізований хоботок.
Цикл розвитку: самка відкладає безпосередньо на поверхню
ґрунту одну личинку (за все життя 6— 12 личинок), яка пере
творюється в лялечку. Через 3—4 тижні виходить імаго. Ж и
виться муха кров’ ю диких і домашніх тварин, а також людини.
Живуть ці мухи у заростях кущів на берегах річок і озер, поб
лизу житла людини.
Епідеміологічне значення: переносник трипаносом ( Тгурапозота Ьгисеі датЬіепсе) — збудника сонної хвороби.
Медична паразитологія __________________________________________4 21
Заходи боротьби та профілактика: у місцях відкладення,
личинок вирубування кущів і дерев. Для знищення дорослих
мух застосовують інсектициди і липучки.
Географічне поширення: західні регіони Африки.
Родина ТаЬапісІае — ґедзі
Ґедзь сірий — ТаЬапиз Ьготіпез
Ґедзь вузьколобий — НуЬотііга зоїзііііаііз
Місце паразитування: поверхня тіла людини і тварин.
Морфоанатомічні особливості: ґедзі — це великі мухи,
довжина тіла яких 8—ЗО мм, на голові є короткі тричленисті
вусики, колючо-ріжучий ротовий апарат, нижньощелепні щу
пики і складні фасеткові очі, забарвлені в синій, зелений та
інші кольори. Крила широкі, прозорі, іноді з темним рисунком.
Ноги помірної довжини. На гомілках середньої пари ніг є ко
роткі щетинки (так зва ні шпори). Черевце сірувате, останні
сегменти черевця самців і самок видозмінені в копулятивні
органи.
Цикл розвитку: розвиток із повним перетворенням. Самка
відкладає 400— 1000 яєць на нижню поверхню листків або
стебел, які ростуть на берегах водойм. Із яєць виходять личин
ки, які 5—6 разів линяють, зариваються у мул і там зимують.
Навесні личинки знову линяють і перетворюються в лялечку,
з якої протягом 5—ЗО днів виходить дозрілий ґедзь. Живлять
ся ґедзі соком рослин, а самки після запліднення мусять на
смоктатися крові, необхідної для розвитку яєць.
Епідеміологічне значення: при укусах самки ґедзів у рану
потрапляє гемолізин, який руйнує еритроцити, з рани довго
тече кров, яку, ймовірно, злизують самці. При укусах ґедзі
переносять збудників туляремії, сибірки, піроплазмозів. У краї
нах Африки вони є переносниками філярії — Ьоа Іоа. Завдають
шкоди тваринництву, в лісотундрі спричиняють до смерті
оленів.
Заходи боротьби та профілактика: знищення личинок
шляхом гідромеліоративних заходів, використання інсектици
дів, а також обробка шерсті тварин пестицидами. Використання
Розділ 6
4 22
природних ворогів — ос, які ловлять ґедзів під час лету, їзд
ців.
Географічне поширення: всі кліматично-географічні зони.
Овод кінський — У а з і г о р к і ї и з і п і е з і і п а ї і з
Овод бичачий — Н і р о с і е г т а Ь о о із
Місце паразитування личинок: поверхня тіла людини і тва
рин.
Морфоанатомічні особливості: оводи відрізняються від ґе
дзів тим, що відкладають свої личинки урани на тілі тварин.
Цикл розвитку: із повним метаморфозом.
Епідеміологічне значення: проковтування людиною з їжею
личинок оводів викликає кишковий міаз, уринальний міаз
розвивається у випадку, коли овод відкладає личинки на бруд
ну білизну, які потім проникають у сечові шляхи. Оводи також
можуть відкладати личинки на волосся людини, які проника
ють у шкіру і викликають свербіж.
Профілактика: використовують ті ж самі заходи, що і для
боротьби з ґедзями.
Географічне поширення: усі географічні зони.
Мокреці — Родина
С е га іо § о п іс Іа е
Місце паразитування: поверхня тіла людини.
Морфоанатомічні особливості: родина включає три роди
СиІісоМе8, Ееріосопорз і Еазіокеїеа. Основна маса кровосисних
мокреців належать до роду Сиіісоідез. Це найдрібніші з літа
ючих кровоносних комах, розміром 1,0—2,5 мм. Голова підіг
нута під тулуб, довгий хоботок, по боках його є довгі вусики,
густо покриті у самців хітиновими волосками, і щупики. Колір
хітину темний. На кінці черевця — добре розвинуті статеві
придатки. На крилах є темні плями.
Цикл розвитку: місцями розмноження мокреців є різні
водойми, вологий ґрунт, лісові підстилки, дупла дерев. Розви
ток із повним перетворенням.
Епідеміологічне значення: самки нападають на людину і тва
рин зранку і ввечері. При масових укусах на шкірі виникають
Медична паразитологія
423
запальні реакції, які супроводжуються сильним свербежем.
Мокреці — механічні переносники збудника туляремії, а у тро
піках — філярій.
Заходи боротьби та профілактика: заходи боротьби такі
ж, які проводяться для знищення комарів.
Географічне поширення: всі материки і всі кліматично-гео
графічні зони, крім Арктики.
Мошки — Родина Зіт иіійае
Мошка —
З іт и ііи т о г п а іи т
Місце паразитування: поверхня тіла людини і тварин (екто
паразит).
Морфоанатомічні особливості. Дрібні комахи, розміром
З—6 мм, колір тіла чорний. На голові у самців фасеткові очі
стикаються, а у самок роздвоєні лобовою смуж кою, вусики
багаточленисті, ротовий апарат колючо-смоктального типу. Від
грудей знизу відходять три пари коротких ніг, а зверху пара
широких безбарвних крил ідзиж чальця. Черевце коротке
і широке, складається із дев’ яти сегментів.
Цикл розвитку: самки відкладають яйця уводу на берегах
річок і струмків (каміння, листки рослин). Кількість личинко
вих стадій коливається від 3 до 6, які в коконі перетворюють
ся на лялечки. З лялечок формується імаго і випливає на по
верхню води.
Епідеміологічне значення: при масовому нападанні у людей
можуть виникати симптоми загального отруєння. Переносник
збудників туляремії та онхоцеркозу.
Заходи боротьби та профілактика: використання інсекти
цидів, а для індивідуального захисту— репеленти.
Географічне поширення: у Сибіру, Поволжі, тропічній Аф
риці та Центральній Америці.
Розділ 6
4 24
Родина Комарі — СиІісШае
Комар звичайний—
С и іе х р ір іе п з
(табл. 6.1)
Таблиця 6.1. Відмінності між звичайним комаром (піскуном)
і малярійним комаром
Немалярійний — Сиіех
Малярійний —
АпорКеІез
Щупики
самки
Щупики коротші, ніж хоботок,
у кілька разів
Довжина щупиків
така сама, як хоботка
Щупики
самця
Довжина щупиків
Довжина щупиків дорівнює
хоботку, без потовщень на кінці дорівнює хоботку
з потовщеннями на
кінці
Щиток
Трилопатевий з трьома групами Напівокруглий
волосків
Положення
в спокої
Паралельно до поверхні води
Під кутом до поверхні
води
Яйця
Без поплавків, є багато яєць,
склеєних між собою
Кожне яйце має з двох
боків поплавки, часто
поодинокі
Дихальний
сифон
личинок
Є дихальний сифон
Немає дихального
сифона
Положення
личинки
У ВОДІ
Під кутом до поверхні води
Паралельно до
поверхні води
Дихальні
трубки
лялечок
Форма вузьких циліндрів
Форма лійок
Відмінності
Морфоанаболічні особливості: самки маленька головка
з великими очима, довгими 15-членистими вусиками, хоботком
і щупиками. Нижня губа утворює хоботок, у якому містяться
верхні та нижні щелепи і язик. З боків основи хоботка розміщені
членисті паличкоподібні щупики, які коротші від хоботка
у 3— 4 рази. На випуклих грудях є прозорі крила без плям,
Медична паразитологія___________________________________________ 4 2 5
дзижчальця у вигляді булавоподібних виростів і три пари дов
гих кінцівок із кігтиками. Черевце дев’ ятичленисте. Тіло
вкрите бурими і жовтими лусочками. На крилах шоста жилка
впадає у п’ яту. Щиток задньогрудей трилопатевий. У самців
вусики вкриті довгими волосками. Щупики довші від хоботка,
всі членики однакової товщини.
Цикл розвитку (рис. 6.26): гладенькі овальні яйця склеєні
по 100— 200 шт. І розташовуються на поверхні води у вигляді
човника. У личинок розрізняють голову, груди і черевце. На
голові є паличкоподібні вусики і ротові органи. Від усіх сег
ментів тіла відходять волоски. Черевце дев’ ятисегментне, від
якого відходить дихальна трубка — сифон. Тіло лялечки скла
дається із потовщеної передньої частини, яка відповідає головогрудям, і членистої задньої частини— черевця у вигляді
коми. У ділянці головогрудей розташовані парні, довгі цилінд
ричні дихальні трубки.
Епідеміологічне значення: преносник віруса японського
енцефаліту.
Географічне поширення: по всій Україні біля водойм.
Комар малярійний— АпорНеІез тасиїіреппіз (табл. 6.1)
Морфоанатомічні особливості: комар малярійний за будо
вою подібний до звичайного, але на крилах має темні плями,
шоста жилка крил не впадає у п’ яту. Кінцівки довші від тіла
в 2,5 раза. Щ иток задньогрудей напівокруглої форми.
У самок однакова довжина щупиків і хоботка, а у самців
щупики закінчуються булавоподібними потовщеннями і також
однакової довжини з хоботком.
Цикл розвитку (рис. 6.26): яйця чорного кольору, поодинокі,
овальні з повітряними камерами по боках, довжиною до 1 мм.
На голові личинки видно складні очі і паличкоподібні вусики.
Груди широкі, черевце дев’ ятисегментне. На грудях і черевці
є пучки великих щетинок. На передостанньому сегменті роз
міщений дихальний отвір, який веде у трахею. У лялечки є
дихальна трубка у вигляді конуса.
Епідеміологічне значення: переносник збудників різних форм
малярії.
Розділ 6
426
Апоркеїез тасиїіреппіз
Сиіех ріріепз
Рис. 6.26. Стадії розвитку комарів роду АпорЬеІез і Сиіех:
1 — яйце; 2 — личинка; 3 — лялечка; 4 — імаго; 5 — ротові при
датки: а — хоботок; б — вусики; в — щупики
Медична паразитологія
427
Географічне поширення: біля водойм. В Україні комарі не
переносять збудника малярії, бо в більшості районів вегетатив
ний період короткий для того, щоб здійснився цикл розвитку
Ріазтодіит.
Родина Метелівкові — РзусНоЛіЛае
М оскіт— РКІеЬоіотиз рараіазіі (рис. 6.27)
Місце перебування: епідерма шкіри лю
дини.
Морфоанатомічні особливості: розміри
тіла 1,5— 3,0 мм, колір жовто-коричневий.
Очі великі, чорного кольору, вусики багаточленисті, колючо-сисний ротовий апа
рат. Тіло і крила вкриті волосками, які
Рис. 6.27. Москіт
надають москітові пухнастого вигляду.
Черевце десятисегментне, на кінці його у самок є сперматека
(сім’яприймач). Крила широкі, загострені на верхівці. Кінцівки
довгі й тонкі. Літає “стрибками” . Кров смокчуть лише самки.
Термін життя — близько одного місяця.
Цикл розвитку, повний метаморфоз, розвиток не пов’язаний
з водою. Самки відкладають яйця два-три рази протягом жит
тя у норах гризунів, у стайнях для худоби, печерах, гніздах
птахів, тріщинах підлоги, вологому і захищеному від сонця
смітті. Личинка кілька разів линяє, перетворюється на лялечку,
а з лялечки виходить ім аго, яке ж ивиться к р о в ’ ю най
різноманітніших тварин, — ящ ірок, гризунів, їжаків. Три
валість розвитку 35—45 днів.
Епідеміологічне значення: переносник лейшманій — збуд
ників дерматотропного і вісцерального лейшманіозів, пропас
ниці папатачі.
Географічне поширення: південні області України.
Заходи боротьби та профілактики: обробка приміщення
і нір гризунів, де водяться москіти, отрутохімікатами, бороть
ба з гризунами і бездомними собаками, своєчасне вивезення
сміття і знезараження смітників.
Лікарські засоби для боротьби з двокрилими: інсектициди,
рекеленти.
4 28
Розділ 6
Ряд Воші — А п о р іи га
Воша головна — Редісиїиз Нитапиз саріїіз (рис. 6.28)
Рис. 6.28. Воші та їхні яйця (гниди): 1 — головна воша (самець
і самка); 2 — одежна воша (самець); 3 — лобкова воша (самка).
Місце перебування: волосяна частина голови.
Морфоанатомічні особливості: тіло сплющене, ромбоподіб
ної форми, розміри 2—4 мм, очі прості, ротовий апарат колю
чо-сисний, колір тіла сірий, вусики короткі й товсті. Сегменти
грудей злиті, немає крил, три пари кінцівок, які закінчують
ся кігтиками. Черевце складається зі 7 сегментів, останній із
них у самки роздвоєний, а у самця заокруглений, видно копулятивний апарат. Вирізки між сегментами черевця глибокі.
Цикл розвитку: неповний метаморфоз, стадія лялечки від
сутня. Самка відкладає щодня 10— 15 яєць, прикріплює їх до
волосинки спеціальним секретом. Яйця називаються гнидами,
вони білого кольору, видовжені, розміром до 1 мм. Розвиток
від яєць до статевої зрілості триває 7— 14 днів. Після линяння
перетворюється на статевозрілу форму. Тривалість життя 1,5 мі
сяця. За цей час головна воша відкладає до 150 гнид.
Епідеміологічне значення: збудник педикульозу, переносник
збудників поворотного тифу (спірохет Обермейєра), які потрап
ляють у порожнину тіла воші і там розмножуються. У слинні
залози збудники не потрапляють, тому при укусі вошею людина
не заражається. Зараження відбувається при розчавлюванні
Отруйні організми. Лікарські рослини______________________________ 429
воші на тілі, коли вивільнені спірохети потрапляють через
подряпини на шкірі (контомінація).
Географічне поширення: по всій Україні.
Заходи боротьби та профілактики: профогляди серед вели
кого скупчення людей (школи-інтернати, літні табори, у вій
ську серед новобранців); забезпечення належних санітарногігієнічних умов, саносвітня робота серед населення.
Воша одежна— Редісиїиз Уіитапиз Нитапиз (рис. 6.28, 2)
Місце перебування: тіло та одяг людини.
Морфоанатомічні особливості: розміри 2,1—4,75 мм, рото
вий апарат колючо-сисний, вусики довгі й тонкі, вирізки на
черевці неглибокі.
Цикл розвитку: неповний метаморфоза, відбувається так,
як у головної воші, за життя самка відкладає 300 гнид. Розви
ток триває до 16 днів.
Епідеміологічне значення: переносник збудників поворотно
го тифу (спірохет Обермейєра) та висипного тифу (рикетсій
Провачека), які інтенсивно розмножуються у клітинах кишко
вого епітелію. При цьому клітини епітелію руйнуються, рикетсії
потрапляють у порожнину кишківника воші, а потім виділя
ються з її випорожненнями та через подряпини на шкірі про
никають у кров. Вони можуть потрапляти у кров також при
розчавлюванні воші на тілі.
Географічне поширення: по всій Україні.
Заходи боротьби та профілактики: особиста гігієна, проф
огляди серед великого скупчення людей (школи-інтернати,
літні табори, у війську серед новобранців); забезпечення на
лежних санітарно-гігієнічних умов, саносвітня робота серед
населення. Для обробки одягу використовують 0,15 % емульсію
карбофосу, 5 % піретрум тощо.
Воша лобкова — Ркіігиз риЬіз (рис. 6.28, 3)
Місце перебування: лобок, борода, вуса, вії (крім волосяної
частини голови).
Розділ 7
430
Морфоанатомічні особливості: тіло вкорочене, завужене
ззаду, не виражена межа між грудьми та черевцем, розмір
1,0— 1,5 мм. Голова має форму видовженого прямокутника.
З боків черевця бородавки з волосками.
Цикл розвитку: неповна метаморфоза. Самка відкладає на
волосся яйця, приклеюючи їх білим секретом. Із гниди вилуп
люється личинка, живиться кров’ ю і після кількох линянь
перетворюється в імаго. Живуть до 1,5 місяця. За життя самка
відкладає 50 яєць.
Епідеміологічне значення: є збудником фтиріазу.
Географічне поширення: по всій Україні.
Заходи боротьби та профілактики: систематичне миття
тіла не рідше одного разу на тиждень з одночасною заміною
білизни (постільної й натільної); зголювання волосся на лоб
ковій ділянці та під пахвами, втирання в пошкоджені ділянки
шкіри 10 % білої ртутної мазі, 5 % борної мазі, лосьйону Нітштоф; попередження тісних контактів із завошивленими людь
ми. Дотримання правил особистої гігієни, санітарно-освітня
робота серед населення.
Ряд Блохи — А р К а п ір іега
Блоха лю дська— Риіех іггііапз (рис. 6.29)
Місце перебування: тіло людини.
:Ж * .
з
Рис. 6.29. Людська блоха: 1 — вусики; 2 — очі; 3 — стилети;
4 — пігідій; 5 — стигми; 6 — задні кінцівки
Отруйні організми. Лікарські рослини______________________________ 431
Морфоанатомічні особливості: тіло коричневого кольору,
сплющене з боків, розміром 0 ,5 — 5,0 мм. На голові є прості очі,
короткі вусики, ротовий апарат колючо-сисний, третя пара
кінцівок добре розвинена (стрибальні). Добре помітні сегменти
грудей, крил немає. Черевце складається із 10 сегментів. На
кінці черевця є орган чуття — пігідій, у самців копулятивний
орган, у самок — сім ’яприймач. Усе тіло вкрите щетинками.
Цикл розвитку: відбувається з повним метаморфозом: яй
це -» личинка —> лялечка -> імаго. Самка відкладає 450 яєць
у норах гризунів, у сухому смітті, у щілинах підлоги в примі
щеннях. Личинки вилуплюються з яєць і живляться речовина
ми, що розкладаються. Мають вигляд білих черв’ ячків, линяють,
перетворюються на лялечок, із яких вилуплюються коричневі
статевозрілі блохи, що нападають на людину. Цикл розвитку
триває 250—270 днів. Тривалість життя до 1,5 року.
Епідеміологічне значення: переносить збудників чуми і ту
ляремії.
Географічне поширення: по всій Україні.
Заходи боротьби та профілактики: дотримання санітарногігієнічних умов у приміщеннях. Замащування щілин у підлозі
між дошками. Боротьба з гризунами у підвалах і з бездомними
котами, які часто проживають на горищах або у підвалах, ви
користання хлорофосу й інших хімічних засобів.
Ряд Напівтвердокрилі— Н е т ір іе га ( Н е іе г о р іе г а )
Блощиця постільна —
С іт е х Іе с іи іа гіи з
Місце паразитування: ліжка, під плінтусами, у щілинах
підлоги
Морфоанатомічні особливості: тіло досягає 4— 5 мм. Самку
від самця можна відрізнити за формою черевця. У самки воно
симетричне, із заднього кінця округле, у самця загострене, має
неправильну форму, помітно серповидний копулятивний орган.
Червоно-коричневого кольору, сплющене в спинно-черевному
напрямку. Біля задніх кінцівок є пахучі залози.
Розд'п 7
432
Цикл розвитку: розвиток із неповним метаморфозом. Сам
ка, після того як насмокталася крові, відкладає кількасот яєць
білого кольору. Кладку яєць можна знайти у ліж ках, на
дерев’ яних, металевих або м ’ яких частинах, на килимах біля
ліжок, на рамах картин. Розвиток із неповним метаморфозом.
Статевозрілі блощиці у пошуках їжі пересуваються в інші при
міщення через плінтуси, вікна, по щілинах у підлозі. Швидкість
пересування 1 м/хв.
Епідеміологічне значення: укус статевозрілої блощиці викли
кає свербіж, набряк, а часом і алергічні явища. Збудників
хвороб не переносить.
Географічне поширення: по всій Україні.
Заходи боротьби та профілактика. Для знищення блощи
ць використовують інсектициди, яйця знищують розчином
оцтової кислоти. У містах, особливо у старих приміщеннях,
обов’ язкові профілактичні огляди меблів, килимків.
Поцілуйковий клоп —
Т гіа іо т а т е£ із іи з
Місце паразитування: нори диких і домашніх гризунів,
гнізда птахів, приміщення для перебування тварин.
Морфоанатомічні особливості: розміри тіла 1,5—3,5 см.
Зверху тіло прикрите складеними крилами. По краях тіла
характерний рисунок. Спереду виступають довгі вусики.
Цикл розвитку: розвиток із неповним перетворенням. На
всіх стадіях розвитку клоп живиться кров’ ю. Нападаючи на
людину кусає переважно у місцях переходу шкіри в слизову
оболонку (біля очей або в губи). Після живлення на місце уку
су виділяє фекалії з трипаносомами, які занурюються в ранки
від укусу.
Епідеміологічне значення: переносник збудника хвороби
Чагаса (Тгірапозота сгигі).
Профілактика: покращення соціально-побутових умов,
обладнання приміщень для тварин, боротьба з гризунами.
Географічне поширення: країни Центральної та Південної
Америки.
Отруйні організми. Лікарські рослини_________
4 33
Ряд тарганові — ВІаііоіЛеа
Тарган чорний — В і а і і а о г і е п і а і і з
Тарган рудий — В і а і і е і і а § е г т а п і с а
Морфоанатомічні особливості. Тіло овальне, видовжене,
сплюснуте у спинно-черевному напрямку. Ротовий апарат гри
зучого типу. Мають дві пари крил, верхні щільні. Швидко
бігають. Живуть у житлах людей.
Цикл розвитку: розвиток із неповним метаморфозом. Сам
ки відкладають до 50 яєць у капсулах, так званих оотеках, які
носять на собі. Із капсул виходять личинки, які за 5—6 місяців
перетворюються на дорослих комах. Живляться таргани хар
човими продуктами, які вживають люди, та різноманітними
відходами.
Епідеміологічне значення: чорні таргани можуть кусати
людей. Таргани механічно переносять на своїх кінцівках бак
терії, найпростіших, їх цисти та яйця гельмінтів. Псують їжу,
залазять через найменші щілини у посуд, поїдають приготова
ну їжу.
Заходи боротьби та профілактика: забезпечення належних
санітарно-гігієнічних умов життя людини, санітарна робота
серед населення, боротьба з тарганами шляхом використання
інсектицидів.
Географічне поширення: по всій Україні.
ЛІТЕРАТУРА
1. Айала Ф.Дж., Кайзер Д. Современная генетика: В 3 т. —
М.: Мир, 1987.
2. Атраментова Л .А. Генетика человека: Учеб. пособ. —
Харьков, 1990.
3. Бажора Ю. І. Імуногенетика. — О.: Чорномор’ я, 2000.
4. Бердишев Г. Д., Криворучко И. Ф. Медицинская гене
тика. — К.: Вища школа, 1990.
5. Биология / Под ред. В. Н. Ярьігина. — М.: Медицина,
1999.
6. Биология / Под ред. К. А. Татаринова. — Л.: Вища шко
ла, 1983.
7. Бочков Н. П. Клиническая генетика. — М.: Медицина,
1997.
8. Бочков Н. П., Чеботарев А. Н. Наследственность чело
века и мутагени внешней средьі. — М.: Медицина, 1989.
9. Бужієвська Т. І. Основи медичної ген ети к и .— К .:
Здоров’я, 2001.
10. Дубинин Н. П. Общая генетика. — М.: Наука, 1986.
11. Захаров А. Ф., Бенюш В. А., Кулешов Н. П. Хромосоми
человека: Атлас. — М.: Медицина, 1982.
12. Збірник задач і вправ з біології / За ред. А. Д. Тимченка. — К.: Вища школа, 1992.
13. Зенгбуш П. Молекулярная и клеточная биология:
В 3 т. — М.: Мир, 1982.
14. Инге Вечтомов С. Г. Введение в молекулярную генети
ку. — М.: Внісшая школа, 1983.
Отруйні організми. Лікарські рослини______________________________ 435
15. Инге-Вечтомов С. Г. Генетика с основами селекции. —
М.: Внісшая школа, 1989.
16. Королев В. А., Кривошеина Г. Н., Полякова 9. Г. Руководство к лабораторньїм занятиям по биологии. — К.: Вища шко
ла, 1986.
17. Ленц В. Медицинская ген ети к а .— М .: Медицина,
1984.
18. Лильин Е. Т., Богомазов Е. А ., Гофман-Кадошников
П.Б. Генетика для врачей. — М.: Медицина, 1990.
19. Лильин Е. Т., Савицкая Т. В., Захарова О. М . Пособие
для изучающих медицинскую и клиническую генетику. — М.:
Медицина, 1996.
20. Мендель Г. О пьітьі над растительньїми гибридами. — М.:
Наука, 1965.
21. Мерфи 3. А., Чейз Г. А. Основні медико-генетического
консулнтирования. — М.: Медицина, 1979.
22. Наследственние болезни / Под ред. Л. О. Бадаляна. —
Ташкент: Медицина, 1980.
23. Никитин Ю. П ., Лисиченко О. В., Коробкова Е. Н.
Клинико-генеалогический метод в медицинской генетике. —
Новосибирск: Наука, 1983.
24. Перспективи медицинской генетики / Под ред. Н. П. Боч
кова. — М.: Медицина, 1982.
25. Пішак В. 17., Бойчук Т. М ., Бажора Ю. І. Клінічна
паразитологія з основами лабораторної діагностики. — Чернів
ці: Мед. академія, 2001.
26. Пішак В. 17., Нечитайло Ю. М . Гелнмінтози у дітей. —
Чернівці: Прут, 2000.
27. Пішак В. 17. Паразитарні хвороби. — Чернівці: Прут,
1998.
28. Плозинский Н.А. Математические методи биологии. —
М.: Изд-во Моск. ун-та, 1978.
29. Пузирев В. 77., Степанов В. А. Патологическая анатомия
генома человека. — Новосибирск: Наука, 1997.
30. Руководство к лабораторним занятиям по биологии /
Под ред. Ю. К. Богоявленского. — М.: Медицина, 1988.
31. Служинська 3. О., Калинюк 17. 17., Служинська О. Б.
Спадковістн людини (хромосомні та генні хвороби). — Л.,
1997.
4 36
Література
32. Слюсарев А. О., Жукова С. В. Біологія. — К.: Вища
школа, 1992.
33. Стент Г., Кзлиндар Р. Молекулярная генетика. — М.:
Мир, 1981.
34. Тимченко А. Д. Биология с основами генетики и паразитологии. — К., 1982.
35. Федченко С. Н., Романюк Б. П. Современньїе методьі
диагностики наследственньїх болезней. Задачи медико-генетического консультирования. — Луганск: Злтон-2, 2000.
36. Фогель Ф., МотульскиА. Генетика человека: В 3 т. — М.:
Мир, 1989.
Навчальне
видання
ВОРОНЕЦЬ Зіновій Дмитрович,
ЧУПАІПКО Оксана Ярославівна,
СЕРГІЄНКО Людмила Михайлівна
та інші автори
БІОЛОГІЯ
Навчальний посібник
Обкладинка О. Куця
Підп. до друку 03.11.2009. Формат 60x90 ‘/іеПапір офс. Гарнітура ЗсЬооІВоокСТТ. Друк офс.
Ум. друк. арк. 27,5. Обл.-вид. арк. 25,1. 3ам.№3835
Видавництво “Знання”
01034, м. Київ, вул. Стрілецька, 28.
Свідоцтво про внесення до Державного реєстру
видавців, виготівників і розповсюджувачів видавничої продукції
ДК № 3596 від 05.10.2009.
Тел.: (044) 234-80-43, 234-23-36
Е-таі1: 8а1ев@2паппіа.сот.иа
М ір:/ /
/ . гпаппіа .с о т .иа
ууугу
В Україні книгу м ож на придбати за адресами:
м. Київ, вул. М. Груш евського, 4, маг. “ Наукова дум ка” ,
тел. (0 4 4 )2 7 8 -0 6 -9 6
м. Київ, вул. Х рещ атик, 44, маг. “ Знання” , тел. (044) 234-22-91
м. Київ, вул. Ярославів Вал, 17а, маг. “ А бзац ” , тел. (04 4 ) 234-42-86
м. Київ, просп. М осковський, 6, маг. “ Будинок книги та медіа” ,
тел. (0 4 4 )4 6 4 -4 9 -7 0
м. Вінниця, вул. Привокзальна, 2 /1 , маг. “ Кобзар” ,
тел. (0432) 61-77-44
м. Д ніпропетровськ, вул. Глінки, 24, ТЦ “ М осковськи й ” ,
тел. (056) 726-53-14
м. Донецьк, вул. Артема, 147А, “ Б удинок книги” ,
тел. (0 6 2 )3 4 3 -8 9 -0 0
м. Ж итом и р, вул. Київська, 1 7 /1 , маг. “ Знання” , тел. (0412) 47-27-52
м. З апоріж ж я, просп. Леніна, 142, маг. “ Спеціальна кн ига” ,
тел. (0612) 13-85-53
м. Івано-Ф ранківськ, Вічовий майдан, 3, маг. “ Сучасна україн ська книга” ,
тел. (03422) 3 -04-60
м. К іровоград, вул. Набережна, 13, маг. “ Книж ковий св іт” ,
т е л .(0 5 2 2 )2 4 -9 4 -6 4
м. Кривий Р іг, пл. Визволення, 1, маг. “ Б укініст” ,
тел. (0564) 9 2 -37-32
м. Л уганськ, вул. Радянська, 58, маг. “ Глобус-книга” ,
тел. (0642) 53-62-30
м. Л уцьк, проси. Волі, 41, маг. “ Знання” , тел. (0332) 77-00-46
м. Львів, просп. Ш евченка, 16, маг. “ Н оти ” , тел. (0322) 61-19-64
м. Львів, просп. Ш евченка, 8, маг. “ У країнська книгарня” ,
тел. (0322) 72-16-30
м. М иколаїв, просп. Леніна, 122, маг. “ К обзар” , тел. (05 1 2 ) 55-20-51
м. Одеса, вул. Буніна, 33, маг. “ Б удинок книги” , тел. (04 8 2 ) 32-17-97
м. Полтава, вул. Ш евченка, 29, маг. “ Будинок книги та медіа” ,
тел. (0 5 3 2 )6 1 -2 6 -7 6
м. Рівне, вул. Соборна, 57, маг. “ С лово” , тел. (0362) 2 6 -94-17
м. Тернопіль, вул. Й. Сліпого, 1, маг. “ Дім книги” ,
тел. (0352) 43-03-71
м. Тернопіль, вул. Ч орновола, 14, маг. “ Книж кова хата” ,
тел. (0352) 52-24-33
м. У ж город, пл. Корятовича, 1, маг. “ К обзар” , тел. (03 1 2 2 ) 3-35-16
м. Х арків, вул. П уш кінська, 74, маг. “ Л ексика” , тел. (0 5 7 ) 717-60-16
м. Х арків, вул. Сумська, 51, маг. “ В оок з” , тел. (057) 7 14-04-70,
714-04-71
м. Х ерсон , вул. Леніна, 1 4 /1 6 , маг. “ Книж ковий ряд” ,
тел. (0 5 5 2 )2 2 -1 4 -5 6
м. Х м ельницький, вул. П одільська, 25, маг. “ К ниж ковий св іт ” ,
тел. (0382) 79-25-59
м. Ч еркаси, вул. Б. Виш невецького, 38, маг. “ Світоч” ,
тел. (0 4 7 2 )3 6 -0 3 -3 7
м. Ч ернівці, просп. Н езалеж ності, 90, маг. “ Будинок книги та медіа” ,
тел. (03722) 3-42-70
м. Ч ернігів, просп. М иру, 45, маг. “ Будинок книги” ,
тел. (0 4 6 2 6 )9 -9 2 -6 2
К ниготорговельним організаціям та оптови м покупцям звертати ся за тел.:
(044) 5 3 7 -6 3 -61 , 537-63-62; ф акс: 235-00-44.
Е -таі1: ва 1 ев@ гп а п п іа.сот.и а
ВИДАВНИЦТВО “ЗНАННЯ” ПРОПОНУЄ
Батлук В. А. Радіаційна екологія: Навч. посіб. — К.: Знан
ня, 2009. — 309 с. — Мова укр. — Формат 70х100У і6Пал. тв.
І8ВМ 978-966-346-707-8
У н а в ч а л ь н о м у п о с іб н и к у р о з г л я н у т о іс т о р ію в и н и к н е н н я р а д іа ц ій н о ї
е к о л о г ії, р о з т л у м а ч е н о , ч о м у р а д іа ц ія є о д н іє ю з н а й с у т т є в іш и х п р о б л е м
л ю д с т в а . П о к а з а н о , в я к и й с п о с і б і за д о п о м о г о ю я к и х п р и л а д ів з д ій с
н ю є т ь с я о ц ін ю в а н н я р а д іа ц ій н о ї о б с т а н о в к и . З о с е р е д ж е н о у в а г у н а о с о б
л и в о с т я х в е д е н н я р о с л и н н и ц т в а і т в а р и н н и ц т в а н а з е м л я х , я к і за зн а л и
д ії р а д іо а к т и в н о г о з а б р у д н е н н я . П о р у ш е н о п и т а н н я щ о д о м о ж л и в о с т і
з б е р е ж е н н я та з а х о р о н е н н я р а д іо а к т и в н и х в ід х о д ів . П р е д с т а в л е н о д о в ід
к о в і м а т е р іа л и , щ о д а ю т ь з м о г у о ц ін и т и н а с л ід к и я д е р н и х в и б у х ів і с п р о г н о з у в а т и р ів е н ь з а б р у д н е н н я с іл ь с ь к о г о с п о д а р с ь к о ї п р о д у к ц і ї н а т е р и
т о р ія х , у р а ж е н и х у р е з у л ь т а т і р у й н у в а н ь а б о а в а р ій А Е С і п ід п р и є м с т в
а т о м н о ї п р о м и с л о в о с т і.
П р и з н а ч е н о д л я с т у д е н т ів е к о л о г іч н и х с п е ц іа л ь н о с т е й , а т а к о ж у с іх ,
х т о ц ік а в и т ь с я п р о б л е м а м и р а д іа ц ій н о ї е к о л о г ії.
Книготорговельним організаціям та оптовим покупцям
звертатися за тел.: (044) 537-63-61, 537-63-62, 234-80-43.
Е-шаіІ: за1е8@2паппіа.сот.иа
ЬІІр://уу\у\у.2 паппіа.сот.иа
\ллллл/.2МАММІА.сот.иа
і
І
Широкий вибір навчальної та ділової літератури
Тел. для довідок: (044) 235-00-44, 234-80-43
—
.
ВИДАВНИЦТВО “ЗНАННЯ” ПРОПОНУЄ
М и ха л іч к о Б. М. Курс загальної хімії. Теоретичні основи:
Навч. посіб. — К.: Знання, 2009. — 548 с. — Мова укр. — Фор
мат 70х100у16- — Пал. тв.
І8ІШ 978-966-346-712-2
У н а в ч а л ь н о м у п о с іб н и к у з у р а х у в а н н я м д о с я г н е н ь с у ч а с н о ї н а у к и
та з а л у ч е н н я м ф а к т о л о г іч н о г о м а те р іа л у н е о р г а н іч н о ї, е л е м е н т о о р г а н іч
н о ї, о р г а н іч н о ї х ім ії і к р и с т а л о х ім ії в и ч е р п н о в и к л а д е н о о с н о в н і п и т а н н я
з а г а л ь н о ї х ім ії: а т о м н о -м о л е к у л я р н е в ч е н н я т а х ім іч н а с т е х іо м е т р ія , п и
т а н н я б у д о в и а т о м ів і м о л е к у л , х ім іч н и х з в ’ я з к і в , к р и с т а л іч н о г о с т а н у
р е ч о в и н , к о о р д и н а ц ій н и х с п о л у к , з а к о н о м і р н о с т і п е р е б іг у х і м і ч н и х р е
а к ц ій , о с о б л и в о с т і в з а є м о д ії р е ч о в и н у р о з ч и н а х , о к и с н о - в ід н о в н і п р о ц е
си т а о с н о в и е л е к т р о х ім ії.
П р и з н а ч е н о д л я с т у д е н т ів у н ів е р с и т е т ів , а т а к о ж м о ж е б у т и к о р и с
н и м а с п ір а н т а м , н а у к о в и м п р а ц ів н и к а м т а в с ім , х т о б а ж а є п о п о в н и т и
з н а н н я з о с н о в х ім ії.
Книготорговельним організаціям та оптовим покупцям
звертатися за тел.: (044) 537-63-61, 537-63-62, 234-80-43.
Е-таіі: ва1ев@2паппіа.сот.иа ЬП,р://\у\у\у.2 пашііа.сот.иа
\лллллл2МАММІА.сот.иа
і
І
Широкий вибір навчальної та ділової літератури
Тел. для довідок: (044) 235-00-44, 234-80-43
і