/




































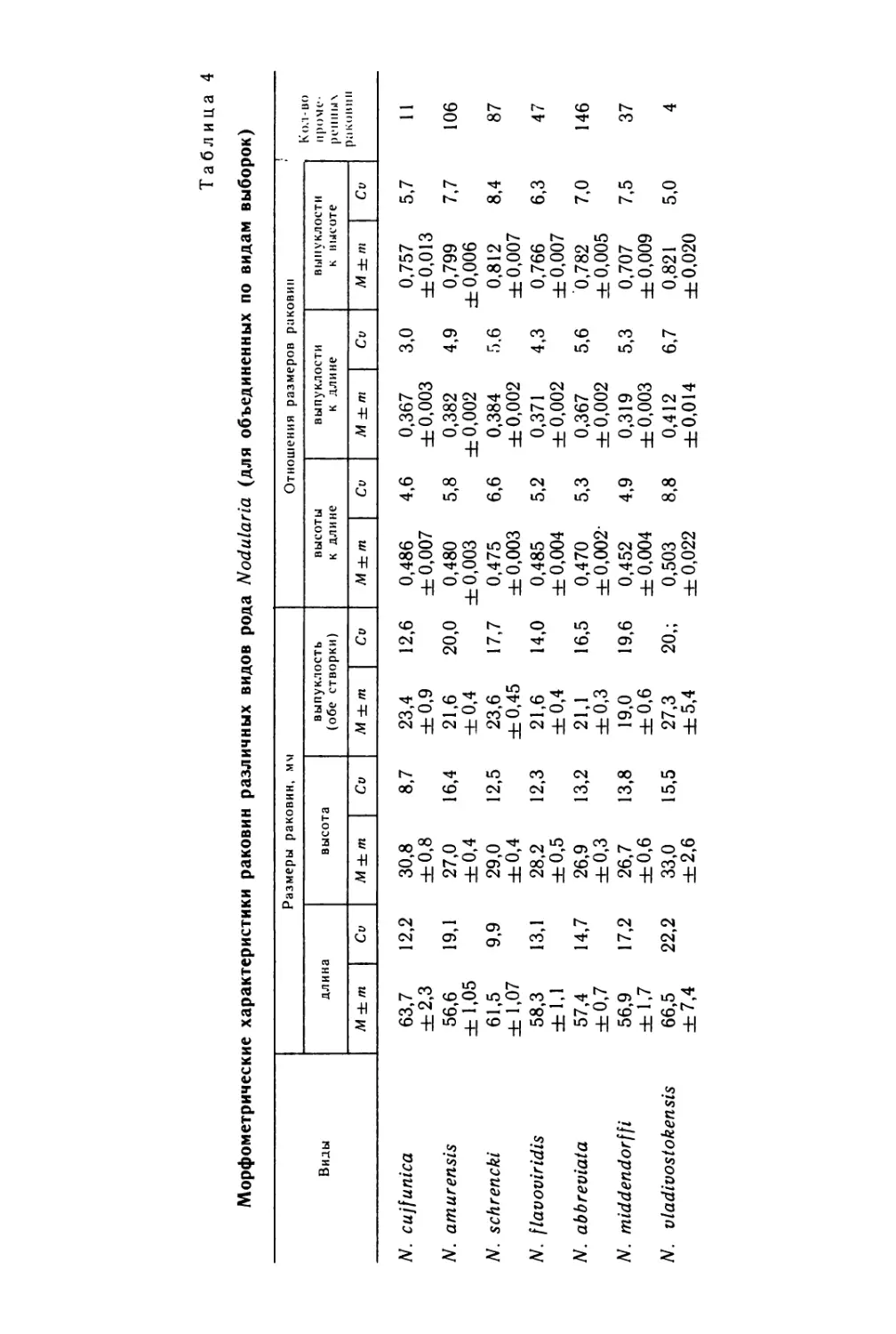



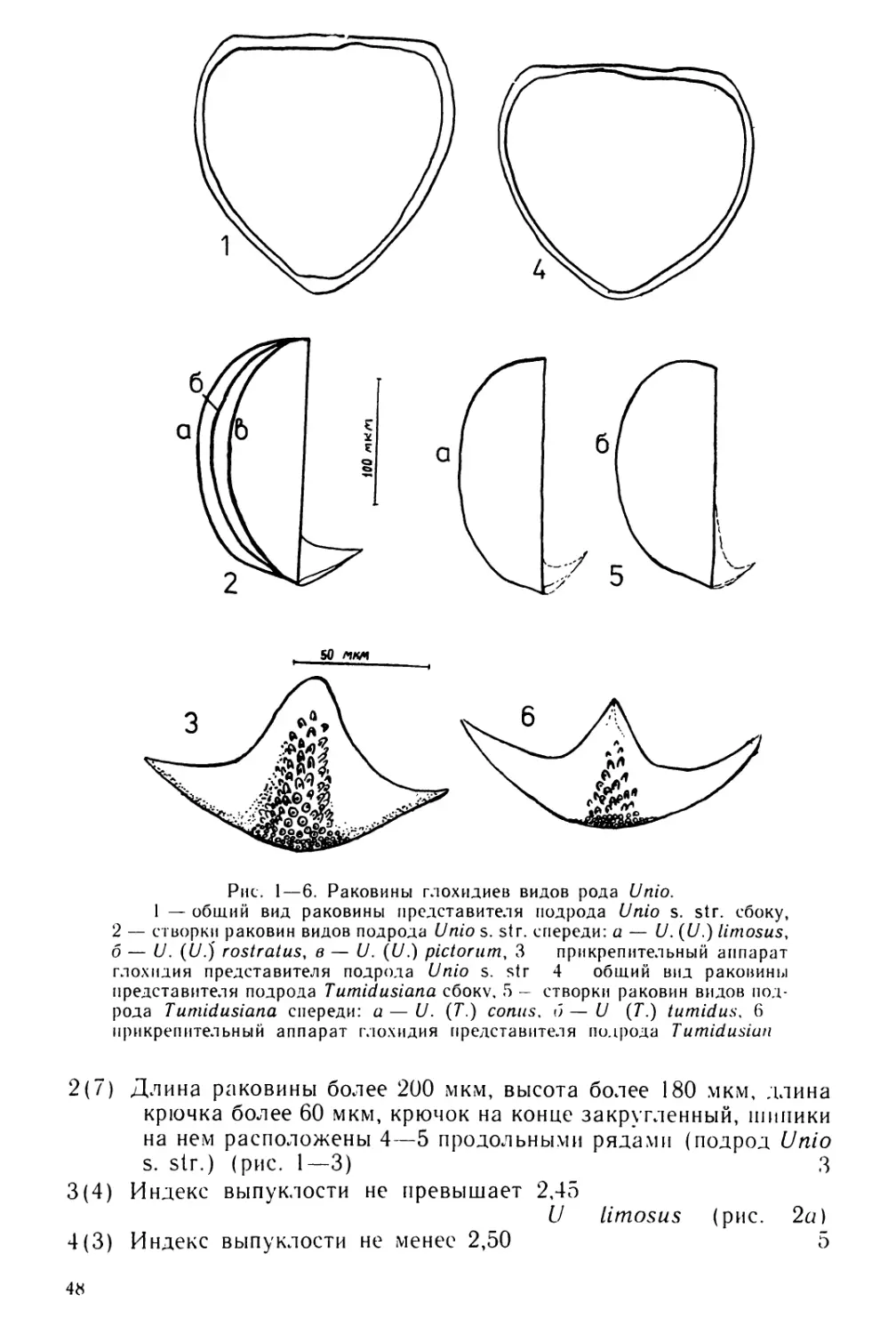























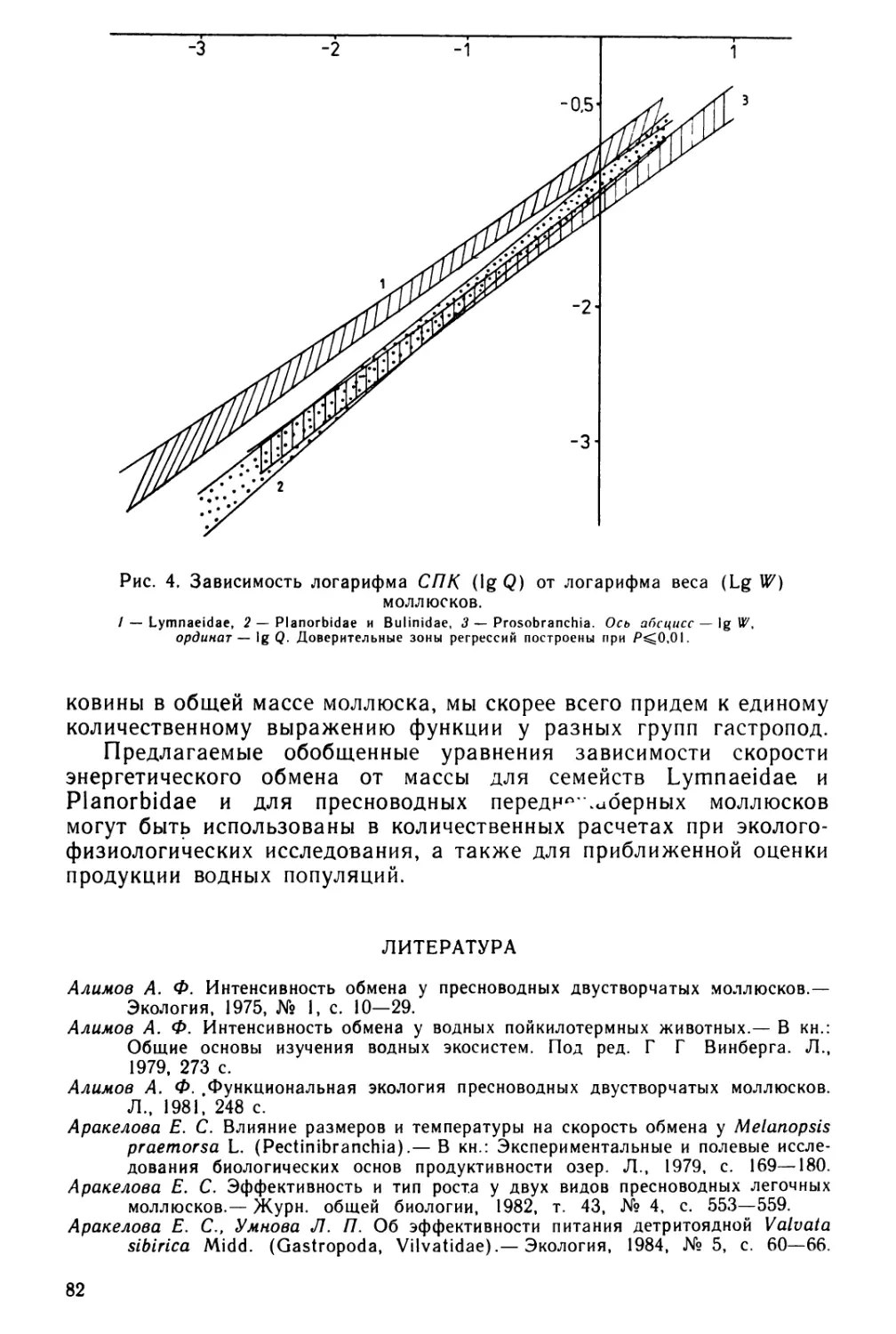








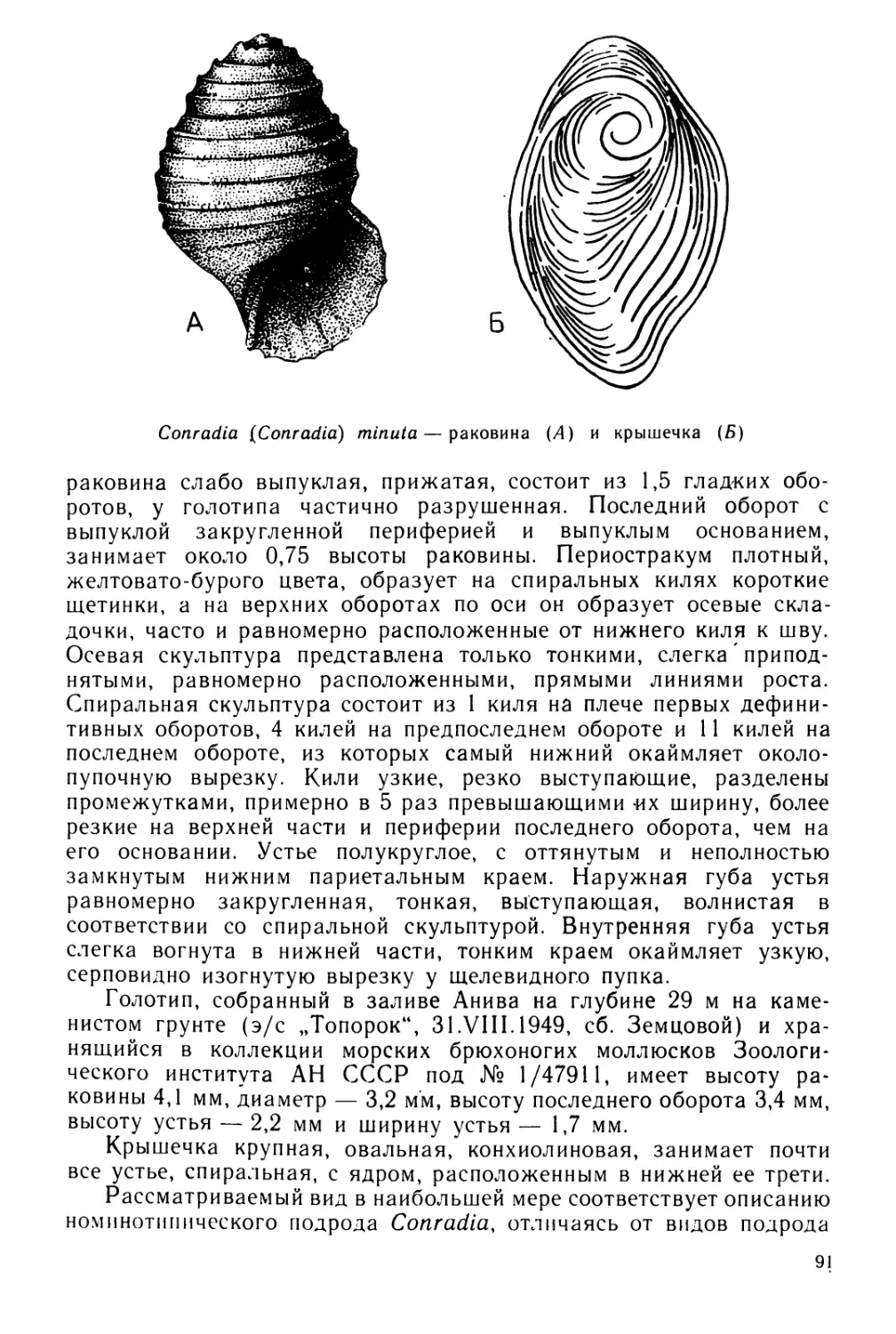








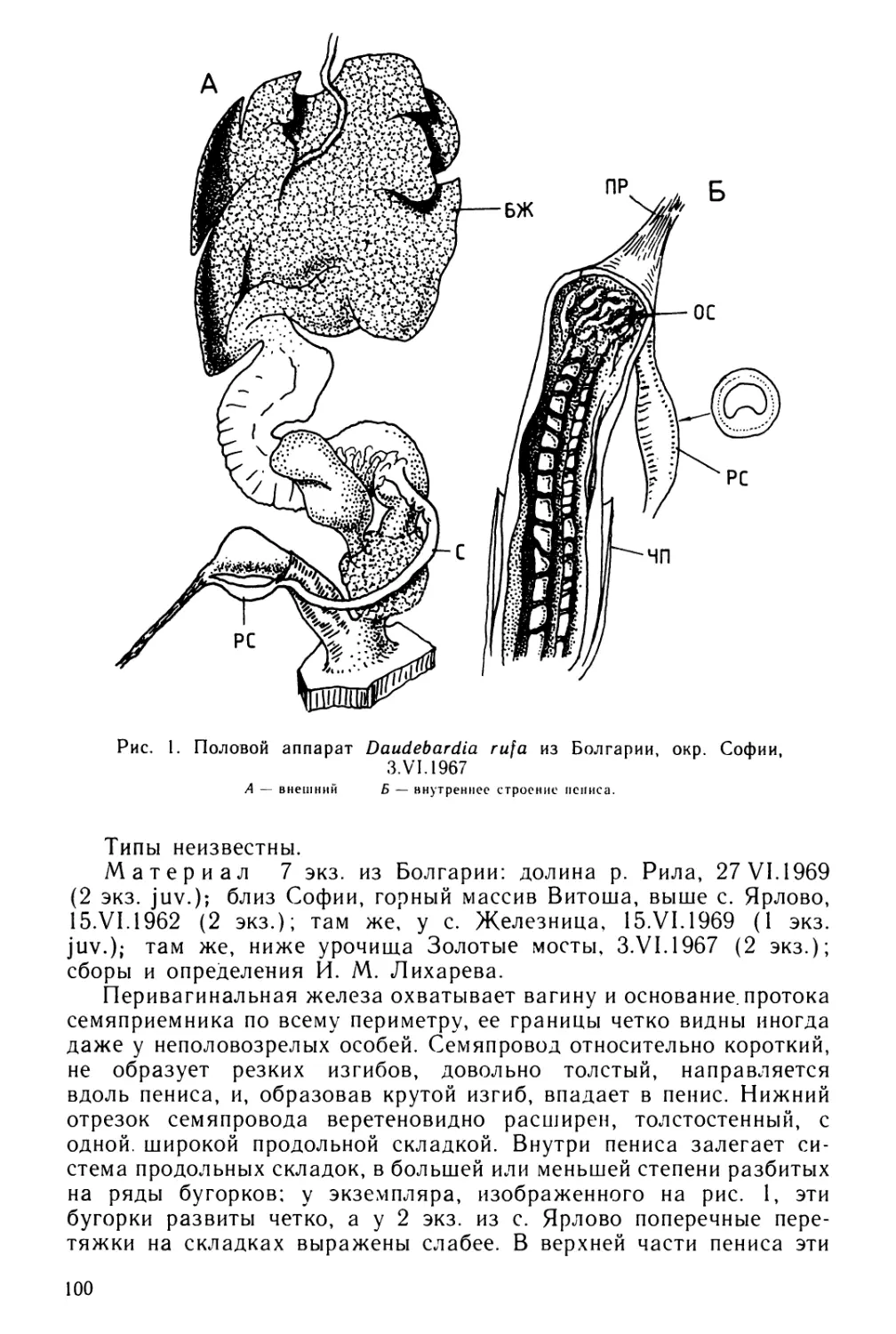

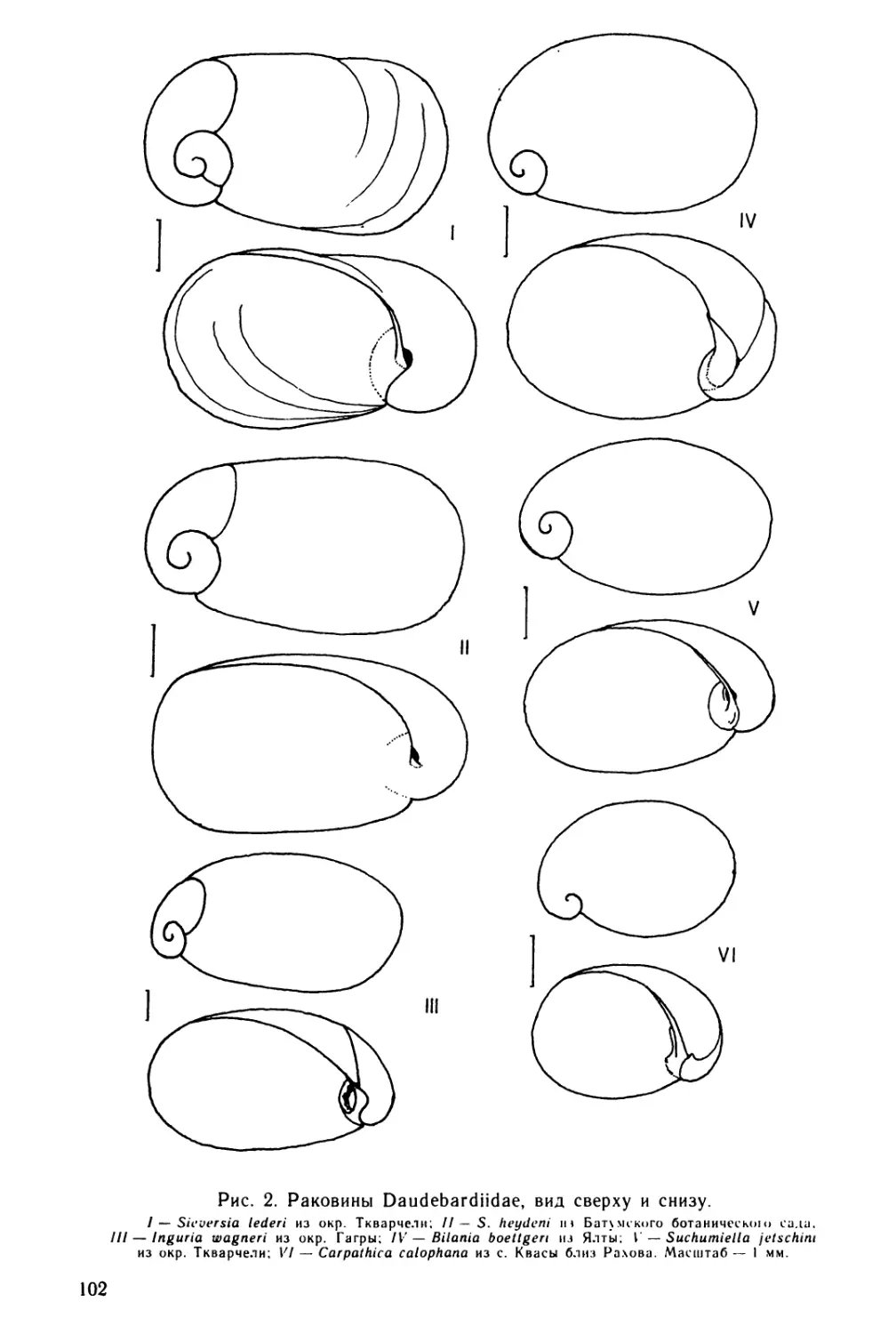
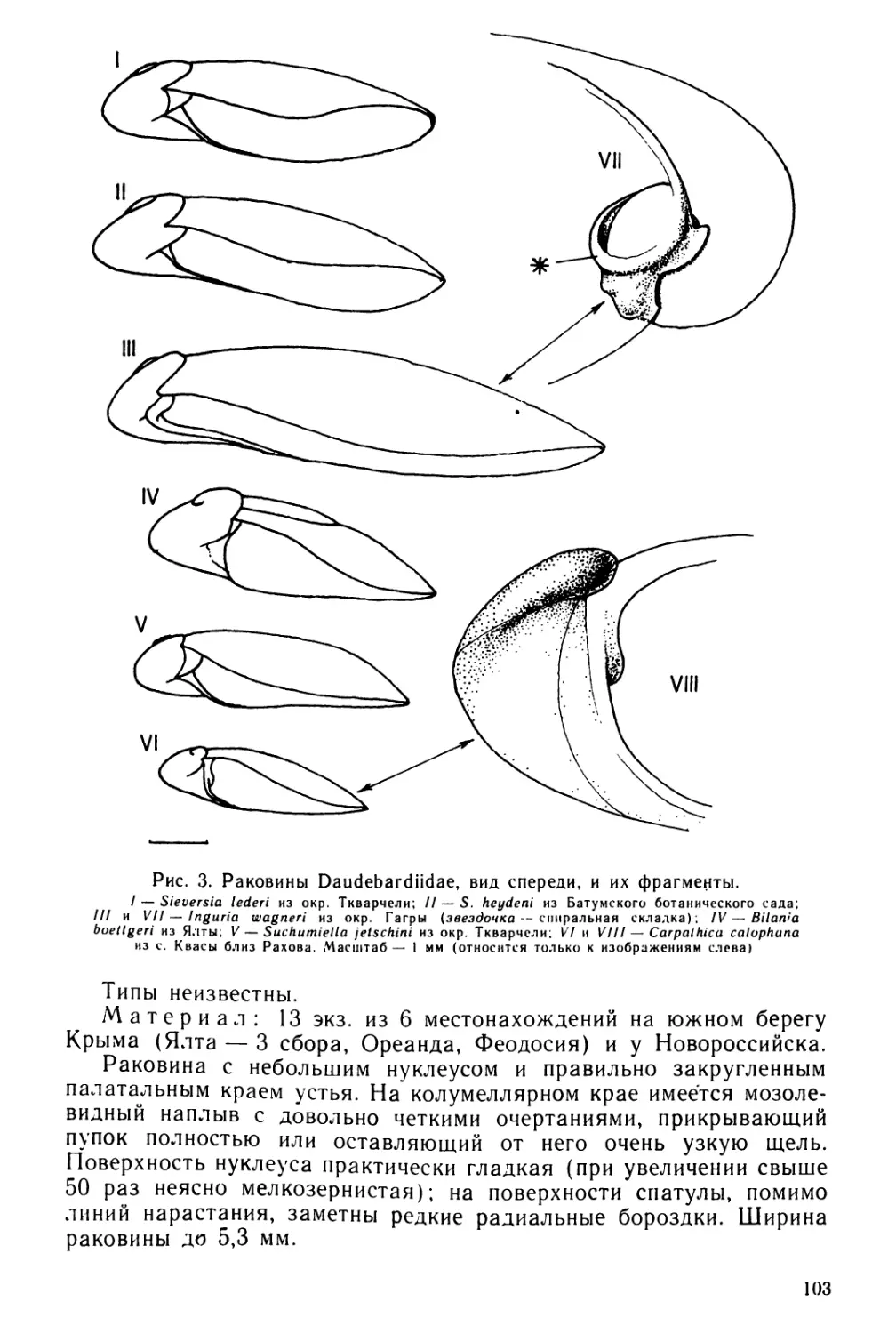
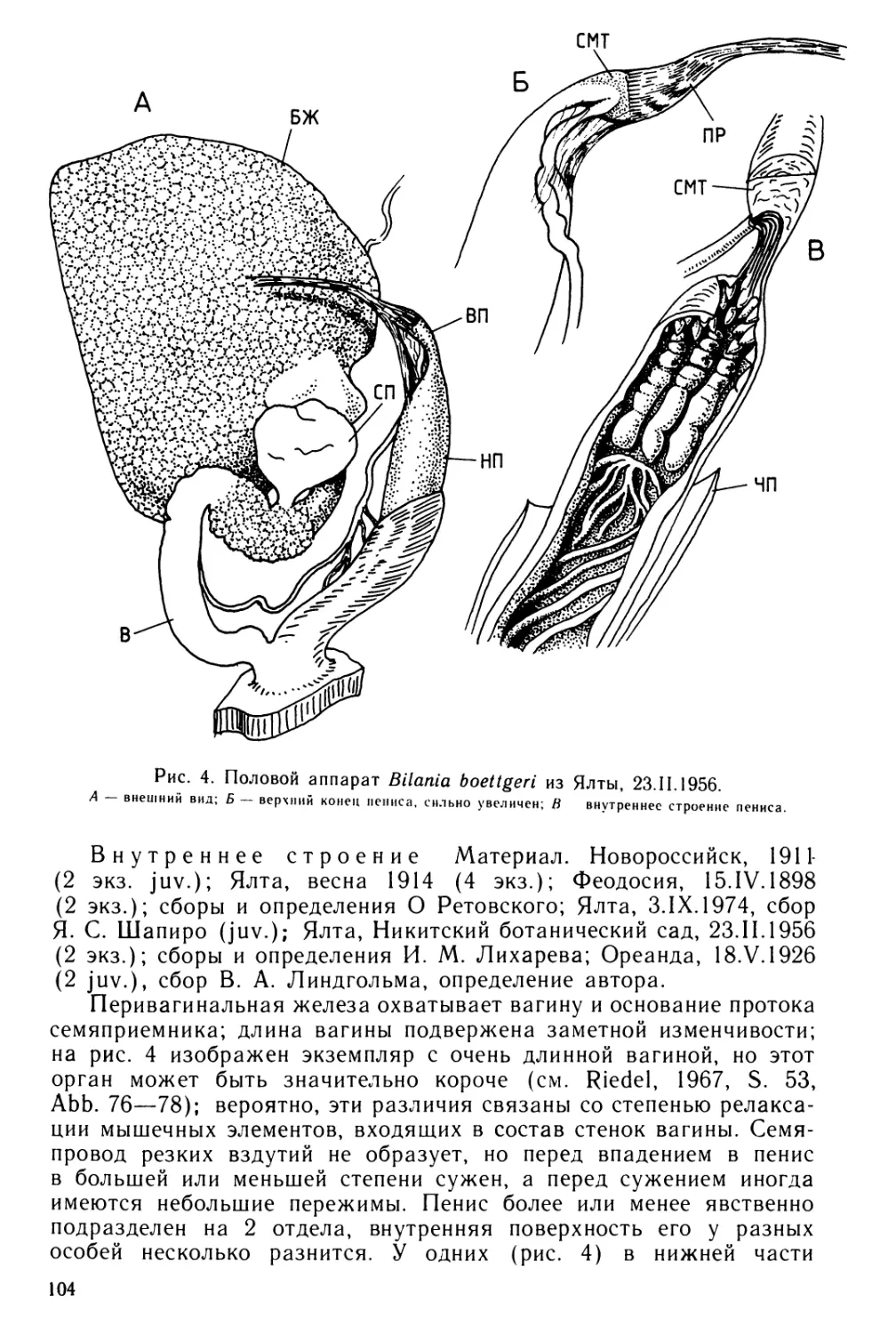

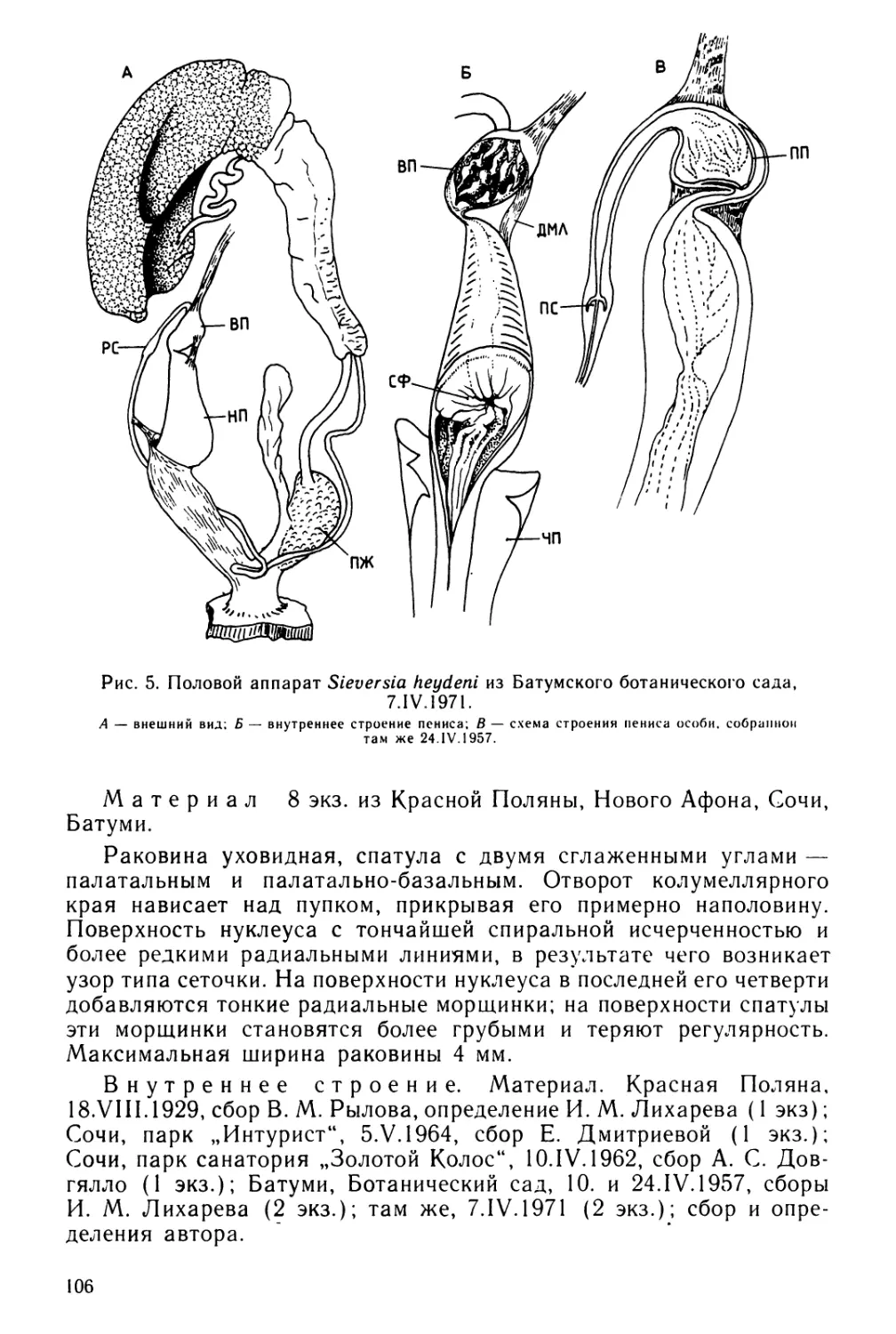

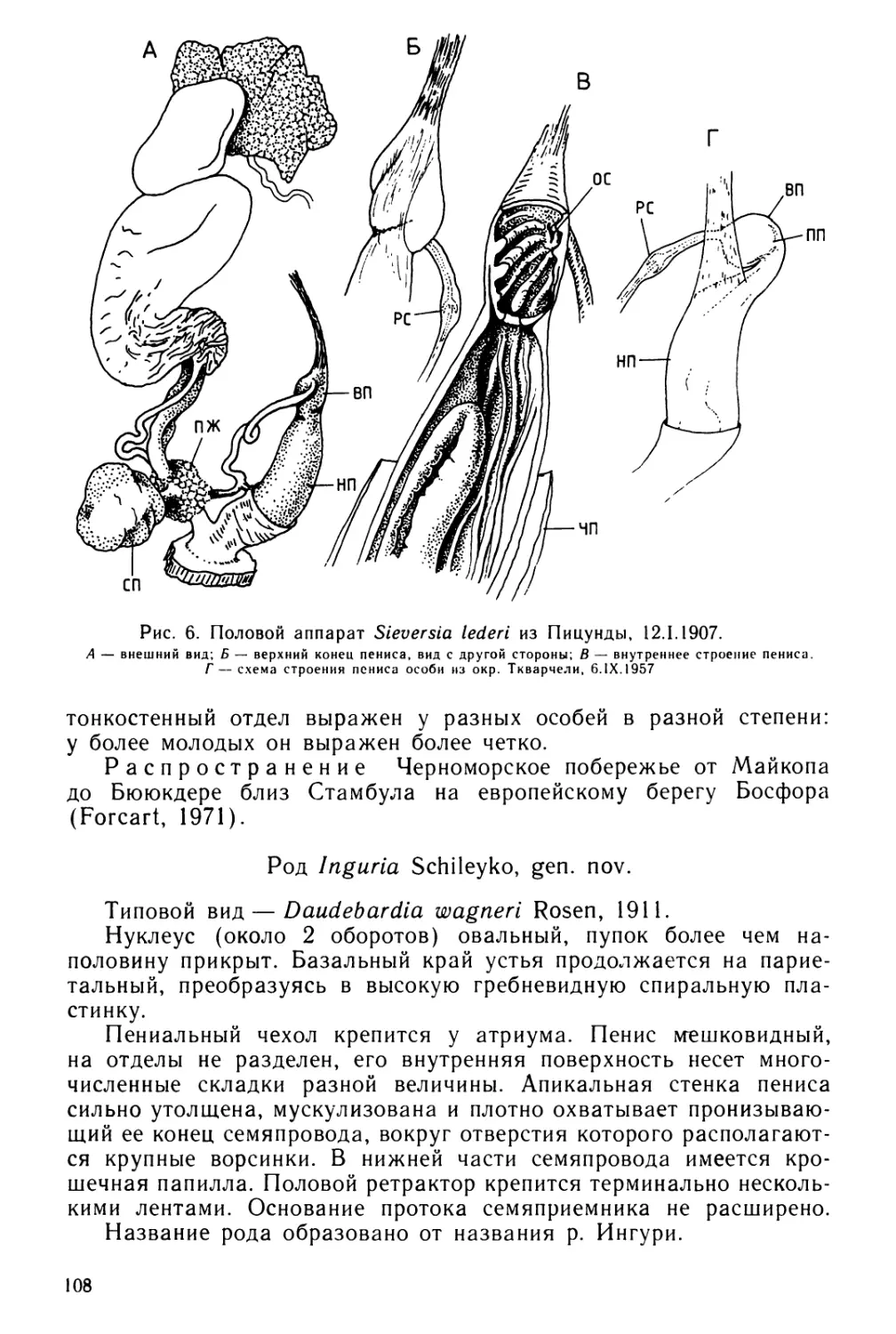














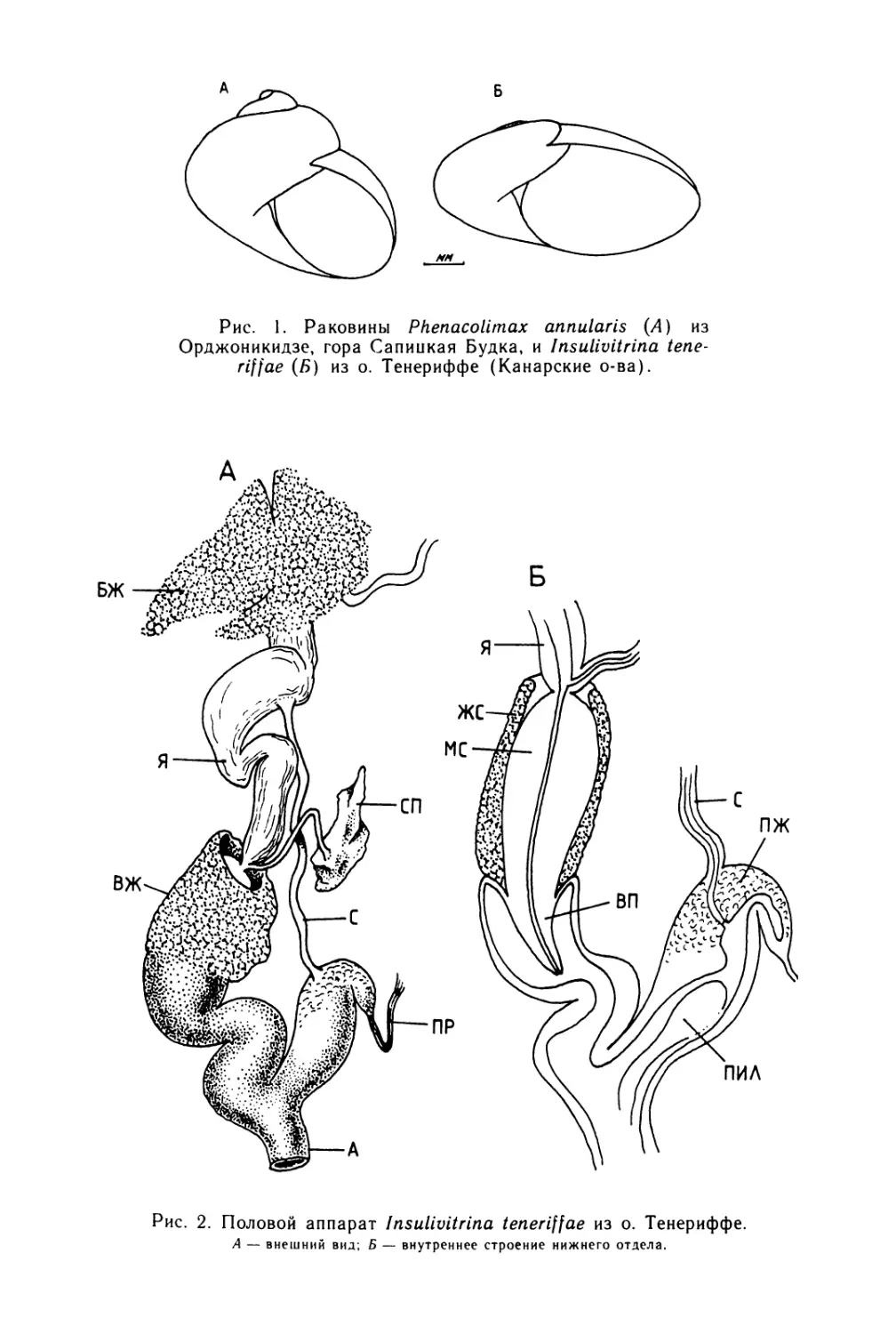

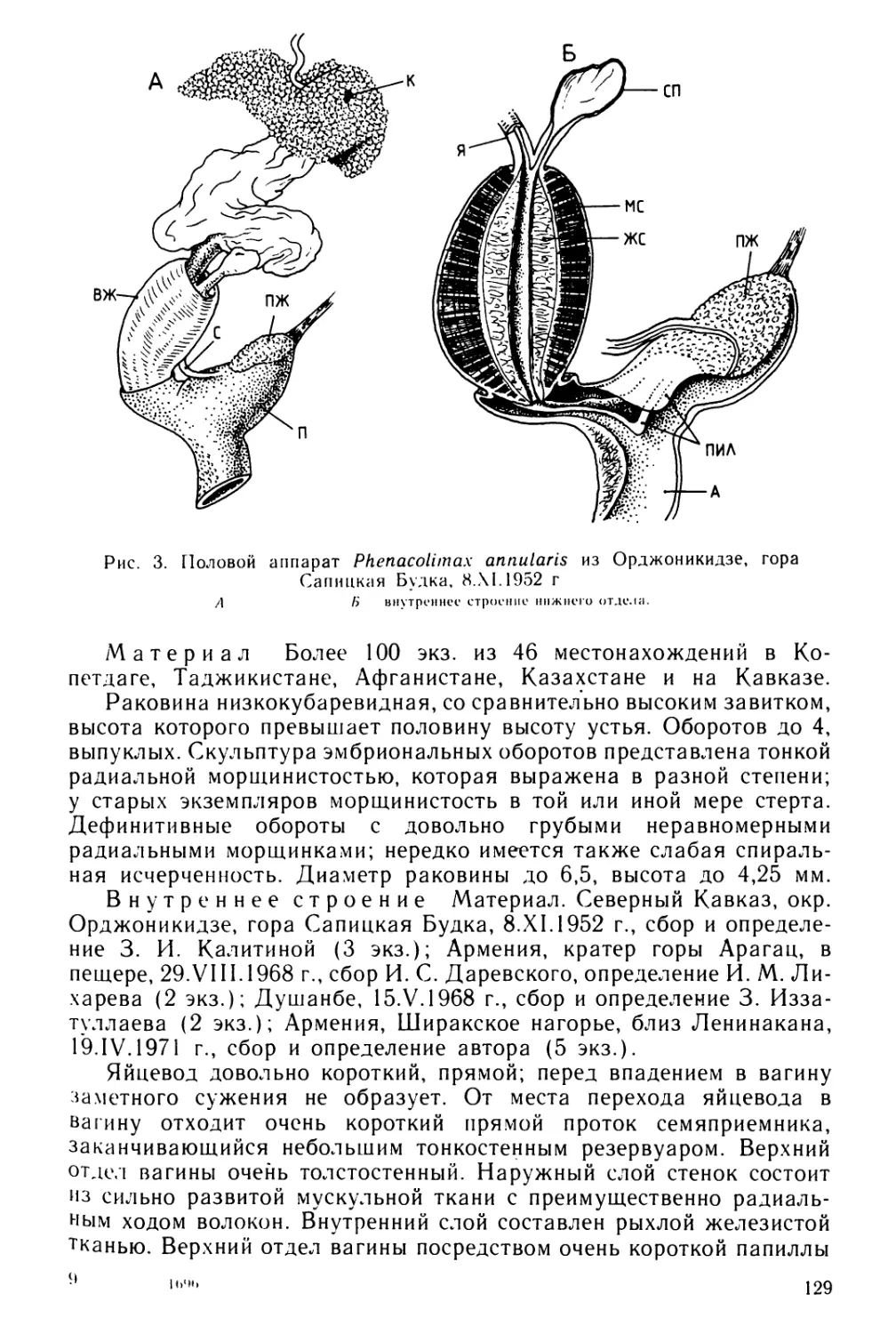

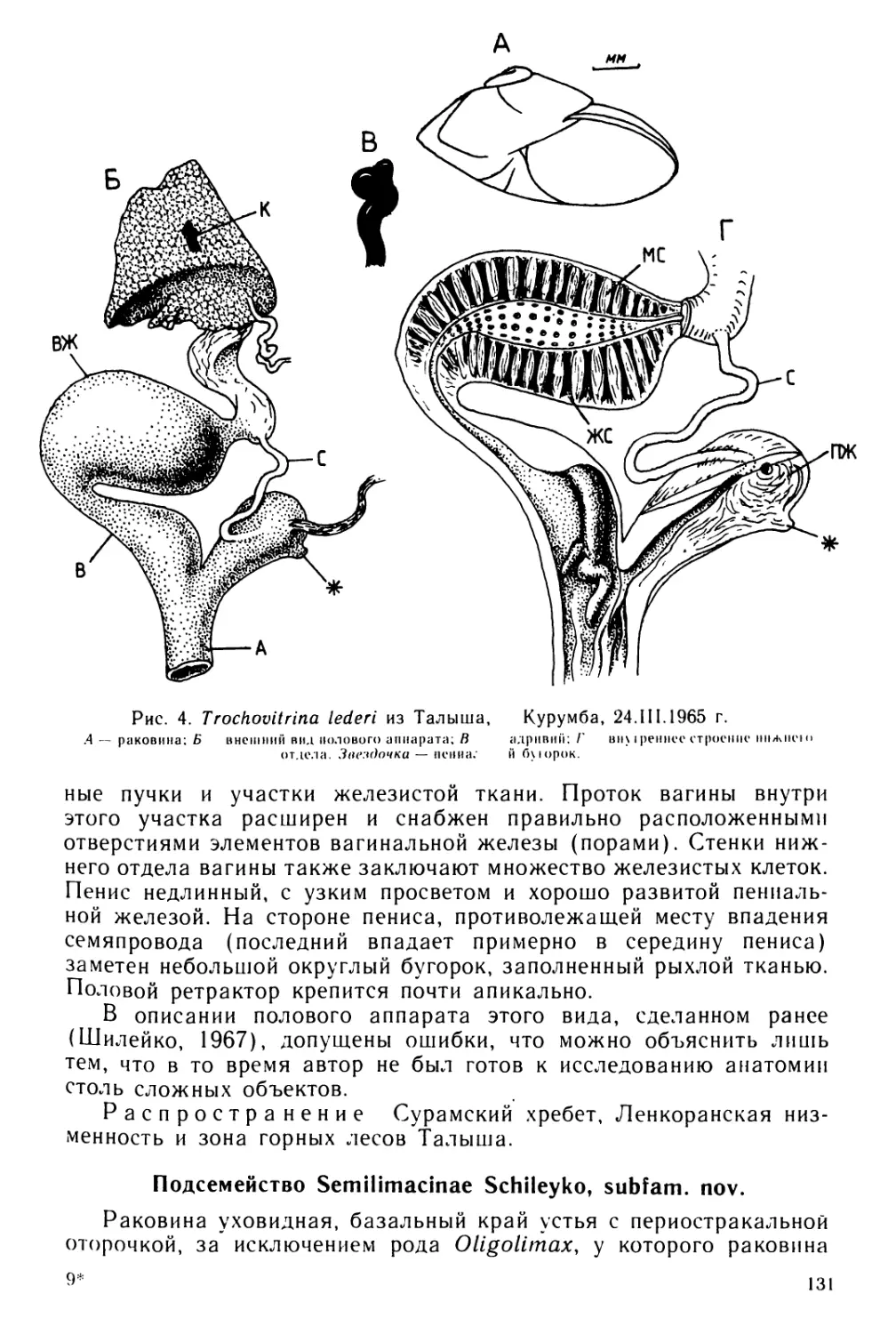


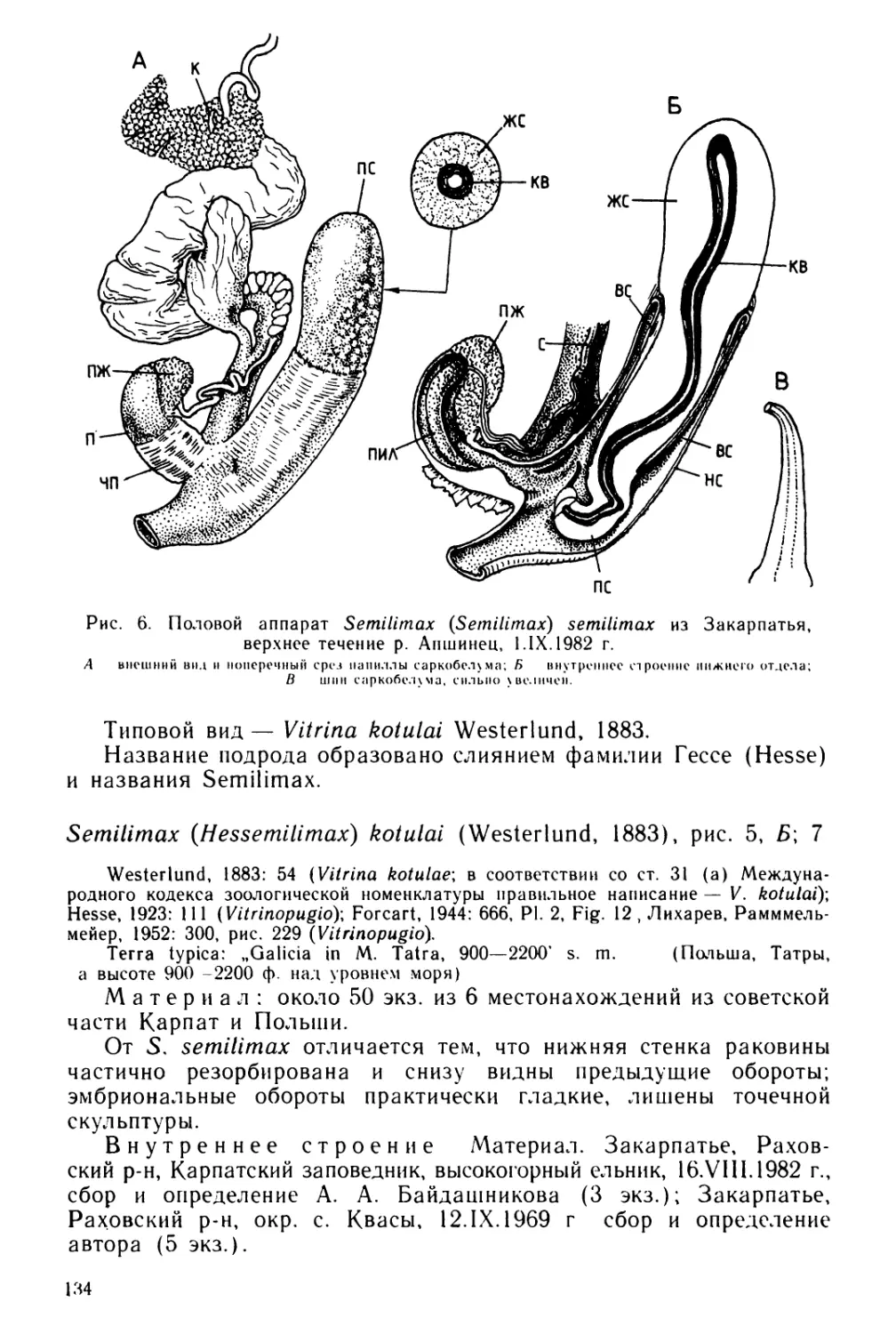
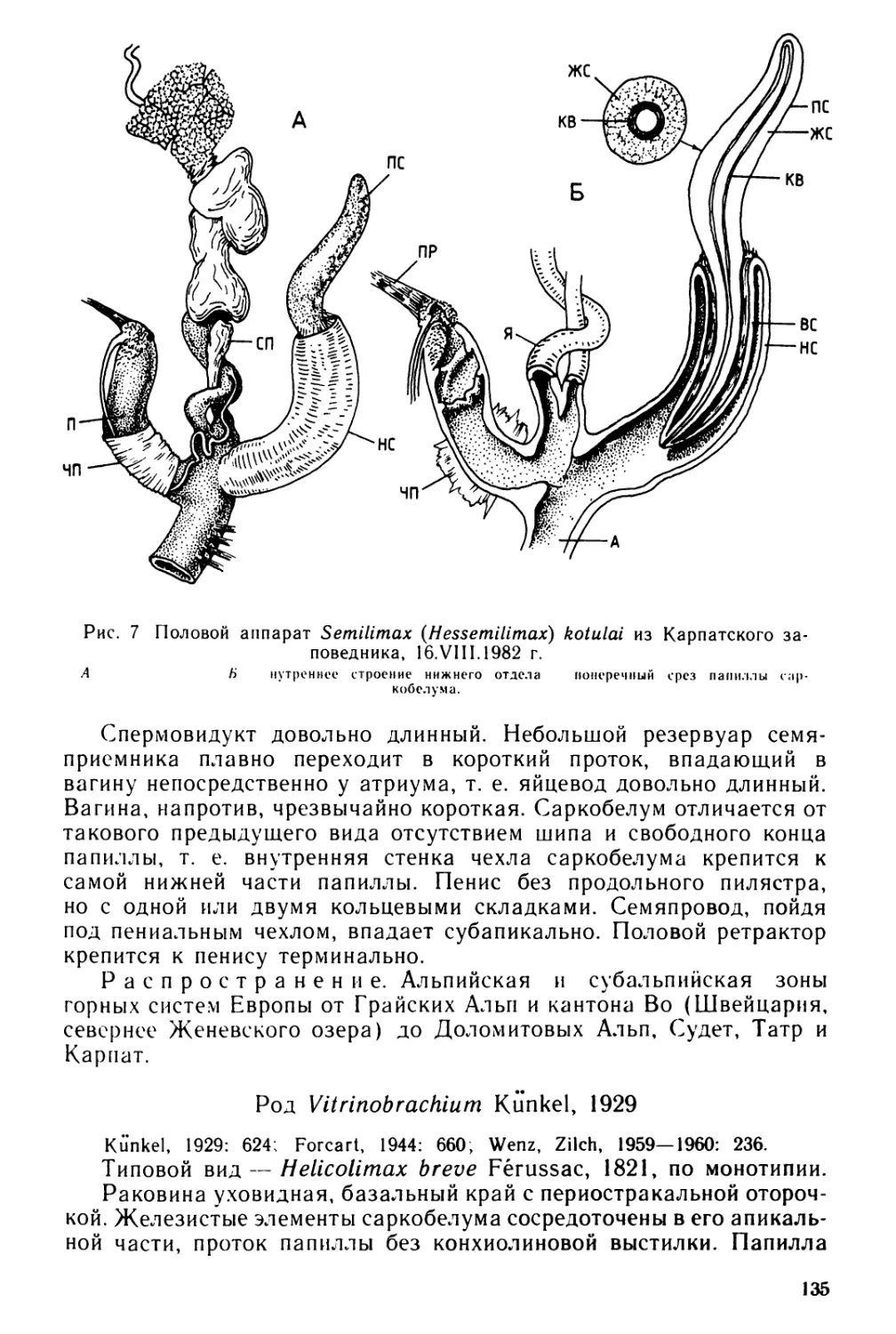

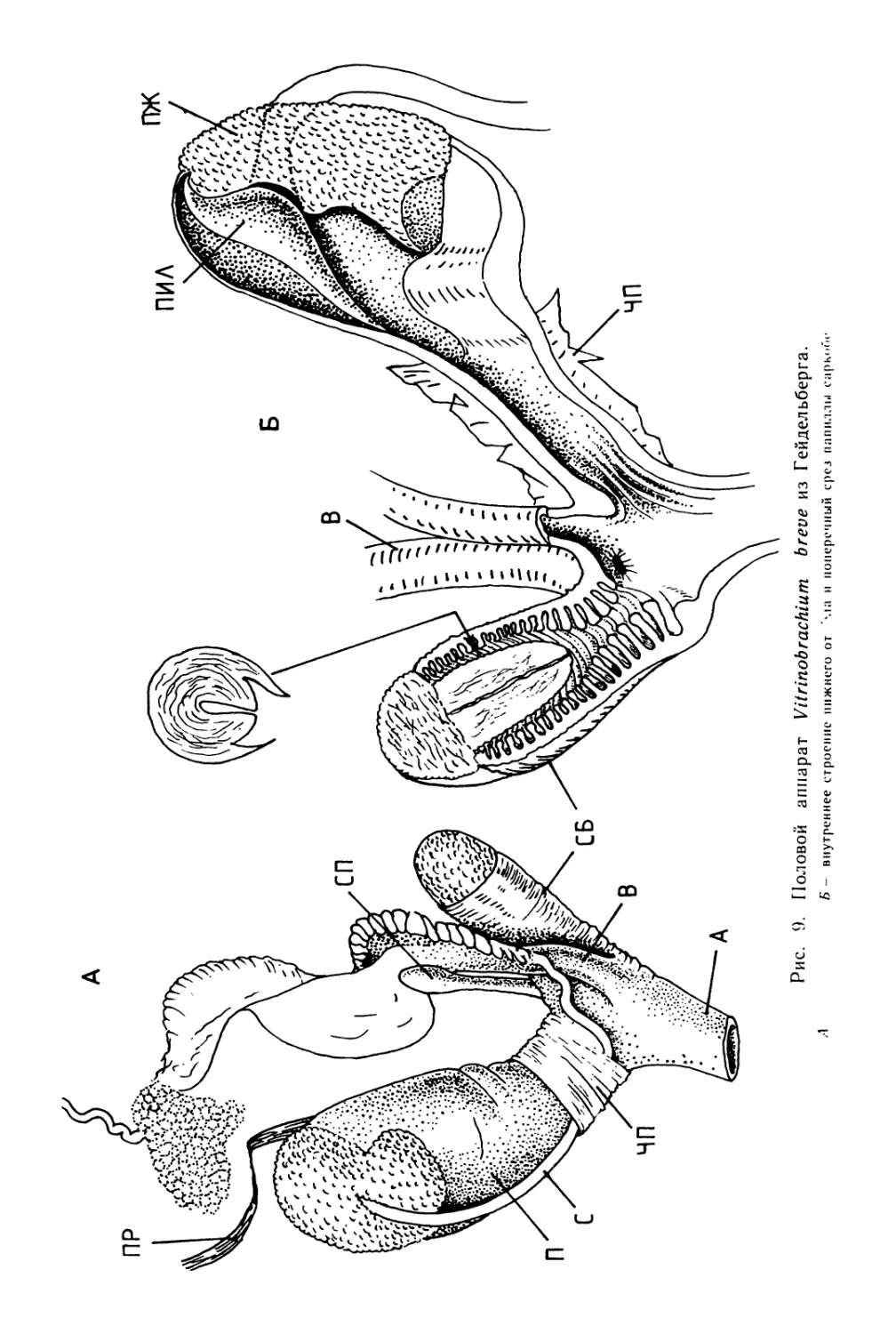


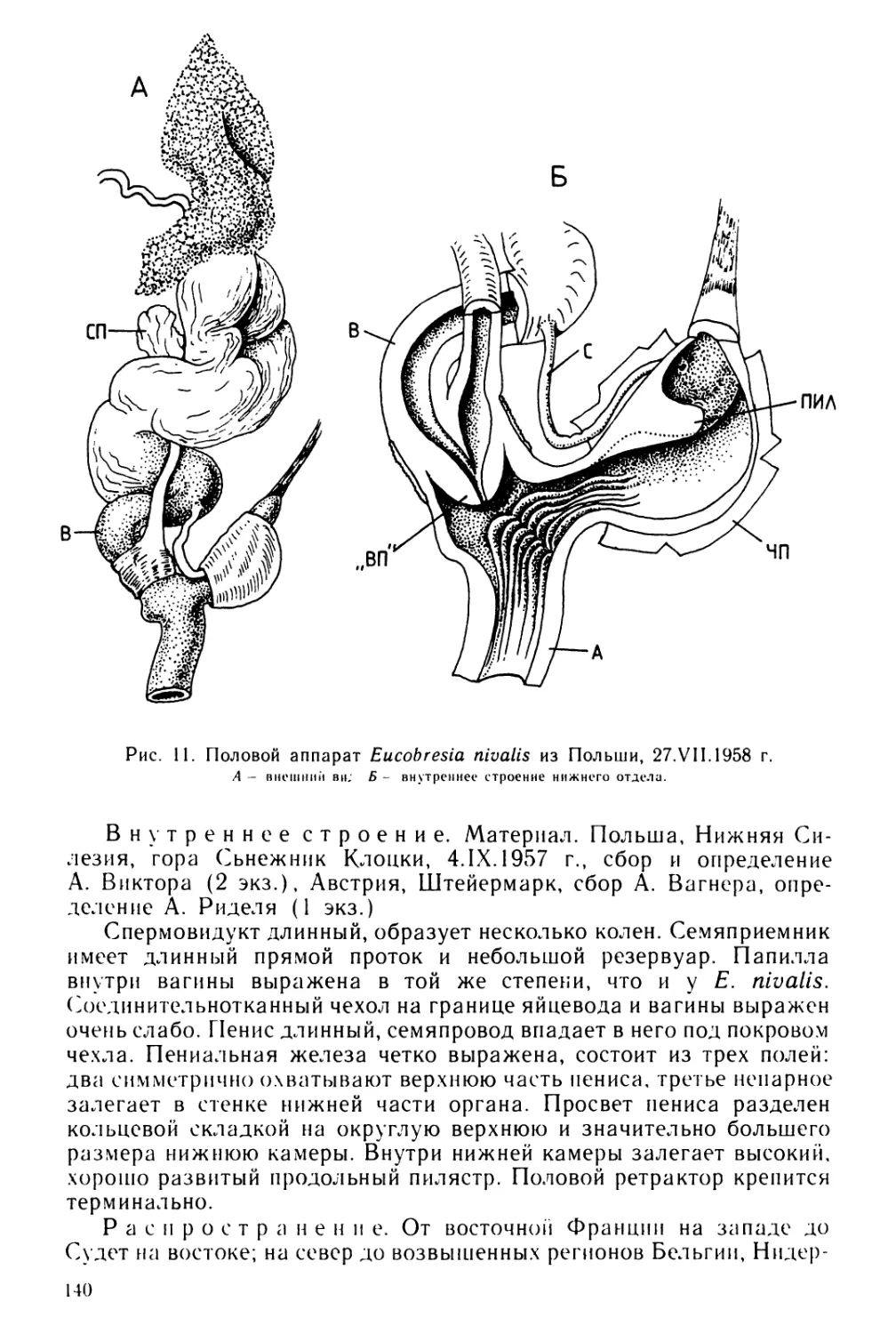
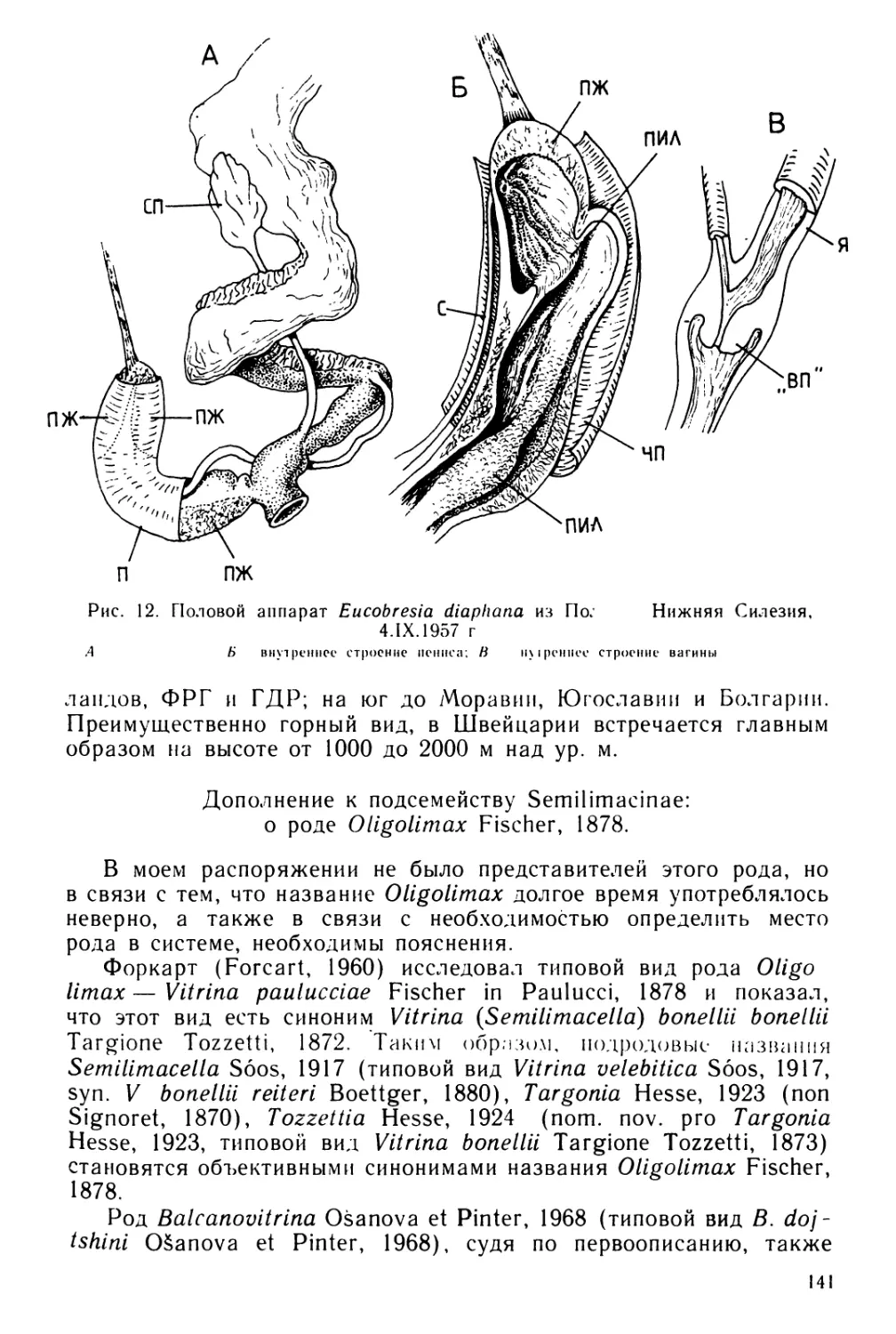






























Text
ISSN 0206— 047
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
МОРФОЛОГИЧЕСКИЕ
И ЭКОЛОГИЧЕСКИЕ
ОСНОВЫ
СИСТЕМАТИКИ МОЛЛЮСКОВ
АКАДЕМИЯ НАУК СССР
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА
Том 148
МОРФОЛОГИЧЕСКИЕ
И ЭКОЛОГИЧЕСКИЕ
ОСНОВЫ
СИСТЕМАТИКИ МОЛЛЮСКОВ
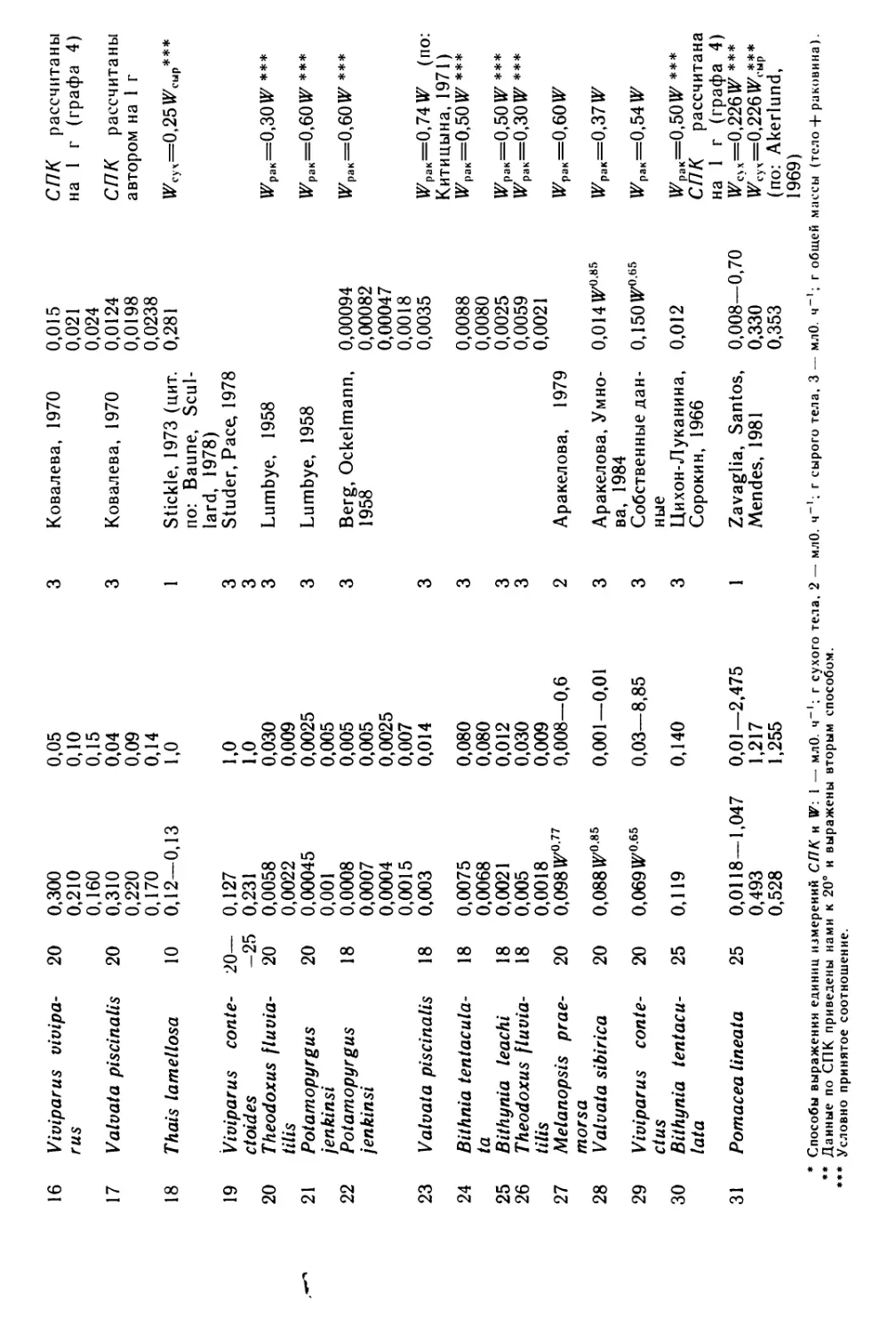
Под редакцией
Я. И. Старобогатова и А. А. Шилейко
ЛЕНИНГРАД
1986
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE; LENINGRAD, 1986, VOL. 148
MORPHOLOGICAL AND ECOLOGICAL BASES
OF THE MOLLUSCAN TAXONOMY
Главный редактор
директор Зоологического института АН СССР
О. А. Скарлато
Редакционная коллегия:
Я. И. Старобогатов (редактор серии), Ю. С. Балашов, Л. Я. Боркин, И. С. Да-
ревский, В. А. Заславский, И. М. Кержнер, М. Г Петрушевская, В. А. Тряпицын,
И. М. Фокин, С. Я. Цалолихин, В. В. Хлебович.
Рецензенты:
И. М. Лихарев, Ю. С. Миничев
Сборник посвящен разносторонним подходам к вопросам систематики и фило-
гении моллюсков. Обсуждается филогения одного из подклассов беспанцирных мол-
люсков, личиночное развитие хитонов и крупных пресноводных двустворчатых —
наяд, морфометрические различия видов наяд, система и филогения двух семейств
наземных моллюсков, интенсивность обмена у брюхоногих моллюсков и экологи-
ческая обусловленность подвидов морских брюхоногих моллюсков. Кроме того, в
статьях сборника описан ряд новых видов и родов брюхоногих и двустворчатых
моллюсков.
АЛ 2005000000—008 с £ л
М “055(02)3-86 Без объяйления © Зоологический институт АН СССР, 1986
ПРЕДИСЛОВИЕ
Настоящий сборник составлен из статей, посвященных морфо-
логии и экологии морских, пресноводных и наземных моллюсков. Эти
исследования непосредственно связаны с разработкой системы четы-
рех классов: панцирных, беспанцирных, двустворчатых и брюхо-
ногих. Наряду со статьями по морфологии и экологии взрослых
животных есть статьи по личиночным формам панцирных (Б. И. Си-
ренко) и пресноводных двустворчатых (Л. А. Антонова). Неболь-
шому, но' крайне своеобразному подклассу беспанцирных мол-
люсков—каудофовеатам.посвящена обстоятельная работа Д. Л. Ива-
нова. Принципиально важна небольшая статья О. А. Скарлато и
Я. И. Старобогатова, поскольку ни одна из существующих систем
индексации элементов замка двустворчатых моллюсков не может
быть признана удовлетворительной. Описания новых видов брюхо-
ногих и двустворчатых моллюсков содержатся в статьях А. Н. Голи-
кова, 3. И. Иззатуллаева и Я- И. Старобогатова. Исследования
А. А. Шилейко содержат материалы по морфологии и соображения
по систематике и филогении древних специализированных семейств
наземных легочных моллюсков. Экологии водных брюхоногих посвя-
щена статья Е. С. Аракеловой. Исследования выполнены сотруд-
никами Зоологического института АН СССР или специалистами из
других учреждений на основе коллекций Зоологического института
АН СССР. Сборник представляет интерес для исследователей, рабо-
тающих в области морфологии, экологии, систематики и филогене-
тики беспозвоночных, а также всех, чья научная деятельность так
или иначе связана с моллюсками.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
Б. И. Сиренко
ЛИЧИНОЧНОЕ РАЗВИТИЕ
ПАНЦИРНЫХ МОЛЛЮСКОВ (POLYPLACOPHORA).
АНАЛИЗ РАБОТ ПО ВИДАМ
С ПЛАНКТОННЫМ ТИПОМ РАЗВИТИЯ ЛИЧИНОК
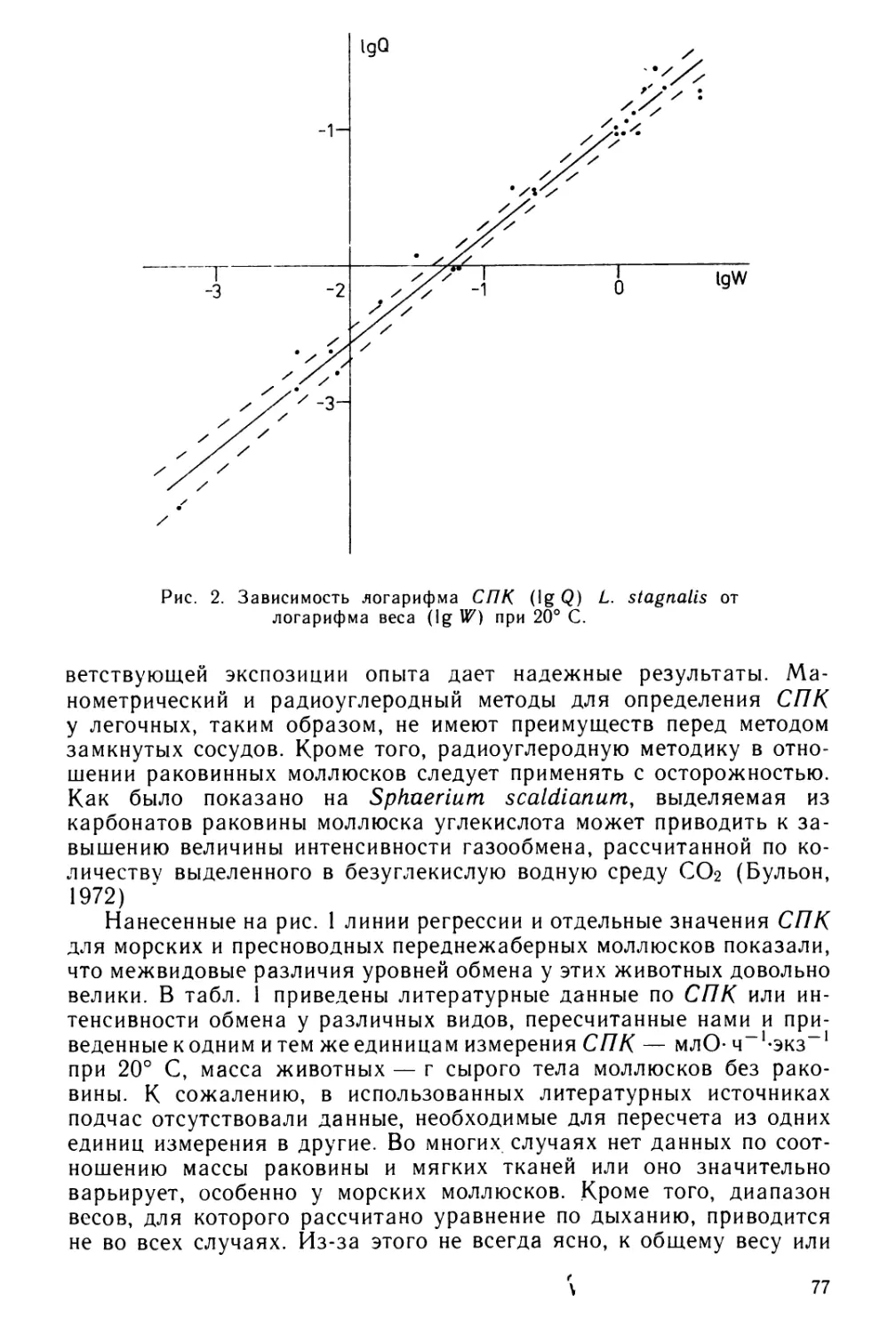
Изучение личиночного развития моллюсков с давних пор при-
влекает биологов, способствуя разрешению многих проблем эмбрио-
логии, морфологии, систематики и экологии. Исследование развития
таких примитивных моллюсков как панцирные позволяет, в частно-
сти, уточнить филогенетические отношения внутри класса Mollusca,
выяснить закономерности закладки различных органов, определить
роль личиночной стадии в распространении видов.
Личиночное развитие хитонов начали изучать в середине прош-
лого столетия, с тех пор только по личинкам с планктонным типом
развития вышло 27 работ (Clark, 1855; Loven, 1856; Kowalevsky,
1879, 1882, 1883; Metcalf, 1892, 1893; Heath, 1899, 1904; Hammar-
sten, Runstrom, 1925; Hull, Risbec, 1930; Hoffmann, 1931; Grave,
1932; Thorson, 1946; Okuda, 1947; Christiansen, 1954; Brewin,
1958; Thorpe, 1962; Nagabhushanam, Gopalakrishna Murti, 1968;
Barnes, Gonor, 1973; Watanabe, Cox, 1975; Haas, Kriesten, Watabe,
1979, 1980; Kniprath, 1980; Soliman, Iskander, 1980, 1982; Rumrill,
Cameron, 1983). Настала пора проанализировать результаты и под-
вести некоторые итоги. Существуют две обзорные работы (Smith,
1966; Hyman, 1967), однако в первой из них рассматриваются в
основном случаи вынашивания яиц и молоди в мантийной полости
хитонов, а также закладка и развитие пластинок раковины у личинок
этих моллюсков. Во второй сводке (Hyman, 1967) использована лишь
половина работ, опубликованных к тому времени, а с момента вы-
хода этой последней сводки появилось 9 статей, содержащих много
новых данных. В литературе на русском языке практически отсут-
ствуют сведения по развитию панцирных моллюсков, если не считать
кратких сообщений в компилятивных работах Б. В. Властова (1940)
и О. М. Ивановой-Казас (1977)
Цель настоящего обзора — познакомить специалистов, интере-
сующихся личиночным развитием хитонов, с библиографией по этому
вопросу, а также представить в сокращенном виде общие законо-
мерности, наблюдающиеся при планктонном типе развития личи-
нок — наиболее распространенном в этом классе моллюсков. Вына-
шивание личинок в мантийной полости этих моллюсков, развитие
в капсулах, отложенных на донный субстрат и случаи живорождения
не обсуждаются и требуют отдельного рассмотрения. Поскольку
дробление яиц хитонов и судьба бластомеров подробно описаны у
4
Ивановой-Казас (1977) эти вопросы также не рассматриваются.
В связи с тем, что названия видов, особенно в работах прошлого
столетия, не всегда были правильными, дается только современное
название вида по каталогу Кааса и Ван Белле (Kaas, Van Belle,
1980).
Основные сведения о развитии, имеющиеся в цитированных
выше работах (в частности, название изученного вида, до какого
возраста он изучен, размеры яиц, хронология развития) приведены
в табл. 1 и 2. При составлении табл. 2 использованы только те работы,
в которых представлены более подробные сведения по хронологии
личиночного развития. За более чем 120-летний период исследований
личиночное развитие изучено у 19 видов хитонов из обоих пред-
ставленных в современной фауне отрядов, причем у 14 до стадии фор-
мирования 7-й и 8-й пластинок. Анализируя имеющиеся данные,
нельзя не отметить высокое качество и непреходящую ценность работ
Ковалевского (Kowalevsky, 1883) и Гаммарстена и Руннстрема
(Hammarsten, Runnstrom, 1925). Об этом свидетельствует то, что
почти в каждой позднее вышедшей работе, используются данные,
полученные этими учеными и заимствуются их точные рисунки.
Необходимо отдать должное также высокому мастерству исполнения
работ Хааса с соавторами (Haas et al., 1979, 1980) и Книпрата
(Kniprath, 1980), почти одновременно исследовавших закономер-
ности закладки пластинок раковины у личинок хитонов.
Общий ход Личиночного развития хитонов с учетом данных, по-
лученных в цитированных работах, выглядит следующим образом.
После оплодотворения яйца (время оплодотворения принимается
за начало отсчета возраста хитонов), в зависимости от запаса
желтка и температуры воды происходит первое дробление яйца. У
тропических видов, таких как Chiton gr anoradiatus, стадия двух
бластомеров наступает через 10 мин (табл. 2), а у большинства ви-
дов умеренных зон океана — через 1—2 ч, стадия четырех бласто-
меров наступает через 20 мин и через 1,5—3 ч соответственно. На
первых двух стадиях дробление равномерное, переходящее затем к
неравномерному по спиральному типу. Через несколько часов обра-
зуется бластула, которая путем инвагинации переходит в гаструлу.
Гаструла уже имеет два ряда клеток с ресничками — будущий прото-
трох. В результате опережающего деления клеток на спинной стороне
зародыша, она расширяется и бластопор постепенно перемещается
на брюшную сторону. Позднее бластопор превращается путем ин-
вагинации в стомодеум. Личинка к этому времени имеет прототрох —
два ряда ресничек, располагающихся посередине и делящих ее на
две половины, верхнюю и нижнюю. Рот помещается сразу же под
прототрохом. На вершине личинка имеет пучок длинных ресничек —
теменной орган. Движения ресничек прототроха замечены у личинок
обычно в возрасте от 5 до 20 ч; на этой стадии трохофоры активно
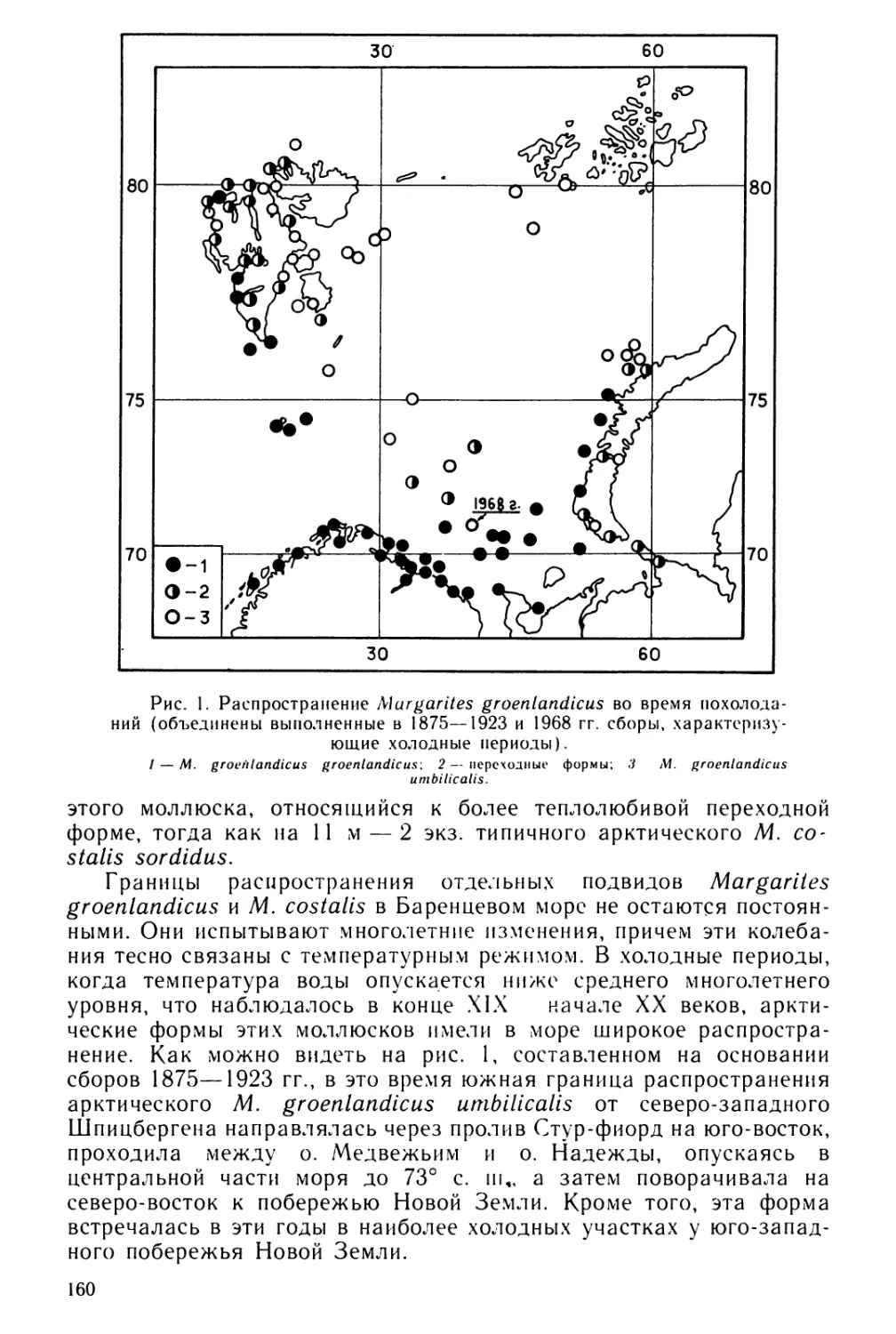
работают прототрохом и через некоторое время, разорвав яйцевую
оболочку, выходят наружу. В норме трохофоры выходят через про-
деланное ими отверстие апикальном концом с теменным органом
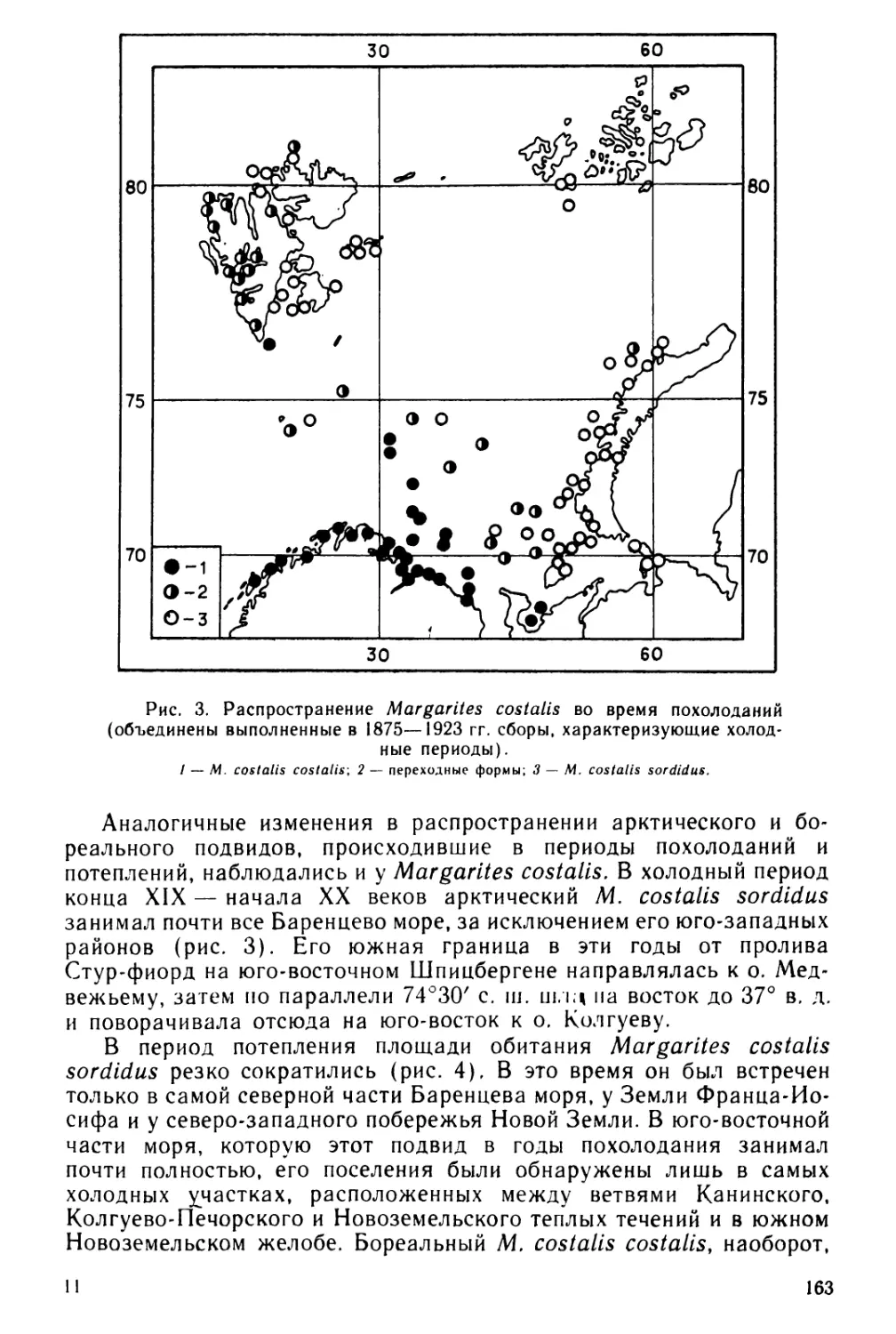
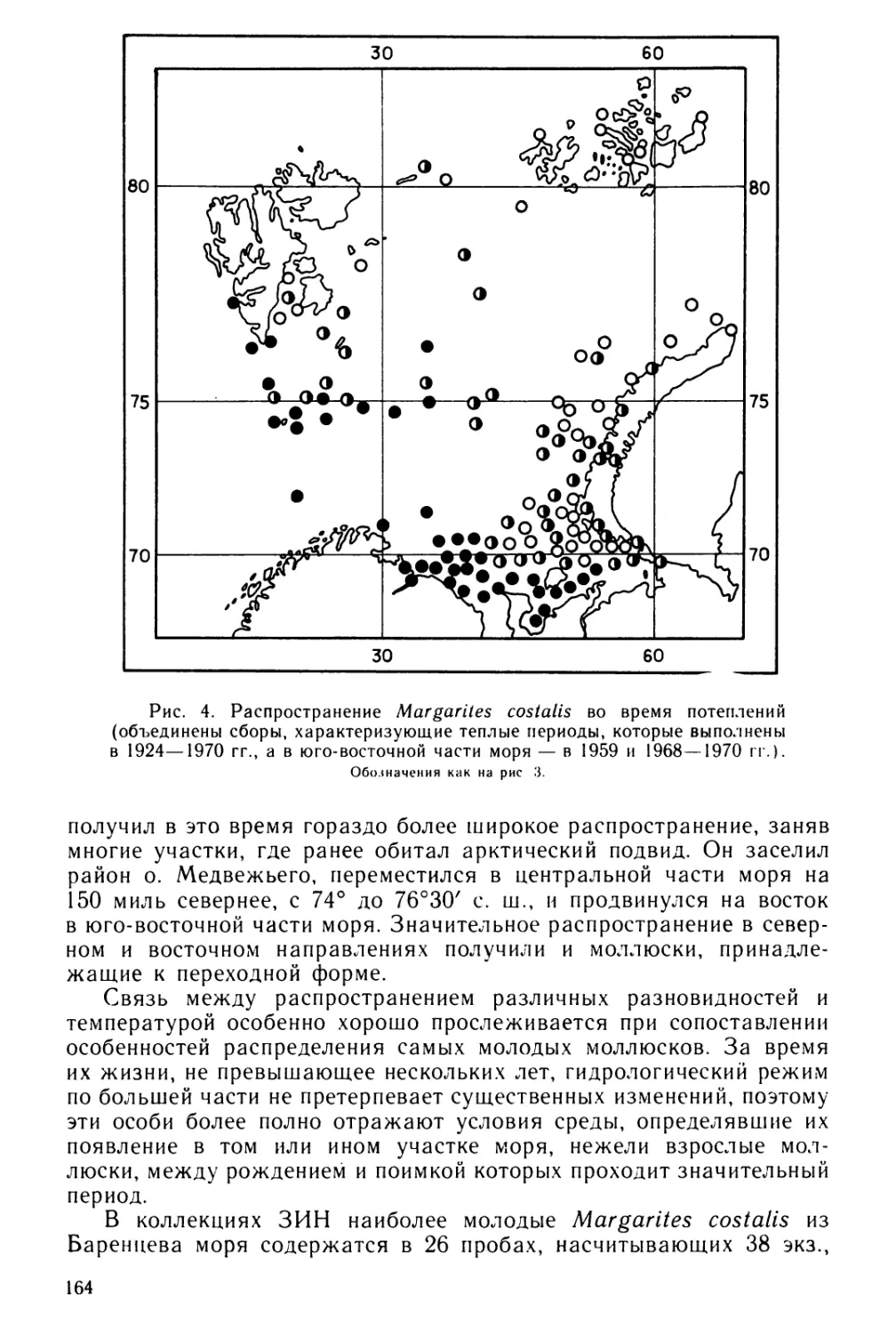
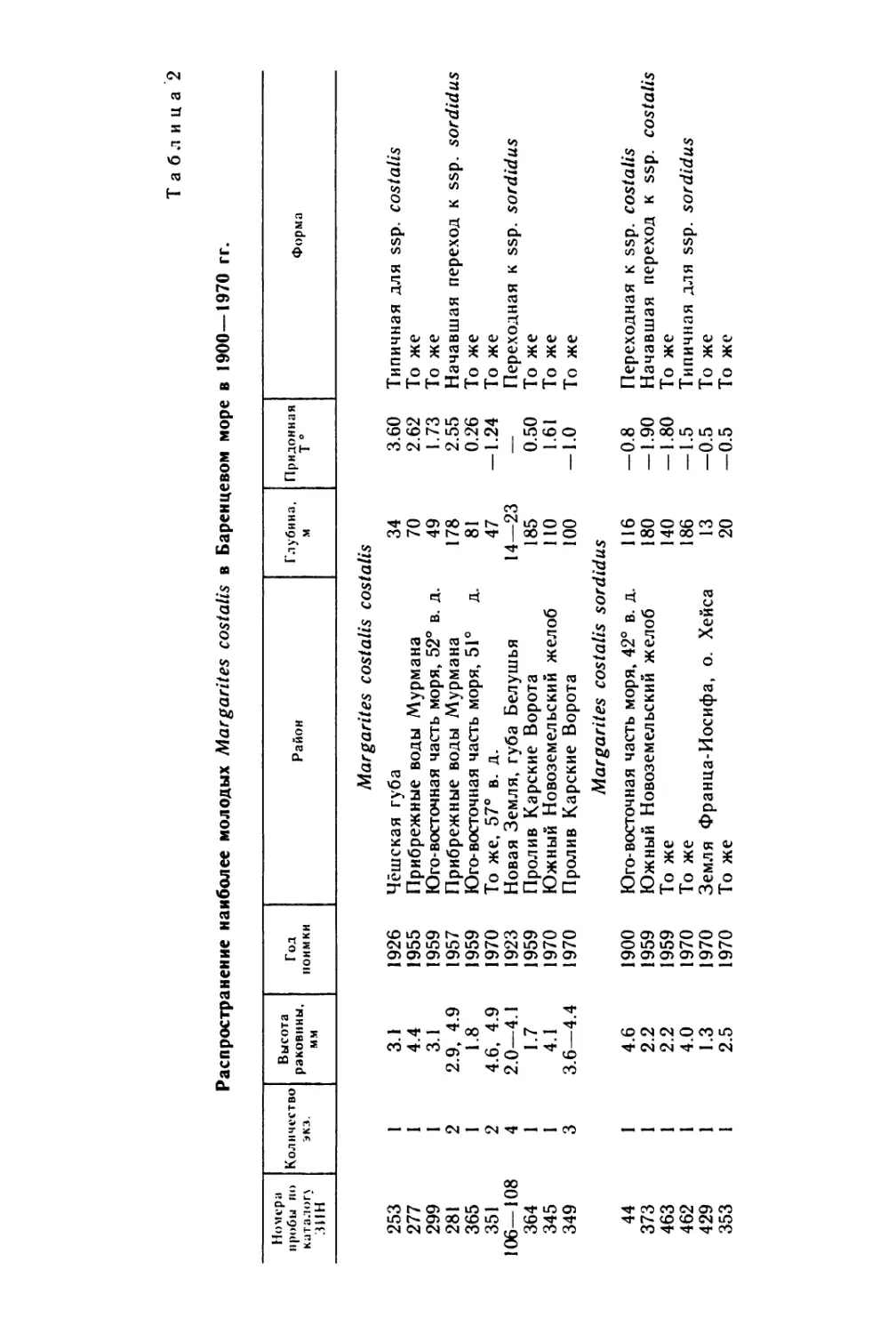
вперед. Возраст, при котором личинки выходят из оболочек, сильно
5
Таблица 1
Изученность личиночного развития хитонов
Вид хитонов
Источник
Leptochiton asellus (Gmelin)
Chaetopleura apiculata (Say)
Ischnochiton acomphus Hull et Risbec
Ischnochiton rissoi (Payraudeau)
Stenoplax magdalenensis (Hinds)
Chiton olivaceus Spengler
Chiton squamosus Linneus
Chiton marmoratus Gmelin
Chiton granoradiatus Leloup
Acanthopleura gemmata (De Blainville)
Lepidochitona cinerea (Linneus)
Lepidochitona corrugata (Reeve)
Mopalia muscosa (Gould)
Mopalia lignosa (Gould)
Mopalia ciliata (Sowerby)
Katharina tunicata (Wood)
Acanthochitona communis (Risso)
Cryptoconchus porosus (Burrow)
Cryptochiton stelleri (Middendorff)
Thorson, 1946(x) *, Cristiansen, 1954(8)
Grave, 1932(8)
Hull, Risbec, 1930(8)
Kniprath, 1980(8)
Heath, 1899(8)
Kowalevsky, 1879, 1882, 1883(x)
Metcalf, 1892(x)
Metcalf, 1892 (x)
Nagabhushanam, Gopalakrishna Murti,
1968(7)
Soliman, Iskander, 1980, 1982(7)
Clark, 1855(8), Loven, 1856(8), Kowalev-
sky, 1879, 1882, 1883(x), Hoffmann,
1931 (x), Thorson, 1946(x), Haas, Kriesten,
Watabe, 1979, 1980(x)
Kowalevsky, 1879, 1882, 1883(8), Kniprath,
1980(8)
Watanabe, Kox, 1975(8)
Watanabe, Kox, 1975(8)
Thorpe, 1962(8)
Rumrill, Gameron, 1983(x)
Hammarsten, Runnstrom, 1925(8)
Brewin, 1958(x)
Okuda, 1947(8)
* В скобках отмечена стадия развития, до которой изучена личинка хитона:
(8) — до формирования 8-й пластинки; (7) — до формирования 7-й пластинки;
(х) — до более ранних стадий.
различается у разных видов — от 8 ч (у Chiton granoradiatus) до
7 дней (у Stenoplax magdalenensis) и находится в положительной
корреляции с диаметром яиц, т. е. запасом питательных веществ
(см. рис.). Личинки большинства изученных видов выходят из обо-
лочек в возрасте 1—3 дня. Личиночные глаза у трохофор начинают
закладываться рано (у Chiton granoradiatus уже в возрасте 5—5,5 ч),
однако большинство исследователей отмечает наличие глаз или в мо-
мент выхода из оболочек, или через 1—2 дня после него. Только у
Lepidochitona cinerea, Chiton granoradiatus, Stenoplax magdalenensis
глаза замечены еще до выхода из оболочек. Сначала трохофоры
большей частью вращаются вокруг собственной оси. Позже дви-
жения их становятся разнообразнее: личинки начинают быстро дви-
гаться горизонтально и наклонно ко дну, то приближаясь, то уда-
ляясь от него, и изредка останавливаются на месте, продолжая
вращаться.
Только что покинувшие оболочки трохофоры имеют слегка оваль-
ную почти шаровидную форму. Постепенно их тело удлиняется,
6
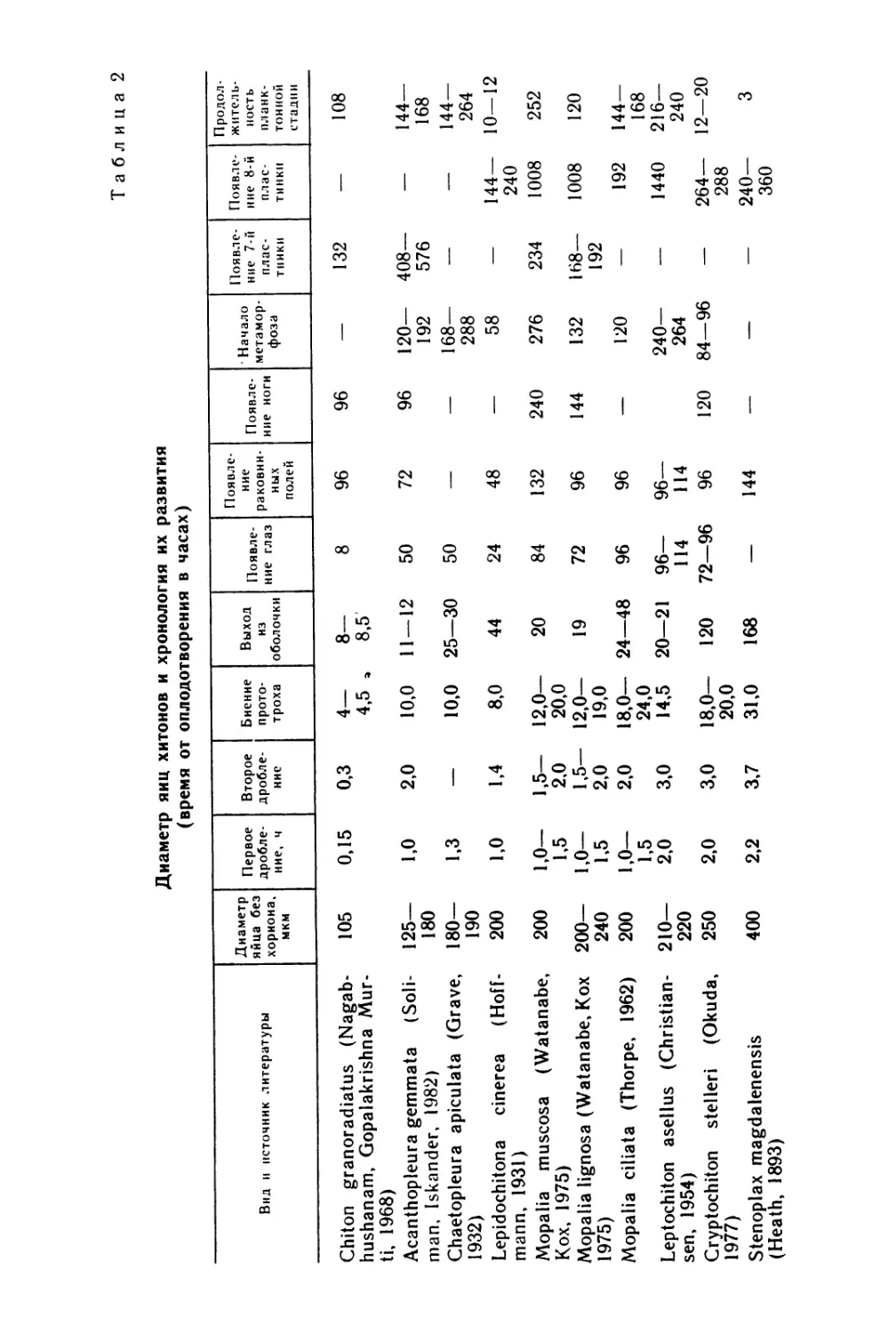
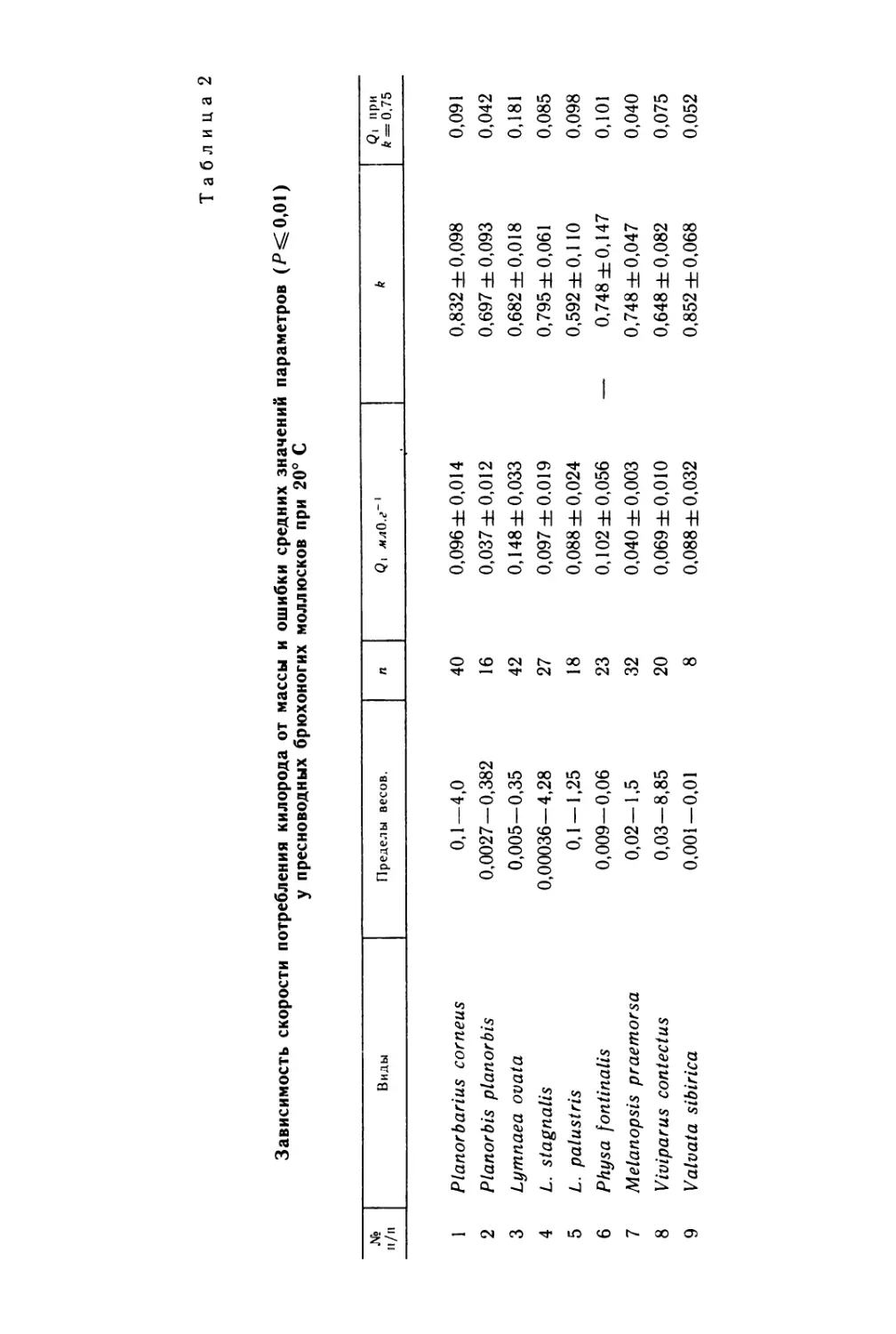
Таблица 2
Диаметр яиц хитонов и хронология их развития
(время от оплодотворения в часах)
Вид и источник литературы Диаметр яйца без хориона, мкм Первое дробле- ние, ч Второе дробле- ние Биение прото- троха Выход из оболочки Появле- ние глаз Появле- ние раковин- ных полей Появле- ние ноги • Начало метамор- фоза Появле- ние 7-й плас- тинки Появле- ние 8-й плас- тинки Продол- житель- ность планк- тонной стадии
Chiton granoradiatus (Nagab- hushanam, Gopalakrishna Mur- ti, 1968) 105 0,15 0,3 4— 4,5 , 8— 8,5 8 96 96 — 132 — 108
Acanthopleura gemmata (Soli- man, Iskander, 19821 125— 180 1,0 2,0 10,0 11 — 12 50 72 96 120— 192 408— 576 — 144— 168
Chaetopleura apiculata (Grave, 1932) 180— 190 1.3 — 10,0 25—30 50 — — 168— 288 — — 144— 264
Lepidochitona cinerea (Hoff- mann, 1931) 200 1,0 1,4 8,0 44 24 48 — 58 — 144— 240 10—12
Mopalia muscosa (Watanabe, Kox, 1975) 200 1,0— 1,5 1,5— 2,0 12,0— 20,0 20 84 132 240 276 234 1008 252
Mopalia lignosa (Watanabe, Kox 1975) 200— 240 1,0— 1,5 1,5— 2,0 12,0— 19,0 19 72 96 144 132 168— 192 1008 120
Mopalia ciliata (Thorpe, 1962) 200 1,0— 1,5 2,0 18,0— 24,0 24—48 96 96 — 120 — 192 144— 168
Leptochiton asellus (Christian- sen, 1954) 210— 220 2,0 3,0 14,5 20—21 96— 114 96— 114 240— 264 — 1440 216— 240
Cryptochiton stelleri (Okuda, 1977) 250 2,0 3,0 18,0— 20,0 120 72—96 96 120 84—96 — 264— 288 12—20
Stenoplax magdalenensis (Heath, 1893) 400 2,2 3,7 31,0 168 — 144 — — — 240— 360 3
ВРЕМЯ ВЫХОДА ЛИЧИНКИ ИЗ ОБО-
ЛОЧКИ ОТ МОМЕНТА ОПЛОДОТВОРЕНИЯ
Зависимость времени выхода личинок из обо-
лочки от диаметра яйца у хитонов разных видов.
Данные взяты из табл. 2
слегка суживается и приобретает удлиненно-овальную форму с более
длинной посттрохальной частью. Одновременно с удлинением тело
трохофор слегка уплощается в дорсовентральном направлении. На
спинной стороне ниже прототроха образуется вначале 6 поперечных
валиков, а позже к ним добавляется еще 1. Между валиками в попе-
речных бороздах формируются раковинные поля. На брюшной сто-
роне начинает обособляться нога. Поперечные валики образуются
обычно на 3—10 день у плавающих личинок и лишь у Stenoplax
magdalenensis и Cryptochiton stelleri — еще у трохофор, находя-
щихся в оболочках. Примерно в то же время начинают появляться
одиночные спикулы, а потом формируется кольцо из спикул, окру-
жающее раковинные поля.
Закладка радулярного мешка происходит довольно рано. У Le-
pidochitona corrugata и Acanthochitona communis радулярный мешок
формируется одновременно с появлением на спине личинки попе-
речных валиков или немного раньше; ко времени образования пер-
вых щитков раковины уже имеются первые пластинки радулы.
В возрасте 3,5—10 дней большинство трохофор претерпевает
метаморфоз. Перед этим плавающие личинки все чаще садятся
задним концом тела на дно и начинают передвигаться, используя
для этого прототрох. Через некоторое время трохофора окончательно
садится на дно и становится малоподвижной или вообще неподвиж-
ной несколько часов. За это время тело ее укорачивается, расши-
ряется и еще более уплощается, она теряет прототрох и теменной
орган; продолжается развитие ноги, мантии со спикулами и появ-
ляются первые известковые пластинки раковины. Как правило, вна-
чале формируется 6—7 первых пластинок. Время нахождения ли-
чинок в планктоне у разных видов сильно различается. Ряд видов
(Lepidochitona cinerea, Cryptochiton stelleri, Acanthochitona com-
munis) имеет короткий планктонный период (от 3 до 24 ч), тогда как
другие (Chaetopleura apiculata, Leptochiton asellus, Mopalia ciliata,
M. lignosa, M. miscosa, Chiton granoradiatus, Acanthopleura gem-
mata) находятся в толще воды от 5 до 7, а некоторые даже до 11
в
дней. Между продолжительностью планктонного периода и диамет-
ром яиц нет четкой зависимости. Однако личинки хитонов с мелкими
яйцами, как правило, дольше находятся в планктоне, а личинки ви-
дов, имеющих наиболее крупные яйца, таких как Stenoplax magdale-
nensis, плавают менее 3 ч. Питаются ли трохофоры в этот период,
пока трудно сказать, поскольку имеется лишь замечание Торсона
(Thorson, 1946) о том, что планктонные личинки Lepidochitona
cinerea и Leptochiton asellus не питаются.
Личинки некоторых видов не садятся на дно и не приступают к
метаморфозу до тех пор, пока не найдут субстраты, на которых
обычно обитают их родители. Так, например, личинки Tonicella
lineata и Katharina tunicata нормально развиваются только после
оседания на корковые известковые водоросли (Barnes, Gonor, 1974;
Rumrill, Cameron, 1983); развитие личинок Mopalia lignosa и M. mus-
cosa продолжается нормально только после оседания на раковины
1иидий, покрытые пленкой микроскопических водорослей, а личинки
Acanthopleura gemmata оседают только на фрагменты водорослей
(Soliman, Iskander, 1982).
В ходе метаморфоза происходит опережающий рост, удлинение
и расширение раковинных полей хитона (заложенных только в пост-
трохальной части), в результате чего раковина и передняя часть
перинотума со спикулами «наползают» на остатки апикальной части
трохофоры — «головной пузырь», и к концу метаморфоза хитон ока-
зывается полностью покрытым сверху раковиной и перинотумом.
В норме каждый щиток раковины закладывается в виде тонкой цель-
ной пластинки, которая растет в дальнейшем за счет добавки из-
вести с краев (Kniprath, 1980). У большинства изученных видов ста-
дия с 7 нормально развитыми щитками раковины наблюдалась на
7—13-й день после оплодотворения. Последний щиток раковины фор-
мировался обычно на 8—10-й день, у некоторых позже (только на
42-й день), а для ряда видов время образования восьмого щитка не
установлено. Сведения по закладке жабр и анального отверстия
очень скудны. Известно, что анус у Mopalia ciliata появляется на
16-й день (Thorpe, 1962), а жабры у М. lignosa и М. muscosa (Wata-
nabe, Сох, 1975) —лишь на 57-й день. Личиночные глаза остаются
некоторое время у молодых хитонов и у М. lignosa и М. muscosa
исчезают в конце второго месяца жизни (Watanabe, Сох, 1975).
ЛИТЕРАТУРА
Бластов Б. В. Класс панцирных моллюсков (Loricata).— В кн.: Руководство по
зоологии, М.—Л., 1940, т 2, с. 286—311.
Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных животных. Тро-
хофорные, щупальцевые, щетинкочелюстные, погонофоры. М., 1977, 312 с.
Barnes J. R., Gonor J J. The larval settling response of the lined chiton Tonicella
lineata.— Mar. BioL, 1973, vol. 20, N 3, p. 259—264.
Brewin B. Note on the development of Cryptoconchus porosus (Burrow).—Trans.
Roy. Soc. N. Z., 1958, vol. 85, N 4, p. 715—716.
Christiansen M. E. The life history of Lepidopleurus asellus (Spengler) (Placophora) .—
Nytt. mag. zool. 1954, vol. 2, p. 52—72.
9
Clark W. On the phenomena of the reproduction of the chitons.— Annals Magazine
Nat. Hist., London, 1855, ser. 2, vol. 16, N 37, p. 446—449.
Grave В. H. Embriology and life history of Chaetopleura apiculata.— Journal of
Morphology, 1932, vol. 54, N 1, p. 153—160.
Hammarsten 0. Runnstr'om J. Die Embriogenie von Acanthochiton.— Zool. Jahrb.
Anat. Jena, 1925, Bd 47, S. 261—318.
Haas W., Kriesten K., Watabe N. Notes on the shell formation in the larvae of the
Placophora (Mollusca).—Biomineralization, 1979, vol. 10, p. 1—8.
Haas №., Kriesten K., Watabe N. Preliminary note on the calcification of the shell plates
in chiton larvae (Placophora).— Meeh. Biominer. Anim. and Plants. Proc. 3rd
Int. Biominer. Symp., Tokyo, 1980, p. 67—72.
Heath H. The development of Ischnochiton.— Zoolog. Jahrb. Anat. u. Ontog., Jena, 1899,
Bd 12, Abth. F, S. 567—656.
Heath H. The larval eye of chitons.— Proc. Acad. Nat. Sci. Philadelphia, 1904, vol. 56,
p. 257—259. м
Hoffmann H. Beitrage zur Kenntnis der Chitonen. I. Uber die Fortpflanzung und
Entwicklung von Trachydermon cinereus L.— Zeitschr. fiir.Morphol. u. OkoL
der Tiere, Berlin, 1931, Bd 20, Heft 4, S. 719—732.
Hull A. F. B., Risbec J. The Loricates of the New Caledonian region (Class Mollusca —
Order Loricata).—Austral. Zool. 1930, vol. 6, part III, p. 277—286.
Hyman L. H. The invertebrates. Volume VI. Mollusca I. McGraw—Hill, N. Y., 1967, 792 p.
Kaas P.t Van Belle R. A. Catalogue of living chitons (Mollusca: Polyplacophora).
Ed. W. Backhuys, Publisher Rotterdam, 1980, 144 p.
Kowalevsky A. O. Uber die Entwicklung der Chitonen.— Zool. Anz., 1879, 2. Jg,
N 37, S. 469—473.
Kowalevsky A. O. Weitere Studien uber die Entwicklung der Chitonen.— Zool. Anz.,
1882, 5. Jg, N 113, S. 307—310.
Kowalevsky A. O. Embryogenie du Chiton polii (Philippi) avec quelques remarques
sur le development des autre Chitons.— Ann. Mus. d'hist. Nat. Marseilles,
Zoologie, 1883, vol. I, N 5, Marseilles, p. 1—46.
Kniprath E. Ontogenetic plate and plate field development in two chitons, Midden-
dorffia and Ischnochiton.— In: W. Roux's Arch. Develop. Biol., 1980, vol. 189,
N 2 p. 97—106.
Loven S. Uber die Entwicklung von Chiton.— Arch, fur Naturg., 1856, Berlin, N 22 (1),
S. 206—210.
Metcalf M. M. Preliminary notes on the embryology of Chiton.— Johns Hopkins
Univ. Circulars, 1892, vol. 11, N 97, p. 79—80.
Metcalf M. M. Contributions to the embryology of Chiton.— Johns Hopkins Univ.
Studies from Biol. Laboratory, 1893, vol. 5, N 4, p. 249—267.
Nagabhushanam R., Gopalakrishna Murti K. Studies on the development of Chiton
granoradiatus Leloup.—Broteria (Ser. Cienc.—Nat.), 1968, vol. 37, N 3—4,
p. 209—214.
Okuda S. Notes on the post-larval development of the giant chiton Cryptochiton
stelleri (Middendorff).— Journ. Fac. Sci. Hokkaido Imper. Univ., 1947, ser. 6,
Zoology, vol. 9, N 3, p. 267—275.
Rumrill S. S., Cameron R. A. Effects of gammaaminobutyric acid on the settlement
of larvae of the black chiton Katarina tunicata.— Mar. Biol., 1983, vol. 12, N 3,
p. 243—247.
Smith A. G. The larval development of Chitons (Amphineura).— Proc. Calif. Acad.
Sci., 1966, vol. 32, p. 433—466.
Soliman G. N., Iskander A. N. The reproduction and development of the common rock
chiton Acanthopleura spiniger (Sowerby) from the northwestern Red Sea.—
Haliotis, 1980, vol. 10, N 2, p. 132.
Soliman G. N.t Iskander A. N. The reproduction and development of the common
rock chiton Acanthopleura spiniger (Sowerby) from the northwestern Red Sea.—
Malacologia, 1982, vol. 22, N 1—2, p. 205—210.
Thorpe S. R. Ir. A preliminary report on spowning and related phenomena in California
chitons.— Veliger, 1962, vol. 4, N 4, p. 202—210.
Thorson G. Reproduction and larval development of Danish marine bottom inver-
tebrates, with special reference to the planktonic larvae on the Sound (Ore-
sund).— Meddeleser fra Kommiss, for Danmarks Fiskeri- og Havundersogelser,
Tie Plankton, 1946, Bd 4, N 1, 523 p.
10
Watanabe J M., Cox L. R. Spawning behavior and larval development in Mopalia
lignosa and Mopalia muscosa (Mollusca: Polyplacophora) in central Cali-
fornia.— Veliger, 1975, vol. 18, Suppl., p. 18—27.
В. I. Sirenko
LARVAL DEVELOPMENT OF POLYPLACOPHORA. ANALYSIS OF STUDIES
OF SPECIES WITH THE PLANCTONIC TYPE OF LARVAL DEVELOPMENT
The general regularities of the planctonic type of the larval development of
chitons are presented on the base of litterature data (27 papers). The larval development
has been studied in 19 species of Polyplacophora during the period more than 120 years,
14 of them having been studied till the formation of 7-th and 8-th valve. The positive
correlation of age of larva in the moment of hatching with the diametr of eggs i. e. the
amount of food material is mentioned.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
Д. Л. Иванов
ФИЛОГЕНИЯ ПОДКЛАССА
CAUDOFOVEATA (MOLLUSCA APLACOPHORA)
В филогенетике и опирающейся на нее эволюционной систематике
для выяснения....родственных связей [в отличие от нумерических
подходов, подразумевающих использование как можно более полных
морфологических данных (Sneath, Sokal, 1973)] основное внимание
уделяется небольшому набору морфологических структур, несущих
наибольшую информацию о родственных взаимоотношениях между
таксонами (Майр, 1971; Юдин, 1974) Кроме того, необходимо учи-
тывать и организацию животных в целом с учетом особенностей
эволюции отдельных систем органов, причем, чем выше таксономи-
ческий ранг группы, тем сильнее возрастает роль указаний на воз-
можные способы и пути эволюционных перестроек различных орга-
нов и аппаратов (Шилейко, 1979).
С точки зрения филогенетической значимости можно выделить
следующие аппараты и структуры, несущие наибольшую информа-
цию о родственных взаимоотношениях между таксонами в подклассе
Caudofoveata: радулярный аппарат, спикульный покров, пищевари-
тельный тракт, головной щиток, внешний вид, кожномускульный
мешок.
Настоящая статья есть результат обработки коллекции Caudo-
foveata из экспедиционных сборов Института океанологии АН СССР
Исследовано 29 видов, принадлежащих 6 родам. Представители
Psilodens и Metachaetoderma в материале отсутствовали.
1. Анализ филогенетически значимых признаков
Радулярный аппарат
Как было показано (Иванов, Цетлин, 1981), радулярные аппа-
раты боконервных моллюсков развиваются на основе слабой и не-
полной дифференциации радулярной мембраны. Зубные пластины за-
частую слабо обособлены от радулярной мембраны, которая лишена
дифференциации по толщине и часто принимает участие в форми-
ровании элементов зубных пластин (см. Scheltema, 1981, Fig. 4,В-С).
Эволюция радулярного аппарата в пределах подкласса проходила
в направлении не столько специализации его отдельных элементов,
сколько модификации и специализации аппарата как целого.
Наиболее близкие по строению радулярные аппараты встречаются
в подклассе Solenogastres (Иванов, Цетлин, 1981). В этой группе
характерен двухрядный полисегментный радулярный аппарат с более
или менее крючковидными зубами. У кораллобионтных Solenogastres
часто появляются серповидные зубные пластины (например, у Geni-
12
Рис. 1. Строение радулы Simrothiella.
/ — ралулярные влагалища (зубные пластины изображены только
в рабочем пименте рал\лы)
toconia, Gymnomenia, Pruvotina) очень похожие на зубные пластины
Scutopus и Psilodens. В ряде случаев у Solenogastres радулярная
мембрана между продольными рядами зубов развита слабо, а у пред-
ставителей рода Simrothiella происходит полное разделение продоль-
ных рядов и формирование их в независимых радулярных влага-
лищах (рис. 1); при этом характер зубных пластин и расположение
продольных рядов относительно одонтофора напоминают картину,
характерную для представителей рода Limifossor (Scheltema, 1981,
Fig- 6,В).
Если предположить, что Caudofoveata произошли от солено-
гастроморфного предка, исходно обитавшего на мягких кораллах и
питавшегося преимущественно полипами, оказывается, что радуляр-
ный аппарат мог быть преадаптирован к микрофагии. Иначе мы не
сможем установить прототип радулярного аппарата Caudofoveata
и объяснить причины столь поразительного сходства в характере
строения этого аппарата в двух группах, сильно различающихся по
стратегии в пищевой специализации.
Наиболее близким к прототипу можно считать двухрядный поли-
сегментный аппарат со слабо дифференцированными серповидными
зубными пластинами с уплощенным основанием, характерный для
рода Psilodens. Дальнейшее преобразование зубных пластин в двух-
рядном полисегментном аппарате шло по пути формирования своеоб-
разных щипцов, которые могли бы не только «хватать» добычу (как
\ Scutopus), но и «откусывать» пищевые фрагменты (как у Limifos-
sor). При этом зубные пластины меняют ориентацию на одонтофоре;
при слабо дифференцированной и тонкой радулярной мембране это
трудно осуществить за счет ее перегиба на передней части субраду-
лярных хрящей (утолщение же радулярной мембраны в данном слу-
13
чае не происходит, так как этот процесс приводил бы к значительному
ограничению подвижности зубных пластин). Изменение ориентации
зубных пластин осуществлялось независимо у Scutopus, с одной сто-
роны, и у Limifossor — с другой. В первом случае это достигалось
изменением характера закладки зубных пластин, а во-втором — пе-
ременой положения радулы относительно субрадулярных хрящей и
расчленением радулярной мембраны в рабочей части на две незави-
симые ленты (рис. 2,7).
Трансформация формы зубных пластин в направлении к Scutopus
приводит к появлению тонкого отгиба, рассеченного на многочислен-
ные треугольные зубцы, и исчезновению уплощенного основания
(рис. 2, 8); в результате зубная пластина закрепляется в радулярной
мембране при помощи специальных выростов (Scheltema, 1981,
Fig. 5, С). В направлении к Limifossor зубные пластины уплощаются
за счет расширения основания зуба, серповидное лезвие загибается
назад и внутрь радулы, преобразуясь в мощный стилетовидный
зубец, а отгиб формирует немногочисленные зубцы на переднем крае
зубной пластины (рис. 2, 7).
При всем разнообразии строения зубных пластин и радулярной
мембраны монофилетичность этой филогенетической ветви может
быть подтверждена наличием дорсального аппроксиматора радуляр-
ных хрящей (Scheltema, 1981), более ни у кого из Caudofoveata не
встречающегося. Попытка гомологизировать верхний аппроксиматор
радулярных хрящей Crystallophrisson (Mu-З у Deimel, 1982) с дор-
сальным аппроксиматором Scutopus несостоятельна, так как верхний
аппроксиматор кристаллофриссонов по происхождению вентраль-
ный и занимает «дорсальное» положение в результате поворота оси
радулы.
На базе двухрядного полисегментного радулярного аппарата
возникает и 3—5-рядный полисегментный аппарат, характерный для
Prochaetoderma и Metachaetoderma. В данной филогенетической
ветви происходит дифференциация центрального ряда зубных пла-
стин и развитие дополнительных образований, располагающихся
латерально от основных (инициальных) зубов. Существенные раз-
личия в характере развития основных элементов радулы связаны
с возникновением у Prochaetoderma челюстей (случай, уникальный
для боконервных).
Поскольку челюсти Prochaetoderma берут на себя основную ме-
ханическую нагрузку по разрушению раковин фораминифер, радула
остается очень изящной и слабо склеротизированной. Радула пред-
ставляет собой серию «пинцетов», при помощи которых прохето-
дермы выбирают органический компонент пищевых объектов. Радула
прохетодерм может высовываться из ротового отверстия и произво-
дить «хватательные» движения (Kovalewsky, 1901), поэтому логично
предположить, что она может также подтягивать к мандибулам ра-
ковины фораминифер.
Основные зубные пластины радулы слегка серповидные, слабо
изогнутые, в центральной части имеется тонкий отгиб, рассеченный
на многочисленные треугольные зубцы. В нижней части зубной пла-
стины имеется мембрановидный придаток (Scheltema, 1981), необ-
14
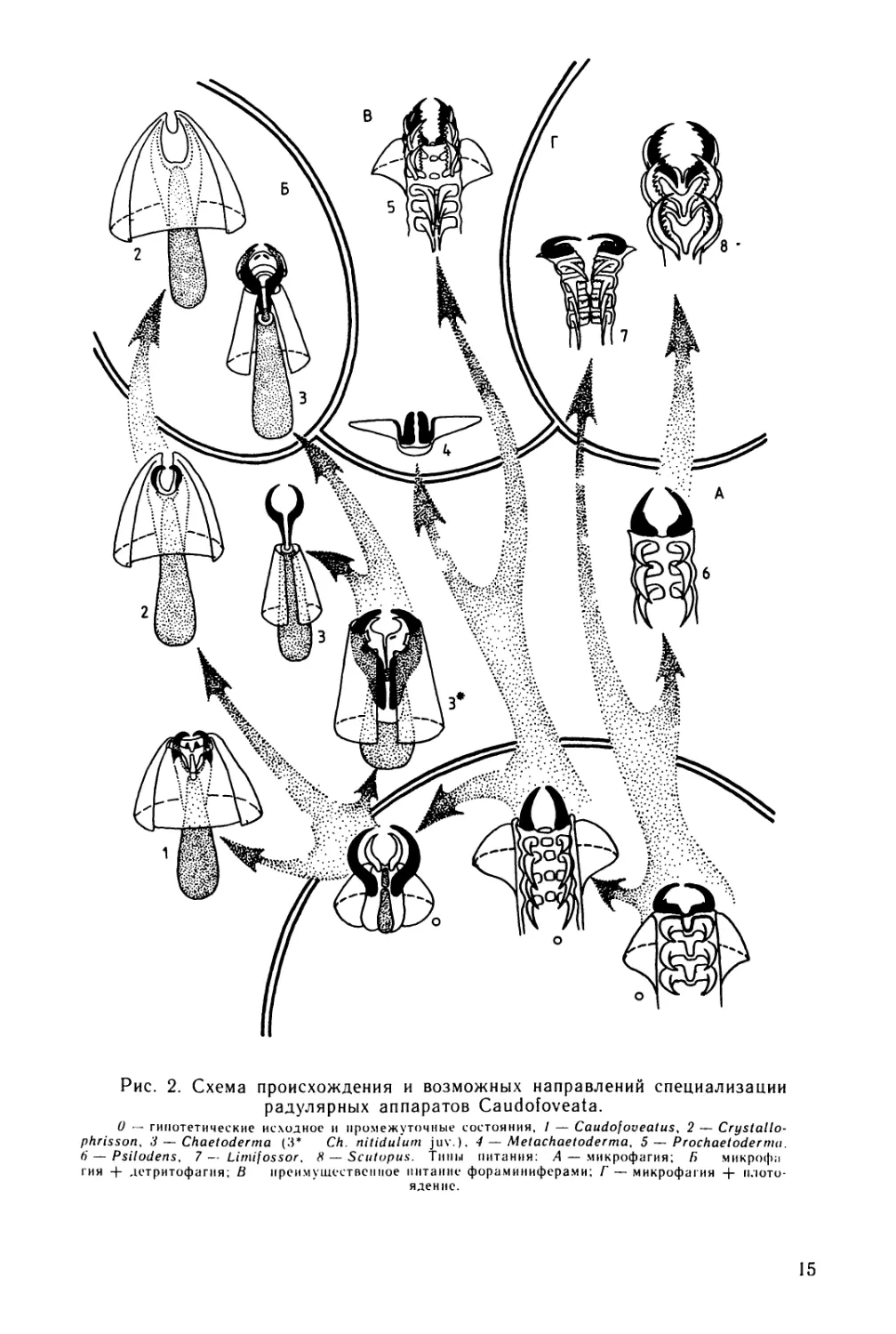
Рис. 2. Схема происхождения и возможных
направлений специализации
радулярных аппаратов Caudofoveata
ГИЯ + детритофагая; В иреямушсетвсш.ое „итак,к- форамнниферамиТ'-^икрофа^ия + яю'то
яденнс.
15
холимый, по-видимому, для переориентации и дополнительной под-
держки зуба, закрепленного в радулярной мембране утонченным
основанием. В нерабочей части радулы зубные пластины залегают
таким образом, что их острия направлены назад и наружу, а в рабо-
чей части они направлены вперед и внутрь радулы. Этот сложный
поворот осуществляется за счет слабого перегиба мембраны на одон-
тофоре и разрастания крыловидного придатка зубной пластины. Для
удержания довольно длинного зуба в вертикальном положении и
передачи усилия на него от мембраны у Prochaetoderma возникают
дополнительные пластинки, отходящие от мембраны около основания
инициальных зубов и охватывающие их с внешней стороны (рис. 2, 5).
Помимо радулярной мембраны, в рабочей части радулы Pro-
chaetoderma латерально от зубных рядов имеется пара крыловидных
кутикулярных выростов, покрывающих верхнюю и переднюю части
одонтофора. Шелтема считает, что это образование гомологично суб-
радулярной мембране (Scheltema, 1981). Однако расположение и
характер развития этого образования наводит на мысль, что оно
гомологично тем участкам глоточной кутикулы, которые у боль-
шинства моллюсков покрывают передне-латеральные участки одонто-
фора (рис. 3) и к субрадулярной мембране отношения не имеют
(см. Kerth, 1983).
Наиболее близким по строению радулярным аппаратом обладают
Metachaetoderma. Но здесь челюсти отсутствуют и основную ме-
ханическую нагрузку несет радула. В результате основные зубные
пластины становятся очень мощными и обладают плоской рабочей
поверхностью. Центральный ряд зубных пластин утолщается и вы-
полняет, по-видимому, функцию естественно-напряженного элемента.
Дополнительные пластинки слабо дифференцированы и выполняют
функции рукояток (или поддержек), к которым крепятся основные
зубы (рис. 2, 4).
Несмотря на взаимную близость этих родов по характеру разви-
тия элементов радулярного аппарата, наличие челюстей и связан-
ные с этим особенности развития зубов у Prochaetoderma ставят
этот род в изолированное положение не только в этой филогенети-
ческой ветви, но и во всем подклассе.
Третье направление перестройки радулярного аппарата привело
к формированию трехрядного моносегментного аппарата (Иванов,
1979), вызвавшего наибольшие затруднения в трактовке его проис-
хождения и установления гомологий слагающих элементов. К этой
филогенетической ветви принадлежат Crystallophrisson, Caudofo-
veatus и Chaetoderma. Указанием на происхождение моносегмент-
ного аппарата от полисегментного служит радулярный аппарат
каудофовеатусов, который, строго говоря, двухсегментный и, если
продолжить направление расположения радулярных сегментов, мы
получим картину, типичную для укладки радулярных сегментов у
большинства моллюсков (рис. 2, /). Кроме того, у Chaetoderma
sterreri (Сальвини-Плавен, 1984) и у Ch. pellucida часто встречается
явление удвоения радулярного аппарата, кбторое.также указывает
на происхождение радулярных аппаратов такого типа от поли-
сегментных.
16
Рис. 3. Строение радулы Cephaiopnda (Loligo gahi).
I — радулярная мембрана,2 — кутикулярная обкладка одонтофора, 3 — раду-
лярное влагалище (радулярные зубы не показаны).
По характеру развития зубных пластин и их прикрепления к осе-
вой пластине как прототип моносегментного радулярного аппарата
можно принять радулярный аппарат, характерный для ювенильных
Chaetoderma (рис. 2,3*): имеется пара крупных серповидных зуб-
ных пластин, рукоятки которых соединены эластичным симфизом
между собой и осевой пластиной. Осевая пластина короткая, го-
раздо короче зубных пластин, и залегает спереди и ниже зубцов
(радулярный симфиз крепится почти в средней ее части). Осевая
пластина — гомолог радулярной мембраны и представляет собой тот
ее участок, который у представителей рода Prochaetoderma диф-
ференцируется в центральный ряд зубных пластин. Радулярный
симфиз, скорее всего, гомологичен тому участку радулярной мембра-
ны, который в полисегментных радулярных аппаратах Caudofoveata
обеспечивает связь зубов между собой.
Существует еще одно образование, характерное для этой фило-
генетической ветви, которому большинство авторов не могли найти
гомологов в радулярных аппаратах остальных моллюсков (Schwable,
1961, Scheltema, 1972; Salvini-Plawen, 1975; Deimel, 1982)—это
радулярный капюшон (Иванов, 1979). По положению, характеру
развития и, отчасти, по функциям, это образование можно считать
гомологом того участка глоточной кутикулы, который у большин-
ства моллюсков покрывает передне-латеральные стенки одонтофора.
Это подтверждается еще и тем, что у Chaetoderma капюшон имеет
характер парного образования (рис. 2, 3). Наличие кутикулярной
выстилки одонтофора сближает моносегментные радулярные аппа-
раты с таковыми Prochaetoderma.
2 Заказ 1696 17
Рис. 4. Схема отличий функционирования радулярных зубов при пинцетном
(Л) и грейферном (Б) способах работы, на примере Chaetoderma.
У представителей рода Chaetoderma от радулярного капюшона
часто отходят своеобразные лентовидные придатки, охватывающие
радулярные зубы и координирующие действия капюшона и зубов
при хватательном акте (сравните с Prochaetoderma). Если рас-
смотреть морфологический ряд Chaetoderma nitidulum—Ch. cros-
sotus—Ch. sagittiferus—Ch. profundus, можно заметить тенденцию
к смещению симфиза из субтерминального к терминальному поло-
жению на вершине осевой пластины и усиление лентовидных при-
датков капюшона (рис. 2, 3).
В пределах Chaetoderma легко видеть два основных направления
специализации радулярного аппарата. В первом — зубы заметно
увеличиваются и получают большую подвижность из-за перемещения
эластичного симфиза в терминальное положение и за счет большей
независимости от радулярного капюшона (например, у Ch. hartmani)
В этом случае радулярный аппарат может фу пщионировать только
как щипцы (Иванов, 1979). Другое направление связано с возник-
новением дополнительных эластичных связок между зубными пла-
стинами, более тесного контакта зубов с радулярным капюшоном
и образованием дополнительных кутикулярных мембраноподобных
пластинок между зубами. Все это позволяет радулярному аппарату
функционировать не только как щипцы, но и наподобие грейферного
ковша (рис. 4).
Радулярные аппараты Crystallophrisson и Caudofoveatus можно
вывести от радулярных аппаратов хетодерматидного типа, еще не
окончательно утративших полисегментность. Их происхождение свя-
зано с иным характером прикрепления зубов к осевой пластине.
У представителей этих групп зубы крепятся не непосредственно
к осевой пластине, а располагаются на вершинах эластичных опор,
которые выполняют функции естественно-напряженных элементов
радулярного аппарата (Иванов, 1979). По всей видимости, они яв-
ляются дериватами радулярной мембраны и гомологичны межзуб-
ному симфизу Chaetoderma. Если в радулярном аппарате хетодер-
матидного типа естественно-напряженный элемент (симфиз) стре-
18
мится развести зубы в стороны, то в радулярном аппарате кристал-
лофриссонидного типа латеральные опоры стремятся их сжать. По-
этому. радулярный капюшон у Crystallophrisson и Caudofoveatus
охватывает зубы целиком и иногда сильно возвышается над раду-
лярными зубами. Поскольку зубные пластины в данном случае уже
не играют исключительной роли при захвате добычи, то наблюда-
ется тенденция к их редукции вплоть до полного исчезновения (на-
пример, у Crystallophrisson simplex).
Радулярный аппарат каудофовеатусов можно считать уклонив-
шимся от основного направления эволюционных перестроек раду-
лярных аппаратов в этой группе, приведших к становлению раду-
лярных аппаратов кристаллофриссонидного типа.
У некоторых Crystallophrisson сильное развитие радулярного
капюшона позволяет перейти от микрофагии к избирательной или
даже неизбирательной детритофагии, так как радулярный аппарат
в данном случае может функционировать не как пинцет, а как грей-
ферный ковш (рис. 2, 2).
Таким образом, можно предполагать, что эволюционные пере-
стройки радулярных аппаратов в подклассе Caudofoveata шли на
основе исходно двухрядного полисегментного аппарата со слабо
дифференцированной радулярной мембраной по трем основным на-
правлениям: 1) сохранение двухрядности и постепенная редукция
центральной части радулярной мембраны и переориентация зубных
пластин (Psilodens, Scutopus, Limifossor)\ 2) дифференциация
центрального ряда зубных пластин и возникновение дополнительных
кутикулярных подставок у инициальных зубов (Prochaetoderma,
Metachaetoderma)', 3) утрата полисегментности и образование ра-
дулярного капюшона (Caudofoveatus, Crystallophrisson, Chaeto-
derma). Положение радулярного аппарата в глотке Prochaetoderma
(Scheltema, 1981, Fig. Зе), характер дифференциации центральной
части радулярной мембраны и кутикулярных элементов радулы в
поперечных сегментах, а также наличие кутикулярной обкладки
одонтофора указывают на тесную связь второго и третьего направ-
лений специализации радулярных аппаратов каудофовеат (рис. 2).
В целом все эволюционные перестройки радулярного аппарата
связаны со специализацией Caudofoveata как микрофагов, однако
в трех «крайних» проявлениях морфологической специализации (Li-
mifossor, Crystallophrisson и Chaetoderma) эти перестройки приво-
дят к полифункциональности радулярных аппаратов, и, следователь-
но, к пищевой «деспециализации» (эврифагии) отдельных предста-
вителей подкласса.
Пищеварительная система
Подробный анализ организации пищеварительной системы
Caudofoveata содержится в статье Шелтемы (Scheltema, 1981), по-
этому в данной работе ограничимся некоторыми замечаниями.
Шелтема выделяет только два основных направления эволюцион-
ных перестроек пищеварительной системы: первое характеризуется
усложнением пищеварительной системы оу Scut opus через Limifossor
2*
19
к Chaetoderma и Crystallophrisson, второе представлено только
Prochaetoderma и характеризуется упрощением пищеварительного
тракта. Однако анализ результатов ее работы (Scheltema, 1981,
Table 1) позволяет предполагать наличие трех направлений специа-
лизации пищеварительной системы, совпадающих с направлениями
специализации • р-адулярного аппарата. Причем, если с Limifossor
и Scutopus прохетодерм сближают только плезиоморфные приз-
наки, то с Chaetoderma и Crystallophrisson их роднит набор апо-
морфных черт организации пищеварительной системы.
Внешний вид
Поскольку переход к роющему образу жизни осуществлялся
постепенно, и вначале Caudofoveata обитали в верхнем слое осадков,
то за исходную можно принять форму тела, характерную для Limi-
fossor Изначально тело не сильно удлиненное, почти цилиндрическое,
без заметной дифференциации на отделы. На вентральной стороне,
вдоль ее средней линии, имелся рудимент ноги в виде продольной
бороздки, лишенной кутикулы и спикул, которая ныне сохранилась
только у некоторых Scutopus (у остальных представителей подкласса
рудименты ноги почти полностью исчезают)
При переходе к обитанию в более глубоких и плотных слоях
осадков тело Caudofoveata удлиняется, приобретая более червеоб-
разный облик; при этом активно участвует в закапывании только
передний участок тела животного, поэтому в ряде случаев передний
участок тела отделяется кольцевой перетяжкой (Crystallophrisson,
Caudofoveatus, Scutopus, некоторые Chaetoderma). В районе пере-
тяжки усиливается кольцевая мускулатура, которая выполняет
функции сфинктера и изолирует головной гемоцель от соматического,
что приводит к интенсификации работы головного участка при
рытье. Кольцевая перетяжка не выражена в тех случаях, когда плохо
обособлен и очень короток отдел пищевода, а печеночный отдел
слегка гипертрофирован (рис. 5, 5). В этих случаях головной участок
гемоцеля легко изолируется от соматического, потому что просвет
между стенкой тела и печенью на большом протяжении очень узок.
Другой фактор, сыгравший значительную роль в формировании
внешнего облика — жаберное дыхание. Поскольку жабры (занимаю-
щие терминальное положение на заднем конце тела) необходимо
удерживать над грунтом, для глубокого зондирования осадков не-
обходимо увеличивать длину тела. В тех случаях, когда печеночный
отдел приближен к головному, удлиняется гоноперикардиальный от-
дел, что приводит к образованию «хвостатой» формы тела (Pro-
chaetoderma и часть Chaetoderma). В тех случаях, когда печеночный
отдел сближается с мантийным, происходит удлинение отдела пище-
вода и дифференцируется кольцевая перетяжка, ограничивающая
головной отдел (все Crystallophrisson). При значительном удлинении
пищевода возникают дополнительные кольцевые перетяжки, создаю-
щие своеобразные «отсеки» тела (рис. 5, 4).
Таким образом, все разнообразие внешнего облика Caudofoveata
определяется различными сочетаниями решений проблемы глубокого
20
Рис. 5. Схема возможных направлений специализации формы тела.
1 Limifossor, 2 — Scutopus, 3 — Caudofoveatus, 4 — Crystallophrisson (4* — форма с дополни-
тельными кольцевыми перетяжками см. текст}, 5 — Chaetoderma, 6 — Prochaetoderma.
зондирования осадков при наличии жаберного дыхания и пери-
стальтического способа движения.
Как видно из сравнительно-морфологических рядов, характери-
зующих возможные направления специализации внешней морфоло-
гии (рис. 5), можно предполагать, что в подклассе Caudofoveata
кольцевая перетяжка, обособляющая головной отдел, возникала
дважды и дважды возникали «хвостатые» формы.
Головной щиток
Головной щиток — слабо мускулизованное образование, располо-
женное около ротового отверстия, покрытое тонкой кутикулой и
лишенное спикульного покрова. Кутикула щитка составляет одно
целое с кутикулой ротовой трубки и сильно отличается от кутикулы,
покрывающей тело (Scheltema, 1981). Это позволяет предполагать,
что изначально головной щиток представлял собой мало дифферен-
цированное околоротовое поле с почти центральным положением
ротового отверстия наподобие буккального «щитка» некоторых Sole-
nogastres (рис. 6).
В строении головных щитков Caudofoveata можно наметить три
основных типа: 1) цельный амфиоральный, 2) цельный подково-
видный, 3) двураздельный.
Цельный амфиоральный щиток с центральным положением рта
ныне не встречается. При амфиоральном щитке имеется тенденция
к смещению ротового отверстия в верхнюю его часть (Scutopus и
некоторые Chaetoderma), при этом кутикула надротовой части ста-
21
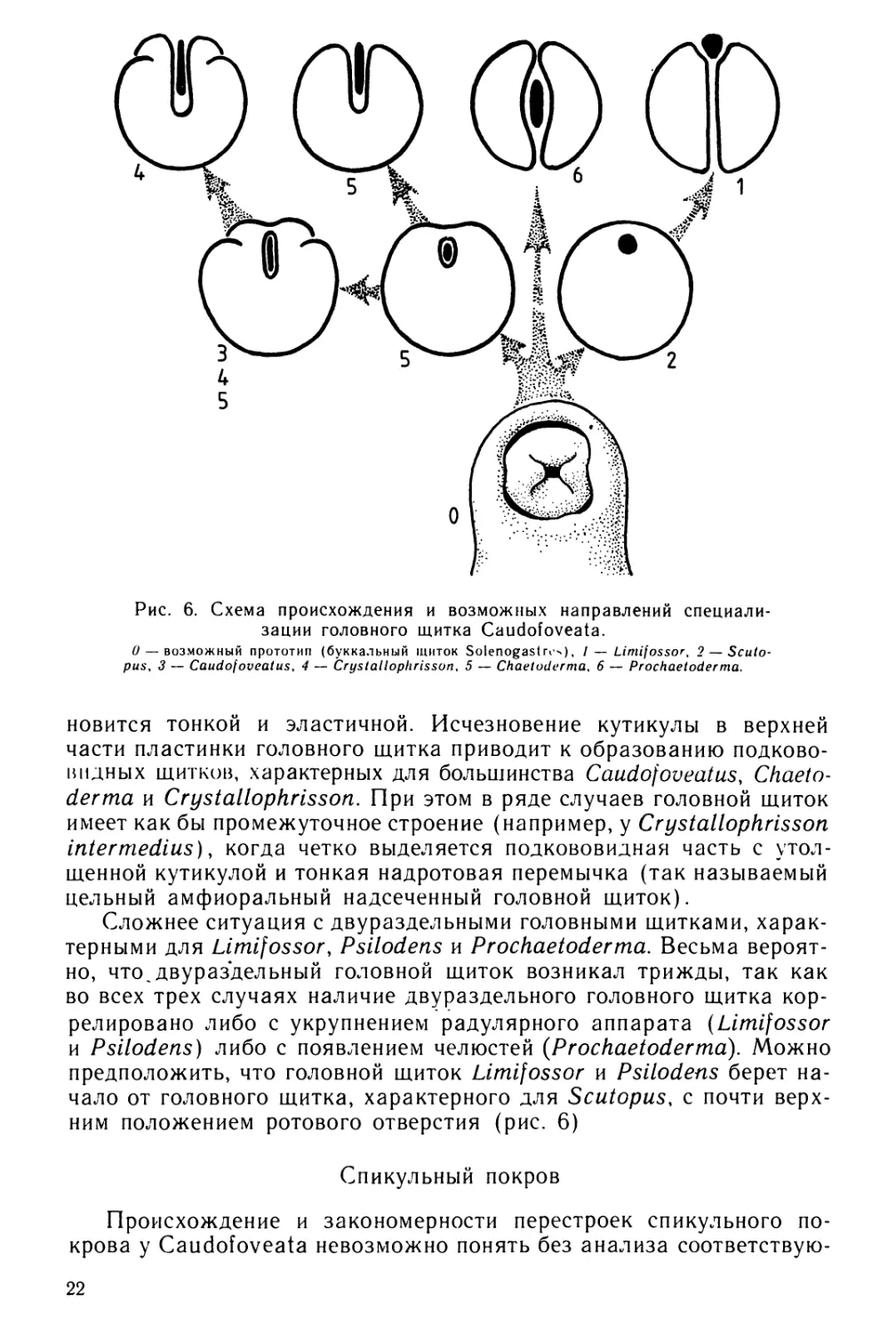
Рис. 6. Схема происхождения и возможных направлений специали-
зации головного щитка Caudofoveata.
0 — возможный прототип (буккальный щиток Solenogastres), / — Limifossor, 2 — Scuto-
pus, 3 — Caudofoveatus, 4 — Crystallophrisson, 5 — Chaetoderma, 6 — Prochaetoderma.
новится тонкой и эластичной. Исчезновение кутикулы в верхней
части пластинки головного щитка приводит к образованию подково-
видных щитков, характерных для большинства Caudofoveatus, Chaeto-
derma и Crystallophrisson. При этом в ряде случаев головной щиток
имеет как бы промежуточное строение (например, у Crystallophrisson
intermedins), когда четко выделяется подкововидная часть с утол-
щенной кутикулой и тонкая надротовая перемычка (так называемый
цельный амфиоральный надсеченный головной щиток).
Сложнее ситуация с двураздельными головными щитками, харак-
терными для Limifossor, Psilodens и Prochaetoderma. Весьма вероят-
но, что~двураздельный головной щиток возникал трижды, так как
во всех трех случаях наличие двураздельного головного щитка кор-
релировано либо с укрупнением радулярного аппарата (Limifossor
и Psilodens) либо с появлением челюстей {Prochaetoderma). Можно
предположить, что головной щиток Limifossor и Psilodens берет на-
чало от головного щитка, характерного для Scutopus, с почти верх-
ним положением ротового отверстия (рис. 6)
Спикульный покров
Происхождение и закономерности перестроек спикульного по-
крова у Caudofoveata невозможно понять без анализа соответствую-
22
щих перестроек спикульного покрова Solenogastres. У последних
основную роль в передвижении играет килевидная нога с ресничным
эпителием. Роль спикульного покрова для большинства соленогаст-
ров состоит в создании наружного опорного скелета, обеспечиваю-
щего постоянство формы тела и необходимую опору для ретракторов
ноги.
У большинства Solenogastres кутикула заметно утолщена и боль-
шая часть спикул залегает в ее «.толще, образуя своеобразный упо-
рядоченный «войлок». Внутрикутикулярные спикулы всегда полые,
с легким S-образным изгибом. Укладка спикул осуществляется
таким образом, что создается более или менее жесткая структура,
способная изгибаться. Помимо внутрикутикулярных спикул, имеются
спикулы, прободающие кутикулу и на большую часть длины тор-
чащие наружу. Как правило, эти спикулы намного крупнее и, воз-
можно, выполняют сенсорные тактильные функции. Кроме пере-
численных видов спикул, практически всегда имеются специализи-
рованные спикулы ножевидной формы без полостей, оформляющие
вентральный желобок ноги.
У некоторых Solenogastres наблюдается более тонкая кутикула
и плоские (иногда ребристые) спикулы, которые укладываются напо-
добие чешуи рыб (Salvini-Plawen, 1967). Это можно связать с
уменьшением размеров тела и с необходимостью увеличения его под-
вижности.
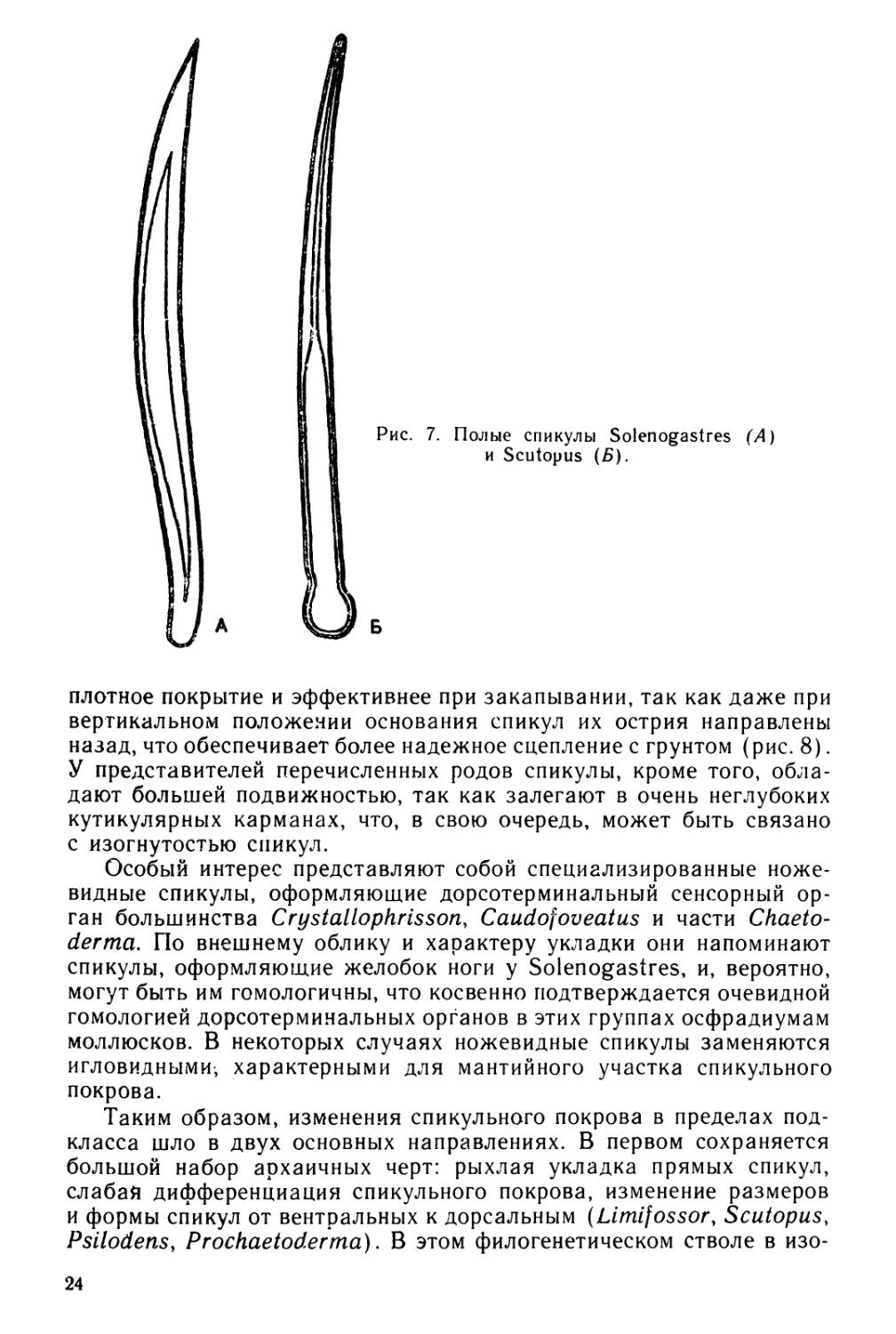
Для Caudofoveata характерны уплощенные бесполостные спику-
лы, напоминающие очертания наконечники стрел или копий. В един-
ственном случае удалось обнаружить у Scutopus (рис. 7) полые
спикулы в мантийном участке спикульного покрова, что, безусловно,
необходимо признать явно плезиоморфным признаком. Кроме того,
у представителей этого рода, так же, как и у Prochaetoderma, наблю-
дается закономерное увеличение длины спикул от вентральных к
дорсальным. У прохетодерм, помимо этого, спикулы располагаются
под значительным углом к оси тела, что, безусловно, можно считать
архаичным признаком.
У Scutopus, Limifossor, Psilodens и Prochaetoderma практически
не наблюдается дифференцировки спикульного покрова по длине
тела. Спикулы у представителей этих родов во всех отделах тела
(за исключением мантийного) однотипны по морфологии и изменя-
ются только их линейные размеры. Кроме того, спикулы в этих груп-
пах не имеют изгиба и укладываются довольно рыхло.
У животных с хорошо выраженными отделами тела (Chaeto-
derma, Crysttallophrisson и Caudofoveatus) наблюдается и значи-
тельная дифференциация спикульного покрова. Практически каждый
отдел имеет спикульный покров, отличающийся наличием специфи-
ческих спикул. Это явление наиболее ярко выражено у ювенильных
форм. В процессе роста происходит унификация спикул и ослабе-
вает дифференциация спикульного покрова на участки. Кроме того,
у представителей этих родов все спикулы имеют более или менее
выраженный изгиб в сагиттальной плоскости.
Из сравнения характера укладки прямых и изогнутых спикул
можно заключить, что изогнутые спикулы могут формировать более
23
Рис. 7. Полые спикулы Solenogastres (А)
и Scutopus (Б).
плотное покрытие и эффективнее при закапывании, так как даже при
вертикальном положении основания спикул их острия направлены
назад, что обеспечивает более надежное сцепление с грунтом (рис. 8).
У представителей перечисленных родов спикулы, кроме того, обла-
дают большей подвижностью, так как залегают в очень неглубоких
кутикулярных карманах, что, в свою очередь, может быть связано
с изогнутостью спикул.
Особый интерес представляют собой специализированные ноже-
видные спикулы, оформляющие дорсотерминальный сенсорный ор-
ган большинства Crystallophrisson, Caudofoveatus и части Chaeto-
derma. По внешнему облику и характеру укладки они напоминают
спикулы, оформляющие желобок ноги у Solenogastres, и, вероятно,
могут быть им гомологичны, что косвенно подтверждается очевидной
гомологией дорсотерминальных органов в этих группах осфрадиумам
моллюсков. В некоторых случаях ножевидные спикулы заменяются
игловидными-, характерными для мантийного участка спикульного
покрова.
Таким образом, изменения спикульного покрова в пределах под-
класса шло в двух основных направлениях. В первом сохраняется
большой набор архаичных черт: рыхлая укладка прямых спикул,
слабая дифференциация спикульного покрова, изменение размеров
и формы спикул от вентральных к дорсальным (Limifossor, Scutopus,
Psilodens, Prochaetoderma). В этом филогенетическом стволе в изо-
24
Рис. 8. А — схема двух основных типов организации кожномускульного
мешка Scutopus (внизу) и Crystallophrisson (вверху).
I ла, 2 — диагонально-кольцевая мускулатура. 3 - продольная мускулатура. 4
ральный кровеносный синус.
Б схема функционирования прямых (/) и изогнутых (2) спикул при
перистальтике.
лированном положении оказывается род Prochaetoderma, где сохра-
няется укладка спикул под углом к оси тела. Во втором — наблюда-
ется заметная дифференциация спикульного покрова по отделам
тела; вторичная его унификация в онтогенезе; более экономичная
и подвижная укладка спикул, возможная в результате их изгиба в
сагиттальной плоскости; сохранение в ряде случаев ножевидных
спикул (Crystallophrisson, Caudofoveatus, Chaetoderma).
Кожно-мускульный мешок
Исходным для Caudofoveata можно предположить кожно-му-
скульный мешок, приближающийся по характеристикам к таковому
Solenogastres. Все отличия в строении кожно-мускульного мешка
у каудофовеат связано с дифференциацией продольной мускулатуры.
Наиболее архаичным кожно-мускульным мешком обладают Scu-
topus. У них отсутствует четкая дифференциация продольных лент;
на вентральной стороне вдоль ее средней линии продольная муску-
латура не развита, а латерально от нее образуются два валика уси-
ленной мускулатуры, что позволяет представителям этого рода скру-
чиваться спиралью (рис. 5, 2).
У представителей родов Limifossor и Prochaetoderma также не
обнаруживается четкой дифференциации продольной мускулатуры
на ленты. Кроме того, у них нет усиленных продольных валиков на
вентральной стороне тела и продольная мускулатура развита почти
гомогенно по всей стенке тела.
25
Co
Рис. 9. Феногра.мма сходства родовых
таксонов Caudofoveata.
Л1 — Metachaetoderma, Рг — Prochaetoder та.
Ch — Chaetoderma, Са — Caudofoveatus, Сг ('гц-
stallophrisson, Sc — Scutopus, Li — Limifossor /
Psilodens (толщина липин пропорциональна чпс.
изоморфных состояний признаков объяснение
тексте).
У Crystallophrisson, Caudofoveatus и Chaetoderma наблюдается
тенденция к усилению продольной мускулатуры в передней части
тела и формирование двух пар лепт (дорсолатеральной и вентро-
латеральной) Наиболее ярко ленты продольной мускулатуры выра-
жены у представителей рода Crystallophrisson в пищеводном отделе.
Таким образом, за исходный тип строения кожно-мускульного
мешка Caudofoveata можно принять такой, который характерен для
рода Scutopus. В специализации кожно-мускульного мешка можно
выделить две основных тенденции: 1) тенденция к гомогенному раз-
витию продольной мускулатуры и 2) дифференциация дорсолате-
ральных и вентролатеральных лент (рис. 8, Л)
2. Филогения и основные направления эволюции
Прежде чем перейти к оценке родственных отношений таксонов
подкласса Caudofoveata, рассмотрим их сходственные отношения.
Для такого анализа построим кольцевую фенограмму сходства
(рис. 9), основанную на подсчете изоморфных состояний 20 призна-
ков, характеризующих следующие 13 черт организации каудофовеат:
1 — характер дифференциации отделов тела; 2 — характер диффе-
ренциации головного отдела; 3 — характер строения головного
щитка; 4 — характер дифференциации продольной мускулатуры;
5 — организация спикульного покрова; 6 — дифференциация раду-
лярной мембраны; 7 — дифференциация зубных пластин радулы;
8 — строение одонгофора; 9 — дифференциация глоточной кутику-
лы; 10 — дифференциация челюстей; 11 —характер дифференциа-
ции среднекишечного отдела пищеварительной системы; 12 — состоя-
ние эпителия пищевода; 13 — сохранение архаичных структур: а) ру-
диментов ноги, б) ножевидных и полых спикул.
С первого взгляда видно разительное отличие Crystallophrisson,
Caudofoveatus и Chaetoderma от Scutopus и Limifossor', Prochaeto-
derma занимают почти промежуточное положение. Два рода (Me-
tachaetoderma и Psilodens) вынесены за пределы кольцевой диаграм-
мы, поскольку эти таксоньг не могут быть пока охарактеризованы
26
полным набором признаков. Тем не менее, имеющиеся данные од-
нозначно определяют большее сходство Metachaetoderma с Pro-
chaetoderma, a Psilodens — со Scutopus и Limifossor.
Теперь необходимо установить — представляют ли эти группы
родов монофилетические таксоны или полученная картина есть ре-
зультат только внешнего сходства.
Scutopus, Psilodens и Limifossor представляют четко очерченную
филогенетическую ветвь, характеризующуюся следующими апо-
морфными признаками: 1 — появление дорсального аппроксиматора
субрадулярных хрящей; 2 — тенденция к разделению мембраны ра-
дулы на две независимые продольные ленты.
Среди этой группы родов скутопусы обладают наибольшим чис-
лом плезиоморфных черт организации, не встречающихся у других
Caudofoveata: примитивно устроенный головной щиток, сохранение
педального отдела центральной нервной системы в полном виде,
сохранение рудиментов ноги, наличие полых спикул. В то же время
у Scutopus происходит дифференциация кольцевой перетяжки, от-
деляющей головной отдел.
По характеру развития радулярной мембраны, форме зубных
пластин и дифференциации отделов тела к Scutopus ближе всего
стоят Psilodens. Limifossor же уклонились как по характеру диф-
ференциации отделов тела, так и по способу развития радулярной
мембраны.
Вторая филогенетическая ветвь представлена Prochaetoderma,
Crystallophrisson, Chaetoderma и Caudofoveatus и может быть оха-
рактеризована следующими апоморфными признаками: 1 —тенден-
ция к четкой дифференциации отделов тела; 2 — дифференциация
в поперечном сегменте радулы пяти слагающих элементов; 3 —
утрата цилиарной выстилки пищевода.
В этой ветви довольно рано уклоняются Prochaetoderma, у пред-
ставителей которых можно отметить появление следующих апоморф-
ных черт организации: 1 —дифференциация парной челюсти в бук-
кальной полости; 2 — формирование двураздельного головного щит-
ка; 3 — утрата тифлозоля; 4 — формирование «хвостатой» формы
тела. При этом прохетодермы сохраняют один специфический плезио-
морфный признак — укладку спикул под углом к оси тела.
К этому же направлению эволюции можно отнести и Metachaeto-
derma, но имеющиеся данные по их организации не позволяют
судить о моменте дивергенции этих двух родов.
Chaetoderma, Crystallophrisson и Caudofoveatus представляют
собой наиболее монолитную (в морфологическом и экологическом
отношениях) группу родов среди подкласса и могут быть охаракте-
ризованы следующим набором апоморфных признаков: 1 — мало-
сегментная радула (1 или 2 сегмента); 2—наличие радулйрного
капюшона; 3 — наличие гастрального щита; 4 — наличие прото-
стиля; 5 — дифференциация двух пар лент продольной мускула-
туры; 6 — спикульный покров сформирован спикулами с сагитталь-
ным изгибом; 7 — появление естественно-напряженных элементов
в радулярном аппарате.
27
В пределах этой филогенетической ветви можно проследить рас-
хождение между Chaetoderma, с одной стороны, и Crystallophrisson
и Caudofoveatus — с другой. Два последних рода характеризуются
удлинением пищевода, становлением подкововидного головного щит-
ка, грифельковидными или коронковидными зубами, связанными
радулярным капюшоном и прикрепленными к осевой пластине ра-
дулы посредством эластичных латеральных опор. Здесь еще сохра-
няется Двухсегментная радула, но зубы уже не связаны непосред-
ственно между собой, а естественно-напряженный элемент стремится
сжать зубные пластины.
В роде Chaetoderma имеется тенденция к удлинению не пище-
водного, а гоноперикардиального отдела и становлению «хвостатой»
формы тела. Зубы серповидной формы связаны между собой и осе-
вой пластиной эластичным симфизом; радулярный капюшон не при-
крепляется непосредственно к зубным пластинам. Естественно-
напряженный элемент радулы (симфиз) стремится развести радуляр-
ные зубы.
В пределах Chaetoderma можно выявить две тенденции в транс-
формации радулярного аппарата. В одном случае зубы перемеща-
ются в терминальное положение на осевой пластине и приобретают
максимальную независимость от радулярного капюшона. Во вто-
ром — появляются дополнительные связки между зубами и медиаль-
ные кутикулярные пластинки, опосредующие действия зубов и раду-
лярного капюшона, что приводит к переходу от пинцетного принципа
функционирования (Иванов, 1979) к грейферному (см. ранее)
На основании проведенного анализа апоморфных состояний при-
знаков построена филогенетическая схема (рис. 10).
В филогении Caudofoveata можно отметить два основных мо-
мента дивергенции. Во-первых, уклонение Scutopus, Psilodens и Limi-
fossor (специализировавшихся, по-видимому, на смешанном пита-
нии: хищничество + плотоядение). Дивергенция этих трех родов
осуществлялась в результате различных стратегий в завоевании
глубоких слоев осадков и различий в переходе от микрофагии к хищ-
ничеству и плотоядению. Во-вторых, расхождение Prochaetoderma и
Metachaetoderma (в связи с их узкой специализацией на питание
фораминиферами), с одной стороны, и Chaetoderma, Crystallophris-
son и Caudofoveatus (пошедших по пути коренной перестройки раду-
лярного аппарата и специализировавшихся на избирательной микро-
фагии) — с другой стороны. Их дальнейшая дивергенция связана
со специализацией радулярного аппарата и с различной стратегией
в завоевании более глубоких слоев осадков.
3. Филогения Caudofoveata и биологический прогресс групп
А. Н. Северцов предложил три признака биологического прогрес-
са: 1) непрерывно возрастающее число особей; 2) увеличение аре-
ала; 3) распадение на соподчиненные таксоны. Из этих трех крите-
риев В. Г Гептнер отдает предпочтение последнему, отмечая, что
«одна из положительных сторон критерия числа видов заключается
28
в том, что он может удерживать от субъективности в суждении о
биологическом прогрессе и от антропоморфизма» (Гептнер, 1965,
с. 1305).
Большое внимание В. Г Гептнер также уделял асимметрии внут-
реннего строения систематических групп, которая, по его мнению,
свидетельствует о наличии в группе более или менее сильного кладо-
генеза. Роды с наибольшим числом видов, очевидно, указывают
главное направление кладогенетической эволюции семейства за не-
давнее прошлое, в настоящее время и перспективу на ближайшее
геологическое будущее. Таким образом, группы, богатые видами,
биологически прогрессивны.
Если под этим углом зрения рассматривать каудофовеат,- оче-
видно, что наиболее биологически прогрессивный род Crystallo-
phrisson: на его долю приходится 57% видового разнообразия под-
класса; вторым по числу видов является род Chaetoderma — 28%;
на долю же остальных 6 родов приходится только 15% видов. Таким
образом, Crystallophrisson и Chaetoderma указывают нам главные
направления кладогенетической эволюции подкласса.
Интересно посмотреть, как связан биологический прогресс груп-
пы с морфологическим. Для этой цели была построена объемная
филогенетическая схема (рис. 11), где по осям оценен биологический
прогресс (выраженный числом видов), морфологический прогресс
(отношение числа апоморфных признаков к суммарному числу ис-
пользованных для анализа признаков) и родственные отношения
(относительное расположение точек дивергенции филогенетических
ветвей)
Из схемы видно, что для Caudofoveata наиболее морфологически
продвинутые таксоны одновременно и наиболее биологически про-
грессивны, за исключением рода Caudofoveatus.
Можно предложить два объяснения этого факта. Во-первых, род
Caudofoveatus описан совсем недавно (Иванов, 1981) и его объем
пока окончательно не установлен. Во-вторых, можно предположить,
что дивергенция Caudofoveatus и Crystallophrisson произошла от-
носительно недавно, и представители последнего рода в ходе эволю-
ции получили дополнительные преимущества, позволяющие успешно
конкурировать с представителями близких по биологии родов, что
в конечном итоге может тормозить биологический прогресс рода
Caudofoveatus.
Можно думать, что основное направление эволюции ориентиро-
вано в сторону становления рода Crystallophrisson. Формирование
у представителей этого рода специфического аппарата добычи пищи
с высокими селективными свойствами (более ни у кого из моллюсков
не отмечавшийся), в сочетании с переходом к перистальтической
локомоции (основанной на существенной перестройке кожно-му-
скульного мешка и спикульного покрова) позволили кристалло-
фриссонам не только обеспечить массовое заселение не свойствен-
ной большинству моллюсков области обитания (толщу илисто-пес-
чэных грунтов), но и позволяют, судя по всему, успешно конкури-
ровать с многощетинковыми червями.
29
о* « 1* Q о V
X^ X x^ ^x^ c==—-
х x XC XC ^><^ x
e>* »• ®?
GD*| (Ю * О 00 “ 00* CD* о
GHZ* GHZ* C_L dZ*
<rl Q‘ Io о 0? о
^xc^ ЩГ ^ьГ? 1PJT
1 * 1 * 1 * nun lllllll lllllll llllllll
1Ж1 &•[ ж 0D x ^x^ ^><^
* * *
4
x ^><^ x
0’1
- jy ^. J* III 1 _* .',,"irTTT/y -li/f j НОШ/МЦ
» ” •* «KB 1^» MBM
2^5)* r-» ~^*
EZ_1 1 C ]| 1 J |l_zd| 1 1* h 1? УюоооооГ
0^0 . (W (^2 r^i * vE/? *
20
19
18
17
16
15
14
13
12
11
10
9
8
7
6
5
4
3
2
1
МОРФОЛ ОГИЧЕСКИЙ ПРОГРЕСС
Рис. 11. Морфобиологический прогресс Caudofoveata.
Обозначение таксонов как на рис. 9 (объяснение в тексте).
ЛИТЕРАТУРА
Гептнер В. Г Структура систематических групп и биологический прогресс.— Зоол.
жури., 1965, ст. 44, вып. 9, с. 1291 —1308.
Иванов Д. Л. Строение и функциональная морфология радулярного аппарата Chaeto-
derma.— Зоол. журн., 1979, т. 58, вып. 9, с. 1302—1306.
Иванов Д. Л. Caudofoveatus tetradens gen. et sp. n. и диагностика таксонов в под-
классе Caudofoveata (Mollusca, Aplacophora).— Зоол. журн., 1981, т. 60, вып. 1,
с. 18—28.
Иванов Д. Л., Цетлин А. Б. Возникновение и эволюция кутикулярного глоточного
вооружения у трохофорных животных с вентральной глоткой.— Зоол. журн.,
1981, т. 60, вып. 10, с. 1445—1454.
Майр Э. Принципы зоологической систематики. М., 1971, с. 1—454.
Сальвини-П лавен Л. Замечания о Chaetoderma и Crystallophrisson (Mollusca, Aculi-
fera).—Зоол. журн., 1984, т. 63, вып. 2, с. 171 — 175.
Рис. 10. Филогения Caudofoveata и распределение состояний признаков (звез-
дочкой отмечены апоморфные состояния).
/ наличие дорсального аппроксиматора субралчлярных хрящей, 2 — дифференциация радулярной
мембраны, 3 — дифференциация отделов тела. 4 — дифференциация продольных рядов радулы. 5 — цили-
ация пищевода, 6 — наличие челюстей, 7 — возникновение сагиттального изгиба спикул, 8 — ножевидиые
спикулы, .9 — протостиль, 10 — гастральный щит, II — кутикулярная обкладка одонтофора, 12— сегмен-
тация радулы, 13 — тифлоюль, 14 - характер дифференциации продольной мускулатуры, /5 — кольцевая
перетяжка головного отдела, 16 состояние головного щитка, 17 положение «печени», /Я—рудименты
ноги, /9 полые спикулы, 20 — дифференциация инициальных зубов радулы (пустые ячейки — первичное
отсутствие структур, перечеркнутые утраченные структуры). Обозначение таксонов как на рис. 9.
31
Шилейко А. А. Система отряда Geophila (=Helicida) (Gastropoda, Pulmonata).
В кн.: Морфология, систематика и филогения моллюсков. Л., 1979, с. 44- 69.
Юдин К. А. О понятии «признак» и уровнях развития систематики животных.--
Тр. ЗИН АН СССР, 1974. т. 53, с. 5-29.
Deimel К. Zur Ableitung des Radulaapparates der Chaetodermatidae (Mollusca,
Caudofoveata).— Z. Zool. Systematik u. Evolutionforsch., 1982, Bd 20, H. 3,
S. 177—187
Kerth K. Radulaapparat und Radulabildung der Moilusken. I. Vergleichende Morpholo-
gie und Ultrastruktur.— Zool. Jb. Anat., 1983, N 110, S. 205- 237
Kovalewsky A. Sur le genre Chaetoderma.— Arch. Zool. exp. (Ser. 3), 1901, p. 261—284.
Salvini-Plawen L. V Neue Scandinavische Aplacophora (Mollusca, Aplacophora).—
Sarsia, 1967, N 27, s. 1—63.
Salvini-Plawen L. V. Mollusca; Caudofoveata — In: Marine Invertebrates of Scandi-
navia, 1975, N 4, p. 1—55.
Scheltema A. H. The radula of the Chaetodermatidae Z. Morphol. Tier. 1972, Bd 72,
s. 361-370.
Scheltema A. H. Comparative morphology of the radulae and alimentary tracts in
the Aplacophora — Malacologia, 1981, vol. 20, N 2, p. 361—383.
Schwable M. Crystallophrisson (=Chaetoderma) hartmani nov. spec., eine neue Apla-
cophora aus dem Ostpacifik — Zool. Anz., 1961, N 166, s. 258—277.
Sneath P H. A., Sokal К. K. Numerical taxonomy: principles and practice of numerical
classification. W H. Freeman, San Francisco, 1973, p. 1—573.
D. L. Ivanov
PHYLOGENY OF THE SUBCLASS CAUDOFOVEATA
(MOLLUSCA APLACOPHORA)
The phylogeny of Caudofoveata is discussed on the base of analyse of external
morphology, epithelial-muscular sac, radular apparatus and another parts of the
alimentary system. It is established that within the subclass Scutopus, Psilodens
and Limifossor are the nearest to ancestral forms. Remaining genera form 2 phylo-
genetic branches: one early separated includes Prochaetoderma and probobly Meta-
chaetoderma and another well specialized and the highest includes Chaetoderma,
Crystalophrisson and Caudofoveatus.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
О. А. Скарлато, Я. И. Старобогатов
ОПЫТ НОВОЙ СИСТЕМЫ ИНДЕКСАЦИИ ЗУБОВ
ЗАМКА ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
Для сокращенного обозначения зубов замка двустворчатых мол-
люсков предложен ряд систем (обзор см. Коробков, 1954, Невес-
ская, 1960, Сох, 1969) Большинство их преследует только названную
цель и лишь наиболее популярная ныне система Бернара — Мунье-
Шальма по замыслу имеет назначение, кроме этого, обозначить
одинаковыми индексами зубы замков разных видов, связанные пре-
емственностью в ходе филогенетического развития («гомологич-
ные») Термин «гомологичные» мы тут заключаем в кавычки, по-
скольку, по нашему мнению, гомологичными могут быть только жи-
вые образования (например, участки мантии, формирующие зубы).
Мы не приводим подробного описания этой системы, поскольку она
достаточно хорошо описана в отечественной литературе (Коробков,
1954, Невесская, 1960). Однако накопленные данные по онтогенети-
ческому развитию замков свидетельствуют, что и система Бернара —
Мунье-Шальма не достигает этой цели (по крайней мере, при срав-
нении систематически далеких родов со сходным по числу зубов
замком)
Ввиду этого мы предприняли попытку построить новую систему,
которая, как и система Бернара — Мунье-Шальма, имела бы задачу
обозначить одинаковыми индексами соответствующие друг другу зу-
бы. Задача предлагаемой заметки — ознакомить малакологов с этой
системой, чтобы она подверглась тщательной проверке на возможно
более широком круге объектов. Это позволит выявить ее недостатки,
в результате чего следует или внести необходимые коррективы, или
в той или иной степени перестроить ее. Предлагаемую систему мы
иллюстрируем несколькими схемами индексации зубов в разных
типах замков, принадлежащих видам разных отрядов Bivalvia
(рис. 1 —14).
Суть предлагаемой системы сводится к следующему.
1. Зубы (равно как и первичные пластинки) передней и задней
половин замка нумеруются независимо; граница между ними — се-
редина прямого спинного края продиссоконха и точки замочного
края раковины на более поздних стадиях онтогенеза, преемственно
связанные с исходной точкой в ходе роста раковины.
2. Первичные пластинки (до их разделения на кардинальные и
латеральные зубы) и соответствующие им зубы прегетеродонтных
и ктенодонтных замков нумеруются снаружи внутрь (т. е. в обратном
по сравнению с системой Бернара — Мунье-Шальма направлении),
в правой створке — нечетными цифрами, в левой — четными, с тем,
однако, чтобы при сомкнутых створках номера зубов образовывали
3 Заказ 1696 33
ПРАВАЯ
рис 1—4. Схемы замка на 3 разных этапах его онтогенетического
развития.
/ _ Gouldia, 2 — Cerastoderma, 3 — Spisula, 4 — Parvilucina.
34
ЛЕВАЯ
5
Рис. 5—8. Схемы замка на 3 разных этапах его онтогенетического развития.
5 — Lorioes, 6 — Euglesa, 7 — Lasea, 8 — Crassinella
3*
35
Рис. 9—14. Схемы замка (рис. 9 и 11 на 3 разных этапах; рис. 11 -
правая створка; рис. 10. 12 — на 2 разных этапах; рис. 12 — правая створка;
рис. 13, 14 — замки взрослых особей.
9 — Cyclocardia, 10 — Plicatula, 11 — Glycymeris, 12 — Malletia, 13 — As-
tarte, 14 — Neotrigonia.
13
14
36
последовательный ряд. К номерам передних первичных пластинок
добавляется буква «а», к номерам задних — «р». Обособленные
или отогнутые части зубов (первичных пластинок) в ктенодонтных
замках обозначаются дополнительными индексами 1 и 2, сдвинутыми
к нижней части буквы.
3. Кардинальные зубы обозначаются добавлением буквы «с»,
латеральные — буквы «1», если из одной первичной пластинки об-
разуется 2 кардинальных зуба, то они обозначаются дополнитель-
ными цифровыми индексами 1 и 2 (снаружи внутрь). Нимфа, обра-
зующаяся за счет части одной из первичных пластинок, на каждой
створке обозначается добавлением буквы «п», другие выступающие
образования, связанные с лигаментом (круры), если они также обра-
зуются за счет первичных пластинок, обозначаются добавлением
букв «сг».
Как видно из прилагаемых схем, замки, обычно считающиеся
астартоидными — замок Cyclocardia (кардитоидный) и Crassinella
(собственно астартоидный) при такой системе индексации довольно
существенно различаются. Замок Cerastoderma (кардиоидный, хотя
обычно считающийся люциноидным) резко отличается от люциноид-
ного замка Parvilucina и более похож на корбикулоидный замок
Gouldia. То же следует сказать и о замке Spisula. Легко видеть,
что черты сходства и различия замков тут более соответствуют
систематическому положению рассматриваемых видов. Различия
замков Loripes и Parvilucina (особенно в отношении кардинальных
зубов правой створки) говорят скорее в пользу сборности семейства
Lucinidae.
Как видно из сказанного, даже первое пробное применение пред-
лагаемой системы индексации открывает новые возможности эволю-
ционной интерпретации замков двустворчатых моллюсков. Можно
предположить, что широкая апробация и надлежащая доработка
новой системы даст новый инструмент для анализа эволюции замков
в пределах класса двустворчатых'моллюсков.
ЛИТЕРАТУРА
Коробков И. А. Справочник и методическое руководство по третичным моллюскам.
Пластинчатожаберные. Л., 1954, 444 с.
Невесская Л. А. Общая характеристика и морфология.— В кн.: Основы палеонто-
логии. Моллюски — панцирные, двустворчатые, лопатоногие. М., 1960, с. 21—47.
Сох L. R. General features of Bivalvia. Treatise on invertebrate paleontology, part N,
vol. 1 (of 3), Mollusca 6, Bivalvia. Lawrence, 1969, p. 2—129.
O. A. Scarlato, Y. I. Starobogatov
TRIAL OF A NEW SYSTEM OF INDEXES FOR BIVALVIAN DENTITION
The proposed system has the next principles. The teeth of the anterior and the
posterior halves of the hinge are numbered independently, the demarcation between
them being the midpoint of the strait dorsal margin of the prodissoconch and those
points in the cardinal margin of the later stages of growth which are connected succes-
sively with the initial point during the growth. Primary lamellae (before their division
to cardinal and lateral teeth) and the teeth of preheterodont hinges respected to
37
them are numbered from outside to inside by the odd number in the right valve and
by the even numbers in the left one in order to form the continuous row of numbers
when the shell being closed. Anterior lamellae are marked by the letter «а», posterior
ones — by the letter «р». Separated or bent part of teeth in ctenodont hinges are
marked by additional sign 1 or 2. The cardinal teeth are designated by addition the letter
«с», lateral ones — by addition the letter «1». When one primary lamella produces
2 cardinal teeth they must be marked as Ci and c2. The nymph formed from the part
of the primary lamella is marked by the addition the letter «n>, and crura — by the
letters «сг». Using of the system is demonstrated on figs 1 —14.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
M. H. Затравкин, A. JI. Лобанов
МОРФОМЕТРИЧЕСКИЕ ХАРАКТЕРИСТИКИ РАКОВИН
ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ РОДА
NODULARIA (UNIONIDAE) ФАУНЫ СССР
И ВОЗМОЖНОСТИ ИХ ИСПОЛЬЗОВАНИЯ
ДЛЯ ИДЕНТИФИКАЦИИ ВИДОВ
И. М. Москвичевой (1973) показано, что на территории СССР
обитает 7 видов рода Nodularia, относимых ранее (Жадин, 1933,
1952) к одному виду Unio douglasiae (Griff, et Pidg.). В работе Моск-
вичевой (1973) приводятся сведения о размерах раковин различных
видов рода из разных мест сбора, в том числе и размеры типовых
материалов.
Цель нашего исследования — сравнение морфометрических ха-
рактеристик раковин различных видов рода Nodularia как из сов-
местных и разных местонахождений, так и видов в целом (независимо
от места сбора), а также изучение возможности использования мор-
фометрических характеристик раковин для идентификации видов
рода Nodularia обитающих в СССР: /V. sujfunica, N. arnurensis,
N. niiddendorffi, N. abbreviata, N. schrencki, N. flavoviridis, N. vladi-
vostokensis.
Материалом для исследования послужили 438 раковин 7 видов из
коллекции Зоологического института АН СССР (Ленинград) и сбо-
ров одного из авторов (М. Н. Затравкина), проведенных в бассейне
Амура в июне—июле 1980 г.
Измерялись штангенциркулем с точностью до 0,1 мм следующие
параметры: длина раковины (/), высота раковины (й). выпуклость
пары створок (В). По полученным промерам для каждого экземпляра
вычислялись отношения высоты раковины к ее длине (й:/), выпук-
лости к длине (В:/), выпуклости к высоте (В:й). Для всех промеров
и их отношений по каждому виду в целом (независимо от места
сбора) и отдельно для каждого места сбора рассчитывались: среднее
арифметическое и его ошибка (M±/7z), коэффициент вариации (Си),
среднее квадратичное отклонение, коэффициенты эксцесса и асим-
метрии. Для сравниваемых видов (в целом независимо от места сбо-
ра и раздельно для разных мест сбора) рассчитывался критерий
Фишера (F) для каждой пары сравниваемых промеров или отношений.
В табл. 1 приведены морфометрические характеристики четырех
наиболее обычных для фауны Дальнего Востока СССР видов из бас-
сейна Амура: N amurensis, N schrencki, N. abbreviaia, N. flavo-
viridis. Как видно из приведенных в табл. 1 цифр, из абсолютных
размеров меньше всего варьирует высота раковины, а из отношений
размеров — отношение высоты раковины к ее длине (для всех видов,
39
Таблица 1
Морфометрические характеристики раковин различных видов рода Nodularia при совместном нахождении (с указанием места сбора)
Вид и место сбора Размеры раковин, мм Отношения размеров Кол-во проме- ренных раковин
высота выпуклость (обе створки) высоты к длине выпуклости к длине ВЫ 11 \ К !'>•’ к высиIV
М ± т | Си М ± т | Cv М ± т | Cv М ± т | Си М ± т | Си М ± т | Cv
N. amurensis Амур, протока Шара- ханда 59,4 ±2,0 8,8 28,0 ±0,8 7,8 23,8 ±1,1 12,8 0,472 ±0,006 3,2 0,399 ±0,009 3,2 0,847 ±0,022 6,7 7
N. schrencki Там же 65,5 ±1,1 13,2 30,3 ±0,4 10,1 24,9 ±0,5 14,6 0,473 ± 0,004 5,8 0,385 ± 0,003 5,7 0,819 ± 0,008 5,7 57
N. flaviviridis Там же 60,4 ±1,1 9,8 29,0 ±0,6 10,1 22,4 ±0,4 9,8 0,480 ±0,003 3,7 0,371 ±0,003 4,4 0,774 ±0,007 4,7 27
N. abbreviate! Там же 57,6 ±1,2 7,2 28,0 ±0,6 7,6 21,1 ±0,4 6,8 0,486 ± 0,004 2,8 0,366 ±0,003 3,2 0,753 ±0,007 3,6 13
N. schrencki Амур у с. Богородское 68,2 ±1,9 7,8 29,3 ±0,6 5,9 6,1 ±0,8 8,3 0,431 ±0,011 7,2 0,383 ±0,011 8,3 0,892 ±0,028 8,8 8
N. abbreviata Там же 60,2 ±0,8 10,7 28,1 ±0,3 8,8 21,9 ±0,4 13,1 0,468 ±0,003 4,5 0,363 ± 0,007 5,5 0,778 6,7 59
N. amurensis Амур у с. Вятское 61,2 ±2,0 15,5 28,8 ±0,7 11,9 23,3 ±0,9 18,0 0,474 ± 0,007 7,1 0,380 ±0,003 4,3 0,807 ±0,017 9,9 23
N. flavoviridis Там же 51,3 ±2,8 18,8 25,4 ±1,3 16,4 18,9 ±1,1 18,9 0,497 ±0,012 7,7 0,368 ±0,004 3,8 0,744 ±0,019 8,6 И
Таблица 2
Вычисленные значения критерия Фишера (при числе степеней свбоды от 1,18 до 1,82)
для видов рода
Виды | N. abbreviata | N. flavoviridis | N. schrencki
ЛЛ amurensis 1 4,78 1,27 0,002
2 17,6 13,4 2,07
3 25,1 17,4 1,3
N. schrencki 1 2,84 1,11
2 10,4 10,2
3 14,5 12,45
N. flavoviridis 1 1,3
2 1,1
3 3,5
Примечание I — отношение высоты
раковины, 3 — отношение выпуклости к высоте
раковины к ее длине, 2 — отношение выпуклости к
раковины.
кроме A. amurensis и A. flavoviridis, собранных в Амуре у с. Вятского;
у этих видов меньше варьирует отношение выпуклости раковины к
ее длине). Из приведенных в табл. 2 значений критерия Фишера, рас-
считанных для N. amurensis, N. schrencki, N. flavoviridis, N. abbre-
viata, собранных совместно в амурской протоке Шараханда, видно,
что по отношению высоты раковины к ее длине достоверно разли-
чаются между собой лишь N. amurensis и N. abbreviata. По признаку
«отношение выпуклости раковины к ее длине» неотличимы друг от
друга пары N schrencki — N. amurensis и N, abbreviata — N. flavo-
viridis', все другие пары достоверно отличимы друг от друга. По
признаку «отношение выпуклости раковины к высоте» все виды до-
стоверно различаются, кроме пары N. amurensis — N. schrencki.
Из приведенных в табл. 2 значений критерия Фишера видно, что
N. schrencki и N. amurensis, собранные совместно в протоке Шара-
ханда, не различаются ни по одному признаку. При анализе значе-
ний критерия Фишера для ЛЛ schrencki и N. abbreviata, собранных
совместно в Амуре у с. Богородского и для N. amurensis и N. flavoviri-
dis, собранных в Амуре у с. Вятского, оказалось, что они достоверно
отличимы друг от друга по всем отношениям (по разным отношениям
Р от 0,95 до 0,999).
Для сравнения всех обитающих в пределах СССР видов (кроме
N. lebedevi, известного лишь по одному экземпляру) были взяты
объединенные выборки для каждого вида независимо от места сбора.
Морфометрические характеристики видов (для объединенных выбо-
рок) приведены в табл. 4. Из всех вычисленных отношений размеров
раковин меньше всего у N. sujfunica, N. amurensis, N. schrencki,
N. flavoviridis варьирует отношение выпуклости раковины к ее длине
(Cv от 3,0 до 5,6%), для N. abbreviata отношение высоты раковины
к ее длине (Cv от 4,9 до 5,3), а для A. vladivostokensis отношение
выпуклости к высоте раковины (Си = 5,0).
По рассчитанным для сравниваемых видов (по объединенным
выборкам) значениям критерия Фишера (табл. 3) видно, что только
41
Таблица 3
Вычисленные значения критерия Фишера (при числе степеней свободы от 1.39 до 1,250)
для объединенных выборок видов
Виды N. vladi- voslokensis A’. inidden- dorjji A' abbre- viata /V flavo- viridis A', schren- cki A’, ainu- rensis
N. sujfunica 1 0,94 20,4 4,24 0,01 1,33 0,56
2 21,5 80,6 0,01 0,44 6,30 6,30
3 6,6 8,1 2,1 0,29 6,7 4,96
N. amurensis 1 2,48 30,6 8,8 1,4 1,25
2 9,4 326,7 36,2 12,53 0,54
3 0,53 67,0 5,5 11,0 2,0
N. schrencki 1 2,87 16,8 1,58 3,9
2 6,11 268,6 37,5 13,7
3 0,07 69,2 13,8 17,1
N. flavoviridis 1 1,54 41,7 13,1
2 21,7 209,9 1,53
3 5,01 28,0 3,2
N. abbreviata 1 6,3 17,1
2 18,4 174,6
3 2,0 55,2
N. middendorffi 1 15,4
2 97,0
3 17,25
Примечание I — отношение высоты
длине раковины; 3 — отношение выпуклости к
раковины к длине раковины, 2
высоте раковины.
шение выпуклости к
пары Af. flavoviridis — N. sujfunica и /V. amurensis — N. schrencki
не различаются ни по одному признаку (отношению). Остальные
виды отличаются друг от друга хотя бы по одному признаку с высо-
кой степенью достоверности. Выше отмечено, что /V. schrencki и
/V. amurensis не различаются морфометрически, и при совместном
нахождении идентифицировать эти виды можно только по другим
признакам, например, по кривизне фронтального сечения створки,
положению точки поверхности створки, наиболее удаленной от
плоскости смыкания створок. Что же касается пары N. sujfunica —
N. flavoviridis, то эти виды не встречаются совместно, так как их
ареалы не накладываются: АЛ flavoviridis — общеамурский вид,
встречающийся также в оз. Ханка, a N. sujfunica — приурочена
то. 1ько к бассейну р. Раздольной. Следует отметить, что среди ис-
следованных видов, обитающих в Амуре, лишь один вид W. midden-
dorffi не отмечен совместно с другими амурскими представителями
рода, но именно он резко отличается от других видов по всем трем
отношениям с высокой степенью достоверности (Р>0,999).
Анализ цифровых данных по морфометрии раковин моллюсков
рода Nodularia (табл. 1,4) показал, что использование морфомет-
рических характеристик для идентификации видов затруднено тем,
что большинство видов достоверно отличимы друг от друга лишь
по 1—2 отношениям, а верхние и нижние пределы индексов могут
накладываться один на другой у разных видов (при статистически
достоверных различиях). Только один вид (ЛЛ middendorffi) легко
42
отличается от всех других видов рода по всем взятым отношениям.
Из сказанного следует, что использовать морфометрические ха-
рактеристики для идентификации видов в большинстве случаев
можно лишь в совокупности с другими конхологическими призна-
ками.
В заключение приводим таблицу для определения видов рода
Nodularia фауна СССР.
1 .(14). Раковина овально-четырехугольная.
2 .(5). Точка поверхности створки раковины, наиболее удаленная от
плоскости смыкания створок (если смотреть спереди), лежит
почти на середине высоты раковины.
3 .(4). Скульптура развита слабо. Макушки смещены ближе к сере-
дине раковины и удалены от ее переднего края на расстояние,
примерно равное 0,30—0,32 ее длины N. amurensis
4 .(3). Скульптура хорошо развита и состоит из коротких часто рас-
положенных валиков. Макушки удалены от переднего края
раковины на расстояние, примерно равное 0,25—0,27 длины
раковины /V. vladivostokensis
5 .(2). Точка раковины наиболее удаленная от плоскости смыкания
створок (если смотреть спереди), лежит заметно выше сере-
дины высоты раковины.
6 .(7). Скульптура раковины хорошо развита в примакушечной ча-
сти. Макушки явно выступают над контуром раковины
W sujfunica
7 .(6). Скульптура раковины развита слабо даже в примакушечной
части. Макушки не выступают над контуром раковины или
выступают крайне слабо.
8 .(9). Макушки раковины смещены к середине раковины и отстоят
от переднего края раковины на расстояние, превышающее
0,33 ее длины N. flavoviridis
9 .(8). Макушки в меньшей степени смещены к середине раковины и
отстоят от ее переднего края на расстояние меньше 0,30 длины
раковины.
10 .(11). Отношение выпуклости раковины к ее длине не более 0,340;
выпуклости к высоте не более 0,740 /V. middendorffi
11 .(10). Отношение выпуклости раковины к ее длине более 0,350,
выпуклости к высоте более 0,750.
12 .(13). Точка поверхности раковины, наиболее удаленная от
плоскости смыкания створок (если смотреть спереди), лежит
выше середины, но ниже границы верхней трети высоты рако-
вины. Макушки отстоят от переднего края раковины на рас-
стояние, примерно равное 0,25—0,26 ее длины .
/V. abbreviata
13 .(12). Точка поверхности раковины, наиболее удаленная от
плоскости смыкания створок (если смотреть спереди), лежит
на границе верхней трети высоты раковины. Макушки отстоят
от переднего края раковины на расстояние, примерно равное
0,28—0,30 ее длины N. schrencki
14 .(1) Раковина удлиненно-эллиптическая N. lebedeui
43
Таблица 4
Морфометрические характеристики раковин различных видов рода Nodularia (для объединенных по видам выборок)
Виды Размеры раковин, мм Отношения размеров раковин Кол-во проме- ренных раковин
длина высота выпуклость (обе створки) высоты к длине выпуклости к длине выпуклости к высоте
М ± т | Си М ± т | Cv М ± т | Си М ± т | Си М±т | Си М ±т | Си
cujfunica 63,7 ±2,3 12,2 30,8 ±0,8 8,7 23,4 ±0,9 12,6 0,486 ±0,007 4,6 0,367 ±0,003 3,0 0,757 ±0,013 5,7 11
amurensis 56,6 ± 1,05 19,1 27,0 ±0,4 16,4 21,6 ±0,4 20,0 0,480 ±0,003 5,8 0,382 ±0,002 4,9 0,799 ±0,006 7,7 106
ЛГ. schrencki 61,5 ± 1,07 9,9 29,0 ±0,4 12,5 23,6 ±0,45 17,7 0,475 ±0,003 6,6 0,384 ±0,002 5.6 0,812 ±0,007 8,4 87
N. flavoviridis 58,3 ±1,1 13,1 28,2 ±0,5 12,3 21,6 ±0,4 14,0 0,485 ±0,004 5,2 0,371 ±0,002 4,3 0,766 ±0,007 6,3 47
N. abbreviata 57,4 ±0,7 14,7 26,9 ±0,3 13,2 21,1 ±0,3 16,5 0,470 ±0,002- 5,3 0,367 ± 0,002 5,6 0,782 ± 0,005 7,0 146
N. middendorffi 56,9 ±1,7 17,2 26,7 ±0,6 13,8 19,0 ±0,6 19,6 0,452 ±0,004 4,9 0,319 ±0,003 5,3 0,707 ±0,009 7,5 37
N. vladivostokensis 66,5 ±7,4 22,2 33,0 ±2,6 15,5 27,3 ±5,4 20,; 0,503 ± 0,022 8,8 0,412 ±0,014 6,7 0,821 ±0,020 5,0 4
ЛИТЕРАТУРА
Жадин В. И. Пресноводные моллюски СССР. Л., 1933, с. 1—232.
Жадин В. И. Моллюски пресных и солоноватых вод СССР.— Определители по
фауне СССР, изд. Зоологическим институтом АН СССР, М.—Л., 1952, т. 46,
с. 1-371.
Москвичева И. М. Наяды (Bivalvia, Unionoidea) бассейна Амура и Приморья.—
Зоол. журн., 1973, т. 52, вып. 10, с. 1458—1471.
М. N. Zatravkin, A. L. Lobanov
MORPHOMETRIC CHARACTERS OF SHELLS OF NODULARIA
(BIVALVIA, UNIONIDAE) FROM USSR AND POSSIBILITY OF USING
THEM FOR SPECIES IDENTIFICATION
Morphometric characters of shells of 7 species of Modularia are studied. The
possibility of using these characters for identification of species are discussed.
ТРУДЫЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
Л. А. Антонова
ВОЗМОЖНОСТИ ОПРЕДЕЛЕНИЯ ЗРЕЛЫХ ГЛОХИДИЕВ
МАССОВЫХ ЕВРОПЕЙСКИХ ВИДОВ UNIONINAE
И ANODONTINAE (BIVALVIA UNIONIDAE)
Глохидии — паразитические личинки пресноводных двустворча-
тых моллюсков надсемейства Unionoidea изучаются уже давно, на-
чиная с работы Ратке (Rathke, 1797), который впервые описал та-
кую личинку у Colletopterum piscinale (Nilss.) (думая, что это особый
организм, паразитирующий в жабрах моллюска) и ввел это название.
Разнообразие формы раковин и прикрепительных аппаратов глохи-
диев позволяют считать, что можно в морфологии этих личинок
отыскать видовые различия. Детальные исследования дали возмож-
ность В. И. Жадину (1938) опубликовать таблицу для определения
видов унионид по глохидиям. Однако за почти полувековой период,
прошедший со времени выхода цитированной выше книги, выясни-
лось, что большинство принимавшихся в ней видов — сборные и
соответствуют родам и подродам в современном понимании. Напри-
мер, Anodonta piscinalis Nilss. соответствует всему роду Colletopte-
rum, Anodonta cygnea (L.) — роду Anodonta, Unto pictorum (L.) —
подроду Unio s. str. a Unio tumidus Phil.— подроду Tumldusiana.
Таким образом, по упомянутой выше таблице можно определить
глохидии только до рода или подрода, и вновь встает вопрос о воз-
можности их видовой идентификации.
Современный список унионид, обитающих в водоемах Европей-
ской части СССР, насчитывает 27 видов (см. Затравкин, 1983). Надо
при этом отметить, что в цитированной работе упоминаются только
24 вида и к ним следует добавить Batavusiana irgizlaica (Lindh.)
с Урала, Colletopterum depressum (Bgt.),4eобнаруженный в дельте
Дуная Т. Л. Алексеенко и С. tumidum (Kust.), найденный в дельте
Волги. Находка последнего вида весьма поучительна. До наших ис-
следований особи этого вида принимались за С. piscinale (Nilss.)
(Пирогов и Старобогатов, 1970), однако отличия в строении рако-
вин глохидиев заставили нас пересмотреть вопрос о видовой при-
надлежности этих особей, и в результате они оказались конспецифич-
ными виду Anodonta tumida, описанному Кюстером (Kuster, 1852)
из Женевского озера.
Изучение глохидиев проводилось в дельте Волги в Астраханском
государственном заповеднике имени В. И. Ленина *. Фауна этого
района насчитывает 14 видов семейства Unionidae: Unio (Unio)
* Пользуюсь случаем принести свою самую сердечную благодарность дирекции
и сотрудникам заповедника за предоставленную возможность работать с унионидами.
46
pictorum (L.), U ((A) rostratus Lam., U. (t7) linwsus Nilss.,
U. (Tumidusiana) tumidus Phil., U. (Г.) conus Spengl., Ano-
donta cygnea (L.), A. zellensis (Gm.), A. stagnates (Gm.),
Colletopterum (Colletopterum) subcirculare (CL), C. (Ponderosiana)
ponderosum „(PL), C. (Piscinaliana) piscinale (Nilss.), C. (Pise.)
tumidum (Kust.), Pseudanodonta complanata (Rossm.), Ps. elongata
(Holl.) т. e. несколько более половины всей фауны Европейской
части СССР. Из этого набора видов удалось изучить глохидии
только у 12 видов; глохидии видов рода Pseudanodonta остались
не изученными в связи с редкостью находок представителей этого
рода в дельте Волги.
Изучали раковину только зрелых инвазионных глохидиев, кото-
рые, будучи извлечены из жабр материнской особи, активно двигали
створками. Для очистки раковины от тканей глохидии помещали в
пенициллиновые склянки в воду на месяц или более, а затем отмы-
вали от остатков мацерированных тканей и хранили в 70% спирте.
Фиксация глохидиев спиртом не рекомендуется: они смыкают створки
и рассмотреть прикрепительный аппарат не представляется возмож-
ным, в то же время такая фиксация сильно затрудняет последующую
мацерацию.
Для изучения глохидиев под оптическим микроскопом их пере-
носят на предметное стекло в каплю глицерина. При медленном сме-
шении спирта с глицерином возникают турбулентные точки, пере-
мещающие и поворачивающие раковины, что позволяет рассмотреть
их со всех сторон. Для удаления прикрепительного аппарата и лига-
мента, что требуется при измерении раковин, последние помещают
на несколько секунд в жавелевую воду.
Описанная методика позволила достаточно подробно изучить
строение раковин глохидиев и прикрепительных аппаратов, пред-
ставленных у унионид крючком, усаженным разного размера шипами.
Для характеристики размеров раковин использовали следующие
промеры: длина — расстояние между передним и задним концами
раковины, измеренное параллельно прямому спинному краю, высо-
та — расстояние по перпендикуляру к спинному краю от него до
вентрального угла створки, выпуклость (толщина) —расстояние
между внешними точками створок по перпендикуляру к плоскости
симметрии, длина крючка — расстояние по прямой от вершины
крючка до вентрального края створки в основании крючка (изме-
ряется в профиль, т. е. при положении створки передним или задним
концом к наблюдателю). Кроме этого, вычислялся индекс выпук-
лости — отношение высоты раковины к половине ее выпуклости (т. е.
фактически к выпуклости одной створки).
Полученные результаты позволили составить следующую таблицу
для определения родов и видов унионид дельты Волги по глохидиям.
1 (10) Раковина маленькая, не более 212 мкм в длину и 190 мкм в вы-
соту, молочно-белая; крючок широкий треугольный (подсе-
мейство Unioninae род Unio) (рис. 1—6) 2
47
Рис. 1—6. Раковины глохидиев видов рода Unio.
1 — общий вид раковины представителя подрода Unio s. str. сбоку,
2 — створки раковин видов подрода Unio s. str. спереди: а — U. (U.) limosus,
б — U. (Uj rostratus, в — U. (U.) pictorum, 3 прикрепительный аппарат
глохидия представителя подрода Unio s. str 4 общий вид раковины
представителя подрода Tumidusiana сбоку, 5 - створки раковин видов под-
рода Tuniidusiana спереди: а — U. (Г.) conus, б — U (Г.) tumidus, б
прикрепительный аппарат глохидия представителя подрода Tumidusian
2(7) Длина раковины более 200 мкм, высота более 180 мкм, длина
крючка более 60 мкм, крючок на конце закругленный, шипики
на нем расположены 4—5 продольными рядами (подрод Unio
s. str.) (рис. 1—3) 3
3(4) Индекс выпуклости не превышает 2,45
U limosus (рис. 2а)
4(3) Индекс выпуклости не менее 2,50 5
48
5(6) Индекс выпуклости не превышает 2,65
U. rostratus (рис. 26)
6(5) Индекс выпуклости не менее 2,70
U. pictorum (рис. 2в)
7(2) Длина раковины не более 190 мкм, высота не более 175 мкм,
длина крючка не более 50 мкм, крючок на конце заостренный,
шипики на нем расположены треугольником в его основании
(подрод Tumidusiana) (рис. 4—6) 8
8(9) Индекс выпуклости не превышает 2,55
U. tumidus (рис. 5а)
9(8) Индекс выпуклости не менее 2,70
U. conus (рис. 56)
10(1) Раковина довольно крупная, не менее 300 мкм в длину и вы-
соту, светло-коричневая, крючок длинный и довольно узкий,
лишь близ основания соединен пленками с вентральным краем
створки (подсемейство Anodontinae (рис. 7—15) 11
11(16) Раковина не очень крупная и несколько вытянутая в про-
дольном направлении: длина ее до 340 мкм, высота до 320 мкм,
вентральный угол раковины не закругленный, крючок длиной
не более 140 мкм, прямой, шипики с крючка заходят на внеш-
нюю поверхность створки (род Anodonta) (рис. 7,
9—11) .12
12(13) Крупные шипы на крючке расположены тремя продольными
рядами, мелкие появляются ближе к основанию крючка,
A. stagnalis (рис. 9)
13(12) Крупные шипы на крючке расположены в один продольный
ряд или не образуют рядов, мелкие разбросаны по всему
крючку 14
14(15) Индекс выпуклости не менее 3,70, на крючке ближе к его
вершине расположен продольный ряд из 4 крупных шипов,
далее к основанию крючка число шипов и продольных рядов
возрастает, высота шипов (если смотреть в профиль) возра-
стает от вершины и основания к средине его длины
A. zellensis (рис. 10)
15(14) Индекс выпуклости не более 3,50, на крючке крупные шипы
расположены в один продольный ряд и только после 6-го шипа
добавляются шипы по сторонам от этого ряда, высота шипов
на всем протяжении крючка примерно одинакова (если смот-
реть в профиль) A. cygnea (рис. 11)
16(11) Раковина очень крупная с примерно одинаковой длиной и
высотой, не меньшей, чем 345 мкм, вентральный угол раковины
закругленный, крючок длиной не менее 150 мкм, изогнутый,
шипики с крючка на внешнюю поверхность створки не заходят
(род Colletopterum) (рис. 8, 12—15) .17
17(18) Крупные шипы идут почти на всем протяжении крючка двумя
продольными рядами и только у основания крючка их число
возрастает, высота шипов (смотреть в профиль) плавно
увеличивается от вершины к основанию крючка и затем резко
падает С. subcirculare (рис. 14)
4
Заказ 1696
49
Рис. 7—8. Раковины глохидиев представителей
подсемейства Anodontinae (сбоку и спереди)
7 — Anodonta, 8 — Colletopterum.
18(17) Крупные шипы на крючке располагаются в один продольный
ряд, по сторонам от которого по мере приближения к основа-
нию крючка добавляются еще шипы, высока шипов примерно
одинакова по всей длине крючка 19
19(20) По сторонам от продольного ряда крупных шипов располага-
ется лишь незначительное число мелких шипиков: на большей
части крючка их не более 3—4 с каждой стороны и лишь у
основания число их возрастает
С. tumidum (рис. 15)
20(19) По сторонам от продольного ряда крупных шипов на всем
протяжении крючка располагается много мелких шипиков
21
21(22) Крупные шипы в привершинной половине крючка располо-
жены строго в один продольный ряд
С. ponderosum (рис. 13)
22(21) Крупные шипы располагаются строго в один продольный ряд
лишь в самой привершинной части крючка, с 4-го по 6-й шип
(считая от вершины крючка) они расположены уже двумя
продольными рядами, далее число продольных рядов еще
возрастает С. piscinale (рис. 12)
50
Рис. 9—11. Прикрепительный аппарат глохидиев видов рода Anodonta
(сбоку и спереди).
9 — A. stagnalis, 10 — A. zellensis, 11 — A. cygnea.
ЛИТЕРАТУРА
Жадин В. И. Сем. Unionidae. Фауна СССР, нов. сер. 18, моллюски IV, 1. М.—Л.
1938, IX + 170 с.
Затравкин М. Н. Unionidae фауны СССР и их роль как промежуточных хозяев
и элиминаторов трематод.— В кн.: Моллюски, систематика, экология и законо-
мерности распространения, сб. 7 Л., 1983, с. 40—44.
Пирогов В. В., Старобогатов Я. И. Моллюски семейства Unionidae Волжской дельты.—
Тр. Астраханск. гос. зап. им. В. И. Ленина, 1970, в. 13, с. 226—248.
4*
51
Рис. 12—15. Прикрепительный аппарат глохидиев видов рода Colletopterum
(сбоку и спереди)
12—С. piscinale, 13—С. ponderosum, 14 — С. subcirculare, 15 — С. tumidum.
52
Raster H. C. Die Gattung Anodonta.— In: Martini und Chemnitz Systematisches
Conchylien-Cabinet, Bd 9, Abt. 1, Nurnberg, 1842, S. 41.—64.
Rathke J. Om Dam-Muslingen.— Skrift. naturhist. Selsk. Cobenhavn, 1797, Bd 4,
N 1, S. 139—179.
L. A. Antonova
ON THE POSSIBILITY OF IDENTIFICATION OF THE RIPE GLOCHIDIA
OF ABUNDANT EUROPEAN SPECIES FROM THE SUBFAMILIES UNIONINAE
AND ANODONTINAE (BIVALVIA UNIONIDAE)
Methods of the optic-microscopical study of glochidian shells is described and
the key for identification of glochidia belonging to 12 species from Volga-delta is
presented.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
3. И. Иззатуллаев, Я. И. Старобогатов
ВИДЫ РОДА ODHNERIPISIDIUM (BIVALVIA PISIDIIDAE)
ФАУНЫ СССР
Группа мелких Pisidiidae с открытым во внутреннюю полость ра-
ковины фиброзным слоем лигамента двадцать с лишним лет назад
была обособлена Ж. Кюйпером (Kuiper, 1962а) в отдельный подрод
Odhneripisidium в пределах обширного рода Pisidium. Кроме
типового вида Р stewarti Prest., он включил сюда Р vincentianum
Woodw., Р prashadi Odhn., Р. annandalei Prash., P parvum Mori,
P. cavernicum Mori (последний с вопросом) и ряд тропических
видов. В том же году (Kuiper, 1962b) он синонимизировал первые
два вида с типовым и добавил Р. dancei Kuip. из Ирана, а в Р. an-
nandalei, описанный с востока Индии, включил все современные
европейские находки моллюсков этой группы (они ограничены Среди-
земноморьем). Несколькими годами позже (Kuiper, 1969) он указы-
вает Р. dancei для востока Казахстана (Семипалатинск, Бухтар-
минск, Усть-Каменогорск). В 1967 г Ю. В. Бутенко (Белякова)
и Я. И. Старобогатов (Бутенко, 1967) описали Р. kungejense But.
et Star, из Кунгей-Алатау. Именно к этому виду, судя по рисунку,
относятся и казахстанские находки Р dancei. В этом же сборнике
Я. И. Старобогатов и Э. А. Стрелецкая (1967) описали еще 4 вида
из северной Азии, рассматривая при этом Odhneripisidium как
самостоятельный род: О. popovae St. et Str., О. janense St. et Str.,
0. indigircense St. et Str., 0. lindholmi St. et Str. и О. kolhy-
mense St. et Str. Позднейшее повторное изучение типовых экземпля-
ров показало, однако, что последние два вида следует относить к
роду Euglesa — у них лигамент не открыт со стороны полости
раковины, а просто очень широкий, занимающий всю замочную
площадку. В 1970 г. С. М. Попова и Я. И. Старобогатов (Попова
и др., 1970) описали еще 5 видов Odhneripisidium из верхне-
неогеновых отложений Чуйской котловины (Алтай). В маленькой
заметке, опубликованной еще в 1961 г., А. Д. Алиев (1961) описывает
находку Р. vincentianum в окрестностях Агдама (Азербайджан-
ская ССР). Позднее Н. Н. Акрамовский (1976) относит найденную
Алиевым форму и сходные с ней экземпляры из Мегри (Армянская
ССР) к О. annandalei. К этому следует добавить, что В. В. Пирогов
и Я. И. Старобогатов (1974), выделяя подсемейство Euglesinae,
отметили, что к нему, кроме родов Euglesa и Neopisidium,
относится и род Odhneripisidium. Таким образом, по литератур-
ным данным род Odhneripisidium насчитывает в водоемах СССР
и в четвертичных и неогеновых отложениях нашей страны 11—14
видов.
54
Целью предлагаемой работы была полная ревизия материалов по
Odhneripisidium из водоемов и неоген-четвертичных отложений
СССР, хранящихся в коллекциях Зоологического института АН СССР
и Института зоологии и паразитологии АН Таджикской ССР, а также
из сборов Е. А. Новикова (Томск). Было изучено несколько сот
экземпляров из Средней Азии, а также из Закавказья и Предкав-
казья, юго-восточного Казахстана, Алтая, Тувинской АССР, Примор-
ского и Хабаровского краев. При этом были обнаружены еще 13
новых для науки видов, а также внесены некоторые уточнения в
систематику видов, описанных ранее. Теперь с территории СССР
известно всего 16 современных видов и 10 обнаруженных пока только
в четвертичных и неогеновых отложениях. В предлагаемой работе мы
приводим описания новых видов и некоторые замечания по ранее
известным. Индексация зубов замка здесь принята та же, что и в
предыдущих работах (например, Пирогов и Старобогатов, 1974).
Голотипы и часть паратипов новых видов хранятся в коллекции
Зоологического института АН СССР (голотипы имеют коллекцион-
ный номер 1 по систематическому каталогу), другая часть парати-
пов хранится в коллекции Института зоологии и паразитологии
АН Таджикской ССР.
Изучение материала заставляет нас делить род Odhneripisi-
dium на 4 подрода.
Подрод Odhneripisidium s. str.
Раковина относительно крупная для рода (длина взрослых осо-
бей 2—3 мм), овально-треугольная, гладкая, покрытая только ли-
ниями роста; зуб За+Зв равномерно изогнут, зуб 2 изогнут, но под
прямым или тупым углом.
Odhneripisidium stewarti (Preston, 1909) (рис. 1)
Экземпляры, точно соответствующие описанию Престона (Pre-
ston, 1909), были собраны на Памире в озерах Зоркуль и Сулукты-
сай (Таджикская ССР) и в оз. Сон-куль (Киргизская ССР). От
О. vincentianum (Woodw.) и О. prashadi (Odhn.) этот вид отлича-
ется заметно меньшей выпуклостью створок и менее утолщенной
задней частью (Зв) зуба За + Зв. Вид, вероятно, ограничен в своем
распространении озерами Памира и Тибета (КНР).
Odhneripisidium prashadi (Odhner, 1937) (рис. 2)
Изучение паратипов этого вида, хранящихся в коллекции Зооло-
гического института АН СССР, и сравнение их с первоописанием
(Odhner, 1937) показало, что он никак не может быть объединен с
О. stewarti, поскольку резко отличается более выпуклой ракови-
ной. От О. vincentianum этот вид отличается почти так же, как и
обсуждавшийся выше О. stewarti, менее изогнутыми кардиналь-
ными зубами. Судя по изученным паратипам и изображениям, при-
веденным Уднером (Odhner, 1937), под этим названием первона-
55
Рис. 1 —10. Раковины видов подрода Odhneripisidium s. str
(вид справа, вид сзади и замок).
1 — О. (О.) steward, 2 — О. (О.) prashadi (паратип, про-
ба 104), 3 — О. (О.) kungejense (паратип О. prashadi, проба 186),
4 — О. (О.) dancei, 5 — О. (О.) chatyrkulense (голотип), 6 —
О. (О.) behningi (голотип), 7—О. (О.) gafurovi (голотип),
8—О. (О.) pamirense (голотип), 9—О. (О.) tolstikovae (голо-
тип—вид слева и сзади; замок правой створки—паратип),
10—О. (О.) kazakhstanicum (голотип; замок левой створки —
паратип).
Масштабные линейки — I мм (меньшая — для изображений раковин, боль-
шая — для изображений замка).
чально была описана из долины Дзачу (Меконга) на юге Цинхая
(КНР) смесь особей О. prashadi (проба 104 в цитируемой работе,
отсюда же происходит и голотип) и О. kungejense (проба 186).
Эти виды, однако, хорошо различаются взаимным расположением
кардинальных зубов на левой створке и общей формой раковины
(рис. 2 и 3). В частности, раковина О. prashadi несколько более
выпуклая, а раковина О. kungejense площе и по контуру более
приближается к треугольнику.
Оба вида распространены в Центральной Азии и сопредельных
районах достаточно широко. Кроме Цинхая (КНР), оба они отмечены
56
из горных районов Таджикской и Киргизской ССР (часто совместно)
и из Алтая — О. kungejense (под названием О. dancei) —
из Семипалатинска, Бухтарминска и Усть-Каменогорска (Kuiper,
1969), О. prashadi — из оз. Теньга в Горном Алтае (материал
Е. А. Новикова).
Odhneripisidium dancei (Kuiper, 1962) (рис. 4)
Экземпляры, достаточно точно соответствующие первоописанию
(только несколько меньших размеров), были обнаружены в Таджик-
ской ССР в родниках на Памире и в окрестностях Куляба и Кир-
гизской ССР в родниках на берегах озер Иссык-куль и Сон-куль,
а также (пустые раковины) в прибрежных отложениях и береговых
выбросах оз. Иссык-куль. От О. kungejense, с которым Кюйпер
объединял этот вид, он отличается более выпуклыми створками
и изогнутым зубом 4а + 4в (рис. 3 и 4). По-видимому, кроме Ирана
(откуда известен из провинций Керманшах и Луристан), вид встре-
чается в среднегорных и высокогорных районах юга и юго-востока
Средней Азии.
Odhneripisidium chatyrkulense Izz. et Star. sp. nov (рис. 5)
Раковина округло-овальная, мало вытянутая (высота составляет
около 0,90 длины), светлая, с широкими умеренно выступающими
макушками, отстоящими от переднего края на расстояние около
0,60 длины. Передний край раковины широко закругленный, задний
еще более широкий и слегка усеченный в средней части, спинной
равномерно закругленный и переходит в передний и задний края без
уголков, брюшной край равномерно и плавно закругленный. Наибо-
лее выступающая точка переднего края помещается несколько ниже
середины высоты раковины. Изменение выпуклости створки с уве-
личением высоты фронтального сечения выражаются следующими
величинами (первая цифра — высота фронтального сечения, вто-
рая— выпуклость одной створки, мм: 1—0,65; 1,5—0,8; 2—0,85;
2,5—0,9. Поверхность створки несет 4—5 резких линий нарастания,
а между ними более тонкие, едва заметные. Замочная площадка до-
вольно широкая. Зуб 2 равномерно изогнутый по всей длине, 4в
довольно короткий, прямой, 4а редуцирован, За и Зв образуют длин-
ный слабо изогнутый зуб, АП и РП почти одинаковой длины, но
последний выступает сильнее, AI длиннее АШ, последний почти
равен PIII. Лигаментная ямка короткая и довольно узкая.
Типовой материал: голотип и 59 паратипов. Голотип имеет эти-
кетку Киргизия, оз. Чатыр-куль, сбор Педашенко, 1906 г. Его раз-
меры (мм): длина раковины — 2,65, высота — 2,4; выпуклость одной
створки — 0,87 Паратипы происходят из того же сбора, а также с
Памира (оз. Каракуль и р. Кара-Джилга), с Нуратинского хребта
(у Джуша, Узбекская ССР) и из окрестностей Ургута (Узбекская
ССР).
57
Типовое местонахождение — оз. Чатыр-куль (Киргизская ССР)
Вид назван по типовому местонахождению.
Замечания Описываемый вид встречается вместе с О. pra-
shadi и О. kungejense, причем никаких переходных форм между
ними не наблюдается. От обоих названных видов он отличается
более вздутой раковиной, широкими макушками и мало изогнутыми
и тонкими кардинальными зубами.
Экология Живет в родниках, родниковых лужах и прибреж-
ных водоемах, у берегов озер на илисто-песчаном и илистом грунте.
Odhneripisidium behningi Izz. et Star. sp. nov. (рис. 6)
Раковина треугольно-овальная, умеренно вытянутая (высота
составляет около 0,8 длины), желтая, с широкими умеренно высту-
пающими макушками, отстоящими от переднего края на расстояние
около 0,8 длины. Передний край неравномерно закругленный, с силь-
но сглаженным углом в средней части, задний край усеченный и
несколько спрямленный в средней части, спинной край относительно
равномерно закруглен, в задний он переходит плавно, а при переходе
в передний образует закругленный тупой угол, брюшной край равно-
мерно закругленный и переходит в передний и задний края без угол-
ков. Наиболее выступающая точка переднего края находится не-
сколько ниже середины высоты раковины. Изменение выпуклости
створки с увеличением высоты фронтального сечения выражаются
следующими величинами (мм): 1—0,5; 1,5—0,65; 2—0,8. Поверх-
ность раковины с 4—5 резкими линиями нарастания и более тонкими
едва заметными между ними. Замочная площадка широкая. Зуб 2
изогнут под прямым углом, причем его задняя часть вдвое короче
передней, 4в прямой, 4а редуцирован, За слегка загнут дорсально
у переднего конца, Зв отогнут вентрально и расширен, АП крупный и
толстый, РП узкий, длинный и низкий, AI массивнее, чем AIII, PI
длиннее, чем РШ, AIII и РШ сравнительно тонкие и короткие. Лига-
ментная ямка короткая и относительно узкая.
Типовой материал: голотип и 32 паратипа. Голотип имеет эти-
кетку оз. Искандер-куль (северный склон Гиссарского хребта),
сбор А. Л. Бенинга, 23.7.1937 г. Его размеры (мм): длина раковины —
2,6; высота — 2,4; выпуклость одной створки — 0,84. Паратипы про-
исходят из того же сбора, что и голотип, а также из оз. Сон-куль
(Киргизская ССР).
Типовое местонахождение — оз. Искандер-куль (Таджикская
ССР).
Вид назван в память советского гидробиолога А. Л. Бенинга.
Замечания По контурам створки вид близок к О. kunge-
jense, но отличается большей выпуклостью, более узкой замочной
площадкой, угловатым передним краем и менее изогнутым зубом
За + 3в.
Экология Обитает в озерах и родниках на илистом грунте,
иногда совместно с О. prashadi на высотах более 2000 м над ур. м.
В конце июля и августе в жабрах встречается молодь в количестве
4—5 экз.
58
Odhneripisidium gafurovi Izz. et Stan sp. noV* (рис. 7)
♦
Раковина округлая, почти шаровидная, мало вытянутая (вы-
сота составляет около 0,9 длины), бледно-желтая, толстостенная,
с широкими мало выступающими макушками, отстоящими от перед-
него края раковины на расстояние около 0,7 длины. Передний край
несколько неравномерно закруглен, так что верхняя его часть менее
выгнута, задний край равномерно закругленный, спинной и брюшной
края равномерно закруглены и только при переходе от спинного к
переднему краю образуется уголок, да и то едва заметный. Наиболее
выступающая точка переднего края расположена несколько ниже
середины высоты раковины. Изменения выпуклости створки с уве-
личением высоты фронтального сечения выражаются следующими
величинами (мм): 1—0,5; 1,5—0,7; 2—0,8. Поверхность раковины с
4—5 резкими, неравномерно расположенными линиями нарастания
и более слабыми, едва заметными линиями между ними. Замочная
площадка умеренно широкая. Зуб 2 плавно изогнут, 4а + 4в образуют
почти прямой зуб, более длинный, чем 2, За + Зв образуют слабо
изогнутый зуб, слегка утолщенный на заднем конце, All и РП почти
одинаковой длины и одинаково выступают, AI заметно длиннее и
толще PI, АШ и PIII очень слабо развиты. Лигаментная ямка срав-
нительно короткая и узкая.
Типовой материал: голотип и 4 + 2 створки паратипов. Голотип
имеет этикетку: Таджикская ССР, восточный Памир, оз. Сулукты-
сай в 9 км от Кызыл-Рабата, сбор А. К. Гафурова 19.7.1974 г. Его
размеры (мм): длина раковины — 2,05; высота 2,0; выпуклость од-
ной створки — 0,75. Паратипы происходят из того же сбора и из
береговых выбросов оз. Кара-куль (Памир).
Типовое местонахождение — оз. Сулукты-сай (Таджикская ССР)
Вид назван в честь советского гидробиолога А. К. Гафурова.
Замечания Вид по общему облику похож на О. prashadi,
но отличается более вздутой и округлой раковиной, более выступаю-
щими макушками и слабо изогнутыми кардинальными зубами.
Экология Обитает в озерах на илистом грунте, на глубинах
1—2 м.
Odhneripisidium pamirense Izz. et Star. sp. nov. (рис. 8)
Раковина округло-треугольная, почти шаровидная, умеренно вы-
тянутая (высота составляет 0,75 длины), бледно- или золотисто-
желтая, толстостенная, с очень широкими выступающими макуш-
ками, отстоящими от переднего края раковины на расстояние около
0,6 длины. Передний край заметно неравномерно закруглен — в
верхней половине он выгнут значительно слабее, чем в нижней, зад-
ний край равномерно закругленный, спинной край равномерно вы-
гнутый и при переходе в передний и задний края образует закруглен-
ные тупые углы, брюшной край равномерно закругленный, перехо-
дящий в передний и задний края без уголков. Наиболее выступаю-
щая точка переднего края лежит на уровне границы нижней трети
высоты раковины, а наиболее выступающая точка заднего края —
59
на уровне середины высоты ракобины. Изменения выпуклости створ-
ки с увеличением высоты фронтального сечения выражаются следую-
щими величинами (мм): 1—0,55; 1,5—0,8; 2—0,9; 2,5—0,95. Поверх-
ность раковины с 3—4 резкими, неравномерно расположенными ли-
ниями нарастания и более слабыми, едва заметными линиями между
ними. Замочная площадка сравнительно широкая. Зуб 2 изогнут от-
носительно слабо, 4в прямой, тогда как 4а полностью редуцирован,
За + Зв — сильно изогнутый зуб, причем За изогнут слабо, а Зв резко
расширен и соединен с ним под углом, ПП и РП относительно ко-
роткие, сильно выступающие, AI и PI длинные, причем AI массивнее,
чем PI, АШ и РШ заметно более короткие. Лигаментная ямка ко-
роткая и относительно узкая.
Типовой материал: голотип и 43 паратипа. Голотип имеет эти-
кетку: Восточный Памир, остаточный водоем в долине р. Кара-
Джилга, сбор И. Э. Эргашбаева 1.9.1972 г. Его размеры (мм): длина
раковины — 2,65, высота — 2,50; выпуклость одной створки — 0,9.
Паратипы происходят из того же сбора, из оз. Кара-куль (выбросы),
оз. Кичик-куль и оз. Сулукты-сай (все три озера находятся на Па-
мире) .
Типовое местонахождение — долина р. Кара-Джилга (Таджик-
ская ССР).
Вид назван по региону, откуда он известен.
Замечания Описываемый вид по характеру замка и лига-
ментной ямки ближе всего к О. gafurovi, от которого отличается
более сильно выступающими макушками и менее изогнутым зубом 2.
От О. kungejense описываемый вид отличается значительно более
вздутой раковиной и менее изогнутыми кардинальными зубами.
Экология Обитает в озерах среди водорослей, на глубинах
около 1—2 м. Во второй половине августа у одной особи встречена
молодь в количестве 5—8 экз. на жабру.
Odhneripisidium tolstikovae Izz. et Star. sp. nov. (рис. 9)
Раковина четырехугольно-овальная, мало вытянутая (высота
составляет 0,86 длины), бесцветная, с мало выступающими макуш-
ками, отстоящими от переднего края раковины на расстояние около
0,6 длины. Передний край неравномерно закруглен — в верхних 2/3
он выгнут более слабо, чем в нижней 1/3, задний край равномерно
закругленный, спинной край равномерно выгнутый, переходит в
передний край плавно, а при переходе в задний край образует за-
кругленный тупой угол, брюшной край равномерно выгнутый и пере-
ходит в передний и задний края без уголков. Наиболее выступаю-
щая точка переднего края лежит на уровне границы нижней четвер-
ти высоты раковины, наиболее выступающая точка заднего края —
на уровне середины высоты раковины. Изменения выпуклости створ-
ки с увеличением высоты фронтального сечения выражаются сле-
дующими величинами (мм): 1—0,5; 1,5—0,65; 2—0,70; 2,5—0,80.
Поверхность раковины покрыта лишь очень тонкими линиями нара-
стания, более резкая линия заметна лишь без края. Замочная пло-
щадка довольно узкая. Зуб 2 изогнут под прямым углом, причем его
60
задняя часть много короче передней, 4а + 4в довольно слабо и равно-
мерно изогнут, причем 4а и 4в примерно одинаковой длины, За + Зв
сильно изогнут, из них Зв много короче и толще, АП и РП длинные,
почти одинаково выступающие, но РП несколько толще, AI и PI
длинные, заметно длиннее, чем AIII и РШ, причем последний наибо-
лее короткий. Лигаментная ямка короткая, но широкая.
Типовой материал: голотип (левая створка) и 10 паратипов (раз-
розненные створки). Голотип имеет этикетку: скважина у устья
р. Или, глубина 20 м, верхнеплиоценочные отложения, сбор Н. В. Тол-
стиковой, 1982 г Его размеры (мм): длина раковины — 2,55; высо-
та — 2,20; выпуклость одной створки — 0,60. Паратипы происходят
из того же сбора.
Типовое местонахождение— верхнеплиоценовые отложения у
устья р. Или (Казахская ССР)
Вид назван в честь советского палеонтолога Н. В. Толстиковой.
Сравнение По форме кардинальных зубов вид ближе всего
к О. kungejense, но отличается менее смещенными назад ма-
кушками и овально-четырехугольной формой раковины, этим же, а
также большей выпуклостью он отличается от О. dancei. От О. pra-
shadi описываемый вид отличается заметно меньшей выпуклостью.
Odhneripisidium kazakhstunicum Izz. et Star. sp. nov. (рис. 10)
Раковина коротко-овальная, мало вытянутая (высота составляет
0,87 длины), бесцветная, с широкими мало выступающими макуш-
ками, отстоящими от переднего края на расстояние, равное 0,6 дли-
ны. Передний край равномерно закругленный, задний также равно-
мерно закруглен, но менее сильно выгнут, спинной край равномерно
выгнут и переходит в задний плавно, а при переходе в передний обра-
зует едва заметный тупой угол, брюшной край равномерно закруглен
и переходит в передний и задний края без уголков. Наиболее вы-
ступающие точки переднего и заднего краев лежат на уровне сере-
дины высоты раковины. Изменения выпуклости створки с увеличе-
нием высоты фронтального сечения выражаются следующими вели-
чинами (мм): 1—0,5; 1,5—0,7; 2—0,75. Поверхность раковины с
4—5 резкими, неравномерно расположенными линиями нарастания,
и более тонкими, едва заметными линиями между ними. Замочная
площадка узкая. Зуб 2 изогнут под прямым углом, причем задняя
часть его очень маленькая, 4а + 4в длинный, умеренно изогнутый,
причем 4а и 4в примерно одинаковы по длине, За+Зв сильно изогнут,
Зв по длине почти равен За, но много толще и имеет треугольную
форму, АП массивный и длиннее, чем столь же толстый РП, AI и
PI примерно одинаковой длины и заметно длиннее, чем AIII и РШ,
последний из них самый короткий. Лигаментная ямка короткая и
довольно узкая.
Типовой материал: голотип (правая створка) и 11 паратипов
(разрозненные створки). Голотип имеет этикетку: скважина у устья
р. Или, глубина 20 м, верхнеплиоценовые отложения, сбор Н. В. Тол-
стиковой, 1982 г. Его размеры (мм): длина раковины — 2,00; высо-
61
та— 1,75, выпуклость одной створки — 0,45. Паратипы происходят
из того же сбора.
Типовое местонахождение — верхнеплиоценовые отложения у
устья р. Или (Казахская ССР)
Вид назван по республике, в пределах которой найден.
Сравнение От О. dancei и О. prashadi описываемый вид
отличается более плоской раковиной, а от О. kungejense к тому же
менее сильно изогнутыми кардинальными зубами. От О. tolstiko-
vae, с которым встречен совместно, вид отличается более округлы-
ми контурами створки и более широкими макушками.
Кроме 11 упомянутых или описанных выше видов, к этому подроду
относятся также О. janense Star, et Str. О. indigircense Star,
et Str. (Старобогатов и Стрелецкая, 1967) и О. paravincentianum
Pop. et Star. (Попова и др., 1970).
Подрод Tuvapisidium Izz. et Star. subg. nov.
Типовой вид: Odhneripisidium popovae Star, et Str., 1967
Раковина наиболее крупная в пределах рода (длина взрослых
особей 2,6—3,3 мм), высокая треугольная, гладкая покрытая только
линиями роста; зуб За + Зв резко изогнут (дугой или под прямым уг-
лом), зуб 2 изогнут под острым углом.
Odhneripisidium terekholicum Izz. et Star. sp. nov. (рис. 11)
Раковина округло-треугольная, мало вытянутая (высота состав-
ляет 0,95 длины) белая, толстостенная, с довольно узкими выступаю-
щими макушками, отстоящими от переднего края на расстояние,
равное 0,65 длины. Передний край выгнут неравномерно, его верх-
няя часть почти прямая, а нижняя резко выгнута, задний край равно-
мерно закругленный, спинной край сравнительно короткий, равно-
мерно выгнутый и при переходе к заднему и переднему краям обра-
зует отчетливые тупые углы, брюшной край равномерно закруг-
ленный и переходит в передний и задний края без уголков. Наиболее
выступающая точка переднего края лежит на уровне границы нижней
четверти высоты раковины, наиболее выступающая точка заднего —
на уровне середины высоты раковины. Изменения выпуклости створ-
ки с увеличением высоты фронтального сечения выражаются сле-
дующими величинами (мм): 1—0,5; 1,5—0,65; 2—0,8; 2,5—0,85.
Поверхность раковины с тонкими линиями нарастания и с 1—2 более
резко выраженными кольцами роста. Замочная площадка довольно
широкая. Зуб 2 изогнут под углом около 45°, 4а + 4в тонкий, прямой,
расположенный наклонно к передне-задней оси раковины. За + 3в об-
разуют подкововидный сильно изогнутый зуб, Зв несколько утолщен
и слегка раздвоен на конце, АП и РП одинаковой длины и высоты,
сильно выступающие, AI и АШ более длинные и мощные, чем PI
и РШ, АШ и РШ короче, чем AI и PI. Лигаментная ямка широкая.
Типовой материал: голотип (правая створка) и 16 паратипов
(разрозненные створки). Голотип и паратипы имеют этикетку: Ту-
62
Рис. 11 —18. Раковины видов подродов Tuvapisidium, Kuiperipisi-
dium и U ssuripisidium (вид справа, вид сзади и замок).
11—0. (Г.) terekholicum (голотип), 12 — О. (Г.) popovae (голотип),
13—О. (Г.) tuvaense (голотип), 14 — О. (/(.) sogdianum (голотип),
15— О. (/(.) polytimeticum (голотип), 16— О. (К.) terekense (голотип),
17 — О. (/<.) issykkulen.se (голотип), 18 — О. (О.) khorense (голотип).
Масштабные линейки — I мм (меньшая — для изображений раковины, большая —
для изображений замка).
винская АССР, оз. Терехоль, сбор С. М. Поповой 8.1963 г. Размеры
голотипа (мм): длина раковины — 2,65; высота — 2,5; выпуклость
одной створки — 0,8.
Типовое местонахождение — оз. Терехоль (Тувинская АССР).
Вид назван .по типовому местонахождению.
Сравнение От О. popovae (рис. 12), с которым описываемый
вид найден совместно, он отличается менее вздутой раковиной, под-
кововидной формой зуба За + Зв (у О. popovae он резко изогнут под
углом несколько меньшим, чем прямой, и Зв раздвоен значительно
сильнее) и относительно более широкой замочной площадкой.
Odhneripisidium tuvaense Izz. et Star. sp. nov. (рис. 13)
Раковина округло-теугольная, мало вытянутая (высота состав-
ляет 0,90 длины), белая, толстостенная, с относительно широкими
выступающими макушками, отстоящими от переднего края на рас-
стояние, равное 0,65 длины. Передний край выгнут неравномерно,
его верхняя часть изогнута более слабо, а нижняя более сильно,
задний край равномерно закругленный; спинной край сравнительно
63
короткий, равномерно выгнутый, плавно без уголков переходящий в
передний и задний края, брюшной край равномерно закругленный,
также без уголков переходящий в передний и задний края. Ниболее
выступающая точка переднего края лежит на уровне границы ниж-
ней трети высоты раковины, наиболее выступающая точка заднего
края — на уровне середины высоты раковины. Изменения выпуклости
створки с увеличением высоты фронтального сечения выражаются
следующими величинами (мм): 1—0,7; 1,5—0,75; 2—0,85; 2,5—0,90.
Поверхность раковины с 3—4 резкими неравномерно расположен-
ными линиями нарастания и более тонкими едва заметными линиями
между ними. Замочная площадка умеренно широкая. Зуб 2 подково-
видно изогнут, почти полукруглый, 4а + 4в изогнут слабо и заметно
утончается к переднему концу, так что неясно представлена тут часть
4а или она редуцирована полностью, зуб За + Зв плавно изогнутый,
утолщенный на обоих концах, причем Зв более сильно утолщен,
но не раздвоен, АН и РП очень мощные и довольно длинные, причем
АП несколько длиннее, AI и PI мощные, примерно одинаковой длины,
причем PI более массивен и несколько отогнут на конце внутрь рако-
вины, АШ и РШ тонкие, почти вдвое короче, чем AI и PI, причем
РШ короче, чем АШ. Лигаментная ямка короткая и широкая.
Типовой материал: голотип (правая створка), 7 паратипов (раз-
розненные створки) и еще 3 паратигга (целые раковины). Голотип
и первые 7 паратипов имеют этикетку: Тувинская АССР, оз. Тере-
холь, сбор С. М. Поповой 8.1963 г. Размеры голотипа (мм): длина
раковины — 2,75; высота — 2,60; выпуклость одной створки — 0,85.
Три остальных паратипа имеют этикетку: Алтайский край, Горно-
Алтайская А. О., Кош-Агачский район, оз. Камышистое близ Кош-
Агача, сбор Е. А. Новикова 1965 г.
Типовое местонахождение — оз. Терехоль (Тувинская АССР).
Вид назван по республике, в пределах которой находится типо-
вое местонахождение.
Сравнение От обоих предыдущих видов подрода, с которыми
он встречен вместе, описываемый вид отличается заметно более ши-
рокими макушками, плавно изогнутым и расширенным на обоих кон-
цах зубом За + Зв, полукруглым зубом 2 и более уплощенной рако-
виной.
Кроме трех описанных или упомянутых выше видов, к этому под-
роду относятся также 4 вида из неогеновых или плейстоценовых
отложений Чуйской котловины Горного Алтая (Попова и др., 1970):
О. altaicum Pop. et St., О. trigon Pop. et St., O. tshuicum V+p.
et St., О racovetzae Pop. et St.
Подрод Kuiperipisidium Izz. et Star. subg. nov.
Типовой вид: Odhneripisidium sogdianum Izz. et Star. sp. nov.
Раковина маленькая (длина до 2,5 мм), овальная, покрытая
только линиями роста, зуб За длинный, почти прямой, а соединенный
с ним зуб Зв короткий, отогнутый вентрально, утолщенный и раз-
двоенный, зуб 2 с короткой отогнутой вентрально задней частью и
длинной прямой передней.
64
Экземпляры первых трех из четырех описываемых ниже видов
этого подрода, судя по опубликованным промерам, были в распоря-
жении Кюйпера (см. Kuiper, 1926b) из Италии, Балканского полу-
острова и Передней Азии. Всех их он отождествил с О. annandalei
(Prash), описанным из Индии. Мы не согласны с таким отождест-
влением, прежде всего потому, что последний вид резко отличается
строением замка (зубы 2 и 4а + 4в прямые и параллельные друг другу)
и заслуживает выделения в самостоятельный подрод.
Odhneripisidium sogdianum Izz. et Star. sp. nov. (рис. 14)
Раковина коротко-овальная, мало вытянутая (высота составляет
0,85 длины), бледно-желтая, тонкостенная, но прочная, с относитель-
но широкими выступающими макушками, отстоящими от перед-
него края на расстояние, равное 0,57 длины. Передний край выгнут
несколько неравномерно, его верхняя треть слегка спрямлена, зад-
ний край равномерно закругленный, спинной край короткий, равно-
мерно выгнутый, не образующий уголков при переходе в передний и
задний края, брюшной край равномерно закругленный, также без
уголков переходящий в передний и задний края. Наиболее высту-
пающие точки переднего и заднего краев лежат почти на уровне се-
редины высоты раковины. Изменения выпуклости створки с увеличе-
нием высоты фронтального сечения выражаются следующими вели-
чинами (мм): 1—0,55; 1,5—0,65; 2—0,80. Поверхность раковины по-
крыта тонкими и довольно частыми линиями нарастания. Замочная
площадка узкая. Зуб 2 изогнут под очень тупым углом, причем
задняя часть его втрое короче передней, 4а+4в изогнут почти так же
и 4в вдвое короче 4а, зуб За прямой, слегка наклоненный вентрально
передним концом, соединенный с ним зуб Зв несколько толще, много
короче и сильно наклонен задним концом вентрально, АП и РП длин-
ные тонкие, более выступающие близ концов, AI и PI длинные, тон-
кие, причем PI заметно длиннее, чем AI, АШ и РШ тонкие маленькие
почти вдвое короче, чем AI и PI. Лигаментная ямка довольно узкая.
Типовой материал: голотип и 16 паратипов. Все они имеют эти-
кетку: Узбекская ССР, северные склоны Туркестанского хребта,
Джизах, Бахмал в роднике, 22.5.1979, сбор 3. И. Иззатуллаева.
Размеры голотипа (мм) длина раковины — 2,0; высота— 1,7; вы-
пуклость одной створки — 0,7
Кюйпер имел 1 экз. этого вида из Италии (Себето близ Неаполя),
материал из коллекции Британского музея естественной истории
№ 196276. Судя по промерам, отдельные экземпляры этого же вида
были найдены А. Д. Алиевым (1961) в ручье и источнике между
Бардой и Агдамом (Азербайджанская ССР).
Типовое местонахождение: Бахмал на северном склоне Турке-
станского хребта (Узбекская ССР).
Вид назван по античному названию оазисных районов Узбеки-
стана — Согд или Согдиана.
Сравнение по общим контурам раковины вид похож на
О. annandalei, но отличается изогнутыми кардинальными зубами
левой створки и заметно более вздутой раковиной.
5
Заказ 1696
65
Экология Живет в родниках на илистом и илисто-песчаном
грунте. Известен пока только из среднегорья.
Odhneripisidium polytimeticum Izz. et Star. sp. nov. (рис. 15)
Раковина овально-четырехугольная, мало вытянутая (высота
составляет 0,97 длины), бледно-желтая, тонкостенная, но прочная, с
относительно широкими умеренно выступающими макушками, от-
стоящими от переднего края на расстояние, равное 0,58 длины. Пе-
редний край выгнут заметно неравномерно — в верхней половине он
почти прямой, в нижней — закругленный, задний край равномерно
закругленный, спинной край слабо выгнутый, образует четкий тупой
угол при переходе к переднему и более закругленный, мало заметный
при переходе к заднему, брюшной край равномерно закругленный,
без уголков переходящий в передний и задний края. Наиболее вы-
ступающая точка переднего края лежит немного выше границы ниж-
ней трети высоты раковины, наиболее выступающая точка заднего
края лежит на уровне середины высоты раковины. Изменения вы-
пуклости створки с увеличением высоты фронтального сечения вы-
ражаются следующими величинами (мм): 1—0,50; 1,5—0,60; 2—0,70;
2,5—0,80. Поверхность раковины покрыта тонкими и довольно часты-
ми линиями нарастания. Замочная площадка узкая, однако не-
сколько шире, чем у предыдущего вида. Зуб 2 в задней части плавно
отогнут вентрально, 4а прямой, параллельный передней части зуба
2, 4в втрое короче, чем 4а, и несколько загнут вентрально, зуб За + 3в
почти прямой, очень слабо изогнутый на переходе от За к Зв, послед-
ний заметно утолщен и слегка раздвоен, АП и РП довольно мощные,
умеренной длины, AI и PI такой же длины как АП и РП, но заметно
тоньше, тогда как АШ и PIII очень тонкие и вдвое короче остальных
латеральных зубов. Лигаментная ямка умеренно узкая.
Типовой материал: голотип и 9 паратипов. Голотип имеет эти-
кетку: Узбекская ССР, Нуратинский хребет лишлак Манишкор у
Джуша, родник, сбор 3. И. Иззатуллаева 14.5.1976 г. Его размеры
(мм): длина раковины — 2,2; высота — 2,1; выпуклость одной створ-
ки — 0,7 Паратипы происходят из того же сбора, из ущелья Ход-
жакон на северном склоне хребта Кугитанг, Туркменская ССР из
родника близ Бахмала, Джизах, северный склон Туркестанского
хребта, Узбекская ССР из родника Чиличор-Чашма, Шартуз, Тад-
жикская ССР, из болотца в пойме близ Мегри, Армянская ССР
В распоряжении Кюйпера были 5 сборов, содержащих этот вид:
1) Италия, Сицилия, Фиуме Чиане, 2) Греция, Эпир, материал из
коллекции Зенкенбергского музея № 153270, 3) оттуда же и из той
же коллекции, № 155885, 4) Иран, Дастгирд, провинция Исфаган,
коллекция Национального музея естественной истории, Париж,
5) Иран, Доулетабад, провинция Керманшах, коллекция того же
музея.
Типовое местонахождение: родник у кишлака Минишкор в Нура-
тинском хребте.
Вид назван по античному названию реки Зеравшан — Поли-
тиметис.
66
Сравнение От предыдущего описываемый вид отличается
менее вздутой раковиной, более резко изогнутым зубом 2 и заметно
менее резко изогнутым, но более сильно утолщенным на заднем
конце зубом За + Зв.
Экология Живет в родниках, а также в лужах и болотцах,
питаемых родниками в среднегорье, реже на равнине.
Odhneripisidium terekense Kazannikov sp. nov. (рис. 16)
Описание вида составлено Е. А. Казанниковым и публикуется
здесь с его любезного согласия.
Раковина овально-четырехугольная, мало вытянутая (высота
составляет 0,90 длины), белая, тонкостенная, но прочная, с довольно
узкими умеренно выступающими макушками, отстоящими от перед-
него края на расстояние, равное 0,60 длины. Передний край выгнут
заметно неравномерно.— верхняя треть его почти прямая, а нижние
2/3 закруглены, задний край равномерно закругленный, спинной
край очень слабо выгнутый и слегка наклоненный назад, в передний
и задний края он переходит с довольно отчетливыми, но закруглен-
ными тупыми уголками, брюшной край равномерно закругленный,
переходящий в передний и задний края без уголков. Наиболее вы-
ступающая точка переднего края лежит немного ниже середины
высоты раковины, наиболее выступающая точка заднего — на уровне
середины высоты раковины. Изменения выпуклости створки с уве-
личением высоты фронтального сечения выражаются следующими
величинами (мм): 1—0,3; 1,5—0,37; 2—0,40. Поверхность раковины
покрыта тонкими и довольно частыми линиями нарастания. Замоч-
ная площадка узкая. Зуб 2 согнут под тупым углом, причем задняя
часть его вдвое короче передней, 4а + 4в слабо согнут, причем обе его
части примерно одинаковой длины, зуб За + Зв согнут под тупым уг-
лом, причем Зв втрое короче и заметно толще, чем За, АП и РП до-
вольно мощные и длинные (на левой створке голотипа, в порядке
аномалии, задних латеральных зубов 2, из которых внешний много
короче и тоньше), AI и PI длинные и тонкие, АШ и РШ вдвое короче,
чем AI и PI и много тоньше (у голотипа в правой створке только
один задний латеральный зуб). Лигаментная ямка узкая и довольно
короткая.
Типовой материал: голотип и свыше 100 паратипов. Голотип
имеет этикетку: Северо-Осетинская АССР, оз. Бекан в окрестностях
с. Карджин, сбор Е. А. Казанникова и Я. И. Старобогатова
26.11.1972 г. Его размеры (мм): длина раковины— 1,55; высота —
1,40; выпуклость одной створки — 0,40. Паратипы происходят из того
же сбора, из родникового ручья у с. Карджин в той же республике,
из родника у Ширванли, Бардинский район Азербайджанской ССР,
из болотца в пойме близ Мегри Армягская ССР, из Кара-су в Талас-
ской долине, Киргизская ССР, из поймы р. Обимазор близ Сари-
хосор, Таджикская ССР, из Бахмала на северном склоне Турке-
станского хребта, Узбекская ССР
В распоряжении Кюйпера были 4 сбора, содержащих этот вид:
1) Греция, о. Корфу, ручей близ Сидари, 2) Сирия, Хомс, р. Оронт
5*
67
близ Эль-Мимас, 3) Иран, окрестности г. Керманшах, коллекция
Национального музея естественной истории, Париж, 4) Иран, Би-
сутун, провинция Керманшах, коллекция того же музея.
Типовое местонахождение: оз. Бекан близ Карджина Северо-
Осетинская АССР
Сравнение От двух предыдущих видов описываемый вид от-
личается еще менее вздутой раковиной, почти равной длиной частей
зуба 4а + 4в и относительно коротким зубом 2.
Экология Живет в родниках, прудах, озерах и речках, но в
последних трех случаях всегда вблизи выхода родников. Молодь в
жабрах до 11 экз. на особь.
Odhneripisidium issykkulense Izz. et Star. sp. nov. (рис. 17)
Раковина овально-четырехугольная, мало вытянутая (высота
составляет 0,90 длины), белая, тонкостенная, но прочная, с довольно
широкими слабо выступающими макушками, отстоящими от перед-
него края на расстояние, равное 0,63 длины. Передний край выгнут
заметно неравномерно, верхняя треть его почти прямая, а нижние
2/3 закруглены, задний край почти равномерно закругленный и лишь
в средней части слегка усеченный, спинной край очень слабо выгну-
тый и почти параллельный продольной оси тела, при переходе к пе-
реднему краю он образует отчетливый тупой угол, а при переходе
к заднему — едва заметный закругленный уголок, брюшной край
равномерно закругленный, переходящий в передний и задний края
без уголков. Наиболее выступающая точка переднего края лежит на
уровне границы нижней трети высоты раковины, наиболее выступаю-
щая точка заднего — слегка ниже середины высоты раковины. Из-
менения выпуклости створки с увеличением высоты фронтального
сечения выражаются следующими величинами (мм): 1—0,25; 1,5—
0,40; 2—0,60; 2,5—0,80. Поверхность раковины покрыта тонкими и
довольно частыми линиями нарастания. Замочная пллощадка узкая.
Зуб 2 плавно дуговидно согнут, 4а + 4в изогнут под тупым углом в
средней части и обе его части примерно одинаковой длины, зуб
За + Зв дуговидный, утолщенный на обоих концах, причем на заднем
заметно сильнее, АП и РП довольно длинные, причем АП заметно
длиннее, чем РП, AI и PI очень длинные и тонкие, а АШ и РШ вдвое
короче их и также очень тонкие. Лигаментная ямка очень узкая и
сравнительно короткая.
Типовой материал: голотип и 8 паратипов. Голотип имеет эти-
кетку: Киргизская ССР, оз. Иссык-Куль, Тонский залив, выбросы
у пос. Кутарга, сбор 3. И. Иззатуллаева 26.7.1980 г. Его размеры
(мм): длина раковины — 1,9; высота — 1,7; выпуклость одной створ-
ки — 0,45. Паратипы происходят из того же сбора, а также с хребта
Хазарати-Шо, Сари-Хосор, пойма р. Оби-Мазор, выбросы, Пянджи-
кент, пойма р. Зеравшан, родник. Оба последних сбора из Таджик-
ской ССР.
Типовое местонахождение: Тонский залив оз. Иссык-Куль и род-
ники на его берегу (Киргизская ССР).
Вид назван по оз. Иссык-Куль.
68
Сравнение Этот вид обладает наиболее уплощенной рако-
виной в пределах подрода, чем и отличается от всех предыдущих
видов. От наиболее сходного с ним О. terekense описываемый
вид отличается более коротким зубом 2, и равномерно изогнутым
зубом За + Зв, утолщенным на обоих концах.
Экология Живет в родниках и в Иссык-Куль, по-видимому,
занесен из какого-то родника. Возможно также, что вид живет в Тон-
ском заливе вблизи выхода родника.
Подрод Ussuripisidium Izz. et Star. subg. nov.
Типовой вид Odhneripisidium khorense Izz. et Star. sp. nov.
Раковина маленькая (длина до 2 мм), овально-четырехуголь-
ная, тонкоребристая. Зубы За + Зв, 4а + 4в и 2 равномерно и довольно
слабо изогнутые.
Odhneripisidium khorense Izz. et Star. sp. nov. (рис. 18)
Раковина овально-четырехугольная, мало вытянутая (высота
доставляет 0,89 длины), серая, тонкостенная, с узкими умеренно
выступающими макушками, отстоящими от переднего края на рас-
стояние, равное 0,70 длины. Передний край выгнут неравномерно,
его верхние 2/3 выгнуты заметно слабее, чем нижняя треть, задний
край равномерно закругленный, спинной край слабо выгнутый, пе-
реходящий в передний и задний края с едва заметными закруглен-
ными уголками, брюшной край довольно слабо выгнутый и плавно
переходит в задний край, а при переходе в передний образует едва
заметный уголок. Наиболее выступающая точка переднего края ле-
жит на уровне границы нижней четверти высоты раковины, наиболее
выступающая точка заднего — на уровне середины высоты раковины.
Изменения выпуклости створки с увеличением высоты фронтального
сечения выражаются следующими величинами (мм): 1—0,45; 1,5—
0,55; 2—0,60. Поверхность раковины покрыта очень тонкими пластин-
чатыми ребрышками и еще более тонкими линиями нарастания,
среди которых выделяются 2—3 более резких. Замочная площадка
узкая. Зуб 2 дуговидно изогнутый, 4а + 4в также плавно изогнут, при-
чем его передний конец находится несколько впереди середины зуба
2, зуб За + Зв изогнут более резко, его передняя часть вдвое длиннее
задней, а последняя сильно утолщена, почти треугольной формы и
слегка раздвоена на заднем конце, АП и РП довольно длинные и
сильно выступающие, причем РП более мощный, чем АП, AI и PI
тонкие и более длинные, чем АП и РП, АШ и РШ очень тонкие; АШ
втрое короче, чем AI, а РШ—лишь вдвое короче чем PI. Лига-
ментная ямка узкая.
Типовой материал: голотип и 1 паратип. Голотип имеет этикетку:
Хабаровский край, р. Хор, родник, сбор И. М. Леванидовой 8.1963 г.
Его размеры (мм): длина раковины— 1,90; высота— 1,70, выпук-
лость одной створки — 0,60. Паратип происходит из родника над
пос. Раздольное Приморского края.
69
Типовое местонахождение: нижнее течение р. Хор (Хабаровский
край).
Вид назван по р. Хор.
Сравнение От всех описанных выше видов отличается прежде
всего тонкой ребристостью поверхности раковины.
Экология Обитатель родников.
ЛИТЕРАТУРА
Акрамовский Н. Н. Моллюски (Mollusca). Фауна Армянской ССР. Ереван, 1976,
268 с.
Алиев А. Д. Новое живое ископаемое.— Природа, 1961, № 2, с.
Бутенко Ю. В. К фауне пресноводных моллюсков южного и юго-восточного Казах-
стана. В кн.: Моллюски и их роль в биоценозах и формировании фаун. Тр. Зоол.
ин-та АН СССР, Т7 42, Л, 1967, с. 205—212.
Пирогов В. В., Старобогатов Я. И. Мелкие двустворчатые моллюски семейства
Pisidiidae ильменя Большой Карабулак в дельте Волги.— Зоол. журн., 1974,
т. 93, вып. 3, с. 325—336.
Попова С. М., Девяткин Е. В., Старобогатов Я. И. Моллюски кызылгирской свиты
Горного Алтая. М., 1970, 95 с.
Старобогатов Я. И., Стрелецкая Э. А. Состав и зоогеографическая характеристика
пресноводной малакофауны Восточной Сибири и севера Дальнего Востока.—
В кн.: Моллюски и их роль в биоценозах и формировании фаун. (Тр. Зоол. ин-та
АН СССР, т. 42), Л., 1967, с. 221—268.
Kuiper J.-G.-J. Note sur la systematique des pisidies.— Journ. de Conchyliol.,
1962a, vol. 102, p. 53—57.
Kuiper J.-G.-J. Etude critique de Pisidium vincentianum.— Bull. Inst. roy. sci.
natur. de Belgique, 1962b, t. 38, N 46, p. 1 — 19.
Kuiper J.-G.-J. Pisidien aus Kasakstan, Sibirien.—Arch. Molluskenk, 1969, Bd 99,
H. 1/2, S. 49—53.
Odhner N. Hj. Pisidium prashadi n. sp., eine neue Hochgebirgsmuschel aus Thibet.—
Arch. Moluskenk., 1937, Bd 69, N 3, S. 117—123.
Preston H. B. Report on a small collection of fresh-water Mollusca (Limnaea
and Pisidium) from Tibet.— Rec. Indian mus., 1909, vol. 3, pt. 2, p. 115—116.
Z. I. Izzatullaev and Y. I. Starobogatov
ODHNERIPISIDIUM SPECIES FROM THE FAUNA OF THE USSR
(BIVALVIA PISIDIIDAE)
The descriptions of 13 new species are presented (among them one has described
by E. A. Kazannikov): O. chatyrkulense, O. behningi, 0. gafurovi, 0. pami-
rense, O. tolstikovae, O. kazakhstanicum (two latter from Neogene deposits),
O. terekholicum, O. tuvaense, O. sogdianum, 0. polytimeticum, O. te-
rekense Kazannikov, O. issykkulense, O. khorense. All species known
from the USSR are divided in 4 subgenera: Ussuripisidium subg. nov. with type
and sole species 0. khorense, Kuiperipisidium subg. nov. with type-species
0. sogdianum and also with 0. polytimeticum, O. terekense, 0. issykku-
lense, Tuvapisidium subg. nov. with type-species O. popovae and also with
0. terekholicum, O. tuvaense and all species described by S. M. Popova
and Y. I. Starobogatov (1970) from the Kyzylgirian beds of Altai mountains (except
O. paravincentianum) and nominotypical subgenus Odhneripisidium s. str. inclu-
ding all remaining species of the genus from the USSR. The distinctions of species
O. stewarti, O. prashadi and O. dancei and their distribution are refined.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
E. С. Аракелова
ИНТЕНСИВНОСТЬ ОБМЕНА У БРЮХОНОГИХ МОЛЛЮСКОВ
К настоящему времени в литературе накоплен огромный материал
по дыханию пойкилотермных животных; в частности, показана связь
между энергетическим обменом водных беспозвоночных и их массой.
Эта связь, как правило, выражается степенной зависимостью и мо-
жет быть описана уравнением типа Q=aWk, где Q — скорость энерге-
тического обмена, выраженная в потреблении кислорода животным
определенного размера за единицу времени (СПК), W — масса жи-
вотного, а и k — коэффициенты уравнения.
При сравнении результатов исследований, посвященных изуче-
нию дыхания животных, видно, что «ни температура, ни какой-либо
из других факторов, влияющих на интенсивность обмена, не могут
идти ни в какое сравнение с размерами тела в отношении амплитуды
вызванных ими различий интенсивности обмена» (Винберг, 1976,
с. 56). Имеются сводки и обобщения по скорости обмена у различных
групп водных пойкилотермных животных, широко используются
уравнения зависимости скорости обмена от массы тела для рако-
образных (Винберг, 1950; Сущеня, 1972), пресноводных двуствор-
чатых моллюсков (Алимов, 1975, 1981), инфузорий (Хлебович, 1974).
свободноживущих плоских и кольчатых червей (Камлюк, 1974). Ши-
рокому кругу вопросов, связанных с дыханием различных водных
беспозвоночных, посвящена обзорная работа (Виленкин, Виленкина,
1979). В отношении пресноводных брюхоногих моллюсков до сих пор
нет работы, обобщающей накопленный экспериментальный материал
и в общем виде описывающей зависимость скорости дыхания от
массы. Оценка интенсивности дыхания пресноводных гастропод по
опытным данным приведена в работе Г Г Винберга и Ю. С. Беляцкой
(1958), в которой исследованы 7 видов легочных и 3 вида передне-
жаберных гастропод. За последние годы опубликованы исследования,
содержащие данные по скорости обмена у пресноводных гастропод,
результаты которых частично обсуждались в работе Кэлоу (Calow,
1975).
Целью данной работы было на основе собственного материала
и литературных данных провести сравнения уровней обмена у ле-
гочных и переднежаберных моллюсков, а также оценить средний
уровень энергетического обмена водных гастропод в пределах всего
класса.
В большинстве работ показателем уровня метаболизма животных
служит скорость потребления ими кислорода, измеряемая на живот-
ных, не ограниченных в свободном перемещении, т. е. когда состояние
покоя перемежается с их спонтанной активностью. Уровень энер-
гетического обмена таких животных, часто обозначаемый как «ру-
71
Рис. 1. Зависимость СПК переднежаберных моллюсков от массы при 20° С.
Обозначения линий регрессий и точек для каждого вида соответствуют порядковым номерам табл. I
Пунктиром проведены линии, обозначающие максимальный и минимальный уровни обмена у брюхоногих
моллюсков (по Винбергу и Беляцкой). Ось абсцисс — UZ, г сырого тела; ось ординат — СПК. млО-ч’1
при 20° С (Q).
тинный» метаболизм, по существу является аналогом стандартного
метаболизма, который и рассматривается нами в данной работе.
На рис. 1 представлены собственные и литературные материалы
по дыханию пресноводных и морских переднежаберных моллюсков.
Эти данные получены при использовании разных методов: мано-
метрического, метода проточной воды, радиоуглеродного, поляро-
графического, метода замкнутых сосудов. Условные границы, в пре-
делах которых укладываются почти все приведенные данные, по-
строены по данным Винберга и Беляцкой (1959) и соответствуют
максимальной (а=0,31 млО- ч-1* г-1 сырого тела при 20°) и минималь-
ной (а=0,05) интенсивности обмена у брюхоногих моллюсков при
£=0,75. Как-видно из рис. 1, лишь в двух случаях линии регрессий
оказались за пределами нижней границы, условно принятой за мини-
мальный уровень обмена у гастропод. Эти линии рассчитаны по урав-
нениям для Nassarius reticulatus (Crisp et al., 1978) и Thais
lapillus (Bayne, S^ullard, 1978). В первом случае в опытах
использовали голодающих животных, во втором — уравнение было
выведено со средним значением степенного коэффициента £=0,511,
тогда как по данным Бэйна и Скалларда пределы колебаний зна-
чений этого коэффициента довольно значительны (£=0,34—0,96)
и зависят от различных условий. Диапазон исследованных весов
72
в обоих случаях составляет 2 порядка величин — 0,0086—0,996 г
для A. reticulatus и 0,003—0,4 г для Т lapillus (веса
рассчитаны для сухих тканей моллюсков), и, надо полагать, что
центры равновесия линий регрессий, которым соответствуют значения
среднего веса ш, находятся в пределах рассматриваемых нами границ.
При сравнении данных табл. 1 видно, что значение коэффи-
циента регрессии k в уравнениях, описывающих скорость энергети-
ческого обмена у разных видов моллюсков, колеблется. Некоторые
авторы считают, что изменчивость k и отклонения его от наиболее
часто встречающейся средней й=0,75 носят адаптивный характер
(Berg, Lumbye, Ockelmann, 1958). Исследования дыхания 12 видов
брюхоногих моллюсков показали, что уравнения зависимости ско-
рости обмена от массы не имеют постоянных параметров и меняются
по сезонам в довольно больших пределах (Berg, Ockelmann, 1959).
Коэффициент регрессии для 12 изученных видов колебался от 0,75
до 1,0. Бранд с соавторами (Brand et al., 1948) объясняют колебания
k различной степенью интеграции органов дыхания, кровеносной
системы, нервной регуляции. Для переднежаберного морского мол-
люска Murex pomum в уравнении зависимости СПК от массы при
нормальных условиях — 27° С и 34%0 — рассчитано высокое значе-
ние й, равное 0,97 (Sander, Moore, 1978). Эти моллюски относятся
к активным хищникам. Переход от седентарного или малоподвиж-
ного придонного образа жизни к высокоподвижному, связанному
с активным плаванием и поиском пищи, некоторые авторы (Бело-
копытин, 1968, 1969; Dykens, Mangum, 1979) считают одной из воз-
можных причин возрастания скорости обменных процессов. Так, в
опытах по дыханию с двустворчатым моллюском Mercenaria mer-
cenaria и двумя видами кальмаров было показано, что при темпе-
ратуре 23° С интенсивность дыхания кальмаров превышает интен-
сивность дыхания двустворчатых моллюсков более чем в 20 раз
(Dykens, Mangum, 1979). Кэлоу (Calow, 1974) на некоторых видах
моллюсков показал, что активность, вызываемая поиском пищи, со-
провождается повышением СПК. Противоположный результат был
получен для Nassarius reticulatus (Crisp, 1979), у которых
активное состояние достигалось введением глицерина, вызывающего
у животных двигательную поисковую реакцию. Было показано, что
активный метаболизм у этих моллюсков не превышает метаболизм
покоя, максимальные его различия не более 20%. Исследуя дыха-
ние литоральных гастропод в связи с воздействием кратковременных
колебаний температуры, Ньюэлл и Норкрафт (Newell, Northcroft,
1967) сделали вывод о том, что колебания СПК вызваны изменениями
уровня активности, которая, в свою очередь, изменяется под прямым
воздействием температуры. На четырех видах средиземноморских
литоральных гастропод показано, что СПК увеличивалась линейно
с увеличением скорости ползания и превышала СПК неактивных
особей в 3 раза (Houlihan, Innes, 1982).
Возможно, что амфибионтная улитка Marisa cornuarietis
(сем. Ampullariidae) отличается наиболее высоким уровнем энер-
гетического обмена (сравнительно с другими пресноводными перед-
нежаберными), благодаря активному образу жизни. Демиан (De-
73
Результаты измерений скорости потребления кислорода у переднежаберных моллюсков
Таблица 1
№ п/п Виды t°c СПК W, г Единицы измерений СПК и массы моллюска * Автор, СПК при 20° С** Примечания
1 Nassarius reticu- latus 16 0,199 HZ0-95 0,0086—0,996 1 Crisp, Davenport, Shumway, 1978 0,069 UZ0-95 UZcyx=0,25UZCbip**
2 Nassarius reticu- lataus, Littorina littorea 20 0,179 HZ0-74 0,002—1,0 2 Zeuthen, 1947 0,192
3 Marisa sp. 30 0,432 1,0 2 Horne, 1979 0,192
4 Thais lapillus — 0,167 HZ0-5" 0,003—0,40 1 Bayne, Scullard, 1978 0,042 UZ° •' летние измерения; UZcyx=0,25UZCbip***
5 6 Littorina irrorata Patella aspera 20 20 20 min 0,308 max 1,323 0,125 UZ0-60 1,0 1,0 1 1 2 Shirley, Denoux, Stickle, 1978 Davies, 1966 min 0,077 max 0,330
7 Patella vulgata 20 20 0,086 HZ0,73 0,093 IT78 — 2 2 Davies, 1966 верхнелитораль- ные нижнелито-
8 Thais lapillus 18 0,082 0,055 1,24 1,24 2 Sandison, 1966 0,065 ральные летом
9 Rapana thomasia- na 20 0,046 UZ078 3—169,2 3 Муравская, Гал- кина, 1979 0,198 HZ0-78 U7Te.la=0,233UZ
10 Valvata piscinalis 23 14 0,66 UZ0-95 0,25 UZ0-98 до 0,031 2 2 Китицына, 1971 » 0,497 UZ0-95 0,390 UZ0-98 UZpaK=0,74UZ
11 Viviparus vivipa- rus — 0,105UZ0,77 0,02—6,00 3 Китицына, 1975 O.175UZ0-77 UZpaK=0,40UZ
12 Marisa cornuari- 20 0,98 UZ0-79 — 1 Akerlund, 1969 0,221 HZ0-79 UZc;x=0,226UZ
etis 20 20 0,773 0,026 3,09 0,044 2 2 » » 0,773 0,026
13 Pila ovata 22 0,39 0,800 1 Coles, 1968 0,416 UZc>x=0,20UZ
14 Cellana tramose- rica 20 20 0,558 UZ0-71 0,246 UZ0,71 — 1 1 Раггу, 1978 » 0,139 Hz0-71 0,061 HZ0-71 UZcyx=0,25UZ
15 Viviparus conte- ctoides 0,117 0,120 0,122 0,089 0,116 0,151 1,14 1,21 1,21 2,45 1,82 1,82 3 Fitch, 1975 СПК у 9 и cf при разных уровнях ак- тивности
16 Viviparus vivipa- 20 0,300 0,05 3 Ковалева, 1970 0,015 СПК рассчитаны
rus 0,210 0,10 0,021 на 1 г (графа 4)
0,160 0,15 0,024
17 Valvata piscinalis 20 0,310 0,04 3 Ковалева, 1970 0,0124 СПК рассчитаны
0,220 0,09 0,0198 автором на 1 г
0,170 0,14 0,0238
18 Thais lamellosa Viviparus conne- 10 0,12—0,13 1,0 I Stickle, 1973 (цит. no: Baune, Scul- lard, 1978) 0,281 U7cy4=0,25U7cup***
19 20— 0,127 1,0 3 Studer, Pace, 1978
ct aides -25 0,231 1,0 3
20 Theodoxus fluvia- 20 0,0058 0,030 3 Lumbye, 1958 U7paK=0,30U7 ***
tilts 0,0022 0,009
21 Potamopyrgus 20 0,00045 0,0025 3 Lumbye, 1958 U7paK=0,60U7 ***
jenkinsi 0,001 0,005
22 Potamopyrgus 18 0,0008 0,005 3 Berg, Ockelmann, 0,00094 UZpaK=0,60U7 ***
jenkinsi 0,0007 0,005 1958 0,00082
0,0004 0,0025 0,00047
0,0015 0,007 0,0018
23 Valvata piscinalis 18 0,003 0,014 3 0,0035 U7paK=0,74U7 (no: Китицына, 1971)
24 Bithnia tentacula- 18 0,0075 0,080 3 0,0088 UZpaK=0,50U7***
ta 0,0068 0,080 0,0080
25 Bithynia leachi 18 0,0021 0,012 3 0,0025 UZpaK=0,50U7 ***
26 Theodoxus fluvia- 18 0,005 0,030 3 0,0059 UZpaK=0,30U7 ***
tilis 0,0018 0,009 0,0021
27 Melanopsis prae- 20 0,098 IV'0,77 0,008—0,6 2 Аракелова, 1979 UZpaK=0,60U?
morsa
28 Valvata sibirica 20 0,088 UP085 0,001—0,01 3 Аракелова, Умно- ва, 1984 0,014 U7085 UZpaK=0,37U7
29 Viviparus conte- 20 0,069 HZ0,65 0,03—8,85 3 Собственные дан- 0,150 IF065 U7paK=0,54U7
ctus ные
30 Bithynia tentacu- 25 0,119 0,140 3 Цихон-Луканина, 0,012 UZpaK=0,50U7 ***
lata Сорокин, 1966 СПК рассчитана на 1 г (графа 4)
31 Pomacea lineata 25 0,0118—1,047 0,01—2,475 1 Zavaglia, Santos, 0,008—0,70 Гсчх=0,226Г ***
0,493 1,217 Mendes, 1981 0,330 и7суч=0,226^*Р**
0,528 1,255 0,353 (по: Akerlund,
1969)
* Способы выражения единиц измерений СПК и W: 1 — млО. ч_|; г сухого тела, 2— млО. ч_|; г сырого тела, 3— млО. ч_|; г общей массы (тело + раковина).
** Данные по СПК приведены нами к 20° и выражены вторым способом.
*** Условно принятое соотношение.
mian, 1965), исследовав дыхательную систему этого вида, показал,
что дыхание осуществляется тремя способами — жаберным, легоч-
ным и сифонально-легочным. Данные Акерлунда (Akerlund, 1969)
по СПК этого вида получены лишь с учетом жаберного дыхания.
Перед опытом воздух из легочного мешка удалялся, животные поме-
щались в респирометр с водой. В течение опыта увеличение двига-
тельной активности моллюсков по сравнению с нормальной не от-
мечалось. Полученная прямая зависимости СПК от массы животных
приближается к линии, условно определяющей верхний уровень ин-
тенсивности обмена у брюхоногих. При сравнении данного вида с
Lymnaea stagnalis оказалось, что уровни энергетического обмена
близки у обоих видов, ведущих сходный активный образ жизни.
Так, для М. cornuarietis по данным автора С77К=0,221 млО- ч_1-г-1
в расчете на сырой вес тела при 20° С. По нашим данным для L.
stagnalis при той же температуре Q=(0,102+0,060) Ц7°’75±004
(рис. 2), а интенсивность обмена, рассчитанная на массу тела без
раковины, составляет 0,261 млО-ч -г'1 (Аракелова, 1979). У амфи-
бионтной улитки Ротасеа lineata не выявлены различия в СПК
в водной и воздушной средах (Zavaglia et al., 1981). СПК определяли
манометрически в аппарате Варбурга при 25° С. Показано, что ско-
рость дыхания моллюсков в воде, осуществляемая ктенидиями, не
отличается от СПК в воздушной среде, когда часть мантии начинает
функционировать как легкое. Это же отмечено и для Р. paludosa
и М. cornuarietis, хотя для второго вида СПК в воде несколько
выше, чем на воздухе (Freiburg, Hazelwood, 1977)
Сравнения скорости потребления кислорода легочных и передне-
жаберных моллюсков, способных поглощать как атмосферный, так и
растворенный кислород, показывает, что в данном случае уровень
энергетического обмена переднежаберных не ниже по сравнению с
легочными. На основе результатов определения СПК у переднежа-
берных и легочных моллюсков, полученных с помощью манометри-
ческого и радиоуглеродного методов (Brand et al., 1948;. Цихон-
Луканина, Сорокин, 1966), было высказано предположение, что уро-
вень энергетического обмена переднежаберных ниже по сравнению
с легочными как более эволюционно прогрессивными. Выбор и при-
менение именно этих методов были продиктованы различиями в
строении органов дыхания этих двух групп моллюсков. В опытах для
легочных моллюсков, многие из которых активные всеядные формы,
полученный уровень энергетического обмена превышал эту же вели-
чину у переднежаберных. Полученные манометрическим методом
значения интенсивности обмена у легочных и переднежаберных
отличались примерно в 3 раза: для легочных а=0,0516, для передне-
жаберных — 0,0163 (а — млО- ч-1т_|) (Brand et al., 1948). Известно,
что легочные моллюски способны заполнять полость легкого водой
и длительное время находиться под водой, не используя атмосферного
воздуха. Учитывая эту способность использовать легочную полость
в зависимости от обстоятельств как жабру или легкое (Павловский,
Лепнева, 1948; Жадин, 1952), для измерения СПК у легочных можно
применять метод замкнутых сосудов, который при определенном
подборе опытных сосудов с учетом размера животного и соот-
76
Рис. 2. Зависимость логарифма СПК (lg Q) L. stagnalis от
логарифма веса (1g W) при 20° С.
ветствующей экспозиции опыта дает надежные результаты. Ма-
нометрический и радиоуглеродный методы для определения СПК
у легочных, таким образом, не имеют преимуществ перед методом
замкнутых сосудов. Кроме того, радиоуглеродную методику в отно-
шении раковинных моллюсков следует применять с осторожностью.
Как было показано на Sphaerium scaldianurn, выделяемая из
карбонатов раковины моллюска углекислота может приводить к за-
вышению величины интенсивности газообмена, рассчитанной по ко-
личеству выделенного в безуглекислую водную среду СО2 (Бульон,
1972)
Нанесенные на рис. 1 линии регрессии и отдельные значения СПК
для морских и пресноводных переднежаберных моллюсков показали,
что межвидовые различия уровней обмена у этих животных довольно
велики. В табл. 1 приведены литературные данные по СПК или ин-
тенсивности обмена у различных видов, пересчитанные нами и при-
веденные к одним и тем же единицам измерения СПК — млО- ч-1-экз-1
при 20° С, масса животных — г сырого тела моллюсков без рако-
вины. К сожалению, в использованных литературных источниках
подчас отсутствовали данные, необходимые для пересчета из одних
единиц измерения в другие. Во многих случаях нет данных по соот-
ношению массы раковины и мягких тканей или оно значительно
варьирует, особенно у морских моллюсков. Кроме того, диапазон
весов, для которого рассчитано уравнение по дыханию, приводится
не во всех случаях. Из-за этого не всегда ясно, к общему весу или
f
\
77
к весу тела относятся данные по СПК. Расчеты осложняются также,
если не известно процентное содержание сухого вещества в сырой
массе моллюска. В некоторых случаях при отсутствии необходимых
для пересчета данных, нами были приняты некоторые допущения:
для перевода единицы сухой массы моллюска в сырую использовано
соотношение U7cyx=0,25 UZcwp, данные по СПК при температуре
опыта, отличающейся от 20° С, приводились к 20° с помощью коэф-
фициента Qio, показывающего изменение скорости процесса при из-
менении температуры на 10° и равного 2,25. При меньших различиях
между температурой опыта t и 20° использовали температурную
поправку q, которую рассчитывали по формуле
<7=2,гб*20-^1 °
Все подобные случаи отмечены в примечании табл. L Именно по этой
причине (из-за недостатка точных сведений) нецелесообразно, по-
видимому, выводить единое уравнение СПК для морских и пресно-
водных переднежаберных.
Для пресноводных видов переднежаберных моллюсков при обоб-
щении собственного материала и литературных данных было рас-
считано уравнение СПК при 20° С, в котором масса моллюсков учи-
тывалась вместе с раковиной:
Q=(0,056±0,022) r0742±0018 (1)
(п=95; г=0,973; Р<0,01).
Для 3 видов из семейства Lymnaeidae по экспериментальным
данным при 20° С получено уравнение зависимости СПК от массы
животных с раковиной:
Q=(0,И5±0,032) Г070,±0’025 (2)
(п=87; г=0,95; Р<0,01).
Для представителей семейства Planorbidae рассчитано уравнение
(рис. 3):
Q=(0,081 ±0,034) №°’83±0’03 (3)
(и=71; г=0,958; Р<0,01),
где, как и в уравнениях (1) и (2), Q — СПК в млО- ч-1, W — масса
моллюска с раковиной в г, п — число измерений, г — коэффициент
корреляции, Р — уровень надежности при (п — 2) степенях свободы.
В полученных нами трех уравнениях относительная ошибка коэф-
фициента а довольно высока и при принятом уровне надежности
(Р^0,01) для пресноводных переднежаберных составляет 39%,
для Lymnaeidae — 28%, для Planorbidae — 42%. Важно, что при
таком широком отклонении от средней величины а в сторону макси-
мального и минимального ее значений, колебания коэффициента й,
определяющего наклон линии регрессии, очень малы во всех трех
случаях и близки к 3%.
78
Рис. 3. Зависимость СПК при 20° С от массы у легочных моллюс-
ков из семейств Planorbidae и Bulinidae.
1 — Planorbis planorbis (Ковалева, 1970; Аракелова, 1982); 2 — Planorbarius
corneas (Borden], 193! no: Calow, 1975; собств. данные). 3 — Biomphalaria alexandrina
(Ishak, Mohamed et al., 1970); 4 — Ancylus fluviatilis (Calow, 1975); 5 — Biomphalaria
glabrata (Brand, Nolan, Mann, 1948): 6 — Biomphalaria glabrata (Lee, Cheng, 1971);
7 — Bulinus truncatus (Ishak, Mohamed et al., 1970); 8— Planorbis contortus
(Calow, 1975).
Для сопоставления и сравнения параметров наших уравнений
необходимо, согласно имеющимся рекомендациям (Алимов, 1979),
привести коэффициенты регрессий к единому значению 0,75, кото-
рый в настоящее время принят как единый показатель степени для
водных беспозвоночных. После соответствующего пересчета коэффи-
циентов уравнения приняли следующий вид:
Lymnaeidae— Q=0,l 17 Г0175 (ш=0,703 г) (4)
Planorbidae — Q=0,070 Г0’75 (ш=0,161 г) (5)
Prosobranchia — Q=0,056 U70,7° (£5=1,0 г) (6)
Как видно из табл. 2, несмотря на значительные межвидовые
различия, значения интенсивности обмена у двух групп моллюсков —
легочных и переднежаберных — отличаются не более чем в 2 раза.
Диапазон весов моллюсков, для которых были рассчитаны уравне-
ния, довольно широк: минимальный и максимальный веса этих жи-
вотных различались на 4 порядка.
При сравнении уровней обмена этих двух групп моллюсков не-
обходимо учитывать, что полученные значения первого коэффициента
79
Таблица 2
Зависимость скорости потребления килорода от массы и ошибки средних значений параметров (Р^0,01)
у пресноводных брюхоногих моллюсков при 20° С
№ п/п Виды Пределы весов. n Qi млО.г 1 k Qi при £ = 0,75
1 Planorbarius corneus 0,1—4,0 40 0,096 ±0,014 0,832 ±0,098 0,091
2 Planorbis planorbis 0,0027-0,382 16 0,037 ±0,012 0,697 ±0,093 0,042
3 Lymnaea ovata 0,005-0,35 42 0,148 ±0,033 0,682 ±0,018 0,181
4 L. stagnalis 0,00036-4,28 27 0,097 ±0.019 0,795 ±0,061 0,085
5 L. palustris 0,1-1,25 18 0,088 ±0,024 0,592 ±0,110 0,098
6 Physa fontinalis 0,009-0,06 23 0,102 ±0,056 — 0,748 ±0,147 0,101
7 Melanopsis praemorsa 0,02-1,5 32 0,040 ±0,003 0,748 ±0,047 0,040
8 Viviparus contectus 0,03-8,85 20 0,069 ±0,010 0,648 ±0,082 0,075
9 Valvata sibirica 0,001-0,01 8 0,088 ±0,032 0,852 ±0,068 0,052
в степенных уравнениях отражают стандартный уровень обмена це-
лого животного с учетом доли раковины. Непосредственно не участ-
вуя в метаболизме, раковина порой составляет значительную долю
от общей массы моллюска, что может отразиться при расчете вели-
чины интенсивности обмена. Так, у многих легочных моллюсков ра-
ковина значительно тоньше, чем у переднежаберных, и составляет
до 5% от общей массы у Lymnaea glutinosa и до 30—35% у дру-
гих прудовиков. У представителей семейства Physidae вес раковины
часто не превышает 10%. Для переднежаберных моллюсков в ли-
тературе приводятся намного более высокие значения этих величин.
Для Rapana thomasiana доля сырого веса тела моллюска от общего
его веса — 23,3% и, вероятно, около 70% приходится на раковину
(Муравская, Галкина, 1979). Соответственно, величина интенсив-
ности обмена значительно повышается, если ее пересчитать на сы-
рой вес тканей. В этом случае она будет равна 0,198 мл О- ч_|-г_|, что
превышает величину интенсивности обмена, рассчитанной на 1 г об-
щей массы, в 4,3 раза. У Valvata piscinalis и Viviparus
viviparus раковина составляет, соответственно, 74 и 40% от общей
массы моллюсков (Китицына, 1971, 1975). По нашим данным, у
Melanopsis praemorsa раковина составляет 60%, у Viviparus
contectoides — 54%, у Valvata sibirica — 37%. Очевидно, что
при столь заметных различиях в соотношении масс тела и раковины
у разных видов моллюсков необходимо учитывать это соотношение
при сравнительной оценке интенсивностей обмена у исследуемых
животных.
Сопоставление уравнений (4), (5) и (6) показало, что величина
интенсивности обмена переднежаберных моллюсков менее чем в 1,5
раза отличается от семейства Planorbidae и в 2 раза ниже, чем у се-
мейства Lymnaeidae. В первом случае доверительные интервалы для
уравнений (5) и (6) взаимно перекрываются на всем интервале ис-
следованных весов (рис. 4). При сравнении уравнений (4) и (6)
выявились достоверные различия в уровнях обмена.
Изложенный в работе фактический материал позволяет придти
к выводу, что различия уровней энергетического обмена легочных
и переднежаберных моллюсков объясняются не эволюционными тен-
денциями и систематическим положением этих групп моллюсков,
а имеют адаптивную природу. Как было показано В. Р Дольником,
„уровень стандратного метаболизма един для всех видов в пределах
крупного таксона (класса)“ (Дольник, 1968, с. 276). При сравнении
средних уровней обмена, определенных для различных семейств
класса Bivalvia, достоверных различий в параметрах уравнений не
обнаружено, что позволило А. Ф. Алимову (1981) вывести единое
для представителей класса Bivalvia уравнение Q =0,094 W0'7'2.
Учитывая сказанное, можно заключить, что различие или сход-
ство в уровне обмена легочных и переднежаберных моллюсков
может быть достоверно установлено только на сравнительном ма-
териале с учетом экологической специфики и морфологических осо-
бенностей моллюсков, в частности, доли метаболически пассивных
образований в виде раковины. Вводя в уравнения дыхания поправки
на активность, связанную с типом питания, и учитывая долю pa-
ts
Заказ 1696
81
Рис. 4. Зависимость логарифма СПК (lg Q) от логарифма веса (Lg W)
моллюсков.
/ — Lymnaeidae, 2 — Planorbidae и Bulinidae, 3 — Prosobranchia. Ось абсцисс — lg W',
ординат — lg Q. Доверительные зоны регрессий построены при Р^О.01.
ковины в общей массе моллюска, мы скорее всего придем к единому
количественному выражению функции у разных групп гастропод.
Предлагаемые обобщенные уравнения зависимости скорости
энергетического обмена от массы для семейств Lymnaeidae и
Planorbidae и для пресноводных переднп\иберных моллюсков
могут быть использованы в количественных расчетах при эколого-
физиологических исследования, а также для приближенной оценки
продукции водных популяций.
ЛИТЕРАТУРА
Алимов А. Ф. Интенсивность обмена у пресноводных двустворчатых моллюсков.—
Экология, 1975, № 1, с. 10—29.
Алимов А. Ф. Интенсивность обмена у водных пойкилотермных животных.— В кн.:
Общие основы изучения водных экосистем. Под ред. Г Г Винберга. Л.,
1979, 273 с.
Алимов А. Ф. .Функциональная экология пресноводных двустворчатых моллюсков.
Л., 1981, 248 с.
Аракелова Е. С. Влияние размеров и температуры на скорость обмена у Melanopsis
praetnorsa L. (Pectinibranchia).— В кн.: Экспериментальные и полевые иссле-
дования биологических основ продуктивности озер. Л., 1979, с. 169—180.
Аракелова Е. С. Эффективность и тип роста у двух видов пресноводных легочных
моллюсков.— Журн. общей биологии, 1982, т. 43, № 4, с. 553—559.
Аракелова Е. С., Умнова Л. П. Об эффективности питания детритоядной Valvata
sibirica Midd. (Gastropoda, Vilvatidae).— Экология, 1984, № 5, с. 60—66.
82
Белокопытин Ю. С. Уровень основного обмена у некоторых морских рыб.— Вопр.
ихтиологии, 1968, т. 8, вып. 2, с. 382—385.
Белокопытин Ю. С. Уровень основного и стандартного обмена у морских рыб
различной подвижности.— Автореф. канд. дис. Л., 1969, 23 с.
Бульон В. В. Опыт применения прибора Ю. И. Сорокина и определения интенсивности
газообмена водных животных радиоуглеродным методом.— Гидробиол.
журн., 1972, т. 8, № 4, с. 91—97.
Виленкин Б. Я., Виленкина М. Н. Дыхание водных беспозвоночных. „Итоги науки
и техники. ВИНИТИ АН СССР. Сер. зоология беспозвоночных", 1979, т. 6,
143 с-
Винберг Г Г Интенсивность обмена и размеры ракообразных.— Журн. общей
биологии, 1950, т. 11, № 2, с. 367—380.
Винберг Г Г Зависимость энергетического обмена от массы тела у водных пойкило-
термных животных.— Журн. общей биологии, 1979, т. 37, № 1, с. 56—70.
Винберг Г Г., Беляцкая Ю. С. Зависимость интенсивности обмена от‘ общих
размеров тела у пресноводных брюхоногих моллюсков.— Бюл. Ин-та биол.
АН БССР за 1957 г., вып. 3, 1958, с. 273—276.
Винберг Г Г., Беляцкая Ю. С. Соотношение интенсивности обмена и веса тела
у пресноводных брюхоногих моллюсков.— Зоол. журн., 1959, т. 38, вып. 8,
с. 1146—1151.
Дольник В. Р. Энергетический обмен и эволюция животных.— Успехи соврем, биол.,
1968, т. 66, № 2, с. 276—293.
Жадин В. И. Моллюски пресных и солоноватых вод СССР. М.—Л., 1952, 376 с.
Камлюк Л. В. Энергетический обмен у свободноживущих плоских и кольчатых
червей и факторы, его определяющие.— Журн. общей биологии, 1974, т. 35,
№ 6, с. 874—885.
Китицына Л. А. Влияние температурного режима водоема-охладителя ТЭС на
интенсивность обмена и размеры моллюска Valvata piscinalis (Muller).—
В кн.: Моллюски. Пути, методы и итоги их изучения. Л., 1971, с. 99—101.
Китицына Л. А. Роль живородки в деструкции органического вещества на участке
Среднего Днепра.— В кн.: Моллюски. Пути, методы и итоги их изучения.
Л., 1975, с. 88—91.
Ковалева А. А. Уровень газообмена у некоторых двустворчатых и брюхоногих
моллюсков.— В кн.: Вопросы физиологической и. популяционной экологии,
вып. 1, Саратов, 1970, с. 31—37
Муравская 3. А., Г алкина О. Н. Интенсивность обмена и состав тела у мидии
Mytilus galloprovincialis и рапаны Rapana thomasiana Grosse.— Биол. моря,
Киев, 1979, № 51, с. 68—77.
Павловский Е. Н., Лепнева С. Г Очерки из жизни пресноводных животных. М.—Л.,
1948, 459 с.
Сущеня Л. М. Интенсивность дыхания ракообразных. Киев, 1972, 196 с.
Хлебович Т В. Интенсивность дыхания у инфузорий разного размера.— Цитология,
1974, т. 16, № 1, с. 103—105.
Цихон-Луканина Е, А., Сокрокин Ю. И. Интенсивность обмена у пресноводных
брюхоногих моллюсков.— В кн.: Планктон и бентос внутренних водоемов.
(Тр. Ин-та биологии внутренних вод, 12 (15), 1966, с. 139—144.
Akerlund G. Oxygen consumption of the ampullarid snail Marisa, cornuarietis L.
in relation to body weight and temperature.— Oikos, 1969, vol. 20, p. 529—533.
Bayne B. L., Scullard C. Rates of oxygen consumption by Thais (Nucella} lapillus
(L).—J. Exp. Mar. Biol, and Ecol., 1978, vol. 32, N 1, p. 97—111.
Berg K-, Lumbye J., Ockelmann K. W. Seasonal and experimental variation of the
oxygen consumption of the limpet Ancylus fluviatilis (O. F. Muller).— J. Exp.
Biol., 1958, vol. 35, p. 43—73.
Berg. K-, Ockelmann K. W The respiration of freshwater snails.— J. Exp. Biol.,
1959, vol. 36, p. 690—708.
Brand von T., Nolan M. O., Mann E. R. Observations on the respiration of Australorbis
glabratus and some other aquatic snails.— Biol, bull., 1948, vol. 95, p. 199—213.
Calow P. Some observations on locomotory strategies and their metabolic effects in
two species of freshwater gastropods, Anculus fluviatilis Mull, and Planorbis
contortus Linn.— Oecologia, 1974, vol. 2, p. 149—161.
6*
83
Calow P. The respiratory strategies of two species of freshwater gastropods (Ancylus
fluviatilis Mull, and Planorbis contortus Linn.) in relation to temperature,
oxygen concentration, body size and season.— Physiol. Zoology, 1975, vol. 48,
N 2, p. 114—129.
Coles G. C. The termination of aestivation in the large freshwater snail Pila ovata
(Ampullariidae) — I. Changes in oxygen uptake.— Comp. Biochem. Physiol.,
1968, vol. 25, p. 517—522.
Crisp M. The effect of activity on the oxygen uptake of Nassarius reticulatus
(Gastropoda, Prosobranchia).— Malacologia, 1979, vol. 18, N 1—2, p. 445—447.
Crisp M., Davenport J., Shumway S. E. Effects of feeding and of chemical stimulation
on the’oxygen uptake of Nassarius reticulatus (Gastropoda! Prosobranchia).—
J. Mar. Biol. Assoc. U. K., 1978, vol. 58, N 2, p. 387—399.
Davies P. S. Physiological ecology of Patella.— I. The effect of body size and
temperature on the metabolic rate.—J. Mar. Biol. Assoc. U. K., 1966, vol. 46,
N 3, p. 647—658.
Demian E. S. The respiratory system and the mechanism of respiration in Marisa
cornuarietis (L.).—Arkiv zool., ser. 2, 1965, vol. 17, p. 539—560.
Dykens J. A., Mangum С. P. The design of cardiac muscle and the mode of
metabolism in molluscs.— Comp. Biochem. and Physiology, 1979, vol. 62, N 3,
p. 549—554.
Fitch D. D. Oxygen consumption in the prosobranch snail Viviparus contectoides
(Mollusca, Gastropoda). I. Effect of weight and activity.—Comp. Biochem.
and Physiol., 1975, vol. A51, N 4, p. 815—820.
Freiburg M. UZ., Hazelwood D. H. Oxygen consumption of two amphibious snails:
Pomacea paludosa and Marisa cornuarietis (Prosobranchia: Ampullariidae).—
Malacologia, 1977, vol. 16, (2), p. 541—548.
Horne F. R. Comparative aspects of aestivating metabolism in the gastropod Marisa.—
Comp. Biochem. and Physiol., 1979, vol. 64A, N 2, p. 309—312.
Houlihan D. F., Innes A. J. Oxygen consumption, crawling speeds, and cost of
transport in four mediterranean intertidal gastropods.— J. Comp. Physiol.,
1982, vol. B147, p. 113—121.
Ishak M. M., Mohamed A. M., .Wafa A. Mousa Аг H., Ayad N. Physiological
studies on Biomphalaria alexandrina and Bulinus truncatus, the snail vectors
of Schistosomiasis. I. Oxygen consumption.— Hydrobiologia, 1970, vol. 35,
p. 333—340.
Lee F. O., Cheng T C. Schistosoma mansoni: respirometric and partial pressure
studies in infected Biomphalaria glabrata.—Exp. Parasitology, 1971, vol. 30,
N 3, p. 393—399.
Lumbye J. The oxygen consumption of Theodoxus fluviatilis (L.) and Potamopyrgus
jenkinsi (Smith) in brackish and fresh water.— Hydrobiologia, 1958, vol. 10,
p. 245—262.
Newell R. G., Northcroft H. R. A re-interpretation of the effect of temperature on
the metabolism of certain marine invertebrates.— J. Zool. (London), 1967,
vol. 151, p. 277—298.
Parry G. D. Effects of growth and temperature acclimation on metabolic rate
in the limpet Cellana tramoserica (Gastropoda: Patellidae).— J. Animal Ecology,
1978, vol. 47, N 2, p. 351—368.
Sander F., Moore E. Comparative respiration in the gastropods Murex pomum and
Strombus pugilis at different temperatures and salinities.— Comp. Biochem.
and Physiol., 1978, vol. A60, N 1, p. 99—105.
Sandison E. E. The oxygen consumption of some interidal gastropods in relation to
zonation.— J. Zoology, 1966, vol. 149, pt. 2, p. 163—173.
Shirley T. C., Denoux G. J., Stickle W B. Seasonal respiration in the marsh
periwinkle, Littorina irrorata.— Biol. Bull., 1978, vol. 154, N 2, p. 322—334.
Stickle W B. The reproductive physiology of the intertidal prosobranch Thais
lamellosd (Gmelin). I. Seasonal changes in the rate of oxygen consumption
and body component indexes.— Biol. Bull., 1973, vol. 144, N 3, p. 511—524.
Studer E. H., Pace G. L. Oxygen consumption in the prosobranch snail Viviparus
contectoides (Mollusca: Gastropoda). IV Effects of dissolved oxygen level,
starvation, density, symbiotic algae, substrate composition and osmotic
pressure.— Comp. Biochem. and Physiol., 1978, vol. A59, N 2, p. 199—203.
84
ZavagUa C. A., Santos Dos, Garcia Mendes E. Oxygen consumption of the amphibious
snail Pomacea Uneata\ influence of weight, sex and environments. Comp.
Biochem. and Physiol., 1981, vol. A69, N 3, p. 595—598.
Zeuthen E. Body size and methabolic rate in the animal kingdom. Comptes rendus
tr. Labor. Carlsberg, Scr. chim., 1947, vol. 26, N 3, p. 1 — 161.
E. S. Arakelova
INTENSITY OF GASTROPODAN METABOLISM
The energetic metabolism of gastropods is evaluated on the base of original
and literary data. The similarity and diversity in the level of metabolism within
the class are stated. The possible causes of these are discussed. It is concluded
that the differences in the level of metabolism between Pulmonata and Prosobranchia
are of adaptive significance.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES ' <!
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
A. H. Голиков
НОВЫЕ ВИДЫ БРЮХОНОГИХ МОЛЛЮСКОВ
ПОДОТРЯДА R1SSOO1DE1
В результате анализа коллекций отряда Littoriniformes в Зоологи-
ческом институте и последних сборов установлено, что в умеренных
водах северо-западной части Тихого океана и в Арктическом океане
этот подотряд представлен, как минимум, 4 семействами, 21 родом
и 62 видами. Из этих видов 4 оказались новыми для науки.
Приводим их описание.
Mohrensternia coronata Golikov, sp. n. (рис. 1)
Раковина относительно крупная, крепкая, толстостенная, из 4,5
отлого-угловатых в верхней части выпуклых оборотов, разделенных
отчетливым вдавленным швом. Периостракум плотно прилегающий
к поверхности раковины, бледно-ржавого цвета. Зародышевая
раковина с 1,5 умеренно-выпуклыми, прижатыми гладкими оборо-
тами. Последний оборот с угловатой периферией и выпуклым
основанием занимает 0,76 высоты раковины. Осевая скульптура
представлена тонкими линиями роста и широкими выступающими
складками, образующими на плече оборотов булавовидные вздутия.
Эти складки не доходят до конца оборотов и не распространяются
на основание раковины, располагаясь с промежутками, примерно
равными им по ширине. На последнем и предпоследнем оборотах
имеется по 15 таких складок. Спиральная скульптура состоит из
низких килей на плече оборотов и на периферии последнего оборота
и из неясной исчерченности. Пупок открытый, щелевидный, огра-
ничен складкой прилежащей части основания раковины. Устье
округло-овальное, слегка выступает у нижнего края, белое внутри.
Голотип, обнаруженный в Охотском море у западных берегов
Камчатки, на глубине около 140 м, на илистом песке (сбор Н. Смир-
нов и А. Бегак на э/с „Командор Беринг“ 18.VIII. 1907), имеет
высоту раковины 3,1 мм, диаметр 2,2 мм, высоту последнего
оборота — 2,6 мм, высоту устья — 1,5 мм и ширину устья — 1,1 мм.
Наличием бульбовидных вздутий, образующих на плече оборотов
подобие короны, и характером спиральной скульптуры новый вид
хорошо отличается от других видов рода.
Единственный экземпляр (голотип) хранится в коллекциях
морских брюхоногих моллюсков ЗИН АН СССР под № 1/42518.
Onoba karica Golikov, sp. п. (рис. 2)
Раковина маленькая, хрупкая, из 5 выпуклых, закругленных
оборотов, разделенных отчетливым прижатым швом. Периостракум
86
Рис. I. Внешний вид раковины Mohre-
sternia coronata Golikov, sp. n.
Рис. 2. Внешний вид раковины Onoba
karica Golikov, sp. n.
тонкий, шелушащийся, светло-желтого цвета. Зародышевая ра-
ковина с 2 выпуклыми, гладкими, закругленными оборотами.
Последний оборот с выпуклой периферией и основанием, занимает
около 2/3 высоты раковины. Осевая скульптура представлена только
тонкими линиями нарастания. Спиральная скульптура состоит из
слегка приподнятых ребер, разделенных промежутками, примерно в
2 раза превышающими их ширину. На последнем обороте, включая
основание, расположено 15 таких ребер, а на предпоследнем обороте
их число сокращается до 8. Пупок закрыт. Устье по*, ги округлое,
с выступающей наружной губой, с сомкнутым краем.
Голотип, обнаруженный в северной части Карского моря на
глубине 300 м, на илистом грунте, при температуре —1,4 и солености
34,92%О (сбор А. В. Иванова, 1934 г.), имеет высоту раковины
3,1 мм, диаметр— 1,5 мм, высоту последнего оборота— 1,9 мм,
высоту устья — 1,2 мм, ширину устья — 0,9 мм.
Вид наиболее сходен с О. aculeus (Gould), от которого отли-
чается существенно более выпуклыми оборотами и меньшим коли-
чеством расставленных (а не сближенных) спиральных ребрышек.
Голотип (единственный экземпляр) хранится в коллекциях
морских брюхоногих моллюсков ЗИН АН СССР под № 1/42541.
Onoba russica Golikov, sp. n. (рис. 3)
Раковина маленькая, хрупкая, из 4,5 умеренно-выпуклых, за-
кругленных оборотов, разделенных тонким вдавленным швом.
87
Периостракум на верхних дефинитивных оборотах хорошс/ развит,
охряно-желтого цвета. Зародышевая раковина небольшая, с 1,5
слабо выпуклыми оборотами. Последний оборот с умеренно-вы-
пуклой периферией и удлиненным основанием занимает 0,64 высоты
раковины. Осевая скульптура представлена только тонкими линиями
роста. Спиральная скульптура состоит из уплощенных ребрышек,
расположенных с промежутками более узкими, чем их ширина.
На последнем обороте, включая основание, расположено около
30 таких ребрышек, а на предпоследнем — 10. Пупок закрыт. Устье
округло-овальное, с тонкой губой. Голотип, обнаруженный в бухте
Русской Восточной Камчатки на глубине 31 м, на песчаном илу
(сбор А. Н. Голикова, 14.VII.1975 г.), имеет высоту раковины
2,7 мм, диаметр— 1,4 мм, высоту последнего оборота— 1,8 мм,
высоту устья — 1 мм и ширину устья — 0,8 мм.
Обнаруженный вид (представленный, вероятно, молодыми
особями) отличается от других видов рода многочисленными
спиральными ребрышками (около 30 на последнем обороте) и
округло-овальной формой устья.
Голотип и паратип хранятся в коллекции морских брюхоногих
моллюсков ЗИН АН СССР, голотип имеет № 1/42551.
Punctulum reticulatum Golikov, sp. n. (рис. 4)
Раковина маленькая, хрупкая, с 4,5 выпуклыми, закругленными
оборотами, разделенными тонким вдавленным швом. Периостракум
тонкий, шелушащийся, бледно-лимонно-желтого цвета. Зароды-
шевая раковина из 1,5 выпуклых закругленных оборотов, покрыта
тончайшей спиральной исчерченностью, составленной 15 нитевид-
ными ребрышками. Последний оборот с закругленной периферией
и выпуклым основанием занимает 0,77 высоты раковины. Осевая
скульптура представлена тонкими линиями роста и узкими осевыми
складками, расположенными с промежутками, примерно в 2 раза
превышающими их ширину, распространяющимися на всю высоту
оборотов, но не заходящими на основание раковины. На последнем
и предпоследнем оборотах расположено по 28 таких складок.
Спиральная скульптура состоит из приподнятых, волнистых на
осевых складках ребрышек, расположенных с промежутками в 2—3
раза более широкими, чем их толщина. На последнем обороте
расположено 10 таких ребрышек, а на предпоследнем обороте их 5.
Пупок не полностью прикрыт отворотом внутренней губы. Устье с
сомкнутым краем, почти округлое, слегка выступает у нижнего
париетального края. Голотип, обнаруженный у берегов Приморья
у залива Рында Японского моря на глубине около 430 м на илистом
с приместью песка грунте (сбор Охрямкина на э/с ,,Россинанте“),
имеет высоту раковины 2,7 мм, диаметр — 2 мм, высоту последнего
оборота — 2,1 мм, высоту устья— 1,4 мм, ширину устья — 2 мм.
Вид наиболее сходен с Р. wyvillethomsoni (Jeffr.), от которого
отличается цветом периостракума, более выступающей скульптурой,
представленной большим числом осевых складок и меньшим числом
спиральных ребер (у Р. wyvillethomsoni их 19 и 18 соответственно).
88
Рис. 3. Внешний вид раковины Onoba
russica Golikov, sp. n.
Рис. 4. Внешний вид раковины Punctu-
lum reticulatum Golikov, sp. n.
Обнаружен в 5 пробах (7 экз.), у берегов южного и среднего
Приморья, на глубинах от 240 до 880 м, на песчано-илистых
и илистых грунтах.
Голотип хранится в коллекции морских брюхоногих моллюсков
ЗИН АН СССР под № 1/36754.
A. N. Golikov
NEW SPECIES OF GASTROPODS FROM THE SUBORDER RISSOOIDEI
In the boreal waters of the Pacific ocean were discovered 3 new species:
Mohrensternia coronata, Onoba russica and Punctulum reticulatum and in northern
part of Kara sea was observed 1 new species — Onoba karica.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
A. H. Голиков, Я. И. Старобогатов
НОВЫЙ ВИД БРЮХОНОГИХ МОЛЛЮСКОВ
CONRADIA M1NUTA ИЗ УМЕРЕННЫХ ВОД
СЕВЕРО-ЗАПАДНОЙ ЧАСТИ ТИХОГО ОКЕАНА
За последние десятилетия в коллекциях Зоологического инсти-
тута АН СССР накопилось несколько экземпляров мелких брюхо-
ногих моллюсков трудно установимой систематической принадлеж-
ности. Все экземпляры были или лишены тела, или сильно высушены,
или полупереварены в желудке конюги Aethia sp. и не пригодны
для изучения мягкого тела. В результате препаровки и вываривания
в щелочи остатков тел удалось выделить лишь несколько крышечек
моллюсков, радулы обнаружить не удалось. По конхологическим
признакам анализируемые моллюски формально нельзя отнести ни
к одному из известных родов, кроме Conradia A. Adams, 1860,
от которого они не отличаются ни одним из краткого перечня
признаков, имеющихся в описании этого рода. Отнесение нашего
вида к роду Conradia и его номинотипическому подроду косвенно
подтверждается еще и тем, что наш вид во многом похож на
единственный изображенный вид подрода Gottiana — С. (G.) cereus
(Wats), но отличается от него, наряду с прочими, признаками,
характеризующими подрод Conradia s. str.
В малакологической литературе род Conradia относят к семейству
Fossaridae. Этому следуем и мы за неимением уточняющих данных.
Однако обращает на себя внимание наличие у нашего вида, равно
как и у С. (G.) cereus, сифональной вырезки, совершенно не свой-
ственной представителям семейства Fossaridae. Это заставляет
подозревать, что этот род не относится ни к семейству Fossaridae,
ни к отряду Calyptraeiformes (=Echinospirida), куда это семейство
входит. Не исключена возможность, что этот представитель отряда
Cerithiiformes (=Entomostoma) и может быть даже близок к се-
мейству Modulidae, что можно будет утверждать или отрицать
только после изучения внутреннего строения и, в первую очередь,
радулы.
Тем не менее, поскольку ни представители фоссарид, ни пред-
ставители модулид не были указаны из холодных и умеренных
вод северного полушария, и виды подрода Conradia s. str. до сих
пор не были изображены, мы публикуем описание и изображение
обнаруженного вида.
Conradia minuta Golikov et Starobogatov, sp. n.
Раковина тонкостенная, овальная, с 4,5 выпуклыми, закруглен-
ными оборотами, разделенными тонким прямым швом. Зародышевая
90
Conradia (Conradia) minuta — раковина (4) и крышечка (Б)
раковина слабо выпуклая, прижатая, состоит из 1,5 гладких обо-
ротов, у голотипа частично разрушенная. Последний оборот с
выпуклой закругленной периферией и выпуклым основанием,
занимает около 0,75 высоты раковины. Периостракум плотный,
желтовато-бурого цвета, образует на спиральных килях короткие
щетинки, а на верхних оборотах по оси он образует осевые скла-
дочки, часто и равномерно расположенные от нижнего киля к шву.
Осевая скульптура представлена только тонкими, слегка'припод-
нятыми, равномерно расположенными, прямыми линиями роста.
Спиральная скульптура состоит из 1 киля на плече первых дефини-
тивных оборотов, 4 килей на предпоследнем обороте и 11 килей на
последнем обороте, из которых самый нижний окаймляет около-
пупочную вырезку. Кили узкие, резко выступающие, разделены
промежутками, примерно в 5 раз превышающими -их ширину, более
резкие на верхней части и периферии последнего оборота, чем на
его основании. Устье полукруглое, с оттянутым и неполностью
замкнутым нижним париетальным краем. Наружная губа устья
равномерно закругленная, тонкая, выступающая, волнистая в
соответствии со спиральной скульптурой. Внутренняя губа устья
слегка вогнута в нижней части, тонким краем окаймляет узкую,
серповидно изогнутую вырезку у щелевидного пупка.
Голотип, собранный в заливе Анива на глубине 29 м на каме-
нистом грунте (э/с „Топорок“, 31.VIII. 1949, сб. Земцовой) и хра-
нящийся в коллекции морских брюхоногих моллюсков Зоологи-
ческого института АН СССР под № 1/47911, имеет высоту ра-
ковины 4,1 мм, диаметр — 3,2 мм, высоту последнего оборота 3,4 мм,
высоту устья — 2,2 мм и ширину устья — 1,7 мм.
Крышечка крупная, овальная, конхиолиновая, занимает почти
все устье, спиральная, с ядром, расположенным в нижней ее трети.
Рассматриваемый вид в наибольшей мере соответствует описанию
номинотипического подрода Conradia, отличаясь от видов подрода
91
Gottiana A. Adams, 1863, наличием щелевидного пупка, ограничен-
ного от основания раковины кильком. От отнесенных к номино-
типическому подроду, Conradia clathrata A. Adams 1860, С. pulchella
A. Adams, 1861, С. tornata A. Adams, 1863, С. doliaris A. Adams,
1863, отмеченных из района Корейского пролива, С. minuta отли-
чается буроватой, тонкостенной, покрытой плотным периостракумом
раковиной и Характером осевой и спиральной скульптуры (у пере-
численных выше видов раковина крепкая, белая, со слабым перио-
стракумом, с осевыми ребрышками и меньшим числом спиральных
элементов скульптуры). От Conradia cinguliferus A. Adams, 1860
(типовой вид рода) из Корейского пролива и С. adamsiana Nevill,
1885 из Персидского залива новый вид отличается большим числом
широко расставленных килей (а не уплощенных ребер) и отсут-
ствием осевых ребрышек.
Тихоокеанский азиатский, преимущественно высокобореальный
вид. Обнаружен в Анивском заливе на глубине 29 м (1 экз.,
возможно, снос) и из желудка конюги Aethia sp., добытой у о. Ионы
в Охотском море (50 экз.). Поскольку конюги собирают пищу
главным образом на мелководье, моллюски, вероятно, обитают в
прибрежных, может быть, лагунных водах.
A. N. Golikov, Ya. I. Starobogatov
NEW GASTROPODAN SPECIES CONRADIA MINUTA
FROM TEMPERATE WATERS OF NORTH-WESTERN PACIFIC
Conradia (Conradia) minuta sp. nov from Okhotsk sea and southern Sakhalin is
described. The picture of shell of the representative of the subgenus is published
in the first time.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
Я. И. Старобогатов
К СИСТЕМАТИКЕ БРЮХОНОГИХ МОЛЛЮСКОВ
РОДА LATIA (GASTROPODA PULMONATA LATIIDAE)
Общепринято считать, что маленькое реликтовое семейство
Latiidae представлено всего одним родом Latia Gray, 1850 и одним
видом L. neritoides Gray, 1850, обитающим в водоемах о. Южного
(Новая Зеландия). Все остальные названия, относящиеся к мол-
люскам этого рода — Crepidula neritoides Recluz, 1851, Pelex latera-
lis Gould, 1852, Latia petitiana Fischer, 1856, L. gassiesiana Fischer,
1856 (Gray, 1850, Recluz, 1851, Gould, 1852, 1856, Fischer, 1856 a, b),
рассматриваются (Suter, 1913) как синонимы L. neritoides.
Несмотря на относительную ограниченность материалов по
новозеландским легочным моллюскам, хранящихся в коллекции
Зоологического института АН СССР, мы получили возможность
подробно проанализировать конхологически все формы Latia с
о. Южного и пришли в результате- к выводу, что этот род там
представлен 3 видами: L. neritoides Gray, 1850 (syn. Crepidula
neritoides Recluz, 1851, Latia petitiana Fischer, 1856) (рис. 1),
L. lateralis (Gould,. 1852) (syn.: L. gassiesiana Fischer, 1856) (рис. 2)
и новым, описываемым ниже видом L. climoi sp. nov. (рис. 3),
названным в честь новозеландского малаколога Ф. Климо (F Climo),
любезно приславшего материал по этому виду. Три перечисленных
вида различаются очень четко относительным возвышением рако-
вины, т. е. отношением расстояния устье — затылок (,,высоты“) к
длине, положением вершины относительно плоскости и контура
устья и формой и положением колумеллярной площадки с ее отрост-
ком, протянутым вдоль правой стороны тела. Эти отличия хорошо
заметны на всех рисунках, сопровождающих первоописания.
Latia climoi sp. nov.
Материал Голотип (№ 1 по систематическому каталогу
ЗИН АН СССР) и 8 паратипов (№ 2), фиксированы спиртом и
снабжены этикеткой: Новая Зеландия, р. Хатт в окрестностях
Веллингтона, сбор Р К. Делла 29 ноября 1963 г., определил как
L. neritoides Ф. Климо. Размеры экземпляров, составляющих ти-
повую серию, представлены в таблице. У всех 9 экз. промерены
только внешние размеры, не требующие удаления тела, а именно,
длина раковины (в таблице обозначена Д), расстояние от плоскости
устья до затылка („высота14 — В), ширина раковины (ZZ/), удаление
вершины от переднего края устья в проекции на продольную ось
устья (Дв), удаление вершины от левого края устья по перпенди-
куляру к продольной оси устья (Шл) и расстояние от вершины до
края устья по хорде наиболее длинного склона (X). Кроме этого,
у голотипа после удаления тела измерены: ширина колумеллярной
93
Рис. 1—3. Раковины видов рода Latia сверху, снизу (со стороны устья)
и справа.
1 — L. neritoides (оз. Токапуна), 2 — L. lateralis (р. Каиварра), 3 —
L. climoi (голотип, р. Хатт).
площадки (вдоль продольной оси устья) — 1,1 мм, длина правого
отросгка колумеллярной площадки — 1,0 мм, ширина открытой части
устья (от переднего края раковины вдоль оси до переднего края
колумеллярной площадки — 6,0 мм и расстояние между передними
концами колумеллярной площадки — 2,9 мм.
Раковина неритоидная, полуяйцевидная, с узкой вершиной, почти
не выступающей за задний контур устья и заметно смещенной
влево от продольной оси тела. Отношение ширины устья к длине
раковины составляет 0,69—0,74, а отношение расстояния от пло-
скости устья до затылка к длине раковины — 0,29—0,35. Вершина
(если смотреть сбоку) расположена на уровне границы нижней
четверти расстояния от плоскости устья до затылка, а самая
верхняя точка затылочной поверхности располагается против сере-
дины длины раковины (в отличие от L. lateralis, где эта точка
Промеры экземпляров типовой серии Latia climoi (мм)
Промеры Д в 1 ш Де | Шл А'
Голотип 7,5 2,5 5,2 7,5 1,5 7,9
Паратипы: 1 Ю,1 3,6 7,5 10,1 10,7 10,7
2 7,0 2,4 5,2 7,0 1,4 7,4
3 7,0 2,4 5,0 7,0 1,5 7,1
4 6,5 2,0 4,6 6,5 1,3 6,6
5 6,8 2,0 4,8 6,8 1,4 7,0
6 6,1 1,9 4,2 6,1 1,3 6,2
7 6,3 2,0 4,5 6,3 1,3 6,2
8 5,5 1,8 3,9 5,5 1,1 5,7
94
располагается над границей привершинной трети длины раковины).
Колумеллярная площадка тонкая, белая, ее правый отросток хорошо
обособлен и заканчивается почти на уровне конца левого выступа
площадки (в отличие от L. neritoides, у которого он заканчивается
заметно позади левого выступа). Правый отросток колумеллярной
площадки длинный, суженный в основании и усеченный на свобод-
ном конце.
Отличия видов рода Latia видны из следующей определительной
таблицы.
1 (2) Отношение расстояния от плоскости устья до затылка к длине
раковины не превышает 0,34 и лишь у самых крупных особей
достигает 0,35, вершина почти не выступает за контур
устья L. climoi
2(1) Отношение расстояния от плоскости устья до затылка к длине
раковины не меньше 0,36, вершина заметно выдается за кон-
тур устья.
3(4) Колумеллярный отросток правой стороны заканчивается далеко
позади левого конца колумеллярной площадки, вентральный
край завитка находится почти на уровне плоскости устья,
наивысшая точка затылка находится против средины длины
устья L. neritoides
4(3) Колумеллярный отросток правой стороны заканчивается незна-
чительно позади левого конца колумеллярной площадки,
вентральный край завитка заметно приподнят над плоскостью
устья, наивысшая точка затылка находится против границы
привершинной трети длины устья L. lateralis
В заключение следует отметить, что раковина представителей
семейства Latiidae (как и близкого к нему семейства Acroloxidae)
левозавитая гиперстрофная, что доказывается положением вершины
и колумеллярной складки (превратившейся здесь в колумеллярный
отросток) по отношению к оси закручивания. В этом смысле ра-
ковины этих двух семейств противоположны раковинам ряда родов
Bulinidae и Planorbidae, которые имеют правозавитую раковину
при левой асимметрии тела.
ЛИТЕРАТУРА
Fischer М. Description de coquilles nouvelles.—J. de conchyliol., 1856a, t. 5,
(ser. 2, t. 1), p. 84—87.
Fischer M. Description d'un nouvelle espece de Latia.— J. de conchyliol., 1856b, t. 5
(ser. 2, t. 1), p. 167—168.
Gould A. A. Mollusca and shells.— U. S. Exploring expedition during the year
1838—1842 under the command of Ch. Wilkes, vol. 12, Philadelphia, 1852,
XV+510 p.
Gould A. A. Atlas. Mollusca and Shells.— U. S. Exploring expedition during the
year 1838—1842 under the command of Ch. Wilkes, vol. 12, Philadelphia,
1856, 16 p. + 52 tab.
Gray J. E. Description of a new genus and several new species of terrestrial
fluviatile and marine molluscous animals inhabiting New Zealand.— Proc.
Zool. soc. London, 1850 (1849), pt. 17, p. 164—169.
95
Recluz M. C. Description de quelques coquilles nouvelles.—J. de conchyliol., 1851,
t. 2, p. 194—216.
Suter H. Manual of the New Zealand Mollusca. Wellington, 1913, XXIV+1120 p.
Y. I. Starobogatov
ON THE TAXONOMY OF THE GASTROPOD MOLLUSCAN
GENUS LATIA (GASTROPODA PULMONATA LATIIDAE)
The description of a new species Latia climoi from the vicinity of Wellington
(New Zealand) and the key for identification of all 3 species of the genus Latia
(L. neritoides Gray, L. lateralis (Gould) and L. climoi sp. n.) recognised by the
author are presented.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
А. А. Шилейко
МАТЕРИАЛЫ ПО АНАТОМИИ И ТАКСОНОМИИ
DAUDEBARDIIDAE (GASTROPODA PULMONATA)
Семейство Daudeuardiidae — четко очерченный небольшой,
таксон, ареал которого охватывает среднюю и юго-восточную Европу,
Италию, Северную Африку (северный Алжир), западную часть
Малой Азии, западный Кавказ и Южный берег Крыма. К настоя-
щему времени известно 24 вида этого семейства, для которых
предложено 13 названий родовой группы, не считая случаев ошибоч-
ного написания и преоккупированных названий (Riedel, 1980).
Внешний вид тела этих хищных полуслизней удивительно
постоянен; раковины членов разных родов и видов порой заметно
различаются, но единственный способ понять структуру таксона
заключается в исследовании анатомии видов, в первую очередь —
полового аппарата.
Крупнейший знаток семейства — профессор А. Ридель — вы-
деляет два рода: Daudebardia (подроды: Daudebardia s. str., Libania,
Cibinid) и Carpathica; при этом он оговаривается, что „синонимизи-
рование кавказских Sieversia и Suchumiella с Daudebardia не
бесспорно, поскольку строение полового аппарата их представи-
телей существенно различается...; отдельные подроды?*— Riedel,
1980, S. 120. В отношении Cibinia Ридель также допускает, что
этот таксон может оказаться самостоятельным родом (op. cit.,
S. 122).
В диагнозах родов и подродов используются в основном 3 комп-
лекса признаков: строение раковины (форма и число оборотов
эмбриональной раковины, степень открытости пупка), место крепле-
ния глоточных ретракторов и особенности внешнего вида дистальной
части мужского отдела полового аппарата. В некоторых случаях
упоминаются отдельные особенности внутреннего строения пениаль-
ной трубки — см. сводную таблицу в работе Риделя (Riedel, 1978,
S. 50-52).
Система семейства в современном представлении изложена
Риделем (Riedel, 1980), где даны диагнозы родов и подродов.
Мною проведено исследование Daudebardiidae, обитающих на
территории СССР (Карпаты — 2 вида, Крым— 1 вид, западный
Кавказ — 4 вида), а также еще 3 видов, собранных И. М. Лихаревым
в Болгарии и любезно предоставленных мне для изучения. Детальный
анализ организации этих видов параллельно с учетом возрастной
изменчивости в деталях строения пениса убедительно показал,
что таксономическая структура семейства значительно сложнее, чем
это принималось до настоящего времени. Об этом же косвенно
свидетельствует и наличие исключений, упоминаемых Риделем в
приводимых им диагнозах родов и подродов.
7
Заказ 1696
97
Прежде чем приступить к изложению полученных материалов
и описаниям таксонов, необходимо уточнить значение некоторых
терминов. Термины „дистальный44 и „проксимальный44 в применении
к участкам полового аппарата употребляются в литературе по-раз-
ному: одни авторы за точку отсчета принимают половой атриум,
другие — гонаду. Думается, что правы вторые, но, во избежание
путаницы, в настоящей работе использованы термины „нижний44
(т. е. расположенный ближе к половому атриуму) и „верхний44
(удаленный от атриума).
Употребление терминов „эпифаллус44 и „бич44 („флагеллум44)
целиком определяется наличием сперматофора: эпифаллус — это
орган, где формируется сперматофор (случаи, когда сперматофоры
формируются в пенисе, как у Trigonochlamydidae — редчайшее
исключение), а бич — придаток, в котором образуется хвостовая
часть сперматофора. В применении к тем группам, члены которых
сперматофора не имеют, употребление термина „эпифаллус44 ограни-
чено: если есть основания считать, что отсутствие, сперматофора —
первичное состояние, то в этом случае термин „эпифаллус44 не-
применим. Если же отсутствие сперматофора — явление вторичное,
то обсуждаемый термин применим для обозначения отдела, гомоло-
гичного настоящему эпифаллусу. Что касается конкретного случая,
то среди Daudebardiidae известен один вид, у которого описан
сперматофор (Riedel, 1973) — это „Carpathica" cretica (Forcart,
1971). Следовательно, вздутый нижний конец семяпровода в данном
случае в полной мере заслуживает наименования эпифаллуса;
независимо от того, есть ли сперматофор у видов Daudebardia
s. str. и Libania, у них имеется отдел, морфологически и топогра-
фически соответствующий эпифаллусу „С.“ cretica, т. е: являющийся
его гомологом. Таким образом, применение этого термина к видам
названных родов вполне закономерно. Что касается членов прочих
родов, то у них имеет место разделение собственно пениса на
2 или 3 отдела, из которых ни один не гомологичен эпифаллусу
упомянутых таксонов, и в этих случаях речь идет об отделах пениса.
По изложенным причинам и слепой конец пениса у видов Carpathica
неверно называть бичем, как это делает Ридель (Riedel, 1980).
Последний термин, подлежащий обсуждению — „папилла44 Здесь
папиллой именуется орган, располагающийся в месте перехода
одной части совокупительного аппарата в другую и принимающий
участие в передаче спермиев. Этот орган может иметь вид трубки
(замкнутая папилла) или желобка (незамкнутая папилла); послед-
ний случай в пределах Daudebardiidae не отмечен. Ридель (Riedel,
1978, 1980) именует папиллами также мелкие сосочкообразные
выросты, нередко встречающиеся на внутренней поверхности
пениальной трубки. Для устранения возможной путаницы эти
маленькие выросты здесь именуются ворсинками.
Семейство Daudebardiidae Kobelt, 1906
Полуслизни Висцеральный мешок и раковина сильно реду-
цированы и сдвинуты к заднему концу тела. Подошва трехраздель-
98
ная. Половое отверстие отодвинуто от основания правого оммато-
фора. На спине располагаются 4 глубоких продольных борозды:
пара прямых затылочных и пара косых боковых. Покровы цефало-
подиума плотные, грубые. Глотка очень сильно развита, парные
глоточные ретракторы отделены от ретракторов омматофоров,
крепятся к столбику или (чаще) к левой стенке тела. Челюсть
развита слабо, слегка полулунно изогнута.
Раковина уховидная, тонкостенная, прижатая, состоит из 1—3
оборотов, гладкая или с тонкой спиральной скульптурой. Граница
между нуклеусом (эмбриональная часть раковины) и спатулой
(дефинитивная часть) выражена слабо. Максимальная длина тела
ползущего животного до 50 мм.
Женский отдел полового аппарата с.перивагинальной железой.
Пенис окутан чехлом, фиксированным только в его нижней части,
внутреннее строение пениса очень разнообразно. Семяприемник
объемистый, с очень коротким протоком, иногда почти сидячий.
Белковая железа обширная, неправильной формы.
Хищники, питаются преимущественно дождевыми червями.
Семейство распространено в западной Палеарктике, преимущест-
венно в восточном Средиземноморье, на Балканском п-ве и на
Карпатах. Северная граница ареала захватывает Подолию, Карпаты,
Польскую Юру, Судеты, Тюрингию, северную Вестфалию, Эльзас,
Англию; южная — южную Италию, Сицилию и Сардинию, северный
Алжир, Крит, Кипр, Израиль; к востоку ареал простирается до
Иракского Курдистана, западного Кавказа и Крыма.
Род Daudebardia Hartmann, 1821
Hartmann, 1821: 41; Clessin, 1878: 98 (Rufina, типовой вид Helix rufa Draparnaud,
1805, по обозначению Forcart, 1950: 108); Westerlund, 1886:4 (подрод Eudaudebardia,
типовой вид Helix rufa Draparnaud, 1805, по обозначению Forcart, 1950: 108);
Лихарев, Раммельмейер, 1952: 290; Riedel, 1980: 119.
Типовой вид — Helix rufa Draparnaud, 1805, по монотипии.
Нуклеус округлый, с медленно нарастающими оборотами, число
которых не менее двух. Пупок полностью или почти полностью
открыт. Пластинок в устье нет. Эмбриональная скульптура в виде
очень слабой спиральной исчерченности.
Чехол пениса крепится у атриума. Пенис на отделы не разделен,
внутри с продольными складками, нередко разбитыми на ряды
бугорков. Семяпровод впадает в пенис почти терминально, под
острым углом, близ пениса образует веретеновидное расширение,
папиллы не заключает. Половой ретрактор крепится к пенису
терминально одной лентой. Основание протока семяприемника не
расширено.
Daudebardia rufa (Draparnaud, 1805), рис. 1
Draparnaud, 1805: 118, t. 8, fig. 26—29 (Helix)-, Westerlund, 1886: 6 [Daude-
bardia naliciensis, fide Riedel, 1978: 23, locus typicus: „Galizien bei Przcmysl“,
типы в коллекции Вестерлунда в Гетеборге, Стокгольме (лектотип) и Зоологическом
ин-те АН СССР, Ленинград]; Riedel, 1978: 23; 1980: 192—193, Abb. 278—283.
Terra typica: Франция.
7=
99
Рис. 1. Половой аппарат Daudebardia rufa из Болгарии, окр. Софии,
3. VI. 1967
А — внешний Б — внутреннее строение пениса.
Типы неизвестны.
Материал 7 экз. из Болгарии: долина р. Рила, 27 VI.1969
(2 экз. juv.); близ Софии, горный массив Витоша, выше с. Ярлово,
15.VI.1962 (2 экз.); там же, у с. Железница, 15.VI.1969 (1 экз.
juv.); там же, ниже урочища Золотые мосты, 3.VI.1967 (2 экз.);
сборы и определения И. М. Лихарева.
Перивагинальная железа охватывает вагину и основание, протока
семяприемника по всему периметру, ее границы четко видны иногда
даже у неполовозрелых особей. Семяпровод относительно короткий,
не образует резких изгибов, довольно толстый, направляется
вдоль пениса, и, образовав крутой изгиб, впадает в пенис. Нижний
отрезок семяпровода веретеновидно расширен, толстостенный, с
одной, широкой продольной складкой. Внутри пениса залегает си-
стема продольных складок, в большей или меньшей степени разбитых
на ряды бугорков; у экземпляра, изображенного на рис. 1, эти
бугорки развиты четко, а у 2 экз. из с. Ярлово поперечные пере-
тяжки на складках выражены слабее. В верхней части пениса эти
100
складки теряют правильность ориентации, но в общем можно за-
метить, что они стягиваются к отверстию семяпровода.
Распространение Юго-восточная и Средняя Европа; на
территории СССР достоверных находок не сделано, но вид отмечен
в Польше близ Пшемысля (под названием D. haliciensis Westerlund,
1886), т. е. в непосредственной близости от нашей границы.
Мною исследован также 1 экз. D. brevipes (Draparnaud, 1805),
собранный И. М. Лихаревым в Болгарии, между Рилой и Пирином.
Вид, как отмечал Ридель (Riedel, 1967, S. 8), очень близок D. rufa
по всем анатомическим признакам, включая внутреннюю фактуру
пениса.
Род Bilania Schileyko, gen. nov.
Типовой вид — Daudebardia boettgeri. Clessin, 1883.
Название представляет собой анаграмму из Libania (Daudebar-
diidae.
Нуклеус (около 1,5 оборотов) овальный. Пупок наполовину
прикрыт наплывом колумеллярного края. Пластинок в устье нет.
Пениальный чехол крепится у атриума. Пенис в общем веретено-
видный. На внутренних его стенках залегают продольные складки,
которые в верхней части становятся более правильными; по мере
приближения к отверстию семяпровода появляются и становятся
все более резкими насечки, в результате чего близ отверстия семя-
провода складки преобразуются в ряды крупных ворсинок. Семя-
провод перед впадением в пенис образует резкий изгиб. Папиллы
в семяпроводе нет, но его нижняя часть заметно гофрирована.
Половой ретрактор крепится терминально к мускулизованным
тканям, в толще которых и происходит впадение семяпровода в
пенис; помимо основной ленты, имеется еще 1—2 дополнительных,
спускающихся ниже. Основание протока семяприемника не рас-
ширено.
Род наиболее близок роду Daudebardia-, отличается значительно
большей относительной длиной пениса, отсутствием веретеновид-
ного расширения в нижней части семяпровода, наличием до-
полнительных лент полового ретрактора, а также мощным разви-
тием ворсинок в верхней части пениса.
Ридель (Riedel, 1978, S. 48) относит типовой вид этого рода к
роду Carpathica на том основании, что у этого вида имеется „флагел-
лум“; этим термином Ридель именует участок мускулизованных
тканей, расположенных в верхней чйсти пениса; однако этот
участок столь резко отличается строением от слепого конца пениса
типового, вида рода Carpathica — С. calophana, что не может быть
сомнений в том, что указанные образования имеют совершенно
различную природу.
Bilania boettgeri (Clessin, 1883), рис. 2, IV; 3, IV; 4.
Clessin, 1883: 38, Taf. 2, Fig. 9, 10 (Daudebardia)-, Лихарев, Раммельмейер,
1952: 293, рис. 222 [D. (Libania)]-, Riedel, 1978: 48, Abb. 76—81 (Carpathica).
Locus typicus: „Strateis“ (Крым, окрестности Ялты).
101
Рис. 2. Раковины Daudebardiidae, вид сверху и снизу.
1 — Sicuersia lederi из окр. Ткварчели; //—5. heydeni in Бат\мского ботаническою сада.
/// — Inguria wagneri из окр. Гагры; IV — Bilania boettgeri из Ялты; V — Suchumiella jetschini
из окр. Ткварчели; VI — Carpathica calophana из с. Квасы близ Рахова. Масштаб — I мм.
102
Рис. 3. Раковины Daudebardiidae, вид спереди, и их фрагменты.
/ — Sieuersia lederi из окр. Ткварчели; // — 5. heydeni из Батумского ботанического сада;
/// и VII — Inguria wagneri из окр. Гагры {звездочка — спиральная складка). IV — Bilan>a
boettgeri из Ялты, V — Suchutniella jetschini из окр. Ткварчели. VI и VIII — Carpalhica calophana
из с. Квасы близ Рахова. Масштаб — I мм (относится только к изображениям слева)
Типы неизвестны.
Материал: 13 экз. из 6 местонахождений на южном берегу
Крыма (Ялта — 3 сбора, Ореанда, Феодосия) и у Новороссийска.
Раковина с небольшим нуклеусом и правильно закругленным
палатальным краем устья. На колумеллярном крае имеется мозоле-
видный наплыв с довольно четкими очертаниями, прикрывающий
пупок полностью или оставляющий от него очень узкую щель.
Поверхность нуклеуса практически гладкая (при увеличении свыше
50 раз неясно мелкозернистая); на поверхности спатулы, помимо
линий нарастания, заметны редкие радиальные бороздки. Ширина
раковины до 5,3 мм.
103
смт
Рис. 4. Половой аппарат Bilania boettgeri из Ялты, 23.11.1956.
А - внешний вид; Б - верхний конец пениса, сильно увеличен; в внутреннее строение пениса
Внутреннее строение Материал. Новороссийск, 1911-
(2 экз. juv.); Ялта, весна 1914 (4 экз.); Феодосия, 15.IV.1898
(2 экз.); сборы и определения О Ретовского; Ялта, 3.IX.1974, сбор
Я. С. Шапиро (juv.); Ялта, Никитский ботанический сад, 23.11.1956
(2 экз.); сборы и определения И. М. Лихарева; Ореанда, 18.V.1926
(2 juv.), сбор В. А. Линдгольма, определение автора.
Перивагинальная железа охватывает вагину и основание протока
семяприемника; длина вагины подвержена заметной изменчивости;
на рис. 4 изображен экземпляр с очень длинной вагиной, но этот
орган может быть значительно короче (см. Riedel, 1967, S. 53,
Abb. 76—78); вероятно, эти различия связаны со степенью релакса-
ции мышечных элементов, входящих в состав стенок вагины. Семя-
провод резких вздутий не образует, но перед впадением в пенис
в большей или меньшей степени сужен, а перед сужением иногда
имеются небольшие пережимы. Пенис более или менее явственно
подразделен на 2 отдела, внутренняя поверхность его у разных
особей несколько разнится. У одних (рис. 4) в нижней части
104
залегают тонкие продольные складки со слегка винтообразной
ориентацией; далее — „узел“ из мелких неправильных складок, вслед
за которым видны четкие короткие мощные пилястры, распадаю-
щиеся ближе к верхнему концу на ряды бугорков. В самой верхней
части эти бугорки преобразуются в крупные ворсинки, заметные
даже без вскрытия пениса сквозь его покровы. У других особей
„узел“ выражен слабее. Верхняя камера, содержащая ворсинки,
очень невелика и отделена от остальной части пениса широкой
вялой перетяжкой. Половой ретрактор крепится к изгибу протока
в месте перехода семяпровода в пенис; надо заметить, что (как
и в роде Illyrica— см. ниже) половой ретрактор в нижней части
превращен в рыхлый соединительнотканно-мускульный тяж, толща
которого пронизана нижней частью протока семяпровода, но, в отли-
чие от видов рода Illyrica, этот тяж очень короткий. Верхняя кромка
пениального чехла связана тонким тяжём с семяпроводом; кроме
того, имеются 1—3 дополнительных тонких ленты, соединяющие
нижнюю часть пениса с верхней; они отходят большей частью
непосредственно от полового ретрактора.
Распространение Горный Крым и окрестности Ново-
российска.
Род Sieversia Kobelt, 1880
Kobelt; 1880, in Iconogr., 7: 28 [Daudebardia (Sieversia)]; Riedel, 1980: 119
[D. (Daudebardia) part.].
Типовой вид — Daudebardia hey deni Boettger, 1879, по монотипии.
Нуклеус (около 2 оборотов) овальный. Пупок не более чем
наполовину прикрыт отворотом колумеллярного края. Пластинок в
устье нет.
Пениальный чехол крепится у атриума. Пенис в общем цилиндри-
ческий, состоит из 2 отделов. Внутри более длинного нижнего отдела
залегают продольные складки, которые примерно на середине этого
отдела образуют подобие сфинктера. Верхний отдел заключает
обширную тонкостенную папиллу со складками внутри; эти складки
могут преобразовываться в ряды ворсинок. В нижней части семя-
провода имеется крошечная папилла. Половой ретрактор крепится
субтерминально. Основание протока семяприемника не расширено.
Sieversia heydeni (Boettger, 1879), рис. 2, //; 3, //; 5.
Boettger, 1879: 3 (Daudebardia); 1880: 112 (Daudebardia), 113 (D. pawlenkoi,
locus typicus: Сурамский хребет, restr. Riedel, 1978), 112 (D. sieversi, locus typicus:
1имотисубани близ Боржоми); 1883: 139 (Daudebardia); Simroth, 1912: 85, 89
n (sic!) ]; Лихарев, Раммельмейер, 1952: 292 [D. Rufina)] ; 293, рис. 220
[D. (Rufina) sieversi]; Riedel, 1970: 35, Fig. 4—8 (Daudebardia); 1978: 15, Abb.
28-36 [D. (Daudebardia)].
Locus typicus: Казбек (указание ошибочно; вероятно, Сурамский хребет —
см. Boettger, 1880, S. 112).
Типы неизвестны.
105
Рис. 5. Половой аппарат Sieversia heydeni из Батумского ботанического сада,
7.IV.1971.
А — внешний вид; Б — внутреннее строение пениса; В — схема строения пениса особи, собранном
там же 24.IV.1957.
Материал 8 экз. из Красной Поляны, Нового Афона, Сочи,
Батуми.
Раковина уховидная, спатула с двумя сглаженными углами —
палатальным и палатально-базальным. Отворот колумеллярного
края нависает над пупком, прикрывая его примерно наполовину.
Поверхность нуклеуса с тончайшей спиральной исчерченностью и
более редкими радиальными линиями, в результате чего возникает
узор типа сеточки. На поверхности нуклеуса в последней его четверти
добавляются тонкие радиальные морщинки; на поверхности спатулы
эти морщинки становятся более грубыми и теряют регулярность.
Максимальная ширина раковины 4 мм.
Внутреннее строение. Материал. Красная Поляна,
18.VIII.1929, сбор В. М. Рылова, определение И. М. Лихарева (1 экз);
Сочи, парк „Интурист44, 5.V. 1964, сбор Е. Дмитриевой (1 экз.);
Сочи, парк санатория „Золотой Колос44, 10.IV. 1962, сбор А. С. Дов-
гялло (1 экз.); Батуми, Ботанический сад, 10. и 24.IV.1957, сборы
И. М. Лихарева (2 экз.); там же, 7.IV.1971 (2 экз.); сбор и опре-
деления автора.
106
Перивагинальная железа верхней частью целиком охватывает
вагину; в нижней части железа выражена лишь на одной сто-
роне вагины, противолежащей семяприемнику. Семяпровод тон-
кий, умеренно извитой; близ впадения в пенис имеется вздутие,
заключающее крошечную папиллу. Нижняя часть пениса (длина ее
составляет не менее 3/4 всей длины органа) толстостенная, внутри
с мощными продольными складками, которые в средней части про-
тока увеличиваются в высоту и формируют в совокупности нечто
вроде сфинктера. Далее пенис резко сужается, толщина его стенок
уменьшается, и этот короткий тонкий и тонкостенный вставочный
отдел переходит в расширенный верхний своеобразного строения.
Полупрозрачные стенки его близ вставочного отдела расслаиваются
и образуют тонкостенную замкнутую папиллу с обширной полостью;
внутри папиллы залегают несколько тонких продольных складок,
гребни которых видоизменены в ряды .крупных остроконечных
ворсинок. Половой ретрактор подходит к месту перехода семяпро-
вода в пенис; перед впадением в пенис семяпровод образует более
или менее ясно выраженный изгиб, края которого стянуты дери-
ватами полового ретрактора. Кроме того, обычно выражена также
лента, соединяющая верхнюю часть пениса с нижней.
Распространение. Приморские районы западного Кав-
каза: к северу до Сочи, к востоку до Сурамского хребта, к западу
до Турции (вилайет Гиресун).
Sieversia lederi (Boettger, 1881), рис. 2, /; 3, /; 6.
Boettger, 1881: 172, Taf. 7, A, В [Daudebardia (Rufina)]-, .1883. 140 [D. (Rufina)]-,
1884: 146 [D. (Rufina)]-, 1889; 5 [D. (Rufina)]’, Reibisch, 1892: 51 (D. rufa det.
err.); A. Wagner, 1906: 181 (D. rufa var. lederi)-, Simroth, 1912: 86, Taf. X, Fig. 44
(D. pontica, locus typicus. Пицунда), 87, Taf. X, Fig. 45 (D. caucasica, locus
typicus: Пицунда); Лихарев, Раммельмейер, 1952: 292. рис. 219 [D. (Rufina)]-,
Riedel, 1878: 19, Abb. 37-39 [D. (Daudebardia)]-, 1980: 120 [D. (Daudebardia)].
Locus typicus: Кутаиси.
Типы неизвестны.
M а т e p и а л. 7 экз. из Красной Поляны, Сочи, Майкопа,
Пицунды, Ткварчели.
Раковина отличается от раковины предыдущего вида отсутствием
элементов радиальной скульптуры на поверхности нуклеуса.
Внутреннее строение. Материал. Окр. Майкопа, запад-
нее р. Белой, 14.IX. 1902,' сбор X. Г Шапошникова, определение
Г Зимрота (1 экз.); Пицунда, 12.1.1907, сбор Казнакова и Шелков-
никова (голотип Daudebardia caucasica Simroth); Сочи, 15.V.1929,
сбор В. А. Линдгольма (1 экз.); Ткварчели, с. Акармара, 6.IX.1957,
сбор Н. Н. Акрамовского (1 экз.); Хоста (Сочи), 20.IX.1956, сбор
И. М. Лихарева (1 экз.); определения автора.
Наиболее постоянное, хотя и трудно обнаруживаемое отличие
от предыдущего вида заключается в том, что внутри верхнего
отдела пениса ворсинки отсутствуют, имеются лишь продольные
складки, стягивающиеся к отверстию семяпровода. Вставочный
107
Рис. 6. Половой аппарат Sieversia lederi из Пицунды, 12.1.1907.
А — внешний вид; Б — верхний конец пениса, вид с другой стороны; В — внутреннее строение пениса.
Г—схема строения пениса особи из окр. Ткварчели, 6.IX.1957
тонкостенный отдел выражен у разных особей в разной степени:
у более молодых он выражен более четко.
Распространение Черноморское побережье от Майкопа
до Бююкдере близ Стамбула на европейскому берегу Босфора
(Forcart, 1971).
Род Inguria Schileyko, gen. nov.
Типовой вид — Daudebardia wagneri Rosen, 1911.
Нуклеус (около 2 оборотов) овальный, пупок более чем на-
половину прикрыт. Базальный край устья продолжается на парие-
тальный, преобразуясь в высокую гребневидную спиральную пла-
стинку.
Пениальный чехол крепится у атриума. Пенис мешковидный,
на отделы не разделен, его внутренняя поверхность несет много-
численные складки разной величины. Апикальная стенка пениса
сильно утолщена, мускулизована и плотно охватывает пронизываю-
щий ее конец семяпровода, вокруг отверстия которого располагают-
ся крупные ворсинки. В нижней части семяпровода имеется кро-
шечная папилла. Половой ретрактор крепится терминально несколь-
кими лентами. Основание протока семяприемника не расширено.
Название рода образовано от названия р. Ингури.
108
Inguria wagneri (Rosen, 1911), рис. 2, /77; 3, 7/7 и V77; 7
Rosen, 1911: 94, Taf. II, Fig. 5, 6 [Daudebardia (Eudaudebardia, sectio Rufina)];
Simroth, 1912: 85, Taf. X, Fig. 43 (Daudebardia); Лихарев, Раммельмейер, 1952:
293, рис. 221 [D. (Rufina]; Riedel, „1978: 15 [D. (Daudebardia) heydeni part.].
Locus typicu.s: „Maikop mit nachster Umgebung“ (Майкоп и ближайшие
окрестности).
Типы неизвестны.
Материал 17 экз. из 6 местонахождений на западном Кав-
казе.
Основное отличие этого вида от других представителей семейства
заключается в наличии сильно развитой пластинки: базальный
край устья при переходе на колумеллярный обособляется и в виде
гребня переходит на париетальный край, изгибаясь по спирали
(рис. 3, VII). В области пупка от этой пластинки отходит языковид-
ный наплыв, более чем наполовину прикрывающий пупок. Другой
наплыв прикрывает часть поверхности париетального края, примы-
кающий к ангулярной области. Скульптура такая же, как у Sieversia
lederi. Диаметр раковины достигает 8,5 мм.
Внутреннее строение. Материал. Майкоп, сбор и опре-
деление О. Розена (1 экз. juv.— синтип?); Гагры, шоссе на Адлер,
8.IV.1911, сбор В. А. Линдгольма (3 экз.); Ткварчели, Акармара,
3.IX. 1957, сбор Н. Н. Акрамовского (2 экз. juv.); Малый Кавказ,
Арсиянский хребет, пер. Годерзи, 2.VII. 1874, сбор И. М. Лихарева
(1 экз.); долина р. Ингури у с. Хаиши, 10.IV.1971 (2 экз.); долина
р. Техури у с. Талери, 28.IV. 1971 (3 экз.); сборы и определения
автора.
Перивагинальная железа охватывает вагину и основание протока
семяприемника. Семяпровод длинный, извитой..Перед впадением его
в пенис диаметр протока внезапно увеличивается и более тонкий
участок „вдвигается** в расширенный; в результате возникает ко-
роткая папилла. Семяпровод впадает апикально, и, пронизывая
толстые сильно мускулизованные ткани апикального конца пениса,
открывается в полость последнего посредством короткой папиллы,
развитой в различной степени; у самых крупных экземпляров (из
окр. Гагры) по краю папиллы имеются крупные конические ворсинки.
Эти ворсинки представляют собой видоизмененные окончания скла-
док, залегающих внутри нижней расширенной части семяпровода.
Пенис мешковидный, широко сообщается с атриумом, внутри него
имеются многочисленные складки преимущественно продольной
ориентации; кроме того, широкая поперечная складка изолирует
полость в верхней части пениса рядом с папиллой. Половой ретрактор
подходит к пенису единой лентой, но в нижней части распадается
на несколько лент, сливающихся концами с тканями покровов
верхней части пениса. Имеются также 1-3 тонких мышечных тяжа,
связывающих между собой участки семяпровода.
Распространение Ареал простирается по западным отро-
гам Большого Кавказа от Майкопа на севере до долины р. Талери на
юге; одна находка сделана на западе Малого Кавказа (Годерзий-
ский перевал на Арсиянском хребте).
109
7 Половой аппарат Inguria wagneri из окр. Гагры, 8.IV. 1911.
А внешний вид; Б внутреннее строение пениса; В го же, схема.
Рис.
Род Suchumiella Н. Wagner, 1952 *
» Н. Wagner, 1945: 53 [Daudebardia (Szuchumiella nom. err.)]-, 1952: 95
(Suhumiella nom. err.), 145; Riedel, 1980: 119 [Daudebardia (Daudebardia) part.).
Типовой вид — Daudebardia (Libania) jetschini A.. Wagner, 1895,
по’обозначению Форкарта (Forcart, 1950, p. 116).
Нуклеус овальный, с медленно нарастающими (не менее 2)
оборотами. Пупок в виде очень узкой щели. Пластинок в устье нет.
Пениальный чехол крепится, заметно отступая от атриума, и
охватывает среднюю треть пениса. Пенис состоит из 3 отделов.
На внутренних стенках округлого нижнего отдела залегают несколь-
ко мощных складок, из которых выделяется одна, рассеченная
глубокими поперечными бороздами. Далее располагается длинный
тонкий отдел, охваченный чехлом; он отделен от нижнего сфинкте-
ром. Верхний отдел расширен, содержит продольные складки, разби-
тые на ряды бугорков; близ отверстия семяпровода эти бугорки пре-
вращаются в ворсинки. Половой ретрактор крепится к семяпроводу
близ его впадения в пенис 1—2 лентами. Семяпровод папиллы не
заключает. Основание протока семяприемника расширено.
Suchumiella jetschini (A. Wagner, 1895), рис. 2, V; 3, V; 8.
A. Wagner, 1895: 618 [Daudebardia (Libania)]; Simroth, 1910: 551, 558, Taf.
VIII, Fig. 51—54 (Daudebardia), 552, 558, Taf. VIII, Fig. 55, 56 (D. kalischewskii,
locus typicus: „Dorf Ashary“, с. Квемо-Ажара в долине p. Кодори, restr. Riedel,
1978); 1912: 87, Taf. X, Fig. 46 (Daudebardia); H. Wagner, 1945: 55 [D. (Szuchumiella)
simrothi, locus typicus: „Umgebung von Suchumi, am Fluss ,,Besla“, окр. Сухуми, restr.
Riedel, 1978]; Лихарев, Раммельмейер, 1952: 294, рис. 223 [D. (Libania)]; Hudec,
Lezhawa, 1969: 120, Abb. 19 [D. (Daudebardia)]; Riedel, 1978: 20, Abb. 40—43
[D. (Daudebardia)]; 1980: 120 [D. (? Daudebardia)].
Locus typicus: ,,Psirsk“ (с. Псирцха у Нового Афона, Абхазия).
Материал 16 экз. из 11 местонахождений на западном
Кавказе.
Поверхность нуклеуса практически лишена скульптуры, сильно
блестит. Поверхность спатулы покрыта тончайшей спиральной
струйчатостью. Спатула удлиненно-овальная, довольно правильной
формы, без сглаженных углов. Пупок наполовину прикрыт, колумел-
лярная мозоль развита в разной степени. Диаметр раковины до
6,8 мм.
Внутреннее строение Материал. С. Лентехи на р. Цхе-
нис-цкали, 4.VIII.1911, сбор Казнакова (1 экз.); гора Апионча,
Цебельда близ Сухуми, 9.IX.1908, сбор Ю. Н. Воронова (2 экз.);
Гудаута, 6.1.1907, сбор Казнакова и Шелковникова (1 экз.); опре-
деления Г Зимрота; с. Ажары (=Квемо-Ажара), 21.VII.1905, сбор
Калишевского (тип Daudebardia kalischewskii Simroth); Военно-Су-
хумская дорога, у Южного приюта, 8.VII.1974,. сбор и определение
И. М. Лихарева (1 экз.); долина р. Кодори выше с. Лета, 8.VII.1974,
* Согласно Международному кодексу зоологической номенклатуры (3-е изд.,
1985, ст. 32) валидным написанием этого родового названия будет Szuchumiella,
произведенное от венгерской транслитерации названия города Сухуми, а дата
опубликования — 1945 г. (прим. ред.).
111
Рис. 8. Половой аппарат Suchumiella jetschini из окр. Сухуми, р. Бесла,
25.VIII.1905.
А — внешний вид; Б — внутреннее строение пениса; В — пенис особи из окр. Ткварчели, 12.IX.1957.
(сильно сокращен). Звездочка — пилястр с поперечными насечками.
сбор я. И. Старобогатова (1 экз.); окр. Ткварчели, 3.IX.1957,
12.IX.1957, 3.IX.1957, сборы Н. Н. Акрамовского (3 экз.); опре-
деления автора.
Перивагинальная железа охватывает только вагину; проток семя-
приемника отходит ниже железы, практически от атриума. Основа-
ние протока семяприемника заметно расширено. Семяпровод длин-
ный, диаметр его на всем протяжении одинаков. Три отдела пениса
всегда выражены четко. Судя по тому, что пениальный чехол
крепится не у атриума, как у прочих Daudebardiidae, а значи-
тельно отступя, нижний отдел пениса есть, вероятно, дериват
атриума. Внутренние стенки этого отдела сильно складчатые; при
переходе в среднюю часть пениса складки увеличиваются и обра-
зуют подобие сфинктера. На растянутом пенисе (рис. 8, Б) одна
из складок, отличающаяся постоянством строения и положения,
несет резкие поперечные насечки и ориентирована почти продольно;
когда пенис сокращен, эта складка приобретает почти поперечную
ориентацию (рис. 8, В). Средний отдел пениса толстостенный, но
очень тонкий, с узким каналом, внутри него видны продольные
складки. Средний отдел переходит в расширенный верхний, заклю-
чающий те же складки. По мере приближения к отверстию семя-
112
провода на этих складках появляются поперечные насечки, которые
все больше углубляются и расширяются, превращая складки в
ряды ворсинок. Половой ретрактор крепится 1—2 лентами к нижней
части семяпровода; от этого места иногда отходят дополнительные
тяжи, уходящие под чехол пениса и прикрепляющиеся к средней
части пениса.
Надо иметь в виду, что пениальная трубка при жесткой фиксации
всегда сокращена, так как при этом сокращается вся мускулатура;
при мягкой же фиксации мускулатура пениса может быть и сокра-
щена, и расслаблена. В последнем случае это бывает тогда, когда
фиксируют еще не полностью умерщвленное животное, которое
способно, например, втянуть щупальца, но уже не в состоянии
мобилизовать мускулатуру покровов.
Распространение Приморские участки Черноморского
побережья Кавказа.
Род 1 llyrica К. Wagner, 1895
A. Wagner, 1895: 621 (Daudebardia, секция Illyrica)\ Riedel, 1980: 122 (Car-
pathica part.).
Типовой вид — Daudebardia (Jllyrica) stussineri A. Wagner, 1895,
по монотипии.
Нуклеус овальный (1 оборот). Пупок запаян или от него остается
очень узкая щель. Пластинок в устье нет. Поверхность нуклеуса
гладкая, спатула несет тонкую радиальную исчерченность с более
выраженными линиями нарастания.
Пениальный чехол крепится у атриума. Пенис состоит из 2 от-
делов. Нижний отдел очень тонкий, длинный, толстостенный, с
очень узким протоком. Верхний отдел расширенный, тонкостенный,
внутри него располагаются 1 или 2 стимулятора. Семяпровод впадает
у вершины пениса, охватывая половой ретрактор; расширений не
образует, папиллы не заключает. Половой ретрактор крепится
терминально одной лентой. Основание протока семяприемника
заметно расширено.
1 llyrica stussineri (A. Wagner, 1895), рис. 9.
A. Wagner, 1895: 624, t. 1, fig. 6, t. 2, fig. 13, t. 5, fig. 36 [Daudebardia
(lllyrica)]; Riedel, 1963: 9, Abb. 14—16 (Carpathica); 1967: 16, Karte 2, Abb. 18—20
(Carpathica); 1978: 46 (Carpathica).
Locus typicus: «Berg „grosser Klek“ bei Musulinski potok nachst Ogulin in
Croatien» (гора Б. Клек, окр. Огулина, Хорватия, Югославия).
Описание и изображение раковины см.: Riedel, 1963: 9, Abb.
14—16; 1967: 16, Abb. 18—20.
Внутреннее строение Материал. Окр. Софии, горный
массив Витоша, 14.VI.1969 (1 экз.); Стара Планина, Петроханский
перевал, 29.VI.1969 (1 экз.); сборы и определения И. М. Лихарева.
Границы перивагинальной железы довольно хорошо заметны:
если половой аппарат ориентирован так, как на рис. 9 (т. е. пенис
направлен от атриума вправо), то железа видна только слева,
8 Заказ 1696 1 13
Рис. 9. Половой аппарат Illyrica stussineri из unp. Софии, 14.VI. 1969.
А — внешний вид; Л' — внутреннее строение верхнего отде.i;i пениса.
охватывая нижнюю часть женского протока и оставляя свободным
основание короткого вздутого протока семяприемника. Семяпровод
тонкий, на всем протяжении равного диаметра, впадает в верхнюю
часть пениса, охватив предварительно основание полового ретрак-
тора. Длина нижней части пениса, охваченной чехлом, зависит от
сокращения: у экземпляра из Стара Планина эта часть почти
вдвое короче, чем у изображенного на рис. 9 из окр. Софии.
Внутри нижней части — тонкие продольные складки, просвет ее
очень узкий, стенки же довольно толстые. За пределами чехла,
ниже концевого вздутия пениса, рельефный узор на внутренних
стенках меняется: здесь залегают короткие, большей частью подково-
видные складки, имеющие преим\ щественно поперечную ориентацию.
Совокупность этих складок образует клапан, способный изолировать
нижний отдел пениса от верхнего. Внутри верхнего отдела имеются
два небольших, но четко выраженных выроста (стимуляторы).
114
Отверстие семяпровода находится в основании языковидного сти-
мулятора, несущего на поверхности множество очень тонких бо-
роздок; на рис. 9, Б стимулятор отогнут для того, чтобы было
видно отверстие семяпровода, но в нормальном положении это
отверстие прикрыто телом стимулятора. Кроме основного языковид-
ного, имеется дополнительный стимулятор цилиндрической формы,
основание которого окружено кольцевыми складками, разбитыми на
ряды бугорков. Половой ретрактор состоит из двух отделов. Верхний
отдел представляет собой тяж обычного строения; нижний отдел
шире и состоит из рыхлой соединительнотканно-мускульной ткани
(полости внутри нет). Является ли этот отдел дериватом пениса
или полового ретрактора, трудно сказать; мне кажется, что первое
предположение более вероятно. Примерно от места впадения семя-
провода, но с противоположной стороны, к верхней кромке пениса
отходит дополнительный мускульный тяж.
Распространение Горный вид- распространен от Южного
Тироля, Венецианских Альп, хребта Караванке до Албании, Сербии
и западной Болгарии (Riedel, 1967).
Помимо этого вида, я исследовал 1 экз. другого представителя
рода — „Carpathica" bielawskii Riedel, 1963, S. 5, Abb. 8—13 из
Болгарии, Странджа, Х.1968, сбор и определение И. М. Лихарева.
Этот вид очень близок /. stussineri и, несомненно, также принад-
лежит роду 1 llyrica\ наиболее существенное качественное различие
заключается в том, что у /. bielawskii в верхней части пениса
имеется только один языковидный стимулятор, а дополнительный
цилиндрический отсутствует.
Род Carpathica A. Wagner, 1895
A. Wagner, 1895: 621 (Daudebardia, секция Carpathica)-, Riedel, 1980: 122.
Типовой вид — Daudebardia (Carpatica') kimakowiczi Wagner,
1895 (syn. Daudebardia calophana Westerlund, 1881).
Нуклеус состоит из 1 —1,5 быстро нарастающих оборотов. Пупок
запаян. На колумеллярном крае имеется вертикальное утолщение.
Пениальный чехол крепится у атриума. Пенис внутри с ветвящи-
мися и аностомозирующими складками. Семяпровод впадает в
середину пениса, перед впадением сильно сужен, папиллы не заклю-
чает. В месте впадения семяпровода в пенис имеется латеральное
утолщение.. Половой ретрактор крепится одной лентой терминально
к слепой части пениса. Основание протока семяприемника, как
правило, расширено.
Carpathica calophana (Westerlund, 1881), рис. 2, VI; 3, VI, VIII; 10.
Westerlund, 1881: 51 [Daudebardia (Libania)]-, 1886: 9 [D. Libania)]-, Лихарев,
ГэвТТэТ^ЬЬ 2762: 277* РИС' ^CarPathica^ Riedel’ l978: 36’ Abb- 61 ~67’
Locus typicus: „Galizia, Przcmysl“ (Польша, Пшемысль).
Типы неизвестны (Стокгольм? Гетеборг?).
Материал 26 экз. из Ивано-Франковской и Закарпатской
областей.
Рис. 10. Половой аппарат Carpathica calophana из с. Квасы близ Рахова,
12—22.IX.1969.
А — внешний вид; Б — внутреннее строение uvinica. Звездочка - латеральное утолщение
» которое впадает семяпровод.
Раковина гетеротрофная, т. е. плоскость, в которой нарастает
первая половина начального оборота, лежит под углом к плоскости
дальнейшего нарастания раковины. Скорость нарастания оборотов
очень велика. Палатальный край правильно закруглен, без признаков
угловатости. Колумеллярный край утолщен и несет развитую в
различной степени вертикальную пластинку. Пупок отсутствует.
Поверхность нуклеуса совершенно гладкая, скульптура спатулы
характерна: здесь отсутствует исчерченность, но имеются тончайшие
продольные штрихи, заметные при увеличении 40—60 раз. Ширина
раковины до 5,0 мм.
Внутреннее строение Материал. Ивано-Франковская
обл., с Яремча, VIII.1963, сбор и определение И. М. Лихарева
(1 экз.); Закарпатская обл., с. Квасы близ Рахова, 12—22.IX.1969,
сбор и определение автора (6 экз.).
Н6
Перивагинальная железа развита в различной степени: у иссле-
дованных мною экземпляров она полностью охватывает вагину и
основание протока семяприемника, но у экземпляра из Бещад
(Восточные Карпаты, Польша) железа сдвинута латерально и
охватывает только основание протока семяприемника, не замыкаясь
вокруг вагины; у экземляра из Румынии, напротив, железа охва-
тывает вагину, оставляя полностью или частично свободным основа-
ние протока семяприемника (Riedel, 1978, S. 38—39, Abb. 62,
65—67). Семяпровод извитой, впадает в утолщение примерно на
середине длины пениса; перед впадением диаметр протока резко
сужается. Пенис цилиндрический или веретеновидный, рельеф по
всей его внутренней поверхности примерно одинаков и состоит
из складок, расположенныл без видимой системы; в нижней части
их ориентация преимущественно продольная, в верхней общая
ориентация отсутствует, одни складки внезапно обрываются, другие
изгибаются, третьи аностомозируют или распадаются на более
мелкие. Отверстие семяпровода окружено хорошо развитой коль-
цевой складкой. Половой ретрактор крепится апикально единой
лентой.
Распространение Карпаты и прилегающая к ним с се-
вера равнина.
Несмотря на то, что в моем распоряжении не было вида, описан-
ного как Pseudolibania (Carpathicd) cretica Forcart, 1950, считаю
необходимым выделить этот вид и близкие формы отдельным родом,
поскольку этот род играет важную роль в филогенетических построе-
ниях. Приводимая характеристика основана на литературных данных
(Simroth, 1891; A. Wagner, 1915*; Riedel, 1968, 1973).
Род Lotharia Schileyko, gen. nov.
Типовой вид — Pseudolibania (Carpathica) cretica Forcart, 1950.
Нуклеус состоит из 1,5 оборотов. Пупок полностью запаян.
Пластинок и складок в устье нет.
Пениальный чехол крепится у атриума. Хорошо выраженный
функционирующий эпифаллус впадает в пенис заметно выше его
середины. Внутри совокупительного аппарата папилл нет. Слепой
конец пениса имеет железистые стенки. В эпифаллусе формируется
гладкостенный заостренно-овальный сперматофор с тонкой хвостовой
нитью. Половой ретрактор крепится терминально к слепому концу
пениса. Основание протока семяприемника не расширено.
Анатомически род формально похож на Carpathica, отличается
наличием хорошо развитого функционирующего эпифаллуса и же-
лезистой выстилкой слепого отдела пениса.
Название рода образовано от имени швейцарского коллеги
д-ра Лотара Форкарта (Lothar Forcart), внесшего большой вклад
в изучение семейства и описавшего типовой вид рода.
?И1^Р?Т и ®агнеР идентифицировали этот вид с Daudebardia sauleyi (Bour-
gmgnat, 1852), живущим на западе Малой Азии и в прибрежных районах среди-
земноморского побережья Аравийского п-ва.
117
Заключительные замечания
Изложенные здесь и известные из литературы факты однозначно
свидетельствуют о том, что разнообразие в строении раковины и,
особенно, совокупительного аппарата внутри семейства настолько
велико, что сведение всего разнообразия форм к четырем таксонам
родового ранга (Daudebardia, Libania, Cibinia, Carpathica) не
адекватно системе семейства и не дает материала для анализа
филогении. В качестве яркого примера можно указать на род
Carpathica sensu Riedel (1980, 10). Отнесенные сюда виды объеди-
няются, в конечном счете, единственным чисто формальным приз-
наком: семяпровод (или эпифаллус) впадает в пенис не апикально.
Следовательно, принимается, что характер впадения семяпровода
(эпифаллуса) — наиболее весомый и стойкий признак, и игнори-
руется множество серьезных различий между группами включаемых
видов. Усложнение таксономической характеристики семейства в
данном случае — единственный способ приблизиться к пониманию
исторических отношений внутри семейства.
Независимо от того, относить ли обсуждаемый таксон в качестве
подсемейства к Zonitidae (Forcart, 1971; Riedel, 1980) или считать
его самостоятельным семейством (Н. Wagner, 1952; Лихарев,
Виктор, 1980), генеалогически он связан с Zonitidae и вместе с
Zonitidae (а также Parmacellidae и Milacidae) составляет надсе-
мейство Zonitoidea. В отличие от родственных, на мой взгляд,
групп, где наблюдается становление морфотипа слизней (Urocycli-
dae, Vitrinidae, Helicarionidae), для Daudebardiidae характерно, что
частичная редукция раковины не является транзитной стадией:
облик полуслизня в этом семействе стабилизировался и своеобразно
модифицированная раковина имеется всегда, без явных признаков
дальнейшей редукции.
Исходя из того, что предками Daudebardiidae были зонитоидные
формы, можно допустить, что наиболее архаичные признаки ра-
ковины заключаются в наличии открытого пупка и сравнительно
многооборотного нуклеуса. Крепление глоточных ретракторов к стол-
бику — примитивный признак и смещение их на стенку тела —
явление вторичное (Forcart, 1950: р. 111). В общем можно констати-
ровать, что процессы уменьшения числа оборотов нуклеуса, сужение
пупка и смещение фарингеальных ретракторов со столбика на
стенку тела проходят параллельно и в ряде случаев независимо
у представителей разных родов.
Что касается совокупительного аппарата, то среди Daudebar-
diidae имеется несколько достаточно резко различающихся вариантов
его строения, и установить наиболее архаичный вариант можно
лишь с помощью некоторых допущений. Первое и наиболее важное
допущение заключается в том, что наличие сперматофора (сле-
довательно, и эпифаллуса) для Zonitoidea — исходная особенность,
на что указывал еще Форкарт (Forcart, 1971, S. 22). В пользу
этого свидетельствует то обстоятельство, что сперматофоры имеются
у представителей (часто немногих) всех семейств Zonitoidea. Более
того, сперматофоры у видов этих семейств имеют общую особен-
на
ность — гладкую поверхность, лишенную ребер, выростов или шипов,
столь характерных для большинства других групп, члены которых
имеют сперматофоры. Если сперматофоры сохранились у всех
Parmacellidae, то у большинства членов трех других семейств они
редуцировались. Среди Daudebardiidae сперматофор описан у един-
ственного вида — Lothario, cretica (см. Riedel, 1973, S. 4—5, Abb. 5).
Это гладкостенная заостренно-овальная капсула с эластичной хво-
стовой нитью (рис. 11, /). Ридель (op. cit., S. 4) не уверен,
формируется ли он в эпифаллусе или во „флагеллу.ме“ (т. е. в слепом
конце пениса); можно не сомневаться, что в эпифаллусе: во-первых,
эпифаллус есть орган, основное назначение которого — формиро-
вание сперматофора; во-вторых, во „флагеллуме“ нет места для
формирования хвостовой нити; в-третьих^ у Oxychilus (Cellariopsis)
orientalis (Zonitidae) на эпифаллусе имеется слепой вырост (це-
кум) — и сперматофор соответственно несет шпору, формирующуюся
в этом цэкуме (см. Riedel, 1980, S. 18*1, Abb. 210); наконец,
на указанном рисунке четко видно, что сперматофор выходит из
эпифаллуса.
Наличие сперматофора — важный, но не единственный довод в
пользу того, чтобы расценивать половой аппарат L. cretica как
наиболее архаичный среди современных Daudebardiidae. Другой
довод заключается в том, что свойственный обсуждаемому виду
облик совокупительного аппарата весьма обычен среди западно-
палеарктических Zonitidae. Вместе с тем закрытый пупок и крепле-
ние глоточных ретракторов к стенке тела не позволяют считать этот
вид предковым в отношении прочих Daudebardiidae. Правильнее
предположить, что эволюция его раковины и мышечного механизма,
обслуживающего глотку, происходила быстрее, чем утрата сперма-
тофора и трансформация элементов совокупительного аппарата.
Судя по всему, обратная картина наблюдается у Daudebardia
s. str.: сохранив примитивные конхологичеСкие черты и архаичное
крепление глоточных ретракторов к столбику, виды этого рода
утратили сперматофор и слепой конец пениса, в результате чего
впадение эпифаллуса в пенис становится почти терминальным
(рис. 11, //). Поскольку половой аппарат вообще более консерва-
тивен, чем раковина, уместно допустить, что гипотетическая исход-
ная форма Daudebardiidae характеризовалась раковиной, свойствен-
ной Daudebardia s. str. и половым аппаратом, свойственным Lotharia.
От этой гипотетической формы оба названных рода развились,
вероятно, независимо. Род Carpathica (в моем понимании) легко
выводится из Lotharia за счет редукции сперматофора и, следова-
тельно, эпифаллуса (рис. 11, /У); вздутие, куда впадает семе-
провод, можно расценить как дериват (рудимент?) эпифаллуса:
оно хорошо развито у С. denticulata и значительно слабее у С. calo-
phana. Сходным образом половой аппарат Libania выводится из
полового аппарата Daudebardia хотя сперматофора у Libania
наверняка нет, у видов этого рода сохраняется, в модифицирован-
ном виде, эпифаллус и терминальное впадение его в пенис
(рис.11, III). Различия между Libania saulcyi и видом, описанным
как Carpathica wirthi Forcart, 1971, сводятся к двум пунктам:
119
Рис. 11. Схемы строения совокупительного аппарата Daudebardiidae и филогенетические связи между некоторыми вариантами.
/ — Lotharia; // — Daudebardia; III — Libania; IV—Carpathica; V—Cibinia; V! — Bilania; VII — Suchumiella; VIII — Sieversia; IX — «Daudebardia* dacica;
X — lllprica: XI — «Daudebardia* wiktori. Сперматофор зачернен, железистые стенки заштрихованы.
у первого вида проток внутри эпифаллуса прямой, и половой ретрак-
тор крепится непосредственно к месту перехода эпифаллуса в пенис,
отмеченному только резким перегибом протока; у второго вида
проток эпифаллуса извит, а от перегиба на границе пениса и
эпифаллуса отходит вначале узкий тяж (Форкарт именует его
флагеллумом), к которому и крепится половой ретрактор. Судя
по изображению (Forcart, 1971, S. 35, Abb. 8), это „флагеллум“
полости не заключает и имеет такое же строение, как у Illyrica
stussineri (см. выше, рис. 9), т. е. является дериватом ретрактора.
Таким образом, оба вида принадлежат одному роду, причем Libania
saulcyi надо считать предковой формой в отношении Libania wirthi.
Вообще организация совокупительного аппарата Lotharia
настолько универсальна, что из нее можно вывести большую часть
вариантов из числа реально наблюдающихся среди Daudebardiidae.
В общем все преобразования этой схемы сводятся к двум основным
путям: 1) смещение места впадения эпифаллуса (или семяпровода)
по пенису до апикального положения и 2) морфологические измене-
ния одного, двух или трех отделов пениса (в полном наборе имеются:
утолщенный нижний отдел; суженный вставочный; расширенный
верхний). Первый путь реализуется во всех линиях развития, кроме
Carpathica (здесь, напротив, место впадения семяпровода несколько
смещается в направлении атриума). Второй путь наиболее акценти-
рован у Cibinia, поскольку преобразованиями здесь затронуты все
три отдела пениса (рис. 11, V). По существу, связь между Lotharia
и Cibinia можно подозревать главным образом потому, что это
единственные роды среди Daudebardiidae, у которых верхний отдел
пениса имеет железистые стенки. Кроме того, если у типового
вида Cibinia — С. transsilvanica— эпифаллус не выражен (Riedel,
1978, S. 33, Abb. 58, 59), to u С. папа он вполне заметен (op. cit.,
S. 34, Abb. 60). Вместе с тем нельзя не видеть и резких различий
между совокупительными аппаратами Lotharia и Cibinia (кроме
ярких внешних различий, наличие ворсинчатой выстилки внутренней
поверхности цилиндрического отдела пениса у С. transsilvanica').
Поэтому о непосредственной преемственности между родами го-
ворить не приходится.
Кавказские роды (Sieversia, Inguria, Suchumiella), а также
крымский род Bilania — древние, аберрантные и сильно специализи-
рованные группы, четко изолированные от остальных Daudebardiidae
(рис. 11, VI, VII, VIII). Если род Suchumiella пошел по пути
разрастания части атриума (судя по месту прикрепления пениаль-
ного чехла, нижний отдел пениса есть дериват полового атриума)
и приобретения более мелких особенностей (рис. 8, Б), то эволюция
видов Sieversia более глубоко затронула верхнюю часть мужского
отдела: наличие двух папилл — внутри семяпровода и внутри
верхнего отдела пениса — свидетельствует о полной и давней утрате
сперматофора и становлении сложных процессов транспорта и на-
коплений аггрегаций спермиев. У родственного рода Inguria в
папиллу пениса превращен нижний конец семяпровода, пронизываю-
щий покровы верхней стенки пениса.
121
В заключение хочу подчеркнуть, что рис. 11 не является филогене-
тической схемой. Здесь представлены схематизированные изобра-
жения основных вариантов строения совокупительного аппарата и
для некоторых из них — предполагаемые связи. Причины, по которым
ряд вариантов строения не включен в цепи предполагаемых связей,
заключаются в своеобразии их внешнего облика и недостаточной
изученности внутренней организации.
Обозначения в иллюстрациях:
БЖ — белковая железа; В — вагина; ВП — верхний отдел пениса; ДМЛ -
дополнительные мускульные ленты: ДСТ — дополнительный стимулятор; НП —
нижний отдел пениса; ОП — основание протока семяприемника; ОС — отверстие
семяпровода; П — пенис; ПЖ — перивагинальная железа; ПП — папилла пениса;
ПР — половой ретрактор; ПС — папилла семяпровода; PC — расширение семяпро-
вода; С — семяпровод; СМТ — соединительнотканно-мускульный тяж; СП — семя-
приемник; СРП — средний отдел пениса; СТ — стимулятор; СФ — сфинктер;
ЧП — чехол пениса.
ЛИТЕРАТУРА
Лихарев И. М., Виктор А. Й. Слизни фауны СССР и сопредельных стран (Gastropoda
terrestria nuda).— В кн.: Фауна СССР, нов. сер., № 122, Моллюски, т. 3,
вып. 5. Л., 1980, с. 1—437.
Лихарев И. М., Раммельмейер Е. С. Наземные моллюски фауны СССР,— В кн.:
Опред. по фауне СССР. Вып. 43. М.-Л., 1952, с. 1—511.
Boettger О. Kaukasische Mollusken gesammelt von Herrn Hans Leder in Paskau.—
Jahrb. dtsch. malak. Ges., 1879, Bd 6, S. 1—42.
Boettger O. Kaukasische Mollusken gesammelt von Herrn Hans Leder, z. Z. in
Tiflis.—Jahrb. dtsch. malak. Ges., 1880, Bd 7, S. 109—161.
Boettger O. Sechstes Verzeichniss transkaukasi^cher, armenischer und nordpersischer
Mollusken aus Sendungen der Herren Hans Leder, z. Z. in Kutais und Dr.
G. Sievers in St. Petersburg.— Jahrb. dtsch. malak. Ges., 1881, Bd 8, S. 167—261.
Boettger O. Siebentes Verzeichniss von Mollusken der Kaukasuslander, nach Sen-
dungen des Hrn. Hans Leder, z. Z. in Helenendorf bei Elisabetpol (Trans-
caucasien).— Jahrb. dtsch. malak. Ges., 1883, Bd 10, S. 135—198.
Boettger O. Liste der von Herrn O. Retowski in Abchasien gesammelten Binnenmol-
lusken.—Ber. senckenb. naturf. Ges., 1884, S. 146—155.
Boettger O. Zehntes Verzeichnis (XII) von Mollusken der Kaukasuslander, nach
Sendungen des Hans Leder, z. Z. in Helenendorf bei Elisabetpol (Transkau-
kasien).— Ber. senckenb. naturf. Ges., 1889, S. 3—37.
Clessin S. Eine mitteldeutsche Daudebardie.— Malak. Bl., 1878, Bd 25, S. 96—99.
Clessin S. Anhang zur Molluskenfauna der Krim.— Malak. Bl., N. F., 1883, Bd 6,
S. 37—52.
Draparnaud J.-P.-R. Histoire naturelle des mollusques terrestres et fluviatiles de
la France. Paris, 1805, p. 1 — 164.
Forcart L. Systematique des mollusques en forme de Daudebardia et revision des
especes d'Anatolie et de Tile de Crete.— J. Conchyl., 1950, vol. 90, p. 107—117
Forcart L. Revision der Daudebardiinae von Vorderasien (excl. Kaukasusgebiet).—
Arch. Moll., 1971, Bd 101, N 1/4, S. 21—38.
Hartmann J. D. W System der Erd:< und Susswasser Gastropoden Europa's.— In:
Sturm. Deutsch., Fauna (6, Wurrner), 1821, Bd 5.
Hudec V Lezhawa G. I. Bemerkungen zur Erforschung der Landmollusken der
Grusinischen Sozialistischen Sowjetrepublik (II).— Sborn. Nar. Muz. v Praze,
1969, vol. 25 B, N 3, S. 93—155.
Kobelt W. Iconographie der Land;, und Susswasser-Mollusken mit vorziiglicher
Beriicksichtigung der europaischen noch nicht abgebildeten Arten. 1880
(1879—1880), Bd 7, S. 1—94.
122
Reibisch P.~ Ueber die bei Gelegenheit einer Reise nach dem Kaukasus gesammelten
Gehauseschnecken.— SB naturf. Ges., Leipzig, 1892, S. 49—58.
Riedel A. Zwei neue Zonitidae (Gastropoda) aus Sudostbulgarien.— Ann. zool.,
Warszawa, 1963, Tom 20, N 22, S. 1 — 13 (473—485).
Riedel A. Daudebardiinae (Gastropoda, Zonitidae) Bulgariens.— Ann. zool.,
Warszawa, 1967, Tom 24, N 8, S. 1—21 (463—483).
Riedel A. Zonitidae (Gastropoda) Kretas.— Ann. zool., Warszawa, 1968, Tom 25,
N 13, S. 1—65 (473—537).
Riedel A. Zonitidae (Gastropoda, Pulmonata) gesammelt von der Niederlandischen
Biologischen Expedition in die Turkei in 1959.— Zool. Meded., 1970, Deel 45,
N 3, S. 25—42.
Riedel A. Eine Zonitiden-Ausbeute (Gastropoda) von den griechischen Inseln.—
Fragm. faun., Warszawa, 1973, Tom 19, N 2, S. 1—6 (21—26).
Riedel A. Kritische Bemerkungen und Erganzungen zur Kenntnis der Subfamilie
Daudebardiinae (Gastropoda, Zonitidae) mit Verzeichnis aller akzeptieren
Arten.— Ann. Zool., Warszawa, 1978, Tom 34, N 8, S. 1—68 (139—206).
Riedel A. Genera Zonitidarum. Rotterdam, 1980, S. 1 — 197
Rosen 0. Die Mollusken Ciskaukasiens und speciell des Kuban-Gebietes.— Ежегод.
Зоол. муз., 1911, т. 16, S. 86—142.
Simroth, H. Die Nacktschnecken der portugiesisch-azorischen Fauna in ihrem
Verhaltniss zu denen der palaarktischen Region uberhaupt.— Nova Acta KsL
Leop.-Carol, dtsch. Akad. Natur., Halle, 1891, Bd 56, N 2, S. 201—424.
Simroth H. Kaukasische und asiatische Limaciden. und Raublungenschnecken.—
Ежегод. Зоол. муз., 1910, т. 15, S. 499—560.
Simroth H. Neue Beitrage zur Kenntnis der kaukasischen Nacktschnecken.— Изв.
Кавказск. муз. 1912, т. 6, S. 1 —140.
Wagner A. Die Arten des Genus Daudebardia Hartmann in Europa und Westasien.—
Denkschr. math.-nat. Cl. kais. Akad. Wiss., 1895, Bd 62, S. 609—626.
Wagner A. Bemerkungen’ zum Genus Daudebardia Hartmann.— NachrbL dtsch.
malak. Ges., 1906, Bd 38, S. 177-186.
Wagner A. Beitrage zur 'Anatomic und Systematik der Stylommatophoren aus dem
Gebiete der Monarchic und der angrenzenden Balkanlander.— Denkschr.
math.-nat. Cl. kais. Akad. Wiss., 1915, Bd 91, S. 429—498.
Wagner H. New systematical researches on carnivorous slugs.— Ann. hist.-nat. Mus.
nat. hung., 1945, Bd 38, p. 53—57.
Wagner H. Die Raublungenschnecken — Gattungen Daudebardia, Testacella und
Poiretia. Budapest, 1952, S. 1—259.
Westerland C. A. Malakologiska bidrag.— (Sfvers. Kongl. Vetensk.-Akad. Forhandl.,
1881, Bd 38, 1 4, p. 35—69.
Westerland C. A. Fauna der in der Palaarctischen Region... I. Fam. Testacellidae,
Glandinidae, Vitrinidae et Leucochroidae. Lund, 1886, S. 1—88.
A. A. Schileyko
SOME DATA ON THE ANATOMY AND THE TAXONOMY
OF DAUDEBARDIIDAE (GASTROPODA PULMONATA)
n Jhue structure of shell and reproductive system are studied in 10 species of
Daudebardiidae, 7 of which living in the U. S. S. R. The* family consists of at least
genera (among them Bilania gen. nov. with type species Daudebardia boettgeri
/n/AS,n’ 18831 in£uria £en- nov- with type species D. wagneri Rosen, 1911 and
na^ia .£en- nov- with type species Pseudolibania cretica Forcart, 1950). The
тЛГ°и ^-va systern of Lotharia is the most archaic having functioning epiphallus,
ine phylogenetic considerations are presented.
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
А. А. Шилейко
СИСТЕМА И ФИЛОГЕНИЯ VITRINIDAE
(GASTROPODA PULMONATA)
Сравнительно небольшое семейство Vitrinidae изучено довольно
детально. Известно строение полового аппарата и радулы боль-
шинства видов, разработана в целом система семейства, предло-
жена морфо-функциональная оценка придаточных органов полового
аппарата, высказан ряд соображений по филогении группы (обзор
см. Forcart, 1944).
Первоначально задача настоящего .исследования заключалась
в осуществлении ревизии видов витринид, обитающих на территории
СССР; однако в ходе работы были выяснены некоторые факты,
позволившие выдвинуть гипотезы относительно исторических связей
как между отдельными группами внутри семейства, так и между
Vitrinidae и другими таксонами.
Фактическую основу работы составили коллекции Зоологического
института АН СССР, Зоологического музея Московского универси-
тета и сборы автора, сделанные в различных регионах нашей
страны; значительная часть этих сборов хранится в коллекциях
двух названных учреждений.
Несколько слов о терминах, использованных в описаниях. Стенки
пениса на той стороне, куда впадает семяпровод, а также апикаль-
ной части пениса, у Vitrinidae сильно утолщены и представляют
собою железу, которая именуется пениальной железой. Вагина у
многих видов имеет утолщенные стенки, также железистые; это
утолщение носит название вагинальной железы. У большинства
видов, имеющих вагинальную железу, нижний ее конец вытянут
в коническую трубку, именуемую вагинальной папиллой.
Наконец, у ряда видов в атриум впадает хорошо развитый
орган, который разные авторы называли по-разному: придаточным
органом (Anhangorgan), рукой оплодотворения (Brachium copula-
torium, Begattungsarm), мешком стрелы (Pfeilsack), любовным кин-
жалом (Liebesdolch), женской половой железой (weiblichen
Genitaldruse), любовной железой (love gland), железой стрелы
(dart gland)
Поскольку этот аксессорный орган аналогичен и гомологичен,
на мой взгляд, такому же органу Ariophantidae и близких групп,
здесь он называется саркобелумом. В схеме саркобелум представ-
ляет собой удлиненное мускулизованное железистое тело (папилла
саркобелума), к которому по периметру крепится трубчатая соеди-
нительнотканная оболочка (чехол саркобелума), сливающаяся на
другом конце с тканями полового атриума. Поскольку папилла
124
саркобелума обычно „вдвинута" в полость чехла, этот чехол напо-
минает не до конца вывернутый палец перчатки; та часть чехла,
которая видна снаружи, именуется здесь внешней стенкой чехла,
а часть, прилегающая к папилле — внутренней стенкой чехла сарко-
белума. Чехол может крепиться к самой нижней части папиллы,
либо отступя от нее — в последнем случае возникает свободный
конец папиллы.
Семейство Vitrinidae Fitzinger, 1833
Улитки или полуслизни (полностью наружная раковина утрачена
только у одного вида — Plutonia atlantica). В соответствии со
степенью редукции раковины висцеральный мешок в разной мере
редуцирован и сдвинут к заднему концу тела. По мере редукции
раковины увеличиваются лопасти мантийного валика, частично при-
крывающие раковину. Подошва трехраздельная. Покровы цефало-
подиума тонкие. Глотка умеренно развита, парные глоточные ре-
тракторы ответвляются от ретракторов омматофоров. Челюсть с
одним срединным выступом.
Раковина от низкоконической до уховидной, очень тонкостенная,
составлена 1,5—3,5 оборотами/гладкая или с тонкой спиральной
струйчатостью; изредка скульптура в виде тонкой радиальной
морщинистости. Эмбриональные обороты гладкие, с радиальными
морщинами или с микроскопическими точками, обычно собранными
в спиральные ряды. Последний оборот преобладает над остальными,
устье большое или очень большое; по мере редукции раковины
базальный край преобразуется в кожистую оторочку (т. е. в этом
участке от раковины остается только периостракум), которая может
резорбироваться. Максимальная длина ползущего животного до
50 мм (у видов, живущих в СССР — не более 20 мм).
Женский отдел полового аппарата с придатками: это либо ваги-
нальная железа, либо саркобелум, заключающий одну или две
папиллы; иногда придаточные органы женского отдела отсутствуют.
Пенис с железой, залегающей в его расширенных стенках, и с
сильно развитым продольным пилястром. Проток семяпровода
пронизывает пениальную железу. Семяприемник развит в различной
степени, у одного рода отсутствует (Тrochovitrina). Белковая железа
сравнительно небольшая, неправильной формы.
Растительноядные животные (исключение — хищные Plutonia).
Семейство распространено в Голарктике, основное разнообразие
в Средней, Западной и Южной Европе, высокогорных районах
Африки (центральной и северной) и островах восточной Атлантики
(Азорские, Канарские, Мадейра, о-ва Зеленого Мыса).
Подсемейство Phenacolimacidas Schileyko, subfam. nov.
Раковина нормально развита (лишь у хищных Plutonia наружная
раковина отсутствует) или слегка уховидная; базальный край может
нести тонкую периостракальную оторочку. Вагина длинная, состоит
125
из двух отделов; верхний отдел заключает хорошо развитую железу,
обычно снабженную папиллой. Саркобелум и чехол пениса от-
сутствуют.
Род Insulivitrina Hesse, 1923
Hesse, 1923: 131, 1924: 226; Forcart, 1944: 654, Wenz, Zilch, 1959-1960: 238
[Phenacolitnax (Insulivitrina)'].
Типовой вид — Helicolitnax lamarcki Ferussac, 1821, по последую-
щему обозначению (Hesse, 1924, S. 226).
Раковина от низко конической до слегка уховидной, базальный
край без периостракальной оторочки или она развита слабо. Перифе-
рия раковины округлая. Вагинальная железа с хорошо развитой
сильно мускулизованной папиллой, имеющей очень узкий проток.
Наружная часть папиллы имеет обкладку из железистой ткани.
Яйцевод длинный или умеренно длинный. Семяприемник с очень
тонким довольно длинным протоком, образующим один резкий
изгиб. Пенис короткий, булавовидный, верхний конец его иногда
сужен, без наружных придатков.
Insulivitrina teneriffae (Quoy et Gaimard, 1832), рис. 1, Б; 2
Quoy, Gaimard, 1832: 142, pl. 3, fig. 4—9 (Vitrina).
Locus typicus: о. Тенериффе (Канарские о-ва).
Гофман (Hoffman, 1929, S. 227) считает этот вид синонимом
типового вида рода. Поскольку в моем материале не было ма-
териала для сравнения, вид здесь обозначается так, как указано
на этикетке: „Vitrina teneriffa, Teneriffa“; дата сбора и фамилия
сборщика не указаны. Материал хранится в Зоологическом му-
зее МГУ.
Раковина низкоконическая, сравнительно твердостенная, устье
без периостракальной оторочки. На эмбриональных оборотах распо-
лагаются очень четкие крупные точковидные вмятины, собранные
в редкие хорошо выраженные ряды. На дефинитивных оборотах —
тонкая четкая неправильная спиральная струйчатость. Максималь-
ный диаметр раковины исследованных экземпляров 6,8, высота 3,4 мм.
Внутреннее строение. Материал. 3 экз. из о. Тенериффе
(Канарские о-ва); дата сбора, фамилия сборщика и определившего
лица не указаны.
Спермовидукт чрезвычайно короткий: семяпровод отходит близ
нижнего края белковой железы. Соответственно яйцевод очень
длинный, образует 1—2 резких изгиба. Резко сужаясь, яйцевод
впадает в расширенный отдел вагины; от этого же места отходит
очень тонкий и довольно длинный проток семяприемника, закан-
чивающийся обширным тонкостенным резервуаром; стенки семя-
приемника у всех исследованных животных были в спавшемся
состоянии. Железистый отдел вагины преобразован в мощную
длинную сильно мускулизованную папиллу, окруженную наподобие
муфты слоем рыхлой железистой ткани. Папилла в той или иной
мере „вдвинута“ в нижний отдел вагины, за счет чего в верхней
126
Рис. 1. Раковины Phenacolimax annularis (Л) из
Орджоникидзе, гора Сапинкая Будка, и Insulivitrina tene-
riffae (Б) из о. Тенериффе (Канарские о-ва).
Рис. 2. Половой аппарат Insulivitrina teneriffae из о. Тенериффе.
А — внешний вид; Б — внутреннее строение нижнего отдела.
части нижнего отдела вагины возникает кольцевой перегиб. Внутрен-
ние стенки нижнего отдела вагины лишены постоянных складок,
в средней части заметно утолщены. Пенис веретеновидный, недлин-
ный; стенки его верхней части заключают пениальную железу,
в толще которой проходит дистальная часть семяпровода. Полость
пениса узкополулунная, щелевидная. Внутри нижней части пениса
имеется один высокий пилястр, сходящий в полости атриума на-нет.
Половой ретрактор крепится к пенису апикально.
Распространение О. Тенериффе (Канарские о-ва).
Род Phenacolimax Stabile, 1859
Stabile, 1859; 422; Bourguignat, 1880: 4 (Gallandia, типовой вид Vitrina
conoidea Martens, 1874, по первоначальному обозначению); Hesse, 1923: 82, 96
(Vitrina, non Draparnaud, 1801, типовой вид Helico-limax major Ferussac, 1807,
по обозначению Hesse, 1923: 82); Forcart, 1944: 651; Лихарев, Раммельмейер,
1952: 296 (Helicolimax part.); Wenz, Zilch, 1959—1960: 237.
Типовой вид — Helico-limax major Ferussac, 1807, по обозначе-
нию Фишера (Fischer in Paulucci, 1878: 24).
Раковина низкоконическая, базальный край устья без периостра-
кальной оторочки. Периферия раковины округлая. Вагинальная
железа с толстостенной папиллой, имеющей сравнительно широкий
проток. Периферический слой папиллы состоит в основном из
радиальных пучков мускульной ткани, внутренний слой — из желе-
зистой. Семяприемник с очень коротким прямым протоком. Пенис
короткий, конический или мешковидный, без придатков.
Phenacolimax annularis (Studer, 1820), рис. 1, А; 3.
Studer, 1820: 86 (Glischrus (Hyalina)]-, Мартенс, 1874: 8, табл. 1, рис. 5 [Vi-
trina (?) conoidea, locus typicus restr. nov.: окрестности Самарканда]; Mousson,
1876: 137 (Helicarion sieversi, locus typicus: Шау-Нахадам у оз. Табацкури, Грузия);
Boettger, 1878: 121 (Vitrina subconica, locus typicus: Казбек); 1879: 392, Taf. 10,
Fig. 4 [V. (Phenacolimax) komarowi, locus typicus: Армения, Кипчаг, Алагез];
Westerlund, 1896: 22 [V (Oligolimax) annularis forma caucasica, locus typicus:
Тбилиси]'; Boettger, 1889: 936, Taf. 27, Fig. 5 [V (Oligolimax) raddei, locus typicus:
Агдаг, Копетдаг]; Forcart, 1944: 658, Abb. 11, Pl. 2, Fig. 8 (Oligolimax)[ Лихарев, Рам-
мельмейер, 1952: 299, рис. 227 [Helicolimax (Oligolimax)]-, Wenz, Zilch, 1959—1960:
237, Abb. 839 [Ph. (Gallandia) conoidea]-, Forcart, 1960: 138 (Nachtrag); Zulch, 1979:
87, Taf. 7, Fig. 10 (фото лектотипа V annularis f. caucasica Westerlund, 1886), Taf. 8,
Fig. 13 (фото лектотипа V raddei Boettger, 1889), 88, Taf. 8, Fig. 14 (фото
лектотипа V. komarowi Boettger, 1879), Taf. 8, Fig. 15 (фото лектотипа V subconica
Boettger, 1878).
Форкарт (Forcart, 1960) справедливо замечает, что если группа
Phenacolimax annularis, ранее ошибочно относимая к подроду
Oligolimax, и заслуживает обособления, то она должна именоваться
Gallandia Bourguignat, 1880 [типовой вид Vitrina conoidea Artens,
1874, syn. Glischrus (Hyalina) annularis Studer, 1820]. Однако
анатомически Helicolimax major Ferussac, 1807 и Glishrus annula-
ris Studer, 1820 настолько близки, что есть все основания считать их
принадлежащими одному роду (подроду), т. е. название Gallandia
есть младший субъективный синоним названия Phenacolimax-
Locus typicus: „In Wallis bey Sitten“ [Швейцария, кантон Вале
(Валлис) ]
128
Рис. 3. Половой аппарат Phenacolimax annularis из Орджоникидзе, гора
Сап и икая Будка, 8. XI. 1952 г
/1 Б внутреннее строение нижнего отдела.
Материал Более 100 экз. из 46 местонахождений в Ко-
петдаге, Таджикистане, Афганистане, Казахстане и на Кавказе.
Раковина низкокубаревидная, со сравнительно высоким завитком,
высота которого превышает половину высоту устья. Оборотов до 4,
выпуклых. Скульптура эмбриональных оборотов представлена тонкой
радиальной морщинистостью, которая выражена в разной степени;
у старых экземпляров морщинистость в той или иной мере стерта.
Дефинитивные обороты с довольно грубыми неравномерными
радиальными морщинками; нередко имеется также слабая спираль-
ная исчерченность. Диаметр раковины до 6,5, высота до 4,25 мм.
Внутреннее строение Материал. Северный Кавказ, окр.
Орджоникидзе, гора Сапицкая Будка, 8.XI. 1952 г., сбор и определе-
ние 3. И. Калитиной (3 экз.); Армения, кратер горы Арагац, в
пещере, 29.VIII.1968 г., сбор И. С. Даревского, определение И. М. Ли-
харева (2 экз.); Душанбе, 15.V.1968 г., сбор и определение 3. Изза-
туллаева (2 экз.); Армения, Ширакское нагорье, близ Ленинакана,
19.IV.1971 г., сбор и определение автора (5 экз.).
Яйцевод довольно короткий, прямой; перед впадением в вагину
заметного сужения не образует. От места перехода яйцевода в
вагину отходит очень короткий прямой проток семяприемника,
заканчивающийся небольшим тонкостенным резервуаром. Верхний
отдел вагины очень толстостенный. Наружный слой стенок состоит
из сильно развитой мускульной ткани с преимущественно радиаль-
ным ходом волокон. Внутренний слой составлен рыхлой железистой
тканью. Верхний отдел вагины посредством очень короткой папиллы
9 ним, ! 29
вдается в полость нижнего отдела, несущего на внутренних стенках
1—2 продольные складки, которые уходят в полость нижнего отдела
пениса и сходят там на нет. Участок стенки полового атриума,
противолежащий пенису, утолщен и содержит железистую ткань.
Пенис короткий, мешковидный или коротко веретеновидный, стенка
его верхнего отдела содержит сильно развитую железу, сквозь
ткани которой проходит проток семяпровода, впадающего в пенис
примерно на его середине. Полость пениса в верхней части на
поперечном разрезе узко полулунная, в нижней части резко расши-
ряется. Половой ретрактор крепится апикально к пениальной железе.
Распространение Преимущественно горный вид: Пиренеи,
Альпы, Апеннины, Карпаты, запад Малой Азии, Крым, восточная
часть Кавказа с Талышом и Эльбрусом, горные системы Средней
Азии.
Род Trochovitrina Boettger, 1880
Boettger, 1880с: 379 Лихарев, Раммельмейер, 1952: 299 [Helicolimax (Trochovi'
Irma)]; Wenz, Zilch, 1959—1960: 237 (syn. ? Gallandia).
Типовой вид — Lampadia lederi Boettger, 1878, по монотипии.
Раковина низкоконическая, базальный край устья без кожистой
оторочки, периферия четко угловатая. Вагина очень длинная, верхняя
часть ее очень толстостенная; в стенках залегают многочисленные
мускульные пучки и железы, открывающиеся крупными порами в
просвет вагины. Яйцевод практически отсутствует. Семяприемник
полностью редуцирован. Пенис коротко булавовидный, утолщенная
верхняя часть его несет маленький вырост.
Trochovitrina lederi (Boettger, 1878), рис. 4.
Boettger, 1878: 121; (Lampadia); -1879: 7, Taf. I, Fig. 2 (Lampadia) 1880b:
379 [Vitrina (Trochovitrina) lederi et var. subcarinata, locus typicus: Ленкорань];
Лихарев, Раммельмейер, 1952: 300, 228 [Helicolitnax (T'c„hovitrina) lederi et var.
subcarinata]; Шилейко, 1967: 28, рис. 4 (T. lederi subcarinata); Zilch, 1979: 87,
Taf. 7, Fig. 11 (Phenacolimax (Gallandia) ledari lederi, фото лектотипа], Taf. 7,
Fig. 12 (Ph. (G.) lederi ? subcarinatus, фото лектотипа].
Locus typicus: Сурамский хребет.
Раковина чечевицеобразная, число оборотов до 3,5. По периферии
проходит четко выраженный угол, иногда киль. Эмбриональные и
дефинитивные обороты шелковисто радиально исчерчены. Диаметр
раковины до 5,75, высота до 4 мм.
Внутреннее строение. Материал. Талыш, с. Курумба в
предгорьях близ Ленкорани, 24.III. 1965 г. (3 экз.); Ленкоранская
низменность, с. Пенсар близ Ленкорани, 17.VI.1964 г. (2 экз.);
сборы и определения автора.
Спермовидукт очень короткий. Поскольку семяприемник отсут-
ствует, судить о длине яйцевода можно косвенно, так как у близких
родов проток семяприемника ответвляется от самой верхней точки
расширенного отдела вагины (вагинальной железы); расстояние
от места отхождения семяпровода до верхнего края вагинальной
железы очень невелико, т. е. яйцевод очень короткий. Стенки расши-
ренного отдела вагины содержат чередующиеся радиальные мускуль-
Рис. 4. Trochovitrina lederi из Талыша,
.4 — раковина; Б внешний вид полового аппарата; В
отдела. Звездочка — пенна/
Курумба, 24.111.1965 г.
адрнвнй; Г вн\i реннее строение ннячнею
й б\юрок.
ные пучки и участки железистой ткани. Проток вагины внутри
этого участка расширен и снабжен правильно расположенными
отверстиями элементов вагинальной железы (порами). Стенки ниж-
него отдела вагины также заключают множество железистых клеток.
Пенис недлинный, с узким просветом и хорошо развитой пениаль-
ной железой. На стороне пениса, противолежащей месту впадения
семяпровода (последний впадает примерно в середину пениса)
заметен небольшой округлый бугорок, заполненный рыхлой тканью.
Половой ретрактор крепится почти апикально.
В описании полового аппарата этого вида, сделанном ранее
(Шилейко, 1967), допущены ошибки, что можно объяснить лишь
тем, что в то время автор не был готов к исследованию анатомии
столь сложных объектов.
Распространение Сурамский хребет, Ленкоранская низ-
менность и зона горных лесов Талыша.
Подсемейство Semilimacinae Schileyko, subfam. nov.
Раковина уховидная, базальный край устья с периостракальной
оторочкой, за исключением рода Oligolimax, у которого раковина
9*
131
нормально развита, а оторочка отсутствует. Вагина короткая, не
разделена на отделы, без вагинальной железы и папиллы; в крайнем
случае, может присутствовать очень короткая папилла, лишенная
железистых элементов и представляющая собой модифицированный
сфинктер. Саркобелум первично имеется (у видов рода Eucobresia
редуцирован), внутри него располагаются одна или две мощные
папилЛы. Пениальный чехол имеется.
Род Semilimax Agassiz, 1845
Agassiz, 1845: 81. Ihering, 1892: 401 (Vitrinopugio, типовой вид Vitrina elongata
Draparnaud, 1805, по обозначению Hesse, 1923: 103)'- Forcart, 1944: 663 Wenz,
Zilch, 1959—1960: 236.
Типовой вид — Helix semilimax Ferussac, 1802, по монотипии
и тавтонимии.
Раковина уховидная, базальный край устья с периостракальной
оболочкой. Стенки папиллы саркобелума железистые, проток па-
пиллы с конхиолиновой выстилкой. Чехол саркобелума крепится к
нижней части папиллы, поэтому большая часть папиллы вне чехла.
За счет того, что папилла вдвинута в чехол, возникают наружная
и внутренняя стенки чехла. Проток семяприемника впадает в вагину.
Папиллы внутри вагины* нет.
Подрод Semilimax s. str.
Базальная стенка раковины не резорбирована. Папилла сарко-
белума завершается коротким пустотелым конхиолиновым шипом.
Семяпровод не связан с пениальным чехлом. Половой ретрактор
отсутствует.
Semilimax (Semilimax) semilimax (Ferussac, 1802), рис. 5, A\ 6.
Ferussac, 1802: 236, pl. 1, fig. А, В, C, D (Helix); Draparnaud, 1805: 120,
pl. 8, fig. 40—42 (Vitrina elongata, terra typica — Франция); Hesse, 1923: 104, Taf. II,
Fig. 11 (V elongata); Mermod, 1930: 106 [Vitrinopugio (Vitrinopugio) elongatus];
Hubendick, 1953: 94, fig. 27 [Vitrina (Vitrinopugio) elongata}; Forcart, 1944: 664
Wenz, Zilch, 1959—1960: 237, Abb. 835.
Locus typicus: bey dem Dorfe Billafingen...“ (Биллафинген близ Бадена,
Швейцария).
Материал около 50 экз. из советской части Карпат и из
Польши.
Уховидная раковина составлена двумя оборотами или чуть
больше. Пупок практически отсутствует. Периостракальная оторочка
базального края устья широкая. Нижняя стенка раковины не резор-
бирована. Поверхность эмбриональной раковины покрыта точками,
сгруппированными в хорошо выраженные спиральные ряды. Диаметр
раковины до 5,0, высота раковины до 2,4 мм.
Внутреннее строение Материал. Закарпатье, Раховский
р-н, Чернотиссянское лесничество, верхнее течение р. Апшинец,
1.IX.1982 г. сбор и определение А. А. Байдашникова -(4 экз.);
Закарпатье, Раховский р-н, близ с. Квасы, 12.IX. 1959 г сбор и
определение автора (7 экз.).
132
Рис. 5. Раковины Sernilimax sernilimax (Л) из Закарпатья, с. Квасы, и S. kotulai
(Б) из того же места.
Вид сверху и снизу.
Спермовидукт довольно длинный. Семяприемник имеет обширный
тонкостенный резервуар и очень короткий проток, впадающий в
вагину чуть ниже места отхождения семяпровода, т. е. яйцевод
чрезвычайно короткий. Вагина длиннее яйцевода не менее чем в
4 раза. Очень большой саркобелум состоит из длинной мощной
папиллы и тонкостенного чехла. Чехол отходит от полового атриума,
поднимается вверх, затем образует резкий кольцевой перегиб, так
как верхний участок его стенки, связанный с папиллой, ввернут
внутрь нижнего — в результате возникают наружная и внутренняя
стенки чехла. Толстые стенки папиллы саркобелума состоят главным
образом из железистой ткани. Канал папиллы имеет конхиолиновую
выстилку и проходит по всей длине органа сверху донизу; открывает-
ся канал на вершине короткого, слегка изогнутого на конце конхиоли-
нового шипа, напоминающего строением иглу шприца. Пенис ко-
роткий, нижняя часть его охвачена хорошо развитым чехлом, а
верхняя (в середину которой впадает семяпровод) содержит пениаль-
ную железу. В полости пениса залегает длинный высокий пилястр.
Половой ретрактор отсутствует.
Распространение Горные системы средней и Западной
Европы, включая советскую часть Карпат.
Подрод Hessemilimax Schileyko, subgen. noy.
Базальная стенка раковины резорбирована. Папилла саркобе-
лума без шипа. Участок семяпровода притянут к пенису пениальным
чехлом. Половой ретрактор имеется.
133
Рис. 6. Половой аппарат Semilimax (Semilimax) semilimax из Закарпатья,
верхнее течение р. Апшинец, 1.IX. 1982 г.
А внешний вид и поперечный срез папиллы саркобелхма; Б внутреннее строение нижнею отдела;
В шин саркобелхма, сильно увеличен.
Типовой вид— Vitrina kotulai Westerlund, 1883.
Название подрода образовано слиянием фамилии Гессе (Hesse)
и названия Semilimax.
Semilimax (Hessemilimax) kotulai (Westerlund, 1883), рис. 5, Б; 7
Westerlund, 1883: 54 (Vitrina kotulae; в соответствии co ст. 31 (а) Междуна-
родного кодекса зоологической номенклатуры правильное написание — V. kotulai);
Hesse, 1923: 111 (Vitrinopugio); Forcart, 1944: 666, Pl. 2, Fig. 12, Лихарев, Рамммель-
мейер, 1952: 300, рис. 229 (Vitrinopugio).
Terra typica: „Galicia in M. Tatra, 900—2200' s. m. (Польша, Татры,
а высоте 900 -2200 ф. над уровнем моря)
Материал: около 50 экз. из 6 местонахождений из советской
части Карпат и Полыни.
От S. semilimax отличается тем, что нижняя стенка раковины
частично резорбирована и снизу видны предыдущие обороты;
эмбриональные обороты практически гладкие, лишены точечной
скульптуры.
Внутреннее строение Материал. Закарпатье, Рахов-
ский р-н, Карпатский заповедник, высокогорный ельник, 16.VI1I.1982 г.,
сбор и определение А. А. Байдашникова (3 экз.); Закарпатье,
Раховский р-н, окр. с. Квасы, 12.IX. 1969 г сбор и определение
автора (5 экз.).
134
Рис. 7 Половой аппарат SemiUmax (Hessemilimax) kotulai из Карпатского за-
поведника, 16.VIII.1982 г.
А Б нутреннсе строение нижнего отдела поперечный срез папиллы сар-
кобелума.
Спермовидукт довольно длинный. Небольшой резервуар семя-
приемника плавно переходит в короткий проток, впадающий в
вагину непосредственно у атриума, т. е. яйцевод довольно длинный.
Вагина, напротив, чрезвычайно короткая. Саркобелум отличается от
такового предыдущего вида отсутствием шипа и свободного конца
папиллы, т. е. внутренняя стенка чехла саркобелума крепится к
самой нижней части папиллы. Пенис без продольного пилястра,
но с одной или двумя кольцевыми складками. Семяпровод, пойдя
под пениальным чехлом, впадает субапикально. Половой ретрактор
крепится к пенису терминально.
Распространение. Альпийская и субальпийская зоны
горных систем Европы от Грайских Альп и кантона Во (Швейцария,
севернее Женевского озера) до Доломитовых Альп, Судет, Татр и
Карпат.
Род Vitrinobrachium Kunkel, 1929
Kunkel, 1929: 624; Forcart, 1944: 660, Wenz, Zilch, 1959—1960: 236.
Типовой вид — Helicolimax breve Ferussac, 1821, по монотипии.
Раковина уховидная, базальный край с периостракальной отороч-
кой. Железистые элементы саркобелума сосредоточены в его апикаль-
ной части, проток папиллы без конхиолиновой выстилки. Папилла
135
Рис. 8. Раковина Vitrinobrachium breve из
Италии, Брианца.
саркобелума целиком помещается внутри чехла, на внутренней
поверхности которого имеются резкие кольцевые складки. Проток
семяприемника впадает в половой атриум. Папиллы внутри вагины
нет.
Vitrinobrachium breve (Ferussac, 1821), рис. 8, 9.
Ferussac, 1819, in Ferussac et Deshayes, 1819—1851, pl. 9, fig. 2 (изображение
без описания); Ferussac, 1821: 21, N 2 (Helicolimax brevis); Mermod, 1930: 108
(Vitrinopugio); Forcart, 1944: 661, Pl. 2, Fig. 10; Wenz, Zilch, 1959—1960: 236,
Abb. 834.
Locus typicus: ,,Billafingen“ (Биллафинген близ Бадена, Швейцария).
Материал: 2 экз. из Гейдельберга.
Уховидная раковина составлена 2—2,25 оборотами. Пупок
закрыт. Поверхность эмбриональных оборотов густо покрыта точ-
ками, группировка точек в ряды не всегда выражена. Периостра-
кальная оторочка базального края устья неширокая. Диаметр
раковины до 5,5, высота до 2,2 мм.
Внутреннее строение. Материал. В Зоологическом му-
зее МГУ хранятся два фиксированных экземпляра с этикеткой
„Гейдельберг“; дата сбора, фамилии коллектора и лица, определив-
шего материал, не указаны. Исследованы оба экз.
Спермовидукт относительно очень длинный, так как семяпровод
отходит близ атриума. Поскольку семяприемник открывается не-
посредственно в половой атриум, вагина отсутствует. Сравнительно
небольшой саркобелум представляет собой орган овальной формы,
вершина которого состоит из железистой ткани. От этой железы
отходит папилла с рыхлыми стенками и очень узким протоком.
Внутренняя поверхность чехла саркобелума покрыта очень резкими
высокими кольцевыми складками, а наружный слой чехла плотный,
мускулизованный, с преимущественно кольцевым ходом волокон.
На поперечном срезе папиллы саркобелума видно, что проток не-
замкнутый, щелевидный; вдоль него проходят два гребня с острыми
кромками, которые, тесно прилегая к поверхности папиллы, способны
136
Рис. 9. Половой аппарат Vitrinobrachium breve из Гейдельберга.
Б — внутреннее строение нижнего от 'ла и поперечный срез папиллы саркобе
полностью изолировать полость папиллы. Пенис массивный, объе-
мистый, верхняя его часть занята хорошо выраженной пениальной
железой. В верхней части полости пениса имеется листовидный
пилястр. Половой ретрактор крепится субапикально.
Распространение. Ареал состоит из двух участков. Пер-
вый участок занимает южную часть Альп, второй — горные системы
юга и центральной части ФРГ и ГДР.
? Род Eucobresia Baker, 1929
Hesse, 1923: 86 (Semilimax, non Agassiz, 1845, типовой вид Vitrina diaphana
Draparnaud, 1805, по обозначению Baker, 1929: 139), Baker, 1929: 139 (nom.
nov. pro Semilimax Hesse, 1923), Forcart, 1944: 641; Лихарев, Раммелымейер,
1952: 298 [Helicolimax (Eucobresia)]’, Wenz, Zilch, 1959—1960: 236.
Типовой вид— Vitrina diaphana Draparnaud, 1805, по первона-
чальному обозначению.
Раковина уховидная, базальный край устья с периостракальной
оторочкой. Саркобелум редуцирован. Проток семяприемника впада-
ет в вагину, внутри которой имеется очень короткая папилла, пред-
ставляющая собой модифицированный сфинктер.
Eucobresia nivalis (Dumont et Mortillet,- 1852), рис. 10, Б; 11.
Dumont, Mortillet, 1852: 209 (Vitrina)', Koch in Hesse, 1923: 92 Taf. 1, Fig. 5
[Phenacolimax (Semilimax) alpina part.]; Forcart, 1944: 644, Abb. 3, 4, Pl. 2, Fig. 4;
1954: 266 (Vitrina kochi auct., non Andreae, 1884, locus typicus — Diluvialsand
von Hangenbieten bei Strassburg): Лихарев, Раммельмейер, 1952: 298 Helicolimax
(Eucobresia) kochi (Andreae, 1884).
Locus typicus: „Anzindaz au-dessus de Bex“ (Швейцария, Бернские Альпы,
долина Роны выше Бе).
Материал: 21 экз. из 5 местонахождений в советской части
Карпат и Польше.
Раковина состоит из 2,5 оборотов. Кожистая (периостракальная)
оторочка неширокая и не доходит до столбика. Точки на поверх-
ности эмбриональных оборотов собраны в ряды, хотя правильность
этих рядов нередко нарушена. Имеется легкая париетальная мозоль.
Пупок отсутствует. Диаметр раковины до 6, высота до 3 мм.
Внутреннее строение. Материал. Польша, Татры, Коше-
лийская долина, 27 VII.1958 г., сбор И. М. Лихарева, определение
А. Риделя (2 экз.); Ивано-Франковская обл., Яремча, VIII. 1963 г
сбор и определение И. М. Лихарева (1 экз.).
Спермовидукт длинный, образует несколько колен. Семяприемник
имеет длинный прямой проток и небольшой тонкостенный резервуар.
Нижняя часть яйцевода и верхняя часть вагины окутаны тонким
соединительнотканным чехлом. Яйцевод в 1,5—2 раза длиннее ва-
гины. Протоки яйцевода и семяприемника, сливаясь, открываются
на вершине короткой сосочкообразной папиллы, лишенной желе-
зистых элементов и напоминающей разросшийся сфинктер. Пенис
довольно короткий, почти по всей длине окутан чехлом, доходящим
почти до места прикрепления полового ретрактора. Очень короткий
семяпровод, входя под чехол близ атриума, впадает в пенис ниже
верхней кромки чехла, т. е. место впадения семяпровода прикрыто
138
Рис. 10. Раковины Eucobresia diaphana (Я) из Польши, Нижняя Силезия,
и Е. nivalis (Б) из Яремчи, Ивано-Франковская обл.
Вид сигр\\
чехлом. Пениальная железа развита слабо. Полость пениса не-
полностью разделена на верхнюю и нижнюю камеры полукольцевой
складкой. Внутри полового атриума имеется серия небольших
параллельных складок, заходящих и в нижнюю камеру пениса.
Половой ретрактор крепится строго терминально.
Распространение. Вид долгое время отождествляли с
Vitrina kochi, описанной из дилювиальных песков близ Страсбурга;
однако, как показал Форкарт (Forcart, 1954), настоящая V kochi
известна только в ископаемом состоянии и не относится к роду
Eucobresia, Поэтому очертить ареал Е, nivalis можно с некоторой
осторожностью. Это в общем альпийский вид, распространенный от
Грайских Альп (Швейцария) на западе до гор Высокий Тауэри (Ав-
стрия) на востоке; Судеты, Карпаты, Татры. Для территории СССР
(Закарпатье) вид впервые отмечен Полевпной (1959).
Eucobresia diaphana (Draparnaud, 1805), рис. 10, А; 12.
Drapamaud, 1805: 120, pl. 8, fig. 38, 39 (Vitrina); Mermod, 1930: 97 [Phenacolimax
(Sernilimax) diaphanus]; Forcart, 1944: 641, Abb. 1, 2, Pl. 2, Fig. 3, Wenz, Zilch,
1959—1960: 237, Abb. 837.
Terra typica: Франция.
Материал: 18 экз. из 7 точек в Австрии, Польше и Болгарии.
Раковина состоит из не более чем двух оборотов. Периостракаль-
ная оторочка базального края устья доходит до столбика. Точки
на эмбриональных оборотах редкие, группировка их в ряды практи-
чески не выражена. Пупок небольшой, круглый. Диаметр раковины
До 6,7, высота до 3,3 мм.
139
Рис. 11. Половой аппарат Eucobresia nivalis из Польши, 27.VII. 1958 г.
А - внешний ви; Б - внутреннее строение нижнего отдела.
Внутреннее строение. Материал. Польша, Нижняя Си-
лезия, гора Сьнежник Клоцки, 4.IX. 1957 г., сбор и определение
А. Виктора (2 экз.), Австрия, Штейермарк, сбор А. Вагнера, опре-
деление А. Риделя (1 экз.)
Спермовидукт длинный, образует несколько колен. Семяприемник
имеет длинный прямой проток и небольшой резервуар. Папилла
внутри вагины выражена в той же степени, что и у Е. nivalis.
Соединительнотканный чехол на границе яйцевода и вагины выражен
очень слабо. Пенис длинный, семяпровод впадает в него под покровом
чехла. Пениальная железа четко выражена, состоит из трех полей:
два симметрично охватывают верхнюю часть пениса, третье непарное
залегает в стенке нижней части органа. Просвет пениса разделен
кольцевой складкой на округлую верхнюю и значительно большего
размера нижнюю камеры. Внутри нижней камеры залегает высокий,
хорошо развитый продольный пилястр. Половой ретрактор крепится
терминально.
Р а с п р о с т р а н е н и е. От восточной Франции на западе до
Судет на востоке; на север до возвышенных регионов Бельгии, Нидер-
140
ландов, ФРГ и ГДР; на юг до Моравии, Югославии и Болгарии.
Преимущественно горный вид, в Швейцарии встречается главным
образом на высоте от 1000 до 2000 м над ур. м.
Дополнение к подсемейству Semilimacinae:
о роде Oligolimax Fischer, 1878.
В моем распоряжении не было представителей этого рода, но
в связи с тем, что название Oligolimax долгое время употреблялось
неверно, а также в связи с необходимостью определить место
рода в системе, необходимы пояснения.
Форкарт (Forcart, 1960) исследовал типовой вид рода Oligo
Umax—Vitrina paulucciae Fischer in Paulucci, 1878 и показал,
что этот вид есть синоним Vitrina (Semilimacella) bonellii bonellii
Targione Tozzetti, 1872. Таким образом, подродовыс названия
Semilimacella S6os, 1917 (типовой вид Vitrina velebitica S6os, 1917,
syn. V bonellii reiteri Boettger, 1880), Targonia Hesse, 1923 (non
Signoret, 1870), Tozzettia Hesse, 1924 (nom. nov. pro Targonia
Hesse, 1923, типовой вид Vitrina bonellii Targione Tozzetti, 1873)
становятся объективными синонимами названия Oligolimax Fischer,
1878.
Род Balcanovitrina Osanova et Pinter, 1968 (типовой вид В. doj-
tshini OSanova et Pinter, 1968), судя по первоописанию, также
141
надлежит свести в синонимы Oligolitnax, что и было сделано Дами-
новым и Лихаревым, 1975, с. 253.
Подсемейство Vitrininae Fitzinger, 1833
Раковина низкоконическая, нормально развитая, базальный
край устья без периостракальной оторочки. Вагина практически
отсутствует, нижняя часть женского отдела внутри без специали-
зированных структур. Саркобелум отсутствует. Чехол пениса либо
охватывает нижнюю часть пениса (Vitrina), либо окутывает пенис
целиком и сливается апикально с половым ретрактором (Calidi-
vitrind).
Род Vitrina Draparnaud, 1801
Draparnaud, 1801 (июль):, 33; Ferussac, 1801 (ноябрь): 390 (Helico Umax,
типовой вид Helix pellucida Muller, 1774, по монотипии); Hesse, 1923: 82 (Phena-
colimax, non Stabile, 1859, типовой вид Helix pellucida, по обозначению Hesse,
1923) Forcart, 1944: 638; Лихарев, Раммельмейер, 1952: 296 (Helicolimax)-, Wenz,
Zilch, 1959—1960: 235.
Типовой вид — Helix pellucida Muller, 1774, по монотипии.
Пениальный чехол муфтой охватывает нижнюю часть пениса и
с половым ретрактором не связан.
Vitrina rugulosa Martens, 1874, рис. 13, Б; 14
Мартенс, 1874: 7 [Vitrina (?) ]; Westerlund, 1886: 22 [Vitrina (Phenacolimax)]-,
1896: 183 [V. (Oligolimax) alexandri, locus typicus — берег оз. Искандеркуль];
Hesse, 1923: 93 [Phenacolimax (Sernilimax)-, Лихарев, Раммельмейер, 1952: 297,
рис. 226 [Helicolimax (Helicolimax)].
Locus typicus: „in montibus Kokandensibus prope Karakasuk" (урочище Kapa-
касук, район Коканда).
Материал Несколько сотен экземпляров из 34 местона-
хождений на Алтае и в Средней Азии.
Раковина с притупленным, слабо выдающимся завитком, высота
которого заметно меньше половины высоты устья. Оборотов 3.
Поверхность эмбрионального оборота покрыта неясной пунктиров-
кой, выраженной в разной степени; группировка точек в ряды
не всегда хорошо заметна. Диаметр раковины до 7, высота до 4 мм.
Внутреннее строение. Материал. Южный Алтай, ущелье
у лога Сары-Алка, 22.VIII.1961 г., сбор и определение К. К. Ува-
лиевой (5 экз.); хребет Петра I, ущ. Нушор, 7.VII. 1968 г., сбор и
определение 3. Иззатуллаева (3 экз.); Гиссарский хребет, перевал
Анзоб, 30.VI. 1956 г., сбор К. Я. Грунина, определение автора
(2 экз.).
Яйцевод короткий, но ясно выражен: проток семяприемника
отходит заметно ниже места отхождения семяпровода. В стенках
самой нижней части яйцевода на стороне, противолежащей семя-
приемнику, залегает выраженный в разной степени участок желе-
зистых тканей. В месте перехода яйцевода в половой атриум имеется
кольцевое утолщение тканей (нечто вроде слабо выраженного
сфинктера), выраженное у разных особей по-разному. Основание
142
Рис. 13. Раковины Vitrina pellucida pellucida из Москов-
ской обл., Белые Колодези на Оке (Л), V rugulosa из Чаткаль-
ского хребта, перевал Чапчима (Б) и V pellucida alaskana
из о. Беринга, Командорские о-ва (В).
протока семяприемника не утолщено. Пенис маленький, внутри него
залегает один мощный округлый в сечении пилястр, в результате
чего просвет пениса очень узкий, полулунной формы. Внутренняя
поверхность вагины лишена постоянных складок.
Распространение Алтай и Средняя Азия.
Vitrina pellucida pellucida (Muller, 1774), рис. 13 A; 15
Muller, 1774: 15 (Helix); C. Pfeiffer, 1821: 47, Taf. 3, Fig. 1 (V beryllina,
locus typicus restr. Forcart, 1955: Bellevnegarten von Cassel); Boettger, 1980a:
115, Taf IV, Fig. 4 (V globosa, locus typicus: Закавказье, Тбатани); Forcart,
1944: 639, Pl. 2, Fig. 1, 2; 1955: 159, Abb. 1, Taf. 12, Fig. 1, 2, 5 (обозначение
неотипа); Лихарев, Раммельмейер, 1952: 297, рис. 225 [Helicolimax (Helicolimax)];
Wenz, Zilch, 1959—1960: 235, Abb. 832.
Locus typicus: „agri Fridrichsdalensis, sen Daniae“ (Фридрихсдаль у Копен-
ганена, Дания).
Материал несколько сотен экземпляров из 76 местонахожде-
ний в европейской части СССР, Закавказье, Средней Азии, на
Урале, в Западной Сибири и Забайкалье.
Раковина с чуть приостренным или притупленным завитком,
высота которого равна половине высоты устья или чуть больше.
Оборотов до 3,75. Эмбриональная скульптура в виде хорошо вы-
раженных точек, четко сгруппированных в ряды. Диаметр раковины
до 6, высота до 3,4 мм.
Внутреннее строение Материал. Байкал, Баргузинский
заповедник, возле устья Б. Речки, 11.IX. 1966 г. (3 экз.); Москов-
ская обл., Белоомут на Оке, 1.V.1960 г. (2 экз.); сборы и определения
Я. И. Старобогатова, определения И. М. Лихарева; Сев. Кавказ,
143
Рис. 14. Половой аппарат Vitrina rugulosa из Южного Алтая, 22.VIII.1961 г.
Л /> BinipvHHee строение нижнею отдела и поперечный срез пениса. Звездочка
железистая ткань.
Орджоникидзе, гора Сапицкая Будка, 8 X1.1952 г., сбор и опреде-
ление 3. И. Калитиной (2 экз.); Московская обл., Пушкино, дер. Ко-
стино, 25.VI. 1958 г., сбор и определение автора (3 экз.).
Место отхождения семяпровода, основание протока семяприем-
ника и место слияния пениса с атриумом находятся примерно на
одном уровне, т. е. как вагина, так и яйцевод практически отсут-
ствуют. Железистых тканей в нижней части женского отдела не
обнаружено. Женский отдел при переходе в атриум лишен коль-
цевых складок. Основание протока семяприемника в той или иной
степени утолщено. Пенис маленький, внутреннее его строение такое
же, как у V rugulosa.
Распространение Палеарктика.
Vitrina nellucida alaskana Dall, 1905, рис. 13 В; 16
Dall, 1905: 37 (I itrina alaskana, nom. nov. pro Vitrina pfeifferi Newcomb,
1861, non Deshayes, 1851), Pilsbry, 1946: 503, fig. 276 (V alaskana); Bequaert,
Miller, 1973: 146, Roth, Lindberg, 1981: 44 (V pellucida part.).
144
Рис. 15. Половой аппарат Vitrina pellucida pellucida из Забайкалья, Баргузин-
ский заповедник, 11.IX.1966 г.
А и поперечный срез пениса с прилегающим семяпроводом; Б — внутреннее строение
нижнего отдела.
Locus typicus: „Carson valley, Churchill Co., Nevada*4 (долина p. Карсон,
шт. Невада, США).
Материал Несколько десятков экземпляров из двух место-
нахождений на о. Беринга, Командорские о-ва.
Раковина отличается от раковины номинативного подвида значи-
тельно более сильно расширенным последним оборотом; скульптура
эмбриональных оборотов (до 1,5) в виде мелких точек, их группи-
ровка в спиральные ряды часто в той или иной мере нарушена.
Высота завитка составляет половину высоты устья или немного
меньше. Диаметр раковины до 10 мм, высота до 6 мм.
Внутреннее строение Материал. О. Беринга (Командор-
ские о-ва), р-н пос. Никольского, VII. 1972 г., сбор А. В. Янковского,
определение автора (3 экз.).
Расстояние между местом отхождения семяпровода и нижней
кромкой пениса примерно равно длине полового атриума; этот
отрезок правильно, вероятно, именовать вагиной, но здесь граница
между яйцеводом и вагиной не может быть точно зафиксирована.
10 Заказ 1696 145
Рис. 16. Половой аппарат Vitrina pellucida
alaskana из о. Беринга, VII. 1972 г.
Внешний вид и поперечн и срез neinn.ii.
Дело в том, что основание протока семяприемника так расширено,
что занимает, по существу, всю длину этого отрезка. Относитель-
ная длина пениса больше, чем у номинативного подвида.
Из описания видно, что между обеими формами, трактуемыми
здесь как подвиды, существуют определенные качественные и коли-
чественные различия. Однако нет уверенности, что отмеченные разли-
чия в устройстве полового аппарата не носят сезонного характера,
поэтому лучше пока эти формами считать подвидами, имея в виду
их значительное сходство и несовпадение ареалов.
Распространение Запад Северной Америки от Мексики до
Аляски и Командорские о-ва. В южных частях ареала живет преиму-
щественно в горах.
Таксономическая структура Vitrinidae
и исторические связи между родами
По поводу системы семейства высказывались многие авторы;
обзор этих взглядов содержится в работе Форкарта (Forcart, 1944).
146
Рассматривая факты, изложенные в систематической части, легко
заметить, что вся совокупность родов, в соответствии с устройством
полового аппарата, четко распадается на 3 группы, что отмечалось
уже давно (Wiegmann, 1886, Simroth, 1889).
Группа 1 Имеется вагинальная железа, обычно снабженная
папиллой (Guerrina, Arabivitrina, Insulivitrina, Phenacolimax,
Trochovitrina, Plutonia}.
Группа 2. Имеется аксессорный орган (саркобелум), отно-
сящийся к женскому отделу (Semilimax, Hessemilimax, Oligolimax,
Vitrinobrachium).
Группа 3 Вагинальная железа и саркобелум отсутствуют
(Vitrina, Calidivitrina).
Род Eucobresia формально принадлежит последней группе, но
заслуживает отдельного рассмотрения.
Среди признаков, используемых в систематике Vitrinidae, обычно
фигурирует такой признак, как степень развития лопастей мантий-
ного валика. В этой связи нужно заметить, что в пределах семейства
наблюдается ярко выраженная тенденция к редукции раковины,
связанная обратной зависимостью со степенью развития мантийных
лопастей: по мере редукции раковины мантийные лопасти все сильнее
разрастаются, частично прикрывая раковину. Поэтому для анализа
взаимоотношений между таксонами достаточно оценивать степень
редукции раковины. Существенно, что редукция раковины у Vitrini-
dae происходила не менее двух раз (в первой и второй из выделенных
групп, которым здесь придается ранг подсемейств); кроме того,
важно, что эволюция раковины и полового аппарата происходит
без взаимной связи. Так, для Phenacolimacinae характерно наличие
нормально развитой раковины, но род Plutonia, сохраняя особен-
ности полового аппарата, полностью утрачивает наружную раковину;
у Semilimacinae раковина исходного типа (нормально развитая)
имеется у единственного рода Oligolimax, половой аппарат которого
обнаруживает черты сильной специализации.
Поскольку тенденция к редукции раковины реализовалась среди
Vitrinidae не раз, анализ эволюции семейства приходится основы-
вать главным образом на путях и способах преобразования по-
лового аппарата.
Для того, чтобы понять генеалогию семейства, в первую очередь
надо установить, какая из групп может быть истолкована как
наиболее примитивная. Зимрот (Simroth, 1889) считает наличие
аксессорного органа архаичным состоянием, во всяком случае,
сравнительно с наличием вагинальной железы. Вигманн (Wiegmann,
1886, S. 75) и Гессе (Hesse, 1923, S. 81), не обсуждая, какое из
состояний более примитивно, считают вагинальную железу и сарко-
белум гомологами. Экхарт (Eckhart, 1914) вообще отрицает связь
между формами, имеющими саркобелум, и формами без саркобелума
и соответственно полагает, что семейство в принимаемом объеме
дифилетично.
Мнение Экхарта будет рассмотрено позже; что же касается
гомологии вагинальной железы и саркобелума, то такая гомология
147
выглядит более чем правдоподобной. Вместе с тем с мнением
Зимрота согласиться никак нельзя, в первую очередь потому, что
невозможно себе представить конкретный ход интеграции саркобе-
лума с тканями вагины, а также функциональный смысл такой
интеграции. Обратный процесс, напротив, выглядит вполне естест-
венным.
Предположение о филогенетическом развитии полового аппарата
выглядит следующим образом (рис. 17). Половой аппарат анце-
стральных форм был простым (типа Vitrina), но концевые его
участки были богато насыщены железистыми тканями — тем исход-
ными материалом, который мог послужить основой для формирова-
ния любых дополнительных органов, придатков и специализирован-
ных желез (рис. 17, /). Эти ткани были развиты как в мужском,
так и в женском отделе. Непосредственные потомки этих предковых
форм — немногочисленные виды рода 1Л7шга(рис. 17, II) и африкан-
ские высокогорные Calidivitrina, пошедшие, по-видимому, по пути
физиологических, а не морфологических перестроек. Многозначи-
тельно в этом контексте обнаружение Хубендиком (Hubendick,
1953, р. 84, fig. 21) железистого участка в стенке вагины у Calidi-
vitrina lactea, который можно трактовать как рудимент вагиналь-
ной железы, хотя Форкарт (Forcart, 1956, р. 115) допускает, что
это железистое образование может быть артефактом. Уместно
также вспомнить, что у V rugulosa в стенке вагины также имеется
участок железистых тканей. Форкарт (Forcart, 1978, р. 325) указы-
вает, что Calidivitrina отличается от Vitrina строением маргиналь-
ных зубов радулы и отсутствием точечной скульптуры. Между тем
имеется еще одно существенное различие: пениальный чехол у
Vitrina охватывает муфтой нижнюю часть пениса и с половым
ретрактором не связан, тогда как у Calidivitrina тонкий мускульный
чехол увеличивается в толщину от атриума к верхнему концу
пениса и продолжается в половой ретрактор (Hubendick, 1953, р. 84).
Если отсутствие точечной скульптуры на эмбриональной раковине —
явление вторичное, то вполне возможно, что сплошной пениальный
чехол надо трактовать как исходную особенность.
В пределах группы 1 (Phenacolimacinae) происходило разраста-
ние и преобразование железистых тканей в том месте, где они
исходно находились, результатом чего стало возникновение не-
большой оформленной вагинальной железы с папиллой (Guerrina)
(рис. 17, III). Дальнейшие этапы развития этой линии (рис. 17,
IV) — последовательность Arabivitrina — Insulivitrina — Phenaco-
Umax. Род Plutonia — наиболее специализированный представитель
подсемейства (да и всего семейства), утративший наружную ра-
ковину и перешедший к хищному образу жизни; вероятно, целе-
сообразно выделение этого монотипического рода в самостоятельное
подсемейство Plutoniinae, как это предлагал Тиле (Thiele, 1931,
S. 600). Род Trochovitrina (рис. 17, V) формально наименее спе-
циализирован (отсутствует папилла вагинальной железы), но отсут-
ствие семяприемника и высокий уровень организации самюй ва-
гинальной железы у Т lederi свидетельствуют об избрании этим
148
Рис. 17. Схема эволюционных преобразований полового аппарата Vitrinidae.
/ — исходный тип строения; // — Vitrina; Ш — Guerrina; IV — Phenacolimax и близкие группы; V — Trochovitrina; VI — гипотетичная форма, у которой ось
вагинальной железы отклонена от оси вагины (начальная стадия обособления саркобелума); VII — Sernilimax; VIII — Vitrinobrachium; IX — Oligolimax; X —
Eucobresia.
родом собственного пути эволюции, связанном, в частности, с утра-
той вагинальной папиллы.
Для того, чтобы принять, что от схемы, соответствующей Phena-
colimacinae (рис. 17, /I/), выводится ветвь, соответствующая ви-
дам с развитым саркобелумом, нужно одно допущение, правомер-
ность которого сама по себе не может вызвать сомнений. Допущение
заключается в том, что ось вагинальной железы несколько отклони-
лась от оси вагины (рис. 17, VI). Основанием для такого допущения
могут служить известные аналогии: у Dyakia striata (Helicarionidae)
семяприемник впадает в саркобелум, причем довольно высоко, т. е.
сильно отступая от атриума. Очевидно, что это положение нс
первично: основание семяприемника должно было „сползти" с ва-
гины до атриума, а затем переместиться вверх по чехлу саркобе-
лума (Van Mol, 1973, р. 217, fig. 4); у Pdpuarion novaguineae
семяприемник впадает в пениальную часть атриума (op. cit., р. 219,
fig. 5) —опять-таки свидетельство того, что взаимное положение
придаточных органов полового аппарата в подотряде Helixina может
меняться в достаточно широком диапазоне.
Если принять допущение о расхождении осей вагинальной же-
лезы и вагины, то образование такого придатка, какой имеется
у Semilimax, будет естественным и закономерным процессом (рис. 17,
VII). Непосредственным производным Semilimax в таком случае
надо считать Vitrinobrachium, у которого железистая часть за-
нимает не всю длину саркобелума, но сосредоточена на слепом
конце' органа (рис. 17, VIII). Происхождение такого саркобелума,
какой имеется у Oligolimax (=Semilimacelld), в деталях реконструи-
ровать трудно, но, вероятно, гомологом папиллы саркобелума
Vitrinobrachium можно считать лишь одну из папилл саркобелума
Oligolimax (большую?); вторая папилла в таком случае будет
новообразованием; представить же формирование двух папилл за
счет расщепления одной трудно как в морфологическом, так и в
функциональном отношениях. Поскольку у Oligolimax железистая
часть занимает апикальный отдел саркобелума (рис. 17, IX), можно
предполагать связь с Vitrinobrachium. Вместе с тем выведению
рода Oligolimax от рода Vitrinobrachium препятствует то обстоя-
тельство, что редукция -раковины Vitrinobrachium зашла далеко,
тогда как виды Oligolimax имеют довольно хорошо развитую ра-
ковину. Поэтому речь может идти не о выведении Oligolimax от
современного V breve, но от его предков, имевших такое же строе-
ние саркобелума, но обладавших еще нормально развитой рако-
виной.
Не исключено и другое предположение: роды Semilimax,
Vitrinobrachium и Oligolimax независимые дериваты гипотети-
ческой формы (прерывистые стрелки на рис. 17).
Остается еще один род Eucobresia, который формально отно-
сится к той же группе, что и Vitrina, поскольку не имеет саркобелума
и вагинальной железы; \ видов рода Eucobresia внутри вагины
имеется короткая папилла, но строение ее резко отлично от той,
которая присутствует \ Phenacolimacinae, поэтому допустимые
150
четыре толкования систематического положения рода: 1) это абер-
рантный член Vitrininae; 2) это представитель Semilimacinae, утра-
тивший саркобелум; 3) это представитель Phenacolimacinae с
дегенерировавшей вагинальной железой, от которой сохранился лишь
рудимент; 4) род составляет отдельное подсемейство, выводимое
непосредственно от гипотетичного предка.
Поскольку у Eucobresia очень хорошо развит пениальный чехол,
предположение о принадлежности рода подсемейству Phenacoli-
macinae приходится отвергнуть. Сближению рода с настоящими
Vitrina мешает место впадения семяпровода в пенис: у Vitrina
он впадает субапикально, а у Eucobresia — латерально, близ сере-
дины пениса; да и само строение пениса у видов этих родов
различается достаточно резко. Для возведения Eucobresia в ранг
самостоятельного подсемейства не „набирается44 достаточно весомых
признаков: ведь род характеризуется главным образом негативными
признаками (нет вагинальной железы, нет папиллы, нет саркобе-
лума). Остается последняя возможность — считать Eucobresia
членом Semilimacinae, утратившим саркобелум (рис. 17, X);- что
касается „папиллы44 внутри вагины, то ее строение не дает основа-
ний к тому, чтобы считать этот орган гомологом вагинальной
железы или вагинальной папиллы Phenacolimacinae: по всей ви-
димости, это увеличенный сфинктер. К тому же раковина видов
Eucobresia чрезвычайно похожа на раковину Semilimax semilimax.
Таким образом, мнение Экхарта (Eckhart, 1914) о дифилети-
ческой природе Vitrinidae несостоятельно. Надо также добавить,
что всех Vitrinidae объединяет еще такой важный (и, по-видимому,
неадаптивный) признак, как наличие яркой и характерной эмбрио-
нальной скульптуры в виде точек, сгруппированных в спиральные
ряды; в отдельных случаях эта скульптура может утрачиваться, но
ее первичность для семейства сомнений не вызывает.
Поскольку строение полового аппарата витринид разнообразно
и часто сложно, понятен интерес к механизму копуляции, который
был исследован рядом авторов (Eckhardt, 1914; Mermod, 1923
Kunkel, 1929, 1933; Gerhardt, 1936; Forcart, 1944, 1949). У тех
видов, которые имеют простое устройство гениталий (Vitrina,
Eucobresia), копуляция осуществляется традиционно: пенис вывора-
чивается и проникает в женские половые пути партнера, передавая
слизистый комок, заключающий спермин. Phenacolimacinae, обла-
дающие вагинальной папиллой, используют ее в качестве органа,
посредством которого происходит всасывание спермиев из пениса
партнера. При этом вагина выворачивается так, что окончанием
ее становится вагинальная папилла; пенис партнера также вы-
ворачивается, и папилла всасывает скопление спермиев почти не-
посредственно из семяпровода. Важную роль в акте насасывания
играет мускулатура, хорошо развитая в стенках вагинальной железы
и папиллы.
Сложнее дело обстоит у Semilimacinae. У Vitrinobrachium
breve, как показал Кюнкель (Kunkel, 1929), при спаривании вначале
выворачивается половой атриум, в результате чего обнажаются
все четыре отверстия (пениса, саркобелума, вагины, протока семя-
151
приемника). Затем у особи, играющей роль самки, выдвигается
папилла саркобелума, которая забирает аггрегации спермиев и
передает их в проток собственного семяприемника; пенис при этом
не выворачивается. Этот механизм не имеет принципиальных отличий
от того, какой имеется у Phenacolimacinae — дополнительное до-
казательство гомологии между вагинальной железой Phenacoli-
macinae и саркобелумом Semilimacinae. У Sernilimax sernilimax,
по Кюнкелю (Kunkel, 1933), пакет спермиев депонируется непосред-
ственно в проток семяприемника посредством выворачивания пениса.
Роль папиллы саркобелума, как полагает Кюнкель, заключается в
фиксации моллюска в определенном положении относительно тела
партнера: конец папиллы, который Кюнкель называет насасываю-
щим сосудом (,,Saugnapf“), присасывается при этом к покровам
партнера. Я не наблюдал спаривания Sernilimax, но сомнительно,
чтобы роль столь сложного устроенного (железистого!) органа,
сводилась к роли присоски, тем более что и представить себе
папиллу саркобелума в этой роли трудно. В самом деле, для
успешного функционирования в качестве присоски необходимо два
свойства: наличие определенного объема, в котором будет происхо-
дить разрежение воздуха, и механизм, который бы это разрежение
создавал. Объемом может служить лишь канал папиллы, а меха-
низмом могли бы стать мышечные волокна, которые расширяли бы
канал, увеличивая в момент присасывания его объем. Если эти
посылки верны, то Кюнкель в трактовке роли саркобелума ошибался,
ибо объем канала слишком мал, а соответствующий мышечный
механизм отсутствует; к тому же конхиолиновая выстилка канала,
несомненно, препятствует изменению его объема, что необходимо в
момент создания присасывающего эффекта.
Функция саркобелума Sernilimax в настоящее время не может
быть удовлетворительно истолкована, хотя бы потому, что стенки
его железистые, но при наличии плотной выстилки неясно, как
секрет этих желез попадает сквозь стенки в просвет канала; а если
не попадает, то каково его значение? Можно предполагать, что
этот секрет служит материалом для конхиолиновой выстилки, но
какова, в таком случае, роль канала, коль скоро в качестве вса-
сывающего органа он действовать не может? Единственное пред-
положение, которое напрашивается в данном случае, заключается в
том, что мышечные элементы, входящие в состав стенок’саркобелума,
способны создать малое разрежение, которого, однако, достаточно
для того, чтобы всосать пакет со спермиями и передать его затем в
семяприемник. Положение свободного конца саркобелума (рис. 6, Б)
таково, что допускает предположение о возможности самооплодотво-
рения у этого вида.
О положении Vitrinidae в отряде Geophila
Анализ этого сложнейшего вопроса составляет предмет отдель-
ного исследования, поэтому здесь уместно лишь краткое изложение
некоторых узловых положений.
152
Общеизвестен так называемый зонитоидный облик раковины,
присущий нескольким семействам Geophila. Этот тип характери-
зуется совокупностью следующих черт. Раковина уплощенная,
тонкостенная, полупрозрачная, с очень гладкой и блестящей по-
верхностью; скульптура постэмбриональных оборотов тонкая, в виде
очень узких спиральных бороздок; дефинитивные признаки (губа,
устьевые зубы), как правило, отсутствуют (если устьевое вооружение
имеется, то оно формируется несколько раз в постэмбриогенезе);
раковина имеет наклонность к редукции.
Такая раковина в целом свойственна следующим семействам:
Zonitidae, Euconulidae, Vitrinidae, Urocyclidae, Ariophantidae,
Helicarionidae, Trochomorphidae и, с оговорками, Gastrodontidae.
У ряда тропических представителей этих семейств раковина порой
отклоняется от описанного типа, но генерализованный тип для
всех перечисленных семейств, несомненно, зонитоидный.
Естественно возникает вопрос: родственны ли между собой пере-
численные семейства? Или: появлялась ли зонитоидная раковина
среди Geophila один раз или несколько? Ответ на этот вопрос
может дать анализ особенностей полового аппарата. Если принять
систему отряда Geophila, предложенную автором ранее (Шилейко,
1979), то легко видеть, что наличие железистых участков на концевых
частях женского отдела полового аппарата или их производных
(саркобелума, стилофоров, слизистых желез, атриальной железы,
перивагинальной железы) есть особенность, свойственная только
членам подотряда Helixina. Уместно вспомнить, что у Achatinina
вообще никаких придатков полового аппарата нет; у Pupillina —
характерного устройства придатки на пенисе (аппендикс, часто
цэкум); у Oleacinina бывают придатки на вагине, но это скорее
исключение, чем правило; у Limaxina придатки если и есть, то
только в мужском отделе. Итак, все перечисленные выше семейства,
для которых характерна зонитоидная раковина, принадлежат
подотряду Helixina.
Необходимо подчеркнуть, что разнообразные и многочисленные
придаточные органы полового аппарата, столь характерные для
Helixina (и только для Helixina) не могли возникнуть без определен-
ных морфологических предпосылок. Такой предпосылкой стало
возникновение и развитие железистых тканей“в'стенках дистальных
отделов гениталий. Следовательно, исходным для Helixina был по-
ловой аппарат, снабженный недифференцированными железистыми
полями, от которого и возникло наблюдаемое ныне разнообразие.
В свете этого положения надо признать правоту Зимрота, который
гомологизировал стилофор Helix с саркобелумом Semilimax (Sim-
roth, 1886). Однако Зимрот пошел дальше и постулировал генеалоги-
ческую связь саркобелума Semilimax с придаточными органами
копулятивного аппарата турбеллярий Rhabdocoelidae, а также
Pteropoda; если мы признаём, что железистые поля, послужившие
основой для формирования придаточных органов Helixina, возникли
только в пределах Geophila,4то с такими широкими гомологиями
согласиться невозможно.
153
Часть групп, имеющих зонитоидную раковину, сохранили прими-
тивное состояние (т. е. сравнительно малодифференцированные
железистые участки) — это преимущественно таксоны, населяющие
умеренные и северные широты (Zonitidae, Daudebardiidae, Vitri-
ninae), африканские представители этих групп (Calidivitrina) живут
только в высокогорье, в условиях отнюдь не тропического климата.
В тропиках от тех же исходных форм развилось колоссальное
разнообразие Ariophantacea sensu Taylor et Sohl, 1962. В пределах
этой группы, на мой взгляд, самые специализированные формы
обладают сложными придатками как в мужском, так и в женском
отделе.
Подотряд Helixina включает три подчиненных таксона — инфра-
отряда: Endodontinia, Helixinia, Zonitinia. Если характеризовать
инфраотряды в целом, то Endodontinia имеют достаточно разно-
образную раковину, но зонитоидный тип здесь практически не
встречается. Половой аппарат простого устройства, причем простота
эта, видимо, первична. В двух других подотрядах легко выделить
две группы семейств, характеризующиеся следующими признаками:
1. Раковина хеликоидная, т. е. толстостенная, часто довольно
грубо скульптурированная, скульптура чаще радиальная; с дефини-
тивными образованиями (самое постоянное и характерное образо-
вание— губа); раковина не обнаруживает наклонности к редукции.
Мужской отдел гениталий обычно с нитевидным бичом (случаи
несомненно вторичной утраты бича в расчет не принимаются),
женский отдел с одним, двумя или четырьмя стилофорами и
несколькими слизистыми железами, функционально связанными со
стилофорами.
Сюда относится 11 семейств, составляющих инфраотряд Heli-
xinia: Humboldtianidae, Helminthoglyptidae, Oreohelicidae, Helicidae,
Bradybaenidae, Hygromiidae, Corillidae, Camaenidae, Ammonitellidae,
Helicodontidae, Sphincterochilidae.
2. Раковина зонитоидная (см. вылие) \ мужской отдел с пениаль-
ной железой или ее дериватами (цэкум, флагеллум); женский отдел
с перивагинальной железой, вагинальной железой, группой паль-
чатых атриальных желез или саркобелумом.
Нетрудно видеть, что' к этой группе (инфраотряд Zonitinia)
относятся семейства, перечисленные в начале данного раздела,
а также полуслизни и слизни: Daudebardiidae, Parmacellidae,
Milacidae.
В предыдущем разделе статьи отмечалось, что эмбриональная
скльптура в виде спиральных рядов точек характерна для Vitrinidae
однако она же встречается и среди перечисленных семейств, в
частности, среди Helicarionidae, Parmacellidae, а для Urocyclidae
такая скульптура вообще очень характерна (Van Mol, 1970, р. 11)
В заключёние, возвращаясь к Vitrinidae, надо заметить следую-
щее. Какой бы ранг ни придавать внутрисемейственным группи-
ровкам в пределах Vitrinidae, очевидно, что разнообразие их внутрен-
него строения значительно больше, чем разнообразие их раковин,
и таксономическая структура семейства сложна. Сам по себе факт
154
большого разнообразия при сравнительно небольшом числе видов
однозначно свидетельствует о значительной древности семейства,
что подтверждается присутствием Provitrina в отложениях нижнего
палеоцена Европы; наличие же многих моно- или олиготипических
таксонов, а также разрывы в ареалах есть признаки реликтовости.
Обозначения в иллюстрациях:
А — половой атриум; БЖ — белковая железа; В — вагина; ВЖ — вагинальная
железа; ВП — вагинальная папилла; ВС—внутренняя стенка саркобелума;
ЖС — железистый слой; К — квадривий; КВ — конхиолиновая выстилка канала
саркобелума; МС — мышечный слой; НС — наружная стенка саркобелума; П —
пенис; ПЖ—пениальная железа; ПИЛ — пилястр; ПР — половой ретрактор;
ПС — папилла саркобелума; С — семяпровод; СБ — саркобелум; СП — семяприем-
ник; ЧП — чехол пениса; Я — яйцевод.
ЛИТЕРАТУРА
Дамянов С., Лихарев И. М. Gastropoda terrestria.— В кн.: Фауна на България.
Т. IV София, 1975, с. 1—425 (болг.).
Лихарев И. М., Раммельмейер Е. С. Наземные моллюски фауны СССР.— В кн.:
Опред. по фауне СССР. Вып. 43. М.; Л., 1952, с. 1—511.
Мартенс Э. Слизняки (Mollusca).— В кн.: А. П. Федченко. Путешествие в Туркестан.
Вып. 1, т. 2, ч. 1. 1874, с. 1—64.
Полевина А. А. К фауне наземных моллюсков Закарпатья.— Докл. и сообщ.
Ужгор. гос. ин-та. Сер. биол., 1959, т. 3, с. 65—68.
Шилейко А. А. Некоторые новые и малоизвестные моллюски Талыша.— Научн.
докл. высш, школы. Биол. науки, 1967, № 4, с. 24—32.
Шилейко А. А. Система и филогения отряда Geophila (=Helicida).— Тр. Зоол. ин-та
АН СССР, 1979, т. 80, с. 44—69.
Agassiz L. Nomina systematica Generum Molluscorum.— Nomenclator Zoologicus
Soludori. T. III. Rostok-Leipzig, 1845 (1842—1896), p. 1—98.
Baker H. B. Nomenclature in the Genus Vitrina.— Nautilus, 1929, vol. 42, p. 137.
Bequaert J. C.y Miller W. B. The molluscs of the arid southwest with an Arizona
check list. Univ. Arizona Press, Tucson, 1973, p. 1—271.
Boettger O. Diagnoses molluscorum novorum a clar. H. Leder in montibus Cauasis
lectorum.—Nachr. Bl. dtsch. malak. Ges., 1878, Bd 10, S. 120—124.
Boettger O. Kaukasische Mollusken gesammelt von Herrn Hans Leder.— Jahrb.
dtsch. malak. Ges., 1879, Bd 6, S. 388—412.
Boettger. O. Kaukasische Mollusken gesammelt von Herrn Hans Leder.— Jahrb. dtsch.
malak. Ges., 1880a, Bd 7, S. 109—150.
Boettger O. Diagnoses molluscorum novorum ab ill. Hans Leder in regione caspia
Talysch dicta lectorum.—Jahrb. dtsch. malak. Ges., 1880b, Bd 7, S. 379—383.
Boettger O. Die Binnenmollusken Transcaspiens und Chorossans.— Zool. Jahrb.,
1889, Abt. 3, Bd 4, S. 925—992.
Bourguignat J. R. Description du nouveau genre Gallandia.— Saint-Germain,
1880, p. 1—8.
Dall W. H. Land and fresh water mollusks of Alaska end adjoining regions.—
Harriman Alaska Series, 1905, vol. 13, p. 1 —171.
Draparnaud J. P. R. Tableau de Mollusques terrestres et fluviatiles de la France.
Paris, 1801, p. 1 — 116.
Draparnaud J. P. R. Historire naturelie des mollusques terrestres et fluviatiles de
la France. Paris, 1805, p. 1 — 164.
Dumont F., Mortillet G. Histoire des Mollusques terrestres et d’eau douce vivants
et fossiles de la Savoie et du Bassin du Leman. Paris et Geneve, 1852,
p. 1-152.
Eckhardt E. Beitrage zur Kenntnis der einheimischen Vitriniden.— Jena. Z. Naturw.,
1914, Bd 51, S. 213—376.
155
Ferussac J. В. L. Expose succinct d’un systeme conchyliologique.— Mem. Soc.
Med. Emul. Paris, 1801, N 4, S. 372—402.
Fkrussac J. B. L. Ueber eine neue Thierart, welche die Gattungen Umax und
Helix mit einander vereinigt, Helix Semilimax.— Naturf. Halle, 1802, Bd 29,
S. 236—241.
Ferussac A. E. J. Tableaux systematiques des animaux mollusques... Tableaux
systematique de la famille Limacpns, Cochleae.— Paris et Londres, 1821,
p. 1-114.
Fkrussac A. E. J., Deshayes G. P. Histoire naturelle generale et particuliere des
Mollusques terrestres et fluviatiles.—Paris, 1819 (1819—1851).
Forcart L. Monographic der schweizerischen Vitrinidae.— Rev. suisse Zool., T. 51,
N 29, 1944, S. 629—678.
Forcart L. Die Erektion der Kopulationsorgan und der Kopulationsmodus von
Phenacolimax major (Fer.).—Arch. Moll., 1949, Bd 77, N 1/6, S. 115—119.
Forcart L. Revision von Vitrina kochi Andreae und Vitrina nivalis (Dumont a.
Mortillet) (Moll., Pulm.).—Verh. Naturf. Ges. Basel, 1954, Bd 65, N 2,
S. 264-269.
Forcart L. Journey to the High Simien (Northern Ethiopia), 1952—3 Three: species of
Phenacolimax (Gastropoda, Vitrinidae), with notes on the taxonomy of the
genus.—Linn. Soc. Journ.— Zool., 1956, vol. 43, N 290, p. 113—122.
Forcart L. Mollusken aus den Abruzzen mit taxonomischen Revisionen und anatomischen
Beschreibungen.— Verh. Naturf. Ges. Basel, 1960, Bd 71, N 1, S. 125—139.
Forcart L. African distribution of genera and subgenera of the family Vitrinidae
(Pulmonata).—J. Conch., 1978, vol. 29, p. 325—327.
Gerhardt U. Weitere Untersuchungen zur Kopulation der Stylommatophoren.—
Z. Morphol. OekoL, .1936, Bd. 30, S. 297—332.
Hesse P. Beitrage zur naheren Kenntnis der Familie Vitrinidae.— Arch. Moll., 1923,
Bd 55, S. 1—25, 81 — 115, 130—145.
Hesse P. Kritische Fragmente.— Arch. Moll., 1924, Bd 56, S. 226—230.
Hoffmann H. Die Vitrinen der Atlantischen Inseln.— Senckenbergiana, 1929,
Bd 11, N 4, S. 218—2235.
Hubendick B. The relationships of the East African Vitrinidae with notes on the
taxonomy of Vitrina.— Ark. Zool., 1953, ser. 2, Bd 6, N 5, p. 83—96.
Ihering H. Morphologic und Systematik des Genitalapparates von Helix.— Z. wiss.
Zool. 1982, Bd 54, S. 386—520.
Kunkel K. Experimented Studie uber Vitrina brevis Fer.— Zool. Jahrb. Physiol.
1929, Bd 46, S. 575—626.
Kunkel K. Vergleichende experimented Studie uber Vitrina elongata Drap. und
Vitrina brevis Fer.— Zool. Jahrb. Physiol., 1933, Bd 52, S. 399—432.
Mermod G. Gasteropodes.— In: Catalogue des Invertebres de la Suisse, 1930,
Fasc. 18, p. 1—583.
Mousson A. Coquilles recueillies par M. le Dr. Sievers dans les contrees Transcau-
casiques.— J. Conchy!., 1876, t. 24, p. 24—51.
Muller O. F. Vermium terrestrium et fluviatilium sen animalium infusoriorum,
helminticorum et testaceorum, non marinorum succincta historia, 2. Havniae
et Lipsiae, 1774, p. J—214.
Osanova N.t Pintkr L. Uber bulgarische Vitrinidae (Gastropoda, Euthyneura).-
Malak. Abhandl. Mus. Dresden, 1968, Bd 2, N 18, S. 243—247
Paulucci M. Materiaux pour servir a 1’etude de la faune malacologique terrestre et
fluviatile de 1’Italie et des Ties. Paris, 1878, p. 1—54.
Pfeiffer C. Systematische Anordnung und Beschreibung deutscher Land- und
Wasser-Schnecken, mit besonderer Rucksicht auf die bisher in Hessen gefun-
denen Arten. Cassel und Berlin, 1821, S. 1 — 134.
Pilsbry H. A. Land Mollusca of North America (North of Mexico).—Acad. Nat. Sci.
Philad., Monogr., 1946, vol. 3, part 2(1), p. 1—520.
Quoy J. R., Gaimard J. P. Zoologie.— In: D’Urville, Voyage de la corvette 1’Astrolabe,
Paris, 1832, vol. 2.
Roth B., Lindberg D. R. Terrestrial Mollusks of Attu, Aleutian Islands, Alaska.—
Arctic, 1981, vol. 34, N 1, p. 43—47
Simroth H. Ueber den Libespfeil der Vitrinen.— Sitz.-Ber. Nat. Ges. Leipzig, 1886,
Bd 12, S. 6-7.
156
Simroth, H. Beitrage zur Kenntnis der Nacktschnecken.— Nova Acta Acad. Leop.,
1889, Bd 54, N 1, S. 1—91.
Studer S. Kurzen Verzeichniss der jetzt in unseren Vaterlande entdeckten Conchy'-
lien.— Meissners Naturw. allg. Anz. schweiz. Ges. Naturw. Bern, 1820, Bd 3,
S. 83—90, 91—94.
Taylor D. W., Sohl N. F. An outline of gastropod classification.— Malacologia,
1962, vol. 1, N 1, p. 7—32.
Thiele J. Handbuch der systematischen Weichtierkunde. Bd I. Jena, 1931, S. 1—778.
Van Mol J.~J. Revision des Urocyclidae (Mollusca, Gastropoda, Pulmonata).—
Ann. Mus. R. Afr. Centr., 1970, N 180, p. 1—234.
Van Mol J.-J. Notes anatomiques sur les Helicarionidae (Mollusques, Gasteropodes,
Pulmones). II. Etudes des genres Pseudaustenia, Dyakia, Helicarion et
comprenant la description de Papuarion, genre nouveau. Discussion sur la
classification des Helicarionidae et les affinites des Urocyclidae.— Ann. Soc.
R. Zool. Belg., 1973, T. 103, fasc. 2—3, p. 209—237.
Wenz W., Zilch A. Euthyneura.— In: Handbuch d. Palaozoologie. Gastropoda, Th. 2,
Berlin, 1959—1960, S. 1—834.
Westerlund C. A. Malakologische Miscellen.—Jahrb. dtsch. malak. Ges., 1883, Bd 10,
S. 51-72.
Westerlund C. A. Fauna der in der palaarctischen Region lebenden Binnenconchylien.
I. Lund, 1886, S. 1—88.
Westerlund C. A.-Neue centralasiatische Mollusken.—Ежегод. Зоол. муз., 1896,
т. I, с. 181 — 198.
Wiegmann F. Der sogenannte Liebespfeil der Vitrinidae.— Jahrb. dtsch. malak. Ges.,
1886, Bd 13, S. 74—95.
Zilch A. Die Typen und Typoide des Natur-Museums Senckenberg, 62: Mollusca:
Zonitacea: Vitrinidae.—Arch. Moll., 1979, Bd 110, N 1/3, S. 81 — 101.
A. A. Schileyko
THE SYSTEM AND THE PHYLOGENY OF VITRINIDAE
(GASTROPODA PULMONATA)
The structure of shell and the reproductive system of 10 species and one
subspecies of Vitrinidae, 7 of which living in the U.S.S.R. are studied. Vitrina
kotulai is separated in subgenus Hessemilimax subgen. nov. There are 3 well definite
variants of structure of the reproductive system within the family which make to
establish 3 subfamilies Phenacolimacinae subfam. nov., Semilimacinae subfam. nov.
and Vitrininae Fitzinger, 1833. Vitrininae and Phenacolimacinae have been evolved
independently from common ancestor, Semilimacinae having been appeared from the
latter during the transformation of vaginal gland to sarcobellum. The family is
connected phylogenetically with Ariophantacea sensu Taylor and Sohl, 1962 + Zonitidae,
Daudebardiidae, Parmacellidae, Milacidae.
ТРУПЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА АН СССР, 1986, Т 148
USSR ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE, LENINGRAD, 1986, VOL. 148
Ю. И. Галкин
ТЕМПЕРАТУРНЫЙ РЕЖИМ И ИЗМЕНЕНИЕ
ПОДВИДОВЫХ КАТЕГОРИЙ У МОЛЛЮСКОВ
MARGARITES GROENLANDICUS И М. COSTALIS
(GASTROPODA: TROCH1DAE)
В работах различных исследователей неоднократно указывалось,
что колебания климата, оказывающие глубокое воздействие на
состояние гидрологического режима, приводят к значительным изме-
нениям в составе и распространении донной фауны. Однако до
сих пор, насколько об этом можно судить по литературе, по-видимому,
не отмечалось, что у некоторых видов, представленных подвидами
различной зоогеографической принадлежности, развитие в направле-
нии той или иной формы определяется температурой и изменяется
на противоположное при перемене температурного режима. Это
явление было отмечено при исследовании многолетних климати-
ческих изменений в распространении в Баренцевом море брюхо-
ногих моллюсков Margarites groenlandicus и М. costalis. Оба этих
моллюска представлены двумя подвидами: бореальными М. groen-
landicus groenlandicus и М. costalis costalis и арктическими
М. groenlandicus umbilicalis и М. costalis sordidus. В процессе их
изучения были обработаны коллекции Зоологического института
АН СССР (ЗИН), в которых М. groenlandicus groenlandicus
представлен 424, М. groenlandicus umbilicalis — 66, М. costalis
costalis — 275 и М. costalis sordidus— 195 пробами, насчитывав-
шими соответственно 1571, 328, 624 и 885 экз. Кроме того, были
использованы литературные источники и постанционные картотеки,
хранящиеся в научных архивах.
Бореальные и арктические подвиды этих моллюсков хорошо
различаются не только по распространению, но и по раковине
(Галкин, 1955), и лишь существование между ними переходных
форм не позволяет считать их отдельными видами. Так, бореальный
Margarites groenlandicus groenlandicus имеет коническую раковину,
покрытую хорошо выраженными спиральными ребрами, и умеренно
широкий пупок, тогда как у арктического М. groenlandicus umbili
calls раковина низкоконическая, поверхность ее, за исключением
первых двух—трех оборотов, гладкая, а пупок широкий и глубокий.
Второй из упомянутых моллюсков представлен бореальным М. co-
stalis costalis, отличающимся сравнительно небольшим числом
спиральных ребер, гребневидной периферией последнего оборота и
уплощенным основанием, и арктическим М. costalis sordidus, у
которого спиральные ребра на оборотах располагаются вдвое чаще,
периферия последнего оборота округлая, а основание выпуклое.
15b
Подвидовые различия хорошо выражены не только у взрослых
индивидов, но и у самых молодых особей, что позволяет определять
систематическую принадлежность и у моллюсков с высотой раковины
порядка 1,5 мм, возраст которых, очевидно, составляет не более
одного года.
Температуры, при которых обитают бореальные и арктические
подвиды этих моллюсков, имеют заметные различия. Средние
придонные температуры, при которых в Баренцевом море встре-
чаются бореальные Margarites groenlandicus groenlandicus и М. co-
stalis costalis составляют соответственно 1,7° и 1,5°, тогда как у
арктических М, groenlandicus umbilicalis и Л4. costalis sordidus
они равняются —1,0° и —0,7° Естественно, что оба первые подвида
населяют прежде всего более теплые южные и западные районы
моря, вторые — северные и восточные участки, а переходные
формы — промежуточную между ними полосу. Но такое распреде-
ление может наблюдаться не только в горизонтальном направлении,
а и по вертикали, причем в последнем случае все эти формы могут
располагаться на ограниченном участке дна. Так, в 1925 г. в губе
Заблудящей, на южном побережье Новой Земли, на глубине от
1 до 16 м были обнаружены все разновидности М. groenlandicus,
возраст которых, судя по размерам раковин, не слишком различался
(табл. 1).
Таблица 1
Систематический состав, количество и размеры раковины
разновидностей Margarites groenlandicus
в губе Заблудящей (Новая Земля) в 1925 г.
Подвид Форма раковины Количество экз. Высота раковины, мм
Margarites groenlandicus gro- enlandicus Типичная для ssp. groenlan- dicus 3 3.4-4.5
Переходная к ssp. umbilicalis 4 3.3-5.4
М. groenlandicus umbilicalis Переходная к ssp. groenlan- dicus 7 2.5-6.0
Типичная для ssp. umbilicalis 10 1.8—7.0
К сожалению, в этой коллекционной пробе были объединены
уловы нескольких драгировок (по крайней мере, четырех, выполнен-
ных на 1, 2,5, 11 и 16 м), вследствие чего отдельные формы нельзя
было точно распределить по глубинам. Но вряд ли можно сомне-
ваться, что особи, принадлежащие к Margarites groenlandicus
groenlandicus, обитали в самых верхних, наиболее прогреваемых
слоях, тогда как индивиды М. groenlandicus umbilicalis распола-
гались на самых больших глубинах, находившихся, однако, всего
лишь в 10—15 м от первых. Это предположение подтверждается
распределением М. costalis sordidus, который также был обнаружен
на этой станции, но уловы двух драг, в которых он был встречен,
были сохранены раздельными. На глубине 2,5 м был пойман 1 экз.
159
Рис. 1. Распространение Margarites groenlandicus во время похолода-
ний (объединены выполненные в 1875—1923 и 1968 гг. сборы, характеризу-
ющие холодные периоды).
/ — М. groenlandicus groenlandicus: 2 — переходные формы; 3 М. groenlandicus
unibilicalis.
этого моллюска, относящийся к более теплолюбивой переходной
форме, тогда как на 11 м — 2 экз. типичного арктического М. со-
stalls sordidus.
Границы распространения отдельных подвидов Margarites
groenlandicus и М. costalis в Баренцевом морс не остаются постоян-
ными. Они испытывают многолетние изменения, причем эти колеба-
ния тесно связаны с температурным режимом. В холодные периоды,
когда температура воды опускается ниже среднего многолетнего
уровня, что наблюдалось в конце XIX начале XX веков, аркти-
ческие формы этих моллюсков имели в море широкое распростра-
нение. Как можно видеть на рис. 1, составленном на основании
сборов 1875—1923 гг., в это время южная граница распространения
арктического М. groenlandicus unibilicalis от северо-западного
Шпицбергена направлялась через пролив Стур-фиорд на юго-восток,
проходила между о. Медвежьим и о. Надежды, опускаясь в
центральной части моря до 73° с. пц, а затем поворачивала на
северо-восток к побережью Новой Земли. Кроме того, эта форма
встречалась в эти годы в наиболее холодных участках у юго-запад-
ного побережья Новой Земли.
160
Рис. 2. Распространение Margarites groenlandicus во время потеплений
(объединены выполненные в 1924—1970 гг. сборы, характеризующие теплые
периоды).
Оботачения как на рис. 1.
В периоды повышенных температур, отраженных сборами
1924—1970 гг., во время „потепления Арктики", южная граница
распространения Margarites groenlandicus umbilicalis оказалась
смещенной далеко на север (рис. 2). В эти годы этот моллюск
не встречался у западного и юго-восточного Шпицбергена; в
центральной части моря, между 30° и 50° в. д., он не отмечался
южнее 77° 15’ с. ш., оказавшись, таким образом, по сравнению с
периодом похолодания на 200 миль севернее, и не был преобладаю-
щим, как ранее, у северо-западного побережья Новой Земли.
Основной формой в Баренцевом море в это время стал бореаль-
ный Margarites groenlandicus groenlandicus. Он встречался вдоль
западного Шпицбергена, в больших количествах обитал в районе
о. Медвежьего и о. Надежды и продвинулся на север в централь-
ной части моря до 76° с. ш., а у Новой Земли — до м. Желания.
При этом М. groenlandicus groenlandicus занял многие районы,
которые прежде служили местом обитания арктического М. groen-
landicus umbilicalis, но которые находились под достаточно сильным
воздействием теплых вод атлантического происхождения — участки
11
Заказ
161
у Шпицбергена и Новой Земли и в центральной части моря.
В других местах арктическая форма была замещена разновидно-
стями, переходными между этими двумя подвидами. Они были
отмечены к северо-востоку от Шпицбергена, куда проходит ветвь
теплого течения, в северо-восточной части моря и даже у Земли
Франца-Иосифа (рис. 2).
Начало перемещения подвидов Margarites groenlandicus в север-
ном направлении относится к 20-м годам, когда стал развиваться
процесс „потепления Арктики" С ним, в частности, можно, очевидно,
связать упомянутое выше обнаружение в 1925 г. в губе Заблудящей
типичной М, groenlandicus groenlandicus и ее формы, переходной к
ssp. unibilicalis, К сожалению, в настоящее время еще не разра-
ботаны методы, с помощью которых можно с достаточной достовер-
ностью определять возраст и, тем самым, время рождения брюхо-
ногих моллюсков, имеющих спирально закрученную раковину. Но
судя по размерам пойманных экземпляров, имевших высоту раковины
всего 3,4—5,4 мм (табл. 1), это были молодые особи, так как
средняя высота М, groenlandicus groenlandicus, встреченных в
юго-восточной части Баренцева моря, составляет 7,0 мм, а макси-
мальная — 10,3 мм. Поэтому, основываясь на размере моллюсков,
можно, по-видимому, предположить, что они появились в губе
Заблудящей в 1920—1922 гг., т. е. в тот период, когда началось
„потепление Арктики", и температуры поднялись выше среднего
многолетнего уровня.
Однако новое положение границ распространения подвидов
Margarites groenlandicus в дальнейшем частично подверглось обрат-
ным изменениям. В 1968 г. арктический М. groenlandicus unibilicalis
вновь был обнаружен в южной части Баренцева моря (рис. 1).
Один небольшой экземпляр этого моллюска был пойман на самой
южной границе Центральной впадины, на глубине 285 м при при-
донной температуре —1,11° Обнаружение здесь этого подвида
можно объяснить двояким образом. С одной ^тороны, в Централь-
ной впадине, принадлежащей к наиболее холодным районам моря,
для которых характерны отрицательные придонные температуры,
даже в период „потепления Арктики" могло сохраняться определен-
ное количество особей, относящихся к арктической форме. В таком
случае один из них и был пойман в 1968 г. Но с другой стороны,
его появление можно связать и с очень сильным похолоданием,
наступившим в Баренцевом море во второй половине 60-х годов.
Поскольку высота раковины пойманного моллюска составляла
5,7 мм, это была молодая особь, так как средняя высота М, groenlan-
dicus unibilicalis из Баренцева моря, представленных в коллекциях
ЗИН, равна 6,4 мм, а максимальная — 17,1 мм, или, по литера-
турным данным (Odhner, 1912),—даже 18,6 мм. Возраст этого
холодолюбивого моллюска насчитывал, очевидно, всего несколько
лет, а время его рождения приходилось, по-видимому, на 1963 или
1965 гг., отличавшиеся низкими температурами. В таком случае в
южной части Баренцева моря арктический М. groenlandicus umbili
calis появлялся дважды: в конце XIX — начале XX веков и в
60-е годы, причем оба раза это совпадало с похолоданиями.
162
Рис. 3. Распространение Margarites costalis во время похолоданий
(объединены выполненные в 1875—1923 гг. сборы, характеризующие холод-
ные периоды).
/ — М. costalis costalis; 2 — переходные формы; 3 — М. costalis sordidus.
Аналогичные изменения в распространении арктического и бо-
реального подвидов, происходившие в периоды похолоданий и
потеплений, наблюдались и у Margarites costalis, В холодный период
конца XIX — начала XX веков арктический М. costalis sordidus
занимал почти все Баренцево море, за исключением его юго-западных
районов (рис. 3). Его южная граница в эти годы от пролива
Стур-фиорд на юго-восточном Шпицбергене направлялась к о. Мед-
вежьему, затем по параллели 74°30' с. ш. ш.ц па восток до 37° в, д.
и поворачивала отсюда на юго-восток к о. Колгуеву.
В период потепления площади обитания Margarites costalis
sordidus резко сократились (рис. 4), В это время он был встречен
только в самой северной части Баренцева моря, у Земли Франца-Ио-
сифа и у северо-западного побережья Новой Земли. В юго-восточной
части моря, которую этот подвид в годы похолодания занимал
почти полностью, его поселения были обнаружены лишь в самых
холодных участках, расположенных между ветвями Канинского,
Колгуево-Печорского и Новоземельского теплых течений и в южном
Новоземельском желобе. Бореальный М, costalis costalis, наоборот,
н
163
Рис. 4. Распространение Margarites costalis во время потеплений
(объединены сборы, характеризующие теплые периоды, которые выполнены
в 1924—1970 гг., а в юго-восточной части моря — в 1959 и 1968—1970 гг.).
Обозначения как на рис 3.
получил в это время гораздо более широкое распространение, заняв
многие участки, где ранее обитал арктический подвид. Он заселил
район о. Медвежьего, переместился в центральной части моря на
150 миль севернее, с 74° до 76°30' с. ш., и продвинулся на восток
в юго-восточной части моря. Значительное распространение в север-
ном и восточном направлениях получили и моллюски, принадле-
жащие к переходной форме.
Связь между распространением различных разновидностей и
температурой особенно хорошо прослеживается при сопоставлении
особенностей распределения самых молодых моллюсков. За время
их жизни, не превышающее нескольких лет, гидрологический режим
по большей части не претерпевает существенных изменений, поэтому
эти особи более полно отражают условия среды, определявшие их
появление в том или ином участке моря, нежели взрослые мол-
люски, между рождением и поимкой которых проходит значительный
период.
В коллекциях ЗИН наиболее молодые Margarites costalis из
Баренцева моря содержатся в 26 пробах, насчитывающих 38 экз.,
164
высота раковины которых составляет 1,3—5,1 мм. Возраст самых
небольших из них, очевидно, равен одному году, а наиболее круп-
ных — вряд ли превышает 4—5 лет. Сведения о большинстве этих
моллюсков приведены в табл. 2.
Как можно видеть из таблицы, молодые типичные особи бореаль-
ного Margarites costalis costalis (пробы № 253, 277), подобно
взрослым индивидам, в течение продолжительного времени обитали
в юго-западных районах Баренцева моря, а на юго-востоке —
только в Чёшской губе, встречаясь на глубинах менее 100 м и при
сравнительно высоких положительных придонных температурах.
Но в 1959 г., во время продолжительного периода потепления,
эта разновидность (№ 299) была найдена гораздо восточнее, в
юго-восточной части моря, на 52° в. д., причем на тех же глубинах
и при тех же температурах, что и более западные представители
этой формы.
Моллюски, находившиеся в самой начальной стадии перехода
к своему арктическому подвиду (№ 281), были отмечены также в
прибрежных водах Мурмана, но на более значительных глубинах,
и в юго-восточной части моря (№ 365), где они находились в более
глубоких и холодных водах, чем типичная разновидность (№ 299).
В 1970 г. они были найдены еще дальше к востоку, в районе Карских
Ворот (№ 351), где они, видимо, появились еще до периода по-
холодания.
Форма, переходная к арктическому ssp. sordidus, и, следова-
тельно, еще менее теплолюбивая, в 1923 г., в начале „потепления
Арктики" была встречена в губе Белушьей, на юго-западном
побережье Новой Земли (№ 106—108). Во время потепления
50—60-х годов она оказалась восточнее, в районе южного Новозе-
мельского желоба и Карских Ворот (№ 364, 345, 349), распола-
гаясь глубже и при более низких температурах, чем предыдущие
разновидности М. costalis costalis.
Подобные же закономерности можно проследить и у арктического
Margarites costalis sordidus. В наиболее теплом западном участке
юго-восточной части моря, на 42° в. д., была встречена только его
наименее холодолюбивая переходная разновидность (№ 44). Формы,
находящиеся в начальной стадии перехода (№ 373, 463) в период
потепления 50-х годов располагались гораздо восточнее, на 50° в. д.,
в южном Новоземельском желобе, при очень низких отрицательных
температурах. Они были найдены здесь на 140—180 м, тогда как
на меньших глубинах и при положительных температурах были
отмечены более теплолюбивые переходные бореальные М. costalis
costalis (№ 345). Типичная разновидность арктического М. costalis
sordidus была встречена лишь на больших глубинах и при очень
низких отрицательных температурах в южном Новоземельском
желобе (№ 462) и у Земли Франца-Иосифа (№ 429, 353), где даже
на мелководьях в течение всего года сохраняется температура
ниже нуля.
Таким образом, в распределении самых молодых Margarites
costalis имеются две закономерности: 1) постепенный переход от
165
Таблица 2
Распространение наиболее молодых Margarites costalis в Баренцевом море в 1900—1970 гг.
Номера пробы ио ката.югх ЗИН ' Количество экз. Высота раковины, мм Год поимки Район Глубина, м Придонная Т ° Форма
Margarites costalis costalis
253 1 3.1 1926 Чешская губа 34 3.60 Типичная для ssp. costalis
277 1 4.4 1955 Прибрежные воды Мурмана 70 2.62 То же
299 1 3.1 1959 Юго-восточная часть моря, 52° в. д. 49 1.73 То же
281 2 2.9, 4.9 1957 Прибрежные воды Мурмана 178 2.55 Начавшая переход к ssp. sordidus
365 1 1.8 1959 Юго-восточная часть моря, 51° д. 81 0.26 То же
351 2 4.6, 4.9 1970 То же, 57° в. д. 47 -1.24 То же
106—108 4 2.0—4.1 1923 Новая Земля, губа Белушья 14—23 — Переходная к ssp. sordidus
364 1 1.7 1959 Пролив Карские Ворота 185 0.50 То же
345 1 4.1 1970 Южный Новоземельский желоб НО 1.61 То же
349 3 3.6—4.4 1970 Пролив Карские Ворота 100 -1.0 То же
Margarites costalis sordidus
44 1 4.6 1900 Юго-восточная часть моря, 42° в. д. 116 -0.8 Переходная к ssp. costalis
373 1 2.2 1959 Южный Новоземельский желоб 180 -1.90 Начавшая переход к ssp. costalis
463 1 2.2 1959 То же 140 -1.80 То же
462 1 4.0 1970 То же 186 -1.5 Типичная для ssp. sordidus
429 1 1.3 1970 Земля Франца-Иосифа, о. Хейса 13 -0.5 То же
353 1 2.5 1970 То же 20 -0.5 То же
бореального Л4. costalis costalis к арктическому М. costalis sordidus
по мере перемещения мест распространения этого моллюска к
востоку, увеличения глубины обитания или понижения температуры;
2) продвижение М. costalis costalis в период потепления в восточном
направлении и замещение им при этом М. costalis sordidus. Обе
эти тенденции, в конечном счете, связаны с температурным ре-
жимом, который, следовательно, является ведущим фактором в
процессе изменения распределения различных зоогеографических
разновидностей.
Рассматривая расширение ареалов бореальных подвидов Marga*
rites groenlandicus и М. costalis, связанное с потеплением, нельзя
не отметить, что у обоих моллюсков оно осуществлялось путем
замещения теплолюбивыми формами арктических разновидностей.
Оба этих вида имеют прямое развитие и не проходят пелагической
эмбриональной стадии (Thorson, 1935). Вследствие этого указанное
явление нельзя отнести за счет особенностей личиночного переноса:
проникновения бореальных личинок во все более отдаленные районы,
где они теперь в связи с повышением температурного режима
могут выжить, тогда как личинки арктических подвидов в силу тех
же причин обречены здесь на гибель. Единственным приемлемым
объяснением процесса подобного замещения является, очевидно,
предположение о зависимости характера развития этих моллюсков
от температуры. Ранее уже указывалось, что отдельные их разно-
видности хорошо различаются по раковине в самом молодом
возрасте. Но, по-видимому, направление развития в сторону той
или иной формы определяется еще во время эмбриогенеза или даже
гаметогенеза. При повышении температуры до определенного пре-
дела оно начинает идти в сторону береального подвида и заканчи-
вается формированием теплолюбивого моллюска. При похолодании,
по всей вероятности, должен наблюдаться обратный процесс, хотя,
надо отметить, из-за ограниченности имевшихся в распоряжении
автора материалов единственным примером, подтверждающим это
предположение было описанное выше появление М. groenlandicus
umbilicalis в 1968 г. в южной части Баренцева моря. Следовательно,
явления, отмеченные у М. groenlandicus и М. costalis, носят обра-
тимый характер и должны, в таком случае, иметь фенотипическую,
а не генотипическую природу. Можно надеяться, что будущие иссле-
дования позволят не только обнаружить указанные закономерности
у других донных беспозвоночных, имеющих аналогичное с М. groen
landicus и М. costalis деление на подвиды разной зоогеографи-
ческой принадлежности, но и дадут возможность подтвердить их
экспериментально.
ЛИТЕРАТУРА
Г алкин Ю. И. Брюхоногие моллюски трохиды дальневосточных и северных морей
СССР (Семейство Trochidae).— Определители по фауне СССР, иэд. Зоологи-
ческим институтом АН СССР, № 57, М.—JT, 1955, 132 с.
167
Odhner N. Northern and arctic invertebrates in the collection of the Swedish State
Museum (Riksmuseum). 5. Prosobranchia. 1. Diotocardia. Kungl. Svenska
Vetenskapsakad. Handl., 1912, Bd. 48, N 1, 93 p., 7 pls.
Thorson G. Studies on the egg-capsules and development of arctic marine prosobranch.
Medd. Gr0nland, 1935, Bd 100, N 5, 71 p.
Yu. I. Galkin
TEMPERATURE AND TRANSFORMATION OF SUBSPECIES
IN MARGARITES GROENLANDICUS
AND M. COSTALIS (GASTROPODA TROCHIDAE)
The relation of distribution of the arctic and boreal subspecies of Margarites
groenlandicus and M. costalis is discussed in connection with warming of the
Arctic. It is established the replacement of Arctic forms by Boreal ones in both
species, which cannot be explained by distribution. It is supposed that the temperature
determins the direction of development to one or another form during the embryo-
genesis or gametogenesis.
РЕФЕРАТЫ
УДК 595.182.4:591.3
Личиночное развитие панцирных моллюсков (Polyplacophora). Анализ работ по
видам с планктонным типом развития личинки. Сиренко Б. И.— В кн.: Морфо-
логические и экологические основы систематики моллюсков. Л., изд. Зоол. ин-та
АН СССР, 1986, с. 4—11. (Тр. Зоол. ин-та АН СССР, т. 148).
На основе анализа литературных данных (27 работ) приводятся общие за-
кономерности, наблюдающиеся при планктонном типе развития личинок хитонов.
За более чем 120-летний период исследований личиночное развитие изучено у 19 видов
панцирных моллюсков, причем у 14 до стадии формирования 7-го и 8-го щитков
раковины. Отмечается положительная корреляция возраста, при котором личинки
выходят из оболочек в воду, с диаметром яиц, т. е. с запасом питательных веществ.
Ил. 1, табл. 2, библ. 32 назв.
УДК 594.312.4
Филогения подкласса Caudofoveata (Mollusca Aplacophora). Иванов Д. Л.—
В кн.: Морфологические и экологические основы систематики моллюсков. Л.,
изд. Зоол. ин-та7 АН СССР, 1986, с. 12—32 (Тр. Зоол. ин-та АН СССР, т. 148).
На основе детального анализа особенностей внешней морфологии, а также
строения кожно-мускульного мешка, радулярного аппарата и других частей пищева-
рительной системы обсуждается филогения подкласса Caudofoveata. Делается вывод,
что наиболее близки к исходным формам Scutopus, Psilodens и Limifossor. Остальные
представители формируют 2 ветви: рано отделившуюся, составленную Prochaetoderma
и, вероятно, Metachaetoderma и высшую, сильно специализированную,составленную
Chaetoderma, Crystallophrisson и Caudofoveatus. Ил. 11, библ. 18 назв.
УДК 594.1
Опыт новой системы индексации зубов замка двустворчатых моллюсков. С кар*
л а то О. А., Старобогато в Я. И.— В кн.: Морфологические и экологические
основы систематики моллюсков. Л., изд. Зоол. ин-та АН СССР, 1986, с. 33—38.
(Тр. Зоол. ин-та АН СССР, т. 148).
Предлагается система индексации, основанная на следующих принципах:
зубы передней и задней половин замка нумеруются независимо; граница между
ними — середина прямого дорсального края продиссоконха или точки замочного
края на более поздних стадиях, преемственно связанные с ней в ходе роста.
Первичные пластинки и зубы прегетеродонтных замков нумеруются снаружи внутрь
нечетными цифрами в правой створке и четными цифрами в левой, так чтобы при
сомкнутых створках ряд чисел был непрерывным. Передние пластинки обозначаются
буквой „а“, задние—„р“ Обособленные или отонутые части зубов ктенодонтных
замков отмечаются добавлением цифры 1. Кардинальные зубы обозначаются до-
бавлением буквы „с“, латеральные — буквы „1“ Если из одной пластинки форми-
руются 2 кардинальных зуба, то они обозначаются как „сГ и „Сг“ Нимфа, если она
формируется за счет первичной пластинки, обозначается добавлением буквы „п“, а
прочие, поддерживающие лигамент образования (круры) — добавлением букв
„сг“ Ил. 14, библ. 3 назв.
УДК 594.141
Морфометрические характеристики раковин двустворчатых моллюсков Nodularia
(Unionidae) фауны СССР и возможности их использования для идентификации
видов. Затравкин М. Н., Лобанов А. Л.— В кн.: Морфологические и
169
экологические основы систематики моллюсков. Л., изд. Зоол. ин-та АН СССР,
1()86. с. 39—45. (Тр. Зоол. ин-та АН СССР, т. 148).
Проведено сравнение морфометрических характеристик раковин 7 видов рода.
Обсуждается возможность использования морфометрических характеристик для
идентификации видов рода. Приведена таблица для определения видов рода, оби-
тающих в водоемах СССР. Табл. 4, библ. 3 назв.
УДК 594.141
Возможности определения зрелых глохидиев массовых европейских видов Unioninae
и Anodontinae (Bivalvia Unlonldae). Антонова Л. А.— В кн.: Морфологи-
ческие и экологические основы систематики моллюсков. Л., изд. Зоол. ин-та
\Н СССР, с. 46—53. (Тр. Зоол. ин-та АН СССР. т. 148).
Описан метод оптико-микроскопического изучения раковин глохидиев и приведе-
на таблица для определения глохидиев 12 видов из дельты Волги. Ил. 15, библ. 5 назв.
УДК 594.153
Виды рода Odhneripisidium (Bivalvia Pisidiidae) фауны СССР. Иззатул-
лаев 3. И., Старобогатов Я. И,—В кн.: Морфологические и экологические
основы систематики моллюсков. Л., изд. Зоол. ин-ia АН СССР, 1986, с. 54—70.
(Тр. Зоол. ин-та АН СССР. т. 148).
Приведено описание 13 новых видов рода Odhneripisidium с территории СССР:
О. chatyrkulense, О. behningi, О. gafurovi, О. pamirense, О. tolstikovae, О. kazakhsta
nicum (два последних вида из неогеновых отложений), О. terekholicum, О. tuvaense,
О. sogdianum, О. polytimeticum, О. terekense Kazannikov, О. issykkulense, О. khorense.
Все известные с территории СССР виды рода сгруппированы в 4 подрода Ussuri-
pisidium subgen. nov. с типовым и единственным видом О. khorense, Kuiperipisidium
subgen. nov. с видами О. sogdianum (типовой вид), О. polytimeticum, О. terekense,
и О. issykkulense, Tuvapisidium subgen. nov. с видами О. popovae (типовой вид),
О. terekholicum, О. tuvaense. Сюда же входят все виды этого рода, описанные
С. М. Поповой и Я." И. Старобогатовым из кызылгирской свиты Горного Алтая,
кроме О. paravincentianum, и номинотипический подрод Odhneripisidium s. str.,
куда входят все остальные известные с территории СССР виды рода. Уточнены
видовые различия и распространение О. steward, О. prashadi, О. dancei. Ил. 18,
библ. 11 назв.
УДК 594.3:591.12
Интенсивность обмена у брюхоногих моллюсков. Аракелова Е. С.—В кн.:
Морфологические и экологические основы систематики моллюсков. Л., изд. Зоол.
ин-та АН СССР, 1986, с. 71-85. (Тр. Зоол. ин-та АН СССР, т. 148).
На основе экспериментального собственного материала и литературных данных
оценивается энергетический обмен гастропод. Выявлены сходство и различия в
уровнях энергетического обмена в пределах класса. Рассматриваются возможные
причины, приводящие к отклонению уровня обмена отдельных видов от среднего
уровня, характерного для класса гастропод. Сделано заключение, что различия
в уровнях обмена которые имеются между двумя подклассами — легочных и передне-
жаберных имеют адаптивную природу. Табл. 2, библ. 54 назв.
УДК 594.32
Новые виды брюхоногих моллюсков подотряда Rissooidei Голиков А. Н. -
В кн.: Морфологические и экологические основы систематики моллюсков. Л.,
изд. Зоол. ин-та АН СССР, 1986, с. 86—89. (Тр. Зоол. ин-та АН СССР, т. 148)
170
В результате ревизии коллекционных фондов Зоологического института АН СССР
по подотряду Rissooidei описаны новые виды Mohrensternia coronata, Onoba russica
и Punctulum reticulatum из умеренных вод северо-западной части Тихого океана и
Onoba karica из северной части Карского моря. Ил. 4.
УДК 594.329
Новый вид брюхоногих моллюсков Conradia minuta из умеренных вод северо-западной
части Тихого океана. Гол и ков А. Н., Старобогатов Я. И.— В кн.: Морфо-
логические и экологические ociroBbi систематики моллюсков. Л., изд. Зоол. ин-га
\Н СССР. 1986, с. 90-92. (Тр. Зоол. ин-та АН СССР, т. 148).
Описание Conradia (Conradia) minuta sp. nov. из Охотского моря и прибрежных
вод Южного Сахалина. Вид этого подрода изображается впервые в мировой литера-
туре. Ил. /.
УДК 594.381.5
К систематике брюхоногих моллюсков рода Latia (Gastropoda Pulmonata Latiidae).
Старобогатов Я. И.— В кн.: Морфологические и экологические основы систе-
матики моллюсков. Л., изд. Зоол. ин-та АН СССР, 93—96. (Тр. Зоол. ин-та
АН СССР, т. 148).
Описан новый вид Latia climoi из окрестностей Веллингтона (Новая Зеландия)
и приведена определительная таблица всех 3 видов рода Latia — L. neritoides
Gray, L. lateralis (Gould) и L. climoi sp. nov., признаваемых автором. Ил. 3,
табл. 1, библ. 7 назв.
УДК 594.382
Материалы по анатомии и таксономии Daudebardiidae (Gastropoda Pulmonata)
Шилейко А. А.— В кн.: Морфологические и экологические основы систематики
моллюсков. Л., изд. Зоол. ин-та АН СССР, 1986, с. 97—123. (Тр. Зоол. ин-та
АН СССР, т. 148).
Исследовано строение раковин и полового аппарата 10 видов Daudebardiidae,
из которых 7 обитают на территории СССР. Таксономическая структура семейства
много сложнее, чем принималось до сих пор: семейство включает не менее 8 родов,
в том числе Bilania gen. nov. (типовой вид Daudebardia boettgeri Glessin, 1883),
Inguria gen. nov. (типовой вид Daudebardia wagneri Rosen, 1911), Lotharia gen. nov.
(типовой вид Pseudolibania cretica Forcart, 1950). Наиболее архаичен половой
аппарат Lotharia) имеющий функционирующий эпифаллус. Приведены соображения
по филогении семейства. Ил. И, библ. 35 назв.
УДК 594.382
Система и филогения Vitrinidae (Gastropoda Pulmonata). Шилейко А. А.—
В кн.: Морфологические и экологические основы систематики моллюсков. Л.,
изд. Зоол. ин-та АН СССР, 1986, с. 124—157. (Тр. Зоол. ии-га АН СССР, т. 148).
Исследовано строение раковин и полового аппарата 10 видов и 1 подвида
Vitrinidae, из которых 7 обитают на территории СССР. Vitrina kotulai выделена
в подрод Hessemilimax subgen. nov. В пределах семейства имеется 3 хорошо
очерченных варианта строения полового аппарата, в соответствии с чем выделено
3 подсемейства: Phenacolimacinae subfam. nov., Semilimacinae subfam. nov., Vitri-
ninae Fitzinger, 1833. От гипотетической предковой формы независимо возникли
Vitrininae и Phenacolimacinae, от последних произошли Semilimacinae за счет
преобразования вагинальной железы в саркобеллум. Семейство филогенетически
связано с Ariophantacea sensu Taylor et Sohl, 1962 + Zonitidae, Daudebardiidae,
Parmacellidae, Milacidae. Ил. 17, библ. 60 назв.
171
УДК 594.325.3:591.543.1
Температурный режим и изменение подвидовых категорий у моллюсков Margarites
groenlandicus и М. costalis (Gastropoda Trochidae). Галкин Ю. И.— В кн.:
Морфологические и экологические основы систематики моллюсков. Л., изд. ин-та
АН СССР, 1986, с. 158—168. (Тр. Зоол. ин-та АН СССР, т. 148).
Обсуждается связь распространения арктических и бореальных подвидов
М. groenlandicus и М. costalis с потеплением Арктики. У обоих видов наблюдается
замещение арктических форм теплолюбивыми, что невозможно объяснить процессами
расселения. Предполагается, что температура определяет направление развития
в сторону той или иной формы еще во время эмбриогенеза или даже гамето-
генеза. Ил. 4, табл. 2, библ. 3 назв.
Содержание
Предисловие .............................................................. 3
Сиренко Б. И. Личиночное развитие панцирных моллюсков (Polyplacophora).
Обзор видов с планктонным типом развития личинок.................... 4
Иванов Д. Л. Филогения подкласса Caudofoveata (Mollusca Aplacophora) . . ' 12
Скарлато О. А., Старобогатов Я. И. Опыт новой системы индексации зубов
замка двустворчатых моллюсков . ’........................................ 33
Затравкин М. И., Лобанов А. Л. Морфометрические характеристики раковин
двустворчатых моллюсков рода Nodularia (Unionidae) фауны СССР
и возможности их использования для идентификации видов рода ... 39
Антонова Л. А. Возможности определения зрелых глохидиев массовых европей-
ских видов Unioninae и Anodontinae (Bivalvia)............................ 46
Иззатуллаев 3. И., Старобогатов Я. И. Виды рода Odhneripisidium (Bivalvia
Pisidiidae) фауны СССР............................................. 54
Аракелова Е. С. Интенсивность обмена у брюхоногих моллюсков.............. 71
Голиков А. И. Новые виды брюхоногих моллюсков подотряда Rissooidei ... 86
Голиков А. Н., Старобогатов Я- И. Новый вид брюхоногих моллюсков Con-
radia minuta из умеренных вод северо-западной части Тихого
океана................................................................... 90
Старобогатов Я. И. К систематике брюхоногих моллюсков рода Latia
(Gastropoda Pulmonata Latiidae).................................... 93
Шилейко А. А. Материалы по анатомии и таксономии Daudebardiidae (Gastro-
poda Pulmonata)................................................... 97
Шилейко А. А. Система и филогения Vitrinidae (Gastropoda Pulmonata) . . 124
Г алкин Ю. И. Температурный режим и изменение подвидовых категорий
у моллюсков Margarites groenlandicus и М. costalis (Gastropoda
Trochidae).........................................................158
Рефераты................................................................ 169
Contents
Preface........................................................................ 3
Sirenko В. I. Larval development of Polyplacophora. Analysis of studies of
species with the planctonic type of larval development................... 4
Ivanov D. L. Phylogeny of the subclass Caudofoveata (Mollusca Aplaco-
phora).................................................................. 12
Scarlato 0, A., Starobogatov Ya. I. Trial of a new system of indexes for
bivalvian dentition..................................................... 33
Zatravkin M. N., Lobanov A. L. Morphometric characters of shells of Nodularia
(Bivalvia Unionidae) from USSR and possibility of using them for
species identification.................................................. 39
Antonova L. A. On the possibility of identification of the ripe glochidia of
abundant European species from the subfamilies Unioninae and
Anodontinae (Bivalvia Unionidae)........................................ 46
Izzatullaev Z. /., Starobogatov Ya. I. Odhneripisidium species from the fauna
of the USSR............................................................. 54
Arakelova E. S. Intensity of gastropodan metabolism........................... 71
Golikov A. N. New species of gastropods from the suborder Rissooidei .... 86
Golikov A. N., Starobogatov Ya. /. New gastropodan species Conradia minuta
from temperate waters of North-Western Pacific.......................... 90
Starobogatov Ya. /. On the taxonomy of the gastropod molluscan genus
Latia (Gastropoda Pulmonata Latiidae)................................... 93
Schileyko A. A. Some data on the anatomy and taxonomy of Daudebardiidae
(Gastropoda Pulmonata).................................................. 97
Schileyko A. A. The system and the phylogeny of Vitrinidae (Gastropoda
Pulmonata) ............................................................ 124
Galkin Yu. I. Temperature and transformation of subspecies in Margarites
groenlandicus and M. costalis (Gastropoda Trochidae)..............158
Abstracts.................................................................... 169
МОРФОЛОГИЧЕСКИЕ И ЭКОЛОГИЧЕСКИЕ
ОСНОВЫ СИСТЕМАТИКИ МОЛЛЮСКОВ
Труды Зоологического института АН СССР
Том 148
Утверждено к печати
редакционно-издательским советом
Зоологического института АН СССР
План 1986 г.
Редактор Т А. Асанович
Художник С. Е. Станкевич
Подписано к печати 25.07.86. М-18803. Формат 60Х90’/|б. Бумага тип.
Гарнитура литер. Печать офсетная. Печ. л. 11, 10,23 усл.-печ. л. 10 уч.-изд.
Тираж 800 экз. Заказ № 1696. Цена 1 р. 50 к.
Зоологический институт АН СССР, 199034, Ленинград, Университетская наб., 1
Типография № 2 Ленуприздата, 191104, Ленинград, Литейный пр., 55