/


















































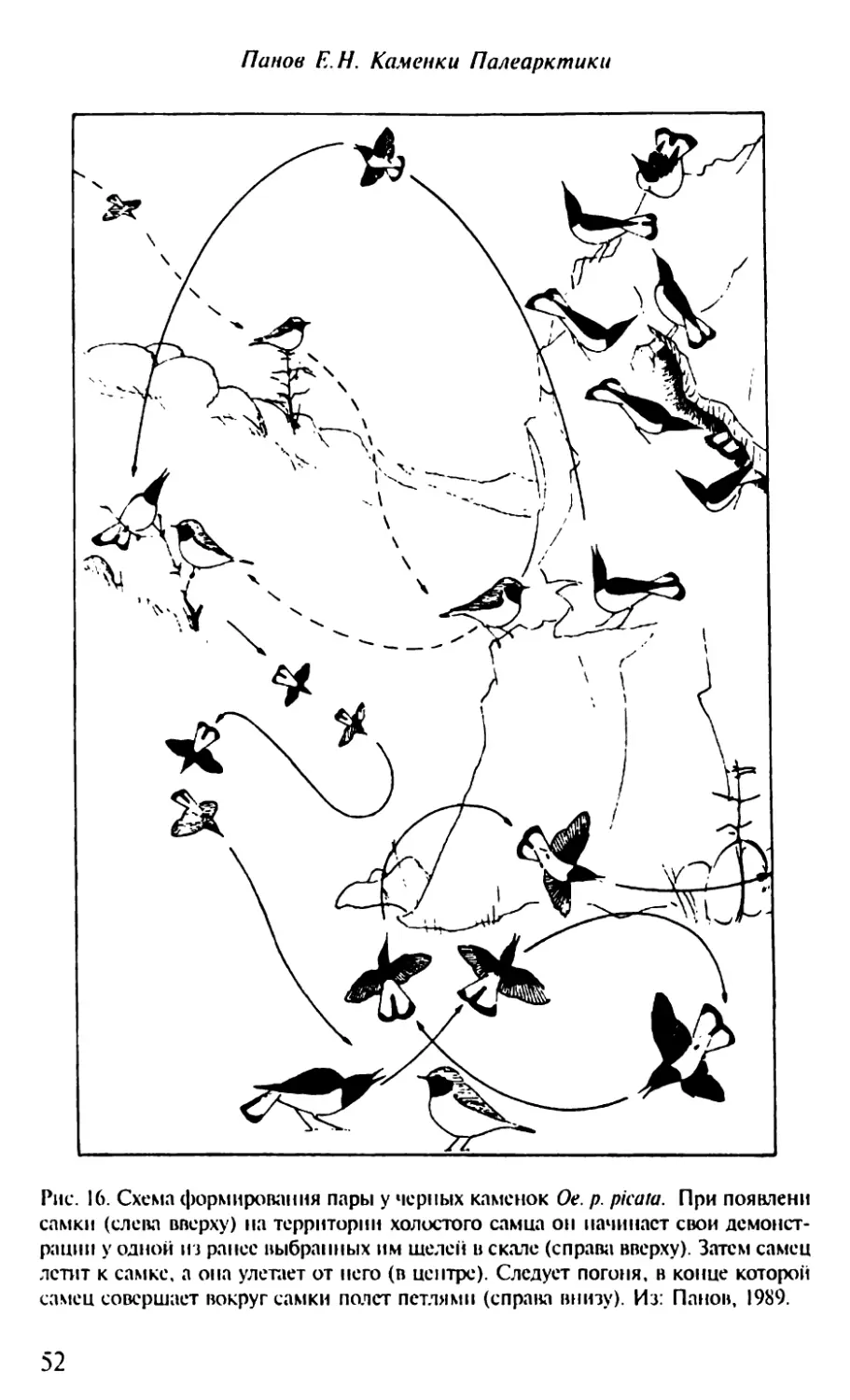




































































































































































































































































































Text
Е. Н. ПАНОВ
КАМЕНКИ
ПАЛЕАРКТИКИ
ЭКОЛОГИЯ, ПОВЕДЕНИЕ, ЭВОЛЮЦИЯ
МОСКВА 1999
Е. Н. ПАНОВ
КАМЕНКИ ПАЛЕАРКТИКИ
ЭКОЛОГИЯ, ПОВЕДЕНИЕ ЭВОЛЮЦИЯ
МОСКВА 1999
Е. Н. Панов. КАМЕНКИ ПАЛЕАРКТИКИ. ЭКОЛОГИЯ, ПОВЕДЕНИЕ, ЭВОЛЮЦИЯ
Издание осуществлено при поддержке Российского фонда фундаментальных исследований по проекту № 98-04-62018
#и
ISBN 5-87317-061-4
Оглавление
Предисловие.......................................................9
Введение.........................................................13
Глава I Некоторые общие сведения о каменках......................18
!.!. Географическое распространение и история расселения ......18
1.2. Некоторые черты морфологии...............................19
Размеры и пропорции........................................19
Окраска....................................................21
Смена нарядов, линька......................................28
Кариотипы..................................................29
1.3. Местообитания............................................31
1.4. Миграции и зимовки.......................................33
1.5. Питание и суточная активность............................34
Объекты питания и кормовое поведение.......................34
Отношение к воде...........................................37
Суточная активность........................................38
1.6. Социальное поведение и гнездовой цикл....................39
Пространственная организация дема..........................39
Выбор территорий самками...................................43
Полигиния..................................................43
Охрана семейной территории.................................44
Взаимоотношения между членами семейной пары................44
Уход за потомством.........................................45
’’Помощники"...............................................46
Территориальность на пролете и на зимовках.................46
1.7. Сигнальное поведение и коммуникация......................47
Структура коммуникативных сигналов.........................47
Саморекламирование самцов и организация взаимодействий при формировании пар.......................................49
Организация взаимодействий при копуляции...................51
Акустическая сигнализация: позывки и песни.................53
1.8. Гнезда и кладки..........................................58
Места расположения гнезд...................................58
Участие партнеров в гнсздостроении.........................59
Конструкция гнездовой постройки............................60
Величина кладки............................................62
Однократное и многократное гнездование .....................62
Окраска яиц................................................63
Насиживание................................................64
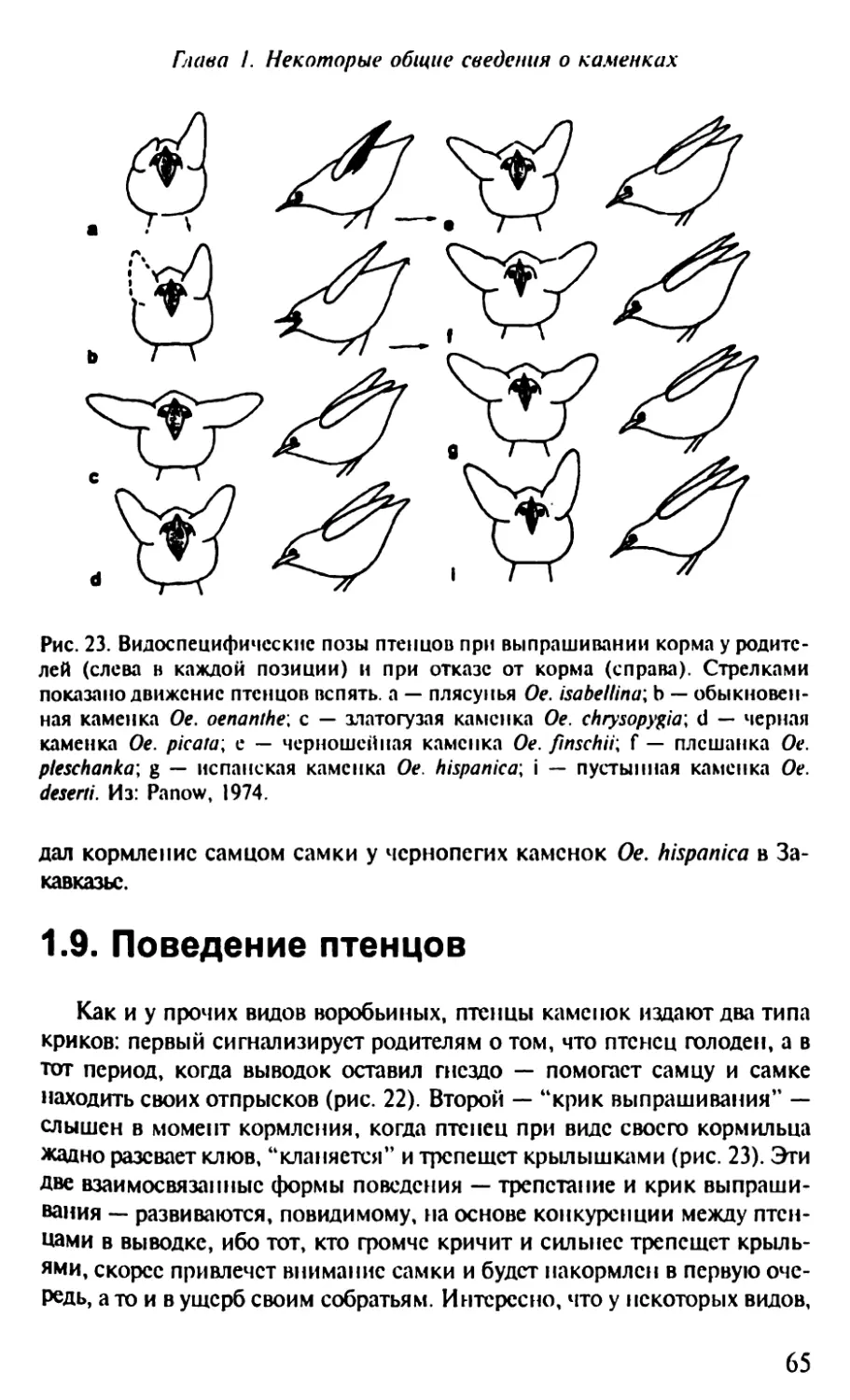
1.9. Поведение птенцов........................................65
1.10. Межвидовые отношения....................................66
Межвидовая территориальность...............................66
Межвидовое "сотрудничество" и гибридизация.................71
1.11. Враги и паразиты........................................73
1.12. Практическое значение...................................74
Панов Е.Н.
Глава 2. Каменка плясунья Oenanthe isabellinaTcmm................76
2.1. Систематические замечания................................76
2.2. Местообитания и возможная история расселения ............78
2.3. Годовой цикл и сроки гнездования.........................81
2.4. Социальное поведение.....................................83
Пространственная и демографическая структура поселений.....83
Территориальное поведение и саморекламирование самцов......85
Формирование пар...........................................88
Гне здострое и ие..........................................89
Копуляция..................................................90
Полигиния .................................................92
2.5. Гнездо, кладка, выводок..................................93
Гнездовые убежища..........................................93
Гнездо.....................................................94
Кладка.....................................................95
Насиживание и репродуктивный успех.........................95
Птенцы.....................................................96
Распадение выводка.........................................97
2.6. Линька и отлет на зимовки................................97
2.7. Биологические заметки....................................98
Глава 3. Обыкновенная каменка Oenanthe oenanthe L...............100
3.1. Систематические замечания...............................102
3.2. Местообитания...........................................103
3.3. Годовой цикл и сроки гнездования........................106
3.4. Социальное поведение....................................108
Пространственная структура поселений......................108
Территориальное поведение и саморекламирование самцов.....108
Формирование пар..........................................110
Развитие отношений между членами пары.....................Ill
3.5. Гнездо,‘кладка, выводок.................................114
Гнездовые убежиша.........................................114
Гнездо....................................................115
Кладка....................................................115
Насиживание и репродуктивный успех........................116
Птенцы....................................................117
Распадение выводка........................................118
3.6. Линька и отлет на зимовки...............................118
3.7. Биологические заметки...................................120
Глава 4. Надвид златогузая каменка: Oenanthe xanthoprymna Hempr. ct Ehr., Oenanthe chrysopygia Hume.121
4.1. Систематические замечания...............................122
4.2. Местообитания...........................................124
4.3. Годовой цикл и сроки гнездования........................125
4.4. Социальное поведение................................... 127
Территориальное поведение.................................127
Саморекламирование самцов.................................129
4
Каменки Палеарктики
Межвидовая территориальность..............................130
Формирование пар......................................... 131
Гнездостросние и конструкция гнезд....................... 132
Развитие отношений между членами пары и копуляция.........135
4.5. Кладка и выводок........................................137
Кладка....................................................137
Птенцы....................................................138
4.7. Линька и отлет на зимовки.............................. 139
4.8. Биологические заметки.................................. 140
Глава 5. Краснопоясничная каменка Oenanthe moesta Licht.........141
5.1. Систематические замечания...............................141
5.2. Местообитания и численность............................ 142
5.3. Годовой цикл и сроки гнездования........................142
5.4. Гнезда и кладки.........................................143
5.5. Биологические заметки...................................143
Глава 6. Черношейная каменка Oenanthe finschii Heug............ 145
6.1. Систематические замечания...............................146
6.2. Местообитания и численность............................ 147
6.3. Годовой цикл и сроки гнездования........................149
6.4. Социальное поведение....................................151
Территориальное поведение и саморекламирование самцов......151
Формирование пар..........................................155
Территориальное поведение самок...........................156
Гнездостросние и конструкция гнезд........................158
Развитие отношений между членами пары и копуляция.........160
6.5. Кладка, выводок.........................................161
Кладка....................................................161
Птенцы, уход за ними со стороны родителей.................162
6.6. Линька и отлет на зимовки...............................164
Глава 7. Траурная каменка Oenanthe lugens Licht.................166
7.1. Систематические замечания...............................166
7.2. Местообитания и численность............................ 169
7.3. Годовой цикл и сроки гнездования........................170
7.4. Гнездо, кладка, выводок.................................171
7.5. Линька, послси(ездовые кочевки, зимовки................ 172
7.6. Особенности поведения.................................. 173
Глава 8. Комплекс черная каменка Oenanthe picata Blyth..........178
8.1. Чем черная каменка интересна для биолога-эволюциониста..179
8.2. Полевые признаки........................................186
8.3. Местообитания и численность............................ 186
8.4. Годовой цикл и сроки гнездования........................189
8.5. Социальное поведение................................... 192
Территориальное поведение и саморекламирование самцов.... 192
Формирование пар..........................................198
5
Панов Е.Н.
Территориальное поведение самок, полигиния............... 198
Филопатрия, персональный состав пар...................201
Гнездостроение и спаривание...............................202
Развитие отношений между членами пары, копуляция..........203
8.6. Кладка, выводок.........................................206
Кладка....................................................206
Репродуктивный успех......................................207
Птенцы....................................................208
8.7. Линька и отлет на зимовки...............................208
8.8. Биологические заметки...................................209
Глава 9. Белочерная каменка Oenanthe atboniger Hume.............213
9.1. Местообитания и численность.............................213
9.2. Гнездовая биология......................................214
9.3. Миграции и зимовки......................................215
9.4. Биологические заметки...................................215
Глава 10. Белохвостая каменка Oenanthe leucopyga Brehm..........217
10.1. Систематические замечания..............................218
10.2. Местообитания н численность............................219
10.3. Годовой цикл и сроки гнездования.......................221
10.4 Социальное поведение....................................222
10.5 Гнездо, кладка, выводок.................................224
10.6. Биологические заметки..................................226
Глава 11. Чернобрюхая каменка Oenanthe leucura Gm...............228
11.1. Систематические замечания..............................228
11.2. Местообитания..........................................229
11.3. Годовой цикл и сроки гнездования.......................229
11.4 Социальное поведение....................................230
11.5 Гнездо, кладка, выводок.................................231
Гнездовые убежиша и конструкция гнезд.....................231
Кладка....................................................233
Репродуктивный успех......................................233
Птенцы....................................................234
11.6. Миграции ..............................................234
11.7. Некоторые особенности поведения........................235
Глава 12. Испанская каменка Oenanthe hispanica L................238
12.1. Систематические замечания..............................240
12.2. Местообитания и численность............................241
12.3. Годовой цикл и сроки гнездования.......................243
12.4. Социальное поведение...................................244
Пространственная структура поселений и территориальное поведение самцов..........................................244
Саморекламирование самцов.................................246
Территориальное поведение самок...........................248
Формирование пар..........................................248
6
Каменки Палеарктики
12.5. Гнездо, кладка, выводок.................................250
Гнездостросние и конструкция гнезд........................250
Развитие отношений между членами пары в преддверии копуляции..................................................253
Кладка.....................................................254
Птенцы.....................................................255
12.6. Биологические заметки...................................256
Глава 13. Плешанка Oenanthe pleschanka Lcpcch....................257
13.1. Систематические замечания...............................259
13.2. Местообитания и численность.............................261
13.3. Годовой цикл и сроки гнездования .......................263
13.4. Социальное поведение....................................264
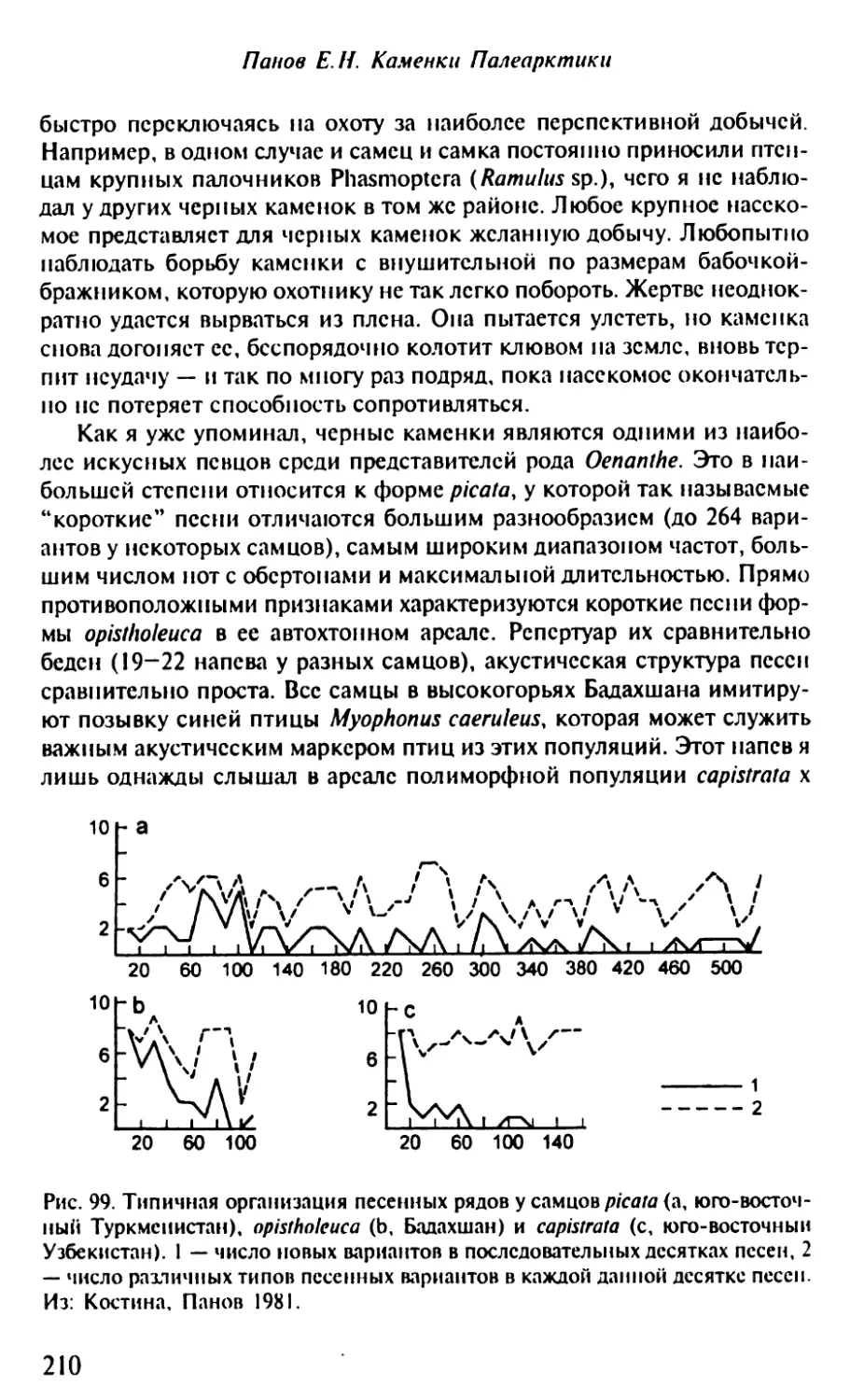
Территориальное поведение и саморекламирование самцов.......264
Формирование пар...........................................267
Развитие отношений между членами пары......................270
13.5. Гнездо, кладка, выводок.................................270
Гнездостроение и конструкция гнезд.........................270
Брачные игры н копуляция...................................271
Кладка.....................................................272
Репродуктивный успех.......................................273
Птенцы.....................................................273
13.6. Линька и отлет на зимовки...............................273
13.7. Биологические заметки...................................274
Глава 14. Гибридные популяции в зонах вторичного контакта испанской каменки Oenanthe hispanica и плешанки Ое. pleschanka.278
14.1. Гибридная популяция Гобустана (восточный Азербайджан)....278
Фенотипический состав, местообитания и численность.........278
Состав гнездящихся пар.....................................282
Сроки и успех гнездования..................................283
Гнезда н кладки............................................283
Особенности поведения......................................284
14.2. Гибридная популяция у южных подножий Главного Кавказского хребта (северо-западный Азербайджан)..............285
14.3. Гибридная популяция Дагестана..........................286
14.4. Гибридная зона в в северном Иране......................287
14.5. Гибридогенныс полиморфные популяции северо-восточного Прикаспия.....................................................288
14.6. Последствия гибридизации и перспективы се углубления.....290
Глава 15. Каменка монашка Oenanthe monacha Tenim.................293
15.1. Местообитания и численность.............................294
15.2. Биология гнездования....................................295
15.3. Миграции и линька.......................................296
15.4. Биологические заметки...................................296
Глава 16. Пустынная каменка Oenanthe deserti Temm................299
7
Панов Е.Н.
16.1. Систематические замечания..............................299
16.2. Местообитания и численность............................300
16.3. Годовой цикл и сроки гнездования.......................303
16.4. Социальное поведение...................................305
Территориальное поведение самцов..........................305
Территориальность самок...................................307
Саморекламирование самцов.................................308
Развитие взаимоотношений между членами пары...............311
16.5. Гнездо, кладка, выводок................................311
Гнездовые убежища..........................................ЗП
Гнездо....................................................312
Кладка....................................................313
Птенцы....................................................314
16.6. Линька и отлет на зимовки..............................315
16.7. Биологические заметки..................................316
Заключение......................................................317
Литература......................................................322
Предметный указатель............................................332
Указатель названий животных.....................................338
8
Каменки Палеарктики
Предисловие
В каждой отрасли биологии существует один или несколько традиционных объектов исследования, именуемых зачастую модельными. Такой объект должен удовлетворять как минимум двум требованиям. Во-первых, основные свойства модельного объекта должны быть таковы, чтобы на его примере можно было решать задачи, актуальные для данной научной дисциплины, и, что особенно важно, проверять справедливость гипотез, сформулированных в рамках существующих здесь теорий. Во-вторых, объект должен быть удобен в работе — то есть, не создавать методических трудностей в дополнение к тем, которых и так немало на пути решения поставленных научных задач.
Классическим примером модельного объекта в биологии может служить мушка-дрозофила. Она очень быстро размножается и тем самым позволяет на сравнительно коротких отрезках времени прослеживать динамику и закономерности преобразования наследственных структур, что, собственно говоря, и есть одна из главных задач генетики. В то же время, особи дрозофилы устраивают специалистов своими размерами — они не слишком малы (что позволяет манипулировать с ними без микроскопа) и не слишком велики, а значит и не требуют больших помещений и запасов корма для своего содержания.
Зоологи, изучающие жизнь диких животных в их естественной обстановке, следуют тем же принципам в выборе объектов своего исследования. Именно поэтому среди десятков тысяч видов живых существ, населяющих нашу планету, более или менее всесторонне и основательно изученными в природе пока что остаются очень немногие. Если говорить о птицах, то среди них можно назвать всего лишь два-три десятка групп близких видов, пристальное изучение которых дало возможность сформулировать ряд важных положений современной эволюционной теории. Среди этих групп можно упомянуть галапагосских дарвиновых вьюрков (дивергенция на основе межвидовой конкуренции), гавайских цветочниц (морфологическая адаптивная радиация), райских птиц (эволюция систем спаривания), колибри (оптимальное кормодобывание), синиц и чаек (географическое видообразование и принцип кольцевых ареалов). Разумеется, приведенный здесь перечень далеко не полон, к тому же он постепенно удлиняется — по мере возникновения в науке новых вопросов и приобщения к традиционным модельным объектам новых, удовлетворяющих тем двум условиям, которые я сформулировал с самого начала.
К числу весьма популярных модельных объектов в орнитологии с достаточным основанием можно отнести и каменок рода Oenanthe. Крупнейшие орнитологи мира — такие в частности, как Э. Штрезс-
9
Панов Е.Н.
манн, Э. Майр, Ч. Вори — посвятили каменкам ряд специальных исследований, затрагивающих как узко специальные вопросы систематики этих птиц, так и более общие, касающиеся, например, природы генетического полиморфизма и его эволюции. Сегодня интерес к каменкам в среде орнитологов нс только не угас, но, как кажется, становится все более пристальным. бДца из причин этого состоит в том, что каменки обитают в открытых ландшафтах (в каменистых предгорьях, в пустынях, на пустошах и в бэдлендах), где за ними легко наблюдать, получая надежно документированные данные относительно самых интимных сторон жизни птиц в природе. Ибо значение модельных объектов в том и состоит, что полученные при их изучении сведения мы можем рассматривать как отражение общезначимых биологических закономерностей, которые, таким образом, позволительно экстраполировать (разумеется, с необходимой осмотрительностью) на достаточно широкий круг других видов, остающихся пока нс исследова-ными с необходимой полнотой.
В современной орнитологической литературе можно найти немало исследований, в которых каменки выступают в качестве модельных объектов для понимания принципов разделения ресурсов между близкими видами при их тесном сосуществовании, приспособления птиц к экстремальным условиям жизни в аридных ландшафтах, стратегий оптимального кормодобывания, закономерностей социальной организации в локальных поселениях, феноменов межвидовой гибридизации и эволюционного происхождения окрасочного полиморфизма, и т. д. В общем, не будет преувеличением сказать, что каменки являются сегодня одной из наиболее детально изученных груп Воробьинообразных птиц. Поэтому они вполне заслуживают того, чтобы их образ жизни, поведение и эволюционная история были обсуждены на страницах еще одной книги о наших пернатых соседях по планете.
Целый ряд обстоятельств, которые будут детально аргументированы в книге, позволяют считать род Oenanthe группой видов, сравнительно молодой в эволюционном масштабе времени. А это значит, что на примере каменок мы можем, применив традиционный сравнительный подход, исследовать начальные и последующие стадии видообразования и дивергенции морфологических, экологических и поведенческих свойств видов в процессе микроэволюции.
Задача монографического описания биологии некого компактного подразделения животного мира — отдельного рода, в данном случае — может быть, в принципе, решена двумя разными способами. Один из них состоит в том, чтобы каждую сторону образа жизни всех видов (например, способы устройства гнезда, особенности миграций либо структуру вокализации) рассматривать в отдельной главе, нарисовав
10
Каменки Палеарктики
им образом обобщенную картину таксономической группы в це-там и путей ее эволюции и дивергенции. Этот подход хорош тем, что Л° позволяет достаточно наглядно показать предполагаемый ход исто-оНчсских преобразований, которыми были затронуты тс или иные фрагменты "биологического портрета" каждого из представителей груп-
Однако, такой подход требует зачастую чрезмерной генерализации исходных данных. Из фрагментов полученной предельно обобщенной картины читателю не всегда легко синтезировать заново целостный образ каждого из тех видов, изучение которых послужило материалом для аналитических интерпретаций. При таком подходе мы безнадежно отдаляемся от живой ткани природы, уходит на задний план идея неповторимости вида как системы, внутри которой все стороны его образа жизни теснейшим образом сопряжены друг с другом и взаимообусловлены. В какой-то степени потерянным оказывается ошушение уникальности вида, которая таится зачастую именно в мельчайших особенностях его экологических предпочтений, социального и сигнального поведения. Эти тонкие нюансы нередко отбрасывают как второстепенные — чтобы нс перегружать деталями конструируемую аналитическую схему эволюции таксона. Имея все это в виду, я предпочел более традиционный подход, суть которого — в подробном описании образа жизни каждого представителя рода как уникальной частицы живого со всеми ее специфическими особенностями, легко ускользающими от глаз поверхностного наблюдателя. Вместе с тем, чтобы облегчить читателю возможность сопоставлений между видами, в первой главе сохранен первый из двух рассмотренных здесь способов представления материала. Иными словами, эта вводная глава представляет собой общий обзор биологии каменок Палеарктики, а в последующих главах каждый, заинтересовавшийся тем или иным вопросом, сможет найти подробные иллюстрации по данной теме в описаниях образа жизни и поведения отдельных видов. Чтобы облегчить возможность
подобных сопоставлений, книга снабжена детализированным предметным указателем.
В книге я попытался также отойти в какой-то степени от сухого академического стиля, ставшего господствующим в современных научных трудах по орнитологии. Эта дисциплина нуждается сейчас в притоке молодых исследователей. Поэтому мне хотелось сделать текст доступным не только для специалистов, но и для тех, кто просто интересуется жизнью природы, будь то школьники старших классов или студенты биологических факультетов. Именно с этой целью содержание многих специальных терминов я оговариваю непосредственно по ходу текста. Мне кажется, что в рамках предложенного подхода описание
11
Панов Е.Н.
поведения пернатых в жанре "Жизни животных" А. Брема может оказаться нс менее полезным в познавательном плане, нежели вполне содержательные рассуждения, изложенные в строгой академической манере. Я надеюсь, что некоторая свобода стиля не повредит научным выводам, предлагаемым мною в этой книге. Так ли это, решать моему читателю.
Я приступил к сравнительному изучению биологии и поведения каменок в 1965 г., в мое первое посещение тогдашней Туркменской ССР в составе зоологической экспедиции новосибирского Института цитологии и генетики, проходившей под руководством Н.Н. Воронцова. В последующие несколько лет благодаря его активности и энтузиазму я имел возможность принять участие в нескольких таких экспедициях, исследованиями которых были охвачены все Среднеазиатские республики бывшего Советского Союза и ряд прилежащих территорий (южное Закавказье, Казахстан, юг Запдной Сибири). В этих регионах, где сосредоточены излюбленые местообитания большинства видов каменок Палеарктики, мне удалось собрать множество ценных сведений по их образу жизни и поведению. Я пользуюсь случаем вспомнить с благодарностью всех своих спутников по этим экспедициям, постоянно деливших со мной радости и невзгоды многомесячных странствий по горам и пустыням. Я имею в виду прежде всего руководителя проекта Н.Н. Воронцов, а также сотрудников руководимой им в те годы Лаборатории эволюции и кариосистематики ИЦИГ СО АНСССР: Е. А. Ляпунову, Н. Ш. Булатову,[А. Д. Базыкина|, А. П. Крюкова, Е. Ю. Иваницкую, и др. В последующие годы значительная часть моих исследований была выполнена в Туркменистане при постоянной дружеской помощи бывшего директора Красноводского заповедника В.И. Васильева и сотрудника заповедника М. Е. Гаузср. В сборе материала во время этих поездок мне помогали мои коллеги — Л. ГО. Зыкова, М. В. Галиченко, В. В. Иваницкий, В. И. Грабовский, Г. Н. Костина, М. Д. Корзухин. В эту книгу вошли материалы двух превосходных многолетних исследований биологии и демографии каменок в гибридных зонах, выполненные С. Ю. Любущснко и А. С. Рубцовым. Во время поездки в Израиль большая помощь в сборе данных по местным видам каменок была оказана автору И. Хохловой и Д. Бланком. Особую признательность хочу выразить Л.Ю. Зыковой, которая проделала огромную работу, связанную с подготовкой рукописи к печати.
Финансирование публикации книги осуществлено при поддержке Российского фонда фундаментальных исследований (грант № 98-04-62018).
Е. Н. Панов
12
Каменки Палеарктики
Введение
Род каменок Oenanthe Vicillot, 1816, относящийся к семейству дроздовых Turdidac и к отряду воробьиных Passeriformes, включает в себя около 20 видов. Читателю может показаться странным, что ему при-подносят эту приблизительную цифру вместо некоего строго установленного числа. Тем не менее, приходится мириться с этим обстоятельством, поскольку точной цифры сейчас никто назвать нс сможет. Род каменок, не в пример многим другим родам птиц, оказался трудным местом систематики. Возможно, что это объясняется тем обстоятельством, что род этот сравнительно молод в эволюционном отношении, и связан целым рядом промежуточных звеньев с другими родственными родами.
Среди мелких африканских дроздовых есть несколько видов, которых систематики не решаются ни включить окончательно в род Oenanthe, ни исключить из него и перенести в другие, близкие роды. Даже один и тот же исследователь может резко менять свое мнение по этому вопросу.Например, известный систематик Рипли (Ripley, 1952, 1964) несколько раз составлял полный список рода Oenanthe. Ниже приведены различия между его списками 1952 и 1964 годов, каждый из которых включает по 18 видов.
Из этой таблицы следует, что разногласия в числе видов, включаемых в состав рода Oenanthe, обусловлены не только расплывчатостью его границ. Во многих случаях неясно также, можем ли мы считать ту или иную форму самостоятельным видом, или только географической расой (подвидом) другого вида. Так, Рипли 1952 году придерживался мнения, что bottae есть всего лишь подвид Ое. isabellina, a lugubris —
Таблица 1. Некоторые разногласия по поводу рода Oenanthe
Ripley, род Mvnnecocichla 1952 род Oenanthe iractrac isabellina + bottae lugenx lugubris Ripley. род Oenanthe 1964 род Cerconiela
bifaxciata bifaxciata ixabe Ilina bottae htgenx + lugubris iractrac
13
Панов Е.Н.
самостоятельный вид. В 1964 г. тот же исследователь выделил bottae в самостоятельный вид, а форму lugubris включил в состав вида Ое. lugens.
В подобных случаях речь идет о так называемых аллопатрических формах, области гнездования которых удалены друг от друга на значительное расстояние. Поэтому о степени родства таких форм мы можем судить, лишь оценивая степень их сходства по размерам, пропорциям, окраске, а также сравнивая особенности их биологии (таких, например, как устройство гнезд, окраска яиц и т.д.) и поведения. К сожалению, детали образа жизни многих популяций каменок юго-западной Азии и Африки до сего времени остаются практически неизученными, так что мы нс имеем достаточного материала для подобных сопоставлений. Поэтому и сегодня остается открытым вопрос о том, можно ли считать самостоятельными видами аравийскую lugentoides (см. Hollomatal., 1988), сомалийскуюрЛ/7//дп (Cramp 1988) и обитающую в западной экваториальной Африке heuglini (Mackworth-Pracd & Grant 1973), или же они являются подвидами в составе видов Ое. lugens, Ое. oenanthe и Ое. bottae. Совсем недавно удалось выяснить, что населяющая о. Кипр популяция плсшанки отличается от всех прочих географических рас этого вида не только меньшими размерами и отсутствием резких различий в окраске самцов и самок, но также совершенно иным звучанием песни (Christensen 1974). На этом основании было решено считать кипрскую плсшанку самостоятельным видом Ое. cypriaca (S\\x\s & Berg 1982, Flint 1995).
При этом исходили из следующих соображений: если бы возникла возможность контакта между популяциямиpleschanka и cypriaca в сезон размножения, самки тех и других нс опознавали бы поющих самцов другой формы как потенциальных половых партнеров, так что гибридизация оказалась бы весьма мало вероятной. А отсутствие гибридизации между двумя данными формами обычно и считают наиболее надежным свидетельством их видовой самостоятельности.
Впрочем, этому четко сформулированному правилу не всегда удастся следовать безоговорочно. Пример тому даст та же самая плешанка в сс взаимоотношениях с испанской каменкой. В отличие от того, что мы видели у только что перечисленных аллопатрических форм (isabelhna-bottae-heuglini, lugens-lugentoides-lugubhs, pleschanca — cypriaca и др.), ареалы плсшанки и испанской каменки частично перекрываются в западном и южном Прикаспии. В этих зонах соприкосновения и перекрывания ареалов (именуемых, соответственно, зонами парапатрии и симпатрии) плешанка и испанская каменка свободно гибридизируют друг с другом. На этом основании некоторые орнитологи (например, Гладков 1954) рассматривали pleschanka и hispanica в качестве подвидов одного вида. Однако в настоящее время практически общепринятой
14
Каменки Палеарктики
стала точка зрения, согласно которой морфобиологическая уникальность каждой из этих форм позволяет считать их самостоятельными видами. Вероятно, такая же судьба ждет в дальнейшем “подвиды” Ое. ^anthoprytnna xanthoprymna и Ое. xanthoprymna chrysopygia, резко различные по окраске и ограниченно гибридизирующие в области соприкосновения их ареалов.
Дополнительные трудности в систематике рода Oenanthe создаст характерное для них явление полиморфизма. В случае так называемого генетического полиморфизма животные данного вида, относящиеся к одной и той же половой и возрастной группе, имеют несколько разных вариантов окраски (как правило, без промежуточных вариантов). Предполагают, что такого рода различия в окраске определяются одним геном или малым числом сцепленных генов. В качестве типичного примера упомянем все ту же плешанку. В некоторых (хотя далеко нс во всех) популяциях этого вида помимо обычного для самцов варианта окраски с черным горлом встречаются единичные бслогорлыс самцы морфы vittata. Такого рода полиморфизм свойственен нескольким видам каменок (см. рис. 1), но поскольку явление это изучено у них совершенно недостаточно, у разных видов он может иметь принципиально различную природу. У одних видов мы можем связывать полиморфизм с процессом мутаций генов (это вероятно в отношении Ое. hispanica), у других он является следствием гибридизации первоначально самостоятельных, различно окрашенных форм (Ое. picata), у третьих полиморфизм может быть трудно отделим от возрастной изменчивости. Последняя характерна для Ое. leucopyga (рис. I) и, возможно, имеет место у южноафриканской Ое. monticola — наиболее полиморфного вида среди всех каменок и одного из наименее изученных. В некоторых популяциях monticola описано до 7 вариантов окраски самцов (Mackworth-Praed & Grant 1963), часть из которых, как я полагаю, могут быть проявлением возрастной изменчивости.
При работе систематика с музейными коллекциями тушек птиц, добытых в ареале некой полиморфной формы, ему зачастую нелегко решить, представляют ли собой разные варианты окраски “генетические морфы”, или же они принадлежат разным подвидам (а то и видам), совместно обитающим в зоне симпатрии. Именно это и произошло в случае с черной каменкой Ое. picata. Сначала было принято думать, что это монотипический (т. е., не распадающийся на подвиды) вид с типичным генетическим полиморфизмом. Присутствие в популяциях Ое. picata грех вариантов окраски самцов объясняли мутациями одного или нескольких генов, определяющих альтернативную — черную либо белую — окраску темени и брюшка (Мауг, Strcscmann 1950). И лишь многолетнее изучение Ое. picata по всему се ареалу в
15
Панов Е.Н.
природных условиях позволило показать, что в действительности этот вид распадается на три подвида, длительная гибридизация между которыми привела к возникновению “полиморфных” популяций. Этот вариант полиморфизма, в отличие от классического генетического (мутационного) полиморфизма, получил название гибридогенного (Panov 1992). Аналогичный случай гибридогенного полиморфизма
Таблица 2.
Перечень "хороших" и "проблемных" видов каменок мировой фауны
мм Имя O&iacib pacn рос i ранения Число подвидов
1 pileala Южная Африка •
2 bottae Экваториальная Африка, Аравня 2
heuitlini* Западная Экваториальная Африка
3 iuibcllina от Малой Алин до северного Китая
4 oenanthe Северо-западная Африка. Европа, северная половина Азии. Гренландия, Лабрадор и Аляска 3 - К»
philhpsi Восточная Африка (Сомали)
6 utnthoprvmna от Закавказья и южного Прикаспня до Таджикистана,Южного Афганистана и северного Пакистана 2>
chryxopygia юго-восточная Турция, западный Иран -
7 mon taolu Южная Африка 4
8 ItHlfUll Северная Африка, Передняя Азия 2
9 fiiixchii Малая Азия, Средняя Азия, северный Иран 2
1« lumens Северная Африка, Ближний Восток. Аравия, Западный Иран 3
lugubris Восточная Африка (Сомали) 3
lutfentoidex Аравия 2
II picata Средняя Азия, Иран, Афганистан 3
12 alboni$er Иран, Афганистан
13 Irucopvftu Северная Африка, Ближний Восток. Аравия 2-3
14 leucura Юго-западная Европа, Северо-западная Африка 2
15 mt mat ha Северо-восточная Африка, Ближний Восток. Аравня, южный Афганистан, юго-западный Пакистан •
16 ple.tthanka Юго-восточная Европа, Передняя Азия. Южная Сибирь, северный Китай 2?
cvpriaca о. Кипр
17 hispanica Южная Европа, Северо-западная Африка. Ближний Восток, Закавказье 2
18 deserti Северная Африка, Азия от Аравин до Памира 3
19 (Mymtet tH H hla') bijasciata Южная Африка
20 (Сен mnela?) tnutnic Южная Африка 2
* Статус форм, нс обозначенных собственными номерами, остается спорным (виды, полувиды либо подвиды)
16
Каменки Палеарктики
описан и в древней зоне гибридизации между Ое. pleschanka и Ое. hispanica северо-западном Казахстане (Панов 1989).
Все сказанное хорошо объясняет, почему перечень видов, включаемых сегодня в род Oenanthe, не может рассматриваться как окончательный. Слишком мало мы знаем сегодня о каменках, населяющих африканский континент и прилежащие к нему районы юго-западной Азии. Прежде чем привести этот перечень, как он видится сегодня, необходимо сделать несколько замечаний по поводу подобных списков вообще. Они преследуют одновременно две цели: первая — дать перечень всех известных видов, вторая — расположить их в порядке родства. И если первая задача сравнительно легко осуществима, то для выполнения второй форма списка мало пригодна. Разные виды имеют разный эволюционный возраст, и нередко несколько видов берут начало от какого-то одного. Могут быть и другие тонкости, которые невозможно передать при простом и стандартном перечислении видов. Именно поэтому списки, составленные разными авторами, неизменно отличаются друг от друга порядком расположения видов. Все сказанное заставляет меня дать сначала примерную схему родственных связей между видами рода Oenanthe (рис. 1), а уже после этого — полный их список с указанием географического распространения (Табл. 2).
Все, что было сказано по поводу трудностей систематики каменок, имеет и другую, пожалуй — более важную сторону. Неясность многих ситуаций, с которыми мы встречаемся при изучении каменок, свидетельствует о том, что процесс эволюции в этой группе еще не закончился, и поэтому мы можем изучать процессы и механизмы эволюции и видообразования “в действии”. С этой точки зрения всестороннее исследование каменок представляет нс только частный зоологический интерес, но может дать многое для развития эволюционного уче!шя, генетики и прочих общебиологических дисциплин.
17
Панов Е.Н. Каменки Палеарктики
Глава 1.
Некоторые общие сведения о каменках
1.1. Географическое распространение и история расселения
Если попытаться проследить историю возникновения и расселения каменок, то она будет выглядеть примерно так. Далекими предками каменок были, невидимому, мелкие лесные дроздовые — нечто среднее между каменкой и горихвосткой. Гнездились они в дуплах деревьев. Такие птицы и сейчас живут в африканских лесах. Относятся они к довольно богатому, объединяющему I7 видов роду Cossypha. Этих птиц англичане называют тем же словом, что и каменок — chats. Характерной чертой их внешнего облика служит рыжий хвост с черным концом и черноватыми средними рулевыми.
Повидимому, птицы такого типа дали начало нескольким другим родам, перешедшим к жизни в местах с разреженной древесной растительностью (например, в саваннах, где обитают многие представители рода Myrmecocichla) или вообще лишенных се. Таковы равнинные полупустыни и пустыни, ставшие типичными местообитаниями большинства видов рода Cercomela и некоторых представителей рода Oenanthe, а также всевозможные каменистые ландшафты от подножий гор до высокогорий. Именно эти последние биотопы с течением времени освоило большинство видов каменок, в связи с чем эта группа птиц и получила свои народные названия в немецком и русском языках.
От своих лесных предков каменки унаследовали привычку гнездиться в закрытых убежищах, в качестве которых равнинные виды каменок приспособились использовать норы грызунов (Ое. pileata, Ое. moesta, Ое. isabellina), а обитатели предгорий и гор — всевозможные пустоты и расселины в глинистых обрывах, в скальных обнажениях и в каменистых осыпях.
Расселение молодых видов, перешедших от жизни в лесах к существованию в открытых ландшафтах, шло двумя основными путями. Одни проникали к югу от зоны африканских тропических лесов, другие — к северу, в область Средиземноморья. Сейчас южнее пояса девственных лесов центральной Африки живут 5 из 9 видов рода Cercomela и только 2-3 из 18-20 видов рода Oenanthe. Эти южноафриканские каменки по сво-
18
Глава /. Некоторые общие сведения о каменках
сму облику нс типичны для рода и являются, невидимому, его древними “осколками”. Основные процессы становления экологического типа каменок и последующего видообразования проходили в области Средиземноморья. Здесь и поныне обитает около половины всех существующих видов каменок.
В последующий период расселения на север и на восток по просторам Европы и Азии большинство видов не смогли порвать со своими исходными местообитаниями — аридными ландшафтами тропиков и субтропиков. Как следует из таблицы 2, основная масса видов нс продвинулась к северу и востоку дальше самых южных районов Европы и горно-пустынных регионов Передней и Средней Азии. Лишь 4 вида расширили свои ареалы далеко за пределы этой обетованной земли каменок. Три из этих четырех видов-первопроходцев (Ое. deserti, Ое. isabellina и Ое. pleschanka) расселились далеко на восток, завоевав новый жизненный плацдарм в крайне негостеприимных горных полупустынях и пустынях Центральной Азии — таких как Такла-Макап, Гоби, Алашань, Ордос. Пустынная каменка Ое. deserti и плясунья Ое. isabellina приспособились к гнездованию на самых высоких плоскогорьях мира — на Памирском плато и на северной окраине Тибетского нагорья (Vaurie, 1972). По мнению Козловой (1975), освоение этих регионов каменками началось более 5 миллионов лет тому назад, когда геологический процесс поднятия высочайших горных плато Центральной Азии находился еще в своих начальных стадиях.
И хотя два из трех названных видов — Ое. isabellina и плешанка Ое. pleschanka в своем движении на восток почти достигли берегов Тихого океана, ни им, ни Ое. deserti нс удалось продвинуться к северу далее 55* с.ш. Этот рубеж миновал четвертый вид из группы первопроходцев — именно, всем нам хорошо известная обыкновенная каменка Ое. oenanthe. Преодолев в силу своей поистине удивительной экологической пластичности широкий лесной пояс Евразии, oenanthe проникла далеко за полярный круг, где успешно освоила материковые тундры и даже суровые арктические ландшафты островов Ледовитого оксана. Болес того, этот бесспорно африканский по происхождению вид двумя независимыми путями — через Исландию и Гренландию и через Берингов пролив — проник в Новый Свет, став там единственным представителем рода Oenanthe.
1.2. Некоторые черты морфологии
Размеры и пропорции. Все каменки стройные, грациозные, сравнительно длинноногие птицы с массой примерно от 15 г до 45 г. Самая мелкая каменка — сомалийская phitlipsi с длиной крыла 79-86
19
Панов Е.Н. Каменки Палеарктики
pileata bottae isabellina oenanthe** phillipst chrysopygia xanthoprymna moesta** Bnschii** I ugens* lugubris** -----picata*
-----alboniger leucopуда leucura monticola**
-----monacha**
deserti** tractrac bifasciala**
Рис. I. Ориентировочная схема филогении рода Oenanthe. Для видов, которым свойственен генетический полиморфизм, приведены оба (или по крайней мере два — как в случае Ое. monticola) варианта окраски самцов. Для Ое. 1еисору%а с таким же вариантом полиморфизма, как у Ое. leucura. показана возрастная изменчивость (молодые особи имеют черную голову). Двумя звездочками обозначены виды, у которых всем популяциям свойственен половой дихроматизм. У видов с од|ioii звездочкой он присутствует не у всех подвидов. Отсутствие звездочки означает, что у вида самцы и самки окрашены одинаково или сходно. Оригинал.
мм, а среди европейских представителей рода — кипрская плешанка cypriaca (крыло 80-85 мм). Общие размеры этих каменок — со щегла или серую мухоловку, то есть они значительно мельче домового воробья. Самая крупная каменка — южноафриканская Ое. monticola (длина крыла до 115 мм), а среди европейских и палеарктических представителей рода — чернобрюхая каменка Ое. leucura массой 38-44 г и величиной примерно с пестрого каменного дрозда. Она заметно уступает в размерах скворцу, вес которого составляет примерно 70-80 г. Рис. I отражает не только различия в окраске, но и относительные размеры разных представи телей рода. Самки обычно мельче самцов, а особи-первогодки мельче птиц более старших возрастных групп.
На рис. 2 порядок расположения палеарктических каменок и одного южноафриканского вида (Ое. monticola) совпадает с уменьшением их общих размеров. Из этой схемы видно, что вес, длина крыла, хвоста и клюва не всегда изменяются про-
порционально. Например, Ое. moesta и Ое. ftnschii — виды, нс совершаю-
щие далеких сезонных миграций, характеризуются опюситслыю короткими крыльями, а Ое. monacha. специализирующаяся на ловле летающих
насекомых, выделяется среди всех прочих видов рода непропорционально длинным клювом и короткими ногами. Напротив, Ое. isabellina. охотя-
20
Глава Г Некоторые общие сведения о каменках
Вид (Подвид) Масса (г) Крыло (мм) Плюсна (мм) Клюв (мм)
leucura 1'1 1 г 1 10 20 30 40 50 1 1 1 1 1 80 90 100 110 120 । । । । 1 20 25 30 35 40 1'1 1-1 1 1 14 16 18 20 22 24
monticola — 1 —
leucopyga
albontgar — — — —
isabellina ' — -—
moesta ? — —
oenanthe leucorhoa —
oenanthe oenanthe — —
finschii —
monacha — — “
picata — ““
deserti oreophila — — — ——
deserti homochroa — — —
lugens persica — — ? —
lugens halophila — — —
xanthoprymna •— — —
hispanica — —— — 1
pleschanka — — —"
cypriaca 9
Рис. 2. Размеры и пропорции каменок Палеарктики и самого крупного представителя рода — африканской Ое. monticola. Оригинал.
щаяся главным образом на земле, обладает самыми длинными цевками.
Можно заметить также, что заметные различия в общих размерах (при сохранении пропорций) могут существовать между разными подвидами одного вида. На примере обыкновенной и пустынной каменок видно, что подвиды, обитающие в более суровых условиях, где выгодны меньшие потери тепла, обладают более крупными размерами. Арктическая Ое. oenanthe leucorhoa имеет массу 25—41 г , тогда как у обитающей в умеренных широтах Ое. oenanthe oenanthe вес обычно нс превышает 29 г. Центральноазиатский подвид пустынной каменки Ое. deserti oreophila, гнездящийся на высокогорных плато, где даже летом ночи довольно холодные, обладает заметно большими размерами, чем равнинная раса Ое. deserti homochroa. Впрочем, даже в тех случаях, когда разные подвиды населяют районы со сходными климатическими условиями, можно видеть тенденцию к увеличением общих размеров в направлении расселения вида из его начального ареала (сравни африканскую, автохтонную Ое. lugens halophila и вторично проникшую на Иранское нагорье Ое. lugens persica).
Окраска. Типичной для каменок принято считать контрастную черно-белую окраску самцов. Однако это справедливо лишь для 12 из примерно 20 видов рода (рис. I). Еще у четырех видов (Ое. moesta, Ое.
21
Панов Е.Н. Каменки Палеарктики
lugubris, Ое. hispanica, Ое. deserti) белый цвет в той или иной степени и на разных участках оперения заменен рыжеватым. Возможны также самые различные комбинации белого и рыжего в брачном наряде самца: так, у окрашенных по одной схеме Ое. lugubris и Ое. moesta самцы первого из этих видов имеют рыжеватые “шапочку”, гузку и рулевые перья, тогда как у самцов второго вида рыжая только гузка, а шапочка и рулевые — белые*. У испанской каменки светлые участки контурного оперения окрашены в рыжевато-бурые тона у самцов западной расы hispanica и почти белые в брачный сезон у восточной расы melanoleuca, в то время как рулевые у тех и у других белые. У Ое. hispanica melanoleuca, как и у ряда других видов с черно-белой окраской самцов (например, у Ое. pleschanca, Ое. finschii, Ое. picata) самцы в свежем перс имеют более или менее выраженный палевый налет на белых участках оперения, который местами сохраняется даже при дальнейшем обнашивании контурных перьев. Чаще всего в палевый цвет при этом остается окрашенным подхвостьс (Ое. finschii, Ое. picata), рыжеватая окраска которого сохраняется на протяжении всего года у самцов Ое. lugens, щеголяющих своим контрастным черно-белым оперением. Изредка рыжеватый налет появляется даже на тех перьях, которые в норме неизменно остаются белыми. Так, у одного из тех птенцов Ое. finschii, которых я выращивал в лаборатории, отрасли не чисто белые, как положено, а чуть желтоватые рулевые. Я убежден, что это атавистический признак, сохранившийся в генотипе вида от его бесконечно Далеких африканских предков, к числу которых могли относиться, в частности, “рыжехвостые” сомалийские Ое. lugubris (рис. I).
Все сказанное склоняет меня к выводу, что у каменок белый цвет в окраске оперения — это эволюционное новообразование, производное от первоначального рыжего цвета, унаследованного родом Oenanthe от родственных ему мелких дроздовых — таких как африканские Cossypha и Cercomela. Мысль о том, что переход от рыжего к белому был очередным шагом в развитии вторичных половых признаков самцов, иллюстрируется следующим любопытным фактом: у Ое. monacha самцы окрашены в сочетание черного и белого, лишенного какой либо примеси желтизны, тогда как самки имеют светлорыжие рулевые. Добавлю к этому, что всевозможные оттенки красноватого и оранжевого в окраске контурного оперения и рулевых широко представлены в очень многих родах дроздовых (зарянки Erithacus, соловьи Luscinia, варакушки Cyanosylvia, горихвостки Phoenicurus, тугайные соловьи Erythropygia, каменные дрозды Monticola, чеканы Saxicola, африканские Thamnolaea, Cossypha, Cercomela
• Здесь и далее подразумевается, что вне зависимости от окраски светлой части рулевых они всегда или почти всегда имеют черные окончания.
22
Глава 1. Некоторые общие сведения о каменках
и многие другие), тогда как чисто белый цвет на этих участках оперения встречается у дроздовых скорее как исключение. Здесь с каменками, больше половины видов которых щеголяют белоснежными отметинами, может соперничать, пожалуй, лишь близкий им африканский род Myrmecocichla. Ряду видов этого рода свойственны ярко-белые “эполеты” на сгибах крыла — признак, сближающий с Myrmecocichla южноафриканскую Ое. monticola (и, в какой-то степени — виды каменок с частично редуцированными “эполетами”: Ое. moesta, Ое. deserti и Ое. bifasciata) (см. рис. 1). Очевидно, в качестве атавизма этот признак проявляется у некоторых самцов Ое. leucopyga и Ое. leucura.
Как можно видеть из того же рис. I, самцы более чем половины изображенных здесь видов сохраняют в своем брачном наряде значительные участки, окрашенные в теплые рыжеватые тона. Если при этом в оперении головы, туловища и крыльев самца отсутствуют бросающиеся в глаза контрастные черные отметины (Ое. isabellina, Ое. (х.) chrysopygia, Ое. tractrac, отчасти — Ое. bottae), то в нем трудно с первого взгляда признать представителя сильного пола. И действительно, у этой группы видов самцы почти нс отличаются внешне от самок своего вида, поскольку оба пола одеты в скромный, покровительственный наряд “самочьего” типа.
Противоположную картину мы видим у ряда контрастно окрашенных каменок (таких, в частности, как Ое. alboniger, Ое. leucopyga, Ое. leucura), у которых самцы и самки также почти неразличимы, но уже по другой причине: здесь самки одеты в праздничный, бросающийся в глаза “самцовый” наряд. И, наконец, в третью группу видов входят такие, у которых одеяние самцов и самок соответствует нашим обычным представлениям о различии облика сильного и прекрасного пола у птиц. У таких видов, как, скажем, Ое. moesta, Ое. monacha, Ое. pleschanka самцы щеголяют контрастным черно-белым нарядом, а самки окрашены в скромные серо-песочные тона.
В чем же причина подобных различий в степени выраженности и в характере полового дихроматизма у разных видов каменок? Считается, что скромная, покровительственная окраска самок полезна в случае их открытого гнездования. Если же самка насиживает в надежном убежище (в дупле, в узкой и глубокой расселине скалы), для ее выживания несущественно, окрашена ли она покровительственно или “вызывающе”. Следуя этой логике, можно было бы допустить, что виды с контрастной окраской самок не так давно (в эволюционном масштабе времени) приспособились к закрытому гнездованию, или же гнездятся в существенно более глубоких и надежных убежищах.
Однако, все эти предположения разлетаются как дым, когда выясняется, что характер полового диморфизма может быть различным у
23
Панов Е.Н. Каменки Палеарктики
Рис. 3. Изменчивость окраски самок черной каменки Ое. picata: a, f — самки формы capistrata; b-e — формы opistholeuca; g~k — формы picata. Предполагается, что у всех подвидов самые светлые самки — это первогодки, самые темные, окрашенные по самцовому типу — самки самых старших возрастных групп. Из: Панов, 1989.
очень близких видов или даже у разных подвидов одного и того же вида. Например, у xanthopryma самцы имеют контрастную окраску, а самки более тусклую, покровительственного типа (хотя некоторые самки почти нс отличимы от самцов); у формы chrysopygia. гибридизирующей с xanthoprymna в зоне контакта, и самцы и самки окрашены по самочьему типу. У всех пяти подвидов Ое. lugens самцы одеты в контрастный черно-белый наряд. Так же окрашены и самки у подвидов Ое. lugens lugens и Ое. /. persica, обитающих в Передней Азии. У африканской расы Ое. I. halophila самки песочно-серые с черноватым горлом, а у южноаравийских подвидов Ое. I. boscaweni и Ое. I. lugentoides они свстло-пссочные без каких-либо темных отметин. К этому следует добавить, что даже в пределах одной популяции окраска самок может сильно варьировать, так что часть из них окрашены в скромные серовато-бурые тона, а у других
24
Глава /. Некоторые общие сведения о каменках
присутствуют отдельные элементы контрастной окраски самцов, причем некоторые самки почти нс отличимы от особей мужского пола. Это явление обычно для многих видов: Ое, oenanthe, Ое. xanthoprymna, Ое. picata picata и Ое. picata opistholeuca (при отсутствии подобной изменчивости у Ое. picata capistrata), Ое. hispanica и Ое. pleschanka (рис. 3) и др. Известно, что у самок птиц гормоны яичника подавляют развитие сам-цового наряда, который самки надевают после кастрации, и что с возрастом продукция этих гормонов у самок может уменьшаться (Завадов-ский, 1922). Поэтому легко предположить, что “пстухопсрыми” у каменок только что перечисленных видов в большинстве случаев являются самки старших возрастов.
Из всего сказанного напрашивается вывод, что окраска каменок в ходе микроэволюции может изменяться сравнительно быстро. Иными словами, она не является достаточно консервативным признаком, так что использование се для выяснение степени родства между видами требует большой осторожности. Например, cypriaca и lugens чрезвычайно сходны друг с другом по окраске самцов и самок и по характеру полового дихроматизма, но, как показывает анализ их поведения, относятся они к совершенно разным ветвям филогенетического древа каменок (рис. I). Причиной же сходства окраски между этими формами служит явление параллельной изменчивости в разных ветвях этого древа, а также у близких видов, имеющих сранительно недавнего общего предка. Свидетельством такого параллелизма служит сходный характер полиморфизма у ряда видов каменок. Например, белобрюхие и чернобрюхие, белоголовые и черноголовые морфы существуют как у видов недавнего общего происхождения (Ое. lugens, Ое. lugubris, Ое. picata), так и у удаленной от них филогенетически южноафриканской Ое. monticola (рис. I). Имея в виду эти и другие подводные камни, ожидающие нас на пути изучения родственных связей между разными видами каменок, схемы их эволюции, построенные на анализе окрасочных признаков (подобные той, что предложены в работах Тай (Туе, 1986, 1989а,Ь), необходимо проверять и уточнять с использованием этологических данных. Эти последние имеют дело с особенностями поведения видов, которое во многих своих аспектах несравненно более консервативно в эволюции, нежели признаки окраски (Lorenz, 1939).
Обсуждая окраску самок, я уже коснулся вскользь вопроса о се приспособительном значении. Вопрос этот до сих пор остается открытым. Не понятны и причины того, почему у этих пустынных, открыто живущих птиц столь часто встречается контрастная черно-белая окраска. Казалось бы, она не должна приносить им никакой пользы, а скорее — только вред. Во-первых, сочетание черного и белого делает птицу очень заметной для хищников (что особенно вредно для самок в
25
Панов Е.Н. Каменки Палеарктики
Рис. 4. Позы каменок, принимающих солнечные ванны: а — Ое. chrysopygia; b — Ое. picata capistrata (самка); c-f — Ое. р. picata; g — Oe.finschii.
период насиживания кладки). Во-вторых, как известно, черные поверхности плохо отражают солнечные лучи, и в условиях аридного климата, под палящим солнцем это может вести к перегреву тела.
Некоторые авторы (Cott, 1947; Mcinertzhagen, 1954) попытались объяснить контрастную окраску каменок предположив, что она играет примерно такую же роль, как отпугивающий (апосематический) черно-желтый рисунок, характерный для многих ядовитых либо несъедобных животных, таких, например, как осы и саламандры. Эти исследователи допускают, что мясо каменок имеет неприятный вкус, о чем и предупреждает хищников контрастная окраска “черно-белых” каменок. Котт проводил опыты, предлагая мясо разных видов птиц шершням Vespa crabro. Среди этих птиц были и четыре представителя контрастно окрашенных
26
Глава /. Некоторые общие сведения о каменках
Рис. 5. Опушение птенца каменок. Точками показаны пуховые птерилии, имеющиеся у других Passerifonnes, но отсутствующие у каменок. По: Ней-фельлт, 1970.
каменок. Оказалось, что мясо трех из НИХ (Ое. leucopyga, Ое. lugens и Ое. monacha) действительно оказалось наименее привлекательным для шершней /“индекс съедобности” составлял для этИХ видов, соответственно, 1, 2 и 4, при максимальном значении 20 для покровительственно, криптичсски окрашенных птиц). Однако мясо четвертого вида, плешанки Ое. pleschanka шершни поедали довольно охотно (индекс съедобности, равный 16). При всем при этом идея о несъедобости мяса каменок для их естественных хищников кажется мало правдоподобной, если учесть, что птенцы этих птиц часо становятся жертвами змей, врановых птиц и млекопитающих (обсуждение см. в работе Gotmark, 1994).
Что касается возможности перегрева, то и здесь нужны экспериментальные данные. О. Митропольский (1980)
сообщает, что в пустыне Кызылкум сплошь черные самцы фенотипа opistholeuca явно страдают от солнца. Мне не приходилось замечать этого. В самые жаркие часы птицы всегда имеют возможность найти укрытие в тени, но иногда они пренебрегают такой возможностью и на самом солнцепеке принимают солнечные ванны (рис. 4).
Однако в аравийских пустынях, где летом дневные температуры часто превышают 40*, обитающие здесь Ое. leucopyga предпочитают укрываться в тени уже около 8.30 утра и оставаться здесь примерно до 16.30. К этому времени жара немного спадает, и птицы отправляются на поиски пропитания (Palfery, 1988). Этот автор за многие часы своих наблюдений лишь однажды, в прохладный январский день, имел возможность видеть каменку, принимавшую солнечную ванну в уютной нише, укрытой от пронизывающего ветра. По моим собственным наблюдениям за этим видом в пустыне Негев в Израиле, в сильную жару птицы несомненно ощущают перегрев: они держат крылья отставленными от туловища и слегка опущенными (чего я никогда не видел у других видов каменок), а клюв часто подолгу остается полуоткрытым (см. рис. 108). Впрочем, в Нсгеве в Дневные часы страдают от зноя и подолгу прячутся в тени не только эти каменки, в оперении которых преобладает черный цвет, но и другие птицы, окрашенные в светлые тона.
27
Панов Е.Н. Каменки Палеарктики
См е н а н а р я д о в, л и н ь к а. Только что вылупившийся из яйца птенец имеет 8 пучков длинного (6—10 мм) эмбрионального пуха: два растут над глазами, два — в плечевой области, два — в бедренной и по одному на затылке и вдоль спины (рис. 5). Присутствием бедренных пуховых птерилий птенцы каменок отличаются от птенцов всех изученных видов чеканов Saxicola и каменных дроздов Monticola (Нейфельдт, 1970). Кроме того, у Ое. oenanthe и Ое. pleschanka обнаружены короткие рудиментарные пушинки над анальным отверстием. Зев окрашен в разные оттенки желтого и имеет неодинаковую форму у разных видов.
За две недели, которые птенец проводит в гнезде, он приобретает свой первый наряд из перьев — так называемый гнездовой наряд. У большинства видов он окрашен в серо-бурые покровительственные тона — иногда без пестрин, так что практически нс отличается от наряда самок (Oe.finschii, Ое. xanthoprymna chrysopygia). чаще же каждое перо имеет темный либо светлый кончик, и птенец выглядит пестрым. Лишь у немногих видов, у которых самки неизменно одеты в контрастный черно-белый "самцовый” наряд (Ое. alboniger, Ое. leucopyga, Ое. leucura). молодые в гнездовом наряде также окрашены контрастно, почти нс отличаясь по внешнему виду от взрослых самок.
Уже через мссяц-полтора после вылета из гнезда птенцы начинают сменять мелкое оперение головы, спины и брюшка. В редких случаях сменяются также крупные перья крыла (чаще — кроющие) и хвоста. После окончания этой линьки молодые оказываются в первом зимнем наряде (мелкое перо новое, крупное осталось от гнездового наряда или сменилось частично).
У большинства видов этот первый зимний наряд сохраняется до будущей весны и с каждым месяцем постепенно становится ярче за счет обнашивания светлых каемок перьев. К началу сезона размножения эти каемки исчезают полностью, и черные участки оперения самцов предстают во всем их великолепии.
Напротив, у Ое. oenanthe, Ое. isabellina, а также у большинства особей Ое. hispanica и Ое. pleschanka первый зимний наряд оказывается промежуточным: он сохраняется лишь до конца зимы, когда происходит новая линька мелкого оперения, после которой птицы надевают свой весенний брачный наряд. Трудно сказать, почему промежуточная предбрачная линька свойственна лишь этим четырем видам. Два из них (Ое. oenanthe и Ое. pleschanka) совершают очень протяженные сезонные миграции, но то же можно сказать в отношении Ое. deserti atrogularis, у которой промежуточная предбрачная линька не описана. Два другие (Ое. isabellina и Ое. hispanica) зимуют сравнительно недалеко от мест гнездования, как и большинство других видов каменок, нс имеющих, в отличие от них, промежуточной линьки.
28
Глава 1. Некоторые общие сведения о каменках
рис. 6. Возрастная изменчивость окраски хвоста у 4 видов каменок: а — Ое. (oenanthe?) phillipsi; b — Ое. р. picata; с — Ое. leucopyga; d — Ое. pleschanka. Во всех случаях слева первогодки, справа — особи старше года. По данным автора (b, d). Cramp, 1988 (с) и Туе, 1986.
Нужно заметить, что у каменок черные окончания перьев хвоста гораздо шире у птиц-первогодков (и у самок), нежели у взрослых самцов (рис. 6). Особенно это бросается в глаза у Ое. leucopyga, у которой первогодки имеют широкую концевую полосу на хвосте, тогда как у взрослых птиц он практически целиком (за исключением центральной пары рулевых) белый (Heim de Balsac, Mayaud, 1962). Таким образом, можно рассматривать широкую черную полосу на хвосте, характерную для deserti, isabellina и ряда других видов (см. рис. I) как признак примитивный.
На вторую осень своей жизни каменки претерпевают полную линьку, в ходе которой сменяется все оперение полностью, а в конце зимы происходит частичная линька мелкого оперения. Линька мелкого оперения начинается обычно с головы, от клюва, после чего распространяется на спину, грудь и брюшко. И первостепенные, и второстепенные маховые сменяются в направлении от внутренних к наружным. Точно также идет и линька хвоста: сначала выпадают средние рулевые, и лишь в последнюю очередь — крайние.
Кариотипы. Для выяснения родственных связей между отдельными видами каменок были исследованы хромосомные наборы 8 видов, обитающих в пределах бывшего Советского Союза. Все кариотипы оказались однотипными, состоящими из 7 пар более крупных хромосом (макрохромосом) и 34 пар мелких микрохромосом (рис. 7). Половые хромосомы, одинаковые в кариотипе самца (две W-хромосо-
29
Панов Е.Н. Каменки Палеарктики
П Ai *Д Л» 4* IX »<
ZZ
XI л.
ZW
С
ZW
А А •• -**
Рис. 7. Кариотипы некоторых видов каменок: черношей->юй Ое. finschii (самец. Копетдаг), черной Ое. picata (самец, Копстдаг), плясуньи Ое. isabellina (самка, Копст-даг), пустынной Ое. deserti (самка, южный Алтай). ZZ, ZW — половые хромосомы (по фотографиям Н.Ш. Булатовой).
мы) и неодинаковые в кариотипе самки (W и Z-xpo-м о с о м ы ) , являются IV (Z) и VII (W) в порядке убывания длины хромосом внутри кариотипа. В итоге кариотип всех каменок состоит из 41 пар хромосом (2N=82). Тот факт, что у всех изученных видов, подчас резко различающихся по внешней морфологии и поведению, кариотипы оказались очень похожими, может свидетельствовать об относительной молодости рода Oenanthe.
По структуре кариотипа каменки довольно сходны с чеканами Saxicola (2N=80) и каменными дроздами Monti со 1а (2N=86), но сильно отличаются от горихвосток Р h о е п i с и г и s (2N=76) и настоящих дроздов рода Turdus (2N = 80) (Bulatova, Panov, 1973; Bulatova, 1981).
30
Глава !. Некоторые общие сведения о каменках
1.3. Местообитания
Подавляющее большинство видов каменок обитает в районах с жарким засушливым климатом. Их излюбленные ландшафты — это приподнятая над уровнем моря каменистая местность, изрезанная неглубокими ушельями или глинистыми оврагами, между которыми возвышаются невысокие холмы. Склоны их покрыты россыпями камней, там и тут видны скалистые обнажения.
Другой тип местообитаний — песчаные и глинистые пустыни, полностью лишенные растительности или же с зарослями кустарников и саксаула Arthrophytum. Нечто подобное евразийским пустыням и полупустыням мы видим в южной Африке. Это так называемые karoo — аридные области с редким малорослым кустарником. Эфемерная травянистая растительность появляется лишь на короткое время после периода дождей. Здесь обитают Ое. pileata и сформировавшаяся в этих местах Ое. monticolagriseiccps(Winterbottom, I960) Немногие виды живут в низкотравянистых степях (среди африканских каменок — Ое. bottae и Ое. bifasciata). Наконец, третий тип местообитаний — это неширокие межгорные котловины с низкой травой, бедные растительностью альпийские луга, выпасы и выгоны.
Среди каменок есть несколько видов, которые могут с равным успехом селиться по крайней мерс, в двух разных типах местообитаний. Так, Oe.Jtnschii и Ое. deserti могут жить и в каменистых предгорьях и в песчаной пустыне, Ое. isabellina — в разных типах пустынь, в межгорных котловинах и на приледниковых лугах. Другие виды более разборчивы: moesta тесно связана с пустынными ландшафтами, Ое. monticola griseiceps — с karoo, Ое. bottae — с сухими степями, Ое. lugens, Ое. lugubris, Ое. leucopyga, Ое. alboniger, Ое. monacha и др. — с каменистыми районами плато и предгорий.
Многие виды каменок проникают высоко в горы, а для некоторых высокогорья стали второй родиной. Например, южноафриканская Ое. monticola nigricauda найдена лишь на склонах горы Моко в Анголе, на высоте около 2600 м. Такие виды, как Ое. isabellina, Ое. chrysopygia, Ое. picata, Ое. hispanica, Ое. pleschanka поднимаются в горы свыше 2000 м над уровнем моря, а Ое. deserti oreophila — почти до 5000 м. Обыкновенная каменка Ое. oenanthe в южной части Евразии — типично высокогорный вид, гнездящийся, например, в Тяньшане до высот 3000-3600 м наду.м. (Губин, Ковшарь, 1985). Когда встречаешь се на усеянных валунами альпийских лугах совсем рядом с ледниками, становится понятным, почему она смогла освоить безбрежные пространства Сибири и проникнуть по обезлесенным долинам рек в приполярные тундры.
31
Панов Е.Н. Каменки Палеарктики
Условия, с которыми вынуждены мириться некоторые виды каменок в их типичных местообитаниях, подчас оказываются чрезвычайно суровыми, поистине экстремальными. В аридных ландшафтах Передней Азии видам, связанным с безбрежными каменистыми пустынями (таким, в частности, как Ое. lugens, Ое. leucopyga, Ое. monacha), приходится противостоять испепеляющему летнему зною, полному отсутствию воды для питья и бедности насекомых-жертв. В высокогорных районах, в особенности в пустынях Центральной Азии с их резко континентальным климатом, к этим неблагоприятным факторам добавляются регулярные ночные похолодания. Во всех подобных местностях каменки оказываются в числе немногих видов мелких воробьиных, способных существовать в столь агрессивной среде. Например, в практически лишенных какой-либо растительности гравийных пустынях Памирского нагорья, часто именуемого “крышей мира” (средние высоты — около 4000 м над уровнем моря), только два вида мелких насекомоядных птиц встречаются в значительных количествах. Это рогатый жаворонок Eremophila alpestris, известный своей терпимостью к экстремальным условиям существования, и пустынная каменка Ое. deserti.
Не менее суровы и нсгостиприимны ландшафты, занимаемые видом-пионером Ое. oenanthe в субарктических и арктических регионах, где его типичными местообитаниями служат как равнинные, так и горные тундры до высот до 1100 м над у. м. Лето здесь короткое и прохладное, начинается оно лишь на рубеже мая и июня. К моменту прилета каменок на места гнездования подвижных насекомых еще очень немного, и птицы первое время вынуждены питаться в основном растительными кормами — например, семенами осок (Портснко, 1973; Кищин-ский, 1980; Данилов, Рыжановский, Рябицсв, 1984). В поисках мест гнездования Ое. oenanthe часто прибегает в тундре к древнему способу, используемому на юге типично равнинными видами каменок: она селится в норах мелких млекопитающих (а иногда — и птиц, таких как глупыши и тупики), где микроклимат, бесспорно, несколько благоприятнее, чем среди нагромождений голых камней. Не избегает Ое. oenanthe на крайнем севере и близости человеческих поселений. Здесь птичкам порой удается устроить пир на свалках отходов либо на трупе павшего домашнего животного. Способность этих каменок использовать любое изменение местности под влиянием деятельности человека в свою пользу выработалось у них в процессе постепенного расселения к северу. И в самом деле, во многих участках своего ареала обыкновенная каменка оказывается почти что синантропным видом, поселяясь в поселках и городах, в искусственных гнезовьях, вывешенных па опушках леса для мухоловок и синиц, в отвалах промышленных разработок и т.д. (Piecre, 1987; Доржиев, Хсртусв, 1992).
32
Глава /. Некоторые общие сведения о каменках
1.4. Миграции и зимовки
В субтропических и тропических районах своего исконного существования местные виды каменок как правило оседлы, и лишь часть особей кочуют во внегнездовой сезон внутри ареала данного вида либо недалеко за его пределами. У обитающих в Средиземноморье и в соседних регионах Ое. moesta, Ое. lugens, Ое. leucopyga, Ое. leucura, Ое. monacha, Ое. deserti deserti многие особи остаются зимовать на своих прежних гнездовых участках, а уже с ноября начинается формирование новых пар, которые начнут готовиться к размножению с первыми проблесками весны (Palfcry, I988; Soler ct al., I995; Shirinai, 1996). Повидимому, все сказанное можно отнести и к почти неизученным видам тропической Африки — таким, например, Kaxphillipsiи lugubris (Smith, 1971).Вболсс северных районах Передней Азии лишь немногие виды гнездящихся здесь каменок придерживаются полуоссдлого образа жизни. Например, в обитающих в Туркменистане, между 35° и 40’ с.ш., Ое. isabellina и Ое. finschii только часть особей (тем большая, чем дальше мы продвигаемся к югу) придерживается зимой своих гнездовых ландшафтов. В основном это самцы. Популяции этих видов из более северных и восточных районов и значительная доля птиц из популяций южного Туркменистана перемещаются на зиму в юго-западную Азию, достигая северо-восточной (Ое.finschii) и экваториальной Африки (Ое. isabellina).
Все прочие виды каменок Европы и умеренных зон Азии — настоящие перелетные птицы, покидающие места гнездования не позже сентября — начала октября. Особенно далекие миграции свойственны тем популяциям, места гнездования которых в результате интенсивного расширения ареалов соответствующих видов оказались локализованными на значительных расстояниях от их первоначальной родины. Например, плешанки Ое. pleschanka из популяций, гнездящихся в восточном Китае, ежегодно дважды покрывают расстояние порядка 8000 км, улетая на зимовки в восточную Африку и возвращаясь весной назад, почти к самому побережью Желтого моря. Еще значительнее расстояния, преодолеваемые обыкновенными каменками Ое. oenanthe, отправляющимися зимовать в экваториальную Африку с Аляски и из Гренландии. Каменка, гнездившаяся на Аляске, чтобы вернуться на то же место весной следующего года, должна проделать два маршрута обшей длиной около 40 000 км. Эти трансконтинентальные перелеты обыкновенные каменки, как и другие дальние мигранты (принадлежащие, например, восточноазиатским популяциям Ое. pleschanka, Ое. isabellina и Ое. deserti), совершают в основном по ночам, останавливаясь на день на кормежку в перевалочных пунктах, отделенных друг от друга сотнями километов. Многие из этих мест отдыха, расположенные в субтропиках
33
Панов Е.Н. Каменки Палеарктики
юго-западной Азии, вполне благоприятны и для зимнего существования обыкновенных каменок. Однако тс, движимые загадочным инстинктом, продолжают из поколения в поколение устремляться на зимние квартиры на свою древнюю африканскую родину.
Очевидно, с течением времени эти архаические связи с древней alma mater постепенно распадаются. Известно, что сегодня немногочисленных Ое. oenanthe можно встретить в зимние месяцы гораздо ближе к местам их гнездования — в Ираке и в Карибском регионе Нового Света (Cramp, 1988). Вероятно, это начало того процесса, который привел к формированию сугубо азиатских зимовок у ряда сравнительно молодых популяций и видов каменок, становление которых происходило уже на Азиатском материке. За его пределы нс выходят в период сезонных миграций такие виды, как Ое. picata и Ое. alboniger. В Индии и юго-восточной Азии зимуют центральноазиатские популяции Ое. deserti oreophila, тогда как гнездящиеся далее к западу расы этого вида продолжают улетать на зиму в северо-восточную Африку (Козлова, 1975).
Очевидно, многие птицы не выдерживают всех трудностей дальних перелетов и гибнут в пути, Однако немало и таких, которые весной возвращаются к предыдущему месту гнездования. Из 9 особей oenanthe, окольцованных на полуострове Ямал в Западной Сибири, далеко за полярным кругом (68’ с.ш.), следующей весной вернулось две — самец и самка, загнездившиеся в 150 м от их прошлогоднего гнезда (Рябицев, 1993). Взрослый самец oenanthe, индивидуально помеченный в высокогорье Заилийского Алатау в Тяньшане в 1978 г., гнездился в 1980 и 1981 гг. в радиусе полукилометра от места первоначальной поимки (Губин, Ковшарь, 1985) Поданным двух названных авторов, на прежние места гнездования возвращается 22-26% взрослых Ое. oenanthe, но ни разу нс были встречены в дальнейшем окольцованные в гнездах птенцы. У среднеазиатской черной каменки Ое. picata, не предпринимающей, в отличие от Ое. oenanthe, столь далеких миграций, возврат взрослых самцов в места гнездования составил 44,3%, взрослых самок — 33,1% и первогодков, окольцованных в гнездах — 12,2% (Любущенко, 1995). В гибридной популяции pleschanka х hispanica в восточном Азербайджане один из самцов-гибридов занимал в период гнездования одну и ту же территорию по крайней мере на протяжении 7 лет (Рубцов, 1995).
1.5. Питание и суточная активность
Объекты питания и кормовое п о в е д е н и е. Многие виды каменок живут в таких условиях, которые не гарантируют им постоянного изобилия пищи. В резко континентальном климате пустыни — особенно высокогорной пустыни, насекомые и прочие бес
34
Глава Г Некоторые общие сведения о каменках
позвоночные периодически исчезают с поверхности почвы; в холодные ночи и жаркие часы дня они прячутся под камни, в норы, уходят глубоко в почву. Особенно неблагоприятны в этом отношении зимние месяцы (для оседлых видов) и ранняя весна. Все это не позволяет каменкам ограничивать свой рацион какой-то определенной группой беспозвоночных, и спектр их питания весьма широк и разнообразен. Так, они могут питаться очень мелкой и очень крупной добычей (Потапова, Панов, 1977), чего обычно нс делают птицы лесов и лугов, в распоряжении которых изобилие насекомых средней величины. Большое место в рационе каменок занимают муравьи — особенно в ранневесенние месяцы. Летом муравьи также составляют важное подспорье. Не исключено, что эти насекомые могут играть роль психологических стимуляторов (наподобие слабых наркотиков у человека).
С другой стороны, каменки часто ловят таких крупных животных, как фаланга, скорпион, египетский таракан, крупных сарапчевых. Птицы поедают такую добычу по частям: они возят и валяют жертву по земле, отрывая от нее по кусочку. Фаланг поедает не только крупная isabellina, но и гораздо более эфемерная Ое. pleschanka. Крупных жуков (например, жуков-чернотелок, диаметром до 15 мм и даже крупнее) Ое. finschii может заглатывать целиком. За полярным кругом Ое. oenanthe — единственный вид насекомоядных воробьиных, не считая сорокопутов, который использует в пищу шмелей Bombus (Успенский, 1969). Жесткие и неперевариваемые части своих жертв каменки выбрасывают через рот в виде погадок.
Некоторые виды каменок (Ое. leucopyga, Ое. leucura, Ое. alboniger, Ое. deserti, Ое. picata) нападают даже на позвоночных, например, на ящу-рок Eremias. С другой стороны, заметное место в их питании осенью занимают плоды. Например, Ое. pleschanka в Памиро-Алае поедает ягоды дикой вишни, а в Румынии — плоды шелковицы. В гибридной популяции pleschanka х hispanica в восточном Азербайджане птицы нередко приносят мелкие ягоды даже птенцам в гнездо. Питание мелкими сочными ягодами Ochradenus baccatus помогает особям Ое. leucopyga в знойной каменистой пустыне Негев восстанавливать запас воды в организме. В местах, постоянно посещаемых туристами в пустыне Негев, местные особи Ое. leucopyga подбирают и поедают либо уносят с собой брошенные людьми кусочки хлеба и фруктов. В периоды бескормицы каменки поедают и сухие семена (Ое. oenanthe, Ое. leucopyga, Ое. leucura и др.). Некоторые мои наблюдения свидетельствуют о том, что во время сильных весенних похолоданий пустынные виды каменок (Ое. finschii) способны подолгу обходиться почти без всякой пищи и оставаться при этом бодрыми и жизнерадостными. Птицы в это время имеют большие запасы подкожного жира.
35
Панов Е.Н. Каменки Палеарктики
Рис. 8. Реакция “разведения крьиьсв” (“движение намерения” — intention movement) у плясуньи Ое. isabeIlina. Оригинал.
Когда каменка занята поисками пищи, она нс ограничивает себя той добычей, которую легко найти на поверхности почвы. Птица постоянно лазает в поры и ниши. Это сопряжено с некоторой опасностью, ибо в норе можно наткнуться на опасного хищника. Каменка отдаст себе в этом отчет, и желание обследовать нору часто борется со страхом перед неизвестностью. Этот внутренний конфликт находит
внешнее выражение в особой позе: птица медленно раскрывает крылья и дела
ет плавные движения вперед (рис. 8). Ту же позу можно наблюдать у всех видов каменок в Момент встречи с крупной незнакомой добычей, да и вообще с любым неизвестным ей предметом (см., например, Smith, 1971). Поэтому описанное движение крыльями наивно толковать как способ “выпугивания” насе
комых из под кустов и из травы, как это делают многие авторы.
У всех известных мне каменок существует еще один способ добывания насекомых — из почвы. Птица роет грунт клювом, ударяя им о землю то с одной, то с другой стороны маятникообразными движениями. Крупную и очень жесткую добычу она долбит клювом сверху — резкими и сильными ударами, наподобие того, как поползень разбивает кедровый орешек, зажатый им в щели коры.
Помимо описанных способов поиска добычи на земле и во всевоз
можных нишах, трещинах скал и пещерках каменки в своих постоянных поисках насекомых нередко используют и другие способы охоты (рис. 9). Они высматривают своих жертв сверху, сидя на камне либо на ветке и при виде ползущего по земле насекомого настигают его быстрым броском сверху, пролетая подчас при этом расстояние до Юм. При отсутствии удобных присад эти птицы иногда высматривают добычу с воздуха, удерживаясь над землей на одном месте быстрыми взмахами крыльев — наподобие охотящейся пустельги Falco finnunculus или зимородка Alcedo atthis. Подобно мухоловкам, каменки с успехом ловят насекомых налету. Особенно часто они охотятся таким образом
36
Глава /. Некоторые общие сведения о каменках
па крылатых особей термитов, в массе покидающих гнездо, где они вывелись, и разлетающихся во все стороны слабым, неровным полетом. У каменок. обитающих в бесплодной каменистой пустыне Негев, существуют и другие способы охогы, практически нс используемые вида ми-обитателями менее суровых ландшафтов. Так, Ое. lugens нередко кормятся здесь насекомыми, которых привлекают цветущие невысокие полукустарнички. Каменка дежурит на валуне, держа в поле зрения несколько кустиков. Заметив жертву, она стремительно, низко над землей проносится по прямой в избранном направлении и схватывает муху или пче-
Время Температура Влажность Ветер
(часы) (°C)
Рис. 9. Частота использование разных способов кормления у обыкновенной каменки Ое. oenanthe при разных погодных условиях (0 а — склевывание с грунта и с низкой растительности, b — выкапывание из грунта, с — ломя налету, d — сзваты-вание при преследовании им землей, е — высматривание с воздуха. Из: Kneis, Liuch, 1983.
лу прямо с кроны куста. Крупные, несколько медлительные Ое. leucopyga высматривают прилетающих на кормежку насекомых снизу, сидя на земле близ основания куста, а
затем влетают в его крону и неуклюже лазают там, схватывая добычу с веток. В Аравии, к этому способу охоты leucopyga прибегает обычно в холодную и в ветренную погоду (Palfcry, 1988).
Отношение к во д е. Создается впечатление, что многие виды
каменок — особенно типично пустынные, могут подолгу обходиться без воды или довольствоваться весьма малыми ее количествами (например, в виде росы или влаги, содержащейся в ягодах). Эта мысль возникает, когда знакомишься с местообитаниями таких видов, как Ое. monacha, Ое. leucopyga, Ое. lugens, Ое. finschii, Ое. deserti, Ое. isabellina, где
37
Панов Е.Н. Каменки Палеарктики
в засушливые летние сезоны порой трудно встретить даже самый маленький водоем. Вместе с тем, когда есть вода, даже наиболее привыкшие к лишениям пустынные виды охотно пьют се в сильный зной. Так поступают, в частности, особи Ое. lugens и Ое. leucopyga, охотно посещают “поилки” в местах сбора туристов в Израиле.
Однако при содержании этих каменок в клетке они пьют много и часто и явно страдают при отсутствии воды. Может быть водный бал-ланс при содержании в искусственных условиях нарушается из-за кормления птиц суррогатами (творог, куриное яйцо, тертая морковь с сухарями).
Молодые Ое. chrysopygia, Ое. deserti, Ое. finschii, взятые из гнезда, в возрасте двух — трех недель интересуются водой, пьют, иногда заходят в плошку с водой, но купаться не пытаются (chrysopygia, deserti) или не решаются (fihschii). Молодые Ое. finschii (иногда — chrysopygia) пытаются “купаться” на сухом месте около плошки с водой. Напротив, молодые isabellina при первом знакомстве с водой залезают в нес и долго, “со знанием дела” купаются. Охотно купаются они и в песке, чего нс делают другие каменки. Живущие в клетках chrysopygia, deserti, finschii практически нс купаются, лишь в очень сильную жару они мочут голову в поилке. Представители других видов купаются при содержании в неволе нерегулярно, от случая к случаю. В естественных условиях Ое. hispanica и Oe.finschН иногда “купаются “ под дождем.
Суточная активность. “Рабочий день” каменки очень длинен. Весной и в начале лета самцы начинают петь еще в глубокой темноте. Интересно, что они поют не только сидя на месте, но и летают с песней на большие расстояния, высоко над землей. Из этого следует, что птицы могут прекрасно ориентироваться в глубоких сумерках, когда глаз человека не способен даже разглядеть певца. Это и не удивительно, если принять во внимание, что каменки совершают свои сезонные миграции в ночное время. В период весеннего пролета именно в ранние утренние часы происходит, по всей видимости, занятие территорий вновь прибывшими самцами, а также формирование брачных пар. Поэтому интенсивная песенная активность самцов-резидентов, заявляющих еще в сумерках свои права на уже освоенные ими земельные наделы и рекламирующих себя самкам более чем оправдана.
С наступлением рассвета самцы начинают кормиться, и возобновляют пение только около 10 часов утра. В середине дня, в самую сильную жару, они обычно мало активны и подолгу могут сидеть неподвижно в тени где-нибудь под навесом камня. Короткий период активности — не столь бурной, как утром, вновь наступает к вечеру. В это время самцы возобновляют пенис. Они посещают свои излюбленные
38
Глава /. Некоторые общие сведения о каменках
песенные посты либо произносят короткие фразы песни, нс прекращая поисков корма. На ночлег птицы устраиваются уже в довольно густых сумерках. Ночуют в глубоких нишах скал или в других закрытых убежищах.
У каменок умеренной зоны спад активности в середине дня выражен не столь резко, как в популяциях, населяющих аридные регионы тропиков и субтропиков. В Субарктике, где в всссннс-летнис месяцы солнце остается над горизонтом день и ночь, местные обыкновенные каменки Ое. oenanthe активны практически на протяжении круглых суток.
Важно заметить, что в начале сезона размножения некоторые виды каменок полупустынных ландшафтов (Ое. picata, Ое. (х.) chtysopygia, Ое. hispanica) часто используют время после захода солнца для брачных игр. В вечерних сумерках нередко происходит и спаривание. К концу дня наблюдается увеличение половой активности и у черношейной каменки Ое. finschii. В ранние утренние часы у всех названных видов эта активность выражена в гораздо меньшей степени.
1.6. Социальное поведение и гнездовой цикл
Пространственная организация д е м а. У оседлых видов каменок по окончании сезона размножения оба члена пары (или только один из них, чаще, невидимому, самец) продолжают держаться в пределах своей гнездовой территории, охраняя се границы — хотя и не столь ревностно, как в брачный период. Ближе к зиме, когда температурные и кормовые условия становятся более благоприятными по сравнению со знойным летом, самцы начинают проявлять признаки повышенной активности. Они все чаще посещают свои любимые песенные посты, извещая округу о своем присутствии и о правах на данную территорию. Начинается восстановление прежних супружеских пар и формирование новых, после чего объединившиеся самец и самка приступают к поискам будущего гнездового убежища. У каменок южных субтропиков все эти события можно наблюдать уже начиная с ноября (например, у аравийской Ое. leucopyga — см. Palfcry, 1988).
У названного и у других оседлых видов права самцов (или пар) на их территории сохраняются из года в год, так что конфликты на почве конкуренции из за пространства редки, они могут возникать лишь при попыток новичков внедриться в уже сложившуюся территориальную структуру. Формирование брачных пар происходит, в общем, асипх-
39
Панов Е.Н. Каменки Палеарктики
Рис. 10. Величина территорий черношейной каменки Ое. finschii в зависимости от времени их занятия (Закавказье): а — нейтральная зона; b — границы территорий; с — гнезда. Из: Панов, 1984.
ронно. В силу всех этих обстоятельств у оседлых видов обычно не приходится наблюдать резких вспышек социальной активности, охватывающих все поселение в целом.
Совершенно иначе обстоит дело у видов перелетных, которые обычно склонны гнездиться в довольно плотных поселениях, внутри которых территории отдельных пар как бы спрессованы друг с друго^ (Дор-жиев, Хсртусв, 1992). Соответственно, площадь территорий в поселении невелика, во много раз уступая размерам гнездовых участков оседлых видов, обитающих в бесплодных аридных местообитаниях тропиков и субтропиков. Если в умеренных зонах средняя площадь территории пары составляет 1-2 га, то в каменистых пустынях Передней Азии эта цифра возрастает до 5-9 га и более (Palfery, 1988 и данные автора по пустыне Негев).
У перелетных видов каменок распределение гнездовых территорий и формирование пар должно происходить в сжатые сроки сразу же по возвращении птиц на места гнездования. Острая конкуренция за право на тот или иной участок местности, как и необходимость без опоздания приобрести полового партнера — все это приводит к тому, что все происходящие события носят бурный и напряженный характер. В общей схеме ход их выглядит следующим образом. Сначала прилетают самцы-пионеры и занимают весьма обширные территории, эффективная охрана границ которых, как вскоре оказывается, не под силу их владельцам. Поэтому самцы, слегка запаздывающие на пролете, селятся здесь же, занимая участки между центрами активности первых поселенцев. Самцов становится все больше, а их территории, соответственно, уменьшаются в размерах. Тем, которые прилетают последними, достаются худшие участки местности, зажатые между активно защищаемыми и лучшими по качеству территориями более ранних поселенцев (рис. 10). Как бы активно нс рекламировал себя хозяин такого неполноценного участка пением и демонстративными полетами, ему либо вообще не удастся привлечь сюда самку, либо это происходит
40
Глава /. Некоторые общие сведения о каменках
лишь тогда, когда на всех соседних участках пары уже образовались. На это может уйти до 25 дней и болсс.
В благоприятных местах гнездования, где поселение неизменно формируется из года в год, уже размножавшиеся здесь самцы в большинстве случаев возвращаются на свои прошлогодние территории. Если собственник участка в момент прилета обнаруживает, что его прошлогодняя территория уже занята прилетевшим ранее самцом, он может смириться с потерей и занять соседний участок, заключающий в себе хорошо известную ему местность. В этом случае оба претендента вступают в непродолжительный конфликт, в ходе которого устанавливается согласованная граница между территориями новоявленных соседей. Однако, прежний собственник участка подчас не склонен идти на подобный компромисс и решительно настроен на то, чтобы отвоевать все, что принадлежало ему в прошлом году. В подобных ситуациях конфликт между обоими претендентами затягивается порой на многие часы. Погони и взаимные Демонстрации порой переходят в жестокую драку, и к концу взаимодействия со-
4000
3000
2000
1000
27 | 3 7 9 11 13 15 17 19
Рис. II. Сроки приобретения самок самцами черных каменок в полиморфной популяции в зависимости от качества территорий самцов: SI — площадь территорий; S2 — площадь вертикальных поверхностей внутри территории (южный Узбекистан); а — самцы capistrata', b — предполагаемые гибриды capistrata х picata, с — opistholeuca: d — evreinowi; е — picata. Из: Панов и Корзухин, I974.
41
Панов Е.Н. Каменки Палеарктики
Рис. 12. Сроки формирования пар в полиморфной популяции испанской каменки Ое. hispanica (а) и время (количество дней) от занятия самцом территории до приобретения им самки (Ь). В позиции b бслогорлыс самцы aurifa обозначены белыми квадратиками, чер-ногорлые самцы stapazina — черными. Слева внизу — самцы-первогодки, окрашенные по самочьему типу (Закавказье). Из: Панов, 1989.
□ □
ПО 15 20
Апрель
перники настолько измождены, что уже почти не в состоянии летать (подробнее см. Панов, 1978). В условиях столь жесткой конкуренции из-за пространства часть самцов вынуждена мириться с участками, непривлекательными для самок (рис. 11), а некоторым первогодкам так и нс удастся занять собственный участок, и они ведут жизнь без
Рис. 13. Интенсивность пения холостого самца Ое. finschii при рекламировании им своей территории (а) и в момент появления на территлории самки (Ь) (Мангышлак). Оригинал.
42
Глава I. Некоторые общие сведения о каменках
домных аутсайдеров в ожидании случайно освободившейся вакансии. Впрочем, если самцу-первогодку удалось занять качественную территорию, он получает перспективу приобрести самку-супругу быстрее, чем самец старше года в роскошном оперении, обладающий “плохим” участком (рис. 12).
Выбор территорий с а м к а м и. С началом прилета самок обстановка в поселении вновь обостряется. В момент появления здесь новоприбывшей она перелетает с территории на территорию с намерением сделать наиболее удачный выбор. Самка ведет себя очень заметно и нередко даже произносит короткие фразы песни. Во время пребывания самки на данном участке его хозяин приходит в экстаз и своим экстравагантным поведением привлекает внимание самцов-соседей (рис. 13). Те зачастую настолько возбуждаются, что склонны игнорировать согласованные границы и вторгаются на территорию счастливчика. Он же пытается изгнать нсзванных пришельцев, а самка тем временем может перелететь на соседний участок, увлекая за собой самца, который из защитника своей территории сразу превращается в нарушителя границ соседних участков. Что касается самки, то она или окончательно останавливается в пределах определенной территории, или вообще покидает поселение, устремляясь на новые поиски. У разных видов каменок описанная картина может варьировать в деталях в зависимости от строгости территориальных запретов (более жестких, скажем, у Ое. finschii и Ое. hispanica и нс столь строгих у Ое. isabellina и Ое. pleschanka) и от общей экспрессивности поведения того или иного вида, а также от таких привходящих факторов, как плотность поселения и степень синхронности его формирования весной (см. Панов, Корзухин, 1974; Панов, 1978, 1989).
Уже гнездившиеся в поселении самки, как и самцы, могут претендовать на свой прошлогодний участок, пытаясь изгнать в момент прилета опередившую ее другую самку. При этом самец-хозяин данной территории порой безучастен и предоставляет самкам самим решать свои проблемы. В такой ситуации конфликт между претендентками может принять столь же затяжной и жестокий характер, как и ранее описанные длительные конфликты самцов. Подчас он продолжается часами и ведет к почти полому обессиливанию одной из самок, которая в конце-концов уступает сопернице и покидает участок. А однажды я наблюдал подобное антагонистическое взаимодействие между самками Ое. hispanica, которое продолжалось весь день и возобновилось на следующее утро.
Полигиния. Иногда самец вмешивается в подобный конфликт самок, принимая, как правило, сторону вновь прибывшей. По отношению к своей первой подруге самец в этих случаях ведет себя довольно
43
Панов Е.Н. Каменки Палеарктики
агрессивно. При таких условиях второй самке удастся закрепиться на территории, и в результате образуется полигиническое трио. Самки, приняв в конце концов соседство друг с другом как неизбежное, делят территорию самца пополам. Каждая кормится в своей зоне, где находится и се гнездо. Впоследствии самец может принимать участие в выкармливании обоих выводков. Подобные полигинные семьи описаны у Ое. isabellina, Ое. picata и Ое. pleschanka (Иваницкий, 1981b; Панов, 1978; Любушенко, 1995). Гораздо реже мы видим у каменок другую форму многоженства — гак называемую политерриториальную полигинию. В этом случае самец имеет две территории в разных местах, и в пределах каждой из них обзаводится супругой. Подобная ситуация описана у Ое. oenanthe (Haartmann, 1969) и у Ое. finschii (Панов, 1978). Понятно, что ожидать ее реальнее всего в разреженных популяциях каменок.
Охрана семейной территории. Как правило, оба члена сформировавшейся пары активно охраняют границы своей общс|й территории. При наблюдениях за видами с резко выраженным половым дихроматизмом хорошо видно, что самцы охраняют территорию от прочих самцов, а самки — от самок. У некоторых видов охрана территории весьма основательна (Ое. finschii, Ое. picata), у других периферические участки соседних территорий могут перекрываться в большей или меньшей степени, и эти зоны перекрывания используются соседями поочередно (Ое. isabellina, Ое. pleschanka). Холостые самцы Ое. hispanica охраняют не только свой земельный надел, но и воздушное пространство высоко над ним, то есть территория у этого вида трехмерна. Впрочем, сразу после образования пары рекламная территория самца резко уменьшается в размерах, а интенсивность ее охраны заметно падает. По существу, некоторые самцы Ое. hispanica охраняют теперь не столько свой первоначальный обширный участок, сколько пространство вокруг самки.
Когда самцы-соседи окончательно выяснили между собой, где именно проходит граница между их участками, они впредь без особых на то оснований стараются не пересекать этот рубеж. Однако время от времени то один, то другой из них прилетает на эту невидимую линию и вызывает соседа на короткий турнир. Продемонстрировав друг другу свое мужество, участники спустя несколько минут покидают место конфронтации, и каждый летит в свои владения. Обычно эти турниры достаточно безобидны, но иногда они выливаются в короткую драку. Самцы словно ищут в этих встречах разрядки своим накопившимся эмоциям.
Взаимотношения между членами семейной п а р ы. На стадиях освоения территории и гнсздостросния супруги чаще всего держатся вместе, хотя у некоторых видов (особенно у Ое. pleschanka) тяготение партнеров друг к другу не столь сильно, так что
44
Глава Г Некоторые общие сведения о каменках
самец нс препятствует самке посещать участки соседних пар. К моменту, когда самка завершает постройку гнезда, оба партнера все чаще проявляют признаки готовности к копуляции. У каменок этот акт каждый раз требует длительной психологической подготовки как со стороны самца, так и самки. Первый своим пением и брачными демонстрациями подолгу пытается привлечь самку в специально отведенное для копуляции место, а та все не решается приблизиться, и даже пойдя на контакт, зачастую нс позволяет самцу сделать садку и довести дело до конца. Эти длительные брачные игры привлекают внимание самцов-соседей, которые прилагают все усилия, чтобы проникнуть на территорию и спариться с самкой, так или иначе обманув бдительность ее супруга. Как правило, эти попытки адюльтера нс приносят успеха пришельцу, которого самец-хозяин свирепо изгоняет прочь.
Когда самка приступает к насиживанию кладки, самец, оказавшись, по существу, в одиночестве, вновь, как и в первые дни после прилета, начинает активно петь и токовать, пытаясь привлечь на свой участок запоздавших на пролете самок. Находясь в состоянии высокой половой активности, он нередко посещает места предыдущих свиданий с самкой и проделывает здесь свои брачные демонстрации в одиночестве, подчас завершая их “спариванием” с камешком, с кусочком глины или с обломанным торчащим стеблем.
Уход за потомством. Вылупление птенцов заставляет самца впервые основательно приступить к выполнению своих родительских обязанностей. В первые дни самка подолгу остается в гнезде, обогревая птенцов, и на самца падает значительная часть забот по их кормлению. Как только молодежь покинет гнездо, самка, если позволяют климатические и погодные условия, сразу же приступает к сооружению нового гнезда для следующего выводка. В первые дни она совмещает строительство с кормлением слетков, но в дальнейшем все реже уделяет им внимание. Когда же самка приступает к насиживанию второй кладки, молодняк оказывается полностью на попечении самца.
Если же пара по тем или иным причинам не собирается выводить потомство вновь, родители нередко делят выводок пополам, так что лишь часть слетков оказывется на попечении самца, тогда как других юнцов кормит самка (Moreno, 1984).
Впрочем, спустя 2-3 недели привязанность родителей к потомству все чаще начинает сменяться проявлениями нетерпимости по отношению к ним, особенно со стороны самца. Интересно, что молодые к этому времени достигают возраста, когда резко меняется их отношение к “приемному родителю”, в качестве которого выступал я, выкармливая юных каменок в клетках. Еще вчера полностью оперившийся юнец охотно брал корм прямо из моей руки, а сегодня он в ужасе шарахается от нес и
45
Панов Е.Н. Каменки Палеарктики
Рис. 14. Элементы сигнального поведения некоторых видов Дроздовых (Turdidae), характерные также для каменок Oenanthe: а, Ь, е — разворачивание перьев хвоста; а,Ь — приподнимание и выдвигание вперед сгибов крыльев; d, j — вытягивание головы вперед и вверх; с, g, h — наклон вперед, часто с распушением контурного оперения; i-k — задирание хвоста; а — Cossypha natalensis: b — Saxicola torquata: c — C. heuglini; d, e — Chaimarromis leucocephala: f — Phoenicurus ochrurosphoenicuroides\ g, h — Monticola saxanlis; i — Myrmecocichla melaena: j — Luscinia svecica: k — Cercofrichasgalactotes. По материалам автора, Farkas, 1969 (a), 1973 (c), Perlo. 1995 (i).
48
Глава /. Некоторые общие сведения о каменках
свойственны практически всем изученным представителям семейства дроздовых, так что вполне естественно обнаружить их и у всех без исключения видов каменок. Сюда относятся, в частности, вертикальная постановка головы (клюв обращен вертикально вверх) и широкая амплитуда движений хвоста вниз и вверх, до вертикального его положения, а также разворачивание рулевых перьев веером (рис. 14). В то же время некоторые ЭДА описаны лишь у немногих видов каменок, а то и вообще у единственного вида — как, например, широкое открывание клюва с демонстрацией партнеру угольно-черного зева у Ое. isabellina.
При обшем сходстве большинства видов каменок в наборах исходных ЭДА и их комбинаций (поз), у разных видов эти компоненты поведения могут использоваться с разной частотой или в неодинаковых ситуациях. Например, разворачивание рулевых веером практически не свойственно Ое. lugens в таких ситуациях, при которых этот ЭДА почти неизменно можно наблюдать у Ое. finschii и Ое. picata. При формировании пар для самцов большинства видов характерна полуверти-кальная или вертикальная поза с поднятым кверху клювом и с широко развернутым хвостом, и лишь у Ое. chrysopygia самец во время таких взаимодействий чаще принимает горизонтальную позу, при которой клюв направлен вперед, а рулевые перья собраны вместе.
Некоторые акты сигнального поведения используются с явно неодинаковой частотой у разных видов каменок. Эффектный “полет петлями” (см. рис. 15, 16, 108) присутствует в коммуникативных репертуарах всех без исключения каменок северной Палеарктики. Самцы совершают этот полет в качестве неизменной прелюдии к акту копуляции (рис. 36, 97). У черношейной каменки Ое. finschii полет петлями лишь в исключительных случаях удается увидеть в иных ситуациях — например, в ходе бурного территориального конфликта двух самцов-соперников. В отличие от названного вида, у плсшанки Ое. pleschanka и испанской каменки Ое. hispanica полст-петлями — вполне обычная акция самцов практически во всех типах их взаимодействий как с самками, так и с самцами своего вида. Самцы нередко проделывают эту демонстрацию даже в отсутствие партнера, “вхолостую”. Болес того, у этих двух видов полет петлями изредка демонстрируют и самки (Панов, 1978: 179).
Не останавливаясь на множестве подобных различий, суть которых читатель сможет уяснить после ознакомления с очерками по каждому виду, можно дать сугубо обобщенную схему использования “зрительных” сигналов в ходе гнездового цикла.
Саморекламирование самцов и организация взаимодействий при формировании п а р. На первых стадиях цикла самец, закрепивший за собой территорию, рскламиру-
49
Панов Е.Н. Каменки Палеарктики
Рис. 15. Амплитуда движения крыльев самца при демонстративном облете территории (левый ряд), траектория демонстративного полета (второй ряд слева) и траектория полета петлями (справа; в позициях с, i показаны по два варианта этой демонстрации, в позиции е — три): а — плясунья Ое. isabe/lina; b — обыкновенная каменка Ое. oenanthe; с — златогузая каменка Ое. chrysopygia; d — черно-шейная каменка Ое. finschii; е — черная каменка Ое. picata; f — плешанка Ое. pleschanka, Ое. hispanica; g — испанская каменка Ое. hispanica; i — пустынная каменка Ое. deserti. Из: Panow, 1974.
50
Глава 1. Некоторые общие сведения о каменках
ет свое присутствие и свои владения с помощью особого демонстративного полета, который у каждого вида имеет свои характерные особенности (рис. 15). В плотных поселениях холостые самцы отвечают на эти воздушные демонстрации своими собственными, так что рекламные полеты соседей часто синхронизированы. Это обстоятельство, вероятно, облегчает обнаружение поселения мигрирующими самками.
При появлении самки в пределах территории холостого самца он в первый момент не делает попыток сблизиться с ней вплотную. Вместо этого самец устремляется к заранее облюбованной им норе или нише, пригодной для устройства гнезда, и демонстрирует здесь экстравагантные брачные позы, то и дело залезая в отверстие и вновь появляясь из него. Когда самка подлетает к месту демонстраций самца, он устремляется к другому потенциальному гнездовому убежищу, где все повторяется сначала. Таким образом самец выступает в качестве гида, знакомя самку со всеми достоинствами своего участка.
Если этот первый этап помолвки прошел гладко, и самка не отправилась на дальнейшие поиски, самец делает первые попытки сблизиться с ней по собственному почину. Самка подпускает самца, но дает ему лишь несколько секунд, чтобы исполнить очередное па брачного танца, отвечающее требованиям ритуала. В следующий момент самка срывается с места, а самец устремляется за ней, стремительно следуя всем изгибам траектории се полета. Когда самка садится или прячется от самца (в нишу либо в гущу куста), тот проделывает вокруг нее уже упоминавшийся, весьма экспрессивный воздушный танец — “полет петлями’* (рис. 16).
Такого рода погони можно наблюдать на протяжении всего времени освоения самкой территории и последующего гнсздостросния. У некоторых видов (Ое. hispanica, Ое. pleschanka, Ое. isabellina) эти погони со временем преобразуются в особый очень медленный совместный полет, во время которого обе птицы подобно двум ночным бабочкам тандемом перемешаются низко над землей. По крайне мере у первых двух из названных видов такие совместные демонстрации происходят обычно в густых вечерних сумерках и заканчиваются, как правило, уже упомянутым воздушным танцем самца (полет петлями). И хотя время близиться к спариванию, эти акции, как правило, не служат прелюдией к копуляции.
Организация взаимодействии при копуляции. Как я уже упоминал, самому спариванию предшествует обычно очень Длительная подготовка. Дело в том, что у каменок, как у видов с низким порогом территориальной агрессивности как самцов, так и самок, прямой телесный контакт между особями сязан, очевидно, с сильным психологическим стрессом. Копуляция происходит в специ-
51
Панов Е.Н. Каменки Палеарктики
Рис. 16. Схема формирования пары у черных каменок Ое. р. picata. При появлени самки (слева вверху) на территории холостого самца он начинает свои демонстрации у одной и з ранее выбранных им шелеп в скале (справа вверху). Затем самец летит к самке, а она улетает от него (в центре). Следует погоня, в конце которой самец совершает вокруг самки полет петлями (справа внизу). Из: Панов, 1989.
52
Глава L Некоторые общие сведения о каменках
алию отведенных для этого местах, куда самка в конце концов прилетает, привлекаемая пением и соответствующими демонстрациями самца. У видов-петрофи-лов роль таких “мест свиданий” выполняют особые небольшие пещерки с плоским полом, где самец появляется, как правило, под вечер, и где он, пытаясь преодолеть нерешительность самки, раз за разом повторяет разнообразные фигуры своего брачного танца. Эту манеру спариваться в укромных, спрятанных от чужих глаз убежищах каменки делят, как и многие другие черты сигнального поведения, со своими близкими и отдаленными родичами из семейства дроздовых — например, с синей птицей Myophonus caeruleus и синим каменным дроздом Monticola solitarius (см. Панов, 1990).
Акустическая с и г н а л и з а ц и я: п о з ы в к и и песни. Помимо описанных средств зрительной сигнализации все каменки используют в общении друг с другом богатый вокальный репертуар. Его
kHz
4
2
0,5 1,0 1,5
2,0 sec
Рис. 17. Основные позывки каменок на примере черношейной каменки Ое. finschii: а — серия коротких свистов (вначале, сигнал N 3) и щелчков (сигнал № 1, две последние посылки; b — разные варианты сигнала № 2 (храпение); с — сигнал № 4 (трещание); d — сигнал № 5 (пикание); е, f — вокализация птенца в ожидании родителей и при выпрашивании у них корма (сигнал № 6, дребезжание).
можно условно подразделить на два класса сигналов: это, во-первых, сравнительно простые по структуре позывки и, во-вторых, гораздо более сложные акустические конструкции — песни. Подразделение это условно потому, что многие типы позывок могут использоваться как сами по себе, так и в составе песенных конструкций. В репертуаре большинства
53
Панов Е.Н. Каменки Палеарктики
хорошо изученных видов каменок насчитывается шесть основных типов позывок. В Табл. 3 в качестве примера приведены акустические характеристики этих сигналов из репертуара Ое. finschii и перечислены основные ситуации, в которых каждый из них может использоваться (см. также рис. 17). Что касается других видов каменок, то у них отличия от обрисованной картины касаются, как правило, лишь частных деталей. Важно, однако заметить, что сигнал № 5 полностью отсутствует по крайней мерс у двух видов — Ое. hispanica и Ое. pleschanka, что вкупе с другими особенностями их вокализации (см. Панов, 1989) говорит о значительной обособленности этой пары близких видов внутри рода Oenanthe. *
Среди песенных конструкций у большинства видов более или менее четко разграничиваются 3-5 типов песен (рис. 18). Это, прежде всего, короткая рекламная песня, которую самец воспроизводит сериями, сидя на том или ином из своих излюбленных постов. Когда по той или иной причине возбуждение самца нарастает, паузы между следующими друг за другом песнями сокращаются, и они постепенно сливаются в рекламную удлиненную песню. Именно она чаще всего воспроизводится во время демонстративного облета самцом своей территории. О повышении половых мотиваций самца может свидетельствовать также подпесня, которая по структуре весьма сходна с агрессивной песней, сопровождающей наиболее напряженные конфронтации самцов. От последней подпесня отличается в основном лишь своим гораздо более тихим звучанием. Наконец, у ряда видов особую категорию составляет брачная песня, с помощью которой готовый к копуляции самец завлекает самку в упоминавшиеся уже “места свиданий”. Впрочем, у большей части видов такая брачная песня чрезвычайно сходна с подпеснсй и агрессивной песней.
Подчас разные виды очень сильно различаются особенностями песенного поведения. У одних видов рекламная песня весьма проста и отличается малым разнообразием вариантов, каждый из которых весьма стереотипен. Примером может служить Ое. deserti. Песенный репертуар других видов поистине неисчерпаем: только у одного самца Ое. р. picata за месяц наблюдений насчитали 264 разных варианта коротких рекламных песен). У этого и у ряда других видов (например, Ое. hispanica и Ое. pleschanka) каждый такой песенный вариант в репертуаре данного самца имеет постоянную структуру и всякий раз воспроизводится певцом практически без изменений. У других видов (например, у Ое. finschii) самцы более склонны к импровизации. У каждого из них в запасе имеется набор фрагментов-нот, комбинируя которые певец постоянно разнообразит свой репертуар рекламных песен (рис. 19; см. Костина, Панов, 1981; Панов, 1989; Грабовский, Панов, 1992).
Если же говорить о других типах песен (удлиненная, подпесня, агрессивная и брачная песни), то здесь принцип импровизации гос-
54
Таблица 3. Репертуар позывок черношейной каменки Oenanthe finschii и их использование в качестве средств коммуникации (из: Панов, 1978, с изменениями).
Название сигнал апо-зывки Структурные признаки Зоны концентрации энергии (кГц) Длина одной посылк-и (мск) Характер использования Как элемент песни
1 Глухой щелчок Шумовой. Одиночный или сериями (также в сериях с сигналом 3) 1.5-5.0 20-50 Территориальные конфликты между самцами и между самками. В вокализации самок при формировании пар. Тревожный сигнал у гнезда (обычно в сочетании с сигналом 3) да
2 Храпение Шумовой. Одиночный, реже -сериями 1.5-5.0 80-450 Патрулирование границ территории самками. Максимальный уровень тревоги у гнезда с больними птенцами нет
3 Короткий свист Тоновый. Обычно сериями (часто -попеременно с сигналом 1) 3.3-5.0 50-80 Гри территориальных конфликтах (самцы). При образовании пар (самцы). После копуляции (самцы). Реакция на хищника. Тревога у гнезда (часто в сочетании с сигналом 1) нет
4 Треск Шумовой. Одиночный или короткими сериями 3.5-5.0 120-160 Гри территориальных конфликтах самок . Гредугреждакхиий сигнал самки, не подпускаюивй к себе самца при образовании пары и в дальнейшем нет
5 Циканье Тоновый. Чянр -сериями 6.5-8.0 50-70 Образование пар (самцы). В преддверии и во время копуляции (самцы). Реже - при территориальных конфликтах (самцы, самки) да
6 Дребезжание Шумовой. Как правило, сериями 2.0-4.0, до 8.0-11.0 200-300 Основной вокальный сигнал птенцов и слетков. Образование пар (самцы) да
Глава /. Некоторые общие сведения о каменках
Панов Е.Н. Каменки Палеарктики
Рис. 18. Основные типы песен каменок на примере Ое. finschii-. а — короткая рекламная песня (advertising song); b — удлиненная песня; с — подпссня; d — брачная (предкопуляционная) песня.
подствуст у всех без исключения видов. Особенно ярко он проявляется у каменок, в вокализации которых заметное место занимают имитации голосов других животных (птиц, лягушек, реже — млекопитающих). Каждый вид имеет излюбленные модели для таких имитаций, так что порой последние оказываются практически постоянным элементом акустического репертуара в той или иной популяции (таков, в частности, "визг” стрижа Apus apus в песнях Ое. hispanica в Закавказье). Наиболее искусные имитаторы среди каменок северной Палеарктики — это Ое. isabellina и Ое. picata. Они с необычайным мастерством подражают звукам, акустические характеристики которых лежат, казалось бы, за пределами вокальных возможностей миниатюрного певца (рис. 20).
56
Глава /. Некоторые общие сведения о каменках
Рис. 19. Организация песни у Ое. finschii. Точками показано присутствие разных нот (1-18) в 25 последовательных песнях одного самца.
2 mnatf 'tuqiiihii ...:.nu ISM .ими Л * Ш -* •»”* ииМ
0,5 1.0 1,5 2.0 2.5
0,5 1,0 1,5 2.0 sec
Рис. 20. Имитации голосов других видов птиц самцом Ое. р. picata: а, Ь. с — голоса сизоворонки Coracias garrulus, ворона Corvus corax и пустельги Falco finnunculus; ар br с( — имитации этих звуков самцами черной каменки.
57
Панов Е.Н. Каменки Палеарктики
Таблица 4. Использование имитаций голосов других видов животных в разных типах песен некоторыми видами каменок Палеарктики.
Вид Подпесня Песня при агрессивных и брачных взаимодействиях Извещающая (рекламная) песня
isabellina +++ + +++
oenanthe ++ + +
(xanthoprymna) chrysopygia + + I +
finschii + + +
picata +++ + ++ ++
hispanica ++ ++ +++
pleschanka ++ + + ++
deserti + + +
+++ имитации используются постоянно, ++ редко, -I- крайне редко.
Наряду с видами-имитаторами среди каменок немало и таких, которые или вообще не склонны копировать звуки окружающего их мира, или делают это крайне редко. Таковы, в частности, Ое. oenanthe, Ое. chrysopygia, Ое. finschii, Ое. iugens и Ое. deserti (Табл. 4).
1.8. Гнезда и кладки
Места расположения гнезд. Почти все каменки устраивают свои гнезда в надежном укрытии — в расселинах между камнями, в щелях скал, в углублениях стенок оврагов. Лишь Ое. tractrac, Ое. deserti и Ое. h. hispanica нередко вьют гнездо прямо на земле — под укрытием камня, кустика или поваленного дерева. Из 32 гнезд последнего вида, найденных в Испании, 29 располагались в неглубоких ямках у оснований густых кустов (Suarez, 1977). Подобное расположение гнезд в виде крайне редких исключений возможно и у других видов (Ое. isabellina,Oe. phillipsi,Oe. pleschanka).Вилы, предпочитающие выровненную местность сильно расчлененному каменистому рельефу (Ое. pileata, Ое. isabellina, Ое. moesta), в качестве укрытий для гнезда используют норы колониальных пустынных грызунов — всевозможных видов песчанок в южных аридных районах, сусликов и пищух — в более северных и восточных сухих степях в ареале Ое. isabellina. Популяции
58
Глава /. Некоторые общие сведения о каменках
Ое. finschii, занимающие каменистые предгорья, устраивают гнезда в различных пустотах скальных обнажений, а те, что вторично перешли к жизни в песчаной пустыне, помешают гнезда в норах песчанок. Названные виды, а среди них в первую очередь Ое. isabellina и Ое. moesfa, нуждаются в период гнездования не просто в каких-нибудь пустотах, где можно спрятать гнездо посреди ровного места. Они явно предпочитают жилые колонии грызунов, в которых недавно использовавшиеся норы находятся сше в хорошем состоянии (Доржиев, Хсртусв, 1992).
К гнездованию в норах млекопитающих очень часто прибегают также некоторые популяции Ое. oenanthe — в частности те, что обитают в суровом климате высокогорий и Субарктики. Например, из 30 гнезд этого вида, найденных в альпийскоми поясе гор Тянь-Шаня, 19 располагались в брошенных норах сурков, а в тундрах северо-восточной Азии гнездовые пары явно концентрируются в поселениях длиннохвостых сусликов Cifellusрлггу'(Кишинский, 1980; Губин, Ковшарь, 1985). Будучи во многих регионах видом-синантропом, обыкновенная каменка вообще необычайно изобретательна в выборе мест для гнезда. Они используют для этого пустоты в штабелях дров, в кучах кирпича, в свалках мусора, под брошенными листами шифера и под крышами строений, скворечники на лесных вырубках и другие самые неожиданные места. Одно гнездо было устроено в камнедробилке, работавшей по 7-8 час. в день при шуме в 120 децибел, причем пара вывела здесь же и второй выводок (Lorck, 1989). Стоит ли после всего этого удивляться успехам обыкновенной каменки в колонизации всей Евразии и части Нового Света?
Участие партнеров в гнездостроен и и. У подавляющего большинства видов каменок все заботы по устройству гнезда ложатся на плечи самки. У видов-петрофилов, предпочитающих для гнездования каменистый субстрат(Ое. chrysopygia, Ое.finschii, Ое. lugens, Ое. picata, Ое. alboniger, Ое. leucopyga) самка начинает постройку гнезда с сооружения платформы из плоских камешков, которые она собирает в радиусе до 50-100 м от гнездовой ниши, тщательно выбирая те, что соответствуют определенной форме и весу. Перебирая клювом камешки и найдя наконец подходящий, самка каждый раз летит с ним к гнездовому убежищу — часто почти вертикально кверху. Я подсчитал, что за 2-3 дня, уходящие на постройку платформы, самка черной каменки Ое. picata проделывает подчас работу величиной порядка 5 кгм. У каменок-петрофилов более крупных размеров суммарные усилия могут быть еще выше. Например, у Ое. leucura некоторые камешки в платформе имеют длину до 6 см и вес до 14 г. Это единственный вид каменок, у которого в сооружсни платформы участвуют, повидимому, оба члена пары (Moreno et al., 1994). У самцов Ое.
59
Панов Е.Н. Каменки Палеарктики
Рис. 21. Использование камешков при сооружении гнезда самками черных каменок: а, b — камешки выполняют свою функцию, предохраняя гнездо от сползания вниз; с, d — необходимость в камешках отсутствует; е — пример иррационального поведения самки; все камешки, принесенные ею на округлую балку, попадали вниз, (d — из: Moreno et al., 1994).
chrysopygia сохра-нились лишь атавистические следы этого поведения. Зачатки других форм гнез^острои-тельной деятельности (например, попытки брать и переносить щепочки, соломинки и пр.) также наблюдаются у самцов этого вида и других видов каменок. Например, самец Ое. picata, очень долго остававшийся холостым, начал обдирать со стеблей растений их тонкую кожицу — точно так же, как это делает самка, добывающая материал для выстилки гнезда.
Конструкция гнездовой постройки. Функция платформы из камешков очевидна в тех случаях, когда структура каменистых обрывов такова, что самке нелегко подыскать гнездовую нишу с горизонтальным полом. В таких случаях приносимые самкой камешки выравнивают поверхность субстрата и предохраняют гнездо от сползания (рис. 21а-с). Однако, поскольку поведение, связанное с постройкой платформы, носит характер слепого инстинкта, самка пытается соорудить платформу и в тех случаях, когда она совершенно бесполезна как опорное сооружение (рис. 2Id,с). Сооружение платформы сохраняется в качестве атавизма у тех популяций каменок-пстрофи-
60
Глава /. Некоторые общие сведения о каменках
лов, которые вторично перешли к гнездованию на ровном мягком грунте — например, у обитающих в глинистых и песчаных пустынях популяций Ое. finschii (Митропольский, 1968b). Повадка закладывать камешки в основание гнезда наблюдается и у Ое. moesta (Shirinai, 1996), которая почти окончательно порвала свои связи с каменистыми ландшафтами и целиком перешла к гнездованию в норах пустынных грызунов.
У видов с относительно мелкими размерами особей, таких как Ое. hispanica и Ое. pleschanka, самки также сооружают помост под гнездом, используя для этого, однако, не камешки, а палочки длиной от 2 до 16 см. В некоторых гнездах hispanica общий вес платформы доходит почти до 100 г (Panow, 1974; Лоскот, 1983). Последний автор также считает, что главная, если не единственная функция такой платформы — опорная, хотя зачастую фундамент сооружается в таких местах, где он совершенно не нужен и обязан лишь слепому врожденному стереотипу. Поэтому мы можем сегодня без колебания отбросить множество гипотез о назначении платформы, связывающих эту конструкцию с потребностями терморегуляции и зашиты от хищников, с адекватным выбором полового партнера при формировании семейных пар и т.д. (см., например, Perrin de Brichambaut, 1989; Moreno ct al., 1994).
Интересно, что у некоторых видов либо подвидов каменок, отнюдь не избегающих каменистых ландшафтов (Ое. oenanthe, Ое. monacha, Ое. deserti oreophila), самки, судя по имеющимся данным, не сооружают специальных платформ, если не считать таковыми нагромождение толстых стеблей, закладываемых в основание гнезда. Не описана эта повадка и у тех каменок, которые изначально связаны с выровненными элементами ландшафта, пусть даже высокогорного — в частности, у Ое. isabellina и у равнинных подвидов Ое. deserti. Самки таких видов, сооружающие гнездо на мягком субстрате, могут, очевидно, слегка углублять грунт в основании гнезда или очищать уже готовую ямку от лишних предметов — растительного мусора и мелких камешков. Так поступают, вероятно, и самки тех видов каменок, которые охотно гнездятся в каменистых ландшафтах (Ое. hispanica — см. Лоскот 1983).
Само по себе гнездо каменок ничем не замечательно. Это открытая сверху чаша, основание которой сооружается из сухой травы. Лоток гнезда самка выстилает более нежными растительными материалами, а У ряда видов — также волосом и перьями. У разных видов размеры гнезда могут быть неодинаковыми. Иногда все сооружение невелико и изящно выполнено (Ое. monacha, Ое. deserti), в других случаях — довольно велико и имеет толстые стенки, особенно в основании гнезда. Таковы, в частности, гнезда Ое. isabellina, гнездящейся в норах грызунов Далеко от входа (до 1,5 м), где субстрат может быть довольно влажным.
61
Панов Е.Н. Каменки Палеарктики
Величина кладки колеблется в зависимости от географической широты и от климатических условий. Виды, ареалы которых целиком лежат в пределах Африки (Ое. tractrac, Ое. monticola,), а также африканские и переднеазиатскис популяции более широко распространенных видов (например, Ое. deserti) имеют небольшую кладку, состоящую из 2-4 яиц, чаше из 3 (Ое. lugubris, Ое. bottae, Ое. tractrac, Ое. leucopyga) У видов умеренной полосы полная кладка содержит4-7 яиц (чаще 5-6). Эта закономерность увеличения числа яиц в кладке с продвижением на север и на восток сохраняется даже при сравнении разных популяций и подвидов одного и того же вида. Например, у Ое. oenanthe в Казахстане большая часть полных кладок содержит по 5 яиц (23 из 43 найденных); кладки из 6 яиц встречаются реже (10), и лишь однажды была обнаружена кладка из 7 яиц. У того же вида в Англии кладки из 5 яиц составляют значительно меньше половины найденных (35%) и уступают в числе более крупным кладкам из 6 (45%), 7 (9%) и 8-9 (менее 2%) яиц. У испанской каменки Ое. hispanica близ южной границы се ареала, в Израиле, численно преобладают кладки из 4 яиц, тогда как у северо-восточной границы ареала этого вида в 13 гнездах из 24 найденных было по 5 яиц и в 9 — по 6. В 36 из 57 гнезд африканской формы Ое. deserti deserti содержались кладки из 4 яиц, а в 14 — из 5, причем это был максимальный размер кладки (средняя се величина 4,1). В Казахстане у местной расы Ое. deserti atrogularis численно преобладают кладки из 6 яиц, а средняя се величина составляет 5,3. У этого вида число яиц в кладке снова уменьшается при продвижении далее к востоку, например, в высокогорья Памира (3600-4500 м над у.м.) с его чрезвычайно суровыми климатическими условиями. Здесь, при бедности кормовой базы, пара Ое. deserti oreophila часто не в состоянии успешно выкормить выводок даже из 5 птенцов, так что наиболее часто встречаются кладки из 4 и из 3 яиц (Потапов, 1966; Сурвилло, 1968; Гаврилов, 1970; Губин, Ковшарь, 1985; Лоскот, 1983; Cramp, 1988; Shirinai, 1996). Общим правилом является уменьшение величины кладки при повторных и при поздних попытках гнездования.
Однократное и многократное гнездование. В районах с благоприятным климатом (раннее начало весны, не слишком жаркое и сухое лето) многие виды каменок склонны в норме гнездиться более одного раза за сезон гнездования. Например, в юго-восточной Испании с ее умеренно-жарким климатом (среднегодовые температуры +10-20") период, когда природные условия не препятствуют гнездованию оседлой Ое. leucura leucura, длится 4 месяца (с начала марта до конца июня). За это время некоторые пары предпринимают до 5 попыток гнездования, из которых 3 могут оказаться успешными (Soler et al., 1995). Два обстоятельства могут воспрепятство
62
Глава Г Некоторые общие сведения о каменках
вать множественному гнездованию каменок в течение одного сезона. Это слишком раннее начало летнего зноя в аридных зонах тропиков и субтропиков, и очень поздняя весна в сочетании с коротким летом в Арктике и Субарктике. Пример первой ситуации мы видим на Аравийском полуострове. Обитающие здесь Ое. leucopyga приступают к гнездо-строению уже в последней декаде января, но даже при этом лишь единичные пары решаются вновь загнсздиться в середине марта, когда угроза надвигающегося знойного лета уже вполне очевидна (Palfcry, 1988). В бесплодной пустыне Негев в Израиле в особенно суровые годы, если зимой не было дождей, местные каменки (в том числе и Ое. leucopyga) вообще могут не приступать к размножению (Paz, 1987).
Прямо противоположный пример дает нам обыкновенная каменка Ое. oenanthe в тех участках ее ареала, которые расположены за полярным кругом. Эти птицы прилетают сюда весной только в самом конце мая — начале июня, а уже в середине августа возможны резкие похолодания. Понятно, что за это время ни одной паре не успеть вырастить более одного выводка, поскольку полный цикл от начала гнсздостро-ения до приобретения слетками самостоятельности занимает 40-50 дней. В более благоприятном климате центральных районов Евразии, например в Забайкалье, более 70% пар обыкновенной каменки выращивают за сезон по два выводка (Доржиев, Хсртусв, 1992).
Окраска яиц внутри рода Oenanthe варьирует от почти чистобелой со слабым голубоватым оттенком до ярко зеленовато-голубой. У многих видов поверх основного фона располагаются мелкие бурые и рыжеватые пятнышки, сосредоточенные в основном “пояском” вокруг тупого конца яйца. Лэк (Lack, 1958) попытался связать межвидовые различия в окраске яиц с характером гнездования того или иного вида. Этот автор предположил, что у видов с гнездами, укрытыми в недоступных убежищах, окраска яиц должна быть белой, поскольку в данном случае кладка невидима для хищников и яйца не нуждаются в более темной покровительственной окраске. У видов, у которых гнездовые убежища неглубоки и содержимое гнезда можно увидеть снаружи, согласно этой гипотезе яйца должны быть более темными, нс столь бросающимися в глаза. Предположение Лэка в применении к каменкам оправдывается лишь частично. Действительно, почти чисто белые яйца мы находим у Ое. isabellina, гнездящейся в глубоких порах мелких млекопитающих, а наиболее “темные”, зеленовато-голубые — У Ое. h. hispanica и Ое. deserti, которые чаще других каменок устраивают гнезда в основании кустика и в других подобных местах. Однако прочие виды каменок, гнездящиеся более или менее однотипно, так что их гнезда скрыты достаточно хорошо, могут иметь как белые яйца, с пестринками либо без них (Ое. xanthoprymna, Ое. monacha), так и ярко-
63
Панов Е.Н. Каменки Палеарктики
Рис. 22. Дивергенция вокальных сигналов птенцов у каменок северной Палеарктики. а — Ое. finschii; b — Ое. picata; с — Ое. oenanthe; d — Ое. deserti; е — Ое. chrysopygia; f — Ое. hispanica; g — Ое. pleschanka; h — Oe. isabellina. Всюду вверху — “маяковая" позывка птенца, извещающая о его присутствии; внизу — вокализация при выпрашивании корма у родителей. Из: Панов, 1978, с изменениями.
голубые (Ое. hispanica melanoleuca, Ое. pleschanka). У большинства же видов окраска яиц очень бледно голубая, причем ее интенсивность сильно варьирует как у каждого данного вида, так, зачастую, и в пределах одной кладки.
Насиживание. Насиживает кладку только самка, которая вплотную приступает к этому обычно после того, как снесет предпоследнее яйцо. Самка сама добывает пропитание на протяжении всех 12-14 дней насиживания. Раз в два—три часа в светлое время суток самка вылетает на кормежку. Она сразу же улетает далеко от гнезда — иногда вообще за пределы территории пары, если кормовые угодья не заняты какой-нибудь другой парой. Например, из поселения Ое. picata, занимающего низкогорья вдоль берегов Каспийского моря, некоторые самки летают кормиться за 150-200 м. на прибойную полосу, где собирают, вероятно, рачков-бокоплавов среди мокрых водорослей. Самец нередко сопровождает самок во время этих прогулок, но не кормит ее в это время, как делают, например, самцы у синиц. Лишь 2-3 раза я наблю-
64
Глава I. Некоторые общие сведения о каменках
Рис. 23. Видоспецифические позы птенцов при выпрашивании корма у родителей (слева в каждой позиции) и при отказе от корма (справа). Стрелками показано движение птенцов вспять, а — плясунья Ое. isabellina; b — обыкновенная каменка Ое. oenanthe, с — златогузая каменка Ое. chrysopygia; d — черная каменка Ое. picata; е — черношейная каменка Ое. finschii; f — плсшанка Ое. pleschanka; g — испанская каменка Ое. hispanica; i — пустынная каменка Ое. deserti. Из: Panow, 1974.
дал кормление самцом самки у чернопегих каменок Ое. hispanica в Закавказье.
1.9. Поведение птенцов
Как и у прочих видов воробьиных, птенцы каменок издают два типа криков: первый сигнализирует родителям о том, что птенец голоден, а в тот период, когда выводок оставил гнездо — помогает самцу и самке находить своих отпрысков (рис. 22). Второй — “крик выпрашивания” — слышен в момент кормления, когда птенец при виде своего кормильца жадно разевает клюв, “кланяется” и трепещет крылышками (рис. 23). Эти Две взаимосвязанные формы поведения — трепетание и крик выпрашивания — развиваются, повидимому, на основе конкуренции между птенцами в выводке, ибо тот, кто громче кричит и сильнее трепещет крыльями, скорее привлечет внимание самки и будет накормлен в первую очередь, а то и в ущерб своим собратьям. Интересно, что у некоторых видов,
65
Панов Е.Н. Каменки Палеарктики
у которых птенцы рассредоточиваются сразу же после вылета из гнезда, “демонстрация трепетания” явно деградирует (например, у Ое. finschii).
У птенцов всех видов каменок есть еще одна интресная демонстрация, извещающая родителей о том, что их отпрыск насытился и больше нс хочет есть. Как правило, эта демонстрация отказа от корма состоит в том, что птенец пригибается к земле и очень мелко трепещет крыльями, приподнятыми над спиной. У Ое. isabellina и Ое. oenanthe сытый птенец пятится назад, особым образом кланяется и подбрасывает кверху одно крыло. Схематическое изображение этих реакций приведено на рис. 23.
Молодежь начинает петь очень рано — уже в первые две недели после вылета из гнезда. Первое время они поют чуть слышно, затем все громче и громче. Сначала развивается подпесня, а уже на се основе формируется типичная территориальная песня. У некоторых видов (например, у finschii) молодые самки поют так же часто и много, как и самцы, у других — самки поют гораздо реже (Ое. oenanthe), у третьих они начинают петь наравне с самцами, но с возрастом поют все меньше (Ое. isabellina).
1.10. Межвидовые отношения
Межвидовая территориальность. Сплошь и рядом мы можем видеть, что несколько видов каменок селятся в непосредственной близости друг подле друга. Например, в предгорьях Копетда-га есть места, где рядом живут Ое. finschii, Ое. picata, Ое. pleschanka и Ое. isabellina; в Закавказье — Ое. finschii, Ое. (х.) chrysopygia, Ое. hispanica, Ое. isabellina', на Алтае — Ое. oenanthe, Ое. pleschanka, Ое. isabellina; в Туве, близ границы России и Монголии — Ое. oenanthe, Ое. pleschanka, Ое. isabellina и Ое. deserti (рис. 24, А). В юго-западном Иране перекрываются гнездовые ареалы Ю видов каменок, и еще один вид (Ое. pleschanka) держится здесь в периоды миграций. Все эти виды могут в разных сочетаниях подолгу существовать бок о бок друг с другом (Cornwallis, 1975).
Многие авторы высказывали мысль, что каменки мало специализированы в отношении питания. Они потребляют широкий спектр кормовых объектов — от очень мелких насекомых (скажем, муравьев) до весьма крупных беспозвоночных — таких, как скорпионы и фаланги, а при случае могут поймать и проглотить небольшую ящерицу. Строение челюстного аппарата каменок разных видов весьма сходно, обеспечивая прежде всего быстроту и большую амплитуду движений надклювья и подклювья друг относительно друга (Потапова, Панов, 1977). Это показывает, что разные виды каменок пошли не по пути специализации к ловле жертв какого-либо определенного размера, а стали
66
Глава /. Некоторые общие сведения о каменках
Рис. 24. Размещение гнезд учетырех видов каменок на экспериментальной площадке в Туве (А), сроки насиживания кладок самками этих видов в том же Регионе (В) и размещение трех видов каменок в Монголии, а — гнезда Ое. pleschanka; b — Ое. oenanthe, с — Ое. isabellina; d — Ое. deserti; е — контуры рельефа; f — скальные массивы; g — пойма реки. А, В — из: Иваниикии, 1980а, С — из. Kneis, Spretke. 1985.
67
Панов Е.Н. Каменки Палеарктики
генералистами. Такой оппортунистический способ питания позволяет каменкам быстро переключаться с одного массового корма на другой при появлении последнего (например, на термитов, обильных в период вылета крылатых особей из гнезд — см. Туе, 1988b). Поэтому обитающие совместно виды каменок часто подолгу существуют, в общем, на одних и тех же группах насекомых. В Туве, например, это жуки (долгоносики и чернотелки) и муравьи. При этом, как показали подсчеты, смешанная популяция четырех видов каменок изымает вдень до 4% от суммарного запаса корма (Иваницкий, 1980а). Иными словами, при совместном обитании нескольких видов, если их популяции характеризуются высокой плотностью, возникает определенная опасность псрсэксплуатации кормовых ресурсов.
Этой возможности, очевидно, в большой степени препятствует явление межвидовой территориальности. Каждая пара охраняет границы своего участка не только против особей того же вида, но изгоняет отсюда также представителей других видов птиц — в первую очередь мелких дроздовых, таких как каменки, чеканы Saxicola и горихвостки Phoenicurus. Чем более сходен пришелец с хозяином территории, тем при прочих равных условиях интенсивнее отпор владельца (рис. 25). Однако разные виды каменок отличаются друг от друга, во-первых размерами и силой особей и, во-вторых — уровнем эмоциональности и агрессивности. Поэтому в местах сосуществования нескольких видов нередко складывается устойчивая межвидовая иерархия. Наример, в тувинской смешанной популяции крупная Ое. isabellina явно доминирует над всеми прочими видами каменок. В местах перекрывания территорий этого и трех других видов за 3 месяца наблюдений отмечено 99 нападений плясуний на особей Ое. oenanthe и 81 — на Ое. deserti. Со стороны Ое. oenanthe — второго по субординации вида, за то же время насчитали 111 нападений на особей Ое. deserti и 11 — на плешанок Ое. pleschanka. Особи Ое. deserti только 5 раз нападали на обыкновенных каменок и 9 раз — на плешанок, занимающих самую низкую ступень в иерархии (Иваницкий, 1980b).
Однако, ситуация может выглядеть и совсем по иному. Флегматичные плешанки, не склонные к межвидовой агрессии, оказываются весьма резистентными к угрозам со стороны особей других видов. В результате они с успехом могут постепенно внедряться на территории доминантного вида и благополучно существовать здесь, попросту игнорируя постоянные нападки первоначальных хозяев этих участков. Именно такую картину можно было наблюдать в восточном Казахстане в период занятия гнездовых участков самцами плешанок (Панов, Иваницкий, 1975а). Все пригодное для гнездования плешанок пространство было давно распределено между гнездящимися парами Ое. finschii — высоко агрессив-
68
Глава /. Некоторые общие сведения о каменках
Рис. 25. Частота нападений (в %%) самцов Ое. finschii на птиц других видов в зависимости от степнени сходства последних с агрессором. Изображены (сверху вниз): Ое. pleschanka, Ое. hispanica, Ое. chrysopygia, Ое. capistrata, Ое. isabellina, Ое. р. picata, Ое. р. opisiholeuca, Phoenicians ochruros, Premophila alpestris, Anthus campestris, Motacilla alba personata. Passer hispaniolensis, Petronia pefronia, Phylloscopus sp., Sitta tephronota, Scotocerca inquieta, Galerida cristata, Ammomanes deserti. Из: Панов, 1989.
ного доминантного вида. Однако вес усилия самцов Ое. finschii воспрепятствовать вселению непрошенных гостей оказались тщетными. Плсшанки попросту улетали от преследовавших их агрессоров и тут же возвращались назад. В конце концов Ое. finschii вынуждены были смириться с присутствием плсшанок, заполонивших их территории,
и почти перестали обращать на них внимание. Произошло то, что у исследователей поведения называется привыканием к стимулу (рис. 26). Аналогичную картину можно наблюдать в южном Закавказье, когда поздно прилетающие с зимовок Ое. hispanica поселяются внутри обширных территорий рано гнездящихся Ое. chrysopygia и Ое. finschii. Иногда первым поселенцам удается удерживать пришельцев на периферии своих участков, но неред-
ко законный владелец нс в состоянии полностью обезопасить от вторжения пришельцев даже центральную зону своей территории.
69
Панов Е.Н. Каменки Палеарктики
Рис. 26. Процесс вселения плешанок Ое. pleschanka в местообитания, полностью освоенные черношейной каменкой Ое. finschii (Мангышлак). А — начальная стадия: самцы pleschanka заняли временные плацдармы (d) в верхней части склонов, откуда начинают колонизовать днише долины. В — общая картина вселения самцов pleschanka на территории пар Ое. finschii. C-F — последовательные этапы захвата плешанками нескольких таких территорий, а — границы территорий finschii; b — сердцевинные зоны этих территорий; с — гнездо finschii; е — самцы pleschanka, вселившиеся до 12 апреля; f — то же, после 12 апреля; g — номера вселяющихся самцов pleschanka. Из: Панов, Иваницкий, 1975а.
70
Глава I. Некоторые общие сведения о каменках
Как мы уже знаем, каменки территориальпы нс только в период гнездования, но и на местах зимовок. Здесь каждая особь (и самцы и самки) удерживает за собой более или менее обширную кормовую территорию. Ее границы хозяин охраняет в первую очередь от особей своего вида, но также старается не допустить сюда и прочих мелких дроздовых. В Кении в январе-марте в конкуренцию из-за жизненного пространства вовлеченны два местные вида каменок (Ое. pileata и Ое. lugubris schalowi) и три свразиатских, проводящих здесь зиму (Ое. isabellina, Ое. oenanthe и Ое. pleschanka), а из других видов мелких дроздовых — местная Myrmecocichla aethiops и зимующие пестрый каменный дрозд Monticola saxatilis и луговой чекан Saxicola rubetra. И здесь, как на местах гнездования в Туве, плсшанку постоянно третируют все названные виды, особенно часто Ое. isabellina и Ое. oenanthe. Как и на местах гнездования, Ое. isabellina оказывается видом-доминантом, а Ое. oenanthe занимает вторую ступень иерархической лестницы. Интересно, что местные виды каменок ведут себя весьма индифферентно по отношению к гостям с севера, разместившимся на их территориях. Если особи Ое. isabellina и Ое. oenanthe выступали в качестве агрессоров в 113 межвидовых взаимодействиях, то Ое. pileata и Ое. lugubris оказались в этой роли всего лишь 9 раз (Leisler, Heine, Siebcnrock, 1983). Это последнее обстоятельство, как и ряд ранее приведенных фактов, наводит на мысль, что явление межвидовой территориальности хотя и может в какой-то степени благоприятствовать более равномерному распределению кормовых ресурсов между сосуществующими видами каменок, но отнюдь не устраняет возможность пищевой конкуренции. Угроза ее минимальна, если суммарный запас кормов избыточен, и если индивидуальные территории достаточно велики, чтобы прокормить не только их хозяев, но и пришлых иждивенцев. Однако, как пишет Смит (Smith, 1971), изучавший взаимоотношения местных и зимующих видов каменок в Эритрее и Марокко, здесь остается пока еще слишком много открытых вопросов.
Межвидовое “сотрудничество” и гибридизация. Иногда межвидовые отношения складываются более необычно: Шмидт (Schmidt, 1968) пишет, что холостые самцы Ое. hispanica и Ое. pleschanka, которые в последние годы все чаше появляются в Венгрии, нс находят самок своего вида и начинают кормить птенцов в гнездах oenanthe, словно бы помогая их родителям На юге Франции в провинции Лозерс самцы весьма многочисленной здесь Ое. oenanthe иногда пытаются ухаживать за самками сравнительно редкой в этих местах Ое. hispanica, а однажды два слетка последнего вида присоединились к семейству обыкновенной каменки и оказались на попечении столь неожиданных приемных родителей (Lovaty, 1985).
71
Панов Е.Н. Каменки Палеарктики
Ранней весной 1973 г. я наблюдал в южном Узбекистане, как самец Ое. finschii, самка которого насиживала кладку, упорно пытался привлечь внимание самки Ое. picata capistrata, строившей в это время собственное гнездо. Самец Ое. finschii раз за разом демонстрировал своей избраннице приглянувшуюся ему норку, то есть вел себя точно так же, как в ситуации формирования пары. Неподалеку от этого места я видел в том же году самца, которого сначала принял за индивида формы capistrata, но затем обнаружил, что оперение спины у него белое, как у типичного Ое. finschii. К сожалению, птицу не удалось поймать и рассмотреть детально, но я нс исключаю мысли, что это мог быть гибрид между названными, довольно близкими видами (см. Панов, 1989: 244-262).
Чрезвычайная редкость подобного рода "случайных гибридов” у каменок (по сравнению с другими родами дроздовых — таких, в частности, как настоящие дрозды рода Turd us) — может указывать на эволюционную давность расхождения большинства видов в роде Oenanthe. Однако внутри рода существуют и такие виды, которые, бесспорно, очень близки генеалогически и зачастую рассматриваются как “неполные виды”, или полувиды (semispecies). Примером могут служить Ое. hispanica и Ое. pleschanka (HafFcr, 1977), из которых вторая, возможно, представляет собой более молодую форму, производную от первой. Они интенсивно гибридизируют во всех местах перекрывания их ареалов, где в результате образуются популяции, почти спошь состоящие из гибридов разных поколений. Окраска этих птиц дает все возможные комбинации признаков исходных видов (Панов, Иваницкий 1975b; Haffer 1977; Лоскот 1982; Панов 1986а, 1989; Грабовский, Панов, Рубцов 1992). Первоначально многие из таких гибридных фенотипов были описаны в качестве самостоятельных видов (например, Saxicola vittata Hemprich et Ehrenberg, 1833 и Saxicola gaddi Sarudny et Loudon, 1904). Аналогичная ситуация существует в западном Иране, где, судя по отрывочным данным, широко гибридизируют формы xanthoprymna и chrysopygia. Эти две формы также могут рассматриваться как semispecies (или как megasubspecies) ввиду резких различий в окраске, несомненно превышающих подвидовой уровень. В качестве megasubspecies следует, вероятно, трактовать формы picata, opistholeuca и capistrata внутри комплекса Ое. picata (Panov, 1992). В обширных зонах гибридизации picata, opistholeuca ncapistrata сформировались весьма своеобразные полиморфные популяции. Аналогичные популяции с устойчивым (сбалансированным) гибридогенным полиморфизмом описаны в древних зонах контакта Ое. hispanica и Ое. pleschanka в северо-западном Казахстане (Панов, Иваницкий, 1975b; Панов, 1989; Рубцов, 1996). Эти и подобные им популяции представляют собой уникальные произведения
72
Глава /. Некоторые общие сведения о каменках
эволюции, нуждающиеся во всемерной охране и в дальнейших углубленных исследованиях. Подробно о них будет рассказано ниже, в очерках, посвященных соответствующим видам каменок.
1.11. Враги и паразиты
Гнезда каменок часто подвергаются нападению многочисленных хищников, желающих поживиться яйцами или птенцами. Если, найдя гнездо каменки, вы хотите, чтобы с ним нс стряслось ничего дурного, лучше не подходите к нему до вылета птенцов. При осмотре гнезда нарушается естественная обстановка вокруг, и сильно возрастает вероятность того, что гнездо будет разорено каким-нибудь мелким млекопитающим вроде ласки, горностая или крысы. Нс исключено, что яйца каменок иногда становятся добычей ежей (Тус, 1980).В столь разных и удаленных друг от друга регионах, как южный Узбекистан и Швеция, значительное число гнезд каменок разоряют лисы. В Узбекистане они откапывают гнезда Ое. picata capistrata и Ое. finschii, спрятанные в горизонтальных промоинах лессовых обрывов, а в Европе от лис гибнут кладки и птенцы Ое. oenanthe (Туе, 1980; Moreno, 1989). В Забайкалье, где последний вид обитает в основном в антропогенном ландшафте, его главным врагом оказываются бродячие собаки (Доржиев, Хертуев, 1992).
В Закавказье гнезда Ое. hispanica, Ое. finschii, Ое. (х.) chrysopygia нередко гибнут от ворон и сорок. Складывается впечатление, что эти птицы (в период массового вылупления птенцов каменок) специально разыскивают их гнезда или выслеживают родителей, доставляющих корм птенцам. Повидимому, немалую роль в разорении гнезд играют и змеи. Во всяком случае, все каменки бурно реагируют на появление змеи: по нескольку птиц, забывающих на время о своей нетерпимости друг к другу, собираются вместе и с громкими тревожными криками трепещутся в воздухе над ненавистным пресмыкающимся.
У подавляющего большинства видов каменок гнезда все же достаточно надежно спрятаны в закрытых убежищах, чтобы предохранить их от паразитирования кукушек. Пожалуй, единственным исключением оказываются популяции испанской каменки Ое. h. hispanica, которые гнездятся в основном на земле под густыми кустами. Кардона (Suarez Cardona, 1976) приводит факты, показывающие возможность паразитирования на этих каменках обыкновенной кукушки Cuculus canorus.
В гнездах каменок подчас можно обнаружить богатую фауну беспозвоночных. Часть из них попросту делят убежище с птицами. Это мелкие пауки, свободноживущие гамазовые клещи, крошечные первичнобескрылые Collembola, муравьи и т.д. Подчас сосуществование
73
Панов Е.Н. Каменки Палеарктики
каменок с беспозвоночными неожиданно влечет за собой трагические последствия для этих птиц. Так, Лоскот (1983) описывст случай, когда крупный паук прикрепил нити своей ловчей воронки к только-что отложенному яйцу испанской каменки Ое. hispanica и загородил этой воронкой вход в гнездо, после чего оно было брошено.
В подстилке гнезд Ое. isabellina, Ое. oenanthe и Ое. pleschanka в Забайкалье, часто встречаются личинки мух рода Protocalliphera. В отличие от этих, в общем безобидных созданий, личинки другого двукрылого (Tripocalliphora lindneri) развиваются под кожей птенцов и живут здесь до тех пор, пока нс уйдут окукливаться в подстилку гнезда. В месте выхода личинки остается сухая язва с черными краями. Обычно птенцы не слишком сильно страдают от этих паразитов, особенно если личинки не поселяются в их ноздрях. Но при высокой зараженности (до 9 личинок на одну особь) возможно истощение и гибель птенцов (Доржисв, Хсртуев, 1992). В южном Алтае на границе России с Монголией этими паразитами поражены гнезда всех птиц-норников, в том числе Ое. isabellina и Ое. deserti.
Среди других паразитов в гнездах каменок Забайкалья Доржиев и Хертуев (1992) обнаружили иксодовых клещей (род Ixodes), 6 видов гамазовых клещей (роды Dermanyssus и Haemogamasus) и 7 видов блох (роды Ceratophyllus и Frontopsylla). Большинство найденых видов имеют широкий круг хозяев, паразитируя как на млекопитающих, так и на птицах, использующих для гнездования норы последних. Поэтому среди каменок Забайкалья наименее богата фауна клещей и блох в гнездах Ое. pleschanka, обычно нс использующей в качестве убежищ норы грызунов. Состав паразитов каменок различен в разных регионах. Например, в Монголии число видов блох в гнездах представителей рода Oenапthe достигает 16 (Кисфср, Крумпал, Лобачев, 1986).
1.12. Практическое значение
В самые последние годы неожиданно выяснилось, что изучение биологии некоторых видов каменок представляет большой интерес для познания природных очагов чумы. Основными хранителями бактерий чумы в природе являются грызуны, особенно некоторые виды песчанок (Gcrbillinae). Чумные бациллы переносятся блохами. Таким образом, те виды каменок, которые гнездятся и кормятся в старых или жилых норах песчанок (Ое. isabellina, Ое. bottae, Ое. finschii, Ое. pileata) могут переносить на себе зараженных чумой блох. В условиях опыта каменку-плясунью удается легко заразить чумой, она болеет и через несколько дней погибает. Блохи грызунов, подсаженные к больной
74
Глава 1. Некоторые общие сведения о каменках
птиис пьют се кровь и затем передают болезнь другим плясуньям, сусликам и тушканчикам (Пейсахис и др., 1970).
Некоторые виды клешей, обитающих в гнездах каменок, известны как переносчики вирусов клещевого энцефалита, туляремии, лихорадки Ку и других природно-очаговых инфекций (Доржиев, Хертуев, 1992).
75
Панов Е.Н. Каменки Палеарктики
Глава 2.
Каменка плясунья, чекан-плясун, Oenanthe /sabe///na Temminck, 1929
Синонимы: Saxicola isabellina Temminck, 1929; Saxicola saltator Menetries, 1832; Saxicola squalida Eversman, 1836; Saxicola saltatrix Shelley, 1872*.
Среди прочих евразийских каменок плясунья стоит несколько особняком. Видимо она очень рано обособилась от общего ствола своих предков (рис. I) и с тех пор существовала в достаточно специфических условиях, которые наложили отпечаток на ее строение, внешний облик и поведение. В сущности, плясунью лишь по традиции можно называть "каменкой”, к камням она имеет косвенное отношение. Это исконный обитатель ровных или слегка всхолмленных ландшафтов типа сухой степи, и присутствие камней в том или ином виде ей совершенно не обязательно. Большую часть своей жизни она проводит на земле, передвигаясь по ее поверхности не прыжками, как большинство прочих каменок, а бегом. Поэтому она обладает длинными и сильными ногами. Не имея обыкновения прятать гнездо среди нагромождений камней или в нишах скал, она устраивает его в большинстве случаев в норах роющих грызунов, от которых существенно зависит в своем распространении.
Плясунья — довольно крупная каменка весом около 30 г., с относительнодлинными заостренными крыльями, сравнительно коротким хвостом и длинными крепкими цевками. В природе плясунью легко узнать по комплексу трех признаков: во-первых, она бегает, высоко держа на ногах свое плотное тело; во-вторых, постоянно покачивает хвостом, наподобие трясогузки Motacilla alba; в-третьих, черная перевязь на конце хвоста широкая. По всем этим признакам она отличается от самок Ое. oenanthe, на которых похожа окраской, а по первому — и от самок Ое. deserti.
2.1. Систематические замечания
На всем протяжении своего ареала — от Передней Азии и Причерноморья до Амура и Хуанхэ (рис. 28) плясунья весьма однотипна и не даст повода подразделять се на подвиды. Правда, Л .А. Портснко (1954) выделяет сибирскую расу Ое. isabellina sibirica на том основании, что
* Здесь и далее синонимы приводятся по Рипли (Ripley, 1965), с некоторыми добавлениями.
76
Глава 2. Каменка плясунья Oenanthe isabellina
птицы из восточных частей ареала несколько темнее и характеризуются большей длиной крыла. Плясуньи из восточных популяций действительно в среднем более длиннокрылы: по данным, 99,8-103,1 мм в Тянь-Шане и Монголии против 96,3 мм в Иране и Афганистане (Cramp 1988). Тем нс менее большинство систематиков сегодня рассматривают плясунью как вид монотипический.
С другой стороны существует точка зрения, что сам вид isabellina есть лишь один из подвидов более широко распространенного вида, включающего в себя (также в качестве под
вида или ряда подвидов) африканских bottae (Ripley, 1952).Не вдаваясь в обсуждение этой проблемы, отмечу лишь, что bottae действительно имеет очень много общего с isabellina в строении, облике и общих чертах биологии, и, по-видимому, близка к их общему предку. Существенно, что bottae населяет области африканских саванн, занимая в период гнездования холмистые плато с редкими акациями либо лишенные древесной растительности, на высотах от 1600 до 4300 м
Рис. 27. Каменка плясунья.
Рис. 28. Ареал плясуньи Ое. isabellina и близких ей африканских каменок (все изображения птиц даны в масштабе). Точками показаны залеты, серым - зимовки Ое. isabellina. Оригинал.
77
Панов Е.Н. Каменки Палеарктики
над уровнем моря (Smith, 1971, Pcrlo, 1995), но нс проникает в настоящие пустыни.
Ряд авторов сближают плясунью также с южноафриканской Ое. pileata. рассматривая эти два вида и Ое. bottae членами одного надвида (superspecies, Formenkreis) (Козлова, 1975, Cramp, 1988).
Для этого, вероятно, есть достаточно оснований (например, сходство песен isabellina и pileata). И если близкое родство между тремя названными видами получит достаточные обоснования в дальнейшем, это станет еще одним аргументом в пользу существенной обособленности плясуньи среди палеарктических каменок. Действительно, окраска pileata не имеет аналогов в роде Oenanthe (рис. 1), как и некоторые особенности демонстраций плясуньи. Манера последней поднимать почти вертикально вверх сложенный либо развернутый веером хвост при стойке “столбиком” (рис. 14i) на редкость сходна с соответствующим поведением представителей рода Myrmecocichla (см. Perlo, 1992, Plate 64, позиция 19), но не описана ни у одного другого из достаточно хорошо изученных видов настоящих каменок. По крайней мерс некоторые виды р. Myrmecocichla — это, как и плясунья, бегающие дроздовые, хорошо приспособленные к жизни в выровненных открытых ландшафтах (см. Cramp 1988: 754-755).
2.2. Местообитания и возможная история расселения
Нс исключено, что некий предковый вид, впоследствии распавшийся на два — bottae и isabellina. принадлежал к числу первых “каменок”, проникших из Африки в Переднюю и Среднюю Азию. Этот предковый вид был приспособлен к жизни в степях и саваннах, и смог проникнуть в новые для него области в тот период, когда его излюбленные ландшафты стали господствовать на месте теперешних североафриканских, аравийских и переднеазиатских пустынь. Это могло случиться в конце миоцена — начале плиоцена, т. с. около 7 млн. лет тому назад. В период последующего увеличения континентальности и аридности климата степные ландшафты вновь сменяются пустынными, причем тем скорее, чем ближе к югу находится местность. Пустыни, возникающие в Аравии и в северной Африке, разделяют ареал предкового вида на две части, которые отныне будут развиваться самостоятельно. Южный фрагмент даст начало bottae, северный — isabellina. Этот непосредственный предок плясуньи в конце плиоцена обитает, по-видимому, в сухих степях юга Средней Азии и постепенно поднимающегося Иранского плато и расселяется в подходящие ландшафты к востоку. В течение четвертичного периода
78
Глава 2. Каменка плясунья Oenanthe isabellina
плясунья начинает осваивать и новые для нес местообитания, которые по своим условиям так или иначе отвечают ее первоначальным потребностям. По мере формирования альпийских лугов, последовавших за интенсивными процессами горообразования, она проникает и сюда, становясь типичным обитателем этих ландшафтов. Сейчас она живет в приледниковых сглаженных участках Тяньшаня, Алтая и Памиро-Алая до высот 2800-3500 м.
Другими местообитаниями плясуньи в высокогорьях оказываются сухие и опустыненные степные участки межгорных котловин и пологих склонов, широкие речные долины и ровные щебнистые полупуст-ни бескрайних плоскогорий восточного Памира и Центральной Азии. Здесь эта каменка гнездится на высотах до 4000 м, а на северо-восточной окраине Тибетского нагорья поднимается местами до 4600 м над у.м. (Козлова, 1975).
В песчаные среднеазиатские пустыни плясунья проникает, очевидно, позже, когда они входят в свою более позднюю фазу и начинают закрепляться в результате вторичного развития травянистой растительности. Сейчас, будучи характерным обитателем Каракумов, Кызылкумов и пустынь равнинного Казахстана, плясунья всюду в этих местах склонна избегать сыпучих песков. В Каракумах она приурочена к местам с плотной почвой — будь то песчаная, закрепленная пустыня, глинистая пустыня, песчано-шсбнистыс участки или сухие склоны останцевых гор (Рустамов, 1954).
Рис. 29. Типичные местообитания плясуньи.
79
Панов Е.Н. Каменки Палеарктики
В Кызылкумах сс любимые места — это плотные, сцементированные пески с редким кустарником, или же такие участки, где полосы глинистой пустыни чередуются с песчаными грядами (Зарудный, 1915). В равнинном Казахстане плясунья изредка селится и в сыпучих, всхолмленных песках с редкой травой и полукустарниками. Не избегает и редких зарослей саксаула Arthrophytum, дюзгена Calligonum pallasii, гребенщика Tamarix и др. (Шнитников, 1949). Каки прочие каменки, плясунья охотно присаживается на ветки одиночных кустов и невысоких деревьев.
Миновав труднодоступный рубеж среднеазиатских пустынь, плясунья получает новые возможности расселения к северу, которые увеличиваются по мере отступания лесов перед степями. Этот процесс идет на наших глазах. На востоке своего ареала плясунья за последние 30 лет значительно продвинулась к северу вдоль восточного побережья Байкала и гнездится сейчас в долине р. Баргузин (Гагина, 1960; Гусев, 1962). На юге Западной Сибири плясунья из степей Минусинской впадины, продвигаясь к северу, в небольшом количестве проникает в лесостепь, где связана с поселениями длиннохвостого суслика Citellus undulatus (Rogacheva, 1992).
В Казахстане плясунья распространилась до 53-54’ с.ш., по долине Волги поднялась к северу на 300-500 км (Гаврилов, 1970). В степях северного Кавказа и Поволжья, куда плясунья проникла, видимо, на протяжении последних 10 тыс. лет, эта птица стала характернейшим элементом ландшафта. Лишь недавно “перешагнув" р. Дон, она к 1960 г. продвинулась на 250 км к западу (Воинственский, I960). Уже в 1973 г. плясунья была в северном Причерноморье весьма многочисленным видом, достигая местами плотности 1,4 пары на 1 га. Здесь она освоила опустыненные участки с сухими глинистыми, глинисто-щебнистыми и песчаными почвами и с разреженной ксерофитной растительностью, образовавшиеся на месте плодородных степей в результате неумеренного выпаса овец. Эрозия почв и опустынивание ландшафта привело к усиленному размножению малого суслика Citellus pygmaeus и тушканчиков AUactaga jaculus, поселения которых оказались прекрасным плацдармом для расселения плясуньи (Панченко, 1976). Здесь плясунья тесно связана с поселениями малого суслика Citelluspygmaeus, в норах которого гнездится. Наконец, в последнее десятилетие плясунья начинает появляться на гнездовье и на Балканском полуострове, куда расселяется из Малой Азии (Kumcrloevc, 1962; Raines, 1962; Нан-лннов, Борисов, 1988). В Греции плясунья селится в норах сусликов — на низких сухих участках с выходами песка, вдоль железнодорожных насыпей, пересекающих бесплодные равнины, и по речным долинам, где устраивает гнезда в дырах песчаных обрывов.
80
Глава 2. Каменка плясунья Oenanthe isabellina
Стоит отметить, что в некоторых регионах плясунья занимает два разных и изолированных друг от друга типа ландшафтов — равнинный и высокогорный. Так обстоит дело в Памиро-Алае, где эта каменка гнездится как на предгорных опустыненных равнинах и в низкогорьях на высоте не более 500-600 м над у.м., так и в альпийском поясе гор под самой снеговой линией, проходящей не ниже 2800 м (Иванов, 1969).
2.3. Годовой цикл и сроки гнездования
Основные зимовки вида расположены в Аравии и северо-восточной Африке, но часть особей зимуют и много севернее, например в Израиле и Ираке. Соответственно, в Израиле начало гнездования приходится на середину марта (Shirinai, 1996). В юго-западном Иране весенний пролет начинается в начале февраля и становится массовым к середине этого месяца. Вероятно, отдельные особи зимуют здесь, как это бывает немногим далее к северу, в южном Туркменистане, где в ноябре — январе можно встретить одиночных особей. Однако в Бадхы-зе, у границы Туркменистана с Афганистаном (около 35е с.ш.), плясуньи полностью перслстны, и формирование гнездовой популяции начинается с середины — конца февраля (Зарудный, 1915; Бельская, 1965а; Иваницкий, 1981). Пролет на островах у юго-восточного побережья Каспийского моря проходит в марте и заканчивается к концу апреля (26.1V). Из северного Ирана птицы откочевывают на самые холодные месяцы — осенний пролет заканчивается в середине ноября, а вновь появляются они уже в конце февраля. Лишь немного раньше (к концу октября) отлетают плясуньи из Туркмении, а весной они возвращаются сюда 19-22 февраля. В некоторые годы одиночки проводят в Туркмении всю зиму — как правило, это самцы, но иногда здесь зимуют и самки. Столь ранний прилет на мссга гнездования обуславливает и ранние сроки размножения плясуньи в Туркмении. В некоторые благоприятные годы гнездостроение наблюдали здесь уже 18-19 и 24 февраля, а вылупление птенцов в трех гнездах — 12 и 18 марта (Бельская, 1965а; Иваницкий, 1978). Однако большая часть южно-туркменской популяции приступает к размножению позже, и, кроме того, многие самки гнездятся дважды в сезон (рис. 30). Весной 1968 г. я видел в окрестностях Ашхабада двух самок, строивших гнезда 14 и 19 марта. На следующий год, в туманную и холодную весну, в этом же месте птицы выбирали убежища для гнезд с 6 марта, но до 20 числа еще нс приступили к строительству.
На островах у юго-восточного побережья Каспийского моря весенний пролет плясуний проходит в марте и заканчивается к концу
81
Панов Е.Н. Каменки Палеарктики
м//////////////////^^^^
а чкшшшяяяшшшу////^^^^
Ь 11№ЯШЯШШШк
»'!“ J. J .—1—>| | ' ' ! | ' Г” "Г1" I I ! ' I I I I I I
10 16 22 28 3 9 15 21 27 3 9
Март Апрель Май
сгаяжиН H2 У/Мм/Л?,
Рис. 30. Сроки размножения отдельных пар и полигиничсских семей Ое. isabellina в поселении этих каменок в Туркменистане. I — гнездостросние, 2 — насиживание, 3 — выкармливание птенцов, а — первые самки в полигиничсских трио, b — вторые самки. Из: Иваницкий 1978.
апреля (26.IV). У северо-восточного побережья Каспийского моря, в западном Казахстане (полуостров Мангышлак, около 44° с.ш.), наиболее ранние даты формирования пар — 20 марта, 3 и 4 апреля, гнсздостросния — 26 марта, вылета птенцов из гнезда — 5 мая. Далее к востоку, по всей территории равнинного Казахстана, начало размножение сдвинуто на несколько более поздние сроки. Массовый вылет молодых из гнезд приходится здесь на самые последние числа мая.
В это время в ешс более восточных районах, в горных степях и полупустынях южного Алтая близ границы России с Монголией, местные плясуньи лишь только приступают к гнездованию. В последней декаде мая здесь наблюдали брачные игры и копуляцию, а со 2 июня начинают попадаться многочисленные нснасиженныс кладки. Массовое вылупление птенцов приходится на вторую декаду июня, к концу которой появляются и первые слетки (17.VI, 22.VI, 28.VI). Второй цикл гнездования отсутствует (Лоскот, 1986; Нсйфсльдт, 1986). Примерно на той же широте (50-52° с.ш ), но в 1500 км восточнее, в Забайкалье, плясуньи появляются на местах гнездования в период между 9 и 15 апреля. Хотя Забайкалье отстоит от мест зимовок далее, чем все уже упомянутые, а весна здесь довольно холодная (среднесуточные температуры на рубеже апреля и мая +2-7°С, с ночными похолоданиями до -5-10, иногда до -20’С), плясуньи начинают гнездиться здесь сравнительно рано. Самая первая дата начала откладки яиц — 19 апреля. Среди самок популяции 26% приступают к яйцекладке в последнюю
82
Глава 2. Каменка плясунья Oenanthe isabellina
пятидневку апреля, и 75% самок заканчивает ее к концу первой декады мая. Массовый вылет птенцов из гнезд заканчивается в целом к середине июня. И хотя лето еще в разгаре, плясуньи не приступают ко второму циклу гнездования, как это делает обитающая здесь же обыкновенная каменка (Доржисв, Хсртуев, 1992).
Примерно в те же сроки, что на южном Алтае и в Забайкалье, размножается плясунья в Монголии. Гнездостроение здесь наблюдали 22 апреля, гнезда с птенцами разного возраста находили 16 и 20 июня. Далее к югу, у северных подступов к Тибетскому нагорью (близ озера Кукунор в Китае) 10 июня встретили уже самостоятельных молодых, а 20 июня обнаружили гнездо с кладкой (Strcscmann, Mcise, Schanwcttcr, 1937; Picchocki et al., 1982).
В европейской части ареала, в южном Закавказье (около 39° с.ш.) в холодную позднюю весну 1974 г. одиночки уже вернулись на места гнездования к 4 марта и начали петь во всю в конце первой декады месяца. Первые супружеские пары наблюдались с 16 марта, но массовый прилет начался лишь десятью днями позднее. В том же районе в теплую раннюю весну 1970 г. гнсздостроение отмечено 11 апреля, а слетки у трех пар, приступивших к гнездованию, очевидно, еще в конце марта, встречены 3 и 4 мая.
На Украине, в районах, освоенных плясуньей совсем недавно (около 47* с.ш.), насиженные кладки встречаются еще в конце мая (25.V, 31.V), но в это же время в большинстве гнезд птенцы уже готовятся к вылету (Панченко, 1976). У восточных рубежей Европы и Азии, в Волжско-Уральском междуречье, примерно на той же широте, прилет плясуний на места гнездования приурочен обычно к концу марта — первой декаде апреля, а вылет птенцов у наиболее рано размножающихся пар — к середине мая (11.V). Массовое появление слетков приходится на конец мая-первые числа июня. Около 35-40% самок гнездятся и выводят птенцов повторно, их кладки можно найти в июле (Гаврилов и др., 1968; Шевченко, 1979).
2.4. Социальное поведение
Пространственная и демографическая структура поселений. Численность плясуньи в разных участках ее ареала и в разных местообитаниях может быть неодинаковой, и даже в одном и том же участке местности подчас довольно сильно меняется по годам. При низкой численности пары плясуний нередко селятся рассредоточен но, на значительном расстоянии одна от другой. Однако, поскольку между самцами в период их прилета на места гнездования существует, по всей видимости, определенное со-
83
Панов Е.Н. Каменки Палеарктики
циалыюс притяжение, они занимают будущие гнездовые территории неподалеку друг от друга. Демонстративные полеты и пение самцов, уже занявших индивидуальные участки, привлекает новых поселенцев, которые стремятся обосноваться здесь же. Они могут заметно потеснить основателей поселения, отторгая часть их владений. В результате складывается довольно плотное поселение, внутри которого крупные территории наиболее конкурентоспособных самцов перемежаются с более скромными наделами тех, что запоздали па пролете (рис. 31). Среди последних преоблада-
л ют, вероятно, самцы-Рис. 31. Размещение территории в поселении Ос. п Т
isabellina в Туркменистане, а — контуры рельефа, b ПСРВОГОДКИ- уркме-— территории самцов-полигамов. с — моногамов, ,,ии величина террито-d — холостых самцов. Из: Иваницкий 1981а. рии в пределах одного поселения может варьировать от 0.7 до 4.2 га, хотя эти цифры весьма приблизительны, по
скольку участки нс имеют четко выраженных границ, так что территории соседей широко (на 20-50 м) перекрываются своими краевыми зонами. В Забайкалье территориальные группировки обычно формируются сравнительно небольшим числом самцов (2-11, чаще 3-5), а их участки менее разнятся друг от друга по величине (1.2-2.0 га).
Дальнейшие события протекают по разному в популяциях, располагающих достаточным временем для того, чтобы завершить два полных цикла гнездования, и теми, у которых длительность размножения лнмитриуется сравнительно коротким летом. В популяциях первого типа (например, в Бадхызе) прилет самок происходит несинхронно, а часть
84
Глава 2. Каменка плясунья Oenanthe isabellina
из них, обосновавшись на окраине поселения, нс торопятся обзавестись супругом и приступить к гнездовым делам. Эти самки (возможно, происходящие из поздних выводков прошлого года) образуют резерв претенденток па более позднее супружество с самцами, которые располагают наиболее крупными территориями. Такие самцы имеют все шансы, обзаведясь одной супругой, в дальнейшем привлечь на свой участок вторую из числа запоздавших па пролете самок либо из контингента местных, "резервных” плясуний. В результате до 30-40% самцов становятся полигамами — при том, что в поселении неизменно присутствуют холостые самцы, владеющие маленькими и непривлекательными для самок участками. Эти холостяки постоянно находятся в состоянии повышенного возбуждения, внося определенный элемент дезорганизации в жизнь всего сообщества.
В популяциях, обитающих в менее благоприятных климатических условиях (например, в Туве и в Забайкалье) прилет самцов и, позже, самок, проходит гораздо более синхронно. Пары формируются без задержки, быстро приступают к размножению и, завершив один цикл гнездования, вскоре покидают поселения, приступая к предмиграци-онным кочевкам (Иваницкий, 1978, 1981а, 1982; Доржиев, Хертусв, 1992).
Территориальное поведение и саморекламирование самцов. Такова общая картина жизни в гнездовых поселениях плясуньи, на фоне которой происходят конфликты между самцами-соседями, формирование супружеских пар и полигиничсс-ких трио, копуляции между половыми партнерами и другие важные события. Мы видели, что для плясуний не характерна жесткая монополизация территориальных владений (чем они отличаются от ряда других видов каменок). Поэтому наряду с многочисленными конфликтами между самцами, возникающими на рубежах их владений, почти столько же или даже больше стычек происходит при попытках самца проникнуть в самый центр территории соседа — например, в момент появления здесь новой самки. В случаях возникновения антагонизма на общей границы соседних территорий самцы принимают весьма экстравагантные позы, медленно передвигаясь шагом параллельно друг Другу в сопровождении особой агрессивной песни (рис. 32а, Ь). Создается впечатление, что противники активно ищут таких встреч, посещая время от времени определенные пункты на обшей границе, где и обмениваются описанными угрозами. Конфликты при залетах соседей па участок более кратковременны и беспорядочны: хозяин стремительно летит к нарушителю границы, а тот обычно сразу же ретируется, преследуемый в воздухе владельцем участка. При обоих типах конфликтов не исключены кратковременные свирепые драки.
85
Панов Е.Н. Каменки Палеарктики
Рис. 32. Некоторые фрагменты сигнального поведения плясуньи, а, b — позы самцов при территориальном конфликте; с, d. е — поведение самца, в вольер которого только что подсажена самка; f, g, 1 — позы самца пр взаимодействиях с самкой около гнездовой норы; i, к — позы самки во время этих взаимодействий; т, п, о — постепенное изменение по ходу гнездостросния “повседневной" позы самца, принимаемой им в присутствии самки; р — позы партнеров перед копуляцией (самец на переднем плане). Из: Panow 1974.
86
Глава 2. Каменка плясунья Oenanthe isabellina
kHz
1 а
2-
*i ч '’i ч I С\ 1\ Л ‘ I
4-
0.5
10 sec
Рис. 33. Сонограмма рекламной песни самца Ое. isabellina. а — d — последовательные фрагменты одной песни. Оригинал.
d
2
J л >
На протяжении всего гнездового сезона самец извещает округу о своем присутствии постоянными демонстративными полетами. Этот полет, за который птица и пл уч ил а свое русское название, весьма своеобразен, резко отличаясь от соответствующих полетов других видов каменок. Развернув веером роскошный черно-белый хвост и очень торопливо работая приспущенными крыльями, совершающими взмахи малой амплитуды, самец медленно, словно бы с большим усилием поднимается под углом вверх, затем на несколько секунд повисает в одной точке, поддерживаемый столь же быстрым биением крыльев, после чего устремляется к земле планирующим либо пики-
рующим полетом (рис. 15а, 36а). Эти воздушные демонстрации сопровождаются звучной и разнообразной песней, подчас включающей в себя имитации голосов разных птиц, млекопитающих (например, сурков), а также позывки самой плясуньи (ювенильную, “тревожную” и Др.) и обрывки агрессивно»! песни, используемо»! при территориальных конфликтах. Заканчивается песня серией лихих звонких посвистов (рис. 33). Особенно часто эти демонстрации проделывают холостые самцы и тс, чьи самки в данный момент насиживают кладки. Самцы нс слишком утруждают себя выкармливанием птенцов, продолжая заниматься саморекламой. Так что даже на этой стадии частота демонстративных полетов довольно высока — до 12 и более в час (Иваницкий, *978). Полеты самца вне всякого сомнения служат важным сигналом
87
Панов Е.Н. Каменки Палеарктики
для самок, готовых обзавестись супругом и участком для выведения
потомства.
Формирование пар. При появлении самки на вакантной территории се хозяин может предпринять попытку сблизиться с ней, демонстрируя ту самую позу, которую мы наблюдаем при конфронтациях самцов (рис. 32а, Ь). Такая тактика нс оправдывает себя: она может привести к столкновению и даже к драке самца и самки, которая вслед за тем скорее всего покинет территорию. Именно это зачастую происходит, если хозяин участка — самец-первогодок (Иваницкий, 1982). Болес зрелые самцы обычно ведут себя иначе, сразу же начиная демонстрировать пришелице потенциальные убежища для гнезда. Самец бежит либо летит особым трепещущим полетом к заранее облюбованной им норке и замирает около нес. Все это время он пост — в основном, короткие варианты песни (рис. 34). Затем самец залезает в нору, издавая позывку птенца. Если самка готова к диалогу, она рань-
ше или позже прилетит сюда же и даже может осмотреть норку изнут-
kHz 4"|а
-jiiiimiHbiiiiiiii
I Т I
I....... I ' " Г 1
0.5 1.0 sec
Рис. 34. Короткие псснин самца Ое. isabellina. Песня в позиции а целиком состоит из “тревожных” сигналов №2.
ри. В этом случае самец выскакивает из отверстия и принимает первоначальную позу. При этом, пока самка находится в норе, он наклоняется к се входу и широко открывает клюв, показывая угольно-черную полость рта (рис. 360.
В первые часы после появления самки самец демонстрирует ей одну или несколько разных нор много раз подряд. При первых встречах около норы партнеры все же не гарантированы от мимолетной драки, инициатором которой может оказаться как самец, так и самка. В промежутках между демонстрациями у нор самец иногда совершает
88
Глава 2. Каменка плясунья Oenanthe isabellina
уже описанные рекламные полеты. После этого он вновь возвращается к норке трепещущим “полетом мотылька” низко над землей. Прежде чем залезть в отверстие, он иногда подбирает с земли травинку или клочок пуха и заносит свою находку внутрь, или же улетает с ней восвояси.
Во время формирования пары на территорию может вторгнуться кто-либо из самцов-соседей. В подобных случаях пришелец устремляется к самке, та издаст угрожающий сигнал № 4 (рис. 35b) и спасается бегством, а хозяин участка несется следом, и все три птицы улетают за пределы территории. Если кому-либо из самцов удается настигнуть самку, oil проделывает возле нес эффектный полет петлями (рис. 36j). Самец-хозяин возвращается на свой участок
1.0 sec d
10-
6-
2
10
6
2
0.5
№*4».... пял
0.5 sec
Рис. 35. Основные позывки плясуньи Ое. isabellina. а — при тревоге у гнезда (чередование сигналов NN 1 и 3), b — сигнал N 4, с — сигнал N 5 (в верхней части сонограммы), d позывка птенца. е — вокализация птенца при выпрашивании корма у родителей. Оригинал.
первым и раз за разом совершает над ним свои рекламные полеты. Спустя некоторое время сюда возвращается и самка, и церемонии показа и осмотра нор возобновляются.
Г нездос троение. Спустя несколько дней (а иногда, например, при плохой погоде — и 2-3 недели) самка приступает к постройке гнезда. Сначала птица вырывает из земли травинки прямо около
входа и заносит их вглубь, а затем начинает носить строительный материал издалека, возвращаясь к норе особым трепещущим полетом, широко развернув хвост. В первый день гнездостросния самец нспрс-
89
Панов Е.Н. Каменки Палеарктики
рывно дежурит около гнезда, и когда самка скрывается в норе, склоняется над отверстием и, широко раскрыв клюв, выкрикивает короткие отрывки песни (или “агрессивный” вариант, или — гораздо реже — более стереотипную короткую песню — рис. 34). В последующие дни самец лишь изредка прилетает к норе издалека, когда увидит самку, вернувшуюся со строительным материалом. Его поведение постепенно меняется в последовательности, изображенной на рис. 32 1-о.
Как мы увидим ниже при описании гнезда, оно помещается в неглубокой ямке, которую, как полагают, выкапывают сами плясуньи. С этим предположением в какой-то мере согласуются следующие наблюдения. Если подсадить самку в большую вольеру к самцу, последний начинает копать песок клювом, ударяя им вправо и влево, как при поисках почвенных насекомых. При этом он раскрыват крылья, держа их в стороны и немного вперед над самой землей, и покачивает сложенным хвостом. Одновременно самец совершает короткие прыжки в стороны. Возможно, это ритуал, используемый при образовании пар (рис. 32с). Нс исключено, что нечто подобное происходит под землей во время пребывания в норе обеих птиц в начале гнездостроения, или же ямку для гнезда самка углубляет сама, разбрасывая землю клювом.
Копуляция. Уже на третий — четвертый день после начала гнездостроения (занимающего обычно 4-7 дней) пара вступает в фазу копуляций. Эта фаза длится 8-12 дней и распадается на два этапа. На первом из них самец всюду следует за самкой, но его поведение довольно флегматично. Самка, напротив, проявляет высокую социальную активность, то и дело прилетая на границу территории и провоцируя здесь конфронтации между самцами-соседями. Ей же принадлежит сейчас инициатива в предкопуляционном поведении. Самка в промежутках между гнездостроснисм перелетает по участку трепещущим “полетом мотылька”, садится неподалеку от самца и продолжает раз за разом мелко трепетать крыльями. Однако, как только самец пытается приблизиться к ней, она слегка отлетает, не переставая провоцировать супруга своим вызывающим поведением. Эта игра в догонялки постепенно переходит в совместный замедленный полет тандемом, при котором самец, словно привязанный за ниточку, следует траектории полета самки низко над землей. Постепенно скорость полета увеличивается, и он перерастает в стремительную погоню, которая в отдельных случаях может завершиться копуляцией (Иваницкий, 1981b).
На втором этапе роли партнеров меняются. Теперь самка становится пассивной и флегматичной, а самец словно оживает, возобновляя свои самые эффектные демонстрации. В это время активность самца сосредоточена на каком-либо возвышении в пределах территории. Здесь он пребывает часами, бегая в разных направлениях по склону холмика.
90
Глава 2. Каменка плясунья Oenanthe isabellina
Самец время от времени спускается к его подножию, забегая здесь в затененные уголки промоин и оврагов. Песня самца в это время чрезвычайно разнообразна, построена по импровизационному типу и произносится либо вполголоса, либо в полную силу. Песня включает в себя завывания, шипение, хриплые свисты, звонкие звуки “цик...-.цик...” (сигнал № 5 — см. Табл. 2, рис. 35с), различные имитации. В промежутках между этими демонстрациями самец совершает рекламные полеты в сопровождении менее богатой звуками и более стереотипной песни.
Самка находится в курсе всех этих событий, по отнюдь не торопится ответить самцу взаимностью. Практически невозможно предугадать, в какой момент она все же примет решение навестить супруга. Самка летит к месту его демонстраций издалека, а последний отрезок
Рис. 36. Поведение самца Ое. isabellina в брачный период. а — рекламный полет, b — трепещущий полет на этапе формирования пары, с — е — элементы поведения в момент образования пары, h, i — то же, в преддверии копуляции, j, к — полет петлями и “валяние” по земле (см. текст). Оригинал.
пути преодолевает бегом. В момент приближения самки самец прихо-
дит в экстаз и совершает вокруг нес эффектный воздушный танец —
91
Панов Е.Н. Каменки Палеарктики
полет петлями. Сев около самки и трепеща развернутыми крыльями, он очень медленно крадется к ней и словно всползает ей на спину (рис. 32р). По окончании коитуса самец стремительно улетает прочь, издавая на лету серии коротких свистов ити...ти...ти.."(сигнал № 3, типичный элемент поведения при тревоге). Следует заметить, что такое благополучное завершение взаимодействия случается нечасто. Примерно в 60% случаев в последний момент оказывается, что либо самка, либо самец нс готовы к копуляции.
Когда самец-хозяин почему-либо отсутствует на холме свиданий, его место может временно занять кто-либо из его соседей. Он ведет себя точно так же, как и владелец территории, и при каждом удобном случае пытается копулировать с самкой отсутствующего хозяина. Таким образом, отсутствие строгого соблюдения территориальных границ у плясуний создает определенные предпосылки к промискуитету.
Полигиния. Я уже упоминал о том, что в некоторых популяциях плясуньи вполне обычны полигинические ячейки (самец и две самки). Учитывая сравнительно небольшие размеры территорий у плясуньи, вселение на участок пары второй самки оказывается довольно драматическим событием. Хотя у плясуньи самки в целом менее агрессивны, чем у ряда других видов каменок (Ое. finschii, Ое. picata), внезапное появление конкурентки в дотоле единоличных владениях первой самки нс может оставить ее равнодушной. Правда, если первая самка находится в завершающей стадии гнездового цикла (кормит взрослеющих птенцов либо слетков), она почти не проявляет агрессивности по отношению к пришел и цс, даже если та обосновывается в каких-то 100 м от гнезда хозяйки участка. Но в тех случаях, когда первая самка еще только строит гнездо либо недавно приступила к насиживанию кладки, она раз за разом пытается свирепо нападать на незванную гостью. Сделать это ей не дает се супруг, который, по человеческим понятиям, вступает в коалицию с пришели-цей. Как только в поле зрения самца, демонстрирующего норы своей новой подруге, попадает его первая супруга, самец бросается в погоню за ней и загоняет ее в какую-нибудь нору, откуда та долго не решается показаться. В одном подобном эпизоде, продолжавшемся 2 ч., самец решительно подавил все 12 попыток первой самки напасть на вторую. Вместе с тем, как только самец оказывается в той части своей территории, которая осталась в распоряжении первой самки, и он и она ведут себя так, словно ничего нс произошло. Они даже могут копулировать спустя 20 мин. после того, как наступила временная пауза в выяснении отношений (Иваницкий, 1978). В конце концов самки делят между собой территорию самца, и каждая избегает появляться на участке соперницы.
92
Глава 2. Каменка плясунья Oenanthe isabellina
Иваницкий в серии своих работ по Ое. isabellina объясняет асинхронность размножения этого вида в Бадхызе разновозрастностыо особей в популяции. Самцов и самок, которые обосновались в поселении, но первое время не принимают участия в гнездовании, этот автор предположительно считает происходящими из поздних прошлогодних выводков. К ним он относит упоминаемых Бельской (1965b) 47% самцов и 33% самок, которые не приступали к размножению в марте, и соответственно, 35 и 10% особей, не начавших гнездиться в апреле. Исходя из этих цифр и из своих наблюдений, Иваницкий высказывает гипотезу, что у самцов окончательное половое созревание запаздывает по сравнению с самками. Эта гипотеза вполне правдоподобна, укладываясь в концепцию биматурантности (Wiley, 1974), но все же остается неподтвержденной, поскольку птицы не были индивидуально помечены, а их возраст остался неизвестен.
2.5. Гнездо, кладка, выводок
Гнездовые убежища. В отличие от прочих наших каменок, плясунья в подавляющем большинстве случаев гнездится в норах грызунов и других мелких млекопитающих, реже — птиц-норников. В Израиле она селится обычно в колониях дневной песчанки Psammomys obessus, в Закавказье характер ее распределения в пространстве определяется размещением колоний песчанки Виноградова Meriones vinogradovi и других видов грызунов (М. persicus, М. blackleri, малоазий-ского суслика Citellusxanthoprymnus). В юго-западной Туркмении 93,8% гнезд располагается в колониях большой песчанки Rhombomis opimus (39,3% — в жилых, 54,5% — в оставленных зверьками колониях). Остальные гнезда располагаются в колониях краснохвостой песчанки Meriones eversmanni (3,1 %) и щурок Merops superciliosa (3,1 %). В пустыне Каракум плясуньи устраивают гнезда преимущественно в норах тонкопалого суслика Spermophilopsis leptodactylus, реже — желтого суслика Citellus fulvus, а в равнинном Казахстане — разных видов тушканчиков Diросйёае.Ввысокогорьях Тянь-Шаня и Алтая эта каменка использует в качестве гнездовых убежищ норы реликтового суслика Citellus relictus, длиннохвостого сурка Marmota caudata и алтайского сурка Marmota baibacina. В горных опустыненных степях плоскогорий южного Алтая большинство гнезд располагались в норах монгольской пищухи Ochotona pallasi. В Забайкалье 74 из 78 гнезд плясуньи помещались в норах длиннохвостого суслика Citellus undulatus, остальные — в убежищах даурской пищухи Ochotona daurica, сибирского тушканчика AUactaga sibirica и тарбагана Marmota sibirica. В европейской части ареала, в Волго-Донских степях, плясунья продвигается к северу вслед за малым сусликом
93
Панов Е.Н. Каменки Палеарктики
30т Ое. isa. О
2°- о
о о
’°' О > Cit und.
+—I I I I r—"I
10 20 30 40 50 60
Cifellus pygmaeus (Шнитников, 1949; Бельская, 1965a; Рустамов, 1954; Бибиков, Бибикова, 1955; Корелов, 1956; Воинственский, 1960; Тарасов, 1975; Панченко, 1976; Лоскот, 1986; Доржиев, Хсртуев, 1992; Shirinai, 1996). Многие исследователи обращали
Рис. 37. Зависимость распределения плясуний Ое. isabellina в Забайкалье от численности сусликов Cifellus undulatus. Цифры — число учтенных особей того и другого вида. По: Доржиев и Хсртуев 1992.
внимание на явную связь распространения плясуньи с тем или иным из перечисленных видов (рис. 16), хотя, как мы видим, эта каменка, расселяясь в новые для
нес регионы, переходит к использованию убежищ наиболее многочисленных здесь млекопитающих-норников. При этом, как показали Доржиев и Хсртуев (1992), плясунья явно предпочитает жилые колонии млекопитающих заброшенным (рис. 37), хотя, возможно, гибель яиц и птенцов выше в первых, чем во вторых (Шевченко, 1979).
Как правило, гнездо располагается очень глубоко — по ходу норы от 95 см до 310 см, в среднем по 22 гнездам в Тянь-Шане — 208 см. В Чуйской степи расстояния от входа норы до гнезда составляли 60-135 см, в северном Причерноморье — 30-145 см. Расстояние от поверхно
сти почвы до гнезда зависит от типичного строения нор тех или иных млекопитающих и колеблется в разных регионах от 10- 75 см (Украина) до 40-120 см (Тянь-Шань). По наблюдениям в Забайкалье, плясуньи предпочитают сравнительно неглубокие норы со слепым Г-образ-ным окончанием, либо зигзагообразные, в изгибы которых не попадает свет. В длинных сквозных норах птицы строят гнездо на расстоянии 130-160 см от входа, а идущий далее отнорок закрывают. Часто гнездо располагается нс прямо по ходу норы, а в коротком боковом ответвлении, дно которого углублено. Предполагается, что и этот “карман”, и ямку птицы выкапывают сами. Так или иначе, гнездо обычно покоится в сферической камере диаметром 12-14 см. (Гаврилов, 1970; Панченко, 1976; Лоскот, 1986; Доржиев, Хертуев, 1992). Изредка плясуньи устраиваются на гнездование в нишах человеческих построек или в кучах камней, а Попов (1959) нашел в Памиро-Алас гнездо
плясуньи, сделанное в ямке на поверхности почвы, под укрытием густой травы и кустика шиповника.
Г н е зд о велико и всегда сильно утеплено, с толстым основанием из сухой растительной трухи, перемешанной с землей. Диаметр гнезда 107-185 мм, высота 60-120 мм, диаметр лотка 61-68 мм, высота 30
94
Глава 2. Каменка плясунья Oenanthe isabellina
mMj глубина лотка 17-30 мм (иногда местами до 75 мм). Обычно гнездо представляет собой плотно сплетенную кострукцию из стеблей злаков, подчас вырванных из земли с корнями (Festuca spp., Роа spp.), перепутанных с большим количеством шерсти, волоса и перьев (Панченко, 1976; Лоскот, 1986; Доржисв, Хсртусв, 1992). Иногда в выстилке гнезда встречаются погадки хищных птиц. Как мы уже видели, строит гнездо только самка.
Кладка содержит4-7яиц (по некоторым сведениям — до 9). По Meinertzhagen (1954), в Сирии кладка состоит из 4 яиц, но в 6 кладках из соседнего Израиля было от 4 до 6 яиц (Shirinai, 1996). Наиболее достоверные данные имеются по Забайкалью: одна полная кладка здесь содержала 4 яйца, 10 — по 5 яиц, 16 — по6и4 — по 7 яиц (ср. 5,74, п = 31; Доржисв, Хсртусв, 1992). По содержимому 14 гнезд с насиженными яйцами или с нсопсрснными птенцами со всего ареала средняя величина кладки составляет 5,29 (4 х I, 5 х 9, 6 х 4). От этих цифр сильно отличаются данные Шевченко (1979) по Волжско-Уральскому междуречью: 6,88 (апрель), 6,03 (май) и 4,90 (июнь). Возможно, на увеличение средних повлияло присутствие в этой выборке кладок из 8 и 9 яиц, нс описанных из других участков ареала. Масса яиц из Чуйской степи 2,7-3,5 г (ср. 3,1, п=15; Лоскот, 1986). Среднее число птенцов на гнездо в высокогорьях Тяиьшаня — 4,9.
Окраска скорлупы яиц бледно-голубоватая, почти белая, с более интенсивным блеском, чем у Ое. oenanthe. Размеры яиц с Украины 22,0-23,5 х 16,0-17,0 (ср. 22,5 х 16,7, n=l I); из Казахстана 20,3-22,9 х 15,3-17,0 (ср. 21,9 х 15,8, п=17); из Чуйской степи (южный Алтай) 21,2-24,1 х 15,1-16,9 (ср. 22,6 х 15,9, n= 18); из Забайкалья 18,2-25,4 х 15,1-18,4 (ср. 21,9 х 16,9, п=124); объединенные выборки по разным регионам: 19,5-25,1 х 15,6-17,3 (ср. 22,3 х 16,6, п=33); 19,0-24,0 х 15,2-17,3 (ср. 21,7 х 16,2, п=100); 22,6 х 16,8 (п=38). По-видимому, географическая изменчивость размеров яиц отсутствует (Hartert, 1910; Пан«-ченко, 1976; Ковшарь, Левин, 1982; Лоскот, 1973 б, 1986 б; Cramp, 1988; Доржисв, Хертусв, 1992).
Насиживание и репродуктивный успех. Эффективность гнездования плясуний различна в разных регионах. В Забайкалье, например, она весьма высока. Неоплодотворенныс яйца составляют 3%, эмбриональная смертность — около 7%, случаев разорения гнезд нс отмечали. Успех вылупления составляет 84,8%, среднее количество слетков на гнездо 4.4 (Доржисв, Хсртусв, 1992). В Волжско-Уральском междуречье доля неоплодотворенных яиц 6,9%, средняя величина выводка перед вылетом 5,3. Гибель гнезд происходит от затопления нор ливнями, она выше в жилых норах грызунов, нежели в оставленных. Птенцы страдают от присутствия муравьев Lasius umbratus.
95
Панов Е.Н. Каменки Палеарктики
Выживают в этом регионе 43% птенцов от числа отложенных яиц (Шевченко, 1979), В Бадхызе — 62% (Бельская 1979). На Украине из 35 гнезд 5 разорили хорьки Mustella eversmanni, причины гибели еще трех гнезд остались невыясненными. Присутствие нс более 2-3 слетков во многих выводках указывает на возможность их значительной гибели на стадии вылета (Панченко, 1996).
Насиживает только самка в течение 12 дней после откладки предпоследнего яйца, 12-15 дней поданным из Туркмении, 13-14 дней в Забайкалье (Бельская, 1965а; Доржиев, Хертуев, 1992).
Птенцы. Эмбриональный пух у птенцов длинный, светлосерый, на голове и на спине он сохраняется до вылета птенцов из гнезда. Зев и валики в углах рта лимонно-желтые. Остроумная методика, разработанная Доржиевым и Хертуевым (1992) для наблюдений за поведением плясуний, кормящих птенцов в глубокой темноте под землей, позволила узнать много любопытного. Прилетая с кормом, взрослая птица продвигается по темному туннелю, держа голову на одном уровне с туловищем и совершая ориентировочные движения клювом из стороны в сторону. Каменка словно ощупывает стенки хода, в чем ей, возможно, помогают щетинки у основания клюва. Птица легко обходит искусственные препятствия, которые исследователи помещали глубоко в норе. Прежде чем сесть на гнездо, плясунья и его ощупывает клювом. Помет птенцов, выпавший из гнезда, родители находят, руководствуясь, возможно, обонянием, после чего уносят из норы. Голодный птенец, встречая кормильца, держит голову с широко раскрытым клювом неподвижно, под небольшим углом по отношению к горизонтали — как раз на том уровне, где должен находиться клюв взрослой птицы. Сытый птенец уходит в тыловую часть гнезда, уступая место впереди своим еще нс накормленным собратьям.
Развитие оперения птенцов идет с отставанием на 1-2 дня по сравнению с птенцами обыкновенной каменки. Молодые в первом наряде пестрые, отличаются от молодых Ое. oenanthe очень светлым, беловатым горлом, почти лишенным пестрин. В возрасте 15 дней (на трое суток позже, чем это происходит у Ое. oenanthe) птенцы выходят из норы, но сидят около ее отверстия и при малейшей опасности ныряют обратно. В это время они могут свободно бродить по разветвленной системе нор, нередко — далеко от гнезда. С этого времени слетки встречают взрослых птиц, прилетающих с кормом, у входа в нору. С наступлением темноты они уходят на ночевку в свою гнездовую нору, и обычно здесь же ночуют их родители. Выводок остается на попечении самца (если самка насиживает вторую кладку) или обоих родителей на протяжении примерно двух недель. Во втором случае самец и самка могут поделить выводок пополам, так что каждый из родителей
96
Глава 2. Каменка плясунья Oenanthe isabellina опекает свою группу слетков. В районах с однократным гнездованием (например, в Туве) выводки распадаются не позже чем через полторы недели после вылета из гнезда, и плясуньи начинают в одиночку широко перемещаться по местности.
Распадение выводка. Родители приступают ко второму циклу размножения или когда молодые еще нс достигли полной самостоятельности, или же дожидаются их полного возмужания. В последнем случае самец нс подпускает своих отпрысков в окрестности вновь строящегося гнезда и свирепо нападает на них. Да и сами молодые уже в месячном возрасте начинают проявлять признаки агрессивности по отношению друг к другу. Если же на территории пары появились слетки второго выводка, сильно подросшие уже юнцы первой генерации явно доминируют над своими младшими собратьями. В возрасте полутора месяцев молодые каменки начинают проявлять первые признаки территориального поведения. Каждая стремится обособиться от себе подобных, занимая небольшой индивидуальный участок, куда по возможности не допускает бывших членов своего выводка и других юных плясуний (Иваницкий, 1982). Эта взаимная нетерпимость, служащая основой территориальности плясуний-первогодков на пролете и на зимовках, ярко проявляется у особей в возрасте около полу-года. Если в клетку к самке этого возраста подсадить самца, она без устали свирепо гоняет его, бьет клювом и вскоре доводит до полного изнеможения.
У молодых птиц (и самцов, и самок) при содержании их в неволе первые робкие попытки пения отмечаются спустя две недели после того момента, как они должны были оставить гнездо. Поначалу “песня” состоит из разнообразных комбинаций позывок типа I и 3 (глухой щелчок и короткий свист — см. Табл. 3 и рис. 34а, Ь), затем постепенно формируется нечто вроде подпесни. Спустя месяц после вылета из гнезда иногда можно наблюдать спонтанный, ничем не спровоцированный короткий и несовершенный еще полет петлями.
2.6. Линька и отлет на зимовки
Линька начинается очень рано, иногда — уже в первой половине июня. У особей данного года рождения контурное оперение сменяется на втором месяце жизни. При содержании их в неволе на пятом месяце начинается новая линька контурного оперения и сменяются рулевые и часть кроющих крыла, а также маховых перьев. У некоторых взрослых особей к середине июня заканчивается нс только линька мелкого оперения, но успевают смениться также маховые перья крыла и руле-
97
Панов Е.Н. Каменки Палеарктики
выс. В эти же сроки может проходить линька молодых первого выводка в Туркмении. Однако в той же Туркмении многие старые птицы и молодые второго выводка линяют и позже, до середины июля, а в равнинном Казахстане — иногда до первой декады сентября. В высокогорных районах Памиро-Алая интенсивно линяющие птицы встречаются на протяжении июля и августа. Зимой (в декабре — феврале) у взрослых особей проходит частичная предбрачная линька контурного оперения и кроющих крыла (Cramp, 1988). К осени зев первогодков из желтого становится красноватым, а к весне — черным.
Отлет на зимовки начинается уже в июле: в Аравии, где плясунья не гнездится, она появляется в конце июля, а массовый пролет приходится на конец августа и сентябрь (Meinertzhagen, 1954). На местах зимовок в северо-восточной Африке первые плясуньи появляются уже в конце сентября, западнее (Чад, Мали) — в октябре. В восточной Африке миграции к югу наблюдаются даже в ноябре (Cramp, 1988). На севере полуострова Сомали, в Эритрее, плясуньи обычны зимой на открытых плато на высотах 2300-2700 м над у.м., а также на песчаных морских побережьях. Иногда держатся на окраинах населенных пунктов. Не исключено, что некоторые особи возвращаются на зимовку на свои прошлогодние участки, которые активно охраняют от других особей своего вида и других видов каменок, и даже упорно атакуют собственное отражение в зеркале. Иногда зимующие плясуньи распевают негромкую подпесню, а при защите территории издают вокальные сигналы №№ 1 и 2 (Smith, 1971; Туе, 1988а).
2.7. Биологические заметки
По характеру питания плясунья не отличается существенным образом от других видов каменок. Спектр кормовых объектов весьма широк — от самых мелких насекомых до пустынных мокриц Hemilepistus (Isopoda), многоножек Myriapoda и скорпионов Scorpiones. В питании в разных регионах и в разные сезоны преобладают те виды беспозвоночных, которые наиболее многочислены в соответствующий период. В Забайкалье и в Чуйской степи это прямокрылые Orthoptera (в последнем регионе — крупные личинки Bryodema sp., Angaractisgebleri). В Туве и в Волжско-Уральском междуречье в изобилии поедают муравьев и жуков (103 вида из 16 семейств, преобладают долгоносики Curculionidae и чернотелки Tenebrionidae), а прямокрылых используют в пишу сравнительно редко, несмотря на их обилие. Изредка плясуньи нападают на мелких ящериц (преимущественно рода Eremias). Я был свидетелем неудачной попытки каменки справиться с круглоголовкой
98
Глава 2. Каменка плясунья Oenanthe isabellina
phrynocephalus sp., но в другой раз птица съела на глазах наблюдателя ящерицу длиной около 7 см. Осенью в желудках птиц попадаются семена. Зимой в питании обычны личинки Tenebrionidae и термиты jsoptera, которых плясуньи, судя по всему, выкапывают клювом из почвы (Бельская, 1965а; Иваницкий, 1980а; Лоскот, 1986; Шевченко, 1986; Доржисв, Хертуев, 1992).
Обитая в норах грызунов и постоянно контактируя с ними, плясунья может оказаться переносчиком и распространителем некоторых особо опасных инфекций. Васильев и Лазарев (1965), обследовавшие гнезда плясуньи в южном Алтае (хребет Сайлюгем), обнаружили в них 6 видов блох, 5 из которых являются типичными паразитами грызунов (Frontopsylla hetera, Fr. baicalis, Fr. data, Neopsylla mana, Rhadinopsylla dahurica, Rh, Uli). Все собранные экземпляры блох имели в кишечнике свежую кровь.
В заключение следует сказать, что Ое. isabellina, судя по многим особенностям ее образа жизни и поведения, существенно уклоняется от всех прочих палеарктических видов рода — в том числе, и от Ое. oenanthe. Поэтому нет достаточных оснований объединять Ое. isabellina и Ое. oenanthe в составе единой ‘‘группы видов”, как это становится принятым в последнее время (см. Cramp, 1988).
99
Панов Е.Н. Каменки Палеарктики
Глава 3.
Обыкновенная каменка, каменка, чекан-попутчик Oenanthe oenanthe Linnaeus, 1758
Синонимы: Motacilla oenanthe Linnaeus, 1758; Motacilla leucorhoa Gmelin, 1789; Motacilla vitijlora Pallas, I8l I; Saxicola rostrata Hemprich et Ehrenberg, 1832; Saxicola libanotica Hemprich et Ehrenberg, 1832; Saxicola oenanthoides Vigors, 1839; Saxicola seebohmi Dixon, 1882; Saxicola phillipsi Shelley, 1885; Saxicola oenanthe argentea Lonnberg, 1909; Oenanthe oenanthe schioeleri Salomonsen, 1927; Oenanthe oenanthe integer. 1950.
Обыкновенная каменка отличается от всех прочих палеарктических видов рода Oenanthe серым цветом спины самцов (рис. 38). Самки евразийских подвидов по общему облику сходны с плясуньей, отличаясь от нес несколько меньшей величиной (масса 22-24 г) и более длинным хвостом, который при постановке птицы “столбиком” касается земли. Кроме того, обыкновенная каменка не столь “большеголова”, как плясунья, клюв сс миниатюрнее, а черная концевая перевязь на хвосте заметно уже (Harris, Shirinai, Christie, 1996). В момент приземления после полета птица I-2 раза мерно поводит хвостом, чуть опуская его и немного мсдлснсс поднимая до уровня туловища. В спокойном состоянии иногда покачивает хвостом с такой же небольшой амплитудой. В состоянии возбуждения резко дергает концами крыльев в стороны и чуть вверх, напоминая этим движением испанскую каменку или плешанку. Как и у плясуньи, полость рта у взрослых особей черная.
Обыкновенная каменка резко выделяется среди прочих видов рода Oenanthe огромной величиной своего ареала. Это пионер среди каменок, единственный вид, который смог порвать с вековыми устоями предков и завоевать новые континенты. Удивительны как разнообразие условий, в которых сейчас существует вид, так и способность его представителей преодолевать гигантскике расстояния в период миграций. Каменки, гнездящиеся на Аляске и зимующие в экваториальной Африке, дважды в год совершают путешествие обшей протяженностью нс менее 20-30 тысяч километров. Пути миграций каменок, выводящих птенцов в Гренландии и на полуострове Лабрадор, вполовину короче, но птицы каждую весну и осень пересекают 4 тыс. км над открытыми водами Атлантического оксана.
100
Глава 3. Обыкновенная каменка Oenanthe oenanthe
В
Рис. 38. Обыкновенная каменка (а — самец, b — самка). Из: Panow 1974.
Виды-одиночки, “выпрыгнувшие”, подобно обыкновенной каменке, из ограниченной области распространения всех своих близких родичей, встречаются во многих родах животных и растений. Ученых всегда интересовал эволюционный возраст таких видов в сопоставлении с возрастом прочих представителей рода. Являются ли эти “выскочки” самыми молодыми или самыми древними представителями данной
101
Панов Е.Н. Каменки Палеарктики
группы видов? На этот счет существуют разные мнения. Что же касается рода Oenanthe, то многие факты свидетельствуют в пользу значительной древности этого вида. В самом деле, наша каменка — это единственный вид рода, который во время миграций регулярно пересекает экватор и зимует в Африке к югу от него. Считается, что пути миграций вида в какой-то мере отражают пути его расселения. Если принять эту точку зрения, то вполне вероятной кажется мысль, что ареал обыкновенной каменки некогда включал в себя обширные пространства Африканского континента. О том же говорит и сушествованис в Африке двух изолированных реликтовых ареалов, населенных каменками, которых обычно считают в той или иной степени близкородственными евразийской обыкновенной каменке.
3.1. Систематические замечания
Эти две африканские формы — именно, seebohmi, населяющую Марокко, и phillipsi, обитающую па Сомалийском полуострове (рис. 39), многие систематики до недавнего вросмсни считали попросту подвидами обыкновенной каменки. В то же время Тай (Тус, 1986, 1989а) склонен думать, что phillipsi — это самостоятельный вид, не состоящий в близком родстве с евразийско-американской Ое. oenanthe. В своих суждениях этот автор основывается на окрасочных признаках, которые, как было сказано раньше (1.2), довольно ненадежны в качестве меры генеалогической близости. Что касается seebohmi, то се Тай называет “зарождающимся видом”, а Крамп (Cramp, 1988) рассматривает в качестве подвида обыкновенной каменки, который обитает в пределах се древнего исконного ареала.
Рис. 39. Ареал Ое. oenanthe (включая форму phillipsi).Точками показаны находки вида за пределами гнездового ареала и стандартных районов зимовки, серым цветом — зимний ареал. Цифры — средняя длина крыла самцов. Оригинал.
102
Глава 3. Обыкновенная каменка Oenanthe oenanthe
Систематика евразийских каменок также вызывает немало споров. Некоторые зоологи выделяют среди них до восьми подвидов (см., например, Портенко, 1954).Рипли (Ripley, 1964) сократил их число до трех: Ое. ое. nivea Weigold,, 1913 (Пиренейский п-ов), Ое. ое. virago Meinertzhagen, 1920 (остров Крит), и Ое. ое. oenanthe Linnaeus, 1758 (остальная часть Евразии и Аляска). Кроме того, четко выделяется еще одна, самая крупная форма — Ое. ое. leucorhoa Gmelin, 1789, населяющая Исландию, Гренландию и северо-восток Северной Америки. Вместе с этим подвидом и двумя африканскими вид, по Рипли, распадается на 6 географических рас.
Однако, позже подвиды nivea, virago, а также переднеазиатскую libanotica и центральноазиатскую argentea объединили под общим названием libanotica (Cramp, 1988), а кавказских rostrata и северосибирских oenanthoides включили в состав подвида oenanthe (Лоскот, 1973а). К последнему подвиду этот автор относит всех каменок, населяющих территорию бывшего Советского Союза — в том числе и южные популяции, прежде относимые к argentea, а ныне — к libanotica. Той же точки зрения придерживается и Степанян (1978), который считает географическую изменчивость обыкновенной каменки не прерывистой, а постепенной (клинальной). В направлении с юго-запада на северо-восток происходит плавное потемнение окраски, увеличение длины крыла (средние для самцов 93,8-96,3 мм в Средиземноморье и 97,5— 99,0 на севере и северо-востоке Сибири) и, возможно, уменьшение длины клюва.
Нечто очень похожее мы видим, двигаясь в ареале обыкновенной каменки от южной Европы на северо-запад — через Фарерские острова и Исландию в сторону Гренландии и Лабрадора. В последних двух регионах обитает крупная, относительно темная Ое. ое. leucorhoa (крыло самцов 105,2 мм, масса 30-35 г), а в Исландии — также довольно длиннокрылая “schioeleri” (102,6 мм), промежуточная по окраске между leucorhoa и более светлыми каменками южной и центральной Европы (Cramp, 1988).
3.2. Местообитания
Область распространения вида охватывает все ландшафтные зоны северного полушария — от высокогорий Атласа в северо-западной Африке, где на высотах 1700-2300 м обитает форма seebohmi, до арктических тундр и суровых побережий Северного Ледовитого Океана. Вместе с тем, местообитания обыкновенной каменки, в общем-то, довольно однотипны, и в большинстве случаев представляют собой низкотравные, в большей или меньшей степени пересеченные участки со скаль-
103
Панов ЕЛ. Каменки Палеарктики
ными обнажениями и
Рис. 40. Типичное местообитание обыкиовс111Юй каменки в среднегорьи Тянь-Шаня.
каменистыми осыпями. Таковы альпийские луга высокогорий, моренные ландшафты средней Европы и тундры Евразии и Северной Америки. В Средней Азии и в Казахстане обыкновенная каменка нередко гнездится в более аридных условиях, в сухих степях и в низкогорных полупустынях — в местах с пересеченным рельефом (часто вдоль обрывистых берегов рек) или же в равнинных районах, где придерживается участков, освоенных человеком: железнодорожных насыпей, заброшенных строений, кладбищ и т. д.
Подобно африканским phillipsi и seebohmi, многие южные популяции евразийских обык
новенных каменок тесно связаны с высокогорьями. В северном Израиле эти птицы наиболее многочисленны на высотах 1800-2300 м над у.м. В хребте Эльбурс каменка обильна в полосе выше 4500 м, в Памиро-Алас нс представляет Г 1кости на высотах 2000-3500 м (иногда до 4 тыс. м), в Тянь-Шане » .сотный предел се распространения составляет около 3 тыс. м. В юж-ой Европе обыкновенная каменка гнездится в Швейцарских Альпах и в Андалузских горах в Испании, поднимаясь до высот 2000-2600 м. (Иванов, 1940; Абдусалямов, 1964; Губин, Ковшарь, 1985; Cramp, 1988; Zamora, 1990; Shirinai, 1996).
Однако во многих южных регионах наряду с высокогорными популяциями Ое. oenanthe вполне обычны также популяции низкогорные (например, во Франции — см. Lovaty, 1985) и равнинные (Тус, 1980; Moreno. 1989). В южном Казахстане и на Алтае каменка распространена от подножий гор до альпийских лугов, до 3000-3300 м пал
104
Глава 3. Обыкновенная каменка Oenanthe oenanthe
уровнем моря (Гаврилов, 1970; Нейфсльдт, 1986). Точно так же в восточной и северной частях ареала некоторые популяции связаны преимущественно с высокогорьями, занимая на гнездовании горные тундры гольцового пояса (хребет Хамар-Дабан к югу от оз. Байкал, запад Корякского нагорья на северо-востоке Сибири), тогда как другие связаны преимущественно с равнинными ландшафтами (центральный и северный Казахстан, Забайкалье). На крайнем северо-востоке Сибири Ое. oenanthe занимет сходные по своему облику местообитания в двух разных высотных поясах — альпийские разнотравные луга и сухие тундры высокогорий, с одной стороны, и зональные (лишайниковые и кустарничковые) тундры речных долин и морских побережий, с другой (Наглов и др., 1968; Кищинский, 1980; Васильченко, 1987; Доржисв, Хсртусв, 1992).
Можно предположить, что в ходе своей эволюции евразийские обыкновенные каменки сначала приспособились к обитанию в высотных поясах гор, что дало им возможность впоследствии, по окончании Ледникового периода, распространиться вслед за отступающим ледником далеко к северу, вплоть до арктических тундр.
Другим фактором, позволившим обыкновенной каменке завоевать громадные пространства Евразии, стала се способность селиться рядом с человеком и обращать в свою пользу плоды его рукотворной, равно как и разрушительной деятельности. В малонаселенных районах каменки используют для гнездования всевозможные сооружения, созданные людьми — от полуразвалившихся строений и временных загонов для скота до каменных оснований столбов в линиях электропередач и железнодорожных путей, предоставляющих этим неприхотливым птицам удобные места гнездования в ровной как стол степи. В восточных и северных регионах России обыкновенная каменка местами стала настоящим синантропным видов — наподобие голубей и воробьев. Например, в Забайкалье не менее 80% популяции обитает в населенных пунктах и рядом с ними. Здесь эти каменки наиболее многочисленны на свалках мусора и промышленных отходов, на строительных площадках, на дровяных складах и т.д., где средняя плотность популяции составляет 15-16 пар/км2, а местами доходит до 30 пар/км2 (Доржиев, Хертусв, 1992). Из южносибирских степей Ое. oenanthe постепенно продвигается к северу, через зону тайги, по долинам крупных рек, используя в качестве плацдармов наступления крупные поселки и города. Например, на отрезке р. Енисей между 63 и 70' с.ш. каменки гнездятся в значительном количестве только в трех больших поселках (Туруханск, Никольское и Усть-Порт), отстоящих друг от Друга на десятки километров. В таежной зоне они селятся также на лесоразработках, устраивая гнезда в кучах срубленных сучьев. В устьях
105
Панов Е.Н. Каменки Палеарктики
сибирских рек, впадающих в Северный Ледовитый оксан, Ое. oenanthe получает доступ к пригодным для нес естественным местообитаниям (береговые обрывы), но и здесь нередко отдает предпочтение свалкам мусора около поселков и буровых вышек (Данилов, Рыжановский, Рябицев 1984, Rogacheva, 1992).
Способность Ое. oenanthe использовать для расширения своего жизненного пространства любые антропогенные изменения природной среды просто поразительна. Приведу лишь два примера. В Бельгии этот вид был широко распространен в начале нашего века. Позже каменки перестали здесь гнездиться, но в 80-х годах появились снова, освоив в качестве мест гнездования шлаковые отвалы металлургических заводов (Pieere, 1987). На юге Восточной Сибири в 1980-1981 гг. 3 пары каменок впервые загнсздились на окраине г. Зея, за пределами своего ареала. В качестве местообитаний они избрали нагромождения железных конструкций, кирпича и бетона на месте строительства Зейской гидроэлектростанции (Ильяшенко, 1986).
3.3. Годовой цикл и сроки гнездования
Обыкновенные каменки начинают покидать свои зимовки в Африке уже в конце января-серсдинс февраля, но часть птиц задерживается здесь до середины мая. На юге Израиля первые весенние мигранты появляются на рубеже первой и второй декады марта (с 12.III), и в эти же даты одиночных особей изредка можно встретить в южном Туркменистане (11.III). Однако заметный весенний пролет обыкновенных каменок в равнинных районах Туркмении и Узбекистана проходит позже, чем у ряда других видов рода (таких, например, как Ое. finschii, Ое. picata и Ое.pleschanka), и более или менее обычными Ое. oenanthe становятся здесь лишь в конце марта — в апреле (в некоторые годы с первых чисел этого месяца, в другие — только в последней его декаде).
Раньше всего — во второй половине марта, обосновываются на местах гнездования равнинные популяции средней и юго-восточной Европы. Например, в Молдавию эти каменки регулярно прилетают весной в период с 23 марта по 4 апреля. Впрочем, ранний прилет у этого вида еще нс означает столь же раннего гнездования. В Молдавии первые гнезда с кладками появляются только 4-7 мая, то есть нс ранее чем через месяц после прилета на места размножения. Та же самая закономерность обнаружена и в равнинных популяциях Уэльса в Великобритании, где весенний прилет приурочен к марту, а также в высокогорных популяциях Швейцарских Альп, где Ое. oenanthe появляется на местах гнездования примерно на месяц позже. В обоих этих
106
Глава 3. Обыкновенной каменка Oenanthe oenanthe
регионах между прилетом и началом откладки яиц проходит около трех недель. В Восточной Англии в разные годы от 15 до 43% пар успевают за лето вырастить по два выводка.
Нс удивительно, что сроки прилета и, особенно — начала гнездования определяются нс только географическими координатами местности и ее удаленностью от мест зимовок, но и также се высотой над уровнем моря. Например, в Забайкалье (около 50'с.ш., ПО' в.д.) каменки прилетают весной сравнительно рано, обычно между 6 и 17 апреля, а первые кладки появляются здесь уже 24 апрсля-5 мая. Значительно западнее и намного южнее, в высокогорьях Тянь-Шаня (43е с.ш., 79е в.д.), весенний прилет падает на вторую половину апреля (12-30.IV), а к откладке яиц каменки приступают не раньше середины мая, т е. примерно на 2 недели позже, чем в Забайкалье. Еще более запаздывает гнездование в горных полупустынях южного Алтая, расположенных примерно на равных расстояниях от Тянь-Шаня и Забайкалья (50е с.ш., 90° в.д.). Видимо, в силу чрезвычайной континентальности климата этого региона (в частности, из-за резких ночных похолоданий) самки нс приступают здесь к откладке яиц раньше последних чисел мая (27.V — 4.VI) В результате у обыкновенных каменок Забайкалья около 70% пар успевают вывести птенцов дважды за лето, в высокогорьях Тянь-Шаня только единичные самки гнездятся второй раз, а в Чуйской степи южного Алтая второй цикл размножения вообще не отмечен (Губин, Ковшарь, 1985; Лоскот, 1986; Доржиев, Хср-туев, 1992).
Наиболее поздно размножаются самые северные популяции обыкновенных каменок Евразии. Прилет Ое. oenanthe на места гнездования на полуострове Ямал за полярным кругом (70' с.ш., 70е в.д.) приходится на конец мая-начало июня (27.V-7.VI). Первые полные кладки появляются здесь только в начале второй декады июня (21.VI). Интересно, что сроки начала гнездования на Ямале практически те же, что и в Корякском нагорье, удаленном почти на 5000 км к востоку (63е с.ш., 170е в.д.). Это значит, что в данном случае столь разная удаленность мест гнездования от зимних квартир никак нс сказывается на сроках начала и окончания гнездового сезона. В последних числах мая (25-26.V) каменки достигают Чукотского полуострова (65* с.ш., 175' з.д.), расположенного уже в западном полушарии. Отсюда рукой подать до Аляски, куда каменки также успевают добраться в мае (Пор-тенко, 1973; Кишинский, 1980; Туе, 1980; Данилов, Рыжановский, Рябицев, 1984; Cramp, 1988).
Продолжительность одного репродуктивного цикла занимает у пары немногим больше 30 дней, а при двух генерациях продолжительность гнездового сезона составляет 80-100 дней (Доржиев, Хертуев, 1992).
107
Панов Е.Н. Каменки Палеарктики
Соответственно, массовый вылет птенцов из гнезд приходится на конец мая — начало июня (первый выводок) и на конец июня — начало июля (второй выводок) в равнинных популяциях Европы; соответственно, на середину июня и на вторую половину июля — в Забайкалье; на конец июня — в южном Алтае и в прилежащих районах Монголии (один выводок); на середину июля — в горных и прибрежных тундрах Заполярья (один выводок).
Отлет на зимовки из районов крайнего севера проходит с середины августа до середины сентября, в южных частях ареала последних пролетных каменок можно изредка встречать на протяжении всего октября (последние встречи в Казахстае — 19 и 25.X) (Гаврилов, 1970; Данилов, Рыжановский, Рябицсв, 1984).
3.4. Социальное поведение
Пространственная структура поселений. На места гнездования первыми прилетают самцы. Там, где условия благоприятны для размножения, они образуют более или менее плотные поселения, включающие в себя несколько примыкающих друг к другу территорий. В антропогенных ландшафтах в степной зоне четко выраженные группировки образуются на мусорных свалках, где в непосредственной близости друг к другу поселяется до 7-8 самцов. Величина территорий зависит от многих факторов — в первую очередь от числа претендентов на данную местность и от того, насколько рано занял свой участок тот или другой самец. Размеры соседних территорий могут быть существенно различными — от 0.8 до 1.9 га, так что в дальнейшем расстояния между гнездами варьируют от 50 до 130 м, составляя в среднем 70 м (Доржисв, Хертуев, 1992).
В местах, нс столь гостеприимных по своим условиям — например, в альпийской зоне Тянь-Шаня на высоте 3000—3500 м, самцы после прилета держатся более рассредоточено. Территории здесь крупнее, их границы нс столь четко выражены, а расстояния между гнездами обычно превышают 100 м, достигая подчас 200 и даже 400 м (Губин, Ков-шарь, 1985). В Швейцарских Альпах (около 2000 м над у.м.) территории также очень велики (5,3— 13,5 га, ср. 8.7 га) по сравнению с равнинными районами Европы (0,9-4,7 га, в среднем 2.3 га в южной Швеции — см. Тус, 1980; Могспо, 1989).
Территориальное поведение и самореклам и-р о в в а н и е самцов. Закрепив за собой индивидуальный участок, самец вскоре начинает активно заявлять о своем присутствии. Рекламируя свой земельный надел, он раз за разом совершает над ним особый демонстративный полет. Мягко и однообразно взмахивая полное-
108
Глава J. Обыкновенная каменка Oenanthe oenanthe
Рис. 41. Некоторые фрагменты сигнального поведения обыкновенной каменки Ое. oenanthe. а, b — позы конфликтующих самцов на стадии занятия территорий; с. d — позы самцов при пограничных конфликтах (высокий уровень агрессивной мотивации); е, f — то же. при низком уровне мотивации; g, i — позы самца в момент формирования пары; k. I. ш — демонстрации самца около гнездового убежища (см. текст). Из: Ра now 1974.
109
Панов Е.Н. Каменки Палеарктики
тыо развернутыми крыльями, птица взлетает вверх под углом 30-40'’, после чего замедленно скользит по наклонной плоскости вниз. В самой высокой точке траектории каменка иногда повисает в воздухе, энергично работая крыльями. Этот типичный полет может отчасти модифицироваться от случая к случаю.
В период распределения территорий и установления их границ конфликты между самцами бывают бурными и продолжительными. Самцы движутся шагом неподалеку друг от друга, обычно в одном и том же направлении, приняв позы, изображенные на рис. 41а, Ь). Они клюют землю, а иногда резко рвут травинки и швыряют их в стороны.
Позже, когда границы участков четко обозначились, живущие по соседству самцы избегают нарушать их, а встречаясь на линии раздела, лишь кратковременно демонстрируют угрожающие позы (рис. 41 d, g, i), после чего каждый улетает к себе. Самки, по всей видимости, не столь агрессивны, как, скажем, у Ое. finschii и Ое. picata. Они способны петь, поделают это очень редко. Иногда самец и самка появляются на территории уже в составе супружеской пары. Обыкновенные каменки так долго продвигаются к местам гнездования, что у них, в принципе, есть время объединиться в семью на пути домой. Описан случай, когда самец и самка вместе опустились на борт судна в Ледовитом оксане под 75" с.ш. Это произошло как раз во время весеннего пролета. Парочка, вероятно, следовала к местам гнездования на о. Врангеля (Berk, 1961; Портенко, 1973; Губин, Ковшарь, 1985).
Формирование пар. Однако чаше самки прилетают нс несколько дней позже самцов, получая возможность сделать выбор между территориями внутри поселения. Самки, которые уже гнездились здесь в прошлые годы, занимают уже знакомые им участки. Владельцем такой территории может оказаться как прежний супруг самки, та и новый самец, если первый нс дожил до очередной весны либо занял новый участок в стороне от прошлогоднего. Известно, что некоторые самцы Ое. oenanthe возвращаются в места прежнего гнездования до 4 лет подряд, а самки — до 5 лет, но совсем нс обязательно, чтобы связи уже знакомых партнеров каждый раз возобновлялись (Губин, Ковшарь, 1985; Cramp, 1988).
При появлении самки на территории, уже занятой самцом, он сразу принимает позу столбиком (рис. 41 е), а затем стремительно летит к ней. Оказавшись рядом с самкой, самец проделывает вокруг нес полет петлями, то взмывая вверх примерно на метр, то почти касаясь земли лапками. Этот танец сопровождается громким пением. Затем следую несколько демонстративных полетов самца; опускаясь на землю, он пробегает несколько шагов с широко развернутым хвостом, который часто бывает опущен и волочится по земле (рис. 41 i).
110
Глава J. Обыкновенная каменка Oenanthe oenanthe
Развмт не отношении м е ж д у членами пар ы. Дальнейшие взаимоотношения партнеров связаны с выбором места для гнезда. Когда самец пытается привлечь самку к тому месту, где есть подходящие ниши и щели, он расхаживает около них, распушив оперение груди и брюшка, широко развернув хвост и запрокинув голову кверху (рис. 41а). В эти минуты самец пост ту самую песню, которую обычно можно слышать во время конфликтов между самцами, оспаривающими право на одну и ту же территорию. В отличите от более короткой и простой песни, сопровождающей демонстративные полеты, это настоящая какофония самых неожиданных звуков: туг и тонкое циканье, и глухое урчание, и резкий стук. Изредка в песню включаются имитации голосов других видов птиц (например, малого зуйка Cha radri us dubius).
Когда самка оказывается, наконец, в избранном ею убежище, самец либо неподвижно сидит рядом, выставив вперед голову по одной линии с горизонтальным туловищем, либо быстро бегает в этой позе, размахивая полностью расправленными крыльями (рис. 41k, m). Если самка долго нс показывается снаружи, самец взлетает и трепещется в воздухе на одном месте, обратив голову к отверстию убежища и немного увеличивая темп работы крыльев. Когда самка показывается из норы, самец подчас ненадолго занимает ее место.
По мере того, как близиться время гнсздостросния, самец часто поджидает подругу у входа в гнездовую камеру. Как только она появляется, пение самца становится громче и эмоциональнее. Он хватает клювом лежащие на земле травинки и резким движение отбрасывает их в сторону. Когда самка подлетает ближе, самец сменяет вертикальную позу на горизонтальную (рис. 41k, I), затем хватает соломинку и забегает с ней в гнездовое убежище. Выбегая оттуда, он снова берет травинку и опять заносит се в отверстие. Так повторяется несколько раз, пока самка нс последует в отверстие за самцом.
На более поздней стадии развития отношений между супругами изредка можно наблюдать другой вариант ухаживания самца за самкой. Кавалер принимает особую горизонтальную позу и, слегка "сгорбившись”, семенит вокруг самки, почти касаясь ее клювом, а та в ответ лихорадочно трепещет чуть отставленными в стороны крыльями. Эта демонстрация самца очень напоминает соответствующее поведение Ое. picata, Ое. finschii и, в какой-то степени, Ос. chrysopygia. Очень похожую позу самец подчас принимает и во время конфронтаций с самцами-соседями (Иваницкий, pers. com.; рис. 42а).
За 24 часа наблюдений за обыкновенными каменками в Туве в период гнсздостросния (с 30 апреля по 10 мая) Иваницкому лишь однажды удалось наблюдать копуляцию. По окончании пограничного
111
Панов Е.Н. Каменки Палеарктики
Рис. 42. Фрагменты поведения Ое. oenanthe в период спаривания, а — “сгорбленная” поза самца (на переднем плане), ухаживающего за самкой, с, d — позы самца при подготовке к копуляции и непосредственно перед садкой на самку, b — угрожающая поза самки при конфликте с самкой Ое. deserti. По зарисовкам В. В. Иваницкого в Туве.
конфликта двух самцов один из них уселся на краю каменного карниза и в течении 15 мин оставался здесь в полной пассивности. Внезапно прилетела самка и села в неглубокую нишу обрыва. Самец устремился к супруге и проделал возле нес весьма эмоциональный полет петлями. Опустившись рядом с самкой, он принял предсовокупитсльную позу и мелкими шажками “заполз” ей на спину. По окончании коитуса самец спрыгнул на землю, и в этот момент самка сделала короткую садку на самца, в ответ на что он второй раз покрыл самку. После этого самец отлетел в сторону и вновь продемонстрировал полет петлями, издавая при этом серию коротких свистов (сигнал № 3) и фрагмент песни (рис. 42с, d, 43а).
Некоторые самцы
контролируют по две территории, разделенные подчас весьма протяженными (до 1400 м) дистанциями, пролегающими через угодья, непригодными для гнездования — такими, например, как сплошной лесной массив либо морская лагуна (Аго, 1968). На каждой территории самец имеет партнера-самку. По данным других авторов, гнезда двух самок, имеющих общего супруга, могут располагаться на расстоянии 75-350 м. Случаи полигинии чаще отмечаются при втором цикле гнездования. Иногда самец кормит птенцов в гнездах обеих своих самок, иногда — только в одном (Cramp, 1988).
112
Eiaea 3. Обыкновенная каменка Oenanthe oenanthe
0,5 sec
Рис. 43. Основные позывки Ое. oenanthe. а — при тревоге у гнезда (чередование сигналов NN 1 и 3), b — сигнал N 4. включенный в песню самца при конфликте его с другим самцом, с — позывка птенца, d — вокализация птенца при выпрашивании корма у родителей. Оригинал
113
Панов Е.Н. Каменки Палеарктики
3.5. Гнездо, кладка, выводок
Гнездовые убежища. Обыкновенные каменки столь же изобретательны в выборе мест для гнезда, как и в своей способности осваивать самые разные местообитания. В горных районах с мягким, теплым климатом они практически неизменно выбирают в качестве мест для гнезда всевозможные пустоты в скальной породе и ниши между камнями (Zamora, 1990). Очевидно, это и есть исходный тип гнездовых убежищ для этого, первично петрофильного вида. Однако в таких же, в принципе, горных местообитаниях, приуроченных к очень большим высотам (3000 м над у.м. и выше), каменки склонны выбирать гнездовые убежища с более устойчивым микроклиматом. Например, в высокогорьях Тянь-Шаня только II из 30 гнезд располагались под и между камнями, а 19 были выстроены в брошенных норах сурков. Можно видеть, что при ухудшении климатических условий обыкновенная каменка начинает пользоваться стратегией видов-норников — таких, скажем, как Ое. isabellina. В норах сусликов обыкновенные каменки гнездятся в альпийской зоне хребта Хамар-Дабан, в норах сурков, сусликов и даурских пищух — в высокогорной Чуйской степи южного Алтая (Губин, Ковшарь, 1985; Лоскот, 1986; Васильченко, 1987).
Эта стратегия особенно оправдывает себя на крайнем севере. Например, в Корякском нагорье Ое. oenanthe почти столь же тесно связана в своем распространении с обитающим в горных тундрах берин-гийским сусликом Citellus parry i. как Ое. isabellina с теми или иными видами песчанок. На полуостровах Таймыр и Ямал за полярным кругом обыкновенные каменки гнездятся в старых норах леммингов, песцов и птиц-норников — например, береговых ласточек Riparia riparia (Кищинский, 1980; Данилов, Рыжановский, Рябицсв, 1984). Впрочем, к гнездованию в норах обыкновенные каменки охотно переходят и в местах с более мягким климатом — например, на о. Скокхолм у юго-западных берегов Великобритании. Здесь 86% гнезд располагаются в норах (кролика Oryctolagus cuniculus, а также морских птиц — глупыша Puffinuspuffinus и тупика Fratercula arctica (Conder, цит. по: Cramp, 1988). Практически все гнезда Ое. oenantheрасполагаются вброшенных норах кроликов и на вересковых пустошах Восточной Англии, поскольку в этих равнинных местностях какие-либо иные гнездовые убежища, пригодные для каменок, попросту отсутствуют (Тус, 1980).
В равнинных и слабо всхолмленных районах Казахстана большинство гнезд располагаются в отверстиях стен жилых или брошенных строений, в углублениях каменных либо саманных оград, в развалинах и на кладбищах. Для синантропных популяций характерно гнездование в ящиках на свалках мусора, в трубах, под досками, листами железа и
114
Глава 3. Обыкновенная каменка Oenanthe oenanthe
шифера, на складах строительных материалов. В деревнях в таежной зоне России каменки гнездятся одиночными изолированными парами в штабелях дров, сложенных рядом с избами. Проникая по вырубкам в тайгу, они иногда занимают под жилье дупла пней и даже скворечники, развешанные на деревьях в разреженном сосновом лесу (Доржисв, Хертуев, 1992).
В убежище гнездо помещается обычно неглубоко, в 5-70 см от входа (в среднем в 30 см в Восточной Англии, 23 см в Тяньшанс, 28 см в Забайкалье, 31 см — в Чуйской степи), даже если за гнездом остается свободное пространство. Крюгер (Kreuger, 1958) нашел на Сомалийском полуострове гнездо формы phillipsi, свитое прямо на земле, под укрытием куста алоэ.
Г н е з д а всех подвидов, от сомалийского до гренландского, совершенно однотипны. Наружный слой сплетен из сухой травы, толстый внутренний — из войлока, волоса, шерсти, перьев и ваты. Если каменкам удастся найти богатый запас утеплительных материалов (например, труп крупной птицы), гнездо может быть свито почти целиком из перьев. Гнезда, выстроенные на мягком грунте, помешаются в неглубоких ямках. Это углубление может быть заполнено толстыми соломинами и древесными прутиками (длиной до 20 см и весом свыше 3 г), составляющими часть толстого основания гнезда. Гнездо выглядит довольно массивным. Его диаметр варьирует от 70 до 240 мм, высота составляет 50-120 мм, диаметр лотка 55-85 мм, его глубина 30-48 мм (Сурвилло, 1968; Левин, Ковшарь, 1985; Лоскот, 1986; Доржиев, Хертуев, 1992).
Строит гнездо только самка, а участие самца в гнездостроен и и носит сугубо ритуальный характер. К постройке гнезда самка приступает не ранее, чем через неделю после образования пары, а при холодной и дождливой погоде этот промежуток подчас превышает 10 дней. Минимальный срок, необходимый самке для постройки гнезда — 4 дня, а на пятый день она уже может отложить первое яйцо (Сурвилло, 1968). В южной Швеции длительность постройки гнезда самкой варьирует от 2 до 7 дней (ср. 3,2 дня) , а первое яйцо может быть отложено как в день завершения работы, так и полторы недели спустя (в среднем — через 2,8 дней) (Moreno, 1989).
Кладка. Полная кладка содержит от 3 до 9 яиц, и се размер увеличивается по направлению с юга на север и с запада на восток за счет увеличения доли крупных кладок. В равнинном Казахстане с его сухим и жарким летом величина кладок распределялась следующим образом: Зх 1,4x7, 5x9, 6x8 (ср. 4,96, п=25). В той же широтной зоне, но высоко в горах Тянь-Шаня, соотношение было следующим: 4 х 3, 5 х 12, 6 х 3, 7 х 1 (ср. 5,00, n= 19), далее к востоку, в Забайкалье: 3x1,
115
Панов Е.Н. Каменки Палеарктики
4 х 4, 5 х 19, 6 х 25, 7 х 4 (ср. 5,75, п=43) и в Сибири у полярного круга 4x2, 5x3, 6x2, 7x4 (ср. 5,73, n=l 1) (Гаврилов, 1970; Кишинский, 1980; Данилов, Рыжановский, Рябицсв, 1984; Губин, Ковшарь, 1985; Доржиев, Хертуев, 1992). В западной Европе средняя величина кладки в Швейцарских Альпах составляет 5.4, на юго-западе Великобритании 5.5-5.8, в южной Швеции в разные годы 6.0-6.4, в Финляндии 6,1 (3 х 2, 5 х 8, 6 х 22, 7 х 15, 8 х I, п=48). В Великобритании величина кладки уменьшается по ходу сезона размножения от 5,7-6,2 в конце апреля до 4.3-4.5 в конце июня. Та же закономерность обнаружена в южной Швеции (Туе, 1980; Cramp, 1988, Moreno, 1989). В Забайкалье, напротив, кладки первого цикла в среднем мельче (5,31, п=29), чем при втором цикле (5,75, п=24). Доржиев и Хсртуев (1992) объясняют это худшими температурными и кормовыми условиями в период первой яйцекладки. У расы leucorhoa нормальная кладка содержит 5-7 яиц, реже встречаются кладки из 4 или 8 яиц, а иногда их бывает 9 в одной кладке.
Размеры яиц: в Великобритании 18,7-24,0 х 14,9-16,8 (ср. 21,0 х 15,8, п=264); в южной Швеции 20,9 х 15,5 (п=31 кладка); в Казахстане 18,8-22,5 х 15,0-17,2 (ср. 20,7 х 15,7, п=47); в высокогорьях Тянь-Шаня 19,7-23,1 х 15,0-16,3 (ср. 21,2 х 15,7, n=l I); в Чуйской степи (южный Алтай) 19,9-22,7 х 15,4-16,9 (ср. 21,5 х 15,9, п=44); в Забайкалье 17,6-23,5 х 14,6-16,1 (ср. 20,8 х 16,1, п=162); на Ямале 20,6-22,1 х 14,6-15,5 (ср. 21,1 х 15,2, п=7); на Чукотке 20,9-21,8 х 14,5-15,8 (ср. 21,4 х 15,2, п=5); на территории бывшего Советского Союза в целом: 20,6 х 15,5 (п=200). У формы seebohmi: 20,5-22,6 х 15,6-16,2 (ср. 21,6 х 15,9, п=10); у формы leucorhoa 21.8 х 16,0 (п=34). Средняя масса яиц составляет 2,6 г в Альпах и в Забайкалье, 2,8 г в южной Швеции и в Чуйской степи, 2,9 г у формы seebohmi. Размеры яиц положительно коррелируют с размерами самок и могут достоверно различаться в одном и том же регионе в разные годы в зависимости от погодных условий. Намечается тенденция к увеличению последнего яйца в кладках, особенно в крупных (Moreno, 1989).
Скорлупа яиц окрашена в бледно-голубой цвет и почти лишена блеска.Единственное известное яйцо формыphillipsi имело блестящую зеленовато-голубую скорлупу с довольно редкими коричневыми пятнышками, напоминая по окраске яйцо дрозда рябинника Turduspilaris (Kreugcr, 1958).
Насиживание и репродуктивный успех. Насиживает только самка. Плотное насиживание начинается после откладки предпоследнего яйца и продолжается в среднем 12-14 дней. В южной Швеции в год с сухой весной продолжительность насиживания была минимальной (12,3±0,8 дней — см. Moreno, 1989). Ваннорман (Bannerman,
116
Глава 3. Обыкновенная каменка Oenanthe oenanthe
1954) считает, что у северной формы leucorhoa самец иногда сменяет самку на несколько часов. В Новой Англии из 98 гнезд 27 (28%) были разорены, 7 (7%) погибли под влиянием активности кроликов и от обвалов гнездовых туннелей. Неразвившиеся яйца (11,1%) содержались в 44,9% кладок, причем они чаще встречались в крупных кладках — в том числе, во всех кладках из 7 яиц. Средний размер выводка в расчете на одну успешную пару составлял здесь в разные годы 4,5-5,9 слетков. Максимальное их число дали кладки из 6 яиц. На юге Великобритании средний размер выводков составляет 4.2 слетка.
Доля яиц, из которых вылупились птенцы, составляет 89-92% в южной Швеции (по благополучным гнездам) и 84,7 % в Забайкалье. В последнем регионе потери обусловлены присутствием в кладках нео-плодотворенных яиц (3,0%), эмбриональной смертностью (4,3%) и разорением гнезд (7,8%). В южной Швеции 15% гнезд были разорены или брошены на стадии насиживания; среди тех, в которых вылупились птенцы, та же судьба постигла 29,2% гнезд. Процент гибели гнезд с птенцами в год с дождливыми весной и летом был в 4-5 раз выше, чем в благоприятные годы. Причинами гибели птенцов стали разорение гнезд и голодание, причем в крупных выводках птенцы чаще гибли от голода. Всего за три года погибло 31 гнездо из 80. В Забайкалье из 194 птенцов погибли только 11 (5,7%). Доля выживших птенцов от числа отложенных яиц составляла в южной Швеции в разные годы 24-49%, в Забайкалье 75,5%, а число слетков на гнездо, соответственно, 1,6-3,0 и 4,2. В высокогорьях Тянь-Шаня из 24 гнезд только 2 были разорены, одно птицы бросили на стадии насиживания и одно — с маленькими птенцами. В общем, гибель гнезд составила 16,7% (Туе, 1980; Cramp, 1988; Moreno, 1989; Губин, Ковшарь, 1985; Доржисв, Хертуев, 1992).
Птенцы вылупляются в течение двух дней. Эмбриональный пух у них темносерый, длиной 11-12 мм, по другим данным — 10 мм на голове и спине и 6 мм на бедренных птсрилиях. с присутствием рудиментарных пушинок на копчике. Уже на XI день птенцы начинают выскакивать из гнезда навстречу родителям, прилетающим с кормом. Окончательно покидают гнездо они обычно на XIV день, по другим данным — на XV-XVI. В своем первом гнездовом наряде слетки выглядят буроватыми и пестрыми. Они способны самостоятельно охотиться уже на VI—VIII день после вылета из гнезда, но обычно находятся на попечении родителей около двух недель.
Птенец, вылупляющийся последним, обычно весит значительно меньше своих собратьев, но по крайней мерс в 13 случаях из 20 эти младшие члены выводка благополучно покидали гнездо одновременно с другими птенцами (Тус, 1980). Впрочем, в Швеции в трех из 7 вы-
117
Панов Е.Н. Каменки Палеарктики
Рис. 44. Слеток обыкновенной каменки. Из: Panow 1974.
водков, судьбу которых детально проследил Морено (Moreno, 1984), младшие птенцы погибли в первый день после вылета из гнезда, вероятно, от голода. Этот автор установил, что родители вообще активнее кормят более крупных птенцов. Спустя 4-7 дней после вылета птенцов самец и самка начинают собирать корм в разных частях своей обшей территории, причем каждый из партнеров увлекает за собой часть выводка, деля его, таким образом, пополам. В начале второй недели после вылета частота кормления отпрысков родителями заметно снижается, и слетки начинают все назойливее преследо
вать самца и самку.
Распадение выводка. У пар, приступающих ко второму циклу размножения, самка иногда начинает насиживать вторую кладку еще до вылета птенцов первого выводка (Тус, 1980). Обычно же самка приступает к постройке второго гнезда сразу же после вылета первого выводка. В этот период основную часть работы по выкармливанию первого выводка берет на себя самец. Спустя 5-6 дней самка уже откладывает первое яйцо. Таким образом, с этого момента вся забота о первом выводке ложится на плечи самца. Но ко времени вылупления птенцов второй генерации самец начинает проявлять признаки агрессивности к своим подопечным. Он не допускает их в окрестности гнезда, где насиживает самка, а в дальнейшем делает попытки нападать на юнцов всюду в пределах своей территории. В этот период самцу удается иногда вступить в связь с новой самкой и стать отцом третьего выводка (Доржиев, Хертуев, 1992). В популяциях, где нормой является один цикл размножения, при гибели гнезда с яйцами самка приступает к новой кладке через 4-5 дней в годы с хорошей весной и через 5-7 дней в неблагоприятные годы. Иногда она делает это уже в первый же день после гибели гнезда (Туе, 1980; Moreno, 1989).
3.6. Линька и отлет на зимовки
Линька проходит примерно в одни и те же сроки по всему ареалу вида. Губин и Ковшарь (1985) обратили внимание на то, что в высокогорьях Тянь-Шаня первыми начинают линять взрослые самцы — еще
118
Глава 3. Обыкновенная каменка Oenanthe oenanthe
в тот период, когда они кормят слетков (14-27.VII). Линька перьев крыла и хвоста настолько интенсивна, что некоторые самцы даже с трудом летают в это время. Самки приступают здесь к линьке гораздо позже, лишь во второй декаде августа, а у молодых линька начинается примерно через месяц после их вылета из гнезд, то есть в середине июля. На крайнем севере молодые заканчивают линьку во II-IH декадах августа. Взрослые птицы сменяют все оперение, тогда как у молодых в это время линяет лишь контурное перо. Однако, известны случаи, когда у молодых в августе происходит полная линька, во время которой сменяются все перья хвоста и значительная часть маховых (Попов, 1959). На зимовках (в январе—марте) взрослые особи претерпевают частичную предбрачную линьку мелкого оперения.
Отлет на зимовки начинается с конца августа. Разгар осеннего пролета приходится на сентябрь. Последние птицы в Среднеазиатских республиках СНГ иногда задерживаются до середины ноября. В период пролета — как весной, так и осенью — эти каменки встречаются иногда разреженными группами до десятка особей. Мсйнсртцхаген (Meinertzhagen, 1954) пишет, что 27 августа он наблюдал в Аравии большое скопление обыкновенных каменок (около 150 птиц), к которым присоединялись все новые и новые, прибывавшие с востока и с севера. Каменки были возбуждены, некоторые взлетали на высоту до 30 м и снова садились на землю. Наконец, уже в вечерних сумерках, вся стая одновременно снялась с места и устремилась к югу.
Птицы подвида leucorhoa из Гренландии и восточной Канады с наступлением осени пускаются в опасное путешествие через Атлантический океан. Пролетных каменок часто видят пассажиры курсирующих здесь судов, и совсем не исключено, что птицам зачастую приходится без остановки преодолевать расстояние в 2400 км над коварной морской пучиной. Они делают привал, достигнув Западной Европы, куда прилетают сильно исхудавшими (вес при отлете с мест гнездования — около 40 г, после трансатлантического перелета — 20-25 г). Дальше этим каменкам предстоит пересечь Средиземное море и значительную часть Африки: зимуют они на западе континента, достигая порой северных берегов Гвинейского залива всего в 5' севернее экватора (Cramp, 1988).
Также в западной Африке, но несколько далее к северу (в Мавритании) находятся, вероятно, и основные зимовки подвида seebohmi. С наступлением осени эти птицы покидают места гнездования в высокогорьях Атласа и спускаются в низменные районы Марокко. Часть особей мигрирует далее к югу, но некоторые остаются зимовать у подножия марокканских гор (Cramp, 1988).
119
Панов Е.Н. Каменки Палеарктики
3.7. Биологические заметки
Как и прочие виды рода, обыкновенная каменка практикует несколько способов поисков пропитания. Была проанализирована частота использования 7 стратегий охоты, среди которых, как оказалось, важное место занимает выкапывание беспозвоночных из грунта (Kneis, Launch, 1983: рис. 9). И все же основной способ добывание корма у обыкновенной каменки — это поиски добычи на земле. Птица движется вперед короткими прыжками, имея радиус обзора около 50 см. Примерно через каждые 70 см каменка замирает секунд на 40, внимательно осматриваясь вокруг (Туе, 1989с). В встренную погоду нередко зависает над землей на высоте 1,5-2 м, высматривая добычу сверху. Среди кормовых объектов преобладают насекомые, наиболее обильные в данное время. В Забайкалье, например, это жуки, в высокогорьях Тянь-Шаня — прямокрылые.
Наблюдения за развитием поведения молодых птиц в неволе показали, что слетки без колебаний пьют и купаются, когда впервые видят воду. В течение двух недель после вылета из гнезда они спят, плотно прижавшись друг к другу, но затем у них внезапно появляется стремление соблюдать индивидуальную дистанцию. В конце третьей недели молодые еще “по-птенцовому” выпрашивают корм у воспитателя, но принимают подношение неохотно, как бы через силу. Уже с первых дней после вылета из гнезда юнцы нередко “летают на месте”, крепко держась пальцами за жердочку и сильно размахивая крыльями. Это поведение сопровождается чмокающими звуками, которые с течением времени перерастают в сигнал № 1. Спустя дней 20 эти упражнения переходят в типичный “полет петлями”. Сигнал № 3, как признак сильного испуга, появляется в лексиконе молодых примерно 10 днями позже, чем сигнал № 1. В это же время юнцы начинают петь. Песня в это время очень тихая, свистовая, отдаленно напоминающая пение славок.
Хотя общая схема сигнального поведения Ое. oenanthe в целом сходна с тем, что мы видим у других изученных в этологическом плане палеарктических каменок-петрофилов — членов picata species-group и Ое. alboniger superspccics, по терминологии Тая (Туе, 1989а), в ее репертуаре присутствуют элементы, нс описанные ни у одного из этих видов. Таковы, например, движения кончиков крыльев вверх, полуверти-кальная поза ‘Толовой вниз”, пробежки с размахиванием крыльев (рис. 41а, b, d, m) и др. Кроме того, как следует из сопоставления этого рисунка с рис. 32 и 36, сигнальное поведение Ое. oenanthe имеет очень мало общего с таковым Ое. isabellina. Это значит, что обыкновенная каменка стоит внутри рода Oenanthe особняком как от последнего вида, так и от всех прочих видов каменок.
120
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
Глава 4.
Надвид златогузая каменка, рыжехвостая каменка Oenanthe xanthoprymna Hemprich et Ehrenberg, 1833 и Oe. chrysopygia Hume, 1871
Синонимы: Форма xanthoprymna'. Saxicola xanthoprymna Hemprichct Ehrcnbeq», 1833; Saxicola cummingi'Whitaker. 1899; Saxicola semenovi Bianchi ct Zarudny, 1900.
Формы chrysopygia и kingi: Drontolaea chrysopygia De Filippi, 1863; Saxicola erythropygia Taylor, 1867; Saxicola kingi Hume, 1871; Saxicola sennaarensis Secbohm. 1881; Saxicola hawkeri Ogilvie-Grant, 1908.
Златогузая каменка относится к числу более мелких представителей рода (рис. 2). Это сравнительно короткокрылая каменка с недлинным хвостом и с относительно короткими и слабыми лапками, весом немногим более 20 г. Крылья у нес широкие и закругленные, а довольно длинный, сильный клюв придаст характерный облик ее силуэту (рис. 45). У взрослых особей полость рта черная. По твердому субстрату златогузая каменка передвигается, как правило, прыжками, и лишь изредка — короткими перебежками. Она охотно садится на кусты и при этом иногда перепрыгивает с ветки на ветку. Птичка постоянно поводит хвостом, довольно быстро опуская его ниже горизонтали и очень плавно приподнимая над уровнем туловища — чаще невысоко, но иногда почти до вертикального положения. “Кланяется”, в отличие от других каменок, сравнительно редко, резко и коротко отво-
дя при этом крылья в стороны и од- > повременно задирая кверху сложен- - • - ' ' Ш
ный либо полуразвернутый хвост. Рис. 45. Златогузая каменка Ое. Нередко в момент возбуждения мяг chrysopygiachrysopygia. Из: Panow 1974.
121
Панов Е.Н. Каменки Палеарктики
ко взмахивает крыльями, приподнимая их либо выставляя вперед. При этом каменка издаст очень характерное "храпение” (сигнал № 2).
На первый взгляд златогузая каменка заметно отличается от других евразийских представителей рода — не только своей окраской, но и общим складом тела, а также посадкой, в которой есть что-то от славковых Sylvidae. Однако по образу жизни и поведению — это типичная каменка, которая близка, по-видимому, к общему предку петрофиль-ных каменок. Рыжеватая окраска хвоста может указывать па давние родственные связи златогузой каменки с лесными предками всего рода Oenanthe.
4.1. Систематические замечания
В пределах вида большинство систематиков выделяют два подвида: Ое. х. xanthoprymna и Ое. х. chrysopygia. Как самцы, так и самки обеих фором резко отличаются друг от друга по окраске. Ареал первой формы тянется от юго-восточных окраин Турции на юго-восток, к горам Загрос. Вторая гнездится севернее и восточнее, занимая северные участки Ирана и Афганистана, а также юг Закавказья и юго-восточный Таджикистан (рис. 46). В зоне соприкосновения ареаловxanthoprymna и chrysopygia в северо-западном Иране эти две формы гибридизируют друг с другом, о чем свидетельствуют находки птиц со всеми промежуточными вариантами окраски, известных под названием “cummingi" (Vauric, 1949; Shirinai, 1996). На этом основании xanthoprymna и chrysopygia были объединены в один вид, вопреки часто высказывавшемуся мнению об их видовой самостоятельности (Зарудный, 1896; Дементьев, 1937; Иванов, 1940). Между тем взаимоотношения рассматриваемых форм совершенно аналогичны ситуации с Ое. hispanica и Ое. pleschanka — с той лишь разницей, что последние несравненно более сходны по окраске, нежели xanthoprymna и chrysopygia. Несмотря на бесспорную генеалогическую близость испанской каменки и плешанки и на весьма интенсивную их гибридизацию в зонах вторичного контакта, эти формы сейчас единогласно признаются самостоятельными видами (или, по крайней мерс, semispecies). Очевидно, что этот прецедент требует аналогичного подхода и по отношению к интересующим нас "зла-тогузым каменкам” xanthoprymna и chrysopygia. которым в этой книге я придаю статус "таксономических видов” (см. Панов, 1993). Я убежден, что после детального ознакомления с образом жизни и с поведением xanthoprymna. о которой нам сейчас практически ничего не известно, этот шаг получит реальное подтверждение и общее признание в среде орнитологов.
122
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
Рис. 46. Ареал надвида Ое. xanthoprymna. а — район гнездования Ое. xanthoprymna, b — то же, Ое. chysopygia chyrsopygia и Ое. chyrsopygia kingi, с — район обитания смешанной популяции Ое. xanthoprymna + Ое. chysopygia, d — зимовки. Оригинал.
Абсолютная неизучснность формы xanthoprymna заставляет меня сосредоточиться в этой главе почти исключительно на “северной злато-гузой каменке” Ое. chrysopygia. Ее географическая изменчивость выражается в весьма очевидном посвстлснии окраски и в увеличении общих размеров особей в направлении с запада на восток (Vaurie, 1949; Степанян, 1971). По сравнения с птицами из Закавказья, каменки из Бадахшана (южный Таджикистан и северный Афганистан) имеют в оперении значительно меньшую примесь рыжеватых тонов, а подхво-стье у последних не рыжее, по почти белое. Длина крыла самцов и самок из Закавказья составляет в среднем 89,3 и 86,7 мм, тогда как соответствующие цифры для бадахшанских каменок — 91,5 и 89,4 мм (Степанян, 1971). На этом основании Степанян предложил выделять два подвида — темный и мелкий западный, который в контексте вышесказанного будет обозначаться здесь как Ое. chrysopygia chrysopygia, и светлый, крупный восточный Ое. chrysopygia kingi.
123
Панов Е.Н. Каменки Палеарктики
4.2. Местобитания и численность
В отличие от многих других представителей рода, златогузая каменка довольно разборчива в выборе местообитаний, причем предпочитает она ландшафты, наиболее суровые и бесплодные с точки зрения человека. Эти каменки живут исключительно в каменистых, сильно пересеченных местностях и никогда нс выходят в равнинные пустыни. Их излюбленный ландшафт — это крутые скалистые склоны с круп-покамепистыми осыпями у их подножий и заваленные валунами узкие каньоны и ущелья (рис. 47). В общем, это типичный горный вид, явно тяготеющий к резко пересеченным участкам ландшафта один из наиболее петрофильных среди всех каменок Палеарктики. В юго-западном Иране златогузая каменка занимает горные ландшафты на высотах 2500-4000 м над у.м. (Cornwallis, 1975: по Cramp, 1988). В Эльбурсе эта каменка не встречается ниже 2000 м над у.м., в Бадахшане предпочитает гнездится по крутым каменистым склонам и террасам вдоль долин горных рек на высотах 2000-2500 м (рис. 48), но не избегает и более суровых высокогорий (до 3600 м над у.м.). Сходные ландшафты Ое. chrysopygia занимает в Афганистане и северо-западном Пакистане.
Рис. 47. Типичные местообитания златогузок каменки Ое. chrysopygia в Закавказье (Нахичевань). В таких ландшафтах гнездятся также отдельные пары испанской каменки Ос. hispanica. Из. Panow 1974.
124
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
Рис. 48. Долина р. Шахдара в окрестностях Хорога (юго-запад Памирского плато). Местообитания златогузой каменки Ое. chrysopygia kingi, черной каменки Ое. picata opistholeuca и плешанки Ое. pleschanka.
Рыжехвостую каменку иногда называют немногочисленной или даже довольно редкой. На самом деле это нс гак. В своих излюбленных местообитаниях она вполне обычна, а то и многочисленна. Например, в юго-восточном Закавказье (окрестности города Джульфа) на километр маршрута можно встретить до десятка пар этого вида. Вдоль русла р. Шахдара в Таджикском Бадахшане численность этих каменок колеблется от 2 до 7 пар ( в среднем 4) на 10 га, а на обширных крупнокаменистых осыпях составляет 1-2 пары на 10 га. По соседству с горными деревушкам-кишлаками, где есть орошаемые участки возделываемых земель, златогузыс каменки подчас образуют более плотные поселения (Абдусалямов, 1973; Лоскот, 1974; Haftcr, 1977; Cramp, 1988). Правда, птицы временами держатся достаточно скрытно, и в силу своей монотонной окраски нс столь часто бросаются в глаза, как контрастно окрашенные “черно-белые” виды каменок.
4.3. Годовой цикл и сроки гнездования
Со своих зимовок в северо-восточной Африке, Аравии и южном Иране эти каменки начинают мигрировать к местам гнездования в
125
Панов Е.Н. Каменки Палеарктики
марте. В Израиле последних зимующих особей видели 17 марта и 11 апреля (Shirinai, 1996) . В Закавказье (Нахичевань) в холодную позднюю весну 1975 г. 15 марта выпал снег, а 17 и 23 марта по ночам замерзала вода. Тем не менее первые самцы появились 7, 8 и 10 марта, а прилет самок и формирование пар я наблюдал 11 и 23 марта и 1 апреля. Самец, занявший территорию 7 марта, приобрел партнершу 23 марта; 2 апреля самка начала строить гнездо и занималась этим по крайней мере до 7 апреля. У другой пары 8 апреля самка уже заканчивала постройку гнезда, 7, 8 и 9 апреля отмечены попытки спаривания, последняя из которых закончилась копуляцией.
В раннюю очень теплую весну 1970 г. в том же районе к 26 марта птицы уже разбились на пары, а 29 марта я наблюдал двух самок, уже приступивших к постройке гнезд. В четырех гнездах, осмотренных 26 апреля, были птенцы примерно одного возраста (нс более пяти дней отроду). В двух других гнездах птенцы такого же возраста обнаружены 4 и 9 мая. Первые выводки молодых, только что покинувших гнезда, встречены 3 и 4 мая.
Очевидно, многие пары подвида chrysopygia гнездятся дважды за сезон. Зарудный (1898) находил в Копетдаге гнезда с ненасижеппыми яйцами в течение всей второй половины мая и первой половины июня. На востоке Турции гнездо с птенцами нашли 8 августа, и в это же время наблюдали выводок с родителями (Cramp, 1988). Митропольский сообщил мне, что он наблюдал в окрестностях г. Джульфа, как самка-мать выводка, состоявшего из совсем короткохвостых слетков, приступила к постройке нового гнезда и однажды пыталась “накормить” проголодавшегося юнца пучком сухой травы. В восточных частях ареала каменки подвида Ое. chrysopygia kingi найдены на местах гнездования в Таджикском Бадахшане (западный Памир) уже 27 марта. У самцов, добытых здесь 31 марта и 14 апреля, семенники были заметно увеличены. Наиболее рання дата гнсздостроения в южной части ареала kingi в Гиндукуше — 9 апреля (Meinertzhagcn, 1938). В горах западного Памира, где снег па перевалах лежит почти до конца мая, златогузыс каменки обычно приступают к гнездованию позже. Птенцов в четырех гнездах я нашел в долине р. [Лахдара в районе пос. Хорог 22 (нсоперсн-ныс), 27 и 31 мая. Однако эти даты относятся, вероятно, к наиболее рано гнездящимся парам. Три гнезда с неполными и нснасижснными кладками обнаружены в том же году в этих местах 10 и 18 июня, гнездо с нсопсренными птенцами 2 июня. Первые слетки отмечены 11 июня, выводок хорошо летающие молодых — 13 июля. По мнению Абдусалямова (1973), хзатогузыс каменки в Бадахшане в норме размножаются один раз в сезон. Однако тот факт, что 18 июня отмечено гнсздостроснис, и что гнезда с яйцами попадаются до 24 июня, зас
126
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
тавляет предположить возможность второго цикла гнездования по крайней мере у части пар.
У Ое. xanthoprymna в горах Загрос (юго-западный Иран) слетков наблюдали с середины июня до середины июля (Cornwallis, 1975 цит.: по Cramp, 1988).
4.4. Социальное поведение
Территориальное поведение. Сразу после прилета на места гнездования самцы занимают обширные территории радиусом до 200-400 м. В первые дни самец обычно молчалив, лишь изредка можно слышать тихую подпесню. Постепенно хозяин участка присматривает несколько господствующих над местностью возвышенных точек, которые впоследствии будут использоваться в качестве песенных постов. Самец все чаще и чаще посещает их, оглашая окрестности громкими мелодичными звуками рекламной песни. Разделенные паузами короткие строфы временами сливаются в длинный вариант песни. Если же проиграть ее запись через динамик магнитофона, певец прилетает к источнику звука и раз за разом воспроизводит здесь песню иного характера, используемую при всевозможных социальных взаимодействиях (рис. 49).
В этот период можно наблюдать конфронтации самцов-соседей в зонах раздела между их территориями. Самцы в характерных позах скачут по земле параллельно друг другу, проделывая движения “ложного кормления” (displacement feeding) и время от времени присаживаясь на низкие кустики (рис. 50а). Следует подчеркнуть, что эти демонстрации не приводят к установлению четких линейных границ. Соседи на протяжении всего гнездового сезона поочередно используют пограничные зоны, где соседние территории широко перекрываются — примерно на 30 м, если участки невелики, и на 100 и более метров, если те достаточно обширны. Кроме того, и ранней весной и в дальнейшем постоянно приходится наблюдать залеты чужаков в срединные зоны территории.
Такие территориальные конфликты, которые очень часто можно наблюдать вконцс марта, трудно интерпретировать однозначно, поскольку самцов и самок порой невозможно распознать по внешнему облику. Однако, создастся впечатление, что при столкновениях соседних пар самцы нападают преимущественно на самцов, а самки — на самок, как это происходит, в частности, у Oe.ftnschiivx Ое. picata. Можно полагать, что владельцы участка определяют пол пришельца по его поведению. Когда в конфликт бывают вовлечены три птицы, две из них выясняют отношения, а третья обычно держится несколько по-
127
Панов Е.Н. Каменки Палеарктики
"1 ' " I | ! | , |
0.5 1.0 sec
Рис. 49. Рахтичныс варианты песен Ое chrysopygia. а — короткая рекламная песня. b — то же, начинающаяся с прерывистого “храпения" (сигнал N I), с — то же. начинающаяся с птенцовой позывки (сигнал N 6), d — короткий фрагмент агрессивной песни. Оригинал.
I28
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
Рис. 50. Поведение самцов Ое. chrysopygia при патрулировании границы между их территориями (а) и поза самца при показе самке (на заднем плане) потенциальных гнездовых убежищ во время формирования пары (Ь). Оригинал.
одаль. Если же их четыре, то они, как правило, распределяются подвое. В одной паре соперников они настроены весьма решительно, так что нередко сцепляются когтями и клубком катаются по земле, нанося друг другу удары клювами. Две другие птицы не решаются перейти к рукопашной схватке и лишь демонстрируют угрожающие позы (рис. 51 а-с) или наблюдают со стороны за дракой своих напарников. Эти конфликты сопровождаются особым угрожающим трещанием (сигнал № 4), и нередко одна или две птицы непрерывно поют во время столкновения. Самки распевают наравне с самцами, хотя их песня обычно более коротка.
Саморекламирование самцов. Самец, окончательно освоившийся на своей территории, подолгу пост, сидя на одном из двух-трех своих песенных постах. Это наиболее возвышенные точки, с которых легко обозреть весь участок. Время от времени самец перелетает с одного поста на другой особым демонстративным полетом. Этот полет наиболее сходен с соответствующими воздушными демонстрациями Ое. picata, но имеет и некоторые характерные особенности. Птица иногда повисает в воздухе почти неподвижно, лишь слегка вздрагивая широко расправленными крыльями. Демонстративный полет златогу-зой каменки сопровождается звучной песней, которая проста пострук-
129
Панов Е.Н. Каменки Палеарктики
Рис. 51. Фрагменты сигнального поведения златогузой каменки, а, Ь, с — позы при территориальных конфликтах; е — демонстрации самца в нише, используемой для копуляций; d — поза самца перед копуляцией; f — взаимодействие членов пары на стадии гнсздостросния (слева самка, справа самец). Из: Panow 1974.
туре (постоянное повторение звучной свистовой фразы с небольшими вариациями), но очень красива и несколько напоминает песню синего каменного дрозда Monticola solitarius. Как в этой песне, так и в других ее типах, о которых будет сказано ниже, полностью отсутствуют элементы имитации. Но самой характерной особенностью песен златогузой каменки является особое храпение, которое птица произносит в минуты наивысшего возбуждения (рис. 49b).
Межвидовая территориальность. Златогузые каменки нетерпимы не только к посторонним особям своего вида. Они свирепо нападают на самцов и самок чернопегой каменки Ое. hispanica, когда те в начале апреля начинают обосновываться прямо на гнездовых уча
130
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
стках златогузых каменок. Первоначальные хозяева постоянно третируют этих пришельцев, и настроены по отношению к ним настолько враждебно, что и самца, и самку златогузой каменки можно поймать в западню на грубо сделанное чучело самца чернопегой каменки.
Формирование пар. Один из самцов, находившийся под постоянным наблюдением с момента прилета (7 марта), оставался холостым до 18 ч. 22 марта. На следующий день в 8.30 я обнаружил на его территорию самку. Скорее всего она была в пути всю ночь и прибыла на место встречи ранним утром 23 марта. Процесс формирования пары продолжался не менее 6 ч. При этом наиболее обычными были два основных типа взаимодействий между партнерами: их сближения по инициативе самца и показ им самке потенциальных убежищ для гнезда.
В первом случае самец летит по направлению к самке с песней, которая временами становится насыщенной разнообразными звуками и напоминает “агрессивную” песню (как при территориальных конфронтациях между самцами). Полет самца перед сближением с самкой отчасти напоминает описанный выше рекламный полет, но движется птица по прямой. Приземлившись около самки, самец принимает горизонтальную позу (рис. 50b). Иногда он при этом разворачивает хвост веером и словно “крадется” мелкими шажками в сторону пришелицы. Та обычно сразу же улетает, а самец преследует ее в стремительной погоне через всю территорию. Такие погони регулярно наблюдаются в первый день помолвки, а в дальнейшем они происходят все реже и реже, хотя их можно видеть на протяжении всего последующего периода гнездостроения.
Демонстрации самцом потенциальных гнездовых убежищ выглядит следующим образом. Самец летит к отверстию в скале, принимает все ту же горизонтальную позу и раз за разом залезает в него и вылезает обратно. При этом он впол-голоса произносит подпесню, а во время пребывания в убежище издает птенцовую позывку (сигнал N 6). Когда самка подлетает сюда, самец устремляется к другому отверстию, и здесь все повторяется сначала. Эти взаимодействия, как и описанные выше, могут оканчиваться погоней самца за самкой.
Во время сближения самца с самкой он иногда проделывает около нее полет дугами. Самец описывает в воздухе короткие дугообразные траектории, взлетая на высоту не более 1 м и касаясь земли (либо на мгновение опускаясь на обе лапки) то справа, то слева от самки. Эта та самая демонстрация, которая в дальнейшем будет неизменно предшествовать копуляции.
После погони самка обычно улетает за пределы территории и отсутствует минут 10-20. Самец же сразу возвращается на свой участок и
131
Панов Е.Н. Каменки Палеарктики
при этом издает продолжительные серии коротких посвистов (сигнал N 3, рис. 52b). В отсутствие самки он несколько раз совершает над своими владениями рекламные полеты с песней.
Холостая самка в момент своего первого появления на территории холостого самца не поет. Некоторые самки при виде летящего к ней самца издают предупреждающее трещание (сигнал № 4), другие самки в подобных ситуациях на редкость молчаливы. Иногда при первых встречах с самцом самка проявляет некоторые признаки агрессивности. Но уже к концу первого дня птицы становятся намного терпимее друг к другу. Они вместе облетают участок, задерживаясь ненадолго около потенциальных гнездовых убежищ, а спустя 2-3 дня могут одновременно посещать пустоты, выбирая окончательное место для гнезда.
Гнездостроение и конструкция гнезд. Места, в которых располагаются гнезда златогузой каменки, довольно разнообразны (рис. 53). Иногда это совершенно недоступные для осмотра узкие щели в каменном монолите, в других случаях — широкие по горизонтали, но узкие по вертикали промежутки между двумя плоскими каменными плитами, лежащими друг на друге. Гнездо может быть выстроено в поре, выкопанной, вероятно, щуркой, в склоне глинистого обрыва. Глубина расположения гнезд варьирует от 20 до 50 см (в среднем по 10 гнездам — 34 см). Гнездо может быть видно снаружи, но по крайней мере 3 из осмотренных гнезд были совершенно недоступны.
Первый этап гнездостросния — это подготовка гнездовой ниши, дно которой самка выравнивает при помощи плоских камешков, которые она в изобилии натаскивает в убежище. Некоторые самки создают таким образом настоящее инженерное сооружение — многослойный настил, выравнивающий дно ниши не менее чем на 10°, почти до строгой горизонтали (рис. 54). Иногда камешками оказываются завалены глубокие щели в дне убежища. Бывает и так, что никакой видимой необходимости в такой работе нет, но самка тем не менее натаскивает много строительного камня, который она складывает не только под будущим гнездом, но и по всей длине прохода — до самого входного отверстия. В отличие от самок черных каменок Ое. picata, которые затрачивают па такого рода подготовку пиши не более полутора дней, некоторые самки златогузой каменки носят камешки полных 4 дня (в одном случае — с 30 марта до середины дня 3 апреля), работая при этом ничуть нс менее интенсивно, чем самки Ое. picata. В одном из гнездовых убежищ златогузой каменки я насчитал 453 камня. Они весили от 3 до 4 г. Так что по самым скромным подсчетам вес всего настила мог составлять немногим менее полутора килограмм. Вероятно, убежище использовалось каменками нс один год, но даже в этом случае затраченная птицами работа производит сильное впечатление.
132
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
Рис. 52. Некоторые позывки . а — при тревоге у гнезда (чередование сигналов NN I и 3), b — фрагмент серии коротких свистов (сипит N 3), издаваемых самцом в ходе формирования пары, с — позывка птенца, d — вокализация птенца при выпрашивании корма у родителей. Оригинал.
133
Панов Е.Н. Каменки Палеарктики
Рис. 53. Расположение гнезд Ое. chrysopygia kingi в горном Бадахшане (d. f — вид сверху). Обратите внимание на отсутствие приносимых самкой камешков у входа в гнездо в позиции “j”. а — камень, b — мягкий грунт. Оригинал.
134
Глава 4. Златогузая каменка
Ое. xanthoprymna и Ое. chrysopygia
Бывает, что самка, затратив много сил и времени на подготовку ниши, позже бросает се по причине полной непригодности. Одна из самок носила камни в узкую шель до тех пор, пока они нс начали вываливаться наружу. За каждым упавшим камнем птица сразу же спускалась по два или по три раза, пытаясь снова водворить его на место — как правило, безуспешно. Вскоре самка оставила эту шель и стала носить камни в нору в глинистой стенке обрыва, метрах в 150 в стороне.
Тот факт, что самцы златогузой каменки также склонны приносить камешки в ниши скал (о чем будет подробнее сказано ниже), наводит на мысль, что этот вид характеризует собой одну из первых исходных стадий в эволюции столь своеобразного гнездост-роитсльного поведения, которое постепенно деградирует в такой последовательности: Ое. leucura— Ое. xanthoprymna— Ое.
picata— Ое. finschii— Ое. pleschanka— Ое. hispanica.
Само гнездо златогузой каменки представляет собой довольно рыхлую чашу, сплетенную из сухой травы и выстланную изнутри лентами луба. Высота гнезда 45-56 мм (70-80 мм), диаметр 110-136 мм (140— 150 мм), диаметр лотка 85-95 мм (60-70 мм), глубина его 32-37 мм (40-50 мм). (Абдусалямов, 1973; цифры в скобках — по данным автора). Если грунт под гнездом мягкий, в нем обычно бывает довольно глубокая ямка, в которой покоится все сооружение. Гнезда, расположенные в узких вертикальных щелях, обычно сильно сплюснуты с боков.
Развитие о т н о ш ени и между членами пары и копуляция. В период после образования пары и во время постройки гнезда самец часто проявляет внимание к самки, причем она сама
Рис. 54. Гнездо хчатогузой каменки на пастиле из плоских камешков (Закавказье). Из: Panow J974.
135
Панов Е.Н. Каменки Палеарктики
нередко инициирует такие взаимодействия. В подобных ситуациях самка поет, после чего летит к самцу. При встрече слышна тихая трслька, и обе птицы принимают своеобразные сгорбленные позы (рис. 51 О-Самец теперь перемещается короткими прыжками вокруг самки, то приближаясь к ней, то несколько увеличивая дистанцию. Обычно он старается приблизиться к супруге сзади, и когда она склонна к продолжению церемонии, то поворачивается несколько раз на месте, пока не обратит хвост в сторону самца. Заканчивается такая встреча, как и у прочих видов каменок, стремительной и беспорядочной погоней в воздухе, во время которой непрерывно слышится тихое “цх вик... цх вик...”. Часто такие взаимодействия приурочены к глубоким вечерним сумеркам.
Эта церемония, во многом аналогичная соответствующим ритуалам у других каменок-петрофилов (см., например, рис. 16), своеобразна в том отношении, что позы самца и самки идентичны. Различия в поведении партнеров состоит лишь в том, что самка в сгорбленной позе иногда трепещет полуразвернугыми крыльями (самцы могут проделывать то же самое во время конфликта с соперником). Сказанное важно в том отношении, что позволяет предположить первичный характер отсутствия полового дихроматизма у златогузой каменки. Известно, что в тех группах птиц, для которых характерно отсутствие различий в окраске между полами, демонстративное поведение самок при формировании пары и на первых этапах ее существования неотличимо от поведения самцов (Lack, 1940). С другой стороны, как уже было сказано, половой дихроматизм отсутствует у представителей родов Cossypha и Cercomela, которые могут стоять у истоков эволюционной истории каменок Oenanthe.
Самец часто посещает самку возле строящегося гнезда, где происходят уже описанные церемонии, иногда — с некоторыми модификациями. Например, самец несколько раз поочередно залезает в гнездовую нишу и выскакивает оттуда, пока самка не последует за ним в убежище. Замечательно то, что помимо собственно гнездовой ниши существует и другая — выбранная самцом, куда он также время от времени приносит камешки, а иногда, посидев с минуту с принесенным камнем возле отверстия, уносит его проочь. Однажды я наблюдал, как самец прилетел с длинной травинкой и передал ее самке.
Эти черты поведения самца я воспринимаю как своеобразный атавизм, указывающий на то, что самцы у ближайших предков каменок принимали участие в гнездостроен и и наряду с самками (см. также Главу 11, посвященную чернобрюхой каменке Ое. leucura). У видов рода Cossypha гнездо строят оба члена пары (Farkas, 1969), у Cercomela familiaris участие самца в постойке гнезда не установлено, но и не отрицается
136
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
(Steyn, 1966). У Cercomela melanura в Израиле, по сообщению Зыковой, самец приносил к гнезду строительный материал наравне с самкой. Под некоторыми гнездами этого вида имеется платформа из камешков (Cramp, 1988)
Самка златогузой каменки затрачивает на сооружение гнезда не менее 5 дней, а вместе с возведением каменного настила — до 8 дней или более. К концу этого периода поведение самца резко меняется. Теперь он часто поет, сидя или бегая по карнизам скалы, над которыми нависают широкие каменные “крыши”. Изменяется и характер песни самца: она становится более разнообразной, включающей в себя звуки разной высоты — от предельно высоких до очень глухих и низких. В эту брачную песню самец часто включает упомянутое уже храпение. Время от времени самец принимает весьма экстравагантную позу: он поднимает расправленные крылья, плавно взмахивает ими вверхи вперед и медленно покачивает широко развернутым хвостом (рис. 52е).
Готовая к спариванию самка прилетает к демонстрирующему таким образом самцу под каменный карниз. При появлении здесь самки самец проделывает вокруг нес неистовый полет петлями, и, приземлившись, трепещет расправленными крыльями (рис. 52d) и словно бы “заползает” на самку. Когда та приступает к насиживанию кладки, самец в первые дни вынужденного одиночества продолжает посещать свои ниши под карнизами и подчас совершает всю церемонию приглашения самки к спариванию “вхолостую”. Все эти действия адресуются теперь какому-либо камешку или кусочку глины, и с таким неодушевленным предметом самец в конце концов и “спаривается”. Я трижды наблюдал подобное поведение одного и того же самца по отношению к двум комкам засохшей глины под его излюбленным карнизом.
4.5. Кладка и выводок
Кладка обычно содержит от 3 до 5 яиц. У формы chrysopygia в Закавказье и северном Иране в двух гнездах кладки состояли из 4 яиц, в четырех — из 5 (ср. 4,67, п=6). В Бадахшане у формы kingi в двух гнездах было по 3 птенца, в трех — 4 птенца, 4 птенца и неоплодотво-ренное яйцо и 5 птенцов, и в четырех — по 5 яиц (ср. 4,44, п=9). При соотношении 3 х 2,4 х 3,5 х 10 средняя величина кладки 4,53 (п=15). Интересно, что две кладки из 3 яиц в Бадахшане отложены, вероятно, одними из первых. Отсутствие кладок, сотоящих более чем из 5 яиц, может быть приспособлением к раннему гнездованию в высокогорьях.
Окраска скорлупы в большинстве кладок чисто белая, поверхность яиц матовая. Иногда яйца белые со слабым голубоватым оттенком. По
137
Панов Е.Н. Каменки Палеарктики
основному фону разбросаны очень мелкие красноватые крапинки, равномерно распределенные по всей поверхности яйца или сосредоточенные вокруг его тупого конца (рис. 55). На яйцах одной кладки из Бадахшана были мелкие голубоватые пятнышки, немногочисленные
и четко очерченные.
Размеры яиц двух кладок из юго-западного Туркменистана 19,6-21,0 х 15,6-16,3 мм, в кладке из Нахичевани (форма chrysopygia) 20,6-22,0 х 14,9-15,2 мм, в среднем 21,1 х 15,0 мм (п=5). В 5 кладках из Бадахшана (слабо дифференцированный подвид kingi) размеры яиц составляли 19,9-23,5 х 15,8-16,1 мм, в среднем 22,0 х 16,0 мм (п=22). По выборке без указания мест находок и количества яиц: 19,0-22,4 х 15,6-16,9 (ср. 20,7 х 16,3, вес 2,95 г (Schonwetter, 1979 цит. по: Cramp, 1988); 21,0 х 16,1 (п=44: Лоскот, 1973b). Обращает на себя внимание очень мелкий размер яиц из Закавказья.
Насиживает, по всей видимости, только самка. Во всяком случае, я нс наблюдал ничего похожего на регулярную смену птиц у гнезда, а
перерывы в насиживании связаны, вероятно, как и у других каменок, с периодическими уходами самки на кормежку. Самец не кормит самку на гнезде, как это происходит, например, у видов рода Cossypha. Лишь однажды я видел, как самец и самка коротко соприкоснулись клювами. Не исключено, что самец передал самке мелкое насекомое.
Может быть, столь редкое и нетипичное для
Рис. 55. Гнездо и кладка хзатогузой каменки Ое. chrysopygia (Закавказье). Из: Panow 1974.
вида поведение есть атавистический след, полученный им от его далеких предков.
/7 т е н ц ы. У только что вылупившихся птенцов кожа желчно-розового цвета. Эмбриональный пух у птенцов из Закавказья серый с теплым оттенком, в Бадахшане — почти белый. На плечевой птерилии эмбриональный пух представлен двумя пучками, рудиментарные пушинки есть на копчиковой птерилии. Клюв желтовато-розовый. валики по бокам его блсдножелтыс, лапы и когти розовые с жсл-
138
Глава 4. Златогузая каменка Ое. xanthoprymna и Ое. chrysopygia
то ваты м оттенком. Зев у птенцов желтый (у взрослых птиц он черный). Глаза открываются на пятый день; на шестой появляются кисточки контурных перьев на груди, брюшке и немного — на плечах. В этот же день становятся видны кисточки маховых. Самка сидит на маленьких птенцах долго, иногда (в холодную и дождливую погоду) до пятого дня после их вылупления. Кормят птенцов оба родителя.
Молодые подвида chrysopygia в первом гнездовом наряде по окраске ничем не отличаются от взрослой самки. По типу окраски гнездовой наряд у этой формы, лишенный пестристости, напоминает гнездовой наряд у Ое. finschii, у которой молодые также окрашены по типу взрослой самки. У молодых в первом гнездовом наряде из Бадахшана бока рыжеватые (как и подвида Ое. chrysopygia chrysopygia), в отличие от взрослых kingi, у которых эти участки оперения почти белые.
В конце второй недели после вылета из гнезда птенцы впервые пытаются петь и самостоятельно брать корм. В это же время у них появляется звуковой сигнал № 1. На 20-ый день некоторые юные самцы поют в клетке все светлое время суток. В месячном возрасте каменки начинают проявлять признаки полового поведения. В одном случае самец принял предкопуляционную позу и сделал садку на другую особь того же возраста. Через 1,5 мсс. после вылета из гнезда у молодых, содержащихся в неволе, пропадает поведение выпрашивания корма. Они начинают бояться воспитателя и при его приближении издают тревожный сигнал № I. В двухмесячном возрасте появляются элементы территориального поведения, иногда самец проделывает короткий полет петлями. В 3 мес. птицы постоянно поют во весь голос. Песня напоминает типичную удлиненную песню с включением в нее уже упоминавшегося характерного храпения. Короткая рекламная песня формируется к началу апреля следующего года.
4.6. Линька и отлет на зимовки
Линька взрослых птиц протекает со второй половины июля до начала сентября. К концу августа заканчивают в целом линьку в зимний наряд и молодые каменки (Иванов, 1969). Молодой самец расы chrysopygia. взятый птенцом из гнезда, сменил все перья хвоста в первую осень своей жизни (август — сентябрь). Смена второстепенных маховых проходила у него с середины октября до середины ноября. На следующий год этот самец начал сменять рулевые в середине июня.
Осенью последние особи chrysopygia могут быть встречены в северном Иране не позднее первых чисел ноября. На местах зимовок в южном Иране Ое. chrysopygia появляются в середине сентября, в Израиле — в тс же сроки (с 12.IX), в Аравии — даже в конце августа. В Эритрее
139
Панов Е.Н. Каменки Палеарктики
они проводят зиму в каменистом пересеченном ландшафте с разреженными насаждениями акации. Примерно в те же сроки проходит осенний пролет Ое. xanthoprymna. хотя часть особей, по-видимому, остается зимовать в нижнем поясе гор в районах своего гнездования (Smith, 1971; Cramp, 1988; Shirinai, 1996).
4.7. Биологические заметки
По способам питания и по составу пищи златогузая каменка ничем не отличается от прочих видов рода. Пропитание они разыскивают на земле, высматривают с присады или “повиснув” на несколько секунд в воздухе, иногда раскапывают почву клювом. С гнездовой территории часто улетают на поиски корма за ее пределы — иногда на расстояние до 700 м. В питании преобладают жуки и муравьи (Лоскот, 1974). Птица может поймать и съесть крупную сколопендру. Единственная наблюдавшаяся попытка напасть на ящурку Eremias закончилась неудачей.
По характеру сигнального поведения Ое. chrysopygia наиболее близка к пстрофильным каменкам Ое. lugens и Ое. finschii. отличаясь от них многими деталями поведения. Это, в первую очередь, своеобразие демонстраций самца при формировании пар (горизонтальная поза со сложенными рулевыми и без приподнимания клюва кверху), а также однотипность поведения супругов в их взаимодействиях в период гнез-достроения.
140
Глава 5. Краснопоясничная каменка Ое. moesta
Глава 5.
Краснопоясничная каменка Oenanthe moesta Lichtenstein, 1823
Синонимы: Saxicola moesta Lichtenstein, IX23; Oenanthe moesta theresae Mcinertzhagen, 1939.
Эта довольно крупная каменка (близкая по общим размерам к особям гренландского подвида обыкновенной каменки Ое. ое. leucorhoa) выделяется среди других палеарктических видов рода своей несколько необычной окраской — именно, значительным распространением у самцов палево-белого цвета на оперении крыльев. Эти беловатые участки оперения бросаются в глаза у сидящей птицы и отличают ее в по
Рис. 56. Самец (на переднем плане) и самка Ое moesta. Оригинал.
лете от всех прочих каменок северного полушария. По общему складу Ое. moesta — это каменка с массивной головой и слегка удлиненным
крепким клювом, высоко держащая тело на довольно длинных, сильных ногах. Во всех этих отношениях она наиболее похожа па черношейную каменку Ое. finschii. Оперение самцов контрастное черно-белое, гузка и основание рулевых перьев оранжево-рыжие, хвост почти целиком черный, без характерного для каменок Т-образного рисунка. Самки по окраске резко отличаются от самцов. Они окрашены в песочные тона, голова и хвост ржаво-рыжие (рис. 56).
5.1. Систематические замечания
Обычно выделяют два подвида — севсроафриканский Ое. т. moesta Lichtenstein, 1 823 и переднеазиатский Ое. т. brooksbanki Mcinertzhagen, I923 (рис. 57). Переднеазиатские птицы несколько крупнее и светлее африканских, но эти различия, по мнению некоторых авторов (Cramp, 1988), недостаточно существенны для выделения названных подвидов.
14I
Панов Е.Н. Каменки Палеарктики
Рис. 57. Ареал Ое. moesta.
5.2. Местобитания и численность
В отличие от златогузой каменки, Ое. moesta — типичная птица равнинных и холмистых ландшафтов. В северной Африке она приурочена к участкам песчаной полупустыни с редким невысоким кустарником, но не избегает холмистой местности с отдельными выходами каменистой породы. В Израиле эта каменка встречается в каменистой пустыне Негев. Всюду в пределах своего ареала она тесно связана с колониями общественных грызунов, в норах которых обычно гнездится. В Израиле это дневные песчанки Psammomys obessus (Shirinai, 1996).
Краснопоясничная каменка нигде не бывает особенно многочисленной. Например, в Марокко при учетах птиц с автомобиля отмечали не более 1-2 этих каменок на 10 км маршрута. Предполагается, что на всей территории Израиля в среднем за год едва ли гнездится более 5-10 пар (Shirinai, 1996). Весьма скудные данные по биологии этого вида получены при наблюдениях за изолированными парами, нс имевшими конспецифичсских соседей.
5.3. Годовой цикл и сроки гнездования
Основная часть популяции, по-видимому, оседла, хотя часть особей, возможно, кочуют во внегнездовой сезон. В Марокко встречали супружеские пары, держащиеся в колониях грызунов, уже в ноябре, но к гнездованию здесь птицы приступают нс ранее февраля. Взрослых каменок, носящих корм птенцам, видели здесь в период с конца марта до конца апреля (Smith, 1971). В Израиле самец из пары, обнаруженной 21 ноября, начал активно петь и изгонять со своей терри
142
Глава 5. Краснопоясничная каменка Ое. moesta
тории каменок других видов начиная со второй педели января. Самка начала строить гнездо 3 февраля, а птенцы вылупились, очевидно, 18 марта. Слетки покинули гнездо 30 марта, и через три недели откочевали из этих мест. Родители приступили ко второму циклу гнездования в мае.
5.4. Гнездо, кладка
Гнездо строит только самка. Судя по беглому замсчснию Ширипаи (Shirinai, 1996), самка приносит в гнездовую нору мелкие камешки, а при постройке гнезда используются, наряду с растительными материалами, шерсть и перья. Гнезда располагаются в нежилых норах млекопитающих на расстоянии до 2 м от входа либо в пустотах другого характера. В кладке 4-5 яиц (Cramp, 1988). Кладка из 4 яиц найдена 12 марта на северо-западе Синайского полуострова. У вышеупомянутой пары в Израиле выводок состоял из 4 слетков (два самца и две самки) (Shirinai, 1996). Яйца беловатые с очень мелкими рыжеватыми точками, расположенными более густо па тупом конце яйца. Размеры яиц подвида moesta: 23,0-24,0 х 15,5-17,0 (ср. 23,4 х 16,3, n=7) (Schdnwettcr, 1979 цит.: по Cramp, 1988). Судя по этим цифрам, яйца относительно более широкие, чем у Ое. xanthoprymna и Ое. finschii.
kHz
4lb
2'г.,'циНМ| М*
Рис. 58. Два последовательных фрагмента “брачной” песни Ое. moesla (а, Ь) и Два фрагмента прсдсовокупитсльной песни Ое. finschii (с, d). а. b — иа: Cramp 1988.
143
Панов Е.Н. Каменки Палеарктики
5.5. Биологические заметки
При поисках пропитания краснопоясничная каменка чаше других видов посещает норы грызунов. При наблюдениях за одной из пар оказалось, что птицы очень часто приносили слеткам скорпионов Scorpio maurus.
Положение этого вида внутри рода Oenanthe вызывает много споров. Тай (Тус, 1989а) сближает Ое. moesta с Ое. xanthopryma (xanthoprymna + chrysopygia). На мой взгляд, для этого нет сколько-нибудь очевидных оснований. Весьма фрагментарные сведения по поведению Ое. moesta (см. Cramp, 1988) наводят на мысль о близости этого вида к Ое. finschii. Судя по описаниям и сонограммам, приведенным в цитированном источнике, обнаруживается определенное сходство между черношейной и краснопоясничной каменками в характеристиках сигнала 1 (“глухой щелчок”) и коротких рекламных песен. Кроме того, “брачная” песня Ое. moesta напоминает прсдсовокупительную песню Ое. finschii монотонностью своего исполнения (рис. 58).
Если самки краснопоясничной каменки действительно начинают гнездостросние с приноса камешков (см. выше), то это указывает на вторичный переход Ое. moesta от петрофильности к гнездованию в равнинных ландшафтах. Первую стадию такого процесса иллюстрируют те популяции Ое. finschii, которые перешли от жизни в каменистых местообитаниях к гнездованию в равнинной пустыне, то есть в тех самых местобитаниях, которые наиболее характерны для Ое. moesta.
144
Глава 6. Черношейная каменка Oenanthe finschii
Глава 6.
Черношейная каменка, чернобокая каменка, белоспинный чекан Oenanthe finschii Heuglin, 1869
Синонимы: Muscicapa melanoleu-ca Giildcnstadt, 1775; Saxicola melano-leuca melanoleuca Hartcrt, 1910; Saxicola finschii Heuglin, 1869; Saxicola erythree Raddc, 1884; Saxicola barne-si Oates, 1890; Saxicola finschii turan-ica Zarudny, 1900; Saxicola finschii neglecta Zarudny, 1900; Saxicola finschii turcomana Loudon, 1903.
Видовое название melanoleuca было неверно присвоено черношейной каменке Хартер-том (Hartcrt, Die Vogel dcr Palaarktischen Fauna, 1910) no
Рис. 59. Черношейная каменка Oe. finschii (самец).
типовому экземпляру Ое. hispanica из Грузии. Это ошибочное название
позже использовал Зарудный (1914).
Рис. 60. Черношейная каменка (самка).
Чуть более короткокрылая и коренастая, чем прочие каменки, одетая мягким пушистым оперением, черношейная каменка выглядит немного крупнее своих истинных размеров (вес се около 25 г). Белая спина и брюшко в сочетании с черными перемычками между горловым пятном и сгибом крыла позволяет легко отличить самца от прочих палеарктических каменок (рис. 59). Спутать его можно лишь с одним вариантом гибридов между Ое. hispanica и Ое. pleschanka (так называемая “белоспинная
145
Панов Е.Н. Каменки Палеарктики
плешанка”), от которых самец черношейной каменки резко отличается голосом и гораздо более флегматичной манерой поведения. Самки обычно выглядят однотонными, светлосерыми (рис. 60). Молодые на первый взгляд неотличимы от самок.
6.1. Систематические замечания
Некоторые авторы (Гладков, 1954; Портенко, I954) ошибочно объединяют этот вид с населяющим Переднюю Азию и Северную Африку Ое. lugens. Неправомерность такого объединения, нс признаваемого большинством систематиков, подтверждается существенными различиями в биологии и поведении этих двух видов. Внутри вида Ое. finschii выделяют сегодня два подвида: Ое. f. finschii и Ое. f. barnesi (рис. 61). Характер географической изменчивости в общем такой же, как у других видов каменок: размеры увеличиваются с запада на восток (в сторону расширения первоначального ареала вида), а в окраске уменьшается примесь рыжеватых тонов. Самцы восточного подвида barnesi (дл и на крыла 91,7-94,3) отличаются от западных Ое. f. finschii (крыло 87,6-89,8 мм) большей белизной брюшка и меньшим развитием рыжеватопалевого оттенка на оперении подхвостья (полная параллель с географической изменчивостью Ое. chrysopygya и, в какой-то степени — Ое. moesta).
Кроме того, по мнению Зарудного (1923), у птиц из северо-восточной части ареала значительно больше развиты белые поля на внутренних опахалах маховых. В ареале западных популяций часто встречаются самки с темносерым или черноватым горлом (около 70%), тогда как на востоке это большая редкость (не более 6%) (Vaurie, 1949).
Рис. 61. Гнездовой ареал Ое. finschii и зимовки (показаны серым; 1 — подвида finschii, 2 — подвида barnesi). Цифрами показана средняя длина крыла самцов. Оригинал.
146
Глава 6. Черношейная каменка Oenanthe finschii
6.2. Местобитания и численность
Наиболее характерные местообитания черношейной каменки — это опустыненные холмы предгорий с отдельными выходами каменистой породы (рис. 62-63). Охотно занимает эта каменка неглубокие каньоны, прорезанные некогда полноводными реками в лессовой толще пустынных низкогорий. Здесь она в изобилии находит удобные убежища для гнезд в многочисленных промоинах и оврагах, соединяющихся с руслами дождевых потоков. Черношсйную каменку нельзя отнести к любителям сильно пересеченной местности, однако она может быть многочисленной в широких известковых каньонах, где, тем не менее, придерживается более выровненных нижних участков долины. Эти птицы обычно нс поднимаются в горы выше 700-800 м. над у.м., но местами их можно встретить и на больших высотах, в горной степи или у нижней границы горного редколесья (Pistaceaspp., Juniperusspp.) вплоть до 1700-1900 м над у.м. (Абдусалямов, 1973; Haffcr, 1977; Shirinai, 1996).
Приверженность вида к относительно мало расчлененным ландшафтам предгорий и низкогорий позволил ему вторично перейти к жизни в закрепленной песчаной пустыне. Вот что о стациях Ое. finschii
Рис. 62. Предгорья центрального Копетдага. Местообитания черношейной каменки Ое. finschii , черной каменки Ое. р. picata и плсшанки Ое. pleschanka.
147
Панов Е.Н. Каменки Палеарктики
Рис. 63. Типичный рельеф пизкогорий: так называемые чинки, по крутым склонам которых гнездятся черношейная каменка Ое. finschii, златогузая каменка Ое. chrysopygia и испанская каменка Ое. hispanica (Закавказье, северный борт долины р. Араке). Из: Panow 1974.
в пустыне Кызыл-Кум писал в свое время Зарудный (1915): “Излюбленные местообитания ... представлены слежавшимися песками с более или менее пышной кустарниковой растительностью; если вдобавок такие пески взметаны особенно высокими буграми и образуют особенно глубокие котловины, так что в общем приобретают довольно пересеченный вид, то нашей птичке нс остается, по-видимому, выбирать ничего лучшего”. В таких и подобных им ландшафтах каменки переходят от гнездования в нишах каменистых и глинистых обрывов к использованию для этой цели нор пустынных грызунов — например, больших песчанок Rhombomys opimus (Митропольский, 1968а). При этом Ое. finschii делит жизненное пространство с плясуньей Ое. isabellina, заметно уступая в численности этому исконному обитателю равнинных ландшафтов.
В южных частях Среднеазиатских республик бывшего Советского Союза популяции обычно разрежены, так что гнездящиеся пары разобщены и не контактируют друг с другом (Панов, Корзухин, 1974). В северных предгорьях Копстдага (Туркменистан) и в Таджикистане численность вида сравнительно невысока. Напротив, в северо-восточ
148
Глава 6. Черношейная каменка Oenanthe finschii
ном Прикаспии (в частности, в приморской части полуострова Мангышлак) черношейные каменки относятся к числу наиболее многочисленных видов, образуя здесь местами довольно плотные поселения (Панов, Иваницкий, 1975а). Столь же плотно селятся они в подходящих местообитаниях в Закавказье (Нахичевань), где группировки, включающие в себя до 10-12 пар отделены друг от друга обширными незанятыми видом угодьями. В этом регионе на 10 км маршрута у подножий скал отмечается 17,4 особей, в каменистых предгорьях — 8,6 особей, в каменистой полынной полупустыне — 4,7 особей (Тарасов, 1975).
Области зимовок расположены сравнительно недалеко от тех районов, где вид выводит потомство. Эти птицы чуть ли нс последними из всех каменок начинают откочевывать к югу, где проводят лишь самое холодное время года, а с первыми признаками весны возвращаются к местам гнездования. Только во второй половине октября черношейные каменки появляются на зимовках в северо-восточном Ираке, а уже впервых числах марта почти поголовно отлетают к северу. Некоторые самцы, а изредка и самки остаются в теплые зимы близ мест гнездования, например, в пустыне Кызылкум, в южных частях Туркменистана и Узбекистана, а в исключительных случаях — даже много севернее, почти у 45* с.ш. (Митропольский, 1968; Степанян, 1970; Абдусалямов, 1973).
6.3. Годовой цикл и сроки гнездования
В тех популяциях, где черношейные каменки частично оседлы, некоторые пары по окончании сезона размножения, возможно, остаются на своих гнездовых территориях до осени, а то и до следующей весны,. В южном Узбекистане на границе с Афганистаном (Термез, 37* с.ш., 67е в.д.) в конце сентября-началс октября 1973 г. в пределах тех самых участков, где каменки в этом году гнездились, я обнаружил, помимо 11 одиночных самцов, также 5 пар, члены которых делили между собой общие территории. На территории одного из одиночных самцов 29 сентября появилась самка, и взаимодействие этих птиц очень напоминало типичное образование пары. По свидетельству Степаняна (1970), в этом районе формирование пар начинается в конце января, причем первыми приобретают самок зимовавшие в одиночку самцы старше года.
К гнездованию некоторые пары приступают здесь уже во второй половине февраля: у двух пар 19-22 марта 1971 г. в гнездах уже подрастали птенцы. Самка в одной из пар, гнездившихся с некоторым запозданием, 15 марта насиживала кладку, а две другие самки 13 и 17 марта были заняты постройкой гнезд. В близкие сроки гнездование
149
Панов Е.Н. Каменки Палеарктики
начинается в годы с теплой весной примерно в 800 км западнее, под 38е с.ш. (Геок-Тспс в окрестностях Ашхабада). Здесь, в северных предгорьях Копстдага, гнездо с маленькими птенцами я нашел 10 марта 1968 г. Самки из двух других пар 12 и 18 марта еше только строили гнезда.
Как видно из этих примеров, даже в самые благоприятные годы размножение разных пар может идти очень неравномерно. По иному обстояли дела в южном Туркменистане в позднюю весну, последовавшую за очень суровой зимой 1969-1970 гг. В течение всего марта стояла холодная сырая погода с сильными туманами и с ночными температурами около 0°С. В первой половине месяца местами лежал еше не растаявший снег,что совершенно необычно для этих районов в марте. Все это сказалось и на численности чернобоких каменок и на сроках их размножения. Немногие пары, образовавшиеся уже в первых числах марта, были распределены гораздо более разрежено, чем в прошлые годы, и не приступали к постройке гнезд до последней декады месяца.
В заметно более поздние сроки события развиваются у северной окраины азиатской части ареала. На полуострове Мангышлак (44* с.ш., 52° в.д.) первые самцы появляются 6-13 марта (Митропольский, 1968а). В 1974 г. ко дню начала наблюдений (15.HI) только один из 9 территориальных самцов успел обзавестись супругой, которая уже строила гнездо. Самцы продолжали прибывать с зимовок и занимать территории вплоть до 22 марта, а массовый прилет самок пришелся на 17-22 марта. Последние эпизоды формирования пар наблюдали 25, 27 и 30 марта. Гнездостросние отмечено 19.111, 26.111, 8.IV и 26.IV. Взрослых птиц, занятых выкармливанием птенцов, видели в период между 24 апреля и 5 мая (8 пар). По данным Митропольского (1968), первых слетков в этом регионе можно встречать начиная с 7-10 мая. В середине мая каменки приступают ко второму циклу гнездования. Наиболее позняя дата вылета птенцов из гнезда — 5 июля.
В европейской части ареала, в Закавказье (Нахичевань, 39° с.ш., 45-46* в.д.) одиночные особи могут быть встречены в середине зимы. В 1975 г., несмотря на позднюю и холодную весну, 26 февраля я наблюдал уже занявшего территорию, поющего холостого самца, а 1 марта обнаружил неподалеку супружескую пару. Массовый прилет самцов происходил 5-9 марта, прилет самок и формирование пар — с 5 по 10 марта. Пролетных самок , пытавшихся внедриться на территории уже сформировавшихся пар, видели 6 и 15 марта.
Самка из пары, образовавшейся до 1 .III, еще насиживала, по-видимому, I4.V (хотя, возможно, у нее уже были маленькие птенцы) Самец, занявший территорию 5.III, приобрел самку 7.III, а 22.111 она строила гнездо. Самец, впервые появившийся 5.III, уже через день
150
Глава 6. Черношейная каменка Oenanthe finschii
обзавелся супругой. Гнездостроенис у этой пары наблюдали 22.III. У другой пары, сформировавшейся 6.III, па следующий день после прилета самца, самка строила гнездо 20-25.III, а 29.111 она носила строительный материал в другую нишу. Самец, занявший участок 9.III, преуспел в супружеских делах в тот же день. Его самка 24.III занималась постройкой гнезда, в котором 16.IV еще были яйца.
В том же районе в очень ранюю, теплую весну 1970 г. гнездо с хорошо оперенными птенцам нашли 8.IV, с маленькими птенцами — 18.IV, вылупление птенцов в двух гнездах произошло II.IV и 17.IV. Самку, строящую гнездо, видели 11 IV, другая насиживала кладку 20.IV. В этом году слетки у двух пар наблюдались 23.IV и 26.IV. Самка в первой из этих пар 23.IV одновременно кормила юнцов и строила новое гнездо. Позже оно было брошено, а в следующем, выстроенном той же самкой, I2.V было 5 яиц. В это время оба супруга еще комили молодых первого выводка, которые, казалось бы, уже могли существовать самостоятельно. Самка из второй пары запимхтась гнсздостроснисм 26.IV.
Кочующие в районе своих гнезд выводки стали обычными с 4-6.V. Вместе с тем, гнездо с только что вылупившимися птенцами найдено I3.V, а с птенцами в недельном возрасте — I4.V. Эти даты относятся, вероятно, уже ко второму циклу гнездования. Самка, кормившая маленьких птенцов 6.V. спустя 10 дней (15.V) строила новое гнездо. Нс исключено, что опа приступила к гнездованию уже третий раз за сезон. О возможности трех репродуктивных циклов у черпошейпых каменок сообщает Бельская (19656).
6.4. Социальное поведение
Территориальное поведен и е и с а м о р е к л а м //-р о в а н и е самцов. В период массового прилета самцов на места гнездования они постоянно вступают в конфликты из-за права на тот или иной участок местности. Наиболее рано прилетающие самцы захватывают территории большего размера и лучшего качества. Те, что запаздывают на пролете, получают возможность запять лишь небольшие участки, как бы вклиниваясь между крупными территориями, уже запятыми в более ранние сроки (см. рис. 10). Появление нового претендента в пределах уже поделенного между самцами жизненного пространства вызывает резкое противодействие со стороны первых поселенцев. Законный хозяин участка раз за разом пытается напасть па нсзвапного гостя. Владелец территории устремляется в сторону пришельца и, приземлившись в 1-3 м от него, словно бы крадется по земле замедленным скользящим шагом в характерной демонстративной позе (рис. 64а, 65-14, /9). Пришелец отвечает ему точно таким же
151
Панов Е.Н. Каменки Палеарктики
поведением. Оба соперника движутся параллельно друг другу, время от времени забегая в щели и пустоты, попадающиеся им на пути. По истечении нескольких минут претендент на участок взлетает, хозяин устремляется за ним, после чего снова повторяются описанные взаимные демонстрации — и так раз за разом, иногда до 6 часов кряду. При сближении оппонентов изредка можно наблюдать полет петлями одного из самцов вокруг другого. Все время самцы непрерывно поют удлиненную песню, чередующуюся с конструкциями типа подпесни. В нее включены особые высокочастотные звуки (“циканьс”, сигнал № 5) и птенцовая позывка (рис. 17).
Я подсчитал, что за время особенно длительного конфликта самцы, вовлеченные в погони друг за другом, находятся в воздухе до 70% времени и пролетают в итоге до 100 км и более. Понятно, что к концу взаимодействия слабейший из них уже почти не в состоянии летать, настолько он обессилен. В результате он либо ретируется, либо вынужден довольствоваться тем малым участком местности, которые ему все же удалось оставить за собой. В плотных поселениях на Мангышлаке, где конкуренция самцов за участки оказывается довольно острой, раз-
Рис. 64. Схема поведения черношейной каменки Oenanthe finschii во время распределения территорий между самцами (а), при формировании пар (Ь) и при копуляции (с). Из: Панов 1989.
152
Глава 6. Черношейная каменка Oenanthe finschii
Рис. 65. Континуум поз Oenanthe finsdiii. Позы NN 15, 19 характерны для конфликтов при распределении территорий между самцами, NN 10, 15 — при патрулировании ими границ территории, NN 12, 13, 17-19 — при взаимодействиях самца и самки во время образования пары и в более поздний период ее существования, NN 4, 5, 8 — на сталии копуляции, NN 19, 20, 25 — непосредственно перед копуляцией. N 9 — агрессивная поза самки, N 24 — ее поза перед копуляцией. Большинство других поз (как и поза N 19) используются самцами в широком спектре ситуаций. Тонкими короткими стрелками показаны резкие прыжки вперед, длинными топкими стрелками — движение шагом, толстыми стрелками -"скользящий” бег. Из: Панов 1978, с изменениями.
меры территорий варьируют от 0,7 до 10 га (в среднем 2,9 га, п=20) (Панов, Иваницкий, 1975а). В Закавказье ширина территорий между противоположными, охраняемыми от соседей участками границ составляет 106-230 м
153
Панов Е.Н. Каменки Палеарктики
После того, как самцы обосновались на индивидуальных участках, они значительную часть времени проводят на песенных постах, распевая и рекламируя свои владения эффектными демонстративными полетами. Птица взмывает высоко в воздух, мерно работая крыльями. Машущий полет периодически сменяется планированием па полностью расправленных крыльях. Самец описывает круги в горизонтальной плоскости, иногда слегка теряя высоту, и его широко развернутый белый хвост сияет в лучах весеннего солнца. Опустившись на песенный пост, самец, нс переставая петь, пробегает несколько шагов в горизонтальной позе с развернутым веером хвостом и с приподнятым кверху клювом. Во время этих демонстраций звучит удлиненный вариант песни, который во время пребывания на песенном посту сменяется короткими рекламными песнями (рис. 18b, а). И тот и другой тип песен, как и у Ое. xanthoprymna, практически лишены имитаций. При этом черношейная каменка, в принципе, способна подражать посторонним звукам, но лишь крайне редко вставляет в песню голоса других птиц.
Время от времени без всякой видимой причины самец устремляется к границе своего участка, и приняв особую угрожающую позу (рис.
Рис. 66. Взаимная стимуляция рекламных полетов у самцов Ое. finschii, занимающих соседние территории. А — оба самца холостые (число полетов 43 и 57). В — штриховкой показаны 35 полетов холостого самца, белыми столбиками — 9 полетов его соседа, самка которого уже начала строить гнездо. Из: Панов 1978.
154
Глава 6. Черношейная каменка Oenanthe finschii
65-10), провоцирует на столкновение того или иного из своих соседей. Если тот склонен принять вызов, он появляется на своей стороне границы, и оба движутся параллельно вдоль нее, угрожая друг другу и проделывая движения ложного кормления. Внезапно может последовать нападение одного из самцов на другого, и тогда возникает короткая драка. За 47 мин. непрерывных наблюдений за поведением одного из холостых самцов он 13,5 мин пребывая на излюбленном песенном посту, 7,5 мин находился в воздухе над своим участком и 5,5 мин конфликтовал то с одним, то с другим из двух своих соседей. Поведение живущих по соседству холостых самцов в той или иной степени согласовано. Они более или менее синхронно совершают рекламные полеты (рис. 66) и склонны выбирать в качестве постоянного спарринг-партнера определенного самца из числа своих соседей, который выступает в своеобразной роли “милого врага" (рис. 67).
Формирование пар. Когда, наконец, на территории внезапно появляется столь долгожданная самка, хозяин участка мгновенно приходит в полный экстаз. Желанная невеста не может остаться незамеченной. Она в момент своего появления проделывает нечто вроде рекламных полетов самца, присаживается в “угрожающей” позе на высо-
Рис. 67. Динамика патрулирования границ территорий у трех соседних самцов Ое. finschii (NN 11, 12 и 13). А — 18.111, В — 19.111, С - 21.111 (Мангышлак). 1 — появление самки на территории холостого самца N 12, 2 — взаимное патрулирование, 3 — провокации самца без ответной реакции соседа. Толщина стрелок дана в масштабе времени. Из: Панов 1978.
155
Панов Е.Н. Каменки Палеарктики
кис валуны и распевает коротенькие песни. Самец в ответ устремляется особым “полетом мотылька” (по прямой, с короткими частыми ударами крыльев) к одной из заранее присмотренных щелей или ниш, и в той самой позе, которая используется во время самых острых территориальных конфликтов, начинает попеременно забегать в отверстие и выбегать из него наружу. Песня и “цик...цик...цик...” звучат непрерывно, а забегая в нишу, самец произносит оттуда также серии птенцовых позывок (рис. 68 а). При звуках “цик...цик...” клюв приоткрывается, и хорошо виден ярко-желтый цвет полости рта, контрастирующий с черным цветом рамфотеки и оперения “лица”.
Если самка готова заключить помолвку, она подлетает к самцу, присаживаясь, однако, не ближе, чем в 1-3 м от него. В ответ тот устремляется к следующей щели, где снова повторяет свой брачный танец — и так раз за разом, подчас на протяжении 15-20 мин. Как и у прочих видов каменок (Ое. isabellina, Ое. xanthoprymna и других, о которых речь пойдет ниже), самец обычно не пытается приблизиться к самке по собственной инициативе. Если же он не выдерживает этого правила и садится рядом с ней, невеста ведет себя откровенно агрессивно и сразу же улетает, а самец устремляется в погоню. Преследуемая самка при первом удобном случае прячется в густой куст или в отверстие скалы, а самец продслываеи вокруг этого места импульсивный полет петлями.
Территориальное поведение самок. Обосновавшись на территории, самка активно защищает ее от других представительниц своего пола, и все се поведение при этом мало чем отличается от поведения супруга. Самка, подобно ему, провоцирует пограничные конфликты с соседками. Если же вдруг появляется новая претендентка на уже освоенную самкой территорию, конфликт между хозяйкой и пришелицей оказывстся столь же длительным и ожесточенным, как и конфронтация самцов в период распределения территорий. Самки гоняются друг за другом, совершают параллельные пробежки скользящим шагом, лазают в пустоты, поют и цикают, а иногда вцепляются друг в друга когтями и ожесточенно клюются, катаясь по земле (рис. 69). Что касается, самца, то он, в общем, игнорирует происходящее, лишь изредка делая ленивую попытку напасть на ту или иную из конкуренток, не делая особой разницы между ними. В конце одного из конфликтов самок, продолжавшегося два с половиной часа, пришлая самка к концу второго часа явно начала терять силы и вынуждена была покинуть место конфликта. Я подозреваю, что ее упорство в попытке отвоевать уже занятую территорию объяснялось тем, что это был ее прошлогодний участок. Что касается победительницы, то она, оставив за собой поле боя, взлетела на песенный пост самца и провозгласила
156
Глава 6. Черношейная каменка Oenanthe finschii
Рис. 68. Поведение самца Ое. finschii при первом появлении самки па его территории (А) и в последующие периоды существования пары. Показано уменьшение экспрессивности демонстраций самца и обеднение его вокального репертуара с течением времени. Сонограммы в позиции А показывают короткую и Удлиненную песни и сигналы NN 5 (циканье) и 6 (видоизмененная птенцовая позывка). Из: Панов 1989.
157
Панов Е.Н. Каменки Палеарктики
Рис. 69. Позы самок Ое. finschii в момент формирования пары и при территориальных конфликтах друг с другом. Из. Панов 1989
свою победу торжествующим, хотя и не слишком мелодичным пением.
Г нездострое-н и е и конструкция гнезд. От момента образования пары до начала постройки первого гнезда может пройти целый месяц, но если позволяют погодные условия, самка приступает к гнездострое-нию в течение недели. Самец не помогает ей в этой работе. Нередко самка, облюбовав ту или иную камеру и уже приступив к гнездостроению, неожиданно находит изъяны в своем выборе и вскоре начинает носить строительный материал в другое убежище. Одна самка даже дважды меняла свой вы
бор, прежде чем выстроить гнездо и отложить в него яйца.
Иногда самец пытается привлечь подругу к какой-нибудь другой нише из числа тех, что он показывал ей во время помолвки, словно желая повторить события того счастливого дня. Однако теперь поведение самца уже далеко не столь эффектно, как в момент формирования пары (рис. 64b, с). Здесь уместно будет сказать, что за несколько лет наблюдений был зафиксирован только один случай (!), когда самка выстроила гнездо в том убежище, которое самец демонстрировал ей при формирования пары.
Места, в которых сооружаются гнезда, достаточно разнообразны. В предгорных районах гнезда помещаются или в глубоких пустотах между камнями (иногда — в щелях монолита, где гнездо совершенно недоступно для хищных птиц и млекопитающих), или в углублениях стенок гли
нистых оврагов, гораздо реже — в норах грызунов на ровных площадках, заросших низкой травой. Очевидно, именно способность использовать убежища последнего типа способствовало расселению вида в песчаные пустыни, где эти птицы гнездятся почти исключительно в норах грызунов (преимущественно — большой песчанки Rhombomys opimus).
158
Глава 6. Черношейная каменка Oenanthe finschii
Рис. 70. Плоские камешки у входа в гнездовое убежите черношейной каменки. Из: Panow 1974.
Гнездо может быть выстроено на разной глубине, обычно в 15-80 сМ от входа, (в среднем по 8 гнездам — в 30 см) но некоторые гнезда располагаются в неглубоких нишах, так что край постройки виден снаружи. Подчас гнездо недоступно для осмотра. Как правило, перед входом в гнездо по ходу тоннеля лежит значительное количество плоских камешков, реже — кусочков костей, стекла, осколков фаянсовой посуды (рис. 70). Митропольский обнаружил в одном убежище 240 камешков общим весом 720 г. Некоторые из них достигали размеров 47 х 32 мм и весили до 10 г. Обычно под самим гнездом можно обнаружить всего 3-4 камешка, тогда как большая часть из них лежит снаружи, у входа в гнездовую камеру, сразу же выдавая присутствие гнезда опытному хищнику. Как справедливо указывает Митропольский (1968), нецелесообразность и демаскирующий характе повадки приносить камни в гнездовой тоннель особенно ярко проявляется при гнездовании черношейных каменок в пустыне, в норах грызунов, где камешки, лежащие на светлом песке, издалека бросаются в глаза.
Так или иначе, самка начинает постройку гнезда именно с приноса камней. Самец нс помогает партнерше, он распевает где-нибудь неподалеку, лишь изредка навещая самку возле строящегося гнезда. Количество камешков, приносимых на место постройки, зависит от индивидуальности самок и от формы самого убежища. Бывают камеры, где камни просто некуда положить (из 47 убежищ, осмотренных Митропольским, таких было два (рис. 71с), но иногда места вполне достаточно, а самка, тем не менее, почему-то нс выкладывает ход камнями. Уже в самом начале работы самка приносит поочередно камешки и соломинки, а вскоре начинает таскать только пучки сухой травы, которую она вырывает клювом из земли нспо-
159
Панов Е.Н. Каменки Палеарктики
Рис. 71. Расположение гнезд Ое. finschii (вид сбоку и сверху). Штриховкой показана каменистая порода, точками — мягкий грунт. Из: Митропольский 1968а.
далеку от места расположения гнезда. На постройку гнезда уходит около недели. Его основа, свитая из сухой травы, обычно выстилается изнутри плотным слоем шерсти, волоса и перьев, но некоторые гнезда бывают лишены такой подстилки.
Развитие о т-ношении между членами пары и копуляция. В период гнсздостросния время от времени можно видеть следующую сцену: самец садится около самки, принимает особую “сгорбленную” позу и начинает расхаживать вокруг супруги мелкими шажками (рис. Ь5-12). Самка почти
не реагирует на эти действия кавалера, и обычно спустя несколько секунд улетает, а тот устремляется за ней, следуя всем поворотам се полета. Стремительная погоня заканчивается в момент приземления
самки, а се преследователь проделывает около нес эмоциональный полет петлями, сопровождаемый возбужденным пением.
Эти взаимодействия весьма сходны с тем, что мы изредка наблюдали у Ое. oenanthe и очень часто — у Ое. chrysopygia (сравни рис. 65-12. 13, 17с рис. 42а и 51 f). К концу гнсздостроения их приходится видеть все реже и реже. Теперь самец уже не пытается приблизиться к самке, которая также явно избегает его, готовясь к яйцекладке. По вечерам
поведение самца выглядит довольно странно: нахохлившись и напоминая черно-белый пушистый шарик, он то и дело отводит крылья в стороны, перелетая время от времени полетом мотылька с одного кустика на другой (рис. 65-4,5,<У). Все это может продолжаться и час, и более. Самец то и дело начинает перебирать оперение клювом, но выглядят эти действия какими-то нервными и беспорядочными. Внезапно самец словно преображается. Оперение теперь приглажено, и птица
160
Глава 6. Черношейная каменка Oenanthe finschii
стремительно залетает в какую-нибудь обширную нишу в скале. Здесь самец начинает перебегать с места на место скользящим шагом, то скрываясь в темном углу ниши, то выскакивая на ее край. Эти демонстрации сопровождаются весьма своеобразной песней, нс похожей ни на что из того, что нам ранее приходилось слышать. Песня звучит как многократно повторяемое, очень тихое и монотонное “тиририрю...ти-ририрю ...тиририрю...” (рис. 18d, 58с, d). Хотя самец, пытаясь таким образом обратить на себя внимание, проводит в нише иногда до получаса, его демонстрации первое время нс производят никакого видимого впечатления на самку. Когда же приближается время откладки яиц, она начинает прилетать на свидания, хотя и теперь такому посещению могут предшествовать весьма длительные демонстрации самца в нише. Но даже явившись на место встречи, самка нс всегда позволяет самцу довести дело до конца. Как оказалось, она готова уступить домоганиям супруга лишь непосредственно во время яйцекладки.
У одной пары я наблюдал посещение самкой супруга, токующего в нише, 26, 28 и 29 марта (25 и 27 марта наблюдения не проводились). Все три посещения закончились копуляцией. 30 марта самка прилетела к самцу, но не позволила ему сделать садку. При инспектировании гнезда наощупь 31 марта выяснилось, что в нем было нс менее 4 яиц. Всего у разных пар я наблюдал 17 прилетов самки в нишу самца. Из них только 6 закончились копуляцией. Десять раз такие свидания происходили в период между 19 ч. 10 мин. и 20 ч. 30 мин., 6 — между 17 ч. 15 мин. и 18 ч. 50 мин., и только один раз — угром (около 11 ч.). Сама копуляция происходит так же, как и у других видов каменок: в момент прилета партнерши самец с торопливой песней проделывает вокруг нее полет петлями, мягко ударяясь грудью о край потолка ниши. При посадке он быстро трепещет широко расправленными крыльями и мелкими шажками “заползает” на спину самки (рис. 65-24, 25). В момент копуляции самец подворачивает свой широко развернутый хвост под гузку самки. При этом он неизменно издает серию высокочастотных “цик...цик,..цик...”. Сразу же после коитуса самец стремительно улетает прочь с песней или, чаще, издавая серию коротких посвистов (сигнал № 3 — см. рис. 17). Детальное описание социального и сигнального поведения Ое. finschii содержится в другой моей работе (Панов, 1978).
6.5. Кладка, выводок
Кладка состоит из 4-6 яиц. По данным Митропольского, на Мангышлаке в гнездах первого цикла численно преобладают кладки из 4 яиц (средняя величина кладки 4,9 яиц, п=26), тогда как кладки второго, летнего цикла крупнее (обычно — 6 яиц, в среднем 5,6, п=6).
161
Панов Е.Н. Каменки Палеарктики
Все кладки, найденные автором в Средней Азии и Закавказье, содержали яйца почти чисто белого, чуть розоватого цвета с небольшим количеством очень мелких красноватых точек, тяготеющих к тупому концу (рис. 72). Размеры их составляли 21,2-22,1 х 16,2-16,7 мм (2 кладки по 5 яиц из Закавказья, в среднем 21,6 х 16,4). По данным Лоскота средние размеры яиц подвида barnesi составляют 21,2 х 16,1 (50 яиц из 9 кладок). Размеры 30 яиц из ареала подвида/ш5сА/7: 19,0-24,1 х 14,9—17,0, в среднем 22,1 х 16,2 (Schonwcttcr, 1979 цит. по: Cramp, 1988).
Птенцы, уход за ними со стороны родителей. Насиживает только самка. Самец не кормит ее, и она время от времени оставляет кладку и улстакт на кормежку. В эти моменты самец может токовать перед пей, а затем сопровождает самку до тех пор, пока та не садится на гнездо. Как правило, в этот период птицы весьма осторожны, и найти гнездо, выследив самку, совсем нелегко.
Вылупление птенцов происходит на 12-13 день после начала насиживания. Пух у новорожденных птенцов очень светлый, серебристо-серый, длиной до 10 мм. Самка выносит скорлупу яиц из гнезда и бросает се на расстоянии в несколько десятков метров в стороне. В течение первых трех — четырех дней она большую часть времени сидит на птенцах, лишь изредка отлучается на кормежку, а возвращаясь, приносит корм и птенцам. Но основная роль в выкармливании потомства в этот период принадлежит самцу.
Следует специально отметить, что самец черношейной каменки, занятый выкармливанием птенцов, не перестает активно петь и охранять территорию. Постоянно можно видеть, как яркая, роскошная птица летит через долину типичным демонстративным полетом — мелко трепеща крыльями, широко развернув хвост распевая в полную силу, несмотря на то, что из клюва се свешивается гусеница либо кобылка, предназначенная на корм птенцам. Нс переставая петь, самец приземляется неподалеку от входа в гнездовую камеру, небрежно оглядывает округу и ныряет внутрь.
По прошествии первых дней, когда птенцы требуют постоянного внимания самки, она уже не сидит на них днем и ночью и предоставляет почти всю заботу о выводке самцу. В это время самку редко увидишь около гнезда — она отдыхает и приходит в себя после почти трех недель сплошного затворничества. Однако, к тому моменту, когда птенцы начинают оперяться, оба родителя кормят их почти на равных началах. Особенно много работы у них впервые дни после вылета молодых из гнезда.
Дело в том, что слетки почти сразу же разбредаются по окрестностям гнездового участка, и каждый облюбовывает для себя постоянное индивидуальное убежище, у входа в которое он греется на солнце и
162
Глава 6. Черношейная каменка Oenanthe finschii
куда прячется при первом же сигнале родителей об опасности. Иногда это убежище имеет один запасной выход или несколько. Все это создает гарантии для выживания если нс всего выводка, то по крайней мере большей его части, но, с другой стороны, усложняет задачу снабжения молодняка кормом для родителей, поскольку слетки держатся подчас на значительном расстоянии друг от друга.
Уже на протяжении первой недели после вылета птенцов первого выводка самка начинает подыскивать место для нового гнезда, и, сделав свой выбор, вновь приступает к строительным работам. Одна и та же камера может быть использована повторно лишь в разные годы (именно за счет этого около наиболее удобных убежищ скапливаются особенно большие запасы каменных плиток), но птицы никогда нс гнездятся дважды подряд в одной и той же нише. В подстилке гнезда после пребывания здесь птенцов обычно скапливается значительное количество личинок мух-кровососок, что не позволяет использовать эту колыбель вторично.
Поведение самки в описываемый период столь же на удивление универсально, как и описанное ранее поведение самца при кормлении птенцов. Самка носит пучки травы для нового гнезда до тех пор, пока нс услышит настойчивого писка какого-либо особенно проголодавшегося слетка. Тогда она бросает гнездовой материал, спешно переключается на ловлю насекомых и несколько раз подряд кормит юнца до полного его насыщения. Тут же самка снова принимается за гнсздостросние. Поведение ее выглядит в это время на редкость рациональным.
По мере того, как самка уделяет все большее внимание постройке нового гнезда, самец все чаще кормит слетков. Когда же самка приступает к насиживанию кладки, на самца полностью ложится забота о первом выводке (рис. 72). В это время слетки выглядят еще совершенно короткохвостыми, так что им еще далеко до приобретения самостоятельности. Впрочем, выводок не распадается еще на протяжении как минимум двух недель, и даже научившись самостоятельно добывать пропитание, молодые держаться разреженной стайкой во главе с самцом, а позже — уже без взрослого руководителя. Примерно через месяц после вылета из гнезда юнцы начинают конфликтовать друг с другом и постепенно откочевывают из окрестностей гнезда.
Птенцы, взятые из гнезда 8.IV в возрасте около 10 дней и помещенные в вольеру, впервые стали подавать голос через 20 дней (сигналы №№ 1 и 3). Один из них 15.V начал тихо и неуверенно петь. Примерно в двухмесячном возрасте молодые птицы (и самцы и самки) громко распевают в клетке в течение всего дня. В этом же возрасте появляются зачатки брачных демонстраций. К этому времени юнцы становятся совершенно нетерпимыми по отношению друг к другу, и
163
Панов Е.Н. Каменки Палеарктики
Рис. 72. Самец черкошейкой каменки Oc.finschi со слетком.
их можно держать теперь только в отдельных клетках. Если же клетки поставить рядом, их обитатели начинают издавать серии звуков “тшок... тшок”, переходящие в громкое “храпение” (сигналы №№ 1 и 2, рис. 17b), а также фрагменты удлиненной песни. Птицы сближаются вплотную и настойчиво стремятся ударить друг друга клювом сквозь прутья клетки.
Недавно оперившиеся птенцы, впервые увидев воду, бывают явно возбуждены, но в воду нс идут, а принимаются “купаться” в песке около плошки с водой. Взрослея, они иногда все же купаются в воде, но по крайней мерс в первые 4 мсс. жизни делают это крайне редко.
6.6. Линька и отлет на зимовки
Линька молодых первого выводка начинается уже в конце июня. К середине августа у них сменяется контурное оперение. У трех самок, взятых птенцами из гнезд в апреле, в течение августа сменились все рулевые, большие кроющие крыла и значительная часть маховых. Очевидно, уже к концу августа (а иногда, возможно, и раньше) молодые самцы могут приобрести контрастный черно-белый наряд, в котором будут щеголять следующей весной.
Перед отлетом на зимовки линяют и взрослые каменки. Перелетные популяции покидают места гнездования в первой половине ноября. На зимовках в Пакистане и юго-западном Иране первые мигранты появляются уже в конце августа, далее к западу и югу, в Израиле — только в начале октября (6.Х). Основной прилет в последнем регионе
164
Глава 6. Черношейная каменка Oenanthe finschii
проходит в течение всего этого месяца (Гаврилов, 1970; Cramp, 1988; Shirinai, 1996).
На зимовках каждая особь, независимо от пола и возраста, удерживает за собой индивидуальную территорию. В юго-западном Иране зимующая популяция включает в себя как пришельцев с севера, так и, вероятно, местных особей. Здесь некоторые самцы занимали один и тот же участок 2 (в одном случае 3) зимы подряд. Территории самцов размещаются преимущественно в местообитаниях лучшего качества и сравнительно невелики здесь (в среднем 3 га). Около 20% самцов и все самки вытеснены в худшие местообитания, где средняя величина территорий составляет 5,8 га у первых и 5,3 га у вторых (Cornwallis, 1975, цит. по: Cramp, 1988).
В Израиле зимующая популяция Ое. finschii оценивается в 3000— 5000 особей. В более благоприятных районах близ побережья Средиземного моря преобладают самцы (55-72%), тогда как в пустыне значительно многочисленнее самки (70-80%). В местах скоплений птицы держатся в среднем на расстоянии около 300 м друг от друга. Иногда каменки встречаются парами (Shirinai, 1996).
Черношейная каменка — это типичнейший представитель “черно-белых0 каменок-пстрофилов, образующих центральное ядро рода Oenanthe. Трудно согласиться с мнением Вори (Vaurie 1949), что Ое. finschii может быть объединена в составе единой “группы видов” с пустынной каменкой Ое. deserti, которая по своим этологическим характеристикам резко уклоняется от видов, составляющих указанное центральное ядро рода (см. Главу 16 и рис. 1).
165
Панов Е.Н. Каменки Палеарктики
Глава 7.
Траурная каменка Oenanthe lugens Lichtenstein, 1823
Синонимы :для Oenanthe lugens s.str.: Saxicola halophilaTristram, 1859; Saxicola lugens Lichtenstein, 1823; Saxicola persica Secbohm, 1881; Oenanthe lugens sarudnvi Sarudny et Harms, 1926;
для “группы подвидов" lugentoides: Saxicola lugentoides Sccbohm, 1881; Oenanthe lugubris boscaweni Bates, 1937;
для “группы подвидов ”lugubris: Saxicola lugubris Rupcll, 1837; Saxicola schalowi Fisher et Reichenow, 1884.
7.1. Систематические замечания
Среди орнитологов нет согласия в том, что именно таится под латинским названием Oenanthe lugens. В тот период, когда в зоологии господствовала так называемая “широкая концепция” вида, было принято объединять все аллопатрические популяции, более или менее сходные морфологически, в некий единый “большой” вид. В это время имя Ое. lugens распространяли на внешне сходных между собой каменок из 4 крупных географических областей, считая, что каждый из этих регионов населяет некий подвид либо группа подвидов траурной каменки (рис. 73). В целом картина выглядела следующим образом:
I. Популяции северной Африки (halophila), Ближнего Востока (lugens) и Иранского нагорья (persica) объединяли в “группу подвидов” lugens.
2. Популяции восточной Африки, распространенные в Эфиопии (lugubris), в Кении и Танзании (schalowi) и на полуострове Сомали (vaurie) объединяли в ‘Труппу подвидов” lugubris.
3. Популяции юго-западной части Аравийского полуострова (lugentoides), южной Аравии (boscaweni), первоначально включенные в “группу подвидов” lugens, позже стали рассматривать как обособленные таксоны, заслуживающие выделения в группу подвидов (или даже в самостоятельный вид) lugentoides.
4. Популяции Иранского нагорья, распространенные далее на северо-восток в Среднюю Азию и объединенные под общим именсм/шс/ш. некоторые орнитологи (Гладков, I954; Портенко, I954) также включали в качестве подвида в состав вида Ое. lugens.
Что касается последнего пункта этого перечня, то здесь просто была допущена ошибка: Ое. finschii и форма persica на обширных простран-
166
Глава 7. Траурная каменка Oenanthe lugens
Рис. 73. Ареал Ое. lugens (а) и близких ему таксонов (b — lugubris, 3 подвида и с — lugen(aides, 2 подвида), d — зимовки. Цифрами показана средняя длина крыла самцов. Оригинал.
ствах Иранского нагорья обитают совместно, нс гибридизируя друг с другом, так что их никак нельзя относить к одному и тому же виду. В отношении же остальных трех “групп подвидов" вопрос о том, насколько они близки друг к другу и должны ли считаться одним видом либо разными видами, до сих пор остается открытым. И в этом плане ничего не изменится до тех пор, пока в руках у орнитологов нс окажутся достаточно подробные сведения о биологии и, особенно, о поведении практически не изученных популяций восточной Африки и Аравии.
Тем нс менее уже сейчас, когда все больший вес приобретает “узкая концепция вида", многие орнитологи склоняются к мысли, что обозначенные выше три группы подвидов (lugens sensu stricto, lugubris и lugentoides) более целесообразно рассматривать в качестве “зарождающихся” либо самостоятельных видов (Vauric, 1950; Hollom el al., 1988; Tye, 1989a; Shirinai, 1996). Этой точки зрения придерживаюсь и я, включая в состав рассматриваемого здесь вида Ое. lugens только три подвида: lugens, halophila и persica. При этом я все же считаю полезным упомянуть в соответствующих местах и о том очень немногом, что известно об образе жизни двух других видов (Ое. lugubris, Ое. lugentoides), стоящих в генеалогическом отношении достаточно близко, по-види-мому, к траурной каменке Ое. lugens.
Траурная каменка по общему складу и пропорциям имеет много общего с черношейной (рис. 74). Средний вес особей — также около 25 г, но клюв немного более длинный и тонкий. От самцов Ое. finschii
167
Панов Е.Н. Каменки Палеарктики
самцы Ое. lugens отличаются черным цветом спины и более яркими рыжеватыми тонами нижних кроющих перьев хвоста. Насыщенность рыжеватого цвета подхвостья увеличивается в направлении с запада на восток в ряду подвидов halophila— lugens — persica. Белые поля на внутренних опахалах первостепенных маховых наименее развиты у расы lugens, а примесь серого на белой “шапочке” наиболее заметна у расы persica. В северной Иордании и южной Сирии обитают сплошь черные каменки с белой гузкой, бело-черным хвостом и с белыми полями на маховых перьях, которых принято считать цветовой морфой Ое. lugens lugens (Wallace, 1983; Cramp, 1988).
У западного подвида halophila некоторые самки по окраске резко отличаются от контрастно окрашенных самцов и сходны с черногор-лыми самками Ое. finschii, другие же в той или иной степени похожи на самцов (см. Vaurie, 1949). У подвида lugens самки окрашены так же, как самцы, у формы persica они чуть тусклее. Резкий половой диморфизм (как у halophila) свойственен восточноафриканским Ое. lugubris и аравийским Ое. lugentoides. Самцы Ое. lugubris отличаются от самцов Ое. lugens тем, что у первых отсутствует белый цвет на маховых перьях, а на хвосте и надхвостье он, как правило, заменен рыжим; цвет шапочки варьирует от светло рыжеватого до темного буро-серого. У этого вида наблюдается полиморфизм самцов: в южной части ареала у части из них брюшко сплошь белое, у других — черное. Самцы Ое. lugentoides, в общем, очень похожи на самцов Ое. lugens (рис. 73).
В пределах вида Ое. lugens размеры птиц увеличиваются с запада на восток. У самой мелкой расы halophila длина крыла самцов составляет в среднем 90,6-91.8 мм, у lugens 93,4-96.1 мм. у persica 95,3-96,6 мм. Восточноафриканские Ое. lugubris отличаются мелкими размерами (средняя длина крыла самцов в Кении 88,4 мм, севернее, в Эритрее и Эфиопии 84,8 мм). У аравийских Ое. lugentoides длина крыла самцов варьирует в пределах от 85 до 90 мм (Meinertzhagcn, 1949; Vauric, 1949; 1950; Mackworth-Pracd, Grant, 1955; Cramp, 1988).
Рис. 74. Траурная каменка Ое. I. lugens. Израиль, цен-тальный Негев.
168
Глава 7. Траурная каменка Oenanthe lugens
7.2. Местобитания и численность
Траурная каменка — это характерный вид каменистых полупустынных и пустынных ландшафтов. Ее местообитания привязаны к аридным, более или менее пересеченным местностям со скалистыми склонами, каменистыми осыпями и сухими ущельями. Растительность здесь зачастую весьма скудная, с бедным покровом из горно-степных травянистых растений и с низкорослой полукустарниковой порослью, сосредоточенной в основном в руслах пересыхающих потоков — так называемых вади. В юго-западной Африке эти каменки гнездятся также среди сухих, безводных глинистых холмов. В Израиле траурная каменка — это наиболее многочисленный вид р. Oenanthe в безотрадной каменистой пустыне Негев (рис. 75). Здесь она избегает лишь наиболее аридных и лишенных растительности участков на обширных выровненных плоскогорьях (Cramp, 1988; Shirinai, 1996).
В юго-западном Иране в местах сосуществования Ое. lugens и Ое. finschii первая вынуждена довольствоваться самыми бесплодными участками каменистых склонов, уступая более богатые растительностью и, соответственно, более кормные угодья у подножий скал своему весьма агрессивному конкуренту. В таких местах только 34% пар Ое. lugens способны удержать за собой территории в оптимальных местообитаниях на границе между травянистым пологим склоном и каменистыми обрывами. Там же, где Ое. finschii отсутствует, в подобных оптимальных местообитаниях сосредоточено уже до 79% пар траурной каменки (Cornwallis, Porter, 1982).
Суровые условия существования траурной каменки обычно не позволяют ей селиться сколько-нибудь плотными группировками. Отдельные пары или полностью разобщены, или образуют весьма разреженные поселения. При учетах с автомобиля, проведенных в Иудейской пустыне и в Негеве (Израиль), сидящих на проводах каменок видели с промежутками в 1-7 км. В Негеве расстояния между гнездами соседних пар составляли в среднем 2,5 км (минимально 0,5 км), а средние размеры территорий — 16,5 га. (Shirinai, 1996). Нетрудно видеть, что плотность популяций Ое. lugens несопоставимо мала по сравнению с тем, что мы видим у каменок в более северных регионах (например, у занимающей сходные местообитания Ое. xanthoprymna).
Траурная каменка обычно связана с предгорьями и низкогорьями, но в юго-западном Иране она поднимается до высот 1700-2700 м над у. м. Местообитания восточноафриканской Ое. lugubris — это каменистые склоны с осыпями и с довольно богатой кустарниковой растительностью на высоте нс ниже 1300 м над у.м. Аравийская Ое. lugentoides обычно занимает сухие каменистые склоны с разреженной кустарни-
169
Панов Е.Н. Каменки Палеарктики
Рис. 75. Местообитания Ое. lugens п пустыне Негев (Израиль).
ковой или древесной растительностью на высотах 1000-3000 м над у. м. (Smith, 1971; Cornwallis, Porter, 1982; Evans, Christensen, Drooks, 1987; Cramp, 1988).
7.3. Годовой цикл и сроки гнездования
В северо-западной Африке и в передней Азии (Израиль, Иордания) значительная часть гнездящихся популяций состоит из оседлых птиц. Например, в Израиле с наступлением сухого сезона из районов гнездования откочевывает в основном возмужавшая к этому времени молодежь, а взрослые птицы остаются до следующего сезона размножения на своих территориях. Самцы начинают активно петь и охранять границы своих владений в феврале, а с середины марта уже в разгаре откладка яиц (Zachai, 1984; Cramp, 1988; Shirinai, 1996). В пустыне Негев в очень сухую, позднюю и холодную весну 1996 г. я наблюдал пару, носившую корм птенцам, 6 апреля. В Египте птицы той же расы lugens приступают к гнездованию даже еше раньше — в феврале (Meincrtzhagen, 1954).
В ареале расы halophila в Марокко в ноябре видели только самцов, но к середине февраля пары уже сформировались. Считают, что откладка яиц начинается в северо-западной Африке с середины марта.
170
Глава 7. Траурная каменка Oenanthe lugens
но каменок, выкармливающих птенцов, видели в Марокко только 20 апреля (Smith 1971).
В популяциях, склонных к оседлости, многие пары гнездятся дважды за сезон. В Израиле начало второго цикла размножения приходится на конец мая-начало июня. Перелетные persica возвращаются с зимовок на места гнездования в западном Иране (около 33 с.ш.) в самом конце февраля и на протяжении марта (29.П-25.III). Птенцов в гнездах здесь находили 6-8 апреля (Zarudny, Harins, 1926). Предполагается, что большинство пар этого подвида гнездятся лишь один раз за сезон — даже в южных частях их ареала, в юго-западном Иране.
7.4. Гнездо, кладка, выводок
Гнезда помещаются довольно глубоко в расселине скалы, под камнем, иногда в норах мелких млекопитающих. Как и у других камснок-пстрофилов, самка, прежде чем приступить к постройке гнезда, выкладывает пол гнездового убежища плоскими камешками до 25-35 мм в поперечнике и весом до 5-7 г. Однако в некоторых случаях камешки в гнездовом убежище отсутствуют. Само гнездо представляет собой неглубокую чашу из сухой травы с более мягкой выстилкой из шерсти, волоса и перьев. Диаметр гнезда — около 110 мм, высота — примерно 50 мм, глубина лотка — 25 мм. Самец нс принимает участия в гнездостроснии. На постройку гнезда самка затрачивает до двух недель.
В кладке бывает от 3 до 6 яиц, чаше 4-5. Соотношение кладок разной величины в северо-западной Африке (подвид halophila): 3 х 4, 4 х 14,5 х 7, 6 х 1 (ср. 4,2, п=26); в Передней Азии (подвид lugens): 3x1,4 х 6, 5 х 3 (ср. 4,2, п=10). Яйца светлого зеленовато-голубого цвета, довольно густо испещренные рыжеватыми пятнышками и точками, наиболее многочисленными вокруг тупого конца яйца. Размеры яиц подвида halophila: 19,0-20,8 х 14,5 х 16,4 (ср. 20,2 х 15,6, п=21). У подвида lugens: 19,6-22,2 х 15,2 х 16,8 (ср. 21,0 х 16,0, п=26). Таким образом, размеры яиц, как и общие размеры особей, увеличиваются в направлении с запада на восток. Это подтверждается и сопоставлением веса яиц у подвидов halophila и lugens: 2,63 г и 2,87 г, соответственно. Насиживает кладку только самка в течение 13-14 дней (Heim de Balsak, Mayaud, 1962, Hue, Etchecopar, 1970, Zachai, 1984, 1989, Cramp, 1988).
Одно из двух гнездах Ое. lugubris vaurie помещалось в отверстии под скалой на глубине 45 см от входа, второе — в 3 м. от земли в нише внутренней стороны стены глинобитной постройки. Оба гнезда были свиты из лоскутьев коры Juniperus procera и выстланы мхом.Вгнсздах было 2 и 3 светлоголубых яйца, в большей или меньшей степени ис-
171
Панов Е.Н. Каменки Палеарктики
остренных красноватыми и сероватыми точками. Яйца в этих гнездах заметно различались размерами: 18,1 — 18,4 х 14,6—15,1 (2) и 19,1—20,7 х 15,5—16,3 (3) (ср. 19,2 х 15,5, n=5) (Krcuger, 1958).
Птенцы покидают гнездо в возрасте 14—15 дней, когда они еще нс способны летать. При средней кладке около 4 яиц до вылета доживают обычно только 2-3 птенца. Слетки делают первые попытки самостоятельно добывать корм на IV день после вылета из гнезда, однако еще долго находятся на попечении родителей. Описаны случаи кормления самками молодых спустя 49 и 55 дней после оставления ими гнезд. Распадение семьи может быть вызвано растущей агрессивностью родителей в отношении молодняка, а также самца по отношению к самке (Cramp, 1988).
7.5. Линька, послегнездовые кочевки, зимовка
Молодые птицы приступают к линьке уже в начале июля. В августе и сентябре практически весь молодняк одет уже в свой первый зимний наряд. Эта частичная линька затрагивает только мелкое оперение и кроющие перья крыла. Изредка сменяются также центральные перья хвоста.
В июле начинаются и кочевки молодежи. В этом и следующем месяцах молодые особи данного года рождения могут быть встречены за 10-30 км от ближайших мест гнездования вида. Взрослые траурные каменки начинают линять даже раньше молодых — иногда уже в первых числах июня. А в конце этого месяца некоторые особи появляются в местах своих традиционных зимовок — например, в долине Нила, куда они откочевывают из бесплодных сухих предгорий. Однако основная масса мигрантов занимают зимние территории в Египте на протяжении августа (Hartley, 1949). В Израиле супружеские пары распадаются в сентябре. Зимующие каменки обосновываются на своих индивидуальных участках в октябре-ноябре, оставаясь здесь до февраля-марта (Shirinai, 1996). Популяции перелетного подвида persica, проводящие свободные от гнездовых дел месяцы в основном на Аравийском полуострове, перемешаются на зимние квартиры в октябре. В южной части ареала этой формы, в юго-западном Иране, в период между серединой ноября и серединой февраля не было встречено ни одной траурной каменки (Cornwallis, 1975, цит. по: по Cramp, 1988).
Зимние местообитания обычно отличаются от гнездовых биотопов менее суровыми условиями. Птицы стремятся обосноваться в угодьях с относительно богатой растительностью. Обычно это подножия гор, где
172
Глава 7. Траурная каменка Oenanthe lugens
в прохладные зимние месяцы температура несколько выше, чем на возвышенностях, и пронизывающие ветры нс столь сильны. Однако в некоторых районах Израиля каменки перемещаются на зиму в горно-степные ландшафты, лежащие выше над уровнем моря, чем их гнездовые мсстобитания (Shirinai, 1996). Нс всегда птицам удается избавится и от изнуряющего зноя. Например, в местах зимовок траурных каменок в Египте дневная температура в тени даже в ноябре доходит до 26*, так что в середине дня каменки зачастую предпочитают проводить время в местах, защищенных от прямых солнечных лучей (Hartley, 1949).
На зимовках птицы поют и охраняют индивидуальные территории от прочих особей своего вида и других видов каменок. Пенис можно слышать на протяжении всего периода между июлем и февралем, однако разгар пения и территориальных конфликтов приходится на сентябрь (Hartley, 1949). Случаи изгнания птиц других видов с территории нередки и в гнездовой сезон. В Израиле в это время нападениям подвергаются в основном пролетные горихвостки Phoenicurus spp. и гнездящиеся в тех же местообитаниях чернохвостки Cercomela melanura.
7.6. Особенности поведения
Траурную каменку обычно сближают с Ое. finschii и Ое. picata. руководствуясь их очевидным сходством по многим признакам внешнего облика (размеры, пропорции, окраска и т.д.). Неоднократно делались даже попытки объединять их друг с другом в разных комбинациях под общими видовыми названиями (Stresemann, 1925; Гладков, 1954; и др ). Тем более неожиданным для меня было обнаружить при наблюдениях за Ое. I. lugens в Израиле, что по ряду особенностей поведения эта каменка резко отличается как от Ое. finschii. так и от Ое. picata, которые в этологическом плане весьма близки друг другу (Панов, 1989).
Первое, что обращает на себя внимание при сравнении траурной каменки с двумя другими названными видами, это весьма малая экспрессивность се сигнального поведения. Наблюдения проводились в разгар сезона размножения (начало апреля), но самцы даже в этот период пели сравнительно мало. Те немногие самцы, которые все же пели вполне активно, очень редко прибегали при этом к воздушным демонстрациям, столь характерным для Oe.finschi и Ое. picata на ранних стадиях гнездового цикла. Например, 30 марта за 4 часа наблюдений (7.30-11.30) один из самых активных самцов Ое. lugens проделал всего лишь 8 рекламных полетов, которые к тому же выглядели мало эмоциональными: птица нс взлетает высоко, лишь иногда разворачивает в полете хвост веером и быстро садится на песенный пост в обыч-
173
Панов Е.Н. Каменки Палеарктики
пой, повседневной позе (для сравнения стоит упомянуть, что холостые самцы Ое. finschii проделывают свои эффектные рекламные полеты с частотой 1,7-2,4 за 5-минутный отрезок времени — см. Панов, 1978). Ни у этого самца, ни у нескольких других ни разу не наблюдались пробежки с широко развернутым хвостом на песенном посту и при посадке по окончании воздушных демонстраций, тогда как у Ое. finschii и Ое. picata такие пробежки — это один из наиболее стандартных компонентов их песенного поведения. Те немногочисленные рекламные полеты, которые мне удалось увидеть, не обладали какими-либо характерными и постоянными видоспецифическими признаками. Создалось впечатление, что у траурной каменки, в отличие от Ос. finschii, Ое. picata и целого ряда других каменок, полет нс ритуализован. Обычно он выглядит скорее как перелет с одного песенного поста на другой, но нс по прямой траектории, а по дугообразной: после взлета самец набирает высоту и остается в воздухе несколько дольше, чем того требует простое перемещение из одной точки в другую.
Редкое использование самцами саморекламирования в полетах нал территорией компенсируется, по-видимому, чрезвычайной громкостью их пения. Записать на магнитофон песню Ое. lugens можно с расстояния по крайней мере вдвое большего, чем того требует запись голосов других видов каменок. К тому же самец во время пения словно привязан к своему очередному песенному посту, оставаясь здесь до 10-15 мин. и долее. Это совершенно нс характерно для поющих самцов Ое. finschii и Ое. picata. У этих двух видов по мерс того, как короткие, разделенные паузами песни начинают сливаться в сплошную, длинную импровизацию, усиливается стремление к двигательной активности. Птица зачастую проделывает при этом короткую пробежку с широко развернутым хвостом, либо сразу взлетает и набирает высоту, после чего садится на тот же либо на другой песенный пост с аналогичной эффектной пробежкой. Ничего похожего я ни разу не наблюла.’! у самцов Ое. lugens. После трансформации коротких песен в удлиненную певец пребывает в неподвижности, оставаясь столь же бесстрастным и невозмутимым, как если бы он просто присел сюда отдохнуть.
Впрочем, сама песня имеет много общего с тем, что мы видели у Ое. finschii. Как и у этого вида, в большинстве песен траурной каменки присутствуют короткие "тщек” или “чрек” (сигнал № 1) (сравни рис. 76 и 18а-с). От Ое. picata траурную каменку отличает более бедный звуковой диапазон и практически полное отсутствие в песнях имитаций других видов птиц. Однако по первому впечатлению очень звучная и приятная па слух песня Ое. lugens более всего напоминает исполнительскую манеру Ое. chrysopygia.
174
Глава 7. Траурная каменка Oenanthe lugens
Рис. 76. Элементы вокализации траурной каменки Ое. /. lugens. а — короткая рекламная песня, Ь — то же, с включением в начале и в конце сигнала N 6 (птенцовая позывка), с — фрагмент удлиненной песни (в полете), d — сигнал N 3 (Израиль). Оригинал.
У холостых самцов Ое. finschii и Ое. picata и утех, которые вынуждены оставаться в одиночестве, пока их самки поглощены насиживанием кладки, постоянно наблюдаются всевозможные демонстрации “вхолостую” (vacuum activity) — вплоть до попыток ухаживать за особями неродственных видов и “копуляций” с неодушевленными предметами. У самцов Ое. lugens какие-либо демонстрации вхолостую практически отсутствуют. За десятки часов наблюдений я лишь однажды наблюдал короткий полет петлями, закончившийся ложной копуляцией с камнем. Совершенно очевидно, что физиологический порог проявления
175
Панов Е.Н. Каменки Палеарктики
экспрессивных форм брачного поведения у этого вида намного выше чем у Ое. finschii и Ое. picata.
То же относится и к сигналам, обслуживающим территориальное поведение. Самца Ое. finschii даже осенью можно заинтересовать грубой имитацией каменки, сделанной наспех из клочка ваты и двух полосок черной бумаги. Хозяин участка сразу же подлетает к "чужаку”, готовый выставить его прочь. Тот же опыт я пытался проделать на территориях двух самцов Ое. lugens, но хозяева участков полностью игнорировали модель (см. также Hartley, 1949: 399). Интересно, что птицы не обращали внимание на подделку даже в том случае, если рядом е ней я помещал магнитофон, воспроизводящий песню самца Ое. lugens. На саму магнитофонную запись (особенно если я проигрывал длинный вариант песни) самцы реагировали с большой готовностью, pas за разом подлетая ко мне и вступая в песенную дуэль с невидимым противником. Однако, демонстрации при этом носили мимолетный характер (рис. 77) и ни разу не сопровождались разворачиванием рулевых, столь характерным в подобных ситуациях для Ое. finschii и Ое. picata. Отсутствие этого элемента демонстраций при конфронтациях самцов, как и общая относительная пассивность соперников, подтверждается также наблюдениями Hartley (1949). Можно предположить, что флегматичность траурной каменки явилась результатом существования в условиях очень низкой плотности популяции, что, в свою очередь, является следствием чрезвычайной бесплодности местообитаний этого вида.
Все изложенные и некоторые другие факты (например, присутствие в репертуаре сигнального поведения “скользящего бега”, упомянутого в предыдущих очерках — см. Bates, 1936) позволяет поместить Ое. lugens в ту же группу видов, куда входят Ое. finschii и Ое. picata. При этом, однако, может оказаться, что родство траурной каменки с этими двумя видами (в особенности с Ое. picata) не является столь тесным, как это казалось ранее. Во всяком случае, вероятно, сейчас нет оснований рассматривать Ое. lugens в качестве возможного прямого предка Ое. picata capistrata, как это сделано в моих более ранних работах (Panov, 1992, и др.).
Удивительная бесстрастность траурной каменки проявлястсяи в се рс-
Рис. 77. Поза самца Ое. lugens в ответ на магнитофонное проигрывание песни того же вида. Стрелкой показаны хпинные прыжки вперед. Ори-
176
Глава 7. Траурная каменка Oenanthe lugens
акции на человека, находящегося в непосредственной близости от гнезда. По наблюдениям за каменками подвида halophila в Алжире, птица порой подпускает в этих ситуациях наблюдателя на 1-2 м от и даже садится в его присутствии на гнездо с кладкой (Jourdain, Wallis, Ratcliff, 1915). В Израиле Ое. I. lugens нс относится к числу доверчивых птиц, но ее поведение у гнезда с птенцами оказалось для меня полным сюрпризом. Я рассчитывал записать на магнитофон сигналы тревоги, но прилетевшая с кормом птица оказалась абсолютно молчаливой. Держа в клюве бабочку, она подолгу сидела па одном месте как вкопанная, и лишь время от времени без каких-либо явных признаков испуга перелетала на другую присяду. В конце концов каменка проглотила бабочку вместе с крыльями и без единого звука улетела прочь. Это наблюдение прекрасно вписывается в сказанное ранее об обшей флегматичности поведения траурных каменок.
Безрадостный характер местообитаний вида в каменистой пустыне Негев делает для него поиски пропитания непростой задачей. Птицы из пары, о которой только что шла речь, неизменно летали за кормом для птенцов за 300 м — из совершенно бесплодного бедленда в долину, где по сухому руслу прежних дождевых потоков тянулась полоска низкорослых полукустарничков. Они находились в цветении и тем привлекали многочисленных летающих насекомых — мух, пчел и бражников. Здесь каменки использовали способ охоты, который я не наблюдал ранее ни у одного другого вида каменок. Птица высматривала добычу, сидя на верхушке валуна, а затем летела низко над землей, мерно работая крыльями, напрямик к кусту, и схватывала насекомое с периферической части кроны. На ровных площадках с разбросанными там и тут небольшими камнями каменки охотятся нссклько иначе. Птица стремительно летит по прямой на высоте около 20 см от земли, преодолевая каждый раз расстояние порядка 5-10 м, и резко падает вниз, увидев на траектории своего движения или несколько в стороне от нес желанную добычу. Иногда эти каменки добывают насекомых из почвы, раскапывая се резкими боковыми движениями клюва. Естественные источники воды практически невозможно найти в Негеве, так что траурные каменки обходятся без питья. Но они охотно пьют из искусственных “поилок” там, где последние сооружены для туристов.
177
Панов Е.Н. Каменки Палеарктики
Глава 8.
Комплекс черная каменка Oenanthe picata Blyth, 1847
Синонимы: Saxicola picata Blyth, 1847; Saxicola capistrata Gould, 1865: Saxicola opistholeuca Strickland, 1849; Saxicola opistholeuca evreinowi Zarudny, I910.
8.1. Чем черная каменка интересна для биолога-эволюциониста
Черная каменка издавна вызывала живой интерес нс только у орнитологов-систематиков, но и у теоретиков-эволюционистов, и ей посвяшсн целый ряд научных исследований. Дело в том, что черную каменку до недавнего времени рассматривали в качестве вида с ярко выраженной дискретной изменчивостью самцов. Было известно три варианта их окраски — I. преимущественно черный, 2. черный с белым брюшком и 3. черный с белым брюшком и белой шапочкой. Предполагали, что эта изменчивость определяется всего лишь одним-двумя генами переменного действия (Mayr, Strcsemann, 1950). Находясь в пеком определенном состоянии в хромосомах одной особи, такой ген обуславливает черную окраску того или иного участка се оперения (например, шапочки); оказываясь у другой особи в ином состоянии, ген делает тот же участок оперения белым. При этом подобные гены альтернативного действия не затрагивают других свойств двух данных организмов, так что те могут быть, в принципе, почти одинаковыми генетически, резко различаясь при этом лишь деталями окраски.
К сожалению, орнитологи, предложившие эту гипотезу генетического полиморфизма для объяснения изменчивости окраски черных каменок, нс знали о существовании одной важной работы, опубликованной русским исследователем Зарули ым еще в 1923 г. В этой статье се автор указывал, что, во-первых, каждый из трех вариантов самцов приурочен к определенной географической области, где могут вообще не встречаться другие варианты; во-вторых, что трем разным вариантам окраски самцов соответствуют также три варианта окраски самок; и, наконец, что в некоторых местах вполне обычны самцы, промежуточные по окраске между теми или иными из трех вариантов, обсуждавшихся Майром и Штрсзсмапном (рис. 78-79).
Суммируя результаты своих исследований, Зарудным счел возможным рассматривать эти три варианта в качестве самостоятельных ви-
178
Глава 8. Черная каменка Oenanthe picata
ов Ое. picata (самцы черные с белым брюшком), Ое. capis-Wta (белые брюшко и тапочка) и Ое. opistholeuca (самцы чер-ныс) — как о,,и бы" ли описаны первоначально другими авторами еше в XIX веке. Самцов, промежуточных по окраске между capistrata и opistholeuca, Зарудный посчитал принадлежащими к подвиду последнего вида и присвоил им название Ое. opistholeuca evreinowi.
Как оказывается сегодня, Зарудныи кое в чем заблуждался, но в целом его точка зрения оказалась много ближе к истине, чем позиция Майра и Штреземан-на. Впрочем, и в подходе последних двух исследователей, как мы увидим ниже, есть определенная доля истины. Результаты многолетних полевых исследова
ние. 78. Ареалы форм Ое. р. picata (а), Ое. р. opistholeuca (с) и полиморфных гибридогенных популяций capistrata х opistholeuca (b). а — высоты ниже 1000 м над у.м., Ь — высоты от 1000 до 3000 м, с — находки picata, d — capistrata, с — opistholeuca, f — примерные границы региона, где самки мономорфны по окраске. Из: Панов 1989, с изменениями.
ний черных каменок, проведенных автором этой книги и его коллегами, позволяют нарисовать следующую гипотетическую схему становления и эволюции черных каменок.
Я предполагаю, что первыми пришельцами с запада (из Передней Азии) в современную область распространения черных каменок были белобрюхие, белоголовые каменки типа capistrata (чрезвычайно сход-
179
Панов Е.Н. Каменки Палеарктики
Рис. 79. Некоторые варианты промежуточной окраски самцов. В левом столбце — разные варианты фенотипа evreinowi. Из: Панов 1989.
ныс по окраске с Ое. lugens). Эти первые поселенцы освоили пустынные пизкогорья северной части среднеазиатского региона. Вторая волна иммигрантов с запада была представлена каменками, расселявшимися на восток по горным ландшафтам южной части того же региона. Обосновавшись здесь, эта первоначально единая популяция со временем распалась на две, которые в дальнейшем эволюционировали независимо друг от друга. Одна из них оказалась приуроченной к Иранскому нагорью и прилежащим к нему горным местностям (picata). Становление второй происходило далее к востоку, в высокогорьях Гиндукуша и примыкающих к нему хребтов (opistholeuca). Со временем обе горные формы начали расселяться как навстречу друг другу, так и на север, в опустыненные низкогорья, занятые формой capistrata. В зонах вторичного контакта началась гибридизация — как это происходит в аналогичных ситуациях контакта близких видов (Ое. xanthoprymna и Ое. chrysopygia, Ое. hispanica и Ое. pleschanka).
В разных зонах контакта и гибридизации события шли неодинаково и имели разные последствия. Некоторые гибридные популяции и по
180
Глава 8. Черная каменка Oenanthe picata
Рис. 80. Соотношение в числе разных фенотипов (%%) в автохтонном ареале формы opistholeuca(1) и в ареале полиморфных популяций capistrata х opistholeuca (3 — 8). 2 — гибридная популяция opistholeuca .г (capistrata л opistholeuca). а — ареал формы opistholeuca. b — стабилизированные гибридогепные популяции capistrata х opistholeuca, с — гибридная популяция opistholeuca х (capistrata х opistholeuca), d — северо-восточный край ареала формы picata. Фенотипы opislholeuca(e), evreinowi(f), opistholeuca ^evreinowi(g), capistrata (h), picata (i), picatax opistholeuca (k). Номера локалитстов: 1 — Таджикский Бадахшан (25), 2 — междуречье Пянджа и Кызылсу (62), 3 — хребет Каратау (14), 4 — хребет Бабатаг (27), 5 — долина р. Шера бал (128), 6 — горы Нуратау (16), 7 — урочише Дарбаза в западных предгорьях Каржантау (192), 8 — остан новые горы пустыни Кызылкум: Актау, Тамдытау, Букантау (129). В скобках число наблюдавшихся самцов. Из: Panov 1992, с изменениями.
сию пору локализованы в пределах сравнительно небольших территорий и характеризуются присутствием в них, наряду с особями трех исходных типов окраски, самых разнообразных промежуточных ее вариантов (рис. 79, 80) Таковы популяции в зонах гибридизации всех трех форм в северо-восточном Афганистане (см. Paludan, 1959) и в юго-западном Таджикистане. Через эти гибридные популяции и по сей день идут встречные потоки генов между автохтонными ареалами родительских форм. Однако, эти генные потоки не настолько сильны, чтобы значительно повлиять на первоначальную генетическую конституцию родительских форм в тех частях их ареалов, которые значи-
181
Панов Е.Н. Каменки Палеарктики
тсльно удалены от гибридных зон. В результате р/сд/д и opistholeuca пока еще сохраняют, в целом, свою генетическую и биологическую специфику и уникальность.
По иному выглядит ситуация в автохтонном ареале формы capistrata. Поток генов на северо-запад из первоначальной зоны се гибридизации с opistholeuca был в свое время настолько интенсивным, что распространился на всю территорию ареала capistrata. В итоге весь этот регион оказался заселенным популяциями гибридного происхождения (capistrata х opistholeuca), в которых, судя по всему, изменчивость окраски самцов обусловлена ныне процессами сбалансированного генетического полиморфизма. Однако эти процессы нс столь просты, как это следовало из модели альтернативного наследования окраски (Mayr, Stresemann, 1950). Наряду с двумя исходными типами окраски самцов, присущими родительским формам, всюду в значительном количестве (10-13%) встречается сильно изменчивый внутри себя третий тип — тот, что был описан Зарудным (1915) под именем evreinowi. Особенно примечательно то обстоятельство, что при весьма широкой изменчивости окраски самцов все самки в ареале гибридогенных популяций capistrata х opistholeuca окрашены совершенно однотипно и по-иному, чем в автохтонном ареале формы opistholeuca (рис. 78b). Возможно, самки сохранили окраску, первоначально свойственную особям этого пола у capistrata. Мономорфизм самок в данном случае определенно говорит о том, что перед нами генетически единая гибридогенная популяция, а нс зона сосуществования двух разных форм.
Таким образом, черная каменка представляет собой нс монотипический вид с повссмссто распространенным генетическим полиморфизмом, а политипический, состоявший первоначально из трех подвидов, среди которых capistrata генеалогически, по-видимому, более удалена от двух других форм (picata и opistholeuca), нежели они друг от друга. Сейчас capistrata уже не существует, как самостоятельная генетическая система: в результате поглотительной гибридизации се первоначальный ареал ныне оказался занятым полиморфной гибридоген-ной формой capistrata х opistholeuca. На стыках ареала этой гибридогенной общности с ареалами двух других подвидов продолжается интенсивная гибридизация. В юго-западном Таджикистане ширина гибридной зоны (capistrata х opistholeuca) х opistholeuca составляет около 100 км (рис. 80, локалитст 2). Из юго-западной части ареала гибридогенных популяций capistrata х opistholeuca идет поток генов в автохтонный ареал формы picata, за счет чего многие особи в ареале последней формы имеют примесь белого в черном оперении головы (рис. 81).
Географическая изменчивость затрагивает не только окраску, но и размерные признаки. Формаpicataотличается наименьшими размера-
182
Глава 8. Черная каменка Oenanthe picata
Рис. 81. Соотношение (%%) самцов “picata” с разной степенью присутствия белого (признак capistrata) в оперении головы (баллы 0 — 7) в северо-восточной части ареала подвида Ое. р. picata (Копетдаг). а — наиболее северные точки находок особей picata в Афганистане, b — наиболее западные находки особей capistrata, с — находки capistrata в северном Афганистане (по Paludan 1959). d — находки capistrata в ареале picata. Номера локалитетов: 1 — Бадхыз (16), 2 — центральный Копетдаг (38), 3 — западный Копетдаг (33), 4 — западные склоны Красноводского плато (57). В скобках число наблюдавшихся самцов. Из: Panov 1992.
ми: средняя длина крыла самцов старше года 90,4 мм (первогодков 88,0 мм), самок 85,7 мм, средний вес, соответственно, 20,1-20,7 г, 19,2-20,4 г и 20,7 г. Длина крыла самцов старше года у формы opistholeuca в автохтонном ареале 92,3 мм (первогодков 90,3 мм), самок 87,1 мм, вес самцов 22,1 г. Длина крыла самцов старше года в автохтонном ареале capistrata 92,9 мм (первогодков 90,5 мм), самок 87,1 мм, вес самцов 23,0 г.
Как мы видим, существует и возрастная изменчивость размеров самцов. Самцы первогодки имеют более длинное первое маховое и более широкую черную перевязь на конце хвоста. У форм picata и opistholeuca изменчивость окраски самок также, как я полагаю, связана с возрастом: самки-первогодки светлые, сходные с самкам capistrata, с возрастом они становятся все более похожими на самцов своего под-
183
Панов Е.Н. Каменки Палеарктики
вида. У самок capistrata на второй год жизни горло из беловатого становится более темным, сероватым (Панов, Грабовский, Любущснко, 1993, Любущенко, 1993; см. рис. 3).
В разных точках автохтонного ареалаpicata самцы с типичным обликом этого подвида составляют от 33 до 87%, самцы с отдельными белыми перышками в оперении головы — 10-33%, самцы липл capistrata
Рис. 82. Черная каменка Ое. р. picata. а — самец, b — самка старшей возрастной группы.
— 0,6-8,3% (рис. 81). Самцы типа opistholeuca встречены всего дважды на крайнем востоке ареала формы picata (Бал-хыз), где возможны залеты птиц с фенотипом opistholeuca из гибридной зоны северного Афганистана. В автохтонном ареале opistholeuca типичные самцы этого подвида составляют от 75 до 100% популяции, самцы picata — 6-10 % (при одном учете их было 5 из 14, т.с. 36%), 5-10% приходится на гибридов opistholeuca а picata и picata х capistrata (единственная находка). Самцы с обликом evreinowi здесь отсутствуют. В зоне контакта всех трех форм в южном Узбекистане на долю самцов capistrata приходится 74-76%, самцы opistholeuca составляют 6-8%. самцыpicata — 3%, самцы типа evreinowi 7-9%, самцы с другими вариантами промежуточной окраски — около 3%. Наконец, в стабилизировавшейся гибридоген ной популяции capistrata х opistholeuca в северном Узбекистане соотношение самцов capistrata : opistho-1енса+evreinowi составляло за 7 последовательных лет 53-69 : 47-31. Здесь среди самцов capistrata с чисто белой шапоч-
184
Fuiea tS. Черная каменка Oenanthe picata
Рис. 83. Изменчивость окраски в комплексе черные каменки, а — Ое. р. capistrata (самец); b— Ое. р opistholeuca,самец совершает демонстративный полет; с — один из вариантов фенотипов, промежуточных между picata и capistrata (самец); d — самка, промежуточная по окраске между capistrata и opistholeuca (все фото — из юго-восточного Узбекистана).
кой встречаются также индивиды с палевым либо “пестрым” теменем, напоминающие по этому признаку самцов Ое. lugubris (Панов, 1986а; Панов, Грабовский, Любущснко 1993; Любушснко, 1995).
Из-за сильной изменчивости самок их подвидовую принадлежность нс всегда легко установить. Так или иначе, в ареале каждой из трех основных групп популяций бесспорно численно преобладают самки соответствующего типа окраски — белобрюхие в ареале picata, грязно-бурые (до черных) — в ареале opistholeuca, песочно-серые — в ареале гибридогенных популяций capistrata х opistholeuca (рис. 78). Разнообра-
185
Панов Е.Н. Каменки Палеарктики
зис окраски самок характерно в основном для гибридных зон на стыках ареалов исходных форм (Панов, 1989).
8.2. Полевые признаки
По общему складу черная каменка, в общем, сходна с черношейной Ое. finschii, отличаясь от нес менее массивной головой и не столь сильными ногами (рис. 82, 83). В спокойном состоянии птица временами слегка покачивает хвостом, очень медленно опуская его немного ниже уровня туловища и чуть быстрее возвращая в исходное положение. При содержании черной каменки в неволе хорошо видно, насколько размерены и изящны все ее движения. В отличие от Ое. finschii и Ое. isabellina она ловко, замедленными, словно “змеиными” движениями перепрыгивает по жердочкам и лазает по стенкам клетки. Будучи озадачена чем-то или испугана, птица совершает глубокий "поклон”, одновременно на мгновение отводя крылья вверх и немного в стороны и опуская хвост, который сразу же слегка разворачивается н поднимается заметно выше горизонтали. Во время питья каменка, сделав очередной глоток, нс поднимает клюв высоко, а держит его почти горизонтально, проделывая челюстями “жующие” движения.
8.3. Местообитания и численность
Основной элемент ландшафта, определяющий распределение черных каменок в пределах их ареала — это присутствие более или менее вертикальных поверхностей, изрезанных всевозможными нишами, щелями и прочими пустотами. Субстрат, из которого сложены такие обрывистые склоны, может быть совершенно различным (каменный монолит, конгломерат, плотно слежавшаяся глина и т. д.), и приурочены эти ландшафты к разным высотам относительно уровня моря — от 400 до 3000 м. Важно лишь, чтобы наряду с обрывистыми поверхностями здесь имелись бы достаточно обширные участки с низким травостоем, где каменки добывают свое пропитание. Присутствует ли в данном месте достаточно богатая древесно-кустарниковая растительность или отсутствует — нс имеет существенного значения, только бы это не были сплошные непроходимые заросли.
В ареале формы capistrata (ныне — гибридогенной популяции capistrata х opistholeuca) каменки гнездятся среди останцовых, сильно разрушенных временем возвышенностей с максимальными высотами около 700 м над уровнем моря посреди закрепленной песчаной пустыни (Кы-зыл-Кум, долина р. Вахш), в каменистых обнажениях невысоких хол-
186
Глава 8. Черная каменка Oenanthe picata
Рис. 84. Лессовые каньоны долины р. Шерабад (юго-восточный Узбекистан). Местообитания гибрилогепной полиморфной популяции Ое. р. capistrata х Ос р. opistholeuca с небольшой примесью фенотипа picata.
Рис. 85. Лессовый псевлокарст по бортам долины р. Шерабад Типичные места гнездования черных каменок фенотипов capistrata, opistholeuca и evreinowi.
187
Панов Е.Н. Каменки Палеарктики
мов предгорий Памиро-Алая, по лессовым каньонам пересыхающих рек на юге Узбекистана и Таджикистана (Потапов, 1959; Панов, Корзухин 1974; Митропольский, 1980; Любущснко, 1995; рис. 84, 85). В таких местообитаниях соседями черных каменок оказываются Ое. finschii, местами также — Ое. pleschanka и Ое isabellina.
Птицы с фенотипом capistrata практически нс встречаются в горах Памиро-Алая выше 1500 м, уступая здесь место форме opistholeuca. господствующей в горах на высотах 2000 м и выше (лишь однажды я нашел здесь гнездящегося самца с обликом гибрида capistrata х picata). Здесь черные каменки гнездятся в одних местообитаниях с Ое. chrysopygia kingi и Ое. pleschanka.
В Иранском нагорье и в прилежащих горных системах Загроса, Эльбурса и Копстдага форма picata распространена от предгорий (соседствуя, например, в Копетдаге с Ое. finschii, Ое. pleschanka и Ое. isabellina) до высот 2100—2400 м (иногда 2700 м) в южном Иране, где она гнездится бок о бок с Ое. с. chrysopygia, и Ое. alboniger, но в целом ниже высокогорных популяций Ое. lugens persica, Ое. oenanthe и Ое. deserti atrogularis (Hafter, 1977; Desfaycs, Praz, 1978; Панов, 1989; рис. 62).
Как следует из сказанного выше, в отличие от capistrata, не изменяющей излюбленным ею низкогорьям, picata и opistholeuca выселялись прежде и охотно выселяются теперь из своих исконных высокогорных ландшафтов в низкогорные местообитания, занятые гибридо-генной популяцией capistrata х opistholeuca. Это приводит к контакту трех исходных форм и к появлению гибридов самой разнообразной окраски. В местах совместного обитания птиц с обликом capistrata, picata, opistholeuca, а также evreinowi и птиц с прочими промежуточными фенотипами, все они занимают одни и те же местообитания, охраняя свои территории друг от друга (рис. 86). Одну и ту же территорию в разные годы могут занимать самцы разных фенотипов. Вместе с тем, самцы со сходными фенотипами (например, opistholeuca либоopistholeuca и evreinowi) подчас селятся внутри смешанной популяции этакими “землячествами”, которые могут существовать в данной точке по нескольку лет, хотя число самцов, удерживающих свои территории внутри такого кластера, и их индивидуальный состав из года в год изменяются (Панов, 1989; Любущенко, 1995).
Обычно черные каменки образуют довольно плотные поселения, причем плотность их тем выше, а размеры территорий, соответственно, тем меньше, чем сильнее расчленен рельеф. В аридных низкогорь-ях юго-западного Туркмсниста с очень скудной травянистой растительностью, выгорающей на солнце уже к началу мая, и при полном отсутствии кустарника и деревьев плотность населения формы picata на скалистых склонах с каменистыми осыпями составляет до 15 парна I кв. км. В Копетдаге и в Бадхызе поселения могут быть гораздо менее
188
Глава 8. Черная каменка Oenanthe picata
ifra Ob EZJc ESid #e ESf L~]g FOh ffli . .1У..29° m
Рис. 86. Размещение территорий самцов capistrata (а) и opistholeuca (b) вдоль лессового каньона в юго-восточном Узбекистане, с — границы тсриторий черных каменок, d — нейтральные зоны, е — пары Ое. finschii, f — русло реки, g — днище каньона, h — овраги глубиной менее 2 м, i — овраги глубиной более 2 м. Из: Панов и Корзухин 1974.
плотными: площадь отдельных территорий составляет здесь от 2 до 3 га (Панов, Грабовский, нсопубл., Панов, 1978). В холмистой полупустыне на границе Узбекистана и Казахстана вдоль склонов грядовых возвышенностей формируются линейные поселения (популяция capistrata х opistholeuca), включающие в себя от 15 до 105 пар при средней величине территорий порядка 0.5-1.0 га (максимально до 3 га) (Любущснко, 1995). Примерно такой же плотностью характеризуются линейные поселения в гибридогенной популяции capistrata х opistholeuca (с примесью отдельных особей picata) по лессовым каньонам южного Узбекистана близ его границы с Афганистаном. Здесь на площади около 2 км2, в 1973 г. размещались территории 38 пар (Панов, Корзухин, 1974; рис. 86). В гибридной зоне (capistrata х opistholeuca) х opistholeuca в междуречье Пянджа и Кызылсу (юго-западный Таджикистан близ границы с Афганистаном) каменки селятся в немногих пригодных для их гнездования местах компактными демами, включающими себя от 6 до 24 пар (в среднем 14 по 7 демам) (Любущснко, 1995). В примыкающих к этому району с востока высокогорьях восточного Памиро-Алая и Бадахшана птицы формы opistholeuca также образуют довольно компактные поселения, но площадь территорий увеличивается по сравнению с приведенными цифрами по мерс увеличения высоты местности, составляя в высотном поясе 2000-2500 м около 2 га.
8.3. Годовой цикл и сроки гнездования
Сроки гнездования черных каменок заметно различаются в разных частях их ареала. Раньше всех приступают к гнездованию южные попу
189
Панов Е.Н. Каменки Палеарктики
ляции в ареале гибридогснсной популяции capistrata х opistholeuca. На юге Узбекистана у границы с Афганистаном (к северу от г. Термез, 37° с.ш., 67е в.д.) даже в поздние весны некоторые птицы успевают заключит брачный союз еще до конца февраля. В 1973 г. самка из пары, сформировавшейся до 27 февраля, начала строить гнездо 6 марта. Однако массовый прилет самок проходил здесь на протяжении всего марта (1-27.111; рис. 11), а гнездостросние я наблюдал в этом году и в другие годы в период между 15 и 30 марта. Слетки появляютеяв здесь с 22-24 апреля, и в эти же дни (24-25.IV) некоторые пары приступают ко второму циклу гнездования (Панов, Корзухин, 1974; Панов, 1989).
Примерно в 400 км. к северу (пос. Дарбаза севернее Ташкента, около 4Г с.ш., 69” в.д.) к 9 марта 1987 г. только 12 из 29 территорий уже были заняты самцами, причем все они оставались пока холостыми. Первые 5 самок появились здесь 11 марта, наиболее рання дата начала гнсздостросния — 15 марта. Однако большинство пар присупают к гнездованию позже. По данным 4-лстних наблюдений откладка яиц у пар с самцами capistrata старше года начинается в среднем 13 апреля (у пар с самцами-первогодками 17 апреля), с самцами opistholeuca 14 (19) апреля, с самцами evreinowi 14 (22) апреля. У некоторых рано размножающихся пар в это время (14-17.IV) самки уже насиживают кладки (Любущенко, 1995). У пары первогодков, окольцованных в этом районе птенцами, при первом цикле гнездования яйцекладка началась 27 апреля, и 5 птенцов первого выводка вылетели из гнезда 28 мая; 4 июня самка отложила яйцо в новое гнездо, в котором 5 птенцов вылупились 20 июня (Матюхин, личн. сообщ.).
Примерно в тс же сроки гнездятся каменки гибридогенных популяций в низкогорьях пустыни Кызылкум (41-43° с.ш., 63-65* в.д.). Здесь гнезда с сильно насиженными кладками и с неоперенными птенцами находили 8-10 мая (и даже 17 мая). Первых слетков видели 11 мая, обычными они становятся с 29-31 мая (Митропольский, 1980; Фотеллср, личн. сообщ.).
В ареале picata популяции, обитающие на той же широте, что и упоминавшиеся выше гибридогенные популяции capistrata х opistholeuca южного Узбекистана, или даже южнее, приступают к гнездованию, тем не менее, почти на месяц позже. Например, в Бадхызс (36° с.ш., 62° в.д.) в раннюю весну 1976 г. холостые самцы заняли территории к 26 марта, но первые самки появились только 27-28 марта, и прилет их продолжался до 7 апреля. В этом году первые даты гнездостросния — 12-13 апреля, в позднюю весну 1977 г.— 5 апреля. Поданным Сухинина (1958) и Бельской (1961) наиболее ранние кладки находили здесь 4, 12 и 13 апреля. В 1976-1977 гг. я находил гнезда с птенцами в период с 24 апреля по 12 мая, слетки отмечались с 5-11 мая (Панов, 1989).
190
Глава 8. Черная каменка Oenanthe picata
Несколько далее к северу, в предгорьях Копетдага близ Ашхабада (38е с.ш., 58° в.д.) в 1969 г. наблюдения начались с 2 марта, но первые два самца picata появились только 6 и 9 марта. С 10 марта их стало заметно больше (в этот день гонады одного из самцов нс превышали в размерах 2 мм). Первый рекламный полет самца я наблюдал 14 марта. В 1968 г. в том же месте самцы были довольно обычны уже вдень начала наблюдений, 8 марта, но лишь один из них активно пел. Первое появление самок в этом районе отмечено в 1968 г. 22 марта, в 1969 г. 15 марта, в 1972 г. прилет самок проходил с 25 марта по 2 апреля. Самая рання дата гнездостроения — 6 апреля 1972 г. На северо-западной границе ареала picata (хребет Большой Бал хан и Красноводский полуостров, около 40е с.ш., 54° в.д.) размножение протекает примерно в тс же сроки, что и в северных предгорьях Копетдага. Здесь в 1985 г. в период с 7 по 16 мая нашли 6 гнезд с кладками (из них две свежие), 10 гнезд с птенцами (в двух шло вылупление, в двух других птенцы были готовы к вылету) и видели 8 выводков (в трех молодые уже хорошо летали). В 1988 г. на Красноводском полуострове 5 и 15 мая обнаружены 2 гнезда с кладками, между 2 и 17 мая — 14 гнезд с птенцами (в одном они только что вылупились). С 28 апреля по 13 мая видели 8 выводков (лишь один, встреченный 2.V, состоял из хорошо летающих молодых).
По данным Степаняна и Степанян (1972), в западном Копетдаге массовый вылет молодежи из гнезд наблюдается 17-20 мая, а 25-27 мая уже встречаются самостоятельные молодые. Вместе с тем, с 15 мая уже встречаются самки с сильно развитыми яичниками, а 21 мая одна из них откладывала яйца. В эти сроки начинается второй цикл гнездования. Гнезда с птенцами находили в Копетдаге в двадцатых числах июня (Ишадов, 1963).
Наиболее поздно протекает размножение в высокогорных популяциях в ареале opistoleuca. Даты прилета каменок в эти районы неизвестны. Наиболее рання дата находки гнезда с яйцами — 18 мая (смешанная пара: самец picata, самка opistholeuca в пос. Калаи-Хумб в долине верхнего течения р. Пяндж). Птенцы должны были вылупиться не ранее чем через неделю. В горах Бадахшана, на больших высотах порядка 2000-2500 м, гнездостроение наблюдали 26-30 мая (одна пара) и 1 июня, гнезда с кладками находили 5 и 11 июня, с птенцами — 7-11 июня, слетков видели 14 июня. Более поздние находки гнезд с яйцами (4 гнезда между 23 июня и 9 июля) и с птенцами (между 15 июня и 19 июля) относятся, вероятно, к повторному гнездованию после потери первого гнезда и ко второму циклу размножения. Слетков отмечали 5 и 27 июля, выводки хорошо летающих молодых — с 13 по 30 июля, самостоятельную молодую каменку видели 17 июля.
191
Панов Е.Н. Каменки Палеарктики
8.5. Социальное поведение
Процесс формирования гнездовых поселений у черных каменок подчиняется той же схеме, что и у Ое. finschii. Поэтому я нс стану повторять сказанное в очерке о последнем виде, а остановлюсь лишь на основных особенностях территориального и брачного поведения черных каменок, а также на некоторых вопросах, нс затронутых ранее.
Территориальное поведение и саморекламирование самцов. Конфронтации между холостыми самцами черных каменок на почве конкуренции из-за территорий редко бывают столь длительными и ожесточенными, как это происходит у Ое. finschii. Обычно конфликтующие самцы нс устраивают погонь в воздухе, а выясняют свои отношения, перебегая неподалеку друг от друга скользящим шагом и периодически залезая в различные пустоты. Птица с пол-минуты остается невидимой, а затем снова “выползает" наружу, устремив клюв кверху и гордо неся над землей свой роскошный широко развернутый хвост (рис. 87а). Эти взаимные демонстрации сопровождаются истинной какофонией звуков — от утробного глухого завывания до тонкого “циканья" при котором, словно вспышки света, становится видимой и снова “гаснет" ярко-желтая полость рта. Иногда самцы неожиданно бросаются друг на друга, и завязывается жестокая драка: птицы сцепляются когтями и, нанося друг другу удары клювами, катаются по земле. Один раз я наблюдал у хозяина территории подобную вспышку гнева — со всеми сопутствующими позами и звуками — в качестве реакции на мертвого самцаpicata, которого я положил на бок (в совершенно неестественной позе) на невысокий кустик посреди охраняемого участка.
Вероятно, подобные острые конфликты происходят в тех случаях, когда самец, вернувшийся на свою прошлогоднюю территорию, застает здесь нового хозяина. Как показали исследования Любущенко (1995), наблюдавшего несколько лет в Дарбазе за каменками, помеченными цветными кольцами, ежегодно на свои прошлогодние территории в среднем возвращается 44% гнездившихся ранее самцов самцов и 33% самок (рис. 88). Суммарно за все годы самцы возвращались на прошлогоднюю территорию 116 раз, и селились при этом нс далее 200 м от нее 23 раза, а в более удаленных частях поселения — всего 12 раз. Для самок эти цифры равны, соответственно, 68, 9 и 12. По наблюдениям Любущснко, прежнему хозяину, как правило, удается отвоевать свои прежние владения у нового хозяина.
Некоторым самцам, замешкавшимся по дороге на места гнездования, остаются в поселении самые никудышные участки, маленькие и лишенные обширных вертикальных поверхностей. Хотя такие самцы
192
Глава 8. Черная каменка Oenanthe picata
Рис. 87. Фрагменты сигнального поведения черных каменок, а — позы конфликтующих самцов на стадии распределения территории; Ь. с — типичное поведение самцов capistrata при пограничных конфликтах; d — то же, у самцов picata: е — взаимодействие самца и самки capistrata в момент формирования пары: f, g — типичные позы самца opistholeuca при формировании пар и в нище, используемой для копуляции; i — поза угрозы у самца capistrata: к — “сгорбленная" поза самца при его взаимодействиях с самкой на сталии гне построения; I — поза самца на сталии подготовки к копуляции (см. рис. 96); m — позы самца (на переднем плане) и самки перед копуляцией. Из* Ра now 1974.
на протяжении всей весны выделяются среди других своим весьма активным пением и обилием рекламных полетов, им обычно так и нс удастся привлечь на свой участок самку (Панов. Корзухин, 1974; рис. 11). Как правило, это самцы-первогодки, которые составляют в поселении резерв холостых особей. При исчезновении хозяина территории кто-либо из этих холостяков занимает его место буквально в течение нескольких часов. Один из взрослых самцов был пойман мной для из-
193
Панов Е.Н. Каменки Палеарктики
Рис. 88. Возврат в поселение особей, окольцованных в предыдущие годы, а — самцы capistrata, b — самцы opistholeuca, с — самки, d — особи- первогодки. Из: Любущсн-ко 1995.
мсрсния и мечения 18 мая. Уже через 2 часа на его участке пел и гонялся за самкой-хозяйкой годовалый самец. Я оставил пойманного самца в клетке и через пару дней выпустил его на прежнюю территорию. Оказавшись на свободе, недавний узник тут же свирепо набросился на узурпатора. Они довольно долго выясняли отношения, но в конце концов первый хозяин все же оставил за собой спорную территорию.
Холостой самец постоянно рекламирует себя и свой участок, совершая эффектные демонстративные полеты. При этом птица, размеренно рабо
тая крыльями, которые совершают взмахи широкой амплитуды, в замедленном темпе поднимается по наклонной траектории высоко в воздух и с пол-минуты красуется на фоне неба, развернув веером хвост и оглашая окрестности звуками ликующей песни (рис. 15с, 16). Каждый самец располагает весьма обширным репертуаром коротких напе-
вов, которые он раз за разом повторяет, сидя на своем песенном посту (рис. 89-91). На начальных этапах сезона гнездования самцы начинают распевать в полный голос минут за 20 до рассвета, летая в полной темноте над своими территориями, и заканчивают свое хоровое
выступление спустя минут десять после появления солнца над горизонтом. Возможно, это наиболее удобное время, чтобы заявить о себе долгожданным невестам, которые, вероятно, совершают свои сезонные миграции по ночам. Если на этот раз помолвка не состоялась, самец с легким сердцем отдаст ранее утро поискам пропитания.
В промежутках между кормежками и пением самцы-соседи время от времени вступают в довольно безобидные турниры на обшей границе своих территорий. Это происходит тем чаше, чем плотнее поселение и, соответственно, меньше размеры земельных наделов. Это именно турниры, а не охрана границ территории в строгом смысле, поскольку конфронтации между данной парой соседей почти неизменно приурочены к одному и тому же короткому отрезку границы, составляющему в лучшем случае одну сотую часть всего ее периметра. Некоторые самцы конфликтуют с соседями охотнее других, некото-
194
Глава 8. Черная каменка Oenanthe picata
Рис. 89. Основные варианты песен самцов Ое. р. capistrata. а — короткие рекламные, b — фрагмент удлиненном песни, с — подпесня, d — фрагмент брачной (прсдсовокупительной) песни. Из: Панов 1989.
рые вообще не склонны к этому. Так, в одном из поселений, где территории 6 холостых самцов занимали площадь около 600 га, за 13 дней я наблюдал 8 турниров-патрулирований. Они были привязаны только к 3 из 8 стыков территорий. Два самца конфликтовали с соседями более активно, а два других вообще не вступали в конфронтации с сосе-
195
Панов Е.Н. Каменки Палеарктики
Рис. 90. Фрагменты песенных репертуаров 4 самцов (А. В. С. D) из ареала Ое. р. picata (Копетдаг) Из: Ианов 19S9.
196
Глава 8. Черная каменка Oenanthe picata
I; Mi /v
***** Ум»ШJIW>
y SMi HI*
U
kHz r6
-4
-2
Рис. 91. Фрагменты несенных репертуаров двух самцов Ое. р. capistrata (Л. В, юго-восточным Узбекистан) п двух самцов Ое. р. opistholeuca (С, D. горнып Бадахшан). Из: Панов 1989, с сокращениями.
дями. Самец, провоцирующий соседа, прилетает па границу особым полетом мотылька, принимает здесь позу угрозы (рис. 87b, с) и ждет, когда появится сосед. Затем оба короткими прыжками движутся параллельно друг другу вдоль границы. Опп вырывают из земли травинки резкими движениями клюва, либо на несколько секунд замирают, плотно прижавшись к земле (рис. 87d). Эта последняя акция, кстати сказать, совершенно нс свойственна самцам Ое. finschii. Если вдоль границы растут деревья, самцы перелетают с кроны на крону парал-
197
Панов Е.Н. Каменки Палеарктики
лсльно друг другу трепещущим замедленным полетом, сильно распушив контурное оперение (Панов, 1978: V. 1).
Формирование пар. Общая схема поведения самцов при формировании супружеских пар у черных каменок в целом сходна с тем, что мы видели у Ое. finschii (см. рис. 16). Основное различие состоит в том, что поведение самцов Ое. picata в этих ситуациях более хаотично. Самец столь же часто демонстрирует будущей супруге привлекательные с его точки зрения щели и ниши, как и сближается с ней — что она воспринимает, как правило, негативно. При первых таких встречах самка издаст сигнал угрозы (рис. 92 с) и нередко проявляет открытую агрессивность по отношению к кавалеру. Иногда она даже пытается ударить самца клювом, что ей подчас удастся сделать. В этом случае самец иногда отвечает тем же, и помолвка начинается с мимолетной драки. Особенно агрессивны самки в гибридогенных популяциях capistrata х opistholeuca, напоминая в этом отношении самок Ое. finschii. В ответ на попытки самца оказаться тет-а-тет с самкой последняя спасается бегством. Самец устремляется за ней, так что сплошь и рядом весь начальный этап формирования пары превращается в назойливое преследование самцом новобрачной, которая только и делает, что пытается освободиться от его навязчивости и приступить к осмотру участка. Иногда серии погонь следуют с некоторыми перерывами на протяжении целого часа, а то и двух часов кряду. Нервозность самца отчасти объясняется нелояльностью самки, которая может одновременно присматриваться к нескольким соседним территориям. Когда самцу кажется, что дело уже сделано и помолвка заключена, самка неожиданно с дерзкой песенкой перелетает во владения соседа, и сразу же начинается сумятица на той территории. Самка же, словно игнорируя самцов, летающих во всех направлениях, поющих без передышки и принимающих эффектные позы, перелетает к следующему соседу либо возвращается на ту территорию, что приглянулась ей при первом осмотре поселения. Если некоторые из здешних самцов уже состоят в супружеских парах, они возбуждены ничуть не меньше холостяков, а их подруги подтягиваются к границам, чтобы защитить свои законные владения от незванной гостьи. Некоторым самцам удастся приобрести подругу лишь с четвертой попытки (Панов, Корзухин, 1974; Панов, 1978).
Территориальное поведение самок, полигиния. Как и у Ое. finschii, самцы защищают свою территорию только от самцов, а самки — от других самок (рис. 93, 94). Однако это не всегда может воспрепятствовать двоеженству самца. В одном случае самецр/ся/я удерживал за собой две территории, разделенные дистанцией около 300 м, и на каждой из них состоял в законном браке. Мне пришлось также наблюдать, как вновь прибывшая самка вторглась на террито-
198
Глава 8. Черная каменка Oenanthe picata
10-1
8-
6-
4-
2- е
0,5
1,0
7— sec
Рис. 92. Основные позывки Ое. р. capistrata. а — при тревоге у гнезда (сигналы N I и N 3), b — разные варианты сигнала N 2 (храпение), с — сигнал N 4 (трещание), d — сигнал N 5 (никакие, в верхней части сонограммы), с — вокализация птенца в ожидании родителей и при выпрашивании у них корма (сигнал N 6, дребезжание).
рию уже сложившейся пары. Первая хозяйка предприняла было попытку изгнать пришелицу, но туг вмешался самец, который встал на защиту последней. События разворачивались точно так же, как это происходит при вселении новой самки на территорию пары у Ое.
199
Панов Е.Н. Каменки Палеарктики
isabellina (см. выше, 2.4). Раз за разом загоняя свою супругу в норку какого-то грызуна, откуда она боялась высунуться, самец полностью сломил се сопротивление. В дальнейшем эта самка все же отложила
яйца в гнездо, которое было почти полностью выстроено к моменту появления соперницы и находилось, по счастливой случайности, на
самой периферии территории, на которую новая самка не претендо
вала. Эта последняя стала хозяйкой большей части территории самца-
Рис. 93. Нападение самки capistrata на чучело самки, выставленное на гнездовой территории агрессора Из:
Рис 94. Самец фенотипа evreinowi , нападающий на самца черной каменки в клетке. Из: Ра now 1974.
хозяина. Точно такую же картину наблюдал Любущснко (1995). В этом случае, к счастью, пришлая самка была ранее окольцована. Опа оказалась прошлогодней хозяйкой оспариваемой территории.
Всего Любушен-ко зафиксировал 5 случаев полигинии в полиморфной популяции capistrata х opistholeиса в Дарбазе В один из сезонов
три самца из 104, гнездившихся в районе наблюдений, имели по 2 самки. В четырех случаях из пяти полигамами стали самцы старше года. Иногда обе самки на территории самца полигама гнездятся почти синхронно, но возможны также самые разные варианты несовпадения всро-
ках гнездования таких самок. На одной
200
Глава 8. Черная каменка Oenanthe picata из территорий вторая самка начала насиживать кладку уже после вылупления птенцов у первой самки. Любушенко считает, что в поселении может существовать резерв холостых самок — наподобие того, что мы видели у самцов.
Филопатрия, персональный состав пар. Как я уже говорил, значительная часть птиц, уже гнездившихся ранее в поселении, на следующий год возвращаются сюда же. Интересно было узнать, восстанавливаются ли при этом прежние супружеские связи. Оказалось, что такие случаи бывают, но они не являются правилом. Из 31 самок, которые гнездились в популяции Дарбазы дважды, на второй год со своим прошлогодним партнером выводили потомство 9 (29%), а 18 самок определенно сменили партнера (в 4 случаях самцов не удалось вовремя пометить, так что можно лишь предполагать, что самки сохранили свои прежние привязанности). Из 5 самок, гнездившихся три года подряд, 2 оставались с одними и теми же партнерами по два года, а на третий загнсздились с другими; одна каждый год гнездилась с новым самцом. Две самки в районе наблюдений выводили птенцов 5 лет подряд. Одна из них в первый сезон пребывала в союзе с одним самцом, три сезона — с другим, а пятый год провела с третьим; вторая два года гнездилась с одним самцом, следующий год — с другим, а четвертый и, возможно, пятый год — с третьим.
При живом прошлогоднем партнере самка может приобрести на следующий год нового, сменив территорию. При этом она подчас полностью меняет свои склонности, выбирая очередного супруга с совершенно иной окраской, чем предыдущий. Например, самка, гнездившаяся один год с черным самцом opistholeuca, на следующий год выбирает в качестве супруга белобрюхого и белоголового capistrata. Подсчеты показывают, что в полиморфной популяции Дарбазы самки, в общем, нс избирательны в отношении окраски партнера, так что вероятность того или иного альянса определяется частотой присутствия разных фенотипов самцов в популяции (Любушенко 1995).
Наряду с птицами, уже гнездившимися ранее в поселении, сюда ежегодно весной прибывают молодые особи прошлого года рождения. В популяции формы picata на Красноводском полуострове в 1990 г. 8 из 9 взрослых самцов определенно гнездились здесь один либо два предыдущих сезона, а остальные 14 (60,9%) были первогодками, из которых лишь один родился в этом поселении. Кроме того, здесь встречена одна самка прошлого года рождения. Среди самок 7 были помечены в 1988 (1) и в 1989 (6) годах, а 10 (58,9%) не имели меток (Панов и Грабовский, нсопубл ). По данным Любушенко (1993), в полиморфной популяции Дарбазы доля самцов-первогодков составляет в разные годы 36,4 — 51,0%, в среднем за 6 лет — 42,7%. При этом значительная
201
Панов Е.Н. Каменки Палеарктики
часть юных новоселов — это каменки, родившиеся в данном поселении. В среднем наследующий год возвращаются 12,2% птиц, помеченных в предыдущий сезон птенцами. Некоторые из них — близкие родственники. Так, известен случай возвращения трех сиблингов из одного и того же выводка. Более того, Любущснко нс обнаружил обмена мечеными особями (в том числе и первогодками) между двумя поселениями, расположенными в 15 км друг от друга.
Как же выбирают себе половых партнеров неопытные юнцы, гнездящиеся впервые? Оказалось, что они чаще, чем можно было бы ожидать, формируют пары со своими сверстниками. Из 15 самок, окольцованных птенцами, 13 в свой первый сезон гнездования объединились в пары с самцами-первогодками, причем 5 пар состояли из птиц, родившихся в данном поселении.
8.6. Гнездостроение и спаривание
Через нсделю-полторы после формирования пары самка приступает к постройке гнезда, на что у нее уходит также не меньше недели. Гнезда располагаются в пустотах самого различного характера, на глубине от 15 до 45 см. Нередко само гнездо и даже его содержимое видно снаружи, если заглянуть в отверстие входа. Наряду с пустотами в каменистых либо глинистых склонах и обрывах, самки изредка могут вить гнездо в дуплах фисташки (Рустамов, 1957) или в слепленных из глины гнездовых камерах большого скального поползня Sitta tephronofa (Рустамов, 1957; Бельская, 1961).
Первым делом самка натаскивает в нишу некоторое количество плоских камешков. Этому делу она целиком посвящает первый день гнсздостросния, а иногда и часть следующего дня. Работает самка весьма интенсивно: на моих глазах одна самка, летавшая от гнездовой ниши вниз, на дно ущелья (за 50-100 м), только за 8 мин. принесла в нишу 18 камней весом от 1 до 18 г. Эта же каменка за 20 мин. вечерней работы и за I ч. 20 мин утренней (с перерывами) подняла на высоту нс менее 10 м по вертикали 70 камней, из которых некоторые были столь велики, что птица нс смогла донести их до места назначения.
Количество камешков варьирует Рис. 95. Расположение гнезд черных от случая к случаю. Их много там, где каменок. Из: Panow 1974. дно убежища неровное и где есть иду-
202
Глава 8. Черная каменка Oenanthe picata
щис вниз вертикальные дыры и щели, которые самка заваливает камнями, выравнивая поверхность. В неглубокие ниши с ровным дном самка обычно приносит немного камней, часть которых оказывается нс в фундаменте гнезда, а в проходе и у выхода. Если вход в гнездо имеет уклон вниз, то наваленные здесь камни создают дополнительное трение, не позволяющее гнезду "съехать” к выходу. Когда самка еще не кончила носить камни, но уже изредка приносит травинки, она тем самым укрепляет основание гнезда в сложенном сю фундаменте из камней (рис. 95). Впрочем, нередко камешки оказываются лежащими не под гнездом, а только перед ним или вокруг него. Впрочем, подчас действия самки выглядят абсолютно бессмысленными. Одна из самок opistholeuca в Бадахшане выбрала в качестве места для гнезда круглую деревянную балку под крышей заброшенного глинобитного сарая. Самка была, видимо, пожилой и многоопытной (судя по черному цвету се оперения, почти не отличимого от наряда самцов), но это не помогло ей. Она раз за разом притаскивала камешки на покатую поверхность балки, и все они без исключения падали вниз, на пол сарая. Закончив свою "работу”, самка свила гнездо прямо на деревянной поверхности балки (см. рис. 21е).
Гнездо самка вьет из сухой травы, выстилая его изнутри более топкими травинками и волокнами, сдираемыми сю с сухих стволов ферулы (Ferula spp.). Иногда в выстилке бывает немного волоса и одно-два пера.
Когда гнездо в целом выстроено и остается лишь немного благоустроить его лоток, наступает время спаривания. Этот процесс довольно длителен и нс прост. Теперь самка в момент приближения к ней самца все чаще принимает особую приглашающую позу (рис. 36m). Что касается самца, то его нередко можно застать в этот период на вполне определенном карнизе скалы, над которым нависает широкая каменная крыша. Это — место любовных свиданий. В распоряжении самца их может быть одно или два, расположенных в разных точках территории, на самых разных расстояниях от гнезда.
Развитие отношений м е ж д у член а м и пар ы, копуляция. В течение первых трех — четырех дней после завершения гнсздостросния партнеры обычно держатся порознь и встречаются лицом к лицу лишь в этих местах свиданий. Инициатива в подготовке свидания чаще исходит от самца, но иногда — и от самки. Обычно перед таким взаимодействием самец пост на каком-либо кустике. Он начинает с многократного повторения короткой извещающей песни, которая затем постепенно переходит в подлесию. В нес самец включает многочисленные и очень точные имитации голосов других птиц (см. рис. 20), среди которых можно назвать сарыча-курганника Buteo rufinus,
203
Панов Е.Н. Каменки Палеарктики
Рис. 96. Поведение самца Ое. р. capistrata при подготовке к копуляции. Объяснения в тексте. Из: Панов 1989.
пустельги Falco tinnunculus, малого зуйка Charadrius dubius, травника Tringa tetanus, черныша Т. ochropus, рябка Pterocles orientalis, домового сыча Athene noctua, стрижей Apus apus и A. melba, зимородка Alcedo atthis, щурок Merops apiastern M. superciliosus, удода Upupa epops, ласточек Hirundo spp. и Delichon urbica, белой трясогузки Motacilla alba, майны Acridotheres tristis, клушицы Pyrrhocorax pyrrhocorax, ворона Corvus corax, большой синицы Parus major, воробьев Passer spp., седоголового щегла Carduelis caniceps и др. Самцы копируют также голоса лягушек и некоторых млекопитающих (например, блеяние ягненка).
Все более и более возбуждаясь, самец сильно распушает оперение и становится похожим на пушистый черно-белый шарик (рис. 87/). Пребывая в этой позе, самец то и дело отводит в сторону крылья, иногда начинает перебирать оперение клювом, но внезапно прекращает эти действия и замедленным трепещущим полетом перелетает на другой кустик. Все это может продолжаться и 10 мин., и более часа (рис. 96). Затем самец устремляется на свой излюбленный карниз и начинает выкрикивать там фразы принципиально иной по структуре песни, включающей в себя своеобразное шипение, завывания, бульканье, лай и прочие звуки, которые совершенно нс вяжутся с обликом этой маленькой птички. И сама песня, и тс позы, которые самец принимает в эти минуты, ничем нс отличаются от того, что мы видели во время
204
Глава 8. Черная каменка Oenanthe picata
наиболее эмоциональных стычек между самцами в период занятия ими гнездовых территорий (рис. 87а, рис. 96, вверху слева).
Если самка в это время находится где-нибудь поблизости и расположена к взаимодействию, она подлетает ближе к карнизу и садится в поле зрения самца. В этот момент последний приходит в полное
Рис. 97. Поведение самца и самки Ое. р. capistrata непосредственно перед копуляцией. Объяснения в тексте. Из: Панов 1989.
неистовство и начинает бегать с песней по карнизу, то выскакивая на его край, то забиваясь в самый
темный угол (рис. 96, вверху слева). Самка взлетает на край карниза, а
самец, не переставая петь, проделывает вокруг нее неистовый полет петлями, поочередно отлетая от ниши и возвращаясь к самке. Хвост самца широко развернут, а амплитуда взмахов крыльев очень мала. Во
время этого полета плоскость скольжения периодически изменяется, и после каждой петли самец мягко ударяется грудью то об пол ниши, то об ее потолок (рис. 97). Затем он усаживается на край карниза, а самка бежит в позе приглашения к спариванию в дальний темный угол ниши. Самец отводит крылья в стороны, очень мелко трепещет ими и плавно взлетает на спину самки (рис. 87 т). Зачастую все это
происходит в густых вечерних сумерках.
Взаимодействие не всегда протекает точно в соответствии с описанной схемой. Иногда самец по несколько раз кряду наведывается на свой карниз, прежде чем здесь появится его супруга. Или же самка прилетает к месту свиданий в тот момент, когда самец пребывает в настроении, не соответствующем случаю. Порой брачные игры почему-либо прерываются, когда они в самом разгаре, и спаривания не происходит. У одной из пар (самец фенотипа evreinowi, самка capistrata), за которой я всл почти непрерывные наблюдения, за 4 дня максимальной сексуальной активности произошло 4 копуляции, но 6 других взаимодействий в месте свиданий окончились ничем. Наиболее гладко проходят первые копуляции, а к концу периода спариваний самка становится агрессивной и в самый последний момент нс подпускает к себе самца.
Материалом для приведенного выше описания поведения птиц при спаривании послужили наблюдения в полиморфной популяции
205
Панов Е.Н. Каменки Палеарктики
capistrata х opistholeuca. Несколько по иному выглядит поведение самцов в популяциях opistholeuca и picata в их автохтонных ареалах. В отличие от самцов из полиморфной популяции capistrata х opistholeuca, самцы из чистых популяций opistholeuca в ожидании самки перелетают по скальной стенке от одного отверстия к другому, не задерживаясь в каждом из них подолгу, не делая длинных перебежек и нс распевая описанной выше брачно-агрессивной песни. Все звуковое сопровождение демонстраций сводится к сериям “цикаюших” звуков (сигнал № 5, рис. 92d). Перелетев к очередному отверстию, самец при посадке словно по инерции делает короткую пробежку с широко развернутым и приподнятым хвостом, затем замирает, прижавшись боком к стенке маленькой ниши (либо скрывается в пей полностью), и произнося свое ритмичное “цик...цик...цик...”. Примерно так же ведут себя непосредственно перед копуляцией самцы в автохтонном ареале picata (Панов, 1978, 1986 б, 1989).
Бывает и так, что за всем происходящим пристально следит самец-сосед либо какой-нибудь бродячий холостяк. Он пытается поймать момент, когда самка окончательно готова к спариванию, а ее самец почему-либо потерял бдительность. В таких ситуациях ловкий проныра быстро залетает в какое-нибудь отверстие на обрыве и оттуда зазывает к себе самку подходящей к случаю брачной серенадой (рис. 89d). Я наблюдал 10 подобных эпизодов, но в 9 из них самец-хозяин во-время замечал неладное и прогонял пришельца. Однако не исключено, что в одном случае узурпатор добился своего. Самка и самец-пришелец находились в глубокой нише, и я нс мог достоверно разглядеть, произошло ли спаривание.
Когда самка приступает к насиживанию, самец нередко демонстрирует описанное брачное поведение перед особями других видов (скальный поползеньSitta tephronota, пустынный жаворонок Ammomanes deserti, скотоцерка Scotocerca inquiera), даже в том случае, если это поющие самцы. Кроме того, у двух самцов opistholeuca и у одного picata я наблюдал весь описанный ритуал, законкившийся “копуляцией” с камнем. Описан также случай спаривания самцаpicata с чучелом самки.
8.6. Кладка, выводок
Кладка состоит обычно из 4-6 яиц (чаще всего из 5). Лишь одна из 60 известных мне кладок содержала 8 яиц, причем в 5 из них зародыши оказались крупнее, чем в 3 других. Птенцы вылупились лишь из двух яиц (Бельская, 1961), так что кладка была либо смешанной (двух разных самок), либо совершенно ненормальной. Соотношение кладок разной величины в ареале формы picata таково: 3 х 1,4 х 2, 5 х 20, 6 х 7
206
Глава 8. Черная каменка Oenanthe picata
(ср. 5,1, п=30); в южной части ареала полиморфной гибридогенной популяции capistrata >• opistholeuca. 5 х 4, 6 х 1 (ср. 5,2, п=5); в северной части того же ареала: 5 х 10, 6 х 6 (ср. 5,38, п=16), по данным Любу-шенко 5,22 (п=9) у пар с самцами capistrata и 5,75 (п=4) у пар с самцами opistholeuca. Все 6 кладок из ареала opistholeuca в высокогорьях Бадахшана содержали по 5 яиц.
У всех трех форм окраска яиц одинакова — от бледно-голубой до беловато-голубой. В пределах одной кладки могут быть яйца различного тона и с разной степенью испсшрснности: некоторые совершенно без рисунка, другие — с большим или меньшим количеством красноватых пятнышек и точек, сосредоточенных гуще на тупом конце.
Размеры яиц из ареала формы picata (юго-западный Туркменистан): 19,1-22,6 х 14,9-16,5 (ср. 20,22 х 15,78, п=34), поданным Заруд-ного (1896) 19,0-20,7 х 14,0-15,5 (ср. 20,05 х 14,98, п= 14). Размеры яиц формыopistholeucaиз Бадахшана: 19,2-22,3 х 14,8-16,4(ср. 20,54х 15,58, п=30), у пары самец picata х самка opistholeuca из ареала той же формы (Дарваз): 20,1-21,2 х 15,9-17,1 (ср. 20,8 х 16,5, п=5). Яйца достоверно крупнее в ареале полиморфной популяции capistrata х opistholeuca. В южной части ее ареала их размеры таковы: 19,3-21,8 х 15,3-16,6 (ср. 20,63 х 16,10, п=26); вес слабонасижснных яиц из этого региона 2,9-3,0 г (п=3), насиженных 2,3-2,5 г (п=4) (Каневский ,1978). На севере ареала тех же популяций размеры яиц 19,0-21,9 х 14,5-16,7 (ср. 20,43 х 16,33, п=37), их вес 2,22-3,02 г., ср. 2,63 г (п=27) (Митропольский, Фоттслср, личи, сообщ.).
Насиживает только самка, самец нс кормит ее на гнезде. Когда самка покидает гнездо и улетает на кормежку, самец иногда пытается сблизиться с ней, но та воспринимает эти попытки негативно и издает в ответ угрожающее трещание (сигнал № 4). Сроки насиживания точно не выяснены. Одна из самок начала насиживать 14 апреля, а птенцы вылупились утром 26 апреля.
Репродуктивный успех. Из 20 гнезд с птенцами, найденных в ареале picata, 5 (20%) содержали погибшие (видимо, нео-плодотворенные) яйца (в четырех гнездах по одному, в необычно большой кладке из 8 яиц — 6). Из 95 яиц нс дали птенцов 10 (10,5%). В 3 гнездах из 20 были найдены также погибшие птенцы (1, 3 и 3, в одном случае со следами травм на голове). Гибель птенцов в гнездах составила 8,2% от числа вылупившихся. Среди 10 гнезд из ареала полиморфной популяции capistrata х opistholeuca погибшие яйца были в 5 (50%) и составляли 9,4% от числа отложенных. Основными врагами черных каменок в период насиживания и выкармливания птенцов являются, вероятно, мелкие змеи в каменистых биотопах и лисы в тех районах, где птицы гнездятся в пустотах глинистых обрывов. Рустамов (1957)
207
Панов Е.Н. Каменки Палеарктики
обнаружил в одном гнезде хорасанскую агаму Laudakia erythrogastra Один из птенцов здесь был уже мертв, другой — при смерти. У обоих на теле и крыльях были кровоподтеки, по всей видимости, от укусов ящерицы.
Птенцы. Только что вылупившиеся птенцы имеют розово-желтую кожу и длинный (до 10 мм) эмбриональный пух серого цвета Зев ярко-желтый, валики в углах рта беловатые. Кормят птенцов самец и самка. По данным Бельской (1961) у птенцов на IV день начинают расходиться веки, на V появляются глазные щели, на VII — кисточки маховых, на VIII глаза уже открыты, на XI день птенцы способны перепархивать. Покидают они гнездо на Х1П-Х1Удснь. Птенцы в гнездовом наряде выглядят пестрыми, отличаясь в этом отношении от одноцветных, светлосерых птенцов Ое. finschii. Средняя величина выводка к моменту вылета составляет в полиморфной популяции Дарбазы у пар с самцами разных фенотипов 4,64-4,76 птенцов (по 82 выводкам)
Самцы полигамы проявляют тревогу в случае приближения наблюдателя к гнездам обеих своих самок и кормят птенцов обоих выводков. В одном случае самец стал кормить птенцов в гнезде на соседней территории после исчезновения се хозяина. Если слеток случайно попадает на участок соседей, его владельцы выкармливают его наравне с молодыми своего выводка. Самка может приступить к новой кладке еще в то время, когда молодые первого выводка нс достигли самостоятельности и остаются на попечении самца (Бельская, 1968).
8.7. Линька и отлет на зимовки
Молодые птицы из северных участков ареала полиморфной популяции, покидающие гнезда в начале мая, приступают к линьке с конца июня. Смена гнездового наряда начинается с контурного оперения и нижних кроющих крыла, позже линяют кроющие второстепенных маховых. У 57 из 87 молодых особей, содержавшихся в неволе, сменялись также некоторые перья из числа третьестепенных маховых, крылышка и хвоста, гораздо реже — отдельные первостепенные и второстепенные маховые и в исключительных случаях — кроющие первостепенных маховых. К концу июля молодые из ранних выводков уже одеты, по существу, в свой первый брачный наряд (Любушенко, 1993). Смена крупных перьев происходит и в природе: 30 июля в Копетдагс поймана молодая птица формы picata, у которой сменялись IV и IX первостепенные маховые. В целом линька молодых и взрослых каменок заканчивается к концу августа (Абдусалямов, 1973).
Отлет с мест гнездования происходит на удивление рано. В последних числах сентября в южном Узбекистане, где в это время стояла
208
Глава 8. Черная каменка Oenanthe picata
Рис. 98. Основыс районы зимовок Ое. picata. а — границы гнездового ареала, b — зимовки picata, с — зимовки capistrata, d — зимовки opistholeuca, с — залет picata, f — залет opistholeuca. Оригинал.
летняя жара и прекрасно чувствовали себя местные Ое. finschii и пролетные Ое. pleschanka, я нс встретил ни одной черной каменки. В центральном Афганистане Палюдан (Paludan, 1959) наблюдал единичных пролетных самок opistholeuca 21 сентября и 9 октября, но к 18 октября каменки полностью исчезли отсюда. Несколько особей picata он видел в долине р. Кабул в разные годы 14 декабря и 19-20 февраля. Одиночный самец opistholeuca встречен на юге Израиля 4 февраля (Shirinai, 1996).
Основные места зимовок черных каменок локализованы на юге Ирана и Афганистана, в Пакистане и в Индии (рис. 98).
8.8. Биологические заметки
Как и другие виды рода, черная каменка использует в пищу весьма широкий спектр кормов. Среди них важное место занимают муравьи, которых птицы могут добыть в достаточном количестве почти в любую погоду. Когда самец в самый ответственный момент подготовки к копуляции, в промежутках между брачными демонстрациями торопливо, словно между делом, склевывает с земли муравьев, создастся впечатление, что поедание этих насекомых в какой-то мерс стимулирует его эмоциональное поведение. Поданным Бельской (1961), в самую жаркую пору лета птицы охотно поедают ягоды (например, ежевики), что, вероятно, восполняет отсутствие питьевой воды. Супружеская пара весьма рационально использует запасы кормов на своей территории,
209
Панов Е.Н. Каменки Палеарктики
быстро переключаясь на охоту за наиболее перспективной добычей. Например, в одном случае и самец и самка постоянно приносили птенцам крупных палочников Phasmoptera (Ramulus sp.), чего я нс наблюдал у других черных каменок в том же районе. Любое крупное насекомое представляет для черных каменок желанную добычу. Любопытно наблюдать борьбу каменки с внушительной по размерам бабочкой-бражником, которую охотнику не так легко побороть. Жертве неоднократно удастся вырваться из плена. Она пытается улететь, но каменка снова догоняет ее, беспорядочно колотит клювом на земле, вновь терпит неудачу — и так по многу раз подряд, пока насекомое окончательно нс потеряет способность сопротивляться.
Как я уже упоминал, черные каменки являются одними из наиболее искусных певцов среди представителей рода Oenanthe. Это в наибольшей степени относится к форме picata, у которой так называемые “короткие” песни отличаются большим разнообразием (до 264 вариантов у некоторых самцов), самым широким диапазоном частот, большим числом нот с обертонами и максимальной длительностью. Прямо противоположными признаками характеризуются короткие песни формы opistholeuca в ее автохтонном ареале. Репертуар их сравнительно беден (19—22 напева у разных самцов), акустическая структура песен сравнительно проста. Все самцы в высокогорьях Бадахшана имитируют позывку синей птицы Myophonus caeruleus, которая может служить важным акустическим маркером птиц из этих популяций. Этот напев я лишь однажды слышал в ареале полиморфной популяции capistrata х
Рис. 99. Типичная организация песенных рядов у самцов picata (а, юго-восточный Туркменистан), opistholeuca (b, Бадахшан) и capistrata (с, юго-восточный Узбекистан). 1 — число новых вариантов в последовательных десятках песен, 2 — число рахпичных типов песенных вариантов в каждой данной десятке песен. Из: Костина, Панов 1981.
210
Глава 8. Черная каменка Oenanthe picata
opistholeuca. что может указывать па крайнюю редкость эмиграции птиц с фенотипом opistholeuca из их автохтонного ареала в ареалы других форм. Песенное поведение каменок в ареале полиморфной популяции capistrata х opistholeuca по всем характеристикам занимает как бы промежуточное положение между тем, что описано для picata и для opistholeuca (Костина, Панов, 1981; рис. 99).
В полиморфной популяции самцы всех фенотипов характеризуются сходным характером песенного поведения. Это и понятно, поскольку самцы, независимо от характера их окраски, принадлежат к единой генетической общности. Это доказано Любущснко (1995) путем прослеживания генеалогий в популяции Дарбазы. Оказалось, что в потомстве самца одного типа окраски могут присутствовать самцы иного фенотипа, причем самцы разных фенотипов подчас принадлежат к одному и тому же выводку (рис. 100). У самок, повидимому, не происходит строгого импринтинга на облик родителей, коль скоро самка подчас выбирает в качестве полового партнера самца с фенотипом иным, чем у ее отца.
Все сказанное выше наводит на мысль, что черные каменки нс вполне укладываются в привычные представления о политипическом виде, в котором все подвиды имеют примерно равную степень генетических различий. Нс исключено, что в данном случае перед нами ситуация, которую Хаффср (Haffcr, 1986) называет “надвидом второго порядка”. Уже было сказано, что комплекс Ое. picata мог образоваться не путем распада первоначального единого вида на три географические расы, а напротив, за счет вторичного объединения (посредством гибридизации) двух первоначально независимых таксонов: монотипического (capistrata) и политипического (picata + opistholeuca). Я допускаю, что эти таксоны могли иметь различное происхождение: capistrata по ряду признаков внешнего облика обнаруживает сходство с Ое. finschii и Ое. lugens. тогда как “горные" черные каменки (picata и opistholeuca) могут вести происхождение от восточноафриканских Ое. lugubris и (или) аравийских Ое. lugentoides. Впрочем, как я уже упоминал, в настоящее время черные каменки столь существенно отличаются по особенностям поведения от траурной каменки Ое. lugens s. str.. что эти два таксона невозможно считать членами одного надвида, как это предлагалось ранее (см. Hall, Moreau, 1970).
Скорее всего, мы никогда нс узнаем доподлинно, как в действительности происходила дивергенция в группе всех названных видов миллионы лет тому назад. Однако мы можем получить новые доводы в пользу или в опровержение той или иной гипотезы, детально изучив поведение всех перечисленных видов и применив к ним быстро развивающиеся методы сравнительно-генетических исследований.
211
Панов Е.Н. Камешки Палеарктики
Рис. 100. Два примера генеалогий в полиморфной гибридогенной популяции capistrataхopistholeuca в южном Казахстане. Самка в разные годы может иметь в качестве полового партнера самцов разных фенотипов. Из: Любушенко 1995
Строго говоря, вторичное объединение двух далеко разошедшихся видов посредством гибридизации — факт достоверно установленный. Как это происходит и какие имеет последствия, мы узнаем ниже, рассмотрев ситуации в зонах вторичного контакта двух других видов ка-менок — плсшанки Ое. pleschanka и чернопегой каменки Ое. hispanica.
212
Глава 9. Бе.ючерная каменка Oenanthe alboniger Ните
Глава 9. Белочерная каменка Oenanthe alboniger Hume
Синоним: Saxicola Alboniger Hume, 1872.
Эта каменка представляет собой как бы увеличенную копию черной каменки подвида picata, с которой неопытный наблюдатель может спутать ее при беглом взгляде. Помимо заметно более крупных размеров (вес ее около 25 г), от черной каменки picata белочерная отличается более массивной головой с несколько удлиненным, сильным клювом и относительно более длинными и толстыми цевками. Черная окраска подбородка и горла нс заходит па верхнюю часть груди, как у черных каменок. Черная “мантия” на спине короче, чем у picata, так что белое над-
хвостье выглядит более “длинным”, 101 Самцы Ое. alboniger и Ое. р - picata в полете. Оригинал.
как бы удлиняя и весь силуэт птицы. р F
Граница между черным и белым на спине проходит нс по прямой линии, а выглядит дугообразной: белое оперение поясницы как бы вклинивается в черное оперение спины (рис. 101). У самцов контраст между глянцевито-черными и снежно-белыми участками оперения очень резок. Самка очень похожа на самца, и различить их можно лишь на небольшом расстоянии, когда становится заметным, что у самки черные участки оперения более тусклые, матовые. Примерно так же выглядят и молодые птицы в гнездовом наряде, у которых черный цвет имеет коричневатый оттенок. Желтый цвет зева птенцов позже не изменяется и остается таким же у взрослых птиц.
9.1. Место обитания и численность
Белочерная каменка — типичный вид аридных пересеченных ландшафтов со скальными выходами и каменистыми осыпями (рис. I02).
2I3
Панов Е.Н. Каменки Палеарктики
Рис. 102. Ареал Ое. alboniger.
На востоке ареала (в Афганистане и в восточном Иране) она делит свое жизненное пространство с Ое. picata и Ое. finschii, причем местами гнездится бок о бок с ними в одних и тех же местообитаниях Однако, в отличие от первого вида она не поднимается в горы выше 1900 м, а второму уступает участки с более сглаженным рельефом. В западных районах области своего распространения бслочер-ная каменка делит об
щие местообитания с каменкой-монашкой Ое. monacha и с траурной каменкой Ое. lugens, придерживаясь в основном подножий гор и низко горий с бедной кустарниковой растительностью. На юге Ирана Ое. a/boniger иногда поднимается выше в горы (до 2300 м над у.м.), но она здесь довольно редка (Desfaycs u. Praz 1978).
В юго-восточном Иране и в соседних районах Афганистана белочерная каменка — вполне обычный вид (Зарудный 1903). То же, вероятно, можно сказать о популяциях юго-западного Ирана, где на 1 кв. км. может разместиться до 6 пар. Как и у других видов каменок, размещение птиц даже в пределах одного региона может быть достаточно неравномерным: более возвышенные над уровнем моря участки с повышенной влажностью и с лучшей растительностью могут прокормить большее число пар, нежели сухие, бесплодные холмы у подножий гор. Соответственно, площадь территорий, занимаемых гнездящимися парами, варьирует в очень широких пределах — от 2.6 до 21 га, составляя в среднем 5.8 га (Cornwallis: по Cramp 1988).
9.2. Гнездовая биология
По сообщению Хью и Эчекопара (Hue, Etchccopar 1970), в Иране самки откладывают яйца в апреле-мае, хотя эти сроки выглядят чересчур поздними для вида, зимующего в местах гнездования либо неподалеку от них. Более правдоподобны сведения о появлении первых
214
Глава 9. Белочерная каменка Oenanthe alboniger Ните
слетков уже во второй половине апреля, при том, что у пар, приступающих к гнездованию позже, птенцы могут покидать гнезда до второй половины мая включительно (данные Cornwallis 1975 для юго-западного Ирана). На юго-востоке Аравийского полуострова (Оман) гнездование начинается, вероятно, очень рано: выводки попадаются уже с конца марта и могут быть встречены до конца апреля.
Гнезда белочерная каменка вьет в расщелинах и во всевозможных пустотах скал. Вход в гнездовую нишу обычно бывает выложен плоскими камешками, причем в некоторых случаях общий вес камней достигает почти килограмма (910 г). Впрочем, такой настил бывает нс во всех гнездовых убежищах. Гнездо свито из сухих стеблей травы с примесью тонких прутиков и небрежно выстлано тонкими травинками, шерстью и перьями. Кладка состоит из 4-5 бледно голубоватых, почти белых яиц — иногда с немногими красноватыми точками. Размеры яиц: 20.3 — 25.0 х 15.5 — 17.1 (ср. 23.0 х 16.3, п = 24). Средний вес яиц 3.25 г (Cramp 1988).
9.3. Миграции и зимовки
Послегнсздовые кочевки молодняка начинаются, по-видимому, уже в конце июня. В октябре белочерные каменки появляются за пределами мест гнездования (западное побережье Персидского залива, некоторые районы Омана), и остаются здесь до февраля включительно. Во время зимовок и самцы и самки удерживают за собой обширные кормовые территории, которые они ревностно охраняют от представителей как своего вида, так и других видов каменок. При этом самки ведут себя ничуть нс менее агрессивно, чем самцы. Самка, помеченная в зимний период (в феврале), на следующий год заняла тот же участок в сентябре (Cornwallis 1975: по Cramp 1988). Складывается впечатление, что все же большая часть особей зимуют на местах гнездования, поскольку настоящего пролета осенью либо в конце зимы никому не удалось наблюдать.
Молодые из ранних выводков успевают перелинять к середине июня, другие еще нс приступают к линьке и на месяц позже. Линька взрослых птиц, проходящая по той же схеме, как и у других каменок, проходит в июне-августе.
9.4. Биологические заметки
Крупные размеры, плотное телосложение и сильный клюв бело-черной каменки, несомненно способствуют расширению се кормовой базы. Птица может справиться с достаточно крупной жертвой, даже
215
Панов Е.Н. Каменки Палеарктики
когда затраты усилий не столь уж оправданы. Например, Зарудный (1903) наблюдал, как белочерная каменка убила ящурку Eremias sp,4 но есть ее не стала. По словам того же автора, эти каменки поедакц даже наиболее крупных представителей саранчовых (Acrididae, Orthoptera). Кормятся они в основном на земле и сравнительно редко пользуются крыльями во время охоты. Вероятно, охотно поедают ягоды, семена которых иногда обнаруживают в желудках этих птиц.
Песню белочерной каменки разные авторы сравнивают с песнями каменок Ое. picata и Ое. lugens, славок Sylvia atricapilla и 5. communis и бюльбюлей Pycnonotus spp. (Cramp 1988).
Положение бслочерной каменки внутри рода Oenanthe трудно обсуждать в виду крайней скудости данных по ее поведению. Однако даже то немногое, что сегодня известно о ней, позволяет почти без колебаний поместить ее в число близких родственников Ое. picata, с одной стороны, и пары близких видов Ое. leucopyga— Oe.leucura, с другой. Впрочем, не имея достаточных сведений об этологических особенностях Ое. alboniger, трудно сказать более определенно, с кем именно ib названных видов она состоит в наиболее близком родстве.
216
Глава 10. Белохвостах каменка Oenanthe leucopyga
Глава 10.
Белохвостая каменка Oenanthe leucopyga Brehm, 1855
Синонимы: Vitiflora leucopyga Brehm, 1855; Dromolaea leucocephala Brehm, 1858; Oenanthe leucopyga aegra Hartcrt, I913.
Белохвостая каменка — это один из наиболее крупных представителей рода (вес около 30 г). Она уступает в размерах лишь чернобрюхой каменке Ое. leucura. От других каменок, щеголяющих белыми “шапочками”, матерые особи белохвостой каменки отличаются меньшим распространением белого на темени, черной окраской брюшка и почти сплошь белым хвостом, лишенным концевой черной полосы (рис. ЮЗ). Взрослые самки почти неотличимы от самцов, только черные участки оперения у них чуть более тусклые. Полость рта у взрослых птиц желтая, как и у птенцов. Птиц-первогодков легко узнать по широкой вершинной полосе на хвосте и по черному темени, на котором концу первого года жизни появляются отдельные белые перышки (рис. 104, 105). Считают, что белую шапочку птицы приобретают после второй зимовки, но, возможно, такая
Рис. ЮЗ. Белохвостая каменка Ое. leucopyga, птица старше года.
смена наряда происходит и позже.
Точка зрения, что изменчивость цвета шапочки есть проявление генетического полиморфизма. сейчас опровергнута. Неверно и то, что черноголовые особи — это самки (Meincrtzhagcn, I93O, Hollom, 1988). Известны гнездящиеся пары, состоящие из двух черноголовых птиц (Palfcry, 1988). Своеобразным проявлением полиморфизма является белое “зеркало” на черном крыле близ его сгиба. Оно образовано верхними кроющими перьями крыла и свойственно лишь очень немногим особям. Однако на крайнем юго-западе арс-
217
Панов Е.Н. Каменки Палеарктики
Рис. 104. Белохвостая каменка Ое leucopyga. птица прошлого года рождения.
Рис. 105. Белохвостая каменка Ое leucopyga, первогодок. Обратите внимание на присутствие отдельных белых перьев на темени
ала, в южном Алжире (нагорье Ахаггар) этот фенотип вполне обычен (из 7 экземпляров в Зоологическом музее в Бонне к нему принадлежали 5: Cramp, 198Х).
10.1. Систематические замечания
В пределах вида выделяют два подвида: leucopyga, занимающую почти всю африканскую часть ареала — к западу до р. Нил, и ernesti. распространенную главным образом в его азиатской части Вторая раса отличается от перво»! несколько большей величиной (средняя длина крыла самцов 112 мм против I06 мм у leucopyga). относительно более мощным клювом и большим ра житием блеска на черных участках оперения. Раса aegra (см. рис. 106) не признана большинством еисюмати-
2IS
Глава 10. Белохвостая каменка Oenanthe leucopyga
ков, но, по мнению Тая (Тус, 1987), это имя может быть сохранено для популяции Сахары (где эти каменки характеризуются мелкими размерами, относительно длинными цевками и тусклой окраской) и ссвсро-
Рис. 106. Ареал Ое. leucopyga. Цифры (по Тус 1987) — средняя длина крыла самцов. Точками обозначены залеты.
восточной Африки. Наиболее резко выделяется среди прочих изолированная популяция из северо-восточной Эфиопии. Здесь живут одни из самых мелких представителей вида (средняя длина крыла около 103 мм), длинноклювыс и “коротконогие”.
10.2. Место обитания и численность
Ареал Ое. leucopyga почти целиком лежит в наиболее знойных районах Африки и юго-западной Азии, ограниченных июльской изотермой 30°, с годовым количеством осадков 25-50 мм. Это единственная из всех каменок, освоившая гористые местности на всем необъятном пространстве пустыни Сахара — от холмов предгорий до высот порядка 3000 м над у.м. Здесь, как и на Аравийском полуострове, типичные местообитания белохвостой каменки — это каменистые террасы, идущие вдоль сухих водотоков и труднопроходимые скалистые склоны, усеянные каменистыми осыпями. Как правило, в таких местах скудный травянистый покров сгорает под палящими лучами солнца уже к началу мая, а древесно-кустарниковая растительность ограничивается низкорослыми кссрофитными видами, отдельные экземпляры которых обычно нс превышает полу-метра в высоту. Считают, что белохвостая каменка — это единственный представитель мелких певчих птиц на всем пространстве низкогорных пустынь Нигера площадью около 160 тысяч кв. км.
Вместе с тем, в Израиле Ое. leucopyga практически отсутствует в наиболее бесплодных, холмистых участках пустыни Негев, уступая эти бэдленды траурной каменке Ое. lugens. Но она становится вполне обычной на юго-востоке и на юге страны, в ущельях депрессии Мертвого Моря и в окрестностях г. Эйлат. Здесь разбросанные тут и там зонтико-
219
Панов Е.Н. Каменки Палеарктики
Рис. 107. Типичное местообитание белохвостой кменки Ое. leucopyga и каменки монашки Ое. monacha на крайнем юге Израиля (Эйлат).
видные акации придают особую суровую красоту безводным долинам и просторным ущельям с красноватыми каменистыми бортами и с нагромождениями валунов (рис. 107). В таких местах Ое. leucopyga обитает совместно с Ое. monacha. На маршруте по одной из таких долин длиной около 6-7 км я отметил 4 мая 3 особи Ое. leucopyga.
Кое-где эта каменка селится рядом с человеком — например, в южном Израиле и в некоторых оазисах Египта (Сива, Дахла) где она теряет свою обычную осторожность и становится на редкость доверчивой. Однако ее не привлекают поселки, где нет каменных строений — например, африканские деревушки в Эритрее с хижинами, построенными из материалов растительного происхождения (Meincrtzhagen, 1930, Smith, 1971, Shirinai 1996). По мнению последнего автора, в окрестности небольших поселков каменок привлекают свалки, где они могут добывать дополнительное пропитание.
В Марокко в некоторых местах пары селятся сравнительно неподалеку друг от друга, образуя разреженные поселения, которые отделены от подобных им “колоний” обширными участками точно таких же угодий, практически нс заселенных этими каменками. В Израиле подобные “групировки” объединяют до 3 пар, причем расстояния между ближайшими гнездами составляют здесь не менее 400 м, а величина территорий варьирует от 10 до 50 га. На Синайском полуострове поселения бывают довольно плотными (средняя величина территорий 0.4 га). В восточной Аравии поселения включают в себя 4-5 пар, плошадь
220
Глава 10. Белохвостая каменка Oenanthe leucopyga примыкающих друг к другу территорий колеблется от 4.8 до 9.1 га (в среднем 6.6 — 6.9 га) (Smith, 1971, Paz 1987, Palfcry, 1988, Shirinai, 1996 ).
10.3. Годовой цикл и сроки гнездования
В северо-западной и северной Африке гнездование проходит в марте и апреле. В испанской Сахаре гнезда с кладками находили I.III, 6.IV и I2.IV, в юго-западном Атласе, около 28" с.ш. (Драа) — с 14 по 23.IV и до 3.V (яйца сильно насижены). Птенцы в гнездах найдены 11 и I9 марта (северный Алжир, северная Ливия) и 26.IV (бывшая испанская Сахара). Молодые, покинувшие гнезда — 2.III и 5.IV (там же), 25.IV — 3.V (юго-западный Атлас). В перечисленных регионах некоторые пары приступают к устройству гнезд в первых числах мая, что указывает на возможность второго цикла гнездования. В конце апреля самцы активно поют, а у некоторых пар птенцы в гнездах могут быть найдены до конца мая. Но уже в первой декаде июня взрослые птицы приступают к линьке.
На Синайском полуострове супружеские пары нс распадаются по окончании брачного сезона и приступают к гнездованию с первыми проблесками весны. Здесь и в соседнем Египте гнезда с кладками попадаются с конца марта, а молодых, оставивших гнезда, видели в последней декаде апреля (22.IV). В Израиле, где члены некоторых пар также остаются в обществе друг друга по окончании сезона размножения, сезон гнездования начинается в конце февраля. Большинство самок откладывают яйца во второй половине марта, а массовый вылет птенцов приходится на середину апреля. В годы с сильными засухами в зимне-весенний период эти каменки вообще не приступают к гнездованию (Paz 1987, Shirinai 1996).
В восточной Аравии самец и самка по окончании очередного сезона гнездования занимают собственные индивидуальные участки, на которых живут в одиночестве до следующей помолвки. Формирование пар начинается в начале ноября, так что те птицы, которым удалось в это время приобрести партнера, проводят в его обществе всю зиму, начиная подыскивать место для гнезда уже с середины декабря. Самки в таких парах заканчивают постройку гнезд в конце января (22.1, 31.1), и первые кладки появляются в середине февраля. Самка в одной из подобных ранних пар умудрилась уже к концу марта отложить вторую кладку, и птенцы этого второго выводка перешли к самостоятельному существованию в середине мая. Разумеется, такой ход событий возможен только при очень благоприятной весне.
221
Панов Е.Н. Каменки Палеарктики
В обычных условиях в январе и даже позже происходит образование пар у тех каменок, которым не посчастливилось найти партнера в более ранние сроки. Соответственно, на месяц и более затягиваются их гнездовые дела. Наиболее поздняя дата окончания постройки гнезда -29 марта. Птенцы оставляют подобные поздние гнезда в конце мая (Palfcry, 1988).
10.4. Социальное поведение
Значительная часть материалов этого очерка базируется на детальных исследованиях биологии белохвостой каменки, проведенных па востоке Аравийского полуострова Джоном Палфсри (Palfcry, 1988).
К концу сезона гнездования молодняк занимает собственные индивидуальные участки по периферии территории своих родителей. Затем и самка начинает все чаще кормиться вне этой территории, и к началу знойных летних дней окончательно покидает ее и устраивается неподалеку, на собственном кормовом участке. В результате всех этих событий каменки рассредотачиваются и на протяжении лета и большей части осени держатся, за редкими исключениями, поодиночке.
Первые признаки возрождения интереса временных отшельников к себе подобным знаменуется возобновлением регулярного пения самцов в середине ноября. Как и у других видов каменок, каждый самец заявляет о своем присутствии с одного или другого из нескольких излюбленных им песенных постов, между которыми время от времени совершает характерный демонстративный полет. С широко разверну-
Рис. 108. Демонстративное поведение Ое. leucopyga. а — рекламный полет самца, b — d — демонстрации на песенном посту, е — поза самца при агрессивном контакте с соперником, f — поза самца при контакте с самкой, g — полет петлями самца вокруг самки. Из: Palfery' I988.
222
Глава 10. Белохвостая каменка Oenanthe leucopyga тым хвостом птица словно скользит по прямой в воздухе, лишь изредка поддерживая этот планирующий полет короткими взмахами крыльев (рис. 108 а). Песни, сопровождающие эту демонстрация — это короткие фразы типа “свии-свии терлитзевии” или “шроо-шроо-сиит-чи-шроо”, разделенные продолжительными паузами. Каждый самец имеет в своем репертуаре значительное число подобных фраз, из которых некоторые характеризуют его индивидуальный репертуар, тогда как другие (например, звучащая наподобие колокольчика “твиилинк-твиилинк-твиилипка") свойственны практически всем самцам данной и других популяций.
Самцы — прекрасные имитаторы, передразнивающие целый ряд видов птиц и других животных, а также проигрываемую в настоящий момент магнитофонную запись своей собственной песни. Среди образцов для имитации каменки часто выбирают звуки, весьма далекие по частотным характеристикам от собственных песен и позывок. Таковы, например, гортанное карканье пустынного ворона Corvus ruficollis, печальный крик обыкновенного канюка Buteo buteo, а также блеяние нубийского горного козла Capra nubiana и перекличка капских даманов Procavia capensis. Нередко хозяева соседних территорий вступают в песенные дуэли, находясь каждый на своем песенном посту, на расстоянии до 200 м друг от друга. Иногда поют, по-видимому, и самки.
Находясь на песенном посту, самец иногда принимает особые демонстративные позы: наклонившись под углом вперед, он широко разворачивает хвост и топчется на месте, поворачиваясь из стороны в сторону либо вокруг вертикальной оси. Либо он словно “проползает” с пол-метра особым скользящим шагом, волоча по земле развернутый веером хвост (рис. 108 Ь, с).
На незваного пришельца, оказавшегося на охраняемой территории, се хозяин решительно устремляется по прямой подчас с расстояния до 300 м и преследует нарушителя границы в воздухе, пока тот нс ретируется. Если же противник отвечает неповиновением, владелец участка, оказавшись рядом с ним, принимает особую “вертикальную” позу (рис. 108 d), после чего погоня возобновляется. Иногда соперники вступают в непродолжительную драку.
В распоряжении Палфери (Palfery, 1988) были лишь фрагментарные наблюдения за взаимоотношениями самцов с самками, и то, что ему удалось увидеть, во многом напоминает поведение других каме-нок-петрофилов, в частности, черной каменки Ое. picata. Члены пары имеют достаточно свободного времени в преддверии гнездостроения, и они по многу раз облетают свой общий участок, осматривая многочисленные здесь щели и ниши в поисках наилучшего гнездового убежища. В этот период самец иногда принимает около самки “горизон-
223
Панов Е.Н. Каменки Палеарктики
тальную” позу и несколько раз семенит вокруг нее по кругу, после чего та улетает, а он устремляется за ней в погоню. Вероятно, это аналог соответствующего поведения у Ое. picata, самец которой принимает в подобной ситуации “сгорбленную” позу и “крадучись”, медленно ходит вокруг подруги (см. рис. 16, 87 к). Очень сходен с “полетом петлями”, столь обычным у всех рассмотренных ранее видов, и описанный Падфсри “танцующий полст“ (dancing flight) самца Ое. leucopyga (рис. 108 g). Самец проделывает его перед своей супругой либо адресует другим самкам, почему либо оказавшимся на его территории. Воздушный танец сопровождается “брачной подпеспей”, которая представляет собой смешение самых невероятных звуков (шипения, бульканья, завывания, хриплых посвистов и т.д.) и более всего напоминает песню скворца Sturnus vulgaris. Однажды отец семейства танцевал таким образом подле дочери, которая всего лишь две недели назад оставила свою колыбель.
Гнездо строит только самка, затрачивая на эту работу иногда несколько недель. Сначала она натаскивает в гнездовое убежище изрядное количество плоских камешков диаметром около 2-3 см и весом 1.5-5 (до 10) г, устилая ими вход в отверстие на расстояние до 20 см. Самка собирает камешки за 100 м от будущего гнезда и летит с каждой находкой по прямой к месту назначения. Камешки присутствовали у входа во все 7 гнезд, найденных Палфери. Общий их вес составил в одном случае 370 г (Valverde, 1957). Самец нс принимает ни малейшего участия ни в сооружении каменного помоста, ни в постройке гнезда.
10.5. Гнездо, кладка, выводок
Гнезда располагаются в щелях скал, в отверстиях в нагромождениях камней, под валунами, в пустотах в стенах домов, на глубине до 75 см от входа. Из 7 гнезд, найденных Палфери, 4 помещались в вертикальных расщелинах скал, два — в пустотах глинистых обрывов и одно — в земляной норе у основания большого валуна. Большинство гнезд недоступно для осмотра. Те, до которых можно было добраться, находились на глубине 50-60 см (до 75 по другим источникам). Все гнездовые убежища располагались в местах, закрытых от солнечных лучей.
Валвсрдс (Valverde, 1957) приводит подробное описание одного гнезда. Его внешний слой был сплетен из лоскутьев коры акации, средний — из стеблей злаков, внутренний — из более тонких травинок, шерсти, тряпичных лоскутьев. Под гнездом были выложены камешки общим весом 370 г. По другим данным, в выстилке гнезда бывают шерсть, волос и перья.
224
Глава Ю. Белохвостая каменка Oenanthe leucopyga
У одной пары, за которой наблюдал Палфсри, в первом выводке было 3 слетка, а во втором 7, что, конечно, редкое исключение для данного вида. В Израиле 13 осмотренных кладок состояли из 2-5 яиц, чаше из 3-4. Распределение величин кладки у формы leucopyga в северной Африке таково: 3x8, 4 х 17, 5x6 (ср. 3.94, п = 31). Есть упоминание об одной или о двух кладках из 6 яиц. Окраска яиц от светлой зеленовато-голубой до голубовато-белой, с небольшим количеством красновато-коричневых точек, сосредоточенных в основном вокруг тупого конца. Размеры яиц формы leucopyga 20.2 — 22.6 х 14.7 — 16.7 (ср. 21.6 х 15.7, вес 3.23 г), формы ernesti 20.2 — 22.6 х 14.7 — 16.7 (ср. 22.0 х 16.6, вес 3.23 г). (Heim de Balsac, Mayaud 1962, Cramp 1988, Shirinai 1996).
Насиживает только самка 13-14 дней, по другим данным — 14-16 (Shirinai 1996), примерно раз в час вылетая на кормежку. В случае повторного гнездования самка строит новое гнездо. В одном случае оно располагалось в 150 м от первого. Ко второй кладке самка приступает через 3-13 дней после вылета первого выводка.
Птенцы остаются в гнезде 13-14 дней. Выводок в течение недели после вылета из гнезда держится неподалеку от него в каком-нибудь нагромождении камней, под которые слетки прячутся при малейшей опасности. Их кормят оба родителя, которые иногда делят выводок на две части, если в нем больше трех-четырех молодых. О своем присутствии птенцы извещают родителей хрипловатым “цвии-цвиии-цвии...” которое переходит в сплошное “зриизриизрии...”, когда самец или самка появляются с кормом. Если самка приступает ко второму циклу гнездования, выводок остается целиком на попечении самца.
Через две недели вольной жизни слетки уже кормятся сами и даже способны подчас поймать насекомое в воздухе. Независимость от родителей молодняк приобретает через 3 недели после вылета из гнезда. Спустя еще неделю взрослые начинают проявлять агрессивность по отношению к своим отпрыскам, особенно если те оказываются в окрестностях нового гнезда, где самка насиживает вторую кладку.
Впрочем, молодые интересуются своими младшими собратьями и даже могут “помогать” родителям в уходе за ними. Палфери наблюдал 16 случаев посещения второго гнезда молодыми первого выводка (не исключено, что во всех случаях это была одна и та же птица). При этом юнцы дважды приносили в клюве насекомых и столько же раз уносили из гнезда помет птенцов. Когда кто-либо из родителей заставал “помощника” в гнезде, все проходило на удивление мирно. По мнению Паза (Paz 1987), в некоторых районах Израиля (например, в Санта Катарина) молодые могут оставаться на территории родителей до следующей весны.
Однако в норме в Израиле молодые расселяются из мест гнездования осенью, но во время кочевок и зимовок остаются в своих типич-
225
Панов Е.Н. Каменки Палеарктики
пых местообитаниях. Многие особи, гнсздяшисся в горах, в октябре спускаются в предгорья. Иногда в это время белохвостых каменок можно встречать за несколько десятков километров от ближайшх мест гнездования (Shirinai 1996). Известны даже залеты в Европу — в южную Испанию, в Италию и даже на Британские острова (Toschi, 1969, Sorgicr. 1978).
10.6. Биологические заметки
На протяжении всего года эти каменки наиболее активны утром примерно до 8.30 и вечером после I6.30. до 8.30 и вечером после 16.30 Жаркие дневные часы они предпочитают проводить в тени, но в начале мая я нередко наблюдал охотящихся особей и в середине дня — около 12-13 час. В это время каменка выглядит так. словно всеми силами пытается избежать перегрева (рис. I09). На протяжении большей части года белохвостая каменка вынуждена обходиться без воды, но она охотно пьет, если предоставляется такая возможность. Дефи щи влаги птицы восполняют, поедая сочные плоды Ochradenus baccate
(Resedacea) (рис. 110). Но при первых каплях дождя птицы прячутся oi него в ближайшее укрытие.
Судя по наблюдениям Валверде (Valverde, 1957), эта каменка еше более всеядна, чем другие представители рода. Наряду со всевозможными насекомыми, от муравьев до крупной саранчи Schistocerca
Рис 109 Ое leucopyga в cnibnyio жару С' кадров видении мки в 11 флюс Оршипап
gregarina. она часто нападает на ящериц (Lacertidae, Gekkonidae), у которых иногда отрывает хвосты. По мнению этого автора, белохвостая каменка но своим хищническим повадкам во многом напоминает сорокопутов. Птицы нередко копают клювом песок, вероятно, в поисках личинок Tenebrionidac. Хотя Палфери пишет, что бе лохвостыс каменки не едя г пойманных ими взросл ы х ж у ко в - ч с р । юте -лок Tenebrionidac, якобы из-за неприятного вкус;)
226
Глава 10. Белохвостая каменка Oenanthe leucopyga
и запаха этих насекомых, я отснял на видеопленку весь процесс длительной “обработки” подобной добычи белохвостой каменкой, которая долго колотила се клювом и деформировала челюстями, после чего проглотила свою жертву. Кроме того, эти жуки были найдены в желудках каменок в Нигере. Около деревень эти птицы поедают всевозможные отбросы, в том числе кусочки мяса. Описан случай поедания каменкой цветов сложноцветного растения Compositac (Palfcry 1988).
Помимо всех тех разнообразных способов охоты, к которым прибегают и все прочие виды ка
менок (высматривание добычи с присад или с воздуха, когда птица зависает в трепещущем полете над землей, поиски насекомых на поверхности почвы или выкапывание их из грунта), у белохвостой каменки есть еще один, весьма своеобразный. Птица садится в редкую крону куста и неуклюже лазает по тонким веточкам, хватая ползающих и перелетающих здесь насекомых.
В целом можно сказать, что Ое. leucopyga — это типичный представитель “черно-белых” каменок-петрофилов, образующих центральное ядро рода Oenanthe.
Рис. 110. Кустарник Ochradenus baccatus, ягодами которого белохвостые каменки утоляют жажду в знойный весен не-лети ни период (Израиль), а — общий вид, b — гроздь ягод.
227
Панов Е.Н. Каменки Палеарктики
Глава 11. Чернобрюхая каменка Oenanthe leucura Gmelin, 1789
Синонимы: Turdus leucurus Gmelin, 1789; Saxicola cachinnans Tcmminck, 1840; Vitiflora leucura Boic, 1822; Sylvia leucura Savi, 1827; Dromolaea leucura Cabanis, 1850; Saxicola leucura Jaubert, 1855; Saxicola syenifica Heuglin, 1869; Saxicola leucurus riggenbachi Hartcrt, 1909.
Рис. 111. Ареал чернобрюхой каменки Ое. leucura.
Эту самую крупную из всех палеарктических каменок (вес се около 40 г) впервые описавший се орнитолог Иоганн Гислин поместил среди настоящих дроздов рода Turdus. Чернобрюхая каменка — птица размером со скворца, которая из-за мощного клюва и толстых цевок кажется еще массивнее, чем есть на самом деле. Черный наряд самцов и самок, кото
рые почти неотличимы внешне, имеет дымчато-коричневый оттенок и при определенном освещении птицы выглядят даже темно-коричневыми. Особенно это свойственно самкам, у которых к тому же бывают неясные пестрины на оперении брюшка. Всем этим чернобрюхие каменки хорошо отличаются от первогодков Ое. leucopyga, густо-черных со слабым голубоватым отливом (Maursberger I987). Молодые в гнездовом наряде очень похожи на родителей. Изредка встречаются особи с частично белыми кроющими крыла — как и у белохвостой каменки Ое. leucopyga (см. выше и Cramp 1988).
11.1. Систематические замечания
Чернобрюхих каменок из европейской части их ареала относят * подвиду leucura, африканских — к расе syenifica (рис. 111). У первых
228
Глава II. Чернобрюхая каменка Oenanthe leucura
менее развиты коричневые оттенки в оперении, концевая черная перевязь на хвосте немного уже, а общие размеры, по-видимому, слегка крупнее. Все эти признаки указывают на более молодой возраст европейских популяций по сравнению с африканскими. Описанный ранее подвид riggenbachi из южного Марокко не признан большинством систематиков.
11.2. Местобитания
Биотоп чернобрюхой каменки — это склоны и ущелья голых скалистых гор и холмов, едва прикрытых скудной травянистой растительностью (Launea fragilis, Asparagus acutifolius, Artemisia sp. и др.). Голые скалы, сухая почва и островки травянистых растений занимают свыше 90% всей площади, и лишь кое-где видны одиночные кусты Nerium oleander и Lycium sp. Низкорослые деревья в таких местах — большая редкость (Richardson I965). Гнездятся эти каменки и вдоль скалистых берегов морского побережья. В горах Атласа на северо-западе Африки эти каменки гнездятся от предгорий до высот около 3000 м над у.м.
Чернобрюхая каменка — это, пожалуй, один из самых пстрофиль-пых видов, теснейшим образом связанный с максимально пересеченным скалистым ландшафтом. В этом отношении вид имеет много общего со златогузой каменкой Ое. chrysopygia и с каменными дроздами рода Monticola.
В пределах своего ареала чернобрюхие каменки размещены неравномерно. Местами они вполне обычны (например, в некоторых районах Триполитании), в других регионах немногочисленны. Во Франции численность вида, возможно, сокращается (Cramp I988).
11.3. Годовой цикл и сроки гнездования
Будучи в целом видом оседлым, чернобрюхая каменка приступает к гнездованию настолько рано, насколько позволяют климатические и погодные условия. На юге ареала, в Алжире и Тунисе, первые кладки можно находить уже в конце февраля. В Сахарском Атласе оперенных птенцов в гнездах видели I6 марта. Однако, поскольку многие пзры склонны выводить птенцов более одного раза за сезон, гнезда с яицами и с птенцами попадаются вплоть до июня включительно. Та же картина наблюдается и в южной Испании: самые ранние кладки мож-1,0 встретить уже с начала марта, но часть пар приступает к размножению только в июне. На юго-западе Франции в ранние весны гнсздос-тРоспис у некоторых пар начинается в самом конце марта. Обычно
229
Панов Е.Н. Каменки Палеарктики
здесь гнезда с кладками находят с середины апреля, но большинство самок приступают к гнездованию на протяжении мая. Возможно и очень позднее гнездование: в одном случае птенцы, недавно покинувшие гнездо, встречены 28 августа (Heim de Balsac, Mayaud 1962, Prodon 1985, Soler ct al. 1995).
11.4. Социальное поведение
Большая часть приведенных здесь сведений заимствована из работы Ричардсона (Richardson I965). В отличие от того, что мы видим у большинства других видов каменок, у чернобрюхой каменки взаимный антагонизм особей на почве территориальных отношений выражен слабо. Возможно, это связано с двумя обстоятельствами. Во-первых, семейные пары сильно рассредоточены. В южной Испании на одну квадратную милю приходится около 7 пар этих каменок, то есть в распоряжении одной пары может быть до 14 га. Среднее расстояние между семью гнездами составляло 363 м, минимальное — 225 м, максимальное 660 м. В юго-западной Франции три гнезда были удалены друг от друга на 400-600 м (Prodon 1985). Понятно, что самец не в состоянии эффективно охранять границы столь обширного участка. Каждая пара проводит большую часть времени внутри более ограниченной зоны, в окрестностях гнезда или гнездовой ниши (нередко используемой парой из года в год), а вблизи границ территорий, которые не очерчены строго, хозяева соседних владений встречаются мирно, как на нейтральной полосе, и, по свидетельству Ричардсона, кормятся нередко группами на расстоянии I — 5 м друг от друга.
Второй фактор, который может объяснить редкость территориальных конфликтов — это оседлость вида. Мы уже видели, что у каменок с жесткой системой территориальности самец наиболее рьяно охраняет границы своей территории до момента формирования пары. Что касается чернобрюхой каменки, как вида оседлого, пары нередко сохраняются из года в год, до гибели одного из партнеров. Впрочем, из этого правила бывают и исключения. По наблюдениям Морено с соавторами (Moreno et al. 1994), в ноябре 1989 г. состав 7 пар оставался тем же, что и летом предыдущеего года, в 4 парах один из партнеров был новым, и на двух территориях прежние самцы-хозяева пребывали в одиночестве. В 4 случаев из 72 птицы меняли своих партнеров на протяжении одного гнездового сезона. Ричардсон, наблюдавший за этими каменками на протяжении полу-года, с октября по апрель, ни разу нс был свидетелем ни пограничных столкновений, ни формирования пар-Самец и самка, живущие в пределах обширного совместного участка» начинают тяготеть друг к другу с начала января (уже 23 декабря ОДИН
230
Глава 11. Чернобрюхая каменка Oenanthe leucura
из самцов начал проявлять интерес к самке и долго преследовал се). В феврале контакты между партнерами становятся все более тесными: 29 января самец и самка вместе ночевали в щели, где позже было выстроено их гнездо. В январе самцы возобновляют пенис, полностью прекращающееся в конце октября, и на протяжении января и февраля поют постоянно. С середины февраля каменки приступают к сооружению гнезд.
В целом социальные отношения у чернобрюхих каменок в высшей степени сходны с тем, что мы видели у белохвостой каменки Ое. leucopyga. Главное различие состоит в том, что последняя размножаются обычно лишь один раз за сезон, тогда как большинство пар у чернобрюхой каменки склонны гнездиться повторно или даже несколько раз подряд. В южной Испании некоторые пары предпринимают до 5 таких попыток за гнездовой сезон длительностью не менее 4 мсс. (с начала марта до июня включительно), и часть из них успевает за весну и лето вырастить 3 выводка. Менее благополучным парам не удается успешно завершить ни одной из этих попыток, и все их усилия заканчиваются ничем (Soler et al. 1995). На юго-западе Франции второй цикл размножения возможен лишь в годы с очень ранней весной, и то лишь у 15% пар (Prodon 1885). Эти различия между двумя близкими видами обусловлены более благоприятным климатом в северных частях ареала Ое. leucura.
11.5. Гнездо, кладка, выводок
Гнездовые убежища и конструкция гнезд. Гнездостроение начинается с благоустройства гнездовой камеры, куда птицы доставляют значительное количество плоских камешков. В отличие от всех прочих видов каменок, у которых этим делом занята только самка (исключение, в какой-то степени, представляет собой златогузая каменка Ое. chrysopygia), у чернобрюхой каменки соответствующее поведение ярко выражено и у самца. Болес того, он первым начинает носить камни в гнездовую нишу и, как полагают, тем самым стимули-РУст самку приступить к гнездовым делам.
Эта первая стадия начинается порой за 3 недели до начала яйцек-ладки, но иногда занимает всего лишь несколько суток. В среднем се продолжительность — 11 дней. На этой стадии лишь у 3 пар из 8, нахо-днв1пихея под наблюдениям, в сборе камней участвовала самка, а самцы проделали около 87% всей работы. Надо сказать, что деятельность эта у еамНов далеко нс столь рациональна, как может показаться с первого П-'Ыла Подводя итог своим наблюдениям. Морено с соавторами ' ,,?по ci al 1994) подсчитали, что в среднем па одну пару при пер
Панов Е.Н. Каменки Палеарктики
вом цикле гнездования только около 33% принесенных камней оказываются в гнездовой нише. Немного меньшее их количество (30%) выпадают из гнездового убежища на землю, а остальные 37% самец приносит не к будущему гнезду, а в совершенно другие отверстия. Похожую картину мы уже видели у Ое. chrysopygia, у которой самец носит камни нс к гнезду, а в приглянувшуюся ему собственную нишу.
Самка начинает активно носить камни лишь в тот момент, когда становится насущной задача реальной постройки гнезда. Она же проделывает и всю работу, связанную с сооружением последнего. Члены пары, в основном, вероятно, самец, нередко приносят все новые и новые камешки до самого начала откладки яиц, но в среднем сбор камешков заканчивается за 4 дня до этого. Результатом является то, что сплетенные из соломы основания большинства гнезд (32 из 37, осмотренных Ричардсоном) оказываются закрепленными в фундаменте из камней.
Большинство камней, приносимых птицами в гнездовую нишу, весят от 3 до 10 г (в среднем 6.8-7.3 г), но некоторые достигают веса 28 г, что превышает половину веса самой птицы (составляющего чуть больше 40 г). Поскольку многие гнездовые убежища используются из года в год, около них скапливаются тысячи камней. В одном месте пол пещерки площадью в 2 кв. м. был сплошь усеян камешками, которых здесь было около 9300 штук. По подсчетам Морено с соавторами (Могепо et al. 1994), каждая пара за сезон переносит немногим более 3 кг камней, причем максимум этой деятельности приходится на период подготовки к первому циклу гнездования. В это время некоторые пары перетаскивают с места на место около 1300-1400 камешков общим весом около I кг.
Изнутри самка выстилает гнездо тонкими травинками, корешками, шерстью, волосом и перьями. В фундаменте наравне с камнями иногда попадаются прутики и палочки длиной до 16 см. Одно из гнезд, осмотренных Ричардсоном, было выстроено на фундаменте, целиком состоявшем из палочек и кусочков стеблей кактуса.
Гнезда располагаются в щелях и нишах скал — иногда на уровне земли, чаще же на высоте от 45 см до 4.5 м (среднее по 32 гнездам — 1.6 м). Каменки почти не используют подходящие для устройства гнезда отверстия на высоте более 3.6 м от земли, хотя они лучше защищены от хищников. Нс исключено, что тем самым они экономят усилия, необходимые для переноса камней на значительную высоту. Вход в гнездо всегда находится в тени, а нередко гнезда помещаются в пустотах в стенах естественных пещер (глубиной до 10 м) или искусственных шурфов, куда вообще проникает очень мало света. На юге Испании эти каменки наиболее охотно гнездятся в нишах рукотворных пе-
232
Глава //. Чернобрюхая каменка Oenanthe leucura
шер, в которых в древности жили люди (Soler et al. 1995). Судя по всему, в большинстве случаев гнездо бывает выстроено недалеко от входа, так что его край виден снаружи. Гнезда довольно велики — около 160 мм в диаметре при поперечнике лотка 70 — 80 мм и глубине его 60 — 70 мм.
Как и у прочих видов каменок, окончание гнсздостросния и начало яйцекладки отделены друг от друга периодом около недели. В южной Испании в одном из двух гнезд, сооружение которых было окончено 12 марта, первое яйцо появилось 21 марта, в другом — 24 марта. Последующие яйца самка откладывает раз в сутки.
Кладка. Полная кладка содержит от 3 до 6 яиц (как исключение, семь). В северо-западной Африке распределение величин кладок таково: 3 х 15, 4 х 67, 5 х 19 (ср. 4.0, п = 101), в юго-западной Франции 3 х 2, 4 х 8, 5 х 9, 6 х 1, 7 х I (ср. 4.57, п = 21). В южной Испании в кладках 3-6 яиц, средняя величина первых кладок 3.99 (п = 87), вторых 4.04 (п = 47), повторных взамен утраченных 4.03 (п = 29). Размеры яиц формы leucura 22.0—26,6 х 16.5—19.4 мм (ср. 25.0 х 17.8 мм, п=65, вес 3,72 г), по другим данным 22.4—27.Ох 17.0—18.7 мм (ср. 24.5 х 17.8 мм (п=62). По описанию Баннермана (Bannerman 1954), окраска яиц — от зеленовато-белой до зеленовато-голубой, с небольшим количеством лилово-красноватых и глубоких фиолетовых пятнышек и точек, сосредоточенных гуще вокруг тупого конца.
Насиживает только самка нс менее 14—15 дней, а в некоторых случаях до 17 — 18, что заметно превышает длительность насиживания У других каменок. У одной из самок период насиживания растянулся на 20-21 день. Значительная изменчивость продолжительности насиживания кладки — характерная и своеобразная черта биологии чернобрюхой каменки. Самка часто сходит с гнезда на кормежку и в дневные часы проводит в гнезде 55 — 65% всего времени (Heim de Balsac, Mayaud 1962, Richardson 1965, Schonwetter, 1979, Prodon 1985, Soler ct al. 1995). Размеры яиц формы leucura 22.0 — 26.6 x 16.5— 19.4 (ср. 25.0 x 17.8, n = 65; вес 3.72 г).
Репродуктивный у с п е х. Г незда часто разоряют всевозможные хищники — по-видимому, змеи (Coluber hippocrepis, Malpolon SP), возможно, ящерицы Lacerta lepida, а также мелкие млекопитающие (садовая соня Eliomys quercinus) и сороки Pica pica. В южной Испании из 175 гнезд 18 (10.3%) были брошены, 34 разорены (19.4%, из ”их 10 гнезд — на стадии насиживания и 13 — с птенцами) и 3 погибли гю неизвестной причине. Отход гнезд первой кладки составляет ^•0%, второй — 20.8%, повторных кладок — 32.4%. В среднем на одно Спешное гнездо приходится 2.1 слетка (1.9 — в первых выводках, 2.5 "" но вторых). За весь сезон пара может вырастить максимально 13
233
Панов Е.Н. Каменки Палеарктики
слетков, в среднем на пару (включая и те из них, которым ни разу не удалось воспитать птенцов) 4.7 слетков. На юге Франции успешными оказались только 10 (58.8%) из 17 гнезд. Здесь среднее число слетков в расчете на “успешное” гнездо составляло 3.5. В случае разорения гнезда с полной кладкой самка может отложить первое яйцо в новое гнездо через 10-11 дней (Richardson 1965, Prodon 1985, Soler et al. 1995).
Успех чаще сопутствует взрослым птицам, чем первогодкам, и старожилам, нежели птицам, недавно обосновавшимся на данной территории. Дело в том, что самки, осваивающие новые для них участки, приступают к откладке яиц в среднем на две недели позже, чем самки-старожилы. Вообще, Солер с соавторами (Soler et al. 1995) пришли к выводу, что успех размножения определяется скорее качеством территории, чем индивидуальными особенностями того или иного самца либо самки. Правда, самцы, сильно пораженные перьевыми вшами, отличались слегка пониженной продуктивностью.
Птенцы.У птенцов эмбриональный пух длинный, серого цвета. Зев желтый, края рта бледножслтыс. В первые дни после вылупления птенцов их кормит главным образом самец, а самка редко оставляет гнездо с молодыми. Вылет их происходит на XIV — XV день. По данным Продона (Prodon 1985), срок пребывания птенцов в гнезде составляет 15—19 дней, в среднем 16.5 дней. Первое время после вылета они держатся около щелей и нор, куда прячутся при малейшей опасности. В это время они способны перелетать на небольшие расстояния, но продолжают эксплуатировать родителей. Спустя неделю-другую молодые вполне могут существовать самостоятельно, но они подчас остаются с родителями в течение месяцев. Так, окольцованного птенца в компании с родителями встретили в на юге Франции в середине зимы (18.XII). В испанских Пиренеях 2 и 3 января видели трех птиц, совместно ночевавших в общем убежище. Нс исключено, что это также были родители со своим отпрыском. Разреженные группы численностью до 6 особей, которые можно иногда встретить в Марокко в ноябре, могут быть не распавшимися еще семьями (Smith 1971, Dorka et al. 1976, Prodon 1985).
11.6. Миграции
Вполне понятно, что по крайней мерс часть первогодков вынуждены расселяться за пределы мест рождения, ио сроки таких кочевок не выяснены. Вероятно, и часть взрослых особей совершает сезонные миграции, покидая на зиму высокогорья и откочевывая к подножиям гор. В некоторых районах (в частности, в провинции Малага на крайнем юге Испании) чернобрюхие каменки не попадаются на глаза в
234
Глава 11. Чернобрюхая каменка Oenanthe leucura
период между ноябрем и февралем. Часть особей в это время оказывается далеко за пределами районов гнездования — например, в Египте и Израиле (6.XI, 2-5.1). Известны также многочисленные залеты в разные районы Европы (конец августа — январь, в одном случае — в июне), к северу до Шотландии и Норвегии (Prodon 1985, Cramp 1988, Shirinai 1996).
11.7. Некоторые особенности поведения
Эти каменки весьма осторожны. Активны они нс только в дневные часы, но и в сумерках, утренних и вечерних. Способы охоты их за насекомыми нс отличаются оттого, что мы видим у других видов рода. Подобно Ое. leucopyga. могут добывать пропитание в кронах густых кустов, где склевывают также ягоды. Охотно поедают плоды аспарагуса (Asparagus acutifolius), барбариса, малины, смилакса (Smilax aspera). мирта, маслины Olea и других кустарников и деревьев. Даже птенцам родители приносят иногда скорпионов и небольших ящериц. С питанием жалящими насекомыми и ядовитыми беспозвоночными (такими, как сколопендры), а также с манерой кормится в кронах колючих кустов иногда связывают особое строение перьев в области клюва, которые напоминают жесткие щетинки со слабым развитием бородок пера.
Брачное поведение, которое, как полагают, предшествует спариванию (хотя я сомневаюсь в этом), весьма своеобразно. Самец пригибается грудью к земле, плавно приподнимает хвост и широко разворачивает его. Оставаясь в этом положении, птица неистово подрагивает хвостом и концами сложенных над ним крыльев, одновременно издавая своеобразное ‘‘хихиканье”. Эта церемония чаще всего происходит недалеко от гнезда (рис. 112). Самка в ответ принимает такую же позу (Kdnig 1966).
Извещающая территориальная песня больше всего напоминает, по общему мнению, песню синего каменного дрозда Monticola solitarius. Короткие полнозвучные строфы нередко заканчиваются шипящими звуками, щебетом или стрекотанием. Продолжительность одной строфы 2 — 4 сек, но иногда песня длинсс, длительностью до 10 — 12 сек. Самцы поют, сидя на своих песенных постах либо во время демонстративного полета, который по описаниям сходен с аналогичным полетом Ое. leucopyga. Вероятно, много общего есть и в структурной организации песен этих двух видов (см. Cramp 1988). Наряду с “короткой” Рекламной песней, в которой отдельные фразы разделены четкими интервалами, самец нередко воспроизводит подпесню, включающую в себя уже упоминавшееся “хихиканье”. Самец воспроизводит эгу пес-
235
Панов Е.Н. Каменки Палеарктики
ню либо пребывая в одиночестве (часто на протяжении всей зимы), либо приглашая самку в гнездовую нишу, куда самец многократно забегает, тут же выскакивая наружу. Иногда при этом самеп и шст птенцовый
Рис. 112. Фрагменты сигнального поведения чер- КРИК 11 >аносит в отвер-нобрюхои каменки Ое. leucura. Из: Kmiig 1966. стие соломинку либо травинку, которую он гут же может вынести нашд. Этим, в сущности, и ограничивается “участие” самца в сооружении колыбели для будущих отпрысков.
Я уже упоминал о том, что Ое. leucura уникальна среди прочих изученных видов каменок в том отношении, что не только самка, но и самец активно участвует в сооружении каменного “помоста” в гнездовом убежище. Морено с соавторами (Moreno et al. 1994) убедительно доказали, что присутствие такого помоста не регулирует температуру в гнезде, нс служит для него защитой против ухудшения погоды и нс предохраняет кладку и птенцов от хищников. Более того, гнезда тех пар, которые “запасали” большее количество камешков, в среднем чаще становились жертвой животных-мародеров. Эти исследователи отрицают, что главная функция помоста состоит в предохранении гнезда от механического смещения (“сползания” к краю ниши), приводя следующие аргументы. Во-первых, как удалось установить, птицы не приносили больше камней в вертикальные щели (где помост необходим в качестве фундамента для гнезда), чем в другие пустоты, в которых гнездо может быть построено и без фундамента. Во-вторых, в некоторых гнездовых убежищах вообще не было камней. В-третьих, каменки приносили камешки и в такие ниши, которые не были впоследствии использованы этими же особями для гнсздостросния. На мой взгляд, ни один из этих аргументов ни в малейшей степени не опровергает “инженерную” функцию помоста. Птица не в состоянии рассуждать подобно человеку и пользоваться своим инстинктом сознательно, отказываясь от его приказаний в тех случаях, где он бесполезен или даже вреден. Когда у птиц нет выбора, и все пригодные ниши требуют для своего использования “достройки” с помощью камешков, инстинкт работает во благо каменок. Если же в распоряжении популяции изобилие ниш с плоским полом, где камешки не нужны вовсе, инстинкт ведет к непроизводительным энергетическим потерям. В сущности, это азы современной теории инстинкта, заложенной
236
Глава 11. Чернобрюхая каменка Oenanthe leucura
сше работами А. Фабра, писавшем о “мудрости и невежестве” инстинкта.
Не убедительны также доводы Морено с соавторами (Moreno et al. 1994) в пользу гипотезы, согласно которой поведение самца в данном случае есть форма “послебрачных половых демонстраций”, выработавшихся в эволюции на основе полового отбора. Идея состоит в том, что самка определяет “качество” самца, ориентируясь на количество переносимых им камней. Правда, некоторые показатели успеха размножения положительно коррелировали с количеством принесенных камней, но при этом общее число слетков, воспитанных парой за год, никак не соотносилось с числом и весом камней в данном гнезде и вокруг него. Добавлю, что в большинстве случаев нс было достоверно известно, какую именно часть камней принес самец, а какую — самка
На мой взгляд, манера самца таскать камни куда надо и куда не надо — это атавизм, унаследованный каменками от тех их далеких предков, у которых оба партнера принимали равное участие в подготовке места для гнезда и в его постройке. У самцов почти всех видов каменок инстинкт “перетаскивания камней” практически полностью деградировал, сохранившись в какой-то мере, помимо Ое. leucura. только у Ое. chrysopygia. где эта форма поведения самца уже никак не связана с гнсздостроснием как таковым. Вероятно, Ое. leucura стоит ближе всех прочих каменок к предковому стволу рода и потому сохранила эту крайне энергоемкую и парадоксальную форму поведения в се более или менее первозданной форме.
Скудные сведения о сигнальном поведении этой каменки, содержащиеся в литературе, нс позволяют проводить продуктивные сравнения с другими видами рода. Довольно очевидно, однако, что чернобрюхая каменка находится в наиболее близком родстве с белохвостой каменкой Ое. leucopyga и вместе с ней принадлежит к ядру рода — мерно-белым пстрофильным каменкам (Ое. picata, Ое. alboniger, Ое. finschii и ДР ).
237
Панов Е.Н. Каменки Палеарктики
Глава 12.
Испанская каменка, чернопегая каменка Oenanthe hispanica Linnaeus, 1758
Синонимы: Motacilla hispanica Linnaeus, 1758; Muscicapa inelanoleuca Goldenstadt, 1775; Saxicola aurita Hemprich et Ehrenberg, 1833; Saxicola xanthomelaena Hemprich et Ehrenberg, 1833; Saxicola eurymelaena Hemprich et Ehrenberg, 1833; Saxicola amphileuca Hemprich et Ehrenberg, 1833; Saxicola gaddi Sarudny et Loudon, 1904; Saxicola albicollis Buturlin, 1909; Saxicola hispanica gaddi Hartcrt, 1910; Saxicola hispanica xanthomelaena Hartcrt, 1910.
Испанская каменка — это типичный представитель небольшой группы видов, заметно выделяющихся среди прочих представителей рода Oenanthe по целому ряду особенностей строения и поведения. Все виды, о которых речь шла до сих пор, по общему складу — скорее “наземные” птицы. Хотя многие из этих каменок прекрасно чувствуют себя в воздухе, в повседневной жизни они значительную часть времени проводят на твердом субстрате, где добывают львиную долю своего пропитания. Несколько иначе ведет себя испанская каменка, ее ближайший родич — плешанка Ое. pleschanka и каменка-монашка Ое. monacha, которая, по моему предположению, относится к этой же группе видов. Все эти птицы гораздо чаще, чем прочие каменки, ловят насекомых налету, напоминая в этом отношении мухоловок. Забавно, что немецкий орнитолог Гюльдснштадт, впервые описавший для науки испанскую каменку, поместил се в род мухоловок Muscicapa. Будучи прекрасно приспособленной для маневренного полета, испанская каменка характеризуется изящным сложением (вес се около 18 г), длинными крыльями со слегка заостренной вершиной и несколько удлиненным хвостом, выполняющим роль руля. Лапки, напротив, сравнительно короткие и слабые, с чем связано, вероятно, нежелание этих каменок передвигаться шагом или бегом, что в определенных ситуациях (в частности, во время агрессивных и брачных демонстраций) столь характерно для всех ранее расмотренных видов Oenanthe. По земле испанская каменка неизменно передвигается короткими прыжками, но предпочитает всегда, когда возможно, пользоваться крыльями.
В природе самцов испанской каменки легко узнать по их характерной контрастной окраске. Что касается самок, то наиболее надежным признаком для их опознавания может служит своеобразная манера их
238
['шва 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
Рис. 113. Ареал испанской каменки Ое. hispanica. а — соотношение (%%) в числе черпогорлых самцов sfapazina и белогорлых aurita, b — ширина черного поля на юрле (мм), с — основные зимовки подвида hispanica, d — то же, подвида melanoleuca (b — по НаПег 1977).
Рис 114. Изменчивость окраски головы самок в популяциях Ое. hispanica "icianoleuca (а. Закавказье), Ос pleschanka (b) н в полиморфной гибридогенной ' 1ЯИИИ полуострова Мангышлак (с. восточный Прикаспии). Из: Ианов 1989
239
Панов Е.Н. Каменки Палеарктики
повседневного поведения. Полет этих птиц стремительный, с частым использованием виражей. При посадке каменка, как правило, делает мимолетное движение грудью вниз и одновременно резко, на мгновение отводит в сторону концы крыльев. Хвост при этом (в отличие от других каменок-пстрофилов) остается на уровне туловища или коротко движется вниз (а нс вверх, как, скажем, у Ое. lugens и Ое. picata). Очень часто этот едва заметный поклон сопровождается глухой, хрипловатой позывкой “зак-зак”. Если другие виды каменок в обыденной жизни очень редко издают позывку этого типа (сигнал N I), то у испанской каменки (и плешанки) она оказывается достаточно постоянным элементом повседневного поведения. Это, в часности, позволяет отличить в поле некоторых гибридов hispanica хpleschanka (так называемых “бслоепинных плешанок”) от чрезвычайно сходных с ними по окраске самцов Ое. finschii. Находясь в спокойном состоянии, испанские каменки медленно покачивают хвостом, поочредно приподнимая его нс выше 10° над уровнем туловища и опуская на на такой же угол ниже горизонтали.
Испанской каменке свойственен генетический полиморфизм окраски. Во всех се популяциях описаны по крайней мере две вариации окраски самцов — черногорлые (морфа stapazina) и бслогорлыс (aurita, или amphileuca). На крайнем западе ареала встречаются также “бело-горлыс” особи со слабым развитием черноты на подбородке, описанные под именем caterinae (рис. 113). Ряд других цветовых вариантов, которые ранее считали морфами испанской каменки (например, melanoleuca, galli и др. — см. Лоскот 1976), в действительности обязаны своим возникновением гибридизации между Ое. hispanica и Ое. pleschanka (Панов и Иваницкий 1975, Haficr 1977, Панов 1986, 1989). Доля чер-ногорлой морфы stapazina возрастает в ареале испанской каменки в направление с запада на восток от 45 — 52% в северо-западной Африке до 80% в Закавказье. Вариации в окраске головы самок указывают на возможность существования генетического полиморфизма и у них, но трудность здесь состоит в том, чтобы разграничить генетическую и возрастную изменчивость (Панов 1989; рис. 114).
12.1. Систематические замечания
В пределах вида выделяют два подвида, которые весьма значительно различаются по окраске и по образу жизни. Светлые участки оперс-ни у самцов западного подвида Ое. h. hispanica окрашены в буровато-палевый цвет, а у восточного подвида они преимущественно белые (палево рыжий налет присутствует у части особей, наиболее выражен он на оперении груди). Чернота на горле занимает меньшую площадь у
240
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica западного подвида и вес большую — по мерс продвижения в ареале на восток (рис. 113). Длина крыла самцов hispanica у западного края ареала подвида (Пиренейский полуостов, южная Франция) составляет в среднем 89.0 мм (п = 21), в восточной части его ареала в Югославии 92.0 мм (п = 21). В ареале nielanoleuca средняя длина крыла самцов также возрастает в направлении с запада на восток, составляя 89.9 мм (п = 9) в Греции и северной Турции, 90.8 мм (п = 19) в юго-западном Иране, 92.2 мм (п = 9) в северном Иране, вблизи гибридной зоны hispanica х pleschanka (Haffer 1977, Cramp 1988, Рубцов 1987). Судя по всему, прослеживается тенденция к увеличению размеров с запада на восток, что в сочетании с параллельной редукцией палевой окраски повторяет изменения, наблюдавшиеся у большинства рассмотренных ранее видов при их расселении в новые участки ареала. Иными словами, это еще один аргумент в пользу гипотезы о расширении в прошлом ареала испанской каменки из западного Средиземноморья (северо-западная Африка и Пиренейский полуостров) в расположенные далее к востоку районы южной Европы и передней Азии.
12.2. Местобитания и численность
Испанская каменка — это типичный обитатель так называемого средиземноморского ланшафта, за что она, собственно, и получила свое немецкое название Mittelmecrstcischmatzcr. Для этого ланшафта характерны резко пересеченный рельеф и сравнительно бедная древесно-кустарниковая растительность. Каменистые склоны гор в среднем высотном поясе (800 — I500 м над у.м.) одеты тонким, местами исчезающим слоем почвы, на котором произрастает угнетенная травянистая растительность (злаки, полыни, зонтичные). В местах с несколько большим количеством подпочвенной влаги растительность более богата и включает в себя разреженные заросли кустарников (мелкоплодная вишня Cerasus microcarpa. миндали Amigdalis spp., жимолость Lonicera iberica, шиповник Rosa spp., держи-дерсво Paliurus spinachristi, гранат Punica grana turn, барбарис Berberis spp. и др.). На каменистых склонах в более высоком поясе гор (1000-2400 м) в таких местах наряду с кустарниками появляются участки низкорослого разреженного леса. В состав древесных пород здесь могут входит в разных сочетаниях дуб Qwe/rw^spp., можжевельники Juniperns spp.и фисташка (в Закавказье, например, Pistacea mutica).
Во всех перечисленных биотопах испанская каменка выбирает ме-ста с максимально расчлененным рельефом — подвергшиеся выветриванию, разрушающиеся выходы скальной породы, стены извилистых каменистых ущелий, крутые террасы в долинах небольших рек и
241
Панов Е.Н. Каменки Палеарктики
ручьев, которые, как правило, полностью или почти полностью пересыхают летом. Хотя этих каменок бесспорно привлекают места с более богатой травянистой и древесно-кустарниковой растительностью, сильно заросших участков они избегают, предпочитая селиться по соседству с ними — на открытых каменистых склонах. Всюду в пределах своего ареала — от гор Атласа в северо-западной Африке до Закавказья — испанская каменка предпочитает в качестве гнездовых местообитаний среднегорья, обычно нс поднимаясь в горы выше 2300, максимум — 2500 м над у.м. (Лоскот 1983, Lovaty 1985, Cramp 1988).
Давние связи испанской каменки с местообитаниями, которым присуща кустарниковая растительность, наиболее очевидны на Пиренейском полуострове, где подавляющее большинство гнезд располагается на земле под густыми кустами (Suarez Cardona 1976). На востоке ареала эти связи, в общем, оказались утраченными, и кустарниковая растительность нс является здесь необходимым для этих каменок элементом ландшафта. Например, в южном Закавказье они во многих районах гнездятся по сухим бесплодным склонам (преимущественно у подножий) невысоких гор, каменистых и глинистых холмов и вдоль неглубоких ущелий (рис. 63). Хотя птицы и не избегают участков низкогорной холмистой полупустыни, они все же предпочитают селиться близ речных долин, арыков и полупсресыхающих ручьев, текущих по дну ущелий. Здесь они очень многочисленны, и территория каждой пары включает в себя не только каменистый либо глинистый склон, в нишах которого каменки устраивают свои гнезда, но и одно-два дерева и участок зеленого луга, изобилующий насекомыми. Плотность на-
Рис. 115. Распределение территорий самцов старше года (а — stapazina, b — aurita) и самцов-первогодков (с — stapazina, d — aurita) в трех типах местообитаний: А — искусственно орошаемый участок с древесно-кустарниковой растительностью, В — сухая долина, С — безводные склоны холмов. Закавказье. Цифры — даты формирования пар.
242
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
селения чернопегой каменки в таких увлажненных угодьях по крайней мере в два-три раза выше, чем на соседних аридных участках (рис. 115).
В благоприятных местообитаниях формируются компактные поселения, где на 30 га может разместиться до 18 пар, а расстояния между гнездами порой сокращаются до 80 — 100 м. (Лоскот 1983). Вероятно, большинство территорий здесь из года в год занимают прошлогодние их владельцы (как мы видели это у Ое. picata). Те же самцы, которым не удастся внедриться между участками их законных хозяев, вынуждены осваивать бесплодные, выжженыс солнцем щебнистые склоны и осыпи — типичные местообитания Ое. chrysopygia и Ое. finschii. И действительно, в таких безрадостных угодьях чаще можно увидеть тусклых самцов-первогодков, своей окраской напоминающих самок. Многим из них удастся привлечь на свой участок самок, но некоторые так и остаются в одиночестве, формируя резервную когорту холостяков.
Иногда испанские каменки гнездятся по окраинам населенных пунктов и даже довольно больших городов. Я наблюдал двух самцов-пср-вогодков и одного старше года, поделивших между собой каменные сооружения средневековой крепости в городе Дербент (Дагестан), которую постоянно посещают множество туристов. На зимовках в восточной Африке (Эритрея) Ое. hispanica занимает места с довольно густыми насаждениями акации, которые, вероятно, непривлекательны для других видов местных и зимующих каменок (Smith 1971).
12.3. Годовой цикл и сроки гнездования
Передвижение зимующих испанских каменок к северу начинается с конца февраля, но основной пролет проходит в марте — апреле. В Израиле пролет самцов, среди которых изредка попадаются и самки, приурочен к последней неделе февраля — третьей неделе марта, а с 21-25 марта начинается массовое перемещение самок, которое продолжается до 10 апреля (Shirinai 1996). В северной Ливии в период между 6 и 9 апреля Станфорд (Stanford 1954) наблюдал сотни этих птиц, а запоздавшие попадались здесь до 15 мая.
В Закавказье первые самцы прилетают на места гнездования в последних числах марта и сразу же занимают наиболее благоприятные территории. В 1970 г. первый самец морфы aurita был встречен в Нахичеванской республике 28 марта, а массовый прилет самцов (преимущественно морфы stapazina) начался 8 апреля. В том же регионе в 1975 первый самец aurita появился 31 марта, но обычными самцы стали только с 3 апреля. Прилет самцов в целом заканчивается здесь к концу апреля, хотя некоторые появляются и позже — до первых чисел мая. Что касается самок, то они, в общем, прилетают заметно позднее.
243
Панов Е.Н. Каменки Палеарктики
Первая самка была встречена в Нахичевани в 1970 г. 7 апреля, массовый прилет начался с 13 апреля, а наиболее поздний срок появления самки и формирования пары — II мая. В 1975 г. появление самок и формирование пар я наблюдал 12 апреля (приобрел самку самец aurita, занявший территорию 8 апреля) и 15 апреля (самки заняли территории двух самцов stapazina). В соответствии с тем, что образование пар растянуто более чем на месяц, сроки размножения столь же растянуты, и в то время, когда в одних гнездах мы уже находим птенцов, в других только начинается откладка яиц. Самые ранние даты вылупления птенцов в 1970 г. в Нахичевани — 7.V, I3.V и I3-14.V. Как раз в это время одна из самок еще только приступила к постройке гнезда (7.V), а у другой пары гнездо было готова, но яйцекладка еще нс началась (8.V).
В целом испанская каменка принадлежит к поздно размножающимся видам. И в южных (северо-западная Африка, Израиль) и в северных частях своего ареала Ое. hispanica приступает к гнездованию не ранее начала — середины апреля. Чем выше в горах расположены места гнездование, тем позже самки начинают откладку яиц. Например, в центральной Испании на высоте около 1000-1300 м над у.м. массовое появление кладок приходится на вторую декаду мая (16 — 20.V), но многие пары начинают гнездиться и позже — вплоть до последней декады июня (Suarez Cardona 1976). Понятно, что при таком положении дел большинство пар размножается лишь один раз в течение лета (Лоскот 1983, Shirinai 1996).
12.4. Социальное поведение
Пространственная структура поселений и территориальное поведение с а м цов.С momci 1та прилета на места гнездования и на протяжении всего сезона размножения каждый самец неустанно охраняет границы своей территории от прочих самцов своего вида. Величина участка может быть неодинакова в разных ландшафтах и зависит главным образом от плотности локального поселения. Вблизи увлажненных участков долин территории соседних самцов как бы “сплюснуты” с боков и тесно прижаты друг к другу своими продольными границами, идущими поперек склона либо ущелья. Протяженность территории вдоль склона составляет нс более 70-I00 м, и здесь границы строго охраняются. На ровные травянистые угодья территории простираются от подножия склона на 200-300 м. Границы между этими периферическими “кормовыми” зонами весьма приблизительны и нс защищаются столь активно. В аридных местообитаниях территории имеют скорее округлые очертания, достигают в
244
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
рис. 116. Элементы сигнального поведения Ое. hispanica nielanoleuca при конфронтации самцов (а, Ь), при патрулировании границ территории (с), при фор-мировнии пар (d-f, g) и в период гпездостроения (h — k). I — поведение самки h период подготовки к копуляции. Из: Ра now 1974, с дополнениями.
245
Панов Е.Н. Каменки Палеарктики
Привлекательный характер некоторых угодий приводит к тому, что плотность населения здесь увеличивается по мерс прилета новых самцов, так что прежние хозяева вынуждены уступать вновь прибывшим часть своих первоначальных владений. Как правило, новые поселенцы втискиваются между территориями пионеров и постепенно расширяют свое жизненное пространство в обе стороны, как бы “расталкивая локтями*' соседей. Лишь в одном известном мне случае новый пришелец сразу же проявил максимум активности и уже в день своего появления в деме вытеснил прежнего хозяина из центра его территории на се периферию. Предшествовавший этому конфликт продолжался почти 6 часов. Большую часть этого времени оба претендента почти непрерывно совершали над оспариваемым участком длительные совместные демонстративные полеты. Когда один из соперников пытался приземлиться, второй стимулировал его к дальнейшим демонстрациям, хотя ни тот ни другой из них не переходили к явно враждебным действиям. Все же турнир закончился дракой, после чего первый хозяин уступил поле боя пришельцу. Потерпевший принадлежа к морфеaurita. Он еше нс приобрел самку, хотя и состоял владельцем территории уже более недели. Пришелец обладал фенотипом stapazina.
После того как территории распределены между самцами и пределы их окончательно установлены, мы наблюдаем лишь кратковременные конфликты между соседями на границах их участков. При встрече самцы обычно распушают оперение брюшка, всплескивают крыльями, клюют землю, вырывают и отбрасывают в сторону травинки, поочередно залезают в ниши обрывов, после чего каждый улетает на свой участок (рис. ! !6 а-с).
Хотя граница между территориями часто проходит по совершенно однородной местности, как-будто бы лишенной сколько-нибудь заметных ориентиров, самец словно видит некую незримую линию, очерчивающую его владения, и почти никогда не пересекает се, даже если хозяина соседнего участка нс видно поблизости. Однако в тот период, когда самка начинает насиживать, некоторые самцы словно получают удовлетворение, нарушая чужие границы и вступая в конфликты с соседями. Так, один из самцов, находившийся под постоянным наблюдением, то и дело залетал на территорию соседнего самца, так что его вторжения стали для того почти привычными. Фактически, настойчивый агрессор превратил значительный фрагмент территории соседа в почти нейтральную зону, где мог находиться подолгу, не встречая активного отпора со стороны хозяина.
Саморекламирование самцов. Каждый самец имеет в пределах своего участка от одного до трех излюбленных песенных постов, расположенных на самых высоких точках местности. Здесь холос
246
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
той самец проводит большую часть времени, распевая громкую песню, которая обычно включает в себя имитации голосов следующих видов птиц: черного стрижа Apus apus, ласточек Hirundo rustica, Delichon urbica и Ptyonoprogne rupestris, щегла Carduelis carduelis, белой трясогузки Motacilla alba, перевозчика Actitis hypoleucus, золотистой щурки Merops apiaster. Наиболее часто каменки имитируют первые четыре вида, причем характерный “визг” стрижа становится практически видоспецифическим признаком песен испанской каменки (рис. 117 с).
Самец то и дело взмывает в воздух и, не переставая петь, совершает длительный облет территории, после чего вновь садится на излюбленное возвышение. Демонстративный облет территории вызывает точно такое же поведение у соседних самцов, и нередко можно видеть, как три-четыре птицы, широко развернув бело-черные хвосты, стремительно носятся в синем небе над своими владениями. Самец нередко взлетает и проделывает эффектый рекламный полет в ответ на появление над его участком стрижа, ласточки, жаворонка, синего каменного дрозда или даже голубя. Особенно часто наблюдаются подобные полеты у холостых самцов. За 26 часов наблюдений у двух холостяков я насчитал 162 демонстративных полета. Стоит отметить, что рекламный полет испанской каменки отличается от повседневного в основном лишь большей стремительностью, обилием виражей и богатым
Рис. 117. Основные типы песен Ое. hispanica melanoleuca. а — тип “стук пишущей машинки”, b — короткая извещающая песня, с — имитация стрижа Apus apus, 11 — имитация горной ласточки Ptyonoprogne rupestris. Из: Панов 1989.
247
Панов Е.Н. Каменки Палеарктики
звуковым сопровождением, то есть он нс столь "ритуализован” как, например, у Ое. isabellina, Ое. picata и Ое. deserti. Так или иначе, можно сказать, что Ое. hispanica — это каменка, для которой воздушное пространство является любимой стихией. И это пространство охраняется от посягательств непрошенных пришельцев столь же ревностно, как и земельный надел внизу. Иными словами, в отличие от всех ранее рассмотренных каменок, в данном случае охраняемая территория не двумерна, а трехмерна. Самцы начинают свои рекламные полеты еще до восхода солнца. Особенно активно они поют практически в полной темноте, которая ничуть не препятствует им перелетать с одного песенного поста на другой.
После формирования брачной пары агрессивность самца заметно снижается (например, он игнорирует теперь другого самца в клетке, поставленной прямо в центре его территории). Много реже появляется он на своих песенных постах и лишь изредка совершает демонстративные облеты участка.
Самцы изредка нападают и на птиц других видов, например, на горихвосток Phoenicurus phoenicurus. При столкновениях с каменками других видов (Ое. chrysopygia и Oe.funschii) самцы чернопегой каменки обычно уступают им без возражений. Даже в тот момент, когда злато-гузая каменка свирепо гоняет самку черношейной, супруг последней нс приходит ей на помощь.
Территориальное поведение самок. Агрессивность самок невысока, и они, как правило, не содействуют самцу, пытающемуся изгнать посторонних самцов-пришельцев. Однако, в первые дни после формирования пары самка иногда нападает на посторонних самок своего вида, нарушающих границы узаконенных владений. При этом самки поют и гоняются друг за другом, но их поведение в таких ситуациях не очень экспрессивно. Впрочем, однажды я оказался свидетелем весьма впечатляющего конфликта двух самок, который продолжался весь день и возобновился на следующее утро. Самки, одна из которых уже несколько дней состояла в браке, гонялись друг за другом, вели себя крайне агрессивно и время от времени вступали в ожесточенные драки в воздухе и на земле. Я думаю, что пришлая самка была хозяйкой данной територии в прошлом году.
Формирование пар. После того, как возвратившийся с мест зимовок самец занял индивидуальный участок, проходит иногда почти целый месяц, прежде чем ему удастся приобрести супругу. Бывает так, что самец, появившийся на месте гнездового поселения одним из первых, долго остается холостым, а только что появившийся кавалер сразу же вступает в брачный союз (рис. 12). Если самцу долго не удается обзавестись подругой, он пребывает в постоянном возбуждении и и ног-
248
r.iaea 12. Испанская каменка, чернопегая каменка Oenanthe hispanica да даже пытается токовать перед птицами других видов, подчас совсем не похожих на каменок (например, перед самкой толстоклювого пустынного снегиря Bucanetesgithagineus).
Когда на участке холостого самца впервые появляется самка, он встречает ее бурным взрывом пения и демонстраций (рис. 116 d-g), чем сразу же привлекает внимание соседей-холостяков. Последние настолько возбуждаются, что забывают о существовании регламентированных границ и нередко вторгаются на участок “молодожена”, пытаясь сблизиться с самкой. Она при этом быстро прячется в щель скалы, а самец тем временем с громкой песней гоняется за пришельцами, вытесняя их со своей территории.
Строго говоря, не самец находит самку, а та, пролетая через владения многих самцов, останавливает свой выбор на одном из них. Очевидно. самка выбирает даже не столько самца, сколько “хорошую” территорию. Качество того или иного участка она оценивает, вероятно. по количеству пустот и ниш, пригодных для устройства гнезда. Поэтому церемония помолвки будущих супругов начинается с того, что самец “ведет” самку по своим владениям и поочередно демонстрирует ей все тс убежища, которые он заранее приметил. Такой осмотр территории не всегда удовлетворяет самку, и в таком случае она покидает самца и отправляется на дальнейшие поиски. На территории одного из самцов в Нахичевани в период с 17 апреля по 3 мая побывало 7 самок, и лишь последняя из них оказалась удовлетворенной инспекцией и осталась здесь гнездиться.
Поскольку перелеты между местами зимовок и районами гнездования происходят по ночам, самки-новоселы нередко появляются в поселении рано утром, еще до восхода солнца. Бесспорно, в подобных ситуациях они находят местность, уже давно поделенную между холостыми самцами, по их хоровому предрассветному пению. Когда же самка по окончании очередного ночного перелета первым делом принимается за поиски корма, и лишь затем отправляснтся на поиски будущего супруга, она может впервые оказаться в поселении самцов и в светлое время суток. Если самка летит издалека, она подчас появляется над поселением на высоте 20-30 м. Дежурящие па своих песенных постах холостые самцы мгновенно замечают се силуэт на Фоне неба и одновременно взмываю в воздух. Наиболее дерзкий из Пих может даже броситься в погоню, и если самка нс имеет ничего против подобной смелости, самцу, вероятно, при счастливом стечении обстоятельств удается “посадить” самку на своей территории. Подобное “сватовство” в воздухе возможно лишь у таких прекрасных летунов, как испанская каменка, и нс описано пока у других представителей рода Oenanthe.
249
Панов Е.Н. Каменки Палеарктики
Когда самка, готовая к формированию пары, путешествует по территориям своих потенциальных супругов, она старается вести себя как можно заметнее и время от времени произносит короткую стереотипную песню. Холостой самец, на территории которого она оказалась, стремительно летит к ней, принимает особую позу (рис. 116 d) и сразу же проделывает вокруг самки серию полетов петлями. Эта демонстрация сопровождается звучной песней, неотъемлемой частью которой служит весьма характерный “стук пишущей машинки” (рис. 117 а). Самка нс дает самцу приблизиться вплотную, она ведет себя агрессивно и даже старается ударить кавалера клювом. Часто она встречает кавалера угрожающей трещанием (сигнал N 4), на которое он отвечает своим обычным “зак-зак” (сигнал N 1), а в моменты наивысшего возбуждения — серией коротких “тик... тик... тик...” (сигнал N 3) (рис. 118). После такого первого знакомства самец взлетает и с песней носится над своим участком, а самка садится на кустик и тоже поет короткую песню. Затем самец устремляется к одной из облюбованых им ниш и, поочередно залезая в нее и выбираясь наружу (рис. 116 e-g), поджидает самку здесь. Когда самка подлетает к самцу, находящемуся возле очередной ниши, он слегка пригибает голову (рис. 117 g, h), издает короткий “стук пишущей машинки” и стрсмительуо перелетает к следующему отверстию, словно бы опасаясь близкого соседства с невестой. Пока самка обследует предыдущее убежище, показанное ей самцом, он продолжает петь, поджидая ее на новом месте, где затем все описанное выше повторяется вновь. Если вся церемония, занимающая с повторениями обычно нс менее часа, прошла гладко, самка остается в пределах территории и спустя несколько дней (от трех до десяти) приступает к постройке гнезда.
12.5. Гнездо, кладка, выводок
Гнездостроение и конструкция гнезд. Окончательный выбор гнездовой ниши оказывается делом весьма ответственным, и здесь от самки требуется немало терпения. Несмотря на кажущееся изобили подходящих мест, подобрать по настоящему удобное и хорошо защищенное отверстие оказывается не столь уж простым делом. На протяжении нескольких дней самка осматривает каждый склон буквально сантиметр за сантиметром. Иногда она настолько поглощена этим занятием, что словно забывает о существовании территориальных рубежей и вторгается на соседний участок, чем повергает самца в страшное волнение. В такие минуты все его поведение направлено на то, чтобы не допустить самку на чужую территорию, и он всясски старается повернуть ее вспять.
250
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
Рис. 118. Основные позывки Ое. hispanica melanoleuca. а — при тревоге у гнезда (сигналы N I и N 3), b — сигнал N 4 (трещание), с — сигнал N 2 (храпение), d. е — вокализация птенца в ожидании родителей и при выпрашивании у них корма (сигнал N 6, дребезжание). Из: Панов 1989.
Нередко самец с самого начала склонен остановить свой выбор на каком-либо определенном убежище, куда он настойчиво приглашает самку. Она же, первоначально разделяя, как будто, решение супруга, Затсм обнаруживает какие-то недостатки и пускается в новые длитель-’•ые поиски. На этой почве между членами пары возможны небольшие
251
Панов Е.Н. Каменки Палеарктики
недоразумения. Самец поджидает подругу около избранной им ниши, то и дело ныряет в нее, а когда самка приближается, принимает эффектную позу и быстро трепещет широко расправленными крыльями (рис. 116 i). Самка, со своей стороны, игнорирует все эти знаки внимания и улсгаст на другой склон ущелья, где вновь принимается за свои поиски.
Места, в которых располагаются гнезда восточного подвида Ое. hispanica melanoleuca, в целом довольно однотипны. Чаще всего это ниша под плоским камнем, лежащем на склоне, вход в которую представляет собой узкую горизонтальную щель. Некоторые гнезда располагаются в горизонтальных расселинах скал. Гнездо выстроено сравнительно близко от входа (в 10-25 см), но оно не видно снаружи, поскольку вход в убежище находится обычно ниже уровня гнезда.
Некоторые гнезда сплетены плотно и искусно, другие — мягкие и рыхлые, а иногда наружный слой вообще отсутствует и есть лишь тонкая выстилка, лежащая в неглубокой ямке. В отличие от гнезд плешан-ки, внутренний слой гнезда обычно содержит мох, тонкие корешки и волос. У входа можно обнаружить некоторое количество толстых палочек, но я ни разу нс находил гнезда, сделанного на настиле, как это бывает у плсшанок. Лоскот (1983) находил в гнездовых нишах от 5 до 182 палочек длиной от 1.5 см до 15.5 см, диаметром 1-13 мм и весом до 3.3 г. Общий вес палочек в гнездах с максимальным их количеством доходил до 50-97 г. В выстилке большинства гнезд, найденные Лоско-том, помимо тонких корешков и растительных волокон, присутствовал волос домашних животных, но лишь в одном гнезде он обнаружил зеленый мох. По данным этого автора, диаметр гнезд — от 90 до 185 мм (ср. 130 мм), высота — от 36 до 78 мм (ср. 56 мм), диаметр лотка 50-68 мм (ср. 61 мм), глубина 30-50 мм (ср. 40 мм).
Совершенно иначе располагаются гнезда подвида Ое. h. hispanica в среднегорьях Пиренейского полуострова. Здесь только 3 гнезда из 32 найденных были спрятаны в нишах скал, остальные покоились в неглубоких земляных ямках у оснований густых кустов (Genista Scorpios, Lavandula sp., Thymus sp., Juniperus communis), под их нависающими кронами. Снаружи к гнезду ведет “туннель” длиной около 10 см, выложенный крупными палочками, а более мелкие прутики располагаются в основании гнезда, в основном вокруг гнездовой постройки (рис. 119). В гнездах было от 8 до 64 палочек длиной 3 — 18 см и диаметром до 12 мм. Внутренний диаметр гнезда — в среднем 7 см, глубина 4 см, толщина стенок — 2 см. Во многих гнездах для выстилки был использован зеленый мох (Suarez Cardona 1976).
Строит гнездо только самка, затрачивая на эту работу около недели. Когда опа летает за строительным материалом, самец обычно сопро-
252
Глава 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
вождаетес. В первые дни после формирования пары он иногда подбирает с земли толстые короткие палочки, но нс относит их к гнезду, а сразу же бросает.
Развитие отношений между членами пары в преддверии копу-л я ци и. Когда гнездо готово, самка начинает демонстрировать перед самцом особую позу (рис. 116/). С приближением стадии копуляции все чаше приходится видеть своеобразные взаимодействия между супругами, происходящие иногда в глубоких вечерних сумерках. Сначала самец, находясь неподалеку от самки, принимает весьма своеобразную позу: он медленно пригибается грудью к земле, широко расправляет крылья и начинает быстро трепетать ими с очень ма
Рнс. 119. Месторасположение (А, В) и устройство гнезд (С) западного подвида испанской каменки Ое. h. hispanica. Из: Suarez Cardona 1977.
л oil амплитудой. Часто он перед началом такой демонстрации взлетает на камешек либо на крошечный кустик, что позволяет ему принять почти вертикальное положение “головой вниз” (рис. 116 i, j). Продолжая трепетать крыльями, самец медленно поворачивается вокруг вертикальной оси, иногда описывая почти замкнутый круг (немногим менее чем 360°). При этом угол наклона туловища то увеличивается, то вновь уменьшается, — до 8 раз за одну демонстрацию длительностью около 45 сек. Затем самец, все также мелко трепеща расправленными крыльями, взлетает и, не изменяя амплитуды взмахов, очень медленно и плавно спускается к самке. Та подпускает его почти вплотную, потом чуть-чуть отбегает, и это повторяется много раз подряд. Затсм
253
Панов Е.Н. Каменки Палеарктики
следует своеобразный совместный полет тандемом: самка летит впереди, очень медленно, с малой амплитудой движения трепещущих крыльев, иногда останавливаясь в воздухе. Самец следует за ней, непрерывно напевая тихую монотонную песню (типа подпесни). Постепенно полет партнеров становится все более ускоренным и наконец переходит в стремительную погоню. Когда самка приземляется, самец в экстазе проделывает вокруг нес эффектный полет петлями, сопровождаемый громкой песней со “стуком пишущей машинки”.
К л а д к а. Средняя величина кладки различна в разных регионах. На юге ареала (например, в Израиле) в кладке бывает от 3 до 5 яиц, чаще всего 4. На Пиренейском полуострове соотношение кладок с разным количеством яиц было таково: 2 х 2, 4 х 3, 5 х 12, 6 х 1 (ср. 4.56, п = 18). В Закавказье среди 9 найденных мной гнезд в 5 было по 5 яиц и в 4 — по 6 (ср. 5.44). По данным Лоскота (1983), соотношение кладок разной величины следующее: 4x2, 5 х 13, 6x9 (ср. 5.29, п = 24; включены данные из гибридных зон). Окраска скорлупы ярко-голубая, часто — с заметным зеленоватым оттенком. В пределах одной кладки яйца могут иметь разную интенсивность окраски фона (до водянисто-голубой) и неодинаковую испещренность (рис. 120). Как правило, красноватые пестрины довольно многочисленны и густо сосредоточены вокруг тупого конца. У подвида Ое. й. hispanica пестрины не столь многочисленны и отчетливы, как у восточной формы melanoleuca.
Размеры яиц подвида hispanica (без указания мест коллсктирова-ния): 17.0 - 21.6 х 14.0 - 16.0 (ср. 19.9 х 15.2, вес 2.46 г, п = 156); подвида melanoleuca: 17.0 — 24.0 х 14.0 — 16.0 (ср. 19.7 х 15.0, вес 2.40 г, п = 150). Размеры яиц hispanicaс Пиренейского полуострова: 18.8 — 21.9 х 14.3 - 15.8 (ср. 20.1 х 15.3, п = 59). Яйца 4 кладок melanoleuca из Закавказья: 18.1 — 22.2 х 14.3 - 15.7 (ср. 19.6 х 15.4, п = 21) (Suarez Cardona 1976, Schdnwcttcr 1979 цит. по: Cramp 1988, данные автора).
Насиживает только самка. От момента от-
„ г . п кладки последнего яйца
Рис. 120. Гнездо и кладка испанском каменки Ое.
hispanica melanoleuca (Закавказье). Из: Ра now 1974. до вылупления птенцов
254
Гшва 12. Испанская каменка, чернопегая каменка Oenanthe hispanica
проходит 12-13 дней. Самец нс кормит насиживающую самку. Лишь однажды я наблюдал, как самец передал что-то самке, когда она вылетела на очередную кормежку. В тот период, когда самка насиживает, самец нередко пытается ухаживать за другими самками.
П т е н ц ы. У недавно вылупившегося птенца кожа розовая, с мясным оттенком. Эмбриональный пух длинный (до 15 мм), темносерый. На бедренных птерилиях он выглядит более светлым. Зев интенсивного желтого цвета, валики в углах рта почти белые. Слетки в гнездовом наряде выглядят коричневато-пестрыми. По наблюдениям Лос-кота (1993), летать, набирая высоту, молодые могут уже через 12 дней после вылета из гнезда, а еще через неделю они способны поймать насекомое в воздухе. Петь молодые самцы начинают на 15 — 17 день вольной жизни, самки — на 25 — 28 день. Пение на первых порах очень немелодичное и состоит главным образом из разных комбинаций сигналов NN 1 и 3, звучание которых птица постепенно начинает всячески видоизменять.
Птенцы, взятые мной из гнезда 13 мая, впервые увидев воду 29 мая, сразу стали купаться. Птица становится в купалку и пьет, делая жующие движения челюстями (как это описано в очерке о черной каменке Ое. picata). Потом садится в воду и проделывает боковые движения крыльями, отставленными в сгибах от туловища. В более старшем возрасте птица при виде воды подчас ведет себя очень своеобразно: она окунает в воду клюв и лоб, проделывая одновременно боковые движения головой, а затем купается в песке около плошки с водой либо в другом конце клетки. Затем каменка “сушит” сухие крылья и совершает движения “ложного кормления”. К середине июня зев юных каменок изменил свой первоначальный желтый цвет, став грязно-розовым. С середины июля молодые приступили к линьке. В это время у них сменилось мелкое оперение на голове и на плечевых партиях, а позже — также большие кроющие крыла. Линька продолжалась до конца августа, когда у самца появилась чернота вокруг глаз и на сгибах крыла, а на спине отросли перья с белыми основаниями. Во второй половине августа у самца сменились также центральные рулевые.
Движение испанских каменок к югу, в направлении мест зимовок, начинается очень рано. Хотя массовый осенний пролет подвида melanoleuca в Передней Азии проходит в основном с середины сентября до середины октября, первые мигранты местами (например, в Израиле) появляются уже с середины июля и становятся вполне обычными с середины августа. В Армении последних птиц видели 7, 20 и 21 сентября (две из них были самцами, пол третьей нс установлен). На своих основных зимовках вдоль южной границы Сахары (Судан, Чад,
255
Панов Е.Н. Каменки Палеарктики
Мали) испанские каменки появляются в сентябре — октябре (Ляй-стер и Соснин 1942, Allouse 1953, Cramp 1988, Shirinai 1996).
12.6. Биологические заметки
Испанская каменка пользуется теми же приемами охоты, что и прочие виды рода, а рацион ее столь же разнообразен, включая в себя кормовые объекты длиной от 3 до 12 мм. Вероятно, чаще других каменок эти птицы приносят птенцам ягоды (в одном из гнезд обнаружено 18 косточек мелкоплодной вишни Cerasus microcarpa). С конца мая значительную часть питания испанских каменок в Закавказье составляют ягоды шелковицы Morns spp. (Лоскот 1983). Интересно было бы выяснить, в какой степени высокая приспособленность Ое. hispanica к стремительному маневренному полету отражается на специфике ее кормового поведения. В отношении ближайшего родича испанской каменки — плсшанки Ое. pleschanka показано, что именно эти качества прекрасного летуна позволяют ей избегать конкуренции с другими видами каменок, не столь часто использующими полет при добывании пропитания (Lcisler ct al. 1983). Поскольку существует мнение (Cornwallis 1975, Лоскот 1983), что кормовые стратегии и спектры питания пле-шанки и испанской каменки чрезвычайно сходны, легко предположить, что аналогичные выводы в дальнейшем будут сделаны и в отношении последнего вида.
256
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
Глава 13.
Плешанка, чекан-плешанка
Oenanthe pleschanka Lepechin, 1770
Синонимы: Motacilla pleschanka Lepechin, 1770; Motacilla leucomela Pallas, 1771; Saxicola vittata Hemprich ct Ehrenberg, 1833; Saxicola niorio Hemprich et Ehrenberg, 1833; Saxicola melanogenys Severtzow, 1872; Saxicola melanotis Severtzow, 1872; Saxicola cypriaca Homcycr, ! 884; Saxicola somalica Sharpe, 1895.
Все то, что было ранее сказано по поводу особенностей строения, повседневного поведения и полевых признаков испанской каменки, в такой же степени относится и к плешанкс. При первом знакомстве с этими двумя видами в глаза бросаются лишь резкие различия в окраске самцов, тогда как распознавание самок плсшанки и испанской каменки в поле затруднено и требует от орнитолога определенного опыта (рис. 121, 122, Н4).
Дело в том, что плешанка и испанская каменка — очень близкие виды, которых одно время многие крупные орнитологи вообще рассматривали в качестве подвидов единого вида (Kleinschmidt 1938, Гладков 1954, Портенко 1954). Затем эта точка зрения была отвергнута на том, якобы, основании, что испанская каменка и плешанка сосушс-
Рис. I21. Передовые самцы плсшанки Ое. pleschanka прилетают на места гнездования, когда там еше случаются сильные снегопады
ствуют в некоторых регионах Турции, западного Ирана и Прикас-пия, где если и гибридизируют друг с другом, то лишь в сдинич-ных случаях (Vauric I949, Ripley 1964, Strescmann, Portcnko. VietinghofT-Scheel 1947, Лоскот 1976, Степанян 1983). Между тем, как оказалось в дальнейшем, гибридизация в зонах перекрывания ареалов плсшанки и испанской каменки чрезвычайно интенсивна и
257
Панов Е.Н. Каменки Палеарктики
Рис. 122. Плсшанка Ое. pleschanka, самка.
соответствует ситуациям так называемого “слияния видов” (по терминологии Майра 1965). При таком положении вещей, если строго следовать пониманию вида как замкнутой генетической системы, следовало бы вернуться к воззрениям Кляпншмидта, Гладкова и Портснко. Однако никто нс делает этого — видимо потому, что биологическая интуиция исследователей оказывается сильнее приверженности формально-логическим схемам.
Интуиция эта подсказывает нам, что плешанка и испанская каменка долгое время эволюционировали независимо друг от друга — как истинные виды, и лишь вторично оказались связанными воеди
но через сравнительно небольшие гибридные популяции. За время не
зависимого существования эти две разные генетические системы накопили множество различий, которые, как выяснилось совсем недавно, касаются нс только окраски самцов и самок, но также отдельных черт строения и, что особенно замечательно — множества особенностей социального и сигнального поведения (Панов 1978, 1989). Отдавая дань формальной логике, некоторые исследователи предлагают считать испанскую каменку и плешанку не “полными” (“хорошими”) видами, а всего лишь полувидами (semispecies), обозначая их, тем не менее, полными видовыми названиями (НаПег 1977) и помещая в таксономических и фаунистических списках рядом друг с другом, что, к сожалению, не всегда делается и сегодня (см. Shirinai 1996).
Между тем, помимо окрасочных признаков (черная, а нс палевая либо белая спина у самцов плсшанки; большее развитие черного на горле самцов плсшанки и на лбу — у самцов испанской каменки) эти виды достоверно отличаются также по длине первого махового пера (оно в среднем длинсс у Ое. hispanica) и цевки (длинсс у Ое. pleschanka)-Кроме того, плсшанки в среднем несколько крупнее испанских каменок (Haffer 1977, Рубцов 1997). Для самок плсшанки, помимо больше-
258
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
Го развития темных тонов на горле и груди, характерны тонкие продольные пестрины на темени и на груди, что в большинстве случаев позволяет установить видовую принадлежность не только “темногор-лых”, по и “белогорлых” самок (Лоскот 1983; см. рис. 114). Различаются испанская каменка и плешанка также особенностями пения (см. ниже) и, вероятно, частотными характеристиками некоторых других вокальных сигналов: у испанской каменки спектр заполнения шумовых сигналов (таких, например, как сигнал N 1) заходит в более низкочастотные области (Панов 1989).
13.1. Систематические замечания
Как и у других ранее рассмотренных видов, у Ое. pleschanka общие размеры и, особенно, длина крыла увеличиваются в направлении путей расселения вида — с запада на восток. Наименьшими размерами характеризуется популяция о. Кипр (ср. длина крыла самцов 84.4 мм). В материковой Евразии западные популяции (до Тянь-Шаня и Алтая на востоке) характеризуются меньшей длиной крыла (87 — 94 мм, ср. 91-92 мм), чем восточные, населяющие юг Восточной Сибири, Монголию и Китай (93 — 102 мм, ср. 96 мм). Предлагалось эти восточные популяции выделить в подвид Ое. pleschanka hendersoni, но по ряду причин большинство систематиков нс признают реальность этого подвида и относят всех материковых плсшанок к форме Ое. р. pleschanka. Что касается кипрской плешанки cypriaca, то се многие склонны рассматривать как самостоятельный вид, поскольку опа отличается от материковых плсшанок не только меньшими размерами и более закругленным крылом, но также слабым развитием полового дихроматизма и особенностями песни. Однако нс все согласны с тем, что этих различий достаточно для выделения кипрской плсшанки в самостоятельный вид (Cramp 1988, Туе 1989а). Я также склоняюсь к этому выводу и буду далее рассматривать кипрскую плсшанку подвидом под именем Ое. р. cypriaca.
Обычно плешанку называют в числе тех видов каменок, которым свойственен полиморфизм окраски. Однако у плсшанки дискретная изменчивость окраски самцов имеет совершенно иной характер, нежели у испанской каменки. У последнего вида два варианта окраски Самцов присутствуют во всех без исключения популяциях, а доли чер-Ногорлого и бслогорлого фенотипов вполне сопоставимы (грубо говоря» От 1:1 до 1:3 в разных регионах). Что касается Ое. pleschanka, то у се бслогорлый фенотип (так называемая vittata) во многих участках
ала вообще никогда и никем нс был встречен (юго-восточная Ев-а’ Кипр, Киргизия, большая часть Казахстана и Монголии, Ал
259
Панов Е.Н. Каменки Палеарктики
тай, Тува). Если взять ареал плешанки в целом, то доля имеющихся в коллекциях самцов vittata составит всего лишь 2.8% (25 vittata на 851 черногорлых самцов — см. Лоскот 1976). Однако находки самцов vittata в ареале плсшанки распределены крайне неравномерно. Белогорлые самцы составляют в некоторые годы до 11% в популяции полуострова Мангышлак, сравнительно часто встречаются на юге Средней Азии (в Туркмении и Узбекистане — до 4-5%) и крайне редки далее к востоку (коллекции содержат всего лишь около 10 экземпляров с огромных пространств южной Сибири, Монголии и Китая) (рис. 123).
Все известные факты заставляют думать, что полиморфизм у Ое. pleschanka возник в результате се гибридизации с Ое. hispanica. Сейчас в ареале плсшанки есть только одна истинно полиморфная популяция, локализованная на полуострове Мангышлак и в соседних регионах северо-западного Казахстана. Как мы увидим из следующей главы (посвященной зонам вторичного контакта Ое. pleschanka и Ое. hispanica). эта популяция в настоящее время полностью изолирована от ареала Ое. hispanica. Иными словами, здесь на протяжении многих тысячелетий шли генетические процессы, со временем преобразовавшие первоначально гибридную общность в популяцию со сбалансированным генетическим полиморфизмом (точно так же, как это происходило в ареале гибридогенной полиморфной популяции Ое. capistrata х
Рис. 123. Ареал плешанки Ое. pleschanka. а — район с высокой концентрацией фенотипа vittata, b — места, где этот фенотип сравнительно обычен, с — единичные находки фенотипа vittata, d — находки плсшанок за пределами ареала вида в гнездовой сезон, е — районы зимовок.
260
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
opistholeuca — см. 8.1). Одной из таких гибридогенных морф стала вариация vittata. Из восточного Прикаспия генетический фактор “бслогор-лости”, берущий начало в геноме испанской каменки, в течение тысячелетий мигрировал по череде поколений в другие, подчас весьма удаленные популяции, где он, естественно, имеет очень низкую концентрацию и проявляет себя исключительно редко (Панов 1989).
13.2. Местобитания и численность
Типичный биотоп плсшанки — это, главным образом, открытые, просшис степной растительностью сухие склоны гор и холмов с отдельными выходами коренной породы и с разбросанными тут и там валунами (рис. 124). В такой обстановке плешанка гнездится отостан-цовых гор пустыни Кызылкум до альпийской зоны западного Пами-ро-Алая и Алтая. Столь же охотно селится плешанка по глинистым и лессовым обрывам берегов Черного, Каспийского и Аральского морей. Плотность населения вида ниже в холмистых сухих степях Казахстана, где эти птицы придерживаются участков пересеченной местности, железнодорожных насыпей и развалин построек человека.
Можно задать вопрос, чем же объясняется бесспорный успех пле-
I24. Местообитания плсшанки Ое. pleschanka в Алтае. Здесь же гнездятся о^Ь1К|1оВениые каменки Ос. oenanthe. Из: Panow 1974.
261
Панов Е.Н. Каменки Палеарктики
шапки в освоении колоссальных пространств Азиатского материка, простирающихся от гор Эльбурса и Копетдага почти до самого побережья Тихого океана. В общем, плешанка во многом использовала те же способы освоения новых стаций, что и Ое. oenanthe. хотя и не достигла при этом столь же ошеломляющего прогресса. Подобно обыкновенной каменке, плешанка приспособилась к гнездованию на равнинах, используя здесь любые фрагменты расчлснененного рельефа (овраги, незначительные по площади выходы горной породы и т.д), а при их отсутствии — всевозможные рукотворные сооружения (временные и заброшенные строения, железнодорожные насыпи и пр.). Во многих местах плешанка стала селиться по окраинам населенных пунктов, устраивая гнезда в отверстиях стен домов и под крышами. Например, в Забайкалье в такой обстановке обитает 3-5% местной популяции. В некоторые годы отдельные пары проникают севернее своего основного ареала, где гнездятся на окраинах больших городов (например, Воронежа, у 52° с.ш. — Воробьев, Лихацкий 1987).
Плешанка начала постепенно заселять и окраины леса. В северном Казахстане и в предгорьях Алтая птицы занимают глинистые и скальные обнажения в разреженных сосновых борах, причем самцы используют в качестве песенных постов вершины высоких сосен. На юге Казахстана, в песчаных пустынях Прибалхашья, они гнездятся по периферии старых лесов из саксаула Arthrophytum spp. В Забайкалье отдельные пары селятся в разреженном сосняке или на полянах в кучах мусора. В Хэнтес (Монголия) этих каменок можно встретить в довольно узких долинах с рощами сосны и лиственницы или с развитой древесно-кустарниковой растительностью вдоль увлажненного днища долины (Шнитников 1949, Гаврилов 1970, Козлова 1975, Доржиев, Хертуев 1992). Наконец, в некоторых регионах Ое. pleschanka стала обычным видом высокогорий. В западном Тянь-Шане эти каменки гнездятся до высот 2800 — 3000 м, а в горах Бадахшана поднимаются даже до 3200 — 3400 м над у. м. В то же время во многих местах, где все условия, казалось бы, благоприятстуют существования плешанки, этот вид, тем не менее, полностью отсутствует (Бадхыз в южной Туркмении, юг Узбекистана и др.).
Чаще всего плешанки селятся отдельными парами на расстоянии 0.5 и более км друг от друга, либо образуют разреженные поселения из 2 — 4 пар, так что каждая пара контролирует участок от 1 до 3 га (в среднем около 2 га). В Туве (крайний юг Восточной Сибири) среднее расстояние между гнездами 126 м, минимальное — 40 м (Иваницкий 1980а, Доржиев, Хертуев 1992). Как это ни удивительно, наибольшая численность и плотность наблюдается в гибридогенной полиморфной популяции Мангышлака, где плсшанки гнездятся по крутым, сильно
262
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
выветренным склонам глубоких известняковых каньонов, по долинам солоноватых ручьев с более сглаженным рельефом и на окраинах населенных пунктов. Здесь в наиболее предпочитаемых местообитаниях на площади порядка 5 га может разместиться до 20 пар (Панов, Иваницкий 1975а). Высокой численностью и плотностью характеризуются также популяции в гибридных зонахpleschanka х hispanica (см. 14.1-14.3).
13.3. Годовой цикл и сроки гнездования
Хотя места гнездования большинства популяций плсшанки отстоят от ее зимних квартир дальше, чем у чернопегой каменки Ое. hispanica, весенний пролет первого из этих двух очень близких видов начинается заметно раньше. Весной первые самцы плешанки появляются в Среднеазиатских республиках в самом начале марта (6 — 12.III — в южном Туркменистане, 2 — 13.III — в южном Казахстане, 4 — I5.III — в предгорьях Памиро-Алая). Однако, это именно одиночки, появление которых непредсказуемо и мало связано с погодными условиями в том или другом году. Например, в южном Узбекистане близ Термеза в раннюю теплую весну 1971 г. я вообще не встречал плсшанок в марте. Один самец и две самки появились одновременно 8 апреля. Здесь же в холодную, дождливую весну I973 г, когда временами еще выпадал снег, в период с 6 по 29 марта встречены 9 самцов. В общем, пролет самцов сильно растянут во времени, и даже в районах африканских зимовок некоторые особи задерживаются до начала апреля. На островах в южной части Каспийского моря пролетные плешанки встречаются с середины апреля до середины мая. В Туркменистане и Узбекистане массовый пролет самцов начинается в последней пятидневке марта, первые самки были встречены 28.111 в предгорьях Нуратау (западнее Ташкента) и 29.111 в Бадхызе (юго-восточный Туркменистан). В этих южных районах, наиболее близких к местам зимовок, холостые самцы бывают обычными иногда еще до начала второй декады апреля (12-13.IV в Копетдаге), хотя отдельные пары могут сформироваться здесь уже к I апреля. Наиболее ранняя дата гнездостроения в этом регионе — 3 апреля. Основная масса плсшанок размножается гораздо позже. Гнездо с незаконченной кладкой найдено 8.V-66, !2.У-82удвух пар были слетки. Самостоятельные молодые встречаются в конце мая. В южном Афганистане, на высотах порядка 2600-2800 м Палюдан (Paindan 1959) нашел гнездо с 6 свежими яйцами 23 мая.
В низкогорьях Забайкалья, примерно в 3700 км восточнее, прилет плсшанок отмечается с Ю-х чисел апреля, массовое появление — в середине этого месяца (15 — 18. IV). Менсе 2% самок приступают к откладке яиц в первой пятидневке мая, 22, 20 и 24% — соответствен -
263
Панов Е.Н. Каменки Палеарктики
но, во второй, третьей и четвертой пятидневках, 3.3% задерживаются с началом гнездовых дел до первой пятидневки июня. Весь цикл от откладки первого яйца до вылета молодых из гнезда продолжается 31-33 дня. Плсшанки размножаются здесь один раз в году (Доржиев, Хер-тусв 1992). В Монголии 5 гнезд с кладками найдены между 6 и 9 июня (в других 4 гнездах в это время были птенцы), в Гоби (Наньшань, 38° с.ш., 100° в.д.) — 27 и 30 июня и 4 июля (нснасижснная кладка) (Strcscmann et al. 1937, Piechocki ct al. 1982, Kneis, Spretke 1985). В высокогорьях Бадахшана (около 2500 м. над у. м.) близ границы Таджикистана с Афганистаном я наблюдал откладку яиц в одном гнезде с 20 по 25 мая, в другом со 2 по 7 июня, в третьем — с 12 по 16 июня.
Весенний пролет расы cypriaca в Израиле проходит, в общем, в те же сроки, что и у номинативного подвида, хотя первые мигрирующие кипрские плешанки появляются здесь почти на месяц раньше — с 17 февраля. На Кипре первые слетки появляются в начале мая (Flint 1995, Shirinai 1996), то есть размножение проходит в тс же сроки, что и у рано гнездящихся пар на той же широте на юге Туркменистана. Кипрская плешанка гнездится дважды в лето.
13.4. Социальное поведение
Территориальное поведение и саморекламирование самцов. Как уже было сказано, многие самцы прилетают на места гнездования задолго до того, как у них появится возможность приступить к выведению потомства. В предгорьях Копетдага весной 1968 г. передовые самцы обосновались на своих территориях с 15 марта, а в позднюю весну 1969 г. — с 6 марта. В оба сезона самки не появлялись вплоть до двадцатых чисел марта (когда наблюдения были прерваны), и все это время самцы пребывали на своих индивидуальных участках в одиночестве, лишь изредка конфликтуя друг с другом на рубежах территорий. В первые дни после прилета они почти не поют и держатся довольно скрытно. В этот период птицы крайне осторожны и проводят большую часть времени в поисках насекомых, которых очень мало на поверхности почвы, так что плешанкам приходится добывать их в основном в нишах и норах скалистых и глинистых обрывов.
Спустя 2-3 дня посе занятия участка самцы начинают петь — сначала изредка, в пол-голоса, затем все чаще и громче. В этот период можно слышать песню двух типов. Одна длительная и разнообразная, включающая в себя характерное стаккато “пишущей машинки”; другая — короткая, стереотипная, включаемая нередко в виде фразы в песню первого типа. Песня материковых плешанок при общем сходстве с песней Ое. hispanica отличается от нес несколько более медлен
264
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
ной манерой исполнения и большей мелодичностью. Эти различия подтверждаются при сравнительном количественном анализе структуры песен плешанки и импанской каменки (рис. 125).
Плешанки склонны имитировать голоса определенных видов птиц,
Рис. 125. Типичные песни материковых плсшанок Ое. р. pleschanka (1 — IV сле-в<ч), формы cypriaca (V и VI слева) и испанской каменки Ое. hispanica melanoleuca (4 песни справа). На диаграмме справа внизу приведены результаты сравнения структуры песен Ое. h. melanoleuca (а — западная Армения, b — восточная Армения) и Ое. pleschanka (с — северное Предкавказье, Ставропольская область, d — северо-западное Причерноморье, Измаил). По: Cramp 1988, Панов 1989, Рубцов 1997.
265
Панов Е.Н. Каменки Палеарктики
kHz 81
Рис. 126. Песни Ое. pleschanka. а — вариант “стук пишущей машинки”, b — имитации малого жаворонка Calandrella cinerea.
которых, как правило, не копирует испанская каменка. В песнях плешанок отсутствуют имитации “визга” стрижа Apus apus, но они часто передразнивают малого жаворонка Calandrella cinerea (рис. 126 b), коноплянок Cannabina spp., а также щегла Carduelis carduelis, желчную овсянку Emberiza bruniceps и желтую трясогузку Motacilla flava. Кроме того, плешанки гораздо реже испанских каменок демонстрируют свое певческое искусство в воздухе. В полиморфной популяции Мангышлака за 49 ч. наблюдений я насчитал у 28 самцов 115 демонстративных полетов (менее чем 0.1 полета на самца в час). По сообщению Иваницкого, в Туве за 32 ч. у 7 самцов он наблюдал 81 полет (0.36 полетов на самца в час). Для сравнения скажу, что только два самца Ое. hispanica за 26 ч. проделали 129 демонстративных полетов (в среднем 2.5 полета на самца в час, то есть в 7-25 раз больше, чем это наблюдалось у плешанок).
Вообще говоря, плешанки по своему темпераменту резко отличаются от испанских каменок, будучи гораздо более флегматичными. Это сказывается как на взаимоотношениях плешанки с другими видами каменок, при которых она выступает в качестве аутсайдера и оппортуниста, так и на всей структуре социальных отношений в популяциях этого вида. Плешанка в известном смысле “асоциальна”. Самцы нс склонны образовывать плотные поселения, а когда обстоятельства заставляют плешанок селиться с высокой плотностью, птицы нс охраняют территорий с четко обозначенными границами, но придержива-
266
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka ются небольших участков обитания, используя широкие зоны перекрывания этих участков наравне с соседями. Залет в самый центр участка соседа — более чем обычное явление у этих каменок. Еще более беззаботны в этом отношении самки, которые у плешанки ведут себя эмансипированно и нс склонны ставить свои планы и перемешения под контроль супруга.
Формирование пар. Упомянутые особенности поведения самок ярко проявляются уже в момент формирования супружеской пары, так что весь процесс помолвки выглядит совершенно иначе, чем у Ое. hispanica. При первом появлении самки на участке самца он. как это обычно происходит у каменок, летит к обрыву и начинает лазать здесь около всевозможных отверстий и щелей. При этом самец не скрывается надолго в пустотах, а мгновенно выбирается наружу, нс меняя все это время своей экстравагантной позы (рис. 127 d, е). Порой он словно замирает на пол-минуты, прижавшись к стенке неглубокой ниши, так что его хорошо видно снаружи. Если поведение самца в целом имеет много общего с тем, что мы видели у других видов каменок (и у Ое. hispanica в частности), то образ действий самки довольно неожидан. Дело в том, что она словно бы игнорирует самца: в одном эпизоде формирования пары самка вообще ни разу нс подлетала к самцу за 2 ч., в другом за час наблюдений сблизилась с ним только один раз, и в третьем подлетала к будущему супругу 3 раза за 50 мин. (всего 4 сближения по инициативе самок почти за 4 ч. наблюдений).
Спустя 10-15 мин. после первого появления самки на участке самец берет инициативу в свои руки и сам начинает раз за разом сближаться с самкой, на что она отвечает угрожающим треском (сигнал N 4 — рис. 128 b ), но не улетает от самца и, таким образом, не стимулирует погони, обычной в таких ситуациях у всех прочих видов каменок. При формировании одной из пар самец за первые 50 мин устремлялся к самке 28 раз, а та, в свою очередь, пыталась неоднократно (47 раз), хотя и довольно вяло, сблизиться с самкой из соседней пары. Последняя, полностью игнорируя угрозы в ее адрес, оставалась все это время на участке молодоженов. Только 6 из 75 наблюдавшихся сближений вылились в погони самца за самкой-”новобрачной”.
Присутствие посторонних особей на участке, где формируется пара — обычное явление у плешанок. В 6 эпизодах помолвки на участки, где происходили события, 14 раз вторгались посторонние самцы и дважды — самки. Пришелец либо пришелица порой остаются в гуще событий До 20 мин. и даже около часа, но это, судя по всему, не так уж волнует хозяев участка. Самец больше занят своей новобрачной, а нсзванный юсть довольно безучастен к вялым угрозам со стороны владельца территории и даже пытается наравне с ним приблизиться к самке. Лишь
267
Панов Е.Н. Каменки Палеарктики
Рис. 127. Основные позы из сигнального репертуара Ое pleschanka. а — при исполнении песни "стук пишущей машинки” (как правило, холостыми самцами), Ь, с — при территориальных конфликтах высокой и низкой интенсивности, соответственно, d, е — при формировании пар, f. g — при посадках во время и после полета петлями, h, i — переходные варианты между позами с, d. I' и демонстрациями сампа (j, к) перед сто замедленным совместным полс-юм с самкой, 1 — в преддверии копуляции, in - повеление самки, ютовой к копуляции (н г Панов 1989)
268
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
Рис. 128. Основные позывки Ое. pleschanka. а — при тревоге у гнезда (сигналы N N 3 и 1), b — сигнал N 4, с — переход сигналов N 1 в сигнал N 2 (последний в серии), d — вокализация птенца (сигнал N 6). Из: Панов 1989.
однажды я стал в такой ситуации свидетелем короткой драки между самцами, причем самка нс обращала никакого внимания на происходящее. Все это странно видеть, зная, насколько эмоциональной была бы реакция участников подобных событий у Ое. hispanica. Там самка не подпускает к себе на короткую дистанцию даже собственного избранника, а при появлении чужого самца сразу же старается спрятаться в густой куст либо в расселину скалы.
Отсутствие резко выраженной агрессивности между самками должно, вероятно, благоприятствовать формированию полигиничсских семей. На Алтае мне удалось найти супружеское трио, включавшее в себя самца и двух самок, которые выстроили гнезда на расстоянии 20 м одно от другого. Обе самки кормили своих птенцов, нс вступая в конфликты друг с другом, а самец помогал той и другой поочередно. Такая ситуация вряд ли возможна у гораздо более агрессивных испанских каменок Ое. hispanica.
Обычно спустя час после своего появления на участке самка начинает самостоятельно осматривать ниши и щели, а самец иногда садит
269
Панов Е.Н. Каменки Палеарктики
ся при этом рядом с ней — уже далеко нс в столь эффектных позах, как в первые минуты помолвки (рис. 127 h, i). Самка осваивается на новом месте очень быстро, и вот уже можно видеть, как молодожены кормятся неподалеку друг от друга, словно они всегда были вместе.
Развитие отношений между членами пары. Момент формирования пары нередко отделен от начала гнездостроения довольно длительным промежутком времени. В приальпийской зоне Гиссарс-кого хребта (западный Памиро-Алай на высоте свыше 2 тыс. м) в конце апреля — первых числах мая температура по ночам падала ниже нуля, а днем нередко шел холодный дождь, иногда — с мокрым снегом или градом. Большинство плешанок в это время уже разбились на пары. Постепенно прибывали и новые, холостые самцы. Некоторые из них пытались обратить на себя внимание самок, которых уже связывали узы брака. Самцы — пришелец и хозяин территории — принимали угрожающие позы (рис. 127 Ь, с), поочередно лазали в отверстия между камнями, а временами сцеплялись когтями и клубком катались по земле.
Члены супружеской пары большую часть времени держались порознь и были поглощены поисками пропитания. Самцы временами принимались петь, и только в редкие более теплые, солнечные дни можно было наблюдать кратковременные “брачные” взаимодействия между супругами. Размножаться в этих местах плсшанки начинают лишь во второй половине мая.
13.5. Гнездо, кладка, выводок
Гн е з д о с троение и конструкция гнезд. Гнезда чаще всего располагаются в неглубоких вертикальных щелях в скале или между камнями. Гнездо самка вьет у самого входа, так что край его виден снаружи. Чаще всего гнездо расположено под навесом, но иногда втиснуто в узкую вертикальную щель и нс имеет укрытия сверху. На полуострове Мангышлак, в гибридогенной полиморфной популяции, гнезда нередко бывают выстроены под плоским камнем, лежащем на склоне (по типу гнезд Ое. hispanica) и в ажурных выветриваниях и “выдувах” белого известнякового монолита. В Забайкалье 35 из 37 найденных гнезд помещались в расщелинах и нишах скал и только 2 — в норах длиннохвостого суслика Citellus undulatus. В норах сурков Marmota sibirica пле-шанки иногда гнездятся в Монголии. Известно гнездо, выстроенное в расщелине ствола саксаула. Как правило, до гнезда легко дотянуться рукой, и лишь в редких случаях оно бывает выстроено в 30 см от входа или глубже. Обычно внутренняя часть гнезда неплохо освещена, и птенцы плсшанки начинают пользоваться зрением раньше, чем это происходит у других видов каменок (Абдрсимов 1981; Доржиев, Хертуев 1992).
270
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
На постройку гнезда самка затрачивает не менее 5 дней (Абдусаля-мов 1964). Некоторые самки начинают гнсздостроение с того, что натаскивают в нишу значительное количество палочек примерно одинаковой толщины и длины. Они служат основанием гнезда, а часть их лежит у входа в убежище. Из семи гнезд, осмотренных мной на Алтае, три были выстроены на таких настилах. Так же выглядело гнездо, найденное Абдусалямовым в Зеравшанском хребте. В некоторых случаях присутствие настила оправдано неровностями пола гнездовой ниши, но в целом поведение, связанное с доставкой палочек в гнездовое убежище носит, вероятно, атавистический характер.
Сама гнездовая постройка состоит из сухих стеблей и листьев злаков, более грубых в наружном слое и более тонких — во внутреннем. Лишь изредка гнездо бывает выстлано шерстью или конским волосом. Некоторые гнезда свиты довольно искусно, другие выглядят скорее неряшливо. Диаметр гнезда 80 — 160 мм (в среднем 130мм), высота 47 — 85 мм (60 мм), глубина 60 — 80 мм (70 мм), толщина стенок 30 — 50 мм (45 мм).
Брачные игры и копуляция. К концу гнездостроения супруги все чаще принимают участие в своеобразных брачных играх, во многом похожих на соответствующие ритуалы испанской каменки. Эти игры чаше всего можно видеть перед самым закатом солнца, а продолжаются они порой в глубоких вечерних сумерках. Самец, пригнувшись грудью к земле и медленно поворачиваясь вокруг вертикальной оси то вправо, то влево, трепещет, словно ночная бабочка, полу-расправлснными крыльями (рис. 127 j, к). Затем он, не меняя темпа и амплитуды движения крыльев, низко над землей летит в сторону самки, а она, не давая ему приблизиться, чуть отлетает в сторону. Постепенно эти демонстрации переходят в совместный замедленный полет “тандемом”, а тот — в стремительную погоню самца за самкой, сопровождаемую крутыми виражами и неожиданными изменениями траектории движения. Наконец самка садится, а самец проделывает вокруг нес полет петлями с песней “на тему пишущей машинки”. На начальной фазе этих демонстраций позы самцов Ое. pleschanka и Ое. hispanica несколько различаются. Самцы плешанок обычно не столь широко разводят в сторону трепещущие крылья, поворачиваются на меньший угол из стороны в сторону и нс так сильно наклоняют туловище вперед (сравни рис. 116 i, j и 127 к). Существуют, вероятно, и другие мелкие отличия. В частности, трепещущие полеты самцов испанской каменки более длительны, и птица в таком полете даже может набирать высоту (Панов 1978).
Описанные брачные игры, вопреки первому впечатлению, не ведут к копуляции. По наблюдениям Иваницкого (неопубл.) она про-
271
Панов Е.Н. Каменки Палеарктики
сходит примерно так же, как и у других видов каменок. Вот как описывает он ход событий. Поздно вечером (в 21.00) самец внезапно принял одну из своих самых эффектных поз (рис. 127 g), которая сразу же сменилась совершенно другой. Самец распушил оперение груди и брюшка, втянул голову в плечи (рис. 127 1) и начал петь монотонный мотив подпесни — нечто вроде “цви...цви...цви...цви...”. Потом он полетел к обрыву, нырнул в нишу скалы, развернув хвост веером, и спел там несколько песен; перелетел в соседнюю нишу, затем кинулся назад в первую — и так 5-6 раз подряд. В одной из этих ниш, как выяснилось, сидела самка. Самец в конце концов сел к ней, проделал длительный полет петлями, покрыл самку и сразу же стремительно улетел прочь, издвая короткие свисты (сигнал N 3).
К л а д к а. Из 11 кладок, найденных Сурвилло (1968) в Зайсанской котловине (восточный Казахстан), 6 содержали по 6 яиц, 4 — по 5 и в одной было 4 яйца (среднее 5.45). Соотношение кладок разного размера в 86 гнездах, найденных на территории бывшего Советского Союза, таково: 3 х 1,4 х 10, 5 х 39, 6 х 36 (ср. 5.28). Отдельная выборка из Забайкалья, характеризующая восточные популяции, дает следующие цифры: Зх 1,4x6, 5 х 15, 6 х 15, 7x2 (ср. 5.28, п = 39). По данным Лоскота (1983) средняя величина кладки 5.31 (включены данные из гибридных зон). В гибридогенной полиморфной популяции Мангышлака кладки разной величины распределялись следующим образом: 4 х 3, 5 х 14, 6 х 18 (ср. 5.43, п = 35; Гаврилов 1970). В западном Тянь-Шане средняя величина ранних кладок 5.2 (п = 13), поздних (повторных и/ или относящихся ко второму циклу гнездования) 3.3 (п = 13) (Митропольский и др., 1981).
Размеры яиц подвида pleschanka в Копетдаге 19.4 — 20.7 х 14.5 — 15.7 (ср. 20.0 х 15.1, п = 22); в Казахстане 18.4 — 20.7 х 14.2 — 15.6 (ср. 19.3 х 14.8, п = 37); в Западном Тянь-Шане в среднем 19.9 х 15.2 (п = 46); в высокогорьях Бадахшана 18.7 — 20.1 х 14.4 — 16.3 (ср. 19.1 х 15.1, п = 14); в Забайкалье 17.8 — 20.6 х 14.2 -15.9 (ср. 19.2 х 14.9, п = 95); в полиморфной популяции Мангышлака 18.6 —20.9 х 14.2-15.6 (ср. 19.6 х 15.0, п = 39) (Зарудныи 1896, Митропольский и др. 1981, Ковшарь, Левин 1982, Доржиев, Хертуев 1992, данные автора). По сборам Лоскота (1983) из разных точек ареала (в том числе из гибридных зон): 17.5 - 21.8 х 14.0 - 16.0 (ср. 19.7 х 15.0, п = 100), вес 2.35 г. То же, по Крампу (Cramp 1988): 17.1 - 21.0 х 13.5 -16.1 (ср. 19.6 х 15.2, п = 75), вес 2.43 г. Вес яиц из Забайкалья 2.30 г. Размеры яиц формы cypriaca: 18.0 — 20.6 — 14.0 — 15.4 (ср. 19.3 х 14.6, п = 55), вес 2.19 г. (Schdnwctter цит. по: Cramp 1988). Длина яиц может сильно варьировать даже в пределах одной кладки.
Скорлупа окрашена в разные оттенки голубого — от светло-голубого до интенсивно бирюзового с примесью зеленого тона; мелкие крас-272
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka
новатыс пятнышки рассеяны по всей поверхности яйца, но располагаются гуше на тупом конце.
Репродуктивный успех. В Забайкалье из 92 отложенных яиц 6 (6.5%) не развились и педали птенцов. Из 77 вылупившихся птенцов погибли 6 (7.8%). Два гнезда (11.1%), построенные в куче строительных материалов и на свалке, были случайно разорены людьми. В нескольких выводках младшие птенцы были затоптаны более старшими. Средняя величина выводка 4.0 (Доржисв, Хсртуев 1992). В Ставропольском крае (северный Кавказ) среднее число птенцов перед вылетом 5.25 (Рубцов 1997).
Птенцы. Маленькие птенцы имеют желтоватую кожу и эмбриональный пух мышино- или темно-серого цвета длиной 7—10 мм Несколько рудиментарных пушинок есть в районе кобчика (у птенцов Ое. hispanica длина пуха достигает 15 мм, рудиментарные копчиковые пушинки не описаны — см. Лоскот 1983). Зев оранжево-желтый, валики в углах рта беловатые и иногда своеобразно изогнуты. Доржисв и Хсртуев (1992) специально подчеркивают, что глаза у птенцов открываются на 4-й день, раньше чем у птенцов Ое. oenanthe. Я также в свое время обратил внимание на удивительно живую реакцию голых еще птенцов плешанки, вынутых из гнезда, на все происходящее вокруг. Они несомненно прекрасно видели и следили глазами, поворачивая голову, за движущимися объектами. На 12-й день птенцы могут вылезать из гнезда и сидеть у входа, на 14-й день они, как правило, окончательно покидают свою колыбель.
В неволе молодые плешанки в месячном возрасте купаются редко и неохотно, а если в жаркую погоду их опрыскать водой из шприца, сидят неподвижно, дрожат и нс пытаются приводить в порядок мокрое оперение. В этом возрасте птицы необычайно молчаливы. При кормлении хватают пинцет на лету, а в возрасте около 18 дн. (считая от момента вылупления) активно едят сами, причем заглатывают иногда очень большие куски (в частности, листьев капусты). Еще через 12 дней юнцы начали издавать тихий сигнал N 1, неуклюжее повторение которого немного похоже на самое начало пения. Впервые одна из молодых птиц запела в возрасте около 65 дней. Незадолго до этого у нес началась линька (I8.VI II). Линька подошла к концу в середине сентября. Помимо мелкого оперения сменились все перья хвоста и третьестепенные маховые.
13.6. Линька и отлет на зимовки
Полная линька взрослых птиц, в ходе которой сменяется все мелкое и крупное оперение, протекает преимущественно в августе, но
273
Панов Е.Н. Каменки Палеарктики
некоторые линяют раньше, с первых чисел июля, а другие позже — в сентябре. Отлет на зимовки начинается в последних числах августа. В северном Иране первые пролетные плешанки появились около 25 августа, а массовый пролет шел с 28 августа по 13 сентября; 24—25 октября довольно часто попадались на глаза только самки и первогодки. Исчезли плешанки после 1 ноября (Зарудный 1986). В Ираке этот вид пролетает в небольшом количестве в конце сентября и в октябре (Allouse 1953). В Израиле пролет происходит в период между 19 сентября и 11 ноября, пик миграций приходится на 5-25 октября (Shirinai 1996). На местах зимовок в Африке эти каменки появляются в сентябре — октябре. В это время они еще попадаются изредка в пределах своего гнездового ареала — например, в Казахстане (13.Х) и в Памиро-Алае (27.Х, 14.XI). Осенние миграции птиц расы cypriaca происходят примерно в то же время, в августе — октябре (Cramp 1988).
На зимовках в Африке плешанка местами многочисленна (например, в Эритрее). Она может быть встречена в это время от среднего пояса гор до высот 2600-2700 м, причем и самцы и самки занимают индивидуальные кормовые территории в самой различной обстановке: в сельскохозяйственных угодьях, отвоевываемых человеком в каменистых местностях, на окраинах городов, в садах, по заросшим кустарником склонам гор и т.д. (Smith 1971, Leisler et al. 1983).
13.7. Биологические заметки
По спектру питания плешанка не отличается явным образом от других сосуществующих с ней видов каменок (Иваницкий 1980а, Дор-жисв, Хертуев 1992). При этом, однако, существует мнение, что при охоте за насекомыми она в большей степени пользуется полетом. Это нс значит, что плешанки чаще, чем другие каменки, ловят добычу в воздухе, хотя такое тоже не исключено: Лэк (Lack 1980) сообщает, что из 100 жертв плсшанок 64 были пойманы на земле, 30 — в воздухе и 6 — в кронах кустов. Наиболее характерный способ охоты плешанок выглядит так: птица перелетает над землей, присаживаясь каждый раз на невысокий кустик либо на травянистые растения, выдерживаю-щиее ее небольшой вес. С этой присады она высматривают добычу на земле или в траве, схватывает ее и перелетает на следующую приезду (Leisler et al. 1983). Такая манера охоты в большей степени свойственна чеканам Saxicola. нежели каменкам. При всем при этом плешанка не упустит и возможности поймать небольшую ящерицу-круглоголовку Phrynocephalus.
Как уже было сказано, Ое. pleschanka и Ое. hispanica — это очень близкие, сестринские виды, из которых первый представляет собой
274
Глава 13. Плешанка, чекан-плешанка Oenanthe pleschanka более молодую ветвь, производную от африканской по происхождени испанской каменки. Отделение “пра-плешанки” от испанской каменки произошло в восточном Средиземноморье, и реликтом периода разделения является, по-видимому, кипрская плешанка. Аргументом в пользу такого предположения может служить тот факт, что песня расы cypriaca по общему принципу организации более сходна с песней Ое. hispanica, чем с песней материковых плешанок (см. рис. 125). Затсм началось расселение плешанки на восток.
Можно предположить, что ареал Ое. pleschanka уже в середине плиоцена включал в себя восточный Прикаспий, а Ое. hispanica к этому времени успела расселиться на восток до Закавказья. В конце Понтического времени (около 6 млн. лет назад) произошло резкое снижение уровня Каспийского моря, и возник сухопутный мост между юго-восточными отрогами Кавказского хребта и северо-западными отрогами Хорасанских гор (Atamuradov 1994). Именно в это время могла произойти первая инвазия Ое. hispanica в ареал Ое. pleschanka. Продуктом их гибридизации стала нынешняя популяция Мангышлака и Устюрта, которая затсм, когда Каспийская впадина вновь заполнилась водой, оказалась отрезанной от ареала испанской каменки,
Плешанка между тем расселялась не только к востоку, но и к северу. Она обогнула с северо-востока Каспийское морс и отсюда стала расширять свой ареал к западу (этот процесс продолжастсяи и сегодня). Тем самым она преградила все пути продвижения на восток испанской каменке. Всюду, где последняя претендовала на земли, уже занятые плсшанкой, начиналась интенсивная гибридизация между этими видами. Как считает Хаффер (Haffcr 1977), вторичный контакт между ними установился в южном Прикаспии, в горах Эльбурса сравнительно недавно, около 13000 — 5000 лет тому назад, когда сухой и холодный климат сменился здесь более теплым и влажным, а суровые горно-степные ландшафты — древесно-кустарниковой растительностью средиземноморского типа. Нс исключено, что примерно в то же самое время произошло расселение испанской каменки на север вдоль западного побережья Каспийского моря. Эта инвазия испанской каменки в районы, уже давно занятые плешанкой, привела к возникновению гибридных зон в северо-восточном Азербайджане и в Дагестане.
275
Панов Е.Н. Каменки Палеарктики
Глава 14.
Гибридные популяции в зонах вторичного контакта Ое. hispanica и Ое. pleschanka
Особи, соединяющие в себе окрасочные признаки испанской каменки и плешанки, были известны орнитологам давно. Однако в то
время их считали представителями новых для науки подвидов или даже
видов, которым присваивали соответствующие латинские названия. Позже такие экземпляры начали трактовать как цветовые морфы одного либо другого вида. Так в орнитологической литературе появилось не
сколько имен, которые ныне используются для обозначения тех или иных гибридных фенотипов (gaddi, Hbyca, iransfuga, vittata). Сторонники
идеи полной генетической замкнутости вида еще до недавнего времени
отрицали возможность гибридного происхождения подобных экземпляров, настаивая на том, что они есть нс что иное, как разные цветовые морфы двух интересущих нас видов (Лоскот 1983, Степанян 1983).
Приверженцы такой точки зрения вынуждены были отказаться от нес, когда оказалось, что изменчивость окраски этих каменок наиболее велика там, где типичные представители обоих видов обитают совместно. При этом выяснилось также, что в подобных “смешанных” популяциях встречаются все мыслимые комбинации ок
Рис. 129. Окрасочные признаки (1—4) Ое. hispanica и Ое. pleschanka и их комбинаторика при гибридизации. Из: Панов и Иваницкий 1975b.
расочных признаков испанской каменки и плешанки, причем эти промежуточные варианты укладываются в непрерывный ряд от типичного фенотипа одного вида до типичного фенотипа другого (Панов, И ван и ц-
276
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
Рис. 130. Зоны вторичного контакта и гибридизации между Ос. hispanica (а) и Ое. pleschanka (b) (вверху) и локализация изученных гибридных популяций (внизу). I — Дагестан, 2 — северо-восточный Азербайджан, 3 — восточный Азербайджан (Гобус-тан), 4 — северный Иран (Эльбурс), 5 — полуостров Мангышлак. Из: Грабовский, Панов, Рубцов 1992.
кий 1975, HafTcr 1977; рис. 129). Сказанное наиболее очевидно в применении к окраске самцов, но то же относится и к самкам, хотя картина здесь не столь наглядна из-за сходства окраски самок того и дру
гого вида.
Такие популяции с чрезвычайно высокой изменчивостью окраски (чью гибридогенную природу теперь уже трудно отрицать) более или менее тщательно изучены к сегодняшнему дню в следующих пяти регионах: горы Эльбурс в северном Иране (HafTcr 1977), урочище Гобу-стан близ г. Баку в восточном Азербайджане (Панов и др. 1992, Рубцов 1997, и др.), окрестности г. Махачкала в Дагестане (Панов 1986), окрестности с. Кахи в северо-западном Азербайджане (Лоскот 1986) и полуостров Мангышлак в северо-восточном Прикаспии (Панов, Иваницкий 1975) (рис. 131). В первых четырех из названных регионов гибридизация идет в настоящее время — за счет постоянного притока сюда особей из популяций обоих родительских видов. В пятом регионе (Мангышлак), как я уже упоминал и как попытаюсь доказать ниже, приток особей возможен только из ареала Ое. pleschanka. Здесь обитают древние гибридогенныс популяции, которые, строго говоря, следует рассматривать в составе вида Ое. pleschanka — как своего рода географическую расу плешанки, полиморфную по окраске самцов и высоко изменчивую по окраске самок (см. ниже).
277
Панов Е.Н, Каменки Палеарктики
Я начну с описания процессов, происходящих в “молодых” гибридных зонах, взяв в качестве примера наиболее детально изученную популяцию Гобустана (Panov et al. 1994). После этого я сопоставлю сказанное о ней с тем, что известно о других гибридных зонах того же характера — в Иране, северо-западном Азербайджане и Дагестане. И завершу этот раздел, коротко рассмотрев своеобразие древней полиморфной популяции Мангышлака, некоторые сведения о которой уже приведены в очерке о Ое. pleschanka (Глава 13).
14.1. Гибридная популяция Гобустана (восточный Азербайджан)
Фенотипический состав, местообитания и численность. Эта популяция занимает склоны одиночно стоящей столовой горы Бсюк-Даш, которые местами выглядят как отвесная каменная стена, а в других участках сравнительно пологи и представляют собой глинистую поверхность с отдельными выходами основной породы и с нагромождениями крупных валунов (рис. 131). Кое-где у подножий горы встречаются угодья с хорошо развитой древесно-кустарниковой растительно-
Рис. 131. Урочище Гобустан (восточный Азербайджан). Местообитания гибридной популяции Ое. hispanica х Ое. pleschanka и черношейной каменки Ое finschii.
278
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
Рис. 132. Фенотипы самцов в гибридной популяции Гобустана. Кружками показаны численные соотношения фенотипов (черный кружок — "pleschanka", белые — "hispanica", серые — “гибриды”)- а — "hispanica aurita", b — "hispanica sfapazina", c — "pleschanka", d — "черногорлый gaddi", e — "gaddi", f — “белоспинная pleschanka", g — "transfuga", h — “черногорлая libyca". Нс обозначены буквами фенотипы, промежуточные между перечисленными. Из: Грабовский, Ианов, Рубцов 1992, с изменениями.
стыо, но в основном склоны выглядят сухими и бесплодными. Здесь на площади около 75 га в разные годы размешается примерно от 50 до 70 пар интересующих нас каменок. Среди самцов наравне с обеими морфами испанской каменки (sfapazina и aurita), обычны типичные самцы плсшанки, а также все переходные варианты между теми и другими. Эти промежуточные фенотипы мы для простоты распределим по 8 классам (gaddi, transfuga, “бслоспинная плешанка” и т.д.). Все 11 основных вариантов окраски самцов показаны на рис. 132. По склонам горы пары с самцами разных фенотипов размещены случайным образом, так что в непосредственном соседстве могут оказаться птицы с любыми вариантами окраски (рис. 133). Селятся здесь эти каменки вместе с менее многочисленными Ое. finschii, которые ведут себя по отношению к ним довольно агрессивно.
279
Панов Е.Н. Каменки Палеарктики
За 6 лет наблюдений доля промежуточных фенотипов, которые бесспорно представляют собой гибридов разных поколений, составила в среднем 55% (рис. 134). Столь высокая концст-рация гибридов в популяции, а также тот факт, что при формировании супружеских пар самки выбирают супругов “наугад” (о чем будет сказано ниже) — все это заставляет усомниться в чистоте крови тех птиц, которые на первый взгляд выглядят как “типичные” плешанки либо испанские каменки. Чтобы прояснить этот вопрос, мы сравнили по ряду признаков местных плешанок и испанских каменок друг с другом и с типичными представителями этих видов из районов, где гибридизация отсутствует. В каче-
Рис. 133. Размещение территорий самцов разных фенотипов в Гобустане в 1989 и 1992 гг. а — “hispanica aurifa". b -“hispanica sfapazina", c — “pleschanka", d — “белоспинная pleschanka", e — “transfuga", f — “gaddF, g — прочие. Из: Рубцов 1997
стве признаков для сравнения были выбраны такие, по которым “чистые” популяции Ое. hispanica и Oe.pleschanka надежно отличаются друг от друга (степень почернения лба и длина первого махового). Результат этйх сопоставлений показан на рис. 135. Оказалось, что местные “испанские каменки” и “плешанки” в гораздо меньшей степени отличаются друг от друга, чем каждая из этих выборок — от представителей того же вида из генетически чистых популяций. Обнаружив это, мы решили и в дальнейшем ставить видовые названия местных птиц в кавычки, подразумевая тем самым, что они с большой степенью вероятности являются не чистокровными, а гибридами от многократных возвратных скрещиваний — таких, например, как [(hispanica xpleschanka) х pleschanka] х pleschanka и т.д. С учетом этого обстоятельства соотноше
280
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
ние самцов “pleschanka" : “гибриды” : “hispanica" составило за все годы 31 : 55 : 14. Изменения фенотипического состава популяции в разные годы показано на рис. 134 (см. Грабовский и др. 1992).
Длина крыла самцов старше года составляет в этой популяции 89.7 мм (все фенотипы, п = 78), самцов-первогодков 87.5 мм (п = 37). Вес самцов 16.9 г (май, п = 18). Длина крыла самок 85.9 мм, вес 18.6 г (май, п = 4). Длина крыла взрослых самцов меньше, чем в гибридной зоне северного Ирана (91.1 - 92.4 по HafTer 1977). Эти различия могут быть обусловлены разными методами измерений, однако малый вес самцов (лежаший у нижнего предела весов Ое. hispanica и Ое. pleschanka в мае — см. Cramp 1988) и мелкие размеры яиц (см. ниже) могут свидетельствовать об уменьшении об-
Рис. 134. Численное соотношение фенотипов в популяции Гобустана в разные годы и за весь период исследований среди самцов и самок, а — “hispanica ”, b — “pleschanka ”, с — “гибриды ”.
ших размеров особей в гибридной популяции Гобустана.
Рис. 135. Сравнение по двум признакам фенотипов “hispanica ”(а) и “pleschanka" (d) из гибридной популяции Гобустана с испанскими каменками (Ь) и плс-шанками (с) из “чистых" аллопатрических популяций. А — длина первого махового относительно кроющих первостепенных маховых (мм), В — развитие черного цвета на лбу (баллы) По данным Рубцова, Грабовского и автора.
281
Панов Е.Н. Каменки Палеарктики
Состав гнездящихся пар. Можно было бы предположить, что самки с фенотипом “hispanica" или “pleschanka" склонны выбирать в качестве супругов самцов, одетых, соответственно, в типичный брачный наряд испанских каменок и плсшанок, или, по крайней мерс, сходных с такими самцами по окраске. Оказалось, однако, что состав пар в большой мере обязан случаю. Скорее всего, самка выбирает не самца, а наиболее приглянувшуюся ей территорию, не обращая внимания на облик се хозяина. Из 84 пар только 12 состояли из партнеров, которых можно было бы отнести к одному и тому же виду (все 12 -“pleschanka" х “pleschanka"). Среди прочих пар 15 были смешанными (“pleschanka" х “hispanica”). а остальные состояли из гибридов (7) либо включали в себя гибрида в качестве одного из родителей (40).
Индивидуальное мечение птиц показало, что они очень привязаны к своим территориям и, если пережили очередную осень и зиму, возвращаются сюда на следующий год. Один из самцов-гибридов гнездился на одном и том же участке не менее 6 лет подряд. Возвращаются на свои прошлогодние территории и самки, что позволило выяснить, есть ли у них какие-либо индивидуальные вкусы при выборе супруга. Под наблюдением было 18 самок, гнездившихся в Гобустане в течение двух последовательных сезонов. Из них 6 вернулись на второй год на территории своих прежних партнеров, то есть их “выбор” был предопределен стремлением осесть на прошлогоднем участке. Прочие самки останавливали свой выбор на том самце, который занял прошлогоднюю территорию самки в этом году, либо на его ближайшем соседе. Так, из 4 самок, гнездившихся в прошлом году с самцами фенотипа “hispanica sfapazina”. все выбирали в дальнейшем в качестве супругов самцов иных типов окраски (позиции с, е,/и g на рис. 132).
В принципе, изменение состава пары возможно даже на протяжении одного сезона, если самец-супруг погибнет по той или иной причине. Дело в том, что ежегодно в популяции присутствует значительно число самцов-первогодков — в среднем за все годы 28%, что близко соответствует их доле в чистых популяциях Ое. hispanica (31.2 — 33.1% по данным Лоскота 1983). Часть из них занимают собственные территории, а некоторые держатся в качестве “сателлитов” на периферии участков взрослых самцов. Хозяин участка постепенно привыкает к присутствию потенциального соперника и почти не обращает на него внимания. Подобно сателлитам, многие территориальные самцы-первогодки отают-ся холостяками. Из 26 таких самцов 10 нс смогли завоевать расположения самок (Рубцов 1995). Так или иначе, первогодки создают значительный резерв самцов, готовыхъ заменить погибшего отца семейства.
Нс раз бывало так, что сразу же после поимки самца из пары (для осмотра, измерений и кольцевания) словно из-под земли на его тер
282
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
ритории появлялся новый самец, и самке нс оставалось ничего другого, как смириться с этой неожиданной сменой супруга. Если пойманного самца на несколько дней оставляли в клетке, пришелец вел себя на освободившейся территории как се законный хозяин, хотя по внешнему виду он зачастую резко отличался от прежнего владельца.
Сроки и успех гнездования. Размножение в этой гибридной популяции происходит практически в те же сроки, что и в ближайшей изученной популяции Ое. hispanica (Нахичевань, 39° с.ш., 46° в.д ), локализованной примерно в 400 км к юго-западу от Гобустана (40° с.ш., 49° 30’ в.д.). В Нахичевани наиболее ранняя дата вылупления птенцов — 13 мая (сразу в трех гнездах). В Гобустане в 16 гнездах (39%) из 41 птенцы вылупились в период с 11 по 15 мая, еще в 12 гнездах (29.3%) — в следующую пятидневку. В 2 самых поздних гнездах вылупление произошло между 9 и 14 июня. Таким образом, размножение в Гобус-танс происходит очень синхронно. Это можно объяснить высокой плотностью популяции, что, как известно, обеспечивает взаимную социальную стимуляцию гнездящихся пар.
Осмотренные гнезда содержали обычно от 4 до 6 яиц. Средняя величина кладки при различных типах скрещиваний колебалась в пределах от 4.9 (в гнездах 31 самок гибридных фенотипов) до 5.5 (21 самка “pleschanka'"), составляя для популяции в целом 5.2 (п = 73). Доля не-развившихся яиц — от 0.27 (30 самок-гибридов) до 0.50 (18 самок "hispanica" и 14 самок “pleschanka”). Число птенцов при разных типах скрещиваний варьировало от 4.3 (6 пар самцы-гибриды х самки “hispanica”) до 5.4 (7 пар “pleschanka” х “pleschanka”), в среднем для всей популяции 4.7 (п = 65). Количество птенцов существенно меньше в поздних гнездах, где вылупление идет с конца мая до середины июня, по сравнению с ранними (3.0 и 4.8, соответственно) (Рубцов 1997).
В один из сезонов 6 гнезд (14.6%) из 41 были разорены, в двух количество птенцов уменьшилось к концу периода выкармливания, в трех обнаружены мертвые птенцы вместе с живыми. В одном из гнезд птенцы умерли от голода: самка, вероятно, погибла, а самец имел вторую самку и кормил преимущественно се птенцов (Рубцов, 1997).
В общем, средняя величина кладки очень близка к тому, что мы видим в ближайшей популяции Ое. hispanica (Нахичевань: 5.4, п = 9; Армения: 5.1, п = 9). Среднее количество птенцов в гнездах в Гобуста-нс меньше, чем в ближайшей чистой популяции Ое. pleschanka (Ставрополье: 5.3, п = 12), но различия статистически недостоверны.
Гнезда и кладки. Расположение гнезд более сходно с тем, что мы видим в популяциях Ое. pleschanka. Они почти неизменно бывают выстроены во всевозможных пустотах скал либо отдельно стоящих крупных валунов. Помещаются гнезда близко от входа, но часто нсдоступ-
283
Панов Е.Н. Каменки Палеарктики
ны для осмотра из-за малых размеров входного отверстия. Размеры яиц: 17.2 — 20.7 х 13.5 — 15.4 (ср. 19.30 х 14.67, п = 56). Яйца заметно мельче, чем в ближайшей популяции Ое. hispanica из Нахичевани (19.6 х 15.4, п = 21) и в популяции Ое. pleschanka из Копетдага ( 20.0 х 15.1, п = 22). Окраска яиц голубая, часто яркая, с бирюзовым оттенком или чисто бирюзовая. Однако, в одной и той же кладке наравне с яйцами голубого цвета разной интенсивности могут оказаться почти белые яйца. Красноватые пестрины и точки сосредоточены гуше на тупом конце.
Особенности поведения. Несмотря на то, что в популяции доля птиц с фенотипом “pleschanka" (31% среди самцов, до 50% в некоторые годы среди самок) намного выше, чем доля особей "hispanica" (11% среди самцов; рис. 134), общий характер исполнения коротких песен
Рис. 136. Результаты сравнения структуры песен самцов из чистых популяций Ое. hispanica (I — Закавказье) и Ое. pleschanka (5, 6 — юго-западный Туркменистан, 7 — Алтай) и из гибридных популяций (2 — Гобустан, 3 — Дагестан, 4 — Мангышлак). А, В — сравнение средних индексов по разным популяциям, С — сравнение напевов самцов с разными фенотипами в гибридных зонах западного Прикаспия (вверху) и Мангышлака (внизу), а — самцы hispanica из чистых популяций Закавказья, b — "hispanica" в гибридных популяциях, с — “гибриды”, d — vittata (ее отсутствие в западном Прикспии показано звездочкой), е — '"pleschanka" в гибридных популяциях, f — Ое. pleschanka из чистых популяций. По: Грабовский и Панов 1992.
284
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
самцами этой популяции ближе к манере пения Ое. hispanica, не отличаясь от него статистически. Все самцы, независимо от их фенотипа, и в том числе окрашенные как типичные плешанки, поют в этой манере (Грабовский, Панов 1992; рис. 136). Этот факт позволяет предположить, что область, в настоящее время занятая гибридной популяцией, первоначально входила в ареал Ое. hispanica. Экспансия “музыкального стиля hispanica" прослеживается далеко на север, в гибридной зоне Дагестана, где фенотип “hispanica" крайне немногочислен (см. ниже), и даже в “чистой” популяции Ое. pleschanka на северном Кавказе (Грабовский, Панов 1992, Рубцов 1997; см. рис. 125). Возможно, эта экспансия в значительной мерс обязана расселению самцов-первогодков из гибридной зоны. Из 120 птенцов, окольцованных в Гобустане, ни один нс был встречен здесь в последующие годы.
Подводя общий итог, можно сказать, что в Гобустане происходит весьма интенсивное перемешивание генофондов испанской каменки и плешанки, так что подавляющее большинство особей, в том числе и фенотипически “чистых”, имеют гибридное происхождение. Генетически чистыми являются, вероятно, лишь немногие иммигранты из соседних популяций Ое. pleschanka и Ое. hispanica. Иными словами, здесь перед нами нс зона симпатрии с ограниченной гибридизацией между родительскими видами, а ситуация их полного слияния.
14.2. Гибридная популяция у южных подножий Главного Кавказского хребта (северо-западный Азербайджан)
Эта популяция локализована примерно в 250 км к северо-западу от Гобустана (41° 20’ с.ш., 47° в.д.). Каменки гнездятся здесь в нижних участках долин рек Курмухчай, Турианчай и Гсокчай, текущих с Главного Кавказского хребта. Лоскот (1983, 1986) подробно исследовал поселение в низовьях р. Курмухчай у с. Кахи, где в 1978 г. на площади порядка 150 га учтена 21 пара, а в 1979 г. на 420 га — 22 пары. Общая численность этого поселения составляет, вероятно, около 30 пар. Соотношение фенотипов в выборке из 53 самцов в целом сходно с тем, что наблюдается в Гобустане (Рис. 137-2).
Гибридная популяция населяет здесь участок долины с каменными дамбами и насыпями, выстроенными для противодействия дождевым потокам. Берега реки заняты редколесьем из тополя, дуба и каркаса (Cellis caucasica, С. glabrata) с разреженным кустарниковым ярусом из облепихи (Hippophoe rhamnoides), боярышника, шиповника, ежевики и т.д. (Лоскот 1983).
285
Панов Е.Н. Каменки Палеарктики
Рис. 137. Соотношение (в %%) разных фенотипов в гибридных зонах западного и южного Прикаспия. I — Дагестан, 2 — северо-восточный Азербайджан, 3 — Гобустап, 4 — Эльбурс (см. рис. 138). Левый столбец — фенотипы ''hispanica aurita*’ (вверху) и "hispanica stapazina’' (внизу), правый столбец — "pleschanka". Данные по популяции 2 — по материалам Лоскота (1986), по популяции 4 — по HafTcr 1977. Из: Панов 1989.
В двух случаях Лоскоту удалось установить, что фенотипы отцов и их сыновей могут существенно отличаться. У пары hispanica aurita х бслогорлая самка один из сыновей выглядел как hispanica stapazina, а другой имел окраску гибрида (вариант е на рис. 132). У пары самец hispanica stapazina х темногорлая самка сын имел фенотип “черногорло-го gaddi" (тот же рис., позиция d). Эти факты служат дополнительным аргументом в пользу предположения, что самец “чистого” фенотипа может в действительности иметь смешанную наследственность.
По существу, это поселение вместе с поселениями соседних речных долин, входит, вероятно, в единый ареал гибридных популяций восточного Закавказья, охватывающий район Гобустана на юге, восточную Грузию на западе и Дагестан на северо-востоке.
14.3. Гибридная популяция Дагестана
В этом регионе наблюдения проводили в ближайших окрестностях г. Махачкала (43° с.ш., 47° 30' в.д.), расположенного примерно в 370 км к северо-западу от Гобустана и в 175 км к северо-востоку от Кахи, у северо-восточных подножий Главного Кавказского хребта. Здесь каменки наиболее многочисленны по скальным выходам вдоль шоссе, ведущего на юго-запад в сторону г. Буйнакска, но селятся также в окружающей холмистой местности с выходами каменистой породы. В отличие от двух только что рассмотренных гибридных популяций здесь в 1985 г. более половины (почти 62%) из учтенных 23^самцов приходилось па фенотип "pleschanka", а самцы с обликом типичной "hispanica" были крайне немногочисленны (Панов I986; рис. 137-/). В 1992 г. их доля оказалась несколько большей (10,0% "stapazina", 16,7% "aurita"
286
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
при 36,7% "pleschanka" и 36,6% гибридных фенотипов, п = 30; Рубцов, 1997).
В 1985 г. в последней декаде апреля (20 — 23.IV) почти все самцы уже были в парах, но самки еще не приступили к гнездостроснию. В 1992 г. в в позднюю дождливую весну вылупление птенцов началось в период с 31 мая по 6 июня. Средняя величина кладки составила 5.3 яйца (п = 11), среднее число птенцов на гнездо 5.0 (п = 8) (Рубцов 1997). Общий стиль исполнения коротких песен у самцов этой популяции промежуточен между манерами их исполнения у Ое. pleschanka и Ое. hispanica, но в целом по этому признаку популяция стоит ближе к последнему виду. Песни самцов с фенотипом "pleschanka" по своим структурным характеристикам в среднем нс отличаются статистически от песен самцов с другими фенотипами (рис. 136).
14.4. Гибридная зона в северном Иране
Эта зона гибридизации локализована в горах Эльбурс. Она протянулась вдоль 36° с.ш. почти на 600 км (рис. 138). Как видно из этого рисунка и рис. 137--#, в центральной части гибридной зоны соотношение фенотипов "pleschanka": гибриды : "hispanica" составляет 20.3 : 62.7 : 17.0. Иными словами, концентрация гибридов здесь максимальна по сравнению со всеми рассмотренными гибридными популяциями, что может указывать на более значительный возраст этой зоны контакта. По мнению Хаффера (Haflcr 1977), ареалы Ое. pleschanka и Ое. hispanica могли войти здесь в контакт около 5-11 тысяч лет тому назад.
Рис. 138. Распределение разных фенотипов в гибридной зоне Эльбурса (северный Иран). Из: HafTer 1977, с изменениями.
287
Панов Е.Н. Каменки Палеарктики
В Эльбурсе интересующие нас каменки обитают от подножий гор до высот 1500 м (местами до 2200 м). Они избегают сухих, бесплодных местностей, предпочитая возделываемые человеком участки долин. Эти каменки часто селятся в значительных количествах вокруг деревень, посещая окраины садов и добывая пропитание на полях и огородах.
14.5. Гибридогенные полиморфные популяции северо-восточного Прикаспия
Ареал интересующих нас популяций занимает около 40-50 тысяч кв. км между 43 и 45° с.ш., простираясь от северо-восточного побережья Каспийского моря до склонов Западного чинка Устюрта (Гаврилов 1970, данные автора). Интересующие нас каменки занимают здесь опустыненные низкогорья, концентрируясь по каменистым склонам немногочисленных солоноводных речушек, а также выходят на холмистые плато и в изрезанные оврагами участки глинистой пустыни с весьма скудным травяным покровом (преимущественно из полыней Artemisia spp. и солянок Salsola spp.). Бедная кустарниковая растительность сосредоточена в основном по долинам и представлена жидкими полосками тамариска (Tamarixspp.) по руслам пересыхающих потоков и отдельными кустами крушины и боярышника по каменистым известняковым склонам. Гнездятся каменки также в заброшенных строениях человека и по окраинам поселков, которые весьма редки в этой суровой местности с ее постоянными сильнейшими ветрами, знойным летом и очень холодной зимой.
В то время как все четыре ранее рассмотренные группы гибридных популяций западного и южного Прикаспия в целом сходны друг с другом, различаясь в основном процентным соотношением общих для них фенотипов, популяции Мангышлака и прилежащих регионов резко отличается от них по своему фенотипическому составу. Главное своеобразие ее состоит в следующем. Во-первых, здесь отсутствует фенотип “hispanica stapazina'\ преобладающий в популяциях испанской каменки Закавказья (где соотношение stapazina : aurita = 3 : 1) и потому вполне обычный в гибридных популяциях западного и южного Прикаспия. Во-вторых, на Мангышлаке появляется новый фенотип vittata (до 11% в некоторые годы), практически отсутствующий в Эльбурсе, Азербайджане и Дагестане. В-третьих, в некоторых поселениях каменок на Мангышлаке (например, в окрестностях пос. Куйбышев) встречаются фенотипы, никем и нигде не описанные ранее — с белой “уздечкой” либо с ожерельем из черных перышек поперек груди (рис. 139). И, наконец, среди самок очень высок процент особей со светлым, желтоватым либо
288
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
почти белым теменем (рис. 114). Доля таких самок составляет в некоторых поселениях около 40%. Они встречаются и в чистых, мономорфных популяциях Ое. pleschanka, но сравнительно редки там, будучи представлены единичными особями. Высокая частота в популяции Мангышлака подобных самок с маскулинизированной окраской отчасти сближает ее с кипрской плсшанкой Ое. р. cypriaca, для которой характерно слабое развитие полового диморфизма.
Тот факт, что в северо-восточном Прикаспии не найден фенотип stapazina, бесспорно свидетельствует об отсутствии в настоящее время обмена особями и генами меж-
vittata
1974 (п=234)
1977 (п=154)
1985 (п=99)
1986 (N=153)
Рис. 139. Соотношение (в %%) разных фенотипов в популяции Мангышлака в разные годы. Левый столбец — "hispanica aurita" правый столбец — "pleschanka". Данные за 1977 г. — по материалам Лос-кота (1986). Из: Панов 1989.
и популяциями hispanica х pleschanka западного и южного Прикаспия. В то же время интересующие нас популяции Мангышлака открыты для широкого обмена генами с популяциями Ое. pleschanka, населяющими Казахстан и Среднюю Азию. Таким образом, присутствующий на Мангышлаке фенотип “hispanica aurita" с точки зрения популяционной генетики представляет собой нс что иное, как один из вариантов окраски (морфу) Ое. pleschanka. Согласно предсказаниям теорем популяционной генетики, концентрация этого фенотипа, как и всех прочих в данной популяции, остается относительно постоянной из поколения в поколение. Исследования показали, что соотношение фснотпов здесь не изменялось сколько-нибудь существенным образом на протяжении по крайней мерс I9 лет (с 1974 по 1993 г. — см. рис. 139 и Рубцов, 1997), и, по всей видимости, было таким же за 27 лет до начала систематических учетов, в 1947 г. (Долгушин 1948,
289
Панов Е.Н. Каменки Палеарктики
Лоскот 1983). Таким образом, здесь перед нами типичный случай сбалансированного генетического полиморфизма, возникшего в результате давней гибридизации между Ое. hispanica и Ое. pleschanka. Один из возможных сценариев этого процесса, происходившего в незапамятные времена, я описал в конце предыдущей главы.
В 1974 г первые самцы появились здесь 6 апреля. Их прилет и занятие ими территорий продолжались до 3 мая. Первая самка встречена 8 апреля, когда началось формирование пар, продолжавшееся до 6 мая. Гнездостроенис у двух пар началось 26 апреля, до 11 мая еще 12 самок приступили к постройке гнезд. Первые кладки были начаты 28 апреля и 1 мая. В 1985 году гнездостроение наблюдали 24-29 мая (3 пары с самцами pleschanka. одна с самцом "aurita” и одна — с самцом gaddi). яйца в гнездах найдены 24 — 29 мая у 5 пар (4 с самцами pleschanka и одна с самцом gaddi). гнезда с птенцами обнаружены между 23 мая и 4 июня (5 пар с самцами pleschanka и одна с самцом vittata). слетков видели 2 июня (самец — белоспинная плешанка) и 4 июня (самец vittata). Не обнаружено явных различий в успехе размножения пар с самцами разных фенотипов (см. Панов 1989).
От молодых гибридных популяций, рассмотренных ранее, полиморфная популяция Мангышлака отличается не только своим фенотипическим составом, но и некоторыми другими особенностями. В частности, если сравнивать популяции Мангышлака и Гобустана, то в первой каменки в среднем крупнее. Это следует не только из сопоставления длины крыла самцов (максимальные значения 94.0 и 91.5, соответственно; средние 90.1 и 89.7), но и из сравнения средних размеров яиц (19.6 х 15.0 мм и 19.3 х 14.7 мм). Репродуктивный успех в популяции Мангышлака существенно выше, чем в Гобустанс, но не отличается от успеха размножения в ближайших чистых популяциях Ое. pleschanka. Например, в ареале последнего вида на северном Кавказе средняя величина кладки составляет 5.4 яйца, в популяции Мангышлака 5.6. Среднее число птенцов в гнезде равно, соответственно, 5.5 и 5.6 (Рубцов 1997).
14.6. Последствия гибридизации и перспективы ее углубления
Каменок с гибридными фенотипами (“белоспинная плешанка”, “gaddi" и др.) орнитологи время от времени обнаруживают в “чистых” популяциях родительских “видов далеко за пределами гибридных зон — иногда на расстоянии до 400-700 км от границ распространения ближайшей гибридной популяции. Их находили в Армении, в Турции,
290
Глава 14. Гибридные популяции Ое. hispanica и Ое. pleschanka
на юге Туркменистана, в Крыму и в Молдавии. Подробный перечень таких залетов содержится в работах Хаффера (Hafier 1977) и Лоскота (1986). Происхождение этих иммигрантов вполне понятно: мы видели, что первогодки, как правило, не возвращаются в свое поселение. Следовательно, они расселяются в другие районы, осуществляя, в частности, перенос “чуждых” генов в чистые популяции родительских видов.
Кроме того, плешанка и испанская каменка продолжают расширять свои ареалы навстречу друг другу. Особенно заметно это в юго-восточной Европе, В частности, в юго-восточной Болгарии, где оба вида каменок гнездятся на скалах по берегу Черного моря, найдены два гибридных самца (Baumgart 1971). Рубцов (1997) наблюдал самца с фенотипом transfuga, который успешно размножался в парс с самкой плешанки, близ границы Болгарии с Молдавией. Много далее к северу, в окрестностях Будапешта в Венгрии, в 1947 — 1963 гг. неоднократно отмечали самцов плешанки и испанской каменки, которые, не находя самок своего вида, вступали во всевозможные контакты с местными обыкновенными каменками и даже пытались кормить их птенцов (Schmidt 1968). Понятно, что в случае появления здесь самок плсшанки либо испанской каменки весьма высокой оказалась бы вероятность формирования гибридных пар. Таким образом в этих районах, а также, возможно, в Турции и Греции, можно в дальнейшем ожидать по меньшей мере случайной гибридизации.
Рис. 140. Схема, иллюстрирующая становление гибридогенного полиморфизма в популяциях Ое. hispanica и Ое. pleschanka за пределами гибридной зоны. Из: Нafter 1977, с изменениями.
291
Панов Е.Н. Каменки Палеарктики
Хаффср (HafTcr 1977) идет еще дальше, предполагая, что полиморфизм Ое. hispanica вообще обязан своим происхождением тысячелетиям обмена генами между этим видом и Ое. pleschanka (рис. 138). По мнению этого автора, испанская каменка первоначально могла быть мономорфной, с единственным типом окраски самцов, соответствующим морфе aurita. Тены чсрногорлости” плешанки, проникая все дальше в ареал испанской каменки, положили здесь начало черногор-лой морфе stapazina. Хаффер аргументирует свою точку зрения, указывая на уменьшение концентрации морфы stapazina в направлении с востока на запад, то есть с увеличением расстояния от ареала плешанки. Он подчеркивает также, что при движении в том же направлении уменьшается и степень развития черного цвета в горловом оперении особей с фенотипом stapazina. ‘Тены белогорлости” испанской каменки, проникая, в свою очередь, в ареал исходно черногорлой плешанки, породили здесь морфу vittata. Весьма высокая концентрация этого фенотипа в популяции Мангышлака, в древней зоне гибридизации двух видов, хорошо укладывается в эту схему (Панов, Иваницкий 1975b). Вся гипотеза выглядит весьма убедительной, и орнитологам остается лишь накапливать новые факты, чтобы подтвердить либо обоснованно опровергнуть ее.
292
Глава 15. Каменка монашка Oenanthe monacha
Глава 15. Каменка монашка Oenanthe monacha Temminck, 1825
Синонимы: Saxicola monacha Temminck, 1825; Saxicola semenowi Bianchii et Zarudny, 1900.
Наряду c Oe. alboniger — это наименее изученный вид среди всех палеарктических каменок. У самца каменки монашки угольно-черная мантия контрастирует с белоснежными шапочкой и брюшком, рулевые перья, за исключением центральных, почти целиком чисто белые. От самцов других каменок с похожим типом окраски (Ое. lugens, Ое. picata capistrata, Ое. pleschanka) самец монашки отличается в первую очередь тем, что черный цвет горла гораздо дальше распространяется вниз и охватывает все оперение груди. Самки этого вида выглядят одноцветно-песочными с желтоватым хвостом, лишенным черной вершинной полосы (рис. I41).
Каменка-монашка выглядит сравнительно крупной птицей, но впечатление это складывается скорее и j-за ее длинного хвоста и непропорционально большого клюва, что дает некоторым авторам основание сравнивать силуэт сидя шеи монашки с очертаниями турок Мcrops spp. Можно признать правомерность такого сравнения, если добавить к сказанному, что сидит монашка обычно почти горизонтально. Среди прочих каменок такая посадка наиболее характерна только для двух видов — Ое. hispanica и Ое. pleschanka. С этими видами монашку роднит также се легкое телосложение — хотя птица выглядит крупной, весит опа всего лишь около 20 г (18 — 23 г). Причина такого несоответствия между кажущейся массивностью монашки и се истинным небольшим весом складывается за счет “вытяпусти” силуэта этой каменки, обяанного се длинному хвосту и крыльям. Для сравнения можно сказать, что пустынная каменка Ое. deserti oreophila имеет такой же вес при длине крыльев 96-104 мм (в среднем 100 мм), тогда как длина кры-
Рис. 141. Каменка-монашка Oenanthi т Самцы — по кадрам видеосъемки автора н Израиле Самка (внизу) — по фот oi рафии Frcihof из: Stobcncr 1989.
293
Панов Е.Н. Каменки Палеарктики
ла Ое. monacha составляет 102 —111 мм (всреднем 106 мм).
Еще одна особенность каменки-монашки — это ее короткие и слабые лапки, которые кажутся еще короче по контрасту с удлиненным, вытянутым силуэтом. Эта черта вкупе с длинным клювом выдает в нашей каменке воз-душного охотника. Иными словами, среди
Рис. 142. Ареал Oenanthe monacha. пустынных певчих птиц
монашка занимает ту же экологическую нишу, что и щурки, в отличие от которых с равным успехом может добывать пропитание не только в воздухе, по и на земле. Укороченные ноги Ое. monacha — это еще одна черта, сближающая ее с парой близких видов Ое. hispanica— Ое. pleschanka.
Каменка монашка — вид монотипический, подвиды внутри него нс выделены (рис. 142).
15.1. Местообитания и численность
Судя по литературным данным, каменка монашка обитает в наиболее аридных и бесплодных каменистых ландшафтах типа бедлендов. В Пакистане, Южном Иране и в Аравии типичные местообитания вида — это низкогорья, пересеченные просторными сухими долинами (вади) и практически лишенные древесно-кустарниковой растительности, которую монашка, якобы, сама избегает (Cramp 1988). По мнению Корнуоллиса (Cornwallis I975), эти каменистые пустыни, простирающиеся от уровня моря до высот порядка 1400 м, из всех каменок способна занимать только монашка. Поскольку эти птицы существенную часть своего пропитания добывают в воздухе, они способны мириться с нагреванием почвы и камней выше 50°, когда наземные насекомые уходят в подземные убежища и становятся недоступными.
Мои наблюдения в Израиле рисуют несколько иную картину. Здесь монашка практически отсутствует в наиболее бесплодных участках пустыни Негев, уступая эти бедленды траурной каменке Ое. lugens. Зато интересующий нас вид становится довольно обычным па юге страны,
294
Глава 15. Каменка монашка Oenanthe monacha
вблизи г. Эйлат, где присутствие в широких каменистых долинах разреженных насаждений древесных акаций с зонтиковидными кронами вызывает в памяти картины африканских саванн (рис. 107). Здесь Ое. monacha делит жизненное пространство с довольно обычной Ое. leucopyga, причем, вероятно, лишь немногим уступает последней в численности. На маршруте по одной из таких долин длиной около 6-7 км я отметил 4 мая три особи Ое. leucopyga и трех самцов Ое. monacha. Следует заметить, что каменки-монашки не только не избегают древесной растительности, но, напротив, очень охотно садятся на верхние ветви самых высоких акаций, оставаясь здесь подчас довольно продолжительное время.
Эти наблюдения расходятся с ранее сделанными подсчетами, согласно которым расстояние между гнездящимися парами в окрестностях Эйлата составлят в среднем 7 км (Shirinai 1996). В южном Иране пара, кормившая птенцов, использовала территорию площадью около I кв. км. В этом же регионе на маршруте длиной 8 км в гнездовое время (май) отметили 5 самцов (Cornwallis, Porter, цит. по: Cramp 1988). Предполагают, что общая численность популяции Израиля составляет 100-200 пар (Shirinai 1996).
15.2. Биология гнездования
Наиболее полные сведения по размножению вида получены в Израиле. Гнездовой сезон продолжается здесь с марта до конца июня. Большинство самок приступают к откладке яиц в самом конце апреля. В качестве убежищ для гнезд используются всевозможные пустоты в каменистых вертикальных поверхностях и между валунами. Гнездо неглубокое, небольших размеров (в среднем 130 мм в диаметре). Материалом для него служит сухая трава, лоток выстлан шерстью и перьями. Величина кладки в 7 найденных гнездах варьировала от 3 до 5 яиц, преобладали кладки из 4 яиц. Цвет скорлупы — очень светлый небесно-голубой, с крошечными рыжеватыми точками, гуще на тупом конце. По Шонветтеру (Schonwcttcr I979), размеры яиц 18.2 -20.5 х 15.7 — 16.0 (ср. 19.3 х 15.9, п = 3: вес 2.6 г.). По другим данным: 2I.6 х 15.6 (Cramp I988). Последний автор намекает, что эти сведения могут и не относиться к Ое. monacha. К сожалению, их нельзя сверить с данными по Израилю, поскольку там измерения делались, вероятно, линейкой (20 х 15 мм — Shirinai 1996).
Продолжительность насиживания составляет 14-15 дней, столько же времени птенцы проводят в гнезде. Слетки появляются в заметных количествах в конце мая — начале июня (Shirinai 1996). По другим регионам сведения еще более скудны. В Иордании брачные игры на
295
Панов Е.Н. Каменки Палеарктики
блюдали в конце апреля, а самца, доставляющего корм птенцам — в конце мая. В Египте слетков встречали в начале июня (Cramp 1988).
15.3. Миграции и линька
Молодые птицы данного года рождения начинают кочевать в Израиле с середины июля. Наиболее часто их встречают за пределами гнездовых местообитаний в августе, реже — до середины сентября. Предполагается, что самцы зимуют в основном на своих территориях или неподалеку от них, тогда как самки предпринимают более протяженные сезонные миграции, проводя холодные месяцы немного далее к югу, на синайском полуострове. Вместе с тем, пара отмечена в гнездовом ареале (Nizanna) 20 декабря (Shirinai 1996).
Полная линька взрослых особей проходит с конца июня до конца августа. Линька молодежи заканчивается к середине сентября. У них сменяется мелкое оперение и часть верхних и нижних кроющих крыла (Cramp 1988).
15.4. Биологические замётки
Крайняя скудость данных по биологии и поведению каменки-монашки отчасти объяснима. Дело в том, что эти птицы выработали своеобразный, экстенсивный способ использования жизненного пространства. Мои наблюдения показывают, что в поисках корма монашки не обшаривают досконально какой-нибудь небольшой участок местности, а все время перемещаются в пределах своей обширной территории на большие расстояния, не задерживаясь нигде подолгу. Обычно каменка появляется совершенно неожиданно на вершине какой-либо из возвышеностсй. окружающих долину. Как правило, псхватаст даже времени, чтобы поймать се в поле зрения бинокля, как она перелетает через долину и сразу оказывается практически недоступной для наблюдений. Полет се стремительный, с неожиданными виражами и крутыми поворотами. Вероятно, каменка нс упускает ни единой возможности схватить подвернувшуюся ей жертву налету, так что она совмещает охоту с перемещениями по огромному кормовому участку. Судя по тому, что в период моих исследований малые щурки Merops orientalis даже в полуденный зной явно не испытывали недостатка в летающих насекомых, эти каменки также могли успешно существовать за счет воздушной охоты. Своих жертв они зачастую ловили на очень большой высоте — до 20-30 м. Кормятся они также па скалистых стенках и на земле — подобно всем прочим видам каменок. Содсржи-
296
Глава 15. Каменка монашка Oenanthe monacha
мое немногих исследованных желудков содержит обычный спектр беспозвоночных, потребляемых каменками (кузнечики, муравьи, жуки, пауки и др.). Полевые наблюдсня за птицами показывают, что они ловят также стрекоз и бабочек.
Оказавшись однажды на территории белохвостой каменки Ое. leucopyga, самец каменки-монашки немедленно подвергся преследованию и, как это уже описано, сразу улетел за гребень хребта. При этом он действовал точно так же, как ведут себя при нападении особей видов-доминантов (таких, как Ое. finschii и Ое. chrysopygia) живущие, фактически, на их территориях плешанки и испанские каменки. Они не делают ни малейших попыток ответить агрессору тем же, а просто продолжают заниматься своими делами, сообразуясь с неожиданными обстоятельствами. Очевидно, такой же оппортунизм свойственен и каменке-монашке. Мои наблюдения лишь подтверждают факт существование иерархических отношений между Ое. monacha и Ое. leucopyga, при которых первая играет роль аутсайдера, а вторая — доминанта (Jennings 1980 цит. по: Cramp, 1988). Известен также случай вытеснения самца монашки с занятой им зимней территории самцом Ое. lugens (этот вид значительно уступает в размерах крупной Ое. leucopyga) (Hartley 1949).
Разумеется, отсутствие сколько-нибудь детализированных данных относительно сигнального поведения Ое. monacha нс позволяет всерьез обсуждать се положение относительно других видов рода Oenanthe. И все же то очень немногое, что нам уже известно, заставляет меня склоняться к мысли о наибольшей близости этого вида к Ое. hispanica и Ое. pleschanka. В пользу этого предположения говорят общие черты строения, склада и повседневного поведения каменки-монашки. Нет ни одного серьезного аргумента в пользу се более тесного родства с предками “группы picata" (с которыми монашку пытается сблизить Тай — см. Тус 1989а), нежели с филумом, к которому принадлежат Ое. hispanica и Ое. pleschanka. Даже тот признак окраски оперения самцов Ое. monacha, который в своем обсуждении упоминает Тай (именно, присутствие серых оторочек на черных перьях спины) характерен также и для самцов Ое. pleschanka.
Только два факта нс вполне согласуются с моей гипотезой. Поданным из работы Cramp (1988), полость рта у взрослых Ое. monacha желтая, как у каменок “группы picata' и других, близких к ним (у плс-шапки и испанской каменки зев у взрослых птиц черный). Песня Ое. monacha, судя по описаниям и по тому, что мне мельком удалось услышать издалека, типично “дроздового” типа и в этом отношении более сходна с напевами picata и близких к ней видов, чем с песнями плешанки и испанской каменки.
297
Панов Е.Н. Каменки Палеарктики
Глава 16.
Пустынная каменка, пустынный чекан Oenanthe deserti Temminck, 1825
Рис. 143. Пустынная каменки Oenanthe deserti oreophila (а — самец, b — самка). Из: Panow I974.
Синонимы: Saxicola deserti? Temminck, 1825; Saxicola montana Koch, 18I6; Saxicola alflbifrons Brandt, 1844; Saxicola atrogularis Blyth, 1847; Saxicola salina Eversmann, 1850; Vitiflora paradoxa Brehm, 1855; Saxicola homochroa Tristram, 1859;
Самец пустынной каменка рыжевато-палевым цветом светлых участков оперения наиболее напоминает черно-горлых испанских каменок западного подвида Ое. h. hispanica var. stapazina, отличаясь от них тем, что черный цвет на горле распространяется на верхнюю часть груди и соединяется с черными сгибами крыла (рис. 143). Однако сходство это чисто поверхностное. Пустынная каменка имеет более плотное телосложение (вес около 20 г) и более длинные цевки, что, очевидно, связано с ее способностью неплохо бегать (испанская каменка почти никогда не использует этот способ передвижения, перемещаясь, как правило, “по-воробиному”, короткими прыжками).
Самки пустынной каменки окрашены в песочно-па
левые тона и по первому впечатлению весьма сходны с самками Ое. oenanthe и Ое.
298
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
isabellina. Наиболее надежный признак для опознавания самок Ое. deserti в поле — это их манера покачивать хвостом наподобие трясогузок. В отличие от Ое. isabellina. которые ведут себя в этом отношении сходным образом, пустынные каменки покачивают хвостом быстрее, с ритмичным изменением скорости движения хвоста, и, кроме того, при каждом покачивании крайние рулевые слегка разводятся в стороны. Сидящая на камне либо на веточке пустынная каменка поводит хвостом вверх и вниз примерно так же, как это делают Ое. finschii и Ое. picata: решительно опускает хвост немного ниже горизонтали и чуть медленнее приподнимает до линии туловища.
Эти первые впечатления при знакомстве с пустынной каменкой сразу же наводят на мысль, что внутри рода Oenanthe она занимает обособленное положение и нс может быть заподозрена a priori в близком родстве с каким-либо из рассмотренных ранее видов.,Весьма своеобразное, “трясогузочье” потряхивания хвостом, о котором будет еще много сказано, во многом напоминает соответствующее поведение птиц рода Cercomela. По описанию Фаркаса (Farkas личн. сообщ.), примерно такие же движения хвоста характерны для южноафриканской Cercomela/ Oenanthe tractrac. Нечто подобное я наблюдал у Cercomela melanura, которая, правда, нс потряхивает хвостом, а ритмично распускает и складывает его.
Другая особенность пустынной каменки, которую просто невозможно нс заметить — это се поразительная доверчивость. К взрослому поющему самцу можно подойти на 3-4 метра, а когда парочка занята брачными играми в районе строящегося гнезда, птицы совершенно игнорируют ваше присутствие и чуть ли не садятся па голову. Смит (Smith 1971) пишет, что самец пустынной каменки, чья зимняя территория включала в себя лагерь экспедиции, постоянно залетал в палатку и хватал кузнечиков, которые бросал ему автор, у самых его ног. Замечу, что многие орнитологи, наблюдавшие за видами Cercomela familiaris, С. tractrac и С. melanura. специально подчеркивали их необычайную доверчивость. Ничего похожего вы не увидите ни у какого другого вида каменок, кроме как у пустынной.
О близости этого вида к роду Cercomela могут свидетельствовать также некоторые особенности окраски пустынной каменки (в частности, почти целиком черный хвост), структура и характер развития песни (см. ниже) и некоторые другие особенности биологии и поведения.
16.1. Систематические замечания
Внутри вида принято выделять четыре подвида: Ое. d. deserti, Ое. d. homochroa, Ое. d. atrogularis и высокогорная Ое. d. oreophila (рис. 144).
299
Панов Е.Н. Каменки Палеарктики
Рис. 144. Ареал пустынной каменки Oenanthe deserti. Точками показаны залеты, серым цветом — зимовки (1 — подвида homochroa. 2 — deserti и atrogularis. 3 — oreophila).
Самыми мелкими размерами характеризуется населяющая северо-западную Африку homochroa (средняя длина крыла самцов 91.2 мм). Размеры птиц увеличиваются с запада на восток, в направлении расселения вида: у переднеазиатских deserti крыло самцов 92.3 мм, у среднеазиатских atrogularis 95.4 мм (вес 19.5-20.7 г), у центральноазиатская oreophila 100.1 мм (вес 23.1 г). У двух восточных подвидов, в отличие от homochroa и deserti. менее развиты рыжеватые тона в окраске мантии и более — на груди, оперение верхней части головы светлее. К востоку увеличиваются белые поля на внутренних опахалах маховых перьев и уменьшается развитие черного на хвосте. Некоторые орнитологи выделяют подвид salina (Средняя Азия и Казахстан), но большинство систематиков не признает его реальности (см. Vaurie 1949, Степанян 1978, Cramp 1988).
Птенцы в гнездовом наряде похожи на самок, отличаясь от них слабой псстристостью.
16.2. Местобитания и численность
Наиболее характерная особенность вида — это его способность селиться в совершенно бесплодных ландшафтах, куда не решается проникнуть ни один другой вид каменок. Таковы, в частности, участки гравийной пустыни в северной Ливии, на плоскогорьях южного Алтая и восточного Памира (рис. I45). Эти места почти безжизненны, практически лишены какой-либо растительности и характеризуются весь-
300
Etaea 16. Пустынная каменка, пустынный чекан Oenanthe deserti
Рис. 145. Местообитания пустынно!! каменки Oenanthe deserti oreophila в высокогорьях восточного Памира.
Рис. 146 Местообитания пустынной каменки Oenanthe deserti atrogularis в песках долины р. Вахш (Таджикистан). Из: Panow 1974.
301
Панов Е.Н. Каменки Палеарктики
ма бедной орнитофауной. Например, на Памирском плато на этих мертвых землях помимо пустынной каменки селится в заметном количестве лишь рогатый жаворонок Eremophila alpestris.
Другой тип местообитаний — это песчаная пустыня, ровная или бугристая, нередко с пышными зарослями тамариска или саксаула (рис. 146). В песчаных пустынях южного Казахстана пустынная каменка проникает в леса из черного саксаула Arthrophytum ammodendron, отдельные деревья которого достигают 80 см. в диаметре и 8 м в высоту. Здесь Ое. deserti и Ое. isabellina обитают на песчаных “полянах”, устраивая гнезда в норах тушканчиков и больших песчанок Rhombomis opimus, а первый вид использует в качестве гнездовых убежищ дупла саксаула либо укромные места под обильным здесь древесным валежником (Шнитников 1949). Гнездится пустынная каменка и в глинистой пустыне, причем в Каракумах она предпочитает именно такие места, тогда как в Кызылкумах и в пустынях Казахстана отдаст предпочтения песчаным участкам. В песках эта каменка обитает и во многих регионах северной Африки и Передней Азии.
Популяции каждого из подвидов могут гнездиться в широком диапазоне высотных поясов — от равнин до высокогорий. Например, каменки расы deserti размножаются в Израиле как в холмистой пустыне Негев, так и высоко в горах (1650-2400 м над у. м.). В южном Иране размножающаяся пара подвида atrogularis встречена на высоте 2950 м.
Рис. 147. Местообитания пустынной каменки Oenanthe deserti atrogularis в Чуйской степи (южный Алтай).
302
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
Популяции того же подвида в южном Алтае и в Туве распространены в гнездовой сезон по самым различным местообитаниям — от каменистой опустыненной степи на подгорных плато (рис. 147) до субальпийского пояса (1500-2000 м над у. м.). Наконец, подвид oreophila предпочитает на Памире области на высотах 3600—4300 м. В благоприятные годы эти каменки доходят до 4550 м, а в Тибете и Ладакс они гнездятся до высот 4950 м (Потапов 1966, Desfayes, Praz 1978, Иваницкий 1980а, Нейфельдт 1986, Shirinai 1996).
Пустынные каменки обычно нс образуют плотных поселений. Типичная для формы atrogularis численность в Туве — от 1 до 10 пар на один кв.км. В наиболее предпочитаемых местообитаниях в этом регионе среднее расстояние между ближайшими гнездами составляет 288 м (минимальное — 168 м). Вероятно, с наибольшей для этого вида плотностью селятся популяции расы deserti в некоторых районах Израиля. Здесь компактные поселения (до 27 пар на площади 6 кв. км. со средними расстояниями между гнездами около 100 м) перемежаются с почти незаселенными угодьями. Довольно высока также плотность в популяциях oreophila, занимающих каменистые террасы вдоль долин горных речек в Восточном Памире. В таких местах на маршруте длиной 9 км можно насчитать до 12-15 пар пустынных каменок (Потапов 1966, Иваницкий 1980а, Shirinai 1996).
16.3. Годовой цикл и сроки гнездования
В южных участках ареала (Африка, Передняя Азия) популяции частично оседлы, а мигранты из числа местных особей начинают возвращаться к местам гнездования уже в конце февраля. В северо-западной Африке гнездо с кладкой нашли уже Ю марта (здесь многие пары, вероятно, успевают вывести птенцов дважды за сезон). В Израиле наиболее ранние сроки начала гнездования — это конец марта, хотя большинство пар приступает к размножению не ранее середины апреля. В Сирии гнездо с 4 свежими яйцами найдено 21 апреля, в восточном Иране гнездо с единственным только что отложенным яйцом — I7 апреля (Sarudny 1903; Meincrtzhagen 1954; Heim de Balsac, Mayaud 1962; Shirinai 1996). С начала марта первые пролетные самцы начинают появляться и много севернее, в Среднеазиатских республиках бывшего Советского Союза: в юго-восточном Туркменистане и в пустыне Каракум (с 6 — 8.III), в Кушке (с 11.III), в южном Казахстане (с II-14.III). Несколько позже прилетают первые самцы в северо-западный Казахстан (Мангышлак, I — 6.IV), когда в этих местах еще случаются заморозки и сильные снежные бураны. В высокогорьях Памира самцы появляются с I6.IV, а самки — с 26.IV (Меклепбурцев I946). Пролет
303
Панов Е.Н. Каменки Палеарктики
продолжается в равнинных районах до начала апреля, а в высокогорьях — еще и в мае.
Несмотря на то, что пустынная каменка рано достигает мест гнездования (опережая такие виды, как Ое. oenanthe, Ое. hispanica и Ое. pleschanka), она размножается в северных частях своего ареала, в общем, позже всех прочих видов каменок. На это обратил внимание еще Митропольский (1968), который пишет, что гнездовой период пустынной каменки на полуострове Мангышлак очень растянут, и что пара едва ли успевает вырастить за лето более одного выводка. В равнинных районах Средней Азии и Казахстана лишь у немногих пар молодые покидают гнезда к началу последней декады мая (17 — 23.V). Из 33 гнезд, найденных в этих районах (между 42 и 48° с. ш.) в мае и в начале июня, только в восьми были птенцы, тогда как прочие содержали кладки. Пять гнезд с кладками найдены в первой декаде мая. 6 — во второй, 7 — в третьей. 4 — в первой декаде июня и одно — во второй декаде этого месяца.
Интересно сопоставить сроки прилета и размножения Ое. deserti и Ое. oenanthe в горных полупустынях южного Алтая, в Чуйской степи на высотах около 2 тыс. м над уровнем моря. В 1969 г. пустынные каменки были здесь многочисленны 5 мая, и некоторые уже сформировали пары. Самки начали подыскивать убежища для гнезд. Одна из них облюбовала подходящее место к 10 мая, но гнездо начала строить нс ранее 17 мая. Все эти дни по ночам было холодно, так что лужи покрывались за ночь слоем льда, который стаивал только часам к II утра. Первые самцы обыкновенной каменки были замечены здесь только 7 и 8 мая; 10 мая я наблюдал образование пары, а 17 и 18 мая две самки уже строили гнезда. На следующий год в 20-х числах июня у многих пар обыкновенной каменки здесь уже были слетки, а другие кормили птенцов в гнездах. Что касается пустынных каменок, то у них большинство самок в это время насиживали, и только 29 июня я встретил кочующий выводок. В 1971 г. в те же сроки у обыкновенных каменок были большие птенцы, и у многих пар молодые уже оставили гнезда. Болыиинстьво самок пустынной каменки еще насиживали кладки, и лишь у одной пары были птенцы.
В том же районе в другие годы самку, строящую гнездо, видели 16 мая, неполные и нснасижспныс кладки находили 5-7 июня, гнезда с насиженными яйцами — 4 и 5 июня. Недавно вылупившихся птенцов отмечали 3 и 16 июня, птенцов перед вылетом — 7 и 14 июня, слетков — с 22 июня по I июля. Пустынные каменки определенно гнездятся здесь только один раз за сезон (Лоскот 1986а, Нсйфельдт 1986). В других северо-восточных участках ареала atrogularis, где условия не столь суровы, как в Чуйской степи, гнездование начинается в более ранние
304
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
сроки. В Туве, на высотах 1600-1700 м над у. м. формирование пар заканчивается к 30 апреля, вылупление птенцов начинается с 25-26 мая. Размножение разных пар Ое. deserti идет здесь наименее синхронно по сравнению с тремя другими местными видами каменок (Ое. oenanthe, Ое. isabellina и Ое. pleschanka), так что некоторые самки насиживают яйца еще в июне и в июле (рис. 24 В). Далее к югу, в Монголии, наиболее ранние гнезда с кладками находили 1 и 12 мая, птенцов в возрасте 3-4 дня — 20 мая (Иваницкий 1980а, Picchocki et al. 1982,).
В высокогорьях Памира первые гнезда с яйцами можно находить с середины мая (12.V, 23.V), начало массовой откладки яиц датируется первыми числами июня, первые выводки встречаются с 25 — 30 июня, а массовый вылет молодых из гнезд происходит впервой декаде июля. В этих районах в конце мая по ночам бывают заморозки, и птенцы пустынной каменки иногда гибнут от переохлаждения. Каменки размножаются здесь один раз в сезон, так что поздние кладки принадлежат, вероятно, парам, у которых первое гнездо по какой-либо причине погибло (Абдусалямов 1961; Потапов 1966).
16.4. Социальное поведение
Будучи обитателем самых бесплодных и бедных в кормовом отношении местностей, пустынные каменки селятся здесь очень разреженно, на расстоянии от нескольких сот метров до километра от пары до пары. В таких условиях границы территорий практически нс выражены, и соседние пары редко контактируют друг с другом. Даже в тот период, когда пара недавно сформировалась, и самка присмотрела подходящее укрытие для гнезда, хозяев участка не всегда легко застать в этом месте. Партнеры посещают его лишь несколько раз в течение дня, а все остальное время проводят в поисках пищи, держась вместе либо порознь. Совершая эти охотничьи экскурсии вдали от гнездового убежища, птицы могут случайно нарушить границу чужого участка, и лишь тогда мы становимся свидетелями территориального конфликта.
Т е р и т о р и а л ь н о е поведение самцов. Столкновения между самцами чаще можно наблюдать в период пролета, когда обосновавшиеся на своих территориях самцы охраняют их от мигрирующих собратьев. В тех местах, где плотность этих каменок выше, конфликты между самцами случаются достаточно регулярно в период формирования пар. Например, в некоторых долинах центрального Памира пара от пары живет на расстоянии 100 — 200 м, и здесь в начале мая между самцами то и дело вспыхивают драки (Потапов 1966). Впрочем, при такой же примерно плотности в Туве Иваницкий (неопубл.) за 49
305
Панов Е.Н. Каменки Палеарктики
Рис. 148. Фрагменты сигнального поведения пустынной каменки Oenanthe deserti oreophila. а, а’ — позы самца при конфликтах на почве конкуренции из-за территорий (и, возможно, при формировании пары); b, с, d — позы самцов при пограничных конфликтах; е. f — конфликт самок на границе их территорий; g, i — поведение самца при взаимодействиях с самкой па стадии консолидации пары. Из: Panow 1974.
ч. наблюдений на стадии гнсздостроепия насчитал всего 19 пограничных конфликтов с участием 6 самцов..
Самцы пустынной каменки пытаются иногда изгонять со своей территории особей других видов каменок, а также горихвосток Phoemcurus
306
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
spp., черноголовых чеканов Saxicola torquata, белых трясогузок Motacilla alba, и других мелких птиц. Впрочем, пустынные каменки гораздо реже нападают на своих соседей из числа прочих видов каменок, нежели сами подвергаются их нападениям. Например, самец Ое. isabellina за 2.5 ч. 24 раза нападал на самца Ое. deserti на территории последнего, и 6 раз — на его самку (Иваницкий 1980 Ь). Пустынные каменки относятся стоически к таким постоянным преследованиям, и только отлетают от агрессора на 2-3 м. Во время столкновений с особями своего вида самцы принимают позы, изображенные на рис. 148 b-d.
Рис. 149. Некоторые варианты рекламной песни Oenanthe deserti (а — с, южный Алтай, Чуйская степь) и двух самцов черпохвостки Cercomela melanura (d, е, Израиль). Оригинал.
Все эти демонстрации сопровождаются покачиванием развернутого или сложенного хвоста, непрерывным либо прерывистым. Если самцы очень возбуждены, они одновременно взлетают навстречу друг другу и сближаются вплотную, стараясь вцепиться в противника когтями. Вслед за
этим начинается длительная погоня в воздухе; когда один из самцов садится, другой пикирует на него с развернутым хвостом, а сидящий на земле вздергивает хвост и резко, многократно отводит крылья в стороны. Бывает, что один из соперников совершает подле другого полет петлями, либо беспорядочно носится низко над землей, проделывая неожиданные виражи и повороты. Самцы конфликтуют или молча, или же поочередно произносят короткую стереотипную песенку (рис. 149 а — с). Во время погони слышны тихие прерывистые звуки, нечто вроде “цх... цик... кха кха...”.
Территориальность самок. Иногда в конфликтах принимают участие и самки, которые молча демонстрируют друг перед другом, нс ввязываясь в ссору своих супругов. Самки распушают
307
Панов Е.Н. Каменки Палеарктики
оперение и мягко припадают грудью к земле около кустика или камня, поворачиваясь при этом на 90°то в одну, то в другую сторону или поочередно залезают в попадающиеся на их пути отверстия (рис. 148 c,f). Бывает и так, что каждый из самцов, прерывая на время конфликт с соседом, старается оттеснить свою самку от границы в свои владения. Конфиктуя друг с другом, самки могут произносить короткую свистовую фразу, отдаленно напоминающую песню самца, но слышал я этот звук только однажды: хозяйка строящегося гнезда возвратилась к нему с очередной порцией строительного материала и застала поблизости другую самку. Бросившись на при шел ину и широко развернув хвост, хозяйка издала звонкое “пейви", а затсм- угрожающий крик (сигнал N 4, звучащий у пустынной каменки более хрипло и гнусаво, чем у других представителей рода).
С а м о р е к л а м и р о в а н и е с а м ц о в. На протяжении всего брачного периода самец постоянно поет на одном из двух-трех камней, которые служат его песенными постами. Песня пустынной каменки имеет очень мало общего с гем, что мы видели у других видов каменок. Она коротка, полностью лишена имитаций и почти неизменно включает в себя своеобразное короткое потрескивание. Из всех известных мне мелких видов дроздовых эта песня по простоте своей структуры наиболее напоминает песню чернохвостки Cercomela melanura (рис. 149 d, е). Каждый самец имеет в своем репертуаре до 7-10 вариантов такой короткой песни, которые он чередует друг с другом, сидя на песенном посту либо совершая весьма характерный демонстративный полет: мерно взмахивая крыльями, птица с песней поворачиват в горизонтальной плоскости то в одну, то в другую сторону, затем резко взмывает вверх, на мгновение повисает на одном месте и сразу же, еле! ка развернув рулевые, пикирует вниз. Затсм вес то же повторяется снова. Во время пике слышна глхчая трескучая трслька (рис. 15 i, 149 b). Приверженность самцов пустынной каменки к этим воздушным демонстрациям довольно велика, и в этом отношении их поведение наиболее сходно с тем, что мы видели у Ое. hispanicka, хотя сам полет у этих двух видов выглядит совершенно по разному. У 6 самцов Ое. deserti Иваницкий насчитал 147 полетов. Самец, которому исследователь посвятил максимум времени (около 20 ч ), проделал за это время 95 полетов (4.8 полета в час).Самка этого самца строила гнездо, так что близилось время копуляций.
Развитие в з а и м о т н о ш е н и й м е ж д у член а м и пары. Самого момента формирования пары мне наблюдать не удалось, но я предполагаю, что оно сопровождается позами, изображенными на рис. 148 a, g. При этих демонстрациях, как и при территориальных конфликтах, самец все время мерно покачивает широко раз-
308
Гмма 16. Пустынная каменка, пустынный чекан Oenanthe deserti
Рис. 150. Элементы поведения пустынной каменки Oenanthe deserti в период гнездостроения и подготовки к копуляции, а — наиболее характерная поза самца в этот период, b — трепетание крыльями в присутствии самки, с — преследование самцом самки, прилетевшей к гнезду со строительным матери* алом, d, е — позы самца перед и во время “копуляции" с камнем. По зарисовкам В. В. Иваницкого (Тува).
верпугым хвостом, иногда проделывает вокруг самки полет петлями. Самка первое время агрессивна, и при попытке самца приблизиться к пей встречает его угрозой клевка. Затем птицы начинают осматривать ниши и норы, пригодные для устройства гнезда. Пока самка осматриваем? убежище изнутри, самец бегает рядом, слегка приподняв голову, покачивая хвостом (рис. 148 а) и произнося в пол-голоса трескучую трельку — ту самую, что сопровождает демонстративный полет. Своеобразие брачного поведения пустынной каменки, во многом непохожего на соответствующие демонстрации у прочих видов каменок, усугубляется также особым движением хвоста вбок (рис. 148 а’)
Когда подходящее место для гнезда выбрано, самка еще долго не приступает к гнездостроению (иногда — до 10 дней, а то и больше), но птицы несколько раз на день посещают это убежище. Иногда они появляются вместе, или же самец прилетает первым и подолгу пост, сидя поодаль и поджидая самку. В это время можно слышать иной гип песни. Она сплошная, протяженная, довольно разнообразная, и вклю
309
Панов Е.Н. Каменки Палеарктики
чает в себя звуки разной высоты, но самым характерным ее элементом оказывается все та же трескучая трелька. Поет эту песню самец обычно в пол-гол ос а. В целом она несравненно беднее, чем соответствующий ей тип удлиненной песни у всех палеарктических каменок.
Когда самка, наконец, после долгого отсутствия возвращается к месту будущего гнезда, она при виде супруга обычно проделывает короткий беспорядочный полет, а затем ныряет в отверстие. Самец подлетает сюда же, пригибает грудь к земле, опускает голову и мелко трепещет слегка отставленными крыльями (рис. 148 i, 150 b). Иногда он при этом наполовину залезает в отверстие, так что наружу торчит только приподнятый хвост. В это время можно слышать короткую звонкую трсльку (гомологи которой есть в репертуаре некоторых других видов каменок) либо тихое глухое “трюв-трюв”. Если же самец поджидает супругу у входа, а она долго остается в убежище, самец взлетает выше, на край обрыва, запевает свою обычную извещающую песню, а иногда хватает клювом камешек и, проделав несколько воздушных эволюций, улетает с ним прочь.
Когда подходит время спаривания, птицы начинают вести себя несколько иначе. Они подолгу кормятся неподалеку друг от друга, перелетая при этом с места на место особым замедленным трепещущим полетом, и при посадке складывают крылья не сразу, а на две-три секунды задерживают их в полурасправлсном положении. Контурное оперение самца сильно распушено, голова втянута в плечи, он всем своим обликом напоминает пушистый шарик (рис. 150 а). Внезапно самец подлетает вплотную к самке, но та не улетает, а лишь отбегает от самца, который торопливо семенит вслед за ней. При этом самка, а порой и самец быстро трепещут слегка отставленными крыльями (рис. 150 Ь, с). Такая “игра в догонялки” подчас продолжается довольно долго, причем самка иногда бегает вокруг кустика, а самец упорно следует за ней. Бывает так, что самка как раз перед началом этих игр набрала очередную порцию шерсти для постройки гнезда, и тогда она “бегает наперегонки” с самцом, держа свою находку в клюве. Лишь однажды я видел, как такие преследования переросли в нечто вроде замедленного совместного полета тандемом, уже описанного ранее в очерках о Ое. isabellina, Ое. hispanica и Ое. pleschanka. При этом самка время от времени издавала предупреждающую трескучую позывку, нечто вроде “жсжсжсжс” (сигнал N 4). Инициатором таких ухаживаний может быть не только самец, но и самка, если она по собственной инициативе подлетает вплотную к кормящемуся самцу.
Взаимодействия супругов в этот период могут выглядеть и несколько иначе. Например, самка проделывает нечто вроде короткого демонстративного полета, садится у подножия обрыва и начинает раз за
310
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
разом залезать в неглубокие отверстия, непрерывно трепеща при этом крыльями. В некоторых случаях и самец проделывает то же самое неподалеку от подруги. Самка особенно интенсивно трепещет крыльями каждый раз, когда она после пребывания в норке появляется на поверхности. В такие моменты самец иногда пытается взлететь на самку, но та издает предупреждающий сигнал и улетает. Подобные попытки садок на самку без длительных предварительных демонстраций самца очень часто наблюдаются у Ое. deserti. но совершенно нс характерны для всех прочих видов каменок. Стоит отметить, что самки пустынной каменки весьма терпимы к такому нарушению индивидуальной дистанции, нс в пример самкам других видов Oenanthe. весьма ревностно оберегающим свое “персональное пространство”.
Иваницкому, подробно описавшему эти брачные игры (нсопубл.), так и не удалось увидеть сам акт спаривания. Однако все говорит о том, что происходит она, в общем, так же, как и у других видов каменок. В момент готовности к спариванию самец устремляется к ближайшей вертикальной поверхности и остается там, без конца повторяя какой-либо из самых простых вариантов своей короткой песни. Затем его поза внезапно меняется: все оперение головы и туловища плотно прижато, клюв приподнят под углом, хвост широко развернут веером. В этой эффектной позе самец начинает перелетать от одной ниши к другой, ненадолго задерживаясь в каждой из них. Если в этот момент сюда прилетает самка, самец проделывает около нее полет петлями и садится в уже описанной позе, природняв развернутый хвост над уровнем туловища и мелко трепеща крыльями. Иваницкому не пришлось видеть сам момент садки, но он наблюдал “ложную копуляцию” самца с камнем, во время которой поза самца ничем существенным нс отличалась от того, что мы видим в аналогичных ситуациях у самцов других видов каменок (рис. 150 d, с).
16.5. Гнездо, кладка, выводок
Г нездовые убежища. Каменки подвида atrogularis чаще всего строят гнезда в горизонтальных норах глинистых стенок оврагов на высоте 0.9 — 2.8 м (обычно 1.0-1.5 м). В качестве гнездовой ниши иногда используются также норы грызунов, дупло саксаула, горизонтально лежащая металлическая труба и т.д. Однако, при отсутствии нор, ниш и прочих укрытий такого рода, эти каменки, в отличие от всех прочих (за исключением Ое. h. hispanica) могут довольствоваться убежищами совершенно иного типа. Так, известны гнезда, свитые под поваленным деревом; под нависшим пластом высохших водорослей, зацепившимся за маленький кустик; в углублении под куртиной зла-
311
Панов Е.Н. Каменки Палеарктики
ка, и в других по-добпых местах. Одно из найден
ных мною гнезд располагалось в неглубокой норке между корнями колючего кустарника, в самой его гуще. Чтобы добраться до этого гнезда, мне пришлось при помощи острого ножа
преодолеть около Рис. 151. Расположение гнезд пустынной каменки. Из: полутора метров Panow 1974. густых, непрохо-
димых кустов (рис. 151). Существенно и то, что даже в том случае, когда выбранная самкой нора достаточно глубока, птица сплошь и рядом строит гнездо у самого ее края, так что постройка хорошо видна снаружи. Из 12 гнезд, найденных Сурвилло (1968). 3 были свиты на расстоянии 5 см от входа, 4 — до 10 см, 4 — до 15 см. и лишь одно — на глубине 35-40 см.
Все сказанное может свидетельствовать о том, что ближайшие предки пустынной каменки гнездились полуоткрыто, наподобие некоторых видов чеканов Saxicola. Подобный же способ расположения гнезд часто наблюдается у разных представителей рода Cercomela (например, у С. schlcgeli. С. sinuate, С. (Ое. fracfrac).
В высокогорьях Памира каменки расы oreophila. подобно большинству других представителей рода, обычно устраивают гнезда в пустотах каменистого субстрата. Из 27 найденных здесь гнезд 11 располагались в щелях и нишах скал, 6 — под камнями на склонах, 3 — в каменных стенах загонов для скота и только 6 - в норах земляных обрывов (как у формы afrogularis) (Потапов 1966).
Г н е j д о обычно выглядит искусно сплетенным и аккуратным. Степки гнезда состоят из трех слоев: наружный, более рыхлый слой содержит грубые стебельки полыни, тсрескспа, лубяные волокна, метелки и листья злаков. Средний слой сплетен из более мягких и нежных стебельков, во внутреннем присутствует значительное количество волоса, шерсти, растительного пуха, реже встречаются мелкие перья. При отсутствии таких мягких материалов их заменяют самые тонкие растительные волокна и корешки. Как правило, внутренний слой очень плот-
312
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti ный и хорошо спрессован, хотя иногда попадаются и небрежно сплетенные гнезда. Размеры гнезд также варьируют. Их диаметр 125 — 180 мм, высота — 50 — 85 мм, диаметр лотка 45 — 75 мм, глубина его — 20 — 50 мм. Наиболее искусно свитые гнезда невелики и имеют толстые, плотные стенки. Под гнездом или около него нс бывает никаких следов платформы из камешков или палочек. Строит гнездо только самка, которая затрачивает на эту работу от 3 — 4 до 10 дней.
Кладка содержит обычно от 4 до 6 яиц, но повторные кладки взамен утерянных могут состоять из 2-3 яиц. Средний размер кладки увеличивается в направлении с юго-запада на северо-восток, но, судя по всему, вторично уменьшается в высокогорьях. Соотношение величины кладок в Северной Африке (раса homochroa): 3 х 7, 4 х 36, 5 х 14 (ср. 4.1, п = 57); в Иране (раса atrogularis) 4x2, 5x5, 6x2 (ср. 5.0, п=9); в равнинных районах восточного Казахстана (раса atrogularis, только ранние кладки): Зх 1, 4 х 1,5x4, 6x6 (ср. 5.25, п = 12); в 500 км восточнее, в Чуйской степи (atrogularis, 2000 м над у.м.): 4 х 3, 5 х 7, 6 х 2 (ср. 4.92, п = 12); в Монголии (atrogularis): 4 х 1, 5 х 2, 6 х 1 (ср. 5.0, п = 4). У высокогорной расы oreophila кладка содержит 3-5 яиц, обычно 4 (Zarudny, Harms 1926, Heim de Balsac, Mayaud 1962, Piechocki, Bolod 1962, Потапов 1966, Сурвилло 1968, Piechocki et al. 1982, Лоскот 1986b, Нейфсльдт 1986). Потапов (1966) приводит свои наблюдения над гнездом Ое. d. oreophila, содержавшим 5 птенцов, которые явно голодали и, несмотря на сравнительно большой возраст, имели плохо развитое оперение. Очевидно, в суровых условиях горных пустынь Памира пара пустынных каменок может успешно выкормить не более 4 птенцов.
Размеры яиц подвида deserti: 18.6 — 21.5 х 14.0 — 16.0 (ср. 19.9 х 14.9, п = 43, вес 2.35 г); подвида atrogularis из Ирана 15.2 — 17.0 х 19.7 — 21.5 (ср. 20.55 х 15.76, п=25); того же подвида из равнинного Казахстана: 18.1 — 21.2 х 14.2 — 16.0 (ср. 19.9 х 15.2, п = 38); того же подвида из Чуйской степи 19.6 — 22.0 х 15.0 — 16.8, п = 24 (ср. 20.8 х 16.1, п = 9); того же подвида из Монголии: 18.1 — 20.0 х 14.3 х 15.4 (ср. 19.1 х 14.9, п = 10, вес 2.17 г); подвида oreophila: 20.0 — 22.4 х 15.0 — 16.0 (ср. 20.8 х 15.5, п = 7). Выборка без указания мест коллсктирования с территории бывшего Советского Союза: 20.6 х 15.6 (п = 64) (Zarudny, Harms 1926, Piechocki, Bolod 1962, Потапов 1966, Сурвилло 1968, Schonwcttcr 1979, Piechocki et al. 1982, Лоскот 1973b, 1986b, Нейфсльдт 1986).
Скорлупа яиц окрашена в яркий голубой цвет с зеленоватым оттенком и более или менее сильно испещрена красноватыми точками и мелкими пятнышками. Встречаются кладки с бледно-голубыми яйцами.
Насиживает самка в течении 13— 14 дней. Насиживание начинается после откладки последнего яйца. В одном случае в высокогорном Памире на откладку 4 яиц самка затратила 8 дней, и вылупление птен-
313
Панов Е.Н. Каменки Палеарктики
Рис. 152. Некоторые позывки Oenanthe deserti. а — при тревоге у гнезда (сигналы 1 и 3), b - другой вариант тревожной позывки, с — позывка птенца, d — вокализация птенца при выпрашивании корма у родителей (Чуйская степь). Оригинал.
нов растянулось на 4 дня (Потапов 1966, Абдусалямов 1973). Самец не кормит насиживающую самку и обычно держится вдали от гнезда.
П т е н ц ы. По распредлению эмбриональсного пуха, светлосерого у птенцов пустынной каменки, этот вид не отличается от прочих видов рода. Зев желтый, ротовые валики белесые. Кормят птенцов оба
314
Глава 16. Пустынная каменка, пустынный чекан Oenanthe deserti
родителя. Гнездо молодые покидают па тринадцатый — четырнадцатый день. Позывки птенцов при выпрашивании ими корма сильно отличаются от соответствующих звуков у прочих видов каменок (рис. 152 с, d). Первый гнездовой наряд выглядит пссочно- либо мышино-серым со слабо заметной пестристостью. Тревожные позывки родителей, которые они издают около гнезда с большими птенцами, весьма своеобразны и напоминают скорее звуковые сигналы горихвосток Phoenicurus.
В неволе самец, взятый из гнезда, впервые начал упражняться в пении в возрасте около 3 недель (считая от момента вылета). Первые опыты состоят в повторении с небольшими вариациями птенцовой позывки (рис. 152 d), которая впоследствии, судя по всему, преобразуется в характерную трескучую трель — этот постоянный элемент песни взрослых самцов (рис. 149 Ь). Еще через три недели птица уже поет нечто вроде подпесни и впервые проделывает демонстрацию “полета петлями”, сопровождая се громким, возбужденным пением. Молодые каменки, живущие в клетке, купаются крайне редко и неохотно, а в сильную жару только мочат в поилке лоб. Когда пьют, очень своеобразным движением “клюют” воду почти отвесно опущенным клювом. Оперение их сохнет очень плохо, и попавшая под дождь каменка чувствует себя явно не в своей тарелке. От других каменок пустынные отличаются поразительной невозмутимостью. К своим сожителям по вольере (каменкам других видов) пустынная каменка подходит без всякого колебания и отнимает у них корм. Если тс пытаются противиться, просто угрожает им, беззвучно открывая клюв. При кормлении, в отличие от других каменок, берет очень мелкие порции корма.
16.6. Линька и отлет на зимовки
Молодые ранних выводков, заканчивающие линьку во взрослый зимний наряд, начинают попадаться уже с первых чисел июля. Самец, взятый птенцом из гнезда 23 июня, почти полностью сменил мелкое оперение в середине сентября. Наиболее ранние сроки линьки взрослых птиц — это вторая половина июня, а в первой декаде августа многие заканчивают линьку, сменив не только мелкое оперение, но также перья крыльев и хвоста.
Послсгнсздовые кочевки начинаются уже в июле, а отлет на зимовки из северных районов ареала — в конце августа-началс сентября. Самые поздние встречи пустынных каменок в Среднеазиатских республиках датируются последними числами октября. В Казахстане последних пустынных каменок видели 18—20.Х. В период осеннего пролета пустынные каменки иногда держатся разреженными группами до
315
Панов Е.Н. Каменки Палеарктики
15 особей и более. Прилет на места зимовок происходит в сентябре-октябре. Местообитания в зимние месяцы столь же разнообразны, как и в гнездовой сезон.
16.7. Биологические заметки
Существующие перечни кормов пустынной каменки не отражают своеобразия кормового поведения этого вида. Наряду с обычными способами добывания корма» общими у пустынной каменки с прочими представителями рода (высматривание насекомых с присады или при зависании низко над землей, выкапывание их клювом из почвы, ловля в воздухе), существуют и другие, которые мне не приходилось видеть у других видов каменок. Например, при массовом вылете комаров Tipulidae птица ловит их быстро одного за другим, набирая в клюв пучками. При этом опа очень быстро летает по ломанной линии над самой землей, совершая резкие броски то в одну, то в другую сторону, либо, пригнувшись грудью к земле и подняв хвост почти вертикально, стремительно скачет по ровной поверхности почвы, время от времени переходя от передвижения прыжками к быстрым перебежкам. Поймав крупное насекомое (например, саранчовое прямокрылое Bryodema), каменка поедает се, отрывая маленькие кусочки. Описанная манера охоты во многом напоминает соответствующее поведение южноафриканской Ое. (Myrmecocichla) bifasciata, с которой Ое. deserti во многом сходна и по окраске (см. Туе 1988b, 1989b).
Все те черты своеобразия пустынной каменки, которые я попытался подчеркнуть в этом очерке, нс позволяют рассматривать ее как близкого родича какого-либо из видов, рассмотренных мною ранее. Строго говоря, по особенностям своего поведения эта каменка в равной степени далеко отстоит и от Ое. finschii, и от Ое. xanthoprymna, с которыми Ое. deserti в свое время пытались сближать разные авторы (Vauric 1949, Cramp 1988, Тус 1989а, и др.). Можно было бы говорить о некоторых чертах сходства между Ое. deserti и Ое. hispanica (Козлова 1975), но и эти два вида столь далеки друг от друга, что не могут быть объединены даже в составе некой единой ‘Труппы видов” (species group).
316
Заключение
Заключение
Как было сказано в предисловии к этой книге, сс автор рассматривает род Oenanthe в качестве благодарного модельного объекта, на примере которого удастся решать задачи, актуальные для современной зоологии и эволюционной теории, проверяя, в частности, справедливость ряда гипотез, сформулированных в рамках последней.
Как объект сравнительно-эволюционных исследований род Oenanthe удобен своей компактностью: большинство его представителей характеризуются значительной униформностью внешней морфологии. Среди них нет видов, которые уклонялись бы очень резко по размерам и пропорциям от некоего среднего типа. Показательно также единообразие кариотипа и эмбрионального птерилозиса у всех видов, исследованных к настоящему времени в отношении этих признаков, показатели которых отделены четким хиатусом от структуры соответствующих признаков у таксономически близких групп (таких, например, как роды Saxicola и Monticola).
С другой стороны, нс исключено, что род каменок не отграничен столь же четко от некоторых африканских родов мелких дроздовых, с которыми каменки, судя по всему, должны иметь общие корни. О размытости границ между родом Oenanthe, с одной стороны, и родами Cercomela и Myrmecocichla, с другой, говорит отсутствии единогласия относительно принадлежности ряда видов мелких африканских дроздовых к тому или иному из названных таксонов.
Компактность рода указывает на его относительную молодость, что позволяет допустить сопоставимость масштабов эволюционного возраста по крайней мерс большинства представителей группы и, таким образом, обещает сопоставимость сравнительных данных и возможность правдоподобных реконструкций сс истории. Следует, однако, заметить, что, говоря о компактности рода каменок, автор нс имеет в виду отсутствия в нем какой-либо морфо-биологическое дифференциации на надвидовом уровне. В действительности, в пределах группы выделяется весьма компактное, гомогенное “центральное ядро” каменок-петрофилов (9-13 видов, по разным оценкам), “сателлитное ядро” (3-4 вида) и неоднородная периферия (7-10 видов, в отношении части которых нс исключена принадлежность к упомянутым выше африканским родам — см. рис. 1).
В пределах обоих ядер, а также “периферии” наравне с далеко ди-вергировавшими “хорошими видами” присутствуют также кластеры форм, очень близких друг другу генеалогически и зачастую рассматриваемых в качестве “неполных видов”, алловидов (allospccics) или полувидов (semispecies).
317
Панов Е.Н. Каменки Палеарктики
Род каменок сформировался на Африканском континенте. Расселение палеарктических представителей рода шло в северном и северо-восточном направлении, в те районы Евразии, где пригодные для каменок местообитания были свободны от потенциальных конкурентов сходного экологического облика
Сравнительный анализ биологии и поведения палеарктических представителей рода позволяет следующим образом резюмировать наиболее существенные результаты проведенной ревизии, которые, вероятно, могут быть экстраполированы на другие подразделения класса птиц и с этой точки зрения имеют определенную предсказательную ценность.
1. При поэтапном освоения расселяющимися видами новых местообитаний последние все более и более отличаются от исходных, оказываясь в конечном итоге весьма далекими по своим свойствам от первоначальных (см. 1.1, 1.3). Многие виды каменок дают пример весьма высокой экологической пластичности, которая позволяет разным географическим популяциям успешно эксплуатировать местообитания с принципиально разными наборами средовых параметров (в предельном случае от горных ландшафтов субтропиков до арктических тундр). Некоторые виды имеют дипоясный характер распространения: часть локальных популяций вида занимают равнинные полупустыни и сухие степи, другие тесно связаны с альпийской зоной высокогорий.
2. Расселение из субтропиков в расположенные севернее регионы»с более суровым климатом и с укороченным периодом, пригодным для гнездования, сопровождалось практически у всех видов увеличением размера кладки (см. 1.8). Можно допустить, что потребность в дополнительных энергетических ресурсах для продукции большего числа яиц послужила стимулом к увеличению размеров особей, которое наблюдается всех видов каменок в направлении экспансии их ареалов
3. Структура ландшафтов, населяемых каменками, не настолько гетерогенна, чтобы в ней четко дифференцировались дискретные местообитания с резко различными свойствами. В большинстве случаев биотопические предпочтения симпатрических видов достаточно единообразны, хотя и могут различаться в деталях. В результате пространственная сегрегация сосуществующих видов выражена слабо либо отсутствует вовсе. В последнем случае формируются смешанные гнездящиеся популяции, образованные особями нескольких видов (до четырех и более). Эта ситуация вполне типична, и наблюдается не только в сезон гнездования, но и на зимовках (см. 1.10).
4. По типу питания каменки являются генералистами, практикующими оппортунистические стратегии кормодобывания. Рационы сим-биотопических видов широко перекрываются (см. 1.5). С учетом этих
318
Заключение
обстоятельств факт существования упомянутых смешанных популяций, характеризующихся к тому же сравнительно высокой плотностью, не вполне укладывается в известное правило Гаузе, тем более что ландшафты, занимаемые этими поливидовыми популяциями, не отличаются высокой продуктивностью.
5. Согласно классическим воззрениям, в подобных условиях должны четко работать механизмы устранения потенциальной межвидовой конкуренции. При отсутствии адаптивной радиации в морфологии челюстного аппарата и, соответственно, дивергенции по пищевым предпочтениям роль такого механизма часто отводят феномену межвидовой территориальности. Показано, однако, что межвидовой антагонизм в смешанных популяциях каменок далеко не всегда гарантирует надежную монополизацию локальных ресурсов собственникам конкретной территории и популяции того вида, к которому они принадлежат (см. 1.10). Благополучие смешанных популяций при отсутствии действенных механизмов устранения пищевой конкуренции заставляет усомниться в реальности последней. Не исключено также, что даже в угодьях с невысокой продуктивностью, каковыми представляются на первый взгляд местообитания каменок, запасы корма высоко избыточны.
6. При сравнении разных популяций одного вида, а также при межвидовых сопоставлениях прослеживаются определенные корреляции между качеством местообитания и величиной гнездовых территорий. Тот факт, что размеры гнездовых территорий в пессимальных местообитаниях (например, в засушливых регионах пустынь северной Африки и прилежащих участков западной Азии) существенно выше, чем в районах с менее агрессивной средой, указывает на важную роль территориального поведения в качестве механизма монополизации кормовых ресурсов.
7. Острая конкуренция из-за пространства, реализуемая средствами территориального поведения, характерна для перелетных популяций, занимающих местообитания сравнительно высокого качества в регионах умеренной зоны, где формируются довольно плотные гнездовые поселения. Из-за краткости периода формирования таких демов, социальные процессы, связанные с распределением гнездовых участков и с образованием брачных пар, протекают весьма динамично. Размеры территорий высоко изменчивы и определяются сроками прилета их обладателей на места гнездования. Формируется резерв нераз-множающихся особей (главным образом среди самцов, а у ряда видов и среди самок), готовых быстро занять освобождающиеся вакантные позиции. В оседлых популяциях, приуроченных к низкопродуктивным аридным местообитаниям южных субтропиков, ресурсы распределе
319
Панов Е.Н. Каменки Палеарктики
ны между членами поселения более равномерно, социальная обстановка гораздо более стабильна и предсказуема — в основном благодаря относительному постоянству состава брачных пар, сохраняющихся из года в год.
8. Между симпатрическими видами в большинстве случаев поддерживается строгая репродуктивная изоляция. Эффективность изолирующих барьеров не может базироваться на механизмах пространственной изоляции в силу отсутствия четкой сегрегации видов по местообитаниям, особенно в ситуациях симбиотопии. Роль хронографической изоляции, вероятно, также ничтожна, поскольку в большинстве случаев близкие симпатрические виды каменок размножаются более или менее синхронно. Таким образом, основным действующим фактором оказывается способность особей выбирать в качестве половых партнеров конспецифических индивидов и избегать половых контактов с представителями других видов (этологическая изоляция).
9. Анализ взаимоотношений между сестринскими алловидами (такими, например, как испанская каменка и плешанка), показывает, что разрушение барьеров этологической изоляции возможно даже при существенном несходстве контактирующих форм в окраске, вокализации и в особенностях социального поведения (Главы 12 и 13).
10. На стыках ареалов таких форм с течением времени формируются популяции, фактически полностью состоящие из гибридов разных поколений. В таких популяциях происходит нивелирование не только признаков внешней морфологии, но также и поведенческих признаков (в частности, видоспецифических песен). Таким образом, картина противоположна той, предсказываемой гипотезой “смещения признаков”, согласно которой в зонах контакта и гибридизации следует ожидать ускоренной дивергенции видовых опознавательных признаков контактирующих форм (Глава 14).
11. Более правдоподобным представляется сценарий превращения гибридогенной популяции в некую высоко интегрированную общность, обладающую мозаикой признаков обоих родительких форм. Иными словами, гибридизация, приводящая к локальному слиянию видов, может оказаться первопричиной становления популяций, которым свойственен сбалансированный генетический полиморфизм (Главы 8 и 14).
12. При относительном единоообразиии признаков внешней морфологии разных представителей рода их сигнальное поведение видов зачастую оказывается принципиально различным. Эти различия отнюдь нс сводятся к несходству в структуре отдельных “сигналов” (вокальных либо оптических, генераторами которых служат видоспсци-фическис паттерны двигательных акций). Различия затрагивают весь
320
Заключение
сложнейший комплекс моторики и вокализации, выступающий в качестве высоко интегрированной системы, уникальной для каждого вида. Неповторимый характер коммуникативной системы вида коренится нс только в особенностях сигнального репертуара, но определяется также правилами “синтаксиса”, которые закономерно трансформируются по ходу коммуникативного процесса (подробнее см. Панов 1978:232-261, 1989: 323-325, 1997: 28-43). Глубокий изоморфизм коммуникативных систем в группах близкородственных видов указывает на высокую консервативность этих поведенческих структур в эволюционном процессе. С этой точки зрения они представляются наиболее надежными критериями для выявления генеалогических отношений между видами, обладая в этом смысле явными преимуществами перед окрасочными признаками, которые, как показано, эволюционируют сравнительно быстро и непредсказуемо (см. 1.2).
В заключение автор обращается к отечественным и зарубежным орнитологам с предложением внести свой вклад во всестороннее изучение биологии каменок вообще и их поведения — в особенности. Для того, чтобы описания поведения, сделанные разными исследователями, можно было продуктивно сравнивать, была предложена методика, разработанная автором именно при работе с интересующей нас группой птиц (см. Панов, 1978, 1989 , Panov, 1996).
321
Панов Е.Н. Каменки Палеарктики
Литература
Абдрсимов Т. (1981): Птицы тугаев и прилежащих пустынь низовьев Аму-Дарьи. Ташкент.
Абдусалямов И. А. (1961): Птицы долины озера Ранг-куль на Памире. — Тр. Инет. зоол. и паразитол. АН Тадж.ССР. Т.П: 1-151.
Абдусалямов И. А. (1964): Птицы горного Зеравшана. Душанбе.
Абдусалямов И. А. (1973): Фауна Таджикской ССР. Т.19. Ч. 2. Птицы. Душанбе.
Бельская Г. С. (1961а): К экологии черной каменки — Тр. Инет. зоол. и паразитол. АН ТССР. Т.7: 43-54.
Бельская Г. С. (19616): Материалы по питанию некоторых видов каменок Туркмении. Изв. АН ТССР. Сер. биол. Т.2:64-73.
Бельская Г. С. (1965а): Экология каменки-плясуньи в Туркмении — Изв. АН ТССР. Сер. биол.Т.З: 76-79.
Бельская Г. С. (19656): О количестве репродуктивных циклов у каменок в Туркмении. — В кн. Новости орнитологии: .23-24. Алма-Ата.
Бибиков Д. И., Бибикова В. А. (1955): К изучению каменки-плясуньи и ее эктопаразитов. — Зоол. журн. 34(2): 399-407.
Васильев Г. И., Лазарева Л. А. (1965): Эктопаразиты гнезд птиц Сайлюгсмского хребта. — В кн. Новости орнитологии: 23-24. Алма-Ата.
Васильченко А. А. (1987): Птицы Хамар-Дабана. Новосибирск.
Воробьев Г. П., Лихацкий Ю. П. (1987): Новые данные по редким птицам Воронежской области. — Орнитология 22: 176-177.
Воинственский М. А. (1960): Птицы степной полосы Европейской части СССР. Киев.
Гаврилов Е. И.(1970): Род Каменка — Oenanthe.— В кн. Птицы Казахстаиа.Т.З: 517-556. Алма-Ата.
Гаврилов Е. И., Наглов В. А., Федосенко А. К., Шевченко В. Л.,Татаринова О. М. (1968): Об орнитофауне Волжско-Уральского междуречья. — Тр. Инет. зоол. АН КазССР. 29: 153-207.
Гагина Т. Н. (1960): Новые данные о распространении птиц в Восточной Сибири — Орнитология 3: 219-225.
Гладков Н. А. (1954): Род Каменки Oenanthe Vicillot. 1816. — В кн. Птицы СССР. Т.6. 493-522. М..
Грабовский В .И., Панов Е. Н. (1992): Конвергенция песен каменки-плешанки Oenanthe pleschanka и испанской каменки Otnanthe hispanica в зонах вторичного контакта. — Зоол. журн. 71(4): 75-84.
Грабовский В. И., Панов Е. Н., Рубцов А. С. (1992): Фенотипический состав и успех размножения в гибридной популяции плешанки Oenanthe pleschanka и испанской каменки О. hispanica в восточном Азербайджане. — Зоол. журн. 71(/): 109-121.
Губин Б. М., Ковшарь А. Ф. (1985): К биологии обыкновенной каменки в высокогорьях Тянь-Шаня. — Орнитология 20: 53-59.
Гусев О. К. Орнитологические исследования на северном Байкале. — Орнитология 5: 149-163.
322
Литература
Данилов Н. Н., Рыжановский В. Н., Рябицсв В. К. (1984): Птицы Ямала. Москва, 1984.
Дементьев Г. П. (1973): Воробьиные. — В кн. С. А. Бутурлин, Г.П. Дементьев. Полный определитель птиц СССР. Т.4. М.-Л
Доржиев Ч. 3., Хертуев В. Н. (1992): Экология каменок в Забайкалье. Улан-Уде.
Завадовский М. (1922): Пол и его признаки. М
Зарудный Н. А. (1896): Орнитологическая фауна Закаспийского края (северной Персии, Закаспийской области, Хивинского оазиса и равнинной Бухары). — Мат к позн. фауны и флоры Российской Империи. Отд. зоол. Т.2: 1-555. М.
Зарудный Н. А. (1903): Птицы восточной Персии. — Зап. Русск. Императ. Географ. Обш. Т. 36. №2: 1-467. М.
Зарудный Н. А. (1915): Птицы пустыни Кызыл-Кум. — Мат к позн. фауны и флоры Российской Империи. Отд. зоол. Т. 14: I- 149. М.
Зарудный А. Н. (1916): Птицы Аральского моря. — Изв. Туркестанск. отд. Русск. Географ. Общ. Т. 12: 1-229. Ташкент.
Зарудный Н. А. (1923): К вопросу о подвидах белоспинного чекана (Saxicola finschii HeugL). — Изв. Туркестанск. отд. Русск. Географ. Обш. Т. 16: 72-81. Ташкент.
Зарудный Н. А. (1923): О некоторых чеканах (Saxicola picata Blyth, S. capistrata Gould, 5. opistholeuca Strick, и 5. evreinovi Zar). — Изв. Туркестанск. отд. Русск. Географ. ОбшТ. 16:65-72. Ташкент.
Зыкова Л. Ю., Панов Е. Н. (1982): Случай межвидовой помощи: черная каменка (Oenanthepicata) выкармливает птенцов каменного воробья (Petroniapetronia). — Зоол. журн. 61(7): 1113-1116.
Иваницкий В. В. (1978): Экологические и поведенческие предпосылки полигинии у каменки-плясуньи Oenanthe isabellina (Turdidae, Aves). — Зоол. журн. 57(/0): 1555-1565.
Иваницкий В. В. (1980а): Межвидовые отношения симпатрических видов каменок (Oenanthe. Turdidae, Passeriformes). I. Экологические предпосылки конкуренции и пространственная структура смешанной популяции. — Зоол. журн. 59(4): 587-597.
Иваницким В. В. (198096): Межвидовые отношения симпатрических видов каменок (Oenanthe, Turdidae, Passeriformes). 2. Поведенческие аспекты сосуществования близких видов. — Зоол. жури. 59(5): 739-749.
Иваницкий В. В. (1981а): Пространственно-этологическая структура популяции каменки-плясуньи (Oenanthe isabellina) в разных частях ареала. — Биол. науки. 7: 53-58.
Иваницкий В. В. (19816): Коммуникация и поведение каменки-плясуньи Oenanthe isabellina (Turdidae, Aves) в период спаривания. — Зоол. журн. 60(Л): 1212-1221.
Иваницкий В. В. (1982): Социальное поведение камсики-плясуньи в период формирования гнездовой популяции. — Зоол. жури. 61 (/): 71-81.
Иванов А. И. (1940): Птицы Таджикистана — Тр. Тадж. базы АН СССР. Т. 10: 1 — 300.
Иванов А. И. (1969): Птицы Памиро-Алая. М.
Ильяшенко В. Ю. (1986): О птицах бассейна верхней Зеи. — В кн. Распрострапенис и биология птиц Альтая и Дальнего Востока. Гр. Зоол. Инет. АН СССР. Т.150: 77-80.
323
Панов Е.Н. Каменки Палеарктики
Ишадов X. (1963): Гнездовая орнитофауна Гсрмаба (Копетдаг) и его окрестностей. — Изв. АН ТуркмССР. Сер. биол. науки. № 1: 73-78.
Каневский В. Я. (1978): Эколого-этологические особенности размножения некоторых видов воробьиных птиц хребта Пянджский Каратау. — Изв. АН ТаджССР. Отд. биол. науки. 71(2): 21-27.
Кейфер М., Крумпал М., Лобачев В. С. (1986): Фауна блох и иксодовых клешей и их медицинское значение в Монголии. — В кн. Природные условия и биологические ресурсы Монгольской Народной республики: 158-160. М.
Кишинский А. А. (1980): Птицы Корякского нагорья. М.
Ковшарь А. Ф.. Левин А. С. (1982): Каталог оологической коллекции Института зоологии КазССР. Алма-Ата.
Козлова Е.В. (1975): Птицы зональных степей и пустынь центральной Азии. Л.
Корелов М. Н. (1956): Материалы к авифауне хребта Кетмень (Тянь-Шань). — Тр. инет. зоол. АН КазССР. Т.6: 109-157.
Костина Г. Н., Панов Е. Н. (1981): Индивидуальная и географическая изменчивость песни у черной каменки Oenanthe picata. — Зоол. журн. 60(9): 1374-1385.
Лоскот В. М. (Loscot V. М) (1973а): Гсограф1чна мпшивктьсвропейсько-аяатських популяцши кам'янки звичайно! Oenanthe oenanthe (L.). — 36ipiuiK праць зоол. муз. 35: 72-77.
Лоскот В. М. (19736): Чеканы и каменки фауны СССР. Автореферат канд. дисс. Киев.
Лоскот В. М. (1976): О систематическом положении черно-пегой каменки и плешанки [Oenanthe hispanica (L.), О. pleschanka (Lepechin)J. — Сб. тр. Зоол. муз. Т. 36:84-89. Киев
Лоскот В. М. (1983): Биология восточной черно-пегой каменки Oenanthe hispanica melanoleuca (Guld.) в СССР. — Тр. Зоол. Инет. АН СССР. Т. 116: 69-107.
Лоскот В. М. (1986а): Фенотипический состав популяции в зонах вторичного контакта чернопегой каменки и плешанки. — В кн. Актуальные проблемы орнитологии: 17-33. М.
Лоскот В. М. (19866): Материалы по птицам окрестностей Ташанты (юго-восточный Алтай). — В кн. Распространение и биологи птиц Алтая и Дальнего Востока. Тр. Зоол. Инет. АН СССР. Т. 150:44-56.
Лоскот В. М., Петрушенко А. А. (1974): О питании каменки рыжехвостой (Oenanthe xanthoprimna Hempr. et Ehr.) в Бадахшане. — Вестник зоол. 5: 59-65.
Любушенко С. Ю. (1995): Популяционная структура и полиморфизм в комплексе “черные каменки*' Oenanthe picata Blyth. 1847 (Aves, Turdidae). Автореферат канд. дисс. М.
Ляйстер А. Ф., Соснин Г. В. (1942): Материалы по орнитофауне Армянской ССР. Ереван.
Майр Э. (1968): Зоологический вид и эволюция. М.
Мекленбурцев Р. Н. Зимующие и пролетные птицы Восточного Памира. — Бюлл. МОИП. 1946. Сер. биол. 51: 87-110.
Митропольский О. В. (1968а): Размножение черношейной каменки на Мангышлаке. — Тр. Инет. зоол. АН КазССР. Т. 29: 67-70.
324
Литература
Митропольский О. В. (19686): О биологии пустынной каменки. — Тр. Инет. зоол. АН КазССР. Т. 29: 64-66.
Митропольский О. В. (1980): Материалы по гнездовой биологии некоторых пустынных птиц низкогорий Кызылкумов. — В кн. Экология и морфология животных: 96-99. Самарканд.
Митропольский О. В.. Третьяков Г.П., Фотгслер Э.П. (1981): Материалы к оологической характеристике птиц Западного Тянь-Шаня. — В кн. Экология некоторых видов млекопитающих и птиц равнин и гор Узбекистана: 69-98. Ташкент
Нейфельдт И. А. (1970): Пуховые птенцы некоторых азиатских птиц. — В кн. Орнитологический сборник. Тр. Зоол. Инет. АН СССР. Т. 47: 111-181.
Нейфельдт И. А. (1986): Из результатов орнитологической экспедиции в юго-восточный Алтай. — В кн. Распространение и биология птиц Алтая и Дальнего Востока. Тр. Зоол. Инет. АН СССР. Т. 150: 7-43.
Панченко С. Г. (1976): О новых находках каменки-плясуньи (Oenanthe isabellina Temm.) на Украине. — Вестник зоол. 6: 24-27.
Панов Е. Н. (1978): Механизмы коммуникации у птиц. М.
Панов Е. Н. (1986а): Новые данные по гибридизации плсшанки (Oenanthe pleschanka) и испанской каменки (О. hispanica). Зоол. журн. 65(//): 1675-1683.
Панов Е. Н. (19866): Поведение как фактор репродуктивной изоляции у птиц. Авторсф. докторской дисс. М.
Панов Е. Н. (1989): Гибридизация и этологическая изоляция у птиц. М.
Панов Е. Н. (1990): Сравнительный анализ поведения синего и пестрого каменного дроздов (Monticola solitarius,M. saxatilis).— Зоол.журн.96(4): 65-77.
Панов Е. Н. (1993): Границы вида и гибридизация у птиц. — В кн. Гибридизация и проблемы вида у позвоночных. Сб. тр. Зоол. музея МГУ. Т. 30: 53-96.
Панов Е. Н., Грабовский В. И., Любушенко С. Ю. (1993): Дивергенция и гибри-догенный полиморфизм в комплексе “черные каменки" Oenanthe picata.— Зоол. жури. 72(#>: 80-95.
Панов Е. Н., Иваницкий В. В. (1975а): Межвидовые территориальные отношения в смешанной популяции чернобокой каменки Oenanthe finschii и каменки плешанки Ое. pleschanka на полуострове Мангышлак. — Зоол. журн. 54(9): 1357-1370.
Панов Е. Н., Иваницкий В. В. (19756): Эволюционные и таксономические отношения между чернопегой каменкой Oenanthe hispanica и каменкой-пле-шанкой О. pleschanka. — Зоол.журн.54( 12): 1860-1872.
Панов Е. Н.. Корзухин М. Д. (1974): Качество территорий и успех самцов при образовании пар у черной каменки Oenanthe picata. — Зоол. журн. 53(5) : 737-746.
Пейсахис Л. А., Стогов И. И., Степанов В. М.. Бибиков Д. И. (1970): Экспериментальное исследование чумы у каменки-плясуньи (Oenanthe isabellina) в связи с ее вероятным значением в природной очаговости этой болезни. — Зоол. журн. 49(//): 1691-1696.
Попов В. А. (1959): Птицы Гиссаро-Каратегина. Душанбе.
Портенко Л.А. Род Oenanthe Vieillot. — В кн. Птицы СССР. Т.З: 158-173. Л.,
325
Панов Е.Н. Каменки Палеарктики
Портснко Л. А. (1973): Птицы Чукотского полуострова и острова Врангеля. Т.2. Л.
Потапов Р. Л. (1959): Очерк летней орнитофауны заповедника “Тигровая балка”. — В кн. Заповедник “Тигровая балка”. Т.1. Тр. Инет. зоол. паразитол. АН ТаджССР. Т. 115: 179-201.
Потапов Р. Л. (1966): Птицы Памира. Тр. Зоол. Инет. АН СССР. Т. 39: 3-119
Потапова Е. Г., Панов Е. Н. (1977): Особенности строения челюстного аппарата каменок рода Oenanthe (Turdidae, Passeriformes) в связи с вопросом о путях снижения конкуренции между близкими видами. — Зоол. журн. 56(5): 743-752.
Рубцов А. С. (1995): Относительная продолжительность жизни самцов в гибридной популяции испанской каменки Oenanthe hispanica и плешанки Oenanthe pleschanka. — Зоол. журн .74(6): 112-121.
Рубцов А. С. (1997): Проблема вида и природная гибридизация у птиц на примере взаимоотношений испанской каменки и плешанки (Oenanthe hispanica L., 1748; Oenanthe pleschanka Lepech., 1770). Автореферат канд. дисс. M.
Рустамов А. К. (1954): Птицы пустыни Кара-Кум. Ашхабад.
Рустамов А. К. (1957): Эколого-зоогеографические заметки о птицах фисташковых насаждений юго-восточной Туркмении. — Зоол. журн. 36(5): 742-751.
Рябицев В. К. (1993): Территориальные отношения и динамика сообществ птиц в Субарктике. Екатеринбург.
Синская Е. Н. (1948): Динамика вида. М.-Л.
Степанян Л. С. (1970): Птицы нижнего Ширабада (междуречье Кугитанга и Байсунского хребта). — Уч. записки МГПИ им. Ленина. Т. 394: 102-150.
Степанян Л. С. (1971): О систематическом положении закавказских и бадах-шанских рыжехвостых каменок (Oenanthe xantoprymna Hemprich et Ehrenderg.). — Научные докл. высш, школы. Биология. 6: 26-29.
Степанян Л. С. (1978): Состав и распределение птиц фауны СССР. Воробьинообразные. М.
Степанян Л. С. (1983): Надвиды и виды-двойники в авифауне СССР. М.
Степанян Л. С., Степанян Е.Н. (1972): Материалы к орнитологической фауне западного Копетдага. — В кн. Фауна и экология животных: 193-213. М.
Сурвилло А. В. (1968): Об экологии каменок южной части Зайсанской котловины. — Тр. инет. зоол. АН КазССР. Т. 29: 71-75.
Сухинин А. Н. (1958): Снегопад в Бадхызс весной 1955 года и его влияние на размножение некоторых видов птиц. — Изв. АН ТуркмССР. Т. 6: 107-110.
Тарасов М. П. (1975): О размещении и численности массовых видов каменок (Oenanthe) в южном Закавказье. — Вестник зоол. 5: 7-10.
Успенский С. М. (1969): Жизнь в высоких широтах (на примере птиц). М.
Шевченко В. Л. (1979): О судьбе кладок, птенцовой смертности и некоторых факторах, определяющих численность каменки-плясуньи в северном При-каспии. — В кн. Экология гнездования птиц и методы се изучения: 236-237. Самарканд.
Шевченко В. Л. (1986): Материалы по питанию каменки-плясуньи в северном Прикаспии. Вестник зоол. 6: 43-46.
Шнитников В. Н. (1949): Птицы Семиречья. М.-Л.
Allouse В. Е. (1953): The avifauna of Iraq. — Irak Nat. Hist. Mus. Publ. 3: 159 pp.
326
Литература
Atamuradov Kh. I. (1994): Paleogeography of Turkmenistan. In: Biogeograpy and Ecology of Turkmenistan: 49-64. Kluwer Acad. Publ.
Aro M.: Kaksi bigamia-tapausta Kitivaskulla (Oenanthe oenanthe). — Omis Fennica. 1968. Vol. 45. P. 16-18.
Bannerman D. A. (1954): The birds of British Isles.. V. 3. Lnd.
Bates G. L. (1936): Birds of Jidda and Central Arabia collected in 1934 and early in 1935, chiefly by Mr. Philby. — Ibis 13: 674-712.
Berck K.-H. (1961): Bcmerkungen zur Biologic des Stcinschmatzers. — Vogelwelt 82: 89-112.
Bulatova N. Sh. (1981): A comparative karyological study of passerine birds. — Acta. sci. Nat. Brno 15(3): 1 -44.
Bulatova N. Sh., Panov E. N. (1973): Comparative analysis of caryotypes of 18 species of the family Turdidae (Aves) — Caryologia 26: 229 — 244.
Christensen S. (1974): Notes on the plumage of the female Cyprus pied wheatear. — Ornis Scand. 5: 47-52.
Cornwallis L. (1975): The comparative ecology of eleven species of wheatear (genus Oenanthe) in S. W. Iran. Ph. D. thesis Oxford University.
Cornwallis L., Porter R.F. (1982): Spring observations of the birds of North Yemen — Sandgrouse 4: 1-36.
Cott H. B. (1947): The edibility of birds: illustrated by five year’s experiments and observations (1941-1946) on the food preferences of the hornet, cat and man; and considered with special reference to the theory of adaptive coloration. — Proc, zool. Soc. London 116: 371-524.
Cramp S., ed. (1988): Family Turdidae: Chats, Thrushes. In: Handbook of the Birds of Europe, the Middle East and North Africa. V. 5.: 585-1025. Oxf., N.Y.
Desfayes M., Praz J. C. (1978): Notes on habitat and distribution of montane birds in southern Iran. — Bonn. Zool. Beitr. 29: 18-37.
Dorka v. V., Dorka U., Haas V.(1976): Gemcinschaftliches Hdhlcnnachtigen beim Trauersteinschmatzer Oenanthe leucura zur Nichtbrutzeit. — Sondcr. Verhandl. Omithol. Gesellschaft Bayern 22: 467-471.
Farkas T. (1969): Notes on the biology and ethology of the Natal robin Cossypha natalensis. — Ibis 111: 281-291.
Farkas T. 1973): Notes on the biology and ethology of the Heuglin's robin Cossypha heuglini. — Ostrich 44: 95-105.
Flint P. (1995): Separation of Cyprus pied wheatear from pied wheatcar. — Brit. Birds 88: 230-241.
Gotmark F. (1994): Are bright birds distasteful? Re-analisys of H. D. Cott data on the edibility of birds. — J. Avian Biol. 25: 184-197.
Grimes L.G. (1976): The occurence of cooperative breeding behaviour in African birds. — Ostrich 47: 1 — 15.
Haartmann L. v. (1969): Nest-sites and evolution of polygamy in European passerine birds. — Omis Fennica 46: 1-12.
Hafter J. (1977): Secondary contact zones of birds in Northern Iran. — Bonn. Zool. Monogr. N 10: 1-64.
Hafter J. (1986): Superspccies and species limits in vertebrates. — Zool. Syst. Evolut.-forsch. 24: 169-190.
327
Панов Е.Н. Каменки Палеарктики
Hall В. Р., Moreau R. М. (1970): An atlas of speciation in African Passerine Birds. — Brit. Mus. (Nat. Hist.), Lnd.
Harris A , H. Shirinai. Christie D. (1996): The MackMillan Birder's Guide to European and Middle Eastern birds. — Lnd., Basingstoke.
Hartcrt E. (1910): Die Vogel der palaarktischen Fauna. Bd. 1. — Berlin.
Hartley P. H. T. (1949): The biology of the mourning chat in winter quarters. — Ibis 91: 393-413.
Heim de Balsac H., Mayaud N. (1962): Les oiseaux du nord-oucst de I’Afrique. — Paris.
Hollom P. A. D.. Porter R. F., Christensen S.. Willis I. (1988): Birds of the Middle East and North Africa. — Callon.
Hue F.. Etchecopar R. D. (1970): Les oiseaux du Prochc et du Moyen Orient de la Mediterranee aux contrcforts de Г Himalaya. — Paris.
Kleinschmidt O. (1938): Katalog meincr ornithologischen Sammlung. — Halle. Saale.
Kncis P., Lauch M. (1983): Untersuchungen zum Nahrungserwerb der Steinschmatzer Oenanthe oenanthe. — Zool. Jb. Physiol. 87: 381-387.
Kncis P., Sprctke T. (1985): Zwischenartliche Aspektc der Besiedlung zweier mongolischcr Bcrghange durch Drossclvdgel der gattung Oenanthe, Monticola und Phoenicurus. — Mitt. zool. Mus. Berl. 61. Suppl.: Ann. Orn. 9: 71-92.
Konig C. (1966): Zur Brutbiologie des Truersteinschnuzers Oenanthe leucura. — Vogelwelt 87: 182-188.
Kreuger R. (1958): Hitherto undiscribed nests and eggs from British Somaliland. — Ibis 100: 278-279.
Kumcrloeve H., M. (1962): Isabell-Steinschmatzcr Oenanthe isabellina (Cretzschmar) in Sudost-Bulgaricn. — J. Ornithol. 103: 108-109.
Kumerloeve H., Kasparek M., Nagel K.-O. (1984): Der Rostbiizel-Steinschmatzer, Oenanthe xanthoprymna (Hemprich & Ehrenberg 1833) als neuer Brutvogel im ostlichcn Anatolien (Tbrkci) — Bonn zool. Bcitr. 35: 97-101.
Lack D. (1958): The significance of the colour of turdinc eggs. — Ibis 100: 145-166.
Lcisler v. B. Heine G.. Siebcnrock K.-H. (1983): Einnischung und interspezifische Territorialitat uberwinternder Steinschmatzer (Oenanthe isabellina, O. oenanthe, O. pleschanka) in Kcnia. — J. Orn 124: 393-413.
Lorenz K. (1939): Vergleichende Verhaltensforschung. — Zool. Anz. (Suppl.) 12:69-102.
Lorek G. (1989): Niezwykla lokalizazaeja gniazd bialorzytki (Oenanthe oenanthe) — Not. ornitol. 30: 104-106 (N 3-4).
Lovaty F. (1985): Statut de traquet oreillard Oenanthe hispanica dans la region de Mcnde (Lozere) — Alauda 53: 310-311.
Mackworth-Pracd C. W., Grant С. H. B. (1963): Birds of the southern third of Africa. African Handbook of Birds, Ser. 2, Bd. 2. — Lnd., N.Y., Toronto.
Mackworth-Praed C. W., Grant С. H. B. (1973): Birds of West Central and Western Africa.- Laugman.
Mayaud N. (1989): Les oiseaux du nord-oucst de I’Afrique notes complemcntaries. — Alauda 57: 10-16.
Mayr E., Strescmann E. (1950): Polymorphism in the chat genus Oenanthe. — Evolution 4: 291-300.
McLachlan G. R., Liverside R., eds (1957): Roberts birds of Southern Africa. — Cape Town
328
Литература
Meincrthagen R. (1930): NicoH's birds of Egypt. V. I. — London.
Mcinerthagcn R. (1938): On the birds of the Northern Afganistan. — Ibis 81: 480-520, 671-717.
Mcinerthagcn R. (1954): Birds of Arabia. — Edinburgh, Lnd.
Mcinerthagcn R. (1949): New races of a courser, woodpecker, swift, lark, wheatear, and serin from Africa. — Bull. Brit. Orn. Club 69: 104-108.
Moore H. J., Boswell C. (1956): Field observations on the birds of Iraq. — Irak Nat.
Hist. Mus. Publ.9/10: 12.
Moreau R. E. (1966): The bird fauna of Africa and its islands. — N. Y., Lnd.
Moreno J. (1984a): Parental care of fledged young, division of labor, and the development of foraging technique in the northern wheatear (Oenanthe oenanthe L.).-Auk 101:741-752.
Moreno J. (1984b): Search strategics of wheatears (Oenanthe oenanthe) and stonechat (Saxicola torquata): adaptive variation in perch height, search time, sally distance and inter-perch move length. J. of Animal Ecology, 33: 147-159.
Moreno J. (1989a): The breeding biology of the wheatear Oenanthe oenanthe in South Sweden during three contrasting years. — J. Orn. 130: 321-334.
Moreno J.(1989b): Energetic constrains on uniparental incubation in the wheatcar Oenanthe oenanthe (L ). — Ardea 77: 107-115.
Moreno, J., Soler M., Moller A.P., Linden M. 1994): The function of stone carrying in the black wheatcar Oenanthe leucura. — Anim. Bchav. 47: 1297-1309.
Nanlinov D., Borisov E. 1988): Izabelovoto kamenarcc (Oenanthe isabellina, Temminck, 1829) — sresci v B'lgaija i nacini za oprcdeljane. — Ornitol. inf. bjul. № 23-24: 67-76.
Palfcry J. (1988): Observation on the behaviour of the white-crowned black wheatear in Eastern Arabia. — Sandgrousc 10: 1-25.
Paludan K. (1959): On the birds of Afganistan. — Vid. medd. Dan. naturhist. foren. 122: 1-332.
Panov(w) E. N. (1974): Die Stcinschmatzerder nordlichcn Palaarktis. Gattung Oenanthe. — Die Ncue Brehm-Bucherei Bd. 482, Wittenberg Luthcrstadt.
Panov E. N. (1992): Emergence of hybridogenous polymorphism in the Oenanthe picata complex. — Bull. British Orn. Club. Centerary Suppl. V. 112A: 237 — 249.
Panov E. N. (1996): Bird comparative ethology. In: Phys. Gen. Biol. Rev. 12: 1-60. — Amsterdam.
Panov E. N., Grabovsky V. I., Rubtsov A. S. 1994): Spatial and temporal dynamics of hybridization by pied wheatcar Oenanthe pleschanka and black-cared wheatcar O. hispanica. — J.Omithol. 135. 361.
Paz U. (1987): The birds of Israel. — London.
Perlo van Ber (1992): Birds of Eastern Africa. — London.
Perrin de Brichambaut S. (1989): Sur la construction du ”mur” du nid du Traquet ricur (Oenanthe I. leucura) — Alauda 57: 226-227. (N 3)
Piechocki R., Bolod A. (1972): Beitrage zur Avifauna der Mongolei. Teil. 2. Passeriformes. — Mitt. Zool. Mus. Berlin 48 (1): 41-175.
Piechocki R., Stubbc, M., Uhlenhaut K., Sumjaa D. (1982): Beitrage zur avifauna dcr Mongolei. Toil. IV. Passeriformes. — Mitt. Zool. Mus. Berlin 58. Suppl.: Ann. Om. 6: 3 - 53.
329
Панов Е.Н. Каменки Палеарктики
Piccre Р. (1987): Nidification du traquet motteux (Oenanthe oenanthe) en Lorraine Beige et dans les regions limitrophes — Aves 24: 105-111.
Portenko L. A., Vietingoff-Schecl E. v. 1971): Oenanthefinschii (Heuglin). In: Stresemann, E., L. A. Portenko, G. Mauersbergcr (Eds). Atlas der Verbreitung Palaarktischen Vogel. — Berlin.
Raines R. J. (1962): The distribution of birds in northeast Greece in summer. — Ibis 104: 490-502.
Richardson, F. (1952): Breeding and feeding habits of the black wheatear Oenanthe leucura in South Spain. — Ibis 107: 1-16.
Ripley S.D.(1952): The trushes.— Postilla 13.
Ripley S.D.( 1964): Subfamily Turdinac.In: Mayr, E., R.A.Paynter, jr.Chcck-list of Birds of the World, Bd. 10. — Cambridge, Mass.
Rogacheva H. (1992): The birds of Central Siberia. — Husum.
Schmidt E. (1968): Uber die Bedeutung der Vorkommen siidlischer Steinschmatzerarten im Karpatenbecken. — Aquila 75: 79-91.
Schdnwettcr U. (1979): Handbuch derOologie. Bd. 2. Berlin.
Shirinai H. (1996): The birds of Israel. Lnd., N.Y.
Sluis R., Berg M. van den (1982): On the specific status of the Cyprus pied wheatear Oenanthe cypriaca. — Ornis Scand. 13: 123-128
Smith K. D. (1971): Notes on Oenanthe species in winter in Africa. — Bird Study 18: 71-79.
Soler M., Moreno J., Moller A. P., Linden M., Soler J. J. 1995): Determinants of reproductive success in a Mediterranean multi-brooded passerine: the black wheatcar Oenanthe leucura. — J. Ornithol. 136: 17-27.
Sorgier R.-C. (1978): Primera cita de la ycbelica (Oenanthe leucopyga) en la Peninsula Iberica. — Donana, Acta Vertebrata 5: 109-110.
Stanford J. K. (1954): A survey of the ornithology of Northern Libya. — Ibis 96: 606-623.
Stein P. (1966): Observations on the breeding biology of the familiar chat Cercomela famitiaris (Stephens). — Ostrich 37: 176-183.
Stobener W. F. (1989): Europaische Singvogel in der Voliere. Steinschmatzer — Oenanthe. — Die Europaische Vogclwelt 7/8. Songcheft: 21-64.
Stresemann E. (1925): Oenanthe opistholeuca (Strickland) — cine melanistische Mutante von Oenanthe picata (Blyth). — Ornit. Monastber. 33: 178-181.
Stresemann E., Meise W, Schonwetter M.(1937): Aves Beickanae. Beitrage zur Ornithologie von Nordwest-Kansu nach den Forschungen von Walter Beik (4-) in den Jahren 1926 - 1933. J. Ornithol. 35, 375 - 576.
Suarez Cardona F. (1977): Nidificacion у puesta la collabra rubia (Oenanthe hispanica). — Ardcola 23: 63-79.
Ticehurst С. B. (1922): Notes on some Indian wheatears. — Ibis 11: 151-158.
Toschi A. (1969): Aviafauna Italiana. — Firenze.
Tye A. (1980): The breeding biology and population size of the wheatear (Oenanthe oenanthe) on the Breckland of East Anglia, with implications for its conservation. - Bull. Ecol. 11:559-569.
Tye A. 1986: Plumage stages, moult, sexual dimorphism, and systematic position of the Somali wheatear Oenanthe philli psi.— Bull. Brit. Orn. Club 106: 104- 111.
330
Литература
Туе А. (1987). Clinal variation and subspeciation in the White-crowned Black Wheatcar Oenanthe leucopyga. — Bull. Brit. Orn. Club 107: 157-165. (4)
Tye A. (1988a): Vocalization and territorial behaviour of wheatears Oenanthe spp. in winter quarters. In: Proc. Sixth Pan-African Ornithol. Congr.: 297-305
Tye A. (1988b): Foraging behaviour and selection of prey by the buffstreaked chat Oenanthe bifasciata. — Ostrich 59: 105-115.
Tye A. (1989a): Superspecies in the genus Oenanthe (Aves, Turdidae). — Bonn. zool. Beitr. 40: 165-182. (N 3-4).
Tye A. (1989b): The systematic position of the buff-streaked chat (Oenanthe/Saxicola bifasciata). — Bull. Brit. Om. Club 109: 53-58.
Tye A. (1989c): A model of search behaviour for the northern wheatear Oenanthe oenanthe (Aves, Turdidae) and other pause-travel predators — Ethology 83: 1-18.
Valverde J. A. (1957): Aves del Sahara Espanol (ctudo ecologico del deserto). — Madrid.
Vaurie Ch. (1949): Notes on the bird genus Oenanthe in Persia, Afganistan and India. — Amer. Mus. Novit. 1425: 1-47.
Vaurie Ch. (1950): Variation in Oenanthe lugubris. — Ibis 92: 540-544.
Vaurie Ch. (1972): Tibet and its birds. Withcrby Lim.
Wallace D. I. M. (1983): The first identification of eastern pied wheatear in Jordan. —
Sandgrouse 5: 1-18
Wiley R. H. (1974): Evolution of social organization and life history among grouse. — Quart. Rev. Biol. 49: 201-227.
Winterbottom J. M. (I960): Zoogeographical affinities of the avifauna of the western Cape Province. — Ibis 102: 383-393.
Zachai G. (1989): Breeding biology in the desert: the mourning wheatear — Israel-Land and Nature 14: 174-181.
Zamora R. (1990): Nest-site selection of the common wheatear in high mountain areas of southeastern Spain — Wilson Bull 102: 178-180.
Zarudnyj N. A., Harms M. 1926): Bemerkungeniibereinige Vogel Persiens. TeiL 4. - J.
Ornithhol. 74: 1-52.
331
Панов Е.Н. Каменки Палеарктики
Предметный указатель
Аллопатрия (см. Виды: алловиды)
трудности в оценке статуса аллопатрических форм 14, 78, 102, 167
Ассортативность скрещиваний
по возрастным признакам 202
по окраске
— отсутствие в гибридных и полиморфных популяциях 42, 201, 211, 212, 282
Виды
алловиды 77- 78. 102, 166, /67, 317
перечни их, методические трудности составления 17
полувиды (semispecies) 72, 122, 258. 317
проблемные 13-15, 77-78, 102, 166
слияние их 258, 285
таксономические 122
Выводки смешанные
одновидовые 208
усыновление молодняка других видов 71
Выводок
механизмы распадения его 45-46, 97, 118
Гибридизация 15-17,320
в результате дисперсии особей в ареал другой формы 182-183, 184, 290-291
в результате встречного расширения ареалов 180, 188, 275
случайная между симпатричсскими видами 72, 291
массовая (см. Виды, слияние их) 285
парапатрическая (см. Гибридные зоны) 122, 257
поглотительная 182
углубление ее 182, 290-291
Гибридные зоны /25, 277-292
Гибридные популяции 180-186, 189-190, 201-202, 211- 212, 277-292,320
динамика фенотипического состава 280, 281, 287, 289 стабилизированные полиморфные (см. Полиморфизм гибридогенный) 72, 184, 260, 277, 288
Гибриды 145
вокализация их 284, 287
дисперсия в ареалы родительских форм 182-183, 290-291
жизнеспособность 281, 282
окраска 122, 129, 132, 180, 182, 183, 185, 281, 286, 291
успех размножения 283, 290
Гнездостросние
332
Предметный указатель
сооружение опорной платформы
- из камешков 59-61, 132-/54, 136-137, 159, 171, 202-203, 215, 224, 231-232
- из палочек 61, 115, 232, 252, 271
— как атавистическая форма повеления у самцов 134, 144, 231-232, 237
— функциональное значение 202-203, 236-237
— деградация данной формы поведения в эволюции 115, 135 Изменчивость
возрастная 15, 24, 28, 29, 183-184, 213, 217, 218, 240 географическая
— полового дихроматизма выраженности 24-25, 102, 123, 168
— размеров тела увеличение в направлении экспансии вида 21, 77, 103, 123, 141, 146, /67, 168, 219, 241, 259, 300. 318 параллельная 25, 146, 241, 289
Имитации акустические (см. Песня: имитаций включение)
Импринтинг на облик родителей
отсутствие его 211
Индивидуальная дистанция 269, 311, 315
как механизм диссоциации выводка 45-46, 97, 139, 163-164 онтогенез формирования 120, 139
трудности преодоления ее
— при копуляции (см. Копуляция: фрустрация при подготовке
к ней) 45
— при формировании пар 88
Коммуникация 47-58, 321
Комфортное поведение
солнечные ванны 26, 27
купание 38, 120, 164, 2556 273, 315
Конкуренция
внутривидовая
— из-за половых партнеров 40, 92
— из-за пространства 165, 246, 319
межвидовая (см. также Территориальность межвидовая) 68, 71, 169,319
Консерватизм гнездовой 34, 41, ПО, 156, 192, 194, 200, 201, 282 Копуляция
вмешательство посторонних особей 45, 206
внебрачные, редкость их 45
психологический стресс при подготовке к ней 45, 51-53, 160, 204-205
с неодушевленными предметами 137, 175, 206, 311
средства маскировки 53, 137, 161
333
Панов Е.Н. Каменки Палеарктики
Кормовое поведение
растительные корма 32, 35, 209, 216, 226-227, 235, 256
оппортунистический характер 35, , 68, 210, 318
стратегии кормодобывания 36-57, 177, 210, 227, 235, 256, 274, 294, 296-297, 316
Местообитания
антропогенные 32, 105-106, 114, 220, 262, 288
дипоясный характер распространения вида 81, 105, 318 однотипность их у близких видов 66, 68-69, 70, 188, 214 сегрегация биотопическая между видами 67, 169, 318 оптимальные и субоптимальные 108, 125, 169, 214, 242-243
Миграции
как указание на пути расселения видов 33-34
ночные 33, 38, 119, 131, 249
Окраска
адаптивное значение 23, 25— 27
апосматическая 26 — 27
изменчивость возрастная 28, 29
— индивидуальная (см. также Полиморфизм) 114
и характер гнездовых убежищ 23
покровительственная (криптическая ) 23
роль в конкуренции между самцами 41, 42, 43
эволюция ее (См. также Изменчивость географическая, параллельная) 25, 321
Оседлость 33. 39
сохранение территорий из года в год 39, 221-222, 230-231
— и постоянство брачных пар 230, 320
факультативная 81, 149
Надвид 78, 121, 166, 211
второго порядка 211
Пар формирование
поведение самок при выборе территории 43, 198
Песни (см, также Позывки)
в гибридных популяциях 284-285
— нивелирование различий в напевах родительских видов 211, 2Я4-285, 287
изменчивость
— в репертуаре особи 54, 196, 197, 210, 223
- ситуационная 42, 161, 174, 195, 203-204, 272, 309-310 имитации в них 56, 57, 58, 203-204, 210, 223, 247, 266 онтогенез 97, 120, 139, 163, 255, 315
самок 66, 97, 250
334
Предметный указатель
структура и организация 54-57, 87, 88, 128, 143, 156, 174, 195,
210-211, 224, 247, 265, 307, 308
типологизация внутри видового репертуара, условность ее 54, 174
Парапатрия (см. Гибридизация парапатрическая)
Погадки 35
Подвиды
уровень морфо-биологических различий 21
Позывки 53-54
как составная часть песен 55, 88, 97, ИЗ, 128, 156, 174, 255, 307, 315
Полигиния 43-44, 82, 92-93, 118, 198-201, 269
политерриториальная 112, 198
самцы-полигамы 85, 200
— роль их в уходе за потомством 112, 208, 269
— роль их при формировании полигинических трио 92, 199-200
Полиморфизм 15-16, 25, 167, 168, 217, 239, 240
генетический 178, 259, 320
как результат мутаций 15-16, 178
гибридогенный 15-16, 72, 182, 240, 259-261, 289-290, 291, 292
и возрастная изменчивость 15, 217
и параллельная изменчивость видов 25, 228
Половой дихроматизм (см. также Изменчивость географическая) 23-25
адаптивное значение, гипотезы 23, 25
отсутствия его разные варианты 23
— и гнездовой наряд 28
— и особенности коммуникативного поведения 136
Помошничество
внутривидовое 46, 225
межвидовое 71
Полувиды (см. Виды)
Признаки
атавистические 21, 22, 228
вторично-половые 22
консервативные 25
примитивные 29
Промискуитет 92
Резерв популяционный 319
самцов 84, 93, 193-194, 282-283
самок 85, 93, 201
Репродуктивный успех
и возраст производителей 234
335
Панов Е.Н. Качен к и Палеарктики
и качество территории 234 факторы снижение его
— голодание птенцов 117, 118, 283
— погодные условия 95, 116-117, 305
— хищничество 73, 97-98, 207, 233
Род
субъективизм в оценке его границ 13, 317 — числа видов в нем 13-15
Сигналы коммуникативные 320-321
акустические (см. также Песни, Позывки) 53-58 гомологичные у разных видов 160, 224 — частота их использования у разных видов 49 оптические как комбинации ЭДА (см.Элементарный двигательный акт) 47-49 переадресованные другим видам 69, 206, 249 различия межвидовые 206, 271 ювенильные 64, 65-66
Синхронизация
размножения в деме 283
рекламного поведения самцов /54, 247
Смещения признаков гипотеза 320
Стимуляция социальная 83-84
как модификатор поведения членов дема 154-155 Температурный режим 26-28
перегрева опасность 27-28, 226
Территориальное поведение 41
конфронтации на границах и эффект “милого врага” 44, 155, 194-198
на зимовках 46
онтогенез 97, 163-164
самок 43, 44, 127, 156, 1258, 198-200, 248, 307-308 Территориальность
и резерв негнездящихся особей 43, 84
круглогодичная при оседлом образе жизни 149
“мягкая”, с перекрыванием участков обитания пар 84, 85, 230, 266-267, 305
пол территориальность 44, 112, 198
Территориальность межвидовая 66-71, 130, 189, 248, 279, 319
и межвидовая иерархия 68, 71 на зимовках 71, 165, 173, 215 функции ее 68, 71
336
Предметный указатель
Территория гнездовая
влияние се качества на успех при формировании пар 41, 84, 249
выбор территории самками 43, 198
изменчивость размеров 40, 83, 84, 108, 153, 220-221, 319
структура, утилизация разных фрагментов сс 230, 244, 305
трехмерная 44, 248
Территория кормовая на зимовках 46. 165
повторное использование в разные годы 215
Филопатрия натальная 34, 201-202
Экологическая пластичность
в выборе местообитаний 32, 147-148, 158, 262
и обитание в антропогенном ландшафте 32, 105-106, 220, 262
и обитание в экстремальной среде 32, 169, 219, 294, 300-302
и расселение видов 19, 32, 318
Экспрессивность поведения
межвидовые различия в высоте порогов реагирования 49, 173-174, 176
Элементарный двигательный акт (ЭДА) 47-49
Этологические признаки
консерватизм их в эволюции 25-49
337
Панов Е.Н. Каменки Палеарктики
Указатель названий животных
МЛЕКОПИТАЮЩИЕ
Кролик Oryctolagus cuniculus 114, 117 Лемминги 14
Лисица обыкновенная Vulpes vulpes 73, 207 Песец Alopex lagopus 14
Песчанки Gerbillinae 74, 114
Виноградова Meriones vinogradovi большая Rhombomis opimus 148, 158, 302 дневная Psammomys obessus 93, 142 краснохвостая Meriones eversmanni персидская M. persicus 93 малоазиатская M. blackleri 93
Пищухи Ochotonidae даурская Ochofona daurica 114 монгольская 0. pallasi
Соня садовая EHomys quercinus 233
Сурки Marmota 114
алтайский M. baibacina 93 длиннохвостый M. caudata 93 тарбаган M. sibirica 93, 270
Суслики 114 бсрингийский Citellus parryi 59, 114 длиннохвостый C. undulafus 94, 270 желтый C. fulvus 93 малоазийский C. xanthoprymnus 93 малый C. pygmaeus 94 реликтовый C.relictus 93 тонкопалый Sperniophilopsis leptodactylus 93
Тушканчики Dipodidac 302
большой Allacfaga jaculus 302 сибирский Allactaga sibirica 93
Хорек Mustella eversmanni 96
ПТИЦЫ
Бюльбюли Pycnonotusspp. 216
Варакушка Cyanosylvia svecica 22, 48
Воробей черногрудый Passer hispaniolensis 69
Глупыш Puffinus puffinus 114
338
Указатель названий животных
Горихвостки Phoenicurus 22, 30, 47, 68
обыкновенная Ph. phoenicurus 248
чернушка Ph. ochruros 48, 69
Горихвостка водяная Chaimarrornis (Thamnolaea) leucocephala 48
Горихвостки чекановые Cossypha 18, 22, 47
белобровая С. heuglini 48
натальская С. natalensis 48
Дроздовые Turdidae 13
Дрозды настоящие Turdus 30, 47, 72, 229
Дрозды каменные Monticola 22, 28, 30, 47, 317
пестрый М. saxatilis 48, 71
синий М. solitarius 53, 130, 235, 247
Жаворонки
рогатый Eremophila alpestris 32, 69, 302
пустынный Ammomanes deserti 69, 206
хохлатый Galerida cristata 69
Зарянки Erithacus 22
Каменки Oenanthe
белохвостая Ое. leucopyga 15, 16, 20, 21, 23, 27, 28, 29, 31, 32, 33, 35, 37, 38, 45, 59, 62, 63,216, 217-227, 228, 231, 235, 237, 295, 297 белочерная Ое. alboniger 16, 20, 21, 23, 28, 31, 34, 35, 59, 120, 188, 213, 216, 237, 293
бледноплечая Ое. (Myrmecocichla, Saxicola) bifasciata 13,16, 20, 22, 31, 316
горная Ое. monticola 15, 16, 19, 20, 21, 25, 31, 62
златогузая северная Ое. (xanthoprymna) chrysopygia 15, 16, 20, 23, 24, 26, 28, 31, 38, 39, 49, 50, 58, 59, 60, 64, 65, 69, 72, 73, 121-140, 143, 144, 146, 148, 154, 156, 160, 169, 174, 180, 188, 229, 232, 237, 243, 248, 297, 316
златогузая южная Ое.(xanthoprymna) xanthoprymna 15, 16, 20, 21, 24, 25, 63, 72, 121-123, 127, 144, 180
испанская Ое. hispanica 14, 15, 16, 17, 20, 21, 22, 25, 28, 29, 38, 39, 42, 43, 49, 50, 51, 54, 58, 61, 62, 63, 64, 65, 69, 71, 72, 73, 122, 124, 135, 145, 148, 180, 212, 238-256, 257, 258, 260, 263, 265, 267, 269, 274-275, 276-292 (гибридизация с плсшанкой), 293, 294, 297, 308, 310, 311, 316
кипрская Ое. (pleschanka) cypriaca 14, 16, 20, 21, 259, 264, 265, 274, 275, 289
краснопоясничная Ое. moesta 16, 18, 19, 20, 21, 22, 23, 31, 33, 58, 59, 61, 141-144, 146
монашка Ое. monacha 16, 20, 21, 22, 23, 27, 31, 32, 33, 37, 61, 63, 214, 220, 238, 293-297
339
Панов Е.Н. Каменки Палеарктики
обыкновенная Ое. oenanthe 14, 16, 19, 20, 21, 25, 28, 31, 32, 33, 34, 35, 37. 39, 44, 46, 50, 58, 59, 61, 62, 63 , 64, 65, 66, 67, 68, 71, 74, 100-120, 160, 188, 261, 298, 305
- форма phillipsi 14, 16, 19, 20, 29, 33, 58. 102, 104, 116
— форма seebohmi 102, 103, 104, 116, 119
плешанка Ое. pleschanka 14, 16, 17, 19, 20, 21, 22, 23, 25, 26, 8, 29, 31. 33, 35, 43, 44, 46, 49, 50, 51, 55, 58, 61, 64, 65, 66, 67, 68, 69, 70, 71, 72 , 74, 106, 122, 125, 135, 145, 180, 188 , 209,212, 239, 240, 241, 256, 257-275, 276-292 (гибридизация с испанской каменкой), 293, 294, 297, 305, 310
- морфа vittata 15, 259, 260, 261, 276, 284, 288, 290, 291, 292 плясунья Ое. isabellina 13, 16, 18, 19. 20, 21, 23, 28, 29, 30, 31, 33, 35, 36, 37, 43, 44, 49, 50 , 51, 56, 58, 59, 61, 63, 64, 65, 66, 67, 68, 69, 71, 74, 77 - 99, 114, 120, 148, 156, 186. 188, 248, 298, 299, 302, 305, 307, 310 пустынная Ое. deserti 16, 19, 20, 21, 22, 23, 29, 30, 31, 32, 33, 34, 35, 37, 38. 50, 54, 58, 61. 62, 63, 64, 65, 66, 67, 68, 74. 188, 248, 293, 298-316
равнинный скромный чекан Ое. (Cercomcla) tractrac 16, 23, 62 рыжегрудая Ое. bottae 13, 14, 20, 23, 31, 62, 74, 77, 78 — форма frenata 77 — форма heuglini 14, 77
траурная Ое. lugens 13, 14, 16, 20, 21, 24, 27, 31, 32. 33, 37, 38, 49, 59, 62, 140, 166-177, 180, 211, 214, 216, 219, 240, 293, 294 — форма lugentoides 14, 16, 166-169, 211
- форма lugubris 13, 16, 20, 22. 25, 31. 33, 46, 62 , 71, 166-169, 171-172, 185, 211
черная Ое. picata 15, 16. 20. 21, 22, 24, 25, 26,27, 29, 30, 34, 35. 39,
41, 44, 46, 49. 50, 52. 54, 56, 57, 58, 59, 60. 66, 69, 72, 73, 106,
ПО, 120, 125, 127, 132, 135, 173-176, 178-212, 213, 214, 216, 223,
224, 237, 240, 243, 248, 260-261, 293, 299
чернолобая Ое. pileata 16, 18, 20, 31, 58, 71, 74
чернобрюхая Ое. leucura 16, 20, 21, 23, 28, 33, 35, 59, 62, 135, 216, 217 черношейная Ое. finschii 16, 19, 20, 21, 22, 26. 28, 30, 31, 33. 35, 37, 38, 39, 40 , 42, 43, 44. 49. 50. 52. 54, 55. 56. 57. 58, 59. 61. 64. 65. 66. 68, 69. 70. 72, 74, 106, ПО. 127, 135, 139, 140, 141, !43. 144. 145-165, 148, 166, 167, 169, 173-176, 186, 188, 189, 192, 197, 198, 209, 211, 214, 237, 240, 243, 248, 278. 279, 297, 299, 302, 316 Копек полевой Anthus campestris 69.
Кукушка обыкновенная Cuculus canorus 73 Ласточки
береговая Riparia riparia 114
горная Ptyonoprogne rupestris 247
340
Указатель названий животных
Поползень скальный большой Sitta tephronota 69. 202, 206
Синяя птица Myophonus caeruleus 53, 210
Славка вертлявая Scotocerca inquieta 69. 206
Снегирь пустынный толстоклювый Bucanetes githagineus 249
Соловьи тугайные Cercotrichas (Erythropygia) 22
С. galactotes 48
Сорока обыкновенная Pica pica 73, 233
Стриж черный Apus apus 56, 247
Трясогузка белая Motacilla alba 69. 307
Тупик Fratercula arctica 114
Чеканы Saxicola 22, 28, 30, 47, 68, 274, 312, 317
луговой 5. rubetra 71
черноголовый S. to rq и ata 48. 307
Чеканы муравьиные Myrmecocichla 13, 16, 18, 23, 47, 317
M. aethiops 71
M. bifasciata 16, 316
M. melaena 48
Чеканы скальные Thamnolaea 22
Чеканы скромные Cercomela 13 , 16, 18, 22 , 23, 47, 136, 299
C. familiaris 136, 299
C melanura 137, 299, 307. 308
C. sinuata 312
C. schlegeli 312
C. (Oenanthe) tractrac 16, 23, 299, 312
Щурки Merops 293
зеленая M. superciliosa93
малая M. orientalis 296
РЕПТИЛИИ
Змеи 73, 207
полоз подковчатый Coluber hypocrepis 233
ящеричная змея Malpolonsp. 233
Ящерицы 66
Агама хорасанская Laudakia erythrogastra 208
Круглоголовки Phrynocephalus spp. 98, 274
Ящерица глазчатая Lacerta lepida 233
Ящурки Eremiasspp. 98, 140, 216
ЧЛЕНИСТОНОГИЕ
Многоножки 98
сколопендры 140, 235
341
Панов Е.Н. Каменки Палеарктики
Насекомые
Блохи 74
Ceratophyllus spp. 73
Frontopsylla spp. 73
F. baicalis 99
F. elata 99
F. hetera 99
Neopsylla mana 99
Rhadinopsylla dahurica 99
R. Uh 99
Двукрылые
комары долгоножки Tipulidac 316
муха-кровососка Tripocalliphora lidneri 74
— Protocallipherasp.74
Жесткокрылые (Жуки) 98, 120, 140
долгоносики Curculionidac 68, 98
чернотелки Tcncbrionidae 68, 98, 99, 226-227
Ногохвостки Collembola 73
Перепончатокрылые
муравей желтый волосистый Lasius umbratus 95
муравьи 35, 66, 68, 73, 98, 140, 209, 226
шмели 35
Прямокрылые 120
палочники Ramulussp. 210
саранча пустынная Schisfocerca gregarina 226
саранчовые Acrididae 216
трещотка Angaractis gebleri 98
— Bryodema sp. 98, 316
Термиты 68, 69
Чешуекрылые (Бабочки)
бражники 210
Паукообразные
клещи 73, 74
— Dermanyssus 74
— Haemogamasus 74
— Ixodes 74
сольпуги (фаланги) 66
Ракообразные
мокрицы пустынные Hemilepistussp. 98
Скорпионы 66, 98, 235
Scorpio maurus 143
342