/















































































































































































































































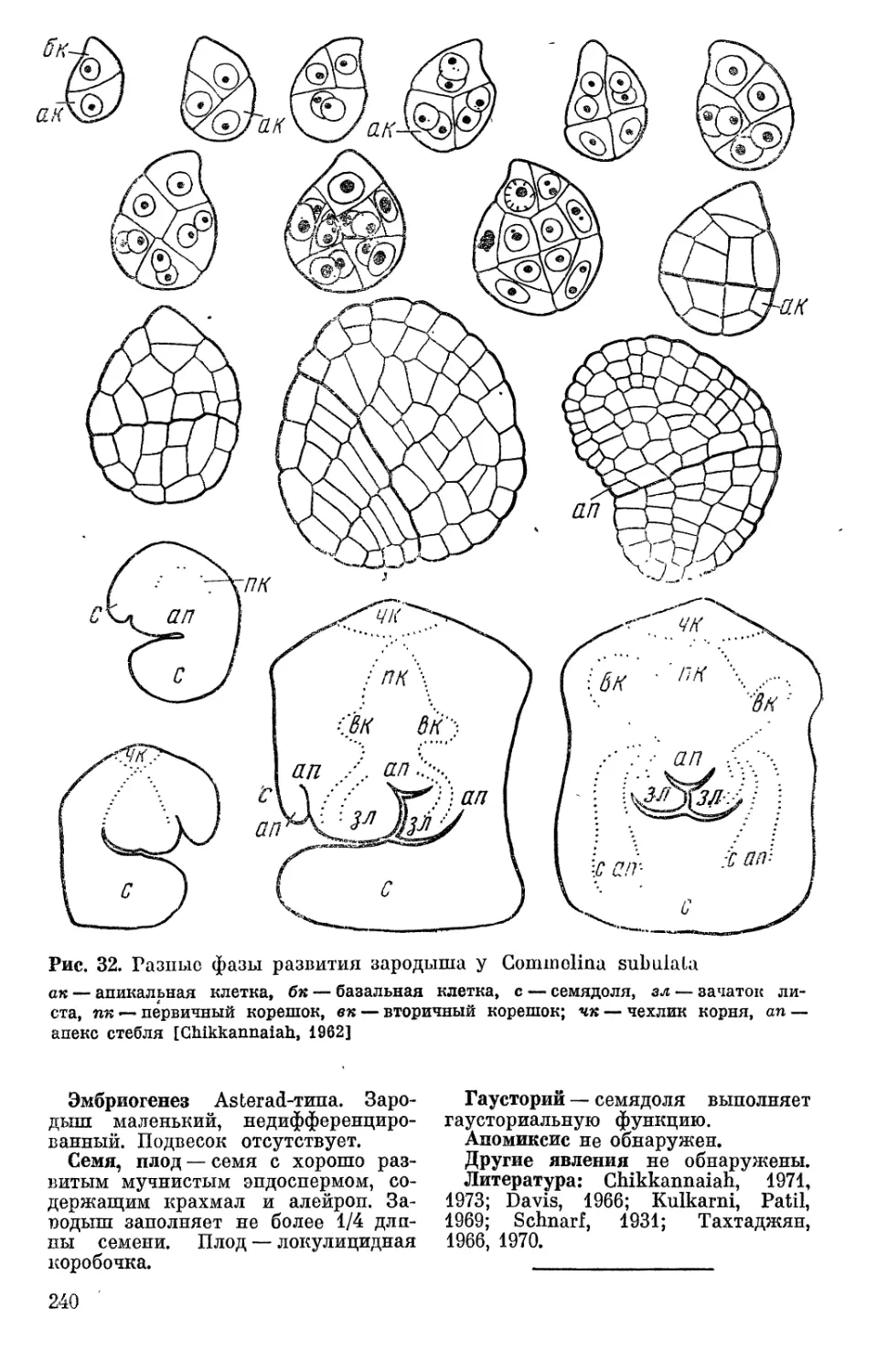




















































































































Text
АКАДЕМИЯ НАУК СССР
ГЛАВНЫЙ БОТАНИЧЕСКИЙ САД
В. A. ПОДДУБНАЯ-АРНОЛЬДИ
ХАРАКТЕРИСТИКА
СЕМЕЙСТВ
ПОКРЫТОСЕМЕННЫХ
РАСТЕНИЙ
ПО
ЦИТОЭМБРИОЛОГИЧЕСКИМ
ПРИЗНАКАМ
ИЗДАТЕЛЬСТВО «НАУКА»
МОСКВА 1982
УДК 582.5/9
Поддубная-Арнольди В. А. Характеристика ее-
мейств покрытосеменных растений по цитоэмбриологическим
признакам. М.: Наука, 1982.
Книга является справочным изданием. В ней обобщены
современные сведения по цитоэмбриологии покрытосеменных
растений. Описания 296 семейств даны по единой схеме в
сравнительном аспекте, что важно при решении вопросов
филогении и систематики растений.
Подобная сводка по сравнительной цитоэмбриологии в
Советском Союзе издается впервые.
Книга рассчитана на цитоэмбриологов, морфологов, си-
стематиков, генетиков и других специалистов.
Ил. 34, список лит. на 79 с.
Ответственный редактор
академик | Н. В. ЦИЦИН |
"OO4ZOO0000 (V0?
вии, oe Нодниеное. Б3 — 27—408—1980
© Издательство «Наука»,
1982 г.
=>
ВВЕДЕНИЕ
В опубликованных нами монографиях 1964 и 1976 гг. были
описаны многообразие цитоэмбриологических признаков покры-
тосеменных растений и закономерности изменений эмбриональ-
ных процессов под влиянием особых условий существования, гиб-
ридизации, полиплоидии, мутагенеза и различных воздействий
химическими и ионизирующими веществами.
В настоящей работе мы попытаемся показать, как цитоэмбрио-
логические признаки характеризуют различные семейства и по-
рядки покрытосеменных и какое это имеет значение для их сис-
тематических и филогепетических взаимоотношений. Цитоэмбрио-
логическую характеристику различных семейств и порядков мы
будем описывать применительно к системе А. Л. Тахтаджяна
[1970], так как она является одной из наиболее хорошо разра-
ботапных отечественных систем. Цитоэмбриологическую тер-
мппологию в настоящей работе мы будем приводить в соответ-
ствий с принятой в наших монографиях 1964 и `1976 гг. Клас-
сификацию тинов образовапия степки пыльника мы даем по Д>-
вис ГРлуйёь 1966[, тинов строения спородермы пыльцевых зерен—
но }рлману | Метан, 1953 Л. А. Цунрияновой [41965] и Л.А. Ву-
прияновой се И. А. Аленшицой [4972]; типов семяпочки — по
М. И. Савченко [4973], типов женского гаметофита — по И. Д. Ро-
манову [4974], типов оплодотворения — по Е. Н. Герасимовой-Не-
вашиной [Gerassimova-Navashina, 41961], типов зародыша — по
Дкогансену [Johansen, 1950], типов семяп — по сведениям
№. С. Смирновой; типов апомиксиса — по В. А. Поддубной-Ар-
польди [1964, 1976].
Типы строения цветков, плодов, способы опыления и размно-
жения мы описываем по работам М. И. Голенкина [1937], Фрик-
cena [Fryxel, 1957], II. Maxemspapu [Maheshwari, 1950, 1964]
и А. Л. Тахтаджяна [1966, 1970].
Окраска зародышей приводится нами из работы М. С. Яков-
лева и Г. Я. Жуковой [1974], а данные о химическом составе
зародышей и эндосперма из работ Леббока ПлЪок, 1892], Ф. Не-
толитекого [ПМею| ку, 1926] и Корнеру [Сотпег, 1976]. Числа
хромосом приведены по Дарлингтону, Уайли [Darlington, Wylie,
1955] и из «Хромосомного атласа» [1969]. Мы, как и другие иссле-
`дователи, считаем, что такие цитоэмбриологические признаки, как
развитие и строение стенки микроспорангия, тапетума, микроспор,
3
мужского гаметофита, семяпочки, макроспор, женского гаметофи-
та, опыление, прохождение пыльцевых трубок в пестике, опло-
дотворение, эндосперма, зародыша, подвеска, гаустория, семени,
плода и апомиксиса, характеризуя определенным образом ту или
иную группу покрытосеменных, являются систематическими
и филогенетическими признаками.
В последнее время все чаще и чаще раздаются голоса в поль-
зу того, что для построения филогенетической системы покрыто-
семенных необходимо привлечь различные методы исследования,
однако на деле это осуществляется довольно редко. В частности,
систематики слабо используют или даже вовсе не используют дан-
ные цитоэмбриологии, которые могут помочь в выяснении система-
тических и филогенетических взаимоотношений между различными
группами и внутри различных групп покрытосеменных. А цито-
эмбриологи, описывая те или иные цитоэмбриологические приз-
наки, обычно не интересуются систематическими и филогенети-
ческими положениями изучаемых ими групп и нередко дают их
неполные цитоэмбриологические характеристики. После опубли-
кования монографий Шнарфа [Schnarf, 19341, 4933] и Дэвис
[Рау1з, 1966] данных по сравнительной цитоэмбриологии покры-
тосеменных, которые можно было бы использовать в целях по-
строения филогенетической системы, становится все больше
и больше. Поэтому, как нам кажется, уже настает время для комп-
лексного применения различных методов исследования, в том
числе и цитоэмбриологического, в целях построения филогенети-
ческой системы покрытосеменных, так как теперь уже совершен-
но ясно, что на основании использования одного какого-нибудь
признака нельзя построить такую систему.
Мы считаем, что цитоэмбриологические признаки являются
важными для систематики и филогении, хотя и относительными,
а не абсолютными, что, впрочем, характерно и для любых дру-
гих признаков. Ни один из вышеуказанных цитоэмбриологиче-
ских признаков не может считаться более важным по сравнению
с остальными. Ноэтому не следует приписывать исключительное
значение одному какому-нибудь из пих и на основании сходства
или различия одного из них судить о систематическом положе-
нии той или иной группы. Необходимо, как правило, учитывать
комплекс различных цитоэмбриологических призпаков, или «эм-
бриологическую диаграмму», по терминологии Шнарфа [Schnarfi,
19331. Однако в некоторых случаях даже с помощью единствен-
ного цитоэмбриологического призпака, особенно такого своеоб-
разного, как, например, тип гаустория, можно сделать очепь
важные выводы для систематики и филогепии. Цитоэмбриологи-
ческие признаки имеют перед другими систематическими призна-
ками то преимущество, что они более копсервативны, так как
в значительной степени не подвержены непосредственному
влиянию внешних условий существования. Это свойство цитоэм-
бриологических признаков позволяет иногда установить цитоэм-
бриологическое сходство и на основании этого связать узами
4
родства виды, роды, семейства растений, внешне морфологически
уже ‘довольно изменившиеся по отношению друг к другу. Однако
большая консервативность цитоэмбриологических признаков, ко-
торая создает им преимущество по сравнению с другими систе-
матическими признаками, вместе с тем и ограничивает их зна-
чение, ‘так как сравнительно малое разнообразие их очень
сглаживает различия между отдельными группами покрытосемен-
ных, вследствие чего морфологически очень отдаленные группы
могут обнаружить сходные черты в своей цитоэмбриологии и, на-
оборот, морфологически сходные группы — отличающиеся черты.
Одни цитоэмбриологические признаки являются первичными,
исходными, другие вторичными, производными. Как правило, те
из них, ‘которые отличаются более сильным развитием, дольше
развиваются и хуже приспособлены к выполнению специальных
функций, являются первичными, исходными признаками. В та-
ким признакам относятся:
1) малое разпообразие основных чисел хромосом;
2) мпогорядная стенка микроспорангия;
3) четыре микроспорангия;
4) двухклеточные, одиночные пыльцевые зерна;
5) паличие большого числа семяпочек внутри одной и той
‚ке завязи;
6) прямая анатропная семяпочка с двумя интегументами;
7) хорошо развитый нуцеллус (крассинуцеллятный);
8) наличие париентальных клеток и нуцеллярного колпачка,
или клювика;
}) многоклегочный женский археспорий;
1) мопоспоричеекний тии (Polygonum-ratt) зародышевого
личико,
ЕР) три эфемерные антиноды;
1..) огсутствие разпого типа гаусториев;
1:2) порогамия;
1^) двойное оплодотворение; большой промежуток времени
между онылением и оплодотворением, а также между оплодотво-
рением, развитием зародыша и эндосперма;
15) зародыш слабо дифференцированный, маленький, прямой,
бесхлорофилльный;
16) семена с эндоспермом;
17) присутствие в семенах перисперма;
18) отсутствие апомиксиса.
Ко вторичным производным признакам относятся:
1) большое разнообразие основных чисел хромосом;
2) стенка микроспарангия с числом рядов не более четырех;
3) микроспорангиев больше или меньше четырех;
4) трехклеточные пыльцевые зерна, соединенные в диады,
тетрады, массулы или поллинии;
5) одна семяпочка внутри (одной и той же) завязи;
6) амфитропная, кампилотропная, цирцинотропная и другие
семяпочки, с одним интегументом или без интегумента;
5
7) нуцеллусе слабо развитый (тенуинуцеллятный), часто
сведенный лишь к одному ряду клеток;
8) отсутствие париетальных клеток и нуцеллярного колпач-
ка, или клювика;
9) одноклеточный женский археспорий;
10) ди- и тетраспорические (Allium, Drusa, Fritillaria, Penaea,
Peperomia, Adoxa um др.) типы зародышевого мешка;
11) наличие более трех долго сохраняющихся антипод;
12) присутствие разного типа гаусториев (макроспоровогто,
зародышева мешка, антиподального, синергидного, эндоспер-
миального, подвесочного и т. д.);
13) халазогамия, мезогамия;
14) одинарное оплодотворение; маленький промежуток време-
ни между опылением и оплодотворением, а также между опло-
дотворением, развитием зародыша и эндосперма;
15) зародыш хорошо дифференцированный, большой, согну-
тый, хлорофиллоносный;
16) семена без эндосперма;
17) отсутствие перисперма;
18) наличие апомиксиса.
Не всегда легко решить, является ли данный цитоэмбриоло-
гический признак первичным, исходным, или вторичным, произ-
водным, и поэтому различные исследователи по-разному оценива-
ют некоторые из них. Это относится, например, к типам тапету-
ма пыльника, семяпочек, эндосперма и образования микроспор.
Давно ведется дискуссия по вопросу о том, какие типы тапету-
ма (секреторный или амебоидный, периплазмодиальный), эндо-
сперма 'нуклеарный, целлюлярный или гелобиальный), семяпо-
чек (обращенная, полуобращенная, прямая или кампилотропная)
и образования микроспор (сукцессивное или симультанное) явля-
ются первичными или вторичными. Мы склопяемся к мнению
тех исследователей, которые считают первичпыми секреторный
тип тапетума в пыльниках, симультанный тип образования мик-
роспор, нуклеарный тип эндосперма и ортотропный тип семяпоч-
ки. Последний тип в процессе эволюции, по-видимому, быстро
исчез, но в онтогенезе он сохранился. После ортотропного типа
наиболее примитивным является анатропный тип семяпочки
с двумя интегументами. Однако для решения этих вопросов тре-
буются дальнейптие исследования.
Некоторые цитоэмбриологические признаки наряду с крайни-
ми имеют и переходные типы. К таким признакам относятся, на,
иример, типы семяпочек, эпдосперма и эндоспермальных гаусто-
puen, которые у одних иредетавителей хоропто развиты, у дру-
их выражены более или менее слабо, и различные способы зало-
жения у пих клеточных перегородок. Существование переход-
ных типов может способствовать сближению разпых семейств,
отличающихся друг от друга противоположными призпаками.
Итак, цитоэмбриологические признаки, подобно морфологиче-
ским, имеют переходы от более простых к более отклоняющим-
6
ся типам, что, однако, в некоторых случаях подвергается дис-
куссии. Эволюция цитоэмбриологических признаков имеет свое
направление, которое часто не совпадает с направлением эволю-
ции во внешней морфологии. Поэтому в более примитивных по
внешним морфологическим признакам группах покрытосеменных
.могут встречаться более прогрессивные цитоэмбриологические
признаки, и наоборот. Кроме того, внутри одной и той же группы
растений одни цитоэмбриологические признаки могут быть при-
митивными, другие прогрессивными. Это указывает, что для эво-
люции как различных цитоэмбриологических, так и морфологиче-
ских признаков характерно наличие гетеробатмии — разноступен-
чатости признаков, и гетерохронии — разновременности прохож-
дения эволюции того или иного признака. Все это создает извест-
ные затруднения и ограничения при попытке использовать дан-
ные цитоэмбриологии в целях систематики и особенно филогении
покрытосеменных; тем не менее направления эволюции морфоло-
гических и цитоэмбриологических признаков нередко совпадают,
и поэтому данными цитоэмбриологии не следует пренебрегать. Так,
например, раздельнолепестности обычно свойственны более при-
митивные исходные, а спайнолепестности — вторичные, более про-
грессивные цитоэмбриологические признаки.
Нам представляется, что в настоящее время в связи с недос-
таточно широкими и всесторонними исследованиями по сравни-
тельной цитоэмбриологии покрытосеменных с помощью этих дан-
ных следует стремиться не столько к поискам первичных и вто-
ричных рядов Форм, выяснению происхождения тех или иных
груиин покрытосеменных, сколько к использованию цитоэмбриоло-
гоческих прианаков ДЛЯ возможно более детальной характери-
стики отдельных групи, в целях более бесснпориого отграничения
остествениых единиц друг от друга. В настоящей мопографии мы
стремились обобщить имеющиеся сведения цитоэмбриологии раз-
личпых семейств и порядков, насколько это возможно при су-
цествующих в настоящее время знаниях, и показать, какие ци-
тоэмбриологические признаки обнаружены в том или ином се-
мействе. Однако мы не занимались выяснением систематических
и филогенетических взаимоотношений внутри семейств, между
различными их подсемействами, трибами, родами и видами, так
как для этого пока не имеется достаточного материала.
При разрешении вопроса, насколько отдельные части системы
однородны и естественны, мы бесспорно должны результаты
сравнительной цитоэмбриологии подкреплять данными, получен-
ными другими методами систематики. Неудачные попытки пост-
роения системы покрытосеменных отчасти объясняются тем, что
данные сравнительной цитоэмбриологии, даже совершенно бес-
спорные, не принимаются во внимание и в основу построения
систем обычно берут не комплекс признаков, а один какой-ни-
будь признак. Ценпый материал для установления родственных
отношений между многими систематическими единицами, полу-
ченный даже при чрезвычайно неполных и случайных цитоэм-
7
бриологических данных, позволяет предположить, что обширные
и планомерные цитоэмбриологические исследования дадут, еще
более ценный материал, и сравнительная цитоэмбриология зай:
мет подобающее ей место при создании филогенетической систе-
мы покрытосеменных растений.
Данные сравнительной цитоэмбриологии, наряду с другими
данными, могут помочь разобраться в некоторых сложных и' за-
путанных вопросах систематики и филогении, поэтому необходи-
мы сравнительно-цитоэмбриологические исследования разных,
‚в особенности сомнительных в систематическом и филогенетиче-
‘ском отношениях, групи покрытосеменных. Эти исследования
должны быть не беспорядочными, а планомерными, и их следу-
ет направить в первую очередь на то, чтобы заполнить имею-
щиеся пробелы и исправить сомнительные или ошибочные дан-
ные. Разумеется, одному и даже нескольким исследователям это
не. под силу, и, так как задача построения филогенетической
системы растений является очень важной, необходимо объедине-
ние усилий многих цитоэмбриологов для ее разрешения.
К цитоэмбриологическому методу, как и к любому другому,
следует относиться критически и делать заключения на основа-
нии исследования достаточно обширного и разнообразного
материала.
В заключение надо отметить, что настоящая монография не
является исчерпывающей. Она обобщает лишь основные данные
по этой дисциплине и показывает, где имеются пробелы и что
требует дальнейших исследований. Нашим намерением является
привлечение внимания цитоэмбриологов к проведению исследова-
ний, с применением данных цитоэмбриологии в целях использо-
вания их для построения филогенетической системы покрытосе-
менных. Несмотря на то что в настоящее время уже накоплел
значительный материал по сравнительной цитоэмбриологии по-
крытосеменных, в этой области науки необходимо сделать еще
очень много.
„При оформлении рукописи и составлении цитоэмбриологиче-
ских характеристик семейств: Asteraceae, Boraginaceae, Diosco-
raceae, Caryophyllaceae, Cucurbitaceae, Fabaceae, Grossulariaceae,
Malvaceae, Nelumbonaceae, Numphaeaceae, Orchidaceae, Oroban-
chaceae, Oxalidaceae, Poaceae, Polygonaceae, Pyrolaceae, Rosace-
ae, Scrophulariaceae, Solanaceae wu Theaceae HamM помогали:
Т. Б. Батыгина, В. А. Верещагина, Л. И. Григорьева, А. Х. Да-
ниелян, А. К. Дзевалтовский, Г. В. Камалова, Л. И. Кострикова,
В. Ю. Мандрик, С. Н. Мовсесян, Н. Л. Мудрова, Г. И. Савина,
С. Б. Салахова, 9. С. Терёхин, Д. Ю. Турсунов, Н. С. и В. Н.
Юрцевы и И. К. Юсупова, которым мы выражаем свою сердеч-
ную благодарность. Кроме того, мы очень благодарны Е.С. Смир-
новой, разрешившей воспользоваться ее пока еще не опуб-
ликованной классификацией типов семян.
ОТДЕЛ МАСМОПОРНОТА (ANGIOSPERMAE)
KJLACC MAGNOLIATAE (DICOTYLEDONES) TIOWKIIACC A. MAGNOLIIDAE
НАДПОРЯДОЕК 1.
MAGNOLIA NAE
ПОРЯДОК 1.
MAGNOLIALES
(ANNONALES),
CEMEHCTBO MAGNOLIACEAE
Основное число хромосом — x= 19.
Цветок обоеполый или очень ред-
ко однополый.
Пыльник — 4А микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 3—4
средних слоя и секреторный дву-
мнотоядерный, двух-трехрядпый та-
летум, Имоются орбикулы (тельца
"Убиша).
Микроспорогенез и микрогамето-
генез. Образование микроспор си-
`’ мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зер-
па двухклеточные, с одной дисталь-
ной бороздой и гладкой экзиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле прямое, образуется внут-
ренним интегументом. Сосудистый
пучок обычно доходит до халазы,
но иногда проникает и во внешний
интегумент. Имеются фуникулярный
обтуратор и гипостаза. Семяножка
короткая. Число семяпочек колеблет-
ся от многих до двух.
Макроепорогенез и макрогамето-
генез. Археспорий одно-многокле-
точный. ПНариетальная ‘ткань мас-
сивная. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок формируется по Polygonum-
типу. Синергиды грушевидные. Име-
ются три эфемерные антиподы. По-
лярные ядра сливаются до опло-.
дотворения. |
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодот-
ворение.
Эндоспермогенез целлюлярный. Эн-
досперм хорошо развитый, но не
руминированный.
Эмбриогенез нерегулярный, ‘т. е.
не соответствующий какому-либо
типу или Опаотад-типа. ’Зародыш
бесхлорофилльный, очень маленький,
с двумя, изредка тремя, семядолями
и корешком. Подвесок зародыша
массивный, имеет 8—10 клеток в
длину и 4—6 клеток в ширину. ©
Семя, плод — зрелое семя запол-
пепо массивпым эндоспермом, ‘CO-
держащим масло и алейрон, малень-
кий зародыш располагается в его
основании, обычно занимая ‘от 1/4
до 1/2 линейной высоты семени.
Кожура семени образуется наруж-
ным интегументом и состоит из ка-
менистого и мясистого слоев, внут-
ренний интегумент — пленчатый.
Плод — спиральная многолистовка
йли реже многоорешек, коробочка
или ягодообразный. Семена при
растрескивании плода свисают на
тонких лигниновых фибриллах.
Гаусторий —у Magnolia obovata
обнаружен эндоспермальный гаусто-
рий в халазальной части зародыше-
вого мешка. Семядоли выполняют
гаусториальную функцию.
Апомикеие не отмечен.
Другие явления не обнаружены,
Литература: ВБапдаг!, 1967, 1971;
Corner, 1976; Davis, 1966; Kapil, Bhan-
dari, 1965; ЗеБпатЁ, 1934; Смирнова,
1977; Тахтаджян, 4966, 1970; Yoshia-
ki, 1964.
ig
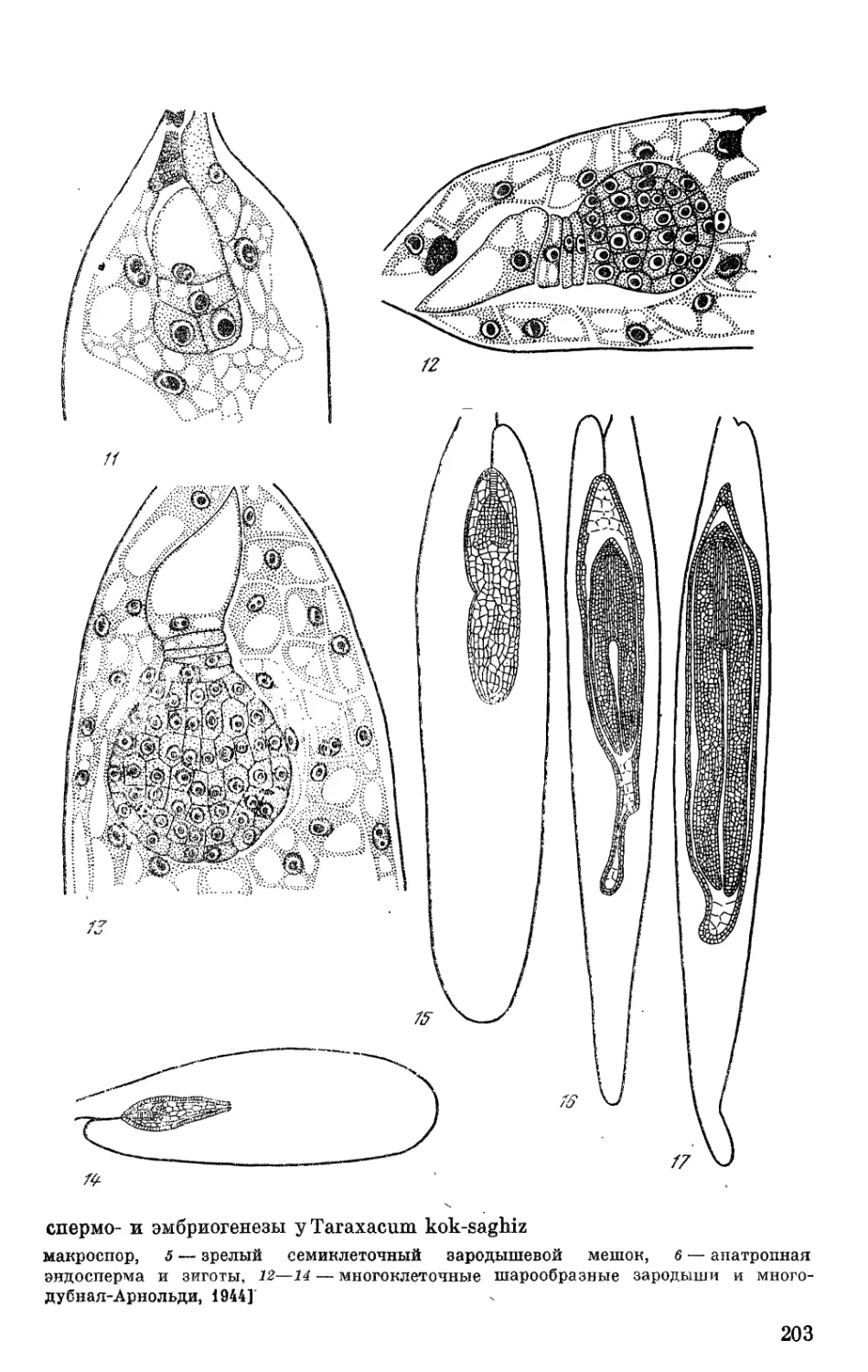
Puc. 1. Muxpocnopanruit m mMukpocnoporenes y Magnolia stellata
1—3 — диаграммы пыльников на различных фазах их развития; а — один микро- спорангий из рис. 1 в увеличенном размере, б — часть микроспорангия из рис. 2 в увеличенном размере; в — часть микроспорангия из рис. 3 в увеличенном разме- ре; 4 — двуядерная клетка тапетума; 5—12 — разные фазы образования микроспор; 13—15 — образование двухклеточных пыльцевых зерен, 16 — зрелое пыльцевое зер-
но [Kapil, Bhandari, 1965]
СЕМЕЙСТВО РЕСЕМЕВТАСЕАЕ
Осповпое число хромосом — х=12.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспоралгия образуется
по базальтому типу и имеет эпидер-
мие, фиброзный эпдотеций, 3
10
средних слоя и секреторный одно-
рядный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточпые, с одной ди-
стальной бороздой.
Prue. 2. Maxpocuoporenes y Magnolia stellata 1—5 — разные фазы развития семяпочки; 6 — перед оплодотворением;: 7 — после оп-
лодотворения [Kapil], Bhandari, 1965]
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле прямое, образуется внут-
ренним иптегументом. Имеется фу-
никулярный обтуратор. Семяпочек
много (30—32).
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальная ткань массивная.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]ухопит-типу. Имеются три эфе-
мерные антиподы,.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Тройное слияние предшествует син-
гамии.
Эндоепермогенез — целлюлярный,
руминированный, содержащий масло.
Эмбриогенез — нерегулярный. За-
родыш очень маленький, бесхлоро-
филльный, обычно с двумя, но часто
с 3—4 семядолями. Нодвесок заро-
дыша короткий, массивный.
Семя, плод — семя с хорошо раз-
11
Рие. 3. Maxpocnoporenes y Magnolia stellata
1 — зачаток семяпочки с тремя археспориальными клетками; 2, 3 — зачаток семя-
почки с тремя спорогенными и париетальными клетками; 4 — ссмяпочка с материн-
ской клеткой макроспор; 5—11 — разные фазы образования и разное расположение
макроспор; 12 — функционирующая и дегенерирующие макроспоры [Kapil, Bhan-
dari, 1965]
Рис. 4. Женский гаметофит у МаспоНа з{еПава. А — два зародышевых мешка в одной и той же семяпочке, Б — восьмиядерный за-
родышевый мешок,
ант — антиподы, я — яйцеклетка,
ные клетки [КарИ, Ввапдат, 1965]
витым эндоспермом. Кожура семени
возникает из наружного интегумен-
та. Внутренний интегумент редуци-
руется до тонкого пленчатого слоя,
Семена при растрескивании плода
свисают на тонких длинных лигни-
новых фибриллах. Плод не растре-
скивающийся, © твердым эндокар-
пием, содержащим каменистые клет-
ки и эфирное масло.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления пе паблюдались.
Литература: Bhandari, 1967a,
1970a, 1971; Corner, 1976; Davis, 1966;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АММОМАСЕАЕ
Основное чиело хромосом — х =
= 7—9,
Цветок обоеполый или редко од-
нополый.
В — семиклеточный
зародышевый мешок. с — синергиды,
дн — дегенерирующие нуцелляр-
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двухдольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—4
эфемерных средних слоя и одноряд-
ный, обычно секреторный, изредка
амебоидный тапетум. Тапетум двух-
четырехъядерный, ядра его клеток
сливаются и становятся полипло-
идными.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное, промежуточное и симуль-
танное. Тетрады микроспор тетраэд-
ральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые
зерна двухклеточные, с одной 60-
роздой на проксимальном конце.
Экзина сетчатая или шиповатая.
Пыльцевые зерпа иногда соедипены
в тетрады.
Семяпочка анатропная, с двумя,
реже с тремя интегументами,
13
ии и»
ГЕТЕ
LY
am —
tit tiie
SCO te
ина
ее
ЕН
ии
ЕН
ЕСН
by —.
Lui
Prue. 5. ImMOpuorenes y Magnolia stellata
1 — зигота; 2, 3 — деление зиготы; 4 — двухклеточ-
ный проэмбрио; 5, 6 — трех-четырехклеточные про-
эмбрио; 7—9 — разные стадии развития зародыша:
10 — зародыш с тремя семядолями; 11 — зрелое семя
y Magnolia ороуаа; с — семядоля, п — проэмбрио,
под — подвесок; з — зародыш, а — эндосперм, ги” —
гипостаза, ви — внутренний интегумент, ме — мясис-
тый и кс — каменистый слой наружного интегумен-
та, сп — сосудистый пучок [Kapil, Bhandari, 1965]
крассинуцеллятная. — Париетальная
ткань массивная. Микропиле прямое,
образуется наружным, внутренним
или обоими интегументами. Имеют-
ся гипостаза и ариллус. Семяножка
короткая. Число семяпочек колеб-
лется от многих до одной.
Макроспорогенез и макрогамето-
генез — археспорий. одно-несколько-
клеточпый. Париетальная ткань
массивная. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро]угопат-типу. Синерги-
ды с крючкообразными выростами.
Чри антиподы, — дДегеперирующие
вскоре после оплодотворения. По-
лярные ядра сливаются до оплодот-
ворения. У Сапапса одога{йа зароды-
шевый мешок богат крахмалом.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление, порогамия.
У Annona sativa оплодотворение
происходит через 2 ч после опыле-
ния, а через 6 ч после опыления
происходит деление первичного яд-
ра эндосперма.
Эндоепермогенез целлюлярный.
Эпдосперм роговой и руминирован-
ный, содержит крахмал. Имеется
периснерм.
мбриогенез Опастад-типа, вариа-
ции ТгИоНат или Г0й1$. Зародыш
очень маленький, бесхлорофилльный,
с двумя семядолями и корешком.
Семя, плод — зрелое семя имеет
массивный эндосперм, содержащий
масло и алейрон. Зародыш распола-
гается в основании семени и обычно
занимает 1/4 или 1/2 его линейной
высоты. Кожура семени образуется
наружным интегументом, а внутрен-
НИЙ интегумент редуцируется.
Нлод ягодообразный, редко деревя-
нистый.
Гаусторий — семядоли при про-
растании семени выполняют гаусто-
риальную функцию.
Апомиксие не обнаружен.
Другие явления — партенокарпия
y Annonasa sativa.
Литература: Bhandari, 1971; De-
vis, 1966; Corner, 1976; Parulekar,
4967, 1970; Schnarf, 1933; Смирнова,
1977; Тахтаджяп, 1966, 1970; Voigt,
1971.
СЕМЕЙСТВО САМЕШ.АСЕАЕ
Основное /число хромосом — х=
— 14, 43.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия состоит из
эпидермиса, фиброзного эндотеция,
двух-трехэфемерных средних слоев
и однорядного, секреторного, двух —
четырехъядерного танпетума. В ре-
зультате слияния ядра тапетума
становятся полиплоидными. Имеют-
ся орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, одноборозд-
ные, соединенные в тетрады.
Семяпочка анатропная или геми-
тропная, с двумя иптегументами,
крассинуцеллятная. Микропиле зиг-
загообразное, образующееся обоими
интегумептами. Имеется ариллус.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточ-
ный. Париетальная ткань массив-
пая. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Роусопит-типу. Синергиды груше-
видные, с фибриллярными аппара-
тами. Три небольшие эфемерные ан-
типоды. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
даппых. .
Эндоспермогенез целлюлярный.
Эндосперм хорошо развит, руми-
пированпый, содержит масло, алеи-
рон.
Эмбриогенез — тип не определен.
Зародыш очень маленький, бесхло-
рофилльный, слегка согнутый, диф-
ференцированный на семядоли и
корешок, содержит масло.
Семя, плод — семя обильно запол-
нено эндоспермом. Зародыш распо-
ложен в основании семени и обычно
пе превышает И/4 его линейной вы-
соты. Кожура семени образуется
впутренним и наружным интегумен-
тами. Плод паракарпная ягода с дву-
мя и больше семепами.
Гаусторий — семядоли выполняют
гаусториальную функцию,
Апомикеис не отмечен.
Другие явления не обнаружены.
Jiurepatypa: Bhandari, 1967b, 1974;
Corner, 1976; Davis, 1966; Смирно-
ва, 1977; Тахтаджян, 1966, 1970; Wil-
son, 1966.
15
СЕМЕЙСТВО МУВТЗТЕАСЕАЕ
Основное чиело хромоеом — х=7.
Цветок однополый. Растения обыч-
но двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средних слоя и однорядный,
секреторный, одноядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор сук-
'цессивное. Тетрады микроспор изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, однобороздные.
Семяпочка — одна анатропная, с
двумя интегументами, крассинуцел-
лятная. Микропиле прямое, образу-
ется обоими интегументами. Имеют-
ся ариллус и гипостаза. Семяножка
короткая.
акроепорогенез и макрогаметоге-
гез — археспорий одноклеточный.
Париетальная ткапь массивная. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Рау-
сопат-типу.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Тройное слияние предшествует син-
гамии. Пыльцевые трубки долго со-
храняются в семяпочке.
Эндоспермогенез — нуклеарный,
эндосперм хорошо развитый, руми-
нированный, содержащий масло или
крахмал, алейрон, кристаллы окса-
лата кальция.
ЭЗмбриогенез — тип не определен.
Зародыш очень маленький, бесхло-
рофилльный, дифференцированный
на две семядоли и корешок, содер-
жит масло.
Семя, плод — зрелое семя с мас-
сивным эндоспермом. Зародыш рас-
положен в основании семени и обыч-
но не превышает 1/4 линейной его
высоты. Семенная кожура образует-
ся из наружного интегумента. Пло-
ды кожисто-мясистые или почти де-
ревянистые.
Гаусторий — семядоли выполняют
таусториальпую функцию.
Апомиксие не обнаружен.
Другие явления не наблюдались.
Литература: Впап@ат, 1967}, 1971;
Corner, 1976; Пау1з, 1966; Смирнова,
1977; Тахтаджяп, 1966, 1970.
СЕМЕЙСТВО УТУТЕКАСЕАЕ
Основное число хромосом — х=
=7, 13.
1)
Цветок ‚обычно обоеполый, редко
однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—4
эфемерных средних слоя и одно-дву-
ядерный секторный или амебоидный
тапетум. Ядра клеток тапетума сли-
ваются и становятся полиплонд-
HBIMH.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изо-
билатеральные. Зрелые, пыльцевые
зерна двухклеточные, соединенные
в тетрады; с одпой дистальной порой
и сетчатой экзиной.
Семяпочка — одна — несколько,
анатропные, крассинуцеллятные с
двумя интегументами. Микропиле
прямое, образуется впутренним ин-
тегументом. Семяножка короткая.
Имеются обтуратор, гипостаза и ну-
целлярный колпачок. Сосудистый
пучок доходит до халазы семя-
почки.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань — массивная.
Тетрады макроспор линейные, 7-
образные. Зародышевый мепюк об-
разуется по Ро!угопат-типу. Синер-
гиды с крючкообразными выростами
и фибриллярпыми аппаратами, Три
маленькие эфемерные антиподы. По-
лярные ядра сливаются до оплодот-
ворение.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Тройное слияние предше-
ствует сингамии. Двойное оплодот-
ворение.
Эндоспермогенез целлюлярный.
Эндосперм хорошо развитый, руми-
нированный, содержит масло.
Эмбриогенез нерегулярный. Заро-
дыш очень маленький, бесхлоро-
филльный, недифференцированный,
по с подвеском, содержащий крах-
мал.
Семя, плод — семя с массивным
эндоспермом. Зародыш обычно не
превьшнает 1/4 липейной высоты се-
мени. Кожура семени образуется
обычно лишь наружным интегумен-
том. Внутренний интегумент pe-
дуцируется. Плод — многолистовка
коробочковидный, ягодообразный.
В ткани етенки плода имеются ка-
менистые клетки и клетки, содержа-
щие эфирное масло и таннин. Пери-
карпий мясистый, состоящий из
14—16 слоев паренхиматозных кле-
ток.
Гауеторий — семядоли выполняют
гаусториальную функцию.
Апомикеис не обнаружен.
Другие явления не наблюдались.
Литература: ВБапдат1, 1967Ъ, 1971; .
Bhandari, Venkataraman, 1968; Bo-
er R. de, Rouman T., 1974; Corner,
1976; Пау1з, 1966; Смирнова, 1977;
Тахтаджян, 1966, 1970.
Из семейства порядка МаспоНа-
les, по классификации А. Л. Тах-
таджяна [1970], два семейства,
a umMeHHo Himantandraceae u Eupo-
таНасеае, в отношении цитоэмбрио-
логии почти не изучены. Поэтому
y Hac пока пет цитоэмбриологиче-
ских данных для определения по-
ложения этих семейств по отноше-
нию друг к другу и к другим се-
мействам МаспоНаез. Однако уже
и сейчас можно отметить, что другие
более или менее подробно исследо-
ванные в отношении цитоэмбриоло-
гии семейства этого порядка харак-
теризуются рядом примитивных ци-
тоэмбриологических признаков, а
именно: малым разнообразием осноз-
ных чисел хромосом, двухклеточным,
одиночным, однобороздиым, мужеким
гаметофитом, большим числом апа-
тропных, крассинуцеллятных, с дву-
мя интегументами семяпочек, мас-
сивной париетальной тканью, Poly-
сопат-типом зародышевого мешка, с
тремя эфемерными антиподами, мас-
сивным руминированным эпдоспер-
мом, очень маленьким, слабо диффе-
ренцированным или вовсе не диффе-
ренцированным зародышем, отсутст-
вием, за исключением МаспоПа obo-
уайа, гаусториев и апомиксиса. Эти
особенности порядка Magnoliales
вполне оправдывают представление
об его примитивпости и расположе-
нии в начале системы покрытосемен-
ных растений. По цитоэмбриологиче-
ским признакам семойства МаспоПа-
ceae wu Degeneriaceae имеют много
общего между собой. Довольно сход-
ны по цитоэмбриологическим призна-
цам и семейства Аппопасеае, Сапе]-
]}асеае и МунзЯсасеае. Несколько
обособленное положение занимает
семейство Winteraceae, xoTA uw оно
имеет некоторые цитоэмбриологиче-
ские признаки, сходные с другими
2 В. А. Поддубная-Арнольди
предотавителями порядка МаопоНа-
]ез. На основании современных зна-
ний невозможно установить фило-
генетические линии происхождения
семейств порядка Махпо!аез, одна-
ко можно предположить, что они
произошли от общих предков и что
порядок этот естественный, объеди-
няющий самостоятельные, но сход-
ные по ряду цитоэмбриологических
признаков семейства.
По мнению А. Л. Тахтаджяна
[1966], Magnoliaceae стоит близко
к Перепеасеае и оба эти семейства
примыкают к Нииащапдагасеае, ко-
торые, как он считает, занимают
промежуточное положение между
МагпоПасеае и Пебепемасеае, с од-
ной стороны, и Аппопасеае — с дру-
rou. KH Himentandraceae, Kak yKa-
зывает А. Л. Тахтаджян, наиболее
близко подходит ЕпротаНасеае. Но
поскольку Himatandraceae um Eupo-
шаНасеае цитоэмбриологически поч-
ти не изучены, решить этот вопрос
в настоящее время нам не пред-
ставляетея возможным. Относитель-
по Аппопасеае, СапеПасеае и Му-
risticaceae мы согласны с А. Л. Тах-
таджяном в том, что эти семейства
довольно тесно связаны между со-
бой. Что касается Winteraceae, KOTO-
рое из всех семейств порядка Мао-
поНа]ез является наиболее специа-
лизированиым, то мы не можем со-
гласиться © Хатчинсоном [Настю -
son, 1959], относившим к Wintera-
сеае pom Degeneria (единственный
представитель Оесепетасеае), сбли-
жая и объединяя тем самым два
цитоэмбриологически отличающиеся
друг от друга семейства. По цито-
эмбриологическим признакам О,еге-
пегасеае, несомпенпо, очень близко
к МаспоНасеае и далеко от Утще-
гасеае, что неоднократно отмечали
индийские цитоэмбриологи [Swamy,
1949; Kapil, Bhandari, 1965; Bhanda-
11, 1967, 1970, 4974 и mp.].
Порядок МаспоНа]ез с давних пор
привлекал внимание индийских ци-
тоэмбриологов, и в результате их
усилий многие семейства этого по-
рядка были более или менее подроб-
но исследованы в отношении цито-
эмбриологии [ЭЗ\уату, 1949; ВВапда-
ri, 1967, 1970, 1974; Kapil, 1964; Paru-
lekar, 1967, 1970; Bhandari, Venka-
{агашаи, 1968; и др.]|. Индийские ци-
тоэмбриологи не — ограничивались
только описанием цитоэмбриологи-
ческой характеристики, а пытались
17
сделать заключение и 0б их фило-
генетических
Они показали, что многие цитоэм-
бриологические признаки Маспойа-
сеае. Песепегасеае и Аппопасеае
сходны, что указывает на их род-
ство, а также отметили, что Сапе]-
]асеае родственно МугзЯсасеае. Ка-
пил, Бхандари [Kapil, Bhandari,
1964] на основании Данных цито-
эмбриологии, морфологии и анато-
мии пришли к заключению, что
Magnoliaceae, Degeneriaceae ua An-
попасеае имеют такие сходные при-
знаки, как секреторный тапетум с
двуядерными клетками, как однобо-
роздные пыльцевые зерна, в кото-
рых генеративные клетки образу-
ются на дистальных концах пыль-
цевых зерен, как анатропные, с дву-
мя интегументами, крассинуцеллят-
ные семяпочки, как Ро|убопат-тип
образования зародышевого мешка,
с тремя эфемерными антиподами,
как целлюлярный эндосперм, что
указывает на их тесное родство.
Вместе с тем по некоторым важным
признакам они отличаются друг от
друга. У МаспоПасеае тетрады мик-
роспор вскоре после их образования
распадаются на отдельные микро-
споры, у ПОегепетасеае тетрады со-
храняются вплоть До образования
экзины и борозд в пыльцевых зер-
пах, У некоторых Аппопасеае тет-
рады микроспор не распадаются на
отдельные микроспоры. Орбикулы
имеются у МаспоПасеае и не имеют-
ca y Degeneriaceae mw Annonaceae.
Руминированный эндосперм имеется
y Degeneriaceae wu Annonaceae, OT
сутствует у МаспоПасеае. Ариллус
имеется у Аппопасеае и отсутствует
у Degeneriaceae и Magnoliaceae.
У МаспоЦасеае основное число хро-
mocom — x= 19, y Degeneriaceae +=
—12, у Аппопасеае х=7, 8 или 9.
Таким образом, хотя эти три семей-
ства тесно связаны друг с другом.
они не могут быть расположены в
одпу филогенетическую — линию.
Можно заключить, что, несмотря на
тесную связь друг с другом этих
семейств, они являются CaMOCTOA-
тельными семействами магнолиаль-
ного комплекса. Что же касается
отнесения рода Оерепеша к семей-
ству У/ищегасеае, то Капил и Бхан-
дари считают это неправильным.
Ресепема отличается от представи-
телей Упуегасеае некоторыми при-
знаками, в том числе строением
18
взаимоотношениях. _
пыльцевых зерен, зародыша, семен-
ной кожуры и т. д. Поэтому отне-
сение ее к самостоятельному семей-
ству Пегепемасеае подтверждается
данными цитоэмбриологии. Постоян-
ное наличие у Ущегасеае пыльце-
вых зерен, соединенных в тетрады,
указывает на большую специализа-
цию и большую подвинутость Win-
feraceae, по сравнению с другими
свойствами Маопо|а|ез.
ПОРЯДОК 2.
LAURALES
СЕМЕЙСТВО МОМУМТАСЕАЕ
Основное число хромосом — х==41.
Цветок обоеполый или однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и секреторный, одно-
рядный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточные, одиночные
или соединены в тетрады, безапер-
турные или реже одно-двубороздные.
Семяпочка анатропная, с одним
или двумя интегументами, красси-
нуцеллятная. Микропиле образуется
обоими интегументами. Имеются ну-
целлярный колпачок, гипостаза и
фуникулярный обтуратор. Семяпо-
чек несколько.
Макроспорогенез и макрогамето-
тенез — археспорий —’одно-многокле-
точный. Париетальная ткань массив-
ная. Тетрады макроспор линейные.
Зародышевые мешки формируются
no Polygonum- или АШат-типам.
Синергиды грушевидные. Клеток
антипод сначала три, потом число
их увеличивается путем деления
до 5—20. Полярные ядра сливают-
ся перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия.
Эндоепорогенез целлюлярный.
Эмбриогенез Азегад-типа. Заро-
дыш маленький, прямой, двусемя-
дольный, бесхлорофилльный. Подве-
сок короткий.
Семя, плод — семя с хорошо раз-
витым маслянистым эндоспермом.
Зародыш располагается в основании
семени и обычно занимает от 1/4 до
1/2 линейной его высоты. Плод —
многоорешек. .
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомикеие — нуцеллярная эмбрио-
НИЯ.
Другие явления — гинодиэция.
Литература: ВВапдаг!, 19676, 1974; .
Corner, 1976; Davis 1966; P. Mahesh-
wari, 1960; Тахтаджян, 1966, 1970;
Sampson; 1969a, b, c; CmumpHora, 1977.
СЕМЕЙСТВО СНГОВАМТНАСЕАЕ
Основное число хромосом — х=7,
15.
Цветок обоеполый или однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, два-
три средних слоя и секреторный дву-
ядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, (трех), четырех- восьмибо-
роздные или безапертурные, с тол-
стой сетчатой экзиной.
Семяпочка одна висячая, орто-
тропная, с двумя, тремя интегумен-
тами, крассинуцеллятная. Третий
интегумепт рудимептарный. Микро-
пиле образустся только внутренним
или обоими интсгументами.
Макроспорогенез и макрогамето-
генез — археспорий одно-, многокле-
точный. Париетальная ткань мас-
сивная. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Ро]угопит-типу. Синергиды груше-
видные. Три антиподы рано дегене-
рируют или долго сохраняются и де-
лятся (образуя до 24 клеток). По-
лярные ядра сливаются до или во
время оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
‚рогамия и двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эндосперм содержит масло.
Эмбриогенез — Onagrad- или СЪе-
nopodiad-tunos. Зародыш в зрелом
семени недоразвит, маленький, бес-
хлорофилльный, содержит масло.
Подвесок короткий, широкий.
Семя, плод — семя с хорошо раз-
витым периспермом и слабо разви-
тым эндоспермом. Величина зароды-
ша не превышает 1/4 осевой длины
2*
семени. Семядоли слабо развиты.
Плод — костянка. Клетки семенной
кожуры и перикарпия содержат
эфирные масла, а клетки интегумен-
тов — таннин.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомикесис пе обнаружен.
Другие явления не наблюдались.
Литература: Согпег, 1976; Davis,
1966; Murty, 1967a, Schnarf, 1931; Сми-
рнова, 1977; Тахтаджян, 1966, 1970.
"СЕМЕЙСТВО САБУСАМТНАСЕАЕ
Основное число хромосом — х=11.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 4—5
средних слоев и секреторный или
амебоидный тапетум.
Микроеспорогенез и микрогаметоге-
нез — образование микроспор cH-
мультанное. Тетрады микроспор тет-
раэдральные или изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, одиночные или изредка сое-
диненные в тетрады (СЬлтопат из
[гаостапз), двухбороздные с сетчатой
экзиной.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
или обоими интегументами. Имеются
пуцеллярный колпачок и гипостаза.
Семяножка короткая, тонкая. Семя-
почек много. Проводящий пучок до-
ходит до халазы.
Макроспорогенез и макрогаметоге-
нез — археспорий многоклеточный.
Периетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок формируется по Ро-
]угопит-типу. Часто образуется не-
сколько зародышевых мешков внут-
ри одной и той же завязи. Как пра-
вило, зародышевый мешок со всеми
своими элементами дегенерирует.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — вви-
ду дегенерации зародышевых меш-
ков оплодотворения нет.
Эндоспермогенез пеллюлярный, 0б-
разующийся автономно, без оплодот-
ворения полярных ядер.
Эмбриогенез — обычно образуются
только нуцеллярные зародыши, со
спирально закрученными семядоля-
MH, бесхлорофилльные, богатые
маслом.
19
Семя, плод — семепа без эндо-
сперма. Зародыш заполняет почти
все семя. Семядоли его выполняют
запасающую функцию. Плод — мно-
гоорешек.
Гауеторий отсутствует.
Апомиксие — обнаружена
лярная эмбриония.
Другие явления — полиэмбриония.
Jiutepatypa: Bhandari, 1967a, 1971;
Corner, 1976; Davis, 1966; Mathur,
1958; Schnarf, 1931; Тахтаджян, 1966,
70.
нудел-
СЕМЕЙСТВО ГАОКАСЕАЕ
Оеновное число хромосом — х==12.
Цветок обычно обоеполый, реже
однополый.
Ныльник — 2—4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и амебоидный или сек-
реторный, дву-, многоядерный та-
петум.
Mux pocnoporenes и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, изобилатеральные, T-
образные и линейные. Зрелые пыль-
цевые зерна двухклеточные, без пор
или иногда однобороздные, экзина
слегка шиповатая.
Семяпочка одна, анатропная,
с двумя инптегументами, крассину-
целлятная. Микропиле образуется
обоими интегумептами. Сосудистый
пучок доходит лишь до халазы.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-, многокле-
точный. Париетальная ткань мас-
сивная. Тетрады макроспор линей-
ные. Зародышевый мешок формиру-
ется по Ро]угопат-типу. У Саззу фа
функционирует много макроспор и
много зародышевых мешков образу-
ется в одпой и той же семяпочке.
Причем опи сильпо удлиняются в
верхней своей части и выступают из
микропиле, достигая семяножки. Си-
нергиды имеют фибриллярные аппа-
раты; антипод обычно три, эфемер-
ные, по ипогда число их увеличива-
отеля; опи представлены в виде ядер
или клеток. Полярные ядра сливатюот-
ся до оплодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия, перекрестное опыление.
Эндоспермогенез нуклеарный или
поллюлярный (у Саззу ал).
20)
Эмбриогенез Piperad-, Asterad-,
Опастад-типов или нерегулярный.
Зародьши крупный, прямой, с двумя
семядолями, почечкой и корешком,
бесхлорофилльный, содержащий
крахмал, масло и мелкие зернышки
алейрона. Зародьши у лавра благо-
родного формируется 3—4 месяца,
а плод 4—0 месяцев. Подвесок ко-
роткий.
Семя, плод — семя без эндосперма.
Зародыш заполняет весь объем зре-
лого семени. Семядоли выполняют
запасающую функцию. Семенная ко-
жура образуется наружным интегу-
ментом, эпидермис которого содер-
жит таннин. Плод ягодообразный
или реже костянка, или сухой не-
раскрывающийся. Перикарпий имеет
наружный эпидермис с клеткаии,
содержащими танпин, 7—9 рядов
паренхимных клеток, один ряд уд-
линенных и внутренний эпидермис
с одревесневшими клетками.
Гаусторий зародышевого мешка.
Апомикеис не найден.
Другие явления — паразитизм.
Jimrepatypa: Bhandari, 1967f; Cor-
ner, 1976; Davis, 1966; Schnarf, 1934;
CmupuHosa, 1977; Souéges et al., 1968;
Тагиева, 1967, 1972; Тахтаджян, 1966,
1970; Терёхин, 1977.
Из 10 семейств, отнесенных
А. Л. Тахтаджяном [1970] к порядку
Гамга]ез, 6 семейств, а именно Апз{-
robaileyaceae, Amborellaceae, Trime-
miaceae, Gamortegaceae, Hernandia-
ceae и Гафог!9асеае, в отношении
цитоэмбриологии почти или вовсе не
изучепы. Остальпые 4 семейства
(Monimiaceae, Chloranthaceae, Caly-
сап(Васеае и Гаптеасеае) хотя и изу-
чены в этом отношении, но недо-
статочно. Несмотря на то, что цито-
эмбриология порядка Laureales uc-
следована очень слабо, все же неко-
торые филогенетические линии на-
мечаются. Семейства Monimiaceae u
Calycanthaceae mo цитоэмбриологи-
ческим признакам сходны. Оба се-
мейства отличаются симультанным
типом микроспорогенеза, двухкле-
точным мужским — гаметофитом,
анатропной с двумя интегументами
крассинуцеллятной семяпочкой, ну-
целлярным колпачком, гипостазой,
многоклеточным женским археспо-
рием, Ро]угопат-типом женского га-
метофита, целлюлярным эндоспер-
мом. Эти же призпаки, за исключе-
нием эндосперма, характерны и для
Гаигасеае. Эндосперм же у Гапга-
сеае нуклеарный. Однако представи-
тели семейств Мот!иласеае и Гапга-
сеае размножаются при помощи
афимиксиса, а Са!усапасеае — при
помощи апомиксиса.
По мнению П. Махешвари и HKa-
numa [Maheshwari, Kapil, 1966] род
Саззуща следует выделить в само-
стоятельное семейство Саззутасеае,
так как он отличается от других
представителей Laureaceae: секре-
торным тапетумом, образованием
большого числа сильно удлиненных
зародышевых мешков и целлюляр-
ным эндоспермом. Другого мипения
придерживается Бхандари [ВБапда-
г1, 1967], который считает, что род
Саззу Ва не должен быть выделен в
самостоятельное семейство Cassyt-
Басеае, что его следует оставить в
семействе Гапгасеае. Вместе с тем
Бхандари считает, что Lauraceae тес-
но примыкает к Мопшасеае, а Са-
lycanthaceae cpa3saHo c Magnoliaceae,
Аппопасеае и Eupamatiaceae. По
сравнению с МаспоПасеае Гапгасеае
1981) подвинутое семейство [Еатз,
1964].
ПОРЯДОК 3.
PIPERALES
СЕМЕЙСТВО ЗАОВОВАСЕАЕ
Основное число хромосом — х==.
= 8, 9, 14.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, сред-
пий слой и секреторный двуядер-
ный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультаннос. Тетрады — микроспор
тетраэдральные, крестообразпые, изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, одноборозд-
ные, с гладкой экзипой.
Семяпочка ортотропная, с двумя
интегументами, крассинуцеллятная,
Микронпиле образуется обоими инте-
гументами. Имеется эндотелий. Се-
мяпочек пемного.
Макроспорогенез и макрогаметоге-
нез — археспорий одпоклеточный. Па-
риетальных клеток два ряда. Заро-
дышевый мешок образуется по Ро-
lygonum-tuny. Сипергиды с крюч-
кообразпыми выростами; три эфе-
мерные антиподы. Полярные ядра
сливаются до оплодотворения.
` Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоепермогенез целлюлярный.
Эндосперм слабо развит. Перисперм
массивный.
Эмбриогенез — Азега4-типа. Заро-
дыш маленький. Подвесок короткий,
однорядный.
Семя, плод — значительная часть
семени заполпепа периспермом, тог-
да как эндосперм по размерам неве-
лик. Величипа зародыша не превы-
шает 1/4 осевой длины семени. Се-
менная кожура твердая. Плод из
нераскрывающихся, немного мяси-
стых, сросшихся у основания одно-
семяпных плодиков.
Гаусторий эндоспермальный (ха-
лазальный). Семядоли выполняют
гаусториальную функцию.
Апомиксие — облигатный апомик-
cuc y Houttuynia cordata.
Другие явления не обнаружены;
Литература: Corner, 1976; Davis,
1966; Murty, 1967b; Schnarf, 19314;
Cuupitona, 1977; Yoshida, 1961;
СЕМЕЙСТВО РТРЕВАСЕАЕ
Основное число хромоеом — х=8,
14, 12, 14, 16.
Цветок обосполый или иногда од-
нополый.
Ныльник — 2—4 микроспозтангия.
Стенка микроспорапгия образуется
по одпопольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секретор-
ный двуядерпый тапетум.
Микроснорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изоби-
латеральные. Зрелые пыльцевые зер-
на двухклеточпые, одпобороздные
или безапертурные, с гладкой эк-
зипой.
Семяпочка одна, ортотропная, с
одним (Ререгопма) или двумя (Pi-
рег) интегументами, крассинуцел-
лятная. Микропиле образуется обои-
21
ми интегументами или
ним внутренпим (Piper).
Макроепорогенез и макрогаметоге-
нез — археспорий одпоклеточный,
иногда несколькоклеточный (Piper).
Париетальных клеток 1—2 ряда. За-
родышевые мешки формируются по
gunam Fritillaria (y Piper), Ререго-
mia, подтипу АсаНрва (у Pepero-
па). Синергиды часто пе дифферен-
цировапы. Антиподы долго сохраня-
ются и даже делятся. У Р!рег обра-
зуется до 100 клеток антипод. По-
лярные ядра сливаются до оплодот-
ворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение.
Эндоспермиогенез — целлюлярный
(y Peperomia), нуклеарный (у Р-
рег). Перисперм хорошо развит, со-
держит крахмал, белок и маело, в
котором растворены пиперин и ку-
барин. Эндосперм слабо развит.
Эмбркогенез Р!регад-типа. Заро-
дыш маленький, слабо развитый, со-
держит масло и алейрон (Р1рег).
Подвесок несколькоклеточный.
Семя, плод — значительная часть
объема семени заполнена периспер-
мом, который содержит масло и
крахмал (у Р1рег) или только масло
(у Ререгопца). Эндосперм по разме-
рам невелик. Величина зародыша
не превышает 1/4 осевой длины се-
мени. Семенная кожура образуется
из внутреннего интегумента, на-
ружный интегумент разрушается.
Плоды ягодообразные или костянко-
образные. Перикарпий дифференци-
руется на 2—3 слоя.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления — обнаружена по-
лиэмбриония у Ререгопла mu Piper.
Литература: Согпег, 1976; Davis,
1966; Гваладзе, 19785; Murty, 1967c;
ЗевпатЕ, 1934; Смирнов, 1977; Тахтад-
жян, 1966, 1970.
только од-
Цитоэмбриологические данные по-
казывают, что семейства Saururaceae,
Piperaceae u Chloranthaceae имеют
сходные признаки, что, возможно,
указывает на их родственные отно-
шения [Могбу, 1967]. Цитологиче-
ские признаки родов Рег и Рере-
топца, настолько различны, что, мо-
жет быть, следовало pox Peperomia
22
выделить в 0с0обое семейство [Миг-
ty, 1967c]. Наличие тетраспориче-
ских зародышевых мешков у пред-
ставителей Р!'регасеае указывает на
большую подвинутость этого семей-
ства по сравнению c Saururaceae.
ПОРЯДОК 4.
ARISTOLOCHIALES
СЕМЕЙСТВО
ARISTOLOCHIACEAE
Основное чиело хромосом — х =
—= 6—9, 12, 13.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный двуядерный тапетум. Име-
ются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор обыч-
но симультанное. Тетрады микроспор
тетраэдральные, изобилатеральные,
крестообразные, линейные, Г-образ-
ные. Зрелые пыльцевые зерна двух-
клеточные, безапертурные или одно-
бороздные, с гладкой или скульштур-
ной экзипой.
Семяпочка анатропная, цирцино-
тропная (у Вгавапна) с двумя инте-
тументами, крассинуцеллятная. Мик-
ропиле образуется внутренним или
обоими интегументами. Сосудистый
пучок доходит до халазы. Имеются
нуцеллярный колпачок и гипостаза.
У Азагаш обнаружена карункула.
Семяпочек много.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевый мешок образует-
ся по Роусопаю-типу. Синергиды
грушевидные, 3—6 антипод; они
эфемерные или долго сохраняются,
крупные.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестпое опыление и самоопыле-
ние. Порогамия. Двойное оплодот-
ворение. .
Эндоспермогенез целлюлярныи,
руминированный (у Арата sili-
quosa).
Эмбриогенез перегулярпый. Заро-
дыш очень маленький слабо диффе-
ренцированный или не дифферен-
цированный (у Азагиот), бесхлоро-
филльный, не превышает 1/4 ли-
нейной высоты семян.
Семя, плод — семя с маслянистым
эндоспермом. Иногда эндосперм со-
держит крахмал. Семядоли при про-
растании семени выполняют гаусто-
риальную функцию. Семепная ко-
жура образуется обоими интегумен-
тами. Плод — полуапокарнная мно-
голистовка (у багата) или обычно
коробочка, или редко нераскрываю-
щийся, односемянный.
Гаусторий — у Asarum clematis
A. Sp. антиподы проявляют гаусто-
риальную активность.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
y Aristolochia bracteata.
Jintepatypa: Bhatnagar, 1967a; Cor-
ner, 1976; Davis, 1966; Periasamy,
1966; Schnarf, 1931; CmupHosa, 41977;
Тахтаджян, 1966, 4970.
Индийский — ученый Батнагар
[ВБабпасаг, 1967] исследовавший ци-
тоэмбриологию семейства Aristoloc-
!асеае, считает, что это семейство
следует сближать с семейством Ап-
попасеае, как предложил Веттштейн
[Wettstein, 1935}, а пе с семейством
Berberidaceae, как это признавал
Хатчинсоп [Ни<Ьтзоп, 1926].
ПОРЯДОК 5.
RAFFLESIALES
СЕМЕЙСТВО КАЕЕГЕЗТАСЕАЕ
Основное число ‘хромосом х=12,
46, 20.
Цветок обычно однополый, редко
обоеполый.
Пыльник — нет данпых.
Тапетум секреторпый.
Микроспорогенез и микрогаметоге-
нез — образовапие микроспор сук-
цессивное. Зрелые пыльцевые зерна
двухклеточные, однобороздные или
безапертурные. ,
Семяпочка анатропная или орто-
тропная, с одним — двумя интегу-
ментами, тенуинуцеллятная. Семя-
почек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
ПНариетальные клетки отсутствуют.
Зародышевый мешок образуется по
Polygonum-tuny. Имеются три хоро-
шо развитые долго сохраняющиеся
антиподы. Полярные ядра сливают-
ся до оплодотворения.
Опыление, прохождение пыльще-
вой трубки, оплодотворение — поро-
гамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный; эн-
досперм хорошо развит, масляни-
стый. Имеется перисперм.
Эмбриогенез — зародыш — малень-
кий, слабо дифференцированный
или не дифференцированный у Ва{-
Пез1а, содержит масло.
Семя, плод — семя с эндоспермом.
Зародыти располагается в его осно-
вапии и занимает обычно от 1/4 до
1/2 линейной высоты семени. Плод
ягодообразный.
Гаусторий — семядоли в начале
прорастапия семепи выполняют га-
усториальную функцию.
Апомиксие не обнаружен.
Другие явления — все представи-
тели семейства паразитные.
Литература: Согпег, 1976; Guzow-
ska, 1964; Ponzi, Pizzolongo, 41976;
Schnarf, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970; Терехин, 1977.
СЕМЕЙСТВО НУОМОВАСЕАЕ
Основное число хромосом — нет
даптых.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка пыльника имеет эпидермис,
эндотеций, 3 средних слоя и секре-
торный тапетум.
Микроспорогенез и микрогаметоге-
Hes — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двухклеточные, двух-трехбороздные,
тетерополярные.
Семяпочка ортотропная, с одним
интегументом, тенуинуцеллятная.
Семяпочек мпого.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальных клеток нет. Зароды-
шевые мешки образуются no Allium-
и Адоха-типам. Синергиды груше-
видные, имеются три скоро дегене-
рирующие антиподы, представлен-
ные не клетками, а ядрами.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
дапных.
23
Эндоспермогенез целлюлярный.
Эмбриогенез Зо]ап4-типа. Зародыш
маленький, слабо дифферепцирован-
ный.
Подвесок длинный, состоит
из 15 клеток, расположенных в
один ряд.
Семя, плод — семя с хорошо раз-
витыми эндоспермом и периспер-
мом, содержащим масло. Плод яго-
дообразный, но с почти деревяни-
стым внешним слоем.
Гаусторий — семядоли при про-
растании выполняют гаусториаль-
ную функцию.
Апомикеие не найден.
Другие явления — все представи-
тели семейства паразитные расте-
НИЯ.
Литература: Сосасс!, 1976; Согпет,
1976; Schnarf, 1931; Тахтаджян, 1966,
1970; Терехин, 1977.
Несмотря на то что семейство
Ва езасеае в отпошепии цитоэм-
бриологии изучено очепь слабо,
Шнарф [Schnarf, 1934] высказал
предположение, что это семейство
имеет родственные отношения с
Анзоосвасеае, с одпой стороны, с
Nymphaeaceae u Ceratophyllaceae — c
другой.
Относительно семейства Нудпога-
сеае, которое цитоэмбриологически
изучено более основательно, чем се-
мейство Rafflesiaceae, Кокучи [Со-
cucci, 1976], довольно детально ис-
следовавший Нудпогасеае, пришел
к заключению, что это семейство
возникло из Аппопасеае и не свя-
зано родственно пи с ВаШезасеае,
пи с Атзюо]осШасеае. Следовательно,
отнесение его к порядку ВаШеза-
]ез может быть ошибочным. Однако
для решения этого вопроса требуют-
ся дальнейшие исследования.
ПОРЯДОК 6.
МУМРНЕАТЕ$
СЕМЕЙСТВО САВОМВАСЕАЕ
Оеновное чиело хромосом — х==12,
40, 52.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций без фиброзных
утолщений и секреторный одно-,
дву-, многоядерный тапетум.
24
Микроепорогенез п микрогаметоге-
нез — образование микроспор си-
м`ультанное. Зрелые пыльцевые зер-
на 'двухклеточные, однобороздные.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная,
микропиле образуется впутренним
интегументом. Семяпочек много.
Имеется гипостаза.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки имеются. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевый мешок образует-
ся по Ро]усопиш-типу и содержит
зерна крахмала. Синергиды с крюч-
кообразными выростами и фибрил-
лярными аппаратами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются до оплодотворепия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворение.
Эндоепермогенез телобиальный,
целлюлярный и пуклеарный.
Эмбриогенез Азегад-типа.
дыш маленький.
Семя, плод — семя с тонким слоем
эндосперма и массивным периспер-
мом. Зародыш расположен в его ос-
новапии и обычно не превышает 1/4
линейной высоты семени. Несмотря
на малую величину, зародыш диф-
ференцирован на семядоли и коре-
шок. Семядоли при прорастании се-
мени выполняют гаусториальную
функцию. Плод переходного типа от
многолистовки к мпогоорешку. Пе-
рикарпий толстый и колючий.
Гаусторий эндоспермальный, рас-
положенный в халазальной части
зародышевого мешка.
Апомикеис пе обнаружен.
Другие явления не наблюдались.
Литература: Ramji, Padmanab-
han, 1965; Davis, 1966; Padmanabhan,
Ramji, 1966; Тахтаджян, 1966, 1970;
Батыгина и др., 1979.
Заро-
СЕМЕЙСТВО МУМРНАЕАСЕАЕ
Оеновное число хромоеом — х = 8,
10, 12, 14, 29, 48, 56, 80.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—5
средних слоев, а также секреторный
или амебоидный, дву-, многоядерный
тапетум, ядра которого могут сли.
ваться и становиться полиплоидны-
ми. Имеются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультапное, ипогда сукцессивное.
Тетрады микроспор тетраэдральные,
изобилатеральные. Зрелые пыльце-
вые зерна двух-, трехклеточные,
одно-, лвух- или трехбороздные.
Семяпочка висячая, анатропная,
с двумя интегументами, крассину-
целлятная. Микропиле образуется
внутренним интегументом. Имеются
нуцеллярный колпачок, ариллус и
гипостаза. Семяпочек много. Cocy-
дистый пучок доходит до халазы.
Семяножка массивная и короткая
или длинная и узкая.
Макроепорогенез и макрогаметоге-
нез — археспорий одпоклеточный.
Один — четыре слоя париетальных
клеток. Тетрады или триады макро-
спор линейные. Зародышевый ме-
шок формируется по Ро]ухопит-ти-
пу и имеет зерна крахмала. Синер-
гиды грушевидные; имеются три
эфемерные антиподы; полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный
(y Nuphar luteum и У1сюма стида-
na), нуклеарный (у Гигуае {огох).
Деление первичного ядра эндоспер-
ма происходит обычио до деления
зиготы. Подвесок присутствует или
отсутствует.
Эмбриогенез Азегад-типа, вариа-
ции Репаеа. Зародыш маленький,
двусемядольный, с почечкой и ко-
решком, содержит крахмал и белок.
Зародыши хлорофиллоносные или
бесхлорофилльные.
Семя, плод — семя со слаборазви-
тым эндоспермом и`массивным ру-
минированным периспермом, содер-
жащим крахмал и белок. Зародыш
расположен в основании семени и
обычно не превышает 1/4 линейной
высоты. Семядоли при прорастании
семени выполияют гаусториальную
функцию. Плод — синкарпная мно-
толистовка.
Гауеторий — халазальная часть
эндосперма превращается в гаусто-
рий.
Апомикеие — обнаружена апоспо-
рия.
Другие явления — полиэмбриония
и клейстогамия.
Литература: Батыгина и др., 1979.
Davis, 1966; Corner, 1976; Khanna,
1964, 1965b, 1967; Li, 1955, Мейер,
1960; Padmanabhan, 1967a, 4970a,
Schnarf, 1934; Schneider, 4978a;
Schneider, Ford, 1978; Sen, 1969;
Cmuprosa, 1977; Тахтаджян, 1966,
1970; Вальцева, Савич, 1965.
СЕМЕЙСТВО
CERATOPHYLLACEAE
Основное число хромосом — х=8,
10.
Цветки однополые, растения одно-
домные.
ПНыльник — число микроспоранги-
CB пе известно.
Стенка микроспорангия имеет
эпидермис, эндотеций, 2 средних
слоя и амебоидный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двухклеточные, однобороздные, без-
апертурные.
Семяпочка — одна ортотропная, C
одним интегументом, крассинуцел-
лятная. Имеется гипостаза.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор линейные, 7-образ-
ные. Зародышевый мешок образует-
ся по Polygonum-Tully и содержит
крахмальпые зерна. Сипергиды гру-
шевидпые; три маленькие, скоро
дегенерирующие антиподы. Поляр-
ные ядра сливаются до оплодотворе-
HUA.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эндосперм содержит крахмал.
Эмбриогенез Азега4-типа. Заро-
дыш двусемядольный, прямой с хо-
рошо развитыми семядолями, почеч-
кой и корешком, хлорофиллоносный,
содержит крахмал и белок. Подвесок
отсутствует.
Семя, плод — семя без эндосперма
и перисперма. Зародыш заполняет
весь объем зрелого семени. Эндо-
сперм полностью израсходуется в
процессе развития зародыша и со-
зревания семени. Семядоли выпол-
няют запасающую функцию. Плод —_
орешек с остающимся стилодием.
Гаусторий не найден.
Апомиксие не обнаружен.
К
qt
Другие явления не наблюдались.
Литература: Согпег, 1976; Davis,
1966; Vijayaraghavan, 1967c; Смир-
нова, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВАВСГАУАСЕАЕ
Основное чиело хромоеом — =
=—47, 18.
Цветок обоеполый.
Пыльник — о числе микроспоран-
гиев нет данных. Стенка микроспо-
рангия имеет фиброзный эндотеций,
два средних слоя и секреторный
одно-многоядерный тапетум, ядра
которого после слияния становятся
полиплоидными.
Микроспорогенез и микрогаметоге-
нез — образование микроспор не
изучено. Зрелые пыльцевые зерна
двухклеточные, однобороздные.
Семяпочка ортотроппая, анатроп-
ная, крассинуцеллятная, с двумя ин-
тегументами. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Зародышевый мешок образуется по
Polygonum-tuny. Имеются три эфе-
мерные антиподы.
Опыление, прохождение пыльце-
вых трубок, оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез Азега4-типа, вариа-
ции Репаеа. Зародыш дифференци-
рован и имеет две семядоли, почеч-
ку и корешок.
Семя, плод — семя с маленьким
зародышем, окруженным скудным
эндоспермом и обильпым периспер-
MoM. Плод — синкарпная многоли-
стовка © остатком трубки венчика.
Гаусторий не найден.
Другие явления не обнаружены.
Литература: Батыгина и др., 1980;
"Гахтаджян, 1966, 4970.
Относительно Вагс]ауасеае цито-
эмбриологические данные до 1980 г.
отсутствовали. Порядок МушрВеаез
неоднократно привлекал к себе вни-
мание многих исследователей [Мей-
ер, 1960; Вальцева, Савич, 1965;
Khanna, 1967; Padmabhan, 1970], oco-
бенно в связи с вопросом о проис-
хождении однодольных.
Исследуя цитоэмбриологию видов
Nuphar и МушрЬава, К. И. Мейер
[1960], а также 0. В. Вальцева и
Е. И. Савич [1965] пришли к заклю-
чению, что по признаку развития
обеих семядолей представители этих
родов не отличаются от типичных
двудольных, но по строению рамт-
]а с ее двумя неодинаково развиты-
ми листочками они очень напоми-
нают однодольные. Сходство в фор-
мировании раша МушрВаеасеае и
однодольных является признаком,
сближающим обе эти группы и вме-
сте с тем лишним доказательством
в пользу происхождения однодоль-
ных от каких-то общих с МутрВаеа-
сеае предков.
Эндлихер [Endlicher, 1841] сбли-
xan Ceratophyllaceae c Chlorantha-
сеае, что по Вижьярагавану [УЦауа-
гаспауап, 1967с] цитоэмбриологиче-
скими данными не подтверждается.
ПОДКЛАСС В. КАМОМСОГТАЕ
НАДПОРЯДОК 1.
RANUNCULANAE
ПОРЯДОК 7.
ILLICIALES
СЕМЕЙСТВО ПЛАДСТАСЕАЕ
Основное число хромосом — х=7.
Цветок обоеполый или однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эни-
26
дермис, фиброзный слой, 2—3 эфемер-
ных средних слоев и секреторный,
двуядерный, двурядный тапетум.
Имеются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые —пыльцевые
зерна двухклеточпые, трехбороздные
с сетчатой экзиной.
Семяпочка анатропная, с двумя
интегумептами, крассинуцеллятная.
Микропиле образуется наружным
интегументом. Семяночек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевый мешок формиру-
ется по Ро]ухопат-типу. Имеются
три эфемерные антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоепермогенез целлюлярный.
Эмбриогенез Asterad-tuna. Заро-
дыш очень маленький, прямой с
двумя семядолями и корешком, бес-
хлорофилльный.
Сея, плод — семя с хорошо раз-
витым маслянистым ондоспермом.
Зародыш располагается в основапии
семени и обычно не превышает 1/4
линейной высоты сего. Плод — цик-
лическая многолистовка.
Гаусеторий — соемядоли при прорас-
тании семени выполняют гаустори-
альпую функцию.
Апомиксие не найден.
Другие явления пе обнаружены.
Литература: Согпег, 1976; Bhan-
dari, 1967e, 1971; Davis, 1966; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970; Vijayaraghavan, 19671.
Г
СЕМЕЙСТВО ЗСНТЗАМОВАСЕАЕ
Основное число хромосом — х=7.
Цветок однополый.
Пыльник — 4 — микроспорапгия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, 1—2 слоя фиброзного эндо-
теция, 2—3 эфемерных средних слоя
и секреторный, дву-, многоядерный,
двурядный тапетум. Ядра тапетума
сливаются и становятся полиплоид-
ными.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные. dpe-
лые пыльцевые зерпа двухклеточ-
пые, трех — шестибороздные с тол-
стой сетчатой экзипой.
Семяпочка апатроппая с двумя ин-
тегументами, крассипуцеллятная.
Микропиле образуется обоими инте-
гументами или только наружным.
Семяпочек много.
Макроепорогенез и макрогаметоге-
нез — археспорий мпогоклеточный.
Париетальных клеток 3—5 рядов.
Тетрады макроспор линейные,
Т-образные. Ипогда образуются
только три макроспоры. Зародыше-
вые мешки формируются по Ро]уг0-
пит и ОепоФега-типам. Синергиды
грушевидпые. Имеются три эфемер-
пые антиподы. Полярные ядра сли-
ваются до оплодотворения.
Опыление, прохождение ныльце-
вых трубок и оплодотворение — по-
рогамия. Между опылением и пер-
вым делением зиготы проходит 4—5
недель.
Эндоспермогенез целлюлярный.
Эмбриогенез Аз{ета4- или Onagrad-
типа. Зародыш маленький, двусемя-
дольный и с корешком. Имеется под-
весок.
Семя, плод — семя с массивным
эпдоспермом, содержащим крахмал
и масло. Зародыш расположен в 0с-
повации семени и обычно не превы-
тает 1/4 линейной высоты семени.
Семенная кожура формируется обо-
ими интегументами. Наружный ин-
тегумент состоит из нескольких сло-
св клеток, а впутренний интегумент
представляет собой тонкий слой де-
геперирующих толстостенных кле-
ток. Плоды из многочисленных одно-
пятисеменных ягодообразных пло-
диков. Степки плода мясистые и со-
держат 14—16 слоев удлиненных
паренхиматозных клеток. Внешний
эпидермис перикарпия содержит
клотки с эфирным маслом.
Гаусторий — семядоли при прорас-
тапии ссмепи выполняют таустори-
альную фупкцию.
Апомиксие пе обнаружен.
Другие явления не наблюдались.
Литература: Bhandari, 1987j, 1970,
1971; Corner, 1976; Davis, 1966; Cmup-
пова, 1977; Тахтаджян, 1966, 1970.
Цитоэмбриологичесие данные под-
тверждают самостоятельность семей-
ства Illiciaceae m выделение ро-
na Illicium из семейства МарпоПа-
сеае в особое семейство [ВБапдат,
19671].
По мнению Имса [Eams, 1961],
xota Schisandraceae um Illiciaceae u
снцепиализированные семейства, од-
пако они имеют теспые отношения
с МаспоНасеае, что указывает на
возможность происхождения всех
этих семейств от общих предков.
21
ПОРЯДОК 8.
NELUMBONALES
СЕМЕЙСТВО МЕГОМВОМАСЕАЕ
Основное число хромосом — х=8.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, три
средних слоя и секреторный много-
ядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зер-
на двухклеточные, трехбороздные.
Наблюдаются сложные пыльцевые
зерна с двумя, тремя клетками.
Семяпочка висячая, анатропная,
с двумя интегумептами, крассину-
целлятная. Микропиле длинное, об-
разующееся только внутренним иц-
тегументом. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий, одно-, четырех-
пятиклеточный. Париетальных кле-
ток 4—5 рядов. Макроспоры липей-
ные. Зародышевый мепток образует-
ся по Polygonum-tuny. Антипод
сначала три, но после оплодотворе-
ния число клеток и ядер в них уве-
личивается в результате митотиче-
ских делений. Полярные ядра сли-
ваются в момент оплодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклсарный.
Эмбриогенез — Аз{егад-типа вариа-
ции Репдеа. Зародыш крупный, пря-
мой, с двумя хорошо развитыми мя-
систыми семядолями, почечкой и
корешком, хлорофиллоносный. Под-
весок с двуядервыми клетками.
Семя, плод — зародыш заполняет
весь объем зрелого семени. Эндо-
сперм почти полностью расходуется
в процессе развития зародыша и
созревания семени. Семядоли вы-
полняют запасающую функцию,
а от эндосперма в семени остаются
лишь малозаметные остатки. Плод —
многоорешек. Семеппая кожура об-
разуется одним наружным интегу-
ментом.
Гауеторий антиподальный, кото-
рый дегенерирует на глобулярпой
стадии развития зародыша.
28
Апомикесие не обпаружен.
Другие явления не наблюдались.
Литература: Батыгина, и др., 1980;
КВаппа, 1965Ъ, 1967; Голенкин, 4937;
Gupta, Ahluwalia, 1977. Gupta, Ra-
ni Ahyja, 1969; Padmanabhan, 1970;
Смирнова, 1977; Тахтаджян, 1070;
Venkateswarlu, Seshavataran, 1964.
На основапии данных цитоэмбрио-
логии индийский цитоэмбриолог
Кханна [КВаппа, 19655] согласился
с предложением Ли [Li, 1965] no
А. Д. Тахтаджяна [1959] выделить
порядок МеаитЪапа]ез с монотипвым
родом МеатЬо из порядка Nymphea-
les mw отнести МешшЪо к самостоя-
тельному семейству Nelumbonaceae.
Другой индийский ученый Падма-
набхан [РадтапаЪЪап, 1970а] также
считает, что предложение выделить
род МецииЪо в особое семейство и
даже отдельный порядок МеиЪо-
па]ез согласуется с данными цито-
эмбриологии, так как по некоторым
признакам: строению ‘пыльцевых зе-
рен, семяпочки, эндосперма зароды-
ша, семени и гаусторий, семейство
Ме!атаБасеае отличается от Мушр-
раеасеае. Однако для окончательного
решения вопроса о правомочности
выделения рода М№еитро в особые
семейство и порядок требуются
дальнейшие исследования [Padma-
nabhan, 1970a], так как семейство
Мейлифопасеае в отношении цито-
эмбриологии исследовано пока недо-
статочно.
ПОРЯДОК 9.
RA NUNCULALES
CEMEMCTBO LARDIZABALACEAE
Основное число хромосом — х=7,
‚8, 15.
Цветок обычно однополый, расте-
ния однодомпые или двудомные.
Пыльник — микроспорангия.
‚ Стенка микроспорапгия имеет эпи-
дермис, фиброзпый эндотеций, 2—3
эфемерных средпих слоя и секретор-
ный двух-пятиядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
‘раздральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трех — шестибороздные.
Семяпочка анатроппая, кампило-
троппая или ортотропная, с двумя
интегументами, крассипуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется нуцелляр-
ный колначок. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одно- изредка
двухклеточпый. Париетальных кле-
ток 3—4 ряда. Тетрады макроспор
линейпые, Т-образные. Зародыше-
вый мешок формируется uo Polygo-
пит-типу. Синергиды грушевидные.
Имеются три долго сохраняющиеся
или эфемерные антиподы. Полярные
ядра сливаются перед оплодотворс-
нием.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип не определен.
Маленький прямой зародыш.
Семя, плод — семя с эндоспермом,
содержащим масло. Зародыш распо-
лагается в основании семени и обыч-
но занимает от 41/4 до 1/2 линейной
высоты семени. Плодики мясистые,
нераскрывающиеся или иногда рас-
крывающиеся вдоль шва.
Гаусторий — в пачале прорастания
семени, ссмядоли выполияют гау-
сториальпую функцию.
Апомикеие по обпаружеп.
Другие явления пе наблюдались.
Литература: Corner, 1976; Davis,
4966; Schnarf, 1931; Смирнова, 1977;
Taxtamnan, 1966, 1970; Vijayaragha-
van, 1967¢.
СЕМЕЙСТВО MENISPERMACEAE
Основное число хромосом — х=
='42, 13.
Цветок однополый, растения обыч-
но двудомные.
Пыльник — 4 микроспорапгия.
Степка микроспорангия образустся
по двудольпому типу и имсет эпи-
дермис, фиброзпый эпдотеций, два
эфемерных средних слоя и секре-
торный двуядерный тапетум. Име-
ются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изо-
билатеральные. Зрелые пыльцевые
зсрна двухклеточные, трехбороздно-
поровые, с крахмальными зернами.
Семяпочка анатропная, гемиана-
тропная-амфитропная, с одним-дву-
мя интегументами, крассинуцеллят-
пая. Микропиле образуется внут-
ренним интегументом. У Ттозрога
cordifolia микропиле зигзагообраз-
ное. Имеется нуцеллярный колиа-
yor. Семяпочек обычно немного,
редко много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Роу-
оопаш-типу. Синергиды с крючкооб-
разпыми выростами и фибриллярны-
ми аппаратами. Три эфемерные или
долго сохраняющиеся антиподы. По-
лярные ядра сливаются до оплодот-
ворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Тройное слияние предшествует син-
тамии.
Эндоспермогенез нуклеарный, ру-
минированный.
Эмбриогенез Опастад-типа (Trifo-
Нао-вариации, у ТШасога гасето-
за). Зародыш довольно большой, со-
гнутый, хоропо дифференцирован-
ный, с смеядолями, гипокотилем и
корешком, развитыми в равной сте-
нени, бесхлорофилльпый.
Семя, плод — семя с эпдоспермом
или без него. Для этого семейства
характерно болышое разнообразие в
соотношениях размеров зародыша и
эндоснерма. Вытянутый согнутый
зародыш располагается по перифе-
рии семени, занимая 3/4 его пери-
метра. Эндосперм сохраняется в
центре семени. Кроме согнутых за-
родышей в этом семействе встреча-
ются и прямые узкие, окруженные
эндоспермом зародыши. Эндосперм
содержит масло и алейроповые зер-
па. Плоды — мпогокостянки или ре-
же многоорезтки.
Гаусторий у одпих видов семядо-
ли выполняют — гаусториальную,
у других запасающую функцию.
Другие явления не обнаружены.
Литература: Согпег, 1976; Davis,
1966; Смирнова,’ 1977; Тахтаджян,
1966, 1970; Тульнова, Машкин, 1971;
Vijayaeaghavan, 1967f.
29
СЕМЕЙСТВО ВАМОМСОГАСЕАЕ
Основное число хромосом — х=
==5—10, 13.
Цветки обоеполые или редко одно-
полые, яркие, обычно энтомофиль-
ные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный или не фиброз-
ный эндотеций 2—4 эфемерных
средних слоя и секреторный одно-,
дву-, многоядерный тапетум. При
слиянии ядра тапетума становятся
полиплоидными. У Апетопорз13 на-
ряду с секреторным описан и аме-
боидный тапетум. Имеются орби-
кулы.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, обычно трех —
пятибороздные, с крахмальными зер-
нами и сетчатой экзиной. Иногда
пыльцевые зерна многобороздные
или многопоровые. Спермии-клетки
хорошо выражены.
Семяпочка висячая. анатропная,
темитропная, с одним — двумя инте-
тументами, красси- или тенуинуцел-
лятная. Микропиле образуется внут-
ренним интегументом. Имеются ну-
целлярный колпачок и гипостаза,
Семяпочек много; одни из них фер-
тильные, другие стерильные. По-
следние недоразвитые; у них может
отсутствовать интегумент, а зароды-
шевые мешки могут образовываться
не по моноспорическому, а по тетра-
спорическому типу.
акроспорогенез и макрогаметоге-
нез — археспорий одно-, дву-, мно-
токлеточный. Париетальные клетки
либо отсутствуют, либо присутству-
ют. Тетрады макроспор линейные,
Т-образные. Иногда — образуются
только три макроспоры. Зародыше-
вые мешки формируются по Ро]у-
gonum-, Allium-, Drusa- a Fritillaria-
типам. Синергиды с крючкообраз-
пыми выростами и фибриллярными
аппаратами. Имеются две, три и боль-
шее число крупных пузыревидных
одно- или многоядерных антипод,
обычно долго сохраняющихся. Ядра
антипод в результате слияния и эн-
домитоза становятся полинлоидны-
ми. Часто под антиподами имеется
30
у
особое возвышение — постамент. По-
лярные ядра сливаются до оплодот-
ворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодотво-
рение премитотического типа. Син-
гамия и тройное слияние происхо-
дят одновременно. У СопзоП4а агуеп-
315 оплодотворение происходит через
6—30 ч после опыления, у Вапапса-
lus роуаоешиз — через 3—19 ч.
Эндоспермогенез нуклеарный.
Клеткообразование в эндосперме
у одних представителей начинается,
с микропилярной, у других с хала-
зальной части зародышевого мешка.
Эмбриогенез Onagrad-, Solanad-,
Caryophyllad-tunmos, у некоторых
представителей эмбриогепез нерегу-
лярный. Зародыш маленький, диф-
ференцированный или недифферен-
цированный, бесхлорофилльный, со-
держит алкалоиды. У Ranunculus fi-
caria зародыш односемядольный,
у других представителей cemMel-
ства — двусемядольный. Подвесок
хоропто развит.
Семя, плод — семя с хорошо раз-
витым маслянистым эндоспермом,
содержащим белок и крахмал. За-
родыш располагается в основании
семени и обычно занимает от 1/4 до
1/2 линейной высоты. У семяпочек
с двумя интегументами семенная
кожура образуется обоими интегу-
ментами, но особенно наружным, ко-.
торый становится твердым у CGaltha
или склероидным у Ади|Певла. У се-
мяпочек с одним интегументом верх-
няя часть клеток наружного эпидер-
миса содержит таннин, нижняя —
густую цитоплазму (Helleborus).
'Плоды — многолистовки или много-
орешки, реже однолистовки или яго-
дообразные.
Гауеторий — семядоли при прорас-
тании семени выполняют гаустори-
альную функцию. |
Апомиксие — нередуцированный
партеногенез и апоспория у некото-
pix BuyoB Ranunculus.
Другие явления — гинодиэция у
Anemone, Ficaria mu Ranuneulus,
клейстогамия. Партенокарпия.
JIurepatypa: Babis Henryk, 1973,
4978; Bensesa H., 1974, 1977; Bhan-
dari, 1965, 1966, 1968; Bhandari, As-
nani, 1968; Bhandari, Ram, 1976;
Bhandari, Vijayaraghavan, 1970;
71
Рие. 6. Микроспорангий и микроспорогенез у Апетопе т1у\аг13
1—5 — диаграммы пыльников (часть пыльников в увеличенном виде: а — из рис. 1,
б — из рис. 2, в — из рис. 3, г — из рис. 4, д — из рис. 5; 6—11— разные стадии
микроспорогенеза, 12—15 — разные стадии образования двухклеточного мужского
гаметофита [Bhandari, 1968]
Рис. 7. Развитие семяпочки у зародышевого мешка у Сетайз gauriana
1 — завязь с фертильными и стерильными семяпочками (а — фертильная, 6, в — стерильные семяпочки); 2—6 — разные фазы развития семяпочек; 7—10 — макро- спорогенез; 11, 12 — дегенерация трех верхних макроспор в тетраде; 13 — двуядер- ный зародышевый мешок; 14, 15 — зрелые зародышевые мешки с одноядерными (14) и двуядерными (15) антиподами [УЦауагапауап, 1962]
32
Рис. 8. Прорастание пыльцы, образование пыльцевых трубок и зароды- шевый мешок у Апетопе петогоза, а также оплодотворение у Апетопе
ranunculoides
А — завязь с фертильными и стерильными семяпочками: Б — прорастание пыльцы
на рыльце;
Brouland, 1969; Corner, 1976; Fish,
1970; Гревцова, Коробова-Семенчен-
ко, 1978; Hiroshi, 1973; 1974d, 1974e;
Jalan, 1968; Kent, 1970; Кордюм,
1959a, b, 1960; 1961; 1963; Ly-Thi-Ba,
1971, 1974; Nogler, 1976; Patel, Cook, .
1972; Picci, 1964; Рудепко, 19614;
Schnarf, 1931; Swamy, 1960; Taxtaz-
жян, 1966, 1970; Татинцева, 1975;
Терзийски, Дасвалова, 1979; Trela-
Sawicka, 1976; Vijayaraghavan, 19675,
1969, 1970; Vijayaraghavan, Bhandari,
4967, 1969, 1970; Vijayaraghavan, Mar-
wash, 1968a, Ь; Жукова, 4965; Иоф-
фе, 1968, 1969, 1971.
3 В.А. Ноддтбная-Арнольди
СЕМЕЙСТВО
CIRCAEASTERACEAE
Основное число хвомосом — х=15.
Цветок обосполый.
Пыльник — 2 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный одноядерный тапетум.
Микроеспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
23
Рис. 8 (продолжение)
В — зародышевый мешок до оплодотворения; Г —‘зародышевый мешок во время
двойного оплодотворения [Иоффье, 1968]
зерна двухклеточные, трехбороздные
с тонкоструйчатой сэкзиной.
Семяночка ортотропная с одним
интегументом, тенуинуцеллятная.
Семяпочек немного.
Макроспорогенез и макрогаметоге-
84
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Polygonum-tuny. Синергиды груше-
видные, Антипод 2—3.
Опыление, прохождение пыльце-
7 ево
..
four
иже
oe
WW S00
35
3%
Рис. 9. Зародышевой мешок
Aconitum пареПаз с гигант-
скими антиподами, клеточным
зародышем из нескольких кле-
ток и многоядерным эндоспер-
MOM и постаментом [Озег-
walder, 1898].
Рие. 10. Эмбриогенез у Ва-
nunculus sceleratus.
ак — апикальная, 6x — базальная
клетки, п — подвесок, з — заро-
дыш, гк — гипофилярный конек;
1-—10 — разные стадии развития
проэмбрио, 11—18.— стадии раз-
вития зародыша [Зоиёсез, 4914]
ной трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эндосперм хорошо развит.
Эмбриогенез Спепоро1ад-типа. За-
родыш маленький, прямой.
Семя, плод — семя —с эндоспер-
мом и периспермом. Плодики —
орешки, покрытые `крючкообразны-
ми волосками.
Гаусторни не пайден.
Апомиксие не обнаружен.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1931; Taxranman, 1965,
1970.
CEMEHCTBO PODOPHYLLACEAE
Основное чиело XpoMocoM — x==6.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Степка микроспорапгия имеет эпи-
дермис, фиброзный эпдотеций, 1—2
эфемерных средпих слоя и секретор-
ный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, ре-
яке крестообразные, зрелые, пыльце-
вые зерна двухклеточные, трехбо-
роздные, с сетчатой экзиной.
Семяпочка анатроппая с двумя
интегументами, красспнупеллятлая.
Микроняле образуется обоими иптс-
гументами. Имоестея пуценлятный
колначок, малопькая гипостаза и
ариллус. Семяпочек пемного или
мпого.
Макроспорогенез и макрогаметоге-
нез — архоспорий одноклеточный.
Тетрады макроспор линейные. Парп-
стальные клетки обычно отсутству-
ют. Зародышевый мешок образуется
но Раусопита-типу и имеет зерпа
крахмала. Синергиды с фибрилляр-.
ными аппаратами. Имеются три
крупные аптиподы. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пет
дапных.
Эндоспермогеопез пуклеарный, хо-
рошо развит.
Эмбриогенез Опастад-типа. Заро-
дыш маленький, прямой, бесхлоро-
филльный.
Семя, плод — семя с массивным,
содержащим масло, эпдоспермом.
Зародыш узкий, вытянутый по оси
семени. Занимает от 1/4 до 3/4 дли-
пы семени. Плоды ягодообразные.
‚реже
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; Эрккенова и др., 1979; Смирно-
ва, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВЕВВЕВТРАСЕАЕ
Основное число хромосом — х=
= 7—8, 10,.14.
Цветок обоеполый.
Ныльник — 4 микроспорапгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзпый эпдотеций. 2—3
эфемерных средних слоя и секре-
торпый многоядерный — тапетум.
Y Berberis arestata ranetym амебо-
HJVITDTH.
Микроепорогенез и микрогаметогс-
пез — образование микроспор обыч-
но симультанное, y Berberis aresta-
(а — сукцессивное. Тетрады микро-
спор изобилатеральные, линейные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, с зернами
крахмала; обычно трехбороздные,
многобороздные с сетчатой
или гладкой сэкзиной.
Семяпочка анатропная, с
иптегументами, крассинупеллятная.
Имеются пуцеллярный колпачок,
ариллус, фупикулярпый обтуратор и
гипостаза. Семяночек песколько —
много.
Макроспорогенез н макрогаметоге-
нез — археспорий одно-многоклеточ-
ный. Париетальные клетки OTCYT-
ствуют или присутствуют. Тетрады
макроспор линейные. Зародышевые
мешки образуются nto Polygonum-
типу и по модифицированпому Ре-
регопла-типу. Синергиды грушевид-
ные. Имеются три крупные пузыре-
видные долго сохраняющиеся анти-
поды. Полярные ядра сливаются до
оплодотворения. У Ериаеднит ар!
пит и Е. ршпашш обнаружен под
аптиподами постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворение.
Сипгамия и тройное слияние проис-
ходят одновременно. Пыльцевые
трубки долго сохраняются в семя-
почке.
Эндоспермогенез нуклеарный. Эн-
досперм масляпистый, содержит
также берберин и алейрон.
двумя
37
Эмбриогенез — Опастай-, Solanad-
типов. Зародыш прямой, дифферен-
цированный, с двумя, иногда тремя
семядолями и довольно длинным ко-
решком, бесхлорофилльный. Содер-
жит масло и алейроп.
Семя, плод — семя с массивным
эндоспермом. Зародыш узкий, вытя-
нутый по оси семени и занимает до
3/4 его длины. Плод — обычно ягода
или орехообразпый. Семенная кожу-
ра образуется из обоих интегумен-
тов; эпидермис наружного интегу-
мента содержит таннин.
Гаусторий синергидпый у Berbe-
ris arestata. Семядоли выполняют
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
y Berberis arestata, B. vulgaris.
Литература: Ва-Гу-ТЬ1, 1976; Сог-
ner, 1976; Руденко, 1964; Зазыт, 1969;
Schnarf, 1974; Смирнова: 1977; Тах-
таджян, 4966, 4970; УЦауагаеВауал,
1967Ъ; Жукова, 1965.
Из 10 семейств порядка Вапа|ез
классификации А. Л. Тахтаджяна
(1970) 4 семейства, а именно: Sar-
_ gentodoxaceae, Glacidiaceae, Hydras-
tidaceae u Nandinaceae в отношении
цитоэмбриологии пс исследованы.
Сравнительные цитоэмбриологиче-
ские данные по другим семействам,
по мнению индийских цитоэмбрио-
логов, показывают, что Гаглхара]а-
сеае не сходно с ВегЬег!Часеае, что
Menispermaceae теспо связано с
Ranunculaceae, a He с МаспоПасеае
[Vijayaraghavan, Bhandar, 1967].
Они считают, что некоторые роды,
oTHOCuMBIe K Ranunculaceae, следо-
вало бы выделить в самостоятельные
семейства (Paeoniaceae, Kingdonia-
ceae, Circaeasteraceae), 10 B TO же
время для выделепия НеПорогасеае
в отдельное семейство нет основа-
ний CO стороны цитоэмбриологии
[Vijayaraghavan, Marwah, 1969].
Вместе с тем они полагают, что Вег-
рег1Ч4асеае в отношении цитоэмбрио-
логии настолько сходпо с Вапипси-
laceae, что включепис обоих этих
семейств в порядок Ranunculales
(Ranales) не вызываст сомпений
[ Vijayaraghavan, 1967c].
Представители порядка Ranuncu-
lates Tak se, Kak mu Magnoleales,
MMeIOT HeMAaO UPWMMUTMBIBIX IMTO-
эмбриологических признаков: двух-
38
клеточный мужской гаметофит, ана-
тронную, крассинуцеллятную семя-
почку с двумя интегументами, ну-
целлярный колпачок, наличие па-
риетальных клеток, Роухопат-тип
зародышевого мешка, эфемерные ан-
типоды, хорошо развитый эндо-
сперм, малепький слабо дифферен-
цированный зародыш, отсутствие
гаустория. Однако вместе с тем
представители Капипси]аез имеют
уже пемало и прогрессивных цито-
эмбриологических признаков: ана-
троиную, тепуинуцеллятную семя-
почку с одним интегументом, на-
блюдается умепьшение числа, а в
ряде случаев и отсутствие парие-
тальных клеток, наличие зародыше-
вых мешков АШиш-, Огаза- и Ре-
peromia-Tullos, наличие крупных
долго сохраняющихся многоядерных
антипод, число которых в некоторых
случаях достигает 11 путем митоти-
ческого деления первоначально трех
антипод, появление склонности к
апомиксису и полиэмбрионии. Все
это свидетельствует о том, что по-
рядок Вапипса[а{ез хотя и является
примитивным, но он все же выше,
чем порядок Маспо|а]ез, и располо-
жение его в системе А. Л. Тахтад-
жяна [1966—1970] также выше по-
рядка МаспоПа!ез кажется нам впол-
пе правильным.
На основании изучения цитоэм-
бриологии некоторых представите-
лей семейства Ranunculaceae
Е. Л. Кордюм [1959] пришла к за-
ключению, что семейство это естест-
венное и что выделение из него ро-
да Paeonia правильно, так Kak
последний род отличается от дру-
гих представителей Ranunculales
рядом цитоэмбриологических при-
знаков, особенно же ценоцитным об-
разованием зародыша [Яковлев,
Иоффе, 1957, 4961].
ПОРЯДОК 10.
PAPAVERALES
СЕМЕЙСТВО РАРАУЕВАСЕАЕ
Основное чиело хромосом — х=
—=5— 41, 19.
Цветок обоеполый, яркий, энтомо-
фильный.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи- от остальных маковых, у Platyste-
дермис, фиброзный эндотеций, 1—3 mon по дапным Г. М. Ильиной
эфемерных средних слоя и секре- [1968], процесс оплодотворения осу-
торный, обычно однорядный, иног- ществляется как по премитотиче-
да дву- трехрядный тапетум. У Р1а- скому, так и по постмитотическому
tystemon тапетум представляет с0- типам. У Рарауег зотиИегит двой-
бой «ложный периплазмодий». Слия- ное оплодотворение происходит че-
ние ядер тапетума приводит к поли- рез 48—72 ч после опыления. Пыль-
плоидизации. цевые трубки сохраняются в заро-
Микроеспорогенез и микрогаметоге- дышевом мешке до образования гло-
нез — образование микроспор си- булярного зародыша.
мультанное. Тетрады микроспор тет- Эндоепермогенез нуклеарный. На-
раэдральные, изобилатеральные, кре- иболсе поздпее образование клеточ-
стообразные. Зрелые пыльцевые зер- ных перегородок в эндосперме на-
на двух-трехклеточные, с зернами блюдается у родов с быстро разви-
крахмала, обычно трехбороздные, вающимися зародышами (Р]абузе-
реже многобороздные или мпогопо- топ, Papaver, Roemeria), наиболее
ровые, с сетчатой или зернистой раннее образование клеточных пе-
сэкзиной. регородок, наоборот, наблюдается у
Семяпочка гемикампилотропная, родов с медлеппо растущими заро-
анатропная, амфитроппая с двумя дышами (Езсвзевоа, Ну]отесап,
интегументами, крассипуцеллятная. Glaucium).
Микропиле у большинства видов зиг- Эмбриогенез — Азегад-, Solanad-,
загообразное. Образуется оно обоими СагуорБуПад-типов. Зародыши пря-
интегументами. Имеются постамент, мые, бесхлорофилльные, либо пол-
нуцеллярный колпачок, гипостаза, ностью дифференцированные, двусе-
эпистаза, плацентарный или фуни- мядольные с хорошо развитыми ко-
кулярный обтураторы, эндотелий, решком и гипокотилем, либо слабо
ариллус. Семяпочек много. дифференцированные, у СотудаЙз за-
Макроспорогенез и макрогаметоге- родыши не дифференцированные.
нез — археспорий одно-многокле- Семя, плод — семя с хорошо раз-
точный. Париетальные клетки от- витым эндоспермом, содержащим
сутствуют или присутствуют. Тетра- масло и алейрон. Зародыш занимает
ды макроспор линейпые или T-o6- от Ч/4 до 3/4 длины семени. Семядо-
разные. Зародышевые мешки обра- ли выполияот гаусториальную функ-
зуются обычно uo Polygonum-, из- цию. Запаспыми питательными ве-
редка по ЕгилШайа-тииам. Послед- ществами семени является масло.
ний тип обнаружен у Р]абузцетоп. В образовании семеппой кожуры
Синергиды с крючкообразными вы- участвуют оба интегумента. Плод —
ростами и фибриллярными аппара- коробочка, стручковидная или OK-
тами. Обычно три и больше круп- руглая, раскрывающаяся порами.
ных долго сохраняющихся антипод, Гауеторий — у растений, имеющих
ства они лежат па постаменте. гуустории эндосперма отсутствуют.
У РИмуетоп, однако, наблюдается В тех случаях, когда антиподы сла-
депрессия антипод с трех до одной. бо развиты или рано дегенерируют,
`дра антипод в результате много наблюдается формирование хала-
кратных эндомитозов становятся зального эндоспермального гаустория
полиплоидами, с гигантскими поли- (Platystemon, Eschscholtzia)
3 °
тепными хромосомами. Полярные Anomuxeue а С,
обычно сливаются до оплодот. moms редуцированный пар-
ворепия. Только у Г]абубетоп по-
лярные ядра сливаются после опло- Другие явления — обнаружена по-
дотворения. У Рарауег rhoeas заро- лиэмбриония и клейстогамия.
дышевой мешок имеет крахмал. Литература: Вего, 1966, 1967; Сот-
Опыление, прохождение пыльце- ner, 1976; Davis, 1966; Gracza, Sar-
вой трубки и оплодотворение —у Капу, 1970; Сапа, Зеа1а, 1976; Ильи-
маковых обнаружено как перекрест- на, 4964, 1964, 1968, 1974, 1976, 1978;
ное опыление, так и самоопыление Poal, Gracza, 1969; Schnarf, 1934;
Порогамия. Двойное оплодотворение Singh, КВап, 1974; Смирнова, 1977;
премитотического типа. В отличие Sulmont, 1968, 1969; 3aenp, 1960.
39
СЕМЕЙСТВО НУРЕСОАСЕАЕ
Основное чиело хромосом — х==
=6, 8.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
средпий слой и амебоидный тапе-
тум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
па двухклеточные, двухбороздпые,
с мелкозернистой сэкзиной.
Семяпочка гемикампилотропная,
с двумя интегументами, крассипу-
целлятная. Микропиле образуется
обоими интегументами. Имеется ну-
целлярный колпачок,
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародьтевый мешок образуется по
Го1угопат-типу. Имеются антиподы
с полинлоидными ядрами и гигаит-
скими политепными хромосомами.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия, двойное оплодотворение.
Эндоеспермогенез пуклеарпый.
Эмбрногенез СагуорруПад-типа. За-
родыш дифферепцированный, дву-
семядольный. В гигантских пузыре-
видных клетках подвеска Нуресоит
обнаружены политепные хромосомы.
Процесс политенизации затухает ко
времени формирования клеточных
перегородок в эндосперме.
Семя, плод. Плод стручковидный,
обычно разламывающийся па члени-
ки, реже раскрывающийся двумя
створками.
Гауеторий подвесочпый.
Апомихсие пе обпаружен.
Другие явления пе паблюдались.
Литература: Ильина, 1964, 1968,
1976; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ЕОМАВТАСЕАЕ
Основное число хромосом — r=
—=6— 8.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
но одподольному типу и имеет эни-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секретор-
ный, Дву-, многоядерный тапетум.
40
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зер-
на двухклеточные, трехбороздные
или рассеянпо-мпогобороздные, с
сетчатой или бугорчатой сэкзиной.
Семяпочка анатропная, кампило-
тропная с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется обоими интегументами.
Имеются фуникулярный обтуратор
и ариллус. Семяпочек две — много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Тетрады макроспор линейные. Па-
риетальные клетки присутствуют.
Зародышевый мешок образуется по
Ро1узопат-типу. Синергиды с фиб-
риллярным аппаратом. Имеются три
крупные антиподы с полиплоидны-
ми ядрами и политенными хромосо-
мами. Полярные ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. Пос-
ле образования пескольких сотен
свободпых ядер между ними возни-
кают клеточпые перегородки.
Эмбриогенез Caryophyllad-runa.
Зародыш бесхлорофилльный, очень
маленький, по дифференцированный
на две семядоли и корешок. Подвс-
сок массивный, мпогоклеточный.
Семя, плод — семя с эндоспермом,
содержащим масло. Зародынт распо-
ложеи в осповании семепи и обычно
пе превыттаст 1/4 линейной высоты
семепи. Плод — стручковидная ко-
робочка, раскрывающаяся двумя
створками или реже односеменной
орох.
Гаусторий — семядоли при прорас-
тании семепи выполняют гаустори-
альпую функцию.
Апомиксие по обпаружен.
Другие явления — обнаружена по-
лиомбриония.
Литература: Corner, 1976; Davis,
1966; Ильина, 1968, 1976; Маркова,
1949; Смирнова, 1977; Тахтаджян,
1966, 1970
Наиболее детально в отношении
цитоэмбриологии представители по-
рядка Рарауега]ез до настоящего
времени исследованы советеким ци-
тоэмбриологом Г.М. Ильиной [1964,
1962, 1963. 1964, 1968, 1976, 1978].
По мнепию Г. М. Ильиной, все три
семейства порядка Рарауега!ез ес-
тественны, родственны друг другу
и обнаруживают тесную связь с по-
рядком Вапипси]аез. Такие прими-
тивные цитоэмбриологические при-
знаки, как секреторный тапетум,
двухклеточные пыльцевые зерна,
большое число анатронных, красси-
нуцеллятных, с двумя интегумен-
тами семяпочек в одной и той же
завязи, мпогоклеточный женский
археспорий, наличие париетальных
клеток, Ро]угопит-тип `зародыше-
вого мешка, порогамия, семена с
эндоспермом, недоразвитие зароды-
ша в зрелом семени п т. д., сбли-
жают представителей порядка Ра-
рауега!е$, особеино семейства Рара-
veraceac, © представителями порял-
ка Вапипси]а[ез, главным образом
семейств Ranunculaceae u_ Berberi-
дасеае. Однако наряду с примитив-
ными цитоэмбрнологическими при-
знаками ирсдетавители порядка
Papaverales имеют и такие про-
грессивные признаки, как амобо-
идный тапетум, трехклеточные
пыльцевые зерна, немного гемикам-
пилотронпных и амфитропных семя-
почек впутри одной ип той же за-
вязи, одноклеточный женский ар-
хеспорий, отсутствие паристальных
клеток, зародьииевый мешок Fritil-
]Лаг1а-типа, присутствие разного ти-
па гаусториев,‚ дифференцирован-
ный зародыш и т. Д., что указыва-
ет па дальнейшую эволюцию пред-
ставителей этого порядка.
Г. М. Ильина считаст, что цито-
эмбриологические данные имеют
одинаковые основания для объеди-
нения трех семейств порядка Рара-
уега1ез в одно семейство Рарауега-
сеае или для выделения трех се-
мейств порядка Рарауега!ез в три
самостоятельные семейства, беру-
щие начало от порядка Вапипеца-
]ез. Возможно, что семейства Рара-
уегасеае, Пуресоассае и Fumaria-
сеае иметшт общее происхождение с
Berberidaceae aw Ranunculaceae
[Ильина, 1968].
Сравнительно - цитоэмбрнологиче -
ские данные, несомненно, указыва-
ют на естественность порядка Ра-
рауега!ез и на наличие в нем при-
митивных и прогрессивных цитоэм-
бриологических признаков.
По цитоэмбрпологическим особен-
ностям Рарауегасеае обнаруживают
близкие родственные связи с Сар-
рагасеае, что, возможно, свидетель
ствует о параллельном развитии их
от общего порядка ое
что отмечает Г. М. Ильина [1968
У большинства видов Рарауега-
сеае микропиле зигзагообразной
формы, как и у резеды, многих ка-
персовых и крестоцветпых [Илья-
на, 1968], что сближает Рарауега-
les c Capparidales.
По наличию ложного периплаз-
модия в тапетуме пыльника, трех-
клеточных пыльцевых 3epeH, Fritil-
]ача-тина зародышевого Moura,
эфемерных аптипод и халазального
эпдоспермального Taycropua Platys-
(шит отличается от других пред-
ставителей порядка Papaverales,
что позволяет, по нашему мнению,
выделить его в особое семейство, а
все остальные семейства порядка
объединить в одно семейство Рара-
уегасеае, поскольку опи цитоэмбри-
ологически очень сходны.
ПОРЯДОК 11.
SARRACENIALES
СЕМЕЙСТВО ЗАВВАСЕМТАСЕАЕ
чиело хромосом — х=
Основное
== 42, 13.
Цветок обосполый.
Ныльник — 4 микроспоравгия.
Стенка микроспорангия имеет эип-
дермис, фиброзный эндотеций, 1—3
средних слоя и секреторный, двуя-
дерный, двух-трехрядный тепетум.
Микроспорогенез и микрогамет)-
генез — образование микроспор си-
мультанное. Тетрады микроспор тс-
траэдральные. Зрелые пыльцевые
зерна двухклеточные, 8—10 борозд-
нопоровые, с меридиональными бо-
роздами и гладкой сэкзиной.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Имеются эндотелий и гипостаза. Се-
мяпочек много.
Микроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро!узопит-типу. Синергиды грушс-
видные. Имеются три эфемерные
антиподы. Полярные ядра слива-
ются до оплодотворения,
41
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. ,
Эндоепермогенез целлюлярный.
Эмбриогенез Caryophyllad-tuma.
Зародыпт маленький, но дифферен-
цированный; имеет две семядоли и
корешок, бесхлорофилльный. Под-
весок семиклеточный, однорядный.
Семя, плод — семя с массивным
эндоспермом, содержащим масло и
алейрон. Зародыш расположен вос-
новании семени и обычно не пре-
вышает 1/4 линейной высоты,
Плод — локулицидная коробочка.
Гаусторий — семядоли при про-
растении семени выполняют гаусто-
риальную функцию.
Апомикеис не обнаружен.
Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; ЭсвпатЁ, 1931; Смирнова, 1977;
Тахтаджян, 1966, 1970.
По своим цитоэмбриологическим
признакам семейство Sarracenia-
сеае во многом сходно с представи-
телями Papaverales u Ranunculales,
что оправдывает расположение $Заг-
гасешасеае вблизи этих порядков.
ПОДКЛАСС С. НАМАМЕНОТРАЕ
НАДПОРЯДОК Ш.
HAMAMELIDA NAE
ПОРЯДОК 12.
TROCHODE NDRALES
СЕМЕЙСТВО
TROCHODENDRACEAE
Основное число хромосом — z =
== 419.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенки — микроспорангия имеют
эпидермис, Ффиброзный эндотеций,
2—3 средних слоя и секреторный
двурядный, двуядерный тапетум.
Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевые
зерна двухклеточные, трохборозд-
ные с сетчатой сэкзиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная,
с разрастающимся халазальным вы-
ростом. Микропиле образуется вну-
тренним интегументом. Сосудистый
пучок доходит до халазы. Семяпо-
чек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Имеются три-четыре ряда парие-
тальных клеток. Зародышевый мсе-
шок образуется по Ро]угопит-ти-
пу, синергиды с Ффибриллярными
аппаратами. Имеются три эфемер-
ные антиподы.
42
Опыление, прохождение пыльце-
вой трубки и бплодотворение —
порогамия, двойное оплодотворение.
Зародышевый мешок после оплодо-
творения сильно удлиняется. Син-
гамия и тройное слияние происхо-
дят одновременно.
Эндоспермогенез целлюлярный,
поскольку первичное ядро эндоспер-
ма располагается либо в центре
зародышевого мешка, либо в хала-
зальной его части, то при первом
делении его закладываются нерав-
ные клетки: болышая микропиляр-
ная и маленькая халазальная. По-
следующие несколько делений —
поперечные.
Эмбриогенез — тип не определен.
Зародыш прямой, бесхлорофилль-
ный, дифференцированный на двое
семядоли и корешок. Имеет масло
и алейрон.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Формирование
семени занимает длительный пери-
од времени — 6—7 месяцев, что ха-
рактерно для примитивных расте-
ний. Запасные питательные вещсе-
ства семени — белок и масло. Ко-
жура семени формируется обоими
интегументами. Плод — синкарпная
многолистовка.
Гаусторий — семядоли при про-
растании семени выполняют гаусто-
риальную функцию.
Апомиксис не обнаружен.
Другие явления не наблюдались.
Jimreparypa: Bhandari, 1967, 1971;
Corner, 1976; Davis, 41966; Taxra-
джян, 1966, 1970; УоНе, 1962, 1965.
Cemelicrno Tetracentraceac, также
относящееся по системе А. Л. Тах-
таджяна [1970] к порядку Тгосво-
dendrales, цитоэмбриологически не
исследовано. М. Д. Иоффе [1962,
1965], которая детально исследова-
ла семейство Тгосводепагасеае, счи-
тает, что это семейство находится
на низком уровне филогенического
развития, так как оно характери-
зуется первичными цитоэмбриоло-
гическими признаками: двухклеточ-
ными пыльцевыми зернами, ана-
тропной, крассинуцеллятной семя-
почкой, с двумя интегументами, на-
личием париетальных клеток, за-
родышевым мешком Ро[угопит-ти-
па, медленным развитием семени и
т. д. По мнению Бхандари [Bhan-
dari, 1967x], pox Trochodendron, 1o-
торый раньше относили к Марпо-
liaceae uuu Hamamelidaceae, cueny-
ет относить к самостоятельному се-
MeiitctBy — Trochodendraceae.
ПОРЯДОК 13.
CERCIDIPHYLLALES
СЕМЕЙСТВО
CERCIDIPHYLLACEAE
Основное
== 19,
Цветок однополый, растение дву-
дольное.
Пыльник — 4 микроспорапгия.
Стенка пыльника не изучена.
Тапетум секреторный, двуядер-
НЫЙ.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трехбороздные,
с тонкой сетчатой сэкзиной.
Семяпочка гемианатропная, с дву-
мя интегументами, крассинуцел-
лятная. Микропиле образуется обо-
ими интегументами. Сосудистый
пучок доходит до оспования нуцел-
луса. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Нариетальных клеток 4—5 рядов.
Тетрады макроспор линейные. За-
родышевый мешок формируется по
Ро]угопит-типу. Синергиды груше-
видные. Имеются три эфемерные
антиподы. Полярные ядра распола-
число хромосом — х =
гаются в халазальной части заро-
дышевого мешка и сливаются до
оплодотворения. Зародышевый ме-
ток после оплодотворения сильно
удлиняется.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
При первом делении первичного
ядра эпдосперма у Cercidiphyllum,
как и у Ттосводеп4гоп, образуются
двс неравные клетки: большая ми-
кропилярная ий маленькая халазаль-
ная. Последующие несколько деле-
ний поперечные.
Эмбриогенез Solanad-runa. Sapo-
ALUM относительно круппый, пря-
мой, бесхлорофилльный, двусемя-
длольпый, с хорошо развитыми по-
чечкой и корешком, с длинным ги-
покотилем.
Семя, плод — семя CO скудным
маслянистым эндоспермом, распо-
ложенным по периферии семени.
Зародыш занимает 3/4 объема се-
мени. Семенная кожура образуется
наружным интегументом. Плод —
однолистовка.
Гаусторий не найден.
Апомиксие не обнаружен.
Литература: ВПапдаг!, 1967c, 1971;
Corner, 1976; Пау15, 1966, 1970; Смир-
нова, 1977; Тахтаджян, 1966, 1970;
Yoffe, 41965.
В отношении цитоэмбриологии
семейство Cercidiphyllaceae сходно
c Trochodendraceae. Оба эти семей-
ства характеризуются одинаковым
основным числом хромосом (х =
== 19), секреторным тапетумом в
пыльниках, симультанным микро-
спорогенезом, двухклеточными трех-
бороздными пыльцевыми зерна-
ми. одноклеточным женским ар-
хеспорием, крассинуцеллятной се-
мяпочкой с двумя интегументами,
паристальной тканью, зародыше-
вым мешком Polygonum-tTuma, jese-
нием первичного ядра эндосперма
в халазальной части зародышевого
мешка, разрастанием зародышевого
мешка после оплодотворения и кле-
точным эндоспермом, первые не-
сколько делений которого проходят
поперечно. Это сходство особенно
своеобразно в развитии эндосперма,
и, возможно, указывает на близко-
родственные отношения между по-
рядками Тгосподеп4гаез и Cercidi-
43
Phyllales [Yoffe, 1965; Bhandari,
1967]. Оба вышеупомянутых поряд-
ка не только сходны по цитоэмбри-
ологии между собой, но и с поряд-
ком МаспоПа|ез, что, по-видимому,
указывает на их родство.
ПОРЯДОК 14.
EUPTELEALES
СЕМЕЙСТВО ЕОРТЕГЕАСЕАЕ
Основное число хромосом — х =
— 44.
‚ Цветок обосполый.
Пыльник — число
гиев равно 4. Степка микросно-
рангия имсет эпидермис, фиброз-
ный эплотеций, 2—3 средних слоя
и одночетырехъядерный секретор-
ный танетум. .
Мнкроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевые
зерна двухклеточные.
Семяночка висячая, анатропная, с
лвумя интегументами, крассипуцел-
лятпая. Микропиле образуется обои-
микроспоран-
ми интегументами. Семяпочек пе-
сколько — много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Нариестальная ткапь массивная.
Тетралы макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]угопиш-типу. Синергиды с фибрил-
лярными аппаратами. Иметотся три
антиподы.
Опыление, прохождение пыльце-
вой трубки, оплодотворение — нет
данных.
Эндоеперм целлюлярный. Первые
песколько делений поперечные, как
y Cercidiphyllum wu Trochodendron.
Эмбриогенез бо|апад-типа.
Семя, плод — семя с обпльным
маслянистым эндоспермом и ма-
леньким зародышем. Нлод состоит
из маленьких крылатых орешков, с
околоплодником.
Гаусторий не найден.
Апомикепе не обнаружен.
Другие явления не наблюдались.
Литература: Endress, 1969; Tax-
таджян, 1966, 1970.
Цитоэмбриологию
единственного
44
Eupteleaceae —
семейства порядка
Еир@еаез — довольно детально
изучил Эндресс {Endress, 1969]. По
ряду цитоэмбриологических при-
знаков, особенно по типу развития
и строения эндосперма Еираеа-
ceae, cxomHo c Trochodendraceae 1
Cercidiphyllaceae, aro, по-видимому,
указывает Ha близкородственные
отношения между порядками ‘Tro-
chondrales, Cercidiphyllales u Eupte-
cales.
ПОРЯДОК 15.
DIDYMELALES
Этот порядок с сего единственным
семейством П!Чушеасеае почти но
изучен в отношении цитоэмбрио-
логий, что не позволяет дать его
цитоэмбрпологическую — характери-
стику.
ПОРЯДОК 16.
HAMAMELIDALES
CEMEMCTBO HAMAMELIDACEAE
Основное чиело хромовом — x =
== 8, 12, 45.
Цветки обоеполые.
Пыльник — обычно 4, no y Exbu-
cklandia populnea yBa MEKpocnopan-
гия. Стенка микроспорангия имеет
эпидермис, Ффиброзный эндотеций,
1—4 эфемерных средних слоя н сек-
реторный, дву-, четырех-, — много-
ядерпый, одно-двурядный тапетум.
Нередко в клетках тапетума наблю-
даются слияние ядер и возникно-
венпе полиплоидных ядер. Имеют-
ся орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор тс-
траэдральные, реже крестообраз-
ные. Зрелые пыльцевые зерна двух-
клеточные, трех-четырехбороздные,
или трех-четырех- и пяти — сс-
миругатные, иногда и. многопоровые
с сетчатой экзиной.
Семяпочка висячая, анатропная,
с двумя питегументами, крассину-
целлятная. Микропиле зигзагооб-
разное; образуется обоими интету-
ментами. Сосудистый пучок дохо-
дит до халазы. Имеется гипостаза.
Семяпочек несколько (до 11).
Рис. 11. Мпикроспорангий и микроспорогепез у ЕхБаскКап а populnea
1, 2 — поперечные срезы пыльников на разных фазах развития (уп — участок пыль-
ника);
5 — полипограмма;
3 — одноядерные пыльцевые зерна;
4 — двухклеточное пыльцевое зерно:
6—9 — деление и слияние ядер в клетках тапетума [Качц!, Ka-
pil, 1974]; а — увеличенная часть пыльника из рис. 1, б — увеличенная часть пыль-
ника из рис. 2; э — эпидермис,
клетки микроспор, ср — средний ряд
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Цариетальная ткань массивная (до
5—10 рядов клеток). Тетрады мак-
роспор липейпые. Зародытевый ме-
шок формируется по Polygonum-
типу. Сивергиды грушевпдные. Три
антиноды либо эфемерные, либо со-
храняются до образования двух —
четырехклеточного эндосперма. Но-
лярные ядра сливаются до оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление у Маш ЧатфЪаг
ai, — эндотелий, т — тапетум, мкм — материнские
styraciflua. — Порогамия. Двойное
оплодотворение. Между опылением
и оплодотворением проходит дли-
тельный период времени (до 5—7
месяцев).
Эндоспермогенез целлюлярный,
нуклеарный. После оплодотворения
первичное ядро эндосперма мигри-
рует в халазальный конец зароды-
тшевого мешка и делится значитель-
но раньше зиготы.
Эмбриогенез Solanad-tuna. Заро-
дыш прямой, крупный, двусемя-
дольный, с хорошо развитыми по-
чечкой, гилокотилем и корешком с
45
Рис. 12. Макроспорогепез и развитие зародышевого мешка у ЕхБасКап@а
populnea
1 — анатропная семяпочка, 2—5 — макроспорогенез; 6—11 — дегенерация верхних
макроспор и развитие нижней макроспоры до четырехъядерной стадии; 12 — семи-
клеточный зародышевый мешок до оплодотворения; 13 — зародышевой мешок пос-
ле оплодотворения; и -— наружный интегумент, м — микропиле, мкм — материн-
ская клетка макроспоры, ви — внутренний интегумент, дм — дегенеративные мак-
роспоры; дс — дегенеративная синергида, з — зигота, ак — антиполиальные клетки,
вя — вторичное ядро, я — яйцеклетка, с — синергиды [Кай1, Кари, 1974]. .
46
4
Рис. 13. Эндоспермогенез и эмбриогенез у ЕхБасНап а роришеа
1 — зародышевый мешок со вторичным ядром; 2, 3 — зародышевые мешки с зиго-
тами, а также с двух- и четырехъядерными эндоспермами; 4 — целлюлярный эндо-
сперм окружает двусемядольный зародыш; 5—10 — стадии развития зародыша от
шарообразной до двусемядольной; а—г — поперечные разрезы зародыша на уров-
нях а—г, обозначенных на рис. 4 [Кац, Карй, 1974]; 0 — клетки эндосперма с за-
пасающими веществами; с — синергизы, я — яйцеклетка, ак — антиподиальные
клетки, з — зигота, вя — вторичные ядра, чк — чехлик корня, см — семядоли, э —
эндосперм
47
чехликом,
рон.
Семя, плод — семя с массивным
эндоспермом. Зародыш занимает
3/4 общего объема семени. Перифе-
рия семени заполнена эндоспермом.
В клетках эндосперма содержатся
масло, алейрон и кристаллы окса-
лата кальция. Плод — коробочка.
Перикарпий плода твердый, экзо-
карпий мясистый.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; Kapil, Kaul, 1969, 1972; Kaul,
1969a, b; Kaul, Kapil, 1974; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970.
содержат масло и алей-
СЕМЕЙСТВО РГАТАМАСЕАЕ
Основное число хромосом — х =
= 7,
Цветки однополые.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и секреторный, дву-
четырехъядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
изобилатеральные. Зрелые пыльце-
вые зерна двухклеточные, трехбо-
роздные.
Семяпочка ортотропная, с двумя
интегументами, крассипуцеллятная.
Микропиле образуется внутренним
интегументом. Семяножка корот-
кая. Семяпочек 1—2.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париестальная ткань массивная. За-
родышевый мешок образустся по
Ро1угопит-типу. Имеются три эфе-
мерные антиподы. .
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Оплодотворение у Р]абапиз проис-
ходит через три недели после опы-
ления.
дндоснермогенез пуклеарцый.
Эмбриогеиез зародынг прямой,
двусемядольцый, бесхлорофилльный.
Семя, плод — семя с тонким сло-
ем эндосперма, —сохрапяющимся
лишь на периферии семепи. Заро-
48
дыш заполняет семя но всей cro
длине. Семядоли выполняют запа-
сающую функцию. Плод — обычно
многоорешек, редко почти многоли-
стовка.
Гаусторий нё найден.
Апомиксие не обнаружен.
Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; Гусейнова, 1976; Капинос, Гу-
сейнова. 1978; ЗебпагЁ, 1931; Смир-
пова, 1977; Тахтаджян, 1966, 1970.
Цитоэмбриология семейства Му-
гофаплпасеае, которое А. Л. Тахта-
джян [1970] также относит к ио-
рядку НашатейЧа]ез, почти не ис-
следована. По цитоэмбриологичс-
ским признакам семейства Ната-
melidaceae wu Р]абапасеае обнару-
кивают сходные черты, что не про-
тиворечит отнесению их к одному
и тому же порядку. Вместе с тем
Наташе бдасеае имеет сходные ци-
тоэмбриологические признаки с Тго-
chodendraceae.
ПОРЯДОК 17.
EUCOMMIALES
СЕМЕЙСТВО ЕОСОММТАСЕАЕ
Основное чиело хромосом — х =
== 17,
Цветок однополый, растение дву-
JJomHOC.
Пыльник — 4 микроспорангия.
Стенка мякросиорангия имеет эпи-
дермпс, двурядный фиброзный эн-
дотеций, 1—2 эфемерных средних
слоя и секротный дву- четырех-
шестиядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор то-
траэдральные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
нопоровые, с толстой двуслойной
экзиной.
Семяпочка анатропная, с одним
интегументом, слабокрассинуцсл-
лятная. Имеется нуцеллярный кол-
пачок. Семяпочек 2.
Макроспорогенез и макрогамего-
генез — археспорий — многоклеточ-
ный. Париетальных клеток 2—3 ря-
да. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Polygonum-tuny. Синергиды груше-
видные. Имеются три антиподы.
Полярные ядра сливаются До онло-
дотворения.
Опыление, прохождение пыльще-
вой трубки п оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип не определен.
Зародыш прямой, крупный, диффе-
ренцированный, двусемядольный, с
хорошо развитым корешком.
Семя, плод — ссмя с бедным эн-
доспермом, который остается в се-
мепи лишь в области корешка и на
сомядолях. Зародыш заполпяет по-
чти весь объем семени. Семядоли
сразу после прорастания семени
выполняют ассимиляцпоииую фуи-
кцию. Плод нераскрываюнишся,
сплюснутый и крылатый, тонкоко-
жистый, односемянной.
Гаусторий но обнаружен.
Апомикеие не найден.
Другие явления не наблюдались.
Литература: Сотпег, 1976; Davis,
1966; Тан-Си-хуа, 1962; Тахтаджян,
1966, 1970.
Систематически напболее чаето
сближают Eucommiaceae c Ulma-
ceac. Однако цитоэмбриологические
признаки этих семейств в значи-
тельной степени отличаются друг от
друга. В то время как Висотииа-
сеае имсот двухклеточный мужской
гамстофит, анатропную слабокрас-
синуцеллятную семяпочку с одним
интегументом, многоклеточный жен-
ский археспорий, Ро[ухопит-тип
зародышевого мешка, целлюляр-
ный эндосперм и прямой зародыш,
Ulmaceae имеет двух-трехклеточ-
пый мужекой гаметофит, анатроп-
ную крассинуцеллятную семяпоч-
ку с двумя интегументами, одно-
клеточный женский — археспорий,
Ро!угопит-, Адоха- и Пгиза-типы
зародышевых мешков, целлюляр-
ный эндосперм, согнутый зародыш.
Однако оба эти семейства исследо-
ваны в отношении цитоэмбриологии
пока не достаточно полно, что не
позволяет сделать более точные вы-
воды о взаимоотношениях этих се-
мейств между собой и с другими
семействами.
4 В. А, Поддубная-Арнольди
ПОРЯДОК 18.
URTICALES
СЕМЕЙСТВО ОГМАСЕАЕ
Основное чиело хромосом — & =
== 10, 11, 14.
Цветок однополый или реже обое-
полый.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2
или 3 эфемерных средних слоя и
секреторный дву- четырехъядерный
_- тапетум.
Микроспорогенез и микрогамето-
генсз — образование микроспор си-
мультанное. 'Тетрады микроспор тс-
зраэдральные, крестообразные. Зре-
лые пыльцевые зерна двух или
трехклеточные. Обычно двух-, трех-
пяти- шести-, реже многопоровые,
с бородавчатой или бугорчатой сэк-
зиной. Спермпи-клетки бедиоплаз-
менные.
Семяпочка одна, анатронпная-гс-
мпанатропная, с двумя интегумен-
тами, крассинуцеллятная. Микро-
пиле образустся только внутрен-
ним или обоими интегументами.
Имеются нуцеллярный колпачок,
гипостаза. фуникулярный обтура-
тор. Сосудистый пучок доходит до
халазы.
Макроспорогеиоз и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 2—5 рядов.
Тетрады макроспор линейные. За-
родышевые мешки образуются по
Polygonum-, Адоха- и Огизза-ти-
пам. Синергиды грушевидные. Три
и более (до 10) антипод. Полярные
ядра сливаются перед оплодотво-
рением.
Опыление, прохождение пыльцс-
вых трубок, оплодотворение — пс-
рекрестное. Халазогамия, мезога-
мия. Двойпос оплодотворение.
Эндоспермогенез нуклеарный, со-
держащий масло.
Эмбриогенез Solanad-, Onagrad-
типов. Зародыш прямой или согну-
Th, бесхлорофилльный, хорошо
дифференцированный (развиты сс-
мядоли, гипокотиль и корешок),
имеются масло и алейрон.
Семя, плод — у представителей
семейства Ulmaceae внутреннее
строение семени очень разнообраз-
но. Кроме семян с прямым узким
49
зародышем, окруженным CO всех
сторон эндоспермом, встречаются
семена, в которых изогнутый заро-
дыш располагается по периферии
семени, занимая 3/4 его перимет-
ра, а эндосперм сохраняется лишь
в центре семени. Вместе с тем
встречаются семена, в которых
массивный изогнутый зародыш 3a-
полняет все семя, а эндосперм пол-
ностью израсходовался в процессе
эмбриогенеза; семядоли у этого ти-
па выполняют запасающую Ффунк-
цию и своими размерами превыша-
ют гипокотиль и корешок. В отли-
чие от этого типа семян встречают-
ся семена, в которых прямой заро-
дыш заполняет все семя, эндосперм
весь израсходовалсея, массивные за-
пасающие семядоли скрывают ос-
тальные части зародыша, с едва
выступающим из них корешком.
Плод — односемянный орех, часто
крылатый или костянка с тонким
мезокарпием.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления — обнаружена
полиэмбриония.
Литература: Corner, 1976; Crété
et al, 1966; Davis, 1966; C. Hasa-
шин, 41951; Shattuck, 1905; Taxta-
джян, 1966, 1970; Schnarf, 1931.
СЕМЕЙСТВО МОВАСЕАЕ
Основное число хромосом — 2 =
= 40, 42—14.
Цветки мелкие, невзрачные, анс-
мофильные или энтомофильные, од-
нополые; растения однодомные или
двудомные.
Пыльник — 4 — микроспоравгия.
Стенка пыльника образуется по од-
нодольному типу и имеет эпидер-
мис, фиброзный эпдотеций, обычно
2, изредка 3—4 эфемерных средних
слоя и секреторный, двух-, четы-
рехъядерный двурядный тапетум.
Ядра в клетках тапетума часто
сливаются и становятся полипло-
идными.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультаниое. Тетрады — микроспор
обычно тотраэдральные, крестооб-
разные, изредка изобилатеральные.
Зрелые пыльцевые зорпа двухкле-
точные, двух-, четырех-, роже пя-
ти — многопоровые, с крахмальны-
50
Семяпочка одна, гемианатропная,
анатропная с мя интегумента-
ми, крассинуцейдлятная. Микропиле
образуется внутренним интегумен-
том. Имеются нуцеллярный кол-
пачок и гипостаза. Сосудистый пу-
чок доходит до халазы.
Макроспорогенез и макрогамето-
генез — археспорий одно-, реже
многоклеточный. Париетальных кле-
ток 5—8 рядов. Тетрады макроспор
линейные, Г-образные. Зародыше-
вые мешки образуются no Polygo-
пиш- или АШит-типам. Синергиды
грушевидные. Имеются три и 60-
лее (до 8) клеток антипод. У мно-
гих представителей Могасеае анти-
поды эфемерные, у некоторых (Е!
CUS) они сохраняются довольно до-
лго. Полярные ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
большинство родов опыляется при
помощи ветра. У Е1сиз опыление
производится при помощи насеко-
мых из группы Нутепорега. У Е!-
cus carica u F. afganistanica napa-
ду с амфимиксисом обнаружен апо-
MUKCHC.
Оплодотворение при амфимикси-
ce y Ficus carica uw F. afganistanica
происходит через 27—30 ч после
опыления. Через 16—18 дней после
опыления зародыш достигает шаро-
видной стадии, а эндосперм ставо-
вится 300—350-клеточным.
Эндоспермогенез нуклеарный, со-
держащий масло. У Е. самса и Е.
afganistanica при апомиксисе BTO-
ричное ядро зародышевого мешка
становится ядром эндосперма, без.
оплодотворения — автоматично. До
поздних стадий эмбриогенеза эндо-
сперм остается в виде свободных
ми зернами и rea экзиной.
у
ядер, обнаруживающих высокую
степень полиплоидии.
Эмбриогенез Азега4-типа. Заро-
дыш обычно двух-, иногда трехсе-
мядольный, прямой или согнутый,
большой или маленький, хлорофил-
лоносный или бесхлорофилльный.
Подвесок короткий. Зародыш имеет
масло и алейрон.
Семя, плод — структура семян
этого семейства разнообразна. Ови
либо с эндоспермом, либо без него.
Зародыш с мясистыми запасающи-
ми семядолями лежит в центре се-
мени, занимая почти весь его объ-
ем, за исключением остатков эндо-
сперма лишь на периферии семени.
Зародыши имеют’ узкие длинные
семядоли, которые‘ превышают об-
щую длину гипокотиля вместе с
корешком. Семенная кожура у Е!
cus, Morus и Втоиззопейа образу-
ется наружным слоем внешнего
интегумента и внутренним слоем
внутреннего интегумента. У Аг®о-
сагриз семенная кожура образуется
внешним интегументом. Плоды ко-
стянковидные, в соплодиях, часто
более или менее сросшиеся между
собой.
Гауеторий не найден. .
Апомиксие —у F. carica u F. af-
ganistanica oOnapyKensl нередуци-
рованный партеногенез, анпогаме-
тия, нуцеллярная эмбриопия и гте-
неративная апоспория.
Другие явления — обнаружены
полиэмбриопия, партенокарпия, ге-
теростилия и вивипария.
Литература: Вгееу, Сгапе, 1965;
Corner, 1976; Crété, 1964; Davis,
4966; Crover, 1969; Johri, Konar,
1956; Kaur, 1967c, 41970c; Kumar,
1978; Mukherjce, 1960; Скокова,
1966; Pomanopa, 1979; Scharf, 1934;
Тахтаджян, 1966, 1970; Замотайлов,
1955.
СЕМЕЙСТВО САММАВАСЕЛЕ
Основное
== 8, 10.
Цветок раздельнополый, растение
однодомное или двудомное.
Ныльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, 2—3 эфемерных
средних слоя и секреторный дву-
ядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
число хромосом — 2 —
траэдральные. Зрелые пыльцевые
зерна двухклеточные, обычно трех-,
реже двух-, четырех-, шестипоро-
вые, с тонкой экзиной.
Семяпочка одна, анатропная, с
двумя интегументами, крассину-
целлятная. Микропиле образуется
внутренним интегументом. Имеется
нуцеллярный ’ колпачок. — Сосуди-
стый пучок доходит до халазы.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 7 рядов. Те-
4*
трады макроспор линейные. Заро-
дышевый мешок Ро[узопит-типа.
Синергиды грушевидные. Имеются
три эфемерные клетки антипод.
Слияние полярных ядер происхо-
дит перед оплодотворением.
Опыление, прохождение пыльцс-
вой трубки и оплодотворение — пе-
екрестное. Порогамия. У Canna-
is saliva двойное оплодотворение
премитотического типа; происходит
оно через 12 ч после опыления.
Эндоспермогенез нуклеарный.
Позднее эндосперм становится кле-
точным, за исключением эндоспер-
мального гаустория, который до
-конца своего существования оста-
ется ценоцитным. Процесс образо-
вапия в ондосперме клеточных пе-
регородок начинается в микропп-
лярной зоне зародышевого мешка
около зародыша, а затем продвига-
ется в халазальную его зову.
Эмбриогенез Опартад-типа. Заро-
дыш еогнутый, двусемядольный,
подвесок двух-четырехклеточный.
Семя, плод — семя с масляни-
стым эндоспермом. Зародыш распо-
ложен в центре семени и со всех
сторон окружен эндоспермом. Дли-
на семядолей превышает общую
длину гипокотиля вместе с кореп-
ком. Плод — орех, покрытый оста-
ющейся чашечкой.
Гаусторий зародышевого мешка
и эндоспермальный (халазальный).
Апомиксисе пе обнаружен.
Другие явления — обнаружена
полиэмбриония у СаппаБ$.
Литература: Corner, 1976; Davis,
1966; Мовап Ваш, 1964; Поддубная-
Арнольди, 1936; Ram Manasi, 1960;
Savelli, 1961; Scharf, 1931; Смирно-
ва, 1977; Сорока, 1975, 1977; Copo-
ка, Жатов, 1971; Тахтаджян, 1966,
1970; Цингер, 189ба, 6.
СЕМЕЙСТВО ОВТТСАСЕАЕ
Основные
= 10—43.
Цветки очень мелкие, почти всо-
гда однополые, но изредка обоепо-
лые (например у Рараата).
Пыльник — нет данных.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные или крестообразные.
Зрелые пыльцевые зерна двухкле-
точные. Обычно двух-, трех-, редко
51
числа хромогом — х =
четырех — шести или многопоровые
с тонкой шиповатой экзинои.
Семяпочка одна — ортотроппая..
анатропная — гемианатропная, с
двумя интегументами, крассину-
целлятная. Микропиле образуется
одним впутренним или обоими ин-
тегументами. Имеются нуцелляр-
ный колпачок и гипостаза.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Нариетальных клеток 2—6 рядов.
Тетрады макроспор линейные. За-
родышевый мешок Ро]угопит-типа.
Синергиды грушевидные. Имеются
три и более клеток антипод. По-
лярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез Азогад-типа. Заро-
дыш бесхлорофилльный, прямой,
двусемядольный, с хорошо разви-
тыми почечкой, гипокотилем и ко-
решком; содержит масло и алеи-
рон; подвесок короткий.
Семя, плод — семя с маслянистым
эндоспермом. Зародыш. занимает
3/4 общего объема семени. Пери-
ферия семени заполнена эндоспер-
Mom. Плод — маленький орех или
костянка, часто с неопадающим
околоцветником.
Гаусеторий эндоспермальный вха-
лазальной части зародышевого мое-
шка, короткий.
Anomukene — нуцеллярная оэмбри-
ония, апогаметия, генеративная
апоспория и — нередуцированный
партепогенез.
Другие явления — полиэмбрио-
uua y Cannabis.
Jiureparypa: Corner, 1976; Davis,
1966; Narayanan, 1968; Schnarf, 1934;
Soueges, 1920; Strasburger, 1910;
Тахтаджян, 1966, 1970.
Предетавители порядка Urticales
имеют много сходных цитоэмбрио-
логических признаков, что, по-види-
мому, указывает на естественность
этого порядка. Наиболее специали-
зировавным и подвинутым являет-
ся семейство ОтгИсасеае, поскольку
оно характеризуется наличием раз-
ных форм апомиксиса.
52
ПОРЯДОК
BARBEYALES
19.
СЕМЕЙСТВО 'ВАВВЕУАСЕАЕ
f
Ilopanox Barbeyales c его един-
ственным семейством цитоэмбрнио-
логически не изучен.
ПОРЯДОК 20.
CASUARINALES
СЕМЕЙСТВО САЗОАВТМАСЕАЕ
Основное число хромосом — х =
—= 11—13.
Цветок одпополый, анемофиль-
ный, растепие однодомноес, реже
двудомпое.
Пыльнник — стенка микроспорап-
rid имест опидермис. фиброзный
эндотедий, офемерный, среднпя
слой и секреторный двуядерный
тапестум. 4 мпкроспорангия.
Микроепорогенез и микрогамето-
генез — тетрады микроспор тетра-
одральные. Зрелые ныльцевые зерпа
двухклеточные (двух-}, трех-(ия-
ти-} поровые, с гладной экзиной.
Спермии — клетки.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная
Микропиле образуется внутренним
интегументом. Семяпочен несколько.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный. Париетальная ткань массив-
ная. Тетрады макроспор линейные.
Зародышевый мешок Polygonum- _
типа. В одной и той же семяпочке
образуется много зародышевых
мешков. Имеются три эфемерные
клетки антипод. Полярные ядра
сливаются в момент оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворенне — ха-
лазогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный. Эн-
досперм имеет алейрон.
Эмбриогенез Опартад-тпна (?).
Зародыш прямой, крупный, двусе-
мядольный, содержит алейрон. Пол-
весок с крупной базальной клет-
цой.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет всо семя.
Массивные запасающие семядоли
скрывают остальные части зароды-
та; конец корешка лишь едва вы-
ступает из них. Плод с кожистым
крылатым выростом, односеменной,
ореховидный, вскрывающийся, как
коробочка, двумя створками.
Гаусторий зародышевого мешка.
Апомиксие не обнаружен.
Другие явления не наблюдались.
Литература: Сотпег, 1976; Davis,
1966; Schnarf, 1934; Shandra, 1969;
Cmupnorna, 1977; Swamy, 1967a; Tax-
таджян, 1966, 1970. *
Порядок Сазиаг!а[о$ с едипетвен-
ным сего семейством одпими иселе-
дователями рассматривается как
примитивный, другими — как реду-
цировавный и подвинутый. Пеко-
торые исследователи относят Сази-
arinales « Protangiospermae [Swa-
my, 1967a]. По паличию халазога-
мии Сазпаттасеае сходно с Веща-
ссае и УТифапдасеае, относящихся
к порядкам Ве аз ип Juglandales.
ПОРЯДОК 21.
FAGALES
СЕМЕЙСТВО FAGACEAE
Осневное число хромосом — х =
= 6, 11—13
Цветки обычно раздельнополыс,
иногда обосполые.
Пыльник — 2 микроспорангия.
Стенка микроспорангия пмеет эпи-
дермис, фиброзный эндотеций, один
средний слой.
Тапетум секреторный,
НЫЙ.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультавпое. Тетрады микроспор те-
траэдральные. Зрелые ныльцевые
зерна двухклеточные, обычно трех-
борозднопоровые, реже с пятью
(четырьмя-), семью бороздновидны-
ми апортурами, содержат крахмал
и масло. Жизнеспособность пыльцы
у лещины (Corylus) сохраняется
до трех месяцев.
Семяпочка анатропная,
интегументами,
двуядер-
с двумя
крассинуцеллятная.
Имеются нуцеллярный колпачок и
гипостазы. Семяпочек обычно 6, ре-
ке 12—18. Сосудистый пучок дохо-
дит до халазы, где он разветвляется.
Макроспорогенез и макрогамето-
генез — одно-, многоклеточный. Па-
риетальная ткань массивная. Тетра-
ды макроспор линейные, Г-образ-
ные. Зародышевый мешок образу-
ется по Роусопиат-типу. Синерги-
ды с фибриллярными аппаратами.
Имеются три и более клеток анти-
под. Полярпые ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление. Порогамия.
Энпдоепермогенез нуклеарный. По-
CTCHCHNO стаповитея клеточным, за
исключением халазальпой части за-
родышевого мешка, в которой па-
ходится большая вакуоль.
Эмбриогенез Опаотад-типа. dapo-
дыш большой. прямой, двусемя-
дольный. бесхлорофилльный, содер-
жит крахмал, масло, алейрон, дру-
зы оксалата кальция. Подвесок ко-
роткий.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Массивные запасающие семядоли
скрывают остальные части зароды-
ша, пз них едва выступает лишь
Kouern rKopenma. Плод — желудь, с
кожистым пли твердым околоплод-
пиком, заключенпый обычно в де-
ревянистую плюску.
Гаусторий не найден.
Апомиксие не обнаружен.
Другие явления — обнаружена
нполиэмбриония у Quercus alba u
Q. rubra.
Литература: Акимова, 1972, 1976;
Corner, 1976; Davis, 1966; Brown,
Мосепзеп, 1972; Кавецкая, 1978; Мо-
розова, 1972; Schnarf, 1931; Смир-
Hopa, 1977; Stairs, 1964; Тахтаджян,
1966, 1970; Тышкевич, Ковшова,
1975а, 6, 1976; Ткаченко, 1969.
По неготорым цитоэмбриологиче-
ским признакам Fagaceae сходно с
Вей]асеае. Однако по числу микро-
спорангиев, степени развития па-
риетальной ткапи, типам прохож-
дения пыльцевых трубок и эмбрио-
генеза они отличаются друг от дру-
га, что оправдывает отнесение этих
семейств к разным порядкам.
53
ПОРЯДОК 22.
BETULALES
СЕМЕЙСТВО ВЕТОГАСЕАЕ
Основное чиело хромосом — 5 =
= 10, 14.
Цветок однополый, растения од-
нодомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, 2—3 средних
слоя. Тапетум секреторпый, 1—3-
ядерный.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трех — семипо-
ровые.
Семяпочка анатропная, с одним
интегументом, крассинуцеллятная.
Семяпочек 3—6.
Макроспорогенез и макрогамето-
генез — археспорий — одно — много-
клеточный. Париетальпых клеток
1—4 ряда. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Роугопит-типу. Синер-
гиды грушевидные. Имеются три
эфемерные или долго сохраняющие-
ся клетки антипод. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление. Халазогамия.
Двойное оплодотворение. У Веа
аа между опылением и оплодотво-
рением проходит 4 недели.
Эндоспермогенез нуклсарный, со-
держащий масло.
Эмбриогенез Азегад-типа. Заро-
дыш большой, прямой, двусемя-
дольный, бесхлорофилльный, содер-
жит масло и алейрон. Семядоли
выполняют ассимиляционную фун-
КЦИЮ.
Семя, плод — семя почти без эн-
досперма. Зародыш заполняет поч-
ти весь объем семени. Эндосперм в
виде тонких пленок остается лишь
в области корешка и на семядолях.
Плод — орех, часто с двумя пере-
пончатыми крыльями.
Гаусторий не обнаружен.
Anomukene — нередуцированный
партеногенез и нуцеллярная эмбри-
ония у Ашиз rugosa.
Другие явления — полиэмбриония
y A. rugosa. Партенокарпия.
Литература: Богданов,
54
Ступов,
1
1970; Corner, 1976: Davis, 1966;
С. Навашин, 1954; Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
1970.
ПОРЯДОК 23.
BALA NOPALES
СЕМЕЙСТВО ВАГАМОРАСЕАЕ
Порядок Ва]апопа!ез с его един-
ственным семейством Balanopaceae
цитоэмбриологически не изучен.
ПОРЯДОК 24.
MYRICALES
СЕМЕЙСТВО МУВТСАСЕАЕ
Основное число хромосом — х =
ыы
—
Цветок обычно однополый, анс-
мофильный.
Пыльник — нет данных.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трехпоровые, с
аспидатными порами и гладкой эт-
зиной.
Семяпочка — ортотропная, с Оол-
ним интегументом, крассинуцел-
лятная. Семяпочек 4—6.
Макроепорогенез и макрогамето-
генез — археспорий одно-двуклеточ-
ный. Парнетальных клеток 4—6 ря-
дов. Тетрады макроспор линейные.
Зародышевый мешок формируется
по Ро]угопит-типу. Синергиды гру-
шевидные. Имеются три эфемер-
ные антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. У видов Мумса между
опылением и оплодотворением про-
ходит 4—5 недель. Двойное оплодо-
творение.
Эндоспермогенез нуклеарный. Об-
разование перегородок между яд-
рами начинается с микропилярной
части зародышевого мешка.
Эмбриогенез — тип не определен.
Зародыш крупный, прямой, диффе-
ренцированный, содержащий крах-
мал. масло и алейрон.
Семя, плод — семя почти без эн-
досперма. Зародыш заполняет по-
чти все семя. Остатки эндосперма
имеются лишь в виде тонких пле-
нок в области корешка и на семя-
долях. Массивные запасающие се-
мядоли скрывают остальные части
зародыша, лишь кончик корешка
едва выступает из них. Эндосперм
содержит масло и алейрон. Плоды
костянковидные.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1931; Смирнова, 1977;
Тахтаджян, 1966, 1970.
ПОРЯДОК 25.
JUGLANDALES
CEMEMCTBO JUGLANDACEAE
Основное число хромосом — 2 =
== 16, 47.
Цветок однополый, растения обыч-
но однодомные, реже двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—4
эфемерных средних слоя и секре-
торный многоядерный, иногда дву-
рядный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор то-
траэдральные, изобилатеральные.
Перед раскрытием пыльников пыль-
цевые зерна двухклеточные; после
освобождения пыльцевых зерен из
пыльников они становятся трехкле-
точными. Спородерма пыльцевых
зерен* трех- четырех — многопоро-
вая. Пыльцевые зерна содержат
капли масла. Спермии-клетки мно-
гоплазменные.
Семяпочка ортотропная, анатроп-
ная с одним интегументом, красси-
нуцеллятная. Имеется нуцелляр-
ный колпачок. Семяпочек одна-че-
тыре.
Макроспорогенез и макрогамето-
генез — археспорий одно-, дву-, мно-
гоклеточный. Париетальных клеток
3—7 рядов. Тетрады макроспор ли-
нейные, Г-образные. Зародышевый
мешок образуется по Polygonum-
типу. Синергиды с крючкообразны-
ми выростами. Имеются три скоро
дегенерирующие антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Халазогамия. Двойное
рение.
Эндоепермогенез нуклеарный. Чис-
ло свободных ядер эндосперма до-
стигает 1000—1500, прежде чем зи-
тота начнет делиться. Позднее меж-
ду ядрами образуются клеточные
перегородки и эндосперм становит-
ся клеточным.
Эмбриогенез Asterad-runa. Заро-
дыш прямой, крупный, бесхлоро-
филльный, содержащий масло и
алейрон. Подвесок одноклеточный.
У Сагуа ПШтоеп51$ зигота начинает!
делиться через 1,5—2 месяца после!
оплодотворения. Полного созрева-
ния плод достигает через 5—6 ме-'
сяцев после опыления.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Массивные запасающие семядоли
скрывают остальные части зароды-
ша, из них едва выступает лишь
кончик корешка. Плод — орех или
костянкообразный.
Гаусторий не обнаружен.
Апомиксие не доказан, но пред-
полагается у видов Juglans.
Другие явления — полиэмбрио-
Hus y Juglans niger.
оплодотво-
Литература: Bouillot, 41968, 1969;
Bouman et al., 1976; Davis, 1966:
Гусейпова, 1977; Дгебуадзе, 1980;
Нее, 1972; Кавецкая, 1964; Солн-
цева, Дгебуадзе, 1974; Schnarf, 1931;
Смирнова, 1977; Тахтаджян, 1966,
1970; Щепотьев, Герасименко, 1970.
Семейство Rhoipteleaceae, кото-
рое по системе А. Л. Тахтаджяна
[1970] также относится к порядку
Juglandales, цитоэмбриологически
не исследовано. По своим цитоэм-
бриологическим признакам Juglan-
daceae во многом сходно с ГРаса-
ceae и ВеШасеае. Поэтому распо-
ложение порядков Juglandales, Fa-
gales u Вей|а]ез вблизи друг дру-
га подтверждается данными цито-
эмбриологии.
ПОРЯДОК 26.
LEIT NERIALES
CEMEMCTBO LEITNERIACEAE
Основное число хромосом — z =
= 16, 17.
55
Цветок однополый, растения дву-
домные.
Пыльник — 4 микроспоравгия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эпдотеций, один
эфемерный средний слой и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сп-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трех- четырехборозднопо-
ровые, с толетой экзиной.
Семяпочка одна, амфитропная, с
двумя интегументами, крассину-
целлятная. Микропиле образуется
обоими интегументами. Имеется ну-
целлярный колпачок.
Макроспорогенез и макрогамето-
генез — архсспорий одноклеточный.
Париетальных клеток 9 рядов. Те-
трады макроспор липейные. Заро-
дышевый мешок образуется по Ро-
]усопит-типу. Синергиды треуголь-
ные, без вакуолей. Имеются три
эфемерные антиподы.
Опыление, прохох:дение пыльце-
вой трубки и оплодотворение — 110-
рогамия.
Эндоспермогенез нуклеарный, эн-
досперм содержит крахмал.
Змбриогенез — тип, не определен.
Зародыш прямой, массивный, дпф-
ференцированный, двусемядольный,
содержащий масло и алейроня. Под-
весок широкий.
Семя, плод — семя © топким сло-
ем эндосперма лишь в области ко-
решка и семядолей. Зародыш за-
полняет почти весь объем ссмени.
Семядоли широкие, листовидные,
выполняют ассимиляционаую Ффун-
кцию сразу же после прорэстания.
Плод костянкообразный.
Гаусторий не найден.
Апомикснс пе обнаружен.
Доугие явления не отмечевы.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1934; Тахтаджян,
1966, 1970.
По некоторым цитоэмбриологиче-
ским признакам Т.еЦпег1асеае сход-
но с Уифапдасеае. Оба сомейства
характеризуются маличием 4 мп-
‹росиорангиев: фиброзного эндоте-
ция, секреториого тапетума, си-
мультаниого микроспорогенеза, двух-
хлеточных пыльцевых зерен, пари-
стальных клеток, зародышевого
мешка Роугопит-типа, нуклеар-
ного эндосперма, крупного прямого
дифференцированного зародыша.
ПОДКЛАСС О. САКУОРНУГГГЛАЕ
НАДПОРЯДОК ТУ.
CARYOPHYLLA NAE
ПОРЯДОК 27.
CARYOPHYLLALES
CEMENCTBO PHYTOLACCACEAE
Основное число хромосом — 5 =
Цветок обоеполый или раздель-
нополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу. Эндотеций
фиброзный. Тапетум секреторный,
дву-, многоядерный. .
Микроспорогенез и микрогамето-
генез — образование микроспор си-
56
мультапное Тетрады тетраэдраль-
ные, кростообразные. Зрелые пыль-
цевые зерна трехклеточные, трех-
бороздные, реже четырех-, много-
бороздные, пли многопоровые.
Семяпочка анакампилотропная,
амфитотроцная, с двумя интегумен-
тами, крассипуцеллятная. Имеется
пуцеллярный клювик, фуникуляр-
ный обтуратор и ариллус. Микро-
пиле образуется внутренним ипте-
гументом. Семяпочек 14—16.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток два ряда.
Тетрады макроспор линейные, Иног-
да вместо тетрады образуется триа-
да макроспор, в результате того
что верхняя клетка диады макро-
спор не делится. Зародышевый ме-
шок образуется to Polygonum-tu-
пу, содержит крахмальные зерна.
Синергады грушевидные; имеются
три клетки антипод, дегенерирую-
щие вскоре после оплодотворения.
Нолярвые ядра сливаются после
оплодотворения одного из ядер.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — по-
рогамия.
Эндоспермогенез нуклеарный.
Поздвее эндосперм становится кле-
точным во всем зародышевом мо-
шке.
Эмбриогенез Caryophyllad- или
Onagrad-runa. Зародыш бесхлоро-
филльный, согнутый, подковообраз-
ный, семядоли складчатые. Подве-
сок Узкий и длинный.
Семя, плод — семя с периспермом,
содержащим крахмал. Зародыш 3a-
нимает весь периметр семени, при
этом семядоли и корешок почти
смыкаются. Семядоли по размеру
гораздо меньше гипокотиля и ко-
решка. Перисперм занимает центр
семени. Семенная кожура образует-
ся из внешнего эпидермиса, внеш-
него интегумента. Плоды — много-
орешки, локулицидные коробочки,
орехи или ягодообразные. В стен-
ках плода и в паренхимных клет-
ках внешнего интегумента содер-
жатся кристаллы оксалата кальция.
Гаусторий не пайдси.
Апомиксие ле обпаружен.
Другие лвлепия ие наблюдались.
Литература: Corner, 19760; Davis,
1966; Карагьозова, 1969; Sclinarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО МУСТАСТМАСЕАЕ
Основное число хромосом — т =
— 10, 43, 47, 29.
Цветок обычно обоеполый, реже
одпополый.
Пыльник — 4 микроспорапгия.
Стенка микроспорапгия образуется
по базальному типу и имост эин-
дермис, Фиброзный эндотеций, 2
средних слоя и секреторпый дву-
ядерный тапетум.
Микроепорогенез н микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
Траэдральные, изобилатеральпые.
Зрелые пыльцевые зерна трехкле-
точные, трехбороздные, четырех-,
многобороздные или многопоровые.
Семяпочка одна, гемианатропная-
анакампилотропная, с одним или
двумя интегумситами, крассинуцел-
лятная. Микропиле образуется внут-
ренним интегументом. Имеется ну-
целлярный колпачок.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 14 рядов. Те-
трады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
lygonum-tuny. После оплодотвоуе-
ния зародышевый мешок сильно
удлиняется и изгибается. Синерги-
ды с крючкообразными выростами.
Три и более антиподы дегснериру-
ют вскоре после оплодотворения.
Опыление, прохождение пыльще-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение, премито-
тического tuna. Y Mirabilis jalapa
развитие зиготы и эндосперма на-
чинастся через 18—20 ч после опы-
ления, а через 15 суток возникает
зрелый зародыш.
Эндоспермогенез нуклеарный, но
затем он становится целлюлярным,
за исключением халазальной части,
где остаются свободные ядра.
Эмбриогенез Азетад-типа. Заро-
дыш хлорофиллоносный, прямой
или согнутый, двусемядольный,
имеет масло и алойрон. Подвесок
массивный, по короткий.
Семя, плод — семя без эндоспер-
Ma, по с периспермом, содержащим
крахмал. Зародыит занимает весь
периметр семепи, при этом семядо-
ли и корешок почти смыкаются
концами. Перисперм заполняет центр
семени. Плод — орешек.
Гаусторий не пайден.
Апомикеие нс обнаружен.
Другие явления — полнэмбриония
и клейстогамия.
Литература: Corner, 1976; Davis,
1966; Дьякова, 1962; Kapil, Vani,
1966; Schnarf, 1934; Тахтаджян,
1966, 1970; Устпнова и др., 1964.
СЕМЕЙСТВО МОГГОСТМАСЕАЕ
Основное число хромосом — х =
— `.
обычно обоеполый, реже
Цветок
однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет энпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и сокре-
торный, дву-, трех-, многоядерный
57
тапетум. В результате слияния
ядер клетки тапетума становятся
полиплоидными. Имеются — орби-
кулы.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные, изо-
билатеральные. Зрелые пыльцевые
зерна трехклеточные, трехборозд-
ные.
Семяпочка анатропная, анакам-
пиллотропная, с двумя иптегумента-
ми, крассинуцеллятная. Микропиле
образуется внутренним интегумен-
том. Имеются нуцеллярный колпа-
YOK, плацентарный и фуникуляр-
ный обтураторы и ариллус. Семя-
почек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 1—2 ряда.
Тетрады макроспор линейные. Т-0б-
разные. Зародышевый мешок обра-
зуется по Роусопашм-типу. После
оплодотворения зародышевый Me-
шок удлиняется и изгибается. Си-
нергиды с крючкообразными выро-
стами и фибриллярными аппарата-
ми. Имеются три скоро дегенери-
рующие или долго сохраняющиеся
клетки антипод. Полярные ядра
сливаются перед оплодотворением
или во время оплодотворения. У
Тнап ета monogyna зародышевый
мешок содержит крахмал.
Эндоспермогенез нуклеарный.
Клеткообразование в эндосперме
начинается с микропилярной части
зародышевого мешка.
Эмбриогенез СагуорвуПа4-, Зо]а-
па4д-типов. Зародыш бесхлорофилль-
ный, согнутый, двусемядольный.
Подвесок массивный.
Семя, плод — семя без эндоспер-
ма, но с периспермом. Зародыш за-
нимает весь периметр семени, при
этом семядоли и корешок почти
смыкаются концами. Семядоли по
размеру гораздо меньше гипокоти-
ля и корешка. Перисперм заполня-
ет центр семени. Плоды сухие, рас-
крывающиеся или редко нераскры-
вающиеся, обычно окруженные ос-
тающейся чашечкой.
Гаусторий зародышевого мешка.
Апомиксис не обнаружен.
Другие явления не наблюдались.
Литература: Пау1з, 1966; Naraya-
na, Lodha, 1972; Siva Rao,1975; Tax-
таджян, 1966, 1970; Смирнова, 1977.
58
СЕМЕЙСТВО АТЛОАСЕАЕ
Основное число хромосом — х =
= 9, 10.
Цветок обычно обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эндотеций, один
средний слой и секреторный дву-,
многоядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна трехклеточ-
ные, с зернами крахмала, трехбо-
розднопоровые или трехбороздные.
Семяпочка анакампилотропная-
кампилотропная © двумя интегу-
ментами и длинной семяножкой,
крассинуцеллятная. Микропиле об-
разуется внутренним интегументом.
Имеются плацентарный обтуратор,
ариллус и пуцеллярный кКлювик.
Сосудистый пучок доходит до ха-
лазы. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий обычно много-
клеточпый, ипогда одпо-двуклеточ-
ный. Имеются париетальные клет-
ки. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Polygonum-Tully и содержит крах-
мальные зерна. Обычно четыре,
иногда три макроспоры. Синергиды
со слабо выраженными крючками.
Имеются три эфемерные антиподы.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — По-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный, по-
зднее становится клеточным.
Эмбриогенез Зо]апад-типа. Заро-
дыш бесхлорофилльный, согнутый,
двусемядольный. Подвесок тонкий
и ДЛИННЫЙ,
Семя, плод — семя без эндоспер-
ма, но с периспермом, содержащим
крахмал. Зародыш занимает весь
периметр семени, при этом семядо-
ли и корешок почти смыкаются
концами. Семядоли по размеру го-
раздо меньше гипокотиля и кореш-
ка. Семенная кожура образуется
двумя интегументами. Клетки на-
ружного интегумента толстостен-
ные и содержат таннин. Перисперм
занимает центр семени. Плод —
обычно локулицидная коробочка,
редко ягода.
Гаусторий зародышевого мешка.
Апомикеис не обнаружен.
Другие явления — полиэмбрио-
ния. Добавочные зародыши образу-
ются из клеток подвеска.
Литература: Согпег, 1976; Davis,
1966; Ly-Thi-Ba, Guignard, 41970;
Dnyansagar, Melkhede, 1963; Рга-
kash, 1964, 1966, 1967а, Ъ, с; Смирнова,
1977; Schnarf, 19341; Тахтаджян,
4966, 1970.
СЕМЕЙСТВО САСТАСЕАЕ
Основное число хромосом — х =
—: 9, 44, 12.
Цветок обоеполый.
Пыльник — 4 микроспорапгия.
Стенка микроспорапгия имеет эпи-
дермис, одно-трехрядпый фиброз-
ный эндотеций, один эфемерный
средний слой и секреторный дву-
многоядерный тапетум. Наблюдает-
ся слияние ядер тапетума и обра-
зование полиплоидных ядер.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевыс
зерна трехклеточные, трехборозд-
ные, 6—15-расселипобороздные. 9—
15-рассеяппопоровые, с шиноватой
или сетчатой экзиной.
Семяпочка кампилотропная, ана-
тропная, гемицирципотропная или
цирцинотропная, с двумя интегу-
ментами, крассинуцеллятная. Мик-
ропиле образуется внутренним ин-
тегументом. Имеются обтуратор, ну-
целлярный колпачок и ариллус. Сс-
мяножка часто длинная. Семяпочек
много.
Макроспорогенез и макрогамето-
генез — археспорий одно- многокле-
точный. Париетальных клеток 1—2
ряда. Тетрады или триады макро-
спор линейные. Зародышевый ме-
шок образустся по Роугопит-типу
и имест крахмальпьте зерна. Синер-
гиды © крючкообразными выроста-
ми и фибриллярными аппаратами.
Имеются три эфемерные клетки ан-
типод. Слияние полярных ядер про-
исходит перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение
промежуточного типа. От опыле-
ния До оплодотворения проходит
от недели до месяца. Пыльцевые
трубки сохраняются в семяпочке и
после оплодотворения.
Эндоспермогенез нуклеарный. Об-
разование клеток в эндосперме про-
текает центробежно.
Эмбриогенез типа поздно диффе-
ренцирующихся зародышей. Зре-
лые зародыши либо хорошо диффе-
ренцированные двусемядольные, ли-
бо недифференцированные, округ-
лой формы, бесхлорофилльные, со-
держат алейрон. Подвесок массив-
ный. Средняя продолжительность
развития половых зародышей око-
-ло 30 суток, нуцеллярных — 15—20
суток.
Семя, плод — семя с периспер-
мом и эндоспермом, содержащими
крахмал. Зародыши заполняют ли-
бо почти все, либо все семя. Семя-
доли составляют менее четверти
всего объема зародыша, или семя-
доли, гипокотиль и корешок соста-
вляют примерно равные по объему
части зародыша. Семенная кожура
образуется обоими интегументами,
наружные и внутренние слои кото-
рых расширяются, утолщаются и
содержат таннин. Плод обычно мя-
систый, ягодообразный, изредка су-
хой.
Гаусторий отсутствует.
Апомиксис — нуцеллярная и ин-
тогументальная омбрионии У неко-
торых видов Opuntia.
Другие явления — полиэмбриония
y Opuntia.
Литература: Согпег, 1976; Davis,
1966; Kapil, Prakash, 1969; Nagaraj,
Nijaligappa, 1973; Haymosa, 1978a,
6; Haymopa, Axopnes, 1978; Schnarf,
1931; Тахтаджян, 1977, 1970; Tiagi,
1967с — 4, 1970а.
СЕМЕЙСТВО РОВТОТАСАСЕАЕ
Основное число хромосом — z =
= 4—8, 10, 11, 19.
Цветок обосполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций; один
эфемерный средний слой и секре-
торный, дву-, многоядерный тапе-
тум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
изобилатеральные, тетраэдральные.
59
Зрелые пыльцевые зерна трохкле-
точные с крахмальными зернами,
трехбороздные, рассеянноборозд-
ные, многопоровые. Спермии клеток
хорошо выражены.
‘Семяпочка анатропная-амфитроп-
ная, с двумя интегументами, крас-
синуцеллятная. Микропиле образу-
ется внутренним — интегументом.
Семяпочек две или много, изредка
одна.
Макроспорогенез и макрогамето-
генез — археспорий одно- двуклеточ-
ный. Париетальных клеток 3—4 ря-
да. Тетрады или триады макроспор
линейные, Г-образные. Зародыше-
вый мешок формируется по Poly-
зопит-типу и имеет крахмальные
зерна. Синергиды имеют крючко-
образные выросты и фибрилляр-
пые аппараты. Три клетки антипод
скоро дегенерируют Полярные
ядра сливаются либо до оплодо-
творения, либо во время оилопо-
творения.
Опыление, прохождение пыльцс-
вой трубки н оплодотворение — нс-
рекрестное опыление и самоопыле-
пие.
Эндоспермогенез нуклеарный. Об-
разование клеток эндосперма начи-
нается с микропилярной части за-
родышевого мешка.
JMOpHorenes Solanad-, Caryophyl-
lad-tunos. Зародыши согнутые, дву-
семядольные или недифференциро-
ванные, бесхлорофилльные. Подве-
сок с двумя или несколькими клет-
ками. Базальная клетка подвеска
сильно увеличенная.
Семя, плод — семя с периспермом,
содержащим крахмал. Зародыш за-
нимает весь периметр семени, при-
чем семядоли и корешок почти смы-
каются концами. Семядоли по раз-
меру гораздо меныне гипокотиля и
корешка. Перисперм занимает центр
семени. Плод — обычно коробочка,
очень редко орех.
Гауеторий не обнаружен.
Апомиксис — редуцированный пар-
теногенез.
Другие явления — полиэмбриония
и клейстогамия. ,
Литература: Corner, 1976; Davis,
1966; Guignard, 1965; Rhan, 1973;
Schnarf, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970; Vignon, Lobé-
gue, 1969b.
60
CEMEHCTBO BASELLACEAE
Основное число xpomocom — x =
== 41, 12.
Цветок обычно обоеполый, реже
однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуотся
Но однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
средний слой и сскреторный много-
ядерный тапетум. ~
Микроеспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна трохкле-
точные, трехбороздные, рассеянно-
бороздные, многобороздные.
Семяпочки одна, базальная, ана-
тропная, амфитропная, с двумя ин-
тсгументами, крассинуцеллятная.
Микронпиле образуется внутренним
интегумеитом, сильно удлиняющим-
ся в микропиле (нитегументальный
клювик), имеются пуцеллярпый
колпачок и маленький ариллус.
Макроспорогенез и макрогамето-
генез — археспорий одно-, изредка
двуклеточный. Париетальная ткань
массивная. Тетрады или триады
макроспор линейные. Зародышевый
мешок образуется no Polygonum-
типу и имеет крахмальные зерна.
Синергиды грушевидные. Имеются
три эфемерные антиподы. Поляр-
ные ядра сливаются перед омлодо-
творением.
Опыление, прохождение пыльце-
вой трубки, оплодотворение — норо-
гамия.
Эндоспермогенез нуклеарный. По-
зднее весь эндосперм становится _
клеточным.
Эмбрногенез — СВепорой}ад-типа.
Зародыш хлорофиллоносный, коль-
цевидный или спиральпо закручен-
ный, Двусемядольпый. —Подвесок
имеет несколько клеток. Базальная
клетка подвеска очень большая,
Семя, плод — семя с мучнистым
периспермом. Зародыш занпмает
весь периметр семени, причем се-
мядоли п корешок почти смыкают-
ся концами. Центр сомепи запол-
нен периспермом. Семенная кожу-
ра образуется обоими интегумента-
ми. Плод но раскрывающийся, окру-
женный остающейся, часто мяси-
стой чашечкой. Оболочка плода мя-
систая и состоит из паренхиматоз-
ных клеток.
Гаусторий не найден.
Апомикеие не обнаружен.
Другие явления — клейстогамия.
Литература: Согпег, 1976; Davis,
1966; Maheshwari D., Pullaiah, 1975;
Pullaiah, 1976; Schnarf, 1931; Cmup-
tropa, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
CARYOPHYLLACEAE
Основное число хромосом — х =
= 6—15, 19.
Цветки обоеполые и раздельно-
полые.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному или однодольному
типам и имеет эпидермис, фиброз-
ный эндотеций, 1—2 эфемерпых
средних слоя и секреторный двуя-
дернпый тапетум.
Микроспорогевез и микрогамето-
генез — образовапие микроспор cH
мтльтанное. Тетрады мпкроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна трехклеточ-
ные, с крахмальными зернами, ча-
тце всего многопоровые с толетои
экзиной. Спермин-клетки © хорошо
выраженной цитоплазмой.
Семяпочка — гемианатропная-кам-
пилотропная, с двумя нитегумеита-
ми, крассипуцеллятная. Микрониле
образуется внутренцниим интегумен-
том. Имеются нуцеллярный клювик
и гиностаза. Семяночек много, He-
сколько, изредка только одна.
Макроепорогенез и макрогамето-
генез — археспорий много-, одно-
двуклеточный. Паристальные клет-
ки имеются. Образуются четыре
или три макроспоры. Тетрады ма-
Kpocnop линейные, Г-образные. За-
родышевый мешок формируется по
Polygonum-tuny. Y Dianthus sinen-
3153 и ОПепе зародышевые мешки
имеют зерна крахмала. Синергиды
с крючкообразными — выростами.
Имеются три эфемериые клетки ап-
тинол. Шолярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — пс-
рекрестное опыление при помощи
насокомых. Порогамия. Двойное
оплодотворение происходит по пре-
митотическому и постмитотическо-
му типам. У Acanthophyllum Кга-
эсвешапЩоуИ оплодотворение про-
исходит через 8—10 ч после опыле-
ния, у А. е@аНиз — через 4—6 ч, у ре-
монтантной гвоздики двойное опло-
дотворение происходит через 40 ч
после опыления.
Эндоспермогенез нуклеарный, по-
зднее становится клеточным в верх-
ней половине зародышевого мешка.
Эмбриогенез Caryophyllad-runa,
вариации баса. Зародыш бесхло-
рофилльный, согнутый, двусемя-
дольный с корешком и гипокоти-
лем. Базальная клетка подвеска
достигает гигантских размеров. У
Acanthophyllum подвесок имеет 6—
8 клеток и сохраняется вплоть до
созревания семени. Зародызи содер-
жит масло и алейрон.
Семя, плод — семя с периспермом,
содержащим крахмал. Зародьши за-
нимает весь пернметр семени, ири-
чем семядоли и корешок почти
смыкаются концами. По размеру
семядоли гораздо меныпе типоко-
тиля и корешка. Перисперм занол-
няет центр семени. Плод — коро-
бочка или орех, реже ягода.
Гаустории зародышевого
и подвесочный.
Апомикене не обнаружен.
Другие явления — полиэмбриония
и клейстогамия. Гинодиэция обна-
ружена у многих представителей
семейства. Партенокарпия.
JIureparypa: Corner, 1976; Chaud-
мопца
hury, Narayana, 1978; Демьянова,
Нокатасва, 1977; Фурсов, Беспалов.
1963; Guignard, 1966a,b; Кордюм.
1963; Lebeque, Vignon, 1968; Гопос-
vialle, 41966. 1967, 1968; Матюнвина,
Турсунов, 1977; Мейер, 1962; Мир-
замурадов, Турсунов, 1971; Pal,
1974; Раь Мишу, 1972; Савинова.
Турсунов, 1966, 1977; Schnarf, 1931;
Тахтаджян, 1966, 1970; Терзийски,
1974; Турсунов, 1966, 1967, 1969,
1972, 1974a.6, 1977; Турсунов, Ма-
тюнина, 1975, 1977, 1978а, 6, 1979а:
Vignon, 1965, 1966Ъ, 1970; V.ignon,
Lebegue, 1966a, 1969a, b, 1970. 1972;
тел, Zenktelter, 1973; Яковлева,
‹ OO,
CEMEMCTBO AMARANTHACEAE
Основное
== 6—11, 17.
Цветки обоеполые или реже по-
лигамные пли двудомные.
Пыльник — 2—4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
G4
чиело хромосом — х ==
Рие. 14. Семяпочка, зародышевой мешок и зародыш Agrostemma githago 1 — семяпочка с зародышевым мешком, 8 — зародышевый мешок с боковым ответ-
влением, в котором располагается вторичное ядро;
3 — семяпочка с двусемядоль-
ным зародышем; 4 — молодой шарообразный проэмбрио с мощным одноклеточным
подвеском [Восеп, 1926]
дермис, фиброзный эпдотеций, один
эфемерный средний слой и секре-
торный двуядерный тапстум. Име-
ются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, с крахмаль-
ными зернами, многопоровые.
Семяпочка анакампилотропная,
кампилотропная или цирцинотроп-
ная, с двумя интегументами, крас-
синуцеллятная. Микропиле образу-
ется либо только внутренним, либо
обоими интегументами. Имеются
нуцеллярный колпачок и фунику-
62
лярный ариллус. Семяпочка одна
или их несколько.
Макроспорогенез и макрогамето-
генез — одно-, многоклеточный. Па-
риетальных клеток 5 рядов. Обра-
зуются три или четыре макроспо-
ры. Тетрады макроспор линейные,
Г-образные. Зародышевый мешок
формируется по РоТубопитм-типу и
имеет крахмальные зерна. Синерги-
ды © крючкообразными выростами
и фибриллярными аппаратами. Име-
ются три и более (до 30—40) скоро
дегенерирующих клеток антипод.
Полярные ядра сливаются до опло-
дотворения. Зародышевый мешок
после оплодотворения удлиняется и
изгибается, принимая подковооб-
Рис. 15. Эмбриогеноз У Sagina procumbens. Са, Со — апикальная и базальная клетки $ Г, ст, ©, п, п’, о, р == последующие ря- ды клеток; 1—13 — разные стадии развития проэмбрио; 14—19 — разные стадии раз- вития и дифференциации двусемядольного зародыша [Бопёсез, 1924]
разную форму, но антиподы остают-
ся на прежнем месте.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия.
Эндоспермогенез нуклеарный.
Клеткообразование в -эндосперме
начинается с микропилярного кон-
ца зародышевого мешка.
Эмбриогенез Chenopodiad-, Sola-
пад-типа. Зародыш согнутый, бес-
хлорофилльный,
двусемядольный, содержащий масло
и алейрон. Подвесок одно- или мно-
горядный, массивный.
Семя, плод — семя с периспер-
мом, содержащим крахмал. Заро-
дыш занимает весь периметр семе-
ни. Семядоли но размеру гораздо
меньше гипокотиля и корешка.
Перисперм заполняет центр семе-
подковообразный.
ни. Плод — обычно орех, редко яго-
да или крыночка.
Гауеторий зародышевого мешка.
Апомикеие — нередуцированный
партеногенез и генеративная апо-
спория.
Другие явления — полиэмбриония.
Jiuteparypa: Chowdhuri, 1965; Cor-
ner, 1976; Davis, 1966; Mendoza,
1973; Pal, 1969; Puri, Singh, 1935;
Schnarf, 1931; Тахтаджян, 1966,
1970; Vignon, Lebegue, 1972.
CEMEKCTBO CHENOPODIACEAE
Основное число хромосом — х =
= 6— e
Цветок обосполый или однополый.
ПНыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
63
дермис, фиброзный эндотеций, один,
два эфемерных средних слоя и сек-
реторный дву-многоядерный тапе-
TyM. TombKo y Chenopodium album
oOnapyxen амебоидный (периплаз-
модий) тапетум, причем ядра cro
претерпевают неоднократные деле-
ния и слияния, в результате чего
становятся полиплоидпыми.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, многопоровые.
Семяпочка обычно одна, анатроп-
ная, кампилотропная, с двумя ин-
тегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяножка длинная.
Имеются нуцеллярный клювик, ин-
тегументальный обтуратор и гипо-
стаза. У гибискусов от 60 до 140
семяпочек.
Макроспорогенез и макрогамето-
генез — археспорий одно-, мпого-
клеточный. Паристальпых клеток
1—3 ряда. Образуются четыре или
три макроспоры, располагающиеся
линейно. Зародышевые мешки фор-
мируются mo Polygonum- Allium-
типам, имеют крахмальные зерна.
После оплодотворения припимают
согнутую форму. Синергиды имеют
рючкообразные выросты. Имеются
три эфемерные или долго сохраня-
ющиеся антиподы. У Ваа уцат1з
наблюдается увеличение числа кле-
ток антипод путем митотического
деления первоначально трех клеток.
Нолярные ядра сливаются перед
оплодотворепием.
Опыленис, прохождение ныльцс-
вой трубки и оплодотворение -- са-
моопыление и перекрестное оныие-
ние. Порогамия. Двойное оплодо-
творение. У гибискусов онлодотво-
рение происходит через 5—10 ч
осле опыления. Семена созревают
на 26-й депь после опыления.
Эндоспермогенез нуклеарный,
Клеткообразование в эндосперме
начинастся с микропилярной части
зародышевого мешка.
JmOpuorenes Chenopodiad-tuma. 3a-
родыши подковообразные или спи-
рально закручепные, хлорофилло-
посные или бесхлорофилльные, дву-
семядольные. Подвссок обычно мас-
сивный. Зародьши содержат масло
и алейрон.
64
-Caryophyllales.
Семя, плод — семя с периспермом,
содержащий крахмал. Зародыпт за:
нимаест весь периметр семени, центр
семени выполнен периспермом. Эн-
Досперм слабо развит. Плод — орех,
окруженный остающейся чашечкой.
Гауеторий зародышевого мешка.
Апомикеие — редуцированный па-
ртеногенез у Вёа ушШзаг1.
Другие явления — полиэмбриония
y Beta rubra u B. vulgaris.
Литература: Согпег, 1976; Davis,
1966; Fossard, 1969; Hindmarsh,
1966: Калягин, 1970, 1974, 1976,
1977: Конычева, 1960, 1961, 1963,
1965, 1968, 1976, 1977, 1979; Коныче-
ва, Кадырова, 1978a,6; Конычева,
Ilesayx, 1974, Maheshwari D., Rao,
1976; Младенцева, 1964; Schnarf,
1931; Петрушина, 1978; Тахтаджян,
1966, 1970; Шевчук, Конычева, 4973;
Wunderlich, 1967b.
3 порядке Caryophyllales cemei-
слва Gyrostemaceac, Bataceae, Te-
tragoniaceac, Didieraceac, Halophy-
laceae u_ Lectorellaccae B oTHOIe-
нии цитоэмбрпологии не исследова-
ны. Остальные семейства изучены
цитоэмбриологически более или мс-
нее детально п обнаруживают нс-
мало сходных признаков, что ука-
зывает на естественность порядка
Такими сходными
признаками у ссмейств в настоя-
ее время являются: 4 микроспо-
рапгия; четырех-пятислойная, стен-
ка пыльпика; симультанный тии
микроспорогенева; — трехклеточные,
прех- миогобороздные пыльцевые
верна с крахмалом; анакампилло-
троипая, красспиуцеллятная © дву-
мя иптоегументами семяпочка; одно-
мпогоклеточпый женский археспо-
рий; наристальные клетки; Polygo-
пит-тии зародьииевого мепга с зер-
иами крахмала; гаусторий зароды-
ICBOTO мешка; пуклеарный эндо-
сиерм; согнутый двусемядольный
зародыш; перисперм. Капил и Пра-
nam [Kapil, Prakash, 1963] ormetm-
ли, что по своим цитоэмбриологиче-
ским признакам Сасасеае связано
с другими семействами Caryophy-
Hales, oco6benno с семействами А1270-
асеае и РогИасасеае, что подтвер-
ждает правильность отнесения Ca-
ctaceae kK порядку СагуорвуПаез.
Тияги и Сахар [Tiagi, Sachar, 1967]
считают, что Сасбасеае является
примитивным семейством раналие-
вого типа. По мнению Махешвари,
Деви и Пулаия [Maheshwari, Pu-
Палав, 1975], данные цитоэмбриоло-
гии Basellaceae подтверждают от-
несение этого семейства к порядку
Caryophyllales.
По pany цитоэмбриологических
признаков семейство Мубфарттасеае
сходно с другими представителями
порядка Caryophyllales, a по эм-
6puorenesy —c Thymelaeaceae, что,
возможно, указывает на родство
этих семейств [ Venkateswarlu,
1947].
Такие признаки, Kak cekpeTop-
ный тапетум, трехклеточное пыль-
цевое зерно, обтуратор, нуцелляр-
ный колпачок, Ро[ухопит-тип за-
родышевого мешка, пуклеарный
эндосперм, Азюгад-тип эмбриогене-
за указывают на родство между
Thymelaeaceae u Nyctaginaceae. Hu
в одном из других семейств Сагу-
ормуПа!ез не обнаружен Asterad-
тип эмбриогенеза. Особенно по это-
му признаку Мубаотасеае отлича-
ется от СагуорвуПа]ез и обнаружи-
вает сходство с ТВуше]аеасеае [Уеп-
kateswarlu, 1947].
B oTnHomenun цитоэмбриологии
Cactaccao сходпо с другими пред-
ставителями — СагуорйуПа]ез. При
сравнении порядка Caryophyllales
с порядками Urlicaleas, Plumbagi-
nales и Ро!угопа!ез, с которыми
некоторые систематики сближа-
ют СагуорвуПа]ез, наибольшее сход-
ство в отношении их цитоэмбрио-
логии обнаружено между порядка-
mu Caryophyllales a Polygonales.
На основании изучения сравни-
тельной эмбриологии мы пришли к
заключению, что Caryophyllales
является одним из наиболее есте-
ственнных порядков покрытосемен-
ных, однако требуются дальнейшие
исследования, чтобы убедиться в
этом.
ПОРЯДОК 28.
POLYGO NALES
СЕМЕЙСТВО РОГУСОМАСЕАЕ
Основное число хромосом — х =
== 7—411, 17.
Цветки обоеполые и однополые:
мужские и женские.
5 В. А. Поддубная-Арнольди
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет
эпидермис, фиброзный эндотеций,
один эфемерный средний слой и
секреторный дву-четырехъядерный
тапетум.
`Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
изобилатеральные, крестообразные,
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, с крахмаль-
ными зернами, трехбороздные, реже
четырех-, многобороздные или мно-
гопоровые. Спермии-клетки бедно-
плазменные.
Семяпочка одна, анатропная, с
двумя интегументами, крассинуцел-
лятная. Микропиле образуется вну-
тренним интегументом. Имеются
гипостазаа и фуникулярный обту-
ратор.
Макроспорогенез и макрогамето-
генез — археспорий одно-многокле-
точный. Париентальных клеток 1—
3 ряда. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок образуется по Ро[ухопит-типу.
Синергиды с крючкообразными вы-
ростами и фибриллярными аппара-
тами. Имеются три обычно эфемер-
ные одноядерные антиподы, но у
Питех они становятся многоядер-
пыми. Иногда антиподы лежат на
постаменте.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодо-
творение премитотического типа.
Эндоспермогенез нуклеарный, эн-
доснерм руминированный, содер-
жит крахмал и алейрон. Образова-
ние клеток в эндосперме начинает-
ся с микропилярной части заро-
дышевого мешка, когда зародыш
становится округлым. У Fagopy-
гит езсшШепит в халазальной ча-
сти зародышевого мешка ядра эн-
досперма остаются свободными.
Эмбриогенез Азега4-типа. Заро-
дыш двусемядольный, бесхлоро-
филльный, согнутый или прямой.
Семядоли складчатые, равных раз-
меров.
Семя, плод — семя с хорошо раз-
витым, мучнистым, руминирован-
ным эндоспермом и слабо развитым
крахмалистым периспермом. У од-
них представителей семейства за-
родьши маленький, располагающий-
65
ся в его основании, обычно занима-
eT of 1/4 no 1/2 линейной высоты
семени. У других зародыш круп-
ный, с широкими семядолями, за-
нимающими 3/4 общего объема се-
мени, с хорошо развитыми почеч-
кой, гипокотилем и корешком. Се-
менная кожура двухслойная, обра-
зующаяся из наружного интегумен-
та. Нлод — орех.
Гаусторий — семядоли при про-
растании семени выполняют гау-
сториальную функцию.
Апомиксие — апоспория,
лярная эмбриония.
Другие явления — партенокарпия,
гетеростилия и клейстогамия.
нуцел-
Литература: Corner, 1976; Davis,
1966; Гваладзе, 1973; Mukherjee,
4972; Neubauer, 1971; Поддубная-
Арнольди, 1948; Rao, Mukherjee,
4973; Camaxopa, 1968, 1973, 1978;
Салахова, Смирнов, 1976, 1977,
1978: Смирнова, 1977; Schnarf, 1931;
Тахтаджян, 4966, 1970.
В отношении цитоомбриологии
порядок Polygonales c его един-
ственным семейством Ро[угопасеае
имеет много общего с представите-
лями порядка Caryophyliales, oco-
бенно с семействами Caryophylia-
ceae, Amaranthaceae u Chenopodia-
сеае.
ПОРЯДОК 29.
PLUMBAGINALES
СЕМЕЙСТВО РЕОМВАСТМАСЕАЕ
Основное число хромосом — z =
—
a (j—
Цветок обоеполый.
Пыльник — 4 микроспоравгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный дву-, многоядерный тапе-
тум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелая пыльца трехклеточная, трех-
пятибороздная, реже шести-рассе-
яннобороздная. Спермии-клетки хо-
рошо выражены.
66
Семяпочка одна, анатропная или
цирцинотропная, с двумя интегу-
ментами, крассинуцеллятная. Мик-
ропиле образуется внутренним ин-
тегументом. Имеется обтуратор.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный,
Париетальных клеток 2—3 ряда.
Зародышевые мешки образуются
m0 Plumbago-, Penaea-, Plumbagel-
la- u Fritillaria- типам. Синергиды
образуются или не образуются.
Клеток или ядер антипод от 0 до
трех.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
обычно перекрестное опыление, из-
редка самоопыленис.
Порогамия — двойное оплодотво-
рение премитотического типа. Ниж-
нее полярное ядро нередко не при-
нимает участия в оплодотворении.
Эндоспермогенез нуклеарный. Пе-
рисперм отсутствует.
Эмбриогенез Зо]апа4-типа.
дыш хлорофиллопосный, прямой,
двусемядольный, содержит масло.
Подвосок массивный, многоклеточ-
НЫЙ.
Семя, плод — зародыш заполняет
почти весь объем семени. Семядоли
выполняют ассимиляционную функ-
цию сразу после прорастания се-
мени. Эндосперм в виде тонких
пленок остается лишь в области
корешка и на семядолях, содержит
крахмал. Плод — обычно орех или
редко раскрывающийся створками
снизу вверх.
Гаусторий отсутствует.
Апомиксие — генеративная
спория.
Другие явления — гиподиэция у
Armeria.
Литература: Boyes, 1939; Bense-
Ba H., 1977; Cass, 1972; Corner, 1976:
Dahlgren, 1916; Davis, 1966; Schnarf,
1931; Смирнова, 1977; Гахтаджян,
1966, 1970; Wunderlich, 1967b.
Заро-
апо-
Некоторые систематики порядок
Ришифавта!ез сближают с поряд-
ком СагуорвуПаез. Однако наличи-
em Plumbago-, Penaea-, Plumbage-
Na- uw Fritillaria-runos зародыше-
вых мешков, а также отсутствием
перисперма при наличии эндоспер-
ма, Р]ашфаста|ез отличается от
Caryophyllales.
ПОРЯДОК 30.
THELIGO NALES
CEMEUCTBO THELIGONACEAE
(CYNOCRAMBACEAE)
Основное число хромосом — 2 ==
— 10, 11.
Цветок однополый, растения од-
нодомные или двудомные.
Пыльник — нет данных о числе
микроспорангиев. Имеется одно-
ядерный секреторный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелая пыльца трех-
клеточная (5)6 (7)-поровая или с
(тремя) (пятью) короткими поро-
видными бороздами.
Семяпочка одна, кампилотропная,
амфитропная, с одним интегумен-
том, тенуинуцеллятная. Имеется
нуцеллярный колпачок.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Имеются три,
двух-четырехъядерные клетки ан-
типод, дегсперирующие после опло-
дотворения. Полярные ядра слива-
ются перед оплодотворением или во:
время пего.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоспермогенез нуклеарный,
вскоре становится клеточным. Об-
ПОДКЛАСС Е.
НАДПОРЯДОК У.
DILLE NIA МАЕ
ПОРЯДОК 31.
DILLE NIALES
СЕМЕЙСТВО ОПЛ.ЕМТАСЕАЕ
Основное число хромосом — 7 =
= 8, 13
Цветок — обычно обоеполый.
5%
разование клеточных перегородок
центробежное.
Эмбриогенез Chenopodiad-runa.
Зародыш согнутый, двусемядоль-
ный, с корешком. Подвесок одно-
двухрядный, девятиклеточный.
Семя, плод — семя с эндоспер-,
мом, содержащим масло и алейрон.
Зародыш располагается в основа-
нии семени и обычно занимает от
1/4 до 1/2 линейной высоты семе-
ни. Плод — костянка.
Гаусторий — семядоли обычно в
начале прорастания семени выпол-
няют гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления не наблюдались.
Литература: Васильченко, 1979;
Kapil, 1966; Kapil et al, 1966a;
Schnarf, 1931; Тахтаджян, 1966,
1970; Wunderlich, 19714.
Систематическое положение The-
ligonaceae подвергается дискуссии.
Капил и Рао [Карй, М. Вао, 1966а]
считали, что данные цитоэмбрио-
логии подтверждают целесообраз-
ность выделения семейства ТБеП-
gonaceae B самостоятельный поря-
док. Ненюков [1939] и Вундерлих
[Wunderlich, 1971] сближают ТЬе|-
gonaceae c Rubiaceae m исключают
его из nopagka Centrospermae или
Caryophyllales в особый порядок
ТБоНсораез. Требуются дальнейшие
исследовапия для выяснения систе-
матического положепия Theligona-
сеае. Свое заключение о близости
Theligonaceae kK Rubiaceae P. ByH-
дерлих [1971] сделала на основании
изучения цитоэмбриологических при-
знаков этих семейств.
DILLENIIDAE
<
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, слаборазвитый фиброзный
эндотеций, 2—3 эфемерных средних
слоя и секреторный двуядерный
тапетум. У Wormia _ suffruticosa
описан амебоидный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
линейные. Зрелые пыльцевые зер-
на двухклеточные, трехборозднопо-
ровые или трехпоровые.
67
“¢
; 1
xp t
а
На (|!
я YAY
a
Oa <
Рие. 16. Нуцеллус семяпочки, пыльник и зародышевый мешок Асгогета
arnottianum
1—4 — макроспорогенез; 5—6 — зародышевые мешки; 7 — микроспориальный с ар-
хеспориальными клетками; 8—9 — микроспорангии с одноядерными пыльцевыми
зернами; 10 — двухклеточное пыльцевое зерно; 11 — микропилярная часть нуцел-
луса семяпочки и зародышевого мешка вскоре после оплодотворения; 12 — зигота;
13, 14 — последовательные фазы развития «зиготической мантии» [З\уашу, Рег!-
samy, 1955]
Семяпочка анатропная-амфитроп- ются нуцеллярный колпачок и арил-
ная, кампилотропная, с двумя ин- лус. Семяпочка одна-много.
тегументами, крассинуцеллятная. Макроспорогенез и макрогамето-
Микропиле зигзагообразное, образу- генез — археспорий одноклеточный.
ется обоими интегументами. Име- Париетальных клеток 5 рядов. Те-
68
фе
ea gee
ыы
>
К АХ
а tie
м : aN к Ч Ц \ Ww “+ 2
ie
ite ig:
трады макроспор линейные, Т-об-
разные. Зародышевый мешок обра-
зуется по Ро]угопит-типу и имеет
крахмальные зерна. Синергиды с
крючкообразными выростами и
фибриллярными аппаратами. Имс-
ются три эфемерпые антиподы. По-
лярные ядра сливаются до оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. За-
ложение клеток между ядрами эн-
досперма начинается с периферии
зародышевого мешка.
£33, Vt hee
DB Я Я и,
a8
Эмбриогенез Onagrad-tuna. Y Ас-
rotrema u Wormia o6onoqka зиготы,
называемая «мантия», массивная с
пыльцеобразными выростами. Заро-
дыш очень маленький, прямой, бес-
хлорофилльный, содержащий мас-
ло и алейрон, с двумя семядолями
и корешком.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Зародыш распо-
лагается в его основании и зани-
мает обычно от 4,4 до 1/2 линей-
ной высоты семени. Плод — одно-
карпная или синкарпная многоли-
стовка, многоорешек или ягодооб-
разный.
69
we
Payette yee RMS
arene eres
Рис. 17. Эмбриогенез у Acrotrema arnottianum.
1—4 — разные стадии развития зародыша и постепенное исчезновение «зиготиче-
ской мантии» [Зуату, Регазату, 1955]
Гаусторий — семядоли в начале
прорастания семени выполняют
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; ЭсппатЁ, 1934; Смирнова, 1977;
Тахтаджян, 1966, 1970; Иоффе, Жу-
кова, 1974.
СЕМЕЙСТВО
CROSSOSOMATACEAE
Основное число хромосом — х =
omer
—
Цветок обоеполый.
Пыльник — стенка микроспоран-
гия имеет эпидермис, 2—3 ряда
клеток фиброзного эндотеция, 2—3
средних слоя и секреторный много-
ядерный, двух-трехрядпый тапетум.
В результате слияния ядра тапету-
ма становятся полиплоидными.
- Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехпоровые, двух (трех)-бо-
розднопоровые с сетчатой экзиной.
Семяпочка кампилотропная (ам-
фитропная после оплодотворения),
с двумя интегументами, крассину-
+
целлятная. Микропиле образуется
обоими интегументами. —Имезлотся
гипостаза и ариллус. Сосудистый
пучок доходит до халазы. Семяно-
чек несколько — много.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный. Париетальная ткань массив-
ная. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Ро]угопит-типу. Три, четыре, шесть
клеток. антипод рано дегенерируют.
После оплодотворения. зародыше-
вый мешок начинает сгибаться и
постепенно принимает подковооб-
разную форму.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Тройное слияние предшествует син-
гамии. Остатки пыльцевых трубок
сохраняются до стадии глобулярно-
го зародыша.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма начина-
ется © микропилярной части заро-
дышевого мешка.
Эмбриогенез Опаотад-типа. Заро-
дыш крупный, согнутый в виде
подковы, двусемядольный.
Семя, плод — семя с эндоспермом.
Зародыш располагается в основа-
нии семени и обычно занимает от
1/4 до 1/2 линейной высоты семе-
ни. Семенная кожура образуется
обоими интегументами. Из клеток
семяножки образуется нитевидный
ариллус, который окружает семя.
Плод — многолистовка.
Гаусторий — семядоли в начал
прорастания семени выполняют
гаусториальную функцию.
Апомиксис не обнаружен.
- Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; Kapil, 1967с, 1970а; Тахта-
джян, 1966, 1970; Уаш, 1966; Смир-
нова, 1977.
Pon Crossosoma Бентам и Гукер
[Bentham, Hooker, 1862—1883] от-
носили к ОШещасеае, но Энглер и
Прандль 11897] высказывались за
необходимость выделения его в 0со-
бый порядок. Хатчинсон [НиюЫт-
son, 1959] ornee Crossosomataceae kK
ПШеп1аез. В этом его поддержали’
Имс [1861] и А. Л. Тахтаджян
[1966]. Капил [КарИ, 19676], кото-
рый детальпо исследовал Crossoso-
та(ассас, считает, 4To poy Crosso-
soma заслуживает выделения его
из ОШешасеае и отнесения его к
самостоятельному семейству, так
как имеет ряд цитоэмбриологиче-
ских признаков, не сходных с та-
ковыми ОПШетасеае, а именно мно-
гоклеточный женский археспорий,
отсутствие «мантии» вокруг зиготы,
крупный, согнутый в виде подковы
зародыш. Вместе с тем по характе-
ру стенки пыльника, тапетума, ми-
кроспорогенеза, семяпочки, зароды-
иезвого мешка и эндосперма Сгоз50-
зоша{асеае имеет сходпые черты с
Dilleniaceae. Поэтому Crossosomata-
сеас следует относить к ОШеюмаез,
а не к Воза!ез или Вапа!ез, как это
делали некоторые ученые [Карй,
1967с]. Порядок ПШен!а]ез, по мне-
нию Капила [КарП, 1967с], тесно
связан © порядком Раеоп1аез. По-
этому расположение этих порядков
вблизи друг к другу, по-видимому,
правильно.
71
с
ПОРЯДОК 32.
PAEONIALES
СЕМЕЙСТВО РАЕОМТАСЕАЕ
Основное число хромосом — 7 =
=5, 7.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, 2—3 слоя фиброзного эндо-
теция, 1—2 слоя средних клеток и
секреторный одно- или двуядерный,
дву-, многорядный тапетум.
Микроеспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехборозднопоровые, с сетча-°
той экзиной и с крахмальными зер-
нами. Спермии-клетки хорошо вы-
ражены.
Семяпочка анатроппая, с двумя
интегументами, крассинуцеллятная.
Микропиле слабо зигзагообразное,
образуется обоими интегументами.
Имеются нуцеллярный колпачок,
гипостаза и ариллус. Семяпочек
много.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный. Париетальных клеток 9 рядов.
Тетрады макроспор линейные, 7-об-
разные. Зародышевый мешок обра-
зуется по Роугопит-типу. В одной
и той же семяпочке может быть
несколько зародышевых мешков,
причем иногда они сливаются. Си-
нергиды имеют крючкообразные
выросты и фибриллярные аппара-
ты. Три клетки антипод сохраняют-
ся, и иногда число антипод увели-
чивается; клетки их имеют одно —
несколько ядер. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение
премитотического типа.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка. В халазальной
части зародышевого мешка ядра
остаются свободными.
Эмбриогенез Раеопад-типа. Этот
своеобразный тип обнаружен толь-
ко У Раеопасеае. Он характеризу-
72
ется тем, что ядро зиготы делится
свободно, образуя многоядерный
циноцит; затем между ядрами ци-
ноцита возникают клеточные пере-
городки. Некоторые перифериче-
ские клетки превращаются в зачат-
ки зародышей. Хотя в семяпочке
может возникнуть несколько заро-
дышей, но только один из них до-
стигает полного развития. Зародыш
бесхлорофилльный, маленький, дву-
семядольный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
капли масла и алейроновые зерна.
Зародыш расположен в основании
семени и обычно занимает от 1/4
до 1/2 линейной высоты семени,
Плод — многолистовка.
Гауеторий — семядоли при прора-
стании семени выполняют гаусто-
риальную функцию.
Апомиксис не обнаружен.
Другие явления не наблюдались.
Литература: Согпег, 1976; Carniel,
1967; Гау!з, 1966; Смирнова, 1977;
Тахтаджян, 1966, 1970; Тао, 19705;
Циунович, 1971; Яковлев, Иоффе,
1957, 1961, 1965; Жгенти, 1974.
Данные морфологии, эмбриоло-
гии, анатомии и цитологии оправ-
дывают выделение рода Раеоша из
Ranunculaceae m oTHecenwe его к
самостоятельному семейству Раео-
п1асеае [Т1а21, 1967]. Ввиду нали-
чия своеобразного типа эмбриоге-
пеза, род Раеоп1а заслуживает вы-
деления его в 0с0бое семейство и
даже порядок [Яковлев, Иоффе,
1957, 1961]. Однако оно имеет мно-
го o6Omero c Ranunculaceae. /
ПОРЯДОК 33.
THEALES
СЕМЕЙСТВО ОСНМАСЕАЕ
Основное число хромосом — х =
== 12, 14.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, эфе-
мерный средний слой и секретор-
ный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
нопоровые, с крахмальными зер-
нами.
Семяночка анатропная, © двумя
интегументами, тенуинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Имеется эндотелий.
Семяпочек одна — много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные, Т-
образные. Зародышевые мешки об-
pasyrotca mo Polygonum- u Allium-
типам и имеют крахмальные зерпа.
Синергиды с крючкообразпыми вы-
ростами и фибриллярпыми аппара-
тами. Имеются три эфемерные ан-
типоды. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка.
Эмбриогенез Опаота4-, СатуорВу-
Пад-типов. Зародыш круппий, иря-
мой или согпутый, двусемядоль-
ный, ‘содержащий масло, крахмал
и алейрон.
Семя, плод — семя без эндоспер-
ма или с эндоспермом, содержащим
масло и алейрон. Зародыш запол-
няет все семя. Массивные запасаю-
щие семядоли скрывают остальные
части зародыша, лишь конец ко-
решка едва выступает из них.
Плод — многокостянка, . паракарп-
ная ягода или коробочка, не рас-
крывающаяся или раскрываю-
щаяся.
Гаусторий не обнаружен.
Апомиксие — поредуцированный
партеногенез, нуцеллярная эмбрио-
ния и апоспория обнаружены у
Ochna multiflora u O. serrulata.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; Narayana, 1975; Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
1970; Vani, 1966.
СЕМЕЙСТВО
DIPTEROCARPACEAE
Основное чиело хромосом — < =
== 6, 7, 10.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, эндотеций без фиброзных
утолщений, 3 эфемерных слоя и
секреторный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерпа двухклеточные, трехборозд-
bie или трехборозднопоровые.
Семяпочка одна, анатропная, с
двумя интегументами, крассинуцел-
лятная. Микропиле — образуется
внутренним интегументом.
Макроспорогенез и макрогаметс-
генез — археспорий одноклеточный.
Периетальных клеток 5 рядов. Те-
трады макроспор линейные, Т-об-
разные. Зародышевый мешок обра-
зуется по Роугопит-типу. Синер-
гиды © крючкообразными выроста-
ми. Имеются три антиподы, дегс-
нерирующие вскоре после оплодо-
творения. Полярные ядра сливают-
ся в центре зародышевого мешка.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклсарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка, и весь эндосперм
становится клеточным, за исключе-
нием Уэета, у которой он остает-
ся свободноядерным.
Эмбриогенез Shorea robusta по
хож на Афегад-тип. Зародыш пря-
мой, крупный, двусемядольный,
хлорофиллоносный, содержащий
крахмал, алейрон и масло. Семядо-
ли массивные, складчатые.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Массивные запасающие семядоли
скрывают остальные части зароды-
ша, лишь кончик корешка едва вы-
ступает из него. Плод сухой, нерас-
крывающийся, односемянный.
Гауеторий антиподальный у Уз-
teria indica.
Апомикеис — обнаружена
поли-
эмбриония у Hopea odorata.
73
Другие явления — обнаружена по-
лиэмбриония у Нореа одогаца.
Литература: Corner, 1976; Davis,
1966; Murty, 1970; Тахтаджян, 1966,
1970; ЗеВпатЁ, 1934; Смирнова, 1977.
СЕМЕЙСТВО ТНЕАСЕАЕ
Основное число хромосом — 2 —
== 15, 21. . ;
Цветок обоеполый, реже однопо
лый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет уши.
дермис, фиброзный эндотеции, 1—
эфемерных средних слоя и секре-
торный дву-четырехъядерный та-
петум.
Микроепорогенез и микрогамето-
генез — образование микроспор CH-
мультанное. Тетрады микроспор те-
траэдфальные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
ные. Спермии-клетки хорошо выра-
жены. Пыльца сохраняет свою жиз-
неспособность преимущественпио в
течение 10 дней.
Семяпочка анатропная, анакам-
пилотропная, с двумя интегумента-
ми, тенуинуцеллятная. Микропиле
образуется внутренним интегумен-
том. Имеется гипостаза. Сосуди-
стый пучок достигает конца наруж-
ного интегумента. Семяпочек He-
сколько — много, изредка одна.
Макроспорогенез и макрогамето-
генез — археспорий одно-, двух“,
трехклеточный. Париетальные клет-
ки отсутствуют. Зародышевый ме-
шок образуется по АШат-типу. Си-
нергиды имеют крючкообразные
выросты. Три антиподы дегенери-
руют после оплодотворения. Поляр-
ные ядра сливаются до оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление y Thea sinen-
313. Порогамия. Двойное оплодотво-
рение постмитотического типа. Син-
гамия и тройное слияние происхо-
дят одновременно. У ТВеа зтепз13
оплодотворение происходит через
48—72 ч после опыления. Между
оплодотворением и эмбриогенезом
у ТЬеа зтетз1$ обнаружен период
покоя в 5—6 месяцев, а полное раз-
витие зародыша осуществляется
через год после опыления.
Эндоспермогенез нуклеарный.
74
Эмбриогенез 3о]апай-типа. Заро-
дыш крупный, прямой или согну-
тый, содержащий масло, алейрон и
кофеин, бесхлорофилльный, с двумя
длинными или крупными широки-
ми семядолями и очень короткими
гипокотилем и корешком. Подвесок
длинный, однорядный.
Семя, плод — семя со скудным
эндоспермом или без него. В пре-
делах семейства наблюдается боль-
шое разнообразие в строении семе-
ни. Семенная кожура образуется
наружным интегументом и являет-
ся толотой и твердой, имеющей
снаружи склереидную зону, а внут-
ри зону клеток, наполненных тан-
нином. Перикарпий склереидный.
Плод — коробочка, иногда ягодооб-
разный или костянкообразный.
Гаусторий не найден.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
обпаружена у нескольких видов
Camellia. Добавочные зародыши
могут возпикать из клеток плацен-
ты, впутреннего интегумента, синер-
гид и путем расщепления яйце-
клетки.
Литература: Ва\а, 1967, 19705;
Согпег, 1976; Пау1з, 1966; Кутубад-
зе, 1974; Мандрик, 1957, 1959, 1960,
1964; Микатадзе, 1973, 1974, 1975;
Микатадзе-Панцулая, 1978, 1979;
Schnarf, 1931; Sethi, Bala, 1965; Tax-
таджян, 1966, 1970.
СЕМЕЙСТВО MARCGRAVIACEAE
Основное
данных.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный слой, 3—4 эфе-
мерных средних слоя и секретор-
ный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехбороздные.
Семяпочка анатропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется эндотелий.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
число хромосом — нет
Тетрады макроспор линейные. da-
родышевый мешок образуется по
Ро!угопит-типу. Три клетки или
ядра антипод дегенерируют вскоре
после их сформирования. Полярные
ядра сливаются до оплодотворения.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип не определен.
Зародыш прямой или согнутый, до-
вольно крупный, с длинными семя-
долями и коротким корешком.
Семя, плод — семя с очень слабо
развитым эндоспермом, содержа-
щим крахмал. Зародыш заполняет
все семя, содержит масло и алей-
рон. Массивные запасающие семя-
доли скрывают остальные части за-
родыша, лишь кончик корешка ед-
ва выступает из них. Семенная ко-
жура образуется эпидермисом na-
ружного покрова и содержит тан-
нин. Плод кожистый, раскрываю-
щийся или не раскрывающийся.
Гаусторий —у Marcgravia обна-
ружен эндоспермальный (микропи-
лярный) гаусторий.
Апомиксие не обнаружен.
Другие явления не наблюдались.
Литература: Corner, 1976; Davis,
1966; Тахтаджян, 1966, 1970; Смир-
нова, 1977.
СЕМЕЙСТВО ВОММЕТТАСЕАЕ
Основное
данных.
Цветок обосполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный слой, один сред-
ний слой и секреторный двуядер-
ный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трех-четырехборозднопоровые.
Семяпочка анатропная, с двумя
число хромосом — ист
интегументами, тенуинуцеллятная,
сильно вытяпутая. Микропиле об-
разуется обоими иптсгументами.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Ро[узопит-типу. Антиподы не клет-
ки, а ядра; они дегенерируют пе-
ред или вскоре после слияния по-
лярных ядер.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. .
Эндоепермогенез нуклеарный.
Эмбриогенез — зародыш прямой с
двумя короткими семядолями и
длинным гипокотилем и корешком.
Подвесок короткий, однорядный.
Семя, плод — семя со слаборазви-
тым эндоспермом или без него. За-
родышт заполняет все или почти все
семя. Плод — септицидная коробоч-
ка.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Pra-
kash, Lau, 1976; Смирнова, 1977;
Тахталжян, 1966, 1970.
CEMEMCTBO CLUSIACEAE
Основное число хромосом — х =
== 7—10, 12, 19.
Цветок обычно однополый, редко
обоеполый.
Пыльник — нет данных,
Микроспорогенез и микрогамето-
генез — пыльцевые зерна трех-, ре-
же двух-, четырех-пятиборозднопо-
ровые, обычно соединенные в те-
трады.
Семяпочка анатропная — геми-
апатроппая, с двумя интегумента-
ми, тенуипуцеллятная. Микропиле
образустся паружпым интегумен-
том. Имоется ариллус. Семяпочка
одна — много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные, 7-об-
разные. Зародышевый мешок обра-
зуется по Ро]угопит-типу. Синер-
гиды с крючкообразными выроста-
ми и фибриллярными аппаратами.
Имеются три эфемерные антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нот
данных.
Эпдоепермогенез нуклеарный.
Эмбриогенез Опаста4-типа. Заро-
Hb прямой, крупный, двусемя-
дольный. Семядоли маленькие или
большие. Корешок короткий или
длинный.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Массивные запасающие семядоли
скрывают остальные части заро-
дыша, лишь кончик корешка едва
75
виден из них. Плод — септицидная
коробочка, костянка или ягода.
Гаусторий не обнаружен.
Апомиксис — нуцеллярная эмбри-
ония У Сагсйма шапооала.
Другие явления — полиэмбриония
у Clusia u Garcinia.
Литература: Corner, 1976; Davis,
1966; Тахтаджян, 1966, 1970; Смир-
нова, 1977.
СЕМЕЙСТВО НУРЕВТСАСЕАЕ
Основное число хромосом — х =
== 7—10, 12.
Цветок обоеполый.
Ныльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный двуядерный тапетум. `
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
нопоровые.
Семяпочка анатропная, с двумя
интегументами, тенуинуцеллятная;
микропиле образуется обоими ин-
тегументами. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопит-типу и имеет крахмаль-
ные зерна. Синергиды грушевид-
ные. Три — семь небольших анти-
под рано дегенерирующих или дол-
го сохраняющихся. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —у
Hypericum calinum перекрестное
опыление. Порогамия. Двойное оп-
лодотворение.
Эндоспермогенез нуклеарный. O6-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка, когда зародыш
уже хорошо развит.
Эмбриогенез Solanad-tuna. B3apo-
дыш прямой или согнутый, хлоро-
филлоносный, двусемядольный.
Имеется подвесок.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя. Се-
76
мядоли короткие, массивные, вы-
полняют запасающую функцию.
Плод — коробочка или ягодообраз-
НЫЙ.
Гаусторий не обнаружен.
Апомикеие — нуцеллярная эмбри-
onua y Hypericum maculatum.
Другие явления —у Hypericum
maculatum обнаружена полиэмбри-
ония. Добавочные зародыши возни-
кают из синергид. Имеется гино-
диэция.
Bugnicourt, 1969,
Литература:
1971 (1972); Corner, 1976; Davis,
' 1966; Gogellin, 1967; Salisbury, 1969;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ЕГАТИМАСЕАЕ
Основное число хромосом — х =
= 9, 10,
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эпдотеций, два
офемерных средних слоя и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
`мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна двух-трех-
клеточные, трехборозднопоровые.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Имеется нуцеллярный
колпачок. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий многоклеточ-
ный. Париетальных клеток 3—4 ря-
да. Тетрады макроспор линейные,
Т-образные. Зародышевый мешок
образуется по Ро[угопию-типу. Си-
нергиды с крючкообразными выро-
стами. Имеются три эфемерные
антиподы. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный. 3a-
ложение клеточных перегородок у
Вегоеа начинается с микропиляр-
ной части зародышевого мешка, у
ЕаИпае —с халазальной.
Эмбриогенез Зо]апай-типа. Заро-
дыш прямой или согнутый, двусе-
мядольный с гипокотилем и кореш-
ком, содержит масло и алейрон.
Подвесок однорядный,
рехклеточный.
Семя, плод — семя без эндоспер-
ма. зародыш вытянут по длинной
оси семени и заполняет весь его
объем. Семядоли, гипокотиль и ко-
решок составляют каждый около
1/3 общей длины зародыша. Плод —
коробочка.
Гауеторий не найден.
Апомиксие не обнаружен.
Другие явления —у НаМпае об-
наружена клейстогамия.
Литература: Согпег, 1976; Рафал,
Singh, 1970; 1971b; Salisbury, 1967;
Schnarf, 1931; CmupnHosa, 1977; Tax-
таджян, 1966.
трех-четы-
В порядке ТВеа!ез мпого либо со-
вершенно пе исследованных в от-
ношении цитоэмбриологии семейств,
либо исследованных настолько сла-
бо, что нет возможности дать им
цитоэмбриологическую — характери-
стику. К таким семействам отно-
сятся: Lophiraceae, Strasburgeria-
ceae, Ancistrocladaceae, Dioncophy-
llaceae, Diegodendraceae, Pentaphy-
lacaceae, » Tetrameristicaceae, Cary-
ocaraceae, Asteropeiaceae, Pellicie-
raceae, Quiinaceae mu Mcdusaceae.
Отсутствие достаточных цитоэм-
бриологических данпых по подав-
ляющему больынинству семейств
(12 из 20) порядка ТВеа1ез очепь
сильно затрудпяет выяспоние си-
стематических и филогенетических
отношений между его представите-
лями. Возможно, что в этот поря-
док включено излишне много се-
мейств и что он является в значи-
тельной степени искусственным.
Необходимы дальнейшие исследо-
вания цитоэмбриологийи представи-
телей порядка ТВеа]ез, чтобы уста-
новить его естественные границы.
ПОРЯДОК 34.
VIOLALES (CISTALES)
CEMEMCTBO FLACOURTIACEAE
Основное число хромосом — х =
= 40—12.
Цветок обоеполый, реже одно-
полый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный двух-четырехъядерпый та-
петум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка висячая, ортотропвая,
анатропная или гемианатронная, с
двумя интегументами, крассинуцел-
лятная. Микропиле зигзагообразное,
образуется обоими интегумелтами.
Имеются гипостаза и ариллус. Се-
мяпочек много, несколько.
Макроспорогенез и макрогамето-
генез — археспорий. Одпо-, иногда
мпогоклеточный. Париетальных кле-
ток 3—7 рядов. Тетрады макроспо
линейные. Зародышевый мешок об-
pasyetca no Polygonum-tuny um ume-
ет крахмал. Зрелый зародышевый
мешок сильно удлиненный. Синер-
гиды имеют крючкообразные выро-
сты и Ффибриллярные аппараты.
Имеются три эфемерные антиподы.
Полярные ядра сливаются перед
оплодотворением или во время оп-
лодотворения.
Опыление, прохождение пыльце-
вой трубки, оплодотворение — нет
дапных.
идоспермогенсз нуклеарный.
)мбриогенез — тип не определен.
Зародыш прямой, двуссмядольный,
с хорошо развитыми почечкой, ги-
покотилем и корешком, бесхлоро-
филльный, содержащий масло и
алеироновые зерна. Подвесок ко-
роткий.
Семя, плод — семя обычно с хо-
рошо развитым эндоспермом. Заро-
дыш занимает 3/4 или всю длину
семени. Периферия семени запол-
нена эндоспермом, содержащим ма-
сло и алейроновые зерна. Семенная
кожура образустся обоими иптегу-
ментами и состоит из пяти слоев:
паружного эпидермиса, гиподермы,
Удлиненных склереид, паренхимы
и внутреннего эпидермиса. Плод —
локулицидная коробочка или ягода,
реже орехообразный.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Dathan, Singh, 1973a; Hell,
1973; Смирнова, 1977; Тахтаджян,
1966, 1970.
ТТ
СЕМЕЙСТВО STACHYURACEAE
Основное число хромосом не уста-
новлено.
Цветок обоеполый или иногда
однополый.
Пыльник — 4 микроспорангия.
Стенка пыльника имеет эпидермис,
фиброзный эндотеций, средний слой
и тапетум.
Микроспорогенез и микрогамето-
генез — нет данных. Зрелые пыль-
цевые зерна двухклеточные, трех-
борозднопоровые.
Семяпочка анатропная, с Двумя
интегументами, крассинуцеллятная.
Микропиле зигзагообразное, обра-
зуется обоими интегументами. Име-
ются нуцеллярный колпачок, ка-
рункула и ариллус. Семяпочек
много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток три ряда. Те-
трады макроспор линейные, Т-об-
разные. Зародышевый мешок обра-
3yetca по Роугопит-типу. Синер-
гиды грушевидные. Имеются три
эфемерные антиподы. Полярные
ядра сливаются до оплодотворения.
Эндоспермогенез нуклеарный.
Эмбриогенез Solanad-, Onagrad-
типов. Зародыш прямой, маленький,
но дифференцированный. Корешок
короче семядолей. Зародыш содер-
жит масло и алейрон.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Плод ягодообраз-
ный, с кожистым околоплодником.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления —у Stachyurus
chinensis отмечена полиэмбриония.
Дополнительные зародыши образу-
ются путем расщепления зиготы и
из синергид.
Литература: Corner, 1976; Davis,
1966; Mathew, Chaphekar, 1977: Tax-
таджян, 1966, 1970; Yoshihiko, 1976c.
СЕМЕЙСТВО УТОГАСЕАЕ
Основное число хромосом — д =
= 6, 10—13.
Цветок обоеполый, изредка поли-
гамный.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—4:
эфемерных средних слоя и секре-
78
торный двуядерный тапетум. Име-
ются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные, изо-
илатеральные. Зрелые пыльцевые
зерна двухклеточные, трех (пяти)-
борозднопоровые. Пыльцевые зерна
прорастают в полости пыльника.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами или только внутрен-
ним. Имеются нуцеллярный колпа-
YOK, двойная гипостаза, постамент
и ариллус. Семяпочек 1—2, много.
Макроспорогенез и макрогамето-
генез — археспорий одно-, изредка
двухклеточный. Париетальных кле-
ток 2—4 ряда. Тетрады макроспор
линейные, Т-образные. Зародыше-
вый мешок образуется по Ро{уро-
пит-типу. Синергиды удлиненные,
с крючкообразными выростами. У
некоторых видов Ую]а (У. одога&а
var. praccox u V. riviniana) y cuHe-
ргид обнаружены фибриллярные
аппараты. Имеются три эфемерные
antTunogpr Y Viola tricolor, o6Hapy-
жена обращенная полярность заро-
дышевого мешка. Полярные ядра
сливаются До оплодотворения или
при оплодотворении.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — са-
моопыление и перекрестное опыле-
ние увидов У101а. Порогамия. Двой-
ное оплодотворение.
Эндоспермогенез нуклеарный. O6-
разование клеточных перегородок
в эндосперме центростремительное.
Эмбриогенез Аз{ега4-типа. Заро-
дыш прямой, крупный, хлорофил-
лоносный, двусемядольный, с хоро-
шо развитыми почечкой, гипокоти-
лем и корешком, содержит масло и
алейрон.
Семя, плод — семя с хорошо раз-
витым мясистым эндоспермом, со-
держащим масло и алейрон. Заро-
дыш занимает 3/4 общего объема
семени. Эндосперм располагается
по периферии семени. Семенная ко-
жура формируется обоими интегу-
ментами; внутренний эпидермис на-
ружного интегумента содержит ок-
салат кальция, в то время как
внутренний эпидермис внутреннего
интегумента склеренхиматозный.
Плод — локулицидная коробочка,
редко ягода или ореховидный,
Апомикеис не обнаружен.
Другие явления —у Viola odora-
{а отмечена клейстогамия.
Литература: Corner, 1976; Davis,
1966; Kuta, 1978; Schnarf, 1931; Singh,
1967a, 1970d; Singh, Gupta, 1967;
Смирнова, 1977; Тахтаджян, 1966,
1970; Верещагина, 1980.
СЕМЕЙСТВО BIXACEAE
Основное число хромосом — 1 =
‚ 8.
Цветок обоеполый.
Пыльник — 8 микроспорангиев.
Стенка микроспорангия имеет эпи-
дермис, слабофиброзный —эндоте-
ций, 1—3 эфемерных средних слоя
секреторный двуядерный тапс-
тум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траздральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехборозднопоровые, с глад-
кой экзиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеются нуцелляр-
ный колпачок, ариллус, слабо раз-
витая гипостаза. Семяпочек мпого.
Макроспорогенез и макрогамето-
хенез — архесторий одноклеточный.
Париетальпых клеток 4—6 рядов.
Тетрады макроспор липейные. За-
родышевый мешок образуется по
Polygonum-tuny и имеет зерна крах-
мала. Синергиды с крючкообразны-
ми выростами и фибриллярными
аппаратами. Имеются три эфемер-
ные антиподы. Полярные ядра сли-
ваются во время оплодотворения.
У Нха отеПапа описан слабо раз-
витый постамент. |
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез пуклеарный.
Позднее весь эндосперм становится
целлюлярным. Эндосперм содержит
крахмал, кристаллы оксалата каль-
ция.
Эмбриогенез нерегулярный. Заро-
дыш прямой, согнутый, крупный,
двусемядольный, бесхлорофилль-
ный, с хорошо развитыми почечкой,
гипокотилем и корешком, содержит
масло и алейрон.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародылт зани-
мает 3/4 общего объема семени. Эн-
досперм заполняет периферию се-
мени. Семенная кожура многослой-
ная, образуется обоими интегумен-
тами. Плод — локулицидная коро-
бочка.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Сворга, Каиг, 1965;
Corner, 1976; Dathan, Singh, 1972а,
b; Davis, 1966; Kaur, 1967a, 1969a,
1970; Singh, 1970; Singh, Gupta,
1967; Смирнова, 1977; Тахтаджяв,
1966, 1970.
СЕМЕЙСТВО СТЭТАСЕАЕ
Основное число хромосом — 5 =
—=5, 8—41.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный двух-четырехъядерный та-
петум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, ° изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, двух-четы-
рехборозднопоровые.
Семяпочка ортотропная, с двумя
интегумептами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Имеются нуцелляр-
ный колпачок, гипостаза и ариллус.
Семяпочек обычно много, изредка
одна-две.
Макроспорогенез и макрогамето-
генез — археспорий многоклеточ-
ный. Париетальных клеток 1—3 ря-
да. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Polygonum-tuny. Синергиды с крюч-
кообразными выростами и фибрил.
лярными аппаратами. Имеются три
Офемерные антиподы.
Нолярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный.
Позднее весь эндосперм становится
целлюлярным.
Эмбриогенез Solanad-runa. 3apo-
дыш прямой, согнутый или сверну-
19
тый, двусемядольный, © хорошо
развитыми почечкой, гипокотилем
и корешком. Содержит масло и алей-
рон. Нодвесок однорядный.
Семя, плод — семя с мучнистым
или хрящевидным эндоспермом, со-
держащим крахмал. Зародыш зани-
мает 3/4 общего объема семени.
Периферия семени заполнена эндо-
спермом. Кожура семени образует-
ся обоими интегументами. Плод —
кожистая или деревянистая коро-
бочка. Перикарпий волосистый, со-
держит большое число кристаллов.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления — клейстогамия.
Полиэмбриония. ,
Литература: Согпег, 1976; Davis,
1966; Kapil, 1967b; Kapil, Maheshwa-
ry, 1965; Maheshwary P, 1966; Poma-
nospa, 1968, 1970; Schnarf, 1934; Tax-
таджян, 41966, 1970c.
В порядке Violales семейство
Cochlospermaceae — цитоэмбриологи-
чески очень слабо изучено, а Рет!-
discaceae u Schyphostegiaceae вовсе
не изучены. По мнению Синга
[Singh, 1967], Violaceae по данным
цитоэмбриологии обнаруживает
большое сходство с Самсасеае, Ci-
staceae, Bixaceae wu Resedaceae.
На основании данных цитоэмбри-
ологии Нхасеае сближают c Cista-
ceae и СосШозрегтабсеае [Kaur,
1970; РаФап, ЗтеВ, 1972]. На осно-
вании данных цитоэмбриологии
Cunr u Tynta [Singh, Gupta, 1967]
высказали предположение о TOM,
что \У!1асеае, Capparidaceae и
Везедасеае — близкородственные се-
мейства и, возможно, возникли от
общего предка, сходного с ныне су-
ществующими Сарраг1дасеае, путем
параллельной эволюции. Однако не-
обходимы дальнейшие исследова-
ния этих семейств, чтобы выяснить
их филогенетические взаимоотно-
шения.
ПОРЯДОК 35.
PASSIFLORALES
CEMEMCTBO PASSIFLORACEAE
Основное число хромосом — х =
= 6, 9, 10.
80
Цветок обычно обоеполый, реже
однополый.
Ныльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя, секретор-
ный многоядерный, одно- или дву-
рядный тапетум. Наблюдается сли-
яние ядер и образование полипло-
идных ядер в клетках тапетума.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, 3—12-борозднопоровые, с
сетчатой экзиной.
Семяпочка анатропная, изредка
ортотропная, с двумя интегумента-
ми, крассинуцеллятная. Семяножка
довольно длинная. Микропиле об-
разуется обоими интегументами,
Имеется ариллус. Сосудистый пучок
доходит до халазы. Семяпочек
много.
Maxkpocnoporenes п макрогамето-
генез — архоспорий одно-, много-
клеточный. Париетальная ткань
массивная (до 20 рядов). Тетрады
макроспор линейпые. Зародышевый
мешок формируется mo Polygonum-
типу. Синергиды с крючкообразны-
ми выростами. Имеются три эфе-
мерные антиподы. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление у видов Раз-
siflora. Порогамия. Двойное опло-
дотворение. Пыльцевые трубки дол-
то сохраняются в семяпочке.
Эндоспермогенез нуклеарный. Эн-
досперм руминированный. Образо-
вание клеток в эндосперме центро-
стремительное. На поздних стади-
ях развития весь зародышевый
мешок заполнен клетками.
Эмбриогенез Опаота4-типа. Заро-
дьши прямой, крупный, двусемя-
дольный, © хорошо развитыми по-
чечкой, гипокотилем и корешком,
бесхлорофилльный, содержащий
алейрон.
Семя, плод — семя с эндоспер-
мом, содержащим масло и алейрон.
Зародыш занимает 3/4 общего объ-
ема семени. Эндосперм располага-
ется по периферии семени. Кожура
семени образуется обоими интегу-
ментами. Внешний эпидермис внут-
реннего интегумента формируется
в склереидный ряд. Плод коробоч-
ка или ягода,
Апомиксис не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Dathan,
Singh, 1969, 1973b; Davis, 1966; Kaur,
19674; Schnarf, 1934; Singh, 1967c,
1970c; CmupHosa, 1977; TaxTapnan,
4966, 1970.
СЕМЕЙСТВО ТОВМЕКАСЕАЕ
Основное число хромосом — х =
= 7, 10.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средних слоя и секре-
торпый дву- многоядерпый танетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультаннос. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехбороздные, с гладкой эк-
зиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Имеются гипостаза и
ариллус. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий — одно-, — Дву-,
многоклеточный. Паристальных кле-
ток 3 ряда. Тетрады макроспор ли-
нейные. Зародышевый мешок фор-
мируется по Polygonum-tuny. Си-
нергиды с крючкообразными выро-
стами. Имеются три эфемерные ан-
типоды. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Пыльцевые трубки долго сохраня-
ются в семяпочке.
Эндоспермогенез нуклеарный. На-
блюдается скопление ядер и цито-
плазмы эндосперма вокруг зароды-
ша и в халазальной части зароды-
шевого мешка. Эндосперм содержит
крахмал.
Эмбриогенез Опастад-типа. Заро-
дыш крупный, прямой, двусемя-'
дольный с хорошо развитыми по-
чечкой, гипокотилем и корешком,
содержит масло и алейрон. Подве-
сок короткий, двухклеточный.
Семя, плод — семя с эндоспер-
мом, содержащим масло и алейрон.
6 В. А. Поддубная-Арнольди
Зародыш занимает 3/4 общего объ-
ема семени. Эндосперм располагает-
ся по периферии семени. Семенная
кожура образуется двумя рядами
наружного интегумента и тремя ря-
дами внутреннего. Плод — локули-
цидная коробочка. Перикарпий па-
ренхиматозный, разделенный на
три зоны.
Гауеторий отсутствует.
Апомикеис не обнаружен.
Другие явления — не отмечены.
Литература: Corner, 1976; Davis,
1966; Kaur, 1967d, 1970d; Mahalin-
gappa, 1975; Смирнова, 41977; Тах-
таджян, 1966, 1970; Vijayaraghavan,
Kaur, 1966.
СЕМЕЙСТВО САВТСАСЕАЕ
Основное число хромосом — х ==
——
——
Цветок обычно однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный слой, один эфе-
мерный средний слой и секретор-
ный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровьыюо с сотчатой экзиной.
Семяпочка апатронная, с двумя
иптогументами, крассинуцеллятная.
Микропило образустся двумя инте-
гументами. Имеются нуцеллярный
колпачок и гипостаза. Сосудистый
пучок Доходит до халазы, где он
принимает зонтиковидную форму.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 8 рядов. Те-
трады макроспор линейные. Заро-
дышевый мешок Ро]усопит-типа.
У видов Самса обнаружены тетра-
скопические зародышевые мешки.
Сипергиды грушевидные с фибрил-
лярпыми аппаратами. Имеются три
офемерные антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление y Carica pa-
рауа. Порогамия. Пыльцевые труб-
ки долго сохраняются в семяпочке.
Эндоепермогенез нуклеарный. Об-
разование клеточных перегородок
в эндосперме центростремительное.
Начинается оно с микропилярной
81
части зародышевого мешка через
79 дней после опыления.
Эмбриогенез нерегулярный. На
основании современных знаний не-
возможно отнести его к какому-ни-
будь известному типу. Зародыш
крупный, прямой, лопатчатый, бес-
хлорофилльный, с хорошо развиты-
ми широкими плоскими семядолями,
почечкой, гипокотилем и кореш-
ком.
Семя, плод — семя сэндоспермом,
содержащим масло. Зародыш зани-
мает 3/4 объема семени. Эндосперм
расположен по периферии семени.
Семенная кожура содержит миро-
зин. Она образуется обоими инте-
гументами и дифференцируется в
саркотесту и эндотесту. Плод мя-
систый, тыквинообразный.
Гауеторий отсутствует.
Апомиксис не обнаружен.
Другие явления — партенокарпия.
Jlureparypa: Corner, 1976; Dathan,
Singh, 1969, 1976; Davis, 1966;
Schnarf, 1931; Singh, 1967a, 1970a;
Смирнова, 1977; Тахтаджян, 1966,
1970.
В порядке РаззШога]ез семейства
Маезвегасеае и АсвВатасеае цито-
эмбриологически не изучены. Срав-
нительная цитоэмбриология пока-
зывает, что Тагпегасеае скорее
сродни РаззШогасеае, чем Гоаза-
сеае, и что отнесение его к поряд-
ку Гоазаез, как это предложил
Хатчинсон, неправильно. По мне-
нию Д. Синга [5тор, 1967], Самса-
сеае по цитоэмбриологии сходно с
Violaceae, Cistaceae, Bixaceae m Re-
зедесеае. Исследованные в отноше-
нии цитоэмбриологии — семейства
Passiflorales обнаруживают значи-
тельное сходство друг с другом,
что может указывать на естествен-
ность этого порядка и правильность
сближения его семейств. Особенно
сходны РаззШогасеае и Turnera-
сеае. Оба семейства характеризу-
ются симультанным микроспороге-
незом, двухклеточными пыльцевы-
ми зернами, анатропными, с двумя
интегументами, крассинуцеллятны-
ми семяпочками, одно-многоклеточ-
ным женским археспорием, парие-
тальными клетками, зародышевы-
ми мешками Ро]угопит-типа, двой-
ным оплодотворением, нуклеарным
эндоспермом, эмбриогенезом Опа-
82
отад-типа, крупным, прямым, хоро-
то дифференцированным зароды-
шем, семенами, содержащими эндо-
сперм.
ПОРЯДОК 36.
CUCURBITALES
CEMENCTBO CUCURBITACEAE
Основное число хромосом — х =
— 7—14.
Цветок — обычно однополый, из-
редка обоеполый. Растения одно-
домные или двудомные.
Пыльник — 2—4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—3
эфемерных средних слоя и секре-
торный дву-трехъядерный тапетум.
Микроепорогенез и микрогамето-
гепез — образование микроспор си-
мультаннос. Тетрады микроспор те-
траэдральныс, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерпа двухклеточные, трех- десяти-
борозднопоровые, или трех-пятипо-
ровые, обычно с гладкой, изредка
с сетчатой экзиной. Апертуры про-
стые или сложные.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле прямое, образуется оно
внутренним интегументом. Имеют-
ся нуцеллярный клювик и гипоста-
за. Сосудистые пучки ветвятся или
не ветвятся, они доходят до халазы
и проникают в наружный интегу-
мент. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий —одно-, дву-,'
трех-, многоклеточный. Париеталь-
ных клеток 5—6 рядов. Тетрады
макроспор обычно линейчые, реже
Т — Г-образные. Зародышевые меш-
ки образуются по Polygonum- и Al-
Пат-типам, содержат крахмал. Си-
нергиды с крючкообразными выро-
стами и фибриллярными аппарата-
ми, Имеются три обычно эфемерные
антиподы. Иногда число антипод
увеличивается до пяти и более. По-
лярные ядра сливаются при опло-
дотворении. В центральной клетке
зародышевого мешка, а иногда и в
яйцеклетке обнаружены крахмаль-
ные зерна.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодо-
творение премитотического типа.
Пыльцевые трубки долго сохраня-
ются в семяпочке, изредка ветвят-
ся. У Сисипз зайуиз оплодотворе-
ние происходит через 24—36 ч по-
сле опыления.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
в эндосперме центростремительное.
Начинается оно с микропилярной
части зародышевого мешка. В эн-
досперме обнаружены ядра с чрез-
вычайно высокой плоидностью
Эмбриогенез Onagrad-, Solanad-
типа. Зародыш прямой, крупный,
лопатчатый, двусемядольный, бес-
хлорофилльный, содержит алейрон
и масло. Подвесок обычно двухкле-
точный.
Семя, плод — семя с очень слабо-
развитым маслянистым эндоспер-
MOM или без него. Зародыш запол-
няет все семя. Массивные запаса-
ющие семядоли скрывают осталь-
ные части зародыша, лишь кончик
корешка едва выступает из них.
Семенная кожура образуется из на-
ружного интегумента и имеет 5 зон:
1) эпидермис, 2) гиподерму, 3) скле-
ренхиму, 4) аэренхиму и 5) хло-
ренхиму или парепхиму. Плод —
ягода или тыквина. Порикарпии со-
стоит из эпикарпия, гиподермы, ме-
зокарпия и эпдокарпия.
Гаусторий энпдоспормальпый, аг-
рессивный. Образуется в халазаль-
ной части эндосперма и значитель-
но различается по длине. У од-
них видов он сравнительно корот-
кий — от 9,90 до 14,90 мм, у других —
очень длинный —от 13,90 до
20,7 мм. Кроме того, у одних видов
гаустории целлюлярные, у других
ценоцитные.
Апомиксис — генеративная — апо-
спория, редуцированный партено-
генез, апогаметия, нуцеллярная эм-
бриония.
Другие явления — полиэмбрио-
ния, партенокарпия.
Литература: Алиева, 1952; Ашур-
метов, 1974а, 6, в; 1975; Ашурметов,
Дзевалтовский, 1975; Ашурметов,
Саидов, 1975; Сворга, Асагуа], 1960;
Chopra, Basu, 41965; Chopra, Seth,
4977; Corner, 1976; Dathan, 1974a, b;
Dathan, Singh, 1971a; Дзевалтов-
ский, 1964, 1962, 1963а, 6, в, 1964,
1965, 4967, 1968, 1969, 1970, 1971,
6*
1973, 1975, 1977; Дзевалтовский и др.,
1973a, 6; Дзевалтовский, Шайтан,
1969; Дзевалтовский, Жалалов, 1976;
Davis, 1966; Mihov, Zagorcheva,
1966; Муратов, 1972; Nikam, 1976;
Саидов и др., 1975; ЗебпатЁ 1934;
Sing, 1967Ъ, е, 1970, 1971—1972;
Singh, Dathan, 1969, 1974a, b,c, 1972,
4973, 1974, 1976; Смирнова, 1977;
Тахтаджян, 1966, 1970; Тиги]а, 1966,
4969, 1971; Turula, Szybowska, 1974;
Чолохян, 1965; Загорчева, 1969,
1972, 1976а, 6; Загорчева, Молхова,
1965, 1976; ЗЖалалов, 1972, 1973,
1974; Жалалов, Дзевалтовский, 1973,
1975, 1977; Жалалов и др., 1977.
Цитоэмбриологические данные
проливают свет на систематическое
положение Cucurbitaceae. Crpoenue
семяпочки, интегументов, сосуди-
стого пучка, эндосперма и семенной
кожуры не сходно с таковыми
Aristolochiaceae, с которым его
сближали некоторые ученые [\и!-
Лет, 1923]. На этом же основании
Campanulaceae, имеющего тенуину-
целлятную семяпочку, с одним ин-
тегументом, целлюлярный —эндо-
сперм с микропилярным и хала-
зальным гаусториями, семя с эндо-
спермом ис семянной кожурой,
имеющей лишь один эпидермис с
несколькими гиподермальными ря-
дами, также по сходно c Cucurbita-
сеае. Пыльцевые зерпа Сатрапи]а-
сеае отличаются от таковых Сиеиг-
bitaceae [Егдшат, 1952]. Сравнение
цитоэмбриологических признаков
Cucurbitaceae с такими семейства-
ми, Kak Begoniaceae, Bixaceae, Са-
ricaceae, Cistaceae, Datiscaceae, Fla-
courtiaceae, Passifloraceae, Turnera-
сеае и У1о]асеае также не обнару-
живает признаков родства между
ними. Пыльцевые sepa Cucurbita-
сеае отличаются от таковых выше-
отмеченных — семейств — [Ег4тап,
1952]. Только с РаззИогасеае они
обнаруживают сходство. СиситЪНа-
сеае отличается цитоэмбриологиче-
ски от всех тех семейств, с кото-
рыми его сближали, за исключени-
ем РаззШогасеае. Цитоэмбриологи-
ческие данные не подтверждают
мнений Мельхиора [1964] и Кронк-
виста \[4968], которые сближали
Cucurbitales c У10]аез и произво-
дили первые из последних. Цито-
эмбриологические данные показы-
83
вают, что СиситЬЦасеае является
высокоразвитым семейством, выде-
ление которого в самостоятельный
порядок и расположение его вблизи
Разз1ота]ез не вызывает сомнения.
ПОРЯДОК 37.
BEGO NIALES
(DATISCALES)
CEMEMCTBO DATISCACEAE
Основное число хромосом — z=
== 11.
Цветок — обычно однополый. Ра-
стения двудомные. `
Пыльник — нет данных.
Микроспорогенез и микрогамето-
генез — нет данных. Зрелые пыль-
цевые зерна двухклеточные.
Семяпочка анатропная с Двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 5 рядов. За-
родышевый мешок образуется по
АШит-типу. Синергиды грушевид-
ные. Имеются три антиподы, деге-
нерирующие после оплодотворения.
Полярные’ ядра сливаются до опло-
дотворения. °
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарпый.
Позднее весь эндосперм становится
целлюлярным.
Эмбриогенез Onagrad-tuna. Заро-
дыш прямой или слегка согпутый,
двусемядольпый, содержащий мас-
ло и алейрон. Подвесок короткий,
малоклеточный.
Семя, плод — семя без эндоспер-
ма или со слаборазвитым эндоспер-
мом, содержащим масло. Зародыш
заполняет все или почти все семя.
Эндосперм, если он имеется, распо-
лагается по периферии семени.
Плод — коробочка.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis
1966; Schnarf, 1934; Тахтаджян,
1966, 1970.
84
СЕМЕЙСТВО ВЕСОМТАСЕАЕ
Основные числа хромосом — Zz =
= 40—15, 48, 24, 24.
Цветок однополый. Растение од-
нодомное.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1, из-
редка 2 эфемерных средних слоя и
секреторный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные. Зрелые пыльцевые
зерна двухклеточные.
Семяпочка анатропная, с Двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальпые клетки в числе двух
образуются, но вскоре дегенериру-
ют. Тетрады макроспор линейные.
Зародынтевый мешок образуется по
Ро]угопит-тпиу. Синпергиды груше-
видире. Три эфемерпые антиподы.
Полярные ядра сливаются перед
оплодотворенпем.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма центро-
стремительное.
Эмбриогенез Опастад-типа. Заро-
дыш прямой или согнутый, двусе-
мядольпый, содержит масло и алей-
pow. Подвесок короткий, малокле-
точный.
Семя, плод — семя без эндоспер=
ма или со слабо развитым эндоспер-
мом. Зародыш заполняет все или
почти все семя. Если имеется эндо-
сперм, то оп запимает периферию
семени. Плод — локулицидная коро-
бочка, или редко ягода.
Гаусторий не найден.
Апомиксие не обнаружен.
Другие явления не обнаружены,
Литература: Corner, 1976; Davis,
1966; Schnarf, 1931; Тахтаджян
1966, 1970.
В отношений цитоэмбриологии
Datiscaceae исследовано слабее, чем
Begoniaceae. Однако все же видно,
что оба эти семейства имеют сход-
ные цитоэмбриологические призна-
ки, что подтверждает правильность
отнесения их к общему порядку.
ПОРЯДОК 38.
CAPPARALES
CEMEMCTBO CAPPARACEAE
Основное число хромосом — z =
= 8—11, 13, 16, 17.
Цветок обоеполый, реже однопо-
лыйЙ.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—4
эфемерных средних слоев и сскрс-
торный, одпо-, дву-, четырехъядер-
ный тапетум. Ядра в клетках тапе-
тума часто сливаются, в результа-
те чего становятся полиплоидными.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна обычно двухклеточные, из-
редка трехклеточные, двух-трех-че-
тырехборозднопоровые, с толстой,
слабо шиповатой окзипой, содер-
KAT крахмальпьо зорпа.
Семяпочка апакампилотропная,
кампилотроиная, с двумя интегумен-
тами, крассипуцеллятпая. Микро-
mune образуется обычно обоими
интегументами, изредка только вну-
тренними (у Сарраг!$). Часто ми-
кропиле зигзагообразное. Имеется
нуцеллярный колпачок. Семяпочек
много, несколько или только 1—2.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный или одноклеточный, изредка
двухклеточный. Париетальных кле-
ток 3—7 рядов. Тетрады макроспор
линейные, Т-образные. Зародыше-
вый мешок образустся по Ро]уго-
пит-типу п имеет крахмальные
зерна. Синергиды с крючкообраз-
ными выростами и фибриллярными
аппаратами. Имеются три малень-
кие эфемерные антиподы, за исклю-
чением Маегиа, у которой антипо-
ды крупные и долго сохраняются.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение. По-
ный, хлоро
рогамия. Двойное оплодотворение
премитотического типа. Сингамия и
тройное слияние протекают одно-
временно.
Эндоспермогенез нуклеарный. При
глобулярном зародыше эндосперм
становится клеточным. Число сво-
бодных ядер эндосперма достигает
4000, прежде чем начнется образо-
вание клеточных перегородок. Име-
ется перисперм.
Эмбриогенез Onagrad-, Solanad-,
Caryophyllad-runos. Зародыш круп-
ный, согнутый, складчатый, хорошо
дифференцированный, двусемядоль-
биллоносный содержит
масло, алейрон, мирозин, ипогда и
крахмал. Подвесок длиппый, одно-
рядный.
Семя, плод — семя без эндоспер-
ма или со скудным эндоспермом,
содержащим алейрон и масло. За-
родыш со своими мясистыми запа-
сающими семядолями располагает-
ся в центре семени и в тех случа-
ях, когда имеется эндосперм, со
всех сторон окружен эндоспермом,
состоящим из одного слоя клеток.
Семенная кожура образуется дву-
мя интегументами; наружный слой
содержит танниноподобные соеди-
нения. Плод — коробочка, стручок
или ягода, иногда орех или ко-
стяпка.
Гаусторий подвесочный.
Аномиксие — пуцоллярная эмбрио-
ius y Capparis frondosa.
Другие явления — гинодиэция У
Cardamine.
Литература: Corner, 1976; Davis,
1966; Fathima, Kumari, 1970; Mitra,
1969; Narayana, 1967a, 1970а; Рта-
sad, 1977; Rao A., 1967b; Rao B. et
al., 1977; Rao P. et al., 1976; Rao P.,
Rao B., 1975; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО МОМКСАСЕАЕ
Основное число хромосом — х =
Цветок обоеполый.
Ныльник — 4—2 микроспоравгия.
Стенка микроспорангия многоряд-
ная. Тапетум секреторный, дву-,
многоядерный. В результате слия-
ния ядра в клетках тапетума ста-
новятся полиплоидными. Имеются
орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
85
крестообразные. Зрелые пыльцевые
зерна двухклеточные, двух-трех-бо-
розднопоровые со слегка шипова-
той экзиной.
Семяпочка анатропная, кампило-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется обоими интегументами.
Оно зигзагообразное. Имеется сла-
боразвитая гипостаза. Семяпочек
много.
Макроспорогенез и макрогамето-
генез — археспорий одно-, изредка
двух-, трехклеточный. Париеталь-
ных клеток 5—8 рядов. Тетрады
макроспор линейные, Т-образные.
Зародышевый мешок образуется по
Ро]угопит-типу. Синергиды с крюч-
кообразными, выростами и фибрил-
лярными аппаратами. Имеются три
эфемерные антиподы. Полярные
ядра сливаются при оплодотворе-
нии.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия, двойное оплодотворепие.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка.
Эмбриогенез Азега4-типа. Заро-
дыш прямой двусемядольный, хло-
рофиллоносный, содержит масло,
алейрон. Подвесок массивный. Се-
мядоли неодинаковой величины.
Семя, плод — семена без эндо-
сперма. Зародыш заполняет все
семя, массивные запасающие семя-
доли скрывают остальные части за-
родыша, лишь кончик корешка ед-
ва выступает из них. Семенная ко-
жура образуется обоими интегумеи-
тами. Внешний слой массивпый со
склереидной тканью, впутренний ре-
дуцирован до тонкого пластипчатого
слоя. Плод — стручковидпая удли-
ненная трехстворчатая коробочка.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; Dutt, 1978b; Marayana, 41967b,
19705; Schnarf, 4934; Смирнова,
1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВВАЗЗТСАСЕАЕ
Основное число хромосом — х =
4—13, 45.
86
Цветок обоеполый, реже однопо-
лый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 4—3
эфемерных средних слоя и секре-
торный, двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, ;‚изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, двух-трех-че-
четырехборозднопоровые. Клетки-
спермии хорошо различимы.
Семяпочка висячая, анатропная,
анакампилотропная, после оплодо-
творения анамфитропная, © дДву-
мя интегументами, тенуи- или
крассинуцеллятная. Микропиле зиг-
загообразное, образуется обоими
интегументами. Имеется эндотелий.
Y Heliophila amplexicaulis o6napy-
жено пеобычайное не только для
Brassicaceae, по и для всех других
семейств покрытосеменных строе-
nue семяпочки. После оплодотво-
рения у Н. ашр]ех1сацИ$ клетки во
внутреннем интегументе сильно
разрастаются, ядра в них делятся,
и они становятся крупными, мно-
гоядерными. В процессе развития
зародыша эти клетки разрушаются,
и к началу дифференциации заро-
дыша от внутреннего интегумента
остается лишь два наружных мел-
коклеточных слоя. Зародышевый
мешок у H. amplexicaulis не изо-
гнут подковообразно, как У других
представителей Вгазз1сасеае. По-ви-
димому, это объясняется отсутстви-
ем в вектральной части семяпочки
Н. атрехсаиз «базального тела»,
свойственного другим представите-
лям Вгазусассае. Семяпочек две,
несколько-много. Сосудистый пучок
доходит до халазы.
Макроспорогенез и макрогамето-
генез — археспорий много-, одно-,
двухклеточный. Париетальные клет-
ки либо присутствуют, либо отсут-
ствуют. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок образуется по Ро]угопит-ти-
пу; яйцеклетка нередко достигает
значительной длины. Синергиды с
крючкообразными выростами и фиб-
риллярными аппаратами. Имеются
три небольшие эфемерные антипо-
ды. Полярные ядра сливаются при
оплодотворении.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление, самоопыле-
ние. Порогамия. Двойное оплодо-
творение премитотического типа.
Эндосперм нуклеарный. Заложе-
ние клеточных перегородок между
свободными ядрами происходит
обычно во время дифференциации
зародыша. У Succowia balearica
число свободных ядер достигает
4000, прежде чем начнется клетко-
образование.
Эмбриогенез Опаота4-типа. Заро-
дыш согнутый, двусемядольный,
хлорофиллоносный, содержащий
алейрон, мирзоин и друзы оксала-
та кальция. Семядоли по своим
размерам в песколько раз превы-
шают гипокотиль и корешок. Под-
весок длинный, многоклеточный, но
одноядерный. У некоторых видов
с гигантской пузыревидной клет-
кой Y основания.
Семя, плод — семя без эндоспер-
ма или с незначительными остат-
ками его, содержащими алейрон.
Зародыш заполняет все или почти
все семя. Семядоли зародыша вы-
полняют запасающую функцию. Се-
менная кожура формируотся обои-
ми интегументами. Плод — коробоч-
ка, стручочек или ятода, ипогда
орех или костянка. Исходным ти-
пом является стручок.
Гаусторий отсутствует.
Апомиксис — нуцеллярная эмбри-
ония, редуцированный партеноге-
нез, псевдогамия при нередуциро-
ванном партеногенезе.
Другие явления — клейстогамия,
полиэмбриония. Партенокарпия.
Литература: Беляева Л., 1963,
1967, 1968; Беляева Л., Фурса, 1978,
1979; Беляева Л. и др., 1977а, 6:
Беляева Л. и др., 1978; Вуоф 1966;
Chaturvedi, Rathore, 1970; Corner,
1976; Davis, 1966; Ilakki, 1974; Wne-
ина, 1962; Кавецкая, 1958, 1960;
Кордюм, 1963; М1зга, 1966; Поляко-
Ba, 1964; Prasad, 1977; Rathore, Singh,
1969; Родионова, 1966a, 6, B, 1967a,
6, 1971a, 6, B, 1972, 1973, 1976, 1978,
1979; Шнайдер, Сильверс, 1974:
Schnarf, 1931; Смирнова, 1977; Со-
колов, 1975; Тахтаджян, 1966, 1970;
Tykarska, 1979; Vaughan, 1974; Яков-
лев, Алимова, 1976.
СЕМЕЙСТВО ВЕЗЕРАСЕАЕ
Основное число хромосом — 5х =
= 6, 7, 10. .
Цветок обоеполый, реже однопо-
лый.
Пыльник — 4 микроспорангия.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
изобилатеральные, тетраэдральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
ные или трехборозднопоровые.
Спермии-клетки хорошо различимы.
Семяпочка анакампилотропная,
анатропная, анамфитропная, с дву-
мя иптегумептами, тенуинуцеллят-
пая или крассинуцеллятная. Ми-
кропиле зигзагообразное; образует-
ся обоими интегументами. Наблю-
дается ослизнение эпидермальных
клеток нуцеллуса. Семяпочек мно-
го, изредка 10—18.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный. Париетальные клетки присут-
ствуют или отсутствуют. Тетрады
макроспор линейные. Зародышевый
мешок образуется по Ро1угопит-ти-
пу и имеет крахмал. Синергиды с
крючкообразными выростами. Име-
ются три эфемерные антиподы. По-
лярные ядра сливаются до оплодо-
творения.
Опылепие, прохождение пыльце-
вой трубки и оплодотворение —- пе-
рекрестное опыление у Reseda lu-
tea u R. odorata. Порогамия. Двой-
ное — оплодотворение. —Пыльцевые
трубки долго сохраняются в семя-
почке.
Эндосперм нуклеарный.
Эмбриогенез Опаотад-типа. Заро-
дыш согнутый, хлорофиллоносный,
двусемядольный, содержит алейрон
и мирозин. Семядоли массивные,
выполняющие запасающую функ-
цию. По размерам семядоли значи-
тельпо больше гипокотиля и ко-
решка.
Семя, плод — семя с эндоспермом
или без него. Зародыш располага-
ется в центре семени. Он либо со
всех сторон окружен эндоспермом,
либо занимает весь объем семени
при отсутствии эндосперма. Плод —
коробочка, ягода или состоит из од-
носеменных плодиков.
Гаусторий отсутствует.
Апомикеис не обнаружен.
Другие явления не отмечены.
87
Литература: Corner,
1966; Schnarf, 1931; Singh, Gupta,
1967; Смирнова, 1977; Тахтаджян,
1966, 1970; Чабан, 1975, 1979; Ча-
бан, Яковлев, 1974; Яковлев, Ча-
бан, 1979.
1976; Davis,
В порядке Сарраг19а]ез семейства
Koelberliniaceae, Pentadiplandraceae,
Tovariaceae wu Emplingiaceae muro-
эмбриологически He исследованы.
Изученные в пастоящее время пред-
ставители Сарраг1да!з имеют мно-
го общих цитоэмбриологических
признаков. Нараяна ([Narayana,
1967] считает, что Capparidaceae
наиболее примитивное семейство в
порядке Сарраг@а1ез и что Мотт-
gaceae тесно связано с Capparida-
сеае. Возможно, что Могшоасеае и
Везедасеае возникли из Capparida-
сеае. Brassicaceae цитоэмбриологи-
чески сходно с Саррагассае и пред-
ставляет собой наивысшую ветвь
Саррага1ез. Это семейство удиви-
тельно гомогенное. Представители
Вгаззсасеае отличаются значитель-
ным сходством по большинству ци-
тоэмбриологических признаков, что
указывает на естественность семей-
ства. Своеобразное строение семя-
почки Heliophila amplexicaulis, по
мнению Т.Б. Родионовой [1972],
подтверждает обособленное поло-
menue H. amplexicaulis Bp сёмействе
Brassicaceae и правильность отне-
сения его к самостоятельной три-
бе — НеНорПеае согласно с систс-
мой Янхена [ТапсвВеп, 1942].
ПОРЯДОК 39.
TAMARICALES
CEMEHMCTBO TAMARICACEAE
Основное число хромоеом — += 12.
Цветок обычно обоеполый.
Пыльник — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средних слоя и секреторный
дву-, трехъядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изо-
билатеральные. Зрелые пыльцевые
88
зерна двухклеточные, (двух), (четы-
рех)-бороздные. Спермии-клетки хо-
рошо различимы.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Сосудистый пучок до-
ходит до халазы. Семяпочек две —
много.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальных клеток один ряд. За-
родышевый мешок у Мумсаша сег-
manica образуется по ЕгиШана-ти-
пу. Что касается видов Tamarix, то,
по мнепию одних исследователей,
у этого рода даже в пределах одного и
того же вида типы зародышевого
мешка различные (Адоха, Огаза, Fri-
ИПана-типы), по мнению других, ви-
ды рода Татагх характеризуются на-
личием dump Fritillaria-rana [Benae-
ва, 1978]. Синергиды широкие, с
короткими крючкообразными выро-
стами, изредка с фибриллярными
аипаратами (Мумсама вотташса).
Аитиподы дегеперируют в момент
оплодотворепия или вскоре после
него. Полярные ядра сливаются при
оплодотворении.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоепермогенез целлюлярный,
нуклеарный.
Эмбриогенез Solanad-tuma. Заро-
дыш прямой или согнутый, двусе-
мядольный, содержащий масло и
алейроп. Семядоли массивные, вы-
полпяющие запасающую функцию.
По размерам семядоли превышают
гипокотиль и корешок.
одпорядпый.
Семя, плод — семя со слабо разви-
тым эндоспермом, содержащим
крахмал или без эндосперма. Заро-
дыш располагастся в цептре семени.
Эндосперм в тех случаях, когда они
имеется, располагается по перифе-
рии семени. Плод — локулицидная
коробочка.
Гауеторий отсутствует.
Другие явления — полиэмбриония.
Литература: Беляева Н., 1977; Сог-
ner, 1976; Davis, 1966; MockosBa, Xpz-
ctos, 1968, 1969; Schnarf, 1931; Cmup-
нова, 1977; Тахтаджян, 1966, 1970.
Подвесок_
‹
СЕМЕЙСТВО ЕОЧООТЕВТАСЕАЕ
Основное число хромосом — х==8.
Цветок обоеполый.
Пыльник — нет данных.
Микроспорогенез и микрогаметоге- .
нез — нет данных. Зрелые пыльце-
вые зерна двухклеточные, трехбо-
розднопоровые.
Семяпочка анатропная, с двумя
интегументами тенуинуцеллятная,
Микропиле образуется внутренним
интегументом. Семяножка короткая.
Имеется эндотелий. Он окружает
зародышевый мешок. Семяпочек
пемного.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-, многокле-
точный, Париетальные клетки отсут-
ствуют. Тетрады макроспор липей-
ные. Зародышевый мешок образуст-
ся по Роугопит-типу. Зародышевый
мешок Т-образной формы. Синерги-
ды грушевидные. Три антиподы де-
генерируют вскоре после оплодотво-
рения. Полярные ядра сливаются во
время оплодотворения.
Опыление, прохождение пыльще-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Тройное слияние предшествует син-
гамии.
Эндоспермогенез целлюлярный.
Эмбриогенез Азегад-типа. Заро-
дыш прямой, двусемядольный, со-
держащий масло и алойроп. Сомя-
доли выполняют запасающую Фупк-
ЦИЮ.
Семя, плод — семя со слаборазви-
тым эндоспермом, содержащим мас-
ло и алейрон. Зародыш заполняет
центр семени. Эндосперм распола-
гается по периферии семени. Плод —
трехстворчатая коробочка.
Гаусторий эндоспермальный (лате-
ральный) и зародышевого мешка.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Соуй, 1967а, 1970а; Тахтаджян,
1966, 1970; Смирпова, 1977.
СЕМЕЙСТВО ЕВАМКЕМТАСГАЕ
Основное чиело хромосом — х==5.
Цветок обоеполый.
Пыльник — число микроспоранги-
ев пе известно. Стенка микроспо-
рангия имеет эпидермис, фиброзный
эндотеций, 2—3 средних слоя и дву-
рядный секреторный двуядерный та-
петум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые
зерна трехклеточные, (двух)- трех-
(четырех)-бороздные, редко ругат-
ные.
Семяпочка анатропная, c Двумя
или одним интегументами, красси-
пуцеллятная. Микропиле образуется
внутренним интегументом. Иногда
образуется рудиментарный третий
интегумент. Семяножка длинная. Се-
мяпочек 2—3 — много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Паристальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопиат-типу и имеет крахмаль-
ные зерна. Синергиды с крючкооб-
разными выростами и фибрилляр-
ными аппаратами. Имеются три
крупные эфемерные антиподы. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Сингамия и тройное слияние проте-
кают одновременно или пеодновре-
мепно. Пыльцевые трубки долго со-
храняются в семяпочкс.
Эндоспермогенез нуклеарный.
Эмбриогепез Зо]апа4-типа. Заро-
дыш маленький, прямой, двусемя-
дольпый. Семядоли, почечка, гипо-
котиль и корешок хорошо развиты.
Подвесок длинный на ранней ста-
дии развития, но у зрелого зароды-
ша отсутствует.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Зародыш занимает 3/4 об-
щего объема семени. Периферия се-
мени заполнена эндоспермом.
Плод — локулицидная коробочка, с
остающейся чашечкой.
Гаусторий — микропилярная часть
энпдосперма — дифференцируется в
двух- или четырехклеточный гаусто-
рий с полиплоидными ядрами.
Anomukene не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; Kapil, 1967; Тахтаджян, 41966,
4970; Walia, Kapil, 1965; Cmmpxosza,
1977.
89
По цитоэмбриологическим данным
Fouquieriaceae правильно относят к
Татат1са!ез, наряду c Tamaricaceae,
так как семейства эти имеют сход-
ные цитоэмбриологические признаки
[Govil, 1967a, 1970a]. Hanua [Kapil,
1967а] считает, что ЕгапКешасеае
не следует относить к порядку Та-
таг са!ез, как это делают Хатчин-
сон и А. Л. Тахтаджян, а его подоб-
но Ветштейну и Рендлю необходимо
включить в порядок Рамеа]ез вбли-
зи с ЕаИпасеае, так как по цито-
эмбриологическим признакам Егап-
Кепласеае больше всего сходно с
Elatinaceae.
ПОРЯДОК 40.
SALICALES
CEMEHCTBO SALICACEAE
Основное чиело хромосом — х=
==42, 19, 24, 22.
Цветок однополый, растения дву-
домные или одподомные.
Ныльник -- 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секретор-
ный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изоби-
латеральные. Зрелые пыльцевые
зерна двухклеточные, безапертурпые
или трехбороздпопоровыс.
Семяпочка анатропная с двумя ин-
тегументами, крассинуцеллятная.
Тетрады макроспор линейные, T-
образные. Имеется эпдотелий. Семя-
почек много, 2—10.
Макроспорогенез и макрогаметоге-
нез — археспорий обычно много-, из-
редка одноклеточный. Париетальных
клеток 2—6 рядов. Тетрады макро-
спор линейные. Зародышевый ме-
шок Ро]усопат-типа. Синергиды ©
крючкообразными выростами и с
фибриллярпыми аппаратами. Имеют-
ся три эфемерпые аптиподы. Поляр-
ные ядра сливаются до оплодотво-
репия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
90
рекрестное опыление у видов Рорл-
11$. Халазогамия, порогамия. У Ро-
раз Ба]затМега от опыления до
созревания семени проходит около
двух месяцев.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эпдосперма центро-
стремительное.
Эмбриогенез Аз{ега4-, Опастад-ти-
пов. Зародыш прямой, хлорофилло-
носный, двусемядольный, содержит
масло и алейрон. Семядоли массив-
ные, выполняющие —запасающую
функцию, скрывают остальные ча-
сти зародыша, лишь кончик кореш-
ка едва выступает из них.
Семя, плод — семя обычно без эн-
досперма. Зародыш заполняет все
семя. Плод двух — четырехстворча-
тая коробочка.
Гауеторий отсутствует.
Апомиксис — редуцированный пар-
теногенез.
Другие явления — полиэмбриовия
у Populus deltoides.
Литература: Барла, 1967, 1968,
1969a, 6; Corner, 1976; Davis, 41966;
Коц, 4964, 1970, 1972 а, 6; Николаева,
1964, 1978; ЗсВпатЁ, 1931; Смирнова,
1977: Тахтаджян, 1966, 1970; Зонова,
1953.
Некоторые систематики сближают
раПсасеае с Ташаг1сасеае. Однако в
отношении цитоэмбриологии, наря-
ду со сходетвом, они существенно
отличаются друг от друга. В то вре-
мя как БаПсасеае имеет зародыше-
вый мешок Ро]угопат-типа, халазо-
гамию, нуклеарный эндосперм, Аз-
{егад-тип эмбриогенеза, Татагсасеае
характеризуются зародышевым меш-
ком Tuna Fritillaria, порогамией,
целлюлярным эндоспермом, эмбрио-
генезом 50]апад-типа. Что же касает-
ся отношения ЗаПсасеае с НасопгИа-
сеае, которые А. Л. Тахтаджян
[1966] сближает друг с другом, то
данные цитоэмбриологии не проти-
воречат этому, так как между Sa-
Нсасеае и Flacourtiaceae больше
сходных цитоэмбриологических при-
зпаков, чем между баПсасеае и Та-
тагсасеае. Однако для решения
этого вопроса требуются дальней-
шие исследования.
s
НАДПОРЯДОЕ УТ.
ERICA NAE
ПОРЯДОК 44.
ERICALES
CEMEMCTBO SAURAUIACEAE
Основное число XpomMocoM — z=
= 11, 28, 48.
Цветок обычно обоеполый.
Пыльник — нет данных.
Микроепорогенез и микрогаметоге-
нез — нет данных. Пыльцевые зер-
на трехборозднопоровые, ипогда
соединены в тетрады.
Семяпочка апатролпая, с одпим
интегументом, тенуинуцеллятная.
Имеется эпдотелий, который окру-
жает нижнюю половину зародыше-
вого мешка. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевой мешок Роугопат-типа.
Синергиды © крючкообразными вы-
ростами. Имеются три эфемерные
антиподы. Нолярные ядра сливатот-
ся перед оплодотворонисм.
Эндоепермогенез целлюлярпый.
Эмбриогеинез --- тии по определел.
Зародыш прямой или слогка согцу-
тый, двусесмядольный с хорошо раз-
витым гипокотилем и корешком.
Подвесок короткий, однорядный.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш зани-
мает от 1/4 до 3/4 длины семени.
Плод — обычно ягода.
Гаусторий — при прорастании се-
мени семядоли выполняют гаусто-
риальную функцию.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Сотгпег, 1976; Davis,
1966; Schnarf, 1931; Смирпова, 1977;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АСТТУТГОТАСЕАЕ
Основное число хромосом — х=58.
Цветок обоеполый или полигам-
ный, растения двудомные.
Пыльник микроспоравгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и секреторный много-
ядерный тапетум. Ядра в клетках
тапетума часто сливаются и стано-
вятся полиплоидными. В стенках
пыльника и связника имеются ра-
фиды.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые зер-
на двухклеточные, трехборозднопо-
ровые, с толстой экзиной.
Семяпочка анатропная © одним
интегументом, тенуинуцеллятная.
Вокруг зародышевого мешка имеет-
ся эндотелий, под зародышевым
мешком гипостаза. В семяпочке
имеется оксалат кальция. Семяпочек
мпого или около 10.
Макроспорогенез и макрогаметоге-
нез — археспорий ’одно-трехклеточ-
ный. Париетальные клетки OTCYT-
ствуют. Тетрады макроспор линей-
ные. Зародышевые мешки обычно
Polygonum-, изредка АШиаш-типов.
Синергиды удлиненные. Имеются
три эфемерные антиподы. Поляр-
ные ядра сливаются до оплодотворе-
ния. Наблюдались случаи наличия
обращенных зародышевых мешков.
пыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия.
дидоспермогепез целлюлярный.
идосперм содержит крахмал.
JmMOpuorenes Solanad-tuna. Заро-
дыш прямой или слогка согнутый,
двусемядольный с хорошо развиты-
ми корешком и гипокотилем, бес-
хлорофилльный, содержащий масло
и алейрон. Подвесок маленький.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Зародыш распола-
тается в центре семени и занимает
от 1/4 до 3/4 длины семени. Семен-
ная кожура содержит таннин.
Плод — ягода или коробочка.
Гаусторий — при прорастании се-
мени сомядоли выполпияют гаустори-
альпую функцию.
Апомиксис не обнаружен.
Другие явления — полиэмбриония
y Actinidia chinensis.
Литература: Согпег, 1976; Davis,
1966; ЭсЬпатЁ, 1931; Смирнова, 1977;
Тахтаджян, 41966, 1970; Vyayaragha-
van, 1965, 1967a, 1970a.
91
СЕМЕЙСТВО СГЕТНКАСЕАЕ
Основное число хромосом — х=8.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средних слоя и секреторный
двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые.
Семяпочка анатропная © одним
интегументом, тенуипуцеллятная.
Две трети нижней части зародыше-
вого мешка окружены эндотелием.
Семяпочек много.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный,
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
\усопит-типу, удлиняется и впедря-
ется в микропиле. Сипергиды удли-
ненные, грушевидпые, достигающие
микропиле. Антипод три, дегенери-
рующих перед оплодотворением.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вых трубок, оплодотворение — по-
рогамия.
Эндоеперм целлюлярный. _
Эмбриогенез Азегад-типа. Заро-
дыш маленький, прямой, бесхлоро-
филльный, двусемядольный, с хоро-
шо развитым гипокотилем и кореш-
ком. Подвесок однорядпый, состоя-
щий из 10 клеток.
Семя, плод — семя с мясистым эн-
доспермом, содержащим алейрон и
масло. Зародыш расположен в цент-
ре семени, занимает 1/4 до 3/4 дли-
ны семени и содержит масло и алей-
рон. Семенная кожура тонкая одно-
рядная. Плод — локулицидная коро-
бочка.
Гаусторий — эндоспермальные га-
устории образуются как в микро-
пилярной, так и халазальной частях
эндосперма. При прорастании семе-
ни семядоли выполняют гаустори-
альную функцию.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
92
1966; Ganapathy, 1967а, 1970а;
ОсВпат?, 1934; Смирнова, 1977; Тах-
таджян, 1966, 4970.
CEMEHCTBO ERICACEAE
Оеновное число хромосом — х =7
8, 11, 12, 19.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций с фиброзными
Утолщениями присутствует или от-
сутствует, 1—3 эфемерных средних
слоя и секреторный двуядерный та-
петум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, соединенные в тетрады, либо
одиночные, обычно трехборозднопо-
ровые.
Семяпочка — апатропная или кам-
иилотропная, с одпим иптсгументом,
тенуинуцеллятная. Иместся эндоте-
лий. Микропиле образуется наруж-
ным интегументом. Семяпочек
много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
lygonum- a Allium-tanam. Ou copep-
жит крахмальные зерна. По мере
развития зародышевый мешок уд-
линяется и внедряется в микропиле,
Яйцеклетка удлипеппая. Синергиды
имеют крючкообразпые выросты.
Антиподы обычно маленькие и эфе-
мерные, их бывает три или две; но
у некоторых видов Геисо ое число
их увеличивается и они долго сохра-
няются. Обычно антиподы дегене-
рируют после оплодотворения. По-
лярные ядра сливаются перед оплс-
дотворением или во время него.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
обычпо самоопыление, изредка пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение промежу-
точного типа, У рододендронов оп-
лодотворение происходит через 10—
13 дней после опыления.
Эндоспермогенез обычно целлю-
лярный, у Rhododendron нуклеар-
НЫЙ.
Эмбриогенез
Зо]апад-типа. Заро-
дыш небольшой, бесхлорофилльный,
прямой, двусемядольный, сначала
богатый крахмалом, позднее имеет
только алейрон и масло. Первое де-
ление зиготы у видов Вододепдгов
происходит через 18—28 ч после оп-
лодотворения. Подвесок имеется.
Семя, плод — семя с хорошо разви-
тым эндоспермом, содержит масло
и протеин. Зародыш расположен в
центре семени и у одних видов за-
нимает от 1/4 до 1/2, а у других от
1/4 no З/А длины семени. Зародыш
со всех сторон окружен эндоспер-
мом. Плод — септицидная или локу-
лицидная коробочка, ягода или ко-
стянка.
Гауеторий — обнаружены микро-
пилярный и халазальный эпдоспер-
мальные гаустории. Эпдоспермаль-
ные гаустории имеют от 2 до 16
гипертрофированпых ядер.
Апомикеие пе обпаружен.
Другие явления — клейстогамия.
Литература: Corner, 1976; Corsi,
1972; Davis, 1966; De Gaetano, 41967;
Eaton, Jamont, 1966; Ganapathy,
19674; Милясва, 14960; Ведду et al.,
1966; Reddy, Narayana, 1966; Robert-
son, McNaughton, 1978; Schnarf, 1931;
Смирнова, 1977; Stushnoff, Palser,
1969; Тахтаджяиц, 1966, 1970; УепКа-
teswarlu, Марезмуаг П., 4973; Ши-
лова, 1962; Якобсон, 1963а, 6, 1969а, 6,
1976; Yamazaki, 1975a, 1979.
СЕМЕЙСТВО ЕРАСВШАСЕАЕ
Оеновное число хромосом — х=4,
6—12.
Цветок обычно обоеполый.
Пыльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 4—3
средних слоя и секреторный двуя-
дерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточпые, сосдипепы в тет-
рады, либо одиночные, трех-четы-
рехборозднопоровые. У Leucopogon
дашрегииз пыльцевые зерпа имеют
пластиды и крахмальные зерна.
Семяпочка анатроппая, с одним
интегументом, тенуинуцеллятная.
Эндотелий окружает зародышевый
мешок в нижней его части. Имеется
фуникулярный обтуратор. Семяпо-
чек несколько — мпого.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
1усопат-типу. Он удлиняется и вра-
стает в микропиле. Синергиды име-
ют фибриллярные аппараты. Обыч-
но двух-трехэфемерные антиподы,
однако у Г. ет1со14ез число их уве-
личивается. Полярные ядра слива-
ются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
_ OMOpHorenes Caryophyllad- unu So-
lanad-tama. Зародыш маленький,
прямой, двусемядольный, с хорошо
развитыми гипокотилем и корешком.
Семя, плод — семя с хорошо разви-
тым мясистым эпдоспермом, содер-
жащим алейрон и масло. Зародыпт
располагается в центре семени, за-
нимает от 1/4 до 3/4 длины семени
и содержит алейрон и масло. Ос-
тальной объем семени заполнен эн-
доспермом. Семенная кожура одно-
или многослойная. — Плод — двух-
створчатая локулицидная коробочка
или костяпка.
Гаусторий — имеются микропиляр-
ные и халазальные эпдоспермальные
гаустории или только микропиляр-
пый. Семядоли выполняют гаусто-
риальпую фупкцию.
Апомиксие пс обпаружен.
Другие явления пе отмечены.
Литература: Сотпег, 1976; Davis,
1966; Сапара фу, 1967 c; Schnarf,
1934; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО РУВОГАСЕАЕ
Основное чиело хромосом — r=8,
11—13.
Цветок обоеполый.
ПНыльник — 4 микроспорангия.
Стенка микроспораигия имеет эпи-
дермис, один средпий слой и секре-
ториый двуядерный тапетум. Эндо-
теций отсутствует.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехборозднопоровые, соединен-
пые в тетрады или одиночные.
Семяпочка анатропная, © одним
интегументом, тенуинуцеллятная.
93
Эндотелий окружает зародышевый
мешок. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро!усопит-типу. Синергиды с широ-
ким основанием. Три довольно долго
функционирующие антиподы. По-
лярные ядра сливаются до оплодот-
ворения.
пыление, прохождение пыльце-
вой трубки и оплодотворение — са-
моопыление. Порогамия. Двойное
оплодотворение.
Эндоепермогенез целлюлярный.
Эмбриогенез СагуорвуПад-типа.
Зародыш маленький, не дифферен-
цированный, бесхлорофилльный, со-
держит алейрон и масло.
Семя, плод — семя с хорошо раз-
витым мясистым эндоспермом, со-
держащим алейрон и масло. Семен-
ная кожура тонкая. Плод — локули-
цидная коробочка.
Гаусторий — имсется слабо разви-
тый эндоспермальный гаусторий в
микропилярной части зародышевого
мешка.
Апомиксиес не обнаружен.
Другие явления — паразитизм.
Литература: Пау!з, 1966; Сапара{-
hy, 1967e; Pyykk6, 1978; Терёхин,
1962а, 6, в, 1963, 19736, 4977; Тахтад-
жян, 1966, 1970.
+
СЕМЕЙСТВО МОМОТВОРАСЕАЕ
Основное число хромосом — х=8.
Цветок обоеполый.
Пыльник — 4 микроспорапгия.
Стенка микроспорангия имсет эпи-
дермис и сокреторный одпоядерный
тапетум. Другие слои ие дифферен-
цированы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточные,
четырех (пяти) -борозднопоровые.
Семяпочка анатропная, с одним
интегумептом, тенуинуцеллятная.
Семяпочек мпого.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный,
Париетальные клетки отсутствуют.
Тетрады макроспор линейные, 7-06-
разные. Зародышевые мешки обра-
зуются по Polygonum и АШит-ти-
94
пам. Имеются три антиподы, сохра-
няющиеся до начала эмбриогенеза.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльще-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Caryophyllad-ruma.
Зародыш маленький, не дифферен-
цированный, бесхлорофилльный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и белок. Плод — локулицид-
ная двух-шестистворчатая коро-
бочка.
Гаусторий — микропилярный и ха-
лазальный, эндоспермальные гаусто-
рии.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Рау!з, 1966; Терехин,
1962а, 6, в; 1963, 1973в, 1977; Тахтад-
man, 1966, 1970.
CEMEAMCTBO CYRILLACEAE
Основное чиело xpomocom — x= 10.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и секреторный дву-
ядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточные,
трехборозднопоровые.
Семяпочка анатропная, с одним
интегументом; тенуинуцеллятная
или слабокрассинуцеллятная: Семя-
ножка довольно длинная. Имеется
эндотелий. Семяпочек 2—145.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-, реже дву-
четырехклеточный. Париетальные
клетки отсутствуют. Тетрады макро-
спор линейные. Зародышевый ме-
шок образуется по Роувопат-типу;
он сильно удлиненный, узкий. Анти-
под — три клетки, они дегенерируют
после оплодотворения. Полярные
ядра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. .
Эндоспермогенез целлюлярный.
Эмбриогенез, возможно, Solanad-
типа. Зародыш маленький, прямой,
двусемядольный, с хорошо развиты-
ми гипокотилем и корешком. Подве-
сок небольшой.
Семя, плод — семя с эндоспермом,
содержащим масло и протеин. За-
родыш занимает от 1/4 до 3/4 длины
семени; остальной объем семени за-
пят эндоспермом. Плод — коробочка
или костянковидная ягода.
Гауеторий — имеются двухклеточ-
ный микропилярный и одноклеточ-
ный халазальный эндоспермальные
гаустории.
Апомиксис — соматическая апо-
chopua y Cliftonia monophylla. Unc-
ло апоспорических зародышевых
мешков колеблется от 1—
Другие явления не обпаружены.
Литература: Согпог, 1976; Davis,
1966; Смирнова, 1977; 'Гахтаджян,
1966, 1970; Vijayaraghavan, 19674,
тв, Ъ, 19705; Vijaraghavan, Dhar,
1978.
СЕМЕЙСТВО ЕМРЕТВАСЕАЕ
Основное число хромосом — х=13.
Цветок однополый или редко обое-
полый. Растения обычно двудомные,
редко однодомные.
Ныльник — 4 микроспорангия.
Стенка микроспораигия имеет DIT
дермис, 2—3 средних слоя и секре-
торный мнпоголядерный танетум. Эн-
дотеций отсутствует.
Микроспорогенез и микрогаметогс-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые зер-
на двухклеточные, трехборозднопо-
ровые. Соединены в тетрады.
Семяпочка анатропная с одним ин-
тегументом, тенуинуцеллятная. Вок-
руг зародышевого мешка имеется
эндотелий. У Етрехгиш имеется обту-
ратор. Семяпочек 2—9.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальпые клетки отсутствуют.
Тетрады макроспор липейные. Заро-
дышевый мешок образустся по Ро-
]усопит-типу. Синергиды грушевид-
ные. Имеются три антиподы, деге-
нерирующие вскоре после оплодотво-
рения. Полярные ядра сливаются
перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Зо]апад-типа. Заро-
дыш прямой, двусемядольный, с хо-
рошо развитыми гипокотилем и ко-
решком, содержит масло и алейрон.
Имеется подвесок.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
злейрон. Зародыш занимает от 1/4
до 3/4 длины семени. Остальной
объем семени занят эндоспермом.
Семенная кожура формируется на-
ружным интегументом. Плод — соч-
пая или сухая костянка.
Гаусторий — обнаружены микро-
пилярный и халазальный эндоспер-
мальные гаустории.
Anomukene пе обнаружен.
Другие явления пс отмечены.
Литература: Согпег, 1976; Davis,
1966; Ganapathy, 1967b, Schnarf, 1931;
Смирнова, 1977; Тахтаджян, 1966,
1970.
CEMEMCTBO GRUBBIACEAE
Основное
данных.
Цветок обоеполый.
Пыльник — нет данных.
Микроепорогенез и микрогаметоге-
нез — нет данпых.
Зрелые ныльцевые зерна трехбо-
роздпопороговые, одипочные.
Семяиочка апатропная, с одним
интегументом, тепуипуцеллятная.
Микрониле длиппое. Имеется эндо-
телий. Семяпочек 2.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]усопат-типу, он длинный и внед-
ряется в микропиле.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных. .
Эндоспермогенез целлюлярныйи.
Эпидермис эпдосперма покрыт тол-
стым слоем кутикулы.
Эмбриогенез — тип пе установлен.
Зародыш прямой, двусемядольный,
с хорошо развитыми гипокотилем и
корешком, содержит масло.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и протеин. Зародыш занимает
от 4/4 до 3/4 длины семени; осталь-
ной объем семени занят эпдоспер-
95
чиело хромосом — HCT
мом. Плод — односемянная костянка.
Гауеторий — имеются микропиляр-
ный и халазальный эндоспермаль-
ные гаустории.
Апомиксис не обнаружен.
Другие явления не отмечены.
Jiurepatypa: Davis, 1966; Fagerlind,
1947; Ram, 1967d, 19704; Смирнова,
1977; Тахтаджян, 1966, 1970.
По данным сравнительной цитоэм-
бриологии Ег!са]ез — довольно моно-
литный, естественный порядок со
значительным числом прогрессив-
ных цитоэмбриологических призна-
ков. Характерными признаками
представителей этого порядка явля-
ются: 4 микроспорангия, четырех-
пятислойная стенка пыльника, сек-
реторный тапетум, симультанный
микроспорогенез, двухклеточные,
трехбороздпопоровкно пыльцевые
зерна, соедипениыс в тетрады, апа-
тропная, тепуинуцеллятная, с одним
интегументом семяпочка, отсутствие
париетальных клеток, одноклеточ-
ный женский археспорий, Ро]уго-
пиш-тип зародышевого мешка, три
эфемерные антиподы, двойное опло-
дотворение, целлюлярный — эндо-
сперм, 30]апад-тип эмбриогенеза, се-
мя с хорошо развитым эндоспермом,
эндоспермальные гаустории в мик-
ропилярной и халазальной частях
зародышевого мешка и отсутствие
апомиксиса. Из всех семейств Е-
са]ез наименее подробпо исследова-
ны баигашасеае u Grubbiaceac.
По даппым цитоэмбриологии Аси-
п1Ф1асеае обпаруживаот сходетво с
Margraviaceae и ТрЬоасеае, поэтому
оно должно быть отнесено к поряд-
ny Theales [Vijayaraghavan, 1967].
По мнению того же автора, Saurauia
не заслуживает ранга самостоятель-
ного семейства и ее следует отнести
к АСИ 91асеае, так как цитоэмбрио-
логически она не отличается от него.
Clethraceae считают примитивным
представителем Ег!саез [Сапара\у,
1967а, 1970а]. В отношении цитоэм-
бриологии СугШасеае сходно с Е!1-
сасеае, и отнесение обоих этих се-
мейств к Е!1са!ез не вызывает сом-
neHua [Vijayaraghavan, 1967d, 1970e].
СугШасеае, как и Ericaceae, xapak-
теризуется семяпочкой с одним ин-
тегументом и скоро исчезающим ну-
96
целлусом, отсутствием париетальных
клеток, Ро]угопат-типом зародыше-
вого мешка, целлюлярным эндоспер-
мом, хорошо развитыми микропи-
лярными и халазальными эндоспер-
мальпыми гаусториями, Зо]апад-ти-
пом эмбриогенеза — признаками,
свойственными Егсасеае. Словом,
цитоэмбриологические признаки ука-
зывают на тесную связь СугШасеае
с Ечсаез и на отсутствие связи его
с Celastrales [Vijayaraghavan, 1967].
Таким образом, отнесение Cyrilla-
сеае к Егса]ез согласуется с данны-
ми цитоэмбриологии [Vijayaragha-
van, Dhar, 1978]. Vaccinium HacTOIBKo
сходен с Енсасеае, что нет необхо-
димости выделять сго в особое се-
mMelictBo Vacciniaceae [Venkateswar-
1а, 1967]. Ерасг!асеае обладает мно-
тими сходными с Емсасеае призна-
ками, хотя и является несколько 60-
лее полвинутым [Ganapathy, 1967].
Цитоэмбриологические данные под-
тверждают отнесение Руго]асеае и
Empotraceao x Ericales [Ganapathy,
1967b]. Monotropaceac lo цитоэм-
бриологическим признакам близко-
родствепно Руго]асеае. На основании
данных цитоэмбриологии включение
СтаЪМасеае в порядок Ег1саез впол-
не правильно [Fagerlind, 1947]. Од-
нако СтаЪЫасеае требует дальней-
ших исследований, так как цито-
эмбриологическая характеристика
его пе полная.
А. Л. Тахтаджян [1966] отмечает,
что Saurauiaceae, Actinidiaceae u
Clethraceae oOHapymupator pay o6-
щих черт с ТВеасеае, что не проти-
ворсчит даппым цитоэмбриологии.
ПОРЯДОК 42.
DIAPE NSIALES
СЕМЕЙСТВО РТАРЕМЗТАСЕАЕ
Основное чиело хромосом — х=6.
Цветок обоеполый.
Пыльник —4 иногда 2 микроепо-
рангия. Стенка — микроспорангия
имеет эпидермис, фиброзный эндо-
теций, 2—3 средних слоя и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — нет данных. Зрелые пыльце-
вые зерпа двухклеточные, одиноч-
ные, трехборозднопоровые, не сое-
диненные в тетрады.
Семяпочка гемианатропная-кампи-
лотропная, с одним интегументом,
тенуипуцеллятная. Эндотелий отсут-
ствуст. Семяпочек несколько-много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальных клеток нет. Тетрады
макроспор линейные. Зародышевый
мешок образуется по Polygonum-ru-
пу. У Барепча синергиды с фибрил-
лярными аппаратами, у ЭКогИа ga-
]а5ТоПа обычно три или до 40 анти-
под, дегенерирующих в начале эм-
бриогепеза. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — ист
данных.
Эндосиермогепез пеллюлярный.
Первая перстородка в эпдосперме
проходит либо поперечно, либо про-
дольно.
Эмбриогенез бо|апад-типа. Заро-
дьни прямой или слегка согнутый,
с хорошо развитыми семядолямн,
тлпокотилем и корешком.
Семя, плод — семя с хорошо разви-
тым эндоспермом, содержащим алей-
рои и масло. Зародыш занимает от
1/4 no 3/4 длины семети; остальной
0бъ»ем семени заполиен эпдоснер-
мом. Плод- трехетворчатая локули-
пидная коробочка.
Гаусторий отсутствует
Апомиксие пе обиаружеин.
Другие явления не отмечены.
Литература: Пау!з, 1966; Palser,
1963; Reynolds, 1968; Schnarf, 1931;
Смирнова, 1977; Тахтаджян, 1966,
1970; Yamazaki, 1966.
Выделение Парепзасеае из Е1-
сшез и отнесепие его в самостоя-
тельный порядок ПО1арепза]ез впол-
не обосповапо со сторопы цитоэм-
бриологии, так как Diapensiaceae or-
личастся от №7сасодо одилочными
пыльневыми зернами, отсулствием
эпдотелия и эндоспермальных ray-
сториев [Рафзег, 1963]. Вместе с тем
| аропзтасеае имеет сходные цитоэм-
брнологические признаки, характер-
hole для ricales, aro cOnmxaet Dia-
pensiales ¢ Lricales,
ТВ. Л. Поддубная-Арнольди
ПОРЯДОК 443.
ЕВЕ МАТЕ$
(DIOSPYRALES)
СЕМЕЙСТВО STYRACACEAE
Основное число хромосом — х=8,
12.
Цветок обычно обоеполый.
Пыльник — 4 микроспорапгия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный двуядерный тапетум.
икроспорогенез и микрогаметоге-
нез — археспорий одиоклеточный.
мультаппос. Тетрады микроспор тет-
раэдральные и изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка анатропная-гемиана-
тропная, с двумя интегументами, те-
нуинуцеллятная. Микропиле обра-
зуется внутренним интегументом.
Семяпочек одна-мнпого.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейпые, 7-об-
разные. Зародышевый мешок обра-
зустся по Polygonum-tuny и имеет
прахмальные зерна. Синергиды силь-
но удлиненные. Имеются три эфе-
морные антиподы. Полярные ядра
сливаются при оплодотворении.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез Зо[эпа4-типа. Заро-
дыш прямой или слегка согнутый,
двусемядольный, с хорошо развиты-
ми почечкой, гипокотилем и кореш-
ком, бесхлорофилльный, содержа-
щий масло и алейрон. У Styrax of-
ИЙстаНз широкий мпогорядный под-
BCCOK.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейроп. Зародыш занимает
3/4 общего объема семепи. Перифе-
рия семени заполнена эндоспермом.
Плод — костянка, локулицидная ко-
робочка или деревянистый нера-
скрывающийся с остающейся ча-
шечкой.
Гаусторий отсутствует.
Апомикеиес не обнаружен.
Другие явления пе отмечены.
97
Литература: Сотпег, 1976; Davis,
1966; Смирнова, 1977; Тахтаджан,
1966, 1970; Yamazaki, 1970a.
СЕМЕЙСТВО ЗУМРГОСАСЕАЕ
Основное
данных.
Цветки обоеполые или изредка
полигамные.
Ныльник — нет данных.
Микроспорогенез и микрогаметоге-
нез — нет данных. |
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка анатропная, с одним
интегументом; тенуинуцеллятная.
Зародышевый мешок окружен эндо-
телием. Семяпочек немного.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]узопат-типу и имеет зерна крах-
мала. Сипергиды удлинеппые, с
крючкообразными выростами. Име-
ются три антиподы, скородегенери-
рующие.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип не определен.
Зародьшш прямой, двусемядольный,
с гипокотилем и корешком.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш pac-
полагается в центре семени и зани-
мает от 1/4 до 14/2 или от 1/4 до 3/4
общего объема семепи. Остальной
объем семени заполняет эпдосперм.
Плод ягодо- или костянкообразный,
с 'остающимися долями чашечки.
Гаусторий но обнаружен.
Апомиксис — инцтегументальная
9mM0puonua y Symplocos klotzschii.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1931; Смирнова, 1977;
Тахтаджян, 1966, 1970; Yamazaki,
1970b, 1972.
чиело хромосом — нет
СЕМЕЙСТВО ЕВЕМАСЕАЕ
Основное чиело хромосом — х=5.
Цветок обычно однополый, реже
обоеполый. Растения обычно дву-
домные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
98
дермис, не фиброзный эндотеций, 2
средних слоя и секреторный много-
ядерный тапетум. |
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые.
Семяпочка апатропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется эндотелий.
Семяножка короткая. Семяпочек
2—32.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопит-типу, он длинный, узкий.
Три маленькие эфемерные анти-
поды.
Опыление, прохождение пыльще-
вой трубки и оплодотворение — пе-
рекрестное опыление у видов 010з-
pyros.
Эндоспермогенез целлюлярный.
Эпдосперм твердый, руминирован-
ный.
Эмбриогенез — тип не определен.
Зародыш маленький прямой. или слег-
ка согнутый, бесхлорофилльный, дву-
семядольный, с корешком и OTHOCH-
тельно длинным гипокотилем. Со-
держит масло, алейрон. <
Семя, плод — семя с`ховошо раз-
витым эндоспермом, содержащим
масло и крахмал. Зародыш занимает
от 1/4 до 4/2 линейной высоты се-
мени. Соменпая кожура тонкая.
Плод — обычно более или менее соч-
пая или кожистая ягода, а иногда
раскрывающийся створками.
Гаусторий не пайден.
Апомикесис пе обнаружен,
Другие явления — у ПО10зругоз у!г-
оп1апа обнаружена полиэмбриония.
Партенокарпия.
Литература: Пау!з, 1966; Мурадов,
1974; Periasamy, 1966; Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
1970; Тутаюк, Мурадов, 1969; Уата-
zaki, 1972.
СЕМЕЙСТВО ЗАРОТАСЕАЕ
Основное
—12, 13.
Цветок обычно обоеполый.
число хромосом — 7=
Пыльник А
_ микроспорангия.
Стенка микроспорангия имеет эпи-’
дермис, фиброзный эндотеций, 3—4
средних слоя и секреторный мното-
ядерный тапетум. В результате
слияния ядра в клетках тапетума
становятся полиплойдными.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изоби-
латеральные. Зрелые пыльцевые
зерна двух-трехклеточные, трех, че-
тырех (пяти-шести) борозднопо-
ровые, с сетчатой экзиной.
Семяпочка анатропная, © одним
интегументом, тенуинуцеллятная,
Семяножка короткая, тонкая. Семя-
почек немного.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейпые, T-
образные. Зародышевый мешок об-
разуется no Polygonum-tTuny. Cunep-
тиды с крючкообразными выростами
и фибриллярными — аппаратами.
Имеются три скородегенерирующие
антиподы, представленные не клет-
ками, а ядрами. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — са-
моопыление и перекрестное опыле-
Hue y Argania sideroxylon.
Эндоспермогенез нуклеарпый, O6-
разование клеточных перегородок
между ядрами эндосперма протекает
центростремительно. Образование
клеток в эндосперме происходит
тогда, когда образуется более 1000
свободных ядер.
Эмбриогенез — зародыш крупный,
прямой или согнутый, двусемядоль-
ный с сетчатым жилкованием, бес-
хлорофилльный, с хорошо развиты-
ми семядолями, почечкой, гипокоти-
лем и корешком. Содержит крахмал,
алейрон и дубильные вещества. Под-
весок массивный, мпогоклеточный.
_ Семя, плод — семя со слаборазви-
тым эндоспермом, содержащим ами-
лоид, масло, алейрон и кристаллы
оксалата кальция. Иногда семя без
эндосперма. Согнутый зародыш рас-
полагается по периферии семени,
занимая 3/4 его периметра, эндо-
сперм сохраняется в центре семени.
Прямой зародыш занимает 3/4 объе-
ма семени, периферия семени запол-
нена эпдоспермом, или прямой за-
7
родыш заполняет почти весь объем
семени, а эндосперм в виде тонких
пленок остается лишь в области ко-
решка и на семядолях. Семенная
кожура толстая, твердая, богатая
таннином. Плод — ягода, редко ко-
жистая и раскрывающаяся..
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Bhatnagar, Gupta,
1970; Davis, 1966; Taxranman, 1966,
1970; Yamazaki, 1971.
В порядке Ефепа]ез семейство Гуз-
зосаграсеае цитоэмбриологически не
исследовано, а семейства Styracaceae
и бутр]осасеае, хотя и исследованы,
но не достаточно. В отношении ци-
тоэмбриологии Ефепасеае и Зароа-
сеае обнаружено значительное не-
сходство, что не подтверждает рас-
положение их в общем порядке.
ПОРЯДОК 44.
PRIMULALES
СЕМЕЙСТВО МУВЗТХАСЕАЕ
Основное число хромосом — х=10,
12, 23.
Цветок обоеполый или однополый.
Растения обычно двудомные.
Пыльник — нет данных.
Микроспорогенез и микрогаметоге-
нез — нет дапных. Зрелые пыльце-
вые зерна двухклеточные, трех (пя-
ти) -борозднопоровые.
Семяпочка анатропная, кампило-
тропная, с одним, двумя интегу-
ментами, тенуинуцеллятная. Микро-
пиле образуется внутренним интегу-
ментом. Зародышевый мешок окру-
жен эпдотелием. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
1усопат-типу.
Эндоспермогенез нуклеарный.
Эмбриогенез Опаотад-типа. Заро-
дыш прямой или слегка согнутый,
двусемядольный, бесхлорофилльный,
с хорошо развитыми гипокотилем и
корешком. Содержит масло и алей-
99
§
рон, иногда крахмал. Подвесок длин-
ный.
Семя, плод — семя с мясистым или
роговым эндоспермом, содержащим
масло и алейрон. Иногда семя без
9 -
‘эндосперма. Зародыш занимает от
1/4 до 3/4 объема семени. Плод —
обычно шарообразная ягода или ко-
стянка.
Гаусторий — семядоли выполняют
гаусториальную: функцию.
Другие явления не обнаружены.
Литература: Сотрег, 1976; Davis,
1966; Rao S., 1972; Schnarf, 1931;
Смирнова, 41977; Тахтаджян, 1966,
1970.
СЕМЕЙСТВО
THEOPHRASTACEAE
Основное число хромоеом — х=18.
Цветок обоеполый.
Пыльник — нет данных.
Микроспорогенез и микрогаметоге-
нез — нет данных. Зрелые пыльце-
вые зерна двухклеточные, трехбо-
розднопоровые.
Семяпочка — анатропная-гемиана-
тропная или кампилотропная с дву-
мя интегументами, тенуинуцелляг-
ная. Микропиле образуется обоими
интегументами. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Си-
пергиды с крючкообразными выро-
стами. Имеются три антиподы, ско-
ро дегенерирующие. Полярные ядра
сливаются перед оплодотворением.
Эндоспермогенез пуклеарный.
Эмбриогенез — тип пе определен.
Зародыш прямой или слегка согну-
тый, содержит масло и `алейрон.
Семя, плод — семя’ с хорошо раз-
витым роговым эндоспермом, содер-
жащим масло. Плод — ягода или од-
носеменная костянка.
Гаусторий не найден.
Anomukene не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; ЗеВпатЁ, 1931; Тахтаджян, 1966,
1970.
СЕМЕЙСТВО РЕТМОГАСЕАЕ
Оеновное число хромосом — х=8,
9, 11—45.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
100
Стенка микроспорангия имеет эпи-
дермис;, фиброзный эндотеций, один
эфемерный средний слой и секре-
‚торный одно-двуядерный тапетум:
Микроспорогенез и микрогаметоге-
нез — образование микроспор си:
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые зёр-
на двухклеточные, обычно трехбо-
розднопоровые, реже трех-десяти-
бороздные.
Семяпочка — анатропная-гемиана-
тропная, кампилотропная, с двумя
интегументами, тенуипуцеллятная.
Микропиле образуется обоими ин-
тегументами. Зародышевый мешок
окружен эндотелием. — Семяпочек
много или немного.
Макроепорогенез и макрогаметоге-
нез — археспорий одно-двухклеточ-
ный. Париетальные клетки OTCYT-
ствуют. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок образуется по Роубопит-тицпу.
Синергиды удлиненные. Имеются
три скоро дегенерирующие антипо-
ды. Полярные ядра сливаются при
оплодотворении,
. Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Самоопыление,
Порогамия. Двойное оплодотворение
Эндоспермогенез нуклеарный.
Y Primula officinalis o6pa3yrorca 70
1000 свободных ядер ’эндосперма,
прежде чем между ядрами начина-
ется образование клеточных перего-
родок. Заложение клеток в эндоспер-
ме начинается с периферии.
Эмбриогенез СагуорвуПа@-типа. 3a-
родыш маленький, прямой, двусемя-
дольный с хорошо развитыми гипо-
котилем и корешком, бесхлоро-
филльный. Подвесок почкообразпый.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
амилоид, масло и алейроп. Зародыш
занимает от 4/4 до 3/4 длины семе-
ни. Остальной объем семени запол-
нен эндоспермом. У зародыша Сус-
]атеп одпа из семядолей меньше
другой. Плод — коробочка или кры-
ночка.
Гаусторий не обнаружен.
Апомиксие — персдуцированный
партеногенез.
Другие явления — полиэмбриония
-y Primula auriculata. Клейстогамия
и диморфная гетеростилия. ,
Литература: Согпег, 1976; Davis,
1966; Dahlgren, 1916; Каграмова,
1972; Пашук, 4969; . Schnarf, 1934;
Смирнова, '1977; Тагиева, 1970; Тах-
таджян, 1966, 41970; Заргатапуат,
Магауапа, 1968.
Несмотря на то что семейства Мут-
sinaceae u Theophrastaceae исследо-
ваны слабее, чем семейство Ретшл-
[асеае, все же видно, что все эти
семейства имеют много общих пито-
эмбриологических признаков, что
указывает на естественность поряд-
ка Рг!пч1ез и на родство этих се-
мейств. Вместе с тем -Primulales
имеет сходные c Ebenales
бриологические признаки, поэтому
расположение их вблизи друг с дру-
гом подтверждается данными цито-
эмбриологии.
НАДПОРЯДОК УИ.
МАТА МАЕ
ПОРЯДОК 45.
MALVALES
СЕМЕЙСТВО ЕГАЕОСАВРАСЕАЕ
Основное число хромосом — х=6
10, 14, 43,
Цветок обычно обоеполый.
Пыльник — 2 микроспорангия.
Стенка микроспорапгия имеет эпи-
дермис, фиброзный эндотеций, 1—2
средпих слоя и секреторный тапе-
тум.
Микрсепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые, с гладкой экзиной.
Семяпочка анатропная, гемиана-
тропная с двумя интегументами,
крассинуцеллятная. Имеются Эндо-
телий и гипостаза. Семяпочек две-
много,
Макроспорогенез и макрогаметоге-
нез — археспорий одно-двуклеточ-
ный. Париетальные клетки имеются.
Зародышевый мешок образуется по
Ро]угопат-типу и имеет крахмаль-
ные зерна. Имеются три скоро деге-
нерирующие антиподы.
Опыление прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия.
Цитоэм- —
Эндоеспермогенез нуклеарный.
Эмбриогенез — тип ‚не определен.
Зародыш прямой, хлорофиллонос-
ный, дифференцированный, бодер-
жит масло и алейроновые зерна. Се-
мядоли толстые.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейроновые зерна. Плод —
коробочка или реже костянковид-
НЫЙ.
Гауеторий отсутствует.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Тахтад-
жян, 1966, 4970.
CEMEVCTBO TILIACEAE
Основное чиело хромосом — х=
== 7—9, 44.
Цветок обычно обоеполый.
Пыльник — 4 микроспорангия.
Стенка`пыльника образуется по ба-
зальному типу и имеет эпидермис,
фиброзный эндотеций, 1—2 эфемер-
ных средних слоя и секреторный
двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образованиб6 микроспор` си-
мультанное. Тетрады микроснпор тет-
раэдральпые. Зрелые пыльцевые зер-
на двухклоточные, трехборозднопо-
ровые,
Семяпочка — апатропная-гемиана-
троппая, с двумя интегументами,
крассипуцеллятпая. Микропиле об-
разуется наружным или внутрен-
ним или обоими интегументами,
иногда зигзагообразное. Имеется ну-
целлярный колпачок, эндотелий и
гипостаза. Семяпочек одна-не-
сколько.
Макроепорогенез и макрогаметоге-
нез — археспорий многоклеточный,
Париетальных клеток 4—8 рядов.
Тетрады макроспор линейные, 7-
образные. Зародышевый мешок - об-
pasyetca mo Polygonum-tuny u co-
держит зоерпа крахмала. Сипергиды
с крючкообразными — выростами,
очень длинные. Имеются три эфе-
мерные антиподы, представленные
не клетками, а ядрами. Полярные
ядра сливаются до оплодотворения.
Y Tilia platyphylla описан поста-
мент. .
Опыление, прохождение пыльце-
вой трубки и оплодотворение — у ви-
дов СогсВогиз самоопыление. Поро-
гамия.
‘101
<
Зидоспермогенез нуклеарный.
Иногда в халазальной части эндо-
сперма скапливаются ядра и цито-
плазма. Эндосперм содержит масло
и алейрон.
Эмбриогенез Опаота4-, Азегад-ти-
пов. Зародыш прямой, двусемядоль-
ный с хорошо развитыми гипоко-
тилем и корешком, хлорофиллонос-
ный, содержит масло и алейрон.
Семядоли расчлененные.
Семя, плод — семепа со скудным
или обильным маслянистым эндо-
спермом. Зародылт занимает от 1/4
до 3/4 длины семени. Остальной
объем семени заполнен эпдоспер-
мом. Плоды сочные или сухие, раз-
ных типов.
Гауеторий отсутствует.
Апомиксис ° — редуцированный
партеногенез.
Другие явления — у Сотебогиз оН-
{отаз обнаружены полиэмбриония и
клейстогамия. Партепокарпия.
- Литература: Сотпег, 1976; Datta,
Mukherjee, 1969; Davis, 1966; Hop-
дюм, 1963; Schnarf, 19381; Смирнова,
1977; Тахтаджяц, 1966, 1970.
СЕМЕЙСТВО ЗСУТОРЕТАГАСЕАЕ
Основное число хромосом — 2 =
=9, 11.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный слой, 1—2 сред-
них слоя и секреторный, одно-
трехъядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образовапие микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые с гладкой окзипой.
Семяпочка анатроппая, © двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется эндотелий.
Семяпочек несколько-много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок очень удли-
ненный; образуется по Ро]ухопат-
типу. Синергиды длинные, с крючко-
образными выростами и фибрилляр-
ными аппаратами. Имеются три
эфемерные антиподы.
Опыление, прохождение пыльце-
102
1
вой трубки и оплодотворение — нег
данных.
Эндоспермогенез — нет данных.
Эмбриогенез — тип не определен.
Зародыш маленький, прямой, двусе-
мядольный © корешком. Семядоли
широкие.
Семя, плод — семя с хорошо раз-
витым, роговым, содержащим мас-
ло, с руминированным или с про-
стым эндоспермом. Зародыш запол-
няет от 1/4 до 3/4 длины семени. °
Плод — деревянистая локулицидная
коробочка или односеменная ко-
стянка.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Смир-
нова, 1977; Тахтаджян, 1966, 1970;
Vijayaraghavan, Dhar, 1976.
СЕМЕЙСТВО ЗТЕВСОТТАСЕАЕ
Основное число хромосом — х=7,
8, 10, 19, 23.
Цветок обоеполый или реже одно-
полый.
Пыльник — 4 микроспорангия.
Стенка пыльника образуется по дву-
дольному типу, за исключением Рге-
tospermum и Э&егсаПа, у которых
она образуется по базальному типу.
Стенка пыльника имеет эпидермис,
фиброзпый эндотеций, один эфемер-
ный средпий слой и обычно секре-
торпый, многоядерный тапетум.
Y Guazuma tomentosa u Waltheria
indica TalleTyM периплазмодиальный.
Микроспорогенез и микрогаметоге-
нез — образование микрбспор си-
мультаппое. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерпа двухклеточные, часто с крах-
мальными зернами, трехборозднопо-
ровые или мибгопоровые, с шипова-
той экзиной.
Семяпочка анатроппая, © двумя
интегументами, крассинуцеллятная.
Микропиле обычно зигзагообразное,
образующееся обоими интегумен-
ramu. Y Buettneria herbacea u He-
|{с‘егез 1зота нуцеллус голый, так
как оба интегумента не развивают-
ся. Имеются нуцеллярный колпачок,
гипостаза и ариллус. Семяпочек
две-много.
Макроспорогенез и макрогаметоге-
нез — археспорий — многоклеточный
(до 10—15 клеток). Париетальных
\
А
клеток 3—6. рядов. Тетрады макро-
спор линейные. Зародышевый мешок
образуется mo Polygonum-tuny и
имеет крахмальные зерна. Синерги-
ды © крючкообразными выростами.
Три антиподы дегенерируют перед
оплодотворением. Полярные ядра
сливаются при оплодотворении.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия и
двойное оплодотворение.
Эндоепермогенез нуклеарный.
У ТЬебЪготаа сасао образование кле-
ток в эндосперме начинается через
50 дней после оплодотворения.
Эмбриогенез Азегад-типа. Заро-
дыш прямой или согнутый, хлоро-
филлоносный, двусемядольный, с
хорошо развитыми почечкой, гипо-
котилем и корешком. У Со{а уега
зародыши содержат кофоип. Пер-
вое делепие зиготы у Theobroma
сасао происходит через 40—50 дней
после оплодотворения. —Подвесок
‚ трехклеточный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло или крахмал. Для этого се-
мейства характерно разнообразие в
соотношениях размеров зародыша и
эндосперма. Согнутый зародыш рас-
полагается по периферии семени,
занимая 3/4 его длины, а эпдосперм
заполняет цептр семени. Прямой
зародыш паходится в центре семе-
ни, занимая 3/4 объема сомени; эп-
досперм при этом располагает-
ся по периферии семени. Плоды ко-
жистые или мясистые, редко де-
ревянистые, раскрывающиеся или
не раскрывающиеся, часто распада-
ющие на отдельные мерикарпии.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления — полиэмбриония,
партенокарпия.
Литература: СБеезтап, 1927; Cor-
ner, 1976; Davis, 1966; Duncan, Todd,
4972; Martinson, 1972; Schnarf, 1934;
_ Тахтаджян, 1966, 4970.
СЕМЕЙСТВО ВОМВАСАСЕАЕ
Основное число хромосом — 5=9,
10, 11.
Цветок обоеполый.
Пыльник — число микроспорангиев
не определено. Стенка микроспо-
рангия имеет эпидермис, фиброзный
эндотеций, 3—4 средних слоя, секре-
`
торный многоядерный тапетум. Яд-
ра клеток тапетума сливаются и ста-
новятся полиплоидными. 'Имеются
орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультаппое. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, трехпоровые
или трехборозднопоровые.
Семяпочка анатропная © двумя
интегументами, крассинуцеллятная.
Микропиле зигзагообразное, образу-
ющееся обоими — интегументами.
Имеются нуцеллярный колпачок, ги-
постаза и ариллус. Семяпочек не-
много.
Макроспорогенез и макрогаметоге-
пез — археспорий — одноклеточный.
Париетальных клеток 8—12 рядов.
Тетрады макроспор линейные, T-06-
разные. Зародышевый мешок образу-
ется по Ро]угопат-типу. Синергиды
с крючкообразными выростами и
фибриллярными аппаратами. Име-
ются три обычно эфемерные анти-
поды. Полярные ядра сливаются
перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
uue y Ceiba pentandra var. indica.
Эндоспермогенез нуклеарный, эн-
досперм содержит масло и крахмал.
Эмбриогенез Азегад-типа. Заро-
дыш двусемядольный с мясистыми,
запасающими складчатыми семядо-
лями, согпутый, хлорофиллоносный.
Семя, плод — семя со слаборазви-
тым эндоспермом или без него. За-
родыш лежит в центре семени, за-
нимая почти весь его объем. Остат-
ки эндосперма сохраняются лишь на
периферии семени. Плод — локули-
цидная коробочка, иногда не рас-
крывающаяся.
Гаусторий — эндоспермальный в
халазальной части зародышевого
мешка,
Апомиксие —у РасШга oleaginea
обнаружепа нуцеллярная эмбриония.
Другие явления не обнаружены.
Литература: Согпег, 1976; Davis,
1966; Пипсап, 1970; Смирнова, 1977:
Тахтаджян, 1966, 1970.
CEMEMCTBO MALVACEAE
Основное число хромосом — х=
=5—13, 15—17, 21, 22.
Цветок обычно обоеполый.
103
Ныльник — 2 микроспорангия.
Стенка микроснорангия образубтся
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секретор-
ный или амебоидный многоядер-
ный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование ‚ микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изред-
ка 7/-образные и линейные. Зрелые
пыльцевые зерна двухклеточные,
многопоровые (or 10 yo 30 пор),
трехборозднопоровые с шиповатой
или изредка гладкой экзиной. Клет-
ки спермиев хорошо выражены.
`Семяпочка — анатропная-кампило-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле зиг-
загообразное, образующееся обоими
интегументами. Имеются пуцелляр-
ный колпачок, гипостаза и фунику-
лярный обтуратор. Семяночек . одна-
много. | |
Макроепорогенез и макрогаметоге-
нез — археспорий одно-, двух-, трох-
многоклеточный. ПЦариетальных кло-
ток 12 рядов. Тетрады макроспор
линейные, Г-образные. Зародышевый
мешок образуется по Ро]убопат-ти-
пу. Синергиды с крючкообразными
выростами и фибриллярными аппа-
ратами. Имеется 3—20 антипод либо
эфемерных, либо долго сохраняю-
щихся. Полярные ядра сливаются
до оплодотворения. :
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекреетное опыление и самоопыле-
ние. Порогамия. Двойное оплодотво-
рение премитотического или проме-
жуточного типов. У хлопчатника
сорта 108-Ф от опыления до опло-
дотворения проходит 24—27 ч, Син-
гамия и тройное слияпие протекают
одновременно. Зигота делится через
16—48 ч после оплодотворения.
Пыльцевые трубки долго сохраняют-
ся в завязи.
Эндоспермогенез нуклеарный. Не-
сколько сот свободных ядер образу-
ется в эндосперме, прежде чем
между ядрами пачнется образование
клеточных перегородок.
Эмбриогенез Азегад-типа. Заро-
дыш прямой или согнутый, с двумя
неравной величины складчатыми се-
мядолями, с хорошо развитыми по-
чечкой, гипбкотилем и корешком,
хлорофиллопосный или бесхлоро-
104
`филльный, содержит’ масло, крахмал
и алейроновые ‘зерна. У Соззур1ат,
кроме того, имеются кристаллы ок-
салата кальция. Подвесок. короткий.
Семя; плод — семя часто с масля-
нистым эндоспермом, иногда без эн-
досперма или со слабыми остатками
его. Зародьши заполняет либо 3/4
объема семени, либо все семя. Пря-
мой зародыш располагается в цент-
ре, а согнутый по периферии семе-
ни. Плод — обычно локулицидная
коробочка или при созревании рас-
падается на мирикарпии; иногда
ягода или крылатки. В условиях
Ташкента для сверхекороспелых
сортов хлопчатника (Соззурпаиа her-
baceum) период от цветения до 60-
зревания тянется около 45 дней,
у среднеспелых (С. hirsutum) — oKO-
ло 60 дней; у позднеспелых (С. Баг-
badense).— около 75 дней. В преде-
лах одного и того же сорта наиболее
быстро развиваются семена в усло-
виях июня — июля, медленнее — в
августе, еще медленнее в сентябре.
Гаусторий отсутствует. ,
Апомиксие — редуцировапныи
партегогенез.
Другие явления — полиэмбриония
и клейстогамия.
Литература: Баранов и gp., 1937;
Беляева Н., 1963, 4964, 1977; Свапага,
Bhatnagar, 1975; Chaudhury, Mellik,
4965; Corner, 1976; Davis, 1966; Dny-
ansagar, Gaoli, 1969; Dunean, Todd,
1972; Johri, Poundir, 1976; Joshi, Wadh-
wani, 1967; Hamamoza, 1966, 1975, 1976,
1978, 1979; Камалова, Ilantemeesa,
1979; - Камалова и др., 1979; Kapil,
Bhatnagar, 1975; Honzrueza, 1960, 1964;
Медведева, 1944; МогМег, 1969, 1972;
Никифоров, 1964, Пантелеева, 1977;
Ramchandani et al., 1966; Романов,
1936, 1937a, 6, 1938, 1939, 1947, -1954,
1957, 1960; Руми, 4960, 1964а, 6, 1966,
1969; Руми, Камалова, 1975; Руми
uw mp., 1976; Schnarf, 1931, Singh B.,
49678, с; Тахтаджян, 1966, 1970;
Умарова, 1977; Усанов, 1968, 1970.
5
В порядке Маузез семейства Заг-
colaenaceae, Rhopalocarpaceae mu Hua-
` сеае цитоэмбриологически не иссле-
довапы. Цитоэмбриологические дан-
ные указывают на сходство Scytope-
talaceae c Elaeocarpaceae, Tiliceae u
Bombacaceae и на отличие их от
Ма|уасеае, который от других се-
мейств Ма[уаез отличается двумя
микроспорангиями; амебоидным та-
петумом, многопоровыми пыльцевы-
ми зёрнами с шиповатой экзиной и
компилотропной семяпочкой [УЦауа-
raghavan, Dhar, 1976]. Ha ocHonanuu
этого вышеупомянутые авторы счи-
тают, что цитоэмбриологические
данные подтверждают мнение Хат-
чинсона [1959] о необходимости от-
несения Ма|уасеае к порядку Ма[-
уа|ез, а всех остальных семейств в
порядок ТШаез. Однако вышеупомя-
нутые семейства исследованы в от-
ношении цитоэмбриологии не до-
статочно полно, чтобы делать такое
заключение. Несомненно, что между
семействами Malvaceae u другими
семействами Ма!уа]ез, наряду с раз-
личиями в отношении цитоэмбрио-
логии имеется немало и сходства,
что сближает все исследованпые се-
мейства этого порядка. Дальнейшие
исследования семейств порядка Ма|-
уа]ез необходимы, так как они по-
могут более точно выявить естест-
венные границы этого порядка.
ПОРЯДОК 46.
EUPHORBIALES
СЕМЕЙСТВО ВОХАСЕАЕ
Основное число хромосом — х=
—40, 14.
Цветок обычпо обоеполый, изред-
ка однополый. Растения чаще одно-
домные.
Ныльник — 4 микроспорангия.
Стенка микроспорапгия имеет эпи-
дермис, фиброзный эндотеций, 1—3
эфемерных средних слоя и секретор-
ный одно-двуядерный тапетум.
Микроеспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, многопоровые, с сетчатой
“ экзиной.
Семяпочка висячая, апатропная,
с двумя интегументами, крассину-
целлятная. Микропиле образуется
внутренним интегументом. Имеются
нуцеллярный колпачок, карункула
и ариллус. Семяпочек 4—8.
акроепорогенез и макрогаметоге-
нез — археспорий однонлеточный.
Париетальных клеток 2—15 рядов.
Тетрады макроспор линейные. За-
родышевый мешок образуется по’
Ро[угопат-типу. Синергиды грушо-
видные и имеют фибриллярные ап-
параты. Три и большее число анти-
под дегенерируют вскоре после оп-
лодотворения. Полярные ядра сли-
ваются до оплодотворения. ,
Опыление, прохождение пыльце-
вой трубки и оплодотворение — Нау-_
мова [1980] считает, что виды Заг-
сососса Утратили половой способ
воспроизведения и перешли на апо-
миктический путь.
Эндоспермогенез нуклеарный У
Simmondsia, целлюлярный у Вз-
xus, Sarcococca ma Pachysandra.
Эмбриогенез Опастад-типа. Заро-
дыш прямой или слегка согну-
тый, двусемядольный с гипокоти-
лем и корешком, бесхлорофилльный,
содержащий масло. Подвесок ко-
роткий.
Семя, плод — семя © хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Иногда семя без
эндосперма. Зародыш занимает от
1/4 до 3/4 объема семени при нали-
чии эндосперма. При отсутствии эн-
досперма зародыш занимает весь
объем семепи. Плод — локулицид-
ная коробочка или костянка.
Гауеторий — у некоторых предета-
вителей семейства семядоли выпол-
пяют гаусториальную функцию.
Апомиксие — нуцеллярная эмбрио-
пия обнаружена у Загсососса ргип-
folia, S. humilis uw S. hookeriana.
Другие явления — у видов Sarco-
сосса обнаружена полиэмбриония.
Литература: Согпег, 1976; Davis,
1966; Меликян, 1968; — Наумова,
~41979a, 6, 1980; Schnarf, 1934; Cmup-
нова, 1977; Тахтаджян, 4966, 1970.
СЕМЕЙСТВО
DAPHNIPHYLLACEAE
Основное число хромосом — х=8.
Цветок однополый, растение дву-
домное.
Пыльник 4 микроспорангия.
Тапетум двух-трехрядный, много-
ядерный, секреторный.
Микроспорогенез и микрогаметоге-
нез — нет данных. Зрелые пыльце-
вые зерна трехборозднопоровые.
Семяпочка висячая, анатройная,
крассинуцеллятная, с двумя интегу-
ментами. Микропиле образуется на-
ружным интегументом. Семяпочек 4,
105
Макроепорогенез и макрогаметотге-
нез — археспорий — одноклеточный.
Зародышевый мешок образуется по
Ро]угопиш-типу. Синергиды имеют
фибриллярные аппараты. Антиподы
эфемерные.
Опыление, прохождение пыльце-
вой трубки, оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип не определен.
Зародыш маленький, прямой, дифде-
ренцированный на семядоли и коре-
шок.
Семя, плод — семя с хорошо раз-
витым маслянистым эндоспермом.
Зародыш не превышает 4/4 линей-
ной высоты семени.
Плод — односеменная костянка.
Гаусторий не обнаружен,
Апомиксие предполагается, но
точно не описан. |
Другие явления — пе отмечены.
Jiurepatypa: Bhatnagar, Meenakshi,
1977; Corner, 1976; Davis, 1966;
Schnarf, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970; Уоз о, 1972.
СЕМЕЙСТВО EUPHORBIACEAE
`Оеновное число хромосом — +=
—6—15.
Цветок однополый. Растения одно-
или двудомные.
Пыльник — обычно 4 микроспоран-
rua, Ho y Acalypha spp., Chrozophora
spp., Euphorbia dracunculoides KE. mic-
rophylla — 2 mukpocnopanrua, Crenka
микроспорангия у ЕпрВогЫа образу-
ется по двудольному типу, у Аса-
1урБа и РВуПаш из — по одподоль-
ному типу. Стенка микроспорангия
имеет эпидермис, фиброзный эндо-
теций, 1—2 эфемерпых средних слоя
и секреторный дву-, трехъядерный
тапетум. Имеются орбикулы. У Аса-
]урва ш@са тапетум амебоидный.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна 2—3-клеточные, 3, 4, 10,
12-поровые или без пор, чаще всего
3-бороздные, встречаются также бо-
розднопоровые. Экзина гладкая или
сетчатая.
Семяпочка висячая, ортотропная,
гемианатропная или анатропная, ©
двумя интегументами, крассинуцел-
лятная. Микропиле обычно образу-
106
ется обоими интегументами, иногда
только одним наружным. Имеются
нуцеллярный колпачок или клювик,
подиум, гипостаза, плацентатный
или плодолистиковый — обтуратор,
каранкула и ариллус. Сосудистый
пучок доходит до халазы. Семяпочек
немного.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-, многокле-
точный. Париетальных клеток не-
сколько рядов.` Тетрады макроспор
линейные или Г-образные. Зароды-
шевые мешки образуются по Poly-
gonum-, Allium-, Penaea-, Plumba-
go-, Drusa-, Fritillaria-, Adoxa-rutam
и подтипу Аса!урва. Зародышевые
мешки имеют крахмальные зерна.
Синергиды с крючкообразными вы-
ростами. Обычно три эфемерные ан-
типоды. Ипогда число антипод уве-
личивается до 4—5. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опылепие, самоопыление.
Двойное оплодотворепие.
Эпдоепермогенез нуклеарный. O6-
разование клеточных перегородок
между свободными ядрами эндо-
сперма начинается на глобулярной
стадии развития зародыша, когда
число ядер эндосперма достигает
100 штук.
omOpuorenes Onagrad-, Solonad-, As-
terad-, Chenopodiad- a Piperad-runos.
Зародыш относительно большой,
прямой или согнутый, двувемядоль-
пый с гипокотилем и корешком,
бесхлорофилльный или хлорофилло-
поспый. Подвесок, одпо-‘или мпогс-
рядный, одно- или многоклеточный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, в котором име-
ются масло, алейроновые зерна и
друзы оксалата кальция. Зародыш,
если он согнутый, располагается по
периферии семепи и занимает 3/4
его периметра, а эндосперм запол-
няет центр семени. Если зародыш
прямой, то он заполняет почти весь
объем семени, а эндосперм распо-
лагается лишь по периферии семе-
ни. Семенная кожура образуется
обоими интегументами. Внешний
эпидермис внутреннего интегумента
дает начало каменистой зоне, кото-
рая в зрелом семени является мно-
тгослойной. Плод — обычно особый
tun (perma) коробочки, реже ягода
или костянка.
ТГаусторий эндоспермальный, до-
стигающий 1000 мм в длину и рас-
полагающийся в халазальной части
эндосперма. |
Апомикеис — нуцеллярная эмбрио-
nua (y Euphorbia 91113) и редуци-
рованный партеногенез.
Другие явления — полиэмбриония
наблюдается часто.
Литература: Bajpai, 1965; Bhatna-
gar, Kapil, 1973; Bhatnagar, Mee-
nakshi, 1977; Bor, Kapil, 1975, 1976;
Corner, 1976; Cesca, 1969; Chopra,
4970; Chopra, Singh, 1969; Dang-van-
Liem, 1960, 1964; Davis, 1966; Dnyan-
sagar, Tiwari, 1966; . Кари,
Кордюм, Заець, 1962, Maki, Mu-
kerjee, 1969; Mukerjee, 1969a, b;
Rao Р., 4967, 4970; Rao P. .et
al, 1975; Rao P., Sarveswara Rao,
‘4976; Schnarf, 1931; Singh B., 1965b;
Sing R., 41968, 1969, 1970а, №,
4972; Singh, Chopra, 1970; Singh R.,
Jain, 1965; Тахтаджян, 1966, 1970;
Tiwary, 1965; Vinghan, 1965; Venka-
*teswarlu, Rao R., 1974, 1975; Venkaes-
varlu et al., 1974; Yoshihiko, 1972.
Cemectsa Simmondsiaceae, Dicha-
petalaceae, Pandaceae, Picrodendra-
ceae и Ахеюжсасеае, включеппые
А. Л. Тахтаджяном [1970] в порядок
Euphorbiales, цитоэмбриологически
не исследованы, а семейство ОарЬп:-
рпуПасеае исследовано очень слабо.
Euphorbiaceae по данным цитоэм-
бриологии — чрезвычайно гетероген-
ное семейство. Однако род В1зВоНа
по всем цитоэмбриологическим при-
знакам сходен с другими предста-
вителями Euphorbiaceae, ostomy он
должен относиться именно к этому
cemeiictsy [Bhatnagar, Kapil, 1973]
и его не следует выделять в особое
семейство.
Ввиду слабой изученности поряд-
ка ЕпрпогЫаез в отношении цито-
эмбриологии, мы не располагаем по-
` ка данными, которые могли бы вне-
сти ясность в вопрос о систематиче-
ском положении включенных в него
семейств. Можно лишь предполо-
жить, что гетерогенность Euphorbia-
сеае, возможно, указывает на поли-
филетическое происхождение его и
на то, что оно имеет несколько ли-
ний эволюции и специализации.
Для решения этого вопроса требуют-
ся дальнейшие исследования.
1960; —
ПОРЯДОК 47.
THYMELACALES
СЕМЕЙСГВО THY MELAEACEAE
Основное число хромоеом — х=9.
Цветок обоеполый, иногда однопо-
лый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия у Газозрроп
ег1серБа\аз образуется по однодоль-
ному Tuny, y Wikstroemia canes-
сепз — по базальному. Стенка пыль-
ника имеет эпидермис, фиброзный
эндотеций, 1—2 эфемерных средних
слоя и секреторпый многоядерный
тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные.
Зрелые пыльцевые зерна трехкле-
точные, часто содержащие крах-
мальные зерна, восьми — шестидеся-
типоровые или трех — шестипоровые
с сетчатой экзиной.
Семяпочка висячая, анатропная,
гемианатропная, с двумя интегумен-
тами, крассинуцеллятная. Микропи-
ле образуется внутренним интегу-
ментом, Имеются пуцеллярный кол-
пачок, обтуратор, гипостаза и арил-
лус. Семяпочек 1—12.
Макроспорогенез и макрогаметоге-
нез — археспорий одпоклеточный.
Париетальных клеток обычно 3—5,
но у Парьпе — 8—10 рядов. Тетрады
микроспор линейные, Г-образные.
Зародышевой мешок образуется по
Ро!усопат-типу. Синергиды с крюз-
кообразными выростами и фибрил-
лярными аппаратами. Y Wikstroe-
и1а сапезсепз 3—6 клеток антипод,
y Daphne cannabina u Thymelaea ar-
vensis KieToK aHntumon 30 um более.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Iloporamua.
Двойное оплодотворение премитоти-
ческого типа.
Эндоепермогенез нуклеарный. 0б-
разование клеточных перегородок
между свободными ядрами начина-
ется © микропилярной части заро-
дышевого мешка.
Эмбриогенез Азегад-типа, вариа-
ции Erodium. Зародыш большой,
прямой, двусемядольный, бесхлоро-
107
филльный, содержит масло, алейрон
и иногда. крахмал. Подвесок корот-
кий или отсутствует,
Семя, плод — семя со слабо- или
‚хорошо развитым эндоспермом или
без него. Зародыш заполняет семя
по. всей его длине. Семядоли выпол-
няют запасающую функцию. Тонкий
слой эндосперма располагается на
периферии семени. Плод нераскры-
вающийся, сухой или сочный, ягодо-
или костянкообразный, изредка ло-
кулицидная коробочка. |
Гаусторий отсутствует.
Апомиксие — нередуцированный
партеногенез и нуцеллярная эмбрио-
ния у Wikstroemia indica var. viri-
diflora.
Другие явления — полиэмбриония
у Daphne cannabina.
Литература: Corner, 1976; Davis,
1966; Тахтаджян, 1966, 1970; Venka-
teswarlu, 19671.
/
7
По мнению некоторых исследовате-
лей, Тушеаеасеае“родетвенно Geis-
solomataceae m Penaeaceae, с одной
стороны, и с СошЬгеасеае — с дру-
гой. Сближение его с Nyctaginaceae,
Geissolomataceae m Penaeaceae, Kak
это предложил Хатчинсон [Hutchin-
зоп, '1926], согласуется с данными
цитоэмбриологии, [Venkateswarlu,
19672]. Данные цитоэмбриологии по-
казывает, что ТБВутеаеасеае имеет
сходные с некоторыми представите-
лями ЕпрВогЫасеае цитоэмбриологи-
ческие признаки. У представителей
обоих семейств имеются: 4 микро-
спорангия, секреторпый — тапетум,
симультанный микроспорогенез, ви-
сячая ° анатропная-гемиапатропная
семяпочка, нуцеллярный колпачок,
гипостаза, обтуратор, ариллус, одно-
клеточный женский археспорий,
массивная париетальная ткань, Ро-
]узопит-тип зародышевого мешка,
три и более антипод, порогамия,
нуклеарный эндосперм, семепа © эн-
доспермом, Азегад-тип эмбриогене-
заа дифференцированный зародьци, -
апомиксис. Однако, в отличие от
Euphorbiaceae, Thymelaeaceae, но
имеют эпдоспермального гаустория.
Требуются дальнейшие исследова-
пия, чтобы выяснить систематиче-
ское положение семейств порядков
Thymelaeales um Euphorbiales.
ПОДКЛАСС Е. КОЗТРАЕ
НАДПОРЯДОЕ УШ.
ВОЗА МАЕ (ВКЛЮЧАЯ
CUNONIALES,
GROSSULARIALES,
PITTOSPORALES)
TOPHHOR 48.
SA XIFRAGALES
ПОДПОРЯДОК 1.
GROSSULARINAE
СЕМЕЙСТВО СОМОМТАСЕАЕ
(ВКЛЮЧАЯ ВАОЕВАСЕАЕ)
Основное число хромосом — х= 12,
15, 16
Цветок обычно обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций 2 эфемерных
.
108
средних слоя и одноядерный секре-
торный тапетум. ее
Микроспорогенез и микрогаметоге-
нез — образование спор симультан-
ное. Тетрады микроспор тетраэдраль-
ные, — изобилатеральные. Зрелые
пыльцевые зорна двухклеточные,
двух- или трехборозднопоровые, ©
гладкой экзипой, содержат крахмал.
Семяпочка апатропная-гемиана-
тропная, с двумя интегументами,
крассипуцеллятная. Микропиле зиг-
загообразное, образующееся обоими
интегументами. Имеются нуцелляр-
ный колпачок, гипостаза и обтура-
тор. Семяпочек две — много.
Макроепорогенез и макрогаметоге-
нез — археспорий одно-дву-много-
клеточный. Париетальных клеток
несколько рядов. Тетрады макроспо
линейные. Зародышевый мешок об-
pasyetca no Polygonum-tuny mu имеет
зерна крахмала. Синергиды груше-
видпые. Имеются три эфемерные ан-
типоды. Полярные ядра. сливаются
до оплодотворения. У Ceratopetalum
‚ Нез — археспорий
ареал описан слабо выраженный
постамент. и
Эндоепермогенез нуклеарпый. Пос-
ле образования большого числа сво-
бодных ядер эндосперм становится
клеточным.
Эмбриогенез — тип не определен,
Зародыш маленький прямой, с хоро-
шо развитыми семядолями, гипоко-
тилем и корешком.
Семя. плод — семя с хорошо
развитым эндоспермом, содержа-
щим масло. Зародыш занимает от
1/4 до 3/4 длины семени. Остальной
объем семепи заполнен эндоспермом.
Семенная кожура образуется двумя _
интегументами. Семядоли выполня-
ют запасающую функцию. Плод —
обычно коробочка, реже — нераскры-
вающийся или костянка,
Гаусторий не найден.
Апомиксис но обнаружен.
Другие явления пе отмечены.
JIutepatypa: Corner, 1976; Davis,
1966; Prakash, McAlister, 1977; Cuup-
нова, 1977; Тахтаджян, 1966, _ 1970.
СЕМЕЙСТВО ЕЗСАТТОМТАСЕАЕ
Основное число хромосом — x=
—44, 12.
Цветок обычно обоеполый, редко
однополый. Растения изредка дву-
домные или полигампые.
Пыльник — нет дапных.
Микроспорогенез и микрогаметоге-
нез — нет данпых о типе микроспо-
рогенеза. Зрелые пыльцевые зерна
трехклеточные, трех-пятибороздно-
поровые или 3—2-поровые.
Семяпочка. анатропная, © одним
или двумя интегументами, тенуину-
целлятная или крассинуцеллятная.
Эндотелий окружает на 2/3 нижнюю
часть зародышевого, мешка. Семяпо-
чек много.
Макроспорогенез и макрогаметоге-
одноклеточный.
Париетальные клетки отсутствуют
y Escallonia m Polyosma, но у Brexia
образуется один ряд их. Тетрады
макроспор линейные. Зародышевый
менток образуется по’ Ро[угопат-ти-
пу. Синергиды удлиненные. Имеются
три скоро дегенерирующие антипо-
ды. Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пылье-
вой трубки и оплодотворение — нет
данных,
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не определен.
Зародыш маленький. ''
Семя, плод — семя с хорошо раз-
витым эндоспермом. Плод -— коро-
бочка или реже костянко- или яго-
дообразный.
Гаусторий не найден.
Апомикеис не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; Тахтаджян, 1966, 4970.
СЕМЕЙСТВО GROSSULARIACEAE
Основное число хромосом — х=8.
Цветок обычно обоеполый, реже
однополый. ,
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, два
средних слоя и секреторный дву-
многоядерный тапетум. У некоторых
видов Ribes тапетум многорядный.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, многопоровые. Спермии-
клетки имеют незначительное коли-
чество цитоплазмы.
Семяпочка. анатропная, с двумя
интегументами, крассинуцеллятная.
Имеются ариллус и гипостаза. Семя-
почек много или пемпого.
Макроспорогенев и макрогаметоге-
нез — археспорий одпо-, изредка
многоклеточный. Париетальных кле-
ток несколько рядов. Тетрады макро-
спор линейные, Г-образные. Зароды-
шевый мешок образуется по Polygo-
пат-типу. Синергиды с крючкообраз-
ными выростами. Имеются две-три
скоро дегенерирующие или долго со-
храняющиеся антиподы. Полярные
ядра сливаются до онлодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опылепие у видов ВШез,
порогамия. Двойное оплодотворение
премитотического типа. У Ribes ni-
сег оплодотворение происходит че-
рез 4 дня после опыления.
Эндоспермогенез целлюлярный, ге-
лобиальный.
Эмбриогенез Caryophyllad-runa,
гариации ЗахИгаса. Зародыш’ ма-
ленький, дифференцированный на
семядоли и корешок, бесхлоро-
филльный.
Семя, плод — семя с эндоспермом,
109
‘
>
содержащим масло. Зародыш распо-
ложен в основании семени и обычно
не превышает 1/4 линейной высоты
семени. Плод р— паракарпная коро-
бочка с остающейся чашечкой.
Гауеторий — семядоли выполняют
таусториальную функцию.
Апомикеис — апогаметия.
Другие явления не отмечены.
Литература: Березенко, Лиферова,
4970; Corner, 1976; Davis, 1966; Ко-
мар, 1965а, 6; Мандрик, 1969 а, 6,
1972, 1976; Мандрик, Андрусяк,
1969 a, 6; Попова, 1965, 1970; Радио-
ненко, 1969, 1970; Радионенко, Лифе-
рова, 1970 6; Семенихина, 1972; Смир-
нова, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО НУРВАМСЕАСЕАЕ
§
Основное число хромоеом — х=13,
14, 17, 18.
Цветок обоеполый.
Пыльник — нет данпых.
Микроспорогенез и микрогаметоге-
нез — микроспорогепез пе изучен.
Зрелые пыльцевые зерпа трехбо-
розднопоровые или изредка трехбо-
роздные, с мелкосетчатой сэкзиной.
Семяпочка апатропная, с одним
интегументом, тенуинуцеллятная.
Имеется эндотелий. Он окружает
нижнюю часть зародышевого меш-
ка. Семяпочек обычно много.
Макроспорогенез и микрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевые мешки образуются по
АШаш- и Адоха-типам. Синергиды
с крючкообразными выростами. Три
антиподы становятся двуядерными;
обычно они скоро дегенерируют. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез 50]опад-типа (?). За-
родыш прямой дифференцирован-
ный, двусемядольный с гипокотилем
и корешком, бесхлорофилльный.
Семя, плод — семя с эндоспермом.
Зародыш занимает от 1/4 до 3/4 дли-
ны семени. Эндосперм заполняет
остальной объем семени. Плод —
септицидная или локулицидная ко-
робочка или редко ягода.
Гаусторий антиподальный (у Ki-
тепоезВота). Семядоля выполняет
гаусториальную функцию.
110
Апомикеисе не обнаружен. .
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Смирнова, ‘1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО РТТОЗРОВАСЕАЕ
Основное чиело хромосом — х=12.
Цветки обычно обоеполые или
иногда полигамные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный, двух-четырехъядерный та-
петум. Ядра клеток тапетума слива-
ются и становятся полиплоидными.
Микроспорогенез и микрогаметоге-
Hes — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
лые пыльцевые зерна трехклеточ-
ные, трехборозднопоровые.
Семяпочка анакампилотропная,
с одним интегументом, тенуинуцел-
лятпая, Имеется обтуратор. Семяпо-
чек мпого, реже песколько.
Макроспорогенез и макрогаметоге-
нез — археспорий одпо-, двух-, мно-
гоклеточный. Париетальпые клетки
отсутствуют. Тетрады — макроспор
линейные, Г-образные. У Morianthus
rigens u Sollya heterophylla muxpo-
пилярная клетка диады макроспор
не делится и вместо тетрады обра-
зуется триада; синергиды с крючко-
образными выростами. Зародышевый
мешок формируется по-Ро]ухопит-
типу. Имеются три эфемерные анти-
поды. Полярные ядра сливаются до
оплодотворепия.
Опылепие, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Тройпое слияние предшествует син-
гамии.
Эндоспермогенез пуклеарный. Пер-
вичное ядро эндосперма делится
раньше зиготы.
- Эмбриогенез Азегад-типа, вариз-
nun Geum. Зародыш очень малень-
кий, но, несмотря на малую величи-
ну, дифференцированный.
Семя, плод — семя с хорошо раз-
витым твердым маслянистым эндо-
спермом. Зародыш расположен в
основании семени и обычно не пре-
вышает 1/4 линейной высоты семени.
Плод — локулицидная коробочка.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Narayana, Sundari, 1977a, Ъ;
Pandey, Saxena, 1977; Rau, 1967;
Schnarf, 1934; Shella, Narayana,
1966; CmupHosa, 1977; Sundari, 1977;
Sundari, Narayana, 1977; Taxramxan,
1966, 1970.
CEMEMCTBO BYBLIDACEAE
Оеновное
данных.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
чиело хромосом — нет
Микроепорогенез и микрогаметоге- _
нез — микроспорогенез не изучен.
Зрелые пыльцевые. зерна трехбо-
розднопоровые с зерпистой экзиной.
Семяпочка анатроппая, с одним
или двумя интегументами, тенуину-
целятная. Микропиле образуется
внутренним интегументом. Имеется
эндотелий.
Макроспорогенез и макрогаметоге-
нез — архесторий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Polygonum-tuny mw содержит крах-
мальные зерна.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
дапных.
Эндоспермогенез поллюлярпиый.
Эмбриогенез — тип пе определен.
Зародыш малопький, прямой, диф-
ференцировапный па семядоли, ги-
покотиль и корешок. Семядоли ко-
роткие, гипокотиль длипный. Подве-
сок. длинный.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш pac-
положен в основании семени и обыч-
но не превышает 1/4 липейной высо-
ты семени. Плод — двух-четырех-
створчатая коробочка.
Гаусеторий — имеются микропиляр-
ный и халазальный эндоспермаль-
ные гаустории. Семядоли выполняют
гаусториальпую фупкцию.
Апомикеис не обпаружен.
Другие явления пе отмечены.
Литература: Corner, 1976; Davis,
4966; Тахтаджян, 1966, 1970; Vani,
1966; Vani, Hardev, 1972.
CEMEMCTBO BRUNIACEAE
Основное число хромосом — х=8.
Цветок обоеполый.
Ныльник — нет данных,
Микроспорогенез и микрогаметоге-
нез — микроспорогенез не изучеп.
Пыльцевые зерна трех или шести-
одиннадцатиборозднопоровые.
Семяпочка анатропная © одним
интегументом, крассинуцеллятная.
Микропиле длинное, нуцеллус ма-
ленький. Семяпочек не много, число
их редко достигает 8.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки имеются (?).
Тетрады макроспор линейные. Заро-
дышевый мешок формируется по
РЫ]узопит-типу и имеет крахмаль-
ные зерна. Синергиды сильно удли-
няются и проникают в микропиле.
Имеются три эфемерные антиподы.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез — нет данных.
Эмбриогенез — тип не определен.
Зародыш очень маленький, прямой,
дифференцированный па семядоли,
‚ гинокотиль и корешок.
Семя, плод — семя с хорошо раз-
витым мясистым эндоспермом. Заро-
дыш занимает от 1/4 до 3/4 длины
семени. Эпдосперм заполняет ос-
тальной объем семени. Плод сухой,
пе раскрывающийся или коробочка.
Гаусторий подвесочный, много-
ядерный. Семядоли выполняют гау-
сториальпую фупкцию.
Апомиксиес не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1934; CmupHosa, 1977;
Тахтаджян, 1966, 1970.
В поднорядке Стоззагтае поряд-
ка Saxifragales семейства ПОау1930-
niaceae, Eucryphiaceae, Paracryphia-
ceae, Crypteroniaceae, BruneNiaceae,
Tribelaceae, Tetracarpaeaceae, IJtea-
ceae, Brexiaceae, Dulongiaceae, Pte-
rostemonaceae, Montiniaceae, Roridu-
]асеае и Аезепозиласеае либо пол-
ностью пе исследованы в отношении
цитоэмбриологии, либо исследованы
настолько слабо, что по имеющимся
данным в настоящее время невоз-
можно дать их цитоэмбриологиче-
скую характеристику. Из остальных
четырех семейств только Стоззи[а-
riaceae исследовано цитоэмбриоло-
тически довольно подробно, а се-
414
MelicTBa Cunoniaceae, Escalloniaceae
и Нудгапоеасеае исследованы в этом
отношении далеко не полно. Такое
слабое исследование цитоэмбриоло-
тии подпорядка Grossularininae He
позволяет на основании цитоэмбрио-
логических признаков судить о род-
ствепных олпошепиях между его
семействами и очертить его естест-
венные границы.
По цитоэмбриологическим призна-
кам род Ваига скорее относится к
Сипотшасеае, чем к Saxifragaceae
[Prakash, McAlister, 1977]. Pittospo-
тасеае сходно с ЕзсаЦотласеае (Sa-
xifragaceae) 10 наличию тенуину-
целлятной, с одним интегументом,
семяпочки, по одноклеточному жен-
скому археспорию, зародышевому
мешку Роугопат-типа, эфемерным
антиподом и нуклеарпому эндоспер-
му. Необходимо дальнейшее изуче-
ние подпорядка Стозза]агтае.
ПОРЯДОК 18.
SA XIFRAGALES
ПОДПОРЯДОК 2.
SA XIFRAGI NEAE
CEMEHMCTBO CRASSULACEAE
Основное число хромоеом — х=13,
15, 17—19, 24.
Цветок обоеполый или очень ред-
ко одпополый.
Ныльник — 4 микроспораптия.
Степка микроснорангия имеет оии-
дермис, фиброзный эидотеций, 1—2
эфемерных средних слоя и сокре-
торнпый одпо-двуядерпый танотум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор cu-
мультанное. Тетрады микроспор тет-
раэдральпые. Зрелые — пыльцевые
зерна двухклеточные, трех-, четы-
рехборозднопоровые, с тонкосетчатой
или гладкой сэкзиной, иногда имеют
крахмальные зерна.
Семяпочка апатропная с двумя
интегументами, крассипуцеллятная.
Микропиле узкое, длинное, образу-
ется обоими иитогумептами или
иногда только паружным иптегу-
ментом. Семяпочек мпого или реже
немного.
Макроспорогевез и макрогаметоге-
112
нез — археспорий одно-многоклеточ-
ный. Париетальных клеток один-не-
сколько рядов. Тетрады макроспор
линейные, ХТ-образные, изобилате-
ральные. Зародышевые мешки обра-
зуются по Polygonum- и АШат-ти-
пам, имеют крахмальные зерна. Си-
нергиды грушевидные. Имеются три
скоро дегенерирующие антиподы.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных. Порогамия. Двойное опло-
дотворение. Тройное слияние проис-
ходит раньше сингамии.
Эндоспермогенез целлюлярный.
Эмбриогенез СагуорвуПа@-типа. За-
родыш маленький или большой,
дифференпцированный на семядоли,
гипокотиль и корешок, бесхлоро-
филльный, содержит масло и алей-
рон.
Семя, плод — семя со слаборазви-
чым ондоспермом или без него.
В первом случае зародыш пе пре-
выцпает 1/4 объема семепи, во вто-
ром — зародьзи заполняет семя по
всей cro длине. Плод — многоли-
стовка, редко коробочковидный.
Гаусторий — обнаружено несколь-
кос типов гаусториев: макроспоро-
вый, эндоспермальный в халазаль-
ной части эндосперма, синергидный,
антиподальный И подвесочный.
У некоторых представителей семя-
доли выполняют гаусториальную
функцию.
Апомиксие не обнаружен.
Другие явления — партенокарпия.
Литература: Commer, 1976; Davis,
1966; Fetre, Lebegue, 1964: Кордюм,
1963; Мандрик, Голышкин, 1973;
Schnarf, 4931; Subramanvam, 1967b, f,
1970; Тахтаджяи, 1966, 1970; Vignon,
Fetré, 1966a, b, 1972.
СЕМЕЙСТВО ЗАМЕКАСАСЕАЕ
Основное число хромосом — z=
6—15, 17, 19, 24.
Цветок обоеполый,
лый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секретор-
ный одно-, дву- четырехъядерный,
обычно одно-, иногда двурядный та-
петум.
Микроспорогенез и микрогаметоге-
редко однопо-
нез — образование микроспор си-
хгультавное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые, с сстчатой экзипой, иног-
да шести-девятипоровые или трех-
бороздные.
Семяпочка висячая, анатроппая,
с двумя или одним интегументом,
крассинуцеллятная или тепуинуцел-
лятная. Микропиле образуется
обоими интегументами или только
одним наружным. Имеется эндоте-
лий. Семяпочек много-несколько.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-многокле-
точный. Париетальных клеток 2—6
рядов. Тетрады макроспор линпей-
ные, Г-образные. Зародышевые меш-
ки образуются mo Polygonum- или
Allium-tumam. Y Astilbe grandis опи
содержат крахмал. Crneprujr ¢
крючкообразными выростами и фиб-
риллярными аппаратами. 3—5 анти-
под либо скоро дегенерирующих,
либо долго сохраняющихся. Поляр-
ные ядра сливаются до оплодотво-
рения. У некоторых представителей
имеются постаменты.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
обычно перекрестное опыление. По-
рогамия. Двойное оплодотворелие
премитотического типа
Эндоспермогенез цоллюолярный, го
лобиалытый, нуклеарный.
Эмбриогенез - - СатуорйуПа@-, Аз-
rad-, Solanad-, um Onagrad-runos. 3a-
родыш прямой, двусемядольный, с
хорошо развитыми гипокотилем и
корешком, бесхлорофилльный, содер-
жит масло, алейрон, иногда крахмал.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло. Зародыш располагается по
оси семени и занимает от 1/4 до 3/4
его длины. Эндосперм занимает ос-
тальной объем семени. Семядоли
иногда выполняют гаусториальную
функцию. Плод — коробочка или
ягода.
Гаусторий аптиподальный, эидо-
спермальный в микропилярпой и
халазальной частях — оэидосперма,
подвесочный.
Апомикесие — нуцеллярная эмбрио-
HUA,
Другие явления — гинодиэция у
Saxifraga.
Jinreparypa: Anantaswami, Sharma,
1967; Arecal et а1., 1970; Blamish,
4967; Biamish, Lin-Shu-Chang, 1965;
ВВ. А. Подду@ная-Арнольди
Coq, 1967; Corner, 1976; Davis, 1966;
Капранова, 1970; Коробова-Семепчен-
ко, 41965, 1966, 1969, 197ба, 6, в, 1977:
Ratnaparkhi, 1973; Rau M., 41967a,
1970b. Rau M., Dehra Dun, 1967b;
Rau M., Sharma, 1967c; Saxena, 1969a,
b, 1970a, b, 1972; Schnarf, 1931; Cuup-
нова, 1977; Тахтаджян, 1966, 1970;
Vignon, Fetre, 1972; Vijayaraghavan,
Ratnaparkhi, 1976.
CEMEMCTBO VAHLIACEAE
Основное число xpoMocoM — x =
Пветок обосполый.
Пыльник — 4 микроспорапгия. Та-
потум секреторный, двуядерный.
Микроспорогенез и микрогаметоге-
иез — образовапие микроспор си-
мультапное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
лые пыльцевые зерна двухклеточ-
ные, трехборозднопоровые.
Семяпочка анатропная с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеются гипостаза и
эндотелий. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одпоклеточный.
Паристальные клетки отсутствуют.
Тетрады макроспор липейпые. Заро-
дышевый мешок образуется по Ро-
Гуропинг типу. Сипоргиды с крючко-
образными выростами. Имеются три
аитиподы, дегенерирующие вскоре
после оплодотворепия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез СагуорвуПад-типа.
Семя, плод — плод — коробочка.
Гаусторий — в халазальной части
эндосперма образуется не агрессив-
ный, двухклеточный гаусторий.
Апомиксие не обпаружен.
Другие явлепия пс отмечены.
Литература: Davis, 1966; Тахтад-
ast, 4966, 1970.
СЕМЕЙСТВО ЕКАМСОАСЕАЕ
Основное число хромосом — х==13.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Микроепорогенез и микрогамстоге-
нез — нет данных.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
113
Микропиле образуется двумя инте-
гументами. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий — однокле гочный.
Париетальных клеток 6 рядов. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Рау-
гопит-типу. Синергиды с крючкооб-
разными выростами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
‘между ядрами эндосперма начинает-
ся с микропилярной части зароды-
шевого мешка. В халазальной части
ядра эндосперма долго сохраняются
в свободном состоянии.
‚ Эмбриогенез — тип не определон.
Зародьии хотя и маленький, но диф-
ференцированппый на семядоли, ги-
покотиль и корешок.
Семя, плод — семя с хорошо раз-
витым масляпистым эпдоспермом.
Зародыш обычно запимает от 1/4 до
1/2 длипы семени. Остальной объем
семени — заполнен эндоспермом.
Плод — локулицидная коробочка.
Гаусторий — семядоли в начало
прорастания выполняют гаустори-
альную функцию.
Апомикеис не обнаружен.
Другие явления не отмечены.
Литература: Пау!з, 1966; Тахтад-
жян, 1966, 1970.
СЕМЕЙСТВО РАВМАЗЗТАСЕАЕ
Основное число хромосом — х==9.
Цветок обоеполый,
Пыльник — 4 микроспорангия,
Стенка микроспорангия имеет эпи-
дермис, фиброзный эпдотеций, 2
средних слоя и секреторный одно-дву-
ядерный тапетум.
Микрсспорогенгз и микрогаметоге-
нез — образование микроспор си-
мультапное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые
гнезда двухклеточные, трехбороздно-
поровые.
Семяпочка — анатропная-кампило-
тропная, с двумя или одним интегу-
ментами. Тенуипуцеллятная. Мик-
ропиле образуется обоими интегу-
ментами. Семяпочек много.
Макроспорогенез и макгогаметоге-
нез — археспорий одноклеточный.
114
i
/ {
1 .
Париетальных клеток нет. Тетрады
макроспор линейные. Зародышевый
мешок образуется по Ро]угопат-ти-
пу. Синергиды с крючкообразными
выростами и фибриллярными аппа-
ратами. Имеются 3—5 крупных
долго — сохраняющихся — антипод.
Полярные ядра сливаются перед оп-
лодотворением.
Опыление, прохождение пыльцз-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез Азегад-, Опаста4-ти-
пов. Зародыш прямой; семядоли вы-
полняют запасающую функцию. Под-
весок состоит из нескольких клеток
или отсутствует.
Семя, плод — семя со слаборазви-
тым маслянистым эндоспермом или
без эндосперма. Зародыш заполняет
семя по всей его длине. Эндосперм,
если оп имеется, то в виде тонкого
слоя клеток располагается по пери-
ферии семепи. Плод — локулицидная
коробочка.
Гаусторий отсутствует.
Апомикесис пе обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; ЗВагта, 1968а, 5; Смирнова, 1977;
Тахтаджян, 1966, 1970.
В подпорядке бахИтастае поряд-
ка БбахИгаса!ез семейства Реп{Пога-
ceae, Cephalotaceae m Eremosynaceae
цитоэмбриологически He исслецева-’
ны. Центральное семейство ЗахИга-
сасеае в отношении цитоэмбриоло-
гии готерогенцо, особенно по харак-
тору эндоспормо- и эмбриогенезов.
Отсутствием париетальных клеток,
разных тинов гаусторий и апомикси-
са Рагпаз\а отличается от предста-
вителой БахИгабасеае, поэтому вы-
деление ее в самостоятельное семей-
ство правильно. Даппые цитоэмбрио-
логии указывают на сходство Сгаз-
зи]асеае с ЗахИгасасеае и на сход-
ство СгаззШасеае с Родозетасеае и
Нудгозасвуасеае [Subramanyam,
1967], так как все эти семейства,
имеют одинаковые цитоэмбриологи-
ческие признаки. Возможно, что в
порядок ЗахИгасаез, особенно в
подпорядок Стаззцагтае, А. Л. Тах-
таджяи включил слишком много се-
мейств, которые не соответствуют
этому порядку и которые следует
отнести к другим порядкам. Р Даль-
’
тгрен [Dahlgren, 1975] некоторые
семейства, отнесенные А. Л. Тахтад-
жяном к порядку бахИгаса|ез, выде-
ляет в порядки Сапоп1айез (Сапотша-
ceae, Iteaceae, Brunelliaceae, Eucryp-
hiaceae, Baueraceae, Brunoniaceae) и
Pittosporales (Pittosporaceae). OnHako
питоэмбриологических данных по по-
рядку бахИтаса!ез и по обоим под-
порядкам (Grassularinae, Saxifragi-
mae) пока еще слишком недостаточ-
но, чтобы делать окончательные вы-
воды о систематическом положении
семейств зтого порядка.
ПОРЯДОК 49.
ROSALES |
СЕМЕЙСТВО ВОЗАСЕАЕ
Основное числе хромосом — х =
= 6—10, 12, 17.
Цветок большей частью обое-
полый.
Пыльник — 4 микроспоравгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный, дву-восьмиядерный тапе-
тум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, ‚Зрелые пыльцевые
зерна двухклеточные, трехпоровые
или многоборозднопоровые.
Семяпочка анатропная-гемиана-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется обоими или только внут-
ренним интегументами. Имеются
нуцеллярный колпачок, фуникуляр-
ный обтуратор и гипостаза. Семя-
почек 1—2, несколько.
Макроспорогенез и макрогамето-
генез — археспорий много- однокле-
точный. Париетальных клеток 9ря-
дов. Тетрады макроспор линейные,
2-образные, Зародышевый мешок
образуется по Polygonum-runy u
имеет крахмальные зерна. Синер-
гиды грушевидные с фибриллярны-
ми аппаратами. Имеются три и бо-
лее эфемерных или долго сохраня-
ющихся антипод. Нолярные ядра
сливаются до оплодотворения или
во время оплодотворения. Наблюда-
ется присутствие нескольких заро-
дышевых мешков в одной и той же
8*
семяпочке, что связано главным об-
разом с наличием многоклеточного
женского археспория и апоспории.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодо-
творение премитотического типа.
Тройное слияние происходит раньше
сингамии.
Эндоепермогенез нуклеарный. О0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярного конца заро-
дышевого мешка. ЗВ халазальной
части зародышевого мешка ядра
эндосперма долго остаются свобод-
ными, Эндосперм содержит масло и
алейрон.
Эмбриогенез Азегад-типа вариа-.
uuu Geum. Зародыш дифференци-
рованный, крупный, прямой, бесхло-
рофилльный, . содержит — алейрон, '
крахмал, друзы оксалата кальция,
эмульсии и амигдалин. Семядоли
выполняют запасающую функцию.
Семя, плод — семя без эндоспер-.
ма или редко со слаборазвитым эн-
доспермом. Зародыш заполняет се-
мя по всей его длине. Если эндо-
сперм имеется, то он располагает-
ся тонким слоем лишь по перифе-
рии семени. Плод — апокарпная или
синкарпная многолистовка, орешек,
коробочка, яблоко или костянка.
аусторий эндоспермальный в ха-
лазальном конце зародышевого ме-
шка.
Апомиксис — редуцированный и
нередуцированный партеногенез, ну-
пеллярная и интегументальная эм-
бриония, генеративная и соматиче-
ская апоспория, у некоторых видов
Rubus, Potentilla a Alchemilla.
Другие явления — полиэмбраония,
при которой дополнительные заро-
дыши образуются из синергид. Пар-
тенокарпия и клейстогамия.
Литература: Березенко, 1969; Бе-
ридзе, 1962, 1964, 1969; Сотпег, 1976;
Czapik, 1962; Davis, 1966; Tpesnosa,
1971а ‚6; Здруйковская-Рихтер, 1970;
Ивановская, 1963; Ивановская, Ор-
лова, 1971; Juguet, Lebegue, 1965;
Каграмова, 1978; — Константинов,
1960; Крылова, 1970; Мандрик, 1976;
Мандрик, Ментковская, 4977, 1978а,
6; Мандрик, Петрус, 1976; Магауа-
na, Sharma H., 1976; Nordborg, 1967;
МипЦатта, РЫррз, 1979; Петрус,
1975; Phelouzot, 41965; Печеницин,
1973; Плиско, 1970; Радионенко,
115
<.) KS
. eon < а PON
Pue. 18. Семяпочка и элемеиты зародышевого мешка Роусопат-типа
у земляпики Ггасайа апапаза (1), вхождение ныльцевой трубки в заро-
дышевый меиюк и двойное оплодотворение у яблока сорта Макинтош
[Радиопенко, 19601 (Б)
Д: а — многоклеточный экецпский археспорий, 6 — яйцевой аппарат, в — полярные
ядра. г-- вторичное ядро зародышевого мешка, д — аптиподальный аппарат [Солн-
цева, 1961, 1965]; Б: а — вхождение пыльцевой трубки, б — двойное оплодотворение
1963, 1964, 1966, 1967, 1968; Саму-
Пыльвик — пет данных о числе
тия, 1971; Schnarf, 1934; Sharma, микроспорангиев. Зрелые пыльце-
Narayana, 1971; Солнцева, 1961, вые зерна трехклеточные, биполяр-
1965а; Спицын, 1966; Сухарева, но-трехщелевые, шести (иногда
1970; Тахтаджян, 1966, 1970; Тор- восьми)-поровые.
ham, 1970; Virdi, Eaton, 1969. Микроспорогенсз и микрогамето-
генез — образование микроспор си-
мультанное.
СЕМЕЙСТВО МЕОВАРАСЕАЕ
Оеновное число хромосом — 1 =
—
—
, 7.
Цветок обоеполый.
116
Семяпочка с двумя интегумента-
ми. Семяпочек от 5 до 20.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Имеется одна париетальная клетка.
Рие. 18 (окопчание)
Зародышевый мептгок образуется по
Polygonum-tuny. Зрелый зародыше-
вый мешок выступает из нуцеллу-
ca. Имеются три эфемерные анти-
поды.
Опыление, прохождение ныльце-
вой трубки н оплодотворение — пет
данных.
Эндоснермогенез — нет данных.
Эмбриогенез — нет данных.
Семя, плод — семя без
сперма.
Гауеторий не найден.
Anomukene пе обнаружен.
Другие явления не отмечены.
Литература: Schnarf, 1931; Taxta-
джян, 1966, 1970.
эндо-
В порядке Rosales семейство
Chrysobalanaceae — цитоэмбриологи-
чески совершенно не исследовано.
а семейство Меигадасеае исследова-
HO в этом отпотиепийи очень слабо.
Хатчиисон ‘[Wulchinson, 1969, 1973],
nan on Л. Л. Тахтаджян [4970], к по-
рядку Rosales относил лишь три
семейства, но в то время как
А. Л. Тахтаджян включил в Воза-
les семейства Rosaceae, Chrysoba-
lanaceae uw Neuradaceae, XaTauncon
относил к этому порядку Rosaceae,
Dichapetalaceae mu Calycanthaceae.
Кронквист ([Cronquist, 1968] sme B
порядок Rosales включил 20 ce-
мейств, так как представителей по-
117
рядков ЗахИтасаез и Fabales on
включил в Rosales. Р. Дальгрен
[Ра степ, 1975] относит к Rosales
7 семейств (Crossomataceae, Rosa-
ceae, Malaceae, Amygdalaceae, Con-
naraceae, Mellanthaceae, Chrysoba-
]апасеае). Такое различное понима-
ние объема порядка Воза!ез указы-
вает на то, что в настоящее время
не накоплено достаточных данных
для установления естественных гра-
ниц этого важного и интересного
порядка.
ПОРЯДОК 50.
FABALES (LEG UMINALES)
СЕМЕЙСТВО МТМОЗАСЕАЕ
Основное число хромосом — х =
— 8, 13, 14, 18.
Цветок обоеполый. -
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных, средних слоя, секретор-
ный одно-многоядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна двух-
трехклеточные, одиночные или сое-
диненные в тетрады и поллинии,
содержат зерна крахмала.
Семяпочка — гемианатропная-ана-
тропная с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется обоими интегументами или
одним наружным. Обычно микропи-
ле зигзагообразные. Семяножка ча-
сто длинная. Имеются нуцеллярный
колпачок и фуникулярпый ариллус.
Макроспорогенез и макрогамето-
генез — археспорий одно- дву- много-
клеточный. Париетальных клеток
2—7 рядов. Тетрады макроспор ли-
нейные, реже Г-образные. Зароды-
шевый мешок образуется по Роу-
сопит-типу и имеет крахмальные
зерна. Синергиды c крючкообраз-
ными выростами и фибриллярны-
ми аппаратами. Имеются три эфе-
мерные или долго сохраняющиеся
антиподы. Полярные ядра сливают-
ся перед оплодотворением или при
оплодотворении.
Опыление, прохождение пыльце-
118
вой трубки и оплодотворение — нет.
данных.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка. В халазальной ч&-
сти зародышевого мешка ядра эндо-
сперма до начала образования у за-
родышей семядолей остаются CBO-
бодными.
Эмбриогенез Опаота4-типа. Заро-
дыш согнутый или прямой, диффе-
ренцированный на семядоли, гипо-
котиль и корешок, хлорофиллонос-
ный. Семядоли выполняют запаса-
ющую функцию. Подвесок отсут-
ствует.
Семя, плод — семя с эндоспермом
или без него. Зародыш располага-
ется в центре семени и в первом
случае со всех сторон окружен эн-
доспермом, во втором он заполняет
весь объем семени. Плод — боб.
Гаусторий эндоспермальный в ха-
лазальной части зародышевого меш-
ка.
Апомиксис не обнаружен.
Другие явления — полиэмбрио-
НИЯ.
Литература: Davis, 1966; Капи-
нос, Керимова, 1972; Schnarf, 1931;
Тахтаджян, 1966, 1970; Vassal, 4968,
71.
СЕМЕЙСТВО CAESALPINIACEAE
Оеновное число хромосом —х =
— 6—12, 44. ‘
Цветок обоеполый или однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 3
эфемерных, реже 6—8 средних сло-
ев и секреторный -многоядерный
тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные, изо-
билатеральные, Г-образные. Зрелые
пыльцевые зерна двухклеточные,
обычно одиночные или соедине-
ны в тетрады, трехбороздноанер-
турные. .”
Семяпочка — анатропная-кампило-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле зиг-
загообразное, образуется двумя ин-
тегументами. Только у Сазза мик-
ропиле образуется одним внутрен-
ним интегументом. Семяножка ча-
сто длинная. Имеются нуцелляр-
ный колпачок и ариллус. Семяпо-
чек до 30.
Макроспорогенез и макрогамето-
генез — археспорий — многоклеточ-
ный, реже одноклеточный. Парие-
тальных клеток 2—4 слоя. Тетрады
макроспор линейные. Зародышевый
мешок образуется по Ро]усопит-
типу. Синергиды с крючкообразны-
ми выростами и фибриллярными
аппаратами. Имеются три эфемер-
ные антиподы. Полярные ядра сли-
ваются до оплодотворения. .
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
начинается с микропилярной части
зародышевого мешка. В халазаль-
ной его части ядра эндоспёрма ос-
таются свободными.
Эмбриогенез Азегад-типа. Заро-
дыш дифференцированный, прямой
или согнутый, хлорофиллоносный.
Семядоли выполняют запасающую
функцию.
Семя, плод — семя без эндоспер-
ма или лишь со следами его. Заро-
дыш располагается в центре семе-
ни и заполняет вось или почти
весь объем семени. Плод — боб.
Гаусторий эндоспермальтый в ха-
лазальной части зародышевого меш-
шка.
`Апомиксис не обнаружен.
Другие явления — полиэмбриония
y Cassia tora.
Литература: Алимджанова, 1979;
Пау1з, 1966; Райкова, Алимджанова,
1978; ВешЪег 1969; Чубирко, 1978;
Тахтаджян, 1966, 1970.
:
СЕМЕЙСТВО ЕАВАСЕАЕ
(PAPILIONACEAE)
Основное число хромосом — zr =
Цветки обоеполые.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—3
эфемерных средних слоя и секре-
торный одно-двуядерный тапетум.
В клетках тапетума обнаружены
кристаллы оксалата кальция. Ядра
в клетках тапетума могут сливать-
ся, в результате чего они становят-
ся полиплоидными.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные,
крестообразные: Зрелые пыльцевые
зерна обычно двухклеточные, оди-
ночные, трехпоровые с гладкой или
сетчатой экзиной, содержат масло.
Спермии-клетки равной или нерав-
ной величины, бедноплазменные.
Семяпочка анатропная, гемиана-
тропная, амфитропная, анакампи-
лотропная, кампилотропная с дву-
мя интегументами, красси- или те-
нуинуцеллятная. Микропиле ` обра-
зуется обоими интегументами, ча-
сто зигзагообразное. Иногда микро-
пиле образуется только одним вну-
тренним или наружным интегумен-
том. Семяножка короткая. Имеются
нуцеллярный колпачок, эндотелий,
обтуратор, гипостаза, карункула и
ариллус. Проводящий пучок дохо-
дит до халазы. Семяпочек от 1 (у
Onobrychis) до 25—30 (y Lotus).
Макроспорогенез и макрогамето-
генез — архесяорий много-однокле-
точный. Париетальных клеток 2—4
ряда. Тетрады макроспор линейные,
Т-образные. Наряду с тетрадами
иногда возникают триады. Зароды-
шевые мошки образуются по Роу-
gonum- u изредка АШит-типам и
MMCIOT крахмальные зерна на всех
стадиях развития. Зародышевый
мешок созревает до раскрытия цве-
тка. Синергиды © крючкообразными
выростами и фибриллярными аппа-
ратами. Имеются три клетки анти-
под, либо эфемерных, либо долго
сохраняющихся. Полярные ядра
сливаются перед оплодотворением.
Иногда полярные ядра не сливают-
ся даже до образования двухкле-
точного зародыша (у палестинской
разновидности Trifolium alexan-
drinum).
Опыление, прохождение пыльце-
вой трубки и оплодотворение — са-
моопыление, перекрестное опыле-
ние, скрытое опыление. Промежу-
ток времени от опыления до опло-
дотворения у разных видов колеб-
лется от 65 до 24 ч. Порогамия.
Двойное оплодотворение премито-
тического, постмитотического или
промежуточного типов, причем
меньший спермий сливается с яйце-
клеткой, больший —с полярными
119
”
ядрами. Тройное слияние
ствует сингамии.
Эндосперм нуклеарный. После
оплодотворения первым — делится
первичное ядро эндосперма. Зало-
жение клеточных перегородок меж-
ду ядрами эндосперма начинается
с микропилярного конца зародыше-
вого мешка, когда зародыш круп-
ный шарообразный. В халазальном
конце зародышевого мешка эндо-
сперм остается ядерным и функци-
онирует как гаусторий. Ядра эндо-
сперма и гаусторий часто являют-
ся высокополиплоидными. Клетки
эндосперма часто содержат хлоро-
пласты.
Эмбриогенез Onagrad-, Asterad-,
бо]апа4- и СагуорпуПа4-типов. Заро-
дыш крупный, хлорофиллоносный,
содержащий масло, белок и крах-
мал, редко кумарин, дифференци-
рованный на Две семядоли, гипоко-
тиль, мощный корешок и слабо раз-
витую почечну. согнулый или пря-
мой. Семядолни выполняюг запасаю-
щую функцию. Подвески очень раз-
нообразны от слабо- до сильно раз-
витых, часто с хлоропластами. Ялра
клеток подвески у Р\|Вазео]из сосс1-
neus вследствие эндомитоза дости-
гают высокой степени полиплоидии.
Семя, плод — зрелое семя без зн-
досперма или со скудными оетат-
предше-
ками его. Зародыш расположен в
центре семени и занимает либо
весь объем семени, либо значи-
тельную часть его. Семенная кожу-
ра образуется из наружного интегу-
мента и состоит из склеренхима-
тозного эпидермиса, равиомерно
утолщенных RECTOR гинодермы и
почти из 10 слоев паренхиминых
клеток. Пекоторые представители ее-
мейства имеюг алкалонды. Ило р —
обычно 606. Эпидермие экзокарния
склерофицированный.
Гаусторий эндоспермальный (ха-
лазальный и латеральный). У мно-
гих видов имеются также зароды-
шевый и подвесочный гаустории.
Гаустории имеют либо клеточное.
либо ценоцитное строение, разнооб-
разны по форме. величине и про-
должительностн существования. Ос-
татки гаусторий сохраняются иног-
да до полной дифференциации ор-
ганов зародыша.
Апомиксис— релуцированный пат-
теногенез, соматическая — апоено-
рия и нуцеллярная эмбриония.
120
Другие явления — клейстогамия.
Полиэмбриония. Добавочные заро-
дыши образуются или из синергид
или при расщеплении зиготы, или
при образовании нескольких заро-
дышевых мешков в одной и той же
семяпочке.
Литература: Аминов, 1967; Anata-
swamy, 1950, 1954, 4953; Ахундова,
1978; Ашурметов, Норматов, 1978;
Атабекова, 1959, 1960, 4962; Атабе-
кова, МЛин-Дзян-син, 1963, 1964;
Behl, Tiagi, 1976, 1977; её а|., 1969;
Chandravadana, 1965; Corner, 1976;
Datta, Choudhary, 1967; Deshpande,
Bhasin, 1974; Deshpande, Urtawal,
1969, 1974; Dmnyansagar, 1967, 1970;
Фодорчук, 1944; Галимова, 1979а, 6,
1978; Гамбарова, 1978; Goursat, 1969;
Hutemo, 1967; Ивашкина, 1969;
Johri, Garg, 1959; Кадырова, 1973,
1974a, 6, 1976; Капинос, Веримова,
1968; Hepumosa, 1970, 1972; Hucenes,
Козлова, 1970; Колев и др., 1972;
Конпычева, 1970а; Кострикова, 1956,
1964, 1968, 1972, 4973а, 6, 1975,
197ба, 6, 1979; Крылова, 1962, 1965;
Кчирко, 1973; Makde, 1971; Манд-
рик, Мотковская, 1977, 1978а, 6;
Мандрик, Петрус, 1976; Marinos,
1970; Маркова, 1947а; МоеМаиеЩьоп,
1976; Mersy et al., 1974; Misra, Sahu,
1969; Nitchell, 1975; Narang, Chand-
га, 1977; Норматов, 1973, 1974, 1977;
Qjchomon, Sangaolu, 1970; Останен-
ко, 1970; Padhye et al. 1969; Pra-
kash, 1960; Prakash, Chan, 1976;
Rembert, 1966, 1967, 1969, 1972, 1977;
Руденко, 1965; Рустамова, 1965;
Salgare, 1973, 1974, 1975; Schnarf,
(934; Sclmept, Nagl, 1970; Iesayx,
Конычева, 1972; Seshavataram,
Ga, by Singh, Shivapuri, 1935;
Shukla, Prakash, 1967; Taxramsan,
1960, 1970; Терзийски, 1969а, 6, в,
170, 19 1а, 6, 1973, 1974, 1978; Тер-
зниски, Ноддубиая-Арнольди, 1973;
Чайкнна, 1960; Чубирко, 1962, 1963,
1964a, 6, 1965, 1968, 1969, 1972, 1976;
Venkareswarlu, Sechavateram, 1971;
Wojciechowska, 1963, 1971, 1972,
1975; Закржевская, Колесников,
1963; Замотайлов, 1958, 1960.
В порядке Рара]ез все семейства
Донольно хорошо исследованы в от-
нонснии цитоэмбриологии. По ци-
тозмбриологическим признакам все
семейства, включенные А. Л. Тах-
таджяном в этот порядок, имеют
много сходных черт. По-видимому,
этот порядок довольпо естествен-
ный. Особенно близки друг другу
семейства Саеза]римасеае и КаБа-
сеае. Вследствие паличия у пред-
ставителей Mimosaceae сложной
пыльцы, что является вторичным
признаком, нам представляется бо-
лее правильным расположение М1
mosaceae не в начале, ав конце
порядка КаЪа!ез. Однако необходи-
мы дальнейнтие исследования, что-
бы выяснить филогенетические свя-
зи между Саеза]римасеае, М!итоза-
ceae u Papilicnaceae [Goursat, 1969].
ПОРЯДОК 51.
CONNARALES
CEMEMCTBO CONNARACEAE
Основное числе хромосом — 7 =
— 13, 14.
Цветок обычно обосполый, реже
однополый, растения редко двудом-
ные.
Пыльник — 4 микроспорангия.
Микроспорогенез и микрогамето-
генез — нет данпых. Зрелые пыль-
цевые зерпа трех (четырех)-апер-
турные (бороздные пли бороздно-
поровые).
Семяпочка гомпапатропная с дву-
мя интегумептами, прассинуицедлят-
ная. Микрониле образуется обоими
интегумоитами. Имсется ариллус.
Семяпочек 2—10.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки имезотся. Те-
трады макроспор линейные, T-00-
разные. Иногда вместо, тетрад обра-
зуются триады. Зародышевый ме-
шок образуется по Рофузопит-типу.
Имеются три скородегенерирующие
‹летки антипод.
Опыленне, прохождение пыльцс-
вой трубки и оплодотворепие — нот
данных.
Эндоспермогслез пуклеарпый.
Клеточные перегородки между яд-
рами эпдосперма начинают возни-
кать после того, как образуется не-
сколько тысяч свободных ядер.
Эмбриогенез — тин не определен.
Заподыш крупный, поямой или со-
гвутый, лифференцированный на две
толстые семядоли, выполняющие за-
пасающую функцию, короткий гипо-
котиль и маленький корешохм; заро-
дыш содержит алейрон и крахмал,
кроме того, имеются дубильные вс-
щества и слизистые клетки.
Семя, плод — семя с обильным
или скудным эндоспермом или без
него. Прямой зародыш располагает-
ся в центре семени и заполняет ли-
бо весь объем семени, либо часть
его, в зависимости от того, насколь-
ко сильно развит эндосперм. Согну-
тый зародыш располагается по пе-
риферии семени, а эндосперм в цен-
тре семени. Плодики обычно одно-
семенные или реже двусеменные,
раскрывающиеся (листовка, реже
бобы) или реже пераскрывающис-
ся (орстикя).
Гаусторий отсутствует.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; Тахтаджян, 1966, 1970.
Некоторые систематики [Cronqu-
ist, 1957; Тахтаджян, 1966] сближа-
ют Connaraceae с Сипошасеае и
Grossulariaceae, что не противоре-
чит данным цитоэмбриологии, таг
как некоторые цитоэмбрпологиче-
ские признаки этих трех семейств
сходны. Однако в отношении цито-
эмбриологии эти семейства изуче-
пы исдостаточно подробно, чтобы
делать околчательные выводы о вза-
имоотношенилх между этими се-
мействами.
ПОРЯДОК 52.
МЕРЕ МТАТЕ$
СЕМЕЙСТВО ОВОЗЕВАСЕАЕ
Основное число хромосом —х =
= 6, 10, 14—16.
Цветок обосполыиц.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по одподольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
средний слой и секреторный мно-
гоядерный тапетум. В результате
слияния ядер в клетках тапетума
образуются полиплоидные ядра.
Y Drosera bifinnata ип D. capensis
описан амебоицный тапетум.
Микроспорогенез и микрогамето-
гснсз —- образовапие микроснор си-
121
мультанное. Тетрады микроспор те-
траэдральные, изобилатеральные.
Зрелые пыльцевые зерна трехкле-
точные или двухклеточные, соеди-
ненные в тетрады или одиночные,
шести-восьмипоровые с шиповатой
окзиной.
Семяпочка анатропная, с двумя
интегументами, красси- или тенуи-
нуцеллятная. Микропиле образует-
ся внутренним интегументом. Во-
круг зародышевого мешка имеется
ложный эндотеций, возникший из
клеток нуцеллуса, а не внутренне-
го интегумента. Семяпочек много
или реже несколько.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 4 ряда. Те-
трады макроспор линейные, T-00-
разные. Зародышевый мешок обра-
зуется по Роусопат-типу. Синер-
гиды с крючкообразными выроста-
ми и фибриллярными аппаратами.
Три антиподы дегеперируют вскоре
после оплодотворения. Полярные
ядра сливаются До оплодотворения.
Опыление, прохождепие пыльце-
вой трубки и оплодотворение — са-
моопыление y Androvandia vesicu-
losa. Порогамия. Двойное оплодо-
творение.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярного конца заро-
дышевого мешка; постепенно заро-
дышевый мешок заполняется клет-
ками эндосперма. Только в хала-
зальной части зародышевого мептка
эндосперм остается свободноядер-
ным, функционируя как гаустории.
Эмбриогелез боапа4- и Сагуорпу-
Пад-типов. У видов Пгозега — Aste-
гад у Попаса шизарша — Опа-
отад-типы. Зародьзи прямой, очень
маленький, слаборазвитый, но диф-
ференцированный на семядоли и
корешок.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Зародыш располагается в
основании семени и обычно не пре-
вышает 4/4 линейной высоты семе-
ни. Плод — локулицидная коро-
бочка.
Гаусторий — семядоли выполняют
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления — клейстогамия.
Полиэмбрионня.
122
Литература: Corner, 1976; Raj,
1965; ЭсппатЁ, 1934; Смирнова, 1977;
Тахтаджян, 1966, 1970; Транков-
ский, 1938.
СЕМЕЙСТВО МЕРЕМТНАСЕАЕ
Основное число хромосом — нет
данных.
Цветок однополый, растения дву-
домные.
Пыльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, средний слой и
секреторный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор те-
траэдральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, соединенные в тетрады с не-
ясно выраженными — апертурами.
Пыльца жизнеспособна в течение
трех дней.
Семяпочка анатропная, с двумя
интегумептами, крассипуцеллятная.
Микропиле образустся двумя ин-
тегументами. Сосудистый пучок до-
ходит до халазы. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одпоклеточный.
Париетальных клеток один ряд. Те-
трады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
lygonum-tany. Синергиды груше-
видные. Имеются три антиподы. По-
лярные ядра сливаются перед онло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
дпдоспермогенез пуклсарный. Ци-
токинез начинается с халазального
конца.
Эмбриогенез Solanad-runa. Заро-
дыш прямой, маленький, но диф-
ференцироваптыяй па две семядоли
и корепток, содержит масло, проте-
ин и крахмал.
Семя, плод — семя с мясистым
эндоспермом, содержащим крахмал.
Зародыш располагается в середине
семени и обычно не превышает 1/4
линейной высоты семени. Семенная
кожура образуется обоими интегу-
ментами. Плод — локулицидная ко-
робочка.
Гаусторий — семядоли выполня-
тот гаусториальную функцию.
Апомикеие не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Li-An-Lan, Prakash, 1973;
Schnarf, 1931; CmupHosa, 1977; Tax-
таджян, 1966, 1970.
Цитоэмбриологические данные по-
казывают, что МереТасеае более
сходно с ПОгозегасеае, чем с Загга-
сетасеае, и что правильнее выде-
лить его из порядка баггасеюа[ез
и расположить в 0с0бом порядке
Netenthales, рядом с Droseraceae
[Lim-Ah-Lan, Prokash, 1973]. Dro-
зегасеае, как wu Nepenthaceae, umect
секреторный тапетум, пыльцевыо
зерна, соединенные в тетрады, ана-
тропные, крассинуцеллятные, с дву-
мя интегументами семяпочки, за-
родышевые мешки Polygonum-tuna,
нуклеарный эндосперм и эмбриоге-
нез Solanad-tuma. В отличие от
МерепВасеае представители Dro-
зегасеае имеют двухклеточные, а
не трехклеточпые, как у Nepenth-
асеае, зрелые пальцевые зерна, что
указывает на болыную примитив-
ность Droseraceae по сравнению с
Nepenthaceae.
ПОРЯДОК 53.
PODOSTEMALES
СЕМЕЙСТВО PODOSTEMACEAL
Основное число хромосом —х=10.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный (двурядный у
Dicraea) эндотеций, 1—2 средних
слоя и секреторный, одно-, дву-
ядерный тапетум. У Zeylanidium
амебоидный тапетум.
Микроспорогенез и микрогаме-
тогенез — образованние микроспор
симультанное или сукцессивпос.
Тетрады микроспор тетраэдральные,
изобилатеральные, кростообразные.
Зрелые пыльцевые зерпа двухклето 1-
ные, одиночные или соединены в ди-
ады, трехборозднопоровые, редко
многопоровые или безапертурные.
Семяпочка анатропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется наружным
интегументом. В результате дезин-
теграции оболочек клеток нуцеллу-
са в центре семяпочки образуется
полость, названная псевдозароды-
шевым мешком. В этой полости
имеется много ядер и густая ци-
топлазма. Образуется так называ-
емый нуцеллярный плазмодий. Се-
мяпочек обычно много, редко 2—4.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Макроспор — 2. Зародышевый ме-
шок четырехъядерный образуется по
Podostemum-nogtuny, АШат-типа.
Некоторые ученые называют этот
тип зародышевого мешка — Dicraea-
или Apinagia-Tumamu. Он имеет
яйцеклетку, две синергиды с крюч-
кообразными выростами и цент-
ральную клетку с одним полярным
ядром. Антиподы отсутствуют. В
зародышевом мешке имеются крах-
мальные зерна. Наблюдаются обра-
щенные зародышевые мешки.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное или одинарное
оплодотворение.
Эндоспермогенез отсутствует.
Эмбрисгенез Зоапа4-типа. Заро-
дыш прямой, бесхлорофилльный,
дифференцированный, двусемядоль-
ный. Содержит масло и крахмал.
Семядоли массивные, выполняющие
запасающую функцию. Подвесок
сильно развитый.
Семя, илод — семя без эндоспер-
ма. Зародылт заполняет все семя.
Семядоли скрывают остальные час-
ти зародыша, лишь кончик кореш-
ка едва выступает из них. Илод —
септицидная коробочка.
Гаусторий подвесочный.
Апомиксис не обнаружен.
Другие явления — клейстогамия.
Литература: Arekal, Nagendan,
1974, 1975a, b, 1977; Battaglia, 1971;
Chopra, Mukkada, 1966; Corner, 1976;
Davis, 1966; Govindappa et al., 1975,
1977; Jager-Ziirn, 1967, Kapil, 1967h,
1970b; Maurilzon, 1933; Mukkada,
1969; Mukkada, Chopra, 1973; Nagen-
dran, 1974; Nagendran, Arekal, 1976;
Nagendran et al. 1977; Nagendran
el al, 1976; Razi, 1966; Schnarf,
1931; Subramanyam, Sreemadhavan,
1969; Смирнова, 1977; Тахтаджян,
1966, 1970.
По мнению некоторых ученых,
цитоэмбриологические признаки Ро-
dostemaceae сходны с таковыми У се-
мейства Стаззи]асеае [Kapil, 1970b].
123
По мнению Maypursona [Mauritzon,
1933], хорошо развитые подвесо-
чные гаустории зародышей Родо-
зетасеае и Сгаззшасеае указыва-
ют на тесное родство этих двух
семейств. Р. Махешвари ([МаБе-
shwari, 1945] отметил, что Родо-
stemaceae — это редуцированное,
безлепестное семейство — производ-
ное Сгаззасеае. Однако наличием
зародышевого мешка Podostemon-
подтипа, АШит-типа, нуцеллярного
плазмодия, отсутствием макроспоро-
вых, антиподальных и эндоспер-
мальных гаусториев Родозетасеае
отличаются от Crassulaceae, что
противоречит их сближению.
НАДПОРЯДОК IX.
МУВТАМАЕ
ПОРЯДОК 54.
MYRTALES
СЕМЕЙСТВО ТУТНВАСЕАЕ
Основное число хромосом —х =5,
6. 8, 9, 15, 25.
Нветок обоеполый.
Пыльник — 4 микроспораягия.
Стенка микроспорангия образуется
но двудольному типу и имеет эпи-
дермис, фиброзный эндотеции,
2 эфемерных средних слоя и секре-
торпый многоядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые, реже © 06—12 чередуюпие
мися поровыми или беспоровыми
бороздами.
Семязочка анатропная, с двуми
интегументами, амфитропная, крас-
си- или тенуинуцеллятная. Име-
ются нуцеллярный колпачок и ги-
постаза. Микропиле образуется обо-
ими интегументами. — Семяпочек
обычно много.
Макроепорогенез и макрогамето-
генез — археспорий одно-мпого-
клеточный. Париетальных клеток
2—7 рядов. Тетрады макроспор ли-
нейные. Зародышевый мепток обра-
зуется по Роугопат-типу и содер-
жит зерна крахмала. У Wodfordia
floribunda зародышевый mention ©
gee
12:
обращенной полярностью. Синерги-
ды с крючкообразными выростами
и фибриллярными аппаратами. Име-
ются три эфемерные антиподы. По-
лярные ядра сливаются до оплодо-
TBopenua. Y Guphca lanceolata onu-
сан постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоепермогенез пуклеарный.
Эмбриогенез Onagrad-tuna. dapo-
дыш прямой или согнутый, круп-
ный, дифференцированный, двусе-
мядольный, содержит масло и алей-
рон. Семядоли выполнняют запаса-
ющую функцию. Подвесок одно-
или многорядный.
Семя, плод — семя без эндоспер-
ма. Плод сухой, обычно коробочка.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления — илейстогамия.
Триморфная retepocrumua y Lyth-
rum.
Литература: Corner, 1976; Davis,
1966; Smith, Tlerr, 1974; | Schnarf,
1931; Тахтаджян, 1966, 1970; Ven-
kateswarlu, 1967c.
СЕМЕЙСТВО ЗОММЕКАТТАСЕАЕ
Основное чиело хромосом — z = 9.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 3—4
эфемерных средних слоя и секре-
торный двуядерный тапетум.
Микроспорогспез и мпкрогамето-
тепез — образование микроспор си-
мультапное. Тотрады микроспор
теграэдральные. Зрелые пыльцевые
верна двухклеточные, трехпоровые
или бороздновидные, с крахмаль-
иыми зерцами. -
Семяпочка анатроиная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется двумя инте-
гументами. Имеется нуцеллярный
колпачок. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одпо-, дву-, мно-
гоклеточный. Паристальных клеток
4—6 рядов. Тетрады макроспор ‚ли-
нейные, ТГ-образиые. Зародышевый
мешок oOpasyerca no Polygonum-
типу и содержит капли масла. Си
ноергиды с крючкообразными вырос-
тами. Имеются три эфемерные ан-
типоды. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльцсе-
вой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворение.
Эндоспермогенез нуклеарный. За-
ложение клеточных — перегородок
между ядрами эндосперма, начина-
ется с микропилярного конца за-
родышевого мешка. Центральная
часть зародышевого мешка не за-
полняется клетками.
Эмбриогенез Опастад-типа. Заро-
дъи прямой или согнутый, хлоро-
филлоносный, маленький или боль-
шой, дифференцированный на се-
мядоли, гипокотиль и корешок.
Семя, плод — семя без эпдоспер-
ма. Зародьши вытянут по длинной
оси семени и заполняет весь cro
объем. Семядоли, гипокотиль и ко-
решок составляют каждый около
1/3 общей длины зародыша. Плод—
коробочка или ягода.
Гаусторий отсутствует.
Апомиксие пе обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Taxranman, 1966, 1970; Schnari,
1934. Смирнова, 1977; УспЕз{ез\ат-
lu, 1967¢.
СЕМЕЙСТВО PUNICACEAE
Основное число хоомосом х-=8.
Цветок обосполый. .
Пыльник — пот данных.
Микроеснорогсенез и микрогамето-
генез — тип микроспорогенеза не
определен. Зрелые пыльцевые зер-
на двухклеточнные, трехбороздно-
поровые.
Семяпочка анатропная, с двумя
интегументами, крассинупеллятная.
Микропиле образуется обоими ин-
тегументами. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Паристальных клеток 3 ряда.
Тотрады макроспор липейные. За-
родьииевый мешок образуется но
Ро!угопат-типу. Синергиды удли-
ненные, обычно без вакуолей. Име-
ются три одноядерные антиподы.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльцсе-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не определен.
Nw
Зародыш крупный, двусемядольный,
прямой, бесхлорофилльный, содер-
жит масло и алейрон. Семядоли
выполняют запасающую функцию.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет весь объем
семени. Семядоли крупнее гипоко-
тиля и корешка. Наружный слой
соменной кожуры сочный, внутрен-
ний роговой. Плод гранатина.
Гаусторий не обнаружен.
Апомиксис ное обнаружен.
Другие явления не обнаружены.
Литература: ‘Сотпег, 1976; Davis,
1966; Смирнова, 1977; Тахтаджян.
1966, 1970; Татевосян, 1968; Vanka-
teswarlu, 19671.
СЕМЕЙСТВО ВНТЛОРНОКАСЕАЕ
Основное число хромосом — & =
—
‚ 9.
Цвстки обоеполые, редко однопо-
лые или полигамные.
Пыльник — нет данных.
Тапетум секреторный.
Микроспорогенез и микрогамето-
генез — образование микроспоо си-
мультанное. Зрелые пыльцевые зер-
на двухклеточнные, трех (четырех) -
бороздпопоровые.
Семяпочка апатропгая-гемпана-
троппая, с двумя интегумеитами,
прасенпупеллятная. Микоопило
обычно зитзагообразное, образую-
цееся обоими интегументами. Од-
nano y Gynotroches mukponume o6-
разуется одиим впнутренчнипим интегу-
ментом. Иногда имеется ариллус.
Семяпочек 2—4 — мпого.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 2 ряда. Тет-
рады макроспор линейные. Заролы-
шевый мешок образуется по Poly-
2опит-типу. Синергиды грушевид-
ные. Имеются три одноядерные ско-
ро дегенерирующие клетки анти-
под. Полярные ядра сливаются пе-
ред оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
дапцпых.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не определен.
Зародыш прямой или согнутый, дву-
семядольный, часто хлорофиллонос-
ный, содержит крахмал. Семядоли
выполняют запасающую функцию.
Подвесок массивный.
Семя, плод — семя обычно с мя-
систым эндоспермом. Зародыш рас-
125
полагается в центре семени и за-
нимает болыпую часть его. Плод —
ягода, реже сухая или мясистая
коробочка, иногда односеменной и
не раскрывающийся.
Гауеторий — у Ceriops найден
эндоспермальный гаусторий в хала-
зальной части зародышевого мешка.
Апомиксис не обнаружен.
Другие явления — у мангровых
представителей трибы В2орвогеае
обнаружена вивипария, так как у
них семена прорастают в плодах
на материнском растении.
Литература: ‘Согпег, 1976; Davis,
joo Тахтаджян, 1966, 1970; Schnarf,
CEMEMCTBO COMBRETACEAE
Основное число хромосом — x =
=7, 14—13. *
Цветок обычно обоеполый, реже
однополый.
Пыльник — 4 микроспорапгия.
Стенка микроспорангия образуетея
по базальному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—3,
изредка 4—6 эфемерных, средних
слоя и секреторный одно-двуядер-
ный, одно-многорядный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые, с тол-
стой гладкой экзиной. ,
Семяпочка висячая, с двумя ин-
тегументами, крассинуцеллятная.
Микропиле, образующееся двумя ин-
тегументами, зигзагообразное. Име-
ются нуцеллярнный колпачок, Py-
никулярный обтуратор и гипостаза.
Семяпочки на тонких длипных се-
мяножках. Семяпочек от 2 до 6.
Макроспорогенез и макрогамето-
генез — археспорий одно-несколь-
коклеточный. Париетальная ткань
массивная. Тетрады макроспор ли-
нейные. Зародышевые мешки обра-
зуются mo Polygonum- u Репаеа-
типам. Синергиды с крючкообраз-
ными выростами и фибриллярными
аппаратами. Три эфемерные анти-
поды обпаружены у Ро{угеа и Тег-
ичтаНа, у Слйега антиподы увели-
чиваются в числе и долго сохраня-
ются. Полярные ядра сливаются
перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
126
рогамия. Двойное оплодотворение.
Эндобпермогенез нуклеарный, но
позднее весь эндосперм становится
клеточным.
Эмбриогенез Азега4-типа, вариа-
ции Erodium. Зародыш крупный
прямой или согнутый, двусемядоль-
ный, бесхлорофилльный или хлоро-
филлоносный, содержит, масло и
алейрон, иногда крахмал. Семядоли
обычно спирально свернутые, они
выполняют запасающую функцию.
Подвесок короткий, двурядный.
Семя, плод — семя без эндоспер-
ма. Зародышг заполняет весь объем
семени. Эндосперм израсходован в
процессе развития зародыша. Пло-
ды кожистые или реже костянко-
образные, односеменные.
Гауеторий отсутствует.
Аномикеие — нуцеллярная эмбри-
Ония.
Другие явления не отмечены.
Литература: ‘Согпег, 1976: Davis,
1966; Dutt, 1978a; Schnarf, 1931; Tax-
таджян, 1966, 1970; Venkateswaliu,
1967a; Venkateswarlu, Rao P., 1972.
СЕМЕЙСТВО МУВТАСЕАЕ
Основное число хромосом — т =
= 6—9, 11.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—
3 эфемерных, средних слоя и сек-
реторный дву- трехъядерный тапе-
тум. Имеются орбикулы. ©.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные и изобилатераль-
ные. Зрелые пыльцевые зерна двух-
клеточные, обычно трехбороздные,
содержащие масло. _ |
Семяпочка анатропная, кампило-
тропная с двумя или с одним (у
видов Eugenia, Syzygium) интегу-
ментами, крассинуцеллятная. Мик-
ропиле обычно образуется обоими
интегументами, но у видов ТЬтур®ю-
шепе и УевПа оно образуется толь-
ко одним внутренним интегументом.
Сосудистый нучок доходит до хала-
зы. Имеются нуцеллярный колпа-
чок и ариллоподобное образование.
Семяпочек много. |
Макроспорогенез и макрогамето-,
генез — археспорий одноклеточный.
Паристальных клеток 2—12 рядов.
N
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по’ Ро-
]угопиш-типу. Синергиды с крюч-
кообразными выростами и фибрил-
‚‹лярными аппаратами. У видов Епса-
]}ураз синергиды своим строением
похожи на яйцеклетки. Имеются
три скоро дегенерирующие антипо-
ды. Полярные ядра сливаются пе-
ред оплодотворением или при опло-
дотворении.
Опыление, прохождение ныльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение. ‘.
Эндоспермогенез нуклеарный. За-
ложение клеточных — перегородок
между ядрами эндосперма начина-
ется с микропилярной части заро-
дышевого мешка‘и постепенно весь
эндосперм становится клеточным.
Эмбриогенез Опаотад-типа. Заро-
дыш крупный, прямой или согну-
тый, двусемядольный, бесхлоро-
филльный или хлорофиллоносный.
Семядоли выполняют запасающую
функцию. Запасные вещества заро-
дыша: алейрон, масло, реже крах-
мал.
Семя, плод — семя без эндоспер-
ма или со слабо развитым эндо-
спермом. Плод — ягода или сухая
хоробочка, очень редко костянкооб-
разный. Зародыш заполняет все
или почти все семя,
Гаусторий отсутствует.
Апомикеие — нуцеллярная и ип-
тегумептальная эмбриония у Euge-
nia jambos u E. hookeri (2—21 за-
родыш в одном и том же семени).
Другие явления не обнаружены.
Литература: Баранов, 1956; Бара-
нов и др., 1955; Corner, 1976; Davis,
1966, 1968b, 1969; Narayanaswami,
Воу, 1960; Полунина, 1957, 1958a, 6,
в, 1959, 1963, 1964; Prakash, 1968a,
b, e, 1969 a—e, g; Sohai, Roy, 1964;. Тахтаджян, 1966, 1970; УепКацез\уаг-
lu, 1967.
СЕМЕЙСТВО МЕГАЗТОМАСЕАЕ
Основное чиело хромоеом — х ==
== 7—12, 14, 17.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций име-
ется или отсутствует, 2—3 эфемер-
ных средних слоя.и секреторный од-
ноядерный двурядный тапетум.
`Микроепорогенез и микрогамето-
генез — образование: микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, обычно с 6
чередующимися поровыми или бес-
поровыми бороздами, с гладкой
сэкзиной.
Семяпочка анатропная, анакам-
пилотропная, с двумя интегумента-
ми, крассиннуцеллятная. Микропи-
ле образуется обоими интогумента-
ми. Имеются гипостаза и обтуратор.
Семяпочек обычно много.
Макроспорогенез и макрогамето-
генез — археспорий много-, одно-
клеточный. Париетальных клеток
2—6 рядов. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро]угопиш-типу. Синер-
гиды с крючкообразными выроста-
ми. Имеются три эфемерные анти-
поды. Полярные ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный, ас-
симилируется зародышем в свобод-
ноядерном состоянии.
Эмбриогенез Onagrad- или Sola-
пад-типа. Зародыиг большой, пря-
мой или согнутый, дифференциро-
ванный, бесхлорофилльный, содер-
жит масло или крахмал. Семядоли,
гипокотиль и корешок хорошо раз-
виты и составляют каждый около
1/3 общей длины зародыша. Подве-
сок многоклеточный, массивный и
короткий.
Семя, плод — семя без эндослер-
ма. Зародыш заполняет весь объем
семени. Плод — локулицидная ко-
робочка или ягода.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Лругие явления — полиэмбриония
y Melastome.
Литература: Corner, 1976; Davis,
1966; Etheridge, Herr, 1968; Cmupuo-
ва, 1977; Schnarf, 1931; Тахтаджян,
1966, 1970; Venkateswarlu, 19674.
СЕМЕЙСТВО OLINIACEAE
Основное число хромосом — пет
данных. * ‘
Цветок — обоеполый.
Пыльник — нет давных о числе
микроспорангиев. Стенка микроспо-
рангия имеет эпидермис, фиброзный
127
—
эндотеций, 2—3, иногда до 7 эфемер-
ных средних слоев и секреторный
(?) одноядерный двурядный тане-
тум. . ..
'’ Микреспорогенез и микрогамето-
генез — нет данных. Зрелые пыль-
цевые зерна трехборозднопоровые.
Семяпочка гемианатропная, с дву-
мя интегументами, крассинуцеллят-
ная. Микропиле образуется двумя
интегументами. Семяпочек 6—15.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 2 ряда. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Ро]у-
опит-типу. Синергиды грушевид-
ные. Имеются три эфемерные анти-
поды.
Оплодотворение, опыление, прохо-
ждение пыльцевых трубок — нет
данных.
Эндоспермогенез нуклеарный. Об-
разуется несколько сот свободных
ядер эндосперма, прежде чем на-
чнется образование клеточных пе-
регородок между ядрами,
Эмбриогенез — нет данных. За-
родыш прямой, заполняет весь
объем семени.
Семя, плод — семя без эндоспер-
ма. Семядоли, гипокотиль и коре-
шок составляют каждый около 1/3
общей длины зародыша. Плод кос-
тянкообразный. -
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Пау13, 1966; Смирно-
ва, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО PENAEACEAE
Основное число хромосом — нет
данных.
Пветок обоеполый.
Ныльник — нет данных.
Микроспорогенез и микрогамето-
генез — нет данных. Зрелые пыль-
цевые зерна трех-четырехбороздно-
поровые.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими инте-
гументами. Семяпочек 8—16.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 5 рядов. За-
родышевый мешок образуется по
Репаеа-типу и является тетраспо-
рическим, 16-ядерным.
128
Опыление, прохождение ‘пыльце-
вой трубки и оплодотворение — нет
данных. ,
Эндоепермогенез нуклеарный. Пе-
регородки между ядрами эндосперма
начинают возникать тогда, когда
зародыш достигает глобулярной
стадии.
Эмбриогенез Asterad-runa. Заро-
дыш прямой, дифференцированный
на две семядоли, гипокотиль и ко-
решок, каждый из которых состав-
ляет около 1/3 общей длины заро-
дыша, содержит масло. |
Семя, плод — семя без эндоспер-
ма. Зародыш занимает весь объем
семени. Плод — локулицидная коро-
бочка с неопадающей чашечкой.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления не отмечены.
Литература: Davis, 1966; Смирно-
ва, 1977; ЭеЪпат[, 1934; Тахтаджян,
1966, 1970.
CEMEHCTBO ONAGRACEAE
Основное число хромосом — х =
== 5—9, 14, 15.
Цветок обоеполый, редко однопо-
лый (у некоторых BupoB Fuchsia).
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и секреторный дву-
ядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование. микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточные, трехапертур-
ные (бороздные, борозднопоровые,
или пароратные), соединенные в тет-
рады или одиночные. У ОепоВега
пыльцевые зерна содержат крахмал.
Семяпочка анатропная, с двумя
‚интегументами, крассинуцеллятная.
Микропиле образуется обоими ин-
тегументами. Имеются нуцелляр-
ный колпачок, гипостаза и ариллус.
Семяпочек много или несколько.
`Макроспорогенез и макрогамето-
генез — археспорий `одно-многокле-
точный. Париетальных клеток 2 ря-
да или париетальная ткань массив-
ная. Тетрады макроспор линейные.
Зародышевый мешок образуется по
ОепоШега-типу и имеет крахмаль-
ные зерна. Синергиды с фибрил-
лярными аппаратами. Антиподы от-
CYTCTBYIOT.
Опыление, прохождение пыльце-
вой трубки и: оплодотворение — пе-
рекреетное‹ опыление,
Hue. Порогамия. Двойное оплодо-
творение премитотического типа.
‘Эндоспермогенез нуклеарный. 0б-
разуется: до 900 свободных ядер эн-
досперма, прежде чем между ними
начнется образование клеточных пе-
регородок. Первичное ядро эндо-
сперма на одно деление опережает
деление зиготы. -
Эмбриогенез Onagrad-tuna. 3apo-
дыш крупный, прямой или согну-
тый, бесхлорофилльный, дифферен-
цированный, двусемядольный, со-
держит масло, алейрон. Семядоли
выполняют запасающую функцию.
Подвесок короткий.
Семя, плод — ссмя без эндоспер-
ма. Зародыш заполняет все семя.
Массивные — семядоли скрывают
остальные части зародыша, лишь
конец корешка едва выступает из
них. Плод — локулицидная коробоч-
ка, орех или редко ягода. .
Гауеторий отсутствует.
Апомиксис — нуцеллярная и ин-
тегументальная эмбриония. Редуци-
рованный партеногенез.
Другие явления — клейстогамия.
Полизмбриония у Taraxia ovata.
Партенокарпия,
итература: Ва, 1969; Bednara,
1977; Corner, 1076; Davis; 1966; Гва-
ладзе, 1978; Гваладве, Гагечиладае,
4977; Гагечиладзао, Гваладае, 1970:
Jaluzot, 1970, 1971; Кордюм, 1983;
Schnarf, 1931; Seshavataram, 1967,
1970; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО ТВКАРАСЕАЕ
Основное число хромовом — z =
= 20, 24.
Цветок обоеполый. .
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотоций, 2—3
эфемерных средних слоя и секре-
торный многоядерный двурядпый
тапетум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные.
релые пыльцевые зерна двухкле-
точные, с хорошо выраженными ме-
ридиональными гребнями, соприка-
сающимися у полюсов, и с тремя
9 В. А. Поддубная-Арнольди
самоопыле-`
экваториальными , ;меридионально
удлиненными порами.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Истинного микропиле нет. Имеются
нуцеллярный клювик и гипостаза.
Семяпочек 2.
Макроспорогенез и макрогамете-
генез — археспорий одноклеточный.
Париетальных клеток несколько ря-
дов. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Ро|усопит-типу. Синергиды груше-
видные. Имеются три эфемерные
антиподы. У Trapa natans описан
постамент.
Опыление, прохождение пылье-
вой трубки и оплодотворение —
двойное оплодотворение. Первичное
ядро эндосперма делится раньше
зиготы.
Эндоепермогенез нуклеарный.
Первичное ядро эндосперма претер-
певает одно- два деления, после
чего ядра эндосперма — дегенери-
руют.
Эмбриогенез — Solanad-tuma. 3a-
родыш с очень неравными двумя
семядолями. Подвесок длинный,
многоклеточный. Ядра подвеска
путем эндомитоза достигают высо-
кого уровня полиплоидии (до 256 п).
Семя, плод — семя без эндоспер-
ма. Плод односемянной, костянко-
образпый, с каменистым эндокарпи-
ом и с 4 или 2 твердыми рогами.
Гаусторий подвесочный.
Апомиксис по обпаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Тахтаджян, 1966, 1970; Тгеа-
Sawinska, 1978.
СЕМЕЙСТВО LECYTHIDACEAE
Основное число хромосом — х =
= 13, 16—18. `
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и амебо-
идный двуядерный тапетум.
Микроспорогенез и микрогамето-
гемез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна —двух-
трехклеточные, трехборозднопоровые
или трехбороздные.
129
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Зародышевый мешок
окружен эндотелием. Семяножка
длинная. Имеется ариллус. Семяпо-
чек одна — много.
Макроспорогенез и макрогамето-
генез — археспорий много-однокле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроспор ли-
нейные, Г-образные. Иногда вместо
тетрады макроспор образуются три-
ады макроспор. Зародышевый ме-
шок образуется по Ро]угопит-типу.
У Napoleana imperialis o6mapyxeHEI
зародышевые мешки с обращенной
полярностью. Синергиды грушевид-
ные. Имеются три эфемерные анти-
поды. Полярные ядра сливаются
перед оплодотворепием или при оп-
лодотворении.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез пуклоарпый.
Эмбриогенез — тии по определен.
Зародыш прямой, крупный, обычно
дифференцированный. Семядоли
массивные, выполняющие запасаю-
щую функцию. Иногда зародыш не-
дифференцированный, состоящий
главным образом из утолщенного
гипокотиля, содержит масло, алей-
рон, крахмал.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет весь объем
семени. Плод костянковидный; яго-
дообразный или коробочка.
Гаусеторий отсутствует.
Апомикеие не обнаружен.
Другие явления пе отмечены,
Литература: Davis, 1966: Тахта-
джян, 1966, 1970; Venkateswarly,
1967.
В порядке Мущаез семейство
АшзорвуНасеае и Репаеасеае цито-
эмбриологически пе исследованы, а
Oliniaceae u Penaeaceae исследовапы
в этом отношепии очепь слабо.
Систематическое положение Гу -
raceae, опрелеленное Энглером,
Гильгом и Веттштейном, по мнению
Капила [КарИ, 1967с], согласуется
с данными сравнительной цитоэм-
бриологии. Sonneratiaceae B oTHO-
шении цитоэмбриологии похоже на
другие семейства Мушаез, особен-
но на Гугасеае \{Venkateswarlu,
130
19672]. Вепкатесварлю [У\УепКафез\аг-
lu, 41967Ё| считает, aro Lythraceae,
Sonneratiaceae, Punicaceae u Mela-
stomataceae nuTosMOpHOoNoruyecky Ha-
столько близки, что их следует от-
носить к общему порядку. СошЪ-
ге@асеае — однородное и естествен-
ное семейство, сходное с другими
представителями Мушаез [УепкКа-
teswarlu, 1967a], за исключением
Ошега, которую следует выделить
в tTpuOy Quieraceae. Lecythidaceae
в отношении цитоэмбриологии не
имеет сходства с другими семей-
ствами Муга]ез, поэтому его не
следует включать в порядок Муйа-
les [Venkateswarlu, 1967b]. Onagra-
сеае в отношении цитоэмбриологии
сходно с Ме]азютасеае, а послед-
нее — с Трутеаеасеае [|АШег,
Herr, 1968]. Мугаез — довольно хо-
рошо исследованный в отношении
цитоэмбриологии порядок и явля-
ется более или менее однородным
и естественным.
Дапные цитоэмбриологии показы-
вают, что Му[асеае — одпородное и
естественное семейство, что исследо-
ванные роды его родственны друг
другу [Полунина, 1963]. Семейства
Ту гасеае, Sonneratiaceae, Punica-
ceae, Oliniaceae, Melastomaceae,
Onagraceae, MHaloragaceae, Hydro-
cariaceae, Myrtaceae и Rhizopho-
тасеае настолько похожи друг
на друга, что их следует от-
нести к одному и тому же по-
рядку [Seshavataram, 1970]. Это
согласуется с мнениями Энглера и
Дильса, Веттштейна и Рендля, но
не согласуется с мпением Хатчин-
сопа, который относил их но к од-
ному, а трем порядкам [Seshavata-
тат, 1970]. Все вышеуказанные ce-
мейства характеризуются следую-
щими цитоэмбриологическими при-
знаками: соекреторным тапетумом,
симультанным образованием микро-
спор, двухклеточным мужским га-
метофитом с тремя порами, ана-
тропной с двумя интегументами
семяпочкой, одноклеточным жен-
ским археспорием, наличием пари-
етальных клеток, зародышевым ме-
шком типа Ро[угопит, нуклеарным
эндоспермом и Опастад-типом эм-
бриогенеза.
Данные цитоэмбриологии подтвер-
ждают правильность выделения из
Опастасеае рода Тгара в самостоя-
тельное семейство ‘Trapaceae [Se-
shavataram, 1967]. Семейство Тгара-
сеае характеризуется Polygonum-
типом зародышевого мешка, 5о]а-
пад-типом эмбриогенеза, наличием
подвесочного гаустория и отсут-
ствием апомиксиса, в то время как
семейство Опартасеае отличается
Оепо{Тега-типом зародьшцевого меш-
ка, Опаогад-типом эмбриогенеза, от-
сутствием подвесочного гаустория и
наличием апомиксиса.
ПОРЯДОК 55.
HIPPURIDALES
(HALORAGALES)
CEMEUCTBO HALORAGACEAE
Основное число хромосом — 5 =
= 6, 7, 12, 16, 17.
Цветок обосполый, чаще однопо-
лый, растения однодомные или по-
лигамные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия формирует-
ся по однодольному типу и имеет
эпидермис, фиброзный эндотеций,
1—2 эфемерных средних слоя и
секреторвый одно- двуядерный та-
нетум.
Микроспорогенез и микрогамсто-
генез — обралование микроснор си-
мультанное. —"'Геграды микроснор
тетраздральные и крестообразные.
Зрелые ныльцевые зерна трехкле-
точные, четырех-нятиноровые с тол-
стой и гладкой экзиной, имеют зер-
на крахмала.
Семяпочка висячая, гемианатроп-
ная-анатропная, с одним, двумя ин-
тегументами, крассинуцеллятная.
Микропиле образуется обоими инте-
гументами. Имеются нуцеллярный
колпачок, гипостаза и обтуратор.
Семяпочек 2—4.
Макроспорогенез и макрогамето-
генез — архоспорий обычно одпо-,
редко дДвух- или трохклеточный.
Париетальных клеток 2--8 ряда.
Тетрады макроспор линейные или
Т-образные. Зародышевый мешок
формируется по Роусопит-типу.
Синергиды имеют фибриллярные
аппараты. Три антиподы дегенери-
руют после оплодотворения. Поляр-
ные ядра сливатотся до оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
9* .
порогамия. Двойное ’оилодотворе-
ние. Сингамия предшествует трой“
ному слиянию. Первичное ядро эн-
досперма делится раньше зиготы.
Эндоспермогенез целлюлярный
или нуклеарный.
Эмбриогенез — Опастай- или Са-
ryophyllad-, sapuanuu Myriophyllun-
типов. Зародыш крупный, прямой
или согнутый, двусемядольный, с
хорошо развитыми гинокотилем и
корешком, бесхлорофилльный, со-
держит масло и алейрон.
Семя, плод — семя с эндоспермом,
содержащим масло, крахмал и алей-
рон. Зародыш занимает от 1/4 до
3/4А длины семени или заполияет
почти вссь объем семени. Семядоли
виолняют Либо гаусториальную,
либо запасаюощие фупкции. Эндо-
сиерм располагается по периферии
семени. Семенная кожура тонкая,
образуется из варужного интегу-
мента. Перикарпий шести-семия-
дерный, твердый благодаря присут-
ствию внутренней склеренхимной
зоны, содержит таннин. Плоды мел-
кие, орешковидные или костянко-
видные.
Гауеторий подвесочный.
Апомиксис не обнаружен.
Другие явления пос отмочепы.
Литература: Ваха, 1967а, 1969а, Ъ,
1970a, Davis, 1966; Kapil, Bawa, 1968;
Nae, 1967; Nagaraj, Nijalingappa,
1967, 1974; Nijalingappa, 1967, 1975;
Schnarf, 1934; Смирнова, 1977; Тах-
таджян, 1966, 1970.
СЕМЕЙСТВО СОММЕВКАСЕАЕ
Основное число хромосом — х =
== 7, 12, 17.
Цветок обоеполый.
Пыльник — нет данных.
Микроспорогенез и микрогамето-
генез — нет данных. Зрелые пыль-
цевые зерпа двухклеточные.
Семяпочка одпа, висячая, ata-
тронная, © Двумя иптегументами,
крассинуцеллятпая.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки имеются. За-
родышевый мешок образуется по
Peperomia-tuny. Имеются шесть
долго сохраняющихся антипод.
Опыление, прохождение пылье-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез целлюлярный.
131
Эмбриогенез — тип не определен.
Зародыш маленький, слабо диффе-
ренцированный на семядоли и ко-
решок, без подвеска.
Семя, плод — семя с эндоспермом.
Зародьши расположен в основании
семени и запимает от 1/4 до 3/4
объема семени. Нлод костянкооб-
разный.
Гаусторий — семядоли выполня-
ют гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не обнаружены.
Литература: Модилевский, 1908;
Зевпатё, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970.
СЕМЕЙСТВО НУРРОВТШАСЕАЕ
Основное чиело xpomMocom— x= 16.
Цеток большей частью обоепойпый.
Ныльник — нет данных.
Микроепорогенез и микрогамето-
тенез — образование микроспор си-
мультаннос. Зрелая пыльца трох-
клеточная.
Семяпочка оцпа, Bioessays, ана-
тропная-гемианатроппая, C одним-
двумя интегументами, тенуи- или
крассинуцеллятная. Имеются ну-
целлярный колпачок и фуникуляр-
ный обтуратор.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. 3a-
родышевый мешок образуется по
Ро]угопит-типу. Синергиды имеют
фибриллярные аппараты. Три анти-
поды дегенерируют после оплодо-
творения.
Опыление, прохождение пыльце-
вых трубок и оплодотворепие —
нет данных.
Эндоспермогепез целлюлярный.
Эмбриогенез Опаигай-тина. Заро-
дыш прямой или согпутый, двусо-
мядольный, содержит масло и алей-
рон.
Семя, плод — ссмя с эндоспермом
или без него. Зародыш располага-
ется в центре семени и заполняет
либо большую часть семени, либо
все ссмя. Плод костянкообразный.
Гаусторий подвесочный.
Апомиксие пе обиаружен.
Другие явления пе обнаружены.
Литература: Ва\уа, 1970; Schnarf,
1931; Тахтаджян, 1966, 1970.
43
На основании цитоэмбриологиче-
ских данных роды Са Шёлеве, Сип-
пега и Hippuris отличаются от
представителей На]огасасеае и дол-
жны быть отнесены к самостоя-
тельным семействам СаШилевасеае,
Gunneraceae uw Hippuridaceae соот-
BeTcTBeHHO [Bawa, 1970a]. Schnarf
[1931] wm Wettstein [1924] pasnuada-
au Haloragaceac, Gunnaraceae u Hip-
puridaceae как самостоятельные се-
мейства, что, по мпению Бава [Ва-
wa, 1970а], согласуется с данными
цитоэмбриологии. Хатчинсон [Ни
chinson, 1959] относил роды Сипше-
та и Нрритз к Натгасасеае, что
Бава [Ва\уа, 1970а] считает непра-
ВИЛЬНЫМ.
НАДПОРЯЛОК Х.
RUTANAE
ПОРЯДОК 56.
RUTALES
CEMEHMCTBO ANACARDIACEAE
Основное чиело хромосом — Z =
— 7, 8, 10, 12, 15.
Цветок обосполый или чаще од-
нополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эндотеции, обы-
чно 2 реже 1—3 эфемерных сред-
них слоя и сокреторный, двуядер-
пый танетум.
Микроспорогенез и микрогамето-
генсз — образование микроспор си-
мультаниое. Теотрады микроспор
тетраэдральнтыюе. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые или трех-восьмиапертур-
ные.
Семяпочка звисячая, анатропная,
с одним или двумя интегументами,
крассинуцеллятная. Микропиле обы-
чно образуется обоими - интегумен-
тами, но у Тох1содепагот — только
одним внутренним. Имеются нуцел-
лярный колпачок, гипостаза и фу-
никулярный обтуратор. Семяпочек
1—5.
Макроспорогенез и макрогамето-
тенез — археспорий одноклеточный.
Париетальная ткань массивная (до
10 рядов). Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро[усопит-типу. Синер-
гиды с крючкообразными выроста-
ми. Три антиподы, дегенерирующие
до оплодотворения. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение. У Р1$асеае
первое деление зиготы происходит
через 6—14 недель после оплодо-
творения.
Эндоепермогенез нуклеарный. За-
ложение клеточных перегородок меж-
ду ядрами эндосперма начинается
с микропилярной части зародыше-
вого мешка и постепенно весь эн-
досперм становится клеточным. Од-
HaKO y Anacardium occidentale xa-
лазальный конец ондосперма оста-
ется свободпоядерпым.
Эмбриогенез Опаотад- или Азе-
га-типов. Зародыш более или ме-
нее согнутый, крупный, двусемя-
дольный, бесхлорофилльный или
хлорофиллоносный, содержит крах-
мал и алейроновые зерна. Семядоли
выполняют запасающую функцию.
Семя, плод — семя со слаборазви-
тым эпдоспермом или без него. За-
родьии лежит в пептре семени и
заполияст НОЧТИ BCCH или BCCh
объем семени. Модвесок ие выра-
жен. Семенная кожура образуется
из обоих питегумоептов и остаТКОВ
нуцеллуса. Плод обычно костянко-
видный.
Гауеторий отсутствует.
Апомиксие — нуцеллярная эмбри-
oHuaA y Mangifera indica.
Другие явления — партенокарпия
полиэмбриония.
Jiurepatypa: Corner, 1976: Davis,
1966; Grundwag, 1976; Grundwag,
Fahn, 1969; Kelkar, 1961; Schnarf,
1931; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВОВЗЕВАСЕАЕ
Основное чиело хромосом — т =
— 11—13.
Цветок обосполый или чаше ол-
нополый, растения двудомные.
Ныльник — 4А микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный одно- двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные. Зрелые пыльцевью
зерна двухклеточные, трехбороэдио-
поровые, с гладкой экзиной.
Семяпочка гемианатропная или
кампилотропная, с двумя или од-
нилг интегументом, крассинуцеллят-
ная. Микропиле образуется внут-
ренним интегументом. Имеется ну-
целлярный колпачок. Семяпочек
2—10.
Макроспорогенез и макрогамето-
генез — археспорий одно-многокло-
точный. Парпетальная ткань мас-
сивная (до 14 рядов). Тетрады за-
кроспор линейные, Т-образные. За-
родьниевый мепток образуется но
Ро]узопию-типу. Синергиды с крюч-
кообразными выростами и фибриин-
лярными аппаратами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются до оплодотворения.
Оныление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Сапагиия
ovatum. Порогамия. Двойное опло-
дотворение. После оплодотворения
У Сагира ршпафа первым делится
первичное ядро эвдосперма.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип пе определоз:.
Зародьыит крупный, прямой или со.
гпутый, двусеемядольный, с хоровю
развитыми типокотилем и KOpert-
KOM, хлорофиллоноспый, содержит
масло и алейрол. Семядоли склад-
чатые, выполыяют занасающую
фупкцию.
Семя, плод — семя без эндосиез-
ма. Зародыш занимает весь объем
семени. Плод одно-пятисеменная
ягода или иногда коробочка.
Гаусторий не обнаружен.
Апомикеие — нуцеллярпая OR -
бриония у Garuga pinnata.
Другие явления не обнаруженн.
Литература: Corner, 1976; Davis.
1966; Chosn, 1970; Gulati, Mathur
1977; Rao, 1967a, 1970a; Srivastava,
1968; Тахтаджяи, 1966, 1970.
СЕМЕЙСТВО ЗТМАКООВАСЕАЕ
Оеновное число хромоеом — х =
= 13—15, 18.
Цветок обоеполый или чаще од-
нополый.
Пыльник — 4 микроспоравгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
фемерных средних слоя и cekpe-
133
торный дву- многоялерный, иногда
двурядный тапетум. Ядра тапетума
часто сливаются и становятся нпо-
липлоидными.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, крестообразные, изо-
билатеральные, 7-образные. Зрелые
пыльцевые зерна двухклеточные,
трехборозднопоровые, с гладкой эк-
зиной.
Семяпочка гемианатропная, ана-
тропная, или кампилотропная, с
двумя или одним интегументом,
крассинуцеллятная. Микропиле зиг-
загообразное, образуется обоими
или одним внутренним интегумен-
том. Имеются нуцеллярный колпа-
чок, гипостаза и плацентарный 66-
туратор. Семяпочек немного (2—75).
Макроспорогенез и макрогамето-
генез — архоспорий ’одно-многокле-
точный. Паристальная ткань мас-
сивная (до 17 рядов). Микроспоры
линейные, Г-образные. Зародьтио-
вый моно образуется мо Polygo-
пит-типу. Синергиды грушевилные,
с фибриллярными аппаратами. Име-
ются три эфемерные антиноды. По-
лярные ядра сливаются до оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндоснерма начина-
ется с периферии и постепенно
весь эндосперм становится клеточ-
ным. Эндосперм содержит масло и
алейроп.
Эмбриогенез Опарга@-тина. Заро-
дыш прямой или согпутый, круп-
ный, двусемядольняй, хлорофилло-
носный, содержит масло, алеироп и
слизистые клетки.
Семя, плод — семя со слабораз-
витым эндоспермом или без него.
Зародыш заполняет семя по всей
его длине. Тонкий слой эндосперма
имеется лишь на самой периферии
семени. Семенная кожура образуст-
ся двумя иптегументами. Плод pac-
падающийся па сухис или костян-
ковидные плодики, или плод — ко-
робочка, крылатка, редко ягода или
костянка. Перикарпий имеет скле-
реидный эндокарпий, молочный ме-
зокарпий и двухслойный эндокар-
nui,
134
Гаусторий эндоспермальный, рас-
положенный в халазальной части
зародышевого мешка у Suriana.
Апомикеис не обнаружен.
Другие явления — полиэмбриония
y Erucea.
Литература: Davis, 1966; Rao D.,
1967b, 1970b; Schnarf, 1931; Смирно-
ва, 1977; Тахтаджяи, 1960, 1970.
СЕМЕЙСТВО ПВОТАСЕАЕ
Основное число хромосом — х =
= 7—13, 17, 19.
Цветок обычно обоеполый, редко
однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный, дву-многоядерный танпетум.
Ядра танетума часто сливаются и
становятся полиплоидпыми.
Мнкроспорогенез и микрогамето-
генсз — образовапие микроспор си-
мультанное. Тетрады — микроспор
тераздральные, крестообразные, изо-
билатеральтыье. Зрелые пыльцевые
зерпа двухклеточные или трехкле-
точные, (двух) трех- шести (вось-
ми)-борозднопоровые.
Семяпочка анатропная-гемиана-
тропная, с двумя, изредка с одним
интегументом, крассинуцеллятная
или тенуинуцеллятная. Микропиле
образустся обоими или только од-
ним внутренним — интегументом.
Имеются нуцеллярный колпачок,
гипостазаа фуникулярный обтура-
тор и ариллус. Семяпочек обычно
исмпого, изредка много. Сосуди-
стый нучок доходит до халазы, где
он ветвитея.
Макроспорогенсз и макрогамето-
генез — архоснорий одно- многокле-
точный. ПНарпетальных клеток 2—
10 рядов. Тетрады макроспор ли-
нейные, Г-образные. Ипогда функ-
ционирует несколько макроспор. За-
родышевый мешок образуется по
Ро]угопит-типу и имеет крахмаль-
ные зерна, часто образуются слож-
ные зародышевые мешки. Синерги-
ды с крючкообразными выростами
и фибриллярными аппаратами. У
Asperula odorata антиподальный ап-
парат сильно развит. Имеются обы-
чно три эфемерные антиподы. По-
лярные ядра сливаются перед опло-
дотворением. Иногда — образуется
несколько зародышевых мешков в
одной и той же семяпочке вслед-
ствие функционирования несколь-
ких макроспор.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворение,
в яйцеклетке — постмитотического
типа, в центральной клетке — пре-
митотического типа. У СИгиз па
sudaidae оплодотворение происхо-
дит через 8 дней после опыления,
У С. чпзШа — через 14 дней. За-
родыит достигает змаровидной ста-
дии через 56 дней, а полной зрело-
сти через 120 дней после опыления.
Эндоспермогенез нуклеарпый. Об-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярного конца за-
родышевого мешка, и постепенно
весь эндосперм становится клеточ-
ным.
Эмбриогенез Onagrad- una Sola-
пад-типа. Зародыш крупный, пря-
мой или согнутый, двусемядоль-
ный, с хорошо развитыми почечкой,
гипокотилем и корешком, бесхло-
рофилльный или хлорофиллонос-
ный. Содержит масло и алейрон,
иногда и крахмал. Подвесок корот-
кий.
Семя, плод — ссмя с эпдоспермом
или O6c3 ного, содержит масло и
алейрон. Зародыпг располагается в
центре или по иериферии семени,
заполняя 3/4 общего объема семе-
ни или все семя. Млоды — ягоды,
в эндокарпии которых содержится
много эфирного масла и лимонкой
кислоты.
Гаусторий агрессивный, эндоспер-
мальный у Аее тагте]ос в хала-
зальной части эндосперма. Анти-
подальный гаусторий у Sherardia
arvensis.
Апомиксис — нуцеллярная oM-
бриония и апоспория y Cilrus, Tri-
phusia mu Xanthoxyllum.
Другие явления — партепокарлия
и полиэмбриония у paylos Citrus
y Ruta graveolens.
Jiurepatypa: Boesewinkel, Bouman,
1978; Corner, 1976; Davis, 1966;
Esan, Soost, 1977; Farooq, Inamud-
din, 1969; Ghosh, 1974a; Horner,
Lersten, 1971; Капанадзе, 4966,
1967, 1968; Кордюм, 1963; Саникидзе,
1967; Смирнова, 1977; —Старшо-
ва, 1974; Старшова, Солнцева, 1973,
1976; Schnarf, 1934; Тахтаджян,
1966, 1970; Yang-Hsu-Jen, 4968.
СЕМЕЙСТВО СМЕОВАСЕАЕ
Основное чиело хромосом — пет
данных.
Цветок обоеполый.
ПНыльник — нет данных.
Микроспорогенез и микрогамето-
генез — по микроспорогенезу нет
данных. Зрелая пыльца трехклеточ-
ная, трехборозднопоровая и четы-
рех-шестиборозднопоровая.
Семяпочка висячая. амфитроп-
ная, с двумя интегументами, крас-
синуцеллятная. Семяпочек немного.
Макроепорогепез и макрогамето-
генез — археспорий одноклеточный.
Данных о париетальных клетках
нет. Тотрады макроспор липейныс.
Зародыптевый мешок образуется по
Роугопит-типу. Синергиды груше-
видные. Имеются три скоро дегене-
рирующие антиподы. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез нуклеарный.
Эмбриогенез Азегад-типа. Заро-
дыш прямой или согнутый с хоро-
шо развитыми семядолями, почс-
чкой, гипокотилем и корешком, бес-
хлорофилльный, подвесок короткий.
Семя, плод — семя с мясистым
эпдоспермом. Согнутый зародыш
располагается по периферии семе-
пи, запимая 3/4 его периметра.
Прямой зародыш располагается в
центре семени. Плод распадается
наз или 4 костянковидных плодика.
Гауеторий не найден.
Апомиксие не обнаружен.
Другие явления не отмечены.
JImreparypa: Davis, 1966; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО MELIACEAE
Осповное число хромосом — х =
= 10, 12, 14, 15, 18.
Цветки обоеполые или полигам-
ные — двудомные.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный одно-двумногоядерный та-
петум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
135
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двух- или трехклеточные,
трех-пятиборозднопоровые с глад-
кой экзиной и большим числом
крахмальных зерен.
Семяпочка анатропная, с двумя,
изредка с одним интегументом,
крассинуцеллятная. Микропиле обы-
чно образуется внутренним интегу-
ментом, но y Dysoxylum гаш о-
тиш — наружным. Имеются вуцел-
лярный колпачок, Фуникулярный
обтуратор и ариллус. Семяпочек
обычно две, реже одна — несколь-
ко или много.
Макроепорогенез и макрогамето-
reves — археспорий одно-многокле-
точный. Париетальных клеток 4—7
рядов. Тетрады макроспор липей-
ные, Г-образные. Зародышевый ме-“
тнок образуется по Ро]усхопит-типу
и имеет крахмальпые зерпа. У не-
‹оторых представителей семейства
каблюдается по поскольку зароды-
шевых мепг:ов в одной и той жо
завязи. Синергиды с кртючкообраз-
ными выростамия и фибриллярпыми
аппаратами. Имеются три эфемер-
ные антиподы. Иногда (y Melia)
антиподы многоядерные. Полярные
ядра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоснермогенез нуклеарный. Об-
разование клеточных перегородок
начинается с микропилярного кон-
ца зародышевого мешка.
Эмбриогенез ОпаотаЯ-типа. Заро-
дыш крупный, прямой, содержит
масло и алейроп, иногда и друзы
оксалата кальция. Бесхлорофилль-
ный или хлорофиллоносный. Семя-
доли выполняют запасающую фуип-
КЦИЮ.
Семя, плод — семя с масляни-
стым эндоспермом или без него. За-
родыш расположен в центре семе-
ни и заполняет почти весь или да-
же весь его объем. Плод — коро-
бочка, ягода или иногда костянка.
Гаусторий отсутствует.
Апомиксие — нуцеллярпая эм-
бриония, апоспория y Lansium
domestica.
Другие явления — полиэмбриония
y Lansium domestica um Azadirachta
indica.
Литература:
1966; Ghosh,
\
Corner, 1976; Davis,
1966; Mestre, 1966;
135
Маг, 1967с, 19705; РгаКаз её а|.,
1977; Вао К., 1972: ЗейпатЁ 1934;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО СОВТАВТАСЕАЕ
Основное чиело хромосом— х==20.
Цветки обоеполые или полигам-
ные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2
средних слоя и секреторный одно-
четырехъядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сг-
мультанное. Тетрады — микроспор
тетраэдральные и крестообразные.
Зрелые пыльцевые зерна двух-
трехклеточные, — трехборозднопоро-
вые. Поры располагаются по мери-
диану.
Семяпочка висячая, анатропная.
с двумя интегументами, крассину-
целлятпая. Микропиле образует-
cH внутренним интегументом. На-
ружиый интоегумент имеет крыло-
образные выросты по обеим сторо-
нам семяножки. Семяпочек 5—10.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 12 рядов.
Тетрады макроспор линейные и
Г-образные. Зародышевый мешок об-
разуется по Polygonum-tuny. Си-
нергиды с крючкообразными выро-
стами. Имеются три эфемерные ан-
типоды. Полярные ядра сливаются
во время оплодотворения.
Олыление, прохождение пыльце-
пой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворение.
)идосиермогенез пуклеарный.
Эмбриогслез Опаста4-типа. Заро-
дыпг прямой, двусемядольный, со-
держит масло. Семядоли выпояня-
10T запасающую функцию.
Семя, плод — семя со скудным
эндоспермом. Зародыш заполняет
семя по всей его длине. От эндо-
сперма сохраняется лишь тонкий
слой на самой периферии семени.
Семенная кожура образуется одним
внутренним или обоими интегумен-
тами. Плод — орех, окруженный мя-
систыми сросшимися лепестками.
Гаусторий отсутствует.
Апомикеис не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Nair, 1967a, 1970; Rau Sharma,
19706; Sharma, 1968a; Смирнова,
1977; Schnarf, 1931; Тахтаджяп,
1966, 1970.
Семейства ]иПтасеае, Родоасеае,
Stylobasclaceae, Aitoniaceae и Ки-
К1асеае, относимые А. Л. Тахтаджя-
ном к порядку Ви\фаез, цитозмбри-
ологически пе исследованы, а Споо-
тасеае исследовано очевь слабо. По
цитоэмбриологии Вигзегасеае похоже
Ha Meliaceae, Simaroubaceae uw Ru-
{асеае [D. Rao, 1967]. Семейство
Вигзегасеае сходно с Эитагоифасеае
по ряду цитоэмбриологических при-
знаков: секреторному — тапетуму
пыльника, фиброзному эндотецию,
симультанному образованию микро-
спор, тетраэдральной тетраде мик-
роспор, трехклеточному, трехборозд-
нопоровому пыльцевому зерну, апа-
тропвой, крассипуцеллятной семя-
почки, с массивной париетальной
тканью, многоклеточному женскому
эрхеспорию, Polygonum-tuny заро-
дышевого мешка, нуклеарному эндо-
сперму и семенной кожуре, образу-
ющейся. обоими интегументами. От
Simaroubaceae Burseraceae отличает-
ся отсутствием долго сохраняюще-
гося средиего слоя тапетума, гино-
стазы, халазального эидоспермаль-
ного гаустория и обтуратора. 'Га-
ким образом, носмотря па цитоэм-
бриологическое сходство между обо-
ими семействами, имеются и разли-
чия, что оправдываст их обособлен-
ное положение. Д. Рао [Вао, 1970]
считает, uTo Burseraceae u Simaro-
ubaceae — близкородственные ce-
мейства и что их следует отнести к
одному и тому же порядку. Мей!-
асеае имеет сходные цитоэмбриоло-
гические признаки c Rutaceae.
Систематическое положение Coria-
т1асеае спорное. Его относили к
Sapindales, payom c Anacardiaceae,
K Celastrales pagom c Salvadoraccae,
K Teribinthales между IJlippocasta-
naceae ww Balsaminaceac, « Ericales
psytom © Emipetracceae mot. jy. Шарма
[Sharma, 1968а|, сравиив цитоэмбри-
ологию Сопатасеае с таковой Ет-
petraceae, АпасагФасеае и За|уа-
doraceae, пришел к заключению
что в отношении дцитоэмбриологии
Созатасеае сходно с Апасаг асеае;
он считает правильным отнесение
этих семейств к одному и тому же
порядку. Носмотря на значитель-
ные пробелы в исследовании цито-
эмбриологии представителей Вща-
les, ero семейства, более или мепос
подробно изученные в настоящее
время, обнаруживают сходные цито-
эмбриологические признаки, что ука-
зывает на естественность этого по-
рядка.
ПОРЯДОК 57.
SAPINDALES
CEMEHCTBO STAPHYLEACEAE
Основное число хромосом — х =
== 42, 13.
Ивстки
ные.
ПНыльник — 4 микроспорангкя.
Стенка микроспорангия имеет эги-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
обосполые или полигам-
зерна двух-трехклеточные, Tpex-
шестибороздпопоровые.
Семяпочка анатроппая, с двумя
пнтсгументами, крассипуцеллятная.
Микроинле образуется обоими ин-
гегументами. Имеются нуцелляр-
ный колпачок и ариллус. Семяпо-
чек обычно 6—10, изредка 1—2.
Макроспорогсиез м макрогамето-
генез — археспорий одпоклеточный.
Париетальвых клеток 3—8 рядов.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро|узопиш-типу. Синергиды с крю-
чкообразными выростами и фибрил-
лярными аппаратами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются: до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотвореснисо.
Эидоенермогеиез пуклеарный.
Эмбриогенез — тип не определен.
Зародыш прямой двусемядольный,
хлорофиллоносный. Семядоли вы-
полняют запасающую функцию.
Семя, плод — семя обычно с хо-
Polo развитым, мясистым эндо-
спермом, содержащим масло или
крахмал. Тонкий слой эндосперма
располагается по периферии семе-
ни. Плод — перепончатая коробочка
или ягодообразный, или состоящий
137
из отдельных вскрывающихся пло-
ДИКОВ.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явление не отмечены.
Литература: Corner, 1976; Davis,
1966; Narayana, 1960; Schnarf, 1934;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АСЕВАСЕАЕ
Оеновное число хромосом — х==43.
Цветки полигамные, растения
двудомные.
Ныльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—3
эфемерных средних слоя и секре-
торный многоядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады —микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трехбороздиые или трехбо-
розднопоровые.
Семяпочка анатроппая-гемиапа-
тропная с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется внутренним интегументом.
Имеется нуцеллярный колпачок.
Семяпочек немного (3—16).
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток несколько ря-
дов. Тетрады макроспор линейные.
Иногда вместо тетрад макроспор
образуются триады макроспор. За-
родышевый мешок образуется по
Polygonum-tuny и имеет зерна
крахмала. Синсргиды с крючкооб-
разными выростами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльцсе-
вой трубки и оплодотворение —
перекрестное опыление и само-
опыление. Порогамия, халазогамия.
Двойное оплодотворение.
Эпдоспермогенез нуклеарный. До
поздней стадии развития эндосперм
остается нуклеарным.
Эмбриогенез Опаста4-типа. Заро-
дыш согнутый, двусемядольный,
хлорофиллоносный, содержащий
алейроновые зерпа и друзы оксала-
та кальция. Семядоли выполняют
запасающую функцию.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Эндосперм полностью израсходует-
138
ся в процессе развития зародыша.
Плод плоский, крылатый, распада-
ющийся на два односеменных оре-
шкообразных плодика.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления — полиэмбриония.
y Acer platanoides.
Литература: Corner, 1976; Davis,
1966; Haskell, Postlethwait, 19714;
Mestre, 1965b; Тахтаджян, 1966, 1970.
CEMENCTBO SAPINDACEAE
Основное число хромосом — х =
== 4141, 12, 15, 16.
Цветки обычно полигамные, ра-
стения двудомные.
Ныльник — 4 микроспорангия.
Стенка пыльника имеет эпидермис,
фиброзный эндотеций, 1—2 эфемер-
пых средних слоя и секреторный
одно-двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тоетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые с тол-
стой экзиной, содержащие крах-
мальные зерна.
Семяпочка висячая.
гемианатропная, с двумя интегу-
ментами, крассинуцеллятная. Ми-
кропиле образуется обоими инте-
гументами или только одним внут-
ренним (у СагФозрегтит). Имеют-
ся ариллус, нуцеллярный колпачок
и плацентарный обтуратор. Семя-
почек пемпого.
Макроспорогснез и макрогамето-
генез — архсспорий одноклеточный.
Париетальных клеток 6—15 рядов.
Гетрады макроспор линейные. За-
родыпевые мешки образуются по
Polygonum- и АНшт-типам. Синер-
гиды имеют крючкообразные выро-
сты. Хотя обычно три антиподы
офемерные, но у Cardiospermum
halicacabum число их увеличивает-
ся до 14 и они становятся много-
ядерными. Полярные ядра слива-
ются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение. Первич-
ный ядро эндосперма делится рань-
ше зиготы.
Эндоспермогенез нуклеарный. Эн-
досперм поглощается, прежде чем
он станет клеточным.
анатропная-
Эмбриогенез Азега4-типа. Заро-
дыш согнутый, двусемядольный,
бесхлорофилльный или хлорофил-
лоносный, содержит крахмал и
масло, масло алейрон, дубильные
вещества, иногда и сапонин. Семядо-
ли выполняют запасающую функ-
цию, они тонкие или толстые, не рав-
ной величины.
Семя, плод — обычно семя без
эндосперма. Зародыш заполняет
обычно все семя. Эндосперм пол-
ностью израсходовался в процессе
развития зародыша. Плод — трех-
гнездная коробочка. Семенная ко-
жура образуется обоими интегумен-
тами.
Гауеторий ондоспермальный, в
микропилярной части зародышево-
го мешка. ,
Апомиксие вес обпаружсн.
Другие явление не отмечены.
Литература: Corner, 1976; Davis,
1966; Karkare-Khushalani, Mulay,
4964; Singh, Shiam, 41977; Тахта-
джян, 1966, 1970.
СЕМЕЙСТВО
НТРРОСАЗТАМАСЕАЕ
Основное число хромосом — х= 20.
Цветок обычпо обосполый, по
верхний, пиогда мужской.
Пыльцик — 4 микроспораигия.
Тапетум сокреторный, многоядер-
ный.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультапное. Тетрады — микроспор
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка анатропная, с двумя
интегументами, обычно крассину-
целлятная, иногда тенуинуцеллят-
ная. Микропиле образуется внут-
ренним интегументом. У Н1рросга-
{еа имеется эндотелий. Семяпочек 6.
Макроспорогенез и макрогамето-
генез — архоспорий многоклеточный.
Париетальных клеток 4 ряда. 'Тет-
рады линейные. Зародышевый ме-
шок образуется по Ро[угопиш-типу.
Синергиды с крючкообразными вы-
ростами и фибриллярными аппара-
тами. Имеются три эфемерные ан-
типоды. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоепермогенез нуклеарный.
Эмбриогенез — тип не определен.
Зародыш крупный, согнутый, дву-
семядольный, хлорофиллоносный,
содержит крахмал, алейрон и ду-
бильные вещества. Семядоли тол-
стые, корешок короткий.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Эндосперм полностью израсходовал-
ся в процессе развития зародыша.
Плод — обычно односеменная локу-
лицидная коробочка.
Гауеторий отсутствует.
Апомикеис не обнаружен.
Другие явления — полиэмбриония.
Литература: Согпег, 1976; Davis,
1966; Taxranman, 1966, 1970; Schnarf,
1934.
СЕМЕЙСТВО МЕШМАКХМТНАСЕАЕ
Основное число хромосом — х =
= 18, 149. -
Цветок обоеполый, реже одпо-
полый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, эндотеций без фиброзных
утолщений, 3—5 эфемерных сред-
них слоев и сокреторпый много-
ядерный тапетум.
Микроепорогенез и микрогаметс-
генсз -— образование микроспор си-
мультанное. Тетрадут — микроспор
тетраэдральные, изобилатеральные.
Зрелые ныльцевые зерна трехклсе-
точные, трехбороздноноровыс.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется одним внут-
ренним или обоими интегументами.
Имеются нуцеллярный колпачок и
ариллус. Семяпочек 8—20, или 4—5.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 6—13 рядов.
Зародышевый мешок образуется по
Ро!узопит-типу. Сипергиды с крю-
чкообразными выростами. Имстотся
три оэфемерные антиподы. Поляр-
ные ядра сливаются до оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез нуклеарный, за-
ложение клеточных — перегородок
между ядрами эндосперма начина-
ется с микропилярного конца заро-
дышевого мешка.
139
Эмбриогенез. Азбегай-типа, вариа-
ции Репаёа. Зародыш маленький,
прямой двусемядольный. „
’_. Семя, плод — семя с хорошо раз-
витым роговым, амилоидным эндо-
спермом. Зародыш располагается в
основании семени, занимая от 1/4
до 3/4 линейной высоты семени.
Плод — коробочка.
аусторий не найден.
Апомикесие не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Mathur, 1977; Schnarf, 1934;
Steyn, 1975; Taxranman, 1966, 1970.
CEMENCTBO SABIACEAE
Основное чиело xpoMocoM — z =
= 42, 16.
Цветки обоеполые или полигам-
ные — двудомные. -
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — микроспорогенез не изучен.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка гемианатропная, с од-
ним интегументом, крассинуцеллят-
ная. Микропиле не образуется. Се-
мяпочек несколько. _
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 4 ряда. Тет-
фады макроспор линейные. Зароды-
певый мешок образуется mo Poly-
зопи-типу. Синергиды с крючко-
образными выростами. Имеются три
эфёмерные антиподы. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных, .
Эндоспермогенез нуклеарный.
Эмбриогенез Опаогад-типа. Заро-
ды крупный, согнутый, двусемя-
дольный. Семядоли выполняют за-
пасающую функцию.
Семя, плод — семя со слабо раз-
витым эндоспермом или без него.
Зародылт заполняет почти все или
даже все семя. Плод сухой, нерас-
крывающийся или костянковидный,
обычно одногнездный.
Гаусторий эндоспермальный.
Апомиксие не обнаружен.
440 -
Другие явления не отмечены.
Литература: Davis, ' 1966; Тахта-
джян, 1966, 1970. `
В порядке Sapindalés семейства
Akaniaceae, Bretschneideraceae и
Сгеуласеае почти или }полностью не
изучены цитоэмбриойогически. По
данным цитоэмбриодогии Асегасеае
cxomHo c Polygalageae u Balsami-
naceae [Mestre, 1965b].
Более или менее полно изучен-
ные семейства Заршаез обнару-
живают сходные цитоэмбриологи-
ческие признаки, что указывает на
естественность этого порядка. Он
характеризуется наличием четырех
макроспорангиев, четырех-пятислой-
ной стенкой пыльника, секретор-
ным тапетумом, симультанным ми-
кроспорогенезом, . двухклеточной,
трехборозднопоровой пыльцой, геми-
анатропной-анатропной, с двумя ин-
тегументами, крассинуцеллятной се-
мяпочкой, наличием нуцеллярного
колпачка и ариллуса, одноклеточ-
ным женским археспориумом, не-
сколькослойной париетальной тка-
нью, Ро]угопит-типом зародышево-
го мешка, эфемерными антиподами,
нуклеарным эндоспермом. По этим
же признакам предетавители Зарш-
да]ез сходны с семействами Вшаез.
ПОРЯДОК 58.
GERA NIALES
СЕМЕЙСТВО ТЛМАСЕАЕ
Основное число хромосом — х =
== 6, 38—16.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный, двух-четырехъядерный та-
петум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, трехборозд-
ные, многобороздные или многопо-, зернами. `
ровые, с крахмальными
Семяпочка анатропная, с двумя
интегументами, красси- или тенуи-‘
нуцеллятная. Микропиле образует
—
ся внутренним интегументом. Име-
отся эндотелий, фуникулярный об-
туратор, карункула и гипостаза.
Семяпочек 40—6 (4).
Макроспорогенез и макрогамето-
генёез — археспорий одно-, редко
двух-многоклетечный. Париетальных
клеток 2—5 рядов. Тетрады макро-
спор линейные. \Зародышевый ме-
шок образуется цо Роубопит-типу
и имеет зерна крахмала. Синерги-
ды с крючкообразными выростами
и фибриллярными аппаратами. Име-
ются три эфемерные антицоды. По-
лярные ядра сливаются до оплодо-
творения. ,
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление. У культур-
ного льна Двойное оплодотворение,
проходит через 5 ч после опыления.
Эндоспермогенез нуклеарный.
Эмбриогенез Зопапад-типа. Заро-
дыш крупный, прямой или согну-
тый, двусемядольный, хлорофилло-
носный, содержащий масло и алей-
рон. Семядоли выполняюТ запасаю-
щую функцию. Подвесок четырех-
девятиклеточный, однорядный.
Семя плод — семя со слаборазви-
тым эндоспермом, содержащим мас-
ло. Зародыш располагается в цент-
ре или по периферии семени и за-
полняет собой почти весь объем се-
мени. Эндосперм в виде тонкого
слоя сохраняется лишь на пери-
ферии семени. Семенная кожура
образуется обоими интегументами,
за исключением Нибопла, у кото-
рой она образуется одним внут-
ренним интегументом. Плод — сеп-
тицидная коробочка или нерас-
крывающийся.
Гауеторий эндоспермальный, об-
разующийся лишь в халазальной
части зародышевого мешка.
Апомиксие—редуцированный пар-
теногенез.
Другие явления — полиэмбриония,
Terepoctunua y Linum.
Литература: Corner, 1976; Davis
66; Дунаева, 1973; Кантор, 1956,
1959, 1964: Модилевский, Дзюбенко,
1966; Магауапа, 19674, 19704; Оста-
пенко, 1970; Поддубная-Арнольди,
1952; Schnarf, 1934; Сизова, 1958а, б
1963; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО JXONANTHACEAE
Основное число хромосом — нет
данных.
?
` мультанное.
Цветок обоеполый или редко од-
нополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет ‘эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и секреторный, двух-.
четырехьядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование’ микроспор си-
Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, трехбороздно-
поровые, с бакулятной экзиной.
Семяпочка анатропная с двумя
интегументами, крассинуцеллятная.
Имеются фуникулярный обтуратор,
ариллусе и гипостаза. Семяпочек
10—2.
Макроспорогенез и макрогамето-
генез — археспорий ’одно-двухкле-
точный. Париетальных клеток 2—3
ряда. Зародьипевый мешок образу-
ется m0; Ро[уропит-типу. Синерги-
ды с крючкообразными выростами'`
и фибриллярными аппаратами. Име-
ются три эфемерные антиподы. По-
лярные ядра`‘сливаются до оплодо-
творения. ,
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез нуклеарный.
Эмбриогенез Зо]апад-типа. Заро-
дыш крупный, прямой, двусемя-
дольный.
Семя, плод — семя со скудным
эндоспермом или без него. Плод —
септицидная или локулицидная но-
робочка или костянка, или кры-
латка.
Гаусторий не найден.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Forman,
1965; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
ERYTHROXYLACEAE
Основное число хромосом— z= 12.
Цветок обычно обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, зрелые пыльцевые
144
зерна трехклеточные, трехбороздно-
поровые.
Семяпочка — анатропная-гемиана-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется внутренним интегументом.
Имеются эндотелий и ариллус. Се-
мяпочек 6—2.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток 2—5 рядов.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро!угопит-типу. Синергиды с фиб-
риллярными аппаратами. Имеются
три эфемерные антиподы. Поляр-
ные ядра сливаются до оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез нуклеарный. Эн-
досперм содержит крахмал, но не
всегда.
Эмбриогенез Зо]апа4-тина. Заро-
дыш крупный, прямой, двусемя-
дольтый, с хорошо развитыми ночеч-
кой, гипокотилем и корешком, хло-
рофиллоносный, содержит алейроп.
Семя, плод — семя с эндоспермом
или редко без эндосперма. Зародыш
занимает 3/4 общего объема семени
или все семя. Эндосперм распола-
гается по периферии семени. Се-
менная кожура образуется обоими
интегументами. Плод костянковид-
ный. Стенка плода дифференциру-
CTCA в эпикарий, мезокарпий и
эндокарпий.
Гаусторий отсутствует.
Апомиксие не обпаружен.
Другие явления - - партенокарния.
Литература: Согпог, 1976; Davis,
1966; Магауапа, 19675, 19706; Rao,
1968; Schnarf, 1931; Смирнова, 1977;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО МАГРТСНТАСЕАЕ
Основное чиело хромосом — z =
= 6, 9—11.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспораптгия образуется
по двудольному тниу и имеет эпи-
дермис, Ффиброзный эндотеций, 2
эфемерных средпих слоя и секре-
торный двух-многоядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
142
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трех-пяти(се-
ми)-борозднопоровые, — олигопорат-
ные или 4—12 (и более)-поратные,
с тонкой экзиной и толстой некзи-
ной, с зернами крахмала.
Семяпочка гемианятропная, с дву-
мя интегументами, крассинуцел-
лятная. Микропиле образуется внут-
ренним или наружпым, или обои-
ми интегументами. Семяпочек не-
сколько.
Макроспорогенез и макрогамето-
генез — археспорий многоклеточ-
ный. Париетальные клетки имеют-
ся. Зародышевые мешки образуют-
ся по Репаеа- и АШит-типам. Име-
ются три эфемерные антиподы. По-
лярные ядра сливаются до оплодо-
„творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
)пдоспермогенез пуклеарный. По-
зднее весь эндосперм, за исключе-
инем таустория, стаповитея KETO
иным. Гаусторий остается свобод-
ноядерным.
Эмбриогенез Зо]апа4д-типа. Заро-
дыш крупный, прямой или согну-
тый, двусемядольный, с хорошо
развитыми гипокотилем и кореш-
ком, бесхлорофилльный, содержит
масло и алейрон. Семядоли выпол-
няют запасающую функцию.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
В процессе развития зародыша эн-
досперм полностью расходуется.
Нлод обычно распадающийся па от-
дольные, часто крылатые орешко-
видные или костянковидные плоди-
ки или реже раскрывающиеся. Ред-
ко плод орех или костянковидный.
Гаусторий эпдоспермальный, об-
разующийся в халазальной части
зародышевого мешка.
Апомиксие — нуцеллярная эмбри-
ония у видов Марша.
Другие явления — партенокарпия
и клейстогамия,
Литература: Corner, 1976: Davis,
1966; Miyashita et al., 1964; Mohini,
Farooq, 1969; Molini et al., 1972—
1974; Schnarf, 1931; Siddiqui, 1968b;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО 2УСОРНУГТАСЕАЕ
Освовнос число хромосом — х =
= 8—13.
Цветок обычно обоеполый, очень
редко однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—8
эфемерных или долго сохраняю-
щихся средних слоя и секреторный
дву-трехъядерный, двухрядный тапе-
тум. Ядра в клетках тапетума час-
то сливаются, в результате чего
образуются полиплоидные ядра.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные,
изобилатеральные. Зрелые пыльце-
вые зерна двух-трехклеточные, трех-
борозднопоровые, реже трехборозд-
ные или многопоровые. Спермии —
клетки многоплазменные.
Семяпочка апатропная, гемитроп-
ная с двумя интегументами, крас-
синуцеллятная. Микропиле образу-
ется внутренним или обоими инте-
гументами. Имеются эндотелий и
гипостаза. Семяпочек 20—25.
Макроспорогенез и макрогамето-
генез — археспорий много-однокле-
точный. Париетальных клеток 1—4
ряда. Тетрады макроспор линей-
ные. Паряду с тетрадами образу-
ются триады. Зародытевый меток
образуется имо Polygontm-rany и
имеет крахмальные зериа. Синперги-
ды © фибриллярными аннаратами.
Имеются три обычно эфемерные
антиподы, но ипогда опи долго со-
храняются. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворение
премитотического типа. У Tribulus
феггез{ 13 слияние гамет происходит
через 15—30 мин после прораста-
ния пыльцы на рыльце.
Эндоепермогенез нуклеарный. Об-
разуется несколько сот свободных
ядер эндосперма, прежде чем меж-
ду ядрами начпут образовываться
клеточные псрегородки. Последние
начинают возникать с микропиляр-
ного конца зародышевого мешка,
и постепенно весь эндосперм ста-
новится клеточным. Эндосперм со-
держит масло и алейрон.
Эмбриогенез Зо]апа4-типа. Заро-
дыш крупный, прямой или согну-
тый, хорошо дифференцированный
на семядоли, гипокотиль и коре-
шок, хлорофиллоносный. Семядоли
выполняют запасающую функцию.
Подвесок однорядный.
Семя, плод — семя с мясистым
или роговым эндоспермом или ре-
же без него. Зародыш заполняет
большую часть объема семени или
весь объем его, в зависимости от
присутствия или отсутствия эидо-
сперма. Семенная кожура образу-
ется обоими интегументами. Плод
обычно — локулицидная или септи-
цидная коробочка или распадаю-
щийся на односеменные мерикар-
MH.
Гаусторий отсутствует.
Апомикеис обнаружена нуцелляр-
ная эмбриония.
Другие явления — полиэмбриония
y Peganum harmala u Zygophyllum
[аБаго. Добавочные зародыши воз-
никают из синергид, клеток нуцел-
луса и подвеска зародыша.
Литература: Беляева, 1972, 1977;
Corner, ‚1976; Davis, 1966; Masand,
1967, 1970; Phatak, 1971; Schnarf,
1934: Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВАГАМТТАСЕАЕ
Основное чиело хромосом — z=9.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
но однодольному типу и имеет эпи-
дермие, фиброзный эпдотеций, два-
три долго сохрапяющихся средних
слоя и сокреторпый двуядерный
тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна двух-
трехклеточные, — трехборозднопоро-
вые, с экзиной более толстой, чем
некзина.
Семяпочка гсмианатропная, с дву-
мя иптегумептами, крассинуцеллят-
пая. Микропиле зигзагообразное,
образуется двумя интегументами.
Эпдотелий окружает зародышевый
мешок. Клетки эндотелия становят-
ся двух-трехъядерными. Семяпочек
обычно 5, реже 10.
Макроспорогенез и макрогамето-
тенез — археспорий одно-четырех-
клеточный. Париетальных клеток
3—4 ряда. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по РоГусопим-типу. Синер-
‘143
гиды с крючкообразными выроста-
ми. Имеются три. эфемерные анти-
поды. Полярные . ядра сливаются
при оплодотворении.
Опыление, прохождение ныльце-
вой трубки и оплодотворение —
нет данных. ,
Эндоспермогенез нуклеарный, но
позднее весь эндосперм становится
целлюлярным.
Эмбриогенез — тип не определен.
_Зародыш прямой, двусемядольный.
’ Семядоли толстые выполняют запа-
сающую функцию.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя,
массивные запасающие семядоли
скрывают остальные части зароды-
ша, лишь кончик корешка едва вы-
ступает из них. Плод — односемен-
пая синкарпная костянка, с очень
толстым эндокарпием, окруженным
сладким маслянистым мезокарпием.
Гаусторий не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis
1966; Смирнова, 1977; Тахтаджян,
1966, 1970.
CEMEMCTBO OXALIDACEAE
Основное число хромосом — z =
= 5—7, 9, 44, 42, 15.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный дву-четырехъядерный тапе-
тум. Ядра в клетках тапетума, час-
то сливаются, в результате чего
ядра становятся полиплоидпыми.
Имеются орбикулы. У клейстогам-
ных цветков фибриллярные утол-
щения в эндотеции не образуются.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные и крестообразные.
Зрелые пыльцевые зерна двух-
трехклеточные, трехбороздные, трех-
борозднопоровые, с тонкосетчатой
или почти гладкой сэкзиной.
Семяпочка анатропная, с двумя
‚интегументами, тенуи- или красси-
нупеллятная. Микропиле образует-
ся обоими интегументами. У В1о-
phyllum наружный — интегумент
длинный, трубчатый. Имеется эндо-
телий. Семяпочек 5 — много.
Макроспорогенез и макрогамето-
144
генез`— археспорий ` одноклеточный.
Париетальных клеток. 72 ‚ряда. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевые мешки образу-
ются по Ро]угопит- -или АШаит-
типам. Синергиды грушевидные, ‚у
ВюорвуПиш с крючкообразными вы-
ростами.. Три антипо дегенериру-
ют вскоре после 9плодотворения.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление и самоопы-
ление у ОхаПз, Порогамия. Двой-
ное оплодотворение.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с периферип зародышевого
мешка. Деление первичного ядра
эндосперма предшествует делению
зиготы.
Эмбриогенез Азега4-типа. Заро-
дыш крупный, прямой, с хорошо
развитыми семядолями, почечкой,
гипокотилем и корешком, хлоро-
филлоносный. Подвесок многокле-
точный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло. Зародыш располагается в
центре семени и занимает 3/4 об-
щего объема его. Периферия семе-
ни заполнена эндоспермом. Оба ин-
тегумента принимают Участие в
формировании семенной кожуры.
Плод — локулицидная коробочка
или редко ягодообразный.
Гауеторий подвесочный.
Апомиксис не обнаружен.
Другие — явления — клейстогамия
и триморфная гетеростилия у Оха-
is.
Литература: Саги!е], 1969; Согипег,
1976; Davis, 1966; Herr et al., 1968;
Krupko, 1971 (1972); Narayana L.,
14967e, 1970e; Schnarf, 1931; Тахта-
mean, 1966, 1970; Veldkamp, 4967;
Bepemaruna, 1965, 1979, 1980; Zenk-
teler, 1965, 1967 (1968).
CEMEUCTBO GERANIACEAE
Основное число хромоеом — х =
== 8—11, 13.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и обычно’ секретор-
пый одно-дву-многоядерный тапетум.
У Егофит и Сегамит обнаружен
амебоидный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные\ — крестообразные.
Зрелые пыльцевые зерна двух-трех-
клеточные, трехборозднопоровые, с
сетчатой экзиной. .
Семяпочка анатропная или кам-
пилотропная или амфитропная с
двумя интегументами, крассинуцел-
лятная. Микропиле формируется
внутренним интегументом. Имеют-
ся нуцеллярный колпачок, обтура-
тор и гипостаза. Семяночек обычно
немного, редко много.
Макроепорогенез и макрогамето-
тенез — археспорий одно-, изредка
многоклеточный. Париетальных клс-
ток’4—6 рядов. Тетрады макроспор
линейные, Т-образные. Зародыше-
вый мешок образуется по Ро]Туро-
пит-типу и имеет зерна крахмала.
Синергиды грушеобразные. Имеют-
ся три обычно скоро дегенерирую-
щие антиподы, но иногда они уве-
личиваются в числе путем митоти-
ческого деления и долго сохраня-
ются. Полярные ядра сливаются
перед оплодотворением.
Опыление, прохождение пыльце-
вой ‘трубки и оплодотворение —
перекрестное опыление и самоопы-
ление. Порогамия. Двойное оплодо-
творение. Пыльцевые трубки сохра-
`няются в течение эмбриогенеза.
Эндоепермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма центро-
бежное. Деление первичного ядра
эндосперма предшествует делению
зиготы. 3
Эмбриогенез Onagrad-, Азегад-,
Зо]апад-типов. Зародыш согнутый;
двусемядольный, хлорофиллонос-
ный, содержит масло и алейрон.
Семядоли складчатые, выполняют
запасающую функцию. Подвосок
мощный, многорядный.
Семя, плод — семя без эндоспер-
ма или реже со слабо развитым эн-
доспермом. Имеется перисперм. За-
роды заполняет все или почти
все семя. Массивные, запасающие
семядоли скрывают остальные час-
ти зародыша, лишь кончик кореш-
ка едва выступает из них. Семен-
ная кожура образуется двумя ин-
10 В. А. Поддубная-Арнольди
тегументами. Плод — коробочка.
Гаусторий подвесочный.
Апомиксие не обнаружен.
Другие явления — гинодиэция у
Erodium u Geranium. о
Литература: Corner, 1976; Davis,
4966; Калягина, 1977а, 6; Kumar,
4977; Narayana, 1967c, 19706; Смир-
нова, 4977; Тахтаджян, 1966, 1970;
Tsai et al., 4973.
СЕМЕЙСТВО ТКОРАЕОГАСЕАЕ
Основное число хромосом — Zz =
o_o
—
‚ Т.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, 1—2 средних
слоя и секреторный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
па двухклеточные, трехборозднопо-
ровые с сетчатой экзиной.
Семяпочка анатропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяпочек 3.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальных клеток нет. Тетрады
макроспор линейные. Зародышевый
мешок образуется по Ро!ухопит-
типу. Синергиды грушевидные. Три
антиподы эфемерные. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез нуклеарный. ‹
Эмбриогенез бо]апад-типа. Заро-
дьзи крупный, прямой, с двумя
массивными семядолями, выполня-
ющими запасающую функцию, хло-
рофиллоносный, , содержит Kpax-
мал, алейрон и мирозин. Сильно,
развитый подвесок. Ядра’ клеток
подвеска у Тгораеот шалаз дости-
гают высокой степени полиплоидии
(до 409 п) вследствие эндомитоза.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Семядоли скрывают остальные час-
ти зародыша, лишь кончик кореш-
ка едва выступает из них. Плоды
распадаются на односеменные ко-
стянковидные, плодики.
Гаусторий подвесочный, очень аг-
рессивный, достигающий плаценты
и проникающий даже в семенную
кожуру и перикарпий.
145
Апомиксие не обнаружен.
Другие явления пе отмечены.
Литература: Corner, 1976; Davis
1966; Narayana, 1967[, 1970f; Schnarf,
1931; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВАТЗАМТМАСЕАЕ
Основное число хромосом — х =
= 6—10.
Цветок обоеполый.
Пыльник — 4 микроспорангия, но
y Impatiens fulva tompxo 2. Стенка
микроспорангия имеет эпидермис,
фиброзный эндотеций, 2—3 эфемер-
ных средних слоя и секреторный
двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, трех-пятибороздные с сет-
чатой сэкзиной.
Семяпочка анатропная с ДВУМЯ
интегументами, телуинуцеллятпая.
Микропиле образустся виутрениим
интегументом. Имеются эндотелий
и гипостаза. Сосудистый пучок до-
ходит до халазы семяпочки. Семя-
почек обычно много.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные, Т-об-
разные. Зародышевые мешки обра-
зуются по Ро]угопит- или АШию-
типу, имеют крахмальные зерна.
Синергиды удлиненные, грушевил-
ные с фибриллярными аппаратами.
Имеются три эфемертияе аптиподы,
Полярные ядра сливаются до онло-
дотворелия.
Опылепие, прохождение пыльце-
вой трубки и оплодотворение —
перскрестное опылеписе и само-
опыление у ГПирайепз Иуа. Поро-
гамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Азегад-типа. Заро-
дыш прямой, с массивными семя-
долями, выполняющими запасаю-
щую функцию, бесхлорофилльный,
содержит амилоид и белок. Подве-
сок короткий.
Семя, плод — семя с очень слабо
развитым эндоспермом или без не-
го. Массивные, запасаютщие семя-
доли скрывают остальные части за-
родыша, лишь кончик корешка едва
выступает из них. Семенная кожу-
145
ра состоит из 2—5 рядов толето-
стенных клеток. Плод — сочная ко-
робочка, раскрывающаяся пятью
створками, реже ягодообразная сеп-
тицидная.
Гауеторий эндоспермальный, ми-
кропилярный и халазальный. Осо-
бенно агрессивный микропилярный
эндоспермальный гаусторий, кото-
рый сильно разветвляется и ветви
которого достигают плаценты. Ха-
лазальный эндоспермальный гау-
сторий развит слабо.
Апомиксис не обнаружен.
Другие явления — клейстогамия у
Impatiens fulva.
Литература: Corner, 1976; Davis,
1966; Narayana, 1965, 1967a, 1970a;
Schnarf, 1934; Смирнова, 1977; Та-
Као, 1968; Тахтаджян, 1966, 1970; Уеп-
kateswarlu, Lakshminarayama, 1957.
«>
СЕМЕЙСТВО ШММАМТНАСЕАЕ
Основное число хромосом — z=).
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, два
эфемерных средних слоя и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, иног-
да изобилатеральные. Зрелые пыль-
цевые зерна двухклеточные, двух-че-
тырехбороздные, с тонкой экзиной.
Семяпочка анатропная с одним
интегументом, тенуинуцеллятная.
Сосудистый пучок доходит до кон-
ца интоегумента. Семяпочек несколь-
ко (2—5).
Макроспорогенез и макрогамето-
генез -- архоспорий многоклеточ-
ный. Паристальиъе клетки отсут-
ствуют. Зародышевые мешки реду-
цированные, тетраспорические Пги-
за- или Адоха-типа, обычно с 5,
реже с 6—7 ядрами. Синергиды
крупные, с крючкообразными вы-
ростами. Изредка наблюдается при-
сутствие 1—2 эфемерных антипод.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Пыльцевые трубки долго сохраня-
ются.
Эндоспермогенез нуклеарный. Эн-
досперм до конца своего существо-
вания остается нуклеарным.
Эмбриогенез Опарга4-типа. Заро-
дыш прямой, двусемядольный, со-
держит алейрон и мирозин. Семя-
доли с вилкообразными выростами
У основания, массивные, выполняю-
щие запасающую функцию, коре-
шок короткий. Подвесок длинный,
однорядный.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Семядоли скрывают остальные час-
ти зародыша, лишь кончик кореш-
ка едва выступает из них. Семен-
ная кожура состоит из 8—10 слоев,
перикарпий состоит из 3—4 слосв.
Плод распадается на односеменные
нераскрывающиеся плодики.
Гаусторий эндоспермальный (ла-
теральный, микропилярный); синер-
ГИиДНныЫЙ.
Апомикеие не обиаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Johri, 1967c, 1970b; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970.
В порядке Сегата]ез семейства
Hugoniaceae, Humiriaceae, Lepido-
botryaceae, Nitrariaceae, Peganaccae,
Hypseocharitaceae, Dirachmaceac, Le-
docarpaceae, Viviaceae wo Bieherstei-
п1асеае очень слабо или почти со-
вершенно не исследованы ЦИТОЙМ -
бриологически. "Таким образом, из
241 семейства, относимых А. Л. 'Гах-
таджяном [1970] к Сегапа!е$, для
10 семейств на основании сущест-
вующих данных невозможно соста-
вить цитоэмбриологические харак-
теристики. Ыпасеае сходно с Егу\-
roxylaceae по строению и развитию
пыльника, мужского , гаметофита,
семяпочки, зародышевого мешка,
эндосперма, зародыша, семени и
плода '[Магауапа, 41970а]. Однако
обтуратор и гипостаза, обнаружен-
ные у Ыпасеае, отсутствуют у Ету-
throxylaceae. B то время как сход-
ство в цитоэмбриологических приз-
наках этих сомейств указывает na
их родство, различия в некоторых
из них оправдывают отнесение их
к двум самостоятельным семейст-
вам [Магауапа, 1970а]. Родство 14-
пасеае с другими семействами Се-
raniales подтверждается данными
цитоэмбриологии. Между Erythro-
xylaceae и другими семействами
Сегат1а]ез также имеется сходство
10*
в отношении цитоэмбриологии, что
указывает на их тесное родство.
Цитоэмбриологические данные но
подтверждают выделения ролов
Ос осозшиз и Тхопапез в oco-
бое семейство Jxonanthaceae, tax
как эти роды очень сходны © Шпа-
сеае и выделять их из этого семей-
ства нет достаточных оснований
[Магауапа, 19704]. Некоторые уче-
ные считают, что по данным цито-
эмбриологии роды Peganum nu Ba-
lanites правильно отнесены к се-
мейству ГугорвуЦПасеае и что это
семейство должно находиться в по-
рядке Geraniales [Masand, 1967].
Масанд '[Masand, 1970] полагает,
uto xota Zygophyllaceae, Balanita-
ссае и ГРерапасеае цитоэмбриологи-
чески HCCKONLKO отличаются друг
от друга, однако эти различия ве
настолько велики, чтобы оба семей-
ства выделить из_ ДугорБуПасеас.
По его мнению, правильнее не вы-
делять роды Ва]апЦез и Ресапишит
в особые семейства, а относить их к
Zygophyllaceae. Oxalidaceae cxonno
c Geraniaceae, Balsaminaceae u Tro-
раео]асеае по следующим цитоэм-
бриологическим признакам: ceKpe-
торному тапетуму пыльников, си-
мультаппому образованию микро-
спор, двухклеточному пыльцевому
зерлу, апатропной с двумя интегу-
ментами семяпочкой, Ро[узопит-ти-
ну зародьпевого мешка, раннему
слиянию полярпых ядер, эфемер-
ным антинодам, порогамии, нукле-
арному эндосперму, Азега4-типу
эмбриогенеза и семени с эндоспер-
мом. Выделение Averrhoa m3 семей-
ства ОхаНдасеае в особое семейство
данные цитоэмбриологии не под-
тверждают [Магауапа, 1970е].
Сегашасеае сходно с ОхаПдассае.
Balsaminaceae и Тгораео]асеае п)
следующим —‘цитоэмбриологическим
признакам: секреторному тапету-
му пыльпиков, симультанному об-
разовапию микроспор, двухклеточ-
пому мужскому гаметофиту, ана-
тропной с двумя интегументами се-
мяпочке, Ро[угопит-типу женского
гаметофита, раннему слиянию по-
лярных ядер, эфемерности антипод,
порогамии, нуклеарному эндоспер-
му и семени с эндоспермом. Вместе
с тем от других семейств Gerania-
1ез, Сегалласеае отличается красси-
нуцеллятной семяпочкой, 2—5 ря-
дами паристальных клеток, нали-
147
чием нуцеллярного жолпачка. Это
семейство может рассматриваться
как более примитивное, чем другие
вышеотмеченные семейства, хотя
сходство в цитоэмбриологических,
цитологических и анатомических
признаках указывает на близкое
родство этих семейств, однако раз-
личия подтверждают отнесение их
к самостоятельным семействам [Ма-
гауапа, 1970].
- Тгорае]асеае в отношении цито-
эмбриологии сходно с ОхаПдасеае,
Balsaminaceae wu Geraniaceae. Оно
отличается от других семейств
лишь наличием агрессивного подве-
сочного гаустория.
Итак, данные цитоэмбриологии
указывают на близкородственные
отношения между Tropaeolaceae,
Oxalidaceae, Geraniaceae u Balsami-
naceae [Narayana, 19706]. Цитоэм-
бриологически Balsaminaceae cxon-
Ho c Oxalidaceae, Geraniaceae u Tro-
раео]асеае, что ‘указывает на их
близкородственные отношения и на
то, что они правильно отнесены к
одному и тому же порядку Сегапл-
аез. Цитоэмбриологические данные
показывают, что включение Lim-
nanthaceae в порядок ` Сегашаез
вряд ли правильно. Это семейство
скорее всего следует выделить в
особый порядок — Шитапаез, так
как от представителей порядка Се-
raniales Limnanthaceae отличается
отсутетвием париетальных клеток
‘и наличием особого тетраспориче-
ского типа зародышевого мешка
[Johri, 1970b].
ПОРЯДОК 59.
POLYGALALES
СЕМЕЙСТВО РОГУСАГАСЕАЕ
Основное чиело хромосом — 5 =
==9,7—12, 45, 47.
Цветок обоеполый.
Ныльник —4 или 2 микроспоран-
гия. Стенка микроспорангия обра-
зуется по двудольному типу и име-
ет эпидермис, фиброзный эндоте-
ций, 1—2 эфемерных средних слоя
и секреторный двуядерный тапе-
тум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
148
>
тетраэдральные, изобилатеральные.
Зрелые ‘`пыльцевые зерна трех-,
двухклеточные, многоборозднопоро-
вые, с тонкой экзиной.
Семяпочка — анатропная-гемиана-
тропная с двумя интегументами
крассинуцеллятная. Микропиле об-
разуется обоими интегументами, но
y Solomonia сашошеп$$ — только
одним наружным. Имеются нуцел-
лярный колпачок, гипостаза и арил-
лус. Семяпочек 1—3, реже 2—46,
Макроепоротенез и макрогаметоге-
нез — археспорий одноклеточный.
Нариетальных клеток 2—10 рядов.
Тетрады макроспор линейные. 3a-
родышевый мешок формируется по
Ро]угопит-типу. Синергиды с крюч-
кообразными выростами и фибрил-
лярными аппаратами. Обычно три
эфемерные антиподы. Реже число их
увеличивается до 7—12 и они долго
сохраняются. Полярные ядра слива-
ются перед оплодотворением или во
время оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез Азега4-типа. Заро-
дыш крупный, прямой, двусемядоль-
пый, с хорошо развитыми почечкой,
гипокотилем и корешком, хлорофил-
лоносный. Подвесок короткий.
Семя, плод — семя с эндоспермом, .
содержит масло, следы крахмала и
алейроновые зерна. Зародыш зани-
мает 3/4 общего объема семени.
Эндосперм занимает периферию с6-
мени. Семенная кожура двуслойная,
образующаяся из наружного интегу-
мента. Плод — септицидная коробоч-
ка, реже орех, иногда крылатый.
Гаусторий не найден.
Апомиксис — обнаружен нереду-
цированный партеногенез у ЕрнтВ!-
zanthes cylindrica.
Другие явления — клейстогамия.
паразитизм.
Литература: Адета, 1966; Corner,
1976; Davis, 1966; Schnarf, 1931; Stee-
nis van, 1968; Смирнова, 1977; Тах-
таджян, 1966, 1970; Терехин, 1977.
Из пяти семейств, относимых
А. Л. Тахтаджяном к порядку Ро]у-
galales, cemelicrsa Trigoniaceae, Voc-
hysiaceae, Krameriaceae a Tremand-
тасеае исследованы цитоэмбриологи-
чески настолько слабо, что по име-
ющимся данным в настоящее время
нельзя составить цитоэмбриологиче-
ские характеристики этих семейств.
Что-касается семейства Роуса]асеае,
то в отношении цитоэмбриелогии оно
обнаруживает сходство с представи-
телями порядка Сегап!а]ез.
НАДПОРЯДОК ХИ.
ARALIA NAE
ПОРЯДОК 60. _
CORNALES (APIALES,
ARALIALES,
UMBELLALES)
СЕМЕЙСТВО МУЗЗАСЕАЕ
Основное число хромосом — х=11.
Цветки полигамные, растения дву-
домные, реже однодомные.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, три
средних слоя и секреторный, дву-
трехъядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
крестообразные, Тетраэдральные.
Зрелые пыльцевые зерна двухклеточ-
ные, трехборозднопоровые, с зерни-
стой сэкзиной. ‚
Семяпочка одна, апатропная, с од-
ним интегументом, тонуинуцеллят-
ная. Имеется нуцеллярный колна-
чок, гипостаза и эндотелий. Сосу-
дистый .- пучок доходит до халазы.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Ро|угопит-типу. Синергиды груше-
видные. Имеются три эфемерные ан-
типоды.
Опыление, прохождение пыльще-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
. Эндоспермогенез нуклеарный.
Эмбриогенез — тип не определен.
Зародыш довольно крупный, прямой,
двусемядольный. Семядоли выпол-
няют ассимиляционную функцию.
Подвесок хорошо развит.
Семя, плод — семя со слаборазви-
тым эндоспермом. Зародыш распо-
лагается в центре семени, а эндо-
сперм по его периферии. В эндоспер-
ме содержатся масло и алейроновые
зерна двух тинов: 1) с друзами ок-
салата кальция и 2) с глобоидами.
Плод — обычно односеменная или
редко двусеменная ‘костянка.
Гаусторий подвесочный.
Апомикеие отсутствует.
Другие явления не обнаружены,
Литература: Corner, 1976; Davis,
1966; Mohana, Rao, 41972a; Schnarf,
1931; Tandon et al., 1971; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО СОВМАСЕАЕ
Основное чиело хромосом — х =
—8— 14.
Цветок обычно обоеполый, реже
однополый.
Пыльник — 4 микроспорантия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
средних слоя и секреторный дву-
трехъядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зер-
на двухклеточные, трехборозднопо-
ровые, с гладкой экзиной. Поры рас-
положены по меридиану.
Семяпочка анатропная, с одним
интегументом, крассинуцеллятная.
Имеются нуцеллярный колпачок,
опдотелий, гипостаза и обтуратор.
Семяпочек 1—4.
Макроспорогенез и макрогаметоге-
нез — археспорий одно — многокле-
точный. Париетальных клеток 3—7
рядов, или они отсутствуют. Тет-
рады макроспор линейные, Г-об-
разные. Зародышевые мешки обра-
зуются m0 Polygonum- u Fritillaria-
типу. Синергиды — грушевидные.
Имеются три антиподы эфемерные
или долго сохраняющиеся. Поляр-
ные ядра сливаются перед оплодот-
зорепием.
‚Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез не определен. Заро-
дыш прямой, двусемядольный, с хо-
рошо развитыми почечкой, гипоко-.
тилем, корешком, хлорофиллоносный.
Семя, плод — семя с мясистым эн-
доспермом, содержащим крахмал и
алейрон. Зародыш располагается в
центре семени и занимает 3/4 его
объема. Эндосперм располагается по
149
периферии семени. Плод — синкарп-
ная костянка или реже ягодообраз-
ный.
Гаусторий отсутствует.
Апомиксие — апоспория,
лярная эмбриония.
Другие явления не обнаружены.
Литература: Сотпег, 1976; Davis,
1966; Chopra. Harjinder, 1965; Kaur,
4967b; 1970b; Mestre, Guignard, 1970;
Schnarf, 1931; CmmpHosa, 1977; Smith,
1973; Тахтаджян, 1966, 1970; УозЬ11-
Ко, 1971, 1976.
нуцел-
СЕМЕЙСТВО САВВУЛСЕЛЕ
Основное число хромосом — х=41.
Цветок однополый, растения дву-
домные.
Пыльник — 4 микроспорапгия.
Стенка- микроспорангия имеет эпи-
дермис, фиброзный эндотеций, одип
офемериый средний слой и секре-
торный одпо-двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образовапие — микроспор си-
мультапноес. Тетрадь микроспор тет-
раэдральпые, крестообразпые. Зре-
лые пыльцевые зерпа двухклеточ-
ные, трех-, реже четырех-пятибо-
роздпопоровые, с крупносетчатой
экзиной и крахмальными зернами.
Семяпочка апатропная с одним-
двумя интегументами, крассинуцел-
лятпая. Имеются пуцеллярный кол-
пачок, фуникулярпый и плацептар-
пый обтураторы и гипостаза. Ce-
мяпочек две. Семяножка длинная.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальных клеток 4—5 рядов.
Тетрады макроспор липейпые. За-
родышевый меитюок образуется но
Ро]угопит-тину. Три антиподы иног-
да подвергаются делению, образуя
5—7 клеток. Яйцеклетка сильно уд-
линяется. Полярные ядра сливают-
ся перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пет
данных.
Эндоспермогенез нуклеарный. Об-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропиллярного копца за-
родышевого мешка, и постепенно
весь эндосперм становится клеточ-
ным.
Эмбриогенез 50]0опа4-типа. Заро-
дыш маленький, прямой, дифферен-
нированный на две семядоли и ко-
150
4
решок. Подвесок длинный,
рядный.
Семя, плод — семя ©е хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. У Саттуа эндосперм
роговой. Зародыш расположен в 0с-
новании семени и обычно занимает
не более 1/4 линейной высоты семе-
ни. Плод одно- или реже двусемен-
ная ягода.
Гауеторий не найден.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
У Саггуа уеакЬ и. Добавочные заро-
дыши образуются из клеток нод-
веска.
Литература: Согпег, 1976; Davis,
1966; Kapil, Mohana Rao, 419665;
Schnarf, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970.
одно-
СЕМЕЙСТВО АГАМСТАСЕАЕ
Основное число хромоеом — х =
==41, 12.
Цветок обосполый.
Ныльник - 4 микросиорангия.
Степка микроспорангия имеет эпи-
дермис, фиброзный эпдотеций, один
офемерпый средпий слой и секре-
торный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — тетрады микроспор изобила-
теральпые. Зрелые пыльцевые зериа
двухклеточные, четырех-пяти-, реже
трехборозднопоровые, с экзиной,
покрытой крупными густо располо-
женными сосочками.
Семяпочка анатропная, с одним
иптегументом, крассинуцеллятная.
Имеются нуцоллярпый колпачок,
эндотелий и гиностаза. Семяпочек
4 ‘)
Макроспорогелез и макрогаметоге-
Hes архоспорий одпоклеточный.
Наристальных клеток 1—2 ряда.
Гетрады макроспор липеойные. Заро-
дышевый мешок образуется по Ро-
1угопат-типу. Синергиды грушевид-
ные. Три ядра антипод дегенериру-
ют, прежде чем образуются клетки,
но иногда наблюдается вторичное
деление и образование до 12 и 60-
лее клеток антипод. Полярные яд-
ра ло оплодотворения не сливаются.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез пуклеарный или
целлюлярный.
Эмбриогенез не определен. Заро-
дыш прямой, хорошо развитый, с
листовидными семядолями и длин-
ным корешком, хлорофиллоносный.
Семя, плод — семя © хорошо раз-
витым эндоспермом, содержащим
сначала крахмал, позднее масло и
алейрон. Плод — односеменная ко-
стянка, увенчанная чашечкой и ди-
ском.
Гауеторий не найден.
Апомиксие — редуцированная апо-
гаметия.
Другие явления не обнаружены.
Литература: Согпег, 1976; Davis,
1966; Eyde, 1968; Kapil et al., 1966b;
Эсвпаг{, 1931; Тахтаджян, 1966, 4970.
СЕМЕЙСТВО АВАМАСЕАЕ
Основное чиело хромосом — x=
—141, 42.
Цветки обосполые, однополые и
полигамные, растепия одно- или
двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный — многоядерный тапетум.
В результате слияния ядра тапету-
ма стаповятся полиплоидными.
Микроеспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточпые, трех- ипогда
двух-четырехборозднопоровые, трех-
борозднопоровые или шестиругат-
ные, с сетчатой экзиной.
Семяпочка висячая, анатропная, с
одним интегументом, обычно крас-
синуцеллятная, изредка тенуинуцел-
лятная. Имеются эндотелий, гипо-
стаза фуникулярный обтуратор.
Наряду с Ффертильными имеются
стерильные семяпочки.
акроспорогенез и макрогаметоге-
нез — археспорий одпоклеточимй.
Париетальпая клетка либо дегепери-
рует, либо делится, образуя ткань,
либо париетальные клетки вовсе но
образуются. Тетрады макроспор ли-
нейные. Иногда вместо тетрады об-
разуется триада макроспор. Зароды-
шевый мешок образуется по Роу-
гопит-типу. Синергиды грушевид-
ные. Имеются три эфемерные анти-
поды, представленные не клетками,
а ядрами. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пере-
крестное опыление и самоопыление.
Эндоспермогенез нуклеарпый.
¥Y Arthrophyllum diversifolium a He-
4ега ВеНх эндосперм руминирован-
НЫЙ.
Эмбриогенез Опастад- или Свепо-
ро1а4-типа. Зародыпшгт маленький,
прямой, но обычно слабо дифферен-
цпированный на две семядоли и ко-
решок, бесхлорофилльный. Иногда
зародыш не дифференцированный.
Семя, плод — семя с эндоспермом,
содержащим белок, алейрон и мас-
ло. Зародыцт располагается в оспо-
вании семени и занимает от 1/4 до
1/2 его объема. Семеппая кожура
топкая, перспопчатая, образующая-
ся из паружного слоя интегумента.
Плод — синкарпная костянка или
ягода, редко вислоплодник.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Артамонова, Иванов,
4969; Богданова, Грушвицкий, 1970;
Сотпег, 1976; Пау1з, 1966; Иванова,
1965; Кордюм, 1966; Вао, 19725; Ре-
riasamy, 1966; Петровская-Баранова,
1959; Schnarf, 1931; Смирпова, 1977;
Тахтаджяп, 1966, 1970.
СЕМЕЙСТВО АРТАСЕАЕ
Основное
—4— 12.
Цветки обычно обоеполыс,
однополые, растения
или двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный дву — многоядерный тапе-
тум. Веледствие слияния, ядра в
клетках тапетума стаповятся поли-
ИЛОИДНыЫМиИ.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультаппое. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, трех-, реже
двух-пятиборозднопоровые, с глад-
кой экзиной.
Семяпочка висячая, анатроппая,
с одним интегументом, обычно те-
нуинуцеллятная. Микропиле очень
длинное. Имеются нуцеллярный кол-
число хромосом — х=
редко
однодомные
151
пачок; гипостаза, фуникулярный об-
туратор / и `эндотелий: Семяпочек
две-одна. Наряду с фертильными
имеются и ‘стерильные семяпочки.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-многоклеточ-
ный. Париетальные клетки отсут-
ствуют. Тетрады макроспор линей-
ные. Зародьышевые мешки образу-
ются по Polygonum-, Allium-, Pe-
паеа- и Огаза-типу. Синергиды ко-
роткие и широкие. Число антипод
варьирует в зависимости от типа
зародышевого мешка. Полярные
ядра сливаются перед оплодотворе-
пием. У Daucus carota, Orlaya gran-
diflora a O. platycerpa описан поста-
мепт.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодотво-
рение. Пыльцевые трубки долго со-
храпяются в семяпочке.
Эндоспермогенез пуклеарпый.
Y Bupleurum tenue формирование
клеточпых перегородок между яд-
рами эндосперма начинается на ста-
дии трех-четырехклеточного заро-
дыша.
Эмбриогенез Зо]апад-типа. Заро-
дыш маленьикй, прямой, слабо диф-
ференцированный на две семядоли
и корешок, бесхлорофилльный.
Семя, плод — семя с эндоспермом,
содержащим белок, алейрон, масло.
Зародыш располагается в основании
семени и занимает от 4/4 до 1/2 ero
объема. Семепная кожура однослой-
пая, образуется из наружного эпи-
дермиса интегумента. Плод — висло-
цлодник или реже костяпка, увепча-
па на верхушке остатком чашечки,
Гаусторий отсутствует.
'Апомикеис — иптегумонтальная
эмбриония.
Другие явления — партенокарнпия.
Литература: Согпег, 1976; Davis,
1966; Jlackanosa, 1977; Dhakre, 1968;
Гамбарова, 1973; Сара, 1967, 1970;
Сара 5. бара М., 1964; Иванова,
41966; Кордюм, 1963, 1964, 1965, 1966,
1967Та, б, в, г, 1968а, 6, в, 1970, 1971;
Кордюм, Веледницкая, 1964; Мезте,
Guignard, 1971; Schnarf, 1931; Смир-
нова, 1977; Sehgal, 1968; Taxtansan,
1966, 1970.
Из представителей Cornales, ce-
мейства Davidiaceae, Aucubaceae,
Melanophyllaceae, Curtisiaceae, Mas-
152
tixiaceae, Griseliniaceae, Taricella-
ceae uw Helwingiaceae цитоэмбриоло-
гически не изучены. Пб данным
цитоэмбриологии род М№узза правиль-
но выделен из Сотпасеае в само-
стоятельное семейство Nyssaceae
[Вао Р., 1972а]. Музза по ряду ци-
тоэмбриологических признаков` от-
личается от Пау11а, которая также
заслуживает ранга самостоятельно-
го семейства Davidiaceae [Rao P.,
1972а]. Систематическое положение
Cornaceae подвергалось постоянной
дискуссии. В настоящее время из
него исключено несколько родов и
отнесено к самостоятельным семей-
ctpam: Alangium «x Alangiaceae, Gar-
rya K Garryaceae, Nyssa k Nyssaceae,
Davidia «x Davidiaceae, Helwingia i
Helwingiaceae u T. J., 4TO согласует-
ся с данными цитоэмбриологии. Вме-
“сте с тем данные цитоэмбриологии
подтверждают — близкородственные
отношения между Araliaceae a Ар!-
aceae. Rao P. [1972a] считает, что
по цитоэмбриологическим данным
АтаПасеае ближе к Арасеае, чем к
Согпасеае, и что оба эти семейства
следует оставить в порядке ОтЬе]-
Ц!егае, а Согпасеае выделить в 0с0-
бый порядок Согпа!ез. Данные цито-
эмбриологии показывают, что се-
мейство ОтЬеШегае, или Ар!асеае,
очепь естественное, монолитное се-
мейство, так как все изученные
представители его отличаются сход-
ными цитоэмбриологическими при-
знаками. Семейство это характери-
зуется наличием прогрессивных ци-
тоэмбриологических признаков. При-
зпаком высокой оргапизации Apia-
сеае является паличие трехклеточ-
пой пыльцы, тенуинуцеллятной с
одпим покровом семяпочки, одно-
клеточного женского археспория,
биспорического и тотраспорическо-
го типов зародышевых мешков.
Представление А. Л. Тахтаджяна
[1960] о том, что Арасеае очень
тесно связано с АгаПасеае, но замет-
но более специализированное, впол-
не согласуется с данными цито-
эмбриологии. Е. Л. Кордюм, наибо-.
лее полно исследовавшая цито.
эмбриологию АгаНасеае и Арласеае,
также считает, что оба эти семей-
ства тесно связапы между собой.
общим происхождением, хотя Ага-
Пасеае — более примитивное семей-
ство, чем Ар1асеае. Вместе с тем она.
полагает, что род Hydrocotyle сле-
дует выделить. из Ар1асеае в/^ само-
стоятельное семейство НудгософУ1а-
сеае’.м равсматривать его как само-
стоятельную линию эволюционного
развития, берущего начало от об-
щих с зонтичными и аралиевыми
предков. Возможно, что Garryaceae
родственно А]апртасеае и Cornaceae,
так как имеет с ними сходные ци-
тоэмбриологические признаки [Ка-
р, Вао, 1966]. В таком случае Gar-
гуасеае следует отнести к порядку
Umbelliferae
1966b].
Род Не\мпола, отпосимый' Хатчин-
coHom [Hutchinson, 1969] к АгаНа-
сеае, по мнению некоторых исследо-
вателей, заслуживает ранга само-
стоятельного семейства Helwingia-
сеае [За®, 1976]. Нам представляет-
ся возможным разделить порядок
Cornales na два порядка: более при-
митивный Согпа]ез и более высоко-
развитый Ар!а|ез.
НАДПОРЯДОЕ ХИ.
СЕТАЗТВА МАЕ
ПОРЯДОК 61.
CELASTRALES
СЕМЕЙСТВО AQUIFOLIACEAE
"” Оеновное число хромосом — х=9,
10, 17.
Цветок — однополый, растения
обычно двудомные.
Пыльник — 4 микроспорангия.
Тапетум секреторный, двуядерный.
Микроепорогенез и микрогаметоге-
нез — микроспорогенез не описан.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка — анатропная-гемиана-
тропная, C одним интегументом,
обычно красси-, иногда тенуинуцел-
лятная. Имеются эндотелий, обтура-
хор и гипостаза. Семяпочек пемного.
акроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальных клеток 1—2 ряда.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]усопат-типу. Синергиды грушевид-
ные. Имеются три долго сохраняю-
щиеся антиподы. Полярные . ядра
сливаются до оплодотворения.
Опыление, прахождение пыльце-
[Kapil, Mohana, Rao, ©
вой трубки и опледотворение — по-
рогамия.
Эндоспермогенез целлюлярный,
Эмбриогенез, — СагуорБуПад-типа.
Зародыш очень маленький, прямой,
‚слабо дифференцированный на две
семядоли и корешок, бесхлоро-
филльный. ,
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Зародыш распола-
гается в основании семени-и зани-
мает не более 1/4 линейной высоты
семени. Весь остальной объем семе-
ни заполнен эндоснермом. Плод —
костянка.
Гауеторий отсутствует.
Апомиксие не обнаружен.
Другие явления не отмечепы.
Литература: Сотгпёт, 1976; Davis,
1966; Herr, 1959; Schnarf, 1931; Cuup-
пова, 4977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ТСАСТМАСЕАЕ
Основное число хромосом — x=
= 10—11.
Цветки обоеполые или реже поли-
тамные, растения однодомные ‚„. или
реже двудомные.
Пыльник — ‘число’ микроспоранги-
ев не установлено. Стенка микро-
спорангия имеет эпидермис, фиб-
ровный эндотеций, 1—2 эфемерных
средних слоя и секреторный пяти-
шестиядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточные, трехборозд-
ные, трехпороратные, безапертур-
ные.
Семяпочка анатропная, © одним
интегументом, обычно красси-, из-
редка тенуинуцеллятная. Семяпочек
Макроепорогенез и макрогаметоге-
нез — археспорий многоклеточный.
Париетальных клеток 5 рядов. Тет-
рады макроспор линейпые. Зароды-
шевый мешок образуется по Ро]у-
сопит-типу и имеет зерна крахма-
ла. Синергиды с крючкообразными
выростами и фибриллярными аппа-
ратами. Имеются три эфемерные
антиподы. Полярные ядра сливаются
до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоепермогенез нуклеарный.
153
Эмбриогенез — тип не определен.
Зародыш маленький, слабо диффе-
ренцированный на две семядоли и
корешок, хлорофиллоносный, пря-
мой или слегка согнутый.
Семя, плод — семя обычно с эндо-
спермом. Зародыш располагается в
осповании семени и занимает от
1/4 до 3/А линейной высоты его.
Плод обычно костянкообразный, од-
носеменной, редко сухой и крыла-
тый.
Гаусторий эндоспермальный, об-
паружен у Nathapodytes Фоейда в
халазальной части зародышевого
мешка.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; Swamy, Ganapathy, 1957; Cump-
нова, 1977; Тахтаджян, 1966, 4970.
СЕМЕЙСТВО ЗАГУАРОКАСЕАЕ
Основное чиело хромоеом — х= 12.
Цветки обосполые, нолигамные
или двудомпыс.
ПНыльник — число микроспорангиев
не определено. Степка микроспо-
рангия имеет эпидермис, фиброзный
эндотеций, 1—2 средних слоя и та-
петум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Зрелые — пыльцевые
зерна двухклеточные, трехбороздно-
поровые.
Семяпочка анатропная с двумя
интегументами, крассинуцеллятная.
Y Salvadora регэса микропиле об-
разуется внутренним, у Атпта бе та-
cantha паружиым иптогумептами.
Семяпочек 2—4.
Макроспорогенез и макрогаметогс-
нез — археспорий одно-, двух- и бо-
лее клеточный. Паристальных кле-
ток 10 рядов. Тетрады макроспор
линейные. Иногда образуются триа-
ды макроспор. Зародышевый мешок
формируется по Polygonum-tury.
Синергиды грушевидные. Имеются
три крупные аптиподы. Полярные
ядра сливаются до или во время
оплодотворепия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нот
данных.
Эндоспермокенез нуклеарный.
Эмбриогенез — тип не определен.
Зародыш крупный, прямой, с двумя
толстыми семядолями, вынполняю-
154
щими запасающую функцию, содер-
жит масло.
Семя, плод — семя без эндосперма.
Зародыш заполняет все семя. Мас-
сивные семядоли скрывают осталь-
ные части зародыша, лишь кончик
корешка едва выступает из них.
Плод — ягода или костянка, в боль-
шинстве случаев односеменной.
Гаусторий отсутствует.
Апомиксис — пуцеллярная
бриония.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; Maheshwari Devi, 1969, 1972;
Смирнова, 1977; Тахтаджян, 1970.
СЕМЕЙСТВО СЕГАЗТВАСЕАЕ
Основное число хромосом — х=8,
12, 23, 40.
. Цветки обоеполые или однополые.
Ныльник — 4—2 микроспоранвгия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
офемерных средних слоя и секре-
торный многоядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образовациие микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные. Зре-
лые пыльцевые зерна двух-трех-
клеточные, трехборозднопоровые, с
сетчатой сэкзиной.
Семяпочка анатропная, с двумя
интегументами, красси- или тенуи-
нуцеллятная. Микропиле образуется
обоими интегументами. Имеются
эндотелий, фуникулярный обтуратор
и ариллус. Семяпочек 2—40, много.
Макроспорогенез и макрогаметоге-
нез — архоспорий одпоклеточный.
Паристальных клеток 1—4 ряда, или
они не образуются. Тетрады макро-
спор линейные, 7-образные. Заро-
дышевый menor формируется по
Polygonum tuny. Синергиды груше-
видные, с крючкообразными выро-
стами у некоторых видов. Три ан-
типоды обычно эфемерные, но y
Gymnosporia spinosa сохраняющие-
ся до оплодотворения. Полярные
ядра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма центро-
стремительное и начинается после
образования около 200 свободных
ядер.
эм-
\
Эмбриогенез 50]апа4- или Сагуор-
руПад-типа. Зародыш круппый, пря-
мой, с хорошо развитыми семядоля-
ми, почечкой, гипокотилем и кореш-
ком, хлорофиллоносный.
Семя, плод — семя © хорошо раз-
витым эндоспермом, содержит мас-
ло, зародыш занимает 3/4 общего
объема семени. Эпдосперм запол-
няет периферию семени. Плод —
локулицидная коробочка, костянка,
крылатка или ягода.
Гаусторий отсутствует.
Апомикесис — нуцеллярная и ин-
тегументальная эмбриония У Се]а${-
rus a Kuonymus.
Другие явления — полиэмбриония
y Celastrus u Euonymus.
Литература: Copeland, 1966; Cor-
ner, 1976; Davis, 1966; Herr, 1959;
Mestre, 1965a; Haymona, 1970a, 9,
49788, 19796; Schnarf, 1931, 1933;
Смирнова, 1977; Тахтаджяи, 1966,
1970.
CEMEMCTBO STACKHOUSIACEAE
Основное число хромосом — х=9,
10, 15.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорапгия имеет эпи-
дермис, фиброзпый эпдотеций, один
эфемерпый средиий слой и секре-
торный двуядерный танетум.
Микроепорогенсз и микрогаметогс-
нез — образование минроспор си-
мультанпое. Тотрады — микросиор
изобилатеральные, тетраэдральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, с зернами
крахмала, трехборозднопоровые с
сетчатой экзиной.
Семяпочка анатропная, с
иптегументами, тенуинуцеллятная.
Микропиле образуется обоими
иптегументами. Имеется гипостаза.
Семяпочек 2—5.
Макроспорогенез и макрогаметоге-
нез — арехспорий одпоклеточный.
Париетальных клеток пет. Тстрады
макроспор линейные, Т-образпые.
Зародышевый мешок образуется по
Ро]угопат-типу. Синергиды груше-
видные. Обычно три, иногда 10—15,
одно- или многоядерных антипод.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия.
Эндоепермогенез нуклеарный. Пос-
двумя
ле образования около 130 свободных
ядер, весь эндосперм стаповится
клеточным.
Эмбриогенез СагуорвуПад-типа.
Зародыш крупный, прямой, с хоро-
шо развитыми толстыми семядоля-
ми, почечкой, гипокотилем и ко-
решком, содержит масло. Подвесок
маленький, короткий.
Семя, плод — семя с эндоспермом,
содержащим крахмал или масло.
Зародыш занимает 3/4 общего объс-
ма семени. Эндосперм заполняет
периферию семени. Наружный эпи-
дермис семенной кожуры содержит
тапнин. Перикарпий дифферепциро-
ван на наружпый топкоклеточный
и впутрепний склерепхиматозный
‚лой Плоды сухие, распадающиеся
па (2) 3—5 мерикарпия. Наружный
опидермис перикарпия сильно кути-
иизирован.
Гаусторий отсутствует.
Апомиксие не обнаружен
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Gulati, 1967d; Schnarf, 1934;
Смирнова, 41977; Тахтаджян, 1966;
1970.
СЕМЕЙСТВО
GEISSOLOMATACEAE
Основное
данных.
Цветок обосполый.
Пыльник — 4 микроспорангия.
Строение стсики микроспорапгия пе
изучено.
Микроепорогенез и микрогаметоге-
нез — типы — микроспорогепеза и
микрогаметогенеза не изучены. Зре-
лые пыльцевые зерна трехбороздно-
поровые, с сетчатой сэкзиной.
Семяпочка апатропная, с двумя
интогументами. Микропиле образу-
ется обоими интегументами. Семя-
ночек 8.
Макроепорогенез и макрогаметоге-
нез — археспории одпоклеточими.
Паристальные клетки имеются. За-
родышевый мешок образуется по
Ро|угопит-типу и имеет крахмаль-
цые зерна. Имеются три эфемерные
аптиподы. Полярные ядра сливают-
ся до оплодотворения. У Се15$0]ота
таготафа описан постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пет
данцых.
Эндоспермогенез — нет данных.
число хромоеом — нет
155
Эмбриогенез — тип пе определен.
Зародыш согнутый, крупный, диф-
ференцированный. Семядоли мас-
сивные, выполпяющие запасающую
фунпкпию.
Семл, плод — семя со скудным эн-
доспермом. Зародыш лежит в цент-
ре ссмепи, занимая почти весь
объем его. Остатки эндосперма со-
храпяются лишь на периферии се-
мени. Плод — коробочка.
Гауеторий не найден.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Dahlgren, Rao, 41969;
Davis, 1966; Schnarf, 1931; CaumpHosa,
. 1977; Тахтаджян, 1966, 1970.
Семейства РуеШтасеае, Siphono-
dontaceae, Goupiaceae u Согупосагра-
сеае, включаемые А. Л. Тахтаджя-
пом [1970] в порядок Celastrales,
цитоэмбриологически пе исследова-
ны, а С61550]отацасеае исследовапо
очень слабо. Цитоэмбриологические
даииые подтверждают родетвештьо
связи АдиЦоПассае и Сфазгасеас,
при этом призпаки первого семси-
ства примитивнее второго [Негг,
1959]. Но мнению Махешвари [Ма-
heswari, Devi, 1972], Salvadoraceae m0
цитоэмбриологии сходно с Се]азйта-
ceae. Stackhousiaceae no цитоэмбрио-
логическим признакам ближе к Се-
lastraceae, uem к Scrophulariaceae,
Lobeliaceae mum Selaginaceae [Gu-
lati, 1967a]. HecmorpaA Ha TO что по-
panox Celastales 8B отношении цито-
эмбриологии исследован пока нелдо-
статочно, все же между отдельпы-
ми его представителями (Ади Пойа-
ceae, Icacinaceac, Salvadoraceae, Cc-
lastraceae nm Stackhousiaceac) имеют-
ся CXOJUIDIO цитоэмбриологические
признаки, что ие противорочит от-
иесению их в обиий порядок. Од-
нако требуются дальнейшие исслс-
дования для определения естествеп-
пых границ Се]азга]ез и выяснения
родственных связей между его пред-
ставителями.
ПОРЯДОК 62.
RHAM NALES
СЕМЕЙСТВО ВНАММАСЕАЕ
Основное чиело хромосом — 7 =
== 10—13.
Цветок обоеполый, реже однопо-
лый,
156
Пыльник — число микроспоранги-
ев не определено. Стенка микроспо-
рангия образуется по базальному
типу и имеет эпидермис, фиброз-
ный эндотеций, 2—3 эфемерных
средних слоя и секреторный двух-
четырехъядерный тапетум. Имеются
орбикулы.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерпа двухкле-
точные, — трехборозднопоровые, с
гладкой экзиной.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллят-
ная. Микропиле образуется внут-
ренним или наружным интегумеп-
том. Имеются нуцеллярный колпа-
чок, гипостаза и эпистаза. Семяпо-
“ чек песколько (5—10).
Макроепорогенез и макрогаметоге-
нез — археспорий одно — многокле-
точпый. Париетальпых клеток 41%
рядов. Тотрады макроспор липей-
ные, Г-образные. Зародьлтевые меш-
‹и образуются wo Polygonum- u Alli-
и1-тинам. Часто образуется He-
сколько зародышевых мешков внут-
ри одной и той же семяпочки. Си-
нергиды с крючкообразными выро-
стами и фибриллярными аппарата-
ми. Имеются три антиподы или бо-
лее (одно-четырехъядерные) скоро
дегенерирующие или долго сохраия-
ющиеся. Полярные ядра сливаются
во время оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворение.
Пыхльцевыюе трубки долго сохраня-
OTC,
Эидоскермо:снез пуклеарный. За-
NOFKCHMUE цклоточных перегородок
между ядрами эпдосперма пачипа-
ется с мпкропилярного копца заро-
дышевого мешка. У Заша и 1тур-
Вуз халазальный копец эндосперма
остается свободноядерпым. Имеется
перисперм.
Эмбриогенез обычно Азегад-, ред-
ко бо]апад-типа. Зародыш крупный,
хлорофиллоносный, прямой или со-
тпутый, дифференцированный па
две семядоли, почечку, гипокотиль
и корешок. Семядоли выполняют
запасающую функцию. |
Семя, плод — семя с маслянистым
эндоспермом или без него. Зародыш
располагается либо в центре семени,
либо по его периферии и завимает
часть объема семепи или зесь его
объем, в зависимости от присут-
ствия или отсутствия эвдоснерма.
Семепная кожура мнпогорядпая, об-
разующаяся из обоих иитегументов.
Плоды костяпковидные, орсхозид-
ные или распадающиеся па мери-
карпии.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления — партенокарпия
и полиэмбриония (у Zizyphus).
Литература: Corner, 1976; Davis,
1966; Эрккенова, 19726; Гагипа, Co-
cuccii, 1971; Nair, 1967d, 1970c; Pau-
кова, Эрккенова, 41973; Schnarf, 1934;
Тахтаджян, 4966, 1970.
СЕМЕЙСТВО VITACKHAL
Основное число хромосом д
10—14, 16, 19, 20.
Цветок обоеполый или одпополый,
растения одпо- или двудомные.
Пыльник 4—2 микроспорангия.
Стенка микроспорангия образуется
по базальному типу и имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средпих слоя и секретор-
ный двух-восьмиядерпый тапетум.
Микроспорогенез и микрогаметогс-
нез — образование микроспор си-
мультанное. Тетралы микросиор тет-
раздральные, изобилатеральные, KPO-
стообразвые. Зрелые иыльцевые зер-
па двухклеточиые, трохбороздионо-
ровые с гладкой сокзипой.
Семяпочка апатроппая, с двумя
интегументами, крассивуцеллятная.
Микропиле образуется внутренним
интегументом. Имеются нуцелляр-
ный колпачок, гипостаза, эпистаза и
обтуратор. Семяпочек несколько (4).
Макроспорогенез и макрогаметоге-
нез — археспорий одно-двухклеточ-
ный. Париетальная ткань массивная.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]угопаш-типу. Сипергиды © крючко-
образными выростами. Имеются три
эфемерные аптиноды. Поляриые яд-
ра сливаются до оплодотворения.
Опыление, прохождение лыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия. Двойное оплодо-
творение. Сингамия и тройное слия-
ние происходит одновременпо. У Vi-
из уаЦега оплодотворение происхо-
дит через 24 ч после опыления,
а деление оплодотворепных жен-
ских ядер — через 24 ч после опло-
дотворения.
Эндоспермогенез нуклеарный,
Позднее эндосперм становится цол-
люлярным и руминированным.
Эмбриогенез Азега-типа. Заро-
дыш маленький, дифференцировав-
ный на две семядоли и корешок,
бесхлорофилльный.
Семя, плод — семя с роговым эн-
доспермом, содержащим — масло,
алейроновые зерна и друзы оксалата
кальция. Зародыпт располагается в
основанпии семени и занимает не
больше 1/4 линейной высоты семе-
ни. Остальной объом семени занпол-
няет эпдосиерм. Оба иптогумепта
участвуют в формировании семеп-
ной кожуры. Плод — мясистая соч-
пая ини сухая ягода.
Гаусторий — зародышевого мешка.
Апомикеис не обнаружен.
Другие явления — партепокарпия.
Гинодиэция у У!5. Клейстогамия.
Литература: Баранов, 1946; Cor-
ner, 1976; Davis, 1966; Маш, 1965,
1967е, 19704; Новикова, 1978; Маг,
Bajaj, 1966; Periasamy, 1962; Самве-
лян, 1973; Симонян, 1965; Симонян,
Самвелян, 1965; ЭсвпатЁ, 1931; Смир-
нова, 1977; Tarnavschi, Petria, 1968;
Гахтаджяли, 1966, 41970; Tradgernxo,
1958, 1961; Якимов и др., 1977, Воло-
совцев, 1967.
ЕМЕЙСТВО ГЕЕАСЕАЕ
Основное число хромосом — нот
данных.
Цветок обоеполый.
Ныльник — число микроспорангиев
не установлено. Стенка микроспо-
рангия имеет эпидермис, фиброзный
эпдотеций, 2 средних слоя и секре-
торный многоядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образовапие микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые — пыльцевые
зерна трехклеточные, трехбороздно-
поровые, с сетчатой сэкзиной.
Семяпочка апатропная, с двумя
иптегументами, крассинуцеллятная.
Микрониле образуется обоими инте-
гументами. Имеются нуцеллярнын
колпачок, обтуратор и гипостаза.
Семяпочек 3—8.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. 3За-
157
родышевый мешок формируется по
Ро]угопит-типу. Синергиды сходны
с яйцеклеткой. Присутствуют три `
эфемерные антиподы. Полярные
ядра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — но-
рогамия. -
Эндоспермогенез нуклеарный, ру-
‘минированный. В клетках эндоспер-
ма имеются друзы оксалата каль-
ция.
Эмбриогенез Азегад-типа. Заро-
Jbl маленький, двусемядольный.
Семя, плод — семя с эндоспермом,
содержащим масло. Плод — ягода.
Гаусторий отсутствует.
Апомикеие не обнаружен.
Другие явления пе отмечены.
Литература: Nair, 1967, 1968,
1970а; Тахтаджян, 1966, 4970.
Все три семейства, относимые
А. Л. Тахтаджяпом к Rhamnales,
довольно хорошо изучены в отпоше-
пии цитоэмбриологии и имеют мпо-
го сходных цитоэмбриологических
признаков, что указывает на род-
ственные отношения этих семейств
и на естественность порядка. ВВают-
пасеае и УЦасеае цитоэмбриологи-
чески сходны между собой по сле-
дующим признакам: строением стен-
ки микроспорангия, двухклеточным,
трехбороздным пыльцевым зерном,
апатропной крассинуцеллятной се-
мяпочки с двумя интегументами,
наличием нуцеллярного колпачка и
гипостазы, Роугопит-типом › заро-
дышевого мешка, пуклеарным эндо-
спермом и Азчетад-типом эмбриоге-
mesa [Nair, 1970a].
Обравовапие семепи у Vitaccac
©ходно с Аппопасеае [Nair, 1967b].
Leea относилась к УЦасеае, по те-
перь выделена в самостоятельное
семейство, что согласуется с данны-
ми цитоэмбриологии [Машт, 19675].
Цитоэмбриологические данные со-
тласуются с отнесением Гееа в са-
мостоятельное семейство Leeaceae,
“так как, несмотря на сходные с \!1-
(асеае цитоэмбриологические при-
знаки, род этот отличается от УЦа-
<сеае отсутствием гаустория зароды-
шевого мешка. Несомненно, что оба
эти семейства тесно родственны и
должны относиться к одному и то-
му же порядку, а именно к ВЗаш-
nales [Nair, 1970a].
158
ПОРЯДОК 63.
OLEALES
СЕМЕЙСТВО ОГЕАСЕАЕ
Основное чиело хромосом — х =
= 11—14, 22, 23, 24.
Цветок обоеполый и однополый,
растения однодомные и двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средпих слоя и секре-
торный многоядерный, иногда дву-
рядный тапетум. Ядра в клетках
тапетума сливаются и становятся
полиплоидными.
Микроспорогенез и микрогаметоге-
нез — образование микроспор
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
`ипогда Г-образные и линейные. Зре-
лые пыльцевые зерна двухклеточ-
ные, обычно трехборозднопоровые,
или трехбороздные с сетчатой сэк-
зипой. Спермии — клетки, яспо вы-
ражоплые. У ясепя жизнеспособ-
ность пыльцы сохрапяется до одно-
го-двух месяцев.
Семяпочка анатропная амфитроп-
пая, с одним интегументом, тенуи-
пуцеллятная. Имеются эндотелий и
гипостаза. Семяпочек 4—2, 8 — много.
Макроспорогенез и макрогаметоге-
нез — археспорий одно много-
клеточный. Цариетальных клеток
нет. Тетрады макроспор линейные,
Г-образные. Зародышевые мешки
образуются по Polygonum- u.Alliim-
типу, содержат крахмал. Синергиды
имеют крючкообразпые ° выросты.
Образуются три эфемерные аптипо-
ды. Полярные ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестпое опыление. Порогамия.
Двойное оплодотворение. У ясеня
от опыления до оплодотворения про-
ходит 2—8 недели. Тройное слияние
и сингамия происходят одновремен-
но. После оплодотворения первым
делится первичное ядро эндосперма.
Эндоспермогенез целлюлярный.
Эмбриогенез 50]апа4- или Сагуор-
ВуПад-типа. Зародыш прямой, дву-
семядольный, бесхлорофилльный,
содержит масло и алейрон. Подве-
сок ДЛИННЫЙ.
Семя, плод — семя с эндоспермом,
>
содержащим масло, ` алейроновые
—
СИ-,
и
м t
А
зерна, кристаллы оксалата кальция.
Зародыш занимает от 1/4 до 3/4
общего объема семени. Кожура се-
мени тонкая. Плод — локулицидная
коробочка, крылатый орех, ягода или
костянка, часто односеменный.
‚ Гауеторий отсутствует.
Апомикеис не обнаружен.
Другие явления — полиэмбриония
y Fraxinus, Jasminum.
Литература: Беляева Л., Фурса,
1978; Беляева Л. и др., 1977Та; Сог-
ner, 1976; Рау1з, 1966; Иванов, Ша-
ренкова, 1969а, 6; Kapil, 1967f; Ka-
ри, Уап1л, 4966; Кулиева, 1966, 1969,
1972; Литвиненко, Шпилевой, 1978;
Maheshwari D., 1975; Mazzolani, 1964,
1963; Николаева 3., 1962a, 6, B, 1963,
1966; Pacini, 1977; Шаренкова, 1969;
Schnarf, 1931; Тахтаджян, 1966,
4970.
По многим цитоэмбриологическим
признакам порядок О]еа]ез сходен с
Celastrales ma Rhamnales. Для пред-
ставителей этих порядков характер-
но наличие: 4 микроспорангиев, че-
тырех-пятислойных стенок микро-
спорангия, секреторного тапетума,
симультанного образования микро-
спор, двухклеточного мужского га-
метофита, анатроппой крассинуцел-
лятной семяпочки с двумя иптегу-
ментами, одноклеточпого женского
археспория, Роубопат-типа sapo-
дышевого ›мешка;, нуклеарпого эпдо-
сперма, семян с эпдоспермом, диф-
ференцированного зародыша.
ПОРЯДОК 64.
SANTALALES
СЕМЕЙСТВО ОГАСАСЕАЕ
Основное чиело хромосом — =
—19, 20.
Цветок обычно обоеполый, рас-
тения очень редко двудомные.
`Пыльник — 4 микроспорапгия.
Стенка микроспорангия имеет эпи-.
дермис, два слоя ' фиброзного эндо-
теция, 2—3 средних слоя и секре-
торный одно — многоядерный, иног-
да двурядный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор. си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двух-трехклеточные, трехбо-
розднопоровые, трехбороздные или
трех-восьмипоровые.
Семяпочка анатропная, недиффе-
ренцированная, с одним или двумя
интегументами. Нуцеллус подверга-
ется редукции, между ним и инте-
гументами исчезают и органичные.
линии. Одпако ошибочно считать,
что у Оасасеае интегументы отсут-
ствуют. Микропиле длинное. Семя-
почек несколько (2—5).
Макроепорогенез и макрогаметоге-
нез — археспорий одно-четырехкле-
точный. Париетальные клетки отсут-
ствуют. Тетрады макроспор линей-
ные, Г-образные. Зародышевые меш-
ки образуются по. Ро]угопат- u Al-
Поп-типу и имеют крахмальные
зерна. Синергиды с крючкообразны-
ми выростами и Ффибриллярными
аппаратами. Имеются три эфемер-
ные антиподы. Полярные ядра сли-
ваются до оплодотворения. Зароды-
шевые мешки сильно удлиняются и
доходят до столбика.
Опыление, прохождение пыльцке-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение..
Эндоспермогенез целлюлярный
или гелобиальпый.
Эмбриогенез — тип но определен.
Зародыш маленький, прямой, диф-
ферепцировапный па две семядоли
и корешок, хлорофиллопосный, со-
держит масло и алейрон. Крахмал
присутствует или отсутствует.
Семя, плод — семя с хорошо раз-
витым эндоспермом, с крахмалом
или без крахмала. Зародыш распо-
лагается в основании семени и за-
нимает от-41/4 до 1/2 его объема.
Семенная кожура отсутствует. Пло-
ды почти всегда односеменные..
обычно костянковидные, иногда за-
ключенные в разросшуюся чашечку..
Гауеторий — эндоспермальный, ха-
лазальный с одним или несколькими
ядрами и зародышевого мешка.
помиксис пе обпаружен.
Другие явления — паразитизм.
Литература: Согпег, 1976; Davis,.
1966; Ram, 1967b, 1970b; Rao K., Shi-
уагапцап, 1978; Смирнова, 1977; Тах--
таджян, 41966, 1970.
СЕМЕЙСТВО ОРПЛАСЕАЕ
Основное
=—10.
число хромосом — 5 =
159:
Цветки обычно обоеполые, реже
двудомные.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фибриллярный ` эндотеций,
1—2 эфомерных средних слоя и сек-
реторпый дву — многоядерный та-
потум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанпое. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна двухклеточпные, трехбороздно-
поровые.
Семяпочка одна, висячая, анатроп-
пая, с одним иптегументом, тенуину-
пеллятная. Нуцеллус сильно реду-
цировап, микропиле очень узкое.
У Аропапага семяпочка не диффе-
ренцированная.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париестальные клетки отсутствуют.
Тетрады marpocnop липейные. За-
родьпиевый мепюк Polygonum -rima
и содержит крахмальпые зерна. Си
нергиды с крючкообразными выро-
стами и фибрилляриыми аппарата-
ми. Имеются три эфемерные антипо-
ды. Полярные ядра сливаются до оп-
лодотворения. Зародышевый мешок
сильно удлиняется, изгибается и до-
стигает семяножки, принимая U-
образную форму.
Опыление и оплодотворение — нет
данпых.
Эндоспермогенез целлюлярный.
Эмбриогенгз — тип не определен.
Зародыш маленький, прямой, диф-
ференцированный па две семядоли
и корешок. Подвесок мпогоклеточ-
ный.
Семя, плод — ссмя с хоропо раз-
витым эплоспермом. Зародыш pac-
полагается в осповании семепи и за-
нимаст от 1/4 до 1/2 его объема. Се-
менная кожура отсутствует. Плод
костяпковидный.
Гаусторий эндоспермальный, в
холазальной части зародышевого
мешка и является одноядерным.
Первичный гаусторий образует вто-
ричный, который ветвится и ветви
которого доходят До сосудистого
пучка. Имсется также гаусторий
зародышевого мешка.
Апомикеис пе обнаружен.
Другие явления — паразитизм.
Литература: Corner, 1976; Davis,
1966; Ват, 1967е, 1970е; Смирнова,
150
1977; Swamy, Daynand, 1963;
таджян, 1966, 1970.
Тах-
СЕМЕЙСТВО
ERYTHROPALACEAE
. Основное число хромосом — пет
дапных.
Цветок обоеполый.
Пыльник — пот дасепых,.
Микроепорогенез и микрогаметоге-
нез — пет данпых. Зрелые пыльце-
выс зерна трехбороздные.
Семяночка анатропная, с одним
интегументом, тепуинуцеллятпая,
Имеется эндотеций. Семяпочек 3—2.
Макроспорогенгз и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор липейпые. За-
родьшшевый мешок образуется по
Ре!угопит-типу и имеет зерна крах-
мала. Синергиды — грушевидные.
Имеются три малепькие антиподы.
Опыление, прохождение пыльце-
пой трубки и оплодотворение — нет
данных.
дидоепермогснез - ист даппых.
‚мориогенез — тии пе определен.
Зародыш маленький, прямой, диф-
ференцированный па семядоли и
корешок.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш pac-
полагастся в осповании семени и
занимает 1/4 его объема. Плод —
костянка.
Гаусторий не найден.
Апомикеие не обнаружен.
Другие явления пе отмечены.
Литература: Davis, 1966; Смирно-
ва, 1977; Гахтаджян, 4966, 1970.
CEMERCTBO SANTALACEAE
Основное чиело хромосом — z= 10,
42,15, 49.
Цветох обосполый или одпополый,
растения однодомные или двудом-
ные.
Пыльники — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
и более эфемерных средпих слоя и
секреторный одно — многоядерный
тапетум. Ядра в клетках тапетума
часто сливаются и становятся поли-
плоидными.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, крестообразные, изо-
билатеральные. Зрелые пыльцевые
зерна двух- трохклеточные, трех-
апертурные с гладкой экзиной.
Апертуры разного типа.
Семяпочка висячая или прямая,
апатропная-гемианатропная, с од-
ним интегументом, тенуинуцеллят-
ная или недифференцированная.
Нуцеллус редуцированный. Микро-
пиле удлиненное. Семяпочек 1—3,
реже 4—5.
Макроеспорогенез и макрогаметоге-
нез — археспорий обычно — много-,
редко одпоклеточный. Париеталь-
ные клетки отсутствуют. Тетрады
макроснор линейные. Зародышевый
мешок обычно образуется по Роу-
gonum-runy, но у Висеуа и Тод-
na—uo АПцим-тину. — Сипергиды
имеют крючкообразпьые выросты и
фибрилляриые аипараты. Сиперги-
ды сильпо удлипяются, достигая 2/3
столбика (1200 мм) и долго сохра-
няются. Имеются три антиподы.
Они либо эфемерные, либо долго со-
храняются. Полярные ядра сливают-
ся перед оплодотворением. Зароды-
шевые мешки сильно удлиняются
и пропикают в столбик, они содер-
Hat крахмальные зерпа.
Опылепиес, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойпое оплодотворенио.
Пыльцевые трубки долго сохраия-
ются впутри семяпочки.
Эндоспермогенез целлолярпый
или гелобиальный (М!4а, Защаата).
Эмбриогенез нерегулярный. Заро-
дыш маленький, прямой, дифферен-
цировапный на две семядоли и ко-
решок, бесхлорофилльный, богатый
маслом и алейроном. Подвесок при-
сутетвует или отсутствует.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
хлорофилл, зародыш располагается в
основании семени, занимает от 1/4
до 1/2 его объема. Перикарпий диф-
ферепцироваи на три зопы: 1) па-
ренпхимпый опикарпий, 2) камепи-
стый мезокарпий и 3) парснхимпый
эндокарпий. У ВисМеуа перикарпий
пе дифференцирован. Плод opexo-
или костянковидный.
Гаусторий синергидный, аптипо-
дальный, зародышевого мешка и эн-
доспермальный в халазальной ча-
сти эндосперма. У То4та последний
очень агрессивный, сильно развет-
вленный, с ядром до 600 мм. У The-
11 В. А. Поддубная-Арнольди
зат ядра гаусториев достигают вы-
сокой степени полиплоидии (42n —
384") в результате эндомитоза.
Апомикеие не обнаружен.
Другие явления — паразитизм, по-
лиэмбриония у Ехосагриз и ВисК]еуа.
Добавочные зародыши образуются
вследствие пролиферации клеток
подвеска зародыша.
JIurepatypa: Bhatnagar S., 1965,
1966, 1967b, 1970; Bhatnagar, Sabhar-
wal, 1966, 1968; Corner, 1976; Davis,
1966; Johri, Agarwal, 1965; Johri,
Bhatnagar, 1969; Raj, 1967, 4970b,
1972; БебпаеЁ, 1931; Смирнова, 1977:
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
MISODENDRACEAE
Основное число хромосом — 7=8,
9, 40, 42, 44.
Цветок обоеполый, растения дву-
домные.
Пыльник —2 микроспорангия.
Микроспорогенез и микрогаметоге-
нез — нет данных. Зрелые пыльце-
вые зерна 4—12-поровые, с шипова-^
той экзиной.
Семяпочка гемианатропная-геми-
кампилотроппая, не дифференциро-
ваппая па пуцеллус и иптегумент,
тепуипуцеллятная. Семяпочек 3.
Макроспорогенез и макрогаметоге-
иез — археспорий одпоклеточный.
Паристальные клетки отсутствуют.
Готрады линейпыс, Т-образные. За-
родышевый мешок образуется по Ро-
]усопит-типу и имеет изогнутую
форму. Синергиды грушевидные,
Имеются три эфемерные антиподы.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез — тип пе определен.
Зародьш маленький, прямой, диф-
ферепцированпый па две сомядоли
и корешок, хлорофиллоносный.
Семя, нлод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Зародыш располагается в
основании семени и занимает от 1/4
до 1/2 его объема. Плод ореховид-
ный, обычно односеменный, реже
двух-трехсеменный. Семенная кожу-
ра отсутствует.
Гаусторий эндоспермальный, рас-
положенный в халазальной части за-
161
родышевого мешка. Он сильно удли-
няется и достигает плаценты.
Апомиксис не обнаружен.
Другие явления — полиэмбриония,
паразитизм.
Литература: Пау1з, 1966; Ram,
1967a, 1970a; Schnarf, 1931; Cmuupno-
ва, 1977; Тахтаджян, 11966, 1970.
CEMEMCTBO LORANTHACEAE
Основное число хромосом — < =
= 8—12. .
Цветок обоеполый или однополый.
Пыльник — 4,2 или 1 микроспо-
рангий. Стенка микроспорангия 00-
разуется по однодольному типу и
имеет эпидермис, фиброзный эндо-
теций, 1—2 эфемерных средних слоя
и‘ секреторный одно-четырехъядер-
НЫЙ, двух-трехрядный тапетум.
У Hyphear тапетум амебоидный.
Имеются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образовапие микроспор си-
мультаппое. Тетрады — микроспор
тетраэдральныс, крестообразиые.
Зрелые пыльцевые зерна двухкло-
точные, трехборозднопоровые или
трехбороздные © крахмальными зер-
нами, с сетчатой или шиповатои эк-
зиной.
Семяпочка — типичных семяпочек
нет. Вместо них из лопастей пла-
центы образуются сосочки (таште-
]0п). Имеется гипостаза.
Макроспорогенез и макрогаметоге-
нез — археспорий много-, одно-дву-
клеточный. Париетальные клетки
отсутствуют. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро]угопит-тину, содер
крахмал. Зародьпиевые метки иря
мые или согиутые, О-образные,
сильно удлипяются, достигая стол-
бика- и даже рыльца. Одии и тот
же зародышевый мешок может до-
стигать 48 мм. В одной и той же
семяпочке может — образоваться
несколько зародышевых мешков.
В этом семействе зародышевые меш-
ки самые длинные для покрытосе-
менных. Синергиды с крючкообраз-
ными выростами и фибриллярными
аппаратами. 3 или 2 антиподы деге-
нерируют на ранней стадии разви-
тия эндосперма. Полярные ядра
сливаются до оплодотворения.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
162
Эндоспермогенез целлюлярный.
В результате слияния эндосперма
нескольких зародышевых мешков
образуется сложный эндосперм.
Эмбриогенез Piperad-tuna. Sapo-
дыш маленький или большой, диф-
ференцированный на семядоли и
корешок, ложнооднодольный, хлоро-
филлоносный, содержит масло, алей-
рон, иногда крахмал. Подвесок очень
длинный.
Семя, плод — семя обычно с хо-
рошо развитым эндоспермом, содер-
жащим масло, алейрон, иногда
крахмал. Зародыш располагается в
основании семени и занимает от
1/4 до 1/2 его объема. Остальной
объем семени заполняет эндосперм.
Плод — ягода, реже костяпковид-
ный, односеменной, реже двух-трех-
семенной. Болышой зародыш распо-
лагается в центре семени, заполняя
семя по всей его длине. Эндосперм
в этом случае занимает периферию
семени. В эндосперме содержится
хлорофилл. Перикарний имеет 4
или о различных зон.
Гауеторий эндоспормальпный в ха-
лазальной части зародышевого ме-
шка. Имеется также и гаусторий
зародышевого мешка.
Апомикеие не обнаружен.
Другие явления — полиэмбриония,
паразитизм.
Литература: Bathnagar, Chandra,
1968; Bhandari, Indira, 1969; Bhan-
dari, Nanda, 1968a, b; Davis, 1966;
Garg, 1959; Johri, Bhatnagar, 1969;
Johri, Prakash, 1965; Johri, Raj,
1969; Prakash, 1968; Raj, 1967, 1970a;
Schnarf, 1931; Tainter, 1968; Taxtaz-
экг, 1966, 1970; Терехин, 41977.
СЕМЕЙСТВО СУМОМОВТАСЕАЕ
Осповное число хромосом — х==12.
Цветки полигамные.
Пыльник — число микроспноранги-
ев не установлено. Стенка микро-
спорангия имеет эпидермис, фиб-
розный эндотеций, один средний
слой и секреторный двуядерный,
одно-двухрядный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трехбороздноно-
ровые с сетчатой экзиной.
Семяпочка одна, висячая, орто-
тгемитропная, анатропная, с одним
массивным интегументом, крассину-
цсллятная. Имеется пеевдонуцелляр-
пый колпачок.
Макроспорогенез и макрогамето-
"енез — археспорий одноклеточный.
Нариетальные клетки отсутствуют.
'Готрада макроспор Т-образная. За-
родышевый мешок образуется по
Polygonum-tuny. Имеются три, ино-
тда больше (до шести), долго со-
храняющиеся антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез целлюлярный.
Первое деление поперечное.
Эмбриогенез нерегулярный. Заро-
дыш недифференцированный, округ-
ло-линзовидной формы. Подвесок
отсутствует.
Семя, плод — семя с эндоспермом.
Плод ореховидный.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления — паразитизм.
Литература: ЗсбпатЕ, 1931; Тахта-
джян, 1966, 1970; Терехин, Никити-
чева, 1975; Терехин, Никитичева,
Яковлев, 1975; Терехин, Яковлев,
Никитичева, 1975.
СЕМЕЙСТВО
BALANOPHORACEATE
Основное чаело хромосом г -
— 1: 15.
Цветки
НОЛОМНЫе
Иильник л микроспорапгия.
Crouka микроснорангия образуется
10 дДвудольному типу и имеет 2
эфемерных субэпидермальных слоя
и секреторный тапетум. Эндотеций
ис дифференцирован.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трех-пятиборозд-
ные или трехмногопоровые.
Семяпочка не дифференцирована
на нуцеллус и интегументы, пред-
ставляет собой сосочкообразный вы-
рост на плаценте. Семяпочек 1—8.
Макроспорогенез и макрогамето-
гепез — археспорий одноклеточный.
Зародьшшевые мешки образуются по
Polygonum- и АШит-типу. Наблю-
даются зародышевые мешки с об-
ратепной полярностью.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
11*
однополые, растепия оД-
Эндоспермогенез целлюлярный.
Эмбриогенез — тип © неопределен-
ной ориентацией деления зиготы.
Зародыш маленький, недифферен-
пированный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Зародыш располагается в
основании семени и занимает 1/4
линейной высоты семени.
Гауеторий эндоспермальный, в
халазальном конце эндосперма.
Апомиксеие — нередуцированный
партеногенез и генеративная апо-
спория у видов Ваапорвога и Не-
losis.
Другие явления — полиэмбриония
у Ва апорбога globosa u _ Helosis
guianensis. Паразитизм.
Литература: Пау!з, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970; Tepé-
хин, Яковлев, 1967.
Из представителей семейств, от-
носимых А. Л. Тахтаджяном [1970]
к Santalales, семейства Schoepfi-
aceae, Octoknemaceae, Cardiopterida-
ceae, Dipentodontaceae u Medusand-
тасеае цитоэмбриологически не изу-
чены, а Erythropalaceae изучено
очень слабо. Цитоэмбриологические
данные показывают, что Olacaceae
и папасеае — семейства, тесно
связанные друг с другом, и выделе-
ние О! асассае в особый порядок
Olacales пе оправдапо [Ram, Mana-
si, 419705]. Фагерлинд [Fagerlind,
1948] считает, что ОрШасеае не
имеет таких особенных признаков,
на основании которых следовало
бы выделить семейство ОрШасеае.
Джори и Батнагар, [Зот1 Впабюта-
gar, 1969] подтвердили мнение Фа-
торлинда и полагают, что ОрШ-
асеае следует оставить в семействе
Santalaceae как трибу ОрШеае,
связывающую трибы Thesieae u
АпТоБо]еае (к первой из них отно-
сится ТВезтат, ко второй — Exocar-
риз). Цитоэмбриологические данные
помогли выяснить истинное система-
тическое положение Ехосагриз и по-
казали, что род этот должен быть от-
песен к Заща[асеае, а не к голосемен-
ным или протангиоспермам, как ду-
мали раньше [ВБабтасаг, 1967Ъ, 1970].
По цитоэмбриологическим данным
выделения из Santalaceae особого
семейства М150о4епдгасеае совершен-
но правильно, хотя это семейство
163
исследовано пока недостаточно
[Ват, Мапаз, 1970а]. Радж [Вау
1967, 1970а] полагает, что подсемей-
ство ‘ У15с014еае- семейства Loran-
{Басеае следует выделить в само-
стоятельное семейство, так как оно
по цитоэмбриологическим данным
отличается от подсемейства Loran-
{Бо1деае. На основании цитоэмбрио-
логического исследования Супото-
riaceae u Balanophoraceae, 9. C. Te-
оёхин и М. С. Яковлев [1975] раз-
личают три семейства Cynomori-
aceae, Balanophoraceae и Helosi-
aceae, oTHo¢éa их к“ баща]а]ез. У
Супотогит, наряду с закономер-
ными для паразитных растений из-
менениями структур зародыша, вы-
ражающимися в редукции органов и
тканей, сохраняются высокодиффе-
ренцированные структуры семяпо-.
чек, что резко отграничивает Супб-
morium ot Balanophoraceae, Helosi-
асеае и других представителей San-
{а]а!ез, хотя и не противоречит от-
несению его к этому порядку.
Помимо семейства СупотоНасеае
паразитизм широко распространен
и среди других семейств порядка
Заща]а]ез. Все’ представители М!50-
dendraceae, Santalaceae, Lorantha-
ceae mw Balanophoraceae являются
паразитными растениями. В семей-
ствах О]асасеае и ОрШасеае также
имеются паразитные растения. Па-
разитизм указывает на родственные
связи между этими семействами и
обусловил У этих растений особен-
ности развития семяпочек, зароды-
шевых мешков, зародышей и эндо-
сперма, выражающиеся в их редук-
ции ов, Bhatnagar, 1969; Tepe-
хин, 1977].
НАДПОРЯДОЕ XIII.
РВОТЕА МАЕ
ПОРЯДОК 65.
ELAEAGNALES
СЕМЕЙСТВО ЕГАЕАСМАСЕАЕ
Основное число хромосом — zr =
= 6, 11, 13, 14.
- Цветок обычно обоеполый, реже
однополый, растения одно- или дву-
домные.
164
Пыльник — число микроспоранги:
ев не определено. Стенка микро-
спорангия имеет эпидермис, - `фиб-
розный эндотеций, один средний
слой и секреторный дву-четырехъ-
ядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор ои-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна трехкле-
точные, трехборозднопоровые.
Семяпочка одна, анатропная,. с-
двумя интегументами, крассинуцел-
лятная. Имеются обтуратор и гипо-
стаза.
Макроспорогенез и макрогамето-
генез — археспорий одно-трехкле-
точный. Париетальных клеток 5—7
рядов. Тетрады макроспор линей-
ные. Зародышевый мешок образует-
ся по Роусопит-типу. Антиподы
эфемерные.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Hippop-
hae rhamnoides. Порогамия.
Эндоспермогенез нуклеарный, но
позднее становится целлюлярным и
руминированным.
Эмбриогенез Аз{ега4-типа. Заро-
дыш крупный, прямой, с утолщен-
ным гипокотилем и двумя мясисты-
ми семядолями, выполняющими за-
пасающую функцию, бесхлорофилль-
ный, содержит алейроновые зер-
на. Отчетливого подвеска нет.
Семя, плод — семя. со скудным
эндоспермом, содержащим _алейро-
новые зерна, или без него. Зародьии
заполняет почти все или даже все
семя. Массивные семядоли скрыва-
ют остальные части зародыша,
лишь кончик корешка едва высту-
пает из них. Кожура семени мас-
сивная, образуется из наружного
интегумента и состоит из 35—40
слоев клеток. Плод — сухой нерас-
крывающийся орешек, заключенный
в мясистую чашечную трубку.
Гаусторий эндоспермальный, об-
разующийся в халазальной части
эндосперма.
Апомиксис не обнаружен.
ругие явления не отмечены.
Литература: Corner, 1976; Davis,
4966; Rau M., 1967a, 1970a; Rau,
Sharma, 1970; Schnarf, 1934; Смир-
нова, 1977; Sharma, 1966; Тахтад-
жян,` 1966, 1970; Wunderlich, 1967a;
Yamazaki, 1975b.
Цитоэмбриологические данные
указывают на более ‘близкое род-
ство Еаеастасеае к ТЬутеНасеае,
чем к ВБашпасеае. Шарма [Shar-
ma, 1966] пришел к заключению, ‚
что Еаеаспасеае — высокоспециа-
лизированное семейство, близкое
'ГрутеНасеае. Тахтаджян [4966] по-
мостил Еаеаспасеае в самостоя-
тольный порядок Еаеаста]ез и рас-
положил его вблизи с Ргоеа|ез, что
пе вызывает возражения, так как
оба эти порядка имеют сходные
цитоэмбриологические признаки.
ПОРЯДОК 66.
PROTEALES
СЕМЕЙСТВО РВОТЕАСЕАЕ
Основное число хромосом — х =
=7, 10, 11, 12, 13, 15.
Цветок обычно обоеполый.
Пыльник — 4 микроспорангия.
Стенка ‘микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный двуядерный тапетум.
икроспорогенев и микрогамето-
генев — образование микроспор си-
мультанное и, сукцессивное. тетра-
ы микроспор тетраэдральные, изо-
илатеральные, крестообразные. Зуе-
лые пыльцевые зерна двухклеточ-
ные, двух-трех-семипоровые © зер-
нами крахмала.
Семяпочка орто-, геми- или ана-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется внутренним интегументом.
Имеются нуцеллярный колпачок,
Эпистаза, гипостаза, постамент или
подиум; Семяпочек много — одна.
Сосудистый пучок разветвленный,
доходит до халазы.
Макроспорогенез и макрогамето-
. генез — археспорий одно-многокле-
точный. Париетальных клеток 10
рядов. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок образуется по Polygonum-rury
и содержит крахмальные зерна.
Синергиды © крючкообразными вы-
ростами и фибриллярными аппара-
тами. Имеются три. эфемерные ан-
типоды. Полярные ядра сливаются
во время оплодотворения. У` Ргфеа
/
‚Пер1осатроп ‘верхняя часть зароды-
шевого мешка внедряется в микро-
пиле.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление и самоопы-
ление. Порогамия. Двойное оплодо-
творение.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между ядрами эндосперма начина-
ется с микропилярного конца за-
родышевого мешка. У Brabejum,
Hakea и Масадапла халазальный
конец эндосперма остабтся свободно-
ядерным и превращается в гаусто-
рий. Имеется перисперм.
_Эмбриогенез Азега4-типа. Заро-
дыш прямой, дифференцированный,
двусемядольный, бесхлорофилль-
ный, содержит масло и алейрон.
Семядоли выполняют запасающую
функцию, Подвесок отсутствует.
Семя, плод — семя без эндоспер-
ма, но с периспермом. Зародыш за-
полняет все семя. Массивные семя-
доли скрывают остальные части за-
родыша, лишь кончик корешка ед-
ва выступает из них. Плод — одно-
листовка, орешек, крылатка или
костянка.
Гаусторий эндоспермальный в ха-
лазально части зародышевого
мешка.
Апомиксис не обнаружен.
пругие явления не отмечены.
итература: Согпег, 1976; Davis,
1966; Haber, 1966; Rao C., 1967a, b,
1968, 1969; Schnarf, 1934: Смирнова,
1977; Тахтаджян, 1966; Уепкаа Вао,
1965, 1967a, b, 1968. ,
В отношении цитоэмбриологии
Ргоеасеае обнаруживают сходетво
c Santalaceae u Loranthaceae [Rao
C., 1967a].
Вместе с тем Ргофеасеае имеет
сходные цитоэмбриологические при-
знаки и © Едеастасеае. Поэтому
сближение всех этих семейств с
цитоэмбриологической точки зрения
вполне оправдано. .
Представители как Еаеаспасеае;
так и Ргобеасеае имеют секретор-
ный тапетум, симультанный. микро-
спорогенез, анатропную с двумя
интегументами крассинуцеллятную
семяпочку, гипостазу, одно — мно-
токлеточный женский археспорий,
зародышевый мешок Ро|усопит-ти-
165
“у
па, три антиподы, порогамию, ну-
клеарный эндосперм с халазальным
гаусторием, эмбриогенез Азега4д-ти-
па, прямой, дифференцированный
зародыш и семя со скудным эндо-
спермом или без него.
ПОДКЛАСС G. ASTERIDAE
НАДПОРЯДОЕ ХГУ.
ТАМТА МАЕ
ПОРЯДОК 67.
DIPSACALES
CEMEMCTBO CAPRIFOLIACEAE
Основное число хромосом — х =
= 8, 9, 10, 16, 18, 19.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспораигия образуется
по двудольпому типу и имест эци-
дермие, фиброзный эидотеций, одип
эфемерный средпий слой и секуе-
торный двуядерный, ипогда амебо-
идный (у ЗатБисиз и УШфигпию),
тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, трехбороздно-
поровые, реже трехбороздные или
трехпоровые.
Семяпочка висячая, анатронпная с
одним интегументом, обычно тенуи-
нуцеллятная, иногда крассипуцел-
лятпал. Имеется эпдотелий. Семя-
почек одпа — много.
Макроепорогенез и макрогамето-
renes — - археспорин ОДИОКЛЕТОЧНыЫй.
Паристальных клеток один ряд.
Тетрады макроспор линейные. За-
родышевые мешки образуются по
Ро]усопит-, АШиат- и Адоха-типу.
Синергиды грушевидные. Три эфе-
мерные антиподы, полярные ядра
сливаются перед оплодотворением.
В зародышевом мешке имеются
крахмальные зерна.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Уфигвит
carlesii. Порогамия, Двойное опло-
дотворение.
Эндоспермогенез целлюлярный, у
Уфигиии юоеепз руминированный
по типу ложной руминации.
166
Эмбриогенез — зародыш образует-
ся по Азегад-типу. Он маленький,
прямой, слабо или хорошо диффе-
ренцированный, бесхлорофилльный.
Подвесочного гаустория нет.
Семя, плод — семя имеет хорошо
развитый эндосперм, содержащий
масло и алейрон и слабо дифферен-
цированный зародыш, или эндо-
сперм заполняет лишь периферию
семени, а хорошо дифференциро-
ванный с широкими плоскими ли-
стовидными семядолями зародыш
заполняет 3/4 общего объема семе-
ни. Плод — коробочка, костянка
или ягода.
Гаусторий отсутствует.
Аномиксие пе обпаружен.
Другие лвления ис отмечены.
Литература: Пау!5, 1966; Moissel,
1941; Periasamy, 1966; Schnarf, 1931;
Смирнова, 1977; Sunesson, 1933;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АРОХАСЕАЕ
Основное число хромосом — х=9.
Цветок обоеполый.
Пыльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, один
офемортый средпий слой и секре-
порный двуядерный танетум. Ипог-
да танотум амоебоидцый.
Микроспорогенез и микрогамето-
генез - образование микроспор си-
мультанное. Тотрадя — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные, трехбороздно-
поровые.
Семяпочка висячая,
с одним интегументом,
целлятная. Семяпочек 3—5.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Адоха-типу. Синергиды грушевилд-
ные. Имеются три антиподы, из ко-
торых одна круппее остальных. По-`
лярные ядра сливаются во время оп-
лодотворения.
анатропная,
тенуину-
Опыление. прохождение пыльце-
вой трубки и оплодотворение —
двойное оплодотворение.
Эндоспермогенез целлюлярный.
Первые несколько делений идут в
продольном, а затем в поперечном
направлениях.
Эмбриогенез Азегад-типа. Заро-
дыш маленький, но дифференциро-
ванный, прямой, с многоядерным
подвеском, содержит крахмал.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
сначала амилодекстрин, а потом
масло и алейрон. Зародыш обычно
не превышает 1/4 линейной высо-
ты семени. Несмотря на малую ве-
личину, зародыш дифференцирован
на две семядоли и корешок. ПлодЫ—
костянка.
Гаусторий — сомядоли при про-
растании семени выполняют гау-
сториальпую фупкцию.
Апомиксие не обнаружен.
Другие явления — у Адоха mo-
schatellina oOHapymena полиэмбри-
ония.
Литература:
1966; Mamenuua,
Малдыбаева, 1980; Schnarf, 1934;
Смирпова, 1977; Шугаева, 1972;
Тахтаджян, 1966, 1970.
Corner, 1976; Davis,
1977; Камелина,
СЕМЕЙСТВО УЛГЩТАМЛАСЕЛЕ
Основное число хромосом -- т ==
—= 7, 8, ®), 11, 16.
Цветок обоспольый или однополый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и амебо-
идный, многоядерный, с ложным
периплазмодием тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроепор
тетраэдральные. Зрелые пыльцевыо
зерна трехклоточпые, трех (четы-
рех)-борозднопоровые. Спермии-кле-
тки бедноплазменныо.
Семяпочка одна, висячая, ана-
тропная с одним интегументом,
тенуинуцеллятная. Имеется эндо-
телий.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
/
Ро!ухопит-типу. Синергиды груше-
видные, без фибриллярного аппа-
pata, обычно с крючкообразиями
выростами. Три антиподы дегене-
рируют после оплодотворения. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Уа]етапа
officinalis. Порогамия. — Двойное
оплодотворение.
Эндоепермогенез целлюлярный.
Эндосперм содержит масло и белок.
Эмбриогенез Asterad-tuma, 3apo-
дыш хорошо дифференцированный,
крупный, прямой, хлорофиллонос-
ный, содержит масло.
Семя, плод — семя со скудным
опдоспермом. Зародыш заполняет
почти все семя. Массивные запаса-
ющие семядоли скрывают осталь-
ные части зародыша, лишь конец
корешка едва выступает из них.
Плод сухой, ореховидный, часто
снабженный крылаткой или летуч-
кой.
Гаусеторий отсутствует.
Апомикеие — редуцированный па-
ртеногенез.
Другие явления — гинодиэция у
Valeriana.
Литература: Corner, 1976; Davis,
1966; Schnarf, 1931; Камелина, 1977,
1978; Камелина, Малдыбаева, 1980;
талии, 1966, 1970; ШуУгаева,
972.
СЕМЕЙСТВО ПОТРУАСАСЕАЕ
Основное число хромосом — х =
= 5, 7, 8, 9, 10, 17.
Цветки обычно обоеполые.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный многоядерпый тапетум.
Микроеспорогснез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна трехкле-
точные, трех- или четырехбороздно-
поровые или иногда трехпоровые.
Семяпочка одна, висячая, ана-
тропная, с одним интегументом, те-
нуинуцеллятная, с хорошо разви-
той проводящей системой. Имеют-
ся эндотелий и гипостаза.
167
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный.
Нариетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопит-типу. Синергиды груше-
видные, без фибриллярных аппара-
тов. Три антиподы, число ядер в
которых иногда увеличивается до
четырех. Полярные ядра сливаются
перед оплодотворением или во вре-
мя ного. В зародышевом мешке
имеются зерна крахмала.
Опыление, прохождение пыльке-
вой трубки и оплодотворение —
перекрестное опыление y Stccisa
ргафеп$15. Порогамия. Двойное опло-
дотворение промежуточного и пре-
митотического типов.
Эндоспермогенез целлюлярный.
Эндосперм содержит масло и алей-
рон.
Эмбриогенез Р!рега4-типа, вариа-
ции Scabiosa. Зародыш прямой,
xopomo дифференцированныяй, хло-
рофиллоноспый, содержит масло и
алейрон.
Семя, плод — CCMA со скудным
эндоспермом, который в виде тон-
ких пленок остается лишь около
корешка и на семядолях. Семядоли
широкие, листовидные, выполняю-
щие ассимиляционную функцию
сразу после прорастания семени.
Плод — орешек, обычно с остаю-
щейся чашечкой.
Гаусторий не найден.
Апомиксис отсутствует.
Другие явления — полиэмбриония.
Гинодиэция у Knautia, Scabiosa и
Succisa. Партенокарпия.
Литература: Corner, 1976; Davis,
1966; Fray, 1976; Maxine, 1929; Ка-
молина, 1976, 1977, 1978, 1979a, 6;
Камелипа, Яковлев, 1974, 1976,
1979a, 6; Kopntom, 1963; Poddubnaja-
Arnoldi, 1934; Comaiuesa, HKamennua,
1976; ЗсппатЁ, 19341; Смирнова, 1977;
Souéges, 1963; Тахтаджян, 1966,
1970; Винокурова, 1959; Vijayaragha-
van, Sarveshwari, 1968.
CEMEMCTBO MORINACEAE
Основное число хромосом — z=
— 17.
Цветок обоеполый.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, Ффиброзный эндотеций, 2
168
средних слоя и секреторный дву-
ядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные. Зрелые пыльцевые
зерна трехклеточные.
Семяпочка одна, висячая, ана-
тропная, с одним интегументом, те-
нуинуцеллятная. Имеется эндоте-
лий.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Синергиды груше-
видные, без фибриллярного аппара-
та. Три клетки антипод делятся,
образуя многоклеточные, подобные
зародышам, образования. Зародыше-
вый мешок имеет крахмальные
зерна.
Опыление, прохождение ныльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
)ндоенермогенсз исллюлярный,
руминированиый по типу ложной
румипации. Эндосперм содержит
белок и масло.
Эмбриогенез — зародыш образует-
ся по Asterad-tuny. On прямой,
крупный, хорошо дифференцирован-
ный, бесхлорофилльный.
Семя, плод — семя с эндоспермом,
содержащим белок и масло. Плод —
орешек.
Гаусторий отсутствует.
Апомиксие отсутствует.
Другие явления по отмечены.
Титература: Винокурова, 1959;
Камелина, 1976, 1977, 1978, 1979а, 6;
Камолила, Яковлев, 1974: Тахта-
Juan, 1966, 1970; Vijayaraghavan,
sarveshwari, 1968.
В порядке Dipsacales Bs настоя-
mee время цитоэмбриологически
наиболее полно исследованы семей-
ства П1рзасасеае и Могтасеае, что
способствовало выяснению система-
тического положения этих семейств.
До сих пор систематическое поло-
жение Могта было спорным: одни
систематики выделяли его в само-
стоятельное семейство Могтасеае,
другие — включали в семейство
О1рзасасеае. Однако О. П. Камели-
на [1976, 1977], которая очень де-
тально исследовала цитоэмбриоло-
гию семейства П1рзасасеае, совер-
шенно обоснованно доказала, что
род Мотпа с его 15 видами заслу-
живает ранга семейства Могтасеае,
так как резко отличается от ПУ1рза-
сасеае образованием и строением
стенки пыльника, антипод, эндо-
сперма и зародыша. Вместе с тем
О. П. Камелина считает, что семей-
ство Могтасеае ближе к Caprifoli-
асеае, а П1рзасасеае ближе K Vale-
rianaceae. /
В отличие от Dipsacaceae — jo-
вольно однородного семейства, Сар-
rifoliaceae reTeporeHHo; тем не ме-
нее имеются некоторые черты сход-
ства между Caprifoliaceae u Mori-
пасеае. У обоих этих семейств се-
креторный двуядерный тапетум в
пыльниках, обильный эпдосперм в
зрелых семепах, эмбриогенез типа
Азсга, хлорофилл в зародышах
отсутствует. Однако характерные
для Могтасеае долго сохраняющие-
ся многоклеточные антиподы, руми-
нированный по типу ложной румина-
ции эндосперм и массивная халаза
семяпочки не свойственны ни Сар-
rifoliaceae, uu Dipsacaceae ua Vale-
rianaceae. Ha оснований этого
О. П. ЦКамелипа [1977] считает, что
СаргИоПасеае по может быть не-
посредственным предком Мотгта-
сеае и что оба эти семейства, веро-
ятнее всего, произошли от какого-
то общего, давио вымершего ипред-
ка. Valerianaceac и Dipsaceae
сближали с семействами Азега-
ceae, Rubiaceae, Calyceraceae, Acan-
thaceae и представителями Tubif-
lorae. XarauHcon [Hutchinson, 1969],
как и другие систематики, сбли-
жал Dipsacaceae с Уэжетапасеае,
но относил оба эти семейства не к
‘Dipsacales, a x Valerianales, наряду
c Calyceraceae, B TO BDeMA Kak
А. Л. Тахтаджян [1970] относил
последнее семейство в особый по-
рядок Са[усесга!ез и располагал ето
вблизи порядка Asterales ¢ ere
ственпым семейством = Asbleraceac.
Цитоэмбриология Са|уссгассеае usy-
чена далеко не полно. Одпако пс-
которые его отличия от Dipsacaccac
видны и по этим неполным дан-
ным. У Са[усегасеае зрелые пыль-
цевые зерна двухклеточные, анти-
поды эфемерные, Зо]апа4-тип эм-
бриогенеза. Азбегасеае отличаются
от П!рзасасеае амебоидным-` тапету-
/
MOM B пыльниках, анатропной пря-
мостоячей семяпочкой, Азюгад-ти-
пом эмбриогенеза, наличием анпти-
подальных и синергидных гаусто-
рий, отсутствием эндосперма в зре-
лых семенах и хлоропластов в за-
родышах, наличием разных типов
апомиксиса. Вместе с Tem y Dipsa-
сасеае и Asteraceae имеются и
сходные черты в эмбриональных
признаках, что может указывать,
однако, не на их родство, а на их
относительно одинаково высокий
уровень развития.
Valerianaceae u Dipsacaceae зна-
чительнно не сходны и c Rubiaceae,
так как последнее семейство, в 07т-
личие от нервых двух, имеет: мно-
гоклеточный женский археспорий,
разнообразные типы зародышевых
мешков, часто многоклеточные ан-
типоды, нуклеарный — эндосперм,
обильный в зрелых семенах, а ино-
гда и руминированный, Зо|апа4-тип
эмбриогенеза; в этом семействе
присутствуют — антиподальные и
подвесочные гаустории, отсутству-
ют хлоропласты в зародышах ит. д.
Все это может указывать на то,
что нет родства между Узечазва-
ceae, Dipsacaceae a Rubiaceae. OT
Acanthaccae и представителей Ту-
biflorae ПО1рзаса]ез отличается дру-
гими типами семяпочек и эмбрио-
геисза, отсутствием подвесочных и
ондоспермальных гаусториев и т. д.
Поэтому отнесение Acanthaceae к
порядку Scrophulariales, a Rubia-
сеае — к порядку СепИапаез, как
это признает А. Л. Тахтаджян
[1970], кажется нам более правиль-
ным. Что касается Ти Йогае, то в
системе А. Л. Тахтаджяна [1970]
такого порядка нет, а входящие в
него семейства отнесены к поряд-
Kam Polemoniales, Scrophulariales a
Lamiales, из которых порядок
Scrophulariales является наиболез
крупным и гетерогенным.
Сравпительпо - цитоэмбриологиче-
ское исследовапие П!рзаса!ез се се-
мействами Capripholiaceae, Adoxa-
ceae, Morinaceae, Valerianaceae u
Dipsacaceae показывает, что в та-
ком составе порядок этот кажется
довольно естественным, так как
все семейства, входящие в него,
имеют много сходных цитоэмбрио-
логических признаков, причем пред-
ставители П1рзаса]ез характеризу-
ются более прогрессивными цито-
169
эмбриологическими признаками по
сравнению с представителями дру-
тих ранее описанных порядков. | .
ПОРЯДОК 68.
GE NTIANALES
CEMEMCTBO LOGANIACEAE
Основное число хромосом — х =
— 4, 6, 7, 8, 11, 12.
Цветок обычно обоеполый.
Пыльник — число микроспоранги-
ев не установлено. Стенка микро-
спорангия имеет эпидермис, фиб-
розный эндотеций (двурядный У
Strychnos potatorum), 2 средних
слоя и секреторный одно-двуряд-
ный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые 'зер-
на двух-трехклеточные, трехборозд-
нопоровые или трех-четырех (шес-
ти) поровые.
Семяпочка гемианатропная-ана-
тропная, с одним массивным инте-
тументом, тенуинуцеллятная. Име-
ются эндотелий и гипостаза. Семя-
почки многочисленные или одино-
чные.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Имеются три ско-
ро дегенерирующие антиподы. По-
лярные ядра сливаются до оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Пыльцевые трубки дол-
го сохраняются. Двойное оплодо-
творение.
Эндоспермогенез нуклбарный, цел-
люлярный.
Эмбриогенез Опаотад-типа. Заро-
дыш прямой, дифференцированный
на`две семядоли, почечку, гипоко-
тиль и корешок, содержит масло и
алейроновые зерна.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло, алейроновые зерна и алко-
лоиды. Зародыш занимает 3/4 об-
щего объема семени. Периферия
семени занята эндоспермом. Плод—
септицидная коробочка.
170
у
Гаусторий эндоспермальный, в
халазальной ‘части зародышевого
мептка.
Апомикеие не обнаружен.
Другие явления не отмечены.
JIureparypa: Bendre, 1975; Corner, .
1976; Davis, 41966; Gulati, 4967b;
эсппатЁ, 1934; Смирнова, 1977; Тах-
таджян, 1966, 1970.
СЕМЕЙСТВО АРОСУМАСЕАЕ
Основное число хромосом — х =
== 8—12, 18, 23.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, слабо выраженный эндоте-
ций, 1—2 средних слоя и секретор-
ный одноядерный тапетум. У Уоа-
сапра 1оей4а фиброзный эндотеций
многосяойный.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное или сукцессивное. Тет-
рады микроспор тетраэдральные,
изобилатеральные, крестообразные.
Зрелые пыльцевые зерна трехкле-
точные, одиночные или соединены
в тетрады или поллинии, трехбо-
розднопоровые или двух-трехпоро-
вые с крахмальными зернами У
некоторых родов, например у Аро-
супит, пыльцевые зерна соединены
в тетрады. x
Семяпочка гемианатропная, ана-
тропная с одним интегументом,
красси- или тенуинуцеллятная.
Имеются ариллус и эндотелий. Ce-
мяпочек несколько — много.
Макроспорогенез и макрогамето-
генез — археспорий обычно одно-,
реже двух-трехклеточный. Парие-
тальные клетки отсутствуют. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевый мешок образу-
ется по Роусопит-типу и содер-
жит крахмальные зерна. Синергиды
се .крючкообразными — выростами.
Имеются три эфемерные антиподы.
Полярные ядра сливаются перед
оплодотворением.
Опыление,. прохождение пыльце-
вой трубки и оплодотворение —
нерекрестное опыление. Порогамия.
Эндоспермогенез нуклеарный, эн-
досперм руминированный.
Эмбриогенез Caryophyllad-tuma.
Зародыш прямой или слегка согну-
THM, дифференцированный, двусе-
мядольный, бесхлорофилльный, со-
держит алейроновые зерна и дру-
зы оксалата кальция.
Семя, плод — семя с эндоспермом,
содержащим масло, алейроновые
зерна, крахмал и друзы оксалата
кальция. Зародыш лежит в центре
семени и занимает 3/4 его объема.
Плод обычно состоит из двух ли-
стовок, реже ягодообразный или
костянка.
Гаусторий отсутствует.
Апомиксис не обнаружен.
Другие явления — полиэмбриония
y Kopsia longifolia m Herium indi-
cum. ВОВ
Литература: Corner, 1976; Davis,
1966; Khan, 1970b; Lamba, 1976;
Latoo, 1969, 1974; Maheshwari D.,
4970, 1971; ЗсБптатЁ, 1931; Смирнова,
1977; Тахтаджян, 1966, 1970.
CEMEMCTBO ASCLEPIADACEAE
Основное число хромосом — х =
== 10, 11, 12.
Цветок обоеполый.
Пыльник —4, 2 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций (мно-
горядный У СатаПита attenuata),
3—9 средних слоев (у Рещайторз
microphylla 3—4 средних ряда с
фиброзными утолщениями) и се-
креторный одпо — мпогоядерный,
обычно одно-, реже дву- или даже
трехрядный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное или сукцессивное. Тет-
рады микроспор линейные, тетра-
эдральные, изобилатеральные. Зре-
лые пыльцевые зерна трехклеточ-
ные, соединенные в тетрады или
поллинии. Спермии-клетки хорошо
выражены. `
Семяпочка — гемианатропная-ана-
тропная, © одним интегументом,
тенуинуцеллятная, Имеется нуцел-
лярный колпачок, Семяпочек обыч-
но много, иногда несколько.
Макроспорогенез и макрогамето-
гепев — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные и Т-
образные. Зародышевый мешок об-
разуется по Роухопит-типу и co-
держит крахмальные зерна. Синер-
гиды грушевидные с фибрилляр-
ными аппаратами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются До оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление. Порогамия.
Двойное оплодотворение. Сингамия
и тройное слияние происходит бо-
лее или менее одновременно. Пыль-
цевые трубки сохраняются в семя-
почке до образования нескольких
ядер эндосперма.
Эндоспермогенез нуклеарный. O6-
разование клеточных перегородок
между ядрами эндосперма центро-
стремительное. Постепенно весь эн-
досперм становится целлюлярным.
Эмбриогенез Зо]апа4-типа, вариа-
gun Physalis П. Зародыш прямой,
двусемядольный, с хорошо разви-
той почечкой, гипокотилем и ко-
решком, хлюрофиллоносный, содер-
жит алейрон.
Семя, плод — семя с эндоспермом,
содержащим масло и алейрон. За-
родыш располагается в центре се-
мени и занимает 3/4 его объема.
Эндосперм располагается по пери-
ферии семени. Плод состоит из
двух листовок. Перикарпий состоит
из трех ясно различимых зон и
очень массивен.
Гаусторий не обнаружен.
Апомиксисе — нуцеллярная эмбри-
ония У видов Ушееюжеиш.
Другие явления — полиэмбриония
у Ушсеюох1сит. Клейстогамия.
JIurepatypa: Bisuas, 1967; Corner,
1976; Davis, 1966; Наизпег, 1976;
Кордюм, 1964а, 6; Maheshwari D.,
Lakshminarayana, 1976, 1977a;
Schnarf, 1931; Смирнова, 1977; Тах-
таджян, 1966, 1970.
CEMEMCTBO GENTIANACEAE
|
Основное число хромосом — х =
= 5, 6, 7, 9, 14, 12, 13, 15, 19.
Цветок обоеполый,
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и секреторный тапе-
тум. У Сапзсога 4еситтелз и Сеп-
Напа deffusa тапетум амебоидный.
Y Cotylanthera ua Exacum эндотеций
без фиброзных утолщений.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
171
Зрелые пыльцевые зерна двух-трех-
клеточные, двух-трех (четырех)-бо-
розднопоровые или редко двух-трех
и даже однопоровые, с сетчатой эк-
зиной.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
У Exacum pumilum имеется эндо-
телий. Семяпочек обычно много.
Макроеспорогенез и макрогамето-
генез — археспорий обычно одно-,
редко двух-трехклеточный. Парие-
тальные клетки отсутствуют. Тэтра-
ды макроспор линейные. Зародыше-
вый мешок образуется по Ро[уоо-
пиш-типу и содержит зерна крах-
мала. Синергиды — грушевидные.
Имеются три обычно скородегене-
рирующие антиподы, но у Swertia
сагоЦпеп$13 наблюдается увеличе-
ние их числа. Полярные ядра сли-
ваются перед оплодотворением. У
На[епла еШрИсеа описан постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
двойпое оплодотворение. Тройное
слияпие происходит более или мс-
нее одновременно.
Эндоепермогенез нуклеарный, цел-
люлярный. Образование клеточных
перегородок между свободными яд-
рами эндосперма центростремитель-
ное.
Эмбриогенез бо]апад-, Asterad-
типов. Зародыши прямые, диффе-
ренцированные или недифференци-
рованпые, бесхлорофилльные или
хлорофиллоносные, содержат масло
и алейрон.
Семя, плод — семя обычно с хо-
рошо развитым эпдоспермом, содер-
жащим масло п алеирон, по ипогда
эпдосперм очень слабо развит. В за-
висимости от степени развития эи-
досперма и зародыпта, последний
занимаст от 1/4 до 3/4 общего объ-
ema семени. Плод — септицидная
коробочка, редко ягодовидный. Ко-
жура семени образуется из наруж-
ного интегумента.
Гаусторий отсутствует.
Апомиксие — нередуцированный
партеногенез, нуцеллярная эмбри-
Ония.
Другие явления —у Swertia an-
gustifolia обнаружена полиэмбрио-
ния. Добавочные зародыши образу-
ются из синергид. Гинодиэция у
Gentiana. Клейстогамия. Парази-
TH3M.
Литература: Corner, 1976; Davis,
172
1966; Gulati, 1967a; Maheshwari D.,
Lakshmanarayana, 1977b; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970; Терехин, 1977: Vijayara-
ghavan, Padmanabhan, 1969c.
CEMEMCTBO MENYANTHACEAE
Основное число хромосом — х =
ee
—
‚ 9.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эопи-
дермис, фиброзный эндотеций. 2—3
эфемерных средних слоя и секре-
торный дву- многоядерный тапе-
тум. В клетках танпетума ядра не-
редко сливаются, в результате чего
они становятся полиплоидными.
Микроепорсгенез и мникрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двух-трех-
клеточиье, трехборозднопоровые.
Семяпочка апатропная, с одним
питегументом, тепуипуцеллятная.
Имеются ондотелий и гипостаза.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады микроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Синергиды груше-
видные. Три антиподы обычно ско-
ро дегенерируют, но иногда долго
существуют. Полярные ядра сли-
ваются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
)идоспермогенсв целлтюлярный.
дидоснерм роговой, амилоидный.
Эмбриогенез Asterad-ruma. 3apo-
дыш маленький, прямой, дифферен-
цированный. Подвесок длинпый.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш рас-
полагается в центре семени и за-
нимает от 1/4 до 3/4 его ` объема.
Плод — коробочка или не раскры-
вающийся.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
y Limnanthemum indicum.
Литература: Davis, 1966; Gulati,
1967c; Schnarf, 1931; Cuupnosa, 1977;
Тахтаджян, 1966, 1970.
CEMEHCTBO RUBIACEAE
Основное число хромосом — х =
= 9—15, 18, 20, 22, 28, 33, 34, 36,
42, 54, 77.
Цветок обычно обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
или 2—3 эфемерных средних слоя
и секреторный одноядерный тапс-
tym. Y Macrosphyra longistyla o6na-
ружен амебоидный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двух-трехклеточные, Tpex-,
много-, двух-четырехборозднопоро-
вые, трехпоровые, редко мпогопоро-
вые или безапертурные. Пыльцевые
зерна обычно одиночные, но ино-
гда соединены в тетрады, как У
Macrosphyra_ longistyla.
Семяпочка анатропная-гемиана-
тропная, с одним интегументом, те-
нуинуцеллятная. Имеются нуцел-
лярный колпачок, обычно фунику-
лярный, изредка плацентарный и
интегумелтальный обтураторы и
эпдотелий. Семяночек много или
HCCKONDICO.
Makpocnoporenes и макрогамето-
генез — архоеспорий обычно мното-,
изредка одпоклеточный. Наристаль-
ные клетки отсутетвуют. Тотрады
макроспор обычно линейныс, изред-
ка Г и Т-образные. Зародышевые
мешки образуются mo Polygonum-,
АШит- и Ререгопла-типу. В заро-
длышевых мешках имеются крах-
мальные зерна. Обнаружены заро-
дышевые мешки с обращенной по-
лярностью. Синергиды с клювооб-
разными выростами. Имеются обы-
чно три эфемерные антиподы, но
у некоторых видов число антипод
увеличивается путем митотического
деления и аитиподальный аппарат
СТАПОВИТСЯ МПОГОКЛОТОЧНыЫмМ. Поляр-
ные ядра сливаются До оплодотво-
репия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
обычно перекрестное — опыление,
редко самоопыление. Порогамия.
Двойное оплодотворение. У ЕВсВаг-
dia scabra зигота иногда начинает
делиться после того. как в эндо-
сперме образуется около 200 ядер.
Эндоспермогенез нуклеарный или
целлюлярный. У Psychotria, Randia
чи Тагеппа эндосперм руминирован-
ный. У СоНеа ага Мса деление пер-
вичного ядра эндосперма происхо-
дит через 21—27 дней после опло-
дотворения. У РзусБойа, ВапФа и
Tarenna эндосперм руминирован-
ный.
Эмбриогенез Solanad-truna. Заро-
дыш прямой, дифференцированный,
двусемядольный, бесхлорофилльный,
содержит крахмал.
Семя, плод — семя обычно с мас-
лянистым эндоспермом, содержа-
щим обычно масло, изредка крах-
мал, реже эндосперм почти отсут-
ствует. Зародыш занимает от 1/4
до 3/4 объема семепи. Плод — локу-
лицидпая или септицидная коро-
бочка, ягода или костянка. Семен-
ная кожура однослойная.
Гауеторий подвесочнпый, антипо-
дальный,
Апомиксиз — редуцированный
партеногенез.
Другие явления — полиэмбриония
у СоНеа агаса. Клейстогамия.
Литература: Андронова, 1972, 1973,
1975, 1977, 1978; Boesewinkel, Bou-
mann, 1978; Corner, 1976; Davis,
1966; Faroog, Inamuddin, 1969; Go-
nal, Lakshmanan, 1977; Конычева,
1976; Periasamy, Parameshwaran,
1965; Rajan et al., 1975; Rao S., Shi-
varamaiah, 1976; Schnarf, 1934; Shi-
varamaiah, 1971; Shivaramaiah, Rao
K., 1977; Shivaramaiah, Rajan, 1973;
Shivaramaiah et al., 1975; Siddiqui
S., Siddiqui Sw., 1968a, b; CmmpuHosa,
1977; Тахтаджян, 1966, 1970; Virdi,
Eaton, 1969; Vishvershwara, 1965;
Vishveshwara, Chinnappa, 1965.
Из представителей, относимых
А. Л. Тахтаджяном к порядку Gen-
tianales, cemeiictsa Desi¢ntainiaceae,
Antoniaceae, Potaliaceae, Plocosper-
maceae, Dialypctalaccae ui Ducke-
odendraccae цитоэмбриологически не
изучены. Азер!адасеае и Аросупа-
ссас сходны цитоэмбриологически
и, по-видимому, родственно тесно
связаны и произошли от общего
предка. Аз ер1адасеае разделяют
на два подсемейства Рег!р]осо14еае
и Супапспо!еае. Хатчинсон поднял
Periplocoideae до уровня самостоя-
тельного семейства, что. по мне-
нию Бисваса [В13\аз, 1967], согла-
суется с данными цитоэмбриологии.
173
Наличие трехклеточных пыльцевых
зерен, соединенных в тетрады, об-
наруженные у представителей Аро-
супасеае и Азферладасеае, сближа-
ет оба эти семейства. Семейство
Menyanthaceae B настоящее время
выделено в самостоятельное семей-
ство, а раньше оно было подсемей-
ством Сепйапасеае, что цитоэмбри-
ологически, по мнению Гулати
[СШай, 196749], оправдано, так как
наличием целлюлярного эндосперма
Мепуащасеае отличаются от дру-
гих представителей Gentianales.
Gentianaceae um Rubiaceae A. JI. Tax-
таджян [1966] сближает с Госап]-
асеае, что не противоречит дан-
ным цитоэмбриологии, так как эти
семейства имеют сходные цитоэм-
бриологические признаки. Все эти
семейства характеризуются наличи-
ем секреторного тапетума, двух-
трехклеточных пыльцевых зерен,
гемианатропной-апатропной, тенуи-
нуцеллятной семяпочки с двумя
интегументами, одпо-многоклеточио-
го женского археспория, зародылие-
вого мешка Ро]усопит-тина, нуклс-
арного эндосперма, хорошо диффе-
ренцированного зародыша, семян с
эндоспермом, отсутствием парие-
тальных клеток. Однако Виасеае
более высоко развитое семейство,
чем Loganiaceae и СепЧапасеае,
так как в нем наряду с Ро]угопит-
типом зародышевого мешка встре-
чаются и другие типы (АШашт и
Peperomia), а также обнаружены
подвесочные и антиподальный гау-
сторий, отсутствующие у Гораз-
асеае и СепНапасеае.
ПОРЯДОК 69.
POLEMONIALES
СЕМЕЙСТВО РОГЕМОМТАСЕАЕ
Основное число хромосом — х =
— 6—9.
Цветок обоеполый.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секре-
торный одно-двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
174
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна двух-трех-
клеточные, трехборозднопоровые,
многобороздные, многопоровые с
гладкой экзиной.
Семяпочка — гемианатропная-ана-
тропная с одним интегументом, те-
нуинуцеллятная. Имеются гипоста-
за, эндотелий и слабо дифференци-
рованный фуникулярный обтуратор.
Семяпочек одна — много.
Макроепорогенез и макрогамето-
генез — археспорий одно-многокле-
точный. Париетальные клетки от-
сутетвуют. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро]усопит-типу. Синер-
гиды грушевидные. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вой ‘трубки и оплодотворение —
перекрестное опыление и самоопы-
ление.
дпдоспермогенез пуклеарный.
)мбриогенез Chenopodiad-runa.
Зародыш круиниый, прямой или
слегка согпутый, дифференцирован-
ный на две семядоли, почечку, ги-
покотиль и корешок, хлорофилло-
носный, содержащий масло и алей-
рон.
‚Семя, плод — семя с херошо раз-
витым или со слабо развитым эндо-
спермом, содержащим масло, алей-
рон и крахмал, семядоли выполня-
ют ассимиляционую функцию. За-
родыш располагается в центре се-
мени и занимает 3/А или почти
весь объем семени. Плод — обычно
локулицидпая коробочка с одним
или многими сомепами. Перикар-
ний твордый.
Гаусторий отсутствует.
Апомикеие ио обнаружен.
Другие явлепия — полиэмбриония
и клейстогамия.
Литература: Corner, 1976; Crété,
1965; Davis, 1966; Govil, 1967c, 1970c;
Kapil et al. 1968; Кордюм, 41963:
Kumar, Jain, 1977; Mapkosa, 1947b;
эсЬпах{, 1934; Смирнова, 1977; Тах-
таджян, 1966; 1970.
СЕМЕЙСТВО CONVOLVULACEAE
Основное число хромосом — х =
= 7, 10, 11, 12, 14, 415.
Цветок обоеполый, редко однопо-
лый, растения иногда двудомные.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и секреторный много-
ядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные,
изобилатеральные. Зрелые пыльце-
вые зерна двух-трехклеточные, трех-
борозднопоровые, — многобороздные
или многопоровые, с шиповатой
или гладкой экзиной.
Семяпочка анатропная, с одним
интегументом, тенуи- или красси-
нуцеллятная. Имеются плацентар-
ный обтуратор и эндотелий. Семя-
почек 14—10.
Макроспорогенез и макрогамето-
генез — археспорий одно-многокле-
точный. Париетальные клетки при-
сутствуют или отсутствуют. Sapo-
дышевый мешок образуется по Ро-
]угопит-типу и имеет крахмальные
зерна. Синергиды сильно удлинен-
ные. Имеются три обычно эфемер-
ные антиподы, но иногда антиподы
дегенерируют после оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестпое опыилепие и самоопы-
ление у видов Гротеа. Порогамия.
Двойное оплодотворенио.
Эндоспермогенез пуклеарный. Пе-
рвичное ядро эндосперма делится
раньше зиготы.
Эмбриогенез Опаста4-, СагуорВу]-
]а4-типов. Зародыш крупный, пря-
мой или согнутый, дифференциро-
ванный на две семядоли, почечку,
гипокотиль и корешок, бесхлоро-
филльный или хлорофиллоносный,
содержит масло и алейрон. Семя-
доли выполняют ассимиляционную
функцию.
Семя, плод — семя с твердым
хрящеватым эндоспермом, содержа-
щим алойроп и масло и крахмал.
Зародыи! заполняет 3/4 или почти
все семя. Плод — обычно локулицид-
ная коробочка, редко пе раскрываю-
щийся и сочный или ореховидный.
Гаусторий подвесочный.
Апомиксие — редуцированный
партеногенез.
Другие явления — полиэмбриония
и клейстогамия.
Литература: Согпег, 1976; Davis,
1966; Fedortschuk, 1932; Govil, 1967b,
1970Ъ, 1974; Kaur, 1969b; Kaur,
Singh, 1970; Momin, 1975; Samuel,
Hillson, 1968; Shan, Hillson, 1968,
1971; Schnarf, 1931; CmupnHosa, 1977;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО СОЗСОТАСЕАЕ
Основное число хромоеом — х =
—= 7, 15.
Цветок обоеполый.
Ныльник — 4 — микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный, дву-четырехъядерный Ta-
петум. Имеются орбикулы.
_Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двух-трехклеточные, трех-че-
тырехбороздные.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Имеется плацентарный обтуратор.
Семяпочек 4.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный,
Париетальные клетки отсутствуют.
Зародышевые мешки образуются по
Го] угопит- и АШию-типам, они со-
держат крахмальные зерна. Синер-
гиды удлиненные. Имеются три
офемерные антиподы. Полярные
ядра сливаются перед оплодотво-
рением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворе-
ние.
Эндоепермогенез нуклеарный. Воз-
никает до 1500 свободных ядер эн-
досперма, прежде чем между ними
начнут образовываться клеточные
перегородки.
Эмбриогенез Caryophyllad-, Sola-
пад-типа. Зародыш скручелпый, не-
дифферсепцироваипый, хлорофилло-
посиьй. Подвесок слабо или сильно
развит.
Семя, плод — семя с эндоспермом,
содержащим алейрон и крахмал.
Зародыш заполняет 3/4 или почти
весь объем семени. Плод — пере-
пончатая или ягодообразная коро-
бочка.
Гаусторий подвесочный,
синер-
TUTE.
175
Апомиксис не обнаружен.
Другие явления — полиэмбриония.
Паразитизм.
Jiurepatypa: Das, Banerji, 1966;
Davis, 1966; Fedortschuk, 1931; Раг-
vathi et al., 1978; Schnarf, 1934; Tia-
21, 1965а; Тахтаджян, 1966, 1970; Те-
рехин, 1977.
СЕМЕЙСТВО
HYDROPHYLLACEAE
Основное число хромосом — 2 =
= 5—13, 19.
Цветок обоеполый.
Пыльник — число микроспоранги-
ев и строение стенки микроспоран-
гия не определено. Тапетум секре-
торный, одноядерный.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на двухклеточные, трехборозднопо-
ровые или шестибороздные.
Семяпочка — анатропная с одним
интегумептом, тенпуипуцеллятиная.
Имеются эндотелии, ариллус и ка-
рункула. Семяпочек много — двс.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Ро|угопит-типу и имеет крахмаль-
ные зерна. Синергиды с фибрилляр-
ными аппаратами. Имеются три
эфемерные антиподы. Полярные
ядра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия.
Эндоспермогенез целлюлярный и
нуклеарный.
Эмбриогенез Зо]апа@-типа.
дыши прямые или согнутые, ма-
леньцие или большие, слабо или
хорошо дифференцированные, дву-
семядольные, бесхлорофилльные,
или хлорофиллоносные. Семядоли
выполняют запасающую или асси-
миляционную функции.
Семя, плод — семя с хорошо или
слабо развитым амилоидным эн-
доспермом. Зародыш заполняет от
1/4 до 3/А длины семени или почти
весь объем семени. лод — обычно
локулицидная коробочка, иногда не
раскрывающийся.
Гаусторий — эндоспермальный,
образуется как в микропилярной,
так и в халазальной частях заро-
дышевого мешка.
176
Заро-
Апомиксиес не обнаружен.
Другие явления не обнаружены.
Литература: Corner, 1976; Davis,
1966; ЗсВпатЁ, 1934; Смирнова, 1977;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ВОВАСТМАСЕАЕ
Основное число хромосом — х =
== 4—13,
Цветок большей частью обоепо-
лый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
торный одно-, дву—многоядерный
тапетум. В клетках тапетума на-
блюдаются слияние ядер и поли-
плоидия.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зорна, трох-двухклоточные, обычно
с крахмалытыми зерпами, трех-мно-
гоборозднопорвые, мпогобороздные
или трехпоровые с гладкой экзиной.
Семяпочка анатропная-гемиана-
тропная, с одним интегумевтом,
красси- или тенуинуцеллятная. Име-
ются фуникулярный обтуратор, эн-
дотелий и гипостаза. Сосудистый
пучок проникает в интегумент и
там разветвляется. Семяпочек 1—4.
Макроспорогенез и макрогамето-
генез — археспорий одно-, двух-,
трехклеточный. Париетальные клет-
ки отсутствуют или присутствуют.
Готрады макроспор липейпые или
7Г-образиые. Зародыневые мешки
образ‘ ются по Polygonum- a Alli-
иш-тину. имеют крахмальные зер-
на. Сипергиды групевидные, ино-
гда с прючкообразными выростами.
Имеются три аптиподы, обычно
эфемерные, но иногда долго сохра-
няются и делятся, образуя до 6
клеток. Полярные ядра сливаются
перед оплодотворением или во вре-
мя него.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
опыление перекрестное и самоопы-
ление. Порогамия. Двойное оплодо-
творение. Y Cynoglossum officinale
оплодотворение происходит через
24—48 ч после опыления.
Эндоепермогенез нуклеарный, ге-
лобиальный, целлюлярный.
Эмбриогенез СПепороа4д-, Азе-
rad-, Solanad- u Onagrad-tuma. 3a-
родьии прямой или согнутый, дву-
семядольный, крупный, бесхлоро-
филльный, содержащий масло и
алейрон. Семядоли выполняют запа-
сающую функцию. Подвесок трех-
четырехклеточный или OTCYTCT-
вует.
Семя, плод — семя с хорошо или
слабо развитым эндоспермом, со-
держащим масло и алейрон. Заро-
дыш занимает 3/4 или почти весь
объем семени. Семенная кожура
имеет эпидермис, содержащий тан-
нин, среднюю зону, состоящую из
паренхимных клеток, и внутрен-
нюю зону — из утолщенных кле-
ток. Плод — костянка с одно-четы-
рехсеменной косточкой или раснада-
ющийся па 4—2 пераскрывающихся
орсховидных членика, редко коро-
бочка.
Гаусторий эндоспермальный, в
микропилярной или халазальной
частях зародышевого мешка.
Апомиксие — образуется от 4 до
12 апоспорических зародышевых
мешков.
Другие явлепия — гиподиэция У
Echium u Myosolis. Mnelicroramun.
Terepoctunus y Pulmonaria offici-
nalis.
Литература: Corner, 1970; Davis,
1966; Fathima, 1966, 1967; Khaleel,
1974, 1975, 1977, 1978; Khanna, 1964;
Malecka, 1975, 1977; Nagaraj, Fathi-
ma, 1967, 1968, 1971; Pal, 41967:
Schnarf, 1931; Tasneem, 1966; Toks,
1976; Тахтаджян, 1966, 1970; Вере-
щагина, 1971, 1975, 1976а, 6, 197За,
6, в, 1979; Верещагина, Бояршино-
а 1979; Верещагина, Ермишина,
СЕМЕЙСТВО ТЕММОАСЕАЕ
Основное число хромосом — х==9.
Цветок обосполый.
Ныльшик — 4 микроспорапгия.
Стенка микроспорапгия имест эпи-
дермис, фиброзный эпдотеций, один
эфемерный средний слой и секре-
торный одно-двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — тип микроспорогенеза не
установлен. Зрелые пыльцевые зер-
на двухклеточные, трех-четырех
(шести) -борозднопоровые.
12 в. А. Поддубная-Арнольди
Семяпочка анатропная,
интегументом,
Семяпочек 12—28.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевый мешок образуется по
Ро!ухопит-типу. Синергиды груше-
видные. Три антиподы сохраняют-
ся до начала эмбриогенеза. Поляр-
ные ядра сливаются до оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет`
данных.
Эндоспермогенез целлюлярный.
JMOpnorenes Solanad-, Caryophyl-
]а4-типа. Зародыш маленький, пе
дифференцированный. Подвесок
сильно редуцированный.
Семя, плод — семя с эндоспермом,
содержащим масло, крахмал и бе-
лок. Зародыш обычно не превыша-
ет 1/4 линейной высоты семени.
Плод — мясистая коробочка.
Гаусторий антиподальный.
Апомикеие не обнаружен.
Другие явления — паразитизм.
JIurepatypa: Davis, 1966; Schnarf,.
1931; Смирнова, 1977; Тахтаджян,
1966, 1970; Терёхин, 1977.
с одним
тенуинуцеллятная.
СЕМЕЙСТВО ТОАЗАСЕАЕ
Основное число хромосом — 5 =
= 8, 9, 14, 13.
Цветок обосполый.
Тильник — лот дапзых.
Микроспорогенез и микрогамето-
генез — тип микроспорогенеза не
выяснен. Зрелые пыльцевые зерна
трехклеточные, — трехборозднопоро-
вые.
Семяпочка — гемианатропная-ана-
тропная с одним интегументом, те-
нуинуцеллятная. Имеется эндоте-
лий. Семяпочек одна — много.
Макроспорогенез и макрогамето-
renes — архоспорий одно-, ипогда
мпогоклоточный. Париетальные клет-
ки отсутствуют. Тетрады Makpo-
спор линейные. Зародышевый ме-
шок образуется по Ро]угопит-типу.
Синергиды грушевидные. Имеются
три антиподы, — дегенерирующие
вскоре после оплодотворения. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
177
Эндоспермогенез целлюлярный.
Эмбриогенез Зо]апай-типа. Заро-
дыш прямой или согнутый, бесхло-
рофилльный, хорошо дифференци-
рованный, имеет две короткие се-
мядоли, почечку, гипокотиль и ко-
решок.
Семя, плод — семя с эндоспермом,
содержащим масло, крахмал и
алейроновые зерна. Согнутый заро-
дыш располагается по периферии
семени и занимает 3/4 его перимет-
ра, а эндосперм находится в цент-
ре семени; прямой зародыш распо-
лагается в центре семени, занимая
3/4 его объема, а периферия семе-
ни заполнена эндоспермом. Плод —
локулицидная или септицидная ко-
робочка или ореховидный.
Гаусторий зародышевого мешка и
эндоспермальный как в микропи-
лярном, так и в халазальном кон-
Wax эндосперма. У Гоаза микропи-
лярный эндоспермальный гаусто-
рий агрессивпый и впедряется в
ткани иптегумента, семяножки, а
иногда и плаценты, у других родов
семейства микропилярпый гаусто-
рий развит слабо и малоактивен.
Халазальный эндоспермальный гау-
сторий одноядерный или многоядер-
ный, разветвленный или неразвет-
вленный.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; Garcia, 1962: Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
1970.
В порядке Polemoniales из 8 сс-
мейств цитоэмбриологически болео
или мепее подробио исследованы 7
‘семейств. Только семейство ПорЮ-
stigmataceae B этом отпошепии
осталось неизученным.
По цитоэмбриологическим призна-
Kam Polemoniaceae сходно с Сопуо]-
vulaceae mM происходит оно, по-ви-
димому, из Сегашасеае [Govil, 1967,
1970]. По мнению Говила, данные
цитоэмбриологии подтверждают
предположение Хатчинсона [4926]
о происхождении Ро]етотасеае из
Geraniaceae.
По данным цитоэмбриологии вы-
деление Сизсщасеае из Convolvu-
laceae правильно, так как цитоэм-
бриологически они отличаются друг
от друга [СоуП, 1967], в то время
178
как у Сопуо[ущШасеае зародыпт диф-
ференцированный, у Сизсщасеае он
недифференцированный. Borragina-
сеае, как показывают данные цито-
эмбриологии, — гетерогенное семей-
ство [Ра], 1967], в нем обнаружены
два типа зародышевых мешков, три
типа эндосперма и четыре типа
эмбриогенеза, что может указывать
на неестественность его.
В отношении систематического
положения Ро]етот1асеае существу-
ют два мнения. Согласно одному
из них Роетошасеае произошли
из Сегашасеае, согласно другому —
из СагуорвуПасеае. Рендль [Веп4@е,
1959] считал, что Polemoniaceae
сходны c Convolvulaceae. Taxta-
джян [1966, 1970] располагает Ро-
]етот1асеае вблизи с Convolvula-
сеае. Рао [Вао, 1940] выводил Ро-
lemoniaceae из Geraniaceae. Polemo-
т1асеаё ближе Kx Geraniaceae, 4em
к Caryophyllaceae [Kapil et al,
1968]. В общем же порядок Рае-
тол!а1е$ имост пекоторысе сходные
цитоэмбриологические признаки с
Gentianales u Scrophulariales.
ПОРЯДОК 70.
SCROPHULARIALES
CEMEMCTBO SOLANACEAE
Основное чибло хромосом — х =
= 6—12, 14, 17, 23, 30.
Цветок обоеполый, редко одно-
полый.
Пыльпик — 4 микроспорангия.
Стенка микроспорапгия образуется
но двудольному типу и только у
Withonia зотпИсга — по базально-
му и имеот эпидермис, фиброзный
ондотеций (за исключением тех ро-
дов, У которых пыльники вскрыва-
ются порами), 1—3 эфемерных сред-
них слоя и секреторный двуядер-
ный тапетум. У ПРабита зтатопшит
был описан амебоидный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады ‚, микроспор
тетраэдральные, крестообразные,
изобилатеральные. Зрелые пыльце-
вые зерна двухклеточные, (двух),
трех-пяти (шести)бороздные, борозд-
нопоровые или иногда безапертур-
ные, содержат крахмал.
Семяпочка — анатропная-гемиана-
тропная, амфитропная, кампилотро-
пная, с одним интегументом, тенуи-
нуцеллятная. Имеются эндотелий
и гипостаза. Семяпочек много или
немного.
Макроепорогенез и макрогамето-
генез — археспорий одно-, реже мно-
гоклеточный. Париетальные клетки
отсутствуют. Тетрады макроспор
линейные, Т-образные. Иногда обра-
зуются триады. Зародышевые меш-
ки образуются mo Polygonum-, Alli-
um- и Adoxa-tuny. B зародышевом
meme Nicotiana silvestris’ o6mapy-
жен крахмал. Синергиды удлинен-
ные или грушевидные, с крючко-
образными выростами и фибрил-
лярными аппаратами. Имеются три
обычно эфемерные или иногда дол-
го сохраняющиеся аптиподы. По-
лярные ядра сливатотся перед опло-
дотворепием. Y Datura metel onu-
сан постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление и самоопы-
ление. Порогамия. Двойное оплодо-
творение. У Lycopersicon esculen-
tum оно происходит через 24 ч
после опыления. У М№Мсойапа rusti-
са оплодотворепие премитотиче-
ского типа.
Эндоспермогепез целлюлярный,
нуклеарный и гелобиальный.
Эмбриогенез Зо!апад , Опарта@-ти-
па. Зародыш прямой, согнутый или
спирально закрученный (y Lyco-
регз1соп), дифференцированный на
две семядоли, гипокотиль и коре-
ток, бесхлорофилльный, имеет мас-
ло и алейроновые зерна. Подвесок
содержит зерна крахмала.
Семя, плод — семя с эндоспермом.
Зародыш располагается в центре или
по периферии семени, занимая 3/4
его объема. В эндосперме имеются
следы крахмала, белка и масла.
Семенная кожура состоит из эпдо-
телия, остатков иптегумептальной
паренхимы и эпидермиса. Плод —
ягода или септицидпая коробочка,
редко костянковидный.
Гаусторий эндоспермальный, сла-
бо выраженный в халазальной час-
ти зародышевого мешка у Schizan-
thus.
Апомикеис редуцированный
партеногенез, интегументальная и
нуцеллярная эмбрионии.
Другие явления — партенокарпия.
42*
-
Полиэембриония у некоторых видов
Lycopersicon, Solanum, Nicotiana mu
Capsicum, Добавочные зародыши
образуются из синергид. Клейсто-
гамия.
Литература: Arecal et al. 1971;
Банникова, 1970; Банникова, OB-
сянникова, 1973; Банникова, Хве-
дынич, 1969, 1973; Cochran, 1970;
Corner, 1976; Даниелян, 1962, 1963а,
6; Davis, 1966; Dharamadhaj, 1978;
Иванова, Орел, 1963; Jos, Singh
1968; Еурвандян, 1965, 1966; Frou-
ssart, 1966a, b; Глущенко, 1962,
1963а, 6, 1964, 1967, 1968а, 6; Гра-
бовская, 1968; МЛудникова, 1970;
Микаелян, 1967a, 6; Морап К..,
1966; Кордюм, 1963; Mohan К.,
Singh, 1969; Молхова, 1963; Мол-
хова, Филева, 1969; Мутафян, 1964,
1966; Оганесян, 1968; Орел, Жакова..
1976; Орлова, 1965; Поддубная-Ар-
нольди и др., 1974; Pacini, Sareatti,
1978; Parveen et al. 41972; Rajan,
1968b, c; Raghuvanshi et al. 1976;
Рыбченко, 1960, 1963а, 6; Захепа её а}!
1969a, b; Schnarf, 1934; Singh D.,
Ргазаа, 1976; Тахтаджян, 1966, 1970;
Vallade, 1970; Варшавина, 1973,
1979; Varghese, 1967, 1970.
CEMEMCTBO NOLANACEAE
Основное число хромосом —х == 12.
Цветок обоеполый.
Иыльиик — вет данпых 0 числе
микроспорапгиев. Стенка микроспо-
рапгия имсет эпидермис, эпдоте-
ций, два средних слоя и секретор-
ный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна двухкле-
точные, трехборозднопоровые.
Семяпочка гемианатропная с од-
ним иптегументом, тепуинуцеллят-
vas. Иместея эпдотелий. Семяпо-
чек 20—5.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Синергиды груше-
видные. Имеются три антиподы, де-
генерирующие вскоре после опло-
дотворения. Полярные ядра слива-
ются до оплодотворения.
179
Опыление, прохождение пныльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез Solanad-runa. Sapo-
дыш согнутый, дифференцирован-
ный на две семядоли, гипокотиль
и корешок. Подвесок длинный.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш pac-
полагается по периферии семени и
занимает 3/4 его периметра. Эндо-
сперм заполняет центр семени.
Плод распадающийся на одно-семи-
‘семенные мерикарпии. Эндокарпий
обычно каменистый.
Гаусторий отсутствует.
Апомикеие не обнаружен.
Другие явления не отмечены.
Jimrepatypa: Davis, 1966; Emil Di
Fulvio, 1969; Schnarf, 1934; Смирно-
ва, 1977; Тахтаджян, 1966, 1970.
CEMEHCTBO BUDDLEJACEAE
Основное число хромосом — т ==
== 15, 19.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Нет данных о строении стенки ми-
кроспорангия.
Микроспорогенез и микрогамето-
тенез — нет данных. Зрелые пыль-
цевые зерна трех-четырехбороздно-
поровые.
Семяпочка гемианатропная с од-
ним интегументом, тенуинуцел-
лятная. Имеются эндотелий и гино-
стаза. Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одпоклеточпый.
Паристальные клетки отсутствуют.
Тоетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny. Синергиды групгс-
видные. Три антиподы существуют
до начала формирования эндоспер-
ма. Полярные ядра сливаются до
оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворение.
ндоспермогенез целлюлярный.
Эмбриогенез Опаота4-типа. Заро-
дыш маленький или большой.
Семя, плод — семя с хорошо
или слабо развитым эндоспермом.
Плод — обычно коробочка, редко
костянка или ягода.
Гаусторий эндоспермальный; ми-
кропилярный гаусторий, сильно
180
разветвленный, двуядерный; хала-
зальный двух-четырехъядерный.
Апомиксие не обнаружен.
Другие явления не обнаружены.
Литература: Пау1з, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
ЭСВОРНОГАВТАСЕАЕ
Основное число хромосом — z+ =
= 6—13, 15—18, 20, 24, 23—75, 30.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций (2
или 3 слоя у Са]сео]ат1$), 1—2 эфе-
мерных средних слоя и секретор-
ный двух-четырех-, иногда много-
ядерный тапетум. Ядра в клетках
тапетума нередко сливаются и ста-
новятся полиплоидными. У Anti-
сВаг!з тапетум многорядный.
Микроспорогенез и микрогамето-
reves — образование микроспор си-
мультанное. 'Тетрады — микроспор
тотраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, двух-семи-
апертурные, двух-четырехбороздные,
трех-шестиборозднопоровые, много-
бороздные, с более или менее сет-
чатой экзиной. У АпИсйаг1$ пыль-
цевые зерна соединены в тетрады.
Спермии-клетки бедноплазменные.
Семяпочка анатропная, гемиана-
тропная, кампилотропная с одним
интегументом, тенуинуцеллятная.
Эндотелий окружает часть или ре-
Me BCCh зародышевый мешок. Име-
ется гипостаза. Семяпочек много.
Макроснорогенез и макрогамето-
генез — архоспорий одпо-, двух-, у
Pedicularis многоклеточный. Пари-
отальные клетки отсутствуют. Тет-
рады макроспор линейные, Т-образ-
ные. Зародышевые мени образу-
ются m0 Polygonum-, Allium-, Dru-
за-типу. У некоторых видов заро-
дышевые мешки удлиняются и вда-
ются в микропиле. Яйцеклетка
очень удлиненная. Синергиды гру-
шевидные; 3 или 2 антиподы, деге-
нерирующие вскоре после оплодо-
творения. Полярные ядра сливают-
ся До или во время оплодотворе-
ния. У Меатругутш ргаепзе ниж-
нее полярное ядро в сплодотворе-
нии не участвует. Зародышевые
мешки содержат крахмальные зерна.
Флыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное оныление и самоопы-
ление. Порогамия. Двойное оплодо-
творение — премитотического или
постмитотического типа.
Эндоспермогенез целлюлярный,
нуклеарный.
Эмбриогенез Onagrad-, Solanad-
или Азегай-типа. Зародыш круп-
ный, прямой или согнутый, диффе-
ренцированный на две семядоли,
почечку, гипокотиль и корешок или
маленький недифференцированный
(У паразитных видов). Семядоли
массивные, выполняющие запасаю-
щую функцию. Семенная кожура
состоит из эндотелия и эпидермиса.
Зародыш бесхлорофилльный или из-
редка хлорофиллоноспый, содержит
алейрон. Подвесок длинный, миого-
клеточпый.
Семя, плод — семя с эндоспермом,
содержащим следы крахмала или
масло, белок и алейрон. Дифферен-
цированный зародыш располагает-
ся по периферии или в цептре се-
мени, занимая 3/4 или почти весь
объем семени. Недифференциро-
ваппый зародыш располагается в
основании семепи и запимает пе
более 1/4 ero липойной высоты.
Плод — обычио коробочка, редко
ягода или костяиковидиый. Пери-
карпий имеет двое зопы: внениною
из тонкостеппых клеток и впутрен-
нюю из толстостенных.
Гаусторий подвесочный, эпдосиер-
мальный. Эндоспермальные гаусто-
рии образуются в халазальной, ми-
кропилярной и латеральной частях
эндосперма и очень разнообразны
по форме, числу ядер и клеток, а
также по степени своей агрессив-
HOCTH.
Апомиксие —
партеногенез.
редуцированный
Другие явления — клейстогамия.`
Партенокарпия. Паразитизм в три-
6ax Rhinanthoideac и Вийпегеас.
Литература: Лфанасьева, 1959,
1964, 1966, 1971, 1976; Афанасьева,
Граханцева, 1975; Афанасьева и др.,
1966; Афанасьева, Окулова, 1971;
Афанасьева и др., 1975; Афанасье-
ва, Юсупова, 1978; Arekal, 1965,
4966; Arekal et al. 1970; Arekal et
al, 1971; Arekal, Swamy, 1974;
Bhandari et al., 1976; Chandra et al.,
1980; Грахапцева, 1978; Greilhuber,
1973; Накк, 1975, 1977; Юсу-
1977; Crété, 1964; Kapil,
Kapoor et al., 1975;
Карлова, 1973, 1975; Кордюм,
1963; Мешкова, Антипова, 1966;
Mestre, Guignard, 1969; Михайлова,
1966; Natesh, Bhandari, 1975; Оку-
лова, 1973; Никитичева, `4966, 1968,
1970, 1971, 1978, 1979а, 6; Никити-
gepa, Tepéxun, 1973; Pal, 1969; Raju,
1973, 1976; Raju, Arekal, 1977;
Schnarf, 1931; Schrock, Palzer, 1967;
Taneja, 1969a; Taxtanman, 1966, 1970;
Терёхин, Никитичева, 1980; Tiagi
B., 1965b, 1966, 1967b, 1968, 1970d;
Tiagi Y., 1968; Tschermak-Woess,
1967; Vijayaraghavan, Ratnaparkhi,
1972; Иоффе, 1976.
пова,
Masad, 1973;
СЕМЕЙСТВО ВТСМОМТАСЕАЕ
Основное число хромосом — х =
— 41, 43, 15, 17—20, 22.
Цветок обоеполый.
ПНыльник — 4, иногда 2 микроспо-
рангия. У В1епоша стенка микро-
спорангия образуется по двудоль-
ному типу, но фиброзных утолще-
ний не имеет. У Огоху|оп образу-
ется 9 слоев и фиброзный эндоте-
ций многорядный. Тапетум секре-
торный или амебоидный, дву-много-
ядерпый. В результате слияния яд-
ра танетума становятся полиплоид-
НЫМИ.
Mukpocnoporenes и микрогамето-
генез — образование микроспор си-
мультаниое. Тотрады микроспор
тетраэдральные. Зрелые пыльцевые
зерна богаты маслом, трехклеточ-
ные, трехборозднопоровые или двух-
двенадцатибороздные, реже без-
апертурные, спирапертурные и дру-
гих типов, обычно с сетчатой сэк-
зиной.
Семяпочка анатропная-гемиана-
тропная, с одним интегументом, те-
нуинуцеллятная. Имеются эндоте-
лий и гипостаза. Семяпочек много.
Макроспорогенез и макрогамето-
генез — архоспорий одпо-двухкле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроснор ли-
нейные, Зародышевый мешок обра-
зуется по Polygonum-tuny. Яйце-
клетки удлиненные. Синергиды с
крючкообразными выростами. Име-
ются три антиподы, либо сохраня-
ющиеся до начала формирования
эндосперма, либо эфемерные. По-
лярные ядра сливаются перед опло-
дотворением.
181
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Tabe-
luia serratifolia m Tecomaria capen-
Sis.
Эндоспермогенез целлюлярный.
Эмбриогенез Опастад-типа. Заро-
дыш прямой, дифференцированный,
двусемядольный, бесхлорофилльный,
содержащий масло и алейрон. Се-
мядоли выполняют —запасающую
функцию. Подвесок длинный.
Семя, плод — семя без эндоспер-
ма или со слабо развитым эндо-
спермом, содержащим алейрон и
масло. Зародыш заполняет все или
почти все семя. Массивные семядо-
ли скрывают остальные части заро-
дыша, лишь кончик корня едва вы-
ступает из них. Плод — двухствор-
чатая коробочка или сочный и пе-
раскрывающийся.
Гаусторий эндоспермальный; ха-
лазальный гаусторий двух-четырех-
клеточный, агрессивияй. У Вепоша
и Огохуоп кроме халазального об-
разуется и микропилярный эпдо-
спермальный гаустории.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Corner, 1976; Davis,
1966; Top6 1975; Lakshmanan, 1968;
Nanda, Gupta, 1971; Райкова, Па-
menko, 1977; Schnarf, 1931; Смирно-
ва, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО РЕРАТТАСЕАЕ
Основное чиело хромоеом — x =
= 8, 13, 14, 15, 48.
Цветок обоеполый.
Пыльник — 4 микроспорапгия.
Стенка микроспораития образуется
по двудольпому типу и имеет DIMI
дермис, 2 слоя фиброзпого эидото-
ция, 2—3 эфемерных средних слоя
и секреторный дву-многоядерный
тапетум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные, Г-образные. Зрелые
пыльцевые зерна трехклеточные,
пяти-тринадцатибороздные, с тон-
кой гладкой экзиной. У Sesamno-
thamnus un Stigmatosiphon noimpre-
вые зерна соединены в тетрады.
Семяпочка висячая, анатропная,
с одним интегументом, тенуинуцел-
лятная. Эндотелий окружает 2/3
182
нижней части зародышевого мешка.
Имеется гипостаза. Семяпочек не-
сколько — много.
Макроспорогенез и макрогамето-
генез — археспорий одно-трехкле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроспор ли-
нейные. Зародышевый мешок обра-
зуется по Ро[узопит-типу. Синер-
гиды удлиненные и грушевидные.
Имеются три эфемерные антиподы.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
самоопыление у Sesamum indicum.
Порогамия.
ндоспермогенез целлюлярный.
Эмбриогенез Опастад-типа. Заро-
дыш прямой, богат маслом и алей-
роном. Подвесок однорядный. Семя-
доли широкие, выполняющие асси-
миляцдионную функцию.
Семя, плод — семя с тонким сло-
CM эпдосперма, содержащим масло
и алейрон. Плод — локулицидная
коробочка или пераскрывающийся.
Гаусторий эпдоспермальный; мик-
ропилярный — двух-четырехклеточ-
пыи, халазальный четырехклеточный
(Sesamum) или четырехъядерный
(Реда ат), агрессивный.
Апомикеис не обнаружен.
Другие явления — не отмечены.
Литература: Corner, 1976; Davis,
1966; Kulkarni, Patashkar, 197 3,
1974; Padmanebhan, 1976; Schnarf,
1931; Смирнова, 1977; Singh S.,
1967Ъ, 19705; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ТКАРЕГГАСЕАЕ
Основное
данных.
Цветок обосполый.
Ныльник — /^ микроспорангия.
Стенка микроспораигия имеет эпи-
дермис, фиброзный эндотеций, сред-
ний слой и секреторный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на трехклеточные, трехборозднопо-
ровые, с тонкой гладкой экзиной.
Семяпочка висячая, -анатропная,
с одним интегументом, тенуинуцел-
лятная. Имеются гипостаза и эндо-
телий. Семяпочек две.
Макроспорогенез и макрогамето-
генез — археспорий одно-трехкле-
точный. Париетальные клетки от-
чиело хромосом — нет
сутствуют. Зародышевый мешок об-
разуется по Polygonum-tuny. Tpu
антиподы сохраняются до оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез целлюлярный.
Эмбриогенез Опаста4-типа. Заро-
дыш прямой.
Семя, плод — семя с0 слабораз-
витым эндоспермом. Плод нерас-
крывающийся, односемянный.
Гауеторий эндоспермальный, в
микропилярной и халазальной ча-
стях зародышевого мешка.
Апомикеие не обнаружен.
Другие явления не отмечены.
Литература: Schnarf, 1934; Taxta-
джян, 1966, 1970.
СЕМЕЙСТВО МАВТУМТАСЕАЕ
Основное чиело хромосом — х =
— 15—16.
Цветок обоеполый.
Пыльник — число микроспоранги-
ев не установлено. Стенка микро-
спорангия имеет эпидермис, фиб-
розный эндотеций, один средний
слой и секреторный двуядерный
тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Теотрады — микроспор
тетраэдральшые, изобилатеральныо,
крестообразные. Зрелые пыльцевые
зерна двух-трехклеточлые, многопо-
ровые (до 15 пор), с толстой сет-
чатой экзиной.
Семяпочка анатропная, Cc одним
интегументом, тенуинуцеллятная.
Имеется эндотелий. Семяпочек мно-
го или немного.
Макроспорогенез и макрогамето-
генез — археспорий одно-двухкле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроспор ли-
нейные. Зародышевий мешок обра-
зуется mo Го!ухопит-типу. Синер-
гиды грушевидные. Имеются три
эфемерные или долго сохрапяю-
щиеся антиподы. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльке-
зой трубки и оплодотворевие —
порогамия. Двойное оплодотворе-
ние. Сингамия и тройное слияние
происходят почти одновременно.
Пыльцевые трубки обычно долго
сохраняются в семяпочке. У Маг-
tynia оплодотворение происходит
через 6 ч после опыления.
Эндоспермогенез целлюлярный.
Образуется несколько сот свобод-
ных ядер эндосперма, прежде чем
между ними начнут появляться
клеточные перегородки.
Эмбриогенез Опаотай-типа. Заро-
дыш прямой, двусемядольный, с0-
держащий масло и алейрон. Семя-
доли выполняют запасающую функ-
ЦИЮ.
Семя, плод — семя почти без эн-
досперма. Зародыш заполняет по-
чти все семя. Массивные сэмядоли
скрывают остальные части зароды-
ша, лишь кончик корешка едва вы-
ступает из них. Семенпая кожура
состоит из тонкостепных клеток
наружного эпидермиса иптогумен-
та, нескольких слоев, дегенериру-
ющих гиподермальных клеток и
внутренней кутикулы. Плод — локу-
лицидная коробочка с мясистым
экзокарпием и деревянистым эндо-
карпием.
Гауеторий эндоспермальный в ха-
лазальной и микропилярной частях
зародышевого мешка.
Апомиксис не обнаружен.
Другие явления не обнаружены.
Литература: Сотпег, 1976; Davis,
1966; Schnarf, 1934; Singh S., 1967a,
1970а; Смирпова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО СЕЗМЕВТАСЕАЕ
Основное число хромосом — х =
= 4, 8—15, 17.
Цветки обоеполые.
Пыльник — число микроспоранги-
ев не установлено. Стенка микро-
спорангия образуется по двудоль-
ному типу и имеет эпидермис, фиб-
розный эндотеций, один эфемерный
средний слой и секреторный много-
ядерный тапетум. Ядра в клетках
тапотума передко сливаются и ста-
повятся полиплоидными.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, крестообразные.
Зрелые пыльцевые зерна двухкле-
точные, (двух) трех(четырех)-бороз-
днопоровые.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Ниж-
183
HAA часть зародышевого мешка
окружена эндотелием. Имеется сла-
боразвитая гипостаза. Семяпочек
много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Polygonum-tuny и имеет зерна
крахмала. Синергиды очень длин-
ные, проникающие в микропиле.
Три эфемерные антиподы. Поляр-
ные ядра сливаются перед оплодо-
творением или во время оплодо-
творения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворе-
ние премитотического типа. Первич-
ное ядро эндосперма начинает де-
литься значительно раньше зиготы.
Эндоспермогенез целлюлярный,
пуклеарный.
Эмбриогепез Опавгай типа. Зиго-
та сильно вытяпутая. Зародынт
прямой или сотнутый, маленький,
бесхлорофилльный, содержит масло,
алейрон, иногда крахмал, диффе-
ренцированный на две неравные
семядоли, гипокотиль, почечку и
корешок. В семядолях и гипокоти-
ле хорошо виден центральный про-
камбиальный тяж. Подвесок четы-
рех-пятиклеточный.
Семя, плод — семя обычно с эн-
доспермом, реже он отсутствует
или выражен очень слабо. В зависи-
мости от развития эндосперма заро-
дыш занимает большую или мень
ую часть семени. При отсутствии
эндосперма зародыш заполияет весь
объем семени. Семядоли выполияют
запасающутю функцию. Илод — хоку-
лицидная или реже сеитицидная ко-
робочка или ягодообразный.
Гауеторий эндоспермальный в ха-
лазальной и микропилярной частях
зародышевого мешка. Халазальный
эндоспермальный гаусторий одно-
двуядерный, микропилярный, двух-
клеточный; первый из них более
агрессивный, чем второй.
Апомиксие пе обнаружен.
Другие явления — клейстогамия.
Литература: АдаНа еф а|., 1971;
Алимова. Яковлев, 1978; Padmana-
bhan, 1966, 1967а; Schnarf, 1934;
Sharma, Vijayaraghavan, 1971; Tax-
таджян, 1966, 1970.
184
СЕМЕЙСТВО ОВОВАМСНАСЕАЕ
Основное число хромосом — z =
= (6), 12, 18, 19, 20.
Цветок обоеполый.
Пыльник — 4—2 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
средних слоя и секреторный двух-,
трех-шестиядерный тапетум. Ядра
в клетках тапетума нередко слива-
ются и становятся полиплоидными.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, трехбороздно-
поровые с гладкой экзиной.
Семяпочка гемианатропная или
анатропная, с одним интегумен-
TOM, тенуинуцеллятная. Имеется
многорядный эндотелий. Семяпо-
чек много. `
Макроепорогенез и макрогамето-
генез — археспорий обычно одно-
клеточный. Чаристальные клетки
отсутствуют. 'Теотрады макроспор
линейные, иногда ГТ-образные. За-
родышевый мешок образуется по
Ро!узопит-типу и имеет крахмаль-
ные зерна. Синергиды с крючкооб-
разными выростами и фибрилляр-
ными аппаратами. Имеются три
долгосохраняющиеся антиподы. По-
лярные ядра сливаются до или в
момент оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия. Двойное оплодотворение.
Силгамия и тройпое слияние про-
исходят одновременно.
„)идоенермогелсз целлюлярный.
‚)мбриогенез Onagrad-, Caryophy-
Па ё-тина. Зародыпт прямой, малень-
кий, недифФференцированный, богат
крахмалом, бесхлорофилльный. Под-
вссок трех-четырехклеточный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло и алейрон. Зародыш распо-
лагается в основании семени и
обычно занимает 41/4 линейной вы-
соты семени. Семенная кожура
твердая, одно-(двух — восьми)-ряд-
ная, одревесневшая с разнообраз-
ными утолщениями. Обычно обра-
зуется из эпидермиса наружного
интегумента и эндотелия. Плод —
локулицидная коробочка. .
Гаусторий эндоспермальный в ми-
кропилярной и халазальной частях
зародышевого мошка. Микропиляр-
ный эндоспермальный гаусторий,
двухклеточный с одним, двумя яд-
рами в каждой клетке, разветвлен-
ный с ветвями, проникающими в
интегумент, агрессивный; халазаль-
ный гаусторий одноклеточный, дву-
ядерный менее активный.
Апомиксие не обнаружен.
Другие явления — широко рас-
пространен облигатный или факуль-
тативный паразитизм.
Литература: Corner, 1976;, Davis,
1966; Iaitly et al, 1968а; Маркова,
1951; Никитичева, 1968, 1970, 1971;
Никитичева, Тербёхин, 1976; Raju,
Chamaich, 1972; Schnarf, 1934: Cmu-
рнова, 1977; Тахтаджян, 1966, 1970;
Терёхин, 1966, 1973a, 6, 1977, 1979;
Терёхин, Никитичева, 1980; ‘Tiagi,
1965c, 1967a, 1970a.
СЕМЕЙСТВО
LENTIBULARIACEAE
Основное число хромосом — х =
= 8, 11, 16.
Цветок обоеполый.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия формирует-
ся по базальпому типу и имеет
эпидермис, фиброзный эпдотеций,
один эфемерный средиий слой и
секреторный двух-четьрехъядеритяи,
одно, редко двурядпый тапетум.
Микроспорогспез и микрогамето-
генез — образовапие микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, трех-много-
апертурные, с крахмальными зер-
нами. Обычно пыльцевые зерна
одиночные, но иногда соединены в
тетрады.
Семяпочка обычно анатропная,
но иногда гемианатропная, орто-
тропная или анакампилотроппая, с
одним иптегумептом, топуипуцел-
лятная. Иместся двух-трехрядный
эндотелий, окружающий цептраль-
ную часть зародышевого мешка.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий обычно однокле-
точный, изредка двух- и даже
трех-четырехклеточный. Париеталь-
ные клетки отсутствуют. Тетрады
макроспор линейные Г и {-образ-
ные и изобилатеральные. Зароды-
шевый мешок образуется по Роу-
оопит-типу и имеет крахмальные
зерна. По мере развития зародыше-
вые мешки удлиняются и достига-
ют плаценты. Синергиды с крючко-
образными выростами. Три антипо-
ды, из которых одна сильно увели-
чивается, не дегенерирует до ран-
них стадий развития эндосперма.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
порогамия, мезогамия. — Двойное
оплодотворение. Зигота представля-
ст собой удлиненную трубкообраз-
ную клетку.
Эндоспермогенез целлюлярный.
Первичное ядро эндосперма делит-
ся значительно раньше зиготы.
Эмбриогенез Onagrad-, Asterad-,
Solanad-, Chenopodiad-tuma. 3apo-
дыш недифференцированный или
дифференцированный, хлорофилло-
носный, содержит алейрон и крах-
мал, либо алейрон и масло.
Семя, плод — семя без эндоспер-
ма или лишь с одним слоем эндо-
сперма. Семенная кожура образу-
ется только из наружного эпидер-
миса иптегумента. Плод — многосе-
меппая раскрывающаяся коробочка
или пераскрывающийся односемян-
ный.
Гаусторий зародышевого мешка,
аптипод и эндоспермальный. Ми-
кропилярпый и халазальный эндо-
спермальтые гаустории агрессив-
ные, особенно первый из них. Он
достигает плаценты.
Апомикеие не обнаружен.
Другие явления — полиэмбриония
y Utricularia flexuosa.
Jiuteparypa: Begum, 1965, 1969;
Corner, 1976; Davis, 1966; Fkaroog,
1965, 1966; Farooq, Bilquis, 1966a, b;
Farooq, Siddiqui, 1966, 1967; Khan,
1967, 1970a; Magqbool, 1965; Rajan,
Kumar, 1974c; Schnarf, 19341; Shiva-
ramiah, 1967; Shivaramiah et al,
1975; Siddiqui, 1968a, b, 1975, 1978a,
Ъ, с, 1979; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО МУОРОВАСЕАЕ
Основное число хромосом— х==54.
Цветок обоеполый.
Пыльник — нет данных по числу
микроспорангиев и строению стен-
ки микроспорангия. Тапетум секре-
торный, дву-трехъядерный.
185
Микроспорогенез и мйкрогамето-
генез — нет данных по типу микро-
спорогенеза. Зрелые пыльцевые зер-
Hd двухклеточные, трех-четырехбо-
розднопоровые.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Имеется эндотелий, который окру-
жает, на 2/3 нижний конец зароды-
шевого мешка. Семяпочек 6—80.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро|угопит-типу. Синергиды груше-
видные. Имеются три небольшие ан-
типоды, дегенерирующие в начале
образования эндосперма. Полярные
ядра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльше-
вой трубки и оплодотворение —
порогамия.
ндоспермогенез целлюлярпый.
Эмбриогенез Опаотад-типа. Заро-
дыш прямой или слегка согнутый,
дифференцированный. Семядоли вы-
полняют ассимиляционную функ-
цию. Подвесок состоит из 7 клеток.
Семя, плод — семя со слабо раз-
витым эндоспермом или без него.
Зародыш заполняет почти весь или
даже весь объем семени. Плод ко-
стянковидный.
Гауеторий — эндоспермальный в
микропилярной и халазальной час-
тях зародышевого мешка. Первый
из них у одних видов многокле-
точный, у других — четырехклеточ-
ный, второй — небольшой, четырех-
клеточный.
Апомиксис не обнаружен.
‚ Другие явления не обнаружены.
JImreparypa: Davis, 1966; Schnarf,
1931; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО GLOBULARIACEAE
Основное число хромосом — х =
— 8, 10. `
Пыльник — нет данных.
Микроспорогенез и микрогамето-
тенез — тип . микроспорогенеза не
установлен. Зрелые пыльцевые зер-
на двухклеточные, (двух) трехбороз-
дноноровые.
Семяпочка одна, анатропная, с
одним интегументом, тенуинуцел-
лятная. Имеется эндотелий.
186
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный,
Париетальные. клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро!узопит-типу. Синергиды груше-
видные. Имеются три антиподы, де-
генерирующие вскоре после опло-
дотворения. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
нет данных.
Эндоспермогенез целлюлярный.
Эмбриогенез Опаргад-типа. Заро-
дыш прямой, дифференцированный
на две семядоли, почечку, гипоко-
тиль и корешок.
Семя, плод — семя с эндоспермом.
Зародыш занимает 83/4 общего объ-
ема семени. Периферия семени за-
полнена эндоспермом. Плод орехо-
обравный, заключенный в остаю-
щуюся чашечку. | `
Гаусторий — агрессивные эндо-’
спермальные гаустории на микро-
пилярном и халазальном концах
зародышевого мешка; оба они вы-
ступают из семяпочек и достигают
стенки завязи.
Апомиксис — соматическая апо-
спория обнаружена у Globularia
vulgaris.
Другие явления не обнаружены.
Литература: Davis, 1966; Corsi,
Garbari, 1971; Schnarf, 1931; Cmup-
нова, 1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АСАМТНАСЕАЕ
Основной число хромосом — х =
= 9—19.
Цветок обоеполый.
ыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и обычно
имеет эпидермис, фиброзный эндо-
теций, 1—3 эфемерных средних
слоя и секреторный двух-четырехъ-
ядерный тапетум. Фиброзный эндо-
теций у некоторых видов отсутству-
ет. В клетках тапетума ядра не-
редко сливаются и становятся по-
липлоидными. Имеются орбикулы.'
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна двухклеточные, с крахмаль-
ными зернами, трех-восьмибороЗд-
+
ные, двух-четырехпоровые, Tpex-
пятиборозднопоровые, с, гладкой эк-
зиной.
Семяпочка анатропная, гемиана-
тропная, амфитропная или кампи-
лотропная с одним интегументом,
тенуинуцеллятная. Имеются энлдо-
телий (за исключением ТВипЬегаа),
гипостаза и фуникулярный обтура-
тор. Семяпочек несколько.
Макроеспорогенез и макрогамето-
генез — археспорий одно-двухкле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроспор обы-
чно линейные, иногда Г-образные.
зародышевый мешок образуется по
Ро]узопит-типу и имеет крахмаль-
ные зерна. Зародышевый мешок
выступает из микропиле и дости-
гает плаценты у Рзеидогатетит
он У-образной формы. Синергиды
грушевидные. Обычно имеются три
эфемерные антиподы, но у АрВе-
landra aurantiaca u Berleria cristata
антиподы подвергаются вторичному
делению, в результате чего число
их увеличивается до 13. Одно по-
лярное ядро сливается со сперми-
ем, а затем с другим полярным
ядром. |
Опыление, прохождение пыльце-
вой трубки и оплодотворение —
перекрестное опыление у Thunber-
gia grandiflora. Порогамия. Двойное
оплодотворение. Сингамия и трой-
ное слияние происходят одновре-
менно.
Эндоспермогенез целлюлярный,
руминированный, нуклеарный у
ТвипЪегоа. Ниже клеточного эндо-
сперма у многих представителей
-Acanthaceae лежит свободноядерный
базальный аппарат. Эндосперм име-
ет масло, алейрон и кристаллы ок-
салата кальция.
Эмбриогенез Onagrad-, Solanad-
типа. Зародыш прямой, крупный,
двусемядольный, с хорошо равиты-
ми почечкой, гипокотилем и кореш-
ком, бесхлорофилльный. Подвесок
варьирует от короткого трехклеточ-
Horo (y Elytraria) yo длинного 60—
70-клеточного (у ЕсЪаНит). Заро-
дыш содержит масло, алейрон и
кристаллы оксалата кальция и ино-
гда крахмал. .
Семя, плод — семя, обычно без
эндосперма или со следами ero.
Плод — обычно локулицидная коро-
бочка, но иногда костянковидный.
Гаусторий подвесочный, зароды-
шевого мешка и эндоспермальный.
У НерВаг1з и ТБипЪего1а халазаль-
ный эндоспермальный гаусторий
отсутствует, у остальных предста-
вителей семейства имеются как
халазальный, так и микропилярный
эндоспермальные гаустории. Оба
гаустория одноклеточные, но с дву-
мя или четырьмя ядрами. Микро-
пилярный эндоспермальный гаусто-
‚ рий агрессивнее халазального. У
Buyos Andrographys, Elytraria, Nap-
Лат Тиз и №5от1а образуются вто-
ричные гаустории из перифериче-
ского слоя эндосперма.
Апомикеис не обнаружен. `
Другие явления — клейстогамия.
Полиэмбриония у Вейема cristata
ua Dipteracanthus рай|из, добавоч-
пые зародыши образуются из кле-
ток подвеска.
Литература: Вазкш У., ВазКш С.,
1971; Bhatnagar, Kallarackal, 1973;
Bhatnagar, Puri, 1970; Corner, 1976;
Davis, 1966; Fathima et al., 1973;
Jaitly, 1969; Karistrom, 1972, 41973,
1974a, b; Lakshmanan, 1968; -Mohan
Ваш, 1960; Sahi, 1966; Schnarf, 1934;
Sell, 1977; Тахтаджян, 1966, 1970;
Wadhi, 1967, 1970; Wall, Long, 1965.
CEMEMCTBO PLANTAGINACEAE
Основное число хромоеом — х =
== 4, 5, 6, 9, 10, 12.
Цветок обычно обоеполый.
Пыльник — стенка микроспоран-
гия имеет эпидермис, фиброзный
эндотеций, один средний слой и се-
креторный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые пыльцевые зерна двухкле-
точные, четырех-четырнадцатипоро-
вые с тонкой гладкой экзиной.
Семяпочка анакампилотропная,
анатропная, с одним интегументом,
тенуинуцеллятная. Вокруг зароды-
шевого мешка образуется эндоте-
лий. Семяпочек одна—несколько.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады. макроспор линейные. За-
родышевый мешок, образуется по
Ро!ухопит-типу. Синергиды груше-
видные, у Р]ащаро шефа —с крю-
чкообразными выростами. Три ан-
типоды дегенерируют вскоре после
187
оплодотворения. Полярпые ядра
сливаются до оплодотворепия.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление, роже само-
опыление у видов Р]ап(аго. Поро-
гамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Опастай-типа. Заро-
дыш прямой или слегка согнутый,
двусемядольный, с хорошо разви-
тыми гипокотилем и корешком, бес-
хлорофилльный, содержит алейрон,
масло, крахмал. Подвесок длинный.
Семя, плод — семя с мясистым
или роговым эндоспермом, содер-
жащим алейрон. Зародыш занимает
от 1/4 до 3/4 длины семени. Семен-
ная кожура двухслойная, образую-
щаяся наружным и внутренним
эпидермисами интегумента. Плод —
перопончатая раскрывающаяся по-
перечной щелью коробочка или
ореховидпый, окруженпый остаю-
щейся чашечкой.
Гаусторий эпдоспермальный в мн-
кропилярной и халазальной чаетях
зародышевого мешка. Микропиляр-
ный гаусторий четырехъядерный,
халазальный — двуядерный. Оба
гаустория агрессивные и существу-
ют до созревания семени. Кроме
Toro, y Plantago шефа образуются
латеральные эндоспермальные гау-
стории. Семядоли выполняют гау-
сториальную функцию.
Апомиксие не обнаружен.
Другие явления — гинодиэция У
Plantago. HKueitcroramua.
Литература: Corner, 1976; Davis,
1966; Kaul, Durani, 1969; Misra,
1967, 1970; Schnarf, 1931; Cuupirona,
1977; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
HYDROSTACHYACEAE
Основное чиело хромосом — <=
= 10, 12.
Цветок однополый, растения обы-
чно двудомные, редко однодомные.
Ныльник — микроспорангия.
О строении стенки микроспорангия
нет данных.
Микроспорогенез и микрогамето-
генез — тип образования микроспор
не изучен. Зрелые пыльцевые зер-
на двухклеточные, безапертурные,
соединенные в тетрады.
Семяпочка анатропная. с одним
188
интегументом,
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные, 7-об-
разные. Зародышевый мешок обра-
зуется по Ро]угопит-типу и имеет
крахмальные зерна. Синергиды гру-
шевидные. Три скоро дегенерирую-
щие антиподы являются не клет-
ками, а ядрами. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эндоспермальные гаусторийи отсут-
ствуют.
Эмбриогенез — тип не установлен.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет весь объем
семени. Зародыш прямой, семядоли
массивные, выполняют запасающую
фупкцию. Плод — септицидная ко-
робочка.
Гаусторий подвесочный у Пуфдго-
stachys imbricatus.
Аномиксис но обпаружсн.
Другие явления не отмечены.
Литература: Davis, 1966; Jiiger-
Zurn, 1965; МаигИтоп, — 19365;
осппагЁ 1934; Тахтаджян, 1966, 1970.
тенуинуцеллятная.
Из 18 семейств системы А. Л. Тах-
таджяна [1970] только два семей-
ства, а именно ВНебласеае и Со]и-
melliaceae, He изучены цитоэмбрио-
логически. Хотя в отношении цито-
эмбриологии Solanaceae исследова-
но недостаточно, тем не мелес уже
и при современных дапиых видно,
что оно сходно с Моапасеае. Сбли-
жение Solanaceae с Сопуо]ущЩасеае
и Погатасеае цитоэмбриологиче-
скими дапными не подтверждается.
Зоапасеае имеет сходные цитоэм-
бриологические признаки с 5сто-
phulariaceae. Вархезе [Varghese,
1970] считает, что оба эти семей-
ства произошли от общего предка.
По мнению Тиаги и Вархезе |Т1а21,
1970; Warghese, 1970], Scrophulari-
асеае близкородственпо Orobancha-
сеае и наиболее правильно тесно
увязывать Scrophulariaceae c Oro-
banchaceae, так как они имеют
сходные цитоэмбриологические при-
знаки.
Некоторые ученые подсемейство.
Rhinanthoideae семейства Зсгорпи-
]ат1асеае относят к Асапасеае, что,
по мнению Тиаги [Т1а51 1970], не
согласуется с даппыми цитоэмбрио-
логии. РедаПасеае сближали с 5сго-
phulariaceae um Bignoniaceae. Эрдт-
ман [Erdtman, 1952] считал, что
Sesamum и РедаНит ближе Kk Big-
пошасеае, а ТгареПа ближе к Scro-
phulariaceae. OpyTmMan {[Erdtman,
1952] выделил ТгареПа в особое се-
мейство ТгареПасеае, что поддержал
и Сингх [Singh, 1960, 1970]. По цито-
эмбриологическим признакам Маг-
{упасеае сходно с РейаПасеае, В15-
noniaceae au Scrophulariaceae. On-
nako Cuurx [Singh, 1970] считает,
ato Martyniaceae Oumxe к Bignoni-
асеае, чем к РедаПасеае.
Одни ученые сближали Oroban-
chaceae с Сезпегасеас, другие с
Scrophulariaccae. Ilo миспию Тиа-
ги [Tiagi, 1970], Orobanchaceae mo
цитоэмбриологическим — признакам
сходно с подсемейством ВЫташ®о-
ideae семейства Scrophulariaceae,
так как оно имеет как паразитиче-
ских, так и непаразитических пред-
ставителей. На основании обшир-
ных и разносторонних исследова-
ний Терёхин и Никитичева [1980]
пришли к заключенито, что Огофап-
chaceae Onusku c Scrophulariaceae,
а не c Gesneriaccae. Эти авторы
убедительно показали, что паразит-
ные норичниковые (триб Rhinan-
theae m Buchnecreac) и заразиховые
сходны между собой но миогим при-
знакам, что подтверждает их тес-
ные родственные отношения. Склон-
ность к паразитизму этих таксонов
- свидетельствует об общности их
происхождения [Терёхин, Никити-
чева, '1980]. Глизик ([Glisic, 1929]
высказала предположение O TOM,
aro Orobanchaceae u Scrophulariaceae
по строению эндосперма настолько
сходны, что их можно было бы
объединить в одпо семейство. Кап
[Khan, 1970] отметил, что цитоэм-
бриологические данные ие проти-
Bopeyat cOnmaxennio Lentibulariaccac
c Scrophulariaceae.
Сравнение цитоэмбриологических
признаков Р]ащасотасеае с таковы-
mu Acanthaceae, Labiatae, Oleaceae,
Polemoniaceae, Primulaceae, Rubia-
ceae, Scrophulariaceae u Verbena-
сеае побудило одного из индийских
ученых [Mirsa, 1970] высказать
предположение, что Р1ащаотшасеае
более близкородственно Scrophula-
гасеае, чем другим семействам, и
поэтому его положение в порядке
Scrophulariales вполне оправдано.
Tpuoy Selagineae семейства Scro-
риа масеае некоторые систематики
выделяют в самостоятельное семей-
ство Зеас1тасеае, что не подтвержда-
ется данными цитоэмбриологии. Оба
семейства настолько сходны по ци-
тоэмбриологическим признакам, что
нет достаточных оснований для их
разъединения. В общем порядок
Scrophulariales, necmMoTp— на свою
гетерогенность, довольно естествен-
ный и между некоторыми его се-
мействами намечаются родственные
связи. Особенно характерпо для по-
рядка Эсгорва]ата]с3 является нали-
чис опдоспермальных гаусториев,
разпой степени развития и разной
формы.
ПОРЯДОК 71.
ГАМТАГЕЗ
СЕМЕЙСТВО УЕВВЕМАСЕАЕ
Основное число хромосом — z=
= N—12, 15, 17, 19.
Цветки обычно обосполые,
полигампые.
Иыльник — 4 — микроспорангия.
Степка микроскорангия обычно об-
разустея но двудольпому, но у Шр-
pia nodiflora 110 одподольпому ти-
пам и имеет эпидермис, фиброзный
эндотеций, один средний слой и
секреторный двух-четырехъядерный
тапетум. Имеются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор см-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зер-
на двух-трехклеточные, с крахмаль-
ными зернами (двух)трех(пяти)-бо-
роздные, шостиругатпыс или трех-
бороздиопоровые.
Семяпочка анатропная, с одним
иптегументом, тенуинуцеллятная.
Имеются эндотелий и ариллус. Семя-
почек 2—5.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопит-типу и имеет крахмаль-
редко
189
ные зерна. ‚Синергиды с крючкооб-
разными и клювиковидными выро-
©стами. Клеток антипод три, но иног-
да число их увеличивается до 12.
Антиподы обычно эфемерные, но
иногда долго сохраняются. Поляр-
ные ядра сливаются перед оплодот-
ворением или в момент оплодотво-
рения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление у СШегодепдгоп
tomentosum. Порогамия. Двойное
оплодотворение. Пыльцевая трубка
сохраняется в зародышевом мешке
до полного развития семени.
Эндоспермогенез целлюлярный.
Первое деление ядра эндосперма
опережает деление зиготы.
Эмбриогенез Опастад-типа у Уег-
bena и Solanad-runa y Avicennia. 3a-
родыш прямой или согнутый, диф-
ференцированный на две семядоли,
почечку, гипокотиль и корешок,
бесхлорофилльный, содержащий мас-
ло и алейроновые зерна. Подвесок
удлиненный.
Семя, плод — семя с эндоспермом,
содержащим масло и алейроновые
зерна или без эндосперма. Зародыш
занимает 3/4 или почти весь и даже
есь объем семени, в зависимости
от степени развития или отсутствия
энпосперма. Плод — обычно костян-
K или состоит из четырех орешко-
образных мерикарпиев или двух-
чететоехстворчатая коробочка. =
Г ‚еторий эндоспермальный в мик-
ропилярной и халазальной частях
‚эндосперма. Микропилярный, эндо-
сперм ъный гаусторий двух- или
четырехклеточный, халазальный дву-
или од ядерный. Микропилярный
таустории более агрессивный, чем
халазальный, особенно агрессивен
у Амсепт . Ядра халазального гаус-
тория становятся полиплоидными,
достигая 12 путем эндомитоза. Яд-
ра микропилярного гаустория и соб-
ственно эндосперма остаются три-
плоидными.
Апомиксие не обнаружен. |
Другие явлензы — у Avicennia об-
наружена вивипария.
итература: “Со ег, 1976; Davis,
1966; Fathima, Nalini, 1972; Jaitly et
а1., 19684; Ка]ежзКа, 1977; Карагьо-
sopa, Jlorod@erosa, 1967; Mukherjee,
4969c: Padmanabhan, 1967b, 19705;
Schnarf, 1931; Tohda, 1971; Taxrap-
man, 1966, 1970.
190
СЕМЕЙСТВО ТАМТАСЕАЕ `
Основное число хромосом — х =
= 6—10, 12, 13, 15, 47, 19.
Цветки обычно обоеполые или
редко полигамные.
Пыльник — 4 микроспорангия.
Стенка пыльника образуется по дву“
дольному типу и имеет эпидермис,
фиброзный эндотеций, один — два
эфемерных средних слоя и секре-
торный дву — многоядерный тапе-
тум. У клейстогамных цветков La-
mium ашрехсаи!е эндотеций не
имеет фиброзных утолщений.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные, кре-
стообразные. Зрелые пыльцевые зёр-
на двух-, трехклеточные, трехмери-
диональнобороздные или шести-, ре-
же четырехбороздные. Пыльцевые
зерна с тремя бороздами высевают-
ся из пыльников на двухклеточной
стадии, тогда как шестибороздные
пыльцевые зерпа высеиваются на
трехклеточной стадии.
Семяпочка — гемианатропная-ана-
тропная, кампилотропная, с одним
интегумептом, тенуинуцеллятная.
Имеются гипостаза, эндотеций и фу-
никулярный обтуратор. Семяпочек
2—4. Сосудистый пучок доходит до
основания зародышевого мешка.
Макроепорогенез и макрогаметоге-
нез — археспорий одно-, редко двух-.
трехклеточный. Париетальные клет-
ки отсутствуют, Тетрады макроспо
линейные. Зародышевый мешок об-
разуется по"Ро]усопит-типу и имеет
крахмальпые зерна. Синергиды с
крючкообразпыми выростами и фиб-
риллярпыми аппаратами. Антипод
обычно три, и только y Physostegia
угоймапа число их увеличивается пу-
тем деления. Хотя обычно антиноды
дегенерируют до оплодотворения, но
иногда они дегенерируют и после
оплодотворения. Изредка антиподы
формируются в гаусториальные
структуры. Полярные ядра сливают-
ся перед опледотворением или во
время оплодотворения.
`У Clerodendron thomsonae onucaH
постамент. ‘
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление. I[loporamus.
Двойное оплодотворение премитоти-
7
\
`
4
ческого типа. Сингамия происходит
намного раньше тройного слияния.
У некоторых видов Гауапда]а опло-
дотворение наблюдается через 9—
46 ч после опыления.
Эндоспермогенез целлюлярный.
Эмбриогенез Опаота4-типа, вариа-
ции Мепфа. Зародыш прямой или
согнутый, дифференцированный,
двусемядольный, с хорошо развиты-
ми гипокотилем и корешком, бес-
хлорофилльный, содержащий масло,
алейроновые зерна. Семядоли мас-
сивные, выполняющие запасающую
функцию. Подвесок длинный.
Семя, плод — семя ‚со слабо разви-
тым эндоспермом, содержащим мас-
ло и алейроновые зерна или без не-
го. Зародыш заполняет почти весь
или даже весь объем семепи. Плод
из четырех, реже, вследствие недо-
развития, из трех — одного орешков.
Редко плод с мясистым околоплод-
ником. .
Гаусторий — эпдоспермальный в
микропилярной и халазальной час-
тях зародышевого мешка. Микро-
пилярный эндоспермальный гаусто-
рий болеё агрессивный, чем хала-
зальный. Он дву- или многоядерный
(до 19 ядер), разветвленный, внед-
ряется в иптегумент, халазальный
гаусторий редуцированный, двуядер-
ный. Наблюдается полиплоидизация
ядер эндоспермальных гаусториев,
которая осуществляетея эндомито-
зом.
Апомиксис пе обнаружен.
Другие явления — цинодиэция у
многих представителей семейства.
Клейстогамия у Гашшт атр]ех1сал-
]ез. Партенокарпия.
Литература: Адмиральская, 1960а,
6; Corner, 1976; Пау1з, 1977; Дзеваль-
товский, Полищук, 1974, 1975: Илие-
ва, 1965; Тау, 1966, 1968, 1969а, Ъ,
1974, Тау её а1., 1968Ъ, с; Кандела-
ки, Кобахидве, 1971, 1977; Kapil, Va-
ni, 1966; Кобахидве, 1975, 1977; Ко-
бахидзе, Кобаснидзе, 1978: Ковален-
ко, 1971; Кордюм, 1963; Курбанов,
41971, 1973, 41978; Mortier, Lebegue,
1966; Полищук, 1972, 1973; Дзевал-
товский, 1971а, 6; БеВпагЁ, 1931; Шо-
феристова, 1970, 1973a, 6, 1978;
Singh В., 1967; Тахтаджян, 4966,
1970; Tasneem, Nalini, 1972; Bepema-
wana, Manauuna, 1974; Wunderlich,
a.
СЕМЕЙСТВО РНВУМАСЕАЕ
Основное число хромосом — х = 7.
Цветок обоеполый.
Пыльник — нет данных.
Микроепорогенез и микрогаметоге-
нез — тип микроспорогенеза не ус--
тановлен. Зрелые пыльцевые зерна
трехклеточные, трехбороздные.
Семяпочка одна, анатропная, орто-
тропная, с одним интегументом те-
нуинуцеллятная. Имеется гипостаза.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по Ро-
усопат-типу и содержит крахмаль-
ные зерна. Синергиды с фибрилляр-
ными аппаратами. Три антиподы де-
генерируют вскоре после оплодотво-
рения. Полярные ядра сливаются
после оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Эмбриогенез 50]апай-типа. Заро-
дыш прямой,- двусемядольный © ги-
покотилем и корешком. Семядоли
массивные, выполняют запасающую
функцию, по своим размерам пре-
вышают гипокотиль и корешок. Под-
весок длинный.
Семя, плод — семя без эндосперма..
Зародыш заполняет вёсь объем се-
мени. Плод — односеменный орех.
Гаусторий эндоспермальный в ха-
лазальном конце зародышевого меш--
ка. Он четырехклеточный, достигаю-
щий гипостазы.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Согпег, 1976; Davis,
1966; Тахтаджян, 1966, 1970.
\
СЕМЕЙСТВО САЦШЛТЕТСНАСЕАЕ
Основное чиело хромосом — х = 3,
5, 6, 8.
Цветок однополый, растения одно-
домные.
Пыльник — число микроспорангиев
и строение стенки микроспорангия
не изучепы. Тапетум секреторный,
многоядерный. °
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Пыльцевые зерна трех-
клеточные, трех (четырех)-бороздные
или безапертурные.
194
Семяпочка висячая, анатропная, ©
одним интегументом, тенуипуцеллят-
ная. Имеется хорошо развитый эн-
дотелий. Семяпочек 4.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальпые клетки отсутствуют.
"Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
1усопит-типу. Синергиды грушевид-
ные. Три эфемерные антиподы. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — са-
моопыление и перокрестное опыле-
uue y Callitriche hamulata. Ilopora-
мия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Опастад-типа. Заро-
дыш прямой, дифференцированный
на семядоли, гипокотиль и корешок,
содержит масло и алейрон. Семядо-
ли выполняют —ассимиляционную
функцип. Подвесок длинный.
Семя, плод — семя с мясистым эп-
доспермом, содержащим масло и
алейрон. Зародыш заполпяет почти
весь объем семени. Плод распадает-
ся на 2 или 4 мерикарпия.
Гаусторий эндоспермальный в мик-
ропилярном и халазальном концах
эндосперма. Микропилярный эндо-
спермальный гаусторий двух-четы-
рехклеточный, агрессивный, внедря-
ющийся в микропиле и доходящий
до семяножки. Халазальный гаусто-
рий значительно слабее развит.
Апомиксие не обнаружен.
Другие явления — полиэмбриония.
Литература: Согрег, 1976; Davis,
1966; Schnarf, 1931; Тахтаджяп, 1966,
970.
Данные цитоэмбриологии показы-
вают, что род А\м1сепп1а не следует
отделять от УегЬепасеае и распо-
лагать его в самостоятельном семей-
стве Ау1сепшасеае, так как он схо-
ден с другими представителями Уег-
benaceae [Padmanabhan, 1970].
Цитоэмбриологические признаки
Lamiaceae сходны с Уетепасеае,
Orobanchaceae m Scrophulariaceae, 8
то время как Boraginaceae имеет
с ними лишь поверхностное сходство
[Singh, 1970]. Hanwaue y подавляю-
ero большипства представителей
Тапта!ез трехклеточных пыльцевых
зерен, тенуинуцеллятной с одним
192
‚нез — образование
интегументом семяпочки, однокле-
точного женского археспория, эндо-
телия и эндоспермальных гаусторий,
отсутствие париетальных клеток го-
ворят о естественности и подвинуто-
сти порядка Гап1аез. Вместе с тем
наличие сходных цитоэмбриологиче-
ских признаков, в особенности эндо-
спермальных гаусторий, сближает
порядок Гапуаез с порядком Scrop-
hulariales.
НАДПОРЯДОК ХУ.
АЗТЕВА МАЕ
ПОРЯДОК 72.
САМРА NULALES
СВМЕЙСТВО CAMPANULACEAE
Основное
(3-15, 47.
Цветок обосполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, 1—2
эфемерных средних слоя и секретор-
ный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
чиело хромосом — х =
‚лые пыльцевые зерна двухклеточ-
ные, с крахмальными зернами, ше-
сти-десятибороздные, трех (двух)-бо-
розднопоровые, трехпоровые или
мпогопоровые с гладкой или шипо-
ватой окзиной.
Семляночка анатроппая, с одним
интегументом, топуипуцеллятная.
Иместея хорошо развитый эндоте-
лий. Семяпочек мпого.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-многоклеточ-
ный. Париетальные клетки отсут-
ствуют. Тетрады макроспор линей-
ные. Зародышевый мешок образует-
ся по Ро[угопат-типу и содержит
крахмальные зерна. Синергиды с
крючкообразными выростами и фиб-
риллярными аппаратами. Присут-
ствуют три эфемерные антиподы.
Полярные ядра сливаются перед оп-
лодотворенисм.
Опыление, прохождение ныльце-
вой трубки и оплодотворение -—- пе-
рокрестное опыление. ’Порогамия.
Двойное оплодотворение. У видов
Помоштала оплодотворение происхо-
дит через 17—19 дней после опыле-
ния. Сипгамия и тройное слияние
происходят одновременно.
идоспермогенез целлюлярный.
)мбриогенез Solanad-tuna. Заро-
дыш маленький, прямой, двуссмя-
дольный, бесхлорофилльный. Зигота
сильно удлиненная. Подвесок длин-
ный, нитчатый.
Семя, плод — семя с эндоспермом,
содержащим масло, крахмал и алей-
рон. Эпидермис семянной кожуры
имеет характерные утолщения.
Плод — коробочка, иногда ягодооб-
разный.
Гаусторий эпдоспермальный в ха-
лазальной и микропилярпой частях
зародынтевого метика. Оба гаустория
имоют 1, 2 или 4 клоток. Халазаль-
пый эпдоспермальный гаусторий Me-
пос агрессивпый, чем микропиляр-
ный.
Апомиксие не обнаружен.
Другие явления — полиэмбриония
у Тазопе и УаШетЪегяа. Гинодиэ-
gaa y Adenophora. Клейстогамия,
Партенокарпия.
Литература: ВеНгап, 1970; Corner,
1976; Davis, 1966; Mrdelska, 1975; Ka-
pil, Vijayaraghavan, 1965; Kaplan,
1968, 1969a, Ь, 1970; Кордюм, 1963;
Longoviallo, 1906, 1968a, b, 1969:
Schnarf, 1931; Subramanyam, 1967a, с,
1970a, с; 'Гахтаджян, 1966, 4970; Уца-
yaraghavan, Malik, 1972; Want, 1963.
СЕМЕЙСТВО ГОВЕМАСЕАЕ
Оеновное число хромосом — х = 7,
9, 14, 13.
Цветок обоеполый.
Пыльник — 4А микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секретор-
пый двуядорпый таиетум.
Микроепорогенез и микрогаметогс-
нез -— образование микроснор cH
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
лые пыльцевые зерна двухклеточ-
ные, с крахмальными зернами, трех-
борозднопоровые с крупнозернистой
сэкзиной.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Эндотелий окружает 2/3 нижней ча-
13 В. А. Поддубная-Арнольди
сти зародышевого мешка. Семяпочек
много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок формируется по Ро-
]угопат-типу и имеет крахмальные
зерна. Синергиды длинные, с фиб-
риллярными аппаратами. Три анти-
поды обычно эфемерные, но иногда
сохраняются до начала образования
эндосперма. Полярные ядра сливают-
ся в момент оплодотворения.
Опыление, прохождение пыльце-
вой трубки и оплодотворение. Пе-
рекрестное опыление у Lobelia ful-
сепз. Порогамия.:
Эндоспермогенез целлюлярный.
Эмбриогенез Зо]апа4-типа. Заро-
дыш прямой, маленький, но диффе-
репцированный на две семядоли и
корешок, бесхлорофилльный. Зигота
очень сильно удлиненная.
Семя, плод — семя с обильным мя-
систым эндоспермом, содержащим
масло или крахмал. Зародыш pac-
полагается в основании семени и
занимает от 4/4 до 1/2 линейной вы-
соты его. Плод — ягода или коро-
бочка.
Гаусторий эпдоспермальный в
обоих концах зародышевого мешка.
Микропилярный и халазальный эн-
доспермальные гаустории двухкле-
точные.
Аномиксие пе обнаружен.
Другие явления — полиэмбриония
У видов ГоБеЙа. Паразитизм.
JIuteparypa: Beltran, 1970; Davis,
1966; Kaplan, 1969a, b; Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
1970; Терёхин, 1977.
СЕМЕЙСТВО SPHENOCLEACEAE
Основное
данных.
Цветок обосполый.
Ныльник — число микроспорангиев
неизвестно. Стенка микроспорангия
имеет эпидермис, фиброзный эндоте-
ций, один средний слой и секретор-
пый двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады — микроспор
обычно тетраэдральные, изредка изо-
билатеральные. Зрелые пыльцевые
зерна трехклеточные, трехбороздно-
поровые.
число хромосом — нет
193
Семяпочка анатропная, е одним
интегументом, тенуинуцеллятная.
Эндотелий хорошо развит. Семяпо-
чек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Синергиды удлиненные. Антиподы
дегенерируют во время развития эн-
досперма.
Опыление, прохождение пыльке-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Опастад-типа. Заро-
дыш маленький, прямой, дифферен-
цированный на две семядоли, гипо-
котиль и корешок. Подвесок корот-
кий.
Семя, плод — семя со слаборазви-
тым эндоспермом. Зародыш заполня-
ет почти весь объем семени. Кожура
семени образуется из трех наруж-
ных слоев интегумента. Плод —
двухгнездная, переиопчатая кры-
ночка.
Гауеторий — эпдоспермальный в
обоих концах зародышевого мешка.
Микропилярный и халазальпый эн-
доспермальные гаустории четырех-
клеточные, агрессивные.
Апомиксие не обнаружен.
Другие явления не отмечены.
JIurepatypa: Corner, 1976; Subra-
manyam, 19674, 19709. Тахтаджян,
1966, 1970.
CEMEKMCTBO STYLIDIACEAE
Основное число хромосом — r= 15.
Цветок обоеполый или однополый.
Пыльник — 4 — микроспорапгия.
Стенка микроспорапгия образустся
по двудольпому типу и имеет эни-
дермис, фиброзный эпдотеций, 1—2
эфемерпый средний слой и секретор-
ный двуядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые зер-
па трехклеточные, с крахмальными
зернами, трех-шестибороздные.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Эндотелий хорошо развит. Семяпочек
много.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-, изредка
двухклеточный. Париетальные клет-
ки отсутствуют. Тетрады макроспор
194
линейные. Зародышевый мешок об-
разуется по Ро]угопат-типу и имеет
зерна крахмала. Синергиды очень
удлиненные, с крючкообразными
выростами и фибриллярными аппа-
ратами. Три антиподы обычно деге-
нерируют после оплодотворения.
Полярные ядра сливаются перед оп-
лодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез Зо]апа4-типа. Заро-
дыш прямой, маленький, но диффе-
ренцированный на две семядоли и
корешок.
Семя, плод — семя с эндоспермом.
Зародыш располагается в основании
семени и обычно не превышает 1/4
линейной высоты семени. Плод —
септицидная коробочка, иногда пе
раскрывающийся.
Гаусторий эндоспермальный в 0бо-
их копцах эндосперма; как микро-
пилярный ондоспермальный гаусто-
рий, так и халазальный являются
двухклеточными.
Аномиксие по обнаружен.
Другие явления — не обнаружены.
Литература: Согпег, 1976; Davis,
1966; Philipson W., Philipson M., 1973;
Schnarf, 1931; Subramanyam, 1967e,
1970е; Смирнова, 1977; Тахтаджян,
1966, 1970.
СЕМЕЙСТВО СООБЕМТАСЕАЕ
Основное число хромосом — z = 7,
8, 9. °
Цветок обосполый.
Тыльник - 4 микроспорангия.
Стенка микроспорапгия образуется
но двудольному типу и имеет эпи-
дормис, фиброзный опдотеций, 1—2
средних слоя и секроторпый много-
ядерный тапстум. Ядра в клетках
тапетума сливаются и становятся по-
липлоидными.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
лые пыльцевые зерна двухклеточ-
ные трехборозднопоровые. У Ге-
зспепаМа пыльцевые зерна соеди-
нены в тетрады.
Семяпочка анатропная, с одним
интегументом, тенуинуцеллятная.
Эндотелий присутствует, но он пе
окружает конец зародышевого меш-
ви. Имостся гипостаза.
Макроспорогенез и макрогаметоге-
мез - - археспорий одноклеточный.
Нариестальные клетки отсутствуют.
'Готрады макроспор линейные. За-
родьикевый мешок образуется по Ро-
Гукопит-типу. Синергиды сильно уд-
чинеппые, с фибриллярными аппа-
ратами. Три антиподы дегенерируют
вскоре после оплодотворепия. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез целлюлярный.
Эмбриогенез 30]апад-типа. Заро-
дыш прямой, маленький, по диффос-
ренцированный на две семядоли и
корешок, содержит масло.
емя, плод — COMA C хорошо раз-
витым эпдоспермом, содержащим
масло. Зародыш располагается в ос-
повании семени и обычно занимает
от 1/4 до 3/4 линейной его высоты.
Плод — обычно коробочка, реже ко-
стянко- и ореховидный.
Гаусторий отсутствует.
Апомиксис не обпаружен.
Другие явления пс обпаружены.
JIntepatypa: Carolin, 1966; Corner,
1976; Davis, 1966, Rosen, 1946;
Schnarf, 1931; Ganpuona, 1977; Tax-
таджян, 1966, 1970, УцауатаТауай
Malik, 1972.
CEMEACTBO BRUNONIACKAE
Основное число хромосом — z = 8.
Цветок обоеполый.
Ныльник — нет данных.
Микроспорогенез и микрогаметоге-
нез — тип микроспорогенеза не вы-
яснен. Зрелые пыльцевые зерна
двухклеточные, — трехборозднопоро-
вые.
Семяпочка одпа, апатропная, ©
одпим иптегумеитом, топуипуцел-
лятпая. Имеется эпдотелий.
Макроспорогенез и макрогаметогс-
нез — архоспорий одпоклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор липейные. За-
родышевый мешок формируется по
Ро]угопит-типу. Синергиды сильно
Удлипенпые, проникающие в микро-
пило. Имеются три эфемерные анти-
моды. Полярные ядра сливаются пе-
ред оплодотворением.
Опнылешие, прохождение пыльце-
13*
вой трубки и оплодотворение — пет
данных.
Эндоспермогенез — нот данпых.
Эмбриогенез — тип не определен.
Зародыш прямой, двусемядольный,
содержит масло. Семядоли выполня-
ют запасающую функцию. Подвесок
массивный.
Семя, плод — семя без эндосперма
Зародыш заполняет все семя. Мас-
сивные семядоли скрывают осталь-
пые части зародыша, лишь кончик
корешка едва выступает из них.
Плод — орех, заключенный в трубку
чашечки.
Гаусторий подвесочный.
Апомиксие пе обнаружен.
Другие явления пс обпаружены.
Литература: Пау!з, 1966; Тахтад-
man, 1966, 1970; Rosen, 1946; Смир-
нова, 1977.
Данпых по цитоэмбрпологии Допа-
Насеае нет, остальные семейства по-
рядка Сатрап]а]ез более или менее
полно исследованы в этом отноше-
нии. По цитоэмбриологическим при-
знакам Сатрапи[асеае очень сходны
с ГоБеНасеае и отнесение ux B 06-
щий порядок, как это сделали Ветт-
штейи [1935] п Хатчинсоп [Hutchin-
50п, 1959], вполне оправдано [Зига-
manyam, 1967, 1970]. Pom Sphenoclea
был выделен в самостоятельное се-
mevcTBo Sphenocleaceae и располо-
жен между РЬую]ассасеае u Primu-
Jaceae [Airy, Shaw, 1948]. Одиако
Субраманиам [Subramanyam, 1967.
1970] считает это неправильным.
Sphenocleaceae обнаруживает боль-
moe сходство с Сатрапи]асеае по
цитоэмбриологическим — признакам,
поэтому это семейство должно стоять
вблизи Сатрап]асеае в одном с ним
порядке [Subramaniam, 1967, 1970].
Род Pentaphragma также выделен
в самостоятельное семейство Реп-
taphragmataceae [Airy, Shaw, 1948],
что, но мпению Субраманиама [5иБ-
татапуат, 1967, 1970], неправильно,
так как по цитоэмбриологическим
признакам pon Pentaphragma на-
столько сходен с Сатшрапа]асеае, что
нет оснований для выделения его в
самостоятельное семейство.
Cy6pamanuam [Subramanyam, 1967,
1970] cauraet, uro Stylidiaceae o6Ha-
руживает тесное сходство и родство
се ГоБеНасеае. Кроме того, по не-
которым цитоэмбриологическим при-
195
sHakam Stylidiaceae cxoquo c Astera-
сеае. Goodeniaceae по своим цито-
эмбриологическим признакам сходно
с другими семействами Сашрапл]а-
]ез ‘и, несомненно, является закон-
ным членом этого порядка. Из всех
- изученных в отпошении цитоэмбрио-
логии семейств Campanulales nan-
менее подробно исследовано семей-
ство Вгапотасеае; ‘тем не менее и
это семейство имеет сходные цито-
эмбриологические признаки. харак-
терные для Сатрап]аез. Мы счи-
таем, что в общем Campanulales ес-
тественный порядок и расположен-
лые в нем семейства более или ме-
нее близки друг другу.
ПОРЯДОК 73.
CALYCERALES
СЕМЕЙСТВО САГУСЕВАСЕАЕ
Основное чиело хромосом — < = 8,
`45, 24.
Цветок обычно обоеполый.
Ныльник — число микроспорангиев
и строение стенки микроспорангия
не изучены. Тапетум секреторный.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Зрелые пыльцевые зер-
на трехклеточные, трехборозднопоро-
вые.
Семяпочка одна, висячая, анатроп-
ная, с одним иптегументом, тенуи-
нуцеллятная. Микропиле образуется
обоими интегументами. Семяножка
короткая;
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный,
Париетальные клетки отсутствуют.
'Гетрады макроспор линейные. За-
одышевый мешок образуется по Ро-
ухгопиш-типу. Синергиды удлинен-
ные, с клювиком. Имеются три эфе-
мерные антиподы. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез целлюлярный.
Первые два деления поперечные. за-
тем наступают продольные деления.
Эмбриогенез Solanad-tuna. Заро-
eI согнутый, дифференцирован-
ный, с хорошо развитыми гипокоти-
лем и корешком. Семядоли сверну-
тые.
196
Семя, плод — семя с хорошо или
слабо развитым эндоепермом, со-
держащим масло и алейроновые: зер-
на. Зародыш располагается в центре
семени и со всех сторон окружен
эндоспермом. Плод семянковидный,
с остающейся чашечкой.
Гаусторий отсутствует. .
Апомиксис не обнаружен.
Другие явления — полиэмбриония.
Jluteparypa: Davis, 1966; Schnarf,
4931; Тахтаджян, 1966, 1970.
Са]усёгасеае раньше относили к
порядку Сашрапи]а]ез, но теперь оно
выделено в самостоятельный поря-
док Са{усега]ез, что согласуется с
данными цитоэмбриологии. Са[усега-
]ез по ряду цитоэмбриологических
признаков (трехклеточным пыльце-
вым зернам, висячей семяпочкой,
отсутствию эндоспермальных гаусто-
риев) отличается от Сашрапа]а]ез и
заслуживает ранга порядка.“Вместе с
тем цитоэмбриологические признаки
Са]усегасеае во многом сходны с та-
ковыми Афегасеае. Существенным
отличием в цитоэмбриологии обоих
семейств является то, что у Са]усе-
гасеае семяпочка висячая, у Азега-
сеае — прямостоячая. Наличие вися-
чей семяпочки, наряду с другими ци-
тоэмбриологическими — признаками,
сближает Calyceraceae с Dipsaca-
сеае. Некоторые систематики отно-
сили СаГусегасеае вместе с П1рзаса-
сеае к порядку ВиМа|ез. Однако от-
сутствие эндоспермальных гауето-
риев сближает Са[усегасеае с Соойе-
niaceae. По-видимому, Calycerales
правильно. расположить между Сат-
panulales m Asterales, Tak как OH
имеет сходные цитоэмбриологические
признаки как с первым, так и со
вторым порядками. Однако, ввиду
недостаточной изученности Са[усега-
]ез, требуются дальнейшие исследо-
вания.
ПОРЯДОК 74.
ASTERALES
CEMEMCTBO ASTERACEAE
Основное число хромосом — х =
= 2—49, 23—28. 30.
‚ Цветки обоеполые, реже однопо-
лые мужские и женские, а также
бесплодные.
Ныльник —4, 2 микроспорангия.
=
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и амебоид-
пый дву — многоядерный тапетум.
В клетках тапетума наблюдается об-
разование полиплоидных ядер раз-
пой ‘величины и формы путем слия-
пия ядер и эндомитоза. Часто обра-
зуется истинный периплазмодий.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
крестообразные. Зрелые ‚ пыльцевые
зерна трехклеточные, трехбороздно-
поровые, реже многобороздные или
трехпоровые, с шиповатой экзиной.
Спермии-клетки бедноплазменные.
Ныльцевые зерна содержат масло.
Семяпочка одна, прямостоящая,
анатропная с одпим интегументом,
тенуинуцеллятная, Сосудистый пучок
обычно доходит до конца интегумен-
та. У Carthamus tinctorius иногда
образуется несколько семяпочек.
Имеются однорядный или многоряд-
ный, одноядерный или несколько-
ядерный эндотелий, а также фуни-
кулярный обтуратор. Нуцеллус силь-
но редуцированный, однорядный.
акроспорогенез и макрогаметоге-
нез — археспорий обычно — одно-,
двух-, иногда мпогоклеточный. Па-
риетальные клетки отсутствуют, Тет-
рады макроспор линейные, Т-образ-
ные. Зародышевые мешки образуют-
ся обычно по Ро]уропит-, реже по
Allium-, Peperomia-, Drusa-, Fritilla-
ria- mw даже Адоха-типу. Синергиды
с крючкообразными выростами и
фибриллярными аппаратами. Анти-
под 2, 3, несколько, много, одно-дву-
ядерных или многоядерных (до 60),
обычно долго сохраняющихся. По-
лярные ядра сливаются перед опло-
дотворением.
Опыление, прохождение пыльце-
вой трубки и опледотворение — опы-
ление перекрестпое или самоопыле-
лие, Порогамия. Двойное оплодотво-
решие премитотического типа. Трой-
ное слияние происходит раньше син-
тамии или одновремепно. В редких
случаях сингамия предшествует
тройному слиянию. Оплодотворение
y Taraxacum kok-saghyz происходит
через 15—20 минут после опыления,
а развитие зародыша и эндосперма
через 6—10 дней после оплодотворе-
пия.
4
~
2
Эндоспермогенез нуклеарный, цел-
люлярный.
Эмбриогенез Азегад-типа, вариа-
ции 5епес1о. Зародыш прямой, круп-
ный, дифференцированный на две
семядоли, гипокотиль и корешок,
бесхлорофилльный, содержит масло.
Семядоли выполняют запасающую
функцию.
Семя, плод — семя без эндосперма.
Зародыш располагается в центре ‘се-
‘мени и заполняет весь его объем.
Массивные семядоли скрывают 0с-
тальные части зародыша, лишь кон-
чик корешка едва выступает из них.
Подвесок нитчатый, слабо развитый,
имеет 3—8 клеток. Семенная кожура
образуется из нескольких наружных
слоев интегумента. Плод — семянка.
Перикарпий зрелой семянки имеет
эпидермис, гиподерму и несколько
слоев паренхиматозных клеток. °
Гаусторий антиподальный, синер-
ГИДНЫЙ.
Апомикеие — редуцированный и
нередуцированный — партеногенезы,
генеративная и соматическая апос-
пории, семигамия (гемигамия).
Другие явления — полиэмбриония.
Добавочные зародыши образуются
из синергид, антипод и-клеток эндо-
телия. Гинодиэция у Амепияа, С!:г-
sium, Serratula, Taraxacum и Н1ега-
стат. Клейстогамия.
Литература: Adamkiewiez, 1966;
Ahlstrand, 1978, 41979a, b, c; Ander-
son, 1970; Bauer, 1964, 1965; Bense-
ва Н., 197ба, 6, 1977; Бенецкая, 1952,
11954; Bijok, 1964; Bijok et al., 1977;
Богомолова, 1959; Bobek, Kavacik,
4974; Chennaveeraiah, Patil, 1974;
Chopra, 1969a, b; Corner, 1976; Davis,
1964, 1966, 1967, 1968a, b; Deshpande,
1967, 1970; Дзюбенко, 1965; Eliasson,
1971, 1972, 1974; Gerassimova, 1933;
Герасимова-Навашина, 1947, 1955;
Герасимова-Навашина и др., 4973;
СВозВ, 19745; Гусейнова, Ахундова.
4978; Хведынич, 1972; Хохлов, 1968;
Но\же, 1969, 1975; Юрукова-Грънчаро-
ва, -1978, 1979; Юрцев, Юрцева, 1974;
Капипос, 4975; Касымова, 1966: Kaul
et al., 1975; Khan, Khotoon, 1969a, b, с;
Kirsten, 1970; Kokwesa, 1932; Koscin-
ska, 1980; Houpraesa, 19706; Kordijum,
ВоЩо, 1962; Кострюкова, 19646: Лит-
виненко, Дзевалтовский, 1972; Ма-
heshwari Devi, 1963; Maheshwari D.,
Pullaiah, 1976a, b; Maleckka, 1967,
1971; Мандрик, 1976; Маркова, 1946;
Maugini, 1967; Mestre, 1964; Misra,
197
Рие. 19. Мигроспорогенез, спермиогенез и развитие зародышевого мешка y Crepis tectorum А — симультанное образование тетрады микроспор; Б — разные стадии образова-
ния спермиев; В — разные стадии развития зародышевого мешка [Gerassimova-Na- vashina, 1957]
198
Рие. 19 (окончание)
4972; Mitra, 1974; Мовсесян, 4964,
1968, 1970, 1973, 1979; Мовсесяи, Бог-
дасаряи, 1966; Мовсесяп, Каспаряп,
1972; Магауап, ЗВаКиг, 1969; Низа-
мова, 1973; Obuchowska, 1969; Онкго
Видаяти, 41973; Padamabhan, 1968;
Pandey, Singh, 1976, 1978; IInucxo,
1970, 1971; Поддубная-Арнольди, 1926,
1927, 1934, 1933а, 6, 1947, 1948, 1951,
1958а, 1964, 1976; Поддубная-Арноль-
ди, Дианова, 1937; Росегз, 1969; Роз-
liani, 1966; Попова, 1979; Pullaiah,
1976, 1979а, b, Rajan, 1968a, b, 1972,
4974a, b; Rao, Sudarik, Narayana,
1979; Richard, 1964; Schnarf, 1934;
Sharma, Murty, 1977, 1978; Schgal,
1966; Skalinska, 1973; Skvarla et al.,
1977; Smith, 1969; Conunesa, 1971,
1973a, 6, 1976, 1978; Tatuunesa, 4/971,
19756; Тахтаджян, 1966, 1970; Чубир-
ко, 1978; Устинова, 1955, 1967, 1968,
1970, 1974; Valentine, Richard, 1967,
1973, 1976; Видаяти, Поддубная-Ар-
нольди, 1971; ГаЪтзКа, 1977.
О систематическом положепии Аз-
[сгасеао ведотся большая дискуссия.
iro cOnmnanu c Cucurbitaceae, Ado-
xaceae, Campanulaceae, Lobeliaceac,
Calyceraceae, Valerianaceae, Rubia-
сеае, О1рзасасеае и т. д. Считают, что
Азегасеае происходит из Сатрапл-
laceae или ГоБеПасеае. В последнее
время происхождение Asteraceae
стали выводить us Rubiaceae — Dip-
sacaceae.
Цитоэмбриологические признаки
показывают, что Азегасеае высоко-
развитое, гомогепное семейство и
разбивать его на несколько семейств,
как ото предлагают пекоторые си-
стематики, было бы неправильно.
Сближение Asteraceae c Cucurbita-
сеае и Adoxaceae не согласуется с
данными цитоэмбриологии, так как
по строению микроспорангия, тапе-
тума, пыльцевого зерна, семяпочки,
гаусториев, зародыша и т. д. опи
`не сходны. Семейство Азегасеае по
цитоэмбриологическим признакам от-
личается также от Campanulaceac,
199
А
Рис. 20. Зародышевые мешки и оплодотворение у Crepis teclorum
А — поздние стадии развития зародышевых мешков (а, д — слияние полярных ядер
и образование вторичного ядра); (б—г — увеличение числа антипод путем митоза);
Б — премитотический тип двойного оплодотворения у Сгер1$ сарШаг!$ [@аегаззйтота,
1933; Gerassimova-Navashina, 1951]
200
Рис. 20 (окончанис)
Goodeniaceac, Stylidiaceae nu Calyco-
тасеае. Наоборот, Азюогасеае по ря-
ду цитоэмбриологических призпаков
cxomno c Dipsacaceac, Valerianaceac
wu Rubiaccac. Ojmiako u3 этого не сле-
yet, uTo Dipsacaceae, Valerianaceae,
Rubiaceae и Азегасеае надо распо-
ложить в одном и том же порядке,
uto Asteraceae возникло из одного
из этих семейств. Наряду со сход-
ством эти семейства имеют и разли-
чия, поэтому правильнее всего Аз-
{егасеае выделить в особый порядок
Asterales, Dipsacaceae u Valeriana-
ceae ocTaBuTh B MOpayKe Dipsacales,
a Rubiaceao n Gentianales.
Пулайя [Pullaiah, 1976], изучив-
ипий дотально 10 tTpnO Asteraceae,
категорически утверждает, uro Aste-
гасеае по цитоэмбриологическим при-
знакам сходно c Dipsacaceae u Vale-
т1апасеае, и считает их предками
АЗ{егасеае. Однако выводить Aste-
raceac m3 Rubiaceae-Dipsacaceae, ue-
смотря па пекоторое сходство в их
цитоомбриологии, преждевременно.
Но исключено, что П1рзасасеае и
Азегассас представляют собой при-
мер параллельной эволюции, что ови,
относясь к разным ветвям системы
и ное являясь родственными друг
другу, достигли примерно одинако-
во высокого уровня эволюции. Одна-
ко данные цитоэмбриологии под-
тверждают, несомненно, что А$ега-
сеае заслуживает отпесения его к
самостоятельному порядку.
Махешвари Деви [Maheshwari,
1963], исследовав трибу НеНап®еае
семейства Азегасеае, пришла к за-
ключению, что данные цитоэмбрио-
логии не дают материала для разли-
чепия триб сомейства Азегасеае и
HC позволяют судить о происхождо-
нии как отдельных триб, так и всего
сомейства в целом. Мы тоже счита-
ем, что говорить о происхождении
как всего семейства А$егасеае, так
и отдельных его триб преждевремен-
но, так как для этого пока нет до-
статочных данных. Однако уже и
сейчас ясно, uTo Asteraceae BBICOKO-
развитое, естественное, широко рас-
пространенное и многочисленное се-
мейство, безусловно заслуживающее
ранга самостоятельного порядка.
201
I 1a
Рис. 21. Микро- и макроспорогенезы, микро- и макрогаметогенезы, эндо
1, 2 — микроспорогенез, 3 — зрелое пыльцевое зерно со спермиями, 4 — тетрада
зародышевым, мешком, 7 — 11 — первые деления ядер
семяпочка CO зрелым
клеточный эндосперм, 15—17 — разные стадии дифференциации зародыша [Под
202
ИО
cy
к)
Rd
СХ
гы
aon
НН
НН
if ESE |
Я
АСЕ
СИМ
arated
we НЙ
ИЕ
УМН Be
ИЯ a
BEAT si
WEA “At
area ess
КИС Е
В ем
М
Aes
“,
спермо- и эмбриогенезы у 'Тагахасаш kok-saghiz
макроепор, 5 — зрелый семивклеточный зародышевой мешок, 6 — анатропная
эндосперма и зиготы, 12—14 — многоклеточные шарообразные зародыши и много-
дубная-Арнольди, 1944] 5
203
ни ne
ТЕР
а
cH
“ah
of
204
= ©) 8 weet
ga ОН SN
me y ее > 3
TMM
ma es . , 3
я WL
f
aceae
ическое (6) размноже-
миксиса у представителей Азег
апо
Рис. 22. Разные типы
+
А — амфимиктичное (а) у Chondriila ambigua u naprenorenert
ние у СпопагШа асапТо!ер1з [Поддубная-Арнольди, 1976]
Б — замена мейоза митозом при образовании нередуцированного зародышевого
В — половые и апоспорические зародышевые
.
>
1935]
MemlKu y Helianthus annuus [YctuHosBa
’
мешка y Antennaria [Bergman
1973], Г — разные стадии гемигамии (семи-
)
1973]
ramun) y Rudbeckia laciniata (cn — cnepmMuu) [Comnmesa,
205
КЛАСС ГИЛАТАЕ (МОМОСОТУГЕРОМЕЗ) ПОДКЛАСС А. АЛ$ЗМГЛАЕ
НАДПОРЯДОК 1.
ALISMA NAE
ПОРЯДОК 75.
АТЛЗМАТЕ$ (ВКЛЮЧАЯ
BUTOMALES)
СЕМЕЙСТВО ВОТОМАСЕАЕ
Основное число хромосом — х =7,
8, 10, 14, 43. .
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
но однодольному типу и имеет эпи-
дермис, фиброзпый эидотеции, одип
эфемерный средпий слой и амобоид-
ный, © истинным периплазмодиумом
тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, изобилатеральные, Т-
‘образные, линейные. Зрелые пыль-
цевые зерна трехклеточные, однобо-
роздные, с сетчатой сэкзиной.
Семяночка анатропная, © двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется нуцеллярный
колпачок. Семяпочек много.
Макроепорогенез и макрогаметоге-
нез — археспорий ’одно-двухклоточ-
ный. Париетальпые клетки отсут-
ствуют. Тетрады макросиор линей-
ные и Г-образпые. Зародышевые
менки образуются по Ро]узопаш- и
А]Шиат-типам. Синергиды с крючко-
образными выростами и фибрилляр-
ными аппаратами. Антипод 3—0.
Наблюдается тенденция к уменьше-
нию числа ядер в зародышевом меш-
ке от 8 до 5. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — пе-
рекрестное опыление у В\юта$ и-
феПавиз. Порогамия. Двойпое опло-
потворение премитотического типа.
Тройное слияние происходит раньше
сингамии или одновременно. В ред-
ких случаях сингамия предшествует
206
тройному слиянию. У Вшошиз пй-
beallatus Джори [Johri, 1963] наблю-
дал прорастание пыльцы непосрел-
ственно на семяпочке.
Эндоспермогенез гелобиальный,
Меньшая халазальная камера оста-
ется одноядерной и дегенерирует,
большая микропилярная становится
многоядерной и формирует собствен-
но эндосперм. В некоторых семяпоч-
ках ядра эндосперма становятся по-
липлоидпыми и достигают бп и да-
же 12м.
Эмбриогенез Caryophyllad-tuna.
Зародыш прямой, дифференцирован-
ный, односемядольный, бесхлоро-
филльный, содержащий крахмал.
Подвесок с одной гигантской клет-
кой.
Семя, плод — семя без эпдосперма.
Зародьии заполияет вес семя. Семен-
пая кожура трехрядная, образую-
щаяся двумя иптегументами. Плод —
многолистовка.
Гаусторий не обнаружен.
Апомиксие не обнаружен.
Другие spuenua—y Limnocharis
обнаружена полиэмбриония.
Литература: Davis, 1966; Johri,
1967, 1970а; Schnarf, 1931; Савит,
1969; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО АШЗМАСЕАЕ
Основное число хромосом — х=5—
9, 41—13. .
Цветки обычно обосполые или ред-
KO однополые или полигамные.
Ныльник -- 4 микроспорангия.
Стеика микроспорантия образуется
но одподольиому типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и амебоид-
ный, одноядерный, с типичным пе-
риплазмодием тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сукцес-
сивное. Тетрады микроспор тетраэд-
ральные, изобилатеральные. Зрелые
пыльцевые зерна трехклеточные,
двух-трехпоровые, с гладкой шипо-
ватой или струйчатой сэкзиной.
Семяпочка анатропная, амфитроп-
ная с двумя или одним интегумен-
тами, тенуинуцеллятная. Микропиле
образуется внутренним интегумен-
Рис. 23. Микроспорогенез и микрогаметогенез у барЩама зай оПа
А — образование микроспор по сукцессивному типу, Б — продольный срез участка пыльника с периплазмодиальным тапетумом и отдельными микроспорами, В — раз- ные стадии образования трехклеточного пыльцевого зерна [Савич, 1967]
Рие. 24. Развитие зародыша и подвеска у Alisma plantagouaquatica (1—86, 7, 9} wy Sagiltaria sagittifolia (6—S) 1—6 — разные стадии развития проэмбрио и подвеска; 7, 8 — образование семядоли и Маша; 9 — сформированный зародыш [Савич, 1967]
том. Имеется гипостаза. Семяпочек
несколько — много.
Макроепорогенез и макрогаметоге-
нез — археспорий одно-многоклеточ-
ный. Париетальные клетки отсут-
ствуют. Зародышевый мешок обра-
зуется по АШат-тину. Синергиды ©
208
крючкообразными выростами и фиб-
риллярными аппаратами. Одна — три
скоро дегенерирующие антиподы.
Наблюдается тенденция к уменьше-
нию числа ядер в зародышевом меш-.
ке от 8 до 5. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение
премитотического типа. Сингамия и
тройное слияние обычно протекают
одновременно.
Эндоспермогенез гелобиальный или
нуклеарный. Меньшая халазальная
камера скоро дегенерирует, в боль-
шей микропилярной образуется не-
сколько свободных ядер, прежде чем
в ней начнется образование клеток.
Эмбриогенез СагуорвуПад-типа, ва-
риации ЗасЦата. Зародьии согнутый,
подковообразный, дифференцирован-
ный, бесхлорофилльный. Содержит
масло и крахмал. Подвесок с одной
гигантской пузыревидной клеткой.
Семя; плод — семя без эндосперма.
Зародыш заполняет все семя. Семсен-
ная кожура четырехслойная, образу-
ющаяся двумя интегумептами. На-
ружный эпидермис наружного ипте-
гумента образует утолщения. Плод
многоорешек. Внутренний эпидермис
внутреннего интегумента утолщается
и кутинизируется.
Гаусторий отсутствует.
Апомикеисе не обнаружен.
Другие явления — полиэмбриония
y Sagittaria отапутеа. Добавочные
зародыши образуются из синергид.
Клейстогамия.
Литература: Davis, 1966; Frey,
4966; Johri, 1967a, 1970a; Кудряшов,
4964; Hyapamos, Casuy, 1963; Ly-thi-
Ba, Guignard, 1976; Canuu, 1964, 1967,
1969; ЗеппатЕ, 1931; Тахтаджяп, 1966,
1970.
Семейство ГАлшпосфагЦасеае, отно-
симое А. Л. Тахтаджяном [1970] и
А. Кронквистом [1968] « Alismales,
по мнению Джори [Т0011, 1970в], не
заслуживает ранга самостоятельного
семейства. Он считает, что на осно-
вании данных цитоэмбриологии к
АПзтасеае кроме АПзта следует от-
носить Ну@гос]с1з, Limnocharis, Bu-
tomopsis u Tenagocharis, a « Butlo-
maceae TombKo Butomus, что, по ого
мнению, подтверждастся данными
морфологии, анатомии, кариологии
и палинологии. ГлтпосрагЦКасеае на-
столько слабо изучено в отношении
цитоэмбриологии, что нет возможно-
сти представить его систематическое
положепие.
Семейства АНзшасеае и Виютасеае
имеют между собой много сходства
14 В. Л. Поддубная-Арнольди
по цитоэмбриологическим призна-
кам, что подтверждает включение их
в общий порядок АНзшаез. Общие
цитоэмбриологические признаки для
АНзтасеае и Вщотасеае следующие:
четыре микроспорангия, однодольно-
го типа четырехслойная стенка мик-
роспорангия, амебоидный с типич-
ным периплазмодиумом тапетум,
сукцессивный тип образования мик-
роспор, трехклеточные пыльцевые
зерна, анатропная, с двумя интегу-
ментами, тенуинуцеллятная семя-
почка, одноклеточный женский архе-
спорий, АШаш-тип зародышевого
мешка с тенденцией к редукции
ядер (от 8 до 5), отсутствие парие-
тальных клеток, порогамия, двойное
оплодотворение. гелобиальный эндо-
‚перм. Семена без эндосперма, эмб-
риотеиез СагуорБуПад-типа.
ПОРЯДОК 76.
HYDROCHARITALES
СЕМЕЙСТВО
HYDROCHARITACEAE
Основное чиело xXpoMocOM — 1 =
== 7-42.
Цветок обоеполый, чаще однопо-
лый, растения чаще двудомные.
Ныльник — 4, 2 микроспорангия.
Стенка микросипорапгил образуется
но одподольпому типу и имеет
опидермис, фиброзный эндотеций,
два средних слоя и амебоидный,
одноядерный, с типичным периплаз-
модием тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор
сукцессивное. Тетрады микроспор
изобилатеральные, крестообразные,
линейпые. Зрелые пыльцевые зерна
двух-трехклеточные, однобороздные
или бозапертурные, нитевидные.
У УаШ5пема пыльцевые зерпа без
экзины. Сисрмии-клетки хорошо вы-
ражены.
Семяпочка анатропная или орто-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется внутренним интегументом.
Имеется нуцеллярный колпачок. Се-
мяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Нариетальная ткань имеется. Тетра-
209
ды макроспор линейные, Г-образные,
тетраэдральные. Зародышевый Me-
шок образуется по Polygonum-tuny.
Синергиды © крючкообразными вы-
ростами и фибриллярными аппара-
тами. Три антиподы, дегенерирую-
щие в начале эмбриогенеза. Поляр-
ные ядра сливаются перед оплодот-
ворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — гид-
рофильное. Порогамия. Двойное оп-
лодотворение.
Эндоспермогенез гелобиальный.
В микропилярной камере сначала
идет свободное деление ядер, а за-
тем между ядрами образуются кле-
точные перегородки. Халазальная ка-
мера одноядерная, приобретающая
иногда гаусториальную активность.
Эмбриогенез СагуорйуПад-типа, ва-
риации За Щана. Зародыш диффе-
ренцированный, односемядольный,
крупный, прямой, бесхлорофилль-
ный, содержит масло и крахмал.
Семя, плод — семя без эндосперма.
Зародыш заполняет всс семя. Кожу-
ра семени хорошо развита. Плод
ягодообразный.
Гаусторий отсутствует.
Апомиксие не обнаружен.
Другие явления — клейстогамия у
ОНеПа оуаШоПа, полиэмбриония у
Nechamandra alternifolia.
JImtepatypa: Davis, 1966; Lakshma-
nan, 19656, 1967b, 1970a; Schnarf,
1931; Тахтаджян, 1966, 1970.
Индийский цитоэмбриолог Лакш-
манан [Lakshmanan, 1970] считает,
что семейство НуагосВагЦасоае ис-
обходимо отнести к порядку Пео-
Ыа]ез. Гго пос следует сближать с
семействами Вигтащасосае, Огс19да-
ceae uu Butomaceae, kak это делали
некоторые систематики. По своим ци-
тоэмбриологическим признакам Нуд-
госвагНа!ез очень сходен с АНзта[ез,
что не противоречит расположению
их вблизи друг друга. Однако Hyd-
госвагйасеае наличием «нитевидных»
пыльцевых зерен без экзины с од-
ной бороздой или без апертур отли-
чается от представителей АПзта]ез,
что оправдывает отнесение Нуйго-
свагНасеае в особый порядок Hyd-
rocharitales.
210
ПОРЯДОК 77.
NAJADALES
СЕМЕЙСТВО
SCHEUCHZERIACEAE
Основное число хромоеом — x= 11.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 4—2
средних слоя и амебоидный, одно-
рядный, превращающийся в типич-
ный периплазмодий тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Микроспоры соединены в
диады. Зрелые пыльцевые зерна
двух-трехклеточные, безапертурные
с сетчатой экзиной.
Семяпочка апатропная, © двумя
иптегументами, крассинуцеллятная.
Микропиле образуется внутренним
иптегументом. Семяпочек немного
(12—6).
Макроспорогенез и макрогаметоге-
нез — археспорий одпоклеточный,
3—4 слоя париетальпых клеток. Тет-
рада макроспор линейная или Т-об-
разная. Зародышевый мешок образу-
ется по Ро]угопат-типу. Синергиды
грушевидные. Имеются три эфемер-
ных ядра антипод. Клеток антипод
не образуется. Полярные ядра сли-
ваются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез гелобиальный.
В микропилярпой клетке эндосперма
спачала происходит свободное деле-
ние ядер, а затем между ядрами
возникают клеточпые перегородки.
Халазальпая клетка остается одно-
ядерной и пе обпаруживает гаусто-
риальной активности.
Эмбриогенез Caryophyllad-runa?
Зародыш крупный, дифференциро-
ванный, односемядольный, хлоро-
филлоносный, содержит крахмал и
масло. Подвесок из одной крупной
пузыревидной клетки.
Семя, плод — семя без эндосперма
или со слаборазвитым эндоспермом,
содержащим крахмал и алейрон. За-
родыш заполняет все семя. Кожура
семени тонкая, образующаяся толь-
ко из одного внешнего интегумента.
Плод многолистовка.
Гаусторий не найдеп.
Апомиксис не обнаружен.
Другие явления — полиэмбриония.
Литература: Davis, 1966; Lakshma-
nan, 1967f; 1970e; Canuy, 1969; Schnarf,
1934; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО
JUNCAGINACEAE
Основное число хромосом — z = 6,
8, 9, 12, 45.
Цветок обоеполый или однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет
эпидермис, эндотеций, 2 эфемерных
средних слоя и амебоидный одно-
ядерный, развивающийся в пери-
плазмодий тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивпое. Тетрады микроспор изо-
билатеральпые. Зрелые пыльцевые
зерна двухклеточные, безапертур-
ные, с сетчатой экзиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяпочек 6—4.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-многоклеточ-
ный. 5—6 слоев паристальпых кле-
ток. Тетрады макроспор липейпые,
Т-образные. Зародышевый мешок об-
разуетея по Ро]усопат-типу. У Tri-
glochin maritimum 3 azrunojt де-
лятся, образуя до 14 клеток, и сохра-
няются до начала эмбриогенеза. По-
лярные ядра сливаются после опло-
дотворения одного из ядер.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоепермогенез нуклеарный.
Эмбриогенез СагуорпуПад-типа, ва-
риации ЗасИата. Зародыш прямой,
дифференцированный, односемядоль-
НЫЙ.
Семя, плод — семя без эндосперма,
зародыш заполняет все семя. Кожу-
ра семепи образуется из паружного
слоя эпидермиса впешиего иптегу-
мента и из впутреннего слоя эпидер-
миса — внутренного интегумента.
Плод — многолистовка.
Гаусторий не обнаружен.
Другие явления — полиэмбриония
y Triglochin palustre.
JIurepatypa: Davis, 1966; Lakhsma-
nan, 1967e, 1970b; Taxtayman, 1966,
14*
СЕМЕЙСТВО
АРОМОСЕТОМАСЕАЕ
Основное число хромосом — х = 8.
Цветок обычно обоеполый, редко
одпополый, растения обычно одно-
домные, редко двудомные.
Пыльник — 4 — микроспорангия..
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и амебоид-
ный, одноядерный, превращающийся
в периплазмодий тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное или симультанное. Тетра-
ды микроспор тетраэздральные, кре-
стообразные. Зрелые пыльцевые
зерна трехклеточные, однобороздные,
с сетчатой экзиной.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяпочек 2—8.
Макроеспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Имеются 2—3 слоя париетальных
клеток. Тетрады макроспор линей-
пые. Зародышевый мешок образует-
сл по Ро]усопаш-типу. Синергиды
грушевидпые. Присутствуют три ан-
типоды, дегенерирующие в начале
эмбриогенеза. Полярные ядра сли-
ваются перед оплодотворением.
Опыление, прохождение пылье-
вой трубки и оплодотворение — нет
Даппых.
Эндоспермогенез гелобиальный..
Микропилярная клетка эндосперма
крупная, и в ней образуются свобод-
ные ядра; халазальная клетка ма-
ленькая, и ядро ее делится редко.
Эмбриогенез СагуорвуПад-типа. За-
родыш прямой, дифференцирован-
ный, односемядольный, хлорофилло-
носный, богат крахмалом. Подвесок
с пузыревидной базальной клеткой.
Семя, плод — семя без эндосперма.
Зародьше заполняет все семя,
ИНлод — мпоголистовка.
Гауеторий отсутствует.
Апомиксйе не обнаружен.
Другие явления не обнаружены.
JIutepatypa: Davis, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970.
CEMEMCTBO ZOSTERACEAE
Основное число хромосом — х = 6.
Цветки однополые, растепия одно-
домные или двудомные.
21%
Пыльник — число микроспоранги-
cB и строение стенки микроспоран-
гия не изучены. Тапетум амебоид-
ный.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
трехклеточные «питевидные», удли-
ненные до 2000 мк, без экзины.
Семяпочка одна, ортотропная, с
двумя интегументами, крассинуцел-
лятная. Имеются гипостаза и нуцел-
лярный колпачок. Сосудистый пучох
доходит до халазы.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]усопат-типу. Синергиды грушевид-
ные. Три антиподы дегенерируют в
начале эмбриогенеза. Полярные яд-
ра сливаются перед оплодотворе-
нием. У Гозега шайта хорошо вы-
ражен постамент.
Опыление, прохождение пыльце-
вой трубки и оплодотворение -— ист
данных.
Эндоспермогенез нуклеарный, ге-
лобиальный.
Эмбриогенез Опаста4-типа. Заро-
дыш прямой, дифференцированный,
односемядольный. Базальная клетка
подвеска очень крупная.
Семя, плод — семя без эндосперма.
Зародыш заполняет все семя.
Гаусторий не найден.
Апомиксие не обнаружен.
Другие явления не обнаружены.
Литература: Davis, 1966; Кудря-
шов, Савич, 1969; Савич, 1969; Тах-
таджян, 1966, 1970.
СЕМЕЙСТВО
РОТАМОСЕТОМАСЕАЕ
Основное число хромосом — х = 6,
7, 8, 10, 13, 15.
Цветок обоеполый.
Пыльник — 1 или 4 микроспоран-
rua. Стенка микроспорангия имеет
эпидермис, фиброзный эндотеций,
1—2 эфемерных средних слоя и
амебоидный, двухрядный, одноядер-
ный, развивающийся в истинный
периплазмодий тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные. Зрелые пыльцевые
212
зерна трехклеточные, безапертур-
ные, с тонкой сетчатой экзиной или
без нее, «нитевидные».
Семяпочка ортотропная, анатроп-
ная, амфитроппая или кампи-
лотропная, с двумя инптегумента-
ми, крассинуцеллятная. Микропиле
образуется внутренним интегумен-
том. Имеется гипостаза. Семяпочек 4.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор липейные или T-
образные. Иногда вместо тетрад мак-
роспор образуются триады. Зароды-
шевый мешок образуется по Ро]у-
gonum-Tuny. Синергиды широкие у
основания. Имеются три скоро де-
генерирующие антиподы.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез гелобиальный,
нуклеарный. При гелобиальном эн-
досперме в микропилярной камере
ядра спачала свободпые, затем меж-
ду пими возникают клеточпые пе-
регородки. В халазальной камере
ядро пе делится и дегенерирует,
а вслед за пим дегенерирует и
клетка.
Эмбриогенез Caryophyllad-runa.
Зародыш крупный, согнутый в виде
подковы, дифференцированный, од-
носемядольный, бесхлорофилльный,
содержит крахмал и масло.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Нлод из четырех или меньше ореш-
ков, или костяпок.
Гауеторий пе пайден.
Апомиксис пс обнаружен.
Другие явлепия пе обнаружены.
JIureparypa: Davis, 1966; Lakshma-
nan, 19670, 1970d; Schnarf, 1934; Ca-
вич, 1969; Тахтаджян, 1966, 4970,
СЕМЕЙСТВО ВКОРРТАСЕАЕ
Основное число хромосом — х = 8.
Цветок обоеполый, маленький,
анемофильпый.
Ныльник — тапетум амебоидный.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелая пыльца трехкле-
точпая, с тремя утолщенными ме-
стами и сетчатой сэкзиной.
Семяпочка кампилотропная, с дву-
мя интегументами, ’крассинуцел-
лятная. Микропиле образуется обои-
~
ми интегументами. Семяпочек
сколько.
Макроспорогенез и макрогаметогев-
нез — археспории одноклеточный.
Один слой париетальных клеток.
Тетрады макроспор линейные или
Т-образные. Зародышевый мешок
образуется по Polygonum-tuny.
Имеются три эфемерные антиподы.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохожедние пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез гелобиальный.
Эмбриогенез СатуорвуПад-типа.
Зародыш дифференцированный, од-
носемядольный. Подвесок однокле-
точный, пузыревидный.
Семя, плод — семя без эпдоспер-
ма. Зародыш заполняет все семя.
Плод из очень асимметричных ко-
стянок.
Гаусторий не найден.
Аномиксис не обнаружен.
Другие явления не отмечены.
Литература: Пау!з, 1966; Савич,
1968а, 6, 1969; Тахтаджян, 1966, 1970.
не-
СЕМЕЙСТВО 2АММСНЕШМАСЕАЕ
Основное чиело хромосом --х = 6,
17, 12.
Цветок обосполый или одпонпо-
лый, растения одно- или двудомиые.
Пыльник — 4
микроспорапгия.
Стенка микроспорапгия имсет эпи-
дермис, эндотеций, один средний
слой и амебоидный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двух-трехклеточные, безапертурные
или иногда с бороздновидной апер-
турой, с тонкой экзиной или без
нее, с зернами крахмала.
Семяпочка ортотропная или ана-
тропная, с двумя интегументами,
крассинуцеллятпая. Чаще всего 3—4
семяпочки, по может быть 9—1.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Париетальная ткань массивная. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Poly-
гопиш-типу. Три клетки антипод
притерпевают последующие деле-
ния, образуя 4—5 клеток. Полярные
ядра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — нет
данных.
Эндоспермогенез гелобиальный.
Эмбриогенез Caryophyllad-runa.
Зародыш согнутый в виде подковы,
дифференцированный. Базальная
клетка трехклеточного однорядного
подвеска, крупная, пузыревидная.
Семя, плод — семя без эндоспер-
ма. Зародыш заполняет все семя.
Кожура семени тонкая, образую-
щаяся почти полностью из наруж-
ного интегумента. Плод состоит из
нераскрывающихся костянковидных
плодиков.
Гаусторий пе пайден.
Апомиксис пе обпаружен.
Другие явления — пс обнаружены.
Литература: Davis, 1966; Lakshma-
nan, 1965a, b; 1967d, 1970f; Miceli Den-
хоп, 1978; Савич, 1969; Schnarf,
1934; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО МАЗАРАСЕАЕ
Основное число хромосом — х =
eran
— |
?
Цветок — однополый, растения
обычпо одподомные, редко двудом-
IbIC.
Пильник — 4 микроспорангия.
Стенка микроспорангия образуется
но редуцировапному типу и имеет
эпидермис, эпдотеций без фиброз-
HBIX утолщений и амебоидный, од-
поядерный, превращающийся в пе-
риплазмодий тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Микроспоры тетраэдраль-
ные, изобилатеральные, линейные и
Т-образные. Зрелые пыльцевые зер-
на техклеточные, апертуры и эк-
зина отсутствуют, имеются зерна
крахмала.
Семяпочка одна, анатропная, с
двумя интегументами, крассинуцел-
лятпая. Микропиле образуется внут-
рештим иптегумоптом.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
1—2 слоя париетальных клеток.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]угопит-типу. Синергиды грушевид-
ные. Три антиподы дегенерируют в
начале эмбриогенеза. Полярные яд-
ра сливаются во время оплодотво-
рения,
213
Опыление, прохождение пыльце-
вой трубки и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный. O6-
разование клеточных перегородок
начинается с микропилярного по-
люса зародышевого мешка.
Эмбриогенез Caryophyllad-ruma,
вариации Зас1ата. Зародыш диф-
ференцированный, прямой, односе-
мядольный, хлорофиллоносный, со-
держит масло и крахмал. Базальная
клетка подвеска сильно удлиняется.
Семя, плод — семя без эндосперма.
Зародыш заполняет все семя. Ко-
жура семени образуется из наруж-
ного и внутреннего слоев внешнего
интегумента и становится твердой.
Плод не раскрывающийся с тонким,
но мясистым околоплодником.
Гаусторий подвесочный.
Апомикеие не обнаружен.
Другие явления не отмечены.
JImreparypa: Davis, 1966; Lakshma-
nan, 1965, 1967d, 1970c; Schnarf, 1934;
Лодкипа, 1976; Тахтаджян. 1966,
1970.
ПОДКЛАСС
НАДПОРЯДОК П.
LILIA NAE
ПОРЯДОК 78.
TRIURIDALES
CEMEHMCTBO TRIURIDACEAK
Основное число хромосом — х=14,
22, 24.
Цветок обычно однополый, иногда
обоеполый, растения одно- или дву-
домные.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, не фиброзный эндотеций,
эфемерный средний слой и секре-
торный (?) тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, изобилатеральные.
Зрелые пыльцевые зерна трехкле-
214
В порядке Ма]адаез по семействам
Роз1доп1асеае и Сутойосеасеае ци-
тоэмбриологических данных нет.
Jlaxkmmanan [Lakshmanan, 1970] по-
лагает, что ЗеВеисЬте1асеае и Лал-
caginaceae, XOTA и самостоятельные,
но близкородственные семейства,
так как они имеют много сходства
в отношении цитоэмбриологии. Кро-
ме того, он считает, aTo Ruppia ua
Роашореюоп настолько сильно от-
личаются друг от друга, что отнесе-
ние их к разпым семействам Rup-
piaceae um Potamogetonaceae вполне
правильно. Он считает также, что
Najadaceae и Zannichelaceae род-
ственны друг другу, так как
они имеют много общего в отно-
шении цитоэмбриологии. По его
мнению [1967], Ма]дадасеае — высо-
коподвинутое семейство. Порядок
Ма}ада]ез, соответствующий поряд-
ку Неюае классификаций преж-
них ‚авторов, в общем состоит из’
семейств, цитоэмбриологические
признаки которых во мпогом сход-
ны, что ло противоречит объедине-
иию этих семейств в общем порядке.
B. LILITDAE
точные, однобороздные или безбо-
роздные.
Семяпочка анатропная, с двумя
интегумептами, тенуинуцеллятная.
Микропиле образуется внутренним
иптегументом. Семяпочек 6—50.
Макроспорогенез и макрогаметоге-
нез — архоспорий одноклоточный,
ПНаристальные клетки отсутствуют.
'Тотрады макроспор линейпые. За-
родышевый мешок образуется по
Ро]угопит-типу. Сипергиды груше-
видные. Имеются три пебольшие
эфемерные антиподы. Полярные яд-
ра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вых трубок и оплодотворение —
двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен;
маленький, прямой, недифференци-
рованный зародыш. Подвесок двух-
клеточный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
белок. Плодики с толстым около-
плодником,
OHO.
Гаусторий не найден.
Апомиксие пе обнаружен.
Другие явления — все представи-
тели семейства паразитические рас-
тения.
Литература: Оау!з, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970; Тере-
хин, 1977.
раскрывающимся про-
А. Л. Тахтаджян [41970] высказал
предположение, что порядок Triu-
г1а]ез имеет общее происхождение
с порядком АПзша]ез, что противо-
речит данным цитоэмбриологии, так
как представители этих порядков
‘отличаются друг от друга некото
рыми важными цитоэмбриологиче
скими признаками. В то время как
представители АП5та|о5$ характери-
зуются наличием АШат-типа заро-
дышевого мешка, гелобиальным эн-
доспермом и семенами без эндоспер-
ма, представители Тгци1Ча]ез имеют
Ро]усопит-тип зародышевого меш-
ка, нуклеарный эндосперм и семена
с хорошо развитым эндоспермом.
Кроме того, у представителей Тга-
т1Ча!ез птироко распрострапеи пара-
зитизм, отсутствующий у предста-
вителей АНзта]ез. Однако оба эти
порядка, особенно Тгии14а103 изучое
ны в отношении цитоэмбриологии
пока недостаточно, поэтому делать
окончательные выводы о взаимоот
ношениях этих порядков прежде-
временно.
ПОРЯДОК 79.
LILIALES
СЕМЕЙСТВО ГИЛАСЕАЕ
Основное чиело хромосом — х =
= 3—28, 30.
Цветок обычно обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2—3
эфемерных средних слоя и секре-
торный дву-многоядерный тапетум.
Имеются орбикулы.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное или симультанное. Тет-
рады микроспор изобилатеральные,
крестообразные, тетраэдральные, ли-
нейные. Зрелые пыльцевые зерна
двухклеточные, обычно одноборозд-
ные, с гладкой мелкосетчатой экзи-
пой с крахмальными — зернами.
Спермии-клетки бедно- или много-
плазменные.
Семяпочка анатропная, тгемиана-
тропная или ортотропная, с двумя
интегументами, красси- или тенуи-
пуцеллятная. Микропиле образуется
внутренним интегументом. Имеются
нуцеллярный колпачок, гипостаза,
ариллус, постамент, интегументаль-
ный обтуратор и эндотелий. Cocy-
дистый пучок доходит до халазы.
Семяпочек песколько — много.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-многокле-
точпый. Париетальные клетки при-
сутствуют или их нет. Тетрады мак-
роспор линейные, Г-образные. Иног-
да вместо тетрады образуется триа-
да макроспор. Зародышевые мешки
образуются mo Polygonum-, Allium-,
Fritillaria-, Adoxa-, Tulipa tetraphyl-
lea-, Eriostemones-, Огиза-типу и под-
типу С№июоща. Синергиды © крюч-
кообразпыми выростами и фибрил-
лярными аппаратами, — одно-трех-
мпогоядерпые антиподы. Число ан-
типод может увеличиваться с 3 до
141 и долго сохраняться. Антиподы
офемерные или долго сохраняются.
Иногда под антиподами имеется
постамент. Полярные ядра сливают-
ся до или во время оплодотворения.
В некоторых случаях нижнее по-
лярное ядро в оплодотворении не
участвует. Наблюдается депрессия
халазального конца зародышевого
мешка. У некоторых представителей
семейства в зародышевых мешках
имеется крахмал.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
pekpectnoe опыление. Порогамия.
Двойное оплодотворение премитоти-
ческого и постмитотического типов.
y Allium schoenoprasum двойное
оплодотворепие происходит через
23—24 ч после опыления. Первое де-
ление зиготы начинается на 4—5-й
день после опыления.
Эндоспермогенез гелобиальный,
нуклеарный. Первичное ядро эндо-
сперма делится раньше зиготы.
Описана полиплоидизация халазаль-
ных ядер эндосперма путем эндо-
митоза.
215
.. .
Ul
246
LM
Puc. 25. Микроспорогенез, микрогаметогенез и семяпочки у представителей
cemelicTBa Liliaceae
А — микроспорогенез сукцессивного типа у ТАПаш regale [Gerassimova-Navashina,
1957] Б — микрогаметогенез и сингамия у Lilium regale [Cooper, 1936] (1— двух-
клеточное пыльцевое зерно; 2 — пыльцевая трубка с генеративной клеткой и веге-
тативным ядром; $3, 4 — метаморфозы в пыльцевых трубках; 5, 6 — анафазы в пыль-
цевых трубках; 7, 8 — зрелые спермии и вегетативное ядро в пыльцевой трубке);
9 — вхождение пыльцевой трубки в зародышевый мешок и слияние одного из спер-
миев с яйцеклеткой; В — поперечный разрез завязи с семяпочками со зрелым за-
родышевым мешком (ла — яйцевой аппарат, пя — полярные ядра, ан — антиподы,
ни — наружный, ви — внутренний интегументы, м — микропиле, о — обтуратор,
с — семяпочка, см — семяножка, н — нуцеллус, э7 — эпидермис нуцеллуса, х — ха-
.лаза, пр — проводящий пучок) [Савченко, Комар, 1965]
Эмбриогенез Onagrad-, Asterad-,
гументальная эмбриония, партеноге-
Caryophyllad-, Chenopodiad-, Зо1а-
нез, апогаметия и апоспория.
па4-типа. Зародыш прямой, односе-
мядольный, слабо или хорошо диф-
ференцированный, бесхлорофилль-
ный, содержит крахмал.
Семя, плод — семена с хорошо раз-
витым эндоспермом, содержащим
масло, алейрон, иногда и крахмал.
Кожура семени образуется из на-
ружного интегумента, внутренний
интегумент при развитии семени
разрушается. Семядоля выполняет
гаусториальную функцию. Плод —
обычно коробочка, редко — ягода.
Гаусторий зародышевого мешка
и подвесочный.
Апомиксие — нуцеллярная и инте-
Другие явления — вивипария. По-
лиэмбриония. Добавочные зародыши
образуются из сипергид. Клейсто-
гамия.
Литература: Баранов, Поддубная,
1925; Berg, 1960, 1978; Беляева Н..,
1975, 1977; Cave, 1966, 1967, 1974,
1975; Davis, 1966; Dutt, 41967a,
1970a, {; Dutt, Rao P., 1975; TamO6a-
рова, 1972, 1975, 1978; Гваладзе, 1960,
1961, 1962, 1963, 1965, 1969a, 6, 1970,
1973, 1978а; Герасимова-Навашина,
Батыгина, 1958, 1964; Гусейнова,
1967; Капинос, 4960а, 6, 1964, 1972,
1978; Капинос. Гусейнова, 1967,
1968, 1969, 1972; Капинос, Кулиева,
217
GIL.
Рие. 26. Микро- и макроспорогенезы, а также развитие женского гамето-
muta y Phormium tenax
218
Ид}
ar”
y
cat
aie
as
12 rete
illaria-
it
ия зародышевого мешка Ег
3) развит
1
ica [Bamb
Рие. 27. Разные стадии (1—
ioni. 1928]
ich
ia pers
illar
it
Tuna y Fr
<—
ТГ enen
а:
oS eos
= Es
BEEPS
D> Spy Od
но
= i
© gy AO
ино
оннен
ЗЕЯ
Fa mee
Зоны
НЯ | <
НН. ©
© >.
о [ms
2 4
x Men
RAM
=~
rw
|
2
7—11 — разные стадии образования тетрады макроспор
12—14 — разные стадии развития зародышевого мешка
16 — микропилярная часть семяпочки с яйцевым аппара
6 — семяпочка
.
9
семяпочки с антиподальным аппаратом и вторичным ядро
1 == Диакинез в материнской клетке микроспор
ного типа
219
[866т ‘ениичь
= ‘внишечен-еяомиов эт]
имимонэж 9 змеи XUMWOWAWN
эиннико —Я ‘WHHOHOIH я
yewed xmyowAW эинэжиио
-идп — У ‘(иим@эпо — U9
‹апиа@энио ИочЧэн odry
~— low ‘апизбэнио — 22'2)
0119 |
18 90$ А ounedoaLor
-oruo эониоз ‘35 ‘эиа
HW OZ
.`
pene, 86
iy
253
|
DOS
РУ.
СС
Sy
aS
о
he
ot
a0
sees
оч Г
Ls dey
sou
sas
EP
<.)
ее
wy
= eA Se
' 7? „м 4
SN `
NS
et
> we .S oe ae af. fe „ - » „ *
Рис. 29. Двойное оплодотворение (А) и эмбриогенез (Б, В) у АШит пшапз
[Стожарова, 1973]
224.
ii
NNO ns
AIT
BRT)
Ам
СНА УЗ
PON ea
ДАК - -
я 4 33; Sood КА А
} bel 7 СА] als) ea >
SOS ia BEI FINED
CISA LIAM DLN НСО
44 Ale wp: rs] д PX 3 4 A Г
ЕР. ЕКО РАЕН А
ur te 99% POS AS So, PS ete | ОСЬ
СЕНО
АЗ A tq} Yet red) CTS
НИИ
GES м т ene Ue VOR LL Se АНЯ
РЯ р; 7 Е Г." TE A
ee eee ht
ея
ре
a CTS I г
ОА» Си ЧО Куча
(4
$4
i
К
о
|
_
КАРА ne os or me
АСЯ
Sp АХ
[Сто-
tans
Рис. 30. Поздние фазы (А —Е) развития зародьниа у АШиаш пла
жарова, 1973]
222
1972; Karagiosowa, 1970; Hoéaxuzse,
1965, 1967, 1970; Кострюкова, 1961а,
Kaul ct al. 1976; Mahabale, Inam-
дат, 1975; Мосепзеп, 1970; Мирками-
лов, 1969, 1972, 1977а, 6; Наумова,
Яковлев, 4975а, 6; Pahuja, Kumar,
4970; Петрова Т., 1977; Печени-
цын, 1965a, 6; 1972a, 6; Романо-
ва, 1938, 1939, 1945, 1957, 1965, 1971;
Poros, Петрова, 41977; Шпилевой
um yp., 1978; Schnarf, 1934; Соколов,
4968a, 6, B, 1969; Sokolowska-Kul-
ezycka, 1973, 1977, 1978; Солнцева,
Ворсобина, 41978; Стожарова, 1973;
Стожарова, Поддубная-Арнольди,
1977; бисШага её а1|., 1969; Тагиева,
1970; Takeuchi, Yasuyoshi, 1970;
Тахтаджян, 1966, 1970; Ворсобина,
Солнцева, 1979а, 6; Ворсобина, 1973;
Yasuyoshi, 1970; Zabinska, 1972.
СЕМЕЙСТВО
XANTHORRHOEACEAE
Основное число хромосом — х==141.
Цветок обычно однополый, редко
обоеполый.
Пыльник — нет данных.
Микроепорогенез и микрогаметоге-
нез—о числе микроспорангиев и
строении стопки пыльпика нот дап-
ных. Пыльцоевые зерна обычно од-
нобороздные.
Семяпочка апатронная, © двумя
интегументами, крассипуцоллятная.
Микропиле образуется впутренним
интегументом. Имсетея нуцелляр-
ный колпачок. Семяпочек одна -
много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный,
три ряда париетальпых клеток. За-
родышевый мешок образуется по
Ро|угопат-типу. Синергиды груше-
видные, с фибриллярными аппара-
тами. Имеются три клетки антипод.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение --- нет
данных.
Эндоспермогенез — ист данпых.
Эмбриогенез — тил не определен.
Зародыш прямой.
емя, плод р— семя, с довольно
твердым роговидным эндоспермом.
Плод — локулицидная коробочка
или односеменный орех.
Гауеторий не найден.
Апомикесис не обнаружен.
Другие явления не отмечены.
Литература: Пау1з, 1966; Тахтад-
жян, 1966, 1970.
СЕМЕЙСТВО НАЕМОРОКВАСЕАЕ
Основное чиело хромосом — 7 =
= 4—8.
Цветок обоеполый.
Пыльник — нет данных о числе
микроспорангиев. Стенка микроспо-
рангия ‘четырехслойная. `Тапетум
амебоидный или секреторный дву-
ядерный.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное или симультанное. Тетра-
ды микроспор тетраэдральные, изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, одпоборозд-
ные, с оперкулумом или без него.
Семяпочка гемитропная, орто-
тропная или анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими инте-
гументами. Имеется нуцеллярный
колпачок. Семяпочек несколько —
мпого.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный,
2—4 ряда париетальпых клеток.
Тотрады макроспор линейные, Г-
образные. Зародышевый мешок об-
разуется по Ро]ухорят-типу. Синер-
гиды грушевидные. Имеются три
скоро дегенерирующие антиподы.
Нолярные ядра сливаются перед
онлодотворепием.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пет
данных.
Эндоспермогенез гелобиальный,
пуклеарный.
Эмбриогенез Asterad-runa. Заро-
дыш маленький, прямой.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Плод — локулицидная ко-
робочка.
Гаусторий эпдоспермальпый (ха-
лазальный). Халазальпая клетка ге-
лобиальпого эндосперма становится
мпогоядерной или разделяется на
четыре клетки и проявляет слабую
гаусториальную активность. Микро-
пилярпая клетка эндосперма после
периода свободного деления ядер
разделяется на много клеток.
Другие явления не обнаружены.
Литература: Davis, 1966; Dutt,
1967c, 1970c; Lakshmi, 1979; Taxraz-
man, 1966, 1970.
223.
СЕМЕЙСТВО НУРОХТАСЕАЕ
Основное число хромосом — 5 = 6,
9, 41.
Цветок обоеполый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—4
средних слоя и амебоидный с пе-
риплазмодием или секреторный
многоядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, тетраэдральные. Зре-
лые пыльцевые зерна двухклеточные,
однобороздные.
Семяпочка гемианатропная, ана-
тропная, кампилотропная, с двумя
интегументами, красси- или тенуи-
нуцеллятная. Микропиле зигзагооб-
разное, образуется либо обоими
интегументами, или только BHYT-
ренним. Семяпочек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одпоклоточный,
Париетальные клетки присутствуют
или их нет. Тетрады макроспор ли-
нейные, Г-образные. Зародышевые
мешки образуются mo Polygonum-
и АШию-типу. Синергиды с фибрил-
лярными аппаратами. Обычно три
эфемерные антиподы, но иногда
число их увеличивается, и они долго
сохраняются. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пылье-
вых трубок и оплодотворение — по-
рогамия.
Эндоспермогенез
нуклеарный.
Эмбриогенез Азегад-типа, вариа-
ции Anthericum. Sapoppum малопь-
кий, прямой, недифферопцирован-
ный.
Семя, плод — ссмя с хорошо раз-
витым эндоспермом. Семенная ко-
жура образуется из обоих интегу-
ментов. Плод — коробочка или соч-
ный, нераскрывающийся.
Гаусторий — халазальная клетка
гелобиального эндосперма становит-
ся гаусторием.
Апомиксис не обнаружен.
Другие явления не обнаружены.
JIurepatypa: Davis, 1966; Arecal,
1967; Паи, 19674, 19704; Тахтаджян,
1966, 1970.
гелобиальный,
224
CEMEHCTBO VELLOZIACEAE
Основное число хромосом — нот
данных.
Цветок обоеполый.
Ныльник — число микроспоранги-
ев неизвестно. Стенка микроспоран-
гия состоит из эпидермиса, эндоте-
ция, 1—3 средних слоя и секретор-
ного одноядерного тапетума.
Микроепорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, одно-дисталь-
нобороздные. У УеПома compacta
ныльцевые зерпа соединены в тет-
рады.
Семяпочка анатропная, с двумя
интегументами, тенуинуцеллятная.
Имеются фуникулярный обтуратор
и эндотелий. Семяпочек много.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный,
Париетальные клетки отсутствуют,
Зародышевый мешок образуется по
Ро[угопат-типу. Аптиподы много-
ядерные.
Опыление, прохождение пылье-
вых трубок и оплодотворение — нет
данных.
Эндоепермогенез нуклеарный. Пе-
риферический слой эпдосперма со-
держит крахмал и белок.
Эмбриогенез — тип не определен.
Зародыш маленький, прямой.
Семя, плод — семя с хорошо раз-
витым и довольно твердым эндо-
спермом. Плод — коробочка.
Гаусторий энтиподальный, клет-
ки гаустория многоядерные. Семя-
доля выполняет гаусториальную
фупкцию.
Апомиксие но обпаружен.
Другие явления пе обнаружены.
Литература: Davis, 1966; Dutt,
1967e, 19700; Monezes, 1976; Schnarf,
1931; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО PHILESIACEAE
Основное чиело хромосом — нет
данных.
Цветок обоеполый.
Пыльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций и сек-
реторный (?) тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелые цыльцевые з6р-
на двухклеточные, обычно однобо-
роздные, реже безапертурные.
Семяпочка кампилотропная, с дву-
мя интегументами, крассинуцеллят-
ная. Микропиле образуется внут-
реппим интегументом. Семяпочек
много или немного.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Два слоя париетальных клеток. За-
рольшевый мешок образуется по
Ро]угопат-типу. Синергиды груше-
видные. Имеются три крупные ан-
типоды.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нег
данных,
Эндоспермогенез — нет данных.
Эмбриогенез — нет данных.
Семя, плод — плод ягода.
Гаусторий — нст даппых.
Anomukene — пост даппых,
Другие явления — пет даппых.
Литература: Davis, 1966; Тахтад-
жян, 1966, 1970.
СЕМЕЙСТВО ТЕСОРНП.АЕАСЕАЕ
Основное чиело хромосом — х==12.
Пветок обоеполый.
Пыльник — пет даппых о числе
микроспорапгиев и строспий стопки
микроспораигия. Танотум секро-
торный (?), двуядерпми.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдралытые. Зрелые ныльцевьо зер-
на двухклеточные, однобороздные.
Борозда оперкулятная.
емяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими инте-
тументами или только внутренним.
Имеются нуцеллярный колпачок и
фуникулярный обтуратор. Семяпо-
чек много.
Макроепорогенез и макрогаметоге-
нез — археспорий одпоклеточный,
1—2 слоя париетальпых клеток. Тот-
рады макроспор липейные. Ипогда
образуются триады. Зародышевый
мешок образуется по Ро]узопат-ти-
пу. Синергиды грушевидные. Три
антиподы либо скоро дегенерируют,
либо сохраняются до начала обра-
зования эндосперма. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
зых трубок и оплодотворение — нет
данных.
15 в. л. Поддубная-Арнольди
Эвдоспермогенез нуклеарный,
позднее становится целлюлярным,
за исключением халазальпого гаус-
тория, который остается свободно —
ядерным.
Эмбриогенез — тип не известен. За-
родыш довольно крупный диффе-
ренцированный, односемянный, пря-
мой,
Семя, плод — семя с хорошо раз-
витым эндоспермом. Плод — локули-
цидная коробочка.
Гауеторий зародышевого
эндоспермальный
Семядоля выполняет
ную функцию.
Апомиксие — нуцеллярпал
риония.
Другие явления — полиэмбриония
y Cyanella capensis.
Jlareparypa: Davis, 1966; Тахтад-
жян, 1966, 1970.
мешка,
(халазальный).
таусториаль-
эмб-
СЕМЕЙСТВО СУАМАЗТВАСЕАЕ
Основное число хромосом — z =
= 11, 42,
Цветок обоеполый.
Ныльник — число микроспорангиез
не известно. Степка микроспорангия
имеет эпидермис, эндотеций, 1—4
средпих слоя и секреторпый много-
ядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивпое, симультапное. Тетрады
микроспор тетраэдральпые, изоби-
латеральные. Зрелые пыльцовые зер-
на двухклеточпые, однобороздные,
иногда трехщелевые.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная,
Имеется плацентарный обтуратор.
Микропиле образуется внутренним
интегументом. Семяпочек 6.
Макроепорогенез и макрогаметоге-
нез — археспорий одноклеточный,
Париетальные клетки имеются. За-
родышевый мешок образуется по
Ро]увопит-типу. Антиподы малень-
кис, эфемерпые.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен,
Зародыш крупный, дифференциро-
ванный и односемядольный,
Семя, плод — семя без эндосперма,
но спериспермом, содержащим крах-
мал, Плод обычно односемянный.
225
Гаусторий не обнаружен.
Апомиксие — нуцеллярная
риония.
Другие явления не обнаружены.
Литература: Davis, 1966; Dutt,
1967Ъ, 19706; ЗсЬпатЁ, 1934; Тахтад-
жян, 1966, 1970.
эмб-
СЕМЕЙСТВО АЗРАВАСАСЕАЕ
Основное число хромосом — х==10.
Цветок обоеполый или однополый,
растения обычно однодомные, реже
двудомные.
Пыльник — число микроспоранги-
ев не известно. Стенка микроспо-
рангия имеет эпидермис, фиброз-
ный эндотеций, один средний слой
и секреторный ядерный тапетум.
Микроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, ГТ-образные, кресто-
образные и изобилатеральные. Зре-
лые пыльцевые зерна двухклеточ-
ные, с зернами крахмала, однобо-
роздные или борозды более или ме-
нее редуцированы, с гладкой эк-
зиной.
Семяпочка анатропная, гемитроп-
ная, с двумя интегументами, крас-
синуцеллятная. Микропиле образу-
ется внутренним интегументом. Се-
мяпочек 6.
Макроспорогенез и макрогаметоге-
нез — археспорий одно-двуклеточ-
ный. Париетальные клетки имеются.
Тетрады макроспор линейные и 7-
образные. Зародышевый мешок об-
разуется по Polygonum-tuny. Cu-
нергиды с крючкообразными выро-
стами и фибриллярными аппарата-
ми. Антиподы крупные.
Опыление. прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез — нет данных.
Эмбриогенез — тип не известен.
По нашим данным, зародыш круп-
ный, дифференцированный, односе-
мядольный, бесхлорофилльный.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Плод — ягода.
Гауеторий зародышевого мешка.
Апомикеие не обнаружен.
Другие явления не обнаружены.
Литература: Mahabale, Inamdar,
4975; Lazarte, Palser, 1979; Schnarf,
4931; Venkateswarlu, Raju, 1958; Tax-
таджян, 1970. ,
226
СЕМЕЙСТВО ЗТЕМОМАСЕАЕ
Оеновное число хромосом — x = 7.
Цветок обоеполый.
Пыльник —`4 микроспорангия.
Строение стенки микроспорангия не
изучено.
Микроепорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двухклеточные, однобороздные.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Имеется ариллус. Семяпочек две —
много.
Макроспорогенез и макрогаметоге-
нез — зародышевый мешок обуазу-
ется по Ро]угопат-типу.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен.
Зародый маленький,
емя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Плод — коробочка, рас-
крывающаяся двумя створками.
Гаусторий не найден.
Апомиксис не обнаружен.
Другие явления не обнаружены.
Литература: ЭсВпатЁ, 1934; Тах-
таджян, 1966, 1970.
СЕМЕЙСТВО DIOSCOREACEAE
Основное число хромосом — х =
‘='40, 42.
Цветок однополый.
Ныльник — 4 — микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, 2 эфе-
мерных средних слоя и секреторный
двуядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные, изобилатеральные. Зре-
лые пыльцевые зерна двухклеточ-
ные, однобороздные, двух (трех) -бо-
роздные или четырехпоровые. Пыль-
ца очень быстро теряет свою жиз-
неспособность (через час).
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется двумя инте-
гументами. Имеются гипостаза ип
ариллус. Семяпочек несколько —
много.
Макроспорогенез и макрогаметоге-
нев — археспорий одноклеточный.
Носколько слоев париетальных кле-
ток. Тетрады макроспор линейные.
Зародышевый мешок образуется по
Ро]узопат-типу. Синергиды с крюч-
кообразными Выростами. Имеются
три скородегенерирующие антипо-
ды. Полярные ядра сливаются пе-
ред оплодотворением.
Опыление, прохождение пыльце-
вых трубок ‘и оплодотворение — по-
рогамия. Двойное оплодотворение
премитотического типа. У ПГ1озсогеа
опо происходит через двое суток
после опыления.
Эндоспермогенез нуклеарный. 0б-
разование клеточных перегородок
между свободными ядрами начина-
стся с халазального полюса заро-
дышевого мешка.
Эмбриогенез Азега4-типа, Заро-
дыш маленький, слабо дифференци-
роваппый, односемядольный, бес-
хлорофилльный. Подвесок широкий,
многоклеточный. Зародыш развива-
стся очень медленно (80—90 дней).
Семя, плод — семя с роговидным
эпдоспермом. Плод — коробочка (ча-
сто крылатая) или редко ягода.
Гаусторий — семядоля выполняет
гаусториальную функцию,
Апомикеис пе обнаружен,
Другие явления не обнаружены.
Jlwrepatypa: Davis, 1966; Schnarf,
1934; Тахтаджян, 1966, 1970; Ta-
keuchi, Kimura, 1968, 1974, 1972; FOp-
цев, Юрцева, 1970, 4973, 1975, 1978;
Yasuyoshi, 1971, 1972; Yasuydshi, Ki-
mura, 1968.
СЕМЕЙСТВО ТАССАСЕАЕ
Основное число хромосом — х==45.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по двудольному типу и имеет эпи-
дермис, эндотеций, 2 эфемерных
средних слоя и секреторный дву-
ядерный тапетум.
икроспорогенез и микрогаметоге-
нез — образование микроспор си-
мультанное. Тетрады микроспор тет-
раэдральные. Зрелые пыльцевые зер-
на двухклеточные, однобороздные.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется корункула.
Семяпочек много.
15*
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Один ряд париетальных клеток. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Ро]уво-
пит-типу. Синергиды грушевидные.
Имеются три эфемерные антиподы,
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не определен,
Зародыш маленький, прямой, одно-
семядольный, содержит крахмал.
Семя, плод — семя с хорощю раз-
витым роговым эндоспермом, содер-
жащим масло и алейрон. Плод —
ягода или локулицидная коробочка,
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомикеие не обнаружен.
Другие явления не обнаружены
Литература: Пау1з, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО РОМТЕРЕВТАСЕАЕ
Основное число хромосом — х = 8,
44, 15, 26.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Строение стенки микроспорангия не
изучено. Тапетум амебоидный, дву-
ядерный.
икроспорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна `двухклеточ-
ные, с 1—2(3) дистальными бороз-
дами.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется двумя инте-
гументами. Семяпочек много, реже
одна.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Один слой париетальных клеток.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
lygonum-tuny. Синергиды с фиб-
риллярными аппаратами. Имеются
три антиподы в виде ядер или
клеток, эфемерны. Полярные, ядра
сливаются перед оплодотворением.
У Наегат Вега’ Нтозит и Рощеде-
та сот4аба антиподы лежат на по-
стаментах.
227
Опыление, прохождение ныльце-
вых трубок и оплодотворение — пе-
рекрестное опыление у Рощедега
cordata. Tloporamua. Двойное опло-
дотворение.
Эндоспермогенез гелобиальный.
В обеих камерах сначала образуют-
ся свободные ядра, а затем между
ядрами возникают клеточные пере-
городки.
Эмбриогенез Аз{егай-типа. Заро-
дыш прямой, — дифференцирован-
ный, односемядольный, содержит
масло. Подвесок короткий.
Семя, плод — семя с эндоспермом,
содержащим крахмал. Плод — мно-
тосемянная, локулицидная коробоч-
ка, односемянный или ореховидный.
Гауеторий у Monochoria spp. Мик-
ропилярная клетка гелобиального
эндосперма имеет два боковых гаус-
тория. Семядоля выполняет гаусто-
риальную функцию.
Апомикеиес не обнаружен.
Другие явления не отмечены.
Читература: Пау1з, 1966; Schnarf,
1931; Тахтаджян, 1966, 4970.
СЕМЕЙСТВО РНП.УРВАСЕАЕ
Основное чиело хромосом — 5 =
== 8, 47.
Цветок обоеполый.
Пыльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 2—4
эфемерных средних слоя и секре-
торный двуядерный тапетум.
Микроепорогенез и микрогаметоге-
нез — образование микроспор сук-
цессивное. Тетрады микроспор ‘тет-
раздральные, изобилатеральныо. Зре-
лые пыльцевые зерпа двухклеточ-
ные, обычно однобороздные, с сет-
чатой экзиной. Пыльцевые зерна
соединены в тетрады.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеются карункула
и фуникулярный обтуратор. Семя-
почек много.
Макроспорогенез и макрогаметоге-
нез — археспорий одноклеточный.
Олин слой париетальных клеток.
Тетрады макроспор линейные. Заро-
хышевый мешок образуется по Ро-
}усопаш-типу. Синергиды с крючко-
образными выростами. Имеются три
эфемерные антиподы. Полярные яд-
228
ра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльдце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Сингамия и тройное слияние обычно
протекают одновременно, реже син-
гамия предшествует тройному слия-
нию.
Эндоспемрогенез гелобиальный.
Обе камеры сначала многоядерные,
а позднее между ядрами возникают
клеточные перегородки.
Эмбриогенез Onagrad-tuna. Philyd-
rum- ula Juncus-Bapuanun. Зародыш
прямой, односемядольный.
Семя, плод — семя с мясистым эн-
доспермом, содержащим крахмал.
Плод — локулицидная коробочка.
Семенная кожура образуется из
двух интегументов.
Гауеторий — семядоля выполняет
гаусториальную функцию.
Апомиксис не обнаружен. ,
Другие явления не обнаружены.
JIutepatypa: Davis, 1966; Kapil,
4967g¢; Kapil, Walia,
anu, 1966, 1970.
1965; Тахтад-
Из 17 семейств порядка Г Па|ез,
согласно классификации А. Л. Тах-
таджяна [1970], три семейства,
a umenHo Aphyllanthaceae, Peterman-
niaceae u Smilacaceae, NOUTH HIM BOB-
се не исследованы цитоэмбриологи-
чески. Остальные семейства порядка
исследованы более или менее под-
робно, но еще далеко не достаточно,
особенно слабо изучены семейства
Xanthorrhoeaceae и РЬПезасеае. По-
рядок ТПаоз очепь гетерогенный.
Особонпо готорогеппо семейство Ш-
Пасоао, в котором обнаружено не-
сколько разпых типов микроспоро-
генеза, семяпочок, Makpocmopore-
незаа зародышевых мешков, эн-
доспермо- и эмбриогенезов, апомик-
сиса и т. д. Гетерогенность, возмож-
но, указывают на то, что в этом по-
рядке и внутри некоторых его се-
мейств (как, например, ГлПасеае)
объединены неродственные таксо-
ны. Тем не менее между некоторы-
ми семействами обнаружено сход-
ство в цитоэмбриологических при-
знаках. Так, например, Наетодога-
сеае в отношении цитоэмбриологии
наиболее близко к Hypoxidaceae
[Dutt, 419704]. По мнению Дутта
[Рай, 1970], включение АтагуШ9а-
свае в семейство Т1Насеае правиль-
110, так как цитоэмбриологически
семейства эти очень сходны.
Xatanucon [Hutchinson, 1959] can-
тал, что Суапа гит и СуапеНа род-
ственны друг другу и их следует от-
нести к ТесорбуПасеае. Однако Дутт
[Рай, 1970] пришел к заключению,
что, так как Суапазтасеае и Те-
сорвуПасеае отличаются друг от
друга по цитоэмбриологическим при-
знакам, их следует считать само-
стоятельными семействами и распо-
лагать вблизи с Dioscoreaceae u
Тассасеае. УеПожма сходна с Атагу|-
|1дасеае, но по данным морфологии
больше заслуживает ранга само-
стоятельного семейства [Dutt, 1970].
РЬПуйгасеае обнаруживает большее
сходство с Роп(едемасеае и Вготе-
Насеас, чом c Commelinaceae u Or-
chidaceae.
Нам представляется, что порядок
АПа]ез в целом и его главное се-
мейство ГЛПасеае следовало бы сде-
лать менее обширными. Хотя се-
мейства АтагуШдасеае и Асатасеае
по цитоэмбриологическим призна-
кам очень сходны c Liliaceae, тем
не менее их лучше сохрапить как
самостоятельные семейства, так как
они отличаются от ТлНасеае по ряду
морфологических и экологических
признаков.
ПОРЯДОК 80.
IRIDALES
CEMEMCTBO IRIDACEAE
Оеновное число хромосом — х=
= 3—18, 22, 25.
Цветок обоеполый.
Ныльник — 4 микроспоравгия.
Стенка микроспорангия имеет эпи-
дермис, средний слой и секреторный
гапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
изобилатеральные, тетраэдральные.
Зрелые пыльцевые зерна двухкле-
точные, обычно однобороздные, из-
редка с двумя дистальными бороз-
дами.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеется обтуратор.
Обычно семяпочек много, редко не-
сколько или одна.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный,
один, несколько слоев париетальных
клеток. Тетрады макроспор линей-
ные, Г-образные. Зародышевый ме-
шок образуется no Polygonum-tury.
Синергиды © крючкообразными вы-
ростами и фибриллярными аппара-
тами. Имеются три актиподы, деге-
нерирующие в начале развития эн-
досперма, Нолярные ядра сливают-.
ся перед оплодотворением. У неко-
торых представителей семейства
обнаружены антиподальные поста-
менты.
Опыление, прохождение пыльцке-
вых трубок и оплодотворение — пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез Азегай-, Caryophyl-
lad-runa. Зародыш дифференциро-
ванный, бесхлорофилльный, содер-
жит маело и алейрон. Подвесок ко-
роткий, многорядный.
Семя, плод — семя с роговым эн-
доспермом. Семенная кожура обра-
зуется в основном наружным инте-
гументом. Нлод — локулицидная ко-
робочка.
Гаусторий не обнаружен.
Апомиксие — партеногенез.
Другие явления — полиэмбриония
и клейстогамия.
Литература: Davis, 1966; Кордюм,
1963; Lakshmanan, Philip, 19714,
19735; Савченко, Комар, 1065;
Schnarf, 1931; Steyn, 4973a; Schnarf,
1931; Тахтаджян, 4966, 1970; Яков-
лев, Соколовская, 1978.
СЕМЕЙСТВО ВОЕКМАММТАСЕАЕ
Основное
— 6, 8.
Цветок обоеполый.
Ныльник — число микроспоранги-
ев не известно. Стенка микроспоран-
гия имеет эпидермис, эндотеций,
один средний слой и секреторный
одно-двуядерный тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двух, трехклеточные, однобороздпные
или однопоровые, иногда безапер-
турные.
чиело хромосом — Z7 =
229
Семяпочка аНатропная, с двумя
интегументами, ‘тенуинуцеллятная.
Семяпочек много.
Макроспорогенез и макрогамето-
генез — археспорий одно-, двухкле-
точный. Париетальные клетки от-
сутствуют. Тетрады макроспор ли-
нейные. Зародышевые, мешки обра-
зуются по Ро[угопит- и АШам-ти-
пу. Синергиды грушевидные. Име-
ются три маленькие эфемерные ан-
типоды. У ТЫзима атег1сапа анти-
поды присутствуют в форме не кле-
TOK, а ядер. Полярные ядра слива-
ются перед оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез гелобиальный.
Эндосперм содержит амилодекстрин.
Эмбриогенез Опастад-, Азегад-ти-
па. Развитие зародыша останавли-
вается на четырех, десятиклеточ-
ной стадиях, однако после опадения
семени на субстрат продолжается
его развитие. Зародыш недиффереп-
цированный.
Семя, плод — семя со слабо разви-
тым эндоспермом или без него.
Плод — коробочка, часто трехкры-
лая.
Гаусторий — в халазальной части
эндосперма образуется дву- или че-
тырехъядерный гаусторий.
Апомиксис — нередуцированный
партеногенез, апогаметия, генера-
тивная апоспория у Burmannia
coelestis m Thismia javanica.
Другие явления — полиэмбриония.
Клейстогамия у Вигшапта. Пара-
зитизм.
Литература: АгеКа], Ramaswamy,
1973; Davis, 1966; Schnarf, 1934; Ra-
maswamy, 1967, 1970; Тахтаджян,
1966, 1970; Терёхин, 1977.
Из четырех семейств, относимых
А. Л. Тахтаджяном к порядку Iri-
4а]ез, семейства Geosiridaceae au
Corsiaceae цитоэмбриологически не
изучены. А. Л. Тахтаджян сближает
'Burmanniaceae c Iridaceae, ofHaKko
первое семейство отличается от вто-
рого рядом существенных цитоэм-
бриологических признаков, а имен-
но: сукцессивным, а не симультан-
ным типом микроспорогенеза, гело-
биальным, а не нуклеарным типом
эндосперма, недифференцированным
зародышем, присутствием эндоспер-
230
мального гаустория.в халазальной
части зародышевого мешка. Для вы-
яснения систематического положе-
ния Burmanniaceae — необходимы
дальнейшие исследования.
ПОРЯДОК &.
ZINGIBERALES
CEMEMCTBO STRELITZIACEAE
Основное чиело XpomocoM — z =
= 7, 8, 11, 12.
Цветок обоеполый.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 3—6
эфемерных средних слоев и секре-
торный двух-трехрядный тапетум,
Микроспорогенез и микрогамето-
генез —‘образование макроспор сук-
цессивное. Тетрады микроспор T-
образные, липейные. Зрелые пыль:
цевые зерпа двухклеточпые, без-
апертурные.
Семяпочка апатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется обоими инте-
гументами. Имеется ариялус. Семя-
почек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Один слой париетальных клеток.
Тетрады макроспор линейные, T-
образные. Зародышевый мешок об-
разуется по Ро[угопит-типу. Синер-
гиды грушевидные. Имеются три
скоро дегенерирующие антиподы.
Полярные ядра сливаются перед
оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение пе-
рекрестное опыление у Утешла ге-
ginae.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен.
Зародыш маленький, прямой, удли-
ненный, дифференцированный с од-
ной массивной семядолей. Подвесок
короткий.
Семя, плод — семя с хорошо раз-
витым мучнистым периспермом и
остатками эндосперма. Плод — яоку-
лицидная коробочка. .
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Davis, 1966; Martin,
1070; Philip, #968; Тахтаджян, 1966,
1070; Venter, 1976a, b.
СЕМЕЙСТВО МОЗАСЕАЕ
‚
Основное число хромосом — х =
кз 7, 90—13.
Цветок однополый или обоеполый.
Пыльник — число микроспоранги-
св и строение стенки микроспоран-
гия не изучены. Танетум секретор-
ный.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
трехклеточные, безапертурные, с тон-
кой экзиной и толстой интиной.
Семяпочка ”анатропная, © двумя
интегументами, крассинуцеллятная,
Микропиле образуется обоими ин-
тегументами, Семяпочек мпого, Име-
ется ариллус.
акроспорогенез и макрогамето-
генез — археспорий одноклеточный,
один — два слоя париетальных кле-
ток. Тетрады макроспор линейные,
Т-образные и /-образные. Зароды-
шевый мешок образуется по Ро]у-
гопат-типу. Синергиды грушевид-
ные. Имеются три скородегенери-
рующие антиподы, представленные
ядрами или клетками. У Маза сос-
сшеа антиподы лежат на поста-
менте.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление у видов Млза.
Порогамия. Тройное слияние осу-
ществляется вблизи антипод.
Эндоспермогенез нуклеарный.
- У Musa аспшшаё эндосперм стано-
вится клеточным через 55 дней пос-
ле опыления.
Эмбриогенез — тип не определен.
Зародыш прямой, маленький, но
дифференцированный, с одной семя-
долей. Подвесок короткий.
Семя, плод — семя с хорошо раз-
витым мучнистым периспермом и
остатками эндосперма, содержащи-
ми крахмал. Плод ягодообразный,
мясистый, толстокожий, выделенный
в особый тип, называемый «Банан».
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления — полиэмбрио-
ния. Партенокарпия у видов Муза.
Литература: Пау1з, 1966; ЗеВпатЬ
1934; Тахтаджян, 1966, 1970.
CEMEMCTBO ZINGIBERACEAE
Оеновное число хромосом — х=9,
44, 42, 16, 17, 24, 24, 26. -
Цветок обычно обоеполый.
- Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, 4—5
средних слоя и секреторный дву-
ядерный тапетум. У Атотишм deal-
рат и №со]а1а абториграгеа тапе-
тум амебоидный.
Микроепорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, линейные, ГТ-образ-
ные, тетраэдральные. Зрелые пыль-
цевые зерна двухклеточные, одно-
бороздные или безапертурные с топ-
кой экзиной, содержат крахмал.
Семяпочка анатропная, изредка
ортотропная, с двумя интегумента-
ми, крассинуцеллятная. Микрониле
образуется внутренним интегумеп-
том. Имеются эпистаза, гипостаза,
карункула, ариллус, нуцеллярный
колпачок и постамент. Семяпочек
много. °
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Один ряд париетальных клеток.
Тетрады макроспор линейные. Обыч-
но зародышевый мешок образуется
no Polygonum-tuny, no у Costus 06-
паружен Адоха-тип. Синергиды с
крючкообразными выростами. Име-
ются три эфемерные антиподы, По-
лярные ядра, располагаясь вблизи
халазальной части зародышевого
мешка, сливаются перед оплодотво-
рением. У некоторых представите-
лей семейства имеются более или
менее хорошо выраженные антипо-
дальные постаменты.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление. Порогамия.
Двойное оплодотворение. Сингамия
и тройное слияние происходят син-
хронно.
Эндоспермогене гелобиальный.
Халазальная камера эндосперма де-
генерирует на двуядерной стадии.
В микропилярной камере деление
ядер продолжается до начала омб-
риогенеза, после чего между ядрами
начинают возникать клеточные пе-
регородки.
Эмбриогенез Азега4-, Caryophyl-
]а4-типа. Зародыш прямой, малень-
кий, односемядольный, дифференци-
231
‚рованный, бесхлброфилльный. Под-
весок, не образуется. =
Семя, плод — семя с хорошо раз-
витым периспермом и остатками эн-
досперма, ‘содержащими крахмал,
масло, алейрон и кристаллы оксала-
та кальция. Плод — коробочка, реже
ягодообразный.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления — полиэмбриония
у Недусит аси тайии. Добавоч-
ные зародыши возникают из синер-
Гид.
Jlurepatypa: Davis, 1966; Panchaks-
harappa, 1966, 1967, 1970; Philip,
1968; Sachar, Arora, 1963; Schnarf,
19384; Тахтаджян, 1966, 1970.
СЕМЕЙСТВО CANNACEAE
Основное чиело хромосом — х=9.
Цветок обоеполый.
Пыльник — число микроспоранги-
ев и строение стенки микроспоран-
тия не изучены. Тапетум амебоид-
ный © истинным периплазмодием.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное, сукцессивное. Зрелые
пыльцевые зерна двухклеточные,
безапертурные с шиповатой экзи-
ной. .
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Имеются нуцелляр-
ный колпачок и гипостаза. Семяпо-
чек много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Один ряд париетальных клеток,
Тетрады макроспор линейные, За-
родышевый мешок образуется по
Ро]узопит-типу. Синергиды груше-
видные. Имеются три антиподы,
представленные не клетками, а яд-
рами. Полярные ядра находятся
вблизи антипод, сливаются перед
оплодотворением. У Canna _ indica
под антиподами обнаружен поста-
мент.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — но-
рогамия. Двойное оплодотворение.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен.
Зародыш прямой, маленький, одно-
семядольный, содержит крахмал,
232 -
масло,
развит.
Семя, плод — семя с хорошо раз-
витым, твердым периспермом и тон-
ким слоем эндосперма. Зародыш за-
нимает от 1/4 до 1/2 осевой длины
семени. Плод — раскрывающаяся ко-
робочка, иногда не раскрываю-
щийся.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не отмечены.
Jinteparypa: Davis, 1966; Schnarf,
1931; Тахтаджян, 1966, 1970.
алейрон. ‘Подвесок слабо
2
СЕМЕЙСТВО MARANTHACEAE
Основное 4nCIO xpomMocoM — z=4,
6, 9, 44—48.
Цветок обоеполый. .
Пыльник — число микроспоранги-
ев и фтроение стенки микроспоран-
гия не изучены. °
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
двухклеточные безапертурные с
тонкой и гладкой экзиной.
Семяпочка одна, анатропная или
анакампилотропная, с двумя интегу-
ментами, крассинуцеллятная. Мик-
ропиле ‘образуется обоими интегу-
ментами или только одним внут-
ренним. Имеется ариллус.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Цариетальные клетки имеются. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Роу-
вопаш-типу. Синергиды грушевид-
ные. Имеются три антиподы, либо
эфемерные (у Thalia), mu60 coxpa-
HALOTCA до оплодотворения (у Ма-
rantha). Наблюдается увеличение
числа антипод (у Саа Теа). Поляр-
ные ядра сливаются перед оплодот-
ворением.
Опылениге, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия.
Эндоспермогенез — нуклеарный.
Эмбриогенез СВепороа4-типа. За-
родыш маленький, согнутый,. диффе-
ренцирбванный, бесхлорофилльный.
Подвесок слабо развит.
Семя, плод — семя © хорошо раз-
витым периспермом при отсутствии
эндосперма. Зародыш занимает OT
114 до 4/2 осевой длины семени.
Плод — коробочка или ягодообраз-
ный. -
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксие не обнаружен.
Другие явления не отмечены.
Литература: Пау!з, 1966; Schnarf,
1934; Тахтаджян, 1966, 1970.
Семейства НеНсотласеае, Созасеае
и Го\тласеае, относимые А. Л. Тах-
таджяном к порядку Zingiberales,
цитоэмбриологически не исследова-
ны. Подсемейства Zingiberoideae u
Costoideae семейства Zingiberaceae
настолько различны, что подсемей-
ство Созвю14еае следует выделить в
самостоятельное семейство * Costa-
ceae [Panchaksharappa, 1970].
Несмотря на в общем недостаточ-
но полное исследование цитоэмбрио-
логии Zingiberales, Bce me уже и
сейчас видно, что некоторые его се-
мейства характеризуются сходными
цитоэмбриологическими признаками.
те! ласеае и Миазасеае характери-
зуются секреторным тапетумом, сук-
цессивным микроспорогенезом, двух-
клеточными безапертурными пыль-
цевыми зернами, анатропной, с дву-
мя интегументами, крассинуцеллят-
пой семяпочкой, ариллусом, одно-
клеточным женским археспорием,
наличием париетальных клеток, Ро-
]усопит-типом зародышевого меш-
ка, нуклеарным эндоспермом и диф-
ференцированным взародышем. Йт-
giberaceae otamaaetca or Strelitzia-
сеае и Мизасеае наличием не нук-
леарного, а гелобиального эндоспер-
ма, при сходстве в остальных цито-
эмбриологических признаках. Can-
пасеае имеет сходные цитоэмбрио-
логические признаки с Marantha-
соае.
'
ПОРЯДОК 82.
ORCHIDALES
CEMEMCTBO ORCHIDACEAE
Основное число хромосом — < =
om OT 7 ДО 22, 24, 25.
Цветок обоеполый, редко однопо-
лый, растения однодомные или дву-
домные.
'’ интегументом.
Пыльник —.4 микроспорангия.
Стенка. микроспорангия ‘образуется
по однодольному типу и имеет эпи-
дермис, фиброзный — эндотеций,
один — два эфемерных средних слоя
ий одно-двуядерный секреторный та-
петум. :
Микроспорогенез и микрогамето-
генез — образование микроспор, си-
мультанное. Тетрады микроспор пзо-
билатеральные, крестообразные, тет-
раэдральные, линейные, Г-образные.
Зрелые пыльцевые зерна двухкле-
точные. У орхидей подсемейства
Cypripedioideae (Diandrae) пыльце-
вые’ зерна одиночные, у представи-
телей подсемейства Orchidoideae
(Monandrae) они соединены в тет-
рады, массочки, поллинии, содер-
жат масло. Спородерма с одно-дву-
дистальными бороздами и тонкой эк-
зиной. Спермии — клетки бедноплаз-
менные.
Семяпочка анатропная, тенуину-
целлятная, с 1—2 интегументами;
микропиле образуется внутренним
‚ Семяпочки — очень
мелкие, число их очень большое (от
нескольких тысяч до нескольких
миллионов). Семяпочки образуются
после опыления.
Макроспорогенез и макрогамето-
генез — археспорий обычно одно-
клеточный, париетальные клетки
отсутствуют. Тетрады макроспор ли-
нейные, иногда вместо тетрад обра-
зуются триады макроспор. У Сурн-
ред о19еае зародышевый мешок об-
asyetca no Allium-rumy, y Orchidoi-
veae —no Polygonum-tuny. [lua op-
хидных характерна редукция числа
ядер в зародышевых мешках от 8
до 4. Антиподы представлены’ ядра-
ми или клетками. Синергиды груше-
видные, с фибриллярными аппара-
тами. Имеются 1—3 эфемерные ан-
типоды. У некоторых орхидей (на-
пример; у Саап а) ядра антипод
сливаются с полярными ядрами,
образуя полярно-антиподальную
группу ядер.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — op-
хидеи хорошо приспособлены к пе-
рекрестному опылению, но у них
встречается самоопыление и клей-
стогамия. К моменту опыления се-
мяпочки обычно представляют со-
бой недифференцированные бугор-
ки. Порогамия. Оплодотворение обыч-
но двойное, постмитотического, из-
233 .
редка премитотического типа, но
иногда одинарное, вследствие отсут-
ствия слияния спермия со вторич-
ным ядром зародышевого мешка.
Между опылением и оплодотворе-
нием, а также между оплодотворе-
нием и делением зиготы промежут-
ки времени различные, иногда очень
большие.
Эндоспермогенез нуклеарный, но
возникает лишь от 2 до 16 ядер эн-
досперма. Нередко вторичное ядро
эндосперма или первичное ядро
эндосперма дегенерируют вскоре
после возникновения.
Эмбриогенез Опаога4д-, Азегад-ти-
па. Зародыши очень маленькие,
обычно недифференцированные,
бесхлорофилльные или хлорофилло-
носные, округлой или овальной фор-
мы, но иногда с семядолями (у В1е-
‘tia зН1айа), содержат масло и бе-
лок.
Семя, плод — семена многочиелен-
ные, очень мелкие, без эндосперма,
обычно с чрезвычайно маленькими
недифференцированными зародыша-
ми, с прозрачной пленчатой одно-
рядной семенной кожурой и воздуш-
ной полостью вокруг — зародыша.
. Дифференциация зародышей проис-
ходит во время прорастания семян.
Семенная кожура образуется из
наружного слоя наружного инте-
гумента. На начальных фазах раз-
вития проростки орхидей имеют
своеобразный вид. Их принято на-
зывать протокормами или зароды-
шевыми клубнями. Плод — коробоч-
ка, вскрывающаяся тремя-шестью
створками, редко ягодообразный,
Перикарпий состоит из 10—43 слоев
паренхимных клеток.
Гаусторий — нередко имеются
очень сильно развитые подвесочные
таустории, которые — сохраняются
даже при прорастании семян
(у Саеуа). Иногда подвесок от-
сутствует.
Апомикеие редуцированный, не-
редуцированный партеногенез, апо-
таметия, интегументальная эмбрио-
ния.
Другие явления — широко распро-
странена нолиэмбриония. Дополни-
тельные зародыши особенно часто
образуются из синергид. Парази-
TH3M.
Hurepatypa: Abe K., 1967, 1968a, b,
1971a, b, 1976, 1977; Ansari, 41977;
Afzelius, 1966; Bapanos, 1925; Cha-
rdard, 1969; Cocucci, 1969; Cocucci,
Jensen, 1969; Davis, 1966; Dieter,
1977; Ekanthappa, Arekal, 1977; Hi-
roshi, 1967, 1968, 1971, 1974a, b, ¢;
Jeyanayaghy, Rao A., 1966; Kimura,
1968, 1971; Kim-Lang-Huynh, 1976;
Mohana Rao, Sood, 1979a, b, c; Mo-
дилевский, 1918, Nagaraja et al.,
4967; Поддубная-Арнольди, 19586,
1960а, 6, в, 1964, 1964, 1967, 41978;
Поддубная-Арнольди, Селезнева,
1957, 1961; Prakash, 1973b, 1967; Ca-
вина, 1964, 1965a, 6, 1974, 1972a, 6,
1973, 1974, 1975, 1978, 1979; Sagawa
Israel, 1964; Schnarf, 1931; Swami-
nanthan, 1967; Swamy, 1967b; Tax-
Taman, (1966, 1970; Tepéxun, 1966,
1973, 4977; Терёхин, Камелина, 1969,
1972; Valmayor, Sagawa, 1966; Val-
mayor et al., 1976; Veyret, 1967, 1975,
nee; Yoshihiko, 1974, 1976b; Young,
Изучение цитоэмбриологии семей-
ства орхидных показывает, что
семейство это высокоспециализиро-
ванное, стоящее на высокой ступени
эволюции, и по своему свогобразию
и объему вполне обосновано выде-
лено в особый порядок ОгсМЧаез.
A. Л. Тахтаджян [1966, 1970] cun-
тает, что CemMeucTBO Orchidaceae o6-
наруживает наибольшие родствен-
ные связи с семейством Нурох!а-
сеае, порядка ТаПа]ез. Однако для
решения вопроса о родственных
связях Orchidaceae требуются даль-
нейщие исследования. Существова-
ние значительных различий в цито-
эмбриологии Сурйрефо14еае и Ог-
сЬ14о14еае, по мнению Свами [5\а-
шу, 19675], дает достаточно основа-
ний разделить орхидные на два са-
мостоятельных семейства — Cypripe-
dioideae (Diandrae) u Orchidoideae
(Monandrae).
ПОДКЛАСС С. СОММЕММТАЕ'
НАДПОРЯДОК П1.
ЗО МСА МАЕ
ПОРЯДОК 83.
JUNCALES
СЕМЕЙСТВО JUNCACEAE
Основное число хромосом — 2=3,
6, 8, 9, 10, 15, 17.
Цветок, как правило, обоеполый,
редко однополый, растения обычно
однодомные, редко двудомные.
Пыльник микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, 1—2 слоя фиброзного эндо-
теция, одии — средний слой и сек-
реторный одно — двуядерный тапе-
тум.
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады — микроспор
тетраэдральные, изобилатеральные.
Зрелые, пыльцевые зерна трехкле-
точные, одпобороздные. Пыльцевые
зерна обычно соединены в тетрады.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Семяпочек много.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный,
1—2 ряда париетальных клеток.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
]усопит-типу. Синергиды крупные.
Имеются три маленькие антиподы,
сохраняющиеся до начала эмбрио-
генеза. Полярные ядра сливаются
перед оплодотворением.
Опыление, прохождение пылье-
вых трубок и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние у видов Luzula.
Эндоспермогенез гелобиальный.
Микропилярпая камера крупная,
и в пей после свободного делецпия
ядра между ядрами возникают кле-
точные перегородки. Халазальная
камера дегенерирует, имея от 1 до
8 ядер.
Эмбриогенез Опастад-типа. Sapo-
дыш прямой, маленький, слабо или
хорошо дифференцированный, без-
хлорофилльный. Подвесок короткий,
двухклеточный.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Зародыш не превышает
1/4 осевой длины семени. Плод — ло-
кулицидная коробочка.
Гауеторий — семядоля выполняет
гаусториальную функцию.
Апомикейс не обнаружен.
Другие явления — клейстогамия
y Juncus.
Литература: Ва Кола, Brukarc-
zyk, 1965; Davis, 1966; Juguet, 1969;
Schnarf, 1934; Shah, 1963; Taxraz-
man, 1966, 1970.
Из двух семейств, относимых
А. Л. Тахтаджяном [1970] в порядок
Тапса[ез, семейство ТБигиасеае ци-
тоэмбриологически не исследовано.
По строению пыльцевых зерен и ти-
пу развития зародыша Juncaceae
приближается к Сурегасеае. У Лапа-
сасеае, как и у Сурегасеае, из че-
тырех микроспор три микроспоры
дегенерируют, в результате чего об-
разуются псевдомонады или крип-
тотетрады.
ПОРЯДОК 84.
CYPERALES
СЕМЕЙСТВО СУРЕКАСЕАЕ
Основное чиело хромосом — х = 5,
6, 8—15, 47, 19, 24—23. ,
Цветок обоеполый или однополый,
невзрачный, обычно анемофильный,
редко энтомофильный. Растения
обычно однодомные, редко двудом-
ные. `
Пыльник — 4 микроспорангия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзпый эндотелий, 1—2
эфемерных средних слоя и секре-
торный одно-двуядерный тапетум.
Имеются орбикулы. ,
Микроспорогенез и микрогамето-
генез — образование микроспор си-
мультанное. Тетрады микроспор
тетраэдральные, крестообразные,
изобилатеральные. Из четырех мик-
роспор три дегенерируют, в резуль-
тате чего образуются псевдомонады
или криптотетрады. Зрелые пыльце-
235
^^.
вые зерна трехклеточные, одно — че-
тырехпоровые. Сперми-клетки бед-
ноплазменные.
Семяпочка одна, анатропная, с
двумя интегументами. крассинуцел-
лятная. Микропиле образуется внут-
ренним интегументом. Имеются фу-
никулярный обтуратор и гипостаза.
Макроепорогенез и макрогамето-
генез — археспорий одноклеточный,
4 слоя париетальных клеток. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Роу-
г0опип-типу. Синергиды с крючко-
образными выростами. Имеются три
эфемерные антиподы. Полярные яд-
ра сливаются перед оплодотворе-
нием.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление у Carex palu-
доза. Порогамия. Двойное оплодот-
ворение. Сингамия и тройное слия-
ние протекают синхронно. Первич-
ное ядро эндосперма делится рань-
ше зиготы.
Эндоспермогенез пуклеарный,
В некоторых клетках оэндосиерма
обнаружено 6—8 ядер, отмечены
случаи слияния их.
Эмбриогенез Опастад-типа, вариа-
ция Лшсаз. Зародыш маленький,
односемядольный, хорошо или слабо
дифференцированный, бесхлоро-
илльный, содержащий масло.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло, крахмал и белок. Зародыш
не превышает 1/4 длины семени.
Кожура семени двуслойная, образу-
ется обоими интегументами. Плод
орехообразный, ‘нераскрывающийся.
Гауеторий — семядоля выполняст
гаусториальную фупкцию.
Апомикеие по обпаружен.
Другие явления — полиэмбриопия.
Гинодиэция у Eriophorum.
urepatypa: Carniel, 1971; Cass,
1970; Creziska, 1964; Davis, 1966;
Dunajska, 1965; Juguet, 1966a, b,
1968, 1969a, b, 1970a, b, 1972; Juguet,
Lebegue, 1966; Kamdar, 1969a, b, c;
Khanna, 1965a, b; Murty, Kumar,
1967d; Nagaraj, Nijalingappa, 1968,
1972, 1973а, 1979; Nijalingappa, Teja-
vathi, 1977; Padhye, 1968, 1969, 1971;
Padhye, Chaube, 1969; Padhye et al.,
1969; Padhye, Geed, Bhasin, 1969;
Padhye, Geed, Kaur, 1969; Padhye,
Kasture, 1970; Padhye, Yyer, 1969;
Schnarf, 1931; Shah, 1965; Shah, Nee-
236
fakandan, 1971; Suquet, 1970; Tarar,
Mukherjee, 1969; Тахтаджян, 1966,
1970; Tiwari, 1968, 1969; Tonepam,
1980; Тодераш, Чеботарь, 1978; Оп-
tawale, Makde, 1971; Untawale, Pad-
hye, 1969a, b.
В отношении цитоэмбриологии Су-
регасеае обнаруживает сходство ©
Juncaceae [Эвай, 1967]. В прежних
классификациях семейства Cypera-
ceae u Gramineae (Poaceae) относи-
ли к одному и тому же порядку
Стаплиа]ез, что He согласуется с
данными цитоэмбриологии, так как
эти семейства имеют разные цито-
эмбриологические признаки. Оба се-
мейства настолько сильно отлича-
ются друг от друга, что отнесение
их к разным порядкам — Сурега{ез и
Розез, как это делают современные
систематики, совершенно правильно
и согласуется с данными цито-
эмбриологии. Семейство Сурегасеае
настолько обширное и своеобразное,
YTO оно вполне заслуживаст ранга
порядка.
НАДПОРЯДОК
СОММЕММАМАЕ `
ПОРЯДОК $55.
BROMELIALES
СЕМЕЙСТВО ВВОМЕШМАСЕАЕ
Основное число хромосом — х=14,
16, 17, 18, 24, 25, 27, 28.
Цветок обосполый, редко функ-
циопально однополый.
Цыльник - - число микроспоранги-
ев и стенка микроспораигия пс изу-
чены. Тапетум секреторпыйи.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна двух-трехкле-
точные, одно-бороздные или дву-
многоапертурные с сетчатой экзи-
ной, имеют крахмальные зерна.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяпочек много.
Макроспорогенез и макрогамето-
'спез — археспорий одноклеточный.
Париетальные клетки имеются. Тет-
рады макроспор линейные. Зароды-
шовыи мешок образуется по Ро]у-
попит-типу. Синергиды грушевид-
ные. Имеются три маленькие скоро
доетеперирующие антиподы. Поляр-
ные ядра сливаются перед оплодот-
ворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез гелобиальный.
Эмбриогенез Asterad-tuma. Sapo-
дыш маленький, односемядольный,
слабо дифференцированный, бесхло-
рофилльный, содержит масло и про-
теин. Подвесок короткий, многоряд-
пый.
Семя, плод — семя с хорошо раз-
витым мучпистым эпдоспермом, со-
держащим масло, алейроп и крах-
мал. Зародыш не превышает 1/4
длины семени. Плод — коробочка
или ягода.
Гауеторий — семядоля выполняет
гаусториальную функцию.
Аномикеис не обнаружен.
Другие явления — полиэмбриония
y Tillansia juncea.
JInreparypa: Davis, 1966; Lakshma-
nan, 1967a; Schnarf, 1931; Тахтал-
жян, 1966, 1970.
Тахтаджян [4966, 1970], как и дру-
гие систематики, производит Вхго-
meliaceae or Liliaceae, что пе про-
тиворечит данным цитоэмбриологии,
так как оба эти семейства характе-
ризуются сходными признаками
(сукцессивным микроспорогенезом,
анатропной с двумя интегументами
семяпочкой, присутствием парие-
тальных клеток, одноклеточным
женским apxecuopuem, Polygonum-
типом зародышевого мешка, гелоби-
альным эндоспермом, семенами с
эндоспермом, Азегад-типом эмбрио-
генеза и т. д.).
ПОРЯДОК 86.
COMMELINALES
CEMEHMCTBO COMMELINACEAE
OcHOBHOe число хромосом — х =
=: 5—10, 12—15, 18, 49.
Цветок обоеполый или однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, эфе-
мерный средний слой и амебоид-
ный с периплазмодием тапетум.
Микроспорогенез и микрогамето-
veHes — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, однобороздные, с шиповатой
экзиной, имеют крахмальные зерна.
Семяпочка ортотропная, гемиана-
тропная, с двумя интегументами,
крассинуцеллятная. Микропиле об-
разуется обоими интегументами.
Имеются нуцеллярный колпачок,
эпистаза, гипостаза и ариллус. Со-
судистый пучок доходит до халазы,
Семяпочка одна или их несколько.
Макроспорогенез и макрогамето-
генез — археспорий одно-двухкле-
точный. Париетальные клетки име-
ются, но скоро дегенерируют. Мак-
роспоры линейные, Т-образные. За-
родышевые мешки образуются по
Ро]угопит-, АШат- или Адоха-ти-
пам. Синергиды с крючкообразны-
ми выростами. Имеются три анти-
поды, обычно эфемерпые, но иногда
долго сохраняются. Полярные ядра
сливаются перед оплодотворением.
Опыление, прохождение пыльце-
вой трубки и оплодотворение — са-
моопыление у Commelina bengha-
lensis u Floscopa spp.
Эндоспермогепез нуклеарпый. O6-
разование клеточпых перегородок
начинается с микропилярного по-
люса зародышевого мешка, за ис-
ключением халазального полюса,
который остается свободноядерным.
Эмбриогенез Азегад-типа. Заро-
дыш маленький, слабо дифференци-
рованный, односемядольный, бес-
хлорофилльный. Подвесок отсут-
ствует.
Семя, плод — семя с хорошо раз-
витым мучнистым эпдоспермом, со-
держащим крахмал. Зародыш за-
полняет не более 1/4 длины семени.
Семенная кожура формируется обо-
ими интегументами. Плод — обыч-
но локулицидная коробочка, редко
сочный, нераскрывающийся.
Гаусторий — семядоля выполняет
гаусториальную функцию. Некото-
рые виды Суапойз имеют в хала-
зальных частях зародышевых меш-
ков эндоспермальные гаустории.
Апомиксие не обнаружен.
237
Рис. 34. Пыльник, микроспорогенез и мужской гаметофит Commelj ~ } р р у y Co elina su bulata 1—7 — участки пыльников с разпыми фазами развития микросиор и пыльцевых зе-
рен; 8—./11 — микроспорогенез сукцессивного тина; JZ 15—24 — разные фазы развития макроспор и зародышевого мешка;
цевом зерисе;
пт — пыльцевая трубка, 21 — эпидермис, с — сипергиды, 3 — -
‘14 — первое деление в пыль-
зигота, вя — вторичное
ядро зародышевого мешка, ант — антиподы [Chikkannaiah, 1963]
Другие явления — клейстогамия
y Commelina.
Jiutepatypa: Chikkannaiah 1964—
1965а, b, 1974, 41978; Chikkannaiah,
Hemarddi, 1979; Davis, 1966; Kulkar-
ni, Patil, 1969; Lakshmanan, 1977;
Mepham, Lane, 1969; Schnarf, 1931;
Тахтаджяп, 1966, 1970.
СЕМЕЙСТВО ХУВТШАСЕАЕ
Основное чиело хромосом — х = 9,
13, 16; 47.
Цветок обоеполый.
238
Пыльник — 4 — микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций без фиброзных
утолщений, один эфемерный сред-
ний слой и секреторный одно — дву-
ядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, однобороздные или безапер-
турные.
Семяпочка ортотропная, с двумя
иптогумеитами, Микропило образуется обоими иите-
тепуинуцеллятиная.
гументами. Семяночек
много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Зародышевые мешки образуются по
Ро!угопат- или АШат-типу. Синер-
гиды с крючкообразными выростами.
Имеются три обычно скоро дегене-
рирующие антиподы, представлен-
ные не клетками, а ядрами. Поляр-
иссколько —
ные ядра сливаются перед оплодот-
ворением.
Опыление, прохождение пыльце-
вых трубок и оплодотгосение — по-
рогамия. Двойное оплодотворение.
Эндоепермогенез нуклеарпый. Об-
разование клеточных перегородок
начинается с микропилярного полю-
са зародышевого мешка. Первые
клетки эндосперма одноядерные,
позднее в халазальной части эндо-
сперма возникают клетки с несколь-
кими ядрами.
239
Рие. 32. Разпыс фазы развития зародыша y Comunclina subulata ак — апикальная клетка, бк — базальная клетка, с — семядоля, зл — зачаток ли- ста, пк — первичный корешок, вк — вторичный корешок; чк — чехлик корня, ап — анекс стебля [Chikkannaiah, 1962]
Эмбриогенез Азегад-типа. Заро-
дыш маленький, недифференциро-
ванный. Подвесок отсутствует.
Семя, плод — семя с хорошо раз-
витым мучнистым эпдоспермом, со-
держащим крахмал и алейроп. За-
родыш заполняет не более 1/4 длп-
пы семени. Плод — локулицидная
коробочка.
240
Гауеторий — семядоля выполняет
гаусториальную функцию.
Апомикеие не обнаружен.
Другие явления не обнаружены.
JiIurepatypa: Chikkannaiah, 1974,
1973; Davis, 1966; Kulkarni, Patil,
1969; Schnarf, 1934; Тахтаджян,
1966, 1970.
Из четырех семейств, относимых
А. Л. Тахтаджяном к порядку Сот-
иеПоез, семейства Мауасеае и
Нараеасеае цитоэмбриологически
но исследованы. Соштейпасеае и
Xyridaceae имеют сходные цитоэмб-
риологические признаки, а именно:
сукцессивный микроспорогенез,
двухклеточные пыльцевые — зерна,
ортотропную крассинуцеллятную се-
мяпочку с двумя интегументами, од-
токлеточный женский археспорий,
Polygonum-, АШаш-типы — зароды-
шевого мешка, нуклеарный эндо-
сперм, семена с хорошо развитым
мучнистым эндоспермом, эмбриоге-
цез А\ега4-типа и т. д. Это обуслов-
ливает расположение их в общем
порядке.
ПОРЯДОК 87.
ЕВТОСА ОТАТЕ$
СЕМЕЙСТВО ЕВТОСАОТАСЕАЕ
Основное чиело хромосом — vz =
== 8, 9.
Цветок однополый.
Ныльник — 4 микроспорапгия.
Тапетум секреторный.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Зрелые пыльцевые зерна
трехклеточные, © одной-пескольки-
ми узкими апертурами (спирапер-
турные) и мелкошиповатой экзи-
пой.
Семяпочка ортотропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется внутренним
интегументом. Семяпочек 3—2.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. Заро-
дышевый мешок образуется по Ро-
lygonum-tuny. Синергиды груше-
видпые. Имеются три эфемерные
антиподы. Полярпые ядра сливают-
ся перед оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. Двойное оплодотворение.
Эндоспермогенез телобиальный,
нуклеарный.
Эмбриогенез Азегад-типа. Заро-
дыш маленький, слабодифференци-
рованный, односемядольный.
16 в. А. Поддубная-Арнольди
Семя, плод — семя с хорошо раз-
витым мучнистым эндоспермом, со-
держащим крахмал. Зародыш запи-
мает не более 1/4 длины семени,
Плод — локулицидная коробочка с
пленчатым околоплодником.
Гауеторий — семядоля выполняет
гаусториальную функцию.
Апомикеис ne обнаружен.
Другие явления — полиэмбриония
y Eriocaulon septangulare.
Jiureparypa: Begum: 1968;° Davis,
4966; Maqbool, 1966, 1968; Schnarf,
1934; Тахтаджян, 1966, 1970.
Цитоэмбриологические признаки
Eriocaulales B значительной степе-
ии сходпы с таковыми Commelina-
les, что подтверждает правильность
расположепия их рядом друг с
другом.
ПОРЯДОК 88.
RESTIO NALES
СЕМЕЙСТВО ВЕЗТТОМАСЕАЕ
Основное чиело хромоеом — х==7,
9, 44—13.
Цветок обоеполый или однополый.
Ныльник — 2 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эпдотеций, один
офемерный средний слой и ceKpe-
торпый (?) одноядерпый тапетум.
Микроепорогенез и микрогамето-
генез — зрелые пыльцевые зерна
трехклеточные, однобороздные,
обычно граминоидные.
Семяпочка ортотропная, с двумя
интегументами, тенуинуцеллятная.
Микропиле образуется обоими ин-
тегумептами. Имеется — гипостаза.
Семяцпочек 3—2,
Макроепорогенез и макрогамето-
гешез — археспорий одноклеточный.
Паристальные клетки отсутствуют.
Зародышевый мешок образуется по
Polygonum-tuly uw имеет крахмаль-
пые зерна. Синергиды имеют крюч-
кообразные выросты и фибрилляр-
пые аппараты. Имеются три анти-
поды обычно эфемерные, но ипогда
делятся до образования 15 клеток.
Полярные ядра сливаются перед
оплодотворением.
241
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный.
Эмбриогенез — тип не известен.
Зародыш маленький, дифференци-
рованный, односемядольный.
Семя, плод — семя с хорошо раз-
витым мучнистым эндоспермом. За-
родыш занимает пе более 1/4 дли-
ны семени. Плод — орешек или ло-
кулицидная коробочка.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксис не найден.
Другие явления не обнаружены.
Литература: Davis, 1966; Голен-
кин, 1937; Miodziahowski, 1963; Tax-
таджян, 1966, 4970.
СЕМЕЙСТВО
CENTROLEPIDACEAE
Основное число хромосом — х =
= 40, 13.
Цветок однополый или 0боспо-
лый, растение одподомное.
Пыльник — 2 микроспорапгия.
Стенка микроспорангия образуется
по однодольному типу и имеет эпи-
дермис, фиброзный эндотеций, один
средний слой и секреторный дву-
ядерный тапетум. Имеются орби-
кулы.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор ли-
нейные, Г-образные, изобилатераль-
ные. Зрелые пыльцевые зерна трех-
клеточные, однобороздные, с крах-
матьпыми зернами.
Семяпочка висячая, ортотроппая,
с двумя инптогумептлами, топуину-
целлятпая. Микрониле образуется
обоими иптегумептами. Cemanouci
немпого.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Париетальные клетки отсутствуют.
Тетрады макроспор линейные. За-
родышевый мешок образуется по
Ро]угопит-типу и имеет зерна крах-`
мала. Три антиподы сохраняются
до оплодотворения. Полярные яд-
ра сливаются до оплодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный.
Эмбриогенез Опаста4-типа, вариа-
242
ции Juncus. Зародыш маленький,
слабо дифференцированный, односе-
мядольный, содержит масло.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
крахмал. Плод — коробочка или
орешек. Околоплодник плепчатый.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомикеис не найден.
Другие явления не обнаружены.
Jintepatypa: Davis, 1966; Prakash,
1969f; Schnarf, 1934; Tonenxun, 1937;
Тахтаджян, 1966, 1970.
Кроме вышеописанных двух Ce-
мейств к порядку Restionales
А. Л. Тахтаджян относит также се-
metictsa Flagellariaceae u Hanguana-
сеае, но они цитоэмбриологически
почти не изучены. Хотя Везйопасеае
исследовано в отношении цитоэмб-
риолокии недостаточно, однако уже
и сейчас видно, что по многим
цитоэмбриологическим признакам
ото семейство сходно с Сешхоер\9а-
ceac. Общими для пих признаками
являются: 2 микроспорангия, че-
тырехслойная стенка пыльника, сек-
реторный тапетум, ортотропная ©
двумя интегументами тенуинуцел-
лятная семяпочка, одноклеточный
женский археспорий, Роухопит-тип
зародышевого мешка с крахмаль-
ными зернами, нуклеарный эндо-
сперм, семя с хорошо развитым
эндоспермом. Таким образом, дан-
ные цитоэмбриологии подтверждают
отнесение ВезНопасеае и Centrole-
р1Часеае к одному и тому же по-
рядку.
ПОРЯДОК 859.
POALES (GRAMINALES)
СЕМЕЙСТВО РОАСЕДЕ
(GRAMINACEAE)
Основное число хромосом — 5 = 2,
4—15, 17, 19, 20, 23.
Цветок сильно редуцированный,
обычно обоеполый, редко однопо-
лый.
Ныльник — 4 микроспорангия.
Развитие стенки микроспорангия
идет по однодольному типу и имеет
эпидермис, фиброзный эндотеций,
1—2 эфемерных средних слоя и сек-
реторный двуядерный или одно-
ядерный тапетум. Обнаружены ор-
бикулы.
Микроепорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор обыч-
но изобилатеральные, иногда Т-об-
разные, линейные. Зрелые пыльце-
вые зерна одиночные, трехклеточ-
ные, обычно с одной порой, крах-
малсодержащие. В зрелых пыльце-
вых зернах спермии-клетки бедно-
плазменные, более или менее сильно
вытянутые. Пыльца злаков быстро
теряет свою жизнеспособность.
Обычно она сохраняет ее не более
трех дней.
Семяпочка одна, гемианатропная
или анакампилотроппая, с двумя
иптегумептами, крассипуцеллятная,
с сильно укорочеппой сомяножкой.
Последняя практически отсутствует
и заменена образованием, называе-
мым плацентохалазой. Микропиле
прямое, образующееся обычно внут-
ренним, но изредка одним наруж-
ным или обоими интегументами. Co-
судистые пучки доходят до халазы.
Имеются пуцеллярпый колпачок,
иптегумептальный обтуратор м ги-
постаза.
Макроспорогенез и макрогамето-
генез — археспорий ’одноклеточный.
Париетальные клетки OTCYyTCTBYIOT.
Тетрады макроспор линейные, 7'-06-
разные. Зародышевый мешок обра-
зуется по Ро]угопаю-типу. У пеко-
торых представителей семейства он
имеет крахмал. Синергиды с крюч-
кообразными выростами и фибрил-
лярными аппаратами. Полярные яд-
ра сливаются перед или во время
оплодотворения. Первоначально об-
разуются три клетки антипод, но
позднее число их более или менее
сильно увеличивается и у некото-
рых злаков достигаст болышной ве-
личины. Так, папример, у заза рап/-
сшайа обпаружено до 300 антицод.
У пшеницы, ржи, кукурузы и дру-
гих злаков их значительно мепьше
(до 40—60). Антиподы обычно долго
сохраняются. Ядра антипод нередко
‚бывают крупными, гипертрофиро-
ванными, содержащими политен-
ные хромосомы. Клетки антипод
могут иметь более одного ядра, и
ядра могут сливаться. Наблюдается
наличие многих зародышевых мепг-
13%
ков в ОДНОЙ и ТОЙ же семяпочке.
Это объясняется либо тем, что мо-
жет функционировать более одпой
макроспоры, либо тем, что по-
сколько клеток нуцеллуса могут
дать апоспорические зародышевые
мешки.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — са-
моопыление, перекрестное опыле-
ние, клейстогамия и гейтеногамия.
Порогамия. Оплодотворение двойное,
премитотического типа. При благо-
приятных внешних условиях про-
растание пыльцы разпых видов пшс-
ницы происходит через 5—15 мин
после опыления. Через 6—7 ч после
опыления образуются два ядра эп-
досперма, через 48 ч после ошыле-
ния, когда эпдосперм имест около
100 свободных ядер, а зародыш 4—6
клеток, между ядрами эндосперма
начинают закладываться клеточные
перегородки. Через 6—7 дпей после
опыления зародышевый мешок пол-
ностью заполняется эндоспермом.
Через 20—25 дней после опыления
зародыш достигает полного разви-
тия и своеобразно дифференциро-
ван. Сингамия и тройное слияние
происходят одповремепно.
Эндоспермогенез нуклеарного ти-
па. Однако после большего или
меньшего числа делений свободных”
ядер между ними возникают кле-
точные перегородки, и эндосперм
стаповится целлюлярным. Эндо-
сперм содержит запасные питатель-
ные вещества в виде крахмала, мас-
ла и белков. Периферический слой
эндосперма дифференцируется в
алейроновый слой.
Эмбриогенез — зародыш бесхлоро-
филльный, своеобразного типа, свой-
ственного только злакам. Этот тип
можно назвать Роад-типом. Подве-
сок зародыша развит очень слабо.
В зрелом состоянии зародыш имеет
такие органы, как щиток (семядо-
ля), колеоптиле (влагалищный
лист), почечку (перышко) первые
2—4 листа, зачаток стебля, эпи-
бласт, корешок, чехлик и колеразу.
Плод, семя — зерновка заполнена
мучнистым эпдоспермом. Зародыш
мал по отношению к объему зер-
новки. Он занимает не более 1/4
общей ее длины, но сильно диффе-
ренцирован и функционально высо-
коспециализирован. Зародыш пря-
мой, прилегающий сбоку к эндо-
243
сперму. Перикарпий образуется из
всех слоев завязи. Редко плод оре-
хообразный, костянка, ягода или
мешочек.
Гаусторий — щиток (семядоля)
выполняет гаусториальную Ффунк-
ЦИЮ.
Апомиксис — редуцированпый и
нередуцированный партеногенез, ге-
неративная и соматическая апо-
спория, апогаметия и нуцеллярная
эмбриония.
Другие явления — часто наблюда-
ются полиэмбриония и клейстога-
мия. Гейтеногамия у кукурузы. Ви-
випария у Роа. Гинодиэция у Сог-
taderia, Pennisetum a Poa.
Литература: Аминова, 1969; Баты-
гина, 1961, 1962а, 6, 1968, 4969, 1971,
1974а, 6; Батыгина, Фрейберг, 1979;
Батыгина, Маметьева, 1979; Сефгав,
1979; Chatterji, Pillai, 1970; Chikkan-
naiah, Mahalingappa, 1975, 1976; Iu-
mun, 1978; Христов, Крушева, 1968;
Христов, Москова, 1966, 1972; Хри-
стов, Терзийски, 11979; Das, 1969;
Davis, 1966; Deshpande, 1976; Desh-
pande, Raju, 1976; Emery, Guy, 1979;
Фрумкина, Колесников, 1969; Фур-
сов, Беспалов, 1963; Glidhill, 4967;
Главацкая, Кабы, 1976; Guignard,
Мезте, 1970, 1974; Сара, 1968; Хве-
дынич, 1969; отт, Атьерсаокахг, 1976;
Kapil, Bhatnagar, 1975; Kopmiom, 1970;
Коробова, 1961, 1962; Kubien, 1968,
1970a, b; Lee, Salunkhe, 1968; Лит-
виненко, 1970; Лихова, 1967; МаЪа-
lingappa, 1978; Maze, Bohm, 1970,
1973, 1974, 1977; Maze et al. 1970:
Модилевский, Бейлис, 1937; Моди-
левский и др., 1958; Моватеф, Со-
и14, 1966; Москова, 1969, (976: Мо-
скова, Мандилова, 1970; Мошкович,
YeOotapp, 1976; Muniyamma, 1969,
1976; Nagabhushana, Rao Sinde, 1976;
Цетрова, 1978; РЫШрзоп, 4977, 1978;
Поддубная-Арнольди, 1979; Поддуб-
ная-Арнольди, Иванова, 1970; Под-
дубная-Арнольди, Пащенко, 1962;
Поддубная-Арнольди и др. 1968:
Purcell, 1965; Reddy, Cruz, 1969a, b:
Pomanos, 1966, 1970, 1972; Ross, Wil-
зоп, 1969; Савченко, Комар, 1065;
Савченко, Петрова, 4973; Saran, De
Wet, 1969; Schnarf, 1931; Shatamma,
1976; Shantamma, Narayana, 1977:
244
Seshvatharam, Satyamurtii, 1976;
Singh D., 14965; Солнцева, Левков-
ский, 1978; Солнцева и др., 1977;
Солнцева, Яковлев, 1964, 1977; Та-
тинцева, 1968а, 6, 1965; Тахтаджян,
1966, 1970; Чеботарь, 1970, 1972; Тер-
зийски, Христов, 1973; Untawale,
Deshpande, 1969; Устинова, 1960,
1962, 1967; Venkateswarlu, 1965; Be-
peutaruua, 1976a, Vijendra, Sreeran-
gasamy, 1978; Weimarck, 1967a, b;
Woodland, 1964; Hrosmes, 1948, 1950,
1969, 1970; Яковлев, Солнцева, 1965;
Зотова, Филатова, 1968; Зуева, 1976.
Цитоэмбриологические признаки
хотя и дополняют характеристику
Роасеае, однако не позволяют раз-
решить вопрос о его филогенетиче-
ских связях. Систематики выводили
Poaceae us Liliaceae, сближали с
Сурегасеае, производили от Junca- .
ceae, Bromeliaceae, Rapateaceae,
Thurniaceae, Restionaceae u Flagel-
lariaceac. По мнению А. Л. Тахтад-
жяна [1966, 1970], Poaceae пред-
ставляют собой конечный этап в
линии анемофильной эволюции од-
нодольных, начинающейся от Сот-
шеНпа]ез, и произошли непосред-
ственно от ВезНопа]ез, возможно от
каких-то вымерших ЕасеПатасеае.
М. С. Яковлев, на основании орга-
низации эндосперма и структуры
зародыша считает, что Везйопасеае
и НасеПатасеае если и не являются
непосредственными предками Роа-
сеае, то во всяком случае филоге-
нетически весьма близки и имеют
ряд общих признаков в строении
эпдосперма и зародыша. По цито-
эмбриологическим признакам ТАНа-
сеае, Суротасеао, Juncaceae u Brome-
Насеае более или мепес сильно от-
личаются от Роасеае, поэтому для
сближения их нет оснований. Цито-
эмбриология Rapoteaceae, Thurnia-
ceae, Restionaceae u Flagellariaceae
исследована недостаточно, чтобы
сделать окончательное суждение об
их отношении к Роасеае. Таким об-
разом, вопрос о филогенетических
связях Роасеае требует дальнейших
исследований.
ПОДКЛАСС D. ARECIDAE
ПАДПОРЯДОК У.
АВЕСА МАЕ
ПОРЯДОК 90.
ARECALES
CEMEHCTBO ARECACEAE,
Основное число хромосом — х=8,
12, 18, 14, 16, 48.
Цветок обоеполый, чаще однопо-
лый, растения обычно однодомные,
реже двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорапгия имест эпи-
дермис, фиброзный эпдотеций, 2—6
эфемерных средних слоев и секре-
торный двурядный двуядерный та-
петум.
Микроепорогенез и микрогамето-
генез — образование микроспор си-
мультанное или сукцессивное. Тет-
рады микроспор изобилатеральные,
тетраэдральные, Т-образпые. Зрелые
пыльцевые зерна двухклеточпыс,
однобороздные, с трохщелевой 60-
роздой или реже с двудистальными
бороздами.
Семяпочка анатроппая-гемиала-
тропная, с двумя иптегументами,
крассинуцеллятпая. Микропиле об-
разуется обоими иптегументами.
Имеются нуцеллярный колпачок,
эндотелий и постамент. Семяпочек
обычно три.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Обычно 1—2, редко до 6 париеталь-
ных слоев. Тетрады макроспор ли-
нейные. Зародышевые мешки обра-
зуются по Ро!угопиат- или АШат-
типу. Синергиды грушевидные. Ан-
типоды либо эфеморные, либо со-
храняются до пачала эмбриогенеза.
Антиподы представляют собой пе
клетки, а ядра. Полярные ядра сли-
ваются до оплодотворения. У неко-
торых представителей семейства ан-
типоды лежат на постаменте.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление и самоопыле-
ние. Порогамия.
Эндоспермогенез нуклеарный.
Эмбриогенез Опаотад- u Asterad-
типа. Зародыш маленький, односе-
мядольный, бесхлорофилльный.
Семя, плод — семя с хорошо раз-
витым роговым или хрящевым,
иногда руминированным эндоспер-
мом, содержащим масло, алейроно-
вые зерна и кристаллы оксалата
кальция. Зародыш занимает от 1/4
до 4/4 общей длины семени. Плод —
сухая или мясистая синкариная ко-
стянка, реже ягодообразный.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомикеис — редуцированный пар-
тепогепез.
Другие явления — полиэмбриония
y Phoenix canariensis u Pinanga
patula.
Литература: Biradar, 1968; Bira-
dar, Mahabale, 1968; Kulkarni, Ma-
habale, 1974; Mahabale, Biradar, 1968;
Schnarf, 1931; Смирнова, 1977; Во-
Бег(зоп, 197ба, Ъ, с; Тахтаджян, 1966,
1970; Vallade, 1966, 1969.
По мнению А. Л. Тахтаджяна
[1966, 1970], Arecales, возможно,
произошел от ближайших апокарп-
ных предков ЫПайез, что, может
быть, пе лишено осповапий, так как
между обоими этими порядками
имеются сходные цитоэмбриологи-
ческие признаки. Среди гетероген-
ного семейства ГАПасеае обнаружены
такие представители, которые име-
ют сходные цитоэмбриологические
признаки с Атесасеае, а именно:
А микроспорангия, секреторный та-
петум, сукцессивный микроспороге-
нез, двухклеточные пыльцевые зер-
па, анатропную или гемитропную
крассипуцеллятпую семяпочку с
двумя иптегументами, пуцеллярный
колпачок, эпдотелий, постамент,
одпоклеточный женский археспо-
рий, париетальные клетки, зароды-
шевые мешки АШат- или Ро]уо-
пит-типа, нуклеарный эндосперм,
эмбриогенез Опабта4- или Азега4-
типа, семядоли, выполняющие га-
усториальную функцию, семя с хо-
рошо развитым эндоспермом, апо-
миксис и полиэмбрионию.
245
Рие. 33. Части пыльников на разных фазах развития у НурВаепе 1т@1са 1—5 — заложение и развитие археспориальных клеток; 6, 7 — одно- и двуядерные
клетки тапетума; 8 — часть пыльника,
ст — секреторным тапетумом, дегенерирую-
щим средним слоем и с одноядерными пыльцевыми зернами; 9 — часть пыльника с фиброзным эндотецием, дегенерирующим средним слоем, секреторным тапетумом и двухклеточными пыльцевыми зернами; 10--21 — разные фазы развития микроспор; 22—27 — разные фазы развития мужского гаметофита [Mahabale, Chennaveeraiah,
1957]
ПОРЯДОК 91.
CYCLANTHALES
СЕМЕЙСТВО СУСГАМТНАСЕАЕ
Основное число хромосом — х==9,
15, 16.
Цветок однополый, растение од-
нодомное.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндотеций, один
эфемерный средний слой и секре-
246
торный двух-четырехъядерный тапе-
тум. Имеются орбикулы.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные, крестообразные. Зре-
лые пыльцевые зерна двухклеточ-
ные, однобороздные.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле, образующееся —обои-
ми интегументами, зигзагообразное.
Имеется нуцеллярный колпачок. Се-
мяпочек много.
+ al Vay
hay ate
“I | № № us
Mahe tp
Uti
АЯ
. 416% hgh oe , 41077072 и» 5% < 427
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
2 слоя париетальных клеток. Тет-
рады макроспор линейные. Зароды-
шевый мешок образуется по Рау-
сопит-типу. Синергиды грушевид-
ные. Антиподы разнообразны по
своей жизнеспособности. Число ан-
типод равно трем; они представляют
собой не клетки, а ядра. Полярные
ядра сливаются перед оплодотво-
рением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез гелобиальный.
247
й и
“=o
ис
о
» ana
т; =>
Рис. 34. Макроспорогенез и развитие зародышевого мешка НурВаепа ша
1—7 — разные фазы макроспорогенеза; 8—20 — разные фазы развития биспоричес-
кого зародышевого мешка Allium —- tuna [Mahabale, Chennaveeraiah, 1957]
248
Эмбриогенез — тип не определен.
Зародыш маленький, прямой, диф-
ференцированный, односемядоль-
ный.
Семя, плод — семя с хорошо раз-
витым эндоспермом. Зародыш зани-
мает от 1/4 до 4/4 длины семени.
Плод сочный, состоящий из отдель-
ных или сросшихся ягодообразных
плодиков. :
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомикеис не найден.
Другие явления не обнаружены.
Литература: ПОау1з, 1966; Смирно-
ва, 1977; Тахтаджян, 1966, 1970.
Цитоэмбриологически семейство
Cyclanthaceae изучено педостаточно
полно, чтобы судить 0б ого взаимо-
отпоптениях с другими семействами
и о правильности выделепия его в
самостоятельный порядок — Суб]ап{-
ales.
ПОРЯДОК 92.
ARALES
СЕМЕЙСТВО АВАСЕЛЕ
Основное число хромосом — т =
— 7—9, 41—47, 22.
Цветок обоеполый или однополый,
растения обычно однодомнысе, редко
двудомные.
Пыльник —2 или 4 микроспоран-
гия. Стенка микроспорангия имеет
эпидермис, фиброзный эндотеций,
1, 2 и болыне эфемерных средних
слоев, амебоидный — двух-четырех-
рядный, превращающийся в истин-
ный периплазмодий тапетум.
Микроепорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, изобилаторальные,
крестообразные. Зрелые пыльцевые
зерна двух- или трехклеточные, с
крахмальными зернами, одноборозд-
ные, двух-четырехдистально-борозд-
ные, трех-четырехпоровые, безапер-
турные и др. Пыльцевые зерна иног-
да соединены в тетрады.
Семяпочка ортотропнйя, гемиана-
тропная или анатропная, с двумя
интегументами, крассинуцеллятная
или изредка тенуинуцеллятная.
Микропиле образуется только внут-
ренним или обоими интегументами,
Имеются нуцеллярный колпачок,
типостаза, постамент, эндотелий, об-
тураторы плодолистиковый, плацент-
арный и фуникулярный. Семяпочек
одна — много.
Макроспорогенез и макрогамето-
генез — археспорий одно-, изредка
многоклеточный. Париетальные клет-
ки отсутствуют. Тетрады макроспо
линейные. Зародышевые мешки об-
pasyrotca no Polygonum- u Allium-
типу. Они имеют крахмальные зер-
на. Синергиды грушевидные. Три
обычно эфемерные антиподы, но
иногда они значительно увеличи-
ваются или делятся. Полярные ядра
сливаются до оплодотворения.
У Асогиз са]атиз под аптиподами
имеется постамент.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опыление, самоопыле-
ние. Порогамия. Двойное оплодотво-
рение.
Эндоспермогенез целлюлярный.
Эмбриогенез Caryophyllad-ruma.
Зародыш крупный, прямой, диффе-
ренцированный, одпосемядольный,
хлорофиллопосный или безхлоро-
филльный, содержащий масло и
алейрон.
Семя, плод — семя с хорошо раз-
витым эндоспермом, содержащим
масло. Зародыш располагается в
центре семени и занимает от 1/2 до
3/4 его длины. Плод — ягода или
кожистый раскрывающийся, одно-
многосемянный.
Гауеторий — семядоля выполняет
таусториальную функцию. Халазаль-
ная часть эндосперма обнаруживает
гаусториальную активность.
Апомиксие — нуцеллярная эмбри-
Ония.
Другие явления — полиэмбриония
У Bugos Spathiphyllum. Busunapmza.
Литература: Davis, 1966; Kapil,
1967а; Мапя-Черней, 1978; РашсКег,
1965; ЭсБпатЁ 1931; Смирнова, 1977;
Тахтаджян, 1966, 1970.
СЕМЕЙСТВО ГЕММАСЕАЕ
Основное число хромосом — х=20,
21, 22, 32.
Цветок однополый.
Пыльник —2 или 4 микроспоран-
гия. Стенка микроспорангия обра-
зуется по редуцированному типу
249
и имеет эпидермис, фиброзный эн-
дотеций и амебоидный, с периплаз-
модием, одноядерный тапетум. Сред-
ние слои клеток не образуются.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор тет-
раэдральные, изобилатеральные,
крестообразные. Зрелые пыльцевые
зерна трехклеточные, однопоровые,
шиповатые, с крахмальными зер-
нами.
Семяпочка ортотропная или ана-
тропная с двумя интегументами,
крассинуцеллятная. После оплодот-
ворения семяпочка становится ге-
мианатропной. Микропиле обычно
образуется внутренним интегумен-
том, но иногда обоими. Имеются
нуцеллярный колпачок и гипостаза.
Семяпочек 4—7.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
2—3 слоя париетальных клеток. За-
родышевые мешки образуются по
Ро]угопаит- и АШиш-типу. Синерги-
ды грушевидные. Имеются три эфе-
мерные антиподы, по ипогда опи
сохраняются до начала эмбриогене-
за. Полярные ядра сливаются до оп-
лодотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия.
Эндоепермогенез целлюлярный.
Эмбриогенез Опаста4-, Азжегад-ти-
па. Зародыш крупный, прямой,
дифференцированный, односемя-
дольный, содержит крахмал. Под-
весок короткий.
Семя, плод — семя с эндоспермом,
содержащим крахмал. Зародыш рас-
положен в центре семепи и зани-
мает от 1/2 до 3/4 длипы семепи.
Плод мешочкообразный.
Гаусторий — халазальная часть
эндосперма превращается в гаусто-
рий. Семядоля выполняет гаустори-
альную функцию.
Апомиксие не найден.
Другие явления не обнаружены.
Литература: ПОау1з, 1966; Kapil,
1967е; Лодкина, 1976; Maheshwari S.,
Kapil, 1963a, b; Maheshwari S., Ma-
heshwari N., 1963; Schnarf, 1934;
Смирнова, 1977; Тахтаджян, 1966,
Данные цитоэмбриологии показы-
вают, что Агасеае и Гетпасеае —
близкородственные семейства, Tak
как по многим цитоэмбриологиче-
ским признакам они обнаруживают
сходство [КарЦ, 1967е].
Представители обоих семейств ха-
рактеризуются наличием 2—4 мик-
роспорангиев, амебоидного (пери-
плазмодиального} тапетума, сукцес-
сивного типа микроспорогенеза, трех-
клеточных пыльцевых зерен, орто-
тропной или анатропной крассину-
целлятной с Двумя интегументами
семяпочки, нуцеллярного колпачка
и гипостазы, одноклеточного жен-
ckoro apxecnopua, Polygonum- или
Allium-runa зародышевых мешков,
порогамии, двойного оплодотворе-
ния, целлюлярного эндосперма с га-
усторием в халазальной его части,
крупного прямого дифференциро-
ванного зародыша и семян без эп-
досперма.
ПОРЯДОК 983.
PANDA NALES
CEMEMCTBO PANDANACEAE
Основное число xpomMocoM — z=
== 30, 32.
Цветок однополый, растения всог-
да двудомные.
Пыльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, 2 средних слоя
и секреторный или амебоидный
двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивиое. 'Гетрады микроспор кре-
стообразиые, изобилатеральные. Зре-
лые пныльцевые зерпа двухклеточ.
ные, однопоровые.
Семяпочка анатропная, © двумя
интегументами, крассинуцеллятная.
Микропиле образуется обычно одним
внутренним, но иногда обоими ин-
тегументами. Имеются нуцеллярный
колпачок и гипостаза. Семяпочек
одна — много.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный,
имеются 3—4 ряда париетальных
клеток. Тетрады макроспор линей-
ные, —Г-образные. Зародышевые
мешки образуются по Роусопит- u
АШиат-типу. Первоначальное число
ядер в зародышевом мешке рав-
но 8, позднее значительно увели-
чивается. Антипод три и более (до
200). Пояярные ядра сливаются пе-
ред оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — по-
рогамия. ,
Эндоспермогенез нуклеарный.
В эндосперме образуется большое
число свободных ядер, прежде чем
между ними возникают клеточные
перегородки. Последние возникают
центробежно.
Эмбриогенез — тип не установлен.
Зародыш маленький, прямой, не-
дифференцированный.
Семя, плод — семя с мясистым эн-
доспермом, богатым маслом и алеи-
роном. Зародьии запимает от 1/4 до
4/4 плины семени. Плод ягодообраз-
ный или костянка, или более или
менее деревянистый, образующий
соплодие.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомикеие не найден.
Другие явления не обнаружены.
Jiutepatypa: Bidyut Ganguly, 1976;
Chean, Stone, 1975; Davis, 1966;
Schnarf, 1931; CmupHosa 1977; Tax-
таджян, 1966, 1970.
Рапдапа1ез имеет сходные с Сус-
Лат аез цитоэмбриологические при-
знаки (4 микроспорангия, сукцес-
сивный микроспорогенез, двухкле-
точные пыльцевые зерна, анатроп-
ную крассинуцеллятную семяпочку
с двумя интегументами, однокле-
точный женский археспорий. Ро-
|угопат-тин зародышевого мешка,
семена с эндоспермом), что оправ-
дывает расположение этих порядков
вблизи друг с другом.
ПОРЯДОК 94.
TYPHALES
СЕМЕЙСТВО ЗРАВСАМТАСЕАЕ
Основное число хромосом — х=15.
Цветок однополый, растение одно-
домное.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, эндотеций, один средний
слой и амебоидный многоядер-
ный, образующий периплазмодий
тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
цессивное. Тетрады микроспор изо-
билатеральные. Зрелые пыльцевые
зерна двухклеточные, однопоровид-
ные, с однопоровидной апертурой.
Семяпочка анатропная, с двумя
интегументами, крассинуцеллятная.
Семяпочек 1—2.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный.
Имеются париетальные клетки. За-
родышевый мешок образуется по
Ро[угопат-типу. Синергиды груше-
видные. Три антиподы делятся, в
результате чего образуется ком-
плекс не менее чем из 150 клеток.
Полярные ядра сливаются до опло-
дотворения.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — нет
данных.
Эндоспермогенез нуклеарный или
гелобиальный (?). Халазальная ка-
мера настолько маленькая, что ее
легко не заметить.
Эмбриогенез Опаста4-типа. Заро-
дыш прямой, дифференцированный,
односемядольный, — бесхлорофилль-
ный, имеет крахмал и алейрон.
Семя, плод — семя с хорошо раз-
витым мучнистым — эндоспермом.
Имеется перисперм, содержащий
крахмал и алейрон. Плод сухой,
нераскрывающийся, с губчатым эв-
зокарпием и твердым роговым эндо-
карпием.
Гаусторий — семядоля выполняет
гаусториальную функцию.
Апомиксис не обнаружен.
Другие явления не отмечены.
Jluteparypa: Asplund, 1968, 1973;
Пау!з, 1966; Глущенко, 1968; ЭсВпатё,
1931; Смирнова, 1977; Тахтаджян,
4966, 1970.
СЕМЕЙСТВО ТУРНАСЕАЕ
Оеновное число хромосом — х=15,
Цветок однополый.
Ныльник — 4 микроспорангия.
Стенка микроспорангия имеет эпи-
дермис, фиброзный эндоТтеций, один
эфемерный средний слой и амебо-
идный двуядерный тапетум.
Микроспорогенез и микрогамето-
генез — образование микроспор сук-
251
цессивное. Тетрады микроспор тет-
раэдральные, изобилатеральные, T-
образные, линейные. Зрелые пыль-
цевые зерна двухклеточные, с одно-
поровидной апертурой, одиночные,
либо соединены в тетрады.
Семяпочка одна, анатропная, с
двумя интегументами, крассинуцел-
лятная. Микропиле образуется внут-
ренним интегументом. Имеется об-
туратор.
Макроспорогенез и макрогамето-
генез — археспорий одноклеточный,
\—2 слоя париетальных клеток. Тет-
рады макроспор линейные, Г-образ-
ные. Зародышевый мешок образует-
ся по Polygonum-tuny. Синергиды
грушевидные. Имеются три антипо-
ды, дегенерирующие вскоре после
оплодотворения. Полярные ядра сли-
ваются перед оплодотворением.
Опыление, прохождение пыльце-
вых трубок и оплодотворение — пе-
рекрестное опылепие у видов ТурВа.
Эндоспермогенез нуклсарпый или
телобиальный (?). Халазальпаля
часть эндосперма становится или пе
становится клеточной.
Эмбриогенез Азегад-типа. Заро-
дыш прямой, дифференцированный,
сдносемядольный, — бесхлорофилль-
ный, содержит крахмал, масло и
алейрон.
Семя, плод — семя с хорошо раз-
витым мучнистым эндоспермом и
тонким слоем перисперма, содержа-
щим крахмал, масло и алейрон. За-
родыш занимает от 1/4 до 4/4 дли-
ны зародыша. Плод маленький, су-
хой, с остающимся столбиком.
Гауеторий — семядоля выполняет
гаусториальную функцию.
Апомикеис не обнаружен.
Другие явления не отмечены.
Литература: Asplund, 1972, 1973;
Davis, 1966; Schnarf, 1934; CarmpHosa,
1977; Тахтаджян, 1966, 1970.
Семейства Зрагоашасеае и ТурВа-
ceae по многим цитоэмбриологиче-
ским признакам настолько сходны,
что их можно считать близкорол-
ственными семействами и отнесе-
ние их к одному и тому же поряд-
ny Typhales не вызывает сомнений.
Оба семейства имеют по 4 микро-
спорангия, четырехслойные стенки
пыльников, двухклеточные, однопо-
ровые пыльцевые зерна, анатрон-
ные, крассинуцеллятные с двумя
интегументами семяпочки, однокле-
точные женские археспории, непо-
средственно становящиеся материн-
скими клетками зародышевых меш-
ков Tuna Polygonum, нуклеарный
или гелобиальный типы эндосперма,
прямые, дифференцированные, од-
носемядольные, бесхлорофилльные
зародыши, семена с мучнистым эн-
доспермом и периснермом, семядо-
ли, выполняющие гаусториальную
функцию.
ЗАКЛЮЧЕНИЕ
Настоящий обзор известных до последнего времени материа-
лов по цитоэмбриологии покрытосеменных, сделанный нами при-
менительно к системе А, Л. Тахтаджяна [1970]! по 20 цитоэмбри-
ологическим признакам, показывает, что еще имеются огромные
пробелы в исследовании цитоэмбриологических признаков многих
семейств и порядков, даже самых крупных широко распростра-
ненных и важпых из них, не говоря уже о второстепенных и
редко встречающихся, что особенно относится к тропическим и
субтропическим представителям. Из 438 семейств покрытосемен-
ных, согласно системе А. Л. Тахтаджяна, 140 семейств почти
или вовсе не исследованы в отношении цитоэмбриологии. Кроме
того, некоторые из данных по цитоэмбриологии являются непол-
ными, сомнительными и даже неправильными. Однако и сейчас
видно, что сходство или различие в цитоэмбриологических приз-
наках может дать ценный материал для характеристики, провер-
ки и установления систематических и филогенетических взаимо-
отношений как между более круппыми (отдел, подотдел, класс,
подкласс), так и между более мелкими (порядок, семейство, род
и вид) таксономическими единицами, причем каждый цитоэмбри-
ологический признак в различных случаях может характеризо-
вать единицы разного таксономического значения. В настоящей
монографии мы остановились главным образом на общей харак-
теристике семейств и порядков, оставляя в стороне подсемей-
ства, трибы, роды, виды.
Учитывая современное состояние сравнительной цитоэмбри-
ологии покрытосеменных, которое пока не позволяет проследить
линий эволюции различных семейств и порядков, попытаемся в
заключении показать, какое значение имеют эти данные для
разрешения таких важных вопросов, как:
1) однородны ли покрытосеменные по своему происхождению;
2) каковы взаимоотпошения между покрытосеменными и го-
лосеменными;
3) какие семейства или порядки покрытосеменных являются
примитивными, исходными; какие высокоорганизованными, вто-
ричными;
4) каковы взаимоотношения между однодольными и двудоль-
ными, между раздельнолепестными и спайнолепестными;
1 В 1980г. появилась новая система А. Л. Тахтаджяна, [Так а]ап А. Ё.
Outline of the classification of flowering plants (Magnoliophyta).— Bot. rev.,
1980, vol. 46, М 3, р. 225—259], которую мы: уже не смогли использовать.
253
5) каковы взаимоотношения внутри разных порядков и меж-
ду разными порядками.
Для всех покрытосеменных, как двудольных, так и однодоль-
ных, характерны сходные типы мужского и женского гаметофи-
тов, оплодотворения, развития и строения эндосперма. Следова-
тельно, цитоэмбриологические признаки покрытосеменных одно-
родвы и располагаются в границах одного и того же ясно выра-
женного основного типа, что указывает на их монофилетическое
происхождение. Среди всех доказательств в пользу их монофиле-
тического происхождения наиболее важными и убедительными,
по нашему мнению, являются те из них, которые дает нам срав-
нительная цитоэмбриология, показавшая своеобразие мужского
и женского гаметофитов, двойного оплодотворения и полинлоид-
ного эндосперма покрытосеменных, что невозможно представить
себе как многократное возникновение этих признаков. Представ-
ление о том, что покрытосеменные являются однородными, т. е.
что они возникли монофилетично, не ново. Наоборот, более новы-
ми являются представления ряда ученых о полифилетическом
происхождении покрытосеменных, которые считают, что выше-
отмечеппые цитоэмбриологические признаки могли возникать
много раз, в разных местах и в разное время. Впрочем, обе точ-
ки зрения были бы примиримы, если иризпать, что свособразные
цитоэмбриологические признаки возникли очень давно у далеких
предков покрытосеменных, от которых впоследствии путем эво-
люции в разных направлениях возникло все многообразие этого
отдела высших растений.
Данные сравнительной цитоэмбриологии пока недостаточны
для разрешения вопроса о происхождении покрытосеменных.
Однако они вскрывают специфические особенности различных
отделов высших растений и указывают на то, что между покрыто-
семенными и остальными отделами высших растений существует
довольно большой разрыв. Несмотря на существование глубокой
пропасти между покрытосеменными и голосеменными, между
этими отделами в отношении цитоэмбриологии все же больше
сходства, чем между покрытосемениыми и остальными отделами
высших растений, что может указывать па родство, хотя и очень
отдаленное, покрытосеменных с голосеменными, Можно предпо-
ложить, что покрытосеменные возникли от общих с голосемен-
ными предков, либо какие-то древние уже вымершие голосемен-
ные дали пачало покрытосеменным.
Цитоэмбриологические исследования высших растений,
вскрывшие специфические особенности каждого из отделов и
подотделов, легли в основу различных систематических и фило-
генетических построений и подкрепили представления, касаю-
щиеся эволюции растительного мира от форм низших к высшим.
В результате этих исследований установлены чередование гаме-
тофита и спорофита, редукция гаметофита и высокое развитие
спорофита, наличие антеридиев и архегониев у более примитив-
ных из них и редукция или даже исчезновение их у более высо-
254
воразвитых растений. При выяснении вопроса о происхождении
покрытосеменных немаловажное значение имеет вопрос о проис-
хождении и развитии мужского и женского гаметофитов, а так-
же характера оплодотворения.
Что касается мужского гаметофита покрытосеменных, то, по
мнению некоторых исследователей, выведение его из гаметофита
толосеменных не представляет особых трудностей, хотя по срав-
нению с последними он сильно редуцирован. Одни ученые счи-
тают, что мужской гаметофит покрытосеменных произошел из
мужского гаметофита каких-то голосеменных предков в резуль-
тате упрощения и утраты клеток заростка и числа сперматозои-
дов, представляя собой только один антеридий с двумя спермия-
ми. Другие исследователи вегетативную клетку мужского гаме-
тофита покрытосеменных рассматривают как клетку заростка,
генеративную же — как сильно редуцированный антеридий, имс-
ющий всего лишь два спермия. Третьи считают, что мужской
гаметофит покрытосемепных пастолько упрощен, что антеридии
в нем перестали образовываться. В отношении женского гамето-
фита все обстоит значительно сложнее. В настоящее время име-
ется несколько гипотез происхождения зародышевого мешка по-
крытосеменных, однако ни ‘одна из них не может считаться
достаточно обоснованной. Наиболее разработанными являются
две основные гипотезы: 1) архегональная и 2) гнетовая.
1. Архегональная гипотеза впервые была высказана Поршем
[Ротзсв, 1907], который исходил из предпосылки, что обе синер-
гиды, с одной стороны, и яйцеклетка с верхним полярным ядром,
с другой, являются сестринскими парами. Согласно этой гипотезе
микропилярная четверка зародышевого мешка покрытосеменных
представляет собой фупкционирующий архегоний, а халазаль-
ная — нефупкционирующий. Порш [1907] считал, что зародыше-
вый мешок покрытосеменных возник путем редукции из женп-
ского гаметофита каких-то голосеменных, возможно хвойных, и
состоит из 1,2 или 4 архегониев без тканей заростка. Гипотеза
Порша привлекла к себе большое внимание, и ее разделяли мно-
гие исследователи [МПзз0п, 1941; Змату, 1946; и др.]. Однако
время от времени против нее высказывались возражения. -
Серьезными возражениями против архегональной гипотезы
происхождения зародышевого мешка покрытосеменных является
невозможность объяснить на ее основании строение би- и тетра-
спорических зародышевых мешков, отделение «брюшных каналь-
цевых клеток» (т. е. полярных ядер), наличие верхнего и нижне-
го «архегониев» (т. е. яйцеклетки с синергидами и трех анти-
под), расположение «брюшной канальцевой клетки» (т. е. поляр-
ных ядер) в центре зародышевого мешка и слияние их друг с
другом, а также образование эндосперма в результате оплодот-
ворения.
По мнению ряда ученых [Fagerlind, 1944; P. Maheshwari,
1948: Романов, 1945; Герасимова-Навашина, 1955, 1958; Под-
дубпая-Арнольди, 1964, 1976; и др.], гипотеза Порша должна
255
быть отклонена, по той причине, что накопилось много
данных, не подтверждающих ее. Фагерлинд [КасегЦ!и49, 1944],
И. Д. Романов [1945], Е. Н. Герасимова-Навашина [14955] и дру-
гие исследователи считают, что для объяснения строения заро-
дышевого мешка покрытосеменных нет необходимости прибегать
к сомнительным гомологиям с архегониями голосеменных, так
как его легко объяснить теми закономерностями развития, кото-
рые ему свойственны. ЦП. Махешвари [Мавез\уаг!, 1948] отметил,
‚ что образование архегониев связано с клеточной фазой, а в заро-
дышевом мешке покрытосеменных наблюдается свободное деле-
ние ядер, указывающее скорее всего на то, что и покрытосемен-
ные, и их ископаемые предки никогда не имели архегониев. На
отсутствие архегониев у покрытосеменных указывает и Свами
[Swamy, 1974].
Еще менее обоснованной, чем архегониальная гипотеза Пот-
ша, является архегониальная гипотеза Шюргофа [Schiirhoff, 1926],
которая базируется на представлении о том, что одна синергида
является сестринской клеткой яйцеклетки, другая — верхнего
полярного ядра. По мнению Шюргофа, микропилярная четверка
представляет собой два архегония, Халазальная же четверка —
заросток. Одпн архегопий состоит из сипергиды и яйцеклетки,
другой — из синергиды и верхпего полярпого ядра, причем синер-
гиды Шюргоф рассматривает как брюшные канальцевые клетки.
«Яйцеклетки» обоих архегониев оплодотворяются, давая начало
зародышу и эндосперму. Гипотеза Шюргофа находится в проти-
воречии с существующими в настоящее время фактами, так как
базируется на неправильном представлении. На самом деле не
синергида и яйцеклетка, а обе синергиды, с одной стороны, и яйце-
клетка с верхним полярным ядром, с другой, являются сестрин-
скими. Гипотеза Шюргофа, как не согласующаяся с современ-
ными представлениями о развитии и строении зародышевого
мешка покрытосеменных, не получила широкого признания.
В настоящее время архегониальпая теория появилась в новой
интерпретации. Фавр-Дюшартр [Favre-Duchartre, 1969, 1970,
1971, 1974, 1975], выводя зародышевой мешок покрытосеменных
из женского гаметофита голосемеппых, рассматривает антиподы
как вегетативные клетки заростка, а остальные клетки — как
архегонии, причем он признает наличие трех архегопиев, из ко-
торых один одноядерный (яйцеклетка), а два другие двуядерные
(синергида -- верхнее полярное ядро и синергида -- нижнее по-
лярное ядро). Это представление также не соответствует действи-
тельности, так как синергиды и полярные ядра не являются се-
стринскими.
2. Гнетовая гипотеза особенно четко выкристаллизовалась
на основе данных по цитоэмбриологии Спейии, полученных Фа-
repmunyom [Fagerlind, 1947]. Позднее эту гипотезу развивали
Il. Maxemsapn (P. Maheshwari, 1948, 1950) m Barransa [Battag-
Па, 19514]. Базируется она на представлении о том, что в зароды-
евом мешке все ядра равноценны и являются потенциальными
256
'аметами-яйцеклетками, из которых каждая может превратиться
в зародыш.
Внервые гнетовая гипотеза, или «теория эквивалентности»,
как ес некоторые называют, была высказана В. Гофмейстером
| 1561], Э. Страсбургером [ЗёгазБитесег, 1884] и С. Г. Навашиным
14900].
Сравнивая покрытосеменные с Спезез, исследователи, при-
знающие гнетовую гипотезу происхождения покрытосеменных,
указывают следующие общие между покрытосеменными и Gneta-
1е$ признаки, а именно: 1) варьирование числа ядер макроспор,
принимающих участие в развитии женского гаметофита и в 0б-
разовании моно-, би- и тетраспорических зародышевых мешков
(пи у одного из других порядков голосеменных такого сходства
с покрытосеменными не обнаружено); 2) тенденция к уменьше-
иню числа ядер в зародышевом мешке, которая сильнее выра-
спа у покрытосементых; 3) отсутствие архегониев; 4) слабое
развитие эндосперма до оплодотворепия и интенсивное его раз-
витие ноело оплодотворения. 3 обоих случаях при этом наблю-
дается слияние ядер эпдосиерма. Фагерлинд [КасегИю@, 1948],
высказывая мнение, что между покрытосеменными uw Gnetum
существует тесная генетическая связь, считает, что 1) полярные
ядра в зародышевом мешке покрытосеменных являются остатка-
ми свободных ядер женского гаметофита Спеб и раппих фаз
развития женского гаметофита других голосеменных; 2) цент-
ральная вакуоль зародышевого мешка покрытосеменных гомоло-
гична таковой у Спели и других голосеменных; 3) клетки зароды-
шевого мешка покрытосеменных гомологичны клеткам перифери-
ческого слоя женского гаметофита Слет, причем яйцеклетка —
фертильная клетка периферического слоя, антиподальные же
клетки покрытосеменных соответствуют нижней части женского
гаметофита; 4) эндосперм покрытосеменных — это гаметофит,
который стимулируется к дальнейшему развитию слиянием со
спермием.
Исследователи, которые придерживаются гнетовой гипо-
тезы, считают, однако, что современные представители Спе-
tum He являются тем типом, из которого возникли покрытосемен-
ные, и что как те, так и другие возникли от общих предков и
развивались параллельно друг другу. Исходя из гнетовой гипо-
тезы, зародышевый мепюк покрытосеменных пе ирямо возпик
из зародышевого мена Спейит, а редукция зародышевого меш-
ка обеих групи шла параллельно друг другу и в обоих случаях
привела к полной потере архегопиев — условия, при котором все
ядра верхней части зародышевого мешка могут рассматриваться
как гомологи женских гамет. По мнению П. Махешвари [Маъе-
shwari, 1948], гнетовая гипотеза из всех ныне существующих
теорий происхождения зародышевого мешка покрытосеменных
является паиболее привлекательной и могла бы стать рабочей
теорией, полезной для стимуляции дальнейших исследований по
этой проблеме. Однако гнетовая гипотеза не может преодолеть
17 В. А. Поддубная-Арнольди 257
}
некоторые трудности. Так, например, исходя из этой гипо-
тезы не удается объяснить возникновение эндосперма покрыто-
семенных после оплодотворения, а также присутствие синергид
в зародышевом мешке покрытосеменных. Таким образом, и гне-.
товая гипотеза происхождения зародышевого мешка покрытосе-
менных далеко не совершенна и оставляет открытым вопрос о
филогенетической близости покрытосеменных и Спеаез.
Особое положение занимает гипотеза происхождения зароды-
шевото мешка покрытосеменных, предложенная Е. Н. Герасимо-
вой-Навашиной [1954, 1956, 1958]. Исходя из предпосылки, что
основой возникновения зародышевого мешка являются общие
для любой клетки закономерности делений, Е. Н. Герасимова-На-
вашина пришла к заключению, что зародышевый мешок покры-
тосеменных — это не что иное, как сильно сокращенный в ходе
исторического развития гаметофит, гомологичный гаметофиту
голосеменных, но в силу сокращения периода своего развития
лишенный архегониев. Из-за ускорения развития и упрощения
яйцеклетка покрытосеменных не является, по мнению Е. Н. Гера-
симовой-Наваптиной, гомологом яйцеклетки голосеменных, хотя
обе эти клетки по функциям сходны “между собой. Синергиды,
антиподы и центральную клетку зародышевого мешка Е. Н. Ге-
расимова-Навашина [41958] рассматривает как клетки женского
заростка и утверждает, что нет достаточных оснований видеть
в зародышевом мешке покрытосеменных специфическую особен-
ность этой группы растений, которая могла бы возникнуть толь-
ко однажды и в одном месте. Вследствие этого Е. Н. Герасимова-
Навашина, в отличие от других цитоэмбриологов, склонна к отри-
цанию строго монофилетического происхождения покрытосемен-
ных и высказывается за их политопно-монофилетическое проис-
хождение.
Перечисленными представлениями о происхождении женско-
го гаметофита покрытосеменных не исчерпываются все попытки,
сделанные в этом направлении. Мы привели здесь лишь наиболее
известные из них. Какая из точек зрения на происхождение за-
родышевого мешка покрытосеменных окажется правильной — по-
кажут время и дальнейшие исследования.
При решении вопроса о происхождении покрытосеменных
немалое значение играет и роль двойного оплодотворения и по-
липлоидного эндосперма. Е. Н. Герасимова-Навашина [1947]
предложила митотическую гипотезу двойного оплодотворения,
которая заключается в том, что благодаря имеющейся в зароды-
шевом мешке митотической полярности и отсутствию у спермиев
митотического покоя к моменту оплодотворения, спермии, попав
в митотическое поле, отходят к полюсам, как сестринские груп-
пы хромосом, где и встречаются с ядрами яйцеклетки и цент-
ральной клетки. Таким образом, механизм двойного оплодотворе-
ния Е. Н. Герасимова-Навашина объясняет, исходя из представ-
ления об общих закономерностях митоза. По ее мнению, двойное
оплодотворение, как и зародышевый мешок, не представляет со-
258 .
бой ничего нового, присущего лишь покрытосеменным, а есть
проявление общих закономерностей, имеющихся в основе разви-
тия любой клетки. Она считает, что возникновение двойного оп-
лодотворения, как и зародышевого мешка, — необходимое след-
ствие дальнейшего упрощения строения гаметофитов покрыто-
соменных, особенно женского.
В чем биологическая сущность двойного оплодотворения?
Слияние одного из спермиев с ядром яйцеклетки не вызывает
сомпений — это истинный акт оплодотворения. Слияние же дру-
гого спермия со вторичным ядром зародышевого мешка объяс-
пяют по-разному. С. Г. Навашин [1898] рассматривал его как
обычный половой акт и признавал, что первичное ядро: эндоспер-
ма равноценно зиготе, а самый эндосперм приравнивал к видоиз-
мененному зародышу, предназначенному для питания зародыша,
образующегося из яйцеклетки. В противоположность С. Г. Нава-
тину некоторые исследователи, в том числе и Страсбургер [5%таз-
Бигоег, 1884], рассматривали слияние ядер при образовании эндо-
сперма не как истинный половой акт или «генеративное оплодо-
творение», а как «вегетативное оплодотворение».
По мнению Бринка и Купера [ВгшК, Соорег, 1947], двойное
оплодотворение является как бы компенсацией крайней редук-
ции женского гаметофита и повышает активность эндосперма,
давая этой ткани физиологические преимущества гибридизации.
Некоторые исследователи считают, что возникновение у покрыто-
семенных эпдосперма — особого питательного материала, обога-
щенпого двойственной наследственностью, возможно, значитель-
чо повысило жизнеспособиость и приспособленность покрытосе-
менных, что обусловило их победу в современную геологическую
эпоху.
На основе данных электронно-микроскопического исследова-
ния кукурузы А. А. Чеботарь [1970, 1972] высказал гомеоста-
тическую гипотезу двойного оплодотворения. Согласно этой гипо-
тезе, вследствие редукции хромосом и особого морфолого-физио-
логического состояния мужские и женские гаметы характеризу-
ются нарушенным гомеостазом, который восстанавливается при
двойном оплодотворении. А. А. Чеботарь считает, что двойное
оплодотворение выводит мужские и женские гаметы из создав-
шегося «тупика» и восстанавливает нарушенный гомеостаз. В ре-
зультате этого зигота отличается более уравновешенным генети-
теским и физиологическим гомеостазом. Эта гипотеза пока еще
педостаточпо развита и обоснована.
По представлению Н. С. Беляевой [1977] в зародышевом меш-
ке при двойном оплодотворении происходит следующее: пыль-
цевая трубка проникает в одну из синергид и с силой вскрыва-
ется в ней. Образующейся струей спермии выносятся за пределы
синергиды. При вскрытии пыльцевой трубки проибходит разру-
шение лизосом и освобождение ферментов, которые лизируют
цитоплазму спермиев. Из синергиды спермии выходят в виде
голых ядер. Передний ‘из них попадает в зону притяжения яйце-
47* 259
клетки и приходит в контакт с ее мембраной, после чего притя-
гающее действие прекращается. Спермий растворяет мембрану,
проникает в яйцеклетку и благодаря наличию направленных
токов цитоилазмы приходит в контакт с ядром яйцеклетки. Вто-
рой спермий, не испытывая притяжения током цитоплазмы, пе-
реносится к центральному ядру. После их объединения движение
цитоплазмы в центральной клетке меняется. Н. С. Беляева под-
держивает мнение тех ученых, которые считают, что генератив-
ные элементы и окружающая их цитоплазма находятся в тесном
взаимодействии. Несмотря на существование ряда точек зрения
о сущности двойного оплодотворения, проблема эта остается не
решенной и требует дальнейших исследований.
В течение многих лет идет дискуссия о том, какую группу
покрытосеменных следует считать исходной, какую следует помес-
тить в начале системы. Долгое время одни ученые считали исход-
ной группой МопосМату4еае или Атеп егае, другие — Polycar-
picae, согласпо системам Ветштейна [Wetstein, 1924] u Qurnepa
[Engler, 1931]. Oqnaxo moaqHee On1n0 mpusHaHo, что обе эти груп-
ны неодпородпы, искусствениы, и их разделили на ряд самостоя-
тельных порядков, что вполне согласуется и с длавпыми цитоэм-
бриологии. Л. А. Куприянова [4964, 1965] относит к Мопоса-
пу4деае (АтепИЁегае) следующие порядки: 1) ЗаПсаез, 2) Огы-
cales, 3) Batidales, 4) Balanopales, 5) Eucommiales, 6) Myricales,
7) Juglandales, 8) Casuarinales, 9) Betuales, 10) Fagales, 11) Gar-
ryales, 12) Leitneriales, 13) Julianiales, 14) Rhoipteleales.
На основании данных палинологии она считает, что Атеп-
tiferae He может рассматриваться как монофилетическая группа
и что порядки, включенные в эту группу, находятся на разных
эволюционных уровнях. Пыльцевые зерна большинства растений
этих порядков по строению спородермы относятся к весьма под-
винутым эквивалентно-апертурным типам, наиболее широко рас-
пространенным среди пыльцы высокооргапизоваппых двудоль-
ных. «Атен [егас», по дапным Т, Л. Куприяновой [4964, 1965],
охватывают почти все тины апертур двудольных, что наглядно
указывает па отсутствие гепетической одпородности в этой груп-
пе и на ее связь со многими порядками двудольных растений,
a umenHo c Hamamelidales, Ranales, Caryophyllales, Euphorbiales,
Proteales, Araliales u Заршдаез. Однако сама Л. А. Куприянова
признает, что одни из этих связей установлены ею с полной уве-
ренностью, в то время как другие в некоторой степени проблема-
тичны. Таким образом, в этом направлении необходимы дальней-
шие исследования.
То, что Amentiferae Ha протяжении многих десятков лет
объединялись в одну группу, по мнению Л. А. Куприяновой
[1964, 1965] объясняется развитием у них конвергентных черт,
выработавшихся в процессе эволюции, в связи с приспособлени-
ем к ветроопылению. Энглер, Ветштейн и многие другие исследо-
260
ватели рассматривали однопокровость цветков как первичный
исходный признак и соответственно этому помещали представи-
телей «МопосМатуйдеае» («АтепИ{егае») в начале системы по-
крытосеменных. Галлир, Бесси, Хатчинсон, А. А. Гроссгейм,
A. Л. Тахтаджян и другие исследователи помещали Мопос|-
1атудеае (АтепЁ{егае) в системе довольно высоко, как про-
изводные молодые группы. В настоящее время большинство си-
стематиков, морфологов, анатомов и цитоэмбриологов разделяют
вторую точку зрения и в начале системы покрытосеменных ста-
вят порядки МаспоНаез, Вапа!ез или Капипси|аез наряду с
Annonales, Nympheales, Sarraceniales u Aristolochiales, Ha Kotopsie
делят Polycarpicae.
Данные питоэмбриологии не противоречат тому, чтобы поря-
док МаспоПа]ез считать исходным и располагать его в начале
системы, как это сделали А. А. Гроссгейм [1945], А. Л. Тахтал-
жян [1959, 1966 и 1970], Xarauucon [Hutchinson, 1959, 1969] u
др. Onuako Xaranucou cuutaet, 4ro ot Magnoliales mpoucxonar
только древесные растения, травянистые же — из Вапа[ез, в то
время как А. Л. Тахтаджян и другие в основу всех покрытосе-
менных помещают лишь один порядок — МаспоНаез. Несмогря
па то, что в порядке МаспоПа]ез семейства Епратайсеае и Н!-
шащапагасеае почти не исследованы в отношении цитоэмбриоло-
гии, все же можно сказать, что семейства этого порядка харак-
теризуются примитивными цитоэмбриологическими призпаками,
а именно: малым разпообразием чисел хромосом, двухклеточным
мужеким гаметофитом, апатропной крассинуцеллятной семяпоч-
кой с двумя интегументами, наличием массивного комплекса па-
риетальных клеток, Ро]угопат-типом зародышевого мешка с тре-
мя эфемерными антиподами, массивным руминированным хоро-
шо развитым эндоспермом, очепь малепьким слабо дифферен-
цированным зародышем, отсутствием апомиксиса и т. д. Эти 0со-
бенпости порядка МаспоПа!ез оправдывают представление
06 его примитивности, и расположение его в начале систе-
мы покрытосеменных не вызывает противоречия со стороны
цитоэмбриологов. Порядок Вапа!ез, или Ranunculales, также
имеет немало примитивных цитоэмбриологических признаков:
пебольшое разнообразие чисел хромосом, двухклеточный муж-
ской гаметофит, анатропную крассинуцеллятную семяпочку
с двумя интегументами, пуцеллярлый колпачок, наличие
париетальных клеток, Роугопат-тии зародьипевого мешка, хо-
ролло развитый эндоснерм, маленький слабо развитый зародыш,
отсутствие гаусторий. Вместе с тем у представителей этого по-
рядка уже имеется немало прогрессивных цитоэмбриологических
признаков, особенно в семействе Вапипсу[асеае. И этим призна-
кам относятся: значительное разнообразие чисел хромосом, тен-
денция к образованию анатропной тенуинуцеллятной семяпочки
с одним интегументом, уменьшение числа, а в ряде случаев и
исчезновение париетальных клеток, наличие зародышевых меш-
ков АШат-, Огаза- и Ререгопта-типов, наличие крунных долго
264
сохраняющихся. многоядерных антипод, число которых иногда
увеличивается путем митотического деления первоначально трех
клеток, довольно хорошо дифференцированного зародыша, появ-
ление склонности к апомиксису и полиэмбрионии. Все это свиде-
тельствует о том, что порядок Вапапсиез, хотя и является
примитивным, все же выше порядка МаспоПа!ез и расположение
его в системе А. Л. Тахтаджяна [14966, 1970] выше порядка Мас-
поаез, но вблизи его, представляется нам правильным. Однако
как среди вышеупомянутых, так и других порядков покрытосе-
менных в настоящее время не обнаружено цитоэмбриологических
типов, которые могли бы быть каким-нибудь образом связаны с
голосеменными и которые выходили бы за рамки изменчивости
покрытосеменных и являлись бы промежуточными между обоими
типами, характерными каждому из них. Поэтому вопрос о том,
являются ли порядки МаспоПа]ез или Вапа[ез действительно са-
‘мыми примитивными исходными, не может пока считаться окон-
чательно решенным. Некоторые исследователи, в том числе и
читоэмбриологи [РасегЦп@, 1948], высказывают такую точку зре-
ния, согласно которой как Monochlamydeae unu Amentiferae,
так и Роусагрсае являются древними, примитивными груп-
пами, произошедшими от еще более примитивных, ныне уже вы-
‘мерших покрытосеменных и развивавшихся параллельно друг
друту.
Для окончательного решения вопроса о том, какую и сколько
трупп покрытосеменных поставить как исходные в начале систе-
мы, требуются дальнейптие исследования, поскольку существую-
цих данных по этой проблеме пока еще недостаточно.
При рассмотрении вопроса о том, возникли ли однодольные
параллельно с двудольными, или однодольные возникли из дву-
дольных, или наоборот — двудольные из однодольных, данные
цитоэмбриологии скорее свидетельствуют в пользу того, что
©днодольные произошли из двудольных и именно из порядка
'МутрВаеаез, так как имеют с ним сходные цитоэмбриологиче-
ские признаки. В отношении цитоэмбриологии глубоких различий,
‘за исключением строения зародыша, между однодольными и дву-
дольными растениями не имеется. Да и различие в строении
зародышей одно- и двудольных сказывается лишь на более по-
здних стадиях развития. Развитие зародышей у представителей
обоих классов на начальных стадиях протекает настолько сходно,
что, по мнению ряда ученых, нет необходимости выделять особые
типы развития и строения зародышей для одно- и двудольных.
Джоганеен [1950] привел 6 типов развития и строения зародышей,
общих для одно- и двудольных растений. Сходство в развитии
и строении мужского и женского гаметофитов, в характере опло-
дотворения, развития и строения зародыша, эндосперма и ран-
них стадий у одно- и двудольных подтверждает, как нам кажет-
ся, монофилетическое происхождение покрытосеменных. ‚
262
Согласно представлениям-А. Л. Тахтаджяна [4945, 1948, 1961,
1170], М. С. Яковлева [4946], Л. В. Кудряшова [1964, 19661,
|. П. Солнцевой и М. С. Яковлева [41964], а также Т. Б. Баты-
гиной [1974], односемядольный зародыш возник из двусемядоль-
ного, но не путем срастания двух семядолей в одну или недораз-
пития одной из двух семядолей, как думали некоторые другие.
исследователи, а путем задержки делений одних клеток при ин--
тоцсивпом делении других, что привело к нарушению симмет-
рии, изменению всего онтогенеза зародыша и образованию одной
не боковой, как у двудольных, а конечной семядоли. Tak Kak
односемядольный зародыш — явление вторичное, то однодольные,.
следовательно, произошли из двудольных, а не наоборот, что.
согласуется с мнением ряда современных систематиков.
Не только в отношении морфологии, но и в отношении цито-
эмбриологии разделение двудольных на подклассы — раздельно-
лепестных и спайнолепестных,— является искусственными соз-
дает лишние трудности для определения систематического поло-
жения отдельных групп покрытосеменных, так как нередко за-
ставляет объединять несходные и разъединять сходные в отно-
шении цитоэмбриологии семейства и порядки. Поэтому и со сто-
роны цитоэмбриологии раздельнолепестные и спайнолепестные
Цитоэмбриологические признаки раз-
дельнолепестных представителей
Цитоэмбриологические признаки спай-
нолепестных представителей
—
Семяпочка анатропная, крассинуце-
лятная с двумя интегументами (исклю-
чение представляет собой семейство
Umbelliferae, rge семяпочка тенуинуцел-
лятная с одним интегументом)
Нуцеллус большой, мощно развитый,
не исчезающий вплоть до оплодотворе-
ния, а иногда и дольше
Париетальные клетки перед образо-
ванием макроспор присутствуют. Число
„их обычно колеблется от одной до не-
скольких. Иногда число их очень вели-
ко и они образуют массивную ткань. Ну-
целлярный колпачок обычно более или
менее хорошо выражен
Эндотелий вокруг зародышевого мешка
отсутствует (исключением является се-
мейство Оше!Шегае, у представителей
которых он обнаружен)
Эндосперм чаще всего гелобиальный,
реже нуклеарный и очень редко целлю-
лярный. Наряду с`эндоспермом иногда
образуется и перисперм. Семена обычно
с эндоспермом, а иногда и с периспер-
мом или только с периспермом
Семяпочек и семян в одной и той же
завязи много
Мужской гаметофит чаще всего двух-
клеточный
Тапстум в пыльниках обычно амебо-
идный (периплазмодиальный)
Зародыши небольших размеров, пря-
мые, бесхлорофилльные
Семяпочка анатропная, тенуинуцеллят-
ная с одним интегументом (исключение
представляют собой порядки Рг/и]а]ез.
и Cucurbitales, y которых семяпочки
крассинуцеллятные с двумя интегумен-
тами)
Нуцеллус слаборазвитый, скоро ис-
чезающий, нередко сведенный к одно-
му эпидермальному слою клеток, окру-
жающих единственную клетку оурхеспо-
рия
Париетальные клетки отсутствуют,
благодаря чему клетки археспория непо-
средственно превращаются в материн-
ские клетки макроспор. Нуцеллярный
колпачок обычно отсутствует
Эндотелий присутствует, состоит из
более или менее крупных ›таблитчатых
клеток, образующихся из внутреннего.
слоя интегумента, и окружает большую
или меньшую часть зародышевого меш-
ка или весь зародышевый мешок
Эндосперм чаще всего целлюлярный,
реже науклеарный и совсем уедко ге-
лобиальный. Перисперм отсутствует. Ce-
мена обычно без эндосперма и без пе-
рисперма
Семяпочек и семян в одной и той же
завязи обычно немного, часто всего по
одному
Мужской гаметофит чаще всего, трех-
клеточный
Тапетум в пыльниках обычно секуре-
торный
Зародыши крупные, нередко согнутые
хлорофиллоносные
263
тредставители, сходные по другим признакам, следует рассмат-
ривать вместе, как это сделали А. А. Гросстгейм [1945], А. Л. Тах-
таджян [1966, 1970] и др. Однако в большинстве случаев спайно-
лепестные характеризуются более высокой ступенью развития
по сравнению с раздельнолепестными не только в отношении
морфологии, но и со стороны цитоэмбриологии, так как имеют
более прогрессивные цитоэмбриологические признаки (см. с. 263).
После выхода в свет обзоров Шнарфа [ЗсЪпатЁ, 1934] и Да-
вис [Дау!з, 1966] по эмбриологии покрытосеменных, которые в
значительной степени способствовали дальнейшему развитию
работ по сравнительной эмбриологии, одни семейства не подвер-
гались исследованию, другие изучались в единичных случаях,
‘третьи — неоднократно, четвертые — многократно. Особенно ин-
тенсивно изучались и исследуются семейства Ranunculaceae,
Fabaceae, Scrophulariaceae, Asteraceae, Liliaceae, Orchidaceae u
Poaceae.
Представителям раздельнолепестных обычно свойственны
более примитивные, а спайнолепестным более прогрессивные
цитоэмбриологические признаки. Хотя эволюции морфологиче-
‘ских и цитоэмбриологических признаков часто идут независимо
друг от друга, однако нередко паиравление их совпадает, и мы
видим, что морфологически более развитые группы имеют и бо-
лее развитые цитоэмбриологические признаки, а морфологически
менее развитые группы имеют и цитоэмбриологические признаки
менее развитые.
Изучение раздельно- и спайнолепестных вместе очень удоб-
но, так как позволяет наметить переход целого ряда представи-
телей раздельнолепестных в спайнолецестные.
Рассмотрение цитоэмбриологии различных порядков и се-
мейств покрытосеменных растений в соответствии с системой
А. Л. Тахтаджяна [1970] показываст, что из 438 семейств, имею-
щихся в этой системе, 140, т. е. примерно 1/3 семейств, не изу-
чены или очень слабо изучены в отношении цитоэмбриологии.
Особенно это касается тропических и субтропических растений.
Такие порядки, имеющие по одному семейству, как барбеаез,
Didymelales, Barbeyales, Balanopales, Myricales mu другие, цитоэм-
бриологически совершенно не исследованы. Некоторые многосе-
моейственные порядки, например Тнезез, СагуорвуПа[ез, ЕмрВо-
rbiales, Saxifragales, Rutales, Geraniales, Gentianales, Liliales 1
другие, исследованы цитоэмбриологически лишь наполовину или
даже того меньше. Так, из 20 семейств, относимых А. Л. Тахтад-
жяном [1970] к порядку ТВеамез, данные по цитоэмбриологии
известны лишь для 8 семейств, а из 8 семейств порядка Еарпог-
biales цитоэмбриологически изучены лишь два семейства — Bu-
хасеае и ЕарпогЫасеае. Вместе с тем имеется немало порядков с
большим или меньшим числом семейств, которые в отношении
цитоэмбриологии изучены более или менее подробно. К, таким
264
порядкам относятся Magnoliales, Ranunculales, Рарауега]сз,
Рисола]ез, Сасатрйаез, Мушаез, ВБашпа!ез, П1рзасаез, Рое-
moniales, Asterales, Orchidales, Poales u ap. Особенно следуст
огметить mops70K Scrophulariales, us 18 cemeiicts KoTOporo TOALKO
лля двух семейств, а именно a Retziaceae n Calumellaceac,
нитоэмбриологических данных не имеется.
Среди тех порядков и семейств, которые более или менее под-
робно исследованы в отношении цитоэмбриологии, не обнаружено
таких семейств и порядков, которые имели бы либо только при-
митивные, первичные, либо только высокоразвитые, вторичные,
цитоэмбриологические признаки. Как правило, в одном и том же
порядке или семействе наряду с первичными встречаются и вто-
ричные цитоэмбриологические признаки. Однако первичные
питоэмбриологические признаки в большем числе наблюдаются
в более примитивных порядках и семействах, а вторичные — в
более высокоразвитых. Такие примитивные порядки, как Маспо-
liales, Ranuncules ип Гапта|е$, характеризуются первичвыми
цитоэмбриологическими призпаками, а такие высокоразвитые
порядки, как Азбега!ез, П1рзасайез и Роа|ез — вторичными. Много
порядков и семейств покрытосеменных (в этом отношении) ло
настоящего времени цитоэмбриологически исследованы недоста-
точно полно. Все это сильно затрудняет применение цитоэмбрио-
логического метода в целях систематики и особенно в целях
филогении. И все же, песмотря на неполноту, отрывочиость,
а иногда и недостаточную точность цитоэмбриологических данных,
кое-какие обобщения можно сделать и теперь. Некоторые поряд-
Ru, Hampumep Caryophyllales, Cucurbitales, Lamiales, Droserales,
Asterales, Orchidales, Poales u apyrue, uMeloT CXOAHBIC B OTHOMEHNH
цитоэмбриологии семейства и, по-видимому, объединяя действи-
тельно родственные семейства, роды, виды, являются естествен-
ными. С другой стороны, такие порядки, Kak Euphorbiales u
Liliales, ocoOeHHo cemelicrsa Euphorbiaceae nu Liliaceae, mo muto-
эмбриологическим признакам наиболее гетерогенные из всех
других порядков и семейств покрытосеменных, возможно, явля-
ются искусственными, и их следовало бы разделить на ряд само-
стоятельных порядков и семейств. По-видимому, те порядки и
семейства, в которых цитоэмбриологические признаки однородны
и в большей степепи примитивны, более древние, чем те, в кото-
рых эти признаки неодпородны и более высоко развиты. Возмож-
по, что разпообразие типов развития впутри одного и того же
порядка или семейства указывает на молодость их, на интенсив-
ное течение в них формообразовательных процессов.
Большое значение данные ‘сравнительной цитоэмбриологии
покрытосеменных могли бы иметь для выяснения вопроса о том,
в какой мере принятые в настоящее время порядки и семейства
могут считаться естественными. Цитоэмбриологические исследо-
вания внутри различных порядков покрытосеменных дают боль-
265>
той материал для установления систематических отношений
между многими семействами, родами и даже видами.
Сходство в развитии цитоэмбриологических признаков у раз-
ных порядков и семейств может свидетельствовать о родственных
отношениях между ними и естественности объединения их. Раз-
личие в развитии цитоэмбриологических признаков у разных
порядков и семейств может указывать либо на разные ступени
развития внутри родственных линий, либо на ошибочность дан-
ных, либо на искусственное объединение’ неродственных групп,
что должно определяться в каждом случае отдельно путем тща-
тельного исследования возможно более обширного и разнообраз-
ного материала. При сравнительно цитоэмбриологических иссле-
дованиях разных семейств, входящих в тот или иной порядок,
невозможно ожидать сходства всех цитоэмбриологических при-
знаков. Поэтому было бы неправильно делать заключение о род-
стве только в тех случаях, когда цитоэмбриологические признаки
разных семейств и порядков обнаруживают полное сходство. При
использовании цитоэмбриологических признаков в целях систе-
матики необходимо учитывать, что в, процессе развития той или
иной группы один или несколько цитоэмбриологических призна-
ков могли подвергнуться изменению, могли в процессе дальней-
шего развития подняться на иную, более высокую ступень раз-
BUTHA.
Некоторые ученые [МачтИхоп, 1939] считают, что если в од-
ном и том же семействе или порядке будут обнаружены разные
цитоэмбриологические признаки, то нельзя на основании этого
разделить эти таксоны, прежде чем не будет произведено иссле-
ъдование большого материала, ибо не исключена возможность,
что отклоняющиеся признаки могут оказаться вершинами разви-
тия внутри одной и той же родственной линии и будут связаны
постепенными переходами. По мнению ряда ученых, при сравни-
тельно цитоэмбриологических исследованиях необходимо прини-
мать во внимание не только возможность развития того или
иного цитоэмбриологического признака, но и влияние на него
экологических условий. Из двух близкородственных видов одного
‘и того же рода один вид, приспособленный к особым условиям
существования, может иметь совершенно иные цитоэмбриологиче-
ские признаки. Так, например, необычайные условия существова-
ния паразитов, сапрофитов, эпифитов и мангровых растений
наложили отпечаток на строение генеративных органов и способ
размножения у этих растений.
Имеется ряд очень важных данных, указывающих на влияние
экологических факторов (у ОгоБапеВасеае, Ругоасеае, Мопото-
paceae, Podostemaceae, Orchidaceae, Crassulaceae, Margraviaceae
и др.), которыми нельзя пренебрегать.
| Хотя существующие системы покрытосеменных [Гроссгейм,
41945; Тахтаджян, 1970; Cronquist, 1968; Hatchinson, 1959, 1969;
Dalhgren, 1975 и др.] более совернтенны по сравнению со стары-
ми системами [Engler, 1931; Wettstein, 19241 m Bo многом под-
266
творждены данными цитоэмбриологии, однако, учитывая состоя-
нио наших знаний, ни одна из них-не является бесспорной, фило-
"опетической. Возможно, что разграничение порядков, а в ряде
‹‘лучаев семейст, и расположение их относительно друг друга у
покрытосеменных подвергнется более или менее значительным
изменениям, когда комплексному методу исследования, в том
числе и сравнительной цитоэмбриологии, будет уделено больше...
вцимания, когда цитоэмбриологические исследования позволят
заполнить те пробелы, ‘которые имеются в настоящее время и
которые так мешают широким обобщениям при построении
филогенетической системы покрытосеменных.
Некоторые изменения в классификации покрытосеменных
можно было бы сделать уже и сейчас, если принять во внимание
известные в настоящее время данные по цитоэмбриологии пок-
рытосеменных растений. Необходимо подчеркнуть, что система-
тическое и филогенетическое значение цитоэмбриологических при-
знаков зависит не только от качества и масштабов исследования,
по и от самого объекта.
Рассмотрение данных сравнительного изучения цитоэмбри-
ологии ‘покрытосеменных показывает, что в этом направлении
в настоящее время сделано еще очень мало и что совершенно
необходимы дальнейшие исследования.
ЛИТЕРАТУРА '
Адмиральская С. А. Стерильность
перечной мяты (МепшТа р1регЦа
Е.) и ее преодоление: Автореф.
дис. ... канд. биол. наук. Л.: БИН
1960а: 17c.
Адмиральская С. А. Оплодотворе-
ние, развитие зародыша и эндо-
сперма у полиплоидной формы
перечной мяты (Меп а ррегца
Г.). ДАН CCCP, 19606, 13, 4:
919—921.
Акимова А. А. В вопросу эмбрио-
логии мужского цветка лецины
обыкновенной. Горьк. Гос. sen.
ин-т, 1972, 144: 22—24.
Акимова А. А. Цитоэмбриологиче-
ское исследование лещины обык-
новепной. — В кн.: Морфогенез
деревьев и кустарников. Горький,
1976: 36—57.
Алиева А. Г. Наблюдения над рос-
том пыльцевых трубок у арбу-
зов.— В кн.: Сборник научных ра-
бот по бахчовым культурам. Таш-
кепт; УзССР, Госиздат, 1952.
Алимджанова Д. Р. К биоморфоло-
гии кассии (пыльца).— Узб. биол.
журн. АН УзССР, 1979, 6:63—65.
Алимова Г. К., Яковлев М. С. ВК эм-
бриологии Streptocarpus rexil
Lindl. (Gesneriaceaec).— B xn.: VIT
Всесоюз. симпоз. по эмбриол.
раст. дев: Наукова думка, 1978,
Аминова М. Ш. Особенности разви-
тия пыльцы как систематический
признак семейства Gramineac:
Автореф. дис. ... канд. биол.
наук. Л., 1969. 24 с.
Аминов Н. Х. Микроскопическое и
субмикроскопическое строение эн-
досперма бобовых (Vicia faba L.):
Автореф. дис. канд. ... наук. JL,
1967. 25 с.
Андронова Н. Н. В эмбриологии
Fadogia cienkowskii Schweinf.—
bot. *ypu., 1972, 57, 8: 826—831.
Андронова Н. Н. К эмбриологии Ос-
todon setosum Hiern. (Rubia-
сеае).— Бот. журн., 1973, 58, 9:
1371—1377.
Андронова Н. Н. Спорогенез и раз-
витие гаметофитов y Macrosphyra
longistylia (DC) Hook. fil, (Ru-
biaceae Juss, Gardeniae DC).—
Бюл. МОИП. Отд. биол., 1975, 80,
2: 68—76.
Андроноса Н. П. О строении семя-
почки У Rubiaceae Juss. — Бот.
журн., 1977, 62, 10: 1461—1469.
Андропова Н. Н. Спорогенез и раз-
витие гаметофитов у Asperula,
odorata L. (Rubiaceae Juss. Ga-
lieae Dumort).— Bot. sxyps., 1978,
63, 3: 404—410.
Apramonoea J. H., Иванов М. А.
К эмбриологии акантопанакса си-
дячецветкового Acanthopanax ses-
silflorum (Rupr. et Maxim.)
Sum.— Mss. 6uon.-reorp. HAM nupu
Иркут. yu-re, 1969, 23, 3:284—
292.
Атабекова А. И. К изучению эм-
бриологий гороха. — Изв. ТСХА,
1960, 33 : 177—182.
Лтабекова Л. П., Лин-Пзян-син. Эм-
бриология люпипа.— В кн.: Лю-
пин. М.: 1962: 97—113.
Атабекова А. И., /Лин-Цзян-син.
Сравнительно - эмбриологические
исследования рода Биршиз (To-
ити.) — Изв. ТОХА. 1963, 2: 219—
221. -
Атабекова А. И., Лин-Цзян-син. Эм-
бриология у люпина. — Изв.
ТСХА., 1964, 4: 38—41.
Афанасьева Н. Г. Микроспорогенез
и некоторые вопросы филоге-
: В списке приводятся работы, опубликованные главным образом в период с 1965 по 1979 г., а также не включенные ранее в книги В. А. Поддубной-Ар- нольди [1964, 1976] и Г. Дэвис [Оау1з, 1966].
268
нии поричниковых (Scrophulari-
асеае).— Науч. докл. высш. шко-
лы. Биол. науки, 1959, 4: 414—122.
Афанасъева Н. Г. Микроспорогенез
м некоторые вопросы филогении
норичниковых.— В кн.: Морфоге-
нез растений. М.: Изд-во МТУ,
1961, 2: 502—506.
Лфанасьева Н. Г. Цитоэмбриологи-
ческое исследование вероники
альпийской (Veronica alpina L.).—
Науч. докл. высш. школы. Биол.
пауки, 1966, 1: 446—119.
Афанасьева Н. Г. Эндоспермальные
гаустории и их значение для по-
знания рода Veronica L.— Бот.
жхурн., 4974, 56, 2: 215—228.
Афанасьева Н. Г. Эмбриология ве-
роник (р. Уегошса Г.). Казань:
Казан. ун-т, 1976 : 144 с.
Афанасьева Н. Г., Граханцева Л. Ш.
К эмбриологии Редсатз КаиЕ-
mannii Pinzc. (Scrophulariaceae).—
Бот. журн., 1975, 60, 6: 777—787.
Афанасьева Н. Г., Михайлова Е. И.,
Мешкова Л. В. Цитоэмбриологи-
ческое исследование вероники
альпийской (Уетопса alpina L.).—
Науч. докл. высш. школы. Биол.
науки, 1966, 1: 146—119.
Афанасьева Н. Г., Окулова С. М.,
Процесс оплодотворения у веро-
nuk (Veronica зр.).— Бот. журн.,
1974, 56, 7: 1013—1119.
Афанасьева Н. Г., Окулова С. М.
Граханцева Л. Ш., Юсупова И. К.
Об оплодотворении у Scrophula-
г1асеае.— Бот. журн., 1975, 60, 12:
1770—1776.
Афанасьева Н. Г., Юсупова И. К.
Эмбриология рода марьянник —
Melampyrum L. (Scrophulariaceae
Lindl—B кн. VII Веесоюз.
симпоз. по эмбриол. раст. Киев:
Наукова думка, 1978, 2:6—7.
Ахундова Е. С. Формирование жен-
ского гаметофита у вязеля чир-
канского.— В Kin: VII Веесоюз.
симпоз. по эмбриол. раст. Киев:
Наукова думка, 1978, 3:4—5.
Ашурметов А. А. Микроспорапгий,
микроспорогенез и развитие га-
метофита у Мео аотозИз Рапо.—
Узб. биол. журн., 4974а, 2: 68—71.
Ашурметов А. А. Развитие пыльни-
ка, микроспорогенез и формиро-
вание пыльцы у Втуоша melano-
сагра Ма. — В кн.: Морфобиол.
особенности дикорастущих расте-
ний Узбекистана. Ташкент: Фан,
19740: 48—24.
Ашурметов А. А. Биология цвете-
ния и цитоэмбриология некото-
рых узбекистанских представите-
лей дынь и брионии (Ме!о Adans
Bryonia Г.): Автореф. дис. ...
канд. биол. наук. Ташкент, 1974в.
28 с.
Ашурметов А. А., Дзевальтов-
ский А. К. КарологчнЕ дослдже-
ния динь Узбек!стану.— Укр. бот.
журн., 1975, 34, 4:522—524.
Ашурметов А. А., Норматов Б. А.
Развитие женской генеративной
сферы у видов рода СЛусуггЫ та
Г.— В кн.: УП Всесоюзный сим-
позиум по эмбриологии растений.
Киев: Наукова думка, 1978, с.
2:78.
Ашурметов А. А., Саидов Д. К.
Строение семяпочки и развитие
женского гаметофита у дикой
сорно-полевой дыни. — ДАН
УзбсСсСР, 1975, 5: 54—56.
Баиникова В. П., Овсянникова О. В.
Цитоэмбриологические особенно-
сти развития семян (М№МсоНапа
гизИса Г.).— Сб. пауч. исслед. ра-
бот ВНИИ табака и махорки,
1973, 160—161 : 142—120.
Банникова В. П., Хведынич О. А.
Ранняя дифференциация ядер в
зародышевых мешках М№сойа-
па.— В кн.: Цитология и генети-
ка. Киев: 1968, 3, 3: 195—201.
Баннкова В. П., Хвединич О. А.
Процесс заплднения у №соИапа
rustica L.— Укр. журн., 1973, 30,
5:668—670.
Баранов П. А. О редукции женско-
го полового поколения в семей-
стве Отс Часеае.— Бюл. Средне-
аз. ун-та, 1925, 10: 181—195.
Баранов П. А. История эмбриоло-
гии растений. Л.: Изд-во AH CCCP,
1955. 439 с.
Барашов П. А. Строение виноград-
ной лозы.— В кн.: Ампелография
СССР. М.: М-во вкус. пром-сти
CCCP, 1946.
Баранов П. А. Колеориза у мирто-
вых.— В кн.: Академику В. Н. Су-
качеву к 75-летию со дня рожде-
ния. М.: Изд-во АН СССР, 1956.
Баранов П. А., Баранова Е. А., По-
лунина Н. Н. Интересная особен-
ность эмбриогенеза эвкалипта.—
Бот. журн., 1955, 40, 1:99—102.
Баранов П. А., Поддубная ВБ. А. К
эмбриологии Туркестанских ама-
риллисовых [Опрегиа зеуегхоуй
269
B. Fedtsch. и [хюШюп tataricum
(Pall.) НегЪ.].— Бюл. Среднеаз.
ун-та, 1925, 11 :4—14.
Баранов П. А., Райкова И. А., Ро-
манов И. Д. Строение и развитие
хлопчатника. Атлас. М. JL:
ОГИЗ — Изогиз, 1937.
Барна М. М. Особливост! розвитку
аночного гаметоф!та у трипло-
1дно1 осики (Роршиз tremula L.
var. gigas Munt.).— Укр. бот.
mypu., 1967, 24, 6: 42—49.
Барна М. М. Ембролопчне дослид-
жения топол!: шрам!дально1 (Ро-
pulus pyramidalis Roz).— Бот.
журн., 1969, 26, 1:93—100.
Барна Н. И. Морфогенез генератив-
ных органов и эмбриональное
развитие осины. — В кн.: Лесо-
водство и агролесомелиорация.
Респ. Межвед. темат. науч. сб.
1968, 15: 126—135.
Барна Н. Н. Цитоэмбриологическое
исследование некоторых видов
рода Роршиз Т.. в связи с гибри-
дизацией: Автореф. дис. ... канд.
биол. наук. Киев: 1969. 24 с.
Батыгина Т. Б. Некоторые данные
по цитологии процесса оплодо-
творения у пшениц.— В кн.: Мор-
фогенез растений. М.: Изд-во
МГУ, 1961, 2: 334—340.
Батыгина Т. Б. Микроспорогенез и
развитие пыльцевого зерна у
пшеницы.— ДАН СССР, 1962а, 142,
5: 1205—1208.
Батыгина Т. Б. Процесс оплодотво-
рения у пшениц.— Тр. Бот. ин-та
АН СССР, 19626. Сер. 7, 5: 260—
293.
Батыгина Т. Б. О возможности вы-
деления нового типа эмбриогеис-
за Angiospermae. — ДАП CCCP,
1968, 181, 6: 1499—1502.
Батыгина Т. Б. Эмбриология ише-
ницы. Л.: Колос, 1974а. 207 с.
Батыгина Т. Б. Эмбриология пше-
ницы: Автореф. дис. ... докт.
биол. наук. Л.: 19746, 54 с.
Батыгина Т. Б., Васильева В. Е.,
Маметьева Т. Б. Проблемы мор-
dorenesa in vivo u in vitro. Om-
бриогенез покрытосеменных pa-
стений.— Бот. журн., 1978, 68, 1:
87—111.
Батыгина Т. Б., Кравцова Т. И..,
Шамаров И. И. ВК сравнительной
эмбриологии представителей по-
panna Nymphaeales u Nelumbona-
les— Bot. журн., 1980, 65, 9:
1071—1087.
270
Батыгина Т. Б., Маметьева Т. Б.
К эмбриологии рода Роа L.—B
кн.: Актуальные проблемы эмбри-
ологии покрытосеменных расте-
ний. М.; Л.: Наука, 1979.
Батыгина Т. Б., Фрейберг Т. Е. По-
лиэмбриония у Poa pratensis L.
(Роасеае).— Бот. журн., 1979, 64,
6: 793—804.
Беляева Л. Ю. Развиток чолов!ч0-
го та жщночого гаметофутв, за-
пиления та заплднепия у катра-
ну аблсанського (Crambe abyssi-
nica Hochst.).— Yup. Oor. журн.,
1963, 20, 6: 24—31.
Беляева Л. Ю. Биология цветения и
цитоэмбриология катрава абис-
синского: Автореф. дис. ... канд.
биол. наук. Одесса, 1967, с. 20.
Беляева Л. Ю. Развиток чолов!1чого
Ta яаночого гаметофеив жовтуш-
ника лакф!олетового (Erysimum
cheiranthoides L.). — Укр. бот.
mypH., 1968, 25, 5:54—559.
Беляева Л. Ю., Фурса М. С. Разви-
ток пиляка, пилкових зерси т за-
родкових мпипкв Syrenia cana
(Pill et Mitt) Neilr.— Bor. sxypn.,
1978, 35, 3: 241—245.
Беляева Л. Ю., Фурса М. С. Форму-
вания чоломчих 1 жночих
структур кытка Berteroa incana
(L.) РС — Укр. бот. журн., 1979,
36, 6:574—577.
Беляева Л. Е., Фурса Н. С., Смыч-
ко Л. А. Эмбриология Зугеша са-
na (Pill. et Mitt.) Neilr.— Bor.
журн., 1972а, 62, 10: 1453—1461.
Беляева Л. Ю., Фирса М. С., Смы-
ченко А. О. EmOpionoriune досл!-
ждення Lepidium perfoliatum
L.— Yup. Oor. atypu., 19776, 34, 3:
207-262, 336.
Beaneea Л. Ю, Чайна К. А., Фур-
са М. С. Заилтднения т развиток
пастики y Diplotaxis tenuifolia
Д. С.— Укр. бот. журн., 1978, 35, 5.
Беляева Н. С. К вопросу оплодо-
творения у хлопчатника. — Изв.
АН ТССР. Сер. биол. паук, 1963,
5: 39—45.
Беляева Н. С. Процесс оплодотво-
рения у хлопчатника (Соззуртат
barbadense L.): Автореф. дис. ..
канд биол. наук. Ашхабад: 1964,
16 с
Беляева Н. С. К эмбриологии Ти-
bulus terrestris L. II. Оплодотво-
рение.— Изв. АН ТССР. Сер. биол.
наук., 1972, 4: 3—9.
Беляева Н. С. Развитие зародыше-
вого мешка и оплодотворение у
дельфиниума. — Изв. АН ТССР,
1974, 2: 3—12.
Беляева Н. С. Разнокачественность
спермиев у некоторых покрыто-
семенных и ее роль в процессе
двойного оплодотворения. — Изв.
АН ТССР. Сер. биол. наук., 1975,
3:3—8.
Беляева Н. С. Эмбриологическое
a3syienvne Calendula persica C. A.
Меу. Развитие женского гамето-
фита.— Изв. АН ТССР. Сер. биол.
наук, 1976a, 1: 42—47.
Беляева Н. С. Эмбриологическое
нзучение Calendula persica C. A.
Меу. Оплодотворение. — Изв. АН
ТССР. Сер. биол. наук, 19766,
2: 63—67.
Беляева Н. С. Развитие зародыше-
вого мешка и оплодотворепие у
покрытосеменных: Дис. ... д-ра
биол. наук. Л.: 1977. 288 с.
Бенецкая Г. К. Оплодотворение и
омбриогенез у подсолнечника при
различных способах опыления.—
Изв. АН ApmCCP, 1952, 5, 7:
17—33.
Бенецкая Г. К. Оплодотворение и
первые фазы эмбриогенеза у под-
солнечника. — Изв. АН АрмССР,
1954, 7, 12: 7—17.
Березенко Н. П. Цо вивчения м1-
кроспорогенезу, чолов!1чого гаме-
тоф!та та ембр!огенезу в садово!
суниц (Fragaria grandiflora
Ehrh.).— Укр. бот. журн., 1969, 26,
А; 81—89.
Березенко Н. П., Лиферова В. В.
Некоторые наблюдения над эм-
бриологией крыжовника (Grossu-
]ат1а гесПпа4а).— Укр. бот. журн.,
1970, 27, 3 : 318—324.
Беридзе Р. К. Развитие женского
гаметофита у рода Fragaria L.—
Тр. Тбилис. бот. ин-та АН ГССР,
1962, 22: 215—226.
Беридзе Р. К. К изучению эмбрио-
логии рода Егагата Т.— Тр. Тби-
лис. бот. ин-та АН ГССР, 1964,
23 : 235—239.
Беридзе Р. К. Полиэмбриония у
земляники.— Тр. Ин-та ботаники.
АН ГСООР, 1969, 26, Сер. Культ.
флора, 3: 102—108.
Богданова В. М., Грушвицкий И. В.
О размерах зародыша в семенах
аралиевых советского Дальнего
Востока — В кн.: Растительные
ресурсы. М.: Наука, 1970, 5:
523—527.
Богомолова Т. Д. Оплодотворение и
развитие семени цикория (СлеБо-
rium Intybus).— Jloxn. TCXA, 1959,
46 : 227—232.
Богданов П. Л., Ступов В. И. Пар-
тенокарпия и апомиксис у бе-
рез.— В кн.: Апомиксис и селек-
ция. М.: Наука, 1970: 242—246.
Вальцева О. В., Савич Е. И. О раз-
витий зародынта y Nymphaea can-
dida Prest uN. tetragona Georgi.—
Бот. журн., 1965, 50, 9: 4323—
1326.
Васильченко И. Т. О работе С. С. Не-
нюкова «Систематическое поло-
жение семейства ТВе|оопасеае».—
Бот. журн., 1979, 64, 8 : 1206—1207.
Варшанина Т. П. Полиэмбриония y
овощных пасленовых культур и
лука.— Бюл. Всесоюз. ин-та, 4973,
33 : 90—98.
Варшанина Т. П. Изучение полиэм-
брионии У томата, перца и бак-
лажана (Solanaceae L., Lycopersi-
con esculentum Mill. Capsicum
annuum LL. Solanum Melongena
Г.): Автореф. дис. ... канд. биол.
паук. Л., 1979. 23 с.
Верещагина В. А. Экология цвете-
пия и опыление Oxalis acetosel-
la Г. Бот. журн., 1965, 50, 8:
1078—1091.
Верещагина В. А. Микроспорогенез
у гинодиэцичного вида Echium
уЩраге Т.— Бот. журн., 1974, 56,
8 : 1006—1043.
Верещагина В. А. Об опылении не-
которых гинодиэцичных видов
Echium.— B ku.: Экология опыле-
ния. Пермь: Гос. ун-т, 1975, 1:
109—412.
Верещагина В. А. Цитоэмбриологи-
ческое изучение клейстогамии у
Рогалика хенке (СегаюсШоа ha-
епкеапа С. Ртгез1).— В кн.: Эколо-
гия опыления. Пермь: Гос. ун-т,
1976а, 2: 82—89.
Берещагина В. А. Некоторые эмбри-
ологические особенности гиноди-
эцичных видов Ес (Boragi-
пасеае).— Бот. журн., 19766, 61,
11: 1582—1589.
Верещагина В. А. Цитоэмбриология
разных половых форм цветков у
гинодиэцичных растений.— В кн.:
УП Всесоюзн. симпоз. но эмбри-
ол. раст. Киев: Наукова думка,
1978а, 2: 12—13.
Верещагина В. А. Изучение гино-
271
quoynuma y Echium amoenum Fisch.
её Меу. П. Развитие пыльников
и пыльцы в обоеполых и функ-
ционально женских клетках. —
Экол. опыления. Пермь: Гос. ун-т,
19786 : 98—104.
Верещагина В. А. Изучение гино-
диэции у Echium amoenum (Bo-
raginaceae).— Бот. журн., 1978s,
63, 2: 183—190.
Верещагина В. А. Изучение гино-
диэции y Echium amoenum Fisch.
et Mey. III. Зародышевый мешок
и эндосперм. — Экол. опыления
(Пермь), Гос. ун-т, 1979, 4: 3—12.
Верещагина В.А. Эмбриологическая
характеристика семейства Oxali-
daceae R. Brown.— Buon. науки,
изд-во МГУ. 1980а, 3:52-—58.
Верещагина В. А. Эмбриология не-
которых клейстогамных форм.—
Бот. журн., 19806, 65, 8 : 1047—
1156.
Верещагина В. А., Бояршинова О.Н.
К эмбриологии разных половых
форм цветков душицы обыкно-
BeHHow (Origanum vulgare 1.).—
Экол. опыления (Пермь), 1979, 4:
13—24.
Верещагина В. А., Ермишина 0. М.
Изучение мейоза в микроспоро-
цитах у половых форм цветков
Echium russicum У. F. Gml. (Bo-
raginaceaec).— Bor. smypiu., 1915,
D8, 12: 1760—1764.
Верещагина В. А., Малацина Л. И.
О гинодиэции душицы обыкно-
венной (Ог1еапит уШраге Т..).—
Науч. докл. высш. школы. Биол.
науки, 1974, 6: 51—56.
Видаяти O., Поддубная-Арноль-
ди В. А. Цитоэмбриологическое
исследовапие Calendula officina-
13 1.—В ки.: Морфология цвет-
ковых растений. М.: Паука, 1971:
15—84.
Винокурова Л. В. Палинологические
данные к систематике семейств
Dipsacaceae и Morinaceae.— IIpo-
6nemer Goranunu, 1959, 4: 54—76.
Волосовцев В. Д. Биология цвете-
ния и цитоэмбриология бессемян-
ных сортов винограда: Автореф.
дис. ... канд. наук. Ташкент,
1967.
Ворсобина Л. И. Процесс оплодо-
творения и ранний эмбриогенез
У некоторых амариллисовых (р.
Zephyranthes Herb., Sternbergia
Wald. её КИ.) в связи с вопроса-
ми семенного воспроизведения.
272
Капд. дис. биол. наук. Л., 1973.
Ворсобина Л. И., Солнцева М. П.
Развитие гелобиального эндеспер-
ма в семействе амариллисовых у
родов Zephyranthes ui Sternber-
gia.— В кн.: Актуальные вопросы
эмбриологии покрытосеменных. Л.:
Наука, 1979а: 75—81.
Ворсобина Л. И., Солнцева М. П.
Особенности цветения и процес-
са оплодотворения у некоторых
амариллисовых.— В кн.: Актуаль-
ные вопросы эмбриологии покры-
тоссменных. Л.: Наука, 19796:
23—38. .
Гагечиладзе М. П., Гваладзе Г. Е.
К изучению эмбриологии Опаста-
ceae.— Сообщ. АН ГССР, 1970, 60,
3: 677—680.
Галимова К. А. Некоторые данные
о развитии женской геператив-
ной сферы у солодки. — В ки.:
Эколого-биологические особенно-
сти и хозяйственно-ценные при-
знаки солодки в культуре. Таш-
кент: Фап, 1975а : 38—40.
Галимова К. Л. О микроспорогене-
зе и развитии мужского гамето-
фита у солодок толой и ураль-
ской.— В кн.: Эколого биологиче-
ские особенности и хозяйствен-
ные признаки солодки в культуре.
Tamkeut: Man, 19756: 35—37.
Галимова Р. А. Развитие мужской
геперативной сферы У солодки.—
В кн.: УП Веесоюз. симпоз. по
эмбриол. раст. Киев: Наукова
думка, 1978, 3:12—13.
Гамбарова ГР. К. Эмбриологические
особенности гиаципта римского
(Н. готапиз 1.) в условиях Ап-
тшерона.— В кн.: Цитоэмбриологи-
ческие исследовапия раститель-
пой Флоры Азербайджана. Баку:
ОЛМ, 1972: 112—120.
Гамбирова Р. К. Особенности стро-
ения мужеского цветка и иекото-
рые цитоэмбриологические дан-
ные борщевика жесткого и бор-
щевика Сосновского в Азербай-
джане (Пиркули). ВИНИТИ, Ба-
ку, 1973, № 554—73, Ден.., с. 6.
Гамбарова Р. К. Эмбриологические
особенности мерендеры трехстол-
биковой (Merendera trigyna) B
условиях Апшерона АзССР. Ин-т
ботаники АН АзССР. Баку: 1975:
Гамбарова Р. К. Эмбриологические
особенности Нуаст из готапиз
(Liliaceae) в условиях Апшеро-
па.— В кн.: Тез. докл. УГ делегат.
Всесоюз. бот. 0-ва, Л.: Наука,
1978а : 14—75.
Гамбарова Р. К. Сравнительно-эм-
бриологическое исследовапие Ас-
трагала ложносладколистного и
Астрагала нутового в Азербай-
джане (Куба-Кусары). — В кн.:
УП Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 19786,
3: 11—12.
Гваладзе Г. Е. Эмбриология АШит
schoenoprasum Г. — Сообщ. АН
ГССР, 1960, 25, 3:321— 334.
Гваладзе Г. Е. К изучению эмбри-
ологии рода Allium L.— Сообщ.
АН ГССР, 1964, 26, 2: 193—200.
Гваладзе Г. Е. Развитие генератив-
ных органов и эмбриогенез у не-
которых видов pona Allium L.:
Автореф. дис. капд. биол.
наук. Тбилиси, 1962.
Гваладзе Г. Е. В изучению полиэм-
брионии рода Аим. — Сообщ.
ГССР, 1963, 2: 393—398.
Гваладзе Г. FE. Вивипария и спо-
собность к генеративному раз-
множению у АШаш sativum L—
Сообщ. АН ГССР, 1965, 40, 2: 421—
427.
Гваладзе Г. Е. К сравнительно-эм-
бриологическому изучению амфи-
миксиса и апомиксиса в роде Al-
lium ТГ.— Тр. Ин-та ботаники АН
ГССР, 1969а, 25, Сер. Культ. фло-
ра, 3: 34—43.
Гваладзе Г. Е. ВК изучению энло-
сперма в роде АШиют Г.— Сообщ.
АН ГССР, 19696, 53, 3:681—684.
Гваладзе Г. Е. Формы апомиксиса
в роде АШим.— В кн.: Апомик-
сис и селекция. М.: Наука, 1970а:
155—159.
Гваладзе Г. Е. В изучению поли-
эмбрионии у АШишм schoenopra-
зит Г.— Сообщ. АН ГССР, 19706,
58 : 185—188.
Гваладзе Г. Е. Сравнительпо-эомбри-
ологическое исследование зароды-
шевых меитков типов Polygonum,
Oenothera, Allium, Frilillaria y
некоторых растений: Автореф.
дис. ... д-ра биол. наук. Тбилиси,
1973.
Гваладзе Г. Е. Халазальное поляр-
ное ядро центральной клетки за-
родьшшевого мешка покрытосе-
менных. Тбилиси: Мецниереба,
1976 : 138.
Гваладзе Г. Е. В эмбриологии рода
Сасеа. — Вестн. Груз. бот. о-ва
18 В. А. Поддубная-Арнольди -
АН ГОСР, 1978a, 7Т:127—146.
Гваладзе Г. Е. Зародышевый ме-
шок типа Peperomia.— B xu.: VII
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 19786,
3:14—415.
Гваладзе Г. Е., Гагечиладзе М. И.
К эмбриологии онагровых.— Изв.
ГССР. Сер. биол., 1977, 3,6: 514—
519.
Герасимова-Навашина Е. Н. О по-
ведении спермиев в пыльцевой
трубке y Crepis.— ДАН СССР,
1947, 57, 3: 285—288.
Герасимова-Навашина Е. Н. Развп-
тие зародышевого мешка, двой-
ное оплодотворение и вопрос о
происхождении покрытосемен-
ных.— Бот. журн. 1954, 39, 5:
655—680.
Герасимова-Навашина Е. Н. Двон-
ное оплодотворение покрытосс-
менных, его природа и происхох:-
депие: Автореф. дис. д-ра
биол. наук. Л. 1955: 55 с.
Герасимова-Навашина Е. Н. Неко-
торые цитологические закономер-
ности процесса оплодотворения у
растений.— В кн.: Проблемы со-
временной эмбриологии. Изд-во
Ленингр. гос. ун-та, 1956: 48—57.
Герасимова-Навашина Е. Н. О га-
метофите и об основных чертах
развития и функционирования
воспроизводящих элементов у по-
крытосеменных растений.— В кн.:
Проблемы ботапики, 1958, т. ПЕ
125—167.
Герасимова-Навашина Е. Н. Эмбри-
ологические наблюдения над Ага-
chis hypogaca ТГ.. — Бот. журн.
1959, 44, 10: 1453—1466.
Герасимова-Навашина Е. Н. Цвой-
ное оплодотворение покрытосс-
менных и некоторые его теорети-
ческие аспекты.— В кн.: Пробле-
мы эмбриологии. Киев: Наукова
думка, 1971: 113—152.
Герасимова-Павашина Е. Н. Ноко-
торыс данные об обусловленно-
сти строения зародышей покры-
тосеменных растений. — Бот.
журн., 1972, 57, 4: 441—457.
Герасимова-Навашина Е. Н. Двой-
ное оплодотворение и митоз.—
В кн.: УП Всесотоз. симпоз. по эм-
бриол. раст. Киев: Наукова дум-
ка, 1978, 3: 15—17.
Герасимова-Навашина Е. Н., Баты-
гина Т. Б. Процесе оплодотворе-
nua y Scilla sibirica Andr.— Bor.
273
журн. 1958, 43, 17:959—989.
Герасимова-Навашина Е. Н., Баты-
гина Г. Б. Некоторые дополни-
тельные данные о половом аппа-
рате и о процессе двойного опло-
дотворения у покрытосеменных
растений (на примере ЭеШа sibi-
rica Andr.).— В кн.: Проблемы
современной эмбриологии. M.:
МГУ, 4964: 86—99.
Герасимова-Навашина Е. Н., Гуля-
ев В. А., Васильцова Т. М. Неко-
торые данные об ультраструкту-
ре клеток зародышевого мешка
Crepis capillaris (L.) Wall.—
Бюлл. Глав. бот. сада AH CCCP,
1973, 89: 14—19.
Главацкая Т. П., Кабыш Н. А. В
эмбриологии пырея сизого Арто-
pyron glaucum (Desf) Roem et
Schult.— B xu.: Warponyroma wpe-
тения и морфогенез монокарпи-
ческих побегов травянистых по-
ликарпических растений. Сверд-
ловск: Гос. уп-т, 1976 : 96—102.
Глущенко Г. И. К биологии цвете-
ния и эмбриологии скополии
карниолийской.— Бот. журн., 1962,
47, 1: 1017—1025.
Глущенко Г. И. Эмбриологическое
исследование начальных этапов
развития скополии карниолий-
ской.— Бот. журн., 1963а, 48, 11:
1680—1686.
Глущенко Г. И. Некоторые особен-
ности в эмбриологии скополии
карниолийской (Scopolia сагшо-
lica).— Изв. АН СССР. Сер. биол.,
19636, 5 : 724—734.
Глущенко Г. И. М!кроскошчне до-
слджения скополи карнолськот
(Scopolia carniolica Jacg) ta rima-
лайско! (5. lurida Dun in DC).
У звязку з питапиями стериль-
ности.— Укр. бот. журн., 1964, 21,
4:53—61.
Глущенко Г. И. Ембр!олойчне до-
слджения скополй пмалайско!
(Scopolia lurida Dun.) Berpomenoii
в ycnopax Киева.— Укр. бот.
журн., 1967, 24, 3:26—32.
Глущенко Г. И. Биология и эмбри-
ология паслена птичьего. — Укр.
бот. журн., 1968a, 25, 1:89—94..
Глущенко Г. И., Кордюм Е. Л. Осо-
бенности формирования репро-
дуктивных органов у Зрагоапиии
знирех На4$.— С.-х. биол., 19686,
3, 4: 564—571.
Голенкин М. И. Курс высших ра-
стений. М.; Л.: 1937: 598 с.
274
Горб Л. К. Биология некоторых ви-
дов рода Саёара Эсар. в услови-
ях западных областей УССР:
Автореф. дис. канд. биол.
наук. Львов: Гос. ун-т, 4975: 20с.
Грабовская С. Р. Развитие много-
клеточного археспория в семя-
почках видов рода Capsicum
Тоиги.— Тр. по прикл. бот., ген.
и селекции. 1968, 40, 1:65—73.
Граханцева Л. Ш. Особенности ми-
кроспорогенеза мытника кауфмана
(Pedicularis kaufmannii Pinzg.)—
В кн.: УП Всесоюзн. симпоз. по
эмбриол. раст. Киев: Наукова дум-
Ka, 1978, 1: 21—23.
Гревцова Н. А. Некоторые данные
о строении семяпочек яблони и
груши.— В кн.: Морфология цвет-
ковых растений. М.; Наука, 1971а:
64—74.
Гревцова Н. А. О полиэмбрионии
яблони и груши.— Вестн. МГУ,
19516, 2: 47—52.
Гревцова Н. А. Сравнительно-эм-
бриологическое исследование не-
которых продставителей родов
Машз МШ и Ругиз Г.: Автореф.
дис. канд. биол. наук. M.,
1974. 23 с.
Гревцова Н. А. Сравнительно-эм-
бриологическое исследование яб-
лони и груши в связи с их систе-
матическим положением.— В кн.:
Материалы 5-го Моск. совещ. по
филогении раст. М.: Наука, 1976:
40—42.
Гревцова Н. А., Коробова-Семенчен-
ко Л. В. К эмбриологии воронца
заостренного.— В кн.: УП Весесо-
юз. симпоз. по эмбриол. раст.
Киев: Наукова думка, 1978, 3:
20—22.
Грушвицкий П. В. Подземное про-
растапие и фупкции семядолей.—
Бот. журн., 1963, 48, 6:906—915.
Гуляев В. А. Применение сравни-
тельно-эмбриологического метода
К систематике рода СИитиШиз
Schrad.— Тр. плодоовощ. ин-та
им. И. В. Мичурина, 1955, 8:
331—334.
Гуляев В. А. К описанию процесса
оплодотворения у СИгии$ vul-
garis Schrad. — Bron. Всесоюз.
ин-та растениеводства, 1961, 9:
21—30.
Гуляев В. А. О типе зародышевого
мешка. Praecitrullus fistulosus
(Sticus) Pang.— [AH CCCP, 1962,
146, 5: 1216—1219:
yineo B. A, Сравнительная эмбри-
опогия СаситоКасеае и се значе-
ные для систематики семейства.—
Бот. журн., 1963, 48, 1:80—85.
‘усейнова С. О. Цитоэмбриологиче-
ское исследование — некоторых
древних и редких кавказских ви-
дов семейства лилейных: ABTO-
реф. дис. ... канд. биол. наук.
Баку, 1967. 29 с.
'усейнова С. О. К цитоэмбриоло-
run Platanaceae.— Бюл. Глав. бот.
сада АН СССР. 1976, вып. 102:
07—71.
Гусейнова С. О. Микроспорогенез у
лапины — Pterocarya pterocarpa
(Michx) Kunth (Juglandaccae).—
Бот. журн., 1977, 62, 11:1647-—1652.
Гусейнова С. 0., Ахундова Е. С. Об
апомиксисе в родах тысячелист-
пик, ромашка, пупавка в семей-
стве СотрозКае. — В ки.: УП
Всесоюз. симиоз. по эмбриол.
раст. 1978, Киев: Наукова думка,
3:22—23.
Даниелян А. Х. Эмбриологическое
изучение процесса оплодотворе-
ния табака в условиях района
им. Hamo.— Изв. АН ApmCCP.
Сер. биол. наук., 1962, 15, 7:67
70.
Даниелян А. Х. Мегаспорогенез и
микроспорогенез табака в усло-
виях Армянской ССР.— Изв. АН
АрмССР. Сер. биол. наук., 1963За,
16, 1:87—90.
Даниелян А. Х. Эмбриологическое
исследование табака в условиях
некоторых районов Армянской
ССР: Автореф. дис. ... канд. биол.
паук. Ереван: Гос. ун-т, 1963: 24 с.
Дгебуадзе Н. В. Особенности репро-
дуктивных оргапов пекана иллий-
ского.— В кн.: 15 региональная
научная сессия Совета ботаниче-
ских садов Закавказья по интро-
дукции и акклиматизации суб-
тропических растений. Сухуми,
1979 : 68—69.
Демьянова Е. И., Покатаева П. В.
Некоторые дапные о гинодиэции
y Dianthus acicularis Fisch. (Са-
гуорвуПасеае).— Бот. журн., 1977,
62, 10: 1469—1479.
Дзевальтовський А. К. Вплив disio-
логично активных речовин на
р1ет, розвиток 1 морфологю гене-
ративных орган1в деяких пред-
ставникв родини гарбузовых.—
УКР. бот. журн., 4962, 19, 6:143—
«-
13*
Дзевалтовский А. К. Особенности
развития женского гаметофита у
триплоидной формы арбуза Аса-
хи Ямато.— Цитология и reneru-
ка, 1969, 3, 1: 7—16.
Дзевальтовський А. К. Морфолоти-
чн1 1 цитоембрологчнЕ особли-
вост: розвитку пормальних та
схильных До партенокарпи orip-
к1в.— Укр. бот. журн., 1964, 18,
1: 33—41.
Дзевальтовський А. К. Особливост1
розвитку жночного гаметофита у
Momordica charantia L.— Укр. бот.
mypH., 1963a, 20, 2: 53—60.
Дзевалтовский А. К. Развитие жен-
ского гаметофита у Benincasa
hispida Thunb.— B кн.: Питанис,
физпология, цитоэмбриология и
флора. Киев, АН УССР. 19656,
60—72.
Дзевальтовський А. К. Цитоембрло-
лог1чн!: досллджения деяких пред-
ставпикКв родипи гарбузових.—
Укр. бот. журн., 1963в, 20, 3:
16—79.
Дзевальтовський А. К. До ембро-
логи огрка — прекача (Ecballium
elatum (L.) А. Rich.— Yup. 6or.
жури., 1965, 22, 4:15—22.
Дзевалтовский А. К. Эмбриологи-
ческие исследовация представи-
телей семейства тыквенных.— Ци-
тология и генетика. Киев: 1967,
2:32—441.
Дзевалтовский А. К. Апоспория у
триплоидного арбуза. — В кн.:
Тез. П совещ. по проблемам апо-
миксиса у растений и животных.
Новосибирск. 1968: 20—24.
Дзевальтовський А. К. Розвиток
нормальних та партенокаршчних
плодв у ряду 1нтродукованих
представник!1в родини гарбузо-
вих.— В кн.: Интрод. та акл!мат.
рослин на Укр. ЮМев: Наукова
думка, 1970: 140—149.
Дзевалтовский А. К. Некоторые осо-
бепности развития женского га-
метофита у огурца (Cucumis sa-
Чуиз [.), обусловленные воздей-
ствием колхицина.— Цитология и
генетика, 1974, 5, 1:23—28.
Дзевалтовский А. К. Естественная
и стимулятивная апоспория y
представителей семейства тыквен-
ных.— В кн.: Проблемы апомик-
сиса у растений и животных. Но-
восибирск: Наука, 1973: 96—105.
Дзевалтовский А. К. Цитоэмбрио-
логические исследования поли-
275
плоидных форм культивируемых
тыквенных. — В кн. Тез. ХИ
Междунар. бот. конг. Л.: Наука,
1975, 1: 248.
Дзевальтовський А. К. Особливост!
цитоембруологИ експерименталь-
ных тетраплойив. СИгиПиз уШоа-
ris Schard.— Укр. бот. журн.,
1977, 34, 1:13—18.
Дзевалтовский А. Е. Эмбриология
губоцветных (Labiatae Juss.).—
Bot. *ypa., 1970, 64, 1: 100—111.
Дзевальтовський А. К., Жалалов Г.
До ембрюологи Меюо ма scabra
№ ид. Укр. бот. журн., 1976, 33,
3: 287—900.
Дзевальтовський А.К., Муратов 3. М.,
Жалалов Г. ЦДеяк1 дант до вивче-
пия розвитку голов1чо!: генератив-
но1 сфери у антильского ог1рка
(Cucumis anguria L.).—3y6. Bor.
сади. Вуз1 в YPCP наупй нар.
rocu. Hies: Вища школа, 1973а:
84—94.
Дзевалтовский А.К., Муратов 3.М.,
Жалалов Г. Явление апоспории у
некоторых представителей семеи-
ства СисигЬЦасеае. — Узб. биол.
жжурн., 19736, 5 : 35—37.
Дзевалтовский А.К., Муратов 3.М..,
Жалалов Г. Апомиксис у пред-
ставителей семейства тыквенных
Momordica charantia Г.— ДАН
УзССР, 1973в, 5 : 70—74.
Дзевальтовський А.К., Помщук В.С.
Дослиджения розвитку ж1ночно!
генеративно! сфери у деяких 1н-
тродукованних представникв ро-
дини губоцытних.— В кн.: Бота-
н1чн! сади вуз1в УРСР — наущ 1
народному господарству. 1974.
Дзевалтовский А. К., Полищук В.С.
Исследовапие ‘особенностей раз-
вития элемоптов женской гепера-
тивной сферы У шалфея мускат-
Horo (Salvia sclarea L.).—B ku:
Материалы науч. конф. «Новые
культуры в народном хозяйстве
и медицине». Киев, 1975.
Дзевальтовський А.К., Шайтан И.М.
Випадки апоспорного утворения
зародкових мпикмв у триплодно!
форми кавуна асах! ямато (Аса-
xi iamato).— Yup. bot. »ypa., 1969,
26, 1: 84—92.
Дзюбенко Л. К. Особливост! розвит-
ку чолов1чого та яЧночого гаме-
тоф1в топинамбура (НеПап лаз
tuberosa Г.).— Укр. бот. журн.
1965, 22, 1:43—5$3.
Дунаева Г. В. Цитоэмбриологиче-
276
ские исследования У льна-долгун-
ца. — Тр. ВНИИ льна, 1975, 41:
11—19.
Дьякова М. И. К изучению процес-
са оплодотворения У Mirabilis
jalapa L.— Бюл. МОИП. Отд.
биол., 1962, 67, 3: 145.
Ервандян С. Г. Макроспорогенез и
развитие женского гаметофита у
картофеля. — Изв. АН АрмССР.
cep. биол. наук., 1965, 18, 14 : 76—
1.
Ервандян С. Г. Цитоэмбриологиче-
ские исследования картофеля в
разных условиях возделывания в
Армянской ССР: Автореф. дис. ...
канд. наук. Ереван. 1966. 23 с.
Жадовский А. Э. К эмбриологии ро-
да ЕритЬтащез Г.— Биол. журн..,
1941, 2, 1:29—50.
ЯЖадовский А. 3. Типы развития
пыльцы и их филогенетическое
значение.— Тр. по прикл. бот.,
ген. и сел., 1925а, 14, 5.
Жадовский А. 9. Типы развития
зародышевых мешков у покры-
тосемепных растепий. — Журн.
Гусск. бот. о-ва, 192560, 10:3—4.
ЯЖалалов Г. Микроспорогенез и раз-
витие мужского гаметофита у
Momordica charantia.— Науч. тр.
Ташкент. ун-та, 1972, 398: 48—50.
Жалалов Г. К вопросу изучения
развития элементов мужской гене-
ративной сферы у бенинказы
волосистой [Benincasa hispida
(Thunb) Cogn]. Hayy. rp. Таш-
кент. ун-та, 1973, 439: 273—276.
Жалалов Г. Биология иветения
и цитоэмбриология у некоторых
представителей семейства тыквен-
ных: Автореф. дис. канд.
биол. паук. Ташкеит.: Гос. ун-т.
1974. 22 ¢.
Шалалов Г., Дзсвалтовский А. К.
К эмбриологии эхипоцистиса ши-
поватого Echinocystis lobata
(Michx) Torr. et A. Grey.— Науч.
тр. Ташкент. ун-та, 1973 : 289—294.
Жалалов Г., Дзевалтовский А. К.
К цитоэмбриологии антильского
огурца (Cucumis anguria L.).—
Узб. биол. журн., 1975, 3:41—43.
Жалалов Г., Дзевалтовский А. Е.
Развитие и строение некоторых
элементов мужской генеративной
сферы у мелотрии повислой.—
Узб. биол. журн., 1977, 2:65—67.
Жалалов Г., Дзевалтовский А. К..,
Шпилевой Б. Е. Формирование и
строение некоторых элементов
мужекой генеративной сферы у
представителя семейства тыквен-
ных.— Узб. биол. журн., 1977, 5:
17—80.
Жгенти Л. П. Цитоэмбриология не-
которых кавказских видов рода
Paeonia L.: Автореф. дис. ...
канд. биол. наук. Тбилиси, 1974:
17 с.
Жукова П. Г. Развитие семяпочки
и макроспорогенез у Ancmone
crinita Juz.— Бот. журн., 1965, 50,
3 : 378—381.
Заець В. О. Развитие мужского и
женского гаметофитов мака снот-
BopHoro (Papaver somniferum L.)
и эшольции калифорнийской
(Eschscholtzia californica Cham.).—
УКР. бот. журн., 1965, 22, 1:54—
Закржевская А.М., Колесников С.М.
Особенпости двойного оплодотво-
репия у вигны. — Изв. АН
МолдССР. Сер. биол. и хим. на-
ук., 1969, 5:72.
Замотайлов С. С. Эмбриология ин-
жира при разных способах опы-
ления.— Изв. АН СССР, 1955, 2:
103—120.
Замотайлов С. С. Эмбриологические
исследовапия арахиса: Автореф.
ДИС. канд. биол. наук. М.:
1958 : 44 с.
Замотайлов С. С. Эмбриология ара-
хиса.—В кн.: Научно-исследова-
тельские работы по масличным и
эфиромасличным культурам. М.:
1960: 192—206.
Эдруйковская-Рихтер А. И. Явление
полиэмбрионии у персика. — Тр.
Кит. бот. сада, 1970, 46: 473—
77.
Зонова Е. С. К эмбриологии неко-
торых представителей семейства
VMBOBHIX (Salicaceae Lindl.): Asro-
реф. дис. ... канд. биол. наук. Л.,
1953; 46 с.
Зотова Г. С., Филатова И. А. Мега-
спорогенез и женский гамстофит
у проса поссвпого.— В ки.: Апо-
миксис и цитоэмбриология расте-
ний. Саратов. ун-т, 1968 : 416—129.
Зуева Г. В. Оплодотворение у двух
видов рода Аторесигиз Т..— В кн.:
Матер. по экол. и физиол. раст.
Уральской флоры. Свердловск:
Гос. ун-т. 1976; 64—71. .
Иванова И. А. Развитие зародыша
B cemMeHax Acanthopanax sessili-
florum (Rupr. et Maxim) Seem.
перед UX прорастанием. — Бюл.
Глав. бот. сада АН СССР, 1965,
59 : 36—44.
Иванова И. А. Биология прораста-
ния семян борщевика Сосновско-
го.— Бюл. Глав. бот. сада АН
СССР, 1966, 63 : 66—73.
Иванова К. В., Орел Л. И. Некото-
рые биологические особенности и
эомбриологическая характеристика
Solanum guinense L.— Tp. по
прикл. бот., ген. и сел. 1963, 35,
2: 146—150.
Иванов М. А., Шаренкова Ё#Е. А. К
цитоэмбриологии сирени. Сирень
венгерская (Syringa josikaea
Тае4.) и сирень волосистая (Sy-
ringa villosa Wahl.).— Mas, Buon.
геогр. н-и ин-та при Иркут. ун-те,
1969а, 23, 3: 19—34.
Иванов М. А., Шаренкова Е. А.
К цитоэмбриологии сирени. Т. Си-
репь обыкновенная (Зугшеа vul-
garis Г[.).— Изв. Биол.-геогр. н.-и.
ин-та при Иркут. ун-те, 19696, 23,
3:3—18.
Ивановская Е. В. Об эмбриологии
яблони.— Науч. докл. высш. шко-
лы. Биол. науки, 1963, 1:106—113.
Ивановская Е. В., Орлова И. Н.
Данные по изменчивости яблони
(оэмбриологич. — исследование). —
Науч. докл. высш. школы Биол,
науки., 1964, 1: 182—188.
Ивашкина Л. А. Некоторые особен-
ности оплодотворения и формиро-
вания зародыша кормовых бобов
в условиях Ленинградской обла-
сти. Уч. зап, Перм. гос. пед.
ин-та, 1969, 68 : 109—114.
Ильина Г. М. Эмбриологическое ис-
следование Papaver somniferum
L.— Бюл. МОИП. Отд. биол. 1964,
66, 4: 13—25.
Ильина Г. М. Эмбриологическое ис-
следование горчицы Brassica jun-
cea (L.) Czern.— Becta. Моск.
ун-та. Сер. биол. и почвовед..,
1962, 67, 1: 34—45.
Ильина Г. М. Развитие пыльцы и
зародышевого мешка у Нуресо-
um procumbens L.— box. MOU.
Отд. биол., 1964, 69, 2: 130—134.
Ильина Г. М. Сравнительню-эмбри-
ологическое исследование семей-
ства маковых (Рарауегасеае В.
Juss), B CBA3H © положением его
в системе покрытосеменных: Азв-
тореф. дис. ... канд. биол. наук.
М.: Гос. ун-т, 1968: 24 с.
Ильина Г. М. Эмбриология Р1айузе-
mon californicus Benth. B cBasu
277
с положением его в системе ма-
ковых. — В кн.: Морфология по-
крытосеменных растений. М.: На-
ука, 1971: 10—33.
Ильина Г. М. Эмбриология маковых
(Papaveraceae B. Juss) B связи с
филогенией. — В кн.: Материалы
5-го Моск. совещ. по филогении
раст. 1976. М.: Наука, 1976:66—69.
Ильина Г. М. Особенности эмбрио-
логии мачка в связи с полюжени-
ем его в системе маковых.— В
кн.: Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 1968,
3: 30—34.
Иоффе М. Д. К эмбриологии Тга-
chodendron aralioides Sieb. et
Jucc.— Tp. Bor. wu-ra, 1962, Cep.
7, 9: 250—259.
Иоффе М. Д. В эмбриологии Тга-
chodendron aralioides Sieb. et
Тисс.— (Развитие зародыша и эн-
досперма). — В кн.: Морфология
цветка и репродуктивный процесс
у покрытосеменных. М.; Л.: Нау-
ка, 1965: 177—188.
Иоффе М. Д. Процесс оплодотворе-
ния У Апетопе петотоза 1. и
Anemone ranunculoides Г. — Бот.
журн., 1968, 53, 6: 891—905.
Иоффе М. Д. Развитие эндосперма
и зародыша у Апетопе nemorosa
L.— Bor. журн., 1971, 56, 414:
1582—1592.
Иоффе М. Д. Особенности двойного
оплодотворения в роде Ме|атру-
гит L. (Scrophulariaceae) B cBas-
зи с формированием диплоидного
эндосперма.— Бот. журн., 1976, 61,
11:1515—1530.
Иоффе М. Д., Жукова Г. Я. Моди-
фикация оболочки зиготы в про-
цессе се развития в роде ОШета
(Dilleniaceae).— Бот. журн., 1974,
59:40, 1409—4416.
Кавецкая А. А. Цитоэмбриологиче-
ские исследования рапса: Авто-
реф. дис. ... канд. биол. наук.
Киев. 1960: 16 с.
Кавецкая А. А. Формирование и
жизнеспособность пыльцы у ле-
щипы.— В кн.: УП Всосоюз. сим-
поз. по эмбриол. раст. Киев.: На-
укова думка, 1978, 3: 32—33.
Кавецька Г. О. Розвиток чолов1чого
гаметофита ор1ха волоського (Тио-
lans regia Г.).— Укр. бот. журн.
1964, 21, 1:52—57.
Кавецька Г. О. Ембрюлоя озимого
ршака. — Укр. бот. журн., 1958,
15, 1:37—47.
278
Каграманова Ф. В. Сравнительно-
эмбриологическое исследование
первоцветов разноцветного и Во-
ронова.— В кн.: Цитоэмбриологи-
ческое исследование растений
флоры Азербайджана. Баку: ЭЛМ,
1972: 82—92.
Каграманова Ф. В. Некоторые во-
просы эмбриологии кизильника
кистецветного.— В кн.: УП Весе-
союз. симпоз. по эмбриол. раст.
Киев: Наукова думка, 1978, 2:
21—23.
Кадырова Р. У. Развитие семяпоч-
ки и зародышевого мешка у эс-
парцета ферганского.— В кн.: Мор-
фогенез растений аридной зоны.
Ташкент: Фан, 1973: 77—80.
Кадырова Р. У. Сравнительно-эм-
бриологические исследования ма-
кросиорогенеза и развития жен-
cKkoro ramMero@uta Onobrychis fer-
ganica (Sirjaev) Grossh. u O. amo-
ena М. Рор. её Ууе4. — В кн.:
Морфологические особенности ди-
корастущих растений Узбокиста-
на. Ташкепт: Фан, 1974а: 53—60.
Кадырова Р. У. Биология цветения
и цитоэмбриология некоторых
многолетних эспарцетов: Автореф.
дис. ... канд. биол. наук. Таш-
кент, 19746: 18с.
Кадырова Р. У. Развитие зародыша
и эндосперма у Опофгус 1$ Ёегра-
nica (Sirjaev) Grossh—B xa. Би-
ологические и морфологические
особенности полезных растений
Узбекистана. Ташкент: Фан, 1976:
102—104.
Кадырова Р. У. К эмбриологии не-
которых видов рода Onobrychis
АЧапз.— В кн.: Биологические и
структурные особенности полез-
ных растений Узбекистана. Таш-
кент: Фан, 1979: 105—144.
Калягина Л. Г. Макроспорогенез и
развитие женского гаметофита у
некоторых видов Geranium L.—
Узб. биол. журн., 1977а, 1: 60—63.
Калягина Л. Г. Микроспорогенез и
развитие мужского гамотофита у
некоторых видов герани.— В кн.:
Биологические и структурные о0со-
бенности полезных растений Уз-
бекистана. Ташкент: Фан, 19776:
81-85.
Калягин Ю. Н. Микроспорогенез и
развитие мужсного гаметофита у
КосМа prostrata (L.) Schrad.—
В кн.: Материалы по структуре
и функциональным особенностям
полезных дикорастущих растений
Узбекистана. Ташкент: Фан, 1970:
66—72.
Калягин Ю. Н. Биология и цитоэм-
бриология двух подвидов изеня—
Kochia_ prostrata (L.) Schrad.:
Автореф. дис.... канд. биол. наук.
Ташкент, 41974: 24с.
Калягин Ю. Н. Макроспорогенез,
формирование женского гамето-
фита и зародыша у Kochia pro-
strata (L.) Schrad.— B xu.: Buono-
тические и морфологические осо-
бенности полезных растений Уз-
бекистана. Ташкент: Фан, 1976:
88—93.
Калягин Ю. Н. Формирование ми-
кроспорангия ‘у функционально
женских цветков изеня.— В кн.:
Вопросы ботанических исследова-
ний в Узбекистане. Ташкент:
Фан, 1977 : 62—63.
Камалова Г. В. Морфологические
и гистохимические особенности
пыльцевых зерен видов мальво-
вых.— В кн.: Вопросы генетики и
эмбриологии хлопчатника. Таш-
кент, 1966.
Камалова Г. В. Явление несовме-
стимости при отдаленной гибри-
дизации в семействе Мауасеае.
Ташкент: Фан, 1975. 142c.
Камалова Г. В. О макроспорогенезе
у Hibiscus manihot L.— Y36. 6non.
mypH., 1976, 2: 56—58.
Камалова Г. В. Цитоэмбриология
видов Сепиесома Cav. (Malva-
сеае).— В кн.: УП Всесоюз. сим-
поз. по эмбриол. раст. Киев. Нау-
кова думка, 1978, 2: 23—24.
Камалова Г. В. О развитии женско-
го археспория y Malvaceac.— Y30.
биол. журн., 1979, 5: 63—66.
Камалова Г. В., Пантелеева А. П.
Оплодотворение и эмбриогенез у
Hibiscus manihot—B кн.: Гене-
тика и цитоэмбриология хлопчат-
ника. Ташкент: Фан, 1979: 57—60.
Камалова Г. В., Серебрякова J. E.,
Пантелеева А. П. Микроспороге-
нез и развитие мужского гамето-
muta y Althea broussonetifolia
Iljin——B ka.: Генетика и цитоэм-
бриология хлопчатника. Ташкент:
Фан, 1979: 46—52.
Йамелина О. П. О сравнительно-эм-
бриологическом исследовании се-
мейств ПО!рзасасеае и Morina-
сеае. — В кн.: Материалы 5-го
Моск. совещ. по филогении` раст.
1976. М.: Наука, 1976: 71—73.
Камелина 0. П. —Сравнительно-
эмбриологическое исследование
представителей П/1рзасассае и Мо-
гпасезе: Автореф. дис. ... канд.
биол. наук. Л., 1977.
Камелина О. П. Развитие зароды-
шевого мешка и дифференциации
его элементов B poze Scabiosa L.
(сем. П!рзасасеае) — В кн.: УП
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 1978,
3: 33—35.
Камелина О. П. Развитие клеточно-
го эндосперма в семействах ПО1р-
засасеае и Могтасеае.— В кн.:
Актуальные вопросы эмбриологии
покрытосеменных. Ji.: Наука,
1979а: 69—74.
Камелина О. П. Особенности эмбри-
отенеза у представителей семейств
П1рзасасеае и Могтасеае.— В кн.:
Актуальные вопросы эмбриологии
покрытосеменных. —Л.: Наука,
19796: 82—89.
Камелина 0. П., Малдыбекова К. С.
Развитие пыльника и пыльцевого
зерна y Valeriana (Valeriana-
сеае) — Бот. журн., 1980, 65, 5:
640—646.
Камелина О. П., Яковлев М. С. Раз-
витие зародышевого мешка в ро-
де Могша Г.— Бот. журн., 1974,
59, 14: 1609—1617.
Камелина О. П., Яковлев М. С. Раз-
витие пыльника и микрогамето-
генез у представителей сем. П!р-
sacaceae и Morinaceae. — bor,
журн., 1976, 64, 7 : 982—945.
Канделаки Г. В., Кобахидзе Л. А.
К эмбриологии базилика огород-
ного.— В кн.: Материалы У Все-
союз. совещ. по эмбриол. раст.
Кишинев, 1971.
Канделаки Г. В., Кобахидзе Л. А.
К изучению зародышевого мешка
и двойного оплодотворения у 68-
зилика огородного (Ocimum Ъа-
silicum Г.) — Сообщ. АН ГССР,
1977, 86, 1: 177—180.
Кантор Т. С. Сраввительно-эмбрио-
логическое исследование культур-
ного льна и некоторых его диких
родичей: Автореф. дис. ... канд.
биол. наук. М., 1956. 15е.
Кантор Т. С. Сравнительно-эмбрио-
логическое исследование культур-
ного и некоторых ‘диких видов
льна.— В кн.: Эмбриологические
исследования покрытосеменных.
М.: Изд-во АН CCCP, 1959, 6:
170—190.
279
Кантор Т. С. Сравнительный эмбри-
огенез некоторых видов льна в
связи C их филогенетическими
взаимоотношениями.— В кн.: Мор-
фогенез растений. Изд-во МГУ.
1964 : 444—444. `
Капанадзе И. С. Явление многоза-
родышевости у померанцевых.—
Сообщ. АН ГССР, 1966, 43, 2: 447.
Капанадзе И. С. Оплодотворение и
образование добавочных эмбрио-
нов У цитрусовых.— Тр. Сухум.
опыт. ст. субтроп. культур., 1967,
1:189—196.
Капанадзе И. С. Образование п раз-
витие адвентивных эмбриоидов
цитрусовых.— В кн.: Материалы
УП Всес. симпоз. по эмбриол.
раст. Киев: Наукова думка, 1968:
88—89.
Капранова Н. ПН. Тапетум пыльника
и микроспорогенез у некоторых
видов и садовых гибридов рода
PhiJadelphus 1. — Науч. докл.
высш. школы. Биол. пауки, 1970,
6: 56—60.
Капинос Г. Е. Эмбриологическое
исследование культурных пред-
ставителей p. Narcissus L.— Тр.
Ин-та ботан. АН АзССР, 1960а,
22 : 5—16.
Капинос Г. Е. Цветение, опыление
и gMOpHonorua Sternbergia lutea
(Г.) Ker-Gawl. u S. fischeriana
(Herb.) Roem.— Бот. журн., 19606,
45, 7: 1044—1055.
Капинос Г. Е. Биология цветения
и эмбриология крупноцветных
штернбергий. В кн.: Пробл. сов-
ременной эмбриологии. М.: Изд-во
МГУ, 1964: 137—143.
Капинос Г. Е. Морфолого-эмбриоло-
гическое исследовапие пролески
Гогенакера (Scilla hohenackeri
Schischk.). Baxy, BUNMUTH, 1972,
№ 41. 72—76, Деп., с. 24.
Капиное Г. Е. Эмбриологический
очерк Calendula persica C. A.
Меу.— Бюл. Глав. бот. сада АН
СССР. 1975, 96 : 38—46.
Капинос Г. Е. О цитоэмбриологиче-
ских исследованиях реликтов и
эндемов.— В кн.: Материалы УП
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка. 1978,
ч. 3.: 35—86.
Капинос Г. Е. Сравнительное эмбри-
ологическое исследование пред-
ставителей рода Scilla. — Бюл.
Глав. бот. сада АН СССР, 1980,
145 : 989—105.
280
Капинос Г. Е., ГРусейцова С. 0.
О прорастании пыльцы в муж-
ском гаметофите некоторых ви-
дов иглицы (Ruscus Tourn). —
Hioxn. AH AsCCP, 1967, 23, 7: 76.
Капинос Г. Е., Гусейнова С. О. Ис-
следование репродуктивного ан-
парата неноторых древних расте-
ний Закавказья в связи с вопро-
сом о степени их реликтовости и
эволюционной адаптацией.— В кн.:
Матер. Всесоюз. симпоз. по эм-
бриол. раст. Киев: Наукова дум-
ка, 1968: 89—94.
Капинос Г. Е., Гусейнова С. 0. О
сексуализации цветка и эмбрио-
логических особенностях Danaé
racemosa Moench. — Докл. АН
АзССР. 1969, 25, 5:58—58.
Капинос Г. Е., Гусейнова С. О. Ма-
териалы к цитоэмбриологии рода
Rhinopetalum Fisch. — ВИНИТИ,
1972, № 4428—4472, Деп. с. 11.
Капинос Г. С., Гусейнова Н. А. Ми-
кросиорогенез и развитие муй-
ского гаметофита У платана.—
Бюл. Глав. бот. сада ЛИ СССР,
1978, 109: 69—73.
Капинос Г. Е., Керимова А. А. Ми-
кроспорогенез и развитие муж-
ских гамет у чингиля серебристо-
го и дрока испанского.— Изв. АН
АзССР. Сер. биол. наук, 1968, 4:
22—28.
Капинос Г. Е., Керимова А. А. О
диморфизме и функциях цветков
и соцветия Albizzia julibrissin
Durazz.— Bot. журн., 1972, 57,12:
1600—1609.
Капинос Г. Е., Кулиева Н. А. Эм-
бриологический очерк поденежни-
ка каспийского [Galanthus caspi-
cus (Rupr.) A. Grossh.].— Bo xum.:
Цитоэмбриологические исследова-
ния растений флоры Азербайджа-
на. Баку: ЭЛМ, 1972: 1241—1530.
Карлова А. А. Развитие стенки
пыльцика, микроспорогенез и
формирование мужского гамето-
фита у четырех видов рода П!01-
talis.— Hayy. докл. высш. школы,
1973, 8:55—57.
Карлова А. А. Сравнительно-эмбрио-
логическое исследование некото-
рых видов норичниковых (Scro-
ри\анасеае Тизз.): Автореф. дис.
. канд. биол. наук. Свердловск:
Ин-т экологии растений и живот-
ных. 1975. 22 с.
Касымова Х. Н. Биология цветения
и плодоношения декоративных ви-
дов сложпоцветных (Емоегой сац]-
teri P. E. a E. speciosus D. C.): As-
тореф. дис... канд. биол. наук. Таш-
кент, 1966 : 3З—25.
Kachidze N. [Kaxudse H. T.|. Kary-
ologische Studien tiber die Fami-
lie der Dipsacaceae.— Planta, 1929,
7, 4, 482—505.
Керимова А. А. Биология цветения
и цитоэмбриологические особен-
ности некоторых бобовоцветных:
Автореф. дис... канд. биол. на-
ук. Баку, 1970. 27 с.
Керимова А. А. Мегаспорогенез и
развитие женского гаметофита
чингиля серебристого. — В кн.:
Цитоэмбриологические исследова-
ния растительной флоры Азер-
байджана. Баку: ЭЛМ, 1972, 103.
Киселев С. Д., Козлова И. В. Раз-
витие женского гаметофита у
Vicia villosa Roth.— 3an. Jlenuurp.
с.-х. ин-та, 1970, 139, 1:128—134.
Кобахидзе Л. А. В эмбриологии Ш-
lium szovitsianum Fisch. et Lall.—
Сообщ. AH ГССР, 1965, 37, 3:
661—667.
Кобатидзе Л. А. Цитоэмбриология
некоторых кавказских видов рода
Lilium Г.:Автореф. дис. ... канд.
биол. наук. Тбилиси, 1967. 30с.
Кобахидзе Л. А. Апомиксис у эн-
демичных для Кавказа видов
лилий.— В кн.: Апомиксис и се-
лекция. М.: Наука, 4970: 160—164.
Кобахидзе Л. А. В изучению неко-
торых процессов, влияющих на
формирование эндосперма и эн-
доспермальных гаусториев у ба-
зилика огородного (Оспиит Ба-
silicum Г.).— Сообщ. AH ГССР,
1975, 80: 137—140.
Кобахидзе Л. А. Цитоэмбриология
wa6bepa cayosoro (Satureja horten-
sis L.).— Mss. AH ГССР. Сер.
биол., 1977, 3, 5:415—424.
Побахидзе Л. А., Кобаснидзе 5. Я.
К эмбриологическому изучению
некоторых представителей рода
Пеютиса Г.— В ки.: УП Зсесоюзн.
симпоз. по эмбриол. раст. Киев:
Наукова думка, 1978: 39—40.
Коваленко В. А. Формирование тка-
ней пыльника и пыльцевых зерен
у лаванды настоящей.— В кн.:
Материалы У Всесоюз. совещ. по
эмбриол. раст. Кишинев, 1971:
718—179. `
Кокиево Е. И. История развития
женского гаметофита у Partheni-
um argentatum С. («Guayle»).—
Бот. журн. СССР, 1932, 171, 1:72.
Комар Г. А. Ариллусы, их природа,
строение и функции.— Бот. журн.,
1965а, 50, 5: 715—724.
Комар Г. А. О структуре ариллуса
некоторых представителей семей-
ства крыжовниковых (Grossulari-
асеае).— В кн.: Морфология цвет-
ка и репродуктивный процесс
у покрытосеменных растений. М.;
Л.: Наука, 19656, 114—130.
Константинов А. В. Эмбриология
некоторых сортов яблони.— Изв.
АН СССР. Сер. биол., 1960, 2:
256—264.
Конычева В. И. К эмбриологии ги-
бискусов.— В кн.: Вопросы био-
логии и краевой медицины. Тап-
кент, 4960: 34—38.
Конычева В. И. Биология цветения
и плодоношения некоторых видов
и гибридов гибискусов: Автореф.
дис. ... канд. биол. наук. Таш-
кент, 1964 : 20 с.
Конычева В. В биологии цветения
и эмбриологии Salsola_ rigida
Ра!].— Узб. биол. журн., 1963, 6:
43—48.
Конычева В. И. К биологии цвете-
ния и эмбриологии Salsola gem-
тазсепз Ра|.— Узб. биол. журн.,
1965, 5: 58—68.
Конычева В. И. Биология и эмбри-
ология напофитона ежового. —
Узб. биол. журн., 1968, 5:18—79.
Конычева В. И. В биологии цвете-
ния и эмбриологии Азтара! из
villosisimus Bge. wu A. unifoliola-
{15 Вое.—В ки.: Материалы по
структуре и функциональным
особенностям полезных растений.
Узбекистана. Ташкент: Фан, 1970а:
52—65.
Конычева В. И. О развитии зароды-
ша и семени полыни — Ацепляа
curanica Krasch. uw A. diffusa.—
В кн.: Материалы по структуре и
функциопальпым особенностям
полезных дикорастущих растений
Узбекистана. Ташкент: Фан, 19706:
71—94.
Еонычева В. И. Строение зароды-
шевого мешка у некоторых маре-
вых в тычиночную фазу цвете-
ния.— В кн.: Биологические и
морфологические особенности по-
лезных растепий Узбекистана.
Ташкент: Фан, 1976: 93—104.
Конычева В. И. О цветении и эм-
6puonoruu Salsola richteri Kar.—
В кн.: Биологические и структур-
281
ные особенности полезных рас-
тений Узбекистана. Ташкент:
Фан, 1977, 85—94.
Конычева В. И. Развитие микроспо-
рангия и пыльцы У многолетних
солянок.— В кн.: Биологические
и структурные особенности по-
лезных растений Узбекистана.
1979: 84—86.
Конычева В. И., Кадырова Р. У.
К эмбриологии СВепороасеае.—
В кн.: Тез. докл. УТ делегат. Все-
союз. бот. о-ва, 1978а : 79—80.
Конычева В. И., Кадырова Р. У.
Эмбриологические особенности He-
которых представителей семейст-
ва Chenopodiaceae.—B кн.: УП
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 19786,
2: 26—28.
Конычева В. И., Шевчук Л. П. Эм-
бриология терескена Eurotia ce-
га(ю14ез.— В кн.: Морфологиче-
ские особенности дикорастущих
растений Узбекистапа. Ташкент:
Фан, 1974: 115—124.
Кордюм Е. Л. Сравнительню-омбрио-
логическое исследование семей-
ства лютиковых: Автореф. дис ...
канд. биол. наук. 4959а : 16c.
Кордюм 9. Л. Порйвпяльно-ембрю-
логичне досл1джения родини жов-
тецевних — Ranunculaceae D. C.—
Укр. бот. журн., 19596, 16, 1:
32—43.
Кордюм Е. Л. Процесс запиления —
запл1днения у деяких представ-
ник!в родини жовтецевних.— Укр.
бот. журн., 1960, 17, 6:61—67.
Кордюм Е. Л. Сравнительно-эмбрио-
логическое исследование сем. лю-
тиковых.— В кн.: Морфогенез ра-
стений. М.: Изд-во МГУ, 1964а:
473—477.
Кордюм Е. „Л. И вопросу полиэм-
брионии у ластовня локарствен-
ного.— Укр. бот. журн., 19616, 18,
3: 48—54.
Кордюм Е. Л. Микроспорогенез и
особенности развития тапетума у
некоторых видов рода ластовня
(Vincetoxicum Moench.).— Yxp. 607.
журн., 1961n, 18, 5: 6—14.
Кордюм 9. Л. До питания про по-
ттирення прикладнот партенокар-
пи у рослин з сухими плодами.—
Укр. бот. журн., 1963, 20, 1:24.
Кордюм Е. Л. Особенности микро-
спорогенеза у бутия ароматного
(Chaerophyllum aromaticum). —
Укр. бот. журн., 4964, 21,5: 49—52.
282
Кордюм Е. Л. Особенности макро-
спорогенеза володушки кругло-
листной — Bupleurum rotundifoli-
um L.—B кн.: Цитология и гене-
тика. Киев: Наукова думка, 1965:
140—144.
Кордюм Е. Л. Морфология алейро-
новых зерен HCKOTOPLIX видов
аралиевых и зоптичных.— В кн.:
Цитология и генетика. Киев: 1966,
2:89—102.
Кордюм 9. J. flo emOpionorii Hip-
pomarathrum caspium (DC) Grossh.
(родина ОшеШ\Ъегае).— Укр. бот.
журн., 1967а, 24, 3:33—40.
Кордюм Е. Л. Особенности разви-
тия стенки микроспорангия и за-
родышевого мешка y Crithmum
maritimum Г. (семейство Отае]-
Шегае).— В кн.: Цитология и ге-
нетика. Киев: Наукова думка,
19676, 14, 1:67—75.
Кордюм Е. Л. До ембр1ологи Соео-
pleurum gmelini (DC) LDB.—
Укр. бот. журн., 1967p, 24, 2:
24—31.
Кордюм Е. Л. Цитоэмбриология се-
мейства зоптичных. Киев: Науко-
ва думка, 1967г: 176 с.
Кордюм Е. Л. Особенности разви-
тия мужского гаметофита зонти-
чных.— В кн.: Материалы Всесо-
юз. симпоз. по эмбриол. раст. Ви-
ев: Наукова думка, 1968а: 100—
104. -
Кордюм Е. `Л. Сравпительная эм-
бриология и цитология видов зон-
тичных в связи с их филогенией
и эволюцией: Автореф. дис. ..
д-ра биол. наук. Л.: 19686.
Кордюм Е. Л. Эмбриология и кар-
диология зоптичных в связи с
филогенией.— В кп.: Морфология
высших растений. М. Наука,
1968в: 27—86.
Пордюм Е. ЛП. Апомиксис в роде
Гоа Г.—В кн.: Апомиксис и се-
лекция. М.: Наука, 1970, 144—149.
Кордюм Е. Л. Значение эмбриоло-
гии для решения вопросов систе-
матики и филогении покрытосе-
менных растений.— В кн.: Про-
блемы ботаники. Киев: 1971:
196—215.
Кордюм Е. Л. Политопно-монофиле-
тическая концепция происхожде-
ния покрытосеменных в свете эм-
бриологических данных. — Тр.
МОИП. Отд. биол., 1974, 54:37—49.
Кордюм Е. Л. Эволюционная цито-
омбриология покрытосеменных ра-
стений. Киев: Наукова думка,
1978 : 220c.
Кордюм Е. Л., Beaeduuyraa J. J.
Особенности развития тапетума
пыльника и микроспорогенез у
ряда представителей Umbellife-
тае. — Бот. журн., 1964, 49, 11:
1609—4615.
Кордюм Е. Л., Глущенко Г. И. Ци-
тоэмбриологические аспекты про-
блемы пола покрытосеменных.
Киев: Наукова думка, 1976: 198.
Кордюм 9. A., Заэць В. А. Цо ем-
бр1ологИ молочаю щербиковидно-
го (Euphorbia peplis L.).— Yxp.
бот. журн., 1962, 19, 5: 42—48.
Коробова С. Н. Эмбриологическое
исследование кукурузы. Автореф.
дис. канд. биол. наук. Л.: 1964.
Коробова С. Н. В эмбриологии ку-
курузы.— Тр. Бот. ин-та AH
СССР, 1962. Сер. Т, 5: 294—344.
Коробова-Семенченко Л. В. К эм-
бриологии представителей семей-
ства БахИгасасеае. Т. Мегаспоро-
тенез и развитие женского гаме-
тофита у Вегоета стаззМНоПа (Г..)
Fritsch wu Chrysosplenium alterni-
folium L.— Hayy. докл. выс.
школы. Биол. пауки, 1965, 4:
101—107.
Коробова-Семенченко Л. В. К эм-
бриологии сем. камнеломковых
(Saxifragaceae). 1. Развитие эн-
досперма, зародыша и формиро-
вание семени Бадана толстолист-
ного [Bergenia crassifolia (L.)
ЕгИзсв].— Бюл. МОИП. Отд. биол.
1966, 74, 2: 64—69.
ЕКоробова-Семенченко Л. В. К эм-
бриологии семейства камнеломко-
вых. ПЛ. Развитие эндосперма и
зародыша у Chysospelenium al-
ternifolium Г.— Вестн. МГУ, 1969,
2:54—58.
Коробова-Семенченко Л. В. К эм-
бриологии семейства Saxifraga-
сеае. ТУ. Мегаспорогепез, разви-
тие женского гаметофита, эндо-
сперма и зародыта у Saxifraga
moschata У’аП.— В кн.: Морфоло-
гия цветковых растений. М.: На-
ука, 197ба : 55—63.
Коробова-Семенченко Л. В. Эмбрио-
логические особенности Chrysos-
решит alternifolium Г. и их
эволюционное значение. — В кн.:
Материалы 5-го Моск. совещ. по
Филотении раст. М.: Наука, 19766:
Коробова-Семенченко Л. В. В эм-
бриологии семейства Saxifraga-
сеае. УТ. Микропилярные гаусто-
рии и ранние стадии развития
зародыша у Согузозретит аКег-
nifollum Г.— Вестн. МГУ. Сер.
биол. почвовед., 19768, 5: 92—94.
Коробова-Семенченко „Л. В. В эм-
бриологии семейства Saxifraga-
сеае. УП. Отклонения при раз-
витии эндосперма у Вегоепа сгаз-
sifolia (L.) Fritsch.— Bectu. МГУ.
Сер. биол., 1977, 2:81—84.
Кострикова Л. Н. Эмбриологическое
исследование рода Vicia.— Вести.
МГУ, 1956, 2: 59—80.
Кострикова Л. Н. О развитии заро-
дыша и эндосперма язвенника
многолистного.— Вестн. МГУ. Сер.
УТ. Биол. почвовед., 1964, 6: 54—
57.
Кострикова Л. Н. К эмбриологии
Robinia pseudoacacia Г.— Вестн.
МГУ. Биол., почвовед., 1968, 4:
63—67.
Кострикова Л. Н. К эмбриологии
Amorpha~ glabra Оез3Ё. — Вестн.
МГУ. Биол., почвовед., 1972, 4:
142—114.
Кострикова Л. Н. О полиэмбрионии
У мотыльковых. — Науч. докл.
высш. школы. Биол. науки, 1973За,
12: 57—60.
Кострикова Л. Н. Хлоропласты за-
родыша и эндосперма некоторых
представителей — мотыльковых. —
Бюл. Глав. бот. сада АН СССР,
19736, 89: 29—35.
Кострикова JI. H. К эмбриологии
рода Amorpha L. B связи с его
положением в семействе бобо-
вых.— Бюл. Глав. бот. сада АН
СССР, 1975, 96 : 33—37.
Костривова Л. Н. Развитие зароды-
ша, эндосперма и семени у 50-
phora flavescens Ait. (Papiliona-
сеае — Sophoreae).— Haya. — докл.
высш. школы. Биол. вауки, 1978а,
7: 88—914.
Кострикова Л. Н. Эмбриологическое
исследование софоры желтеющей
(сем. мотыльковых).— В кн.: УП
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 19786,
2: 29—30.
Кострикова Л. Н. К эмбриологии
Sophora flavescens -Ait. (Papilio-
naceae — Sophoreae). — Вестн.
Моск. ун-та, 1979, 2:26—30.
Кострикова Л. Н. Полиэмбриония у
Sophora flavescens Ait. (Papilio-
283
пасеае).— Вестн. МГУ. Сер. биол..,
1980, 1:37—38.
Кострюкова К. Ю. К эмбриологии
Lilium martagon 1.— Изв. АН
АрмССР биол. науки, 1961а, 14,
1:3—16.
Кострюкова К. Ю. Оплодотворение
y Heliopsis helianthoides Sweet.—
Журн. общ. биол., 19616, 22, 1: 58.
Koy 3. П. Развитие зародышевого
мешка тополя белого.— В сб.: Се-
лекция, интродукция и семено-
водство древесных лесных пород.
Киев: Урожай, 1964: 60—63.
Коц 3. П. Цитоэмбриологическое
изучение тополей.— В кн.: Лес-
ная генетика, селекция и сомено-
водство. Петрозаводск: Карелия,
1970 : 33—38.
Koy 3. П. Развиток набниевых за-
чатк1в та эмночого археспоря в
род! Роршаз Т.— Укр. бот. журн.,
1972а, 29—14: 19—24.
Коц 3. П. Развиток я1ночо! катки
Tononi cH30i (Populus pruinosa
Schrenk. subg. turanga Bege.).—
Укр. бот. журн., 19726, 29, 2:
228—232:
Крылова В. В. Полиэмбриония в
семействе бобовых.— В кн.: Био-
логия оплодотворения и гетеро-
зис культурпых растений. Киши-
нев: Штиинца, 1962, 41: 140—145.
Крылова В. В. Биология цветения
и опыления фасоли.— В кн.: Био-
логия оплодотворения и гетеро-
зис культурных растений. Киши-
нев: Картя Молдовеняскэ, 1965,
3: 126—185.
Крылова В. В. Апоспория и поли-
эмбриония у яблони. В кн.:
Апомиксис и селекция. М.: Пау-
ка, 1970: 120—126.
Кудряшов Л. В. Происхождепие од-
носемядольпости (па примере
Неюмае) — Бот. журн., 1964, 49,
4: 473—484.
Кудряшов Л. В., Савич Е. И. Неко-
торые данные к эмбриологии
Alisma plantago aquatica L.— Бюл.
МОИП. Отд. биол., 1968, 68, 4:
50—63.
Кудряшов Л. В., Савич Е. И. Эм-
бриология Zostera шагша Г. в
связи с вопросами о морфологи-
ческой природе семядоли у одно-
дольных.— В кн.: Морфология выс-
pax растений. М.: Наука, 1968:
—16.
Кулиева Н. А. Формирование ре-
продуктивных органов и цитоэм-
284
бриологические особенности ма-
слины европейской на Апшероне:
Автореф. дис. ... канд. биол. на-
ук. Баку, 1966 : 20с.
Кулиева ПН. А. Формирование семя-
почки, мегаспорогенез и разви-
тие женского гаметофита масли-
ны езропейской на Апшероне.—
Изв. АН АзССР. Сер. биол. наук,
1969, 3: 40—14.
Кулиева Н. А. Развитие женской
сферы цветка маслины европей-
ской на Апшероне.— В кн.: Цито-
эмбриологические — исследования
растительной флоры Азербайджа-
па. Баку: ЭЛМ, 1972: 74—84.
Куприянова Л. А. Палинологиче-
ские данные к систематике и
филогении сережкоцветных (Ате-
ntiferae): Автореф. дис. ... д-ра
биол. наук. Л.: 1964. 29с.
Куприянова Л. А. Палинология се-
режкоцветных. М.; Наука,
1965.
Куприянова Л. А., Алешина Л. А.
Пыльца и споры растений флоры
свропейской части СССР. М.; Л.:
Паука, 1972.
Курбанов 9. А. Микроспорогенез.
(Thymus collinus M. B.).—B xn:
Материалы У Всесоюз. совещ. по
эмбриол. раст. Кишинев, 1974.
Курбанов 9. А. Цитоэмбриология
чебреца в условиях Апшерона.—
кн.: Материалы Всесоюзного
симпозиума «Половой процесс и
эмбриогенез растений». М.: 1973.
Курбанов 9. А. Микроспорогенез и
оплодотворение зизифоры Бибер-
штейна. — В кн.: УП Всесоюз.
симпоз. по эмбриол. раст. Киев:
Паукова думка, 1978, 3: 50—54.
Кутубидзе В. В. Явлопие полиэм-
брионии в чайном растении.—
Субтроп. культуры, 1974, 4:45—48.
Литвиненко С. П. Цитоэмбриоло-
гическое и гистохимическое ис-
следование костра безостото (Bro-
mus inermis Leyss): Автореф.
дис. ... канд. биол. наук. Киев,
1970. 28 с.
Литвиненко Л. М., Дзевалтов-
ский А. К. К эмбриологии дипло-
идной и тетраплоидной эмилии
пылающей (Emilia flammea
Сазз).— Укр. бот. журн., 1972, 29,
5: 614—618.
Литвиненко Н. М., Шпилевой Б. Е.
Микроспорогенез и развитие муж-
ского гамотофита у сирени китай-
ской.— В кн.: УП Beecows. сим-
поз. по эмбриол. раст., Киев, 1978,
1:31—32.
Лодкина М. М. Своеобразные чер-
ты развития пыльцевых мешков
у некоторых видов Najadaceae
Juss wu Lemnaceae S. Gray.— Бот.
журн., 1976, 61, 11:1536—1545.
Лудникова Л. А., Янушевич 9. Б.
Партенокарпия у томатов.— Ви-
шинев: Картя Молдавеняскэ, 1970,
97 с., 1964, 3: 3—18.
Мандрик В. Ю. Макроспорогенез и
развитие женского гаметофита у
чайного растения. — Науч. зан.
Ужгородск. ун-та. Ботапика, 1957,
23 : 69—88.
ЛМандрик В. Ю. Микроспорогенез и
развитие мужского гаметофита у
чайного растения в условиях За-
карпатья.— Науков1 зап. Ужго-
родьск. ун-та, 1959, 41: 271—278.
Мандрик В. Ю. Эмбриология чайно-
го растения в условиях Закар-
патья: Автореф. дис. ... канд. би-
ол. наук. Ужгород, 1960
Мандрик В. Ю. К эмбриологии чай-
ного растения в условиях Закар-
патья.— В кн.: Проблемы совре-
менной эмбриологии. М.: Моск.
ун-т, 1964, 148—152.
Мандрик В. Ю. Различия в разви-
тии женской генеративной сферы
смородины (ВШез Г.) и крыжов-
ника (Grossularia МШ.).—В кн.:
Вопросы охраны природы Кар-
ат. Ужгород: Карпаты, 41969а:
—57
Мандрик В. Ю. Макроспорогенез и
развитие женского гаметофита у
крыжовника (Grossularia reclinata
Mill.) и альпийской смородины
(Ribes alpinum L.).— Бюл. МОИП.
Отд. биол., 19696, 74, 5: 101—108.
Мандрик В. FO. Эмбриологическое
исследование Ribes nigrum L,,
развитие женской генеративной
сферы, оплодотворение, развитие
эндосперма и зародыша. — Бот.
журн., 1972, 57, 2:234—244.
Мандрик В. Ю. Развиток наночних
репродуктивних структур на за-
пл1днения у деяких вид! в роду
смородини (Вез Г.).— Укр. бот.
журн., 1976a, 33:600—603, 670.
Мандрик В. Ю. Ембрологичне до-
слджения деяких видв роду
Alchemilla Г.— Укр. бот журн.
19766, 33, 5:477—480.
Мандрик В. Ю., Андрусяк М. А.
Изучение пыльцевых трубок и
спермиогенеза у крыжовника
Grossularia reclinata МШ. и cmo-
родины Ribes alpinum L. (mopdo-
логические и гистохимические ис-
следования).— Бот. журн., 1969а,
54, 7:1142—1447.
Мандрик В. Ю., Андрусяк М. А. Ци-
тоэмбриологическое исследование
микроспорогенеза крыжовника
Grossularia reclinata Mill. a cmo-
родины Ribes alpinum L.— Bor.
журн., 19696, 54, 9: 1388—1396.
Мандрик В. Ю., Голышкин Л. В. Эм-
бриологическое исследование не-
которых видов семейства Сгаззи-
laceae.— Bot. журн., 1973, 58, 2:
263—272.
Мандрик В. Ю., Ментковская Е. А.
Цитоэмбриологическое исследова-
ние некоторых популяций Poten-
tilla erecta (L.) Hampe (Rosaceae)
в украинских Карпатах (микро-
спорогенез, дифференциация ce-
мяпочек и развитие зародышево-
го мешка).— Бот. журн., 1977, 62,
7: 1062—1063.
Мандрик В. Ю., Ментковская ЕЁ. А.
Оплодотворение и апомиксис y
Potentilla erecta (L.) Hampe.—
В кн.: VII Всесоюз. симпоз. по
эмбриол. раст. Киев: Наукова дум-
ка. 19783а, 2: 34—36.
Мандрик В. Ю., Ментковская Е. А.
Цитоэмбриологическое исследова-
ние некоторых популяций Poten-
tilla erecta (Г.) Нашре. Оплодо-
творение, развитие эндосперма и
зародыша.— Бот. журн., 19786, 63,
9: 1326—1334.
Мандрик В. Ю., Петрус Ю. Ю. Апо-
миксис и половой процесс у не-
которых видов рода Rubus L—
В кн.: Апомиксис и его использо-
вание в селекции. М.: Колос,
1976 : 898—100.
Маня-Черней. Е. Н. Анатомическое
и морфологическое исследование
соцветий некоторых видов семей-
ства Агасеае.— Бот. журн., 41978,
63, 4:510—522.
Маркова Л. Г. Эмбриологический
анализ гваюлы и родственных ей
видов.— Бот. журн., 1946, 34, 1:
: 19—26.
Маркова Л. Г. К эмбриологии Рое-
monium соегщеит.— Изв. Зап.-
Сибир. филиала АН СССР. Сер.
биол., 1947а, 2, 1:9—19.
Маркова Л. Г. Материалы по эм-
бриологии некоторых представи-
телей семейства Leguminosae.—
Бот. журн., 19476, 29, 5: 220—231.
285
Маркова Л. Г. В эмбриологии Со-
rydalis bracteata L.— Bion. MONITIL
Отд. биол., 1949, 54, 1:77—83.
Маркова „Л. Г. К эмбриологии ред-
кого северного паразита бошма-
кия.— Бюл. МОИП. Отд. биол.,
1954, 56, 4:55—64.
Матюнина Т. Е., Турсунов Ж. Ю.
Развитие и строение стенки пыль-
ника у некоторых сапонинонос-
ных растений Узбекистана.— В
кн.: Вопросы ботанических иссле-
дований в Узбекистане. Тез. докл.
Г Реси. конф. молодых ученых.
Ташкент: Фан 1977 : 71—72.
Медведева Г. Б Эмбриологический
очерк кенафа.— Бот. журн., 1944,
29, 6.
Мейер К. И. К эмбриологии Мир-
har luteum Sm.— Brn. MOMIL
Отд. биол., 1960, 65, 6: 48—58.
Мейер К. И. Эмбриология Согопата
Поз-сисиН.— Бот. журн., 1962, 47:
: 827—852.
Меликян А. П. Положение семейств
Buxaceae mw Simmondsiaccae Bcn-
стемо.-; Бот. журн., 1968, 53, 8:
: 1043—1047.
Мешкова Л. 3., Антипова С. М. Ци-
тоэмбриологическое исследование
вероники сибирской (Уеготса 31-
Ыгса Г.).— В кн.: Аспирантские
работы. Казанск. ун-та. Казань.
ч. Г. 1966: 39—48.
Микаелян С. Г. Цитоэмбриологиче-
ское исследование баклажана в
условиях возделывания в Армян-
ской ССР: Автореф. дис. ... канд.
биол. наук. Ереван: Гос. ун-т,
1967a. 19с.
Микаелян С. Г. Микроспорогенез и
микрогаметогенез у баклажана.—
Buon журп. Армении, 19676, 20,
8 : 95—100.
Микатадзе Ц. А. Микроспоро- и га-
метогенез у некоторых форм чай-
ного растения, разводимого в Гру-
зии.— Журн. субтроп, культуры,
1973, 5:163—165.
Микатадзе Ц. А. В цитоэмбриологи-
ческому исследованию некоторых
форм Теа sinensis ТГ.— В кн.:
УПТ Научная конференция моло-
дых ученых, посвященная 250-ле-
тию АН СССР. Тбилиси, 1974, (на
груз. яз.).
Микатадзе Ц. А. Макроспорогенез
и развитие женского гаметофита
у некоторых форм ТЬеа $з10епз$1$
L.— Сообщ. АН ГССР, 1975, 18,3:
: 677—680.
286
Микатадзе-Поанцулая Ц. А. К изу-
чению процесса оплодотворения и
эмбриогенеза у некоторых рас-
пространенных в Грузии форм
Thea sinensis L—B xa.: VII Bce-
союз. симпоз. по эмбриол. раст.
Киев: Наукова думка, 1978, 3:
: 57-59.
Микатадзе-Панцулая Ц. А. Биология
и цитоэмбриология некоторых с0-
ртов и форм чая, разводимых в
Грузии: Автореф. дис. ... канд.
биол. наук. Тбилиси, 1979. 24с.
Миляева 9. Л. Эмбриологическое
исследование земляничного дере-
Ba (Arbutus andrachne L.).— Ha-
уч. докл. высш. школы. Биол. на-
уки, 1960, 4: 129—134.
Миркамилов М. А. Развитие жен-
ского гаметофита у АШиш аНа-
tunense.— Узб. биол. науки. Таш-
кент: 1969, 6: 33—36.
Миркамилов М. А. Полиэмбриония
У видов рода АШиш (секция Mo-
Нит оп.) — Докл. АН УзССР,
1972, 6:48—49.
Миркамилов М. А. Микроспорогенез
и развитие мужского гаметофита
У видов АШиют Г.— В кин.: Вопро-
сы ботанических исследований в
Узбекистане. Ташкент: Фан, 1977а
83—85.
Миркамилов М. А. Опыление и оп-
лодотворение у нокоторых средне-
азиатских видов лука.— Вкн.: Во-
просы ботанических исследова-
ний в Узбекистане. Ташкепт:
Фан, 19776: 81—83.
Михайлова Е. И. К эмбриологии ве-
роники Линдлеана (Veronica lin-
Ффеуапа).— В кн.: Аспирантские
работы. Казанск. уп-та. Казань,
т. Г. 1966: 49--58.
Младенцева М. С. Изучение эмбри-
OJIOCHH илодообразование у хме-
ля в связи с выясиениеом зозмож-
ности партеногепеза: Автореф.
дис. ... канд. биол. наук. М. 1964.
17с.
Мовсесян С. Н. Процесс оплодотво-
peuua y Rudbeckia maxima.— Изв.
AH ApmCCP 1964, 17, 2:
Мовсесян С. Н. Сравнительно-цито-
эмбриологическое исследование
некоторых видов рудбекий.— В
кн.: Матер. Всес. симпоз. по эм-
бриол. раст. Киев: Наукова дум-
ка, 1968.
Мовсесян С. Н. Цитоэмбриологиче-
ское исследование апомиксиса у
представителей рода ВиаесМа.—
В кн.: Апомиксис и селекция. М.:
Наука, 1970: 426—134.
Мовсесян С. Н. Отклонения в раз-
витии женского гаметофита у
рудбекии.— В кн.: Эмбриология
покрытосеменных растений. Ки-
шинев: Штиинца, 1973.
Мовсесян С. Н. К характеристике
типов зародышевого мешка Вид-
beckia triloba (L.) (Asteraceae).—
Биология (Ереван), 1979, Т:42—
49
Мовсесян С. Н., Багдасарян А. М.
Поведение антиподального аппа-
pata Rudbeckia speciosa. Биол.
журн. Армении, 1966, 19, 7:47—
52
Мовсесян С. Н., Каспарова И. П.
Цитоэмбриологические особенно-
сти рудбекии трехлопастной.— В
win: «Tes. докл. УП Всесоюзн.
симп. по эмбриол. раст. 1978, 3:
56—57.
Модилевский Я. С. ВК эмбриологии
Gunnera chinensis.— Вег. dtsch.
bot. Ges., 1908, 26: 550—556.
Модилевский Я. С. К эмбриологии
u wutonorua Neottia nidus avis
(L.) Rich.— 3an. Hues. о-ва есте-
ствоиспыт., 4918, 26:1—55.
Модилевський Я. С. Запидниения у
ячменю (Hordeum sativum) в
звязку з вивченням особливостей
‚заптднення ‘у покритонаснних
рослин.— Бот. журн. АН УССР,
1953, 140, 2: 13—31.
Модилевский Я. С. К вопросу о про-
исхождении зародышевого мешка
покрытосеменных растений.— Бот.
журн., 1953, 38, 1:31—48.
Модилевский Я. С. Цитоэмбриоло-
гия высших растений (Современ-
ное состояние проблемы). Киев:
АН УССР, 1963. 370с.
Модилевський Я. С., Betiaic P. A.
До emO6pionorii i nuronorii ume-
Hui. I EmMOpiorenes numenuni Biz
археспор1я до s8aponKa.— Журн.
Ин-ту бот. АН УРСР, 1937, 13, 44:
: 127—144.
Модилевський Я. С., Дзюбенко Л.К.
]лекитимне запиления готеро-
стильного льону KPYWHOKBITKOBO-
ro (Linum grandiflorum Desf.). y
звязку з застосуваниям ф1з1оло-
чно активних реговин.— Укр.
бот. журн., 1966, 23, 1:31—40.
Модилевский Я. С., Оксиюк П. Ф..,
Худяк М. И., Дзюбенко J. K.,
Бейлис-Выровая Р. А. Цитоэм-
бриология основных хлебных зла-
ков. Киев: Изд-во АН УССР, 1958.
336с.
Морозова Е. Л. Морфотенез соцве-
тий бука. Тр. Кишипев. с-х
ин-та, 1972, 93: 77—84.
Москова Р. Д. Апомиксис у Во&11о-
chloa ischaemum Г.—В кн.
Апомиксис и его использование
в селекции. М.: Колос, 1976:
101—147.
Мошкович А. М., Чеботарь А. А.
Рожь (Кариология, цитоэмбрио-
логия, цитогенетика). Кишинев:
Штиинца, 1976. 92е.
Мурадов А. ТГ. Развитие репродук-
тивных органов и ранний эмбри-
огенез хурмы восточной (Diospy-
ros Kaki L.) в условиях занадной
зопы Азербайджана. Автореф.
дис. ... канд. биол. наук. Баку,
1974. ЗА с.
Муратов 3. М. Цитоэмбриологиче-
ское исследование дыпь.— Науч.
тр. Ташкент. ун-та, 1972, 398:
46—47.
Мутафян Е. М. Мегаспорогенез и
образование зародышевого мешка
у перца.— Изв. АН АрмССР. Сер.
биол. наук., 1964, 17:95—99.
Навашин С. Г. Избранные труды.
М; Л.: Изд-во АН СССР, 1954а.
364 с.
Навашин С. Г. О цветневой трубке
У вяза.— Изб. труды. М.; Л.: Изд-
во AH CCCP. 19516 : 3—364.
Наумова Т. Н. Полиэмбриония у
Euonymus macropteria a E. pla-
nipes (Koehne) Koehne.— bor.
журн., 1970а, 55, 9: 1270—1284.
Наумова Т. Н. Развитие половых и
апомиктических зародышей у бе-
ресклета европейского (ЕКиопу-
mus еигораеа Г.).— Бот. журн.,
19706, 55, 5: 638—648.
Наумова Т. Н. Особенности разви-
тия нуцеллярной ткани и нуцел-
лярная полиэмбриония у ОрипИа
elata (Cactaceae).— Bot. журн.,
1978a, 638, 3: 344—355.
Наумова Т. Н. Эмбриогенез поло-
вых и нуцеллярных зародышей у
Opuntia (Cactaceae).— B uu.: Tes.
докл. УГ делегат. Всесоюз. бот.
о-ва. 19786: 87—88.
Наумова Т. Н. Половой процесс, ну-
целлярная и интегументальная
эмбриония У представителей се-
мейств Се]азтасеае В. ВВ. и Са-
ctaceae Лиз3.— В кн.: УП Всесоюз.
симпоз. по эмбриол. Киев: Науко-
ва думка, 1978в, 3:59—61.
287
Наумова Т. Н. Семяпочка, макро-
спорогенез, микро- и макрогаме-
тогенез у Sarcocca humilis Hort.—
Бот. журн., 1979а, 64, 5: 635—
645.
Наумова Т. Н. Эмбриология пред-
ставителей сем. Се|азтасеае. Зна-
чение интегументальной эмбрио-
нии в эволюции рода Ецопу-
mus.— B ka.: Актуальные вопро-
сы эмбриологии покрытосемен-
ных растений. М; Л.: Наука,
19796 : 46—57.
Наумова Т. Н. Нуцеллярная поли-
эмбриония в роде Загсосса (Bu-
хасеае) Бот. журн., 1980, 65, 2:
230—240.
Паумова Т. Н., Яковлев М. С. Ад-
вентивная эмбриония У покрыто-
семенных растений.— Бот. журн..,
1972, 57, 8: 1006—1024.
Наумова Т. Н., Яковлев М. С. Ма-
KporametoreHes Trillium camschat-
сепзе Ker-Gawl.— bot. — журн.,
1975а, 60, 3: 342—350.
Наумова Т. Н., Яковлев М. С. ГРаз-
витие ‚эмбриональных структур
Trillium camschatcense Ker-Gawl.
после опыления.— Бот. журн.,
19756, 60, 5: 627—635.
Наумова Т. Н., Яковлев М. С. По-
ловое воспроизведение у ОрипИа
ficus-indica (Cactaceae).— Bor.
журн., 1978, 63, 4:548—556.
Низамова Х. Развитие зародыша и
эндосперма у двух видов эриге-
рон (мелколенестник).— Уч. зап.
Душанбин. гос. пед. ин-та. 1973,
87 : 90—93.
Нивитичева 3. И. К эмбриологии
Lathraea squamaria L. (Scrophu-
lariaceae).— Бот. журн., 1966, 51,
14: 1607—1617.
Никитичева 3. И. Развитие пыль-
пика и микроспорогепез у неко-
торых представителей Scrophula-
riaceae и Orobanchaceaec.— Бот.
журн., 1968, 53, 12:1704А—1715.
Никитичева 3. И. Эмбриология па-
разитных представителей — се-
мейств Scrophulariaceac u Ого-
фапсрасеае в связи с их образом
жизни: Автореф. дис... канд. биол.
паук. Л. БИН АН СССР, 1970:
21 с.
Никитичева 3. И. Эмбриогенез не-
которых паразитирующих видов
из семейств Scrophulariaceae u
Orobanchaceae.— Бот. журн. 1971,
56, 1:93—105.
288
Пикитичева 3. И. Морфогенетиче-
ские корреляции репродуктив-
ных и эмбриональных структур в
семействе Scrophulariaceae.— В
кн.: УП Всесоюз. симпоз. по эм-
бриол. раст. Киев: Наукова дум-
ка, 1978, 2: 39—40.
Никитичева 3. И. Развитие зароды-
ша, эндосперма и семени у пара-
зитных представителей Scrophu-
]ат1асеае.— В кн.: Актуальные во-
просы эмбриологии покрытосе-
менных. Л.: Наука, 1979а: 130—
157.
Никитичева 3. И. О взаимосвязях в
развитии эндосперма и ткани се-
мязачатка у видов семейств Эсго-
phulariaceae wu Orobanchaceae.—
Бот. журн., 19796, 64, 14: 1567—
1581.
Никитичева 3. И., Терёхин 2. C.
К oemO6puonorun Tozzia alpina L.
(Scrophulariaceae).— bot. журн.,
1973, 58, 5: 680—691.
Никитичева 3. И., Терёхин 29. C.
Развитие семепи и проростка
Orobanche раШаога Уппи. et
Grab. (Orobanchaceae).— Бот. журн.
1976, 61, 6: 690—700.
Никифоров Ю. „/Л. Развитие муж-
ского гаметофита хлопчатника.—
Изв. АН ТССР. Сер. биол. наук,
1964, 5: 54—67.
Пиколаева 3. В. Микроспорогенез у
ясеней.— Бот. журн., 1962a, 47,
9 : 1333—4338.
Николаева 3. В. Пыльца ясеней
(морфология, жизнеспособность).
Татикент. Гос. ун-т им. В. И. Ле-
нина, Ташкент, 19626, 204: 115—
427, ~
Пиколаева 3. В. Морфофизиологи-
ческие особеппости прорастания
пыльцы у ясеней.— ДАН CCCP,
1962в, 116, 4 : 221—224.
Николаева 3. В. Морфогенез гене-
ративной сферы и биология цве-
тения ясеней в условиях Таш-
кепта: Автореф. дис. канд.
биол. наук. Ташкент, Гос. ун-т,
1963.
Николаева 3. В. Развитие женского
гаметофита, семени и плода ясе-
ней Fraxinus.— Бот. журн., 1966,
51, 2: 288—291.
Николаева Е. С. К эмбриологии то-
поля Populus balsamifera L.—
Бот. журн., 1964, 49, 11: 1644—
1649.
Николаева Е. С. К эмбриологии не-
которых представителей семей-
ства ивовых (ЗаПсасеае 4па].).—
В кн. УП Всесоюз. симпоз. по
омбриол. раст. Киев: Наукова
думка, 1978, 2:40—42.
Новикова В. М. Эмбриогенез неко-
торых раносозревающих сортов
винограда.— В кн.: УП Всесоюз.
симпоз. по эмбриол. раст. Киёв:
Наукова думка, 1978, 3:61—62.
Норметов Б. А. Макроспорогенез,
развитие женского гаметофита и
эмбриогенез у эспарцета красиво-
ro.— В кн.: Морфогенез растений
аридной зоны Ташкент: Фан,
1973 : 88—87.
Норматов Б. Строение микроспо-
рангия и макроспорогенез у эс-
парцета мелкоцветущего Опогу-
chis micrantha Зеобтепк.— В кн.:
Морфобиологические особенности
дикорастущих растений Узбеки-
стана. Ташкент: Фан, 1974: 66—
74.
Норматов Б. Микроспорогенез у од-
нолетних эспарцетов.— В кн.: Во-
просы ботанических исследова-
ний в Узбекистане.— В кн.: Тез.
докл. Т Респ. конф. молодых уче-
ных. Ташкент: Фан, 1977 : 77—18.
Оганесян М. Г. Рост пыльцевых
трубок и спермиогенез у томата.—
Биол. журн. Армении, 1958, 21,
3:52—58.
Окулова С. М. Эмбриологические ис-
следования некоторых видов ве-
роник: Автореф. дис. канд. биол.
наук. Казань, 1973. 23 с.
Онкго Видаяти. Цитоэмбриология
некоторых декоративных сложно-
цветных: Автореф. дис.. канд.
биол. наук. М., 1973.
Орел Л. И., Жакова М. А. Циффе-
ренциация стенок пыльника и ди-
морфизм тапетума томата Гусо-
persicon esculentum Mill. (Solan-
ceae).— Бот. журн., 1976, 614, 12:
1720—1729. `
Орлова И. Н. Развитие зародышево-
го мешка у некоторых видов кар-
тофеля.— Тр. по прикл. бот., ген.
и сел., 1965, 37, 3:143—152.
Остапенко Е. К. Цитоэмбриологиче-
ское исследование некоторых
представителей ‘семейства бобо-
BEIx (Leguminosae Juss.) m льно-
вых (Linaceae Dumort.) в связи
с накоплением запасных пита-
тельных веществ: Автореф. дис...
канд. биол. наук. Киев, 1970. 24 с.
Пантелеева А. П. Микроспорогенез
19 В. А. Поддубная-Арнольди
и развитие мужского гаметофита
y Althea broussonetifolia— В кн.:
Вопросы ботанических исследова-
ний. В Узбекистане. Ташкент:
Фан, 1977: 85—86.
Пашув Х. Т. Развиток чолов1чого
та ж1ночного гаметоф!т1в у соль-
данели угорсько! (З01ЧапеПа hun-
сана ЗиК.) — Укр. бот. журн.,
1969. 26, 4: 76—81.
Петрова Л. Р. Некоторые данные
эмбриологии бамбуков.— В кн.:
Тез. докл. УП Всесоюз. симпоз.
по эмбриол. раст. Киев: Наукова
думка, 1978, 3:67—68.
Нетрова Т. Ф. Цитоэмбриология ли-
лейных. Подсемейство ТлПо1еае.
М.: Наука, 1977. 246 с.
Петровская-Баранова Т. П. Эмбрио-
логическое исследование женьше-
ня.— Тр. Глав. бот. сада AH CCCP,
1959, 6: 214—225.
Петрус Ю. Ю. Цитоэмбриологиче-
ское исследование некоторых ви-
дов ежевик (Rubus Г.): Автореф.
дис. ... канд. биол. наук. Киев,
1975.
Петрушина М. П. К эмбриологии у
Beta trigina W. et K.—B xa: VII
Всесоюзный симпозиум по эмбри-
ологии растений. Киев. Наукова
думка, 1978, 2:44—45.
Неченицын В. П. Морфогенез и эм-
бриогенез некоторых видов рода
Тира Г.: Автореф. дис. ... канд.
наук. Ташкент 41965а. 19 с.
Печеницын В. П. В эмбриологии
тюльпана Введенского. АН УзССР,
Buon. журн. 19656, 2:53—57.
Печеницын В. П. Двойное оплодо-
творение у видов Тира c Fritil-
laria-THIIOM зародышевого меш-
ка.— Бот. журн., 1972а, 51, 5:
465—469.
Печеницын В. П. Развитие зароды-
шевого мешка Fritillaria-runa y
некоторых среднеазиатских ви-
дов ТиПра.— Бот. журн., 19726,
57, 2:221—229.
Печеницын В. П. Автономный апо-
миксис у интродуцированных ви-
дов рода Сгааегиз.— В кн. Ин-
тродукция и акклиматизация ра-
стений. Ташкент, Фан, 1973,
10: 131—137.
Нлиско М. А. Эмбриологическое
изучение апомиксиса у предста-
вителей Возасеае и СотрозЦцае.—
В кн.: Апомиксис и селекция. М.:
Наука, 1970: 134—141.
289
Ллиско М. А. Электронно-микроско-
пическое исследование особенно-
стей мегагаметогенеза у Calen-
да оотаНз Г.— Бот. журн.,
1974, 56, 5: 582—588.
Лоддубная В. В эмбриологии и ци-
тологии рода — Сагациз.— Бюл.
МОИП. Отд. биол., 1926, 35—3—4.
Поддубная-Арнольди В. А. Срав-
нительно-эмбриологическое иссле-
дование некоторых нормальных
в половом отношении видов и ги-
бридов Тагахасит.— Бюл. МОИП.
Отд. биол., 1947, 52, 1:81—90.
Поддубная-Арнольди В. А. Срав-
нительно-эмбриологическое иссле-
дование диплоидных и тетрапло-
идных форм гречихи.— Бот. журн.,
1948, 33, 2: 181—194.
Поддубная-Арнольди В. А. Значе-
ние эмбриологических исследова-
ний высших растений для систе-
матики.— Усп. соврем. биол.,
1951, 32, 3(6): 352—392.
Поддубная-Ариольди В. А. Иссле-
дование зародышей у ипокрытосс-
менных растений в живом состо-
янии.— Бюл. Главн. бот. сада
AH CCCP, 1952, 14: 3—12.
Поддубная-Арнольди В. А. Значе-
ние эмбриологических исследова-
ний для построения филогенети-
ческой системы покрытосеменных
растений. В кн.: Проблемы бо-
таники. М.: Изд-во АН CCCP.,
1958а, 3: 196—247.
Поддубная-Арнольди В. А. Иссле-
дование процесса оплодотворения
У некоторых покрытосеменных
растений на живом материале.—
Бот. журн., 19586, 43, 2: 178—193.
Поддубная-Арнольди В. А. Матери-
алы к истории исследования про-
цесса оплодотворения у покрыто-
семенных растений.— Тр. Глав.
бот. сада АН СССР, 1959, 6:7—
Поддубная-Арнольди В. А. Полиэм-
бриония у орхидей.— Бюл. Глав.
бот. сада АН СССР, 1960а, 36:
56—61.
Поддубная-Арнольди В. А. О под-
весочном гаустории зародышей в
семенах орхидей.— Бюл. Глав.
бот. сада АН СССР, 19606, 37:
47—52.
Лоддубная-Арнольди В. А. Пласти-
ды в клетках генеративных орга-
нов орхидей.— Бюл. Глав. бот. са-
да АН СССР, 1960в, 39: 54—61.
290
Поддубная-Арнольди В. А. Разви-
тие, современное состояние и пер-
спективы эмбриологии покрыто-
семенных растений.— В кн.: `Мор-
фогенез растений. 1964: 31 : 35.
Поддубная-Арнольди В. А. Общая
эмбриология покрытосеменных
растений. М.: Наука, 1964a.
482 с.
Поддубная-Арнольди В. А. Сравни-
тельно-эмбриологическое исслело-
вание представителей семейства
орхидных.— Бюл. Глав. бот. сала
АН СССР, 19646, 54: 51—62.
Поддубная-Арнольди В. А., Кудря-
шов Л. В. Успехи эмбриологии
покрытосеменных растений в
СССР.— Науч. докл. высш. шко-
лы. Биол. науки, 1967, 7:7—11.
Поддубная-Арнольди В. А. Микро-
спорогенез и мужской гаметофит
покрытосеменных — растений.— В
кн.: Проблемы эмбриологии. Киев:
Наукова думка, 1968: 26—72.
Поддубная-Арнольди В. А. Цости-
жения и перспективы эмбриоло-
гии покрытосеменных растений в
ХХ в.—В ки.: Материалы Всесо-
юзного симпоз., посвященного 75-
летию открытия С. Г. Наваши-
ным двойного оплодотворения у
покрытосеменных растений. М.,
1973а: 184—185.
Поддубная-Арнольди В. А. Е исто-
рии цитоэмбриологии растений.—
В кн.: Из истории биологии. М.,
19736 : 5—40.
Поддубная-Арнольди В. А. Цитоэм-
бриология покрытосеменных ра-
стений. М.: Наука, 1976—507.
Поддубная-Арнольди В. А. Цитоэм-
бриологическая характеристика
семейства = Orchidaceae.— В кн.:
УП Всосоюз. симпоз. но эмбриол.
раст. Киев: Паукова думка: 41978,
2:45—48.
Поддубная-Арнольди В. А. Цитоэм-
бриологическая — характеристика
семейства злаковых.— Бюл. Глав.
бот. сада АН СССР, 1979, 109:
57—60.
Поддубная-Арнольди В. А., Диано-
ва В. И. Характер размножения
некоторых каучуконосных и не-
каучуконосных видов рода Тага-
хасит.— Бот. журн., 1937, 22, 3:
2671—2595.
Поддубная-Арнольди В. А., Ивано-
ва Й. А. Проращивание пыльцы
некоторых злаков на искусствен-
ной питательной среде.— В кн.:
Отдаленная гибридизация и по-
липлоидия. М.: Наука. 1970: 199—
206.
Поддубная-Арнольди В. А., Луне-
ва М. 3., Джалилова Х. Х. Мор-
фологичесное и цитоэмбриоло-
гическое исследование некото-
рых видов и гибридов №сойа-
па.— Бот. журн., 1974, 59, 6:
874—883.
Поддубная-Арнольди В. А., Пащен-
ко 3. М. Исследование мужского
гаметофита 27еа шауз.—В кн.:
Морфогенез кукурузы. М.: МГУ,
1962 : 162—174.
Поддубная-Арнольди В. А., Селез-
нева В. А. Орхидеи и их культу-
ра. М.: Изд-во АН СССР, 1957.
176 с.
Поддубная-Арнольди В. А., Селез-
нева В. А. Орхидеи из семян.—
Цветоводство, 1961, 7:6—7.
Поддубная-Арнольди В. А, Стеши-
на Н. А., Сосновец А. А. Матери-
ал к биологии цветения и раз-
множения — оЭсоттопега _— tau-sag-
hyz.— Bor. журн., 1934, 19, 4:
338—366.
Поддубная-Арнольди В. А., Ци-
цин Н. В., Махалип М. А. Микро-
спорогенез и спермиогенез у ди-
плоидной и тетраплоидной ози-
мой ржи Вятки московской.—В
кн.: Генетични изследования в
памет на Дончо Костов. София,
1968 : 68—67.
Полицук В. С. Ембрологчн! досл!
джения шавелй лкареькой (Sal-
via officinalis L.) и щавели му-
скатно! (3. зсатеа Т.).— Укр. бот.
журн., 1972, 29, 1:25—30.
Полищук В. С. Цитоэмбриологиче-
ские исследования майорана са-
дового (Majorana hortensis Mo-
ench.) B cBA3H с его интродукци-
ей и акклиматизацией на Украи-
не.— Изв. АН МолдССЬР. Сер. биол.
и хим., 1973, 1:81—82.
Полицув ВБ. С., Дзевалтовський А.К.
До ембрюлогИ лаванди колосово!
(Lavandula spica Т.).— Укр. бот.
журн., 1971а, 28, 6:718—1781.
Полищук В.С., Дзевалтовский А.ЁЕ.,
Эмбриологическое — исследование
некоторых представителей семей-
ства губоцветных.— Цитология и
генетика. Киев, 19716, 5, 3.
Полунина Н. Н. Материалы по био-
логии цветения и эмбриологии
19*
фейхоа.— Бюл. Гл. Gor.
AH CCCP, 41957, 29: 60—71.
Полунина Н. Н. Биология цветения
и эмбриология некоторых пред-
ставителей семейства миртовых.
Автореф. дис. ... канд. биол. на-
ук. М., 1958а: 16 с.
Полунина Н. Н. Биология цветения
и эмбриология СаШзетоп ]апсео-
latus Sweet.— Bot. журн., 19586,
43, 8: 1169—1178.
Полунина Н. Н. К биологии цвете-
ния и эмбриологии Евгении мир-
толистной (Eugenia mzyrtifolia
Sims.).— Bron. Глав. бот. сада
АН СССР, 1958в, 30: 59—65.
Полунина Н. Н. Биология цветения
и эмбриология эвкалипта.— Тр.
Глав. бот. сада АН СССР, 1959,
6: 191—210.
Полунина Н. Н. Сравнительно-эм-
бриологическое исследование не-
которых представителей семей-
ства миртовых.— Бюл. Глав. бот.
сада АН СССР, 1963, 49:82—90.
Полунина Н. Н. Сравнительное изу-
чение эмбриологии, биологии цве-
тения и плодоношения фейхоа в
различных условиях произраста-
ния.— В кн.: Проблемы современ-
ной эмбриологии. М.: МГУ, 1964:
153—158.
Полякова Т. Ф. Развитие мужского
и женского гаметофита у Arabi-
4ор51з ШаПапа Г.— В кн.: Иссле-
дования по генетике. ЛГУ, 1964,
2: 125—133.
Попова А. Ф. Закладания та раз-
виток генеративных орган1в у чор-
но! смородины (ВШез шота L.).—
Укр. бот. журн., 1965, 22, 6: 83—
сада,
Попова А. Ф. Цитоэмбриологиче-
ское изучение представителей ро-
mos Ribes um СтоззщШата в связи с
гибридизацией: Автореф. uc.
канд. биол. наук. Киев, 1970 : 24 с.
Попова Л. К. Развитие женского
гаметофита у ястребинки волоса-
tou (Hieracium pilosella L.).—
Изв. АН МолдССР. Сер. биол. и
хим. наук. 1979, 2:81—82.
Радионенко А. Я. Некоторые на-
блюдения над эмбриологией аб-
рикоса обыкновенного 8 (Armeni-
ca vulgaris).— Yap. 607. журн.,
1963, 6: 15—23.
Радюненко А. Я. О развитии жен-
ского гаметофита у яблони (Ma-
lus domestica).— Укр. бот. журн.,
1964, 21, 4: 62—70.
291
Радионенко А. Я. Двойное оплодо-
творение у яблони.— ДАН СССР,
1966, 168, 5: 1208—1241.
Радионенко А. Я. Исследование по-
лового процесса у яблони.— Ци-
тология и генетика. Киев. 1967,
1, 5:26—34.
Радионенко А. Я. Исследование
процесса оплодотворения у ябло-
Hu Malus domestica Borkh.— Бот.
mypu., 1968, 53, 8: 41101—1110.
Радионенко А. Я. Исследование оп-
лодотворения черной смородины
(Ribes nigrum Г.).— Цитология и
генетика. Киев. 1969, 3, 1: 36—45.
Радионенко А. Я. Процесс оплодо-
творения у ВЪез nigrum L.—
Бот. журн., 1970, 55, 6:807—813.
Радионенко А. Я., Лиферова В. В.
Цитоэмбриологическое изучение
смородины черной (Ribes_ ni-
grum).— Укр. бот. журн., 1970, 27,
1:90—97.
Райкова И. А., Алимджанова Д.Р.
Микроспорогенез и развитие муж-
ского гаметофита у кассий, ин-
тродуцированиых в ‘Tamkenre.—
В кн. УП Всесоюз. симпоз. по
эмбриол. pact. Hares. Наукова
думка, 1978, 3:75—17.
Райкова И. А., Нащенко 3. М.Е ци-
тоэмбриологии недзвецкой.— В
кн.: Интродукция и акклимати-
зация растений. Ташкент: Фан,
1977, 14: 142—120.
Райкова И. А., Эрккенова Е. М. Ге-
неративная сфера в функциональ-
но мужскнх и функционально
женских цветках жестеров.— В
кн.: Половой процесс и эмбрио-
генез растений. М.: Наука, 1973:
193—494.
Родионова Г. Б. Мегаспорогенез у
Незрег1$ steveniana DC.— Beetu.
МГУ. Сер. УГ 196ба, 5: 08—72.
Родионова Г. Б. Относительно эм-
бриогенеза Тлтама аппиа L.—
Бот. журн., 19666, 51: 1506—1541.
Родионова Г. Б. Эмбриологическое
исследование Егиса зайуа Гашт.—
Вестн. МГУ. Сер. VI, 19663, 1:
61—68.
Родионова Г. Б. Сравнительно-эм-
бриологическое исследование сс-
мейства крестоцветных (Crucife-
rae Juss): Автореф. дис. ... канд.
биол. наук. М. 1967.
Родионова Г. Б. Оплодотворение у
Hesperis steveniana DC.— Hay.
докл. высш. школы. Биол. науки,
1967, 12:73—78.
292
Родионова Г. Б. Эмбриологическое
исследование Hesperis steveniana
рС.— В кн.: Морфология высших
растений. М.: Наука, 1971а: 34—
54.
Родионова Г. РБ. Эмбриогенез Глта-
тг1а аппиа Г.— Вестн. МГУ. Биол.,
почвовед., 19716, 4: 59—64.
Родионова Г. Б. Эмбриологическое
развитие Егузииит pannonicum
Сгап(2.— Вестн. МГУ, 1974в, 5 : 52—
57.
Родионова Р. Б. Развитие семяпоч-
ки и интегументов у НеПорПа
amplexicaulis L—bBor. журн.,
1972, 57, 9: 4453—1161.
Родионова Г. Б. Эмбриологическое
passutue Heliophila amplexicau-
lis L. B связи с положением B CII-
стеме крестоцветных (Cruciferae
]155.).— В кн.: Эмбриология по-
крытосеменных растений. Киши-
нев: Штиинца, 1973: 70—84.
Родионова Г. Б. Эмбриологическое
развитие Macropodium nivale
(Pall.) B. BR. m3 семейства кре-
стодветных.-—- В ки.: Рост расте-
ний и пути cro регулирования.
М., 1976 : 126—139.
Родионова Г. Б. Особенности раз-
вития и основные пути эволюции
в эмбриологии крестоцветных.—
В кн.: УП Всесоюз. симпоз. по
эмбриол. раст. Киев: Наукова
думка, 1978, 3:79—81.
Родионова Г. Б. Эмбриология резу-
хи кавказской. Бюл. Глав. бот.
сада AH CCCP, 1979, 114: 98—106.
Романов И. Д. Развитие зародыше-
вого мешка в роде Gossypium.—
В кн.: Вопросы цитологии эмбри-
ологии и анатомии хлопчатника.
Ташкент: СоюзПИХИ. 1936: 17—
36
Романова Г. С. Цитоэмбриологиче-
ские исследования пекоторых
видов ладонника (Cistus L.): Ar-
тореф. дис. ... канд. биол. наук.
М. 1968 : 47 с.
Романова Г. С. Цитоэмбриологиче-
ские исследования некоторых ви-
дов цистуса (С13и1$).— Тр. Никит.
бот. сада, 4970, 46: 184—193.
Романова Г. С. Эмбриология инду-
цированного апомиксиса у Е!сиз
Г.— Бюл. Глав. бот. сада АН СССР,
вып. 143, 1979, 143:84—90.
Романов И. Д. Развитие зародыша
и эндосперма.— В атласе: Строе-
ние и развитие хлопчатника. М:
Л: ОГИЗ—ИЗОГИЗ, 1937а.
Ломанов И. Д. Развитие семяпочки,
зародышевого мешка, оплодотво-
рение.— В атласе: Строение и раз-
витие хлопчатника. М; Л.: ОГИЗ—
ИЗОГИЗ, 19376.
Романов И. Д. Новая форма заро-
дышевого мешка РОгиза-типа у Т.
tetraphylla и Т. ostrovskiana.—
ДАН СССР, 1938, 149, 1:143—115.
Романов И. Д. Две новые формы
зародышевого мешка в роде Ти-
lipa.— JAH CCCP, 1939, 22, 3:
140—142.
Романов И. Д. Эволюция зародыше-
вого мешка в группе Tulipeae
семейства Liliaceae.— Бюл. Сред-
неаз. ун-та, 1945, 23.
Романов И. Д. Стерильность семя-
почек и поведение пыльцевых
трубок в завязях межвидовых ги-
бридов хлопчатника.— Изв. АН
УзССР, 1947, 1.
Романов И. Д. Эмбриологические
исследования хлопчатника. Сре-
днеаз. ун-т, 1954 : 3—58.
Романов И. Д. Зародышевый меток
в рода ТиНра.— ДАН СССР, 1957,
115, 5: 1025—1027.
Романов И. Д. Развитие семяпочек
и зародышевого мешка.— В кн.:
Хлопчатник. Ташкент, 1960, 3.
Романов И. Д. Опыт анализа неко-
торых особенностей развития за-
родышевого Memka Fritillaria-ru-
па.— Бот. журн., 1965, 50, 9:
1276—1287.
Романов И. Д. Специфические осо-
бенности развития пыльцы зла-
ков. ДАН CCCP, 1966, 169, 2:
456—459.
Романов И. Д. Особенности разви-
тия пыльцы злаков и значение их
для некоторых генеративных ис-
следований.— Генетика, 1970, 6,
10; 14—24.
Романов И. Д. Типы развития за-
родышевого мешка покрытосе-
менных.— В сб.: Пробл. эмбриоло-
гии. Киев, Наукова думка, 1974:
712—143.
Ротов Р. А., Петрова Т. Ф. Особен-
ности развития зародышевого
Memka Notholirion thomsonianum
(Кое) Stapf.— Bron. Глав. бот.
сада АН СССР, 1977, 106:96—100.
Рыбченко О. И. Цитоэмбриология
развития партенокарпических
плодов у томатов.— Автореф. дис.
канд. биол. наук. Киев, 1960 : 24 с.
Рыбченко 0. И. Цитоэмбриологиче-
ское исследование женского га-
метофита в семействе паслено-
вых.— Укр. бот. журн., 1963а, 29,
5: 33—43.
Рыбченко 0. И. Цитоэмбриологиче-
ское исследование, развитие муж-
ского гаметофита в сем. паслено-
вых.— Укр. бот. журн., 19636, 20,
6, : 3—14.
Руденко Ф. Е. Сперматогенезис и
оплодотворение у экзота магонии
(Mahonia aquifolium L.).— Бюл.
Моск. о-ва испыт. прир. Отд. би-
om, 1964a, 66, 1:133—137.
Руденко Ф. Е. Цитологическое ис-
следование мужских гамет у
представителей семейства люти-
ковых.— Бюл. Глав. бот. сада АН
CCCP, 19646, 42: 57—66.
Руденко Ф. Е. Развитие мужских
гамет у представителей РарШо-
пасеае.— Бот. журн., 1965, 590, 11:
1575—1578.
Руми В. А. Развитие пыльцы и
мужского гаметофита хлопчатни-
ка.— В кн.: Хлопчатник. Ташкент,
1960, 3.
Руми В. А. Процесс образования
генеративной клетки у хлопчат-
ника.— Узб. биол. журн., 1964а, 1;
18—82.
Руми В. А. О сперматогепезе у хлоп-
чатника.— Узб. биол. ›курн., 19646.
4: 11—76.
Руми В. А. Морфология пыльцы че-
тырех видов хлопчатника.— В кн.:
Вопросы генетики и эмбриологии
хлопчатника. 1966: 25—33.
Руми В. А. Эмбриолотия хлопчат-
ника. Ташкент, Фан, 1969,
200с.
Руми В. А., Камалова Г. В. О хло-
рофиллоносности зародышей в се-
мействе Malvaceae.— ДАН УзССР,
1975, 4: 63—64.
Руми В. А., Камалова Г. В., Сере-
брякова Л. Е. О микроспорогене-
se y Hibiscus manihot L.— Цитоло-
rua wu renetuna, 1976, 10, 2: 107—
140.
Рустамова Д. М. Биология цвете-
ния и цитоэмбриология сои в ус-
ловиях Ташкентского оазиса: Ав-
тореф. дис. ... канд. наук. Таш-
кент, 1965: 19.
Савина Г. И. Оплодотворение у Су-
pripedium calceolus L.— Bor. sypt.,
1964, 49, 9: 1817—1322.
Савина Г. И. Развитие половых эле-
ментов и процесс оплодотворения
у некоторых видов Огсвуз.— Бот.
журн., 1965а, 50, 1:96—102.
293
Савина Г. И. Оплодотворение у ор-
хидных (ОтгсЬ9асеае) — Автореф.
Дис. канд. биол. наук. Л. 19656.
Савина Г. И. Развитие зародыше-
вого мешка y Ophrys insectifera
Г.— Бот. журн., 1972а, 57, 1: 118—
123
Савина Г. И. Процесс оплодотворе-
nua y Ophrys insectifera L.— Бот.
mypH., 19726, 57, 3: 382—392.
Савина Г. И. Оплодотворение y op-
хидных.— В кн.: Материалы Все-
союз. симпоз. «Половой процесс
и эмбриогенез растений». М.:
1973.
Савина Г. И. К эмбриологии орхид-
ных, исследование Cephalanthera
longifolia (L.) Fritsch—B_ ka.:
Тез. ХИП Междунар. бот. конгр.
Л. Наука, 1975.
Савина Г. И. К эмбриологии орхид-
ных. Развитие Сутпо4дета cono-
рзеа (Т.) В. Вг.—В кн.: УП Вее-
союзн. симпоз. по эмбриол. раст.
Киев: Наукова думка, 1978, 3: 89.
Савина Г. И. Особенности эмбрио-
логии семейства Orchidaceae.—
В кн.: Актуальные вопросы эм-
бриологии покрытосеменных. Л.:
Наука. 1979: 19—28.
Савинова Г. Ю., Турсунов J. BO.
Процессы оплодотворения у не-
которых представителей рода
Acanthophyllum Сата.— В кн.: Во-
просы ботанических исследова-
ний в Узбекистане. Ташкент: Фан,
1977 : 73—74.
Савич Е. И. К эмбриологии семей-
ства АПзшабасеае.— В кн.: Мор-
фология растений. М.: Наука,
1967 : 45—61.
Савич Е. И. Мейоз и мужской га-
метофит у Вирма maritima L.—
В кн.: Тезисы симпозиума, посвя-
щенного 70-летию со дпя откры-
тия двойного — оплодотворения
С. Г. Навашиным. Киев : 1968а.
Савич Е. И. Эмбриологическое изу-
чение Вирра шагШта L.— B ku.:
Морфология растений. М.: Нау-
ка, 19686.
Савич Е. И. Эмбриологическое ис-
следование Helobiae: Автореф.
ДИС. канд. биол. наук. М.
1969 : 24 с.
Савченко М. И. Морфология семя-
почки покрытосеменных расте-
ний. М.: Наука, 109 с.
Савченко М. И., Комар Г. А. Мор-
фология семяпочек некоторых
однодольных.— В кн. Морфоло-
294
THA цветка и репродуктивный
процесс у покрытосеменных ра-
стений. М.; Л.: Наука, 1965 :174—
113.
Савченко М. И., Петрова Л. Р. Мор-
фология семяпочки ячменя Ног
deum vulgare L. и некоторые осо-
бенности ее — развития.— Бот.
журн., 1963, 48, 11: 1623—1638.
Саидов Д. Е., Ашурметов А. А.,
Дзевалтовский А. Н. Семяпочка
и женский гаметофит у пере-
ступня черноплодного.— ДАН
УзССР, 1975, 6:45—А47.
Самвелян Г. Е. Цитоэмбриологиче-
ское исследование начальных и
поздних стадий эмбриогенеза не-
которых сортов винограда (Уз
ушШега), возделываемых в Усло-
виях АрмССР: Автореф. дис. ...
канд. биол. наук. Ереван: Гос.
ун-т, 1973.
Салахова Г. Б. О некоторых осо-
бенностях микроспорогенеза и
развития мужского гаметофита у
конского щавеля (Витех confer-
(13 \УШа.) в связи © неполным
диморфизмом его цветков.— В кн.:
Краткие сообщения. — Казань:
1968 : 35—37.
Салахова Г. Б. Микроспорогенез и
мужской гаметофит Виштех соп-
fertus Willd. 8 cBazu c сексуали-
зацией его особей.— В кн.: Поло-
вой процесс и эмбриогенез расте-
ний М.: Наука, 1973 : 207—208.
Салахова Г. Б. Макроспорогенез,
женский гаметофит и оплодотво-
рение Rumex confertus Willd. sb
связи с особенностями ето сек-
суализации.— В кн.: VII Beeco-
юз. симпоз. по эмбриол. раст. Ки-
ев: Наукова думка, 1978, 1:48—
-
90.
Салахова Г. Б., Смирнов Л. Г. Не-
которые особеппости строения
цветков конского щавеля в свР-
3H C формирующимся половым
полиморфизмом.— В кн.: Матери-
алы У Моск. совещ. по филоге-
нии растений. М.: Наука, 1976:
149—151.
Салахова Г. Б., Смирнов А. Г. 0 не-
которых аномалиях в строении
цветков у конского щавеля.— В
кн.: Аспирантские работы. Ва-
зань, 1978: 24—30.
Самушия М. Некоторые вопросы
развития женсного гаметофита у
абрикоса.— Тр. Ин-та сад-ва, ви-
иод., виногр-ва ГССР, 19714, 19—
20 : 132—140.
Гапикидзе Д. Д. Макроспорогенез и
развитие женского гаметофита у
растения лимона.— Тр. Груз. ин-
та субтроп., 1967, 11: 339—344.
(‘еменитина Л В. Микроспороге-
нез и морфология пыльцы У ви-
дов смородины ЁВШФез ]фапсхехзки
А. Pojark. и ВЪез УШозим
Wall.— Изв. АН 'ТаджССР. Отд.
биол. наук, 1972, 1:81—83.
Сизова М. А. Изучение оплодотво-
рения у растений льна.— Тр. по
прикл. бот., ген. и сел., 1958а,
31 :61—81.
Сизова М. А Процесс оплодотворе-
ния У льна Linum usitatissi-
mum.— Tp. по прикл. бот., ген. и
сел., 1958в, 33 : 155—172.
Сизова М. А. Процесс оплодотворе-
ния у тетраплоидных форм льна
Linum usilatissimum pr. elonga-
ta Бот. журн., 1963, 48: 537—
5
Симонян Е. Г. Макроспорогенез и
развитие женского гаметофита у
винограда.— Изв. с.-х. наук (Ере-
ван), 1965, 40.
Симонян Е. Г., Самвелян Г. Е. Ци-
тоэмбриологическое исследование
винограда (У. ушЦега).— Изв.
АН АрмССР. Биол. науки, 1965,
18, 9:12—22.
Скокова А. А. Цитоэмбриология
бессемянных сортов шелковицы:
Автореф. дис. ... канд. наук. Та-
шкент, 1966. 24 с.
Смирнова Е. С. К морфогенезу се-
MAH покрытосеменных — расте-
ний.— Докл. TCXA, 1961, 72:
268—272.
Смирнова Е. С. Морфологические
типы семян однодольных расте-
ний.— Бюл. Глав. бот. сада АН
СССР, 1964, 55: 71—81.
Соколов И. Д. Развитие семяпочки
репчатого лука (АШит сера
L.).— Изв. АН ТССР. Сер. биол.,
1968а, 4:21—27.
Соколов И. Л. Цитоэмбриологиче-
ские исследования культурных
видов лука (АШит сера Ё., A.
porrum L. uw A. fistulosum L.):
Автореф. дис. ... канд. биол. на-
ук. Л.: ВИР, 49686.
Соколов И. Д. Развитие зародыше-
вого мешка репчатого лука АШ-
ит сера Г.— Изв. АН ТОСР. Сер.
биол., 1968в, 1:87—89.
Соколов И. Д. Макроспорогенез у
некоторых видов Allium.— Bor.
журн., 1969, 54, 2: 202—207.
Соколов Г. Д. Розвиток HacinHoro
зачатка У Arabidopsis thaliana
(L.) НеупЬ.— Укр. бот. журн.,
1975, 32, 4:518—522.
Соколов Д. Д., Алимов Н. Х., Ро-
жанов И. Д. Цитология эндоспер-
ма цветковых растений. Киев;
Донецк: Вища школа, 1979.
Солнцева М. П. Некоторые данные
о процессе оплодотворения у
Fragaria ananasa Duch.— Bor.
журн., 1964, 46, 3:371—377.
Солнцева М. П. О развитии много-
клеточного археспория у земля-
ники.— В кн.: Морфология цвет-
ка и репродуктивный процесс у
покрытосеменных растений. М.;
Л.: 1965а : 189—204.
Солнцева М. П. К биологии цвете-
ния и эмбриологии ковылей.—
Tp. бот. ин-та AH CCCP, 19656,
Сер. 3, 47: 185—150.
Солнцева М. П. Особенности цве-
тения и эмбриологии ковылей.—
Уч. зап. Ульянов. гос. пед. ин-та,
1968a, 23, 3: 29—40.
Солнцева М. П. О поведении спер-
миев в зародышевом мешке
Rudbeckia laciniata—B кн.: Ма-
териалы УП Всесоюз. симп. по
эмбриол. раст. Киев: Наукова
думка, 19686.
Солнцева М. П. Основы эмбриоло-
гинеской классификации апомик-
сиса покрытосеменных.— В кн.:
Апомиксис и селекция. М.: Нау-
ка, 1970:87—100.
Солнцева М. П. О полярности заро-
дышевого мешка и морфологии
зародыша Rudbeckia laciniata L.—
В кн.: Материалы У Всесоюз. со-
вещ. по эмбриол. раст. Кишинев:
Штиинца, 1974: 168—169.
Солниева М. П. Типы развития за-
родышевого мешка и морфоло-
rus зародыша Видфесма ]асимафа
Т.— В кн.: Эмбриология покрыто-
семенных растений. Кишинев:
Штиинца, 1973а : 47—57.
Солнцева М. П. Семигамия и опло-
дотворение у Rudbeckia lacinia-
ta Г.— Бот. журн., 19736, 58, 9:
1264—1277.
Солнцева М. П. Гемигамия и ее
проявление у растений.— В кн.:
Апомиксис и его использование
в селекции. М.: Колос, 1976: 73—
81.
295
Солнцева М. П., Ворсобина Л. И.
Гелобиальный тип эндосперма в
родах Zephyranthes u Sternbergia
(Amaryllidaceae)— B кн. Tes.
докл. УГ делегат. съезда Bee-
союз. бот. о-ва. Л.: Наука, 1978:
92—93.
Солнцева М. П., Дгебуадзе Н. В.
К эмбриологии нпекана Сатуа 1-
linoensis (Wangenheim) K. Koch
(Juglandaceae).— Бот. журн., 1974,
59, 12: 1794—1804.
Солнцева М. П., Камелина О. П.
Двойное оплодотворение и заро-
дыш у Cephalaria gigantea
(Ledsb.), Bobr. (Dipsacaceae).—
Бот. журн., 1976, 64, 2: 254—263.
Солнцева М. П., Левковский В. П.
Эмбриологическая характеристи-
ка некоторых арктических зла-
ков. В кн.: УП Всесоюз. сим-
поз. по эмбриол. раст. Киев: На-
укова думка, 1978, 3:96—97.
Солнцева М. П., Левковский В. П.,
Кишечникова Т. И. К эмбриоло-
гии мятлика (Роа Г..) о. Врангс-
ля.— ДАН СССР, 1977, 237, 1.
Солнцева М. П., Яковлев М. С.
Предпосылки развития однодоль-
ности в раннем эмбриогенезе у
ковылей.— Бот. журн., 1964, 49,
5: 625—633.
Сорока В. П. Сравнительное цито-
эмбриологическое исследование
двудомной и однодомной конопли
(Cannabis sativa L.): Автореф.
ДИС. канд. биол. наук. До-
нецк, 1977.
Сорока В. П., Жатов А. И. Некото-
рые особенности мужского гаме-
тофита у однодомной и двудом-
ной конопли.— Бюл. Глав. 00т.
сада АН СССР, 1974, 78: 67—69.
Спицын И. П. Эмбриологические пс-
следования вишни, черешни и
вишне-черешневых гибридов сред-
ней полосы Союза. Тамбов: Изд-во
«Тамбовская правда», 1966 : 34 с.
Старшова Н. П. Особенности эколо-
гии семенного размножения бар-
хата амурского (Phellodendron
amurense Rupr.) в среднем По-
‚ волжье: Автореф. дис. ... канд.
биол. наук. Пермь: Гос. ун-т,
1974.
Старшова Н. П., Солнцева М. П.
К характеристике зародышевого
мешка Phellodendron amurense
Вирг.— Бот. журн., 1973, 58, Ч:
1644—1654.
296
Старшова Н. П., Солнцева Н.П,
Солнцева М. П. Процесс оплодо-
творения у бархата амурского
Phellodendron amurense Rupr.—
ДАН CCCP. 1976, 228, 6: 1485—
1488.
Стожарова И. А. Цитоэмбриологи-
ческое исследование АШишт nu-
фапз Г.: Автореф. дис. канд. биол.
наук. М.: Гос. ун-т, 1973: 24 с.
Стожарова И.А., Поддубная-Арноль-
ди В. А. Роль синергид при оп-
лодотворении, эмбрио- и эндо-.
спермогенезе у АШиш nutans L.-—
Бюл. Глав. бот. сада АН СССР,
1977, 105: 70—78.
Сухарева Н. Б. Элементы апомик-
сиса у земляники.— В кн.: Апо-
миксис и селекция. М.; Наука,
1970 : 116—120.
Тагиева О. А. Развитие репродук-
тивных органов и цитоэмбриоло-
гические особенности лавра благо-
родного на Апшероне: Автореф.
ДИС. канд. биол. наук, 1967:
20 с.
Т'агиева О. Л. Микроспорогенез ци-
кламена изящного (Суатет @е-
хапз 1.). Баку: Ин-т ботан. АН
АзССР, 1970 : 6.
Тагиева О. А. Мегаспорогенез лав-
ра благородного.— В кн.: Цито-
эмбриологические — исследования
растительной флоры Азербайджа-
на. Баку: ЭЛМ, 1972: 58—66.
Татевосян В. Б. Микроспорогепез и
мегаспорогенез граната в усло-
виях Араратской равнины Армян-
ской ССР.— Биол. журн. Арме-
Hun, 1968, 21, 3:66—71.
Татевосян В. Б. Цитоэмбриологиче-
ские особенности закладки и диф-
ферснциации гспосративных орга-
тов траната (Punica granatuni
1..): Автореф. дис. ... канд. биол.
паук. №реван, 1969 :27 с.
ТГатинцева С. С. Развитие мужских
репродуктивных органов у фер-
тильной и стерильной форм сор-
ro (Sorghum caffrorum Г.): Авто-
реф. дис. ... канд. биол. наук. Л.,
1968а.
Татинцева С. С. Спермотенез в
пыльцевом зерне сорго (ЗотеВит
саЙгогит Г.).— Изв. АН ТССР.
Сер. биол., 19686, 3:26—34.
Татинцева С. С. Развитие мужско-
го гаметофита топинамбура (Не-
lianthus фиБегозиз).— Изв. AH
ТССР. Сер. биол., 1971, 1:14—2.
Татинцева С. С. Развитие пыльце-
вого зерна и образование спер-
миев в пыльцевых трубках Ое]-
phinium camptocarpum Fisch. et
Меу.— Изв. АН ТЕСР. Сер. биол.,
1975а, 3: 9—14.
Гатинцева С. С. Образование и раз-
витие спермиев у некоторых по-
крытосеменных. 1. Роасеае (Ога-
mineae); If Asteraceae (Composi-
{ае).— Бот. журн., 19756, 60, 12:
: 1746—1752.
Гахтаджян А. Л. Система и фило-
гения цветковых растепий. М.;
Л Наука, 1966: 608 с.
Гахтаджян А. Л. Происхождение и
расселение цветковых растений.
Л.: Наука, 1970. 145 с.
Терёхин 9. С. Эмбриология груша-
нок и вертляниц в связи с их
биологией и систематическим по-
ложением. Автореф. дис. канд.
биол. наук. Л.: 1962а : 19 с.
Терёхин 9. С. О развитии зароды-
шей у некоторых Pyroleae-Mono-
tropoideae.— Bor. журн., 19626,
47, 12:1811—1816.
Терёхин 9. С. Развитие эндосперма
и характер роста зиготы у евро-
пейских грушанок.— Бот. журн..,
1962в, 47, 2:254—258.
Tepéxun 2. С. Развитие семяпочки
и женского гаметофита у Руго-
leae и Мопотореае.— Бот. журн..,
1963, 48, 3: 406—444.
Герёхин 9. С. Эмбриология ОтоЪап-
chaceae. I. Phelypaea coccinea
(М. В.) рог.— Бот. журн. AH
СССР, 1966, 51, 9: 1601—1607.
Tepéxun 9. С. Эмбриология Огофап-
сВасеае. 11. К эмбриологии и эко-
логии Phacellanthus tubiflorus
Sieb. et Zucc.— Bot. mypx., 1973,
58, 12 :1775—1778.
Герёхин 9. С. Закономерности эво-
люционных преобразований эм-
бриональных и репродуктивных
структур паразитных Angiosper-
тае. Автореф. дис. д-ра биол. па-
ук. Л., 1973.
Герётии I. С. Паразитные цветко-
вые растения. Л.: Маука, 1977:
219 с.
Герёхин 9. С. Жизпеппые формы
зародышей цветковых растений. —
В кн: УП Всесотюз. симпюз. по
эмбриол. раст. Киев: Наукова
думка, 2:55—56.
fepéxux 9. C. Orobanchaceae. III
Mannagettaea htimmeli H. Smith.—
Бот. журн., 1979, 64, 10:1390—
4400.
Терёхин 9. С., Камелина О. П. Эн-
досперм Отс асеае (к вопросу
о редукции}.— Бот. журн. АП
СССР, 1969, 54, 5: 657—666.
Терёхин 93. С., Камелина О. П. Эн-
досперм Habenaria linearifolia
Maxim u Neottia nidus-avis (L.)
Rich.— Bot. sypnH., 1972, 57, 411:
1373—1383.
Терёхин 9. С., Никитичева 3. И.,
Яковлев М. С. Развитие семени,
зародыша и эпдосперма Cynomo-
rium songaricum Вирг. (Супошо-
т1асеае).— Бот. журн., 1975, 60,
44 : 1603—1613.
Терётин 9. С., Никитина 3. И. Ото-
Бапсвасеае. Онтогенез и филоге-
нез (в печати).
Tepéxun 9. С., Яковлев М. С. Эм-
бриология Ва!апорпогасеае (К во-
просу о гомологиях «цветка» Ва-
lanophora) — Bor. журн., 1967,
92, 6: 745—758.
Герёхин 9. С., Яковлев М. С., Ни-
китичева 3. И. Развитие микро-
спорангия, пыльцевых зерен, се-
мяпочки и зародышевого мешка
y Cynomorium songaricum Rupr.
(Cynomoriaceae).— bor. IKYPH.,
1975, 60, 2: 453—162.
Терзийски Д. П. Цитоэмбриологиче-
ское исследование Опоп1$ со]ит-
пае АП. Бюл. Гл. бот. сада АП
CCCP, 1969, 74 : 22—30.
Герзийски Д. П. Сравнительно ци-
тоэмбриологическое изучение не-
которых представителей РарШо-
пасеае Giss. (-Fabaceae Lindl.):
Канд. дисс. Гл. бот. сад АН СССР,
М., 1970.
Герзийски Д. П., Поддубная-Ар-
нольди В. А. Цитоэмбриологиче-
ское исследование люцерны ро-
допской (Medicago rhodopaea
Уе!.).— Бюл. Гл. бот. сада АН
CCCP, 41973, 89: 20—29.
Тихомиров В. Н. O68 отраженин
пекоторых особенностей эволю-
ции покрытосеменных в фило-
генетической системе.— Бюлл.
МОИП. Отд. биол., 1972, 77 (3):
73—87.
Ткаченко Г. В. Биология цветения
и оплодотворения винограда в
Закарпатье: Автореф дис. ... д-ра
биол. наук. М., 1958 :50 с.
Ткаченко Г. В. Некоторые данпые
о процессе оплодотворения у ви-
нограда.— В кн.: Морфогенез ра-
стений. M.: МГУ, 1961, 2: 371-—
74.
297
Ткаченко П. И. В эмбриологии ро-
буроидных дубов Кавказа. В с6б.:
работ аспирантов Краснодар. Гос.
пед. ин-т, 1969, 4:42—50.
Тодераш Л. Г. Вариологическое и
цитоэмбриологическое исследова-
ние ocok Молдавии.— Автореф.
дис. ... канд. биол. наук. Киши-
нев, 1980 : 28 с.
Тодераш Л. Г., Чеботарь А. А.
Мейоз и развитие пыльцевото
зерна у некоторых видов Сагех
(Сирегасеае).— В кн.: Тез. докл.
УТ делегат. Всесоюз. бот. о-ва,
1978 : 94.
Транковский Д. А. Спермиогенез и
оплодотворение у росянки (Dro-
зега).- Бюл. МОИН. Отд. биол.,
1938, 41, 1: 104—112.
Тульнова Н. Н., Машкин С. И. Ми-
кроспорогенез у Schizandra chi-
пепз15.— Тр. Воронеж. ун-та, 1971,
78 : 102—105.
Турсунов Д. Ю. Особенности цве-
тения и плодоношения Gypsophy-
la paniculata В кн.: Материалы
по филогении и экологии Сред-
ней Азии. Ташкент: ФАН, 1966:
100—104.
Турсунов Д. Ю. Микроспорогенез и
спермиогенез у мыльнянки ле-
карственной.— ДАН УзССР, 1972,
10:52—54.
Турсунов Д. Ю. Биология цветения
и цитоэмбриология видов «мыль-
ного корня»: Автореф. дис. ..
канд. биол. наук. Ташкент: Гос.
ун-т, 1969. 18 с.
Турсунов Д. Ю. Развитие женско-
го гаметофита у мыльнянки ле-
карственной.— ДАН УзССР, 1972,
3:60—62.
Турсунов Jl. FO. Семяпочка и
женский гаметофит у Gypso-
phila paniculata Г. и С. tri-
chotoma Wend—B_ xu.: Mop-
фобиологические особенности ди-
корастущих растений — Узбеки-
стана. Ташкент: ФАН 1974а,
141—115.
Турсунов Д. Ю. Развитие женской
генеративной сферы у колючели-
стника качимовидного Асащюрву-
llum gypsophiloides Rgl_—B ku.:
Морфобиологические особенности
дикорастущих растений Узбеки-
стана. Ташкент: ФАН, 197406,
108—141.
Турсунов Д. Ю. К эмбриологии ро-
да Acanthophyium С. А. М.—
В кн.: Материалы совещания по
298
филогении центросеменных. М.:
1977 : 58—60.
Турсунов Д. Ю., Матюнина Т. Е,
Микроспорогенез и развитие муж-
ского гаметофита у АсапФорву-
lum krascheninnikovii Schischk.
и А. еаНиз Вое.— В кн.: Перспе-
ктивы сапониносных растений и
опыт их культуры. Ташкент’
ФАН, 1975 :78—83.
Турсунов Д. Ю., Матюнина Т. Е.
Развитие женской генеративной
сферы У колючелистника Кра-
шенинникова Асап{ВорвуШа kra-
scheninnikovii a колючелистника
высокого A. elatius Bge.—B Ka.°
Биологические и структурные
особенности полезных растений
Узбекистана. Тазикент;: ФАН,
1977 : 109—113.
Турсунов Д. Ю., Матюнина Т. Е.
Сравнительная цитоэмбриология
видов Acanthophyllum (Caryophy-
Цасеае).--В кн.: Тез. докл.
делегат. Всесоюз. бот.
1978а : 94—55.
Гурсунов Д. Ю., Матюнина Т. Е.
О строении пыльника у некото-
рых представителей рода Асап-
thophyllum C. A. M.— B кн.: Эко-
лого-биологические — особенности
важнейших сырьевых растений в
культуре. Ташкент: 197860 :172—
7
Турсунов Д. Ю., Матюнина Т. Е.
Развитие мужской генеративной
сферы Acanthophyllum — subgla-
brum Schischk. uw Acanthophyllum
leiostegivm Vved—B xuu.: Биоло-
гические и структурные особен-
ности полезных растений Узбеки-
стала. Ташкент: Фан, 1979а : 87—
91.
Турсунов Д. 10, Матюнина Т.Е.,
Савинова Г. Ю. Сравнительно-эм-
бриологическое исследование важ-
нейших представителей сапони-
ноносных растений рода АсапФо-
phyllum С. А. Меу—В кн.: УП
Всесоюз. симпоз. по эмбриол.
раст. Киев: Наукова думка, 1978,
2:57—59.
Турсунов Д. Ю., Савинова Г. Ю.
Эндоспермо-эмбриогенез у колю-
челистника железистого.— В кн.:
Биологические и структурные
особенности полезных растений
Узбекистана. Ташкент: Фан,
19796 : 101—104.
Турсунов Д. Ю., Савинова Г. Ю.
Процесс оплодотворения у Асап-
о-ва,
thophyllum elatius Bge. u A. kra-
scheninniikovii Schischk.—B «ka:
Биологические и структурные
особенности полезных растений
Узбекистана. Ташкент: Фан.
1977 : 105—109.
Лутаюк В. Х., Мурадов А. Т. Раз-
витие тычинок и микроспороге-
нез у восточной хурмы.— Уч. зан.
Азерб. с-х ин-та. Сер. агр., 196%,
4:10—13.
Poukeeuu Г. Л., Koewoea РГ. А.
Формирование женского гамето-
фита и первые этапы эмбриогене-
за бука.— Лесоведение, 1975а, 4:
93—97.
Лышкевич ГР. Ji, Ковшова РГ. А.
Эмбриологическое изучение бу-
ка.— Цитология и генетика, 19756,
9, 4: 361—362.
Гышкевич Г. Л., Ковшова Г. А.
Микроспорогенез и развитие муж-
ского raMeTo@uta Fagus sylvatica
Т.— Бот. журн., 1976, 61, 6: 890—
894.
Умарова Т. В. К эмбриологии крас-
ной махровой штокрозы.— В кн.:
Вопросы ботанических исследо-
ваний в Узбекистане. Ташкент.:
Гос. ун-т 1977 : 78—80.
Усанов М. О темпе процесса опло-
дотворения у хлопчатника сорта
108-Ф.— Узб. биол. журн., 1968.
5: 77—78.
Усанов М. О развитии зародыша
хлопчатника.— ДАН УзССР, 1970,
6: 54—55.
Устинова Е. И. Явление апоспории
у подсолнечника.— ДАН CCCP,
1955, 100, 6: 1163—1166.
Устинова Е. И. Цитоэмбриологиче-
ское исследование зародышевого
мешка и процесса оплодотворе-
ния кукурузы.— Журн. общ.
биол., 1960, 21, 4: 261—269.
Устинова Е. И. Цитоэмбриологиче-
ское исследование зародышевого
мешка процесса оплодотворения
и эмбриогонеза у кукурузы.—
В кн.: Морфология кукурузы. М.:
МГУ, 1962: 241—265.
Устинова Е. И. Цитоэмбриологиче-
ские исследования подсолнечни-
ка и кукурузы: Автореф. дис.
д-ра биол. наук. М., 1967:
36 с.
Устинова Е. И. Апомиксис у под-
солнечника.— В кн.: Апомиксис и
селекция. М.: Наука, 1970: 140—
16.
Устинова Е. И., Дьякова М. И., Ше-
дрина Р. Н. Развитие зародыша и
эндосперма у Маз ]аара Г.
при ограниченном 'опылении.—
Науч. докл. высш. школы. Биол.
науки, 1961, 3: 173—179.
Федорчук В. Ф. Развитие и строе-
ние семяпочек и семян красного
клевера (ТгНоНиш pratense) M.:
ТСХА, 1935, 4:1—38.
Фрумкина А. П., Колесников С. М.
Некоторые особенности оплодо-
творения у пшеницы.— Изв. АН
МолдССР. Сер. биол и хим. наук,
1969, 5: 72—73.
Фурсов В. И., Беспаев С. Б. Неко-
торые цитоэмбриологические дан-
ные колючелистника качимовид-
ного.— Тр. Алма-ат. бот. сада АН
КазССР, 1963, 7:121—124.
Хазова И. И. Биология цветения и
эмбриология некоторых культур-
ных видов сорго в условиях Уз-
бекистана: Автореф. дис. ...
канд. биол. наук. Л.: 1970: 25 с.
Хазова И. И. К эмбриологии куль-
турных видов сорго.— Бот. журн.,
1970, 55, 1—6: 93—102.
Хведбинич О. А. Процесс подейного
заплднения в ячменю (Ногдеию
vulgare Г.).— Укр. бот. журн.,
1969, 26, 5:120—123.
Хведынич 0. А. Процесс оплодотво-
рения и ранние стадии развития
зародыша и эндосперма при от-
даленной гибридизации. Автореф.
дис. канд. биол. наук. Киев, 1972:
24 с.
Хохлов С. С. Апомиксис: классифи-
кация и распространение у по-
крытосеменных растений.— В кн.:
Усп. совр. генет. М.: Наука, 1967,
43—105.
Хохлов С. С. Закономерности рас-
пространения апомиксиса в се-
мействе сложноцветных.— Уч. зап.
Ульяновского roc. пед. ин-та,
1968, 23, 3:57—66.
Хромосомные числа цветковых рас-
тений, Л.: Наука, 1969.
Цингер Н. К истории развития ко-
нопли.— тр. СПб. о-ва естество-
испыт., 1896ба, 27, 4.
Цингер Н. К морфологии сем. ко-
ноплевых.— Тр. СПб. о-ва есте-
ствоисныт., 18966, 27, 7—8.
Циунович 0. Д. Спермиогенез уне-
которых представителей рода
Раеота и их гибридов.— Вестн.
МГУ. Биол. и почвовед., 1974, 2:
114—146.
Цицин Н. РВ. Отдаленная гибриди-
299
зация растений. М.: Наука, 1978.
Цицин Н. В. Многолетняя пшени-
ца. М.: Наука, 1978 :288 с.
Цицин Н. В., Поддубная-Арноль-
ди В. А. Материалы к биологии
цветения многолетних форм пше-
нично-пырейных — гибридов. — В
кн.: Отдаленная гибридизация
в семействе злаковых. М.: 1958:
99—414.
Чабан И. А. Электронно-микроско-
пическое исследование эндоспер-
ма y Reseda lutea L—B xku.: An-
туальные вопросы эмбриологии
покрытосеменных. Л. Наука,
1979 : 58—69.
Чабан И. А. Эмбриогенез и эндо-
спермогенез Reseda lutea Г. и
Chelidonium maius L. no данным
световой и электронной микро-
скопии. Автореф. ... дис. канд.
биол. наук. Л., 1975: 26 с.
Чабан И. А., Яковлев М. С. Эмбри-
ология Reseda lutea L. I. Mera-
спорогенез и развитие зародыше-
вого мешка. Бот. жури, 1974,
59,1 : 24—37.
Чайкина К. В. Цитологоэмбриологи-
ческое изучение клевера открыто-
зевого.— Сообщ. Моск. отд. Всес.
бот. о-ва, 1960, 1:97—102.
Чеботарь А. А. Цитоэмбриологиче-
ское и электронно-микроскопиче-
ское исследование кукурузы: Ав-
тореф. ... дис. д-ра биол. наук.
Кишинев, 1970.: 57 с.
Чеботарь А. А. Эмбриология куку-
рузы. Кишинев: Штиинца, 1972.
Чолалян Д. П. Данные цитоэмбри-
ологических исследований неко-
торых представителей семейства
тыквенных в условиях Арарат-
ской равпипы АрмССР.— Уч. зап.
Ерсв. ун-та, 1965, 97 : 3—9.
Чубрко М. 1. Микроспорогенез и
развитие мужского гаметофита у
Vicia villosa Во .— Наук. зап.
Ужгород. ун-та, 1962, 49:10—
17
Чубрко М. М. Макроспорогенез и
развитие женского гаметофита
клевера ползучего (Trifolium re-
репз Г.) — Укр. бот. журн., 1963,
20, 2: 47—52.
Чубирко М. М. Эмбриологическое
исследование . некоторых кормо-
вых бобовых:. Автореф. ... дисс.
канд. биол. наук. М. 1964a:
22 с.
Чубрко М. М. Эмбриогенез ледвян-
ца рогатого (Lotus corniculatus
300
Г.).— Укр.
3:271—35.
Чубирко М. М. Микроспорогенез и
развитие мужского гаметофита у
KAeBepa Trifolium repens L.— Бот.
журн., 1965, 50, 41: 1578—1584.
Чубирко М. М. Развитие мужского
гаметофита лядвенца рогатого.—
Бюл. МОИП, Отд. биол., 1968, 13,
6: 106—112.
Чубирко М. М. Формирование се-
мян клевера ползучего.— В кн.:
Вопр. охраны природы Карпат.
Ужгород: Карпаты, 1969:58—68.
Чубирко М. М. Формирование пе-
стика, семяпочек и макроспороге-
бот. журн., 19646, 21,
Hes лядвенца poraroro.— Бюл.
МОИП. Отд. биол., 1972, 77, 2:
125—134.
Чубрко М. М. Особмвост1 запиле-
ния Та подвйне заптднения у
лядвенця poratoro (Lotus corni-
сШафиз Г.).— Охорона, вивчения,
та збагачения рослин сыту. Мж-
д. наук 36. 1976, 3:32—42.
Чубирко М. М. Эмбриологическое
necnenopaune Gleditschia triacant-
hus (Caesalpinaceac).— В кн.: Тез.
докл. УГ делегат. Всесоюз. бот.
о-ва, 1978а : 97.
Чубирко М. М. Эмбриологическое
исследование арники горной.—
В кн.: УП Всесоюз. симпоз. по
эмбриол. раст. Киев: Наукова
думка, 19786, 2:62—64.
Шаренкова Е. А. Биология цвете-
ния, опыления и цитоэмбриологи-
ческое исследование некоторых
видов сирени в условиях Прибай-
калья: Автореф. ... дис. канд. би-
ол наук. Иркутск: Гос. Ун-т.
1969, 49 с.
Шевчук JT. П., Копычева В. И.
К омбриологии ОпоЪгусВ1$ отап-
dis Тарзку и O._ chorossanica
Bge.—B кн.: Морфологические
структурные особенности кормо-
вых растений Узбекистана. Таш-
кент: Фан, 1972: 85—94.
Шевчук Л. П., Конычева В. И. Ми-
кроспорангий, микроспорогенез и
развитие мужского гаметофита у
саксаула безлистного.— В кн.:
Морфогенез раст. аридной зоны.
Ташкент: Фан, 1973: 55—60.
Шилова Н. В. Строение зародыша
и проростка в подсемействе Ап-
dromedoideae (Drude) Е. Busch.—
Бот журн., 1962, 47, 3 :344—353.
Пнайдер Т., Сильверс Анте-Пээп.
Полиэмбриония У рапса.— Изв.
АН ЭССР. Биол.,
267—270.
Шоферистова Е. Г. Некоторые во-
просы эмбриологии у лаванды.—
В кн.: Материалы У конф. моло-
дых ученых бот. садов Украины
и Молдавии. Киев: 1970.
Шоферистова Е. Г. Спорогенез и
развитие гаметофитов лаванды.—
Матер. У конф. молодых ученых
бот. садов Украины и Молдавии.
Киев: 1973а.
Шоферистова Е. Г. Особенности
формирования тенеративной сфе-
ры лавандина.— В кн.: Материа-
лы Всесоюз. симпоз. «Половой
процесс и эмбриогеноз растений».
М., 1973.
Шоферистова Е. Г. Цитоэмбриоло-
гическое исследование некоторых
представителей Lavandula Г. в
связи с задачами селекции: Ав-
тореф. ... дис. канд. биол. наук.
Кишинев, 1978.
Шпаков А. М. Особенности цито-
эмбриологии некоторых водных
цветковых растений: Автореф....
дис. канд. биол. наук, Кишинев,
1974 ; 18 с.
Шпилевой Б. Е., Ященко Н. If,
Литвиненко Н. М. Формирование
зародышевого мешка у Тира
turkestanica Rgl.—B xu.: VII Bee-
1974, 23, 3:
союз. совещ по эмбриол. раст.
Киев: Наукова думка, 1978,
63—65.
Штэпа И. С. Морфология пыльцы
рода Сгзшт МШ. и близких ро-
дов трибы Cynareae (Composi-
tae).— Tp. Тбилис. бот. ин-та,
1960, 24 : 81—126.
Шугаева Е. В. Нарушение микро-
спорогенеза у эксперименталь-
ных полиплоидов валерьяны бле-
стящей.— Генетика, 1972, 8, 7:
Щепотьев Ф. J., Герасименко А. Г.
Об апомиксисе у грецкого оре-
ха.—В кн.: Апомиксис и селек-
ция. М.: Наука, 1970: 232—238.
Эрккенова Е. М. Микроспорогенез
y Zizyphus jujuba МШ.— Изв.
ТаджССР. Отд. биол. наук, 1972a,
1:32—35.
Эрккенова Е. М. Развитие генера-
тивной сферы унаби и жестеров
в связи C обильным опадением
цветков и низким качеством де.
MAH унаби: Автореф. ..
канд. наук. Ташкент, 19726: с,
Эрккенова Е. М., Николаева 3. В.,
Тетуева 3. Ш. 'Микроспорогенез и
развитие мужского гаметофита у
леонтицы — эвермана (Leontice
eversmannii Bge). Узб. биол.
журн., 1979, 2:61—64.
Юрцев 'В. Н., Юрцеза НЙ. С. Цито-
эмбриологическое изучение дио-
скореи кавказской (Пюзсогеа сан-
сазха Шрэк У Докл. TCXA,
1973, 195: 187 198.
Юрцез В. Н., Юрцева Н. С. Эмбри-
ологическое изучение крестовни-
ка ромболистного.— Докл. ТСХА,
1974, 204: 213—218.
Юрцева Н. С., Юрцев В. Н. Эмбри-
ологическое изучение диоскореи
ниппонской.— 0б. научн. работ
БНИИ лекарственных растений,
1975, 7: 143—441.
Юрцева Н. С., Юрцев В. Н. Цито-
эмбриологическое изучение П103-
согеа Че{ю14еа.— В кн.: Тез. докл.
VI делегат. Всесоюз. Бот. о-ва.
1978 : 96.
Юрцева Н. С., Юрцев В. Н., Даки-
личев Л. С. Микроспорогенез и
развитие мужского гаметофита у
диоскореи кавказской.— В кн.:
Поиски новых источников биоло-
гически активных веществ. М.:
1970, 1:46—49. ,
Юсупова И. К. Цитоэмбриологиче-
ские исследования некоторых.
представителей семейства норич-
никовых: Автореф. ... дис. канд.
биол. наук. М., 1977.
Якимов /. М., "Литвак A. H., Ba-
лан Ю. Г., Малтабар T. B. Atnac
по эмбриологии винограда. Киши-
нев: Штиинца, 1977, 103 с.
Якобсон „Л. Я. Развитие пыльчи-
ков у рододендронов.— Изв. АН
ИТССР, 1968a, 12 (257): 101:
Якобсон Л. М. Спермиогенез у ВБо-
dodendron mucronulatum Turcz.—
Науч. докл. высш. школы. Биол.
науки, 19686, 6.
Якобсон Л. Я. Образование и раз-
витие спермиев y Rhododendron
luteum Sweet—B кн.: Вопросы
биологии. Рига: Знание, 1969а:
—46.
Якобсон Л. Я. Цитоэмбриологиче-
ские исследования некоторых ви-
дов рододендрона (Ввододепагоп
Т.): Автореф. ... дис. канд. биол.
наук. Рига, 19696 : 16 с.
Якобсон Л. Я. Эмбриологическиь
исследования рододендронов.—
Изв. АН ЛатвССР, 1976, 11: 84—
96.
301
Яковлева Г. В. Оплодотворение у
ремонтантной гвоздики.— В кн.:
УП Всесоюзный симпоз. по эм-
бриол. раст. Киев: Наукова дум-
ка, 1978, 2:69—70.
Яковлев М. С. Морфологические ти-
пы зародыша и филогения зла-
ков. ДАН АрмССР, 1948, 8, 3:
127—134.
Яковлев М. С. Эмбриогенез и его
значение для филогении расте-
ний. М.; Л.: 1960: 40 с.
Яковлев М. С. Основные задачи в
области изучения эмбриогенеза. —
В кн.: Проблемы эмбриологии.
Киев: Наукова думка, 1971:
152—170.
Яковлев М. С. Основные типы эм-
брионального развития высших
растений.— В кн.: Эмбриология
покрытосеменных растений. Ки-
шинев. 1973.
Яковлев М. С. Гаметогенез, зароды-
шевый мешок и пыльцевое зерно
(к проблеме происхождения An-
giospermae).— Бот. журн., 1974,
59, 12: 1721—4727.
Яковлев М. С., Алимова Г. ЕК. Эм-
бриогенез у Arabidopsis thaliana
(L.) Heynh. (Cruciferae).— Бот.
журн., 1976, 61, 1:12—24.
Яковлев М. С., Иоффе М. Д. Осо-
бенности эмбриогенеза рода Рае-
onia Г.— Бот. журн., 1957, 42,
10 : 1494.
Яковлев М. С., Иоффе М. Д. ЦДаль-
нейшее изучение нового типа
эмбриогенеза покрытосеменных.—
Бот. журн., 1961, 46, 10:1402—
4424,
Яковлев М. С., Иоффе М. Д. Эмбри-
ология некоторых представителей
рода Раеопла Г.— В кн.: Мюорфо-
логия цветка и репродуктивный
процесс у покрытосеменных ра-
стений. М.; Л.: Наука, 1965:
140—176.
Яковлев М. С., Жукова Г. Я. По-
крытосеменные растения с зеле-
ным и бесцветным зародышем.
Л.: Наука, 1973 : 100 с.
Яковлев М. С., Соколовская Т. Б.
К эмбриологии Iris pseudacorus
Г.— Матер. сессии УП ЗВеесоюз.
симп. по эмбриол. раст. Киев: На-
укова думка, 1978, 3: 144.
Яковлев М. С., Солнцева М. П. Не-
которые вопросы — морфологии
цветка и эмбриологии ковылей.—
В кн.: Морфология цветка и ре-
продуктивный процесс у покры-
302
тосеменных pacteHuw. M.; JL:
Наука, 1965: 61—73.
Яковлев М. С., Чабан И. А. Ультра-
структурные аспекты эмбриогене-
3a Reseda lutea L.—B xa: Axty-
альные вопросы эмбриологии но-
крытосеменных. Л.: Наука, 1979:
: 96—104.
Abe K. Contributions to the embryolo-
gy of family Orchidaceae. I. Deve-
lopment of the embryo sac in Cym-
bidium goeringii Reich. f.— Sci.
Repts Tohoku Univ. Ser. 4, 1967,
34, 1: 79—82.
Abe K. Contributions to the embryo-
logy of the family Orchidaceae. П.
Development of the embryo sac in
Pogonia japonica Reichb. F.— Sci.
Repts Tohoku Univ. Ser. 4, 1968a,
34, 1: 59—65.
Abe K. Contributions to the embryolo-
gy of the family Orchidaceae. III.
Development of the embryo sac in
Cremastra appen. diculata.— Sci.
Repts Tohoku Univ. Ser. 4, 1968b,
34, 2—4: 119—123.
Abe K. Contributions to the embryo-
logy of the family Orchidaccae (Ble-
lilla striata).— Sci. Repts Téhoku
Univ. Ser. 4, 1971a, 35, 4: 243—218.
Abe K. Contribution to the embryolo-
gy of the family Orchidaceae. De-
velopment of the embryo sac in Ore-
orchis patens.— Sci. Repts Tohoku
Univ. Ser. 4, 1971b, 35, 4: 249—224.
Abe K. Contributions to the embryolo-
gy of the family Orchidaceae. VI.
Development of the embryo sac of
15 species of orchids.— Sci. Repts
Tohoku Univ. Ser. 4, 172a, 36, 3:
135—178.
Abe K. Contributions to the embryo-
logy of the family Orchidaceae. VII.
A comparative study of the orchid
embryo sac.--Sci. Repls Téhoku
Univ. Ser. 4, 1972b, 36, 3: 179—201.
Abe K. A reinvestigation of the de-
velopment of the embryo sac in
Gastrodia elata Blume (Orchida-
ceae).— Ann. Bot. 1976, 40, 4165:
99—102.
Abe K. Development of the embryo
sac in Amitostigma kinoshitae (Ma-
kina) Schltr (Orchidaceae).— Ann.
Bot., 1977, 44, 174: 897—899.
Adamkiewicz E. Studia cyteembriolo-
giczne nad zdziczalym gatunkiem
Chrysanthemum parthenium (L.)
Bernh.— Zesz. nauk Wyzszej szkoly
roen. Olsztynie, 1966, 22, 14: 109—4122.
Adatia R. D., Sharma Y. B., Vijayara-
chavan M. R. Studies in the Gesne-
riaceae. I. Morphology and Embry-
ology of Platystemma_ violoides
Wall.— Bot. Notiser, 1971, 124, 4:
25—38.
Даета F. A review of the herbaceus
species of Polygala in Malesia.—
Blumea, 1966, 14: 253—276.
Afzelius K. Cleistogamy in Pleurotha-
llis procumbens Lindl.— Acta hor-
ti berg., 1966, 20, 5: 313—317.
Ahlstrand L. Embryology of Ursinia
(Compositae).— Bot. Notiser, 1978,
431, 4: 487—496.
Ahlstrand L. Embryology of Arcto-
teae — Arctotinae (Compositae).—
Bot. Notiser, 1979a, 132, 14: 109—116.
Ahlstarand L. Embryology of Arctoti-
deae — Gorteriinae (Compositae).—
Bot. Notiser, 1979b, 132, 3: 381—376.
Ahlistarand L. Embryology of Arctoti-
deae — Gundeliinae (Compositae).—
Bot. Notiser, 1979c, 132, 3: 377—380.
Airy Shaw H. K. Additions in the flo-
ra of Borneo and other Malay Is-
land. The Pentaphragmataceae of
the Oxford University Expedition
of Sarawak.— Bull. Misc. Inf. Kew,
1941: 233—286.
Airy Shaw H. K. Sphenocleaceae —
Flora Malesiana, 1948, Ser. 4, 4:
27—28.
Anantaswamy R. The suspensor haus-
toria of some species Crotolaria.—
Linnean New Ser., 1950, 14, 56.
Anantaswamy R. The endosperm in
some of the Papilionaceae.— Phyto-
morphology, 1951, 1, 8—4: 153—158.
Anantaswamy R. Some observations
on the endosperm in Papiliona-
ceae.— Phytomorphology, 1953, 3, 3:
209—222.
Anantaswamy R. M., Sharma V. K.
Early embryologeny in Saxifraga
L.— Phytomorphology P. Maheshwa-
ri Mem. vol., 1967, 17: 252—255.
Anderson L. C. Embryology of Chry-
sothamnus.— Modrono, 1970, 20, 7:
337—342.
Ansari R. Observations on the Ocur-
rence of polyembryony in two spe-
cies of orchids.— Curr. Sci. (India),
1977, 46, 17: 607.
Arekal G. D. Embryology of Mimulus
ringens.— Bot. Gaz., 1965, 126, 1:
58—66.
Arekal G. D. Embryology of Veronica
serpyllifolia L.— Proc. Indian Acad.
Sci., 1966, 64B, 5: 241—257.
Arekai G. D. Contribution to the em-
bryology of Hypoxis aurea Lour.—
J. Indian Bot. Sci., 1967, 46, 2—3:
192—198.
Arekal G. D., Girijamma 5., Swa-
my S. N. Contribution to the embry-
ology of Lindernia hyssopioides (L.)
Haines.— Proc. Indian Acad. Sci.,
1970, 728, 6: 224—235.
Arekal С. Р., Мавепагап С. В. А991-
tional notes on Farmeria indica Wil-
lis (Podostemaceae).— Proc. Indian
Acad. Sci., 1974, 80: 226—228.
Arekal G. D., Nagendran C. R. Emb-
ryosac of Hydrobryopsis sessilis
(Podostemaceae)— origin, organiza-
tion and significance. Bot. Notiser,
1975a, 128, 3: 332—338.
Arekal G. D., Nagendran C. R. Is the-
re a Podostemum type of embryo
sac in the genus Farmeria? — Cuto-
logia, 1975b, 28, 2: 229—235.
Arekal G. D., Nagendran C. R. The
female gametophyte in two Indian
genera of Fristichoideae (Podostema-
ceae)— a reinvestigation.— Proc. In-
dian Acad. Sci., 1977, 86, 43, 5:
287—294.
Arekal G. D., Ramaswamy S. N. Em-
bryology of Burmania pusilla (Wall
ex Miers) THW and its taxonomic
status.— Beitr. Biol. Pflanz., 1973,
49, 1: 35—45.
Arekal G. D., Rajeshwari S., Swa-
my S. N. Contribution to the embry-
ology of Scoparia dulcis L.— Bot.
Notiser, 1974, 124: 237—248.
Arekal G. D., Swamy S. N. R. The
endosperm organisation in Microcar-
paea R. Br. (Scrophulariaceae).—
Ст. Sci. (India), 1974, 43, 3: 87—
Asplund I, Embryological studies in
the genera Sparganium and Ty-
pha.— Sven. bot. tidskr., 1968, 62, 2:
410—442.
Asplund I. Embryological studies in
the genus Typha.— Sven. bot.
tidskr., 1972, 66, 1: 1—17.
Asplund I. Embryological studies in
the genus Sparganium.— Sven. bot.
tidskr., 1973, 67, 2: 177—200.
Atabekowa A. I. Cytologische Unter-
suchungen an der Kulturerbste Pi-
sum sativum.— Biol. Zbl., 1959, 78,
3: 424—438.
Babis H. General description of a ma-
ture ovule and embryo sac, early
stages of development of the endo-
sperm and of embryo development
in some species of Delphinium L.—
Genet. pol., 1973, 14, 1: 29—36.
Babis H. Early stages of embryo de-
303
velopment in Delphinium panicula-
tum Host.— Acta Soc. bot. pol., 1978,
47, 4: 359—362.
Bajpai P. N. Studies on the vegetaive
growth and development of male and
femal gemetophyte in Emblica of-
ficinalis Gaertn.— Agra Univ. J. Res.
Sci., 1965, 14: 467—187.
Bala B. S. Theaceae: Seminar Compa-
rative Embryology of Angioserms.
New Delhi, 1967: 36.
Bali P. N. A contribution to the emb-
ryology of Oenothera rhombipetala
Nutt. Proc. 56th Indian Sci. Congr.
Bombay, 1969, 4: 33—34.
Ba-Ly-Thi. Embryogénie des_ berbéri-
~ dacées. Développement de l’embuon
*“¢hez le Mahonia aquifolium Nutt.—
Paris. C. r. Acad. Sci., 1976, D282, 5:
445—441.
BannikowaW.P. [Банникова В. П.].
Development features of female ge-
metophyte of Nicotiana. Ann.
Univ. Reims, 1970, 6/7: 73—77.
Bartkowia E., Brukarczyk K. Megas-
porogenesis, development of the em-
bryo sac of endosperm in Juncus
subnodulosus Schrank. and Juncus
tenuis Wild.— Bull. Soc. amis. Sci,
et lettres, Poznan, 1965, D, 6: 79—97.
Baskin J. M., Baskin C. C. Polyem-
bryony in Ruellia humilis Nutt.—
Bull. Torrey Bot. Club, 1971, 98:
‹ 4109—4140.
The bases of Angiosperm phylogeny.—
Ann. Missuri Bot. Garden, 1975, 62,
3: 621—646.
Battaglia E. The embryo sac of Podo-
stemaceae — an interpretation.— Ca-
ryologia, 1971, 24, 1: 403—420.
Вауста Т. В. [Батыгина Г. Б.].
On the possibility of separation of
a new type of embryogenesis in An-
giospermae.— Rev. cytol. ct biol.
véget., 1969, 32, 3/4: 335—341.
Batygina T. В. [Батыгина Т. Б.]|.
Some peculiar features of macro-
sporogenesis and development of
the female gametophyte in wheat.—
Ann. Univ. Reims, 1971, 9, 14: 46—
50.
Bauer S. Cytological and embryologi-
cal studies in the genus Inula L.—
Acta biol. cracov. Ser. bot. 1964
(1965), 7, 2: 147—130.
Ваша S. B. Theaceae.— In: Seminar
on Comparative Embryology Angi-
osperms. New Delhi, 1967a: 36.
Ваша 5. В. Haloragaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 79.
304
Bawa S. B. Embryological studies on
the Haloragidaceae. II Laurember-
gia brevipes.— Proc. Nat. Inst. Sci.
India, 1969a, B35, 4: 273—290.
Bawa S. B. Embryological studies on
the Haloragidaceae. III Myriophyl-
lum DC. Beitr. Biol. Pflanz., 1969b,
49, 3: 447—464.
Bawa S. B. Haloragaceae: Symp. on
Comparative embryology of Angio-
sperins. New Delhi, 1970a, 44: 226—
Bawa S. B. Theaceae: Symp. on Com-
parative Embryology of Angio-
sperms. New Delhi, 1970b, 4:
75—77.
Bednara J. Female gametophyte and
pollen tube of Epilobium palustre
L.— Acta Soc. bot. pol., 1977, 46, 4:
603—615.
Begum M. Studies on the embryology
of Utricularia graminifolia.— Curr.
Sci. (India), 1965, 34, 11: 355—356.
Begum M. Sporogenesis and the deve-
lopment of gametophytes in Erio-
caulon quinquangulare L.— Curr.
Sci. (India), 1966, 35: 262—263.
Begum M. Embryological studies in
Kriocaulon quinquangulare L.—
Proc. Indian Acad. Sci., 1968, 67B,
4: 148—156.
Begum M. A note on the development
of endosperm and embryo in Utri-
сапа graminifolia Vahl.— Curr.
Sei. (India), 1969, 38, 18: 445—446.
Behl H. M., Tiagi B. Seed development
in Fabaceae (Phaseoleae and Vi-
cieae). Embryology of crop plants:
Indo-Soviet Symp. Delhi, 4976: 4.
Behl H. M., Tiagi B. Development of
seed in Rhynchosia, Cajanus and
Atylosia (Subtribe Cajaneae).— In-
dian Acad. Sci., 41977, 86B, 5:
303 —310.
Beltran S. C. Embryology of Isotoma
patraee.— Austral. J. Bol., 1970, 18,
2: 213—224.
Bendre A. M. Studies in the family
Loganiaceae. II. Embryology of
Buddleia and Strychnos.— J. Indian
Bot. Soc., 1975, 54, 3—4: 272—279.
Bentham G., Hooker J. D. Genera
plantarum. London, 1862—1883,
vol, I—III.
Berg R. Y. Ovary, ovule and endo-
sperm of Calochortus amabilis.
With remarks on the taxonomic po-
sition of Calochortus.— Nytt. mag.
bot., 1960, 6: 1489—206.
Berg R. Y. Seed-dispersal of Dendro-
mecon: its ecological, evolutionary
and taxonomic significance.— Amer,
J. Bot., 1966. 53: 64—73.
Berg R. Y. Megagametogenesis and se-
ed development in Dendromecon ri-
gida (Papaveraceae).— Phytomor-
phology, 1967, 17, 1—4: 223—233.
Berg R. Y. Development of ovule, em-
bryo sac and endosperm in Brodiea
(Liliales).— Norw. J. Bot., 1978, 25,
1: 4—7.
Bessey C. E. The phylogenetic taxono-
my of flowering plants.— Ann. Miss.
Bot. Gard., 1915, 2: 109—164.
Bhandari N. N. Studies in the family
Ranunculaceae. VIII. Variation in
the development of the embryo sac
of Anemone vitifolia Buch. Ham. ex
р. C.— Phytomorphology, 1965, 15,
3: 285—291. -
Bhandari N. N. Studies in the family
Ranunculaceae. IX. Embryology of
Adonis Dill. ex Linn.— Phytomor-
phology, 1966, 16, 4: 578—587.
Bhandari N. N. Calycathaceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967a: 20.
Bhandari N. N. Canellaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 46.
Bhandari N. N. Cercidiphyllaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967c: 23.
Bhandari N. N. Degeneriaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967d: 12.
Bhandari N. N. Illiciaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967e: 48.
Bhandari N. N. Lauraceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967f: 24.
Bhandari N. N. Magnoliaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967g: 11.
Bhandari N. N. Monimiaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967h: 19.
Bhandari N. N. Myristicaceae: Semi-
nar comparative Embryology of An-
giosperms. New Delhi, 1967i: 45.
Bhandari N. N. Schisandraceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967j: 17.
Bhandari N. N. Trochodendraceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967k: 22.
Bhandari N. N. Winteraceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 19671: 413.
‘Bhandari N. N. Studies in the family
Ranunculaceae. X. Embryology of
20 В. A. Поддубная-Арнольди
Апетопе L.— Phytomorphology.
1968, 18, 4: 487—497.
Bhandari N. N. Degeneriaceae: Зушр.
on Comparative Embryology of An-
giosperms. New Delhi. 1970a, 41:
Bhandari N. N. Schisandraceae: Symp.
on Comparative Embryology of An-
giosperms. New Delhi. 1970b, 41:
42—44.
Bhandari N. N. Embryology of the
Magnoliales and comments on _ the-
ir relationships.— J. Arnold Arbore-
tum, 1974, 52, 4: 1—32; 285—304.
Bhandari N. N., Asnani S. Studies in
the family Ranunculaceae. XI. Mor-
phology and embryology of Cerato-
cephalus falcatus Pers.— Beitr. Bi-
ol. Pflanz., 1968, 45, 2: 271—290.
Bhandari N. N., Bouman F., Natesh S.
Ovule ontogeny and seed coat.
structure of Scrophularia himalen-
sis Royle.— Bot. Jahrb. Syst. Pflan-
zengesch. und Pflanzengeogr., 1976,
95, 4: 535—548.
Bhandari N. N., Chaube R. K. Studies.
in the Viscaceae: morphology and.
embryology of Karthalsella opuntia
Merr. Seminar on Morphology ana-
tomy and embryology of land plants.
Delhi, 1969: 59—60.
Brhandari N. N., Indira K. Studies in
the Viscaceae. 1V. Embryology of
Eubrachion (Hook et Arn.) Engl.—
Bot. Notiser, 1969, 122, 2: 183—203.
Bhandari N. N., Nanda K. Studies in
Viscaceae. I. Morphology and em-
bryology of the Indian dwarf mist-
letoe.— Arceuthobium minutissi-
mum.— Phytomorphology, 1968a, 18,
4: 435—450.
Bhandari N. N., Nanda K. Studies in
Viscaceae. II. A reinvestigation of
the female gametophyte — Arceu-
thobium douglasii— Amer. J. Bot.,.
1968Ъ, 55: 1028—1080.
Bhandari N. N., Ram K., Natesh S. On-
togeny, Cytology and Histochemist-
ry of anther tapetum in relation to
pollen development in Nigella da-
mascena.— Phytomorphology, 1976,
26, 1: 46—59.
Bhandari N. N., Venkataraman R.
Embryology of Drimys winteri.— J.
Arnold Arboretum, 1968, 49, 4:
509—524.
Bhandari N. N., Vijayaraghavan M. R.
Studies in the family Ranuncula-
ceae. XII. Embryology of Aquile-
gia vulgaris.— Beitr. Biol. Pflanz.,
1970, 46: 337—354.
305
Bhatnagar S. P. Morphological and
embryological studies in the family
Santalaceae. VII. Exocarpus bidwil-
lii Hook F.— Proc. Inst. Sci. India,
1965, ЗАВ, 1—2: 34—44.
Bhatnagar S. P. Microsporogenesis
and male gametophyte in Santalum
L.— Proc. Nat. Inst. Sci. India, 1966,
32B: 47—52.
Bhatnagar S. P. Aristolochiaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967a: 33.
Bhatnagar S. P. Santalaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 7.
Bhatnagar S. P. Santalaceae. Symp.
on Comparative Embryology of An-
giosperms. New Delhi. 1970, 41:
15—18.
Bhatnagar S. P., Chandra S. Endo-
sperm in Pseittacanthus.— Curr, Sci.
(India), 1968. 37, 24: 704—706.
Bhatnagar S. P., Gupta M. Morpholo-
gy and Embryology of Mimusops
elegans L.— Bot. Notiser, 1970, 123,
4: 455—473.
Bhatnagar S. P., Kallarackal J. Mor-
phology and embryology of a plie-
tesial species (Phlebophyllum Kun-
thianum).— Phytomorphology, 1973,
23, 3/4: 205—211.
Bhatnagar A. K., Kapil R. N. Bischo-
fia javanica—its relationship
with §Euphorbiaceae.— Phytomor-
‘ phology, 1973, 23, 3/4: 264—267.
Bhatnagar A. K., Meenakshi G. Affi-
nities of Daphniphyllum — palyno-
logical approch.— Phytomorpholo-
gy, 1977, 24, 14: 92—97.
Bhatnagar S. P., Puri S. Marphology
and embryology of Justica betonica
Linn.— Osterr. bot. Z., 1970, 118,
55—74.
Bhatnagar S P., Sabharwal G. Vema-
le gametophyte and endosperm of
Iodina rhombifolia Hook, Arn.—
Phytomorphology, 1966, 16, 4: 588—
594.
Bhatnagar S. P., Sabharwal G. Mor-
phology and embryology of Iodina
rhombifolia Hook, Arn.— Beitr. Bi-
ol. Pflanz., 1968, 45, 3: 465—479.
Bhojwani S. S. Morphogenetic behavi-
our of mature endosperm of Croton
bonplandianum Baill. in culture.—
Phytomorphology, 1966, 16: 349—
353.
Bhojwani S. S. Morphogenesis of em-
bryo in a parasitic angiosperm Exo-
carpus cupressiformis.— Curr. Sci.,
1969, 38: 6—8.
306
Bidyut Ganguly. Some observations on
the flower of Freycinetia funicula-
re.— Sci. and Cult. 1976, 42, 5:
270—272.
Bijok K. Cytoembryological studies ox
species Chrysanthemum subcorym-
bosum (Schur) Beck. and Chrysan-
themum corymbosum L.— Zesz. na-
uk. Wyzszej szkoly roln. Olsztynie,
1964, 18, 1: 124—140.
Bijok K. Studia cyto-embriologiczne
nad gatunkiem Crambe maritima
L.— Zesz. nauk. Wyzszej szkoly
гот. Olsztynie, 1966, 22, 14: 123—4136.
Bijok K., Pawlak T., Krenska B. Emb-
ryological studies on Anthemis tin-
ctoria L.— Acta Soc. bot. pol., 1977,
46, 4: 647—657.
Biology and chemistry of the Compo-
sitae I. L. etec., 1977.
Biradar N. V. Studies on Palms: em-
bryology of Phoenix pusilla Gaertn.,
P, acaulis Buch. and P. reclinata
Jacq.— Proc. Indian Acad. Sci., 1968a,
67B, 4: 165—173.
Biradar N. V., Mahabale T. S. Studies
on palms: embryology of Phoenix
robusta Hook.— Proc. Indian Akad.
Sci., 1968b, 68B, 1: 1—8.
Biswas I. Asclepiadaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 91.
Blamish K. I. A Pasific coast saxifra-
ge with 10 pairs of chromosomes:
meiosis, development of female ga-
metophyte and seed production.—
Canad, J. Bot., 1967b, 45: 1797—1801.
Biamisch K. f., Lin-shu-Chang. Ferti-
lization and seed development in Sa-
xifraga integrifolia Hook.— Cahad.
J. Bot., 4965, 43, 7: 861—865.
Bobek J., Kovdcik K. A. On the via-
bility of pollen in sunflower (He-
lianthus annuus L.).— Sci. Agric.
bohemosl., 1974, 6, 4: 7—12. .
Boer R. de, Bouman F, Integumetary
studies in the Polycarpicae. III.
Drymis winteri (Winteraceae).—
Acta bot. neerl., 1974, 23, 4: 149—27.
Boesewinkel F. D., Bouman F. Deve-
lopment of ovule and testa in Ruta-
ceae. II. The unitegmic and pachy-
chalazae seed of Glycosmis C. F. ar-
borea (Rotb.) D. C.— Acta bot. ne-
erl., 1978, 27, 1: 69—78.
Bouillot J. Ver investigastion de fe-
male gametogenesis chez Juglans re-
gia L.—C. r. Acad. sci., 1968, D266,
12: 1397—1403.
Bouillot J. Individualisation progres-
sive des cellules du sac embryon-
naire chez le Juglans regia.— Rev.
cytol. et biol. végét., 1969, 32, 3—4:
203—208.
Bor J., Kapil R. N. Euphorbia genicu-
lata — ovule to seed.— Acta bot. ne-
er., 1975, 24, 3—4.
Bor J., Kapil R. N. Anatropy and on-
togeny of the bitegmic ovule in
Chrozophora А. Н. 1. Jussiew
(Euphorbiaceae).— Acta bot. neerl.,
4976, 25, 6: 385—400.
Boys J. W. Development of the emb-
ryo sac of Plumbagella micrantha.—
Amer. J. Bot., 1939, 26: 539—547.
Bradley M. V., Crane J. C. Supernu-
merary ovule development and par-
thenocarpy in Ficus carica L.— Phy-
tomorphology, 1965, 15, 4: 85—92.
Brewbaker J. L: The distribution and
phylogenetic significance of binuc-
leate and trinucleate pollen grains
in the angiosperms.—Amer. J. Bot.,
1967, 54, 9: 1069—1083.
Brouland M. Recherches cytologiques
sur la fécondation chez quelques
Ranunculacées: Caltha palustris L.,
Anemone nemorosa L., Ranunculus
acris L., Helleborus foetidus L.—
Rev. cytol. et biol. végét., 1969, 32,
3—4: 309—345.
Brown В. С., Мовепзеп Н. Г. Late
ovule and early embryo develop-
ment in Quercus gambelii.— Amer.
J. Bot., 4972, 59, 3: 341—316.
Bugnicourt M. Polyembryonie chez
VHypericum tetrapterum Fries (Hy-
pericum acutum Moench.).— Bull.
Soc. bot. France, 1969, 116: 495—504.
Bugnicourt M. Embryogénie des hy-
pericacées. Development de l’embry-
on chez lHypericum tetrapterum
Fries.— Bull. Soc. bot. France, 1974,
(1972), 118, 5—6: 349—334.
Buss P. A., Ir. Galen Donald F., Ler-
sten Nels R. Pollen and tapetum in
Desmodium glutinosum and D. illi-
noense (Papilionoideae, Legumino-
sae).— Amer. J. Bot., 1969, 56, 40:
1203—1208.
Carniel K. Uber die Embryobildung in
der Gattung Paeonia.— Osterr. bot.
Z., 1967, 114, 1: 4—19.
Carniel K. Beitrage zur Entwicklungs-
geschichte des Antherentapetum in
der Gattung Oxalis. II. Oxalis ace-
tosella.— Osterr. bot. Z., 1969, 147,
2: 201—204.
Carniel K. Zur kenntnis der Ubisch-
Когрег von Heleocharis palustris.—
Osterr. bot. Z., 41974, £19, 4—5:
496—502.
20*
Carolin R. C. Seeds and fruits of the
Goodeniaceae.— Proc. Linn. Soc. N.
S. W., 1966, 91: 58—83.
Cass D. D. Occurrence and develop-
ment of a filiform apparatus in the
egg of Plumbago capensis.— Amer.
J. Bot., 1972, 59: 279—283.
Cass D. D., Jensen W. A. Fertilizati-
on in Barley.— Amer. J. Bot., 1970,
17: 233—239.
Cave M. S. The female gametophytes
of Lapageria rosea and Philesia ma-
gellanica.— Gayana Bot., 41966, 15:
25—34.
Cave M. S. The megagametophyte ot
Androcymbium (Liliaceae).— Phy-
fomorphology, 1967, 17, 1—4: 233—
Cave M. S. Female gametophytes of
Chlorogalum and Schenolirion (Ha-
stingsia).— Phytomorphology, 1974,
24, 1—2: 56—60.
Cave М. 5. Embryological studies in
Stypandra (Liliaceae).— Phytomor-
phology, 1975, 25, 1: 95—99.
Cebrat J. Embryologiczne Badania nad
diploidaenym Zytem (Secale cerea-
le).— Hodowla Roslin Aklimatyza-
cja i Nasiennictwo, 1979, 23, 5:
293—310.
Cesca G. Cytological and embryologi-
cal studies in the genus Euphorbia
epithymoides L.— Proc. Nat. Inst.
Sci. India, 1969, B35, 2: 139—4152.
Chandra S. Post-pollination develop-
ment of wheat caryopsis. Embryolo-
gy of crop plants: Indo-Soviet Symp.
Delhi, 1976: 46—47.
Chandra §. Pollen morphological cont-
ribution to the segregation of a new
genus Gymnostoma from Casuarina-
ceae: Proc. 56th Indian Sci. Congr.
Bombay, 1969, 3: 342—343.
Chandra S., Bhainagar S. P. Seed de-
velopment of Abelmoschus esculen-
tus (Malvaceae).— Plant Syst. and
Evol, 1975, 423, 4: 255—262.
Chandra S., Singh R. P., Pandey A. K.
Embryological studies in Места
parasitica Rich. var. chitrakutensis
Rau. (Scrophulariaceae).— Flora.
1980, 169, 1: 111—120.
Chandravadana P. A note on the em-
bryology of Gliricidia sepium.—
Current Sci., 1965, 34: 185—186.
Chardard R. Aspects de quelques or-
chidacées.— Rev. Gytol. et biol. vé-
gét., 1969, 32, 1—2: 67—86.
Chatterji A. K., Pillai G. K. Apomixis
in Pennisetum pedicellatum Trin.—
Sei. and Cult., 4970, 36, 12: 667—669.
307
Chaudhuri S. К., Маше N. Pollen
morphological studies of the order
Malvales.— Bull. bot. Soc. Negalor,
1965, 19: 32—36.
Chandhury J. P., Narayana H. S. Stu-
dies on the development and struc-
ture of seed in the Caryophyllaceae.
I. Spergula arvensis L.—J. Indian
Bot. Soc., 1978, 57, 1: 46—52.
Chaturvedi S. N., Rathare R. P. Mor-
phological studies in Barbarea vul-
garis R. Br.—J. Indian Bot. Soc.,
1970, 49, 4—4: 58—63.
Chean C. H., Stone B. C. Embryo зас
and microsporangium development
in Pandanus (Pandanaceae).— Phy-
tomorphology, 1975, 25, 2: 228—238.
Cheesman E. E. Fertilization and emb-
ryology in Theobroma cacao L.—
Ann. Bot. London, 1927, 41: 107—
126.
Chennaveeraiah M. S., Раш В. М.
Apomixis in Blumea.— Phytomor-
phology, 1971, 241, 1: 71—76.
Chikkannaiah P. S. A contribution to
the life history of Floscopa зсап-
dens Lour.— J. Karnatak Univ. Sci.,
1964—1965, 9—10: 140—122.
Chikkannaiah P. S. Studies in Tinan-
На fugax Scheidw.— J. Karnatak
Univ. Sci, 1964—4965, 9—10:
123—137.
Chikkannaiah P. S. Post-fertilization
studies in Aneilena paniculatum
Wall.— Proc. Indian Acad. Sci,
1971, 73B, 2: 96—104.
Chikkannaiah P. S. Development of
endosperm, embryo and seed coat
in Amischophacellus axillaris L.—
Proc. Indian Acad. Sci., 1973, 77B,
4: 166—175.
Chikkannaiah P. S., Нетатаа4 В.
A note on the megasporogenesis and
endosperm of some Commelina-
ceae.— Curr. Sci. (India), 1979, 48,
8: 363.
Chikkannaiah P. S., Mahalingap-
ра М. 5. Antipodal cells in some
members of Gramineae.— Curr. Sci.
(Indian), 1975, 44, 4: 22—23.
Chikkannaiah P. 5., Mahalingap-
pa M. S. Embryological studies in
Eleusine. Embryology of crop
plants: Indo-Soviet Symp. Delhi,
1976: 7.
Chopra R. N., Agarwal G. S. The fe-
male gametophyte of Benincasa ce-
rifera Savi.— Bot. Notiser, 1960, 113,
2: 192—202.
Chopra R. N., Basu B. Female gameto-
308
phyte and endosperm of some mem-
bers of the Cucurbitaceae.— Phyto-
morphology, 1965, 15, 3: 248—223.
Chopra R. N., Harjinder K. Some as-
pects. of the embryology of Cor-
nus.— Phytomorphology, 41965, 15,
4: 353—859.
Chopra R. N., Kaur H. Embryology of
Bixa orellana.— Phytomorphology,
1965, 152: 211—244.
Chopra R. N., Mukkada A. J. Gameto-
genesis and pseudo-embryo sac in
Indotristicha ramosissima (Wight)
von Royen.—Phytomorphology, 1966,
16, 2: 182—188.
Chopra R. N., Seth P. N. Some aspects
of endosperm development in Cucur-
bitaceae.— Phytomorphology, 1977,
27, 4: 112—115.
Chopra 5. Morphological studies of
Callendula officinalis L.— Plant
science, 1969a, 4: 126—129.
Chopra S. Development of the female
gametophyte in niger (Guizotia ab-
yssinica Cass). Curr. Sci. (India),
1969b, 38, 6: 392—393.
Chopra S. Development of female ga-
metophyte in Sopium § setiferum
Roxb.— Curr. Sci. (India). 1970, 39:
17—18.
Chopra S., Singh R. P. Structure and
development of seed in Phyllantuus
urinaria L.— J. India Bot. Soc., 1969,
48, 83—4: 242—216.
Chowdhuri N. The antipodals in Ama-
pantiaceae.— Sci. and Cult. 41965,
20: 611.
Христов М., Крушева Р. Цитоембрио-
логични проучвания на три вида
от рода Кезбаса Т..— Генетика и се-
лекция (НРБ), 1968, 1, 3: 233—250.
Христов М., Москова Р. Морфологич-
ни и цитоембриологични проучва-
Ish върху формироването и раз-
витието па съцветисто при Расфу-
lis glomerata L.— Hayau. rp. Brrcm.
селскостоп. ин-т Г. Димитров. Аг-
рон. dak. Сер. растениеводство,
1966, 18: 137—147.
Христов М., Москова Р. Апомиксисът
и полиэмбрионията при ВофосвВ-
loa ischaemum L. (Andropogon
ischemum ТГ. Кепо.).— Генетика и
селекция (НРБ), 1972, 5, 1:71—
86.
Христов М., Терзийски Д. Върху
произхода на близничните расте-
ния при някои апомикто рамзножа-
ващи се видове от рода Роа L.
София: Фитология, Българска
Академия на науките, 1979; 13: 3—
14.
Cocuccit A. E. Embriologia de orquide-
as. La megaspora de Epidendrum
scutella.— Kurtziana, 1969, 5: 7—214.
Coceuci A. E. Some suggestions on
the evolution of gametophytes of
higher plants.— Phytomorphology,
1973, 23, 1—2: 109—124.
Cocucci A. E. Estudies en el genero
Prosopanche (Hydnoraceae). 3. Em-
priologia.— Kurtziana, 1976, 9: 19—
Cocucci A., Jensen W. Orchid embry-
ology: megagametophyte of Epiden-
drum scutella following fertilizati-
on.— Amer. J. Bot., 1969, 56, 6:
629—640.
Сосйтап Н. Г. ' Pollen-tube growth
through the style of pimiento pep-
рег (Capsicum frutescens L.).—
Hort. Sci., 1970, 4: 50.
Copeland H. F. Morphology and emb-
ryology of Euonymus japonica.—
Phytomorphology, 1966, 16, 3: 326—
334.
Cog Cl. La mégasporogenése chez le
Saxifraga cochlearis Reichb.— Rev.
cytol. et biol. végét., 1967, 30, 3:
207—243. `
Corner E. J. H. The seeds of Dicoty-
ledons. Cambridge, 1976, 1.
Corsi G. The suspensor of Erica sati-
va Miller during embryogenesis in
vitro.— Giorn. bot. ital., 1972, 106,
1: 44—54.
Corsi G., Garberi F. Aspetti citotasso-
nomici ed embryologici del contin-
gente endemico apueno. II. Globu-
laria incanescens Viv.— Atti Soc.
tosc. sci. natur., 1971, 78: 46—54.
Crété P. Embryogenie des Moraceae.
Developpement de l’embryon chez
VDimerocarpus brenieri Gagner.—
С. г. Acad. sci., 1964, 259D, 22:
4102—4105.
Crété P. Embryogenie des Polemonia-
ceac. Developpement lembryon
chez Navarretia squarrosa Hoou.—
С. г. Acad. sci., 1965, 260D, 21:
5617—5619.
Crété P., Quignard J. L., Mestre J. C.
Embryogenie des Ulmacées. Deve-
loppement de l’embryon chez ГОГ
mus campestris L.— C. г. Acad. sci.,
1966, 2620, 9: 986—988.
Cronguist A. Outline of a new system
of families and orders of flowerling
plants.— Bull. Jar. bot. Brux., 1957,
27: 13—40.
Cronguist A. The evolution and classi-
fication of flowering plants. L::
Edinburgh, 1968.
Czapik R. Embryological studies in the
genus Potentilla. 2. Potentilla are-
naria.— Acta Biol. Cracov., 1962, 5:
29—42.
Czapik R. Apomiksja w systemach
klasyfikacyjnych rozmnazania An-
giospermae.— Wiad. bot., 1977, 21,
4: 239—248.
Dahigren K. V. Zytologische und em-
bryologische Studien ther die Rei-
hen Primulales and Plumbaginales.—
Kgl. Sven. vetenskap. handl, 1916,
568: 1—80.
Dahigren R. A system of classificati-
on of the angiosperms to be used to
demonstrate the distribution of
characters.— Bot, Notiser, 1975a, 428,
1: 119—447.
Dahlgren R. Current topies. The dist-
ribution of characters within an an-
giosperms. I. Some embryological
characters.— Bot. Notiser, 1975b, 123,
4: 181—197.
Dahlgren R., Rao V. S. A study of
the family Geissolomataceae.— Bot.
Notiser, 1969, 122, 2: 207—232.
Dang-van-Liem IL. Embryogénie de
euphorbiacées. Developpement de
Yembryon chez Mercurialis peren-
nis L—C, r. Acad. зс1., 1960, 2500,
19: 3247—3219.
Dang-van-Liem L. Embryogénie des
euphorbiacées. Developpement de
VYembryon chez le Phyllanthus gran-
difolius L—C. r. Acad. sci., 1961,
252D, 3: 443—445.
Даскалова Ц. Образоване на семе и
плод при Зезей гит У. К.—
Фитология (НРБ), 1977, 7: 54—74.
Das L. D. A note on twin megaspores
and embryo sac in Saccharum.—
Curr. Sci. (India), 1969, 38: 248—
249.
Das L. D., Banerji M. L. Pollen mor-
phology of a new species of Cuscu-
ta.— Curr. Sci., 1966, 35: 405—106.
Darlingion C. D., Wylie AP. Chro-
mosome atlas of flowering plants
London. George Allen and Unwin
Ltd., 1955: 520 p.
Dathan A. S. Embryology and seed de-
velopment of Dactyliandra welwi-
tschii Hook. F.— Proc. Indian Acad.
Sci, 1974а, В80, 5: 207—215.
Dathan A. S. Embryology of Meloth-
ria leucocarpa (Bl) Congn.— Sci.
and Cult., 4974b, 40, 3: 119—122
Dathan A. S., Singh D. Female game-
tophyte and seed of Carica canda-
309
marensis Hook f.—In: Proc. 56th
Indian Sci. Congr. Bombay, 1969,
3 (4): 394—395.
Dathan A. S., Singh D. Development
of female gametophyte and seed of
Tacsonia mollissima HBK.— In:
Proc. 56th Indian Sci. Congr. Bom-
bay, 1969, 3 (4): 389—390.
Dathan A. S., Singh D. Development
and structure of female gametophy-
te and seed of Bergia odorata Ed-
gew.— In: Proc. 57th Indian Sci.
Congr. Bombay, 1970, 3 (4): 254—
205.
Dathan A. S., Singh D. Morphology
and embryology of Marah macrocar-
pa Greene.— Proc. Indian Acad. Sci.,
1971a, 73B, 5: 240—250.
Daihan A. S., Singh D. Embryology
and seed development in Bergia
L.— J. Indian Bot. Soc., 1971b, 50:
362—370.
Dathan A. S., Singh D. Female game-
tophyte and endosperm of Hydno-
carpus laurifolia (Dennst) Sleu-
mer.— Curr. Sci. (India), 1972a, 414,
17: 647—649.
Dathan A. S., Singh D. Development
of embryo sac and seed in Bixa L.
and Cochlospermum Kunth.— J. In-
dian Bot. Soc. 4972b, 54, 3—4,
254—266.
Dathan A. S., Singh D. Structure and
development of ovule and seed in
Flacourtia indica (Burn. F.) Mer-
rill.— Proc. Indian Nat. Acad., 1973a,
B39, 2: 172—179.
Dathan A. S., Singh D. Development
and structure of seed in Tacsonia
Juss. and Passiflora L.— Proc. Indi-
an Acad. Sci., 1973b, 77B, 1: 5—48.
Dathan A. S. R., Singh D. Embryolo-
gical studies papaya and mountain
papaya. Embryology of crop
plants.— Indo-Soviet. Symp., Delhi,
1976: 8—9.
Dathan A. S. R., Singh D. Embryology
and seed structure of cultivated Cu-
curbits. Embriology of crop plants:
Indo-Soviet. Symp. Delhi, 1976: 9.
Datta R. M., Choudhary P. C. Compa-
rative studies on the rate of pollen
tube growth of some species of Cro-
talaria and Sosbania.— Biologia,
Bratislawa, 1967, 22: 34—38.
Datta R. M.. Mukherjee P. Comparati-
ve studies on the rate of the pollen
tube growth of two wild jute spe-
cies (Corchorus urticaefolium W.,
A. and C. siamensis Craib).— Beitr.
Biol. Pflanz., 1969, 46: 155—168.
310
Davis G. L. Embryological studies in
the Compositae. IV. Sporogenesis, ga-
metogenesis and embryology in
Brachycome ciliaris (Labell) Less.—
Austral. J. Bot., 1964, 12, 2: 1442—15]
Davis G. L. Systematic embryology of
the angiosperms. N. Y. ete., 1966:
505 р.
Davis G. L. Apomixis in the Composi-
tae.— Phytomorphology, 4967, 17.
1—4: 270—277.
Davis G. L. Apomixis and abnormal
anther development in Calotis lap-
pulacea Benth. (Compositae).—
Austral J. Bot. 1968a, 16, 4: 1—47.
Davis G. L. Floral morphology and
the development of Gametophytes
in Eucalyptus melliodora A. Cunn.—
Austral J. Bot., 1968b, 16, 4: 19—35.
Davis G. L. Floral morphology and the
development of the gametophytes in
Kucalyptus stellata Sieb.— Austral
J. Bot., 1969, 17, 2: 177—190.
Deshpande P. K. Morphology of the
endosperm in Caesulia axillaris
Roxb.- Curr. Sei. (India). 1960, 29,
2: 56—57.
Deshpande P. K. Compositae: Seminar
on comparative embryology of an-
giosperm. New Delhi, 1967: 110.
Deshpande P. K. Compositae. Sympo-
sium on comparative embryology of
angiosperms. New Delhi. 1970, 4:
325—333.
Deshpande P. K. Development of em-
bryo and endosperm in Eragrostis
uniolodes (Poaceae).— Plant Syst.
апа Еуо]., 1976, 125, 4: 258—259.
Deshpande P. K., Bhasin R. K. Embry-
ological studies in Phaseolus aconi-
tifolins Jacq. Obs.— Bot. Gaz., 1974,
135, 2: 104—113
Deshpande P. K, Raju P. S. G. Deve-
Jopment of Caryopsis Triticum. Em-
bryology of crops plants: Indo-Sovi-
et. Symp. Delhi, 1976: 10.
Deshpande P. K., Untawale A. G. Stu-
dies in the Leguminosae. III. Male
and female gametophytes of Eryth-
rina suberosa Roxb: Proc. 56th In-
dian Sci. Congr. Bombay, 1969, 3:
155—159.
Deshpande P. K., Untawale A. G. De-
velopment of seed and fruit in In-
digofera enneaphylla L.— Bot. Gaz..,
1971, 432: 96—102.
Dexheimer J. Une nouvelle interpreta-
tion du sac embryonnaire des an-
giosperms.— Bull. Acad. et Soc. lor-
rain. sci., 1976, 15, 2: 79—87.
Dhakre J. S. Morphological studies in
the order Umbelliflorae — Umbel-
liferac. II. Anethum graveolens L.—
J. Indian Bot. Soc., 1968, 47, 1—2:
13—22.
Dharamadhaj P., Prakash N. Develop-
ment of the anther and ovule in
Capsicum L.— Austral Bot. 1978,
26: 433—440.
Dieter R. Sobre a reproducao em. Ma-
xillaria brasiliensis Brieg. et Yllg.
e M. cleistogama Brieg. et Yllg.
(Orchidaceae).— Rev. brasil. biol.,
1977, 37, 2: 267—279.
Dnyansagar V. R. Leguminosae: Se-
minar comparative embryology of
angiosperms. New Delhi, 1967: 41.
Dnyansagar V. R. Leguminosae: Sym- —
posium on comparative embryology
of angiosperms. New Delhi. 1970,
41: 93—108.
Dnyansagar V- R., Gaoli H. P. Embry-
ology of Thespesia lampas Dalz. and
Gibsson and T. macrophylla Blu-
me.— J. Biol. Sci. 1969, 12, 1:
33—43.
Dnyansagar V. R., Malkhede S. R. De-
velopment of the seed of Trianthe-
ma portulacastrum L.— Proc. Indi-
an Acad. Sci., 1963, B57, 6: 343—355.
Dnyansagar V. R., Tiwari H. N. Deve-
lopment of seed of Euphorbia ro-
thiana Spreng.— J. Biol. Sci., 1966,
9, 1—2: 1—7.
Dundjska L. Embryo sac development
of Carex praecox Schreib.— Acta
Soc. bot pol., 1965, 34, 2: 264—272.
Duncan E. J. Ovule and embryo onto-
genesis in Bombacopsis glabra. A.
Robyns.— Ann. Bot., 1970, 34, 136:
671—683.
. Duncan E. J, Todd A. W. Structure
of the mature embryo of Theobro-
ma cacao L.— Ann. Bot., 1972, 36,
148: 939—945.
Рин В. 5. Amaryllidaceae: Seminar
comparative embryology of angios-
perms. New Delhi, 1967a: 121.
Рин В. 5. Cyanastraceac: Seminar
comparative embryology of angio-
sperms. New Delhi, 1967b: 120.
Dutt B. S. Haemodoraceae: Seminar
comparative embryology of angio-
sperms. New Delhi, 1967c: 119.
Рин В. 5. НурохЯасеае: Seminar
comparative embryology of angio-
sperms. New Delhi, 1967d: 122.
Dutt B. S. Velloziaceae: Seminar com-
parative embryology of angiosperms.
New Delhi, 1967c, 3: 123.
Dutt B. S. Amaryllidaceae: Symposi-
um on comparative embryology of
angiosperms. New Delhi, 1970a, 41:
365—367.
Dutt B. S. Cyanastraceae: Symposium
on comparative embryology of an-
giosperms. New Delhi, 1970b, 41:
362—364.
Dutt B. S. Haemodoraceae: Symposi-
um on comparative embryology of
angiosperms. New Delhi, 1970c, 41:
358—361.
Dutt B. S. Hypoxidaceae: Symposium
on comparative embryology of an-
giosperms. New Delhi, 1970d, 41:
368—372.
Dutt B. S. M. Velloziaceae: Symposi-
um on comparative embryology of
angiosperms. New Delhi, 1970e, 41:
373—314.
Dutt B. S. Embryology of Pancrati-
um longiflorum.— Phytomorphology,
19701, 20, 1: 1—5.
Dutt R. S. Embryology in Gyrocarpus
americanus Jacq.— Curr. Sci. (In-
dia), 1978a, 47, 17: 636.
Dutt B. S. Embryo development in
Moringa concanensis Nimmo.—
Curr. Sci. (India), 1978b, 47, 18: 6983.
Dutt B. S. Anther in Moringa conca-
nensis Nimmo.— Curr. Sci. (India),
1978c, 47, 16: 589.
Dutt B. S. Fruit and seed of Gyrocar-
pus amerikanensis Jacq.— Curr. Sci.
(India), 1979a, 48, 13: 594—596.
Dutt B. S. Ovule and seed of Moringa
concanensis Nimmo.—Curr. Sci.
(India), 1979, 48, 14: 652—654.,
Dutt B. S., Rao P. S. Embryo sac de-
velopment in Scilla peruviana L.—
Curr. Sci. (India), 1975, 44, 3: 94.
Eames A. J. Morphology of the angio-
sperms. New York; Toronto; Lon-
don, 1961.
Eaton G. W., Jamont A. Megagameto-
genése of Vaccinium corymbosum
L.— Canad. J. Bot. 1966, 44, 5:
112—714.
Ekanthappa K. Q., Arekal G. D. A con-
tribution to the embryology of Cir-
rhopetalum fimbriatum Lindle.— In-
dian Acad. Sci., 1977, 86B, 4: 211—
216.
Eliasson U. Studies in Galapagos
plants. X. The genus Leucocarpus.—
Sven. bot. tidskr., 1971, 65: 246—277.
Eliasson U. Studies in Galapagos
plants. XI Embryology of Macraea
laricifolia Hook. f. (Compositae).—
Sven. bot. tidskr., 1972, 66, 1: 43—47.
Eliasson U. Studies in Galapagos
plants. XIV. The genus Scalesia
Arn.— Opera bot., 1974, 36: 4—117.
311
Emery W. H., Guy M. N. Reproducti-
on and embryo development in Te-
xas Wildrice (Zizania texana
Hitche.).— Bull. Torrey Bot. Club.,
1979, 106, 1: 29—34.
Emil Di Fulvio Teresa. Embryologia
de Nolana paradoxa (Nolanaceae).—
Kurtziana, 1969, 5: 39—54.
Endlichers S. Enchiridion botanicum.
Lipsiae, Vienne, 1844.
Endress P. Gesichtpunkte zur syste-
matischen Stellung der Fuptelaceae
(Magnoliales).— Ber. Schweiz. Bot.
Ges., 1969, 79: 229—278.
Engler A., Prantl K. Die naturliche
Pflanzenfamilien. Leipzig, 1934.
Engler A., Prantl K. Die naturlichen
Pflanzenfamilien. Leipzig, 1895—
1915: 20 p.
Erdtman G. Pollen morphology and
plant taxonomy Angiosperms.— In:
Chronica Bot. Waltham. Mass., 1952.
Erdelska O. Pre-fertilization develop-
ment of ovule of Jasione montana
L.— Phytomorphology, 1975, 25, 4:
16—81.
Езап А., 500$: В. К. Адуепйуе ешЪ-
ryogenesis in Citrus and its relati-
on to polination and fertilization.—
Amer. J. Bot., 1977, 64, 6: 607—614.
Etheridge A. L., Herr J. M. The deve-
lopment of the ovule and megaga-
metophyte in Rhexia mariana.— Ca-
nad. J. Bot., 1968, 46: 133—139.
Eyde R. H. Flowers, fruits and phy-
logeny of Alangiaceae.— J. Arnold
Arboretum, 1968, 49: 4167—192.
Fagerlind F. Die systematische Stel-
lung der Familie Grubbiaceae.—
Sven. Bot. tidskr., 1947, 44: 345—
320.
Faroog T. Studies in Lentibulariaceac.
3. The embryology of Utricularia
uliginosa.— Phytomorphology, 1965,
15: 121—131.
Farooq M. Studies in the Lentibulari-
aceae. 4. The embryology of Utri-
cularia striatula Sm.— J. Indian Bot.
ос., 1966, 45, 1: 41—18.
Farooq M., Bilquis S. Studies in the
Lentibulariaceae. 7. The embryoge-
ny in Utricularia scandens Benj.—
Beitr. Biol. Pflanz., 1966a, 42, 4:
127—134.
Farooq M.,Bilquis 5. Studies in the
Lentibulariaceae. 8. The life histo-
ry of Uttuaria scandens Beni.—
Beitr. Biol. Pflanz., 1966b, 42: 3638—
371.
Faroog M., Siddiqui S. Studies in the
Lentibulariaceae. 5. The deve lop-
312
ment of Utricularia vulgaris var.
americana. A. Gray.— New Phytol,
-1966, 1: 50—53.
Ратоод М., Siddiqui S. Studies in the
Lentibulariaceae. 6. The embryolo-
gy of Utricularia stellaris Lin. f.—
J. Indian Bot. Soc. 1967, 46, 1:
31—46.
Farooq M., Ynamuddin M. The emb-
ryology of Oldenlandia nudicaulis
Roth.— J. Ind. Bot. Soc., 1969, 48,
1—2: 166—173.
Fathima T. Sporogenesis and the dc-
velopment of gametophytes in Cor-
dia alba L—Curr. Sci. (India).
1966, 35, 3: 73—74.
Fathima T. Embryological studies in
Trichodesma zeylanicum R. Br.—
Curr. Sci., 1967, 36: 53.
Fathima T., Boraiah G. Cytoembryo-
logy of Thunbergia mysorensis T.
Anders.— Proc. Indian Nat. Sci.
Аса4., 1973, В, 39, 2: 157—174.
Fathima T., Kumari P. К. Embryolo-
gical studies in Capparidaceae. J.
Sporogenesis and the development
of Gamctophytes in Cleome viscosa
Linn.— Proc. Indian Acad. Sci., 1970,
72, B, 5: 207—2145.
Fathima T., Nalini A. S. Embryology
of Lantana aculeata L. var. nivea
Bailg.— Curr. Sci. (India), 1972, 41,
13: 491—494.
Favre-Duchartre M. Expose d'intro-
duction du Calloque sur les aspects
cytologique de la reproduction se-
xuée des plantes ovulées.— Rev. Cy-
tol. et biol. végét., 1969, 32. |
Favre-Duchartre M. Des ovules aux
grains. Paris, 1970.
Favre-Ducharire M. Interpretation de
la reproduction sexuée des angio-
spormes; la lumier d’organisations
gamclophytique eb embryonnaires
d’aulres Archegoniales.— Phyto-
morphology, 1974, 241, 4: 353—361.
Favre-Ducharire M. Aspects phylogé-
nitiques de la double fécondation
des spermaphytes.— Bull. Assoc.
prof. biol. et geol., 1974, 64, 243:
267—275.
Favre-Duchartre M. Interprétation de
albumen et des éléments des sacs
embryonnaires selon divers autec-
urs.— Bull. Soc. bot. France, 41976,
123, 1—2: 17—32.
Редотзсйик У. РГ. [Федорчук В.Ф].
Embryologische Untersuchung von
Cuscuta monogyna und Cuscuta epi-
thymum.— Planta 1931, 14: 94—111.
Fedorischuk V. F. [®edopuyrx B.®}.
Entwicklung und Bau des manlichen
Gametophytes bei den Arten der
Convolvulaceae-Gattung |§Ouamoc-
lit.— Planta, 1932, 16: 554—574.
Fétré J., Lebegue A. Embryogénie des
Crassulacées. Developpement de
Yembryon chez Sedum spectabile
Boreau.— C. r. Acad. sci. Paris,
1964, 258: 5035—5088.
Fish RB. K. Megagametophyte in Cle-
matis and its taxonomic and phylo-
genetic implication.— Phytomorpho-
logy, 1970, 20: 347—327.
Forman L. L. A new genus of Ixonan-
thaceae with notes on the family.—
Kew Bull, 1965, 19: 517—526.
Forman L. L. The reinstatement of
Galeria Zoll. et Mor. and Microdes-
mis Hook f. in the Pandaceae.—
Kew Bull., 1966, 20: 309.
Forman L. L. A synopsis of Galeria
Zool. et Mor. (Pandaccae).— Kew
Bull., 1971, 26: 153—165.
Fossard R. A. de. Development and
histochemistry of the endothecium
in the anthers of in vitro grown
Chenopodium rubrum L.— Bot. Gaz.,
1969, 130: 10—22.
Frey Г. Studia embriologiczne nad
Alisma lanceolatum With.— Acta bi-
ol. cracov. Ser. bot., 1966, 9, 2:
125—436.
Frey L. Embryological studies ап
Scabiosa lucida Will. and S. ochro-
leuca L. in Poland.— Acta biol. cra-
cov. Ser. bot., 1976 (41977), 19, 2:
173—179.
Fryzele P. A. Mode of reproduction of
higher Plants.— Bot. Rev., 1957, 23,
3: 435—238.
Froussart N. Cytoembryological inves-
tigation of sexual propagation of So-
Janum nigrum.— Ann. Univ. Assoc.
reg. etude et rech. sci. 1966a, 4,
3: 149—423.
Froussart N. Cytologycal investigation
of femal gametophyte development
bei Solanum nigrum L.— C. r. Acad.
sci, 1966b, D263, 3: 236—238.
Gaetano de A. Osservazioni sull emb-
riologia di una specie rara di Erica:
Е. sicula Guss.—Giorn. bot. ital.,
1967, 101, 1: 33—39.
Ganapathy P. S. Clethraceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 82.
Ganapathy P. S. Empetraceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967b: 85.
Ganapathy P. S. Epacridaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967c: 86.
Ganapathy P. S. Ericaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967d: 84.
Ganapathy P. S. Pyrolaceae: Seminar
Comparative Embryology of Angio
sperms. New Delhi, 1967e: 83.
Ganapathy P. S. Clethraceae: Sympo-
sium on Comparative Embryology
of Angiosperms. New Delhi, 1970a,
41: 239—240.
Ganapathy P. S., Palser B. F. Studies
of floral morphology in the Erica-
les. VII. Embryology in the Phyl-
lodoceae.— Bot. Gaz., 1964, 125, 4:
280—297.
Garcia V. Embryological studies in the
Loasaceae. Development of Endo-
sperm in Blumenheachia hieronymi
Urb.— Phytomorphology, 1962, 412,
3: 307—342.
Garg S. Morphologycal and embryo-
logical studies in the Loranthaceae:
Thesis for the degree of Philos.
Univ. Delhi, 1959.
Сегазятота Е.М. [Герасимова E. H.}
Fertilization in Crepis сарШаги$.
Cellule. 1933, 42.
Gerassimova-Navashina H. [Гераси-
мова-Навашина E. H.| Fertilizati-
on and events leading up fertilili-
zation and their bearing of the ori-
gin of Angiosperms.— Phytomorpho-
logy, 11, 1/2: 139—146.
Shash R. B. Studies in the family
Meliaceae. I. Development of the fe-
male gametophyte of Aphenamixis
polystachya (Wael.).— Beitr. Biol.
Pflanz., 1966, 42, 1: 133—138.
Chosh R. B. Studies on the embryolo-
gy of Burseraceae — embryology of
Garuga pinnata Roxb.— Broteria.
Ciénc. Natur., 1970, 39, 1—2: 17—23.
Ghosh R. B. Embryological studies in
the family Rutaceae. I. Megasporo-
genesis and the development of the
female gametophyte in Ravenia
spectabilis Eng.— Bot. Gaz., 1974a,
135, 2: 89—93.
Ghosh R. B. A contribution to the em-
bryology of Eupatorium odoratum
L. together with a discussion on its
interrelationship.— Brotéria. Ciénc.
natur, 1974b, 43, 3—4: 103—117.
Gledhill D. Embryo sac formation in
African Axonopus species.— Phyto-
morphology, 1967, 17, 1—4: 214—
2
Gogellin A. J. A revision of the genus
Cratoxylon Bl. (Guttiferae).— Blu-
mea, 1967, 15: 453 —479.
313
Gonal V., Lakshmanan K. K. Embryo-
logical studies in the Rubiaceae. Or-
ganogenesis and microgametogene-
sis in Morinda citrifolia L.— Curr.
Sci (India) 1977, 46, 12: 423—
424.
Goursat M. J. Recherches sur l’embryo-
logenie des Papilionacées.— Univ.
Paris, fac. pharmac. Sér. E, 1969,
190, 190: 3—187.
Govil C. M. Fouquieriaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 93.
Govil C. M. Convolvulaceae. Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 94.
Govil C. M. Polemoniaceae. Seminar
Comparative Embryology of Angio-
sperms. New. Delhi, 1967c: 92.
Govil C. M. Fouquieriaceae. Symposi-
um on Comparative Embryology of
Angiosperms. New Delhi, 1970a, 41:
244—245.
Govil C. M. Convolvulaceae. Symposi-
um on Comparative Embryology of
Angiosperms. New Delhi, 1970b, 41:
246—249.
Govil C. M. Polemoniaceae. Symposi- |
um on Comparative Embryology of
Angiosperms. New Delhi, 1970c, 44:
241—243.
Govil C. M. Morphological studies in
the family Convolvulaceae: 1. De-
velopment and structure of the se-
ed-coat.— J. Indian Bot. Soc., 1971,
50: 32—38.
Govindappa D., Arekal G. D., Nagend-
ran C. R. Embryo sac of Hydrobry-
opsis sessilis (Podostemaceae): ori-
gin, organization and significance.—
Bot. Notiser, 1975, 128.
Govindappa D. A., Nagendran C. R.
Female gametophyte of Zeylanidi-
um (Podostmaceac): A_ clarificati-
on.— Phytomorpholy, 1977, 27, 2:
423—129.
Gracza P., Sarkany S. Organisation
und histogenetische Korrelationen
der Samenanlage von Papaver som-
niferum L.— Acta bot. Acad. sci.
hung., 1970, 16, 1—2: 37—957.
Greilhuber J. Uber die Entwicklung
des Embryosacks von Melampyrum
and Parentucellia latifolia.— Os-
terr. bot. Z., 1973, 124: 81—97.
Grezicka W. Megasporogenesis, ovule
and embryo sac development in Ca-
rex aristata R. Br. var. cujavica.—
Acta Soc. bot. pol. 1964, 33, 2:
307—322.
Grover H. Development of the female
314
gametophyte in some Ficus speci-
es.— J. Indian Bot. Soc., 1969, 48.
3—4: 334—338.
Grundwag M. Embryology and fruit
development in four species of Pi-
stacia L. (Anacardiaceae).—Bot. J.
Linnean Soc., 1976, 73, 4: 355—370.
Grundwag M., Fahn A. The relation
of embryology to the low seed set
in Pistacia vera (Anacardiaceae).—
ry tomorphology, 1969, 19: 225—
Guignard J. L. Embryogenie des por-
tulacacées. Developpement de l’emb-
ryon chez le Talinum patens
Willd.— C. r. Acad. sci., 1965, 261,
25: 5599—5601.
Guignard J. L. Embryogenesis du Ca-
ryophyllaceae. Le developpement de
Yembryon chez le Gypsophila ее-
gans Bieb.— C. r. Acad. Sci., Paris,
1966a, D262, 26: 2718—2721.
Guignard J. L. Embryogenesis du Ca-
ryophyllaceae. Le developpement de
Vembryon chez le Cerastium glome-
ratum Thuill.— C. r. Acad. sci. Pa-
ris, 1966Ъ, 0263. 12: 856—859.
Guignard J. L. Embryogénie et classi-
fication embryogénique.— Bull. Soc.
bot. France, 1975, 122, 7—8: 281—
294,
Guignard J. L., Mestre J. C. Liembry-
on des graminées.— Phytomorpho-
logy, 1970, 20, 2.
Gulati N. Gentianaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967a: 89.
Gulati N. Loganiaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967b: 88.
Gulati N. Menyanthaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967c¢: 90.
Gulati N. Stackhousiaceae: Seminar
Comparative Kimbryology of Angio-
sperms. New Delhi, 1967d: 57.
Gulati N., Mathur S. Embryology and
taxonomy of Filicium decipiens.—
Phytomorphology, 1977, 27, 3:
261—267.
Gunn C. R., Seldin J. Seeds and fruits
of North American Papaveraceae.
Wash.: Agr. Res. Serv. US Dep. Agr.,
1976, 1—96.
Gupta P. K. Observations on degree
of apospory in three members of
Andropogoneae.— Curr. Sci., 1968,
37: 295—296.
Сирю 5. С. Umbelliferae: Seminar
Comparative Embryology of Angi-
osperms. New Delhi, 1967: 81.
Сира 5. С. Umbelliferae: Symposium
on Comparative Embryology of An-
giosperms, 1970, 414: 233—238.
Gupta S. C., Gupta M. Some observa-
tion on the embryology of Bupleu-
rum tenue Buch. Ham ex D. Don.—
Curr. Sci., 1964, 33, 18: 563—564.
Gupta S. C., Rani Ahyja. Embryology
and systematic position of Nelumbo
nucifera: Seminar on morphology,
anatomy and embryology of land
plants. Delhi, 1969: 63—64.
Guzowska I. Reinvestigation of emb-
ryo sac development, fertilization
and early embryogeny in Cytinus
hypocistus L.— Acta Soc. bot. pol.,
1964, 33, 1: 157—166.
Haber J. M. Comparative anatomy and
morphology of the flowers and in-
florescences of the Proteaceae III.
Some African taxa.— Phytomorpho-
logy, 1966, 16, 4: 490—527.
Hakki M. I. Embryologische und mor-
phologische Beobachtungen an
Succowia balearica (L.) Medik.
(Brassicaceae).— Bot. Jahrb. Syst.
Pflanzengesch. und Pflanzengeogr.,
1974, 94, 3: 360—382.
Hakki M. I. Embryologische und mor-
phologische Untersuchungen an
Pflanzen aus Westindien. I. Zur Em-
bryologie von Capraria biflora Г.
(Scrophulariaceae).— Bot. Jahrb.
Syst. Pflanzengesch. und Pflanzen-
geogr., 1975, 96, 1—4: 125—153.
Hakki M. I. Embryologische und mor-
phologische Untersuchungen an
Pflanzen aus Stidafrika. I. Uber die
Embryologie, Morphologie und Sys-
tematische Zugehorigkeit von Der-
matobotrys saudersii Bous.— Bot.
Jahrb. Syst. Pflanzengesch. und
Pflanzengeogr., 1977, 98, 1: 98—
149
Haskell D. A., Postlethwait S. N.
Structure and histogenesis of the
embryo of Acer saccharinum. I. Em-
bryo sac and proembryo.— Amer. J.
Bot., 1971, 58: 595—603.
Hausner G. Embryogenese und Nucel-
lar — Polyembryonie bei Cy-
nanchum/Vincetoxicum — Arten.—
Beitr. Biol. Pflanz., 1976, 52, 1—8:
401—126.
Heel van W. A. Anatomical and onto-
genetic investigations on the mor-
phology of the flowers and fruit of
Scyphostegia borneensis Stapf (Scy-
phostegiaceae).— Blumea, 4967, 15:
107—125.
Heel van W. A. Flowers and fruits in
Flacourtiaceae: 1. Scaphocalyx spa-
thacea.— Blumea, 1973, 24: 259—
279.
Heel van W. A., Bouman F. Note on
the early development of the integu-
ment in some Juglandaceae.— Blu-
шеа, 1972, 20: 155—159.
Herr J. M. Embryological evidence for
the relationship of Aquifoliaceae to
Celastraceae.— Virginia J. Sci., 1959,
10: 259.
Ilerr J. M., Dowd Marybruce L. Deve-
lopment of the ovule and megaga-
metophyte in Oxalis corniculata L.—
Phytomorphology, 1968, 18, 14: 48—
53
Hindmarsh G. J. An embryological
study of five species of Bassia All.
(Chenopodiaceae).— Proc. Linn. Soc.
N. S. W., 1966, 90: 274—289.
Hiroshi T. An embryological study of
Hetaeria shikokiana, a saprophytic
orchid in Japan.— Sci. Repts Toho-
ku Univ. Ser. 4, 1967, 33, 2: 83—95.
Hiroshi T. Development of the emb-
ryo of Bletilla striata.— Sci. Repts
Tohoku Univ. Ser. 4, 1968, 34, 2+—4:
125—131.
Hiroshi T. Development of embryo of
Orchis aristata.— Sci. Repts Toho-
ku Univ. Ser. 4, 1971, 35, 4: 239—
243.
Hiroshi T. Embryo sac development
in Trautvetteria japonica Sieb. et
Zucc. (Ranunculaceae).— Sci. Repts
Téhoky Univ. Ser. 4, 1973, 36, 4:
255—259.
Hiroshi T. Development of the embryo
of Leucanorcnis japonica.— Sci.
Repts Téhoku Univ. Ser. 4, 1974a,
37, 1: 89—93.
Hiroshi T. Development of the emb-
ryo of Pogonia (Orchidaceae) .— Sci.
Repts Tohoku Univ. Ser. 4, 1974b,
37, 1: 89—93.
Hiroshi T. Development of the emb-
tyo of Gymnadenia cucullata.— Sci.
Repts Tohoku Univ. Ser. 4, 1974c,
37, 1: 71—77.
Hiroshi T. The embryology of Cimici-
fuga acerina Sieb. et Zucc., C., tana-
ka (Ranunculaceae).— Sci. Repts
Tohoku Univ. Ser. 4, 1974d, 37, 4:
55—64.
Hiroshi T. The embryology of Anemo-
nopsis macrophylla Sieb. et Zucc.
(Ranunculaceae).— Sci. Repts То0-
hoku Univ. Ser. 4, 1974e, 37, 1:
63—69.
Horner H. T., Lersten N. R. Microspo-
rogenesis in Citrus limon (Ruta-
315
ceae).— Amer. J. Bot., 1974, 58:
12—19.
Howe P. D. The female gametophyte
of Machaeranthera pattersonii (As-
ter pattersonii) and of M. tanaceti-
folia— Amer. J. Bot, 1969, 56:
641—645.
Howe P. D. The female gametophyte
of three species of Grindelia and
Prionopsis ciliata (Compositae).—
Amer. J. Bot., 1975, 62, 3: 273—279.
Hu-s-y. Studies in polyembryony of
Hosta coerulea Tratt. 1. Observati-
ons on the embryogenese and de-
velopment on polyembryony.— Ac-
ta bot. sinica, 1960, 9, 2: 147—452.
Hutchinson J. The families of flowe-
ring plants. London, 1926, 2937,
I—II.
Hutchinson J. The families of flowe-
ring plants. 2nd ed. London, 1959,
I—II
Hutchinson J. Evolution and Phyloge-
ny of Flowering Plants. L.; N. Y.:
Acad. Press, 1969: 717.
Hutchinson J. The families of flowe-
ring plants. 3rd ed. Oxford: Claren-
don Press, 1973.
Hutomo W. On the embryology of Mu-
cuna bennetti ЕР. Muell.— Ann. bo-
gor., 1967, 4, 3: 2014—211.
Илиева Стоянка. Изследвания
върху цыфтежа, опрашването и
оплождането на ба]\м1а зФагеа Г..—
Растениевъдни науки. София, 1965,
2, 10: 31—46.
Jaéger-Ziirn I. Zur Frage der systema-
tischen Stellung der Hydrostachya-
ceae.— Ost. Bot. Zeitschr., 1965, 112:
621—639.
Jéger-Ziirn I. Embryologische Unter-
suchungen an vier Podostomace-
en.— Ost. Bot. Z., 1967, 114: 20—45.
Jaitly S. C. Development of seeds and
fruits in Anisomeles indica (Linn.)
O. Kze.— Phytomorphology, 1966,
16: 430—436.
Jaitly S. C. Le développement de l’al-
bumen chez les genres Salvia, Hyp-
tis, Ocimum, Pogostemon, Mentha
et Leucas.— Bull. Soc. bot. France,
1968 (1969), 145, 7—8: 373—378.
Jaitly S. C. Morphology and embryolo-
gy of Justicia betonica Linn: Symp.
Morphology, Anatomy and Embry-
ology of Land Plants. 1969a, 62—63.
Jaitly S. C. Structure and development
of seed and fruit in certain mem-
bers of the Labiatae: Symp. Morpho-
logy, Anatomy and Embryology of
Land Plants, 1969b, 64—65.
316
Jaitly S. C. Studies in the Labiatae. I.
Structure and development of seed
and fruit in two species of Salvia
L.— J. Indian Bot. Soc., 1971, 50, 2:
182—188.
Jaitly S. C., Guignard J. L., Mest-
re J.C. Embryogenie des Orobancha-
сбез. Le developpement de l’emb-
ryon chez le Lathrea clandestina
Тат.— С. г. Асад. sci., 1968a, D267,
2: 170—172.
Jaitly S. C., Guignard J. L., Mest-
re J. C. Embryologenie de labiées.
Le developpement de lembryon
chez Scutellaria columnae Host.—
C. r. Acad. sci., 4968b, D267, 21:
1747—1719.
7аНу 5. С.. Guignard J. Г., Mest-
re J. C. Embryogenie de labiées. De-
veloppement de l’embryon chez le
Mentha aquatica L.— Bull. bot.
France, 1968¢ (1969), 115, 7—8:
369—372.
Jatly S. C., Guignard J. L., Mest-
re J. C. Embryogenie des Verbena-
cécs. Developpement de l’embryon
chez le Caryopteres mastacanthus
Schau.—C. rv. Acad. sci. Paris,
1968d, D267: 59—61.
Jalan S. Some further observation on
the embryology of Caltha palustris
L.— Current Sci. (India), 1968, 37,
21: 621—623.
Jalouzot M. F. Mise en évidence de
parois callosique au cors de la me-
gasporogénese et de l’oogénés d'Oeno-
thera biennis.— C. R. Acad. de Paris,
1970, D, 270: 347—319.
Jalouzot M. F. Aspects ultrastructuraux
de la megasporogenese d’Oenothera
lamarckiana et rapport avec les de-
pots callosiques observes.— Annal.
de ’Univer. Reims., 1971, 9, 1: 36—
45.
Janchen Г. Das System der Crucife-
ren.— Osterr. bot. Z., 1942, 91: 1—28.
Jeyanayaghy S., Rao A. N. Flower and
seed development in Bromheadia
finlaysoniana.— Bull. Torrey Bot.
Club, 1966, 93, 2: 97—108.
Тойт В. М. Alismataceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 1414.
Johri B. М. Butomaceae: ‘Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 142.
Johri B. M. Limnanthaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967c: 43.
Johri B. M. Angiosperm embryology
and taxonomy: Symp. on Newer
Trends in Taxonomy, 1967, 34:
263—268.
Johri B. M. Alismataceae and Butoma-
сеае: Symp. on Comparative Emb-
ryology of Angiosperms. New Del-
hi, 1970a, 44, 334—335.
Johri B. M. Limnanthaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970b, 41:
410—113.
Johri B. M., Agarwal S. Morphology
and embryological studies in the
family Santalaceae. VIII Quincha-
malium ihilense Lam.— Phytomor-
phology, 1965, 15, 4: 361—372.
Johri B. M., Ambegaokar K. B. Seed
development in Triticales. Embryo-
logy of crop plants: Indo-Soviet
Symp. Delhi, 1976: 15—16.
Johri B. M., Bhatnagar S. P. Endo-
sperm in the Santalales.— Rev. Cy-
to]. et biol. végét., 1969, 32, 34:
353—369.
Johri B. M., Garg 5. Development of
endosperm haustoria in some Legu-
minosae.— Phytomorphology, 1959,
9: 34—46.
Johri B. M., Konar R. N. The floral
morphology and embryology of Fi-
cus religiosa.— Phytomorphology,
1956, 6: 97—111.
Johri B. M., Prakash S. Morphological
and embryological studies in the fa-
mily Loranthaceae. 11. Tapinostem-
ma acacia.— Phytomorphology,
1965, 15: 150—158.
Johri B. M., Pundir N. S. Embryology
studies in cultivated Cottons. Emb-
ryology of crop plants: Indo-Soviet
Symp. Delhi, 1976: 16—17.
Johri B. M., Raj B. Morphological and
_. embryological studies in the fami-
ly Loranthaceae. XII Moquiniella
rubra (Spreng f.). Balle.— Osterr.
bot. Z., 1969, 116, 1—5: 475—485.
Johri B. M., Vasil I. K. The embryo-
logy of Ehretia laevis Roxb. Phyto-
morphology, 1956, 6, 2: 134—143.
Jos J. S., Singh S. P. Gametophyte de-
velopment and embryogeny in the
genus Nicotiana. J. Indian Bot.
Soc., 1968, 47, 1—2: 147—128.
Joshi A. S., Wadhwani M., Johri B. M.
Morphological and embryological
studies of Gossypium L.— Proc. Nat.
Inst. Sci. India, 1967, 33B: 37—93.
Юрукова-Грънчарова П. Д. Развитие
на женския гаметофит на два вида
от род Leontodon L. София, Българ-
ска инадемия на науките, 1978, 9:
2—69.
Юрукова-Грънчарова П. Д. Сравни-
тельно ембриолого — таксономиче-
ское проучкане на Български
представители от подсемейство
Cichorioideae Kitam. на семейство
Asteraceae Dumort. (Compositae
giseke). Канд. дисс. София, 1979:
с.
Juguet M. Embryogénie des Cypera-
cées. Developpement de l’embryor
chez Scirpus maritimus.—C. _r.
са. sci., 1966, 2263, 22: 1710—
1713.
Тихие: M. Polyembryonie chez le Ca-
тех arenaria L.— Bull. Soc. bot.
France, 1966 (1967), 113, 9: 448—
452.
Juguet M. Polyembryonie chez le Kob-
resia bellardii— Bull. Soc. bot.
France, 1967 (1968), 114, 7—8: 277—
279.
Juguet M. Develppement de l’embryon
chez TEriophorum angustifolium
Roth. (Cyperaceae).— Bull. Soc. bot.
Егапсе, 1969а, 116, 3—4: 137—143.
Juguet M. Embryogénie des juncacé-
es. Developpement de l’embryon
chez le Luzula pilosa (L.) Willd.—
С. г. Асад. sci, 1969b, D268, 25:
3036—3039.
Juguet M. Embryogenie des cyperacé-
es developpement de lembryon
chez le Cyperus fuscus L.—C. г.
Acad. sci., 1970, D271, 6: 580—583.
Juguet М. Developpement de l’embry-
on chez guelques Cyperacees’ afri-
caines.— Adansonia, 1970, 10, 2:
271—288.
Juguet M. Observations embryogeni-
ques chez les Cyperacées et les jun-
cacées: C. r. 93e Congr. nat. Soc.
savantes, Tours, 1968, Sec. Sci. 3.
Paris, 1972: 547—572.
Juguet M., Lebegue A. Embryogénie
des Cyperacées. Developpement de
Vembryon chez le Carex arenaria
Т.— С. г. Acad. sci., 1966, D262, 17:
1844—1841.
Kajewska E. Studies of Verbena offi-
cinalis L.— Acta biol. cracov. Ser.
bot., 1977, 20, 1—2: 41—49.
Kamdar H. M. Gametophytes of Scir-
pus articulatus L: Proc. 56th Indian
Sci. Congr. Bombay, 1969a, 3: 369.
Kamdar H. M. Male und female game-
tophytes of Fimbristylis diphylla
(Retz.) Vahl: Proc. 56th Indian Sci.
Congr. Bombay, 1969b, 3: 369.
Kamdar H. M. Studies in the genus.
Cyperus L.: Proc. 56th Indian Sci.
Congr. Bombay, 1969c, 3: 396.
17
Kapil R. N. Embryology of Acalypha
Linn.— Phytomorphology, 1960, 19,
2: 174—184.
Kapil R. N. Embryology and systema-
tic position of Theligonum Linn.—
Proc. Nat. Inst. Sci. India, 1966, B,
32: 218—232.
Kapil R. N. Araceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967a: 126.
Kapil R. N. Cistaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967b: 66.
Kapil R. N. Crossosomataceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967c: 34.
Kapil R. N. Frankeniaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967d: 68.
Kapil R. N. Lemnaceae:Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967e: 4127.
Kapil R. N. Oleaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967f: 87.
Кари В. № Philydraceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967g: 125.
Kapil R. N. Podostemaceae: Seminar
comparative Embryology of Angio-
sperms. New Delhi, 1967h: 42.
Kapil R. N. Crossosomataceae: Symp.
on Comparative Embryology of An-
giosperms, 1970a, 44: 63—68.
Kapil R. N. Podostemaceae: Symposi-
um on Comparative Embryology of
Angiosperms, 1970b, 41: 104—109.
Kapil R. N. Contemporary spectacle in
Angiosperm Embryology.— Acta
bot. indica, 1974, 2: 79—106.
Kapil R. N., Bawa S. B. Embryologi-
cal studies on the Haloragidaceae.
I. Haloragis colensoi Skottsb.— Bot.
Notiser, 1968, 124, 1: 11—28.
Kapil R. N., Bhandari N. N. Morpholo-
gy and Embryology of Magnolia.
Dill. ex Linn.— Proc. Nat. Inst. Sci.
India, 1965, 30, 5—6: 245—262.
Kapil R. N., Bhatnagar A. K. A fresh
look at the process of double ferti-
lization in Angiosperms.— Phyto-
morphology, 1975, 23, 3: 334—368.
Кари В. N., Kaul U. Embryological
studies in the Hamamelidaceae:
Proc. 56th Indian Sci. Congr. Bom-
bay, 1969, 3: 395.
Kapil R. N., Kaul U. Embryologically
little known taxon Parrotiopsis
jacquemontiana.— Phytomorphology,
4972, 22, 3—4: 234—245,
318
Kapil R. N.,. Maheshwari P. Embryo-
logy of Helianthemum vulgare Ga-
ertn.— Phytomorphology, 1965, 414:
547—557.
Кари В. N., Mohana Rao P. R. Emb-
ryology and systematic position of
Theligonum Linn.— Proc. nat. Inst.
Sci. India, 1966a, B32: 218—232.
Kapil R. N., Mohana Rao P. R. Studi-
es of the Garriaceae. II. Embryolo-
gy and systematic position of Gar-
ria douglas Lindley.— Phytomorpho-
logy, 1966b, 16, 4: 564—578.
Kapil R. N., Prakash N. Embryology
of Cereus jamacarn and Ferocactus
wislizeni and comments on the Sys-
tematic position of the Cactaceae.—
Bot. Notiser, 1969, 122, 3: 409—426.
Kapil R. N., Rustagi P. N., Venkatata-
raman R. A contribution to the em-
bryology of Polemoniaceae.— Phy-
tomorphology, 1968, 18, 3: 403—412.
Kapil R. N., Vani R. S. Nyctanthes
arbor — tristis Linn: embryology
and relationships.— Phytomorpho-
logy, 1966, 16, 4: 553—563.
Kapil R. N., Venkatataraman R. Emb-
ryology of Gilia capilata Sims. Proc.
soth Indian Sci. Congr. Benares,
1968, 3: 345.
Kapil R. N., Vijayaraghavan M. Е.
Embryology of Pentaphragma hors-
fieldii, with a discussion on the sy-
stematic position of the genus.—
Phytomorphology, 1965, 15: 98—
102.
Kapil R. N., Walia Karvita. The emb-
ryology of Philydrum lanuginosum
Banks ex Gaerth. and the systema-
tic position of the Philydraceae.
Beitr. Biol. Pflanz., 1965, 41, 3:
381—404.
Kaplan О. В WVistogenesis of the and-
roecium and gynoccium in Downin-
gia bacigalupii.— Amer. Т. Bot.,
1968, 55, 8: 933—950.
Kaplan D. R. Sporogenesis and game-
togenesis in Downingia (Campanu-
laceae; Lobeliodeae).— Bull. Torrey
Bot. Club., 1969a, 96, 4: 418—434
Карп 0. В. Seed development
in Downingia.— Phytomorphology,
1969b, 19, 3: 253—278.
Kapoor T., Parulekar N. K., Vijayara-
ghavan M. R. Contribution to the
Embryology of Celsia coromandeli-
ana Vahl. With a discussion on its
affinities with Verbascum thapsus
L.— Bot. Notiser., 41975, 128, 4:
438—449.
Карагьозова М. Д. Цитоемриологично
изучаване на Еаесю1еега ригра-
rea Kuntze.— Изв. Бот. ин-т Boar,
АН, 1969, 19: 117—126.
Karagisowa M, D. Besonderheiten bei
der doppelten Befruchtung und die
Embryogenese von Galanthus niva-
lis— JJAH 1970, 23, 6: 745—718.
Карагьозова М. Д., Логофетова В.
Цитоембриологично изучавне на
Сегодепдгоп Шотзопае ВаН.—
Изв. Бот. ин-т Бълг. АН, 1967, 11%:
187—198.
Karkare-Khushalani Ishwari, Mu-
ву В. N. Studies in Sapindaceae I.
Embryology of Dadonaea_ viscosa.
Phyton, 1964, 41, 1—2: 83—92.
Karlstrém Per-Olof. Embryological
studies in the Acanthaceae. I. The
genus Ruellia L.— Sven. Bot. Tidskr.,
1972, 66, 4: 303—313.
Karlstrém Per-Olof. Embryological
studies in the Acanthaceae. II. The
genera Pseuderanthemum Radek,,
Ruspolia Lindau, Eranthemum ТГ.
and Lankesteria Lindl.— Sven. bot.
Tidskr., 1973, 67, 3: 257—280.
Karlsirém Per-Olof. Embryological
studies in the Acanthaceae. III. The
genera Barleria L. and Crobbea
Harv.— Sven. bot. tidskr., 1974a, 68,
2: 121—135.
Karlstrém Per-Olof. Embryological
studies in the Acanthaceae. IV. The
genera Asystasia Bl. and Chamaran-
themum Nees.— Sven. bot. tidskr.,
1974Ъ, 68, 3: 315—328.
Kaul U. Morphology and embryology
of Exbucklandia populnea (Hama-
melidaceae): Seminar on Morpholo-
gy, anatomy and embryology of
land plants, 1969a: 66—67.
Kaul V. Endosperm in Parrotiopsis
jacquemontiana.— Phytomorphology,
19695,19, 2: 197—199.
Kaul V., Dathan A. S., Singh D. Em-
bryological studies on the genus
Sonchus L.—J. Indian Bot. Soc.,
1975, 54, 3—4: 238—245.
Kaul U., Kapil R. N. Exbucklandia
populnea — from flower to fruit.—
Phytomorphology, 1974, 24, 3—4:
247—228.
Каиг Н. В1хасеае: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967a: 67.
Kaur H. Cornaceae: Seminar Compara-
tive Embryology of Angiosperms.
New Delhi, 4967b: 80.
Kaur H, Moraceae: Seminar on Com-
parative Embryology of Angio-
sperms. New Delhi, 1967c: 2.
Kaur D, Turneraceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967d: 64.
Kaur H. Embryology investigations on
Bixa orellana L.— Proc. Nat. Inst.
Sci. India., 1969a. 35, 6: 487—506.
Kaur H. Stucture and Development
of seed in Ipomoea obscura Ker.—
Gawl.— J. Indian Bot. Soc., 1969b,
48, 3—4: 346—351.
Kaur H. Bixaceae: Symp. on Compara-
tive Embryology of Amgiosperms.
1970a, 44: 205—207.
Kaur H, Cornaceae: Symp. on Compa-
rative Embryology of Angiosperms.
New Delhi, 1970b, 44: 230—232.
Kaur H. Moraceae: Symp. on Compa-
rative Embryology of Angiosperms.
New Delhi, 1970c, 1441: 1—3.
Kaur D. Turneraceae: Symp. on Com-
parative Embryology of Angio-
sperms, 1970, 41: 194—198.
Kaur H., Singh R. P. Structure and
development of seeds in three Ipo-
moea species.— J. Indian Bot. Soc.,
1970e, 49, 1—4: 168—174.
Kelkar S. S. The development of en-
desperm and embryo in Lannea co-
romandelica (Houtt) Merr.—J.
Univ. Bombay, 1961, 29, B: 1—5.
Khaleel T. F. Embryology of Cyno-
glossum denticulatum D. C.— Bot.
Notiser, 1974, 127, 2: 193—210.
Khaleel T. F. Embryology of Cordia
alba L.— Bot. Gaz., 1975, 136, 4,
380—387.
Khaleel T. F. Embryology of Tricho-
desma (Boraginaceae).— Bot. Noti-
зег., 1977, 130, 4: 441—452.
Khaleel fT. F. Embryology of Heliot-
ropium scabrum and H. strigosum
(Boraginaceae).— Plant Syst. and
Еуо]., 1978, 129, 1—2: 45—62.
Khan A. M. Development of ovule and
female gametophyte in Talinum pa-
tens Willd.— Curr. Sei. (India), 1973.
42, 10: 364—365.
Khan R. Lentibulariaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 4967: 104.
Khan R. Lentibulariaceae: Symp. on
Comparative Embryology of Angio-
sperms, 1970a, 44: 290—297.
Khan P. S. Structure and development
of seed coat and fruit wall in Ca-
tharanthus pusillus (Murr) С.
Don.— Proc. Nat. Acad. Sci. India,
1970b, 40B: 24—24.
Khan R., Khatoon F, Embryology of
319
the Compositac, tribe Inuleae. I.
Gnaphalium purpureum L. and G.
indicum L.: Seminar on morpholo-
gy, anatomy and embryology of
lands plants. Delhi, 1969a: 67—68.
Khan R., Khatoon F. Development of
the female and male gametophytes
in Gnaphalium purpureum L.: Proc.
56th Indian Sci. Congr. Bombay,
1969b, 3: 378.
Khan R., Sharma H. P. Embryology
- Of the Compositae, triba Asteraceae.
I. Brachycome iberidifolia Benth,
and Erigeron linifolius Willd: Semi-
nar on morphology anatomy and
embryology of land plants, Delhi,
1969: 68—69.
Khanna P. Embryology of Trichodes-
ma amplexicaule.— Bull. Torey bot.
Club., 1964, 94: 105—144.
Khanna P. A. A contribution to the
embryology of Cyperus rotundus L.,
Scirpus mucranatus L. and Hyllin-
ga melanospora Nees.— Canad. J.
Во%,, 1965а, 43, 12: 1539—1547.
Khanna P. Morphological and embryo-
logical studies in Nymphaeaceac.
II. Brasenia schreberei Gmel. and
Nelumbo nucifera Gaertn.— Austral.
J. Bot., 1965b, 13: 379—387.
Khanna P. Morphological and embryo-
logical studies in Nymphaeaceae.
III. Victoria cruziana D’arb and Ny-
mphaea stellata Willd.— Bot. Mag.
Tokyo, 1967, 80, 950: 305—342.
Kim-Lang-Huynh. Quelques phenomé-
nes de polarité du pollen Orchida-
ceae.— Bull. Soc. bot. suisse, 1976,
86, 3—4: 115—128.
Kimura C. On the embryo sac of Cyp-
ripedium debile Reichb. f.— Sci.
Repts Tohoku Univ. Ser. 4, 1968,
34, 4: 67—TA4.
Kimura C. Embryological studies of
Galeola septentrionalis Reichg.—
Sci. Repts Tohoku Univ. Ser. 4,
1971, 35: 253—258.
Kirsten E. Embryological investiga-
tions in Arnica alpina from Green-
land.— Sven. bot. tidsskr, 1970, 65,
3: 225—244.
КЖолев И. fl, Седмакова-Личан-
ска Е. Някои ембриологични 060-
бенности на Trifolium arvense
L., T. preslianum Boiss. и Т. smy-
rnaeum Boiss.— Генетика и селек-
ция (НРБ), 1972, 5, 2:167—172.
Koscinska M. Embryo and endosperm
in Rudbeckia bicolor Nutt.— Acta
biologica Cracovensia. Ser. Botanica,
1980, 22, 4: 73—79.
320
Kordyum Е. 2. {Кордюм Е. Л.] Раг-
ticularités du developpement du
reproduction sexuelle chez des Om-
belliferes.— Ann. Univ. Reims, 1970,
6/7: 195—200.
Kordijum E. L., Boiko A. P. [Kop-
дюм Е. Л., Бойко А. П.]. Оп Ще
embryology of Gerbera anandria
Schultz.— Dokl. Akad. Nauk Ukrai-
па, 1962, 8: 1109—1142.
Кош А. К., Durani P. К. Cytokinesis
and microspore formation in Plan-
tago lanceolata L.— Curr. Sci, 1969,
38: 524—522.
Koul A. K., Wafai B. A., Wakhlu A. K.
Studies on the genus Gagea. III.
Sporogenesis, early embryogeny and
endosperm development in hexa-
ploid Gagea_ stipitata.— Phytomor-
phology, 1976, 26, 3: 255—263.
Krupko 5. Some embryological and
cytological characteristics of two
varieties of Oxalis cernua
Thurnb. var. sericea and var
compressa from South Africa.—
Bull. Soc. Amis sci. et lett. Pozn.,
1971 (1972), 1, 12—43: 164-496.
Krupko 5. Megasporogenesis and de-
velopment of the embryo sac in the
Palestine variety of Trifolium ale-
xandrinum L.— Acta Soc. bot. pol.,
1973, 12, 4: 647—625.
Kubier E. Cytological processes du-
ring the development of antipodals
in Ammophila arenaria Link.— Ac-
ta biol. cracov. Ser. bot., 1968, 414,
di: 21—29.
Kubier E. Embryological studies in
Ammophila arenaria. Link.— Acta
biol crac. Ser. bot., 1970a, 13, 4:
1—10.
Kubiet E. Cytological and embryolo-
gical studies in Ammophila baltica
(Fligge) Link.— Acta biol. crac.
Ser. bot., 1970b, 13, 2: 99—110.
Kulkarni K. M., Mahabale T. S. Stu-
dies on Palms: embryology of Li-
vistona chinensis R. BR.— Proc. In-
dian Acad. Sci., 1974, 80, 4: 1—17.
Kulkarni A. R., Pataskar P. G. Game-
togenesis in Sesamum laciniatum
Klein ex Willd.— J. Biol. Sci., 4973—
1974, 16—17, 1—2: 86—91.
Kulkarni A. R., Patil V. D. On the em-
bryology of Cyanonotis cucullata
Kunth.— Curr. Sci. (India), 1969, 38,
9: 220—221.
Kumar A. Studies in Geraniales. V.
The embryology of some indian spe-
cies of Feranium L.— J. Indian Bot.
Soc., 1977, 56, 4: 328—334.
Kumar 5. Embryological studies on
Indian species of Morus. I. Micro-
sporogenesis.— Curr. Sci. (India),
1978, 47, 2: 807—808.
Kumar S., Jain D. K. Microsporoge-
nesis in Gillia capitata Sims.—
Curr. Sci (India), 1977, 46, 21: 749—
750.
Kuta E. Cyto-embryological studies on
the species of the Viola L. genus,
Nomimium Ging, Section from the
territiry of Poland.— Fragmenta
floristica et geobotanica, 1978, 24,
4: 23—91.
Laguna I. G., Cocucci A. E. El ova-
rio el ovula y el megagametofito
de Colletia spinosissima (Rham-
naceae).— Kurtziana, 1971, 6: 53—
Lakshmanan K. K. Note on the endo-
sperm formation in Zannichellia pa-
lustris L.— Fyton, 1965a, 22, 1: 48—
14.
Lakshmanan K. K. Embryological stu-
dies in the Hydrocharitaceae. IV.
Post-fertilization development in
Hydrilla verticillata Royle.— Fyton,
1965Ъ, 22, 1: 45—50.
Lakshmanan K. K. Embryological stu-
dies in the Bromeliaceae. I. Lindma-
nia pendulifera (C. H. Wright)
Stapf.— Proc. Indian Acad. Sci.,
1967a, 65B, 2: 49—55.
Lakshmanan K. K. Hydrocharitaceae:
Symp. on Comparative Embryology
of Angiosperms. 1967b: 113.
Lakshmanan К. К. Juncaginaceae:
Seminar Comparalive Embryology
of Angiosperms New Delhi. 1967c:
118
Lakshmanan K. K. Najadaceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967d:
115
Lakshmanan K. K. Potamogetonaceae:
Seminar Comparative Embryology
Angiosperms. New Delhi, 1967e: 416.
Lakshmanan K. K. Scheuchzeriaceae:
Seminar Comparalive Embryology
of Angiosperms. New Delhi, 1967f:
114.
Lakshmanan K. K. Zannichelliaceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi, 1967g:
147.
Lakshmanan K. K. Preliminary note
on the embryology of Diplanthera
uninervis Aschers.— Curr. Sci., 1968,
37: 534—535.
Lakshmanan K. K. Hydrocharitaceae:
21. А. Поддубная-Арнольди
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970a,
41: 336—341.
Гакзртапап К. К. Juncaginaceae:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970b,
41: 344—347.
Lakshmanan K. K. Najadaceae: Symp.
Comparative Embryology of Angio-
sperms. New Delhi, 1970c, 44: 354—
57.
Lakshmanan К. K. Potamogetona-
ceae: Symp. on Comparative Emb-
ryology of Angiosperms. New Del-
hi, 1970d, 44: 348—351.
Lakshmanan K. K. Scheuchzcriaceae:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970e,
441: 342—348.
Гакзйтапап К. К. Zannichelliaceae:
Symp. on Comparative Embryology
of Angiosperms, New Delhi, 1970f,
44: 352—358.
Lakshmanan K. K. Studies on the de-
velopment of Commelina bengha-
lensis L. I. Zygote to globular pro-
embryo.— Proc. Indian Acad. Sci.,
1977, 86B, 3: 167—174.
Lakshmanan K. K., Philip V. J. A con-
tribution to the embryology of Iri-
daceae.— Proc. Indian Acad. Sci.,
1971—1973 B, 3: 410—116.
Lakshmi N. A contribution to the em-
bryology of Sansivieria zeylanica
Wield.— Curr. Sci. (India), 1979, 48,
9: 440—412.
Lamba L. C. Microsporogenesis and
male gametophyte in Rauwolfia
serpenlina (L.) Benth. ex Kurz.—
Curr. Sci. (India), 1976, 45, 10:
387—388.
Latoo C. S. Embryology of Hollarrhe-
na antidysentcrica Wall.— In: Proc.
56th Indian Sci. Congr. Bombay,
4969, 3: 379—380.
Latoo C. S. Morphology and embryolo-
gy of Nollarrhena antidysenterica
Wall.— Bot. Gaz., 1974, 135, 3: 173—
180.
Lazarte J. E., Palser B F. Morpho-
logy, vascular anatomy and emb-
ryology of pistillate and stamina-
te flowers Asparagus officina-
lis— Amer. J. Bot., 1979, 66, 7:
153—764.
Lebégue A, Nouveaux aspects d'une
classification embryologigue.— Bull.
Soc. bot. France, i966, 113, 3: 87—
98.
Lebégue A., Vignon F. Contribution a
321
Vembryogénie des Caryophyllaceac.
Le développement de l’embryon chez
le Melandrium album (Mill) Gar-
cke. (Lychnis alba).—C. r. 93¢e
Congr. nat. Soc. savantes, Tours,
toe Sec. sci. Paris, 1972, t. 3: 485—
507.
Lebégue A., Vignon J. Embryologie
des Saxifragacées. Développement
de Yembryon et formation de l’albu-
men chez le Saxifraga aizoides L.—
C. r. Acad. sci. Paris, 1966, D262, 2.
Lee C. Y., Salundkh D. K. Observati-
on on degree of apospory in three
members of Andropogoneae.— Curr.
Sci., 1968, 37: 295—297.
Li-Ah-Lan, Prakash N. Life history of
Nepenthes gracilis— Malaysian J.
Sci., 1973, 2(A): 45—53.
Li Hi. Classification and phylogeny of
Nymphaeaceace and allied famili-
es.— Am. Midl. Nat., 1955, 54: 33—
AA,
Longevialle M. Embryogenie des Ca-
ryophyllaceae. Developpement de
Vembryon chez PArenaria controver-
sa Boiss—-C. r. Acad. sci. Paris,
1966a, D263: 1205—1208.
Longevialle M. Embryogenie des Cam-
panulacées. Développement de l’emb-
ryon chez lAdenophora bulleyana
Diels (Fisch.).— C. r. Acad. sci. Pa-
ris, 1966b, D262: 1527—1529.
Longevialle M Embryogénie des Cam-
panulacées. Développement de l’emb-
ryon chez Wahlenbergia hederacea
Reichb.— C. г. Acad. sci. Paris,
1968a, D267: 946—949.
Longevialle M. Embryogenie des Cam-
panulacées. Développement de Рет-
bryon chez le Specularia speculum
L. (DC).—C. r. Acad. sci. Paris,
1968b, D266: 461—464.
Longevialle M. Embryogénie des Ca-
ryophyllacées. Developpement de
Vembryon chez le Silene otites
Sm.— C. r. Acad. sci. Paris, 1968c,
0266, 20: 2071—2074.
Longevialle M. Embryogenie des Cam-
panulacées. Developpement de Гет-
bryon chez Specularia castellana
Lang — Acad. sci Paris, 1969, D269,
8: 838—840.
Longevialle M. Embryogenie des Ca-
ryophyllacées. Developpement de
Vembryon chez le Cucubalus bacci-
fer L.— C. r. Acad. sci. Paris, 1967,
0264: 268—271.
Lubbock J. A. Contribution to our
Knowledges of Secdlings. London,
1892.
322
Luxova M. Fertilization of Barley
(Hordeum distichum L.).— Biologia
plantarum (Praha), 1967, 9, 4:
304—307.
Ly-Thi-Ba. Embryogénie des ranuncu-
lacées. Developpement de l’embryon
chez le Cimicifuga europaea
Schips.— C. r. Acad. sci. Paris, 1974,
0272, 9, 1248—1254.
Ly-Thi-Ba. Embryogénie comparée et
phylogénie des Helléborées.— Rev.
обп. Воё., 1974, 81: 154—194.
Ly-Thi-Ba, Guignard J. Embryogénie
des aizoacées. Developpement de
Vembryon chez le Mollugo nudica-
ulis—C. r. Acad. sci, Paris, 1970,
D271, 12: 980—982.
Ly-Thi-Ba, Guignard J. L. Embryogé-
nie des Alismatacées. Devcloppe-
ment de l’embryon chez, le Dama-
sonium stellatum Thub—C. г.
Acad. sci. Paris, 41976, D282, 6:
549—552.
Mahabale T. S., Birader N. V. Studies
on palms: Embryology of Phoenix
sylvestris Roxb.— Proc. Indian
Acad. Sei, 1968, 67B, 2: 77—96.
Mahabale 7. S., Inamdar A. C. Mor-
phology and embryology of Aspara-
gus virgatus Baker.— Proc. In-
dian Acad. Sci., 1975, 81B, 4: 162—
169.
Mahalingappa M. S. Anther and male
gametophyte development in Tur-
nera ulmifolia Linn. (var. angusti-
folia Willd.).— Curr. Sci. (India),
1975, 44, 17: 640—641
Mahalingappa M. S. Microsporogene-
sis and male gametophyte in €yno-
don dactylon.— Curr. Sci. (India),
1978, АТ, 16: 594—596.
Maheshwari D. H. Embryological stu-
dies in СотрозЦае. ТУ. НеНав-
theae.— Proc. Indian Acad. Sei.
1963, B58, 5: 274- -290,
Maheshwari Devi HW. Embryology and
systematic position of Salvadora-
ceae.— In: Symp. Morphology, ana-
tomy and embryology of land
plants, 1969—72—73.
Maheshwari Devi H. Embryology of
Rauwolfia serpentina and Catharan-
thus roseus.— Curr. Sci., 1970, 39:
376—377.
Maheshwari Devi H. Embryology of
Apocynaceae. I. Plumiereae.— J. In-
dian Bot. Soc., 1971, 50, 1: 74—85.
Maheshwari Devi H. Salvadoraceae.
A study of its embryology and sy-
stematics —J. Indian Bot. Soc.
1972, 51, 1: 56—62.
Maheshwari Devi H. Embryology of
Jasminus and its bearing on the
Composition of Oleaceae.— Acta bot.
indian., 1975, 3, 1: 52—64.
Maheshwari Devi H., Lakshminaraya-
na K. Embryology of Oxystelma es-
culentum. Embryology of crop
plants: Indo-Soviet Symposium. Del-
Бу, 1976: 20—21.
Maheshwari Devi H., Lakshminaraya-
na K. Embryology of Oxystelma es-
culentum.— Phytomorphology, 1977a,
27, 1: 59—67.
Maheshwari Devi H., Lakshmanaraya-
na K. Embryological studies in Gen-
tianaceae.— J. Indian Bot. Soc., 1977b,
56, 2: 182—188.
Maheswari Devi H., Narayana L. K.
Embryology of two cultivars of Ne-
rium indicum Mill.— Curr. Sci. (In-
dia), 1975, 44, 17: 641—642.
Maheshwari Devi, Pullaiah T. Life
history of Basella rubra L. and taxo-
nomic status of the family Basella-
ceae.— J. Indian Bot. Soc., 1975, 54:
154—166.
Maheshwari Devi H., Pullaiah T. Emb-
ryology investigation in the Melam-
podinae I. Melampodium divarica-
+ tum.— Phytomorphology, 1976a, 21,
S: 77—86.
Maheshwari D. H., Pullaiah T. Emb-
ryology of Niger and sunflower.
Embryology of crop plants: Indo-So-
viet Symposium. Delhi, 1976b, 21—
22
Майезшат Ш. Н., Вата Вао М. 5.
Embryology of Suaeda maritima
(L.) Dumort.— Curr. Sci. (India),
1976, 45, 10: 383—385.
Maheshwari P. Embryology of angio-
sperms as a field of research.— Na-
ture, 1945, 156: 354—355.
Maheshwari P. The angiosperm emb-
ryo sac.— Bot. Rev., 1948, 14: 1—56.
Maheshwari P. An introduction to the
em oryclogy of Angiosperms. N. Y.,
50.
Maheshwari P. Embryology in relati-
on to taxonomy.— Vistas Bot., 1923,
4: 55—97.
Maheshwari P., Kapil R. N. Some
indian contribution to the em-
bryology of Angiosperms.— Phy-
tomerphology, 1966, 16, 3: 239—
294.
Maheshwari R. Embryology of Heli-
anthemum vulgare Gaerth.— Phy-
tomorphology, 1964 (1965), 14, 4:
547—557.
Майезйшат 5. С., Кари В. N. Mor-
21*
phological and embryological stu-
dies on the Lemnaceae. 1. The flo-
ral structure and gametophytes of
Lemna paucicostata.— Amer. J. Bot.,
1963a, 50: 677—686.
Maheshwari S. C., Kapil R. N. Morpho-
logical and embryological Studies
on the Lemnaceae. 2. The endo-
sperm and embryo of Lemna pauci-
costata.— Amer. J. Bot., 1963b, 50:
907—914.
Maheshwari S., Maheshwari N. The
female gametophyte endosperm and
embryo of Spiridela polyrrhiza.—
Beitr. Z. Biol. Pflanzen. dtsch., 1963,
39: 379—388.
Maheshwari J. K. Cleistogamy in An-
giosperms.— In: Proc. Summer
School. Bot. Darjeeling., 1966. 145—
155.
McNaughton Judith E. Embryology of
Erythrina caffra Thunb. sporogene-
sis and gametogenesis.— J. S. Afr.
Bot., 1976, 42, 4: 395—400.
Makde K. H. Gamethophytes and fer-
tilization of Indegofera pulchella
Roxb.— J. Indian Bot. Soc., 1971, 50,
4: 308—311.
Make K. H., Mukherjee P. K. Study of
seed coat and fruit wall in Eupho-
rbia heterophylla Linn.— In: Proc. °
56th Indian Sci. Congr. Bombay,
1969, 3: 362.
Malecka J. Cytoembryological studies
in the Taraxacum scanicum Dt.—
Acta biol. cracov. Ser. bot., 1967, 10,
2: 195—206.
Malecka J. Cyto-taxonomical and emb-
ryological investigation on a natu-
ral hybrid between Taraxacum kok-
saghiz Rod. and T. officinale Web.
and their parent species.— Acta bi-
ol. cracov. Ser. bot., 1971, 14, 2:
179—197.
Malecka J. The course of differenti-
ation of the endosperm of Echium
vulgare L.— Acta biol. cracov. Ser.
Ъо+., 1975, 18: 13—26.
Malecka J. The course of differenti-
ation of the endosperm of Lithospe-
rmum arvense.— Acta biol. cracov.
Ser. bot., 1977, 20: 34—40.
Magqbool B. Studies on the embryolo-
gy of Utricularia graminifolia
Vahl.— Curr. Sci., 1965, 34: 355—
306.
Marinos N. G. Embryogenesis of the
pea (Pisum ‘sativum). I. The cyto-
logical environment of the develo-
ping embryo.— Proloplazma, 1970,
10, 3/4: 261—279.
323
Martin F. W. The stigmatice cxudate
of Strelizia— Phyton (B. Aires),
1970, 27: 47—51.
Martinson V. Polyembryony in Theo-
broma cacao L.— Ann. Bot., 1972,
36, 148: 947—954.
Masand P. Zygophyllaceae. Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967: 47.
Masand P. Zygophyllaceae: Sympo-
sium on comparative embryolo-
gy of Angiosperms., 1970, 41:
123—126.
Mathew C. J. Development of male
and female gametophytes in Camel-
lia sasangua.— Phytomorphology,
1978, 28, 3: 262—269.
Mathew C. J, Chaphekar М. Develop-
ment of female gametophyte and
embryology in Stachyurus chinen-
sis.— Phytomorphology, 1977, 27,
4: 68—79.
Mathur S. L. Development of the ma-
le and female gametophytes of Ca-
lycanthus fertilis— Walt. Proc. Nat. -
Inst. Sei. India, 1968, B34, 6:
323—329.
Mathur 5. L. Embryology of Melian-
thus major L. I. Microsporangium
and pollen..—Curr. Sci. (India),
1977, 46, 23: 820—821.
Maugini E. Sul’embriologia di Leon-
topodium nivale (Ten) Huet ex
Hand. Mazz.—G. bot. ital., 1967,
101, 3: 185-—187.
Mauriizon Jf. Uber die systematische
Stellung der Familien Hydrostachy-
aceae und Podostemonaceae.— Bot.
Notiser, 1933: 172—180.
Maury G. Differents types de polyem-
bryonie chez quelques dipterocar-
pacées Asiatiques.— Bull. Soc. hist.
natur. Toulouse, 1970 (1971), 106,
3—4: 282—288.
Maze J. Bohm Г. R. Comparative
embryology of Stipa elmeri (Gra-
mineae).— Canad. J. Bot. 19713,
51, 1: 235—247.
Maze J., Bohm I. R. Comparative
embryology of Stipa elmeri.—
Canad. J. Bot., 41974, 92: 365 —
379.
Maze J., Bohm L. R. Embryology of
Festuca microstachys (Grami-
neae).— Canad. J. Bot., 41977, 59,
13 : 4768—4782.
Maze Т, Войт Г. R., Mehlenba-
cher L. E. Embryo sac and early
ovule development in Oryzop-
sig miliacea and Stipa torlilis.—
Canad. J. Bot., 1970, 48, 4: 27.
324
Mazzolani G. Osservazioni sulli svi-
luppo del gametofito femminile in
Olea europaea L.— Ann. bot., 1964,
27, 1: 140—119.
Mehlenbacher L. E. Floret develop-
ment, embryology and systema-
tic position of Oryzopsis hendero-
ni (Gramineae).— Canad. J. Bot.,
1970, 48: 1741—1758.
Melchior H. A. Епфегз Sylla-
bus der Planzenfamilien. Berlin,
1964.
Mendoza Sister Rosalinda C.° Embry-
ogenesis in Amaranthus spinosus
L. and Amaranthus viridis Г. Phi-
lipp.— J. Sei, 1973, 103, 4: 243—
268.
Menezes N. L. Megasporogénese, me-
gagametogénese e embryogénese em
Velloziaceae.— Bol. bot. inst. bei-
osci., 1976, 4: 41—59.
Mepham R. H., Lane G. R. Formati-
on and development of the tape-
tal periplasmodium in Tradescan-
tia bracteata.— Protoplasma, 1969,
68, 1/2: 175—192.
Mercy 5. Т., Kakar S. N., Varghe-
5е Т. М. Mmbryology of Cicer arie-
tinum and C. soongaricum — Bull.
Torrey Bot. Club., 1974, 101, 4:
26—30.
Mestre J. C. Recherches d’embryo-
génie comparée: les raports phylo-
génétiques des composées: Thés.
doct. Fac. pharmacie Univ. Paris,
1964, 399 : 1—137.
Mestre J. C. Embryogenie chez Ce-
lastraceae. Developpement Гет-
bryon chez Evonimus eurcopeus.—
C. r. Acad. sci. Paris; 1965a, D260,
12 : 3453—3456.
Mestre J. C. Embryogénie des acéra-
cees. Development de Vembryon
chez VAcer pseudoplatanus Г.— С.
г. Acad. sci. Paris, 1965b, D261,
25 : 5602—5604.
Mestre J. C. Embryogénie des meli-
acées. Le developpement de l’em-
bryon chez le Cedrela sinensis
L—C. r. Acad. sci. Paris, 1966,
D262, 10 : 1074—1076.
Mestre J. C. La_ signification phy-
logenétique de lEmbryoge-
nie.— Rev. gen. bot, 1967, 74,
213—322.
Mestre J. С, Guignard J. L. Em-
bryogénie des scrofulariacées. Le
developpement de VTembryoa chez
le Zaluzianskya capensis Walp.—
CG. r. Acad. sci. Paris, 1969, D268,
26 : 3170—3172.
Mestre J. C., Guignard J. L. Embry-
ogénie des cornacées. Developpe-
ment de lembryon chez Aucuba
Acad.
japonica ТВопЪ.— С. т.
561. Рагз, 1970, 0274, 16: 1358—
1360.
Mestre J. C., Guignard J. L. Embry-
ogenie des umbelliferes. Deve-
loppement de VTembryon chez le
Daucus carota—L. C. т. Асад.
sci. Paris. 1974, D272, 10: 1383—
1386.
Miceli P., Renzoni G. C. Alethenia
filiformis Petit subsp. filiformis
(Zannichelliaceae): analisi morfolo-
gica ed embriologica.— Atti. Soc.
tosc. Sci. Natur. Mem., 1978, B,
85 : 233—249.
Mihov A., Zagorchaeva L. A _ case
of apogamy in Cucumis sativus
L.— Comptes Rendus de lAcade-
mie Bulgare des Sciences, 41966,
19, 6: 527—529.
Miodzianowski F. The structure and
the later stages of development in
the embryosac of Thamnochortus
fruticosus Berg (Restionaceae).—
Bull. Soc. amis. sci. et lett. Poz-
nan, 1963 (1964) D, 4: 3—11.
Misra R. C. Morphological studies on
Sisymbrium irio L—J. Indian
Bot. Soc., 1966, 45: 14—23.
Misra R. C. Plantaginaceae. Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967:
105
Misra R. C. Plantaginaceae. Sympo-
sium on comparative embryology
of Angiosperms. New Delhi, 1970,
41 : 298—305.
Misra 5. Floral morphology of the
family Compositae. IV Tribe Ver-
nonieae — Vernonia anthelminti-
ca.— Bot. Mag (Tokyo), 1972, 85:
: 187—199.
Misra R. C., Sahu R. C. Morphologi-
cal studies in Phaseolus I. Embry-
ology and _ seed structure in Pha-
seolus aureus.— In: Proc. 56th In-
dian Sci. Congr. Bombay, 1969,
3: 39.
Mitchell J. P. Megasporogenesis and
microsporogenesis in Vicia faba.—
Canad. J. Bot., 1975, 53, 2: 2804—
2812.
Mitra K. Pollen morphology in rela-
tion to taxonomy in some Indian
Capparidaceae.— In: Proc. 56th In-
clan Sci. Congr. Bombay, 1969,
3: 34.
Mitra J. N. A contribution to the
embryology of some Compositae.—
J. Indian Bot. Soc., 1974, 26 : 105—
123.
Miyashita В. К., Nakasone I. Y.,
Lamoureux С. Н. Reproductive
morphology of Acerola (Malpighia
glabra L.).— Techn. Bull. Haw. Agr.
Exp. Stat. Univ. Haw., 1964, 63:
31
Mogensen П. L. Megagametophyte
development in Agave Parryi.—
Phytomorphology, 1970, 20, 1: 16—
22
Mohamed A. K., Gould F. W. Bio-
systematic studies in the Boteloua
curtipendula Complx. V. Megaspo-
rogenesis and embryo sac develop-
ment.— Amer. J. Bot., 1966, 53:
166—169.
Mohana Rao P.R., Sood S. K. Life
history of Saturium nepalense
(Orchidaceae).— Norw. J. Bot.,
1979a, 26: 285—294.
Mohana Rao P. R., Sood S. K. Em-
bryology of Habenaria densa (Or-
chidaceae).— Bot. Notiser, 1979b,
132 : 145—148.
Mohana Rao P. R., Sood S. K. De-
velopment of the Embryo in Her-
minium angustifolium Benth.—
Ann. Bot. 1979c, 43 : 661—663.
Mohan K. M. The ovule and embryo
sac development in Browallia de-
missa L.— Proc. Indian Acad. Sci.,
1966, 64B, 1: 26—31.
Mohan K. M. Morphological studies
in Solanaceae. II. Morphology,
development and structure of seed
of Browallia demissa L.— Proc.
Nat. Inst. Sci. India, 1968, B34, 3:
142—148. .
Mohan K. M., Singh B. Morphologi-
cal studies in Solanaceae. III.
Structure and development of seed
of Solanum torvum Sw.— J. Indian
Bot. Soc. 1969, 48, 3/4:338 —
345,
Mohan Ram H. Y., Ravindra Nath.
The morphology and embryology
of Cannabis sativa L.— Phytomor-
phology, 1964, 14, 3: 414—429.
Mohini Munshi, Mond Faroog. The
embryology of Stigmaphyllon peri-
plocaefolium A. Juss. Seminar on
morphology, anatomy and embryo-
logy of land plants. Delhi, 1969:
73—14. ,
Mohini Munshi, Tabat Wasseem, Fa-
roog M. The embryology of Stig-
maphyllum periplocaefolium. A.
Juss. and the cause of its sterili-
320
ty.— Hort. 1972—1974, 9:
55—57.
Moissel E. Vergleichende embryologi-
sche Studien uber die Familie der
Caprifoliaceae.— Osterr. bot. Zeit.,
1941, 90: 153—212.
Молхова Е. Проучване върху фор-
мирането на ембриосака, опраш-
ването и оплождането при Сарз1-
cum annuum L.— Изв. инст. расте-
ниевод., 1963, 17: 19—56.
.Молхова E., Филева М. Проучвапе
върху развитисто на зародыша и
ендосперма в различно плоидни
видове и межувидови хибриди в
роде Сарясит.— Генетика и се-
лекция (ПРБ), 1969, 2, 2:77—85.
Мотт А. В. Оп Embryology formu-
lae of some Convolvulaccae mem-
bers.— Sci. and Cult., 1975, 44, 7:
318—319.
.Mortier J. Embryogenie des malva-
cées. Developpement de l’embryon
chez JVAlthaea officinalis.— Bull.
Soc. bot. France, 1969 (1970), 116,
9: 505—509.
Mortier J. Developpement de l'em-
bryon chez quelques malvinées
indigénes.— In: С. г. 93е Congr.
nat. soc. Savantes. Tours, 1969.
Sec. Sci. Paris, 1972, t. 3:509—
520.
Mortier J., Lebegue A. Embryogénie
des Labiées. Développement de
VYembryon chez le Ballota foetida
Lamk.— CG. г. Acad. sci. Paris,
1966, D262, 1: 87—91.
„Москова Р. Морфолого-апатомични,
кариологични и цитоембриоло-
гични проучвания при Koeleria
nitidula Vel. Генетика и селек-
ция (НРБ) 1969, 2, 3: 169—179.
Москова Р., Христов М. А. Върху
ембриологията па разпростране-
ни У нас видове от рода Тата-
их 1. Научни тр. высший с.-х.
ин-т им. Г. Димитрова, Сер. Pa-
стениев., (София), 1968, 20(48):
61—86.
Москова Р., Христов М. А. В эм-
бриологии распространенных в
Болгарии видов рода Tamarix
Г..— Научни тр. Высш. с.-х. ин-та
им. Димитрова. Сер. Растениев.,
(София), 1969, 20: 161—186.
Москова Р., Мандилова С. Цитоем-
бриологично и кариологично про-
Учване upu Panicum miliaceum L.
сорт Мироновско 85.— Генетика
и селекция (НРБ), 1970, 3, 6:
485—491.
Adv.,
326
Mukherjee P. K. Trends of speciali-
zation in Euphorbiaceae. II. The
female gametophyte.— In: Proc.
o6th Indian Sci. Congr. Bombay,
1969a, 3 : 395—396.
Mukherjee P. K. Trends of speciali-
zation in Euphorbiaceae. III. Em-
bryo and endosperm.—In: Proc.
o6th Indian Sci. Congr. Bombay,
1969b, 3: 361.
Mukherjee P. K. Pollen morphology
of some Indian Clerodendrum L.—
In: Proc. 56th Indian Sci. Congr.
Bombay, 1969c, 3: 344.
Mukherjee P. K. Embryological stu-
dies in Polygonum barbatum L.—
J. Indian Bot. Soc., 1972, 51, 2:
106 : 142.
Makherjee S. K. Vivipary in mul-
berry.— Sci. and Cult., 1960, 26,5:
234,
Mukkada A. J. Some aspects of the
morphology, embryology and _ bio-
logy of Terniola zeylanica (Gard-
ner) Tulasne.— New Phytol., 1969,
68, 4: 1145—1158.
Mukkada A. J., Chopra R. N. Post-
fertilization development in Indo-
tristicha ramosissima (Wight.) van
Royen.— New Phytol., 1973, 72,3:
639—646.
Muniyamma M. Occurence of twin
embryo sacs in Themeda cymba-
ria Hack.—In: Proc. 56th Indian
Sci. Congr. Bombay, 1969, 3: 356.
Muniyamma M. A. cytoembryologi-
calstudy of Agrostis pilosula.—
Canad. J. Bot., 1976, 54, 21: 2490—
2496. .
Maniyamma M., Phipps J. B. Cyto-
logical proof. of apomixis in Cra-
taegus (Rosaccae).— Amer. J. Bot.,
1979, 66, 2: 149—155.
Murty Y. S. Chloranthaccae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967a: 32.
Murty УТУ. S. Saururaceae: Semi-
nar Comparative Embryology of
angiosperms. New Delhi, 1967b:
30.
Murty Y. S. Piperaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967c: 31.
Murty Y. S., Kumar V. Development
of the female gametophyte and
embryo in Fimbristylis diphylla
Vahl.— Proc. Indian Acad. Sci,
1967d, 65B, 5: 185—194.
Nagabhushana Rao Sindhe. Repro-
ductive behaviour in Penisetum
squamulatum: Crop. plants. Indo-
Soviet symposium. Delhi, 1976:
41—42.
Nagaraj M., Fathima T. Studies on
the structure and development of
pericarp and seed coat in Rotula
aquatica Lour.— Proc. Indian Acad.
Sei, 1971, 74B, 6: 314—318.
Nagaraj M., Fathima T. Embryologi-
cal studies in Rotula aquatica Lo-
ur.— Proc. Indian Acad. Sci., 1967,
66В, 3: 106—116.
Nagaraj M., Fathima T. A note on
the sporogenesis and gamctogenc-
sis in Adeloceryum.— Current
Sci. (India), 1968, 37, 9 : 265—267.
Nagaraj M., Nijalingappa B. H. Em-
bryological studies in Myriophyl-
lum intermedium DC.— Proc. In-
dian Acad. Sci., 1967, 65B, 2: 210—
220.
Nagaraj M., Nijalingappa B. H. So-
me aspects of embryology in Cy-
perus alopecuroides Rottb.— Curr.
Sci. (India), 1968, 37, 41: 324—325.
Nagaraj M., Nijalingappa B. H. Spo-
rogeneses and development of ga-
metophytes in Hypolytrum latifo-
lium Г. C, Rich.— Curr. Sci. (In-
dia), 1972, 44, 7: 260—261.
Nagaraj M., Nijalingappa B. I. Em-
bryological studies in Cyperus
alopecuroides Rottb.— Proc. Indi-
an Acad. Sci., 1973, 77B, 6: 252—
263.
Nagaraj M., Nijalingappa B. H. Em-
bryological studies in Laurember-
gia hirsuta. Bot. Gaz. 1974, 1:
19—28.
Nagaraj M., Nijalingappa В. Н.,
Tejavathy D. H. A note on the
embryology of Lipocarpha sphace-
lata Kunth. (Cyperaceae).— Curr.
Sci., 1976, 45, 2: 67—69.
Nagaraja Rao A., Shamanna S. Flo-
wer and seed development in
Arundina graminifolia.— Phytomo-
rphology, P. Maheshwari Mem.
Vol. 1967, 17 : 191—300.
Nagendran C. R. Is the embryo sac
of Podostemaceae bisporic? — Curr.
Sci. (India), 1974, 43, 8 : 259—260.
Nagendran C. R., Arekal G. D. Em-
bryo sac of Griffithella hookeria-
na. A_ reinvestigation.— Phytomor-
phology, 1976, 26, 4: 359—363.
Nagendran C., Arekal G., Subrama-
пуат K. Embryo sac studies in
three indian species of Polypleu-
rum (Podostomaceae).— Plant.
Syst. and Evol. 1977, 128, 3—4:
215—226.
Nagendran C. R., Subramanyam К.,
Arekal G. D. Development of the
female gametophyte in Hydrobry-
um griffithii (Podostemaccac).—
Ann. Bot., 1976, 40, 167 :541—513.
Nair N. С. Development of the ovule:
in a species Cissus.— Nature, 1965,
208, 402.
Nag K. K., Nijalingappa B. H. Km-
bryological studics in Myriophyl-
lum intermedium DC.— Proc. Indi-
an Acad. Sci., 1967, 65: 210—220.
Nair М. C. Coriariaceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967a:
54
Nair N. C. Leeaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967b : 60.
Nair N. C. Meliaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967c : 53.
Nair N. C. Rhamnaceae: Seminar
Comparative Embryology of An-
giosperms. New Delhi, 1967d: 58.
Nair N. C. Vitaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967e: 59.
Nair N. C. Contribution to the flo-
ral morphology and embryology
of two species of Leea with a dis-
cussion on the taxonomic position
of the genus.— J. Indian Bot. Soc.,
1968, 47, 3/4: 193—205.
Nair N. C. Leeaceae: Symp. on Com-
parative Embryology of Angio-
sperms, 1970a, 44: 180—184.
Nair N. C. Meliaceae: Symp. on Com-
parative Embryology of Angio-
sperms, 1970b, 44: 154—155.
Nair N. C. Rhamnaceae: Symp. on
Comparative Embryology of An-
giosperms, 1970c, 44: 168—173.
Nair N. C. Vitaceae: Symp. Compa-
rative Embryology of Angiosperms,
1970d : 174—179.
Nair N. C., Bajaj Y. P. Floral mor-
phology and embryology of Cyp-
hostemma setosa (Wall.) Aeston
and a discussion on the taxono-
mic position of the genus Cypho-
stemma.— J. Indian Bot. Soce.,
1966, 45: 108—115.
Nanda K., Gupia S. C. Endothecium
in Pyrostegia venusta.— Curr. Sci.
(India), 19741, АО, 17: 470—471.
Narang A. K., Chandra N. Structu-
re and development of sced coat
in Crotalaria and Tephrosia speci-
es.— Proc. Indian Acad. Sci., 1977,
85B, 5 : 283—291.
327
Матауапа Н. 5. Capparaceae: Semi-
nar Comparative Embryology of
Angiosperms, New Delhi, 1967a:
37
Narayana H. S. Moringaceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967b:
38
Narayana H. S. Capparaceae: Symp.
on Comparative Embryology of An-
giosperms. New Delhi, 1970c, 41:
81—83.
М№агауапа Н. 5., Гоара В. С. Ет-
bryology of Glinus lotoides L.—
Proc. Indian Acad. Sci, 1972,
158, 2:77—85.
Narayana H. S., Prakasa Rao C. G.
Floral morphology and embryolo-
gy of Seetzerria orientalis De-
che.— Phytomorphology, 1963, 43.
1: 197—205.
Narayana H. S., Sharma M. K. Em-
bryology of fruit crops of Rosa-
сеае. Embryology of crop plants:
Indo-Sovict Symp. Delhi, 1976:
28—29.
Narayana L. L. Embryology of Stap-
hyleaceae.— Current. Sci., 1960,
29: 403—404.
Narayana L. £. Contribution to the
embryology of Balsaminaceae.—
J. Jap. Bot., 1965, 40, 4: 104—116.
Narayana Г. Г. Balsaminaceae: Sce-
minar Comparative Embryology
of Angiosperms. New Delhi, 1967a:
55
Narayana Г. L. Erythroxylaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967b:
49
Narayana L. Г. Geraniaceae: Sc-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967c:
45
Narayana LL. Г. WUinaceae: Seminar
Comparative Embryology of An-
giosperms. New Delhi, 1967d: 48.
Narayana Г. LE, Oxalidaceae: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967e, 3:
44
Narayana Г. Г. Тгораео]асеае: Se-
minar Comparative Embryology
of Angiosperms. New Delhi, 1967f:
46.
Narayana L. L. Balsaminaceae: Symp.
on Compartive Embryology of An-
giosperms. New Delhi, 1970a, 41:
158—162.
Narayana Г. Г. Erythroxylaceae:
Symp. on Comparative Embryolo-
328
gy of Angiosperms. New Delhi,
1970Ъ, 41: 133—135.
Narayana L. L. Geraniaceae: Symp.
on Comparative Embryology of
Angiosperms. New Delhi, 1970c,
44 : 147—120.
Narayana ZL, Г. Шпасеае: Зутр. оп
Comparative Embryology of An-
giosperms. New Delhi, 1970d, 41:
127—132.
Narayana L. L. Oxalidaceae: Symp.
on Comparative Embryology of
Angiosperms. New Delhi, 1970e,
44: 114—116.
Narayana L,. L. Tropaeolaceae: Symp.
on Comparative Embryology of
Angiosperms. New Delhi, 1970,
44 : 124—122.
Narayana L. Г. А contribution to
the floral anatomy and embryolo-
gy of Ochnaceae.— Jap. J. Bot.,
1975, 44: 329—336.
Narayana L. L,, Sundari K. T. Em-
bryology of Pittosporaceae I.—
Jap. J. Bot., 1977a, 52: 204—209.
Narayana L. L., Sundari K. T. Em-
bryology of Pittosporaceae II.—
Nat. Acad. Sci., 1977b, 52.
Narayana L. L., Sundari K. T. Em-
bryology of Pittosporaceae III.—
Proc. Indian Acad. Sci., 1978, 87b,
8 : 205—214.
Narayanan C. R. Polyembryony in
Streblus taxoides (Heyne) Kurz.—
J. Indian Bot. Soc., 1968, 47, 3—
4: 354—358.
Narayan K. N., Shukur A. Gameto-
genesis in the genus Blumea.—
In: Proc. 56th Indian Sci. Congr.
Bombay, 1969, 3: 354.
Narayanaswami S., Roy S. K. Em-
bryology of the genus Psidium.—
J. Indian bot. Soc., 1960, 39: 35—
45.
Natesh S., Bhandari N. N. Two ty-
pes of Embryo sac development
in Scrophularia himalensis Roy-
le— Ann. Bot. 1975, 39, 162:
129—732.
Netolitzki F. Anatomie der Angio-
spermen Samen.—In: Pfl. Anat.,
Linsbauer Handb., 1926, 10.
Neubauer B. F. The development of
the achene of Polygonum pensyl-
vanicum: embryo endosperm and
pericarp.— Amer. J. Bot, 1971,
58 : 655—664.
Nijalingappa B. H. Morphology stu-
dies in Myriophyllum intermedi-
um DC.— Curr. Sci., 1967, 36: 104—
105.
Nijalingappa B. H. Embryological
studies in Haloragis micrantha.—
Phytomorphology, 1975, 25, 2:
146—155.
Nijalingappa B. H. Sporogenesis and
gametogenesis in some Сурега-
ceae.— Proc. Indian Acad. Sci,
1976, 838A, 2: 66—72.
Nijalingappa B. H. Post-pollination
studies in some Cyperaceae.—
Proc. Indian. Acad. Sci, 1977,
85B, 6 : 391—398.
Nijalingappa B. H., Devaki N. Em-
bryological studies in Diplacrum
caricinum В. B.—Beitr. Biol.
Pflanz., 1978, 54, 2: 215—225.
Nijalingappa B. H., Tejavathi D. H.
Embryo development in Scirpus
mucronatus L.— Curr. Sci. (India),
1977, 46, 3:84—85.
Nikam E. R. Embryological studies
in Тасепама siceraria Standl.—
Curr. Sci. (India), 1976, 45, 7:
250—253.
Nogler G. A. Apomixis in Ranuncu-
lus auricomus: Indo-Soviet. Sym-
posium. Embryology of crop
plants. Delhi, 1976: 29.
Nordborg G. Embryological studies
in the Sanguisorba minor com-
plex (Rosaceae).— Bot. Notiscr,
1967, 120, 1: 109—119.
Obuchowska В. Вадаша kariologi-
czne nad rozwojem tapetum и ру
nikach Galinsoga parviflora Cav.
Shidia.— Soc. Sci. torun., 1969,
8D, 5/6 : i—16.
Ojchomon O., Sanyaolu M. O. Ovule
formation and embryo develop-
ment in persisting and abortive
fruits of the cowpea Vigna ип-
guiculata (L.) Walp.— Niger. J.
Sci., 1970, 4, 1:341—40.
Paal H., Gracza P. Contributions to
the seed development of field po-
ppy (Papaver rhoeas_ L.).— Acta
agr. Acad. sci. hung., 1969, 18, 3—
4: 387—390.
Pacini E. Sviluppo del gametofito
maschile di Olea eropaea. Osser-
vazioni embriologiche.—G. Bot.
ital., 1977, 1441, 6: 377.
Pacini E., Sareatti G. The repro-
ductive calendar of Lycopersicum
peruvianum Mill— Bull. Soc. bot.
France, 1978, 125, 1—2: 295—299.
Padhye M. D. Male and female га-
metophytes of Eleocharis genicu-
lata Roem. et Schult.— Proc. In-
cian Acad. Sci., 1968, 67B, 2 :54—
Padhye M. D. Studies in Cypera-
ceae. VI Contribution to embryo-
logy of the genus Fimbristylis
Vahl.—In: Proc. 56th Indian
Sci. Congr. Bombay, 1969. 3: 391.
Padhye M. D. Studies in Cyperaceae.
Ill. Life history of Kyllinga bre-
vifolia Rottb. with a brief discus-
sion on systematic position of
Kyllinga.— Bot. Gaz., 1974, 132:
172—179.
Padhye M. D., Chaube S. N. Studies
in Cyperaceae. VIII. Gametophy-
tes and fertilization in Scirpus
corymbosus Heyne.—In: Proc,
56th Indian Sci. Congr. Bombay,
1969, 3: 362—363.
Padhye M. D., Chaube S. N,,
Jyer A. Studies in Cyperaceae.
VIII. Gametophytes and fertiliza-
tion in two members of Cypera-
ceae.— J. Indian Bot. Soc.. 1970,
49, 1—4: 86—92.
Padhye M. D., Geed Г. N., Ka-
иг В. В. Studies in Cyperaceae.
X. Gametophytes and fertilization
in Cyperus squarrosus L.— In:
Proc. 56th Indian Sci. Congr. Bom-
bay, 1969, 3: 363—364.
Padhye M. D., Jyer A. V. Gameto-
phytes of Fimbristylis monosta-
chya (L.) Hassk.— In. 56th Indi-
an Sci. Congr. Bombay, 1969, 3:
363.
Padhye M. D., Kasture S. W. Stu-
dies in Сурегасеае. У. Gameto-
phytes and fertilization in Pycre-
us globosus (All) var. miligiri-
cus (Stead) C. B. ClL—J. Biol.
Sci., 1970. 13, 2:37—42.
Padhye M. D., Yawakar V. V. An-
dankar D. W. The gametophytes
of Alysicarpus hamosus Edgev.—
In: Proc. 56th Indian Sci. Congr.
Bombay, 1969, 3: 364.
Padmanabhan D. The octant of Epi-
Фета carnosum Benth.— Proc.
Indian Acad. Sci., 1966, 64B:
283—292.
Padmanabhan D. Embryogenesis in
Epithema carnosum J.— Indian
Bot. Soc., 1967a, 46, 1: 90—408.
Padmanabhan D. Nymphaeaceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi,
1967b : 28.
Padmanabhan ‘D. Verbenaceac: Se-
ininar Comparative Embryology of
Angiosperms. New Delhi, 1967e:
96.
329
Padmanabhan D. Development from
zygote to seedling in Tridax pro-
cumbens L.—J. Indian Bot. Soc.,
1968, 47, 1—2:94—112.
Padmanabhan D. Nympheaceae:
Symp. on Comparative Embryolo-
gy of Angiosperms. New Delhi,
1970a, 44: 59—62.
Padmanabhan D. Verbenaceae: Symp.
on Comparative Embryology oi
Angiosperms. New Delhi, 1970b,
44 : 250—254.
Padmanabhan D. Embryo develop-
ment in Sesamum indicum: Indo-
Soviet Symposium. Embryology
of crop plants. Delhi, 1976: 30.
Padmanabhan D., Ramji M. V. De-
velopmental studies on Cabomba ca-
roliniana Gray. II. Floral anato-
my and microsporogenesis.— Proc.
Indian Acad. Sci., 1966, 64B, 4:
246—223.
Pahuja A. N., Kumar V. Embryo
sac development, cytology and
systematic position of Clintonia.—
Phytomorphology, 1970, 20, 2:
97— 102.
Pal A. Structure and development
of seed in Celosia cristata L.— J.
Indian Bot. Soc., 1969, 47, 3—4:
381—387.
Pal P. К. Boraginaceae: Seminar
Comparative Embryology of An-
giosperms. New Delhi, 1967: 95.
Pal S. Microsporogenesis of Stella-
ria aquatica (Scop.).— Curr. Sci.
(India), 1974, 43, 6: 192.
Pal S., Murty Y. S. Structure and
development of female gametophy-
te and embryo in Stellaria aqua-
tica Scop.—J. Indian Bot. Soc.,
1972, 51, 83—4 : 232—234.
Palser B. F. Studies of floral mor-
phology in the Ericales. VI. The
Diapensiaceae.— Bot. Gaz. 1963,
124 : 200—219.
Palser B. F. The basis of Angio-
sperm phylogeny: Embryology.—
Ann. Missouri Bot. Gard., 1975,
62, 3: 621—646.
Panchaksharappa M. G. Embryologi-
‘cal studies in some members of
Zingiberaceae. II. Elettaria carda-
monum, Hilchena caulina and
Zingiber macrostachyum.— Phyto-
morphology, 1966, 16, 4: 412—417.
Panchaksharappa M. G. Zingibera-
ceae: Seminar Comparative Em-
bryology of Angiosperms. New
Delhi, 1967 : 129.
380
/
Panchaksharappa M. G. Zingibera-
ceae: Symp. on Comparative Em-
bryology of Angiosperms. 1970,
41: 380—385.
Pandey B. P., Saxena N. P., Struc-
ture of gametophytes in Litho-
phragma affinis Gray.— Acta Bot.
Indica, 1977, 5: 207—209.
Pandey A. K., Singh R. P. Develop-
ment of female gametophyte in
Hymentherum tenuifolium Cass.—
Curr. Sci. (India), 1976, 45, 4 :29-—
30.
Panicker T. K. Female gametophyte
and endosperm in Lagenandra ova-
ta Thw.— Curr. Sci. (India), 1965,
34, 21: 6414—615.
Parulekar N. K. Annonaceac: Semi-
nar Comparative Embryology of
Angiosperms. New Delhi, 1967: 44.
Parulekar N. K. Annonaceae: Symp.
on Comparative Embryology of
Angiosperms, 1970, 41: 38—44.
Parvathi А., Narayana Е. Т,,
Dutt B. S. Embryology of Cuscuta
chinensis Lamk.— Nat. Acad. Sci.
Lett., 1978, 1—12 : 433—435.
Parveen A., Iftikhar N., Khan N. R.
The megasporogenesis and the de-
velopment of embryo sac in Wi-
thania somnifera.— Pakistan J. Sci.
and Ind. Res., 1972, 15, 3: 196--
198.
Patel T. K., Cook C. D. Embryologi-
cal studies on Ranunculus tripar-
titus D. С. (Ranunculaceae).—
Bot. Jahrb. Syst. Pflanzengesch.
und Pflanzengeorg., 1972, 91, 4:
438—461.
Periasamy K. Studies on seeds with
ruminate endosperm. VI. Rumina-
tion in the Araliaceae, Aristolo-
chiaceae, Caprifoliaceac and Ebec-
naceae.— Proc. Indian Acad. Sci.,
1966, 64B, 3: 427—134.
Periasamy K., Parameswaran N. A
contribution to the floral morpho-
logy and embryology of Tarenna
asiatica.— Beitr. Biol. Pflanz.,
1965, 44: 123—138.
Phatak V. G. Embryology of Zygop-
hyllum coccineum L. and Z. faba-
go L.— Proc. Kon. ned. acad. we-
tensch., 1971, 74C, 4:379—397.
Phelouzat R. La gametogenese etla
fertilité dans les fleurs femelles
et les fleurs hermaphrodites du
Poterium sanguisorba L.— Bull.
Soc. Bot. France, 1965, 112, 7—8:
370—378.
Philip V. I. The origin of cotyledon
and epicotyl in Elettaria cardamo-
num.— Austral. J. Bot., 1968, 16:
421—431.
Philipson М. N. Haustorial syner-
gids in Cortaderia (Gramineae).—
N. Z. J. Bot., 1977, 15, 4: 777—779.
Philipson M. N. Apomixis in Corta-
deria jubata (Gramineae).—N. Z.
J. Bot., 1978, 16, 1: 45—59.
Philipson W. R. Ovular morphology
and the major classification of the
dicotyledons.— Bot. J. Linnean
Soc., 1974, 68 : 89—108.
Philipson W. R., Philipson M. N. A
comparison of the embryology of
Forstera L. and Donatia J. R. and
G. Forst.—N. Z. J. Bot., 1973, 11.
3: 449—460.
Pogers S. Some observation on the
reproduction of Tridax procum-
bens L. II. Development of the
embryo sac and embryo.— Cytolo-
gia, 1969, 34, 2: 196—204.
Pogliani M. Ricerche embriologiche
in Anacyclus radiatus Lois.— Ann.
Bot., 1966, 28, 3: 725—729.
Poddubnaja ТУ. [Поддубная В.].
Spermatogenesis bei einigen Com-
positen.— Planta, 1927, 4, 3: 284—
298.
Poddubnaja-Arnoldi ТУ. А. [Поддуб-
ная-Арнольди В. А.]. Ein Ver-
such der Anwendung der embryo-
logischen Methode bei der Losung
einiger syslematischer Fragen.—
Beih. Bot. Zbl., 19341, 48.
Poddubnaja-Arnoldi V. A. {[foddy6-
ная-Арнольди В. A.| Geschlech-
tliche und ungeschlechtliche Fort-
pflanzung bei einigen Chondri-
llaarten.— Planta, 1933a, 19, 4: 46—
86.
Poddubnaja-Arnoldi Т. A. [Поддуб-
ная-Арпольди В. A.|. Kunstliche
Kultur und Zytologische Untersu-
chung des Pollenschlauches von
Senecio platanifolius | Benth.—
Planta, 1933b, 19, 2: 299—304.
Poddubnaja-Arnoldi V. (Поддубния-
Арнольди В.|]. Spermazellen in
der Familie der Dipsacaceac.—
Planta, 1934, 21, 4:381—386.
Poddubnaja-Arnoldi V. [Поддубная-
Арнольди B.|. Beobachtungen
liber die Keimung des Pollens eini-
ger Pflanzen auf kiinstlichen
Nahrboden.— Planta, 1936, 25:
502—529.
Ponzi R., Pizzolongo P. Cytinus hy-
pocistus L. embryogenesis: Ultra-
structural aspects ot megasporo-
genesis and megagametogenesis.—
J. Submicrosc. Cytol., 1976, 8, 4:
327—336.
Poddubnaya-Arnoldi V. A. [Поддуб-
ная-Арнольди В. А.|. Comparati-
ve embryology of the Orchida-
ceae.— Phytomorphology, 1967,
17: 312—320.
Prakash N. The endosperm of Ara-
chis hypogea.— Phytomorphology,
1960, 10: 60—64.
Prakash N. <Aizoaceae and Cacta-
ceae. A study of their embryolo-
gy and Relationships—Ph. D.
thesis University of Delhi, 1966.
Prakash N. Gametogenesis and seed
development in Hereroa hesperan-
tha (Dtr) Dtr. et Schwant (Aizoa-
ceae).— Austral. J. Bot. 1967a.
15 : 425—435.
Prakash N. Life history of Tetrago-
nia tetragonioides (Pall) O. Kunt-
ze.— Austral. J. Bot. 1967b, 415:
413—424.
Prakash N. Aizoaceae—A_ study of
its systematics.— Bot. Notiser.
1967с, 120, 3: 305—323.
Prakash N. The embryology od Kun-
zea capilata Reichb.— Ausiral. J
Bot. 1969a, 17, 41: 97—106.
Prakash М. Seed development in
Leptospermum scoparium Forst.—
Austral. J. Sci., 1969b, 34,9: 3314—
333.
Prakash М№ А contribution to the
life history of Angophora flori-
bunda (SM.) Sweet. (Myrta-
ceae).— Austral. J. Bot., 41969c,
47 : 457—469.
Prakash N. Reproductive develop-
ment in two species of Darwinia
(Myrtaceae).— Austral J.
Bot., 1969d, 47: 245—227.
Prakash N. Some aspects of the li-
fe history of Callistemon citrinus
(Curt) Skeels.— Austral. J. Bot.,
1969e, 17, 4: 407—117.
Prakash N. The floral development
and embryology of Centrolepis
fascicularis. —- Phytomorphology.
4969f, 19, 3: 285—291.
Prakash N. A_ contribution to the
life-history of Angophora floribun-
da (Sm) Sweet. (Myrtaceae).—
Austral. J. Bot., 19699, 17: 457—
469.
Prakash N. A study of the embryo-
logy of Melaleuca.— Malaysian. J.
Sci., 1973a, 2(A) : 81—85.
Prakash N., Aow Lee-Lee. Life hi-
Rudge
334
story of a common Malaysian Or-
chid Spathoglottis plicata.— Phy-
tomorphology, 1973b, 23, 1—2:9—
17
Prakash N., Chan Y. Y. Embryology
of Glicine Max.— Phytomorpholo-
gy, 1976, 26, 3: 302—309.
Prakash N., Lau Y. Y. Morphology
of Ploiarium alternifolium and
the taxonomic position of Ploiari-
um.— Bot. Notiser, 1976, 4129, 3:
219—285.
Prakash N., Lim L.L., Menurung В.
Embryology of Duku and Lang-
sat varieties of Lansium domesti-
cum.— Phytomorphology, 1977, 27,
1:50—559.
Prakash N., McAlister E. J. An em-
bryological study of Bauera capi-
tata with comments on the syste-
matic position of Bauera.— Aus-
tral. J. Bot., 1977, 25, 6: 645—622.
Prasad K. The development and
structure of basal body in the
ovule and seed of certain species
Cruciferac.— Bol. Jahrb. Sysl.
Pflanzengeorg., 1977, 98, 2: 266--
272.
Pullaiah T. Embryological investi-
gations in the Compositae with
some observations on the Basel-
laceae: Thesis for the Degree of
doctor of philosophy. Andra Univ.
Walter (India), 1976: 190.
Pullaiah T. Embryology of Adeno-
stemma, Elephantopus and Verno-
nia (Compositae).— Bot. Ntiser,
1979a, 132, 1:51—56.
Pullaiah T. Studies in the embryo-
logy of Composilac IV. The tribe
Inuleae.— Amer. J. Bot., 1979b,
66, 10: 4419—1127.
Puri V., Singh B. Studies in the fa-
mily Amaranthaceac. J. The life-
history of Digcera arvensis
Torsk.— Proc. Indian Acad. Sci.,
1935, 1, 12 : 893—908.
Purcell C. M. Embryo sac develop-
ment in two accessions of giant
pangola, Digitaria valida Stent.—
J. Agric. Univ. Puerto Rico, 1965,
49, 4: 477—483.
Pyykké M. Embryological and ano-
tomical studies on Finnish speci-
es of the Pyrolaceae.— Ann. bot.
{епп., 1978, 5: 453—165.
Raghuvanshi В. K., Prasad fT.,
Singh О. Embryological studies
in Capsicum. Embryology of crop
plants: Indo-Soviet Symp. Delhi,
1976 : 30—34.
332
>
~ Raj
В. Pollen structure in Drose-
ra.— Sci. Cult., 1965, 34:531.
Raj B. Loranthaceae: Seminar Com-
parative Embryology of Angios-
perms. New Delhi, 1967: 9.
Raj B. Loranthaceae: Symp. on Com-
parative Embryology of Angio-
sperms. 1970a, 44: 22—28.
Raj B. Morphological and embryo-
logical studies in the family San-
talaceae — Buckleya lanceolata
Sieb. et Zucc.— Beitr. Biol. Pflanz.,
1970Ъ, 46, 2: 193—207.
Raj B. Morphological and embryo-
logical studies in the family San-
talaceae. III. Choretrum latirifo-
lium. R. Br.— Osterr. bot. Z., 4972,
120, 1/2: 143—454.
Raja Rajeswari, Devi K., Prakasa
Rao P. S. Embryo development in
Euphorbia peplus L.— Curr. Sci.
(India), 1975, 44, 2: 57—59.
Rajan S. S. Embryological studies in
Compositae. Floral, morphology,
sporogenesis and gametogenesis
in Emilia sonchifolius (Linn.).—
Gurr. Sci. (India), 1968a, 37 : 26—
28.
Rajan S. S. Observations on female
gametogenesis in Synedrella eg-
gplant (Solanum melongena L.).—
Bull. Torrey Bot. Club, 1968b, 97:
146—118.
Rajan 5. 5. Observations on female
gametogenesis in Synedrella по-
difera Gaertn.— Curr. Sci. (In-
dia), 1968c, 37, 113: 385—386.
Rajan §S. S. Embryological studies
in Compositae. III. A contribution
to the Embryology of Blainvillea
rhomboides, Cass (В. latifolia
DC.).— Proc. Indian Acad. Sci.,
1972, 75b, 4: 167—176.
Rajan S. 5. Embryological studies
in Compositae.: IV. A contribution
to the life history of Spilanthes
acmilla Murr. (S. calva Wt. Yc).—
Proc. Indian Acad. Sci., 4974а,
79B, 6 : 267—282.
Rajan 5. 5. Embryological studies
in Compositae. V. On the embry-
ology of Eclipta alba Hassk.—
Ст. Sci. (India), 1974b, 43, 2:
—57.
Rajan S. 5. Micropyle formation in
the ovule as an indicator of pri-
mitiveness in Angiosperms.—
Proc. Indian Acad. Sci., 1976.
84B, 4: 124—127.
Rajan S. S., Kumar Y. D. Embryo-
logical studies in Lentibularia-
}
сеае. I. Floral morphology and
embryclogy of Untricularia smit-
hiana.— Proc. Indian Acad. Sci.,
1974c, 80b, 1: 18—25.
Rajan §&., Shivaramaiah G, Sathya-
nanda N. Embryology studies in
Richardia scabra Linn.— Curr. Sci.
(India), 1975, 44, 4: 1382.
Raju D. Embryological studies in
the tribe Selagineae — Agathelpis
dubia.— Phytomorphology, 1973,
23, 34: 194—205.
Raju D. Gametogenesis, endosperm
development and embryology in
Microcarpaea muscosa (Scrophu-
lariaceae, Gratioleae).— Plant Syst.
and Evolut., 1976, 126, 2 : 209—220.
Raju D., Arekal G. D. Contribution
to the embryology of Melampy-
rum pratense L.— Proc. Indian
Acad. Sci., 1977, 86B, 1: 23—31.
Raju D., Chamaiah H. N. Contribu-
tion to the embryology of Christi-
sonia subacaulis Gardn. (Oroban-
chacecae).— Proc. Indian Acad. Sci.,
1972, 75В, 3: 129—139.
Ram M. Misodendraceae: Seminar
Comparative Embryology of An-
giosperms. New Delhi, 1967a: 8.
Ram M. Olacaceac: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967b: 4.
Ram M. Opiliaceae: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967c : 5.
Ram M. Grubbiaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967d: 6.
Ram M. Mysodendraceae: Symp. on
Comparative Embryology of An-
giosperms. New Delhi, 1970a, 41:
Ram M. Opiliaceae: Symp. on Com-
parative Embryology of Angio-
sperms. New Delhi, 1970b, 41:
4—8.
Ват М. ОрШасеае: Symp. on Com-
parative Embryology of Angio-
sperms. New Delhi, 1970c, 41:
9—12.
Ram M. Grubbiaceac: Symp. on Com-
parative Embryology of Angio-
sperms. New Delhi, 1970d, 41:
` 13—14.
Ramaswamy 5. № Вигтапшасеае:
Seminar Comparalive Embryology
of Angiosperms. New Delhi, 1967:
124.
Ramaswamy 5. М. Вигтащасеае:
Symp. on Comparative Embryolo-
gy of Angiosperms. New Delhi,
1970, 41: 375—379.
Ramchandani 5. P., Joshi A. C., Pun-
dir N. 8S. Seed development in
Gossypium L.— Indian Cotton. J.,
1966, 20: 97—106.
Ramji M. V., Podmanabhan D. Deve-
lopmental studies on Cabomba caro-
liniana Grag. I. Ovule and carpel.—
Proc. Indian Acad. Sci., 1965, 62 B:
215—223.
Rao A. №. Flower and seed develop-
ment in Arundina graminifolia.—
Phytomorphology, 1967а, 17, 1—4:
291—300.
Rao A. N. Gametophytes and sced de-
velopment in pineapple.— Curr. Sci.
(India), 1974, 43, 6: 174—172.
Rao A. V. Embryological studies in
Cleome monophylla L.— Proc. Indi-
an Acad. Sci., 1967. 65B, 6: 249—256.
Rao B. A., Mukherjee P. K. A note on
the embryology of Polygonum glab-
rum Willd.—Curr. Sci. (India),
1973, 42, 7: 250—252.
Rao B. H., Prakasa Rao P. S. Anther
in Capparis decdua (Forsk) Pax.—
Curr. Sci. (India), 1977, 46, 49:
683—684.
Rao C. V. Proteaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967a: 3.
Rao C. V. Evolution of the endosperm
in Proteaceae.— New Phytol., 1967b,
66, 4: 755—768.
Rao C. V. Studies in the Proteaceae.
XI. Morphology, floral anatomy and
embryology of Lambertia Sm. with
a discussion on the taxonomic posi-
tion of the genus.— Proc. Indian
Scad. Sci., 1968, 68B, 14: 11—24.
Rao C.V. Studies inthe Proteaceae. X.
Morphology and embryology of Ha-
kea Schard.—J. Indian Bot. Soc.,
1969, 48, 3—4: 340—321.
Rao D. Burseraceae: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967a: 52.
Rao D. Simaroubaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967b: 51.
Rao D. A contribution to the embryo-
logy of Erytroxylaceae.— Proc. Nat.
Inst. Sci. India, 1968, 38: 53-—65.
Rao D. Burseraceae: Symp. on Com-
parative Embryology of Angio-
sperms. 1970a, 41: 148--150.
Rao D. Simaroubaceae: Symp on Com-
parative Embryology of Angio-
sperms. 1970b, 44: 142—147.
333
Rao К. Gametogenesis and gameto-
phytes in Aglaia roxburghiana
Bedd.— Curr. Sci. (India), 1972, 41,
6: 223—224.
Rao K. S., Shivaramian G. The emb-
ryology of Ximenia americana L.—
Proc. Indian Acad. Sci., 1978, 87B,
2: 23—27.
Rao P. N. Euphorbiaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967: 50.
Rao P. N. Euphorbiaceae: Symp. on
Comparative Embryology of Angio-
sperms. 1970, 44: 136—141.
Rao P. N. Embryology of Excoccaria
agallocha Linn. Seminar on morpho-
logy, anatomy and embryology of
land plants. Delhi, 1969: 75—77.
Rao P. N., Sarceswara Rao Embryo-
logy of Cassava. Embryology of
Crop plants: Indo-Soviet Symposi-
um. Delhi., (976: 34.
Rao P. R. Embryology of Nyssa silva-
tica and systematic consideration of
the familly Nyssaceae.— Phytomor-
phology, 1972a, 22, 1: 8—21.
Rao P. R. Morphology and embryolo-
gy of Tieghemopanax sambucifo-
lius with comments of the affinities
of the family Araliaceae.— Phyto-
morphology, 1972b, 22, 1: 75—87.
Rao P. §., Prakasa Rao B. H. Embryo
development in Cleome aspera L.
(Capparidaceae).— J. Jap. Bot., 1976,
51, 4: 110—117.
Вао Р. 5., Raja Rajeswari Devi K. So-
me embryological features of
Euphorbia vermiculata Raf.— Curr.
Sci. (India), 1974, 43, 8: 257—
259.
Rao P. S., Rao B. H. Anther develop-
ment in Cleome tenella Linn.— Curr.
Sci. (India), 1975, 44, 12: 438—440.
Rao P. S., биадаг К. Т., Naraya-
na L. L. Some embryological featurs
of Emilia flammea Cass.— Curr.
sci. (India), 1979, 48, 3: 129—131.
Rao S. G., Shivaramaiah N., Sathya-
nada N. Reverse polarity in Nauc-
lea orientalis L. (Sarcocephalus cor-
datus Mig.).— Proc. Indian Sci.
1976, 84В, 5: 180—184.
Rao S. K. Studies in Myrsinaceae. I.
A contribution to the embryology
of Maesa dubia Wall.— Proc. India
Acad. 5с1., 1972, 75В, 1: 160—166.
Rathore R. K., Singh R. P. Embryolo-
gical studies in Brassica campes-
tris L. var. Yellow Sarson Prain.—
334
J. Indian Bot. Soc., 1969, 47, 3—4:
341—349.
Ratnaparkhi S. Co-existence of mono-
sporic and bisporic embryo sacs in
Tellima grandiflora.— Phyton, 1973,
15, 1-2: 123—126.
Rau M. A. Elaeagnaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 62.
Ray M. A. Saxifragaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 40.
Rau M. A., Sharma V. K. Early em-
bryology in Saxifraga L.— Phyto-
morphology, 1967c, 17, 1—4: 252—
5
Rau M. A., Sharma V. K. Elaeagna-
ceae: Symp. on Comparative Iimb-
ryology of Angiosperms. New Del-
hi, 1970a, 41: 185—187.
Rau M. A. Saxifragaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1790b, 44: 90—
92.
Rau M. A, Sharma V. K. Coriaria-
ccae: Symp. on Comparative of An-
giosperms. New Delhi, 1970c, 44:
156—157.
Razi B. A. Some observation on the
embryology of Podostemaceae.—
Proc. Autumn. Sch. Bot. Mahabale-
shwar (India), 1966: 303—319.
Reddy K. B., Murthy S., Narayana В.
Male and female gametophytes and
endosperm in Vaccinium leschenaul-
tii W.— Curr. Sci. (India), 1966, 35:
130—131.
Reddy K. B., Narayana R. The female
gametophyte in Rhododendron nigi-
ricum Zenk.— J. Mysore Univ., 1966,
20B: 1—6. ^^
Веа4у Л. 5., Р’Стаз В. Месфапизт of
apomixis in Dichanthium annula-
tum -- Bot Gaz, 1969a, 130, 1: 71—
79.
Reddy P. S., D’Cruz R. Polyembryony
in Dichanthium annulatum (Forssk)
Stapf.— Bot. Gaz., 1969b, 139, 3:
462—165.
Rembert D. H. Megasporogenesis in
Laburnum anagyroides Medic— A
case of bisporic development in
Leguminoseac.— Trans. Kentucky
Acad. Sci., 1966, 27, 3/4: 47—50. |
Rembert D. H. Development of the
ovule and megagametophvie in
Wisteria sinensis.— Bot. Gaz., 1967,
128, 3/4: 223—229.
Rembert D. H. Comparative megaspo-
rogenesis in Papilionaceae.— Amer.
J. Bot., 1969, 56, 6: 584—591.
Remberit D. H. Phylogenetic signifi-
cance of megaspore tetrad pattern
in Leguminales.— Phytomorphology,
1972, 24, 4: 1—9.
Rembert D. H. Ovule ontogeny, mega-
sporgenesis and early gametogene-
sis in Trifolium repens (Papiliona-
ceae).— Amer. J. Bot., 1977, 64, 4:
483—488.
Rembert D. H. Comparative megaspo-
rogenesis in Caesalpiniaceae.— Bot.
Gaz., 1969, 130, 4: 47—52.
Rendle A. B. The classification of flo-
wering Plants. Cambridge, 1952.
Rendle A. B. The classification of flo-
wering plants. Cambridge, 1930,
4938. 1—11.
Reynolds J. D. Morphological studies
in Diapensiaceae. I. Chromosome
number and microspore develop-
ment in Pyxidanthera brevifolia
Wells.— Bull. Torrey Bot. Club.,
1968, 95: 653—656.
Richard A. Studies on embryo sac
and fertilization by some species of
Compositae.— Rev. Cytol. et biol. vé-
gét., 1964, 28, 1: 1—37.
Robertson B. L. Embryology of Jubae-
opsis caffra Becc. 1. Microspororan-
gium, microsporogenesis and micro-
gametogenesis.— J. 5. Afr. Bot.
1976a, 42, 2: 97—108.
Robertson B. L. Embryology of Jubae-
opsis caffra Becc. 2. Megasporangi-
um, Megasporogenesis and megaga-
metogenesis.— J. S. Afr. Bot., 1976b,
42, 3: 173—184.
Robdertson B. L. Embryology of Juba-
eopsis caffra Becc. 3. Endosperm
and embryogeny.— J. S. Afr. Bot.,
1976с, 42, 4: 285—299.
Robertson B. L., McNaughion J. E.
The reproductive potential of Erica
junona Bolus: megasporogenesis and
megagametogenesis.— J. S. Afr. Bot.,
1978, 44, 2: 97—101.
Rodkiewicz В. Embryologia roslin
kwiatowych. Warzawa, 1973: 285.
Romanov I. D. Романов И. Д.|.
Developpement du gametophyte ma-
le chez le Froment (Triticum aesti-
vum L.).— Ann. Univ. Reims, 1870,
6/7: 188—194.
Rosen W. Further notes on the emb-
ryology of the Goodeniaceae.— Acta
Hort. Géteb., 1946, 16: 235—249.
Ross W. M., Wilson J. A. Polvembrio-
ny of Sorghum.— Crap. Sci. 1969,
9, 6: 842—843.
Rutishauser A. Embryologie und Fort-
pflansungsbiologie der Angiosperms.
Wien; New York: Springer-Verlag,
1969: 163.
Sachar R. C., Arora U. Some embryo-
logical aspects of Amomum dealba-
tum und Hedychium acuminatum,
1963, 124: 353—360.
Sagawa J., Israel H. W. Post pollina-
tion ovule development in Dendro-
bium orchids. I. Introduction.— Ca-
ryologia, 1964, 17, 14: 583—64.
Sahai R., Roy S. K. Polyembryonia
in Eugenia heyneana Duthie.— Sci.
and Cult., 1962, 28, 14: 37—38.
Sahi B. G. Embryo sac and _ endo-
sperm in Andrographis paniculata
Nees.— Current. Sci. 1966, 35:
368—369.
Salgare S. A. The megagametophyte
of Sesbania aeguptica Poir. I.—
J. Biol. Sci. 1974, 16—17, 1—2;
108—110.
Salgare S. A. On the megagametophy-
te of Sesbania aculeata Poir. IV.—
Sci. and Cult., 1975, 41, 4: 172.
Salisbury E. J. On the reproduction
and biology of Elatine hexanda
(Lapierre) DC. (Elatinaccae).—
Kew Bull. 1967, 21: 189—149.
Salisbury E. J. A note of fertile seed
production by Hypericum calyci-
num.— Watsonia, 1969, 7: 2%.
Sampson F. B. Studies on the Moni-
miaceae. I. Floral morphology and
gametophyte development of Hedi-
carya arborea J. R. and G. Forst.—
Austral J. Bot., 1969a, 17, 3: 403—
424,
Sampson F. B. Studies on the Moni-
miaceae. II. Floral morphology of
Laurelia novae-zelandiae A. Cunn.
(subfamily Atherospermoideae) .—
oa Zeland J. Bot., 1969b, 7: 214—
240.
Sampson F. B. Studies on the Moni-
miaceae. III. Gametophyte deve-
lopment of Laurelia novae-zelan-
diae A.—Cunn. Austral. J. Bot.,
1969с, 17, 3: 325—439.
Samuel C., Hillson C. J. Developmen-
tal morphology of the magasporan-
gium and embryogeny in Ipomea
reptans Poir. and related species.—
Bot. Gaz., 1968, 129, 2: 150—4156.
Samuel C. K., Hillson C. J. NDevelop-
ment morphology of the microspo-
rangium in [pomea replans Poir.—
Bot. Gaz, 1971, 132, 3: 224—
229.
Saran §. De Wet J. M. A structural
peculiarity observed in the sexual
embryo sacs of Dichanthium in-
339
termedium.— Canad. J. Bot., 1969,
47, 7: 1205—1206.
Saran S., De Wet J. M. The mode of
reproduction in Dichanthium inter-
medium (Gramineae).— Bull. Tor-
rey Bot. Club, 1970, 97: 6—13.
Sastri R. L. Comparative morphology
and phylogeny of the Ranales.—
Biol. Rev., 1969, 44: 291—349.
Sasiri R. L. Floral morphology, emb-
ryology and relationships of Ber-
beridaceae.— Aust. J. Bot., 1979, 17:
69—79.
Sato Y. Development of the embryo
sac of Daphniphyllum macropodum
var. humile (Maxim) Rosenth.—
Sci. Rep. Téhoku Univ., 1972,
Зег. 4 (Ъ10].), 36: 129—133.
байта @. Г. [Савина Г. И.]. Deve-
lopment of the embryo sac in Oph-
ris insectifera L.— Ann. Univ. Re-
ims, 1971, 9, 4.
баргта С. Г. [Савина Г. И.]. Ferti-
lization in Orchidaceae.— In: Fer-
tilization in higher plants/Ed.
H. F. Linskens. Amsterdam, 1974:
197—204.
Saxena N. P. Studies of the family
Saxifragaceae. IJI. A conrtibution
to the morphology and embryology
of Astilbe revularis.— J. Indian Bot.
Soc., 1969a, 48, 3—4; 368—314.
Sarena №. P. Studies in the family
Saxifragaccae. IV. A contribution
to the embryology of Bergenia cillia-
ta (Roule) Raizada— Proc. Indian
Acad. Sci., 1969b, 70, 2: 104—110.
Saxena N. P. Studies in the family
Saxifragaceae. V. Structure and de-
velopment of the gametophytes in
Mitella diphylla L.— J. Indian Bot.
Sci., 1970a, 49, 1—4: 38—41.
Sarena №. P. Studies in the family
Saxifragaceac. VI. Structure and
development in the gametophytle in
Deutzia corymbosa R. Brox. and
Dichroa febrifuga Lour.— Ber.
Schweiz. bot. Ges., 4972 (1971), 81:
91—96.
Sazena N. P. Studies in the family
Saxifragaceae. VII. Floral morpho-
logy and development of gametophy-
tes in Ribes L.— Proc. Indian Acad.
Sci., 1970b, 71B, 5: 213—220.
Saxena T , Singh D. Comparative emb-
ryology and seed structure of Sola-
num nigrum complex: Symp. mor-
phology, anatomy and embryology
of land plants. 1969a, 77—78.
Sarena T., Singh H. Embryology and
secd development in Capsicum an-
336
nuum L. var. acuminata.— In: Proc.
56th Indian Sci. Congr. Bombay,
1969Ъ, 3: 388—389.
Schnarf K. Vergleichende Embryolo-
gie der Angiospermen. B.: Gebriider
Borntraeger, 1931: 354 c.
Schnarf K. Die Bedeutung der emb-
ryologischen Forschung fiir das na-
tiirliche System der Pflanzen.— Bio-
logia Generalis, 1933, 9, 2.
Schneider E. L. Morphologycal studies
of the Nympheaceae. TX. The seed
Barclaya longifolia Wall.— Bot. Gaz.,
1978a, 139, 2: 223—230.
Schneider E. L., Ford E. G. Morpholo-
gical studies of the Nymphaeaceae.
X. The seed of Ordinea purpurea
Dn Ilartog.— Bull. Torr. Bot. Club,
1978b, 105, 3: 192—200.
Schnept E., Ма И’. Оъег einige
Strukturbesonderhciten der Suspen-
sorzellen von Phaseolus vulgaris.—
Protoplasma, 1970. 69, 1: 133—143.
Schrock G. F., Palzer B. F. Floral de-
velopment, anatomy and embryolo-
gy of Collinsia heterophylla with
some noles on ten other species Col-
linsa and on Tonclla tenella.— Bot.
Са27., 1967, 128, 2: 838—104.
Sahi B. G. Embryo sac and endosperm
in Andrographis paniculata Nees.—
Curr. Sci., 1966. 35, 368—369.
Salisbury E. J. On the reproduction
and biology of Elatine hexandra
(Lapierre) DC. (Elatinaceac).— Kew
Bull., 1967, 24: 139—149.
Salisbury E. J. A note on fertile seed
production by IHlypericum calyci-
num.— Watsonia, 1969, 7: 24.
Sastri R. L. Floral morphology, emb-
ryology and relationships of Berbe-
ridaceaec.— Austr. J. bot., 1969, 17:
69—79.
Sato Y. Development of the embryo
sac of Daphniphyllum macropodum
var. humile (Maxim.) Rosenth.—
Sci. Rep. Téhoku Univ. Ser. 4, 1972
(Biol.), 36: 129—133.
Sehgal C. B. Morphological and emb-
ryological studies on Erigeron bo-
nariensis L. (E. linifolium Willd.).—
Beitr. Biol. Pflanz., 1966, 42: 161—
183.
Sele Y. La cleistogamie chez Rucllia
loreniziana Griseb. ct quelques aut-
res Acanthacécs.— Ber. Dtsch. bot.
Ges., 1977, £0, 1/2: 185—147.
Sen S. Comparative embryological
studies of principal genera of Nymp-
haeaceae.— In: Proc. 56th Indian
Sci. Congr., 1969, 3: 358.
Seshavataram V. Onagraceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967: 78.
Seshavataram V. Contribution to the
embryology of the genus Tephrosia
Pers.— In: Proc. 56th Indian Sci.
Congr. Bombay, 1969a, 3: 379.
Seshavataram V. Development of emb-
ryos in Papilionatae: Seminar on
morphology anatomy and embryo-
logy of land plants. Delhi, 41969b:
78—80.
Seshavataram V. Onagraceae: Symp.
on Comparative Embryology of An-
giosperms. 1970, 41: 220—225.
Seshavataram V., Satyamurtii T. ТУ.
Embryology of some forage grasses.
Embryology of crop plants: Indo-
Soviet Symp. Delhi, 1976: 39.
Sethi S., Bala. Structure and develop-
ment of seed in Camellia sinensis
(L.) O. Kuntze.— Proc. nath. Inst.
Sci. India, 1965, B, 34: 24—33.
Shah C. K. The life history of Juncus
bufonius L.—J. Indian Bot. Soc.,
1963, 42, 2: 239—254.
Shah C. K. Embryogeny in some Су-
peraceae.— Phytomorphology, 1965,
15, 1: 4—9.
Shah C. K, Cyperaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967: 128.
Shan S. C., Hillson C. J. Developmen-
tal morphology of the megasporan-
gium and embryology in Ipomea
reptans Poir, and related species.—
Bot. Gaz., 1968, 129: 150—156.
Shan S. C., Hillson C. J. Developmen-
tal morphology of the microsporan-
gium in Ipomoea reptans Poir.—
Bot. Gaz., 1971, 132: 224—229.
Shah C. K., Neelakandan N. Embryo-
geny in some Cyperaceae.— Beitr.
Biol. Pflanz., 1971, 47, 2: 215—227.
Shantamma C. Apomixis in Cenchrus
glaucus. Embryology of crop plants:
Indo-Soviet Symp. Delhi, 1970:
40—41.
Shanthamma C., Narayan K. N. For-
mation of nucellar embryos with
total absence of embryo sacs in two
species of Gramineaec.— Ann. Bot.,
1977, 44, 172: 469—470.
Sharma H. P., Murty Y. S. Embryolo-
gical studies in the Compositae. 3.
Tribe Asteraceae, Brachycome ibe-
ridifolia Benth. A casual apomict.—
a Indian Bot. Soc., 1977, 56, 4: 327—
Sharma H. P., Murty Y. S. Embryo-
logical studies in the Composi-
22 В. А. Поддубная-Арнольди
tae — Astereac. II.— Proc. Indian
Acad. Sci., 1978, 87B, 5: 149—4156.
Sharma M. K., Narayana H. S, Deve-
lopment of male and female gamc-
tophytes in Prinsepia utilis Royle.- -
Curr. Sci. (India), 1971. 49, 1:
7—8.
Sharma V. K. Morphological and emb-
ryological studies of some familics
and genera of disputed systematic
position.— Ph. D. Thesis, Univ. Ag-
ra, 1963.
Sharma V. K. Embryology of Elaeag-
nus conferta Roxb.— Curr. Sci. (In-
dia), 1966, 35, 7: 185—186.
Sharma V. K. Floral morphology, ana-
tomy and embryology of Coriaria
nepalensis Wall. with a discussion
on the inter-relationshops of the
family Coriariaceae.— Phytomorpho-
Тору, 1968а, 118: 143—153.
Sharma V. К. Morphology, floral ana-
tomy and embryology of Parnassia
nubicola Wall.— Phytomorphology,
1968Ъ, 18, 2: 193—204.
Shattuck C. H. A morphological study
of Ulmus americana.— Bot. Gaz.,
1905, 20: 209—223.
Sheela R., Narayana L. L. Embryology
of Pittosporaceae.— Curr. Sci., 1966,
35: 74—75.
Shivaramiah G. Observations on the
floral morphology and embryology
of Utricularia stricticaulis Stapf.—-
Proc. Indian Acad. Sci., 1967, 65B:
56—62.
Shivaramiah G. A contribution to the
embryology of Anotis wightiana
Wall.— Curr. Sci. (India), 1971, 40,
22: 606—607.
Shivaramiah G., Rao K. S. Studies in
Rubiaceae. III. Structure and de-
velopment of ovule seed of Olden-
landia gracilis DC.— Curr. Sci. (In-
dia), 1977, 46, 19: 662—664.
Shivaramaiah G., Rajan S. 5. А cont-
ribution to the embryology of
Oldenlandia umbellata L.— Proc.
Indian Acad. Sci., 1973, 77B, 4: 19—
24.
Shivaramiah G., Rajan S., Rao S.
A contribution to the embryology
of Utricularia scandens Oliver.—
Curr Sci. (India), 1975, 44, 14: 377—
378.
Shukla S. Prakash. On polyembryony
in Cajanus cajan (L.) Millsp.— Sci.
and Cult., 1967, 33; 2: 80—81.
Siddiqui S. A. Embryology of Utri-
cularia reticulata var. parviflora -
Santapau and Utricularia nivea
037
Vahlenum.— Sci. and cult., 1968a,
ЗА, 10: 422—424.
Siddiqui S. A. The microsporangium
and the male gametophyte in Mal-
pighia coccigera L.— Beitr. Biol.
Pflanz., 1968Ъ, 14, 3: 361—364.
Siddiqui 5. A. Studies in the Lenti-
bulariaceae. 7. The development of
Endosperm and Embryo in Utricu-
laria coerulea var. filicaulis Clar-
ke.— Bot. Notiser, 1975, 124 4:
432—445.
Siddiqui S. A. Studies in the Lenti-
bulariaceae. 8. The development of
gametophytes in Utricularia dicho-
toma Labill._— Flora, 1978a, 167, 2:
441—416.
Siddiqui S. A. Studies in the Lentibu-
lariaceae. 9. Pollination fertilization,
endosperm, embryo and seed in
U. dichotoma Labill.— Bot. Jahrb.
syst., 1978b, 100: 237—245.
Siddiqui S. A. Studies in the Lenti-
bulariaceae. 10. The development of
gametophytes in U. cornuta Misch.—
Plant Sci., 1978c, 8: 14—15.
Siddiqui S. A. Lentibulariaceae. 11.
The development of endosperm and
embryo in Utricalaria cornuta
Mixch.— Proc. Indian Acad. Sci.
1979, 88Ъ, 1414, 3: 213—247.
Siddiqui S. A., Siddiqui S. B. Studies
in Rubiaceae. II. A contribution to
the morphology of Borreria sticta
L.— Beitr. Biol. Pflanz., 1968, 44:
353—360.
Siddiqui S. A., Siddiqui B. Studies in
the Rubiaceae. I. A Contribution to
the embryology of Oldenlandia dic-
hotoma Hook. f.—Beitr. Biol.
Pflanz., 1968a, 44, 3: 343—3514.
Singh B. Labiatae: Seminar Compara-
tive Embryology of Angiosperms.
New Delhi. 1967a: 97.
Singh B. The structure and develop-
ment of Abelmoschus moschatus.
Medic seed.— Phytomorphology,
1967b, 17, 1—4: 282—290.
Singh B., Shivapuri T. N. The game-
tophytes of Neptunia oleracea La-
ur.— Proc. Indian Acad. Sci., 1935,
1, 8: 423—434.
Singh D. H. Apomixis in Paspalum. I.
Paspalum dilatatum Poir.— Proc.
Nat. Acad. Sci. India, 1965a, 35B,
2: 229—231.
Singh D. Structure and development
of seeds in Codiaeum variegatum.—
J. Indian Bot. Soc., 1965b, 44:
205—210.
338
Singh D. Caricaceae: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967a: 69.
Singh D. Cucurbitaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967b: 70.
Singh D. Passifloraceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967c: 65.
Singh D. Violaceae: Seminar Compa-
rative Embryology of Angiosperms.
New Delhi, 1967d: 39.
Singh D. Structure and development
of seed coat in Cucurbitaceae: 1.
Seeds of Biswarea Cogn., Edgarica
Clarke and Herpetospermum Hook.
f.— Proc. Indian Acad. Sci., 1967e,
65B: 267—274.
Singh D. Caricaceae: Symp. on Com-
parative Embryology of Angio-
sperms. 1970a, 44: 208—211.
Singh D. Cucurbitaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970b, 44: 242—
219.
Singh D. Passifloraceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970c, 44: 199—
204.
Singh D. Violaceae: Symp. on Compa-
rative Embryology of Angiosperms.
New Delhi, 1970d, 441: 188—193.
Singh D. Structure and development
of seed coat in Cucurbitaceae. П.
Seeds of Luffa.—J. Indian Bot.
Soc., 1971, 50A: 208—215.
Singh D. Structure and development
of seed coat in Cucurbitaceae. VITI.
Seeds of Marah Kell.— Bull. Torrey
Bot. Club., 1972, 99: 239—242.
Singh D., Dathan A. S. Structure and
development of Seed-coat in Cucur-
bitaceae: 5. Seeds of Melothria L.—
In: Proc. 56th Indian Sci. Congr.,
1969, 3 (4): 393.
Singh D., Dathan A. S. Morphology
and embryology of Marah macro-
carpa Greene.— Proc. Indian Acad.
Sci., 1971a, 73B: 241—249.
Singh D., Dathan A. S. Structure and
development of seed-coat in Cucur-
bitaceae; 7. Seeds of Cucumis spp.—
In: Proc. 58th Indian Sci. Congr.,
1974Ъ, 3 (4): 443—444.
Singh D., Dathan A. S. Further studi-
es on the endosperm of the Cucur-
bitaceae.— Plant Sci. 1973, 5:
42—51.
Singh D., Dathan A. S. Structure and
development of seed coat in Cucur-
bitaceae. X. Seeds of Trichosanthes
Linn L.— J. Indian Bot. Soc., 1976,
55, 2—3: 160—168. <.
Singh D., Gupta S. The seeds of the
Violaceae and Resedaceae: A com-
parison.— J. Indian Bot. Soc., 1967,
46, 2—3: 248—256.
Singh D., Prasad T. Embryology and
seed structure of Lycopersicon.
Embryology of crop plants: Indo-
Soviet Symp. Delhi, 1976: 43—44.
Singh R. P. Structure and develop-
ment of seeds in Euphorbiaceae:
Melanthesa rhamnoides Wt.— Beitr.
Biol. Pflanz., 1968, 45, 4: 127—138.
Singh R. P. Structure and develop-
ment of seeds in Euphorbia helios-
copia.— Bot. Mag. Tokyo, 1969, 82,
973: 287—293.
Singh R. P. Structure and develop-
ment of seeds in Putranjiva rox-
burghii Wall—J. Indian Bot. Soc.,
1970a, 49, 1—4: 99—105.
Singh R. P. Structure and develop-
ment of seeds in Euphorbiaceae.
Beitr. Biol. Pflanz., 1970b, 47: 79—90.
Singh В. P. Structure and develop-
ment of seed in Phyllanthus miru-
ri L.— J. Indian Bot. Soc., 1972, 54,
1: 73—77.
Singh R. P., Chopra S. Structure and
development of seeds in Croton
bonplandianum __ Baill.— Phytomor-
phology, 1970, 22, 1: 83—87.
Singh R. P., Jain J. Development of
femaly gametophyte in Euphorbia
pilosa L.— Curr. Sci. (India), 1965,
34, 24: 614—612.
Singh R. P., Khan A. M. Development
of ovule and female gametophyte in
EFomecon chionantha Hance.— Curr.
Sci. (India), 1974, 48, 24: 695—697.
Singh R. P., Shiam R. Microsporoge-
nesis in Litchi chinensis L.— Curr.
Sei. (India), 1977, 46, 17: 643—615.
Singh S. P. Martyniaceae: Seminar of
Comparative Embryology of Angio-
sperms. New Delhi, 1967a, 102.
Singh S. P. Pedaliaceac: Seminar of
Comparative Embryology of Angio-
sperms. New Delhi, 1967b: 101.
Singh S. P. Martyniaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970a, 41:
218—284.
Singh 9. P. Pedaliaceae: Symp. on
Comparative Embryology of Angrio-
sperms. New Delhi, 1970b, 44:
273—277.
Siva Rao B. S. Embryo development
in five species of Mollugo.— Curr.
Sci. (India), 1975, 44, 19: 712—713.
22*
Skalinska M. Further studies in fa-
cultative apomixis of MHieracium
aurantiacum L.— Acta. biol. cracov.
Ser. bot., 1973, 16, 1: 121—133.
Skvarla J. J., Turner B. L., Patel V.C,
Tomb A. S. Pollen morphology in
the Compositae and in morphology-
cally related families. The biology
and chemistry of the Compositae. J.
L. etc.: 1977.
Smith B. B. The use of a new clea-
ring technique for the study of ear-
ly ovule development, megasproge-
nesis and megagametogenesis in
five species of Cornus L.— Amer. J.
Bot., 1973, 60, 4: 322—338.
Smith B. B., Herr J. M. Ovule deve-
lopment, megagametogenesis and
early embryology in Ammania coc-
cinea Rothb.— J. Mitchell Soc., 1971,
87, 4: 192—199.
Smith E. C. Pollen characteristics of
Arnca species of Vernonias.—
J. Arnold Arbor, 1969, 50: 469—477.
Sokolowska-Kulczycka A. Develop-
ment of bisporic embryo sac in Ve-
ratrum lobelianum Bernh.— Acta
biol. cracov. Ser. bot., 1973, 16,
85—98.
Sokolowska-Kulczycka А. Develoyp-
ment of bisporic embryo sac in Ve-
ratrum nigrum L.— Acta biol. cra-
cov. Ser. bot., 1977 (1978), 20, 1—2:
11—24.
бойтега М. Р. [Солнцева М. П.]
Apomixis and hemigamy as one of
its forms.— Proc. Indian Sci. Acad.,
1978, В, 44: 78—90.
Soueges R. Embryogénie des Urtica-
cees. Developpement de l’embryon
chez lUrtica pilulifera.— C. r. Acad.
sci. Paris, 1920, 171: 1009—1011.
Soueges R. Embryogenie des Dipsaca-
cees. Developpement de l’embryon
chez de Dipsacus sylvestris.— C. r.
Acad. sci., Paris, 1963, 256D, 2268—
2273.
Souéges R., Guignard J. L., Mestre J C.
Développement de Vembryon chez
le Laurus nobilis L. (Lauraceac).—
Phytomorphology, 1967, 47, 1, 4:
255—261.
Sporne K. R. The ovule as on indi-
cator of evolutionary status in an-
giosperms.— New Phytol., 1969, 68,
2: 555—566.
Srivastava С. N. Male and female ga-
metophytes and devélopment of the
seed in Bursera delpechiana Po-
iss.— J. Indian Bot. Soc., 1968, 1—2:
53—59.
339
Stairs G. R. Microsporogenesis and
embryogenesis in Quercus.— Bot.
Gaz., 1964, 125, 2: 115—121.
Steenis C. G. Notes on Bredemeyera
(Comosperma).— Acta bot. neerlL,
4968, 17: 377—384.
Steyn E. Embryogenie van Meliant-
hus major L.—J. S. Afr. Bot., 1975,
41, 3: 199—205.
Steyn Е. Nembriologiese ondersoek
van Romulea rosea Eckl. var. ref-
lexa Beg. 1. Die bou, ontwikkeling
en bevrugting van die Saadknop.—
J. S. Afr. Bot., 1973a, 39, 2: 113.
Steyn E. Nembryologiese ondersoek
van Romulea rosea Eckl. var. ref-
lexa Beg. 2. Die ontwikkeling van
die endosperm, embrio en saed-
huid.— J. S. Afr Bot., 1973b, 39, 3:
235—243.
Strasburger E. Uber Befruchtung.--
Bot. Ltg. Alt., 1901, 2: 59.
Strasburger E. Sexuelle und apogame
Fortpflanzung bei der Urticaceen.—
Jahrb. wiss. Bot., 1910, 47: 245—
288.
Stushnoff S., Palser B. F. Embryology
of five Vaccinium taxa including
diploid, tetraploid and hexaploid
species or cultivars.— Phytomorpho-
logy, 1969, 19, 4: 342—331.
Subramanyam K. Campanulaceae. Se-
minar Comparative Embryology of
Angiosperms. New Delhi. 1967a, 106.
Subramanyam K. Crassulaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967b: 39.
Subramanyam K. Pentaphragmatace-
ae: Seminar Comparative Embryo-
logy of Angiosperms. New Delhi,
1967с: 108.
Subramanyam K. Sphenocleaceae Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967d: 107.
Subramanyam K. Stylidiaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967e, 109.
Subramanyam K. Some aspect of the
embryology of Sedum chrysanthum
(Boissier). Raymond-Hamlet. with
a discussion on its systematics po-
sition.— Phytomorphology, 1967f, 17,
1—4: 240—247.
Subramanyam К. Campanulaceae:
Symp. on Comparative Embryology
of Angiosperm. New Delhi, 41970a,
44: 306—312.
Subramanyam K. Crassulaceae: Symp.
on Comparative Embryology of An-
giosperms. New Delhi, 1970b, 41:
—89.
340
Subramanyam K. Pentaphragmatace-
ae: Symp. on Comparative Embryo-
logy of Angiosperms. New Delhi,
1970c, 44: 317—320.
Subramanyam K. Sphaenocleaceae:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970d,
41: 313—346.
Subramanyam K. Stylidiaceae: Symp.
on Comparative Embryology of An-
giosperms. New Delhi, 1970e, 41:
321—324.
Subramanyam K., Narayana Г. Г. Е]о-
ral anatomy and embryology of Pri-
mula floribunda Wall.— Phytomor-
phology, 1968, 18, 1: 105—143.
Subramanyam K., Sreemadhavan C. P.
A conspectus of the families Podo-
stemaceae and Fristichaceae.—
Bull. Bot. Surv. India, 1969, 11,
161—168.
Sugihara Y., Yoshimura H., Unezu N.
Embryo sac development in Dispo-
rum .— Phytomorphology, 1969, 19,
2: 112—144.
Sulmont G. Formation du sac embry-
onnaire chez le Papaver argemo-
ne L.— Bull. Soc. bot. France, 1968
(1969), 115, 3—6: 167—172.
Sundari K. T. Embryology of the Pit-
tosporaceae.— J. Jap. Bot., 1977, 52,
7: 204—209.
Sundari K. T., Narayana L. L. A note
on the embryology of Marianthus
bignonaceus F. Muell.— Curr. Sci.
(India), 1977, 46, 8: 282—283.
Suneson S. Zur Embryologie der Gat-
tung Viburnum.— Bot. Notiser. 1933:
181—194.
Swaminathan D. Embryological studi-
es in Polystachya flavescens (BI.)
J. J. Smith— Curr. Sci. (India),
1967, 36, 14: 384—385.
Swamy B. G. Contribution to the emb-
ryology of Cansjera rheedii.— Phy-
tomorphology, 1960, 10, 4: 397—409.
Swamy 8. G. Casuarinaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967a: 4.
Swamy B. G. Orchidaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967b, 130.
Swamy B. G. Comparative morphology
and phylogeny of the Ranales.—
В101. Веу., 1969, 44: 291—349.
Swamy B. G. On the presumed ancest-
ry of angiosperm embryo sac.— Phy-
tomorphology, 1974, 24, 1—2: 102—
106
Swamy B. G., Dayanand R. The endo-
sperm in Opilia amentacea Roxhb.--
Phytomorphology, 1963, 13, 4: 423—
428
Swamy B. G., Krishnamurthy K. V.
On the so-called Endothelium т
the Monocotyledons.— Phytomorpho-
logy, 1970, 20, 3: 262—269.
Swamy B. G., Krishnamurthy K. V.
The Helobial endosperm: a decen-
nial review.— Phytomorphology,
1973, 23, 1—2: 74—79.
Swamy B. G., Krishnamurty K. V.
Embryo sac ontogenesis in Angio-
sperms and _ elucidation.— Phyto-
morphology, 1975, 25, 4: 12—18.
Tainter F. H. The embryology of Ar-
ceuthobium pusillum.— Canad. J.
Bot., 1968, 46, 12: 1473—41476.
Takao S. A study on the development
of embryo sac in Impatiens texto-
ri.— Bot. Mag. Tokyo, 1968, 81, 960:
310—347.
Takhtadjan А. Г. [Гахтаджян A, J.J].
Die Evolution der Angiospermen.
Jena, 1959.
Tandon, Shiv R, Herr J. M. Embryo-
logical features of taxonomic signi-
ficance in the genus Nyssa.— Ca-
nad. J. Bot., 19741, 49: 505—5144.
Taneja S. Embryogeny in Nemesia
strumosa Benth.— Curr. Sci., 1969a,
38 : 522.
Taneja 5. Embryology, seed and
fruit structure of Hebenstreitia
comosa Hoghst. with discussion of
the systematic position of the fa-
mily Selaginaceae.— J. Indian Bot.
Soc., 1969b, 48, 3—4: 300—310.
Tarar J. L., Mukhrjee P. K. Studies
in the male and female gameto-
hytes of Cyperus globosus All.—
In: Proc. 56th Indian Sci. Congr.
Bombay, 1969, 3: 370.
Tarnavschi I. T., Petria E. Dezvol-
tarea gametofitului masculin la
unii reprezentantiai familici Vita-
ceae.— An. Univ. Bucuresti. Ser.
stint. natur. biol., 1968, 17: 177—
98.
Терзийски Д. П. Цитологично и хи-
стохимично проучване върху/
развитиего на мъжкия гамето-
фит при Уча striata M. B. a V.
melonops 5. $.— Генетика и се-
лекция (НРБ), 1969а, 2, 3: 153—
167.
Терзийски Д. П. Цитоембриологич-
но проучване на ТтМоПил moli-
nieri Ва. Българска Акад. по
науките.— В кн.: Втора нац. конф.
по ботанике. София, 19696: 355—
Терзийски Д. П. Цитоембриологич-
но проучване ua Vicia striata
М. В.— Изв. Oo7. un-ta Boar.
AH, 1974а, 24:81—99.
Герзийски Д. П. Развитие на ендо-
сперма и зародыша при Trigone-
lla gladiata Э%еу.— Генетика и се-
лекция (НРБ), 19716, 4, 5: 343—
351.
Терзийски Д. П. Цитоембриологич-
но проуване ua Trifolium moli-
nieri Balb—-B кн.: Втора нац.
конф. по ботанике. София, 1973:
355—367.
Терзийски Д. П. Върху някои OCO-
бенности на ембриогените про-
цес и характера на полиембрио-
нията при Spergula arvensis L.—
Генетика и селекция (НРБ),
1974, 7, 4:322—330.
Герзийски Д.П. Въерху някои про-
блеми на съвременната расти-
тельна ембриология.— В кн.
Проблеми на бислогията. 1978,
11 : 121—154.
Терзийски Д., Христов М. Цитоем-
бриологично проучване на Festu-
ca pratensis Huds., Alopecurus
pratensis L., Holcus lanatus L. a
Agrostis vulgaris With.— lenern-
ка и селекция (ПРБ), 1975: 6, 1:
45—57.
Терзийски Д. П., Даскалова Ц. Д.
Цитоембриологично проучване па
видов от рода Тпайсиит L. I.
Микроспорогенезис и развитие
на мъжския гаметофит при ТЬаН-
ctrum lucidum L.— Фитология
(HPB), 1979, 13: 53—70.
Гас B. Development of the em-
bryo-sac, and embryogeny in Cus-
cuta lupuliformis.— Curr. Sci. (In-
dia), 1965a, 34, 23: 671—672.
Tiagi B. Development of the seed
and fruit in Melampyrum nemoro-
sum and M. arvense.— Canad. J.
Bot., 1965b, 43: 1511—1521.
Tiagi B. Studics in the family Oro-
banchaceae. VI. Development of
the seed in Conopholis americana
(L. fil.) Wallr.— Acad. bot. hung,,
1965c, 14: 253—261.
Tiagi B. Development on the seed
and fruit in Rhinanthus major
and R. serotinus.— Amer. J. Bot.,
1966, 53: 645—651.
Tiagi ВБ. Orobanchaceae: Seminar
Comparative Embryology of An-
giosperms. New Delhi, 1967a: 4103.
Tiagi B. Scrophulariaceae: Seminar
Comparative Embryology of An-
341
giosperms. New Delhi, 1967b: 99.
Tiagi B. Orobanchaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970a, 41: 282—
289.
Tiagi B. Scrophulariaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi: 1970b, 44: 259—
266.
Tiagi Y. D. Cactaceae: Seminar Com-
parative Embryology of Angio-
sperms. New Delhi, 1967a: 10.
Tiagi Y. D. Contribution to the embry-
ology of the genus Pereskia.— In:
Proc. 54th Indian Sci. Congr., 1967b,
3: 324—325.
Fiagi Y. D. A contribution to the em-
bryology of Veronica longifolia L.—
Acta bot. Acad. sci. hung., 1968, 14,
4—2: 197—217.
Tiagi Y. D. Cactaceae: Symp. on Com-
parative Embryology of Angio-
sperms. New Delhi: 1970a, 41:
29—35.
Та: У. О. Раеощасеае: Зушр. оп
Comparative Embryology of Angio-
sperms. New Delhi: 41970, 41:
53—58.
Tiwari D. K. Embryological studies in
the Cyperaceae. II. Endosperm and
embryo development in Fimbristy-
lis quinquangularis Kunth.— J. In-
dian Bot. Soc., 1968 (1969), 47, 3—
4: 264—271.
Tiwari D. K. Embryological studies in
the Cyperaceae. A contribution to
the embryology of Pycreus sangui-
nolentus Nees.— J. Biol. Sci., 1969,
12, 2: 24—30.
Tiwari H. N. Embryology of Baliosper-
mum axillare Blume.—J. Univ.
Bombay, 1965, 33, 3: 76—88.
Tohda H. Embryological study on
Idesia polycarpa.— Sci. Repts Toho-
ku Univ. Ser. 4, 1971, 35, 4: 231—
237.
Toke E. Endosperms and embryo de-
velopment in Cynoglossum officina-
le L.— Acta biol. cracov. Ser. bot.,
1976, 19, 1: 47—59.
Topham P. B. The histology of seed
development in diploid and tetrapo-
lid raspberries (Rubus idaeus L.).—
Ann. Bot., 1970, 34: 128—135, 187—
145.
Trela-Sawicka L. Further embryologi-
cal studies in the genus Anemone
L. from Poland.— Acta biol. cracov.
Ser. bot., 1976, 19: 883—92.
Trela-Sawinska Z. Embryology stu-
dies Trapa natans L.— Acta biol.
342
cracoy. Ser. bot., 1978, 24—22: 101—
[sai H., Harney P. M., Peterson R. L.
Megasporogenesis in Pelargonium
hortorum.— Canad. J. Bot., 1973, 54,
3: 607—612.
[schermak-Woess E. Uber das zwei-
kernige endopolyploide Anthéren
tapetum von Rhinanthus.— Phyto-
morphology, 1967, 17, 1—4: 188—195.
furala K. Endopolyploidie in Endo-
sperm von Echinocystis lobata.—
Oster. Bot. Zeit, 1966, 113: 235—
furala K. Endopolyploidie and struc-
turs of the nuclei in the endosperm
ol the Cucurbitaceae.— Osterr. Bot.
Zeit., 1969, 116: 95—111.
Turala K. Disturbed mitotic divisions
and endomitoses during the diffe-
rentiation of the endosperm in Ec-
ballium elaterium (Cucurbita-
ceae).— Acta biol. Crac. Ser. Bot.,
1974, 14: 27—35.
Tykarska T. Rape embryogenensis. II.
Development of embryo proper.—
Acta Soc. pol., 1979, 48, 3: 391—421.
Lurala-Szybowska К. Karyological
anatomy of the endosperm of Bryo-
nia dioica and Cucurbita pepo.— Ac-
ta Biol. Crac. Ser. bot., 1974, 17:
75—90.
Tuschnjakova М. М. [Тушняко-
ва М. М.]. Embryologische und
Zytologische Beobachtungen itiber
Listera ovata (Orchidaceae) .— Plan-
ta, 1929, 7, 1: 29—44.
Untawale A. G., Deshpande P. K. Stu-
dies in the Gramineae. I. Male and
‘female gametophytes of Eragrostis
unioloides (Retz.) Nees. Exstends.—
J. Indian Bot. Soc., 1969, 48, 3—4:
386—392.
Untawale A G., Makde K. H. Gameto-
phytes of Cyperus alternifolia
Linn.— Curr. Sei. (India), 1971, 40,
2: 327—328.
Untawale A. G., Padhye M. D. Embry-
ology studies in the Cyperaceae.
III. Sporogenesis and gametophytes
in Scleria stocksiana Boeck.— In:
Proc. 56th Indian Sci. Congr. Bom-
bay, 1969a, 3: 377.
Untawale A. G., Padhye M. D,. Embry-
ological studies in the Cyperaceae.
IV. Male and female gametophytes
of the genus Fimbrystylis Vahl.—
In: Proc. 56th Indian Sci. Congr.
1969Ъ, 3: 377.
Urbanska-Worytkiewiez K. L’agamo-
spermie, systéme de reproduction
important dans la speciation des an-
giosperms.— Bull. Soc. bot. France,
1974 (1975), 124, 9: 329—346.
Vallade J. Developpement de l’embry-
on chez Elaeis guineensis Jacq. in
proces de germination.— C. r. Acad.
sci., 1966, D 262, 9: 989—992.
Vallade J. Organogenese embryonnai-
re chez Elaeis guineensis Jacq.—
Rev. cytol. et biol. végét., 1969, 32,.
3—4: 343—350.
Vailade J. Developpement embryonnai-
re chez Petunia hybrida Hort.— C.
r. Acad. sci., 1970, D 270, 15: 1893—
1896.
Vaimayor H. L., Sagawa Y. Ovule de-
velopment of Broughtonia sangui-
nea R. Br.— Philippine Agr., 1966,
50, 4: 356—363.
Valmayor H., Sorita E, Panklang A.,
Price G. Ovule development in some
Sarcanthinae orchids.— Philippine
Agr., 1976, 60, 5—6: 227—236.
Vani R. Systematic embryology of
Crossosoma, Lophira, Roridula and
Nyctanthes: Ph. D. Theses Univ.
Delhi, 1966.
Vani-Hardev. Systematic embryology
of Roridula gorgonias Planch.—
Beitr Biol. Pflanz., 1972, 48, 3:
339—354.
Varghese T. M. Solanaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967: 98.
Varghese T. M. Solanaceae: Symp. on
Comparative Embryology of Angio-
sperms. New Delhi, 1970, 44: 255—
258.
Vassal J. Grains albumineusées chez
les Acacia.— Trav. lab. forest. Tou-
lose, 1968, 4: 7.
Vassal J. Contribution a l’etude mor-
phologique des graines d’Acacia.—
Bull. Soc. hist. natur. Toulouse,
1974, 107: 191—246.
Vaughan J. G., Rest J. A. Note on the
testa structure of Panda Pierre, Ga-
learia Zoll. et Mor. and Microdesmis
Hook f. (Pandaceae).— Kew Bull.,
1969, 23: 215—248.
Vaughan J. G., Whitehouse J. M. Se-
ed structure and the taxonomy of
Cruciferae.— Bot. J. Linnean Soc.,
1974, 64: 383—409.
Veldkamp J. E. A revision of Sarcothe-
ca Bl. and Dapania Korth (Oxalida-
ceae).— Blumea, 1967, 15: 549—543.
Venkata Rao C. Pollen grains of Pro-
teaceae.— J. Palynol., 1965, 4: 1—9.
Venkata Rao C. Evolution of endo-
sperm in Proteaceae.— New Phytol.,
1967a, 66: 755—768.
Venkata Rao C. Studies in the Protea-
ceae: 8. Morphology, floral anatomy
and embryology of Grevillea R.
Br.— Proc. Nat. Inst. Sci. India,
1967b, 33B: 162—199.
Venkata Rao C. Studies in the Proic-
aceae. XI. Floral morphology, flo-
ral anatomy and embryology of
Lambertia Sm. with a discussion on
the taxonomic position of the ge-
nus.— Proc. Indian Acad. Sci., 1968,
48B: 14—24.
Venkateswarlu J. A contribution to
the embryology of Sonneratiaceae.—
Proc. Indian Acad. Sci., 1937, 5, 3:
206—223.
Venkateswarlu J. Embryology of so-
me Indian grasses.— Curr. Sci. (In-
dia), 1965, 338, 4: 104—106.
Venkateswarlu J. Combretaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967a: 77.
Venkateswarlu J. Lecythidaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967b: 75.
Venkateswarlu J. Lythraceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967c: 71.
Venkateswarlu J. Melastomataceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi, 1967d:
76
Venkateswarlu J. Myrtaceae: Seminar
Comparative Embryology of Angio-
sperms. New Delhi, 1967e: 72.
Venkateswarlu J. Punicaceae: Semi-
nar Comparative Embryology of An-
giosperms. New Delhi, 1967f, 74.
Venkateswarlu J. Sonneratiaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967i: 73.
Venkateswarlu J. Thymelaeaceae: Se-
minar comparative Embryology of
Angiosperms. New Delhi: 1967k: 61.
Venkateswarlu J., Lakshminaraya-
ma Il. A contribution to the embryo-
logy of Hydrocera triflora W. A.—
Phytomorphology, 1957, 7, 2: 194—
20
Venkateswarlu J., Maheshwari Devi H.
An embryological approach to the
taxonomic status of Vacciniacea.—
Proc. Indian Acad. Sci., 1973, 78B,
6: 282—291.
Venkateswarlu J., Raju C. S. Male and
female gametophytes in four speci-
es of Asparagus.— J. Indian Bot.
Soc., 1958, 37, 2: 290—299.
Venkateswarlu J., Rao P. S. Embryo-
343
logical studies in some Combreta-
ceae.— Bot. Notiser., 1972, 125, 2:
161—179.
Venkateswarlu J., Rao P. N. A contri-
bution to the embryology of the tri-
be Hippomaneae of the Euphorbia-
ceae.— J. Indian Bot. Soc., 1975, 54,
1—2: 98—103.
Venkateswarlu J., Rao P. N., Rao S. D.
Occurrence of stylar obturator in
two Euphorbiaceae.— Curr. Sci. (In-
dia), 1974, 43, 4: 128—129.
Venktateswarlu J., Seshavataram YV.
Germination of pollen grains in Ne-
lumbium speciosum.— Curr. Sci.,
1964, 33: 117.
Venkateswarlu J. Seshavataram У.
A contribution of the embryology
of Abrus precatorius L. with re-
marks on its systematic position.—
J. Indian Bot. Soc., 41971, 50, 4:
332—331.
Venter H. A. van de. Microsporogene-
sis and gametogenesis in Strelitzia
regina Ait.— J. S. Afr., 1976a, 42,
1: 25—31
Venter H. A. van de. Notes on the
morphology of the embryo and seed-
ling of Strelitzia reginae Ait.— J. S.
Afr., 1976b, 72, 4: 63—69.
Veyret Y. L’apomixie chez le Cynosor-
chis lilaeina Ridley (Orchidaceae) .—
C. r. Acad. sci. Paris, 1967, D265,
22: 1713—1716.
Veyret Y. Sur le développement des
ovules avant la pollinisation chez
certains orchidacées (Orchideae et
Neottieae).— C. r. Acad. sci., 1975,
D 280, 24: 2761—2763.
Veyret Y. Embryologye du Tylostig-
ma tenellum Schltr et d'un Stenor-
rhynchus sp. (Orchidacées).— Bull.
Soc. bot. France, 1976, 123, 3/4:
137—144.
Vignon-Fétré J. Embryologie des Cras-
sulacées. Formation et evolution de
Valbumen chez le Sedum spectabile
Boreau.—C. r. Acad. sci. Paris,
1966a, D 262: 1848—1851.
Vignon-Fétré J. Embryogénie des Cras-
sulacées. Developpement de l’emb-
ryon chez le Sedum sieboldi Swe-
et.—C. r. Acad. sci. Paris, 1966b,
D 262: 2145—2148.
Vignon-Fétré J. Relations embryogé-
niques entre Crassulaceae et Saxi-
fragaceae.— In: C. r. 93e Congr. nat.
Soc. savantes, Tours. Paris, 1972:
5211-546.
Vignon F. Embryogenie des Caryo-
phyllacéés. Developpement 4е Гет-
344
bryon chez le Lychnis flos — cuculi
L.—C. г. Acad. sci. Paris, 1965,
D 260, 21: 5643—5616.
Vignon F. Embryogénie des Caryo-
phyllacées. Developpement de Гем-
bryon chez le Spergularia margina-
ta Kittel—C. r. Acad. Paris, 1966,
D 263: 505—508.
Vignon F., Lebégue A. Embryologie
des Caryophyllacées. Developpement
de Vembryon chez le Melandrium
rubrum Garcke.— C. r. Acad. sci. Pa-
ris, 1966a, D262: 2149—2152.
Vignon F., Lebégue A. Embryogénie
des Caryophyllacées. Developpement
de l’embryon chez le Cerastium ob-
scurum Chaub.—C. r. Acad. sci.,
1969a, D268, 4: 664—667.
Vignon F., Lebégue A. Recherches
comparatives sur la_ signification
phylogénétique des systémes emb-
ryogéniques appliquées aux Centro-
_ spermae.— In: C. r. 93e Congr. nat.
Soc. savants, Tours. Paris, 41972:
439—483.
Vignon F., Lebégue A. Embryogenie
des portolacacées. Гез pricipaux
stades du developpement de l’emb-
ryon chez l’Anacampseros arachnoi-
des (How.) Sims.— C. r. Acad. sci.,
1969Ъ. 24: 2091—2094.
Vignon Е. L’Embryogenie des Centro-
spermae: Theses présanté a la fa-
culté des Sci. de luniv. d’Amiens,
pour le grade de docteur dés Sci.
naturelle. 1970: 506 p.
Vignon J., Lebégue A. Embryologie
des Saxifragacées. Developpement
de l’embryon et formation de [al-
-bumen chez le Saxifraga aizoides
L—cC. r. Acad. sci. Paris, 1966b,
D 262, 255—258.
Vijayaraghavan N. R. Morphology and
embryology of Actinidia polygama
Franch and Sav. and systematic po-
sition of the family Actinidiaceae.—
Phytomorphology, 1965, 15: 224—
235.
Vijayaraghavan M. R. Actinidiaceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi, 1967a:
35
Vijayaraghavan M. R. Berberidaceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi, 1967b:
25
Vijayaraghavan M. R. Ceratophylla-
ceae: Seminar Comparative Embry-
ology of Angiosperms. New Delhi,
4967с: 29.
Vijayaraghavan M. R. Cyrillaceae: Se-
minar Comparative Embryology of
Angiosperms. New Delhi, 1967d: 56.
Vijayaraghavan M. R. Lardizabala-
ceae: Seminar Comparative Embry-
ology of Angiosperms. New Delhi,
1967e: 26.
Vijayaraghavan M. R. Menisperma-
ceae: Seminar Comparative Embryo-
logy of Angiosperms. New Delhi,
1967f: 27.
Vijayaraghavan M. R. Ranunculaceae:
Seminar Comparative Embryology
of Angiosperms. New Delhi, 1967g:
24.
Vijayaraghavan M. R. Studies in the
family Cyrillaceae. I. Development
of male and female gametophytes
in Cliftonia monophylla (Lam.)
Britton ex Sarg.— Bull. Torrey Bot.
Club., 1969a, 96, 4: 484—489.
Vijayaraghavan M. R. Embryology of
Cyrilla racemiflora L. and affini-
ties of the Cyrillaceae: Seminar on
morphology, anatomy and embryo-
logy of land plants. New Delhi,
19695: 86—87.
Vijayaraghavan M. R. Embryology and
systematic position of Helleborus:
Seminar on morphology, anatomy
and embryology of land plants. New
Delhi, 1969c: 85—86.
Vijayaraghavan M. R. Actinidiaceae:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970a,
41: 69—74. ~
Vijayaraghavan M. В. СугШасеае:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970b,
Al: 163—167.
Vijayaraghavan M. R. Ranunculaceae:
Symp. on Comparative Embryology
of Angiosperms. New Delhi, 1970c,
44: 45—52.
Vijayaraghavan M. R., Bhandari N. N.
Studies in the family Ranuncula-
ceae: Morphology and embryology
of Nigella damascena L.— Phyto-
morphology, 1969, 19: 147—153.
Vijayaraghavan M. R., Bhandari №. М.
Studies in the family Ranuncula-
ceae: Morphology and embryology
of Thalictrum javancum Blume.—
Flora, 1970, 159: 450—458.
Vijayaraghavan M. R., Dhar М. Scy-
topetalum tieghemii — embryologi-
cally unexplored taxon and affini-
ties of the family Scytopetalaceae.—
Puylomorphology, 1976, 26, 1: 16—
Vijayaraghavan M. R., Dhar N. Em-
bryology of Cyrilla and Cliftonia
(Cyrillaceae).— Bot. Notiser, 41978,
131, 1: 127—138.
Vijayaraghavan M. R., Kaur D. Mor-
phology and embryology of Turne-
ra ulmifolia and affinities of the
family Turneraceae.— Phytomorpho-
logy, 1966, 16, 4: 5389—553
Vijayaraghavan M. R., Malik N. Mor-
phology and Embryology of Scaevo-
la frutescens K. and affinities of
the family Goodeniacea.— Bot. No-
tiser, 1972, 125, 3: 241—254.
Vijayaraghavan M. R., Marwah K. N.
Studies in the family Ranuncula-
ceae: morphology and embryology
of Nigella damascena.— Phytomor-
phology, 1969a, 19, 2: 4147—453.
Vijayaraghavan M. R., Marwah K. N.
Studies in the family Ranuncula-
ceae microsporangium, microsporo-
genensis and Ubisch granules in Ni-
gella damascena.— Phytomorpholo-
су, 1969Ъ, 13, 3—4: 203—209.
Vijayaraghavan M. R., Padmana-
bhan N. Morphology and embryolo--
gy of Centaurium ramosissimum
Druce and affinities of the Gentia-
naceae.— Beitr. Biol. Pflanz., 1969c,
46, 1: 15—37.
Vijauaraghavan M. R., Ratnaparkhi 5.
Some aspects of embryology of
Alectra thomsoni.— Phytomorpholo-
gy, 1972, 22, 4: 1—8.
Vijayaraghavan M. R., Ratnaparkhi S.
Morphogenesis and female Game-
tophyte in Saxifragaceae.— In:
Form, structure and Function in
plants. Johri commomeration Volu-
me (Meerut: Sarita Prakashan).
New Delhi, 1976: 215—222. .
Vijayaraghavan М. R., Sarveshwa-
ri G. S. Embryology and Systema-
tic Position of Morina longifolia
Wall.— Bot. Notiser, 1968, 121, 3:
382—402.
Vijendra L. D., Sreerangasamy 9. ПВ.
Megasporogenesis in Oriza sativa 1..
and Rhynchoryza subulata (Ness)
Baill. indicating some taxonomic
significance.— Acta bot. neerl., 1978,
27, 1: 61—68.
Virdi B. V., Eaton G. W. Embryo sac
and early embryo development in
the salmonberry (Rubus spectabi-
lis)— Canad. J. Bot. 1969, 47:
1891—1893.
Vishveshwara 5. С. Embryological
studies in Coffea arabica L.— Tur-
rialba, 1965, 15, 4: 307—316.
Voigt A. Unigue type of angiosperm
345
pollen from the family Annona-
ceae.— Amer. Ass. Adv. Sci., 1974,
1972: 565—567.
Wadhi Mridul. Acanthaceae: Seminar
Comparative Embryology of Angio-
sperm. New Delhi, 1967: 4100.
Wadhi Mridul. Acanthaceae: Sym-
posium on Comparative Embryolo-
gy of Angiosperm. New Delhi, 1970,
44: 267—272.
Walia K., Kapil R. N. Embryology of
Frankenia Lim. with some com-
ments on the systematic position of
the Frankeniaceae.— Bot. Notiser,
1965, 118, 4: 412—429.
Wall W. F., Long R. W. Megasporoge-
nesis and embryo sac development
in Ruellia caroliniensis (Acantha-
ceae). Bull. Torrey Bot. Club, 1965,
92, 5: 372—377.
Want G. Sporogenesis, gametogenen-
sis and embryogeny of Wahlenber-
gia bicolor N. Lothian. Austral. J.
Bot., 1963, 11, 2: 152—167.
Weimark G. Apomixis and Sexuality
in Hierochloé australis and in Swe-
dish H. odorata of different poly-
ploid levels. Bot. Notiser, 1967a, 120,
2: 209—235.
Weimark G. Apomixis in Hierochloé
monticola (Gramineae). Bot. Noti-
ser, 1967b, 120, 4: 448—455.
Walker J. W. Unique type of angio-
sperm pollen from the family An-
nonaceae.— Science, 1971, 172:
565—567.
Werner A., Zenkteler M. The embryo
sac development and fertilization in
Dianthus amurensis Jeck.— Bull.
Soc. Amis. Sci. et lett. Pozn., 1973,
44D: 57—69.
Wettstein R. Handbuch der systemati-
schen Botanik. 3 aufl. Wien; Leip-
zig, 1924.
Wettstein R. Handbuch der Systemati-
schen Botanik. 4 aufl. Wien; Leip-
zig, 1935.
Wojciechowska B. Embryological stu-
dies in the genus Lotus. Pt I. Fer-
tilization and seed devclopment fol-
lowing open—and_ selfpollination
in Lotus corniculatus L.— Genet.
pol., 1963, 4, 14: 53—63.
Wojciechowska B. Embryogenesis in
Ornithopus sp.— Genet. pol., 1971,
42, 1—2: 61—75.
Wojciechowska B. Pollination and
fertilization in Ornithopus sp.— Ge-
пеф. ро]., 1972, 13, 4: 37—52.
Wojciechowska W. Gametogenesis and
pollination processes in Ornithopus
346
pinnatus (Miil.) Druce. in reference
to flower bud development.— Acta
Soc. bot., 1975, 44, 2: 203—215.
Wilson T. K. Comparative morpholo-
gy of the Canellaceae. IV. Floral
morphology and conclusion.— Amer.
J. Bot., 1966, 58: 244—220.
Woodland P. S. The floral morpholo-
gy and embryology of Themeda
australis (R. Br.) Stapf.— Austral.
J. Воё., 1964, 12, 2: 157—172.
Wunderlich R. Ein Vorschung zu einer
natiirlichen Gliderung der Labiaten
auf Grund der Pollenkorner, der Sa-
menentwicklung und des reifen Sa-
men.— Osterr. Bot. Z., 1967a, 144,
4—5: 383—483.
Wunderlich R. Some remarks on the
taxonomic significance of the seed
Coat.— Phytomorphology, 1967b, 17,
4—4: 301—311.
Wunderlich R. Die systematische
Stellung von Theligonum.— Osterr.
Bot. Z., 1974, 119: 329—394.
Yakovlev M. S. [Яковлев М. С.].
Embryogenesis and some problems
of phylogenesis.— Rev. cytol. et bi-
ol. végét., 1969, 32, 3—4: 325—330.
Yakovlev M. S. [Яковлев М. С.].
On the embryology of Melocanna
bambusoides Trn.— Ann. _ bogor.,
1970, 5, 2: 109—415.
Yamazaki T. Tne embryology of Shor-
tia uniflora with a brief review of
the systematic position on Diapen-
siaceae.— Jap. Bot., 1966, 44: 245—
251.
Yamazaki T. Embryological studies in
Ebenales. 1. Styracaceae.— Ibid.,
1970а, 45, 9: 267—273.
Yamazaki T. Embryological studies in
Ebenales. 2. Symplocaceae.— Ibid.,
1970Ъ, 45, 12: 353—358.
Yamazaki T. Embryological studies in
Ebenales. 3. Sapotaceae.— Ibid.,
1971, 46, 6: 161—166.
Yamazaki T. Embryological studies in
Ebenales. 4. Ebenaceae.— Ibid., 1972,
47, 1: 20—28.
Yamazaki T. A system of Gamopeta-
lae based on the embryology.— J.
Fac. Sci. Univ. Tokyo, 1974, Sec. 3,
11, 8/9: 2683—2841.
Yamazaki T. Floral anatomy and em-
bryology of Tripeltaleia paniculata
and T. bracteata (Ericaceae).— Bot.
Mag. Tokyo, 1975a, 88: 267—279.
Yamazaki T. Embryogeny of Elaeag-
nus umbellata Thunb.— J. Jap. Bot.,
1975b, 50, 9: 281—284.
Yang-Hsu-Jen. Fertilization and de-
velopment of embryo on Satsuma
orange (Citrus unshiu Marc.) and
Natsudaidai (C. natsudaidai Haya-
ta).— J. Jap. Soc. Hort. Sci. 1968,
37, 2: 102—108.
Yasuyoshi T. On the embryo sac of a
Japanese species of Smilax.— Sci.
Repts Téhoku Univ. Ser. 4, 1970, 39,
2—3: 170—173.
Yasuyoshi T. Embryo sac development
in Dioscorea izuensis Anahor.— Sci.
Repts Téhoku Univ. Ser. 4, 1971, 39,
4: 225—229.
Yasuyoshi T. Embryo sac development
in Dioscorea japonica Thunb. and
D. quinqueloba Thunb.— Acta phy-
totaxon. et geobot., 1972, 25, 2—3: .
57—60.
Yasuyoshi T., Kimura C. On the em-
bryo sac formation of Dioscorea
nipponica Makino and D. tokoro Ma-
kino.— Sci. Repts Tohoku Univ. Ser.
4, 1968, 34, 2—4: 137—140.
Уойе М. Р. [Иоффе М. Д.] Fertili-
zation and formation of the primary
endosperm nucleus in Anemone ne-
morosa L.— Rev. cytol. et biol. vé-
gét., 1969, 32, 3: 319—328.
Yoshiaki H. The embryology of the fa-
mily Magnoliaceae sens. lat. ИТ. Ме-
gasporogenesis, female gametophy-
te and embryology of Magnolia li-
liiflora Desrousseaux and Michelia
fuscata Blume.— Sci. Repts Tohoku
Univ. Ser. 4, 1964, 30, 2: 89—98.
Yoshida O. Embryologische Studien
tiber die Ordnung Piperales. V. Em-
bryology von Saururus loureiri Dec-
ne.— J. Coll. and Sci. China Univ.,
1961, 3, 3: 314—316.
Yoshihiko S. Embryological study in
Aucuba japonica Thunb. with spe-
cial reference to the unusual deve-
lopment of the embryo sac.— Sci.
Repts Tohoku Univ. Ser. 4, 1974, 4,
201— 206.
Yoshihiko S. Development of the em-
bryo sac of Daphniphyllum macro-
podum var. humile (Maxim). Ro-
senth.— Sci. Repts Tohoku Univ.
Бег. 4, 1972, 36, 3: 129—133.
Yoshihiko S. Embryological studies in
the Japanese Epipactis (Orchida-
ceae).— Sci. Repts. Tohoku Univ.
Зег. 4, 1974, 37, 1: 33—45.
Yoshihiko S. Embryological studies of
some Cornaceous plants.— Sci. Rept
Tohoku Univ. Ser. 4, 1976a, 37, 2:
117—130.
Yoshihiko S. Development of the em-
bryo sac of Cephalanthera longibra-
cteata Blume.— Sci. Repts. Tohoku
. Univ. Ser. 4, 1976b, 37, 2: 189—144.
Yoshihiko S. Embryological studies
on Stachyurus praecox and its va-
riety.— Sci. Repts Tohoku Univ.,
Ser., 4, 1976с, 37, 2: 131—138.
Young D. P. Studies in the British
Epipactis. VI.: Some further notes
on E. phylanthes.— Watsonia, 1962,
5: 136—139.
Zabinzska D. Embryo sac and endo-
sperm development in some represen-
tatives of the group Ornithogalum
gussone? from Poland.— Acta biol.
cracov. Ser. bot., 1972, 15, 2: 177—
190.
Zabiniska D. Embryology of the dio-
ecious species Cirsium arvense (L.)
scop.— Acta biol. cracov. Ser. bot.,
1977, 20, 1—2: 133—146.
Загорчева Л. Цитоэмбриологични из-
следования на геномни мутанти в
Cucumis sativus L. I. Причины за
намаления фертилетет на една ав-
тотетраплоидна форма.— Генетика
и селекция. София, 1969, 2, 2: 99—
113
Загорчева Л. Отклонения в устрой-
ство на женския гаметофит на инп-
цухт — линия в Сисаи1$ зайуиз Г.
и някои особенности на макроспо-
рогенеза в тоза вид.— Генетика и
селекция. София, 1972, 2, 3: 244.
ЭЗагорчева Л. Макроспорогенезис и
MakKporaMeToreHesuc Ha Cucumis sa-
tivus L. uw съетояние на женския
гаметофит и женските цветове.—
Генетика и селекция. София, 1976,
9, 5: 386—399.
Загорчева Л., Молхова Е. Проучкане
на ембриосака, опрашването и оп-
лождането при Cucumis sativus
L.— Градинарска и лозарска нау-
ка. София, 1965, 2, 3: 285—296.
Загорчева Л., Молхова Е. Развитие
и кариология на эндосперма на
диплоидпа и тетраплоидна форма
и на реципрочните им кръстоски в
Cucumis sativus.— [Tenerura Mu ce-
лекция, София, 1976, 9, 4: 303—445.
Zenkteler M. Comparativo embryolo-
gy of three heterostylic races of
Oxalis cernua Thunb.— Bull. amis.
Sci. et lett. Poznan, 1965, 6D: 99—
115.
Zenkteler M. Comparative embryology
of three heterostylic races of Oxalis
cernua Thunb. II. Microsporogenc-
sis and chromosome number.— Bull.
Soc. amis. sci. et lett. Poznan, 1967
(1968), 8D: 103—118.
УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ ТАКСОНОВ
Acanthaceae 169, 186, 189
Aceraceae 138, 140
Achariaceae 82
Actinidiaceae 91, 96
Adoxaceae 166, 169, 205
Aextoxicaceae 107
Agavaceae 229
Aitoniaceae 137
Aizoaceae 58
Akaniaceae 140
Alangiaceae 150, 152, 153
Alismaceae 206, 209
Alismales 206, 209, 210, 215
Alismanae 206
Alismidae 206
Alseuosmiaceae 111
Amaranthaceae 61, 66
Amaryllidaceae 228, 229
Amborellaceae 20
Amentiferae 260, 261
Amygdalaceae 118
Anacardiaceae 132, 137
Ancistrocladaceae 77
Angiospermae 9
Annonaceae 13, 17, 18, 21,
24, 158
Annonales 9
Antoniaceae 173
Anisophyllaceae 130, 228
Aphyllanthaceae 228
Apiaceae 151, 152, 153
Apiales 149, 153
Apocynaceae 170, 173, 174
Aponogetonaceae 211
Aquifoliaceae 153, 156
Araceae 249, 250
Arales 249
Araliaceae 151, 152, 153
Araliales 149
Aralianae 149
Arecaceae 245
Arecales 245
Arecanae 245
Arecidae 245
Aristolochiaceae 22, 23, 24,
83
Aristolochiales 22, 24
Asclepiadaceae 171, 173, 174
Asparagaceae 226
348
Asteraceae 169, 196, 205
Asterales 169, 196
Asteranae 192
Asteridae 166
Asteropeiaceae 77
Aucubaceae 152
Austrobaileyaceae 20
Balanitaceae 143, 147
Balanopaceae 54
Balanopales 54
Balanophoraceae 163, 164
Balsaminaceae 137, 140, 146,
147
Barbeyaceae 52
Barbeyales 52
Barclayaceae 26
Baueraceae 108, 115
Bataceae 64
Basellaceae 60, 65
Begoniaceae 83, 84
Begoniales 84
Berberidaceae 23, 36, 38, 4!
Betulaceae 53, 54, 55
Betulales 53, 54, 55
Biebersteiniaceae 147
Bignoniaceae 181, 189
Bixaceae 79, 80, 83
Bombacaceae 103, 104
Bonnetiaceae 75
Boraginaceae 176, 178, 188,
192
Brassicaceae 86, 88
Bretschneideraceae 140
Вгех!асеае 111
Bromeliaceae 229, 236, 237, `
244
Bromeliales 236
Вгии!асеае 111
Brunoniaceae 115, 195, 196
ВгиреШасеае 111, 115
Buddlejaceae 180
Burmaniaceae 210, 229, 230
Burseraceae 133, 137
Butomaceae 206, 209, 210
Butomales 206
Buxaceae 105
Bybleidaceae i114
Cabombaceae 24
Cactaceae 59, 64, 65
Caesalpiniaceae 118, 121
Callitrichaceae 132, 194
Calycanthaceae 19, 20, 21,
117
Calyceraceae 169,
Calycerales 196
Campanulaceae 83, 192, 195,
205
Campanulales 192, 195, 196
СапеПасеае 15, 17
Cannabaceae 51
Cannaceae 232, 233
Capparaceae 80, 85, 88
Capparales 85, 88
Caprifoliaceae 166, 169
Cardiopteridaceae 163
Caricaceae 80, 81, 82, 83
Caryocaraceae 77
Caryophyllaceae 61, 66, 178
Caryophylales 56, 64, 65, 66,
67
Caryophyllanae 56
Caryophyllidae 56
Cassythaceae 21
Casuarinaceae 52, 53
Casurinales 52, 53
Celastraceae 154, 156
Celastranae 153
Celastrales 96, 137, 153, 156,
159
Centrolepidaceae 242
Cephalotaceae 114
Ceratophyllaceae 25, 26
Cercidiphyllaceae 43
Cercidiphylales 43, 44
Chenopodiaceae 63, 66
Chloranthaceae 19, 20, 22, 26
Chrysobalanaceae 117
Circaeasteraceae 33, 38
Cistaceae 79, 80, 82, 83
Cistales 77
Clethraceae 92, 96
Clusiaceae 75
Cneoraceae 137
Cochlospermaceae 80
Columelliaceae 188
Combretaceae 108, 126
196, 205
Commelinaccae 229, 237, 241
Commelinales 237, 241, 244
Commelinanae 236
Commelinidae 235
Connaraceae 118, 121
Connarales 124
Convolvulaceae 174, 178, 188
Coriariaceae 136, 137
Cornaceae 149, 152, 153
Cornales 149, £32, 153
Corsiaceae 230
Corynocarpaceae 156
Costaceae 233
Crassulaceae 112,
124
Crossosomataceae 71, 118
Crypteroniaceae i141
Cucurbitaceae 82, 83, 205
Cucurbitales 82
Cunoniaceae 108, 112,
124
Cunoniales 108, 415
Curtisiaceae 152
Cuscutaceae 175, 178
Cyanastraceae 225, 229
Cyclanthacese 246, 249
Cyclanthales 246, 249,
Cymodoceaceae 214
Cynanchoideae 173
Cynomoriaceae 162, 164
Сурегасеае 235, 236, 244
Cyperales 235, 236
Cyrillaceae 94, 96
114, 123,
115,
Daphniphyllaceae 105, 107
Datiscaceae 83, 84
Datiscales 84
Davidiaceae 152
Davidsoniaceae itt
Degeneraceae 10, 17, 18
Desfontainiaceae 173
Dialypetalaceae 173
Diapensiaceae 96, 97
Diapensiales 96, 97
Dichapetalaceae 107, 117
Dicotyledones 9
Didieraceae 64
Didymelaceae 44
Didymelales 44
Diegodendraceae 77
Dilleniaceae 67, 71
Dilleniales 67, 74
Dillenianae 67
Dilleniidae 67
Dioncophyllaceae 77
Dioscoreaceae 226, 229
Diospyrales 97
Dipentodontaceae 163
Dipsacaceae 167, 168, 169,
196, 205
Dipsacales 166, 168, 169
Dipterocarpaceae 73
Dirachmaceae 147
Donatiaceae 195
Droseraceae 121, 123
Duckeodendraceae 173
Dulongiaceae i144
Ebenaceae 98, 99
Ebenales 97, 99
Elaeagnaceae 164, 165
Hlaeagnales 164, 165
Elaeocarpaceae 101, 104
Elatinaceae 76, 90
Empetraceae 95, 96, 137
Emplingiaceae 88
Epacridaceae 93, 96
Егетозупасеае 114
Ericaceae 92, 96, 97
Ericales 91, 96, 97, 137
Ericanae 94
Eriocaulaceae 241
Eriocaulales 241
Erythropalaceae 160, 163
Erythroxylaceac 141, t47
Escalloniaceae 109, 112
Eucommiaceae 48, 49
Bucommiales 48
Eucryphiaceae 111, 115
Euphorbiaceae 106, 107, 108
Euphorbiales 105, 107, 108
Eupomatiaceae 17, 24
Eupteleaceae 44
Eupteleales 44
Fabaceae 119, 124
Fabales 118, 120, 124
Fagaceae 53, 55, 119
Fagales 53, 55
Flacourtiaceae 77, 83, 90
Flagellariaceae 77, 242, 244
Fouquieriaceae 88, 89
Francoaceae 113
Frankeniaceae 89, 90
Fumariaceae 40, 41
Garryaceae 150, 152
Geisolomataceae 108, 153,
156
Gentianaceae 171, 174
Gentianales 169, 170, 174
Geosiridaceae 230
Geraniaceae 144, 147, 178
Geraniales 140, 147, 149
Gesneriaceae 183, 189
Glaucidiaceae 38
Globulariaceae 186
Gomortogaceae 20
Goodeniaceae 194, 196, 205
Goupiaceae 156
Graminaceae 236, 242
Graminales 236, 242
Griseliniaceae 152
Greyiaceae 140
Grossulariales 108
Grossulariaceae 109, 112, 124
Grubbiaceae 95, 96
Соппегасеае 131, 132
Gyrostemonaceae 64
Haemodoraceae 223, 224
Halophytaceae 64
Haloragaceae 130, 131, 132
Haloragales 131
Hamamelidaceae 43, 44, 48
Hamamelidae 42
Jiamamelidales 44, 48
Hammelididae 42
Hanguanaceae 242
Hectorellaceae 64
Heliconiaceae 223, 233
Helobiales 210
Helosiaceae 164
Helwingiaceae 152, 153
Hernandiaceae 20
Himantandraceae 17
Hippocastanaceae 137, 139
Hippuridaceae 132
Hippuridales 131
Hoplestigmataceae 178
Наасеае 104
Hugoniaceae 147
Humiriaceae 147
Ilydnoraceae 23, 24
Hiydrangiaceae 110, 112
Hydrastidaceae 338
ITydrocharitaceae 206, 210
Hydrocharitales 205, 210
Hydrophyllaceae 176
Hydrostachyaccae 114,
Hypecoaceae 39, 41
Hypericaceae 76
Hypoxidaceae 224, 228, 234
Hypseocharitaceae 147
188
Icacinaceae 153, 156
Tlliciaceae 26, 27
Illiciales 26
Iridaceae 229, 230
Iridales 229, 230
Цеасеае 111, 115
Ixonanthaceae 141, 147
Juglandaceae 53, 55, 56
Juglandales 53, 56
Juliniaceae 437
Juncaceae 235, 236, 244
Juncales 235
Juncanae 235
Juncaginaceae 211, 214
Kingdoniaceae 38
Kirkiaceae 137
349
Koelberliniaceae 88
Krameriaceae 148
Labiatae 189
Lactoridaceae 20
Lamiaceae 190, 192
Lamiales 169, 189, 192
Lamianae 166
Lardisabalaceae 28, 38
Lauraceae 20, 21
Laurales 18, 20
Lecythidaceae 129, 130
Leeaceae 157, 158
Ledocarpaceae 147
Leguminales 118
Leitneriaceae 55, 56
Leitneriales 55
Lemnaceae 249, 250
Теппоасеае 177
Lentibulariaceae 185, 189
Lepidobotryaceae 147
Liliaceae 215, 228, 229, 244,
245
Liliales 215, 228, 229, 234,
245
Lilianae 24, 2414
Liliidae 244
Limnanthaceae 146, 147, 148
Limnanthales 148
Limnocharitaceae 209
Linaceae 140, 147
Loasaceae 82, 177
Loasaies 82 .
193,
Lobeliaceae 156, 195,
205
Loganiaceae 170, 174
Lophiraceae 77
Loranthaceae 77, 162, 164,
165
Lowiaceae 233
Lythraceae 124, 130
Magnoliaceae 9, 17, 18, 21,
43
Magnoliales 9,
4h
Magnolianae 9
Magnoliatae 9
Magnoliidae 9
Magnoliophyta 9
Malaceae 118
Malesherbiaceae 8&2
Malpighiaceae 142
Malvaceae 103, 104, 105
Malvales 101, 104, i05
Ма!уапае 101
Maranthaceae 232, 233
Marcgraviaceae 74, 96
Martyniaceae 183, 189
Mastixiaceae 152
Mayacaceae 241
17, 18, 38,
350
Medusaceae 77
Medusandraceae 163
Melanophyllaceae 152
Melastomaceae 127, 130
Meliaceae 135, 137
Melianthaceae 139
Menispermaceae 29, 38
Menyanthaceae 172, 174
Mimosaceae 118, 121
Misodendraceae 161, 163, 164
Molluginaceae 57
Monimiaceae 18, 20, 24
Monochlamydeae 260, 264
Monocotyledones 206
Monotropaceae 84, 96
Montiniaceae 111
Moraceae 50
Morinaceae 168, 169
Moringaceae 85
Musaceae 231, 233
Myoporaceae 185
Myricaceae 54
Myricales 54
Myristicaceae 16, 17, 18
Myrothamnaceae 48
Myrsinaceae 99, 101
Myrtaccae 126, 130
Myrtales 124, 130
Myrtanae 124
Najadaceae 213, 214
Najadales 210, 214
Nandinaceae 38
Nelumbonaceae 28
Nelumbonales 28
Nepenthaceae 122, 123
Nepenthales 121, 123
Neuradaccae 116, 117
Nitrariaceae 147
Nolanaceae 179, 188
Nyctaginaceae 57, 65, 108
Nymphaeaceae 24, 26
Nymphaeales 24, 26
Nyssaceae 149, 152
Ochnaceae 72
Octoknemaceae 163
Olacaceae 159, 163, 164
Olacales 163
Oleaceae 158, 189
Oleales 158, 159
Oliniaceae 127, 130
Onagraceae 128, 130, 131
Opiliaceae 130, 159, 163, 164
Orchidaceae 210, 229, 233,
234
Orchidales 233, 234
Orobanchaceae 174, 188, 185,
192
Oxalidaceae 144, 147
Paeoniaceae 38, 72
Paeoniales 71, 72
Papaveraceae 38, 44
Papaverales 38, 40, 41, 42
Pandaceae 107
Pandanaceae 250
Pandanales 250, 251
Papillionaceae 119, 124
Paracryphiaceae 111
Parnassiaceae 114
Passifloraceae 80, 82, 83
Passiflorales 80, 82, 84
Pedaliaceae 182, 189
Peganaceae 147
Pellicieraceae 77
Penaeaceae 108, 128, 130
Pentadiplandraceae 88
Pentaphragmataceae 195
Pentaphyllaceae 77
Penthoraceae 114
Petermanniaceae 228
Peridiscaceae 80
Periplocoideae 173
Phellinaceae 156
Philesiaceae 224, 228
Philydraceae 228, 229
Phrymaceae i9f
Phytolaccaceae 56, 195
Picrodendraceae 107
Pipecraceae 21, 22
Piperales 21
Pittosporaceae 110, 1412, 115
Pittosporales 108, 115
Plantaginaceae 187, 189
Platanaceae 48
Plocospermaceae 173
Plumbaginaceae 66
Plumbaginales 65, 66
Poaceac 236, 242, 244
Poales 242
Podoaceae 137
Podophyllaceae 36
Podostemaceae 114, 123, 124
Podostemales 123
Polemoniaceae 174, 178, 189
Polemoniales 169, 174, 178
Polycarpicae 261, 262
Polygalaceae 140, 148, 149
Polygalales 148
Polygonaceae 65, 66
Polygonales 65, 66
Pontederiaceae 227, 229
Portulacaceae 59, 64
Posidoniaceae 214
Potaliaceae 173
Potamogetonaceae 212, 214
Primulaceae 160, 101, 189,
195
Primulales 99, 104
Proteaceae 165
Proteales 165
Proteanae 164
Pterostemonaceae 111
Punicaceae 125, 130
Pyrolaceae 93, 96
Quiinaceae 77
Quiraceae 130
Rafflesiaceae 23, 24
Rafflesiales 23
Ranales 74, 260, 264
Ranunculaceae 30,
72
Ranunculales 28, 38, 41, 42
Ranunculanae 26
Ranunculidae 26
Rapataceae 241, 244
Resedaceae 80, 82, 87, 88
Restionaceae 241, 242, 244
Restionales 241, 242, 244
Retziaceae 188
Rhamnaceae 156, 158, 165
Rhamnales 156, 158, 159
Rhizophoraceae 125, 130
Rhoipteleaceae 55
Rhopalocarpaceae 104
Roridulaceae tit
Rosaceae 115, 117, 118
Rosales 71, 1415, 117, 118
Rosanae 108
Rosidae 4108
Rubiaceae 67, 169, 173, 174,
189, 205
Rubiales 196
Ruppiaceae 212
Rutaceae 134, 137
Rutanae 132
Rutales 132, 137, 140
38, 41,
Sabiaceae 140
Salicaceae 90
Salicales 90
Salvadoraceae 137, 152, 154,
156
Santalaceae 160, 163, 165
Santalales 159, 163, 164
Sapindaceae 138
Sapindales 137, 140
Sapotaceae 98, 99
Sarcolaenaceae 104
Sargentodoxaceae 38
Заггасеп1асеае 41, 42, 123
Sarraceniales 41, 123
Saurauiaceae 91, 96
Saururaceae 21, 22
Saxifragaceae 112, 114
Saxifragales — Grossularinas
108, 14414, 4415
Saxifragales — Saxifraginae
1412, 1414
Scheuchzeriaceae 210, 214
Schisandraceae 27
Schoepfiaceae 163
Scrophulariaceae
188, 189, 192
Scrophulariales 169, 178, 188,
192
Schyphostegiaceae 80
Scytopetalaceae 102, 104
Selaginaceae 156, 189
Simarouhbaceae 133, 137
Simmondsiaceae 107
Siphonodontaceae 156
Smilaceae 228
Solanaceae 178, 188
Sonneratiaceae 124, 130
Sparganiaceae 251, 252
Sphenocleaceae 193, 195
Stachyuraceae 78
Stackhousiaceae 155, 156
Staphyleaceae 137
Stemonaceae 226
Sterculiaceae 102
Strasburgeriaceae 77
Strelitziaceae 230, 233
Stylidiaceae 194, 195,
205
Stylobaselaceae 137
Styracaceae 97, 99
Symplocaceae 98, 99
156, 180,
196,
Taccaceae 227, 229
Tamaricaceae 88, 89, 90
Tamaricales 88, 89
Taricellaceae 152
Tecophilaceae 225, 229
Tetracarpaeaceae (it
Tetracentraceae 43
Tetragoniaceae 64
Tetrameristicaceae 77
Theaceae 74, 96
Theales 72, 77, 96
Theligonaceae 67
Theligonales 67
Theophrastaceae 100, 101
Thurniaceae 235, 244
Ttymelaeaceae 65, 107, 108,
130, 165
Thymelaeales 107, 108
Tiliaceae 101, 104, 105
Tovariaceae 88
Тгарассае 129, 130, 131
Trapellaceae 182, 189
Tremandraceae 148
Tribelaceae 111
Trigoniaceae 148
Trimeniaceae 20
Triuridaceae 214
Triuridales 214, 215
Trochodendraceae 42, 43, 48
Trochodendrales 42, 43, 44
Tropaelaceae 145, 147
Turneraceae 81, 82
Typhaceae 251, 252
Typhales 251, 252
Ulmaceae 49
Umbelliforae 152, 153
Umbecllales 149
Urticaceae 51, 52
Urticales 49, 52, 65
Vacciniaceae 96
Vahliaceae 113
Valerianaceae 167,
Velloziaceae 224
Verbenaceae 189, 192
Violaceae 78, 80, 83
Violales 77, 80, 83
Vitaceae 157, 158
Viviaceae 147
Vochysiaceae 148
169, 205
Winteraceae 16, 17, 18
Xanthorrhoeaceae 223, 228
Xyridaceae 241
Zannicheliaceae 213, 214
Zingiberaceae 231, 233
Zingiberales 230, 233
Zosteraceae 211
Zygophyllaceae 142, 147
СОДЕРЖАНИЕ
Введение ........... ее. 3
ОТДЕЛ МАСМОШОРНУТА (ANGIOSPERMAE) oe ee 9
9
9
Kuace Magnoliatae (Dicotyledones). . . . . . .
Подкласс А. Маспой4ае. . . . . . © « « +
Подкласс В. ВапипеаИдае ............ 25
Tlomxmace C. Hamamelididae . ....... 2.04.8 42,
Ilogknace D. Caryophyllidae . . . . 1... we ew 56
Подкласс E. Dilleniidae . ..,..... 646 5. « ee 67
Подкласс Е. Rosidae . ..... 2. 6 © «© © s ~~ 4108
Подкласс С. Ачен4ае.............. 1606
Kaace Liliatae (Monocotyledones) .......... £206
Подкласс А. АПзи9ае ............. 206
Подкласс В. И9дае. . , ‚. . «© «6 « $. «6 «a 244
Подкласс С. СотштеЙ9дае............ 235
Подкласс ПО. Агосае . ..... . 2. 6 © ew ew о. 245
Заключение... еее... 9253
Литература ...... . еее в 268
Указатель латинских названий такеонов ...... 348
ВЕРА АЛЕКСЕЕВНА ПОДДУБНАЯ-АРНОЛЬДИ
ХАРАКТЕРИСТИКА СЕМЕЙСТВ ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
ПО ЦИТОЭМБРИОЛОГИЧЕСКИМ ПРИЗНАКАМ
Утверждено к печати Главным ботаническим садом Академии наук СОСР
Редактор издательства Э. И. Николаева. Художник О. Н. Остащева
Художественный редактор Н. Н. Власик. Технический редактор В. Д. Прилепсевая.
Корректоры Д. Ф. Арапова, Ю. Л. Косорыгиин
ИБ № 21079
Сдано в набор 16.01.84. Подписано к печати 04.12.81. Т-28946. Формат 60х90.
Бумага типографская №2. Гарнитура обыкновенная. Печать высокая. Усл. печ.
л. 22. Усл. кр.-отт. 22. Уч.-изд. л. 32,5. Тираж 1150 экз. Зак. 487. Цена 4 р. 20 к.
Издательство «Наука». 117864 ГСП-7, Москва, В-485, Профсоюзная ул., 90
4-я типография издательства «Наука». 630077, Новосибирск-77, Станиславского, 25